










































































































































































































































































































































































































































































































































































































































































































































Author: Чайковский Ю.В.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез развитие органического мира (филогенез) биология эволюция
ISBN: 5-87317-342-7
Year: 2006




Ю.В. Чайковский
голоцен
(геологи-
ческое на-
стоящее)
Nautilus M^acnnus
ЕЁ somella /,. tr0 nus i
P
Neopilina
Nucula
меп
_____125
Наука о
развитии
жизни
трис
______225
ордовик
______
Lingula ftatastenas
юра
карбон
девон
пермь
______275
силур
_______44С .
0,01 bnaspideslLiphistius
плейсто-
ну .5-2
неоген,па-
леоген 65
34C
40C .
j Lepidocaris
Lingula Villebrun-
aster
Pihma Nucula
। 4 Papaeo
Metacnntjs
Pleuroto-^Nautilus»
maria |
кембрий
60C .
докембрий А^ео1а
папортники
голосеменные плауновые
пермь
Danaeopsis
Опыт
теории
эволюции
карбон
Москва ♦♦♦ 2006
3
Ю.В. Чайковский
Наука о развитии жизни
Опыт теории эволюции
В память о жертвах нелепого и
страшного террора начала XXI века —
тысячах убитых, замученных,
искалеченных и обездоленных
Товарищество научных изданий КМК
Москва ❖ 2006
УДК 575.8
ББК 28.02
Ч 151
Чайковский Ю.В. Наука о развитии жизни. Опыт теории эволюции.
Москва: Т-во научных изданий КМК. 2006. 712 с.
Цель книги — показать, что для сохранения жизни и культуры нам нужны не
только призывы жить лучше, нам нужна еще и новая теория эволюции.
В последние годы стали обычны заявления о подходе с «эволюционных» пози-
ций к проблемам космологии, экологии, экономики и техники, к развитию не-
рвной системы и рака. Появились термины «нервный дарвинизм», «экономи-
ческий дарвинизм» и даже «литературный дарвинизм». В них нет ссылок на
эволюционные труды — авторам хватает смутной школьной памяти, что
эволюция идет «по Дарвину». На самом деле все 4 главных тезиса дарвинизма
(непрерывность и ненаправленность изменений, естественный отбор и рас-
хождение от единого предка) оказались не общими законами, а либо частны-
ми правилами с исключениями, либо не действуют вовсе. И, главное, они обес-
печивают лишь изменчивость в пределах близких видов, но не эволюцию.
Книга рассказывает о появлении и развитии жизни, о развитии взглядов учёных
на этот процесс. При необходимости даются параллели с идеями эволюции в
других науках (космологии, геологии, лингвистике, экономике и др.) и в различ-
ных религиях. Автор исходит из той идеи, что новая теория может быть по-
лезна для практики не объяснениями происшедшего и не предсказанием будуще-
го хода эволюции (это мало кому важно), а конкретными рекомендациями —
например, по охране природы и культуры, по медицине и систематике.
От читателя требуется знание школьного курса биологии и, вдобавок, любой
популярной книжки по иммунологии. Книга рассчитана на студентов старших
курсов и аспирантов биологических специальностей, но ряд мест (особенно п. 5-
2 и глава 11) требует некоторого навыка чтения теоретической литературы.
Книга может быть полезна учителям и старшеклассникам школ биологиче-
ского профиля, но также должна быть интересна и всем специалистам, кому
по роду их занятий надо знать новые эволюционные идеи — биологам, меди-
кам, психологам, экологам, инженерам, экономистам и обществоведам.
Рисунков 92. Литература: 147 названий (рекомендуемый список) и более 200
иных названий, упомянутых в тексте.
НА ОБЛОЖКЕ — «живые ископаемые» из книги:
Г. Крумбигель, X. Вальтер. Ископаемые. Пер. с немецкого. М., Мир, 1980.
Все изображенные на верхней строчке обеих таблиц животные и растения живут
ныне, но ископаемых предков имеют далеко не все: штриховые линии показывают,
что ближайшие их “предки” жили сотни миллионов лет назад. Почему так? Можно
ли считать их настоящими предками? См. параграф 9-14, «Лазаревы таксоны»
© Ю.В. Чайковский, текст, иллю-
страции. 2006
© Т-во научных изданий КМК,
ISBN 5-87317-342-7 издание, 2006
3
ОГЛАВЛЕНИЕ
Предисловие.................................................11
ЧАСТЬ 1. РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ ОБ ЭВОЛЮЦИИ 13
Глава 1. Творение или развитие? 13
1-1. Зачем эта книга.......................................14
1-1 * Дарвинизм и эволюция 15
1-2. Что такое эволюция....................................17
1-3. Начало идей эволюции - в религии......................19
1-3* Библия: мир и человек сотворены дважды 21
1-4. Религия и наука о рождении мира.......................23
1-5. Эволюция у первых натуралистов........................24
1-5* Пифагорйцы, Гераклит и Эмпедокл 27
1-6. От софистов до Аристотеля.............................29
1-6* После Сократа 31
1-6** Существенное и побочное (сущность и акциденция) 34
1-7. Изменчивость, отбор и законы познания.................34
1-7* Ранние стоики 37
1-8. Рождение идеи предначертанной эволюции................38
1-9. Богословы, мореплаватели и эволюция...................40
1-10. Хэйл, основатель эволюционизма.......................44
1-11. Человек - модель эволюции............................46
1-12. Дорожки от Хэйла к Дарвину...........................47
1-13. ... и мимо...........................................50
1-14. Лейбниц и Мопертюи. Эволюционная философия...........52
1-15. Морфология и законы эволюции.........................55
1-15* Эволюция и катастрофы 58
1-16. Эразм Дарвин и принцип активности....................59
1-17. Ламарк и идея прогресса..............................62
Глава 2. Победа эволюционной идеи 65
2-1. Научная революция.....................................65
2-2. Немцы выходят на эволюционную сцену...................68
2-3. Параллелизмы и связь всего со всем. Биосфера..........69
2-4. Эволюция романтическая. Окен..........................72
2-5. Развитие зародышей и эволюция. Жоффруа Сент-Илер......75
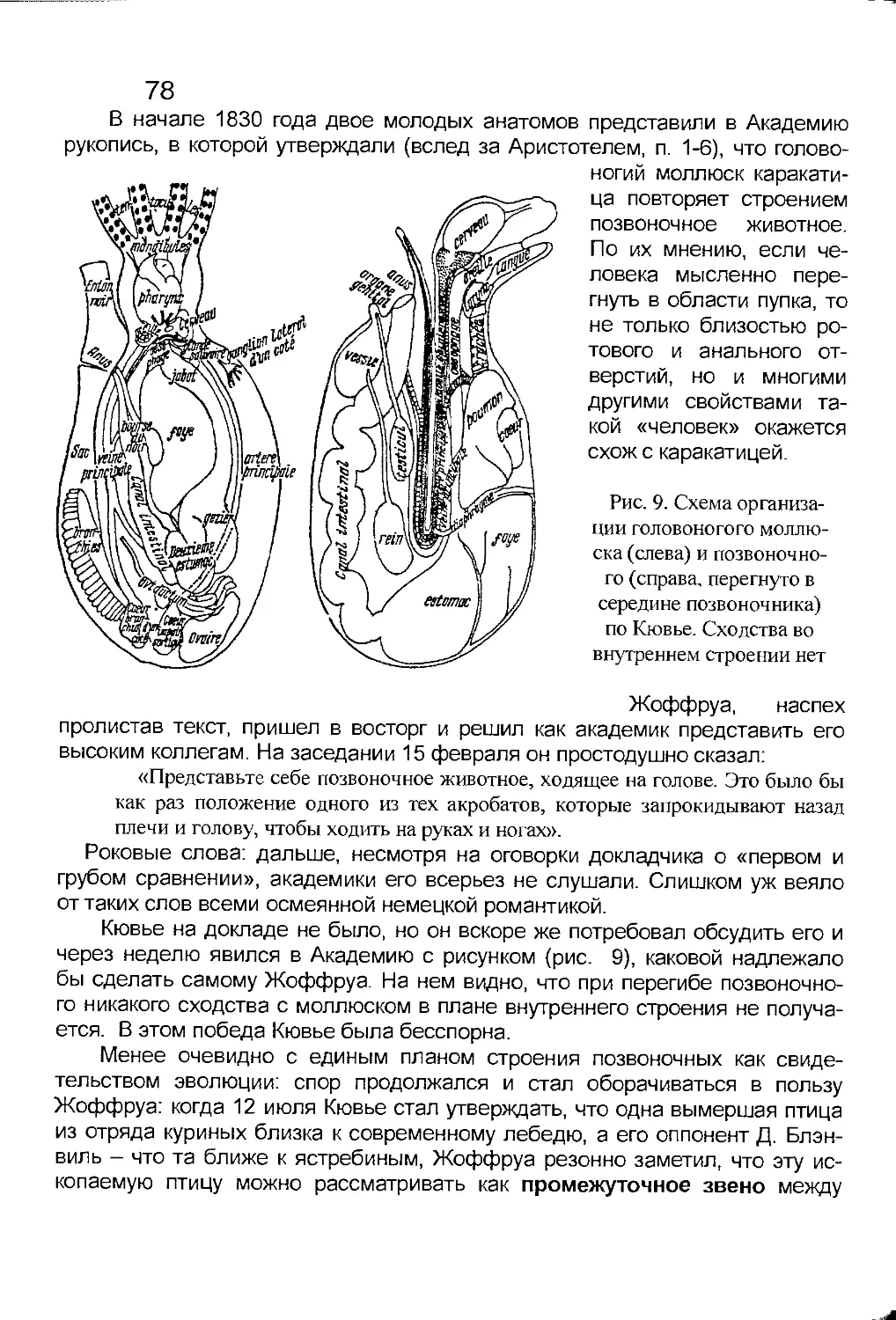
2-6. Знаменитый спор.......................................77
2-7. Что такое жоффруизм...................................79
4
2-7* Гомология 82
2-7** Карл Нэгели и развитие жоффруизма 83
2-8. Молодой Дарвин............................................84
2-9. И острый галльский смысл, и сумрачный германский гений...88
2-10. Пока Дарвин писал «Длинную рукопись».....................90
2-10* Множественный параллелизм 93
2-10** Проблема вида 93
2-11. Первые успехи и неудачи дарвинизма.......................95
2-11* Метод Дарвина и случайность 97
2-12. Дарвинизм без отбора.....................................99
2-13. Дарвин без Мальтуса.....................................100
Глава 3. Наследие Дарвина 102
3-1. Две истории науки........................................102
3-2. Поздний Дарвин...........................................104
3-2* Вынужденные добавления к учению 106
3-3. Зачем половой отбор? И есть ли он в природе?.............108
3-4. «Кошмар Дженкина»........................................111
3-5. Законы наследования. Дарвинизм по Уоллесу................113
3-6. Принцип смены функций. Дарвинизм по Дорну................115
3-7. Диалектика отбора. Дарвинизм по Дарвину..................117
3-7* Насекомые опровергают идею отбора 120
3-7** Тезис Витгенштейна 122
3-8. Активность и морфология. Дарвинизм по Геккелю............124
3-9. Презумпции наследования. Дарвинизм по Вейсману...........129
3-10. Происхождение родов идет не так, как происхождение видов. Дарвинизм
по Копу.................................................132
3-10* Параллельные ряды 133
3-11. Борьба как взаимопомощь. Дарвинизм по Кропоткину........135
3-12. Рождение генетики. Дарвинизм по де-Фризу и Мензбиру.....137
3-13. Российский эволюционизм: мир как целое..................141
3-13*Симбиогенез 142
3-13** Мир гак географическое целое 145
3-14. Мир как целое у Ганса Дриша. Энтелехия..................146
Глава 4. Вокруг советской биологии 148
4-1. Исходные посылки.........................................148
4-2. Московская школа эволюционной генетики. Четвериков.......151
4-3. Системное понимание отбора...............................153
4-4. Эволюция и политика......................................155
4-5. Ученики Четверикова: мутации без отбора..................158
4-6. Ученики Четверикова: творческая роль отбора - что такое?.159
5
4-7. Ученики Четверикова: Балкашина и гомеозис..............160
4-8. Ученики Четверикова: Николай Беляев и генетика чудес.. 162
4-9. Ряды и номогенез. Вклад Вавилова...................... 165
4-10. Берг и рождение номогенеза.............................168
4-10* Не ко времени 171
4-11. Номогенетики-дарвинисты................................173
4-12. Соболев и его «историческая биогенетика»...............175
4-12* Действительно - синтез учений 179
4-13. Новый жоффруизм и винт эволюции........................181
4-13* Перемещение жоффруизма на Запад 184
4-14. Дарвинизмы бывают разные...............................185
4-14* Адаптация ламаркизма к дарвинизму 188
4-14** Крайние варианты, в том числе лысенковщина 189
4-15. Эффект группы..........................................191
4-15* Национальная наука 195
4-16. Биологическое поле.....................................196
4-17. Феномен Любищева.......................................199
4-17* Процесс приспособления — не главный 202
4-17** Связавший несвязанное 205
4-18. Злосчастный опыт Шапошникова..........................208
4-19. Одомашнивание.........................................210
4-20. Закончим с темой отбора...............................211
ЧАСТЬ 2. МЕХАНИЗМЫ ЭВОЛЮЦИИ 214
Глава 5. Принцип активности, развитие особи и иммуногенез214
5-1. Какая нужна теория?....................................214
5-1* Выявление презумпций. “Бритва Оккама” 216
5-1** С историей или без? Каверзные ответы 220
5-1*** Неслучайность «случайных мутаций»: основной постулат СТЭ
давно опровергнут опытами 222
5-2. Принцип активности.....................................224
5-2* Идея активности в эволюционных учениях 226
5-2** Изменения активности. Давление нормы 229
5-3. Познавательные модели эволюционизма....................230
5-3* Идея Творения в дарвинизме и в иммунологии 232
5-4. Молекулярный ламаркизм и иммунология...................235
5-4* Вентребер, старец-пионер 237
5-5. Ламаркизм и новая генетика.............................238
5-5* Генетический текст читается на разных языках. Иносказания 241
5-6. Прыгающие гены и редактирование РНК....................244
6
5-7. Центральный тезис вместо «центральной догмы»............247
5-8. Экспериментальная эволюция. Подход иммунологов..........249
5-8* Не отбор, а расплод 252
5-9. Экспериментальная эволюция. Подходы иных биологов.......254
5-9* Экспериментальная эволюция у бактерий 257
5-10. Экспериментальная эволюция. Модели размножения.........259
5-11. Экспериментальная эволюция. Аршавский, доминанта и стресс.262
5-11* Физиологический стресс (эвстресс) 263
5-11** Патологический стресс (дистресс) 266
5-12. Экспериментальная эволюция. Наследование приобретенных свойств267
5-12* Первичные половые клетки и новый пангенез 269
5-12** Пангенез и стволовые клетки 272
5-13. Генетический поиск и норма.............................274
5-13* Поддержание нормы и ее смена 277
5-14. Новая термодинамика, эволюция и старение...............280
5-14* Биологическое поле и морфогенез 283
5-15. Морфогенез как активность и узнавание. Эпигностика.....285
5-15* С Платоном или без? 289
5-15** Поле, гомеобокс и эволюция 291
5-16. Сравнительный метод и пространство жизни............. 293
5-17. Жоффруизм и давление нормы.............................297
5-18. Случайность в эволюции организмов......................300
5-18* На грани порядка и хаоса 303
Глава 6. Сообщества, уровни и разнообразие 305
6-1. Эволюция организмов и эволюция природы..................305
6-2. Тенденции в эволюции крупных таксонов...................307
6-2* Экологическая прочность 310
6-3. Мейен и наука о разнообразии - диатропика...............312
6-3* Блочность строения и множественный параллелизм 315
6-3** Таксоны, мероны и рефрены. Диасеть 317
6-3*** Грамматика биологии 322
6-4. Наличное разнообразие и эволюционная изменчивость.......325
6-4* Наследуется не свойство, а разнообразие свойств 326
6-4** Транзитивный полиморфизм против идеи отбора 327
6-5. Простые примеры применения диатропики...................328
6-5* Была ли митохондрия симбионтом? 329
6-5** Зачем нужна теплокровность? 331
6-5*** Зачем нужен иммунитет? 332
6-6. Эпигностика, иммунитет и сетевой принцип эволюции.......333
6-6* Эпигностика, мышление и адаптивный иммунитет 336
6-6** Врожденный иммунитет 338
7
6-7. Уровни развития и фракталы живого.......................341
6-7* Наследование фрактала и разнообразия 345
6-7** Фракталы, ДС и катастрофы - три теории или одна? 347
6-8. Сквозная закономерность эволюции.........................348
6-8* Многослойная регуляция 350
6-9. Горизонтальный перенос и генное единство природы........352
6-10. Век генетики...........................................354
6-10* После прочтения генома человека. Тезис Аршавского 356
6-11. Диасеть и структурализм................................359
6-11* Гиперболы Виллиса 362
6-12. Приспособленность. Подбор по Богданову.................365
6-12* Нынешнее состояние проблемы 369
6-13. Эволюция как преобразование разнообразия...............370
6-13* Ниша и таксон 372
6-13** Форма и функция 376
6-14. Эволюция организмов и эволюция сообществ................377
6-14* Образование видов и остальных таксонов 382
6-14** Экосистемная эволюция 383
6-15. Инстинкт и коэволюция. Где мыслящий субъект?...........385
6-15* Ряды окрасок и мимикрия 389
6-15** Коэволюция и платонизм 391
6-16. Избавимся от неоправданных терминов....................393
ЧАСТЬ 3. КАК ЭВОЛЮЦИЯ ШЛА 395
Глава 7. Загадка начала жизни 395
7-1. С чего начать отсчет? Антропный принцип и картина мира..396
7-2. Молодая Земля...........................................398
7-2* Химический отбор 401
7-3. Вода и гель. Тезис Воейкова.............................403
7-4. Сначала биоценоз, затем организмы.......................405
7-5. Биосфера как организм...................................409
7-6. Новая термодинамика: планета делает на себе жизнь.......411
7-7. Жизнь родилась в скороварке?............................412
7-7* Ценозы, но не биосфера 415
7-7** Об энергетике биопоэза 417
7-8. Появление наследственности..............................418
7-8* От РНК к генам. Прогенота 419
7-8** От РНК-ового мира к нашему 422
7-9. Самосборка, хиральность и биопоэз.......................425
7-10. Энергия жизни и ее лучи................................427
8
7-10* Лазеры жизни и новая картина мира 429
7-11. Активность и рождение поля жизни.......................433
7-12. Итог. Проблема четырех А...............................435
Глава 8. Главные черты процесса эволюции 436
8-1. Различные понимания процесса эволюции...................437
8-2. Эволюция и систематика царств...........................441
8-3. Первые бактерии.........................................442
8-4. Эволюция клетки: принцип компенсации....................446
8-5. Эволюция клетки: как появились эвкариоты?...............450
8-5* Диатропика митозов и эволюция 453
8-5** Формирование митохондрий 457
8-6. Часы эволюции: время особи течет вперед и назад.........458
8-7. Принцип блочности в ранней эволюции.....................462
8-7* Блочность эволюции против симбиогенеза 465
8-8. Появление мейоза и многоклеточное™......................468
8-8* Альтернативы многоклеточное™ 470
8-9. Черновики Господа Бога..................................472
8-10. Компенсация как тенденция..............................474
8-11. Тенденции, прогресс и регресс..........................476
8-11 * Пример связи эмбриологии и палеонтологии 480
8-11** Тенденции регресса и рефрены 481
8-12. Эпигностика как двигатель эволюции.....................482
8-12* Диатропика и эпигностика эволюции млекопитающих 485
8-12** Диатропика и эпигностика размножения 488
8-13. Эволюция биосферы......................................489
8-13* Эволюция экосистем 493
8-14. Изменение роли солнечной энергии.......................494
Глава 9. От червя до человека 494
9-1. Рост разнообразия и скоростей...........................495
9-1* В разных временах 498
9-2. Аспекты процесса эволюции...............................499
9-3. Главные изобретения эволюции и их описание..............503
9-4. Преобразование фаун и флор в пространстве. Вклад Мейена.505
9-4* Споры вокруг глобального флорогенеза. Биотогенез 509
9-5. Или прогресс форм, или прогресс функций.................512
9-6. Проблема вымирания......................................515
9-6* Отчего вымерли динозавры? 516
9-7. К диатропике вымираний..................................519
9-7* Статистика вымираний и фракталы 524
9-8. Эволюция и сдвиг материков..............................525
9
9-9. Реальные свидетельства факта эволюции...................527
9-10. Эти живучие крокодилы...................................529
9-11. Блочность и планы строения..............................531
9-12. Блочность и родство групп..............................533
9-12* Происхождение цветковых и гамогетеротопия 535
9-13. Блочность и эволюция позвоночных.......................536
9-13* Преадаптация 538
9-13** Блочность и появление амниот 541
9-14. Блочность и Лазаревы таксоны...........................543
9-15. О происхождении человека...............................544
9-15* Шесть тенденций очеловеченья 548
9-15** Ископаемые мозги 552
9-15*** Дитя жары и мороза 554
9-16. Конец эволюции?........................................554
9-16* Двускоростной механизм эволюции людей 557
9-16** Нынешняя эволюция человека 559
ЧАСТЬ 4. ПРИЛОЖЕНИЯ И ВЫВОДЫ 562
Глава 10. Эволюция и систематика 562
10-1. Система, таксоны и названия............................562
10-1 * Стабильность названий. Тезис Линнея 564
10-1** Смысл иерархии. Тезис Короны 567
10-2. Филогенетика, кладизм и систематика....................568
10-2* Систематика как отражение наличной картины мира 570
10-2** Кладизм 572
10-2*** Какую же систему считать эволюционной? 575
10-3. Альтернативы кладизму и филогениям вообще..............576
10-3* Экоморфологическая систематика 579
10-4. Родство и сущность.....................................580
10-5. Диатропика и систематика...............................582
10-6. Система голосеменных Мейена. Грубая филогения..........584
10-7. Ядро и периферия. Межтаксон............................587
10-8. Логика макросистемы....................................589
10-8* Устойчивость системы. Межцарства 593
10-9. Макросистема эволюционная и нефилогенетическая.........595
10-9* Макросистема как парк 596
10-9** Уровни развития и макросистема 599
10-9*** Единый взгляд на биоразнообразие 601
10-10. Царство человека......................................603
10-11. Диатропические системы меньших уровней................606
10
Глава 11. Абрис теории 608
11-1. Исходные положения....................................608
11-1* Специфика биологии 610
11-2. Стержень теории биологической эволюции................614
11-2* Возникновение нового 617
11-3. Эволюция снизу и сверху...............................619
11-3* Эволюция снизу 620
11-3** Эволюция сверху 626
11-4. О факторах эволюции...................................628
11-4* Факторы, изменяющие организм 630
1 ]-4** факторы, изменяющие отношения между организмами 632
11-5. Акты эволюции.........................................633
Глава 12. Практические рекомендации 635
12-1. Шоры дарвинизма и давление нормы......................635
12-1* Евгеника и эволюция 638
12-1** Поиски остатков внеземной жизни 641
12-2. Загадочный карбон и умеренные креационисты............642
12-3. Анализ рефренов и диатропический прогноз..............645
12-4. Тенденции эволюции и спасение природы.................646
12-5. Как жить в новой биосфере?............................647
12-6. Заключение............................................651
Дополнения 654
Дополнение 1. Юридические презумпции - не пример для учёных 654
Дополнение 2. Лучи Гурвича и биополе 657
Дополнение 3. Аспекты эпигностики 659
Дополнение 4. Фрактал как пример рефрена и рождения новизны 660
Дополнение 5. Свойства рефренов и диасети 666
Дополнение 6. Более тонкие свойства рефренов 671
Дополнение 7. Формирующая причинность 675
Дополнение 8. Активность, компенсация и номогенез 679
Дополнение 9. Параллель мышления и эволюции. Новизна 681
Дополнение 10. Мифы, Библия и эволюция человека 686
Рекомендуемая литература....................................690
Сокращения..................................................695
Указатели 696
Указатель имён..............................................696
Указатель основных терминов.................................708
11
Предисловие
Перед вами новая книга. Хотя в ее основу и положена «Эволюция»
(ЧЭ)7, но понадобилось новое название, поскольку оформился логический
стержень концепции, триединый принцип эволюции: активность - диа-
сеть - смена норм (п. 11-1*). Термином диасеть (диатропическая сеть)
названа рефренная структура разнообразия по С.В. Мейену. И она, и идея
активности, в ЧЭ уже излагались, но теперь мне стало ясно, что усложне-
ние форм действия активности на диасеть - основной принцип эволюции.
Взамен прежней критики дарвинизма, допускавшей, что естественный
отбор как эволюционное явление в природе существует (но не всё может),
упор теперь сделан на демонстрацию отсутствия самого явления. Вме-
сто него использовано реально наблюдаемое явление - подбор по А.А.
Богданову (п. 6-12), не связанный с избирательным размножением и по-
служивший ему основой для первого варианта общей теории систем.
Если идея активности как движущего принципа теряется во тьме веков,
а регулирующий принцип взят мною в форме богдановского подбора, то
идея его реализации для эволюции живого вполне нова и заимствована
мной у зоолога Елены Евгеньевны Коваленко (Петербургский универси-
тет). Это - принцип смены норм, введенный в гл. 5. Смена норм - реаль-
ный акт (не путать с мысленной «сменой адаптивных норм», п. 4-14). А
именно, снятие давления нормы (его частным случаем является стресс)
выступает как механизм, раскрывающий эволюционную изменчивость.
В книге ЧЭ такого стержня не было, и ее было бы лучше в свое время
озаглавить: «Очерки об эволюции жизни» У нее на обложке был изобра-
жен ящер, разительный пример разнообразия, а теперь фото перед тек-
стом книги маркирует новую идейную базу - роль принципа активности.
Итог ряда смен форм активности предстаёт как преобразование раз-
нообразия (организмов и их сообществ). Так было дано и в ЧЭ, но теперь
стали ясны два обстоятельства. 1) Эволюционные новшества представи-
мы как заполнение новых строк диасети, оно происходит через смену
форм активности. 2) Понимание эволюции с позиции триединого принципа
требует новой картины мира, способной отобразить то пространство
жизни, в котором течет эволюция (п. 5-16). Подчеркнута мысль, что наивно
ожидать понимания эволюции и овладения ею, если искать только новые
молекулярные механизмы. Поэтому привлечена новая для теории эволю-
ции методология - метод познавательных моделей (п. 5-3).
Всё это позволило заново поставить и рассмотреть в п. 11-2* проблему
возникновения нового (эмерджентности), обычно избегаемую.
В ЧЭ были плохо изложены и плохо увязаны друг с другом частные
линии изложения. Смысл, очевидный мне, не всегда был понятен тем, с
1 См. список сокращений в конце книги.
12
кем мне пришлось ее обсуждать. Надо было многое изложить подробнее,
а ЧЭ и без того слишком объемиста. Это вынудило что-то изъять - порт-
реты, полемический материал, уже сыгравший свою роль, дублирующие
примеры, уходы в сторону и лишние детали. Исключение сделано лишь
для примеров рефренов: их число даже увеличено. Материал, загромож-
давший изложение или требующий для его понимания знать все выводы
книги, вынесен в Дополнения. Они в основном написаны заново.
Главную экономию дало сокращение критики дарвинизма. Опыт ЧЭ
показал, что она почти не имеет читателя: приверженцы дарвинизма ника-
кой критики не читают (это было мне известно и ранее), а остальным ока-
залось важнее подробное изложение позитивного материала. Критика
иных учений занимала в ЧЭ мало места и сохранена полностью.
Не имея сил писать книгу заново, я выбрал путь изменений текста ЧЭ.
В частности, пришлось сохранить прежний порядок литературных ссылок
(выросший из первой публикации в БПС). Рекомендуется для чтения лите-
ратура, сколько-то доступная в российских библиотеках и вводящая в курс
проблем. Она указана в конце книги, и ссылки на нее даны в квадратных
скобках. Иная литература дана выборочно, прямо в тексте, причем длин-
ные названия сокращены (что указано многоточиями), а статей иногда и
опущены, если это не в ущерб смыслу читаемого. Ссылки на учебники и
справочники приводятся лишь изредка - если это необходимо по сути.
Главные новации: часть 2 переписана и отделена от идеи отбора (он
теперь рассмотрен в части 1). Пункт 3-7* обновлен, стал смысловым цен-
тром главы 3 и исходной точкой главы 5. В п. 6-12 проведена связь разно-
образия с приспособлением. Рождение жизни дано в главе 7 с позиции
гель-ценотической концепции. Параграф «Единая схема эволюции» убран
из главы 6 и развернут в новую главу 11, где сделана попытка наметить
общую теорию. Параграф «Шоры дарвинизма» перенесен из конца главы
3 в начало главы 12 и обрел советующий смысл. Обновлено изложение
эволюционной иммунологии и энергетики, так как в данных отраслях в по-
следние 5 лет достигнут радикальный прорыв.
С этими новациями книга, надеюсь, начнет реализацию многими заяв-
ленную, но никем не проведенную, идею: «эволюция - основа биологии».
Генетик Наталья Александровна Малыгина показала мне, что генетику
надо использовать шире и излагать аккуратнее, чем это было сделано в
ЧЭ; но главное, она внимательно прочла обе книги и сумела сочетать поч-
ти несовместимое: была мне строгим критиком и увлеченным помощни-
ком. Приношу ей глубокую признательность.
Также благодарю всех, кто помогал мне (они названы поименно в пре-
дыдущих работах, а недавние - далее по тексту). Хочется отдельно по-
благодарить тех, кто оказал мне компьютерную помощь: это Е.П. Крюкова
и ее сын Г.А. Базыкин, а также мой коллега по Библиотечному совету
МОИП палеонтолог А.В. Гужов, оформивший новые рисунки.
13
Часть 1. Развитие представлений
об эволюции
Зачем нам теория эволюции живой природы? Почему она вновь и вновь
овладевает умами общества? Принято говорить, что она служит фундамен-
том нынешнего мировоззрения и потому нужна при обсуждении любых об-
щих вопросов. Однако на деле всё не так: эволюцию почтительно поминают
во введениях, чтобы затем никак не принимать ее во внимание. Это и по-
нятно - на фоне сложных нынешних теорий старомодные эволюционные
уверения (например, во всемогуществе отбора полезных мутаций) поража-
ют убогостью мысли и скудостью данных.
Подлинная причина думать об эволюции видится в наше время в дру-
гом: та теория эволюции, которой учат в школе и вузах, говорит лишь о про-
цессах медленных, незаметных глазу, а вокруг мы видим, наоборот, очень
быструю эволюцию, она бьет в глаза и требует к себе внимания.
В последние 70 лет число людей на Земле утроилось, что преобразило
и вскоре еще более преобразит облик планеты. Быстро меняется климат:
отступили ледники, казавшиеся вечными, и обнажили могилы Средневеко-
вья. Появились формы животных, столь живучие, что хоть беги - например,
городские комары, крохотные и ненасытные (причем неизвестно, проникли
они в города или произошли в разных местах заново). Быстро формируется
(особенно в городах) особая природа, приспособленная к тому, чего прежде
никогда не было (например, к ядам и радиации). Многие виды птиц, прежде
перелетные, зимуют теперь в городах. Исчезли такие вечные болезни, как
оспа, и появились новые, столь же грозные.
Живой мир меняется на глазах, и, как выяснено недавно, периоды
очень быстрых изменений в прошлом тоже не раз бывали. Исчезают многие
виды, и попытки их спасать часто беспомощны. Всякий ли вид можно спа-
сти? Можно ли создать новый? В учебнике дарвинизма (и иных старых тео-
рий эволюции) ничего об этом не прочесть.
Словом, нужна теория эволюции, которая опишет процессы быстрые,
медленные и катастрофические, которая поможет нам жить в мире кризиса
и укажет, как из него выйти (если выход вообще есть, что пока неизвестно).
Об этом и пойдет у нас речь - сперва о старых учениях, а затем и о вырас-
тающей сейчас из них новой теории.
Гпава 1. Творение или развитие?
Что такое эволюция? Кто и когда впервые стал думать о ней? Можно ли
считать факт эволюции строго установленным? Каковы движущие силы и
механизмы эволюции? Как фактически шла эволюция? С какого времени
можно считать ее явлением, вполне признанным наукой?
14
Как ни странно, из всех этих вопросов, вроде бы таких привычных, од-
нозначно можно ответить лишь на последний: со времен Чарлза Дарвина.
Даже в самом факте эволюции некоторые ученые снова стали сомне-
ваться, но никто, даже ярые ее противники, не отрицают, что до Дарвина
идея жила в науке на правах лжеучения, а после него стала предметом ши-
рокого исследования и обучения. В этом непреходящая заслуга английского
натуралиста, она останется за ним независимо от того, как пойдет в буду-
щем развитие науки (чтим же мы великого Коперника, хотя давно исповеду-
ем иную космологию). А пойти наука может путями неожиданными.
1-1. Зачем эта книга
Полвека назад, готовясь отметить столетие «Происхождения видов»,
западные историки науки погрузились в архивы, в старые книги и журналы и
пришли к поразительному выводу - Дарвин пренебрег многим из тогдашней
науки, и все крупные натуралисты того времени указывали на это. Неучтен-
ный материал рушил выводы Дарвина, он так и не нашел места в его тру-
дах, но данный факт (вот странность) никак не помешал победе учения.
Его приняли как новую веру. Спорить с верой нет смысла, но начинаю-
щий должен понимать, что он изучает - свод знаний или свод верований.
После юбилея появилось много книг и крупных статей с изумляющими
заглавиями вроде: «Darwin’s metaphor: does nature select?» («Метафора
Дарвина: отбирает ли природа?») или даже: «Evolution without evidence, Ch.
Darwin and the Origin of Species» («Эволюция без доказательства - Дарвин и
Происхождение Видов»), Разумеется, советский читатель не узнал об этом
ничего - ведь дарвинизм был частью коммунистической идеологии (т.е. ве-
ры). Удивительно, что российский не знает до сих пор. Даже в последнем
“академическом” издании «Происхождения видов» (СПб., Наука, 2001) ком-
ментарии игнорируют всю указанную работу дарвиноведов.
Увы, как часто о Дарвине берутся судить, не прочтя его! Сам он, пусть и
не был в рассуждениях безупречен, но всегда был против обращения его
взглядов в догму. Однако догма утвердилась, и с тех пор то и дело можно
прочесть такое: «Согласно Дарвину... движущей силой эволюции является
естественный отбор. Именно он и только он определил те пути, которыми
шло развитие органического мира от самых простых до самых сложных
форм». Если бы прекрасный генетик С.М. Гершензон, написавший в 1984
году эти слова, поискал у Дарвина их подтверждения, он ничего не нашел
бы: Дарвин никогда не считал отбор единственным движущим фактором.
См., например, статьи: «Был ли Дарвин дарвинистом?» (Thuiller Р. Darwin
etait-il darwinienne? // LR, 1982, № 129) и [Чайковский, 1988].
Еще полтораста лет назад великий биолог Луи Пастер писал: «Идеи
превращения видов так легко воспринимаются, может быть, потому, что ос-
вобождают от строгого экспериментирования». В самом деле, эволюцион-
ные упражнения дают биологу отдохнуть от изнурительных норм своей нау-
15
ки и позволяют плыть по течению свободной от их контроля мысли. Трудно
вообразить ученого, который решился бы описать в качестве своих резуль-
татов чужие многократно опубликованные данные, зато легко представить,
как смеялись бы его коллеги. А вот эволюционисты сотнями лет высказы-
вают (как свои собственные!) одни и те же положения, одни и те же на них
возражения, нисколько не смущаясь, когда изредка кто-нибудь укажет на
прискорбные повторы и на отсутствие всякой новизны в суждениях.
Разве еще где-нибудь коллеги допустят утверждать, что некая точка
зрения доказана, если в качестве фактов даются никем не поставленные
эксперименты? А ведь дарвиновы воображаемые олени, убегающие от во-
ображаемых волков, не только были единственным у Дарвина “доказатель-
ством” эффективности отбора у животных, но и до сих пор приводятся как
решительный аргумент в пользу новейшего дарвинизма.
Многие удивятся: разве эффективность естественного отбора не дока-
зана всей биологией? Я тоже 40 лет назад полагал, что доказана, - пока чи-
тал учебники. И даже поставил компьютерный эксперимент, чтобы уточнить
детали вполне очевидной мне теории. Правда, компьютер (тогда слова та-
кого в русском обиходе не было, он звался ЭВМ «Минск-22», не имел даже
экрана, зато занимал целиком небольшой зал) отказался подтверждать
догму (см. далее, п. 5-10), но я решил, что просто не так что-то задал.
Пришлось зарыться в литературу, чтобы понять, где моя ошибка. Тут-то
и выяснилось, что опытов поставлено на удивление мало и все они постав-
лены и истолкованы так, что ответ задан заранее (об этом поговорим в п. 3-
7). Естественно, противники дарвинизма на это указывали, но без успеха -
им просто не отвечали. Никто не ответил и мне, хотя из устных реплик сле-
довало, что статьи с машинными экспериментами заметили многие.
Данный факт меня поразил: ведь по канонам науки следовало либо
найти в них ошибку, либо пересмотреть основы дарвинизма. Тогда мне по
наивности казалось, что дарвинисты озабочены истиной, но позже я прочел
у методологов, что это неверно: как правило, сторонники устаревших воз-
зрений просто охраняют, сознательно или нет, свою догму (подробнее см. п.
3-7**). Тем самым, Пастер и ныне прав.
1-1* Дарвинизм и эволюция
Многие до сих пор уверены, что дарвинизм - то же самое, что теория
эволюции (в американской литературе это общепринято), и что поэтому ему
противостоит креационизм - учение о творении Богом каждого вида в от-
дельности (термин «креационизм» происходит от латинского creatio ex nihilo
- «выбор из ничего»; отсюда и английское creation - творение). В самом де-
ле, сторонники креационизма обычно уверены, что, указывая на изъяны
дарвинизма, ниспровергают эволюционную идею вообще. Это наивно.
Эволюция - явление природы, а дарвинизм - лишь один (притом очень
старый) способ толковать некоторые стороны этого явления; он никогда не
16
давал никакого обоснования эволюции в широком смысле этого слова. И
тот, кто взялся отрицать явление (эволюцию), обязан опровергнуть данные
о самом явлении, а не его старое толкование, удобное для разгрома.
Дарвинизм и креационизм не знают никого, кроме друг друга. Мне уже
случалось писать, что “научный креационизм” - явление вовсе не научное, а
политическое, что он вовсе не противник дарвинизму, а его оборотная сто-
рона («словно цирковые борцы, они нужны друг другу» [Чайковский, 1993, с.
139]), и недаром оба предпочитают только спорить друг с другом, а не изу-
чать нынешнюю науку. За прошедшие годы ситуация еще более проясни-
лась: перегиб в одну сторону всегда рождает перегиб в другую.
Сто лет уверений дарвинистов, что «либо Дарвин, либо творение», ес-
тественно привело верующих к мысли, что изъяны дарвинизма исправимы
лишь возвратом к идее Творения в ее самой примитивной форме. Но и
креационизм ведь бывает разный.
Одно направление (именующее себя «научным креационизмом») отри-
цает эволюцию вообще. Поскольку отрицать изменение органического мира
после всех открытий палеонтологии невозможно, оно объединяет тех, кто
ее не знает или пытается уложить ее в библейские 6 тыс. лет (что, как мы
увидим в п. 12-2, почти то же, но не совсем то же). Такие книги спорят с дар-
винизмом XIX века, и приводить их тут смысла нет.
Второе направление понимает эволюцию как «непрерывное Творение»,
и оно в прошлом много дало науке о природе. Сошлюсь на православный
просветительский сборник:
Той повеле, и создашася. Современные учёные о сотворении мира / ред. А.В.
Гоманьков. Клин (Московская обл.), “Христианская жизнь”, 1999.
К этому направлению близок популяризатор Ариэль Рос (США) в книге:
Рос А. В начале... . Заокский (Тульская обл.), “Источник жизни”, 2002. - Мно-
готочие в заглавии принадлежит оригиналу.
Этих книг мы еще коснемся. Рос не объявляет своей позиции, расска-
зывает многое об эволюции, склоняется к пониманию ее как долгого творе-
ния и допускает, что Вселенная создана до «шести дней творения».
А немецкий учебник (1992 г.) Райнхарда Юнкера и Зигфрида Шерера
(ЮШ) спорит с дарвинизмом середины XX века и даже приводит данные из
биологии и геологии, в самом деле для теории эволюции трудные.
С такими креационистами уже можно беседовать.
В учебнике ЮШ (как увидим далее) много интересных и полезных све-
дений, однако, к сожалению, почти нет ничего из науки последнего полуве-
ка, и спорит он только с вариантами дарвинизма. Огорчают и прямые ошиб-
ки. Так, желая показать, что зачаточные органы нужны сами по себе (а не
являются следами эволюции), авторы пишут: «Остатки крыльев у киви слу-
жат для регулировки равновесия» (ЮШ, с. 124). На самом деле у этой птицы
вообще нет видимых снаружи зачатков крыльев, а сам довод наивен: нали-
чие пользы от органа ничего не говорит против идеи эволюции.
17
За ЮШ пытается следовать российский православный учебник:
Вертъянов С.Ю. Общая биология. Учебник для 10-11 классов общеобразова-
тельных учреждений с преподаванием биологии на православной основе.
Сергиева Лавра, 2005. По благословению Патриарха... Алексия II.
Ошибки ЮШ не только почти все повторены, но и гротескно развиты
(например: «Мы ведь с вами в случае потери равновесия вскидываем рука-
ми - и киви надо же чем-то вскидывать!», с. 202). Автор даже путает ген и
аллель (с. 112). Досадно: ведь это первый в России учебник, который ищет
примирения идеи эволюции с православием. Так, автор назвал (в отличие
от ЮШ) некоторые новые эволюционные взгляды и верно заключил (с. 198):
«Многие серьезные биологи сегодня отделяют эволюционную теорию как науку
о возможных изменениях организмов от реконструкции древа эволюции, призна-
вая последнее лишь предполагаемой историей. Современные эволюционные тео-
рии являются чисто гипотетическими, своеобразной философией в науке».
Увы, «крылья киви», машущие по всему учебнику (подробнее см. Мамонтов
С.Г. // Природа, 2006, № 7), затмевают эти немногие разумные фразы.
Но С.Г Мамонтов, найдя в учебнике море ошибок2, по сути возразил
лишь одной фразой: теория эволюции «покоится на прочном основании» (с.
88). Это неправда, и лишь она будет замечена креационистами, а он
не вправе будет удивляться, когда в школах восстановят «Закон Божий». Да
и не всё ли равно, кто из этих «цирковых борцов» сейчас одержит верх?
На мой взгляд, сегодня важно не заявить, управляет ли Бог эволюцией
или она течет сама по себе (этот вопрос никогда не имел и не будет иметь
приемлемого для всех ответа), а выяснить, как она фактически шла и идет.
То есть надо выявить закономерности эволюции, чтобы жить в ней и, на-
сколько возможно, вписываться в ее ход. До сих пор ни атеисты, ни верую-
щие (разных религий) такой задачи, насколько знаю, не ставили.
1-2. Что такое эволюция
Противников идеи эволюции можно понять: книги по эволюции и впрямь
вызывают оторопь. Так, в американском руководстве читаем:
«Эволюцию можно рассматривать как изменения любого свойства данной
популяции с течением времени. В некоем общефилософском смысле в этом и
2 Его упреки С.Ю. Вертьянову верны, но есть учебник еще хуже: Мамонтов С.Г. Биология
для школьников старших классов и поступающих в вузы. М., Дрофа, 2005. Там происхожде-
ние жизни дано по Ф. Энгельсу и А.И. Опарину плюс опыт Н. Пири (1953 г.), генетика - по Г.
Менделю с упоминанием Т. Моргана (1920-е гг.), теория эволюции - по Дарвину плюс пара
суждений 1930-х гг. (без имен авторов), происхождение человека - по Дарвину и Энгельсу
плюс пара фактов 1930-х гг. При списывании со старых учебников трудно сделать много
ошибок, но Мамонтов их делает: например, путает хромосому и ДНК, не может объяс-
нить ни строения ДНК (нет спирали), ни различий РНК, а вместо описания их рабо-
ты - масса разрозненных деталей (с. 42-44,49). У Вертьянова это сделано лучше.
18
заключается суть эволюции» (О.Солбриг, Д.Солбриг. Популяционная биоло-
гия и эволюция. М.. Мир, 1982, с. 21).
Философия тут ни при чем (любой философ сразу скажет, что смысл
слова «эволюция» здесь не раскрыт, а лишь загнан в слово «изменение»),
но не будем придираться к словам, а прочтем дальше:
«В одном из популярных определений эволюция рассматривается как изме-
нение частоты отдельных генов из поколения в поколение. Это очень точное
определение».
Подобное определение есть и в других книгах. Но если признать его
верным, то креационисты правы - умнее и честнее призвать Бога, чем пы-
таться понять, каким генам надо изменить частоту, чтобы, к примеру, поя-
вился фотосинтез (принято считать, что первые организмы его не имели).
Призыв к Богу вовсе не означает отказа от эволюции. Вот ее определе-
ние, которое дал французский палеонтолог-иезуит Пьер Тейяр де Шарден,
умерший полвека назад:
«Эволюция — что это? Теория? Система? Гипотеза? Нет, но зато нечто боль-
шее: общее условие, которому должны отныне удовлетворять, чтобы быть
осмысленными и истинными, все теории, гипотезы, системы. Свет, осве-
щающий все факты, изгиб, который принимают все линии».
Замечательно, но ведь это - у богослова. А как у учёных? Вот опреде-
ление из советской энциклопедии, благополучно перешедшее в новейшее
российское издание (Российский энц. словарь. М., 2001, т. 2):
«Эволюция (биол.), необратимое ист. развитие живой природы. Определяется
изменчивостью, наследственностью и естеств. отбором организмов. Сопро-
вождается приспособлением их к условиям существования, образованием и
вымиранием видов, преобразованием биогеоценозов и биосферы».
Слова о ценозах и биосфере добавлены в новом издании (веяние вре-
мени), и это хорошо, но в остальном мы видим, как и в советское время, не
определение явления, а декларацию конкретного взгляда на него. Работать
с таким “определением” невозможно.
Однако и у дарвинистов бывали приличные определения. Например,
старый польский учебник «Эволюционизм» Станислава Сковрона (есть рус-
ский перевод, Варшава, 1965) дал такое определение эволюции: это
«в основном необратимый процесс, происходящий во времени, благодаря кото-
рому возникает что-то новое, разнородное, на более более высокой ступени
развития. Говоря об эволюции, мы не имеем в виду действия какой-то таинст-
венной силы, а естественный процесс... Астроном говорит об эволюции плане-
тарных систем и звезд, геолог - об эволюции Земли, а биолог - об эволюции
живых существ... Языковед старается с эволюционных позиций подходить к
вопросам своей специальности, мы говорим об эволюции наших взглядов на
разные проблемы; можно... говорить об эволюции какой-нибудь машины.»
Определение неидеально (я бы уточнил: эволюция живого есть эволю-
ция биосферы, в ходе которой эволюируют входящие в нее сообщества, в
19
свою очередь состоящие из эволюирующих видов), но с ним можно рабо-
тать. Оно ясно говорит, что эволюция живого - частный случай эволюции, и
это очень важно. Еще важнее, что в определении явления нет ни слова о
какой бы то ни было теории, призванной объяснять это явление.
Зато сразу встает вопрос о том, что в биологической эволюции особен-
но, а что обще с другими формами эволюции (космической, языковой и т.д.).
Сковрон его не коснулся, поэтому стоит заметить, что бывают хорошие оп-
ределения эволюции вообще. Так, в дни Дарвина философ Герберт Спен-
сер дал такое определение:
“Эволюция есть интеграция материи и сопутствующее ей рассеяние движе-
ния, причем материя переходит от состояния неопределенной, бессвязной од-
нородности к состоянию определенной, связной разнородности, а сохранен-
ное движение претерпевает параллельные изменения”.
Позже мы увидим, что определение Спенсера актуально для новейшей
эволюционной науки - надо лишь заменить “рассеяние движения” на “рас-
сеяние энергии”. А сейчас поговорим об эволюционизме раннем.
1-3. Начало идей эволюции - в религии
В сущности, эволюционизм (совокупность знаний об эволюции) старше
самой науки. Все древние культуры имели свою космогонию (мифы о рож-
дении мира), пытавшиеся так или
иначе осмыслить появление всего, что
окружает людей. В восточных
космогониях обычным сюжетом было
«мировое яйцо», из которого
вылупился мир. Следы этого мифа
проникли и на Запад (рис. 1).
Рис. 1. Бог - архитектор мира. Миниатюра
из английской Библии XIV века. Вряд ли
художник сознавал, что воспроизводит
идеи древних космогоний: рождение
Вселенной из “мирового яйца” и управ-
ляющие миром законы, которым сами боги
вынуждены подчиняться. От этих законов
у христиан остался только механический
инструмент (циркуль) в руках Творца
В древних космогониях для нас
существенна идея законов природы,
которые даже боги не в праве нарушить. В этом отношении особняком стоит
космогония греков, поскольку только она дала начало науке о мироздании. В
своем завершенном виде она не содержит идеи «мирового яйца» и дошла
20
до нас в поэме Гесиода, жившего в -VII векеЛ Поэма называется «Теого-
ния» («Происхождение богов») и цитируется далее в переводе В. Вересае-
ва. Поэт обращается к музам:
Всё расскажите, - как боги, как наша земля зародилась,
Как беспредельное море явилось и шумные реки,
Звёзды, несущие свет, и широкое небо над нами;
Кто из бессмертных подателей благ от чего зародился,
Как поделили богатства и почести между собою...
И сообщите при этом, что прежде всего зародилось.
Это напоминает программу научного исследования. Вот ответ:
Прежде всего во вселенной Хаос зародился, а следом
Широкогрудая Гея, всеобщий приют безопасный,
Сумрачный Тартар, в земных залегающий недрах глубоких,
И, между вечными всеми богами прекраснейший, Эрос.
Гея - это богиня земли и, одновременно, сама земля; Эрос - бог любви.
Если вдуматься, это важные мысли: сперва появилась первичная субстан-
ция (хаос), т.е. мир мыслится отнюдь не вечным. Первичным духовным на-
чалом названа любовь (тогда как вражда отодвинута в состав младших бо-
жеств). Света и тьмы сперва не было, их породил Хаос:
Черная Ночь и угрюмый Эреб родились из Хаоса.
Ночь же Эфир родила и сияющий День, иль Гемеру...
Далее, первые боги (а с тем и первые сущности) порождены бесполо,
равно как и первые поколения их детей:
Гея же прежде всего родила себе равное ширью
Звёздное небо, Урана, чтоб точно покрыл ее всюду
И чтобы прочным жилищем служил для богов всеблаженных;
Нимф, обитающих в чащах нагорных лесов многотенных.
Также еще родила, ни к кому не всходивши на ложе,
Шумное море бесплодное, Понт. А потом, разделивши
Ложе с Ураном, на свет Океан породила глубокий...
Океан - обтекающая землю кругом река и, одновременно, речной бог.
Каждый бог двулик: природный объект или явление и антропоморфный (по-
добный человеку) бог этого объекта или явления. Половое рождение вто-
рично, и этим сразу задано миропонимание - то самое, которое характерно
поныне. Так, в дарвинизме существует нерешенная проблема: чем выгодно
наличие двух полов? Для сравнения: в китайской космогонии Инь и Ян (жен-
ское и мужское начала) первичны, и этот вопрос встать не мог.
В обильных порождениях Геи нет принципа наследственности: она (а
затем и другие богини) рождала самых немыслимых чудищ. Этот процесс
з
Будем, как принято у историков астрономии, обозначать даты до новой эры в виде
отрицательных.
21
порождений Геи пресёк последний ее сын Кронос, оскопив своего отца Ура-
на, чем прекратил инцест сына с матерью и тем самым положил конец по-
рождению чудищ. Оскопил он его, кстати, серпом - орудием культурного
земледелия, знаменуя начало нового типа и этапа жизни.
Нет в «Теогонии» понимания роли Солнца: свет и тьма - самостоятель-
ные сущности, рожденные задолго до солнца и солнечного бога Гелиоса
(внука Геи и Урана). Появление растений и животных не привлекло внима-
ния греческого космогониста, и в этом его крупный просчет (с нашей точки
зрения) по сравнению с еврейской, финикийской и восточными космогония-
ми. Чья возникла раньше - предмет споров, в которые вступать не будем.
1-3* Библия: мир и человек сотворены дважды
В книге Бытия (первая книга Священного писания евреев и христиан)
читаем, что Бог (его происхождение не рассматривается) сотворил, как и у
греков, сперва землю, затем свет (день первый). Во второй день
“И сказал Бог: да будет твердь посреди воды, и да отделяет она воду от воды.
И создал Бог твердь; и отделил воду, которая под твердью, от воды, которая
над твердью. И стало так. И назвал Бог твердь небом”.
В третий день
“сказал Бог: да соберется вода, которая под небом, в одно место, и да явится
суша. И стало так. И назвал Бог сушу землею, а собрание вод морями”.
Вчитаемся в эти слова, ибо предлагаемая космология совсем отлична
от иных космологий Древности: земной мир подобен водолазному кессону,
где вода снизу и сверху. Модель наглядно объясняет феномен дождя (от-
сюда выражение: «Разверзлись хляби небесные») и делает понятной ле-
генду о Всемирном потопе, но ничего более. Для засушливой Палестины та-
кая космология удивительна (у соседних египтян мифа о Потопе нет). Как
узнали в XX в., она заимствована из шумерского мифа.
Другие народы видели мир иначе. В частности, в иных восточных кос-
могониях отделение земли от неба символизировало разделение женского
и мужского начал. Это видно из того, что обычно небо там является мужчи-
ной, а земля женщиной (у египтян наоборот). Чисто мужская еврейская кос-
могония лишилась этой символики. Подробнее см.: Гоейвс Р., Патай Р. Иу-
дейские мифы. Книга Бытия. М., БСГ Пресс, 2002.
Обычно комментаторы Библии всю тему странного «кессона» обходят,
сразу переходя к творению живого. А именно, в тот же третий день
«И сказал Бог: да произрастит земля зелень, траву, сеющую семя, дерево пло-
довитое, приносящее по роду своему плод, в котором семя его...».
В этом выражении «семя по роду своему» можно видеть намек на
принцип наследственности (который у греков то и дело игнорировался).
И лишь в четвертый день Бог создал небесные светила. Здесь - обыч-
ное для ранних мифов непонимание роли Солнца, уверенность, что оно -
всего лишь сгусток дневного света. Однако тьма уже (в отличие от греков)
22
мыслится как отсутствие света, а не как особая сущность4.
На пятый день Бог творил водных животных, а на шестой - наземных.
Наконец, тоже в шестой день, Бог творит человека.
Суть человека выступает в главе 1 книги Бытия двойной: с одной сто-
роны, к человеку отнесено то же, что и к животным, божье обращение «пло-
дитесь и размножайтесь, и наполняйте землю» и так же, как животным, от-
даны в пищу растения; но с другой, Бог призвал человека обладать землею
и животными (не разрешив, но и не запретив поедать их), а самого его объ-
явил своим образом и подобием - как мужчину, так и женщину.
Однако глава 2 дает иную трактовку творения: «по образу и подобию»
создан мужчина, Адам, он получает в пользование рай, узнаёт от Бога за-
прет на плоды «древа познания добра и зла» и нарекает имена всем птицам
и наземным животным - всё это без женщины. После этого Бог творит жен-
щину (Еву) из ребра человека. Об их размножении сказано лишь после гре-
хопадения: тогда Бог обрек ее на тяжкие роды и подчинение мужу, а его -
на тяжкий труд для пропитания. И обоих, изгнав из рая, лишил бессмертия.
Этот странный разнобой священной истории повлёк огромную литера-
туру и до сих пор влияет на биологию, о чем мы узнаем в конце главы 9.
Но это что! Есть в Библии куда более удивительное место: иной взгляд
на всё творение. В «Притчах Соломоновых» Мудрость говорит:
«Господь имел меня началом пути Своего, прежде созданий Своих, искони...
Я родилась, когда еще не существовали бездны,... когда еще Он не сотворил
ни земли, ни полей, ни начальных пылинок вселенной. Когда Он уготовлял
небеса, я была там. Когда Он проводил круговую черту по лицу бездны [...]
Тогда я была при Нем художницею».
Вдумаемся: история вселенной, начинаемая с мудрости (не сотворен-
ной, а родившейся, как языческие богини), за которой следовали «началь-
ные пылинки вселенной», а вовсе не твердые небо и земля, - вот уж неожи-
данный для нас взгляд древних монотеистов на эволюцию. Все такие тексты
в Библии ясно говорят учёным (в том числе многим верующим ученым), что
Библия, как и все священные книги, несет следы разнородных верований
разных эпох (см. Гоейвс Р., Патай Р.), а вовсе не послана свыше.
Конечно, мыслители думали о рождении мира всюду, в том числе в
странах, где естествознания долго не было. В Ригведе (Индия) читаем:
«Боги пришли после, когда Вселенная уже была создана. Кто же знает, когда
она поднялась из вод? Когда началось творение - быть может, оно само со-
творило себя, а быть может, и нет. Тот, кто... на высочайшем из небес, только
он это знает - а быть может, не знает и он».
См.: Чайковский Ю.В. Доплатонова космология и Коперник // Историко-астрономич.
исследования. Вып. XXX. М., Наука, 2005. Раньше всех Солнце как источник света осозна-
ли. насколько знаю, египтяне. Евреи и греки долго оставались чужды данной идее.
23
Ясно, что «мировое яйцо» не устраивало автора. Как и греков -VII века.
1-4. Религия и наука о рождении мира
В греческих богов давно никто не верит, и мы спокойно можем указы-
вать на различные исторические слои в их мифах. Никого не коробят даже
прямые противоречия - так, у Гесиода в стихе 217 «Теогонии» три Мойры
(богини судьбы) рождены богиней ночи, а в стихе 904 - ее внучатым пле-
мянником Зевсом и его второй женой Фемидой. Ясно, что поэт собирал и
обрабатывал разные мифы. Кстати, в ранних мифах Зевс побаивается
Мойр, а в поздних повелевает ими.
С Библией дело обстоит иначе: она до сих пор - священный текст для
многих, и они отрицают всякие в ней противоречия. Не будем спорить, од-
нако встает вопрос: чему при этом верить, а чему нет?
Еще 2 тыс. лет назад грекоязычный иудей Филон Александрийский, фи-
лософ и богослов, предложил трактовать непонятные и противоречивые
места Библии как иносказание. Его идею отвергли, но она не раз заново
возникала в христианском богословии, и именно из нее в XVII веке родился
конкордизм (от лат. concors - дружный, согласный) - теория, по которой
наука не противоречит священному Писанию, а раскрывает его новый
смысл. Этот смысл якобы не мог прямо быть высказан Богом прежде, чем
люди созрели для понимания. В частности, шесть дней творения конкор-
дизм истолковал как шесть геологических эпох, так что первая глава Книги
Бытия теперь выступала как бы описанием эволюции мира и жизни.
Допустим, говорят скептики (не только атеисты), конкордизм прав, но
как объяснить прямые противоречия - внутри Писания и между Писанием и
наукой? Как нам считать: человек первоначально создан наполнять собой
Землю и повелевать природой (Бытие, гл. 1), или же - жить ограниченным
числом особей в специально уготованном раю (Бытие, гл. 2)? Вопрос стал
особо актуальным в XX веке, когда стало ясно, что завет «наполняйте зем-
лю» человек выполнил и перевыполнил; и актуальность его будет нарас-
тать. Не менее актуален ныне и такой вопрос: человек - это оба пола (как в
главе 1) или только мужчина (как в главе 2)? А как толковать то место Биб-
лии, где прямо сказано, что светила созданы лишь на четвертый день, когда
земля уже зеленела растениями? Параллель с греческим мифом очевидна,
а вот согласование с наукой - отнюдь.
И главное: если Бытописатель изрек для “незрелого” читателя прямую
и очевидную в то время ложь (в Египте и Вавилонии давно знали, что ис-
точник дневного света - Солнце), то верить ли остальному тексту Писания?
Что приличнее для христиан - выдумывать вольные толкования противоре-
чий (их приводит, не соглашаясь с ними, упомянутый в п. 1-1 христианин А.
Рос, с. 352-356) или честно согласиться с противоречием?
«Непонятно, как растения, сотворенные на 3-й "день", могли существовать
без Солнца, сотворенного на 4-й "‘день”» - пишет православный палеобота-
24
ник А.В. Гоманьков в сб. “Той повеле, и создашася" (с. 188).
По мне, именно для авторитета религии лучше и честнее прямо при-
знать, что Писание тоже собрано из мифов разных времен. Однако его сли-
чения, хотя бы с греками, верующие христиане избегают. А как быть с иско-
паемыми, которые явно принадлежат не тем видам организмов, что живут
ныне? Значит ли это для верующего, что Бог творил неоднократно?
Разумеется, спор верующего с неверующим (или иначе верующим)
бессмыслен, и не надо вступать в него. Однако замечу, что возможно всего
4 линии поведения: отрицание науки («научные» креационисты), отрицание
религии (атеисты), отрицание обеих (нищие духом) и конкордизм. Послед-
ний как раз и можно назвать научным (без кавычек) креационизмом. С ним
мы не раз будем иметь дело.
Надо еще добавить, что кроме естествознания есть также история и
филология, и все они ясно показывают, как священные тексты эволюирова-
ли и друг на друга накладывались. В частности, все древние культуры в мо-
мент появления их первых дошедших до нас текстов предстают носителями
широких прав женщин. Отсюда и культы богинь. Следами этого в Библии
являются творение обоих полов людей в главе 1 и наделенная свойствами
богини Мудрость в «Притчах Соломоновых».
Затем, в зрелой Античности, женщины всюду вытесняются из общест-
венной жизни, что сперва ведет к выхолащиванию культа богинь (они ста-
новятся безгласными женами богов), а затем и к победе монотеизма (еди-
нобожия), где Бог мыслится мужчиной. Словом, Писание предстает делом
рук человеческих, и это нисколько не мешает многим ученым веровать, со-
гласно их убеждениям. Подлинный конкордизм и состоит, по-моему, в том,
чтобы признать это обстоятельство.
Если пользоваться нынешним языком, то в главе 1 Книги Бытия человек
понят как биологический вид, особенностью которого (если угодно, тезисом
видового диагноза) является тяга к господству над природой и к познанию, а
также - вера. Этот вид создан после всей прочей природы и поставлен по-
велевать ею. Итог трех тысяч лет господства такого взгляда на мир мы ви-
дим сегодня. В главе 2 Книги Бытия, вероятно написанной много позже, че-
ловек рассмотрен с позиций общественных и хозяйственных.
1-5. Эволюция у первых натуралистов
Сочинений философов, живших прежде Платона, до нас не дошло, и
мы судим о них по отрывочным свидетельствам в трудах других ученых - в
основном, ученых поздней Античности. Почти все сведения о ранних (ранее
Демокрита) мыслителях собраны в книге «Фрагменты...» [Лебедев], которой
мы и воспользуемся. (В цитатах перевод скорректирован по оригиналам и
другим переводам.)
В конце «Фрагментов» приведен небольшой чудом уцелевший текст -
трактат «О седмицах» [Лебедев, гл. 66а], написанный вероятно около -600
25
года, т.е. самый древний греческий прозаический текст. Он описывает
строение мира и притом интересно: мир у него имеет семь частей, Земля
тоже; сложен из семи частей и сам человек, и его голова. Есть и иные се-
мёрки. Будем называть автора трактата гептадором (семёрочником).
Но темы эволюции мы здесь не найдем. Это и понятно: поскольку все
мифы говорили о рождении мира, то естественно, что первые натуралисты
должны были данной темы избегать: желая заменить сказку рассказом, они
нуждались сперва в понимании того, как мир устроен. И у первого известно-
го по имени ученого (то был Фалес из Милета, главного греческого города
на эгейском берегу Малой Азии) нет темы эволюции. Правда, есть тема
первоначала - из чего всё состоит?
Фалес учил, что всё состоит из воды, и есть много разных мнений о том,
как это следует понимать. К нашей теме относятся такие понимания: “геоло-
гическое” («Выпадая в осадок и превращаясь в ил, вода обращается в зем-
лю») и “экологическое” («Фалес, утверждающий, что все рождается из воды,
говорит, что тела следует закапывать, дабы они могли разложиться во вла-
гу») [Лебедев, с. 110]. Иные полагают, что первоначалом у Фалеса была во-
все не обычная вода, а мировая душа (см.: Чайковский Ю.В. Основатели
Милетской школы И Диалог со временем. Альманах интеллектуальной исто-
рии. Выл. 2, М„ УРСС, 2000).
Идея мировой души (независимо от того, был ли ее автором Фалес)
очень важна (она вводит причастность активности человека к активности
мира). Она имела долгую историю и выходы как в религию, так и в науку. В
частности, многие философы полагали, что душа человека есть частичка
мировой души, получаемая, например, с первым вдохом. Вот ее самый ран-
ний след: «Пифагор, Анаксагор, Платон... и Клеанф утверждают, что ум
внедряется извне» [Столяров, с. 182].
Наоборот, Анаксимандр5 (ученик Фалеса, писавший около -550 года)
тему эволюции разработал. Его эволюционную схему изложил историк -I ве-
ка Диодор Сицилийский. При образовании мира
«воздух приобрел непрерывное движение, причем огнистая часть его стек-
лась в самые верхние места, поскольку подобной природе свойственно
устремляться вверх (по этой причине Солнце и прочие множества светил
были вовлечены во всеобщий вихрь), а илистая мутная часть... осела в одно и
то же место в силу тяжести. Непрерывно вращаясь вокруг своей оси и
сбиваясь в комок, она произвела из жидких частиц море, а из более твердых -
землю». «Когда же воссиял огонь Солнца, земля сперва затвердела, а затем,
-----поскольку от нагревания поверхность ее забродила,... возникли гнильцы,
Ранние эволюционисты (до Ч. Дарвина и его круга включительно) здесь и в гл. 2 выде-
лены полужирным курсивом. В остальной части книги им выделены формулировки.
Светлым курсивом по всей книге выделены смысловые акценты и напоминаемые из-
вестные понятия, тогда как прямым полужирным - понятия, используемые далее при
построении теории.
26
от нагревания поверхность ее забродила,... возникли гнильцы, покрытые тон-
кими оболочками, что и теперь еще наблюдается в топях и болотистых мес-
тах... Как только влажные вещества стали живородить от нагревания указан-
ным образом, [они] начали по ночам получать пищу из тумана,... а днем от-
вердевать от жара. Наконец, когда утробные зародыши, вынашиваемые [в пу-
зырях], выросли до зрелого состояния, обожженные оболочки растрескались,
и произошли всевозможные породы животных».
Вот еще свидетельство:
«По мнению Анаксимандра Милетского, из нагретой воды с землей возникли
то ли рыбы, то ли чрезвычайно похожие на рыб животные; в них сложились
люди, причем детеныши удерживались внутри вплоть до зрелости: лишь то-
гда те [утробы рыб] лопнули, и мужчины и женщины, уже способные про-
кормить себя, вышли наружу».
Прошу обратить внимание на подчеркнутые мною слова: первый эво-
люционист, кажется, избежал ловушки, в которую затем попадались едва ли
не все в течении двух тысяч лет: понял, что нельзя считать ныне живущих
рыб предками наземных существ. Они всего лишь близки.
Как видим, эволюционизм Анаксимандра - сквозной, от космоса до че-
ловека. Вот с кого надо бы начинать курсы истории эволюционной идеи!
Анаксимандр открыл грекам совсем новый мир. В вихрях этого мира рожда-
лись и умирали целые космические системы, вспыхивали и гасли мириады
звезд, рождались и вымирали виды животных. Причем всё происходило в
согласии с обыденным опытом - так же, как ветер крутит пыль, как огонь
рвется ввысь, как в гниющей луже появляются «черви», а из них мухи. Одно
обидно - нет и намека на то, что Анаксимандр сам что-то наблюдал. Рассу-
ждения его чисто умозрительны.
Но вскоре нашелся тот, кто подкреплял свои взгляды на историю мира
наблюдениями - уроженец Колофона (близ Милета) Ксенофан. Он более
полувека вел жизнь бродячего певца (рапсода), пока в старости (около -500
г.) его не приняла Элея - греческий город на юго-западе Италии. Ксенофан
высмеял прежних поэтов (Гомера, Гесиода) за наделение богов чертами
людей, в том числе - пороками. Если бы, говорил он, быки или львы могли
рисовать, они изображали бы своих богов похожими на быков или львов
Истины как таковой не существует, а есть лишь различные мнения.
Он мыслил Бога единым, пронизывающим природу, т.е. стал творцом
религиозного течения, именуемого пантеизмом. Разумеется, Бог, разлитый
в природе, ни произойти, ни сотворить природу не мог, т.е. оба мыслились
вечными. Зато Ксенофан говорил об эволюции Земли. По Ксенофану,
«из земли и воды вышла некая мешанина. Кажется, со временем земля осво-
бождается от влаги. Это, как он (Ксенофан - Ю.Ч.) говорил, видно из того,
что ракушки встречаются посреди суши и в горах; говорят, что в Сиракузах...
найдены отпечатки рыбы и тюленей; на острове Парос - отпечаток анчоуса в
глубине камня; и на Мальте - плоские камни с [отпечатками] всякой морской
27
всячины. Это, говорит он, образовалось, когда море все поглотило; затем отпе-
чатанное грязью высохло. Все люди гибли, когда земля, опускаясь в море, ста-
новилась грязью, и рождались вновь, являя такую перемену в каждом мире».
Тем, кто полагает, что для таких выводов не надо много ума, предложу
раздобыть ископаемую раковину и рассмотреть. Она выглядит как истинный
камень (каковым и является: органическое вещество давно замещено в ней
на минеральное), так что первое впечатление - что сходство с живым не
большее, чем, например, у облака, принявшего вдруг форму головы или
цветка. Недаром спор о природе ископаемых продолжался в европейской
науке 2300 лет - от Ксенофана до Ламарка. Много раз ученые, в том числе
и ведущие, объявляли ископаемые остатки растений и животных “игрой
природы”, причем последние примеры относятся уже к нашему времени (п.
9-15**); даже зная о биологической природе ископаемых костей и раковин,
ученый мир то и дело готов объявить “игрой природы” тот или иной образец,
не ложащийся в теорию.
У Анаксимандра и Ксенофана речь шла не о том постепенном превра-
щении одних организмов в другие, которое мы ныне именуем биологической
эволюцией, а об их внезапном появлении. Происхождение человека у Анак-
симандра напоминает выход бабочки из куколки (и, думаю, с него и скопи-
ровано). Но так рассуждали и 2 тыс лет спустя, и вряд ли можно додуматься
до эволюции, минуя данный этап.
Вот слова безвестного мыслителя, поражающие актуальностью. Их со-
хранил упомянутый Филон. В книге «О вечности мира» он писал:
“Тех, кто рассуждает о рождении и гибели мира, обычно вводят в заблужде-
ние четыре явления: неровности земли, отступание моря, разушение всех
элементов мироздания, гибель целых пород сухопутных животных. Для пер-
вого явления они приводят следующее объяснение... Если бы земля не воз-
никла однажды, на ней не были бы заметны никакие возвышенности: все го-
ры стали бы плоскими и все холмы сравнялись бы с поверхностью. При таком
количестве дождей, ежегодно от века выпадающих на землю, все возвышен-
ности непременно исчезли бы... так что всё повсюду имело бы ровный вид. А
сейчас повсеместные неровности и многочисленные горные вершины, дости-
гающие эфирной области, свидетельствуют, что земля существует не вечно”
[Столяров, с. 53-54].
В части 3 мы увидим, что данная мысль оказалась весьма важной при
обсуждении происхождения жизни и ее ранней эволюции. Кого мог цитиро-
вать Филон? Скорее всего, судя по уровню знаний, автор писал после Ксе-
нофана и мог принадлежать безвестному пифагорейцу.
1-5* Пифагорейцы, Гераклит и Эмпедокл
В начале -V века в Южной Италии, в философской школе Пифагора,
развивались все формы тогдашнего знания, причем пифагорейцы хранили
свои достижения в глубокой тайне, так что даже автора знаменитой “теоре-
28
мы Пифагора” мы на самом деле не знаем.
В работах школы пифагорейцев участвовали женщины, в остальной
Элладе уже изгнанные из общественной жизни (прежде роль женщин была
и в Греции весьма заметна, что видно из мифов). Участие женщин и внима-
ние к ним заметно влияло на философию пифагорейцев. Наоборот, полное
невнимание к ним в остальной Элладе во многом определило ущербный
характер греческой биологии, что мы увидим.
Пифагор претендовал на всеведение, за что его ненавидел его млад-
ший современник Гераклит. Вопреки Пифагору, видевшему мир как непод-
вижную гармонию, Гераклит во всем видел движение и потому иногда счи-
тается [Лункевич] как бы эволюционистом. В основе мира он видел огонь,
под которым понимал сразу и жгущее, и творческое начало, но ничего эво-
люционного у него нет. Организующим принципом был для него логос, т.е.
закономерность (и этим он близок ко второму ряду), а движущим - полемос
(вражда, война, спор, склока):
«Должно знать, что полемос общепринят, что он справедлив и что всё возни-
кает через вражду и заимообразно (в ущерб кому-то)» [Лебедев, с. 201].
Этим Гераклит проводил параллель естественных знаний с обществен-
ными, и мы вернемся к данной теме в начале части 2.
Среди ранних пифагорейцев известен врач Алкмеон, которому принад-
лежит первый подход к проблеме наследственности. Он первым из греков
догадался, что человек мыслит не сердцем, а головным мозгом. По его
мнению, сперма образуется в головном мозге как самом главном органе и
по сосудам (очень мало тогда изученным) попадает в половой орган. При
зачатии «воспроизводится пол того из родителей, от кого истекло больше
семени». Мысль для нас нелепа, но она впервые утверждала равное уча-
стие женщины в наследовании свойств организма6, от нее берет начало
идея комбинирования наследственных свойств.
Эту идею блестяще развил сицилийский врач и чародей, поэт и натур-
философ Эмпедокл, писавший в середине -V века. Он развивал учение Ал-
кмеона: «Во время соития часть сердца, очень маленькая и невосприни-
маемая чувствами, исходит от мужчины и часть - от женщины; так и со все-
ми частями тела». И провозглашал:
Но и другое тебе я поведаю: в мире сем тленном
Нет никакого рожденья, как нет и губительной смерти:
Есть лишь смешенье одно с различеньем того, что смешалось.
Что и зовут неразумно рождением темные люди.
& До этого греки простодушно видели в ней лишь почву, засеваемую мужским семенем.
Принцип чисто мужской наследственности не раз возрождался, препятствуя понима-
нию размножения и эволюции. Последний его рецидив имел место в середине XIX ве-
ка, когда прорастание пыльцы на пестике цветка (открыто в 1822 г.) трактовали как
прорастание семени в почве [Баранов, с. 278-281].
29
Значит, Эмпедокл додумался до той мысли, что материя неуничтожима и
что видимые появления и исчезновения тел являются всего лишь измене-
ниями комбинаций одних и тех же частиц (следующее поколение мыслите-
лей назвало их атомами). Но почему их смешение приводит не к бестолко-
вой мешанине, а к той природе, где есть столь многое, в том числе и люди?
Потому, учил Эмпедокл, что подходящее соединяется мировой Любовью
(как у Гесиода). В разные эпохи соотношение любви и ненависти различно,
и в наше (эмпедоклово) время доля любви убывает, но еще превышает до-
лю ненависти. Именно силой любви складывались в целое те первые орга-
низмы, которые были порождены землей.
По Эмпедоклу, «Сначала из как бы беременной земли там и сям роди-
лись отдельные члены, затем они срослись и образовали естество цельного
человека»,
Выросло много голов, затылка лишенных и шеи,
Голые руки блуждали, не знавшие плеч, одиноко
Очи скитались по свету без лбов, им ныне присущих.
А вот другое место: «По словам Эмпедокла, люди родились из земли,
как лебеда». То есть в земле, словно в матке, могут вырастать части тела.
Это уже шаг назад от Алкмеона. Похоже на Анаксимандра, однако у Эмпе-
докла нет анаксимандровой идеи преобразования одного типа организма в
другой. Человек произошел, по Эмпедоклу, путем комбинирования частей,
но не от другого животного, а прямо из земли.
1-6. От софистов до Аристотеля
Пусть рассуждения ранних философов и поражают нас иногда своей
наивностью, но нельзя отрицать, что это был поиск истины. Наоборот, с пе-
ремещением в середине -V века философии в Афины на полвека возобла-
дала иная линия: подбирать доводы в пользу наперед заданного утвержде-
ния. Проводили ее софисты (т.е. “мудрствующие”) - так именовали плат-
ных учителей философии, учивших юношей добиваться положения в обще-
стве. От них пошло словечко “софизм”, означающее нечто похожее на ис-
тину. (Пример: софизм “Рогатый”. То чего ты не терял, то твоё - верно?
Верно. Рогов ты не терял, значит ты рогат.) Вместо философии на полвека
воцарилась риторика - умение красиво строить речь. Такому повороту дел
была историческая причина (росла публичность управления и права), но
для науки настали тяжкие времена - ее стали презирать.
Первый афинский натурфилософ Анаксагор (младший современник
Эмпедокла) пытался рассуждать о чем-то вроде частиц наследственно-
сти. Он учил, что все тела состоят из «семян всех вещей», так что с пищей
(в том числе и растительной) человек получает мельчайшие частички всего
того, из чего сам состоит, - мяса, костей, волос и всего прочего. Правда, ему
еще не приходило в голову, что частицы живого должны чем-то качественно
отличаться от частиц неживой материи, зато он впервые заявил, что части-
30
ца может содержать в себе все свойства целого, а эта мысль была необхо-
димой для рождения идеи наследственных частиц. Анаксагор рассуждал о
строении мира и многом другом. Мир, по его мнению, был устроен неким
Умом, приведшим в порядок мешанину первичных частиц. Подробнее см.
[Лункевич].
Это навело на него гнев афинского общества, оформленный софистами
в гневные обличительные речи. Ему удалось бежать из Афин, однако лет
через 30 те же обвинения были брошены софистами Сократу и привели к
его казни (поскольку бежать он отказался). Хотя Сократ не сказал ничего,
прямо относящегося к эволюции, он для нашей темы весьма важен как ос-
нователь нового способа толковать природу.
Во-первых, произвольным речам натурфилософов и корыстной ритори-
ке софистов он противопоставил важнейший вопрос: “Откуда мы это зна-
ем?”, чем породил критическое направление в философии.
(Он пытался давать всем понятиям строгое определение и потерпел в
этом неудачу. До сих пор многие видят исходный пункт исследования в даче
определений, хотя от этого отказались уже ученики Сократа.)
Он первый исследовал причинность как таковую и пришел к важнейше-
му выводу: каждому факту можно дать два объяснения - механическое и
целевое. В тюрьме он пояснил это ученикам на примере учения Анаксагора,
которым увлекался в молодости. Читая, Сократ увидал,
«что Умом он не пользуется вовсе и не указывает настоящих причин упоря-
доченности вещей, а ссылается на всякие там воздухи, эфиры, воды и множе-
ство других нелепых вещей». «Это всё равно, как если бы кто сперва объявил,
что всеми своими действиями Сократ обязан Уму, а потом, принявшись объ-
яснять причины каждого из них в отдельности, сказал: ... “так как кости сво-
бодно ходят в своих суставах, сухожилия, растягиваясь и напрягаясь, позво-
ляют Сократу сгибать ноги и руки, вот по этой-то причине он и сидит здесь”
... пренебрегши истинными причинами - тем, что раз уж афиняне почли за
лучшее меня осудить, я в свою очередь счел за лучшее сидеть здесь» (Пла-
тон. Федон, 98 b-d).
А во-вторых, Сократ дал свое понимание мира как разумно устроенного
богами для блага человека. Например:
"... как заботливо боги уготовали всё, в чем люди нуждаются”; “прежде всего,
нам нужен свет и... боги даруют нам его... Затем, нам нужен отдых, и боги
даруют нам ночь... а ночью не так ясно всё видно, не потому ли боги зажгли
ночью звёзды, которые показывают нам время ночи?... боги производят нам
из земли пищу, которая нам нужна, и даруют подходящие для этого времена
года... Да разве не очевидно... что и животные рождаются и вскармливаются
ради людей? ” (Ксенофонт. Воспоминания о Сократе, кн. 4, гл. 3).
Тем самым, творчество Сократа явилось рубежом в познании мира. У
Сократа мир устроен благим началом, и надо понять его замысел, чтобы
действовать разумно и нравственно. От него эта мысль перешла в христи-
31
анское богословие, где породила естественное богословие, о котором
речь далее. А оттуда - в нашу науку, где легла в основу дарвинизма и “ан-
тропного принципа” космологии (см. гл. 7, а также: Чайковский Ю.В. Антич-
ная философия как общеобразовательный предмет И ВФ, 2002, № 9). Есте-
ственное богословие формально отвергнуто нынешней биологией (факти-
чески же, как мы увидим, продолжает царить - в дарвинизме), но для -400
года такие мысли были огромным достижением.
Сократ вырос среди софистов, и стиль его бесед с учениками во многом
был софистическим: его вопросы ставили в тупик слушателей. Однако Со-
крат видимо не задавал каверзных вопросов ради позы и самоутверждения,
он задавал их и самому себе. Он отказался бежать из тюрьмы, был казнен и
тем (вот парадокс) одержал над софистами сокрушительную победу: все
возникшие вскоре философские школы были в какой-то мере сократовски-
ми, а о софистах более не слышно несколько веков.
1-6* После Сократа
Самой знаменитой сократовской школой была платонова Академия, и
единственным в Античности, у кого можно найти идею постепенного изме-
нения организмов, был Платон. Но до чего странная то была эволюция!
Подробно описав в конце диалога «Тимей», как боги творили человека
(мужчину), он заявил, что люди, жившие недостойно, в следующем поколе-
нии рождались женщинами, что и побудило богов создать обоим полам ме-
ханизм размножения. Далее,
«дать начало племени птиц пришлось мужам незлобивым, однако легкомыс-
ленным, а именно таким, которые любили умствовать о том, что находится над
землей, но в простоте душевной полагали, будто наивысшая достоверность в
таких вопросах принадлежит зрению» (а не размышлению). «А вот племя сухо-
путных животных произошло из тех, кто был вовсе чужд философии». Еще бо-
лее глупые обратились в пресмыкающихся, а совсем тупые - в водных живот-
ных. Так и повелось, что живые существа «перерождаются друг в друга, меняя
облик по мере убывания или возрастания своего ума или глупости».
Ирония этой аллегории вроде бы очевидна, и она не касалась бы на-
шей (биологической) темы, если бы не два обстоятельства. Во-первых, Пла-
тон повторяет Анаксимандра полным отсутствием идей экологии (растения
даже не упоминаются), чем уступает Библии, где растения созданы в пищу
животным. А во-вторых, размышляя о возможности превращения одних су-
ществ в другие, Платон дал (в диалоге «Государство») печально знамени-
тое предложение - идеальное государство должно вывести породу идеаль-
ных людей путем отбора. Предложению суждена была долгая жизнь Нико-
му не удалось улучшить людей, но замучали многих (см. далее о евгенике).
До идеи постепенного усложнения организмов никто в древности не
додумался. Платон, подробно описав создание мира, всюду говорил об од-
нократных актах творения или переделки; после них шло лишь повторение
32
и ухудшение (деградация). Акты творения совершал Демиург - этим сло-
вом, буквально означающим “мастер, ремесленник”, Платон обозначил
старшего из богов, творившего мир. Очень любопытна та роль, которая от-
ведена Демиургу. Описав творение космоса, Платон задал вопрос:
“взирая на какой первообраз работал тот, кто его устроял, - на... неизменный
или на имевший возникновение? Если космос прекрасен, а его демиург благ,
ясно, что он взирал на вечное. ... Возникши таким, космос был создан по тож-
дественному и неизменному [образцу], постижимому с помощью рассудка и
разума” (Тимей, 29а).
Тут сформулирован главный пункт философии Платона - признание
неизменного мира идей, с которого Демиург лишь считывал вечные истины
мироздания. Это по сути то же самое, что соломонова «Мудрость», бывшая
при творении “художницею” мира (п. 1-3*). Есть и новшество: люди способ-
ны и обязаны познавать мир идей.
Всякая ли вещь имеет идею? В диалоге «Парменид» на вопрос, есть ли
идеи у всякого сора и грязи, платоновский Сократ с некоторым колебанием
отвечает: «Вовсе нет, я полагаю, что такие вещи только таковы, какими мы
их видим. Предположить для них существование идеи было бы слишком
странно» (Парменид, 130 cd). Вернемся к этому в п. 6-15*.
Как и в Библии, у Платона женщины произошли от мужчин. Это против
более ранних религий, где мужчина и женщина сотворены одновременно,
но вполне соответствует эпохе зрелой Античности, когда завершился про-
цесс изгнания женщин из общественной жизни. Есть и важное новшество:
женщина происходит от мужчины не материально (из ребра), а духовно -
путем переселения души, недостойной снова жить в мужском облике.
После Платона пришло понимание того, что из эволюции земных толщ
эволюция организмов сама по себе не следует. Например, великий Аристо-
тель (середина -IV века), воспитанник философской школы Платона и круп-
нейший ученый-энциклопедист древности, эволюцию земных толщ призна-
вал, а организмов - нет.
Зато именно Аристотель впервые задумался о том, как связаны причи-
на и следствие, и поныне мы пользуемся его пониманием причинности. По
Аристотелю, для всякого объекта существует четыре типа причин:
материальная - из чего объект возник (создан),
формальная - форма, или сущность возникающего объекта,
действующая - откуда идет изменение или образование объекта,
целевая - ради чего возникает или существует объект.
Аристотель привёл примеры, вот самый простой: дом сделан из камня и
дерева - вот его материальная причина; мастер-строитель придал мате-
риалу форму дома - эта форма есть его сущность, или формальная причи-
на; деятельность строителей - его действующая (движущая) причина; а на-
значение дома - его целевая причина. Однако в разных сочинениях Аристо-
тель понимал 4 причины различно, и нам надо изложить то понимание, ко-
33
торое нам понадобится далее.
Легко видеть, что причины делятся у Аристотеля на две группы - пер-
вая и третья предметны, а вторая и четвертая понятийны. Предметные
причины просты и наглядны, тогда как понятийные вызывают массу вопро-
сов и порождают споры. Прежде всего, чем замысел (форма, сущность) от-
личается от цели (назначения)? Вопрос труден и неоднозначен, так что
здесь замечу лишь, что дом проектирует архитектор, не обязательно знаю-
щий его будущее назначение, а цель определяет владелец, не обязанный
понимать в строительстве.
Далее, почему цель отнесена к причинам? Ведь обычно мы противопо-
лагаем причину и цель. Тут надо понимать мировоззрение Аристотеля - для
него всё в мире осмысленно, ничего без цели не бывает, а значит, цель так
же служит причиной, как и форма. Однако форма всегда принадлежит са-
мому предмету, тогда как цель может быть и вне предмета (назначение до-
ма определяет заказчик). Человек может ставить себе цель, а Бог ставит
ее себе всегда сам. «Бог есть деятельность» (Метафизика, 1072 Ь 27).
Что касается организмов, то Аристотель видел всего две причины их
появления и существования: материальную и осуществляющую. Осуществ-
ление (энтелехия) являет собой самую важную и трудную часть его учения
о причинах: она может выступать и как форма (самка, по Аристотелю, дает
для потомства материю, а самец - форму), и как действие (душа строит для
себя тело в ходе развития зародыша и ребенка), и как назначение (тело су-
ществует ради души). Всё это, надеюсь, станет понятнее вдумчивым чита-
телям, когда мы займемся в главах 3 и 7 энтелехией в понимании ученых
нашего времени.
Аристотель не был первым, кто исследовал животных (назову хотя бы
Алкмеона и Демокрита), но именно он известен как «отец зоологии», по-
скольку его труды по зоологии - древнейшие, какие до нас дошли. Его «Воз-
никновение животных» - первая книга по эмбриологии, а «О частях живот-
ных» - первая по сравнительной анатомии. Он выявил крупные группы жи-
вотных (звери, птицы, рыбы и т.д.) и выстроил их в ряд, от самых сложных к
самым простым (первый вариант лестницы существ). В отличие от нас, он
не видел в этом свидетельств эволюции.
Наиболее известен его трактат «Исследование животных». Это назва-
ние чаще переводят как «История животных», что неверно: никакой истории
там нет, а есть описание всех известных ему видов животных (около 500); у
греков слово хисториа означало как раз «исследование». Здесь много дан-
ных об изменчивости животных, в том числе и диковинной. Так, дано опи-
сание козла, дававшего молоко, свиньи, имевшей по одному копыту на ноге
и т.п. А в труде «О возникновении животных» Аристотель привел пример,
объяснить который непросто и нам - о женщине, жившей с эфиопом: «Не
дочь ее, а сын ее дочери родился эфиопом», т.е. темнокожим. (Ныне такие
факты объясняют, не очень убедительно, либо плазматической наследст-
34
венностью, либо активацией молчавшего гена.)
1-6** Существенное и побочное (сущность и акциденция)
Для нашей темы наиболее важен трактат «О частях животных». Здесь
Аристотель ясно выразил идею единства строения животных, едва наме-
ченную у Анаксимандра (в Новое время ее назвали теорией единого плана
строения): рыбы, звери и люди устроены сходно. В этом Аристотель усмот-
рел общую сущность данных организмов.
Ныне это всем известно, но мало кто знает, что Аристотель шел много
дальше: отметив (в гл. 9 Книги 4) сходство «фигуры тела» четвероногих и
насекомых, он смело заключил: моллюски тоже устроены так же, но только
если их тело мысленно переломить пополам - так, чтобы рот и задний про-
ход оказались рядом. Как увидим в п. 2-6, этой конструкции суждена была
долгая жизнь и большое влияние на эволюционную науку.
Трактат явился первым опытом классификации животных. Описывая
общие принципы классификации, Аристотель выразил уверенность (в главе
3 книги 1), что «следует производить разделение по тому, что относится к
сущности, а не является само по себе привходящим», т.е. побочным (акци-
денцией). Например, кита надо отнести не к рыбам, а к зверям.
Это важнейшее положение еще помнил в XVIII веке Карл Линней, вели-
кий систематик, но впоследствии оно было забыто, и до сих пор многие
биологи ведут бесцельные споры о конкретных системах организмов, не за-
думываясь ни о существенности признаков, по которым ведется разделение
на группы, ни о сущности самих методов классификации как о чем-то лиш-
нем. Мы займемся этими вопросами в главе 10.
Дело в том, что дарвинизм зиждется на неумении различить сущ-
ность и акциденцию, хотя это умел еще Аристотель. В самом деле, идея
выводить эволюцию из анализа изменчивости осмысленна только тогда, ко-
гда доказано, что достаточно регулярно изменения бывают сущностными;
однако ни один пример изменчивости, приведенный Дарвином и после него,
не затрагивает сути организации живого. Мы вернемся к этому еще не раз.
1-7. Изменчивость, отбор и законы познания
Аристотель был уверен в существовании самозарождения, т.е. в том,
что организмы могут возникать помимо процесса размножения (как, по
Анаксимандру, возникли в жидкой грязи первые животные). Естественно,
что, признавая самозарождение любых животных и растений, нет нужды за-
думываться, возможно ли преобразование одних в другие.
Удивляться господству идеи самозарождения не приходится: с одной
стороны, сам процесс размножения был известен плохо, а с другой, господ-
ствовал такой взгляд на строение веществ, который прямо-таки подталки-
вал к этой идее. А именно, все тела мыслились состоящими всего из четы-
рех стихий, т.е превоэлементов (земля, вода, воздух и огонь), причем тече-
35
ние процессов (в том числе и жизненных) осуществлялось, согласно древ-
ней науке, всего двумя парами оппозиций (противоположных качеств) среды
- парой тепло-холод и парой сухое-влажное. При таком понимании мира
легко было допустить, что в почве могут рождаться сами собой любые орга-
низмы. Другими словами, в такой науке о живом принцип наследственности
не был обязательным.
После Анаксагора прежнее представление о мужском и женском семе-
ни, смесь которых определяет и пол ребенка, и его свойства, можно было
конкретизировать: каждая часть тела посылает свою частичку в половой ор-
ган, где они вместе и образуют “семенную жидкость”. Все свойства ребенка,
в том числе и пол, можно было теперь трактовать просто: от кого из родите-
лей данная семенная частичка попала в зародыш, на того ребенок и похож
данным свойством.
Эту мысль высказывали многие, а наиболее четко - великий врач Гип-
пократ (около -400 г.). При этом взаимоподгонку частичек в единый зародыш
можно было понимать как итог эмпедоклова принципа любви. При этом
очень слабое у Эмпедокла место - «из... земли родились отдельные члены,
затем они срослись» - теперь заменялось на гораздо более сильное: со-
единяться должны не органы, а крохотные частицы живых тел, ответствен-
ные за передачу свойств.
Как ни странно, такое понимание наследственности, важное достиже-
ние античной «генетики», Аристотель подверг убийственной критике. Сде-
лать это было нетрудно - опровергающие примеры лежат на поверхности.
Однако при этом была упущена одна глубокая мысль: наследственные час-
тицы могут по-разному сочетаться, поэтому наследстванность носит комби-
наторный характер. Будучи отвергнуто авторитетом Аристотеля, это мне-
ние, хотя и было известно из трудов Гиппократа, прозябало на задворках
науки вплоть до XVIII века.
У Гиппократа есть интересное описание изменчивости, точнее, того, что
в наше время назвали наследованием приобретенного свойства (НПС):
обычай спеленывать головы младенцам, известный у одного народа, при-
вел к появлению голов характерной формы, которая несколько поколений
сохранялась после отмирания обычая спеленывать (это пример длитель-
ной модификации - см. п. 6-10*). Однако никому тогда не пришло в голову
счесть всё это материалом для появления новых форм организмов, как при-
ходит нам. Зато таковой материал при желании можно увидеть у Эмпедокла
в его «смешении» частей, при котором сила Любви случайным образом со-
единяет порознь выросшие члены. Эмпедокл (лет за 60-70 до Гиппократа)
писал:
Множество стало рождаться двуликих существ и двугрудых,
Твари бычачьей породы с лицом человека являлись...
Женской природы мужчины, с бесплодными членами твари.
Эмпедокл понимал, что одной любви мало, что даже прекрасно собран-
36
ное ею может оказаться ни на что не годным. И он объявил: ни на что не
годные гибли, а годные оставались жить, т.е. происходил (если использо-
вать нынешний термин) их отбор. Эдакий античный дарвинизм.
Первый смутный намек на отбор мы видим уже в самом древнем тексте
- в упомянутом выше трактате «О седмицах». Безвестный милетец (гепта-
дор) писал о частях человека:
«При благоразумии они существуют без страданий; и те, которые всё делают
надлежащим образом, те в течение всей жизни бывают здоровы, полны бод-
рости и живут достаточно. А те, что плохо составлены по ошибкам заботяще-
гося (curantis), наталкиваются на такие тяжкие страдания и такие испытания
своих сил, каких не ожидали», и умирают.
В дни Анаксимандра на другом берегу Эгейского моря, в Мегаре (к се-
веро-западу от Афин) жил другой “дарвинист” - поэт-аристократ Феогнид.
Смена власти аристократов на власть демократов привела его к уверенно-
сти в безнадежной порче человеческой природы - за счет необдуманных
браков лучших с худшими. Вот место из «Элегий» Феогнида:
...Выбираем себе лошадей мы, ослов и баранов
Доброй породы, следим, чтобы давали приплод
Лучшие пары. А замуж ничуть не колеблется лучший
Низкую женщину брать, только б с деньгами была!
Женщина также охотно выходит за низкого мужа,
Был бы богат! Для нее это важнее всего. (...)
Полипаид, не дивись же тому, что порода сограждан
Всё ухудшается: кровь перемешалася в ней.
Через 200 лет именно эту мысль Платон решил обернуть к пользе об-
щества. В утопических мечтах об идеальном государстве он много внима-
ния уделял вопросу создания и сохранения прослойки избранных, которые
могли бы достойно управлять всеми остальными людьми. В диалоге «Госу-
дарство» он прямо заявлял, что
«лучшие мужчины должны большей частью соединяться с лучшими женщи-
нами, а худшие, напротив, с самыми худшими и что потомство лучших... сле-
дует воспитывать, а потомство худших - нет, раз наше стадо должно быть от-
борным».
В XX веке, когда родилась евгеника (идеология улучшения людей с по-
мощью искусственного отбора), Платона признали первым ее теоретиком, и
многие считают до сих пор. Мы вернемся к этому в пп. 4-4 и 10-1, а здесь
отмечу: античная евгеника показывает, сколь хорошо греки владели селек-
цией животных; а из мифов видно, что им также был хорошо известен ин-
цест (браки братьев с сестрами и родителей с детьми), резко осуждаемый
среди людей, но почему-то обычный среди богов (см. п. 9-16*).
Лучше всего наследственность была изучена на домашних растениях.
«Отцом ботаники» признан Теофраст, знаменитый ученик Аристотеля. У не-
го есть много описаний изменений растений под влиянием заботы человека
37
или перемены климата, а также взятых у учителя примеров резких измене-
ний животных (для нас они, по большей части, наивны: например, уверение,
что ястреб-перепелятник на лето превращается в кукушку). В «Исследова-
нии растений» (его тоже часто переводят: «История растений») Теофраст
сравнил их с превращением личинки в насекомое, но, в отличие от Анакси-
мандра, не увидал тут эволюции. И все-таки Теофраст занял достойное ме-
сто в истории эволюционизма - как увидим, через 1500 лет его примеры
были переосмыслены в догадки о происхождении видов.
1-7* Ранние стоики
Во время старости Теофраста, после -300 года, в Афинах открылась
последняя из знаменитых философских школ - школа стоиков. В изучении
природы она дала мало нового и для нашей темы важна своим интересом к
познанию и творчеству. Основатель ее Зенон Китийский7 учил, что
“есть два вида огня: нетворческий (атэхнон), который превращает в себя
свою пищу, и творческий (тэхникон), который умножает и сберегает все,
присутствует в растениях и животных и, таким образом, является природой и
душой”; что “бог пронизывает все вещество и на одном уровне выступает как
ум, на другом - как душа, на третьем - как природа, на четвертом - как
структурное единство” [Столяров, с. 61. 72].
Творческий огонь стоики называли также пневмой. Зенон ввел еще по-
нятие сперматический логос для переосмысления того, что Анаксагор на-
зывал «семенами всех вещей»: новое понятие обозначало не саму мате-
рию, а «порождающую способность всякой первичной пневматической
структуры». «Начало всего, по Зенону, природный и божественный закон»
[Столяров, с. 52, 73].
Эволюции организмов у стоиков нет, но есть нечто более общее. Так,
Клеанф, ученик Зенона, видел причиной всего напряжение:
“заключенное в мировом веществе напряжение (топос) того начала, которое
вечно творит это круговращение и этот распорядок, никогда не прекращается.
Подобно тому как все части чего-то одного в надлежащее время рождаются
из семени, так и части мироздания, к которым относятся существующие жи-
вотные и растения, вырастают в надлежащее время” [Столяров, с. 174].
Здесь нам важна параллель сил, порождающих мироздание и организм.
Ранние стоики прославились еще тем, что ввели понятие: каталепсис - ин-
туитивное ухватывание новой мысли как чего-то целостного [Столяров, с.
25-40]. И тонос, и каталепсис суть разные формы активности, которая тут
впервые выступает в качества порождающей способности. Ее результатом
являются наблюдаемые в мире уровни упорядоченности.
Дарвинизм чужд всему этому кругу мыслей, но для ламаркизма она ис-
ходна, хотя сам Ламарк и не мог знать идей стоиков, поскольку их воззрения
7
Не путать с Зеноном Элейским, автором апорий, жившим лет на 150 раньше.
38
были открыты историками науки позже
Итак, диалектика Платона и логика Аристотеля (общеизвестные основы
западной философии) были дополнены интуицией (что, к сожалению, оста-
валось почти неизвестным до конца XIX века). Нам это понадобится при
анализе эволюционного способа мышления в п. 3-7.
1-8. Рождение идеи предначертанной эволюции
Больше ничего оригинального по нашей теме в рамках античного есте-
ствознания сказано не было. Эволюцию тогда не увидали.
Это может звучать странно - ведь популяризаторы, например, часто
называют великим эволюционистом римского поэта Лукреция (-I век). Одна-
ко внимательные историки всегда признавали, что Лукреций не сказал по
сути о появлении новых организмов ничего сверх Эмпедокла, он лишь под-
робно и талантливо пересказал идею о гибели тех, над кем недостаточно
потрудились любовь и случай. Происхождения одних организмов из других у
него, как и у Эмпедокла, нет. Зато Лукреций смог немного продвинуться в
том, что в XX веке назвали генетикой - он подметил нечто, сходное с рецес-
сивностью:
Может случаться и так, что дети порою бывают
С дедами схожи лицом и на прадедов часто походят.
Ибо нередко отцы в своем собственном теле скрывают
Множество первоначал в смешении многообразном...
Это изумительно как наблюдение, но тут нет и намека на идею истори-
ческого изменения организмов. Подробнее см.: [Берг, с. 49-59].
Раннее христианство и ранний ислам много потрудились, преследуя
«языческую» мудрость и ее носителей. Именно в раннем Средневековьи по-
гибла основная масса античных книг. Оригинальные мысли о живом были
редки крайне. Но тем они для нас интересней.
В IV веке в Каппадокии (область в Малой Азии) был знаменит епископ
Григорий Нисский, признанный позже как один из отцов Восточной церкви.
Он один из немногих в то время отстаивал право научного исследования бо-
гословских вопросов. К ним он относил и толкование творения, изложенного
в Книге Бытия; в труде «Толкование на шестоднев» (на 6 дней творения) он
развил свое понимание развития мира. Вот как описал его историк филосо-
фии Г.Г. Майоров:
«Все вещи и события были созданы богом сразу, в едином вневременном ак-
те, однако только в потенциальной форме, в форме... семян, содержащих в се-
бе скрытую энергию и, так сказать, программу будущего развития. Затем все
это множество семян последовательно, каждое в свое время, естественным
путем и без дополнительного вмешательства бога развивается во все явления,
которые составляли, составляют и будут составлять мир» (Майоров Г.Г. Фор-
мирование средневеков. философии. М., Мысль, 1979).
Это уже близко к эволюции. Легко видеть здесь переосмысление идеи «се-
39
мян всех вещей» Анаксагора и «сперматических логосов» стоиков, а также
развитие идеи Аристотеля об энтелехии (переходе возможности в действи-
тельность). Но нам важнее другое: Бог мыслится здесь не как творец каждой
вещи, а как творец программ, по которым вещи появляются в свой срок и по
своим законам. Как творец предначертанной эволюции.
Впоследствии христианские богословы назвали это миропонимание
теорией вторичных причин, нам же будет удобнее еще более поздний
(XVII век) и более короткий термин - деизм (от лат. Deus - Бог). Он упот-
ребляется для обозначения всех религий и идеологий, в которых Бог созда-
ет законы природы и далее не вмешивается в ход событий. Деистическая
позиция давала возможность верующим ученым XVII века и позже изучать
законы природы. Конечно, сам Григорий не мог видеть себя деистом, но
элемент деизма у него налицо.
Противоположное понимание Бога - как творца каждой вещи (непо-
средственно или через существ, творящих каждый раз по его воле) известно
как теизм (от греч. theos - бог). Это различение (связанное с отличием ха-
рактера латинских мифов от греческих) нам не раз понадобится. Теизм гос-
подствовал в средневековом богословии, и принято считать, что переход
ученых от теизма к деизму в науке Нового времени был шагом к атеизму,
т.е. к отказу от религии.
Около 420 года мысль Григория повторил блаженный Августин, тогда -
епископ в городе Гиппон (север Африки), главный из отцов Западной церк-
ви, в книге «О граде Божием». По-гречески он не читал, но о трудах Григо-
рия знал и тоже, по словам Майорова, видел в шести днях творения «не
временную, а логическую последовательность»; «Как в семенах содержатся
все будущие растения, так и материя мира с самого начала содержит в себе
все, что она может когда-либо произвести на свет». Именно Августин и пе-
ренес эту идею Григория в западный мир.
Августину же принадлежит мысль, что развитие природы спланировано
Богом при творении до конца, что шесть дней творения соответствуют шес-
ти историческим эпохам, и мы живем в шестую, а потому должны готовиться
к концу света.
Читателю, может быть, читать это странно: разве эволюция - это раз-
ворачивание раз и навсегда заданных возможностей? Нет, разумеется, мы
называем эволюцией иное (см. п. 1-2), но первоначально смысл слова зна-
чил именно разворачивание потенций (латинское evolutio означает «разма-
тывание» - например, нити).
В наши дни эволюция не мыслится без приспособления к среде обита-
ния (а дарвинизм даже выводит всю эволюцию нацело из одной-
единственной идеи приспособления), но это не мешает большинству ны-
нешних дарвинистов рассматривать конкретную эволюцию как включение и
выключение генов, а не как создание чего-то принципиально нового. Поэто-
му мы далее не раз вспомним точку зрения Григория Нисского.
40
1-9. Богословы, мореплаватели и эволюция
С крушением греко-римской культуры античная наука погибла. Возник-
нув снова, наука долго витала в рамках толкования Писания. Лишь в XII ве-
ке французский философ и богослов Пьер Абеляр заявил публично, что Пи-
сание может содержать ошибки. Он не отрицал богодухновенности священ-
ных текстов, однако обращал внимание на то, что между небесным автором
и земным читателем неминуемо встает «брат третий» в лице писателей и
переписчиков, искажающих истину. По Абеляру, в случае противоречия ме-
жду священными текстами надо искать истину средствами разума. Естест-
венно, он подвергся церковным гонениям.
Абеляр продолжил шедшую от Филона традицию толкования Библии,
положил начало ее филологическому анализу, сильно укрепил позицию
принципа «Понимаю, чтобы верить», ставшего впоследствии позицией уче-
ных-натуралистов Возрождения [Рассел].
Наиболее известен в плане принципа «Понимаю, чтобы верить» знаме-
нитый богослов и ученый Альберт Великий, монах-доминиканец (XIII век).
Он сделал важный шаг к идее эволюции: отметил самопроизвольную из-
менчивость как способ перехода к иному виду растений: примеры, когда-то
приведенные Теофрастом, он охарактеризовал как трансмутацию одного
вида в другой (термин он взял из алхимии) [Лункевич].
Альберта восхищенные современники именовали Doctor universalis
(всеобщий, целостный), а его современник, монах-францисканец5 Роджер
Бэкон заслужил эпитет Doctor mirabilis (удивительный). Он блестяще про-
должил линию Абеляра по разграничению роли религии и науки в познании
мира и потому значительную часть жизни провел в тюрьмах. Много раз-
мышлял о природе и вере, о методах их познания и пришел к выводу о не-
обходимости различных религий. По его мнению,
“Ничто так не радует природу и волю, как то, что ведет к разнообразию
(diversitas), но причины этого неизвестны” (Bacon R. Opus Majus, IV, 2, 2).
О причинах разнообразия мы и теперь знаем немногим больше. Гово-
рил Роджер и про «умножение видов» (multiplicatio speciorum), но неясно,
имел ли он в виду под видом то же, что и мы. (На эти места из Р. Бэкона
мое внимание любезно обратил историк науки католик И.В. Лупандин.)
Лишь через 500 лет после Абеляра, в либеральной Голландии Борух
Спиноза решился пойти дальше него: заявил, что «не нужно приспосабли-
вать ни Писание к разуму, ни разум к писанию». Но за это еврейская общи-
на отлучила его и изгнала [Лункевич].
О
° В монашеских орденах св. Доминика и св. Франциска состояли почти все известные в
ХШ в. западные ученые. Существенно, что эти ордена были опорой инквизиции: заби-
рая имущество еретиков, они безмерно обогатились, и только в их монастырях процве-
тала наука [Рассел].
41
В XVI веке снова был открыт мир ископаемых организмов, причем лишь
к концу XVII века утвердилась мысль, что они - не «игра природы», не камни
в форме костей или раковин (рис. 2), а остатки древних животных. При этом
никто долгое время не замечал, что среди ископаемых есть формы, отсут-
ствующие в мире нынешних организмов. Это было и невозможно, пока не
был инвентаризован мир организмов нынешних, что было вчерне сделано
Рис. 2. а): “Два рыбоподобных камня” - из книги “Музей металлов" (Улисс Альдрован-
ди, умер в 1605 г.), опубликованной в 1648 г. б): Здесь наоборот: дендрит (проросшая в рас-
щелине скалы кристаллизованная соль) описан как ископаемое растение - из книги “Допо-
топный гербарий” (Иоганн Шойхцер, 1709)
лишь к началу XIX века.
И вдруг эволюционная тематика стала актуальной: мореплаватели об-
наружили в Новом свете неизвестных дотоле животных и неведомую расу
людей, так что встал вопрос: как они пережили Всемирный потоп?
«Удивительно: проблема происхождения впервые поставлена богословами, а
не биологами. Благодаря географическим открытиям, с 1492 года передовые
люди узнавали о большом числе дотоле неизвестных видов животных, так что
старая проблема - как эти животные могли уместиться в Ноевом ковчеге, -
вновь оказалась в центре внимания. Было найдено объяснение: до потопа су-
ществовали не все известные нам сегодня формы, но только ограниченное
число основных групп. Лишь после потопа виды развились из этих групп по
аналогии с тем, как развиваются домашние животные. Так, родилось допуще-
ние, что многие виды диких оленей и диких быков - не настоящие виды, а ва-
42
риации нескольких архетипов9» (Wichler G. Charles Darwin the founder of
evolution and natural selection. Oxford, 1961, p. 3).
Историк науки Джерард Вичлер сослался при этом на работу 1559 года
«Ноев ковчег, его форма и вместимость» Иоганна Бутео, который вычис-
лил, что ковчег не мог вместить все виды известных животных. Замечу, что
этот тезис, по сути эволюционный, напечан ровно за 300 лет до книги Дар-
вина и что далее эволюционная традиция никогда не прерывалась (в том
смысле, что видно знакомство авторов с идеями предшественников), хотя
временами связующая нить становилась столь тонка, что оставалась неза-
метной для современников.
Удивление Вичлера нуждается в пояснении: 370 лет - с того года, когда
Колумб водил напоказ по улицам кастильских городов пленных индейцев, и
до отмены рабства в США после победы Севера в Гражданской войне, за-
падная наука то и дело возвращалась к вопросу о том, являются ли все лю-
ди единым биологическим видом. Научные доводы не удавалось отделить
от эмоций чисто грабительских: завоевателям выгодно было счесть индей-
цев и негров не людьми, а говорящим скотом, и, соответственно, в науке
появилось течение мысли (позже оно получило имя: полигенизм), подби-
равшее аргументы в пользу того, что людей следует относить к нескольким
видам.
Булла римского папы, объявившая в 1512 году всех людей потомками
Адама, долгое время не имела никакого влияния на ход дел в колониях: как
раз католические завоеватели вели себя наиболее жестоко, подчас до не-
лепости. Однако булла послужила важной идейной поддержкой противопо-
ложному течению мысли (моногенизму), видевшему, как видим и мы, во
всех людях членов единого вида. Если понимать вид как потомство единст-
венной пары, то, поскольку никаких индейцев в Писании не упомянуто, по-
лучается, что индейцы произошли от какой-то расы Старого света, и их про-
исхождение оказывалось как-то связанным с расселением. К этой мысли мы
еще вернемся.
В 1575 году Бернар Палисси, художник-керамист и ученый-самоучка,
устроил в Париже выставку ископаемых, где впервые провел их сравнение с
ныне живущими. В 1580 году он высказал в печати вот какую мысль: по-
скольку всё в природе находится «в вечной трансмутации», то многие иско-
паемые остатки рыб и моллюсков относятся к вымершим видам. Термин
«трансмутация» (означавший у алхимиков превращение одних элементов в
другие, в том числе простых металлов в золото) применен здесь, как и у
Альберта Великого, в том смысле, какой мы придаем ныне слову «эволю-
9 Здесь архетип - первоначальный образец (греч.). Этот термин употреблял еще Филон в
значении "прообраз, идея", но здесь он значит еще и “первопредок’'. Впоследствии, на-
чиная с Ч. Дарвина, данное смешение смыслов стало обычным.
43
ция». Вскоре Палисси попал в тюрьму, где и умер.
Еще один путь к эволюции лежал через проведенную Альбертом анало-
гию трансмутации видов с разведением домашних пород. Как ни странно, в
печати ее первым из мирян развил знаменитый «королевский пират», один
из организаторов разгрома англичанами испанской «Непобедимой армады»,
Уолтер Роли (или Рэли, Raleigh). Сидя в тюрьме, он написал книгу «История
мира» (1614), где коснулся и потопа. Развивая идею Бутео, он утверждал
(пересказ Вичлера):
«Ковчег не требовал места для всех видов животных, поскольку некоторые
группы видов развились только после потопа путем скрещивания, как мул
(лошадь с ослом) и гиена (волк с лисой). Сэр Уолтер Роли объяснял, однако,
что затем новые виды стали результатом не только скрещивания, но и прямых
изменений существующих видов. Эти изменения были вызваны, как говорил
Роли, новой окружающей средой; например, европейская дикая кошка разви-
лась из индийской пантеры, и наш черный дрозд изменил цвет и размер в
Вирджинии, тем же путем, каким и люди изменяются в росте и цвете [кожи]
на разных материках... Роли был убежден, что изменения в природе анало-
гичны развитию рас при одомашнении...».
Тема стала популярной, и в 1620 году ее коснулся английский мысли-
тель, в общем от биологии далекий - Френсис Бэкон, политик и философ-
утопист. Он высказал мысль, что виды могут изменяться путем накопления
«ошибок природы», т.е. случайных изменений.
В Италии и Франции богословы традиционно, со Средних веков, стояли
на страже догм, причем в Италии добились в течение XVII века полного раз-
грома науки, которая прежде была лучшей в Европе. Показательна судьба
натурфилософа Луцилио Ванини, который в молодости, избегая гонений,
покинул Италию, но в 1619 году все-таки угодил на костёр инквизиции во
Франции, в Тулузе. За что? Даже его враги не могли объяснить это толком.
Просто за вольнодумство. Нам же важно, что он был, вероятно, первым
итальянским эволюционистом.
В книге (1615 г.), стоившей ему жизни, Ванини повторял Альберта: трак-
товал известные нам замечания Теофраста как свидетельствующие о пре-
вращении вида в вид. Опираясь на подобные авторитеты, он шел дальше:
выстраивал живые существа в порядке их сложности и совершенства, гово-
рил об их (в том числе и человека) происхождении друг от друга. Один из
его диалогов так и назван: «О первоначальном происхождении человека»
(De prima hominis generatione).
После этого не будем удивляться, что крупнейший французский фило-
соф Ренэ Декарт, написав около 1630 года космогонический трактат (где
мир рождался из вихря), даже не пытался печатать его. К нашей теме там
относилась такая мысль:
«Чтобы лучше понять природу растений и животных, гораздо предпочти-
тельнее рассуждать так, будто они постепенно порождены из семени, а не
44
созданы Богом при начале мира... И тогда, поскольку мы будем помнить, что
на самом деле всё это возникло не так, мы изложим природу явлений значи-
тельно лучше».
Цензурный характер выделенной мною оговорки очевиден, однако
цензура проникает ведь не только в тексты гениев, но и в их мысли.
В напечатанной книге («Рассуждение о методе», 1637) Декарт был еще
более осторожен, но и это вызвало бурную реакцию церковников, не только
католических, но и протестантских. На много лет вся дискуссия оказалась
связана с именем Декарта. Для одних выделенная выше его оговорка была
искренним руководством (так, даже еретик Спиноза рассуждал вполне в ее
рамках: «Будем ли мы представлять природу в форме протяжения или в
форме мышления, мы в любом случае найдем один и тот же порядок, одну
и ту же связь причин» - вот первая формулировка параллелизма бытия и
мышления), для других же эволюционный смысл идей Декарта был очеви-
ден. Они-то нам и интересны.
1-10. Хэйл, основатель эволюционизма
Дело оставалось за обобщающим трудом. Он появился в Англии, пере-
жившей революцию. Читатель уже, наверное, не удивится, узнав, что и его
автором был отнюдь не натуралист. Это был юрист, богослов и финансист
Мэтью Хэйл (1609-1676). В 1660 году, будучи ненужным ни республикан-
ским властям (уже), ни королевским (еще), он уехал в свое поместье и напи-
сал натурфилософский трактат «Первоначальное происхождение челове-
ческого рода, рассмотренное и испытанное согласно свету природы». Опуб-
ликован после смерти автора, в Лондоне, в 1677 году.
Вичлер ограничился одной цитатой из Хэйла:
«Мы не должны воображать, что все виды и роды были сотворены в той фор-
ме, в какой мы видим их ныне, - нет, сотворены только те виды и роды, кото-
рые мы зовем архетипами», остальные развились из них под влиянием «мно-
гих обстоятельств».
Влияние книги Роли очевидно. Некоторые из «обстоятельств» Вичлер
затем перечислил, и одного этого, по-моему, вполне достаточно, чтобы ис-
торики эволюционизма постеснялись писать что-либо прежде, чем прочесть
Хэйла самым внимательным образом. Увы, этого не произошло - Хэйла ед-
ва упоминают, и то изредка. Историю эволюционизма принято начинать с
середины XVIII века, с робких намеков Бюффона (о нем поговорим далее).
Почему?
У истории науки свои законы. Как и любой ученый, историк может доко-
паться сам до чего угодно, но далеко не всё может объяснить обществу.
Оно услышит и воспримет лишь то, к чему готово. А принять эволюционное
учение из уст богослова и креациониста оно не готово и ныне.
Да, как ни странно, в книге с «эволюционным» заглавием Хэйл ставил
ту же цель, что когда-то Августин, - доказать истинность книги Бытия и оп-
45
ровергнуть идею вечности мира Аристотеля. Возможно, что Хэйл, повторив
заголовок диалога Ванини, полемизировал со злосчастным еретиком, до-
пускавшим, что человек мог произойти естественным путем. Книга Хэйла
никогда не имела широкого круга читателей: верующие не нуждаются в под-
тверждении своей веры научными аргументами, натуралистам же нужны
другие, небогословские, доводы. А ведь в ней собрано и осмыслено почти
всё то, чем впоследствии стал знаменит Дарвин.
Своих наблюдений Хэйл привел мало, и они тонут в книжной учености,
порою старомодной. Многословие Хэйла подчас изнурительно, некоторые
мысли повторяются десятки раз, а главное поминается походя. Книга на-
правлена против Декарта, однако многое изложено прямо по Декарту. Так,
происхождение Земли объяснено как опускание более массивных частиц к
центру хаотической массы (что, кстати, похоже на нынешнее объяснение),
но этот механизм назван библейским.
Да, всё так, но ведь прошлое не выбирают! Основателем эволюциониз-
ма явился креационист Хэйл. Между прочим, слово «эволюция» упомянуто
им тоже один раз, а ведь это - первое известное ученым применение дан-
ного слова в биологическом смысле. Говоря о человеческом семени, Хэйл
заметил:
“Оно должно в скрытом виде содержать как минимум всё устройство челове-
ческого организма (или же как минимум его абстрактный принцип или об-
раз), в эволюции (evolution) которого должно состоять соединение и форми-
рование человеческого организма” {Hale М. The primitive origination of
Mankind... London, 1677, p. 250).
У Хэйла, как видим, речь идет об индивидуальном развитии, и только
явное сходство с “потенциями” отцов Церкви (см. п. 8) позволяет видеть тут
подход к эволюции в нашем смысле слова. Как и у Декарта, сотворена из
ничего у Хэйла лишь первичная хаотическая материя, остальное развитие
мира он соглашался называть творением лишь «по аналогии». Бог мог бы
построить мир и всё живое мгновенно, но, по Хэйлу, иногда использовал та-
кие методы и «такие времена», которые «в некотором смысле идентичны
естественному процессу». Как видим, в Англии даже богослов и придворный
мог выражаться несколько прямее и смелее, чем независимый французский
философ Декарт.
Эволюция не могла уложиться в шесть дней, но для Хэйла библейская
версия была нерушимой, и вот мы видим у него рассуждения в духе конкор-
дизма (см. п. 1-4), правда - весьма невнятные: исследователям предстоит
выяснить, «в каком порядке и характере времени это было».
У Хэйла предвосхищены многие споры о роли случайности, возникшие
с рождением дарвинизма. Он признавал эволюцию за счет случайных соче-
таний (подобной тому, какая образовала английский язык - «странную
смесь» элементов более древних языков), видел тут необъятный источник
возможностей (ведь разнообразие слов - результат комбинаций букв алфа-
46
вита) и допускал, подобно Альберту, самопроизвольное изменение «семян»
(мы именуем это мутациями). В то же время он решительно отвергал «слу-
чайное соединение атомов» в качестве источника упорядоченности, говоря,
что получение чего-либо упорядоченного таким путем невероятно
(incredible), а потому невозможно. Тут нужно не случайное соединение ато-
мов, а «обычное, естественное и необходимое соединение естественных
причин и действий». (Нынешним языком: случайным может быть акт изме-
нения, но не его механизм, не процесс формообразования - это вполне в
духе номогенеза, учения XX века - см. гл. 4.) И мы сейчас, через триста с
лишним лет, когда механизмы наследственных изменений в значительной
мере выяснены и стала ясна роль самосборки (см. п. 3-13), можем лишь
признать полную правоту Хэйла.
Отдал он дань древнему учению о частицах наследственности («се-
менных молекулах»), из которых якобы сам собой собирается зародыш:
«Форма этих маленьких семенных молекул... есть работа разума, выбора, из-
брания, намерения наиболее разумного и мудрого существа, а никак не про-
стого случая...».
Эти молекулы были, по Хэйлу, первично сотворены, они попадают в
землю, воздух и воду от тел умерших, а затем складываются в низшие ор-
ганизмы. Что же касается высших растений, то тут Хэйл настаивал на на-
следственности:
«говорят: злак может брататься в плевел, и плоды могут выродиться в ходе
культивации... Но во всех этих трансмутациях, как улучшающих, так и ухуд-
шающих, соблюдается принцип семенной специфичности».
Совсем близко Хэйл подошел к дарвинову пониманию изменчивости,
рассуждая о культурных растениях. Сославшись на Бэкона, он убеждал, что
культурные воздействия могут как улучшать, так и ухудшать их свойства.
Вот мысль о случайности эволюционных вариаций:
«Возможно, что виды, ныне различные, были первично одним видом, но при-
обрели некоторые случайные различия с течением времени».
Таким путем он допускал появление новых пород собак, видов врано-
вых и т.п. Он провел параллель между уродствами и эволюционными нова-
циями, очень важную, как мы узнаем далее (п. 1-14), для дальнейшего раз-
вития эволюционной науки.
1-11. Человек - модель эволюции
Как и прежние эволюционисты, главную цель Хэйл видел в объяснении
эволюции человека. И человек же был для него главным фактическим при-
мером эволюции, ибо о нем больше всего известно. Конечно, он не выводил
человека из животных - Адам был для него одним из «архетипов», сотво-
ренных Богом. Понять предполагалось лишь разнообразие рас. Точно так
же понимал он и эволюцию остальных живых существ, только речь шла не о
расах, а о видах.
47
Основной моделью эволюции стало для Хэйла заселение Америки. Со-
брав все сведения о путешествиях в Новый свет до Колумба (а мы-то наив-
но полагаем, что их собрали лишь в XX веке) и допустив наличие в прошлом
сухого пути туда из Азии (это нынешней наукой признано), он пришел к вы-
воду, что отличия тамошних людей и животных вызваны «перерывом в
сношениях» и накоплением изменений у переселенцев. Примерно так писал
впоследствии Дарвин о галапагосских вьюрках.
Более того, в своих рассуждениях о центре творения (Capita
specierum), каковой видел в Азии, Хэйл подошел к основной идее биогео-
графии. От вопросов расселения он, как и многие, естественно перешел к
вопросам роста численности, столь волновавшим англичан в дни детства
Дарвина и столь актуальным ныне. Он прикидывал, сколь быстро неболь-
шая группа может заселить «пустую» страну, чем предвосхитил знаменитое
учение Мальтуса. Беды, снижающие численность людей (эпидемии, голод,
войны и междоусобицы, наводнения, пожары) он трактовал как акты высшей
воли, как «коррективы», избавляющие мир от «перегрузки».
Заодно он сформулировал такие же «коррективы» в отношении низших
животных и отметил, что (хотя те размножаются много быстрее людей) их
численность ограничивается их поеданием другими животными, а также
«равновесием тепла и холода»; тем самым, он оказался одним из творцов
учения о балансе природы - учения, предварявшего нашу экологию. Глядя,
как хищники преследуют жертв, Хэйл утверждал «непрерывное вторжение и
превосходство более сильных, активных и живучих», в чем некоторые исто-
рики видят зачаток идеи борьбы за существование, а с тем - и естественно-
го отбора. Это вряд ли законно, поскольку поскольку здесь нет речи о борь-
бе между близкими формами (например, между двумя хищниками за жертву
или между двумя породами травоядных - за участок). У него нет и того сла-
бого подобия отбора, что было у Эмпедокла и Лукреция.
1-12. Дорожки от Хэйла к Дарвину...
Не продолжая описания взглядов замечательного дилетанта - о них
давно пора написать книгу, - признаем, что первый трактат по биоэволюции
вышел в свет за 125 лет до первой эволюционной книги Ламарка и что его
можно уверенно назвать первой книгой по научному (без кавычек) креацио-
низму. Не имея возможности проследить всю историю от Хэйла до Дарвина,
наметим пунктиром три параллельных пути от первого ко второму - через
анализ 1) накопления изменений и их перемешивания, 2) приспособлений
(«влияние условий» и случайное улучшение), 3) избыточности населения и
баланса со средой.
Все три легко прослеживаются, и неясно одно: что знал о них Дарвин?
Хэйла быстро забыли, но заложенное им учение (научный креационизм)
развивалось и имело блестящее продолжение. Достаточно сказать об из-
вестной книге «Мудрость Бога, явленная в трудах творения» (1691), автор
48
которой, знаменитый ботаник Джон Рэй (Ray), развивал многие мысли Хэй-
ла (например, о балансе). Это течение мысли хорошо было известно Дар-
вину как естественное богословие.
Естественное богословие - учение, трактовавшее все явления мира как
гармонию, говорящую о божьей мудрости, проявленной в ходе творения,
учение, ставившее целью увязать религию с наукой. По сути очень старое
(см. п. 6), оно возникло теперь как реакция христианской мысли на успехи
естествознания, становившегося во второй половине XVII века самостоя-
тельным мировоззрением. Эти успехи породили стремление обосновать
христианскую точку зрения на мир, исходя из успехов науки.
По книге Вильяма Пэйли «Естественное богословие» (1802) Дарвин
учился в колледже и до старости благодарно вспоминал ее. Он мог не знать
имени Хэйла, но знал его мысли. Достаточно сказать, что Пэйли начал с во-
ображаемого примера, который есть у Хэйла (философ, никогда не видев-
ший часов, обнаруживает неизвестно кем сделанные часы и рассуждает -
могла ли их сделать природа?), а всю книгу построил по образцу книги Рэя.
К нашей теме относятся три аспекта естественного богословия - конкор-
дизм, адаптационизм и селекционизм.
О конкордизме мы уже говорили в пп. 1-4 и 1-10. Сам его принцип был
высказан смутно еще Хэйлом, а затем многими уточнен, но для укоренения
эволюционной идеи требовалось большее - показать, что в ходе «творе-
ния» живые организмы действительно изменялись. В 1665 году знаменитый
физик Роберт Гук издал книгу «Микрография», где, в частности, описал
микроскопическое строение раковин - нынешних и ископаемых. Он пришел
к выводу: ископаемые раковины принадлежат не просто организмам, но ор-
ганизмам, каких в нынешних водах не бывает.
Вывод, хотя и повторял идею Палисси столетней давности, был черес-
чур смел: состава фаун никто не знал, на что вскоре же и указал Рэй. Осто-
рожно признав эволюцию в пределах рода, он отрицал возможность выми-
рания какого-либо вида (ибо тем были бы нарушены и природный баланс, и
мудрость Творца) и уверял, что описанные Гуком аммониты (раковинные
головоногие моллюски) еще будут найдены живыми в водах океана. (На са-
мом деле те вымерли в меловом периоде.) Чтобы продолжать спор, пред-
стояло создать две науки - фаунистику и палеонтологию.
Адаптационизм, т.е. учение о приспособленности каждого организма к
своим условиям обитания, был еще старше - рассуждения о пользе тех или
иных органов можно найти у древних греков. Он всегда носил характер сла-
вословия в адрес богов, но в XVII веке открытие множества новых фактов
заставило всерьез задуматься - действительно ли все свойства каждого ор-
ганизма полезны. И если да, то всегда ли - самому владельцу? Сводку до-
водов о полезности дал Рэй, за следующие сто лет тут родилась целая ли-
тература. Вот пример из книги Христиана Шпренгеля «Раскрытая тайна
природы в строении и оплодотворении цветов» (1793):
49
«Премудрый творец природы не создал ни единого волоска без какой-либо
определенной цели», однако «для насекомых было бы мало пользы в том, что
большинство цветов выделяет сок и что сок этот защищен от дождя, если бы
в то же время им не была обеспечена возможность легко находить эту пред-
назначенную им пищу. Природа, которая ничего не делает наполовину... по-
заботилась о том, чтобы насекомые могли уже издали замечать цветы путем
зрения или обоняния...».
Наряду с серьезными описаниями приспособлений, бывали курьезные
- один ботаник уверял, что оболочка некоторых дынь имеет дольчатую
форму для того, чтобы удобнее делить ее за столом; один анатом убеждал,
что человек наделен ягодицами для того, чтобы, удобно усевшись, раз-
мышлять о величии божьем и т.д.
Словом, когда Пэйли сел писать свой учебник, сказано было, казалось,
всё возможное. Однако и он внес свою лепту, сформулировав вероятност-
ный аргумент-, если пользы не видно, можно сделать непроверяемое пред-
положение и назвать его вероятным. Например, он писал:
«А в рыбьем глазу радужка неспособна к сужению. Это большое отличие, ве-
роятный смысл которого тот, что ослабленный водой свет никогда не бывает
слишком резким на сетчатке».
Теперь можно было доказать всё что угодно. Этим и занялось следую-
щее поколение, породившее дарвинизм.
Селекционизм, т.е. утверждение, что изменение видов в природе про-
исходит путем, аналогичным селекции домашних пород человеком, был
разработан меньше всего, но и он присутствовал. Сама аналогия бытовала
в литературе, как мы видели, с XVI века, а наиболее четко выражена в ран-
ней рукописи Чарлза Дарвина (1844 г.):
«Предположим теперь, что некое Существо, одаренное проницательностью,
достаточной, чтобы постигать совершенно недоступные для человека разли-
чия в наружной и внутренней организации и предвидением, простирающимся
на будущие века, сохраняло бы с безошибочной заботливостью и отбирало
бы для какой-нибудь цели потомство организма... я не вижу никакой причи-
ны, почему бы оно не могло создать новую расу» (Ч. Дарвин. Соч., т. 3. М.-Л.,
1939, с. 133).
Это похоже с виду на дарвинизм, но это пока еще - естественное бого-
словие: «Существо» явно наделено божественным сверхразумом. Однако
дело даже не столько в сверхразуме (для отбора он не нужен), сколько в
малозаметном словечке «безошибочно»: в действительности выгодное ка-
чество дает выгоду не безошибочно, а при сочетании (всегда редком) вы-
годных обстоятельств. В другом месте (с. 86) Дарвин заметил, что «природа
не позволяет своей расе портиться от скрещивания с другой расой, и сель-
ские хозяева знают, как трудно это предупредить».
Но как раз природа (если она - не Бог) позволяет спариваться всем, кто
может, и новый вариант не рождается с запретом на встречу со старыми, а
50
значит не может находить себе пару «безошибочно». Для безошибочности
нужно то самое «Существо». Ко времени написания «Происхождения ви-
дов» оно (вместе со сверхразумом) из записей Дарвина исчезло. Чем «Су-
щество» было заменено? Когда мы поймем это (см. гл. 3 и 5), мы поймем,
что такое дарвинизм и чем он отличен от естественного богословия, а чем с
ним схож. Тогда только и можно будет задать вопрос, что можно взять из
дарвинизма для работоспособной теории эволюции.
1-13. ... и мимо
От Хэйла шли дорожки и мимо Дарвина. Прежде всего, Дарвину была
совсем чужда мысль проводить эволюционную идею через всю историю
мироздания, как делали это многие, начиная с Анаксимандра. Вопрос о про-
исхождении жизни тоже не занимал его. Если сравнивать с предшественни-
ками, то эволюционизм Дарвина более похож на взгляды Рэя, признававше-
го «трансмутацию» то ли в рамках рода, то ли вида.
Утверждение эволюционной идеи в умах было невозможно без указа-
ния места, занимаемого историей жизни в истории мира, и младшие совре-
менники Хэйла активно этим занимались. Гук, например, в 1667 году высту-
пал в Лондоне с лекциями «О землетрясениях», в которых учил, что Земля,
подобно организму, стареет, и рассматривал переходы между видами в
качестве следствия эволюции природы как целого.
Существенной тут была мысль Гука о медленных, незаметных наблю-
дателю, изменениях в течении многих веков, усвоенная впоследствии и ла-
маркизмом, и дарвинизмом. Наоборот, рассмотрение Земли как организма
выглядело фантазией и прошло незамеченным. К этой теме мы еще вер-
немся в части 2.
Столь же мало внимания привлекло высказывание английского свя-
щенника Томаса Бернета, описавшего (1681) декартову космогонию и сде-
лавшего вот какой вывод: жизнь должна была зародиться, при содействии
солнечного света, на границе загрязненного (вулканами) воздуха, воды и
плававшей в ней «жирной материи» (мы бы ныне сказали - на границе трех
фаз; как мы узнаем в гл. 7, эта мысль оказалась очень глубокой). Наземные
же организмы, как считал Бернет, появились после частичного высыхания
вод. Если бы не жуткое для нашего глаза название его книги («Священная
история Земли»), его имя, надо полагать, украшало бы все книги по истории
эволюции.
А помните цитату об архетипах в начале п. 10? И у Роли, и у Хэйла под
архетипами понимались те виды, которые были сотворены Богом прямо при
создании мира. Сколько их было и каких именно? Для ответа предстояло
развить научную дисциплину, которая получила имя сравнительной ана-
томии. Начало ее восходит к Аристотелю, а в Новое время первым ее опы-
том было «Исследование природы птиц с их описанием и простодушными
рисунками, взятыми с натуры». Ее написал французский зоолог Пьер Белон
51
(1555). Один из этих «простодушных» (naifs) рисунков предлагаю читателю
разглядеть: тут впервые дано детальное сравнение скелетов человека и
птицы (рис. 3).
Рис. 3. Из книги "Исследование птиц” (Belon, 1555). Гомологичные кости обозначены
одинаковыми буквами и притом почти всюду верно
Разумеется, в те времена никому в голову прийти не могло, что данное
сходство можно истолковать как происхождение людей и птиц от общего
предка. Сходство трактовалось как единство божественного замысла. Через
90 лет единый замысел видели уже в строении всех животных. Так, италь-
янский анатом Аврелий Северино, изучавший самых различных животных
(позвоночных, насекомых, моллюсков), в 1645 году писал:
«Прототип, лежащий в основе всех сотворенных животных, настолько очеви-
ден. что всякий анатом замечает это почти помимо своей воли».
Читая это, можно даже подумать, что автор действительно сравнивал
планы строения очень различных животных и находил в них важное сходст-
во, но это не совсем так. Его познания в строении позвоночных (этого тер-
мина еще не существовало) были ниже, чем у Белона, о котором он вряд ли
знал. А единый прототип он понимал по Платону: образцом творения слу-
жил человек, другие оценивались сравнением с ним, и черты сходств ана-
том искал не столько в строении, сколько в поведении; лиса, например, бы-
ла образцом коварства, осел - глупости и т.д.
52
Зато Северино первым сравнил разные члены, служащие сходной це-
ли: легкие и жабры, руку и хобот, рожки улитки и глаза позвоночных. Позже
такие сходства назвали аналогиями. Отметил и сходства, впоследствии по-
лучившие название гомологий', между шерстью зверей и перьями птиц, ме-
жду конечностями зверей и человека, между рукой и ногой.
Заметим кстати, что в Италии говорить всё это было накладно - хотя
Северино и заявлял об отличии анатомии человека от анатомии животных,
ему пришлось однажды иметь дело с инквизицией.
Впоследствии, в дарвинизме, аналогии были истолкованы как незави-
симые приспособления к одной и той же функции, а гомологии - как следст-
вие происхождения от общего предка. Другие эволюционные теории отка-
зываются от столь простого деления всех сходств; наиболее из них извес-
тен упомянутый выше номогенез.
В 1672 году в Англии мысли о единстве плана строения развил знаме-
нитый врач и натурфилософ Томас Виллис. Анатомируя омара, он пришел к
выводу, что у ракообразных «не кости покрыты мясом, но мясо покрыто кос-
тями». Назвав панцырь наружным скелетом, Виллис стал искать и находить
такие же вывернутые сходства с позвоночными во всём, в том числе и в пе-
редвижении - потому, мол, рак и пятится назад.
Вот сколько сказано до Хэйла об архетипах. Хэйл видел их как первич-
но созданные виды, но другие видели их иначе. В 1678 году в Англии был
напечатан труд немецкого ботаника, математика и философа Иоахима Юн-
га, где архетип понят иначе, по Филону: идея архетипа есть идея поиска
единого во многом. Данный ход мысли имел многих последователей, а в
наше время лёг в основу диатропики (см. гл. 6).
Тем самым, Юнг положил начало тому, что через 160 лет после его
смерти поэт и натурфилософ Иоганн Вольфганг Гёте назвал морфологией
- общей наукой о форме. Морфологию и сравнительную анатомию часто
рассматривают как синоноимы, но это не совсем точно - первая есть теоре-
тический итог второй. Задача морфологии, науки о фундаментальных сход-
ствах, в некотором смысле противоположна задаче систематики - той час-
ти биологии, что изучает различия организмов. Подробно этот вопрос будет
рассмотрен в главе 6, в том числе в п. 6-11 будет дано более точное пони-
мание морфологии по Гёте.
1-14. Лейбниц и Мопертюи. Эволюционная философия
Эволюционисты никогда не исчезали совсем, но их было мало, и они не
печатались. Среди них был великий философ и математик Готфрид Виль-
гельм Лейбниц (1646-1716) который в молодости сам обшарил горы и руд-
ники Южной Германии и Северной Италии с молотком в руке.
Вечно чем-то увлеченный философ написал книгу «Первичная Земля»
(Protogea), проявив завидную эрудицию в вопросе, для него побочном, но не
удосужился издать ее. Она пролежала в рукописи полвека, увидев свет
53
лишь в 1749 году, а первый ее перевод с латыни на современный язык
(французский) появился еще через 110 лет, в год выхода «Происхождения
видов». Но отнюдь не устарел: несмотря на всеобщее признание эволюции
Земли, общество 300 лет (от Бутео и Палисси до Ламарка и Дарвина) обсу-
ждало один и тот же вопрос - происходила ли на самом деле эволюция жи-
вого. Доводы Лейбница в пользу биологической природы ископаемых костей
и раковин всё еще были актуальны, но важнее была постановка вопроса: «И
не вероятно ли, что в больших изменениях, которым подвергался земной
шар, большое число животных было преобразовано?» Он по-прежнему не
имел ответа.
Блестящее предисловие Бертрана Сен-Жермена к французскому из-
данию «Протогеи» служит не только лучшим очерком предыстории палео-
нтологии, но и укоризненным памятником нашему небрежению к ученым
прошлого. Ведь ни современники Дарвина, ни мы с вами, не осведомлены о
трудах большинства предшественников нисколько.
Так, часто пишут, что палеонтология родилась 200 лет назад в трудах
Жоржа Кювье, а она, оказывается, вдвое старше. В этом есть заслуга и
Лейбница, и более ранних авторов, труды которых Кювье использовал, но
не назвал [Чайковский, 2000].
В «Протогее» ясно проведена идея древности жизни и изменения обли-
ка Земли, констатировано вымирание многих видов морских животных, но
ничего более об эволюции организмов не сказано. Примерно в то же время
(1690-е годы) Лейбниц в одном из писем заявлял, что «всё во Вселенной так
связано, что настоящее таит в себе зародыш будущего» - мысль, которую
мы видели еще у Григория Нисского.
Далее Лейбниц сформулировал свой знаменитый «закон непрерывно-
сти», который обычно выражают формулой: «Природа не делает скачков».
Для организмов это значит, что между несходными видами можно указать
цепочку промежуточных и притом слабо различающихся видов.
Тут-то Лейбниц и выразил скупо свое видение эволюции:
«Поскольку, как я думаю, этот закон создает полную непрерывность в плане
последовательности, он создает нечто сходное и в плане одновременности».
Вот проблеск идеи, которая позже породила теорию параллелизма ме-
жду последовательностью ископаемых и разнообразием ныне живущих. О
ней речь будет в конце п. 2-9, пока же приведу место, показывающее, что
Лейбниц распространял закон непрерывности на виды живого:
‘Что касается постепенного перехода видов, то... философы высказали ряд
соображений о пробелах в ряду форм или видов. В природе все совершается
постепенно, в ней нет скачков... Но для красоты природы, требующей раз-
дельных, отчетливых восприятий, необходимы видимость скачков и, так ска-
зать, музыкальные интервалы в явлениях... На каком-нибудь другом небесном
светиле могут быть виды, промежуточные между человеком и животным”
{Лейбниц ГВ. Соч., т. 2, М., Мысль, 1983, с. 486-487).
54
Вот и всё, что есть у него об эволюции. Добавлю: набожный Лейбниц
понимал ее как непрерывное творение. Так понимали тогда почти все.
С 1735 года ботаник Карл Линней, вскоре ставший высшим авторите-
том в систематике, утверждал, что виды организмов неизменны; однако в
поздних работах он перешел на позицию того ограниченного эволюциониз-
ма, который тянулся тоненькой ниточкой от Альберта и Роли. В 1774 году
Линней писал, что
“Бесконечно Сущий сперва, в продвижении от простого к сложному и от мало-
го к многому, при начале растительной жизни, создал столько растений, сколь-
ко есть естественных порядков. Что Он Сам затем растения этих порядков пе-
ремешал путем скрещивания, что появилось столько растений, сколько есть
разных отчетливых родов. Что затем Природа эти родовые растения... переме-
шала между собой и умножила существующие виды, все, какие возможно”.
Это были лишь смутные намеки на идею эволюции, а для утверждения
идеи эволюции в умах ученых нужно было другое - новая идеология. И она
была в середине XVIII века найдена - в аналогии с химией. Точнее, в поня-
тии химического сродства.
В 1744 году в Париж привезли редкостного уродца - негритёнка с белой
кожей, голубыми глазами, рыжими волосами и похожими на лапы животного
руками. Среди глазевших на него обывателей (шутивших, естественно, о
легкомысленном поведении негритянки) был крупный физик Пьер Луи Мо-
пертюи (1698-1759). Он навечно вошел в пантеон математической физики
как автор принципа наименьшего действия.
Учёный быстро понял, что перед ним вовсе не помесь негра с белым, а
именно уродство, т.е. результат неправильного развития. И написал «Рас-
суждение о белом негре».
У Мопертюи поставлены вместе три вопроса: почему дитя похоже на
обоих родителей, как происходит развитие зародыша и откуда берутся
уродства. Он вряд ли знал авторов предыдущего века, писавших про урод-
ства, но, как и все тогда, читал древних авторов. К уверению древних о
смешении мужской и женской “семенных жидкостей” Мопертюи добавил
следующее: плавающие в этих жидкостях частицы соединяются в зародыш
точно так же, как в ходе химической реакции собираются в единое веще-
ство элементы - по законам, аналогичным закону химического сродства.
Свойства «частиц», аналогичные этому сродству, он охарактеризовал как
«нечто аналогичное желанию, неприязни, памяти».
Замечательно, что свои допущения, для нас чисто произвольные, он
полагал выведенными из опытных данных. (Это важно запомнить, ибо через
сто лет по сути то же самое сделал Дарвин.) Согласно Мопертюи, опыт за-
ставляет принять три допущения:
«1. Что в семенной жидкости всякого вида животных есть несметное число
частиц для формирования, путем комбинаций, животных того же вида.
2. Что в семенной жидкости каждого индивида частицы, годные для образо-
55
вания черт, сходных с его собственными, присутствуют в гораздо большем
числе, и к ним он имеет наибольшее сродство, как бы много ни имелось дру-
гих [частиц] для других черт.
3. Что касается того тела, которое формируется из семени каждого животного
вида из частиц, свойственных этому животному, то было бы смелой догадкой
думать, что каждая частица предназначена для своих зародышей (germes)»
(Maupertids P.-L. Oeuvres. T. 2. Lyon, 1756. p. 120).
Конечно, тут гораздо больше взято не из опыта, а из натурфилософии
Античности и Возрождения. Как и многие до него, Мопертюи полагал, что
частицы организма одного вида могут служить для сборки организма друго-
го. В виде примера он приводил кишечных паразитов, появление которых
толковал как самосборку частиц организма-хозяина.
Объяснение для нас наивно, но оно навело физика на один из самых
глубоких вопросов биологии - откуда берется сходство.
Например, почему так похожи земляные черви, личинки насекомых и
глисты? Мопертюи склонен был видеть здесь, выражаясь нынешним язы-
ком, сходство законов формообразования; примерно так по законам химии и
физики растет кристалл (добавлю: при одних условиях углерод кристалли-
зуется в графит, при других - в алмаз, но не в кристалл вроде соли). Для
нас важно, что параллель морфогенеза с химическим сродством явила со-
бой первый намек на принцип активности.
1-15. Морфология и законы эволюции
Законы морфологии хорошо видны там, где они нарушаются, - наруше-
ния сами по себе закономерны. Белый негр, урод среди негров, реализовал
ряд свойств, обычных у европеоидов. Имея совсем немного подобных при-
меров (тут ему могла бы помочь всеми забытая книга Хэйла), Мопертюи
нащупал общее правило, которое морфология XX века сформулировала
так: признак, являющийся уродством у данного таксона (считая расу тоже
таксоном), почти всегда присутствует в качестве нормы у какого-то другого
таксона (правило Кренке - см. далее, п. 4-11).
Чем тяжелее уродство, тем у более далекого вида оно может явиться
нормой. Вот примеры, касащиеся людских дефектов. Если нарушение пиг-
ментации - относительно безобидный дефект - образует признаки другой
расы (белый негр с голубыми глазами), то, к примеру, «заячья губа» (не-
сращение половин верхней губы) обычна у низших обезьян (лемуров), а ру-
ки, похожие на лапы, сходны с конечностями зверей других отрядов. Самые
тяжелые дефекты, из числа совместимых с жизнью, выводят за рамки клас-
са (например, конечностей не имеют змеи) или даже за рамки типа (двуго-
ловыми в норме бывают лишь некоторые черви).
Всего этого Мопертюи не знал, но понял главное: то, чем одни виды от-
личаются от других, могло сперва возникнуть как уродство. Примеры он ви-
дел в работе селекционеров, когда те скрещивают носителей редких ано-
56
малий. Почти все его мысли есть у Хэйла, но установки их сильно различны:
Хэйл предварял естественное богословие, а Мопертюи (отнюдь не чуждый
богословия при трактовке принципа наименьшего действия) страстно спо-
рил с основной посылкой естественного богословия - идеей полезности ка-
ждого свойства. Он отметил (1747 г.), что Исаак Ньютон видел целесооб-
разность (понятую как божественный замысел) в однообразии орбит планет
и строения животных, а сам спрашивал:
«если единообразие, наблюдаемое нами во многом, является доказательст-
вом, то не опровергается ли это доказательство бесконечным разнообразием,
которое мы наблюдаем во многом другом?»
Так, некоторые видят целесообразность в складках жесткой кожи носо-
рога (без них она не могла бы сгибаться), но забывают при этом о черепахе,
панцырь которой в самом деле не сгибается. Тем самым, Мопертюи сопряг
два основных в билогии вопроса: откуда берутся и как связаны сходство и
различие? То есть поставил проблему разнообразия (привлекавшую, как мы
видели, еще Р. Бэкона и Хэйла):
«Настоящий философ не должен позволить ни ослепить себя частями Все-
ленной, где блещет порядок и соответствие, ни поколебать себя частями, в
которых он этого не находит».
Мопертюи здесь близко подошел к пониманию того, что (если пользо-
ваться нашим нынешним языком) морфология и систематика
взаимодополнительны друг другу (в смысле Нильса Бора). См. [Чайковский,
1990, с. 29, 33, 63]. Мы еще будем говорить об этом в главах 6 и 10.
В 1748 году в либеральной Голландии вышла анонимная книга «Тел-
лиамед, или беседы индийского философа с французским миссионером об
отступании моря, образовании суши, происхождении человека и прочем». В
диковинном имени философа читалась фамилия автора: им был Бенуа Де
Малье (De Maillet), давно умерший французский дипломат, востоковед и
геолог-любитель. Книга, написанная около 1716 года, повторяла древнюю
идею о том, что морские животные могли породить наземных, т.е. исходила
из аналогии с превращением личинки в насекомое, что далеко от эволюции
в нашем понимании. Морфологическая догадка Анаксимандра (недаром
ведь выводившего людей из рыб, а не из беспозвоночных) была теперь
разработана: были построены ряды сходств, пусть и фантастические (на-
пример: рыбы - летучие рыбы - птицы).
Самое понятное у Де Малье - нечто вроде отбора:
«Пусть сто миллионов из них (рыб, выброшенных на сушу - Ю. Ч.) погибнет,
будучи неспособными свыкнуться с новой средой; достаточно, если это уда-
стся двоим, чтобы возник новый вид».
Де Малье, признавая образование новшеств за счет скрещивания, дал
свое определение рода: род - множество способных скрещиваться видов.
Остальные натуралисты считали, вслед за Рэем (большинство считает так и
ныне), что скрещивание идет лишь в рамках вида (что неверно).
57
Книга Де Малье (как и труды Мопертюи) попала на рабочий стол круп-
нейшего натуралиста Жоржа Луи Бюффона, одного из основателей фран-
цузской зоологии. Де Малье щедро цитировал предшественников и послу-
жил для Бюффона источником мудрости XVII века, которая сквозит в его
«Естественной истории» (первые тома вышли в 1749 г.) повсюду.
Бюффон руководил Королевским ботаническим садом и музеем при
нем, он наблюдал осаждение осадков из вод и пришел к выводу: наблю-
даемые земные толщи никак не могли образоваться за библейские 6 тысяч
лет, что Земле как минимум 75 тысяч лет. Срок этот нам сейчас кажется
смешным, но геохронология родилась именно с этого подсчета Бюффона, и
ему пришлось даже извиняться перед богословами.
Описывая вид за видом, Бюффон отмечал, наряду с приспособления-
ми, признаки, явно бесполезные. Зачем, к примеру, свинье вполне сформи-
рованные пальцы, которыми она не может воспользоваться?
Тем самым, возникало еще одно сомнение в правоте естественного бо-
гословия, утверждавшего, что Господь сотворил каждый волосок на пользу
его обладателю или же в пользу человеку. Не лучше ли сказать, что пер-
вично создан лишь прототип, или общий план, который затем без конца
варьируется? Бюффон так и утверждал.
Как великий человек Бюффон позволял себе странности - например,
начисто отвергал систематику в духе Линнея. Зато преуспел в морфологии.
Соединив мысли многих (от Белона и Роли до Мопертюи и Де Малье - ни-
кто из них назван не был), он решил, что общность плана строения можно
истолковать как общность происхождения. Если вспомнить, что даже среди
редких тогда эволюционистов царил взгляд Рэя (изменение в рамках вида),
то это - очень смелая мысль (что еще не значит - верная).
Вот пример, говорящий о склон-
ности Бюффона к математике:
«возьмите скелет человека, наклоните
кости таза, укоротите кости бедер, голеней
и рук, удлините таковые ступней и
ладоней, соедините вместе фаланги,
удлините челюсти, сократив лобную кость,
и, наконец, удлините также позвоночник ...
это будет скелет лошади».
Вскоре такие сравнения стали мод-
ны (рис. 4), но это еще не эволюция.
Рис. 4. Сходство опорных аппаратов
коровы и птицы, как его представлял
голландский анатом Питер Кампер на
лекции для студентов (1772 г.)
58
нов. четыре названные им типа
близ Парижа в карьере камень,
Крупнейшие после кончины Бюффона (1788 г.) морфологи Гёте и Кювье
добились блестящих результатов без ссылки на эволюцию. Гёте выдвинул
знаменитую «позвоночную теорию черепа», утверждавшую, что череп по-
звоночных сложен из измененных позвонков (рис. 5). В ней можно видеть
шаг к эволюционной морфологии, если считать, что первые шесть позвон-
ков бесчерепных предков постепенно преобразовались в череп. Но можно
обсуждать это и на языке вариации единого плана (божественного замыс-
ла).
На этом языке и говорил Кювье. Он преобразовал морфологию вве-
дением двух положений - о четырех типах животных и о корреляции орга-
строения: позвоночные, членистые, мягко-
телые (моллюски) и лучистые
(радиальные) - должны были пресечь
разговоры о едином плане строения всех
животных: таковой план можно, по Кювье,
искать лишь в пределах типа.
Рис. 5. Череп человека, представленный в
виде шести воображаемых
преобразованных позвонков (из книги
Карла Кар уса, 1841)
Зато в этих пределах Кювье творил
чудеса. Как-то ему принесли выломанный
из которого торчала пара зубов. Кювье уве-
ренно заявил, что они принадлежат вымершему сумчатому, что и оказалось
в действительности, когда образец был отпрепарирован. Помог тут именно
принцип корреляции органов, гласящий, что далеко не любое сочетание
свойств возможно в рамках одного организма, - такие зубы могли быть
только у сумчатого.
1-15* Эволюция и катастрофы
Смену фаун Кювье объяснял катастрофами земной поверхности, при-
водившими к сменам суши и моря, причем нигде не объяснил, каков был
механизм появления новых форм. Их прямое творение после каждой ката-
строфы он отрицал: «я не говорю, что нужно было новое творение». Он да-
же признавал постепенное преобразование одних форм в другие:
«вариации привели постепенно классы водных животных к их теперешнему
состоянию» (Кювье Ж. Рассуждение о переворотах на поверхности земного
шара. М.-Л., 1937, с. 150, 81).
Эти фразы заставляют включить его в число эволюционистов, хотя это
- отдельные фразы, а знаменит Кювье отрицанием эволюционных работ
своих современников.
59
До Кювье в науке говорили про одну катастрофу - Всемирный потоп,
взятый христианством из восточных мифов. Кювье отверг эту идею, видя на
каждом материке следы не менее трех катастроф, а причину каждой видел
в опускании и подъеме материка: «Это был ряд мелких катастроф и пере-
мен на земном шаре, по всей вероятности довольно кратковременных» (там
же, с. 249).
У Кювье был ученик, знаменитый палеонтолог Альсид Д’Орбиньи. Он
выступил с наиболее радикальной идеей: катастрофы якобы носили все-
мирный характер и приводили к полному исчезновению жизни, каковая воз-
никала каждый раз в ходе нового творения. (Таковых катастроф он насчитал
27, по числу известных ему геологических эпох.) Как бы ни казалось нам это
наивно, надо признать, что идея упорядочивать слои по катастрофам фак-
тически идет от Д’Орбиньи и господствует в геологии до сих пор. Хотя ныне
мы знаем, что жизнь не прерывалась.
Впротивовес Кювье, итальянский поэт и натуралист Джованни Батиста
Брокки в 1814 году высказал ту мысль, что причина смен флор и фаун не в
катастрофах, а в старении таксонов: каждый таксон, как и организм, спер-
ва расцветает, затем старится и, наконец, умирает. Хотя этим и нельзя объ-
яснить синхронность смен, но сама идея старения таксонов популярна по-
ныне и носит название “броккизм”.
Об эволюции Кювье говорил мало и с насмешкой, но жил он в эпоху
стремительных перемен (революция, Наполеон, реставрация), а в такие
времена люди особенно склонны видеть перемены и в природе. И вот, когда
он как-то раз разглядывал зуб динозавра, кто-то сострил: «Кювье ищет свою
переднюю ногу». Эволюционизм властно стучался в дверь науки.
1-16. Эразм Дарвин и принцип активности
Одной морфологии для понимания эволюции мало. Чтобы понять, как
фактически шло преобразование организмов в ходе истории Земли, надо
привлечь палеонтологию, но и ее недостаточно: креационист скажет, что ис-
копаемые лишь иллюстрируют ход последовательного творения, и возра-
зить ему, оставаясь в рамках палеонтологии, нечего. Эволюция останется
предметом веры, пока не будет показано, как изменения могут фактически
совершаться и наследоваться в череде поколений.
Разумеется, сразу решить такую задачу невозможно (как мы узнаем, до
конца она не решена и поныне), но ее надо было хотя бы поставить. Как
обычно в эволюционизме, это сделал не биолог-профессионал, а талантли-
вый дилетант - врач и поэт Эразм Дарвин (1731-1802).
Он заявил о себе поучительной поэмой «Любовь цветов» в духе Гесио-
да и Лукреция, чем изрядно навредил себе и своему делу: публика охотно
раскупала его поэмы, они издавались на разных языках, но ученый мир иг-
норировал и их, и ученые труды их автора. Когда Э. Дарвин все же написал
и издал серьезный научный трактат «Зоономия» (1794-96, Лондон), было
60
поздно: ученые его почти не читали - даже обширную 39-ю главу, ту, где
шла подробная речь об эволюции. А ведь это - пожалуй, главный эволюци-
онный труд XVIII века. Эволюционисты не читают его поныне.
От биологов этот опыт переняли многие историки науки, но, к счастью,
не все. Английский историк науки Питер Баулер так описал его взгляды:
«Эразм Дарвин... был деистом, убежденным, что Бог замыслил живые суще-
ства самоусовершенствующштся во времени. В своем постоянном желании
преодолеть трудности внешнего мира они развивают новые органы посредст-
вом механизма, который... стал знаменит как “'наследование приобретенных
свойств”. Результаты усилий особей наследуются их потомками, так что по
наколении в ходе многих поколений может сформироваться целый новый ор-
ган. По-видимому Дарвин полагал, что всеобщим результатом такого усилия
приспособиться к среде может быть постепенный прогресс жизни в направ-
лении более высоких ступеней организации» {Bowler Р. Evolution. The history
of an idea. Berkeley, 1984, c. 77).
Здесь нужны три замечания. Во-первых, НПС не придумано Э. Дарви-
ном, а заимствовано из древнего общего мнения (см. п. 1-7); зато ему при-
надлежит, как верно отметил Баулер, понимание данного наследования как
фактора эволюции. Во-вторых, тексты Э. Дарвина действительно не дают
уверенности, что он имел ясное представление о прогрессе как итоге эво-
люции, хотя неявно касался его многократно. В-третьих, самоусовершенст-
вование и «постоянное желание преодолеть» явились более общими, чем у
Мопертюи («сродство»), выражениями принципа активности - основного в
нашем дальнейшем исследовании.
Подобно дилетанту Де Малье, Э. Дарвин внимательно штудировал
предшественников, но, в отличие от него, мало их цитировал. Видно его
знакомство с трудами геологов, а также с воззрениями эволюционистов XVII
-XVIII веков. Он знал о параллели развития Земли и жизни, о самозарожде-
нии жизни на границе сред, об изменении аммонитов и их вымирании, об
идее борьбы и параллелях между выживанием в природе и селекцией на
ферме, о происхождении видов растений путем скрещивания первичных
видов, о «семенных молекулах» и случайности вариаций - словом, едва пи
не всё, что было сказано до него об эволюции. Многое он при этом добавил
- например, борьбу между близкими видами. В поэме «Храм природы» он
писал, говоря об активности:
Деревья, травы вверх растут задорно,
За свет и воздух борются упорно.
Ему же принадлежит выражение «миллионы веков» эволюции (вместо
бюффоновых 75 тысяч лет). Но всё это было лишь базой, на которой он
строил свое учение. В цитированной поэме «Храм природы» он в примеча-
нии отметил:
«Один натуралист... считал возможным, что первыми насекомыми были
пыльники и рыльца, каким-то образом отделившиеся от материнского расте-
61
ния (наподобие цветов валлиснерии), и что из них с течением времени разви-
лись другие насекомые, одни из которых получили крылья, другие - плавни-
ки, а третьи - когти, благодаря непрерывным усилиям добыть пищу или
обезопасить себя от повреждения. Однако эта идея не легче для понимания,
чем превращение гусеницы в бабочку».
Кого он имел в виду? Такие сказочные построения, обычные в старину,
давно уже никем из ученых не поминались, но нам важно другое: кто-то еще
до Э. Дарвина видел в активности организма фактор эволюции. Эту мысль
поэт-врач подробно развил. Отсюда и пошел физиологический эволю-
ционизм. В нем главный поставщик новшеств - не слепая случайная из-
менчивость и не предустановленные потенции, а собственная активность
особи как самостоятельный фактор эволюции.
«Многие органы животных образовались в результате трех великих потреб-
ностей: полового чувства, чувства голода и чувства самосохранения» - писал
врач-мыслитель. Не надо думать, что речь идет только о сознательных жела-
ниях. Нет. активность автор видел на всех уровнях жизни, вплоть до микро-
скопических пульсирующих «волокон»: «Особенности форм животных, от-
личающие их друг от друга,... образуются постепенно из сходных живых во-
локон и видоизменяются при воспроизведении».
Итак, еще двести лет назад мыслитель указал на процесс наследствен-
ного изменения: это - изменения «волокон» при «воспроизведении», т.е.
примерно то, что мы называем мутациями. Один из биографов Э. Дарвина
прямо сравнил «волокна» с ДНК, и даже если это натяжка, трудно найти
лучшее сравнение. В последнем издании «Зоономии» (1801) «волокна» не-
сут не только наличные качества, но и способности к развитию (formative
aptitudes), т.е. они несут также и особую форму активности.
Их можно сопоставить с генами как носителями не только элементар-
ных биохимических, но и морфологических, и эмбриологических функций: по
Э. Дарвину, в высших животных «эти волокна и молекулы, плавающие в
циркулирующей крови родителей, собираются в соответственных железах
самца или самки»; их смесь служит основой для порождения зародыша в
матке. Пока еще это в духе Хэйла, Мопертюи и прочих, но вот и новое: «В
одних своих частях зародыш сходен с отцом, в других с матерью - в зави-
симости от количества или активности волокон или молекул в момент их со-
единения». В наших терминах это приблизительно значит: развитие управ-
ляется активностью генов, а эволюция - это изменение ранних стадий раз-
вития зародышей. Насколько знаю, до нашего героя этого не заявлял никто.
«Многие потратили немало сил, пытаясь объяснить законы жизни зако-
нами механики и химии» - сетовал Э. Дарвин; при этом упускали из виду
главное - «жизненный принцип, животворящий дух», которым наделены
даже растения. Организму, учил он, вовсе не нужно понимать цель произ-
водимых им движений, важно совершать сами движения. Если этим удается
избежать неприятности или опасности, оно постепенно станет привычкой, а
62
затем и изменит нужный орган. Так, различные формы птичьих клювов воз-
никли из-за усилий птиц питаться различными видами пищи.
Словом, был дан второй (после Хэйла) синтез знаний об эволюции.
1-17. Ламарк и идея прогресса
В 1798-99 годах в одном из парижских научных журналов появилась по
частям работа врача и натурфилософа Жана Кабаниса «Отношения между
физической и нравственной природой человека», где упоминается Э.Дарвин
и изложена, среди прочего, его идея физиологического эволюционизма. Кое
в чем она развита. Например, Кабанис допускал глубинное, а не только свя-
занное с внешней средой, развитие:
«Организмы могут к тому же без всякой видимой порчи своей природы под-
вергаться глубоким изменениям в своих внутренних положениях; приобре-
тать совсем новую способность (aptitude) к новым впечатлениям и движени-
ям».
Преподавая сам и занимаясь реформой образования, Кабанис хорошо
знал парижскую профессуру и был близок с крупнейшим тогдашним натура-
листом, который сам вскоре стал учить студентов идее эволюции, прежде
ему чуждой. Имя его было Жан-Батист Ламарк (1744-1829).
В 1800 году на лекции Ламарк изложил одно из главных положений Э.
Дарвина - развитие органов под влиянием регулярного упражнения:
«Птица, которую влечет к воде потребность найти добычу, необходимую ей
для поддержания жизни, растопыривает пальцы ног, когда хочет грести и
двигаться по поверхности воды. Благодаря этому кожа, соединяющая пальцы
у их основания, приобретает привычку растягиваться. Так с течением време-
ни образовались те широкие перепонки между пальцами ног, которые мы ви-
дим теперь у уток, гусей».
Э. Дарвин не упомянут (ни тут, ни когда-либо позже), но есть на него
намек: о природе сказано: «Известно, что время для нее не имеет границ и
что поэтому она всегда им располагает». На самом деле «известно» было
тогда совсем иное - что Земле 6 тысяч лет; даже Бюффон, смело удесяти-
ривший этот возраст, отнюдь не полагал его безграничным. О безграничных
«миллионах веков» писал в те годы только Э. Дарвин.
Ламарк был из тех ученых, которым проще придумывать самому, чем
учиться у других. Он и по-французски читал мало, не то что на других язы-
ках, так что нет оснований полагать, что он мог сам читать толстую «Зооно-
мию». Однако он явно ее видел - иначе нельзя понять, почему вышедшая
вскоре «Философия зоологии» Ламарка посвящена тем же темам и скомпо-
нована примерно так же, как «Зоономия», хотя прежде 65-летний Ламарк
ничем подобным не интересовался. Известно, что к физиологической теме
его обратил Кабанис (который в 1802 году издал свою упомянутую выше
работу отдельной книгой). Ламарк не раз ссылался на его книгу, где изложе-
ны идеи «Зоономии».
63
Словом, факт заимствования налицо, и отцом ламаркизма был не Ла-
марк, а Эразм Дарвин, однако Ламарк не был просто плагиатором: он пере-
работал идею физиологической эволюции вполне творчески и добавил к
ней идею прогресса, у Э. Дарвина едва упомянутую (сравнение учений см.
[Лункевич, т. 2]). Да и вряд ли можно вообще говорить о плагиате в то вре-
мя, когда никаких норм цитирования вне экспериментальной науки установ-
лено не было. Подробнее о теме «Э. Дарвин и Ламарк» и вообще о фено-
мене избегания предтеч см. [Чайковский, 2000; 2002].
Ламарк бывал слишком самоуверен. Он утверждал, например:
«Я мог бы доказать, что отнюдь не форма тела или его частей обусловливает
привычки и образ жизни животных, но что, напротив, привычки, образ жизни
и все прочие воздействующие обстоятельства с течением времени создали
форму тела и его частей у животных».
На самом деле доказать это не удалось пока никому, но именно в этой
скупой лекционной фразе заявлено знаменитое утверждение о том, что
функция определяет форму. Есть и противоположное мнение - что форма
определяет функцию. На мой взгляд, обе достаточно однобоки, о чем мы не
раз еще поговорим.
Ламарку принадлежит первый подход к тому, что в наше время получи-
ло имя экологического эволюционизма. В 1802 году он выпустил две
свои первые эволюционные книги: «Гидрогеология» и «Исследования о жи-
вых телах». О них можно прочесть в работе [Чайковский, 2002], а здесь ска-
жу лишь, что «Гидрогеология» впервые рассматривала роль организмов в
образовании земных толщ (эта идея стала популярной лишь в XX веке бла-
годаря работам В.И. Вернадского), а вторая книга явилась как бы первым
наброском главной эволюционной книги Ламарка «Философия зоологии»
(1809); но есть в ней и мысли, позже явным образом не выраженные. На-
пример, только тут Ламарк решился высказаться на самую опасную тему - о
возможном животном происхождении человека.
В отличие от Э. Дарвина, Ламарк с восторгом уподоблял организм ме-
ханизму и в начале «Философии зоологии» появление новых форм объяс-
нил тем, что
«жидкости организма при ускорении их движения преобразуют клеточную
ткань, место своего движения, открывают в ней себе проходы, образуют раз-
ные каналы, наконец создают здесь различные.органы, отвечающие состоя-
нию данной огранизации».
Эти движения и «влияние обстоятельств» - вот «две основные причины
нынешнего состояния различных животных».
Допустим, но как быть с растениями? Туг помогла та мысль Кабаниса,
что активность бывает внешняя и внутренняя. По Ламарку, растения и низ-
шие животные «живут только раздражениями, получаемыми извне», но по
мере усложнения организации всё больше играют роль внутренние движе-
ния и внутренняя активность (кабанисовы «впечатления»):
64
«...Природа, вынужденная вначале заимствовать у окружающей среды возбу-
дительную силу для жизненных движений и действий несовершенных живот-
ных, сумела, всё более и более усложняя животную организацию, перенести
эту силу внутрь самих существ и сумела наконец передать ее в распоряжение
особи».
Тем самым, чем сложнее устроено животное, тем легче ему эволюиро-
вать. Продолжая мысль, Ламарк пришел к своему главному принципу, прин-
ципу градации. (В наше время его называют принципом прогресса, или по-
вышения организации.) До него многие, по сути следуя Платону, писали о
деградации, т.е. об упрощении организации по мере удаления от человека.
В ней видели и путь эволюции: Бюффон, например, писал о деградации
лошади в осла. Ламарк, сам еще будучи в плену платоновой идеи («чем
ближе стоит животная организация к организации человека, тем она совер-
шеннее»), отважился ее переосмыслить: деградацию мы видим всюду лишь
потому, что «берем общий ряд животных в направлении, обратном избран-
ному природой».
На самом деле, по Ламарку, главное - градация (усложнение животных
в силу внутренней активности), и мы наблюдали бы одну ее, если бы не
превратности внешних обстоятельств. Вот ясное выражение данной идеи в
начале главы 6 “Философии зоологии”:
«Если бы природа создала одних только водных животных и если бы эти жи-
вотные — все и всегда - жили в одинаковом климате, в однородной по составу
воде, на одной и той же глубине и т.д. - очевидно, в организации этих су-
ществ наблюдалась бы правильная и равномерная градация».
В реальности же, как полагал Ламарк, градация то и дело прерывает-
ся и искажается процессами приспособления. Вот главное его отличие от
Ч. Дарвина, видевшего в эволюции лишь цепь приспособлений.
Заявив идею прогресса как общую тенденцию организмов, Ламарк
встал перед проблемой - почему существуют одновременно низшие и выс-
шие организмы? Почему низшие не стали высшими? Решил он ее гениаль-
но просто: низшие моложе высших, поскольку низших постоянно поставляет
процесс самозарождения жизни. Цитированный выше Баулер изобразил
этот принцип (совсем в наши дни забытый) наглядной диаграммой (рис. 6).
Пусть жизнь в наше время и не самозарождается, так что ламарково объяс-
нение наличия низших организмов нас сейчас не устраивает, но это никак
не в упрек великому французу.
Как и Э. Дарвин, Ламарк видел основной способ образования приспо-
соблений в активности особей, однако принцип упражнения органов относил
только к высшим животным. А у низших животных и у растений эволюция
состоит, по Ламарку, из одних лишь реакций на изменение среды - в этом
слабое место его учения. Ныне мы знаем, что внутренняя активность даже у
одноклеточных ничуть не ниже, чем у высших. Но это тоже не в упрек.
65
Рис. 6. По Ламарку, прогресс постоянно преобразует низшие формы жизни, возни-
кающие путем самозарождения, в высшие, так что нынешние низшие вовсе не являют-
ся древними (по Bowler, 1984)
Другая его слабость: нет никакого механизма наследования. Гениаль-
ная догадка Э. Дарвина об активности и изменчивости «волокон и молекул»
была напечатана после завершения книги Кабаниса и осталась Ламарку не-
известна. Он смог лишь повторить старую, как мир, установку - что приоб-
ретенные особью улучшения наследуются. Короче, в области анализа на-
следственности у него, в отличие от Э. Дарвина, зияла пустота.
В том же 1809 году, когда на прилавки французских книготорговцев лег-
ла «Философия зоологии», в Англии, в семье покойного уже Эразма Дарви-
на, родился внук Чарлз. По воле отца он сперва учился на врача, но вид
операций приводил его в ужас (наркоза еще не знали), и отец разрешил сы-
ну учиться на священника. Здесь он, между прочим, ознакомился с естест-
венным богословием, которое позже ему очень понадобилось. Но окончить
курс не пришлось: юный Чарлз отплыл в кругосветное плаванье.
А книга Ламарка всё еще лежала на тех же прилавках. Несмотря на
громкое имя автора, эволюция вновь оказалась ненужной. До поры.
Гпава 2. Победа эволюционной идеи
2-1. Научная революция
Итак, даже беглый анализ, проведенный в главе 1, убеждает, что эво-
люционное учение - не детище Ламарка или Дарвина, а мировоззрение, ку-
да более давнее. Притом перечень натуралистов был у нас очень краток -
опущены все, без кого изложение еще сохраняло связность.
Будучи старше самой науки, эволюционная идея время от времени ов-
ладевала умами ученых, хоть и немногих. С середины XVII века можно уже
говорить об эволюционном учении, обоснованном фактическим материалом
66
(пусть и отрывочным). Дважды, у Хэйла и Э. Дарвина, мы видим даже син-
тез различных воззрений, причем у обоих эволюция организмов выступала
как часть эволюции мира (их синтез был более общ, чем у Ламарка и у Ч.
Дарвина.) Но в целом общество оставалось на позициях креационизма, о
твердыню которого неизменно разбивались разрозненные волны эволюци-
онного натиска. Эволюционизм оставался чем-то вроде лженауки, но все-
таки, излагая его историю, приходится с начала XIX века говорить уже не
столько об отдельных людях, сколько о целых направлениях мысли.
Как мы уже знаем, отнюдь не всё, до чего ученый может додуматься, он
способен объяснить согражданам. Так, толкование наблюдаемой изменчи-
вости как причины и итога эволюции, ведет начало от французского монаха-
математика Бутео, но во Франции не прижилось, а развитие получило через
65 лет в Англии, в книге мореплавателя Роли, где и легло позже в основу
дарвинизма. Наоборот, идея физиологического эволюционизма (где первым
и главным фактором выступает собственная активность особи), сформули-
рованная английским врачом Э. Дарвином, не получила в Англии развития,
зато нашла себе подлинную родину во Франции, где развилась под именем
ламаркизма.
Словом, если национальность и место жизни ученого-новатора мало
влияют на его открытия, то относительно массы средних ученых, опреде-
ляющих взгляды эпохи, этого никак не скажешь. Очень даже влияют.
Хоть и твердят нам, что наука не знает национальных границ, но опыт
истории, в том числе истории эволюционизма, говорит иное. Там, где наука
задевает мировоззрение, она всегда одета в яркие национальные одежды.
Позже, когда мировоззрение сложится и станет единым для почти всех, ко-
гда научная работа приобретет характер выяснения деталей, тогда только и
воцарится на время то, что американский методолог Томас Кун назвал в
1963 году нормальной наукой. Ее и можно считать безнациональной.
При своем появлении дарвинизм был вполне национальным учением.
Сравнив учение Дарвина со взглядами английских философов, квалифици-
рованный современник Дарвина заключил: это -
“учение чисто английское, включающее в себя не только все особенности на-
правления английского ума, но и все свойства английского духа” (Данилев-
ский Н.Я. Дарвинизм. СПб., 1885, т. 1, ч. 2, с. 478).
Близкую мыль видим и в наши дни:
“Западная биология сформировалась под доминирующим влиянием англо-
американской, крайне прагматичной пуританской культуры. Здесь из двух
выбирают то, что лежит ближе” [Гродницкий, с. 84].
Мы вернемся к данной теме в п. 4-17, а здесь замечу, что при рождении
дарвинизм в целом был творческим (зовущим к поиску нового) учением. На-
оборот, типичной нормальной наукой является СТЭ (“синтетическая теория
эволюции”), т.е. нынешний вариант неодарвинизма (кавычки означают, что,
на нынешний взгляд, никакого синтеза в ней нет, как нет и теории эволюции,
67
а есть лишь популяционная генетика 1930-х годов). Она с первых лет встала
в глухую оборону к фактам. Мы убедимся в этом не раз, а специально во-
прос освещен в книге [Назаров, 2005].
Смену одного состояния нормальной науки на другое Кун назвал науч-
ной революцией. Крупной научной революцией в биологии была победа
эволюционной идеи в середине XIX века, малой - смена в 1930-х годах ста-
рого дарвинизма на СТЭ. Сейчас нам пора узнать еще об одной мало из-
вестной научной революции.
В середине XVIII века стены Парижской Академии наук сотрясала рас-
тянувшаяся на много лет дискуссия преформистов и эпигенетиков. Пер-
вые отрицали развитие зародыша в яйце - по их мнению, все части орга-
низма предсуществуют в зародыше и лишь увеличиваются в размерах по
мере роста Вместилищем зародыша считалось либо яйцо, либо спермий,
но в обоих случаях приверженцы преформизма оказывались перед двумя
главными вопросами: почему дитя похоже на обоих родителей и где нахо-
дятся зародыши будущих зародышей?
Неубедительность их ответа бросалась в глаза даже современникам:
преформисты просто-напросто признавали главным родителем либо отца,
либо мать, что же касается вопроса о будущих поколениях, то предполага-
лось, что в зародыше заложен будущий зародыш, в нем следующий и так
далее до конца времен. (Отражением подобных взглядов, присущих, как по-
лагают этнографы, нашим далеким предкам, можно считать игрушку - мат-
решку.)
Эпигенетики, наоборот, придерживались мнения, что зародыша как та-
кового заранее нет, а есть лишь «идея» будущего организма. Они одержали
верх после того как в 1759 году Каспар Фридрих Вольф (германский врач и
эмбриолог, долго работавший в России) доказал прямыми наблюдениями,
что растущий зародыш усложняет свое строение, а не просто увеличивает-
ся в размере.
Дискуссия длилась до начала XIX в., хотя решающее слово, в сущности,
было произнесено еще до Вольфа. Его произнес Мопертюи, о чем у нас шла
речь в главе 1: предсуществуют не зародыши, а частицы наследственности.
Его не услышали, но и Вольф с его опытными доказательствами был не
сразу услышан, хотя принять их было куда легче, чем отвлеченные рассуж-
дения Мопертюи.
В год смерти Мопертюи (1759) в своей диссертации Вольф высказал
следующую мысль: сходство между различными видами вызвано тем, что
организмы образуются в ходе онтогенеза посредством одних и тех же
сил. В 1781 году эту мысль развил анатом Иоганн Фридрих Блюменбах,
введя понятие формообразующей силы (nisus formativus) - направляю-
щей силы, свойственной всем организмам и ответственной, в частности, за
развитие их зародышей. Только после этого идея эпигенеза вытеснила
идею преформации, и можно говорить о состоявшейся научной революции.
68
И это тоже закономерно - научное сообщество переходит к новому ви-
дению явлений не тогда, когда публикуется решающий факт, а тогда,
когда оно готово принять объясняющую его теорию. Нам не раз придется
вспомнить это обстоятельство, поскольку в наше время уже опубликованы
как десятки фактов, требующих новой теории эволюции, так и их теоретиче-
ские обоснования, но общество еще не высказалось о том, какую (или ка-
кие) из них следует принять.
С Блюменбахом на сцену эволюционизма вышла новая интеллектуаль-
ная сила - немецкая натурфилософия.
2-2. Немцы выходят на эволюционную сцену
В 1784 году Иоганн Гердер (Herder) издал первый том своих «Идей к
философии истории человечества»; он был старшим другом великого Гёте
(Goethe). Относительно самой эволюции организмов Гердер был еще осто-
рожнее Бюффона (которого часто упоминал), он избегал прямо говорить о
физической связи между видами, но вполне ясно описывал изменения, про-
исходившие в царстве животных во времени. Например:
«если возрастать должны были воля и и некое подобие разумности в животном,
то голова его стала наполняться мозгом и увеличиваться в размерах; три глав-
ные части тела образуют теперь некую пропорцию между собой, тогда как
раньше не было никаких пропорций, как, например, у насекомых, червей...».
Перед нами та форма эволюционизма, которую со времен Лейбница
называют непрерывным творением.
Бюффонову мысль о едином плане строения Гердер смело развил:
«... даже еще весьма неразвитые существа по внутреннему строению некото-
рых главных частей очень напоминают человека. Уже земноводные удаляют-
ся от такого основного типа, и еще более далеки от него птицы, рыбы и насе-
комые, живущие в воде существа - эти последние постепенно теряются среди
растительных образований и окаменелостей». Для Гердера «вполне вероятно,
что и в морских существах, и в растениях, а может быть, и в тех существах,
которые мы называем неодушевленными, заложены одни и те же основы, за-
датки органического строения... Вполне возможно, что для взгляда вечного
Существа, который усматривает единую взаимосвязь всего, форма льдинки,
вокруг которой складывается снежинка, составляет аналогию к образованию
эмбриона в чреве матери».
Тут легко усматриваются идеи отцов Церкви (п. 1-8), но важнее, что
Бюффон соединен с Мопертюи: единый план прочерчен не только через всё
живое, но и через неживое (в гл. 6 нечто сходное назовем диасетью). Есть
и новое: кристаллизация заявлена как аналогия образованию зародыша.
Это позволило Гердеру развить и идею Вольфа - Блюменбаха:
«...Природа, склоняясь к бесконечному разнообразию живых существ, по-
видимому все живое на Земле создала по единой протоплазме (Hauptplasma)
огранического строения».
69
Пусть прямого признания эволюции тут и нет, но именно отсюда ведет
начало наша уверенность в едином корне всего живого.
Наиболее проницательные исследователи уже тогда видели, что рас-
суждения о силах, движущих развитие зародышей, указывают на какой-то
весьма общий закон живой природы. У историка науки Джозефа Нидхэма
находим замечательное суждение английского эмбриолога Джона Хэнтера,
написанное в 1786 году:
«Если бы мы расположили животных в ряды от менее совершенных до наи-
более совершенных, мы, вероятно, нашли бы, что несовершенное животное
соответствует некоторой ступени развития самого совершенного».
Это и был первый подход к данному закону. К одному из основных за-
конов биологии развития, к закону, о котором спорят поныне.
Однако для развития этой мысли в теорию потребовался иной, неанг-
лийский, коллективный ум. Параллель между индивидуальным и историче-
ским развитием была разработана в Германии.
2-3. Параллелизмы и связь всего со всем. Биосфера
В 1790 году Иммануил Кант развил идею, которая вошла в историю
науки как идея параллелизма сил. Его тезис звучит так: силы, через кото-
рые материя воздействует, суть те же, посредством которых она су-
ществует. Легко понять, что мысль эта относилась ко всему на свете и
продолжала Спинозу и Лейбница.
С зоологией Канта увязал, пусть и туманно, немецкий зоолог Карл
Фридрих Кильмейер (у него учился великий Кювье) в своей знаменитой ре-
чи 1793 года в Штутгарте. Она называлась «О взаимосвязи органических
сил в ряду организмов»). В своей речи он выразил тезис Канта так:___
«Сила, благодаря которой происходит развитие индивидуума, тождествен-
______на силе, которая производит различные организации на Земле».__
Эту мысль развил в 1808 году анатом Фридрих Тидеман:
«Следя за метаморфозом лягушек, я пришел к заключению, что они во время
развития последовательно проходят организацию червей, моллюсков, рыб и
только под конец становятся лягушками».
Здесь конкретизировано утверждение Хэнтера (пусть и неверное фак-
тически: черви и моллюски развиваются по пути первичноротых, а позво-
ночные - по пути вторичноротых, поэтому единого ряда на деле тут нет),
иначе говоря, фиксирован онто-морфологический параллелизм, т.е. па-
раллель между развитием организма и повышением организации вдоль
морфологического ряда «от низших к высшим».
Естественно, эрудиты напомнили, что нечто похожее говорил еще Ари-
стотель во второй книге «Возникновения животных»:
«Ведь не одновременно возникает животное и человек или животное и ло-
шадь; то же относится и к другим животным: завершение наступает напосле-
70
док, и то, что составляет особенность каждой особи, является завершением
развития.»
Об эволюции в тех немецких построениях речи еще не было, но она не-
избежно должна была пойти - ведь и осторожный Кильмейер на лекциях
для студентов и в письмах друзьям (где выражался смелее, чем в речи пе-
ред начальством) прямо говорил о параллели между развитием одного ор-
ганизма и последовательным появлением организмов на Земле. Например,
в 1801 году он в письме к Кювье писал:
“полагаю, что различия между ископаемыми животными и ныне еще живу-
щими на Земле объясняются не столько... исчезновением видов, а скорее из-
менением созидательных сил... идущих параллельно с революциями нашей
Земли”. И далее: “Возможно, что эти законы тождественны с теми законами,
по которым еще теперь возникают мимолетные и остающиеся вариететы...
тюльпанов и гвоздик” {Канаев И. И. Кильмейер. Л., Наука, 1974, с. 43-44).
Отсюда (т е. от тезиса Канта - Кильмейера) и пошло общеизвестное
достижение немецкого эволюционизма - теория параллелизма индиви-
дуального и исторического развития. (Она была упомянута в п.1-9 и поз-
же привела к “биогенетическому закону” - см. п. 2-7).
Замечу, что наблюдение за «вариететами» (изменчивостью) цветков
очень похоже на то, чем через полвека стал знаменит Ч. Дарвин.
Взаимосвязь явлений была тогда отмечена натурфилософами и в дру-
гом: в том, что ныне именуют глобальной экологией. Гердер писал:
«Земля способна была создавать органические строения - растения, а потом
животных и человека»; «животные - старшие братья людей. Людей еще не
было, а животные были, и позднее, куда бы ни приходили люди, местность
была уже занята...».
Здесь Гердер едва ли не первый заявил (выражаясь нашим языком),
что эволюция - процесс экологический. Если до него все эволюционисты (и,
что греха таить, почти все после него) поминали среду обитания лишь во
фразах вроде «среда влияет», то у Гердера среда сама эволюирует. Кое-
что у Г ордера звучит так же, как до него у Хэйла и после него у Дарвина:
«Каждый вид заботится лишь о себе, как будто он один на белом свете, а на
самом деле рядом с ним другой вид, который ограничивает его поле
деятельности».
Но Гердер был мастер поворачивать вопрос неожиданно:
«И лишь в таком соотношении противопоставленных друг другу пород на-
шлось у творящей Природы средство сохранить целое» - закончил он приве-
денную мысль. А вот и совсем недарвинская установка: «Если Природа бро-
сит живое существо, кто же позаботится о нем, - ведь весь мир сотворенных
существ вечно воюет... друг с другом!»
То есть одной борьбы недостаточно, нужна еще некая позитивная
активность природы. Напомню, что всё это было сказано до Э. Дарвина,
увидевшего фактор эволюции в активности каждой особи.
71
Итак, природа активна, она сама творит. Но как именно? Гердер начал с
утверждения в духе естественного богословия:
«Всякой области Земли чудесным образом дано Природой все необходимое
для нее: и то, что растение даст, но и то, что оно притянет к себе и возьмет»,
но затем стал развивать отнюдь не веру в чудо, а уверенность в
физиологическом единстве природы:
«растения впитывают как раз горючее вещество, флогистон, убивающее жи-
вотных и во всех животных телах способствующее гниению! Замечено было,
что это полезное свое дело, очищение воздуха, растения производят не с по-
мощью тепла, а с помощью света...».
Различие ролей света и тепла понимали тогда немногие, и Гердер был сре-
ди них. На всем этом построен первый эскиз экологической эволюции:
«...Меня не огорчает, что погибли виды крупных животных. Погиб мамонт, но
погибли и великаны; другим было прежде соотношение между родами живых
существ. А теперь мы видим на Земле равновесие, причем не только в целом,
на всей Земле, но и очевидное равновесие в каждой отдельной стране и части
света. Культура может еще изгнать животных из той или иной страны, но едва
ли может совершенно искоренить целую их породу, - по крайней мере, такого
не случалось еще ни в одной большой части Земли; а с другой стороны, разве
не приходится природе вместо животных хищных и диких питать все больше
животных прирученных? Итак, на Земле... в ее теперешнем состоянии, не вы-
мерло ни одного живого рода, хотя я и не сомневаюсь, что раньше, когда Зем-
ля была совсем другой, могли жить на ней и совершенно другие живые суще-
ства и что позже, когда природа или искусство совершенно изменят Землю,
сложится на ней и другое соотношение между живыми существами.»
Увы, нынешный взгляд не столь светел. Через 36 лет после Гердера
старый слепой Ламарк оценил возможности природы весьма мрачно, от-
нюдь не по Гердеру (и, замечу, забегая вперед, не по Вернадскому), зато
реалистично (ЛамаркЖ.-Б. Избр. произв., т. 2. М., 1959, с. 442):
«Человек, ослепленный эгоизмом, становится недостаточно предусмотри-
тельным даже в том, что касается его собственных интересов: вследствие сво-
ей склонности извлекать наслаждение из всего, что находит... вследствие без-
заботного отношения к будущему и равнодушия к себе подобным, он сам как
бы способствует уничтожению средств к самосохранению и тем самым - ис-
треблению своего вида. Ради минутной прихоти он уничтожает полезные рас-
тения, защищающие почву, что влечет за собой ее бесплодие и высыхание ис-
точников, вытесняет обитавших вблизи них животных, находивших здесь
средства к существованию, так что обширные пространства земли, некогда
очень плодородные и густо населенные разного рода живыми существами,
превращаются в обнаженные, бесплодные и необитаемые пустыни. Подчиня-
ясь своим страстям, не обращая внимания ни на какие указания опыта, он на-
ходится в состоянии постоянной войны с себе подобными... вследствие чего
народности, весьма многочисленные в прошлом, мало-помалу исчезают с лица
72
земли. Можно, пожалуй, сказать, что назначение человека как бы заключается
в том, чтобы уничтожить свой род, предварительно сделав земной шар непри-
годным для обитания.»
Ламарк, как видим, оказался неожиданно весьма близок к нам, к нашим
мрачным прогнозам. Его подчас именуют основателем экологического эво-
люционизма, хотя, как видим, он должен делить таковой титул с немецкими
натурфилософами.
2-4. Эволюция романтическая. Окен
Среди них был знаменит Лоренц Окен (1779-1851). В 1805 году он из-
дал книгу «Зарождение», где, кстати, развил Гердера:
«Подобно тому как все животные находятся в соответствии друг с другом и
всякие изменения в их численности есть... переход пищевых веществ из одно-
го состояния в другое, так и в растительном мире все организмы также нахо-
дятся в соответствии друг с другом... Но не только растительный мир нахо-
дится в указанном равновесии, но, как сказано выше, он находится в таком же
соответствии с животным миром».
Эволюция живого в целом выступала у Окена как процесс перераспре-
деления живого вещества (что было огромным достижением: ведь другие
трансформисты рассуждали тогда лишь об изменении данного вида в за-
данной среде). Отсюда следовала та (неверная) идея, которую веком позже
В.И. Вернадский назовет (со ссылкой, заметьте, на Окена) сохранением жи-
вого вещества, идея постоянства массы биосферы.
«Зарождение» Окена удивительно уже титульным листом, где изобра-
жена пара сплетенных змей, кусающих себя за хвосты (рис. 7). В наше вре-
мя некоторые склонны видеть тут предвосхищение ДНК, но мне больше
нравится то мнение, что кусающая себя змея - древнеегипетский символ
вечности, а сплетенная пара символизировала для Окена переплетение
ролей полов в зарождении организма [Райков, 1969, с. 28].
Историки науки до сих пор спорят - была ли книга Окена умозрением
или обобщением фактов. В самом деле, его поразительные утверждения,
что «всякое живое тело (буквально Fleisch, т.е. мясо) состоит из первичных
зверьков (Urtierchen) и что «семя соединяется при оплодотворении с жен-
ским пузырьком», являются как бы заявкой на клеточную теорию.
Эти утверждения заявлены как факты, однако первые известные нам на
сей счет достоверные данные опубликованы куда позже: яйцо млекопитаю-
щего впервые описал в 1827 году крупнейший эмбриолог Карл Бэр (позже, в
России, он стал известен как зоолог-эволюционист и географ Карл Макси-
мович Бэр), а проникновение спермия в яйцеклетку установлено еще много
позже, в 1840 году; что же касается самой клеточной теории, то о ней тоже
стали писать гораздо позже книги Окена, лишь с 1838 года. Откуда же Окен
мог взять всё это?
73
Рис. 7. Титульный лист “Зарождения”. Бамберг (Бавария), 1805
На мой взгляд, перед нами
обычный пример исторического
заблуждения: мы привыкли вести
историю всякого учения от трудов
его “основателей”, которых при-
знали основателями их совре-
менники, а не от тех работ, где
данные мысли действительно
были высказаны впервые.
«Зарождение» Окена являет
редкий и похвальный пример тру-
да молодого натурфилософа, по-
нимавшего, что с предметом раз-
мышлений следует ознакомиться
самому, глазами и руками. Сам
Окен опытных открытий не сде-
лал, но конечно же слышал раз-
говоры зоологов о всяческих но-
вых наблюдениях, а его блестя-
щая интуиция и огромная начи-
танность позволили ему высказы-
вать то, что у других позже стало
открытиями. К сожалению, он не
сообщил нам, чьими замечаниями
(возможно устными) он восполь-
зовался, но так поступает, увы,
большинство ученых.
Об эволюции Окен тогда еще не говорил, но для нее очень важны два
его тезиса из «Зарождения».
Первый тезис. Развитие зародыша представлено у него как наращива-
ние числа клеток, что делало более понятной всю теорию Вольфа. Лишь
много позже, в 1830 и 1832 годах, был описан сам факт размножения клеток
делением (Вермель Е.М. История учения о клетке. М., Наука, 1970, с. 134). А
понимание того, что ранние стадии эмбриогенеза (развития зародыша) дей-
ствительно состоят в актах деления и перемещения клеток, пришло еще
позже, в 1850-е годы [Баранов; Лункевич, т. 2].
Второй тезис. Окен модифицировал схему Мопертюи: если у того раз-
витие трактовалось как самосборка частиц наследственности, то у Окена -
как самосборка «инфузорий» (клеток). Отрицая общепринятую тогда идею
зарождения живого из неживого, он был уверен, что организм, состоя из
«инфузорий», после смерти разлагается вновь на отдельные «инфузории».
74
По сути, этим он развивал древние идеи Эмпедокла, но разнообразие орга-
низмов (животных и растений) впервые понималось как итог разнообразия
комбинаций составляющих их клеток.
Эволюция самих организмов в «Зарождении» подана, как и у Гердера,
коротко и туманно:
«Первое и постоянное стремление инфузорий - образовывать растения. Это
ведет к образованию водорослей и грибов, а последние опять распадаются,
пока им не удастся превратиться в настоящее растение» и т.д.
Реакция коллег на эту романтику была отрицательна. Критичнее всех
был крупнейший тогда философ Георг Гегель. В параграфе 249 “Философии
природы” (1817) он прямо отверг эволюцию в нашем ее понимании:
“Мыслительное рассмотрение должно воздержаться от такого рода туманных
... представлений, как например представление о так называемом происхож-
дении растений и животных из воды или представление о происхождении бо-
лее развитыж животных из низших”. “На природу следует смотреть как на
систему ступеней, происходящих с необходимостью одна из другой... но этот
процесс не следует представлять себе так, будто одна ступень естественно
возникает из другой... Метаморфоза свойственна только понятию как таково-
му”. Позже Гегель добавил: “Представление, будто роды развиваются посте-
пенно во времени, является совершенно бессодержательным”.
Страннно? И да, и нет. Да, с нашей нынешней позиции всякое развитие
протекает во времени, но Гегель, вслед за Спинозой, признавал соответст-
вие бытия и мышления, а потому не спешил видеть процесс всюду, где вид-
но простое и сложное. Это мы понимаем простое раньше сложного, в при-
роде же, созданной Богом, всё иначе - полагал Гегель. То же, как мы виде-
ли в п. 2, раньше говорил Гердер.
Вопреки Ламарку (которого он знал и цитировал), Гегель был уверен,
что ряд животных форм можно “читать” как от низших к высшим, так и в об-
ратном направлении. “Во всякой системе наиболее абстрактное является
первым членом ... Но это не значит, что из водного животного естественным
образом возникло земное...”. К этим воззрениям нам не раз еще придется
обращаться.
Впрочем, романтиков всё это не остановило. С 1817 году Окен издавал
в Веймаре журнал «Isis», где в 1819 году поместил свою статью «Появление
первых людей». Здесь с истинным философским бесстрашием он довел до
логического завершения (каковое обычно есть абсурд) античную идею, ко-
торую можно обозначить как самосборку зародыша: по Окену, первые чело-
веческие эмбрионы сами собой формировались в морской слизи (рис. 8;
подробнее см. [Райков, 1969] и ЧЭ).
В 1825 году зоолог Мартин Ратке доказал, что зародыши птиц имеют
что-то вроде жабер, и это в каком-то смысле подтверждало фантазии Оке-
на, будто первые дети до двух лет дышали жабрами. Даже прямо наталки-
вало на мысль, что предки птиц были рыбами.
75
Вот тут-то и стали горячие головы утверждать, что «зародыш птицы
проходит через класс рыб», повторяя кратко прежний путь эволюции. Вид-
нейшим из таких авторов был опять-таки Окен.
Рис. 8. Так представлял себе Окен первич-
ный эмбрион человека, плававший в море
(как у Анаксимандра)
Великий Бэр в 1828 году предос-
терегал от поспешных выводов: по его
данным, зародыш птицы вовсе не про-
ходит через класс рыб, а просто заро-
дыши рыбы и птицы сперва сходны, а
потом развиваются по-разному; и в ча-
стности, жаберные щели попросту ха-
рактерны для зародышей всех позво-
ночных, но лишь у рыб развиваются в
жабры. В этом состоит закон сходства зародышей Бэра:
“Общее в каждой достаточно крупной животной группе формируется раньше,
чем специальное”, причем “зародыш высшей животной формы никогда не
бывает подобен другой животной форме (взрослой!), а лишь ее зародышу”, и
“чем дальше погружаемся мы в глубь развития (эмбрионального), тем более
находим сходства даже у очень различных животных” [Лункевич, т. 2, с. 381].
С тех пор эмбриология признает это как факт (точнее см. п. 4-13).
Горячих голов это не остудило, и в 1831 году Окен писал в своем
«Учебнике натурфилософии»:
«Животные совершенствуются мало-помалу, прибавляя один орган к друго-
му, совершенно так же, как усложняется тело отдельного животного. Царство
животных совершенствуется путем увеличения числа органов».
Постепенно эта мысль, обрастая фактами, превратилась в упомянутое
выше учение о параллелизме индивидуального и исторического развития
организмов. Оставалось добавить один пункт - что именно через изменения
пути развития зародыша происходит эволюция видов, - и новая теория об-
рела бы цельность. Однако немцы этого шага не сделали, его сделал
француз, ученик Ламарка.
2-5. Развитие зародышей и эволюция. Жоффруа Сент-Илер
В 1802 году Ламарк опубликовал книгу «Исследования организации жи-
вых тел», где, как и на лекциях, обращался к эволюции (подробнее см. [Чай-
ковский, 2002]). Тут опять звучали модные за рубежом мысли - на сей раз
немецкие. (Хотя в чтении иностранной литературы его заподозрить трудно,
но легко допустить новую беседу с Кабанисом.) Теперь его занимала связь
сил, движущих эволюцию, с силами, формирующими сам организм. Мысль
76
его была проста: поскольку «привычка упражнять» орган приводит не только
к его росту и совершенствованию, но и к изменению до неузнаваемости, то
приобретение новых органов можно, в принципе, понять, если допустить,
что зародыш тоже живет и упражняет свои задатки. Эта идея Ламарка легко
связывается с идеей самозарождения - в сущности, по Ламарку, организм
каждый раз заново самозарождается в ходе развития яйца [Нибхэ/и]. Идея
уже по тем временам была наивна.
В одной рукописи Кювье пародировал Ламарка: «Привычка жевать, на-
пример, привела через несколько поколений к появлению зубов; привычка
гулять - к появлению ног». Кювье видел здесь логический абсурд: следст-
вие стоит у Ламарка впереди действующей причины.
Лет полтораста ирония Кювье почти всем казалась уместной. Но только
- почти. Ламаркисты считали и считают, что надо говорить не о «привычке
гулять», а о необходимости ползать - она действительно преобразует плав-
ники в ласты и лапы, если у данных организмов есть к тому задатки. Что это
значит - задатки, мы узнаем позже, в главе 6 (когда займемся эволюцион-
ными тенденциями), а здесь заметим, что ирония Кювье может быть на-
правлена и против дарвинизма - ведь там тоже следствие (появление ново-
го органа) стоит впереди причины (гибели тех, кто им не обладает). Так что
смеяться над Ламарком не стоит, лучше вспомнить девиз биолога и натур-
философа А.А. Любищева:
если не хочешь быть осмеян потомками, никогда не смейся над предками.
У Ламарка нет еще прямой связи эволюции с изменением условий за-
родышевого развития - на нее указал младший коллега Ламарка, столь же
блестящий и увлекающийся натуралист Этьен Жоффруа Сент-Илер
(1772-1844; Жоффруа - не второе имя, а первая фамилия).
В том же 1802 году (год выхода книги Ламарка) он прославился тем, что
добыл в египетской экспедиции ценнейшие образцы животных и, главное,
смог отстоять их в казалось бы безнадежной борьбе с англичанами.
Когда брошенный Бонапартом на произвол судьбы французский корпус
сдался англичанам, те потребовали сдать им все научные материалы. Уве-
рения ученых, что в чужих руках еще не описанные находки бессмысленны
и погибнут для науки, были напрасны, и тогда Жоффруа в отчаяньи заявил,
что сам уничтожит всё. Уничтожит прежде, чем англичане, ведущие перего-
воры о капитуляции, успеют вызвать отряд солдат, который мог бы захва-
тить находки силой. Англичане уступили.
В наш лживый век трудно поверить, что они не только уступили, но и
выполнили уговор: ученые и находки в целости отбыли во Францию. Что ни
говори, 200 лет назад люди были несколько иные.
К нашей теме относятся привезенные в Париж мумии священных жи-
вотных: по ним зоологи впервые точно установили, что за 2-3 тыс. лет не
произошло сколько-то заметной эволюции данных видов. Кювье торжество-
77
вал: он получил в руки недостававший ему аргумент в пользу неизменности
видов и принялся за свою знаменитую теорию катастроф.
Доводов Ламарка о недостаточности срока (вот где послужили ему
эразмовы «миллионы веков» - увы, только ему) никто не слушал, и каждый
остался при своих взглядах.
А Жоффруа должен был задуматься: так есть ли на самом деле эволю-
ция? Долгие годы эволюция казалось бы не занимала его: как и немцы, он
накапливал знания о фундаментальных сходствах животных, не касаясь
причин. Сами эти данные были великолепны, что мы узнаем далее. Но
сперва ему предстояло еще лет 20 ссориться с Кювье, другом молодости.
2-6. Знаменитый спор
В 1820 году Жоффруа сделал в Парижской академии наук доклад о
едином плане строения позвоночных и насекомых. Не подозревая, что по-
вторяет Т. Виллиса (о котором мыговорили в п. 1-13), он смело заявил, что
членистоногие устроены в принципе так же, как позвоночные. В частности,
«насекомые живут внутри своего спинного хребта, подобно тому, как мол-
люски живут внутри своей раковины».
Кювье после доклада заметил презрительно: «Нам говорили глупости».
Сам он придерживался теории четырех типов (п. 1-15), которую разрабо-
тал, во многом опираясь на ранние работы того же Жоффруа. По Кювье,
единый план строения возможен лишь в пределах одного типа. А Жоффруа
одной фразой свел три типа к единому плану.
Впрочем, легко сказать: «Как моллюски внутри раковины» - ведь если
взять двустворчатых, то их раковина не делится на сегменты, так что даже
поверхностного сходства с другими типами тут нет. А в общем строении тем
более: если позвоночное и насекомое действительно похожи по общему
строению (голова с глазами и ртом, органы выделения и размножения на
другом конце туловища, конечности по бокам туловища, и т.д.), то моллюск
устроен совсем иначе. И десять лет эта лихая формулировка Жоффруа не
имела продолжения, а вельможный Кювье презрительно от дискуссий
уклонялся.
В декабре 1829 году умер Ламарк - нищий, слепой и почти всеми бро-
шенный. Над его могилой от имени профессоров выступил Жоффруа. Оце-
нив заслуги покойного, он резюмировал: неудачно выбирая порой частные
доказательства, Ламарк не имел равных «в установлении общих принци-
пов». Кювье откликнулся сдержанным похвальным некрологом, где упирал
на фактологические книги покойного Ламарка - «Флору Франции», «Естест-
венную историю беспозвоночных» и т.п., но не на его теории.
А через семь недель между двумя академиками начался бой, о ходе и
исходе которого спорят поныне, причем не только историки науки.
78
В начале 1830 года двое молодых анатомов представили в Академию
рукопись, в которой утверждали (вслед за Аристотелем, п. 1-6), что голово-
ногий моллюск каракати-
ца повторяет строением
позвоночное животное.
По их мнению, если че-
ловека мысленно пере-
гнуть в области пупка, то
не только близостью ро-
тового и анального от-
верстий, но и многими
другими свойствами та-
кой «человек» окажется
схож с каракатицей.
Рис. 9. Схема организа-
ции головоногого моллю-
ска (слева) и позвоночно-
го (справа, перегнуто в
середине позвоночника)
по Кювье. Сходства во
внутреннем строении нет
Жоффруа, наспех
пролистав текст, пришел в восторг и решил как академик представить его
высоким коллегам. На заседании 15 февраля он простодушно сказал:
«Представьте себе позвоночное животное, ходящее на голове. Это было бы
как раз положение одного из тех акробатов, которые запрокидывают назад
плечи и голову, чтобы ходить на руках и ногах».
Роковые слова: дальше, несмотря на оговорки докладчика о «первом и
грубом сравнении», академики его всерьез не слушали. Слишком уж веяло
от таких слов всеми осмеянной немецкой романтикой.
Кювье на докладе не было, но он вскоре же потребовал обсудить его и
через неделю явился в Академию с рисунком (рис. 9), каковой надлежало
бы сделать самому Жоффруа. На нем видно, что при перегибе позвоночно-
го никакого сходства с моллюском в плане внутреннего строения не получа-
ется. В этом победа Кювье была бесспорна.
Менее очевидно с единым планом строения позвоночных как свиде-
тельством эволюции: спор продолжался и стал оборачиваться в пользу
Жоффруа: когда 12 июля Кювье стал утверждать, что одна вымершая птица
из отряда куриных близка к современному лебедю, а его оппонент Д. Блэн-
виль - что та ближе к ястребиным, Жоффруа резонно заметил, что эту ис-
копаемую птицу можно рассматривать как промежуточное звено между
79
отрядами. Кювье (всегда отрицавший наличие переходных форм) так за-
нервничал, что прибег к административному аргументу: в открытом заседа-
нии критика неуместна.
Замечу, что столь резкий ход для вельможного Кювье отнюдь не был
характерен, и лишь после потери всех четверых детей (последняя дочь,
любимая Клементина, умерла в 1828 г.) стал он раздражителен. У него бы-
ли в тот день особые причины нервничать: началась Июльская революция,
а он входил в состав структур власти.
Но вскоре на престол взошел новый король, который дал барону Кювье
еще один титул - пэра Франции. Кювье уживался с любой властью.
Заседания Академии возобновились в октябре, и теперь Жоффруа сам
делал доклад о переходных формах. Кювье готов был ответить, но его
июльский аргумент неожиданно был обернут против него же - президент
Академии заявил, что если господин барон намерен дискутировать с пре-
дыдущим докладчиком, то надо организовать закрытое заседание. Кювье
уклонился - теперь акция президента выглядела насмешкой. На том дис-
куссия и кончилась. В вопросе о промежуточных формах, столь важном для
эволюционизма, победа скорее осталась за Жоффруа, однако общество за-
помнило только зимнюю победу Кювье [Амлинский].
Жоффруа, увидав, сколь трудно доказать эволюцию, исходя из ее ре-
зультатов, решил доказать ее, исходя из механизмов. Он серьезно взялся
за почти забытую «Философию зоологии» Ламарка и вскоре представил в
Академию обширный доклад «О степени влияния окружающего мира на ви-
доизменение животных форм...», опубликованный в 1833 году. Увы, реакции
Кювье не последовало: пока доклад ждал печати, тот в расцвете сил и на
вершине карьеры (только что стал министром) неожиданно умер. А доклад
стоил ответа - более, чем всё написанное Этьеном Жоффруа об изменяе-
мости организмов. В нем ламаркизм соединен с немецкой натурфилософи-
ей и развит настолько, что в 1915 году Н.А. Холодковский, зоолог, поэт и пе-
реводчик, предлагал даже ввести термин жоффруизм.
2-7. Что такое жоффруизм
Опираясь на глубокие морфологические исследования, свои и Кювье,
Жоффруа превратил в теорию то, что до него было набором голых и не-
связных идей. По словам Райкова, «Природа, по Сент-Илеру, как бы штам-
пует животных по одному идеальному образцу, а разнообразие в их внеш-
нем устройстве зависит от разнообразных влияний со стороны внешних ус-
ловий». Отличие от Ламарка здесь во-первых - в признании «идеального
образца» (его, как мы знаем, называли еще и архетипом), а во-вторых - в
том, что внешние влияния действуют не на взрослый организм, а на расту-
щий, или даже на его зародыш. Жоффруа писал в докладе:
«Если вы хотите узнать, почему возникает сдавленная и полуэллиптическая
форма головы у лягушки... то взрослое животное не даст вам удовлетвори-
80
тельного ответа». Однако, будучи головастиком, «лягушка имела в известной
степени организацию рыбы, поскольку ее органы дыхания помещались под
затылком и состояли из больших жабер», так что части ее черепа «стойко со-
храняют расхождение в ширину, вызванное вклиниванием жабер между по-
кровами ушной области» (цит. по (Амлинский, с. 326]).
Это чисто механическое объяснение было в то время очень убедитель-
ным. Тот же процесс Жоффруа видел в эволюции:
«Постарайтесь, освободившись от иллюзий и предрассудков прежних времен,
отдать себе отчет в последовательности явлений различия в развитии существа,
прошедшего все фазы своей жизни: перед вами в сжатом виде будет в некото-
рых отношениях... последовательность явлений различия, порожденных одно
другим. Обратить свое внимание на это - значит обратить испытующий взор
на картины великих эпох мира» (там же, с. 329).
Это - первая ясная формулировка того самого, что Эрнст Геккель в
1872 году назвал основным биогенетическим законом.
У Жоффруа было новым еще и то, что изменение влияний среды рас-
сматривалось как основной фактор эволюции. В основе концепции лежала
аналогия с уродствами: ссылаясь как на споры времен Мопертюи и Вольфа,
так и на новейшие опыты, где у зародышей искусственно вызывались урод-
ства, Жоффруа убеждал, что новое эволюционное приобретение вида мо-
жет быть закреплением массового уродства. А массовые уродства могут
возникать при ухудшениях условий среды, порождающих ненормальное
развитие зародышей. Ярче всего это Жоффруа показал на воображаемом
примере эволюции крота.
Если Ламарк приводил утерю кротом глаз как пример «неупотребления
органа», то Жоффруа приложил к нему «закон компенсации». Закон восхо-
дит к Аристотелю:
«Природа везде, взяв с одного места, отдает другой части» и «Общих и мно-
гих средств защиты природа, однако, не дала одному и тому же животному»
{Аристотель. О частях животных. II, 14; III, 2).
Жоффруа развил этот принцип в духе физиологии питания. Вот как из-
ложил дело его биограф:
«Морда крота, употребляемая им на то, чтобы рыть землю, служит для весьма
трудной работы; поэтому она сильно увеличивается в размере, и вместе с ней
растут и все соседние части, особенно весь орган обоняния... за счет того, что
другой, примыкающий к нему орган подвергается самой тяжелой атрофии:
именно глаза и испытывают это вредное воздействие» [Алишнский, с. 333].
Замечу от себя, что атрофия значит по-гречески отсутствие питания;
этим держится тут вся трактовка редукции глаз: они, мол, в ходе роста орга-
низма остаются без питания, тогда как чрезмерному питанию (гипертро-
фии) подвергается нос.
Жоффруа был уверен, что подобными чисто механическими
построениями можно объяснить всё то, чем крот отличается от других
зверей. В этом состоял его ответ на главное возражение противников
81
этом состоял его ответ на главное возражение противников эволюции, ут-
верждавших, что в природе нет переходных форм: их, по Жоффруа, и не
должно быть, если виды образуются путем резких уклонений (variations
brusques), вызванных изменениями среды, и их массового наследования.
Но в вопросе наследования Жоффруа не продвинулся дальше Ламарка.
Зато он ответил на другой важный вопрос: каков сам изменяющий ме-
ханизм? По Жоффруа - это дыхание, тот воздух, каким мы дышим:
«Изменения, неощутимые от одного века к другому, в конце концов... сумми-
руются; вследствие этого дыхание для некоторых систем органов становится
затруднительным и, наконец, невозможным». Под дыханием имеется в виду
снабжение кислородом каждого органа. «Тогда животное создает для себя
ставшее ему необходимым для существования новое устройство, улучшающее
или изменяющее легочные клетки, в которых происходит дыхание. Возник-
шие изменения - полезные или гибельные - распространяются и оказывают
влияние на всю остальную организацию животного».
С помощью этой мысли он объяснил, как ему казалось, весь процесс
превращения рептилии в птицу. Мысль была сумбурна, и ее не заметили
(но именно она оказалась близкой к основной идее российской школы фи-
зиологического эволюционизма, возникшей в начале XX века, - мы узнаем
об этом в главе 4). Впрочем, главное следовало дальше.
Историки науки крайне редко отмечают эту фразу (исключение соста-
вил И.Е. Амлинский), следующую в докладе за вышеприведенной:
«Если эти изменения приводят к вредным последствиям, то жи-
вотные, у которых они возникают, гибнут и заменяются другими
животными со слегка измененными формами, притом изменен-
ными таким образом, что они соответствуют новым условиям».
А ведь фраза - эпохальная: прочтя ее в феврале 1838 года, Ч. Дарвин
стал дарвинистом. О гибели неприспособленных писали многие, начиная с
древности, ново же утверждение, что «слегка измененные» вытесняют
прежних. (У других эволюционистов более приспособленными всегда счи-
тались сразу носители готовых новых приспособлений.) Вот этот-то тезис
использовал через пять лет Дарвин для первого эскиза теории отбора.
Однако у Жоффруа отбору подвергаются (и в этом суть жоффруизма)
не случайные уклонения, а результаты целесообразной реакции зародыша
на изменения среды. Есть, правда, и важная общая черта: и Жоффруа, и
Дарвин ввели в оборот данное положение, даже не пытаясь его хоть чем-то
обосновать. И если вытеснение неприспособленных приспособленными
действительно наблюдается, то «слегка измененные формы» к этому, как
оказывается, абсолютно неспособны, о чем мы еще узнаем в части 2. То
есть Жоффруа указал ложный ход мысли Дарвину, которому ученые в ос-
новной своей массе следовали полтора века.
82
2-7* Гомология
Зато важнейшим достижением Жоффруа явилась идея гомологии.
Еще в 1807 году он удивил самого Кювье, убедительно показав, что черепа
зверя и рыбы состоят из костей, в некотором смысле одинаковых. Это каза-
лось невозможным: в рыбьем черепе костей гораздо больше - куда, хотя
бы, деть четыре кости жаберной крышки? Однако Жоффруа нашел соответ-
ствие всему; в частности - и этим четырем костям: им соответствуют (ныне
это знает любой зоолог) 4 косточки слухового аппарата млекопитающих. Как
же он доказал это? Оказалось, что соответствие очевидно, если рассматри-
вать развитие зародышей. Тут-то и открылся путь, по которому Жоффруа
впоследствии пришел к своей знаменитой теории: эволюцию движет изме-
нение способов развития зародыша, а те следуют за изменениями условий
среды, в которой живут организмы.
В 1822 году Жоффруа ввел 4 понятия: равновесие (balancernent) орга-
нов, связи (connexions) органов, органы-гомологи (homologues) и сродство
(affinite) частей к целому [Амлинский, с. 174]. О них будет речь в п. 5-17.
Слова «гомология» у него нет, но идея принадлежит ему, что признал
ведущий палеонтолог Ричард Оуэн, “английский Кювье”, автор термина «го-
мология» [Амлинский, с. 178]. Оуэн различал три типа гомологии.
(1) Частной гомологией он именовал «органы у разных животных при
всевозможных вариациях их формы и функции». Ее примеры известны: ру-
ка и крыло, волос и перо, слюнная и ядовитая железы змей и т.д. Этой го-
мологии противостоит аналогия: исполнение сходной функции органом ино-
го происхождения (ядовитая железа во рту змеи и на коже лягушки).
(2) Общей гомологией Оуэн именовал указание на положение части в
целом. Все скелеты позвоночных он рассматривал как вариации единого
архетипа (идеального скелета), а все его кости - как вариации частей иде-
ального (архетипического) позвонка. Следуя Окену и Карусу, он полагал че-
реп состоящим из нескольких поясов, каждый из которых являет собой ви-
доизменение идеального позвонка, и таким же позвонком считал пояс рёбер
(реальный позвонок с парой рёбер и элементом грудины), а также пояс ко-
нечностей. В этой мысленной конструкции общей гомологией обладали
элементы, происшедшие из одной и той же части идеального позвонка. Так,
Оуэн выделял в реальном позвонке пару гемапофизов (щитков, ограждаю-
щих кровеносную трубку), и указывал их видоизменения в остальных по-
звонках (поясах): часть нижней челюсти, коракоид (клювовидная кость) пле-
чевого пояса, пара реальных гемапофизов, подвздошная кость - все они, по
Оуэну, связаны друг с другом общей гомологией.
(3) Серийной гомологией он называл тот вариант общей гомологии, ко-
гда гомологичные элементы несут сходные функции. Так, одну серию со-
ставляют позвонки, другую - фаланги пальцев, третью - конечности. Под-
робнее см. [Лункееич, т. 2; Канаев, 1963; Чайковский, 1990].
83
Всё это выглядело запуганно и надуманно, от чего можно легко отка-
заться, что и было сделано с победой дарвинизма. Тем удивительнее, что
через 120 лет биологи вынуждены были всё это вспомнить (см. п. 6-5). За-
мечу, что у Оуэна архетип понят как нечто интуитивное.
2-7** Карл Нэгели и развитие жоффруизма
В том же духе, что и Жоффруа, писал германо-швейцарский ботаник
Карл Нэгели (1817-1891):
«Как потомки прежних индивидов несколько от них отличаются, так должны
отклоняться от прежних и их зародыши. Это должно делать изменение стой-
ким; и это изменение не может вести ни к чему иному, нежели вырождение
вида или переход его в другой вид... Впрочем, так ведут себя далеко не все
индивиды данного вида. Как у самшита среди желтой листвы зеленеет одна-
единственная ветвь... так при переходе от одних геологических периодов у
другим индивиды одного-единственного вида могут послужить основой для
многих других [видов] и оставить после себя способные к развитию зароды-
ши» (С. Nageli. Die Individualitat in der Natur mit vorzuglicher Beriicksichtigung
des Pflanzenreichs. Zurich, 1856, S. 210-211).
Мысль Нэгели замечательна - только у него налицо ясное понимание
двух обстоятельств: во-первых, зародыш изменяется не сам по себе, а от-
ражает изменение родительского организма, и именно этот факт делает из-
менение эволюционным; во-вторых же, для эволюции достаточно успешно-
го изменения немногих, тех, кто способен к развитию. В отличие от обычно-
го ламаркизма и жоффруизма, здесь отведена важная роль случайности
(какой именно вид окажется способным к развитию, не предопределено).
Зародыши изменяются не все, а лишь некоторые, и под влиянием не внеш-
ней среды, в которой живет материнский организм, а - внутренней среды, в
которой развивается зародыш.
По Нэгели, вопрос о возможности происхождения вида из вида весьма
важен. Не называя никого из прежних эволюционистов (опять “избегание
предтеч” [Чайковский, 2000]), он отметил, что новые виды происходят из
прежних путем быстрых перестроек (мысль Шпренгера - см. далее, п. 3-12)
и что новые виды бывают сложнее прежних в силу стремления живого к
развитию (мысль Ламарка). Механизм эволюции он видел в изменении раз-
вития зародышей (мысль Жоффруа), что фактически наблюдаемое образо-
вание новых рас являет очевидную аналогию предполагаемому образова-
нию новых видов (мысль, вскоре высказанная Ч. Дарвином). Первым из
профессиональных биологов Нэгели заявил, что в эволюции принципиальна
роль случайности. Он высказал любопытное соображение: поскольку из-
вестны растения, размножающиеся вегетативно в течение многих поколе-
ний, а затем вдруг являющие половой процесс, то и в отношении эволюци-
онного развития можно допустить резкую неоднородность во времени. При-
чину он видел в обоих случаях одну и ту же - скрытое изменение внутренне-
84
го строения под покровом внешней неизменности:
«Высказывание, что вид оставался неизменным в течение ... исторического
времени, не следует поэтому понимать так, что он находился в абсолютном
спокойствии, а всего лишь так, что его изменения не пересекали определен-
ных границ. Как и вообще любое природное явление, вид не может оставаться
в полном спокойствии» (С. Nageli. Die Individualist..., S. 209).
Здесь и признание самопроизвольности эволюционного процесса (в ду-
хе молодого Дарвина, писавшего об этом в записной книжке 1837 г.), и до-
пущение скрытых от глаза изменений, легшее, например, через 82 года в
основу учения Шмальгаузена о стабилизирующем отборе. Наконец, сразу
после работы Вейсмана (1883) Нэгели понял (1884), что в клетке есть осо-
бое наследственное вещество, идиоплазма. Поэтому вполне можно счи-
тать его одним из основателей современного эволюционизма, и ныне
наука возвращается как раз к Нэгели.
Но вернемся в дни Жоффруа. Его доклад, как и книга Ламарка, был
встречен молчанием. Не сразу заметил его и Чарлз Дарвин (1809-1882), но
по веской причине - он плыл тогда в океане.
2-8. Молодой Дарвин
Неизвестно, когда мысль о превращении видов впервые посетила мо-
лодого Дарвина. Вряд ли это случилось в Эдинбурге, где он учился на вра-
ча, или в Кембридже, куда он, послушный воле отца, перешел учиться на
священника. Скорее это произошло в путешествии, куда его пригласил в ка-
честве натуралиста капитан военно-топографического корабля «Бигль» Ро-
берт Фиц-Рой. Осенью 1835 года Дарвин, находясь на Галапагосских остро-
вах (что к западу от Эквадора), сделал интересную запись в блокноте. Изу-
чая местную птичку - пересмешника, он сравнил экземпляры с четырех ост-
ровов, убедился в их отличии друг от друга и от материковых форм и за-
ключил: «Зоология архипелагов вполне заслуживает исследования, ибо та-
кого рода факты могут подорвать неизменность видов».
Это - первая его запись на тему эволюции, и говорит она немного, по-
скольку гораздо больше можно было прочесть у Ламарка, о котором юный
Дарвин узнал еще в Эдинбурге. У Ламарка было целое учение об эволюции,
тогда как запись Дарвина всего только подвергала робкому сомнению догму
о постоянстве видов.
Через два года, разбирая в Лондоне путевые заметки, Дарвин ощутил
вдруг потребность отложить бумаги и взять с полки том «Зоономии» Эразма
Дарвина, своего давно умершего деда. Известно это и потому, что он любил
расчеркивать читаемое, и потому, что он вел «записные книжки о трансму-
тации видов», первую из которых так и назвал - Зоономия.
Только живые тела поддерживают свое существование путем смены
поколений, и недаром проницательный Эразм связал идею продолжения
рода с идеей преобразования видов; как личинка порождает бабочку, а го-
85
ловастик - лягушку, так и один вид может порождать другой вид. Только
превращение вида в вид (как писали и дед, и Ламарк, да и сам Чарлз видел
на своих материалах) происходит очень и очень медленно. Как же это про-
исходит? Здесь Эразм и Ламарк были единодушны: животные активно при-
способляются к среде, развивая упражнением те органы и свойства, кото-
рые им в это время нужны. Почему так?
Потому, что организмам присуще некое особое свойство - стремление к
совершенствованию. Чарлзу это было не по душе, да и нужды в таких до-
пущениях он не видел, так как имел уже счастье считать себя учеником
Чарлза Лайеля. «Геологическая наука бесконечно обязана Лайелю, больше,
я думаю, чем кому-либо другому на свете» - писал Дарвин в старости.
Лайель писал о формировании горных пород путем медленного дейст-
вия самых обычных причин (осаждение ила на дно, выветривание), а Дар-
вин решил искать эволюцию в медленном накоплении самых обычных из-
менений организмов. Какое же изменение - самое обычное при смене поко-
лений? Огромные коллекции Дарвина молчали, поскольку в них нигде не
зафиксирован этот важнейший акт - смена поколений. На помощь пришел
пример из ботаники: «Семена растений... производят много форм, между
тем как новые особи, произведенные почками, все одинаковы» - записал
он. Это суждение было в те годы довольно обычно.
Так, русский садовод Вианор Беликов писал, что при пересадке в новые
условия растение «так сказать, перерождается сообразно условиям местно-
го климата» (Беликов В.В. И Русский земледелец, 1838, ч. 1, № 1-3, с. 59).
Вскоре, в 1840 году, он добавил, излагая западные взгляды, что достичь
стойкого итога можно только половым размножением, семенами:
“Неделимые (особи - Ю. Ч.) выращиваемые из семян обязаны различием сво-
им этому смешению при оплодотворении”. “Оплодотворение в царстве рас-
тительном, как и в животном, есть исключительное начало жизни... - эта
жизненная способность в обоих царствах уничтожается”, если нет оплодо-
творения; хотя вегетативное размножение и возможно, но “при каждом во-
зобновлении это обновление юности становится короче, ... сила роста умень-
шается” (цит. по [Баранов, с. 275-276]).
Здесь сказано больше, чем у Ч. Дарвина: выделенные мною слова гла-
сят, что Беликов отметил то, мимо чего прошел Дарвин - жизненную актив-
ность как изменяющий фактор.
С приведенной выше заметки Дарвина, сделанной в июле 1837 года,
началась, можно сказать, жизнь его учения: эволюционную изменчивость,
т.е. материал для медленного накапливания, поставляют «семена» - поло-
вой процесс, соединение свойств родителей.
И тут коллекции заговорили! Различие галапагосских пересмешников
вызвано, решил юный Дарвин, тем, что птицы разных островов давно не
спариваются друг с другом; еще больше их различие с птицами континента,
поскольку контакт с ними потерян давным-давно, еще при образовании ар-
86
хипелага. Чем дальше места обитания (т.е. дольше изоляция), тем больше
различие; сперва это - разновидности, а затем и разные виды.
Итак, причиной появления новых видов можно считать изоляцию. Но
это, так сказать, причина пассивная, дающая условия, а что эволюцию дви-
жет? Об этом пока не сказано, но уже в первой книжке Чарлза есть такая
мимоходная запись. Описав постепенную смену флоры при подъеме в горы,
Дарвин задал вопрос: «Как объяснить это при помощи закона малых разли-
чий, производящих более плодовитое потомство?» Вот первое упоминание
Дарвином идеи естественного отбора.
Точнее, у самого Дарвина оно первым не было: легко видеть, что фраза
является ссылкой на прежний текст, и как раз предыдущий листок из книжки
вырезан. Что там цитировалось? Про обработку каких-либо фактических
данных (на двух маленьких страничках размашистым почерком) и речи быть
не может, зато сразу назван «закон». Назвать законом собственную голую
догадку молодой неуверенный в себе Дарвин тоже явно не мог. Очевидно,
что «закон» заимствован откуда-то, где уже был сформулирован, и тут осо-
бо гадать не приходится - у Жоффруа, многократно упоминаемого в этой и
других записных книжках Дарвина.
Помните? У Жоффруа неуспешные животные «заменяются другими
животными со слегка измененными формами» (п. 2-7). Как и у французского
предтечи, у Дарвина здесь нет даже намека на роль перенаселения и кон-
куренции в процедуре отбора70. Однако вскоре ему на глаза попалась книга
«Очерк о населении» покойного Томаса Мальтуса, священника и профессо-
ра политэкономии. По Мальтусу, общество, как и природа, живет постоян-
ной войной всех против всех, и лишь немногие благоденствуют.
Люди, писал Мальтус, размножаются с ростом времени «в геометриче-
ской прогрессии», и любые ресурсы когда-то окажутся исчерпанными. Об
эволюции он не писал, но это и не требовалось. Дарвин додумал сам и за-
писал в книжке, что фактический рост ограничивается возможностью данной
среды прокормить данное число особей. Это приводит в действие механизм
приспособления к требованиям среды: корм достается лучшим.
С этого дня, 28 сентября 1838 года, повел свою хронологию дарвинизм
- идея естественного отбора приняла специфически дарвинов вид. Впро-
чем, «день рождения» дарвинизма остался в тени: многим казалось зазор-
ным выводить “хорошее” учение Дарвина из “плохой” доктрины Мальтуса,
служившей как бы обоснованием злого поверья «человек человеку - волк».
Увы, саму книгу Мальтуса почти никто в руки не брал, и Дарвин не стал ис-
ключением: в его личной библиотеке первый том Мальтуса не был (как всё
остальное, что читал Дарвин) расчёркан, второй же просто остался нераз- 10 *
10 О них, как и о малых изменениях, писал английский лесовод Патрик Мэтью в 1831 г.
(Соболь С.Л. // История биологии. Наук, вып. 9. М., 1962), но Дарвин не знал об этом.
87
резанным. А жаль - книга очень богата фактами, в том числе и опровер-
гающими «геометрическую прогрессию». Точнее, Мальтус на нее опирался,
говоря о Северной Америке, но отвергал ее для Индии и Китая, население
которых считал постоянным.
К счастью, теперь мы знаем, что «геометрическая прогрессия размно-
жения» - наивное заблуждение Мальтуса. Она равно плохо описывает и
общество, и природу. Так что можно обсуждать проблему спокойно, чисто
исторически, как в главе 1 мы обсуждали древние мифы.
Всякий процесс свободного размножения должен быть вовремя оста-
новлен - иначе беда. Двести лет назад (1798 г.) об этом как раз и заявил
Мальтус, книга которого привела тогда в ужас английское общество, а через
40 лет вызвала восторг молодого Дарвина. В ужас привело предложение то-
го, что ныне именуется планированием семьи, а восторг был вызван уве-
ренностью, что избыточное размножение движет эволюцию.
Лишь недавно выяснено, что животные «планируют семью» куда более
строго, чем предлагали мальтузианцы. Свободного размножения нет даже у
растений и микробов. Те немногие примеры неограниченного роста числа
особей, какие приводил сам Мальтус и какие лежат в основе построений
дарвинизма, являются редкими и краткими исключениями. Мы вернемся к
этим вопросам в конце главы, а здесь заметим, что как избыточность раз-
множения, так и изменчивость суть формы активности. От нее, как видим,
никуда не деться.
От записных книжек Дарвина до публикации «Происхождения видов»
прошло 20 лет. За это время Дарвин стал известным ученым, лучшим в ми-
ре специалистом по усоногим ракам, но по эволюции не опубликовал ниче-
го, да и писал мало. Правда, в 1838-1844 годах написал три очерка, а в 1857
году сел писать «Длинную рукопись», но за 12 лет (1845-1856) не записал
почти ничего. Почему?
В ноябре 1844 года в Лондоне вышла в свет анонимная книга «Свиде-
тельства естественной истории творения». Ученый мир ее презрительно от-
верг, несмотря на ее громкий успех у публики, и Дарвин, болезненно чувчт-
вительный к молве, побоялся той же участи.
Вскоре выяснилось, что автором был писатель и издатель Роберт Чем-
берс. Вот уже в третий раз в Англии, и притом опять неспециалистом (каки-
ми были Хэйл и Эразм Дарвин), давался синтез эволюционных воззрений.
Эволюционизм Чемберса тоже был сквозной (от космоса до обществ) и
включал новейшие идеи - например, немецкую теорию параллелизма. Был
популярно описан фактический ход развития организмов, вплоть до появле-
ния позвоночных животных и цветковых растений.
Дарвин прочел «Свидетельства» и сделал много заметок. Например:
«Идея, что рыба переходит в пресмыкающееся, чудовищна». Если вспом-
нить, что тогда пресмыкающихся объединяли в один класс с земноводными
(которых нынешняя наука выводит из двоякодышащих рыб), то станет ясно
88
- Дарвин не был тогда готов признать факт эволюции в полном объеме. Это
важно запомнить, поскольку эволюции в полном объеме нет, по сути дела, и
в «Происхождении видов».
2-9. И острый галльский смысл, и сумрачный германский гений
Нигде Дарвин не описывал того, как, собственно, шла по его мнению
конкретная история земной жизни, как появлялись и исчезали те или иные
группы организмов. Об этом, опираясь на успехи палеонтологии, писали
другие - как популяризаторы вроде Чемберса, так и ученые типа Жоффруа.
Сам Этьен Жоффруа Сент-Илер в июне 1844 года умер (как и Ламарк -
слепой, почти всеми забытый и лишь в гробу удостоившийся горячих при-
знаний, столь нужных ему раньше), но его влияние с годами росло - осо-
бенно в России, в Москве, в трудах Карла Рулье.
Луи Шарль Рулье, несмотря на французское имя, данное ему отцом,
Франсуа Рулье, нижегородским ремесленником (видимо осевшим в России
при отступлении армии Наполеона), был человеком вполне русского воспи-
тания и культуры. Едва начав преподавать, Рулье стал именовать себя
Карлом Францевичем и все популярные статьи (а также учебники) писал по-
русски. Именно его яркий и точный русский язык позволил ему стать первым
пропагандистом эволюции в России (чем он сильно отличался от самого
знамениго эволюциониста России, Бэра, который прожил в России 64 года,
но до конца дней не писал по-русски). В стране, где за любое отклонение от
официальной доктрины можно было тогда, при Николае I, не только ли-
шиться работы, но и угодить в ссылку, такая пропаганда дорого стоила и
была едва ли не важнее, чем собственные эволюционные идеи.
А у Рулье и они были. Можно уверенно сказать, что трудами его и его
школы был создан русский эволюционизм - столь же своеобразный, как
эволюционизм английский, французский и немецкий. (Эволюционисты были
в России и до него, но либо неоригинальные, либо заезжавшие в Россию
ненадолго.) С 1845 года Рулье в лекциях и публикациях подробно излагал
сведения о палеонтологических основаниях эволюции, как раз в то время
обильно поступавшие с Запада. Вот образец его стиля:
«В продолжение второго периода органической жизни (мезозойской эры -
Ю. Ч.) на земле явился в растениях новый тип постепенного развития, тип пре-
лестной роскоши растительности в тропической полосе - пальм и вообще рас-
тений односеменодольных. Прямой, не ветвящийся или маловетвящийся,
ствол, несущий листья на одинаковой высоте, в виде кольца; листья однооб-
разные на обеих поверхностях, с нервами параллельными; цветки, разделенные
обыкновенно на три доли, - вот этот тип, к которому относятся наши травы,
лилейные растения и др. В животных же не явилось ни одной новой основной
формы (типа - Ю. Ч), продолжали только существовать предшествовавшие,
изменяясь, конечно, в видах и часто в родах, и явился новый класс позвоноч-
ных - гады [класс амфибий, затем класс рептилий - Ю. Чф предуготовленный
89
первым периодом. Гады же в свою очередь распались на новые формы, слу-
жившие предвестниками последовавших за ними различных порядков нынеш-
них гад (голокожих, или лягушек; ящериц, змей и черепах), птиц и зверей.»
Рулье начисто отвергал всякие родословные древа:
«Эта мысль, занимавшая еще недавно естествоиспытателей-поэтов, совершенно
отвергнута наукою. Тому немного времени, как представляли все животное и
растительное царство в виде двух деревьев, подымающихся из недр морских и
постепенно разрастающихся на воздухе: корни дерева показывали на то, что не-
совершеннейшие животные и растения живут в воде, а разветвление сучьев - на
мнимое постепенное преобразование одного класса животного и растения в
другие, которые тем были совершеннее, чем более удалялись от ствола».
К этой мысли мы не раз вернемся. Рулье отвергал древа потому, что
отрицал саму возможность существования одиночного вида растений или
животных - как из экологических («ни одно органическое существо не живет
само по себе; каждое вызывается к жизни и живет только постольку, по-
скольку находится во взаимодействии с относительно внешним его ми-
ром»), так и из палеонтологических соображений:
«История же земли, насколько она доступна человеку, - геология свидетель-
ствует, что в древнейших толщах, в которых в первый раз встречают живот-
ных, их находят вдруг из всех классов беспозвоночных, к которым по мере
приближения [к поверхности] земной коры прибавляются постепенно классы
позвоночных».
Механизм эволюции Рулье излагал по Ламарку и Жоффруа. Новым
здесь было отношение к изменчивости: вместо деления изменений на внут-
ренне присущие (прогрессивные) и вызванные (упражнением органов и
прямым влиянием среды на зародыши), молодой московский профессор
предпочитал говорить, что всякое изменение «есть результат взаимодейст-
вия двух факторов - сил образовательных животного и внешних деятелей»,
т.е. взаимодействия внутренней и внешней природы организма. Рулье на-
зывал это «законом двойственных отношений», мы же в таких случаях при-
выкли говорить о системном характере эволюции, т.е. о том, что отделить
эволюцию данного вида от эволюции всей природы нельзя.
Признавая, что вид может вымереть от неспособности приспособиться
ко внешним условиям, Рулье был далек от мысли все сводить на конкурен-
цию. В заметке о зубре (1848 г.) он отметил, что тот склонен к вымиранию
несмотря на строгую охрану, и сделал афористический вывод в духе Брок-
ки: «Как отдельная особь животного живет кратковременно, так и целый вид
(порода) животных имеет свои начало и конец во времени жизни на земле».
Впоследствии эта идея тоже стала одной из основных в ламаркизме. Нако-
нец, у Рулье мы находим одну из самых ранних (1852 г.) формулировок
тройного параллелизма (высшего тогда достижения теории параллелизма):
«...главным законом населения нашей планеты растениями и животными есть
та же последовательность в постепенном изменении форм и перехождении из
90
одной среды в другую, которому следуют животные и ныне в отдельном сво-
ем развитии... Здесь два ясные параллельные ряда - один доисторических жи-
вотных, другой - представляемый развитием нынешних животных. Это гене-
тический ряд. К ним должно прибавить еще и третий, представляемый вполне
развитыми нынешними животными... которые ясно размещаются подобным
же рядом совершенствующихся форм» (К.Ф.Рулье. Избранные биологические
произведения. М.-Л., 1954, с. 161).
А что это значит - совершенствование?
2-10. Пока Дарвин писал «Длинную рукопись»
Что рыба проще человека - всем интуитивно ясно, но можно ли утвер-
ждать, что, например, лягушка проще ящерицы? Конечно, у ящерицы слож-
нее устроено сердце и есть амнион (оболочка, препятствующая высыханию
яйца), но зато у лягушки есть три способа дыхания - жаберное, легочное и
кожное. Чтобы не вести бесконечные споры, нужно общее правило, крите-
рий высоты организации. Такой критерий предложил Бэр: выше организо-
ваны те, у кого можно указать большее число различных (по строению и
функции) органов. В 1828 году ему удалось показать, что четыре типа Кю-
вье являются четырьмя типами онтогенеза, а значит в каждом типе можно
стало выстраивать организмы в ряды еще и по числу различимых стадий
развития зародышей.
Подход Бэра довел в 1853 году до совершенства немецкий зоолог и па-
леонтолог Генрих Брони. Он предложил критерий высоты организации,
наиболее годный для работы: морфологический прогресс он характеризо-
вал не только через дифференцированность и специализацию частей тела,
но и через симметрию тела, уменьшение числа гомологичных органов, кон-
центрацию органов в пределах тела (например, головной мозг у высших
вместо распределенных по телу ганглиев у низших) и их проникновение в
глубь тела, а также - через усложнение онтогенеза.
Исследуя вымершие и нынешние организмы, именно Брони выявил
фактические прогрессивные линии их развития, чем заложил, в сущности,
основы эволюционизма. Тем самым он встал перед трудной проблемой: его
огромный материал ясно говорил об эволюции, тогда как сама идея эволю-
ции считалась в его кругу ненаучной фантазией.
Профессионально он принадлежал морфологической школе Кювье, но
собственные работы и принадлежность (душой) немецкой культуре вели его
в объятия натурфилософии, которую та школа строго порицала. Личный
выход Брони нашел в том, что взялся описать с единой теоретической по-
зиции весь фактический ход развития жизни на Земле (данные об этом бы-
ли уже весьма обширны), однако сам факт порождения одним видом друго-
го отрицал, считая эволюцию непрерывным творением, как когда-то сделал
Лейбниц. Иными словами, Брони как натурфилософ избрал путь Лейбница в
ущерб Окену.
91
Его позиция импонировала кювьеровской школе, и в 1857 году Брони
получил премию Парижской Академии наук за обширную книгу «Исследова-
ния законов развития органического мира во время образования земных
оболочек». Написана она тяжело и беспорядочно - несмотря на просьбу
французских академиков облегчить стиль, Брони ухитрился дать неудобо-
читаемый текст даже по-французски; немецкий же вариант местами вообще
неподъемен. Ее мало читали и вскоре забыли.
Однако «Исследования» Бройна - одна из самых значительных книг той
поры, итог немецкого додарвинского эволюционизма. Без нее нельзя уяс-
нить ни причины успеха «Происхождения видов», ни развития эволюцио-
низма вообще. Мы привыкли считать, что представление о постепенном
развитии жизни вошло в науку с Дарвином, но ведь в его книгах нет ни опи-
сания истории флор и фаун, ни анализа морфофизиологической эволюции
каких-либо групп организмов, ни глобальных обстоятельств (связи эволю-
ции жизни с составом атмосферы, животных с растениями и т. п.), ни много-
го другого. Все это мы видим в трудах Бронна.
(Его выводами, кстати, широко пользовался и Рулье. Увы, как раз «Ис-
следований» Бронна он увидать не успел - в расцвете сил он в 1858 году
внезапно умер, нанеся страшный урон и своей семье и московской эволю-
ционной школе.)
Вот «Первый фундаментальный закон» Бронна (формулировка его
столь длинна, что я вынужден дать ее в выдержках):
«Различные организмы следовали во времени и пространстве в стольких типах
и количествах, какие соответствовали внешним условиям», причем «появле-
ние двух органических царств было одновременно», «первичное население
соответствовало теплому климату без смены времен года», а «все последова-
тельные обновления населения достигались уничтожением прежних видов и
последовательным появлением новых видов, без постепенных переходов»
(H.G. Broun. Untersuchungen uber die Entwickelungsgesetze der organischen Welt
wahrend der Bildungszeit unzerer Erdoberflache. Stuttgart, 1858, S. 483-484).
Добавлю, что среди сил, истребляющих прежние виды, Брони допускал
и вытеснение худших вариантов лучшими - как у Дарвина.
А вот «Второй фундаментальный закон»:
«Наряду с первым, явно существует положительный и независимый закон тво-
рения, даваемый нам в простоте и упорядоченности всех, как одновременных,
так и последовательных обновлений органического мира.»
Запомним про упорядоченность (о ней мы будем говорить в ч. 6). Суть
же данного закона в том, что наряду с внешними условиями (которые, буду-
чи чисто негативным фактором, могли только ограничивать «действия по
плану творения»), необходимо некоторое активное начало, формирующее
как организмы, так и их отношения. В этих рамках Брони отмечал
«постоянное равновесие между растениями и животными, наземными и водными
животными, травоядными и хищными - в каждом творении; и всё это делается
92
гораздо точнее, чем можно ожидать действием одних внешних условий, которые
могут разрушать, но не создавать ... Систематическое и прогрессивное развитие,
как и закон, который им управляет, суть факты, в которых невозможно дольше за-
блуждаться. Тем не менее, мы не должны представлять это прогрессивное разви-
тие как состоящее в первичном появлении одних лишь Phytozoa»,
т.е. животнорастений (ныне этот термин забыт, вместо этого говорят о низ-
ших тканевых животных). И далее:
«Высшие подцарства [типы - Ю.Ч.] были сотворены друг за другом быстро,
тогда как классы и отряды, впервые появившись, были представлены форма-
ми, постепенно восходящими» (там же, S. 487-488).
Как Брони понимал феномен «восхождения» (прогресса), мы уже знаем,
а теперь он решил обсудить еще и его механизм. Допуская конкуренцию как
одну из причин вымирания, Брони связывал появление новых форм не с
конкуренцией, а с действием особой силы природы. “Та же сила, которая
первоначально создала организмы, ... действовала в течение всей геологи-
ческой истории, вплоть до появления человека. При этом нигде не обнару-
живается постепенного превращения прежних видов в новые; наоборот, но-
вые виды всюду возникают без участия прежних” (там же, S. 80). Не надо
думать, что Брони понимал эту силу как постоянное вмешательство божьей
воли - нет, он понимал ее именно как особую силу, аналогичную гравита-
ции, химическому сродству и т.п.
Брони ввел замечательное понятие, закон террипетного развития (по-
французски terripete - нечто устремленное к земле): тип морфологически
«стоит тем выше, чем более преобладают в нем наземные классы и отря-
ды». Брони пояснял: в ходе истории чисто морские формы становились ли-
торальными (живущими в зоне прилива), затем береговыми, просто назем-
ными и, наконец, горными. Такую последовательность форм Брони назвал
террипетной и обнаружил, что вдоль нее наблюдается морфологический
прогресс, т.е. усложнение строения.
В пояснение приведу пример: плавник преобразуется в ласт, ласт - в
лапу и т.д.; тело приподымается на все более длинных ногах, затем прини-
мает наклонное (а потом и вертикальное) положение, а передние конечно-
сти обращаются в крылья или в руки; при этом наблюдается усложнение
мозга позвоночных. Данным законом мы воспользуемся в частях 3 и 4.
Брони не признал, даже намеком, что террипетный ряд может быть ге-
нетическим, но в конце «Исследований» сделал робкий подарок эволюцио-
нистам: «...мы не знаем, какую роль взял на себя Творец в определении
систематического порядка организмов». То есть - Бог не обязательно сам
творил каждый вид.
Некоторые палеонтологи писали об эволюции более прямо. Так, швей-
царский палеоботаник Освальд Геер в 1859 году опубликовал свою теорию
перечеканки (die Umpragungstheorie), согласно которой современная флора
93
произошла из флоры третичного периода77 сразу, после долгого времени
постоянства. Видно влияние катастрофизма Кювье, но “перечеканка” одного
типа строения в другой означала эволюцию, притом в широком смысле (не
только видов, но и флор как целого).
2-10* Множественный параллелизм
Мы уже знаем про тройной параллелизм (Рулье, п. 2-9), но через 5 лет,
как раз в разгар работы Дарвина над «Длинной рукописью», в Америке был
заявлен более сильный результат: геолог и палеонтолог Жан-Луи Агассиц
(приехавший из Швейцарии) писал о пятерном параллелизме. Он назвал 5
типов рядов:
1) “структурная градация” (усложнение строения) в геологической по-
следовательности появления организмов;
2) сходство между взрослыми формами разных таксонов системы;
3) сходство эмбрионов со взрослыми формами иных таксонов;
4) “структурная градация” среди ныне живущих видов;
5) географическое распространение.
Рулье затронул пункты 1, 3 и 4, а пункты 2 и 5 нам надо пояснить. Вто-
рой прост: сходные ряды мы видим, например, между плацентарными и
сумчатыми млекопитающими. Пятый сложнее, о нем Агассиц писал:
“Почти в каждом классе имеются тропические семейства, и они-то, как пра-
вило, высшие в своих классах” (Agassiz L. Contributions to the natural history of
USA. Pt 1. Boston, 1857, p. 121).
Он приводил примеры. В частности: обезьяны живут в тропиках и раз-
витием своим ближе всего к человеку. О ведущей роли тропических форм в
эволюции мы узнаем в п. 9-4, а здесь замечу, что эволюцию Агассиц отри-
цал. Можно и далее подсчитывать, кто сколько параллельных рядов взял
для сопоставления, но мы будем прямо говорить о множественном парал-
лелизме (п. 6-3*).
2-10** Проблема вида
Дарвин знал работы Бронна, Геера и Агассица, ценил их как знатоков
фактов, но преобразование флор и фаун не занимало его нисколько. Его
интересовала лишь «.проблема вида», т.е. процесс накопления внутривидо-
вой изменчивости и возможность образования таким путем нового вида. Ее
четко обозначил в 1852 году английский философ Герберт Спенсер в не-
большой статье «Гипотеза развития». Там изложена идея единого процес-
са эволюции (от инфузории до человека), названо ее причиной приспо-
собление, впервые термин эволюция применен в нынешнем смысле, как
И Тогда геологические слои делили на первичные (ныне палеозойские), вторичные
(ныне мезозойские), третичные (ныне весь кайнозой без антропогена) и четвертичные
(ныне антропогеновые). О Геере см.: Давиташвили Л.Ш. История эволюционной па-
леонтологии от Дарвина до наших дней. М.-Л., Изд. АН, 1948.
94
обозначение процесса исторического преобразования организмов. Но ни-
чего не сказано о движущей силе эволюции (рус. перевод в кн.: Теория
развития. СПб., 1904).
Вскоре о проблеме стали писать чуть ли не все научные журналы, ка-
савшиеся биологии, и я ограничусь одной цитатой. В 1857 году ученик Ру-
лье, молодой зоолог Н.А. Северцов, поместил в одном парижском журнале
статью по классификации хищных, где, между прочим, писал:
«Индивидуальные вариации ведут к образованию видов, создают последние,
делаясь наследственными признаками. Нельзя еще решить, происходит ли это
на основе принципа развития, в силу изменений, свойственных самому орга-
низму и независимых от внешних влияний (принцип градации Ламарка -
Ю.Ч.) ... или же принцип этих изменений заключается в разнообразном влия-
нии окружающей среды» (цит. по [Райков, т. 4, с. 39]).
Дарвин тоже не мог решить этот вопрос, но склонялся к тому, что при-
чина первичных изменений в некотором смысле случайна. Статьи Север-
цева он не заметил, но и без нее знал, что надо спешить, поскольку такие
статьи попадались ему регулярно. Весь 1857 год и весну следующего он пи-
сал так называемую «Длинную рукопись» - текст, в котором собирался, на-
конец-то, подробно и обоснованно изложить свои взгляды.
За 13 лет перерыва сильно изменилась аргументация Дарвина: если
первый его очерк (1838 г.) был комментарием к одному богословскому трак-
тату, прочитанному тогда Дарвином, а два других (1842, 1844) сами были в
некотором смысле богословскими трактатами (оттуда и взято приведенное
мною в п. 1-12 рассуждение Дарвина о всевидящем Существе), - то «Длин-
ная рукопись» уже очень похожа на первое издание «Происхождения ви-
дов», где богословская аргументация почти исчезла (в те годы “естествен-
ное богословие” вообще уходило из английской научной и общественной
мысли), уступая место ссылкам на приспособление, случайность наследст-
венных изменений и на Мальтуса.
Дарвин написал уже полторы тысячи страниц, когда труд его был вдруг
прерван. В феврале 1858 года малоизвестный тогда зоолог Альфред Уол-
лес (1823-1913) послал ему из Индонезии рукопись, в которой изложил ту
же, что у Дарвина, схему образования нового вида. И даже применил Маль-
тусов термин «борьба за существование», которого у Дарвина до тех пор ни
в одном тексте не было. Уоллес простодушно просил именитого натурали-
ста представить его рукопись в какой-нибудь журнал.
В мае Дарвин получил пакет Уоллеса, и настал бурный трагический ме-
сяц: прозрения, сомнения в своей роли, болезнь детей (один умер), советы
друзей скорее публиковать идею и, наконец, написание и отсылка в Лондон
(вместе со статьей Уоллеса) своего «Извлечения», где много взято у Уолле-
са, но имя которого там не упомянуто. (Об этом можно прочесть немного в
ЧЭ и, подробнее, в книге: Brooks J.L. Just before the Origin. N.Y., 1984; а так-
же в моей рецензии на нее, ВИЕТ, 1988, № 2.) Не будем судить строго.
J
95
2-11- Первые успехи и неудачи дарвинизма
Уже в августе статьи Дарвина и Уоллеса появились в лондонском
«Журнале Линнеевского общества». Лед сломан: впервые об эволюции за-
говорило респектабельное ученое общество. Более того, статьи заметили.
Сам Оуэн, председательствуя осенью на съезде Британской ассоциации
содействия развитию наук, высказался о статьях положительно. Заметил,
правда, что ископаемый материал не позволяет делать таких смелых выво-
дов, как у Дарвина, но тот и сам знает, что доказательства надо еще искать
и искать. Главное - сэр Ричард не счел это чепухой!
В январе 1859 года откликнулся журнал «Zoologist». Критик прочел ста-
тьи с интересом, но считает выводы поспешными. В мартовском номере
снова отклик. Этот критик прямо отверг идею естественного отбора: не убе-
ждает его аналогия с искусственной селекцией. Вскоре пришел текст речи
Сэмюэла Хоутона, президента Ирландского геологического общества, кото-
рый говорил, уже довольно грубо, что естественный отбор - не научная, а
натурфилософская идея, противоречащая всему естестврзнанию; правда,
он не возражал против самой идеи изменчивости, но «Против этого нечего
возразить, кроме того, что это не ново».
Весной Оуэн издал книжку «О классификации и географическом рас-
пределении млекопитающих», половину которой составили два приложения
об эволюции. Дарвин назван не был, но сэр Ричард спорил явно с ним. В
первом приложении он без лишней скромности формулировал «величай-
шее обобщение», сравнимое с законами Ньютона и состоящее в том, что в
природе есть разумное начало, снабжающее организмы не только нужными
приспособлениями, но и нужной изменчивостью. Второе приложение иссле-
довало эту изменчивость; вывод был тот, что она не затрагивает сущност-
ных свойств организмов. Например, ни у собак, ни у приматов она никогда
не может привести к изменению ни зубной формулы, ни точек крепления
мышц, ни принципов строения черепа (Owen Fl. On the classification and
geographical distribution of the mammals... London, 1859, c. 100, 103). Фактиче-
ски этим Оуэн предостерегал от подмены понятий, от смешения сути и ак-
циденции (см. п. 1-6**).
Как видим, уже первые критики возражали то же, что новейшие
критики возражают сейчас. Не читать друг друга и сотни раз повторять
один и тот же довод стало традицией эволюционизма (не только дарвиниз-
ма). Дарвин редко отвечал (а когда отвечал, то не по сути, но лишь по дета-
лям), и оставлять критику без ответа по-существу (или вообще без ответа)
тоже стало эволюционной традицией.
Книга Дарвина «О происхождении видов путем естественного отбора,
или сохранение удачных пород в борьбе за жизнь» вышла в свет в Лондоне
в ноябре 1859 года и сразу оказалась в центре внимания прессы. Первая
рецензия появилась в научно-популярном журнале «Athenaeum» еще за два
96
дня до выпуска тиража в свет, носила скандальный характер (в частности,
вполне лживо утверждала, что книга посвящена происхождению человека
от обезьяны), вызвала ажиотаж и привела к тому, что тираж был сразу рас-
куплен случайными покупателями, а ученым достались лишь те две дюжины
экземпляров, что разослал им сам Дарвин.
Подробнее обо всей этой ранней истории дарвинизма см.: Чайковский
Ю.В. Перед выходом «Происхождения видов» // ВИЕТ, 1981, № 4.
Дарвин сразу стал получать массу писем, и они обескураживали впе-
чатлительного автора. Биогеограф Хьюит Уотсон был в восторге: «Вы вели-
чайший революционер в естественной истории нашего века, если не всех
веков», зато старый адмирал Фиц-Рой, друг молодости Дарвина - в ярости:
«Вы едва ли читали труды нынешних авторитетов, кроме тех обрывков, ко-
торые могли обратить себе на пользу». Кому верить?
Вскоре же в печати появился термин дарвинизм. Его пустил в оборот в
анонимном обзоре молодой друг Дарвина зоолог Томас Гексли. Признав,
что у Дарвина мало доказательств и много ошибок в рассуждениях, он тем
не менее заключил статью восторженно:
«У Коперника орбиты - круги, но за ним шли Кеплер и Ньютон... Что из того,
что орбита Дарвинизма окажется не вполне круговой?» ([Huxley Т.Н.] Darwin он
the Origin of Species // Westminster Review, New Ser., 1860, v. XVII, № 2, p. 569).
Оуэн, так дружелюбно принявший журнальные заметки Дарвина и Уол-
леса, теперь, в рецензии на книгу Дарвина, заметил язвительно:
«М-р Дарвин редко цитирует труды своих предшественников, из которых он,
можно полагать, более вывел свои идеи происхождения видов, чем из фено-
менов распределения обитателей Южной Америки».
Сперва Дарвин отмалчивался, потом пробовал всё признавать в печа-
ти, затем опять перестал отвечать на упреки, в том числе важные. Только из
его писем коллегам мы знаем, что некоторые из критиков его интересовали,
и притом всерьез.
Например, геолог А.А. Кейзерлинг - единственный в России, кому
Дарвин послал первое издание, в письме Дарвину говорил не о своем при-
оритете, а о вещах более серьезных. Признавая отбор как причину приспо-
собленности видов ко внешним условиям, он, однако, заметил Дарвину: ви-
ды изменяются слишком регулярно, словно компоненты химических реак-
ций. Он хотел сказать, что отбор играет ту же роль в биологии, что внешние
условия реакций - в химии (набор реактивов, температура, давление и т.п.),
но что у Дарвина нет иного движущего начала, тогда как в химии есть еще
внутренние законы - например, химическое сродство (водород стремится
соединиться с кислородом, но не с металлом); в биологии же такие законы,
по Кейзерлингу, предстояло еще найти [Райков, т. 4, с. 641]. По сути он по-
вторял Бронна. Об этом писали тогда многие: теории Дарвина не хватает
внутреннего активного фактора, какого-то механизма, поставляющего но-
вые варианты организмов.
97
2-11* Метод Дарвина и случайность
У Дарвина эти новые варианты мыслились так: дети непохожи на роди-
телей, пусть и немного, но буквально по всем признакам; варианты возни-
кают всякие, но только те из них, которые чем-то облегчают их обладателям
борьбу за жизнь, сохраняются - они обеспечивают повышенную вероят-
ность оставления потомства. (То, что такая изменчивость есть тип активно-
сти, было ясно, как мы видели, Оуэну, но никак не Дарвину, видевшему тут
аналог броунова движения частиц.)
«Всякие» надо было понимать только в том смысле, что новый признак
появляется независимо от того, нужен он или нет. Проще говоря (а читатель
обычно любит то, что проще), новый вариант возникает случайно, и лишь
взаимодействие организма со средой определит, полезен ли он. Дарвин
возражал против этого упрощенного понимания:
(«Я до сих пор иногда говорил так, как будто изменения... были делом случая.
Это, конечно, совершенно неверное выражение, но оно служит для показа
нашего незнания причины каждого конкретного изменения»),
но теперь с огорчением видел, что многие видят его учение как
«нагромождение случайностей». Как быть? Подчеркивать впредь, что
перемены в строении организмов идут по каким-то четким (как химические
реакции), пусть и непонятным, законам? Дарвин пробовал делать так, но
въедливые критики зашумели, что этим Дарвин отказывается от принципа
естественного отбора: выходит, что облик организма определяется прежде
всего теми возможностями, какие предоставляются законами изменчивости,
отбор же выступает не автором, а лишь цензором (позже из этой мысли
вырос номогенез). Вот Дарвину и приходилось говорить, что первичные
изменения не направленны, а случайны.
Некоторые критики шли еще дальше: если эволюция невозможна без
направленных изменений, то не проще ли сказать, что ею управляет Все-
вышний, задавая изменчивость? В самом деле, наблюдая лишь тот факт,
что все особи различны, вообще нельзя доказать, что виды произошли друг
от друга, а не были прямо созданы такими, какими мы их видим. То, что они
именно произошли, было догадкой.
Дарвин не раз указывал на это, но не решался прямо сказать (а, воз-
можно, даже и подумать), что этим он идет вразрез с логикой тогдашней
науки. Эта его новая методология (в XX веке ее назовут гипотетико-
дедукт иеной) была едва ли не главным, что определило судьбу книги, но
выяснилось это лишь через сто лет (Ghiselin М.Т. The triumph of Darwinian
method. Berkeley, 1969).
Дарвину оставалось надеяться, что новое поколение натуралистов при-
дет в науку, уже зная его логику как само собой разумеющуюся:
«Если суждено моим взглядам когда-нибудь стать общепринятыми, то это
произойдет тогда, когда подрастут молодые люди и заменят старых работай-
98
ков, когда молодые люди найдут, что они могут группировать факты и нахо-
дить новые пути исследований лучше, руководствуясь понятием о естествен-
ном происхождении, чем понятием о творении».
(Отбор, как видим, тут не назван.) Так и произошло. Даже больше: но-
вое поколение приняло не только давнишнее «понятие о естественном про-
исхождении», но и новое - о естественном отборе малых случайных укло-
нений. Что же касается самой проблемы случайности, то она так и застыла
в дарвинизме в том виде, как ее оставил Дарвин: хотя каждое наследствен-
ное изменение в каждой своей стадии происходит по вполне определенным
законам (это ясно показала молекулярная генетика), однако проследить все
взаимосвязи между этими стадиями и их причинами не удается, так что
дарвинизм поныне утверждает, что наследственные изменения случайны. О
случайности мы поговорим в п. 5-18.
Летом 1860 года вышел немецкий перевод книги, сделанный Бренном.
В послесловии Брони задал самые простые и каверзные вопросы. Почему
отбору приписано формирование начальных стадий сложного приспособле-
ния, если пользы можно ожидать лишь от достаточно поздних стадий, когда
новая функция хоть в какой-то мере уже действует? Почему отбор измене-
ний, направленных во все стороны, приводит не к мешанине признаков, а к
тем видам, которые мы наблюдаем? Как формируются явно бесполезные
признаки, вроде зубного рисунка?
И главное: даже если допустить, что начальные и промежуточные ста-
дии формирования полезных качеств чем-то полезны и могли отбираться,
то каждая такая стадия должна вытеснять предыдущую и вытесняться по-
следующей; где следы этого процесса? В ископаемом материале их, по
уверению Бренна, нет^2. То же вскоре заявили все ведущие палеонтологи, и
Дарвин был в растерянности.
Как он был рад, когда в 1861 году ему сообщили, что в Бюллетене
МОИП появилось по-французски краткое сообщение: московский геолог
Герман Траутшольд говорил о промежуточных формах, представляющих
переход от видов к видам и тем поддерживающих точку зрения Дарвина. В
очередном издании книги Дарвин сослался на это французское резюме.
Сам доклад Траутшольда был на немецком языке, которым Дарвин
владел слабо, и читать доклад не стал. А зря: в нем было не совсем то. Ав-
тор был осторожен: расположив раковины аммонитов, найденные им в юр-
ских отложениях Подмосковья, в ряд, показавший постепенные переходы от
вида к виду, он заметил, что промежуточные формы могут быть как пере-
ходными в смысле Дарвина, так и одновременно жившими вариантами, и
даже гибридами. (Позже Траутшольду удалось отнести некоторые формы к
различным горизонтам и тем подтвердить эволюцию, но свидетельств вы-
17
О месте Бронна в науке об эволюции см. Чайковский Ю.В. // Природа, 1984, № 7.
99
теснения одних другими он в этом не увидел, и, приняв идею эволюции,
идею отбора отверг.) Дарвину данное сообщение осталось неизвестным
(подробнее см. [Чайковский, 1989, с. 132]). Невольно опять вышло то, что
Фиц-Рой назвал «обрывками себе на пользу».
2-12. Дарвинизм без отбора
Казалось бы, дарвинизм без идеи отбора немыслим, но это не так. Сам
Дарвин не раз заявлял, что главное для него - продемонстрировать факт
постепенного преобразования вида в вид, а в 1863 году писал даже: «Мне
лично естественный отбор, конечно, очень дорог, но это мне кажется совсем
незначительным в сравнении с вопросом о творении или изменении». Да и
«предшественники» наперебой твердили, что Дарвин их повторяет, хотя у
большинства и намека не было на отбор.
До Дарвина более двухсот авторов писали об эволюции, некоторые ох-
ватывали проблему гораздо шире, чем он, а некоторые рассматривали и
отбор, но услышан и признан обществом был именно Дарвин. Почему?
Уверения, что Дарвин доказал свои идеи фактами, всякий может прове-
рить сам, почитав его книгу. Фактов там много, но касаются они изменчиво-
сти, а не отбора. Есть параграф «Примеры действия естественного отбо-
ра», где даны «два воображаемых примера»: волки разной быстроногое™ и
цветки разной сладости. Более реальных примеров отбора в трудах Дарви-
на нет, в том числе и в Длинной рукописи (содержащей, как считалось до ее
опубликования в 1974 году, все недостающие аргументы). Успех дарвиниз-
ма был явно вызван чем-то другим.
Отбор вообще мало интересовал большинство читателей (это видно
хотя бы из того, что никто из ранних переводчиков не сохранил дарвинского
смысла слова «отбор»; см. ЧЭ, с. 81). Он был неким символом, как бы фор-
мулой, которой пользуются, не интересуясь, верна ли она.
Мнение самого Дарвина мало кого интересует: почти все пишут «Дар-
вин в 1859 г. сказал...», а затем цитируют шестое издание (1872), хотя это
разные книги с очень разными выводами.
Просто для победы идеи эволюции настало время, и обществу оказа-
лось достаточно того, что известный натуралист объявил, что знает меха-
низм этого явления. Обществу, в котором тогда быстро нарастали движения
за социальное переустройство, нужна была сама эволюция, сама идея
борьбы, а не факты и подробности из биологии.
Часто пишут, что в России учение Дарвина было принято очень благо-
желательно. Действительно, ни один ранний рецензент, кроме православ-
ных богословов, не отверг саму идею эволюции. За этим фактом, однако,
скрывалось весьма сдержанное отношение к естественному отбору и почти
полное неприятие мальтузианской идеи [Чайковский, 1989].
Ученик Рулье зоолог С.А. Усов в статье «Зубр» (январь 1859 года, т.е.
еще до книги Дарвина) развивал мысли учителя о вымирании. Усов пре-
100
красно описал конкуренцию самцов за самку: могучие старые самцы легко
отгоняют молодых от самок, хотя сами для размножения уже непригодны, и
это сильно снижает численность стада. Позже, в книге «Зубр» (1865), со-
глашаясь во всем с Дарвином, тем не менее он выказал понимание борьбы
за существование в духе Рулье:
«Как особь истрачивается в борьбе за существование, теряет возможность
реагировать... и умирает, так точно и вид вымирает, если не находит в себе
довольно сил, чтобы противоборствовать влияниям внешней среды».
Зубр, по его мнению, как раз и «не находит»: это выражается и в неле-
пой борьбе самцов, и в низкой плодовитости самок. Спасти вид может
скрещивание с бизоном, не проявляющим подобных свойств. (Как показы-
вает вековой опыт, Усов оказался прав, см. п. 12-4.) Тем самым, Усов ясно
отграничил естественный отбор от объективной склонности вида к выми-
ранию. Запомним это. Заодно обратим внимание и на другое: естественный
отбор не может (в отличие от искусственного) помешать тенденциям, в чем
мы не раз еще убедимся.
2-13. Дарвин без Мальтуса
Другой ученик Рулье, ботаник С.А. Рачинский, первый переводчик
«Происхождения видов», описывая отбор в статье «Цветы и насекомые»
(1863), вообще обошелся без ссылок на конкуренцию. По Рулье, эволюция
начинается там, где внешние воздействия «разрывают целость образова-
ния» (мы бы сказали: нарушают связь вида с его экосистемой), и Рачинский
тоже в строении каждого существа видел «и элементы, служащие для со-
хранения вида, и элементы, служебные прочим живым существам». Это
достаточно далеко от Дарвина (с его конкурентной схемой эволюции), но
Рачинский был уверен, что излагает именно его:
«Из многочисленного потомства всякого поколения органических существ
выживает лишь малейшая доля, выживают лишь те представители, которые
вполне и до последней подробности приспособлены ко всему строю внешней
природы»; она поступает «как искусный заводчик, дорожащий чистотою кро-
ви в своем стаде. Она строго подбирает в каждом поколении те особи, кото-
рые она предназначает для продолжения племени: поэтому Дарвин и назвал
этот процесс естественным подбором родичей (natural selection)».
Почти излишне напоминать, что это не дарвинизм. Дарвин утверждал
подобное в ранних очерках (п. 1-12), а в книге говорил лишь о шансах оста-
вить потомство при свободном спаривании; речь у него шла не об отдель-
ном поколении и не о приспособленности каждого поколения «до последней
подробности», а (выражаясь нашим языком) лишь о сдвиге средних значе-
ний за много поколений. Рачинский повторял давно забытого Гердера и мо-
лодого Дарвина, чьи очерки не были тогда известны никому.
Ход мыслей, характерный для Рулье и его школы, был тогда повторен
русскими авторами многократно. Мальтузианству тут места не было: эле-
101
ментарным актом эволюции мыслилось не вытеснение конкурента из обще-
го местообитания, а выявление некоего смутно осознаваемого зачатка це-
лостности, как бы зачатка подходящей экосистемы. В России надолго воца-
рилось умонастроение «Дарвин без Мальтуса» (см. D.P.Todes. Darwin with-
out Malthus. The struggle for existence in Russian evolutionary thought. N.Y.,
1989), не вполне оставленное поныне. Оно-то и запомнилось обществу как
быстрое принятие дарвинизма Россией. Реально же дело было иначе. Пер-
вый наш переводчик Мальтуса публицист П.А. Бибиков в предисловии к пе-
реводу (1865 г.) писал:
‘‘Все животные и растительные породы ведут двоякого рода борьбу: обуслов-
ленную конкуренцией и нисколько от нее не зависящую”, и “чем сильнее
борьба между ними, не зависящая от конкуренции, тем слабее борьба, вызы-
ваемая конкуренцией”.
Тем самым, утверждение Мальтуса он предлагал заменить на такое:
“Все животные и растительные породы стремятся к размножению до числа,
допускаемого количеством пищи, но никогда не могут достигнуть его по
причине других, неблагоприятных для их размножения условий”.
Он привел много примеров, и среди них такой: катастрофическое раз-
множение сусликов на юге России прекратилось не от того, что был съеден
весь хлеб, а от повального их заражения глистами. И вообще, заключал Би-
биков, земельные угодья обычно могут прокормить много большее населе-
ние, чем фактически наблюдаемое.
Через 90 лет морфолог и паразитолог В.Н. Беклемишев высказал почти
то же: по его мнению, паразиты служат важной частью биоценоза, регули-
рующей численности животных на приемлемых для его устой-чивости уров-
нях, чего не может система “хищник-жертва” (Беклемишев В.Н. Возбудители
болезней как члены биоценозов//ЗЖ, 1956, № 12).
На Западе из труда Мальтуса большинством пишущих был сделан сме-
лый, но необоснованный вывод - что всякая популяция сама по себе стре-
мится к неограниченному росту, пресекаемому лишь нехваткой ресурсов
(прежде всего, пищи). В недавнем исследовании биофизика В.Л. Воейкова
показано, что в природе таковой рост наблюдается только в патологических
популяциях вроде раковой опухоли или нашествий саранчи (Воейков В.Л.
Правда Дарвина и ложь дарвинизма // Человек, 1997, № 3. - По-моему, за-
главие неудачно, но оно дано редактором).
Иными словами, «мальтузианская» схема безудержного размножения,
ограниченного только борьбой за внешний ресурс, описывает не эволюцию,
а редкие патологии вроде нашествия саранчи, кончающиеся гибелью попу-
ляции, подверженной «мальтузианской» страсти. (Воейков сравнил «маль-
тузианский» рост популяции с раковой опухолью.)
Наоборот, в эволюции, как и в онтогенезе, всякий рост численности
(молекул, клеток, организмов) подчинен той системе, частью которой объект
является. Дарвин совершил логическую ошибку, когда писал:
102
«Нет ни одного исключения из правила, по которому любое органическое су-
щество численно возрастает... с такой большой скоростью, что, не подвергай-
ся оно истреблению, потомство одной пары очень скоро заняло бы всю зем-
лю»; слоны плодятся медленно, но и для них «по истечении 740-750 лет от
одной пары получилось бы около 19 миллионов живых слонов».
Нет, обычно лишь обилие семян и спермиев, а у них нет тех органов, по ко-
торым (как полагал Дарвин) идет сравнение особей. Наоборот, обилие
взрослых потомков, друг с другом конкурирующих - редкость, «жи-
вых слонов» всегда мало, а эволюируют они быстрее всех (см. п. 9-1).
Далее, все виды, не только животные, но и растительные и даже бакте-
риальные, обладают эффективными средствами регулирования своего
размножения, ослабляющими и даже сводящими на нет конкуренцию внут-
ри вида. Все нормальные популяции, начиная с молекулярного уровня, об-
ладают жесткими механизмами регуляции своей численности, а все не об-
ладающие («мальтузианские») быстро гибнут.
Не следует думать, что саморегуляция численности - приобретение
высших форм жизни. Нет, к ней способны даже бактерии, вырабатывающие
для этого целое семейство особых веществ. Это бактериоцины - короткие
пептидные цепочки (20-70 аминокислот), синтезируемые специально для
подавления численности, своей и чужой (Котельникова Е.А., Гельфанд
М.С. Выработка бактериоцинов... // ЭГ, 758).
Есть и особый механизм выживания популяции микробов за счет пре-
кращения размножения при действии ядов - персистентность: малая
часть популяции реагирует на антибиотик или иной яд не гибелью, а пре-
кращением всякой активности; яды, действующие на какой-либо биохими-
ческий процесс, бессильны против пассивной клетки, и она возобновляет
активность после вымывания яда из среды (Льюис К. Персистирующие
клетки... // Биохимия, 2005, № 2).
Высшие организмы регулируют свою численность многими способами,
вплоть до пропуска сезонов размножения и рассасывания беременности.
Мальтус этого не мог знать, а мальтузианцы (дарвинисты) не хотят видеть.
Гпава 3. Наследие Дарвина
3-1. Две истории науки
С появлением дарвинизма идея эволюции стала респектабельной и,
главное, преподаваемой. На языке методологов данный факт именуют так:
эволюционизм был до Дарвина лишь феноменом когнитивной, т.е. позна-
вательной истории науки, а теперь стал еще и феноменом социальной ее
истории. До Дарвина эволюция была делом отдельных ученых, а теперь
стала делом общества, если угодно - лицом эпохи. Переплетение двух ис-
торий и породило дарвинизм. О роли обеих историй в эволюционизме XX
века см. [Колчинский, с. 29-38].
103
Социальная история науки часто отстает от когнитивной на 100-200 лет
- ведь наука в целом куда консервативнее, чем мы думаем. Она гораздо
ближе к обществу, которому она служит, чем к своим передовым личностям.
Более того, наука склонна запоминать в качестве первопроходцев отнюдь
не героев когнитивной, а деятелей социальной ее истории (не тех, кто ска-
зал нечто совсем новое, а тех, кто был впервые услышан обществом). Та-
ким деятелем и стал, помимо своей воли, Дарвин.
Как мы знаем из предыдущего, все основные идеи Дарвина высказаны
до него; более того, бывали более широкие, чем у него, обобщающие кар-
тины эволюции. Дарвин оказался в ряду основателей лишь в чисто соци-
альном аспекте (не сказав ничего по-существу нового, первый сумел быть
услышанным всеми), а Нэгели, наоборот - в чисто когнитивном: первую
цельную теорию эволюции мы видим у него, и она всеми забыта.
Все лавры достались одному Дарвину, хотя всерьез у него рассмотрен
один-единственный вопрос - о внутривидовой изменчивости, после чего
высказана гипотеза о происхождении рас (разновидностей) путем накопле-
ния мелких изменений. Об остальной эволюции сказано гораздо меньше,
чем у предшественников, причем отдельные высказывания не всегда увяза-
ны, а описания реальной эволюции нету вовсе. Если бы Чемберс и Брони не
описали до Дарвина конкретный ход эволюции, то его отвлеченные рассуж-
дения о “проблеме вида” никак не смогли бы восприниматься учеными в ка-
честве базы эволюционизма.
Проблема не в приоритете, а в сути дела: всё, что не вошло в дарви-
низм из прежнего эволюционизма, оказалось забыто. Как мы видели, до
Дарвина существовали целые эволюционные школы, например, герман-
ская, и сказали они об эволюции гораздо больше, чем Дарвин, но для даль-
нейшей науки почти всё это пропало. Восстановить историю эволюциониз-
ма - значит, восстановить путь к пониманию самой эволюции.
О какой именно истории должна идти речь - о когнитивной или о соци-
альной? Конечно об обеих. Но их не надо путать: не научившись отделять
когнитивное от социального, мы не поймем в дарвинизме ничего. Излагае-
мые ниже итоги истории дарвинизма, основанные на исследованиях, прове-
денных историками в последние полвека, исходят из такого разграничения.
Сам Дарвин, после первого опьянения всеобщим признанием, стал
оценивать свою доктрину невысоко и затем просто отошел от нее (что хо-
рошо видно из пятитомника его писем). Последние 10 лет жизни Дарвин
почти не касался своего детища, о чем писал Уоллесу: «Я взялся за преж-
нюю работу по ботанике, а все теории забросил». Эволюционисты, для ко-
торых он был духовным отцом, обращались к нему с вопросами, но он отве-
чал редко, купо и не по сути.
Данные факты тщательно скрываются от широкой публики - как у нас,
так и на Западе. С когнитивной позиции и эта фальсификация истории нау-
ки, и это равнодушие читателей удивительны, но для социальной истории
104
науки тут всё обычно - учение живет само по себе, вне связи с поведением
ее основателя. Наоборот, облик самого основателя преображается в соот-
ветствии с нуждами учения. Тем более понятно это в отношении дарвиниз-
ма, сразу ставшего аргументом в политических спорах. Основатель должен
быть мудр и всеведущ, и Дарвин стал им (см. п. 1-1), хотя почти не бывал в
библиотеках и даже свою личную знал плохо, а к теориям вкуса не имел.
3-2. Поздний Дарвин
После появления «Происхождения видов» Дарвин жил и активно рабо-
тал еще 22 года, причем создал основную часть им написанного. Понять,
какие его труды естественно отнести к поздним, нетрудно. Достаточно вник-
нуть в список изданий «Происхождения видов», и грань легко видна: за
1859-1862 годы вышло 6 оригинальных (т.е. содержавших уникальную ав-
торскую правку) изданий - в Англии, США, Гемании и Франции; потом 4 года
их не было вовсе, после чего вышло еще 3 оригинальных издания, все в
Лондоне (1866, 1869, 1872). Перерыв и породил позднего Дарвина: вместо
отдельных переделок, спешно вставляемых в каждое ближайшее издание,
он стал не торопясь превращать книгу, прежде заявленную как «извлече-
ние» (то ли из «Длинной рукописи», то ли из будущего трактата), в основной
труд жизни. Который так и остался памфлетом.
Видимо, именно тогда Дарвин понял, что обещанный им обоснованный
труд по эволюции написан не будет. Длинная рукопись осталась в том виде,
как ее бросил автор в злополучном июне 1858 года. Когда она была наконец
в 1974 году опубликована, ученых ждало разочарование: это оказалось во-
все не то всеобъемлюще обоснованное исследование, на какое дарвинисты
сто лет ссылались как на реальность, а, как бы нулевое издание «Происхо-
ждения видов», более многословное и содержащее ссылки на литературу,
но не на недостающие (и обещанные!) факты.
Что же произошло с Дарвином за четыре года? Ну, например, в 1862
году он ознакомился, наконец, с той трактовкой прогресса, какую дал Брони
(см. п. 2-10), с почтением отозвался о ней в своей книге об орхидеях, но в
«Происхождении видов» не сказал на эту тему, казалось бы, ничего. Надо
очень внимательно сравнить тексты разных изданий «Происхождения» и
выяснить по письмам обстановку во время их подготовки, чтобы понять, на-
сколько слабым считал теперь Дарвин свое понимание прогресса и на-
сколько Брони повлиял на него.
В первом издании Дарвин уверенно заявлял:
«по моей теории, более недавние формы должны быть организованы выше,
чем более древние; ведь новый вид образован за счет наличия некоего пре-
имущества в борьбе за жизнь. Если бы в примерно сходных климатах эоце-
новые обитатели какой-то части мира вступили в соревнование с нынешни-
ми, то эоценовая фауна или флора непременно была бы побеждена и истреб-
лена; так же и мезозойская - эоценовой, а палеозойская - мезозойской. Я не
105
сомневаюсь в этом процессе усовершенствования... но я не знаю способа
проверить этот вид прогресса».
Однако через полгода, в американском и немецком изданиях, послед-
няя фраза данной цитаты заменена на совсем иную концовку:
«Так ли на самом деле? Значительное большинство палеонтологов ответило
бы утвердительно; но по своему слабому (imperfect) суждению я могу... со-
гласиться с этим лишь в некоторой ограниченной мере. Тем не менее, можно
предположить, что будущие геологические исследования представят более
веские доказательства» .
Вскоре «значительное большинство палеонтологов» воспротивилось
такой трактовке, и абзац снова пришлось менять. Дарвину осталось опе-
реться на себя самого, и слово imperfect исчезло; а после ознакомления с
прогрессом по Бронну исчезла вся концовка. Вместо нее в издании 1866 го-
да читаем:
«Я не могу, после прочтения обсуждения этого вопроса Лайелем, Бронном и
Гукером, рассматривать данное заключение как вполне доказанное, хотя и
считаю его высоко вероятным».
Разумеется, никакие будущие исследования не дадут нам информации
о том, как соревновались бы животные прошлого с нынешними, и Дарвин
был прав, изъяв фразу о будущих доказательствах. Рассуждение о сорев-
новании - плод воображения и сделанный из воображаемого наблюдения
вывод - обычная логическая ошибка, именуемая в учебниках логики как
petitio principii (подмена основания): утверждение, которое надо доказать,
само принято в ходе доказательства как исходное.
Критики не раз ему на такие ошибки указывали, а Дарвин отвечал
обычно не изменением по сути, но лишь смягчением формулировок. Крити-
ков это не устраивало, и порой Дарвин просто возвращался к раннему тек-
сту. Так и в вопросе о прогрессе: в последнем издании ссылки на возраже-
ния Лайеля, Бронна и Гукера нет, зато вновь видим:
«Значительное большинство палеонтологов ответило бы утвердительно; и
видимо этот ответ надо признать верным, хотя и трудно это доказать» (гл. 11,
п. «О степени развития древних форм»).
Как видим, критика попросту игнорирована Дарвином.
13
Правка этих двух изданий очень важна для понимания становления и сути дарви-
низма. К сожалению, на нее никто не обращает внимания. Даже благожелательный
рецензент книги ЧЭ, похвалив мой исторический обзор, заключил, несмотря на при-
веденную цитату, что “Происхождение видов” «при жизни автора выдержало 6 изда-
ний» (Миркин Б.М. II УСБ, 2005. № 2), имея в виду лондонские. Изданий было гораз-
до больше, в том числе 9 несли уникальную правку. Вместе с «Длинной рукописью»
(опубликована в 1974 г.) имеем 10 вариантов и можем убедиться, насколько станов-
ление дарвинизма было социальным и психологическим феноменом, шедшим во
многом вразрез с развитием тогдашней биологии (Чайковский, 1988].
106
В 1868 году вышла в свет самая толстая его книга - двухтомное «Изме-
нение животных и растений при одомашнивании». Она обстоятельно, с на-
учным аппаратом (рисунки, таблицы и ссылки на литературу) разбирала те-
му первой главы «Происхождения видов» и выглядела как начало обещан-
ного Дарвином трактата. Ожидание его было столь велико, что первый рус-
ский перевод «Изменения» так и был назван: «Происхождение видов. Отдел
I». Но никаких других отделов не появилось.
Книга имела большое значение для селекционеров, но собственно к
эволюции добавила мало. Как позже писал сын Дарвина, Френсис,
«Она не избегла враждебной критики: так, например, говорили, что публика
терпеливо ждала pieces justificatives (франц.; здесь: решительных аргументов
- Ю. Ч.) г. Дарвина, а по прошествии восьми лет всё, что получилось, свелось
к целой куче подробностей...».
Критики были правы лишь отчасти: анализ изменчивости дал начало
многим идеям, в том числе, в наши дни, идее давления нормы (см. гл. 5).
3-2* Вынужденные добавления к учению
В частности, именно в «Изменении» Дарвин обнародовал свою «вре-
менную гипотезу пангенеза», а мы, россияне, лучше всех знаем, что нет ни-
чего постояннее, чем временное. Пангенезом (т.е., по гречески, всеобщим
порождением) он назвал тот античный принцип, о котором мы говорили в
главе 1: каждая часть тела посылает свою частицу в орган размножения,
где частицы вместе и образуют «семенную жидкость». Этот принцип вводи-
ли многие, в том числе Хэйл, Мопертюи, Бюффон и Э. Дарвин, но лишь внук
последнего дал название и самому принципу, и частицам (назвав их гемму-
лами). Гипотеза понадобилась Ч. Дарвину для ответа на множество каверз-
ных вопросов (например: почему одно и то же растение можно получить и
от семени, и от почки, и от корневой отводки?), но он сам стеснялся ее, и
современники дружно ее отвергли. Причем ни одна сторона не знала, что
повторяет аргументы тысячелетней давности.
Однако как раз эта гипотеза может пережить всё, Дарвином написан-
ное; физиолог-ламаркист И.А. Аршавский любил говорить, что одного «Пан-
генеза» достаточно, чтобы увековечить имя Дарвина. Мало того, что в споре
с идеей пангенеза родилась генетика (см. п. 3-12), но для высших животных
ее через сто лет отчасти подтвердили (см. гл. 5).
Иных глав «Происхождения видов» Дарвин подобным образом разви-
вать не стал, зато выпустил в 1871 году толстую книгу «Происхождение че-
ловека и половой отбор». Во Введении он пояснил свой замысел: поскольку
идея естественного происхождения видов (но не естественного от-бора) уже
победила идею независимых творений, однако еще ни разу всерьез не рас-
смотрена для какого-либо вида, то настало время приложить эту идею к
конкретному виду, в качестве которого автор избрал человека.
лв
107
Это была правда, но не вся правда и даже не главная ее часть. Главная
же, по-моему, состояла в том, что общество ждало от Дарвина именно тео-
рии происхождения человека, а ее, даже при самом мягком отношении к ло-
гике, из идеи естественного отбора вывести не удавалось (см. п. 3-3). В
«Происхождении видов» о человеке практически речи не было, и вот Дар-
вин наконец-то решился прямо высказаться относительно «обезьяньей ро-
дословной» человека. Сама по себе эволюция сразу отошла на второй план
- по крайней мере для простого читателя.
Как уже говорилось в п. 2-12, все ранние переводчики и большинство
ранних критиков понимали естественный отбор как активный сознатель-
ный выбор. В ответ Дарвин в 1861 году вписал в начало главы «Естествен-
ный отбор» разъяснение: «В буквальном смысле слова естественный отбор
- неправильное название (misnomer)», а в 1869 году усилил мысль:
«...естественный отбор - ложный (false) термин». Если так, то надо было
предложить другой термин, правильный, который не вводил бы читате-
ля в заблуждение, но Дарвин ограничился напоминанием, что и в других
науках не все термины удачны.
Видимо, дело было в неясности Дарвину самого описываемого явле-
ния, и заменой терминов помочь делу было нельзя. Не написан обещанный
Дарвином доказательный трактат никем и поныне. Обычно книги по теории
дарвинизма построены примерно по образцу «Происхождения видов»: мно-
го говорится об изменчивости окраски и прочих акциденций (побочных при-
знаков, п. 1-6**), гораздо меньше о конкуренции74, совсем мало о ходе эво-
люции (он излагается скороговоркой, без связи с остальным текстом), почти
ничего об изменчивости существенных свойств и ничего о сущности отбора.
Даже отбора этих самых акциденций.
Даются мнимые примеры и для них пишутся уравнения. Реальных при-
меров постепенного преобразования под действием избирательной раз-
множаемости нет. Приводятся единичные примеры либо постепенной эво-
люции без всякого анализа отбора, либо мгновенного (в одну мутацию) по-
лезного приобретения. Дарвин знал такие примеры (их тогда именовали
спортами) и справедливо, с позиции теории естественного отбора, отрицал
их роль в эволюции: ведь всякий спорт свидетельствует о наличии какого-то
закона изменчивости, который, тем самым, и творит эволюцию. Спорты не
помогали теории Дарвина, они ее заменяли. 14 * * * * *
14 Это понятие, основное у Дарвина, почти не исследовано им и выпало из нынешнего
дарвинизма, оставив после себя лишь “принцип конкурентного исключения Гаузе”,
гласящий, что два экологически идентичных вида не могут сосуществовать. “Принцип”
широко используется, но не подтвержден ничем. Сам Г.Ф. Гаузе дал 70 лет назад лишь
вольные толкования ничего не говоривших опытов. Исследования показали, что в при-
роде этот “принцип” (в изученных случаях) места не имеет [Гиляров].
108
3-3. Зачем половой отбор? И есть ли он в природе?
Половым отбором в дарвинизме именуется преимущественное раз-
множение самцов с теми качествами, которые позволяют победить в борьбе
за самку. Тема появилась еще в первом издании «Происхождения видов» и
тогда уже вызвала упрек Бронна. У Дарвина говорилось, что уродливые от-
ростки у некоторых голубиных и куриных нельзя объяснить никаким отбо-
ром, так как их «мы не можем считать ни полезными в борьбе, ни привлека-
тельными для самок». Это место Брони комментировал на с. 95 своего пе-
ревода «Происхождения видов» иронически: «Но как можем мы судить, что
у жениха привлекательно в глазах курицы или горлицы!»
Дарвин ответил, как обычно - изъял сам пример, но ничего не изменил
в теории. Наоборот, положил ее в основу учения о происхождении человека
(1871). Бронна давно не было в живых, но наиболее въедливый критик, не-
мецкий ботаник Альберт Виганд, заявил, что сама идея полового отбора до-
водит всю теорию Дарвина до смешного:
“Если бы целью было показать, до чего смешна теория отбора, то это нельзя
было бы сделать удачнее, чем при помощи тех доведенных до крайности аб-
сурдов. которые Дарвин, не щадя себя, производит на свет половым отбо-
ром”. И тут же пояснил, что надо бы не только собирать “богатый материал
для теории любви”, но и посвятить “некоторые размышления самой теории”
(Wigand A. Der Darwinismus und die Naturforschung Newtons und Cuviers. Bd 1.
Braunschweig, 1874, S. 180).
В самом деле, книга Дарвина «Происхождение человека и половой от-
бор» удивляет: почему столь фундаментальный акт, как происхождение че-
ловека, поставлен вровень с такой частностью, как половой отбор?
Попробуем понять. Как уже сказано, происхождение человека не уда-
лось объяснить идеей обычного отбора. Точнее, можно было говорить о
преобразовании скелета и тому подобном, но как быть с мышлением? Никто
не брался объяснить, как за счет конкуренции диких предков человека поя-
вилась, например, способность породить математику. Даже Уоллес, соавтор
дарвинизма, отказался признать происхождение интеллекта “по Дарвину”,
мотивируя отказ тем, что жители джунглей, по его наблюдениям, не глупее
«среднего члена наших ученых обществ».
Тут Дарвин и решил в помощь естественному отбору добавить половой
отбор. Поскольку это его воззрение часто упрощается, сводится к афоризму
“Дуры отобрали умников”, приведу его собственные слова. Исходным у Дар-
вина было античное убеждение, что мужчина умнее женщины:
«Главное различие в умственных способностях обоих полов проявляется в
том, что мужчина во всем, за что берется, достигает совершенства, не-
достижимого для женщины». Правда, «у женщин способность интуиции,
быстрое восприятие и, может быть, даже подражание выражены резче, чем у
109
мужчин, но... некоторые из этих свойств характеризуют низшие расы, а сле-
довательно - прошлое или низшее состояние цивилизации».
Опять - подмена основания рассуждения. И опять самое интересное,
происхождение такого чуда, как интуиция, не привлекло Дарвина вовсе. Он
продолжал строить схему эволюции ума попросту:
«Но для того, чтобы избегать неприятелей или успешно нападать на них, для
того, чтобы ловить диких животных, выделывать оружие, необходима по-
мощь высших умственных способностей...»
Вот в чем загвоздка: ум для Дарвина - лишь инструмент в борьбе за
пищу, потому интуиция неинтересна. (Уоллес был глубже, недоумевая, как
борьба могла породить математические способности или мораль.) Далее:
«Эти же способности, равно как и предыдущие, развились у мужчины ча-
стью путем полового отбора, т.е. путем борьбы между соперничающими
мужчинами, частью - путем естественного отбора, т.е. успеха в общей борь-
бе за жизнь. А так как в обоих случаях борьба происходила в зрелом возрас-
те, то приобретенные этим путем особенности должны были передаваться
мужским потомкам полнее, чем женским».
Это странное суждение дано без каких-либо попыток подтверждения,
зато есть столь же странное пояснение: по Дарвину, существует
«стремление признаков, приобретенных в позднюю пору жизни тем или дру-
гим полом, передаваться тому же полу в соответствующем возрасте».
Вот, по сути, и вся “теория” (Ч. Дарвин. Соч., т. 5, М.-Л., 1953, с. 608-
610). Нагромождение голых деклараций. Можно только удивляться, что
дарвинисты продолжают ссылаться на Дарвина как на автора теории проис-
хождения человека. Объяснением, на мой взгляд, служит то обстоятельство
(ставшее известным философам науки через сто лет), что учёные в массе
своей видят труд классика прошлого как священный текст, не ища в нем ни
фактов, ни логики. Мы вернемся к этому вопросу в п. 3-7**.
Гексли, ближайший друг Дарвина, в молодости сам пустивший в оборот
словечко «дарвинизм», завоевавшее науку, писал в старости:
«Единственное, что смягчает мой пессимизм, свидетельствуя о благости
Творца мира, - это моя способность наслаждаться театром и музыкой. Не ви-
жу, чем она могла быть полезна в борьбе за существование».
Это не помешало ему остаться в истории науки в качестве ярого дарвиниста
(недоброжелатели звали и зовут его «бульдогом Дарвина»).
Что касается сути полового отбора, то Любищев объяснил ее так:
«...дарвинизм не может обойтись без признания принципов, которые он
обычно громогласно отвергает», «и теория полового отбора ... вводит на-
стоящий, хотя и бессознательный, целеполагаюший фактор, притом дейст-
вующий в одном направлении в течение многих поколений» [Любищев. 1982,
с. 157].
Он имел в виду, что идея отбора свойств самцов самками предполагает
способность самок из поколения в поколение выбирать самца посредством
110
одного и того же критерия, например, красоты^5 А это противоречит дарви-
низму и характерно как раз для религиозных ветвей эволюционизма.
Данный пункт понимается с трудом, поэтому поясню: в отличие от есте-
ственного отбора, который все признают хотя бы в той форме, что неспо-
собное жить не живет, половой отбор не очевиден ни в какой форме. “Дока-
зательством” полового отбора всегда служат примеры избирательного по-
ведения (например, выбора самца самкой), но никогда даже не ставится во-
прос: как проверить, приводят ли такие акты к наследственным изменени-
ям? Из того явного факта, что самцы некоторых видов бьют друг
друга, а некоторые самки предпочитают некоторых самцов, еще не
следует, что в итоге вид изменяется.
Можно ли узнать, изменяются ли они от этого на самом деле?
Если естественный отбор хотя бы пытались обнаружить и даже изме-
рить (как увидим, без успеха для дарвинизма), то половой отбор был и ос-
тался уловкой для словесного наполнения теоретических пустот.
А ведь еще Усов (п. 2-12) отмечал, что борьба самцов за самку может
мешать выживаемости вида. Орнитолог Е.Н. Панов подробно описывает
бои самцов токующих птиц и даже признаёт, что самка производит активный
выбор самца для спаривания, но не видит в акте победы материала для
эволюции. В самом деле, самка выбирает вовсе не драчуна, а того, кто из-
бегал драки; это почти всегда «ветеран тока», так что шанс на оставление
потомства имеет самец, из года в год тихо идущий к статусу ветерана. Ос-
новная же масса самцов лишь организует социальную структуру популяции.
А главное, токует всего 1% видов птиц (и еще меньшая часть иных живот-
ных, например, головоногих [Несис, с. 13]), причем не видно никакой связи
феномена токования со свойствами вида [Панов, с. 222-234].
То же видно на двух самых ясных примерах самцовой конкуренции:
спермиев за оплодотворение яйцеклетки и самонесовместимости пыльцы (у
растений с перекрестным опылением). Оба приводятся как примеры поло-
вого отбора («скрытый женский выбор» - Skogsmyr /., Lankinen Е. Sexual
selection: ап evolutionary force in plants? // BR, 2002, № 4, p. 556), однако на
самом деле в обоих примерах акт оплодотворения никак не связан с качест-
вом генов “победившей" гаметы. В самом деле: даже если допустить, что
она произвела оплодотворение в силу своих лучших качеств (а это еще на-
*5 Критерий недавно подвергся проверке, и Любищев вроде бы оказался прав: швед С.
Джирланда привел “доказательство того, что куры тоже обладают чувством прекрасно-
го”: они выбирали те же картинки, какие считают красивыми люди. Так это или нет. но
швед получил шуточную премию в США за “работу, поражающую своей бессмыслен-
ностью” (Антинобелевские премии 2003 года // Природа, 2004, № 4). А ведь Брони и
Дарвин спорили об этом всерьез; Любищев видел эстетическую целесообразность важ-
ной для теоретической биологии [Линник Ю.В. // Системность..., с. 82-85]. Выходит,
что опять «громогласно отвергли» непонятное за то лишь, что оно непонятно.
111
до доказать, чего никогда не делается), то эти качества касаются только са-
мой процедуры оплодотворения, а не свойств будущего организма.
Тем самым, о половом отборе говорить нет смысла. Различие
полов играет роль в эволюции, но совсем иную (см. далее, п. 9-16*).
К словам Любищева могу добавить (что он и сам делал не раз): если
признак вреден (например, огромные рога оленя), то самки, выбирающие
таких самцов, будут (согласно логике дарвинизма) уничтожены обычным от-
бором. Мысленное выключение этого отбора и означает признание направ-
ляющего фактора. Он служит как бы буксиром, движущим корабль дарвин-
ской эволюции там, где его парусов не может наполнить ветер внешней
среды. Посмотрим, кто как вводил направляющий принцип. Начнем с крити-
ка, чью правоту признал (это бывало редко) сам Дарвин.
3-4. «Кошмар Дженкина»
Статья английского инженера Флеминга Дженкина появилась в июнь-
ской книжке журнала «North British Review» за 1867 год под заголовком,
столь же скромным, сколь и вызывающим: «Происхождение видов». Автор
не страдал недостатком самоуверенности: о чем он только не писал - об
архитектуре, о санитарии, о финансах, и нужно признать, что ему часто уда-
валось схватить суть проблемы. То, что предпринял Дженкин, можно на-
звать первой успешной попыткой логического анализа дарвинизма.
Не будем, писал Дженкин, обращать внимание на недостатки фактиче-
ского обоснования идеи естественного отбора; допустим, что все примеры,
которые приводит или предполагает Дарвин, правильны, и проследим, что
из них следует. Может ли естественный отбор выбирать новые качества и
скрещивать разновидности так же, как это делает селекционер? Да и сами
возможности человека увеличивать различия между породами - разве без-
граничны? По Дарвину, естественный отбор отличается от искусственного
только тем, что действует медленнее; но почему же надо считать, что есте-
ственный отбор может сделать то, чего селекционеру никогда не удавалось
достичь? Если, скажем, за шестьдесят лет можно вывести новую породу го-
лубя, то разве из этого следует, что за какое-нибудь время из голубя можно
вывести дрозда? (А тем более, добавлю от себя, вывести птицу из пресмы-
кающегося.)
Только дикарь, продолжал Дженкин, глядя, как ядро вылетает из пушки,
может решить, что в конце концов оно долетит до звезд. И скорость ядра, и
размах наследственных вариаций стремительно убывают по мере удаления
от исходной точки. Поэтому нет никаких оснований ожидать, что подходя-
щие изменения будут накапливаться.
Дженкин отнюдь не первый писал про это и был отнюдь не самым язви-
тельным. Так, еще в 1860 году епископ Сэмюэл Вильберфорс, самый тогда
известный критик Дарвина, вопрошал: «Допустимо ли считать, что удачные
вариации репы стремятся стать людьми?» Но сарказм лишь мешает пони-
112
мать суть дела, и яркая статья Вилберфорса (основанная на работах Оуэна)
осталась почти без внимания.
Главный пункт у Дженкина - поглощающее влияние скрещивания.
Предположим, что появился вариант, более удачный, чем существующие
нормальные организмы данного вида. Спрашивается: с кем ему скрещи-
ваться? Если вокруг имеются лишь нормальные особи, то шансов передать
полезное новшество нет: уже через несколько поколений оно будет «засо-
сано болотом» обычных организмов. Следовательно, никакая уникальная
вариация (ни один спорт) не может иметь значения для эволюции. Сущест-
венны только массовые отклонения индивидов от нормы. А они, массовые
отклонения, как уже говорилось, никогда не выводят новую разновидность
за рамки существующего вида.
Остается предположить одно из двух: либо новая вариация не должна
теряться при скрещиваниях, либо она должна возникнуть сразу у значи-
тельного процента особей. Однако, заметил Дженкин, обе гипотезы отрица-
ют суть дарвинского учения. Первая противоречит наследственности, какой
она выглядит в природе и как ее описывает сам Дарвин; вторая же, если ее
допустить, приводит к порочному кругу: чтобы распространиться, новая
вариация уже должна быть достаточно распространенной.
Таково было содержание первой половины статьи Дженкина. Во второй
половине он, среди прочего, развил интересную мысль, что трудности раз-
личения видов еще не являются доказательством их происхождения от об-
щего предка. (Дженкин по сути призывал не путать феномен эволюции с
феноменами нынешнего разнообразия, а также не путать сущность с акци-
денцией -см. конец п. 1-6.)
В своих возражениях Дженкин не был первым, но он был первым, кто
сумел связать их воедино и привлечь внимание Дарвина.
«Он может выдумать - читал Дарвин о самом себе - вереницы предков, чье
бытие ничем не доказано; он выставит против них армию воображаемых вра-
гов,.. и растянуть прошлое до бесконечности ему тоже ничего не стоит; при
таких способностях можно изобрести какие угодно организмы, сослаться на
любые обстоятельства и с их помощью обойти любые затруднения».
Дарвин отметил на полях: «Хорошая насмешка». Далее шло:
«Чувствуя трудность иметь дело с противником со столь обширной фантази-
ей... доверимся таким аргументам, которые он по крайней мере не сможет
разбить простыми усилиями воображения».
И дальше следовало то, что нам уже известно: отбор мог бы быть эф-
фективным лишь при условии, если какое-то уклонение, и притом значи-
тельное, возникнет сразу у большого числа особей; но в действительности
такие отклонения редки и, значит, неизбежно будут поглощены «болотом»
нормальных особей.
Перечитав это место, Дарвин представил себе конкретный пример - так
ему было легче рассуждать. Пусть у невинной птички вылупился птенец с
113
кривым хищным клювом, которым удобно рвать мясо; ведь ему, этому птен-
цу, когда он вырастет, придется искать себе подругу среди птиц с прямыми
клювами, так что у детей клюв будет уже не такой кривой, у внука - еще
прямее. В конце концов - и очень скоро - новое приобретение исчезнет. И
Дарвин написал другу: «Доводы Дженкина меня убедили». В истории дар-
винизма настал критический момент.
Дарвин признал, что спорты не должны играть роли в образовании но-
вых видов, и по этому поводу в очередном (пятом) издании «Происхождения
видов» появился новый примирительный абзац. Теперь Дарвин был вынуж-
ден принять в качестве основы для своей теории массовые вариации осо-
бей, а следовательно, и признать их общеизвестные свойства: они разно-
родны, хаотичны, незначительны (не являются готовыми приспособления-
ми), не очень-то стойко закрепляются в потомстве, а главное, изменения
происходят не одновременно. Следовательно, надо допустить существова-
ние каких-то механизмов (внешних или внутренних), синхронизирующих
изменчивость и закрепляющих удачные приобретения у потомков^ (ис-
пользуя нашу прежнюю аналогию - надо прицепить буксир). Эти механизмы
автор «Происхождения», скрепя сердце, заимствовал из несимпатичной ему
теории Ламарка: влияние климата, упражнения органов и т.д. Катастрофи-
ческий крен дарвинского судна удалось на время выправить, но какою це-
ной! Вместо желанного механистического курса корабль эволюционной тео-
рии поплыл в бермудский треугольник непонятных законов наследования.
3-5. Законы наследования. Дарвинизм по Уоллесу
Вскоре оказалось, что Уоллес предложил, к посрамлению Дженкина,
рассуждение, которое разбивало противника именно «простыми усилиями
воображения». Он заявил, что любая вариация представляет собой
«или увеличение или уменьшение органа, или его усиление, или ослабление;
поэтому все изменения могут быть разделены на две группы; усиливающие и
ослабляющие деятельность животного, иначе говоря, полезные и вредные».
Дальше всё получалось просто: если полезных уклонений в среднем
около половины, то всегда найдется, что отбирать Здесь Уоллес совершал
очередную подмену основания, т.е. исходил из посылки, которая сама по
себе нуждалась в доказательстве: что от всякой нормы регулярно возникает
отклонение в нужную для приспособления сторону и сразу же помогает ор-
ганизму выжить. (Добавлю: как тут не вспомнить анекдотический вопрос -
почему за переполненным автобусом часто следует пустой? Ответ: потому
что вероятность попасть в пустой автобус равна нулю - по определению.
Логика та же.)
16 В отличие от естественного отбора, так никем и не обнаруженного, эти
механизмы в наше время найдены - см. главу 5.
114
Легко заметить, что в рассуждениях Уоллеса речь шла вовсе не обо
«всех изменениях», а лишь об изменении каких-то пропорций; его вариант
дарвинизма не годился для объяснения, откуда берутся новые качества. Как
видим, Уоллес предпочел выбросить за борт почти весь груз, лишь бы не
менять курс корабля. Что и говорить, легкость мысли досадная, но не надо
думать, что соавтор дарвинизма был так уж прост. Это вскоре и выясни-
лось.
Шторм не утих, и скоро корабль дарвинизма опять оказался на грани
крушения. В ноябре 1870 году еженедельник «Nature» поместил письмо бо-
таника Альфреда Беннета под названием «Теория естественного отбора с
математической точки зрения». Математики в этом письме было не больше,
чем у Дженкина: несколько арифметических действий. Но если Дженкин пе-
ремножал дроби, чтобы показать, как быстро «засасывается» уникальный
признак, то Беннета интересовал другой вопрос. Вряд ли отдельная вариа-
ция, рассуждал он, принесет пользу организму, особенно если речь идет о
возникновении нового органа или необычной функции. Допустим, что для
получения полезного качества требуется 10 последовательных стадий, при-
чем на каждой стадии признак может изменяться по крайней мере двадца-
тью способами; в таком случае для обнаружения полезного качества потре-
буется перебрать не менее 10 триллионов особей. (Для примера Беннет
рассматривал подражание одной бабочки другой по рисунку крыльев, при-
чем считал, что его оценки даже занижены.) Пусть, продолжал он, числен-
ность популяции не превышает миллиона особей. Тогда для образования
нового качества понадобится по меньшей мере 10 миллионов поколений, и
в течение всего этого времени отбор не сможет быть полезным. Вывод?
Полная неэффективность отбора как фактора эволюции.
Неизвестно, читал ли Беннет Дженкина (критики дарвинизма всегда
плохо читали друг друга и еще хуже цитировали), но в принципе речь шла о
близких темах (и само возражение Беннета уже было высказано вскользь
Дженкином). Дарвин, как обычно, не удостоил нового оппонента прямым от-
ветом, зато Альфред Беннет и Альфред Уоллес яростно сцепились на стра-
ницах «Nature». Уоллес заявил:
«Каждое успешное поколение под влиянием наследственности получит все
большее преимущество, так что шанс воспроизвести нужную вариацию будет
все выше».
На это Беннет резонно возразил, что Уоллес противоречит своей соб-
ственной теории: выходит, что преимущество (рост вероятности дать по-
томство) может проявляться вне всякой связи с отбором! Тем самым в тео-
рию вводится какой-то новый фактор - наследственность (который, добав-
лю, и служит «буксиром»). При чем же здесь отбор? Как ни странно. Уоллес
торжественно согласился с этой трактовкой: да, новый фактор есть принцип
наследственности (principle of heredity), дарвинизм не может его игнориро-
вать, и Беннет может-де не утруждать себя «математической демонстраци-
115
ей». Диспут продолжался, но главное было уже сказано: эволюционная тео-
рия не смогла обойтись без признания определенной направленности на-
следственных вариаций.
Позже (1889 г.) этот принцип уточнил двоюродный брат Дарвина, ан-
трополог и статистик Френсис Гальтон - он сформулировал “law of ancestral
heredity” {закон предковой наследственности), который, в частности, гла-
сил о «соединении свойств предков в детях»: если оба родителя уклоняют-
ся от нормы в одну сторону, то потомство уклоняется сильнее, чем каждый
родитель. Это хорошо известно селекционерам.
Но много ли он дает эволюции? Ведь главное - понять, как свойство
возникает в зачатке и существует до того, как станет полезным, а не как оно,
почему-то оказавшись сразу у обоих родителей, усиливается. На это четко
указал еще один критик Дарвина, зоолог Сент-Джордж Майварт. Он повто-
рил возражение Беннета и заметил в виде примера:
«Низшие позвоночные совершенно лишены ног... Каким образом можно объ-
яснить сохранение и развитие первых зачатков конечностей, если эти зачат-
ки... были лишены функций»?
Дарвин уделил Майварту в последней редакции «Происхождения ви-
дов» основную часть главы 7, но ответа по-существу так и не дал. И если бы
не стычка двух Альфредов, мы не видели бы сейчас так отчетливо тот но-
вый подводный камень, на который наскочил дарвинский корабль: кроме
неопределенных вариаций, отбора и наследственности, нужен еще один
фактор - организатор начальных стадий. Сойти с этого камня можно было
опять-таки лишь с помощью “буксира” - нового теоретического принципа.
3-6. Принцип смены функций. Дарвинизм по Дорну
Дарвин был лучшим в мире знатоком усоногих раков, но в обоснование
своего учения об отборе на них не сослался: хорошо известный материал
поддавался трактовке хуже, чем воображаемые олени, убегающие от вооб-
ражаемых волков. И едва дарвинисты взялись за усоногих, выводы пошли
удивляющие.
В 1867 году молодой немецкий зоолог Антон Дорн (1840-1909), востор-
женный почитатель и Бэра, и Дарвина, захотел примирить учения обоих.
Его удивили те усоногие, которые, паразитируя на акулах, потеряли весь
животный облик, обратившись в сосущие мешки с половыми железами; а в
то же время их личинки вполне развиты и подвижны, что и позволяет отне-
сти паразитов к усоногим. У Дарвина он нашел подробнейшее их описание,
но не объяснение столь странной эволюции. Понятно, что паразитам почти
все ненужно, но ведь они не конкурируют друг с другом за отсуствие органа,
так чем же направлялось столь радикальное упрощение? Дорн задумался:
что движет усложнением и упрощением животных? Он предположил, что
это - образ жизни, а тот был для низших почти не изучен.
116
Дорн решил построить зоологическую станцию. Буквально выцарапав
деньги у богатого отца (тот и без того финансировал множество научных за-
тей), он купил в Неаполе участок в приморском парке и почти построил ее,
но денег нехватило, чиновники, мягко говоря, не помогали, и Дорн воззвал к
светилам науки о помощи. Столь разные люди, как Лайель, Дарвин и Бэр,
обратились к правительствам, сами собрали какие-то деньги, и вот в 1874
году станция была открыта. Вскоре она стала всемирно известной, и именно
там получены были многие важнейшие данные для эволюционных идей
конца XIX века.
Вернувшись к научной работе в 1875 году, Дорн прочел Майварта, не
был удовлетворен ответами Дарвина и предложил собственный ответ: но-
вый орган развивается не из «зачатка», поначалу всегда бесполезного, а из
иного органа, ранее нёсшего иную функцию (для ползания могли первона-
чально служить плавники и т.п.) Здесь первичным актом эволюции был при-
знан акт смены поведения особи (попытка использовать прежний орган для
новой цели), т.е. налицо возврат к Ламарку, хоть его имя названо и не было.
Дорн сформулировал общий принцип:
«Преобразование органа происходит путем последовательной смены функ-
ций, носителем которых остается тот же орган... Побочная функция посте-
пенно становится главной, вся функция в целом становится иной, и следстви-
ем всего процесса становится преобразование органа» (Л. Дорн. Происхожде-
ние позвоночных животных и принцип смены функций. М.-Л., 1937, с. 160).
Проявляя большую изобретательность в мысленных перемещениях,
видоизменениях и изменении функций органов, Дорн брался объяснить чуть
ли не все акты эволюции животных, но вставал вопрос: если из всего может
возникнуть всё, то почему же образуются вполне определенные организмы,
а не мешанина свойств? Тут он и обращался к Бэру.
В 1861 году Бэр удивленно описывал один вид слепой рыбы, живущей в
темноте, мальки которой имеют нормальные глаза, но взрослея, не просто
слепнут, а утрачивают глазные яблоки. Зачем бы? Бэр отвечал: вероятно,
так экономятся жизненные силы. См. [Чайковский, 1990, с. 168]; позже воз-
ник механоламаркизм, который добавил: глаз нужен в онтогенезе, и его ут-
рата возможна по завершении роста черепа (ЮШ, 126).
Наблюдая похожее у некоторых усоногих, Дорн задался более общим
вопросом: почему одни группы животных прогрессируют, а другие дегради-
руют? Если Дарвин неуверенно склонялся в данном вопросе к каким-то
влияниям среды, то Дорн не видел в этом ответа и следовал за Бэром - де-
ло в жизненных силах; как одни дети могут хорошо учиться, а другие нет, так
одни группы имеют «способнось к усовершенствованию», а другие нет. Та-
кая «способность» и пресекает мешанину (т.е., в наших прежних терминах,
служит «буксиром»).
Это ламаркизм (он, кстати, тоже признаёт постепенность), он был по-
дан Дорном как развитие дарвинизма и как таковое дарвинистами воспри-
117
нят. Как увидим, многие новшества дарвинизм до сих пор умеет объяснять
только по Дорну, причем принцип Дорна понимается очень широко (Кокшай-
ский Н.В. Филогенетический перенос функций И ЗЖ, 1997, № 8). Если же и
по Дорну объяснение требует признания скачков, то честнее будет признать
что никакого объяснения становления новых приобретений нет. Не получив
решения, проблема Бэра-Дорна была забыта. Мы вернемся к ней в п. 5-13*.
3-7. Диалектика отбора. Дарвинизм по Дарвину
Едва Дарвин провозгласил половой отбор фактором, способным объ-
яснять то, чего не объясняет отбор естественный, возникли, как отмечено в
п. 3, проблемы совсем неожиданные - философские. Сам Дарвин избегал
философии («У меня не метафизическая голова» - писал он), и даже у
Дженкина увидал только конкретные примеры, но вовсе не заметил логиче-
ского анализа учения.
Никто из окружения Дарвина и слушать не хотел нескольких критиков,
заметивших, что отсутствие философского анализа может оказаться роко-
вым для всей теории. Один из таких критиков, философ и историк науки
Джордж Генри Луэс (Lewes; это популярное в то время имя писали тогда по-
русски «Льюис»), состоял с Дарвином в переписке, и Дарвин охотно обсуж-
дал его конкретные возражения, но общее возражение о необходимости
философского анализа даже ни разу не упомянул.
Казалось бы, зачем нужен философ, если обсуждаются конкретные
факты? Однако, как мы узнаем в п. 3-10, Луэс ставил вопросы, живо обсуж-
даемые поныне, так что изложу “философский упрек” подробнее.
Когда 2500 лет назад греки задумались над проблемой: откуда мы зна-
ем, что наше мнение верно, - ответов было предложено немного, и, как ни
странно, новых с тех пор по сути не добавилось. Вот они:
1. То, что сказано в священном тексте (или: оракулом), истинно;
2. Мое мнение истинно, ибо я умнее моих противников (или: я опытнее,
моими устами говорит божество, я выражаю мнение большинства);
3. Утверждение истинно, если оно доказано, т.е. логически следует из
очевидных посылок (или: из трактовки доступных наблюдению фактов);
4. Истинно то, из чего следуют проверяемые факты;
5. Истинно то, что получено диалектическим рассуждением;
6. Истина ухватывается интуитивно, а рассуждение ее подтверждает.
Пятый тезис фактически был применен Дарвином и потому является
для нас тут основным. Когда рост культуры сделал недостаточными два
первых ответа, родилась идея доказывать свою правоту тем, что позже по-
лучило имя логики. Однако любое логическое построение из чего-то исхо-
дит, а верно ли это исходное? Здесь возможно либо сослаться на авторитет
(ответы 1 и 2), либо заявить об очевидности предпосылок или наблюдаемых
фактов (ответы 3 и 4), либо признать, что истины как таковой не существует,
а есть более или менее удачные мнения (это полагал еще Ксенофан).
118
Так было до Платона. Платон предложил анализировать сами предпо-
сылки, анализировать до тех пор, пока анализ не убедит нас, что они верны
(в том смысле, что сами следуют из всего, что о них известно и сказано) или
ложны. Такой путь доказательства он назвал диалектическим и полагал,
что этот путь ведет к беспредпосылочному знанию. Так построены его глав-
ные диалоги, они кончаются общим согласием. Но оно обманчиво: платонов
диалог - не беседа реальных людей, а имитация беседы: все участники
идут к цели, заранее заданной автором, и произносят лишь то, что подтвер-
ждает позицию автора. Спора нет и в помине [Рассел].
Однако в способе Платона есть, как мы теперь знаем, смысл: хотя ис-
ходным понятиям (их именуют категориями) нельзя дать определений, но
их можно объяснить друг через друга (это именуется категориальным ана-
лизом), что и позволяет людям о чем бы то ни было говорить.
В наше время (в отличие от Платона) основной признак верности пред-
посылок видят не в их наглядной очевидности, а в том, что сделанные из
них выводы подтверждаются на практике (ответ 4); такой способ проверки
предосылок именуется гипотетико-дедуктивным методом. Как сказано в
п. 2-11, в эволюционизме он связан как раз с именем Дарвина. Метод быва-
ет очень удобен (например, в физике он породил теорию квантов), но опа-
сен: как известно из логики, от ложной посылки верными рассуждениями
можно прийти к верным выводам. Поэтому ложность посылки можно поко-
лениями не замечать - пока из нее не будут получены (и, главное, признаны
сообществом как таковые) выводы, противоречащие практике.
Уже Аристотель, ученик Платона, усомнился в правомерности диалек-
тического метода. Молодой Аристотель не увидел у учителя обоснования
знания, а увидел подмену основания (о ней см. п. 3-2). Старый философ
был обескуражен и в поздних диалогах ушел от диалектики, не предложив
ничего достойного взамен, точнее, предпочтя диалектике миф (в диалоге
«Тимей» он прямо аттестовал свои эволюционные взгляды как миф).
Аристотель искал истину иначе, в форме доказательства (до него оно
было развито лишь в геометрии), и для этого изобрел логику. После него
ранние стоики (п. 1-7) открыли еще однин путь познания - каталепсис (ин-
туитивное ухватывание сути, ответ 6). Ухваченная суть нуждается в обосно-
вании, и оно может быть дано как в форме подтверждения (диалектика), так
и в форме доказательства (логика). Все эти формы поиска истины в ряде
философских школ (начиная с софистов) подменяла риторика - удобные
фразы, заменяющие истину (см.: Чайковский Ю.В. // ВФ, 2002, № 9).
В Новое время термин «диалектика» получил у Гегеля иной смысл, но
нас здесь касается только платонова диалектика, поскольку именно ею
пользовался Дарвин при обосновании идеи отбора. Подробное описание
явлений (которое он не отличал от анализа понятий) создавало у него уве-
ренность, что он дал исчерпывающее беспредпосылочное знание. Критики,
наоборот, видели у Дарвина лишь «кучу подробностей» и риторику.
119
В XX веке философы науки признали: копить доводы в свою пользу -
самообман, видимость обоснования; не бывает беспредпосылочного зна-
ния; всё, что остается ученому - выявлять свои предпосылки и открыто их
формулировать именно как предпосылки, а не как нечто истинное. После
такой процедуры часто возникает новое, более скептическое отношение к
предпосылкам, а затем и желание “пошевелить" их, т.е. посмотреть, как бу-
дут объясняться следствия, если изменить предпосылки.
Дарвин, если заходил в тупик, тоже пробовал выявлять свои предпо-
сылки и даже “шевелить" их. Так появились пангенез и ламарковы факторы.
Но его роковой философский просчет был в том, что делал он это лишь в
форме оговорок, не проводя через логическую структуру учения.
Например, поняв, что некоторые свойства организмов нельзя объяс-
нить как результат отбора, он только для них, как для исключений, допускал
возможность иных факторов, но не исследовал далее эти факторы, а про-
должал приводить примеры в пользу отбора. Это хорошо описал Н.Я. Дани-
левский (натуралист, философ и деятель хозяйства), наш самый обстоя-
тельный критик дарвинизма:
«Нельзя сказать, чтобы он их (Дарвин трудности - Ю. Ч.) совершенно упустил
из виду, - он и сам... то в одном, то в другом месте скажет об них несколько
слов, совершенно ничего впрочем не разъясняющих, или упоминает о возра-
жениях, сделанных другими, признает за ними некоторую силу; но затем все
остается по-старому, и он продолжает свои выводы и доводы, как будто этих
возражений, им нисколько не опровергнутых, вовсе и не существовало» (Н.Я.
Данилевский. Дарвинизм. Критическое исследование. СПб., 1885, т. 1, ч. 2, с. 3).
Развивая мысль Виганда, Данилевский писал, что искусственный отбор
- «хитро устроенная машина», изобретенная человеком и не встречающая-
ся в природе, как и, например, паровая машина (ч. 1, с. 231). Тем самым,
параллель естественного отбора с искусственным, основная метафора
Дарвина, ничего не дает. Мы вернемся к этому вопросу в п. 5-8*.
Позже критики выражались и резче. Так, Теодор Рузвельт, президент
США (1901-1909) и натуралист-любитель, писал, что дарвинисты не столько
доказывают свои воззрения, сколько «упражняют красноречие», ибо всяко-
му признаку можно при желании придумать пользу (ЧЭ, с. 98).
На мой взгляд, истина рождается только интуитивно и должна прове-
ряться практикой, а не убеждениями. Логика необходима при ее обоснова-
нии, но - лишь для выявления логических ошибок. Иными словами, сама
логика истины не дает, но от ложных ходов мысли уберегать может.
Дарвин в 1867 году был в гостях у Генри Бэйтса, недавно выпустившего
книгу «Натуралист на Амазонке», где впервые излагались доводы в пользу
того, что яркие раскраски тропических животных - результат отбора. Зашел
разговор о красно-черной расцветке гусеницы, кормящейся на зеленом лис-
те - чем это ей полезно? Ведь другие гусеницы зелены. «Вы бы лучше
спросили Уоллеса» - сказал Бэйтс, что Дарвин вскоре и сделал. Уоллес (о
120
силе воображения которого мы уже знаем) ответил, что сам видел белых
бабочек на зеленом фоне и склонен предположить, что те предупреждают
контрастной расцветкой о своей несъедобности. Дарвин был в восторге:
«Никогда не слыхал ничего более остроумного... Факт белой моли великоле-
пен; кровь загорается, когда видишь, что истинность теории почти доказана»
- писал он Уоллесу.
Ободренный Уоллес положил подобные рассуждения в основу учения
(в его книге «Дарвинизм» четыре главы из пятнадцати посвящены окраске),
Бэйтс же решил иначе: из второго издания своей книги изъял все рассужде-
ния об отборе расцветок. Кто прав?
3-7* Насекомые опровергают идею отбора
«Почти доказана» была не «истинность теории», а правдоподобность
частного объяснения частного примера. То есть был предложен еще один
довод в пользу своей правоты. Такой довод бесполезен в науке: он убеди-
телен для тех, кто и без него думает «как надо», а для думающих иначе вы-
глядит шуткой. Последние приводят свои аргументы, которые тоже не про-
изводят впечатления на тех, кто не готов к ним заранее. Так, довод Уоллеса
об упреждающей окраске был отвергнут в той самой дискуссии на страни-
цах «Nature», о которой уже шла речь.
А именно, в декабре 1870 года в «Nature» энтомолог Сэмюэл Скёддер
недоумевал: гусеница гротескной формы и угрожающей окраски буквально
кишит пожирающими ее паразитами, тогда как гусеницы близкого вида для
них несъедобны; почему оказался эффективным отбор столь сложной фор-
мы и расцветки, если судьбу решают не птицы, а паразиты, съедающие
99,9% потомства на стадии яиц, гусениц и куколок?
Признаюсь, меня этот вопрос в свое время тоже ошарашил; но для ве-
рующих в отбор он просто неинтересен. Оппонентам оказалось куда инте-
реснее, что Скёддер допустил мелкую ошибку: он не удержался от сарказма
(«Изучая 15 лет бабочек, я не видал ни одной в птичьем клюве»), - они ста-
ли радостно возражать: мол другие видали. А суть дела обсуждать не стали,
ибо эта суть была для них убийственна.
Дело в том, что Скёддер указал на общий для насекомых факт: на од-
ной из стадий размножения они могут подвергаться почти полному истреб-
лению. В те годы было уже известно о тропических термитах - общест-
венных насекомых, которые перед спариванием теряют крылья и, беспо-
мощные, становятся пищей для многих видов [Метерлинк, с. 334]. Лишь
около одной пары на тысячу ускользает от гибели, а это значит, что самою
природой из века в век ставится очень жёсткий селекционный опыт: если
есть вариации съедобности и они могут отбираться, то несъедобные долж-
ны вытеснить остальных. Но этого не происходит, следовательно, никакого
«отбора на несъедобность» здесь нет, и идея отбора опровергнута
121
прямым массовым наблюдением. Нет никаких оснований предполагать
отбор в менее жёстких ситуациях.
Разумеется, в «Дарвинизме» Уоллеса о термитах ни слова нет. Дарвин
помянул смертный полет термитов лишь однажды: «иногда два самца пре-
следуют одну самку» (Соч., т. 5. М., 1953, с. 382), а о сути умолчал. Правда,
в 6-м издании «Происхождения видов» он признал эту трудность (что нет
отбора при общем выедании), но тут же закончил: «тем не менее из числа
тех, которые выживут, наиболее приспособленные особи... будут размно-
жаться в большем числе». Насколько в большем (можно ли эту разницу за-
метить за все время жизни вида?), Дарвин обсуждать не стал.
Это был уже просто уход от обсуждения. К сожалению, такой приём
стал традицией дарвинизма - приводить довод в пользу своего убеждения и
успокаиваться. Это даже не диалектика, это уже риторика.
Смысл массового выедания мы нам станет ясен позже (пп. 6-1, 6-2*, 6-
14**), а пока обсудим полевые и лабораторные опыты по выеданию насеко-
мых разных расцветок птицами, ставившие целью доказать эффективность
отбора. Вот самый яркий пример. Николас Тинберген, нобелевский лауреат,
исследовал поедание птицами гусениц Он выбрал гусеницу пяденицы,
имитирующую сухой сучок, причем
“случай наиболее утонченной маскировки мы выбрали отнюдь не случайно:
если столь точная имитация сучков в мельчайших деталях возникла благода-
ря естественному отбору, значит, можно найти и тех, кто этот отбор произво-
дил. ... Возражения против теории естественного отбора в применении к по-
добным явлениям нередко сводятся всего лишь к ссылкам на то, что птицы и
другие животные вряд ли могут обладать необходимой для этого способно-
стью замечать и правильно оценивать мельчайшие детали”. Он был убежден в
силе их способностей, “но одного убеждения было мало - его следовало под-
крепить самой жесткой экспериментальной проверкой” {Тинберген Н. Осы,
птицы, люди. М., Мир, 1970, с. 146).
«Самая жесткая проверка» свелась к тому, что птицу сойку, одну-
единственную особь, выпустили на площадку, где было много сучков и гусе-
ниц: птица прыгала по ней минут 20, пока не наступила на гусеницу. Та изо-
гнулась и тут же была склёвана, а больше сойка ничего найти не сумела.
Тинберген счел, без повтора и контроля, опыт завершенным, ибо «мы сра-
зу же получили положительный результат». Но коллега Тинбергена продол-
жил исследование и выяснил иное: птицы тратят от 7 до 40 минут на поиск
первой гусеницы, однако «после обнаружения первой гусеницы остальные
почти всегда отыскивались очень быстро» (с. 149). Словом, сойка в “опыте”
Тинбергена была просто бестолковым экземпляром.
Ситуация вполне обычная: опыт ставился для подтверждения теории и
потому был воспринят как ее подтверждение. Заметьте, если бы реальность
отбора не была заранее постулирована, то и вопрос был бы иной: влияет ли
поиск первой гусеницы на выживание их популяции? Факты Скёддера дают
122
основание полагать, что ответ отрицателен, но Тинберген предпочел отве-
чать на другой вопрос: на тот, где заранее знал ответ, - об остроте зрения
птиц А ведь знал (судя по ссылке на возражения), что скептики типа Скёд-
дера существуют. Среди них был и знаменитый тогда французский эволю-
ционист Люсьен Кэно, на основе фактов полагавший, что даже самая со-
вершенная маскирующая окраска не может существенно снижать поедае-
мость хищниками [Назаров, 1974, с. 186].
Пусть скептики и редки, но отвечать надо именно на их неприятные во-
просы, а не на свои, удобные. Откуда такая методическая слепота? В инте-
ресной книге Хью Котта (ЧЭ, с. 98) находим такой ответ:
«Многие нападки на дарвинизм вышли из лабораторий и кабинетов... Здраво
рассуждая, внимание, уделяемое возражениям, преуменьшающим интенсив-
ность борьбы за существование, следует соразмерять с объемом полевого
опыта ученого, выступающего с этими возражениями».
Почему полевого опыта надо требовать именно от выступающего с воз-
ражениями? А если с подтверждением (как Тинберген, проведший свой
“опыт” в лаборатории), то полевой опыт не нужен? Не лучше ли задуматься
(в кабинете, разумеется) о том, что в опыте получено? И хотя бы вспомнить
студенческий практикум и не прерывать опыт, едва он дал желаемый ре-
зультат? Но фраза Котта, сейчас нелепая, имела в дни Тинбергена большой
успех. К счастью, не у всех. Например Любищев, ведущий полевой энтомо-
лог, аттестовал приспособительную окраску как наивный маскарад, который
хищники легко разгадывают.
Поскольку до сих пор серьезных примеров отбора никто не привёл,
можно подвести итог. В «доказательство» эффективности естественного
отбора вот уже полтора века приводятся только диалектические уверения в
своей правоте, тогда как осмысление конкретных («полевых») данных гово-
рит против. Как такая слабая идея могла господствовать?
Любищев любил говорить, что отказ от философствования - худшая из
философий: ведь всяк исходит из неких общих установок, и лучше делать
это явно, нежели утверждать, что обходишься без философии. По Любище-
ву, в его время в науке, особенно в генетике, царил мизологизм (отвраще-
ние к размышлению - термин Платона) [Любищев, 2004, с. 4].
Оценил философию и Дарвин, но - в самом конце жизни. В феврале
1882 года, прочтя «О частях животных» Аристотеля, он писал: «Моими бо-
гами, хотя и очень по-разному, были Линней и Кювье, но они просто школь-
ники по сравнению со стариком Аристотелем». Читать аристотелеву критику
платоновой диалектики было поздно: в апреле Дарвин умер.
3-7** Тезис Витгенштейна
К середине XX века философы науки убедились: для сообщества
ученых никакой факт не имеет значения, если нет объясняющей его
схемы [Кудрин, с. 22; Мейен и др., с. 121]. Сделано это было, в основном,
123
на материале физики, а у биологов и даже философов биологии до сих пор
еще обычно убеждение, что науку движут именно факты и что дарвинизм
победил в силу его соответствия фактам. Это странно, если вспомнить, что
основное понятие, естественный отбор, не имело, как признавал сам Дар-
вин, фактической базы и не имеет, как увидим, до сих пор.
В 1930-х годах австрийский философ Людвиг Витгенштейн (1889-1951)
прояснил этот вопрос, и философ Зинаида Сокулер резюмирует его так:
“Каждая научная теория каким-то образом свидетельствует о мире, но если
попытаться точно сформулировать это свидетельство, то выйдет философская
бессмыслица или ложь” {Сокулер З.А. Людвиг Витгенштейн и его место в фи-
лософии XX века. Долгопрудный (Моск, обл.), 1994, с. 57).
Это выглядит преувеличением, но в нашем случае так оно и есть: в са-
мом факте, что неспособные жить погибают, никто не сомневается, и тео-
рия Дарвина “свидетельствует" об этом. Однако из названного факта еще
ничего не следует для теории эволюции, ибо его признают и креационисты
(видящие в нем свидетельство Божией мудрости). У всех, кто пытался уточ-
нить понятие отбора настолько, чтобы им можно было пользоваться для по-
строения теории, возникали сомнения в реальности отбора; а математиче-
ски точное понятие отбора оказывается вообще ни к чему в природе не при-
ложимо (анализ см. [Чайковский, 2004, с. 167]). Так что Тинберген просто
спутал отбор как наглядную реальность и как фактор эволюции', из того,
что неспособные жить не живут, не следует, что способные жить эволюиру-
ют (последнее надо еще доказать - см. п. 5-8*).
Наглядность отбора, воспринятая как его неопровержимость, делает
для его адепта невозможным какой-либо акт проверки роли отбора как фак-
тора эволюции. Вот этот вывод в изложении Сокулер (с. 158):
“Когда мы придаем предложению статус неопровержимо достоверного, мы
тем самым, как показывает Витгенштейн, начинаем употреблять его как пра-
вило (соответствующей языковой игры) и на его основе оцениваем все другие
предложения. ... Одно и то же предложение может выступать в одних ситуа-
циях как доступное экспериментальной проверке, а в других - как правило для
проверки иных предложений. Но есть предложения, которые настолько закре-
пились в функции правил, что вошли в структуру некоторой языковой игры и
приобрели логический характер. ... Они предшествуют всякому определению
истинности и соответствия реальности”.
Здесь высказан известный тезис Витгенштейна:_______________________
утверждение, нуждавшееся в проверке, становится само правилом для про-
________верки иных предложений и для признания фактов истинными._______
Мысль высказана вне связи с отбором, а выглядит как сказанная прямо
о нем. Следовательно, Витгенштейн подметил общую закономерность по-
знания. Через 30 лет ее выявил Кун в рамках своей концепции нормальной
науки (о ней см. п. 2-1). Суть ее в том, что научное сообщество как целое
124
весьма консервативно и тщательно охраняет основные положения науки от
проверок, пока накопившиеся противоречия не опрокинут систему взглядов
и не приведут к очередной научной революции.
3-8. Активность и морфология. Дарвинизм по Геккелю
Но чем же на самом деле, в природе, достигается столь часто и явно
наблюдаемая приспособленность организма к своей среде? Пора сказать
об общем механизме - активном приспособлении. Оказывается, даже ярые
дарвинисты без этого ламаркова принципа не обходились, только данную
сторону их трудов поминать не принято.
В 1866 году появилась замечательная «Общая морфология» Эрнста
Геккеля (1834-1919), которого вскоре прозвали «немецким Дарвином», а сам
дарвинизм обрел в Германии вторую родину. В этой книге была восстанов-
лена традиция сквозного рассмотрения эволюции, когда развитие жизни
выступает как итог эволюции Вселенной. В предисловии Геккель рассмат-
ривал свою деятельность как борьбу,
“исход которой не может вызывать сомнений после Чарлза Дарвина, который
семь лет назад нашел ключ к твердыне и благодаря своей достойной восхище-
ния селекционной теории создал из теории происхождения (Descendenz-
Theorie), установленной Вольфгангом Гёте и Жаном Ламарком, победоносное
завоевательное оружие”.
Фраза очень характерна - и романтизмом, и оптимизмом, и установкой
на борьбу, и грубыми ошибками. «Теория происхождения» была выдвинута
до Гёте, он вовсе не развивал ее (хоть и выражал к ней симпатию), и Дар-
вин ни в каком смысле не исходил из Гёте; наоборот, положив в основу из-
менчивость, он заявил позицию, противоположную морфологии Гёте.
Сам Гёте работал в той же традиции, что и Кювье, которого Геккель
счел креационистом [Колчинский, с. 59]. Что касается теории Ламарка, то из
нее Дарвин действительно исходил (хотя отрицал это), и слова Геккеля
имеют основополагающий смысл: он предлагал читателю не столько дарви-
низм, сколько дополненный идеей отбора ламаркизм.
Главным у Ламарка он видел идею приспособления за счет активно-
сти особи, и материалом для отбора полагал результат такой активности,
т.е. унаследованное направленное приспособление. Акт выживания взят у
Дарвина, но механизм, поставляющий материал для такового выживания (в
наших терминах, «буксир») - у Ламарка и Жоффруа: «Все свойства орга-
низмов приобретаются ими или в силу наследования, или в силу приспо-
собляемости». Тем самым, геккелеву триаду надо было бы выражать сло-
вами. приспособление, наследование, отбор.
Такое понимание сути дарвинизма шло от Бронна и было для Германии
характерно. Людвиг Плате, последователь Геккеля, даже называл это «чис-
тым дарвинизмом» (в пику "неодарвинизму", который признаёт лишь нена-
правленные вариации). Историк науки Эм. Радль писал, что Геккелю
125
«свойственны два слова, приспособление (Anpassung) и наследование (Verer-
bung)», причем «через первое слово обозначается изменчивость вместе со
всеми ее причинами и следствиями», так что «под приспособлением понима-
ется возникновение новых форм, а под наследованием - их фиксация. Так
Геккель двумя словами ухватил все содержание дарвинских работ...» {Radi Е.
Geschichte der biologischen Theorien. T. 2. Leipzig, 1909, S. 278).
Вскоре, с развитием генетики, мысль Геккеля оказалась забыта. С его
именем связывают сверхпростую формулировку дарвинизма: триаду «на-
следственность, изменчивость, отбор». Наследственность - это сходство
детей с родителями, изменчивость - случайное отклонение свойств детей
от свойств родителей, отбор - выживание только удачных отклонений. Как
видим, половой процесс, с которого начал Дарвин (см. п. 2-8), и активность
исчезли из сверхпростой схемы.
У социальной истории науки свои законы, и понемногу первая форму-
лировка (самого Геккеля) под перьями последователей превращалась во
вторую. Так, в 1923 году наш блестящий генетик, подробнее других изло-
живший геккелево понимание приспособления как первичного фактора эво-
люции, тем не менее резюмировал его мысль совсем упрощенно:
«В результате взаимодействия наследственности и изменчивости и происхо-
дит расхождение признаков... Благодаря борьбе за существование возникает
естественный подбор...» [Филитенко. с. 69].
Иными словами, социальная история пошла в сторону, противополож-
ную когнитивной, или, говоря проще, общество усвоило идею в той форме,
которая фактического обоснования не имела. И винить тут некого - общест-
во хотело слышать так, и никто из говоривших иначе не был услышан, пока
не настало их время. Запомним это и вернемся к Геккелю.
У Дарвина вопросы морфологии затронуты лишь мельком, в самом об-
щем виде, и многие морфологи (включая Бэра, Бренна и Оуэна) заявили,
что дарвинский механизм (отбор мелких изменений) не объясняет сути эво-
люции, т.е. процессов, связанных с изменением типа организации. Поворо-
том в судьбах морфологии как раз и явилась «Общая морфология».
В ней Геккель развил новую дисциплину, проморфологию, учение о
формах организмов. Это была блестящая попытка завершить линию Гёте -
Кювье - Бэра - Бронна. Главной была идея классифицировать организмы,
исходя из сходства форм их тел, а сходство понимать, прежде всего, в тер-
минах симметрии и соотношения положений органов. Он выявил много раз-
личных типов симметрии организмов, и два из них сохранились до сих пор -
радиальные и двусторонне симметричные организмы. Различие типов сим-
метрии Геккель считал фундаментальным.
Архетипы (о них мы говорили в гл. 1, пп. 9, 10, 11, 13) Геккель отожде-
ствил с предковыми формами, отчего система организмов приняла форму
филогенетического древа, т.е. родословной, ветвящейся от предка к по-
томкам. Он выводил все группы организмов из амеб, т.е. вместо неизмен-
126
ных типов вводил идею единства типа, идя в этом даже дальше Жоффруа,
поскольку относил к этому древу и все растения. Сложное учение Оуэна о
гомологии (п. 2-7*) обратилось в очень простое: частная гомология была ис-
толкована как унаследование от общего предка, а остальные типы гомоло-
гии игнорированы. Причину сходства руки и ноги Геккель видел в симмет-
рии. (Когда в XX в. симметрия выпала из анализа, эту гомологию, как и об-
щую гомологию, просто перестали упоминать.)
На учение Геккеля отозвался старый Бэр. Допуская идею эволюции, он
резко возражал против смешения морфологического сходства с историче-
ским родством (в частности, архетипа с предком). Для него это смешение -
«разрастающийся сорняк в вопросе о трансмутации», т.е. об эволюции.
Выводить разнообразие из единой предковой формы Бэр считал нена-
учным, в чем, кстати, был близок Дарвину, который свою схему постепенной
эволюции никогда не пытался приложить к становлению какого-либо архе-
типа, как бы этот термин ни понимать. Более того, в последнем издании
«Происхождения видов» архетип определен по Оуэну и Бэру, а не по Гек-
келю: «Архетип. - Идеальная первичная форма, по которой кажутся органи-
зованными все существа той или иной группы» (Ч.Дарвин. Соч., т. 3, М.-Л.,
1939, с. 667). Кстати, из последнего “академического” издания «Происхож-
дения видов» (СПб., 1991; 2001) это определение, как и весь Словарь тер-
минов, изъято. Не будем вдаваться в спор о том, стоит ли делать из Дарви-
на большего дарвиниста, чем он был в действительности.
Система Геккеля стала образцом на целое столетие, и в этом она впол-
не следовала духу Дарвина: ведь Дарвин считал факт эволюции Ав Б дока-
занным, когда между А и Б удавалось найти ряд переходных форм. И до сих
пор многие уверены, что эволюция данной группы понята, если указаны ли-
нии от воображаемого предка ко всем наблюдаемым потомкам.
Однако есть и другая точка зрения, гласящая: указание пути преобра-
зования ничего не говорит о механизмах преобразования. Ее высказывали
еще в дни Геккеля, в начале XX века она почти господствовала [Филипчен-
ко, с. 75-76], но затем биологи вновь увлеклись филогениями. Задача пони-
мания механизмов эволюции едва начинает ставиться в наше время (о чем
пойдет речь в части 2) и будет в нашем рассказе главной.
С середины XX века отношение к Геккелю стало меняться: от его родо-
словного древа ничего не осталось, зато вновь ожила проморфология, чту-
щая его как основателя [Беклемишев, т. 1, с. 22]. Она служит одной из основ
номогенеза, о котором далее у нас будет речь не раз.
Весь XX век шли споры о том, как понимать морфологию. В начале ве-
ка столкнулись два мнения: «Морфологично то, что еще не может быть по-
нято физиологически» (К. Гёбель) и противоположное: «Морфологично то,
что вообще нельзя понять физиологически» (В. Тролль) - см. [Канаев, 1966,
с. 133]. Как мы увидим, это противостояние бытует до сих пор и, видимо,
оно вечно В частности, физиологи уверены, что гомологическое сходство
127
свидетельствует о какой-то неведомой (быть может, прежней) пользе, а
морфологи указывают примеры, когда сходство, всеми признаваемое гомо-
логическим (например, нелепо сложные плавники некоторых рыб и ихтио-
завров) возникало заново и явно без всякой пользы (см. п. 9-10). И до сих
пор учебники дают гомологию по Геккелю.
В «Общей морфологии» Геккель описал и знаменитый “Основной био-
генетический закон”. Он ввел его в такой форме: «Онтогения являет собой
короткое и быстрое повторение (рекапитуляцию) филогении»77. Вместо
развития зародыша по заданным природой самого развтия законам, тут
предполагается, что организмы тащат за собой сотни миллионов лет свое-
образный "исторический хвост”. Легко видеть, что здесь переосмыслен в
механическом духе тезис Канта - Кильмейера (п. 2-3).
Видеть-то легко, а вот понять ход мысли Геккеля трудно (если вообще
можно). Ведь если налицо бесполезный “хвост”, почему (должен бы заду-
маться Геккель) он не отсечен естественным отбором?
Геккель об этом не задумался, но это сделали, как и следовало ожи-
дать, креационисты наших дней:
“Бессмысленные ходы в развитии, которые указывают нам только на филоге-
нез, должны быть исключены под воздействием отбора, так как они предпо-
лагают бесполезный расход времени и материала и, следовательно, являются
селективно отрицательными” (ЮШ, с. 130).
На самом деле организмы никакого “хвоста” не тащат: более тщатель-
ный анализ ранних стадий развития показал, что повторение пути - лишь
кажущееся. На самом деле онтогенез не повторяет историю, а история есть
последовательность онтогенезов (см. далее, п. 4-13). Увы, “закон” Геккеля
был частично обоснован поддельными рисунками, что вызвало скандал и
что позже признал сам Геккель (ЮШ, с. 128). Это не мешает им кочевать из
учебника в учебник в качестве подлинных черт онтогенеза.
Наконец, в «Общей морфологии» Геккель ввел в научный оборот слово
экология. Если у Дарвина всё взаимодействие особей сводилось к борьбе
за существование (остальное свелось к оговоркам, ни к чему не обязываю-
щим), то у Геккеля в самом деле есть азы экологического подхода. Так, он
обратил внимание на удивительный параллелизм сумчатых и плацентарных
млекопитающих (рис. 10) и объяснил его сходством условий существования
соответствующих животных.
17
Филогения, в понимании Геккеля, - историческое развитие, при котором процесс эво-
люции представляется в форме родословного древа. Есть другое, более широкое значе-
ние термина (почти синоним эволюции), которым мы пользоваться не будем.
128
/о
Рис. 10. Параллелизм сумчатых и плацентарных (из Raven, Johnson, 1989)
129
К старости Геккель разочаровался в своей схеме и, несмотря на ее ог-
ромную популярность, развил именно отвергнутую в то время обществом
идею всеобщей активности (о которой у нас пойдет речь в гл. 5):
«по Геккелю... уже атомам следует приписать простейшую фому чувствова-
ния и стремления, т.е. “душу” самого примитивного качества, а у организмов
имеются самые различные категории души - “душа клетки”, “душа союза
клеток”, “душа ткани”, “нервная душа” и т.д.» [Фшипченко, с. 76].
Как Дарвин до него и многие после него, Геккель знаменит отнюдь не
тем, чем в старости хотел бы. То же, как увидим, было с Вейсманом.
3-9. Презумпции наследования. Дарвинизм по Вейсману
Презумпция - это тезис, который признается истинным без доказатель-
ства до тех пор, пока не будет доказана его ложность. Широко известна
правовая идея - презумпция невиновности, подозреваемого надо считать
невиновным, пока суд не признает его виновным. См. Дополнение 1.
Когда скептики дружно заявили Дарвину, что наглядность идеи отбора
не может заменить доказательств реальности отбора как фактора эволю-
ции, он стал защищать идею отбора именно как презумпцию: не раз он
заявлял, что единственный пример свойства, необъяснимого отбором,
явился бы «сильнейшим ударом» всему учению.
Такие примеры указывали многие: еще Теофраст писал о бесполезно
огромных рогах оленя, а в дни Дарвина нашли ископаемого ирландского
оленя с рогами непомерными, явно вредными. Затем много примеров обна-
ружил сам Дарвин (в том числе самый важный, ум человека - п. 3-3), но
предпочел не менять учение, а искать каждому такому случаю отдельное
оправдание. Это, как мы знаем, всегда сделать можно.
Особенно преуспел в этом Уоллес. Даже про ирландского оленя Уоллес
нашел, что сказать: гигантские рога, оказывается, хоть потеряли роль «по-
лового вооружения» и стали цепляться за ветки, но зато стали защищать
некоторые точки тела при беге сквозь те же ветки. Интересно - сколько, по
мнению Уоллеса, от этого родилось добавочных оленят за всё время жизни
всех оленей? Если бы Уоллес задал себе этот вопрос, ему, полагаю, стало
бы ясно, что пользу от рогов он толкует не по Дарвину, а по Ламарку. Одна-
ко такие вопросы в дарвинизме задавать не принято.
До конца дней Уоллес призывал считать отбор всемогущим, пока не бу-
дет доказано противное. Напомню: он пришел к выводу, что ум отбором (ни
естественным, ни половым) не создан, но презумпцию не сменил.
Легко видеть, что «принцип наследственности» (п. 3-5) был введен Уол-
лесом тоже как презумпция - после того, как Беннет загнал его в угол. Мож-
но вспомнить тут и тезис Витгенштейна: утверждение, нуждавшееся в про-
верке, становится само правилом для проверки иных предложений, т.е. (до-
бавлю) нормой или презумпцией. Уоллесу же принадлежит еще одна пре-
130
зумпция дарвинизма: считать каждый признак полезным - либо по извест-
ной причине, либо по неизвестной.
В наше время порою раздается призыв решать эволюционные вопросы
так же, как в суде, т.е. признавать данное учение (обычно это какая-то часть
дарвинизма) истинным, пока не доказана его ложность.
Опыт показывает, что тот ученый, который принял какую-то презумп-
цию, уже не ищет истину, а копит доводы в свою пользу. Истину ищут дру-
гие. Мы уже говорили, что люди, встретясь с опровержением своих взгля-
дов, склонны не менять их, а игнорировать само опровержение или, в луч-
шем случае, искать в нем слабые места, а найдя хоть одно, успокаиваться
(«чего хочется, в то и верится» - говорил по этому поводу Данилевский).
Так, например, поступали Жоффруа (пп. 2-2, 2-6) и Дарвин (п. 3-6). Наобо-
рот, призыв М.Д. Голубовского (в сборнике ЭБ) отказаться от “демона авто-
ритетов” есть по сути признание неприемлемости презумпций в науч-
ном поиске. См. Дополнение 1.
Ученый, принимая презумпцию, заданную его окружением, теряет плю-
сы индивидуального мышления и должен понимать, что перестает быть са-
мостоятельным теоретиком. Уверять себя: «я сменю убеждение, как только
мне предъявят убедительные доводы» - наивный самообман. Сменить
убеждение он может, только если готов его сменить. Только тогда доводы
полезны. Иначе он будет лишь искать контрдоводы. А обычный ученый (т.е.
работающий в “нормальной науке”, см. п. 1) бывает готов к смене только
вместе с обществом. Получается порочный круг, выход из которого произ-
водится всем обществом лавинообразно и с большим опозданием, причем
не по осознании причин, а под влиянием повода.
Мне видится верной та мысль английского сборника (1859: Entering in
age of crisis. Bloomington, 1959), что победу дарвинизма определило общее
падение религиозности в 1850-х годах, а книга Дарвина лишь дала повод.
Столетиями эволюцию признавали единицы и вдруг, в течение полуго-
да, стало признавать большинство, хотя Дарвин не привел ни одного нового
факта, да и из прежде известных взял малую часть - феномен изменчиво-
сти. Идея эволюции стала вдруг презумпцией для общества потому, что ему
был Дарвином дан повод (голословное, но наглядное объяснение - отбор
полезных наследуемых вариаций) и оно к этому моменту созрело для такого
повода (статьи о “проблеме вида” появлялись в западной научной периоди-
ке 1858-59 годах едва ли не каждый месяц). А созрело общество потому, что
религиозность проходила тогда на Западе через свой очередной минимум.
Дарвин, как и все до него, был уверен, что приобретенные особью при
жизни свойства могут наследоваться. При каких условиях НПС на деле про-
исходит, ни он, ни кто другой объяснить не мог. НПС имело тогда статус
общепризнанного постулата (но не презумпции).
Приведя в «Изменении» огромное число примеров как наследования,
так и его отсутствия, Дарвин пришел к вполне верному выводу: НПС на-
131
блюдается, но нерегулярно. Некоторые его выводы очень важны - на-
пример: 1) из всех традиций уродования людского тела только обрезание
(если, добавлю, проводится мальчикам во младенчестве) производит на-
следственный эффект, пусть нерегулярно и неполно, но достаточно часто;
2) в других случаях НПС происходит часто лишь тогда, когда «передается
именно болезненное состояние нервной системы»; 3) принято считать, что
сеянцы яблонь, груш и слив вырастают дичками, но на деле всё сложнее: у
них очень высока изменчивость, но дичками большинство сеянцев считать
нельзя (Дарвин Ч. Изменение..., гл. 12).
Казалось бы, эволюционисты должны были броситься изучать НПС, но
вышло иначе. То было время становления математической статистики, ко-
гда идеология “надежных фактов” уступала место идеологии “статистиче-
ской достоверности”. НПС не удовлетворяло ни той, ни другой и было обре-
чено на столетнее забвение. Тот факт, что наибольший интерес для пони-
мания процесса могут представлять редкие события, не имеющие опреде-
ленной частоты (возникающие на грани порядка и хаоса - см. далее, п. 5-
18*), едва начинает осознаваться в наше время.
В 1883 году немецкий зоолог Август Вейсман (1834-1914) заявил, что
НПС - постулат, не имеющий обоснования. Он указал, что для передачи по-
томству изменение должно попасть в половую клетку, тогда как проявляется
оно в соматических (неполовых) клетках. Он сформулировал на сей счет
презумпцию, которая легла в основу неодарвинизма: надо отрицать НПС,
поскольку «до сих пор нет ни одного факта, который действительно доказы-
вал бы, что приобретенные свойства могут передаваться по наследству».
То есть сменил постулат наследования на противоположную презумпцию,
что никак нельзя назвать хорошим научным приемом.
В терминах логики: отрицание квантора всеобщности всегда дает кван-
тор существования (от отрицания), но не квантор всеобщности. Вейсман за-
конно мог утверждать лишь, что существуют акты ненаследования и что они
преобладают. Он же сменил при отрицании один квантор всеобщности на
другой. Логику в те годы изучали в гимназиях, так что ошибка была очевид-
на. Однако смена установки (поводом для которой послужила книга Вейс-
мана) в обществе ученых произошла, и ошибку никто из принявших пре-
зумпцию не замечал. Даже когда на нее указал сам Вейсман - см. далее.
Обычно презумпции защищаются именно с данной логической ошибкой - с
подменой кванторов.
Доказав в опыте, что не наследуются лёгкие травмы (он рубил мышам
хвосты в течение ряда поколений), Вейсман по аналогии заключил, что не
наследуются и никакие прочие изменения. И хотя он ясно понимал, что
имеет дело с аналогией, его последователи стали писать и, как ни странно,
пишут до сих пор, что невозможность НПС доказана.
В действительности опытные факты, как собранные во времена Дарви-
на, так и новейшие, ясно говорят, что свойства, приобретенные особью при
132
жизни (в основном, в детстве), наследуются, но не всегда и не все (новей-
шие ссылки см. [Назаров, 2005, гл. 14]).
Однако бессмертна заслуга Вейсмана в том, что он первый заявил: по-
ловые клетки защищены от влияния внешней среды (“барьер Вейсмана"
[Стил и др.]). Этим был положен конец наивным построениям (таким, на-
пример, какое ярко описано у Пушкина: «И не диво, что бела / Мать бере-
менна сидела / Да на снег лишь и глядела»).
Позже Вейсман понял, что был неправ, считая барьер непроницаемым,
и указал возможный путь изменения наследственного материала при жизни
- патологическое воздействие на половые клетки [Филипченко]. Это было
великолепное прозрение (предвиденье патологического стресса), однако
оно опережало век, и тут Вейсмана не слушали. Правда, бывали исключе-
ния: у Любищева в письме (1945 г.) читаем:
“ заслугой Вейсмана, по-моему, является не его логичность, а правильный
упор на то, что наследственная субстанция гораздо более независима от
внешних факторов, чем это было принято думать до него. Но он, конечно,
сделал грубую ошибку, пытаясь абсолютизировать независимость, от чего он,
впрочем, почти отказался к концу жизни” [Любящее - Гурвич, с. 174].
Презумпция Вейсмана овладела ученым миром и только одна из его
наследия запомнилась. То был тупик, из которого биология лишь через сто
лет начала выбираться - мы узнаем об этом в части 2.
3-10. Происхождение родов идет не так, как происхождение
видов. Дарвинизм по Копу
Дарвин не дал ископаемых иллюстраций эволюции, ограничился увере-
нием, что ископаемая летопись слишком неполна. Ведущие палеонтологи с
этим не согласились и вообще встретили дарвинизм холодно. Но уже в 1868
году молодой американский палеонтолог Эдвард Коп (1840-1897) выступил
в поддержку Дарвина - с одной, правда, важной оговоркой: естественным
отбором можно объяснить происхождение видов и ничего более.
Сын богатого торговца, Коп обратил свое состояние на организацию
экспедиций, чем встал у начала не только нового типа научных исследова-
ний (тогда же начал свои раскопки Трои богач Шлиман, а Дорн стал строить
свою станцию), но и планомерного палеонтологического изучения США,
ставших одной из наиболее обследованных стран. Он был самым знамени-
тым палеонтологом США, «американским Кювье», и облик многих ископае-
мых мы знаем именно по реконструкциям Копа. Но, в отличие от Кювье, он
стал знаменитым эволюционистом. Его чтут как основателя психоламар-
кизма - течения мысли (близкого Дорну), которое ценит у Ламарка более
всего идею волевого акта особи. О психоламаркизме см. [Филипченко, гл. 7],
мы же коснемся взглядов Копа на отбор.
Сравнивая видовые и родовые признаки, он пришел к выводу, что лишь
первые можно рассматривать как результат отбора, а в образовании вторых
133
его роль мала. Еьце в 1864 году Спенсер охарактеризовал естественный от-
бор как «переживание наиболее приспособленного» (the survival of the
fittnest). Теперь же (1871 г.) Коп призвал отличать переживание от происхо-
ждения: по его мнению, отбор говорит многое о первом, но мало о втором.
Он писал, что эволюционные силы можно разделить на производящие
(originative) и направляющие (directive), причем отбор является только на-
правляющим фактором, тогда как еще предстоит «выяснить причины проис-
хождения наиболее приспособленного (origin of the fittnest)».
Чем выше таксономическое значение признака, тем более в его фор-
мировании видна роль производящих сил, и легче всего это увидеть на от-
личии происхождения родов от происхождения видов: есть признаки видо-
вые и признаки родовые, они эволюируют независимо, так что родовые
(требующие данной силы) могут изменяться без изменения видовых.
В этом Копу следовал немецкий палеонтолог Густав Штейнман, кото-
рый положил идею сохранения свойств низших таксонов при смене высших
в основу схемы эволюции. По Штейнману, основные группы не вымирают, а
изменяют таксономическое положение. Среди млекопитающих
«древнетретичные амблиподы (предки копытных - Ю.Ч.)... продолжают су-
ществовать ныне в виде гиппопотама и моржа, современные дельфины про-
исходят от ихтиозавров, кашалоты - от плезиозавров, беззубые киты - от
Thallatosauria» (Steinmann G., 1908, цит. по: Давиташвили Л.Ш. История эво-
люционной палеонтологии... Изд. АН СССР, 1948, с. 250).
Коп и Штейнман увидели черты того, что в п. 6-3 названо диасетью.
В 1875 году мысль, близкую «производящей силе» Копа, выразил Луэс
в Англии, назвавший акты появления принципиально нового словом эмерд-
женты. В отличие от Копа, термин Луэса прижился, и мы не раз еще упомя-
нем эмерджентную теорию эволюции.
3-10* Параллельные ряды
Идея независимой эволюции видовых и родовых признаков привела
Копа к необходимости дать начало еще одному эволюционному течению -
теории гомологических рядов. Еще Дарвин приводил примеры поразитель-
ных параллелей в наследственной изменчивости организмов. Например,
персики бывают бархатистые и гладкокожие, причем
«Гладкий персик - потомок бархатистого; разновидности бархатистых и
гладких персиков представляют замечательные параллели: у плодов белая,
красная или желтая мякоть, косточки то не отделяются, то отделяются, цветки
крупны или мелки, листья пильчаты или зубчаты... Надо отметить, что при-
знаки каждой разновидности гладкого персика получились не от соответст-
вующей разновидности бархатистого» (Ч. Дарвин. Изменения..., гл. 26, и.
«Аналогичная или параллельная изменчивость»).
Дарвин видел тут затруднение для теории, но не придал ему веса:
134
«Признаки, происходящие исключительно в силу аналогичных изменений,
будут, по всей вероятности, несущественного свойства, потому что сохране-
ние всех функционально важных признаков будет определяться естествен-
ным отбором» (Ч. Дарвин. Происхождение видов, гл. 5).
Легко видеть обычную у него подмену основания (п. 3-2): ведь именно тот
факт, что важные признаки определяются отбором, надо проверить.
Это и проделал Коп. Он пришел к выводу, что параллели наблюдаются
даже в таких важных свойствах, которые приписать их отбору нельзя, ибо в
эволюционном ряду такое свойство может как расти, так и убывать. В числе
примеров он привел параллельные ряды ископаемых лягушек, у которых
явно полезные свойства (перепонки для плаванья, диски для прилипанья к
субстрату и т.п.) в разном историческом порядке сочетались с такими явно
бесполезными свойствами, как недоразвитие грудины или рта (Давиташви-
ли Л.Ш. История эволюцион. палеонтологии... М.-Л., 1948, с. 176-177). Коп
видел причину параллелизма так: сходство между родами (членами данного
ряда) обязано своим происхождением наследственности, а их сходство с
членами другого ряда «происходит от тождественных эволюционных влия-
ний», т.е. от общности законов эволюции.
В качестве самого наглядного примера параллельных рядов Коп указы-
вал на млекопитающих - сумчатых и плацентарных. Саму эту параллель,
как мы знаем, до него описал Геккель, приписавший ее, однако, действию
отбора. Доводы в пользу этого допущения были чисто эмоциональны (как
всегда при упоминании роли отбора), поэтому Коп искал иного обоснования.
Он провел аналогию между параллельными рядами животных и уже из-
вестными тогда в химии «гомологическими рядами» углеводородов (спир-
тов, эфиров, меркаптанов). Тем самым, его смело можно отнести к предше-
ственникам номогенеза, о котором пойдет речь в главе 4.
Итак, Копу (и, насколько знаю, только ему) довелось встать у истоков
сразу трех эволюционных традиций. Однако палеонтологи помнят его преж-
де всего как автора «принципа неспециализированного», который гласит,
что предками будущих групп служат вовсе не высшие представители групп
прежних, а их примитивные (неспециализированные) члены. Поясню: нет
смысла искать предка человека среди гоминид (высших обезьян), он мог
быть только среди низших обезьян, еще не специализировавшихся к дре-
весной жизни. А специализированные формы неспособны к существенной
эволюции, если верить Копу.
Идея красива и подтверждается многими примерами, но неясно, откуда
неспециализированные всякий раз берутся. Это уже совсем другая пробле-
ма, в дни Копа еще не родившаяся (согласно излагаемой в данной книге
концепции, массовую деспециализацию производит снятие давления нор-
мы, например, стресс - об этом пойдет речь в главе 5).
135
3-11. Борьба как взаимопомощь. Дарвинизм по Кропоткину
В 1890 году в Лондоне независимо появились сразу две работы - кем-
бриджского экономиста Альфреда Маршалла и русского эмигранта П.А.
Кропоткина, теоретика анархизма. Каждая из работ пыталась в своей науке
поколебать мнение о благотворности конкуренции. Маршалл убеждал, что
победа в рыночной конкуренции происходит не столько в силу лучшего ка-
чества или меньшей себестоимости изделий, сколько в силу «удачного на-
чала деятельности» фирмы (ныне это хорошо известно, только теперь гово-
рят более общо - в силу случайности, не имеющей отношения к свойствам
продукции - см. далее). Через сто лет выяснилось, что это обстоятельство
рушит базовую аналогию дарвинизма.
А Петр Алексеевич Кропоткин (1842-1921) показал: ни перенаселение,
ни связанная с ним конкуренция не являются в природе общим правилом.
Так, в холодных странах никакого перенаселения нет, животные гибнут не в
борьбе друг с другом, а от иных, не зависящих от плотности индивидов фак-
торов, и все же эволюция вполне идет.
Кропоткин напоминал, что петербургский зоолог К.Ф. Кесслер в 1880 го-
ду говорил на Съезде русских естествоиспытателей:
«Я ведь не отрицаю борьбы за существование, но только утверждаю, что про-
грессивному развитию... не столько соответствует взаимная борьба, сколько
взаимная помощь... Всем органическим телам присущи две коренные потреб-
ности - потребность питания и потребность размножения. Потребность пита-
ния ведет их к борьбе за существование и к взаимном}' истреблению друг
друга, а потребность размножения ведет их к сближению между собой и к
взаимной помощи друг другу».
Сам Кропоткин сожалел, что Дарвин, признав роль сотрудничества,
«не подверг более строгому исследованию сравнительную важность и относи-
тельную распространенность двух форм борьбы» и что, признав (в гл. 6 «Про-
исхождения видов») уход от конкуренции в качестве самого обычного способа
действия естественного отбора, не связал его с основной идеей учения (П.А.
Кропоткин. Взаимная помощь как фактор эволюции. Харьков, 1919, с. 14. 68).
Успех дарвинизма на Западе Кропоткин связывал с тем, что западные зоо-
логи обычно путешествуют в тропиках, где видят перенаселение; русские же
зоологи чаще ездят в Сибирь и на Север, потому мальтузианство у них не в
ходу. То есть продолжал традицию «Дарвин без Мальтуса» (п. 2-12).
Аргументация слаба (как у Дарвина, примеры в свою пользу без попыт-
ки сравнительного анализа), но надо сделать два замечания. Во-первых,
если в тропиках регулярно действует фактор, отсутствующий или слабый
вне тропиков, то эволюция в тропиках должна идти в целом по-особенному.
Так отчасти и оказалось (см. п. 9-4, «Глобальный флорогенез»). Во-вторых,
русские эволюционисты подметили главное - не все акты взаимодействия
особей, важные для эволюции, управляются конкуренцией.
136
“Уход от конкуренции” - вот главное, на мой взгляд, что отметил и при-
нял Кропоткин в дарвинизме (в отличие от последователей Дарвина, не за-
метивших этого фактора). А XX век ознаменовался признанием того, что и в
экономике главным движущим фактором является не конкуренция, а попыт-
ка уйти от нее плюс самоорганизация', они рождают к жизни корпорацию как
основной тип организации производства. Как выяснил экономист Роналд
Коуз (США) в 1932 году, внутри корпорации правит не конкуренция, а ад-
министрация, друг же с другом корпорации не столько конкурируют, сколько
делят сферы влияния. Именно данный тип организации крупного производ-
ства и означает в развитых странах так называемую рыночную экономику.
Эпитет “рыночная” тут столь переосмыслен, что честнее приискать иной тер-
мин. В науке это термин “корпоративная экономика”. Она явила ряд важных
сходств с экономикой социализма, и в 1967 г. экономист Джон Гэлбрейт (США)
выступил с теорией “нового индустриального общества”, идущего на смену ка-
питализму и социализму. Добавлю: главной машиной при этом стал компьютер.
Но в публицистике и в политике до сих пор царит идиологема “рыночная эко-
номика”, противополагаемая идеологеме “плановая экономика”, хотя таковой
экономики у нас давно нет: при Хрущеве возник, а при Брежневе расцвел, в тер-
минах американских советологов, «административный рынок». Он, а не свире-
пая плановая экономика, привел к «застою». Суть его в том, что хозяйством и
культурой правят чиновники, меняющие один ресурс (“дефицит”) на другие.
Поскольку дарвинизм и рыночная экономика издавна служат друг другу моде-
лями, предлагаю прочесть статьи: Артур У.Б. Механизмы положительной обрат-
ной связи в экономике // ВМН, 1990, № 4; Гэлбрейт Дж.К. Фасад и скрытая за
ним истина // ВМН, 1991, № 8; [Чайковский, 1993; 2004, п. 5-3.2]. К теме биоэво-
люции ближе всего тот вывод экономистов, что конкуренция - феномен положи-
тельной обратной связи, т.е. дестабилизирующий фактор. Поэтому “одна из тех-
нологий в конечном итоге вытеснит все остальные... Однако, во-первых, нельзя
заранее предсказать, какая технология победит, а во-вторых, победившая техно-
логия может и не оказаться потенциально лучшей из тех, между которыми про-
исходила конкуренция” (Доси Дж., Нельсон Р., Введение в эволюционную эко-
номическую теорию // Вестник молодых ученых. Эконом, науки. 1999, № 1, с.
95). Этот феномен тоже именуется отбором (там же, с. 87), хоть и не связан с
избирательной смертностью (ср. п. 6-12). Рыночная идеология близка к дарви-
низму еще и тем, что смешивает сущность и акциденцию (п. 1-6**): если худший
товар действительно хотя бы иногда вытесняется с рынка лучшим (хотя гораздо
чаше хороший товар вытесняется дешевым), то никто не показал, что появление
новшеств бывает хотя бы иногда следствием конкуренции. Ее роль как движу-
щей силы развития общества додумывается. В главе 11 мы узнаем, что конку-
ренция - условный, а не причинный фактор. На примерах успешного взаимодей-
ствия фирм видно преобладание договора над актами конкуренции.
По Кропоткину, роль взаимопомощи возрастает по мере усложнения
животной организации, становясь постепенно основным фактором, веду-
137
щим эволюцию. Отсюда взял начало социабилизм - направление эволюци-
онной мысли, видящее в социальной организации организмов и даже их
клеток основную движущую силу эволюции. (Новейшим примером служит
обширная книга [Панов], к сожалению, не упоминающая Кропоткина.) На-
оборот, роль конкуренции, по Кропоткину, падает.
Но без конкуренции лучшие не вытесняют худших, так что дарвинизм
лишается единственного (пусть и воображаемого) движущего принципа.
Кропоткин не заметил этого - он не строил своей теории, он был уверен, что
дарвинизм нуждается лишь в единственном уточнении, в добавлении идеи
взаимопомощи. (Подробнее см.: Чайковский Ю.В. Князь Кропоткин: револю-
ция и эволюция // ВИЕТ, 2003, № 3.)
3-12. Рождение генетики. Дарвинизм по де-Фризу и Мензбиру
Ранее мы видели, что в дни Дарвина многие ученые, приветствуя его
учение, пытались ввести в него внешний движущий принцип, «буксир». Од-
нако нынешний дарвинизм, как известно, никакого «буксира» не признаёт и
потому отвергает все факты, его требующие. Как это вышло?
Как уже сказано, Уоллес видел движущий принцип в наследственности.
Позже, в «Дарвинизме», он ни слова не сказал об этом, поскольку принял
презумпцию Вейсмана. Столь странные шаги были в эволюционизме часты
и даже неизбежны, пока не было теории наследственности.
Принято считать, что такая теория, генетика, родилась в 1865 году, с
законами Менделя. Однако почему их почти никто не замечал 35 лет?
Их цитировали, но не понимали. Как показала эстонская исследова-
тельница Майе Реммель (Вальт), с ними просто нечего было делать при по-
нятийном аппарате тогдашней биологии, исходившей из понимания наслед-
ственного материала как чего-то непрерывно текучего, связанного со сме-
шением каких-то не вполне ясно выразимых жидкостей (Valt М. Mendel ja
Darwin И Eesti Loodus, 1972, July; резюме русское).
Дарвин сам признавал, что не имеет на сей счет четких представлений,
тем не менее именно он в 1868 году ввел в научный оборот понятие на-
следственной частицы (геммулы). Оставалось соединить Менделя с Дарви-
ном в единой концепции. Но чтобы понять роль дискретного (частиц) в био-
логии непрерывного, понадобился гений, и его пришлось ждать. Им стал
голландский ботаник Гуго де-Фриз (1848-1935).
Он опубликовал в 1889 году книгу «Внутриклеточный пангенез», само
название которой ясно говорит о ее сути: дарвиновы геммулы навели его на
ту мысль, что существуют частицы наследственности (у де-Фриза: пангены),
но что мигрировать они должны не по всему телу, а лишь внутри клетки (Де-
Фриз Г. Избр. произв. М., 1932).
До него уже понимали, что ядро содержит материал наследственности
(в качестве ее носителей даже называли хромосомы), а цитоплазма их реа-
лизует. Однако проницательный де-Фриз пошел дальше:
138
«И чтобы отдать себе отчет во всех явлениях, надо для каждой наследствен-
ной особенности принять отдельную частицу... Эти пангены невидимо малы,
однако они - совсем другого порядка, чем химические молекулы и и их бес-
численные соединения; они должны расти, размножаться и распределяться по
всем или почти всем клеткам организма при делении клеток. Они неактивны
(латентны) или активны, но размножаться могут в обоих состояниях. Будучи
преимущественно латентны в клетках зародышевого пути, они развивают
обычно высокую активность в соматических клетках. И именно так, что у
высших организмов не все пангены в одной клетке достигают активности, но
в каждой одна или же несколько небольших групп пангенов достигают гос-
подства и придают клетке ее характер».
В этом отрывке - программа развития генетики, но дальше текст еще
удивительнее: де-Фриз смело заявил, что хлоропласты («хроматофоры»)
должны иметь свои пангены; что в эволюции клетки был момент, когда со-
вместное нахождение активных и неактивных пангенов в цитоплазме стало
мешать работе клетки, и тогда возникло ядро, которое являет собой «прак-
тическое разделение труда»; что ядро содержит все пангены организма, а
цитоплазма - только «те пангены, которые ей нужны для деятельности».
Увы, тут намечены не только взлеты будущей науки, но и ее тупики. По
де-Фризу, частицы наследственности несут «наследственные особенности»,
но нет ни слова об основных, общих для всех клеток, функциях, и такое по-
нимание наследственности надолго стало всеобщим: неявно считалось, что
организм буквально сложен из своих характерных признаков; а его целост-
ность если и изучалась, то вне генетики.
По де-Фризу, «однажды вышедшие из ядра пангены уже не могут быть
допущены обратно» ни в это, ни в иное ядро. В части 2 мы узнаем другое:
обратный поток генов исчезающе слаб по сравнению с прямым, но он-то и
движет эволюцию.
В те годы де-Фриз исследовал «мутации» - резкие наследуемые изме-
нения свойств. Большинство из них ныне именуется макромутациями, но
среди них были и те, наследование которых подчиняется соотношениям
Менделя. Как и другие ботаники, де-Фриз отметил их, но ему еще и повезло:
в начале 1900 года ему показали статью Менделя. А он был не из тех, кому
надо объяснять дважды. Он сразу понял, что дискретные частицы - не ред-
кость и что мутация - качественное изменение «пангена».
Плод открытия явно перезрел: едва появилась краткая заметка де-
Фриза (апрель), как два немецких ботаника заявили о похожих результатах
и успели их в том же году опубликовать (ходила даже легенда, что все трое
переоткрыли Менделя независимо, пока ее не опроверг историк науки М.Д.
Голубовский). Генетика родилась.
Де-Фриз дал свое понимание эволюции - мутационизм'. один вид пере-
ходит в другой скачком, в одну мутацию. Теперь «буксиром» эволюции ста-
новился механизм мутирования: он определял, что из чего делается.
139
Собственно говоря, ничего нового открыто не было, просто все обрати-
ли, наконец, внимание на то, что прежде лишь мельком поминали как до-
садную помеху. Еще в 1590 году немецкий аптекарь Филипп Стефан Шпрен-
гер разослал известным ботаникам семена новой разновидности чистотела,
неожиданно выросшей в его саду. Разновидность Шпренгера выросла в го-
товом виде из семени обычного чистотела и оказалась устойчивой, т.е. со-
храняла отличия при размножении семенами. Ее стали кое-где культивиро-
вать и описали как подвид', она проявляла, как следует особому подвиду,
способность преимущественного выживания в некоторых особых условиях
(на остатках стен), тогда как на грядках уступала обычному чистотелу. На-
лицо элементарный акт эволюции (рис. 11). Он получил имя “гетерогенез”
(от греч. гетерос - другой), поскольку здесь одно порождает прямо другое
(С.И. Коркинский. Гетерогенезис и эволюция // Записки Имп. Академии наук.
Физ.-мат. отд., т. 11, № 2,
1899).
Рис. 11. Чистотел Chelidonium
majus: слева типичная форма (из
книги D. Chabraeo. 1678); справа
форма foliis quernis, т.е. дуболи-
стная; она открыта Шпренгером
в 1590 г. (из книги С. Bauhin,
1620, где описаны обе формы,
но дан рисунок только второй)
Дарвину пример
Шпренгера остался неиз-
вестен, однако он знал
несколько других (такова
плакучая разновидность
туи; описание и рисунки
этой туи привел Данилев-
ский в своем «Дарвиниз-
ме» как пример эволюции не по Дарвину - рис. 12), но не стал менять своей
презумпции: эволюция идет путем отбора мелких вариаций - как не сме-
нило ее и общество. Эту презумпцию удалось сменить де-Фризу: в 1901 го-
ду он заявил, что вид переходит в вид не по Дарвину, а путем мутации; и хо-
тя перехода в одну мутацию наука не признала, но воцарилась презумпция:
вид образуется путем мутаций.
Уже в 1902 году московский зоолог Михаил Александрович Мензбир
(1855-1935) произнес в МОИП речь «Мнимый кризис дарвинизма» (Русская
мысль, 1902, № 11), где сочетал Дарвина с де-Фризом. По Мензбиру, Дар-
вин «совершенно логически приходит к заключению, что тот путь, которым
140
происходят породы домашних животных, должен иметь место и по отноше-
нию к происхождению пород диких животных», но вместо искусственной се-
лекции действует мальтусова схема: перенаселение рождает борьбу за су-
ществование. Этот процесс «настолько прост и ясен, что против него воз-
ражать нечего», причем он равно действует в отношении происхождения
видов, родов и вплоть до «типов, общих животному и растительному царст-
ву». Так был заложен нынешний дарвинизм.
Конечно, Мензбир знал, что данная схема вызывает массу возражений,
но, как видим, отверг само их существование. Такой научный фанатизм до-
саден, ибо всегда рождает переупро-
щенные теории. Однако он хорошо от-
ражал ожидания общества, и мало кто
хотел видеть примитивизм нового дар-
винизма.
Рис. 12. Туя восточная (Thuja orientalis) из книги
Н.Я. Данилевского. 1885 (где дано старое родо-
вое название Biota). Сверху: типичная форма;
снизу: плакучая форма (pendula)
По Мензбиру, мутациями следует
называть не только крупные измене-
ния, порождающие разновидности и
виды, но и любые хорошо наследуе-
мые скачкообразные изменения; он
был уверен, что мутация, как и дарвин-
ская вариация, не создает ничего со-
всем нового, но откуда берется новое,
не сказал. Вскоре такое понимание
эволюционной изменчивости стало
общим (см. п. 4-6), и эволюционные ис-
следования дарвинистов сосредоточи-
лись на анализе судьбы мутаций в по-
пуляциях. Остальное либо выражалось на этом языке, либо выпадало из
круга эволюционных исследований.
Через 40 лет после речи Мензбира, с появлением книги Джулиана
Хаксли «Эволюция, новый синтез», данное понимание дарвинизма стало
почти всеобщим и получило название СТЭ. (Ее излагают все учебники, в
том числе ЮШ; о ее связях с остальным дарвинизмом см. [Воронцов; Гоод-
ницкий; Зусмановский; Назаров, 2005]; о ее пренебрежительном отношении
к внезапным крупным эволюционным изменениям см. [Колчинский].)
Итак, было решено, что в мутациях реализуется дарвинская идея из-
менчивости и что прежние споры вокруг дарвинизма можно забыть. Однако
141
ни одного примера той последовательной замены небольших улучшений,
какую молодой Дарвин нашел в форме идеи в труде Жоффруа (но не нашел
в природе) и какую с полным правом назвал «законом малых различий,
производящих более плодовитое потомство» (п. 2-8), обнаружить так и не
удалось.Начиная с Мензбира, дарвинисты считают все подобные трудности
просто несуществующими.
Если знать историю, то легко видна логическая дыра: мутацию (то есть
стойкое скачкообразное изменение наследственности, «спорт») в кругу Дар-
вина не раз обсуждали, но никому тогда в голову не приходило сказать, что
один вопрос снимает остальные. Им не приходило в голову отвергнуть все
движущие принципы, не предложив взамен ничего. Все ученые в XIX веке,
включая даже Вейсмана, чувствовали необходимость «буксира». Наоборот,
преподаватели (а профессор Мензбир стал даже первым советским ректо-
ром Московского университета), журналисты и проповедники ее не чувство-
вали. И их читатели и слушатели, став профессорами, обратили дарвинизм
в наивную догму.
3-13. Российский эволюционизм: мир как целое
Как мы видели в главе 2, в России еще до рождения дарвинизма утвер-
дились две эволюционные школы - Бэра в Петербурге и Рулье в Москве.
Тогда они мало отличались от западных: Бэр нес в Россию немецкий эво-
люционизм с его симпатиями к сравнительной анатомии, эмбриологии и на-
турфилософии, а Рулье исходил из французского жоффруизма. В 1858 году
начинающий петербургский зоолог Н.Н. Страхов писал:
«Вообще дело не в том, как произошли формы и могут ли они переходить од-
на в другую, а в том, как они существуют в своем разнообразии» (Страхов
Н.Н. О методе наук наблюдательных. // Журнал Министерства народи, про-
свещения, 1858, № 1).
Читая его, диву даешься: до выступления Дарвина и Уоллеса еще пол-
года, а Страхов пишет так, будто ход и исход боев за эволюционную идею
ему уже сообщён: главное для эволюциониста, полагал он, - понять, как в
каждый данный момент организовано разнообразие живого.
Впоследствии Страхов не раз пояснял эту свою мысль: если теорией
эволюции считать выявление родословных древ, то она мало что дает для
понимания живого, в том числе - и его истории Среди свойств организмов
есть те, которые взял за исходные Дарвин (размножение и наследствен-
ность), и те, на которые он едва обратил внимание (рост и развитие орга-
низма, взаимодействие органов и организмов). Дарвинизм не объяснил «со-
держания и разнообразия животной и растительной жизни», не ответил на
вопрос: почему организмы имеют эти свойства, а не другие? Поэтому нача-
ла Дарвина «недостаточны для предмета, теорию которого он задумал по-
строить», а натуралисты не заметили этой «скудости начал».
142
Почему? Потому, отвечал Страхов, что «им нужно было не объяснение
дела, а какая-нибудь теория, поскорее нужен был новый авторитет.» По
Страхову, переворот в науке произошел под влиянием вненаучных факто-
ров, которые Страхов объединил под именем «европейского нигилизма»
(Н.Н.Страхов. Борьба с Западом в нашей литературе. Изд. 3-е, кн. 2, Киев,
1897, с. 266, 269).
Мы уже говорили, что дарвинизм победил как явление социальное, не
взявшее почти ничего из биологических новинок своей эпохи, и Страхов
один из первых понял это. В России Страхов отнюдь не был в таких мыслях
одинок. Данилевский (ученик Бэра и духовный учитель Страхова) писал в
конце «Дарвинизма», что «для успеха необходимо появиться своевремен-
но», что «чего хочется, тому верится», что дарвинизм порожден английским
способом мышления.
Ведущий физиолог растений А.С. Фаминцын, сурово критиковавший
Данилевского за желание возразить буквально на каждое слово Дарвина,
все же признал: проблема эволюции Дарвином не решена и дело не столько
в фактах, сколько в готовности Дарвина толковать их в свою пользу. Факти-
чески именно Фаминцын поставил вопрос о презумпциях.
Сторонники и противники идеи эволюции обладали одним и тем же на-
бором фактов, но - разными презумпциями. Так, Виганд, полагавший, что
эволюции в обозримом прошлом не было (она могла быть лишь в «перво-
начальном периоде»), не видел в географическом распределении видов
даже довода в пользу эволюции, тогда как Спенсер включил его в число
прямых доказательств эволюции. Победа досталась тем, чью презумпцию
поддержали околонаучные круги.
В сущности, напомню, научная революция по Куну (п. 2-1) есть смена
презумпций основной массой ученых. Сейчас это - азы науковедения, но
тогда самого науковедения не было.
I' Приведя презумпцию Виганда (надо признавать виды неизменяемыми,
пока фактические данные не убедят нас в противном) и признав, что ни од-
ного твердого факта перехода вида в вид нет, Фаминцын тем не менее зая-
вил: «Мы обязаны иметь в виду обе эти возможности».
Вот установка, хоть и трудная в исполнении (попробуй-ка давать одну
цену и милым душе фактам, и неприятным), но куда более плодотворная,
чем презумпции. Виганд ведь так и умер, не не найдя никаких доводов в
пользу эволюции, хоть и был близок к открытию важного эволюционного за-
кона. А Фаминцын, теми же фактами владея и к той же школе ботаников
принадлежа, нашел. (О презумпциях см. Дополнение 1.)
'I 3-13* Симбиогенез
Фаминцын был одним из тех, кто открыл в 1860-х годах поразительный
факт: лишайник является симбиозом гриба и водоросли. В этом долгом
утомительном открытии участвовали многие, но, по-видимому, из них лишь
143
Фаминцын увидал тут возможный общий принцип эволюции: как лишайник
составлен из гриба и водоросли, так и всякий организм, по Фаминцыну, со-
ставлен из клеток, а всякая клетка - из «наипростейших жизненных еди-
ниц», т.е. внутриклеточных органелл^. И онтогенез, и эволюция предстают
при этом как процесс самосборки.
Главным аргументом послужил Фаминцыну тот факт, что некоторые ор-
ганеллы размножаются делением: до конца XIX века было установлено, что
делением размножаются хромосомы, хлоропласты, а у некоторых однокле-
точных - еще клеточное ядро и ядрышко.
Фаминцын счел хлоропласты зелеными бактериями, живущими внутри
растительной клетки и до конца дней
«старался выделить из зеленых растений... более простой организм, снабжен-
ный хлоропластом и способный продолжать жить и размножаться вне расте-
ния» (А.С.Фаминцын. О роли симбиоза в эволюции организмов // Записки
Имп. Академии Наук, VIII серия, 1907, т. XX. № 3. с. 4).
Замечу, что Виганд тогда же считал бактериями также и митохондрии.
Несмотря на тщетность опытов петербуржца Фаминцына, у него был
страстный конкурент, казанский биолог К.С. Мережковский. Он дал новому
учению термин «симбиогенез» и попытался выявить его суть. Он заявил,
что в каждой клетке имеются «две плазмы»: микоидная (бактериальная и
грибная) и амебоидная (растительная и животная). Первая химически вы-
нослива, не требует ни кислорода, ни органической пищи, зато неподвижна,
тогда как вторая подвижна. Хлоропласты он едва помянул, на Фаминцына
не сослался вовсе (тот отплатил ему тем же), зато построил схему происхо-
ждения живого (рис. 13).
Цитологи, найдя массу противоречий ее с данными строения клетки,
вскоре дружно ее отвергли. Идея складывать организмы из порознь возник-
ших частей стара, как мир: ее высказывал еще Эмпедокл (п. 1-5). И любо-
пытно напомнить, за что Аристотель отверг ее - за то, что она ничуть не
проясняет, каким образом сложенный из фрагментов организм оказывается
целостным. Но этого не проясняет и идея симбиогенеза.
Так что же, зря трудился полвека Фаминцын? Отнюдь. Он внес свою
лепту в понимание эволюции мира как целого, если у Дарвина каждый ор-
ганизм борется со всем миром, и потому вопрос о целостности мира и об
его эволюции как целого даже поставлен всерьез быть не может, то у Фа-
минцына предложен в качестве элементарного противоположный принцип -
сотрудничество (точнее, его частный случай - симбиоз).
Говоря нынешним языком, речь шла о самоорганизации.
18
До него в 1861 г. австрийский физиолог Эрнст Брюкке писал, что клетка - механизм,
состоящий из меньших механизмов.
144
Рис. 13. Симбиогенез по Мережковскому, 1909:1, царство грибов (сим-
биоза нет); II. царство растений (двойной симбиоз — ядро и хроматофоры,
т.е. хлоропласты); III. царство животных (одинарный симбиоз — ядро).
Митохондрии не привлекли внимания автора
О сходном думал и Страхов: для дарвиниста
«употребление органа... случайная целесообразность, не имеющая отношения
к внутреннему развитию организма, истинный же телеолог (т.е. эволюцио-
нист - Ю. Ч.) видит здесь то, как организм стремится осуществить свою об-
щую цель, видит ответ самостроющегося существа на внешние возбуждения
и обстоятельства. Т.о. изучение целесообразностей становится изучением ор-
ганического творчества». Это творчество и ныне продолжается: «...великая
тайна создания мира совершается перед нами до сих пор». (НН.Страхов.
Мир как целое. СПб., 1892, с. XVII, 89).
Конечно, он повторял немецких натурфилософов, начиная с Блюмен-
баха (п. 2-1), но, по-моему, подчеркнутые мною два слова вновь показыва-
145
к т, сколь замечательна была биологическая интуиция автора: об эволюции
щк самоорганизации заговорили только в наши дни; зато об эволюции как
творчестве уже через 15 лет написал французский философ-эволюционист
Анри Бергсон, о чем мы узнаем в главе 6 А о мире как целом речь вновь
пошла вскоре же.
3-13** Мир гак географическое целое
Об этом писал еще немецкий путешественник и натурфилософ Алек-
сандр Гумбольдт в своей знаменитой «Географии растений» (1807): по его
наблюдениям, изменение облика планеты меняло и распределение расте-
ний. В эволюционном плане о связи облика Земли с обликом ее обитателей
говорил в 1830 году минералог Рудольф Герман в торжественном заседа-
нии МОИП. По его мнению, катастрофы (в смысле Д’Орбиньи) порождались
। актами разогрева земных недр; после охлаждения
“этот необъятный океан сам оживлял себя (beiebte sich) всеми видами мор-
ских животных”, которые “формировались до высшего совершенства своих
форм. Путем непрестанной борьбы элементов они, однако, были распростра-
няемы (wurde ilberdect) по еще несвязным частям нашей каменной оболочки”
(Hermann J.R. // Actes de la seance publique extraordinaire... // Bulletin Soc. Imp.
Nat. Moscou, 1830, № 2, p. 241).
Тут, как и у последующих немецких натурфилософов, борьба - фактор
не эволюции, а расселения. Отметим также и «еще несвязные части»:
жизнь не сразу стала биосферой (если тут позволителен наш термин).
С победой эволюционизма тема единства живого и неживого миров
стала всё чаще обсуждаться, и к концу века геолог В.П. Амалицкий (Варша-
ва) подвел (тоже в торжественном собрании) итог западных воззрений:
“главные моменты в развитии организованного (живого - Ю.Ч.) мира, на ос-
новании которых земную кору делят на группы..., совпадают с главными мо-
ментами в развитии лика земли. Но кроме того и самое направление этого
развития идет параллельно с развитием организованного мира: ... дифферен-
цировка земного рельефа может быть поставлена в связь с усложнением со
временем условий горизонтального или географического распределения ор-
ганизмов на земной поверхности, а увеличение материков — с интенсивным
развитием континентальной органической жизни, начиная со второй полови-
ны палеозойской эры. Наконец, знаменательно также совпадение моментов
наиболее энергичного эволюционного движения в земном рельефе с интен-
сивною эволюцией среди организованного мира. Продолжительные периоды
спокойного изменения рельефа земной коры... сменявшиеся периодами ин-
тенсивного проявления горообразовательных процессов, должны быть по-
ставлены в параллель с периодами спокойного последовательного развития
организованного мира, прерываемого короткими промежутками интенсивной
эволюции” (Амалицкий В.П. О геологич. развитии организмов и земного
рельефа // Варшавские университ. изв., 1896, N6, с. 26).
146
Как видим, вполне выражена связь био- и геоэволюции с географиче-
ским распределением видов, а также идея плавной эволюции, перемежае-
мой скачками. К этому стоит добавить смутную догадку Амалицкого о пере-
мещении материков:
“В переживаемую нами эпоху первичный облик земли совершенно преобра-
зился: два первично обособленные полярные материковые пространства, вы-
тянутые в западно-восточном направлении, через последовательный ряд пре-
образований придвигаются друг к другу и дают две разнородно составленные
нынешние материковые массы, вытянутые в обратном направлении к пер-
вичным материкам” (там же, с. 26).
Хотя на Западе тогда об этом писали более конкретно - см. Суворов
А.И. История мобилизма в геотектонике. М., Наука, 1994 (Труды ГИН, вып.
494), нам важно завершение Амалицким раннего варианта темы «мир как
целое», к которой мы вернемся при обсуждении картины мира в п. 6-14.
3-14. Мир как целое у Ганса Дриша. Энтелехия
В 1905 году немецкий эмбриолог и философ Ганс Дриш (1867-1941) вы-
ступил с книгой «Витализм», где связал целостность мира с наличием в нем
цели. (Витализм — взгдяд на мир, при котором феномен жизни - нечто
данное, что необходимо исследовать, но что не следует пытаться опреде-
лить, поскольку всякое определение - это попытка объяснить феномен че-
рез какие-то иные, более понятные термины, а для жизни таких терминов
быть не может.) Аргументом в пользу целостности мира была для Дриша
гармония, т.е. само наличие жизни и человека в ней.
“Мы не колеблемся признать, что... природа как таковая существует ради оп-
ределенной цели, и вряд ли возможно представить себе эту цель не с чисто
антропоморфической точки зрения. Мы признаём, другими словами, неорга-
ническую природу в ее главных свойствах целесообразной для существова-
ния организмов и человека”. Против этого взгляда “выдвигаются соображе-
ния, представляющие собой нечто вроде дарвинизма ... Гармония между ор-
ганическим и неорганическим мирами есть будто бы лишь результат пережи-
вания тех жизненных форм, которые оказались чисто случайно приспособ-
ленными к окружающей среде. Но это возражение не может уничтожить са-
мого факта, что природа существует в таком виде, что одна ее часть целесо-
образна для другой” (Дриш Г. Витализм. М., 1915, с. 273-274).
Это объективное свойство мира (пригодность к жизни) позже названо
“антропным принципом”, а затем, в другой связи, “гипотезой Геи”, о чем мы
узнаем в главе 7. В отличие от Страхова, Дриш не был уверен в реальности
феномена эволюции («нам покамест ничего неизвестно относительно объе-
диняющего исторического начала или исторической “эволюции”»), однако
обращал внимание, как и Дарвин в молодости, на факт размножения как на
ключевой. Только если для Дарвина размножение представляло интерес
147
лишь как источник изменчивости, то для Дриша главным здесь было «пред-
ставление о сверхъиндивидуальной цели».
Эта цель требовала для своего осуществления того фактора, который
выше мы характеризовали как “буксир", причем Дриш полагал этот фактор
всеобщим - действующим и в индивидуальном развитии, и во Вселенной.
Называл его Дриш словом «энтелехия», взятым у Аристотеля. Как и Ари-
стотель, он использовал его не всегда одинаково, но можно все-таки грубо
сказать так: если Аристотель под энтелехией имел в виду процедуру пере-
хода возможности в действительность, то Дриш - некий действующий агент,
производящий эту процедуру, причем не во всех ее аспектах, а лишь в од-
ном, неэнергетическом. Точнее, энтелехия не противоречит действиям си-
лы и энергии, но и не следует из них:
“Не будучи энергией, энтелехия не может создавать сама разности интенсивно-
стей, но ... энтелехия в состоянии приостанавливать те реакции, которые по
условиям системы могли бы возможны и состоялись бы, если бы не вмеша-
тельство энтелехии” (с. 259). Или: “Представление об энтелехии ... напоминает
ставшую знаменитой фикцию Максвелла о “демонах”, могущих вмешательст-
вом в распределение молекул достичь перехода тепла из места низшей темпе-
ратуры к месту более высокой”, но если у Максвелла была фикция, то тот же
эффект “осуществляется в полной реальности энтелехией” (с. 260). Наконец,
различие между' силой и энтелехией в следующем: “В первом случае мы поль-
зуемся категорией причинности, во втором - индивидуальности” (с. 272),
т.е. энтелехия у Дриша ответственна за свободу воли.
Энтелехию долго шельмовали как идеалистическое понятие, но затем
оно было востребовано наукой, что прекрасно изложено у С. В. Мейена, па-
леоботаника и философа [Мейен, 1977]. В п. 5-2 мы увидим, что “энтелехия”
Дриша может быть поглощена понятием активности, что она - одна из
форм активности живого и тем самым не выпадает из общего строя науки.
А здесь следует заметить, что представления о наличии энтелехии воз-
никли у Дриша при эмбриологическом исследовании: ему удалось, разделив
пополам начинавший развиваться зародыш, получить два нормальных за-
родыша, из чего он сделал вывод, что развитие есть процесс целенаправ-
ленный, а не механический. Получение одного итога различными путями
именуется эквифинальностью. Лучше всего о ней сказал в 1924 году Ганс
Шпеман, немецкий эмбриолог:
“Эмбриональный зачаток выглядит так, как будто бы он был построен доми-
нирующей силой из оказавшегося налицо материала, без внимания к его про-
исхождению...” (цит. по [Любищев, 2004, с. 102]).
Активность зародыша здесь на первом плане, и потому это много глубже,
чем возникшая позже трактовка развития как включения и выключения ге-
нов. Мы вернемся к данной теме в главе 5.
148
Гпава 4. Вокруг советской биологии
Теперь нам надо погрузиться в перипетии советской науки, где познава-
тельные аспекты порой совершенно заслонялись социальными.
Зачем, казалось бы, ворошить столь старое и давно преодоленное? За-
тем, что преодолено не всё и даже, на мой взгляд, не основное. Не только у
нас, но и на Западе, как ни странно, до сих пор ссыпаются на эпоху террора
в советской науке как на причину поразительного отставания эволюциониз-
ма от остальной биологии последнего полувека (например: [Стил и др., с.
30]). Только сейчас мы узнаём, что
“корифеи нашей отечественной физиологии ... склонялись к признанию ла-
маркистских принципов, лежащих в основе эволюции” (Аршавский И.А. Мы
все равноправны друг перед другом. Рига, 2002, с. 72),
что уверения советских идеологов о полной победе дарвинизма были не-
правдой. О ламаркизме речь у нас пойдет позже, в части 2, а пока не будем
забывать, что даже “лысенковщина”, которую обычно подают как уродливую
форму ламаркизма, именовала сама себя «творческим дарвинизмом». Та-
ков был язык эпохи.
4-1. Исходные посылки
Когда в 1900 году началось развитие генетики, эволюционисты понача-
лу разделились на три лагеря.
Одни сочли, что дарвинизму пришел конец, поскольку появилась более
простая и обоснованная теория - мутационизм, согласно которой новый
вид возникает из прежнего скачкообразно, за счет мутации (такая теория
имеет и другое название - сальтационизм, от лат. saltus - прыжок), причем
отбору остается только одна функция, а именно отбраковка неудачных му-
таций. Напомню, что, по Дарвину, отбор работает не с крупными скачкооб-
разными изменениями (улучшение организма за их счет Дарвин полагал не-
вероятным), а с последовательными мелкими изменениями отдельных при-
знаков. Основателем мутационизма был, как уже говорилось, де-Фриз. Он
полагал, что время от времени данный вид входит в «мутационный пери-
од», в течение которого происходит массовый переход потомков в новый
вид, и даже указал конкретный вид цветковых растений - ослинник
(Oenothera lamarkiana), находящийся, по его мнению, в данном периоде и
регулярно (с частотой выше 1/100) дающий особей новых видов (например,
Oenothera gigas - рис. 14).
Другие, наоборот, отвергли мутационизм как ничего не объясняющий и
продолжали держаться ортодоксального дарвинизма. К ним принадлежал
патриарх дарвинизма Уоллес. К сожалению, это стало традицией: дарви-
низм сперва яростно отвергает почти все научные открытия как излишние,
чтобы впоследствии, когда они все-таки утвердятся в науке, объявить их
своею составной частью.
149
Рис. 14. Мутация по Де Фризу: справа Oenothera lamarkiana, слева Ое. gigas
Третьи же, очень
немногие, признали сра-
зу, что мутации - то са-
мое, чего не доставало
дарвинизму. Наиболее
четко эту позицию зая-
вил, как уже сказано в п.
3-12, Мензбир в том же
1902 году. От мутацио-
низма ее радикально от-
личала уверенность, что
эволюцию движут не
«видовые мутации»,
скачком дающие новый
вид, а небольшие скачки,
слегка изменяющие от-
дельные признаки. Это и
был генетический дар-
винизм.
Что касается ослин-
ника, то пример был действительно красив: скачком появлялся новый вид.
Правда, пример был не вполне нов (мы говорили в главе 3 о сальтациях у
чистотела и туи), но теперь был возможен генетический анализ. Вид Ое.
gigas - тетраплоид, т.е. произошел путем удвоения набора хромосом. По-
скольку эволюция как целое не может идти таким путем, теория быстро по-
теряла привлекательность. Как водится, при этом потеряли интерес и оста-
лись без ответа важнейшие вопросы, поставленные опытами де-Фриза. В
частности, до сих пор никто не знает, как и почему возникают «мутационные
периоды». Хотя их вновь и вновь обнаруживали (например. Р.П. Берг).
К сожалению, сторонники генетического дарвинизма не видели тех
трудностей, какие встают при их собственном подходе к эволюции. Так, им
казалось, что ни о каком «поглощающем влиянии скрещивания» (которое
Дженкин доказал Дарвину - см. п. 3-4) нет и речи: каждый ген неделим и не
может «измельчать», сколько ни сменится поколений. По этому поводу фи-
зиолог растений К.А. Тимирязев в 1910 году писал: «Кошмар Дженкина, ис-
портивший столько крови Дарвину, рассеивается без следа».
Так ли уж без следа? Если бы Тимирязев и другие (в том числе все яро-
стные защитники генетического дарвинизма) обратились к статье Дженкина,
они увидели бы, что неделимый ген предсказан в ней как одна из логических
возможностей. Вот это место:
150
«Мы вправе предположить, что потомок спорта неуклонно воспроизводит
удачное свойство без измельчания. Тогда новый вариант может заменить
прежний вид. Но наследственность, которую признает такая теория, так от-
лична от ежедневно наблюдаемой, что ее можно назвать теорией последова-
тельных творений; она не соответствует дарвинской теории».
Почему же проницательный инженер заключил, что таким путем нельзя
«воспроизвести удачное свойство»? Почему он решил что такой вариант
радикально отличается от «ежедневно наблюдаемой наследственности»?
Почему, наконец, за эту возможность не ухватился сам Дарвин?
Все дело в словечке «неуклонно» (faithfully), которое здесь означает (в
нынешних терминах): признак должен определяться одним, и притом доми-
нантным, геном, да и в этом случае, чтобы не затеряться, признак должен
обеспечить своим обладателям плодовитость, намного превосходящую
плодовитость прочих особей - ведь каждый гетерозиготный ген передается
только половине детей. Да, Дженкин был прав, отрицая эволюционную роль
уникальной вариации, и Дарвин справедливо соглашался с ним
Однако именно генетика в одном смысле помогла решить спор Дарвина
с Дженкином, в смысле, о каком те не могли и подозревать: оказалось, что
природа не может отбирать отдельные генотипы по той причине, что каж-
дый из них, однажды возникнув, практически никогда не появляется
вновь. Процесс полового размножения перемешивает гены, и на суд отбо-
ра может быть представлено только все многообразие генотипов сразу - ге-
нофонд популяции. Уже одно это лишало смысла идею отбора.
Так что и Дарвин, и Дженкин напрасно сосредоточили внимание на
свойствах отдельного организма и на борьбе близких особей: будут они бо-
роться или нет, рано или поздно (если, конечно, они оставят потомство) им
предстоит перемешать свои гены. Поэтому борьба индивидов ни сильным,
ни слабым фактором эволюции явиться не может. Если борьба за жизнь и
может играть роль в эволюции, то лишь борьба между теми группами, кото-
рые не смешивают своих генов. Они-то и выступают как эволюционные ин-
дивидуальности - своего рода сверхорганизмы. Это в сущности и утвер-
ждает СТЭ, когда называет эволюционной единицей популяцию. Вот почему
сам термин «борьба за существование» из СТЭ понемногу исчез, оставшись
лишь в восхваляющих фразах о Дарвине.
Хотя в п. 3-7* уже сказано, что нет никаких доказательств действия ес-
тественного отбора в природе, о нем придется еще поговорить, поскольку
вокруг него вертелись все эволюционные споры XX века.
Помните «математическое» возражение Беннета - Майварта (п. 3-5)?
Когда возникла генетика, ему дали действительно математическую форму:
в 1915 году появились расчеты английского математика Г. Нортона, где бы-
ла дана оценка времени, потребного для замены худшего гена лучшим.
Оказалось (как легко догадаться), что судьбы рецессивного и доминантного
аллелей резко различны.
151
Если полезное новшество возникает в форме рецессивного гена, то на-
до долго ждать, пока оно хоть немного распространится в популяции: даже
аллель, несущий признак, повышающий вероятность выживания сразу на
50%, достигнет, по Нортону, частоты гомозигот в 0,01% численности попу-
ляции, лишь через 2 тыс, поколений после возникновения. Если же повы-
шение вероятности выживания равно 10%, то ждать придется почти 20 тыс.
поколений, а если 1%, то - целых 200 тысяч. Это и естественно: вероят-
ность встречи двух носителей поначалу весьма мала. Зато потом дело пой-
дет куда быстрее: три указанных аллеля завладеют всей популяцией (точ-
нее, составят 99,9% ее численности) через 120, через тысячу и через 10
тыс. поколений соответственно.
У доминантных аллелей наоборот: полезный признак легко овладевает
популяцией, зато неспособен за реальное время вытеснить рецессивную
исходную норму: даже при подавляющем превосходстве (рост вероятности
выживания на 50%) надо ждать 4 тыс. поколений, пока частота прежней
нормы упадет до 0,1% в гетерозиготах. При вероятностях 10% и 1% ждать
надо 20 тыс. и 200 тыс. поколений соответственно.
Вывод Нортона был прост: таким механизмом ничего, кроме сдвига час-
тот, достичь невозможно, а следовательно, модель отбора отдельных мута-
ция никуда негодна, и на самом деле эволюция идет иначе. Подумайте: 20
тыс. поколений - это же почти расстояние от нас до питекантропа. По Нор-
тону, эволюция может идти лишь крупными скачками (см. [Бабков, с. 30]).
Вывод Нортона никогда никем опровергнут не был, его попросту игнориро-
вали. Запомним это.
4-2. Московская школа эволюционной генетики. Четвериков
Генетический дарвинизм господствовал всю советскую эпоху. В знаме-
нитой статье «О некоторых моментах эволюционного процесса с точки зре-
ния современной генетики» (Журнал эксперим. биологии, 1926) москов-
ский^ зоолог Сергей Сергеевич Четвериков (1880-1959) рассмотрел вопрос
о мутациях и их отборе. Говоря о судьбе единичного мутанта, он признал,
что вероятность спаривания двух одинаковых мутантов ничтожна, что по-
этому его мутация действительно будет «засосана» болотом исходных осо-
бей (т.е. признал возражение Дженкина, рассмотренное выше), однако вы-
вод сделал совсем другой, чем у Дженкина:
«Но судьба ее будет все же совершенно иная, чем это мыслилось прежним
эволюционистам. Геновариация (мутация - Ю.Ч.~) не погибнет, не растворит-
ся в массе нормальных особей. Она будет существовать в гетерозиготном со-
стоянии, из поколения в поколение оставаясь скрытой от глаз...».
19
Далее город будет указываться только для российских (советских) ученых, работав-
ших вне Москвы.
152
Но так ли это на самом деле? Позже сам Четвериков признал [Бабков,
с. 38], что для эволюции важны лишь массовые мутации, т.е. согласился,
как и Дарвин, и де-Фриз, с Дженкином. Более этой темы Четвериков не
касался, так что его рассуждение о гетерозиготах - возражение лишь на
одну фразу Дженкина. Тот шотландский журнал, где печатался Дженкин, в
Россию не попал. Сложный и тонкий анализ, данный Дженкином и
приведший в смущение Дарвина, остался Четверикову неизвестен.
Дженкин полагал, что анализ наличного разнообразия не есть анализ
эволюции, и в этом был прав (см. ч. 2). Не ведая о данном ходе мысли (поч-
ти не известном тогда на Западе и почти исчезнувшем из русской литерату-
ры со смертью Страхова), Четвериков ориентировал собранную им в 1919-
1924 годах группу блестящих молодых генетиков как раз на анализ генети-
ческого разнообразия ныне живущих организмов.
Был начат анализ разнообразия дрозофил Подмосковья, который трак-
товался именно как анализ малых шагов эволюции.
Сперва надо было осознать вывод Нортона. Однако Четвериков, при-
ведя его численные данные, ни словом не обмолвился о том выводе, какой
из них был сделан самим Нортоном. Наоборот, он декларировал:
«Таблица Нортона учит нас тому, что всякий эволюционный процесс, вызван-
ный отбором, безразлично для доминантных или рецессивных форм, всегда
идет до конца, до полной замены менее приспособленной формы более при-
способленной. Она же учит нас тому, что отбор подхватывает и окончательно
закрепляет каждое, даже самое незначительное улучшение организма».
Вновь вспомним Данилевского: чего хочется, в то и верится. Но в дан-
ном случае получилась прямая ложь.
Что же имел в виду Четвериков? А вот что: он изменил само понимание
эволюции. Он назвал эволюцией процесс изменения частот генов. Правда,
он отметил, что процесс распространения рецессива почти невозможно на-
чать, а доминанта - закончить, но вывод и тут сделал самый оптимистиче-
ский для дарвинизма. Вот откуда идет отмеченная мною в начале книги
традиция называть эволюцией не появление новых форм, а сдвиг генных
частот. Она пресекла лет на 70 почти всю эволюционную мысль.
В действительности Четвериков не был столь прост, как может пока-
заться, и в той же статье начал рушить свою благостную схему. Как бы вто-
ря Рузвельту (см. п. 3-7), Четвериков писал:
«Систематика знает тысячи примеров, где виды различаются не адаптивны-
ми, а безразличными (в биологическом смысле) признаками, и стараться по-
дыскивать им всем адаптивное значение является столь же малопроизводи-
тельной, как и неблагодарной работой, где подчас не знаешь, чему больше
удивляться - бесконечному ли остроумию самих авторов или их вере в неог-
раниченную наивность читателей».
Его правоту легко видеть на рис. 15: пользы у такой изощренной особенно-
сти шовной линии искать нечего.
153
Рис. 15. Раковина аммонита
Philloceras (юрский период). В одном
месте оболочка раковины счищена,
чтобы показать сутуру (шовную ли-
нию). Характер сутуры является глав-
ным при различении видов аммони-
тов. но вряд ли кто-нибудь станет ут-
верждать. что различие этих причуд-
ливых завитков полезно в борьбе за
жизнь
Сам он делил эволюцию
на два процесса - происхожде-
ние видов, к отбору не относя-
щееся («Истинным источником... происхождения видов является не отбор, а
изоляция»), и прогрессивную эволюцию, причиной которой «является борь-
ба за существование и вытекающий из нее естественный отбор». Как обыч-
но, второй процесс никаким примером не пояснен, и мне остается напом-
нить, что, Коп решал тот же вопрос прямо противоположно (п. 3-10): проис-
хождение вида полагал обязанным отбору, а прогресс - нет.
Причину этого прискорбного разнобоя понять легко: как мы видели в п.
3-7* и увидим далее, в природе естественный отбор не действует, так что о
нем можно говорить всё что угодно. Рассмотрим вопрос подробнее.
4-3. Системное понимание отбора
Напомнив позицию Уоллеса (от всякой нормы возможно отклонение как
в сторону ухудшения, так и в сторону улучшения данного свойства - п. 3-5),
Четвериков в той же статье выразил уверенность:
«Любая вновь возникающая геновариация может оказаться по отношению к
отбираемому признаку либо его «усилителем», либо его «ослабителем». В
случае «усилителя» отбор ее подхватит и в последующих поколениях распро-
странит этот ген на все население, усилит отбираемый признак. Таким обра-
зом, действие отбора не прекращается с переходом отбираемого признака в
гомозиготное состояние, а продолжается... Отбирая один признак, один ген,
отбор косвенно отбирает и определенную, наиболее благоприятную для про-
явления данного признака генотипическую среду - генотип».
Читатель может спросить: чем это лучше схоластики Уоллеса, которую
сами дарвинисты сто лет не вспоминают? По-моему, это интересно (хоть и
биологически бессодержательно), поскольку показывает, что вся расплыв-
чатая фразеология изменчивости «по Дарвину» будет использована и
впредь, что открытие дискретных генов не будет мешать громоздить туман-
ные фразы (ну хотя бы, что значат слова «отбор ее подхватит», если при-
знаётся, что отбор действует не на мутацию, а на весь генотип?). Объясни-
154
тельные возможности дарвинизма расширены за счет ввода как бы гене-
тических понятий («усилитель» и т.п.). Они опять заменят дарвинистам от-
сутствие примеров реального отбора и реальной эволюции и позволят по-
прежнему игнорировать критику Дженкина и всё остальное.
Через полвека после статьи Четверикова такое понимание отбора было
названо системным, но тогда оно осталось никем не замечено как ненуж-
ное - ни сторонникам, ни противникам.
Впрочем, вскоре (1931 г. и позже) немецкий генетик Рихард Гольд-
шмидт, прежде дарвинист, заявил, исследуя популяции насекомых, что на-
копление изменчивости не ведет к видообразованию и что отбор не движет
эволюцию, а может лишь уничтожать неудачных мутантов. Постепенно он
пришел к теории системности, но не к системному отбору (которого никто не
наблюдал ни до, ни после), а к системным мутациям (он их называл мак-
ромутациями), о которых речь пойдет далее (п. 5-5*).
Однако почти никем не было замечено куда как большее: в том же 1926
году вышла книга, напомнившая, что никакие разговоры о естествен-
ном отборе вообще не нужны для биологии, поскольку ничего по-
добного естественному отбору в природе просто нет. В ней фран-
цузский драматург и натурфилософ Морис Метерлинк (родом из Бельгии),
автор знаменитой у нас «Синей птицы», писал в изумлении о половом поко-
лении тропических термитов (мы говорили о нем в п. 3-7*):
«не многих существ природа так плохо вооружила для борьбы за существо-
вание. У него (термита - Ю. Ч.) нет ни пчелиного жала, ни замечательной хи-
тиновой брони муравья - его самого непримиримого врага. Как правило, у не-
го нет крыльев; но если даже он ими обладает, то они, словно бы в насмешку,
даются ему лишь для того, чтобы долететь до гекатомбы. Он тяжел, непово-
ротлив и не может избежать опасности бегством..., он беззащитен перед все-
ми, кто в мире птиц, рептилий и насекомых жаден до его сочной плоти» [Ме-
терлинк, с. 278].
Гекатомба - это массовая могила. Метерлинк имел в виду тот факт, что
крылатые особи теряют крылья при образовании пары и тут же подвергают-
ся почти полному истреблению хищниками. Как сказано в п. 3-7*, гибель
крылатых термитов показывает полную бессмысленность принципа естест-
венного отбора. И если уж он бессилен в создании простейшего колоссаль-
но выгодного признака (несъедобности), то говорить о системном естест-
венном отборе сложных конструкций просто несерьезно. (Зато можно гово-
рить о системном отборе в иных смыслах: пп. 4-8, 6-14, 6-15.)
Если бы не слова «для борьбы за существование», мы не смогли бы
понять, имел ли Метерлинк в виду дарвинизм. Он сомневался в самом фак-
те эволюции, однако, как ни странно, приветствовал занятие ею:
«Я знаю все то, что можно возразить против теории трансформизма... Не
нужно никогда вверяться текущим истинам эпохи... Многое свидетельствует
о том, что она ложна; но, пока верят в ее истинность, - она полезна, ибо под-
155
держивает бодрость духа и побуждает к новым исследованиям. С первого
взгляда может показаться, что было бы лучше вместо всех этих замысловатых
предположений объявить во всеуслышание глубокую истину о нашем ном не-
ведении. Но эта истина была бы полезна только в том случае, если бы было
доказано, что мы и впредь ничего не узнаем... Мы созданы таким образом,
что, исходя из заблуждений, можем достигнуть наивысшего увлечения» [Ме-
терлинк, с. 238-239].
Вряд ли вера в ложную теорию так уж полезна (она побуждает к ложным
действиям, а мир мстит, и это многих делает несчастными), но насчет новых
исследований и увлечения Метерлинк явно был прав. А уж «бодрость духа»
была тогда (как и в наши дни) нужна ой как. Просто чтобы хоть продолжать
жить, а тем более работать.
4-4. Эволюция и политика
Увы, Четверикову продолжить исследования не пришлось. В начале
1929 года он был под нелепым предлогом арестован. Его начальник биолог
Н.К. Кольцов бросился к высокому начальству, ему помог писатель А.М.
Горький, и их ждал успех - Четвериков был всего лишь сослан. Но в Москву
он никогда больше не вернулся, и группа его распалась. Об этом, как и о
многом другом, можно прочесть в статье: В.В. Бабков. Н.К. Кольцов: борьба
за автономию науки и поиски поддержки власти // ВИЕТ. 1989, № 3.
Но советская репрессивная машина, выросшая из царской охранки, окрепшая в
годы Гражданской войны и на политических процессах 1920-х годов, в целом не
была тогда брошена против ученых. Весной 1929 года она получила неслыхан-
ное по размерам и опасности задание: уничтожить крестьянство как отдельный
класс. Согласно негласной доктрине тогдашнего социализма, в СССР полагалось
остаться всего двум классам - пролетариату и бюрократии. Крестьянству надле-
жало стать частью первого, а ученым, как и всей интеллигенции - распределить-
ся между первым и вторым. Террор обрушился сперва на крестьянство, а основ-
ная масса ученых продолжала пока работать.
Наша тема - биологическая эволюция, а не политика, но некоторых на-
учных течений и выводов просто нельзя понять, не погрузившись в тогдаш-
нюю обстановку. В п. 3-5 мы упоминали имя Гальтона, а сейчас пора ска-
зать, что он был основателем печально известного направления в дарви-
низме, которому дал имя - евгеника (от лат. eugeneus - благородный, наи-
лучший). Евгеника утверждает, что человеческую природу можно улучшить
посредством селекции (искусственного отбора). Мысль такая высказыва-
лась не раз в древности (Феогнид, Платон - см. п. 1-7), а затем многими
утопистами, так что дарвинизм лишь подвел под нее «научную» базу (поче-
му тут уместны иронические кавычки, мы узнаем в главе 6, когда познако-
мимся с началами диатропики и итогами евгеники).
На горе себе и сотрудникам, Кольцов развивал тот вариант евгеники,
который коммунистическая идеология признавала буржуазным: отрицая
156
НПС, он видел задачу евгеники только в организации размножения (или
устранения от него) определенных пар. Наоборот, «пролетарская евгеника»
делала упор на воспитание - в уверенности, что удачно воспитанные поро-
дят удачное потомство. Понятно, что с установлением открытого противо-
стояния СССР и Германии (где «буржуазная» евгеника свирепствовала)
Кольцов и его институт были обречены.
Однако дарвинизм не только продолжал существовать в СССР, но и
стал правящей идеологией коммунистов, что вроде бы странно - ведь его
исповедовал Запад. Разве идея медленного накопления случайных вариа-
ций могла нравиться преобразователям мира, крушившим старое и мол-
ниеносно воздвигавшим новое? Вряд ли. Проблема не изучена, но могу
процитировать Любищева: он отмечал важный факт-
«выдвижение марксизмом на первый план наиболее примитивных потребно-
стей, которые в действительности лишь ограничивают рамки развития (по-
добно естественному отбору), но отнюдь не являются движущими причинами
развития общества» (Любищев А.А. Понятие эволюции и кризис эволюцио-
низма. // Известия Биол. ин-та при Пермск. ун-те. 1925, т. 4, вып. 4, с. 151). (В
издании [Любищев, 1982] это место, и только оно, искажено).
Генетик и историк науки И.А. Захаров, наоборот, полагает, что истоки
бед генетики «не следует искать в области идеологии», что виной был про-
вал надежд коммунистов на коллективизацию сельского хозяйства:
«инициаторы экспериментов признавать свою ответственность за провалы
не желали и стремились найти “козлов отпущения”, например, на первом эта-
пе - кулаков-вредителей. Подходящим “козлом отпущения” была и сельско-
хозяйственная наука, которая... “отгораживается от нужд” и т.п. ... генетика
представлялась не как фундаментальная, а как сельскохозяйственная наука»
Бурное развитие привело ее к упадку: «в 1929 г. при образовании Академии
сельскохозяйственных наук в ее составе был 1 институт, а через 5 лет их чис-
ло перевалило за 100... Скороспелые выдвиженцы , подчас энергичные и
честолюбивые, ... составили армию Т.Д. Лысенко, которую он возглавил и
двинул против “буржуазных ученых”» (Захаров И.А. Генетика в XX веке.
Очерки по истории. М.„ 2003, с. 20).
Это верно, но это пол-правды. Надо уметь отделять беды страны и нау-
ки вообще от особых бед генетики и эволюционизма.
Выдвиженцы свирепствовали во всех отраслях жизни и были прямым следст-
вием Октябрьской революции, которая сделала чиновничество единственным
Поясню Захарова. Выдвиженец - центральная фигура кадровой политики коммуни-
стов. С первых и до последних дней своей власти они зорко выслеживали активных и
способных молодых людей в деревнях и на заводах с целью выдвижения их на руково-
дящую работу (в этом состояла одна из основных функций комсомола). Народ оставал-
ся без возможных вождей, а партия .могла называть себя рабоче-крестьянской.
I
157
социально активным классом (его роль в жизни России была непомерна и преж-
де). Этот факт тщательно скрывался: оно никогда не рассматривалось не только
как класс, но и вообще как особая часть населения. (Первым про “новый класс”
написал в 1957 г. югославский политик и публицист Милован Джилас, но он на-
зывал новым классом компартию, а не чиновничество; на самом деле оно старо,
как само государство, а компартия - лишь его новый инструмент.) Напрочь иг-
норировав разделение властей, вожди революции породили жуткий террор, в ко-
тором погибли сами, и новое рабство (как при Петре I - он тоже возвысил чи-
новничество и возродил угасавшее тогда крепостное право). После Сталина раз-
деление властей вновь возникло: различные аппараты (партия, госбезопасность,
армия, технократия, юстиция и другие, помельче) не вполне подчинялись друг
другу (при внешнем господстве партии), что, на мой взгляд, и дало возможность
недолгого процветания. Для дальнейшего нам важно, что разделение властей -
исторически первый способ говорить про многослойную регуляцию (см. п. 6-8*).
Разделение властей было попрано в 1979 г. вторжением советских войск в Аф-
ганистан, полной военизацией экономики и поголовной мобилизацией студентов
(о чем не было даже решения Политбюро партии). Генералы взяли верх над ос-
тальными чиновниками, что и повело власть к распаду. Это - общее.
Что касается особых бед генетики и эволюционизма, то приведу свои
догадки. Большевики были равнодушны к эволюции вообще и к дарвинизму
в частности - их занимала не эволюция, а революция. Но победив, они, как
и все победители, нуждались в новой идеологии и важную часть ее увидали
в биологической теории, основанной на идее всепроникающей борьбы.
Именно эта идея привлекала в дарвинизме Маркса и Энгельса, хотя оба
смеялись над тягой Дарвина видеть в природе буржуазное общество (под-
робнее см.: Чайковский Ю.В. История науки и обучение науке (на примере
дарвинизма) // ВИЕТ, 1987, № 2).
Коммунисты сталинской поры уже не нуждались в революционной
идеологии. Им нужна была именно эволюция путем борьбы, с тем услови-
ем, что в идеологии речи быть не могло ни о далеких целях, требующих ты-
сяч поколений, ни о борьбе на уничтожение среди своих (как раз она и шла
внутри когорты прежних соратников, но публично отрицалась).
Первое условие обеспечивал дарвинизм по Геккелю (с НПС и упражне-
нием органов), что вскоре было сформулировано в партийной литературе
(см.: Б.М. Завадовский. Дарвинизм и марксизм. М.-Л., 1926). А второе усло-
вие обеспечил в 1930-х годах выдвиженец-агроном Лысенко: он предложил
свой «творческий дарвинизм», где не было ни случайных мутаций, ни внут-
ривидовой борьбы. За этот вариант дарвинизма компартия держалась до
падения власти Хрущева (1964 г.).
Вот почему эволюционисты работали под большим, чем остальные
биологи, идеологическим прессом, но до воцарения Лысенко (1948 г.) все-
таки могли работать, что и дает нам материал для рассказа. В основе сле-
дующих четырех параграфов лежит материал книги [Бабков].
158
4-5. Ученики Четверикова: мутации без отбора
Летом 1926 года ученики Четверикова П.Ф. Рокицкий и С.М. Гершензон
привезли с Кавказа 239 особей Drosophila melanogaster, потомство которых
(151 тыс. мух в двух поколениях) дотошно исследовала вся лаборатория. У
239 особей было найдено 33 различных гена, по которым они были гетеро-
зиготны, причем всего зафиксировано 632 гетерозиготных рецессивных ге-
на, т.е. каждый из них в среднем встретился 19 раз, а на исходную особь их
пришлось по два-три. Разумеется, обнаружена была лишь малая часть при-
знаков (легко видимые при просмотре).
Так рухнуло представление о некоем «диком типе»: несмотря на внеш-
нее сходство, двух генетически одинаковых особей в популяции не бывает
(позже это подтверждалось на самых разных объектах). Еще более замеча-
тельно было распределение 33-х обсчитанных генов: один был найден у
50% особей, другой у 40%, третий у 28%, четвертый у 11%, еще два - у 7%
каждый, причем всем им
«едва ли можно приписать какое-либо биологическое значение», ибо они оп-
ределяли различие в числе щетинок, жилковании крыла и т.п.; поэтому при-
чина их широкого распространения в популяции «остается пока для нас неиз-
вестной» (Четвериков С.С. Проблемы общей биологии и генетики. Новоси-
бирск, 1983, с. 225).
Основная масса гетерозигот встретилась по 1-2 раза (т.е. менее 1% ис-
ходных особей), и среди них были явно вредные. Ни одного случая вариа-
ции гена, который можно было хотя бы предположительно счесть улуч-
шающим исходный генотип, найдено не было. Это не помешало Четверико-
ву сделать свои выводы. По его убеждению, для эволюции имеют значение
не единичные мутации, с которыми «никакого дела не сделать», а тот «гро-
мадный запас наследственной изменчивости, из которого отбор может чер-
пать... когда необходимо» (там же, с. 226).
Тем самым, он занял ту же позицию, что за 60 лет до него Дарвин, в от-
вет на возражение Дженкина признавший эволюционную роль массовых, а
не единичных вариаций. Новым было то, что вывод делался на основании
численных данных, и данные эти относились как раз к признакам, по мне-
нию самого Четверикова не явившимся результатом отбора.
«Четверикову прежде всего было важно утвердить общие принципы исследо-
вания: совместимость генетики и дарвинизма... Экспериментальные же ре-
зультаты были важны лишь как иллюстрация» [Бабков, с. 40].
Мы уже не раз видели, как обманчива подобная цель и как опасно ви-
деть в опытных данных иллюстрацию своих убеждений.
Забегая вперед, отмечу диатропическую деталь: различие частот в 7
раз (7% и 50%) между признаками, которые не имеют приспособительных
отличий, говорит о том, что изменчивость должна быть чем-то упорядочена.
Чем именно? Четвериковцы ответили: прежними случайными колебаниями
159
численности (обычная для дарвинизма отговорка, уход от неприятного во-
проса). Этому в книге [Бабков] посвящена глава «Генетико-автоматические
процессы». В те же годы был дан и другой ответ, о котором скажу в п. 6-11*,
когда речь пойдет о гиперболах Виллиса.
4-6. Ученики Четверикова: творческая роль отбора - что такое?
Вопросы конкретного действия отбора у самого Четверикова совсем не
были разработаны, и ученики смело взялись за них. Сперва Б.Л. Астауров
описал мутацию tetraptera (четырехкрылость), при
которой у мухи появляется на месте жужжальца
крыло второй пары, давным давно всем отрядом
J f двукрылых утерянной (рис. 16).
у y'ff Рис. 16. Мутация tetraptera дрозофилы (рисунок
Е.И. Балкашиной, 1929 г.)
Он отметил феномен генетической неста-
бильности: мутация tetraptera возникает исче-
зающе редко, зато возникнув, может проявляться у потомства с самыми
различными частотами (различия в десятки раз). Притом различия возника-
ют из-за различий условий среды: на холоду частота tetraptera не превыша-
ет 1%, а в тепле достигает 35%. Естественно возникает мысль, что мутация
может делать неустойчивым какой-то ген, который включает, в зависимости
от условий, в каких развивается зародыш, определенный механизм разви-
|тия особи. Это дает возможность ввести в схему эволюции влияние внеш-
ней среды, причем ввести без НПС; в этих терминах генетическая неста-
бильность смогла стать предметом интереса эволюционистов в годы без-
раздельного господства неодарвинизма. (Замечу, что прямое влияние сре-
ды на развитие зародыша - аргумент жоффруизма).
Отбор создает тут не саму отбираемую конструкцию, а лишь условия
для ее реализации. Отсюда всего шаг до введения понятия творческой ро-
ли отбора. О ней говорил еще Дарвин, а яснее всех - Уоллес, когда писал,
без всяких аргументов: «Каждое преуспевающее поколение под влиянием
наследственности получает все большее преимущество, так что шанс вос-
произвести нужную вариацию будет все выше».
Другими словами, ими было сделано допущение, что преуспевающие
(те, кто выжил в прошлом) получают некие новые возможности для выжива-
ния в будущем. Каковы эти возможности, оставалось неясно, и с рождением
генетики вопрос обострился. А именно, один из первых генетиков, датский
ботаник Вильгельм Иоганнсен (он, кстати, ввел сам термин «ген») ясно по-
казал на своих объектах, что никакой новой изменчивости отбор не создает,
отбор (притом только искусственный) может выбрать лучшее из
наличного, но не более.
160
С ним спорили, и этим в 1932 - 1940 годах занимался Рокицкий, в опы-
тах которого велся отбор (искусственный - это обстоятельство никем тогда
не принималось во внимание) дрозофил на увеличение и на уменьшение
числа грудных щетинок при условии рентгеновского облучения. Выявилось
три группы данных. В первой даже долгий отбор никакой новой изменчиво-
сти не породил; во второй после 30 поколений безрезультатного отбора из-
менчивость вдруг резко возросла, дала материал для отбора, а затем вновь
упала; в третьей изменчивость возрастала каждые 5-10 поколений, что тоже
обеспечило длительный успех отбора. Рокицкий истолковал эту картину по
Четверикову, через генотипическую среду:
«Первое наследственное изменение, действительно новое, вскрывает целый
ряд других, фактически уже бывших, но прежде бесполезных для отбора».
Конечно, это было лишь толкование. Не будем вступать в спор толкова-
телей, это бессмысленно. Заметим лучше, что Рокицкий дал именно то по-
нимание «творческой роли отбора», которое господствует в дарвинизме по-
ныне: отбор является творческим фактором эволюции не потому, что изме-
няет изменчивость в нужную сторону, а потому, что изменений, «фактически
уже бывших», достаточно для объяснения всей будущей эволюции. То есть
дарвинизм вернулся в IV век, к верованию Григория Нисского.
Тем самым, вопрос о подлинно новом даже не ставился. Кроме того,
все примеры “творческой роли отбора” относятся к акциденциям (побочным
свойствам - п. 1-6**) и даже если их признать, эволюцию не объясняют. От-
сюда и пошло то понимание дарвинизма, которое стало господствующим и
которое критики вовсе не относят к эволюционным учениям. Наоборот, они
называют СТЭ «смягченным фиксизмом», т.е. скрытым способом отрицания
эволюции (подробнее см.: ЧЭ, с. 178-179) Вот почему в книгах по СТЭ так
много разговоров про щетинки и совсем ничего - про саму эволюцию: сто-
ронникам СТЭ просто нечего о ней сказать.
4-7. Ученики Четверикова: Балкашина и гомеозис
Если у насекомого оторвать усик, на этом месте может вырасти беспо-
мощная лапка. Такая же лапка сама вырастала на месте аристы (ветвистого
стерженька на конце усика) у потомства одной из кавказских дрозофил, изу-
ченных четвериковцами. Среди них была замечательная исследовательни-
ца Елизавета Ивановна Балкашина (1899-1981), и она взялась за анализ
этого явления. Она назвала данную мутацию термином aristopedia, под-
черкнув этим направление замены - аристы на ножку.
Появление одного органа на месте другого именуют гетероморфозом
или гомеозисом. Гетероморфоз по-гречески значит иначе сформирован-
ный, а гомеозис (термин ввел Вильям Бэтсон, один из основателей генети-
ки, в 1894 г.) - уподобление. Как видим, оно может быть и прижизненным, и
наследственно обусловленным. Исследуя развитие органа, Балкашина ус-
тановила, что в обоих случаях формирование идет сходно. Явление оказа-
161
лось общим у членистоногих. Балкашина суммировала литературу почти за
сто лет, так что в 1928 году смогла резюмировать:
«Ряд гомеозисов, возникших вследствие изменения наследственной субстан-
ции и развивающихся в организме в течение его эмбриональной жизни, явля-
ется параллельным ряду совершенно аналогичных гомеотических изменений,
происшедших путем регенерации». Таким образом, «силы в организме, опре-
деляющие развитие органа, действуют одинаково как при внешнем воздейст-
вии (экстирпация [вырывание - Ю.Ч.] органа), так и при внутреннем воздей-
ствии (генотипическая изменчивость)» (цит. по [Бабков, с. 86]).
Иными словами, отмечен параллелизм между прижизненными и на-
следственными изменениями. В сущности, это - продолжение той вековеч-
ной идеи, которой мы касались в главе 2 в связи с именами Вольфа, Киль-
мейера и других и которая привела Геккеля (уже в дни Дарвина) к «биогене-
тическому закону». Только речь у Балкашиной шла не о том, что потомок
невесть зачем и почему повторяет своим зародышем историю взрослых
предков (по Геккелю), а о единстве законов формообразования (по Киль-
мейеру). Впервые было показано, как именно мутация меняет строение ор-
ганизма: в данном случае она переключает онтогенез с одного пути на дру-
гой, притом - на уже существующий в том смысле, что для него у организма
есть программа.
Результат сам по себе замечательный, но Балкашина сделала больше:
она определила место гена, чья мутация вызывает данный гомеозис, на
хромосоме и заметила, что там же в тесном соседстве располагаются три
другие (все известные тогда у дрозофил) гомеозисных гена, в том числе и
исследованный Астауровым ген, давший мутацию tetraptera. Открытые поз-
же другие гомеозисные гены оказались там же. Стало понятно, что тут кро-
ется какой-то эволюционный смысл.
Гольдшмидт быстро оценил эти открытия и положил их в основу своей
теории эволюции путем системных мутаций. Теория (опубликованная им в
1940 г. в США, после эмиграции из фашистской Германии) постулировала,
что виды и другие таксоны образуются скачком (сальтационно), причем от-
личие от де-Фриза состояло в конкретизации путей: системная мутация ме-
няет хромосому как целое и в особенности - гомеозисную область. Как мы
узнаем далее (п. 5-5*), он оказался целиком прав.
Формируемый таким путем новый организм Гольдшмидт нарек обнаде-
живающим уродом (hopeful monster), подчеркнув этим, что вопрос о его бу-
дущем решает отбор в понимании де-Фриза, т.е. как апробация готовой це-
лостности (но не в понимании Дарвина).
Развитием идей Балкашиной сделали себе имя и некоторые другие
биологи, забыв ее саму. Зато ее не обошли вниманием советские органы:
она была, как почти все четвериковцы, арестована. Сосланная в Восточный
Казахстан, жила там с дочерью до конца дней. Работала энтомологом на
санэпидстанции, но в науку не вернулась. С другими обошлись куда хуже.
162
(Это и многое другое мне любезно сообщил историк науки В.В. Бабков.) Всё
это было еще до начала лысенковщины, так что корень бед надо искать не в
ней. Наоборот, сама лысенковщина выросла из того глубокого корня, что
повсюду подымал выдвиженцев (п. 4-4), губя специалистов.
4-8. Ученики Четверикова: Николай Беляев и генетика чудес
Николай Константинович Беляев (1899-1937) с детства любил бабочек
и, поступив в Московский университет, сблизился с лучшим тогда в России
специалистом по бабочкам - Четвериковым. Под его руководством Беляев,
как и все, изучал дрозофил, но после ареста учителя вернулся к бабочкам,
став вскоре ведущим в мире знатоком их генетики.
В те годы бурного развития генетики многие были уверены, что она и
только она дает биологии подлинную базу, и искали, в частности, эволюци-
онное родство в сходстве хромосомного аппарата. Беляев отдал дань этой
идеологии - например, поддержал гипотезу об общем предке бабочек
(Lepidoptera) и ручейников (Trichoptera) на том основании, что в обоих отря-
дах (и, среди насекомых, только в них) самки гетерозиготны по половым
хромосомам (система WZ, в отличие от обычной для большинства организ-
мов системы XY, где гетерозиготны самцы).
С этим доводом трудно спорить, но Беляев предостерег от увлечения
хромосомной систематикой, напомнив, что сходство хромосомного аппарата
у таксономически близких (в классическом смысле, т.е. по совокупности
свойств) групп далеко не всеобще: даже число хромосом, будучи приблизи-
тельно постоянно у некоторых крупных групп, у других групп сильно варьи-
рует в пределах рода, а иногда (например у бабочек рода Biston из семей-
ства пядениц) даже в пределах вида.
Самое полезное для страны, что мог сделать тогда лепидоптеролог
(специалист по бабочкам) - заняться шелководством, что Беляев и сделал,
сперва в Ташкенте, а затем в Тифлисе. (Тем же занялся Кольцов в Москве,
а позже и ссыльный Четвериков в Горьком.) В считанные годы советским
генетикам удалось, несмотря на репрессивную обстановку, создать пригод-
ные для местных условий породы шелкопрядов. Тогда, до эры синтетики,
парашютный шелк был важным стратегическим материалом, и даже ссыль-
ный Четвериков получил орден.
А вот острый на язык Беляев был в 1937 году арестован и вскоре рас-
стрелян. Его работы успешно продолжил В.П. Эфроимсон (позже просла-
вившийся и в медицинской генетике), которому дали даже стать доктором
наук, но потом (в 1948 г.) все же дали 10 лет лагерей.
Блестящие, прерванные в самом расцвете работы Беляева важны для
нашей темы: он впервые показал практически, что невозможно отбирать
«полезные признаки», если те входят в единый комплекс (отбор на хорошие
коконы приводит к снижению жизнеспособности, скорости размножения, и
т.д.), зато можно легко и быстро отбирать сами комплексы. То есть идея
163
системного отбора (п. 4-3) вполне разумна, но только если отбор - искусст-
венный. Тогда и это никем замечено не было.
Как же удается получать хорошие породы? Словами Бабкова:
«Ближайший практический вывод, который и сделал Эфроимсон, - требова-
ние выводить и испытывать породы в провокационных условиях», т.е. в сре-
де, похожей на «неблагоприятные условия промышленной выкормки».
Конечно, и ламаркисты, и жоффруисты скажут, что этим шелководы
признали влияние среды на наследственность, однако сами дарвинисты со-
чли достаточным вновь сослаться на сложность отбора.Так и живем.
Другой эпизод касается открытия молекулярной основы наследствен-
ности. О загадочной природе вещества наследственности писали многие; из
упомянутых ранее эволюционистов XIX века назову хотя бы Спенсера и
Геккеля, проводивших аналогию предполагаемого копирования наследст-
венных признаков с кристаллизацией. По словам историка, обследовавшего
этот вопрос, работ на эти темы было полторы сотни (АЛ. Пилипенко. Про-
блема информационных молекул и матричного биосинтеза в XIX - первой
трети XX века. И ВИЕТ, 1988, № 2), и последним в списке (по порядку, но ни-
как не по значению) назван Кольцов.
Кольцов на съезде зоологов (1927 г.) напомнил, что вопрос имеет
большую литературу, и сделал странный для нас шаг - отверг гипотезу, по
которой веществом наследственности является нуклеиновая кислота. Для
него было несомненно, что этим веществом может быть только белковая
нить и что хромосома составлена из двух одинаковых нитей2Л Его речь на
съезде важна нам своими эволюционными репликами. Вот одна из них:
«Конечно, в основе процесса эволюции белковых молекул наблюдается неко-
торая закономерность ... И если у одного вида млекопитающих замена мети-
ловой группы ионом водорода в определенной аминокислоте вызывает, по-
ложим, альбинотическую мутацию, то естественно, что такая же мутация мо-
жет появиться независимо в параллельных рядах близких видов, может и по-
вторно возникнуть у того же вида. Однако... хромосомные молекулы настоль-
ко сложны (допуская сантильоны изомеров), что из всех возможных комби-
наций до сих пор в течение жалких 1 000 миллинов лет существования Земли
21
Н.В. Тимофеев-Ресовский, ученик Кольцова и Четверикова, позже заменил одинако-
вые нити на комплементарные (как позитив и негатив), чем ввел идею конвариантно-
сти. “Возможно, самый важный вклад Н.В. в науку - концепция конвариантной редуп-
ликации” (Шнолъ С.Э. Н.В. Тимофеев-Ресовский // БПС. 1996, № 45, с. 8). Н.В.. рабо-
тая в 1925-1945 гг. в Германии, оказался одним из основателей военной радиобиоло-
гии. Поэтому вполне понятны и даже неизбежны “устрашающие легенды о его про-
шлой деятельности в Германии" (Чесноков В.А. Николай Владимирович Тимофеев-
Ресовский // БШ, 2000, № 6, с. 21). В 1945 г. Н.В. попал в советский лагерь, где стал
инвалидом: затем был ведущим радиобиологом. Его эволюционные взгляды легли в
основу СТЭ и изложены в БЖ, 1958, № 3.
164
осуществлена лишь ничтожно малая часть их» (Кольцов Н.К. Организация
клетки. М.-Л., 1936, с. 490).
Тут мы подошли к третьему эпизоду. Смысл реплики в том, что «неко-
торая закономерность» эволюционного процесса не только призна-ется, но
и прямо связывается с параллельными рядами наследственной изменчиво-
сти, о которых у нас шла уже речь в п. 3-10; однако отмечено и то, что ряды
эти не могут сами по себе объяснять закономерности эволюции, поскольку
природой «осуществлена лишь ничтожно малая часть» возможных вариан-
тов, а потому надо объяснить появление именно наблюдаемых форм, а не
каких-то еще. И Кольцов пояснял:
«Без дарвиновского принципа естественного отбора и отметания неприспо-
собленных фенотипов белковые молекулы находились бы до сих пор в самом
начале своей эволюции... А вместе с исчезнувшей молекулой уносятся без-
возвратно и квадрильоны комбинаций, которые, согласно законам номогене-
за, могли бы возникнуть в ходе дальнейшей эволюции».
Надо пояснить сантильоны, квадрильоны и номогенез. Сантильон - это
единица с тридцатью нулями, квадрильон - с пятнадцатью, т.е. корень
квадратный из сантильона. Столь огромное число наследственных комби-
наций означает, что изменчивость предполагалась у Кольцова такая, какую
постулировал Дарвин, - случайная ненаправленная.
Но всегда ли так на деле? Четвериковцы задумывались над этим, и од-
ного из них, Гершензона, ждал успех: в 1939 году он опубликовал в «Докла-
дах АН СССР» заметку «Вызывание направленных мутаций у Drosophila
melanogaster». Вряд ли автор тогда подозревал, что она рушит его собст-
венный взгляд на мир. В опытах Гершензона почти смертельные дозы те-
лячьей ДНК вызывали у дрозофил направленные мутации', мутировали
очень немногие гены и в очень немногих направлениях (иногда даже в од-
ном-единственном), зато часто. Вместе с линией исследований трансфор-
мации бактерий, шедшей от Ф. Гриффита (см. п. 5-12), она открывала новую
эру в эволюционизме.
В 1940 году генетик Ю.Я. Керкис (Ленинград) дал общее понимание му-
тации как следствия нарушения нормальной работы клетки (Керкис Ю.Я.
Физиологические изменения в клетке как причина мутационного процесса И
УСБ, 1940, № 1). В нынешних терминах, речь шла про стресс, явление,
описанное под этим именем в 1936 году Гансом Селье, но фактически во-
шедшее в оборот гораздо позже. О стрессе мы будем говорить в главе 5, а
сейчас надо напомнить более ранние работы:
«Мёллер (Genetics, 1928, July) в своих исследованиях действия температуры
на количество мутаций установил, что... чувствительным к температуре явля-
ется главным образом период созревания, т.е. образования полноценных по-
ловых клеток». Вообще, «все данные экспериментальной генетики говорят за
то, что мутации вызываются повреждением живой материи, т.е. такими дей-
ствиями, которые лежат за пределами физиологического приспособления»
165
(Бауэр Э.С. Теоретическая биология. M.-JL, Изд. Всесоюз. Ин-та эксперимен-
тальной медицины, 1935, с. 196-197).
Как видим, явление влияния стресса на мутации было хорошо известно
задолго до появления самого понятия «стресс».
4-9. Ряды и номогенез. Вклад Вавилова
А что такое номогенез, который у Кольцова как бы извлекает корень из
изменчивости? Номогенез - это эволюция на основе закономерностей.
Реплика Кольцова - видимо последнее сочувственное упоминание учения,
которое затем полвека разрешалось только поносить, не разъясняя сути, а
лучше всего - не упоминать вовсе. Номогенез для Кольцова - несомненный
эволюционный факт, указывающий на механизмы, поставляющие материал
для отбора. Номогенез, по Кольцову, дан нам в наблюдении параллельных
рядов изменчивости. Поступим и мы так же: сперва поговорим о рядах, а
через них уже перейдем к номогенезу.
Как уже говорилось в главе 3, сейчас модно говорить о презумпции
объяснений «по Дарвину»; мы-де согласны, что дарвинизм не всё объясня-
ет, и пользуемся им только до тех пор, пока нам не докажут, что он неверен.
Сторонники презумпции могут признать негодность какого-то своего приме-
ра, но не видят причин менять точку зрения. Сходна позиция ламаркистов:
если нельзя придумать, как можно развить путем упражнения какое-то свой-
ство (например маскирующую окраску), то признаётся трудность только в
этом конкретном случае.
Совсем иной оказывается позиция номогенетиков: их исходный пункт
(параллельные ряды - п. 3-10*) реально наблюдаем, а не додуман. Это ко-
нечно не значит, что у них нет презумпций, но все-таки их учение можно на-
чать не с презумпции, а с осмысления фактов. Ведь веских примеров отбо-
ра вообще нет, а ряды наблюдаются во всем.
В п. 4-7 приведена цитата Балкашиной о рядах формообразований, а в
п. 3-10, шла речь о рядах сходных (но таксономически далеких) организмов.
С рядами сталкивается любой систематик, но относятся к ним систематики
различно. Напомню хотя бы, что паралелизм между сумчатыми и плацен-
тарными (в обоих подклассах есть свои хищные, свои грызуны, свои насе-
комоядные и т.д., а среди хищных - свои медведи, свои собаки и т.д.) Гек-
кель объяснял независимым приспособлением к сходным условиям жизни,
а Коп - сходством законов формообразования.
Читатель спросит: как на самом деле? Отвечу: на самом деле каждый выбирает
то объяснение, какое ему понятнее. Есть, правда, несколько примеров, гораздо
проще объясняемых с одной позиции, чем с другой, но простота объяснения нико-
гда противников убедить не может (зачем вводить новое, если мне нравится ста-
рое?). И даже если с одной из позиций нет никакого объяснения какому-то классу
фактов, ее сторонники могут сослаться на неясные прошлые обстоятельства или на
непременные будущие открытия. Со времен Спинозы (XVII в.) такой довод име-
166
нуется asylum ignorantiae (лат.: убежище невежества). Но как тогда вообще новое
входит в науку? В точности этого никто не знает, и господствуют два объяснения.
Первое (его исповедует марксизм) гласит: «Практика - критерий истины».
При всей внешней очевидности, оно далеко и от истины, и от практики: мы уже
видели и не раз увидим, что общество может признавать «практикой» отнюдь не
факты, а мифы, рекламу и даже анекдоты. Наоборот, достоверно установленные
факты могут очень долго оставаться в полном небрежении.
Второе объяснение дал Кун (см. п. 2-1): в рамках нормальной науки новое
входит быстро и сравнительно безболезненно, а всё остальное копится в ожида-
нии очередной научной революции. Как отмечено в п. 3-13, ей сопутствует смена
презумпций. Долг ученого видится мне в том, чтобы выискивать и осмысливать
новое, связывать его со старым и прилагать к практике, а не ждать научной рево-
люции, т.е. не следовать консервативному большинству. Иногда феномен нор-
мальной науки может напрочь блокировать развитие науки (см. пп. 5-3 и 12-1).
Ряд одинаковых покровительственных окрасок (белый медведь, белая
куропатка, заяц-беляк и т.д.) естественно считать рядом независимых при-
способлений - неважно, по Дарвину, по Ламарку или еще как-то. Подобные
примеры кочуют из книги в книгу, но никогда не соседствуют в них с рядами,
которые не удается связать с приспособлением. Таковы, например, ряды
сходных форм раковин (самых разных моллюсков и даже - одноклеточных
фораминифер) или ряды листьев.
Обыкновенный остролистный клён и платан столь сходны контуром
листа, что новичок их путает. Этот клён так и называется - платановидный,
Acer platanoides. Аналогично, обыкновенный платан именуется клёнолист-
ным: Platanus acerifolia.
Чем такой лист может быть полезен дереву, никто до сих пор не приду-
мал, а обычное «когда-нибудь узнаем» всё меньше удовлетворяет сомне-
вающихся. В самом деле, вопрос задают уже триста лет (с тех пор, как ес-
тественное богословие занялось придумыванием назначения всем ясно ви-
димым особенностям организмов - см. п. 1-12), а ответа нет ни для каких
контуров и, насколько знаю, его никто даже не ищет. Зато сами контуры вы-
страиваются в великолепные параллельные ряды.
Возьмем, к примеру, всевозможные клёны (120 видов) и расположим
их листья в горизонтальный ряд - от остролистного клёна (кончики листа
не просто остры, но снабжены иголочками) через контуры простой пяти-
пальчатый, размытый пятипальчатый, округлый и яйцевидный и т.д., до
ланцетовидного. Под этим рядом расположим ряд листьев различных
смородин (150 видов) - он будет похожим, хотя на месте первого члена
ряда будет пропуск: кажется, нет смородины с иголочками на концах лис-
та. Ниже можно поместить ряд листьев семейства аралиевых (в него вхо-
дят, в частности, плющи, листья которых сходны с листьями смородин) -
картина будет примерно та же (рис. 17).
167
Рис. 17. Пример размывания “кленового” контура листа:
клён остролистный, платан восточный, смородина Макси-
мовича, клён ложноплатановый (внизу справа), плющ
Есть клёны со сложным
листом, состоящим из пяти
ланцетовидных листочков, но
есть такой лист и среди ара-
лиевых - у женьшеня. Можно
добавить и другие ряды, на-
пример, ряд платанов; но пла-
танов мало (10 видов), поэтому
в их ряду много пробелов. При
всём том, остроконечные фор-
мы образуют свою вертикаль-
ную колонку, округлые - свою,
яйцевидные - свою, сложноли-
стные - свою и т.д.
Вам это что-нибудь напоми-
нает? Многим напоминает пе-
риодическую таблицу химиче-
ских элементов.
Как и в химии, иногда с по-
мощью рядов удается даже сделать прогноз (указать еще не открытый вид),
что в биологии казалось бы немыслимо. Первый такой прогноз удался пет-
роградскому ботанику Николаю Ивановичу Вавилову (1887-1943). Сравнив
разные виды пшеницы и ржи, он
обнаружил полный параллелизм,
полный настолько, что отсутствие
одного вида ржи - безлигульная
пшеница была известна на Пами-
ре, а безлигульной ржи не было -
побудило его в ближайшей поезд-
ке на Памир искать безлигульную
рожь. Вавилов поехал на Памир и
нашел ее (рис. 18).
Рис. 18. Стебель без лигулы и с
лигулой (левая пара — пшеница,
правая — рожь)
Недаром, когда доклад Вавилова в Саратове (1920 г.) вызвал овацию,
кто-то сказал: «Это биологи чествуют своего Менделеева». Наконец-то.
168
Ведь ряды описывали еще до Дарвина, и аналогию их с химией провел еще
Коп. Однако в химии есть параметры, естественно упорядочивающие клетки
таблиц (например, химические элементы упорядочены атомным весом), а в
биологии ничего такого нет, поэтому трудно понять, как воспользоваться
рядами для описания эволюции.
И всё же эволюционная концепция, осмысливающая ряды биологических
форм, была построена. Она и получила название, упомянутое в п. 4-8 - но-
могенез. Фактически сторонники номогенеза поменяли презумпцию:
если дарвинизм и ламаркизм полагают всякое сходство либо свиде-
тельством общего происхождения, либо результатом независимого приспо-
собления, то номогенез утверждает, что всякое сходство - результат общ-
______________ности законов развития, если не доказано иное._______
Попробуй-ка докажи! Да не друг другу, а противникам!
4-10. Берг и рождение номогенеза
Идея о том, что изменчивость как-то упорядочена и что виды, изменя-
ясь, занимают место в какой-то огромной многомерной таблице, полезна,
хотя саму таблицу редко удается изобразить (ситуация, обычная в матема-
тике; там есть теория матриц, изучающая свойства таблиц произвольного
размера). То, что бабочки двух разных видов имеют одинаковый рисунок
(мимикрия), объясняется не их подражанием друг другу («подражатель» и
«образец» иногда живут по разные стороны океана), а всего лишь принад-
лежностью обоих видов к сходным рядам изменчивости. Об этом как раз и
сказал Вавилов в 1920 году на саратовском съезде и сделал эволюционный
вывод [Вавилов, с. 19-20]:
“Явления мимикрии, подражание одних видов и родов другим по форме и ок-
раске, несомненно во многих случаях представляют ... повторение тождест-
венного цикла изменчивости у различных семейств и родов, и не представля-
ют какого-либо исключения, иллюстрирующего роль отбора в создании
форм, как это склонны были предполагать дарвинисты, а общую для органи-
зованного мира повторность форм изменчивости”.
И далее:
“Явления мимикрии общи всем классам и семействам, и те необычайно рази-
тельные ее формы, которые открыты, например, у бабочек, являются лучшей
иллюстрацией закона тождества рядов генотипической изменчивости. ... При-
рода кристаллизует формы в определенные системы ... Природа оказывается
бессильной до бесконечности разнообразить виды и роды и производит неред-
ко аналогичные ... формы у разнообразных родов, семейств и даже порядков”.
Как видим, дарвинизм отнесен к прошлому, что было разумно, хоть и
преждевременно. Позже Вавилов никогда не выражался об эволюции столь
прямо (и потому основателем номогенеза стал не он), зато в 1935 году чётко
отделил феномен рядов от фактов генетики:
169
‘В случаях параллелизма отдаленных семейств, классов, конечно не может
быть речи о тождественных генах даже для сходных внешне признаков”
[Вавилов, с. 92].
К сожалению, многие писали и пишут, что Вавилов рассматривал свои
ряды как генетические. Нет, как видим, они у него номогенетические.
। Основателем номогенеза стал петроградский географ и зоолог Лев Се-
менович Берг (1876-1950), который 1922 году опубликовал книгу «Номоге-
нез, или эволюция на основе закономерностей». Принципиальным новше-
ством был у Берга внимательный анализ основ учений. Он выявил то об-
щее, что есть у всех эволюционных учений [Берг, с. 73, 88]:
1) все они исходят из наличия организмов как некоей данности (проис-
хождение живого ни одно учение даже не пытается рассматривать);
2) организмы обладают целесообразностью, и это тоже не обсуждается
(«Рассмотрение вопроса, почему живое отличается свойством реагировать
целесообразно..., относится уже к философии природы»);
3) эволюция в основном (а у некоторых авторов целиком) «основана на
развертывании уже имеющихся налицо задатков».
Ламарк не объяснил, как организмы приобрели активность, Дарвин -
откуда взялась избыточность размножения, они это ввели как аксиомы. По-
няв всё это, Берг стал строить свою теорию тоже на аксиомах.
Номогенез Берга сложился из пяти эволюционных идей: это шедшая от
Мопертюи и Кейзерлинга аналогия с химией, порожденные ею гомологиче-
ские ряды Копа - Вавилова, о которых мы уже знаем, и три идеи, для нас
новые: ортогенез, преадаптация и пророческие фазы.
Ортогенезом называют длительную тенденцию какой-либо группы к
эволюции в некотором определенном направлении. Термин сделал извест-
ным в 1897 году немецкий зоолог Теодор Эймер [Полое; Филипченко]. Он
обосновал концепцию на бабочках, но она верна для самых различных
групп организмов (такова эволюция ног предков лошади от трехпальцевых к
однопальцевым). Ортогенез, начавшись как нечто полезное для организмов,
подчас продолжается далеко за пределы какой-либо пользы.
Еще Аристотель в «Частях животных» писал, что рога оленя чрезмерны,
а в Новое время палеонтологи нашли в Ирландии ископаемого оленя с та-
кими огромными рогами (рис. 19), что об их пользе говорить не стоило. Са-
мые рьяные сторонники отбора сумели предложить лишь объяснения того,
как эти рога совместимы с идеей отбора (хоть и невыгодны), но это вышло
столь вычурно, что даже Конрад Уоддингтон, всё на свете легко объясняв-
ший отбором, признал в 1939 году - рога ирландского оленя теорией отбора
необъяснимы (подробнее см.: Симпсон Дж.Г Темпы и формы эволюции. М.,
1948, с. 261).
Столь же непомерны спинные шипы некоторых наземных ископаемых
яЩеров, шея изображенного на обложке ЧЭ ящера, клыки саблезубого тигра
и многое другое. Так вот, Берг постулировал, что ортогенез - одна из основ-
170
ных закономерностей эволюции. Он же по-
стулировал, что общей закономерностью
живого является целесообразность уст-
ройства и поведения организмов, и заявил,
что это он не сам придумал, а заимствовал
у Ламарка и Дарвина.
Рис. 19. Ископаемый олень (точнее, лось)
Megaceros (плейстоцен, отдел четвертич. пе-
риода) с непомерными рогами, явно затруд-
нявшими жизнь его обладателю
В самом деле - разве изменчивость ор-
ганизмов и их стремление к избыточному
размножению, постулированные Дарви-
ном, сами по себе более понятны, чем
стремление рогов избыточно расти?
Аналогично ввел Берг в свое учение и две другие идеи.
Преадаптацией называют такое изменение организма, которое оказы-
вается полезным не в тех условиях, в каких данный вид жил в момент воз-
никновения изменения, а в тех, в какие попал позже (Кэно, 1911 г. [Назаров,
1974]). В виде примера приведу сложный членистый скелет конечности,
спрятанный внутри плавника кистепёрой рыбы. Его сложность являет ра-
зительный контраст с простотой выполняемой функции (рис. 20). Он всерьез
понадобился гораздо позже, наземным животным и при этом (вот удиви-
тельно) сильно упростился (см. далее, п. 8-11*). Преадаптацию часто рас-
сматривают как довод в пользу ортогенеза [Попов, с. 147].
Пророческой фазой эволюции геолог и палеонтолог А.П. Павлов на-
Рис. 20. Скелет плавника кистепёрой рыбы
(девонский период)
звал в 1901 году ситуацию,
когда зародыш или молодая
особь предка обнаруживает
признак, обычый у взрослых
потомков. Пример еще в 1828
году привёл Бэр: «У голова-
стика есть настоящий клюв,
как у птицы». Здесь развитие
особи не повторяет эволюцию
(как в «биогенетическом зако-
не» Геккеля), а, наоборот,
предваряет ее.
171
Итак, известные до него закономерностей Берг свел воедино в основ-
ные постулаты эволюции. В качестве шестой идеи он заимствовал у Стра-
хова (о нем см. п. 3-13) философское отрицание дарвинизма, которое до-
полнил аналогичным отрицанием ламаркизма. На этой основе он смог уве-
ренно заявить, что в ходе эволюции организмы перестраиваются согласно
собственным определенным законам, а роль внешней среды осуществляет-
ся как отбором, так и прямым «влиянием ландшафта».
Бергу возражали, что его теория не объяснение, а лишь новое имя для
непонятного. Это верно лишь отчасти: всякая новая теория начинает с груп-
пировки фактов и выявления правил, а понимание возникает позже. Ведь и
Дарвин не мог ни сказать о природе изменчивости, ни дать реальные при-
меры отбора, но его идею критики Берга считали понятной.
4-10* Не ко времени
На упомянутом съезде зоологов (1927 г.) Берг обсудил интересный
пример: он рассказал о редком виде миног с необычным расположением зу-
бов, причем аналогичное расположение зубов иногда в качестве уклонения
(на отдельных особях) наблюдается у других, широко известных видов.
Этим замечанием он мимоходом вторгся во владения французской школы
тератологического эволюционизма (о ней см. [Назаров, 1974]), шедшей от
Жоффруа и видевшей эволюцию в форме превращения индивидуальных
уродств в видовую норму.
Главное там (но не у Жоффруа) отрицание приспособительного харак-
тера эволюции. Так, генетик Эмиль Гийено в 1921 году заявлял, что многие
виды являют собой уродства, «едва совместимые с жизнью», а отнюдь не
приспособления [Назаров, 1974, с. 206].
Нечто похожее не раз уже предлагалось. Первым к этой теме, насколь-
ко знаю, подошел в 1913 году Бэтсон в Англии: по его мнению (цитирую по
[Берг, с. 179]):_____________________________________________________
«животные существуют не благодаря своим признакам, но вопреки им, и тот
факт, что животные продолжают существовать, доказывает лишь то, что в
________целом баланс их свойств склоняется несколько в их пользу».____
К этому тезису Бэтсона мы еще не раз обратимся.
На упомянутом съезде Берг заключил:
«Такие единичные уклонения не имеют эволюционного значения, но когда
процесс подобного новообразования захватит значительную массу особей,
мы сразу получим новый вид... Вот этим именно способом и идет образова-
ние новых форм - путем закономерного развертывания скрытых (рецессив-
ных) признаков, а не путем отбора случайных вариаций. Никаких случайных
вариаций нет - все вариации предопределены и могут быть уложены в гомо-
логические ряды Вавилова; отбор же лишь распределяет организмы по зем-
ной поверхности, играя роль географического фактора».
Берг имел в виду мнение Вавилова, который писал:
172
«Мутации, идущие как бы случайно в разных направлениях, при объединении
их обнаруживают общий закон» [Вавилов, с. 97],
Через три года, в статье «Воздействие географического ландшафта на
культурные растения и животных», Берг изложил пример с миногами почти
дословно, но цитированной мною фразы там нет. Время наступило иное.
То была его последняя эволюционная работа, хотя он прожил еще два-
дцать лет и издал еще семьсот работ. Время сколько-то свободных речей
кончилось, и вавиловские ряды больше как основу номогенеза никто не по-
минал. Дело тут было отнюдь не только в советской политике - она лишь
выбрала угодный ей вариант дарвинизма (по Геккелю), назвав его «мичу-
ринской биологией», и силой утвердила его монополию в СССР, - но сам по
себе дарвинизм стал господствовать повсеместно, надолго оттеснив даже
во Франции обычный для нее ламаркизм. Гомологические ряды (как и весь
номогенез) оказались не ко двору. На Западе их поминали уважительно, но
все реже и реже, так что понемногу о них забыли и после Второй мировой
войны стали открывать (в который раз) заново.
Почти все примеры можно толковать и так и эдак, но не все. Разве
сходство жилкований и их вариаций у крыла насекомого и у листа растения
связано с их общим предком? Если таковой и был, то заведомо не имел ни
крыла, ни листа. Да и плотва весьма далека от летучих рыб.
И вот в 1924 году ленинградский генетик Ю.А. Филипченко прямо зая-
вил, что кроме параллелизмов, вызванных сходством генов, есть и другие
(он назвал их анатомическими и гистологическими), известные у генети-
чески очень далеких видов. Добавлю еще параллелизм наследственных и
ненаследственных изменений, упомянутый выше, в п. 4-7 (подробнее см.
[Чайковский, 1990, с. 32, 107]). Их лучше объединить единым термином
морфологический параллелизм, который в 1923 году предложил Любищев в
письме Бергу (ЛЧ, 1996, с. 10).
В нем он высказал целый ряд возражений Бергу, в некоторых из кото-
рых взял под защиту дарвинизм, чего позже не делал и что, тем самым, го-
ворит о долгом его пути к своей эволюционной позиции. Для нашей темы
интереснее всего его утверждение:
“самые интересные факты, Вами приводимые, свидетельствуют о закономерно-
сти в многообразии, а не о развитии организмов”, и это потому, что “многооб-
разие организмов не может быть истолковано на эволюционном языке” (с. 13).
Этот намек Любищева стал понятен через полвека, когда появился но-
вый вариант номогенеза, в котором эволюция есть процесс заполнения кле-
ток абстрактной таблицы (мы узнаем об этом в гл. 6).
Из данного утверждения Любищева следует, в частности, что систе-
матика не должна быть эволюционной. К этой теме мы обратимся в
главе 10. В конце письма Любищев высказал свое понимание номогенеза:
законы эволюции следует искать не столько в форме предписаний, сколько
в форме «того, что не может произойти». Предложенное им тут понимание
173
закона как запрета обычно для точных наук, но в биологии сформулировать
запрет непросто. Сам он ограничился примером:
‘удлинение плавников у черепах никогда не производится путем гиперфалан-
гии (излишних пальцев - Ю. Ч.\ точно так же, как удлинение шеи млекопитаю-
щих или удлинение усиков у жуков-усачей”.
Поясню: у жирафа, как и у человека, и у кита, и у мыши, всего 7 шейных
позвонков. Это - общее для млекопитающих правило: удлинение шеи идет
за счет удлинения позвонков. Но бывают очень редкие исключения (у ле-
нивца 9 шейных позвонков), и их наличие говорит о том, что налицо не за-
прет, а тенденция. Тенденциями мы займемся в главе 6.
В смысле социальной истории науки Берг сделал для эволюционизма
то же, что до него сделали Ламарк и Дарвин: дал новому учению основателя
и первую книгу. Притом, номогенез был замечен и сохранился в науке, но
лишь на правах маргинального (не входящего в научную норму своей эпохи)
учения. Как и ламаркизм, он почитается в основном на родине основателя
(хотя ламаркизм и известен во всем мире).
И это отнюдь не худший вариант - жоффруизм (п. 2-17) до сих пор ни
одному обществу учёных не служил флагом, хотя обоснован фактами пол-
нее, а механизмы его куда нагляднее, чем у других учений.
4-11. Номогенетики-дарвинисты
В 1930-х годах ботаник Н.П. Кренке на огромном материале высказал
(развивая Вавилова) более сильное, чем у Копа, Вавилова и Гийено, утвер-
ждение. признак, являющийся уродством у данного вида, как правило
нормален у какого-то иного вида. Он же сформулировал «закон родст-
венной изменчивости»: если виды близки, то близка и их изменчивость.
Смысл обоих утверждений виден на таком примере. Обычно у миндаля
плод сухой, а у персика - окруженный мясистым околоплодником, но изред-
ка бывает наоборот; эта пара видов проявляет целый ряд подобных сходств
в своей изменчивости. То же мы видим у многих близких видов, на что и
указал Кренке. Например, волк может быть рыжим, а лиса - серой.
Хотя о выгодности уродств и речи быть не может, однако Кренке видел
причину рядов изменчивости только по Дарвину, т.е. считал, что они вызва-
ны не сходством неизвестных законов формообразования, а сходством ге-
нов, унаследованных от общего предка (в силу их предполагаемой прежней
выгодности), и их мутациями (почему-то не отсекаемыми отбором).
Важно отметить, что за развитие главного козыря номогенеза (ряды)
взялся Кренке, крайний дарвинист, которого никак не заподозришь в симпа-
тиях к номогенезу. Его идеи были востребованы через 40 лет, что мы узна-
ем в главе 6. Зато Кренке сразу же дал советским биологам урок в другом:
если называть номогенез дарвинизмом (неважно - искренне или для обхода
цензуры), то можно печататься без помех. То же и тогда же показал зоолог
А.Ф. Котс - у него это были ряды окрасок тетеревов.
174
Искать материал, подтверждающий правила Кренке, утомительно, по-
скольку подавляющее большинство особей почти одинаково, а особи, за-
метно отличные от нормы, очень редки. Легче всего это делать на тех же
листьях растений и на насекомых, труднее - на мелких грызунах, а для бо-
лее крупных животных это почти немыслимо - трудно получить в распоря-
жение достаточное число экземпляров.
И всё же трое российских орнитологов сумели, живя в разное время,
представить нужный материал - ряды окрасок тетеревов. Для этого им
пришлось совершить научный подвиг: в общей сложности 90 лет ходить по
базарам и скупать подряд все необычно окрашенные экземпляры. Просмот-
рев несколько миллионов тушек, они выбрали сотни полторы «аберрантов»,
составивших коллекцию Дарвиновского музея в Москве. Последний из тро-
их, основатель музея, расположил их в ряды - это и был Котс.
Эпиграфом своего труда он взял слова Мензбира, мимоходом брошен-
ные тем в молодости: «Спорные вопросы разрешаются только тогда, когда
их перестают прятать». Сам Мензбир, имея массу иных достоинств, не жил
под этим лозунгом - вспомним, как он «прятал» в 1902 году все спорные во-
просы, на какие не имел ответа (п. 3-12), а вот Котс явно им пользовался:
будучи дарвинистом, смело погрузился в изучение рядов - как делали до
него далёкие от дарвинизма Коп, Соболев, Вавилов и Берг.
Упорядоченность уродливых окрасок тетеревов оказалась высокой: у
самцов она уложилась всего в 9 форм. Это можно считать морфологиче-
ским параллелизмом по Филипченко, так как о приспособлении речи тут ид-
ти не может: ведь формы были уродствами. Подтвердился закон Кренке.
сходство с близким видом (глухарь) было наибольшим, с куропаткой мень-
ше, а с далеким видом (фазан) - наименьшим. Котс восхищался:
«каскады форм, скрывающиеся под однородным с виду оперением птицы, --
как не вяжутся они с былым наивным представлением о постоянстве облика
живых существ! Эта изменчивость в потенции понятна лишь как отголосок
прошлого или как вестник будущих возможных изменений» (А.Ф. Котс. О
гомологических рядах в окраске оперения Tetraonidae и Phasianidae // Памяти
акад. М.А. Мензбира. М.-Л., 1937, с. 227).
Видимо, «изменчивость в потенции» должна была напомнить о сослан-
ном Четверикове. О номогенезе же говорить было в те дни нельзя совсем.
Но, как ни относиться к номогенезу, трудно счесть его цельной теорией
эволюции - прежде всего из-за дефекта, общего с дарвинизмом: невнима-
ния к поведению особи и физиологии. И там и тут мы видим на сей счет
лишь отдельные оговорки, но не анализ сути дела. В остальном, и там и тут
организм не живет, а лишь предъявляет среде свои свойства, зара-
нее заданные, так ли уж важно, что происхождение их мыслится по-
разному? Словом, номогенез - лишь аспект будущей теории.
Зато, как ни относиться к ламаркизму и жоффруизму (о нем см. п. 2.6),
надо признать, что в этих учениях жизнь особи - в центре внимания
175
4-12. Соболев и его «историческая биогенетика»
Яркую картину эволюции как процесса, одновременно морфологическо-
го, физиологического и экологического, развернул в те годы палеонтолог
Дмитрий Николаевич Соболев (1872-1949). К тому же только у него одного
чувствуется понимание истории - прежние взгляды приводятся им не для
красоты и не для показа эрудиции, а с целью понять, что такое эволюция. И
только у него мы находим попытку создать действительно теорию, т.е. огля-
деть с единой позиции эволюцию всю, а не те лишь ее черты, что милы са-
мому автору.
Он опубликовал диссертацию по филогении гониатитов^- (гониатиты
- отряд подкласса аммонитов, вымерших головоногих моллюсков - получи-
ли свое название за угловатость
шовной линии - рис. 21.) Защитить ее не
удалось, формально коллеги возмути-
лись обилием цитат из Лукреция, но ре-
альную причину Соболев видел «в ере-
тическом характере» своих идей; Собо-
лев предложил синтез того главного, что
нашел в ламаркизме и дарвинизме, с
идеями Копа (т.е. с будущим номогене-
зом). У Ламарка он взял принцип града-
ции (прогресса), у Дарвина - изъятия не-
пригодных (отбора как причины вымира-
ния), а у Копа - идею рядов и принцип
независимой эволюции видовых и родо-
вых признаков.
Рис. 21. Раковина гониатита (девонский период).
Внизу показана ее простая угловатая сутура (ср.
с очень сложной сутурой на рис. 15)
Начал Соболев с того, что определил предмет исследований;
«многие формы мало отличаются одна от другой... В сети многочисленных пере-
плетающихся вариаций не всегда легко уловить расплывающиеся границы “вида”.
Но если вместо цельных форм мы будем рассматривать их признаки как самостоя-
тельный объект исследования, мы легко заметим, что пределы вариаций этих при-
знаков не широки. Небольшое число типов формы... в разнообразных сочетаниях
повторяется у различных форм» (с. 7-8).
Тем самым, предлагалось вести исследование эволюции форм, отвлека-
ясь и от наличной систематики, и от “происхождения видов”.
27
z Соболев Д. Наброски по филогении гониатитов. Варшава, 1913 (также: Известия Вар-
шавского политехнич. института. 1914. № 1. с той же нумерацией страниц).
176
Посылая Соболеву свой саратовский доклад, Вавилов писал:
«На днях от проф. Л.С. Берга я узнал, что в вашей работе о филогении гониа-
титов развиваются мысли, аналогичные тем, к каким пришли мы, работая с
растениями. Книгу вашу я прочитал, и действительно ваш... закон параллель-
ного развития в сущности весьма близок к тому, что я назвал законом гомо-
логических рядов».
Близость, конечно, есть, но у Соболева сказано больше: на ископаемых
описана реальная эволюция, а не одна нынешняя изменчивость, как у Ва-
вилова. Соболев разделял признаки на комбинационные (такова форма ра-
ковины), такие же, как позже у Вавилова, а также градационные (такова су-
тура) - те, которые определяют постепенную эволюцию группы, и мутаци-
онные (такова всякая перемена в онтогенезе, см. далее) - каких в материа-
ле Вавилова нет.
До войны Соболев работал в Варшаве, был учеником Амалицкого, а за-
несенный войной в Харьков, стал профессором и с 1915 года читал лекции
- как для студентов, так и для публики. Из них родилась его теория, опубли-
кованная в 1924-28 годах и вскоре забытая. Как и Берг, больше об эволюции
Соболев не высказывался, избежал репрессий и заведовал в Харьковском
университете кафедрой геологии до конца дней (1949 г.). Подробнее см.
Колчинский Э.И И ВИЕТ, 1995, № 3.
Как когда-то Теофраст, он видел в процессах, которыми учебники “дока-
зывают" эволюцию, не создание новизны, а лишь продолжение онтогенеза,
который всеми признан как процесс запрограммированный (эволюция в
первичном значении этого слова, т.е. разматывание нити). Эволюция для
Соболева - единый процесс «органического роста», идущий как при разви-
тии особи (онтогенезе), так и при историческом развитии (филогенезе). «Ор-
ганический рост» протекает в череде поколений, так что каждое поколение,
вырастая, повторяет онтогенез своих родителей не в точности, а с ничтож-
ными изменениями. Идею эту Соболев взял у Нэгели. Соболеву она была
важна тем, что отрицала за обычными постепенными историческими изме-
нениями новизну, создание чего-то совсем нового.
Подлинно новое возникает, по Соболеву, скачком и являет собой не
эволюцию, а биогенетику. И свою теорию исторического развития жизни
Соболев назвал исторической биогенетикой-^. Его обоснование биогене-
тики шло с трех сторон. Во-первых, все достоверно известные примеры
появления новых видов являют собою, начиная с чистотела Шпренгера (п.
3-12), примеры скачков. Во-вторых, история флор и фаун является чере-
дованием периодов скачков и периодов постоянства, о чем писал еще Геер
(поэтому в ископаемой летописи почти нет переходных форм). Его поддер-
живали многие палеонтологи, но их не слушали. Большинство держалось
Соболев Д. Начала исторической биогенетики. Харьков-Симферополь, 1924.
177
позиции Дарвина, в пользу которой есть несколько примеров медленного
постепенного увеличения какого-то признака.
Дарвинизм легко мирится с отсутствием ископаемых переходных форм,
для него гораздо важнее те редкие примеры, когда ископаемые переходные
формы есть и становятся в “непрерывный” ряд.
Наиболее известный и важный из них - эволюция североамериканских
предков лошади. С начала кайнозойской эры известен отряд Condylarthra,
который считается предковым и для копытных, и для грызунов, и даже ки-
тов. От него до нынешней лошади выстроен ряд, местами весьма деталь-
ный. Парадным примером в дарвинизме является, как в дни Соболева, так и
ныне, эволюция предков лошади от трехпалых к однопалым, фундамен-
тально описанная Владимиром Ковалевским в 1873 году.
Оппоненты возражали: такие примеры единичны, а 95% видов появля-
ется сразу, скачком, причем в слоях отличной сохранности. Их не слушали,
пока в 1972 году группа американских палеонтологов (Стивен Гулд и др.) не
переломила общественное мнение: вдруг большинство согласилось, что
часто таксон образуется быстро и затем мало меняется. Вместо «перече-
канки», термина давно забытого Геера, в обиход вошли термины «прерыви-
стое равновесие», или, что то же, пунктуализм (о нем см. п. 9-1).
Описание эволюции лошади Соболев шутливо называл «парадной ло-
шадью эволюционной теории», где парадный эффект достигнут за счет
смешения понятий: он писал, что начальные члены ряда
«не могут быть включены в непосредственную родословную лошади, так как
другие признаки, кроме ног, оказываются у различных членов этого ряда раз-
личными».
Указав «предков» лошади по Ковалевскому, Соболев иронизировал:
«палеотерий давно уже вымер, не претерпев изменения, когда появился пер-
вый анхитерий, а этот в свою очередь исчез, тоже не изменившись, прежде
чем был замещен внезапно появившимся гиппарионом. Таким образом, «ро-
дословная» обращается в простое изображение процесса, каким трехпалая
ступня могла превратиться в однопалую» (Д. Соболев. Начала.... с. 135).
Как увидим в п. 9-12, данный пример Соболева оказался ошибочным,
но зато тут поднят важный вопрос: всюду, едва исследователь начинает
строить конкретный ряд от третичного зверя к современному, выясняется,
что он следит только за избранными им признаками - за скелетом пальцев,
за рисунком зубов, за «утолщением костей, поддерживающих рога» и т.п.
(т.е. за семофилогениями, см. п. 9-12), но не за преобразованием целых ор-
ганизмов. Выходит, что сам факт эволюции вне сомнения, но все конкрет-
ные пути сомнительны. Почему так?
Соболев отвечал: потому, что в организмах на самом деле не содер-
жится достаточной информации о путях их прежних преобразований. Мысль
не была новой (ее высказывал еще Мопертюи), но понималась с трудом и
немногими. И если большинство биологов, отклонив чужой ряд как недосто-
178
верный, выстраивают свой, не более достоверный и не более долговечный,
то меньшинство (и Соболев с ними) ищет иного принципа упорядочения ис-
копаемых, без детальных родословных.
Изменчивость ископаемых сходна с изменчивостью ныне живущих:
реализованы различные варианты, их можно выстраивать в ряды и назы-
вать эти ряды эволюционными, но чаще всего это будет лишь исповедани-
ем своей веры. Если в случае «органического роста», когда ничего нового
не образуется, Соболев соглашался верить и в постепенность, и в скачки, то
принципиальную новацию он соглашался принять исключительно в форме
скачка, ибо промежуточные формы, «не обладая должной устойчивостью,
не существовали» (Начала..., с. 148). Это в-третьих.
Тут до Соболева всё зависело от презумпции исследователя: одному
достаточно пары “непрерывных рядов", чтобы счесть, что и у остальных ко-
гда-нибудь найдут переходные формы (а потому скачков не бывает), друго-
му же эта пара - неинтересное исключение из общего закона, закона скач-
ков. Но Соболев глянул с третьей стороны: и то и другое есть, и то и другое
важно, суть же спора в том, кто как читает материал. (Мы вернемся к этому
вопросу в п. 9-12.) Но как же появляется нечто новое?
Соболев видел главный механизм новаций так же, как Жоффруа, через
изменение онтогенеза, причем основывался на двух главных группах фак-
тов: параллелизм таксономических различий с уродствами (как Мопертюи и
многие после него) и предварение эволюции онтогенезом (пророческие фа-
зы Павлова).
Соболев привел примеры массового характера таких новаций, и один
из них так прост, что я кратко изложу его. У всех аммонитов есть сифон -
канал, проходящий через все камеры раковины и позволяющий регулиро-
вать ее плавучесть. Канал закручивается вместе с раковиной, обычно по
наружной стороне ее полости. В конце девонского периода вдруг появилось
много видов аммонитов, у которых сифон шел по внутренней стороне рако-
вины - это климении (названы по имени мифической Климении, дочери
Океана). Данная особенность столь фундаментальна и постоянна, что их
выделили в особый отряд и, тем самым, сочли происшедшими от общего
предка. Однако Соболев описал 12 таких рядов позднедевонских гониати-
тов, что последним членом каждого ряда можно поставить какую-то климе-
нию - единственное ее отличие от остальных членов ряда в том, что сифон
у нее идет по внутренней, а не по внешней стороне полости камер. По-
скольку известно, что у некоторых аммонитов сифон в раннем детстве пе-
реходил с внутренней стороны полости на внешнюю, Соболев объяснил по-
явление климений как скачок хода онтогенеза в каждом из этих рядов в от-
дельности. Если так, то отряд климений ввести можно, но он ничего не ска-
жет о родстве. Откуда взялась синхронность появления климений? Все они
появились в позднем девоне и в нем же вымерли. (Внутренний сифон - му-
тационный признак.)
179
Это стоит выразить как тезис Соболева-.
Объектом эволюции служит не вид, а совокупность свойств, характер-
_____________ная для рассматриваемой группы и ее эпохи._____________
Тут мы подходим к самой оригинальной части исторической биогенети-
ки-к теории регулярных смен флор и фаун. Теория была забыта (хотя са-
мого Соболева и его «Биогенетику» многие знают и чтут), и лишь недавно
историки науки вновь открыли ее [Колчинский; Попов]. Почему забыта, мож-
но лишь гадать - потому ли, что развита была в серии популярных статей,
или потому, что опережала остальную науку на лет на 60.
4-12* Действительно - синтез учений
Вопреки всем канонам, Соболев отрицал тесную связь мира животных с
миром растений: еще в 1915 году он обращал внимание на то, что даже
глобальные смены флор не вели к сменам фаун. Как бы ни менялись рас-
тения, основная масса животных приспосабливалась к этому, не вымирая.
По Соболеву, их массовые вымирания связаны исключительно с геологиче-
скими катастрофами - горообразованием и ростом активности вулканов,
переполнявшим атмосферу углекислым газом. Гипоксия (нехватка кислоро-
да) делала животных чувствительными к другим факторам среды.
(В отрицании связи смены флор и фаун он ошибся, так, при распро-
странении цветковых сильно изменился мир насекомых. См. п. 8-13.)
В периоды угнетения животных растительность, по Соболеву, наоборот,
расцветала - в силу обилия СО2 и связанного с этим потепления климата. С
ослаблением вулканизма и усилением растительности состав атмосферы
восстанавливался за несколько миллионов лет, что влекло новый рост раз-
нообразия животных, а с тем и их численности. Выедание растительности
усиливало выветривание и размывание горных пород и почв, в частности -
снос карбонатов в осадочные породы, ведший к еще меньшей концентрации
СС>2 в атмосфере и к похолоданию. Начиналось массовое вымирание рас-
тений. Это вело к синхронному снижению биомассы животных, но не их раз-
нообразия, которое изменялось медленно - до следующего акта горообра-
зования. Тем самым, Соболев изящно объяснил загадочное несовпадение
вымирания фаун и флор.
По Соболеву, «земное население» (мы теперь говорим: биосфера) -
единый организм, онтогенез которого основан на актах эволюции таксонов.
Если так, то таксоны должны, как у Брокки, стареть и умирать, а физиологи-
ей этого организма должна служить экология, что мы в его труде «Нача-
ла...» и видим:
«...земное население в каждый момент истории земли представляет собою не
случайный аггрегат различных форм, но стройное целое, связанное бесчис-
ленными узами... части его находятся в состоянии подвижного равновесия
(Еленкин) между собою и с окружающей средой» (с. 196).
180
Как видим, у Соболева вместо отбора - ссылка на «подвижное равно-
весие» по Еленкину, каковое надо разъяснить.
Ботаник А.А. Еленкин выдвинул принцип взаимодействия организмов,
частным случаем которого были борьба и взаимопомощь. Изучая лишайни-
ки, он пришел к выводу, что симбиоз - это «взаимный паразитизм» и потому
быстро распадается, едва паразитизм перестает быть взаимным. Аналогич-
ную картину дает симбиоз грибных нитей с корнями деревьев или бактерий
с корнями бобовых, и не надо следовать Фаминцыну и Мережковскому в их
попытках видеть в симбиозе главный путь эволюции:
«Я считаю, что первозданная клетка... не нуждалась в симбиозе с другими
чуждыми ей организмами... Скорее всего эволюция организмов и симбиоз
суть параллельные, но не гомологичные ряды развития жизни на земле, ибо
между обоими рядами очень мало внутреннего сходства» (А.А. Еленкин. За-
кон подвижного равновесия... // Известия Главного ботанич. сада РСФСР.
Петроград, 1921, т. XX, № 2, с. 87).
В основу своей теории Еленкин положил философскую идею Спенсера,
по которой любой процесс эволюции идет, словами Еленкина,
«через переходное состояние уравновешенных движений (как в планетной
системе) и уравновешенных отправлений (как в живом теле)». Разложению
одного объекта сопутствует развитие другого, и результатом явится «под-
вижное равновесие всего сущего», при котором «нет борьбы за существова-
ние и нет взаимопомощи. Есть только функциональные отношения между ор-
ганизмами и внешней средой».
Добавлю: это похоже на химическую систему: нагреешь - идет распад,
охладишь - в ней же идет синтез.
В этом - различие организма и надорганизма (симбиоза):
«жизнь индивидуального организма в большей степени регулируется внут-
ренними законами, определяющими его развитие и жизнь, чем внешними
факторами; наоборот, жизнь симбиоза в большей степени регулируется
внешними факторами, т.е. окружающей средой, чем внутренними, т.е. воз-
действием симбионтов друг на друга». Поэтому симбиоз «вполне подчиняет-
ся закону подвижного рав-новесия, тогда как жизнь индивидуального орга-
низма регулируется этим законом лишь отчасти» (там же, с. 88).
Вот Соболев и решил, считая сообщество надорганизмом, что его мож-
но подчинить «закону подвижного равновесия». Отсюда, как видно ныне,
всего лишь шаг до эволюционной термодинамики, о которой речь пойдет в
главе 5. Сам Соболев предпочитал экономическую аналогию:
«не все получаемое тратится на покрытие издержек производства, остается
излишек, идущий на расширение производства и на улучшение орудий труда»
(1927 г.; цит. по [Колчинский. с. 308]).
В наши дни данная схема эволюции гораздо легче понимаема, чем при
Соболеве: благодаря популярности концепции биосферы Вернадского (то-
же, кстати, непонятого при жизни) мы легко оперируем глобальными круге-
181
воротами и равновесием. Соболев почтительно ссылался на Вернадского, а
тот не упоминал его - возможно, не знал брошюр провинциального профес-
сора. Так или иначе, ничего похожего на глобальную эволюцию в духе Со-
болева у Вернадского нет: в целом его анализ остался на уровне балансов,
единых во все времена. А у Соболева биосфера развивается. Поэтому он
будет весьма полезен нам, когда мы займемся в главе 6 построением еди-
ной теории.
4-13. Новый жоффруизм и винт эволюции
В 1859 году, наряду с огромным скачком вперед (победа эволюционной
идеи), произошла и огромная утрата: по сути был отброшен почти весь опыт
франко-германского эволюционизма с его вниманием к активности особи, к
плану строения и к развитию зародышей, с его признанием медленных и
быстрых изменений. Став лицом к английскому пониманию эволюции (уп-
рощенному, а потому легкому для понимания), ученый мир на сто лет от-
вернулся от куда более сложных и полезных идей.
Они, если и бывали кем-то поддержаны, не встречали сочувствия и
снова забывались. Так, в 1864 году немецкий зоолог Фриц Мюллер в эмо-
циональной статье «За Дарвина» ясно высказался о рекапитуляции (крат-
ком повторении филогенеза в онтогенезе):
«В короткий промежуток времени нескольких недель или месяцев сменяю-
щиеся формы зародышей и личинок дают нам более или менее верную кар-
тину тех изменений, благодаря которым в течение многих тысячелетий вид
достиг своего настоящего состояния».
Это всем известно, и Мюллер вошел в историю дарвинизма. Однако в
той же статье есть и совсем иное, не “за Дарвина”, а скорее “за Нэгели”. Он
признал роль изменений онтогенеза в эволюции:
«как правило, преимущество, заключающееся в численности потомства, да-
леко не может сравниться с преимуществом, заключающемся в большем со-
вершенстве молоди» (Ф. Мюллер, Э. Геккель. Основной биогенетический за-
кон. М.-Л., 1940, с. 153),
- а это уже шаг в сторону Нэгели. Замечен он был нескоро.
Так, крупнейший специалист по эмбриологическому эволюционизму
Алексей Николаевич Северцов (сын Николая Алексеевича Северцева, упо-
мянутого в главе 3, и учитель И.И. Шмальгаузена) в докладе «Эволюция и
эмбриология» (1910 г.) полностью отрицал позицию Нэгели, поскольку пря-
мо отождествил эволюционизм с дарвинизмом и пропагандировал «основ-
ной биогенетический закон Мюллера - Геккеля», согласно которому разви-
тие зародыша лишь кратко повторяет эволюцию. В этом Северцов зашел
так далеко, что даже утверждал:
«Онтоненез есть функция филогенеза, т.е. ... без понимания законов эволюции
мы не можем понять и законов индивидуального развития, скажу больше, за-
конов жизни».
182
Позже он изменил позицию, а в конце жизни даже признал, что «пови-
нен» в пренебрежении ко второй части вывода Мюллера (А.Н. Северное.
Собр. соч., т. 5. М.-Л., 1949, с. 374). Однако слов Нэгели тогда уже никто не
помнил. Ныне, как уже говорилось в главе 2 и будет рассмотрено далее,
наука возвращается к догеккелевым взглядам.
Роль А.Н. Северцова в понимании эволюции как перестройки онтогене-
за огромна: если до него связь эволюции и онтогенеза в основном служила
предметом восхищения, то он основательно исследовал вопрос и разъяс-
нил суть дела. Обосновав идею Мюллера о двух типах изменений онтогене-
за (уклонение от прежнего пути и надставка к прежнему онтогенезу), Север-
цев указал третий - архаллаксис (греч: начальное изменение), т.е. измене-
ние онтогенеза данного органа с самого появления его закладки в зароды-
ше. В этом случае о рекапитуляции говорить нельзя, а значит "основной
биогенетический закон” тут вообще не действует.
Рис. 22. Зародыши геккона (слева) и ужа на одном и том же этапе развития.
Уж обгоняет геккона по числу позвонков, а геккон ужа — развитием головы
Вот простой и ясный пример архаллаксиса: если сравнить зародыши
двух пресмыкающихся - геккона и ужа, то легко видеть, что с довольно ран-
них стадий развития уж (змея) демонстрирует большее количество сомитов
(основных сегментов тела), в том числе позвонков и ребер, чем геккон
(ящерица). Этот архаллаксис определяет всё дальнейшее развитие: напри-
мер, уж отстает от геккона в развитии головы (рис. 22).
Как крайний пример архаллаксиса приведу различие животных в типе
дробления - оказывается, на самых ранних стадиях развитие может резко
различаться у таксономически близких видов (рис. 23).
Здесь очевидно нарушение и “основного биогенетического закона”, и
упомянутого в п. 2-4 закона Бэра. Зато можно говорить об эквифинальности
по Дришу (п. 3-14). В части 3 мы увидим, что изменение типа дробления
важно для понимания эволюции млекопитающих. А пока видим, что разно-
образие дроблений рушит всю идеологию Геккеля: вместо «биогенетическо-
го закона» лучше говорить про «узел сходств», т.е. про единообразное раз-
витие зародышей всех позвоночных, начиная со стадии нейрулы и кончая
стадиями закладки осевых органов (подробнее см. п. 5-17).
183
к к л м н
шей. Соболев пришел
Рис. 23. Различие способов дробления
яйца круглых червей. Eustron-gylides
excisus (А - 3) имеет два варианта, схо-
дящиеся к общей картине на стадии 12
бластомеров; у Trichocephalus trichurus
(И - Н) путь единствен; как видим, он
приводит к совсем другой картине ста-
дии 12 бластомеров. Если бы класси-
фикация на самом деле проводилась
(как часто говорят) по способам разви-
тия, эти виды попали бы в разные фи-
лумы (типы)
Свое понимание связи онто- и
филогенеза позвоночных Северцов
в 1912-1936 годах развил в извест-
ную теорию филэмбриогенеза, где
главным механизмом эволюции бы-
ло как раз преобразование зароды-
к близким выводам на моллюсках, но напомню, что
родоначальником данного направления мысли был Жоффруа.
В теории филэмбриогенеза выявлено, кроме архаллаксиса, еще не-
сколько «модусов», т.е. способов, которыми смена онтогенеза порождает
смену форм организмов (эволюцию). Самый, на мой взгляд, удивительный -
неотения, т.е. размножение в невзрослой (например, личиночной) стадии.
Так, амбистома и аксолотль - два вида хвостатых амфибий, однако аксо-
лотль является личинкой амбистомы, способной к размножению. (Сам тер-
мин «неотения» старше: еще в 1885 г. его ввел немецкий эмбриолог и ан-
трополог Юлиус Кольман.) Поскольку у личинки органы недоразвиты, она
идеальна на роль «неспециализированного» (п. 3-10*) и тем самым, неоте-
нию можно рассматривать как способ поворачивать направление эволюции.
Мы обратимся к этому в п. 8-6.
К сожалению, в школе Северцова связь эволюции и онтогенеза изуча-
лась и изучается исключительно как преобразование форм. В последние
годы творчества Северцова расцвело совсем иное направление - физиоло-
гия развития, именовавшая себя сперва “механикой развития”; она искала
понимания физиологической природы развития зародышей. (О ней см.:
Светлов П.Г. Физиология (механика) развития. Л., Наука, 1978.) Северцов в
своей последней книге (изданной в 1939 г. посмертно) выразил восхищение
ее успехами, однако отметил с огорчением, что она совершенно игнорирует
тему эволюции, и высказал надежду, что «наступит время, когда механика
развития вплотную подойдет к вопросам эволюции» (Северцов А.Н. Собр.
соч„ т. 5. М„ 1949, с. 350).
184
4-13* Перемещение жоффруизма на Запад
Ждать пришлось недолго. Примерно тогда же, когда Северцов писал в
Москве эти слова, в Лиссабоне, на Международном съезде зоологов (1935
год) французский эмбриолог Поль Вентребер сделал доклад «Физиологиче-
ский эпигенез». Он утверждал, что эмбриология остается, как в XVII-XVIII
веках, преформистской наукой (см. пп. 1-14; 2-1), и предлагал обратить ее в
эпигенетическую посредством физиологии развития.
Как мы знаем из п. 3-14, зародыш проявляет при развитии черты
целостности. Шпеману, сформулировавшему в 1921 году это положение
(вслед за Дришем и другими), принадлежит замечательный результат: если
рассечь зародыш на две части, развиваться дальше будет только та,
которой достанется определенный участок - организатор. Шпеман был
уверен, что организатор запрограммирован генетически, и искал то
вещество, выделение которого является организующим сигналом. Поиски
привели к разочарованию: эту роль могли играть очень многие вещества, и
природа “того самого” вещества оставалась загадкой.
Вентребер предложил разрубить гордиев узел: поиск такого вещества -
ненужное наследие преформизма. Возможность запустить организатор са-
мыми различными веществами означает, что организатор - не структура и
не мишень, а «сцепление функций». По Вентреберу, онтогенез - это после-
довательность сцепленных между собой процессов и порождаемых ими
структур, причем сцепление процессов (он говорил - функций) ничем не
преформировано и являет собой «физиологический эпигенез», дающий ко-
манды «морфологическому эпигенезу». Отсюда следовал вывод, что и
строение организма не преформировано, а создается постепенно, так что
каждая стадия задана предыдущей (в частности, организатор появляется на
соответствующей стадии развития зародыша).
Об эволюции Вентребер тогда речи не вел, но вскоре занялся именно
ею (см. п. 5-4). Затем, уже в разгар Мировой войны, в оккупированном нем-
цами Париже вышла книга палеонтолога и историка науки Раймона Фюрона,
который расставил смысловые акценты. Изложив кратко позицию Вентре-
бера, Фюрон резюмировал: если онтогенез является переплетением взаи-
мозависимых процессов, то вмешательство самых разных факторов может
изменить его ход, а это - эволюция (Furon R. La paleontologie. Paris, 1943, p.
99-101). Оставалось выяснить, что это за факторы. Мы займемся ими в п. 5-
11. А здесь замечу следующее.
Для жоффруизма принципиально указание на то, что изменение разви-
тия зародышей происходит одновременно у значительной части самок дан-
ного вида под влиянием неблагоприятных изменений среды обитания. Эту
мысль подробно назвил в 1949 году бельгийский эмбриолог Альбер Дальк.
Резкую однократную перестройку зародыша он называл онтомутацией.
Как и Гольдшмидт, он полагал, что она вызвана реорганизацией хромосом и
185
окружающей их цитоплазмы, однако придавал больше, чем тот, значения
изменениям генов [Назаров, 2005]. Доказать это в те годы было невозмож-
но, и оба ученых подверглись жестоким нападкам со стороны СТЭ, что за-
тормозило эволюционную науку на 30 лет - первая работа В.Н. Стегния на
данную тему появилась в 1979 г. (см. п. 5-5*).
Это, впрочем, характерно для всех форм дарвинизма, что мы уже виде-
ли, и еще увидим: все попытки раскрыть реальные, а не воображаемые, ме-
ханизмы эволюции сперва грубо отвергаются как ненаучные на - том осно-
вании, что и так всё ясно. Затем, когда сопротивление выглядит уже смеш-
ным, прежде отвергаемые взгляды либо объявляются составной частью
дарвинизма, либо (что чаще) просто перестают упоминаться.
подчеркнув этим, что
Рис. 24. Хологенез; сле-
ва - позвоночного (W.
Zimmermann, 1953),
справа - цветкового (из
учебника: Г.17. Яковлев и
В.А. Челомбитъко. Бо-
таника. М., 1990)
В 1953 году немец-
кий палеоботаник Валь-
тер Циммерман придал
концепции связи онтоге-
неза с эволюцией на-
глядную графическую
форму винтовой линии
(рис. 24); само эволюци-
онное явление он назвал
словом хологения (от
греч. холос - цельный),
онто- и филогенез являют собою единый процесс,
трансиндивидуальный онтогенез. Как увидим в п. 5-17, хологения лежит в
основе нынешнего понимания сущностной эволюции организмов.
4-14. Дарвинизмы бывают разные
Чуть раньше книги Фюрона, в 1942 году, тоже в разгар войны, в Лондоне
вышел «Новый синтез» Джулиана Хаксли, обозначивший формальное рож-
дение СТЭ. Изложив в главе 1 идею естественного отбора и кое-что о ее ис-
тории, в главе 2 он высказал свое отношение к эволюции в целом: прогрес-
сивно то, что отобрано, и прогрессивные линии эволюции это «линии адап-
тивной радиации». Далее, как обычно в дарвинизме, почти вся книга изла-
гает мнения о роли отбора в формировании подвидов и видов, а в конце две
186
небольших главы бегло говорят об остальной эволюции - что она должна
идти так же. В этом смысле СТЭ явилась обычным дарвинизмом. В осталь-
ном в ней завершена традиция, которую со времен Вейсмана называли
неодарвинизмом и которую Филипченко назвал в 1923 году «левым крылом
дарвинизма»: она нацело свела изменчивость к случайным мутациям и слу-
чайным рекомбинациям.
В отличие от последующих классиков СТЭ, Хаксли еще был озабочен
каверзными вопросами, знал о ламаркизме, номогенезе и прочем, даже по-
началу изучал зародыши. Любопытно его рассуждение об органическом
отборе. Идею такого отбора высказал в 1895 году американский дарвинист
Джеймс Болдуин для объяснения удивительного параллелизма (уже помя-
нутого в п. 7) прижизненной и наследственной изменчивости. Наблюдается
он весьма часто. Так, "загар" наследствен у негров, а нам надо загорать ка-
ждый год заново. Вот Болдуин и допустил, что приспособление может воз-
никать при жизни, позволять организму сразу же выжить, а затем отбор по-
степенно выработает соответствующее наследственное изменение. Такой
отбор он назвал органическим, другие называли его совпадающим, но никто
не исследовал, возможен ли такой отбор на самом деле. (Впрочем, никто не
исследовал на деле никакой отбор.)
С появлением генетики стали говорить, что органический отбор
«заменяет модификации мутациями». Хаксли привел пример:
«Известно, что головная и платяная вошь человека так различны строением,
что им даны разные названия. Если, однако, головную содержать на теле, она
обращается в платяную за 4 поколения... Эти два типа постоянно общаются
путем миграции. Видимо, мы наблюдаем здесь первую фазу органического
отбора, когда основная часть различий еще модификационна (прижизненна -
Ю. Ч.). Но если отбор обеспечит лучшее приспособление, это приведет к бы-
строму расхождению наследственности этих форм».
Уоддингтон назвал (тоже в 1942 г.) органический отбор генетической
ассимиляцией, поскольку генотип как бы усваивает то, что возникло вне его.
Позже этот ход мысли был в СТЭ отвергнут. Так, американский зоолог
Эрнст Майр, классик СТЭ, суммируя мнения коллег, писал:
«Некоторые утверждают, что ненаследственные изменения подготавливают
почву для эквивалентных мутаций, однако для такого утверждения нет доста-
точных оснований. Способность фенотипа изменяться под непосредственным
воздействием внешней среды без мутирования сильно снижает давление от-
бора»; «введение отдельного термина “генетическая ассимиляция’" (Уоддинг-
тон) для обозначения накопления таких генов посредством отбора лишь за-
туманивает предмет обсуждения» {Майр Э. Зоологический вид и эволюция.
М., Мир, 1968, с. 128,484).
В самом деле: если уж организм поведенчески приспособился и выжил,
то полезным мутациям отобраться не проще (как думали Хаксли и другие), а
труднее. Беда в том, что своего объяснения самого факта параллелизма
187
прижизненной и наследственной изменчивости СТЭ не предлагает А факт
до сих пор служит опорой ламаркизма и номогенеза. Ссылки см. в моей ста-
тье в сб. [Кудрин].
В том же 1942 году вышла книга московско-киевского (в то время эва-
куированного вместе с другими академиками в Казахстан) зоолога И.И.
Шмальгаузена «Организм как целое в индивидуальном и историческом раз-
витии» (первое неполное ее издание появилось в 1938 г.).
В 30-е годы в СССР быстро укоренялась при поддержке власти гекке-
лева трактовка дарвинизма («правое крыло дарвинизма» по Филипченко),
вскоре перешедшая в «творческий дарвинизм» Лысенко, начисто отрицав-
ший внутривидовой отбор, зато целиком признававший НПС. Но Шмальгау-
зен сумел устоять, так сказать, в рядах “левых” и даже стать классиком на-
рождавшейся СТЭ. Это тем более удивительно, что суть дела была у него
местами вполне “правая”:
«Мы видим, что непосредственное модифицирующее влияние измененной сре-
ды, а также «упражнение» и «неупражнение» органов может привести и, ко-
нечно, приводит в новой обстановке к установлению новых, исторически нико-
гда не существовавших форм... Из семейства белок суслики живут на полях и
роют норы, белки перешли к лазанью по деревьям, а летяги лазают на деревьях
и перепархивают с ветки на ветку. Различная функция конечностей в этих слу-
чаях явно их преобразует... Функциональные изменения здесь должны играть
ведущую роль... Прямые наблюдения над прирожденным дефектом передних
конечностей у молодой собаки, благодаря которому она вынуждена была пере-
двигаться на одних задних конечностях, показали очень крупные изменения в
строении последних, в положении таза и т.п.» (Шмальгаузен И. И. Организм как
целое в индивидуальном и историческом развитии. М., Наука, 1982, с. 157).
В этом поразительном ламаркистском признании последняя фраза ну-
ждается в пояснении: при постоянном перемещении на задних лапах собака
вырастает похожей на тушканчика или на кенгуру; и хотя такая форма тела
не наследуется, но встает вопрос: если столь сложное изменение достигну-
то в ходе одного онтогенеза, то естественно вспомнить Жоффруа и его
идею эволюции путем смены онтогенезов. Не шла ли реальная эволюция
прямоходящих зверей по Болдуину? Мы вернемся к этой теме в п. 5-11.
Еще в 1940 году Шмальгаузен основал, взамен прекращенных журна-
лов Кольцова, «Журнал общей биологии» (ЖОБ), где сразу поместил три
своих программных статьи. Третья называлась «Изменчивость и смена
адаптивных норм в процессе эволюции» и завершала то, что выше мы рас-
смотрели как системное понимание отбора. Отбор, оказывается, сравнивает
не полезные признаки, а целостные функциональные системы, так что его
итогом является переживание тех, чья «норма» наиболее «адаптивна». От
критиков не могло укрыться, что это - не дарвинизм:
«...тогда придется сделать вывод, что ведущим фактором в развитии организ-
мов является какой-то фактор, связывающий в одну единую систему весь ор-
л
188
ганизм вплоть до мелочей, как например рисунки на пальцах; получается
номогенез» [Любищев, 1982, с. 108].
Конечно, подобных слов тогда печатать было нельзя - это был бы пря-
мой донос, но Любищев тогда работал у Шмальгаузена в Киеве и в какой-то
форме должен был это шефу сказать, ибо мыслей своих никогда не скры-
вал. Так или иначе, но больше Шмальгаузен об этом прямо не высказывал-
ся, хотя саму идею пропагандировал до конца жизни. Она видна и в упомя-
нутой его книге; переход к лазанию или прямохождению - это смена «адап-
тивных норм» (вопрос: адаптивны ли они на самом деле? - никогда не ста-
вится). Замечу: если нужная для этого перестройка тела фактически идет
прижизненно, то ее унаследование надо как-то объяснять, а не полагать
(как позже у Майра), что проблемы тут нет. Вот Шмальгаузен и показал, как
на языке дарвинизма “объяснять", не ища понимания.
Для понимания тут нужен материал части 2 (см. Дополнение 6).
4-14* Адаптация ламаркизма к дарвинизму
Если результат упражнения наследуется, это ламаркизм, но если ска-
зать, что он стабилизируется (а “упражнение” употребить в кавычках), это
будет всего лишь введение нового термина. Шмальгаузен ввел этот термин
- стабилизирующий отбор (естественный отбор, который «идет на основе
селекционного преимущества самой приспособленной нормы перед всеми
отрицательными от нее уклонениями» - Шмальгаузен И.И., 1982, с. 167).
Одно из преимуществ «самой приспособленной» состоит, по Шмалыаузену,
в автономизации развития особи от условий внешней среды. Суть ее в
том, что она наследственно воспроизводит то, что «менее приспособлен-
ные» реализуют каждый раз заново, поведенчески и физиологически. (По-
ясню; негр в Африке более приспособлен, чем белый, который вынужден
заново загорать.) Если так, то превращение ненаследственного в наследст-
венное есть селективный механизм, что и требовалось доказать.
Давно отмечено, что тут совершены две грубых подмены. 1) Термин
«стабилизирующий отбор» введен для обозначения вполне реального, но
мало интересного явления - преимущественной гибели тех особей, которые
несут уклонение от нормы, - а используется для обозначения совсем иного
явления, весьма важного для теории, но никем не наблюдавшегося: авто-
номизации развития (преимущественной гибели тех, кто выглядит и устро-
ен точно так же, но у кого этой автономизации нет). 2) Применена подмена
основания - логическая ошибка, о которой говорилось в п. 3-2: обещано
доказательство наличия нового типа отбора, а на деле этот тип
постулирован. На это позже и указал Майр.
Неужели Шмальгаузен, эрудит и интеллектуал, не увидал столь оче-
видных ошибок? Думаю, он всё видел, но не имел иного пути поведать миру
свою идею: то, что сперва найдено индивидом в ответ на изменение среды,
затем реализуется наследственно, автономно (независимо от среды). Так,
189
хождение на задних лапах возможно для четвероногого и даже может, как
мы видели, приводить его к «двуногому» облику, но эволюционным дости-
жением становится лишь тогда, когда детеныш прямо развивается по «дву-
ногой» программе. Идея автономизации путем отбора была, по-моему, по-
следней крупной идеей в дарвинизме.
Крупная - не значит верная в смысле подтвержденное™ фактами. Ни-
каких фактов в данной “теории” нет, стабилизирующий отбор как автономи-
зирующий фактор был введен в форме постулата и таковым остался. Уточ-
ню: стабилизация есть факт, такой же факт, как борьба между самцами, по-
влекшая идею полового отбора (п. 3-3), но связь этих явлений с отбором
ненаправленных наследственных вариаций никем не отмечена. В “опытах
по изучению стабилизирующего отбора” (например: Зеликман А.Л. И ЖОБ,
1946, № 4; Берг Р.Л. // БЖ, 1958, № 1) наблюдалась стабилизация призна-
ков, а отбор и автономизация додумывались.
Самую доходчивую характеристику я услыхал лет 25 назад от энтомо-
лога Ю.А. Захваткина: «Вся теория стабилизирующего отбора - теория
адаптации. Адаптации ламаркизма к дарвинизму». Подробнее см. мою ста-
тью: Чайковский Ю.В. Стабилизирующий отбор, или святость веры И [Куд-
рин}, - где доводы Шмальгаузена характеризованы как сознательная паро-
дия, ясно говорящая запретную правду тем, кто владеет материалом и спо-
собен читать между строк. Увы, пародия стала священным текстом.
Запад эту идею Шмальгаузена игнорирует.
Для дальнейшего очень важно иметь в виду, что стабилизирующий от-
бор - такая же фикция, как и остальные формы отбора, рассмотренные на-
ми ранее (естественный и половой). Важно по той причине, что в россий-
ском неодарвинизме бытует мнение, будто во всех формах отбора можно
сомневаться, но не в стабилизирующем, ибо он «очевиден». На самом деле
сторонниками этой точки зрения обоснованы лишь два факта:
1) повышенная смертность организмов, очень сильно уклонившихся от
нормы, т.е. уродов. Никакой стабилизации уцелевших из этого не следует:
никто не смог, да и не пробовал, показать в опыте, что гибель одних улуч-
шает других (это, впрочем, относится к любому отбору);
2) стабилизация физиологических и поведенческих свойств организмов,
попавших в новые условия. Но она протекает столь быстро (иногда - в 1-2
поколения, см. пп. 4-18, 4-19), что ни о каком отборе говорить нельзя.
Объединение этих абсолютно различных по сути явлений одним тер-
мином и создало иллюзию, что наследственный стабилизирующий процесс
может быть следствием отсева уклонений от нормы.
4-14** Крайние варианты, в том числе лысенковщина
В 1943 году, пока в Европе бушевала война с фашизмом, двое бежав-
ших от фашизма ученых, Макс Дельбрюк (из Германии) и Сальвадор Лурия
(из Италии), поставили в США эксперимент, обеспечивший победу левому
190
(по Филипченко) крылу дарвинизма. Они подвергали колонию бактерий дей-
ствию смертельной дозы вируса и убедились, что выживают лишь те из них.
которые в этот момент уже имеют к вирусу устойчивость. Сама же устойчи-
вость возникала у отдельных членов колонии случайным образом. Вот как
писал широко известный учебник:
«как рождение генетики датируют 1865 г., когда появилась статья Менделя,
так и рождение генетики бактерий можно датировать 1943 г., когда Лурия и
Дельбрюк опубликовали статью, озаглавленную «Мутации, в результате ко-
торых бактерии, чувствительные к вирусу, приобретают устойчивость к не-
му». Лурия и Дельбрюк... впервые показали, как следует ставить опыты, как
обрабатывать результаты этих опытов и, что самое главное, как следует рас-
суждать, чтобы получить осмысленные и однозначные результаты»
(Г.Стент. Молекулярная генетика. М., Мир, 1974. с. 133).
В самом деле, оба вирусолога показали идеальное владение методикой
СТЭ (заложенной Мензбиром и Четвериковым), в частности, полным отри-
цанием неугодных фактов, и поставили опыт так, чтобы ничего нежелатель-
ного произойти не смогло. «Однозначные результаты» были, если говорить
прямо, методическим подлогом; но их приняли на ура.
Если внимательно рассмотреть опыты, то окажется, что сама их поста-
новка лишала возможности получить что-либо, кроме «доказательства»
случайности мутаций. В частности, в них всегда использовались столь вы-
сокие концентрации инфекций и ядов, что исключалась возможность фи-
зиологического приспособления, хотя вроде бы ставилась цель - проверить,
идет ли таковое приспособление и носит ли оно наследственный характер.
Позже Лурия и Дельбрюк были увенчаны нобелевской премией за ра-
боты по строению вирусов, но запомнились обществу именно как авторы
названного опыта. Сам он явился последним решительным аргументом в
пользу СТЭ, согласно которой единственным движущим фактором эволю-
ции является естественный отбор случайных ненаправленных вариаций; а
главным видом вариаций были сочтены ненаправленные мутации. Ни опы-
ты советских генетиков, ни появившиеся вскоре сходные данные А. Буавэна
(Франция) ситуацию изменить не смогли. (Сам же опыт Лурии - Дельбрюка
был опровергнут лишь через 45 лет - см. п. 5-1 ***.)
Пока на Западе утверждалась власть «левого крыла дарвинизма», со-
ветская биология изнывала под властью его «правого крыла». В 1948 году в
Москве прошла печально знаменитая Августовская сессия ВАСХНИЛ (Все-
союзная Академия сельскохозяйственных наук имени Ленина; ее сперва
возглавлял Вавилов, а с 1938 года - Лысенко), официально утвердившая
победу Лысенко над генетиками. Победа была в том, что его поддержал
Сталин. Лысенковщине посвящена огромная литература (см. например:
[Воронцов; Колчинский]), и мне остается заметить, что на сессии крайне
правый вариант дарвинизма праздновал победу над крайне левым. Там не
было слышно никаких других течений эволюционизма.
191
Однако Лысенко безраздельно правил всего 4 года и 4 месяца. В де-
кабре 1952 года по указанию Сталина против Лысенко выступили сразу два
журнала - БЖ и Бюллетень МОИП. Оба тогда редактировал ботаник В.Н.
Сукачев, яркий ученый и порядочный человек, чье имя было использовано
партийным аппаратом для прикрытия очередной идеологической кампании
(обзор дискуссии см.: БЖ, 1954, N№ 1, 2). Она могла повлечь увольнения и
аресты, и Лысенко лучше всех знал это. Он даже не пытался спорить, но тут
его спасла смерть Сталина. (Подробнее см.: Ю.В.Чайковский. О классиках и
комментаторах // ВИЕТ. 1992, № 4.)
Еще 10 лет, при Хрущеве, Лысенко задавал тон в советской биологии,
но уже не был в ней единственным хозяином, и генетика стала оживать. В
июле 1953 года Любищев уже отмечал:
“В настоящее время многие практические ошибки исправлены, но зажим в
области теории продолжает существовать, хотя имеются некоторые симпто-
мы ослабления” (Любищев А.А. В защиту науки. Л., Наука, 1991, с. 26).
Итак, безраздельная монополия Лысенко длилась всего 52 месяца. А
вот монополия его противников длится более 40 лет и лишь с падением со-
ветской цензуры перестала быть безраздельной. Могут сказать: «И слава
Богу!», но не будем спешить - любая монополия для науки гибельна.
Будучи как ученый слаб, предвзят и даже просто лжив, Лысенко старал-
ся произвести впечатление серьезного ученого обширными цитатами из от-
вергаемых им трудов, иногда воспроизводя небольшие статьи противников
почти целиком. Его «Агробиология» печаталась огромными тиражами и во-
лей-неволей служила просвещению масс.
Наоборот, с 1960-х годов установилась прискорбная традиция не знать
об эволюции ничего, кроме своих собственных взглядов. Исключение со-
ставляли очень немногие, например, Любищев и Мейен.
Влияние Лысенко на генетику и эволюционизм длилось целых 30 лет
(1935-1964), так что тогда выросло целое поколение ученых, видевших в
нем лидера. Эти ученые получали результаты, в том числе и интересные,
однако с изгнанием Лысенко попавшие под запрет. Попробуйте найти в ру-
ководствах по биологии что-нибудь о вегетативной гибридизации, которой
стал знаменит И.В. Мичурин. Пусть Лысенко и раздул ее роль в садовом се-
лекционном деле, но само явление существует и очень важно для понима-
ния морфогенеза растений. Подробно о ней см: [Любищев, 2006].
Иначе тогда (в 1960-е годы) вряд ли и быть могло, но давно пора с этим
кончать, пора учиться отличать научные аргументы от политических - иначе
мы вперед не двинемся Вот один пример.
4-15. Эффект группы
Термин эффект группы ввели французские зоологи Пьер-Поль Грассэ
и Реми Шовен в 1944 году, хотя сам феномен был описан ранее. У них он
означал «психофизиологическую реакцию отдельной особи на присутствие
192
других особей своего вида». Наиболее он известен и поразителен у саран-
чи: одиночные особи пассивны и безвредны, соединенные же в стаю -
опаснейший агрессивный враг полей.
По сути не менее поразителен тот факт, что одиночная особь пчелы не
живет и трех суток. В коллективе же пчёлы совершают чудеса, в том числе
проявляют разумное (не запрограммированное) поведение. Так, пчёлы вы-
таскивают иглу, воткнутую в сот, а если это не удаётся, перестраивают сот
так, чтобы замуровать ее в стенку (Панов Е.Н. Бегство от одиночества. М.,
Лазурь, 2001, с. 501). Далеко не целиком запрограммировано и их поведе-
ние при роении (там же, с. 480).
Можно отнести к эффекту группы и большинство фактов, собранных
Кропоткиным. Ленинградский ботаник Ю.В. Титов распространил понятие
“эффект группы” на растения [Титов]. В этой книге есть список литературы,
в том числе иностранной; об эффекте группы см. также: БЖ, 1972, № 5, с.
434; БЖ, 1975, № 9, с. 1237; 1351.) Титов писал:
“Механизмы взаимоотношений у растений и животных существенно разли-
чаются У последних, особенно у высших, преобладают информационные
формы связи между особями. Однако эффекты взаимодействия между расте-
ниями и между животными ... в основных чертах сходны. Поэтому определе-
ние эффекта группы у растений можно пока принять аналогичным ... - как ре-
зультат их взаимодействия, проявляющийся в отличительных особенностях
жизнедеятельности растений в группе по сравнению с изолированными друг
от друга растениями” [Титов, с. 16].
Этот факт сходства итогов взаимодействия совершенно несходных ор-
ганизмов загадочен и будет еще предметом нашего внимания, а здесь надо
сказать о самом эффекте группы. Особи одного вида далеко не всегда кон-
курируют - даже если есть острая нехватка ресурса. Об этом писали давно.
Вот очень яркий пример.
В 1953 году в Бюллетене МОИП (№ 2) появилась статья лесовода Ю.П.
Бялловича «К вопросу внутривидовых и межвидовых взаимоотношений» с
анализом уникального эксперимента, поставленного войной. Семена раз-
личных видов деревьев были посеяны на прямоугольных делянках чрез-
мерно густо, в расчете на прореживание, но война спутала планы, и дерев-
ца 10 лет росли сами по себе. Почти все делянки приняли характерную
форму «ванн» - равно угнетенные деревца одного роста по всей делянке
(«дно») и резкое повышение роста на краях, независимо от того, была ли
рядом поляна или же «ванна» другого вида. Что-то вроде стражи по грани-
цам. (Как тут не вспомнить Кропоткина.)
Этот факт, для практики явно важный, есть важный частный случай
“эффекта группы”, точнее, краевой эффект [Титов, с. 70]. Из-за отсутствия
информации Титов ничего не знал о статье Бялловича (хотя Юрий Петро-
вич, самый крупный ученик Сукачева, даже заслужил у коллег прозвище
«Юпитер»), как ныне мало кто помнит книгу Титова.
193
Бяллович не смог объяснить этот феномен обычными теориями и сде-
лал вывод - популяция являет собою некое функциональное единство, по-
этому конкуренция между особями одного вида выражена гораздо слабее,
чем между особями разных видов: «конкуренция преобладает на стыках»,
«взаимопомощь же ей постоянно сопутствует». Другими словами, особи од-
ного вида поддерживают друг друга и направляют свои силы на преодоле-
ние экспансии особей чужого вида.
В статье нет речи ни о Лысенко, ни об отрицании внутривидовой борь-
бы, но ее «правый» смысл был очевиден, а о Кропоткине никто тогда не
помнил. Редактор Сукачев был умен, честен и либерален, он не отклонил
статью ученика, но не удержался от упреков автору в «недостаточно обос-
нованных выводах» и в «телеологии» (т.е. в признании стремления вида к
некоей цели). Какой вывод из данных фактов был бы более обоснован, он
не сказал. Казалось бы, очевидно: Бяллович переоткрыл подвижное равно-
весие Еленкина. Но за 32 года после статьи Еленкина (и за 11 лет после его
самоубийства) о нем тоже забыли.
В 1959 году Бюллетень МОИП посвятил номер столетию «Происхожде-
ния видов». Там Сукачев, наряду с обычными юбилейными статьями, по-
местил свою необычную статью о борьбе за существование. Сперва заяв-
лено, что понятия «борьба за существование» и «взаимопомощь» неудачны
(добавлю: удачным было бы взаимодействие) и что Дарвин идею борьбы
за существование ничем фактически не обосновал. Автор решил навести
здесь порядок и получил замечательные результаты.
В опыте с ивами двух видов им было выяснено, что
«дарвиновское положение о более напряженной внутривидовой конкуренции
не имеет места» (Сукачев В.Н. Новые данные по экспериментальному изучению
взаимоотношения растений // Бюллетень МОИП, отд. биол.. 1959, № 4, с. 38).
Дело в том, что ни расы, ни виды нельзя поделить на лучшие и худшие:
например, при тесной посадке форма А вытесняет форму В, а при разре-
женной - наоборот. Добавлю: поэтому одна форма другую никогда целиком
не вытеснит; а это значит, что, как и в расчетах Нортона, борьба за суще-
ствование оказывается фактором экологии, но не эволюции. При
всём показном почтении к памяти Сукачева, этот его вывод, противореча-
щий дарвинизму, упоминать не принято.
При чрезмерной густоте посадки растения не могут проявить все свои
потенции (это значит, добавлю я, что отсутствует или резко ослаблена воз-
можность выявлять полезные вариации). Например, семена, собранные с
буйно разросшихся особей, сами дают растения с лучшими семенами, но
выявить это при тесной посадке невозможно, поскольку нет буйно разрос-
шихся особей (с. 45).
Кроме приведенных Сукачевым, есть и другие факты. Некоторые дере-
вья, например ель и бук, в молодости склонны образовывать мертвопок-
ровные сообщества', деревца стоят столь густо, что под ними ни травинки
194
не растет; до стадии размножения доживает едва ли одна особь из сотни,
зато вся биомасса данной площадки достается единственному виду. Особи
не конкурируют друг с другом, а отвоевывают у других видов площадку для
одной-единственной особи, которая даст потомство. Детальное исследова-
ние ёлочек показало, что еще до смыкания крон загущение участка вызыва-
ет, как и следовало ожидать, «резкое снижение прироста деревьев по высо-
те и и диаметру ствола», причем деревца начинают резко различаться на
господствующие и угнетенные. Это понять легко.
Но дальше идет неожиданное: «После смыкания крон... наблюдается
некоторое увеличение прироста деревьев в высоту и по диаметру». На рис.
25 показана динамика этого процесса: момент смыкания крон - наихудший
для популяции (рост и толщина падают втрое), но затем средние по ней па-
раметры вновь растут: средняя толщина на треть, а средний рост даже
вдвое. Ботаники, описавшие этот факт, назвали его “сменой внутривидовой
конкуренции на взаимную
толерантность” [Титов, с.
73].
Рис. 25. Изменение прироста
ели (в % от прироста свободно
растущих) в ходе смыкания
крон: 1 - по высоте, 2 - по диа-
метру (из [Титов, с. 73])
Хотя почти все особи тут
угнетены, но конкуренции в
ее привычной форме тут нет,
и “борьба за существование”
выражается в сотрудничест-
ве угнетателей и угнетенных. Это обеспечивает полную монополию данного
вида на данной площадке, поэтому для дарвиновой схемы расо- и видооб-
разования места нет. Со временем останутся лишь крупные особи, но и они
не конкурируют, ибо в юности разделили между собой участки (благодаря
массе угнетенных особей, затем погибших).
Нет господства конкуренции и в межвидовых отношениях:
“Пространственно-временная мозаичность и обособление группировок растений
отражают комплементарную природу большинства естественных сообществ.
Комплементарные сообщества в отличие от конкурентных состоят из видов ...
дополняющих друг друга” [Титов, с. 12].
Наконец, растения, как и животные, размножаются отнюдь не по Маль-
тусу: в случае неблагоприятных условий многолетники перестают плодоно-
сить, двухлетники нередко идут по тому же пути (становятся многолетника-
ми), и только однолетники ведут себя двояко - у одних видов основная мае-
195
са престает размножаться (т.е. популяция почти вымирает), а у других дис-
комфорт вызывает ускорение вегетации и плодоношения [Титов, с. 78-80].
Последнее можно счесть мальтузианством, тогда как всё вместе - эффек-
том группы.
Об этом в стандартной экологии, опирающейся на дарвинизм, можно
прочесть не больше, чем у Лысенко - о генах. Но нам важно не уязвить оп-
понента, а понять причину замалчивания. Редкие упоминания эффекта
группы наводят на мысль, что авторы просто не понимают, о чем пишут.
Например, подают это явление как оптимизацию физиологических процес-
сов или как один из механизмов регуляции численности популяции.
Для объяснения эффекта группы было предложено «фитогенное поле»,
природу которого признали концентрационной и электрохимической для
почвы, а для воздуха - электромагнитной. Тип взаимодействия растений
охарактеризован как информационный, а не энергетический, причем уста-
новлено, что растения одной популяции выравниваются по всем сущест-
венным параметрам (размеры, сроки роста и т.д.), чем предположительно
обеспечивается выравнивание полей.
Однако чем и как производится различение растениями своего поля от
чужого, не сказано. В п 4-16 мы поговорим про биологическое поле, а пока
заметим, что к эффекту группы следует отнести и столь же загадочный фе-
номен синхронного плодоношения яблонь и других плодовых деревьев: все
знают, что яблони цветут через год, но никто не знает, почему они это де-
лают сразу почти все по всей округе.
В п. 6-15 эффект группы будет сопоставлен с другими загадочными яв-
лениями, общим в которых является феномен узнавания.
4-15* Национальная наука
Шовен, соавтор эффекта группы, видел причину непонимания эффекта
в национальном характере науки:
“любое животное реагирует на приближение сородичей, изменяя не только
характер своего поведения, но и физиологию... Это так называемые эффекты
группы, которыми особенно интересуются французские исследователи. Круг
их интересов лежит довольно далеко от интересов американских ученых, ко-
торые выращивают обширнейшие популяции вообще всех насекомых - вре-
дителей зерна и подсчитывают их число - подход чисто статистический”
(Шовен Р. Мир насекомых. М., 1970, с. 90).
Здесь стоит напомнить и ту давнюю мысль, что ранний дарвинизм -
чисто английское явление (п. 3-7). Мысль Шовена верна, но мне видится
более важной другая причина, им тоже упомянутая.
По Шовену, эффектом группы демонстрируется «не дарвиновская, по
меньшей мере не предусмотренная Дарвином, система регуляции, посколь-
ку тут не имеется в виду ни отбор, ни борьба за существование» (с. 96-97).
Точнее, эффект группы говорит против мальтусовой схемы, а она в те годы
196
царила в США безраздельно, да и в других странах преобладала. Наоборот,
во Франции теоретическая мысль до 1980-х годов во многом держалась ла-
маркизма с его вниманием к активности особей.
В п. 2-1 мы уже говорили о национальном характере науки. На эту тему
есть прекрасная брошюра: Линник Ю.В. Русская биология. Петрозаводск,
Музей космич. искусства им. Рериха, 2001.
Автор, поэт и натурфилософ, живущий в Карелии, опирается на идущую
от Еленкина традицию «без малейшего шовинистического оттенка» выяв-
лять национальную компоненту науки. Он вспоминает, что, по Еленкину
(1909 г.), немецкая биология «устремлена к единству», английская ориенти-
рована на «трезвый позитивизм, чуждый метафизики», французская содер-
жит «элемент фантастичности», а русской свойственна (в пересказе Линни-
ка) взаимодополнительность механицизма и витализма. Почти век спустя
Линник пытается придать этой “русской биологии” платонический смысл
(идея реальнее, чем ее частные воплощения), причем ссылается на многих
из цитированных выше (и ниже) наших биологов. При всей ее многоплано-
вости, русская компонента биологии определяется, по Линнику, вниманием
к симметрии и красоте природы. Далее мы не раз встретимся с таким ходом
мысли.
Основная часть эволюционистов работает в безликом безнациональ-
ном стиле, но запоминаются не они, а те немногие, кто имел собственное
лицо; лицо это обычно несет черты национальной науки. Например, в Рос-
сии всегда был силен жоффруизм, а на Западе он до недавних пор был со-
всем слаб (даже термина для него там нет). В 1930-е годы эмбриолог И.И.
Ежиков высказал идею эволюции насекомых, где ведущую роль играли уд-
линение и сокращение эмбриональных стадий развития. На этой основе
зоолог А.А. Захваткин развил концепцию эмбрионизации (удлинения заро-
дышевых стадий) для всех животных:
“эмбрионизация является ведущей линией, столбовой дорогой прогрессивно-
го развития также и у позвоночных”; она охраняет зародыш “от преобразую-
щего действия естественного отбора”, создает “явления, описываемые под на-
званием биогенетического закона” (Захваткин А.А. Сб. научных работ. М-
1953, с. 376-377).
К той же традиции можно отнести прекрасную работу Н.Ю. Сахаровой
по сравнительной эмбриологии (п. 8-12*). Мы вернемся к теме эмбриониза-
ции в п. 5-11*.
4-16. Биологическое поле
В п. 4-13 у нас была речь о том, что Шпеман искал вещество, органи-
зующее развитие зародыша, и не нашел его. Вентребер делал из этого вы-
вод, что развитие ничем не преформировано, а сам Шпеман решил иначе:
развитие преформировано, но преформирующий агент - не вещество, а
биологическое поле. Это понятие разные биологи вводили по-разному,
197
общим же было то, что таким образом хотели описать феномен целостно-
сти организма, оставаясь в рамках материализма.
Первым, насколько знаю, понятие поля ввел в 1910 году немецкий ци-
толог Теодор Бовери. Для него поле было чем-то механическим, и он пы-
тался понять его, крутя зародыши на центрифуге или сдавливая их покров-
ным стеклом. Вскоре (1917) американский эмбриолог Чарлз Чайлд ввел по-
нятие градиента, близкое понятию поля. По Чайлду, существует некая ска-
лярная величина, характеризующая физиологическую активность как заро-
дыша, так и взрослого животного, она убывает вдоль каждой его оси сим-
метрии - это и есть градиент. Его легко наблюдать на низших животных: у
них головная часть легче включает внешний краситель и она же первая
умирает (“рыба гниет с головы”). Ленинградский эмбриолог П.Г. Светлов ре-
зюмировал:
“физиологический осевой градиент - это и есть свойство, которое обеспечи-
вает целостность организма и связь между отдельными его частями” (Свет-
лов П.Г. Физиология (механика) развития. Л., 1978, т. 1, с. 232).
(У него же о Бовери: т. 2, с. 16.) От этих положений будем двигаться.
1) Для Шпемана биополе было полем химических концентраций. Это
направление мысли понемногу превратилось в поиск веществ, вызывающих
деление соответствующих клеток зародыша. Замечательного результата
тут в 1936 году достиг эмбриолог Г.В. Лопашов: помещая убитые спиртом
зачатки глаза лягушки в гасгрулу тритона, он наблюдал формирование за-
чатка глаза; поскольку мертвый зачаток развиваться не мог, налицо ясное
свидетельство химического воздействия (индукция).
За последующие 60 лет было выяснено, что эти вещества суть короткие
белки. Индукция существенна в начальной части эмбриогенеза, которая
«заканчивается возникновением терминальных клеточных типов, из которых
в основном и состоит тело животных»; у позвоночных этих типов клеток око-
ло двухсот; в последующих процессах «индуцирующие факторы не участ-
вуют» (Лопашов Г.В., Земчихина В.Н. Основные факторы и периоды индук-
ционных процессов в развитии животных // УСБ, 2000, № 6). К сожалению
(для нашей темы), линия работ Шпемана - Лопашова выродилась в чисто
химическую, и о поле как факторе целостности в ее рамках не вспоминают.
Лопашов установил «закон минимальной массы», нужной для индукции,
провел пересадку клеточных ядер, что привело в наши дни к успехам кло-
нирования животных (Корочкин Л.И. Георгий Викторович Лопашов. К 90-
летию // ЭГ, 1167).
2) Другая линия породила теорию биополя как организатора целостно-
сти. В 1922 году петроградский биолог А.Г. Гурвич ввел идею поля чисто
Формально:
“Участие каждой клетки в эмбриогенезе определяется не свойствами самой
клетки и не взаимодействием с соседними клетками, а общим для большого
клеточного комплекса ... фактором. ... Такая формулировка участия этого
198
надклеточного фактора исчерпывает для нас всю его сущность и реальность”
(Гурвич А.Г. Теория биологического поля. М., 1944, с. 11).
Такое понимание поля позволило описать некоторые стадии некоторых
процессов эмбриогенеза и в нескольких случаях даже записать формулы,
близкие к реальности (с. 14-21), но разочаровало автора.
3) В 1930-х годах Гурвич обнаружил в опыте митогенетические лучи,
т.е. ультрафиолетовые лучи, вызывающие митоз. Это побудило его в 1944
году изменить взгляды и ввести биополе как клеточное (в противополож-
ность прежнему его пониманию как надклеточного), причем электромагнит-
ное излучение он понимал как одну из форм его проявления. Прежнее поле
понималось теперь как совокупность клеточных, и это вполне естественно
для физиолога: если зародыш состоит из клеток, то его поле должно состо-
ять из клеточных полей. Напомню, что для объяснения эффекта группы бо-
таники позже вводили фитогенное поле (п. 4-15), и они тоже видели его
электромагнитную форму как частный случай.
Новое видение биополя позволило Гурвичу
“исходя из единого принципа непротиворечиво объяснить глубинные меха-
низмы основных жизненных проявлений: морфогенеза, обмена веществ, кле-
точного деления, наследственности, изменчивости, процессов их реализации.
Многие следствия теории биологического поля подтверждаются эксперимен-
тально” [Воейков, 1998, с. 301].
С удивительной проницательностью Гурвич при этом описал явления,
которые в наши дни были открыты как диссипативные структуры (о них см.
далее, п. 5-14) и как внутриклеточный лазер (Гурвич А.Г. Принципы анали-
тической биологии и теории клеточных полей. М., 1991, с. 5, 218).
Однако даже близкие друзья А. Г. Гурвича отказались принять новый
вариант поля. Так, в 1945 году Любищев писал Гурвичу:
“Никак не пойму: каким образом клеточные поля, не связанные никаким об-
щим надклеточным фактором, могут образовать закономерное распростране-
ние системы...” [Любищев - Гурвич, с. 167].
А в 1957 году Любищев вспоминал:
“В разговорах с Беклемишевым и Смирновым мы все трое пришли к едино-
душному суждению, что причиной отказа от старого понимания поля являет-
ся именно то, что к морфологии у Александра Гавриловича нет настоящего
интереса” (там же, с. 43).
Удивительнее, что от нового понимания поля отказался еще один друг
Любищева, эмбриолог Светлов, человек физиологической ориентации: по
его мнению, как раз старые взгляды Гурвича «имеют наибольший эмбрио-
логический интерес» (Светлов П.Г, т. 1, с. 240).
В 1989 году, через 35 лет после кончины Гурвича, германский физик
Фриц-Альберт Попп установил для одного типа биополя его природу - это
оказался ультрафиолетовый лазер ничтожной мощности. Этим биополю
был, наконец, придан научный статус, и сомнения в его существовании поч-
199
тй прекратились (их продолжают лишь те, кто не знает новых работ). Но на
смену упрекам пришло молчание, и это вновь напоминает нам, что науку
движут далеко не только факты, а если затронуто мировоззрение, то - от-
нюдь не факты. См. Дополнение 2.
4) Вопрос о природе поля не может быть решен без обращения к при-
роде силы или сил, образующих поле. Здесь огромную роль сыграл прие-
хавший в Москву из Венгрии биофизик Эрвин Бауэр, хотя он и не говорил
сам про поле. По Бауэру, главное отличие живого состоит в способности на-
капливать химическую энергию не только в форме химических связей, но и
в форме изменения структур и, прежде всего, конфигурации молекул. (Воз-
никшая вскоре квантовая химия позволила уточнить мысль Бауэра: речь
должна идти не столько о разных конфигурациях, сколько о разных энерге-
тических уровнях. Именно они определяют специфику биополей [Воейков,
1998].) Здесь Бауэр нам интересен тем, что, как установлено в наши дни,
энергией поля Гурвича естественно считать структурную энер-
гию Бауэра, о чем мы узнаем в главах 5 и 7.
Бауэр связал мутабильность с нарушением физиологии организма, чем
опередил идею генетического поиска (о ней см.п. 5-13), разделил понятия
жизни и размножения, чем мы воспользуемся в гл. 7 (Бауэр Э.С. Теоретиче-
ская биология. М.-Л., 1935, с. 197, 138) и мн. др. Он возражал против биохи-
миков, полагавших, что «существует столько ферментов, сколько они нахо-
дят реакций»; они «еще вынуждены предположить существование чудесно-
го, гармонического совместного действия самых различных ферментов» (с.
102). Гурвич также полагал, что «все внутриклеточные ферменты просто
обломки единого действующего комплекса» [Любищев - Гурвич, с. 179].
Этот взгляд еще ждет своего исследователя.
В 1937 году Бауэр был арестован и расстрелян, книга же его попала под
запрет. Но главная причина долгого замалчивания его взглядов была в том,
что они намного опережали эпоху. Так было всегда со всеми, но в ту эпоху
это заметно особенно. Похожа судьба мыслей Любищева.
4-17. Феномен Любищева
Александр Александрович Любищев (1890-1972), как и все еретики, по-
сле 1930 года не мог работать в теории и ушел в практику, став лучшим зна-
током проблемы насекомых-вредителей. Его работы перевернули всю про-
блематику борьбы с вредителями посевов (Наумов Р.В. И [Системность...]).
Одна деталь: интуитивно очевидно, что падение урожайности должно моно-
тонно расти с ростом доли объеденных листьев, но Любищев показал иное
- падение урожая начинается при поражении более четверти площади ли-
стьев, а до этого он даже растет за счет интенсификации фотосинтеза в по-
врежденных листьях.
Из работ Любищева по насекомым-вредителям выяснилось, между
прочим, что подлинным бичом земледелия чаще оказываются не на-
200
секомые, а чиновники. Разумеется, такой вывод не мог остаться без вни-
мания, и Любищев, как он сам говорил, «едва не угодил на казенные харчи».
Хотя его статья (1933) и была использована для изменения методов борьбы
с вредителями, сам он был в 1937 году уволен и чуть было не был аресто-
ван как вредитель (Смирнов Е.С. К 75-летию А.А. Любищева // ЭнО, 1966, №
1). Спасло его то, что его главный гонитель сам был арестован и Любищев
стал выглядеть жертвой “врага народа”.
Не имея выходов в печать, Любищев много писал. В 1946 году, живя и
работая в Киргизии, он написал большую статью «Проблема целесообраз-
ности», где показал общенаучный (а не только биологический) характер
проблемы, провел (с помощью четырех причин Аристотеля - см. п. 1-6)
группировку видов целесообразности и пришел к странному в те годы выво-
ду, что проблема является важной для биологии, но не главной.
Советую всем прочесть статью, а здесь приведу из нее пример указа-
ния на загадочное. Все говорят, что яркий цвет и сладкий вкус плодов нуж-
ны растению, привлекая опылителей; но яркий и сладкий арбуз покрыт
крепкой кожурой защитного цвета, а вот ряд диких арбузов - сладкие, кис-
лые, горькие и ядовитые. «Наличие очень сходных плодов с прямо противо-
положными свойствами как будто не соответствует их приспособительной
роли» [Любищев, 1982, с. 164]. В главе 6 мы узнаем, что такие ряды рушат
всю прежнюю теорию эволюции и ведут к новой науке.
Любищев щедро дарил мысли и в письмах. В 1949 году, в письме другу
Б.С. Кузину, зоологу и мыслителю, он дал шутливую, в строительных тер-
минах, характеристику своего понимания основной тройки учений: дарви-
низм для него - «теоретический свинарник ... считающий, что все в природе
движется чистым свинством, борьбой за существование и размножение», а
ламаркизм - «дворец физкультуры», в котором идет «активная эволюция
организмов». Номогенез, к которому примыкал сам Любищев, охарактери-
зован как «храм истины, красоты и закона».
Больше всего Любищев запомнился как блестящий критик, считавший
долгом выискивать и разбивать не слабые (как это обычно), а самые силь-
ные места критикуемых учений. Этим он продолжил (кажется, первый в би-
ологии после Данилевского) линию Сократа - Канта, зовущую ученых по-
стоянно задавать себе вопрос: «Откуда я знаю то, во что верю?» Затем эту
линию блестяще развил Мейен, о чем узнаем в главе 6. Если бы критикуе-
мые ученые (в эволюционизме это были дарвинисты) включились с ними в
серьезный спор, им пришлось бы задавать себе Сократов вопрос ежечасно,
но никто не включился. Подробнее см.: Салихов М.В. А.А. Любищев об «об-
щепринятых» постулатах науки и философии. Ульяновск, 2001.
Он справедливо призывал, следуя Платону, освещать проблему со всех
сторон («мы должны бороться ... со всякой монополией: гегемония одного
направления есть отказ от подлинной диалектики» [Любищев, 1982, с. 244]),
и возразить можно одно - это лучше именовать не диалектикой (данный
201
термин Сократа ныне слишком связан с Гегелем), а методом множествен-
ных гипотез. Данный метод сто лет назад ввел геолог и палеонтолог Т.
Чемберлин, который видел изъян науки в господстве «метода ведущей тео-
рии» и предлагал допускать к одновременному использованию более одной
рабочей гипотезы [Мейен// Системность..., с. 20].
Что касается самой диалектики, то свойственное советской эпохе по-
нимание ее по Платону (собирание доводов в свою пользу, п. 3-7), сильно,
на мой взгляд, мешало Любищеву. Смею допустить, что именно оно сни-
зило его успех в положительной теоретической деятельности. (По выясне-
нии сути дела из множества рабочих гипотез надо либо выбрать одну, ли-
бо провести их синтез, что у Любищева редко.) В 1925 году он писал:
“вместо четырех сформулированных Соболевым самостоятельных законов
(наследственности, органического роста, обратимости и прерывности) воз-
можно сформулировать один закон диалектического развития организмов”
[Любищев, 1982, с. 145],
но потом за 47 лет жизни не сдвинулся в этом направлении ни на шаг. При-
чин вижу две: 1) на самом деле диалектика - не закон развития природы, а
набор правил мышления, и 2) она противоположна всякому поиску истины,
хотя полезна для поиска доводов. Подробнее см. (Чайковский Ю.В. // ВФ,
2002, № 9, с. 170).
Летом 1953 года Любищев направил властям письмо против лысенков-
щины, это вызвало скандал, повлекший увольнение его в 1955 году на пен-
сию. Другие в таких случаях приходили в тоску, а он сел писать главные
свои труды. Он более всех прояснил логику систематики (см. гл. 10) и пока-
зал, что эволюция никак не сводится к цепи “приспособлений организма к
среде”, что в ней столь же важны законы преобразования форм.
В том же 1953 году в Англии была раскрыта двуспиральная структура ДНК, в
1961-66 годах в США - генетический код (соответствие нуклеотидных трипле-
тов двадцатке аминокислот), и последние 40 лет XX века прошли под знаком
господства молекулярной биологии. Успехи в понимании работы микроструктур
живого были поразительны, но столь же поразительно полное отсутствие успе-
хов в эволюционной теории. Наоборот, она откатилась назад, с позиций основа-
телей СТЭ (Московская школа, Шмальгаузен, Хаксли) к примитивным пред-
ставлениям начала века (в духе Мензбира), едва прикрытым новой терминологи-
ей, часто применяемой без понимания.
Эксперимент стал чудовищно сложен, а потому и дорог, и центром науки, ес-
тественно, стала самая богатая страна, США. Это наложилось на общее господ-
ство США после Второй Мировой войны. Английский язык стал языком науки
настолько, что западные научные издания почти сплошь перешли на английский
(удержалась, насколько знаю, одна Франция, перешедшая на английский лишь в
наши дни), и англоязычные авторы стали в массе терять знание иностранных
языков, а с тем и теоретическую культуру (появились также советские авторы,
избегавшие цитировать русскую научную литературу).
202
Ныне 85% статей писаны по-английски. И, хота всего 31% их написан в США,
но 2/3 ссылок в американских статьях приходится на американские же статьи; в
них почти нет ссылок на статьи из России и Китая, даже писанные по-английски.
Ни в одной стране нет такого, и науковеды США видят в этом превосходство
науки США (Макрусова В.А. Цитируемость российских публикаций в мировой
научной литературе // ВРАН, 2003, № 4). По-моему же, это говорит о самоуве-
ренной самоизоляции. Напомню, что иностранных ссылок почти не было и в со-
ветской позднесталинской науке.
Популярное американское руководство провозглашало: «В настоящее время
генетика играет роль краеугольного камня в общей структуре биологических на-
ук. Все чаще проблемы... физиологии, биохимии, медицины, экологии и эволю-
ции возникают как продолжение современных генетических идей... Наш подход
к проблеме действия генов будет простым и непосредственным: действие гена
мы будем отождествлять с образованием и функционированием белка, предпо-
лагая при этом, что все другие формы проявления гена являются следствием
этих процессов» (Хартман Ф., Саскайнд 3. Действие гена. М., 1966, с. 11). Как
увидим далее, этот путь оказался тупиковым.
Что касается эволюции, то беда усугублялась забвением того, что основные
мысли по теории эволюции содержались тогда отнюдь не в английских текстах,
а у немецких и французских морфологов.
Примитивизм коснулся даже блестящих умов. Так, английский биофизик
Джон Бернал, в блеске ума которого мы убедимся в главе 7, провозгласил в 1967
году такую максиму: “Жизнь перестает быть таинством и практически становит-
ся чем-то вроде головоломки ... Жизнь есть частичная, непрерывная, прогрессив-
ная, многообразная и взаимодействующая со средой самореализация потенци-
альных возможностей электронных состояний атомов” (цит. по [Мейен и др.,
1977]). На мой взгляд, с тем же правом можно сказать, что творчество поэта -
лишь реализация возможностей алфавита. Это не столько ложно, сколько неин-
тересно: отсюда не следует реальных результатов. В годы этой примитивизации
и стали появляться в печати статьи Любищева.
4-17* Процесс приспособления — не главный
После 32-х лет теоретического молчания (не считая работ по биомет-
рии), Любищев вновь заявил о себе как теоретике биологии. Живя с 1950
года в Ульяновске, он в 1962 году опубликовал в Москве статью «Понятие
сравнительной анатомии» (перепечатана в сб. [Любищев, 1982]).
Главное в ней для нашей темы: если до Дарвина зоология была осно-
вана на сравнительной анатомии, то по воцарении дарвинизма интерес к
ней у большинства биологов пропал. Она, как предмет преподавания и ис-
следования, сохранилась лишь в качестве набора свидетельств в пользу
эволюции. Если Гёте ставил перед морфологией «задачу раскрытия зако-
нов органических форм», то само их наличие стало сомнительным. Орга-
низм стали рассматривать как сумму приспособлений.
203
Знание конкретных законов органических форм уступило место рас-
плывчатым объяснениям полезности каждого свойства (добавлю: про-
изошел возврат к основной идее естественного богословия). «Удовле-
творенность расплывчатым объяснением притупляет желание искать зако-
ны природы, что и случилось» - писал Любищев. Призывая вернуться к
сравнительной анатомии как основе биологии, он признал, что с утвержде-
нием идеи эволюции «радикально изменились основные постулаты морфо-
логии». Он привел 7 пар новых постулатов, сопоставив каждому новому
старый. Я изложу их со своими (петит) пояснениями позиции Любищева.
1. Историзм: главная причина сходства организмов - их родство. Прежний
постулат: сходство вызвано сходством формообразующих сил (Кильмейер, Бэр).
2. Акгуализм Лайеля (формы всегда образовывались по тем же законам,
что и ныне) против, например, прежнего катастрофизма Кювье.
3. Монизм в таксономии: таксоны всех рангов образуются тем же путем,
что и виды. Иной постулат: даже виды и роды образуются различно (Коп).
4. Скрытый телеологизм (фактическое признание цели в любом акте эво-
люции, сочетаемое с заявлениями об отсутствии у нее цели): «все органы,
части и даже признаки получают рациональное объяснение через указание
их служебной роли». Сам Любищев полагал главной не пользу, а гармонию.
5. Аддитивный характер организма (фактическое рассмотрение организма
как суммы признаков, сочетаемое с декларацией его целостности) против
принципа корреляции Кювье, на деле работавшего с целостностями.
6. Пассивность эволюции: организм изменяется лишь в силу условий сре-
ды. Против - идея номогенеза, видная уже у ранних авторов (Де Малье и др.).
7. Неповторимость эволюции. Этот принцип выдвинул бельгийский палео-
нтолог Луи Долло (1893 г.), он слабо обоснован, и против выступали многие,
начиная с Копа. Любищев полагал, что «закон Долло основывается во мно-
гих случаях на появлении сходных образований на иной морфологической
основе» (ЛЧ, 1996, с. 12). В наше время вместо “закона Долло” предложен “закон
неидентичности сходных структур” [Серавин // ЭБ, с. 89]; по сути это совпадает с
позицией Любищева, о которой пойдет речь в пп. 6-13* и 9-14.
Данная семерка относится не только к морфологии, но и к другим дис-
циплинам, так что собственно морфологической статью делает рассужде-
ние о гомологии. Любищев привел определение Оуэна (гомология - сход-
ство по положению, аналогия - по функции) и отметил: “Историческая мор-
фология заменила сходство по положению сходством в силу общего проис-
хождения”, но вышла путаница; например, рука гомологична крылу, но, в
новом понимании, не гомологична ноге. Основную часть статьи занимает
анализ пониманий и приложений понятия гомологии.
Выводы каждый ее читатель должен делать сам, мое же мнение таково:
ныне “историческая морфология” выглядит набором противоречащих друг
Другу презумпций. Жаль, что статью Любищева почти никто за 40 лет не ис-
пользовал. Ее почти не заметили, и вряд ли могло быть иначе: она сетует
204
на низкую культуру биологов, а требует для понимания как раз высокой
морфологической культуры. Но Любищев прожил еще 10 лет и успел напи-
сать более доступные работы, позволившие ему стать весьма известным
среди теоретиков самых разных школ. Заговорили о «феномене Любище-
ва» [Мейен и др., 1977].
Главный тезис Любищева звучит для нас странно. Он таков:__________
_______________Основой природы является не польза, а гармония.________
Для Любищева даже «приспособление есть частный случай гармониче-
ского строения» [Любищев, 1982, с. 66]. Отсюда некоторые выводят общее
понятие эстетической целесообразности:
«Если “утилитарная” целесообразность - это соответствие... целям жизне-
деятельности, то под “эстетической” целесообразностью мы понимаем (в ду-
хе любищевской и кантовской точек зрения) - соответствие более возвышен-
ному целому (или целостности), о конечной цели которого у нас нет пред-
ставления» (Гуркин Б.А. Эстетика целесообразности // ЛЧ, 1992, с. 9).
См. также 6-15**. Сперва кажется странным: разве не очевидно, что ка-
ждый орган служит своей цели? Да, служит, но из этого вовсе не следует,
что он возник в ходе процесса приспособления к ней (это надо доказывать,
а не постулировать). Об этом писал еще Бэтсон (п. 4-10), а Любищев в руко-
писи 1946 года двинулся дальше. Например:
“возникновение узкой кормовой специализации никакой пользы животному
не приносит, но эта специализация развивается, так сказать, в силу инерции,
по привычке... и может стать фатальной. По-моему, это не адаптация, а то,
что удобно назвать рутинизацией.” И далее: “Я вовсе не уверен, что такой
классический случай, как образование ...копытных животных, есть действи-
тельно приспособление к быстрому бегу.” Вернее, что копыто лошади —
“следствие однообразных беговых движений в силу рутинизации, а не адап-
тации” [Любищев, 1982, с. 171-172].
В самом деле: не все копытные быстро бегают, лучшие бегуны — не
копытные, а гепарды, и вне бега копыто лошади малоудобно.
Как видим, к идее гармонии (номогенез) добавлено упражнение (ламар-
кизм), но, что еще важнее, полезность отодвинута на второй план. При этом
Любищев отметил, что физика стала бурно расти тогда, когда отка-
залась от целевого объяснения в пользу причинного. Мы увидим в п.
6-12, что биологии в самом деле придется учесть этот опыт.
Каково же тогда место естественного отбора в эволюции? Любищев,
вслед за Бергом, полагал, что отбор, не будучи вообще движущим факто-
ром эволюции, тем не менее, может играть роль «цензора» (отсекать не-
жизнеспособные формы) и «квартирмейстера», т.е. распределять виды по
зонам обитания. Здесь приходится возразить Любищеву.
Откуда он это знал, если ни одного примера не привел? Как видим, он
совершил ту же ошибку, за которую так порицал дарвинистов - ввел отбор
205
как непроверяемый гипотетический фактор. Как мы уже знаем, цензором
отбор не может служить ни в какой мере (пп. 3-7*, 4-3), а роль квартирмей-
стера весьма сомнительна. Ведь сам Любищев приводил примеры несоот-
ветствия между строением организма и его зоной обитания (оляпка, т.е. во-
дяной воробей, ныряет и бегает по дну, имея совсем сухопутный облик, и
многое другое). Во всяком случае, впредь до проведения опыта, который
покажет, что какой-то вид изменил свою зону обитания в результате раз-
личия размножаемостей разных ненаправленных вариаций, говорить об от-
боре как квартирмейстере нельзя.
В главе 6 мы узнаем, что разнообразие видов и условий их существо-
вания столь просто не связаны, и узнаем о другом «квартирмейстере».
Вообще же, своей теории эволюции Любищев не оставил; главная при-
чина этого (помимо пристрастия к диалектике) видна из его слов:
“Для меня центральная, наиболее интересная проблема биологии - многооб-
разие органических форм. К физиологии я весьма равнодушен” [Любищев —
Гурвич, с. 31].
Без физиологии и экологии теорию эволюции не построить, и, как видно
из его мимоходных замечаний, он понимал это. Зато его трактовка понятия
эволюции остаётся самой удачной до сих пор. Термин «эволюция» он он
определил через через 5 противоположений:
эволюция как трансформизм (отрицание постоянства);
эволюция как преформизм (отрицание новообразования);
эволюция как постепенное развитие (отрицание скачков);
эволюция как восхождение, прогресс (в противоположность нис-
хождению, т.е. эманации, или регрессу);
эволюция как необратимый процесс (в противоположность обра-
тимому развитию, инволюции} [Любищев, 1982, с. 11].
По Любищеву, основная часть споров об эволюции беспредметна, ибо
участники говорят о ней в разных смыслах. А основная часть “доказа-
тельств” эволюции на поверку оказывается примерами эманации и инволю-
ции. К этому стоит добавить, что к доказательствам эволюции обычно отно-
сят различие ныне живущих организмов (ланцетник - предок рыб и т.п.), то-
гда как все ныне живущие виды прошли одинаково длинный эволюционный
путь, поэтому никто из них никому не предок.
4-17** Связавший несвязанное
Самое, на мой взгляд, важное у Любищева - метод рассуждения. Он
противоположен “бритве Оккама” (см. п. 5-1*), презумпциям и вообще вся-
кой косности мышления. В основе его лежит то, что хочется назвать чтени-
ем поперёк, т.е. умение при чтении любой литературы видеть параллели
прочитанного со всем, что ему известно, и быть готовым встать на новую
точку зрения. Такое умение ведет к возможности давать противоречащие
примеры ко всем непродуманным утверждениям, а главное - отказаться от
206
своего убеждения, как только оно будет аттестовано самим мыслящим в ка-
честве догмы. Метод был усвоен Мейеном и, насколько удалось, мною.
Любищев спрашивал: «Что лучше - неверная гипотеза или никакой?» в
его ответе корень его расхождений не только с СТЭ, но и с большинством
ученых. Большинство ученых предпочитает избегать гипотез, выходящих за
привычные рамки, т.е. по сути отвечает: “лучше никакой”.
Меньшинство поступает наоборот: оно видит в выдвижении гипотез
смысл своей теоретической деятельности и ведет на их основе бесконеч-
ные бесполезные споры друг с другом.
А сам Любищев отвечал разумнее - что неверная гипотеза лучше, чем
ничего, если она является действительно теоретической конструкцией, под-
лежащей проверке (прежде всего, самим ее автором), но хуже, чем ничего,
если исходит из догмы, не подлежащей проверке, и выдвинута в ее под-
держку [Любищев, 1975, с. 210]. Для пояснения тезиса он приводил приме-
ры, когда невиновные бывали осуждены просто потому, что в поле зрения
судьи не было конкурирующей версии [Любищев, 1982, с. 244].
В 1960-е годы он обратился к самым общим вопросам. Тут ему помог-
ла установка, которую он еще в 1927 году выразил в виде шутки: «Будучи
беспринципным в выборе цитат, я приведу определение блаженного Авгу-
стина...» (Любищев А.А. Понятие номогенеза // Природа, 1973, № 10, с. 43).
Это значит, что он был готов черпать мудрость из любого источника, какой
считал достойным, тогда как почти все ученые избегают упоминать чуждых
им авторов (даже если расшаркиваются при встрече). Любищев читал и ци-
тировал всех, кого считал интересными, в том числе тех, кто избегал его.
Можно спорить с каждой мыслью Любищева, но нельзя спорить с тем
фактом, что он читал своих противников и находил у них полезное. Наобо-
рот, никто из известных мне дарвинистов не показал знания трудов Люби-
щева (и вообще ни одного из “чужих” направлений). Хотя многие его упоми-
нают, но всегда не по делу и без понимания сути. В итоге для будущего Лю-
бищев оказался полезнее, чем те, кто считал его чудаком и неудачником.
За полгода до смерти Любищев наиболее четко выразил свое понима-
ние номогенеза в рецензии на книгу М.С. Гилярова «Закономерности при-
способлений членистоногих к жизни на суше» (М., 1970). Там он отметил:
«Гиляров прямо подходит к номогенетическому пониманию сходств и к ор-
тогенезу (с. 238): “Направляют эволюцию членистоногих общие принципы
организации типа, выработавшиеся еще в прежней среде обитания...”». И
добавил от себя: “нельзя отрицать наличия сходств, связанных с общностью
происхождения, но значение этих сходств все время отступает перед следст-
вием сходства формообразующих сил” (Любищев А.А. И ТПЭ-3).
Любищев соединил номогенез с ламаркизмом:
“сходное решение приспособлений при переходе на сушу является следстви-
ем не общности предков..., а следствием закономерной реакции изменяюще-
гося организма на требования новой среды” (там же).
207
Он пояснил это примером эволюции трахейного дыхания. Таков его метод:
соединять то, что для других несоединимо, и показывать соединимость.
Идеи Любищева стали за 30 лет после его смерти известны у нас всем,
кто хотел их знать, и сыграли огромную роль в повышении общей культуры
наших эволюционистов - даже тех, кто с ним не соглашался. Это очень
важно: любищевский клан, к которому считаю за честь принадлежать и я,
пропагандирует его идеи не потому, что со всеми из них мы все согласны, а
потому, что считает их общекультурным наследием.
Например, мне чужда любищевская идея примата формы над функцией
и следующее из нее почти полное равнодушие Любищева к физиологии,
экологии и этологии. Однако эта идея высказана им впротивовес тоже со-
мнительной идее примата функции над формой. В основе у Любищева ле-
жит уверенность в том, что форма - самодовлеющая сущность, не своди-
мая к физиологии и экологии (см. анализ: Марасов А.Н. Понятие феноме-
нальности в контексте проблемы реальности или об органической форме
жизни И ЛЧ, 2002), а об этой сущности важно знать всем. По-моему, форма
и функция взаимодополнительны.
Чужда мне и симпатия Любищева к диалектике Платона, из которой вы-
текала неприязнь к “линии Демокрита”, изложенная в работе «Линии Демок-
рита и Платона в истории культуры»24. Ярким примером “линии Демокрита”
служит как раз «Тимей» Платона, что делает для меня всю концепцию “двух
линий” совсем слабой. Сам автор отошел от нее: бросил рукопись на полу-
слове и позже (он писал еще 8 лет) не разрабатывал этих “двух линий”, а в
предсмертной работе писал прямо о трех линиях:
“Упомянув два направления, теоретическое Пифагора и доктринальное Ари-
стотеля, стоит сказать о третьем великом имени в античной философии - Де-
мокрите” (Любищев А.А. К классификации эволюционных теорий // Пробле-
мы эволюции, т. 4. Новосибирск, Наука, 1975, с. 214).
Поясню: Любищев причислял Платона к линии Пифагора. Как видим,
теперь этой линии он противопоставил прежде всего Аристотеля, а Демок-
рит поставлен с ними как бы в один ряд «великих».
В отличие от меня, Марасов (ЛЧ, 1994, с. 18; ЛЧ, 1999, с. 56) видит в
диалектике Любищева важное для биологии достижение мысли, и это вовсе
не мешает нам общаться.
Кстати, роль Платона отнюдь не сводится к диалектике: его “мир идей”
очень существен в систематике, и Любищев был единственным, у кого мы
об этом в 1960-е годы узнавали. О платонизме приходится вспомнить и при
анализе роли математики в материальном мире, что мы увидим в п. 6-7.
Любищев А.А. Линии Демокрита и Платона в истории культуры. М., Электрика, 1997
(ред. Б.И. Кудрин, с указателем и приложениями). Есть и другое издание: СПб., “Але-
тейя”, 2000 (ред. Р.Г. Баранцев, ввод, статья Ю.А. Шрейдера).
208
Думаю, именно благодаря Любищеву и его школе наша эволюционная
мысль никогда не замыкалась в тесных рамках СТЭ и была лучше, чем за-
падная, подготовлена к восприятию появившихся в конце XX века новых
эволюционных идей. К сожалению, за рубежом Любищева почти не знают.
4-18. Злосчастный опыт Шапошникова
Почти как с Любищевым, советская биология поступила с другим клас-
сиком, хотя вёл он себя совсем иначе и еретиком не был. Ленинградский
зоолог Георгий Христофорович Шапошников (1915-1997) поставил в 1957
году близ Майкопа (сев. Кавказ) опыт, который справедливо называют клас-
сическим. Изложу его суть, опуская всё, без чего смысл еще виден.
Шапошников работал с тлями рода Dysaphis. Эти тли - монофаги, т.е.
питаются листьями растений только одного вида. В течение лета они произ-
водят 15-18 бесполых (точнее, партеногенетических - см. ЧЭ, с. 195, сноска)
поколений, давая одно половое лишь осенью, перед зимовкой. Шапошников
сажал бесполых тлей на растение, для данного вида почти или вовсе негод-
ное, и тли быстро эволюировали. Точнее, взято три вида тлей: а, в и с, жи-
вущих на растениях видов А, В и С соотетственно. Вид а на растении вида
С не мог жить вовсе, но мог кое-как жить на растении вида В, куда его и са-
жали. Сперва падали численность и размеры, зато сильно росли изменчи-
вость и смертность. В 9-10-м поколениях произошел
“скачкообразный необратимый переход в новое адаптивное состояние с... рез-
ким снижением изменчивости, прекращением естественного отбора, ... повы-
шением плодовитости; в 8-10-м поколениях возникли существенные морфоло-
гические различия между исходной формой и образовавшейся новой, уже не-
способной жить на старом хозяине” (Шапошников Г.Х. Морфологическая ди-
вергенция и конвергенция в эксперименте с тлями... // ЭнО, 1965, № 1, с. 23).
Выделенные мною слова звучат странно, но означают лишь то, что ав-
тор понимал под естественным отбором попросту рост смертности, дос-
тигавшей в некоторых поколениях 75%. Никакого отбора (избирательного
размножения носителей определенных качеств) он не исследовал.
Хотя возможность у Шапошникова была: одна из сотен личинок вида а
выжила прямо на виде С (ЭнО, 1961, с. 745). Идеология дарвинизма тре-
бовала тщательно обмерить эту тлю (насколько сходна с видом с), поста-
раться получить расплод от нее и бросить силы на отыскание случаев по-
вторного выживания особей а на растениях С, но об этом не сказано. Ав-
тора привлекало именно то, что он именовал «воспитанием» новой фор-
мы, как нечто реально наблюдаемое в виде закономерности.
Сама методика была направлена на поддержку гибнущих, т.е. препят-
ствовала отбору: в самые трудные для тлей моменты автор возвращал их
на сутки на исходное растение «для восстановления иссякающих жизнен-
ных сил» (Шапошников Г.Х. // ЭнО, 1961, № 4, с. 745). Это - ремарка в духе
ламаркизма. Но, насколько знаю, ламаркистом он себя не считал.
209
С 11-го поколения тлей сажали на растение вида С, и теперь они уже не
гибли сплошь, а дожили до осени, до появления полового поколения, дав
новую форму; она не могла скрещиваться с исходным видом а, зато давала
плодовитое потомство с видом с. Иными словами, Шапошников получил но-
вый вид животных за одно лето!
Сам он говорил осторожно: это новая «видовая форма». Внешне она
несла черты как вида с, так и вида Ь. А это, как мы узнаем далее, - блоч-
ность, элемент номогенеза. Но автор предпочел не заглядывать вглубь, а
всего лишь расширить понятие вида так, чтобы в его опыте границы вида не
нарушались (Шапошников Г.Х. И ЭнО, 1966, № 1, с. 30).
Открытие нуждалось в генетическом анализе, но вместо этого опыт был
прекращен на самом интересном месте: никто так и не знает, плодовито ли
потомство полового поколения. Автор прожил еще сорок лет, но не смог ни
продолжить опыт, ни даже опубликовать сделанное. (Насколько знаю из
разговоров, он, сменив вид насекомых и ужесточив условия опыта - выше
95% смертности в отдельных поколениях, - сумел преодолеть барьер не
только вида, но и рода - тут уж изменением определения вида делу не по-
мочь.) Вместо помощи коллег, двадцать лет Шапошников отражал их обви-
нения -доказывал, что он не ламаркист.
Пытался защищаться в печати, пробуя без ереси объяснить свои ере-
Ееские результаты. В частности, допускал особые формы отбора: сверх-
зтрая эволюция якобы вызвана неким «дестабилизирующим отбором»
алогия со стабилизирующим отбором, каковой всякую изменчивость
уменьшает) и даже «отбором на раскрытие потенций» (Г.Х. Шапошников.
Динамика клонов, популяций и видов и эволюция //ЖОБ, 1978, № 1). Но та-
кого «раскрытия» бесполой тле по теории не полагалось, а одного полового
поколения для отбора явно мало, и о ней скоро забыли.
Суть же гонений в том, что Шапошников на своих тлях получил запрет-
ное “наследование упражнения органов” по Ламарку и запретную “расша-
танную наследственность” по Мичурину (этот термин использовал Лысенко,
и с тех пор исследовать само явление считается неприличным).
Одним из немногих Шапошникова тогда поддержал Мейен:
“Намечается следующая логическая связь. Продукты метаболизма как-то
влияют на генетический код. Тогда изменение этих продуктов должно вести к
изменению таких влияний, а значит, к перестройке... самого кода. Почему
же... не предположить, хотя бы в виде рабочей гипотезы, что опыты Шапош-
никова знакомят нас как раз с таким явлением” (Мейен С.В. Закон? Есть за-
кон? // Знание — сила, 1974, № 9, с. 9).
Как позже выяснилось, Мейен был прав (п. 5-13). Еще десять лет опыт Ша-
пошникова не поминали, а затем, забыв его суть, стали поминать как дока-
зательство эффективности отбора. Сам автор в 1983 году был уволен из
Зоологического института на пенсию, что для ученого такого ранга было
Редкостью, и, говорят, до конца дней жил в обиде на родной институт.
210
Оптимист скажет: ведь не расстреляли же (как отца Шапошникова) и
даже не посадили! Но таких возможностей у гонителей Любищева и Ша-
пошникова не было. Ате, что были, использованы, увы, полностью.
4-19. Одомашнивание
Доместикация (одомашнивание) была исходным пунктом у Дарвина, и
ей он посвятил самый свой объемистый труд (см. п. 3-2). В ходе доместика-
ции изменчивость возрастает, причем в самых разных направлениях. Уже
сам Дарвин отмечал, что изменчивость иногда бывает упорядоченной, а в
XX веке это оказалось общим правилом: выявлены параллельные ряды из-
менчивости, как у Вавилова. Так,
“на основании построения параллельных признаков по рядам мы можем для
собак разных пород предсказывать нахождение... признаков, до сих пор неиз-
вестных” (Боголюбский С.Н. Происхождение и преобразование домашних
животных. М., Советская наука, 1959, с. 533).
И в самом деле, полувековые опыты по доместикации американской
серебристо-черной лисицы (их начали в 1948 г. в Эстонии, а с 1960-х ведут
в Новосибирске), показали ясную параллель с собаками. Селекцию вели на
“домашнее” поведение лисят, а впридачу получили у них целый комплекс
собачьих черт: висячие уши, хвост кольцом, размножение вне сезонов, ко-
роткие морды, собачьи типы окраса шерсти (пегость, подпалость). Послед-
нее досадно, поскольку снижает ценность шкурок. Феномен так и назвали:
«особачивание лисиц».
Но вместо рассмотрения идеи Вавилова (явного, как у С.Н. Боголюбско-
го, поиска в новом ряду свойств, известных по старым рядам) генетики ог-
раничились тем, что ввели новый тип отбора: рост изменчивости описали
как итог дестабилизирующего отбора (Беляев Д.К. Дестабилизирующий от-
бор... // Природа, 1979, № 1). Как и в случае со стабилизирующим отбором,
никаких данных о собственно отборе нет, да их никто и не искал.
Факты говорят, что изменчивость (дестабилизация) растет сразу (у де-
тей тех особей, с которых начата доместикация) и массово, а это значит, что
ни о каком отборе говорить нет смысла. Замечательно и то, что сходные
процессы наблюдаются при доместикации самых разных животных: собаки,
овцы, свиньи, коровы, лошади и кошки обнаруживают уменьшение мозга,
пегость и др., часто — висячие уши (Бердников В.А. Основные факторы
макроэволюции. Новосибирск, Наука, 1990, с. 28-30).
В новой статье новосибирских исследовательниц (Трут Л.Н., Плюснина
И.З., Оськина И.Н. Эксперимент по доместикации лисиц и дискуссионные
вопросы эволюции собак // Генетика, 2004, № 6) уже нет “дестабилизирую-
щего отбора”, и это отрадно. Вместо этого сказано, что собачьи черты лисят
возникают как при селекции на нужное поведение, так и без нее, только в
сто раз реже; что мутациями и выщеплением гомозигот их объяснить нель-
зя; что в одном помёте бывает целый спектр изменений.
211
Здесь, как и у Шапошникова, можно говорить про синтез ламаркизма с
номогенезом. Это станет ясным по ознакомлении с материалом Части 2.
4-20. Закончим с темой отбора
Идея отбора сходна с идеей теплорода: обе наглядно толковали мно-
гое, но в природе этих явлений не оказалось. Рассверливание пушки нагре-
вало ее тем больше, чем меньше оставалось в ней металла, откуда физик
Бенджамин Румфорд (Англия, 1798) понял, что тепло - не вещество, а итог
движения. Но физики верили в теплород еще полвека, читающая публика -
сто лет; тепло названо веществом в словаре В.И. Даля (вплоть до 1913 г.).
Обнаружив, что принцип отбора давно опровергнут наблюдением за
крылатыми термитами (пп. 3-7*, 4-3), я был ошеломлён: как же так? Ведь те,
кто почти не размножается, на самом деле (думал я) должны уступать место
тем, кто размножается успешно. А этого у крылатых термитов нет, и никто
об этом вот уже 80 лет не вспоминает. Метерлинк сильно упростил картину,
но это не в укор ему - потрясающие факты надо было высветить. К сожале-
нию, он остался одинок. Все, с кем я говорил о термитах, уходили от обсуж-
дения сути дела, предпочитая ссылаться на побочные обстоятельства.
Известные мне книги обходят данный вопрос начисто, а устные реплики, ка-
кие я слышал, по сути все одинаковы: термиты - общественные насекомые, у
них всё не так; они содержат симбионтов в кишечнике, их изменчивость ограни-
чена выживанием симбионтов; несъедобность - дело сложное. Почему отбор
должен идти на несъедобность? А вдруг как раз съедобность виду выгодна? Ведь
массовое выедание известно у многих видов. Вот окуни вообще питаются своей
молодью. Может быть, кал хищников, объевшихся термитами, удобряет почву
вокруг термитника и способствует его процветанию? (Трудно поверить, но этот
вопрос действительно был мне задан, притом умным благожелательным собе-
седником, ибо он видит в идее отбора презумпцию - см. п. 3-9). И так далее.
Но надо не уходить от каверзного вопроса, а тщательно обдумать его.
Общественная организация термитов похожа на таковую у многих видов
муравьев и пчёл, но такого выедания у них нет. Подобная расточительность
известна у некоторых рыб (вкусная икра) и растений (вкусные семена) и она
тоже входит в противоречие с идеей отбора (см. пп. 6-1,6-2*).
Можно взять руководство по насекомым (например, т. 3 «Жизни живот-
ных») и прочесть о тех тропических термитах, которые живут весь год в за-
купоренных термитниках, бесполые, слепые и бескрылые; откуда раз в году
вылетает зрячее крылатое половое поколение (каста) - самцы и самки. Это
похоже на пчелиный рой, но есть три принципиальных отличия.
Во-первых, самок много, во-вторых, в момент образования пар все осо-
би обламывают себе крылья и тут же становятся жертвой множества хищ-
ников, а в-третьих, термиты неуклюжи, очень привлекательны как пища,
легко заметны и совсем беззащитны, поэтому выедаются мгновенно. Лишь
менее одной пары на тысячу ускользает от гибели.
212
Значит, как сказано в п. 3-7*, самою природой из века в век ставится се-
лекционный опыт: если существуют, как гласит дарвинизм, вариации всех
свойств, то среди них должны быть вариации съедобности (в широком
смысле: колючий покров, яд, отпугивающий запах или вкус и т.д.) и они
должны отбираться, а это значит, что несъедобные должны вытеснить съе-
добных. Такова логика дарвинизма, на которой построено всё учение.
Однако несъедобных не возникает. Столь же просто пресечь отпадание
крыльев, но вместо этого налицо, наоборот, специальные приспособления
для их обламывания (см. п. 6-14**). Итак, схема отбора здесь не действует.
Опыт природы поставлен идеально: тысяча видов (сотни термитников в
каждом) дает достаточный материал для реализации изменчивости. Часто-
ты мутаций известны (одна на миллион особей и выше - по каждому из-
менчивому признаку), численности тоже (у некоторых видов - миллион по-
ловых особей на термитник, сотни термитников у каждого вида), эффектив-
ность отбора - практически 100% (съедобные гибнут практически все, но
успешная пара создает целую колонию). Поскольку несъедобность может
быть достигнута многими, в том числе и очень простыми способами, она
должна возникать часто и распространяться быстро (менее чем за 100 по-
колений). И тот, кто утверждает, что этот мощный фактор (отбор) чем-то на-
чисто блокирован, должен указать блокирующий фактор и показать, почему
он еще более мощен. Или молчать, ибо у него нет теории.
Но если отбор не действует в идеальных условиях, то может ли он дей-
ствовать хоть где-то? Действительно биологический (без отговорок) ответ
будет дан в п. 6-14**, а пока замечу, что если отбор бессилен в простейшей
идеальной для него ситуации, то ссылаться на него нет оснований.
Можно бы сказать, что механизм отбора у крылатых каст термитов вы-
ключен (и искать выключатель), если бы имелся хоть один аккуратный при-
мер того, что механизм существует, что он у кого-то бывает включён.
Дарвинистам надо было рассмотреть аккуратно, с числами и графи-
ками, хотя бы один пример эволюционного приобретения, приписываемо-
го отбору, чтобы убедиться, что оно произошло именно под действием
отбора ненаправленных вариаций. Для этого надо каталогизировать из-
менчивость и измерять размножаемость каждого варианта (из каталога) в
каждом поколении. Но этого никем сделано не было.
Полутора веков бесплодного ожидания такого примера достаточно, что-
бы усомниться, что он вообще возможен. Пока и поскольку его не имеется,
вернее будет признать: фактов естественного отбора не существу-
ет, есть лишь факты приспособленности и их толкование в тер-
минах отбора. Более того, в описанных выше рамках естественный отбор
- опровергнутая гипотеза. Почему же она продолжает царить?
В п. 5-3 мы узнаем, что она - феномен не нынешней биологии, а дав-
ней социальной психологии. Именно по старым внебиологическим при-
213
чинам в дарвинизме налицо набор элементарных логических ошибок, об-
щий для всех его форм. В отношении отбора действительность такова:
1) Из самого факта гибели одних организмов еще не следует ничего о
том, как будет (если будет) протекать эволюция других, выживших.
2) Как заметил Нортон (п. 4-1), избирательная размножаемость никого
не способна вытеснить начисто (мы вернемся к этому явлению в п. 6-2*).
3) Даже если полное вытеснение в одной популяции произошло (в СТЭ
это называют случайной фиксацией), из этого еще не следует, что успешная
популяция сохранит (а тем более усилит) свою отличительную особенность
в ряду поколений. Это сохранение надо доказывать опытом.
4) Естественный отбор не может идти по схеме искусственного, о чем
мы не раз упоминали и о чем будет подробно сказано в п. 5-8*.
5) Сами дарвинисты признают, что естественный отбор является меха-
низмом обратной связи, т.е. регулятором. Но регулятор бессмыслен без
двигателя, т.е. без механизма прямых связей (тема “буксира”, см. главу 3).
Все немногочисленные «доказательства естественного отбора» демон-
стрируют либо искусственный отбор, либо просто сдвиг генных частот. В ос-
тальных же случаях термином «естественный отбор» пользуются для обо-
значения всего лишь такого изменения, которому удается придумать полез-
ность. Нужны не толкования, а наблюдения, опыты и теория.
Критики дарвинизма привели много примеров эволюции без отбора и
даже против отбора. Они сформулировали общее теоретическое положе-
ние: отбор в принципе неспособен вести прогрессивную эволюцию (услож-
нение организмов), но может служить для второстепенных эволюционных
актов (выбраковка негодных, простые очень эффективные улучшения).
Наиболее четко эта линия (как теперь понятно, тоже чисто умозритель-
ная) проводена в книге [Любищев]. В данном ключе написаны и мои преж-
ние работы, в том числе книга ЧЭ, где введено умозрительное понятие
«квант селекции». Увы, эта линия оказалась ложной: идеальным квантом
селекции должна бы быть несъедобность термитов, а ее нет. Нет и других
реальных примеров естественного отбора, зато есть опровержения. Это за-
ставляет отказаться от естественного отбора как фактора эволюции.
Разумеется, неспособное жить не живет, но это нельзя счесть причи-
ной какого-то процесса. Подробнее см.: Чайковский Ю.В. IIЛЧ, 2006.
Отказываясь от идеи естественного отбора, мы избавляем себя от нуж-
ды вести бесконечные списки толкований и бесконечные споры с толкова-
телями. Однако при этом мы лишаемся основного инструмента объ-
яснения, гласящего: «полезное выживает потому, что лучше раз-
множается». Пусть его очевидность и обманчива, пусть принцип вообще
ложен, но что ему противопоставить?
Словом, принцип отбора должен быть не просто отброшен за непригод-
ностью, а заменен на иной, годный для построения теории. Таковым пред-
ставляется принцип активности., который мы рассмотрим в главе 5.
214
Часть 2. Механизмы эволюции
Эволюционизм последних сорока лет лучше излагать не в истори-
ческом порядке, а как нынешнюю науку, лишь иногда углубляясь в историю,
например, чтобы увидать точку ветвления, где сделан тупиковый выбор.
Всё сказанное в предыдущих главах должно было, по-моему, убедить
читателя в том, что философские принципы нельзя игнорировать, что они
присутствуют неизбежно в любой теории, а потому должны быть сформули-
рованы в явном виде. Теперь следует добавить: они должны быть (что го-
раздо труднее) выдержаны затем в ходе всего изложения.
Таких попыток в отношении теории эволюции в целом мне неизвестно,
и есть опасность, совершая ее в первый раз, сменить традицию “как бы
объяснения” (дарвинизм) на концепцию того же качества. Как этого избе-
жать? С такого вопроса мы и начнем разговор о механизмах эволюции.
Все несуразности дарвинизма можно было терпеть, пока эволюцион-
ное учение просто удовлетворяло нашу потребность объяснять и обслужи-
вало правящую идеологию. Но нынче - экологический кризис, он требует
принятия срочных мер и, соответственно, реально работающей теории эво-
люции. Меры, которые ныне принимаются, помогают мало. Точнее, пока что
они помогли в одном - показали, что мы, человечество, вмешались в про-
цесс эволюции и она оказалась совсем не такой, как нас учили.
В главе 5 мы рассмотрим новые взгляды на эволюцию, касающиеся
строения организмов, а в главе 6 - относящиеся к сообществам и иным
множествам организмов, к их разнообразию.
Гпава 5. Принцип активности, развитие особи и
иммуногенез
5-1. Какая нужна теория?
Часто пишут, что наука должна давать объяснения и предсказания, что
качество теории определяется полнотой объяснений и выполняемостью
предсказаний. Однако это не вполне так.
Самое полное и безукоризненное объяснение явлениям эволюции дает
религия («Бог так творил»), но наукой это никто, включая верующих, не счи-
тает. Предсказания? Нет, дело тоже не в них: ни историческая наука, ни
дарвинизм не дают предсказаний, и объясняют они лишь малую часть
известного, но считаются науками (в отношении первой это общепризнано).
Наоборот, масса важных и успешных предсказаний (например, земле-
трясений) отвергается наукой из-за отсутствия объяснений, приемлемых
для ученых среднего интеллекта и высокого положения (а именно они опре-
деляют стандарты науки).
215
Мнение, что теория должна верно предсказывать ход событий, заим-
ствовано из небесной механики XVII века (в нынешней отнюдь не всё так
просто: уже в проблеме трех тел возможны хаотические решения) и к тео-
рии эволюции не относится. Дело не столько даже в том, что живая природа
слишком сложна, сколько в том, что предсказания на самом деле мало кого
интересуют. То, что будет через 2 года - не эволюция, а то, что будет даже
через 20 лет, берутся предсказывать слишком многие, не боясь ответствен-
ности. Что уж тогда говорить о долгих процессах!
И как оценить, оправдался ли прогноз? Какие-то параллели реального и
предсказанного всегда есть. Для одних Нострадамус дал прогнозы, которые
сейчас, через 450 лет, блестяще сбываются, а другие видят у него наме-
ренно туманные фразы, которые можно толковать и так, и эдак.
Не отрицая роли объяснений и предсказаний вообще, хочу всё же глав-
ную роль отвести иному - это описание и рекомендации.
Описать объект или явление - значит не только указать их свойства, но
и поместить их в ряд (в ряды) сходных, а также указать на отличия от иных
объектов и явлений. Этого всерьез почти нигде не делают.
Далее, общество гораздо больше ждет от нас практических советов (как
быть, что делать, чего не делать), чем предсказаний. Более того, если совет
дан в катастрофической ситуации и ставил целью избежать бед, т.е. дан на
основе мрачного прогноза, то само исполнение совета ведет к неисполне-
нию (или неполному исполнению) прогноза. Таковы, например, верные про-
гнозы войн и эпидемий, позволявшие пресечь опасный ход событий. Иными
словами, совет оказывается тем лучше, чем хуже оправдался прогноз, на
котором он был основан. Этого тоже почти никто не учитывает.
Полагаю, что дарвинизм был привлекателен не только своими обман-
чиво очевидными объяснениями (всегда без анализа рядов), но и своими
удобными советами. Они все оказались ложными (см. п. 4-20), и ждать да-
лее нет ни возможности, ни смысла. Мое мнение: дарвинизм ныне теряет
почву прежде всего потому, что многие перестают верить его советам.
Однако неуживчивые люди, уверенные, что «миром движет Гераклитово нача-
ло», т.е. вражда, склока (п. 1-5*), были и будут всегда. Люди такого типа всегда
берут верх в эпоху смут. Нам надо осознать, что это никак не связано с уровнем
их знаний, что это их этический изъян: они строят (пусть и без умысла) «теоре-
тический свинарник» (п. 4-17), а такая «теория» неминуемо обращает нас в сви-
ней и даже хуже. Пусть мы не можем от таких людей избавиться, пусть они есть
и всегда будут наверху, но не надо считать их советы благом.
Если ужасы, дарованные нам дарвинизмом, неизбежны, то теория
Должна доказать эту неизбежность и дать совет, как с ними ужиться. Одна-
ко, к счастью, все советы дарвинизма чисто эмоциональны, а единственный
процесс, им реально рассмотренный (изменение частот генов в популяциях)
не имеет отношения к эволюции (см. например: [Гоодницкий, п. 2.5; Наза-
ров, 2005]). Осознание этого открывает, как мне представляется, широкое
216
поле для опытов по созданию теории эволюции. Повторю, она должна ста-
вить целью не прогноз, а обоснованную рекомендацию.
Разумеется, теория должна быть обоснована фактами. Одни уверены,
что Дарвин дал теорию, содержавшую обобщение фактов, другие - что у
него вообще нет фактов, относящихся к сути дела, что его учение голослов-
но. Кто прав? Ясно, что сами понятия факта и обоснования понимаются
разными учеными по-разному и что именно здесь нужен анализ - что счи-
тать достоверным фактом (см. начало гл. 7). Мы уже знаем от Витгенштей-
на, что обоснованность теории не может быть выяснена в рамках самой
теории (п. 3-7**), но можно ли ее выяснить вообще?
Ситуацию несколько прояснил в 1934 году молодой австрийский мето-
долог Карл Поппер, разделивший всё научное знание, грубо говоря, на
теории и доктрины. Теория допускает проверку, которая в принципе может
привести к опровержению теории, тогда как доктрина такой проверки не до-
пускает: любой опыт может быть истолкован в ее пользу - достаточно лишь
придумать неожиданнное его толкование или изменить понимание какого-
то из используемых терминов.
(К доктринам у Поппера отнесены марксизм, фрейдизм и дарвинизм.)
До Поппера примерно ту же мысль высказал Любищев, но не развил ее
[Любищев, 2004, с. 97]. Лишь в конце жизни Любищев высказался на ту же
тему более определенно и опять - в духе Поппера:
“истинная роль философии в науке” состоит “в разработке систем постула-
тов, которая не может быть ни доказана, ни опровергнута, но которая может
служить базой для конкретных гипотез и теорий, могущих быть опровергну-
тыми” [Любищев, 1982, с. 130].
Тут уже видно влияние Поппера и австрийского логика Курта Гёделя.
Согласно Попперу и Любищеву, никакое количество доводов в свою
пользу не ведет к теории. Добавлю: накопление их - основа той диалек-
тики, о которой у нас шла речь в п. 3-7. Зато оно ведет к укреплению своей
веры, и не зря Августин писал о диалектике: «Это она меня научила, что все
приведенные положения, которыми я пользовался, истинны». Вот что род-
нит дарвинистов, богословов и креационистов — диалектика.
Когда Любищеву говорили, что в пользу критикуемой им идеи собран
“Монблан фактов”, он отвечал, что против нее можно собрать “Гималаи
фактов” и указывал подчас на целые блоки таких фактов. Но никогда не
подсчитывал баланс “за” и “против”, а всегда предлагал выяснить суть про-
тиворечий, ибо доводы в свою пользу не ведут к теории.
К ней может привести лишь чёткое разграничение философских
постулатов, личных симпатий, аналогий и реально добытых фактов.
5-1* Выявление презумпций. “Бритва Оккама”
Ранее (п. 3-9) уже говорилось, что эволюционные постулаты обычно
вводятся неявно, в форме презумпций. Недавно палеонтолог К.Ю. Еськов
217
издал учебник (о котором пойдет речь в части 3), где провозгласил их осно-
вой теории эволюции. Методология действительно должна быть ясно обо-
значена, и спасибо Еськову, отважившемуся сделать это. Однако моя точка
зрения противоположна: всякая презумпция мешает поиску истины.
Там, где она вводится, поиск истины часто прекращается вообще. Поясню
свою мысль примером из самой книги Еськова.
В качестве основной презумпции эволюционизма Еськов на с. 19 при-
вел «принцип актуализма», введенный Лайелем в 1830 году и утверждаю-
щий, что в прошлом эволюция шла по тем же законам, что ныне.
“И пускай, к примеру, в докембрии существовали экосистемы, не имеющие
современных аналогов, но камень-то, надо думать, и тогда падал с ускорени-
ем 9,8 м/с2, вода замерзала при нуле градусов Цельсия, а молекула хлоро-
филла исправно поглощала кванты света”.
В действительности, цитата говорит прямо против актуализма:
1) Если сказано, что прежние экосистемы ныне невозможны, то этим
признано, что для них актуализм не работает, и надо понять причину этого.
Дело как раз в изменении действия законов природы. Хотя сами законы
физики за время биоэволюции, вернее всего, не изменялись, но, вопреки
Еськову, они в разные эпохи действовали по-разному.
2) Так, вода при начале жизни замерзала отнюдь не при нуле, а при ми-
нус 20°С и ниже, поскольку давление ранней атмосферы было гораздо вы-
ше нынешнего, и именно поэтому смогла возникнуть жизнь (см. гл. 7).
3) Вообще, лёд, мощная сила эволюции, отсутствовал в биосфере мно-
го раз, и в эти эпохи эволюция экосистем текла иначе.
4) Согласно различным концепциям расширяющейся Земли, ускорение
свободного падения в ходе эволюции уменьшалось, и это изменяло ее ход.
Независимо от того, верны эти теории или нет, нельзя делать вид, что их не
существует - можно оказаться вне науки. Дело в том, что они берутся объ-
яснять факты, о которых Еськов просто молчит. Так, с позиции актуализма
пока не удается объяснить наличие леса на Северном полюсе былых эпох,
тогда как указанные концепции кое-что предлагают (п. 9-7).
5) Сам Лайель видел основной геологический процесс в деятельности
вод (накопление осадков и размывание берегов) и полагал, что так было
всегда (на принципе актуализма основан и дарвинизм). Однако катастро-
фисты (Кювье и др.), видели главный геологический фактор в ином - в ред-
ких катастрофах, преображавших облик планеты. В наше время роль ката-
строф никто не отрицает, а некоторые признают их одним из главных явле-
ний эволюции [Катастрофы...], т.е. отрицают актуализм Лайеля.
Не странно ли, что Еськов, зная всё сказанное, ограничился столь
слабыми примерами актуализма? Нет, не странно: презумпцию никто ни-
когда не обосновывает, ее полагается лишь пояснять - неважно чем. Хотя
Еськов и признал ограниченность актуализма, но не объяснил, в каких
218
случаях от этой презумпции следует отказываться и чем ее в принципе
можно заменить. Вообще, вне “методологической” главы он своей
методологии не проводит.
Как уже мог заметить читатель, моя установка иная: выявлять принци-
пы всюду по тексту, где это нужно, и отказываться от тех, которые не
работают. Подробнее см. [Чайковский, 1990; 1993].
Но отказываться трудно. В книге палеонтолога А.П. Расницына [Расни-
цын] принципы тоже выявлены, обсуждены и даже положены в основу рас-
суждения (научная честность автора достойна восхищения), но часто про-
тиворечат его же фактам, и их принятие для него самого сомнительно (одну
из его презумпций разберем чуть далее). Это не мешает ему пользоваться
презумпциями как истинами, что, напомню, делал и Дарвин.
Вековой успех дарвинизма был бы невозможен, если бы не презумпция:
«отбор малых ненаправленных вариаций есть ведущий фактор эволюции».
Вместо поиска ее доказательств (их у Дарвина, как он сам признавал, нет),
дарвинизм оставил, как и прокламирует метод презумпций, опровегать ее
оппонентам. Они это не раз проделывали (с особым блеском - Любищев),
но их доводов дарвинисты не читали, заявляя, что им и так все трудности
известны. Итог этой лености тоже известен (п. 4-20).
Для оправдания своей лености дарвинисты приняли еще и установку:
«Не умножай сущностей сверх необходимого минимума».
Это - известная “бритва Оккама". Она имеет массу сторонников и про-
тивников. Еськов и Расницын приняли ее, Любищев и Мейен относились к
ней негативно (противополагая ей требование всестороннего анализа про-
блемы, п. 4-17**); а математик и философ Ю.А. Шрейдер любил даже гово-
рить, что___________________________________________________________
_____________бритва Оккама годна лишь для духовной кастрации._______
Этот тезис Шрейдера верен в том смысле, что автор обязан обдумы-
вать свою идею всесторонне, в том числе и с разных сущностных позиций;
только тогда в ней обнаружатся изъяны. Однако “бритва Оккама” полезна
при компоновке материала - для публикации и преподавания.
Кстати, Вильям Оккам, логик XIV века, на самом деле говорил осторож-
нее, чем его позднейшие толкователи: «Не следует делать большими сред-
ствами то, что можно сделать меньшими» [Рассел].
Такой умеренный тезис вполне можно принять: ведь новая сущность
вовсе не обязательно есть “большее средство”. Часто бывает даже наобо-
рот: попытка оставаться в рамках прежде принятых сущностей заставляет
нагромождать допущения, а новая сущность как раз и есть то малое, что
бывает надо добавить к прежним истинам, чтобы перестать громоздить
произвольные допущения и начать делать дело.
Знаменитый ученик Оккама, логик Жан Буридан (которому тоже при-
писывают то, чего он прямо нигде не написал - дилемму “Буриданов осёл")
219
блестяще развил мысль учителя: ввел в оборот почти всеми забытую ан-
тичную идею вращения Земли; то была новая сущность, должная заменить
нагромождение прежних - движений светил вокруг Земли.
Оккам пояснял, что его тезис означает прежде всего недопустимость
введения гипотетических понятий, не поддающихся проверке опытом (отче-
го и стал предтечей эмпиризма). Выходит, что всевозможные типы отбора,
щедро вводимые в СТЭ, ни один из которых не был наблюдаем, как раз и
подлежат, если следовать Оккаму, отсечению. Кстати, Тимофеев-Ресовский
так и делал: он не раз говорил (не знаю, писал ли), что естественный отбор
един, а все вводимые его частные формы, включая стабилизирующий от-
бор, - лишь способ выражаться. Его классическое изложение основ СТЭ
(БЖ, 1958, № 3) построено именно так.
Он был прав - в том плане, что нагромождение форм отбора лишь мас-
кирует недостачу нужных сущностей. Так, отсутствие в дарвинизме меха-
низма прогресса заменяется “отбором на высшую организацию”; отсутствие
идеи сотрудничества - “групповым отбором” и “ценотическим отбором”; от-
сутствие идеи самоорганизации - “стабилизирующим отбором” и т.д. Не-
давно появился термин “замещающий отбор", снявший для его привержен-
цев даже проблему появления человеческого ума: если природа отбирает
организмы, то человек - мысли. К нашей теме прямо относится “мобили-
зующий отбор” (Бердников В.А. Основные факторы макроэволюции. Ново-
сибирск, Наука, 1990, гл. 6), призванный заменить отсутствующее в дарви-
низме понятие активности.
Начав отсекать “лишние” сущности на том лишь основании, что без них
проще, неодарвинизм отнюдь не добился простоты, зато отсек почти всю
биологию как лишнее. А потому и не дал рекомендаций.
Важно отметить, что “бритва Оккама” тоже - презумпция: никто ведь не
отказывается от введения новых сущностей вообще; мол если понадобится,
то введем. Однако еще Мопертюи отмечал, что только в конце исследова-
ния становится ясно, какой набор сущностей явился для него необходимым
(см. [Чайковский, 1990, с. 29]); и уверение Еськова, что основу европейского
научного мышления являет “бритва Оккама”, просто ложно. Проблемы тео-
рии познания столь просто, увы, не решаются.
Если уж требуется познавательный афоризм, то лучше такой:
«Великий физик Максвелл как-то сказал: из всех гипотез ... выбирайте ту,
которая не пресекает дальнейшего размышления...» [Берг, с. 92].
А идея презумпции как раз и призвана пресекать. Такова и главная пре-
зумпция Расницына: «сходство следует считать унаследованным, пока и
поскольку не доказано обратное». Сам он знает, что она неверна:
«независимо возникшее сходство, вещь гораздо более обычная, чем мы привык-
ли думать, и она особенно обычна у близко родственных форм»; у далеких то-
же: известно множество примеров «независимого достижения определенных
уровней организации—уровней амфибий, рептилий, млекопитающих и птиц».
220
И всё же он, как ни странно, верит в нее:
«Тем не менее я думаю, что в основе... параллельной эволюции лежит... од-
нажды приобретенная особенность» [Раснш/ын, с. 32-33].
Оснований не приведено, и мне остается напомнить про ряды Копа -
Вавилова. Как увидим далее, они ведут совсем не туда, куда ведет пре-
зумпция родства. Подробнее см. Мейен С.В. Путь к новому синтезу, или Ку-
да ведут гомологические ряды? И Знание - сила, 1972, № 8 (перепечатано в
книге: Мейен С.В. Листья на камне. М., ГЕОС, 2001).
Хотелось бы обойтись вовсе без презумпций, но не выходит. Остается
выявлять собственные презумпции и отказываться ото всех, от каких можно.
К примеру, я в молодости верил в положения дарвинизма, и только работы
Любищева открыли мне, что эти положения суть презумпции. После этого,
далеко не сразу, я стал проверять их, и они рассыпались.
Но столь же тщательно надо исследовать сами суждения, в ходе кото-
рых рассыпается дарвинизм (нет ли среди них тоже неявных презумпций?),
чтобы не начать строить догму того же качества. По-моему, ничто так не
тормозит науку, как вера в возможность уложить факты в уже заданную тео-
рию, вера в силу презумпций.
В данной книге сознательно принята всего одна-единственная презумп-
ция - презумпция рационального объяснения явлений. То есть, не от-
рицая того, что некоторые положения эволюционизма могут по сути носить
иррациональный (мистический) характер, я исхожу из возможности их пони-
мания в рациональных терминах, пока мне не докажут противоположное.
Без нее я не вижу возможности строить какую-либо эволюционную теорию,
кроме теории непрерывного творения, которая, кстати, удовлетворяет
“бритве Оккама” еще лучше, чем дарвинизм.
5-1** С историей или без? Каверзные ответы
Ламарк впервые выступил с эволюционной идеей на лекции в мае 1800
года, и ровно через 200 лет, весной 2000 года, в том же Париже появилась
книга АЕ. Ее авторы (космолог, палеонтолог и экономист) возродили дав-
нюю традицию сквозного описания эволюции - от космоса до общества - и
уложились в 350 страниц текста. Каждый написал свой раздел, а затем все
трое написали раздел о единой (фрактальной) закономерности трех форм
эволюции. Согласен я не со всем, что там сказано, но она - единственная
известная мне нынешняя работа, в которой рассуждения о механизмах со-
единены с описанием фактического хода биоэволюции.
Биологический раздел тут подчеркнуто внеисторичен, причем его автор,
палеонтолог Жан Шалин, почти не называет прежних имен и ни с кем не
спорит. Не последовать ли этому примеру? Как хорошо не ввязываться по
каждому вопросу в спор со сторонниками различных взглядов, а просто из-
лагать нынешнюю науку, как она мне видится. Так писали многие, но я ре-
шил сохранить свой обычный стиль: выводить каждую нынешнюю тему из
221
работ прошлого и показывать сложное к ней отношение в разные времена.
Первая эволюционная статья Спенсера (1852 г.) построена как спор с креа-
ционистами. Сейчас нам это излишне, но тогда было неизбежно.
Надеюсь, скоро книги вроде АЕ будут широко читаться и переводиться
и мой способ изложения окажется архаичным. Пока это, увы, далеко не так.
У Владимира Высоцкого автобиографический стих кончен фразой:
А мы всё ставим каверзный ответ
И не находим нужного вопроса.
В этом суть - чтобы задавать нужные вопросы, надо уметь видеть не-
годность каверзного ответа и прямо признавать, что ответа нет.
Само состояние эволюционной науки требует повсюду указывать раз-
ные позиции. Если этого не делать, многие читатели будут удивлены - за-
чем все эти сложности, если всё можно объяснить гораздо проще? Да, мож-
но, но такие объяснения (дарвинизм и креационизм) никуда не ведут.
Пока учебники убаюкивают ученика уверениями, что всё сущее про-
изошло «под действием отбора» или «по воле божьей» (что по сути то же) и
пока он видит в этом ответ на все вопросы, в том числе и на еще не постав-
ленные, приходится то и дело разъяснять, что это - каверзный ответ.
Впрочем, религиозная позиция все-таки продуктивнее: давая универ-
сальный и потому ненаучный ответ на вопрос «почему?», она, в отличие от
дарвинизма, оставляет открытым вопрос «как?». Это дает возможность ве-
рующим исследовать механизмы эволюции, притом любые.
В отношении механизмов эволюции организмов (а не их сообществ) до
сих пор предложено всего 4 принципиально различные концепции - ламар-
кизм, жоффруизм, дарвинизм и номогенез (приведены в порядке выхода
главных работ Ламарка, Жоффруа Сент-Илера, Дарвина и Берга.)
Из этой четверки терминов на Западе знают дарвинизм и ламаркизм,
причем второй понимают узко, как НПС (к сожалению, такое понимание от-
части проникло и в нашу литературу). Вместо двух других Запад применяет
много разных терминов, обозначающих частные концепции. Так, словом
ортогенез называют и изменение под прямым действием среды (это эле-
мент жоффруизма), и развитие в прямом, неизменном, направлении (эле-
мент номогенеза), и эволюцию к предначертанной цели (элемент богослов-
ского эволюционизма Тейяра), а это ведет к путанице. Отрицание жесткой
зависимости эволюции организма от среды обитания выражается на Западе
термином автогенез, но в этой концепции нет, в отличие от номогенеза,
речи о структуре разнообразия. Наличие всей четверки в понятийном аппа-
рате сильно облегчит нам работу.
Сложилась досадная традиция противопоставлять концепции, так то
выпадает из поля зрения их идейное сходство. Например, ламаркизм и дар-
винизм многие считают антагонистами, хотя их объединяет допущение не-
прямого действия окружающей среды на свойства организмов. В ламар-
кизме это - влияние упражнения органа на его развитие, а в дарвинизме -
222
выживание более приспособленных. Оба учения ввели это своё исходныое
допущение как чисто умозрительный постулат. Аналогично, ламаркизм и
номогенез близки принятием положения о самодовлеющем характере про-
гресса (тогда как дарвинизм едва касается его, полагая его одним из след-
ствий приспособления к среде2-5); а дарвинизм и номогенез одинаковы в
том, что игнорируют активность особи - она в этих учениях не живет, а лишь
предъявляет свои заданные качества.
Истории уже посвящена часть 1, и можно в одном плане считать крити-
ку оконченной: поскольку отсутствие естественного отбора в природе
показано с той строгостью, какая принята в других разделах биологии, да-
лее идея отбора рассматривается только в порядке сопоставления учений.
Тот единственный известный мне реальный процесс избирательного раз-
множения, который может влиять на ход эволюции, далее будет описан под
названием «расплод» (п. 5-8*), чтобы не путать его с вымышленным явле-
нием - естественным отбором.
5-1*** Неслучайность «случайных мутаций»: основной постулат
СТЭ давно опровергнут опытами
Видеть в мутациях основной источник эволюционной изменчивости ста-
ли после того, как в 1932 году американский генетик Томас Хант Морган, в
прошлом эмбриолог, а в скором будущем нобелевский лауреат, провозгла-
сил (вопреки фактам), что «генетика заменила эмбриологию и внесла поря-
док в изучение эволюции» [Гилберт и др., с. 325]. До этого главным факто-
ром эволюции обычно полагали изменение зародышей, которое можно бы-
ло толковать (а можно и не толковать) как вызванное мутацией гена.
Но каковы сами мутации? В СТЭ их принято считать возникающими
беспорядочно и меняющими организм в любых направлениях, хотя давно
показано, что это не так (пп. 4-8, 4-14**). Еще в 1931 году германский генетик
Виктор Йоллос привел, длинный (6 позиций) ряд аллелей гена цвета глаз
дрозофил, в котором вызванные теплом мутации могут следовать только в
одном порядке - от дикого типа к white (белому) [Полое, с. 72].
В 1988 году американский генетик Джон Кэйрнс (с сотрудниками) поста-
вил опыт, сходный с опытом Лурии - Дельбрюка и пришел к выводу, ничто
тогда доказано не было. Кэйрнс смог лишь воскликнуть: «Поразительно,
сколь мало обоснованным было общепринятое мнение».
Действительно, общее мнение основывалось просто на априорной уве-
ренности ученых, что их мировоззрение безупречно. Опровергнутые Кэйрн-
сом опыты ставили, в сущности, лишь одну цель - собрать факты в пользу
царившего мировоззрения. А у Кэйрнса смертельные дозы были уменьше-
75
Н.В. Тимофеев-Ресовский в своей последней эволюционной статье (Природа, 1980, № 9)
признал, что дарвинизм следует дополнить принципом прогресса, но он остался в этом
среди дарвинистов одинок.
223
ubi до стрессовых, и сразу же обнаружились акты наследуемого приспо-
собления бактерий: кроме случайно возникших мутантов, нашлись и такие,
которые образовались в ответ на новый внешний фактор. Описание этих и
подобных опытов см. [Зусмановский; Назаров, 2005].
Исключив мутантов, экспериментаторы (Кэйрнс и соавторы)
“нерастущие клетки оставляли в условиях углеводного голодания. Сначала му-
танты отмирали. Но спустя неделю и более наблюдался новый рост за счет
вспышки реверсий именно в гене lacZ (без которых был невозможен рост в лак-
тозной среде — Ю. Ч). Как будто клетки в условиях жесткого стресса, не делясь
(!), вели генетический поиск и адаптивно меняли свой геном” (Голубовский М.Д.
Неканонические наследственные изменения // Природа, 2001, № 8, с. 8).
О генетическом поиске речь пойдет ниже, в п. 5-13.
В 1990 году генетик Барри Холл (США) получил удивительный резуль-
тат на двойных мутантах, т.е. при наличии сразу двух дефектов, прекра-
щающих рост на данной среде. Об этом пишет Голубовский:
“Выжить могли только особи, у которых реверсии возникали одновременно в
двух триптофановых генах. Частота таких особей была в 100 млн раз выше,
чем ожидалось при простом вероятностном совпадении мутаций в двух генах.
Холл... показал, что они возникают и у дрожжей, т.е. у эвкариот2'5”.
Иными словами, мутации у бактерий у оказались случайными, но на-
правленными. На дарвинистов всё это не произвело никакого впечатления;
они этого просто не стали читать (в чем я не раз убеждался в беседах). За-
чем знать лишнее?
Вскоре такие же результаты были получены и на более сложных орга-
низмах, и всюду оказывалось, что дело в физиологии. Об этом см. ЧЭ, с.
177 и [Назаров, 2005, гл. 14]. Однако, после короткой вспышки интереса
многих журналов (опыты были многократно подтверждены), всё вернулось к
прежнему, словно экспериментального опровержения СТЭ и не было.
Как отметил 90-летний Гершензон, учебники наивно продолжают уве-
рять, что мутации случайны (С.М. Гершензон. Избирательность мутагенного
действия ДНК и других полинуклеотидов // ЖСБ, 1996, № 6). В чем тут де-
ло? Давайте снова внимательно прочтем слова Стента: «...самое главное,
как следует рассуждать, чтобы получить...» - дело в них. Как мы теперь ви-
дим, результаты были заданы заранее, заданы всей тогдашней атмосферой
науки, и для социального успеха надо было рассуждать так, чтобы получить
именно их и ничего более.
2б
Эвкариоты, т.е. "собственно ядерные" - организмы, чьи клетки имеют ядро (в отличие
от прокариот, т.е. доядерных организмов, бактерий в широком смысле). Это русское на-
писание согласно с греческой и русской фонетикой. Греческий дифтонг ей- читается как
эв- (эвглена, эвкалипт, эврика, эвтрофия и т.д.) или как ев- (Евангелие, Евгений. Европа,
евнух и т.д.). Написанию же "эукариоты" нет русских аналогов, оно противоречит рус-
ской фонетике, и его трудно признать грамотным.
224
Разумеется, иные ученые получали тогда и другие результаты, но уче-
ный мир не принимал их [Голубовский И ЭБ], да и сейчас плохо принимает.
Так в науке было, есть и (полагаю) будет всегда. Единственное, что остает-
ся - пытаться сократить время, нужное обществу для усвоения нового. Для
этого мало быть способным и добросовестным, надо хоть немного знать
теорию научного познания, уметь выявлять недопустимые презумпции и от-
казываться от них. Тут главная роль выпадает историкам науки, философам
и учителям. Увы, до сих пор они занимаются, в основном, иным - насажда-
ют в качестве установленных фактов презумпции, привычные обществу.
Выяснив свою методологическую позицию, можно перейти к выявлению
своих теоретических положений. Прежде всего, нужно ясно указывать свои
постулаты и их источники. Затем, ни в коем случае нельзя замалчивать не-
приятные результаты. И, наконец, свои проблемы надо сопоставлять с по-
хожими проблемами других наук. Всё это вместе понадобится нам при опи-
сании понятия, основного для дальнейшего - активности.
5-2. Принцип активности
Мы видели, что самые разные эволюционисты, от Мопертюи до Кей-
зерлинга, пытаясь нащупать движущий эволюцию фактор, сравнивали его
с химическим сродством-, что появление дарвинизма не только не сняло
проблему движущего фактора (“буксира"), но даже обострило ее. Она была
не решена, а просто забыта со смертью современников Дарвина, как был
забыт весь прежний эволюционизм, но ныне ученый мир снова обращается
к эволюционным проблемам, и следует воспользоваться опытом прошлого.
Мне очевидно, что в основе возможности развития мира лежит некото-
рое свойство мира, действующее на всех его уровнях, некоторое мирообра-
зующее начало, более общее, нежели сродство частей к целому. Ему не раз
давали название (такова, например, воля Шопенгауэра), и я решил выбрать
самое общее и нейтральное слово - активность27. Ее нельзя ни из чего ло-
гически вывести, ее можно только ввести как постулат, интуитивно обоб-
щающий единое впечатление от природы. Нельзя и дать данному началу
определения, поскольку это - первичное понятие, такое же, как пространст-
во или время. Зато его можно и нужно обсудить, т.е. выразить через другие
основные понятия, тоже данные нам интуитивно.
Через всю историю европейской мысли, от мифа (Эрос), пантеизма
Ксенофана, Бога как деятельности у Аристотеля и мирового напряжения у
27 От лат. activus — действенный, деятельный. Слово активность в русском языке (как и
англ, activity, и франц, activite) многозначно, означая не только рассматриваемое здесь ми-
рообразующее начало, но и рабочую деятельность людей. Поэтому нужно пояснение. Та-
ковым для вводимого термина активность мне видится немецкое Vtirksamkeit. Оно обра-
зовано от прилагательного wirksam (действующий) и (в отличие от немецких Aktivitat и
Tatlgkeit) означает прежде всего активность как действующую силу и деятельное свойство.
225
Клеанфа, проходит та мысль, что мир существует и развивается за счет
пронизывающей его активности. В частности, физика как наука о неживой
природе приняла свою классическую форму в XVIII веке именно тогда, когда
признала свои собственные типы активности (сила, поле, энергия), отделив
их от феномена активности живого. В наше время физика строится на том,
что всякая сила есть итог действия соответствующего поля. Сила тяжести
есть итог действия гравитационного поля, а наиболее заметная механиче-
ская сила является совокупным итогом действия электромагнитных полей.
Поле часто принимает форму сродства (гравитация сближает всё, что
имеет массу, электростатическое поле сближает отрицательный и положи-
тельный заряды). Борьба физики Ньютона (поле) с физикой Декарта (вихри
частиц) была борьбой за право видеть активность материи как таковую.
Противники идеи поля отказывались признать гравитацию, говоря, что
‘тело не может действовать там, где его нет”, но сторонники Ньютона побе-
дили - тогда, когда понятие поля стало привычным. А привычным оно стало
потому, что позволяло получать практические результаты, и тем постепенно
привлекло ученых. (Наоборот, вихревая физика Декарта была в то время
способна лишь на произвольные толкования, как ныне дарвинизм.)
Через 200 лет то же повторилось с полями, вводимыми в теории кван-
тов. У физиков бытует афоризм: «квантовую физику нельзя понять, к ней
надо привыкнуть». Привыкнув, физики ею овладели, и полагаю, та же
участь ждет идею активности живого.
Существенно, что химия тоже ввела свои типы активности (валент-
ность, химический потенциал и др.), которые вроде бы могут быть выведены
из физических типов активности, но фактически эта задача решена лишь
для простейших примеров. А в общем случае она так и осталась благим по-
желанием, и проще сказать, что с охлаждением материи до планетных тем-
ператур возникли химические типы активности.
Аналогично с активностью живой материи: задачу ее выведения из фи-
зико-химической активности ставить можно (что и проделал на своем круге
задач В.Л. Воейков - см. гл. 7), однако на деле почти все свойства активно-
сти живого приходится рассматривать как самостоятельные.
Ныне общепринято, что по мере охлаждения Вселенной появлялись
новые типы полей и сил и что это усложняло Вселенную: с понижением
температуры до той, при которой кварки и глюоны могут сливаться в прото-
ны и нейтроны, началась атомная эволюция; когда атомы смогли соеди-
няться в молекулы, началась химическая эволюция. Повышение (с ростом
массы) давления внутри небесного тела приводит к его эволюции в звезду
или в планету с ее геологической эволюцией.
Область жизненных переменных находится внутри области химических,
поэтому начало биологической эволюции связано не с охлаждением мате-
рии и не со снижением давлений, а с ее усложнением - усложнением ве-
ществ, росте их разнообразия и усложнением их потоков в рамках планеты.
226
В физике переход вещества в качественно иное состояние именуется
фазовым переходом. Всем известны переход газа в жидкость и жидкости в
твердое тело, но соединение кварков и глюонов в нейтрон - тоже фазовый
переход (Райордэн М., Зэйц У. Первые микросекунды // ВМН, 2006, № 8).
Образование частиц в потоке плазмы, атомов - в потоке частиц, массы пу-
зырьков - в жидкости, деление клетки и т.д. - примеры локальных фазовых
переходов. А расслоение планеты на кору, мантию и ядро - глобальный
фазовый переход. Любой из них означает усложнение формы активности.
Это ведет к пониманию эволюции как расширения набора форм актив-
ности, достигнутых в фазовых переходах. Сперва это - активность космиче-
ских полей, затем частиц, веществ и их планетных потоков, ряд специфиче-
ски жизненных активностей. Последний сам удлиняется (так, у многоклеточ-
ных есть своя активность - активность клеточной дифференцировки). Нако-
нец, эволюция обществ и изделий - явный итог людской активности.
5-2* Идея активности в эволюционных учениях
Феномен активности живого известен всем культурам в форме призна-
ния души организмов. Начиная с ботаника Цезальпина (XVI в.) говорят и о
душе растения. Можно даже говорить о душе микроба, но - не вируса, ибо
он неактивен25, и нет оснований видеть в нем организм (подробнее см. гла-
ву 7). Начиная с Мопертюи, активность живого сопоставляют с химической
активностью, а начиная с Эразма Дарвина, все эволюционные концепции
так или иначе рассматривают активность как фактор эволюции.
Ламаркизм основан на двух типах активности: это активность особи и
стремление к прогрессу; является ли он активностью молекул, клеток, осо-
бей или таксонов - не говорится, но ясно, что в наших терминах акт про-
гресса таксона или экосистемы есть фазовый переход.
Цитолог растений В.Я. Александров (Ленинград) яснеее других понял,
что все задачи познания внутриклеточных, включая макромолекулярные,
процессов являют собой проблему поведенческой активности. Он даже
ввел в 1970 году (УСБ, т. 69, № 2) термин цитоэтология (наука о клеточном
поведении) [Александров]. Тогда эволюция - такая смена поведений микро-
структур, при которой меняются наследственные свойства организмов.
Жоффруизм, понимаемый в смысле п. 4-13, вводит собственную ак-
тивность зародыша как фактор эволюции. В работе возрастных физиологов
(Аршавский И.А., Немец М.Г. // Успехи физиол. наук, 1996, № 1) основной ее
механизм видится им как реализация доминанты (о ней см. далее, п. 5-11),
т.е. допускается, что зародыш ведет себя как активное целое. Иногда доми-
нанту связывают с потребностями развивающегося организма:
2Я
Вот иная позиция: “Вирусы... не проявляют свойств жизни. Но, попадая информацион-
но-энергетическое поле живых организмов, проявля[ют] свою активность, инициирован-
ную единой потребностью: существовать, размножаясь” (Зусмановский, с. 18].
227
“Выстраивается схема процесса самоорганизации: потребность — генетиче-
ская программа структурогенеза, потребность — генетическая программа
функциогенеза. [...] При одновременной актуализации ряда потребностей,
важнейшая из них формирует доминанту как первый акт поиска способа ее
удовлетворения” [Зусмановский, с. 112, 118].
Очевидно, что здесь потребность понята и как сама активность, и как
цель, ею достигаемая. Мы будем стараться этого смешения избегать.
Дарвинизм ввел активность в виде двух идей: активность особи - как
стремление к избыточному размножению (отсюда борьба особей за дефи-
цитный ресурс, а это тоже активность) и изменчивость наследственного
материала, не имеющая определенного направления (случайная). Обе по-
нимаются очень узко: они напоминают не гравитацию и не валентность, а
молекулы газа, бьющиеся в стенки сосуда. Однако уже Гольдшмидт ввел
понимание активности наследственного вещества в более прямом смысле:
“ген есть нечто, присутствие чего в половых клетках заботится о том (dafiir
sorgt), чтобы в организме возникла определенная особенность”; вскоре, в
1920 году, Гольдшмидт уточнил: ген — это “определенное количество опре-
деленного активного вещества” [Любищев, 2004, с. 38-39].
Номогенез достаточно равнодушен к идее активности, но все-таки у
Берга находим замечательную мысль:
“Какая причина заставляет организм изменяться в определенном направле-
нии, это пока для нас скрыто... Но замечательно, что организм обладает спо-
собностью активно приспособляться к среде, обнаруживая при этом как бы
присутствие некоего внутреннего регулирующего принципа”, причем “разви-
тие идет, нередко вопреки внешним условиям, в определенном направлении в
силу внутренних конституционных причин, связанных с химическим строе-
нием протоплазмы” [Берг, с. 135].
Если ни одна концепция не обходится без идеи активности, то разумно
ввести ее в явной форме, чтобы сделать из нее все нужные следствия.
Объекты склонны соединяться друг с другом, образуя при этом нечто
новое (т.е. образуя эмердженты по Луэсу). Эту тягу друг к другу для образо-
вания качественно нового отмечали многие. Напомню лишь то, о чем упо-
миналось ранее: принцип любви Гесиода и Эмпедокла, принцип сродства
Мопертюи (о сродстве в данном смысле см. [Чайковский, 1990, п. 9.3]) и
принцип взаимопомощи Кропоткина. В XX веке применяли и другие назва-
ния (например, социабильность). Биология лишь вступает на путь, пройден-
ный физикой и химией, на путь осмысления усложнения форм активности.
В 1907 году французский философ-эволюционист Анри Бергсон, из-
вестный ламаркист, назвал всякую активность живого объекта жизненным
порывом (elan vital). Термин «активность» предпочтительнее, ибо подчерки-
вает общность феномена, его господство на всех уровнях организованности
~ от элементарных частиц, через физику и химию, через жизнь и культуру,
До космоса. На мой взгляд, введение активности как первичного свойства
228
позволяет понять появление новизны (эмерджентность) как появление
новой формы активности, неизбежное при том усложнении материи, какое
происходило в ходе действия прежних форм активности.
Другое дело - считать ли активность живого особой сущностью или пы-
таться выводить ее из чего-то. В главе 7 мы увидим, что некоторые свойст-
ва активности живого можно вывести из химии, но не все. Есть и противопо-
ложная точка зрения: активность как живого, так и неживого, вытекает из
более общего свойства природы. Это свойство физик и натурфилософ Джо-
ханнес Бёрджерс (США) называл концептуальной активностью и полагал
его первичным; даже материя и поле для него вторичны. Благодаря такой
активности нарушается принцип причинности (точнее, добавляется целе-
вая причина Аристотеля):
«Каждое событие включает как эффекты прошлых ситуаций, так и предвиде-
ние будущих возможностей» (Burgers J.M. Causality and anticipation П Science,
1975, vol. 189, № 4198, p. 197).
Обычная физика, где принцип причинности работает, служит Бёрджер-
су таким же приближением к физике концептуальной активности, как физика
Ньютона - к физике Эйнштейна. За следующие 40 лет физика сделала важ-
ные шаги по пути отказа от привычного понимания материи (в пользу пони-
мания ее как взаимодействия полей, а затем и «темной энергии», являю-
щейся чистой активностью космоса), тогда как биологи надеются понять
эволюцию в рамках физики XVII века (когда в ней не было полей, а до-
пускался к рассмотрению лишь контакт частиц) до сих пор. Поэтому нам
стоит посмотреть, что же следует из идеи Бёрджерса. Сам он писал:
“Информацию или множество инструкций, зашифрованных в ДНК клеточно-
го ядра, можно скорее рассматривать как множество стратегий, чем как мно-
жество фиксированных программ. Эти стратегии обладают некоторой пла-
стичностью и могут адаптироваться к обстоятельствам... При их реализации
вступает в силу различающая, или концептуальная активность... Стратегии
могут вступать в дело одновременно или раздельно под влиянием старших
(master) стратегий. Поэтому должны быть формы концептуальной активности
разных уровней”. И далее: “Я допускаю, что можно рассматривать мутации
прежде всего как изменение стратегии. Это изменение может вызываться
перемещением определенных групп в ДНК и ферментах и, если оно не дест-
руктивно, оказаться подходящим” (Burgers J.M. Experience and conceptual
activity. Cambridge (Mass.), 1965, p. 175-176,179).
Как видим, процесс приспособления (адаптация) здесь выступает то-
же как форма активности (т.е., в сущности, по Ламарку).
Во многом Бёрджерс оказался прав: активность многоуровнева (п. 5-14),
а мутации являются, в основном, транспозициями (п. 5-6). Его интуиции (он
был физик-гидродинамик) биолог может лишь позавидовать.
229
5-2** Изменения активности. Давление нормы
Итак, будем понимать эволюцию как усложнение форм активности. Од-
ни скажут - активности материи, другие - активности материи и духа, третьи
(среди них был знаменит К. Бэр) - как возрастание роли и доли активности
духа. Не будем спорить. Главное, задан новый вектор исследования.
Но есть опасность - загнать всё непонятное в термин «активность», за-
менив тем самым исследование природы на игру словами (как это про-
изошло в дарвинизме). Выход видится мне в том, чтобы
1) чётко оговорить, что в эволюции не является активностью. Такова
упорядоченность материи, т.е. тот материал, на который действует актив-
ность. О типах упорядоченности речь пойдет в главе 6;
2) следовать Бергу, указавшему на «регулирующий принцип» (см. вы-
ше, а также далее, п. 6-12), и выявить регулирующие эволюцию принципы.
Они должны быть (в отличие от активности, вводимой как постулат) не по-
стулатами и не презумпциями, а осмыслением биологических фактов. В ос-
новном, это факты, которые прежде замалчивались или (что по сути то же)
объяснялись особыми формами отбора, специально для этого вводимыми.
Первое правило активности живого назовем давлением нормы. Суть
его в том, что в нормальном состоянии (когда объект соответствует своей
функции, своему назначению) изменчивость особей низка, а в ненормаль-
ном - высока. Мы рассмотрим этот феномен в п. 5-13, после введения поня-
тия стресса. И увидим, что снятие давления нормы - основной акт, запус-
кающий эволюционное изменение.
Второе правило - эволюцию движет избыток активности. Ее исся-
кание, наоборот, ведет к застою, а затем к вымиранию (в этом состоит брок-
кизм, п. 1-15*). Данным правилом регулируется ход эволюции и, в частности,
эмерджентность (см. выше). Последняя, видимо, поглощает всю избыточ-
ную активность вида, отчего новые формы сперва малочисленны.
Третье правило - это принцип компенсации (п. 2-7), который будет
рассмотрен в главе 8. Он регулирует распределение активностей по видам.
Суть его вовсе не в том, что активность достаётся всем поровну (некоторым
ее достаётся совсем мало), а в том, что ни один организм не получает ее в
таком избытке, чтобы развивать все доступные ему качества.
Бергсон говорил: «эволюция есть творчество». Но творчество есть выс-
шая форма активности, и в любищевском клане эту мысль Бергсона не раз
вспоминали. Зоолог Б.С. Кузин, давний друг Любищева, признавался;
“не могу представить никакого порядка, никакого космоса, возникшего без
участия творческого начала” {Кузин Б.С. Из писем И ВФ, 1992, № 5, с. 182).
Р Г. Баранцев, глава питерских любищевцев, уверен:
“Творческие способности живого существа проявляются через его активность.
Принцип активности играет ведущую роль в учении Ухтомского о доминанте”
[...] “Внутренний механизм упорядочения, каким бы он ни был, поддерживает-
230
ся, должно быть, потенциалом творческой активности живого вещества” (Ба-
ранцев Р.Г. Имманентные проблемы синергетики // ВФ, 2002, № 9, с. 96,98).
Нам надо наполнить эти философские тезы биологическим содержани-
ем, для чего сперва придется узнать кое-что о доминанте (п. 5-11), о новой
термодинамике и новой биологии развития (п. 5-14 и далее).
5-3. Познавательные модели эволюционизма
Для обозначения приёмов упорядочения и истолкования разнообразно-
го материала сообществом учёных методолог А. П. Огурцов в 1980 году ввел
понятие познавательной модели. Познавательная модель (ПМ) - это на-
бор приёмов и утверждений, которые данному учёному (учёным) настолько
наглядны и очевидны, что через них принято объяснять (к ним сводить, ими
моделировать) все основные факты и понятия. Огурцов видит ПМ как «ба-
зисную метафору». Подробнее см. работы [Чайковский, 1990; 1993] и осо-
бенно [Чайковский, 2004], где методу ПМ посвящена целая глава.
Для усвоения основной консервативной частью общества теория долж-
на соответствовать той ПМ, какая принята в обществе в данное время. Ино-
гда сразу господствуют две конкурирующие ПМ, но вряд ли более.
До рождения науки Нового времени в обществе преобладала этико-
эстетическая ПМ, видевшая природу как храм. Затем появилась знаковая
(первая научная) ПМ, видевшая природу как текст или как шифр', после
нее - механическая ПМ, видевшая природу как часы. В дни Дарвина ее вы-
тесняла статистическая ПМ, видевшая природу как совокупность весов и
бухгалтерских балансов. Именно она привела к господству кинетической
теории газов и рыночной идеологии (конкуренция с целью сдвига баланса в
свою пользу). В наше время ее сменяет системная ПМ. Она видит природу
и общество как целостность, более общую, нежели часы или весы, что
влечет призывы к созданию системной теории эволюции. Системная ПМ
склонна моделировать мир организмом (например, представлять себе и
клетку организма, и всю биосферу как организм).
Ныне статистическая и системная ПМ конкурируют, но уже нарождается
диатропическая познавательная модель, ставящая во главу угла феномен
разнообразия. О ней мы будем говорить в главе 6.
При господстве знаковой ПМ возникли зачатки номогенеза, при механи-
ческой возникли ламаркизм и жоффруизм, при статистической - дарвинизм,
при системной - физиологический и экологический эволюционизм. Как уви-
дим в главе 6, диатропическая ПМ дала новую жизнь номогенезу.
Рассмотрение всеобщей роли феномена активности само собой ставит
вопрос: какова будет следующая ПМ?
Поскольку ПМ по определению есть феномен социальный, то сейчас
нельзя сказать, какая из нынешних идей, не поддерживаемая еще никаким
общественным слоем, сможет стать будущей ПМ. Однако новую ПМ следу-
ет ожидать, если ни одна из существующих не указывает пути к решению
231
какой-то фундаментальной проблемы. Именно такой проблемой является
проблема эмерджентности эволюции (п. 3-10) и вообще всякой новизны.
Поэтому новую ПМ, нужную для ее усвоения, придется обозначить, пусть и
рискуя дать совсем ложный прогноз.
Вероятно, это будет активностная ПМ, видящая мир как обретающий
в ходе эволюции всё более и более сложные формы активности. Проще го-
воря, я надеюсь, что излагаемая концепция эволюции будет признана об-
ществом. Подробнее см. Дополнение 9.
Итак, сегодня можно говорить о семи ПМ - одной донаучной и шести
научных, последняя из которых лишь предполагается. Выпишем их:
0) этико-эстетическая (религиозная, донаучная);
1) знаковая (в прежних работах: семиотическая, схоластическая);
2) механическая;
3) статистическая (балансовая);
4) системная (организмическая);
5) диатропическая;
6) активностная (в прежних работах: креативно-пропенсивная).
Легко видеть, что чётные ПМ тяготеют к целостности восприятия (син-
тезу), а нечётные - к расчленяющему знанию (анализу, элементности). С
исторической точки зрения это понятно: маятник общественной моды посто-
янно колеблется от предпочтения целого к предпочтению деталей и обрат-
но. Некоторое сходство всех чётных ПМ ведёт к тому, что мы легко заимст-
вуем понятия для конструируемой будущей шестой ПМ из арсенала ранне-
го, в том числе донаучного знания. Такова и идея активности.
Каждая ПМ используется своими сторонниками для объяснения всего
на свете, однако устаревшая ПМ тормозит науку, а новая ПМ, наоборот, от-
крывает путь новым теориям. Так, всем известна польза от параллельной
деятельности многих на одном участке, однако статистическая ПМ видит эту
пользу в конкуренции, системная - в многослойной регуляции (о ней см. п.
6-8*), а диатропическая - в разнообразии самом по себе. Легко предсказать,
что активностная ПМ истолкует ее как синергию (рост совместной активно-
сти), повторяя этим нулевую ПМ. Ведь еще Гесиод дал в поэме «Труды и
дни» двойную характеристику Эриде (борьбе, конкуренции):
Знай же, что две существует различных Эриды на свете,
А не одна лишь всего. С одобреньем отнесся б разумный
К первой. Другая достойна упреков. И духом различны:
Эта - свирепые войны и злую вражду вызывает...
Первая раньше второй рождена многосумрачной ночью...
Эта способна понудить к труду и ленивого даже;
Видит ленивец, что рядом другой близ него богатеет,
Станет и сам торопиться...
Вот эта Эрида для смертных полезна.
Замечу, что тема конкуренции и ее благотворной (или разрушительной)
232
роли проходит через всю историю культуры и каждой эпохой принимается
как открытие. Не лучше ли сперва прочесть Гесиода и понять, что природа
соревнования действительно бывает различна?
Каждая ПМ имеет свой резон, но беда в том, что ее сторонники видят в
ней набор презумпций. Так, статистическая ПМ исходит из презумпции од-
нородного изотропного пространства (события, о связи которых ничего не-
известно, полагаются независимыми - см. [Чайковский, 1990, с. 210]), тогда
как действительная задача науки - поиск еще не известных зависимостей.
В отдельных науках могут господствовать ПМ, в целом уже ушедшие из
научной моды. До недавних пор генетика развивалась в понятийных рамках
знаковой ПМ, поскольку рассматривала наследственность в качестве гене-
тического текста. Возникновение науки в знаковой форме обычно (вспомним
пифагорейцев (п. 1-5*) и средневековую схоластику), но уход генетики из
этих узких рамок затянулся и еще далеко не осознан основной массой гене-
тиков, не говоря уж о работниках психологии, медицины, сельского хозяйст-
ва и природопользования, до сих пор ищущих гены отдельных свойств.
В настоящее время генетика освоила механическую ПМ (всевозможные
механические модели действия макромолекул, рибосом и т.п.) и статисти-
ческую ПМ (популяционная генетика, подсчеты частот генов, их мутаций, их
средней давности и т.п.), понемногу осваивает системную ПМ (это началось
с открытия оперонной регуляции) и даже диатропическую (это началось с
отказа от формулы «один ген - один фермент»). Однако царит в ней по-
прежнему знаковая ПМ, и это, как увидим в п. 5-5, неудобно. Тем более, ес-
ли принять законность новых (пятой и шестой) ПМ.
5-3* Идея Творения в дарвинизме и в иммунологии
Итак, внимательный анализ побуждал многих ученых вводить в дарви-
низм некий “буксир” и некоторые авторы прямо называли его активно-
стью. В XX веке про активность надолго забыли, полагая, что успехи гене-
тики решили или решат все эволюционные проблемы в чисто механических
понятиях дарвинизма. На мой взгляд, эта линия потерпела полный крах, и
основная причина этого - устаревшая ПМ. Все достижения биологии по-
следнего полувека получены помимо дарвинизма, а часто и в противоречии
с ним. Таков вопрос о понимании иммуногенеза, т.е. прижизненного обрете-
ния устойчивости к заразе (о врожденном иммунитете см. п. 6-5***).
Дарвинисты разрабатывали селективную теорию иммуногенеза, а их
противники - его инструктивную теорию. Обе возникли в 1900-х годах
{Т.И.Ульянкина. Зарождение иммунологии. М., Наука, 1994, гл. 6).
Автором селективной теории был Пауль Эрлих, основатель иммунохи-
мии. Он писал: «Физиологические аналоги антител должны существовать
заблаговременно в организме и его клетке», а иммуногенез состоит в пере-
делке аналогов в сами антитела и в усиленной выработке этих веществ.
Дальнейшее развитие науки показало правоту данного мнения, но на этом
233
пути не было ответа на вопрос, откуда «аналоги антител» берутся. Эрлиха,
разумеется, упрекнули в преформизме, т.е. в признании Творения.
Инструктивная теория, наоборот, видела антиген как матрицу, на кото-
рой как-то формируется антитело. Автор ее - Карл Ландштейнер, один из
творцов иммуногенетики. Она привела в свою пользу решающий факт: ан-
титела вырабатываются даже на синтетические антигены, в природе от-
сутствующие. Как возможен иммунитет к ним, если не путем распознавания
антигена? Но царил дарвинизм, и решающий факт оставили без внимания.
Оба, Ландштейнер и Эрлих, позже были увенчаны нобелевскими пре-
миями, но проблема от этого, естественно, с места не сдвинулась. Иммуно-
логи 1950-х годов прямо повторили ход мысли генетиков, которые видели в
эволюции лишь отбор полезных генов, возникших независимо от их пользы.
Вот данная мысль у иммунолога Питера Медавара, нобелевского лауреата:
«рождение “новой иммунологии”, как ее назвали, фактически датируется при-
знанием теории... что антиген просто выявляет способность, уже ранее сущест-
вовавшую». Точно так же, «процесс “обучения” бактерий - это эволюционный
процесс, сводящийся к естественному отбору форм, уже обладавших новой
способностью» {МедаварП., Медовар Л. Наука о живом. М., 1983).
Как видим, Лурия и Дельбрюк повторены точно.
Странно: ведь выработка антител на синтетические антигены была Ме-
давару известна. Но факт во внимание принят не был, и, более того, была
признана истинной селекционная теория - только на том основании, что ин-
структивная напоминала о ламаркизме (о чем у Медаваров сказано тоже).
Такое “понимание” эволюции на полвека закрыло путь к выяснению
становления иммунитета, о чем мы будем говорить далее подробно.
В самом же эволюционизме данное “понимание” бытует до сих пор. Вот
новейший пример. Петербургский зоолог В.В. Хлебович вспоминает старую
идею Болдуина, по которой полезные свойства возникают в виде прижиз-
ненных модификаций, а затем уж становятся наследственными (п. 4-14). Мы
уже знаем тот вывод СТЭ, что она противоречит идее отбора, и Хлебович
признал этот вывод логичным, но, как обычно в дарвинизме, привёл иное
мнение, чем и успокоился. Его в тот момент занимал не отбор, а само появ-
ление признака, и этим его статья нам интересна. Он пишет:
«Модификационная изменчивость связана с регуляцией активности генов [...]
Так называемую наследственную фиксацию признака, который ранее прояв-
лялся как модификационный, легче всего объяснить выпадением (дезактиваци-
ей) альтернативной программы» {Хлебович В.В. II Природа, 2006, № 7, с. 28).
Причем тут «легче»? Откуда в эволюции берутся сами «альтернатив-
ные программы»? Откуда гены, чья активность меняется? Автор ничего,
кроме ссылки на «эксперимент природы» с удвоением числа хромосом, на-
звать не смог. Позиция выглядит ненаучно, но она отнюдь не нова в науке.
На ней стоят противники эволюции (креационисты): у них все свойства
организмов, в том числе изменчивость, заранее заданы Богом при Творе-
234
нии, они обеспечивают частные приспособления сотворённых видов, но не
эволюцию. Так, напомню, понимал изменение видов сам Дарвин в молодо-
сти, а позже критики его книги дружно указали ему, что это - не эволюция.
Дарвин ничего по сути не ответил, а затем всё это было забыто - на сто лет
(п. 1 -1). Неужели креационизм вновь общепризнан? И да, и нет.
Да, ибо приведенные цитаты из Медавара и Хлебовича (а таких фраз -
многие тысячи) - являются примерами так называемого смягченного креа-
ционизма (п. 4-6) в духе Альберта Великого, вполне могут быть включены в
умеренный вариант «Закона Божия», что подчас учителями и делается.
Нет, ибо из таких фраз ничего для науки не следует. Это видно, кстати,
и из статьи Хлебовича: он даже не пытается увязать их с реальной генети-
кой, а множество приведенных в статье интереснейших фактов не имеет
никакого отношения к этим цитатам. Эволюционная наука развивается на
совсем ином интеллектуальном поле, что и постараюсь показать далее.
А пока, забегая вперед, отмечу обычный безрадостный факт: новое
знание во многих отраслях биологии добывается (с большим опозданием и
очень обрывочное) при яростном отрицании всякой возможности новизны. И
связано это, как увидим, именно с господством дарвинизма, отрицающего
активный поиск на генетическом уровне. Например, факт синтеза антител
путем распознавания антигенов многими отрицается до сих пор. Даже авто-
ры, признающие синтез антител «по принципу прямого слепка», могут в сле-
дующем же абзаце заявить, что сами они надеются на успех поиска
«механизмов клональной селекции» [Марков, Куликов, 2006, с. 140].
Как обычно, нормальная (по Куну) наука отчаянно сопротивляется
всему новому, если оно грозит ей самой. Это - отличная иллюстрация к
феномену давления нормы: нормальная наука есть та самая норма, которая
подавляет изменчивость, и ее снятие (научная революция) родит новизну.
Спрашивается - откуда же тогда колоссальный и быстрый прогресс
науки XX века, той же генетики? Ответ неожиданно прост: прогресс огромен
там, где не задевал мировоззрения, а там, где задевал, его не было вовсе.
Не следует думать, что иммунология и эволюционизм - что-то особен-
ное. Нет, точно то же можно наблюдать в разных науках, в том числе самых
что ни на есть прикладных. Не в “века мракобесия”, а в XIX - XXI веках вра-
чи предпочитали и предпочитают массами убивать и калечить пациентов,
нежели брать пример с коллег, применяющих методы вполне успешные, но
противоречащие привычному мировоззрению и господствующей ПМ.
Поразительно чудовищные примеры я нашел у историка медицины Л.С.
Салямона. Так, в середине XIX века в европейских клиниках свирепствовала
родильная горячка, уносившая порой до 30% рожениц. Молодой венгерский
врач Игнац Земмельвейс установил причину - грязные руки акушеров (руки
было принято мыть не до, а после “грязной” работы по приему родов; даже
если перед этим акушер вскрывал труп). В 1847 году Земмельвейс доложил
свой результат: мытье рук перед родовспоможением снижает смертность ро-
235
жениц в десятки раз - но был отвергнут коллегами. Он отчаянно боролся за
спасение пациенток и умер в 1865 году в психбольнице. Салямон писал:
“Наука движется как бы спиной к будущему; она пятится вперед и позволяет
нам обозревать пройденную дорогу. Тот, кто движется быстрее и обгоняет
своих современников, выпадает из поля зрения” (Салямон Л.С. О некоторых
факторах, определяющих восприятие нового слова в науке // Научное откры-
тие и его восприятие. М., Наука, 1971, с. 113).
Эти слова: наука пятится вперед - мы не раз припомним. Чудовищ-
ная практика продолжается поныне (примеры будут далее). Но всё же про-
видцы бывают. Из упомянутых ранее эволюционистов, это наш Страхов,
немец Нэгели и француз Вентребер. К последнему пора вернуться.
5-4. Молекулярный ламаркизм и иммунология
В 1937 году Вентребер вышел на пенсию и удалился в свой уютный до-
мик у Средиземного моря, как бы зная, что судьба дарит ему вторую жизнь -
28 лет размышлений на новую для него тему, об эволюции. В саду росли
мандарины, неподалеку были коллеги (академическая морская лаборато-
рия), через год из Парижа пришла весть, что он избран в академики, и даже
начавшаяся еще через год Мировая война не слишком изменила ход жизни
старого ученого. Еще через 10 лет, когда коллеги в перипетиях войны уже
его подзабыли, он прибыл в Париж и сделал в Академии наук блестящий
доклад со странным названием: «Химический ламаркизм» (Wintrebert Р. Le
lamarckisme chimique И CR, 1949, t. 228, № 13). Химии как таковой в нем не
было, и в наше время его назвали бы иначе - “Молекулярный ламаркизм”.
Вентребер повторил здесь то положение ламаркизма, что прогресс
«господствует над адаптацией» (т.е. служит основной чертой эволюции), и
согласился, что открытие мутаций стало решительным шагом в понимании
адаптации, но счел, что «интерпретация экспериментальных мутаций тре-
бует размышления». Он поделил их на три категории - 1) вызванные физи-
ческими и химическими мутагенами, 2) управляемые и 3) обязанные изме-
нению климата. По его убеждению,
«первые, полученные вне всякого естественного воздействия, являются пря-
мыми увечьями самых уязвимых генов, реализующими случайные ва-риации,
и в целом регрессивными. Лишенные эволюционного значения, они породили
у дрозофилы подлинный “двор чудес” в рамках этого вида».
Термином «двор чудес» («соиг des miracles» - кур дэ миракль) в сред-
невековом Париже называли притон нищих, где можно было встретить все
виды уродств, природных и сфабрикованных. Этой параллелью Вентребер
подчеркивал (замечу: не вполне законно), что спектр искусственных мута-
ций не дает материала для эволюции. Гораздо более перспективными он
счел вторую категорию мутаций, управляемые (dirigees).
Они, по его мнению, возникают в силу естественных воздействий - как
химических веществ (внешних и внутренних), так и чужих генов. Вентребер
236
сослался на совсем новые (1948 г.) результаты генетика А. Буавэна: тот вы-
зывал у одних бактерий мутации, обрабатывая их раствором ДНК других
бактерий. (Аналогичной работы Гершензона на дрозофилах он не знал.) Это
позволило Вентреберу высказать пророческий тезис:___________________
«организм может вписать ген в свою наследственность... и это легче
_____________происходит у низших организмов, чем у высших»._________
Сославшись еще на данные по мутациям у низших грибов, Вентребер
сформулировал свой тезис и иначе: имеющая эволюционный смысл мута-
ция - это не поломка гена, а скорее «прививка гена», подобная тем привив-
кам, какими пользуются растениеводы. Она должна быть вызвана воздейст-
вием природного агента и состоит в приспособлении аппарата наследст-
венности к этому агенту. (Замечу, забегая вперед, что это похоже на тот ме-
ханизм иммуногенеза, который узнали через 35 лет.)
Третью категорию (мутации, обязанные изменению климата) Вентребер
счел единственной, относящейся к традиционному ламаркизму, поскольку
она вызвана естественными изменениями жизненных условий.
Перечислив всё это, он заявил, что может снять два главных возраже-
ния против ламаркизма. Первое носит экспериментальный характер: при-
обретенные признаки не проявляют наследуемости. Он отвечал: нет ничего
удивительного в том, что изменения, происшедшие во взрослом родителе,
не достаются детям - ведь его репродуктивная сфера уже сформирована.
Надо изучать другие изменения - те, что идут при формировании организма
(он не сказал, что это одна из основных идей Э. Жоффруа Сент-Илера).
Второе возражение умозрительно: тело организма неспособно влиять на
собственную репродуктивную сферу. Тут Вентребер сослался на собствен-
ные эмбриологические работы, показавшие, что зародыш не так уж изоли-
рован от посторонних воздействий.
Итак, Вентребер решительно покинул «двор чудес» - область тех мута-
ций, с которыми имела дело генетика, и занялся подлинными, в его глазах,
эволюционными изменениями, которые называл «биологическими мута-
циями». Он напомнил, что главное в ламаркизме - активность особи, и зая-
вил, что организм активно производит собственные мутации. Подробнее см.
Аронова Е.А. Неумирающий ламаркизм... И БПС, 1997, №№ 41,42.
Как эмбриолог, Вентребер мог бы прямо выйти на эволюционную про-
блематику через описание изменений в развитии зародышей, но он предпо-
чел путь, которым не ходил до него, кажется, никто - представил в качестве
основного способа эволюции иммуногенез, т.е. выработку организмом ре-
акции на чужое вещество. Вне эволюции иммунитет и наследственность со-
поставляли давно (раннюю литературу см.: [Любищев, 2004, с. 91]). Эволю-
цию с иммуногенезом тоже связывали до Вентребера многие, но то были
иммунологи, и они всегда ставили противоположную ему цель - понять им-
муногенез, взяв за данное определенную эволюционную концепцию.
237
5-4* Вентребер, старец-пионер
Вентребер смело допустил, что у иммуногенеза и наследственности
есть что-то общее, и приступил к описанию собственного понимания мута-
ции, т.е. к сути своего химического ламаркизма, который «ищет опору в об-
щих и самых элементарных свойствах живого и находит ее в реакциях им-
мунизации». Вентребер предложил следующую схему приспособления ор-
ганизма к новой среде: если какой-то орган перестает справляться со своей
работой, он вынужден работать в ненормальном режиме, в нем начинается
производство какого-то вещества, вредного для организма, и это вещество
служит сигналом для поиска адаптации.
Мы теперь хорошо знакомы с таким пониманием механизма запуска
адаптации: это концепция, согласно которой процедуру адаптации запуска-
ет стресс (а запускающие саму механику стресса вещества - стрессоры).
Австрийский физиолог Ганс Селье развивал концепцию стресса с 1936 года,
но вряд ли о ней Вентребер знал. Фактически он отождествил стрессоры с
антигенами и решил, что иммунный процесс, ими запущенный, ведет к пе-
рестройке того гена, который ответствен за работу данного органа.
Здесь нетрудно заметить также то понимание работы генетической сис-
темы, которое позже получило название «.оперонная регуляция» (француз-
ские генетики Франсуа Жакоб и др., 1960). Как это происходит, Вентребер,
разумеется, сказать не мог - ведь тогда, в 1949 году, сами генетики далеко
еще не достигли согласия даже в том, что является веществом наследст-
венности - ДНК или белок. Однако Вентребер не только уверенно встал на
сторону приверженцев ДНК, но и дал набросок предполагаемого механизма
ее генетического действия: нуклеиновая кислота, находясь в цитоплазме,
как-то узнает о строении антигена; а сохраняет она это знание, делая его
наследственным, когда попадает в клеточное ядро. Об этом генетики стали
говорить лишь в 1960-е годы.
Мысли своего доклада Вентребер уточнял в докладах и статьях еще
три года, а затем на целых десять лет снова замолчал - писал книгу. За это
время была расшифрована двуспиральная природа ДНК, поставлена и ре-
шена проблема генетического аминокислотного кода и открыт оперон - ос-
новной элемент генетической регуляции у бактерий; но книга Вентребера
«Живое, творец своей эволюции» (1962) оказалась вовсе не архаизмом, но
наоборот, вполне актуальной.
Читая книгу, никак не скажешь, что автору 95 лет. Сперва (с. 3) он про-
вел ту идею, что живое есть двойственное единство “организм-среда" и «ре-
зервуар связей и запросов, которые могут быть удовлетворены», в чем по
сути повторял Спенсера и шел впараллель с нашим физиологом П.К. Ано-
хиным. Затем он уточнил прежние свои мысли:
1) повреждение гена ведет не к адаптивной реакции, а к ухудшению ра-
боты организма, к дефициту какой-то функции, и ее надо восстановить (в
238
наше время это называют генетическим поиском, см. п. 5-13);
2) сильное воздействие среды может привести к общему повреждению
гена, что делает дальнейшее развитие организма по прежнему пути невоз-
можным; тогда генетическая система реализует иной путь, например атави-
стический (возврат признаков предка);
3) поэтому мутагенез при сильном дискомфорте может не иметь ничего
общего с обычно изучаемым мутагенезом и даже приводить к появлению
нового вида (с. 131-132). Легко видеть, что тут он повторял сальтационистов
(см. п. 4-1), но важнее то, что он увидел, возможно, первый, два различных
стресса, о чем у нас будет речь ниже.
Примечательно: тогда началось неумеренное увлечение генетикой как
основой и даже сутью всей биологии (см. п. 4-17), но Вентребер заявлял со-
всем иное:
“Ген есть продукт протоплазмы. Составленный из ДНК и нуклеопротеина, он
... ничто иное, как продукт, сотворенный живым [веществом]. Тем самым, он
- его делегат в хромосомах, гормональная субстанция, состоящая в резерве и
используемая, когда надо” (Wintrebert Р. Le vivant createur de son evolution.
Paris, 1962, c. 137-138).
К сожалению, в книге не видно следов того, что автор прежде был эм-
бриологом и за это стал академиком. Теперь, как обычно у ламаркистов и
дарвинистов, он едва поминал развитие зародыша, предпочитая от половых
клеток прямо переходить ко взрослому организму, и считал форму следст-
вием функции. Ничего серьезного сказать об эволюции как последователь-
ности онтогенезов (ср. п. 4-13) он не мог по той простой причине, что считал
теперь онтогенез не самодостаточным процессом (как считал в 1935 г.), а
лишь кратким повторением (рекапитуляцией) филогенеза.
Главное однако, что Вентребер вернул ламаркизм в ряды современных
ему учений. Сперва тот ожил во Франции, затем увял там, но в 1980-х годах
дал ростки в англоязычных странах. Нас же интересует не судьба доктрин, а
понимание феномена эволюции, от кого бы оно ни исходило.
5-5. Ламаркизм и новая генетика
Под влиянием Вентребера к ламаркизму обратился крупнейший тогда
во Франции зоолог Грассэ. Ему принадлежит чеканный тезис ламаркизма,
прокламирующий активность:
___________«Жить, значит реагировать, а отнюдь - не быть жертвой»___
(Grasse Р.-Р. (-’evolution du vivant. Paris, 1973, p. 351). Он отметил и феномен
направленности эволюции, причем, в отличие от англоязычной традиции,
увязал его с параллелизмом:
“Палеонтология обнаруживает, что линии, исходящие от общего корня мате-
ринской формы, проявляют совершенно одинаковые тенденции к реализации
определенной формы... но в неравной степени” (с. 400).
239
Если Вентребера феномен направленности занимал лишь в рамках ор-
тогенеза, то Грассэ явно имел в виду параллелизм в духе Копа - Вавилова.
К тезису Вентребера о работе гена Грассэ добавил:
“внесение информации в геном есть операция, отличная от ее приобретения;
эти операции следуют друг за другом, а не осуществляются одновременно. ...
Возможно, что производство информации идет медленно и продолжается
очень много поколений ... Словом, ДНК регистрирует и стабилизирует эво-
люцию, но не творит ее” (с. 401).
Этим он отметил, что грань между ламаркизмом и дарвинизмом прохо-
дит вовсе не там, где тогда считали почти все, т.е. не по линии НПС, а по
линии активности особи вообще и активной записи информации в геном в
частности. Одним из первых Грассэ указал конкретные механизмы записи
(трансформацию ДНК, обратную транскрипцию и др.) в качестве факторов
эволюции, против чего дарвинисты тогда грубо протестовали.
Вообще, надо отметить, что в те годы мысли ламаркистов отвергались
без обсуждения как ненаучные. Теперь, когда они, в основном, приняты к
обсуждению и даже рассматриваются некоторыми как часть дарвинизма
(такова идея Вентребера об отсутствии эволюционной роли у большинства
типов мутаций), нельзя забывать, что дарвинизм яростно сопротивлялся им
всем без исключения и что это - обычная черта дарвинизма, быть может,
неотъемлемая. Отринув все открытия генетики (кроме одного: генетического
полиморфизма популяций) при их появлении, он не может, по-моему, счи-
таться научной дисциплиной, хотя сто лет назад ею был.
Сейчас он выполняет одну лишь полезную функцию: тормозя новое, ог-
раждает общество от скороспелых решений. Однако и ее следует выпол-
нять грамотно, на что дарвинизм уже почти негоден, ибо не выходит за по-
нятийные рамки генетики 1930-х годов. Даже серьезный научный креацио-
низм выполняет эту функцию лучше. Такова книга ЮШ, часто старомодная
и даже наивная, но указывающая на многие радикальные трудности в пони-
мании эволюции, чего дарвинизм избегает в принципе.
Генетик и историк науки МД. Голубовский в своих работах увлекатель-
но рассказывает, с каким трудом и опозданием внедрялись в сознание и в
практику биологов самые важные идеи генетики - от законов Менделя до
новейших открытий молекулярного ламаркизма. Его вывод:
“задержка, примерно 25-30 лет, в признании многих выдающихся открытий -
вполне закономерное явление в развитии науки” {Голубовский М.Д. Личност-
ное знание и парадоксы истории генетики // Природа, 1997, № 7, с. 68).
Это верно, но это половина правды. Судьба открытия радикально зави-
сит от того, насколько оно задевает правящую идеологию. Если не задева-
ет, то усвоение может быть мгновенным, и вся история генетики тому при-
мер: ее успехи за сто лет огромны и молниеносны. Но если открытие заде-
вает презумпции (например, дарвинизма), неприятие может продолжаться
сто лет и больше.
240
В качестве примера приведу ту тему, которой занимался сам Голубов-
ский, эпигенетическую наследственность. Еще в 1895 году Болдуин (см.
п. 4-14) высказал догадку, что полезное свойство возникает в ненаследст-
венной форме, а затем становится наследственным; позже ее высказывали
многие, но она отвергалась - дарвинисты видели тут вариант НПС. Разви-
тие молекулярной генетики в 1970-х годах указало на конкретный механизм
краткосрочного и не затрагивающего строение гена наследования: наследо-
ваться может не только структура гена, но и его состояние (активен или
нет); когда оно изменится, наследование пропадает.
Обычный путь дезактивации гена - метилирование некоторых нуклео-
тидных оснований в ДНК; фермент метилаза узнаёт нужное основание при
репликации и добавляет метильную группу в дочернюю цепь ДНК, так что
дезактивация наследуется, пока метил не будет удален. (Эпигенетическая
схема признана, в форме метилирования, действительно с опозданием на
25 лет, однако если считать от Болдуина, то получится 100 лет, а если
учесть, что учебники СТЭ молчат о ней поныне, то еще ждать и ждать.)
Дезактивация при этом необязательна, и такое метилирование очень
старо; достаточно напомнить, что этим путем родилась ДНК: тимин - это
метилированный урацил (подробнее см. гл. 7). Урацил может быть метили-
рован в разных местах, и один из вариантов (оксиметилурацил) служит вме-
сто тимина у многих панцирных жгутиконосцев (Dinoflagellata). Данный факт
открыт еще в 1973 году и затем подтвержден на нескольких видах (Galleron
C.//Orig. Life, 1984, vol. 13, N 3-4), но игнорируется до сих пор.
Приведя в сборнике ЭБ ряд фактов такого игнорирования, Голубовский,
к сожалению, не сказал, что речь обычно идет о молекулярном ламаркизме.
То же видим у биокибернетика С.Н. Гринченко: вводя два главных постулата
Ламарка (активность особи и усложнение организации), он ни Ламарка, ни
ламаркизм не назвал [Гоинченко, с. 24].
Пусть для укоренения нового в массовом сознании в самом деле по-
лезно избегать одиозных терминов, но для нашей темы (понять эволюцию)
надо называть вещи своими именами, дабы стало ясно, чем из прошлого
пора воспользоваться. Если мы будем, как прежде, ждать, пока дарвинизм
объявит нынешние достижения своей составной частью, то не только поте-
ряем время, но и останемся в неведении, куда двигаться дальше, ибо дар-
винизм, отрицая феномен активности как эволюционный, никуда не зовет
(подробнее см. [Чайковский, 2002]). Он очень плохо умеет даже “пятиться
вперед” (п. 5-3*), как это делают остальные науки.
В качестве недавнего примера напомню историю с интронами - уча-
стками ДНК, которые ничего не кодируют, в отличие от экзонов, между ко-
торыми интроны вставлены (для вырезания интронов и сшивки экзонов
служит особая процедура — сплайсинг). Интроны открыты в 1977 году.
Кроме них, загадкой являются многочисленные и многократные (до ста ты-
сяч раз в одном геноме) повторы коротких некодирующих последовательно-
241
стей ДНК. Повторы и интроны были объявлены “мусорной ДНК” (вот явная
издержка господства знаковой ПМ в генетике), и дарвинизм вскоре же раз-
вил доктрину «эгоистического гена», который якобы паразитирует внутри ге-
нома, где может размножаться безудержно, как саранча.
Однако тогда же было выяснено, что интроны и повторы служат для ре-
гуляции; появилась идея, что они нужны для эволюции: отделяя экзоны друг
от друга, они облегчают перебор комбинаций, и уже первые бактерии долж-
ны были иметь их и передать эвкариотам; позже многие бактерии могли их
утратить (Gilbert W. Why genes in pieces? // Nature, 1978, № 5645).
Затем роль повторов ДНК была осознана как задающая ряд черт онто-
генеза и эволюции [Галимов, 2001, с. 147-150; Корочкин, с. 17]. Наконец,
было выяснено, что интроны служат не только для компоновки генов, но и
как резерв генетического материала: иногда интрон включается в состав эк-
зона, причем кодирует активную часть белка и даже обеспечивает ему до-
бавочную функцию (Doherty J.K. е.а. И PNAS, 1999, vol. 96, р. 10869).
Конфуз с “мусорной ДНК” поучителен: причиной его была не какая-то
частная модель, а обычная для дарвинизма вера в то, что суть устройства
мира уже понята и потому всё непонятное можно счесть мусором. Мир же,
как мы не раз увидим, устроен удивительно и в главном еще непонятен.
5-5* Генетический текст читается на разных языках. Иносказания
Самое радикальное утверждение о повторах гласит, что избытка ДНК
вообще нет, а есть иной, кроме 4-х-нуклеотидного, вид информации:
“ДНК располагает иным, помимо уотсон-криковского механизма, способом
хранения и передачи информации. Оба процесса... могут взаимодействовать, а
могут быть практически независимы” (Акифьев А.П., Потапенко А.И. И Гене-
тика, 2001, № 11, с. 1455).
В качестве такого способа авторами названа упомянутая выше эпигене-
тика. Она поначалу кажется не имеющей отношения к эволюции - ведь тут
не создается новых генетических текстов. Но если видеть текст не только в
самой цепочке нуклеотидных пар, но, например, еще и в ряду сидящих на
ней метильных групп, то мы видим, что новый текст при метилировании
создается. Можно ли на этом языке записать нечто новое? Код нового белка
записать нельзя, но ведь эволюция эвкариот и не мыслится ныне в терми-
нах новых белков. Новое у них - это прежде всего новая расстановка бел-
ков, а это как раз и делается проще всего путем расстановки и снятия ме-
тильных групп (другие способы см. п. 5-6).
Информация содержится на ДНК не только в химической, но и в физи-
ческой форме (более общо: не только в форме текстов). Для этих целей
удобны как раз длинные повторы однотипных участков, главная часть ДНК.
Вероятно, что всё, касающееся пространственных форм (о них далее, п. 5-
15), наследуется именно так. В этом плане интересны соображения о “вол-
новом геноме” (ГаряевП.П. Волновой геном. М., 1993, 1997):
242
“Наши хромосомы реализуют программу строительства организма из яйце-
клетки через биологические, фотонные и акустические, поля. Внутри яйце-
клетки предварительно создается образ будущего организма,... генетический
аппарат проявляет свои потенции через голографическую память” (цит. ц0
[Зусмановский, с.32-33]).
По-моему, это интересно как иносказание. Голография (тип записи
изображения, при котором можно по малой части плоской записи восстано-
вить трехмерное изображение всего объекта), даже если принять, что она
тут возможна, мало чем полезна, так как требуется не воссоздать изобра-
жение, а создать сам живой объект и притом не сразу, а постепенно, путем
роста и развития (тезис Икскюля - см. далее, п. 5-15). Лучше говорить (на
сегодня, и тоже иносказательно) о фрактальном росте (см. далее, п. 6-7),
при котором малая часть также бывает подобна целому.
Таких иносказаний ныне в науке много, ими принято пренебрегать, но в
них есть глубокий смысл: все вместе их авторы ярко очерчивают круг неве-
домого (как прежде блистательно очерчивали, например, Берг и Вентре-
бер), чего вовсе не видно в академической науке. Собрав целый спектр но-
вых высказываний такого рода, А.Г. Зусмановский подвел итог:
“Если все действительно так... то генетический код - это ключ к блокам ин-
формации, которая... несет сведения обо всей совокупности параметров объ-
екта, закодированного в геноме”. Имеются “свободные последовательности
ДНК для фиксации благоприобретенной информации”; “сам по себе акт дуп-
ликации генов не создает новой информации, но подготавливает соответст-
вующий субстрат для кодирования на нем информации... необходимой для
конкретных конструкторских задач” [Зусмановский, с. 33, 35, 181].
К сожалению, ничего не сказано о количестве такой информации: вол-
новые и полевые иносказания предлагают каналы передачи, в тысячи раз
менее мощные, чем нужно для построения органов. См. Дополнение 2.
Однако запись генетической информации сразу на нескольких языках
наводит на мысль, что есть более высокий уровень записи и чтения
информации, нежели известный нам; он пособен понимать ее цельно, а не
только “буква за буквой”. Эту мысль подтверждает такой пример.
Исследовав хозяйственно полезного мутанта (барана с особо мясистым
задом и его потомство, наследовавшее это свойство непонятным образом),
бельгийский генетик Мишель Жорж (с сотрудниками) нашел, что для полу-
чения желаемого потомства нужны одновременно
«обычный белок-кодирующий ген, один или более генов, кодирующих только РНК,
два эпигенетических фактора (метилируемых участка ДНК - Ю. Ч.) и в заклю-
чение - точковая мутация, замена А на G “посреди генной пустыни, за 30 тыс.
пар нуклеотидов до ближайшего гена”» (Гиббс У. «Теневая» часть генома: за
пределами ДНК И ВМН, 2004, № 3, с. 66),
причем эта сложная совокупность действует согласованно, цельно, хоть и
разобщена в геноме (правда, вся помещается в одной хромосоме).
243
С первых лет генетики были известны хромосомные мутации, т.е. скач-
кообразные наследуемые изменения структуры хромосом. О них много пи-
сали, но роль их оставалась неясной до тех пор, пока информацию рас-
сматривали как текст, записанный четырехбуквенным нуклеотидным кодом
(вот наглядный пример того, как устаревшая ПМ мешает познанию).
В настоящее время известно, что основная масса эвкариотной ДНК
служит не для кодирования нуклеотидных текстов, а для пространственной
организации хромосомы - как при функционировании неделящейся клетки
(Петрова Н.В., Яровая О.В., Разин С.В. Специфическая пространственная
организация хромосом... поддерживается ядерным матриксом И Доклады
АН, 2006, т. 406, № 1), так и при ее делении (Поляков В.Ю. и др. Структурно-
функциональная модель митотической хромосомы И Биохимия, 2006, № 1).
Это наводит на ряд мыслей о том, что хромосомные мутации, меняя строе-
ние и ориентацию генетического материала, должны играть не меньшую
роль в эволюции, нежели мутации в генах.
Подтверждением служат классические опыты цитогенетика В.Н. Стегния
(Томск), показавшие ход видообразования у двукрылых насекомых. Еще не
зная большей части нынешних данных внутриядерной цитогенетики, Стег-
ний сумел показать, что видообразование у эвкариот вызвано перестройкой
ядерных мембран. Она связана с перестройкой хромосомного аппарата, ко-
торая не затрагивает самих генов, но меняет пространственную организа-
цию интерфазных (вне митоза) хромосом. Вид образуется скачком, причем
из вида, чья зона обитания сузилась. Исходный вид должен иметь высокий
эволюционный потенциал (сложное понятие; в частности, требуется слабое
крепление хромосом к ядерной мембране).
Существенно, что у Стегния отрицается роль полиморфизма в видобра-
зовании: по исследованным свойствам хромосом все особи данного вида
(всего изучено 7 видов малярийных комаров) идентичны. Затем сходные
результаты были получены на дрозофилах (Стегний В.Н. Архитектоника
генома, системные мутации и эволюция. Новосибирск, НГУ, 1993).
Причины скачков Стегний видит в холодовом стрессе и жёстком инбри-
динге29, каковой неизбежен при падении численности-?0. Автор указал, что
развивает идеи макромутаций Гольдшмидта и Далька (системные по сути).
В терминах п. 5-3: он отказался от знаковой ПМ как от искажающей матери-
20 В отличие от большинства авторов, сводящих инбридинг к росту доли гомозиготных
। енов, Стегний отмечает, что «жёсткий инбридинг», т.е. спаривание братьев с сестрами и
родителей с детьми, порождает высокую изменчивость всех типов, особенно в генера-
। ивных клетках. Этот важный для эволюции вопрос почти не изучен.
' ° А.М. Словников любезно обратил мое внимание на тот факт, что инбридинг при сни-
жении численности приближает процедуру естественного размножения к процедуре
искусственной селекции. В этом свете данные Стегния обретают особый эволюционный
смысл. См. далее, п. 5-8*
244
ал и рассмотрел всю картину в рамках системной ПМ. Мы вернемся к теме
генетических языков в п. 6-7, при обсуждении генетического кодирования
программ роста в онтогенезе.
Следует говорить об усложнении не только структур, но и сил. Дело в
том, что информационные процессы в живом загадочны, между прочим, в
энергетическом смысле: направление биологического процесса часто про-
тивоположно потоку энергии в нем. Иными словами, сигнал ничтожной силы
вызывает мощную реакцию (такова замена нуклеотида, если влечет пере-
мену в организме; таковы все акты поведения [Александров]), подчас много-
кратную. В неживой природе похожие процессы (автокатализ, фазовый пе-
реход, сход лавины) всегда однонаправленны и для повторения требуют
восстановить объект.
Итак, на иных языках, вероятно, записана та информация, о которой се-
годня можно лишь фантазировать. Если мы прямо признаем, что новое на-
до искать и осмысливать, а не отвергать и не сводить (ссылаясь на “бритву
Оккама”) к старым понятиям, то тут же увидим, что новые открытия молеку-
лярной генетики выстраиваются в единую стройную картину. Вот два таких
открытия, и оба связаны с активностью в смысле п. 5-2.
5-6. Прыгающие гены и редактирование РНК
В 1951 году Барбара Мак-Клинток (США) открыла на кукурузе гены, ко-
торые не имеют определенного места на хромосоме. Хотя она была одним
из самых известных в мире генетиков, ее открытие не принималось сообще-
ством лет 20, поскольку противоречило уверенности, что каждый ген распо-
ложен на хромосоме жестко и может менять место только в ходе инверсии и
транслокации. (Издержки устаревших ПМ - знаковой и механической.) Лишь
к 1977 году было выяснено, что “прыгающие гены" есть у организмов всех
царств - у бактерий, грибов, растений и животных (LR, 1977, № 81, с. 784).
Генетическую частицу, способную к миграции, именуют транспозоном.
Обычно она содержит несколько генов, однако мигрировать может и мень-
ший генетический элемент.
То, что обычно называют мутацией, чаще всего (у дрозофил на 80%)
являет собой итог транспозиций. Частота этих актов сходна с частотой
классических “мутаций” и бывает от одной миллионной до одной десятой
[Геном..., с. 55-56, 87]. То есть тоже никаким числом не выражается.
Об эволюционной роли транспозонов пишет Голубовский. В его экспе-
рименте 1977 года транспозиционная “мутация” в раннем онтогенезе дро-
зофилы привела к стопроцентному появлению уродливого потомства:
“Сие чудо, когда все потомство одного самца несло мутацию-уродца, снимает
обычное возражение Р. Гольдшмидту, а с кем же будет размножаться макро-
мутант? С самим собой...”.
Голубовский видит тут генетический довод в пользу афоризма палеонтолога
Отто Шиндевольфа: “Первая птица вылетела из яйца амфибии” {Голубов-
245
ский М.Д. Век генетики: эволюция идей и методов. СПб., 2000, с. 201).
Сейчас о транспозонах мы знаем больше. Наиболее изучены рет-
ротранспозоны (продукты обратной транскрипции РНК в ДНК):
«Мутагенез, вызываемый ретротранспозонами, имеет ряд особенностей.
Инсерции (вставки - Ю.Ч.) ретротранспозонов происходят не случайно по
длине хромосом. Характерно наличие “горячих” и “холодных” сайтов, склон-
ность к встраиванию в определенные гены и в премоторные области генов...
Возникающие мутации обычно нестабильны и продолжают мутировать в
иные аллельные состояния и ревертировать к норме» (Андрианов Б.В. Инду-
цированная ретротранспозиция // УСБ, 2005, № 1, с. 87).
Мы вернемся к транспозонам после знакомства с явлением стресса.
Оказываясь в новом месте, пен иначе работает сам и изменяет работу
соседей, так что мобильность генов - фактор эволюции. Механизм ее по-
началу был совсем непонятен, и его сочли случайным, как прежде сочли
случайными мутации. Но в обоих случаях правильнее говорить о законо-
мерностях (переработки генетических текстов), до сих пор слишком слож-
ных для уяснения.
Ведущий генетический инженер первой волны Филип Курильский
(Франция) писал о “генетическом гении”, т.е. механизме разумного перебора
вариантов {Kourilsky Р. Le genie genetique // LR, 1980, № 110). Идея под-
твердилась открытием в 1980-х годах механизмов альтернативного сплай-
синга и редактирования РНК.
Альтернативный сплайсинг - сшивание гена из экзонов (при удале-
нии интронов) разными способами. Наличие такого явления означает, что,
читая один и тот же генетический текст, можно прочесть разные смыслы.
Это заставляет отказаться от самых привычных объяснений (см. п. 6-12).
Редактирование РНК (the RNA-editing) — феномен направленного (не
вполне случайного) изменения текста РНК в ходе транскрипции или после
нее, но до сплайсинга. Редактированию подвергаются как информацион-
ные, так и рибосомные, и транспортные РНК многих организмов. Феномен
известен для ядерных, митохондриальных и хлоропластных генетических
систем, как для экзонов, так и для интронов, а для прокариотных генетиче-
ских систем его возможность пока-только обсуждается. Но если учесть, что
митохондрии и хлоропласты похожи своей генетикой на прокариот и что сам
феномен редактирования РНК рассматривается как один из самых древних,
то наличие его и у прокариот более чем вероятно. Редактированию подвер-
гаются также РНК многих вирусов (Дейчман А.М. Редактирование РНК. М.,
2001, с. 4-8, 102). Открытие данного феномена важно не только само по се-
бе, но и для понимания того, насколько мы мало знаем о переработке гене-
тической информации и сколь неоправданно легко многие списывают непо-
нятное на случайное.
Изложу основное для нас ее суждение подробно, опуская, однако,
ссылки на обширную литературу.
246
Редактирование РНК - более тонкая процедура, чем те, какими обычно
объясняют возникновение новой генетической информации (рекомбинация,
дупликация, транспозиция и др.), так как обеспечивает точную замену не-
большого (вплоть до точечного) фрагмента генетического текста. Ученые
признают редактирование РНК загадочным, поскольку неясны ни его цель,
ни, тем более, селективные преимущества редактируемых сайтов. Меха-
низм этого многошагового «фермент-каскадного процесса» непонятен, как
неясны выбор сайта для редактирования и запускающая процедура. Благо-
даря редактированию (и, добавлю, не только ему) нарушается тот принцип
генетики, согласно которому один ген ответствен за синтез только одного
белка. Непонятно,
«почему клетки часто предпочитают постоянно содержать и запускать энер-
гоемкие “редактирующие машины” (в том числе для редактирования мРНК-
транскриптов), вместо того, чтобы однократно... внести нуклеотидные изме-
нения в сами гены».
Неясно, существует ли единый контроль за одновременно идущими ак-
тами редактирования. Влияет ли редактирование РНК на тексты ДНК? На-
конец, если редактирование исправляет ошибки в РНК, то зачем в ДНК
сохраняются “ошибочные” тексты? (Дейчман А.М., с. 4-18,103, 104).
Ответ на последние два вопроса по-моему очевиден: обратная транс-
крипция (см. п. 5-7) наверняка, пусть и редко, переводит редактированные
тексты в ДНК. Тексты, подлежащие редактированию, вовсе не все ошибоч-
ны, им достаточно быть всего лишь не соответствующими моменту. Само
же наличие механизма быстрого направленного изменения генетической
информации открывает огромные возможности как для прижизненных из-
менений, так и для эволюции. В сущности, прижизненное и эволюционное
смыкаются, и мы должны с благодарностью вспомнить Эразма Дарвина,
Ламарка и Жоффруа, особенно - принцип активности.
По-моему, открытие феномена редактирования РНК заполняет давнюю
брешь в понимании эволюции как преобразования генетических текстов.
Хорошо известна параллель между таким преобразованием и записью но-
вой мысли. Математик А.Н. Паршин (ВФ, 2000, № 6, с. 109), изображая дан-
ную параллель в табличной форме, положил транскрипцию и трансляцию
аналогом логического вывода, что явная натяжка, поскольку в этих процес-
сах текст изменяется лишь в порядке ошибки. Наоборот, редактирование
изменяет тексты РНК в порядке нормальной работы и тем делает аналогию
мышления с созданием новых генетических текстов законной.
Очевидно, что здесь налицо активность генетической системы, сход-
ная с мышлением. Загадочность обоих явлений значительно уменьшится,
если допустить, что редактирование РНК выступает в качестве акта
внутриклеточного мышления. Допущение сможет быть принято как
факт или отвергнуто, когда будет выяснено, в чем конкретно состоит суть
привносимой редактированием информации (уже сейчас ясно, что процесс
247
общезначим, поскольку основная масса ДНК транскрибируется, но не участ-
вует в кодировании белков). Мы воспользуемся этим допущением в пп. 5-
13* и 6-15*, а сейчас посмотрим, как феномен редактирования изменяет
наши основные представления об эволюционной генетике самой по себе.
5-7. Центральный тезис вместо «центральной догмы»
Вскоре после понимания наследственной роли ДНК надолго утверди-
лась идея, что ДНК является «главной молекулой»: она может служить мат-
рицей как для себя, так и для РНК, а РНК - для себя и для белка, тогда как
ни белок, ни иные молекулы не могут служить матрицами ни для себя, ни
для нуклеиновых кислот. Это - центральная догма молекулярной биоло-
гии, ее выдвинул Френсис Крик в 1958 году. Идею в 1970 году пришлось
подправить - была открыта обратная транскрипция (синтез ДНК на РНК-
овой матрице), которую используют РНК-овые вирусы, чтобы встраиваться
в геном хозяина. Однако в остальном идея однонаправленного потока ин-
формации еще лет двадцать оставалась незыблемой.
Верна ли она? Если понимать ее узко, как запрет «обратной трансля-
ции», т.е. синтеза РНК на белковой матрице, то, вернее всего, р,а31. Но
обычно данную “догму" понимают широко - как главенство ДНК в биологии и
как невозможность передачи наследственной информации иначе, нежели
на нуклеиновых матрицах. А это уже неверно.
Легко видеть, что “догма” в широком понимании (а мы будем говорить
только о нем) - утвердилась в умах как новая формулировка отрицания
НПС, т.е. как форма отрицания дарвинизмом собственной активности ге-
нома. Ситуацию четко обозначил в 1968 году английский биоматематик-
дарвинист Джон Мейнард-Смит:
«Огромное достоинство центральной догмы состоит в том, что она наконец
прояснила, что должен делать ламаркист: он должен ее опровергнуть».
Другими словами, центральная догма - презумпция, и опровержением ее
откровенно предложено заняться тем, кто в нее не верит. К сожалению,
среди ламаркистов не нашлось молекулярных генетиков, поэтому задачу
пришлось решать людям, от проблем эволюции далеким.
Первые два удара по догме нанес американский вирусолог Дейвид Бал-
тимор. В 1970 году он был одним из троих, открывших обратную транскрип-
цию, и ему же посчастливилось в 1982 году оказаться в паре, открывшей
нематричный синтез ДНК. Он обнаружен в системе иммуногенеза (Alt
F.W., Baltimore D. И PNAS, 1982, vol. 79, p. 4118-4122). Оказывается, в ходе
синтеза гена, кодирующего иммуноглобулин (белок, атакующий чужеродное
31 Впрочем, обратная трансляция вполне могла существовать в прошлом (см.
п. 7-8**), а некоторые молекулярные биологи (например, А.М. Дейчман) допус-
кают ее наличие и в современных организмах.
248
тело), происходит сшивка фрагментов прежних генов, причем в точке сшив-
ки в текст ДНК встраивается небольшой (в опыте Альта и Балтимора: коди-
рующий до 8 аминокислот) фрагмент, никакой матрицей не кодируемый и
синтезируемый (как и встраиваемый) чисто ферментативно. Подробнее см.
популярную статью [Чайковский, 2003].
Третий удар нанесло открытие наследственности без нуклеиновых ки-
слот, от белка к белку. К 1973 году было уже известно, что центриоль (ор-
ганелла внутри клеточной органеллы - центросомы, организующая полюс
деления клетки), образуется от материнской центриоли без участия нуклеи-
новых кислот (Фрей-Вислинг А. Сравнительная органеллография цитоплаз-
мы. М., Мир, 1976, с. 106). Однако данный факт оставался без всякого вни-
мания лет 25, пока не были открыты прионы - белки, передающие инфек-
цию (до этого считалось, что инфекцию могут передавать только микробы и
вирусы). Лишь теперь стало ясно, что в клетке
“существуют две категории матричных процессов: копирование последова-
тельностей ДНК и РНК и копирование конформаций... некоторых белков. Сво-
его рода матричное воспроизведение конформаций имеет место в таких про-
цессах, как образование структур цитоскелета-^2..., воспроизведение ядерной
мембраны... Эти механизмы могут лежать в основе так называемой эпигенети-
ческой наследственности/изменчивости” (Инге-Вечтомов С.Г Прионы дрож-
жей и центральная догма молекулярной биологии // ВРАН, 2000, № 4, с. 305).
Главным же ударом по догме стало раскрытие феномена редактирова-
ния РНК: если теперь и можно говорить о “главной молекуле”, то ею оказа-
лась РНК. Она играет центральную роль во всех клеточных процессах, в
том числе и в качестве опорной структуры (в рибосомах), и в записи новой
информации на ДНК.
В то же время ДНК инертна и выполняет только функцию хранения и
передачи наследственной информации. Ее, кстати, РНК тоже умеет выпол-
нять (в РНК-вирусах), но в наше время выполняет редко: именно из-за от-
сутствия инертности она - плохой хранитель. Этот факт будет для нас ва-
жен в главе 7, при обсуждении происхождения жизни.
От «центральной догмы» не осталось ничего вне ее самого узкого
смысла (запрета на обратную трансляцию), а в таком понимании она пере-
стала быть центральной - независимо от того, существует ли обратная
трансляция в природе или нет.
Редактирование РНК (и альтернативный сплайсинг как частный случай)
может рассматриваться одновременно и в качестве активности генома, и
в качестве акта внутриклеточного мышления.
Учтя сказанное в пп. 5-4 и 5-6, в качестве центрального тезиса молеку-
лярной биологии, но никак не ее догмы (ведь хорошо известно, что любые
J Имеются в виду процессы вроде активного роста микротрубочек (см. п. 5-15).
249
догмы губят науку), примем следующий тезис (рис. 26):
Главное назначение белка - обеспечение функций (ферменты) и всевоз-
можных структур. Главное назначение РНК - обеспечение функций (синтез
других РНК, ДНК и белков; геномная цензура), структур (в рибосоме) и из-
менение наследственной информации. Главное назначение ДНК-хране-
_____________ние и передача наследственной информации._____________
Есть и менее общие роли данных макромолекул: белок может переда-
вать инфекцию (прион), РНК может быть ферментом (рибозим), ДНК служит
опорной и ориентирующей структурой в эвкариотной хромосоме.
Рис. 26. Центральный тезис молекулярной биологии. Сплошные стрелки
означают копирование матриц, а штриховые - ферментативные связи
Предложенная форма
центрального тезиса пред-
ставляется мне умеренной
- есть гораздо более жёст-
кая позиция. Ее вот уже лет
40 прокламирует биолог
широкого профиля Барри
Коммонер (США), уверен-
ный, что белок играет не
меньшую роль в наследст-
венности, чем ДНК. Многие
годы он был одинок (и ушел
в охрану природы), но сей-
час и к его молекулярным
взглядам стали прислуши-
ваться. См. например:
Commoner В. Unravelling the secret of life: DNA self-duplication, the basic pre-
cept of biotechnology, is denied //Gene Watch, 2003, Mai - June.
К этому можно добавить, что 2/3 регуляторов метаболизма - не белки, а
РНК (Маттик Дж. Тайна программирования сложных организмов И ВМН,
2005, № 1, с. 30).
5-8. Экспериментальная эволюция. Подход иммунологов
Рассмотрим подробнее самый изученный тип альтернативного сплай-
синга. Он наблюдается в иммуногенезе.
Наконец-то сдвинулся с мертвой точки давний спор сторонников селек-
тивной и инструктивной теорий: обе оказались по-своему правы. Выявление
антител (иммуноглобулинов), пригодных для борьбы с данным антигеном, и
их размножение действительно текут приблизительно так, как утверждает
первая, однако их гены не предсуществуют, а формируются в ответ на по-
250
явление антигена, и формируются отнюдь не только путем мутаций. Стано-
вится все более очевидно, что информация о строении антигена каким-то
образом используется при формировании антитела, а если так, то надо го-
ворить о собственной активности генома.
Теплокровные имеют все те виды иммунитета, что у растений и у дру-
гих животных, плюс ТВ-систему, т.е. совершенный адаптивный (приобре-
тенный) иммунитет. Она состоит из двух подсистем - одна генерирует Т-
клетки, связывающие антигены на своей поверхности (имеется в различ-
ных формах у всех позвоночных), а другая генерирует В-клетки, синтези-
рующие иммуноглобулины (есть в развитом виде только у теплокровных).
Общее назначение иммунной системы видится как надзор за целостностью
(п. 5-15), а в отношении борьбы с заразой надо знать, что Т-система борется
с вирусами, а В-система - с бактериями.
Иммуноглобулины различаются вариабельной частью, она меняется от
клетки к клетке и ответственна за то, что каждая клетка поражает один тип
антигена. Рассмотрим один аспект работы одной подсистемы - синтез ва-
риабельной части легкой цепи иммуноглобулина в В-клетке. Дело идет так
(см. например [Стил и др.]).
У зародыша млекопитающих совсем немного генов иммуноглобулинов
- около сотни. В ходе развития организма их разнообразие каждый раз соз-
дается заново (точно так же, как создается любой орган) создается путем
комбинирования фрагментов существующих генов. Но этого разнообразия
оказывается мало, поэтому конкретное антитело обычно не выбирается из
наличных, а создается в ответ на конкретную заразу (на антиген). В стрес-
совой ситуации, какую создает вторжение антигена, включается механизм
перестройки иммуноглобулиновых генов: по каким-то не вполне еще понят-
ным правилам генетическая система режет и сшивает фрагменты ге-
нов до тех пор, пока не найдет приемлемый вариант - тот, что синтезирует
антитело, которое реагирует с вторгшимся антигеном. Найденный вариант
клонируется (размножается из единственного родоначального экземпляра;
клон - это бесполое потомство единственной особи).
Можно ли сказать, что она режет и сшивает случайно? Нет, нельзя
(точнее см. далее, п. 5-18). Проблему стараются обходить, но уже сейчас
одно надо прямо признать: тут налицо активность генома.
Механизм комбинаций работает, но грубо: поставляет антитела, связы-
вающие антигены, но довольно слабо. Поэтому нужен еще один механизм -
соматический гипермутагенез, который включается после создания нуж-
ной комбинации фрагментов. Заключается он в том, что при клонировании
гены найденного варианта мутируют с огромной частотой (тут каждый ты-
сячный нуклеотид заменяется, тогда как самопроизвольный точковый мута-
генез в 100 млн раз менее интенсивен), так что порождается масса чуть от-
личных антител, различающихся одной аминокислотой или двумя, чем и
достигается точная подгонка антитела к антигену. Конечный вариант снова
251
клонируется в качестве нормы (вот самый яркий пример давления нормы)
и запоминается генетической системой, т.е. наследуется на время жизни
особи (а иногда и долее).
В этом, грубо говоря, состоит «генетический принцип обеспечения раз-
нообразия антител», за открытие которого получил в 1987 году нобелевскую
премию Сусуму Тонегава-?-? (работа начата в Швейцарии, завершена в
США). Представляя лауреата, шведский иммунолог Ханс Вигзел сказал:
“Когда Тонегава проводил свои опыты, царило убеждение, что каждый белок,
каждая полипептидная цепь управляется своим геном, в отношении один к
одному. Но в то же время подсчет числа генов, кодирующих белки, в хромо-
сомах человека дает число порядка ста тысяч генов. Они должны обеспечи-
вать все белки тела — для гемоглобина эритроцитов, для пигмента в наших
глазах и т.д. Только меньшая часть, быть может один процент, вероятно мо-
жет быть использована для порождения антител. Около тысячи генов, спо-
собных породить миллиарды различных форм антител? Задача казалась не-
разрешимой”. И далее: “Тонегава ... открыл нечто совсем новое и революци-
онное в генетике.” (Les Prix Nobel 1987. Stockholm, 1988, с. 25.)
Суть открытия в том, что ген может быть активно переделан в
цитоплазме: возникшие при перестройках (собственно механизм Тонегавы)
фрагменты сшиваются, причем с нематричными вставками (механизм Альта
- Балтимора), затем успешный вариант точно подгоняется к антигену (ме-
ханизм гипермутагенеза), клонируется и запоминается (соматическое на-
следование). (Позже, в 1995 г., был открыт еще один этап - замена отдель-
ных фрагментов [Стил и др., с. 112].)
Словом, гены антител образуются не за счет мутаций, как думали
прежде, а путем четырехступенчатого процесса активности, в кото-
ром лишь одну ступень можно назвать мутагенезом, и то в особом
смысле: он направлен - в том плане, что происходит только в нужных
генах и в малом числе направлений, зато с неимоверной частотой.
Воздав должное такой разумности природы, Вигзел заметил:
“Каждую минуту наше тело производит миллионы белых кровяных телец,
лейкоцитов. В каждом из них идет гибридизация ДНК, приводящая к созда-
нию ее собственных, уникальных антител. Те, что не будут выявлены, быстро
погибнут. Если, однако, они вступят в контакт с подходящими внешними
структурами, они будут вознаграждены, допущены к размножению и прожи-
вут долго. После большой случайностной генной лотереи естественный отбор
поддержит победителей...”. (Подчеркнуто мною.)
Сам Тонегава провел параллель с эволюцией еще более прямо:
33 Великолепное открытие, за которое Тонегава получил премию, далеко не це-
ликом принадлежит ему. Подробнее см. ЧЭ, с. 190-191. Увы, это норма. В от-
ношении открытия спирали ДНК см.: Андреева Н.С. II Природа, 2006, № 8.
252
“Подобно организмам в экосистеме, эти лимфоциты - субъекты отбора анти-
генами, и приспособленнейший будет выживать. И... иммунную систему ин-
дивида можно видеть как своего рода Дарвинов микрокосм” (Les Prix Nobel
1987. Stockholm, 1988, с. 223).
Параллель с эволюцией налицо, и можно сказать больше: иммуногенез
выступает как эволюционный эксперимент, как модель эволюции путем ак-
тивности генома. Но надо выяснить, при чем тут Дарвин.
5-8* Не отбор, а расплод
«Допущены к размножению» - термин селекционеров, он относится не к
естественному, а к искусственному отбору, и это вовсе не оговорка Вигзе-
ла или Тонегавы, а самая суть дела: оказывается, иммуногенез схож не с
тем отбором, на котором Дарвин построил свое понимание эволюции, а с
другим, искусственным, который был привлечен им лишь для аналогии.
Выходит, что Данилевский (п. 3-7) был прав, указывая на теорию есте-
ственного отбора как на подмену понятий. Но прошло сто лет, и оказалось,
что он был неправ в другом - когда заявлял, что в природе нет аналога ис-
кусственному отбору. Аналог не только есть, но именно он работает эффек-
тивно - это отбор антител.
Данная подмена трудно уловима. Даже вдумчивые биологи подчас
склонны считать всякий отбор, идущий в природе, естественным по Дарви-
ну, поскольку-де искусственный не может происходить в природе по опре-
делению. Однако сам Дарвин прекрасно видел разницу в механизмах дей-
ствия этих отборов и в 4-й главе «Происхождения видов» писал:
“При методическом отборе животновод или растениевод отбирает с некото-
рой определенной целью, и если допустить свободное скрещивание особей,
его труд будет совершенно потерян”.
К этому надо добавить: селекционер сам назначает высокую размно-
жаемость избранным обладателям желаемого свойства, тогда как естест-
венный отбор по определению действует лишь на то свойство, которое дает
размножаемость более высокую, чем у других. Кроме того, вспомним, что
искусственный отбор действует наверняка (см. п. 1-12).
Этими свойствами естественный отбор не обладает, и Дарвин данную
брешь в своем учении видел, но полагал простодушно, что большие време-
на и численности могут восполнить все недостатки, хотя это ниоткуда не
следовало. Наоборот, все эффективные процедуры случайного поиска по-
строены принципиально иначе, чем избирательная размножаемость [Гоин-
ченко, гл. 4.4]. С этой позиции иммуногенез и надо исследовать.
«Дарвинов микрокосм», как он изложен и в «Происхождении видов», и в
СТЭ - это мир редких малых вариаций, последовательно вытесняющих
друг друга в борьбе за дефицитный ресурс. Данного явления никто никогда
не видел, и сказанное в п. 4-20 позволяет считать, что не увидит.
Иммунногенез же - реальность, и в нем всё течет иначе, не по Дарвину.
253
Сигнал к размножению дает клетке ее антитело, связавшееся с антиге-
ном, причем ни конкуренции за ресурсы, ни сравнения клеток по выживае-
мости тут нет: словно селекционер на ферме, иммунная система колос-
сально размножает тех, кто несет желаемый признак, и вовсе не допускает к
размножению остальных. Малые вариации являют здесь только одну из
ступеней изменчивости, тонкую подстройку, и отнюдь не являются редкими.
Еще в дни Дарвина были ученые, желавшие видеть в схеме Дарвина
процедуру активного выбора, что показали все ранние переводы (француз-
ское election, итальянское elezione, немецкое Zuchtwahl, русское подбор ро-
дичей и т.п. - см. ЧЭ, с. 81). В XX веке дарвинизм эту точку зрения отверг и с
тех пор во всех своих формах отрицает эволюцию путем активного выбора,
а вот иммунологи странным образом признают факт активного выбора дар-
винизмом. Попробуем выяснить причину этой странности.
Селекционизм первоначально появился в эволюционной литературе
как утверждение, что изменение видов в природе происходит путем, анало-
гичным селекции домашних пород человеком (божество активно отбирает).
Эта мысль, заимствованная из естественного богословия, четко выражена
в ранней рукописи Дарвина (см. п. 1-12), и только в этом смысле можно го-
ворить про “Дарвинов микрокосм” иммуногенеза. Но в самом дарвинизме
эта параллель исчезла, уступив место статистической схеме (медленный
сдвиг средних характеристик популяции под действием конкуренции за де-
фицитный ресурс). Ничего такого при синтезе антител нет.
Скорее тут Тонегаве надо бы сказать не «Дарвинов микрокосм» (не
имел же он в виду ранний очерк Дарвина, известный лишь дарвиноведам), а
«микрокосм Эмпедокла и Клеанфа» (см. пп. 1-5 и 1-7).
Дело, однако, не в образном выражении, а в уяснении того факта, что
иммуногенетика, если ее рассматривать как модель эволюции, заставляет
принять идею активного выбора, и сразу в двух смыслах:
1) он действует на итог направленной изменчивости и
2) не связан рамками размножаемости (он сам задает их).
На первый смысл указал еще Геккель (в этом состоял тогдашний ламарко-
дарвинизм), на второй - Виганд и Данилевский.
Предложена «Двухэтапная схема макроэволюции», в точности повторяющая
два этапа синтеза антител - комбинацию блоков и точную подгонку мутагенезом
{Тиходеев О.Н. Молекулярные механизмы макроэволюции // ЖОБ, 2005, № 1),
но автор не подтвердил ее фактами и не провел параллели с иммуногенезом.
Рассмотренному типу активности следует дать такое название, которое
исключит путаницу. Название не должно содержать слова «отбор»
(selection), поскольку слишком прочна его связь с дарвинизмом, а исходное
значение (сознательная сельскохозяйственная процедура) утеряно.
Не желая вводить неологизм и не найдя лучшего варианта, предлагаю
старинный термин расплод - в том значении, которое дает ему словарь
В.И. Даля: отглагольное существительное от глагола расплодить', тем са-
254
мым, оно исходно означает процедуру получения массового потомства от
избранного варианта. В этом значении оно соответствует латинскому слову
proseminatic?4 и английскому the breeding.
(Кроме этого, в быту слово расплод применяется еще и как итог дан-
ной процедуры, т.е. как множество молодых особей; точно так же, как слово
приспособление означает и процесс, и его итог. Далее в тексте слово рас-
плод будет означать только процедуру, а не ее итог.)
Перечислю свойства расплода как процедуры:
1) производится активный выбор того свойства, которое подлежит изби-
рательному размножению;
2) особям, обладающим данным свойством, обеспечивается размноже-
ние, тогда как остальные особи от него устраняются;
3) если размножение половое, то желательным свойством (свойствами)
должны обладать оба родителя (нужен активный подбор пары);
4) Все эти три процесса направленны, случайность минимальна.
И селекция на ферме, и опыт Агаева (см. ниже, п. 5-9), и завершение ак-
та иммуногенеза, это - точно установленные акты расплода.
Возможно, что быстрая эволюция в опыте Шапошникова (п. 4-18) тоже
включала акт расплода (напомню, учёному не пришлось завершить опыт).
Наоборот, если изменчивость не имеет нужного для эволюции направ-
ления, избирательная смертность ничего изменить не может (самый яркий
пример: половое поколение термитов). Одна из причин в том, что ненаправ-
ленная изменчивость не может преодолеть «барьер Нортона», т.е. изъять
вредный признак нацело (пп. 4-1,6-4**) ни при какой смертности.
5-9. Экспериментальная эволюция. Подходы иных биологов
Итак, иммунология выступила как экспериментальный полигон эволю-
ции. Впараллель работали и другие “полигоны”. Вот некоторые.
1) Индустриальный меланизм. В Англии, в середине XIX века, когда
лишайники, не выдержав загрязненного воздуха, стали вымирать, а березо-
вые стволы потемнели от гари, неожиданно потемнели и бабочки березо-
вые пяденицы (Biston betularia), чья окраска имитирует лишайник.
Первую меланистическую, то есть темную, форму бабочки поймали в
Манчестере в 1848 году, а всего 70 лет спустя она преобладала во многих
индустриальных областях Европы и Америки. Впервые на глазах ученых
возникло приспособление, для которого были точно известны начальное и
конечное состояния, скорость распространения и внешняя причина. Было
установлено, что птицы чаще склевывают бабочек, отличающихся по цвету
от их обычного фона, что темный цвет - результат мутации одного гена, что
мутация эта изредка появляется во всех популяциях березовых пядениц, но
широко распространяется только там, где темный цвет оказывается выгод-
34 Впервые употреблено М. Хэйлом (в англ, форме proseminatiori) на с. 119 его книги.
255
ним. Естественно, пример меланизма березовой пяденицы бытует в книгах
по дарвинизму как пример «эволюции по Дарвину».
Однако скептики не находят в истории с пяденицей подтверждения
дарвинизму. В примере с пяденицей главное для ламаркиста - активное
стремление бабочек приспособиться к новой среде (здесь: выбрать нужный
фон), для дарвиниста - в независимости меланистических мутаций от усло-
вий среды, а для номогенетика - в том, что из необозримого множества по-
тенциально возможных мутаций природа достаточно часто извлекает при-
способительную, меланизм (для проявления меланизма нужно, чтобы в
нужное время в нужных клетках шел синтез черного пигмента).
Не менее важно, что меланизм не создает никакого нового рисунка, а
лишь разрушает прежний, заменяя его черным пятном. Наконец, как и пред-
сказал Нортон (п. 4-1), отбор пядениц лишь сдвигает долю гена, но не может
сравнять ее ни с нулем, ни с единицей. Т.е. отбор даже в этом случае не
привел к акту эволюции [Зусмановский, с. 139].
И все-таки пример с березовой пяденицей - блестящий пример эволю-
ции. Подумайте: какой смысл в темном наряде тому, кто рожден сидеть на
светлом фоне? Однако выяснилось, что темные бабочки так же боятся
светлого фона, как светлые - темного. Бабочка сама выбирает себе фон,
сообразуя его со своей раскраской. Но это - активность, а ее как раз и ут-
верждают ламаркисты в качестве фактора эволюции. Без этой поведенче-
ской активности «полезная мутация» была бы бесполезна.
Это справедливо и в отношении растений: еще Дарвин обращал вни-
мание на важную роль, которую играют в жизни растений различные движе-
ния (в поисках опоры, света, влаги). Иногда, как и в случае термитов, иде-
альный опыт с растением ставит сама природа, заставляя его проявить
подлинную изобретательность55. Приспособительную роль играют различ-
ные изменения физиологии организма при попадании его в новые условия.
Словом, живое активно приспосабливает себя, каждая особь пытается при-
строить к делу даже свое уродство, и если это ей удается, то она - «обна-
деживающий урод» (термин Гольдшмидта, п. 4-7).
Поэтому, в согласии с тезисом ламаркизма (п. 5-5),
поведение особи оказывается в самом деле эволюционным
________________фактором, обеспечивающим приспособление.______________
2) Опыты Кэйрнса. Кэйрнс (п. 5-2) не сам придумал свою схему опыта
- выращивание организмов в непригодных для них средах имело давнюю
Изумительный пример поведенческой адаптации: обыкновенная сосна, растущая на пес-
ке в зоне ураганов, изогнула от середины ствола вниз пару ветвей, причем эти ветви
вросли в песок (Агафонов Б.П. И Природа, 2005, № 2). См. фото в начале книги. На отбор
тут нельзя сослаться хотя бы потому, что объяснить надо реакцию особи в данном поко-
лении, а не в их ряду. Впрочем, в этом смысле загадочен любой онтогенез.
256
историю. Идею высказал еще Дарвин. В «Происхождении видов» есть пункт
«Акклиматизация», где выражена надежда, что превратить теплолюбивые
турецкие бобы в холодостойкие сможет тот, кто попробует
«высевать свои бобы в течение по крайней мере двадцати поколений так рано,
чтобы основная их часть погибала от мороза... соберет затем семена с выжив-
ших экземпляров, тщательно избегая случайных скрещиваний».
Опять видим, что в отбор случайностей Дарвин не очень верил.
3) Яровизация. Давно известно, что если озимую пшеницу многократно
высевать весной, она становится яровой. К сожалению, на яровизации сде-
лал карьеру Лысенко, и потому заниматься ею стало непрестижно и даже
опасно, но все-таки дагестанский агроботаник М.Г. Агаев, позже работавший
близ Ленинграда, сумел доказать, что при яровизации, как и при озимиза-
ции, происходит настоящая экспериментальная эволюция: за 10-15 поколе-
ний образуется новая стойкая «видовая форма»; он избегал говорить о но-
вых видах, указывая, что дарвинизм отрицает быстрое видообразование
([Агаев]; основные его мысли можно найти также: БЖ, 1965, № 6; 1967, № 7;
1968, № 1; 1969, № 9; 1970, № 1). Замечательно, что его объекты - автога-
мы (размножаются самоопылением, т.е. клонально).
Наиболее нам интересно то, как именно получалась «видовая форма».
Так, в опытах с яровой пшеницей сорта Диамант видим, что сперва выжи-
ваемость при осенних посевах очень низка (1-8%) и необычных особей нет,
но на 7-й год вдруг сразу выжило 98,7% особей и среди них более половины
оказались необычны строением. Выживаемость осталась очень высокой и
далее, т.е. озимая форма выведена, зато доля необычных особей упала на
8-й год до 6% и на 11-й до 1,5% [Агаев, табл. 73]. Новая форма сразу най-
дена почти всеми, кто сумел, в одно (7-е) поколение.
Не будучи генетиком, Агаев не мог выявить наследственный механизм
изученной им эволюции. Не будем винить его за отсутствие генетического
анализа - привлечь генетиков к своей еретической работе он вряд ли в то
время мог. Скорее наоборот - лучше было не привлекать их внимания. У
Агаева есть мимолетная ссылка на аналогичные работы Шапошникова с
тлями, о которых мы уже говорили в п. 4-18 - они тоже остались без генети-
ческого анализа, зато вызвали санкции.
4) Клоны тлей. До Шапошникова с тлями работал ламаркист Е.С.
Смирнов и получил НПС, но лишь на 15 поколений (так называемые дли-
тельные модификации - см. п. 6-10*). Шапошников (ЭнО, 1965, № 1, с. 22)
полагал, что если в опыт Смирнова ввести высокую смертность, то можно
получить стойкое изменение строения тлей.
Прав ли был Шапошников, неясно, так как его опыт не был завершен,
но похоже, что Смирнов имел дело просто со снятием давления нормы, а
Шапошников добавил суровый стресс (о нем см. п. 5-11**) и наблюдал, что
«до 7-го поколения включительно жизненность тлей была крайне неустой-
чивой» (Шапошников Г.Х. И ЭнО, 1961, № 4, с. 745).
257
Позже опыты были повторены на другом роде тлей и дали результаты,
вовсе несовместимые ни с каким отбором, ибо весь процесс приспособле-
ния проходил за 2 (два!) бесполых поколения при ничтожных численностях:
десятки начальных особей в каждом варианте и всего две сотни конечных,
во всех вариантах вместе (Сапунов В.Б. Адаптация к перемене экологиче-
ских условий и фенотипическая изменчивость тлей // ЖОБ, 1983, № 3; во-
преки заглавию, речь шла и о генетической изменчивости тоже).
Через 45 лет после первых статей о тлях проблема дошла до Запада.
Сходные результаты получены в США еще на одном виде тлей, и уровень
аргументации остался примерно тот же: быстрая перестройка объяснена
отбором, а источник столь обширной изменчивости связывается с единст-
венным половым поколением (S. Wia, A.J. Shaw. Short-term evolution in the
size and shape of pea aphids // Evolution (USA). 1996, № 1). Тут признано, что
«механизмы пока неясны», и это само по себе - успех: прежде было ясно
всё. Мы вернемся к данному вопросу в п. 5-13*.
Осталось незамеченным, что как раз клон для такого быстрого
процесса и нужен. Все примеры сверхбыстрой эволюции относятся к слу-
чаям резкого ограничения процессов перемешивания генов - либо к кло-
нальному размножению, либо к тесному инбридингу (в том числе инцесту).
К сожалению, в традиции дарвинизма принято игнорировать роль кло-
на и инбридинга в эволюции - на том основании, что они снижают изменчи-
вость, а инцест, к тому же, снижает численность и вообще ухудшает свойст-
ва потомства (в среднем). Но в природе живут не средние величины, а жи-
вые существа, и тесный инбридинг, как знают селекционеры, необходим при
создании породы - в качестве этапа. Мы вернемся к этому в п. 9-16*.
5-9* Экспериментальная эволюция у бактерий
Немало было опытов и на бактериях. В сущности, они велись по той же
схеме - на непригодной для питания среде почти все клоны вымирают, зато
оставшиеся быстро эволюируют. Но, в отличие от опытов ботаников и зоо-
логов, здесь проводился детальный генетический анализ. В 1976 году груп-
па английских биохимиков во главе с Б. Хартли опубликовала изящный ре-
зультат: бактерию Klebsiella aerogenes, способную расти на рибитоле, нау-
чили расти на ксилитоле и выяснили природу ее эволюции. Рибитол и кси-
литол - два пятичленных спирта, оптические изомеры, различные положе-
нием одной ОН-группы (рис. 27). Эволюция бактерии состояла в выработке
нового фермента (ксилитолдегидрогеназы) на базе прежнего, рибитолде-
гидрогеназы. Отличие их - в одной аминокислоте в активном центре фер-
мента, и для получения новой формы нужно каждый раз около 100 поколе-
ний в хемостате, содержащем сто миллиардов бактерий, т.е. один мутант
приходился на 10 триллионов бактерий (если код новой аминокислоты от-
личается от кода прежней двумя нуклеотидами). Однажды такой опыт удал-
ся без применения мутагенов и еще в четырех случаях - при обработке
258
слабым мутагеном. Такие результаты отмечены серьезным научным креа-
ционизмом (ЮШ, с. 43-47) как показывающие предел эволюционных воз-
можностей процедуры отбора мутантов.
СН2ОН I сн.оь I * Рис. 27. Пятиуглеродные спирты: ксилитол
сно® сно® (слева) и рибитол. Кружком обведен водо- род, отрываемый при первом шаге ассими-
но сн сн он ляции молекулы бактерией
сн он сн он Вскоре (1978 г.) появился обстоя-
сн2он I сн2он тельный обзор американского биохимика Тай Тэ By (Т.Т. Wu), растившего бактерии
на ксилитоле, где подведен итог «экспе-
риментальной эволюции бактерий»: путем замены нуклеотидов можно дос-
тичь только замены одной-единственной аминокислоты в белке; таким спо-
собом можно слегка изменить активный центр фермента, но и только; более
сложные изменения белка требуют других средств. Например, By сравнил
три сходные дегидрогеназы, у которых различие состоит в том, что их ак-
тивные центры расположены в разных участках белковой цепи, и заключил,
что это различие не могло быть следствием точкового мутагенеза. То есть
бактериологи, как и иммунологи, могут объяснить отбором ненаправленных
мутаций лишь тонкую подстройку. Литературу см. [Чайковский, 1990; 2002].
Разумеется, слова «могут объяснить» вовсе не означают, что так оно в
природе и есть. Как раз аналогия с иммуногенезом (где происходит расплод
избранных иммунной системой вариантов, но не сравнение выживаемостей)
наводит на мысль, что у бактерий суть дела тоже не в отборе.
Например, главную роль в выживании популяции бактерий зачастую иг-
рает биоплёнка (тонкое скопление бактерий на какой-нибудь поверхности).
Немногие особи плёнки (от одной на сто до одной на сто тысяч), выживаю-
щие не за счет полезной мутации, а за счет прекращения активности, тем
самым переживают недолгую катастрофу без приспособления к новым ус-
ловиям. (Это похоже на эффект группы у растений, п. 4-15.) Выживание
биоплёнок - один из главных типов лекарственной устойчивости бактерий
(Льюис К. Персистирующие клетки... И Биохимия, 2005, № 2).
Перемещение активного центра по белковой цепи в наше время при-
вычно, поскольку хорошо известны механизмы перемещения фрагментов
ДНК (транспозоны). Называть такое перемещение мутацией, дело вкуса, но
надо понимать, что при этом полностью меняется смысл слова. В частно-
сти, нет пока никаких оснований называть перемещения случайными - кро-
ме той традиции, которая полагает случайным всё, что еще не понято.
Идея замены источника пищи с целью наблюдения эволюции вытекает
из принципа смены функций Дорна (п. 3-6). В итоге выяснено, на что спо-
собны мутации (с учетом принципа Дорна): если нет гипермутагенеза, то
259
для простейшего полезного изменения необходимы (это не означает, что
достаточны) триллионы особей. Даже для бактерий такие числа многоваты,
а для многоклеточных просто нереальны. Ни один вид крупных животных не
имел триллиона особей за всю свою историю, а эволюируют они, как ни па-
радоксально, намного быстрее мелких (см. п. 9-1).
Эволюция явно требует направленных изменений, и они вскоре были
найдены. В 1984 году было описано появление у бактерии нового фермен-
та, разлагающего нейлон, т.е. вещество, отсутствующее в природе. Фер-
мент образовался не путем серии мутаций, а скачком: в фермент-
предшественник добавлена вставка и обрезаны концы (Ohno S. // PNAS,
1984, vol. 81, р. 2421, цит. по [Зусмановский]).
5-10. Экспериментальная эволюция. Модели размножения
Математики (начиная с Нортона, п. 4-2) взялись заполнить ту брешь в
дарвинизме, что создалась из-за отсутствия доказательств (и даже наблю-
дений) эффективности отбора малых вариаций. Английский статистик Ро-
налд Фишер в 1922 году показал, что слабо выгодная мутация имеет неко-
торую вероятность прожить неограниченно долго.
Статья задала тон огромной литературе по математическому дарви-
низму, поэтому в параграфе 5-10 книги ЧЭ она была рассмотрена. Там рас-
сказано, что исходные допущения Фишера лишены биологического смысла,
а его основной вывод вовсе не означает возможности эволюции путем сла-
бовыгодных мутаций. Далее там же рассмотрены мои статьи по выживанию
мутантного клона, вынуждающие отказаться от отбора как ведущего факто-
ра эволюции. Всё это теперь потеряло актуальность после выяснения того
обстоятельства, что сама идея естественного отбора однозначно опровер-
гается фактами (см. п. 4-20). Единственное, что, пожалуй, стоит отметить,
это - катастрофический отбор.
В 1979 году был поставлен компьютерный эксперимент, где клон пона-
чалу имел миллион особей и за время 100-120 поколений уменьшал свою
численность по разным правилам, задаваемым различными видами функ-
ций s(t)<0, где s(t) - селективная ценность особи, a t - номер поколения
[Маленков, Чайковский]. Отрицательность всех вариантов s(t) как раз и опи-
сывала ситуацию катастрофы. Мы решили считать близкими по селектив-
ной ценности такие клоны, у которых средние по времени значения s разли-
чаются не более, чем на 2%. Большинство таких клонов имело в опыте
близкие судьбы, но удалось найти режим, в котором они хорошо различа-
лись. А именно, удалось подобрать такие s(t) и такие моменты времени Т,
что если прекратить катастрофу при t = Т, то один клон оказывается почти
вымершим (живо лишь несколько особей), а другой - почти в целости (живо
больше половины от начального миллиона).
Вот он, решили мы, долгожданный отбор малых вариаций - тот, в кото-
рый верил Дарвин и в который еще не утеряли тогда веры мы.
260
Радость длилась недолго. Поскольку роль отбора не была для нас пре-
зумпцией, а от бритвы Оккама нас уберег тезис Шрейдера, мы решили вы-
яснить, как же должны конкретно различаться селективные ценности, чтобы
отбор их уверенно различал. И скоро обнаружили много таких вариантов,
когда клон с большей средней s вымирал, а клон с меньшей средней s пе-
реживал катастрофу и сохранял значительную численность. Разумеется,
если средние s различались достаточно сильно (в опыте: более чем на 5%),
то большей s всегда соответствовало большее значение численности после
катастрофы, но при малых различиях селективных ценностей различия в
судьбе клона определялись не ими.
Наш результат легко представить наглядным графиком (рис. 28). По го-
ризонтальной оси отложим значения s, а по вертикальной - значения веро-
ятности выживания p(s). В модели Фишера p=2s, и он был уверен, что в лю-
бых случаях p(s) растет с ростом s монотонно. Но это не вполне так: в на-
шем опыте точки не ложатся ни на какую кривую, а занимают S-образную
область шириной около 5% в середине. Линией она кажется лишь издалека,
а это значит, что при росте среднего числа потомков на особь можно
говорить о росте вероятности выживания лишь как о грубом при-
ближении. У нас условия нарочно подобраны для различения мутантов, а в
жизни следует ожидать гораздо худшего их различения, т.е. ширины, много
большей, чем 5%.
В самом деле, в жизни, в самых различных природных ситуациях, мы
видим: грубая зависимость судьбы организма от его приспособленности, как
правило, очевидна, но нет никаких оснований говорить о роли мелких раз-
личий. Подробнее см. [Чайковский, 2004, гл. 9]. Более того, есть много при-
меров, когда прекрасно выживают даже очень плохо приспособленные ор-
ганизмы (п. 6-2*). Организмы могут нести самые чудовищные изъяны, ком-
пенсируя их чудовищной плодовитостью или даже организуя расплод осо-
бей, предназначенных видом на уничтожение в пользу экосистемы (из них в
пп. 3-7* и 4-20 были рассмотрены термиты).
Вот еще одна модель размножения, по-моему, полезная. Для распро-
странения вида удобны быстро размножающиеся организмы, а для его
формирования - долго живущие. Первые реализуют, на языке, принятом у
экологов, r-стратегию, а вторые К-стратегию.
«Стратегия-r предполагает бурное размножение, короткую продолжитель-
ность жизни при высокой активности... и быструю смену поколений. Эта
стратегия полезна для переломных моментов эволюции. [...] Стратегия-/^
предполагает низкий темп размножения и длительную жизнь особей. Она
создает условия для “благоденствия популяции” при сравнительно стабиль-
ных условиях» (Зусмановский А.Г. Потребностно-информационная теория
эволюции. Ульяновск, 2004, с. 42; ссылка на статью В.П. Скулачева).
Многочисленные модельные расчеты привели ученых к мысли, что г-
стратегия предпочтительна при первичном расселении таксона, а К-
261
стратегия - при его длительной эволюции в уже освоенной экологической
нише. Палеонтология дает тому подтверждения (см. п. 8-11).
Рис. 28. Катастрофический отбор (компьютерный эксперимент). Типичная
.уд-диаграмма, показывающая выживание различных клонов в режиме ката-
строфы: каждая точка обозначает вероятность выживания p(s) для клона, за-
даваемого своей функцией s(t). Любая пара точек, в которой левая располо-
жена выше правой (например, пара, выделенная стрелками), демонстрирует
тот факт, что при большей размножаемости возможна меньшая выживае-
мость, а это противоречит концепции естественного отбора в ее общеприня-
той форме (по [Маленков, Чайковский})
Очевидно, что вид, сочетающий преимущества г- и ^стратегий, должен
эволюировать быстро и эффективно. Таким видом вероятно оказался чело-
век, о чем будет речь в п. 9-16*.
Важно отличать эту реально различаемую пару явлений от воображае-
мой пары “г-отбор и К-отбор”, бытующей в экологии. Вообще надо заметить,
что, отделяя успешность эволюции от эффективности размножения, мы от-
крываем целый спектр возможностей, скрытых от дарвинизма. Мы уже
встречались и еще встретимся с инбридингом как фактором эволюции, а
ведь он всегда вызван снижением численности, а затем сам к нему ведет (в
силу отбраковки вредных гомозигот).
Механизм распространения нового свойства вовсе не состоит в “давле-
нии отбора”, т.е. в вытеснении новыми особями прежних за счет большей
размножаемости. Есть гораздо более эффективный механизм - внутриге-
номный. Для его действия достаточно, если нужный комплекс генов регу-
лярно копируется в гомологичную хромосому транспозоном и, тем самым,
распространяется среди половых клеток одной особи, т.е. подвергается го-
ризонтальному переносу™. Такой комплекс внешне похож на пресловутый
36
° То есть переносу генетического материала помимо процесса размножения. См. п. 6-9.
262
“эгоистичный ген” (см. п. 5-5), но несёт он отнюдь не “мусор” и размножается
отнюдь не безудержно. Он может успешно размножаться даже при дли-
тельном снижении численности его носителей, что в рамках идеи отбора
немыслимо.
5-11. Экспериментальная эволюция. Аршавский, доминанта и
стресс
Надо понять, сколько полезных свойств вид может воспринять одно-
временно. В дарвинизме считается, что отбор может отбирать сразу всё,
что угодно данному дарвинисту, но проверить это никто из них не пытался.
Наоборот, в известной мне физиологической модели, подкрепленной опы-
тами, приобретения могут происходить и фиксироваться лишь по одному.
Физиолог Илья Аркадьевич Аршавский (1903-1996) писал, что в дарви-
низме «на долю эволюционной физиологии выпадает задача лишь регист-
рировать эволюционные преобразования, но не раскрывать природу их воз-
никновения» [Аршавский, с. 248]. Сам он строил чисто физиологическую
картину эволюции и главным эволюционистом считал Ламарка. Исходным
понятием была у него метаболическая спираль. Суть ее такова.
Метаболизм принято делить на два класса процессов - анаболизм, при
котором характерно удлинение ковалентных цепочек и колец молекул плюс
поглощение энергии; и катаболизм, при котором цепочки укорачиваются, а
энергия выделяется. Оба класса тесно сопряжены в любом организме, но в
продуцентах, естественно, преобладает анаболизм.
Казалось бы, в других организмах должен преобладать катаболизм
(ведь они живут за счет разрушения молекул, синтезированных продуцен-
тами), но дело обстоит сложнее. Организм может, к примеру, проглотив 3 г
пищи, 1 г лизировать и за счет полученной при этом энергии включить ос-
тальные 2 г в состав своего тела. В смысле судьбы всей пищи это редукция
(взято 3 а, усвоено только 2 а), но в смысле эффективности метаболизма
это - превышение анаболической фазы над катаболической вдвое.
Аршавский рисовал процесс, в котором анаболизм и катаболизм равны
по величине, в форме движения по окружности, а превышение анаболизма
над катаболизмом - в форме расходящейся спирали. Он полагал это пре-
вышение основной эволюционной характеристикой организма, так что эво-
люционный прогресс наблюдается лишь в группах с высоким превышением
(Аршавский И.А. К обоснованию принципов и задач эволюционной физиоло-
гии // Журнал эволюционной биохимии и физиологии. 1985, № 2). До него
похожее писал Соболев (п. 4-12*).
Это превышение видится мне как материальная основа той активно-
сти, о которой на разных языках говорили столь многие. В терминах новой
термодинамики (п. 5-14) эти взгляды получат у нас развитие.
Нам понадобится понятие доминанты - той формы активности, которую
с 1911 года исследовал петербургский физиолог А.А. Ухтомский.
263
По нынешнему определению, доминанта - “господствующая группа
нервных центров, направляющая поведение организма в данной ситуации
на решение наиболее важной для него задачи”, причем введение этого по-
нятия в физиологию означало признание активности субъекта в качестве
исходного понятия. Роль его аналогична (в терминах 1920-х гг.) роли теле-
фонистки на телефонной станции, соединяющей искомого с ищущим (Кос-
тандов Э.А. Психофизиология установки и теория доминанты // Российский
физиология, журнал, 2000, № 8, с. 929).
В широком смысле доминанта - это установка особи на то, чтобы де-
лать одно дело всеми возможными силами, когда остальные функции вы-
падают из сферы ее активности и, у высших, - из ее внимания. Так, если ей
больно, она стремится избежать боли, поневоле оставив иные дела. Осо-
бенно это существенно, как мы увидим далее, в стрессовых ситуациях: ме-
ханизм доминанты позволяет до предела напрячь ту функцию, от которой
зависит выживание - например, усилить синтез фермента, которого нехва-
тает для усвоения непривычной пищи; при этом, как оказывается, ген фер-
мента активно мутирует.
Благодаря доминанте, в катастрофической ситуации все организмы
испытываются по одному-единственному блоку, и те особи, что прошли ис-
пытание успешно, могут осуществить расплод. Именно так происходит в
конце иммуногенеза, когда бурному размножению подвергается тот единст-
венный пулл клеток, который синтезирует нужное антитело. Что касаестя
эволюции, то на сегодня нам остается, в ожидании опытов, только рассуж-
дать по аналогии. Например, таким расплодом мне представляется раз-
множение тлей, научившихся питаться на новом растении (опыт Шапошни-
кова, п. 4-18). Но о том, как приобретения становятся наследственными, из-
вестно мало; литературу см. [Назаров, 2005, с. 367-368].
5-11* Физиологический стресс (эвстресс)
Аршавский, ученик Ухтомского, до последних дней своей динной жизни
считал долгом напоминать о ведущей роли доминанты и стресса.
Собака, лишенная передних лап, вырастает похожей на кенгуру, а зна-
чит, изменение образа жизни может порождать облик, характерный для со-
всем иных животных, даже - иного отряда. Насколько это наблюдение мож-
но признать общим? Аршавский выращивал животных в несвойственных им
условиях и смотрел за изменением их облика.
Наиболее наглядны опыты с крольчатами. Кролик похож на зайца, но
впечатление обманчиво, недаром их относят к разным родам. Кролик мало-
подвижен и при опасности спасается в нору, а заяц - бегством. Это отража-
ется в их внешнем облике, и в их физиологических параметрах. Аршавский
заставлял крольчат подолгу плавать, и они, вырастая, обретали заячий об-
лик (рис. 29) - настолько, что не могли спариваться с обычными кроликами.
264
Рис. 29. Кролики: снизу - обычный,
сверху - зайцеобразный (выросший в
условиях физиологического стресса)
(по: [Аршавский, 1982])
Менялась в сторону заячьей
и их физиология (например,
параметры крови). Налицо
адаптивная модификация, т.е.
ненаследственное полезное
изменение. Здесь мы вновь
видим параллелизм наследст-
венного и прижизненного, на
что указывали Балкашина (п.
4-7) и многие другие.
Вот к чему ведет избыточная двигательная активность. По Аршавскому,
именно она движет эволюцию: взросление в состоянии регулярного уме-
ренного стресса изменяет физиологию, а через нее строение.
В 1960-х годах советские физиологи дружно заявили, что под словом
«стресс» у Селье скрываются два совсем разных процесса: физиологиче-
ский (умеренный) и патологический стрессы. Первый - основа процессов
приспособления и развития, а второй - основная причина болезней.
Пример с тем же холодом: все мы знаем, что на холоду можно просту-
диться и заболеть, но краткое погружение в холодную (а иногда и в ледя-
ную) воду - лучший и простейший способ закалки организма. Холодными
ваннами Аршавский буквально творил чудеса: не только делал хронически
больных детей здоровыми, но и выращивал, как мы видели, животных в ка-
ком-то смысле иного вида.
Конечно, все эти изменения ненаследственны, и крольчонок, родив-
шийся от “зайца", вырастет, если его не купать, обычным кроликом, но разве
не удивительно, что столь сильное различие получено в одно поколение,
одним лишь различием способов воспитания? Выходит, что “заячьи” черты
в кролике уже есть, их надо только выявить (это Аршавский сумел) и на-
следственно закрепить - этого Аршавский, будучи физиологом, а не генети-
ком, даже не пробовал, ибо для этого нужна другая лаборатория, другие
методы, другая квалификация сотрудников.
Чуть позже в Ростове Л.Х. Гаркави и ее коллеги выявили в физиологи-
ческом стрессе две стадии, которые были названы активацией и трени-
ровкой. Различие их прежде всего в том, что стадия тренировки доступна
всем, тогда как активация более полезна и действенна для молодого орга-
низма. Она-то и ответственна за механизм холодового закаливания по Ар-
шавскому (Ааркави Л.Х., Квакина Е.Б., Уколова М.А. Адаптационные реак-
ции и резистентность организма. Ростов н/Д., 1979).
265
Вижу недоумение читателей - если изменения ненаследственны, при-
чем тут эволюция? В сущности, вопрос - не к физиологу, а к генетику, и ес-
ли генетикам неинтересно, почему преобразование организма в опыте так
похоже на его изменение в ходе эволюции, то тем хуже для них. Аршавский
не мог ставить генетических опытов (в 1980 г. его непостижимым образом
уволили на пенсию из Института нормальной физиологии, тем самым пре-
кратив даже его обычные физиологические опыты), но вопрос его, конечно,
занимал, и в 1985 году он изложил следующие свои соображения (Аршав-
ский И.А. К обоснованию принципов и задач эволюционной физиологии //
Журнал эволюц. биохимии и физиологии. 1985, № 2).
1. В «теории стабилизирующего отбора» Шмальгаузена (п. 4-14*) отби-
раются не мутации, а адаптивные модификации. Как они становятся на-
следственными, никто не объяснил, а суть как раз тут. Для ее понимания
прежде всего важна старая идея эмбрионизации А.А. Захваткина (п. 4-15*).
Аршавский указывал: благодаря эмбрионизации оказывается, что наследо-
вание состоит в постепенном сдвиге приобретенного свойства на всё более
и более ранние стадии развития, пока оно не станет врожденным (т.е. появ-
ляющимся еще у эмбриона), а затем и наследственным.
В пояснение мысли Аршавского сошлюсь на зоолога и палеонтолога
М.А. Шишкина. Он привел примеры эмбрионизации у позвоночных:
«образование мозолей у млекопитающих и бегающих птиц есть обычная
адаптивная реакция кожи на трение. Однако в участках, подвергающихся
трению постоянно... утолщение эпидермиса наступает еще в эмбриональный
период» (М.А. Шишкин, закономерности эволюции онтогенеза // Соврем, па-
леонтология. М., Недра, 1988, т. 2, с. 176).
Такие примеры, давние козыри ламаркизма, Шишкин счел фактическим
подтверждением идеи Шмальгаузена. Это верно если понимать ее как пе-
ресказ ламаркизма. У Шишкина оригинально (и Аршавским отмечено) иное
- его понимание наследуемости.
2. Для Шишкина НПС - ложная проблема: новые свойства, возникаю-
щие прижизненно, отличаются от наследственных только тем, что потомок
может как реализовать их, так и нет; если же он теряет способность выбора,
то мы говорим, что один из вариантов развития стабилизировался, т.е.
унаследовался (Любищев называл этот феномен рутинизацией, п. 4-17*).
По Шишкину, наследование - не приобретение новой информации, а ее
утрата, приобретение же происходит раньше, при становлении адаптивной
модификации. (Это, добавлю, утверждают все формы ламаркизма.) Аршав-
ский и другие физиологи указали на роль стресса: стресс, нарушая обычную
изоляцию половых клеток, может делать прижизненное наследственным.
3. Для такого наследования надо, чтобы найденная информация попала
в половые клетки. В связи с этим Аршавский предлагал вспомнить старую
теорию пангенеза, которой объяснял наследование еще Дарвин. Последуем
совету Аршавского позже, а пока закончим со стрессом.
266
5-11** Патологический стресс (дистресс)
Изменение наследственных свойств тоже оказалось связано со стрес-
сом, только с другим стрессом. Если организм подвергать чрезмерным на-
грузкам или почти смертельным дозам яда, то наступает реакция, которую
Аршавский назвал патологическим стрессом. При нем нарушается нор-
мальное развитие, и если он достаточно продолжителен, то подопытные ор-
ганизмы в массе гибнут (стадия истощения по Селье). При этом, между про-
чим, рушатся многие физиологические барьеры, и в том числе те, что огра-
ждают наследственный материал от внешних воздействий («барьер Вейс-
мана», п. 3-9). Патологический стресс ведет к многочисленным наследст-
венным изменениям т.е. среди немногих выживших велика доля “мутантов”
Подавляющая часть их несет уродства, портящие организм, но небольшая
часть особей несет те же самые изменения 37, какие наблюдались (в качест-
ве ненаследственных) при физиологическом стрессе. То есть стресс может
играть ключевую роль не только в жизни данного поколения, но и в эволю-
ции. Для микроорганизмов это уже не просто возможность, а эксперимен-
тально доказанный в последние годы факт.
Для высших организмов генетический анализ итогов стресса мне неиз-
вестен, но имеются косвенные свидетельства наследственной адаптации к
ледяной воде. Так, магаданские физиологи, изучавшие закалку людей ле-
дяной водой, пишут, что
“уровень изменений системы крови, рассматриваемый в качестве адаптивного,
не достигает у пришлого населения [уровня], присущего коренным жителям,
даже после 10 лет пребывания на Севере и после 5 лет применения закаливаю-
щих процедур. Это указывает на решающую роль генетического фактора” (Бе-
лобородов ГС., Белобородов А.Г. Эволюционные аспекты изучения закалива-
ния в условиях Крайнего Северо-востока России // ТПЭ-3, с. 30).
Узнав от советских коллег про две формы стресса, Селье уточнил свое
учение. Русских терминов он не принял, введя вместо них короткие слова
eustress и distress (эвстресс и дистресс).
О том, что патологический стресс прорывает барьер наследственности
(«барьер Вейсмана» [Стил и ф.]), теперь пишут многие, в том числе в свя-
зи с иммунитетом. Стресс оказался общим явлением. Он описан и у расте-
ний, у которых выявлено вещество, синтез которого ответствен за стресс -
абсцизовая кислота. Она определяет тот удивительный факт, что растения
одинаково отвечают и на нехватку воды, и на избыток солей; ее нехватка
37
Загадочный для дарвинизма факт данного параллелизма (мутации часто повторяют
прижизненную изменчивость) побуждал некоторых дарвинистов вводить особые фор-
мы отбора (совпадающий, органический, стабилизирующий и т.п. - п. 4-14). Основная
же масса дарвинистов попросту отрицала сам феномен такого параллелизма. С пози-
ции диатропики, излагаемой в гл. 6, он естествен.
267
вызывает прорастание семян на материнском растении и многое другое. Ей
был посвящен блок статей (ФР, 1994, № 5). Нам надо выявить то, что объе-
диняет все формы стресса, включая бытовые (стресс как “гонка по жизни”).
Общее и главное, что никакой стресс нельзя разъять на части: всякий
стресс - цельная реакция цельной системы (например, организма).
5-12. Экспериментальная эволюция. Наследование приобре-
тенных свойств
После триумфа опыта Лурии - Дельбрюка и воцарения «центральной
догмы» говорить про НПС стало опасно, да и просто неприлично. Исследо-
вания на многоклеточных были практически прекращены-^. Однако в одной
отрасли исследования продолжались, пусть и без упоминания запретных
терминов, продолжались как практическое медицинское направление. Речь
идет о генетической трансформации бактерий.
Еще в 1928 году английский бактериолог Фредерик Гриффит обнаружил
странный феномен: в ходе эпидемии пневмонии один пневмококк уступал
место другому. Гриффит верно предположил, что происходит превращение
(трансформация) бактерий (Серов О.Л. Перенос генов в соматические и
половые клетки. Новосибирск, Наука, 1985, с. 7). Замечу, что фактически им
наблюдалась эволюция бактерий в природе. Вскоре ее начали исследо-
вать и экспериментально.
К 1960 году была довольно хорошо изучена инфекционная наследствен-
ность бактерий, т.е. наследуемое их изменение (например, превращение
безопасных в опасные) посредством эписом - генетических элементов, спо-
собных передаваться между бактериями и встраиваться в их геномы (Жакоб
Ф., Вольман Э. Пол и генетика бактерий. М., ИЛ, 1962).
Вскоре выяснились и другие механизмы горизонтального переноса ге-
нов, и все они привлекли внимание думающих как возможный фактор эво-
люции. Дело в том, что происходят они примерно с той же частотой, что и
мутации (1СГ5 - 1СГ8 на клетку, см. Серов О.Л., с. 17, 57), но сразу обеспе-
чивают новый генетический текст, а с тем и новое наследуемое свойство.
Разрозненные данные нуждались в обобщении, и его предпринял в 1979
году Стил (о нем см. п. 5-8*). Исходная мысль его была парадоксальна: хотя
«центральная догма» (п. 5-7) ничем не доказана и мешает пониманию
эволюции, но путь к новой теории лежит через уяснение сути и статуса этой
В том числе исследования Гершензона на дрозофилах. На одноклеточных эвкариотах
(инфузориях) НПС успешно исследовал Йоллос (Назаров, 2005]. В работах 1913-1939 гг.
он развил учение о длительных модификациях - нестойком наследовании, служащем ша-
гом к обычному наследованию (Jollos V. Grundbegriffe der Vererbungslehre, inbesondere
Mutation, Dauermodifikation, Modifikation // Handbuch der Vererbungswissenschaft. Bd IV.
Berlin, 1939). Из фашистской Германии уехал в США, но не смог там работать и в 1941 г.
покончил с собой. Насколько знаю, с его смертью данный круг работ прекратился.
268
люции, но путь к новой теории лежит через уяснение сути и статуса этой
догмы. Поскольку она - молекулярный аналог идеи зародышевой плазмы
Вейсмана, то начать надо с него:
«Моя точка зрения такова: основной блок любого понимания наследования
приобретенных признаков можно найти прямо в доктрине Вейсмана».
Стил был прав: именно Вейсман в 1892 году писал:
«Корень наследственных изменений должен лежать глубже (чем в слиянии
половых клеток родителей - Ю.Ч.} и заключаться в прямом воздействии
внешних влияний на биофоры» (т.е. на единицы наследственности).
В чем же состоит такое воздействие? Этого, разумеется, Вейсман не
знал и, в стиле натурфилософов прошлого (например, К.-Ф. Вольфа), напи-
сал: «Начало изменения основывается на малых неправильностях в пита-
нии зародышевой плазмы». Тут Стил смог дать важное уточнение: «пита-
ние» наследственного материала состоит в усвоении им генетического ма-
териала некоторых вирусов.
Конечно, сводить эволюцию, даже только приспособительную, к одному
лишь НПС наивно, зато такое сужение задачи позволило Стилу вчерне ре-
шить ее. По его мнению, зародышевая плазма действительно играет веду-
щую роль в эволюции, и, в отличие от Вейсмана, мы сейчас можем утвер-
ждать, что ее фрагменты передаются между органами и организмами с по-
мощью вирусов и прочих форм горизонтального переноса.
Стил (с соавтором) получил НПС в опыте на мышах, пользуясь тем
фактом, что новорожденные самцы линии А, обработанные лимфоцитами
линии В, обретают совместимость с тканями линии В (за открытие этого
факта получил нобелевскую премию Медавар). Исследователи два года
упорно скрещивали таких самцов с обычными самками линии А и определя-
ли совместимость получаемого потомства к тканям линии В. “Положитель-
ная передача была непостоянной, появляясь с большой частотой у одного-
двух из десяти самцов” [Стил и др., с. 154].
Эти данные взялся проверить Медавар, отрицавший возможность НПС.
Он тоже получил несколько случаев НПС, однако не счел это доказательст-
вом реальности феномена НПС, поскольку случаи наблюдались редко. Это
был нечестный приём, поскольку сам он отстаивал гораздо более редкую
процедуру - случайные мутации, которые, кстати, тоже не проявляют устой-
чивости частот. Но что мог возразить стажёр лауреату?
Медавар потребовал от Стила прекратить опыты, Стил был вынужден
вернуться в Австралию и был подавлен. Даже там для зачисления его в
штат университета потребовалась помощь Поппера: философ оказался
единственным, кто встал на защиту свободы ученого и научного поиска.
Подробнее о работах и судьбе Стила см.: Аронова Е.А. Карл Поппер,
“наука по Попперу” и дискуссии о ламаркизме... // ВИЕТ, 2002, № 4.
Вообще, возражение о нерегулярности НПС предъявляется во всех
случаях, когда против его реальности возразить нечего. Как обычно, против
269
неприятной теории хорош любой довод. Но ведь НПС и должно наблю-
даться не всегда - это ясно из самой природы явления: горизонтальный
перенос заведомо не может происходить регулярно (с ним борется особый
вид иммунитета, “геномные цензоры” - см. далее, п. 6-6). Жаль, что в по-
следующие годы аналогичные опыты (они очень трудоёмки) не были по-
ставлены на других объектах и с другими приобретаемыми иммунными
свойствами.
Тем не менее, сам факт НПС в последующие годы был показан - спер-
ва на бактериях (опыты Кэйрнса, п. 5-1***), а затем на самых разных объек-
тах [Назаров, 2005, с. 367-368],
5-12* Первичные половые клетки и новый пангенез
Казалось бы, те клетки, которые будут у взрослого организма порож-
дать гаметы (спермин и яйцеклетки), должны формироваться у зародыша
вместе с его гонадами (половыми железами), но на самом деле они дела-
ются у большинства животных где угодно, только не там. Гонады сперва
пусты, а затем заселяются пришедшими извне половыми клетками (рис. 30).
«Сага о зародышевом пути» - так названа глава об этом в руководстве
[Гилберт, т. 3]. Словно викинги, авторы саг, немыслимым для нас образом
находившие
бескрайней
Исландию
в
Рис. 30. Первичные по-
ловые клетки (выделены
темным) гидроида Clava
sqummata, мигриру-
ющие в половую железу
(слева) и за-селившие ее
(справа)
Атлантике, первичные половые
клетки (ППК), сперва
рассеянные по зароды-
шу, находят гонаду.
В 1880 году немецкий эмбриолог Мориц Нуссбаум ввел термин «заро-
дышевый путь» (die Keimbahn) для обозначения того, что ныне именуется
каналом передачи наследственной информации от предков потомкам. Для
передачи требуются клетки, чья наследственная информация никак не по-
вреждена. По-видимому, это должны быть клетки, не затронутые процесса-
ми специализации, тогда как почти все клетки организма (в том числе и
клетки тканей половых желез) специализированы. Нуссбаум постулировал,
что те клетки, которые ответственны за наследственность, не участвуют в
процессах специализации, а лишь размножаются друг от друга, т.е. полно-
стью изолированы от всех процессов развития и выполняют единственную
Функцию - служат источником развития для других клеток.
270
Вскоре Вейсман уточнил этот постулат и положил его в основу своей
теории двух плазм, по которой половые клетки образуют особую субстан-
цию (ее он, вслед за Нэгели, назвал идиоплазмой), недоступную внешним
влияниям. Ныне этот факт именуют «барьером Вейсмана» (п. 3-9). Осталь-
ные клетки организма (соматоплазма) формируются из половых и не пере-
дают свои изменения потомкам.
Далее произошло нечто странное: с одной стороны, теория Вейсмана
легла в основу неодарвинизма, а с другой - была отвергнута генетикой,
вторым краеугольным камнем неодарвинизма. Отвергнута потому, что со-
матические клетки почти всегда содержат те же хромосомы, что и в половых
клетках. Неясно, на чем держался тогда неодарвинизм, кроме как на уве-
ренности в своей правоте. Зато она была в избытке, поскольку поддержива-
лась симпатиями общества и политиками разных направлений.
Затем теорию Вейсмана отвергло и большинство эмбриологов, в том
числе Вентребер. Но слишком простые решения редко бывают верными.
В том же 1962 году, когда вышла главная книга Вентребера, француз-
ский эмбриолог Этьен Вольф собрал в Париже международную конферен-
цию по половым клеткам, показавшую, что вопрос куда сложнее, что теория
двух плазм имеет глубокий смысл, хотя и отличный от вейсмановского. Тру-
ды конференции переведены в сборнике [Происхождение...].
Непрерывный зародышевый путь, который Вейсман считал общим
свойством живой материи, нашелся не у всех: его нет ни у растений, ни у
самых низших животных (губок и некоторых других) - там в качестве поло-
вой может выступить любая клетка. Однако у большинства животных поло-
вые клетки действительно развиваются особым путем, и нельзя понять ос-
новную тенденцию эволюции животных, не поняв свойств зародышевого пу-
ти. В частности, ни одно животное, имеющее мозг, без него не обходится. О
тенденциях мы будем говорить в главе 6.
ППК - единственный источник будущих яйцеклеток и спермиев - всегда
имеют полный набор хромосом, тогда как все остальные клетки (соматиче-
ские) имеют таковой набор лишь почти всегда: у некоторых низших живот-
ных (например, аскарид) хромосомы соматических клеток теряют фрагмен-
ты, а у комара рода Miastor некоторые хромосомы соматических клеток про-
сто уничтожаются. Хотя такие примеры и редки, они ясно говорят, что «эво-
люция по Вейсману» вполне возможна. Столь же важно, что ППК имеют со-
всем особую цитоплазму.
Как происходит миграция ППК? Зачем она нужна? По первому вопросу
можно сказать, что они используют обычные для онтогенеза способы ми-
грации клеток [Гилберт], но второй пока точного ответа не имеет (мы вер-
немся к нему в п. 5-14*). На Конференции у Вольфа отмечено, что ППК
«систематически избегают участия в превращениях, которые претерпевают
соматические клетки, причем это продолжается до момента, когда гоноциты
заселяют территории будущих половых желез» [Происхождение..., с. 201].
271
Происходит это не так уж рано: на рис. 31 видно, что ППК находятся еще
вне зародыша, когда уже реализован основной план строения позвоночного.
Изоляция ППК от действия эмбриональных тканей действительно важна
для сохранения основных свойств генотипа, которые следует передать по-
томству, но эволюции она лишь препятствует. Зато мигрируют ППК в пото-
ке жидкостей, окружающих соматические клетки (у птиц - плывут в кровото-
ке, у зверей - ползут по тканям), и могут подвергаться всевозможным влия-
ниям, которые позже затруднены или даже невозможны. Например, могут
заражаться вирусами. О роли вирусов в передаче наследственной инфор-
мации в 1962 году еще почти не знали, зато многое знали про иммунитет, и
один докладчик выразил удивление:
«первичные гоноциты как бы иммунизированы против индукционных воздей-
ствий со стороны окружающих их тканей» (с. 183).
Он хотел этим сказать, что ППК не реагируют на окружающие клетки,
если только это не клетки гонад (конечный пункт миграции). С этим мы еще
столкнемся не раз: онтогенез удобно понимать
/ в терминах иммунитета. И в этом мне видится
@ ) ключ к построению теории эволюции.
I * . Рис. 31. Первичные половые клетки проникают в
U тело зародыша цыпленка: 1 - достигшие половой
( \ ку /лЦт складки, 2 - в кровеносной системе
JI \/ Теперь мы знаем, что ППК избегают далеко не
------------всяких контактов: яйцеклетка взаимодействует с
окружающей ее средой и получает, в частности,
генетическую информацию (см. [Зусмановский], пункт «Включение новой
информации в ооцит»). В свете новых данных ясно, что она получает от
самца не только хромосомы спермия, как думали прежде, но и информаци-
онную РНК (Rassoulzadegan М. е.а. И Nature, 2006, vol. 441, р. 469).
Тем самым, пангенез вполне возможен прямо путем передачи отдель-
ных плазматических генов в яйцеклетку. Но есть и иные пути.
По прибытии в гонаду ППК подвергаются отбраковке, чем-то похожей на
иммунное отторжение. Так, в яичнике зародыша девочки они сперва бурно
размножаются, а затем основная масса их гибнет (рис. 32).
Успешные поступают на долгое хранение: в профазе мейоза они ждут,
пока девочка станет женщиной и ППК поодиночке породят яйцеклетки [Гил-
берт, т. 3, с. 273]. Все эти годы ППК стареют (вот почему здоровых детей
чаще рожают молодые матери). Вероятно, их старение связано с порчей
вторичной (метилирование) и третичной (укладка) структур ДНК.
Наоборот, популяция ППК мужчины порождает спермин ежечасно, так
что спермин стареют гораздо медленнее яйцеклеток (в силу «обнуления»
при мейозе, как мы узнаем в п. 8-8).
272
Рис. 32. Количество половых клеток в яичнике женщины. Как видим, оно после
завершения миграции возрастает у зародыша раз в 12, а затем во столько же раз
падает к семи годам. Зачем бы это?
5-12** Пангенез и стволовые клетки
Все, кто говорил о пангенезе (начиная с Алкмеона), имели в виду мигра-
цию частиц наследственности в половой орган. С появлением клеточной
теории стали говорить о миграции клеток, несущих оные частицы. Таковыми
являются ППК, но какую они могут нести новую (т.е. эволюционную) инфор-
мацию? Вот если бы они мигрировали из уже работающих органов! Однако
мигрирующие клетки есть во всех органах. Прежде всего, это фибробласты
(клетки соединительной ткани животных, ответственные за регенерацию) и
стволовые клетки. О последних пишут:
"’Стволовые клетки представляют собой загадочный и еще мало понятный
феномен. От них в буквальном смысле зависит вся наша жизнь. Стволовая
клетка - это клетка, способная к интенсивной пролиферации (последователь-
ным делениям - Ю.Ч.)... Так, одна стволовая клетка может дать начало клону,
содержащему миллионы дифференцированных клеток, [или] небольшому
числу стволовых клеток... Кроветворные стволовые клетки млекопитающих
мигрируют в костный мозг, где сохраняются в течение всей жизни животно-
го. Стволовые клетки имеются и для таких непрерывно возобновляющихся
тканей, как эпидермис, а также для возобновления числа спермиев в семенни-
ках... стволовые клетки скелетной мускулатуры существуют, вероятно, в те-
чение развития плода” [Гилберт, т. 1, с. 212].
Теоретики возлагают на них огромные надежды. Ведущий французский
журнал «Comptes Rendus Biologies» посвятил им особый номер, где, в част-
273
пости, патриарх французской генетики Жакоб в статье «Мир стволовых кле-
ток» писал о том, что у нас выше названо активностью. Он резюмировал:
«Фазу разобщения, характерную для двухсот лет биологии, сменяет фаза вос-
соединения (reconstruction) живого». Например, «Эмбриональные СК могут са-
ми себя дифференцировать (se differencier), ибо, впрыснутые в эмбрион, они
участвуют в создании мыши, давая химеру, ткани которой, в том числе зароды-
шевый путь, могут содержать впрыснутый генотип». Далее, «они могут диффе-
ренцироваться (se differencier): как in vivo, после впрыскивания в во взрослую
мышь, у которой они образуют мелкие тельца, “эмбриоиды”, содержащие де-
риваты (необычные варианты - Ю. Ч.) трех зародышевых листков, так и in vitro,
где они могут за несколько дней произвести тот или иной тип клеток, из кото-
рых состоит взрослая мышь. Дифференциация обращается на тот или иной тип
клеток комбинацией факторов роста, которые открыты недавно» {Jacob F. Le
monde des cellules souches // CR Biol., 2002, t. 325, № 10, p. 999, 1000).
Нейл Тейз (США) выдвинул «новый принцип клеточной пластичности»,
призванный заменить прежний «принцип однонаправленного развития»:
«Любая клетка, содержащая цельный геном - без делений, дупликаций и пе-
регруппировок - может стать клеткой любого другого типа» Theise N.D. New
principles of cell plasticity (там же, с. 1040).
А французский референт обобщил обоих (Жакоба и Тейза):
«клетки одного органа, становящиеся клетками другого, преодолевают даже
барьер между первичными зародышевыми листками эмбриона. Эти результа-
ты... подтверждают, что однонаправленных путей дифференцировки в дейст-
вительности не существует» (там же, с. 1039).
Хочу особо обратить внимание на то, что ППК - это СК зародышевого
пути. СК есть во всех органах, в том числе в головном мозге, и естествен
вопрос: не могут ли одни СК контактировать с другими и не могут ли СК од-
ного типа занимать место других? Какие здесь нужны опыты?
Опытов нечего и ждать, пока общество не видит самой проблемы. Если
Аршавскому было достаточно наличных данных, чтобы принять пангенез
как факт, то многим другим достаточно “центральной догмы”, чтобы ни о
чем таком не думать. Третьи ставят изящные опыты и получают яркие ре-
зультаты, но не видят их связи с пангенезом, поскольку не знают о самой
этой идее. Так, нейробиологи из Сингапура Гэвин Доуи и Чжи-Чэн Сяо
«скрестили здоровых самок мышей с самцами, у которых благодаря гене-
тической модификации все клетки производят белок, флуоресцирующий
зеленым светом. Исследователи обнаружили зеленые клетки плода в мозге
матерей, которые превращались в нечто очень похожее на нейроны...»
(Ребенок в голове И ВМН, 2006, № 3. с. 11).
Здесь установлен факт миграции СК из плода в мозг и преодоление ими
физиологических барьеров. Работ по этой теме много на различных живот-
ных. Осталось обнаружить обратный поток, что может быть быстро сделано,
если только будет поставлена такая цель.
274
К сожалению (для эволюционизма) работы по СК ставят почти исключи-
тельно медицинские цели, и их вывод разочаровывает: наилучшие резуль-
таты дают опыты по использованию собственных СК организма (ибо избе-
гают проблем иммунного отторжения), зато у них
«высока частота генетических мутаций, делающих невозможным их исполь-
зование для лечения» {Пальцев М.А. и др. Перспективы использования ство-
ловых клеток в медицине И ВРАН, 2006, № 2, с. 101).
Это нам и важно: СК, перемещаясь по своему организму, активно мути-
руют. Этого вполне можно было ожидать в силу снятия давления нормы, но
ясно (независимо от способа объяснения данного явления), что СК, если
они могут попадать в гонады, представят собою материал для эволюции.
Словом, данные в пользу пангенеза есть, но они разрознены и предсто-
ит их сборка в целое. Приведу лишь некоторые соображения.
1) новые свойства и даже облик организма могут достигаться особью
без перемен в наследственности (опыты Аршавского),
2) тем самым, в организме таятся разные возможные облики,
3) надо узнать, где эти облики таятся (прежде говорили: «предсуще-
ствуют») - в геноме (ядерном или внеядерном) или же вне его (в конструк-
ции организма),
4) существует движение генов внутри клетки, внутри организма и ме-
жду ними (горизонтальный перенос),
5) существует движение клеток по организму (миграция), изредка пре-
одолевающее физиологические барьеры,
6) существует категория клеток (СК), сохраняющая способность об-
ращаться в клетки любых типов; к ним относятся и ППК,
7) мы не знаем, как черты нового облика становятся (если становятся)
генами, не знаем и обратного - как гены порождают облик, а без этого пан-
генез не понять;
8) но мы знаем один пример того, как ген приводится в соответствие с
обликом его продукта - в иммуногенезе;
9) ППК избегают зон, где делается общий облик организма, зато по-
падают в гонаду из зон, где закладываются органы.
Последнее наводит на мысль, что морфогенез управляется биополем
(п. 4-16) и что ППК избегают действия общего поля зародыша (о нем см.
ниже, п. 5-15), но не частных полей. Если так, то пангенез будет понят не
ранее, чем морфогенез. К последнему мы обратимся в п. 5-15, но прежде
нам надо ввести ряд новых понятий.
5-13. Генетический поиск и норма
«Экспериментальной эволюцией» ясно показано, что эволюцию,, вы-
званную стрессовыми изменениями среды, наблюдать можно.
Само слово “выгодная” по отношению к единичной мутации почти все-
гда лишено смысла, а чисто случайное совпадение в одной особи несколь-
275
них нужных мутаций невероятно и, главное, неповторимо. Встает вопрос о
механизме, порождающем новые генетические тексты. Ему в 1976 году бы-
ло мною дано название «генетический поиск»39.
Генетическим поиском будем далее называть режим работы гене-
тической системы, в котором изготовляются новые генетические тек-
сты. Простейшей формой генетического поиска является повышение мута-
бильности при дистрессе. Его молекулярный механизм был описан как
инерционный мутагенез [Чайковский, 1976, с. 157]. Затем стали встречать-
ся и иные термины. Таков, например, термин «мутации как проявление по-
исковой активности» [Гринченко, с. 151]: он подчеркивает, что генетический
поиск— пример активности, а именно, активность на генном уровне.
Механизмов поиска известно несколько, и все начинают работу с того,
что активность генетической системы данной группы организмов переклю-
чается с поддержания нормы на создание нестабильности. Затем идет
противоположный процесс - поиск новой нормы.
Если это происходит в катастрофических условиях, то почти все осо-
би погибают, зато немногие оставшиеся подвергаются, в силу длительного
дистресса, массовым генетическим изменениям, среди которых есть и те,
которые фиксируют в генотипе полезные изменения организмов. Некоторые
из них бывают найдены прижизненно - например, по Шапошникову или Ар-
шавскому. В последнем случае дистресс видимо позволяет унаследоваться
изменениям, найденным при стрессе физиологическом, но надо признать,
что взаимосвязь двух форм стресса в эволюции неясна.
Как показывает 60-летний опыт, мутации статистически направленны.
Наиболее эффектно это видно в иммуногенезе. Чуть более сложный вари-
ант генетического поиска - направленный «адаптивный мутагенез»:
«Было показано (ссылка на Кэйрнса - Ю. Ч.), что по крайней мере у бактерий
существует механизм, обеспечивающий приобретение ими в условиях селек-
ции преимущественно адаптивных мутаций. Этот термин получил название
“адаптивный мутагенез”» (Бабынин Э.В. //Генетика, 2004, № 5, с. 581).
Существуют более сложные формы поиска, они основаны на редакти-
ровании РНК и транспозиции мобильных генов. Один класс последних (рет-
ротранспозоны - п. 5-6) изучен, и можно сделать некоторые выводы. Пере-
мещаются не структурные гены, а регуляторы, и их очень много:
«структурные гены действительно не перемещаются, но... существует целая
система подвижных элементов, составляющая в сумме 10-30% всего генома у
животных и до 50% у растений». В итоге, «эволюционное значение рет-
ротранспозонов несомненно связано с их способностью вызывать мутации.
Выявление принципиальной возможности... перемещения ретротранспозонов
В качестве самых ранних авторов тогда мною были указаны Э.С. Бауэр и Ю.Я. Керкис.
Лучше было бы начать с работ Ф. Гриффита (1928) и других по трансформации бактерий,
поскольку с них началось понимание факта направленности изменчивости. См. п. 5-12.
276
под действием экологических факторов позволило предложить гипотезу, объ-
ясняющую быструю эволюцию дрозофил. Согласно этой гипотезе, критиче-
ские стрессовые условия существования приводят к вспышкам индуцирован-
ных транспозиций, которые создают генное разнообразие у потомства немно-
гих выживших особей.» Это должно происходить не у всех особей, а лишь v
имеющих гены контроля транспозиции в нужном состоянии; у них частота
наблюдаемых мутаций должна «повышаться на один-два порядка по сравнению
с особями, имеющими нормальные контролирующие транспозицию аллели и
почти неизменную частоту спонтанных мутаций в любых условиях» (Андриа-
нов Б.В. Индуцированная ретротранспозиция // УСБ, 2005, № 1, с. 78, 87).
Напомню, поток таких мутаций случаен, но направлен. Теперь можно
говорить о механизмах, поставляющих эволюционную изменчивость.
Если мутагенез можно сравнить с бросанием несимметричной играль-
ной кости, а редактирование РНК - с выводом теоремы, то ретротранспози-
цию - с разумным экспериментом, исход которого неизвестен заранее.
Благодаря стрессу, доминанте и генетическому поиску, объектом эво-
люционных экспериментов служат не признаки, а целостные конструкции.
Итоги экспериментов могут подвергаться расплоду. Механизмы запуска
расплода известны (например, для иммуногенеза), но не исследованы.
Термин «генетический поиск» входит в научный оборот только в по-
следние годы [Голубовский // ЭБ; Зусмановский; Назаров, 2005], однако по-
существу он в своей простейшей форме (влияние стресса на мутабиль-
ность) общепризнан уже лет 20. Например, «инерционный мутагенез» был
использован в книге: Жученко А.А., Король А.Б. Рекомбинация в эволюции и
селекции. М., Наука, 1985, на с. 218-223.
Опыты Шапошникова с тлями (п. 4-18) служат ясным примером генети-
ческого поиска. Вместо бессмысленного “дестабилизирующего отбора” (де-
стабилизация носит тут массовый характер уже в первом дочернем поколе-
нии), можно уверенно говорить о том, что тли, оказавшиеся на негодном для
них растении, теряют состояние нормы и повышают изменчивость. Они жи-
вут и размножаются в состоянии дистресса, что вызывает высокую смерт-
ность и высокую статистически направленную изменчивость выживших.
Сейчас выясняется, что особенность тлей тут ни при чем. Точно так
же повели себя в условиях экологической катастрофы озёрные лягушки
Rana ridibunda: почти потеряв биотоп около воды, они за 8 лет выросли (ут-
роили вес) и приблизились по физиологии и поведению к жабам (Иноземцев
А.А. Озерная лягушка сильно выросла И Природа, 2002, № 12). То же отме-
чено в других терпящих бедствие биотопах (так изменение аральских рыб
аттестуют как видообразование). К сожалению, генетический анализ ни в
одном из таких случаев не проведен, но генетический поиск во всех подоб-
ных ситуациях достаточно очевиден, хотя бы на внехромосомном уровне.
Есть работы, где стресс рассматривается как физиологический фактор
эволюции. См. например, обзоры австралийского эколога Питера Парсонса:
277
parsons P.A. Stress, extinctions and evolutionary change: from living organisms
to fossils // BR, 1993, № 2; Environments and evolution: interaction between
stress, resourse inadequacy and energetic efficiency // BR, 2005, № 4. Но заме-
чу, что стресс - процесс, протекающий в ходе жизни особи, а эволюционные
события, фиксируемые палеонтологией, текут в миллионы раз медленнее.
О стрессе можно тут говорить лишь в порядке метафоры, а нам интересны
реальные процессы. Какой процесс может действовать на изменчивость по-
добно стрессу, но - в течение очень многих поколений, мы не знаем.
Насколько многих? Здесь предела нет. Долго считалось, что генетиче-
ский код вообще неизменен (универсален), а оказалось, что он изменчив во
всем, только меняется крайне редко. Зато может различаться у близких ви-
дов радикально: есть жгутиковые у которых тимин частично заменен на
оксиметилурацил (п. 5-5); а есть род грибов (Candida), где кодон CUG явля-
ется самой изменчивой частью генома - у некоторых видов рода вместо
лейцина он кодирует серин во многих, но не во всех генах {Gomes А.С. и
др. И МБ, 2006, № 4). Механизм этой изменчивости неизвестен, но полагают,
что он действует миллионы лет. Можно ли тут видеть генетический поиск?
По-моему, о генетическом поиске имеет смысл говорить всюду, где на-
лицо направленная (пусть и случайная) изменчивость, что бывает, как уви-
дим ниже, далеко не только при стрессе.
5-13* Поддержание нормы и ее смена
При всем значении стресса для запуска генетического поиска, дело да-
леко не только в стрессе. Биологи указывают и на иные способы восстанов-
ления утраченного гомеостаза (состояния, обеспечивающего нормальную
работу системы), в том числе на роль стволовых клеток:
“В течение многих лет исследователи пытаются понять, каким образом орга-
низм поддерживает гомеостаз, как восстанавливается после повреждения...
Стволовые клетки (СК) занимают центральное место в клеточном гомеостазе
организма прежде всего потому, что их основной функцией является воспол-
нение естественной потери клеток” (Мусина Р.А., Егоров Е.Е., Белявский А.В.
Стволовые клетки... // МБ, 2004, № 4).
Возможно, СК в самом деле играют решающую роль в востановлении
утраченного гомеостаза там, где они есть, но ведь СК - атрибут тканевых
организмов, тогда как гомеостаз умеют соблюдать все организмы, в том
числе бактерии. Внутри клетки роль агентов поддержания гомеостаза игра-
ют вновь синтезируемые белки. Гомеостаз внутри вида поддерживается ме-
ханизмами регуляции размножения. Кроме них, гомеостаз экосистемы под-
держивается путем замены одних видов другими.
Виден некий общий принцип, и “центральное место” должен занимать
он, а конкретные механизмы гомеостаза лучше рассматривать как средства
его реализации. Таким принципом видится мне поддержание нормы. На-
зовём так реально наблюдаемую форму активности (в отличие от вообра-
278
жаемых: отбора, оптимизации и пр.). Феномен поддержания нормы шире
чем гомеостаз, под которым обычно понимают сохранение существенных
характеристик системы. Ведь если нормальную работу системы не удается
обеспечить сохранением каких-то свойств и значений в нужной области, то
ее приходится поддерживать их активной сменой. Например, обычно жи-
вотные утоляют голод усилением прежней активности, но при иссякании ис-
точника пищи вынуждены менять место обитания.
Поддержанием нормы являются и генетический поиск, и поздняя ми-
грация ППК в гонады (п. 5-12), и автодиагностика (см. далее, п. 6-8), и обо-
стрение слуха при слепоте, и смена старого вожака стаи на молодого, и сук-
цессия экосистемы (о ней см. далее, пп. 8-13*, 11-2, 11-4**).
Дарвинизм видит акт эволюции как постепенную смену признаков, а
прочие учения рассматривают эволюцию прямо как смену старой нормы на
новую (некоторые дарвинисты смену норм тоже называют в терминах отбо-
ра, но это, по существу допускаемых средств, не дарвинизм - см. п. 4-14).
Поэтому механизм распада прежней нормы интересен как механизм за-
пуска эволюционной изменчивости. Выше таковым механизмом выступал у
нас как стресс, и это верно, но слишком узко - столь же узко, как видеть в
СК общий механизм поддержания нормы. Дело в том, что изменчивость
резко возрастает при любом распаде нормы, в том числе и при несомнен-
ном отсутствии признаков стресса. Вот примеры.
1) Удивительно, но факт: в пользу дарвинизма приводятся примеры
усиления эволюции при ослаблении отбора. В изолированных экосистемах
(мелкие острова и озёра) известно много резко отличных и даже уродливых
форм. Так, в молодом озере (ему 10 тыс. лет) два рода рыб семейства кар-
повых строением нижней челюсти вышли
“за рамки всего громадного семейства карповых”, а у рыб более старых озёр
уродливые зубы вышли “за пределы... всего класса костистых рыб” {Шварц
С.С. Экологические закономерности эволюции. М.. 1980, с. 249).
Эколог С.С. Шварц толковал это как следствие слабой конкуренции в
тех неполных экосистемах, какие обычны для изолятов. Если так, то усло-
вия жизни облегчены и это противоположно стрессу, а итог тот же: рост
изменчивости по многим свойствам. То же мы увидим в глобальном флоро-
генезе (п. 9-4).
2) Доместикация (одомашнивание) ведет к росту изменчивости как при
стрессе, так и вне его. Так, наиболее спокойные лисята проявляют наи-
большую изменчивость (многие свойства, обычные для собак, п. 4-19).
3) В опыте с тлями (п. 4-18) направленная изменчивость могла расти
после окончания стресса, в условиях привыкания: тли
“после длительного воспитания в лаборатории в Ленинграде значительно чаше
стали питаться и в течение некоторого времени размножаться на чужих хозяе-
вах, чем это наблюдалось в первый период воспитания в районе Майкопа в ус-
ловиях более близких к естественным” {Шапошников Г.Х. И ЭнО, 1965, № 4).
279
4) Давно отмечена удивительная эволюция в темноте: животные теря-
ют окраску и глаза, причем процесс направлен: у зародыша глаза есть, но
затем исчезают (п. 3-6). Верно отмечено, что здесь «новая среда отменяет
требование к функции» (Бердников В.А. Основные факторы макроэволю-
ции. Новосибирск, 1990, с. 31), но объяснение дано наивное — «уменьше-
ние мутационного груза». Примерно тот же уровень бездумного объяснения
видим и в западной литературе. Например: Wi/kens Н., Strecker U. Conver-
gent evolution of the cavefish... // BJ, 2003, v. 80, № 4.
5) Активация иммунитета при заражении вирусом протекает в условиях
стресса, но ее же можно вызвать интерфероном или ударной дозой аскор-
биновой кислоты. При этом симптомов стресса не наблюдается.
6) Последний пример совсем загадочен:
“По Коху (1951), мутирование ослабляется под влиянием неблагоприятных
условий жизни и усиливается при благоприятных” (Боголюбский С.Н. Проис-
хождение и преобразование домашних животных. М., 1959, с. 221).
Словом, если понимать норму как нечто устоявшееся, как “гомеостаз
особи и среды”, то можно сказать, что изменчивость растет при любом зна-
чительном уклонении от нормы (даже в сторону улучшения жизни), в том
числе и длительном (очень много поколений). Наоборот, поддержание нор-
мы достигается низкой изменчивостью. Причину столь странного явления
можно сегодня обозначить лишь в самом общем виде, но и этого достаточ-
но, чтобы получить нужный вектор для намечающейся теории.
Начнем с того, что норма системы формируется из тех состояний ее
подсистем, какие нужны для системы, а не из тех, какие они принимают са-
ми по себе, когда ничто их не связывает. Пример: сердце бьется само по
себе (в покоящемся организме) медленнее, чем при обеспечении нормаль-
ной работы организма; если стая бежит с единой (и в этом смысле нор-
мальной) скоростью, то разные особи прилагают к этому разные усилия и
сердца их бьются различно.
На этой основе зоолог М.М. Завадовский когда-то разработал идею, по
которой оптимум организма складывается из неоптимальных состояний ор-
ганов (Завадовский М.М. Противоречивое взаимодействие между органами.
М., Изд. МГУ, 1941). Идея противоречит догме рыночной экономики (где
благо общества складывается из благ конкурентов) и ныне игнорируется.
Агроботаник Г.Н. Зайцев распространил идею на культурные растения:
“в любых условиях жизни растение путем процессов гомеостазиса поддержи-
вает лишь одно состояние функций - оптимальное для данного их сочетания
как между собой, так и с факторами внешней среды. Это представляет собой
одно из самых удивительных явлений природы ...” (Зайцев Г.Н. Оптимум и
норма в интродукции растений. М., Наука, 1983, с. 242).
Феномен нормы в самом деле удивителен, но он вытекает из более
общего (и еще более удивительного) феномена активности (сродства, жиз-
ненной силы, жизненного порыва и т.д.), из того загадочного факта, что ак-
280
тивность сама собой распределяется по системе (например, по организму)
так, что образуется норма. Ссылка на отбор ничего тут не дает (даже если
признать, что он есть) потому, что надо понять, как норма е данный момент
осуществляется. Например, как и почему сейчас растет микротрубочка (см.
ниже, п. 5-15), обеспечивая нормальный митоз?
Если состояние системы перестало быть физиологической нормой, то у
подсистем нет ни причин, ни стимулов его поддерживать, а потому каждая
может его изменить в пользу собственной (подсистемной) нормы. Но это не
дает нормы целому, и неизбежен ее поиск (рост изменчивости).
Противоположный феномен (падение изменчивости при достижении
нормы) имеет смысл назвать давлением нормы (см. п. 5-2). Падение из-
менчивости может быть связанным с ростом жизнеспособности, о чем будет
сказано в п. 5-17. Можно сказать, что давление нормы - это доминанта,
выходящая за пределы физиологии в область морфологии. С этой
позиции доминанта и давление нормы - единый тип активности.
Мысль о давлении нормы возникла у меня при чтении статьи Е.Е. Кова-
ленко {Коваленко Е.Е. Эффект нормы признака и его теоретическое значе-
ние // Эволюционная биология: история и теория. Вып 2. СПб., 2003). Хотя в
ее работе данного понятия в явном виде нет, но есть положение: «Норма
признака экранирует остальные его состояния, сокращая реальное разно-
образие», и оно обосновано обширным материалом по норме и аномалиям
крестца лягушек. Как происходит смена норм, мы узнаем позже, после вве-
дения нужных понятий (п. 6-8). Пока же скажу немного о той физике, которая
позволяет говорить о системах, а с тем и об их нормах.
5-14. Новая термодинамика, эволюция и старение
Старая термодинамика, которую по традиции изучают в вузах (ее те-
перь иногда именуют более правильно: термостатика), изучает вообра-
жаемые бесконечно медленные процессы, текущие в изолированных сис-
темах, находящихся вблизи от теплового равновесия. Только для них верны
ее законы, из которых знаменит «второй принцип», гласящий, что все сис-
темы стремятся к тепловому равновесию, т.е. что структуры могут сами со-
бой лишь распадаться. Не раз отмечено: дарвинизм выражает ту же точку
зрения на мир, что и «второй принцип» (см. например: Галимов Э.М. И Гео-
химия, 2005, № 5, с. 470). В терминах п. 5-3 можно сказать, что оба выража-
ли статистическую ПМ и вместе с нею теряют популярность.
Еще К.Э. Циолковский в 1919 году отметил, что «второй принцип» оче-
видным образом неверен, если в системе есть силовое поле', напри-мер,
гравитационное поле собирает космическую пыль в небесные тела, т.е. вся
Вселенная (ее наука рассматривает как изолированную систему) сама со-
бой эволюирует в сторону создания структур. В хорошем приближении
является изолированной и Солнечная система, а в ней гравитация зажгла
звезду и разогрела земные недра, чем создала условия для жизни. Итак,
281
«второй принцип», хорошо работающий в некоторых условиях (например, в
реакциях, текущих вблизи химического равновесия), надо осторожно прила-
гать там, где есть какие-либо поля. Дело в том, что поле активно, а «вто-
рой принцип» описывает объекты неактивные.
Никаких полей в биологии до недавнего времени не признавалось, и
«второй принцип» вовсю прилагался к живому. Естественно, вставал во-
прос: почему живое в целом не деградирует, а развивается? Первое, что
пришло в головы - что дело в открытости биологических систем: если живой
объект изолировать, он погибнет, т.е. придет к тепловому равновесию. От-
вет поверхностный - прежде чем погибнуть, изолированный объект может
долго жить и даже развиваться (куколка насекомого), а значит процессы те-
кут там всё это время против «второго принципа», и непонятно, что их в это
время движет. Более содержательным оказался другой ответ: дело в том,
насколько система удалена от равновесия.
Биосфера находится в потоке солнечной энергии и в потоках из земных
недр, благодаря чему в ней идет преобразование одних структур в другие.
Она сильно удалена от химического равновесия (т.е. описывается нелиней-
ными уравнениями). Столь же далек от него всякий живой организм: даже
если он в данный момент не растет, не движется, не размножается и не
стареет, все равно в нем непрерывно идет обмен веществ, т.е. преобразо-
вание одних веществ в другие. Наоборот, умерший организм понемногу
стремится к химическому и тепловому равновесию с окружающей средой,
т.е. его термодинамика постепенно становится термостатикой.
Естественно, тут встает вопрос о старении. Известные мне гипотезы
механизмов старения делятся на две группы: одна признаёт процедуру ста-
рения генетически запрограммированной, другая отрицает эту идею и вы-
водит старение из всевозможных ошибок функционирования, т.е. по сути -
из «второго принципа». В обеих группах есть гипотезы (их большинство),
выводящие старение высших организмов из старения их клеток, и такие, ко-
торые видят в старении организма особый целостный процесс. Мне более
интересны те, что увязывают программу и целостность, поскольку более яв-
ственно связаны с новой термодинамикой, рассматривающей систему как
таковую, а не как набор частей. Аналогия: государство стареет и гибнет не
оттого, что стареют его граждане, а по своим причинам.
Каждая гипотеза старения объясняет свой круг фактов и вне его бес-
сильна. Поэтому интересна заметка геронтолога О. В. Квитко (Минск), фак-
тически приложившего к проблеме старения идею снятия давления нормы.
По его мнению, процесс старения сдерживается у растущего организма
«развитийной доминантой», т.е. установкой всех механизмов на одну цель.
Она тормозит остальные процессы, в том числе, ведущие к старению, и ее
снятие ведет к их проявлению (Квитко О.В. Развитийная доминанта как ме-
ханизм антистарения // Геронтология и гериатрия. Альманах Ин-та геронто-
логии РАМН. М., 2005). Мы вернемся к теме старения в п. 8-6.
282
Если система достаточно далека от равновесия, то в ней, оказывается
могут сами собой образовываться упорядоченные структуры. Бельгийский
физхимик Илья Пригожин назвал их диссипативными структурами (ДС)
подчеркивая этим, что они существуют за счет диссипации (рассеяния) во-
вне получаемых веществ и энергии. Простейшая ДС возникает в ванне с
водой, когда вынута пробка: при достаточно мощном стоке вода закручива-
ется, водоворот имеет свои простые онтогенез (воронка формируется по-
степенно) и память (закрутившись влево, не сможет закрутиться вправо, ес-
ли не разрушить всю структуру). Он вбирает в себя всё, что плавает вокруг,
и в этом смысле сам создает силовое поле.
Никакая ДС невозможна без движения, она существует как побочный
результат какого-то потока. Школа Пригожина сумела показать значитель-
ную общность феномена ДС (они образуются в самых разных физических и
химических процессах) и провести яркие аналогии с биологией.
Утверждается, что всякий организм (да и всякое сообщество, вплоть до
биосферы) существует и развивается за счет текущих через него веществ и
энергии, за счет их диссипации. Феномен ДС - главный в новой термодина-
мике, он показывает, что в далеких от равновесия системах возможна са-
моорганизация. Это и есть основа термодинамического эволюционизма.
Однако она возможна не только в рамках термодинамики, но и в рамках
механики. Примером служит солитон (уединенная волна). Он тоже, как и
ДС, является примером самоорганизации там, где ее вроде быть не может,
и есть попытка описать самые различные биологические объекты как соли-
тоны (Петухов С.В. Биосолитоны - тайна живого вещества. М., 1999). Об-
разование солитона - фазовый переход. О солитонах см. пп. 6-14** и 7-6.
«Второй принцип», понимаемый как распад структур, может (но не обя-
зан - вспомним Циолковского) действовать в любых системах, и это не ме-
шает возникновению в каких-то частях системы порядка и сложности.
Если до рождения науки природу воспринимали как творение божье, то
наука поначалу восприняла и природу, и живой организм как огромный ме-
ханизм (чаще всего как очень сложные часы) и с тем смогла найти первые
ее законы; в XIX веке и природу, и организм стали понимать как тепловую
машину, и это тоже позволило многое понять. Так вот, понимание организма
как ДС позволяет понять еще кое-что, и этим аналогия полезна. Она на-
столько же выше аналогии с тепловой машиной, насколько та была выше
аналогии с часами. Тем самым, новая термодинамика дает обоснование
(пока весьма абстрактное) самоорганизации в ходе эволюции.
В биохимических процессах самоорганизация возможна и без образо-
вания ДС - за счет превышения анаболизма над сопряженным с ним ката-
болизмом (п. 5-11). Как полагает геохимик Э.М. Галимов, системы «в кото-
рых протекают параллельно и сопряжено неравновесные процессы, реаги-
руют на нарушение стационарности производством упорядочения», причем
нелинейности не требуется (Геохимия, 2005, № 5, с. 473).
283
Вероятно, в близком будущем ДС, солитон и сопряжение станут част-
ными случаями более общей теории самоорганизации. Она будет настолько
же выше теории ДС, насколько та выше термостатики. Она должна вклю-
чить в себя и когерентную логику (о ней см. [Чайковский, 1993, с. 133-134]), и
синергетику (теорию самоорганизации, конкурирующую с новой термоди-
намикой). Мы вернемся к этой теме в п. 6-7**.
Сказанное в этом и предыдущем параграфах можно резюмировать так:
материал для эволюции поставляет не только направленная случайная
изменчивость, но и самоорганизация, причем обе возникают при
снятии давления прежней нормы.
5-14* Биологическое поле и морфогенез
Неизбежно образование не только самих ДС, но и структур из них, при-
чем каждая такая структура сама выступает как ДС более высокого уровня.
При образовании составных ДС какая-то из структур низшего уровня высту-
пает в качестве затравки для ДС высшего уровня. Подробнее см. [Чайков-
ский, 1990, с. 201]. Хороший пример затравки дает эмбриогенез, что мы
сейчас и увидим.
В наше время поле чаще всего понимают по раннему Гурвичу - как ме-
тафору для описания целостности системы управления онтогенезом (п. 4-
16). Например, эмбриолог В.Г. Черданцев дал такое определение поля:
“Мы можем варьировать механизмы развития одной и той же структуры, с
тем, чтобы отыскать их инвариантные характеристики, которые и определяют
морфогенетическое поле системы”.
Там же отмечено, что если в опыте из зародыша удален организатор
(см. п. 4-13), то развитие может пойти по пути, свойственному животным бо-
лее низкой ступени развития: например, развитие зародыша амфибии - по
пути, характерному для зародышей низших хордовых (бесчерепных и даже
асцидий) (Черданцев В.Г. Морфогенетические поля и эволюция // Вопросы
эволюции онтогенеза. М., МОИП, 1985, с. 106, 107, 111).
Тем самым, организатор играет роль упомянутой термодинамической
затравки. Более того, этим показано, что эволюция действительно
может идти по Жоффруа.
Эмбриологи видят развитие как преобразование формы зародыша, а
физиологи, наоборот, обращают на его форму мало внимания и склонны
трактовать онтогенез как смену способов жить. Для физиолога зародыш -
тоже организм, только живущий в особой среде (внутри яйца или внутри ут-
робы матери). С этой точки зрения Аршавский уравнивал поле (которое по-
нимал по позднему Гурвичу) с доминантой (п. 5-11): по его мнению,
“то, что Чайлд (1941) обозначал физиологическим градиентом, а А.Г. Гурвич
(1944) - эмбриональным полем, в физиологическом смысле является меха-
низмом доминанты”; при этом “с момента возникновения организма в виде
зиготы одновременно возникает механизм (принцип доминанты), обеспечи-
284
вающий не только процессы регуляции, но и интеграцию его как целостной
системы ... т.е сохранение целостности развивающегося организма на всем
протяжении его индивидуального развития” [Аршавский, с. 56].
Такое уравнивание поля и доминанты натянуто, однако некоторый
смысл для нас имеет. Насколько понимаю, доминантой зиготы (оплодотво-
ренной яйцеклетки) Аршавский считал электрическое поле, наблюдаемое
между ее полюсами, «периэлектронический контраст» (там же). Далее поле
усложняется, чем, по Аршавскому, и движется онтогенез. С возникновением
нервной системы доминанта обретает обычный для физиолога облик, и оп-
ределяет структуру поведения в течение всей жизни (и, добавлю, далее с
морфогенетическим полем не связана).
Разнородные суждения прежних биологов о поле можно резюмировать
словами цитированного в п. 4-16 Светлова: хотя теория поля «наталкивает-
ся на очень большие трудности», однако «как образ мысли она прочно во-
шла в обиход механики развития» (т. 1, с. 243). Поясню: смысл теории био-
поля не столько в реальных таблицах и формулах, сколько е общем взгляде
на процесс формообразования. Сам же Светлов дал и примеры реальных
картин эмбриональных полей. Они интересны, указывают на параллели
процессов, но не дают подходов к пониманию их специфики (почему из го-
рошины вырастает именно горох и т.п.).
В недавнее время, в связи с новыми открытиями, теория биополя вновь
привлекла внимание в нескольких планах сразу.
1) Как сказано (п. 4-16), была открыта лазерная природа митогенетиче-
ских лучей Гурвича (см. Дополнение 2).
2) При начале развития куриного зародыша подзародышевая часть
желтка заметно излучает ультрафиолет (Белоусов Л.В., Попп Ф.-А., Казако-
ва Н.И. // Онтогенез, 1997, № 5). Как видим, ППК “бегут от греха подальше”,
чтобы излучение не побудило их делиться и не испортило наследственную
информацию.
3) Поле, организующее пространственную структуру зародышей, зада-
ется концентрациями: изменение концентрации одного белка задает место
появления и направление развития конечности или пера. (Лопашов и Зем-
чихина - см. п. 4-16 - уточняют: на стадии индукции действуют короткие
белки-индукгоры, способные вызывать появление новых клеточных типов, а
позже - длинные белки-матрицы, способные только стимулировать деление
клеток уже существующего типа.)
Даже противник идеи поля эмбриогенетик Л. И. Корочкин писал:
“Прообраз будущего организма складывается под влиянием продуктов генов
материнского организма, передаваемых в яйцеклетку из материнских питаю-
щих клеток. Эти клетки расположены на поверхности яйцеклетки неравно-
мерно, и потому их продукты распределяются в ней по градиентам, создавая
полярность будущего организма и размечая его головной и хвостовой концы”
[Геном..., с. 95].
285
Но где градиент, там и поле - в данном случае, по Корочкину, поле кон-
центраций (а по Аршавскому, как мы видели, электрическое). В более слож-
ных случаях приходится искать более сложные поля (см. гл. 7).
4) Открыты гены, управляющие основными полями морфогенеза. Поля
оказались сходно устроенными у самых разных животных. Для эволюции
это главное. Об этом можно прочесть в американском обзоре [Гилберт и
др.]. Эмбриолог Скотт Гилберт - тот самый, про «Сагу о зародышевом пути»
которого говорилось в п. 5-12. Другим соавтором обзора является эмбрио-
лог Рудолф Рэфф, соавтор книги [Рэфф, Кофмен].
В подлиннике обзор назван: «Воссоединение (Resynthezing) эволюци-
онной биологии и биологии развития». Авторы уверены, что СТЭ не была
синтезом; наоборот, она стала отказом от основных достижений эволюцио-
низма начала XX века, затем забытых. На мой взгляд, до нового синтеза и в
этом обзоре еще далеко, но стремление к нему есть. Например:
«Подобно тому как клетка (а не ее геном) служит единицей органической
структуры и функции, морфогенетическое поле (а не гены или клетки) пред-
ставляется основной единицей онтогенеза, и его изменение влечет за собой
изменения в эволюции».
Авторы обзора читают по-немецки, что радикально расширяет их умст-
венный горизонт по сравнению с большинством коллег, но все-таки замеча-
тельная работа Балкашиной о генах гомеозиса, т.е. переделки одних орга-
нов в другие (п. 4-7), осталась им неизвестна. Иначе их простодушное заяв-
ление: «Гомеозисные явления оставались на обочине эволюционной теории
с тех лор, как в 1894 г. их описал Бэтсон» - просто нелепо: Елизавета Ива-
новна выяснила (и описала по-немецки), что гены гомеозиса расположены
компактно на одной хромосоме дрозофилы (см. п. 5-15**). Мы вернемся к
данному обзору в п. 5-17.
5-15. Морфогенез как активность и узнавание. Эпигностика
Считается, что можно понять морфогенез, представив его как склады-
вание клеток в нужном порядке. Тогда проблема морфогенеза сведётся к
двум вопросам: как клетка узнаёт, где она находится, и как, узнав это, она
обретает нужную форму и функцию? Или: как клетки взаимодействуют?
Приблизительный ответ дал иммунолог Джералд Эдельман (США), ко-
гда в 1980-х годах развил теорию топобиологии. Под ней он понимает
«биологические взаимодействия, определяемые пространственным располо-
жением. Такие взаимодействия ярко проявляются в развивающемся зароды-
ше. Их исследование дает ключ [и] к разгадке происхождения иммунной сис-
темы» {Эдельман Дж. Топобиология // ВМН, 1989, № 7, с. 24).
По Эдельману, положение клетки в эмбрионе и, тем самым, структуру
растущей ткани, определяют открытые им молекулы избирательной кле-
точной адгезии (прилипания), расположенные на поверхности клетки. Эти
молекулы оказались сходными с иммуноглобулинами, и Эдельман заклю-
286
чил, что адаптивный иммунитет тоже развился из механизма избирательной
адгезии. Она имеется даже у одноклеточных и позволяет им отличать клет-
ки своей колонии от чужих. Словом, избирательная адгезия есть у всех, он-
тогенез - у многих, а адаптивный иммунитет - у немногих.
Изменение типов адгезии должно вызывать перемену в пространствен-
ном расположении клеток, т.е. изменение процесса морфогенеза. Если так,
то топобиология увязывает в одно целое эволюционизм, эмбриологию и
иммунологию, причем увязывает не на филогенетической («какой организм
от какого?»), а на механизменной («как?») основе. Она станет еще содержа-
тельнее, если включить в нее понятие биополя, о котором шла речь в главе
5 и что мы сделаем ниже.
Тем самым, Эдельман указал общий механизм активности на клеточ-
ном уровне (избирательную адгезию). Его частные случаи - столь разные
явления, как форма клеточной колонии, морфогенез и иммунитет.
Поразительным примером активности в морфогенезе служит рост ней-
рона: его верхушка (конус роста) покрыта шипиками, адгезивные поверхно-
сти которых ощупывают соседние клетки, словно вибриссы кота [Албертс и
др., т. 3, с. 351]. В этой своей поисковой активности нейрон выступает как
индивид. То же (на меньшем масштабе) можно сказать про активность рас-
тущей микротрубочки (см. ниже).
Всю концепцию молекулярного и клеточного узнавания можно назвать
словом эпигностика (от греч. эпигнозис - знакомство, узнавание). Оно
представляется мне более точным, чем топобиология (это слово, кстати, не
прижилось). Иммунология, редактирование РНК (п. 5-6), геномные цензоры
(п. 6-6) и многое другое выступают в этом аспекте как разделы эпигностики
(см. Дополнение 3). 1
Эдельман хотел дать подход к отсутствующей до сих пор теории мор-
фогенеза, и выразил общую точку зрения, понимая морфогенез как укладку
клеток, подобную укладке камешков в мозаику. Но так ли это? Можно ли по-
нимать онтогенез многоклеточного организма как череду клеточных деле-
ний? Почти все эмбриологи ответят “Да”, однако это ошибка: во-первых,
есть несколько ярких примеров неклеточного онтогенеза; во-вторых,
сам клеточный онтогенез не сводится к укладке клеток.
1) Параллелизм между прокариотной и эвкариотной тканями: миксобак-
терии (прокариоты) и миксомицеты (эвкариоты) образуют сходные структу-
ры - крупные плодовые тела, на которых вырастают “грибочки” со шляпка-
ми. При этом плодовое тело миксобакгерии многоклеточно, а у миксомицета
является огромной многоядерной клеткой, т.е. не может быть результатом
клеточной дифференцировки.
2) Сходные формы расчленения листовой пластинки известны как у
многоклеточных (папоротников и семенных), так и у гигантских одноклеточ-
ных - таковы, например, сифоновые водоросли. Их вернее будет назвать
цитоидами (подробнее о них см. п. 8-8*).
287
3) Покровная ткань плоских червей (тегумент) тоже являет собой еди-
ный цитоид (о чем будет речь в п. 8-4).
4) У зиготы кольчатого червя исследователь подавил деление клеток, и
при этом, удивительным образом, наблюдал фрагмент онтогенеза:
“в отсутствие клеток происходили многие события раннего развития. Цито-
плазма зиготы делилась на определенные области,... светлая цитоплазма, ло-
кализованная в анимальной области яйца, перемещалась книзу над вегетатив-
ными участками, напоминая эпиболию клеток (движение клеточных пластов,
рождающее ткань — Ю. Ч.) анимального полушария при нормальном развитии.
Это движение наблюдалось точно в то время, когда должна была происходить
эпиболия у нормального зародыша” [Гилберт, т. 1, с. 111].
5) У дрозофил органы формируются из компартментов {компартмент
- участок зародыша, попав в который, клон клеток не может его покинуть),
не совпадающих с органами: например, из одного компартмента формиру-
ется передняя часть крыла и часть груди, а из другого - задняя часть крыла
и другая часть груди. Уже это удивительно, но 30 лет назад группа испан-
ских эмбриологов (Garcia-Bellido А. е.а.) к тому же показала, что внутри ком-
партмента клеточные клоны располагаются самым прихотливым образом, и
видно: каждый клон, разрастаясь, занимает доставшееся ему место, кото-
рое меняется от опыта к опыту; можно даже взять быстро растущий клон, и
он всё равно не перейдет границ компартмента, хотя бы и окруженный мед-
ленно растущими клонами ([Гилберт, т. 3, с. 140]; [Маресин, с. 117]). Одни-
ми взаимодействиями соседних клеток этого не объяснить.
6) У высших растений онтогенез идет не путем наращивания клеточных
пластов, а иначе; об этом у нас речь пойдет в п. 9-2.
Все вместе данные примеры показывают, что, вопреки уверениям
большинства, онтогенез задан не на языке деления клеток, а в тер-
минах преобразования формы зародыша как целого (о чём 80 лет назад
писал Любищев [2004]). Примерно тогда же проблему хорошо оценил не-
мецкий физиолог и биолог-теоретик Якоб Икскюль:
“Механизм любой машины, например, часов, всегда строится по центростре-
мительной модели. Это значит, что сначала изготавливают все части часов -
стрелки, пружинки, колесики - а затем их монтируют... Напротив, развитие
животного... центробежно, начиная с зародыша. Сначала возникает гаструла,
в которой появляются зачатки, которые затем развиваются в органы... Части
собираются по совершенно противоположным принципам, но в согласии с
одним и тем же планом” (UexkiillJ., цит. по [Том, с. 151]).
Это - замечательный (хоть и мало известный) тезис Икскюля:_______
_______онтогенез - не самосборка клеток, а синтез сложного из простого.
Он явственно противостоит общепринятым тезисам Окена (п. 2-4). Со-
гласно тезису Икскюля, онтогенез - вовсе не последовательность клеточных
Делений, он есть создание формы, которую чаще всего заполняет множест-
288
во клеток, но ее могут заполнять и иные структуры - гигантская клетка, кость
и т.д. Морфогенез надо понимать как образование формы, задаваемой це-
лостностью организма и заполняемой либо клетками (если они имеются),
либо неклеточными массами. Эту еще малопонятную целостность и назы-
вают биополем (см. ниже). Процесс формообразования много сложнее, чем
последовательность актов укладки клеток «по Эдельману». Какой же меха-
низм создаёт форму?
Проблема до сих пор не решена даже на уровне онтогенеза самой
клетки. Хотя он подробно расписан в руководстве (Албертс и др., т. 2) и хо-
тя ясно, что главную роль тут играет активность микротрубочек, но на
указанную тему там есть всего одна фраза. Описывая генетический про-
цесс, определяющий сборку микротрубочек из молекул белка (тубулина),
руководство отмечает:
“сборка молекул тубулина регулируется таким образом, что многие микро-
трубочки направляются к определенным участкам клетки. Еще не вполне яс-
но, как это достигается...” [Албертс и др., т. 2, с. 308].
На самом деле здесь царит полная неясность, причина ее давно указана:
“знание химических превращений веществ в клетке без знания целенаправлен-
ных движений внутриклеточных структур не может привести нас к понима-
нию...”. “Если бы... проблема программирования и регуляции работы генов в
эмбриональном развитии была бы разрешена, то ... осталось бы еще неразга-
данным, какими механизмами обеспечивается закономерная пространственная
расстановка клеток, свойственная данному организму” [Александров, с. 23, 32].
Здесь проблема осуществления (каким образом линейный ген управ-
ляет созданием пространственной формы [Любищев, 2004]) выступает как
проблема понимания активного поведения микроструктур, как проблема
цитоэтологии (п. 5-2). На их поведение указывал и математик Ренэ Том
(Франция): «Молекулы ощупывают, ощипывают, скручивают и разрывают
друг друга как живые существа. Не стоит этому удивляться» [Том, с. 120-
121]. Не стоит потому, что теория катастроф описывает и большое, и малое.
Подходов к решению проблемы осуществления мне не известно, но
важно уже то, что онтогенез можно видеть как совместную проблему цито-
этологии и эпигностики. Единственная известная мне попытка конкрети-
зации этой проблемы — теория биополя. (Напомню, что поле введено фи-
зиками как форма активности материи.) Биополе принято либо отрицать,
либо упоминать как нечто из истории, либо просто не упоминать. Однако на
самом деле никто из пишущих о биологии развития не обходится
без идеи поля. Вопрос разобран в послесловии к книге [Любищев, 2004].
Добавлю только, что недавно А.М. Оловников, строя модель онтогене-
за, снова назвал «позиционную информацию» (он местами пишет: «морфо-
генетическое поле») единственным (притом непознанным и никак не уточ-
няемым) источником сведений о трехмерных структурах зародыша или ор-
ганизма (Оловников А.М. И Клиническая геронтология, 2005, № 1).
289
Важное уточнение в идею поля вносит книга: Черданцев В.Г. Морфоге-
нез и эволюция. М., КМК, 2003 - поле следует рассматривать не как пози-
ционное, а как динамическое, т.е. как зависящее не только от положения
пластов клеток, но и от скоростей деления клеток.
Идея поля имеет не только конкретно научный, но и мировоззренческий
смысл: победа идеи поля (Ньютон) над идеей механического контакта (Де-
карт) позволила 330 лет назад создать теоретическую физику. А в XX веке
на основе идеи поля физики вновь вернулись к Декарту (к идее виртуальных
частиц, заполняющих пространство). См. Дополнение 3.
На основе п. 5-14 можно предварительно сказать, что онтогенез - не
клональный рост, а последовательность ростов разноуровневых ДС. Учтя
сказанное, рассмотрим идею эволюции как смены онтогенезов.
5-15* С Платоном или без?
Кроме указанных в п. 5-14*, были открыты гены, запускающие форми-
рование глаза, сердца и конечности. Один и тот же ген начинает работу по
закладке данного органа - как у насекомых, так и у позвоночных [Гилберт и
др.]. Пишут, что этим доказано единство их происхождения, но ведь данные
органы устроены у насекомых и позвоночных совсем различно.
Те, кто видит здесь общего предка, вряд ли сознают, что стоят на по-
зиции платонизма: у фасеточного глаза насекомого и линзового глаза по-
звоночного общей является только идея зрения, у внешнего скелета насе-
комого и внутреннего скелета позвоночного - идея подвижного каркаса, а у
их сердец - идея прокачки жидкости. Материальные реализации совсем
различны и от общего предка идти не могут. Значит ли это, что от “общего
предка” унаследована идея?
Любищев отвечал положительно. Точнее, он был уверен, что про нали-
чие общего предка мы обычно ничего не знаем, однако сам факт реализа-
ции одной и той же идеи в разных эволюционных ветвях признавал. Более
того, он в 1925 году видел путь к успеху именно тут:
“Единственно серьезным возражением против платонизма в морфологии яв-
ляется чрезвычайная трудность этого направления” [Любищев, 2004, с. 124].
Что изменилось за 80 лет? Тогдашние примеры общности идеи (при
различии материальных реализаций) обрели конкретность: доказано, что
разные материальные процессы, реализующие одну и ту же идею, часто за-
пускаются одним и тем же геном. В данных случаях ген маркирует идею,
а вовсе не кодирует ее материальную реализацию. Платонизм, да и только.
Как биологу отнестись к этому?
Нужно отметить, что математики стоят перед аналогичной проблемой
давно и, в основном, склоняются к платонизму. Рене Том [Том, с. 215] вспо-
минал платоническую мысль математика Шарля Эрмита:
“кажется, что числа существуют вне нас и заставляют нас себя признать с той
же необходимостью, той же неизбежностью, что и натрий или калий”,
290
и прилагал данное умонастроение к биологии:
“...в биологии аналогичным образом существуют формальные структуры,
имеющие геометрическую природу, которые предписывают динамике само-
воспроизводства в заданной среде единственную возможную форму”.
С тремя оговорками (структуры могут иметь не только геометрическую,
но и алгебраическую природу, не только в пространстве, но и во времени и
задавать не только единственную форму, но и некоторое разнообразие
форм) эти слова Тома вполне можно принять. Об этом гласят, кстати, все
аргументы номогенеза. Вот новейший, из эволюционной иммунологии:
«примечательное противоречие: несмотря на отсутствие явных гомологов ря-
да ключевых ряда ключевых компонентов адаптивной иммунной системы по-
звоночных, сама способность к адаптивному иммунному ответу (у беспозво-
ночных - Ю. Ч.) хорошо развита». «По-видимому, многие задачи, стоящие пе-
ред иммунной системой, решались в ходе эволюции разных групп разными
способами» [Марков, Куликов, 2006, с. 142].
Важнейшую поддержку платонизм получил в последние 30 лет от тео-
рии фракталов (см. далее, п. 6-7). Эти объекты возникают при решении
простейших уравнений и поражают своей замысловатой сложностью и раз-
нообразием. Стоит чуть изменить один из параметров уравнения, и компью-
тер рисует совсем новую удивительную картину. Ни математик, ни авторы
компьютера и его программ не имели никакого представления о том, что по-
лучится при данном значении параметров. Само их различие при замене
ничтожно, и ее никак нельзя счесть породившей ту огромную информацию,
которая заключена в открывшейся дивной картине (см. Дополнение 4).
То, что смутно ощущали математики прежде, теперь стало наглядно
очевидно: строя математику, мы лишь открываем шкатулку с драгоценно-
стями, которые лежали в ней ранее. Ситуация не нова: с тем же чувством
мы впервые глядим в микроскоп и в телескоп. Поскольку в них мы разгля-
дываем то, что явно сделано до нас, то становится легче понять, что так же
могут быть сделаны и многие объекты математики.
Большинство биологов слышать не хочет о платонизме, но при этом не-
которые проблемы не может даже назвать. Таковы проблемы эмерджентно-
сти (п. 3-10) и осуществления (см. начало параграфа). Постановку послед-
ней проблемы обычно заменяет ссылка на «каскады генов, последователь-
но реализующих, в конце концов, программу специфической клеточной
дифференцировки» [Корочкин П.И. И Геном..., с. 93]. Иногда еще ссылаются
на «позиционную информацию», но это незаконно: на самом деле она вовсе
не позиционна, а динамична (п. 5-14*), и источник динамики надо узнать.
Как и почему «клеточная дифференцировка» принимает форму органа,
который работает, никогда не говорится. Кстати, дифференцировка вовсе
не обязательна - см. выше примеры неклеточного онтогенеза.
Если один ген запускает разные онтогенезы, это похоже отнюдь не на
выполнение конкретного приказа, а на реализацию архитектором заказа,
291
данного ему заказчиком в самых общих выражениях. Будто в прошлом
предки совсем разных таксонов получили один и тот же “заказ в письменной
форме”, и каждый реализовал его как сумел, творчески. Надо ли признать
тут творчество в смысле Страхова - Бергсона?
5-15** Поле, гомеобокс и эволюция
Таким “заказом в письменной форме” выглядят гомеобоксы. А именно,
установлено, что все гомеозисные гены содержат на З’-конце участок в 180
пар нуклеотидов, который был выявлен в 1985 году и назван гомеобоксом
(он кодирует гомеодомен, полипептид в 60 аминокислот), почти одинаковый
у самых разных животных, в частности - у насекомых и позвоночных (фор-
мулы пяти гомеодоменов см. [Гилберт, т. 3, с. 155]). У тех и других эти гены
ответственны за создание передне-задней оси [Гилберт и др., с. 333; Гоод-
ницкий, с. 87]. Многим казалось, что филогенетика (учение об общих пред-
ках) празднует победу: ведь совпадение генов принято понимать как родст-
во. Но природа, как всегда, оказалась сложнее наших теорий.
Генетики, склонные трактовать онтогенез как простой итог работы ге-
нов, а эволюцию - как накопление и отбор случайных мутаций, стали рас-
суждать, как водится, про общего предка всех животных, но тут вышел кон-
фуз: вскоре гомеобокс был открыт у грибов и высших растений:
“гомеобокс растений значительно отличается от аналогичной последовательно-
сти животных и грибов; имеющееся сходство тем не менее свидетельствует о
том, что они произошли от одного гена, имевшегося у одноклеточного сущест-
ва, которое явилось общим предком всех организмов и существовало миллиард
лет назад. Какова была исходная роль этого гена, остается только гадать”
- писал в редакционной заметке журнал «Scientific American» (ВМН, 1991,
№ 8, с. 23). Придется возразить: гадать можно о том, существовал ли
предковый ген миллиард лет назад, но оснований признавать его носителя
«общим предком всех организмов» нет. Нет оснований признавать, что об-
щий предок у растений и животных был эвкариотом40. Если же допустить,
что гомеобокс был уже у бактерии, то он не мог отвечать за морфологию
эвкариот и тем более - органов многоклеточных. Куда проще видеть в го-
меобоксе маркёр, передаваемый, например, горизонтальным переносом.
В любом случае, гомеобокс управляет чем-то весьма общим, нужным
грибу, кукурузе, червю, мухе и человеку. Тут общностью задачи ничего не
объяснишь: если передне-задняя ось у низшего червя есть, то ничего похо-
жего на сердце и конечность нет. Почти излишне напоминать, что у насеко-
мых и позвоночных совершенно различны строение органов и их формиро-
вание в онтогенезе. В частности, скелет ноги у насекомого наружный, а у
позвоночного внутренний. И если филогенетик все-таки скажет, что гены,
соответствующие сердцу, глазу или ноге, у червя были, то этим он откажет-
40 Их биохимическую общность лучше всего объясняет идея прогеноты (см. гл. 7).
292
ся от привычного понимания эволюции, он присоединится к Григорию Нис-
скому (п. 1-8) и последующим богословам, понимавшим эволюцию как раз-
ворачивание заданных Богом изначальных потенций.
Так видят эволюцию многие, в том числе ведущие ученые. Например,
Корочкин приводил (в качестве эволюционного) пример: гомеозисная мута-
ция изменила тип сегментации туловища дрозофилы так, что один сегмент
стал похож на сегмент туловища многоножки, т.е. далекого предка [Геном...,
с. 86]. Но это - возврат назад, инволюция (по Любищеву, п. 4-17*), а не эво-
люция, не создание нового.
Можно признать горизонтальный перенос гена гомеобокса. Это проще
для понимания, ибо “всего лишь” рушит идею родословных древ. Но сам по
себе он ничего не разъясняет. Пусть сам гомеобокс мог быть создан заново
или перенесен, но каким образом его новый “хозяин” смог им воспользо-
ваться, если его онтогенез устроен по иному принципу?
И мы приходим к необходимости рассмотреть “платонический” вариант:
гомеобокс не запускает стадию онтогенеза, а лишь маркирует ген,
запускающий изменение морфогенетического поля.
Тут нам неожиданно помогает идея редактирования РНК (п.5-6). Стран-
но и даже неправдоподобно, но факт - редактирование имитирует
схему общего происхождения. Оно приводит
“к возрастанию гомологичности отдельных белков различных организмов и, та-
ким образом, способствует поддержанию их консервативности” {Тоусон Е.Л.
Редактирование РНК в различ. генетич. системах // ЖОБ, 2004, № 1, с. 59).
Если так, то не надо искать в родстве причину сходства отдельных ге-
нов (см. п. 5-16). В частности, не надо толковать сходство гомеобоксов об-
щим происхождением от неведомой бактерии. Если приведенный вывод о
редактировании РНК окажется общим, то вся геносистематика (см. далее,
п. 10-3) станет беспредметной. Как и почему редактирование сближает ге-
нетические тексты различного происхождения, неизвестно, но уже сейчас
очевидно, что это вид номогенеза: сходное возникает независимо.
Теории онтогенеза нет и в обозримом будущем не предвидится. Но, хо-
тя механизм онтогенеза непонятен, но на уровне его как явления эволюция
в форме перестройки онтогенезов очевидна, чем и воспользуемся.
Если поле признать как факт, принесет ли это пользу для понимания
эволюции? Можно ли сказать, что в эволюции имеет место искривление то-
го поля, в котором протекает развитие зародыша? В книге [Рэфф, Кофмен]
дан рисунок из книги английского биоматематика Вентворта Д’Арси Томпсо-
на (1917): искривляя координатную сетку рисунка рыбы одного вида, можно
получить рисунок рыбы другого, очень далекого, вида (рис. 33).
Естественна мысль, что зародыш второго вида развивался в искрив-
ленном силовом поле. Напомню: на качественном языке похожее говорил
еще Бюффон. В этом смысле идея поля уже в такой форме работает.
293
Рис. 33. Преобразование контуров организмов по Д’Арси Томпсону
Но если одну видовую форму можно переделать в другую «простым
преобразованием системы координат», значит форма подчиняется своим
законам (законам биополя), а не только приспособлением к среде. После
книги Д’Арси Томпсона стало ясно, что «невозможно всерьез заниматься
проблемами формообразования без четкого понимания пространственных
свойств изучаемых объектов» [Маресин, с. 3].
До сих пор мы говорили о развитии форм. Однако онтогенез состоит в
активном развитии не только формы, но организма, способного жить, т.е.
функционировать. Реализация этого процесса развития тоже протекает как
активность, как жизнь зародыша. Пусть он и не живет во внешней среде,
но он активно живет в том смысле, что строит сам себя, дышит и питается.
Поэтому строить отдельную теорию морфогенеза - то же, что строить макет
телевизора, игнорируя электромагнитное поле.
Это понимал Аршавский и призывал строить теорию развития на фено-
мене активности зародыша [Аршавский]; но зато его не занимал онтогенез
сам по себе (как и почему возникает особь именно данного вида). Здесь еще
вся работа впереди: в частности, предстоит выяснить, что тут означают
давление нормы и искривление поля.
5-16. Сравнительный метод и пространство жизни
Сравнительный метод в эволюционизм ввел астроном Вильям Гершель
в 1789 году: он сравнил звездное небо с садом, где можно одновременно
294
видеть все стадии развития растений (ростки, цветы, плоды, засыхание) и
тем самым понять ход жизни одной особи. Метод прижился в науке, несмот-
ря на то, что Гершель оказался неправ: туманности исследованные им, яв-
ляют собой, как позже выяснилось, скопления множества звезд, тогда как
сам Гершель был уверен, что исследует стадии формирования одной звез-
ды из космической пыли [Чайковский, 1994, с. 206]. Причина данной ошибки
теперь нам понятна: в природе часто большое подобно малому (см. п. 6-7).
Сравнительный метод служит одной из основ всякого эволюционизма -
см.: Раутиан А.С. Апология сравнительного метода // Любищев и проблемы
формы, эволюции и систематики организмов. М., МОИП и ЦСИ, 2003 (Цено-
логич. Исследования, вып. 23). Но, как показал первый же опыт его приме-
нения Гершелем, метод таит опасность ложных толкований.
К этой общей опасности нынешняя биология добавляет свою специфи-
ческую трудность: в наше время принято утверждать, что все ныне живущие
организмы имеют один и тот же эволюционный возраст, поскольку ведут
свою историю от единого акта возникновения жизни.
Ламарк и Дарвин могли применять сравнительный метод вполне закон-
но, поскольку в их время считалось, что жизнь непрестанно возникает вновь
(концепция самопроизвольного зарождения) и, следовательно, нынешние
черви считались младше рыб, рыбы младше зверей и т.д.
В наше время не так: наука гласит, что жизнь заново не возникает. Но
почему тогда в морях до сих пор плавают рыбы, а не только дельфины, по-
чему рыбы не выползли все на сушу, чтобы стать наземными животными?
Экологический взгляд на мир подсказал удобный ответ: каждый организм
приспособлен к своей экологической нише, а она определяет его облик. На-
пример, чтобы зарываться в грунт, лучше всего быть червем, чтобы пла-
вать, лучше всего быть рыбой, и пока существует ниша, она должна быть
занята соответствующим организмом.
Ответ подозрительно прост (зарываются в грунт и насекомые, и позво-
ночные; плавают и змеи, и дельфины), но в целом звучит приемлемо.
Однако иногда глобальная экология задает трудные вопросы. Так, во
многих тропических океанских водах до 80% продукции приходится на зеле-
ную бактерию Prochlorococcus marinus (Пиневич А.В. и др. И ФР, 2000, № 5,
с. 732). Почему она, процветая и будучи (согласно идеологии сравнительно-
го метода) предком зеленых водорослей, не стала за миллиард лет сама
зеленой водорослью? Или: почему в тайге основную массу составляют не
цветковые, а более примитивные деревья - хвойные?
Исторически сравнительный метод в биологии зародился вместе с эво-
люционной идеей: в понимании Анаксимандра рыбы породили людей в силу
того, что вода - общий (по Фалесу) источник жизни, а среди водных живот-
ных его привлекли своим сходством с людьми именно рыбы. В более позд-
них фантазиях это были дельфины (отсюда легенды о русалках). Однако
реальной основой нашей эволюционной картины (четвероногие произошли
295
от рыб) явился не сравнительный метод, а тот палеонтологический факт,
что остатки рыб присутствуют в земных толщах, не содержащих остатков
зверей, в том числе и дельфинов. Затем были открыты переходные формы
от рыб к амфибиям (см. далее, п. 9-13). Именно эти факты позволили вы-
брать одну из двух картин, допускаемых сравнительным методом.
И так всегда: сам по себе сравнительный метод не дает истории.
Историю дает палеонтология, но ископаемые организмы представлены
почти исключительно формами - скелетами, раковинами и отпечатками.
Поэтому почти всё то, что мы знаем о функциях древних организмов, добы-
то сравнительным методом: если установлено, что, к примеру, иммунитет
рептилий проще иммунитета теплокровных, то делается вывод, что слож-
ные формы иммунитета возникли вместе с теплокровностью.
А почему? Какие доводы мешают счесть, что те древние рыбы, которые
были нашими прямыми предками, обладали нашей формой иммунитета или
чем-то вроде? Быть может, они потому и смогли обратиться в наземных жи-
вотных, что сложный иммунитет помогал им жить в новых средах? Или на-
оборот: почему не допустить, что первые млекопитающие вымерли как раз
из-за отсутствия сложного иммунитета нынешнего типа?
И так - во всём. Например, что мешает допустить наличие нынешнего
фотосинтеза (такого же, как у высших растений) у какой-нибудь древней
бактерии или, наоборот, отказать древней бактерии в умении столь же со-
вершенно, как нынешние, реплицировать ДНК?
Общий ответ мне известен один - если какое-то свойство обще для ны-
не живущих членов группы, то его можно допустить и у ее вымерших чле-
нов. Мейен называл эту процедуру типологической экстраполяцией, а
Кювье принципом корреляции (п. 1-15). Но он говорил только о наблюдае-
мых формах (которые связывал с ненаблюдаемыми функциями) и то ока-
зался не во всем прав, что мы узнаем в главе 9 в связи с халикотериями.
Что же касается функций, не отражающихся на форме (иммунитет, цветное
зрение и т.п.), то жёстко увязывать их с формой вообще нет оснований.
Почему же мы так делаем? Думаю, основная причина типологических
экстраполяций кроется в силе той презумпции, которую именуют принципом
актуализма (пп. 4-17*, 5-1*): проще всего думать, что всё неизвестное орга-
низовано так же, как известное (в частности, прошлое - как нынешнее), пока
кто-то не докажет противного. Как я старался показать ранее (пп. 3-9, 5-1, 5-
Ю), науку движет, в широком плане, именно этот “кто-то”, от презумпций от-
казавшийся. Так что поищем более серьезного ответа.
Для меня он естествен в рамках номогенеза: поскольку эволюция есть
процесс смены целостностей, то реализуется не всё что попало, реализу-
ются только целостности; их набор ограничен, поэтому в истории вновь и
вновь являются одни и те же принципиальные конструкции, различающиеся
лишь теми признаками, которые на целостность не влияют (акциденциями).
Те свойства, которые входят в синдром (совокупность свойств, совместно
296
характеризующих явление) группы, должны были присутствовать и у древ-
них членов данной группы - вот идеология номогенеза. Ее косвенно под-
держивает идеология новой термодинамики, в свете которой легче осознать
два факта: что целостность живых объектов регулярно образуется сама, по
законам самоорганизации, еще мало понятным, но несомненно сущест-
вующим, и что только уже сложившиеся новые конструкции (клеток или мно-
гоклеточных организмов) могут служить объектами расплода.
Впрочем, это лишь толкование философских постулатов номогенеза, и
нельзя пользоваться им как фактическим обоснованием последнего. На-
оборот, ряды Копа - Вавилова таким обоснованием являются.
С позиции эпигностики (п. 5-15), у прежних рыб естественно предпола-
гать тот же тип иммунитета, что характерен для нынешних рыб. Но это -
толкование. Превратить его в достоверный факт может либо эксперимент,
либо свидетельство палеонтологии.
В отношении иммунитета палеонтология что-то говорит крайне редко,
но и это бывает. В 1989 году у азиатских верблюдов обнаружили особен-
ность строения антител (кроме обычных, у них есть еще мелкие нанотела),
и ее вскоре же нашли у американских верблюдов - лам (см. далее, п. 6-
5***). Унаследована ли эта особенность от общего предка или возникла не-
зависимо? Ответ дала палеонтология: предки верблюдов возникли в Се-
верной Америке, откуда расселились, эволюируя, в Южную Америку (ламы)
и Евразию. Можно достаточно определенно заключить, что вымершие севе-
роамериканские предки верблюдов уже имели нанотела.
Но нет палеонтологических данных (фактов) для утверждения (а срав-
нительный метод утверждает), что, например, нынешние кистепёрые рыбы
действительно произошли от кистепёрых мелового периода. (В этом утвер-
ждении нет и нужды, см. л. 9-14.)
Как ни странно, господство сравнительного метода - следствие стати-
стической ПМ. Она исходит из той (старой, притом по существу физической)
картины мира41, где мир мыслится однородным по всем направлениям,
включая время. Эта ПМ породила в физике мощное течение, именуемое эр-
годической теорией, которая исходит из идеи сущностного сходства (если
не тождества) тенденций во времени (т.е. процессов) с тенденциями в про-
странстве. Эргодическая теория эффективна при описании газов, но вряд
ли годна для описания каких-либо структур, поскольку работает только там,
где есть перемешивание (подробнее см. [Чайковский, 2004, с. 222]).
41 Термин в близком, но немного ином смысле ввел биолог-теоретик Людвиг Берталанфи
(Австрия). Отметив, что в начале XX в. мир видели физически, он заключил: «Но сегодня
во всех науках выступают проблемы, описываемые через понятия целостности, органи-
зации и формы, т.е. радикально связанные с биологией. В этом... важный вклад биологии
в нынешнюю картину мира» (Bertalanffy L. Das biologische Weltbild. Bem, 1949, S. 11).
297
Не видно оснований приравнивать структуру во времени структуре в про-
странстве, но именно это делает сравнительный метод, не давая тому ника-
кого обоснования (наоборот, эргодическая теория его для газов дает, хотя до
этого 90 лет была гипотезой). Обоснованием может служить диатропика, о
которой пойдет речь в главе 6. Связанные с нею картина мира и ПМ приведут
к другому сравнительному методу, в котором сходство никак не связывается с
общим происхождением. Последнее, если оно для данных групп предполага-
ется, должно быть доказано - методами, о которых пойдет речь в части 3.
Такая картина мира необходима, чтобы увидеть то пространство
жизни, в котором реально течет эволюция. Речь должна идти о том про-
странстве, в котором действует активность. Пока вопрос совсем не разрабо-
тан, можно лишь воспользоваться аналогией с физикой и космологией.
Там, в отличие от биологии, идеи эволюции быстро сменяются, но всё
же видна тенденция: по мере охлаждения и расширения Вселенной чере-
дуются фазовые переходы (п. 5-2), и каждый порождает более сложную
форму активности материи. Сперва царило излучение, а ныне - темная
энергия, так что на нашу форму (вещество планет) приходится 0,03% энер-
гии мира (Чернин А.Д. // Природа, 2006, Ns 10). В рамках этой формы фазо-
вый переход породил жизнь, она тоже являет собой ряд параллельных це-
пей фазовых переходов, и эти параллели - пища сравнительному методу.
5-17. Жоффруизм и давление нормы
Много лет назад зоолог В.В. Малахов вполне убедительно показал:
«Хордовые являются перевернутыми вторичноротыми», поскольку их
брюшная сторона гомологична спинной стороне других вторичноротых (Ма-
лахов В.В. Проблема основного плана строения в различных группах вто-
ричноротых животных //ЖСБ, 1977, № 4, с. 485). Тем самым, хордовые от-
личаются ото всех остальных вторичноротых способом онтогенеза.
Малахов показал, что перевертыванием зародыша в ходе развития
можно объяснить своеобразие хордовых среди вторичноротых и тем сохра-
нить принцип единства этого раздела животного царства. Статья имела в те
годы важное значение как воскрешение идеи плана строения в обстановке,
когда молекулярные методы стали вытеснять морфологию, вытеснять, не-
смотря на нелепость получаемых результатов.
В связи с открытием сходных формообразующих генов у позвоночных и
насекомых (см. далее, п. 5-15), эмбриолог Л.В. Белоусов (внук Гурвича) пи-
шет: поскольку на спинной стороне зародышей позвоночных работают гены,
весьма сходные с теми, которые выполняют аналогичную работу на брюш-
ной стороне членистоногих, то подтвердились
«взгляда! Жоффруа Сент-Илера о позвоночном как об “инвертированном беспо-
звоночном”, высказанные 160 лет тому назад исходя из сравнительноанатомиче-
ских соображений и известные... в виде почти анекдотического примера заблуж-
дений далекого прошлого» (Белоусов Л.В. // Онтогенез, 1997, № 6, с. 471).
298
To же пишет Шапин (АЕ, 178). Словом, общий план строения типов жи-
вотных (п. 2-6) оказался реальностью, и жоффруизм можно включить в
число серьезных эволюционных концепций. Белоусов напоминает нам
об одном из принципов Жоффруа - это
«принцип коннексий - связей, основанных на дофункциональном, стрктурно-
композиционном сходстве зачатков. Сент-Илер стремился, хотя еще и в смут-
ной форме, выразить идею о том, что между различными органами могут
быть связи, основанные не на их последующем функционировании, а на зако-
нах их формообразования» [Белоусов, с. 335].
Принцип не вполне понят до сих пор: если эмбриологи склонны видеть
онтогенез как чистое становление формы, то физиологи (из названных
мною ранее - Аршавский и Зусмановский) - как чистое становление функ-
ций. Попытку сближения этих позиций впервые осуществила (насколько
знаю) книга Жоффруа «Анатомическая философия»: там сказано, что заро-
дыш развивается по законам формы, но в рамках, задаваемых его физиоло-
гией, и эволюцию направляет сдвиг этих рамок.
Вообще, идеи Жоффруа перечислены в п. 2-7*, а теперь обсудим их с
позиций ПМ. Поразительным образом, великий француз рассуждал сразу в
рамках четырех ПМ: представление организма как равновесия органов от-
носилось к третьей (статистической) ПМ, принцип коннексий - к четвертой
(системной), гомологи - к пятой (диатролической), а сродство - к шестой
(активностной) ПМ. Естественно, что его не понимали тогда и до сих пор об-
виняют в схоластике (первая ПМ) и механицизме (вторая ПМ).
Да он и сам не мог четко выразить свою мысль и теперь нуждается в
толковании, а толкований мы стараемся избегать. Единственное, что могу
сказать определенно - он пытался уловить целостность, ускользавшую от
понимания, а столь широкие обобщения осмысленны (как мы видим ныне) в
области идей единого глобального эволюционизма (см. п. 6-14), тогда как
Жоффруа мог рассуждать только в пределах эволюции организмов.
В наше время вспомнили сто раз осмеянную гомологию по Жоффруа -
Оуэну (см. п. 2-7*). Вспомнили в связи с удивительным сходством строения
глобинов, т.е. свернутых в шар белковых молекул. Их частную гомологию
(сходство одноименных глобинов у разных видов организмов) теперь назвали
ортологией, а серийную (сходство глобинов одного организма) - паралогией.
Но, главное, вновь заговорили про общую гомологию: “Общая гомоло-
гия разных глобинов отражает отношение их структуры к особой функции” -
читаем в обзоре [Гилберт и др.]. Авторы поясняют:
«По Оуэну (Owen, 1848), цепь идентичных позвонков образовалась в резуль-
тате “вегетативного повторения” архетипического позвонка. Каждый из этих
позвонков мог подвергаться “независимой модификации” в соответствии со
своим местоположением. Согласно гипотезе Бриттена и Коэна (Britten, Kohen,
1968), тандемные семейства идентичных последовательностей ДНК возника-
ют в результате “скачкообразной репликации”» [Гилберт и др., с. 332].
I
299
Мне остается пояснить: сходство глобинов (например, гемоглобина и
миоглобина) действительно похоже на сходство “позвонков” по Оуэну (на-
пример, плечевого и тазового поясов). И напомнить, что сам Оуэн видел в
таком сходстве вариацию идеи по Платону, но не эволюцию.
Идея общей гомологии пришлась ныне весьма кстати, поскольку биоло-
гия столкнулась неожиданно с явным платонизмом:
“В сердце насекомых и позвоночных едва ли можно найти что-либо общее, кроме
способности перекачивать жидкости. Тем не менее их объединяет экспрессия од-
ного и того же гена”; “органы... структурно аналогичные, могут быть сформиро-
ваны гомологичными процессами! Мы вступаем в новооткрытую, захватываю-
щую сферу гомологии - в сферу гомологии процесса” (Гилберт и др., с. 334].
Впрочем, новизна не в этом: гомология и прежде имела дело с процес-
сами, но их не полагалось замечать42. А проницательный морфолог еще
давно сказал, что организм есть морфопроцесс [Беклемишев, т. 1, с. 11].
Новизна в том, что общее видят там, где раньше видели только частности,
что (по Белоусову) идет поиск инвариантов развития.
Но ведь эволюция - тоже морфопроцесс. Тем самым, параллелизм эво-
люции и индивидуального развития - не метафора натурфилософов, а объ-
ективное сходство процессов. Поэтому гомологизация процессов позволяет
продвинуться в понимании “основного биогенетического закона” (о нем см.
пп. 2-7, 4-7, 4-13). В дарвинизме он принят за истину, но не имеет никакого
объяснения. Удивительно, но факт: А.С. Северцов (многолетний завка-
федрой дарвинизма) вообще не нашел в литературе «обсуждения причин
рекапитуляций, т.е. филогенетически длительного сохранения в онтогене-
зе потомков анцестральных (предковых - Ю.Ч.) признаков» (Северцов А.С.
О причинах рекапитуляции И ЗЖ, 2004, № 10, с. 1187). Он верно отметил,
что с позиции дарвинизма отбор должен бы отсечь излишние свойства,
поэтому рекапитуляция вредных и бесполезных свойств непонятна; она
нуждается в особом механизме. И, добавлю, он начинает проясняться.
А именно, онтогенез обладает своими законами, законами самооргани-
зации; их общая суть состоит в том, что онтогенез может идти к одной
и той же цели различными способами (это эквифинальность, но, в от-
личие от опыта Дриша, на далеких друг от друга видах) и в разных эво-
люционных ветвях проявляется в различных формах. Это могут быть эм-
брионизации по А.А. Захваткину, рекапитуляции по Ф. Мюллеру или проро-
ческие стадии по А.П. Павлову. По-видимому, там, где ослаблено давление
нормы, мы можем видеть всё разнообразие способов онтогенеза.
42 т.
1ак, деление высших животных на первично- и вторичноротых основано на различии
процессов их раннего онтогенеза На самых ранних стадиях (дробление, см. п. 4-13)
первичноротые могут вести себя очень различно, и это не мешает признать их единой
группой - ввиду общности последующей части процесса онтогенеза, образующей
«узел сходства». То же самое можно сказать о вторичноротых [Белоусов, с. 193, 340].
300
Есть примеры, когда жоффруизм (понимаемый как прямое действие
изменений среды на изменение онтогенеза) общепризнан и основан на уче-
те давления нормы (пусть и без данного термина). Пример известен, в част-
ности, всем собаководам: у породистого щенка отнимать зажатую его зуба-
ми вещь путем ее вытягивания запрещено (положено - путем разжимания
челюстей), чтобы не испортить прикус; дело в том, что по прикусу эксперты
судят об отсутствии вырождения. В наших терминах: разнообразие прику-
сов у щенков данной родительской пары говорит о том, что норма породы
тут не выдержана. Сходное установлено в эволюции людей:
“аномалии челюсти (неправильный прикус...) относительно новы для евро-
пейских популяций. В хорошо сохранившихся скелетах людей, живших в XV
и XVI вв., неправильные прикусы почти не встречаются” (Гилберт С.Ф.
Экологическая биология развития... // Онтогенез, 2004, № 6, с. 434).
Массовый неверный прикус объясняют наивно - как итог кормления де-
тей мягкой пищей; однако как раз в прошлом их в массе кормили молоком
лет до двух-трех. Вернее иное - перед нами эволюционный эффект: эко-
этологический сдвиг в жизни людей и резкое падение детской смертности
повлекли ослабление давления прежней нормы.
Для понимания давления нормы надо понять механизм онтогенеза, а он
пока не раскрыт. Генетик В.А. Струнников в опытах на шелкопряде показал
«снижение изменчивости по мере повышения жизнеспособности» и выска-
зал мысль, что главным здесь является снижение числа тех ошибок онтоге-
неза, которые носят случайный характер, не связанный ни с генетикой раз-
вития, ни со средой обитания (Струнников В.А. Природа гетерозиса... М.,
Наука, 1994, с.29-38). Мы подошли к проблеме случайности.
5-18. Случайность в эволюции организмов
Ранее мы не раз уже касались вопроса о том, что именно можно в эво-
люции полагать случайным и в каком смысле. К сожалению, большинство
исследователей уверено, что всякой случайности можно приписать некото-
рую вероятность4^ и далее оперировать с нею. Исключения редки чрезвы-
чайно, и тем отраднее встречать их. Например, у Шалина:
«Существует ли статистический закон для появления этих (эволюционных -
Ю.Ч.) крупных скачков, или они обязаны целиком “случаю”, который подра-
зумевается непредсказуемым?» (АЕ, 273).
Здесь «статистический закон» (повторность событий, позволяющая вы-
числять вероятности) справедливо противопоставлен полной непредска-
зуемости. Мне уже приходилось писать, что случайности бывают разными и
требуют разных подходов, что в природе преобладают как раз те, которые
По А.Н. Колмогорову, случайное явление, происходящее с некой вероятностью (т.е.
имеющее устойчивую частоту), именуется стохастическим. См. [Чайковский, 2004, с.
20-21]. Эволюционные события обычно нестохастичны (см. далее, п. 6-11*).
301
вероятностного подхода не допускают, поскольку более хаотичны, нежели
те, что имеют вероятности (устойчивые частоты) [Чайковский, 2004]. Тако-
вы, в частности, и все «крупные скачки».
Есть также много данных, говорящих о том, что элементарные события,
поставляющие изменчивость, являют собой более сложную случайность,
чем полагали прежде. Например,
“разные гены демонстрируют разное нестохастическое использование кодо-
нов, и гены с высоким уровнем экспрессии отличаются максимальным сме-
щением от стохастического использования кодонов”. При этом “вопреки
ожиданиям экспериментаторов, наиболее часто используемые кодоны и соот-
ветствующие им тРНК при трансляции допускают большее количество оши-
бок по сравнению с редкими кодонами” {Малахов М.П., Семененко В.Е. Ис-
пользование кодонов в генах цианобактерии... // ФР, 1994, № 2, с. 163).
Выходит, что используемое реже используется аккуратнее.
Более того, сама редкость применения какого-то действия может спа-
сать от вымирания: так, если начать редко размножаться, можно выжить в
самых гибельных условиях (п. 2-13). Если таким путем выживает патоген-
ный микроб, то немногих выживших клеток бывает достаточно, чтобы свести
на нет все усилия врачей и естественной эволюции.
Такая случайность (редкое выживание) эволюцию тормозит. Однако
бывает и другая случайность, которая эволюцию движет.
В теме биологической случайности важно, что ею часто именуют актив-
ность. Всякая активность есть свободный выбор, а в классификации слу-
чайностей [Чайковский, 2004] свободный выбор выступает как одна из форм
случайности. Вот сравнительно простой пример такой случайности из эм-
бриологии дрозофил. В п. 5-15 мы говорили о прихотливом распределении
клонов внутри компартмента. При этом
“поведение клеточных клонов в данной ткани изменяется от зародыша к заро-
дышу, т.е. явно идентичные клоны в двух разных мухах дают начало разным
участкам одной и той же структуры. Из этих данных был сделан вывод, что
клональность сама по себе не является единственным параметром, определяю-
щим организацию структур развивающегося организма” [Маресин, с. 117].
Что же еще является, не сказано, а это как раз и есть активность: клоны
активно занимают места, допускаемые их средой, т.е. (можно полагать) ак-
тивностью поля зародыша. При этом выбор между разными вариантами
свободен: он не следует из логики процесса, т.е. выглядит случайно.
Самое интересное насчет случайности обнаружилось в механизме син-
теза иммуноглобулинов. На первом этапе синтеза гена антитела идет ком-
бинирование блоков. Если бы механизм Тонегавы перебирал одну за другой
все возможные комбинации фрагментов, то, как показывает расчет, он на-
работал бы в одном организме мыши 3 млн различных антител [Стил и др.
с. 111]. Но возможных антигенов, по Вигзелу - миллиарды, и нет никакой га-
рантии, что среди созданных были бы те самые антитела, какие в данное
302
время нужны. Поэтому процесс идет иначе: выбирает одни варианты много
чаще других, делает “болванку” нужного антитела и доводит ее до кондиции
путем гипермутагенеза.
Суть процесса пока непонятна, и иммунологи характеризуют ее как не-
кую сложную неравномерную случайность - это выражение применил То-
негава в нобелевской лекции. Через 12 лет в книге [Стил и др.] на эту тему
был приведен следующий интересный цифровой материал.
Разнообразие антител на стадии комбинирования достигается комби-
нированием разнотипных участков, обычно именуемых буквами V, D и J.
Точнее, в каждом иммуноглобулине комбинируются элементы из следующе-
го набора: 100 V-элементов, 20 D-элементов и 4 J-элемента. Поскольку ос-
новной вклад в создание разнообразия вносят V-элементы, можно бы ожи-
дать, что они будут очень отличны друг от друга. Однако оказывается на-
оборот - они почти не отличаются друг от друга. Это похоже на алфавит -
ведь разные буквы одного алфавита могут очень мало отличаться друг от
друга и тем самым вызывать затруднения у иностранца, но прекрасно вы-
полнять свою функцию.
Удивительнее, что «около половины V-элементов никогда не участвуют
в образовании антитела», а реальное одновременное разнообразие анти-
тел - отнюдь не 3 млн, но меньше 10 тыс. [Стил и др., с. 111-112].
Все стадии процесса авторы называют случайными, но, как видим, слу-
чайность эта ничуть не похожа на стандартные (как в теории вероятностей)
явления типа бросаний игральной кости. И легко понять, почему: стандарт-
ная случайность дает в среднем то же, что и сплошной перебор вариантов,
а он был бы тут бесполезен.
В самом деле, механизм Тонегавы перебирает не нуклеотиды, а со-
ставленные из них блоки. Образно говоря - не буквы, а слоги, слова и даже
фразы. Если перебор букв вполне реален, хотя и дает осмысленные слова
редко (можно поочередно заменять в данном слове каждую букву и смот-
реть, осмысленно ли получившееся слово), то перебор слогов весьма за-
труднен (осмысленные слоги складываются в осмысленные слова исче-
зающе редко), а перебор слов просто невозможен, поскольку их в любом
языке более ста тысяч, а словопар - многие миллиарды. Никто не мыслит
путем перебора слов, и вот мы видим, что механизм Тонегавы тоже осуще-
ствляет не перебор, а какую-то более сложную процедуру. Пусть смысл ее
нам пока непонятен, но итог ее работы таков.
У мыши одновременно имеется всего 50 млн экземпляров В-
лимфоцитов, причем каждый синтезирует лишь один тип антител, а деление
лимфоцита занимает более 5 часов. При стохастическом распределении
типов антител (а их 3 миллиона) по клеткам каждый тип будет представлен
всего несколькими (менее 20) экземплярами, причем их клонирование не
сможет поспеть за размножением инфицирующих бактерий (деление у ко-
торых занимает меньше часа).
303
Природа избрала иную стратегию. До недавних пор считалось, что в
ответ на инфекцию иммунная система начинает случайный направленный
четырехступенчатый поиск (п. 5-8), что налицо очень сложная случайность
(о которой говорил Тонегава). Ее свойства совсем не изучались, хотя без
этого нельзя решить, зависит ли судьба больного от случайности (найдено
ли вовремя нужное антитело).
Первое, чего тут следовало ожидать - огромного разброса сроков им-
мунного ответа, а этого нет. Первичная иммунная реакция наступает сразу,
а на полный ответ (через несколько суток) время тратится, в случае бакте-
риальной инфекции, на создание «зародышевых центров» (Сидорова Е.В. //
УСБ, 2006, № 3), т.е., так сказать, “фабрик антител”. Если случайный поиск и
идет, то он занимает очень мало времени по сравнению с остальными про-
цессами. Вернее всего, что никакого поиска редкого варианта здесь нет, что
нужный вариант бывает найден сразу многими тысячами клеток (поэтому
разбросы усредняются, а пул компетентных В-клеток достаточно велик),
следовательно, он достаточно типичен.
Но если очень многие лимфоциты находят один и тот же вариант анти-
тела, то налицо клеточный номогенез. Выяснение данного вопроса ради-
кально повлияет на развитие и идей номогенеза, и иммунологии.
В принципе можно бы допустить здесь горизонтальный перенос найден-
ной генетической информации между лимфоцитами, но ее никто не наблю-
дал (зато наблюдают перенос сигнальных молекул). К тому же известные
процессы горизонтального переноса весьма нерегулярны (идут без опреде-
ленной частоты), тогда как иммунный ответ достаточно регулярен.
5-18* На грани порядка и хаоса
Главная эволюционная мысль книги [Чайковский, 2004] состоит в сле-
дующем: к эволюции способны не детерминированные и не стохастические
объекты, а системы на грани порядка и хаоса.Начало исследованиям дан-
ной темы положил американский биокибернетик Стюарт Кауфман. Его ком-
пьютерная модель совершенно абстрактна, т.е. не имитирует никакого био-
логического объекта, а лишь демонстрирует роль необычной случайности.
Изложить суть его теории здесь невозможно, так что каждый, кому тема ин-
тересна, должен прочесть статью [Кауфман] сам. Я же могу привести оттуда
лишь несколько мыслей. Вот, из начала его статьи:
«Математические модели могут изменить представления биологов об истоках
упорядоченности в эволюции.... Дарвину и в голову не могла прийти мысль о
существовании самоорганизации - недавно открытого природного свойства,
присущего некоторым сложным системам. [...] Хаос, как бы он ни был инте-
ресен, - это лишь часть поведения сложных систем. Существует также не
поддающееся интуитивному осознанию явление, которое можно было бы на-
звать антихаосом. Оно выражается в том, что некоторые весьма беспорядоч-
ные системы спонтанно “кристаллизуются”, приобретая высокую степень
304
упорядоченности. Я полагаю, что антихаос играет важную роль в биологиче-
ском развитии и эволюции».
В предложенной им схеме эволюции очевиден (если использовать на-
ши термины) номогенез: Кауфман показал, что число устойчивых вари-
антов развития больших систем весьма мало. Очевиден и жоффруизм -
эволюция есть смена режимов онтогенеза. Если же в ходе “кристаллизации”
возникнет нечто совсем новое, то налицо эмерджентность.
Основная идея Кауфмана состоит в том, что сложные системы можно
поделить на два исходных класса - “твердые” и “газообразные”, т.е. на упо-
рядоченные и хаотические, причем между классами возможны переходы
(система может как обрести жесткую структуру, так и утратить ее); именно
при таком переходе система и может совершить акт эволюции, т.е. качест-
венно измениться. Такой переход лучше всего сравнить с прямым перехо-
дом твердого тела в газ (возгонка) и обратно.
Добавлю: яркую аналогию дает также осаждение кристаллов льда из
воздуха, образующее снежинки и морозные узоры на стекле, поражающие
своим разнообразием. Особенно интересны морозные узоры, поскольку они
рождают крупные картины, в которых не видно никакого хаоса, а видно об-
щее для каждого рисунка правило. Видно и сходство с биологическими объ-
ектами, прежде всего с листьями:
“Один из рисунков квалифицированный ботаник принял за фотографию чер-
тополоха, в других - очевидное сходство с... листьями пальмы, корневищами
растения с отходящими от него листьями, подобием мхов” (Любищев А.А.
Морозные узоры на стёклах // Знание - сила, 1973, № 7).
Любищев видел биологический смысл данных сходств в том, что узоры
допускают только одну трактовку: их формируют «законы образования
твердых тел из пара», а не приспособление к среде, не история и т.п.; это
вело его к выводу, что и в образовании живых форм играют роль законы об-
разования форм как таковых. В п. 6-7 мы увидим подтверждение этому.
К сожалению, ни Любищев, ни Кауфман не вспомнили в данной связи о
морфогенетическом поле. А ведь связь напрашивается: хотя закон образо-
вания кристаллов льда един, каждый рисунок характерен своей индивиду-
альностью. Чем она задаётся? Удобно сказать: своим полем, конкретный
вид которого задан в зародыше рисунка, причем в качестве зародыша вы-
ступает центр кристаллизации. Можно сказать, что тип данного центра
случаен. Наоборот, рост (онтогенез) узора задан.Переход от одного типа
рисунка к другому (он случаен) напоминает эволюцию.
Конкретный биологический пример перехода от одного устойчивого со-
стояния к другому Кауфман видит в феномене клеточной дифференциров-
ки. Малое число устойчивых состояний, упомянутое выше, он трактует как
малое число различных типов клеток организма.
Огромный класс явлений “на грани порядка и хаоса” представляют так-
же фракталы, о которых пойдет речь в п. 6-7 и которые вновь заставят нас
305
обратиться к теме «С Платоном или без?», затронутой в п. 5-15*. Пока же
позволю себе обратить внимание читателя на то, что многие виды роста -
например, морозного узора на стекле, клона клеток многоклеточного заро-
дыша - можно описать как фрактальный рост.
Глава 6. Сообщества, уровни и разнообразие
Рассмотрев эволюцию в ее обычном с древности понимании, т.е. как
появление новых форм организмов, пора рассмотреть ту эволюцию, о кото-
рой обычно говорят только вскользь: с одной стороны, это появление новых
уровней организации в рамках отдельных организмов, с другой - эволю-
ция сообществ организмов. До этого нужно познакомиться с теорией раз-
нообразия, которая должна обозначить те пути, движение по которым (как
в онтогенезе, так и в эволюции) обеспечивается активностью. Затем при-
дется вернуться к эволюционному значению эпигностики (п. 5-15) как спо-
соба реализции онтогенетической активности.
6-1. Эволюция организмов и эволюция природы
Всякий организм входит в несколько сообществ - в свою популяцию, в
свою пищевую цепь (или цепи), вступает в конкурентные, кооперативные
или симбиотические отношения с особями иных видов, паразитирует или
кормит паразитов. Долго считалось, что всё окружение вида можно выра-
зить словом «среда» и описывать эволюцию как приспособление каждого
вида к своей среде. Но теперь почти всем ясно, что это не так - хотя бы по-
тому, что среда сама изменяется гораздо быстрее, чем идет эволюция дан-
ного вида. В самом деле: стоит всего лишь упасть численности вида, слу-
жившего основным источником пищи (а это подчас происходит в считанные
дни), и питавшийся им вид вынужден полностью изменить образ жизни. То
же самое возможно при резкой смене климатических факторов.
Учение о виде, приспосабливающемся к своей среде, на котором зиж-
дятся и дарвинизм, и ламаркизм, и жоффруизм, становится просто неинте-
ресным. Разумнее говорить о коэволюции (совместной эволюции) видов, а
ее требования совсем не те, что «приспособление к среде».
Коэволюция не только не дает виду времени на постепенную выработку
нужных качеств (в силу быстрой смены обстановки), но и не позволяет вы-
явить отдельные эволюирующие объекты. Нужна теория эволюции сооб-
ществ, но до последнего времени ее не было: эволюция экосистем рас-
сматривалась как процесс, состоящий в замене одних видов другими (без-
различно, каким путем появляющимися), а не как цельный процесс измене-
ния экосистемы путем изменения входящих в нее видов.
Из того факта, что вид задаётся своим местом в экосистеме (экологиче-
ской нишей), прямо следует, что разные виды должны эволюировать совсем
по-разному. Старая идея «приспособления к среде» исходила из некоего
306
смутного представления о том, что каждая особь должна стараться избе
жать гибели и оставить как можно больше потомства.
На самом деле мир устроен иначе, и если вид служит основой питания
другим, то особь лучше всего приспособлена к тому, чтобы быть съеденной
Такова половая каста тропических термитов, чья сплошная гибель застави-
ла меня полностью пересмотреть всю концепцию отбора (п. 4-20). Именно
такие виды наиболее стойки к невзгодам, на что впервые обратил внимание
К. Бэр (см. п. 6-2*). Наоборот, млени, убегающие от волков (пример отбора
по Дарвину), вымирают столь же легко, сколь и преследующие их волки.
Дарвин обратил внимание на то, что два самца термитов гнались за од-
ной самкой (п. 3-7*), но упустил куда более важное: победитель и самка бы-
ли тут же съедены, для чего обладали особым приспособлением - их кры-
лья обламываются до спаривания, а их тела сочны и вкусны для огромного
числа видов. При роении размножение грубо принесено в жертву съедобно-
сти, и термиты имеют особое приспособление для размножения без роения,
внутри термитника. Главное же назначение роения состоит в исполнении
термитником его экологической функции - в доставке на поверхность земли
той биомассы, которую термитник добыл в глубине субстрата.
Это - пример приспособительной коэволюции, в котором явно нет
смысла выделять термитов как отдельный приспосабливающийся объект.
Глобальный кризис наглядно показывает нам, что в природе всё со
всем связано. Первая мысль, какая приходит тут в голову - вообразить всю
живую природу как некий огромный организм, приспосабливающийся к са-
мому себе (так же, как, например, всякий человек приспосабливается к сво-
им болезням) и к природе неживой. Такой видел Землю еще 400 лет назад
Иоганн Кеплер, великий астрсном-мистик, затем Гук, а еще 120 лет спустя -
шотландец Джеймс Гёттон, один из основателей нынешней геологии. В
1788 году он писал, что геология - нечто вроде физиологии Земли. С тех
пор появилась новая наука - экология, которую можно назвать физиологией
биосферы, и теперь у Гёттона много последователей. Таким был Соболев
(п. 4-12*), затем - В.Н. Беклемишев, назвавший земной организм геомери-
дой (1928 г.). Вскоре Тейяр писал: «С самого начала клеточная туманность,
несмотря на свое внутреннее множество, необходимо представляла собой
своего рода рассеянный суперорганизм».
В последнее время у Гёттона вновь появились последователи. В част-
ности, в 1996 году в Оксфорде основано Общество геофизиологии (см. да-
лее, п. 7-5). Более всех «геофизиологов» известен английский физик и на-
турфилософ Джеймс Лавлок; для него человечество - лишь один из органов
земного организма (которому он дал имя греческой богини Геи), и что Гея
может, если возникнет опасность для нее самой, себе этот орган ампутиро-
вать, т.е. уничтожить человечество.
Это выглядит забавно, но здравая мысль тут есть: пора перестать гля-
деть на природу как на сцену для разыгрываемого нами спектакля, пора по-
307
пять, что человечество - лишь один из актёров, которому, быть может,
предстоит сойти со сцены задолго до конца спектакля.
Вспомним концепцию Вернадского - в ней эволюция выглядит тоже как
смена актёров (видов) на постоянной сцене (биосфере), по сути не меняю-
щейся в течение миллиардов лет, несмотря на смену актёров. Правда, для
Еювека Вернадский делал исключение - предполагал, что человечество
даст для себя новую земную оболочку - ноосферу (сферу разума, тер-
мин ввел Тейяр в 1927 г.), как бы натянутую на биосферу, чем сделает себя
таким же вечным, как биосфера.
Вернадский, по-моему, ошибался: человечество не создает ноосферу, а
уничтожает биосферу, так что «натягивать» желанную новую оболочку (ноо-
сферу) скоро будет просто не на что. Да и биосфера, как мы увидим в части
3, не раз изменяла и свой размер, и облик. А вокруг себя мы видим ее ката-
строфически быстро меняющейся. По-моему, пора понять, что это тоже -
эволюция, притом более актуальная, чем «происхождение видов».
Если концепцию биосферы можно счесть теорией, то “увы, этого нельзя
сказать о концепции перехода биосферы в ноосферу” (Левит Г.С. Критиче-
ский взгляд на ноосферу В. И. Вернадского // Природа, 2000, № 5, с. 74). По-
пробуем построить более реалистичную схему эволюции живой природы
как целого, не забывая, конечно, особой роли человека в ней.
Первое, что надо заметить, это многоуровневость коэволюции: цельна
не только биосфера, но в некоторой мере и каждая экосистема, и популя-
ция, и каждый организм, и орган, и ткань, и клетка - словом, всё, что актив-
но. Можно ли всё это рассмотреть с единой точки зрения? Увы, пока что это
ни у кого еще не получилось. Начнем с упорядочения точек зрения.
6-2. Тенденции в эволюции крупных таксонов
Ламарк, будучи увлечен эволюцией высших животных, счел основным
фактором, движущим их эволюцию, сознательную волю, а о низших живот-
ных, как и о растениях, заметил лишь, что причиною их развития являются
прямые влияния внешней среды. Можно сказать, что высшим животным он
приписал собственную активность, тогда как источник активности низших
поместил вне их, в среду их обитания. Но вставал вопрос, Ламарком не за-
тронутый: если этой активности достаточно для эволюции, то зачем высшим
организмам иметь еще 4т6-то? Затем, можем мы сказать теперь, чтобы идти
обоим путем эволюции.
Зато Ламарк отметил очень важное обстоятельство: процесс эволюции
основан на двух различных тенденциях - к развитию (усложнению, прогрес-
СУ - называйте как хотите) и к приспособлению. Вообще, тенденцией мы
будем называть всякую нечеткую (т.е нарушаемую на отдельных примерах)
закономерность, ясно видную лишь на массовом материале.
Так, монета проявляет тенденцию падать гербом примерно в половине
бросаний; и тот факт, что к северу лиственный лес сменяется хвойным, и
308
утверждение «украинцы хорошо поют» тоже указывают на тенденции, и ес_
ли для дарвинистов приспособление к среде обитания - непреложное ос-
новное свойство всякой эволюции организмов, то для ламаркистов приспо-
собление - лишь одна из эволюционных тенденций, маскирующая главную
фому активности - тенденцию к прогрессу.
Кто-то сказал: «Эволюция подобна фонтану - движение струи вверх
направляется внутренним импульсом, а падение воды вниз - приспособле-
нием к обстоятельствам». Этой позиции придерживался и Бергсон (п. 5-2)
он же был первым, обратившим внимание на противоположные тенденции
царств. В 1907 году он отмечал:
«нет ни одного ясно выраженного признака, который отличал бы растение от
животного», и заключал: группу лучше всего определять «не тем, что она об-
ладает известными признаками, но тенденцией к усилению этих признаков»
[Бергсон, с. 127].
Поясню: для животных наиболее характерна подвижность, но есть и
неподвижные животные - например, кораллы; для растений наиболее ха-
рактерен фотосинтез, но есть растения-паразиты без оного; самое харак-
терное для царства грибов - разложение теп отмерших растений и живот-
ных, однако есть небольшая группа хищных грибов, питающихся живыми
почвенными червями. Такие нетипичные группы относят к их царству по со-
вокупности сходств: грибы имеют “ткань” в форме мицелия; растение- пара-
зит заразиха не имеет фотосинтеза, но имеет цветок. О систематике царств
мы будем говорить в главе 10.
Есть тенденции царств, характеризующие развитие особи. Например:
“По-ввдимому, общим в животном мире правилом является первичность дли-
тельного метаморфоза и его постепенная утрата в ходе эволюции” (Тихомирова
А.Л. Перестройка онтогенеза как механизм эволюции насекомых. М., 1991, с. 98).
А вот тенденция личинок подтипа позвоночных:
“Переход от примитивного развития бесчерепных с ресничными свободно-
плавающими личинками типа диплеврулы к появлению личинок, все более
сходных со взрослыми формами, затем к утрате личиночных стадий как тако-
вых и, наконец, к живорождению” (там же, с. 100).
У позвоночных видна также тенденция к замене жаберного и кожного
дыхания на легочное и к заселению суши (террипетность по Бронну); зако-
номерность очевидна, но не всеобща - некоторые четвероногие вернулись
в воду. Можно добавить, что для царства животных характерна тенденция к
раздельнополости (чем выше организованы животные, тем меньше среди
них доля гермафродитных видов), а в царстве растений - наоборот.
Есть и тенденции, общие в различных царствах. «Существует пред-
ставление, что разные царства эвкариот имели разных предков» (Дымсиич
Г.М. И Природа, 2002, № 6, с. 60), однако все эвкариотные клетки устроены в
принципе одинаково. Механизмы клеточного деления (митоз, мейоз) почти
одинаковы у высших животных и высших растений (общая тенденция). На-
309
оборот, у одноклеточных и у грибов разнообразие митозов и мейозов очень
велико (противоположная тенденция).
Сходства, независимо приобретенные в ходе эволюции, именуются
конвергенциями. Сходство митозов растений и животных - типичная кон-
вергенция. Дарвинизм трактует конвергенцию как приспособление к сход-
ным условиям, ничем это не подтверждая. В отношении митозов это осо-
бенно странно, поскольку сходство наблюдается решительно во всех сре-
дах и при всех типах строения, тогда как огромное разнообразие митозов
мы видим в одной-единственной среде и для одного уровня организации - у
планктона (свободное плавание одноклеточных у поверхности моря). По-
моему, тут надо говорить о тенденциях митоза как таковых.
Дарвинизм формально не признает тенденций: в его логической основе
лежит стремление организмов к избыточному размножению, не имеющее
исключений, и вытекающая из него борьба за жизнь, а перечисленные выше
тенденции просто не рассматриваются. Фактически же дарвинизм, как и все
учения, основан на признании тенденций. Так, существование феномена
рудиментов, т.е. органов, потерявших функцию и сохраняющихся вопреки
их бесполезности (а иногда и вредности), превращает феномен отбора из
всеобщего закона в тенденцию - даже если допустить, что он действует.
Далее слово “тенденция” мы будем в основном прилагать к тенденции,
текущей во времени (это тенденция-процесс). Нарастание или уменьше-
ние какой-то черты организма в ходе эволюции наводит на мысль о наличии
механизма, но поиск его всегда затруднен именно наличием исключений.
Причина того, что закономерности обычно наблюдаются в форме тенден-
ций, достаточно очевидна: каждый объект и каждый процесс подвергается
многим одновременным воздействиям, которые не связаны друг с другом,
те. случайны в отношении друг друга. Здесь понятие тенденции призвано
выявить существенную часть процесса.
Многие обращали внимание на то, что объяснить тенденцию как цепь
гриспособлений не удается: некоторые ее этапы обычно бесполезны или
Даже вредны. Сам Дарвин приводил как пример эволюцию цветка, заходя-
щую много дальше, чем требует приспособление к опылению. Тут он фак-
тически вел речь именно о тенденциях-процессах.
Позже было найдено много примеров, когда тенденция прямо ведет к
гибели. Таков ирландский олень (пп. 3-9, 4-10) - последний этап тенденции
семейства оленей к росту рогов. Гигантизм здесь затронул один орган, и
Этот пример аллометрического (непропорционального) роста частей орга-
низма довольно обычен в качестве примера явной неадаптивности.
Так, в небольшом триасовом (200 млн лет назад) море южной Европы
*Ил удивительный морской ящер Tanystropheus, «один из самых причудли-
ЕЫх среди архозавров (а может быть, и вообще среди пресмыкающихся) ...
второго называют жирафошеим ящером» (Бюргин Т. и др. И ВМН, 1989, №
8)- Имея туловище и лапы вполне наземного облика, он жил в воде - об
310
этом судят по тому, что на суше он не смог бы даже ползать: из общей длц_
ны тела (4,5 м) на туловище пришлось всего 18%, остальное составили
хвост и, главное, шея. Она была почти втрое длиннее туловища и включала
всего 9 позвонков (рис. см.: ВМН, 1989, № 8, с. 45-46, а также на обложке
ЧЭ). Столь гигантские позвонки не давали ящеру возможности поднять го-
лову, хоть та и была крохотной. Удивительно, что танистрофеусы были мно-
гочисленны и водились долго. Придумывать этому облику пользу швейцар-
ские палеонтологи, к их чести, не хотят:
“Трудно допустить, что непомерно огромная шея давала Tanystropheus какое
бы то ни было преимущество. Напротив, по нашему мнению, эти животные
просуществовали миллионы лет не благодаря, а вопреки своим длинным ше-
ям” (там же, с. 46).
Как видим, тезис Бэтсона (п. 4-10*) повторен дословно. Его повторяли
многие - все, кто был способен критически видеть свой материал. Но хватит
сто лет повторять тезис, пора его осмыслить.
Налицо одна из последних стадий тенденции наземных ящеров к удли-
нению тела. Напомню: в п. 4-10* шла речь о длинных шеях зверей и малом
числе позвонков в них. Ту же тенденцию видим у наземных ящеров: даже
гигантский ящер диплодок (юрский период) с длиннейшей шеей имел всего
12 шейных позвонков. Теперь мы видим случай, когда жестокая тенденция
буквально загнала ящера в воду. Его потомкам предстоял выбор - либо
приспособиться к тенденции, развив подходящие частные свойства, либо
противопоставить ей другую тенденцию, либо вымереть.
Среди морских ящеров мезозоя мы видим широкий спектр реализаций
таких возможностей [Кэрролл, т. 2]; он наводит на мысль, что пагубная тен-
денция к удлинению тела без роста числа позвонков переставала действо-
вать лишь с утратой наземного облика, т.е. в ходе реализации новой тен-
денции. Так, морской ящер эласмозавр (меловой период) имел 76 шейных
позвонков.
6-2* Экологическая прочность
Осмысление тезиса Бэтсона видится мне в следующем: эволюционные
тенденции действуют сами по себе, многие из них надолго снижают жизне-
способность организмов, но не могут истребить их, пока у тех не исчерпан
запас их экологической прочности. Этим термином будем обозначать
способность индивида успешно переносить ухудшение жизни (условий как
внешних, так и внутренних). Понятие экологической прочности первона-
чально понадобилось мне при анализе экологической концепции Бэра:
«Характерен его (Бэра - Ю.Ч.) шутливый афоризм: на вопрос дамы, зачем
нужны эти несносные комары, он ответил: “чтобы нам побольше иметь рыбы в
наших пресных водоемах”. Он отмечал огромный запас прочности экосистем-
Поэтому в огромной плодовитости рыб он не увидел эволюционного фактора-
гибнут не худшие генотипы, а неуспевшие вырасти экземпляры, причем высо-
311
кая поедаемость вида не является мерой его неуспеха, а наоборот, является ме-
рой его экологической прочности и перспективности (такой вид быстро разрас-
тается при снятии трофического давления); наоборот, вид, не служащий пищей,
легко вытесняется из экосистемы - например, все крупные хищники» {Чайков-
ский Ю.В. Экологическая основа эволюционизма у Бэра и Дарвина // Доклады
МОИП, Общая биология. I полугодие 1978 г. М., МГУ, 1980, с. 87).
Экологическая прочность позволяет организмам на вредное воздейст-
вие (среды или собственной изменчивости) отвечать сперва сменой пове-
дения, затем ухудшением состояния (исхуданием, подсыханием и т.п.), за-
тем снижением и даже временным прекращением размножения (но не утра-
той размножаемости). Лишь за пределами экологической прочности особи
еид отвечает на вредное воздействие заметным ростом смертности.
Однако и рост смертности еще не исчерпывает экологической прочно-
сти вида: многочисленный вид может во много раз уменьшить свою числен-
ность без риска вымереть. Риск вымереть появляется только тогда, когда
размножаемость не восстанавливается в благоприятных условиях (как
t/ы это видим ныне у зубра, панды, стерха и других подобных видов).
Если у вида высока экологическая прочность, он может нести, без рис-
ка вымереть, вредные свойства. А сколь велика экологическая прочность
многих животных, видно из способности инвалидов к нормальной жизни:
У крабов «сильно развита способность в самопроизвольному отбрасыванию
конечностей... Иногда краб теряет таким образом до шести конечностей. Как
ему удается выжить в таком случае, совершенно непонятно, особенно, когда
утрачены обе клешни» {Павлов В.Я. Жизнеописание краба камчатского. М.,
ВНИРО, 2003, с. 54). Или: «Судя по всему, не бедствуют в море и рыбы с раз-
нообразными врожденными уродствами» {Торканов А.М., Орлов А.М. Укло-
нившиеся от рыбьих стандартов // Природа, 2005, № 8, с. 22).
Примерно то же видим у ящериц. У них
“может быть утрачено до 6 пальцев... такой уровень травматизма едва ли может
быть фактором, снижающим жизнеспособность... Агамы с травмированными
конечностями не уступают полноценным особям в скорости и легкости пере-
движения” {Панов Е.Н., Зыкова Л.Ю. Горные агамы Евразии. М., 2003, с. 224).
Как и у кистепёрых (рис. 20), тут сложность конечностей превосходит
Рабочую потребность, и говорить об их приспособленности вообще (и пу-
тем отбора особенно) несерьезно. Их явно формирует нечто иное.
А вот совсем поразительный пример: у пятнистой гиены
“оба пола внешне вообще неотличимы... Копуляция происходит через... канал в
клиторе и поэтому вообще чрезвычайно затруднена. Через него же (т.е. через
клитор) происходит и деторождение. Все это устройство поражает своей неце-
лесообразностью” {Шмальгаузен И. И. Факторы эволюции. М., 1968, с. 136).
Хотя тут и можно произносить разные слова об особой форме отбора
(что Шмальгаузен и сделал), но очевидно, что обычный отбор должен был
Отсечь эту нелепость - если бы на самом деле он существовал.
312
С позиции отбора факт резкого ухудшения функции немыслим, но с по-
зиции экологической прочности он понятен: поскольку пятнистая гиена на-
много превосходит другие три вида семейства гиеновых функционально
(она крупнее, сильнее и активнее их, опасна даже для старых львов), то она
смогла выжить со своими изъянами (и то - в очень узком субтропическом
ареале), выжить вопреки дефекту строения, по Бэтсону.
В реальной жизни все виды имеют ненулевую (а большинство - весьма
заметную) экологическую прочность, и потому в них могут протекать, без
риска вымирания, процессы, снижающие жизнеспособность. Такой вид и
реализует свою эволюционную тенденцию. Нашу главную задачу можно те-
перь выразить так. нужно выявлять тенденции и искать механизмы
их осуществления, не ища для каждой из них адаптивного смысла. Хотя
у многих этот смысл налицо, но навязывать его всем неосмотрительно.
Многие группы организмов демонстрируют приспособление к своим
вредным тенденциям - как поведенческое (уход в щадящую зону обитания,
переход к неудобному, но приемлемому образу жизни), так и морфо-
физиологическое (изменение или утрата органов, усвоение новой пищи и
т.п.). Намек на подобное понимание тенденции есть у Вентребера:
“Полные надежды уроды Гольдшмидта, несчастным образом произведенные
природой, полностью реализовали возникшие позже приспособления ... Безногие
позвоночные имели время реадаптироваться. Они заменили, к примеру, четверо-
ногое передвижение на извивание или ползание” (Wintrebert Р. Le vivant..., р. 90).
Идея экологической прочности в чем-то противоположна «принципу
адаптивной поисковой оптимизации» [Гринченко]. Его приверженцы созна-
тельно пытаются преодолеть неуспех «теории слепого поиска» (т.е. дарви-
низма) введением такого постулата: всякая живая система стремится мак-
симизировать какую-то энергетическую функцию.
Сам факт попытки строго описать активность радует, но, по-моему,
постулат о существовании оптимизации противоречит фактам: во-первых,
примеры оптимальности организмов редки на фоне примеров неоптималь-
ности; во-вторых, даже если допустить, что организмы оптимальны по како-
му-то ненаблюдаемому параметру, их оптимальности противоречит фено-
мен разнообразия - одна и та же функция (например, полёт) выполняется
разными организмами с очень различной эффективностью. А в-третьих, оп-
тимальности противоречит экологическая прочность. Поэтому данной идеей
Гринченко мы далее пользоваться не будем.
Экологическую прочность нельзя выразить уравнениями, но можно -
неравенствами, ограничивающими область допустимого разнообразия.
6-3. Мейен и наука о разнообразии - диатропика /
Теперь на время покинем проблематику активности, чтобы борисовать
то пространство, в котором активность реализуется, ту упорядоченность, ко-
торая была уже не раз упомянута, в том числе в п. 5-2**.
313
Воробей сер - это маскирующая окраска, снегирь ярок - он привлекает
самку, синицы (тут ярки оба пола) опознают по расцветке свой вид. Всё это
ежедневно рассказывается в школах и вузах, и ученики слушают доверчиво,
хотя на деле тут сплошные вопросы: почему снегирю не нужно маскиро-
ваться, почему синица не требует особой расцветки самца, откуда мы узна-
ли, что причина расцветок связана именно с половым поведением? И во-
обще, стоят ли за этими словами опыты и размышления?
Ведь хорошо известно, что многие птицы (например, пингвины) и мно-
гие головоногие моллюски вообще не отличают самца от самки до самого
начала совокупления и тем не менее прекрасно размножаются.
Если такой «объяснительный» ряд, какой приведен выше, продолжать
излагать достаточно долго, ученики начинают смеяться. Даже попадись они
очень доверчивые и серьезные, смех непременно вызывает пример с ту-
рухтанами. У этих куликов голова каждого самца раскрашена на свой лад,
так что в определителе птиц указывается не сама расцветка головы турух-
тана, а факт ее разнообразия. И вот один западный орнитолог догадался:
самцы, оказывается, преодолевают таким путем безразличие самок, кото-
рые у этого вида плохо возбудимы - только присутствие сразу многих раз-
ных кавалеров заставляет самку уступить ухаживаниям одного из них. Под-
робнее см. [Любищев, 1982, с. 157-159]. Это и вызывает смех.
Объяснение поражает произвольностью, но чем оно хуже предыдущих,
взятых из респектабельных учебников? Ни одно из них не проверено опы-
том, и даже не поставлен вопрос, можно ли данные утверждения как-то про-
верить. Это даже не презумпции, это метод софистов (п. 1-6), благополучно
переживший тысячелетия. Пусть для воцарения дарвинизма была в XIX ве-
ке социально-психологическая причина (становление статистической ПМ),
но нельзя же следовать ей вечно.
Если мы позволим себе перебирать любые доводы, пока не найдем
подходящий к данному примеру (это - диалектика по Платону, усвоенная
марксизмом), то лишимся всякого критерия истинности и можем доказать
всё что угодно. Над такими приемами критики смеялись всегда (п. 3-7). Но
критиковать легко, а что предложить взамен?
Сергей Викторович Мейен (1935-1987), палеоботаник, эволюционист и
философ науки, в 1970-х годах сдвинул вековую дискуссию с мертвой точки
- вместо вопроса «зачем?» решил начать с вопроса «у кого?», т.е. с сопос-
тавления разнообразия свойств и разнообразия их обладателей.
Спенсер считал разнообразие причиной устойчивости сообществ; Бэт-
сон отрицал полезность отдельных свойств; Коп и Вавилов выстраивали
сходства в ряды, а Кренке даже пробовал ряды упорядочивать; Любищев
указывал, что разнообразие свойств плохо согласуется с идеей полезности
всякого свойства и с идеей эволюции.
На этой основе Мейен стал строить теорию разнообразия. Уже после
его безвременной кончины эта теория стала основной частью диатропи-
314
ки44 Диатропика является, на мой взгляд, не столько отдельной теорией
сколько аспектом всякой теории, имеющей дело с разнородными объекта-
ми. Для сравнения: во многих науках есть морфология (учение о строении
объекта из частей) - и в геологии, и в биологии, и в филологии; она являет-
ся аспектом данных наук и, как увидим, входит в состав диатропики.
Разнообразие множества - это совокупность отношений сходства и
различия между элементами данного множества [Чайковский, 1990, с. 6].
Так, разнообразием гена «цвет глаз человека» является перечень всех
известных расцветок радужки (задаваемый либо словесным описанием, ли-
бо физическими характеристиками, либо картинками) с указанием аллеля
(аллелей), приводящего к каждой данной расцветке. Разнообразие конечно-
стей позвоночных - это перечень возможных качественно различных конеч-
ностей (плавник, ласт, лапа для ползания, лапа для хватания, нога, крыло,
рука и т.д.) с указанием положения (непарная, передняя и т.д.) и степени
развития (зачаток, пятипалая, однокопытная и т.д.). Разнообразие жизнен-
ных форм растений - это их перечень (водоросль, трава, полукустарник,
кустарник, лиана, дерево и т.д.) плюс перечень степени развития каждой
формы. Он может быть задан через отношение сходства с иными группами
(например, для таллома водоросли: одноклеточный, ценоцитный, подобный
траве, подобный кусту и т.д.).
Даже сам факт осознания феномена разнообразия бывает очень поле-
зен. Так, есть много примеров и яркой окраски самцов, и их драк за самку, и
выбора самками самцов, но нет смысла строить на них “теорию полового
отбора”, поскольку есть много (гораздо больше) противоположных приме-
ров, когда ничего этого нет, а эволюция идет точно так же. Теория должна
описывать оба типа примеров, иначе она представит интерес только для
своих приверженцев.
Есть науки, метод которых прямо построен на отрицании разнообразия
(в силу их приверженности “бритве Оккама”), например физика: она ищет
общие законы поведения своих объектов, а их разнообразие выносит за
рамки исследования как нечто случайное (подробнее см. [Чайковский, 1990,
с. 7; он же, 2004, с. 122-123]). Но и она в последнее время понемногу скло-
няется к признанию феномена разнообразия. А в химии, биологии и психо-
логии разнообразие является первым, бьющим в глаза феноменом.
44 Диатропика (от греч. диатропос - разнообразный), наука о законах организации раз-
нообразия. Рассмотрена в книге [Чайковский, 1990]. Взгляд на мир, в основе которого
лежит идея разнообразия, назван в п. 5-2** диатропической ПМ. Сам Мейен пользо-
вался термином ‘'типология” (что неудачно, так как порождало недоразумения). Этот
термин был первоначально использован и мною [Чайковский, 1983]. Эта повесть может
служить популярным введением в тему разнообразия. См. также подборку: Из научно-
го наследия С.В. Мейена: к истории создания основ эволюционной диатропики И [Эво-
люция флор..., с. 106-135] и мою статью в ВФ [Чайковский, 2006]. /
315
Многие социальные проблемы порождены тем, что общество требует
сто всех своих членов единого поведения, хотя очень люди различны.
Главный вид наказания - лишение свободы - калечит одних и вообще не слу-
жит наказанием для других (многие преступники пытаются попасть в колонию
снова); призыва в армию одни боятся, как казни, а другие желают, как приклю-
чения; одни не могут без чтения, другие уверены, что всякий читающий где-то
учится; одни ищут развлечений (вина, наркотиков, зрелищ, игр и т.д.), другие
любят только делать дело; одни стремятся к выгоде, другие к власти, третьи к
покою; одним важно общее благо, другим личное, третьи любят дискомфорт.
Список можно продолжать бесконечно, и это всем известно, однако почти не
принимается во внимание. Особенно страдают непониманием разнообразия все-
возможные утопии, но элемент утопизма есть, по-моему, в любой социальной
программе, которую устроители творят на свой вкус. Так нам был навязан со-
циализм, а теперь - “рыночные реформы”. Разумный выход видится мне в том,
чтобы не искать единого для всех правила, обязательного априори.
Но мало указать на разнообразие, надо увидеть в нем закономерности,
структуру. Первой частью диатропики стал мероно-таксономический ана-
лиз Мейена, которым мы сейчас займемся. Второй частью - анализ законо-
мерности и случайности разнообразия [Чайковский, 1983; 2004], о чем шла
речь в п. 5-18 и еще пойдет не раз, а третьей - учение о ядре и периферии,
восходящее к Бэру (о чем пойдет речь в гл. 10).
Сразу предупрежу - тот, кто предпочитает мыслить посредством “брит-
вы Оккама”, может дальше не читать, поскольку диатропика начинает с от-
каза от презумпции простых решений.
6-3* Блочность строения и множественный параллелизм
Давно замечено, что биоразнообразие составлено из ограниченного
числа компонент. Тот факт, что организмы как бы собраны из таких компо-
нент, описан как блочность строения животных (Мужчинкин В.Ф. // ЖСБ,
1978, № 5), «мозаичность структурных трансформаций» (Воробьева Э.И. //
УСБ, 1994, № 6), «модульность» строения растений (Корона В.В. И ЖСБ,
2002, № 3), «приобретение и потеря модулей» как путь эволюции мобиль-
ных элементов генома (Капи П., Мэзонот К. // ЭГ, 719), «перетасовка гене-
тических модулей» (Тиходеев О.Н. //ЖСБ, 2005, № 1) и т.д.
Простые примеры блоков: гены, клетки, ткани, позвонки, листья, гриб и
водоросль в лишайнике, правая и левая руки. Чуть посложнее: клёнолист-
ный платан и платанолистный клён, равно как ясень и ясенелистный клён,
имеют в каком-то смысле общие блоки, строящие сходный лист. Точнее см.
в части 3, здесь же замечу, что блочность свидетельствует о номогенезе, а
точнее, реализует тезис Соболева (п. 4-12*).
Легче всего видны блоки, из которых составлены белки. На рис. 34 при-
ведены два основных блока, из которых состоят ферменты.
316
Рис. 34. А - два основных бел-
ковых блока: альфа-спираль и
бэта-лист; Б - участие их в
формировании белка (фермент
лизоцим).
Высказана также гипотеза
модульной организации сред-
ней части эмбриогенеза позво-
ночных (Bininda-Emonds O.R.P.
е. а. И Proc. Roy. Soc. Lond. Ser.
В, 2003, vol. 270, p. 341). Если
так, то из модулей могут быть
составлены не
только формы, но и процессы.
Признав блочность, надо
искать законы ее упорядоче-
ния, и первое, что видно - на-
личие одних и тех же блоков
позволяет строить ряды.
Исходным для диатропики является понятие «ряд», она оперирует им так
же, как другие науки - понятием «факт».
Ряд - та простейшая закономерность, какую удается выявить в иссле-
дуемом разнообразии. Если ставить целью поиск закономерностей, то луч-
ше всего начинать анализ именно с рядов, а не с единичных фактов. Не-
сколько утрируя, Мейен сказал: «единично взятый факт может служить под-
тверждением любой теории» (МС, 294). Чаще всего разнообразие предста-
ет как множество пересекающихся рядов.
Окраска турухтана изменчива? Это не объект интереса диатропики. А
вот то, что каждый индивид окрашен по-своему у турухтанов, у ядовитых ля-
гушек Центральной Америки (ВМН, 1983, № 4, с. 78), у жирафов, у подвя-
зочных змей, - это ряд, объект диатропики. Пусть у нас нет таким фактам
объяснения - не беда. Каждая наука начинается с группировки фактов, с
составления рядов, а диатропика должна показать, как с рядами работать.
В этом смысле роль диатропики похожа на роль статистики, выражае-
мую ходячим афоризмом: «Статистике часто принадлежит первое слово, но
последнее - никогда» (т.е. статистика указывает на закономерности, но не
статистике надлежит раскрывать их механизмы). Различие же в том, что
главный метод статистики - усреднение, а диатропики - обобщение.
Ряд может быть задан разными способами. Первый, самый простой:
общим свойством - например, ряд зеленых стульев: из множества ступь-
317
ев мысленно извлечены зеленые. Второй: способом построения - ряд
гростых чисел, алфавитный порядок слов, строй солдат по росту, ряд ще-
лочных металлов, лестница существ Аристотеля. Третий: путем сопостав-
ления с другим рядом - например, алфавитному ряду английских слов
англо-русский словарь сопоставляет по смыслу ряд русских слов; ряду ста-
дий зародыша цыпленка можно сопоставить ряд стадий зародыша щенка
1'ли малька рыбы. Третий способ наиболее важен нам, поскольку здесь
фиксируется феномен параллелизма, о котором мы много говорили в главе
2' упорядочивая объекты одним способом, мы то и дело обнаруживаем упо-
рядоченность по какому-то другому способу. Таковы в англо-русском слова-
ре латинизмы (основная причина сходств английского и русского слов); или:
упорядочивая зародыши различных видов животных по номерам дня фик-
сации, получаем ряды, между которыми видно сходство, тем более полное,
чем меньше номер дня фиксации (закон Бэра).
Основной прием диатропики - сопоставление рядов. Самым четким (но
редким) видом параллелизма является периодичность. Например, в пе-
риодической системе химических элементов сами элементы упорядочены в
строки по заряду ядра, а сходства реализуются в форме столбцов (таковы,
например, щелочные металлы). Известны периодические системы и в био-
логии, из которых наиболее, на мой взгляд, удачна система метаболических
путей (см. карту с брошюрой: Малыгин А.Г Карта метаболических путей
(периодическая). М., Наука, 1976; позже улучшена тем же автором). О пе-
риодических системах организмов см. главу 10.
Химическая таблица плоская, а почти все таблицы живого многомерны.
Это видно и на клёнах: названия многих из них указывают на сходство с де-
ревьями иных семейств - платановидный, граболистный, ясенелистный,
лавровый. Плоская “кленовая” таблица (п. 4-9) как бы пересекает стоящие
поперек нее плоские таблицы прочих семейств.
Если Агассиц обнаружил множественность параллелизма, если Коп и
Вавилов составляли детальные ряды и параллели, Филипченко отделил ге-
нетические ряды от прочих, а Кренке и Котс добавили закономерности внут-
ри каждого ряда, то Мейен занялся объединениями направленных рядов и
таблицами, при этом получающимися.
Ссылки можно найти в работах [Чайковский, 1990; 2004, гл. 8; 2006].
6-3** Таксоны, мероны и рефрены. Диасеть
Исходной при построении диатропики Мейену послужила та давно за-
бытая идея Страхова (о нем см. гл. 3), что сравнительная анатомия класси-
фицирует части тела так же, как систематика - целые организмы. Для сис-
тематики исходным является понятие таксона (вид, род и т.д.), основной
процедурой - выявление таксонов.
По аналогии с понятием таксона Мейен ввел понятие «мерой», т.е.
«класс частей». Организм имеет органы, свойства, признаки, а таксон имеет
318
мероны. Например, таксон «позвоночные» имеет мерой «парные конечно-
сти», этот мерой принимает у разных позвононых различные значения:
плавники, ласты, лапы, крылья, ноги, руки. В таксоне «рыбы» мерой «пар-
ные конечности» принимает только значения «парные плавники», а также (у
кистепёрых рыб) - «ласты» (ласт отличается от плавника наличием сложно-
го скелета и мускулатуры). Можно сказать и иначе: меньший таксон имеет
меньший мерой: таксон «рыбы» имеет мерой «парные конечности» только в
форме «парные плавники и ласты».
Для склонных к математике приведу другое определение: «Мероны, по
Мейену, - это классы эквивалентности, порождаемые отношением гомоло-
гии» (МС, 73). Поскольку гомология органа задается положением его в архе-
типе (целостностью организма), то здесь тем самым подчеркнуто, что мерой
не имеет смысла вне архетипа. По Мейену,
“гомологизированные, т.е. расклассифицированные и ставшие меронами
части организмов данного таксона в сумме составляют архетип таксона”
[Мейен, 1978, с. 497].
Определение мерона и архетипа друг через друга означает, что архе-
тип - понятие интуитивное (ср. п. 2-7*)- Слова «в сумме» означают, что
понятие принадлежит нечётной (пятой) ПМ. Наоборот, архетип по Оуэну
(см. ниже) характеризует целостность и пригоден для чётных ПМ.
Мейен полагал, что меронами могут быть как органы (формы) любых
организмов, так и их свойства (функции). В виде примера напомню соци-
альную организацию термитов и голых землекопов (см. далее, п. 6-5).
По Мейену, всякая новая система организмов исходит из какой-то
прежней, хотя исследователь часто думает, что строит ее самостоятельно.
А в самом начале была одна интуиция - она позволяла различать животных
и растения, зверей и насекомых, а заодно и многое другое, например - лапы
и крылья. Всё это Мейен шутливо называл «кухонной морфологией». Кста-
ти, еще в пору мифов было понято, что руке каким-то образом соответству-
ет не только крыло птицы, но и лапка насекомого, и даже ветвь дерева. В
наш век гомеобоксов (п. 5-15) эти параллели стали выглядеть вполне серь-
езно, но как бы всё это подчинить общим правилам или хотя бы тенденци-
ям? Тут Мейен высказал три простых важных тезиса.
Первый тезис: классификация любых объектов состоит из двух проце-
дур - выявления таксонов, т.е. групп объектов (акты выявления и описания
таксонов вместе образуют таксономию, ее еще называют систематикой)
и выявления меронов (акты выявления и описания меронов вместе образу-
ют мерономию - это основа морфологии). Существенно, что построение
классификации всегда состоит в попеременном обращении, то к таксоно-
мии, то к мерономии: первоначальная «кухонная морфология» позволила
Аристотелю построить первую систематику животных, на основе которой
была (уже в годы Возрождения) построена первая научная морфология (ар-
хетипы и планы строения, о которых шла речь в гл. 1), позволившая в XVIJJ
319
веке построить научную систематику животных и так далее45.
Второй тезис: у разных таксонов общие мероны (при переходе от одно-
го таксона к другому всегда видно совпадение некоторых меронов). Так, пе-
реходя от птиц к млекопитающим, видим соответствие большинства костей.
Близкие классы различаются в основном значением меронов, а не переч-
нем меронов. Самое замечательное, что от таксона к таксону повторяются
не только сами мероны, но и правила их преобразования. Мейен как бота-
ник любил примеры с рядами расчленения листьев, где преобразованием
является усложнение края листовой пластинки. Один пример мы рассмат-
ривали в п. 4-9: в разных семействах цветковых наблюдается весь ряд
форм листа, от сложной остролистной до простой яйцевидной.
В приведенном примере последовательность наблюдается от таксона к
таксону, но ее же можно видеть в рамках одной особи: на одном дереве
обыкновенного клёна или обыкновенного платана можно обнаружить одно и
то же разнообразие форм листьев - от совсем простого до совсем кленово-
го (см. рисунок: Маслова Н.П. и др. И ПЖ, 2005, № 4). «Эту повторяющуюся,
подчиненную одному правилу преобразования последовательность состоя-
ний мерона назовем рефреном» [Мейен, 1978, с. 501].
Третий тезис: рефрены лежат в основе структуры любого разнообра-
зия, в том числе биологического. Столь парадоксальному утверждению
дал иллюстрацию еще Агассиц, заявивший, что расстояния между орбитами
планет и листьями на стебле подчиняются общему правилу (Agassiz L., с.
130 - см. п. 2-10*). Именно в этой универсальности понятия рефрена Мейен
видел его пользу. Благодаря ей можно говорить, что наблюдаемое разнооб-
разие не хаотично, а образует единую диатропическую сеть (диасеть); что
эволюцию можно рассматривать как достаточно закономерное преобразо-
вание диасети. Этим как бы завершена идея Гердера (п. 2-2), а диасеть вы-
ступает тем пространством жизни, о котором мы говорили в п. 5-16.
Сам Мейен не дал ни рисунков, ни таблиц рефренов, ограничился примерами
из тонкой морфологии голосеменных растений, и не указал даже, что имеет в ви-
ду под «общим правилом». Это было одной из причин неуспеха его идеи. Пер-
вый сколько-то наглядный мой пример (конечности позвоночных, см. далее)
Мейен видел и ничего не возразил, поэтому далее я буду им пользоваться.
Мы будем понимать общее правило образования рефрена широко: как
всякую тенденцию (п. 6-2) изменения мерона в рамках множественного па-
раллелизма. То есть сравнивать можно любые ряды сходств - животных и
Растений, нынешних и вымерших, диких и домашних, взрослых и зароды-
45 с _ _ „ ,
Ьсть два способа определять мероны без попеременной (итеративной) процедуры.
Первый предложил еще Гёте: выявлять естественные части тела, глядя на его онтогенез
(см. ниже, п. 11), а второй - ученик Мейена А.В. Гоманьков: мерой является геометри-
чески заметным функциональным элементом (МС, 74). Оба способа удобны, но неуни-
версальны (например, не выявят такой мерой, как система ТВ-иммунитета).
320
шей, особей и их частей. Лишь бы сравнению подвергалось не менее трех
объектов и наблюдалось не менее двух параллелизмов (рефрен выражался
бы таблицей размером не меньше 3#3), имеющих направление (на рис. 35Д
это - усложнение листа слева направо и сверху вниз).
Рис. 35. Простейшие примеры рефренов у растений
А - рефрен мерона «расчленение листовой пластинки»
Можно отнести к реф-
ренам и одиночный па-
раллелизм, если налицо
два чётких ряда с ростом
общего параметра. Таков
параллелизм щелочных и
щелочноземельных
металлов (таблица раз-
мером 6#2), где наблю-
дается рост активности с
ростом атомного веса в
таблице размером 4#2.
Похож параллелизм
плавников у кистепёрых и
двоякодышащих рыб. Но
главный интерес являют
большие таблицы с
малым числом пустых
клеток (таковы рассмат-
риваемые ниже рефрены, изображенные на рис. 35Б и 36).
Откуда в природе берутся сходства, образующие рефрены? Дарвинизм
списывает их либо на унаследование от общего предка (так, все виды рода
«дрозофила» считаются при такой трактовке происшедшими от единого
предкового вида мух), либо на независимое приспособление (так, считается,
что весь инфракласс плацентарных, повторивший строением соответст-
вующие формы инфракласса сумчатых, приспособился к сходным условиям
существования). Доказательств не приводится, и предлагается принять эти
утверждения на веру.
Допустим, но чем объяснить сходство никак не используемых органов у
таксономически далеких видов? Например, амфибия сирен и рептилия хи-
рот обе обладают совершенно бесполезными крохотными «ручонками» и
притом только передними. Кстати, их бесполезность видна тоже из сопос-
тавления рядов - другие животные в тех же условиях просто избавлены от
конечностей (червяги, змеи, безногие ящерицы).
321
Б - фрагмент рефрена мерена «таллом водоросли» (по: Дьяков, 2000). Че-
тыре типа окраски маркируют четыре (из существующих десяти) типов во-
дорослей - хлорофиты, ксантофиты, хризофиты и родофиты (багрянки).
Усложнение наблюдается от коккоидной формы вверх и вниз. Жгутиковых
багрянок не существует
Наконец, если во всем искать пользу, то чем объяснить сходство не толь-
ко самих свойств, но и их преобразований? Например, может быть и можно
искать пользу от каждого типа расположения листьев на стебле (пока что ее
не нашли), но заведомо никакому виду нет пользы от того, что типы листо-
расположения разных видов растений укладываются как целое в ряд Фибо-
наччи (Мейен С.В. Нетривиальная биология //ЖОБ, 1990, №1, с. 6). Этот
пример мы поставим далее в ряд с другими примерами целостности, каж-
дый из которых сам по себе непонятен (п. 6-13**).
Откуда берутся рефрены, можно пока лишь предполагать. Мое мнение
будет дано после введения фракталов в п. 6-7. Рефренную упорядочен-
ность разнообразия, которую короче назвать упорядоченностью диасети,
Мейен называл типологической упорядоченностью и полагал ее одним из
основных свойств природы - неживой, живой и социальной. Замечу: общую
диасеть для всего на свете (в природе и обществе) вводить вряд ли стоит,
322
лучше (содержательнее) вводить свою собственную диасеть в каждой от-
расли знания.
Сходство структур Мейен объяснял не общностью происхождения, а
общностью законов формы (не путать с общностью путей формообразова-
ния, которой может и не быть) и любил повторять старое изречение:
“Кубы, в которые кристаллизуется поваренная соль, никто не будет рассмат-
ривать с точки зрения общего происхождения... от одного куба”.
Добавлю: разные вещества могут образовывать одинаковые кристаллы,
а одно вещество - разные, чем и объясняется феномен простейших хими-
ческих рефренов. Самые наглядные примеры упорядоченности дают в био-
логии периодические таблицы систематики (см. гл. 10). Возможно, в буду-
щем найдут законы, их определяющие, как узнали в начале XX века закон
заполнения электронных уровней, определяющий свойства (рефрен) уже
давно известной тогда таблицы Менделеева. См. Дополнение 5.
6-3*** Грамматика биологии
Диатропика изучает любые разнообразия, и исходный материал дала
Мейену грамматика. Самые ясные примеры рефренов она дает в виде пра-
вил спряжения и склонения. Очевидно, что слова «бал» и «пол» преобразу-
ются по общему правилу (все падежные окончания совпадают), так что нуж-
но запоминать не каждую падежную форму данного слова, а лишь тот реф-
рен (тип склонения), к которому оно относится. В книге [Чайковский, 1990] в
главе 2 приведены грамматический рефрен (русские падежные окончания),
фонетический (совпадение окончаний морфологически далеких русских
слов) и два зоологических (параллелизм внешних обликов воробьиных и
повторность форм конечностей, о которой сказано ниже).
К сожалению, в биологии ничего похожего на грамматику пока нет, и ка-
ждую видовую форму и форму каждого органа приходится описывать и за-
учивать отдельно. А ведь материал явно проявляет регулярность, и Мейен
искал аналогии ботаники с грамматикой.
Его примеры требуют более сложного языка, чем принят в данной кни-
ге, и я предпочту более простые. Такова таблица форм листовой пластинки,
где рефрен «типы расчленения листа» очевиден (рис. 35А).
Аналогичен у низших растений рефрен, в который выстроены формы
талломов водорослей (рис. 35Б), где «вместо филогенетического дерева
возникает сеть наподобие периодической системы Менделеева» (из книги:
Дьяков Ю.Т. Введение в альгологию и микологию. М., МГУ, 2000, с. 10). В
этом же учебнике на с. 12 дана таблица рефрена форм для актиномицетов
и грибов). Об этих рефренах см. также пп. 8-6,10-8.
А вот пример из зоологии. Возьмем те же конечности позвоночных: все
знают, что плавники есть у рыбы и у кита, что плавники, лапы и крылья мо-
гут быть слабо развиты (зачаточны), но мало кто догадывается, насколько
глубоки тут параллели. В действительности, во всех пяти главных классах
323
(костные рыбы, амфибии, рептилии, птицы, звери) виден один и тот же
рефрен - от полного отсутствия одной или обеих пар конечностей, через
зачаток или слабо развитую пару, через полноценный плавник или ласт, ла-
пу и планирующую поверхность - к органу активного полета, крылу. Правда,
некоторых вариантов не бывает: нет и никогда не было амфибий с крылья-
ми и птиц без задних конечностей - но в остальном параллелизм удиви-
тельно полон (рис. 36).
Рис. 36. Рефрен мерона «парные конечности позвоночных». Усложнение об-
щей организации наблюдается сверху вниз, а конечностей - слева направо
значения мерона |
класс отсутствие или неви- димый сна- ружи зача- ток развита только сла- бая перед- няя пара многофа- ланговые плавники или ласты орган для
ползания планирую- щего полёта активного полёта
костные рыбы мурена китовидка, угорь кистепёрые морской нетопырь летучие рыбы клинобрю- шка
амфибии червяга сирен некоторые ископаемые саламанд- ры яванская ля гушка не бывает
рептилии змеи и безно гие ящерицы хирот морские черепахи черепахи, крокодилы ногокрыл (ископае- мое) летающие ящеры (ископае- мые)
птицы киви (нет пе- редней па- ры) не бывает пингвины поползень, стрижи совиный попугай большинст- во видов
звери киты, ламан тин (нет зад ней пары) КИТЫ ламантин (передние) калан (задние) кроты, тюлени шерсто- крыл летучие мыши
Он дан упрощенно: все признаки выражены в полной мере. На самом
Деле каждую клетку можно заменить целым рядом - от полного выражения
До почти полного “размывания” признака. Если таблицу рефрена предста-
вить лежащей на столе, то ряд степеней выражения будет выглядеть верти-
кальным столбиком, стоящим на каждой клетке таблицы. Такой столбик для
листьев был дан на рис. 17 (там два последних вида помещены рядом с
первыми тремя). Он показывает “размывание” пальчато-лопастного листо-
вого контура; подробнее см. Чайковский Ю.В. И МС, 61.
Для пяти основных классов позвоночных получаем 5 параллельных ря-
324
дов, в которых видно сходство не только отдельных свойств, но их комплек-
сов. Например, киты не имеют брюшных плавников, и не имеет их малень-
кая глубоководная рыбка китовидка (Cetomimus) - черная, с голой кожей,
огромными головой и ртом, с малюсенькими глазками - ну прямо крохотная
копия гренландского кита, бороздящего поверхность вод. Назвать это неза-
висимыми приспособлениями - значит не сказать ничего, ибо условия жизни
тут различны предельно. И придумать пользу от «ручонок» сирена и хирота
никто даже не пробовал - слишком очевидно было бы «упражнение красно-
речия». См. Дополнение 6.
Тенденции, выявленные Мейеном на ботаническом материале, побуди-
ли его ввести понятие рефрена: вместо того, чтобы давать каждой различи-
мой стадии преобразования (например, стадии размывания контура листа
46,
или редукции лапьги) видовое имя, он предлагал давать единое описание
преобразования мерона как характеристику таксона. Этим он продолжал
мысль Вавилова:
“в законе Вавилова... обращается внимание на повторность отдельных при-
знаков от таксона к таксону, но опушена повторность в правилах преобразо-
вания варьирующих признаков”, причем “запомнить порознь все разновидно-
сти и относящиеся к ним самостоятельные термины невозможно (это все рав-
но, что запоминать все склонения каждого существительного, не познако-
мившись с правилами склонения” [Мейен, 1978, с. 500-501].
Уточню: вместо «все склонения» здесь надо читать «все падежные
формы». О падежах надо еще иметь в виду, что в языках, где их много, они
не просто перечисляются, но и упорядочиваются в таблицы^7. В биологиче-
ской грамматике нередко нужно как раз упорядочение. Зоологический при-
мер “падежей” см.: Дополнение 6.
Диасеть фактически давно использовалась в определителях, состав-
ленных по схеме Балковского (Банковский Б.Е. Цифровой полигамический
ключ для определения растений // БЖ, 1960, № 1; он же. Теза, антитеза и и
ряд признаков в диагностике растений // БЖ, 1960, № 11). Первым по этой
схеме был построен полевой определитель: Н.А. Бородина и др. Деревья и
кустарники СССР. М., 1966. Пользоваться им просто и удобно: вместо уто-
мительного выбора антитез “по ключу" (когда каждый ваш неверный выбор
исключает возможность правильного определения, но вы не узнаёте этого)
Замечу для систематиков: в описанном выше случае с лапами перечень стадий их ре-
дукции у разных родов характеризует структуру семейства.
Например, сериальные локативы дагестанских языков: каждая серия из трех падежей
сооответствует вопросам откуда, где, куда (сверху, наверху, наверх; сбоку, на боку, на-
бок и т.д.) — всего до восьми серий. Кроме них, падежная система этих языков включает
обычные индо-европейские падежи — именительный, родительный, эргативный и т.д.,
полагаемые в один ряд (Магометов А. А. Табасаранский язык. Тбилиси, 1965).
г
325
здесь надо просто найти у растения нужные признаки (состояния меронов) и
записать их цифровые значения, указанные в таблице признаков, а затем
найти по полученному шифру нужный вид. Отсутствие или неверный выбор
одного признака мало что меняет, поскольку по таблице легко сравнивать
близкие шифры. Вот пример из Введения к данному определителю:
“Обнаружив, что листья и веточки расположены супротивно, а на ветвях от-
сутствуют колючки, запишите число 3; светло-бурой глубокопродольно-
морщинистой коре соответствует число 3, а молодые побеги, зелено-
чернеющие от железистых бородавочек, обозначены числом 8. Добавив двой-
ку, так как листья определяемого кустарника простые, яйцевидные, голые или
снизу по жилкам опушенные, получаем набор цифр 3382. Определить, какие
были соцветия или цветки, осенью довольно трудно, поэтому переходим к
рассмотрению плодов, поставив 2 прочерка вместо цифр, соответствующих
цветкам. Плоды определяемого кустарника — 4-лопастные коробочки с семе-
нами, окруженными оранжево-красными мясистыми присемянниками (кро-
вельками); это соответствует индексу 16. Окончательный шифр 3382-16 со-
ответствует в таблице бересклету бородавчатому”.
Причину удобства поясняет приведенный выше тезис: «У разных таксо-
нов общие мероны», в силу которого большая часть ходов в обычном опре-
делителе являет собой повторы. Мы вернемся к этому в п. 10-3.
6-4. Наличное разнообразие и эволюционная изменчивость
Исходное допущение дарвинизма - что различие между организмами
одного вида есть фактор эволюции (эволюционная изменчивость) - оказа-
лось принципиальной ошибкой. На это указывал еще Дженкин (и многие то-
гда же), но обманчивая наглядность сто лет не давала увидеть тот факт, что
различие между особями к эволюции не ведет.
Как мы видели ранее, Коп и Четвериков полагали, что внутривидовая и
надвидовая эволюция — принципиально различные процессы; Коржинский,
Де-Фриз, Гольдшмидт и их последователи показывали, что вид отличается
от вида комплексом свойств, отнюдь не разложимыми на признаки, какие
могут появляться порознь. Можно вспомнить и ту идею Дж. Виллиса (1922
год) [Чайковский, 1990; Назаров, 2005], что вид образуется за счет одно-
кратной видовой мутации, а потому эволюция на уровне вида и выше не
имеет никакого отношения к популяционным событиям.
Никто из них услышан не был. Причина этого видится мне в господстве
статистической ПМ, точнее - эргодической идеи (см. п. 5-16).
В 1969-1972-х годах генетик Ю.П. Алтухов (Владивосток) и соавторы по-
казали, что у многих рыб особи одного вида различаются одними белками, а
вид от вида — другими (что подтвердили литературные данные по другим
позвоночным и дрозофилам) К 1981 году было установлено, что лишь 11 %
белков обладают полиморфизмом, выявляемым элекгофоретически (Алту-
хов Ю.П. Генетические процессы в популяциях. М., Академкнига, 2004, с.
326
256)45. На этой узкой базе был сделан вывод, что основная масса белков
“дарвиновской” изменчивостью не обладает и что (словами историка науки)
“видообразование — продукт качественно иных реорганизаций генетического
материала, нежели мутации, лежащие в основе (внутривидового — Ю.Ч.) по-
лиморфизма” [Назаров, 2005, с. 136].
Попытки Алтухова и соавторов осмыслить данное обстоятельство с по-
зиций дарвинизма не дали, на мой взгляд, ничего. Наоборот, работы Стег-
ния (тоже отрицающего роль полиморфизма в видообразовании и в опыте с
двукрылыми насекомыми показавшего мономорфизм видообразующих
свойств хромосомного аппарата, п. 5-5*), вполне раскрыли номогенетиче-
ский механизм видообразования (для данных объектов).
Эти и подобные исследования вплотную подвели к феномену, вскоре
описанному Мейеном. Достаточно было понять, что процесс адаптации не
связан с полиморфизмом, как открылось замечательное явление.
6-4* Наследуется не свойство, а разнообразие свойств
Речь идет про транзитивный полиморфизм (транпол). Понятие ввел
Мейен [1978, с. 505], но сам феномен описал (никак не назвав) еще Дарвин
(п. 3-10): гладкокожие персики повторили своим разнообразием всё разно-
образие сортов бархатистых, хотя все гладкокожие выведены из одного му-
танта, появившегося в культуре бархатистых. Через этот один экземпляр
разнообразие бархатистых “переехало” к гладкокожим, совершило “тран-
зит”. Глядя на эти два ряда сортов, нельзя узнать, какой из гладкокожих сор-
тов был исходным, однако ясно, что он содержал в своей наследственности
не только свои качества, но и весь ряд возможных вариаций, т.е. будущее
разнообразие, и неважно, какой вариант послужил для “транзита”.
Другой пример дает доместикация (п. 4-19): одомашнивая лису, сибир-
ские генетики получили у нее комплекс собачьих свойств. Понятно: селек-
ционер работает с тем же рефреном, что и природа, и оба имеют дело с
транзитивным полиморфизмом.
И транпол, и другие законы разнообразия видны в самых различных
науках, всем там известны, но воспринимаются с трудом. Поскольку человек
сам производит отдельные действия и следит за отдельными объектами, то
и эволюцию принято видеть как появление и изменение отдельных свойств
у отдельных видов - особенно грешат этим ламаркизм и дарвинизм. Нам
трудно усвоить, что природа действует иначе, что природа оперирует с
разнообразиями свойств, взятыми целиком.
Вот пример. В п. 4-11 упоминался «закон родственной изменчивости»
Кренке (это, как теперь видно, частный случай транпола), и был приведен
пример: ряд персиков и ряд миндалей сходны, но если у персика нормален
ДО
Исследован был полиморфизм белков по одному признаку — электрофоретической
подвижности, т.е. по суммарному электрическому дипольному моменту.
327
мясистый плод, а сухой уродлив, то у миндаля - наоборот. Эволюция близ-
ких видов выступает здесь как движение вдоль одного и того же рефрена
(спектр возможных свойств плода у персиков и миндалей одинаков), но в
разые стороны; увидеть это трудно, поскольку частое в одном таксоне очень
редко в другом. Персики и миндали - две нормы на одном рефренном поле
(на одном участке диасети), и эволюция состоит здесь в смене норм на од-
ном и том же поле. См. Дополнение 6.
6-4** Транзитивный полиморфизм против идеи отбора
Самые яркие и известные примеры транпола касаются бесплодия:
свойство организма, ведущее к бесплодию, само по себе не передается по-
томству, и вроде бы очевидно, что оно должно встречаться исчезающе ред-
ко, с частотой мутаций или еще реже. Однако интуиция нас обманывает: та-
кие свойства вполне обычны. Например, полный гомосексуализм поражает
4% мужчин, и лишь ничтожная часть гомосексуалов размножается (путем
искусственных приемов), однако эти 4% воспроизводятся - в каждом поко-
лении заново. Около 1 % людей несут прочие генетические дефекты, приво-
дящие к стерильности. Это значит, что ненаследуемое свойство передается
потомству за счет транзита через полноценных индивидов, и помешать та-
кому положению дел выбраковка не может.
Впечатляют и обратные примеры: так, одна на 10 тыс. женщин рожает
легко и сверхбыстро, но это свойство не стало частым, хотя смертность от
родов была мощным селективным фактором в ходе всей истории человече-
ства. Мнимый отбор и тут оказался бессилен перед реальным транполом.
Это, кстати заставляет усомниться и в идеях евгеники (см. далее, п. 12-1*).
В п. 3-3 говорилось, что идею полового отбора никто не пытался прове-
рить опытом, а теперь можно назвать реальный механизм сохранения по-
ловых различий: половая структура популяции воспроизводится целиком
путем транпола вне связи с размножаемостью разных вариантов.
Если выстроить примеры драк самцов в ряд, то станет ясно, что их био-
логический смысл состоит отнюдь не в том, что драчливые лучше размно-
жаются. Помню, Тимофеев-Ресовский смеясь говорил на одной из своих
лекций по эволюции: «Пока два два петуха дерутся, третий топчет кур». Это
оказалось довольно общим правилом для многих видов.
Так, наш лучший специалист по головоногим К.Н. Несис хотя и писал,
что самцы множества животных в брачный период борются из-за самок и
что «побеждают в этой борьбе, естественно, самые крупные и агрессивные»
- однако затем начисто разбил своё утверждение: пока крупные самцы де-
рутся, мелкий оплодотворяет кладку [Несис, с. 98-99].
Это, добавлю, могло бы вести к измельчанию популяции, если бы не
транпол. Крупный и мелкий, драчливый и тихий, яркий и невзрачный - все
они представлены в разнообразии вариантов, которое воспроизводится не-
зависимо от того, кто кого оплодотворит.
328
Такое же пиратское оплодотворение известно у некоторых рыб, а не-
давно описано у горной популяции обыкновенной травяной лягушки Rana
temporaria (Семенов Д.В. Пиратское оплодотворение // Природа, 2006, № 4).
Опять видим ряды, и встает вопрос: могут ли поведенческие ряды образо-
вать рефрен, подобный рефренам структурным? Об этом см. п. 6-5.
Биологический смысл борьбы самцов вовсе не сводится к ответу на во-
прос “зачем?”, он включает и ответы на вопросы “как?” и “почему?”. Если
вид экологически достаточно прочен, самцы могут драться и вопреки пользе
вида (это, увы, прекрасно показали дуэлянты прошлых веков). А большин-
ство видов вообще прекрасно обходится без этих драк.
(Роль двуполости вообще малопонятна - для размножаемости гермаф-
родитизм вдвое выгоднее - и мы немного проясним ее в п. 9-16*.)
Впрочем, природа, по всей видимости, обходится вообще без отбора, о
чем у нас шла речь не раз, и пора объяснить, в чем тут дело.
Строго говоря, объяснять здесь должны дарвинисты - на каком основа-
нии голословная идея была без проверки принята наукой. Но они этого не
сделали (презумпция^.), и приходится объяснять противоположное.
Как мы видели на термитах, самая жёсткая отбраковка одних не
влечёт сама по себе никакого изменения других. Если изменчивость
выживших не направлена в определенную сторону, то они воспроизводят
прежнюю изменчивость - в этом и состоит транпол. Если же изменчивость
получила направление (А.Г. Зусмановский называет это новой потребно-
стью, а можно сказать и иначе: например, о давлении новой нормы), то
именно этим и будет определяться эволюционный акт. Если такая процеду-
ра идет в форме массового размножения нового варианта при подавлении
размножения прежних, то имеет место расплод (п. 5-8*).
Скучно повторять, но сделаю это еще раз: расплод отличается от есте-
ственного отбора тем, что подавляюще повышенное размножение данного
варианта вызвано не его высокой размножаемостью (наоборот, она может
быть очень низка, как мы не раз увидим - например, в п. 12-1), а работой
всей размножающей системы, пресекающей иные варианты.
6-5. Простые примеры применения диатропики
Закономерности, отмечаемые диатропикой, не раскрывают сути явле-
ний сами по себе - в силу самой ее природы как грамматики науки. Грамма-
тика - вспомогательный прием анализа текстов, не касающийся их смысла.
Это не упрек диатропике: то же относится ко любой процедуре формального
анализа (статистика тоже сама по себе не раскрывает механизмов, но
весьма полезна). Как и всякая грамматика, диатропика нужна при любом
анализе, помогая обозревать не отдельные факты, а их совокупности.
В п. 5-17 мы видели, что возрождается понимание единства всех жи-
вотных по Жоффруа. И даже всех организмов. С пониманием гомеобокса
как маркера (п. 5-15**) стало ясно, что это единство не сводится к родству, а
329
это значит, что рефрены приходится считать реальностью, не выводимой ни
из общности происхождения, ни из сходства онтогенезов. Мы получаем
первый набросок ответа на трудный старый вопрос о сути феномена сход-
ства (о природе общей гомологии по Оуэну).
Известные мне прежние ответы - грубо материалистический (родство
по Геккелю) и грубо идеалистический (общая идея по Платону), не были
вразумительны и, по-моему, просто выражали веру их авторов.
Теперь же очевидно, что сходные структуры и функции могут дости-
гаться различными путями в силу упорядоченности диасети (п. 6-3**).
Данный феномен указывает на некую очень общую закономерность, част-
ные случаи которых давно известны как правила грамматики (во всех язы-
ках), как периодический закон и гомологические ряды в химии и другие при-
меры. К ним выше были добавлены примеры рисунков 35 и 36.
Великолепный поведенческий рефрен образуют общественные насе-
комые - пчелы, муравьи и термиты, причем термиты весьма далеки от пчел
и муравьев как таксономически (принадлежат к другому подклассу), так и
экологически (разлагают древесину, пользуясь кишечными симбионтами).
Однако поведенческий рефрен не выглядит у насекомых самостоятель-
ным, ибо маскируется морфо-функциональными рефренами. Поэтому край-
не важен (и удивителен) такой факт: в тот же самый рефрен, что поведение
общественных насекомых, укладывается и поведение млекопитающего. Та-
ков один вид подземных грызунов - это голый землекоп Heterocephalus gla-
ber. Как показала зоолог Дженнифер Ярвис, известная южноафриканская
исследовательница, семья этих зверьков, которых она изучала около 30
лет, образует сверх-организм (иногда до 300 особей), похожий на те, какие
хорошо известны у общественных насекомых. Вплоть до наличия одной-
единственной плодущей самки, что, казалось бы, для млекопитающих не-
возможно (The biology of naked mole-rat. Princeton Univ. Press, 1991).
Причину их сходств с термитами многие биологи видят в сходстве под-
земных условий жизни, однако это мнение не выдерживает диатропической
проверки: все остальные подземные звери сохраняют индивидуальность, а
среди термитов и других чисто общественных насекомых далеко не все
подземны (некоторые термиты даже живут в кронах деревьев).
Мне представляется, что подземные условия всего лишь запустили вид
по данному пути развития (одному из возможных), далее работает сам реф-
рен. Нет оснований видеть в отдельных его чертах приспособление.
Вот еще три простых примера того, как полезно само признание фено-
мена разнообразия, даже без обращения к аппарату диатропики.
6-5* Была ли митохондрия симбионтом?
Вот уже 35 лет пишут, что митохондрия - потомок бактерии, когда-то
поселившейся внутри клетки. Единственный ясный довод в пользу этого
мнения - многие (но далеко не все) митохондрии похожи на бактерий.
330
Однако анализ разнообразия митохондрий говорит иное. Давно извест-
но, что есть митохондрия, совсем непохожая на бактерию - кинетопласт
некоторых жгутиковых одноклеточных. Митохондрий в клетке всегда много,
тогда как кинетопласт всегда один. Он крупнее их, всегда расположен около
основания жгутика и, очевидным образом, обеспечивает его энергией. Он
“не имеет сходства с бактериями... Единый хондриом трипаносомид во много
раз крупнее любой бактерии, а по трубчатой, иногда сильно разветвленной
форме не имеет с ними ничего общего”. Он “находится на ином, более высо-
ком, чем прокариотный, уровне организации. Существование такой систе-
мы... отменяет обязательность прокариотного уровня организации митохонд-
рий вообще и делает их происхождение от бактерий маловероятным” (Калли-
никова В.Д. Клеточная органелла кинетопласт. Л., Наука, 1977, с. 103).
Вскоре были открыты другие совсем не похожие на бактерий митохонд-
рии, нитевидные; им в 1978 году даже дали название «митохондриальный
ретикулум», т.е. сеть. В наше время стало ясно, что они гораздо обычнее в
природе, чем те митохондрии, что похожи на бактерий:
“Оказалось, что мелкие округлые органеллы (описываемые как митохондрии
- Ю. Ч.)... как правило, представляют собой лишь срезы протяженных ните-
видных структур... В.П. Скулачев предположил, что митохондрии могут слу-
жить электрическими кабелями, доставляя энергию в виде трансмембранного
потенциала из одной части клетки в другую. Эта гипотеза была подтвержде-
на...” (Черняк В.В. и dp. И Биохимия, 2005, № 2, с. 294).
Далее, обычный сперматозоид имеет митохондрию, которая кольцом
облегает осевую струну жгутика; оно собрано путем объединения обычных
митохондрий в ходе сперматогенеза [Албертс и др., т. 3, с. 38; Александ-
ров, с. 11]. Это ясно указывает место митохондрии в клетке как подчиненное
ядру. Наконец, есть митохондрии, в том числе и у нас с вами, которые име-
ют совсем не ту энергетику, что бактерии: в качестве основного продукта
производят не АТФ, а АФК - это митохондрии нейтрофилов (белых кровя-
ных телец). Данный факт мне любезно сообщил В.Л. Воейков.
К тому же у многих одноклеточных митохондриальная ДНК не замкнута
(в отличие от бактериальной) в кольцо; у животных, грибов и жгутиковых ге-
номы митохондрий содержат отличие в генетическом коде: триптофан коди-
руется иначе, чем в ядерных и прокариотных генах (Одинцова М.С., Юрина
Н.П. Геном митохондрий и протистов // ЭГ, 773). Это и многое другое гово-
рит о различном происхождении митохондрий разных организмов и плохо
согласуются с гипотезой об их бактериальном прошлом.
Словом, сходства митохондрии с бактерией, послужившие основой
“теории” - всего лишь самообман поверхностного взгляда на разрозненные
элементы разнообразия. Взгляд же с позиции диатропики всё расставляет
по местам: митохондрия - типичный гибридный орган [Чайковский, 2006,
с. 101], т.е. орган, эволюционно происшедший от наложения сменявшихся
онтогенетических программ. Всё это нам понадобится в п. 8-7*.
331
6-5** Зачем нужна теплокровность?
Учебники и руководства избегают этого вопроса, иногда лишь мельком
замечая, что теплокровность повышает уровень обмена веществ. Однако
гигантский кальмар достигает размеров лошади за два года [Несис, с. 23].
Клод Бернар говорил: «Постоянство внутренней среды организма есть ус-
ловие свободной жизни», и этот афоризм иногда тоже приводят в пользу
теплокровности. Но и это сомнительно: холоднокровные могут насквозь
промерзать и затем оживать, что для теплокровных исключено. Да и не все-
гда постоянна их температура - однопроходные (утконос, ехидна и пр.)
миллионы лет живут с весьма непостоянной температурой тела.
Биохимик С.Э. Шноль вывел теплокровность из роста скорости распро-
странения нервного импульса, ибо рептилии медлительны (Шноль С.Э. Фи-
зико-химические факторы биологической эволюции. М., 1979, с. 214). Да,
рептилии цепенеют на холоду, но как обстоит дело в целом?
Во-первых, рептилии вполне преуспевают: звери хоть и преобладают,
но рептилий не вытесняют. Во-вторых, в жарких странах у рептилий со ско-
ростями всё вроде в порядке, но почему-то и там преобладают звери. В-
третьих, в скорости ли нервного импульса дело? Многие амфибии, рыбы и
моллюски, в отличие от рептилий, сохраняют высокую подвижность даже
при нулевой температуре. Огромная скорость реакции отмечена у головоно-
гих [Несис, с. 82, 99] и у хищных растений (Природа, 2006, № 3, с. 78). В-
четвертых, теплокровные тратят всемеро^9 больше энергии, так что обяза-
тельная теплокровность - тяжкий груз, что-то вроде рогов ирландского
оленя. В-пятых, согревание тела очень просто физиологически (снижение
к.п.д. окислительного фосфорилирования) и, в частности, наблюдается у
многих рептилий при насиживании яиц и у многих растений.
Из растений лучше всех греются аронниковые (до 30 градусов выше среды),
но используют тепло по-разному: наш аронник, цветущий рано, до появления
листьев, согревает себя на холоду; а американская вонючая капуста Symplocar-
pus, цветущая летом, нагревается для имитации гниющего мяса и привлекает
этим мух. Источник их нагрева прост: прямой перевод дыхания в тепло, как при
гниении. Наоборот, у теплокровных процесс сложен: энергия запасается в АТФ
или протонном потенциале, а затем лишь рассеивается в тепло, и зачем такая
растрата, неизвестно (Берцова Ю.В. и др. И Биохимия, 2004, № 5, с. 716-717).
Кстати, неизвестно и то, зачем ароннику нужно цвести раньше других.
Зачем понадобилась перестройка физиологии, приведшая к теплокров-
ности? Обычная точка зрения при проверке разнообразием становится
весьма сомнительной. А что советует диатропика? Да очень просто: надо, 49
49
При одинаковой массе тела энерготраты теплокровных в 6-8 раз превышают траты хо-
лоднокровных (Иванов К.П. И ЖОБ, 1990, № 1). К этому добавлю: наше тело надо и со-
гревать. и охлаждать.
332
как уже говорилось, начинать не с вопроса «зачем?», а с вопроса «у кого?».
Обязательная теплокровность наблюдается у всех птиц и зверей, и притом
только у них, так же как и развитой адаптивный иммунитет. То есть налицо
свойство разнообразия животных - теплокровность и ТВ-система жестко
связаны. Естественна мысль, что причина появления теплокровности прямо
связана с эволюцией иммунитета.
Вернее всего, что теплокровность является частной формой проявле-
ния того общего повышения активности, какое нужно для высших стадий
прогрессивной эволюции. Мы вернемся к этому вопросу в п. 8-12*.
6-5*** Зачем нужен иммунитет?
Странный вроде бы вопрос - иммунитет нужен против заразы. Однако
многие иммунологи отмечают, что для борьбы с заразой достаточно того
иммунитета, что есть у растений и низших животных. Например:
«у более 98% [видов] многоклеточных организмов нет адаптивного иммуни-
тета с Т- и В- лимфоцитами... Тем не менее, они не только успешно справля-
ются с инфекцией, но и не страдают от онкологических и аллергических бо-
лезней» [Лебедев, Понякина, с. 5].
Против повседневных инфекций прекрасно работают три давно из-
вестных механизма: 1) общий для всех «принцип несъедобности» (часто
организму бывает достаточно чуть-чуть изменить одну категорию своих
макромолекул, чтобы лишить инфицирующий вирус или бактерию способ-
ности размножаться в нем); 2) общий для всех растений феномен синтеза
ядов и антибиотиков и 3) общий для всех животных фагоцитоз (поедание
лимфоцитами посторонних клеток). К ним надо добавить: 4) недавно откры-
тые механизмы, о которых пойдет речь в п. 6-6**. Против редких инфекций
еще проще: обычно бывает достаточно повысить рождаемость.
Зачем же нужен сложнейший ТВ-иммунитет теплокровных, о котором
шла речь в главе 5? Опять анализ разнообразия делает сомнительной при-
вычную трактовку. Поэтому напомню, что «Биологический энциклопедиче-
ский словарь» (1986) определил иммунитет как «способность организма за-
щищать собственную целостность и биологическую индивидуальность», а
борьбу с инфекциями назвал «частным проявлением иммунитета». Так
предложил понимать его еще австралийский иммунолог Ф. Макферлейн
Бернет в 1962 году. Будем так понимать иммунитет и мы.
В книгах по иммунологии обычно описывается лишь адаптивный имму-
нитет теплокровных (ТВ-система), и едва упоминается всеобщий механизм
- конститутивный (врожденный) иммунитет. (О нем см. например: [Ру-
мянцев; О’Нилл; Лебедев, Понякина].) Он таит не меньшие загадки, чем
адаптивный, и борьбу со многими видами заразы эффективен именно он.
К примеру, врожденный иммунитет к малярии, какой имеют коренные
жители тех тропических районов, где она обычна, основан на том, что у них
синтезируется модифицированный гемоглобин. Замена всего лишь одной
333
аминокислоты в его молекуле исключает возможность использования такого
гемоглобина малярийным плазмодием [Румянцев, с. 104]. Ту же роль играют
мутации, меняющие структуру клеточных стенок.
Иногда врожденный иммунитет называют молекулярным, а адаптивный
клеточным, что неверно: молекулярный может обладать некоторой способ-
ностью к адаптации (например, у растений), а клеточный - быть врожден-
ным (например, фагоцитоз). Примечательно, что ТВ-системы разных таксо-
нов могут различаться как раз на молекулярном уровне.
Так, верблюды имеют, вдобавок к обычным антителам, еще нанотела
(вдесятеро меньшие и в то же время более эффективные). Это - явное дос-
тижение иммунитета, и им обладают также южноамериканские ламы (Гиббс
У. Нанотела // ВМН, 2005, № 11). Поскольку верблюды и ламы разошлись 20
млн лет назад, свойство достаточно старо. Странно: громоздкий иммунитет
одинаков у мышей и людей, а возможен куда более экономный, верблюжий.
Это - к вопросу о любимой многими оптимальности. Есть ли она?
Растения имеют и конститутивный иммунитет, и элементы адаптивного
(но не клеточного) иммунитета. Они могут распознавать чужеродные белки
и клетки, синтезировать антибиотики в ответ на инфекцию и убивать ее
вместе с собственными зараженными клетками (аналог фагоцитоза, но без
фагоцитов). Самонесовместимость (невозможность самоопыления у рас-
тения) - тоже разновидность иммунитета, это как бы автотолерантность
наизнанку (автотолерантностью называется отсутствие иммунной реакции
теплокровного животного на его собственные белки). При этом
“самонесовместимость занимает у растений примерно такое же место, как
раздельнополость у животных” (Суриков И.М. Несовместимость и эмбрио-
нальная стерильность растений. М., Агропромиздат, 1991, с. 9).
Явления внешне противоположны, тем не менее оба обеспечивают
принудительную рекомбинацию. Самонесовместимость растений ясно пока-
зывает, что их иммунитет носит индивидуальный характер. Она являет ана-
логию с клеточным иммунитетом животных, с их комплексом гистосовме-
стимости (Skogsmyr /., Lankinen Е. И BR, 2002, № 4, р. 554). Самонесовмес-
тимость бывает и у некоторых животных (например, у пустынных дрозофил
[Марков, Куликов, 2006, с. 135]).
Однако известно, что плодовые деревья размножают прививкой и что
иммунная система этому не мешает. Почему растения допускают срастание
тканей разных видов и даже близких родов (например, груши, яблони и ря-
бины)? Очевидно, дело не в простоте и сложности, а в различии функций
эпигностических механизмов (см. п. 5-15) у растений и у животных.
6-6. Эпигностика, иммунитет и сетевой принцип эволюции
Появление простого иммунитета животных довольно понятно: фагоци-
тоз принято рассматривать как приспособление древней способности сво-
бодных клеток (питание) к новому делу - борьбе с заразой. Просто и со-
334
гласно с принципом смены функций Дорна (п. 3-6). Аналогично Эдельман
понял появление адаптивного иммунитета теплокровных (п. 5-15). Сейчас
выясняется, что элементы адаптивного иммунитета есть у всех или
почти всех эвкариотных организмов [Марков, Куликов, 2006]).
Но зачем адаптивный иммунитет выстраивается в каждом поколении
заново? Может быть, именно его формирование в каждом онтогенезе и
нужно? С позиции эпигностики (п. 5-15) ответ положителен: в новых руково-
дствах формирование зародыша подается как совокупность актов клеточно-
го узнавания. Интересно, что при изложении эмбриологам оказался удобен
как раз термин из иммунологии - компетентность. Например:
“Способность отвечать специфическим образом на данный стимул называет-
ся компетенцией” [Гилберт, т. 2, с. 62]. Она возникает и утрачивается: на-
пример, “об эктодерме гаструлы можно сказать, что она компетентна к вос-
приятию индукционных стимулов. Эта компетенция ... приобретается на ста-
диях позднего дробления и утрачивается на стадиях поздней гаструлы” [Гил-
берт, т. 3, с. 52].
Назначение системы узнавания с самого начала жизни не сводилось к
борьбе с заразой, а служило для контроля целостности орга-
низма и вида [Марков, Куликов, 2006]. Прекрасный пример
простого контроля онтогенеза приведен еще в «Номогенезе»
Берга: паразитируя на растениях, насекомые откладывают
яйца в их ткани; растения защищаются от паразитов,
окружая чужеродное тело оболочкой - галлом, причем
форма галла вполне специфична для растения; на хвойном
это - недоразвитая шишка (рис. 37). В этой эпигностической
реакции растения мы видим сразу акт как иммунной защиты,
так и онтогенеза.
Рис. 37. Галл на сосне принимает форму
сосновой шишки
С простым конститутивным иммунитетом нас бы, надо полагать, попро-
сту не было. Обоснованию данной мысли было уделено место в ЧЭ (тоже в
п. 6-6) и в статье [Чайковский, 2003], а мы вернемся к эпигностике.
Датский иммунолог Нильс Ерне предложил в 1974 году сетевую тео-
рию иммунитета, согласно которой иммунная система млекопитающего
являет собой не просто набор клонов, а единый самоподдерживающийся
механизм, в котором каждый клон вплетен в единую сеть. Вплетен как «сни-
зу» (каждая клонообразующая клетка отличается от остальных одним им-
муноглобулином, а он получен, как упоминалось в п. 5-8, перетасовкой на-
бора генов, общего для всех иммуноглобулинов организма), так и «сверху»:
едва численность клона становится высока, он сам подвергается атаке им-
335
мунной системы. Здесь нет иерархии (последовательного подчинения)50, а
есть именно сеть - всякого кто-то ест.
Через 30 лет стало ясно, что особенности млекопитающих тут несуще-
ственны (врожденный иммунитет животных и растений в принципе способен
на то же самое - см. далее), но в остальном Ерне был прав. Естественно
рассмотреть с подобной точки зрения и эволюцию. Если прежде она пред-
ставлялась как набор «клонов» (родословных древ), то открытие диасети и
горизонтального переноса заставляют рассматривать ее как заполнение се-
ти (многомерной таблицы). К данной теме мы вернемся не раз.
Иммунная система похожа на экосистему, и естественно, что в иммуно-
логии развернулись те же споры о роли борьбы и взаимопомощи.
В 1988 году в Новосибирске вышел сборник [Проблемы...], где напом-
нены старые и поставлены новые вопросы. Основная их часть вращается
вокруг загадки: зачем теплокровному организму столь сложный и ему само-
му опасный механизм, готовый в любой момент “съесть поедом” и себя, и
хозяина? Ответ новосибирские иммунологи дали по-разному.
Г.З. Шубинский вспомнил Ерне и других классиков иммунологии, счи-
тавших иммунитет регулятором онтогенеза, поддерживающим целостность
организма. (Добавлю: это и есть эпигностический взгляд.)
В.А. Козлов решил, что организму выгодно держать некоторые свои
ткани на грани канцерогенеза, чтобы пользоваться продуктами их почти
безудержного синтеза - среди них много полезных организму веществ.
Только теплокровные могут позволить себе такую роскошь, но то и дело
расплачиваются за нее раком - если иммунная система не удержала эти
очень активные ткани под контролем.
Остальные организмы вынуждены искать подобные вещества в мире
микроорганизмов (впрочем, данный источник всеобщ: «Эта микрофлора
приносит организму большую пользу: синтезирует необходимые для орга-
низма витамины, регулирует перистальтику кишечника и т.д.» [Лебедев, По-
някина, с. 8]). Организм трактуется экологически, как сообщество.
Можно сказать, что иммунитет ведет себя тут как фермер, пасущий
стадо быков, готовых, если он зазевается, растоптать и его, и всю родню;
зато ферма всегда имеет и мясо, и шкуры, и навоз, и много прочего.
О.К. Баранов обратил внимание на сходство организации иммунной и
нервной систем и допустил, что сперва произошли просто чувствительные
клетки, а из них - клетки нервной и иммунной систем. Он спрашивал:
«не использует ли нервная система молекулярно-генетические структуры и
механизмы, подобные установленным у иммунной системы?» [Проблемы...].
о0 Иерархия в первичном значении (от греч. иерос — священный, архэ — старшинство,
власть) означает систему управления, в котором (как в церковном управлении) ни один
подчиненный не имеет двух или более непосредственных начальников. Изображается
Древом. Другое значение слова (многоуровневость) далее в книге не используется.
336
Думаю, да. В самом деле, у всех млекопитающих мы видим вполне раз-
витой иммунитет, тогда как мозг достиг у них наибольшего развития лишь в
последнюю очередь - у человека. Зная, что иммунная система хорошо обу-
чается и генетически запоминает найденное, вполне можно допустить, что в
ходе антропогенеза мозгу было чему у нее поучиться.
Решения указанных проблем нет до сих пор, хотя многие считают, что
именно изъяны нашего типа иммунитета ведут к появлению новых заразных
болезней, а они способны даже погубить человечество. Например,
“Анализ показал, что важнейшая причина возникновения и широкого распро-
странения новых инфекционных болезней главным образом состоит в недос-
таточно эффективном развитии (эволюции) иммунной системы человека и ее
физиологической регуляции” (Иванов К.П. Традиционные и новые проблемы
физиологии // Успехи физиология, наук, 2004, № 3, с. 94),
поэтому очевидно, что сборник [Проблемы...] отнюдь не устарел. Наоборот,
экологическое (сетевое) понимание коэволюции еще более сблизилось с
иммунным и физиологическим:
«Поддержание оптимального состава микробного биоценоза организма [тоже]
является частью функции иммунной системы» (Лебедев К.А., Понякина И.Д.
Иммунофизиология эпителиальных клеток и образраспознающие рецепторы
// Физиология человека, 2006, № 2, с. 123).
Это понуждает нас заново вернуться к эпигностике (п. 5-15).
6-6* Эпигностика, мышление и адаптивный иммунитет
Пожалуй самое сложное, что произвела эволюция - человеческий мозг.
Он является генетическим парадоксом - устроен в соответствии с наслед-
ственной программой (наследуются даже черты характера), но не может
быть задан ею, ибо генов в миллионы раз меньше, чем связей клеток в моз-
гу. Однако ведь точно так же устроена и ТВ-система, а всякая параллель -
пища для диатропики.
Ерне в своей нобелевской лекции (1984 г.) уже проводил эту параллель:
иммунитет - аналог рассудочной деятельности, и в основе обоих лежит по-
рождающая грамматика51. Сделаю следующие замечания.
1) Редактирование РНК (п. 5-6) по-видимому относится к тому же кругу
явлений, поскольку явно не является бессмысленной.
2) Иммуногенез и онтогенез можно видеть как аналоги мышления, но
без свободы воли (тут Хомский может быть к месту), но видеть порождаю-
щую грамматику основой мышления, значит игнорировать главное в мыш-
лении: свободу воли. Тем не менее, во всех трех на основе малого числа
генов возникает огромное разнообразие структур, и очевидно, что во всех
Термин «прождающая грамматика» ввел около 1955 г. лингвист Ноам Хомский
(США), полагавший грамматику не просто набором правил для построения речи, но и
орудием выражения мыслей. У других лингвистов роль грамматики скромнее.
337
действует некое формирующее начало, причем наследуется именно оно, а
не сами структуры. Мы узнаем про это начало в п. 6-7.
3) Процесс эволюции выглядит скорее как обладающий свободой воли
(так, у Бергсона эволюционное новшество подобно изобретению), т.е. бо-
лее сходный с мышлением, чем с онтогенезом и иммунитетом. Дарвинизм
утверждает случайный характер эволюции, но это - лишь способ невнятно
говорить о том же. Подробнее см. [Чайковский, 2004, с. 219].
4) Недавно параллель адаптивного иммунитета и мышления получила
развитие. Описаны «иммунные синапсы», т.е. зоны контакта клеток ТВ-
системы, которые оказались принципиально сходны с синапсами - зонами
соединения нейронов. Обмен информацией между иммуноцитами напоми-
нает деловую беседу, и английский иммунолог Дэниел Дэвис, много сде-
лавший для понимания явления, пишет: когда
«стало ясно, что мыслительный процесс, осязание или ответ организма на по-
падание... патогена опосредуются сходными белковыми структурами, иссле-
дование механизма иммунной реакции приобретает новый смысл» (Дэвис Д.
Загадочные иммунные синапсы // ВМН, 2006, № 5, с. 26).
Главная особенность ТВ-системы теперь видится в том, что она может
реагировать на отсутствие рецептора, а не только на его наличие (там же,
с. 30). ТВ-система и мышление - это два аспекта эпигностики (п. 5-15).
На мой взгляд, теплокровность нужна просто потому, что обе эти сис-
темы эффективны именно в данном интервале температур.
5) Если Козлов прав, что иммунитет теплокровных держит организм на
грани канцерогенеза, то аналогичная грань должна быть и в мозге. Она до-
вольно очевидна: безумие. Итальянский статистик Чезаре Ломброзо напи-
сал 120 лет назад книгу «Гениальность и помешательство». Да, эти черты
связаны: за способность к творчеству человечество платит постоянную
дань: самые способные пребывают в околокритическом режиме, они то и
дело срываются в безумие. В сущности, сама эволюция состоит из ряда пе-
реходов: через эту грань живое, едва возникнув, стало безудержно (можно
сказать - по Мальтусу) размножаться, и затем всякое новое ее достижение
знаменовалось таким разрастанием. Каждое из них так или иначе доходило
До своей грани и либо гибло, либо вписывалось в свою систему, задавав-
шую всем компонентам определенные границы.
Оренбургский социолог и эволюционист В.С. Жданов (о нем см. [Чай-
ковский, 1993]) убежден, что эволюция на самых разных уровнях (геосфера,
биосфера, общество, вид, особь и, особенно, сознание индивида) во всех
своих существенных чертах определяется такими переходами через грань
Устойчивости. Такой переход он именует словом “опрокид". Это - один из
примеров катастрофы по Рене Тому [Том] и, как увидим далее, одно из
следствий фрактальности диасети.
Вероятно, тот околокритический режим, в котором иммунная (вернее,
эпигностическая) система млекопитающих умеет держать организм, как раз
338
и нужен для онтогенеза нервной системы (делает возможным высшее раз-
витие мозга). Мозг поэтому тоже живет в околокритическом режиме. Сумеет
ли вид, владеющий этим эволюционным достижением, вписаться в свою сис-
тему (биосферу) или погибнет - либо подобно стае саранчи (по Воейкову),
либо подобно безумцу (по Ломброзо), либо подобно ампутированному органу
(по Лавлоку), покажет время. Мы вернемся к этому вопросу в части 4.
6-6** Врожденный иммунитет
Врожденный иммунитет (п. 6-5***) есть у всех эвкариот (а возможно, и у
прокариот). Недавно он снова стал привлекать общее внимание; ему посвя-
тили спецвыпуски французский журнал “CR Biologies” (2004, № 6) и австра-
лийский “Immunology and Cell Biology” (2005, № 3). Затем стали появляться и
популярные обзоры [Лебедев, Понякина; О’Нилл]. Главные для нас их вы-
воды таковы:
1) врожденный иммунитет гораздо сложнее, чем до сих пор думали, хо-
тя и много проще адаптивного; он достаточно общ у всех эвкариот (хотя у
растений и одноклеточных отсутствует фагоцитоз);
2) он является первой линией обороны, как от бактериальных, так и от
вирусных инфекций;
3) он служит у теплокровных для запуска ТВ-системы (именно врожден-
ную систему включает интерферон, известный как то лекарство, которое
действует только в первые полсуток болезни - ср. п. 5-13*);
4) он служит и для контроля строения организма:
«первичная роль врожденной иммунной системы - саморегуляция.., а защит-
ная вторична» (Gordon S. Pathogen recognition or homeostasis? ... // CR Biol.,
2004, № 6, c.606-607).
О ТВ-системе это писали и раньше. Итак, врожденный иммунитет тоже
является частью эпигностической системы (рис. 38).
Именно врожденный иммунитет производит образраспознающие ре-
цепторы - белки семейства TLR (Toll-like receptors; от англ, toll - звонить);
они есть на всех клетках иммунной системы, включая эпителиальные. У са-
мых разных организмов они узнают бактериальную заразу:
«Многие из компонент у самых разных микроорганизмов одинаковы, и по-
этому от всех известных патогенов достаточно десяти TLR. Врожденным им-
мунитетом обладает не только человек. Это очень древняя система, которая
есть почти у всех исследованных организмов: насекомых, морских звезд, даф-
ний и т.д. Круглые черви используют единственный имеющийся у них TLR,
чтобы вовремя обнаружить в среде вредные бактерии и избежать встречи с
ними. TLR в изобилии имеется и у растений. Так, у... сорняка арабидопсиса
число TLR превышает 200» [О’Нилл, с. 36-37].
Обилие «рецепторов» у одной клетки наводит на мысль, что это вовсе
не отдельные рецепторы, а рецепторные функции комплексов, несущих бо-
лее общие функции. Так и есть:
339
«Toll-белок участвует в эмбриональном развитии дрозофилы, а именно, регу-
лирует дорзо-вентральную (спина-брюшко) полярность насекомого» [Лебе-
дев, Понякина, с. 6].
врожденный иммунитет
адаптивный иммунитет
Рис. 38. Упрощенная схема двойной системы иммунитета Вторая часть имеется
только у теплокровных (по [Лебедев, Понякина], с пояснениями)
слева: TLR - Toll-like receptors, молекулярные комплексы, активация которых
оповещает организм об инфекции; хемокины - молекулы, синтезируемые клетка-
ми эпителия для активации клеток врожденного иммунитета; нейтрофил и мак-
рофаг - два вида фагоцитов; дендритная клетка - ветвистая клетка захватываю-
щая антигены с целью их презентации (разложения на фрагменты и выставления
их на поверхность клетки для распознавания) клеткам ТВ-системы; моноцит -
предшественник макрофага (тоже спопобна к презентации антигенов);
справа: цитокины - семейство молекул, активирующих ТВ-систему и служа-
щих для «бесед» иммуноцитов (п. 6-6*); В-лимфоцит — иммунная клетка, произ-
водящая антитела; наивный - лимфоцит, еще не встретившийй антигена; цито-
токсический - Т-лимфоцит, созревший для ликвидации клеток
В свою очередь, действие этого белка высвобождает белок NF-kB, ко-
торый служит фактором транскрипции как у дрозофилы, так и у млеко-
питающих. И снова видим такую же многофункциональность:
«Транскрипционный фактор NF-kB млекопитающих — центральный регуля-
тор воспаления и иммунитета, принимает также участие в процессах апопто-
за, онкогенеза, роста, дифференциации» (Дьяков Ю. Т. На пути к общей тео-
рии иммунитета // ЖОБ, 2005, № 6, с. 454).
340
Напомню, что еще Гурвич и Бауэр (п. 4-16) полагали: не следует всякую
активность воспринимать как работу отдельной, специально для этого соз-
данной молекулы. С данной позиции легче понять, почему, например, у дро-
зофил в создании столь простой вещи, как пробка после спаривания (mating
plug), участвуют около ста белков [Марков, Куликов, 2006, с. 135] - это вовсе
не белки, специально созданные для данной функции.
Очевидно, что в геноме и в клетке нет места для сотни белков для каж-
дой функции. Данные факты указывают на ту целостность, без которой не-
возможно понять феномен целесообразности (см. п. 6-12). Словом,
эволюция идет не столько путем создания новых белковых блоков, сколько
путем их комбинирования для новых целей (полифункциональность).
Участвует иммунитет и в феномене давления нормы. В частности, ра~
бота врожденной и адаптивной иммунных систем против заразы может но-
сить стрессовый характер. Для растений это подчеркивает книга фитофи-
зиолога И.А. Тарчевского (Казань). Хотя слово «иммунитет» используется в
ней только в плане борьбы с заразой, зато отмечена общность «сигнальных
систем» всех организмов (про- и эвкариот), связанность этих систем в еди-
ную сеть, роль их в онтогенезе и их эволюция. Например:
«интермедиаты эволюционно более “молодых” сигнальных систем могут
оказывать губительное действие не только на клетки патогенов, но и на клет-
ки растения-хозяина, приводя их к апоптозу, что препятствует распростране-
нию патогенов из мест инфицирования в другие части растения» [Тарчев-
ский И.А. Сигнальные системы клеток растений. М., 2002, с. 208).
Поясню: апоптоз (запрограммированное самоубийство клетки) рас-
смотрен тут как элемент иммунитета, причем иммунитета растений. Как и у
животных, мы видим его усложнение в эволюции. В отношении эволюции
особо интересна РНК-интерференция - недавно открытый у самых разных
эвкариот эпигностический механизм, использующий короткие двухцепочеч-
ные РНК, который борется с горизонтальным переносом:
“Когда в клетку попадает... опасный для нее ген, система заставляет его за-
молчать, распознавая и уничтожая кодируемую им РНК и не трогая мРНК
других клеток”. Когда чужих генов нет, она переключается на “плановое
вычеркивание” генов из генома, причем “это необходимо для правильного
развития организма - образования различных типов клеток” (Ло Н., Бартел
Д. Геномные цензоры И ВМН, 2003, № 11, с. 31-36).
Выключение генов посредством двухцепочечных РНК также
«позволяет растениям бороться с привнесенными извне генами, что обеспе-
чивает своего рода иммунную защиту на уровне нуклеиновых кислот» [Кле-
нов М.С., Гвоздев В.А. Формирование гетерохроматина: роль коротких РНК
и метилирования ДНК // Биохимия, 2005, № 11, с. 1450).
Интересно, что в растениях именно такая защита выполняет основную
функцию антивирусного иммунитета, поскольку 95% растительных вирусов
341
являются РНК-овыми (Chowdhury D., Novina C.D. II Immunology and Cell Biol-
ogy, 2005, № 3). У теплокровных, напомню, это делают Т-клетки.
Словом, открытие «геномных цензоров» и прочих чудес врожденного
иммунитета позволяет взглянуть на весь процесс развития (и в онтогенезе,
и в эволюции) как на иммунный (точнее, эпигностический) - новые ткани и
органы обычно возникают не за счет появления новых белков, а за счет ре-
гуляции на уровне РНК (матричных, цензорных и др.), которая приводит к
избирательному торможению белковых синтезов.
Чудо эпигностики являет и процесс мейоза, о чем будет речь в п. 8-8.
В итоге можно сказать, что у всех организмов схема имунитета
«принципиально отличается от схемы, предложенной Ф. Бернетом, переме-
щая акцент управления имунными реакциями с Т- и В-лимфоцитов на клетки
врожденного иммунитета» (Лебедев К.А., Понякина И.Д. И Физиология чело-
века, 2006, № 2, с. 123).
Итак, на грани веков в иммунологии свершилась новая революция, в
итоге которой стало ясно, что иммунитет служит всеобщим фактором эво-
люции. Напомню, что так полагал еще Вентребер.
6-7. Уровни развития и фракталы живого
В ходе эволюции, в разных ветвях живого, вновь и вновь текут сходные
процессы и образуются сходные уровни развития. Они замечательны тем,
что часто не удается увидеть их связи ни со сложностью условий, ни с ус-
пешностью проживания в них. Функцию разложения подчас равным образом
осуществляют грибы (эвкариоты) и бактерии (прокариоты); сложнейшие из
цветковых, орхидеи, зачастую очень капризны и потому редки; сумчатые
Америки успешно живут рядом с плацентарными, а сумчатые Австралии —
нет. Примеры можно приводить без конца.
У растений и животных независимо развился почти одинаковый аппарат
клеточного деления, хотя у одноклеточных они весьма различны; у первич-
но- и вторичноротых независимо развились многие одинаковые механизмы
(так, очень сходны глаза позвоночных, головоногих и некоторых пауков);
сходны иммунитеты птиц и зверей, сходны психика зверей и головоногих и
т.п. Все такие ряды сходств вместе образуют диасеть.
Это говорит о том, что основные свойства организмов определя-
ются не их родословными, а тем уровнем, до которого они дошли в
эволюции - не так уж важно, каким путем. Словом, заданное Геккелем по-
нимание эволюционизма как выяснения родословных не слишком интерес-
но (не говоря уж о том, что, в свете данных о горизонтальном переносе, и
недостижимо). Интереснее, по-моему, понять механизмы эволюции - тем
более, что механизмами можно будет воспользоваться для спасения при-
роды, о чем пойдет речь в главе 11, а родословными - нельзя.
Сходство сложного комплекса свойств принято объяснять как унасле-
дованное от общего предка, но, например, общим предком зверей и птиц
342
были рептилии, не имевшие (как принято считать) ТВ-системы. Как можно
унаследовать уровень развития от предка, не имевшего этого уровня, ибо
стоявшего на более низком уровне? Казалось бы, нельзя наследовать то,
чего нет, но это не вполне так. Можно получить в дар или в наследство “зо-
лотой ключик” от дверцы, за которой сам даритель не был. Об одной такой
теории и поговорим. Это - теория фрактального роста.
В каждом поколении уровень развития многоклеточного строится зано-
во, ибо всякий онтогенез начитается с одной клетки. (Часто говорят, что он-
тогенез есть последовательность клеточных делений, но в п. 5-15 мы виде-
ли, что это не всегда так и что лучше характеризовать онтогенез как рост
последовательно возникающих структур, в том числе ДС.)
Взгляните на рис. 39. Что на нем изображено?
Рис. 39. Схема ветвящегося
процесса эволюции не зави-
сит от масштаба процесса.
Дарвин иллюстрировал ею
образование рас и подвидов
Превращение вида в
род, рода в семейство или
семейства в отряд? А мо-
жет быть, дифференциров-
ка клеток зародыша? Или
размножение клона иммун-
ных клеток? На самом деле
схема взята из «Происхож-
дения видов» и рисует рас-
хождение признаков в ходе
борьбы за существование;
но не зная масштаба, по-
нять это невозможно. И не-
даром многие читатели
Дарвина восприняли ее как
иллюстра-цию дивергент-
ной (расходящейся) эво-
люции вообще. Тут эволю-
ция мыслится самоподобной (одинаковой на всех масштабах) для рас, ви-
дов, родов и т.д. В математике самоподобные (масштабноинвариантные)
объекты описывают фракталами.
Если не гнаться за строгостью, то фрактал - это нелинейная (не опи-
сываемая никаким линейным уравнением) структура, у которой часть уст-
роена в каком-то смысле так же, как вся структура. (Самоподобные линей-
343
ные структуры - прямая и плоскость - неинтересны.) Фрактальность нашего
мира, если не считать смутную догадку в «Трактате о седмицах» (п. 1-5),
впервые отметил 300 лет назад Лейбниц: он писал образно:
«всякую часть материи можно представить наподобие сада, полного расте-
ний, и пруда, полного рыб. Но каждая ветвь растения, каждый член животно-
го, каждая капля его соков есть опять такой же сад или такой же пруд».
На рис. 39 “фрактал” получен путем последовательных ветвлений, но
фракталы можно получать и иначе - и путем разрывов, и путем изломов
{fractus по-латыни - сломанный, разбитый). Пример ломанного фрактала -
береговая линия на карте: так, глядя на береговую линию норвежского или
таймырского фьорда, нельзя сказать, каков масштаб карты (по другим эле-
ментам карты это сделать можно). Если выбрать маленький фрагмент бере-
говой линии и «рассмотреть его в лупу» (т.е. на более подробной карте), то
берег будет выглядеть точно так же (в форме столь же ломаной линии),
причем подобную операцию можно последовательно проводить много раз.
Математически говоря, о фрактале идет речь тогда, когда операцию
можно проводить бесконечное число раз. Но так может быть только в иде-
альной модели. В природе так не бывает. На практике всякое увеличение и
уменьшение масштаба имеет предел: глядя на капилляр в микроскоп, уви-
дишь не еще меньшие сосуды, а стенку сосуда; и наоборот - неограничен-
ное увеличение численности, т.е. рост фрактала-древа, на практике ограни-
чено либо ведет клон к гибели. Если быть строгими, то надо говорить о
пред-фракталах, однако мы, как и все прикладники, будем далее называть
их фракталами. На практике процесс конечен, однако в теории, как всегда в
математике, удобнее иметь дело с абстракцией - считать процесс умень-
шения или увеличения бесконечным (таким же путем, как суммы заменяют-
ся в математике рядами и интегралами). Только тогда для фракталов можно
выводить формулы и, в частности - вычислять их размерности.
Пример с картой удобен, чтобы увидеть основное свойство фракталов:
дробную размерность. Обычно мы имеем дело с целыми размерностями
(у точки - ноль, у линии - единица, у поверхности - двойка, у объемного те-
ла - тройка, у процесса роста тела во времени - четверка и т.д.), и вот ока-
зывается, что фрактальные размерности заполняют промежутки между ни-
ми. Так, если вписывать в обычную кривую (например, в окружность) много-
угольники (например: квадрат, 8-угольник, 16-угольник и т.д.), то суммарная
Длина сторон многоугольника с ростом числа его сторон растет, но стремит-
ся к конечному пределу - длине кривой. Если же вписывать ломаную линию
в линию берега, то будет иное: длина ломаной растет с детализацией карты
беспредельно. Если счесть (в идеальной модели!) процесс детализации
бесконечным, то длина ломаной (одномерная величина) тоже будет беско-
нечной, хотя площадь карты (двумерная величина) конечна; данный факт
Удобно описать так: кривая имеет размерность больше единицы, но меньше
Двух. Это и есть дробная размерность.
344
Много разных фракталов можно указать в организме: глядя на ветвя-
щиеся бронхи в легком или на сплетение кровеносных сосудов (рис. 40), то-
же нельзя сказать, каков масштаб рисунка.
Рис. 40. Масштабная инвариантность сети кровеносных сосудов
Дробная размерность позволяет фракталу служить связью структур не-
соизмеримых. Так, тело наше трехмерно, а пронизывающие его капилляры
можно считать одномерными нитями. Как подвести кровь к каждой клетке
трехмерного тела посредством одномерных нитей? С помощью фракталь-
ной сети сосудов, имеющей размерность между двумя и тремя, и капилля-
ров, имеющих размерность между единицей и двойкой. Можно сказать бо-
лее общо: процесс заполнения пространства линиями (фрактальный рост)
дает природе возможность устанавливать связи между пространствами
разных размерностей. В частности - между физиологическими, морфологи-
ческими и экологическими процессами.
345
В системе кровеносных сосудов можно видеть и сращение двух ветвя-
щихся фракталов (артериального и венозного), и новый тип фрактала -
сеть, где важны не ветвление, не изломы и не разрывы, а переплетения.
Это особо интересно тем, что допускает аналогию и с мозгом, и с экосисте-
мой, и с иммунитетом по Ерне. В частности, весьма вероятно, что
мозг столь сложен при столь малом числе генов потому, что в генах
записаны не его связи, а параметры фракталообразующих правил._
Примеры плоских фрактальных форм (в том числе контур спиральной
раковины моллюска) см. [Галимов, 2001, с. 198-204].
Возможно, что феномен самоподобия в биологии шире, чем феномен
фрактала. Для демонстрации значения самоподобия как такового отмечу
поразительные факты одинакового влияния самых разных агентов в самых
разных дозах на самые разные биопроцессы. Если Селье ввел стресс как
сходный ответ на воздействие самых разных агентов, то теперь видно, что
одну и ту же реакцию вызывают и самые разные дозы одного и того же
агента, причем различие доз порой составляет 13 порядков, т.е. 10 трил-
лионов раз. Таковы многие вредные лекарства. См. Дополнение 2.
Подобие большого малому являют и рефренные таблицы - как в живой,
так и в неживой природе. Так, если сравнить рефрен мерона «конечности
позвоночных» (рис. 36) и рефрены химических элементов (Периодическую
систему), то и тут и там видна так называемая вторичная периодичность: в
первом случае это - рефрен «пальцы», а во втором - рефрен «лантаноиды
и актиноиды» (это сказал мне покойный историк химии В.П. Мельников).
6-7* Наследование фрактала и разнообразия
Поскольку для наследования каждого свойства явно не хватает генов,
то ясно, что наследоваться могут лишь общие правила, возможно, содер-
жащие в себе какой-то элемент случайности. Таковыми мне видятся фрак-
талообразующие правила. Именно они, на мой взгляд, и создают все уровни
развития - по крайней мере, выше одноклеточного.
В п. 6-4* шла речь про транпол (странное наследование - не свойств и
не признаков, а их разнообразия). Так вот, наследование фракгалообра-
зующего правила и есть, видимо, механизм передачи разнообразия. В п. 5-
18* говорилось о свойстве морозных узоров - давать крупные рисунки там,
где не видно никакого макроскопического правила. Теперь его можно на-
звать: это правило фрактального роста кристалла льда на плоскости; на-
чавшись по данному правилу, рисунок будет продолжаться в соответствии с
ним. Литературу см. [Чайковский, 1993, с. 111-114; 2004, с. 122-130].
Советую полистать любую иллюстрированную книгу про фракталы, на-
пример: Пайтген Х.-О., Рихтер П.Х. Красота фракталов. М., Мир, 1993. Там
легко убедиться, что простым правилом фракгалообразования можно зада-
вать изумительно сложные фигуры и тела, а совсем простым изменением
такого правила (параметра фрактала) - кардинально их изменять.
346
Постепенно изменяя параметр, на некоторых фрактальных множествах
можно получать длинные серии качественно различных картин, причем
смена картин бывает совершенно непредсказуема и в данном смысле слу-
чайна, хотя ни правило построения фрактала, ни уравнение изменения его
параметров никакой случайности не содержат. Это открывает путь к пони-
манию природы диасети. См. Дополнение 4.
Возможность рождать целые миры заменой одного параметра наводит
на мысль, что в вечном споре о природе математической истины правы сто-
ронники Платона (п. 5-15*), для которых эта истина существует вне матери-
ального мира. Не будучи сам платоником, в одном, данном, пункте я вынуж-
ден признать их правоту, и понятно это стало мне на фракталах.
«Да пропади вся Вселенная, а теорема Пифагора останется верна» -
любил повторять Шрейдер. Это - позиция объективного идеалиста. Субъек-
тивный же идеалист скажет, что теорема возникла вместе с познавшим ее
субъектом, а материалист - что она родилась вместе с первым прямоуголь-
ным треугольником. Каждый выскажет свои “аргументы” (т.е. пояснит своё
убеждение), и советую биологу-практику пропустить всё это мимо ушей.
Другое дело фракталы - здесь понять суть дела биологу необходимо.
Если в математике роль познающего ума обычно столь велика, что
можно допустить (а можно и не допускать) появление истин именно в нем,
то в преобразовании фрактальных картин ум практически не участвует: их
рисует компьютерная программа, автор которой ничего не мог знать о них -
хотя бы потому, что их бесконечно много и они качественно различны. И
уравнение, которое дал программисту математик, может быть просто до
убогости - его можно объяснить даже подростку, чуждому математики (та-
ково уравнение, которое рассматривает Дополнение 4).
Наоборот, сами картины сложны и красивы ошеломляюще. Можно вы-
разить загадочность ситуации и в терминах информации: в уравнении ее
десятки бит, тогда как в каждой картинке - многие тысячи, а качественно
различных картинок - бесконечность. Откуда информация взялась? Единст-
венной разумным видится мне вывод, что она не создается, а читается.
Иными словами, эти картины существуют и всегда существовали вне
нас, а мы только сейчас научились их видеть. Точно так же, сами по себе,
существуют и таблица Менделеева (которую мы лишь дописываем, созда-
вая трансурановые элементы), и рефренные таблицы Мейена.
Надо честно признать: это - платонизм по Любищеву. Даже те, кто яро-
стно протестует против платонизма, в данном пункте смогли до сих пор
лишь выразить своё эмоциональное неприятие. Что делать, мне это тоже
было много лет неприятно, но надо уметь принимать неприятное.
Признав здесь правоту платонизма (т.е. признав данный факт, извест-
ный в теории познания еще с Античности), можно начинать строить долго-
жданную теорию онтогенеза. Строить ее должен эмбриолог-математик, мне
же остается сделать ряд замечаний.
347
Фрактальная структура растет из малого зародыша (способна удовле-
творить тезису Икскюля - п. 5-15), ее рост идет по собственным (морфо-
функциональным, а не генетическим) законам, так что кодировать надо
только переключение режимов роста. На сегодня фрактал - единственное
известное мне теоретическое средство, которым можно описать перенос
больших количеств информации (изменение онтогенеза) через маломощ-
ный канал (наследование), а фрактальный рост - единственная надежда
понять механизм онтогенеза трехмерных форм, ныне в лучшем случае сво-
димый к иносказанию «голограмма» (п. 5-5*). Поэтому будущая теория онто-
генеза включит в себя, смею предсказать, теорию фракталов.
На мысль о фрактале наводит и названная в п. 6-1 многоуровневость
коэволюции: вероятно, эволюция не имеет никакого собственного характер-
ного масштаба в пространстве и времени, т.е. течет сразу в большом и в
малом, так что одни и те же по форме эволюционные процессы могут течь и
в рамках вида, и на уровне класса; и очень медленно (за миллионы лет), и
очень быстро (за считанные поколения, как у Шапошникова), и даже в жизни
особи (таков иммуногенез).
Кроме фракталов-структур, которые существуют в пространстве и лишь
разворачиваются во времени, есть частотные фракталы, существующие
только во времени. Они выражаются в подобии больших интервалов вре-
мени малым (пример см. в п. 6-14). О частотных фракталах в эволюции см.
(Gisiger Т. // BR, 2001, № 2, с. 167). Основная черта каждого из них - отсут-
ствие какой-либо собственной характерной частоты. Когда все возможные
частоты реализуются равномерно, то налицо “белый шум" (полный хаос) ,
но если редкие частоты преобладают, то ищи временной фрактал. Это по-
надобится нам при анализе массовых вымираний в гл. 9.
Авторы АЕ видят в частотных фракталах общую базу теории эволюции,
в том числе биологической, и показывают это, вычисляя моменты появле-
ния крупных таксонов; такие моменты хорошо совпадают с данными реаль-
ной эволюции (АЕ, 295-317), но это не слишком интересно, так как исполь-
зуемое уравнение содержит 4 произвольно задаваемых параметра. Единст-
венное, что видится мне в АЕ серьезным предсказанием, это утверждение,
нто расхождение путей эволюции людей и человекообразных обезьян про-
изошло 10,5 млн лет назад (АЕ, 329). Кстати, ископаемых свидетельств
Данного этапа развития приматов до сих пор нет.
6-7** Фракталы, ДС и катастрофы - три теории или одна?
В АЕ упомянута концепция физика Диого Куэйрос-Конде (Париж):
«Согласно этому автору, эволюция видов не только прерывиста (punctuee) и
фрактальна, но и могла быть основой необозримой турбулентности, содер-
жащей случайную часть и когерентные структуры, той турбулентности, кото-
рая связана с незначительными флуктуациями живого 3,5 миллиарда лет на-
зад в недрах почти однородной биосферы и которая породила, согласно
348
принципу сохранения потока энтропии, сложные организмы, населяющие
нашу планету» (АЕ, 327).
Подробнее см.: Queiros-Conde D. Principe de conservation du flux de
I’entropie pour revolution des especes // CR, Ser. II, 2000, t. 330, № 6). К сожа-
лению, изложение этой концепции выходит за рамки моих возможностей, и
мне остается обратить внимание математически и биофизически квалифи-
цированного читателя на тот факт, что искать природу диасети (рефренной
структуры) надо, на мой взгляд, именно здесь. См. Дополнение 4.
Речь идет о соединении трех физических подходов: турбулентность и
когерентные структуры - это из области стохастической динамики и теории
катастроф, а сохранение потока энтропии - из новой термодинамики. Со-
единение позволяет, на мой взгляд, указать источник активности живого -
он, абстрактно говоря, есть следствие физики сложных систем - точно так
же, как активность неживого есть следствие физики веществ. Добавлен и
математический факт: при реализации фрактала появляется как бы из ниче-
го новая информация. Поэтому именно тут можно ожидать создания единой
теории, которая включит в себя теорию эмерджентности (см. п. 3-10), ту
самую, без которой эволюционизм всегда уступал креационизму.
Если рассматривать эволюцию организмов как перестройку их онтоге-
незов, а каждый онтогенез понимать как фрактальный рост совокупности
ДС, причем акты роста переключаются локальными катастрофами (по То-
му), то открывается необозримое поле эволюционных исследований.
Теперь можно, наконец, выразить то общее, что есть у эволюционных,
экологических и организменных структур, в том числе у мозга и иммунной
системы: их формирование - некое подобие фрактального роста структур;
его результат в целом закономерен, но в деталях не предопределен. Рас-
считывать подобные структуры мы еще не умеем (а возможно и никогда не
сумеем), но польза от них очевидна немалая: они уже сейчас помогают нам
увидеть одну из общих закономерностей развития живого мира.
6-8. Сквозная закономерность эволюции
Почти каждый фрактальный фрагмент строится и соединяется с дру-
гими при содействии некоторого элемента случайности, но не может быть
назван чисто случайным, поскольку встраивается в единую структуру, тоже
в целом неслучайную. Элемент случайности на всех уровнях весьма суще-
ствен - желающему рекомендую статьи: [Голдбергер и др.] (откуда взят рис.
40) и Чайковский Ю.В. Ступени случайности и эволюция IIВФ, 1996, № 9.
Всякое развитие можно рассматривать как многокомпонентный фрак-
тальный рост, а результат развития - как многофрактальную структуру. Тот
удивительный факт, что в итоге развития образуется нечто работающее,
был в главе 5 обозначен как итог активности и давления нормы. Далее, в
п. 6-12, этот факт будет рассмотрен формально, а здесь замечу только, что
феномен взаимной подстройки фрактален в следующем смысле: система с
349
ее нормой состоит из блоков (подсистем), каждый из которых имеет свою
норму, сам состоит из меньших блоков со своими нормами и т.д. При этом
норма системы отнюдь не складывается из норм блоков (п. 5-13*).
Вообще же, можно говорить о подстройке подсистем к системе и утверждать,
что она носит характер компромисса подсистем. Данную процедуру уже 40 лет
(Norris K.S., Lowe С.Н. // Ecology, 1964, № 3) неудачно именуют адаптивным
компромиссом. Адаптивный компромисс недавно назван метафорой [Расницын,
с. 13], что, по-моему, и выражает суть дела: его, как и разные формы отбора, ни-
кто даже не пытался реально обнаружить. Палеонтолог А.В. Марков (2003) на-
звал интересующее нас явление более удачно - регуляторным компромиссом
(см. [Гринченко, с. ПО]). Как понятие кибернетики регуляторный компромисс
вполне реален: он есть норма в излагаемом выше и ниже понимании.
Эволюция, понимаемая таким образом, - не итог отдельных приспо-
соблений видов к своим средам, а единый системогенез (приспособление
целого к самому себе). Примерно так приспособливается сама к себе
мысль, когда из смутной становится ясной, и эта аналогия не мною приду-
мана (Бергсон, Ерне и др.). Каждый блок создан системой фрактальных
ростов и потому сам является переплетением фрактальных фрагментов.
Блоки тоже проникают друг в друга и тем реализуют связь всего со всем.
Это особенно хорошо видно в явлениях, на которых основана восточная
медицина - например, иглоукалывание: укол в заданную точку ушной рако-
вины помогает исцелить определенную болезнь внутреннего органа. Сово-
купность таких фактов позволила группе московских биофизиков говорить о
наличии у человека системы автодиагностики, которая собирает и обра-
батывает сведения, поступающие от органов, и “ставит диагноз”; так что то-
чечная терапия выступает как коррекция данной системы (Чернавский Д.С.,
Карп В.П., Родштат И.В. Биофизическая концепция аутодиагностической
системы человека И Биофизика, 2004, № 4).
Единый блочный системогенез - вот сквозная закономерность
эволюции. В ее рамках реализуется и упомянутый в п. 6-7** акт возникно-
вения нового (эмерджентности). Ее же нередко описывают как смену адап-
тивных компромиссов, но не будем вводить новую метафору, а лучше при-
влечем теорию нормы. Ей посвящена глава в книге [Чайковский, 1990], где
есть и ссылки на литературу. Среди приведенных там пониманий нормы
нам тут удобно такое: нормально то состояние способной к поиску системы,
в котором она выполняет свою функцию, без поиска нового состояния.
Каковы свойства такого состояния? Параметры, задающие поведение
системы, можно разделить на управляемые (их надо “держать в норме”) и
Управляющие (их можно изменять для сохранения значений управляемых).
Так, температура в термостате (управляемый параметр) почти постоянна за
счет управляющих скачков тока в нагревателе.
350
6-8* Многослойная регуляция
Системолог В.Н. Новосельцев в 1973 году показал: если параллельных
связей (переменных величин, влияющих на один и тот же параметр) в сис-
теме много (обычно хватает пяти), то в устойчивом состоянии ее параметры
оказываются равноправными - управляющие изменяются столь же мало,
сколь и управляемые. Таков термостат с грубым и точным нагревателями, с
грубым и точным охладителями и с подвижной теплоизоляцией, т.е. имею-
щий 5 регуляторов (подробнее см. [Чайковский, 1990]).
Когда нагреватель служит отрицательной (стабилизирующей систему)
обратной связью, тогда включенный впараллель с ним охладитель - поло-
жительной (дестабилизирующей) обратной связью. Первая необходима, а
вторая желательна - как усилитель и ускоритель поиска новой нормы.
Состояние равноправия (симметрии) параметров и есть здесь нор-
мальное для системы (замечу, что это - отнюдь не определение всякой
нормы, а свойство данного типа нормы). При смене среды симметрия нару-
шается: управляемые сохраняются за счет смены состояний управляющих.
Если число последних уменьшить, их значения начнут прыгать, чтобы со-
хранить первые “в норме” (хотя это уже не столько норма системы, сколько
ее патология). См. [Чайковский, 1990, с. 129].
Если так, то акт эволюции как акт смены норм происходит не в
ходе приспособления, а при снятии давления нормы (т.е. при разруше-
нии множества связей, которые до этого не удавалось разделить на управ-
ляемые и управляющие), когда открывается возможность для действия ак-
тивности (в данном случае - внутренних законов развития).
Словом, можно считать, что в норме система не ведет поиска, т.е. име-
ет минимум изменчивости. Таков итог многослойной регуляции.
В этих терминах эволюционное приспособление состоит в подстройке
каждого вида не только к среде52, но и к самому себе, к своим возмож-
ностям и к своему "внутреннему импульсу” или “жизненному порыву” (см. п.
6-2). Способность к такой двойной подстройке - это обобщенная адаптив-
ность [Гринченко, с. 23-24]. Слова «к самому себе» можно выразить и ина-
че: «удовлетворяя тенденциям своего таксона». Сами тенденции состоят в
направленном движении вдоль некоторого рефрена и могут не быть адап-
тивными даже в обобщенном смысле.
Например, утеря змеями конечностей выглядит приспособлением вовсе
не к ползанию как освоению какой-то заранее выгодной экологической ни-
52
Поскольку среда может меняться гораздо быстрее, чем идет эволюция вида, приспо-
собление к среде эффективно лишь посредством смены функционирования (в том чис-
ле поведения) его особей. Что касается собственно эволюции (изменения строения ор-
ганов), то она возможна как путем приспособления (адаптации), так и путем преадап-
тации Гп. 4-10) и даже вообще не быть приспособительной (гигантские рога).
351
ши, а к своей, увы, безрадостной тенденции - непомерному росту числа по-
звонков Она образует небольшой рефрен (у рыб, амфибий и рептилий) и
оставляет ее обладателям всего одну возможность двигаться - извиваясь. К
ней они приспособились, избавясь от бесполезных (при столь длинном те-
ле) конечностей. Это коэволюция в рамках одного организма.
Итак, устойчива система с многослойной регуляцией, причем она ус-
тойчива, когда ее параметры симметричны (регуляторы не отличаются по
своему поведению от регулируемых). Многослойность природных систем
обеспечивается той “связью всего со всем”, о которой шла речь в п. 2-3.
При нарушении симметрии параметров (регуляторов и регулируемых)
система эволюирует к новому устойчивому (симметричному) состоянию. Бу-
дет ли это прогресс (усложнение системы и обретение ею новых возможно-
стей) или регресс, заранее сказать невозможно.
Зато можно сказать уверенно (и Новосельцевым было сказано), что
многослойная регуляция в устойчивом состоянии обеспечивает постоянство
потоков через систему, не требуя уставок (уставка - значение регулируемо-
го параметра, устанавливаемое в искусственной системе оператором; тако-
во в термостате значение требуемой постоянной температуры).
Так, организм человека является термостатом, но, насколько знаю, ни-
какого органа, задающего пресловутые “36 и 6” нет: нормальная температу-
ра тела не задается, а вытекает как итог баланса потоков веществ и энергии
через тело. Наоборот, повышенная при болезни температура не бывает по-
стоянной. Это обстоятельство (отсутствие уставок) фундаментально, так как
избавляет природу от проблемы, известной с древности как проблема «кто
будет сторожить сторожей?».
В недавнее время выяснено, что именно так работает генетическая
система: в ней всякую функцию осуществляет своя генная сеть (совокуп-
ность функционально связанных генов), в которой налицо “связь всего со
всем”, т.е. многослойная регуляция. При этом обычно (нормально)
“наличие в генных сетях регуляторных контуров с положительными и отрица-
тельными обратными связями... Благодаря этим двум типам регуляторных кон-
туров возможно поддержание определенного функционального состояния ген-
ной сети или ее переход в другой режим функционирования, в том числе и под
влиянием... среды” (Колчанов НА. и др. Генные сети // МБ, 2000, № 4, с. 543).
Для теории эволюции важно то, что понятие многослойной регуляции
позволяет описывать с единой позиции эволюцию на разных уровнях.
Устойчивость государства тоже понимается в наше время как многослойная
регуляция, хотя была осознана в прошлом иначе - как проблема разделения вла-
стей (п. 4-4), т.е. как проблема баланса (“сдержек и противовесов”). Лучше она
видна как задача регулирования: власть не должна быть вся в одном учреждении,
в одном контуре управления. Иными словами, проблема осознана в рамках ста-
тистической ПМ, но эффективно описана в системной (п. 5-3). Иногда ее пыта-
ются описать в рамках диатропической ПМ (призывы к плюрализму).
352
Сказанное выше позволяет увидеть жизнь как совокупность разно-
масштабных циклов:
1) на молекулярном - рождение и гибель активных форм кислоро-
да и циклы АТФ - АДФ (мы обсудим это в гл. 7);
2) на макромолекулярном - циклы многослойной генной регуляции,
обмен веществ;
3) на субклеточном - например, рост и распад микротрубочек;
4) на клеточном - деление, дифференцировка, апоптоз (п. 6-6**);
5) на ткане-органном - регенерация, циклы активности желёз; цик-
лы многослойной регуляции параметров организма;
6) на организменном - рождение, онтогенез (включая метаморфоз
и смену “хозяев” паразитом), размножение, старение, смерть;
7) на экологическом - сукцессия экосистем (далее, п. 8-13*);
8) на уровне эволюции таксонов - прерывистая эволюция (пунк-
туализм - п. 4-12), “старение таксонов” (броккизм - п. 1-15*).
9) на биосферном - чередование теплых и холодных биосфер (но
смена флор и фаун как правило циклом не является).
Образно выражаясь, эту совокупность циклов можно охарактеризовать
как следствие фрактальной структуры диасети. Последние два уровня и оз-
начают биологическую эволюцию. Хотя данный “фрактал” един, но “физика”
(форма активности), лежащая в основе работы каждого уровня - своя.
Это еще в 1932 году фактически понимал Берталанфи, когда отмечал,
что на каждом уровне организации материи (от электронов до организмов)
«надо ожидать» свою «специфическую закономерность», верную лишь на
данном уровне (Вермель Е.М. История учения о клетке. М., 1970, с. 206).
6-9. Горизонтальный перенос и генное единство природы
Вертикальный перенос - передача наследственной информации потом-
кам, а горизонтальный перенос - передача ее особям того же поколения.
Открытие горизонтального переноса в корне изменило всю эволюционную
проблематику: во-первых, информация может, оказывается, распростра-
няться гораздо быстрее, чем идет размножение, а во-вторых, она может пе-
редаваться между разными таксонами. Вся биосфера выступает как единый
эволюирующий объект не только в том общеизвестном смысле, что всё со
всем связано экологически, но и в новом смысле: в принципе всё со
всем может быть связано генетически без родства.
Возможно встраивание в эвкариотный геном не только вирусов, но и
значительных фрагментов бактериальной ДНК (для бактерии Agrobacterium
см.: Ли А., Тинланд Б. Интеграция Т-ДНК в в геном растений... // ФР, 2000, №
3). Более того, есть мнение, что можно говорить как о реальности и о гори-
зонтальном переносе между высшими животными - в данном случае между
рептилией и млекопитающим (Kordis D., Gubensek F. // PNAS, 1998, vol. 95,
p. 10704-10709). Генетик B.A. Гвоздев добавляет [Геном..., с. 70]:
353
“поедание организмов друг другом может лежать в основе горизонтального пере-
носа, поскольку показано, что ДНК переваривается не до конца, и отдельные моле-
кулы могут попадать из кишечника в клетку и в ядро, а затем... в хромосому”.
Мы уверенно можем включить горизонтальный перенос в число факто-
ров эволюции и даже назвать основной его механизм - это перемещение
транспозонов (п. 5-6) между организмами. Однако не стоит видеть в гори-
зонтальном переносе панацею. Как сказано в п. 5-13, полезные фрагменты
надо сделать, куда-то направить, соединить в систему и заставить новую
целостность работать. То есть горизонтальный перенос не работает без
фактора (или факторов), обеспечивающих целостность.
Сейчас Запад начал осмысливать эволюцию с учетом горизонтального
переноса, но не надо забывать, что это с 1976 года делал киевский генетик
В.А. Кордюм. Он положил это в основу схемы эволюции:
«Развитие генетической инженерии продемонстрировало фактически неогра-
ниченные возможности объединения фрагментов любых геномов [...] Каза-
лось, что такой путь in vivo вряд ли возможен. И тем не менее в лаборатории
Коэна безупречно корректными опытами показали, что... генетическая инже-
нерия в клетках не только возможна, но и осуществляется с относительно вы-
сокой вероятностью, измеряемой тысячными долями процента» {В.А. Кор-
дюм. Эволюция и биосфера. Киев, Наукова думка, 1982, с. 94-95).
Итак, с частотой, близкой к частоте мутаций, происходит обмен фраг-
ментами геное53. То есть изменчивость случайна, но не беспорядочна.
Здесь важны: 1) деление биосферы на два уровня - прокариотный,
служащий источником новых генов, и эвкариотный, потребляющий генети-
ческую информацию для построения сложных структур, причем у эвкариот
«реализуемая в них информация мутационно дорабатывается примени-
тельно к новому молекулярному окружению» (там же, с. 135).
2) новый взгляд на размножение. Низший уровень: для макромолекулы
явный предок - кодирующая ее матрица, но есть неявные предки: молекулы
и процессы, вершащие вставки и вырезки. Уровень бесполого размножения:
одноклеточные размножаются бесполо (один явный предок), но имеют по-
ловой процесс - обмен плазмидами (внутриклеточными органеллами, име-
ющими оболочку и короткую ДНК внутри нее), их поражают вирусы, способ-
ные передавать фрагменты ДНК (п. 5-12). Уровень полового размножения: у
каждого два родителя, но есть и горизонтальный перенос - например, об-
мен вирусами (инфекция), в том числе и между членами далеких таксонов.
К сожалению, ни у Кордюма, ни позже не сказано о том, как появляются
осмысленные конструкции - всё то, на что указывают ламаркизм, жоффру-
изм и номогенез. То есть об активности генетической системы.
53
В п. 5-12 мы уже видели, что направленное наследование происходит достаточно ред-
ко и потому его путают с мутациями.
354
И все-таки горизонтальный перенос понемногу был принят как фактор
эволюции всеми. Соединение генетического поиска с горизонтальным пере-
носом, т.е. активное включение организмом посторонней ДНК, открывает
огромные эволюционные возможности. Становится ясно, почему про- и эв-
кариоты устроены столь различно: первые обеспечивают связь всего со
всем и потому должны быть генетически открыты максимально, насколько
это совместимо с сохранением родовой (даже не всегда видовой) индиви-
дуальности, тогда как вторые обладают сложным онтогенезом и потому вы-
нуждены отделяться друг от друга (и тем более от прокариот) барьерами
несовместимое™, носящей иммунный (точнее, эпигностический) характер.
6-10. Век генетики
Ушедший XX век был веком генетики. Начавшись вместе с нею, он
прошел под флагом ее господства в массовом сознании, причем не только
биологов. Успехи ее огромны, но для понимания эволюции происшедшее
«генетическое переосмысление эволюции» [Гилберт и др., с. 325] принес-
ло, на мой взгляд, лишь вред: было забыто, что эволюируют не гены сами
по себе, а организмы и их сообщества, что их эволюция лишь отображает-
ся в изменении генома.
В 1900 году три биолога (первым был де-Фриз) заявили о переоткрытии
законов Менделя, и с этого момента ведет свою историю генетика. Летом
2000 года три научных центра заявили о расшифровке генома человека,
чем зафиксировали главный результат, достигнутый к столетнему юбилею
генетики. Фактически этим было обозначено, что главный приоритет науки
конца века реализован и что дальше нужны приоритеты новые.
Надежды на эту расшифровку были огромны, хотя скептики и говорили,
что ожидания напрасны. Так, в сущности, и оказалось: ни одна из крупных
проблем биологии и смежных наук с места не сдвинулась - набор генетиче-
ских текстов сам по себе ничего не говорит. Тут самое время вспомнить, что
на это еще 80 лет назад указывал Любищев в своей поразительной, хоть и
небольшой (125 с.), книжке [Любищев, 2004], написанной тогда, когда едва
начал обсуждаться вопрос, является ли вообще ген текстом.
В основе книжки лежит та мысль Гурвича (1910 г.), что менделева гене-
тика являет собою преформизм, тогда как эмбриология демонстрирует эпи-
генез (ср. п. 1-14), и их соединение («проблема осуществления гена») -
труднейшая проблема биологии. Любищев видел возможность решить ее с
помощью представлений Гурвича о биологическом поле, но полвека Люби-
щева не слушали, ибо считалось, что никаких полей нет, а генетика сама
решит все проблемы биологии. Увы, ни «проблему осуществления», ни
проблем эволюции так решить не удалось.
Когда в 1961 году был расшифрован генетический код синтеза белка, то
(как сказано в п. 4-17) многим стало казаться, что генетика, понимаемая как
управление синтезом белков, - сама суть биологии. Лет 20 такая идеология
355
вела к блестящим успехам, и (хотя во многих областях, например в эколо-
гии и психологии, она дала совсем мало) понятно желание счесть генетиче-
ский язык общим, в том числе и описывающим эволюцию, а генетический
код универсальным. Однако код изменчив буквально во всём - как по соста-
ву сахаров (у фагов бывает глюкоза - Кордюм В.А., с. 59) и кодонов (давно
известны минорные основания - Ленинджер А. Биохимия. М., Мир, 1974, с.
284, 298; вероятно, они играли роль в происхождении жизни - см. п. 7-8**, а
одно из них поныне может заменять тимин - п. 5-5), так и по их смыслу (ко-
дон может читаться по-разному, п. 5-13; в особенности это часто у митохон-
дрий, п. 6-5*). Чего-то в проблеме кода мы явно еще не понимаем.
Вершиной генетического подхода к эволюции (и признаком его заката)
была, на мой взгляд, книга [Рэфф, Кофмен]. Ее авторы заявляли, что «эво-
люцию следует рассматривать как результат изменений в генах, регули-
рующих онтогенез». Решить эту задачу им не удалось, зато в книге постав-
лено много интересных вопросов, в том числе старых; ни один из них ответа
тоже не получил, но сами вопросы прелюбопытны, и некоторые я приведу.
Еще в 1874 году, за четверть века до рождения генетики, немецкий эм-
бриолог Вильгельм Гис недоумевал, почему «единственным допустимым
объяснением развития живых существ считается наследственность», а че-
рез 20 лет Вильгельм Ру уточнил - дело в механике. По его мнению,
«все крайне разнообразные структуры многоклеточных организмов можно
свести к... росту клеток, их исчезновению, делению, миграции, активному
формированию, элиминации и качественному метаморфозу».
Ру основал новую дисциплину - экспериментальную эмбриологию, ко-
торую самонадеянно назвал механикой развития (приверженность уста-
ревшей уже тогда механической ПМ) и полагал ее будущим ключом к эво-
люции. С расцветом генетики это было надолго забыто.
Не менее интересны в книге Рэффа и Кофмена мысли эмбриологов,
направленные против учения Геккеля. Так, в 1895 году замечательную
мысль о родословных высказал американский эмбриолог С. Уитмен:
«Нам ничего не дает понимание того, что глаза потому у нас есть, что они
имелись у наших предков. Если наши глаза похожи на их глаза, то это объяс-
няется не генеалогическими связями, а тем, что развитие молекулярной осно-
вы зачатков этих глаз происходило в сходных условиях».
С приходом генетики эти мысли тоже были забыты: все увлеклись ро-
дословными. А ведь это очень странно, поскольку поразительное сходство
глаза осьминога, паука-скакунчика и птицы общими генами не объяснишь.
По-моему, тут надо для начала привлечь не генетику, а диатропику.
Из новых вопросов, поставленных в книге Рэффа и Кофмена, приведу
«С-парадокс»: почему близкие виды могут обладать совсем различным ко-
личеством ДНК, а далекие - сходным? Авторы подробно рассказали, что
более 90% ДНК высших организмов вроде бы ни для чего не используется,
но не смогли сказать о смысле такой чудовищной избыточности ничего,
356
кроме обычного у дарвинистов - что она-де вероятно повышает эволюци-
онную “пластичность”. Но если так, почему избыточность у саламандры и у
лилий в десятки раз превышает человеческую, а у весьма “пластичных” бак-
терий часто отсутствует вовсе? Этого вопроса как раз и не задано.
За прошедшие годы он не раз ставился. Ответа не получено, но осоз-
нана сложная природа С-парадокса (Gregory Т.Н. Variation across amphibian
species in the size of the nuclear genome supports a pluralistic, hierarhical ap-
proach to the C-value enigma // BJ, 2003, v. 79, № 2). Ясно лишь, что наш
обычный подход к ДНК как к тексту не всегда эффективен.
В 1980-е годы эволюционная генетика зашла в тупик, осознаный лишь
в последнее время. Суть его давно предсказал Любищев, говоривший, что
основной прогресс в эволюционизме всегда связан с «преодолением пре-
тензий на универсальность». Генетика хотела объяснить всё из самой се-
бя, но объяснялось не всё. Как бы ни восхищаться успехами генетики, на-
до задавать себе вопросы: каков круг приложимости вновь открытых зако-
номерностей? Что осталось вне его? Их слишком долго не ставили. Те-
перь их начали ставить, и выяснилось, что «вне» осталось очень много.
Грань веков отмечена пониманием этого. Так, Голубовский в сборнике
[ЭБ] подвел итоги некоторым спорам о наследственности. Во-первых, ока-
зывается, наследственность не всегда связана с нуклеиновыми кислота-
ми: прион, переносчик “болезни бешеных коров”, является белком. Во-
вторых, эволюционная изменчивость связана не только с мутациями и ре-
комбинациями, но и с направленным переносом генетического материала
(транспозоны и т.п.), на что, как мы выше видели, прежде безуспешно ука-
зывали многие. В-третьих, сами мутации, главный козырь СТЭ, оказались
зависимыми от условий существования организма. И в-четвертых, приоб-
ретенные свойства оказались кое-когда наследуемыми (см. гл. 5).
6-10* После прочтения генома человека. Тезис Аршавского
Главное прозрение связано именно с полным прочтением генома чело-
века. Получив ворох распечаток, покрытых непонятыми текстами, генетики
наконец стали понимать, что без помощи третируемых “традиционных био-
логов” им не обойтись. Изложу мою надежду на новый статус генетики.
Теперь на ведущее место, прежде занятое генетикой, явно претендует
иммунология: см. беседу с иммунологом В.А. Черешневым «Иммунология -
королева биологических наук» (ВРАН, 2002, № 5). В самом деле, иммунитет
оказался столь же старым и вездесущим, как наследственность (Черешнев:
«Многие ключевые механизмы иммунитета стары, как сама жизнь»)54, да и
сама иммунология подчас указывает путь генетике: вспомним, что открытие
иммунолога Тонегавы было вполне справедливо представлено как “рево-
54 По-моему, лучше говорить о более общей теории (эпигностике), частью которой явит-
ся иммунология (см. п. 5-15).
357
люция в генетике". Но дело не только в иммуногенетике.
Зоолог Д.Л. Гродницкий (Красноярск) выразил уверенность, что
“эволюция требует объяснения в терминах, которые описывают не столько
геном, сколько весь организм в ходе его индивидуального развития и взаи-
модействия со средой обитания. Иными словами, эволюция сопровождается
генетическими изменениями, но не сводится к ним” [Гродницкий, с. 148].
Мысль не нова, ее высказавали Шмальгаузен (1939), Вентребер (1962)
и многие другие, но декларативно. Иначе поступил Аршавский.
И.А. Аршавский был известен как один из основателей возрастной фи-
зиологии, борец за здоровое воспитание детей. В п. 5-11* сказано об их за-
калке, но столь же упорно он добивался грудного кормления с момента ро-
ждения (и добился: уже после его смерти оно разрешено Минздравом).
Для нашей темы главное, что своими опытами, о которых шла речь в п.
5-11, он указал, как конкретизировать декларации о связи генетики и эволю-
ционизма: если крольчонка можно, не меняя его генотип, вырастить похо-
жим на зайца, то эти различия их - не в различии генов. То есть
организм сам ищет и находит нужное изменение своей физиологии,
которому затем подыскивается генетическое выражение; геном является
_____________объектом активности развивающегося организма._____________
Вот этот тезис Аршавского в Заключении его книги:
“Ведущим и первичным в эволюционных преобразованиях является прежде
всего сам организм — его способность осуществлять... адаптивную модифи-
кацию, адекватно соответствующую характеру новых меняющихся условий
среды... в соответствующие возрастные периоды. Именно сам организм явля-
ется субъектом или демиургом эволюции.” Очевидно, что “индивидуальные
адаптации могут возглавить процесс эволюции при условии, что возниющие
модификации сменяются и закрепляются соответствующими мутационными
изменениями” [Аршавский, с. 249-251].
Через 7 лет он уточнил (курсив мой):
«Согласно принятым представлениям геном - субъект, определяющий инди-
видуальное развитие в соответствии с закодированной в нем программой.
Данные наших многолетних исследований позволили прийти квыводу, что
геном является объектом, исполнителем (клавишами), деятельность которого
регулируется через соответствующие эффекторы (гормоны, метаболиты, ме-
диаторы) развивающимся фенотипом» (Аршавский И.А. О физиологических
механизмах преобразования индивидуального развития при одомашнивании И
Сельскохозяйственная биология. 1989, № 6, с. 105).
В этом он был близок к Грассэ (п. 5-5). Поясню: тезис «геном является
объектом» гласит об активном генетическом поиске, который вовсе не сво-
дится к мутациям. Тут стоит отметить, что Аршавский, как и другие физио-
логи, преувеличивал значение адаптации и напрочь игнорировал законы
формы (отсюда, можно полагать, и сомнительная аналогия с клавишами).
358
Доказать тезиса он не мог, и его не слушали, а зря: в те же годы имму-
нологи показали, как активно делаются нужные организму в данный момент
гены (гл. 5, пп. 7, 8); а Шмальгаузен еще в 1939 году приводил список при-
жизненных модификаций, постепенно заменяемых наследственными
(Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.,
Наука, 1983, с. 62). До него это показал Йоллос (п. 5-12, сноска).
В наше время механизм длительных модификаций во многом прояснен
(как и у Йоллоса, на одноклеточных эвкариотах, но не на морфологических
свойствах, как у него, а на биохимических). Как пишет Голубовский, если
подвергать клетку несмертельному яду, то идет
«генетический поиск», в результате которого устойчивость у яду возрастает
от поколения к поколению. При удалении яда из среды устойчивость падает с
поколениями постепенно (это и есть длительная модификация), но «если от-
бор (искусственный - Ю. Ч.) продолжался достаточно долго, часть амплико-
нов (копий гена - Ю. Ч) встраивалась в хромосому», т.е. устойчивость к яду
наследовалась стойко (Голубовский М.Д. И Природа, 2001, №№ 8-9).
Если данный механизм окажется достаточно общим, то тезис Аршав-
ского верен. Однако это еще надо доказать.
Тем не менее, тезис Аршавского уже сейчас позволяет ставить задачи.
В п. 5-15 мы видели странное сходство генов при радикальном различии он-
тогенезов. Идея общего предка здесь ничего не дала, и “каскады генов”,
включающихся друг за другом, процессов развития не прояснили, а тезис
Аршавского оказывается полезным: с его позиции гомеозисные гены сами
по себе не управляют развитием, а лишь включают его этапы.
Но таким знанием о генетике развития наука обладала еще 80 лет на-
зад, т.е. налицо познавательный тупик. Мы как бы копаемся с паяльником и
вольтметром в компьютере, думая, что это лишь очень сложный осцилло-
граф, и наблюдаемые каскады напряжений мало что говорят. Нужен новый
руководящий принцип, новая философия. Суть ее видится мне так.
Очевидно, что генетика развития должна осознать себя как раздел био-
логии развития, поскольку как раздел генетики (такой ее понял Морган) она
зашла в тупик. Пробным камнем должно стать уяснение акта формиро-
вания клетки как акта онтогенеза (пространственного морфогенеза).
Без этого понимание происхождения жизни и ее эволюции вперед не сдви-
нутся (до сих пор, насколько знаю, формирование клетки тоже изучено лишь
с позиции “каскада генов”, а не с пространственной).
С позиции тезиса Аршавского, общие гены говорят не про общего пред-
ка, а про общую задачу, решаемую организмами разных таксонов.
Тут эволюционизм может помочь генетике: в ней есть лишь идея обра-
ботки информации, тогда как он постоянно ставит вопрос - откуда инфор-
мация взялась? Или, образно говоря: генетика видит развитие только как
прорастание семени (по Григорию Нисскому), а в эволюционизме есть, кро-
ме этой идеи, еще идея развития как изобретения (творчества по Страхову
359
- Бергсону). Последняя видится мне как идея активности, конкретизирован-
ная концепцией давления нормы (п. 5-13).
С этой позиции возможно понимание таких явлений, как индуцирован-
ная транспозиция (пп. 5-6, 5-13*), вопрос об информационной сути которой
не ставился. Но гораздо важнее, по-моему, более общее обстоятельство:
винт эволюции (п. 4-13), постоянно крутящийся перед глазами эволюциони-
ста, выпал из поля зрения генетиков. Проницательный Шапошников в свое
время заметил;
“каждый год тли как бы заново осваивают своего вторичного хозяина, посте-
пенно все больше и больше специализируясь на нем и тем самым сужая свою
приспособляемость” (Шапошников Г.Х. И ЭнО, 1961, № 4, с. 751).
Другими словами, последние шаги эволюции еще не попали в постоян-
ную наследственную программу тлей и в каждом онтогенезе работают зано-
во. Генетика весьма далека от описания таких тем, и мне остается напом-
нить, что в феномене редактирования РНК (п. 5-6) удивляет тоже “работа
заново”. И тут, и там видится реализация тезиса Аршавского.
Мы видели, что генетический анализ не проводился, если грозил дать
“не те" результаты (пп. 5-10, 5-11), и примеры можно множить сколько угод-
но. Чтобы начать генетический анализ эволюции, надо покончить с диктатом
века генетики. Признать, что она - лишь раздел биологии.
6-11. Диасеть и структурализм
Завершение века генетики побуждает ученых вновь принять во внима-
ние, что кроме исторических сходств, порождаемых общностью происхож-
дения и функций, есть еще и сходства, в которых законы разнообразия вы-
ступают сами по себе, в чистом виде. Это - сходства диатролические, они
говорят о наличии общности на уровне законов разнообразия, еще слабо
известных, но уже теперь ясно указывающих на ограниченность спектра
возможных форм. К этому надо привыкать: в других науках умеют строить
теорию явлений, основа которых неизвестна (гравитация, психика).
Лучшей иллюстрацией различия исторических и диатропических
свойств была и остается система химических элементов. Будучи сама
рефренной таблицей, чуждой всякой эволюции, она позволяет поставить
вопрос о происхождении элементов, который во всех теориях решается
одинаково: происхождение их явилось заполнением клеток таблицы, объек-
тивно существовавшей всегда. Теории могут различаться только способом
и порядком заполнения клеток (историческими свойствами), тогда как вся
диатропика элементов задана таблицей и никакого отношения к истории не
имеет. Самая первая теория утверждала (как, кстати, и в биологии), что все
химические элементы произошли из водорода путем последовательного
усложнения (добавления протонов и нейтронов в ядро), но она не смогла
объяснить наблюдаемого разнообразия изотопов и была отвергнута, усту-
пив место (как и в биологии) теориям блочным.
360
Понимание эволюции как преобразования разнообразия, как движения
вдоль рядов, задаваемых самой структурой этого разнообразия (а отнюдь
не средой), иногда обозначают как «структурализм в эволюции».
Основателем структурализма (но не автором термина) может считаться
поэт и морфолог Гёте. Он был чужд идее эволюции, зато стал первым, кто
перешел с обычной анатомической точки зрения на морфологическую; он
мысленно расчленял растение не на те части, которые видны прямо (ко-
рень, стебель, лист, цветок, плод), а на части, «отвечающие его природе».
Такими он счел части, выявляемые в ходе развития растения, в том числе
развития уродств. Гёте писал:
“Мы можем одинаково хорошо сказать, что тычинка является сжавшимся ле-
пестком и что лепесток - это тычинка в состоянии расширения”.
На этом языке он хотел познать то, что ныне именуется структурным
инвариантом (анализ см. Корона В.В. О сходстве и различиях морфологиче-
ских концепций Линнея и Гете //ЖОБ, 2002, № 3).
В 1808 году Гумбольдт, в молодости друживший с Гёте, писал о сходст-
ве элементов культуры у очень далеких (исторически, географически, куль-
турно) народов и полагал, что сходство
“имеет скорее психологические причины и зависит от внутренней природы
наших умственных способностей, чем доказывает тождественность происхо-
ждения народов или древние связи между ними” (Гумбольдт А. Картины
природы. М., 1959, с. 87).
Понимание того, что сходство само по себе вовсе не обязано говорить о
родстве, выражено у Гумбольдта вполне ясно, что для начала XIX века уди-
вительно (основная масса ученых далека от такого понимания поныне).
Другим ранним примером структурализма явилась теория химического
полиморфизма Вернадского, тоже отделившая сходство от родства: в ней
утверждалась «близость форм полиморфных тел, хотя бы и принадлежа-
щих разным системам» (Вернадский В.И. О полиморфизме как общем свой-
стве материи // Ученые записки Моск, унив-та, 1892, выл. 9, с. 17).
В 1917 году примерно то же, что Гёте на растениях, сделал Д’Арси
Томпсон на контурах животных (пп. 5-15, 5-17). Предпринятое вскоре описа-
ние параллельных рядов в эволюционных исследованиях Берга и Соболева
тоже означало шаг к выявлению структурных инвариантов.
Термин структурализм возник в 1930-е годы в гуманитарных науках и
в 1950-е годы привел к трактовке социальной эволюции как реализации од-
них и тех же рядов в различных культурах. Ленинградский филолог В.Я.
Пропп, исследуя изоморфизм сказок, прямо указывал на связь своего мето-
да с морфологией растений Гёте (Пропп В.Я. Морфология сказки. Л., 1928).
Ведущий структуралист, французский этнолог Клод Леви-Строс, сравнивая
мифы разных стран, в 1958 году писал, что его интересует «не универсаль-
ность функции, которая далеко не очевидна... но разнообразие...». В этом
разнообразии он искал инварианты:
361
“Мы хотим лишь извлечь из огромного числа разнообразных данных опыта
превышающих возможности наших наблюдений и описаний, такие посто-
янные величины, которые повторяются в иных местах и в иные времена” (Ле-
ви-Строе К. Структурная антропология. М., 1985, с. 412,21, 77).
В недавнее время мы видим идею структурализма в «автоэволюции»
шведского цитолога (испанца по происхождению) Антонио Лима-де-Фариа.
[Пима]. При всех ее недостатках (автор почти незнаком, как он сам признал
в личных беседах, с эволюционной наукой и потому спорит, словно бого-
слов, с одним только нынешним дарвинизмом), книга являет собой обшир-
ный свод вопросов, на которые в дарвинизме нет ответа. Лима стал строить
свое эволюционное учение, не подозревая, что повторяет простейшие логи-
ческие ходы структурализма вообще и номогенеза в частности.
Ближе всех из известных мне натуралистов к структурализму гуманита-
риев приблизились блестящие (увы, умершие в расцвете сил) ботаники С.В.
Мейен и В.В. Корона (им обоим посвящена книга ЧЭ). Они прямо сопостав-
ляли структурные инварианты растений с грамматическими правилами в
языках, что, как показано в п. 6-3**, работает и на зоологических рядах. Ин-
вариантом структуры у Мейена служит архетип (МС, 44); если понимать ар-
хетип не только как план строения, но и как план развития, то видим сбли-
жение с морфологией Гёте.
Другим инвариантом структуры является модуль в концепции модуль-
ной организации. Согласно этой концепции все организмы делятся на уни-
тарные и модульные, т.е. состоящие из однотипных частей (модулей). К мо-
дульным относятся все колониальные животные, все высшие грибы (то, что
в обиходе именуют словом «гриб», т.е. плодовое тело, есть модуль на об-
щей грибнице, тоже состоящей из однотипных нитей) и большинство расте-
ний; ведь обычно растение состоит из множества ветвей, листьев, цветков и
пр. Всё это - модули. Для модульной концепции характерно
“использование новых понятий специально для обозначения общих призна-
ков в конструкции организмов вне зависимости от их систематического по-
ложения и анатомического сходства” (Марфенин Н.Н. Концепция модульной
организации в развитии//ЖОБ, 1999, № 1, с. 8).
Само наличие таких сходств - одно из проявлений номогенеза.
“Блочность” строения организмов (п. 6-3*) - перефразировка структура-
лизма, позволяющая выйти за узкие рамки генетического языка. Словом,
термином «структурализм в эволюции» можно характеризовать ту часть
эволюционной диатропики, которая выявляет структурные инварианты
(рефрены) и описывает эволюцию как движение по диасети (заполнение ее
клеток живыми таксонами и и освобождение - вымершими).
Родившись в биологии, структурализм ушел из нее и заново входит в
нее медленно, с трудом, мало задевая англоязычные страны. Просмотр
эволюционного журнала «BJ» выявил полное отсутствие темы рядов Копа -
Вавилова (простейшей иллюстрации структурализма), причем даже там, где
362
она бросается в глаза и даже у авторов, видящих негодность традиции вы-
думывать полезность каждому признаку. Особенно это странно в серии ста-
тей о гнездовом паразитизме кукушек. Например, в статье (Soler J.J. е.а. //
BJ, 2003, v. 79, № 4) приведен параллелизм окрасок яиц кукушек и их
жертв, сказано, что он вряд ли вызван пользой, но в качестве причины опять
предложены одни лишь случайные мутации. См. п. 12-1.
В англоязычных странах обычно понимают структурализм просто как
анализ структур, а не как выявление инвариантов структур. Так, американ-
ский профессор биологии Стэнли Сэлт в докладе, где назвал себя структу-
ралистом, на самом деле ограничился на сей счет фразой «организмы сами
по себе являются структурами», да еще упомянул ДО термодинамики, кото-
рые «стабилизируются генетической информацией». Это не структурализм,
это просто описание всем известных предметов в модных терминах. Зато
именно у Сэлта мы находим ясное указание на платоновский источник
структурализма (о чем обычно молчат):
“Продолжаете ли вы в духе Платона считать, что структуры существуют где-
то в мире независимо от того, обнаружены они или нет, ... вероятно зависит
от вашего типа личности” (Salthe S. Development and evolution as aspects of
self-organisation // Theory of Evolution - In Need of a New Synthesis? Tampere,
Finland, 1993, pp. 6-7, 9).
В самом деле, рефренная таблица предполагается существующей вся
целиком, независимо от того, какая ее часть на сегодня реализована в
природе, точно так же как таблица химических элементов существовала вся
еще до синтеза трансурановых элементов (ср. п. 6-7*). Однако одни ученые
признают это, а другие отрицают.
Для нашей темы структурализм важен в двух планах: во-первых, он за-
дает, образно говоря, ту систему координат, в которой эволюционное изме-
нение можно описывать как движение вдоль рефренной таблицы (см. да-
лее, п. 6-13); а во-вторых, позволяет понять место эволюционизма среди
наук и тем самым - его возможные цели и средства. Мы уже говорили о том,
что идея эволюции выросла из мифа, а анализ мифов - первая задача
структурализма по Леви-Стросу. Есть мнение, что наука вообще вся вырос-
ла из мифов, и даже - что она поныне состоит из них (так, например, считал
Поппер). Насчет всей науки это, видимо, лишь ошарашивающая метафора,
не претендующая на серьезный анализ, а вот насчет эволюционизма так
оно, на мой (и Поппера) взгляд, и есть. Ламаркизм и дарвинизм как ми-
фологемы рассмотрены в работах: [Чайковский, 1994] и ЧЭ, с. 267-268.
6-11* Гиперболы Виллиса
Единственная в структурализме тема, имевшая хоть какое-то распро-
странение в биологии англоязычных стран, это квази-гиперболические рас-
пределения (открыла их в XIX веке итальянская финансовая статистика, а в
XX веке их стали открывать повсюду - см. статью Б.И. Кудрина [Кудрин]).
363
В 1918 году английский ботаник Джон Кристофер Виллис, изучая свой
«Словарь цветковых», обнаружил, что в каждом крупном семействе более
трети родов - одновидовые. Странно: роды для того и вводятся, чтобы объ-
единять сходные виды, почему же так много одновидовых?
Всего в его Словаре описан 12571 род, из которых 4853 (т.е. 38,6%) со-
держат по одному виду. Двухвидовых родов оказалось 1632 (12,9%), трех-
видовых - 921 (7,3%) и так далее - чем больше видов в роде, тем меньше
таких родов. Точнее, распределение родов по числу входящих в них видов
грубо выражается убывающей кривой, похожей на гиперболу.
Зато в каждом семействе оказался род-гигант (более сотни видов), а
два рода цветковых (астрагал и крестовник) содержали более полутора ты-
сяч видов каждый. То же самое Виллис обнаружил и на животных. Это уж
совсем странно - почему не разбить их на меньшие роды? Однако ничего
не выйдет: пробовали, разбиение оказывалось несуразным и отвергалось.
Графики, касающиеся цветковых, некоторых отрядов насекомых и от-
ряда рукокрылых, а также литературу см. [Чайковский, 2004].
Хвосты этих кривых нерегулярны, а середина всегда содержит горб, по-
этому их лучше называть квази-гиперболами. Они обладают общим инва-
риантом: 35-40% родов имеет по одному виду, 12-17% родов - по два вида,
7-11% - по три вида и т.д., тогда как основная масса видов состоит в немно-
гих крупных родах, и непременно найдется род-гигант.
Сходным оказалось распределение родов по семействам: так, цветко-
вых ныне насчитывают 474 семейства, из которых 170 - однородовые
(36%). Хотя видно, что примерно таково распределение и среди высших
таксонов, но явление это до сих пор не изучено.
Поскольку квази-гиперболы являют собой инвариант, они относятся к
структурализму. Высказывалось мнение, что кривые Виллиса являют собой
не биологический, а психологический феномен: отражают не структуру био-
логического разнообразия, а способ классифицирования. Однако квази-
гиперболы обычны и в тех отраслях знания, где о влиянии психики исследо-
вателей говорить невозможно: были открыты аналогичные виллисовым
распределения слов по их длине, букв и слов по их встречаемости в тек-
сте55, двигателей по их мощности, химических элементов по их обилию в
природе, мутаций по их частоте и т.д. — все они тоже распределены при-
близительно гиперболически.
Поэтому квази-гиперболы приходится принимать как природную зако-
номерность. Они встречаются всюду, где налицо сложная система с не-
жесткими связями, и как раз таковы все эволюирующие объекты. Эволю-
ционные квази-гиперболы являют собой элемент нового номогенеза.
™ На этом основании отрицал всякую роль распределений Виллиса американский гене-
тик растений, классик СТЭ Джордж Стеббинс (Stebbins G.L. И Columbia Biol. Ser., 1950,
№ 16). То есть загадочная общность была ему безразлична именно в силу ее общности.
364
Виллис высказал в 1922 году уверенность, что нашел общий закон эво-
люции: распределение видов по родам есть итог процесса, определяемого
“видовой мутацией”, т.е. появления (с заданной частотой) нового вида, и
“родовой мутацией”, т.е. распада вида (тоже с заданной частотой) на два
рода. Это было в духе де-Фриза.
Модель данного процесса предложил приятель Виллиса математик Га-
ролд Юл в 1924 году: это ветвящийся процесс появления видов и родов.
Полученные модельные гиперболы Юла хорошо легли на опытные данные
(см. [Чайковский, 2004]), но биологи отказались даже рассматривать ту
идею, по которой эволюция таксона не зависит от числа особей в нем.
Это - к вопросу о том, какую роль в науке играют факт и теория. Учёный
мир дружно отверг и сами факты, и объяснявшую их теорию, поскольку они
противоречили нарождавшемуся в те годы популяционному мировоззрению
(а оно вытекало из статистической ПМ - см. п. 5-3).
С появлением теории фракталов оказалось, что ветвящийся процесс
хорошо описывается как фрактальный рост (п. 6-7). В частности, в лога-
рифмическом масштабе гипербола предстает как прямая, тангенс угла на-
клона которой есть размерность фрактала; для модели Юла она равна от-
ношению частоты “видовой мутации” к частоте “родовой мутации”.
Квази-гиперболы Виллиса носят не только таксономический, но и эко-
логический смысл. Виллис обнаружил квази-гиперболическое распределе-
ние видов по их численности, а также тот факт, что размер ареала (зоны
распространения) рода растет с ростом числа видов в нем.
В западной литературе квази-гиперболы обычно именуют степенными
законами (power laws). Канадский физик Томас Гизигер заявляет, что, ос-
тавляя в стороне споры об их природе, можно «высказать следующую гипо-
тезу: степенные законы хорошо представляют статистику эволюции видов,
имеющую место на Земле» (Gisiger Т. И BR, 2001, № 2, с. 189). Например,
так распределены ископаемые роды по длительности их жизни.
Уточненю: да, квази-гиперболы Виллиса верно описывают ход эволю-
ции, если понимать ее как фрактальный рост множества таксонов, но вряд
ли они говорят что-то о ее механизме, поскольку подобные статистические
кривые слишком широко распространены в самых разных науках. Виллис не
открыл нового закона биоэволюции, зато оказался одним из тех, кто открыл
новую статистическую закономерность и, более того, - новый класс случай-
ностей.56. Квази-гиперболы очерчивают круг сквозной закономерности эво-
56 Мир случайных явлений делится, в основном, на два типа: имеющие устойчивые часто-
ты, а с тем и вероятности (их условно именуют “гауссовыми”), и не имеющие. Квази-
гиперболические распределения относятся ко второму типу и характерны для явлений,
имеющих историю (п. 5-18). И если мы, встретив в опыте “гауссово” распределение, не
делаем никакого вывода о природе объекта (слишком они обычны у самых различных
объектов), то не стоит рассуждать о природе явления и при виде квази-гиперболы.
365
люции (п. 6-8): если СТЭ работала с отдельными признаками, подчиняющи-
мися обычной (“гауссовой”) статистике и не имеющими отношения к эволю-
ции (п. 6-4), то квази-гиперболы, видимо, имеют. Обнаружив на некотором
множестве “степенную” статистику, мы получаем довод в пользу того, что
множество не собрано как попало, а сложилось в ходе какой-то эволюции.
Но - не довод в пользу какого-либо конкретного ее механизма.
6-12. Приспособленность. Подбор по Богданову
Ну хорошо, скажет благосклонный читатель, разнообразие проявляет
некоторую упорядоченность, не всегда объяснимую через приспособление.
Пусть даже и были правы те, кто (от Ламарка до Любищева) полагал при-
способление второстепенной чертой эволюции, но откуда все-таки берется
сама приспособленность? Со времен Дарвина ее было принято объяснять
как итог естественного отбора, но это объяснение лопнуло.
Попробуем вывести ее из идеи реализации разнообразия.
Прежде всего замечу, что приспособленность - частный случай целесо-
образности. Рене Том в 1972 году отнес ее к великим проблемам биологии,
причем попытку объяснить ее как итог отбора назвал «вялым и совершенно
непроверяемым аргументом» и признал, что проблема «бросает разуму вы-
зов, который заслуживает применения других средств» [Том, с. 207]. Из та-
ких средств* он назвал лишь ту двойственность причинного и целевого опи-
саний, на которую указал еще Сократ (п. 1-6); это важно но недостаточно.
Если идти по данному пути, то начать надо с того, что целесообразность
- частный случай целостности, которая вовсе не обязательно является
итогом процесса приспособления Например, та целостность организма, что
дает возможность вести иглоукалывание (п. 6-8), никем еще не выводилась
из идеи процесса приспособления, однако целесообразна с позиции враче-
вания. Вряд ли стоит пытаться вывести из данной идеи и ту целостность,
которая выступает в кодировании одним геном сразу нескольких белков (п.
5-6) или в работе Toll-белка одновременно в эмбриогенезе и в распознава-
нии опасной бактерии (п. 6-6**). Поэтому начинать понимание приспособ-
ленности надо не с процесса приспособления, а с целостности.
Есть традиция, идущая от Рачинского (п. 2-13) - называть естествен-
ным отбором самоподстройку. К ней принадлежали многие. Например, Ру (о
нем см. п. 6-10) рассматривал эмбриогенез как борьбу и «внутренний от-
бор» частей зародыша. Термин неудачен, но суть достойна внимания: в хо-
де каждого онтогенеза заново происходит подстройка частей к целому.
В 1913-1928 годах традицию продолжил основатель тектологии
(«Всеобщей организационной науки») Александр Александрович Богданов
(настоящая фамилия: Малиновский, 1873-1928), большевик и утопист, фи-
лософ и ученый-врач, погибший в ходе опыта над собой.
Он хотел описать и объяснить феномен организованности (мы теперь
говорим - системности), причем начал с замечания:
366
«Первые попытки точно определить, что такое организация, привели к идее
целесообразности. Понятие организации относилось тогда, конечно, только к
живым существам» (Богданов А.А. Тектология. Всеобщая организационная
наука, т. 1 [1925]. М., Экономика, 1989, с. 112).
Это как раз то самое, что нужно и нам. Богданов отрицал (в отличие от
Любищева) пользу понимания целесообразности как гармонии. В основу
своей концепции он положил идею активности, причем
«организационное исследование совершенно одинаково оперирует и с актив-
ностями человеческими, и с иными активностями или “энергиями”, свойст-
венными другим живым существам и, наконец, процессам неорганической
природы» (там же, с. 118).
Для описания судеб систем он ввел понятие «подбор» (по-моему, у не-
го оно близко латинскому слову delectus и английским the choice и the self-
assembling). По Богданову, подбор действует в любой системе, причем ес-
тественный отбор является тем его частным случаем, когда имеются раз-
множение и наследственость. Богданов был уверен, что специфика данного
частного случая особой роли не играет (с. 190). Его уверенность была осно-
вана на анализе разнообразия систем, показавшем, что все системы ведут
себя некоторым образом сходно. Если так, то и целесообразность есть об-
щее свойство систем (или хотя бы некоторого класса систем), не зависящее
от того, есть ли в данной системе размножение. Состоит же она в том, что
всякая система самосогласованна (подстроена сама к себе).
Читать это трудно, поначалу кажется, что всё это - заурядная игра сло-
вами, попытка заменить понимание введением новых названий (как в дар-
винизме). И изложение у Богданова сходно с дарвинским: огромное число
разнородных слабых примеров, нет никаких рядов данных; хотя многосло-
вие изнурительно, но внятных формулировок и ссылок на литературу почти
нет. В частности, видно незнакомство с трудами Мальтуса и Дарвина: Бо-
гданов отверг первого и признал второго, просто следуя моде.
Поэтому 16 лет назад, когда моя аспирантка Симона Пустильник, готовя
диссертацию о Богданове, советовала мне обратить внимание на его «под-
бор» (ее вариант англ, перевода - tektological selection), меня это не трону-
ло. А в подборе оказалась суть дела, и теперь я ей глубоко признателен.
Действительно, если все системы, от атомов до скоплений галактик,
включая живые, социальные и технические, проявляют самосогласован-
ность, то причину этого следует искать общую. Ее Богданов выводил из
трех посылок: активность всех форм материи, наличие формирующего
принципа и регулирующего принципа. Последний и есть подбор.
Это разграничение, отсутствующее в дарвинизме, делает концепцию
Богданова содержательной, и нам надо сперва рассмотреть его форми-
рующий принцип. Состоит он в следующем.
В системах всех уровней удается выявить три главных свойства: это ин-
грессия, эгрессия и дегрессия. Как могу, объясню их.
367
Ингрессия - это соединение элементов в цепь, древо или сеть. Направ-
ление в каком происходят акты соединения, не играет роли (но, добавлю,
взаимодействие их может быть направленным).
Эгрессия - это наличие в системе какой-то центральной части.
Дегрессия - это наличие у системы скелетных структур. Такой струк-
турой у организма являютя скелет и кожа, у коллектива - занимаемое им
помещение и объединяющая его идея, у государства - его институты и
идеология. Самой общей дегрессией является следующая пара: «простран-
ство и время - скелет живого опыта» (т. 2, с. 135).
Связь этих трех системных свойств при создании организованности та-
кова: «Ингрессия собирает организуемое содержание, эгрессия его концен-
трирует, дегрессия фиксирует» (т. 2, с. 152).
Наличие этого общего инварианта систем говорит о том, что системы
всех уровней образуют рефрен. Теперь вернемся к приспособлению.
Феномен приспособления настолько общ, что ему следует искать общее
обоснование, вне частных учений и прежде всего - вне ламаркизма и дар-
винизма. Эту мысль высказывали многие. Из ранних генетиков, близких
эволюционизму, ее яснее всех высказал Филипченко:
«Процесс эволюции организмов необъясним ни так называемыми факторами
Ламарка, ни подбором (здесь: естественным отбором - Ю. Ч.), а является од-
ной из коренных особенностей живых существ... Допуская возможность све-
дения отдельных жизненных процессов на чисто механические причины, мы
едва ли будем в состоянии объяснить последними жизнь какого-либо орга-
низма’в ее целом» (цит. по: Бабков В.В. // ВИЕТ, 2006, № 1, с. 174).
Признание «коренных особенностей» означало новый (для начала XX
века) взгляд на мир, а он требовал новой ПМ. Для Богданова и Филипченко,
работавших в обстановке господства статистической ПМ, это могла быть,
разумеется, только системная ПМ, едва начавшая входить в научную прак-
тику. В ней можно хотя бы обрисовать проблему самоорганизации (которой
не видно в рамках механики и статистики). Обычно этот шаг видят у Берта-
ланфи, в его «Общей теорией систем» (1940-е гг.), но он понимал систем-
ность как прежде всего биологический феномен (см. [Пуст ил ьн и к]), тогда
как Богданов (еще до него) видел ее всюду во Вселенной и фактически уже
предлагал новые ПМ: изучал разнообразие и указывал на активность.
В биологии всякую целесообразность принято считать итогом процесса
приспособления, но, насколько я понимаю Богданова, у него целесообраз-
ность - один из итогов активности материи на всех уровнях. Если к этому
добавить отмеченную вскоре Циолковским (п. 5-14) самоорганизацию Все-
ленной под действием полей, то получим, что_________________________
________самоорганизация - основное свойство природы на всех уровнях.
Данное положение явным образом противоречит представлениям тер-
мостатики о пассивности природы. Сам же Богданов двинулся дальше: он
368
выявил то, что через 15 лет после его гибели было названо контуром об-
ратной связи. За это его чтут системологи, однако он сделал большее -
ввел понятие двойного взаимного регулирования (бирегулятора):
«бирегулятор есть такая система, для которой не нужно регулятора извне, по-
тому что она сама себя регулирует» (Богданов. Тектология, т. 2, с. 350).
Этот вывод был им скорее прочувствован, нежели обоснован, но после
появления многослойной регуляции Новосельцева (п. 6-8*) бирегулятор
можно уверенно принять. Далее, об активности примерно то же, что и он,
утверждал Бёрджерс (п. 5-2*). А наиболее близки к Богданову идеи, которые
в 1970-х годах высказал Эрих Янч, системолог и натурфилософ (переехал в
США из Австрии); он во многом определил взгляды известной школы При-
гожина (п. 5-14), которая, к сожалению, очень редко его упоминает. Для Ян-
ча феномен ДС являлся лишь частной (хотя и самой в то время яркой) ил-
люстрацией всеобщего феномена самоорганизации.
Янч полагал, что понятие приспособления надо переосмыслить: оно не
может протекать под прямым воздействием среды, поскольку последняя
слишком изменчива (мы об этом говорили в п. 6-1). По Янчу, эволюционный
процесс, который выглядит как движение к заранее выбранной цели, может
в действительности не быть таковым, поскольку протекает не в нашем
трехмерном пространстве, а в многомерном системном континууме, где ко-
ординатами являются температуры, концентрации, связи и т.п. В таком про-
странстве данный процесс может быть обусловлен не целью, а действую-
щей причиной (очевидна параллель с Бёрджерсом - п. 5-2).
Янч, как и Любищев, видел в приспособлении не цель эволюции, а ее
средство, аспект целесообразности, которую надо оценивать не для одного
момента (как в ламаркизме и дарвинизме), а интегрально, за всё время эво-
люции - от биопоэза до наших дней.
Подробнее о взглядах Янча и их дальнейшем развитии можно прочесть
в работах: Chaikovsky Yu.V. То the evolutionary thermodynamics // Lectures in
theoretical biology. Tallinn, “Valgus”, 1988; а также [Чайковский, 1990].
Эту линию рассуждений, идущую от Богданова, суммируем так:
организмы приспособлены к среде обитания не столько вследствие ее
влияния на них, сколько в силу принадлежности всех организмов одному и
_________________________тому же цельному миру.________________________
Для многих ученых данный вывод слишком абстрактен или вообще не-
научен. Однако вновь обратим внимание на то, что у науки три различные
цели: описание, объяснение и рекомендация (п. 5-1). Если первая цель дос-
тигнута и обеспечила третью, то данной наукой можно пользоваться. При-
том ею могут пользоваться все, хотя объяснения многих могут и не устраи-
вать. Для одних вывод (данный в рамке) разумен сам по себе, другие могут
искать его обоснование в механических или идеальных принципах. Такие
ситуации в науке возникали не раз и со временем становились привычными.
369
Например, еще 320 лет назад почти всем было непонятно, как Луна
сцеплена с Землей, ибо идея гравитационного поля отрицалось почти все-
ми. А сейчас оно очевидно, и в нем нет для нас никакой сложности, хотя в
понимании природы гравитации продвинуться и не удалось.
То же было при рождении квантовой физики (п. 5-2) и теории относи-
тельности, когда выяснилось, что принятое понимание строения действия
(т.е. активности) и пространства-времени пригодно лишь для обыденных
масштабов длин, масс и скоростей. Вне их прежняя механическая ПМ не
работает, ее пришлось сменить на системную (и атом, и Вселенная оказа-
лись системами). Ясно, что системой является и любой биологический объ-
ект от макромолекулы до биосферы, однако их понимание придет, вернее
всего, в рамках акгивностной ПМ (п. 5-3), где предпочтительное направле-
ние столь же естественно, как независимость в статистической ПМ.
6-12* Нынешнее состояние проблемы
Ныне идею Богданова продолжает у нас группа ученых (последователи
Шмальгаузена), которая фактически отрицает отбор в обычном смысле
(основанный на избирательной размножаемости), но нигде не обращает на
этот факт внимания. Ее лидер Шишкин (см. п. 5-11*) пишет:
«Материал отбора (пространство аберраций нормы) в шмальгаузеновской
теории не случаен, но статистически детерминирован свойствами системы
развития; действие отбора заключается в преобразовании свойств системы в
сторону стабильного воспроизведения сохраненной аберрации... Отбор в этих
представлениях есть механизм поиска системой нового равновесия взамен ут-
раченного; ... отбор есть лишь биологическое выражение универсального ме-
ханизма преобразования открытых систем путем последовательных циклов
коррекции их состояния».
Далее автор напоминает об основной мысли Шмальгаузена:
«он утверждал, что поддерживая определенный фенотип, реализуемый гене-
тически неидентичными особями, отбор тем самым преобразует генотип их
потомства в сторону все более устойчивого воспроизведения данного феноти-
па... и не заметить ее (мысли - Ю.Ч.) отличия от традиционного понимания
отбора, казалось бы, невозможно» (Шишкин М.А. И Памяти Сергея Викторо-
вича Мейена (к 70-летию со дня рождения). Труды Международной палеобо-
танич. Конференции. М., ГЕОС, 2005, с. 41,43).
Нет (в отличие от концепции самого Шмальгаузена) и намека на отбор
как избирательную размножаемость. По-моему, автору лучше бы отказаться
от термина «отбор» и взять иное слово (как мне пришлось ввести в п. 5-8*
термин «расплод»). Тогда ему не пришлось бы сетовать, что читатели его
не понимают. Здесь подойдет, по-моему, именно богдановский «подбор».
Поведенческое приспособление всегда идет вне смены поколений, но в
биологии немало также примеров структурного приспособления без раз-
множения - приспособления, которое аналогично наследственному. И они
370
показательны: опыт Аршавского, антитела к синтетическим антигенам, ре-
парация ДНК в неделящихся клетках. В последнее время говорят о едином
механизме репарации белков и нуклеиновых кислот, причем называют акт
основной реакции репарации отбором - фактически, это значит, что отбор
понимается в том же смысле, что «подбор» у Богданова, т.е. без размноже-
ния (Беда Н.В., Недоспасов А.А. // Биоорганич. химия, 2006, № 1, с. 20).
Возможность единого механизма репарации столь различных молекул
говорит о наличии весьма общего свойства биосистем. Таковым является,
прежде всего, фрактальная структура клеточной воды. Она служит «строи-
тельными лесами» всякого биосинтеза, долго не привлекавшими внимания
биологов (Бульенков Н.А. И Биофизика, 2005, № 5, п. 3.2).
Возвращается в науку и внутренний отбор в смысле Ру:
«системы отбора очень эффективно борются и с мутациями, которые появля-
ются в половых клетках, именно поэтому их так трудно зафиксировать»; «от-
бор наверняка существует и на стадии мейоза»; «Создается впечатление, что в
новых экологических условиях интенсивность естественного отбора растет.
Мутантные особи при этом не возникают или не выживают»; «адаптация идет
не путем возникновения и накопления новых, мутантных, вариантов генов, а
за счет поиска подходящих для новых условий адаптивных комбинаций»
(Глазко В. И. Чернобыль 20 лет спустя // Природа, 2006, № 5).
Поясню: все упомянутые автором явления происходят либо в одном
(первом) поколении, либо (последняя фраза) проявляются не постепенно, а
сразу - в 26-м поколении. Пафос статьи украинского эколога В.И. Глазко - в
приспособлении организмов к высокой радиации путем самоподстройки, т.е.
отбором у него тоже именуется подбор по Богданову57.
Сюда же относятся и опережающее отражение (см. далее, п. 6-14) и
общее понимание отбора системологом и энергетиком Б.И. Кудриным [Куд-
рин, с. 23-26], и западные: «эволюционная экономическая теория» (п. 3-11),
«замещающий отбор» (п. 5-1*) и т.п. Словом, нынешнее понимание отбора
«скорее соответствует тому, что имел в виду Богданов: отбор вне смены поко-
лений и борьбы за существование, отбор в контексте системного глобального
эволюционизма, рассматривающего мир как единую систему, как взаиморазви-
тие и взаимоприспособление мирового организма» [Пустилышк, с. 30].
6-13. Эволюция как преобразование разнообразия
С точки зрения диатропики процесс эволюции состоит в заполнении
рефренных таблиц, а они существуют, как и все законы природы, самостоя-
5' К сожалению, большинство эволюционистов попросту обходит проблему приспособле-
ния. Достаточно сказать, что в последней сводке (Назаров, 2005] она вообще при изложе-
нии новых учений не упоминается. Как увидим в п. 7-1, сходная проблема встает в космо-
логии, где нет ни размножения, ни наследственности в биологическом смысле этих слов, а
о приспособленности говорить приходится (антропный принцип).
371
тельно, независимо от наличия или отсутствия конкретных видов (точно так
же как теорема Пифагора была справедлива еще до того, как кто-то начер-
тил первый прямоугольный треугольник, и вся таблица Менделеева была
верна ранее, чем был начат синтез трансурановых элементов, ср. п. 6-7*);
но таблица редко бывает заполнена целиком, и какие ее клетки заполнятся
ныне, а какие останутся ждать пустыми, никто сказать не может. В этом
прежде всего и состоит случайный компонент эволюции.
Пусть рефренные таблицы мы целиком выписать не можем, но сама
идея понимать разнообразие как таблицы многое проясняет.
Вот пример: поразительная картина спаривания некоторых глубоковод-
ных кальмаров - самка, созревая, перестает питаться, почти теряет муску-
латуру и начисто теряет все щупальца, а самец, пользуясь ее беспомощно-
стью, распарывает ее кожу и вкладывает в раны свои сперматофоры; после
нереста рыхлая самка всплывает умирать на поверхность, а плотный самец
падает умирать на дно. Можно придумать всему этому какую-то пользу, но
это вряд ли разумно: у большинства видов кальмаров
«самка до самого нереста сохраняет прекрасный аппетит и остается мускули-
стой... Неужели самка дегенерирует только для того, чтобы не оказать сопро-
тивления самцу, раздирающему ей внутренности? ... Мудрость жестокой
любви кальмаров я так и не уразумел (пока?)» (Несис К.Н. Жестокая любовь
кальмаров // Природа, 1997, № 10, с. 103; [Несис, с. 60]).
Оптимистическое «пока» (изъятое из книжного варианта) вполне оправ-
дано: первый шаг к пониманию сделан - автор отказался от придумывания
«мудрости» (от «упражнения красноречия» по Рузвельту), предпочтя срав-
нить типы спаривания разных кальмаров. Рефрен мерона «тип спаривания»
огромен, и публике обычно показывают либо благостные картинки, чем-то
похожие на образцовую любовь людей, либо курьезы - поедание самца
самкой. Дальние закоулки рефрена, где расположены позиции с общим на-
званием «самец калечит самку», освещать не принято. Их можно выделить
в отдельный небольшой рефрен «самец калечит самку».
Еще Усов (о нем см. п. 2-12) описывал жуткую любовь зубров: могучий
старый самец, уже неспособный оплодотворять, отгоняет молодых самцов
от самки и, взгромоздясь на нее, ломает ей хребет огромным своим весом.
Усов счел это свидетельством склонности вида к вымиранию (и оказался
прав в том смысле, что зубр вскоре в дикой природе вымер, несмотря на
все меры охраны), но ведь искалечить самку пытаются быки многих видов.
Разница лишь в том, что виды, у которых эта склонность не наносит чрез-
мерного вреда воспроизводству, выживают, а зубр не смог.
Кстати, сам Несис провел параллель между кальмарами и обычаями у
людей некоторых тропических племён наносить увечья невестам [Несис, с.
59]; этим он как раз и пояснил рефрен «самец калечит самку».
Словом, смысл отдельных клеток рефренной таблицы можно понять
лишь тогда, когда известна достаточно большая часть диасети - иначе эво-
372
люция выглядит нелепой и беспорядочной. А если таблица перед глазами,
то открывается удивительная картина: всякий вид занимает свое место, и
кальмары не исключение. Живое предстает экологическим, физиологиче-
ским и морфологическим единством. О последнем для растений писал Гёте,
для животных - Бюффон, Жоффруа и Тейяр, а выявление Мейеном фе-
номена диасети позволило говорить о морфологическом единстве
природы как целого. Мы легко понимаем, что всякий вид занимает свое
место в экосистеме (а именно - экологическую нишу), но оказывается, что и
его строение задано не менее жёстко.
Примерно то же можно сказать о его функциях. Ульяновский зоолог и
физиолог А.Г. Зусмановский в своей недавней книге развил ту мысль (в
иных терминах то же утверждает тезис Аршавского), что полезные измене-
ния развития обязаны не случайным мутациям, а приспособительным реак-
циям, которые становятся наследственными в ходе генетического поиска
[Зусмановский]. О генетическом поиске см. пп. 5-10, 6-9. У Зусмановского
вид задается своими потребностями58, которые определяют специфику
вида в физиологическом смысле; эволюция при этом предстает как появле-
ние и удовлетворение новых потребностей.
Можно сказать, что вид занимает свою нишу (нишу вообще), которая
состоит из частных ниш. Если в экосистеме вид занимает экологическую
нишу, то в системе возможных форм, заданных рефренами своих меронов,
он занимает морфологическую нишу, а в системе возможных потребностей
- функциональную нишу. Рассмотрим это подробнее.
6-13* Ниша и таксон
Для понимания морфониши сравним ее с эконишей. Во-первых, говорят
об эконише вида - о совокупности условий среды, в которой вид может су-
ществовать; а во-вторых, говорят про общую эконишу всех видов, сходных
по условиям обитания (например: златокрот занимает в Южной Африке
эконишу нашего крота, колибри занимают в Южной Америке эконишу круп-
ных бабочек). Второе понимание дает нам нужную аналогию: морфониша -
это совокупность сходных черт строения. Сирен занял в классе амфибий ту
же морфонишу, что хирот - в классе рептилий; близкие (сходные) морфо-
ниши имеют кит и китовидка, лист и сидящая на нем листовидка.
Общность морфониш (как и экониш) нельзя вывести из идеи родства.
Она в каком-то смысле аналогична сходству форм, как его понимает термо-
динамика (п. 5-14): сходные ДС порождают сходное строение. Иногда такое
СО
Он использует это понятие по П.В. Симонову (1987): «Потребность есть специфиче-
ская “сущностная” сила живых организмов, обеспечившая их связь с внешней средой
для самосохранения и саморазвития, источник активности в окружающей среде» (Зус-
мановский А.Г. Потребности и эволюция // ЛЧ, 1999, с. 89). О потребностях как факто-
ре эволюции писали Э. Дарвин, Ламарк, Кропоткин (пп. 1-16, 1-17, 3-11) и др.
373
сходство может быть связано с горизонтальным переносом генов (напри-
мер, от растения насекомому), но тут сходные гены задают не общие при-
знаки, а сходную пространственную организацию (сходные поля), так что
сам по себе перенос еще ничего не разъясняет. Вернее признать, что нали-
цо диасеть и допустить, что она - самобытная сущность, столь же харак-
терная для биологии, сколь и для химии (п. 6-3**).
Аналогично, функциониша есть совокупность потребностей и средств
их удовлетворения. Пример Зусмановского: перенесение бактерии с одной
среды на другую (п. 5-9) создает новую потребность, а выработка нового
фермента удовлетворяет ее. Одну потребность можно удовлетворить раз-
личными способами - так, дыхание могут осуществлять жабры, кожа, тра-
хеи и легкие; энергетику могут обеспечить как гликолиз, так и дыхание
(окислительное фосфорилирование), и гашение АФК (о нем см. гл. 7).
Четких границ между эко- и функционишей провести нельзя, но разли-
чие этих понятий очевидно на типичных примерах. Так, различие способов
пищеварения у грибов (наружное) и у подавляющего большинства животных
(внутреннее) есть одновременно экологическое и физиологическое; наруж-
ное пищеварение морских звезд и внутреннее - других морских животных,
глотающих ту же пищу, которую морские звезды обволакивают, различают-
ся чисто физиологически; а различие грибов, способных и неспособных
разлагать древесину, чисто экологическое.
Еще сложнее эта граница у растений, где «естественно применение по-
нятия “жизненная форма”, а не “экологическая ниша”, различающихся как
взгляд изнутри и снаружи» [Заварзин Г.А. // Природа, 1990, № 4, с. 119]. Та-
кой подход становится обычным и в зоологии:
«жизненная форма - обобщающая морфоэкологическая характеристика жи-
вотного, дающая представление об облике организма как целого, его месте и
функциональной роли в экосистеме... Понятие жизненной формы много-
аспектно и основало на широком распространении конвергенций и паралле-
лизмов в живом мире, так что неродственные организмы одного таксона мо-
гут иметь одну и ту же жизненную форму достаточно высокого ранга» (Несис
К.Н., Нигматуллин Ч.М. Жизненная форма... // ЖОБ, 2003, № 3, с. 227).
Поясню: здесь жизненная форма понимается как ряд форм и функций,
проходимый организмом в ходе жизни (например, личинка и взрослая фор-
ма или ряд форм паразита в разных «хозяевах»). Далее, «неродственные
организмы одного таксона» означают, что авторы допускают нефилогенети-
ческую систему (см. гл. 10), а «достаточно высокий ранг» - что единая жиз-
ненная форма объединяет разные виды.
Здесь нам удобно будет понятие “ниша вообще”, которое строится на
основе понятия жизненной формы - как «взгляд снаружи» на жизненную
форму. Оно-то и позволит говорить о единстве живой природы вообще. Де-
ло в том, что это единство принимает порой странные и трудно выразимые
формы. Так, давно известен, но почти забыт тот факт, что
1
374
«класс птиц как целое охотится за классом насекомых как целым, невзирая на
окраску, ядовитость и прочие защитные качества конкретных видов на-
секомых». И хотя «ни одна защита не дает преимуществ», однако «мимикри-
рующих насекомых едят, в основном, те виды птиц, которые специализирова-
ны в их поиске; если бы не мимикрия, эти виды стали бы общей добычей и
были бы съедены целиком» [Чайковский, 1990, с. 132-133].
В те (1930-е) годы, когда факт был описан, он был истолкован (и вряд
ли могло быть иначе) в понятиях третьей (статистической) ПМ:
«Если... гибель от хищников пропорциональна численности жертв, то это
лишь показывает, что в природе существует известное равновесие между раз-
личными типами жертв и что разные типы защитных приспособлений при-
мерно одинаково охраняют их обладателей» (Котт X. Приспособительная
окраска животных. М., ИЛ, 1950, с. 198).
Однако в настоящее время он требует экосистемного описания.
Единую нишу в некотором смысле может занимать и целый класс жи-
вотных. Так в п. 8-11 мы узнаем, что класс Bryozoa (мшанки) может вести
себя как целое не только в данный момент, но и в ходе длительной (мил-
лионы лет) эволюции - если являет собой морфоэкологическое единство
(обладает единой жизненной формой). Здесь целостность трактуется тоже с
позиции четвертой (системной) ПМ.
Однако суть дела видится мне в том, что в таких задачах для понима-
ния явления требуется пятая (диатропическая) ПМ. А именно, все разроз-
ненные примеры целостности следует выстроить в ряды, чтобы увидеть
рефрены целостности. Тогда, быть может, удастся понять, как целостные
конструкции преобразуются. См. п. 6-15.
Есть вопрос, который без понятия “ниша вообще” выглядит безнадеж-
ным: почему продолжают существовать “низшие” организмы? Иначе: поче-
му в ходе эволюции одни группы являют неуклонное повышение уровня ор-
ганизации, а другие так и остаются “внизу”?
Ламарк видел ответ в постоянном порождении низших форм жизни из
неживого (см. п. 1-17), и через 140 лет Тейяр вторил ему, делая лишь ого-
ворку: порождение в наше время не наблюдается. Дарвин видел ответ в от-
сутствии потребности к усложнению у организмов, живущих в простых усло-
виях (мы бы сказали - в постоянном наличии экологических ниш для “низ-
ших” форм жизни). Оба ответа ныне надо признать негодными: жизнь не
возникает в наши дни и, надо полагать, не возникает уже как минимум 2
млрд лет; а самые простые из бактерий (молликуты, они же микоплазмы)
могут занимать поразительно сложные среды.
Например, местообитание и свойства одной из молликут вызвали изум-
ление у группы исследовавших их казанских бактериологов:
“Отсутствие клеточной стенки, рекдукция генома и ограниченные метаболи-
ческие возможности не являются для этих бактерий существенным препятст-
вием в преодолении разнообразных защитных систем высших организмов и
375
выживании в неблагоприятных условиях. Уникальным видом микоплазм с
точки зрения адаптивных способностей является Acholeoplasma laidlavii — вез-
десущая микоплазма, обнаруживаемая в почве, компосте, сточных водах, кле-
точных культурах, тканях человека, животных и растений”, причем причины
их процветания неизвестны (Чернов В.М. и др. Адаптация микоплазм... // Док-
лады АН, 2004, т. 396, № 3).
Столь же существенно, что в одной эконише мы часто видим тварей
самой разной высоты организации. Так, одну эконишу могут занимать бак-
терии и плесени, водоросли и водные цветковые, рыбы и морские звери,
колибри и бабочки. Почему, хотя бы, крупные водоросли не вытеснены
цветковыми, растущими тут же?
Ответ видится мне в том, что клетки рефренной таблицы заполняются
вновь и вновь, но не за счет новых актов происхождения жизни, а за счет
непрестанных переходов как вверх (прогресс), так и вниз (регресс) по шкале
уровней организации. До недавних пор этот ответ был чисто умозритель-
ным, и вместо него в науке господствовал противоположный и столь же
умозрительный “закон Долло”, гласивший, что эволюция необратима (п. 4-
17*). Новый анализ вопроса см. [Попов, с. 138-139], где “закон Долло" рас-
сматривается как аргумент в пользу ортогенеза.
Есть работа, утверждающая, что многие (если не все) низшие много-
клеточные несут в своих геномах следы происхождения от более высоко
развитых форм: совокупные данные по строению 18S РНК и по морфологи-
ческим признакам «свидетельствуют о многократном системном регрессе в
эволюции Metazoa» (Алешин В.В., Петров Н.Б. Молекулярные свидетель-
ства регресса в эволюции многоклеточных животных // ЖОБ, 2002, № 3, с.
197). Хотя доводы авторов шатки, поскольку сами по себе сравнительные
данные никогда не говорят прямо об эволюции, но факт повторности появ-
ления сходных таксонов засвидетельствован палеонтологически (см. п. 9-
14) и тоже противоречит “закону Долло”.
В свете сказанного разумнее будет принять, что морфониши и функ-
циониши заполняются столь же регулярно, как экониши. Хотя и там и
тут возможны пропуски: до появления человека пустовала экониша потре-
бителей свободного углерода (каменного угля); более двухсот лет пустует
функциониша морских коров, т.е. морских зверей, потребляющих заросли
крупных водорослей (различие в том, что водоросли так или иначе разлага-
ются, а уголь без людей выпадает из биологического круговорота). До сих
пор пустует морфониша змееобразных зверей.
Дарвин уделил морфологии лишь несколько самых общих фраз, и эта
традиция сохраняется в руководствах по дарвинизму до сих пор. Учебники
морфологии постепенно обратились, благодаря дарвинизму, в интересные,
но бессвязные рассказы о разных органах (крайний пример: Дзержинский
Ф.Я. Сравнительная анатомия позвоночных животных. М., МГУ, 1998). По-
нятие морфониши призвано изменить ситуацию.
376
Ниши заполняются в ходе эволюции, и все три типа частных ниш игра-
ют примерно равные роли. Природа оперирует не отдельными объектами, а
целыми разнообразиями (п. 6-4), и видимо поэтому почти никакие ниши не
остаются навсегда пустыми (без этого “почти” в диатропике почти никогда
не обойтись). Чем больше видов взято для учета, тем полнее параллельные
ряды и составленные из них рефренные таблицы меронов. В любое время
некоторые частные ниши пустуют, но с учетом вымерших групп рефренные
таблицы довольно полны. Такой полнотой воспользовался Вавилов, когда,
обнаружив отсутствие одной-единственной формы ржи в паре параллель-
ных рядов рожь - пшеница, решил заполнить эту пустую клетку (морфонишу
безлигульной ржи) и сумел это сделать (п. 4-9).
Палеонтолог В.В. Жерихин видел сообщество видов через экониши, ко-
торые те занимают (см. далее, п. 11-3**, сноска). Можно с этим согласиться,
но только если иметь в виду нишу вообще.
6-13** Форма и функция
Мейен придумал рефренные таблицы, изучая морфологический мате-
риал. Он полагал, что на их языке можно рассуждать и о физиологии, и об
экологии, но это верно лишь отчасти: да, рефрены для эко- и физиониш
рисовать можно, но суть дела при этом обычно остается за кадром.
Американский эколог Роберт Уиттекер, будучи далек от веры в уни-
версальность схемы древ для форм, решительно возражал против попыток
описать эволюцию сообществ на языке эволюции форм и таксонов:
«1. Настоящее и прошлое сообществ не упорядочиваются экологами в эволю-
ционные древа так, как в некоторых случаях удается распределить сущест-
вующие и вымершие виды. 2. Эволюция сообществ не может быть положена
в основу... их классификации, как она положена (насколько это возможно) в
основу таксономии организмов. 3. Сообщества не отбираются как целые с их
видами... 4. Необходима осторожность при обсуждении эволюции признаков
сообществ как процесса, обособленного от эволюции их видов» (Уиттекер Р.
Сообщества и экосистемы. М., Прогресс, 1980, с. 319).
Это вызвано тем, что в экосистеме
«Объединения одних видов с другими по преимуществу слабы и изменчивы;
эволюция сообщества подобна сетке в том смысле, что в ходе эволюции виды
различным образом комбинируются... в сообществах» (там же, с. 324).
Уиттекер еще не знал, что данный феномен (блочность эволюции, п. 6-
3*) уже обсуждается и в связи с эволюцией форм, но он был прав в том, что
видам гораздо проще, чем органам, поменять связи друг с другом. А это
значит, что сами связи упорядочиваются плохо и потому не могут дать хо-
роших рефренов. Однако яркие параллели известны и тут (например, роль
бабочек и птичек колибри в опылении цветов). Еще ярче параллели в орга-
низации внутривидовых сообществ. Таково похожее на термитник сообще-
ство особей у грызуна по имени «голый землекоп» (п. 6-5).
1
377
Любищев и Мейен считали форму самодовлеющей сущностью, для фи-
зиологов Аршавского и Зусмановского форма задается функцией, а мате-
матик Том дал нечто примиряющее [Том, с. 154-155]:
“главное в генетическом обеспечении животного заключается не в его мор-
фологии — большинство анатомических деталей второстепенны - но скорее в
механике гомеостаза, физиологической регуляции. Именно благодаря этим
механизмам животное может противостоять неизбежным изменениям сре-
ды... Мы придерживаемся здесь ламарковского взгляда... что функция созда-
ет орган или, точнее, что образование органа является результатом конфлик-
та между элементарным функциональным полем и первичной органической
материей, которая ему противостоит и навязывает ему генетически предопре-
деленные способы осуществления”; “наследуются на самом деле верхние ог-
раничения вариаций физиологической активности взрослой особи”.
В наших терминах это значит: функция создает орган не тем, что творит
его сама, а лишь тем, что задает процедуру давления нормы, чем вводит в
оборот новые клетки диасети; а уж они определят новые формы.
Пример: трава дает место ее поедателю, но им могут стать и птица, и
зверь, и тля, и улитка, и гриб, причем форма каждого может быть выбрана
лишь из вариантов, задаваемых его местом в диасети. Не лист, а диасеть
определяет, что зверь будет жевать его зубами, птица - перетирать камня-
ми в желудке, а улитка и гриб - переваривать наружно. И уж явно не лист
задает форму позвоночного, резко отличную от формы улитки.
Словом, форма и функция не сводятся друг к другу, хоть и зависят друг
от друга, поскольку являются двумя сторонами одной реальности - целост-
ности всякой системы. Иногда эта целостность очевидна (клетка, организм,
экосистема), но иногда загадочна: что побуждает виды самых разных таксо-
нов вести себя согласованно? Листорасположения разных видов выстраи-
ваются в ряд Фибоначчи, термиты приспособлены для почти полного их по-
едания, класс птиц согласованно охотится за классом насекомых и т.д. Этот
ряд загадок мы сильно удлиним в п. 6-15.
6-14. Эволюция организмов и эволюция сообществ
Вид имеет эконишу и может, если она меняется не слишком сильно, к
ней приспособиться, но уже о роде это можно сказать не всегда: попробуйте
описать «нишу» рода Canis, куда входят волк, шакалы и все домашние со-
баки; или рода Solanum, где находятся картофель и баклажан. Недаром бо-
таники предпочитают говорить не об эконишах, а о жизненных формах (о
них см. п. 6-13). Из этого следует, что эволюция высших таксонов не может
течь так же, как видовая. Всё чаще пишут, что только в рамках микроэволю-
ции (т.е. внутривидовой эволюции) есть смысл говорить о приспособлении
вида к среде. Двое петербургских биологов отметили, что
«дарвинизм является только разделом более общей теории развития био-
сферы, и поэтому нельзя требовать, чтобы все факты, касающиеся биологиче-
378
ской эволюции, объяснялись в его рамках» (Старобогатов Я.И., Левченко
В.Ф. Эксцентрическая концепция макроэволюции // ЖОБ, 1993, № 4).
По их мнению, макроэволюция обычно запускается актом распа-
да прежней экосистемы и завершается созданием новой экосистемы с
новым набором таксонов. Позже (сб. ЭБ, с. 40-41) они добавили (фактиче-
ски продолжая мысль Копа): главную роль в экосистемах играют хорошо
адаптированные виды (“специалисты”), имеющие большую численность, но
материал для эволюции представляют не они, а малочисленные виды, не
адаптированные ни к какой узкой нише (‘Тенералисты”)59. Тем самым эво-
люция идет путем смены ведущих таксонов - прежние Тенералисты” дают
начало новым “специалистам”, которые широко распространяются.
Добавлю, что новые ‘Тенералисты” появляются с распадом экосистем
за счет снятия давления нормы.
Если дарвинизм исходил из уверенности, что эволюция на всех уровнях
течет одинаково (потому макроэволюция - лишь замедленная микроэволю-
ция, набор актов микроэволюции), если ламаркизм утверждал, что новое
образуется в силу тенденции к развитию, а учения вроде исторической био-
генетики Соболева видели в смене таксонов полный аналог смены поколе-
ний, то во второй половине века положение стало меняться.
Немецкий палеонтолог Отто Шиндевольф (который еще в 1950 году
был уверен, как Брокки и Соболев, что всякий таксон проходит, в силу внут-
ренних причин, стадии юности, зрелости, старости, смерти) в 1963 году вы-
ступил со статьей «Неокатастрофизм?». Там на огромном ископаемом ма-
териале он показал, что причины эволюции бывают не только внутренние,
но и внешние, что наибольшее количество новых таксонов высокого ранга
появляется сразу же после вымирания предшественников (рис. 41). Всё это
Шиндевольф истолковал как заполнение экологических ниш, опустевших в
силу катастроф, разрушивших прежние экосистемы. Причину древних эко-
логических кризисов он видел в космических излучениях (Ивановский А.Б.
Палеонтология и теория эволюции. Новосибирск, Наука, 1976, с. 14, 49).
Ныне можно привести много иных причин (см. далее, п. 9-7).
Поскольку вновь возникают не прежние виды, а новые (обычно, сложнее
организованные), то они и организуются по-новому, в новые экосистемы.
Новые и старые таксоны пронизывают друг друга неповторимо сложной се-
тью взаимодействий, которая с каждой новой катастрофой еще усложняет-
ся, т.е. эволюирует. Биосфера, если изобразить ее схемой, похожа на
59 С иной позиции нечто сходное увидал Бауэр: “Материал для эволюции поставляют не
победители в борьбе за существование, а побежденные” (Бауэр Э.С. Теоретическая
биология. М.-Л., ВИЭМ, 1935, с. 201). Напомню, что Стегний тоже указал на малочис-
ленность как на необходимый компонент видообразования (п. 5-5*).
379
Перкъ Триас
— FusuLlnLda
Streptelasmatina ColumnorUna Astrocoentina Fungilna Paulina Trlloblta Eurypterlda Beyrich Lida Leperdltilda Thysanura Paloeodlctyopterv r'Jsgasecoptera Protohemlptera Orthoptera Untonacea Carditacea Cardiacea Myacea Ostreacea BeUerophontacea Platycercrtacea SubuUtacea PateUacea Trochacea Littorlnacea Csrithlacea Natlcocea Solenochillda 6onujtLti.no
—
PhyUocerattna Trepostomata Cryptostbmata DalmaneTlacea Productacea Blastoidea Inadunata FlexiblUa Camerota Articulata Rhachrtoml Stereospondyl. Ichthyosaurla Sauropterygla Rhynchoceph. Squamata Archosaurla Pelycosaurta J Ictldosaurla
—
Isoptera
Aptianip-t era
Strepsiptera
Dretssenslacea
Rudlstae
Euomphalacea
Trochonemat.
Nerineacea
Pyramldellacea
Atlarrtacea
Lamellarlacea
Splratellacea
Phyllocerattna
Lytoceratlna
Ammonltlna
Belem raided
Pycnodontoldea
Aspldorhynch.
Urodela
Ichthyosaurla
Saurepterygla
Mesosuchla
Pterosaurta
Theropoda
Sauropoda
OmlthletHa
Odonthognathas
Pnlaeognathae
coenolestoldra
Inseotivora
Chlrqptera
Taeruodorrtla
Primates
Carnivora
Strenta
Condylarthra
LLtepterna
NdtounguUrta
Perisscdactylo
Arttodactyla
Ejferrtata
Rodentla,
Рис. 41. Вымирание крупных групп животных на границах геологических эр (сле-
ва - между палеозоем и мезозоем, справа - между мезозоем и кайнозоем)
фрактальную сеть нервов или сосудов. Опять параллель биосферы с орга-
низмом и большого с малым, медленного с быстрым. Таких параллелей
много. Например, можно рассмотреть заселение древних экосистем после
крупных вымираний с заселением нынешних городов птицами, прежде не-
городскими. Там - миллионы лет и континенты, тут - десятки лет и неболь-
шие долины. Но застройка прежних угодий - тоже катастрофа (для птиц). И
тут и там налицо снятие давления нормы, и закономерности выявились
примерно одни и те же.
380
Правда, птицы не сумели за столь короткое время стать новыми видами
по строению, но по образу жизни успели стать даже новыми семействами и
даже отрядами: голуби научились сидеть на ветках, перелетные стали
оседлыми, а самые умные (врановые) освоили новые способы охоты и пи-
тания - ловят рыбу, словно чайки, и клюют рябину вися вниз головой, слов-
но попугаи. Главное же - городская фауна ведет себя как экосистема, и, в
частности, старые европейские города, подобно сложившейся природе, со-
противляются новым пришельцам, а новые (американские) не могут. Разу-
меется, резко возросла изменчивость организмов и стало много уродливых
особей (Вахрушев А.А., Раутиан А.С. Исторический подход к экологии со-
обществ//ЖОБ, 1993, № 5).
Запомним это: эволюция может течь очень быстро. Город от города
может сильно отличаться по разнообразию птиц, и авторы резонно пишут:
можно, конечно, искать особую причину для каждого города, но это нера-
зумно, ибо «в целом ситуация удивительно аналогичная дарвиновской не-
определенной изменчивости». Не спорю и добавлю: отказ объяснять каж-
дый факт его особой причиной - первый шаг к диатропике, призывающей
начинать не с объяснения, а с упорядочения.
В согласии с п. 6-7 замечу: если в подобии большого малому (в мас-
штабной инвариантности) видеть фрактал, то тут должен быть фрактал
временной. Такие фракталы известны в физиологии - например, сердечные
ритмы [Голдбергер и др.]. В связи с этим сошлюсь на обзор «Масштабная
инвариантность в биологии: совпадение или след общего механизма?», где,
в частности, сделан вывод: «Экосистемы представляются в высокой степе-
ни масштабноинвариантными» (Gisiger Т. // BR, 2001, № 2, с. 204). Следова-
тельно, эволюцию биосферы надо представлять не как взаимодействие за-
данных объектов (экосистем), а как единый процесс взаимопроникновения,
сходный с прорастанием кровеносной системы сквозь все ткани организма
(рис. 40), т.е. как пространственно-временной фрактал.
Это дает новую картину мира (п. 5-16): если в п. 6-3** было сказано,
что разнообразие увязано в диасеть, если в п. 6-8 мы видели, что в орга-
низме всё увязано со всем, причем подчас самым немыслимым с позиции
физиологии образом, то теперь можно добавить: всё живое связано в еди-
ную сеть - как в организме и в биосфере, так и в структуре разнообразия ор-
ганизмов и их сообществ. Соответственно, эволюция предстаёт как услож-
нение этой сети. Тем самым, продолжена идея эволюции мира как целого
(Гердер, п. 2-2; Страхов, п. 3-13; Дриш, п. 3-14; Мейен, п. 6-3** и другие).
Взгляды сверху (с позиции эволюции биосферы) и снизу (с позиции
жизни организма и эволюции вида) попробовал сочетать Зусмановский в
докладе: «На пути к новому синтезу» (ЛЧ, 2000). Отправным пунктом ему
послужил «новый синтез» Гилберта, Опица и Рэффа (см. п. 5-14*, 5-17).
Главной особенностью поведения высших животных он считает ту, ко-
торую физиолог П.К. Анохин в 1962 году назвал опережающим отражением
381
действительности. Зусмановский провел параллель между таким пове-
денческим актом и преадаптацией, т.е. эволюционным актом. По Зусманов-
скому, вид эволюирует не сам по себе, а как часть биосферы:
“Онтогенез биосферы обусловлен эволюцией организмов на основе создания
предшествующими формами жизни среды обитания для последующих
форм”, причем “организмы, непосредственно участвующие в изменении па-
раметров биосферы, первыми это испытывают на себе и в недрах своих гено-
типов и фенотипов формируют предпосылки (преадаптации) к возникнове-
нию новых вариантов изменчивости. Действует механизм опережающего от-
ражения действительности... Функциональные системы Анохина, вероятно,
эквивалентны морфогенетическим полям в фазе генетического поиска” (ЛЧ,
2000). См. также книгу [Зусмановский].
Легко видеть, что «в недрах генотипов и фенотипов» автором фактиче-
ски подмечен богдановский подбор, что в корне отлично от дарвинизма, и
мне пришлось (октябрь 2006 г.) обратиться к автору с вопросом: почему у
него на обложке заявлена правота Дарвина? Александр Григорьевич отве-
тил, что имел в виду медленное постепенное видообразование.
Еще от дарвинизма Зусмановский взял «непрогнозируемость эволюци-
онно значимых потребностей» (можно бы сказать более общо: случайность
как одну из причин изменчивости), от ламаркизма - НПС и активность особи
как факторы эволюции, от номогенеза - гомеозис по Балкашиной и поле по
Гурвичу. Он упомянул и «эмерджентность эволюции» (о ней см. п. 3-10).
Это взгляд снизу. Взгляд же сверху - в том, что каждый вид должен
вписаться в биосферу, состоящую из таких же, как он, видов (а не «приспо-
собиться к среде», которая при этом мыслится заданной). От себя (и, до-
бавлю, Бёрджерса) он ввел в свою модель ту идею, что биоинформация
возникает «в опережающем режиме» и «предопределяет свойства организ-
мов». Этим, на мой взгляд, Зусмановский пришел к подбору по Богданову.
Синтез у Зусмановского состоит, прежде всего, в осознании того, что
каждая концепция отражает свой аспект (угол зрения) процесса эволюци-
ии, а вовсе не описывает (как многие думают) какие-то отдельные явления.
Это радикально отлично от тех попыток синтеза, где в основу положен дар-
винизм, к которому добавлен какой-то еще принцип. У Гилберта с соавтора-
ми и у Гродницкого данный принцип - изменение развития зародыша как
фактор эволюции, т.е. идея Этьена Жоффруа.
По-моему, подходом Зусмановского принцип синтеза исчерпывается, и
дальше нужны шаги в новых направлениях, еще никем не пройденных. Пер-
вым таким шагом видится указанная выше сквозная закономерность эволю-
ции (п. 6-8). В частности, примеры действия во вред себе и во благо целому
образуют внемасштабные ряды на всех уровнях, и теория должна учесть
это. Сюда же следует отнести и все примеры морфологического единства
(п. 6-13). Других шагов потребуют эволюция инстинкта (п. 6-15), эмерджент-
ная эволюция (п. 11-2*) и филоценогенетика (п. 11-3**).
382
6-14* Образование видов и остальных таксонов
Аккуратные примеры эволюции (начиная со Шпренгера, п. 3-12, и кон-
чая Стегнием, п. 5-5*) суть примеры видообразования. Из этого вовсе не
следует, что род и прочие таксоны образуются путем серии видообразова-
ний. Коп (п. 3-10), Аршавский (п. 5-11) и Виллис (п. 6-11*) предлагали свои
механизмы родообразования, а симбиогенез (п. 3-13*) - даже образования
высших таксонов. Однако модель видообразования нужна любой теории.
Вид от вида отличается связным устойчивым чётко различимым набо-
ром свойств - таково единственное известное мне практически работающее
подобие определения вида. Кроме него, существует простое и внешне кра-
сивое “определение”: вид - это множество организмов, плодовитых при
спаривании внутри множества и не дающих плодовитого потомства при
спаривании с членами иных множеств. На практике им пользоваться почти
никогда не удаётся, поскольку существуют межвидовые и даже (изредка)
плодовитые межродовые гибриды, а сама процедура спаривания для по-
давляющего большинства исследуемых пар не проводилась (для вымерших
и бесполых видов она и невозможна). Однако обычно в качестве процесса
видообразования рассматривают именно возникновение барьера для скре-
щивания популяций исследуемого вида организмов.
Новейшая модель видообразования [Марков, Куликов, 2006] вводит
данный барьер как затруднение в процедуре узнавания партнёра, т.е. (в
наших терминах) как барьер эпигностический (п. 5-15). А именно, узнава-
ние мыслится в ней как оценка генетической близости партнёра сперва в
поведенческом и иммунном (свойства спермы) смыслах, затем в цитогене-
тическом (соединение пронуклеусов) и, наконец, в собственно генетическом
(спаривание гомологичных хромосом) смыслах. Авторы модели упоминают
без ссылок опыты по быстрой, в 10-15 поколений (вероятно - это опыт Ша-
пошникова) потере скрещиваемости и заключают, что
«существует некий оптимум генетического сходства партнеров, при котором
вероятность успешного размножения максимальна». Она «характерна... для
некого промежуточного уровня генетической близости партнеров, который,
вероятно, может смещаться в ... зависимости от условий». «В группах, при-
спосабливающихся к стрессовому фактору, появляется тенденция к эндога-
мии — предпочтению в качестве половых партнеров особей из групп, подвер-
гавшихся воздействию того же самого фактора». «Напротив, в благоприятных
условиях оптимум, вероятно, может смещаться в сторону меньшего сходст-
ва». «Обе системы (поведенческая и иммунная - Ю. Ч.), по-видимому, могут
способствовать видообразованию, обеспечивая в некоторых случаях автома-
тическое зарождение у эндогамии (здесь: инбридинга - Ю. Ч.) у групп особей,
по тем или иным признакам (поведенческим или биохимическим) уклонив-
шихся от других представителей вида» [Марков, Куликов, 2006, с. 135-136;
143, 145].
383
Авторы уверены, что здесь имеет место подбор (в отличие от отбора, упо-
минаемого в остальных местах статьи) и что он ведет к новому виду:
«Опыт выведения новых пород... путем целенаправленного подбора брачных
пар (в частности, с чередованием близкородственных и отдаленных скрещи-
ваний) свидетельствует о том, что такой подбор является мощным средством
быстрого формообразования». Аналогично, в природе «предпочтение отдает-
ся особям с промежуточным (не слишком высоким и не слишком низким)
уровнем сходства, но в определенных ситуациях оптимум сходства, по-
видимому, может смещаться (с. 145).
Не входя в обсуждение фактической обоснованности модели, замечу,
что она согласуется с пунктуализмом (п. 4-12) и с данными Стегния о роли
инбридинга (п. 5-5*). Если заменить стресс на более общее снятие давле-
ния нормы (п. 5-2**), то модель станет достаточно общей.
Авторы уверены, что она приложима к любым организмам, имеющим
половой процесс, т.е. могла возникнуть «на заре жизни». Не спорю, но нам
важно иметь в виду, что фактически она описывает только становление ре-
продуктивного барьера, а что именно будет изолировано от прежней нормы
(новый вид, род или выше), ею не описывается. Ранг нового таксона зависит
от другого процесса - смены норм (п. 5-13*). В опыте Аршавского это новый
род, а о механизме образования высших таксонов можно пока лишь гадать.
Например - допустить, что он един, что
«при некоторых чрезвычайных обстоятельствах, когда в геном интегрируется
P-элемент (транспозон - Ю.Ч.), происходит... реорганизация генома, что и
может привести к порождению новых таксонов» (Корочкин Л.И. Параллелиз-
мы... // Молекулярные механизмы генетич. процессов. М., 1985, с. 143).
Единственное, что видится мне несомненно общим - необходимость ка-
тастрофического падения численности вида, рождающего новый таксон;
возможно, вплоть до потомства единственной пораженной транспозоном
беременной самки, как в опыте Голубовского (п. 5-6).
6-14** Экосистемная эволюция
Во всех явлениях эволюции общей является проблема целостности:
целостность как факт несомненна, однако непонятно, в чем состоит меха-
низм ее действия. Рассмотрим экологическую сторону этого явления.
Ярким примером служит эволюция термитов (п. 4-20). Почему более
жизнеспособные мутанты термитов (а они не могли ни разу не возникнуть в
описанных условиях, идеальных для этого) не распространяются? По-чему
не появляются, то тут, то там, термиты, несъедобные или сохраняющие
крылья на время, достаточное, чтобы улететь с места побоища?
Легко заметить, что половая каста специально и сложно приспособлена
для того, чтобы быть выедаемой: обламывание крыльев обеспечено и их
строением, и поведением (активными усилиями) их обладателей; все воз-
можные средства защиты, имеющиеся у иных особей, у крылатых особей
384
изъяты; они ярки, вкусны для всех и неповоротливы. Вдобавок, у многих ви-
дов термитов есть параллельный способ размножения - без выедания: бес-
крылые слепые половые особи (Брайен М. Общественные насекомые. Эко-
логия и поведение. М., Мир, 1986).
Напрашивается мысль, что эволюцией термитов управляет какой-то
внешний механизм. Нетрудно догадаться, что он - экосистемный.
В пп. 4-20 и 6-5 уже было отмечено, что феномен выедания половой
касты вероятно связан не с общественной организацией (обязательной у
всех термитов), а с экологической функцией некоторых термитов. Она же
вполне ясна: так обеспечивается баланс экосистемы: термитник выбра-
сывает в окружающую среду биомассу, полученную при потреблении под-
земной пищи, и это единственный поток органики из почвы наружу:
«Образование почвы в тропиках, перемешивание ее слоев, круговорот ве-
ществ в тропическом лесу - процессы, определяемые деятельностью терми-
тов. Других почвенных животных в тропических лесах часто и не бывает, но
термиты кишат» (Гиляров М.С. Отряд Термиты // Жизнь животных, т. 3, М.,
Просвещение, 1969, с. 209).
У термитов, живущих на свежем воздухе, ничего такого не наблюдает-
ся, хотя крылья их тоже обламываются, как и у муравьев, усилиями самих
их обладателей (Брайен М., п. 12.2).
Если одной выжившей пары на много тысяч съеденных пар хватает для
существования вида, то нужды в появлении несъедобности нет. Термитник
живет неопределенно долго, иногда столетиями, и в размножении (в смыс-
ле появления других термитников) всё это время не нуждается.
Наоборот: если бы возникла несъедобная популяция, она распростра-
нилась бы, словно стая саранчи, и подорвала базу собственного существо-
вания. Тем самым, термитник вписан в экосистему и является итогом ее
эволюции. Можно, в духе дарвинизма, ввести понятие «экосистемный от-
бор», но надо понимать при этом, что он не направляется избирательной
размножаемостью термитов, а сам управляет ею.
Мы подошли к основной теме: что движет эволюцию сообществ?
Разумеется, введение еще одного типа отбора - самообман, никуда не
ведущий тупик мысли. Однако то упорство, с каким дарвинисты весь XX век
вводили эти типы, заставляет спросить: нет ли в этом ходе мысли какого-то
рационального зерна? И оно, по-моему, есть - они неявно имели в виду бо-
гдановский подбор, который действует независимо от процесса избиратель-
ной размножаемости (п. 6-12). Особенно явствен данный ход мысли в смене
адаптивных норм по Шмальгаузену, где надо было бы «всего лишь» убрать
слово адаптивных (п. 6-4*).
Адаптивна ли норма, никто не знает, да и сам этот вопрос почти никто
не ставит - единственное известное мне исключение принадлежит Мейену
[Системность...]. Зато сам феномен смены норм хорошо известен (п. 5-13*),
и здесь приведу только один пример на экосистемном уровне.
385
Петербургский энтомолог О. В. Ковалев описал процесс распростране-
ния жука-листоеда Zygogramma suturalis, ввезенного из Америки на Кавказ
для борьбы с сорняком амброзией. Распространение жука имело форму уе-
диненной бегущей волны (УПВ) т.е. солитона. Она
«характерна необычной концентрацией насекомых в узкой полосе - до 5000
особей на 1 м2. УПВ очень близка к автоволне горения: в посевах некоторых
культур, засоренных амброзией... в тылу волны амброзия полностью уничто-
жалась. Это напоминало распространение степного пожара», причем «были
постоянны форма и скорость, а также начальный импульс при достижении
некоторой критической плотности, т.е. характерные особенности автосолито-
нов». При этом «формирование УПВ в ряду поколений вызвало появление ле-
тающих форм в пятом поколении после интродукции (вселения - Ю. Ч.). Этот
вид не только не обладал способностью к полету, но и не имел... развитых
мышечных структур для полета. Новая форм полностью заменила предшест-
вующую на всем ареале».
Замечательно, что этот акт экспериментальной и одновременно
природной эволюции сопровождался снятием давления нормы:
«на Северном Кавказе среди 10 млн особей (одного подвида листоеда - Ю. Ч.),
составляющих одну волну, можно было обнаружить не только все формы из-
менчивости, свойственные обоим подвидам, но и обнаружить наборы фенов,
которые никогда не могут проявиться на родине вида» (Ковалев О. В. Универ-
сальная модель эволюции... // ЭнО, 1994, № 4, с. 758-759).
Итак, экосистема может побудить вид к тому же, что делал Шапошников
в опыте (п. 4-18). Жаль, что и тут мы не видим генетического анализа.
6-15. Инстинкт и коэволюция. Где мыслящий субъект?
Метерлинк, признавая поведение насекомых почти нацело инстинктив-
ным, сделал исключение для общественных насекомых. У них
«строгий порядок непрестанно приводил бы к неразрешимым и гибельным
конфликтам... и здесь же, как и внутри нас, очень трудно найти зыбкую грань
между инстинктом и разумом. Это тем более трудно, что обе способности, на-
верное, имеют одно и то же происхождение».
Приведя примеры бессмысленной работы инстинктов, он заключил:
«Мы обнаруживаем внутри себя те же чередующиеся примеры разумности и
глупости... Лихорадка... - всего лишь защитная реакция нашего организма,
состоящая из тысячи изобретательных и сложных видов помощи. Прежде чем
мы нашли способ нейтрализовать или регулировать ее избыток, обычно она
уносила жизнь пациента быстрее болезни, в которой боролась» [Метерлинк, с.
385-386].
Словом, для него мышление, инстинкт и работа органов (и клеток) яв-
ляли собой три стороны одного сознательного процесса.
Очень странно умение паразитов управлять поведением своих “хозяев”
(жертв). В 1961^1962 годах в Германии и Англии появились сообщения, что
386
личинки паразитических плоских червей заставляют зараженных ими насе-
комых (муравьев и мух) вести себя так, чтобы их съели те животные, внутри
которых личинки могут развиться во взрослые особи. Вскоре из других стран
подтвердили, что это правда. Так, личинка ланцетовидного сосальщика
Dicrocoelium lancetatum, попав в муравья, мигрирует в его подглоточный
ганглий, после чего эти муравьи
«вползают на растения, окружающие муравейник... прикрепляются челюстя-
ми на верхушке растений... и переходят в состояние оцепенения... По мере
понижения температуры количество оцепеневших муравьев постепенно уве-
личивается, достигая максимума к закату солнца» (Анохин И.А. // Доклады
АН СССР, 1966, т. 166, № 3).
Такие муравьи часто поедаются пасущимся скотом, который иначе поч-
ти не имеет шансов проглотить муравья.
Ученый мир этому не верил, что и понятно: разве можно признать, что
сложный поведенческий акт (миграцию в определенный орган жертвы) осу-
ществляет примитивнейший объект - даже не плоский червь, а его еще го-
раздо проще устроенная личинка? Но затем паразитолог Симона Эллюи
(Франция) назвала около 20 таких примеров. Сама она ставила опыты с
рачком-бокоплавом Gammarus lacustris, который вел себя не менее зага-
дочно: зараженный личинкой скребня Polymorphus marilis, он менял отноше-
ние к свету - вместо темных, выбирал светлые места и быстро поедался
водными птицами. Эллюи назвала такое поведение сумасшествием (la tete
a I’envers - буквально: голова наизнанку), а свою статью озаглавила:
«Манипуляция поведением промежуточного хозяина» (Helluy S. Un mode de
favorisation de la transmission parasitaire: la manipulation du comportement de
I’hote intermediare // Revue d’Ecologie, 1983, v. 38, № 2).
Конечно, можно сказать, что прежде паразит использовал разных “хозя-
ев" (тогда его странствия не выглядели столь удивительно), и назвать даль-
нейшее сужение его экологической базы рутинизацией (о ней см. п. 4-17*).
Так сделано в книге [Чайковский, 1990, с. 123], но этим объяснено лишь за-
вершение загадочного процесса. А его становление (выбор паразитом стра-
тегии) требует иного подхода.
Вот пример, где рутинизацией не объяснить ничего: мелкие бродячие
(не строящие ловчих сетей) пауки рода Tidarren интересны тем, что
“у взрослых самцов не два, как положено, а только один палытус (копулятив-
ный орган). Дело в том, что перед заключительной линькой паук, прежде чем
стать взрослым, ампутирует [себе] один из пальпусов” (Михайлов КГ Каст-
рация и половой каннибализм у пауков И Природа, 2002, № 5, с. 81).
Второй пальпус отламывается в ходе копуляции, за счет согласованных
действий самца и самки, и продолжает ритмичные движения, пока паучиха
съедает кастрированного самца.
Фантастика? Да, но не больше, чем всё остальное. Разве эксплуатация
одноклеточными иммунитета высшего животного (п. 8-12) не фантастика? А
387
примеры целостности, перечисленные в конце п. 6-13**? А термиты, обла-
мывающие себе крылья, чтобы быть съеденными (п. 6-14**)?
Мне видится перспективным построение ряда из примеров, сходных по
типу своей непонятности. Паразит, управляющий иммунитетом жертвы или
поведением жертвы, сходны в том, что оба становятся в один ряд с десят-
ками иных примеров, которые никогда не попадают в учебники. А параллель
иммунитетов растений и теплокровных (п. 6-5**) поразительно схожа с па-
раллелью эффекта группы у растений и у животных (п. 4-15). Такой парал-
лелизм параллелей для диасети достаточно обычен.
Сложнее с оторванным копулирующим органом: хотя параллель с дру-
гими оторванными, но работающими органами найти можно60, но единичная
параллель - еще не ряд. Проводя единичные параллели, можно доказать
всё что угодно. (На это указывает системолог Ю.А. Урманцев, многого до-
бившийся анализом параллельных рядов. Литературу см.: ЧЭ, с. 278.)
В таких случаях надо искать новые параллели, чтобы составить ряд. И
он есть: фороциты (клетки-переносчики) губок, кишечнополостных и иных
низших животных доставляют спермин к яйцеклеткам, зародыши — к мес-
там питания и многое другое [Александров, с. 32-41].
Самую удивительную параллель с пауками рода Tidarren являют голо-
воногие рода Argonauta: и тут и там самка съедает отслужившего самца в то
время, когда его оторвавшийся половой орган (у головоногих это изменён-
ная рука) активно выполняет свою работу [Несис, с. 172].
Обще в этих примерах то, что изощренное поведение проявляют не
только “безголовые” пауки, но личинки и даже клетки, явно не имеющие ни-
чего вроде мозга. Поэтому причину приходится искать в чем-то неизвест-
ном. Где именно? Обычная бездумная ссылка на отбор ничего тут не дает
даже с позиции дарвинизма, поскольку сперва надо понять, какой механизм
обеспечивает данное поведение в данном поколении, тогда как отбор при-
влекают для объяснения хода событий в череде поколений.
Мозг гусеницы слишком примитивен, а у руки аргонавта или у личинки
сосальщика его нет вовсе. Но ведь личинка все-таки действует, и ее дейст-
вие как-то (чем-то) направляется. Пусть это и запрограммировано, но где?
Тут надо вспомнить, что умеют действовать даже бактерии, вспомнить об их
поразительной способности активно приспосабливаться генетически.
Представление о бактериях, пассивно ждущих нужных мутаций, оказалось
ложным. Оказывается, они тоже проявляют генетическую активность:
60 Таковы клептокнидии (греч. украденные капсулы): некоторые голожаберные моллю-
ски (например, Aeolis) используют стрекательные капсулы съеденных полипов, причем
капсула не выстреливает в кишечнике моллюска, но непостижимым образом выносит-
ся из его кишечника в его «кожу», где и становится в рабочее положение [Беклемишев,
т. 2, с. 33]. А оторванные органы являют параллель с ядами съеденных муравьев, пере-
ходящими в кожу ядовитых лягушек, не отравляя их (Природа, 2006,.№ 7, с. 30).
388
“концепция постоянства структуры, организации и экспрессии бактериальной
хромосомы претерпела существенные изменения... адаптационные процессы
у бактерий связаны с непрерывной реорганизацией генетических систем”
(МомынапиевК.Т., Говорун В.М. //Генетика, 2001, № 9, с. 1176).
Эволюция смыкается с поведением особи, и естественно вспомнить
всем известные загадки: например, многие камбалы, будучи положены на
шахматную доску нижней стороной тела, изображают ее на своей верхней
стороне. Всё что можно сказать - это тоже эпигностика (п. 5-15), как и ре-
дактирование РНК (п. 5-6), и иммуногенез, которые, по Ерне, являются ак-
тами внутриклеточного мышления. О таком мышлении пишет и цитолог Г.
Альбрехт-Бюлер (Albrecht-Bueler G. // PNAS, 1992, vol. 89, р. 8288-8292; он
же // Cell Motility and the Cytoskeleton, 1998, vol. 40, p. 183-192). Если замы-
сел записан внутри особи, то налицо инстинкт - явление, никого не удив-
ляющее, но тоже по сути загадочное (особенно в отношении бактерий). А
может быть замысел записан вне особи? Тогда генетическая система особи
играет роль приемника внешних приказов.
Данный вариант наиболее фантастичен, но он и наиболее перспекти-
вен, поскольку открывает возможность понимания поведения, полезного не
самому объекту (он при этом обычно гибнет), а системе, в которую он вхо-
дит. Вполне естественна аналогия: нас ведь не удивляет, что клетки, ответ-
ственные за иммунитет, совершают сложные действия и жертвуют собой на
благо организма, частью которого являются. До сих пор для явлений данно-
го круга ни одна теория эволюции не предложила ничего, если не считать
идею “группового отбора”, которая рассыпается даже с позиции дарвинизма
при попытке оценки потребного времени и числа особей.
Сюда же стоит отнести и загадочную ориентацию животных по све-
тилам. Пусть о ее механизмах и спорят, но нельзя отрицать, что
«штурманское искусство членистоногих по своей сложности нисколько не ус-
тупает “навигационным расчетам” перелетных птиц» (Мазохин-Поршня-ков
ГА. Астрономическая ориентация членистоногих // ЭнО, 1961, № 4).
Об эволюционном значении проблемы звёздной ориентации птиц см.
[Чайковский, 1983]. Сходство проблематики в том, что источник разумного
поведения и тут и там ищут в мозгу животного, тогда как сходно ведут себя
и субъекты, мозга не имеющие.
Американский нейробиолог Роберт Сапольски взялся подвести итог. К
примерам вроде приведенных выше, он добавил неожиданные, хотя и всем
известные: вирус гриппа заставляет больного чихать и тем самым распро-
странять паразита, а вирус бешенства с той же целью заставляет своего
“хозяина” (жертву) кусаться. Ни о каком поведении вируса говорить невоз-
можно, а итог тот же. В чем же дело? Сапольски склонен видеть изъян в са-
мой нашей картине мира (о ней шла речь в п. 5-16):
«В сознании многих из нас глубоко укоренилось представление о “направ-
ленном” и “прогрессивном” характере эволюции: беспозвоночные животные
389
примитивнее позвоночных, млекопитающие - самые совершенные позвоноч-
ные, приматы - наиболее продвинутые млекопитающие. Что бы ни бубнил я
на своих лекциях, на эту удочку попались некоторые из моих лучших студен-
тов... Итак, помните: нас окружают существа, способные управлять работой
головного мозга. Микроскопические... организмы, обладающие большей мо-
щью и властью, чем диктаторы и даже нейробиологи» (Сапольски Р. Тарака-
ны в голове // ВМН, 2003, № 6, с. 67).
Он лишь повторил Метерлинка, и нам надо думать дальше самим.
Некую трудность являет “разумный” вирус: он не может действовать,
разносится пассивно, а итог разумен. Может быть, не надо искать смысла в
активности клеток? Нет, надо: они делают то, чего вирус не может.
Так, Вильгельм Хоорст и Гернот Графе (ФРГ) выяснили, что первая ли-
чинка сосальщика, попавшая в брюшную полость муравья, всегда направ-
ляется в его подглоточный ганглий и там гибнет (изредка так делают две
личинки), а остальные остаются в брюшной полости и могут оставить по-
томство (Hohorst W., Graefe G. Ameisen: obligatorische Zwischenwirte des
Lanzettengels (Dicrocoelium dendriticum) // Naturwissenschaften, 1961, Bd 48,
Heft 7, S. 229). Здесь - как у семенных растений (о них см. далее, п. 8-12**):
на попадание в цель тратится огромное число личинок (семян), ибо лучшего
природа не нашла (или так ей нужно - для баланса экосистемы), зато каж-
дая попавшая в цель личинка действует предельно экономно.
Непонятностей очень много, но напомню простой факт: акт мышления
нам тоже непонятен, однако его никто не отрицает. Считается, что это по-
нимание и ненужно для познания эволюции «низших», поскольку они не
мыслят, но ведь «низшие» оказались не столько низшими, сколько в иную
сторону развитыми. Как и мы, они реализуют замыслы, и для уяснения эво-
люции надо сперва понять, как и чем мы сами думаем. Однако до этого
очень далеко, а эволюцию надо как-то понимать уже сейчас.
Мне видятся три пути: либо возможно мышление на внутрикле-
точном уровне (нечто вроде редактирования РНК), либо мыслит субъ-
ект, находящийся вне этих объектов (Гея или нечто более общее), ли-
бо здесь имеет место не мышление, а более общее явление - соот-
ветствие объекта системе по Богданову (п. 6-12).
6-15* Ряды окрасок и мимикрия
На мой взгляд, ни одна из старых теорий эволюции, развитых на основе
изучения одних внешних и внутренних форм, не описывает реального хода
эволюции именно потому, что на самом деле формы не развиваются одни,
без функций и окружения. Даже саму эволюцию форм при этом понять не
удается, что видно из полной неспособности прежнего эволюционизма по-
нять закономерности окраски и прежде всего - мимикрии.
Очевидна тенденция: огромное множество видов, видимо большинст-
во, окрашены сходно с их основным фоном обитания. Это обстоятельство
390
нуждается в объяснении, но никак не методом софистов и Уоллеса, а каким-
то общим и точным методом. Другими словами, нужно строить не отдель-
ные толкования (каждое из них пасует перед вопросом: «а почему у других
не так, почему сходные виды в сходных условиях решают те же задачи со-
всем по-другому?»), а единую теоретическую схему.
Первое, что бросается в глаза - если виды в сходных условиях могут
как обладать покровительной окраской, так и не обладать, значит, она не
обязательна, т.е. можно выжить и без нее. Как сказано в п. 6-2*, все виды
обладают какой-то экологической прочностью, позволяющей им иметь
вредные свойства. Одним из них можно считать вычурную окраску.
Далее, сходные окраски образуют ряды столь правильные, что Вавилов
называл их циклами изменчивости. Как мы видели в главе 4, ряды легли в
основу номогенеза. Для нового номогенеза ним следует добавить более
общую идею блочности (мозаичности) процесса эволюции (п. 6-3*). Она
вполне применима к рисункам на крыльях многих бабочек:
“Крыловой рисунок имеет модульную природу, т.е. развивается и представ-
лен в конкретных семействах определенными блоками элементов”, причем
соответствующие области крыла “видимо, соответствуют морфогенетиче-
ским полям генов развития крылового рисунка” {Артемьева Е.А. Крыловой
рисунок чешуекрылых как комплексный признак // ЛЧ, 2002).
Блочность проясняет идею циклов Вавилова, но ничем не помогает в
понимании гусеницы, похожей на сухой сучок. Вавилов был уверен:
“Особую категорию, не имеющую никакого отношения к закону гомологиче-
ских рядов и вообще к параллельной изменчивости, составляют случаи ми-
микрии организмов к внешней среде, форме листьев, коре деревьев, лишай-
ников и т.д.” [Вавилов, с. 94].
В наше время этот тип мимикрии именуют словом «мимезия». Хотя слу-
чаи мимезии тоже выстраиваются в ряды, однако Вавилов был прав в том,
что мимезия стоит особняком: подражание налицо, но не видно, как свести
его к законам формы. Засохший сучок не является ни функциональным, ни
морфологическим единством (вряд ли даже платоники примут, что он имеет
собственную идею в смысле Платона, которую может считывать гусеница).
По-моему, для понимания мимезии следует дополнить вавиловский номоге-
нез новым пангенезом (п. 5-12*).
Лучше всех понятна мимезия у птиц: у них ППК плывут по зародышу в
кровотоке (а значит, можно допустить достижение гонады любой мигрирую-
щей клеткой), и у них же видна быстрая адаптивная эволюция - как поведе-
ния, так и расцветок. Например, расцветка яиц кукушки имитирует расцвет-
ки 150-ти видов птиц, в чьи гнёзда она кидает яйца, причем новые (имити-
рующие) цвета нанесены у кукушки на скорлупу поверх исконного (голубого),
и это наводит на мысль, что расцветки унаследованы нестойко [Чайковский,
1990, с. 121, 126]. Показано, что самки кукушки активно выбирают гнёзда то-
го вида, чью расцветку наследственно имитируют их яйца, причем наследо-
391
ванне рисунка скорлупы кодировано в непарной половой (W) хромосоме
(Alives J.M., Moller А.Р. И BJ, 2004, v. 82, № 1). Тем самым, самцы не участ-
вуют в наследовании расцветки яиц, она наследуется клонально. Поэтому
тут и стоит искать пангенез.
В недавнее время у млекопитающих установлен сходный факт - попа-
дание СК в головной мозг (вероятно, с кровотоком, см. п. 5-12**), что откры-
вает широкие возможности для исследования пангенеза и у них.
В более сложных случаях уместно дополнить номогенез и пангенез ла-
маркизмом. Ведь мимезия имеет всем известный функциональный аналог:
многие животные меняют окраску (и тем приспособляются к фону) прямо
при жизни (загар у людей, камбала на шахматной доске и т.д.).
К.Н. Несис поражался брачной игрой некоторых головоногих: мелкий
самец “переодевается женщиной” (принимает яркую брачную окраску сам-
ки), легко достигает кладки, ревниво охраняемой крупным самцом, и опло-
дотворяет ее [Несис, с. 98-99]. Вклад его в потомство мал (основная часть
яиц оплодотворена хозяином), но важно другое: половой диморфизм окра-
ски может меняться, словно одежда, усилием воли. Само же «пиратское»
поведение, при всей его экзотике, берется из рефрена (п. 6-4**).
Ранее мне случилось разбирать феномен мимезии [Чайковский, 1990, с.
116-119], и вывод был такой: если признать прижизненную смену рисунка
понятной, то мимезия тоже понятна - как акт рутинизации (п. 4-17*). Само
же понимание прижизненной смены рисунка упирается в понимание психи-
ки, а она может, как мы видели, быть даже внутри клетки.
В итоге видим: поисковая активность особи копирует чужой рисунок, а
затем ее генетическая активность делает рисунок наследственным. И всё
это происходит в рамках, задаваемых диасетью.
6-15** Коэволюция и платонизм
Пора подвести итог разговору о целостности. Она пронизывает мир жи-
вого на всех уровнях и во всех направлениях и всюду задает загадки. По-
пытка выбирать из них самые простые, оставляя остальные без внимания,
приводит науку в тупик. Остается пробовать решать их совместно, и стано-
вится ясно, что как процедура решения, так и ее результат будут сильно от-
личаться от того, что предлагалось ранее.
Целостность видят по-разному. Довольно обычно понимание ее как
первичного свойств систем - так было еще у Богданова и (для живого) у
Берга. Понимают ее и как полезность, и как красоту, и как итог процесса
приспособления. В биологии традиционно преобладает последнее, однако
необходимо заметить, что обоснований нет: ни для одной системы показать
ее становление через процесс приспособления не удается без грубых не-
проверяемых допущений. Поэтому важны все способы понимать.
Понимание системы как полезной (целесообразной) привело Богданова
к понятию подбора (п. 6-12). Что же касается красоты, то, согласно брошюре
392
Линника (о ней см. п. 4-15*), «форма самоценна независимо от своих адап-
тивных приложений. Красота выше пользы». Пусть слово “независимо”
слишком категорично (некоторая связь просто очевидна), но Линник прав в
том, что сведение красоты к одной лишь пользе натянуто. В своей «Книге
трав» (Петрозаводск, 1986) Линник высказался подробнее:
“Интереснейший момент: там, где появляется взаимность, там рождается
прекрасное ... Красота мира во многом обусловлена цветущими растениями...
Явление красоты многопланово. Но все-таки в этом явлении можно выделить
два основных слоя. Первый - красота конструктивная, структурная: она — в
законах упорядоченности вещества, она - в целесообразности живых форм...
Но как бы поверх этого слоя - на его основе и благодаря ему - существует и
другой вид прекрасного. Образован он информационными процессами”. Они
более понятны у высших животных, но и “у растений - казалось бы, про-
стейших существ — тоже возникли информационные структуры! Причем по
своему разнообразию, по декоративным достоинствам они часто не знают се-
бе равных. Необходимо понять величие содеянного растениями: они создали
сложнейшую службу связи” (с. 360-362).
Линник имел в виду не только связь цветков и насекомых, но и влияние
мира растений на мир людей. «Служба связи» мыслится, насколько пони-
маю, на уровне мира идей Платона: активность мира растений восприни-
мается (и миром насекомых, и миром людей) в цепом, как идея.
Такая же активность отмечена для мира насекомых-паразитов: посколь-
ку разные виды парализуют своих жертв различно, но все демонстрируют
поразительное знание анатомии, то в ходе эволюции может передаваться
лишь «общая тема»: «парализовать, не убивая» [Бергсон, с. 183]. По сути,
речь здесь идет о концептуальной активности (п. 5-2).
Наиболее эффектна и в то же время загадочна эта активность у обще-
ственных насекомых. Метерлинк спрашивал об улье и термитнике:
«Кто здесь правит? Кто отдает приказы, предвидит будущее, строит планы,
восстанавливает равновесие, управляет и осуждает на смерть? ... При малей-
шем нападении тревога распространяется, словно пламя; организуется оборо-
на, аккуратно и методично производится срочный ремонт» [Метерлинк, с.
350, 352].
Отнести всё это на счет инстинкта нельзя, даже если счесть, что сам
феномен инстинкта понятен (Метерлинк это справедливо отрицал), ибо и
улей, и муравейник, и термитник нередко ведут себя рационально даже в
необычных, созданных экспериментатором, условиях (см. п. 4-15).
И Бергсон, и Богданов, и Берг, и Метерлинк, и Любищев, и Бёрджерс, и
Линник являют нам варианты платонизма (см. п. 5-15*). Платонизм принято
отрицать (в самом деле, он малопонятен, а многих, в том числе и меня, еще
и отталкивает эмоционально), но взамен предложить нечего. (Подробнее
см. Дополнение 7.) Если же его принять к рассмотрению, то эволюция
предстает как активное движение вдоль диасети.
393
6-16. Избавимся от неоправданных терминов
Многие читатели, дочитав часть 2, удивятся: а где популяции и их ге-
нофонд, где дивергенция и конвергенция, ароморфозы и алломорфозы,
симпатрическое и аллопатрическое видообразование? Неужели автор всё
это отрицает? На подобные вопросы Кордюм отвечал в конце своей книги
так: все эти термины ввести можно, а некоторые даже полезно, но «нельзя
подменять терминами о процессе механизмы самого процесса».
Да, часто термин служит самогипнозу. Так, А.Н. Северцов именовал
важные акты эволюции ароморфозами; в его школе самый важный аро-
морфоз именуют ключевым (развивая идею Майра о ключевых признаках).
«Ключевой ароморфоз - это морфофизиологический признак или свойство,
обретение которого... изменяет взаимоотношение его функциональных под-
систем» (Современная палеонтология. М., 1988, т. 2, с. 104).
Автор этих слов зоолог А.С. Раутиан привел примеры:
«Ключевым ароморфозом в становлении амфибий было, по-видимому, при-
крепление конечностей к осевому скелету независимо от головы», у птиц -
«наличие оперения», а у млекопитающих - «оптимизация силовой обработки
пищи во рту... а также, вероятно, волосяной покров».
По данной идеологии, внешне маловажный признак ведет к перестрой-
ке организма. Объяснения (даже с позиции отбора) тут нет, поэтому даже не
все дарвинисты могут данный взгляд принять. Например,
“можно только удивляться тому, что Северцов... относит к числу ароморфо-
зов изменения в строении зубов, органов дыхания и кровообращения, кожи и
т.д., но не упоминает изменений в структуре и функции репродуктивной сис-
темы” (Иванова-Казас О.М. Эволюционная эмбриология животных. СПб.,
Наука, 1995, с. 292).
Можно добавить иммунитет и многое другое. На мой взгляд, лучше во-
обще избегать термина «ароморфоз» как неинформативного, поскольку
эволюция никак не может идти путем замены свойств (и тем более призна-
ков), она всегда идет путем смены одной работающей (под)системы на дру-
гую. Появление признака может лишь служить маркёром новой системы, но
не более, а для школы Северцова ароморфоз - отнюдь не маркёр.
Так, сплошной покров шерсти или перьев маркирует систему тепло-
кровности, но объектом расплода служит вся система, и ароморфозом ло-
гичнее было бы, по-моему, назвать саму теплокровность.
Трудность, вынудившую ввести понятие ароморфоза, легко увидеть:
«Телевизор имеет несколько программ (дискретных состояний). Управле-
ние... осуществляется одной ручкой, хотя на самом деле в это время изменя-
ется функция (состояние) многих транзисторных элементов. Если бы на-
страивать нужно было каждый из п элементов... число переключений вы-
росло бы до 2" и пользоваться таким прибором практически было бы невоз-
можно. Отсюда следует, что... условием оперативного и эффективного кон-
394
троля системы является свертка информации» (Веселова Т.В., Веселовский
В.А., ЧернавскийД.С. Стресс у растений. М., МГУ, 1993, с. 132).
Словом, вместо ароморфоза лучше говорить о свёртке информации.
Бывает, что некое свойство организмов непрерывно изменяется при
расселении вида (так, часто особи одного вида мельче в южных популяци-
ях, чем в северных). Это так называемая клина. Она является несомненной
тенденцией, но в дарвинизме тенденции отрицаются (п. 6-2), и феномен
клины здесь “объяснили” тем, что ввели термин клинальный отбор. Таких
форм отбора придумано более двадцати. Самый важный из них - стабили-
зирующий отбор, и мы видели (п. 4-14*), что за этим термином стоит точка
зрения (явившая собой ламаркизм, выраженный в дозволенных тогда тер-
минах), но нет никакого механизма. То же можно сказать про «дестабилизи-
рующий отбор» (п. 4-18). От них надо избавиться безусловно. Но гораздо
важнее решить, надо ли отказаться от самого термина «естественный от-
бор». Примеров его действия как фактора эволюции в природе нет, и мы
видели в п. 4-20, что признание его таковым фактором - итог логической
ошибки и неумения слушать критиков.
Однако для генетика, занятого селекцией полезных хозяйству мутаций,
вызванных искусственно (химически и т.п.), часто выглядит нелепой мысль,
что эволюцию нельзя понять в терминах отбора полезных мутаций. Из на-
званных ранее такими были Четвериков, Тимофеев-Ресовский и Эфроим-
сон. Наоборот, Вентребер (как многие эмбриологи) и Любищев (как многие
систематики) полагали искусственные мутации ни на что не годными, над
идеей же отбора смеялись. Цитолог Лима-де-Фариа вообще назвал свою
концепцию «Эволюция без отбора» [Лима].
Можно ли примирить эти позиции? По-моему, можно, если учесть три
обстоятельства. Первое: сторонники селекции мутантов в данном случае на
самом деле описывают вовсе не естественный отбор (как полагают), а ис-
ключительно факты расплода (п. 5-8*). Второе: то, что они именуют мута-
циями и относят к случайным изменениям в ДНК, при анализе оказывается
физиологическим процессом. В частности, химический мутагенез эф-
фективен не в смертельных, а в стрессовых дозах, т.е. при снятии давления
нормы. Третье: распространяя термин «естественный отбор» с наблюдае-
мых полезных мутаций на никем не наблюдавшиеся преобразования строе-
ния и функций организмов и их сообществ, сторонники дарвинизма хоть и
совершают логическую ошибку, но, как ни странно, часто описывают вполне
реальное явление, правда, не имеющее отношения к естественному отбору
(в смысле избирательной размножаемости) - подбор по Богданову.
Западные работы, насколько они мне известны, всё еще крутятся в
сфере понятия отбора, однако проблематика сама ведет их к теме богда-
новского подбора (пример рассмотрим в п. 12-1). Прежде всего, к ней ведут
факты эпигностики (п. 5-15), показывающие, как подбор реально действует в
пространственных и временных рамках одной особи.
395
Часть 3. Как эволюция шла
Из части 2 мы узнали, что за последние полвека изменилось понимание
задач эволюционизма. Процесс эволюции выглядит не просто как постепен-
ная замена одних свойств или таксонов другими, а как преобразование эко-
систем и иных сообществ, влекущее за собой замену видов. Такой взгляд
радикально меняет представления об эволюции органического мира, и пре-
жде всего - о возможных путях происхождения жизни.
Гпава 7. Загадка начала жизни
Рождение жизни из неживого, самое загадочное (после рождения Все-
ленной) событие, именуют термином биопоэз (от греч. биос - жизнь и по-
иэзис - создание), или менее удачным - абиогенез. С позиции, развивае-
мой в данной книге, надо понять, как появилась та форма активности, какая
специфична для живого. Вопросов встает много, и для начала выделю три.
1) Чего ждать от теории биопоэза? На мой взгляд, она должна стать ба-
зой эволюционизма. Без нее он обходит главные события, списывая их на
некие туманные обстоятельства неописуемо далекого прошлого; в том да-
леке якобы сложилось всё принципиальное, все то, что далее лишь разви-
вается, эволюирует, но более не возникает.
В частности, биопоэз породил биоэнергетику и наследственность, и ес-
ли эволюционизм принимает их как данность, то без понимания остается их
становление, а потому эволюционные концепции обречены говорить лишь о
второстепенном, о «происхождении видов», что мы и наблюдаем.
Такие концепции трактуют развитие «как продукт внешних причин, которые,
для того чтобы сделать их более наглядными, помещаются в темную глубину ве-
ков» - шутил Гегель (Философия природы, п. 249). Он шутил, а вот нобелевский
лауреат Джеймс Уотсон без всяких шуток отнёс к «бестолковой публике» тех
ученых, которые «тратят время и силы на бессмысленную полемику о возникно-
вении жизни или о том, как установить достоверность научного факта». Естест-
венно, что «самым славным событием в биологии со времен книги Дарвина» он
счёл свою с Френсисом Криком модель двуспиральной ДНК {Уотсон Дж. Д.
Двойная спираль. М., Мир, 1969, с. 58, 148). Модель Уотсона, Крика и (напомню
достоверный факт, см. сноску к п. 5-8) Розалинды Франклин действительно ве-
ликолепна, но предлагаю читателю вообразить, что это открытие немного запо-
здало (ведь Уотсон признал, что искал просто «нобелевскую тему», а потому мог
взять любую), что сперва были открыты синтез белка на РНК-матрице и жидкок-
ристаллическая структура воды, определяющая форму спиралей ДНК и белков.
Тогда ДНК не была бы объявлена «главной молекулой», не родилась бы «цен-
тральная догма», которую Уотсон считал достоверным фактом и которая рухну-
396
ла еще при его жизни (на мой взгляд, она принесла биологии больше вреда, чем
лысенковщина). А «самое славное событие» встало бы в один ряд со всеми,
«главной молекулой» вероятно была бы названа РНК, и никто не рассматривал
бы биопоэз как рождение ДНК в «первичном бульоне». Словом, понимать «бес-
толковую публику» бывает иной раз важнее, чем простодушно искать «нобелев-
скую тему». Запомним это в связи с ролью идеи биопоэза в эволюционизме.
2) Неясно, в чем состоит сама специфика активности живого:
«Свести по отдельности любое из жизненных явлений проявлений к физико-
химическим процессам по-видимому можно, а вот вывести из физики и химии
неживого их комплекс - целостный, активный и направленный структуриро-
ванный процесс - нельзя» [Воейков, 1998, с. 287].
Эта трудность знакома математикам как невыводимость и известна как
проблема Гёделя, но биофизики и биохимики долго надеялись понять био-
поэз именно путем мысленного воссоздания физико-химических условий и
процессов, в которых из неживого должно родиться живое. Удалось полу-
чить некоторые вещества и структуры, но они не были живыми.
3) Жизнь возникла на Земле однажды, и прямых свидетельств этого со-
бытия нет, а наука умеет работать только с воспроизводимыми актами.
Посмотрим, как и в какой мере эти трудности преодолеваются.
7-1. С чего начать отсчет? Антропный принцип и картина мира
Большинство эволюционистов (начиная с первого известного по имени,
Анаксимандра) вплетали свои мысли об эволюции живого в общую схему
развития мира, этаких, как Дарвин, не делавший этого, было мало.
В космологии есть понятие «антропный принцип», фактически идущее
от Сократа; суть его в утверждении, что эволюция мира с самого начала
шла таким образом, чтобы в определенный момент в ней мог появиться
мыслящий наблюдатель. Так проще всего дать объяснение удивительному
соответствию значений мировых констант - скорости света, массы протона,
заряда электрона и т.д.: если бы хоть одна из них имела несколько иное
значение, во Вселенной не было бы не то что людей, не было бы даже ато-
мов. Этот принцип позволяет понимать эволюцию трояко.
С одной стороны - как выражение креационизма, т.е. как точку зрения,
привлекательную для верующих: первичное согласование констант дано
Богом, предвидевшим появление людей.
С другой стороны - как естественный процесс, детерминированный
своим концом (как гигантскую преадаптацию). Речь фактически идет о са-
моорганизации и о богдановском подборе (п. 6-12).
С третьей - как итог некоего отбора: из всех мыслимых вселенных че-
ловек живет в той единственной, в которой он смог произойти. А это уже со-
звучно дарвинизму, в рамках которого тоже принято говорить: данное свой-
ство организмов существует потому, что не имевшие его не смогли жить.
Физик Пол Девис (США), пропагандист «антропного принципа», признал, что
397
данная (эволюционная) версия идейно близка дарвинизму. По его мнению,
Вселенная отобрана из «ансамбля» вселенных, а всякий успешно живущий
организм - из «ансамбля» возможных организмов. Фактически речь идет о
счастливом совпадении редчайших случайностей.
Девис привел и критику в адрес принципа, в частности, такую:
«Географ, убежденный в предопределенности всего сущего», мог бы счесть
реку Миссисипи специально сотворенной, поскольку она «очень удобно под-
ходит ко всем пристаням и проходит под всеми мостами» (Девис П. Суперси-
ла. М., 1989, с. 260).
Конечно, река возникла раньше, а технические сооружения - позже и по
другим законам. Не так ли следует подходить и к развитию космоса? Пока
не было частиц, не было и их констант, а когда (по мере охлаждения Все-
ленной) частицы стали возникать, у них обнаружились те свойства (выра-
жаемые в частности, константами), которые вписывались в уже возникший
мир, т.е. возникли те частицы, которые могли существовать.
При объяснении биопоэза возникают такие же трудности: в самой про-
стой клетке все удивительно подогнано друг к другу, равно как подогнаны
друг к другу и разные клетки (организмы и части многоклеточного организ-
ма). Не менее удивляет взаимоподогнанность частей биосферы. И о том,
что все это создано для человека, говорил еще Сократ (п. 1-6).
Обратите внимание: антропный принцип космологами (и то не всеми)
взят на веру лишь для того, чтобы объяснить соотношение констант. Они
вовсе не предлагают обращаться к нему всякий раз, как что-то неясно, а
биологи-дарвинисты обращаются к конечной причине (отбору) постоянно -
всякий раз, когда иной причины не видно.
Антропный принцип часто признают потому, что не видят альтернативы
ему. Более научной мне видится линия рассуждений, отвергающая антроп-
ный принцип в первой форме как излишний, - во всяком случае, пока не ис-
черпаны все остальные аргументы (в этом состоит презумпция рациональ-
ного объяснения явлений, п. 5-1*). Вторую форму можно признать рацио-
нальной, если принять целевую причину Аристотеля в качестве допустимой
в рассуждении (п. 5-2). О третьей можно сказать то же, что о любом отборе:
если речь идет о некоем общем системообразующем факторе, для понима-
ния которого удобна новая картина мира с фазовыми переходами (п. 5-16) и
новая ПМ (п. 6-12), то это - тоже вторая форма; если же речь идет о совпа-
дении, то это не наука - по крайней мере, в понимаемом мною смысле.
Здесь вряд ли когда-нибудь окажется возможным полное взаимопони-
мание, и мне остается признать, что научным для меня выглядит только
второй подход, источник которого можно видеть в принципе наименьшего
действия Мопертюи, а в XX веке - в идее подбора по Богданову.
Трудности, побудившие космологов ввести «антропный принцип», а
биологов - подразумевать его, одинаковы во всех науках и вызваны тем,
что процесса эволюции не видно, а его результаты поражают своей согла-
398
сованностью. И все же одна особая трудность у космологов действительно
имеется: соответствие констант выглядит чудесным потому, что Вселенную
не во что “погрузить”, ее нельзя рассмотреть как часть чего-то, чтобы вы-
вести ее законы из свойств этого “чего-то”. Но к нашей теме данная труд-
ность не относится, если полагать жизнь порождением Земли, как Землю -
порождением космоса. Приняв эволюцию космоса (с ее цепью фазовых пе-
реходов - см. п. 5-2) как нечто данное, мы можем начать распутывать клу-
бок загадок рождения жизни. Оно тоже предстает как фазовый переход.
7-2. Молодая Земля
Марсианская пустыня несет явные следы прежнего течения по ней бур-
ных вод. Куда же они делись? В настоящее время у Марса не только нет
океана, но почти нет и атмосферы, и считается, что миллиард лет назад они
были сорваны с планеты в результате падения крупного астероида. Такие
катастрофы, видимо, случались тогда и с Землей, но наша планета нара-
щивала себе и новую атмосферу, и новый океан, а Марс так и остался го-
лым. Объясняют это тем, что газы и вода поступают на поверхность плане-
ты из недр, а марсианские недра давно мертвы - чем меньше планета, тем
быстрее она «вымерзает».
Наш Мировой океан, кажущийся очень старым, в одном смысле вечно
молод: у него всегда новое дно. Возраст самой старой известной нам океа-
нической коры (на западе Тихого океана, под километровым слоем осадков)
не выше 200 млн лет, тогда как самые древние известные породы континен-
тальной коры (на юго-западе Гренландии) в 19 раз старше - им, как полага-
ют, 3 млрд 850 млн лет. Согласно концепции дрейфа континентов, океан-
ское дно обновляется в ходе спрединга (растекания): из недр новые массы
горных пород непрерывно поступают в срединноокеанические хребты. Они
расползаются, а старая океаническая кора, как полагают, задвигается под
кору континентов. Подробнее см. п. 9-8. Есть и другие концепции, но и они
признают, что океаническая кора молода.
Всем известен «круговорот воды в природе», необходимый для под-
держания жизни, и столь же нужны круговороты всех важных для нее эле-
ментов, - например, фосфора («именно наличие фосфора ограничивает
объем биоты»; поэтому «в эпохи активного вулканизма возникают условия
для расцвета биоты» [Галимов, 2005, с. 477]).
Однако у нас на глазах фосфор мигрирует лишь в одну сторону - вы-
мывается (а теперь и добывается человеком) из горных пород в виде солей,
включается в жизненные процессы, затем сносится в моря и океаны, где
оседает на дно с телами отмерших организмов. Назад из океана возвраща-
ется всего около 1 % фосфора, в основном доставляемого морскими птица-
ми в виде гуано. Круговорот замыкается только через медленные глубинные
процессы (когда захороненные осадочные породы возвращаются на по-
верхность), и без горообразования на континентах жизнь долго не продлит-
399
ся. Даже углерод, куда менее дефицитный, - и тот постоянно поступает из
вулканов в форме углекислого газа.
Всё это привело ученых к формулировке «геохимического принципа со-
хранения жизни», планета может быть обитаема, пока активны ее
недра (Ронов А.Б. Стратисфера, или осадочная оболочка Земли. М., 1993,
с. 133). Активность недр важна не меньше, чем активность Солнца. Чтобы
продолжалась жизнь, Земля сама должна быть в указанном смысле живой.
Это означает, что Лавлок и другие ученые, серьезно считающие Землю ор-
ганизмом (гипотеза Геи, пп. 6-1, 6-15), заслуживают полного внимания. Для
них феномен организованности биосферы - не набор случайностей, а ре-
зультат ее (Гей) собственной активности.
В условиях, похожих на первичные, найдено много способов получения
основных «кирпичиков» живого - аминокислот, сахаров (в том числе и рибо-
зы - компонента РНК), нуклеотидных оснований (пиримидинов и пуринов).
Многие необходимые для жизни молекулы найдены даже в вулканах и ме-
теоритах. Неорганические компоненты (прежде всего, фосфаты) тоже были
в молодой Земле. Но что происходило на ней дальше?
Пионер исследований биопоэза биохимик А.И. Опарин видел задачу в
построении чего-то похожего на первичную клетку, плававшую в «первич-
ном бульоне», содержавшем органические молекулы. Он был уверен, что
именно в ней природой найдены все механизмы первичной жизни. Еще в
1920-х годах его внимание привлекли коацерваты - микроскопические кол-
лоидные капли, содержащие полимеры и плавающие в воде. Он изучал их
более полувека. Коацерваты могут окружать себя простыми оболочками и,
достигнув предельного размера, распадаться на несколько дочерних. Су-
щественно, что концентрация полимеров в коацервате может быть выше,
чем в среде, в 10 тысяч раз.
В 1950-е годы биохимик Сидней Фокс (США) показал, что в качестве по-
лимеров можно использовать протеиноиды - цепочки аминокислот, которые
он получал, нагревая смесь сухих аминокислот. Растворяя их затем в воде и
нагревая крепкий раствор в
автоклаве до температуры выше
130°С (запомним это!), он тоже
получал микрокапли, в которых
наблюдались некоторые фермен-
тативные свойства. У капель возни-
кали оболочки, и при охлаждении
происходило «деление клеток» (рис.
42). Однако оно прекращалось по
исчерпании запаса протеинои-
Рис. 42. Срез делящейся микросферы под
электронным микроскопом (по Фоксу)
400
дов, а пополняться сам собой их запас уже не мог, поскольку для этого не-
обходимы совсем иные условия - перегретой (130°С) жидкости.
Впоследствии в русле идей Опарина - Фокса было высказано много важ-
ных мыслей. Например, кристаллохимик Э.Я. Костецкий (Владивосток) и его
коллеги видят биопоэз как последовательность синтезов макромолекул
(белков и нуклеиновых кислот) на апатитовых матрицах; на них же и проис-
ходил, по их мнению, и синтез первых клеточных оболочек, поскольку мине-
рал апатит обладает подходящими стереохимическими свойствами (Кос-
тецкий Э.Я. ПУК. эволюц. биохимии и физиологии, 1999, № 3).
Однако для «биопоэза по Фоксу» нужно сочетание предельно различ-
ных условий: для синтеза аминокислот - влажная атмосфера (из метана,
аммиака, углекислого газа и паров воды) с электрическими разрядами (гро-
зами), для синтеза протеиноидов - сухой порошок аминокислот, для синтеза
микросфер - перегретая жидкость, а для их деления - вода при «комнат-
ной» температуре. Как их совместить?
Земная кора охладилась ниже 100°С около 4 млрд лет назад, и океан
еще 800 млн лет был теплее 90°С, а самая древняя известная осадочная
порода (3,85 млрд лет) уже несет следы жизни (подробнее об этом погово-
рим в п. 7-4). Выходит, жизнь возникла всего за 200 млн лет и притом в ки-
пятке! Странно, ибо на дальнейшую эволюцию клетки ушло 2 млрд лет.
Тут можно вспомнить старое предположение - что жизнь не возникла
на Земле, а была занесена извне, и что там, где она действительно сама
возникла, биопоэз мог идти миллиарды лет. Однако возможность этого со-
мнительна, поскольку, как сейчас считают, самой Вселенной всего 14 млрд
лет, а жизнь не может возникнуть около звезды первого или второго поколе-
ния - из-за отсутствия тяжелых химических элементов. Солнце же принад-
лежит третьему поколению звезд.
«Появление земных форм жизни нельзя считать задержавшимся, поскольку
не исключено, что они относятся к числу самых первых во Вселенной. Дело в
том, что... возникновение и развитие многоклеточных форм жизни возможно
лишь в системах звезд со средней массой, относящихся к третьему и после-
дующим поколениям» (Аллен Дж., Нельсон М. Космические биосферы. М.,
Прогресс, 1991, с. 60).
Далее, как бы ни попала жизнь на Землю извне, такая гипотеза не объ-
ясняет, как жизнь возникла где-то (см. п. 12-1**). Остается добавить, что по-
ка нет разработанной теории биопоэза, не следует удивляться и его скоро-
сти. Может быть, биопоэз и должен осуществляться быстро?
Сторонники Геи обратили внимание на удивительную особенность зем-
ной атмосферы: по законам химии азот, основной ее газ, должен бы окис-
литься (за счет грозовых разрядов и квантов света) и перейти в нитраты
океана, а кислород, сильнейший окислитель, вообще не должен был поя-
виться в заметных количествах. А концентрация кислорода в атмосфере
401
(20%) близка к предельной: при 25% всё, что может гореть, сгорело бы. Та-
ких примеров можно привести множество, и антропным принципом ничего
не объясняется - сопряжены не только константы, но и постоянно идущие
«не туда» процессы. Тут нужна авторегуляция.
Если же признать Землю активной (живой), то биопоэз выступает как
самый заметный акт взросления нашей планеты, а отсутствие его в наше
время - как выход Земли из фертильного возраста. Конечно, скептик может
заметить, что Земля - не организм, ибо не размножается, что она породила
не другую планету, а собственную часть (биосферу). Однако не всё живое
размножается. Например, не размножаются рабочие пчелы.
В мифе у древних греков Гея, богиня Земли, тоже порождала не другой
мир, а части уже существующего мира - того мира, который благодаря это-
му стал пригоден для жизни людей. С позиции гипотезы Геи проблема био-
поэза ставится совсем по-новому: необходимо понять факт появления
не первой клетки, а первой биосферы.
7-2* Химический отбор
В этом отношении полезно вспомнить «ретроградную концепцию» эво-
люции биохимических систем, которую давным-давно предложил американ-
ский биохимик Норман Горовиц (N.H. Horowitz. On the evolution of biochemical
synthesis // PNAS, 1945, vol. 31, № 6).
«Горовиц... предположил, что метаболические системы эволюционировали,
так сказать, задом наперед. Если в наше время какая-нибудь метаболическая
последовательность ведет, например, от вещества А к веществу Б и далее к
веществам В, Г и Д, то можно думать, что в самом начале возникла потреб-
ность в веществе Д и самой древней реакцией была реакция получения Д из
Г, а Г было сырьем, запас которого имелся в окружающей среде. Лишь после
того, как этот запас начал иссякать, возникало давление отбора в пользу спо-
собности получать Д из другого сырья, т.е. из В. Затем недостаток В породил
конкуренцию» и т.д. Так «медленно и постепенно эволюционировала... в об-
ратном порядке, вся метаболическая цепь» (Дикерсон Р. Химическая эволю-
ция и происхождение жизни // Эволюция. М., Мир, 1981, с. 101).
Жаль, что ни Горовиц, ни Дикерсон не пробовали поставить опыт, что-
бы выяснить, как на самом деле действует «давление отбора», да еще при
отсутствии какой-либо наследственности. Но это упрёк не им, а всей эпохе.
Опять: ссылка на отбор появляется тогда, когда нечего сказать о механизме
явления. К схеме Горовица мы вернемся в п. 7-4.
На самом деле возможна лишь та реакция, какая идет в сторону хими-
ческого равновесия (или его аналога, если система в принципе неравновес-
на). Отсутствие интереса к этому, обычное для объяснений “по Опарину”,
делает их несерьезными. Любищев в одном из писем назвал подход Опари-
на принципом импробабилизма, так как в нем делается ставка на регуляр-
ность событий, практически невероятных, тот же принцип был мною назван
402
“принципом слонопотама” - в честь медвежонка Винни-Пуха, который, ус-
лыхав о существовании неведомого зверя слоно-потама, вырыл у своего
домика яму и стал ждать, пока слонопотам туда свалится [Чайковский, 2004,
с. 230]. Почти вся литература по биопоэзу основана на этом “принципе”, что
досадно, зато избавляет от нужды излагать ее.
Надо не уповать на чудо, а строить теорию. В 1960-х годах шведские
ученые - химик Л. Силлен и геолог М. Руттен - сделали к ней важный шаг:
отвергли концепцию “первичного бульона” как химически невероятную. В
самом деле, нужные для биопоэза реакции - это те, которые должны быть
направлены против химического равновесия, а, следовательно, их продукты
в “бульоне” не могут накапливаться ни за какое время. Поэтому Силлен и
Руттен предложили сменить познавательную установку: не надеяться на
последовательность исчезающе редких удач, а выяснить, при каких усло-
виях нужные реакции могут протекать устойчиво и были ли такие
условия на молодой Земле. Нужным условиям удовлетворил не
«бульон», а поток. В нем может идти любой синтез, если его продукт уно-
сится (или еще как-то предохраняется) от распада [Руттен, с. 297].
Но как это могло достигаться? Первый шаг к ответу на этот вопрос дан
в книге химика А.П. Руденко «Теория развития открытых каталитических
систем» (М., МГУ, 1969). В ней показано, что в потоке веществ и энергии
возможно саморазвитие, причем сам собою развивается не продукт, а ката-
лизатор. Оказывается, активность катализаторов может повышаться в про-
цессе их функционирования, и поток веществ преимущественно перераба-
тывается этими, улучшенными катализаторами. Более активные из них об-
разуются вполне понятным путем - за счет их «избирательного отравле-
ния», т.е. сужения спектра ускоряемых реакций. Теперь катализатор стано-
вится как бы узким, но ценным специалистом, и прежние катализаторы вы-
тесняются им из соответствующих химических потоков веществ.
Это явление Руденко назвал естественным отбором, но мы будем ис-
пользовать устоявшийся к нашему времени термин «химический отбор».
Суть идеи химического отбора такова: ресурс вовлекается в основном в
тот процесс, который его быстрее использует, как бы много ресурса ни бы-
ло. Если «органический» процесс оказывался таким, он занимал место «не-
органического». Реакцию, в которой происходит эволюция катализатора,
Руденко назвал базисной. Тем самым, в анализе биопоэза акцент был пе-
ренесен с привычного вопроса «что синтезировано?» на вопрос «как синте-
зировано?», и процесс рождения жизни предстал как эволюция катализато-
ров неорганических «базисных реакций».
К сожалению, сам Руденко не указал, какие из реакций биопоэза следу-
ет считать базисными, и, тем более, не обратил внимание ни на какие про-
цессы, кроме катализа. Поэтому на фоне появившейся вскоре новой термо-
динамики, давшей более общую теорию открытых систем (в ней показано,
что в открытых системах структуры могут возникать и усложняться сами со-
403
бой), частное достижение Руденко осталось почти без внимания. Мало кто
заметил, что без его теории дальше двигаться невозможно: именно он опи-
сал один из необходимых конкретных механизмов самоорганизации - меха-
низм порождения затравок, нужных для формирования любой термодина-
мической структуры (п. 5-14).
Что касается теории химического отбора, то лавры достались немецко-
му химику Манфреду Эйгену, выступившему с похожей идеей в 1971 году.
Но Эйген ввел отбор как аксиому, взятую из дарвинизма, а у Руденко отбор
- реальное явление в реакции с изменяющимся катализатором.
Биохимик и историк науки К.Б. Серебровская (Ж. физ. хим. 1979, т. 80,
с. 108) указала на то, что базисными реакциями по Руденко могут считаться
биологические стадии обратимой реакции разложения-синтеза воды. Тем
самым, она дала его теории новую жизнь. Теорию Руденко вновь использо-
вал (к сожалению, без ссылки на него) химик В.Н. Пармон (Новосибирск):
электроразряды и прочие «сильные воздействия» на «бульон» могли играть
роль «мутаций», скачком изменяющих катализатор базисной реакции [Пар-
мон, с. 980], а вовсе не роль сдвига точки равновесия, как думали прежде.
7-3. Вода и гель. Тезис Воейкова
Владимир Леонидович Воейков (р. в 1946 г.), биофизик с химическим
мышлением, неожиданно для себя пришел к выводу, что подход Опарина
содержит гораздо больше ценного, чем думали в последние полвека. Разу-
меется, речь не о «принципе слонопотама» (п. 7-2*), а о том, что, как оказы-
вается, в «первичном бульоне» действительно могли идти многие реакции
биопоэза. Прежде всего это могли быть реакции поликонденсации (полиме-
ризации с затратой энергии и выделением воды), источником энергии для
которых служит механическое движение воды. При движении ее через
сверхтонкие поры идет ее диссоциация, и гидроксилы образуют перекись
водорода в неожиданно больших (свыше 1%) концентрациях; она и служит
окислителем. Часть перекиси разлагается на О2 и Н2.
Для необратимости этих реакций требуется сток продуктов. При поли-
конденсации он достигается сменой условий среды; а при разложении пере-
киси О2 и Н2 уходят в атмосферу, где О2 остается внизу и служит основным
окислителем (Voeikov V.L. Reactive oxygen species, water, photon, and life //
Rivista di Biologia / Biology Forum 94,2001).
Поликонденсация является одной из форм первичной самоорганиза-
ции, возможные механизмы которой Воейков рассмотрел в своей доктор-
ской диссертации (Биофак МГУ, 2003).
Однако проблемы биопоэза как целого этим, разумеется, не решаются:
надо еще понять, как и почему полимеры могут собираться в то, что нужно
для жизни. Ленинградские физиологи Д.Н. Насонов (ученик Ухтомского) и
А.С. Трошин (ученик Насонова), а вскоре и Гилберт Линг (прибыл в США из
Китая), разработали в середине XX века концепцию клетки, во многом про-
404
тиворечившую общепринятым взглядам. Главное для нас в ней то, что клет-
ка - не раствор, удерживаемый ее оболочкой, а желеобразная структура
(гель), активность которой и определяет работу клетки.
В настоящее время эта теория67 весьма продвинута и дает понимание
многих вопросов цитологии. Основой работы всех клеточных механизмов
(транспорт ионов через границу клетки, деление клетки, расхождение хро-
мосом и т.д.) признаётся локальный фазовый переход.
Если признать, что полость клетки - не раствор, а гель, то меняется вся
проблематика биопоэза: вместо праздных размышлений о том, как из моле-
кул “бульона” мог сам собой сложиться первый набор с нужными для дан-
ной модели биопоэза качествами, ставится довольно реальная задача - по-
нять, как был устроен нужный для рождения жизни гелевый комплекс.
Его не следует представлять себе как клетку и лучше называть эоби-
онтом (этот термин в 1953 г. предложил Н. Пири).
Первая трудность биопоэза, которая отпадает в концепции геля: нуж-
ные концентрации веществ и их ионов задаются не оболочкой эобионта, а
самой его структурой. Никакие «насосы» для начала жизни не нужны.
Вторая трудность - как первые белки и нуклеиновые кислоты сложи-
лись в нужные спиральные конструкции - отпадает при уяснении того фак-
та, что спирали задаются квазикристаллической структурой воды.
Главное - вода проявляет ту самую активность, на которой зиждится
всё живое. Проявляет сразу в двух совсем различных формах: во-первых,
структура воды определяет пространственную структуру макромолекул и
организует их взаимодействие, а во-вторых, вода служит источником и но-
сителем активных форм кислорода (АФК) - таково общее обозначение для
частиц, содержащих кислород с неспаренным электроном (гидроксил, пере-
кись водорода, озон, 0'2 и др.).
Гашение АФК, достигаемое путем спаривания двух неспаренных элек-
тронов при соединении двух свободных радикалов, является, по Воейкову,
основным и исторически первым источником энергии жизни (АТФ появилась
позже - см. п. 7-7**). АФК всё время возникают и тут же исчезают - либо ис-
пользуются в реакции метаболизма, либо, если таковой потребности в дан-
ный момент в данном месте нет, просто гасятся; причем для гашения в
клетках всех организмов есть особые механизмы.
Такой процесс рождения и гибели АФК напоминает мне флуктуации
квантового вакуума (Воейков с этой аналогией согласился).
01 Так именует свое построение американский физхимик Джералд Поллак (Pollack G.H.
Cells, gels and engines of life; a new, unified approach to cell function. Seattle (Washington),
2001; готовится русское издание под ред. В.Л. Воейкова). На самом деле речь идет об од-
ном аспекте будущей теории; рассмотрена абстрактная клетка; разнообразие клеток (на-
пример, способов деления) игнорировано, и неясно, как его в эту концепцию включить.
Слишком упрощены роль мембраны и ранняя эволюция клетки.
405
Главным окисляемым субстратом биохимии является сильно структу-
рированная вода, продуктом окисления - слабо структурированная вода, а
источником энергии - гашение АФК. Акт структуризации воды есть акт нако-
пления энергии, акт ее деструктуризации высвобождает энергию для био-
химической реакции. Можно сказать, что именно включение данного про-
цесса в реакции геохимического круговорота, повлекшие усложнение ве-
ществ, знаменовало переход химической активности в биохимическую.
Подробнее см.: [Воейков, 2005]. Если вспомнить, что дыханием именуется
окисление субстратов с целью метаболизма, то тезис Воейкова________
[ «Жизнь есть дыхание воды»
- вполне можно принять. Разумеется, это не определение жизни, а указание
на первый и главный биоэнергетический процесс, а также на главное на-
правление поисков решения загадки рождения жизни.
Начнем с того, что коацерват является крохотной порцией водного геля,
но гель может заполнять и крупную структуру (например, лужу). Если доба-
вить, что над водой, в воде и в геле изобилуют АФК, то, как увидим, про-
блема начальных стадий биопоэза значительно упрощается.
7-4. Сначала биоценоз, затем организмы
Подлинно интересную теорию биопоэза начали строить те, кто рас-
сматривал проблему не в рамках “бульона", а в рамках “гипотезы Геи”. Она
основана на утверждении, что жизнь возникла сразу в форме простого био-
ценоза, лишь позднее разделившегося на отдельные особи и виды; что сна-
чала на планете были только геохимические круговороты веществ, затем
возникали одна за другой метаболические реакции - сразу в форме звень-
ев, которые встраивались в прежние круговороты, - а уже потом возникли
наследственность, отдельные особи и размножение. Каждый из первых ша-
гов биопоэза (замена неорганического катализатора той или иной геохими-
ческой реакции на протеиноид) был связан с химическим отбором. Аппарата
наследования на этой стадии еще не требовалось.
Зато требовался аппарат, активно вовлекающий нужные компоненты в
нужную конфигурацию. Это и был, как полагают, водный гель.
Гель-ценотическая концепция (ГЦК) - это модель биопоэза, в которой
водный гель рассмотрен как активная среда формирования первичного ме-
таболизма (включавшегося в неорганические круговороты), т.е. среда, поз-
же разделившаяся на первые организмы (клетки).
Примером включения органического звена в геохимический круговорот
является круговорот серы, изображенный на рис. 43: он существует и вне
жизни, но основной поток серы из морских вод в горные породы в наше
время осуществляют организмы (бактерии Desulphovibrio и др.), которые
восстанавливают сульфаты в сульфиды.
Отличить биогенные сульфиды можно по тому, что в них на несколько
процентов повышено содержание тяжелого изотопа серы с атомным весом
406
34 - за счет того, что биологическая реакция идет другим (многоступенча-
тым) путем.
Исток ГЦК можно видеть в «Биогеохимических очерках» В.И. Вернад-
ского (1940), где написано:
«Появление жизни при создании биосферы должно было произойти не в виде
появления одного какого-нибудь вида, а виде их совокупности, отвечающей
геохимическим функциям жизни. Должны были сразу появиться биоценозы».
Но вопрос о том, как это могло реально получиться, Вернадского не за-
нимал. Первым шагом в конкретизации ГЦК можно считать упомянутую
«ретроградную концепцию» Горовица: он показал, как в принципе органиче-
ский процесс мог встроиться в геохимический круговорот, а именно: в по-
рядке, обратном обычному порядку протекания химических реакций.
Следующий, после Горовица, шаг к ГЦК сделал в 1957 году английский
физик, натурфи-
лософ и историк
науки Джон Бер-
нал - в Москве,
на Международ-
ном симпозиуме
«Возникновение
жизни». В док-
ладе [Бернал] он
обобщил разроз-
ненные высказы-
вания предшест-
венников и выс-
казал следую-
щие тезисы,
ределившие про-
блематику
полвека:
Море
Vх Юя
Атмосфера
ВяпровоД вынос Зрыю
Рига г
I
Осадочные
породы,в ос-
новиом сланцы
2,7 хЮ*
Мвтамарфи-
чеааге и извер-
женные породы
7x10®
Окисленное _
Восаланомшная
Эвапориты
5x10»
Преаям
боды
ЗхЮв
оп-
на
Метаморфизм
Й
Рис. 43. Круговорот серы. Справа от ломаной линии сера представлена веществами, в ко-
торых тяжелого изотопа серы мало, слева - в которых много. Выше прерывистой линии
преобладание сульфатов, ниже - сульфидов. Эвапориты - породы, образовавшиеся за счет
высыхания растворов. 1 и 2 - потоки из недр, не связанные с жизнью. 3, 4 и 5 - потоки, в
которых преобладает биогенная сера. Масса серы дана в тоннах (по: [Руттен])
407
1) Если «первичный бульон» и существовал, то возникает проблема - как
в нем обеспечивались нужные концентрации нужных наборов веществ (это
тот вопрос, который позже разработали Силлен и Руттен);
2) Вероятнее всего, «бульона» никогда не было, а образование мономе-
ров осуществлялось параллельно с их полимеризацией;
3) Для этого были удобны мелкие водоемы на грани высыхания, где в ка-
честве катализаторов, заменявших ферменты, служили глины (уточню: сре-
дами реакций в таких водоемах могли быть гели);
4) В ту эпоху «ничего похожего на обособленный организм возникнуть не
могло, а могли существовать лишь беспорядочные образования довольно
постоянного состава, в которых метаболизм поддерживался некоторое вре-
мя и которые более напоминали холодное пламя, чем организмы»;
5) Такие «субвитальные единицы» могли иметь меняющиеся границы и
сливаться друг с другом, причем несовместимые разрушались, а у подхо-
дящих друг другу «диапазон их биохимической активности расширялся»;
6) Сначала это были «зоны» («субвитальные территории»), занимавшие
целые равнины, покрытые влажным субстратом (Бернал называл его илом
и почвой, что неверно - ничего подобного до возникновения жизни не суще-
ствовало, зато мог существовать гель);
7) Появление разнообразных полимеров привело к возникновению мем-
бран, а это позволило субвитальным единицам локализоваться в коацер-
ватных каплях и других микрообъектах. Они не имели аппарата наследст-
венности и были не протоклетками, а эобионтами.
8) Появление наследственности неясно, но важны 2 опытных факта:
во-первых, «два дополнительных синтетических полирибонуклеотида (один с
группой пурина, а другой с группой пиримидина), каждый в отдельности не-
правильно скрученные, соединились in vitro... в плотную двойную спираль,
аналогичную спирали природной ДНК». Во-вторых, нуклеиновая кислота
может служить матрицей только для точно ей соответствующей, тогда как
один «фермент может может способствовать синтезу многих, если не всех,
нуклеиновых кислот».
Последний пункт надо пояснить. Бернал указал тут на три обстоятель-
ства: (1) РНК может самопроизвольно скручиваться в двойную спираль, (2)
РНК может копировать только саму себя, (3) белок ничего сам не копирует,
но может катализировать копирование любых полимеров.
Не пытаясь решать проблему «взаимной связи между синтезом нуклеи-
новой кислоты и белка» (она не вполне решена и ныне), Бернал указал са-
мое главное в проблеме появления наследственности: первым наследст-
венным веществом удобно считать такую РНК (а, заметим, не ДНК), которая
могла принимать нужную форму сама, без белков, тогда как первой функци-
ей белка наследственной системы должна была служить РНК-полимеразная
активность. Момент появления этой «взаимной связи» Бернал и полагал
моментом рождения жизни.
408
Итак, по Берналу, рождению жизни предшествовало появление об-
ширных субвитальных зон и, затем, обособление внутри каждой из них
многих крохотных эобионтов; лишь после этого появилась наследствен-
ность. До ее возникновения процессы протекали сами собой, поскольку не
были уникальны (как в обычных химических процессах). Удивляться высо-
кой скорости этой стадии биопоэза не надо - медленно осуществляется
лишь то, для чего требуется многократное ожидание маловероятных актов.
В 1970 году к ГЦК еще на шаг подошел биолог М.М. Камшилов, рабо-
тавший после сталинской ссылки в Институте водных проблем (пос. Борок
Ярославской обл.). По Камшилову, если сначала возник метаболизм, а за-
тем клетки и их размножение, то сразу должно было появиться много раз-
ных одноклеточных видов - когда первичные биоценозы расчленялись на
отдельные организмы. Он сочувствовал натурфилософскому мнению Вер-
надского о том, что жизнь всегда существовала в виде биосферы.
Но бактериолог ГА. Заварзин, столь же ревностный почитатель Вер-
надского, тем не менее с 1970-х годов возражет против первичности био-
сферы. По его мнению, о биосфере как целом нет никаких сведений до тех
пор, пока в геологической летописи не появились цианеи (синезеленые во-
доросли). Жизнь же появилась много раньше.
Поясню: следами жизни считается изменение соотношения изотопов (в
осадочных породах) и состава атмосферы; и то и другое выясняется изуче-
нием осадочных пород исследуемой эпохи. Так вот, появление цианей (они
выделяют при фотосинтезе кислород) привело к накоплению кислорода в
атмосфере, каковой стал влиять на развитие жизни около двух млрд лет на-
зад. Наоборот, у молодой Земли была совсем иная атмосфера, почти или
вовсе без кислорода (о двух вариантах ранней атмосферы см. [Самуилов]).
Это ясно из того, что осадочные породы, погребенные в то время, содержат
недоокисленное (двухвалентное) железо.
В подземных водах его много и сейчас, но оно сразу же окисляется на
воздухе: читатель мог сам видеть, как льющаяся из скважины прозрачная
вода быстро мутнеет - растворимые соли двухвалентного железа окисля-
ются воздухом в плохорастворимые соли трехвалентного.
Но если кислорода в атмосфере не было, то почему древнейший след
жизни оказался окисленной породой - бурым железняком? Ответ Заварзи-
ным предложен простой: первые организмы составляли биоценоз, который
был тесно сомкнутым, похожим на нынешний лишайник; окислитель (кисло-
род) в атмосферу не выходил. Заварзин также обратил внимание на важный
факт: в горячих источниках близ нынешних вулканов обнаружено множество
различных литотрофных (питающихся неорганикой) бактерий - и высказал
предположение, что они-то и были первичными организмами. К этой мысли
мы еще вернемся.
Наконец, по Заварзину, всё, чего не мог синтезировать первый орга-
низм, он получал из геосферы (идея Горовица).
409
Изотопный анализ показал, что самые древние осадочные породы (те
самые, из юго-западе Гренландии) несут следы жизни - их изотопный со-
став таков, какой обычен у остатков жизнедеятельности. Однако в этих по-
родах нет никаких свидетельств наличия отдельных организмов (клеток). По
последним данным (LEG, 12), древнейший гренландский осадочный слой
имеет возраст 3,85 млрд лет и уже содержит изотопный сдвиг, а древней-
шие породы, содержащие нечто похожее на бактериальные клетки, - 3,4-3,5
млрд лет (Австралия и Южная Африка), т.е. возможно, что бактерии на 400
млн лет моложе жизни. Этим косвенно подтверждается та идея, что в самом
деле жизнь возникла до клеток, в форме системы биохимических реак-
ций. Мы вернемся к этой теме в п. 7-8*.
7-5. Биосфера как организм
Итак, жизнь, возможно, возникла в форме биоценоза. Но биоценоз - не
биосфера: биоценозы зависят друг от друга, а биосфера одна. Быть может,
и она родилась раньше, чем организмы? Такую биосферу, не поделенную
на организмы, изобразил Станислав Лем в фантастическом романе «Соля-
рис». У него биосфера планеты представлена одним гигантским разумом,
внимательно исследующим прибывших землян. Не так ли выглядела и пер-
вичная биосфера Земли? Думаю, что не вполне так.
Первоначально Лавлок рассматривал свою «Гею» (п. 7-2) как нечто по-
хожее на Солярис. При всей ее экстравагантности, идея вовсе не абсурдна:
пусть биосфера состоит из других организмов, но это не мешает рассматри-
вать и ее саму как организм - ведь другие «составные» организмы хорошо
известны - таковыми можно считать и пчелиный улей, и начиненного зеле-
ными водорослями моллюска, и, конечно, лишайники.
Более того, наш собственный организм тоже в некотором смысле со-
ставной: он состоит из органов, они - из тканей, которые, в свою очередь,
состоят из клеток. А сами клетки могут быть организмами - при подходящих
условиях опыта они долго живут вне организма («культура ткани»).
Иное дело, какими свойствами составной организм может обладать, а
какими нет. Сначала Лавлок наделял его даром предвидения - в ожидании
будущих бед «Гея» якобы устроила на своей поверхности подходящие слои
(атмосферу, гидросферу и биосферу), подобно тому, как человек одевает
себя одеждой:
«Гипотеза Геи постулирует, что физические условия на поверхности Земли, в ат-
мосфере и океане - как прежние, так и нынешние, активно созданы жизнью са-
мой для себя... Это противоположно обычным представлениям, по которым
жизнь приспособилась к условиям планеты и они произошли своими отдельным
путями» (Lovelock J.E. Gaia: A new look of life on Earth. Oxford, 1979, c. 152).
Но затем, под воздействием критики, Лавлок в книге «Возрасты Геи»
ослабил ее предполагаемые возможности, сведя их к способности поддер-
живать ее собственный гомеостаз. Организмом считается уже не планета, а
410
биосфера, и нервной системой ему служит царство животных. Головного
мозга нет, поэтому, образно говоря, Гея из “человека” обратилась в живот-
ное вроде медузы.
Казалось бы, “головным мозгом" Геи можно считать человечество (в ду-
хе «ноосферы», как ее понимал Вернадский), но Лавлок счел иначе - по его
мнению, человечество противостоит Гее, и она чуть ли не готова к ампута-
ции этого больного органа; во всяком случае, Лавлок уверен, что она пере-
живет «ядерную зиму», как больной - ампутацию (Lovelock J.E. The ages of
Gaja. Oxford, 1989, c. 171-177). Хотя Гея теперь лишилась у Лавлока дара
предвидения, но в ней по-прежнему
«эволюция организмов и эволюция окружающей их среды тесно связаны в
единый процесс. Его естественное свойство - саморегуляция».
Многие биологи стали бурно возражать, поскольку
«большинству биологов все-таки трудно принять даже идею глобального
гомеостаза. Дело в том, что почти невозможно представить себе переходное
звено от локальных эффектов к изменениям в масштабе планеты. Организмы,
изменяющие свою среду в интересах потомства, конечно, получают за счет
этого преимущество: вот почему птицы, например, строят гнезда. Но трудно
понять, каким образом эта чисто дарвинистская «семейственность» может
действовать в масштабе многих поколений и на континентальных расстояни-
ях» - писал в редакционной статье журнал «Scientific American» (перевод:
ВМН, 1990, № 2, с. 62).
Можно выразиться сильнее: не просто трудно, а невозможно. В преды-
дущих частях мы не раз убеждались, что дарвинизм бессилен, едва лишь
встают вопросы целостности. Однако биосфера, ни с чем не конкурируя,
миллиарды лет поддерживает свои основные параметры - состав атмо-
сферы, вод - это ли не свидетельство целостности?
Нам надо выяснить, когда эта целостность возникла, ибо первые цено-
зы, как уже сказано, биосферы не образовывали. Бернал простодушно по-
мещал эобионты в ил и в почву, но это, как мы уже говорили, - образования,
сделанные организмами. А как выглядела поверхность суши до появления
организмов? За последние 30 лет ученые пришли к согласию в том, что она
состояла из ровных, почти горизонтальных поверхностей примерно на уров-
не океана и почти вертикальных скал. Как писал Руттен, биопоэз, вероятно,
протекал в период между актами горообразования, а в такое время «мате-
рики, как правило, стачиваются эрозией до уровня моря». Напомню: это по-
нимали и античные натурфилософы (п. 1-5).
Выветривание и размыв (эрозия) происходили на юной Земле несрав-
ненно быстрее, чем ныне: при отсутствии почв ничто не сдерживало буйст-
ва ветров и водных потоков. Ровная суша изобиловала мелкими водоема-
ми, то и дело высыхавшими, а затем снова растекавшимися. Это означает,
что противоположные условия, нужные для синтеза мономеров и для их по-
лимеризации (“биопоэз по Фоксу”, п. 7-2), чередовались.
411
Столь же важен был повсеместный контакт трех фаз - твердой, жидкой
и газовой. Это соединение трех фаз, сохранявшееся многие миллионы лет,
называют эквилибросферой (слоем равновесия). Название неудачно: ведь
на самом деле биопоэз требовал регулярного чередования противополож-
ных условий. Но сам контакт трех фаз был долгим, и введение понятия эк-
вилибросферы проясняет многие вопросы биопоэза.
Вспомним, что первый ученый, рассуждавший о естественном возник-
новении жизни (Т. Бернет, п. 1-13) говорил именно о границе трех фаз.
«Бульон» появился только в концепциях XX века, и, видимо, там и останет-
ся. (Впрочем, и в «бульоне» есть граница фаз - это оболочки коацерватов.)
Аллен и Нельсон (их книга упомянута в п. 7-2), пишут:
«Формирование сравнительно устойчивой эквилибросферы (твердые вещест-
ва - жидкости - газы) происходит в первые несколько сотен миллионов лет ее
существования».
Жизнь и поныне основана на процессах, идущих на границах фаз, толь-
ко теперь эти границы, в основном, пролегают внутри клетки.
7-6. Новая термодинамика: планета делает на себе жизнь
Наша главная задача - показать, что биопоэз был не скоплением сча-
стливых случайностей, а термодинамически оправданным явлением. В п. 5-
14 было сказано, что новая термодинамика, в отличие от прежней (т.е. от
термостатики), показала неизбежность образования и усложнения структур
в любой системе, в которой существует поток вещества и энергии, в доста-
точной мере неравновесный.
Диссипативной структурой (ДС) является и пламя, и организм, и биоце-
ноз, и вся биосфера (видимо, только в этом смысле она - как Солярис.)
Могут возразить, что такие ДС, как водоворот или пламя, не являются
самостоятельными предметами - их, например, нельзя передвинуть, не
двигая их субстрат, а любой организм можно из его субстрата хотя бы на
время вынуть. Поэтому вроде бы его нельзя сравнивать с живыми сущест-
вами. Но живое тоже нельзя произвольно сдвинуть в том пространстве, в
котором оно выступает как ДС - например, в пространстве концентраций
внутриклеточных веществ. Хотя они могут “гулять” сами, как “гуляет” по ван-
не водяная воронка. Важно понять, что слова «живое, понимаемое как ДС»
относятся не к веществу организма (оно может и не измениться, если орга-
низм убить), а к тому, что делает его живым, т.е. к процессам.
С точки зрения термодинамики, жизнь - гигантская ДС, состоящая из
множества меньших ДС, те состоят из еще меньших и т.д.; в этом смысле
жизнь организована иерархически. Земля как планета не перестанет суще-
ствовать, когда за счет ее охлаждения потоки из недр остановятся; но био-
сфера без этих потоков умрет.
Неотъемлемой частью жизни является процесс обмена (вернее сказать
- поток) веществ: и биосфера, и организм, и клетка создают и поддержива-
1
412
ют свою структуру, непрерывно разрушая нечто внешнее и выбрасывая
свои отходы наружу. Земля пока еще достаточно горяча, в ней существуют
потоки веществ, обеспечивающие ее магнитное поле, движения земной ко-
ры и выход веществ из недр (вулканы, источники первичных газов и вод и
т.п.). Эти потоки обеспечивают существование множества различных ДС,
наиболее заметны из которых две - подвижная кора и биосфера. Земля
обеспечивает биосферу потоками веществ, тогда как потоками энергии ее
обеспечивают не только Земля, но еще Солнце и космос.
Концепция жизни как иерархии диссипативных структур взята мною из
более общей теории «самоорганизующейся Вселенной» (Jantsch Е. The self-
organizing universe. Oxford - N.Y., 1980). Добавлю, что ее автор Янч (о нем
см. п. 6-12) понимал термин ДС расширительно; по-моему, термин «соли-
тон» (в смысле Петухова - п. 5-14) был бы не хужеЛ
С изложенной точки зрения, жизнь порождена Землей (точнее, ее пото-
ками), и в этом смысле саму Землю можно признать живой. Более того,
биосфера жива лишь постольку, поскольку жива Земля - в п. 3 мы подошли
уже к тому же выводу с позиций геологии. Жизнь могла родиться только то-
гда, когда Земля находилась в соответствующем возрасте, и потому загла-
вие книги Лавлока «Возрасты Геи» вполне оправданно.
Никакая ДС не может образовываться постепенно, любая ДС (как и вся-
кая целостность) возникает скачком. Этим обстоятельством воспользовался
дальневосточный геолог В.Н. Компаниченко. Он отметил диссипативный ха-
рактер всех процессов биопоэза и выдвинул на этом основании гипотезу,
состоящую в утверждении геологически мгновенного («революционного»)
становления первых живых систем, противопоставленную эволюционной
гипотезе Опарина. Каждая стадия биопоэза характеризовалась появлением
новых типов ДС (Компаниченко В.Н. Возникновение жизни в глубинах гидро-
термальных систем. Хабаровск, 1996).
В заключение замечу, что с позиций термодинамики биопоэз выгоден
для планеты: если термодинамически выгодно образование различных ДС,
а не их разрушение, то выгодно и накрыть их все вместе единой оболочкой,
замедляющей общее разрушение. Такую оболочку совместно создают био-
сфера и созданная ею атмосфера. (Как сказано в п. 7-4, кислородную атмо-
сферу создали цианеи.)
7-7. Жизнь родилась в скороварке?
Как мы уже видели, юная Земля, видимо, не имела суши в нынешнем
смысле: если не считать вертикальных скал, то вместо суши тогда была,
вероятно, чуть ли не сплошная эквилибросфера: во-первых, то и дело вы-
сыхающие и вновь заливаемые отмели (см. п. 4), а, во-вторых, бьющие по-
всюду из земли горячие источники, поставляющие материал на эти отмели.
Первые, еще «не совсем живые» структуры (эобионты) следует искать в
щадящих условиях. Но что ими считать? В эпоху биопоэза океан был кис-
413
лым (а не соленым) кипятком, дули метановые и аммиачные ветры при вы-
соких давлениях. Понятно, что условия, комфортные для нас, для первых
организмов были бы гибельными.
Если добавить, что для образования ДС необходимы устойчивые пото-
ки подходящих веществ, то окажется, что лучшей колыбели жизни, чем го-
рячие источники, нечего и искать. Об этих источниках как о поставщиках
требуемых веществ думали давно, но лишь лет 30 назад стали искать жизнь
прямо в них. И нашли нечто изумительное.
Источники горячих вод на суше и на дне океанов наделены своей осо-
бой жизнью -сообществами бактерий, неспособными жить при обычных для
нас температурах. Сначала они были описаны как особое приспособление к
жару, но затем стало ясно, что именно эти бактерии являются самыми
древними. Большинство из них относят к обширной группе архебактерий,
т.е. древних бактерий. Следовательно, вернее признать, что не они приспо-
собились к жаре, а вся остальная современная жизнь приспособились к
обитанию в условиях «холода».
Гидротерма (или гидротермаль) - это выброс горячих газов, растворов
и взвесей из разломов океанического дна, чаще всего - в срединноокеани-
ческих хребтах, т.е. в зонах спрединга (п. 7-2). Их температура часто намно-
го превышает 100°С, но вода в них не кипит, поскольку при огромных глу-
бинных давлениях ее точка кипения может превышать 300°С. Среди гидро-
терм наиболее эффектны и
поэтому более известны «черные
курильщики» - выбросы, в которых
преобладает пирит (дисульфид
железа), образующий огромные
клубы черной взвеси.
Вокруг выброса формируется
труба твердых осадков высотою до
20 м, а рядом часто располагается
«белый курильщик» из светлых
взвесей, не образующих труб, и в
результате всего этого гидротерма
в свете прожектора выглядит как
сказочный подводный завод (рис.
44).
Рис. 44. Схема гидротермы на дне океана (по [Лобье])
Каждый курильщик существует всего 50-100 лет - до тех пор, пока ак-
тивен выброс, и все это время окружен совсем необычным биоценозом.
Известны литотрофные бактерии (п. 4), живущие в устьях гидротерм и
имеющие оптимум роста при 110°С, как в скороварке. Они восстанавливают
414
углекислый газ и серу, окисляя водород и выделяя метан и сероводород,
чем обеспечивается включение основных химических элементов в биологи-
ческие круговороты. При этом
«нет оснований предполагать, что бактериальные обитатели реликтовых со-
обществ сколько-нибудь серьезно изменились со времени своего возникнове-
ния» {Сергеев В.Н, Холл Э.Х., Заварзин ГА. Первые три миллиарда лет жиз-
ни: от прокариот к эвкариотам // Природа, 1996, № 6, с. 55, 57).
Последнее утверждение следует уточнить: имеется в виду сохранение
древних экологических и физиологических функций микробов, о конкретных
же биохимических реакциях ископаемых микроорганизмов мы почти ничего
не знаем. Если признать, что первые живые организмы были столь же био-
химически сложны и совершенны, как современные, это значит - признать
творение, а не эволюцию.
Читатель может удивиться: сначала утверждалось, что биопоэз проте-
кал в высыхающих лужах, а теперь - что на дне океана. Но противоречия в
этом нет - «условия скороварки» тогда вполне уживались с границей фаз
(эквилибросферой): тогдашняя атмосфера из углекислого газа обеспечива-
ла давление во много раз выше современного (как сейчас на Венере), а
гребни “срединноокеанических” хребтов торчали из еще неглубокого тогда
горячего океана, полного отмелей, плавно переходивших в сушу. Органиче-
ские вещества легко засасывались с боков в наземные термальные источ-
ники или, смытые в океан, - в гидротермы. Тем самым, термальные источ-
ники тоже включались в глобальные круговороты, что понемногу создало
условия для превращения незамкнутых ценозов в замкнутые, т.е. в малень-
кие копии биосферы.
Жизнь гидротерм подробно и увлекательно описал французский зоолог
Люсьен Лобье, один из первых ее исследователей. Среди гидротермных
сообществ существуют такие, которые только из недр получают необходи-
мые им вещества; по-видимому, они представляют собой биоценозы, спо-
собные жить без остальной биосферы [Лобье]. Поясню сказанное.
Основная часть биосферы держится на противоположных реакциях:
фотосинтез: СО2 + Н2О + свет = углевод + О2
дыхание: углевод + 02 = СО2 + Н2О + энергия.
Но это возможно лишь потому, что уже есть биосфера, обеспечившая
кислородную атмосферу, а также есть системы фотосинтеза и дыхания.
Чтобы создать биосферу, надо было включить в органический круговорот
первую порцию органических веществ, а для этого нужны литотрофы (по-
гречески это значит - камнееды, хотя на самом деле основной пищей этим
бактериям служат газы: Н2, СО2 и т.п.); они являются первичными проду-
центами, поскольку именно они делают органику из неорганики. Вот не-
сколько литотрофных реакций (выбраны те, что идут в бактериях гидротерм,
причем не любые, а те, что обеспечивают свой биоценоз энергией):
415
образование метана: 4Н2 + СО2 = СН4 + 2Н2О,
образование сероводорода: Н2 + S = H2S, (1)
образование дисульфида натрия: 5Н2 + №2803 = 2Na2S + ЗН2О.
Получив запас энергии в форме восстановителя, бактерия приступает к
хемосинтезу (аналогу фотосинтеза), например делает глюкозу:
3H2S + 6СО2 + 6Н2О = 3H2SO4 + С6Н-| 2С>6- (2)
(Окислителем тут служит не кислород, а, как ни странно, углекислый
газ.) Этой глюкозой пользуется и сама бактерия, и тот, кто ее съест. Но
пользуется не путем дыхания (свободного кислорода вокруг нет), а путем
гликолиза. Когда сделанное таким путем органическое вещество будет вы-
брошено из гидротермы током кипятка в обычную воду, содержащую рас-
творенный кислород, тогда оно может быть использовано окончательно
(съевший аэроб превратит его в СО2 и Н2О).
Гидротерма могла, в силу сказанного выше, оказаться и на поверхности
вод. Как пишет Заварзин, по мере затухания активности нынешней гидро-
термы в ее выбросах начинает преобладать СО2, а среди ее обитателей -
фототрофы. То есть, жизнь за счет недр переходит в жизнь за счет света и
атмосферы, и условия гидротермы переходят в условия мата.
Роль фотосинтетиков в первых матах должны были играть не цианеи
(появившиеся много позже), а фотобактерии с более простым (бескисло-
родным) фотосинтезом. Из ныне живущих таковы, например, пурпурные
серные бактерии, в которых на свету образуются, вместо сахара и кислоро-
да, формальдегид и серная кислота (ср. с реакцией (2)):
2 СО2 + H2S + 2Н2О = 2СН2О + H2SO4.
Как такой фотосинтез превратился в обычный кислородный фотосинтез
(общий цианеям и растениям), неизвестно (замечу: вновь мы видим повтор-
ное использование старой идеи на новом субстрате - ср п. 6-15**). К сча-
стью, для понимания биопоэза это знать и не нужно. Достаточно того, что
путь от эобионта до подобия биосферы вчерне удалось наметить.
Подробнее о становлении фотосинтеза см. [Самуилов]. В частности,
там сказано: «Скорее всего, фотосинтез формировался как мозаичный про-
цесс», т.е. по-разному в разных группах организмов. Однако у всех высших
(растений) он практически одинаков, так что налицо эквифинальность, но
не онтогенетическая (п. 3-14), а эволюционная. То же мы увидим не раз.
7-7* Ценозы, но не биосфера
Так и хочется сказать, что каждый первичный ценоз - маленькая био-
сфера, но это не так: он не замкнут, не утилизирует своих отходов, а выбра-
сывает их наружу. Таким образом, и понять его возникновение проще, и
роль его в дальнейшей эволюции очевидна - создание базы для остальной,
более сложной и разнообразной жизни, для будущей биосферы. Лишь по
416
накоплении органики жизнь смогла позволить себе такую роскошь, как жи-
вотных, самостоятельно ищущих свою пищу, и круговороты снова стали за-
мыкаться. До этого каждый организм был гораздо теснее, чем ныне, связан
со своим источником питания, и каждое животное, как космонавт на чужой
планете, носило с собой свою «среду обитания». И так же недалеко могло
уползти от «источника жизнеобеспечения» (гидротермы).
Интересно, что в такой экосистеме почти отсутствует привычное нам
поедание крупными организмами мелких: вместо этого большинство круп-
ных животных разводят внутри себя бактерий, поставляющих им пищу. Та-
ковы рифтии (тип погонофор) - огромные, до полутора метров, внешне по-
хожие на кольчатых червей, но с султаном щупалец. Щупальца яркокрасны,
но любоваться ими некому - вокруг полная тьма.
Как и все погонофоры, рифтии не имеют никакой пищеварительной сис-
темы, но если остальные погонофоры, вероятно, сорбенты (всасывают пи-
щу всем телом), то для рифтий установлено иное: они обладают полостью,
наполненной автотрофными (усваивающими СО2) бактериями, которые в
данном случае играют роль растений. Разница состоит лишь в том, что вме-
сто света источником энергии им служит H2S, вероятно произведенный ли-
тотрофом в гидротерме согласно указанной выше реакции (1) и использо-
ванный внутри рифтии согласно реакции (2).
Самое интересное в том, что растения, обычно именуемые продуцен-
тами, в действительности являются лишь вторичными продуцентами: они
возвращают в биологический круговорот тот углерод, который выпал из не-
го, когда другие организмы, разрушая органику, испустили его в форме уг-
лекислого газа. Включить в органический круговорот неорганику (азот, серу
и т.п.) они неспособны, да и СО2 из недр включают в органику не они, а ли-
тотрофы. Однако термин «продуценты» настолько прижился к растениям,
что менять его поздно (подробнее см. п. 8-2).
Множество следов древнейшей жизни сохранилось в форме строма-
толитов - холмов, выраставших на дне прежних мелких водоемов с пере-
менной соленостью. Строматолит (греч.: «ковровый камень») - окаменев-
ший циано-бактериальный мат, т.е. слоистая структура, образованная
биоценозом прокариот. Живой мат - это многослойный (до 2 см) «ковер» из
живых прокариотических клеток. Он покрывает слои, созданные клетками
уже отмерших поколений прокариот и неорганическими осадками.
Роль растений в таком мате играют цианеи: они занимают верхние
миллиметры «ковра» (куда попадает свет и где производится кислород), а
остальные бактерии живут за их счет. Такие маты образуются сами и обра-
зуют строматолиты до сих пор: их можно наблюдать в жарком климате в
очень соленых лагунах морей и озер, например, в озере Балхаш. Для них
характерна смена увлажнения и высыхания, как в эквилибросфере.
В отличие от гидротермы мат - как бы маленькая биосфера со своими
417
замкнутыми круговоротами кислорода и органических веществ; такой «оа-
зис», в принципе, мог существовать и в безжизненной среде (Заварзин Г.А.
Бактерии и состав атмосферы. М., 1984). Добавлю, что так мог внешне вы-
глядеть и первый организм-биоценоз, еще не разделенный на особи. Хочет-
ся понять, как он мог, в принципе, возникнуть. Об этом узнаем далее, в п. 7-
8. Но сперва нужно сказать кое-что об энергетике.
7-7** Об энергетике биопоэза
Прежде всего был нужен носитель энергии для жизненных процессов. У
всех организмов таковым служит АТФ (аденозинтрифосфат) - об этом мож-
но прочесть в любом курсе биохимии. Однако его самопроизвольное появ-
ление непонятно, и вернее будет принять, что основную энергию (энергию
возбуждения молекул) поставлял (и поставляет ныне) не он, а другой меха-
низм - гашение активных форм кислорода (АФК) - см. п. 7-3.
АФК имеются в воде, и никаких особых механизмов для их включения в
процессы биопоэза не требовалось. Данное обстоятельство мне любезно
разъяснил В.Л. Воейков в марте 2006 года. По его мнению, АТФ служит
вспомогательным источником энергии - там, где нужны мелкие порции
энергии. Он мог возникнуть позже, в ходе направленного биохимического
процесса (до сих пор его появление объясняли самопроизвольным накоп-
лением, т.е. на основе“принципа слонопотама”, п. 7-2*).
Компоненты АТФ (аденин, рибоза и фосфаты) действительно могли на-
капливаться химически. Аденин - одно из четырех азотистых оснований
(основы нуклеиновых кислот). Все четыре могут, в соединении с рибозой и
фосфатами, служить для переноса энергии, но только АТФ является пере-
носчиком универсальным. Можно допустить, что
“первичная распространенность аденина, по сравнению с другими основа-
ниями, была обусловлена, очевидно, тем, что он гораздо легче синтезируется
абиогенно... путем последовательной полимеризации молекулы HCN” [Гали-
мов, 2001, с. 96].
Это возможно, но несущественно. Для биопоэза важно другое - нали-
чие потока, обеспечивавшего накопление аденина и АТФ в первичных сре-
дах жизни. Никаких моделей такого потока мне не известно, в литературе на
сей счет царит “принцип слонопотама”. Наоборот, АФК достаточно обильны
в воде сами по себе. С их помощью, полагаю, могли получать нужное на-
правление первичные реакции, в том числе - синтеза аденина, АТФ и т.п.
Биохимик В.П. Скулачев отметил, что аденин лучше поглощает ультра-
фиолет. По его мнению, первые организмы в качестве источника энергии
использовали свет Солнца, но не тот видимый, каким пользуются растения,
а ультрафиолет; он и превращал тогда АДФ в АТФ. Затем
«древние клетки должны были переключиться на... видимый свет». «Замена
опасного ультрафиолетового излучения на безопасный видимый свет могла
быть тем признаком, который лег в основу естественного отбора» (Скулачев
418
В.П. Эволюция биологических механизмов запасания энергии И Соросовский
образовательный журнал, 1997, № 5, с. 15).
Однако, во-первых, в ходе биопоэза Солнце вряд ли было источником
ультрафиолета - оно, по всей вероятности, едва излучало, притом только в
красной части спектра. В таких условиях основную энергию для биопоэза
поставляли недра Земли, как мы это видим до сих пор в гидротермах. (За-
мечу: если первая АТФ возникла и работала “по Скулачеву”, то источником
ультрафиолета могли быть грозы. АТФ могла попадать в водоемы и, всасы-
ваясь в приповерхностные гидротермы, участвовать в биопоэзе.)
А во-вторых, переход фотосинтеза от ультрафиолета к видимому свету
означал бы смену всей биохимии процесса. Если такая смена имела место,
то налицо использование старой идеи на новом материале - то самое, что в
п. 6-15** было аттестовано как платонизм. Моя попытка указать Владимиру
Петровичу на эту философскую трудность (январь 2004 г., личная беседа)
успеха не имела - он выразил уверенность, что времени на отбор случай-
ных мутаций было достаточно. Но, замечу, если это допущение принять, то
ультрафиолетовая стадия не поясняет в становлении фотосинтеза ничего.
И как раз с аденином оказалось непросто: в предполагаемых первичных
средах он нестоек [Галимов, 2001, с. 100]. Почему же вездесущ именно он?
Ответа нет, и разные авторы независимо предлагают преодолеть эту и
сходные неясности ссылкой на закрепившийся результат прежнего отбора
случайных мутаций. (Точно так же Ф. Крик толковал «универсальность гене-
тического кода» - феномен, которого попросту не оказалось - см. п. 5-13.)
Вообще замечу: ни один автор не апеллирует к естественному
отбору, пока понимает исследуемый механизм. Ссылка на отбор все-
гда служит палочкой-выручалочкой из непонятностей.
7-8. Появление наследственности
Итак, первый биоценоз являл собой не набор организмов, а систему
биохимических реакций и не был замкнут. Любопытно, что древнейший жиз-
ненный слой (Гренландия, 3,85 млрд лет) имеет изотопный сдвиг (п. 7-4),
самый мощный из известных (Заварзин Г.А. Лекции по природоведческой
микробиологии. М., 2004, с. 317-319). По-моему, это значит, что жизнь до ор-
ганизмов включала какие-то реакции, ныне не идущие, или необычное соот-
ношение реакций. Каковы были эти реакции, неясно.
До сих пор мы рассматривали метаболические аспекты происхождения
жизни: как именно могли работать первые биохимические системы. Но все
достигнутые ими успехи надо было еще научиться наследовать. Вероятно,
первая система реакций как раз и не была унаследована.
Вспомним, что для копирования ДНК надо сперва расплести двойную
спираль, затем с каждой половинки сделать копию, учитывая то, что на-
правления чтения у двух цепей ДНК противоположны, и снова заплести две
двуспиральные системы. Что касается синтеза белка, то поскольку на самой
419
ДНК белок синтезироваться не может, с ее молекулы следует снять копию в
виде матричной РНК (мРНК), на основе которой и будет синтезироваться
белок - с помощью совокупности транспортных РНК (тРНК). И в каждой ста-
дии участвуют свои ферменты.
А вот для копирования мРНК необходим лишь один фермент - РНК-
полимераза. Аденин, входящий в АТФ, тоже включает рибозу (убрав из АДФ
еще один фосфат, получим рибонуклеотид). Цепи рибонуклеотидов могут
удлиняться сами собой, без ферментов (реакция Четверина [Спирин, 2005,
с. 26]). Так не с РНК ли началась наследственность?
Вопрос ставился не раз, например, Берналом (п. 7-4), однако был
праздным до тех пор, пока считалось, что биохимическую реакцию катали-
зировать могут только белки. Но в 1981 году установлено, что РНК может
выступать как фермент (его назвали «рибозим»). Была высказана догадка,
что в биопоэзе РНК осуществляла как наследственную, так и каталитиче-
скую функции, и в 1985 году даже появился термин «РНК-овый мир».
А в 1986 году предложена общая схема становления ранней наследст-
венности (Darnell J.E., Doolittle W.F. И PNAS, 1986, v. 83, p. 1721), которая
изложена в руководстве: Сингер М., Берг П. Гены и геномы (М., Мир, 1998,
т. 2, Введение). Суть ее вкратце такова.
7-8* От РНК к генам. Прогенота
Для хранения и передачи наследственной информации РНК малопри-
годна. Для этого нужна макромолекула, не участвующая (как участвует РНК)
в жизни клетки, зато обеспечивающая надежность информации, а значит, ее
избыточность. Таковой явилась двуспиральная ДНК, и схема эволюции
должна объяснить, каким образом в РНК-овом мире появилась ДНК-овая
наследственность.
Главным в схеме явилось допущение, что синтез ДНК на РНК-овой мат-
рице, известный как обратная транскрипция, имел место при биопоэзе и что
для этого служила «активность наподобие активности обратной транскрип-
тазы» (рис. 45). Долгое время этому допущению противоречило отсутствие
обратной транскрипции у прокариот, однако недавно у паразитической бак-
терии Wolbachia (о ней см. п. 8-3) обнаружены инактивированные гены об-
ратной транскриптазы. Это делает схему реальной.
Наследственность, по данной схеме, возникла еще до появления пер-
вых организмов (клеток). Ее носителем явилась прогенота - первая систе-
ма, в которой обмен веществ и информации отделен от наследственности.
Авторы сочли, что еще в РНК-овом мире появились основные способы об-
работки генетической информации - не только транскрипция РНК в РНК, но
и сплайсинг, и экзон-интронное строение (и, добавлю, редактирование РНК).
Словом, появление ДНК с ее репарацией, рекомбинацией и репликацией
вероятно было итогом того, что мы выше (п. 6-15) упоминали как “внутри-
клеточное мышление”, причем оно возникло в прогеноте.
420
В этом допущении ук-
репляет и недавний спец-
выпуск журнала «Биохи-
мия» (2005, № 5), по-
священный 70-летию Б.Ф.
Ванюшина. Сам юбиляр
напоминает, что метили-
рование («второй язык
генетики», см. п. 5-5*) мо-
жет протекать без фер-
ментов, что тимин
(отличающий код ДНК от
более древнего кода РНК)
есть всего лишь
метилированный урацил,
что РНК может
направлять процедуру
метилирования ДНК, и
много иных интересных
вещей, относящихся к
ранней эволюции. Из
обзоров этого номера мы
узнаём, что метилирова-
ние, которое много лет
считалось просто одним
из механизмов регуляции
развития особи, на самом
деле есть один из самых
общих информационных
языков жизни.
Рис. 45. Прогенота в гипотетической схеме биопоэза (по: Сингер и
Берг, 1998). Изображение прокариот и эвкариот на одном уровне
отнюдь не означает одновременности их происхождения
При чтении этого спецвыпуска более всего поражает тот факт, что чу-
жеродная ДНК может не уничтожаться иммунной системой, а маркироваться
путем метилирования и в такой форме включаться в геном. Зачем? Похоже
на то, что генетические системы могут, подобно библиофилам, копить тек-
сты «про запас», чтобы затем «почитать на досуге». Читают они вполне
сознательно, делают при этом вполне сознательные «пометки на полях» и
ставят «секретные коды». Опять, как и в пп. 5-6, 5-7, 6-15, видим внутрикле-
точное мышление. Сколь рано оно появилось, непонятно.
421
Прогенота, согласно излагаемой гипотезе, произвела первую клетку
(замечу, что эта стадия до сих пор тоже совсем непонятна), причем клетки
довольно быстро оказались двух типов: первый сохранил интроны, и в нем
произошли эвкариоты, тогда как у второго интроны почти или вовсе утра-
тились, что привело к иному типу эволюции (прокариоты) - клетка оста-
лась сравнительно простой, зато быстро делящейся, легко порождающей
новые типы белков и открытой для горизонтального переноса.
У всех прокариот ДНК замкнута в кольцо, а у эвкариот - линейная. Как
обратил внимание биохимик-теоретик А.М. Оловников, это достоинство эв-
кариот (оно облегчает компоновку хромосомы из блоков) обернулось для
них радикальным изъяном: хромосома, а за ней и вся клетка, стареет от
неполной репликации ее концов. Мы вернемся к данному вопросу в п. 8-6, а
здесь замечу, что это - заведомо не единственная причина старения и что
бактерии стареют тоже.
В прогеноте же должны были возникнуть вирусы. Вероятно, они воз-
никли до клеток - как простейшие компактные переносчики и хранители ге-
нетической информации. Из этого вовсе не следует, что они были в каком-
то смысле предшественниками прокариот: вирус не является организмом, и
принцип его организации не использован в прокариотах. Он использован,
возможно, в некоторых бактериальных органеллах.
Зато вирусный принцип компактной укладки двуспиральной ДНК ис-
пользован в эвкариотной клетке, он лежит в основе строения хромосомы.
По всей видимости, в прогеноте возникли также вирусы с двуспираль-
ной РНК, осуществляющей функцию "геномных цензоров” (п. 6-6**) и тем
самым давшие эвкариотной клетке возможность существовать в потоке про-
кариотных генов, не теряя индивидуальности. Если так, то первая эпигно-
стическая функция возникла еще в прогеноте и ограничивала горизонталь-
ный перенос генов.
Как долго жила прогенота, сказать нельзя. Еще недавно можно было
отметить, что биохимические следы жизни примерно на 700 млн лет стар-
ше, чем самые древние из обнаруженных клеток и что первые следы прока-
риот и эвкариот почти одинаково стары (около 2,7 млрд лет), но экологиче-
ски важными эвкариоты стали лишь еще через миллиард лет (Федонкин
М.А. Изменение доступности металлов и эвкариотизация... И Труды ГИН,
вып. 565. М., ГЕОС, 2004). Однако в настоящее время структуры, похожие
на бактерий, обнаружены во всех осадочных породах, начиная с возраста
3,8 млрд лет (микропалеонтолог А.Ю. Розанов, в ответ на мой вопрос на его
докладе в апреле 2006 г., прямо сказал: «во всех»), так что для прогеноты
собственное время жизни пока не выявляется.
Это еще не значит, что ее вообще не было. Во-первых, она могла оста-
вить след о себе в более древних породах, а во-вторых, гипотеза прогеноты
вовсе не требует, чтобы та была мощным глобальным объектом, сохра-
няющимся в геологической летописи. Она могла быть просто лужей.
422
7-8** От РНК-ового мира к нашему
Рибозимы несравненно менее разнообразны, чем белки, поскольку бел-
ки формируются из 20 разных аминокислот, тогда как РНК - всего из 4
нуклеотидов двух типов: основаниями двух нуклеотидов являются пури-
ны, т.е. аденин и гуанин, а двух других - пиримидины, т.е. цитозин и
урацил. (Напомню: нуклеотидом называется азотистое основание плюс
звено сахаро-фосфатной цепи.) Казалось, что преодолеть малое разно-
образие нуклеотидов невозможно, и с идеей РНК-ового мира следует
расстаться. Но из тупика уже найдено, как минимум, два выхода, причем
оба согласуются с концепцией прогеноты.
Рис. 46. Минорные основания, возможно служившие для абиогенеза. Слева урацил (без
одного водорода); он обратимо образует 5-гидроксиметилурацил - основу множества
минорных оснований, являющихся аналогами обычньгх аминокислот. Далее по часо-
вой стрелке: тимин (аналог аланина), затем изображены аналоги цистеина, аргинина,
тирозина, триптофана, гистидина; в самом низу рисунка - основание нуклеотида, слу-
жащее аналогом сразу трех аминокислот (глютамина, аспарагина, глицина); последним
изображен аналог лизина (по: Робертсону и Миллеру)
Майкл Робертсон и
Стенли Миллер (США)
напомнили, что в
условиях, имитирующих
преджизнь, можно
получать не только 4
обычных, но и ряд
минорных (редких)
оснований. Среди них
интересен 5-оксиметил-
урацил: его можно
получить, соединяя ура-
цил с формальдегидом.
При 100°С (вот она, гид-
ротерма!) 5-оксиметил-
урацил может порож-
дать целый спектр
минорных оснований (рис. 46). Интересны они тем, что каждое служит сте-
реохимическим аналогом одной из аминокислот, т.е. имеет сходные размер
и форму и может участвовать в тех же реакциях (Robertson М.Р., Miller S.L.
Prebiotic synthesis of 5-substituted uracils: a bridge between the RNA world and
the DNA-protein world// Science, vol. 268, 5 May 1995).
Итак, согласно концепции «РНК-ового мира» механизм синтеза белка
возник при биопоэзе следующим образом: сначала рибозимы (тогда гораздо
423
более разнообразные, чем теперь) эволюировали сами по себе, встраива-
ясь в геохимические круговороты наравне с протеиноидами, тоже состояв-
шими из большего, чем теперь, числа типов аминокислот. Те рибозимы, что
катализировали собственный синтез, стали предками генетического аппара-
та, а те, что катализировали синтез протеиноидов - предками аппарата син-
теза белка. Каким образом эти два класса начали взаимодействовать, до
сих пор неясно, и потому надо сказать о втором выходе из тупика.
Международный журнал «Origins of life and evolution of the biosphere»
(Нидерланды) дал в номере 3-5 за 1996 год подборку статей, в которой ав-
торы из разных стран на все лады заявляли о негодности концепции РНК-
ового мира, а завершила подборку статья А.Д. Альтштейна «Происхождение
протоклеток», где сказано, что есть выход лучше: РНК-ового мира никогда
не было, а с самого начала существовали прогены - гипотетические струк-
турные единицы, на которых шел одновременный синтез полинуклеотидов и
соответствующих им полипептидов; проген располагался на мембране, а
работа его состояла в подборе аминокислот, стереохимически соответст-
вующих определенным нуклеотидным триплетам. Клетка, полагает Альт-
штейн, возникла тогда, когда мембрана научилась расти и замыкать собою
полость, содержащую проген. Идея, как видим, близка прежней требовани-
ем стереохимического соответствия, но не требует от рибозимов разнооб-
разия ферментативных свойств.
Обе концепции решил сблизить японский генный инженер Масуями
Нашимото: он полагает, что в эпоху РНК-ового мира был период, когда РНК
и белки играли симметричные друг другу роли, когда наряду с примитивной
трансляцией существовала примитивная обратная трансляция, т.е. проце-
дура формирования транспортных РНК для аминокислот. По мнению На-
шимото, обратная трансляция опасна (как нарушающая принцип наследст-
венности) и вскоре исчезла (Nashimoto М. The RNA/Protein symmetry hy-
pothesis: experimental support for reverse translation of primitive protein // Jour-
nal Theoret. Biol., 2001, vol. 209, p. 181-187). На данную работу мое внимание
любезно обратил А.М. Дейчман (который сам полагает, что обратная транс-
ляция малых фрагментов существует поныне).
С позиции Нашимото понятно, почему из 150 аминокислот только 20
имеют свои тРНК: для остальных аминокислот просто не хватило времени
их выработать до окончания «периода симметрии».
Конечно, все это очень сыро и не раз еще будет пересмотрено, но важ-
но тем, что показывает разрешимость проблемы [Спирин, 2001].
Вероятно, что аппарат наследственности впервые образовался в фор-
ме одноцепочечной РНК (как у некоторых современных вирусов). Вирус - не
организм, он не имеет своего метаболизма и не может самостоятельно раз-
множаться, а первый организм должен был уметь и то, и другое. Жизнь на-
чалась явно не с вирусов, но их изучение полезно диатропически: показы-
вает, сколь различно может быть устроен механизм наследственности. Ви-
424
рус отличается от примитивнейшего организма так же, как дискета с не-
сколькими маловажными программами - от простейшего компьютера (бо-
лее вольно говоря, как компьютерный вирус от компьютера), но именно на
вирусах видно разнообразие «программных языков» жизни, т.е. способов
записи наследственной информации. У вирусов она может храниться в че-
тырех формах - в РНК или ДНК, в одноцепочечной или двуспиральной (хотя
каждый вирус использует только одну форму).
В случае одноцепочечной РНК информацию можно использовать пря-
мо, без расплетения нитей и процессов трансляции. Поэтому естественно
считать, что такой способ был исторически первым. Но он же и самый нена-
дежный: такую молекулу легко разрушить любым ферментом, и всякое по-
вреждение необратимо (поскольку пары для сравнения нет).
Двуспиральная форма РНК удобнее тем, что «несъедобна» для фер-
ментов, зато она труднее для считывания. В такой форме (одиночные РНК
для метаболизма, двуспиральные для наследственности) жизнь, вроде бы,
вполне возможна, и возникает вопрос: почему мы ее не наблюдаем? Ведь
все организмы (вирусы - не организмы) имеют наследственный аппарат,
основанный на двуспиральной ДНК и одноцепочечной РНК. Исключение яв-
ляют “геномные цензоры”, использующие короткие двуспиральные РНК (п.
6-6), но она используется отнюдь не для наследования.
У двуспиральной ДНК замечательных свойств много (например, она хо-
рошо вписывается в кристаллическую структуру воды и потому даже разры-
вы обеих цепей могут устраняться), но сейчас нам важно понять ее место в
процессе биопоэза. Напомним, что согласно ГЦК (п. 4), эобионты - первич-
ные самостоятельные формы жизни - выделились из субвитальных зон.
Этому должно было предшествовать появление хотя бы самой примитивной
наследственности, иначе эобионты были бы бесполезны. Следовательно,
наследственность была нужна еще в субвитальной зоне, где многочислен-
ные РНК были вовлечены в круговороты, а потому неминуемо подвергались
уничтожению; наряду с массовой, подлежала уничтожению уникальная РНК,
кодировавшая едва найденные «изобретения». Отличить нужное было не-
кому. Тут и помог иной тип нуклеиновой кислоты.
Изъятие одного атома кислорода из каждого звена сахаро-фосфатной
цепочки, превращающее рибозу в дезоксирибозу и, соответственно, РНК в
ДНК, позволяет двойной спирали скручиваться более компактно - спираль
ДНК может обладать как обычной формой (A-форма), аналогичной спирали
РНК, так и «сверхскрученной» В-формой, недоступной ферментам.
Возможно, что именно это различие послужило к разделению функций:
РНК - для работы с информацией, для ее неограниченного копирования и
редактирования, ДНК - для ее хранения. Считывать ДНК могла более слож-
ная, более поздняя и более медленная система. Из нее, вероятно, позже
появилась прогенота. Конкретнее о биопоэзе сказать пока нечего, зато в са-
мое последнее время начал выясняться тот факт, что и в наше время РНК
425
служит не менее важным метаболитом, чем белок: лишь 30% генов (транс-
крибируемых участков ДНК, успешно прошедших сплайсинг) кодируют бел-
ки, а остальные используются прямо в форме РНК - как регуляторы (см. ко-
нец п. 5-7). Мы и ныне живем в РНК-овом мире.
7-9. Самосборка, хиральность и биопоэз
Из сказанного создается впечатление, что достаточно мономерам со-
ставить цепочку, и они смогут функционировать. Так ли это? В некоторых
ситуациях - да. Многие полимеры сами принимают форму, пригодную для
функционирования {Финкельштейн А.В., Бадретдинов А.Я. // МБ, 1997, №
3). Это один из простых вариантов важнейшего жизненного принципа - уже
не раз упомянутого принципа активности, точнее, его варианта, самосборки.
Самосборка - самое общее явление природы, прекрасный пример бо-
гдановского подбора, идущего без размножения. Даже образование атома
из элементарных частиц - это самосборка.
О том, какие физические процессы могли быть при этом использованы,
отчасти можно судить по аналогии:
«в абиогенных условиях в пегматитовых жилах образовались углеродистые те-
ла, имеющие формы... микроорганизмов... Н.П. Юшкин рассматривает про-
цесс образования таких тел как абиогенный синтез протоорганизмов из наност-
руктур; Г.А. Заварзин говорит об общем пространстве геометрических возмож-
ностей тел такого размера, уточняя, что не всё, что выглядит как бактерии, яв-
ляется таковым». «Если эти материалы будут признаны за микрофоссилии (т.е.
ископаемыми микробами - Ю. Ч.), то происхождение жизни следует рассматри-
вать как космический феномен. Разумеется, микрофоссилии не могут служить
переносчиками жизни*^, но вполне могут свидетельствовать о жизни как кос-
мическом явлении» {Заварзин Г.А. И ВРАН, 2006, № 2, с. 166].
Самосборка макромолекул так же удивительна, как и соответствие бел-
ков нуклеиновым кислотам, и напрашивается мысль, что в основе обоих за-
кономерностей лежит какой-то общий закон.
Один из подходов к нему намечен в круге работ, которые провел кри-
сталлохимик Н.А. Бульенков, показавший, что феномен самоорганизации
молекул базируется на глубинных свойствах вещества - хиральности моле-
кул и фрактальном росте макромолекул.
Молекулы живого вещества составлены в основном (а нуклеиновые ки-
слоты и альфа-спирали белков - исключительно) асимметрично: из левых
(вращающих в растворе плоскость поляризации света против часовой
стрелки) аминокислот и правых сахаров. Это - хиральность живого. По-
скольку молекула белка является спиралью, она должна быть целиком за-
62 Видимо, Заварзин имеет в виду тот факт, что жизнь может существовать (а
значит и перемещаться) только в форме биоценоза. Подробнее см. п. 12-1*.
426
кручена в одну сторону. По Бульенкову, хиральная молекула белка может
“ввинчиваться” в жидкокристаллическую структуру воды. Причина хираль-
ности сахаров была тогда неясна, зато Бульенков показал, что взаимодей-
ствие ДНК с водой носит характер фрактального роста (Бульенков Н.А. И
Кристаллография, 1988, №2 и последующие его работы). Вскоре было по-
казано, что хиральность двух сахаров (рибозы и дезоксирибозы) необходи-
ма для образования двойной спирали.
Для появления системы белок-полинуклеотидного соответствия тре-
бовалось накопление мономеров определенной хиральности;
«Решение общей проблемы происхождения жизни распадается на ряд част-
ных проблем». К ним «относятся абиогенез (здесь: накопление органических
молекул - Ю.Ч.), разделение фаз и образование дискретных эволюциони-
рующих систем, возникновение матричного синтеза, генетического кода и
двух фундаментальных асимметрий - ионной и хиральной» (Яковенко Л.В.
Возникновение ионной и хиральной асимметрий в предбиологической эво-
люции. Автореф-т докт. дисс., Биофак МГУ, 2006, с. 1-2).
По Яковенко, успех достигнут только в области абиогенеза, тогда как
остальные проблемы находятся на стадии выявления молекулярных запре-
тов. Замечу на будущее; проблема возникновения аппарата клеточного де-
ления ни тут, ни где-либо даже не упомянута - видимо, всем достаточно
факта самопроизвольного деления капель по Фоксу (рис. 42).
Другой подход состоит в том, что для понимания самосборки нужно
предложить новую картину мира (п. 5-16) и, соответственно, новую ПМ.
В самом деле, почему самосборка нас удивляет? Потому, что исходным
состоянием материи принято полагать отсутствие всякой организованности,
однородность и хаос - как наиболее вероятное состояние. Это - следствие
статистической ПМ (п. 5-3), царившей в науке XIX века и породившей «вто-
рой принцип термодинамики». Ее ограниченность давно стала очевидной (п.
5-14). Приняв активность в качестве движущего принципа на всех уровнях
бытия, мы должны быть готовы всюду встретить самопроизвольный рост
организованности, в том числе и самосборку макромолекул. С позиции ак-
тивностной (шестой, будущей) ПМ только это и можно счесть естественным.
Если так, то самосборку надо описывать, но вовсе не надо чем-то оправды-
вать - кроме как тем, что так устроен мир. В сущности, это и есть подход Бо-
гданова, в этом - новая картина мира.
Однако способность к самосборке отнюдь не является всеобщей. От-
крытие прионов (п. 5-7) показало, что белок, собирающийся сам собой в по-
лезную структуру, может под влиянием другого белка изменить свою форму
и функцию и стать гибельным для организма. Сперва прионы считались
инфекционным агентом млекопитающих, но позже оказались вездесущими.
Генетик С.Г. Инге-Вечтомов (Петербург), исследующий прионы дрожжей,
пишет, что прионы являются «частным случаем общебиологического меха-
низма, лежащего в основе эпигенетического наследования» (Инге-
427
Вечтомов С.Г. И ВРАН, 2000, № 4, с. 305). Суть механизма в том, что белок
может быть матрицей для белка (см. п. 5-7).
7-10. Энергия жизни и ее лучи
И всё же, несмотря на самосборку, переход к живому из неживого ни в
природе, ни в лаборатории не наблюдается. Что-то явно не учтено. Дело в
той трудноуловимой сущности жизни, которую Бергсон называл «жизнен-
ным порывом». К ней же в 1957 году (на упомянутом симпозиуме в Москве)
подошел Горовиц: живое, сказал он, обладает «импульсом к развитию».
Никто пока не знает, что это такое и как описать «порыв» или «им-
пульс» в терминах естествознания. Ясно только, что в ходе биопоэза воз-
никла качественно новая форма активности материи. Настолько (добав-
лю) новая, что многие предпочитают говорить уже не «активность материи»,
а «дух», «духовное». Весь XX век эволюционисты старались об этом не ду-
мать, а немногих думавших упрекали в ненаучности. Но теперь, когда кон-
туры проблемы обозначились довольно четко (в основном, в связи с рабо-
тами по биопоэзу, в которых биологию безуспешно пытаются вывести из
физики и химии), не замечать ее уже просто невозможно.
Организм часто сравнивают с машиной. Однако машина не обновляет
вещество, из которого состоит, а организм делает это непрерывно. Вот на
столе прибор - я выключу его и забуду о нем, а когда понадобится, включу
снова. Но вот цветок в горшке или птичка в клетке - о них надо помнить всё
время. Если это и машины, то очень странные - всё время требуют горюче-
го, даже когда не работают. На что? На «борьбу с энтропией»?
Но почему этого не требует прибор? Данный пункт труден, и многие не
видят самого вопроса. Ведь организм может долго быть заморожен и затем
ожить. С другой стороны, прибор может от хранения заржаветь.
Однако давайте сравним не крайности, а сущности. В морозильнике ля-
гушка подобна выключенному прибору, это верно; в данное время она не-
живая. Оживает она при оттаивании и при этом как бы включается. Однако
включенный прибор продолжает ржаветь (если не прогревается) и тратит
энергию не на поддержание себя, а на работу. При этом никакое вещество
втекать в него не обязано, как не обязано вытекать и другое.
У меня есть старинные фамильные часы, недавно я завел их, и они по-
шли. Ни одна молекула в них не была при этом заменена. Лягушку же надо,
отогрев, накормить и убрать ее испражнения. (Известное положение эколо-
гии: ни один организм не может жить в собственных отходах.)
В чем же различие часов и лягушки? Бауэр первый заявил (1935 г.), что
живое, кроме обычной для любых машин энергии, обладает еще и своей
особой «структурной энергией», которую он уже тогда (!) связывал с возбу-
жденным состоянием молекул. Что это за энергия, объясняет Воейков:
«Молекулярный субстрат живых организмов находится в состоянии, весьма
далеком от термодинамического равновесия, вследствие высокой заселенно-
428
сти верхних электронных уровней (т.е. вследствие возбужденного состояния
молекул - Ю. Ч.)... В этом случае энергия не диссипируется через тепловой
канал, а затрачивается на осуществление биологических функций» (Воейков
В.Л. О структурно-энергетической специфичности живого состояния // Соз-
нание и физическая реальность, 1996, №4).
Поясню сказанное. Структурная энергия - это энергия возбуждения
биомолекул. Ее кванты, как правило, намного больше тех, какие может пре-
доставить АТФ. Только возбужденные молекулы могут вступать в те реак-
ции, которые делают живое живым, т.е. способным к развитию и к поведе-
нию. Именно благодаря запасу структурной энергии молекула (и организм)
могут совершать то, что удивляет физиков и химиков - реагировать на сиг-
налы, т.е. совершать крупные перемены под действием малых воздействий.
Такова реакция на запах: две-три молекулы могут заставить зверя взять
след, т.е. радикально изменить поведение.
Материальная основа сигнала может быть вообще не видна: например,
жест или воспоминание. Это отнюдь не только у высших животных: так же
загадочно, с физической точки зрения, поведение многих ферментов, на-
пример - репликаз, которые обратимо меняют направление своего движе-
ния по матрице в ответ на сигналы, энергетически ничтожные.
Неживая система тоже может радикально измениться за счет ничтожно-
го воздействия, но всегда необратимо. Говорят, что снежная лавина может
сойти от громкого крика; да, это возможно, но вернуть ее назад невозможно,
а на новую потребуются годы накопления снега.
Возбужденные молекулы бывают и в неживых системах, но там каждая
быстро и самопроизвольно переходит в исходное, невозбужденное состоя-
ние (на нижний электронный уровень), излучая квант энергии. На этом осно-
ваны, например, все способы получения света.
Живое вещество ведет себя иначе: излучает не сразу и очень неболь-
шую часть структурной энергии, поэтому может довольно долго сохранять
ее для полезной работы (движения, развития, мышления). Но поскольку из-
лучение все-таки имеет место, то запас возбужденных молекул надо возоб-
новлять. Это требует циклов, которые можно осуществлять двумя путями -
либо возбуждать прежние молекулы, либо менять их на поступающие извне
уже возбужденные молекулы (так сказать, либо заряжать“аккумуляторы”,
либо менять “севшие батарейки” на свежие).
Природа использует оба способа. Например, в основе химии зрения
лежит цикл, состоящий в 1) регулярных переходах светочувствительных
молекул в возбужденное состояние под действием света, 2) столь же регу-
лярном переводе этого возбуждения в нервный импульс и 3) возвращении
молекул в светочувствительное состояние. Такой цикл (“аккумулятор”) воз-
можен лишь потому, что светочувствительные молекулы погружены в сис-
тему, снабжающую их энергией и работающую по второму (“севшие бата-
рейки”) способу. Живое как целое не умеет “питаться от розетки”.
429
Для поддержания вещества в живом состоянии нужно непрерывно син-
тезировать новые (возбужденные) молекулы, а прежние расщеплять и уда-
лять. Это невыгодно - кроме энергии, непрерывно расходуется вещество.
Но такова жизнь, иной мы не знаем, и вряд ли она возможна.
В термодинамике это связано с тем, что всякая ДС требует для своего
существования поток вещества, а не только энергии. Однако одного этого
для понимания биоэнергетики мало - надо еще понять квантовохимическое
обстоятельство: если вспомогательные реакции (сближение веществ, рас-
плетание макромолекул и т.п.) требуют малых квантов и потому идут с по-
мощью гидролиза АТФ, то перевод в возбужденное состояние требует
большого кванта, каковой высвобождается иначе - гашением АФК.
Фермент чаще всего изготовляет молекулу в возбужденном состоянии.
В том и состоит механизм действия фермента как катализатора: всякий ка-
тализатор служит для преодоления энергетического барьера химической
реакции, и фермент делает это радикальным методом - молекула находит-
ся на высоком энергетическом уровне уже в момент ее синтеза.
7-10* Лазеры жизни и новая картина мира
Излучение живого вещества крайне слабо и достигается «лазерной»
конструкцией: много молекул вместе излучают общий квант. В такой коопе-
рации (являющей собою одну из форм сродства) Воейков и видит специфи-
ку живого. Иногда нужен квант высокой энергии (ультрафиолетовый), на-
пример, для запуска митоза (лучи Гурвича, см. п. 4-16), и он всегда бывает
редким: множество квантов малой энергии, полученных молекулярным ком-
плексом в ходе метаболизма, порождают один квант высокой энергии.
Но есть и другой (в биопоэзе он, видимо, был основным) путь получения
большого кванта - погасить свободный радикал АФК, возникший сразу как
целое - например, за счет механохимических свойств воды (п. 7-3) - т.е.
спарить неспаренный электрон. В обоих случаях стоит вспомнить с благо-
дарностью про «холодное пламя» Бернала.
Можно утверждать, что «жизненный порыв» на молекулярном уровне
получил первое описание в форме структурной энергии, создающей самые
малые ДС организма (внутриклеточные потоки постоянно заменяемых “ба-
тареек"). Можно и сказать иначе: структурная энергия обеспечила появле-
ние в природе новой - биологической - формы активности.
Наконец, есть третий путь получения большого (притом - неограничен-
но большого) кванта: взять его из внешнего излучения. Об этом пути надо
сказать подробнее. Лет 40 назад радиобиологи неожиданно для себя обна-
ружили, что малые дозы радиации полезны для всех организмов, ибо дей-
ствуют принципиально иначе, чем большие (возбуждая электроны и почти
не ионизируя вещество). А еще лет через 20 стало выясняться, что сверх-
малые ее дозы для жизни просто необходимы. Выясняя механизм данного
явления, патриарх радиобиологии А.М. Кузин (опираясь на работы коллег в
430
России, США и Франции) пришел к выводу, что действие радиации на живое
вещество в корне отлично от действия на неживое:
«Как известно, в молекулах при возбуждении электроны... в ничтожные доли
секунды (10-2 - 10"3 с) обусловливают реакцию возбужденной молекулы или,
возвращаясь на нормальный уровень, излучают кванты... В живых организ-
мах возникают значительно более сложные процессы при возбуждении поли-
меров, находящихся в конденсированном состоянии. Возбужденный электрон
такой молекулы делокализуется в огромном электронном облаке... образуя
нелинейные устойчивые вихревые сгустки энергии, подобные в макромире
солитонам, получившие название поляритоны^. Время их жизни уже не доли
секунды,а часы» [Кузин, с. 41].
Поляритоны, медленно распадаясь, испускают в ультрафиолетовом
диапазоне синхронные лучи, чем обеспечивают вторичное биогенное излу-
чение, наблюдаемое у всех организмов; его частным случаем являются, по
всей видимости, лучи Гурвича (там же). См. Дополнение 2.
По Кузину, эти «лучи жизни» обеспечивают механизм целостности - то
самое биополе, которое есть у всех организмов [Кузин, с. 58-62]. Добавлю:
они же, вероятно, обеспечивают энергией те (видимо, очень немногие) био-
химические акты, для которых нехватает мощности иных квантов.
К сожалению, Кузин ничего не сказал о буйной жизни в гидротермах -
неужели ничтожных концентраций обычных радиоактивных изотопов там
достаточно? Или там есть свой источник жёстких излучений? Вероятно,
речь может идти об источниках радоновых вод.
В предисловии к посмертной книге Кузина почти прямо сказано, что у
него речь шла о новой картине мира. Обсудим этот вопрос.
Кроме всего, на что указывал Бернал (п. 7-4), акт перехода неживого в
живое (химии в биохимию) состоит в появлении нового пути синтеза необ-
ходимых молекул - того пути, на котором они становятся возбужденными и
организованными в молекулярные ансамбли. Теперь мы знаем - эти ан-
самбли характерны тем, что излучают синхронно, как лазер.
Важно, что их удалось наблюдать в неживой системе, в растворе ами-
нокислот. Воейков показал, что в этих растворах молекулы синхронно всту-
пают в реакции полимеризации (а не добавляются к полимеру с одного кон-
ца), синхронно излучают (Воейков В.Л. Докт. дисс., Биофак МГУ, 2002). Увы,
этот круг работ (из многих его участников выше названы Ф.-А. Попп, А.М.
Кузин и В.Л. Воейков, а Дополнении 2 будет назван А.В. Будаговский) вы-
глядит островком в океане старомодной биологии, основанной на физике
XVII века, пусть иногда и применяющей квантовые и иные новые термины.
Можно сказать, что Воейков пытается протянуть связь между химиче-
ской и биологической формами активности. Для обнаружения этой связи
03 О солитонах см. п. 5-13*. Поляритон - одна из квазичастиц в квантовой физике систем.
431
необходимо, чтобы сама химия обладала подходящей познавательной ба-
зой. Предлагаемая мною в данной книге идея эволюции как последователь-
ного усложнения форм активности является картиной мира (о картинах ми-
ра см. пп. 5-16, 6-14, 6-15, а также недавнюю статью: Чайковский Ю.В. Кар-
тины мира и познавательные модели // Экология и жизнь, 2006, № 4). Ее в
разные времена держались некоторые мыслители, но она никогда не овла-
девала обществом, не становилась познавательной моделью. В п. 5-3 я
предположил, что активностная ПМ в будущем может появиться. Из от-
дельных наук зачатки ее лучше всего видны в химии.
Идея активности в форме идеи сродства присутствовала в химии все-
гда, но лишь с утверждением атомизма (вторая половина XIX века) она
смогла из натурфилософского принципа начать обращаться в основу кон-
кретных теорий - теорий химических реакций. Различные ПМ чередовались
при этом в своем обычном порядке. (При изложении я опираюсь на факты,
заимствованные из трех вышедших томов (задумано было шесть) «Всеоб-
щей истории химии» - М., Наука, 1980-1983, ред. Ю.И. Соловьев.)
До Ньютона сродство понимали чисто знаково (первая ПМ): что с чем
может соединяться. Ньютон понимал его как аналог гравитации, а другие -
как аналог электростатики (вторая ПМ). Затем наступил период исследова-
ния балансов реакций (третья ПМ). Переход к системности (четвертой ПМ)
можно видеть в том, что многовалентный атом перестали рассматривать как
набор одновалентных (т.е. субатомов), а признали, что молекула как систе-
ма определяет свойства входящих в нее атомов и, стало быть, в разных
молекулах один и тот же атом может иметь разную валентность.
Замечательно, что системная (четвертая) ПМ завоевала химию практи-
чески одновременно с диатропической (пятой) в те годы (3-я четверть XIX
века), когда в целом наука едва осваивала четвертую ПМ при господстве
третьей. Были предложены гомологические ряды углеводородов (позже по-
служившие Копу, а затем Вавилову аналогом для гомологических рядов в
биологии) и самая красивая диатропическая схема - периодическая систе-
ма Менделеева. Причина столь быстрого продвижения видится мне в том,
что разнообразие веществ является для химиии главным объектом иссле-
дования. Вскоре начала развиваться и собственно химическая концепция
активности - различные теории элементарных актов реакций.
Первая из них - теория соударений - появилась в начале XX века и но-
сила примитивно механическую форму (считалось, что химическую связь
образуют те атомы, энергия столкновения которых преодолевает энергети-
ческий барьер реакции). Однако в 1935 году Генри Эйринг (США) придал
теории реакций чисто активностную форму, введя понятие активированно-
го комплекса - промежуточного возбужденного состояния (конфигурации)
группы атомов, в котором и происходит элементарный акт реакции. После-
дующие 70 лет развития квантовой химии являют собой, на мой взгляд,
процедуру понимания действия этого комплекса.
432
Любопытно, как этот комплекс, в сущности один и тот же, разные авто-
ры трактуют в понятиях той ПМ, какая им близка эмоционально. В недавней
статье двух биофизиков видим чисто механическую трактовку:
«центральная идея: ферменты... являются молекулярными машинами - конст-
рукциями, молекулярная механика которых производит химические реакции и
синтез биомолекул. Энергия такой машины, по Блюменфельду, создается в не-
равновесной конформации... именно она является макроэргом, первичным но-
сителем, который затем преобразует “механическую” энергию в химическую»
(Бучаченко АЛ, Кузнецов ДА. Магнитный изотопный эффект магния - ключ к
механохимии фосфорилирующих ферментов... // МБ, 2006, № 1, с. 12).
Однако действие этих машин «остается загадкой», и вот в 2002 году
биофизик Г. Остер привлек статистическую ПМ: «Остер назвал их дарви-
новскими броуновскими машинами» (там же, с. 13). Разумеется, понимания
больше не стало, и вот сами авторы неосознанно склоняются к системной
ПМ: в исследуемом ими процессе перенос электрона от АДФ к иону магния,
«ключевая стадия реакции», происходить вроде бы не может:
«Эта реакция сразу вызывает протест, она невероятна и, действительно, в
обычных условиях не происходит. Но именно в этой реакции объединяются
молекулярная механика и химия» (там же, с. 14),
т.е. она возможна лишь как системный эффект в ферментном комплексе.
В чем же общий смысл открытого ими «ключа»? Авторы выстраивают
ряд сходных реакций (как бы готовясь применить пятую ПМ), но опять пы-
таются всё свести к механике, и общий смысл остется (для меня) тайной. На
мой взгляд, механическая модель тут полезна не более, чем была полезна
планетарная модель атома при становлении квантовой теории спектров.
Видимо, прав Бульенков, полагая, что понимание принципа действия
«мотор-доменов» белков, в том числе ферментов, требует не выяснения
новых деталей, а новой ПМ (Бульенков Н.А. Роль модульного дизай на в
изучении процессов системной самоорганизации И Биофизика, 2005, № 5).
Он не называет ее, но в беседе со мной сказал, что она - активностная.
Многие натурфилософы утверждали, что вписать биопоэз в общую кар-
тину мира можно только путем смены самой картины мира, и, по всей види-
мости, они тоже правы. Другое дело - какую картину мира принять. Можно
делать это, оставаясь в рамках привычной рациональной науки и лишь ме-
няя исходные пункты рассуждений. Таковы, например, позиции Янча и Во-
ейкова. Расширяя эти рамки, мы в данной книге пользуемся единым прин-
ципом активности, так что биопоэз предстает как вырастание биологической
формы активности из химической.
Описанный Воейковым раствор выглядит живым, но с важным ограни-
чениями: образующийся полимер неустойчив и вне активного раствора бы-
стро распадается, а реакция полимеризации затухает через несколько ча-
сов. Живая система, наоборот, умеет сама себя сохранять. Следовательно,
в растворе аминокислот возникает не сама жизнь, а лишь некое поле жиз-
433
ни полимера, в пределах которого идет полимеризация. Замечательно, что
Воейков, начав с представлений витализма (что жизнь - первичное понятие,
ни из чего не выводимое), пришел к модели, позволяющей вывести разби-
раемое им свойство жизни из неживого вещества (в этом и состоит тезис
Воейкова). Тут мы переходим от химических форм активности к биологиче-
ским - в рамках активностной картины мира.
7-11. Активность и рождение поля жизни
Можно ли увязать глобальные круговороты, самосборку, наследствен-
ность, структурную энергию живого и способность к развитию (от субвиталь-
ной зоны до биосферы и от эобионта до многоклеточного) в единый эволю-
ционный процесс? Если не увязывать все эти процессы друг с другом, а по-
давать их порознь (как традиционно и делается, например, во всех учебни-
ках), то создается впечатление о наличии цепи удивительных совпадений,
что прямо ведет к антропному принципу в третьем смысле (п. 7-1), т.е. в по-
знавательный тупик. Если же пытаться увязать их между собой, то необхо-
дима связующая идея, и в качестве нее не раз предлагалась идея жизнен-
ного поля, называемая по-разному.
Напомню, что поле - одна из форм активности материи. Понимая про-
грессивную эволюцию как усложнение форм активности, мы должны искать
различные, всё более сложные, типы полей. Поле есть причина сродства
(в п. 5-2 мы видели это на примере гравитации), и встает вопрос: можно ли
всякому сродству сопоставить какое-то поле?
Биопоэз предстает как процедура встраивания органических систем в
неорганические потоки, где первые явно проявляли сродство ко вторым.
Сам акт рождения организмов из первичного ценоза был усилением сродст-
ва - теснее сопряженные части ценоза становились одним организмом.
Откуда сродство возникает? Наглядный пример - формирование ДС, и
для наблюдателя оно часто выступает как самосборка. Так, ДС, образую-
щаяся в водовороте (воронка), затягивает в себя плавающие предметы, и
тем самым они оказываются рядом, т.е. проявляют сродство, подобное гра-
витации, именно благодаря ДС. Однако нет оснований полагать некоторые
поля (например, гравитационное) итогом действия какой-либо ДС.
Формами сродства являются и симбиоз (например, гриба и водоросли в
лишайнике), и колониальность, и многоклеточность. Симбиоз выступает как
первичное явление, ослабевающее в ходе эволюции настолько, что при
изучении высших организмов воспринимается как особое приспособление.
С появлением колониальности (и тем более многоклеточное™) стал необ-
ходим апоптоз (п. 6-6**) - такое же средство замыкания круговорота, как и
симбиоз. Ведь апоптоз - это “тихий уход из жизни, малозаметный для окру-
жающих клеток, не причиняющий им ущерба”, тогда как некроз (простое
умирание клетки, часто с гниением) - это “взрыв, когда содежимое клетки...
изливается в межклеточный раствор” [Самуилов, с. 305].
434
Апоптоз - такой же итог сродства частей к целому, как фагоцитоз. То же
можно сказать о выедании половой касты термитов другими животными для
замыкания экосистемного круговорота (п. 6-14**). Остаётся снова напом-
нить, что сродство - тоже форма активности.
Здесь интересна позиция Галимова. Он рассматривает поле лишь как
молекулярное, зато дает нам понять разницу между биохимией и молеку-
лярной биологией: вторая вступает в права там, где надо рассматривать
индивидуальные судьбы молекул, т.е. там, где не работает закон действия
масс. Там, по-видимому, и следует говорить о появлении простейшего био-
логического типа активности. Это - поле крупных молекул:
«Присутствие молекулы способствует синтезу в ее силовом поле аналогичной
молекулярной структуры. Это - предпосылка к самокопированию... Неболь-
шие молекулы слишком подвижны. Они не создают в данном участке про-
странства стабильного упорядоченного поля, в силовом рельефе которого
может происходить формирование молекулярной реплики». И позже: «Упо-
рядочение происходит не на уровне вещества, а на уровне индивидуальных
молекул», причем важно «наличие механизма копирования возникшей моле-
кулярной структуры» [Галимов, 2005, с.60,474].
Причинность и здесь работает, хоть и не на уровне химии, а на уровне
физики субмикромира. Но, поскольку ныне так удается описать далеко не
всё, то у некоторых ученых возникает желание усомниться в справедливо-
сти самих наших приемов мышления, даже в умении увязывать причину и
следствие, т.е. в привычной рациональной науке. Применение понятия поля
может оказаться эффективно и здесь. Например, если физические (про-
стые) поля вызывают взаимодействие молекул в пространстве, то не суще-
ствует ли принципиально более сложное поле, создающее взаимодействие
тел во времени, меняя связь причины со следствием?
Этому вопросу посвящена «гипотеза формирующей причинности» анг-
лийского биохимика и натурфилософа Руперта Шелдрейка (1981). Его точка
зрения может показаться сумасшедшей (а может быть, таковой и является),
но она имеет давнюю философскую традицию и ставит вопросы, которые
иначе никто пока поставить не умеет. См. Дополнение 7.
Шелдрейк видит в ДНК не текст, а резонатор внешних сигналов, гене-
рируемых неким «морфическим» (т.е. формообразующим) полем. Теперь
так пишут многие, например, П.П. Гаряев)6^. Замечательно то, что Воейков
независимо пришел к тому же пониманию наследственности:
Разумеется, сторонники этой точки зрения не отрицают, что ДНК может служить тек-
стом для РНК и, тем самым, для белков. Речь о другом: нет никаких данных о том, что
в ДНК записано хоть что-то о форме, т.е. о пространственных отношениях внутри
клетки, между клетками, между тканями и т.д. Гены гомеобокса (п. 5-15) регулируют
поле, но не образуют его. Добавлю, что основная часть ДНК эвкариот (интроны и по-
вторы, см. п. 5-5) вообще не служит текстом для РНК. Но для чего-то служит.
435
«клеточные ядра действительно выступают в роли центров клеточных полей.
И благодаря исключительными свойствам хроматина (ДНК хромосом -
Ю. Ч.), они могут устойчиво сохранять колоссальный объем информации, а
также обмениваться ею с другими... полями. Если это так, то открывается аб-
солютно новый подход к объяснению природы наследственности и изменчи-
вости» {Воейков В.Л. И Знание - сила, 1996, № 4, с. 54-55).
Зусмановский ввел общее понятие «функционального поля», включаю-
щее поле потребностей [Зусмановский, с. 141]. Жизнь, с этой точки зрения,
возникла тогда, когда активность приняла форму функционального поля
жизни. К этому следует добавить: жизнь возникла тогда, когда проявились
ее основные атрибуты: активностью стало управлять давление нормы (п. 5-
2**), энергетикой - метаболическая спираль (п. 5-11), а прогенота распалась
на отдельные организмы (клетки) со сменой поколений.
К этому надо добавить следующее: А.М. Кузин, открывший вторичное
биогенное излучение (п. 7-10*), допускал, что
«именно в момент образования в “первичном бульоне” коацерватных капель,
перехода к протополимеров из растворенного в конденсированное состояние,
под действием атомной радиации... в них возникают вторичные биогенные
излучения», которые «образуют в ней своеобразное, ранее не существовавшее
электромагнитное поле, несущее информацию от одного полимера к другому,
объединяющее многочисленные полимеры и другие компоненты коацерват-
ной капли в единое целое, в простейший живой протоорганизм» [Кузин, с. 68].
Соединяя выраженные выше позиции, можно сказать, что «функцио-
нальное поле» черпает энергию не только из метаболизма - ему с самого
начала была необходима (в ничтожных количествах) еще и высококачест-
венная энергия радиационного фона. Таково на сегодня понимание поля,
порожденного жизнью и одновременно породившего жизнь.
7-12. Итог. Проблема четырех R
Итак, первые биоценозы возникли до первых организмов и затем обра-
зовали первую биосферу, поля которой с тех пор, видимо, и управляют эво-
люцией организмов; жизнь возникла путем поэтапного, но геологически бы-
строго замещения неорганических звеньев геохимических цепей на органи-
ческие (химических форм активности на биологические). Первые биоценозы
использовали потоки веществ и энергии из земных недр, что привело к соз-
данию сообществ, которые в настоящее время известны как гидротермаль-
ные. Эти ценозы сначала не составляли единой системы (биосферы), но
создавали для будущей биосферы материал - органические вещества и
атмосферу. Первые крупные организмы существовали в тесном симбиозе с
мелкими, т.е. также являли собой ценозы (п. 7-7).
Разумеется, эти положения не окончательны и, наверное, еще не раз
будут пересмотрены. Но загадка рождения жизни видимо прояснилась в од-
ном: решение не упирается в проблему появления всего сразу - мы уверен-
436
но делим биопоэз на последовательные стадии, что впервые наметил Бер-
нал (п. 7-4). В наши дни можно изложить ГЦК гораздо убедительнее, чем это
сделано в его блестящем докладе.
Пусть мы не понимаем, что такое биополе, но хотя бы вопрос постав-
лен. А есть проблемы, по сути и не поставленные. Их по меньшей мере две:
как возникло деление клеток и как возник аппарат наследственной изменчи-
вости? (Мы сразу видим изменчивость как рациональную активность.) Ника-
ких серьезных работ на сей счет мне не попадалось.
Первая клетка могла обходиться без оболочки - ее целостность обес-
печивал гель. Можно надеяться, что вскоре кто-то предложит модель появ-
ления замкнутой клеточной оболочки, синтез и распад которой управляются
находящимися внутри нее генами. Наоборот, вторая проблема представля-
ется мне на сегодня неприступной.
Ее можно обозначить как проблему четырех R - рекомбинации, репа-
рации и репликации ДНК и редактирования РНК65. Все рассуждения о “мире
РНК” приближают лишь к пониманию возникновения транскрипции РНК, ко-
торая при этом выступает как исторически первая процедура чтения генети-
ческого текста. Репарация (починка) ДНК видится на сегодня неотъемлемой
частью генетической системы. Репликация ДНК несравненно сложнее, даже
в ее простейшей форме “катящегося кольца”, чем транскрипция РНК, и по-
нять ее становление никто, насколько знаю, не пытался.
Но гораздо хуже то, что таковое понимание вряд ли само по себе что-то
даст для проблемы биопоэза, поскольку первые три R выглядят единым
функциональным целым, а четвертое R тоже грозит оказаться его частью.
(Только система репарации, возможно, возникла до системы репликации -
как система починки аппарата транскрипции.) В этой ее части проблема
биопоэза остается проблемой “как же всё появилось сразу?”
Выходит, что неприступность проблемы просто отодвинулась. Мы как
бы взяли крепость, но, поднявшись на крепостную стену, увидали внутри
нее поражающий своей неприступностью акрополь. Овладеть им можно,
полагаю, лишь с позиций новой картины мира и новой ПМ (пп. 7-9, 7-10*).
Гпава 8. Гпавные черты процесса эволюции
Рождение жизни ныне принято видеть как появление в первых биоцено-
зах (прогенотах раннего архея66) первых отдельных организмов, т.е. клеток.
Мы будем придерживаться хронологии, данной на рис. 47, в рамках которой
65 Три R, касающиеся ДНК, звучат в известных мне языках сходно (латинская основа), и
“редактирование” тоже звучит сходно в разных языках, однако в английском слово the
redaction почти вытеснено словом the edit, откуда и термин the RNA-editing.
66 Далее прямым полужирным шрифтом выделено первое упоминание геологических
эпох и периодов. См. рис. 47.
437
ранние группы появлялись в следующем порядке (в миллиардах лет назад)
[LEG, 12; Заварзин, 2006]:
Первые следы жизни (изотопный сдвиг) 3,85
первые бактерии 3,8
цианеи (синезеленые водоросли) 3,5
одноклеточные эвкариоты 3,0 -2,7
грибы 2,4-2,2
многоклеточные растения 2,2-1,9
многоклеточные животные 1,8-1,5
целоматы (животные, имеющие полость) 1,4-1,0
Эра Период Время, млн лет назад
КАЙНОЗОЙ ЧЕТВЕРТИЧНЫЙ 1,7 67 137 195 230 285 350 405 440 500 570 680 1700 2700
ТРЕТИЧНЫЙ
МЕЗОЗОЙ МЕЛОВОЙ
ЮРСКИЙ
ТРИАСОВЫЙ
ПАЛЕОЗОЙ ПЕРМСКИЙ
КАМЕННОУГОЛЬНЫЙ
ДЕВОНСКИЙ
СИЛУРИЙСКИЙ
ОРДОВИКСКИЙ
КЕМБРИЙСКИЙ
ПРОТЕРОЗОЙ ВЕНДСКИЙ
РИФЕЙ
КАРЕЛИЯ
АРХЕЙ
Последние К ООО лет
/^Плейстоцен
| Плиоцен
] Миоцен
J § I-——----------
g g. 1 Олигоцен
Й- । Эоцен
I—----------------
I Палеоцен
1.7млн
10 млн
25 МЛН
ТЯ млн
49 млн
67 млн
Рис. 47. Геохронологическая таблица.
Внизу более крупным планом
изображено членение кайнозоя
Около 700 млн лет назад появились
организмы, оставлявшие после себя
твердые остатки (раковины, скелеты),
составляющие основной предмет
изучения палеонтологии. На эту тему
есть множество хороших книг, напри-
мер [Ивахненко, Корабельников',
Кэрролл; Мейен, 1981]. Поэтому
здесь мы лишь посмотрим, как новое
знание о механизмах помогает
понять некоторые ключевые события
и процессы эволюции.
8-1. Различные понимания
процесса эволюции
Для самых ранних эволюционистов
(их принято называть трансфор-
мистами) понять эволюцию -
значило всего лишь одно: убедиться,
что организмы в ходе истории Земли
изменялись. Для первых геологов
задача состояла уже в большем - по-
нять, кто за кем жил.
Вероятно Эразм Дарвин был
первым, кто поставил вопрос о
причинах эволюции: у него главные
из них - огромное время и собственная активность живого. Этим он дап
начало как учению о факторах (элементарных движущих силах) эволюции,
438
так и тому конкретному пониманию этих факторов, которое известно нам
под именем ламаркизма. Теперь понять эволюцию значило - узнать ее ме-
ханизмы. Тех, кто ищет их в работе самого организма, будем называть фи-
зиологами. К сожалению, эволюционисты-физиологи, много думая о меха-
низмах эволюции, мало озабочены изучением ее процесса.
Ламарк увязал идею Эразма с данными геологии и объявил активность
живого основным фактором эволюции высших животных; растения же и
низшие животные у Ламарка изменяются прямо под влиянием среды. Чарлз
Дарвин уточнил таковое влияние (среда отсекает негодные случайные ук-
лонения) и отбросил активность особи как нечто лишнее (на самом деле
признав активность в форме избыточности размножения и изменчивости).
Изменил он и роль времени: хотя время эволюции мыслится в дарвинизме
(как и в ламаркизме) неограниченным, но само по себе оно тут ничего не
значит, и если условия жизни не меняются, организмы должны оставаться
неизменными, ибо дарвинизм целиком (ламаркизм лишь частично) видит
эволюцию как цепь приспо-соблений организма к среде.
Парадокс в том, что задачу трансформистов решило лишь появление
дарвинизма: факт изменения организмов в ходе истории Земли был при-
знан наукой и обществом. Поэтому и процесс, и механизм эволюции стали
понимать по Дарвину - как происхождение видов.
Для происхождения прочих таксонов (родов, семейств и т.д.) при таком
строе мысли ни особого процесса, ни особого механизма не предлагается.
Молчаливо предполагается, что все эти таксоны образуются в принципе так
же, как и виды, т.е. путем многократных видообразований. Таким образом,
видообразование выступает как основной акт эволюции. Далеко не все с
этим согласны, и в следующем пункте мы к данному вопросу вернемся.
Геккель объединил Ламарка с Дарвином (активное приспособление и
отбор) и дал новый способ понимания эволюции: понять ее - значит указать
генеалогию (какой таксон от какого произошел). Так появились филогене-
тики, понимающие эволюционную науку как построение родословных древ.
Их большинство, к ним относятся все дарвинисты, но в науке бытуют и дру-
гие подходы. Кратко говоря, они сводятся к поиску не родственных свя-
зей, а механизмов эволюции. (Это - главная тема книги [Лима].)
Кроме приспособления, в эволюции важны такие явления, как прогресс
и разнообразие. Эти три феномена друг к другу не сводятся, хотя дарви-
низм считает иначе. Понятие прогресса ввел Ламарк, а в основу морфоло-
гического учения его положили Бэр и Брони (п. 2-10). Затем Берг, основа-
тель номогенеза, вновь сделал прогресс главным эволюционным понятием.
Дарвин прогрессом почти не интересовался, тогда как Ламарка почти не
занимало разнообразие, а Берга - приспособление. Легко догадаться, что
реального понимания эволюции организмов можно ожидать только в рамках
теории, которая рассмотрит все три феномена вместе (и рассмотрит, кроме
эволюции таксонов, эволюцию сообществ, но о ней речь пойдет особо).
439
Пренебрегая физиологией, номогенез оказался односторонним, однако
блестящий вклад его видится мне в том, что ясно показано - эволюция со-
стоит в постепенном заполнении рефренных таблиц. Порядок заполнения
клеток таблицы (ход эволюции) может быть различным. Для случая рефре-
на «Типы расчленения листа», рассмотренного в п. 6-3***, Мейен предлагал
рисунок, показывавший невозможность выяснения конкретного пути, каким
был достигнут данный тип сложного листа (рис. 48). В таких случаях фило-
гению построить нельзя, и справедлив вопрос Б.И. Кудрина:
“не ставят ли палеонтологи нереальную задачу: восстановить родство по иско-
паемым остаткам, которые утратили идеальную сторону информации, неиз-
бежную при инфор-
мационных процес-
сах?” {Кудрин, с. 39].
Рис. 48. Возможные
пути эволюции типов
расчленения листа (по:
Мейен, 1973). Одного
итога можно достичь
разными путями
Другой подход обы-
чен для физиологов.
К ним, кроме ламар-
кистов в указанном
смысле (сторонни-
ков собственной
активности особи как ведущего фактора эволюции) и жоффруистов (видя-
щих основной фактор в изменении зародышей под влиянием среды), надо
отнести и тех иммунологов, которые видят в иммуногенезе теплокровных
модель эволюции. Главных заслуг физиологов я вижу две: на уровне орга-
низмов они указали схему приспособления, объяснившую, как приспособле-
ние возникает («свойства приобретаются поочередно»), а на уровне моле-
кулярном - схему, объяснившую, как приспособление наследуется: при
стрессе возникает генетический поиск; он лучше всего изучен в иммуногене-
зе. Однако есть и более общая (чем стрессовая) физиологическая схема
эволюции - расширение активности при снятии давления нормы.
Третий подход к эволюции предлагают физики:
«Наблюдаемый мир соткан из взаимодействий, все ими пронизано, все ими
движется. Материя - это вещество плюс поля взаимодействий. Вещество со-
стоит из молекул, молекулы - из атомов, атомы - из элементарных частиц,
элементарные частицы представляют собой сгустки полей взаимодействий,
так что вещество - это те же поля взаимодействий, только “сгустившиеся” и
440
приобретшие благодаря этому новые свойства» (Хайтун С.Д. Социум против
человека. Законы социальной эволюции. М., УРСС, 2006, с. 37).
Мы уже воспользовались данным подходом, когда вооружились поня-
тием активности. Не будем, однако, думать, что им всё решается.
Есть четвертый подход, холизм (от греч. холос - целый). Холисты - те,
для кого понять эволюцию - значит осознать целостность эволюционного
процесса, его системность. Конечно, о целостности (системности) говорят
все, но приверженцы этого подхода кладут ее в основу всех своих рассуж-
дений, тогда как остальные (большинство биологов) до сих пор рассуждают
о приспособлении вида к изменениям среды (что с холистической точки
зрения бессмысленно). Идею целостности настойчиво разъяснял Любищев
(см. Марасов А.Н. Единство целого при свободе частей - метаидея А.А. Лю-
бищева // ЛЧ, 1992), но тогда еще не настал ее час.
Она и ныне понимается медленно и трудно. Хотя еще Геер, Соболев,
Шиндевольф, Мейен и некоторые другие оперировали с эволюцией как с
преобразованием флор и фаун, хотя экологи давно говорят об эволюции как
о преобразовании экосистем, однако многим удобнее видеть два разных
процесса: первичный - эволюцию таксонов, и вторичный, вытекающий из
нее - перестройку экосистем. Почему вытекающий? Да потому, говорят эти
ученые, что эволюция таксонов - цепь приспособлений организмов к среде,
а эволюция экосистемы из этих приспособлений якобы и состоит.
Но легко видеть, что это не так. Во-первых, среда может меняться
мгновенно (при затоплении, вселении хищника, гибели источника пищи и
т.п.), а если вид должен приспособиться к тому, что меняется быстрее, чем
он сам, то называть его приспособление первичным нет смысла. А во-
вторых, таксон от таксона лишь изредка отличается приспособительными
признаками (и на этом строится систематика - см. гл. 10). Нужно искать дру-
гой, целостный, подход к пониманию процесса эволюции.
Повторяя многих мыслителей, Тейяр де Шарден более полувека назад
заметил: физика, изучая элементарные взаимодействия частиц, видит мир
как скрепляемый ими, т.е. снизу (так называемый редукционизм). Однако
«Более полное наблюдение за движениями мира вынудит нас мало-помалу
перевернуть эту перспективу, то есть открыть, что вещи держатся и держат
друг друга лишь в силу сложности, сверху» [Тейяр, с. 45].
Прошло полвека, и теперь физики сами видят мир «сверху», т.е. третий
и четвертый подходы соединились. Как биосфера, так и каждая ее экоси-
стема непрерывно приспосабливается сама к себе (подобно тому как при-
спосабливается сам к себе ребенок и как приспосабливается каждый из нас
к своим болезням). Это - примерно то, что называл подбором Богданов.
Кроме процесса приспособления к себе, экосистемная эволюция вклю-
чает поиск и заселение организмами новых экологических ниш. Такая эво-
люция носит характер фрактального роста (п. 6-14), что ведет, в частности,
к подобию медленных процессов быстрым.
441
Чаще всего холисты выступают в эволюционной литературе как гло-
бальные экологи, но бывают и далекие от экологии, например виталисты.
Пятый подход - финалисты, т.е. те, кого занимают не столько родо-
словные и механизмы эволюции, сколько тот финал, к которому она, по их
убеждению, должна прийти (о финализме см. [Назаров, 2005]). Первым фи-
налистом можно назвать Августина (п. 1-8), считавшего, что мир вступил в
свою последнюю историческую эпоху, за которой последует Страшный суд.
У него было много последователей; из них в данной книге мы говорили о
Тейяре. Он видел эволюцию как процесс, направленный к заранее наме-
ченной Богом (но отнюдь не страшной) конечной цели.
По сути, описание процесса эволюции зиждется на двух блоках данных
- палеонтологических и диатропических. Палеонтология указывает, в ка-
кой последовательности и в каких условиях жили организмы прошлого, ее
данные наиболее объективны и надежны, но касаются лишь некоторых
свойств некоторых вымерших организмов. Диатропика, наоборот, сравнива-
ет любые организмы по любым свойствам, и потому сравнительный метод
служит основным источником сведений о функциях древних организмов, но
он бывает обманчив (п. 5-16). Нужно сочетать оба блока, а они, увы, часто
друг другу противоречат.
Итак, есть много способов понимать процесс эволюции. Что касается ее
механизмов, то они, насколько они мне понятны, собраны в части 2.
8-2. Эволюция и систематика царств
Ныне систематику трактуют эволюционно: таксоном мы считаем группу
родственных организмов. Так, собаки больше похожи друг на друга, нежели
другие псовые (например, песцы), псовые - нежели другие хищные и так
далее. Общность эту объясняют как происхождение от общего предка: пре-
док вида «собака» жил позже, чем общий предок семейства псовых, тот -
позже, чем общий предок отряда хищных и т.д.
Казалось бы, проще всего проследить фактический ход эволюции, гля-
дя на систему организмов - сперва появились царства (точнее, первые ви-
ды, от которых мы ведем историю царств), затем типы (точнее, виды одно-
го царства, принадлежащие к разным типам), за ними классы и т.д. Долгое
время ученые так и описывали эволюцию, но это оказался тупик. Ведь вся-
кая эволюционная система построена на конкретном понимании эволюции,
так что ничего нового об эволюции, глядя на нее, не узнаешь. Систему надо
строить впараллель с познанием эволюции - вот мысль, которой мы будем
следовать. (Есть иная точка зрения - что систематику надо строить незави-
симо от теории эволюции; ее защищали Любищев и Корона, см. гл. 10.)
Эволюцию таксонов не выше типа (отдела) можно понимать как пере-
делку планов строения (скачком или постепенно), а эволюцию царств так
представить нельзя. О связи эволюции и систематики царств речь пойдет в
главе 10, здесь же замечу только, что нам предстоит понять, как прогенота
442
смогла породить царства. Это и будет эволюционная систематика царств.
Самих царств мы положим, как почти всеми принято, четыре: бактерии,
растения, грибы и животные. Иногда добавляют пятое царство, объеди-
няя в него всех одноклеточных эвкариот, но его не удается построить фак-
тически. Подробнее см. главу 10.
Это деление исходит из эколого-морфологического понимания царст-
ва. Уиттекер (о нем см. п. 6-13) разделил живой мир на два надцарства по
принципу строения клетки: про- и эвкариоты. Всех прокариот он полагал
единым царством, а эвкариот делил на три царства по способу питания:
1) Растения - это продуценты (автотрофы; создают органику).
2) Грибы - сорбенты (всасывают пищу, подобно бактериям); они же
являются редуцентами (разлагают органику).
3) Животные - консументы (поедают пищу; преобразуют органику без
качественного повышения или понижения ее химической сложности). Они
же являются фаготрофами, т. е. механически захватывают пищу.
Такое членение естественно продолжает идею Линнея, отмечавшего,
что растения производят, а животные потребляют. Мы этому последуем.
После Уиттекера выяснено еще одно фундаментальное обстоятельст-
во: среди прокариот имеются первичные продуценты, делающие органику
из неорганики земных недр (это - литотрофы), тогда как растения лишь
замыкают круговорот органики и, тем самым, служат вторичными проду-
центами (п. 7-7). Среди прокариот имеются также и редуценты, т.е. орга-
нику разлагают грибы и бактерии.
Итак, имеются четыре главные трофические функции - первич-
ные продуценты (литотрофы), вторичные продуценты (хемо- и фо-
тосинтетики), консументы (животные) и редуценты (грибы и редуци-
рующие бактерии). Прокариоты выполняют три функции: первичных проду-
центов, вторичных продуцентов и редуцентов (это гнилостные бактерии и
прокариотные аналоги грибов). Эвкариоты тоже выполняют три функции, но
иные: вторичных продуцентов (растения, т.е фотосинтетики), консументов
(животные) и редуцентов (грибы). До появления животных биологические
круговороты уже существовали.
Естествен вопрос: если так, то почему мы не делим прокариот тоже на
три царства? Ответ будет дан в главе 10, после введения понятий ядра и
периферии таксонов. Пока же замечу, что систематику прокариот вообще
невозможно строить на общих с остальными организмами основаниях, по-
скольку в силу горизонтального переноса генов, который у них регулярен,
«геном прокариот мозаичен» [Заварзин, 2006, с. 167].
8-3. Первые бактерии
О том, как появились первые клетки, ныне ничего не известно, вопрос
почти не обсуждается, и могу сказать об этом совсем немногое. Наиболее
полная работа (Martin W., Russel M.J. On the origin of cells: a hypothesis for the
443
evolutionary transitions from abiotic geochemistry to chemoautotrophic procaryo-
tes, and from procaryotes to nucleated cells // Philosoph. Trans. Roy. Soc. Lon-
don, Ser. B, 2003, vol. 358, p. 59-85) признаёт, что существует, но не обсуж-
дается, проблема возникновения клеточной мембраны, однако сама обсуж-
дает только вопрос о происхождении на ней окислительно-
восстановительной биохимии и мембранных липидов. Генетическая сторона
дела (каким образом геном оказался внутри оболочки и стал управлять ее
синтезом и делением) оставлена без обсуждения.
Физиолог Ю.В. Наточин (Петербург) предлагает различать два термина
- протобионт и протоклетка (второй отличен от первого наличием (ци-
топлазматической мембраны, т.е. клеточной оболочки). Он пишет:
«Ни в одной работе не удалось найти гипотез или обсуждения вопроса о вре-
мени и условиях возникновения плазматической мембраны» {Наточин Ю.В.
Роль ионов натрия как стимула в эволюции... // ПЖ, 2005, № 4, с. 20).
Еще, как минимум, одна работа, как видим, была, но в целом он прав.
«Вопрос, который необходимо обсудить - была ли на начальном этапе у про-
токлетки плазматическая мембрана со способностью селективного отношения
к отдельным ионам. Существенно обсудить вопрос, когда эта мембрана воз-
никла, что послужило стимулом к ее появлению» (там же).
По-моему, еще существеннее сперва обсудить вопрос: как мембрана
могла возникнуть? Как протобионт стал протоклеткой? Наточин его не по-
ставил, сразу перейдя к необходимым свойствам мембраны (проницае-
мость, транспорт ионов). Но если мы примем (вслед за Насоновым и в от-
личие от Наточина) во внимание, что основа клетки - гель (а не раствор), то
проблемы проницаемости и транспорта отойдут на второй план, поскольку
гель сам может избирательно адсорбировать молекулы.
Единственная известная мне модель образования оболочки исходит из
идеи клетки как раствора, однако содержит важное положение: условия для
создания первых оболочек возможны в микробрызгах. Когда океан доста-
точно остыл, а атмосферное давление упало67, то в игру смогли вступить
поверхностные явления, невозможные в прежней “скороварке".
При низкой влажности воздуха тонкий поверхностный слой воды (0,1-
0,3 мм) охлаждается примерно на 1 градус, а «градиент температуры в тон-
ком слое может вызвать термодиффузные процессы, приводящие к разде-
лению ионов и других компонентов», характерному для внутриклеточной
среды. Эту среду локализуют и фиксируют микробрызги (полости размером
с клетку, окруженные гидрофобной оболочкой {Яковенко Л.В., Твердиспов
В.А. И Биофизика, 2003, № 6). В таких условиях на поверхности океана
вполне могла идти завершающая стадия биопоэза - появление клеток, но,
_67 Возможно, что плотная первичная атмосфера была сорвана в ходе космической ката-
строфы, как это, по всей вероятности, произошло с Марсом. См. [Резанов].
444
повторяю, непонятно, каким образом внутри “клетки” мог оказаться весь ге-
ном, притом управляющий ее делением. Биофизик Л.В. Яковенко (как и все
прочие) проблему появления управляемого деления клетки не упоминает,
однако увязывает физику тонкого поверхностного слоя с тем, что
«в структуре воды экспериментально обнаружены гигантские гетерофазные
кластеры... обладающие отличными от континуальной воды физико-
химическими свойствами» {ЯковенкоЛ.В. Автореферат..., с. 35).
Замечу, что слово гетерофазные здесь фактически означает гель, и,
следовательно, обе теории (п. 7-3) смыкаются. По-моему, надо сопрячь де-
ление капли по Фоксу с биофизикой капли по Яковенко и теорией геля.
Что касается непонятности феномена управляемого деления, то она
связана с гораздо более общей нехваткой - отсутствием теории онтогенеза:
мы фактически не знаем почти ничего о том, как гены управляют построени-
ем тел в пространстве (п. 5-15). Единственной фактической зацепкой на се-
годня служит сообщение, что у паразитической бактерии Wolbachia оболоч-
ку видимо строит ее хозяин (об этой удивительной бактерии см.: [Марков,
Захаров]). Данный факт ничего не доказывает, но наводит на мысль, что
первую оболочку могла строить прогенота вокруг одной из своих протокле-
ток; что управление оболочкой было отдано геному протоклетки позже.
Первую клетку принято представлять как предельно простую бактерию.
Вероятно, что первые бактерии обитали в редких оазисах жизни, которыми
были гидротермы. Когда они завладели океаном, неизвестно, зато есть сви-
детельство того, что через почти 1,5 млрд лет, т.е. 2,4 млрд лет назад (то
был Карелии) бактерии формировали первые почвы суши.
Сам же океан в то время составлял, как полагают, около трети нынеш-
него по объему и имел среднюю глубину меньше километра (ныне она рав-
на 3170 м). Неясно, когда он приобрел нынешний объем и солевой состав
(Клиге Р.К. и др. История гидросферы. М_, 1998, с. 59, 114).
Хотя всё тут весьма проблематично, но одно несомненно - что первые
бактерии были гораздо проще нынешних. Это ясно хотя бы из того, что все
нынешние бактерии имеют одинаково сложную систему репликации ДНК,
транскрипции и прочего «клеточного хозяйства».
Как же тогда выглядели первые микробы? Тут разнообразие нынешних
микроорганизмов оказывается нам полезно, но не для поиска среди них
предка, а в другом плане: оно дает понять, каково самое простое строе-
ние, какое достаточно для существования.
Самыми мелкими и самыми просто устроенными из ныне живущих бак-
терий являются молликуты (микоплазмы)65. Слово “молликуты” означает
Размер: 0,3-0,8 микрона. В 1999 г. описаны еще меньшие микробы, нанобактерии,
размером 0,1-0,2 микрона. Они бывают паразитами, но известны и в гидротермах, и в
океане. Нанобактерии очень важны для понимания начала жизни, но молекуляное строе-
ние их еще не изучено {Спирин А.С. И ПЖ, 2005, № 4, с. 25).
445
“мягкокожие" - они не имеют клеточной стенки, так что от внешней среды их
отделяет только цитоплазматическая мембрана.) Они также не имеют ни
жгутиков, ни органелл внутри клетки, а метаболизм их предельно прост (в
частности, вся энергетика сводится к гликолизу). Молликуты имеют самый
короткий геном. Он едва обеспечивает самый простой метаболизм, да и то
- в достаточно богатых средах, лишь втрое превышает геном самого круп-
ного вируса и втрое-пятеро короче чем у бактерии Escherichia coli (кишечная
палочка). Это тоже удобно для аналогии, если первые бактерии жили в про-
странстве прогеноты “на всем готовом”.
Молликут известно всего 7 или 8 родов, из которых 6 - паразиты, ино-
гда нестрогие; а один род термоплазма живет свободно в умеренно горячих
источниках и в молекулярном смысле близок к архебактериям.
Сочетание трех качеств термоплазмы - предельно простая для непара-
зита конструкция, умение жить самостоятельно вблизи гидротерм (а они в
гл. 7 описаны как возможная колыбель жизни), отсутствие аппарата под-
вижности*^9 и сходство с остальными архебактериями - все это делает тер-
моплазму похожей на первичный организм.
Иные говорят даже, что она в самом деле дожила до наших дней со
времен начала жизни, но это невозможно. Во-первых, она - аэроб, т.е. при-
способлена жить в кислородной атмосфере, каковой в начале жизни, как
полагают, не было. Во-вторых, с тех пор она должна была прожить более
триллиона поколений, и ей потребовался бы фантастически точный меха-
низм копирования, чтобы за это время не измениться до неузнаваемости; от
первичного организма такого ждать не приходится.
Предки термоплазмы наверняка эволюировали и, в частности, оставили
ей в наследство развитую систему синтеза РНК, ДНК и белков. Кроме того, в
силу горизонтального переноса предками термоплазмы могли быть (и, пола-
гаю, были) очень различные прокариоты.
По идеологии нового номогенеза, эволюция состоит в заполнении объ-
ективно существующей диасети, которая задает принципиальные возмож-
ности строения (план строения каждого таксона) и возможную изменчивость
в их рамках. Клетка диасети, задающая облик и функции термоплазмы, мог-
ла вновь и вновь заполняться (путем изменений других, близких, бактерий),
причем одно и тоже приобретение могло достигаться разными путями. При
таком понимании эволюции вполне естественно, что очень физиологически
и экологически различные организмы имеют сходное строение.
Первые организмы были, вероятно, похожи на нынешних молликут и
дали начало остальным прокариотам. Эвкариоты, вероятно, появились поз-
же прокариот (хотя, по всей видимости, произошли тоже в прогеноте), раз-
делив биосферу на два надцарства (мы узнаем о них в гл. 10).
® Иногда им приписывают наличие жгутика, но это - явная и грубая ошибка.
446
До сих пор эти два типа клеток занимают два взаимно дополнительных
места в биосфере, что мы видели в п. 6-9. Очень раннее обособление над-
царств видно из того, что они независимо обрели многие аппараты и, в ча-
стности, механизмы подвижности. Прокариоты могут как скользить по по-
верхности, так и обладать жгутиками. Их жгутики имеют в основе белок
флагеллин и электрохимическую энергетику; они не имеют ничего общего
со жгутиками эвкариот, построенных на микротрубочках, сделанных из бел-
ка тубулина и берущих энергию от АТФ (Метлина А.Л. Жгутики бактерий...
// Биохимия, 2004, № 11). Аппарат наследственности молликут виден в ос-
нове “хромосом” бактерий и органелл эвкариот (митохондрий и хлоропла-
стов - см. пп. 6-5*, 8-6*). Наоборот, ядерные хромосомы эвкариот устроены
по принципу плотной упаковки, характерной для вирусов (вирусы известны
уже у бактерий).
Из того факта, что эвкариоты выглядят очень древними, нельзя делать
вывод, что они равновозрастны прокариотам. Вернее, что эвкариотный
принцип основан на использовании достижений прокариотного, уже вырабо-
тавшего те биохимические механизмы, без которых жизнь невозможна. Мы
вернемся к этим вопросам в п. 8-5 и п. 8-7*.
Ныне общепризнано, что
«прокариоты играли и играют главную роль в биосфере Земли. Огромное их
разнообразие в сочетании с устойчивостью прокариотных систем заставляют
предполагать особые принципы эволюции в мире прокариот: горизонталь-
ный перенос генов, формирующих сетевую картину эволюции, и адаптивная
динамика сообществ создают единую систему, причем наиболее тесные
функциональные связи наблюдаются как раз между филогенетически дале-
кими прокариотами» [Заварзин, 2006, с. 166].
Всё это верно, но стоит добавить: сетевую картину эволюции структура
диасети обеспечивает и при отсутствии горизонтального переноса.
8-4. Эволюция клетки: принцип компенсации
Как сказано выше, для архебактерий характерно усложнение молеку-
лярных структур в ущерб усложнению строения клетки. По всей видимости,
так было всегда - одни преуспевали в чем-то одном, другие в другом. В
этом состоит принцип компенсации Аристотеля (п. 2-7). Некоторые молеку-
лярные новшества архебактерий (например, экзон-интронная структура ге-
на) нашли место в организации эвкариот, которым, в свою очередь, при-
шлось дорого платить за свою сложность: они сильно отстают от прокариот
биохимически. Есть сведения, что вычленение интронов из цепи ДНК воз-
можно без участия белков; если так, то оно может быть крайне старо - даже
атрибутом РНК-ового мира [Гапимов, 2001, с. 149].
Поразительный пример компенсации: паразитическая бактерия Wolba-
chia столь проста устройством, что ее свойства используют для реконструк-
ции происхождения жизни (пп. 7-8*, 8-3), но ее поведение сложнее, чем у
447
любых иных клеток: она обладает удивительной системой шифров. Из мно-
жества ее свойств отмечу одно. Вольбахия заражает самок насекомых и че-
рез них размножается. Заражать таким образом самцов она не умеет, зато
умеет заражать их совсем иным образом: ставит на их геном метку, которая
позволяет им эффективно спариваться только с заражеными самками. На-
оборот, все зиготы, полученные от спаривания зараженных самцов с неза-
раженными самками, гибнут [Марков, Захаров].
Эта вычурная агрессия против «чужих» (незараженных) самок выгля-
дит избыточной активностью, использующей чрезмерно развитый (в ущерб
всему остальному) эпигностический аппарат. Пример хорошо вписывается в
несколько рефренов, но не становится от этого менее удивительным. Пред-
лагаю читателю сопоставить его со столь же загадочными примерами избы-
точной активности, приведенными в Дополнении 7.
Временем широкого распространения эвкариот принято считать сред-
ний рифей, а в позднем рифее (почти миллиард лет назад) морские мелко-
водья были уже полны многоклеточных эвкариот - бесскелетных животных,
известных нам по следам их жизнедеятельности (норки, следы перемеще-
ния по сырому грунту), а также водорослей, известных по их отпечаткам.
Вероятно, появление многоклеточных связано с ростом концентрации ки-
слорода в атмосфере и в воде, поскольку переход от брожения к дыханию
позволил организмам расходовать пищу экономно, а потому иметь клетки,
не соседствующие с внешней средой. Однако пока Ог было еще мало (в
воздухе - менее 1%), организмы вынужденно оставались тонкими пластин-
ками. Вероятно, некоторые напоминали плоских червей, но имели ли они
основные их качества (двустороннюю симметрию, кишечник и т.д.), сказать
невозможно. Довольно определенно говорят лишь, что в позднем рифее
были губки. Растительные же остатки представлены, в основном, стромато-
литами (п. 7-7) мелководья.
Лучше известен следующий период, венд. В море появились организ-
мы, способные попасть в ископаемую летопись: с накоплением О2 менялась
биохимия организмов - водоросли стали выделять известь, и животные
стали достаточно плотными, чтобы оставлять отпечатки. Быстрая эволюция
была, как считают, связана с резким ухудшением условий жизни. На границе
рифея и венда материки вероятно сместились так, что Южный полюс ока-
зался в середине Африки, а Северный - в Скандинавии; если так, то это вы-
звало оледенение суши, а ледники на суше влекут понижение уровня океа-
на и рост его солености. На уцелевших для жизни мелководьях стало не-
уютно и тесно, что означает стрессовые условия жизни. Как сказано в главе
5, такие условия способствуют быстрой эволюции немногих выживающих.
Напомню, что длящийся миллионы поколений стресс - всего лишь ме-
тафора: длительный стресс гибелен для особи. Поэтому лучше говорить
про снятие давления нормы, лишь начало которого имело форму стресса.
448
По самой своей сущности оно безвредно и может действовать долго -
впредь до установления новой нормы.
Термин «прогресс» биологи прилагают в основном к формам строения,
полагая, что эвкариот прогрессивнее прокариота, многоклеточный - одно-
клеточного и т.д. Но с позиции принципа компенсации всё не так. В частно-
сти, биохимически бактерии прогрессивнее нас. Принято считать, что они
давным-давно не усложняются. Даже рассудительный Шалин смог напи-
сать, что бактерии,
“живущие в нашем кишечнике, сохраняют способы жизни, в точности иден-
тичные приобретенным три миллиарда лет назад. Эти вполне адаптированные
бактерии не подверглись существенным изменениям” (АЕ, 165).
Неверно: они выживают в кислородной атмосфере, которой всего 2
млрд лет. Более того, они могут жить в таком кишечнике, какого не было
еще даже в мезозое, так что нет оснований считать их самих старше. И во-
обще, большинство родов ныне живущих бактерий несет следы
приспособления к эвкариотам.
Нынешние молликуты - итог эволюции и несут следы упрощения: так,
геном Mycoplasma genitalium, самый короткий среди всех организмов (580
тыс. пар нуклеотидов), целиком содержится в геноме М. pneumoninae (Мо-
мыналиев К.Т., Говорун В.М. Механизмы генетической нестабильности мол-
ликут (микоплазм) // Генетика, 2001, № 9) - ясно, что первый произошел из
второго путем утрат. Остальные прокариотные генетические механизмы у
молликут имеются.
Полагаю, что прокариоты потратили на свое биохимическое усложне-
ние как раз те 2 млрд лет, которые у эвкариот ушли на эволюцию форм.
Некоторое представление о том, как могла идти эта биохимическая
эволюция, дает исследование нынешней эволюции лекарственной устойчи-
вости болезнетворных бактерий: она идет быстро, направленно и комплекс-
но (сразу по нескольким направлениям); горизонтальный перенос при этом
регулярен, но не част, а обычные мутации выполняют лишь роль тонкой
подстройки [Прозоров, 2000, с. 522, 523, 526], как и в иммуногенезе. Веду-
щим механизмом, как и в остальной эволюции, мне здесь представляется
редактирование РНК.
Феномен диасети приводит к тому, что сходные формы появляются
вновь и вновь, и у нас нет гарантии, что постоянство ископаемых форм не
таит усложнения биохимических процессов. А процессы внутри клетки столь
сложны, что на их отработку явно нужно было огромное время.
Что касается нынешних одноклеточных эвкариот, то их эволюция со-
стояла в усложнении не столько биохимии, сколько геномной организации и
клеточного строения. Стоит глянуть в микроскоп на инфузорию, чтобы по-
нять, что ее единственная клетка устроена много сложнее, чем любая из
клеток высших. Иногда ее строение именуют даже не одноклеточным, а
сверхклеточным (п. 8-8*).
449
Другой тип сверхсложности - у низших многоклеточных. Всякий, кто
рассматривал в микроскоп губок, должен был удивиться, насколько их клет-
ки и скелеты неожиданно сложны. И это не исключение. В 1925 году гисто-
лог А.А. Заварзин (о его племяннике, бактериологе Г.А. Заварзине, мы гово-
рили не раз), работавший тогда в Перми, отмечал, что не обнаруживается
«никакой взаимосвязи между высотой организации животного и его гистоло-
гической структурой, а гораздо чаще совсем наоборот». Он привел несколько
примеров: «удивительные мышцы червей и более простые элементы моллю-
сков, насекомых и позвоночных», а также «в высшей степени сложные струк-
туры Protozoa и относительно просто организованные клетки Metazoa» (За-
варзин А.А. Труды по теории параллелизма и эволюционной динамике тканей.
Л., Наука, 1986, с. 17).
Для примера неожиданно сложной ткани приведу тегумент (покров-
ную ткань) плоских червей. Он тоже являет собою более чем клетку, но со-
всем не такую, как одноклеточные: это гигантское многоядерное образова-
ние (цитоид, п. 8-8*) без границ между ядрами. Наружная сторона тегумен-
та являет собой сплошной слой цитоплазмы без ядер, ядра же находятся в
выростах, соединенных с ним каналами, проходящими через слой, содер-
жащий мускулатуру (рис. 49). Те 2 млрд лет, которые наши предки потрати-
ли на создание костей, кровообращения, иммунитета, нервной системы и
прочего, низшие животные извели на усложнение клеток, тканей и т.п. Всё
это - частные случаи упомянутого принципа компенсации.
Рис. 49. Покровная ткань плоских червей: а) турбеллярий, б) трематод, в) цестод. Ее ус-
ложнение сопровождается упрощением всей остальной структуры червя (по Догелю)
450
Приходится отказаться от привычного со времен Ламарка изложения
хода эволюции «от низших к высшим»: Ламарк имел право рассматривать
эволюционный прогресс как приближение к человеку, но мы, зная микро-
биологию и гистологию, такого права не имеем. Эволюция “низших” изоби-
лует актами прогресса: таково, например, усложнение тегумента. Оно про-
текает в рамках реализации рефрена «тегумент», достаточно хорошо вид-
ного при сравнении плоских и круглых (вместе со скребнями) червей. Факти-
ческое описание данного рефрена см.: Никишин В.П. Эволюция покровных
тканей паразитических червей И ТПЭ-3; он же. Необычайная организация
покровной ткани скребней И Природа, 2003, N 11.
Что касается структурной сложности прокариот, то она тоже повыси-
лась по сравнению с первичным организмом коллосально, и стало это по-
нятно в последние лет двадцать. Если полвека назад микробиологи говори-
ли про бактерию как про “мешок с ферментами”, а 30 лет назад видели ее
как “мешок с генами”, то сейчас она предстала сложным структурным един-
ством. Единственная “хромосома” бактерии прежде казалась просто коль-
цевой двуспиральной нитью, хоть и было странно - как она умещается в
клетке, превосходя ее длиной в тысячу раз. Теперь известно, что бактери-
альная ДНК очень сложно и неравномерно скручена, что ее упаковка кон-
тролируется особыми белками и что отдельные ее петли удерживаются от
перепутывания с помощью нитей РНК (Прозоров А.А. Геном бактерий //
Микробиология, 1998, № 4).
Вспомним еще, что синтез белка ведут рибосомы, что подвижность бак-
терии обеспечивают жгутики или клеточная стенка, что энергетика (точнее,
окислительное фосфорилирование) сосредоточена в мембранах, а половой
процесс (неотличимый у прокариот от горизонтального переноса) осущест-
вляют плазмиды. Словом, прокариоты сложны очень.
8-5. Эволюция клетки: как появились эвкариоты?
И все-таки возможности прокариотной организации ограничены. У ки-
шечной палочки кольцевая ДНК закручена на полторы тысячи оборотов.
Более длинная двойная нить по всей вероятности начнет запутываться. По-
этому прокариотная клетка не бывает слишком сложно устроена, и, что еще
важнее, из таких клеток нельзя построить сложный организм.
Подавляющее большинство прокариот одноклеточны, а многоклеточ-
ные прокариоты - не более чем простые кустики или грибочки. Многокле-
точная сложность - удел эвкариот. Как только эволюция перешла от поиска
новых генов к поиску новых структур (комбинаций прежних генов), прогено-
те (эту гипотезу см.: п. 7-8*) пришлось менять и принцип записи генетиче-
ской информации, и принцип устройства клетки.
Во-первых, исчерпал себя принцип записи информации на единствен-
ной кольцевой двуспиральной ДНК, характерной для прокариот. В рамках
прокариота можно было только увеличить число колец двуспиральных ДНК
451
(нынешние бактерии с двумя и тремя кольцами ДНК известны). Вероятно, в
клетке сперва появились цепочки различных двуспиральных ДНК - прооб-
раз будущих негомологичных хромосом.
Во-вторых, с появлением данного новшества стали преобладать более
короткие, чем прежде, цепочки ДНК, как у молликут или плазмид. Вернее
всего, процесс начался с очень простых прокариот, т.е. эвкариоты не мно-
гим младше, чем прокариоты и, вероятно, ведут начало из прогеноты.
В-третьих, комбинирование как основной путь новаций заставило при-
роду отказаться от оперонной регуляции активности генов и перейти к
сплайсингу (сшиванию экзонов), в том числе альтернативному (п. 5-7).
В-четвертых, резко увеличилось количество белков, связанных с ДНК, а
это значит, что возросла роль новых способов записи информации, осно-
ванных не на последовательности оснований (п. 5-5*). Это, видимо, было
главным для последующего становления многоклеточное™.
В-пятых, возник новый способ крепления ДНК к мембране - такой, что-
бы разводить в разные стороны не только целые геномы, но и отдельные
кольца. Такой способ найден опять-таки у некоторых молликут: единствен-
ное кольцо ДНК крепится к бляшке, именуемой «полярным диском», причем
диск может делиться на два, и новые диски могут расползаться по мембра-
не без деления самой клетки.
В-шестых, в клетке появился цитоскелет - система из жёстких микро-
трубочек и сократимых микронитей.
В-седьмых, для дальнейшей эволюции надо было мембрану, несущую
кольца ДНК, отделить от той мембраны, что несет энергетическую (обо-
лочка митохондрии) и защитную (клеточная оболочка) функции. Очевидно,
что в основе всех трех оболочек лежала конструкция бактериальной мем-
браны. Замкнутая энергетическая мембрана (митохондрия) у многих (но
далеко не у всех) клеток напоминает по форме бактерию (см. пп. 6-5*, 8-
7*). Замкнутая оболочка, несущая хромосомы, именуется ядром клетки.
Последнее эволюционное достижение было обнаружено первым и лег-
ло в основу разделения всех организмов на ядерные (эвкариоты) и безъя-
дерные (прокариоты). Эволюцию прокариотного механизма деления в эвка-
риотный представить себе нетрудно (рис. 50). Однако в целом схему изме-
нения прокариота в эвкариот никто предложить не может.
И, прежде всего, никто не может сказать ничего о том, как появилось
ядро (наличные идеи на сей счет собраны в недавнем обзоре [Марков, Ку-
ликов, 2005]; они целиком ограничены вопросом «от кого?», не касаются во-
проса «как?» и, на мой взгляд, не продвигают проблему ни на шаг).
Всякий нынешний эвкариот имеет синдром эвкариотности - набор
свойств, который по частям не встречается: либо он целиком есть, либо его
вообще нет (Чайковский Ю.В. Генетическая интеграция клеточных структур
как фактор эволюции //ЖОБ, 1977, № 6).
452
Рис. 50. Эволюция процедуры клеточного деления от прокариот к эвкариотам
по И.Б. Райкову (а, б, в - стадии деления клетки; А, Б, В, Г - стадии эволюции).
Прошу обратить внимание, что таблица имеет ту же форму, что и приведенные
ранее (гл. 6) рефренные таблицы
К названным семи его свойствам можно добавить и другие. Так, веду-
щий английский специалист по эволюции одноклеточных Томас Кавалье-
Смит назвал 22 таких свойства, в том числе пространственное разобщение
транскрипции (в ядре) и трансляции (в цитоплазме), а также новый тип ме-
ханической активности, наряду с активностью растущих мембран, обеспе-
чивающих развитие и подвижность прокариот, появился новый феномен -
микротрубочка, активно растущая, словно стебелёк, и ставшая основой не
только цитоскелета, но еще реснички и жгутика. Он выразил уверенность,
что эвкариотная организация произошла в едином акте «квантовой эволю-
ции» (Cavalier-Smith Т. The origin and early evolution of the eucaryotic cell // Mo-
lecular and Cell, aspects Microb. evol.; 32nd Symp., Edinburgh, 1981, pp. 33-84).
За следующие четверть века предложена, по существу, лишь одна ги-
потеза - прогеноты: по ней, синдром эвкариотности не вырос в каком-то
прокариоте, а возник, как до этого сами прокариоты, в прогеноте, которая
использовала все достижения, найденные ею в предыдущей эволюции про-
кариот, включая их вирусы. Синдром возникал неоднократно: очевидно, что
растительная, грибная и животная клетки, несмотря на фундаментальное
453
единство их строения, произошли независимо (см. п. 6-2). Об этом ясно го-
ворит также и колоссальное разнообразие их митохондрий, многие из кото-
рых совершенно непохожи на бактерии.
Так или иначе, эвкариоты повторили формой прокариотную диасеть, а
затем эволюировали за ее пределы. Организация хромосомного аппарата
эвкариот дает такие возможности, какие прокариотам недоступны (см. рабо-
ты Стегния, п. 5-5*). А главное, у всех эвкариот есть особый механизм кле-
точного деления - митоз и структуры, его обеспечивающие.
8-5* Диатропика митозов и эволюция
В ходе обычного митоза сперва становятся видимыми (спирализуются)
хромосомы, которые до этого дуплицируются, затем возникает веретено,
состоящее из нитей (микротрубочек), соединяющих хромосомы с противо-
положными полюсами ядра, потом ядерная оболочка исчезает, а хромосо-
мы выстраиваются в два ряда и вскоре расходятся, влекомые нитями, к
противоположным вершинам веретена. Наконец, вокруг каждого нового на-
бора хромосом вырастает новая ядерная оболочка, и хромосомы деспира-
лизуются. При этом обычно делится и сама клетка.
Если бы данная картина была у всех одинакова, о ее происхождении
сказать было бы нечего. Но кариология (наука о клеточных ядрах) выясни-
ла, что у одноклеточных любая из указанных стадий может поодиночке от-
сутствовать, а митоз как целое всё же работает. Сводку ранних знаний об
этом дал ленинградский кариолог И.Б. Райков (сын историка науки Б.Е. Рай-
кова, цитированного в части 1) в великолепной книге: Райков И.Б. Кариоло-
гия простейших. Л., Наука, 1967. (Единственный, на мой взгляд, ее недоста-
ток - отсутствие данных о грибах, что мешает пониманию темы.)
Ныне мы знаем, что у многих одноклеточных ядерная оболочка не ис-
чезает, а делится, так что митоз протекает внутри ядра; при этом веретено
может быть как внутри ядра, так и снаружи (тогда нити проходят сквозь по-
ры в оболочке). У многих веретено возникает с одного бока ядра, и лишь в
ходе митоза вершины веретена расходятся к полюсам ядра (рис. 51).
Рис. 51. Пример митоза,
радикально отличного от
митозов, приводимых в
учебниках цитологии:
деление ядра в ходе
митоза в гифах гриба
Fusarium oxysporum.
В частности, здесь митоз
протекает без растворе-
ния ядерной оболочки
454
В учебнике (Мюллер Э., Лёффлер В. Микология. М., Мир, 1995, с. 40)
этот митоз приведен как типичный для грибов. Но в действительности мито-
зы грибов весьма разнообразны. Например, у многих базидиомицетов
ядерная оболочка растворяется частично (так называемые “полярные окна”
— там же, с. 41). У некоторых веретено почти незаметно (нитей немного) и,
видимо, расхождением хромосом управляет не оно, а ядерная оболочка. У
одноклеточной жгутиковой водоросли эвглены внутриядерным митозом, ве-
роятно, управляет ядрышко (точнее см. [Райков И.Б., 1986, с. 42]).
Естественно, возникло желание использовать эти различия для уясне-
ния происхождения митоза, причем желание возникло в самой простой
форме - многим захотелось найти среди ныне живущих одноклеточных
предка, чей митоз первичен. Такую группу нашли и торжественно назвали
(1965 г.) мезокариотами, т.е. промежуточноядерными - ими были объявле-
ны жгутиковые водоросли пиррофиты (они же у зоологов: динофлагеллаты,
т.е. панцырные жгутиконосцы). Хромосомы у них видны всегда, ядерная
оболочка не исчезает, а делится, словно бактерия.
Однако вскоре от идеи
“мезокариот” пришлось
отказаться, поскольку митоз
пиррофит оказался не
проще, а сложнее типичного
(его описание см.: Дьяков
Ю.Т. Введение в альгологию
и микологию. М., МГУ, 2000,
с.33-34).
Рис. 52. Эволюция митоза по
И.Б. Райкову, 1986. Тип А
(хромосомы разъединяются с
помощью ядерной мембраны)
можно считать самым ранним.
Типы митоза, обведенные рам-
кой, возникли явно позже дру-
гих; тип Г характерен для рас-
тений, тип Ж - для животных.
Остальные типы могли появ-
ляться в различной
последовательности
Веретено некоторых
пиррофит тоже сложное:
вне ядра, натянуто между
его полюсов. И наоборот:
есть жгутиковые водоросли
455
криптомонады с очень простым (“примитивным”) митозом, но протекающим
при растворении оболочки ядра, и красные водоросли (багрянки) со вполне
развитым (почти типичным) митозом, но внутри ядра. То же видно и с дру-
гими “примитивными” чертами. Кого же счесть предком?
В 1986 году, после 20 лет безуспешных попыток построить филему70 71 * *
митозов, И.Б. Райков предложил схему эволюции митозов, отрицающую
филогению (рис. 52). Самое интересное в ней - сходство со схемой Мейена,
приведенной на рис. 48. И тут и там имело место заполнение рефренной
таблицы, т.е. номогенез заявил свои права и в кариологии (хотя сам Игорь
Борисович рассуждал при этом как дарвинист [Райков, 1986]).
Очевидно принципиальное сходство этого рисунка с рис. 48. В самом
деле, предка искать нет смысла: первый эвкариот жил 2 млрд лет назад и
следов не оставил, так как не имел ничего, что сохраняется в ископаемом
виде. Наоборот, клеточные оболочки пиррофит хорошо сохраняются и из-
вестны с кембрия, т.е. пиррофиты слишком молоды для предка эвкариот.
Имеет смысл лишь пробовать мысленно собрать минимальный рабо-
тающий механизм митоза (что и сделал Райков), а затем уж искать све-
дения о дальнейшем усложнении митотических механизмов, по возможно-
сти увязывая их с геохронологией77. Но нет смысла указывать на какой-
либо митоз как на предковый. Нынешние митозы все обладают тем свойст-
вом, что их обладатели могут жить в нынешних средах, тогда как от первич-
ного митоза этого свойства ждать не приходится.
Эффективность митотических механизмов очень различна: при нор-
мальной температуре митоз разных форм может занимать от пяти минут
(обычный митоз в бурно растущей ткани) до двух часов (митоз эвглены).
Это не мешает широкому распространению самых разных форм. По-
этому нет оснований считать, что типичный митоз победил в ка-
кой-то прежней конкуренции, лучше вспомнить принцип компенсации
(никому не дано всё сразу). Одним из его проявлений как раз и является то,
что клетки низших зачастую устроены сложнее, чем у высших.
Конечно, вариации не могут быть любыми: они заполняют клетки своей
рефренной таблицы и этим указывают нам спектр былых возможностей.
Каждая из них могла в прежней эволюции встретиться, если организм, кото-
рому она принадлежала, мог существовать. Учтя это, попробуем очертить
ста-новление эвкариотности и, прежде всего, митоза
70
Филема- филогенетическая (в форме древа) схема эволюции.
71
Такие попытки делались давно - например, реконструкция становления митоза зеле-
ных водорослей с помощью данных о митозах пиррофит, эвглен и грибов (Pickett-
Heaps J.D. Evolution of mitosis and the eukaryotic condition // BioSystems, 1974, № 1). Bee
они распадались при получении новых данных и никем не вспоминаются
456
При внутриядерном митозе ядро делится подобно бактерии; значит, тут
мог работать бактериальный механизм; он же мог обеспечить расхождение
хромосом до возникновения веретена; хромосома пиррофит сходна с пуч-
ком бактериальных ДНК, следовательно, она демонстрирует принципиаль-
ную возможность сборки из различных бактериальных «хромосом». Далее,
обычная плотная упаковка ДНК могла быть позаимствована у вирусов - они
есть у бактерий, поэтому могли возникнуть ранее эвкариотности. Что же ка-
сается веретена, то вопрос его происхождения упирается в появление мик-
ротрубочек, а оно непонятно.
В остальном можно мысленно собрать ядерную организацию из эле-
ментов доядерной, не предполагая таинственного одновременного появле-
ния всего сразу. Всё сказанное получено после отказа от филогенетической
идеи, как ее альтернатива. Подробности и литературу см. [Чайковский,
1990, с. 145-146, 192-193]. Немногие более новые обзоры указаны в статьях:
Микрюков К.А. Необычный тип митоза у ризоподы ... И ЗЖ, 1998, № 11; он
же. Система и филогения солнечников... И ЗЖ, 2000, № 8.
К сожалению, в 1990-х годах поток исследований разнообразия митозов
почти иссяк, хотя внятного их итога высказано не было. Причина видится
мне в том же, что и в эволюционизме вообще: филему построить не уда-
лось, а иной идеи не искали (увы, обычные «шоры дарвинизма» - см. да-
лее, п. 12-1). По-моему, внятный итог ныне состоит в том, что описаны почти
все клетки той рефренной таблицы, которую можно назвать «митотические
фигуры». Вообще, на вопрос «как?» можно, в отношении одноклеточных,
ответить нечто вразумительное, лишь отказавшись отвечать на вопрос «от
кого?» (отказавшись от идеи филогении). В этом и состоит первый шаг
диатропического подхода к эволюции.
Возможен он в отношении любых таксонов, но для одноклеточных осо-
бенно эффектен, поскольку традиционные методы не дают тут просто ниче-
го. Разумеется, в будущем наш нынешний ответ на вопрос «как?» покажется
примитивным: ведь мы не очень-то понимаем, каким образом старые части
оказались пригодны для нового целого - вместо этого мы произносим слова
«сродство» и «самосборка» (п. 7-9). Но ведь и в химии сродство обрело чет-
кий смысл не сразу.
Встает вопрос: почему митозы одноклеточных разнообразны, тогда как
все тканевые имеют единственный тип митоза (с небольшими вариациями)?
Обычный ответ (одноклеточные старше и сохранили древние типы митоза)
наивен - все организмы имеют равно длинную историю. Наивна и мысль,
что митоз тканевых победил в конкуренции с иными митозами: в таком слу-
чае митоз у всех тканевых был бы просто одинаков, а это неверно - прими-
тивные черты митоза есть и у некоторых тканевых. Например, у некоторых
насекомых расхождением хромосом управляет клеточная мембрана (Пост
X. Физиология клетки. М., Мир, 1975, с. 658). И уж совсем наивна мысль, что
все тканевые унаследовали свой тип митоза от “общего предка” - тканевые
457
никем не признаются за монофилетический72 таксон; не считаются таковы-
ми ни животные, ни растения.
Единственным разумным допущением видится мне следующее: при-
знать однообразие митозов тканевых как итог давления нормы. Мы видим
(хоть и не можем ныне объяснить, почему), что типичный митоз оказался
нормален для тканевого организма (для растений и животных)7-?. Немногие
вариации, наблюдаемые в митозах тканевых, реализуют некоторые позиции
диасети митозов одноклеточных, и вполне возможно, что они затрудняют
нормальную работу клетки, но не выводят ее за рамки экологической проч-
ности данного тканевого вида. Для низших (нетканевых) ни этой нормы, ни,
следовательно, ее давления нет.
8-5** Формирование митохондрий
Не менее, чем появление митоза, для становления эвкариот было важ-
но появление отдельных энергетических органелл - митохондрий.
Дело в том, что энергетика - отрасль грязная не только в экономике, но
и в физиологии. Как мы знаем из главы 7, первичным и главным источником
энергии живого были, вероятно, АФК, тогда как гликолиз и, тем более, окис-
лительное фосфорилирование появились, по всей видимости, позже.
АФК весьма агрессивны, а потому должны немедленно либо использо-
ваться, либо гаситься - иначе они рушат всё окружающее, например, вызы-
вают массовые мутации. Рост интенсивности энергетики потребовал изоли-
ровать «электростанции» клетки от остальных ее механизмов, что и было
достигнуто появлением митохондрий - органелл, имеющих небольшое чис-
ло собственных генов, весьма, как и следовало ожидать, мутабильных в си-
лу соседства с мощным источником АФК.
Существуют две гипотезы происхождения митохондрий - симбиотиче-
ская и автономная. Первую давно выдвинули сторонники симбиогенеза, по-
лагавшие митохондрию бывшей бактерией (пп. 3-13*, 6-5*), а вторую пред-
ложил Кавалье-Смит в работах 1975-1981 годов. Он допустил, что комплекс
ферментов окислительного фосфорилирования обособился от остальных
структур клетки на особой внутриклеточной мембране, в качестве которой
могла быть использована плазмида (п. 6-9).
77
Монофилия - происхождение от единого общего предка. Изображается филемой. Так-
сон монофилетичен, если все его подтаксоны имеют одного и того же предка.
'J Грибы имеют различные типы митоза, но их не относят к тканевым. Во-видимому, ти-
пичный митоз тканевых необходим для интеграции клеток в ткань. Основное отличие
митоза животных - они имеют центриоль (см. п. 5-7), а у высших растений ее нет. Она,
вероятно, обеспечивает более высокую степень интеграции: если у растения новый ор-
ганизм может вырасти почти из любой клетки, то у животных — только из зиготы и
бластомера, которые, кстати, тоже не имеют центриолей (Ткемаладзе Дж.В., Чичинадзе
К.Н. Центриолярные механизмы... //Биохимия, 2005, № 11, с. 1579).
458
Утверждение об однократном бактериальном происхождении митохон-
дрии, которое «подкрепляется молекулярными данными, указывающими на
монофилетическое происхождение митохондрий всех эвкариот» [Марков,
Куликов, 2005, с. 3], вряд ли верно. Оно игнорирует все факты, кроме по-
верхностного сходства. Что касается «молекулярных данных», то они не
только малоинформативны в силу их вольной трактовки, но и весьма нена-
дежны - дают втрое меньший возраст эвкариот, нежели данные палеонто-
логии. Литературу см.: Наточин Ю.В. // ПЖ, 2005, № 4, с. 20.
В отличие от симбиотической, автономная гипотеза ничему не противо- 1
речит, а в ее пользу говорит многое; например, полное несходство многих
митохондрий с бактериями и полная неспособность митохондрий что-либо
делать самостоятельно (без ядерных генов). Главное же - нет нужды ни до-
пускать многократное независимое укоренение прокариота в прокариоте
(никто такого укоренения не наблюдал), ни громоздить произвольные допу-
щения, как, например, сделано в работе [Марков, Куликов, 2005].
Замечу, что другая органелла, имеющая собственные гены - пластида
(хлоропласт), более автономна, нежели митохондрия, и еще Кавалье-Смит
(1981) допускал, что некоторые пластиды действительно произошли от
цианей (сине-зеленых водорослей), о чем будет речь далее (п. 8-7*). Там же
он обратил внимание на то, что разговоры о симбиотическом происхожде-
нии органелл ни на шаг не приближают нас к пониманию становления син-
дрома эвкариотности и, прежде всего, появления ядра и цитоскелета.
8-6. Часы эволюции: время особи течет вперед и назад
Принято считать, что феномен смерти появился вместе с многоклеточ-
ностью и что одноклеточные в идеале бессмертны, поскольку могут неогра-
ниченно размножаться делением. Но это не вполне так: делящийся клон
рано или поздно вырождается (по крайней мере у эвкариот), и для его со-
хранения нужно хотя бы изредка перемежать деление половым процессом.
Почему клон стареет? Почему время не начинает свой отсчет для каждой
клетки с нуля? Тут не обойти вопрос - что такое время?
Дать ему общее определение не удается, поскольку нет понятий, ему
предшествующих (Кант называл время первой интуицией сознания). Един-
ственное, чему тут люди научились - время измерять, т.е. сравнивать с эта-
лонами. Для эталона годится любой процесс, который неограниченно сам
собою повторяется и всякий раз течет одинаково. Определить, что значит
«одинаково», тоже не удается, но интуитивно мы такие процессы находить
умеем (прежде всего, смена суток, месяцев и лет), и первое их свойство -
течь всегда в одну сторону: солнце ни при каких обстоятельствах не пойдет
с запада на восток. Время как таковое - та основная величина, от которой
зависит и ход эволюции, и облик планеты.
Мы привыкли, что время всегда течет в одну сторону. Разумеется,
«первая интуиция сознания» такова, но стоит придумать времени какую-
459
либо формулировку, как дело усложняется: у придуманного времени можно
поменять знак. Например, физики школы Пригожина любят говорить (силь-
но упрощая природу), что направление времени задается необратимостью
самопроизвольных процессов: Земля остывает, камень падает вниз, железо
из ископаемых пород рассеивается, и время при этом возрастает. О таких
процессах говорят: «энтропия растет». Противоположные процессы (печка
нагревается, снаряд летит вверх, железо скапливается на свалках металло-
лома) можно назвать текущими в обратном направлении, в сторону умень-
шения общефизического времени. А как в биологии?
Тут естественным является направление течения жизни организмов,
которое знаменуется делением клеток (включая одноклеточных): клетки де-
лятся, растут и снова делятся, чем и задается направление жизненного
времени. Клон клеток при этом неминуемо стареет. Но один процесс течет
наоборот - половой процесс, когда клетки соединяются. Именно при нем
происходит, говоря языком техники, «обнуление всех счётчиков», так что
новый организм начинает жизнь “с нуля”, с младенчества. Тем самым, жиз-
ненное время становится циклическим.
Механизмы «обнуления» еще малопонятны. Самый простой и понятный
связан с актом репликации хромосомы: каждый акт сопровождается недо-
репликацией небольшой части концевого участка, так что дочерняя хромо-
сома становится чуть короче материнской. Однако никакой значащий ген от
этого не страдает, поскольку концевой участок хромосомы (теломера) ге-
нов не содержит. Он-то и считает биологическое время. «Обнуление» со-
стоит в присоединении новой теломеры. Идею высказал в 1971 году Слов-
ников, а позже укорочение теломеры было установлено в опыте.
К сожалению, проблема старения клетки так просто не решается: ста-
реют, хотя и за большее число поколений, даже бактерии. Им тоже время от
времени необходим половой процесс (у одноклеточных он не связан с раз-
множением). Очевидно, что есть и какой-то другой механизм «обнуления
счётчиков» при половом процессе.
Нам важно запомнить: не с делением клеток, а с изобретением поло-
вого процесса жизнь получила потенциальное бессмертие.
Самое простое “движение назад во времени” проявляют те же организ-
мы, что обнаруживают самую простую многоклеточность (в отличие от ко-
лониальных, многоклеточными называют организмы, образующие осо-
бые многоклеточные органы размножения); эти организмы - миксоформы,
т.е. миксобактерии в царстве прокариот, а также миксомицеты (слизевики,
слизевые грибы) и акразиевые в царстве грибов. Принадлежа разным цар-
ствам, виды этих типов ведут себя очень сходно (параллелизм) в том смыс-
ле, что индивиды часть жизни проводят как одноклеточные, а затем спол-
заются в колонию, на которой вырастают плодовые тела - яркие “грибочки".
У миксобактерий они малы (меньше 1 мм) и вырастают на псевдоплазмо-
дии, т.е. на особом теле, в котором отдельные клетки сохраняются. Наобо-
460
рот, миксомицеты образуют истинный плазмодий - единое бесклеточное
многоядерное тело с ладонь размером и “грибочками” до 1 см (рис. 53). Тем
самым, миксомицет производит (когда сползаются его одноклеточные час-
ти) возврат в биологическом времени целиком - до слияния клеток вклю-
чительно. К сожалению, миксоформы в ископаемом виде не сохраняются, и
мы не знаем момента их появления.
Рис. 53. Цикл развития миксоформ различных царств: а) у миксобактерий, б) у миксо-
мицетов, где он протекает от стадии А (выход амебоидной клетки из споровой клетки)
до стадии Н (формирование “грибочка”). Движение “назад во времени” происходит от
стадии К к стадии Л
Размножение
Они - простейший пример особого
плана строения организмов, который
состоит из последовательности планов
строения всех стадий. Этот план не
умещается в пространстве, но может
быть описан в пространстве-времени.
Такой план строения именуют
морфопроцессом [Беклемишев, т. 1, с.
11], или хронотопом (от греч. хронос -
время и топос - место). С таким планом
строения мы еще встретимся.
Параллелизм хронотопов прокариотных
и эвкариотных миксоформ - яркий
пример номогенеза.
Возврат во времени выступает у
миксоформ как самостоятельное яв-
ление, которое видно тут лучше, чем в
половом процессе. Возможно, что оно
возникло сперва у миксоформ, а затем
было использовано для полового
процесса, но возможно и наоборот. Так
или иначе, но кроме привычного нам
времени, всегда текущего в одну
сторону, есть и другое, циклическое (ср.
п. 6-8*). К циклам мы вернемся в п. 8-8,
а пока замечу, что каждый цикл есть
функциональный блок, но не в
пространстве, а во времени.
Еще Вернадский принимал «гипотезу, что пространство внутри живого
вещества есть иное, чем внутри косных естественных тел..., что время вы-
ражается в нем полярным вектором» (Вернадский В.И. Проблемы биогео-
461
химии. М., Наука, 1980, с. 81). То есть - что время многомерно. Если так, то
для уяснения сути жизни нужны новые математика и физика - мысль, до сих
пор чуждая биологии. В связи с этим Мейен заметил:
“сейчас носятся с Вернадским, с его биосферой и ноосферой. Но ведь это -
самое неинтересное в его творчестве... Но его концепцию времени и прост-
ранства ни физики, ни философы в упор не видят. Там надо уже крепко шеве-
лить мозгами” (Мейен С.В. Из заметок... // [Эволюция флор..., с. 131]).
Нежелание или неумение «шевелить мозгами» привело к таким стран-
ным идеям, как идея омоложения с помощью таблеток, замедляющих мета-
болизм (таковы, например, антиоксиданты). Из сказанного ясно, что омо-
ложения при этом не наступит. Для омоложения надо не замедлять процес-
сы, а менять их суть. Например, “обнулять счётчики”.
Любой счётчик так или иначе считает время, т.е. являет собою часы. Нет
сомнений в том, что такие часы управляют онтогенезом, но есть в эволю-
ционизме линия мысли, идущая от Брокки и полагающая, что такие же часы
управляют эволюцией, так что вымирание столь же неизбежно, как и смерть
особи (идея цикличности в эволюции). На самом деле
“одни группы стареют и вымирают, другие, хотя и очень старые, не выми-
рают, а некоторые даже процветают, так что проблема, по-видимому, не
имеет однозначного решения” [Назаров, 2005, с. 199].
Уточню: групп-долгожителей мало, причем возрасты ныне живущих родов
ложатся на гиперболу74 - так же, как у Виллиса ложились количества видов
в родах и численности видов. Налицо система, так что «однозначное реше-
ние» возможно, но только если оно опишет всю ее, а не отдельные факты
порознь.
Приняв, что решение (механизм эволюции, приводящий к гипербо-
лическому распределению сроков жизни родов) в принципе существует,
можно понять, что хотел сказать Шалин, когда писал:
«Видимо, эволюция управляется множеством маленьких “часов жизни”, кото-
рые определяют и контролируют различные этапы развития. Но надо уточнить,
что существует определенная степень свободы развития, которая соответствует
эпигенетическим феноменам». Работа часов изменяется мутациями: “Мутации
регуляторных генов управляют также изменениями последовательности собы-
тий или скорости развития. Тем самым, новации могут быть вовлечены в водо-
ворот этих изменений развития. Тогда они усиливаются ускорениями и гипер-
морфозами или ослабляются замедлениями и гипоморфозами” (АЕ, 180).
Поясню: Шалин полагает, что как гиперморфоз (переразвитие — тако-
вы, например, рога ирландского оленя, рис. 19), так и гипоморфоз (недо-
развитие -такова неотения, п. 4-13), являются результатом изменения хода
74 РаупД., Стэнли С., Основы палеонтологии, М., Мир, 1974, с. 264. На графике 78% ро-
дов двустворчатых моллюсков младше 25 млн лет, 5% старше 350 млн лет.
462
часов онтогенеза. Из этого допущения, в частности, следует (АЕ, 173), что
направление эволюции можно, в принципе, поворачивать простым измене-
нием хода часов онтогенеза - достаточно одновременного перехода многих
особей данного вида к неотении.
Наконец, несколько слов о старении высшего организма. Спор двух то-
чек зрения (старение - итог либо действия программы, либо накопления
ошибок - п. 5-14) обретает новый смысл в аспекте диатропики: механизмы
старения, вероятно, образуют рефрен. Его крайние позиции дают примеры
в пользу обеих точек зрения, но в целом справедливы обе. Так, у высших
растений есть группы с явно выраженной программой старения и смерти
(одно- и двухлетники), аналог которым у высших животных являют члени-
стоногие, головоногие и рыбы, гибнущие после акта размножения.
У людей феномен программы обнаружен в форме очень редкой болез-
ни - это прогерия (когда ребёнок быстро становится стариком). Редкость
прогерии не дает ей (к счастью) повлиять на статистику старения, что слу-
жит для многих основанием отрицать генетическую программу старения во-
обще (Гаврилов Л.А., Гаврилова Н.С. Биология продолжительности жизни.
М., Наука, 1991). На самом деле прогерия есть следствие правила Кренке
(п. 4-11), и ее наличие ясно говорит, что программа старения у людей суще-
ствует, но замаскирована другими явлениями.
8-7. Принцип блочности в ранней эволюции
Нам пора узнать, что блочность является одним из главных принципов
эволюции вообще. Начать с того, что феномен наследственности демонст-
рирует блочность сразу на нескольких уровнях. Во-первых, ген состоит из
блоков, каждый из которых кодирует один домен. Во-вторых, прокариотные
гены объединены в функциональные блоки - опероны. В-третьих, процесс
изменения генов - блочный (замены нуклеотидов и их цепочек, вставки, вы-
падения, инверсии, транспозиции, изменение числа копий). В-четвертых,
кроме данных (обязательных), есть тип блоков, без которого жизнь, по-
видимому, возможна - это интроны (п. 5-5).
Появление эвкариотной организации отмечено новыми типами блоков -
хромосомами, различными комплексами генов, сменившими опероны, и
различными органеллами. Затем блоками стали сами эвкариотные клетки.
В 1925 году гистолог А.А. Заварзин, обнаружив, что ткани самых разных
организмов устроены сходно, не пошел по стопам симбиогенетиков (не стал
искать предков и утверждать, что ткани раньше были организмами или чем-
то вроде), а заявил про «параллелизм структур как основной принцип мор-
фологии» и что параллелизм - одно из центральных понятий номогенеза.
Такой номогенез (он идет от Соболева и развит Мейеном) понимает эволю-
цию как перебор различных комбинаций блоков.
В общем виде идею эволюции как комбинации блоков высказал ленин-
градский физиолог А.М. Уголев. Сам он занимался пищеварением и сумел
463
показать, что вся эволюция пищеварения представима в виде смены ком-
бинаций функциональных блоков, появившихся очень рано - в эпоху гос-
подства одноклеточных и самых простых многоклеточных.
Блоки (он их называл технологиями) Уголев видел всюду-
«на уровне биосферы, отдельных биогеоценозов, популяций, организма, орга-
на, клетки, а также на субклеточном уровне. Точно так же эволюцию следует
рассматривать как эволюцию естественных технологий, а происхождение
жизни - как происхождение естественных технологий» [Уголев, с. 465].
Тему технологий затрагивал еще Ламарк [Чайковский, 2002, с. 442].
Основные блоки появились очень рано и после этого новые блоки по-
являются редко. Самый первый и самый общий функциональный блок поя-
вился, по Уголеву, еще в ходе биопоэза - то была «пристеночная орга-
нелла». Этим термином он обозначил структуру, обеспечивающую клетке
отрицательный электрический потенциал относительно внешней среды.
«Сущность нашей гипотезы заключается в том, что в ограниченных водоемах
могла образоваться пристеночная органелла, источником энергии для кото-
рой служил поток ионов». Этот поток мог возникать в регулярно пересыхаю-
щих водоемах, на дне которых неизбежно формировались многослойные
макромолекулярные пленки, похожие на мембраны клеток. «Можно ожидать,
что таким путем могут возникать макроэрги типа полифосфатов и АТФ»
[Уголев, с. 431-432].
Поясню: одна из главных задач мембраны - обеспечение энергетики
клетки. Эта энергетика носит электрохимический характер - клетка заря-
жена отрицательно по отношению к среде, т.е. получается гальванический
элемент, питающий клетку. Простейший электрогенный процесс - выведе-
ние наружу ионов водорода. Он изображен на рис. 54 так, как ныне приня-
то, однако замечу, что внутренность клетки - не раствор, а гель (п. 7-3).
Это, естественно, не мешает признать роль электрохимии.
Рис. 54. Пристеночная органелла (по: Шлегель, 1987, упрощено)
464
Так или иначе, в клетке работает механизм, возможый в неживой
природе, хотя в организмах поток протонов идет в силу окислительных
биохимических процессов в мембране, а у предшественника жизни он шел,
по Уголеву, за счет намокания и высыхания неорганических плёнок, меняв-
ших при этом проницаемость (вновь обращу внимание на роль границы
фаз). В принципе это реально: известно, что проницаемость мембран для
ионов меняется вследствие изменения концентрации ионов кальция внутри
мембраны при скручивании и распрямлении волокон «щелевых соедине-
ний» (Страйер П. Биохимия. Том 3. М., Мир, 1985).
Первично такие волокна могли быть неорганическими. А вот как первый
организм оказался одет замкнутой мембранной полостью, как сопряг деле-
ние ее и генома и как сменил ненаследственные структуры (в том числе
электрогенные мембраны) на наследственные, пока непонятно, о чем уже
говорилось в конце главы 7.
Подходы Для описания фактического хода эволюции удобно фиксиро-
вать появление новых морфологических (Мейен) и функциональных (Уго-
лев) блоков и показывать, какие новые возможности для комбинирования
открывались с появлением каждого из них. Это содержательнее, нагляд-
нее и проще для обучения, чем принятые до сих пор принципы (со-
гласно которым объяснить эволюцию - значит найти каждому виду предка,
а каждому свойству - пользу для выживания).
Уголев ввел еще один принцип эволюции, принцип цикличности:
«На всех уровнях организации (субклеточном, клеточном, органном, орга-
низменном, популяционном, экосистемном и планетарном) биологические
системы частично или полностью циклизированы. Так, очевидна циклизация
окислительных процессов на уровне трикарбоновых кислот (цикл Кребса -
Ю. Ч.)... Возможно, наиболее сложным и совершенным выражением циркуля-
ции молекул, надмолекулярных комплексов и клеток в организме служит
кровообращение... По всей вероятности, циклизация входит в число важней-
ших принципов, обеспечивающих высокую экономичность и эффективность
биологических систем благодаря многократному использованию одних и тех
же структур» [Уголев, с. 458-459].
Остается добавить, что именно в форме циклов (круговоротов) жизнь
возникла и что системой циклов является обмен веществ (м. гл. 7). Соеди-
нение идей блочности и цикличности («новый функционализм» по Уголеву) -
вот нынешнее понимание эволюции как единого целостного процесса.
Предлагаю способ рассказа ученикам о ходе эволюции: фиксировать появ-
_______ление новых циклов - как внутри особи, так и между особями.____
Первым циклическим процессом на уровне организма явилась смена
поколений одноклеточных, т.е. их размножение, вторым - половой процесс,
третьим - многоступенчатая репликация (при переходе к эвкариотности),
четвертым - многоклеточность (смена клеточных поколений в тканях), пя-
465
тым - смена поколений многоклеточных, шестым - появление циклов в жиз-
ни многоклеточных организмов (суточных, месячных, сезонных).
При наследовании молекулярных блоков возможна радикальная смена
их функций. Кроме гомеобокса, приведу такой пример:
«По-видимому, многие сигнально-регуляторные домены, общие для бакте-
рий и эвкариот, у первых выполняли синэкологические функции (обеспече-
ние взаимодействия с другими компонентами прокариотного сообщества), а
у вторых стали использоваться для согласования работы клеточных орга-
нелл и отдельных клеток» [Марков, Куликов, 2005, с. 3].
Схсдные примеры есть и в их новом обзоре [Марков, Куликов, 2006].
8-7* Блочность эволюции против симбиогенеза
Идея эволюции живого в форме комбинирования функциональных бло-
ков впервые, не считая фантазий Эмпедокла, была разработана в учении о
симбиогенезе (см. пп. 3-13*, 6-9). То была слишком грубая идея - попытка
видеть в качестве блоков целые организмы: как гриб и водоросль соедини-
лись в рамках лишайника, так и другие органы и органеллы якобы прежде
были организмами (бактериями). Тогда научное сообщество эту идею от-
вергло. Однако через 60 лет, в 1969 году, цитолог Линн Маргелис (США)
возродила эти взгляды на основе новых данных. Строение некоторых мито-
хондрий и хлоропластов оказалось кое в чем похоже на бактериальное. На-
пример,они имеют свою ДНК (она похожа на бактериальную).
Маргелис назвала свою идею теорией происхождения эвкариот, но что
касается главного - происхождения ядра и цитоскелета, фактически не
предложила ничего (вопрос подробно разобран в книге: Серавин П.Н. Про-
стейшие... Л., Наука, 1984). Она лишь допустила, что веретеном деления
тоже управляет бывший прокариот.Эта гипотеза вскоре была опровергнута
(митотический аппарат основан, как и эвкариотный жгутик, на микротрубоч-
ках, но не имеет ДНК), что признала в 1973 году сама Маргелис.
После этого разговор о симбиогенезе как пути становления эвкариот
утратил смысл. Вскоре потерял смысл и разговор о симбиотическом про-
исхождении митохондрий: сходств с бактериями оказалось меньше, чем от-
личий, а у некоторых нет вовсе сходств (п. 6-5*). Сходство ДНК органелл и
бактерий - не довод: ведь плазмиды никто не считает бактериями.
Вся наследственность органелл закодирована вперемешку - в них и в
ядре: каждый фермент собирается из ядерных и цитоплазматических субъ-
единиц, причем в самой митохондрии нет вовсе генов транскрипции и
трансляции и почти нет генов рибосомных РНК. Даже белки, из которых со-
стоят мембраны митохондрий, в основном кодируются ядерным геномом
(см. Побежимова Т.П., Войников В.К. Импорт белков в митохондрии И ФР,
2000, № 1). Из прочих макромолекул здесь синтезируются только “самые
жирные” (выражение В.П. Скулачева) субъединицы, которые затруднитель-
но транспортировать через оболочку. Вот зачем сюда вынесено несколько
466
генов из ядра (а заодно множество повторов и несколько интронов). Право-
та Кавалье-Смита (п. 8-5**) выясняется всё более и более.
Однако общество приняло идею симбиогенеза “на ура”, и Маргелис в
1975 году вернулась к ней, никак этого не объяснив (литературу см. [Чай-
ковский, 1990]). Когда было накоплено много новых опровергающих эту
идею данных, для спасения идеи симбиогенеза было придумано странное
допущение - что эволюция органелл состояла в упорной раздаче их генов
ядру. По-моему, естественнее вспомнить, что митохондрии содержат много
ядовитых АФК, и сказать, что дыхание эвкариотной клетки выступает как
“грязная технология”, вынесенная в отдельный структурный блок, обла-
дающий своим “подсобным хозяйством”.
Гораздо сложнее вопрос о происхождении хлоропластов. Если мито-
хондрии содержат всего около 3% собственных генов (не дублирующих
ядерные гены), то хлоропласты - около 20%, причем многие из них бывают
более автономны, чем митохондрии7-5. Однако и они весьма отличны от тех
бактерий, которых прочили им в предки7<5_ Вопрос пока открыт.
Остальные органеллы не проявляют никаких или почти никаких симбио-
тических свойств. Прежде всего - идея симбиогенеза ничего не может ска-
зать о появлении цитоскелета.
Позже симбиогенез не раз подвергался убийственной критике, и ныне
многие западные ученые считают его «поэтической фантазией» (УСБ, 2000,
№ 6, с. 620). Ни одно возражение не опровергнуто и даже не рассмотрено
его сторонниками. Ситуация целиком повторила историю с естественным
отбором: адепты симбиогенеза не спорят с тем, что в их учении много про-
тиворечий и мало фактов; они в нее попросту верят, о противоречиях не
упоминая и недостающих фактов не ища. Если прежде сторонников симбио-
генеза упрекали в голословности, то теперь презумпции поменялись, и
уже от противников симбиогенеза требуют доказать, что митохондрии не
могли быть симбионтами, а доказательств не читают.
Содержавшееся в идее симбиогенеза верное соображение о много-
кратном использовании в ходе эволюции одних и тех же блоков уже
давно предлагалось использовать прямо, без натяжек, в форме идеи гене-
75
' Наблюдается спектр промежуточных объектов: от самостоятельного симбионта-
фотосинтетика (цианеи) через полусамостоятельного симбионта (цианеллу) до хлоро-
пласта, столь же зависимого от ядерного аппарата клетки, как митохондрия (Масюк
Н.П. Цианеллы...//Альгология, 1997, №3).
76 Таковыми долго считались прохлорофиты'. они сочетают прокариотное строение с эв-
кариотным типом хлорофиллов. Их называли “прокариотными зелеными водоросля-
ми” (Wither N. W. е.а. И PNAS, 1978, vol. 75, № 5, р. 2301). Но теперь признано, что “ан-
тенны Oxiphotobacteria и пластид имеют независимое эволюционное происхождение”
(Пиневич А. В. и др. Прохлорофиты двадцать лет спустя // ФР, 2000, № 5, с. 730).
467
тической интеграции {Чайковский Ю.В. Генетическая интеграция клеточ-
ных структур как фактор эволюции //ЖСБ, 1977, № 6).
Более ста лет говорили, что ядро тоже произошло из бактерии, приводя
в пользу этого всего один сильный довод: оно, словно эндосимбионт, окру-
жено двумя оболочками - своей и клеточной - и один слабый: у некоторых
одноклеточных ядро делится, словно клетка. Однако долгое время счита-
лось, что прокариот в прокариоте жить не умеет (в прокариотах находи-
ли только вирусы и плазмиды, а они организмами не являются), и это слу-
жило главным доводом против симбиогенетической природы ядра.
Но появилось сообщение, что насекомое мучнистый червец может не-
сти внутри себя паразитическую бактерию, в которой живет другая бактерия
(von Dohlen C.D. е.а. И Nature, 2001, v. 412, р. 433), и это использовано для
модели симбиотической природы ядра (Martin W., Russel M.J., 2003 - см. п.
8-3). Авторы озабочены лишь биохимической стороной дела, а не структур-
ной (например, пишут, что в бактериях есть белок тубулин, но не пишут о
происхождении микротрубочек, состоящих из тубулина), и вся эвкариотная
часть их статьи состоит в перечислении бактериальных функциональных
блоков, которые могли быть использованы при формировании первой эвка-
риотной клетки. Основная их мысль - гипотеза, по которой прокариот, посе-
лившийся в прокариоте, стал обмениваться генами с «хозяином».
Наоборот, группа французских биологов взяла за основу не гипотезу
симбиогенеза, а реальный факт: синтрофизм (соединение бактерий разных
классов в колонию для питания общим субстратом). Центральная клетка ко-
лонии (по их мнению, архебакгерия) обрела функцию ядра, а окружающие -
функции митохондрий и цитоплазмы. Общую генетическую систему посте-
пенно создал горизонтальный перенос (Selosse М.-А. Quels ancetres pour
nos cellules? // LR, 2003, № 362, pp. 48-52). Это гипотетическое построение
похоже на колониальную теорию происхождения многоклеточных (см. ниже,
п. 8-8) и вполне укладывается в рамки блочного понимания эволюции. К со-
жалению, становление синдрома эвкариотности в обеих этих работах, как,
впрочем, и во всех других, выпало из внимания. Зато обе укрепляют пози-
цию блочного понимания эволюции клетки.
Обнаружение прокариота в паразитическом прокариоте ставит новый
вопрос: может ли прокариот жить в свободном прокариоте? Если да, то
можно, наконец, строить модель становления эвкариотности путем сери-
ального эндосимбиоза (гипотеза, которой воспользовалась Маргелис, когда
лопнула идея симбиогенеза, но у которой до сих пор не было основы).
Если нет, если прокариот может жить только в прокариоте, стабилизи-
рованном его внешней средой, то укрепляется гипотеза происхождения эв-
кариот в прогеноте - она-то и была внешней стабилизирующей средой. Ес-
ли не может нынешний прокариот, тем менее был способен прежний.
Судя по всему, ядро, митохондрии и хлоропласты произошли путем
блочной эволюции с участием горизонтального переноса. Вероятно, их
468
внешнее сходство с бактериями имеет чисто диатропическую (рефренную)
природу: например, для реализации биоэнергетической и фотосинтетиче-
ской “машин" всюду реализуются сходные формы.
Поэтому, продолжая п. 8-3, можно заключить: вероятно, первые эвка-
риоты появились на заре жизни и использовали генетический аппарат про-
геноты дважды: сходный с прокариотным - в органеллах, а компактную (ви-
русную) упаковку ДНК - в ядре. При этом возник генетический аппарат ново-
го типа: геном ядра, уложенный в хромосомы и управляющий фрагментар-
ными геномами органелл, имеющими единственную молекулу ДНК.
Энергетика дыхания вероятно была заимствована у прокариот, но никак
не в форме целых бактерий. Вопрос о пути появления цитоскелета остается
открытым во всех концепциях. Недавний обзор [Марков, Куликов, 2005] при
обсуждении появления цитоскелета ограничен, как и при обсуждении появ-
ления ядра, вопросом «от кого?», однако содержит две ценных подробно-
сти: 1) энергетику сборки микротрубочек обслуживает не АТФ, а ГТФ, что го-
ворит о большой обособленности (древности?) процесса; 2) у бацилл (бак-
терий в форме палочек) существует «нечто вроде прокариотического цито-
скелета», сходного с цитоскелетом эвкариот как структурно, так и функцио-
нально, но - на совсем иной белковой основе [там же, с. 10]. Опять налицо
заимствование идеи, а не веществ (аргумент платоников, ср. п. 5-15*).
8-8. Появление мейоза и многоклеточное™
Каким механизмом реализуется комбинирование блоков? У прокариот и
одноклеточных эвкариот он достаточно очевиден: горизонтальный перенос.
Тут комбинироваться может всё что угодно - в этом и плюс, и минус. Плюс в
том, что огромен выбор возможностей; а минус в том самом, что не дает му-
тациям стать заметным фактором эволюции: вреда от беспорядочных ком-
бинаций больше, чем пользы. Их надо как-то ограничить, и тем жестче, чем
крупнее организм - по той простой причине, что численность организмов
падает с ростом их размеров.
У эвкариот горизонтальный перенос почти нацело вытеснен другим ме-
ханизмом; это - рекомбинация, т.е. обмен генами и их частями внутри одно-
го генома. Она есть у всех организмов, но лишь у «высших» берет на себя
роль организатора, поскольку их эволюция состоит почти целиком в пере-
группировке прежних генов. Рекомбинация разных фрагментов происходит с
разными скоростями, и
«Имеются серьезные основания считать, что рекомбинация обычно подавлена
именно в тех участках хромосом, где расположены... блоки коадаптирован-
ных генов» (Жученко А.А., Король А.Б. Рекомбинация в эволюции и селекции.
М., Наука, 1985, с. 225).
Иными словами, рекомбинация сделана так, что сохраняет блоч-
ность конструкций. Налицо еще один пример активности, точнее -
пример узнавания. Как это получается, до сих пор неясно. Зато известно,
469
как в ходе эволюции улучшался механизм рекомбинации. У прокариота
фрагмент единственной «хромосомы» может рекомбинировать с любым
фрагментом его плазмиды, а у эвкариота хромосом несколько, и рекомби-
нируют, в основном, гомологичные участки гомологичных хромосом. Реком-
бинация происходит исключительно в тот момент деления клетки, когда го-
мологичные хромосомы спарены.
Ее осуществляет специальный механизм - кроссинговер. Он изредка
работает в ходе митоза и в таком случае служит важным поставщиком из-
менчивости (у многих низших грибов и водорослей). У большинства же эв-
кариот для обеспечения рекомбинаций важнее другой тип деления клетки -
мейоз (при нем образуются половые клетки, гаметы), при котором кроссин-
говер происходит в тысячи раз чаще и поставляет почти всю изменчивость.
Многоклеточных эвкариотных организмов, не имеющих настоящего мейоза,
видимо, не бывает.
Видимо, именно мейоз, сделав процедуру кроссинговера регулярной,
сделал вид устойчивой единицей эволюционного процесса. Вероятно, что
сама возможность образования нового вида путем скрещивания связана с
нарушением запрета на спаривание несходных хромосом [Бородин и др.,
2002]. Возможно, что это и есть механизм действия пунктуализма (п. 5-1).
“Мейоз начинается с того, что гомологичные (парные) хромосомы разыс-
кивают друг друга. Как они это делают, до сих пор непонятно”, но “грубое
распознавание может быть достигнуто простым совмещением” таких хромо-
сом. “Тонкое опознание начинается с того, что в ДНК мейотических хромо-
сом возникают множественные двунитевые разрывы. Свободные концы разо-
рванных ДНК привлекают на себя специфические ферменты, с помощью ко-
торых они внедряются в неповрежденные районы ДНК других хромосом (не
спрашивайте нас, откуда они знают, что это - ДНК других хромосом, ответа
не знает никто). [...] Прежде, чем перейти на следующую стадию мейоза,
клетка сама себя проверяет... Все ли хромосомы спарены? Не осталось ли не-
спаренных участков, незалеченных разрывов ДНК? ... Если хоть что-то не так,
мейотическая клетка, как истинный самурай, делает себе харакири - перехо-
дит в апоптоз” [Бородин и др., 2005, с. 36-37].
Пусть механизмы и неясны, однако здесь нам достаточно того, что все
эти акты чудесного узнавания суть проявления активности (точнее, эпигно-
стики) и тем сходны с иммунитетом, что описанный контроль есть то самое
давление нормы, снятие которого повышает изменчивость.
О том, как из одноклеточных произошли многоклеточные, высказано
много гипотез (Иванов А.В. Происхождение многоклеточных животных. Л.,
Наука, 1968), и споры идут до сих пор. Одно нынче очевидно: этот процесс
происходил 1,5-2,4 млрд лет назад во многих группах одноклеточных па-
раллельно и многократно (номогенез). При этом в наше время ясно, что в
основе многоклеточное™ лежит избирательная адгезия (прилипание) кле-
точных оболочек друг к другу (п. 5-15).
470
Это вроде бы нехитрое «изобретение» липкой поверхности было со-
вершено еще у прокариот. Адгезия постепенно стала очень сложной. При-
липание превратилось в узнавание клетками друг друга, что и породило всё
то разнообразие организмов, которое мы видим вокруг себя.
Вообще, для понимания феномена многоклеточное™ необходимо
сравнить его у прокариот и эвкариот. Многоклеточные прокариоты известны
давно (кустики цианей, грибочки миксобактерий и т.п.), а в настоящее время
становится ясно, что все вообще бактерии склонны образовывать многокле-
точные структуры. Ограничусь одним примером. Многие бактерии умеют
формировать биоплёнку (п. 5-9*), которая сама умеет формировать не-
большую популяцию персистеров (неактивных клеток, способных пережить
катастрофу). Если рассматривать колонию бактерий как организм, то перси-
стер выглядит как его орган размножения и, в то же время, как аналог ство-
ловой клетки эвкариотного организма.
Возможно, первое многоклеточное выросло из колонии одноклеточных.
К такой мысли ведет анализ колоний, обладающих основными свойствами
многоклеточных организмов: отдельные клетки берут здесь на себя функ-
ции органов, и можно даже говорить о зародыше (Иванов А.В., с. 31-34). На-
следственное разделение их функций предстает как рутинизация (п. 5-11*) и
тем самым понятна, но, замечу, первые клетки сразу же должны были нау-
читься узнавать “своих”, т.е. сразу был необходим простой иммунитет.
Проще представить себе, что сперва появились многоядерные плазмо-
дии, которые постепенно разгораживались стенками; тем более, что такой
процесс у одноклеточных известен. Тогда становление конъюгации (слия-
ния гамет) и миксоформ тоже становится понятнее - как поворачивание
вспять процесса становления многоклеточное™.
Мое мнение: многоклеточное™ на самом деле развивалась разными
путями. На такой вывод наводит сравнение низших многоклеточных, напри-
мер, водорослей: один и тот же тип строения (кустик) можно видеть и в
форме ассоциации одноклеточных диатомей, и в форме многоядерного зе-
леного плазмодия и в форме многоклеточной багрянки (Сапожников Ф.В.
Деревья диатомового мира // Природа, 2002, № 12).
8-8* Альтернативы многоклеточное™
Группы, не ставшие многоклеточными, вовсе не все остановились в
развитии. Много раз высказывалась та мысль, что инфузорий нельзя отно-
сить к одноклеточным, что они реализовали другой, сверхклеточный ва-
риант развития живого, вариант, когда клетка столь усложняется, что ее не
стоит называть клеткой. Достаточно сказать, что любая инфузория имеет
как минимум два ядра с совершенно различными функциями: одно ответст-
венно за наследственность, а другое - за метаболизм. (Почему тут не гово-
рят про симбиогенез? Потому, что сам симбиогенез ничего не умеет сказать
про ядро.) Замечательно, что в первом ядре ДНК не метилирована и при
471
этом не транскрибируется (служит только для репликации), тогда как во
втором наоборот: метилирована и служит только для транскрипции (Хатт-
ман С. И Биохимия, 2005, № 5, с. 673).
Некоторые сверхклеточные обладают такими приспособлениями, какие
у многоклеточных именуются органами (см. рис. 90 в конце книги):
«Ротовое отверстие инфузорий ведет в глотку... Затем следует аналог средней
кишки, которая обособлена от остальной цитоплазмы специальн ыми скелет-
ными элементами. Далее располагается задняя “кишка”, заканчивающаяся
анальным отверстием» (Серавин Л.Н. Пути эволюции протистов И Протесты.
Часть 1. Руководство по зоологии. СПб., Наука, 2000, с. 140).
Есть и крупные «сверхклеточные» организмы (например, плазмодии
миксомицетов). Плазмодий мог играть для первых многоклеточных ту же
роль, что прогенота - для первых клеток: сперва могла возникнуть много-
ядерная организация, а затем уж многоклеточность.
Мне уже случилось заметить, что о настоящей многоклеточное™ есть
смысл говорить только тогда, когда налицо не колония сходных клеток, а
разноклеточность (для разных функций организм использует различные
клетки - Чайковский Ю.В. Опыт эко-физиологической макросистемы // Ме-
тоды исследования в экологии и этологии. Пущино, 1986, с. 23). Петербург-
ский протистолог Л.Н. Серавин тоже счел главным появление
«разноклеточности (дифференцированности) вегетативных клеток в пределах
одного и того же организма» [Серавин Л.И., с. 144),
что произошло независимо у различных водорослей, у высших растений, у
губок, у прочих животных. Напомнив о сверхклеточных, Серавин пишет:
«Слоевище сифоновой водоросли Caulerpa prolifera достигает в длину пол-
метра и более. Оно сильно расчленено и по внешнему облику подобно выс-
шему растению. Стелющиеся по грунту ризомы - аналоги стеблей - много-
кратно ветвятся. Местами вниз от них отходят корнеподобные ризоиды, а
вверх - побеги, которые часто имеют вид листоподобных пластинок; при этом
тело каулерпы не разделено на клетки» (там же, с. 140).
Он предложил для многоядерных организмов, не поделенных на клетки,
понятие цитоид. В цитоидах достигается макроскопическая (видимая про-
стым глазом) форма в двух или трех измерениях, без процедуры деления
клеток. Учтя это, имеем такие уровни клеточной организации:
моноцит - одноклеточное (например, жгутиковое);
гиперцит - сверхклеточное (например, инфузория);
гомоцит - равноклеточное (например, колония вольвокс);
цитоид - многоядерное без клеточных перегородок (например, си-
фон, плазмодий, тегумент - см. п. 8-4); может иметь многоклеточные ор-
ганы размножения или быть частью многоклеточного организма;
гетероцит - разноклеточное (настоящее многоклеточное).
Все уровни появлялись в эволюции многократно, выявляя этим соответ-
ствующую часть (или, лучше сказать, сторону) диасети. Для нее характерно,
472
что клетки настоящего многоклеточного организма проще, чем гиперцит или
цитоид (в этом тоже - принцип компенсации).
Кроме указанных форм клеток, существует еще нить - клетка или ее
часть, достигающая макроскопического размера в одном измерении. Воз-
можна у одноклеточных (такова вертикальная ось зеленой водоросли
Acetabularia), но главный интерес представляют гифы грибов и аксоны (от-
ростки нервных клеток) высших животных. С помощью нитей достигается
проникновение одних структур в другие.
8-9. Черновики Господа Бога
В венде надежно известны красные водоросли (багрянки) и зеленые
водоросли. Их разнообразие начало заполнять собой ту самую рефренную
таблицу (рис. 35Б), которую мы наблюдаем до сих пор.
С животными сложнее: здесь типы не только появлялись, но и исчеза-
ли. От конца венда хорошо сохранилась эдиакарская фауна. Она была най-
дена в 1947 году на плато Эдиакар в южной Австралии, а затем - на всех
материках. В ней мы видим жизнь не только в форме известных нам типов,
но и нигде позже не виданную - новые типы (т.е. совсем особые планы
строения) тогда появлялись и исчезали; в дальнейшей истории (позже
среднего кембрия) типы уже не вымирали, и эдиакарскую фауну называют
«опытами Природы» или «черновиками Господа Бога» [Еськов, с. 120, 126].
Имеется в виду не всеведущий Бог христиан, которому опыты ни к чему, а
Бог в понимании Гегеля, т.е. осознающий себя в творении.
Чем и как питались тогда животные? Сказать почти нечего, но тот факт,
что нет свидетельств наличия у них внутренних органов, наводит на мысль,
что они еще, подобно организмам гидротерм, жили за счет эндосимбионтов.
Однако симбионты были теперь не литотрофами, а фотосинтетиками.
Вендских образцов найдены десятки тысяч, и среди них ни одного повреж-
денного - тела уходили в захоронение целиком. Из этого делают вывод, что
не было еще не только хищников, но и трупоедов. Разлагали органику пря-
мо редуценты, т.е. бактерии и первые низшие грибы.
В венде из известных типов животных описаны кишечнополостные (их
отпечатки преобладают), кольчатые черви, примитивные членистоногие и
столь же примитивные моллюски. Более простые типы, вроде плоских чер-
вей, должны были существовать тоже, но они ископаемых форм не остав-
ляют. В конце венда впервые проявился гигантизм форм (некоторые меду-
зы достигали 40 см), за которым последовало, как всегда, вымирание соот-
ветствующих таксонов.
Так или иначе, в венде жизнь освоила два главных типа симметрии
строения тела - радиальную и двустороннюю - и соответствующие планы
строения. Кроме того, найдены отпечатки, которые не удалось отнести ни к
одному из известных типов. Это и есть “черновики”.
473
Таково странное животное Helicoplacus (т.е. «спирально пластинча-
тый»), жившее в начале кембрия, чем-то похожее на морского ежа, но осу-
ществившее невиданный ни до, ни после тип криволинейной (по Д.В. На-
ливкину) спиральной симметрии (рис. 55 а).
б
Рис. 55. “Черновики Господа Бога’’ из нижнего кембрия: а) гелиотакус, длина
5 см; б) аномалокарис, один из первых хищников, длина до 45 см. Эти типы жи-
вотной организации никогда позже не встречались. Ссылки см. ЧЭ, с. 348
Наоборот, кембрий стал временем становления привычной нам жизни.
Основная вендская фауна вымерла, и ее быстро сменила совсем иная.
Дальнейший рост концентрации кислорода (результат фотосинтеза) и
фосфора (тогда была очередная стадия горообразования) привел к появле-
нию организмов с твердыми скелетами [Хайн, 1994 с. 71]. Все организмы
стали служить кому-то пищей, т.е. экосистемы стали похожими на нынеш-
ние, и биосфера замкнулась. Радикально возросло разнообразие организ-
мов. В частности, появились хищники (рис. 55 б). От кембрия сохранились
остатки всех типов животных, которые вообще попадают в летопись. Жизнь
протекала почти только в морях, зато в них она процветала. В частности, в
мелких водах в одном метре ото дна сложились многоярусные экосистемы,
напоминающие в миниатюре нынешний влажный тропический лес (Рожнов
С. В. И ПЖ, 1993, № 3, с. 127).
Тип хордовых появился в раннем кембрии, древнейшие их находки
имеют возраст 530 млн лет и напоминают нынешнего ланцетника (рис. 56).
Вероятно, они произошли от кольчатых червей, но в таком случае им при-
шлось, вопреки “биогенетическому закону”, сменить тип эмбрионального
развития (черви первичнороты, а хордовые вторичнороты).
Случайно ли появление нашего собственного типа или мы можем быть
уверены, что эволюция пришла к нему закономерно? Шалин пишет:
“Некоторые считают, что должно было существовать около сотни планов ор-
ганизации, из которых большинство исчезло”; при этом он уверен: “К сча-
стью для нас, план организации Pikaia, предка... позвоночных, избежал пер-
474
вичного истребления, иначе никогда не было бы ни рыб, ни амфибий, ни реп-
тилий, ни зверей, ни людей. На чем держится наше бытие - на случайности
эволюции” (АЕ, 163, 166).
Позволю себе иную точку зрения, диатропическую: тип хордовых мог
возникать многократно. Недавние находки в Китае (рис. 56), на 20 млн
старшие, чем найденная в Канаде Pikaia, эту мысль подтверждают: они на-
много старше, однако намного сложнее, чем Pikaia.
Это самые древние хордовые из когда-либо и где-либо найденных. 1 — передний
конец тела, 2 — глаз, 3 — ротовая полость, 4 —передний жаберный сосуд, 5 — мозг, 6 —
жаберные щели, 7 — зндостилъ, 8 — зубовидные структуры, 9 — спинная аорта, 10 — глотка,
11 — брюшная аорта, 12 — спинной плавник, 13 — сердце, 14 — пищевод, 15 — нервный тяж,
16 — половые железы, 17 — желудок, 18 — хорда, 19 — брюшной плавник, 20 — миосепты, 21 —
кишка, 22 — анальное отверстие, 23 — миомеры.
Рис. 56. Первые хордовые. 530 млн лет назад. Китай (подробнее: ЧЭ. с. 349)
В позднем кембрии в типе моллюсков появился их высший класс, голо-
воногие, которые в ордовике тоже стали проявлять гигантизм, и многие
вымерли. Тогда же на суше уже имелась почва, а это означает, что были и
растения, не оставившие о себе следов, вероятно, наземные водоросли, и
грибы. Именно их, полагал Мейен, можно считать предками наземной рас-
тительности, тогда как раньше считалось, что высшие растения формиро-
вались только в море [Еськов, с. 156-160]. Если так, то высшие растения
первыми совершили тот путь “обратно в воду”, который всем известен по
ихтиозаврам, китам и прочим позвоночным.
Черви не попадают в ископаемые, но в почвах ордовика видны следы
их ползания. Дальнейшая эволюция - отдельная тема, мы коснемся ее в
главе 9. А здесь перечислю некоторые общие эволюционные тенденции.
Как сказано в главе 6, выявление тенденций - основная задача диатропиче-
ской теории эволюции.
8-10. Компенсация как тенденция
Тенденция - это правило, верное в целом, но не на всех объектах (п. 6-
2). Одной из общих тенденций всего живого выступает, среди прочего,
1
475
принцип компенсации Аристотеля (п. 2-7): одни преуспели в одном, а другие
в другом (хотя одни - во многом, а другие - нет). Как будто каждой эволю-
ционной ветви при ее появлении был дан какой-то «аванс» (кому большой,
кому малый), которым каждая воспользовалась по-своему. Бергсон назвал
этот аванс «жизненным порывом» и первый заявил, что царства растений и
животных задаются своими основными тенденциями (п. 6-2).
Нас будут в основном занимать тенденции-процессы (см. с. 230). На
что тратили жизненный порыв те группы, которые не преуспели в усложне-
нии строения своего тела? Например, «низшие» многоклеточные? В целом
их жизненный порыв шел на усложнение тканей (и отчасти клеток) в ущерб
усложнению плана строения, но у многих «низших» усложнялся также и
план строения; попробуем это увидеть.
Тип плоских червей включает 5 классов, из них один - турбеллярии
(ресничные черви) в основном состоит из видов, живущих свободно, имею-
щих глаза и кишечник; остальные классы - чисто паразитические. И если
трематоды (сосальщики) не имеют глаз, но имеют кишечник, то цестоды
(ленточные черви) обходятся даже без него. Кто от кого произошел? Обще-
принято, что первыми были турбеллярии, а упрощение остальных - приспо-
собление к «более простым» условиям существования паразитов. Если так,
то внешние формы плоских червей упростились, зато ткани усложнились,
как мы уже видели на рис. 49. (Существенно, что самый упрощенный червь
обладает самым сложным тегументом.) Строение тегумента надо включить
в план строения типа, ибо он характерен именно для этого типа, и тогда на-
лицо усложнение плана у паразитов.
Паразит имеет еще и сложный цикл развития со своим планом строе-
ния каждой стадии, т.е. у него радикально усложнился хронотоп (совокуп-
ный план строения, п. 8-6). Тем самым, с плоскими червями родилась еще
одна тенденция: усложнение плана строения через появление циклов раз-
вития (что является усложнением экологической ниши). Произошло это не
раньше ордовика, когда появились первые рыбы. До этого кишечным пара-
зитам просто негде было сформироваться.
Итак, плоские черви являются «низшими» лишь в очень узком смысле,
в смысле простоты внешней формы тела. То же касается многих других
групп, и встает вопрос: насколько принцип компенсации всеобщ?
Как мы видели на сложнейших тканях (тегументах) просто устроенных
животных, или на орхидных, где есть сложнейшие цветки, но нет главного
приобретения цветковых (двойного оплодотворения), - явление компенса-
ции распространено весьма; таких примеров множество: см. Дополнение 8.
Палеонтологи А.В. Марков и Е.Б. Наймарк приводят пример тенденции,
которая вроде бы противостоит компенсации - это специализация: таксон,
вставший на путь специализации, может утратить всё, чем обладал, не по-
лучив взамен ничего, кроме быстрого роста численности и шанса на бы-
строе вымирание. Однако рост численности и есть искомая компенсация.
I
476
По мнению Маркова и Наймарк, палеонтологические данные показы-
вают, что
«повышение устойчивости к одному фактору повышает вероятность того, что
организм станет более устойчивым и ко всем прочим факторам»77, поэтому
«специализация и генерализация представляют собой две основные макроэво-
люпионные стратегии... Эволюция таксона, раз пойдя по одному из этих пу-
тей, с большой вероятностью будет продолжать по нему идти (ортогенез)»
Марков А.В., Наймарк Е.Б. Количественные закономерности макроэволюции.
Опыт... системного подхода. М., ГЕОС, 1998, с. 256, 260).
Обратимся к их книге за другими примерами тенденций.
8-11. Тенденции, прогресс и регресс
Одна из таких тенденций - отмеченное еще Бюффоном, Жоффруа и
Тейяром морфологическое единство основных групп животных. В наше
время ему придан новый смысл:
«многие надвидовые таксоны эволюционируют как целостные системы,
причем эта целостность обусловлена прежде всего тем, что таксоны часто
обладают экологической определенностью, т.е. в той или иной степени сов-
падают с понятием жизненной формы... В соответствии с этим на ранних
этапах эволюции таксона происходит бурная дивергенция» {Марков А.В.,
Наймарк Е.Б. Количественные закономерности макроэволюции. Опыт...
системного подхода. М., ГЕОС, 1998, с. 204).
Под «бурной дивергенцией» имеется в виду такая, когда преобладают
роды тех отрядов, которые реализуют r-стратегию (см. п. 5-10). Впоследст-
вии, по ее завершении, начинают преобладать роды отрядов с К-
стратегией, а носители г-стратегии вымирают (рис. 57). Тенденция состоит
здесь в преобладании данной стратегии в данном отряде.
Поясню: «надвидовые таксоны» выявляются исключительно по морфо-
логическим, а не по экологическим признакам, так что их преобладание в
коллекциях никак не следует из принципа приспособления каждого вида к
своей среде и скорее выглядит как акт номогенеза: крупный таксон (в дан-
ном случае класс мшанок) выступает как единый субъект эволюции. И сам
феномен тенденции выступает как один из аспектов номогенеза.
В той же книге читаем цитату из Шиндевольфа:
«При вымирании родов организмов окружающая среда играет предельно по-
верхностную роль... Ветер опрокидывает лишь гнилое дерево»
и соответствующий вывод авторов {Марков А.В., Наймарк Е.Б., с. 227):
«Эволюция биоты - не слепок с развития косной материи, с движения мате-
риков и и колебания климата, а самостоятельный природный процесс».
'7 Фактически авторы ведут речь о таксоне с высоким «авансом» активности.
477
Рис. 57. г- и /С-стратегии в эволюции палеозойских мшанок: 1 — быстро эво-
люирующие и быстро вымирающие роды (5 отрядов); 2 - медленно эволюи-
рующие роды (6 отрядов), которые, в основном, и реализуют возможности
класса. По оси абсцисс - геологические ярусы (от раннего ордовика до позд-
ней перми), по оси ординат - число родов (в указанных отрядах). По: Марков
В.А., НаймаркЕ.Б., 1998, с. 220
Здесь отмечена важная тенденция: хотя связь массового вымирания
таксонов с катастрофами неживой природы очевидна, основную причину его
надо искать в самих этих таксонах, в их старении (броккизм, п. 1-15*), а не
во внешних влияниях. (Что касается аналогии с ветром, то Шиндевольфу
следовало бы добавить: ураган валит и очень крепкие деревья. Это пона-
добится нам в п. 9-7.) В той же книге находим длинные, никаким материа-
лом не обоснованные, рассказы о конкуренции как причине эволюции. Им
резко противоречат конкретные данные о том, что реальна не конкуренция,
а расцвет одних после гибели других (например, с. 219, 251).
Не имея возможности рассказать про все важные тенденции эволюции,
приведу в качестве очень общего и важного примера группу тенденций, воз-
никших при выходе жизни на сушу. Этому предшествовало изменение атмо-
сферы. В кембрии в ней было раз в 20 больше СО2, чем ныне, а значит,
был силён парниковый эффект.
Едва в начале кембрия стаяли ледники (материки видимо отошли от
полюсов), а рост вулканизма повысил содержание СО2, как установился
жаркий климат и растения стали бурно потреблять СО2. Столь же бурно они
478
выделяли 02, концентрация которого к концу кембрия почти равнялась ны-
нешней. Возник озоновый экран, позволивший организмам овладеть сушей.
Хотя следы первых наземных растений (вероятно водорослей) известны
еще из протерозоя, но лишь из силура известны высшие растения, которые
образовали наземные (прибрежные) сообщества; а в девоне в этих сооб-
ществах стали существенны грибы. Потребление СО2 еще больше возрос-
ло, и в карбоне (каменноугольном периоде) его осталось в атмосфере мало
- почти столько, сколько ныне.
Бурное развитие растительности, ведшее к накоплению каменного угля,
происходило только в тогдашней тропической зоне, тогда как вне ее климат
был холодный, и на полюсах образовались большие полярные шапки.
Похоже на нынешний климат, но неясно, за счет чего тогда шло мощное
угленакопление. Болотистые тропические леса карбона резко отличны от
торфяных болот нашего времени. В частности, в них очевидны следы
регулярных катастроф, когда целые лесные массивы сносились паводками
и деревья погребались вместе с корнями. О причинах карбоновых
катастроф будет немного сказано в главе 11, здесь же стоит заметить:
вероятно, массовое захоронение неокисленной органики повлекло избыток
свободного кислорода в воздухе, отчего стали на время возможны
гигантские насекомые [Еськов, с. 185].
Животные, оставляющие ископаемые остатки, вышли на сушу еще в
конце ордовика (членистоногие) и в силуре (моллюски); а с конца девона
известны первые наземные позвоночные (амфибии). Их предками, как
считают, были девонские саркоптеригии (греч. мясопёрые) - так именуют
подкласс костистых рыб, чьи мясистые плавники содержат сложный скелет.
Подкласс состоит из двух отрядов: двоякодышащие и кистепёрые. О том,
кто из них был предком амфибий, идут бесконечные споры.
В карбоне появились рептилии; они, начиная с перми, царили на суше.
Затем, в мезозое, установилась так называемая теплая биосфера; в
частности, не было ледников. С триаса, самые интересные тенденции
наблюдаются именно на суше.
Первая связана с террипетностью. По «закону террипетного
развития» (п. 2-10), у группы животных строение тем сложнее, чем более
преобладают в ней наземные подгруппы. (Закон довольно ясно виден и у
растений: так, цветковые устроены сложнее всех, а они почти нацело -
наземная группа; проще же всех устроены водоросли - почти нацело
водные группы.) Высшие таксоны животных можно разделить на более
террипетные, т.е. в основном наземные, и менее террипетные. Так, тип
иглокожих не имеет наземных форм, тип моллюсков имеет их мало, тип
хордовых имеет больше наземных, чем водных, а в типе членистоногих
подавляющее большинство - наземные насекомые. Высшая степень
террипетности - подъем головы, а затем и туловища, обращение лап в руки
или крылья.
479
Замечательно, что террипетность косвенно связана с усложнением
мозга. Известно, что большие полушария позвоночных произошли от
обонятельных зон мозга рыб и разрастались по мере продвижения таксонов
на сушу и по суше. То же описано недавно у брюхоногих моллюсков,
единственного класса этого типа, проявляющего террипетность (легочные
улитки заселили сушу вплоть до гор): переход на сушу влечет усложнение
обоняния и вместе с ним - появление нового отдела мозга; при
возвращении в воду эти органы вновь упрощаются (Зайцева О.В. //
Российский физиология, журнал, 2000, № 8).
Вторая тенденция связана цефализацией, увеличением головы в
ходе эволюции многих групп животных. Понятие цефализации ввел
американский геолог Джеймс Дана в 1852 году. Степень цефализации у
различных групп весьма различна: у столь высокоразвитой (и
высокотеррилетной) группы, как пауки, головы, строго говоря, нет вообще, а
у насекомых есть и у мух весьма велика. На поведении это сказывается
весьма заметно, хоть и весьма неожиданно: головастую муху очень трудно
поймать, а мелкоголового комара (из того же отряда двукрылых) - проще
простого. Головаста и медоносная пчела (отряд перепончатокрылых) с ее
правильными сотами и замысловатым поведением.
Легко собрать подобные примеры и сделать вывод о строгой связи
интеллекта с цефализацией. Но вот перед нами “безмозглые” пауки с их
ловчими сетями и, порою, совершенно невообразимым поведением (п. 6-15)
и умные осьминоги, чей мозг вообще не в голове, а вокруг пищевода.
Поэтому ограничимся, как и Дана, позвоночными. Весьма террипетны, но
мало цефализованы птицы, а у зверей развиты и террипетность, и
цефализация, но в разной мере: так, максимальный подъем головы видим у
жирафа, но цефализация у него минимальна.
Обе эти тенденции друг от друга независимы, и те таксоны, которые
проявляют их совместно, выступают в качестве самых прогрессивных.
Достаточно заметить, что обе тенденции в высокой степени свойственны
приматам, а из них особенно - человеку. Можно сказать, что и ламарково
стремление к прогрессу - тенденция, наиболее явная у групп животных, где
сочетаются террипетность и цефализация.
Третью тенденцию сформулировал Коп (о нем см. п. 3-10) и назвал
филогенетическим ростом: в эволюции многих таксонов животных наблю-
дается увеличение размеров тела. Часто оно кончается гигантизмом (кро-
ме уже упомянутых медуз венда и головоногих ордовика, стоит указать
стрекоз карбона - до 90 см, юрских динозавров - до 25 м и нынешних ки-
тов), за которым следует вымирание. Предел гигантизму кладут физиологи-
ческие причины: для насекомых с их трахейным дыханием это - дециметры,
для наземных позвоночных - несколько метров (большего не дает гравита-
ция), тогда как в океане живет наибольшее из животных всех времен, ны-
нешний синий кит (а отнюдь не динозавр); он достигает 33-х м. Нам эта
480
тенденция важна потому, что высший интеллект достигается у крупных (но
вовсе не самых крупных) животных.
Известно много примеров филогенетического роста и для отдельных
органов - например, рогов у оленей, числа позвонков у змей, и предел тут
тоже кладет физиология (рога мешают передвигаться).
Этой общей тенденции противостоит частная, филогенетическая кар-
ликовость: почему-то некоторые группы вдруг начинают мельчать. Самым
удивительным примером считается карликовость прежних слонов; наи-
меньший, Elephas falconeri, живший 500 тыс. лет назад на островах Среди-
земного моря, был ростом с пони [Рэфф, Кофмен, с. 64].
8-11* Пример связи эмбриологии и палеонтологии
Интересно появление конечностей позвоночных. Шалин пишет:
“Конечности всегда формируются в три последовательных фазы: (I) сперва
плечевая и бедренная кости, (II) затем лучевая и локтевая, равно как большая и
малая берцовые кости, (III) наконец, рука и нога. У рыб третьей фазы, т.е. появ-
ления рук и ног, нет. Это, тем самым, морфологическое приобретение четверо-
ногих. Парные конечности у саркоптеридиевых рыб выражены единственной
костью на плечевом и бедренном поясах, и изучение ископаемых форм показа-
ло, что бедренная и плечевая кости появились первыми, вероятно, в верхнем
силуре”; а в девоне появились кости второй и третьей фаз. “В этом случае име-
ет место согласие между последовательностями онтогенетической и историче-
ской: онтогенез в самом деле повторяет картину филогенеза” (АЕ, 177-178).
Э.И. Воробьева заключает: история кистепёрых была «генеральной ре-
петицией выхода на сушу», поскольку у них то и дело появлялись черты бу-
дущих амфибий. Так, ряд черт среднего уха, органа слуха в воздушной сре-
де, появился ранее самого органа, до выхода в данную среду, и
“обычно отмечается преобладание особенностей исходной организации, в ко-
торую как бы вкраплены отдельные черты вышестоящей группы. В то же
время промежуточные признаки... практически отсутствуют” (Воробьева Э.И.
//УСБ, 1994, №6, с. 648).
Более наглядный пример, уже помянутый в п. 4-10: внутри плавников
кистеперых сформировалась система пальцев, даже более сложная, чем у
четвероногих: восемь пальцев с зачатком девятого (Laurin М. И LEG, 30).
Напомню, что поиск промежуточных состояний был главной установкой
Дарвина, более важной для него, чем даже отбор. С данной установкой
можно теперь расстаться: эволюция шла иначе. Вместо постепенных пере-
ходов мы повсюду видим блочность эволюции. Она выражалась, например,
в случаях «появления у кистеперых признаков, свойственных амфибиям,
ихтиозаврам, териодонтам и млекопитающим» - пишет Воробьева.
В этом, напомню, состояли пророческие стадии Павлова. Проще
говоря, можно проследить изменение органов при выходе на сушу, но
нельзя проследить линию “предок-потомок”. Мы вернемся к этому в главе
10.
481
Вскоре был совершен громадный прорыв в понимании морфогенеза -
открыты гены, ответственные за формирование планов строения как орга-
низма в целом, так и основных органов (п. 5-15). Перечислив различные ге-
ны, последовательно формирующие план строения конечности позвоночно-
го, Шалин подытожил:
“Итак, здесь не функция порождает орган и не приспособление постепенно
преобразует орган под действием естественного отбора, а скачкообразное по-
явление органа оправдывает (autorise) новую функцию. Но имеется обяза-
тельное условие, накладываемое средой, - что новая структура не явится
крупной помехой организму ” (АЕ, 178).
В целом это вполне соответствует изложенному в части 2 пониманию
эволюции, в том числе идее экологической прочности.
8-11** Тенденции регресса и рефрены
Причин тенденций мы не знаем, но это не мешает выявлять их и ими
пользоваться. (Аналогично, химики XIX века, не зная ничего о строении
атома, подметили важные тенденции - например, возрастание щелочной
активности с ростом атомного веса в двух параллельных рядах: щелочных и
щелочноземельных металлов; это помогло и практике, и созданию теории
атома.) Для описания эволюции необходимо знать тенденции хотя бы пото-
му, что всякий таксон приспосабливается не только к своей среде,
но и к своим тенденциям. Так, змеи научились ползать, когда их тенден-
ция (рост числа позвонков) не оставила им другой возможности. Тем же са-
мым легко объяснить возвращение в воду целых отрядов позвоночных:
многие из них ничего, по всей видимости, в воде не приобрели, но они по-
просту не могли существовать нигде более.
Такое понимание тенденций открывает путь и к пониманию регресса,
т.е. упрощения строения и утраты органов, важных для жизни. Вместо вы-
думывания пользы от подобных утрат, лучше посмотрим на утраты как на
тенденции, к которым их обладатели приспособились, кто как сумел. Самое
обычное - утрата зрения, а затем и самих глаз. Что процесс этот может, как
и всякая утрата, идти очень быстро, понял еще Бэр, когда узнал о слепой
рыбе, живущей в геологически молодом водоеме (п. 3-6). Однако на трило-
битах палеозоя отмечена очень медленная утрата глаз - за 3-15 млн лет.
Вот рис. 58, взятый из обзора: Thierry J., Marchand D. Les invertebres
fossiles et (’illustration de revolution... // LEG, 28. Авторы видят в “быстрой” (3
млн лет) утрате глаз пунктуализм, а в медленной (15 млн лет) - градуализм
(обычную дарвинову эволюцию). Но оба процесса очень медленны и потому
требуют иной трактовки: сходство трех картин порождает вопрос: а может
быть, авторы лишь конструируют воображаемый процесс?
В самом деле, подпись к французскому оригиналу рисунка 58 гласит:
“Эволюционные тенденции, ведущие к редукции вплоть до утраты глаз
трилобитов ордовика и девона. Избранные примеры не обязательно являют
482
эволюционную линию, они иллюстрируют тенденции выстроенных в ряд
изменений строения головных щитов с течением времени” -
и это побуждает к положительному ответу. Что же тогда изображено?
Рис. 58. Эволюционные
тенденции утраты глаз у
трилобитов.
А - семейство Phacopi-
dae (франский и фамен-
ский ярусы девона);
В - виды рода Ота-
thops (аренигский и
лланвирнский ярусы
ордовика);
С - виды рода Tropido-
coryphines (франский ярус
девона);
М.а. - миллионы лет.
Фактически изображен
фрагмент рефрена
“степени выраженности
глаз у трилобитов”
Авторы замечают,
что похожие процессы
известны у иных
организмов - юрских
Cryphops
acuticeps
Diemops
griffilhides
Trimerocephalus
mastomphthalmus
Phacops
granuiatus
Famennien
Frasnien
tendance Evolutive
(duree, environ 15 M. a.)
O. nicholsoni
O. llanvemensis
O. alavus
О bomi
Lianvirn
Arenig
tendance Evolutive
(dur6e, environ
Frasnien
Frasnien
Pterocoryphe
languedociana
Pterocoryphe
progrediens
Pteroparia
coumiacensis
Pterocoryphe
ocidata
tendance Evolutive
(durfc, environ 3 M. a.)
аммонитов. Действительно, еще Траутшольд показывал, что гипотетиче-
ские “эволюционные” ряды юрских аммонитов аналогичны рядам заведомо
одновременно живших форм (п. 2-11**).
Здесь, при анализе трилобитов, недостает как раз сравнения с одно-
временно жившими формами. Процессы В и С идут в рамках рода и потому
могут быть названы эволюцией (из вида в вид), тогда как в строке А взято по
одному виду из разных родов, значит реальной эволюции тут не выявлено,
а лишь очерчено разнообразие вариантов. Словом, изображен фрагмент
диасети, точнее - временной рефрен. Вероятно, что в той же диасети мож-
но найти и аналогичный пространственный рефрен.
8-12. Эпигностика как двигатель эволюции
В терминах тенденций можно ставить и решать новые эволюционные
вопросы. Так, мы знаем из главы 6, что высшее развитие психики состоя-
лось у тех, кто имел наиболее развитой иммунитет. Но почему птицы не
достигли тех же высот психики, что дельфины и высшие приматы? Ответ
очевиден: у них маловат мозг; а это - итог малой цефализации всего класса
483
птиц, включая нелетающих. Кстати, летающие зверьки все принадлежат к
плацентарным, и беременные самки вполне могут летать. Значит, нельзя
объяснить яйцерождение птиц, исходя из экономии полетного веса.
Малый мозг птиц тоже нельзя связать с полетом - хотя бы потому, что
мозг мал и у нелетающих птиц (страусы, пингвины), причем группы эти дос-
таточно древние, и времени на эволюцию мозга у них было больше, чем
ушло, например, на эволюцию гоминид вплоть до человека.
Дело в различии тенденций. У всех птиц мозг устроен хоть и сложнее,
чем у рептилий, однако качественно отличен от мозга зверей: новая кора
(неокортекс, основа разумной деятельности млекопитающих), едва наме-
ченная у некоторых рептилий, у птиц хорошо выражена, но разумную дея-
тельность ведет не она, а свойственное только им тело гиперстриатум,
нарост над полосатым телом [Ромер, Парсонс, т. 2, с. 308]. Последнее, кор-
пус стриатум, есть и у других четвероногих, оно ответственно за движения.
У птиц (как, кстати, и у сумчатых) отсутствует мозолистое тело, соеди-
няющее два полушария у плацентарных. Можно сказать, что мышление
птиц-тоже своего рода “черновик Господа Бога” (ср. п. 8-9).
Отставание дельфинов от приматов выглядит как следствие отсутствия
террипетности (китообразные потеряли ее, вернувшись в воду), и примерно
то же можно сказать про эволюционный тупик, в котором оказались голово-
ногие: большой “головной” мозг (вокруг пищевода) при полном отсутствии
террипетности у всего класса (они не живут даже в водах слабой солено-
сти). Можно сказать, что по сходной причине мы умом обогнали слона: мозг
у него больше, но террипетность ниже - сила тяжести не дала его предкам
реализовать подъем головы.
У всех теплокровных имеется ТВ-иммунитет, но у птиц В-система раз-
вита гораздо слабее, чем у зверей. Здесь разнообразие антител
“связано с иным типом организации иммуноглобулиновых генов и происхо-
дит в характерном только для птиц месте - в расположенной у клоаки фабри-
циевой сумке. В локусе легких цепей у курицы имеется один V-ген, который
вначале перестраивается и соединяется с одним комплексом J-C” (Ройт А.,
Бростофф Дж., МейлД. Иммунология. М., Мир, 2000, с. 290).
Если это будет показано для всего класса птиц, то у них налицо окажет-
ся тупик развития ТВ-системы. Не в этом ли кроется причина их слабой це-
фализации? Сама постановка этого вопроса открывает тему эпигностики как
фактора эволюции.
В самом деле, свое от чужого умеют отличать даже бактерии: свои
клетки образуют единую колонию, и именно по форме колоний принято оп-
ределять самые древние бактерии (возрастом до 2 млрд лет); чужие клетки,
наоборот, подвергаются атаке. Тем более должны были с самого начала
уметь узнавать друг друга эвкариотные клетки. Дело в том, что избиратель-
ная адгезия (п. 8-8), возникшая как приспособление одноклеточных к объе-
динению со своими и питанию чужими, у многоклеточных стала средством
484
для построения тканей и органов. Для этого ей пришлось стать более раз-
нообразной: у клеток каждой ткани закон адгезии свой. Но этот закон от
особи к особи не меняется, а вот у клеток иммунной системы он у каждой
особи свой, да к тому же то и дело меняется.
Лучше всего решает задачи узнавания ТВ-иммунитет. Но как средство
против заразы он во многом уступает иммунитету врожденному. Он как бы
специально создан, чтобы зараза к нему подстраивалась. В самом деле,
даже самые просто устроенные бактерии (молликуты) обманывают иммун-
ную систему своих жертв: почти 5% их крохотного генома занял повторяю-
щийся ген белка адгезина, и за счет рекомбинаций между его копиями адге-
зии то и дело меняет свою антигенную структуру, «что помогает преодолеть
иммунный ответ хозяина». Более сложные паразитические бактерии реша-
ют даже собственные иммунные задачи - могут «связывать и поглощать
при генетической трансформации лишь ДНК из клеток того же вида» (Про-
зоров А. А. // Микробиология, 1998, №4, с. 445-446).
А паразитические эвкариоты прямо-таки творят иммунные чудеса: неко-
торые трипаносомы еженедельно меняют свои антигенные качества
синхронно, так что вся популяция, живущая в данном организме, имеет одну
и ту же антигенную структуру оболочки (Организация генома. М., 1989, с.
124). Это говорит о быстром и эффективном горизонтальном переносе
между ними, но не только о нем. Чтобы приноровиться к сверхвариабельной
иммунной системе хозяина, паразит должен быть столь же вариабелен.
Если признать, что адаптивный иммунитет - приобретение позднее, то
останется признать, что паразиты усвоили вариабельность от хозяев
(вероятно, тоже путем горизонтального переноса). Это (как и многое другое)
говорит о продолжающейся эволюции одноклеточных. Можно сказать, что
примитивнейший паразит (молликут) только потому и дожил до наших дней,
что нашел эпигностический ключ к иммунитету высших.
Пора задать вопрос: когда какой вид эпигностики возник? Видимо, все
прокариоты обладают противовирусным иммунитетом. Кроме того,
«Обмен генетическим материалом у прокариот в большинстве случаев проис-
ходит только между близкородственными штаммами, хотя в последнее время
чаще обсуждается как раз менее типичное явление - генетический обмен между
неродственными формами. У прокариот есть несколько способов обмена гене-
тическим материалом, прежде всего, конъюгация (“половой процесс”)-.- Перед
началом конъюгации бактерии вступают в контакт припомощи специальных
отростков - пилей. При этом, по-видимому, происходит оценка партнерами
друг друга» [Марков, Куликов, 2006, с. 132-133].
На этой основе авторы делают заключение, что система распознавания
возникла «на заре жизни», точнее, в наших терминах, в прогеноте. Вряд ли
это можно счесть обоснованным: с тем же правом можно утверждать, что
люди всех времен ходили в тканях фабричного производства - ведь такие
ткани можно нынче встретить у самых отсталых племён.
485
Мое мнение: древность свойства вытекает не из всеобщности его рас-
пространения, а из невозможности без него жить (как, видимо, нельзя жить
без «четырех R», см. п. 7-12) и наличия модели раннего появления свойст-
ва. Исходя из допущения, что всякий организм обходится тем иммунитетом,
какой ему необходим для создания и поддержания своей целостности, мож-
но полагать, что горизонтальный перенос возник вместе с появлением бак-
терий, РНК-интерференция (п. 6-6**) - вместе с эвкариотностью, а эпигно-
стика активных поверхностей - вместе с многоклеточностью. Замечу, что у
самых низших многоклеточных имеются элементы тканевой несовместимо-
сти (Ройт А. и др., Иммунология. М., 2000, гл. 15).
С этой позиции иммунитеты кольчатых червей, боконервных мол-
люсков, улиток и осьминогов должны быть очень различны, но есть ли ра-
боты на эту тему, мне неизвестно. Зато известно, что у самых примитивных
нынешних хордовых (оболочники) уже есть зачаток адаптивного иммунитета
(Du Pasquier /..Innate immunity in early chordates and the appearence of
adaptive immunity // CR Biol., 2004, № 6). Естественно допустить, что им об-
ладали и их кембрийские предки, но проверить это невозможно.
Эпигностический подход к эволюции позволяет продвинуться и в пони-
мании процесса еидообразоеания. Как известно, высшие организмы раз-
множаются половым путем (высшие животные только им). Принято считать,
что спаривание гомологичных хромосом в мейозе требует их почти полного
сходства, однако недавние исследования на грызунах открыли совсем иную
картину: существуют виды, у которых могут спариваться (гомологичными
участками) хромосомы, очень различные по длине и составу. (Эффективно
скрещиваются и организмы с различным числом хромосом - в таком случае
спариваются обычные хромосомы со слившимися.)
Привычное генетике требование сходства гомологичных хромосом ока-
залось не геометрическим, а чисто функциональным запретом, похожим на
иммунную несовместимость. Для описания вида, в пределах которого число
хромосом может сильно меняться при сохранении скрещиваемости, авторы
даже применили иммунологический термин - толерантность (этого вида
грызунов) к хромосомным перестройкам [Бородин и др., 2002]. Создается
впечатление, что у большинства видов, включая человека, данный запрет,
старый, как мейоз, служит для стабилизации вида, а его снятие открывает
путь к видообразованию, это может оказаться искомым механизмом дей-
ствия прерывистого равновесия. К сожалению, явление еще мало изучено.
8-12* Диатропика и эпигностика эволюции млекопитающих
Если не считать рептилий с предположительными чертами теплокров-
ности, известных с конца карбона, то теплокровные (птицы и однопроход-
ные звери) появились в конце триаса и развивались весь юрский период.
Но ископаемые остатки ничего не говорят о том, каков был их иммунитет,
поэтому палеонтологический анализ тут надо дополнить диатропическим.
486
Сумчатые - не предки плацентарных, а отдельная ветвь зверей, оба
эти подкласса появились приблизительно в одно время (ранний мел, т.е.
меловой период), так что иммунитеты сумчатых и плацентарных, вероятно,
развивались независимо. С детства мы привыкли считать сумку сумчатых
забавным, но очень полезным приспособлением, не задумываясь: приспо-
соблением к чему? Гигантская кенгуру рожает детеныша легче грамма; воз-
можно, что это недоразвитие эмбриона как раз ведет к низкому развитию
мозга (слабой цефализации) сумчатых. Не отсюда ли отставание сумчатых
от плацентарных, по крайней мере австралийских? Ведь появление плацен-
тарных в Австралии привело к вытеснению сумчатых. В Южной Америке
сумчатые (опоссумы и т.п.) хотя и благоденствуют, но занимают только ни-
ши низко цефализованных животных (грызунов в широком смысле).
Есть ли в раннем рождении польза для вида? Как когда. Вспомним, что
обычный тип приспособления - к самому себе, к своим тенденциям. Вот к
своему иммунитету приспособились и теплокровные: если иммунная систе-
ма самки отторгает белки самца, то эмбрион надо от нее защитить.
Известно, что живорождение возникало многократно - у позвоноч-
ных и беспозвоночных (Иванова-Казас О.М. Эволюционая эмбриология жи-
вотных. СПб., Наука, 1995), но у холоднокровных оно не вызывает иммун-
ных проблем и, надо полагать, не вызывало в прошлом. Совсем иначе у те-
плокровных: едва в триасе теплокровность предположительно появилась,
как мы видим изменение статуса живорождения: класс птиц начисто его ли-
шен (в отличие ото всех других классов позвоночных), причем нет его и у
нелетающих птиц; нет живорождения и у первых млекопитающих (однопро-
ходных, т.е. яйцекладущих), а сумчатые рожают ранее, чем можно ожидать
отторжения плода, полагает Шалин (АЕ, 150).
Наоборот, плацентарные, как известно, окружают ранний зародыш им-
мунным барьером - трофобластом, отделяющим плод от иммунных атак
организма матери. (Трофобласт - слой клеток, который затем переходит в
плаценту.) Канадский палеонтолог Роберт Кэрролл пишет:
«Свидетельств того, как возник трофобласт, нет. Определить его наличие или
отсутствие по скелетным остаткам мезозойских млекопитающих также не-
возможно. Кайнозойские находки уже демонстрируют наличие эмбрионов
внутри материнского организма, однако исходная радиация (появление и рас-
сселение новых таксонов - Ю. Ч.) плацентарных млекопитающих произошла
намного раньше» [Кэрролл, т. 3, с. 6].
Кэрролл предположил, что трофобласт был уже у первых плацентар-
ных - именно в силу его иммунной необходимости. Если так, то надо при-
знать, что ТВ-система возникла примерно вместе с теплокровностью, а
время от первых однопроходных до первых плацентарных (вероятно, весь
юрский период) - это поиск пути преодолеть отторжение плода, пути к тро-
фобласту. С этой точки зрения, иммунитет птиц оказался тоже достаточен
для отторжения плода, но пути к трофобласту они не нашли.
487
Эмбриология недавно подтвердила данный ход мысли. Известно, что
плацентарные звери развиваются необычно: после укоренения в матке за-
родыш делается (из трех клеток) заново и именно он превращается в новую
особь. Остальные клетки образуют трофобласт [Сахарова]. Недавно уда-
лось добавить к этому важный факт: первичный зародыш не может сам ни
во что развиться потому, что клетки в нем связаны слишком слабо:
“Из-за слабых адгезивных свойств поверхностей бластомеров и перемещений
относительно друг друга эти клетки располагаются хаотически, причем в раз-
ных эмбрионах по-своему” [Сахарова, с. 30].
Эмбриологи удивляются, но с эпигностической позиции тут всё ясно:
мощная ТВ-система не допустит наличия никаких чужеродных клеток, кроме
самых безликих (с «лицом заурядным, как бельевая пуговица» - говорил
писатель С.Д. Довлатов о сотруднике госбезопасности). Отсутствие специ-
фических поверхностных рецепторов позволяет клетке выжить, но не дает
ей стать частью зародыша, и только после формирования трофобласта мо-
жет начаться собственно онтогенез, основанный на клетках с высоко изби-
рательной адгезией поверхностей.
Замечательны параллели, проведенные Н.Ю. Сахаровой:
1) между низшим (кишечнополостные) и высшим уровнями животных:
“дробление у гидромедуз асинхронное и хаотическое, бластомеры способны
перемещаться и формировать цепочки, а также отделяться друг от друга, да-
вач начало полноценным особям. Почти все так же, как у млекопитающих!”;
более того, “при дроблении зародышей млекопитающих бластомеры образу-
ют крестообразную фигуру... крест, явно напоминающий расположение спор
при палинтомическом делении колониальных жгутиковых” (с. 31).
2) между зверьми и наездниками (паразитическими насекомыми):
“У наездников морула^... превращается в паразитическую личинку, полу-
чающую питательные вещества из организма хозяина через оболочку - тро-
фамнион (аналог трофобласта - Ю.Ч.). Из нескольких соматических клеток
тела личинки формируются группы эмбриональных клеток, дающие начало
новым личинкам” (с. 33).
3) между двумя высшими уровнями организмов:
“У млекопитающих и цветковых растений в развивающемся зародыше фор-
мируются ткани, которые обеспечивают поступление питательных веществ от
матери. У [плацентарных -Ю.Ч.] млекопитающих это... трофобласт, а у цвет-
ковых растений - эндосперм” (с. 36).
Есть и другие параллели. Наиболее удивительна плацента (орган свя-
зи матери с плодом), являющая параллель между низшими уровнями жи-
7©
Стадия зародыша, аналогичная бластуле. Сам факт наличия морулы у низших (неко-
торые губки, черви и др.) и высших (некоторые членистоногие и почти все млекопи-
тающие) — параллелизм, демонстрирующий важную часть диасети.
488
вотных (например, онихофоры, или первичнотрахейные — п. 9-3) и высши-
ми (живородящие рыбы и амфибии, а также почти все млекопитающие).
Вместе они рисуют общую картину: диасеть имеет единую основу у
всех организмов, на ней выстроены диасети отдельных царств, на каждой
из них строится меньшая диасеть и т.д. (фрактальный принцип). С этой точ-
ки зрения понятна позиция Сахаровой, видящей в морфогенезе млекопи-
тающих черты колонии жгутиковых, эмбриогенеза гидромедуз (тип дробле-
ния), некоторых насекомых (трофобласт) и цветковых (новый зародыш из
обычных клеток, т.е. не из ППК).
8-12** Диатропика и эпигностика размножения
В девоне появились семенные растения, и в карбоне они стали преоб-
ладать. Семя - отличное средство расселения, но нам интереснее другое
их приобретение: пыльцевое зерно. Самих пыльцевых зёрен нужно много,
но каждое проросшее зерно используется предельно экономно: прорастая в
женской шишке или в женском цветке79 (или женской части цветка) оно ис-
пускает ровно два спермия; один оплодотворяет яйцеклетку, а второй либо
отмирает (у голосеменных), либо оплодотворяет диплоидную клетку (двой-
ное оплодотворение покрытосеменных - время появления: нижний мел).
(Такое экономное оплодотворение семенных справедливо сравнивают
с заботой о потомстве у высших животных. Особенно эффективно оно у
цветковых: двойное оплодотворение приводит к триплоидному эндоспер-
му, который дает возможность очень быстрого развития плода и семени.
Лучше всего это видно на Крайнем Севере, где хвойных нет вовсе, а цвет-
ковые иной раз успевают провести весь цикл жизни за 10 дней - при почти
нулевой суточной температуре.)
Поразителен контраст с оплодотворением у высших животных: все они,
включая людей, производят не два спермия, а миллионы, что выглядит при
внутреннем оплодотворении нелепым излишеством. Столь же нелепа по-
пытка объяснить его как поле для конкуренции спермиев. В самом деле: при
этом в принципе возможна только конкуренция за качество самих спермиев,
но не за качество будущего организма (как многие молчаливо допускают).
Мне известно лишь одно объяснение: высокая концентрация спермиев
позволяет преодолеть женский иммунный барьер. Цель активности спермия
вполне понятна у низших организмов (он ведет поиск объекта оплодотворе-
ния), и вполне естествен тот факт, что спермин семенных неактивны: здесь
поиск ведет другая структура - прорастающее пыльцевое зерно.
7Q
Иногда считают шишку цветком (‘‘Женская шишка рассматривается чаще как соцве-
тие, а не как единичный цветок” - Крюссман Г. Хвойные породы / Пер. с немец. М.,
Лесная промышленность, 1986, с. 16), но при этом термин “цветковые” несёт иной
смысл, чем обычно (подчеркивая сходство всех семенных). Далее мы будем понимать
“цветковые” как синоним термина “покрытосеменные”, хотя это и неточно.
489
Однако у высших животных активный поиск ведет самец, но и у них ус-
тановлена высокая поисковая активность спермиев: головка спермия по-
крыта белками адгезии (п. 5-15) и сходна в этом плане с конусом роста ней-
рона (с помощью которого нейрон активно ищет путь для своего роста - п.
5-15). Статья об этом сходстве так и названа: «Спермин - нейрон с хво-
стом» (Meizel S. The sperm, a neuron with a tail: “neuronal” receptors in
mammalian sperm // BR, 2004, № 4).
Суть же в том, что в обоих случаях использован удивительный меха-
низм активного поиска, действующий на макромолекулярном уровне; он
обеспечивает как рост нервной ткани, так и оплодотворение. Еще удиви-
тельнее, что специфика данного (субклеточного и макромолекулярного)
уровня тут ни при чем: ту же поисковую функцию выполняет и самец, ищу-
щий самку, и отделившееся от тела самца щупальце головоногого; ее же мы
видим и в любом процессе роста, и в ходе мейоза (п. 8-8) и во многом дру-
гом. Налицо поисковая активность как таковая, и она использует всё, что ей
оказалось нужно и удобно. Мы вернемся к этому в п. 8-15, после знакомства
с иными формами активности.
8-13. Эволюция биосферы
Неверно думать, что во все эпохи биосфера имела хотя бы одинаковый
объем. И она порой состояла из очень непривычных нам экосистем. Бывали
эпохи теплые и холодные, с большим или меньшим процентом суши, с
большим или меньшим разнообразием видов. Переход между эпохами яв-
лял собою прогрессивную эволюцию экосистем, что, в сущности, понимал
еще Брони (п. 2-10) до рождения экологии.
Сегодня можно говорить про четыре биосферы, последовательно воз-
никавшие в ходе эволюции. Каждая возникала ввиду появления новой гос-
подствующей категории организмов, и первые три из них длились, вероятно,
примерно по миллиарду лет.
1) Хемосфера произошла из прогеноты и являла собой множество док-
леточных зон, в которых развивались прокариоты. Пищевой базой служили
литотрофы (п. 7-7); основной путь эволюции - включение органических
звеньев в геохимические круговороты. Хемосфера сложилась из первых
очагов жизни и обрела свойства биосферы лишь тогда, когда разрослась
достаточно, чтобы стать планетным явлением, т.е. завладела океаном (су-
ша была безжизненна). Видимо, в ней были обычны предельно короткие
круговороты за счет симбиоза.
В океане она осталась до сих пор (в гидротермах) и очень слабо связа-
на с остальной жизнью, но теперь она состоит из отдельных организмов.
Недавние исследования обнаружили, что остатки этой биосферы распро-
страняются еще и на полкилометра в глубь океанского дна, и некоторые ис-
следователи полагают, что условия жизни там не изменились со времен
биопоэза (Summit М., Barros J.A. И PNAS, 2001, vol. 98, № 5).
490
2) По мере охлаждения земной поверхности (в эпоху рождения жизни
поток тепла из недр был вчетверо выше нынешнего) хемосферу сменяла
прокариотная фотосфера. В ней пищевой базой были фотоавтотрофы; ос-
новной путь эволюции - усложнение биохимии. Она тоже начала с коротких
круговоротов (вспомним циано-бактериальные маты), но затем, наоборот,
стала удлинять цепи круговоротов, чем изменила облик планеты - напри-
мер, создала кислородную атмосферу. В ней, по всей видимости, возникли
основные нынешние механизмы - про- и эвкариотная организации клетки, а
также апоптоз (п. 6-6**), но прокариоты долго еще царили.
3) Затем возникла кариосфера (господство эвкариот; основной путь
эволюции - усложнение структур при прежней биохимии), и прокариоты бы-
ли отодвинуты на периферию биосферы, где господствуют поныне (напри-
мер, в земной и океанической коре), а главными продуцентами выступили
зеленые растения. Длинные круговороты стали преобладать: между проду-
центами и редуцентами (п. 7-7) появились консументы (животные), так что
теперь нормальная пищевая цепь имеет 5 звеньев, а иногда и 7. Кариосфе-
ра выглядит как собственно биосфера, и Г.А. Заварзин именует ее неосфе-
рой, но на ее базе уже вырастает следующая.
4) Антропосфера - ныне возникающая биосфера будущего; основные
круговороты в ней будут рождены людьми, а основным путем эволюции ор-
ганизмов будет создание искусственных пород при сокращении числа есте-
ственных видов. Ламарк, а за ним Вернадский, были правы, указывая на то,
что человечество становится геологической силой. Человек уже сейчас
уничтожает одни виды и создает другие, уничтожает одни ископаемые, соз-
давая другие (свалки отходов), и изменяет круговороты - как удлиняя их в
пространстве (глобальные перевозки), так и укорачивая геохимические цепи
(сжигая лес и т.п.). Антропосфера - та неприглядная реальность, которая
вырастает на месте, где Тейяр и Вернадский полагали увидеть прекрасную
ноосферу. Мы вернемся к этой теме в части 4.
Каждая следующая биосфера вырастает на базе предыдущей и прояв-
ляет новую активность: забирает основную часть тех глобальных функций,
какие может исполнить сама. А.Г. Пономаренко выделил ряд важных био-
сферных перестроек [Пономаренко]. Перечислю самые существенные.
(1) Появление многоклеточности. Хотя многоклеточные прокариоты
известны (грибочки, кустики), но Пономаренко прав, говоря, что у эвкариот
многоклеточность видна “в качестве генеральной линии” и что
“способность увеличивать размеры была реализована эвкариотами в вендской
фауне, состоявшей из необыкновенно крупных организмов, но низко разно-
образных и одинаковых по всей Земле”.
Резкий рост разнообразия был в кембрии. В частности, тогда появились ор-
ганизмы с твердым скелетом и первые членистоногие (трилобиты).
К словам Пономаренко добавлю: геохимически облик биосферы опре-
деляют одноклеточные прокариоты, а в иных смыслах (таксономически,
491
морфологически и поведенчески) ее облик создают многоклеточные эвка-
риоты. В самом деле, у прокариот многоклеточные виды составляют около
1%, а среди эвкариот, наоборот, лишь около 1% видов одноклеточны Про-
изошла реверсия разнообразия (об этом явлении у нас пойдет речь в гла-
ве 10). Да и самих эвкариотных видов более 2 млн - это раз в 500 больше,
чем прокариотных.
(2) Освоение толщи вод и дна. Раннюю жизнь Пономаренко видит, в
основном, в самом поверхностном слое вод, объясняя это высокой мутно-
стью воды. С появлением в кембрии искусных фильтраторов мелкий мате-
риал «быстро отфильтровывается, упаковывается в фекальные пеллеты и
отправляется на дно», что ухудшает условия жизни на поверхности, но де-
лает воду прозрачнее и снабжает органикой дно. Появились илоеды.
Добавлю: дно было освоено раньше, в венде, водорослями и их поеда-
телями (Бурзин М.Б. Эволюция бентосной растительности в позднем до-
кембрии // ПЖ, 2001, № 5). Однако до сих пор жизнь, в основном, приуроче-
на к небольшим глубинам: океанские глубины и дно почти пустынны: в пер-
вых мало органики, а до второго почти не достигает кислород. Исключение
составляют окрестности гидротерм, являющие собой как бы особую био-
сферу (хемосферу) на дне океанов.
(3) Выход жизни на сушу. Пономаренко вновь поднял древний вопрос о
том, что такое море и суша в архее (ср. пп. 1 -5 и 7-5). Он тоже пришел к вы-
воду, что без наземной растительности ландшафт состоял из почти верти-
кальных скал и почти горизонтальных низких равнин.
“Приливные и штормовые волны могли проходить по этим равнинам на
большие расстояния, и береговая черта практически отсутствовала. Водорос-
лево-бактериальные маты могли занимать всю эту равнину, при этом они су-
ществовали не в море и не на суше”.
Добавлю: в таких условиях лучше говорить даже не о выходе жизни на
сушу, а о создании самой суши жизнью. В ходе протерозоя “маты” вероятно
и создали на суше первую почву. Сперва жизнь на суше являла собой тон-
кую пленку, состоявшую из плесеней и наземных водорослей; сгнивая, они
образовали почву, в которой жили черви и членистоногие.
(4) Появление вертикальной зональности наземной жизни. В середине
силура (420 млн лет назад) на суше появились высшие растения (так име-
нуют всё, что сложнее водорослей); первым из них была куксония. В ниж-
нем девоне, с появлением растений с вертикальными стеблями (первой
была риния, Rhynia - рис. 59) возникли ярусы: низкие растения могли рас-
полагаться под высокими. Вертикальный стебель потребовал (в терминах
Уголева) новой естественной технологии - способа подачи водного раство-
ра против силы тяжести.
Добавлю, что механизм этой технологии не выяснен до сих пор (модель
механического насоса и обзор других моделей см.: Кундт В., Робник М.
Водные помпы в корнях растений // ФР, 1998, № 2). Зато ясно, что
492
“Появление листьев, вынесенных в воздушную среду, и корневой системы,
способной использовать подземную воду, - крупнейшее событие в истории
биосферы” {Заварзин Г.А. // Природа, 1990, № 4, с. 119).
Рис. 59. Риния — первое наземное растение с
вертикальным стеблем, нижний девон
В карбоне появились многоярусные леса, давшие
начало залежам каменного угля. Вертикальные
стволы требуют мощных корней, а корни препят-
ствуют размыву почвы. До появления лесов биосфе-
ра не могла иметь нынешней биомассы, и прав
Пономаренко, говоря, что идею Вернадского о пос-
тоянстве количества живого вещества трудно
принять. Добавлю: в наше время больше 90% массы
биосферы заключено в наземных растениях - в
основном, это стволы деревьев. Основной кислород
стало производить болото.
(5) Катастрофы в океане и на суше. История жизни изобилует массо-
выми вымираниями, и некоторые из них носили характер биосферных ката-
строф. Прежде всего это касается вымираний на границе геологических эр:
венд-кембрийское, пермо-триасовое и мел-палеоценовое.
Самым загадочным Пономаренко счел пермо-триасовое вымирание:
“Граница между палеозоем и мезозоем характеризуется самым сильным в ис-
тории Земли падением разнообразия морской биоты... Число семейств мор-
ских организмов уменьшилось более чем вдвое, тогда как... при знаменитом
вымирании между мезозоем и кайнозоем - только шестая часть”. Загадоч-
ность в том, что “гигантскому вымиранию на море не соответствует сколько-
нибудь значительное изменение органического мира суши”.
Добавлю: изменения на суше тоже были значительны, только вырази-
лись в ином: по Мейену, тогда стерлись границы флор, что означает распад
экосистем суши, хотя количество таксонов растений снизилось мало. Ги-
бель именно морской фауны естественно связать с отравлением вод океа-
на, а это вернее всего было итогом растрескивания океанической коры. Оно
не раз происходило, на чем основаны концепции расширяющейся Земли (
мы упоминали их в п. 5-1* и поговорим о них подробнее в п. 9-7).
(6) С начала триаса морская и наземная эволюция шли по-разному:
«Анализ изменений разнообразия позволяет рассматривать мезо-
кайнозойскую историю морской биоты как достаточно единый этап».
На суше, наоборот, было еще знаменитое вымирание динозавров и
других животных в конце мелового периода, и причиной его Пономаренко
назвал распространение цветковых растений. На самом деле они распро-
странились на 30 млн лет раньше, в середине мела - тогда произошла
493
быстрая смена растительных эр (переход от мезофита к кайнофиту, за
30 млн лет до смены мезозоя на кайнозой), вызвавшая свою катастрофу.
Именно поэтому Жерихин выделял два периода - ранний мел и поздний
мел. По Жерихину, при этой смене эр
“уцелевшие от вымирания мезофитные группы - папоротники и хвойные —
меняются, становятся похожими на современные”. “В конце раннего мела об-
наруживаются древнейшие известные плацентарные... Становятся многочис-
ленны птицы. В середине мела появляются первые змеи. Резко меняется со-
став насекомых. Среди них, например, появляются муравьи, становятся
обильными и разнообразными термиты и бабочки. В море появляется множе-
ство групп костистых рыб... Переживают меловую катастрофу динозавры. Их
час еще не пробил... Но... они сильно изменились, особенно растительнояд-
ные” {Жерихин В.В. Избранные труды... М., КМК, 2003, с. 59).
8-13* Эволюция экосистем
Биосфера состоит из экосистем и эволюирует путем их смены. Как пра-
вило, экосистема в ходе своего формирования переживает сукцессию. Сук-
цессия - это обычная плавная перестройка экосистемы; такова смена озера
болотом, а затем смена болота лесом. Сама по себе такая смена - не эво-
люция, а онтогенез экосистемы. Устойчивое состояние экосистемы име-
нуется климаксом, и в приведенном примере - это лес (способный сущест-
вовать много дольше, чем озеро и болото). Сукцессия не считается актом
эволюции, но ее сходство с онтогенезом организма признаётся далеко не
всеми. Для большинства биологов данное
«сходство не является значительным. В сукцессии виды сменяют друг друга в
совершенно различных направлениях, в большой степени зависящих от слу-
чайностей распространения диаспор (отделившихся популяций - Ю.Ч.) ви-
дов, а не в соответствии с генетически управляемым... развитием организма.
Признаки климакса определяются не унаследованной схемой, а условиями
окружающей среды и видами, которые в состоянии прижиться» друг к другу
{Уиттекер Р. Сообщества и экосистемы. М., 1980, с. 320).
Жерихин, наоборот, полагал сходство онтогенеза и сукцессии весьма
глубоким и важным для понимания эволюции экосистем (см. п. 11-2). Это
различие позиций маркирует собой весьма существенное различие понима-
ний эволюции: для учёных круга Жерихина экосистема - реальный субъект
эволюции, определяющий эволюцию входящих в нее видов. Наоборот, для
большинства экосистема - лишь способ описания взаимодействия видов в
природе, а эволюция видов рассматривается порознь, как приспособление
каждого из них к среде обитания (п. 6-1).
Экосистема эволюирует не путем размножения, а преобразованием од-
ной экосистемы в другую, т.е. путем замены одного сукцессионного ряда на
другой. Например, болото может высохнуть и превратиться не в лес, а в луг
или участок степи. Мы вернемся к этому в главе 11.
494
8-14. Изменение роли солнечной энергии
Многие считают Солнце единственным для жизни источником энергии.
Однако нет сомнений, что в ходе эволюции роль тепла недр (главного при
рождении жизни) падала, а роль тепла Солнца росла. Постепенно актив-
ность живого в основном переключилась (2 млрд лет назад) с преобразова-
ния химической энергии недр на преобразование солнечной энергии.
В изменениях солнечной активности можно видеть один из факторов
эволюции, и есть ученые, видящие тут возможность объяснить катастрофы
прошлого (например, грандиозные вымирания, о которых пойдет речь в гл.
9), для чего оснований, по-моему, нет. Но совсем отрицать данный фактор
было бы неразумно: активность Солнца изменяется с периодом около 11
лет, и еще в 1920-х годах биофизик А.Л. Чижевский показал статистическую
связь жизненных процессов с этим циклом. Воейков уверен, что повышен-
ная солнечная активность играет роль универсального «внешнего раздра-
жителя», запускающего ветвящиеся процессы в живых системах [Воейков,
1998, с. 304-305]. Сам по себе период в 11 лет слишком короток, чтобы уви-
деть его связь с эволюцией, но именно его краткость позволила четко уста-
новить различие между причиной изменения в экосистеме и ее поводом.
Ясный анализ этого провел Р.В. Наумов, ближайший ученик Любищева:
за 123 года отмечено 11 эпидемических вспышек численности вредных лес-
ных насекомых, причем максимум численности всегда приходился на мини-
мум солнечной активности. Связь очевидна, но пик активности Солнца лишь
запускает вспышку численности, проявляющуюся через 5-6 лет, тогда как ее
характер (главный вид неумеренно размножающихся насекомых, площади
поражения и т.п.) определяется состоянием биоценоза (Наумов Р.В. Влия-
ние солнечной активности на поражаемость лесонасаждений... // Экология
насекомых и их охрана. Межвуз. сб. науч, трудов. Ульяновск, 1990). Тот
факт, что космический фактор лишь запускает подвижку в ценозе, но не ве-
дет ее, понадобится нам в главе 9.
Подробнее см.: Чайковский Ю.В. Рэм Владимирович и теоретическая
биология И ЛЧ, 2003. Важно и изменение солнечного спектра, возможно,
сыгравшее роль в становлении фотосинтеза [Кондратьев, Федченко].
Гпава 9. От червя до человека
Поскольку жизнь, вероятно, возникла раньше, чем организмы, первые
ее шаги приходится описывать сверху, с позиции биосферы, что мы и сде-
лали в главе 7. В главе 8 был добавлен взгляд снизу, с позиции организма,
и он будет играть всё большую роль по мере нашего движения вверх по
геохронологической шкале, т.е. по мере появления более сложных организ-
мов. С момента появления тканевых в протерозое биосфера приняла прак-
тически нынешнюю кислородную атмосферу и нынешний биохимический
495
облик, поэтому с фанерозоя50 взгляд снизу становится для нас опреде-
ляющим: изменения биосферы будут занимать нас редко - в основном, в
связи с глобальными катастрофами.
Займемся эволюцией тканевых. Тканью называется компактное мно-
жество однородных клеток, специализированных для определенной функ-
ции. Тканевым принято считать организм, в котором выявляется несколько
различных тканей, а если, например, налицо лишь слой генеративных кле-
ток и оболочка, их окружающая, его к тканевым не относят.
9-1. Рост разнообразия и скоростей
Одна из главных черт реверсии разнообразия (л. 8-13) состоит в том,
что основную массу таксонов составляют тканевые. И вообще, чем выше
уровень организации, тем выше разнообразие таксонов, если только
рассматривать всех имеющих центральную нервную систему (и позвоноч-
ных, и насекомых, и головоногих) как один уровень. Но если дробить этот
уровень, то мы увидим, что разнообразие таксонов опять падает: тепло-
кровных меньше, чем прочих, а культуру создал вообще один вид.
Жизнь началась с роста биохимического разнообразия, который в ри-
фее сменился ростом разнообразия форм и функций организмов, причем
тех, которые имеют низкое биохимическое разнообразие. В течение фане-
розоя разнообразие таксонов неуклонно росло: от начала кембрия до плей-
стоцена число одновременно живших ископаемых семейств выросло почти
впятеро (Журавлев А.Ю. Количественная палеонтология: кривое разнообра-
зие // Природа, 2002, № 9, с. 51).
Рост разнообразия таксонов прерывался массовыми вымираниями на
границах многих (но не всех) геологических периодов. Замечу, что сами гра-
ницы периодов были введены по смене руководящих ископаемых и, тем са-
мым, приурочены к вымираниям. Нам же данный факт важен тем, что сви-
детельствует о надежности летописи: если сам по себе рост разнообразия
вверх по разрезу можно связать с лучшей сохранностью более молодых ос-
татков, то синхронные падения говорят о том, что наличных данных доста-
точно для выявления тенденций.
Скорость процесса эволюции в целом довольно постоянна, если рас-
сматривать очень большие периоды времени: эволюция прогеноты, бакте-
рий, одноклеточных эвкариот (почти в одно время с бактериями), многокле-
точных (до появления хордовых) заняли примерно по миллиарду лет каж-
дая, хордовых вдвое меньше. В пределах фанерозоя мы тоже не видим
единой тенденции к ускорению эволюции, но видим ее в отдельных ветвях.
Так, с палеоцена растет скорость эволюции плацентарных.
on
Фанерозой (т.е. хорошо документированную эпоху) традиционно видят с кембрия, но
ныне в него уже можно, с оговорками, включить и венд. См. табл. рис. 47.
496
Во все эпохи виды сменяли друг друга примерно с одной и той же ско-
ростью: вид жил от 500 тыс. до 10 млн лет, род - в 3-4 раза дольше вида, а
семейство - в 3-4 раза дольше рода. Бывала и быстрая смена родов (за
200-500 тыс. лет): она известна как в палеозое (аммониты), так и в кайнозое
(млекопитающие). Переход вида в вид шел, согласно общепринятым взгля-
дам, на порядок быстрее, за 20-50 тыс. лет. Тем самым, вид довольно быст-
ро формируется и затем долго не меняется.
Но насколько быстро? Палеонтолог Стивен Гулд (США) затронул важ-
ный вопрос о переходных формах: их известно слишком мало, чтобы стро-
ить на них учение о процессах перехода вида в вид, но главное не в их ко-
личестве, а в их сущности - каждая форма является жизнеспособным орга-
низмом, а вовсе не стадией. Так что никакое число переходных форм не
приближает нас к ответу на вопрос Майварта (п. 3-5) - как зарождается по-
лезное приобретение в случаях, когда бессилен принцип Дорна (п. 3-6).
По Гулду, ответ содержится в изменениях зародыша, т.е. в жоффруиз-
ме (этого термина у него нет). Он писал:
«Никто так и не смог решить старую (1871 г.) проблему Майварта о стадиях
зарождения полезных приспособлений. Законченная челюсть - предмет вос-
торга для инженеров; те же кости работали не менее успешно для поддержа-
ния жаберных дуг у бесчелюстных предков. Но можете ли вы построить по-
следовательный ряд жизнеспособных промежуточных форм? (Нет надобности
говорить, что никаких ископаемых промежуточных форм не существует.) Ка-
кая польза была бы от костей, отделившихся от жабр, но еще слишком далеко
расположенных, чтобы работать как рот? ... Почему кости не могут передви-
нуться на новое место сразу, в результате небольшого генетического измене-
ния, сильно влияющего на морфологию организма в период раннего разви-
тия? ... Сравнительно недавно появились работы, в которых защищается
пунктуационный (прерывистый) характер происхождения крупных групп по-
средством малых генетических изменений, сильно влияющих на онтогенез»
[Катастрофы..., с. 32-33].
Согласно пунктуализму Гулда и Элдриджа (1972), образование вида
обычно занимает всего десятки и сотни поколений, после чего следует дол-
гий стазис, т.е. период в миллионы поколений, во время которого таксон не
подвержен заметным изменениям; видо- и родообразование идут впарал-
лель и с близкими скоростями (как у Копа и Виллиса), тогда как по схеме
дарвинизма второе должно идти в тысячи раз медленнее первого.
Эти взгляды пунктуалистов содержали мало нового, но они вошли в
учебники, и вот уже их противники обороняются, а их мало слушают. Как
верно отметил В.И. Назаров [2005, с. 384],
“В немалой степени это явилось следствием энергичной пропаганды и рекла-
мирования этой теории ее создателями во всех средствах массовой информа-
ции. Возможно, ради ее упрочения... Гулд и Элдридж пошли на те компро-
миссы со СТЭ, которые противоречат существу пунктуализма”.
497
(Имеется в виду их заявление, что пунктуализм лишь дополняет СТЭ.)
Начинающему кажется, что в эволюционизме противостоят СТЭ и пунк-
туализм; однако на деле последний являет собой лишь иную, чем СТЭ,
форму дарвинизма, улучшенного введением двух старых идей - Жоффруа
и Геера. Ее заслуга видится мне в демонстрации того факта, что разные
таксоны появляются по-разному, и что ни одна теория этого обстоя-
тельства не охватывает. Но никакой своей теории тут нет.
Палеонтолог А.П. Расницын привел следующие данные о скоростях:
“если генетический подход87 к эволюции справедлив, то ее скорость у млеко-
питающих... должна быть минимальной, особенно у наиболее крупных из
них, у одноклеточных - максимальной... В действительности же, как показы-
вают многочисленные палеонтологические данные, эти отношения имеют об-
ратный характер... Так, возраст (в миллионах лет) полусовременной фауны (в
которой половина видов современные, половина - вымершие) у крупных
млекопитающих (хоботные и копытные) 0.2, у мелких 0.5, у птиц и рыб 0.7, у
насекомых 3-7, у моллюсков 3.5-5, у диатомовых водорослей 15. Время полу-
вымирания (по аналогии с полураспадом, время вымирания половины исход-
ного числа видов) в тех же единицах наименьшее у слонов (0.18), а в среднем
у млекопитающих 0.54, у костистых рыб 3.5, у... диатомовых водорослей 5.5,
у динофлагеллат 9. На старших таксономических уровнях... возраст полусо-
временной фауны для родов млекопитающих 4 млн лет, птиц 10, рептилий 20,
рыб 30-50, насекомых 40, моллюсков 60, фораминифер 230” {Расницын, с. 8].
В отношении слонов эта скорость особенно поражает: ни один их вид по
чисто экологическим соображениям ни в одном поколении не мог иметь да-
же миллиона особей (включая и карликовые виды, ибо они жили по остро-
вам), сами поколения сменяются у них очень медленно (половая зрелость
до 20 лет, долгая беременность, низкая плодовитость), а скорость эволюции
огромна: за 2 млн лет сменялись роды. Кроме крупных животных, быстро и
значительно эволюируют животные в изолятах (п. 5-13*).
Быстрая эволюция млекопитающих сочетается с их очень высокой эко-
логической прочностью (п. 6-2*): в окрестностях Чернобыльской АЭС ока-
залось, что радиационная катастрофа снизила численность почвенных бес-
позвоночных во много раз (иногда в 30 раз), тогда как численность грызунов
даже возросла (за счет роста числа овуляций), хотя физиологическое со-
стояние их было угнетенным (Пельгунов А.Н. Паразиты и паразитарные
системы в радиационных биоценозах. М., Наука, 2005).
При такой экологической прочности эволюция не может идти за счет
избирательной смертности и размножаемости: она шла бы (даже если до- 81 * *
81 О какой генетике идет речь, какие генетические данные имеются в виду, Расницын не
уточнил. Видимо, он имел в виду концепцию популяционной генетики, где изменчивость
мыслится растущей с численностью. Как мы видели в части 2. эволюция в опыте на самом
деле идет очень быстро (за несколько поколений), если число особей очень мало.
498
пустить, что вообще шла бы) куда медленнее, чем у беспозвоночных, а она
идет намного быстрее. Очевидно, что сама избирательная смертность яв-
ляется не причиной эволюции (как полагает дарвинизм), а следствием неко-
торого направленного процесса; в п. 5-8* он был назван расплодом.
Как устроен расплод организмов вне лаборатории, мы не знаем, однако
имеем указующую параллель в расплоде клеток: в завершающей стадии
иммуногенеза загадка скоростей подозрительно похожа - каждый лимфоцит
делится впятеро медленнее, чем бактерия, а весь пулл лимфоцитов своей
численностью быстро подавляет колонию опасных бактерий (п. 5-18). И тут
и там первичным изменением охвачены сразу многие (особи, клетки), т.е.
видим атрибут целостности - синхронность.
О том, что новые варианты могут быстро распространяться именно в
малых популяциях, мы теперь знаем - например, от Голубовского (п. 5-6).
У растений скорости иные: крупнейшие организмы (гигантские деревья)
эволюируют медленно. Самое крупное дерево, секвойя, известно как род с
юры (150 млн лет назад); оно же и самый долгоживущий организм: живет
более 3 тыс. лет и всё это время она наводняет окрестность семенами не-
изменного генома. Инерция такого долгожительства огромна, хоть плодови-
тость старых особей и мала.
Словом, эволюция разных групп шла различно, и можно сказать, в тер-
минах п. 8-6, что они как бы жили е разных временах.
9-1* В разных временах
Встает вопрос о роли времени как такового.
“В основе эволюции растений и всего живого лежит закон о непрерывности
движения. Первым, хотя и менее энергичным фактором является само время.
Даже при условии сохранения в течение очень долгого того же климата и... тех
же действующих факторов, эволюция все же продолжается, хотя и замедлен-
ным темпом” (Криштофович А.Н. Палеоботаника. Л., Нефтетоп., 1956, с. 438).
В этом замечательном учебнике самое для меня удивительное — пара-
граф «Факторы эволюции», не упоминающий отбора, зато содержащий при-
веденную цитату. Мысль высказывалась и прежде, в том числе юным Дар-
вином, который затем, однако, всю жизнь утверждал иное — что в постоян-
ной среде меняться незачем, если приспособление достигнуто.
Примеров в пользу времени как самостоятельного фактора эволюции
приводили много. Киты и слоны в последние 2 млн лет изменялись вовсю, а
насекомые и цветковые были почти неизменны. Даже на великие оледене-
ния они откликались, в основном, лишь сменой расселения - следователь-
но, снятие давления нормы здесь явно не работает.
Признание времени отдельным фактором эволюции противоположно
актуализму Лайеля (п. 5-1*), и наш автор не побоялся сказать это:
“Насколько энергичны были древние факторы эволюции, видно из того, что
даже такое грандиозное явление, как четвертичное оледенение, не внесло ни-
499
чего существенно нового... в растительность земного шара, которая уже за-
долго до этого явления приобрела свой современный не только родовой, но в
основном и видовой состав” (Криштофович А.Н., с. 439).
Но если не актуализм, то что? Есть ли здесь какие-то правила?
Самое старое из них дал броккизм (пп. 1-15*, 8-11): каждый таксон,
словно особь, проживает свою жизнь и умирает. То есть живет в своем соб-
ственном времени. Пока оно не подошло к концу, внешние катастрофы ему
не опаснее, чем юноше простуда. Идея вызывает недоверие тем, что нет
указаний ни на какой механизм, способный “изменять ход часов” - не осо-
бей (как в п. 8-6), а всего таксона. Возможно, мы поймем его вместе с меха-
низмом синхронного расплода, который обеспечивает быструю эволюцию.
9-2. Аспекты процесса эволюции
К сожалению, единой (сквозной, всесторонней) теории процесса эволю-
ции пока нет, и нам остается описать разные его аспекты, а затем рассмот-
реть несколько важных явлений - в нескольких аспектах каждое.
Самый привычный аспект - таксономический, когда процесс эволю-
ции рассматривается как последовательность появления и вымирания так-
сонов (типов, классов, отрядов, семейств, родов и видов). Он освещен во
многих хороших книгах и мы будем касаться его лишь попутно, пока же за-
метим, что высшие таксоны сформировались очень давно: надцарства - в
среднем рифее, с появлением эвкариот; эвкариотные царства (растения,
грибы и животные) появились в венде. Правда, эдиакарская “фауна” венда
(п. 8-9) вряд ли состояла из животных; вернее, что эти довольно крупные ор-
ганизмы были осмотрофами, т.е. по форме были близки к животным или
растениям, а по физиологии - к грибам. Однако с ее вымиранием в конце
венда крупные организмы окончательно распределились по трем царствам,
оставив промежуточную организацию некоторым микробам и симбиотиче-
ским организмам вроде лишайников (о межцарствах, т.е. о группах орга-
низмов промежуточной организации, речь пойдет в гл. 10).
Царства будем делить на филумы (тип, или филум, в зоологии - то же,
что отдел в ботанике). Филум (тип) задается своим планом строения. Фи-
лумы животных, известные из палеонтологии, не вымирают и не появляют-
ся с раннего ордовика (последними появились мшанки). Если добавить, что
высший тип грибов, настоящие грибы (Eumycota), известен с позднего вен-
да, что высший тип животных (хордовые) известен с раннего кембрия, а
высший тип растений (семенные) - с девона, то надо признать: фанерозой
почти весь прошел на таксономических уровнях ниже филума (типа)^.
8?
Некоторые филумы (миксомицеты. плоские и круглые черви и т.д.) вообще неизвест-
ны в ископаемом виде. Большинство ботаников считает голосеменные и цветковые
разными отделами; но построены они по единому плану, поэтому мы будем говорить о
едином филуме семенных.
500
Эволюция в рамках филума, заданных планами строения, оказывается дос-
тупной диатропическому анализу, как он очерчен в главе 6.
Тем самым, в рамках филума всегда возможен морфологический ас-
пект, когда эволюция рассматривается как усложнение или упрощение
форм организмов. (Часто он возможен и вне филума, как мы видели, на-
пример, в п. 8-12**). В этом аспекте нашу тему лучше всего начать с упоми-
нания того, что первые тканевые организмы известны с венда. В то время
существовали, кроме эдиакарской фауны, и несомненные тканевые живот-
ные. Самое древнее на сегодня - червь пармия (Parmia). Он найден в венд-
ских отложениях южного Тимана (невысокий кряж в Коми) и внешней фор-
мой был похож на кольчатого червя (Бурзин М.Б., Гниловская М.Б. Какими
были древнейшие животные // Природа, 1999, № 11).
К сожалению, появление ранних уровней в эволюции плохо документи-
ровано: до венда ничего ясного нет, а в венде мы уже видим вторичнополо-
стных в виде червя Parmia, который тем самым оказывается возникшим од-
новременно с двуслойными, что сомнительно. Зато с кембрия ископаемый
материал ясно указывает эволюцию форм: в трех филумах животных (мол-
люски, членистоногие и хордовые) независимо наблюда-ется усложнение
организации вторичнополостных - рост подвижности, усложнение пищева-
рения, кровообращения и нервной системы.
Часто говорят о повышении уровня сложности (уровня организации) как
о самостоятельном процессе, не связанном с рамками таксономии. Для
эволюции животных эти уровни таковы (рис. 60): одноклеточные (Protozoa),
пластинчатые (Placozoa), простые тканевые (губки), двуслойные или лучи-
стые (Radiata), простые трехслойные или бесполостные (Scolecida), первич-
нополостные (Pseudocoelomata), вторичнополостные, или целомические
(Coelomata).
Тканевый уровень четко виден и у растений, тогда как у грибов настоя-
щих тканей нет, роль ткани выполняет плектенхима - подобие ткани, со-
стоящее не из клеток, а из переплетенных гиф.
Следующий уровень сложности - наличие нервной системы; он достиг-
нут только внутри царства животных. Здесь надо перейти к иному аспекту
эволюции - физиологическому.
Хотя систематики и морфологи часто говорят, что учитывают в своих
эволюци-онных построениях физиологию, сами физиологи склонны читать
иначе - что обычные эволюцион-ные схемы лишь называют среди прочих
признаков физиологические, но не раскрывают сути физиологической эво-
люции, т.е. становления функций. Отметим две из этих функций.
Нервная система взяла на себя основную часть текущей работы по ре-
гуляции жизнедеятельности, до этого выполнявшейся только генетической
системой; это позволило сосредоточить гораздо более значительную долю
работы последней на ее эволюционной функции - генетическом поиске
[Зусмановский, с. 113]. К этому наблюдению стоит добавить, что оно отно-
501
сится только к выс-
шим животным, тогда
как у всех организмов,
включая растения,
есть еще и гумораль-
ный уровень, также
освобождающий гене-
тическую систему от
части краткосрочных
забот (замечание Н.А.
Малыгиной).
Рис. 60. Становление
тканевой организации
животных.
А — тканеобразный
слой, окружающий
центральную полость;
Б — два слоя тканей,
внутренняя окружает
кишечник;
В — три слоя тканей,
средний образует как бы
стержень тела;
Г — три слоя тканей
плюс первичная полость
тела;
Д — то же плюс
вторичная полость тела
(по: Валентайн Дж., 1981)
Почти то же можно
сказать про иммуни-
тет. Он тоже есть у
cnidaria (актании, медузы, кораллы kinorhyncha (киноркнхи)
Б
PLATYHELM1NTHES (пЛОСКИб Чвреи)
всех организмов (по крайне мере, у эвкариот), но у растений и низших жи-
вотных - лишь врожденный. При этом
«растения, возможно, изобрели специализированный механизм для поддержа-
ния быстрой эволюции генов устойчивости (иммунного ответа - Ю.Ч.). Это
контрастирует с механизмом распознавания не-себя у млекопитающих, при ко-
тором к разнообразию антител приводят соматические события» (Шамрай С.Н.
Гены устойчивости растений // ЖОБ, 2003, № 3, с. 208).
Надо уточнить: здесь предполагается, что как изощренный иммунитет
цветковых, изменяющийся (полагает харьковский генетик Сергей Шамрай)
только в ряду поколений, так и еще более изощренный иммунитет тепло-
502
кровных, изменяющийся при жизни особи, произошли от общего предкового
примитивного иммунитета. Так оно или нет, не знаю, но очевидно, что кле-
точный иммунитет животных взял на себя эпигностическую работу, до этого
выполнявшуюся основной генетической системой организма.
Среди высших животных можно выделить еще более высокий уровень
организации, который Зусмановский называет «знанием о собственном су-
ществовании». Для этого уровня необходимо (но вряд ли достаточно) нали-
чие центральной нервной системы. Обычно ее связывают с головным моз-
гом, но центральный мозг головоногого моллюска, расположенный вокруг
пищевода, вряд ли можно назвать головным.
Возможно, именно по указанным трем причинам высшие животные эво-
люируют быстрее всех Однако особо быстрая эволюция самых крупных из
них (п. 9-1) имеет иную природу.
Физиологический аспект можно, в свою очередь, рассматривать на раз-
ных уровнях, причем в последние полвека преобладает молекулярный уро-
вень. Хотя самые удивительные открытия последних лет (например, редак-
тирование РНК) являются молекулярными, но я стараюсь вести целостный
рассказ, и не забывать иных уровней. В частности, объединяя взгляды Ар-
шавского (п. 5-11) и Уголева (пп. 8-7, 8-8), можно сказать, что
______неполная замкнутость циклов - это движущий фактор эволюции.___
С этим добавлением известная концепция биосферных циклов Вернад-
ского становится эволюционной.
Об экологическом аспекте эволюции сейчас говорят очень много, но
до сих пор не решен исходный вопрос: образуются ли таксоны одновремен-
но с экосистемами или же экосистемы складываются из уже имеющихся ви-
дов? В данной книге развивается первая точка зрения.
В качестве примера второй точки зрения укажу работу: Чернов Ю.И.
Эволюционная экология... // УСБ, 1996, № 3.
Есть и третья точка зрения - что экосистемы первичны и являют собой
основную реальность. Ее выражают концепция биосферы Вернадского, а
также афоризм: «Эволюционный процесс состоит в том, что отдельные по-
пуляции приходят и уходят из фитоценоза» (Веселова Т.В., Веселовский В.А.,
ЧернавскийД.С. Стресс у растений. М., МГУ, 1993, с. 134).
Труднее всего говорить об этологическом (поведенческом) аспекте
эволюции, поскольку о поведении ископаемых судят почти исключительно
по аналогии с нынешними организмами, что мало интересно. Но бывают
редкие яркие исключения. Так, восточноазиатский верхнемеловой ящер с
перьями, обнаруженный на гнезде с яйцами, получил имя Oviraptor (лат.
“яйцекрад”), однако более детальное обследование находки привело к вы-
воду, что он погиб за высиживанием, т.е. обладал теплокровностью и про-
являл заботу о потомстве (Lange-Barde В. Temoin celebre... // LEG, 27).
503
У растений в определении уровня организации примерно ту же роль,
какую животных играет полость (coeloma), играет побег (cormus). Усложне-
ние строения побеговых растений пошло по пути модульной организации (п.
6-3*), так что прогрессивная эволюция растений являет собой в основном
комбинирование модулей. При этом сопоставлять высшие растения разум-
но не с высшими животными, а с колониальными, где хорошо видна мо-
дульная организация, и виден ясный параллелизм форм при полном разли-
чии выполняемых ими функций. Морфолог растений А.К. Тимонин (Биофак
МГУ) рассматривает данный факт как свидетельство наличия у формы соб-
ственных законов, не сводимых к функциям (Тимонин А.К. Роль морфоло-
гии в ботанике // Гомологии в ботанике: опыт и рефлексия. СПб., СПб союз
ученых, 2001, с. 10-17). О том же у нас шла речь в п. 4-17 и в конце п. 6-3**.
Далее мы познакомимся еще с двумя аспектами эволюции - географи-
ческим (понимание процесса эволюции через расселение таксонов) и био-
сферным (эволюция как единое преобразование биосферы).
9-3. Главные изобретения эволюции и их описание
Пономаренко (ПЖ, 1998, № 4) описал такую черту эволюции: каждый ее
важный акт состоял в одновременном изобретении^ различных элементов
новой организации разными таксонами. Так прошли, по его мнению, «эвка-
риотизация» (становление ядерной организации клетки), а также «метазои-
зация и метафитизация» (становление многоклеточное™ у животных и у
растений), «артроподизация и тетраподизация» (становление главных типов
организации наземных животных) и, наконец, «ангиоспермизация» (появле-
ние цветковых). Для последней характерно «возникновение и распростра-
нение среди голосеменных отдельных черт строения, соединение которых в
дальнейшем дало покрытосеменных» [Пономаренко,с. 23].
Отмеченное им явление Пономаренко никак не назвал. Я пока обозначу
его как “зация". У всех этих процессов он видит следующую общую суть
(Пономаренко А.Г. Артроподизация и ее экологические последствия // Эко-
системные перестройки и биосфера. Вып. 6. М., 2004, с. 8):
а) процесс не только таксономический, а всебиосферный;
б) он начинается созданием подходящей экосистемы и завершается
осуществлением всех основных экологических потенций данной зации;
в) может возникать независимо и неоднократно;
г) идёт самоускоряясь.
В силу (в) всякая зация - рефрен, но важнее, что все вместе они обра-
зуют рефрен высшего порядка (рефрен рефренов). Феномен является част-
ным случаем более общего, известного как блочность эволюции (п. 6-3*).
Изобретенные элементы становились блоками новой организации. Если в
83 Не ставлю здесь кавычек, поскольку не могу указать отличия данного процесса от
изобретения в его обычном смысле.
504
отношении тетраподизации этот процесс появления и сборки блоков до-
вольно прост и сводится к формуле:
кистепёрые рыбы —> амфибии —> рептилии —> птицы и звери
(см. ниже, п. 9-13), то артроподизация (как и сколько раз появлялись члени-
стоногие) шла сложнее: независимо и многократно у членистоногих появля-
лись многие главные черты - например, трахейное дыхание.
Оно впервые появляется у первичнотрахейных (онихофор) в кембрии,
отсутствует у трилобитов и низших хелицеровых, дышащих жабрами, и
вновь появляется в девоне у пауков (вытесняя лёгочное дыхание) и у насе-
комых - как основной способ дыхания. Еще Любищев удивлялся:
“Поразительно, что первичнотрахейные при крайней примитивности и древ-
ности своей организации в известном смысле показывают пример далеко
идущей эволюции... от яйцерождения до совершенного живорождения с об-
разованием плаценты... Нечто подобное показывают и древние... скорпионы”
{Любищев А.А. // ТПЭ-3, с. 8).
Вот, замечу, еще один из примеров реализации одной и той же идеи на
разных материальных основах (п. 5-15*). И это тоже номогенез.
Любищев соглашался с М.С. Гиляровым, что
“сходство трахейной системы определяется не общностью происхождения, а
сходными условиями и принципами функционирования”,
и делал из этого и подобных фактов вывод:
“гипотеза монофилетичности членистоногих даже в узких рамках подрывает-
ся на каждом шагу и держится только в силу консерватизма”; “объединение в
типы должно происходить не по признаку выведения от общего предка, а по
принципу общего плана строения” (ТПЭ-3, с. 7, 9).
Если для Пономаренко главное - из чего?, то других больше занимает
вопрос: как? Такова, например, статья: Старобогатов Я.И. Принцип основ-
ных компонентов тела... // ЗЖ, 2000, N№ 1, 2. «Принцип основных компонен-
тов тела» - это, в сущности, признание того же, что у Пономаренко, - неза-
висимого развития частей. Главным в усложнении строения животного ав-
тор считал усложнение строения целома (выявлено 5 уровней его развития,
почти не связанные с уровнями остальных систем органов). Конечно, это
чересчур односторонне.
Принять уровень развития целома за уровень организации всего орга-
низма нельзя, но попытка Старобогатова выглядит разумной, если принять,
что, в сущности, он вел речь о меронах и архетипе и дал ряд значений ме-
рена «строение целома».
Других меронов он не рассмотрел, но мы можем сделать это сами. Ес-
ли, например, взять усложнение нервной системы и выстроить группы по
этому признаку в ряд, то оба ряда вместе дадут плоскую таблицу, т.е. дву-
мерный фрагмент рефренной таблицы. Перечислив таким образом все су-
щественные мероны, характерные для типов, мы получим таблицу разнооб-
505
разия архетипов животного царства. (Если взято N меронов, то получится N-
мерная таблица.) О дальнейшем усложнении тел организмов см. [Чайков-
ский, 1990, с. 154].
Приняв, что основные изобретения (уровни организации) самих орга-
низмов можно в принципе описать и упорядочить как комбинации блоков,
обратимся к эволюции организации совокупностей организмов.
9-4. Преобразование фаун и флор в пространстве. Вклад Мейена
Еще Уоллес отметил, что реликты (остатки древних групп) склонны за-
нимать окраины прежних ареалов своих групп. Лучше всего тенденция Уол-
леса видна на лесах: лиственные леса заняли теплые страны, а хвойные
(относятся к голосеменным) оттеснены на север, до самой границы лесной
зоны. (В Южном полушарии для зоны тайги нет подходящей суши.) То же
внутри класса хвойных: тайгу формируют сосновые (более древние), а ки-
парисовые (более молодые и сложнее устроенные) остались в теплых стра-
нах (из них до российских широт дотянули только можжевельники).
Голосеменные реликгны не только тем, что старше цветковых и проще
устроены (не имеют основных структур цветка и двойного оплодотворения),
но и тем, что утратили былое разнообразие (см. далее, п. 10-6).
Затем многие обращали внимание, что крупные группы распространя-
лись в обе стороны от экватора (биполярно), что новые их формы постоянно
оттесняют древних представителей групп в более холодные зоны. В 1941
году ленинградский ботаник Е.В. Вульф указал на тропическую родину цвет-
ковых (Игнатьев И.А. // [Эволюция флор..., с. 17]).
Затем Берг привел много примеров биполярности видов средних широт
и объяснял ее расселением видов с севера на юг в ледниковую эпоху, когда
в тропиках похолодало; но он же признал, что так объясняется не всё: есть
несколько биполярных видов, живущих в Арктике и в Антарктике, а им эква-
тор не пересечь; некоторые таксоны рыб разошлись биполярно ранее по-
следней ледниковой эпохи (Берг Л.С. Климат и жизнь. М., 1947). В данной
книге Берг умолчал о запретном в те годы номогенезе, но отметил, что Дана
видел в биполярности повторное видообразование. А это, замечу, и есть
номогенез. Сам Берг в 1930 году тоже прибег к идее повторного видообра-
зования для объяснения тождества двух видов миног из очень далёких рек
- Венгрии и Китая [Берг, 1977, с. 344]. К сожалению, в тогдашних исследо-
ваниях почти нет данных об ископаемых.
Аналогично, самые примитивные по строению птицы (пингвины и гага-
ры) живут в Антарктике и Арктике, хотя произошли в более низких широтах.
Таких примеров, подтверждающих Уоллеса, много, но есть и противополож-
ные: на Крайнем Севере мы видим полное отсутствие хвойных - древесные
породы тундры представлены карликовыми березками, ивами и т.п. Поляр-
ный воробей (пуночка) дальше всех залетает на острова Ледовитого океана,
а воробьиные - один из самых развитых отрядов птиц.
506
Словом, для выводов нужны не примеры, а сплошной анализ всего
массива фактов с единой точки зрения. 250 лет назад, когда системы раз-
ных авторов резко разошлись друг с другом, понадобился гений Линнея,
чтобы единым взором охватить всё известное тогда разнообразие организ-
мов и предложить единую их систему. С тех пор число известных видов вы-
росло раз в сто, да и задача стоит посложнее - о каждом таксоне теперь
надо знать, когда и где он возник, как расселялся и когда вымер. Для всех
видов это немыслимо.
К счастью, для первого шага это оказалось и ненужно. В середине 1980-
х эту задачу в первом приближении решил Мейен для наземных растений.
Он ограничил круг рассмотрения таксонами не ниже семейства и исключил
из анализа мхи и наземные водоросли. В остальном анализ был задуман
полным - не обойти вниманием ни одно семейство, начиная с девона. Увы,
первые же статьи на эту тему он писал смертельно больным и смог едва
обозначить открытую им тенденцию. Но все-таки, на мой взгляд, его вывод
можно (как и открытие хемосферы, п. 8-7) назвать самым ярким в классиче-
ской (немолекулярной) биологии наших дней: Мейеном пунктирно описан
глобальный флорогенез, т.е. процесс эволюции и одновременного с нею
расселения флор (географически компактных групп растений) по суше из
тропиков в высокие широты. Вот его суть.
Жизнь расселялась по суше медленно: в позднем силуре были устой-
чиво заселены лишь побережья океанов. Материки освоены жизнью в дево-
не. Никаких зон у девонской флоры не замечено, что соответствует данным
о жарком климате на всех девонских континентах. Наоборот, начиная с кар-
бона, крупные таксоны растений формируются в экваториальной зоне и
расселяются к северу и к югу, мало при этом эволюируя. Точнее, ни одного
класса высших растений не возникло вне жаркой зоны, а порядков (отрядов)
возникло всего два. И Мейен с полным правом говорил:
«Экваториальная зона сейчас представляется главным источником таксоно-
мического состава растительного покрова континентов и местом всех “экспе-
риментов”, касающихся общей организации... Отсюда таксоны расселялись в
более высокие широты, подвергаясь по мере движения к полюсам все менее и
менее значимым эволюционным преобразованиям» {Мейен С.В. Флорогенез и
эволюция растений // Природа, 1986, № 11, с. 56).
Свой вывод он изобразил в виде схемы (рис. 61), а названную черту эволю-
ции назвал фитоспредингом (растеканием растений).
Главное даже не сам флорогенез, а окончательное понимание того
факта, что историю нельзя понять без географии. Мейен мечтал уви-
деть эволюцию каждой группы, “накладывая филогению на карту”, т.е. увя-
зывая связи между предками и потомками со связями между положением
тех захоронений, в которых эти предки и потомки найдены.
507
Рис. 61. Глобальный флорогенез по Мейену. 1 - южные (гондванские) внетро-
пические области; 2 - тропические; 3 - северные внетропические; 4 - флористи-
ческие области не обнаружены. Видно, что почти все ветви исходят из тропи-
ческой зоны. На врезке - карта климатических поясов позднего палеозоя, как
они видны при нынешнем расположении материков
Вместо привычного филогенетического древа он предлагал рассматри-
вать флородрево (схему последовательного происхождения флор) и успел
эскизно описать его нижнюю часть: связь флор с девона до середины мела.
Подробнее он смог показать один элемент, касавшийся одного позднепале-
озойского класса голосеменных (Природа, 1986, № 11, см. цветные схемы
на с. 52 и 53 журнала). Суть его в том, что в самом конце карбона от тропи-
ческого порядка Peltaspermales класса Ginkgoopsida отошла ветвь, рассе-
лившаяся в ближайшую внетропическую область (в Печорскую провинцию),
а в средней перми ушедшая еще дальше от тропиков. При этом образовы-
вались всё новые и новые семейства и только один порядок образовался в
зоне, где бывали в ту эпоху морозы. (То были арбериевые из голосеменных
Южного полушария, они возникли в карбоне и вымерли в триасе.) В уме-
ренной зоне изредка образуются семейства, а в Арктике возникло всего 2-3
рода. (рис. 62).
508
Рис. 62. Флородрево класса гингкоопсид отдела голосеменных. Как видим, порядок
арбериевых возник вне тропиков из тропической предковой формы (по Мейену)
Поясню мысль Мейена: Печорская провинция располагалась в карбоне
и перми по широте между нынешними устьем Печоры и средним течением
реки Урал и по долготе между нынешней Волгой и Уральскими горами. Она
была вне тропиков и отделялась мелким Казанским морем от остальной Ев-
ропы (где тогда был тропический климат), а море постепенно отступало.
Позже палеоботаник М.А. Ахметьев, зав. лабораторией в ГЙН, которой
прежде руководил Мейен, достроил флородрево и сделал его более де-
тальным, причем сама идея флородрева осталась вне сомнений (Ахметьев
М.А. Географическая дифференциация позднемеловых и кайнозойских
флор... //Стратиграфия. Геологич. корреляция, 1999, № 3).
По Мейену, в мягких климатических условиях эволюция
“отпускает тормоза”, а “чем дальше от экватора, тем чаще и сильнее работают
тормоза... Здесь мало возможностей для эволюционных экспериментов, отбор
сразу пресекает смелые попытки... Очевидно, в тропиках отбор преследует
лишь чрезмерные вольности” (Природа, 1986, № 11, с. 57).
509
Важная мысль: где меньше смертность, там богаче эволюция.
Главный довод в ее пользу высказан еще до Мейена: в тропиках гораздо
полнее видны ряды Копа - Вавилова. А это явный номогенез.
Иными словами, каждый вид обладает запасом экологической прочно-
сти (не при каждом ухудшении условий вымирает), и во влажных тропиках
этот запас оказывается выше за счет того, что организмы находятся далеко
от голодной и холодной смерти, а также от высыхания. Здесь могут возни-
кать и существовать многие (а в высоких широтах немногие) варианты, т.е.
частично снято давление нормы, и как раз в таких условиях оказывается ре-
альным появление нового.
Зато умеренно холодные страны позволяют многим таксонам гораздо
дольше жить. Вспомним, что в тайге до сих пор царят хвойные, чей макси-
мум разнообразия миновал еще в мезозойских тропиках.
9-4* Споры вокруг глобального флорогенеза. Биотогенез
СТЭ видела в высоком тропическом разнообразии итог жёсткого отбора
вследствие жёсткой конкуренции, вызванной перенаселением. Тут в самом
деле налицо прямое противоречие с идеей экологической прочности. Кто
прав? Многочисленные в середине XX века опыты по конкуренции не дали
никаких эволюционных выводов, зато наблюдения за эволюцией при пони-
женной конкуренции (п. 5-13*) дали те же результаты, что у Мейена. Так что
в данном пункте вернее, что прав он — в рамках номогенеза.
Чисто палеоботаническое возражение Мейену высказал его ученик И.А.
Игнатьев. Он пишет о концепции флорогенеза:
“С точки зрения критериев... Поппера, эта концепция находится на грани науки,
поскольку' обосновывается массой подтверждающих фактов и не принимает во
внимание опровергающие примеры... До сих пор не показано ни одного конкрет-
ного примера формирования и последующей миграции надродового таксона
высших растений в соответствии с концепцией фитоспрединга” {Игнатьев И.А.
Биогеографические модели и эволюция растительного покрова... // МС, 169).
Возражение убийственно, но, к моей радости, верно лишь отчасти. Один
пример есть - приведенный выше фрагмент флородрева. Другой пример
мне удалось найти (с любезной помощью самого Игнатьева) в посмертных
материалах Мейена. Скажу о нем чуть подробнее.
Хвойные появились в Печорской провинции в конце ранней перми в той
области, которая ныне располагается восточнее средней Камы (она отме-
чена мною на карте кружком). Тогда это было северо-восточное побережье
Казанского моря, а его противоположный юго-западный берег был границей
тропической зоны. По мере отступания моря остальная часть Печорской
провинции, как видим на составленной Мейеном карте (рис. 63), заселялась
хвойными, причем расселение сопровождалось эволюцией - нигде нет тех
же видов, что в самых ранних захоронениях (Meyen S.V. // Review of
Palaeobotany and Palynology, 1997, № 96), отмеченных кружком.
510
Рис. 63. Распространение и эволюция ранних хвойных в Печорской области ранней
перми. Слева на таблице показаны позднепермские ярусы (татарский, казанский и
уфимский) ярусы и раннепермский (кунгурский) ярус, в середине - места находок, обо-
значенных на карте, справа - виды ранних хвойных. Как видим, более поздние виды
расходятся дугой на север, запад и юг от двух самых древних кунгурских видов (обве-
денных кружком)
WESTERN ANGARALAND
| Tatarian | 1 Upper I Vyatsky Aristovo, Titovo, Viled, Kalinovka, Luptyug, Dor Sashinia aristovensis, S. borealis, Quadrocladus dvinensis, 0. borealis, Dvincstrobus sagittalis, Geinitzia subangarica, Geinitzia sp. SVM—1
Severodvinsky
Isady, Aleksandrcvka Dvincstrobus sagittalis, Quadrocladus schweitzeri, Geinitzia sp. SVM—2
Kargala Mines Kzyl—Bairak "UHmannia" Steirophyllum biarmica, lanceolatum "Waichia" foliosa
Lower
। Kazanian ' Upper
Kityak Pseudovoitzia fcomuta
Sluda Quadrocladus (al. Steirophyllum} komiensis
Lower
Vym-1, Vym—2 Timanostrobus muravievii Conchoiepis harrisii
с <0 Е 5 Sheshminsky Solikamsky
Shapkina Kungurodendron ? sp. SVM—1 Cyparissidium entsovae
с .S Irensky Chekarda—1 Chekarda—2 Kungurodendron sharovii, Cyparissidium appressum, Taxodiella bardaeana
с Filippovsky Krutaya Katushka, Krasnaya Glinka, Matveevo—1 Kungurodendron sharovii, Cyparissidium appressum, Taxodiella bardaeana
Shapkina
Vym—1, Vym—2
Sluda
Vi led
Aristovo
Isady
Titovo
Dor
Luptyug
Kalirovka
Krutaya Katushka, Krasnaya
Glinka, Matveevo—1
Chekarda—1, Chekarda—2
Kityak
Kzyl—Bairak
Aleksandrcvka
Kargala Mines
Разумеется, двух
примеров мало для
теории. Но перед нами не
теория, а концепция, в
которой речь может идти
лишь о тенденции. Как
раз тенденции - основ-
ной объект интереса диа-
тропики. Для принятия
флорогенеза в качестве
модели эволюции надо
убедиться в том, что его
подтверждений хотя бы
на порядок больше, чем
опровержений. К сожале-
нию, ныне такие работы,
насколько знаю, почти
все ведутся в масштабах,
не позволяющих сделать
никакого общего вывода.
Однако одна такая ра-
бота выполнена, и ре-
зультат ее оказался не-
ожидан. Ю.В. Мосейчик
(ученица Игнатьева, ГИН)
приводит данные о том,
что в середине раннего
карбона (335 млн лет
назад) в каждом
полушарии существовала
не одна, а три зоны, в
которых в массе порож-
дались новые крупные
таксоны высших
растений - эти зоны
511
были в тропиках, в субтропиках и в умеренной зоне- Вместо единого (по
Мейену) фитоспрединга, шедшего от экватора к полюсам,
«вероятно имел место сложный механизм расселения из центров происхож-
дения в пределах трех географических “колец эволюции”» (Мосейчик Ю.В.
Номогенетический характер эволюции растений (на примере флоры раннего
карбона) // ЛЧ, 2005).
Если этот результат подтвердится на более общем материале, то кон-
цепцию флорогенеза придется радикально уточнить (принять в каждом по-
лушарии три широтных зоны фитоспрединга), зато она обретет подлинно
диатропический облик: всюду таксоны будут порождаться в рамках одной и
той же диасети, что, в сущности, и показала Мосейчик.
В каждой зоне эволюция носила характер чёткого номогенеза: на раз-
ных континентах происходили
“параллельные изменения в строении листьев разных таксономических групп”,
а у “репродуктивно усложненных плауновидных происходит параллельное
голосеменным формирование семяподобных структур” (там же).
А как дела с животными? Можно ли выявить фауногенез и зооспре-
динг (растекание животных из низких широт в высокие)? Что зооспрединг
никак не может быть всеобщим явлением, видно на примере тех же пингви-
нов. Их разнообразие невелико (15 нынешних и 36 ископаемых видов), но
строение столь отлично от других птиц, что орнитологи выделяют их в осо-
бый надотряд. Пингвины обитают, в основном, в Антарктике, а в более низ-
кие широты расселились вдоль холодных течений (один вид, галапагосский
пингвин, живет даже у экватора). Самые древние из известных пингвинов
жили в миоцене в умеренном климате Новой Зеландии; это значит, что они
не просто живут в высоких широтах, но и распро-страняются в низкие, и при
этом эволюируют. Это - против идеи зооспрединга, но как в целом?
Зоолог Ю.И. Чернов собрал, следуя ботанической идее Мейена, данные
о зооспрединге. Картина вышла пестрая, но тенденция видна и тут. Состоит
она в следующем: в целом разнообразие таксонов животных падает с уда-
лением от экватора и так же падает доля “продвинутых” (недавно возникших
и сложнее устроенных). Самые “продвинутые” (например, попугаи среди
птиц) исчезают уже в субтропиках. Арктические же и антарктические таксо-
ны представляют «нижнюю половину филемы» (примитивные формы), при-
чем возникли они в более низких широтах (Чернов Ю.И. Филогенетический
уровень и географическое распределение таксонов // ЗЖ, 1988, № 10).
Кое в чем зоологический материал аналогичен ботаническим данным
Мейена более конкретно. Зональность расселения тетрапод (четвероногих)
в девоне так же не выражена, и разнообразие их так же ничтожно; зато в
карбоне, едва появились различные климатические зоны, эволюция четве-
роногих выступает как расселение от экватора:
“Фауна тетрапод карбона представлена только в Северной Америке и Запад-
ной Европе, что, очевидно, связано с расположением этих стран в тропиче-
512
ской зоне” (Каландадзе Н.Н.. Раутиан А.С. Юрский экологический кризис
сообщества наземных тетрапол... // Проблемы доантропоген. эволюции био-
сферы. Вып. 3. М., 1993, с. 61).
Любопытно для сравнения: если состарившиеся формы наземных рас-
тений вытеснялись в высокие широты, то морских животных - «в глубины
океана» (Жураелее А.Ю. // Природа, 2002, № 9, с. 53).
Едва ли фауногенез удастся выразить в столь же общих правилах, как
флорогенез, и наверняка не удастся выразить в столь же простых; но это не
умаляет общей роли идеи флорогенеза: ведь и законы Менделя мало на
чем видны точно, но они выражают самую простую и общую генетическую
закономерность и потому легли в основу всей генетики. Точно так же, закон
Ома безупречно работает всюду, но попробуйте открыть его в телевизоре,
где его он маскируется множеством иных законов.
Ныне ископаемого материала хватает только для самых грубых эскизов
процесса биотогенеза (термин Мейена для обозначения фауно- и флоро-
генеза вместе), но и они весьма полезны.
Так, сумчатые возникли в раннемеловых тропиках Северной Америки в
формах, близких к опоссумам, в позднем мелу расселились по Америке и
Европе, а позже (в эоцене) даже попали в Антарктику. (В Африке и Азии их
никогда не было, а в Австралии жили и живут совсем другие сумчатые.)
Сумчатых обычно описывают как единую родственную группу, но стоит на-
ложить список их родов (живых и вымерших) на карту, как родство рушится:
подотряд опоссумов (в широком смысле, т.е. с крысовидными) известен
только в Америке и (ископаемые) в Европе и Антарктиде, а остальные сум-
чатые -только в Австралии и близ нее [Кэрролл, т. 3, с. 210].
Знаток американской фауны Джордж Симпсон не видел возможности
миграции сумчатых ни в Австралию, ни из Австралии и считал вопрос об их
происхождении таинственным (Симпсон Дж. Г. Великолепная изоляция. М.,
Мир, 1983). По-моему, особой тайны тут нет: в обоих регионах могли неза-
висимо возникать близкие клетки общего рефрена, на что фактически и
указывали еще Дана и Берг.
9-5. Или прогресс форм, или прогресс функций
Почему вне тропиков оказываются именно “примитивные” формы? Тут
можно вспомнить Бергсона: группы потому и получают статус примитивных,
что их «жизненный порыв» израсходован не на усложнение формы, а на
физиологию, на приспособление к трудным условиям. Это - частный случай
принципа компенсации (гл. 8). В понимании этого - суть диатропического
подхода к эволюции. Подход вполне нов и сильно отличен от того привыч-
ного, когда изучаются формы, а функции лишь упоминаются. Вот два при-
мера различий старого и нового подходов.
1) Семейство ятрышниковых (орхидных) часто называют вершиной эво-
люции, поскольку их цветки поражают своей красотой и сложностью. Про
513
красоту в дарвинизме (как и в марксизме) говорить не умеют: не то умона-
строение, не тот кругозор. А о причине удивительной сложности цветков ор-
хидей говорят всегда одно - они «высоко и разнообразно специализирова-
ны для опыления насекомыми» (Энц. словарь «Биология». М., 1983). Да,
многие опыляются очень малым числом видов насекомых, но что тут умест-
нее - «для» или «вследствие»? А может быть, «вопреки»? Судите сами.
Странная это “вершина эволюции”: орхидеи не только не имеют эндос-
перма, но из всех цветковых только они утратили двойное оплодотворе-
ние, отчего их размножение протекает так же медленно, как у голосемен-
ных: от опыления до оплодотворения - полгода, до зрелого плода - год или
два. Опыление насекомыми слишком усложнено и идет плохо - многие ор-
хидеи спасаются ветром и даже просто бесполым размножением. Их “высо-
кая специализация” (а вернее - очень узкая, патологически узкая, ниша)
видна и во многом другом: они не могут существовать без симбиоза с гри-
бами (без гриба семя орхидеи вообще не прорастает), многие потеряли фо-
тосинтез и многие из них (что вовсе странно) могут жить на одном-
единственном виде растений, некоторые могут выжить только в союзе (под
охраной) муравьев. Словом, орхидеи недаром всегда служили доводом
против дарвинизма и в пользу ортогенеза [Попов, с. 146].
Да, орхидные - самое большое по числу видов семейство растений (до
35 тыс. видов), и в целом распространено едва ли не по всей суше. Но како-
ва численность его видов в природе? Вот свидетельство экологов:
“Представители семейства ятрышниковых очень редки, поэтому вопрос о
полном запрещении их сбора поставлен” {Лавренова ГВ. Травы, дарующие
здоровье. Донецк, Донеччина, 1994, с. 421).
Редкость северных орхидей отметил и Линник. Естествен вопрос: к че-
му орхидеи приспособились? Разве что к цветочному рынку.
И все-таки в каком-то смысле орхидеи действительно выше всех.
Вспомним, что выражать успех численностью принято только в дарвинизме.
Великий Бэр, наоборот, полагал, что численность вида определяется вовсе
не успехом в борьбе за жизнь, а его местом в «экономии природы».
Очевидно, что орхидеи занимают какое-то особое в ней место. Здесь
нужна новая философия. Для начала приведу слова Линника:
“Семейству орхидных не чужды поиски. Орхидеи смело отходят от традиций,
смело создают новое” {Линник Ю.В. Книга трав. Петрозаводск, «Карелия»,
1986, с. 262).
Как видим, не просто приписано активное поведение травам - активно
ведет себя их семейство. Что это может значить? Буквально или нет?
Еще сто лет назад Бергсон писал:
“когда видишь, с какой уверенностью и точностью используют вьющиеся
растения свои усики, какой поразительный комплекс приемов применяют ор-
хидеи, комбинируя их, чтобы оплодотвориться при помощи насекомых, то
как не думать при этом об инстинктах?” [Бергсон, с. 181].
514
Инстинкты - свойство животных, и Бергсон уверял:
“было бы столь же нелепо отказывать в сознании животному, поскольку оно
не имеет мозга, как заявлять, что оно не способно питаться, поскольку не
имеет желудка. На самом же деле нервная система, подобно другим систе-
мам, зародилась вследствие разделения труда”( с. 130).
Он имел в виду то, что животные подвижны, зато растения усваивают
неорганику. К сожалению, он связывал сознание («самопроизвольность»)
только с механической подвижностью в трехмерном пространстве:
“Движениям же растений не свойственны ни частота, ни разнообразие дви-
жений животных... Если у них и проявляется в исключительных случаях
смутная самопроизвольность, то кажется, что видишь перед собой случайное
пробуждение обычно дремлющей активности” (с. 129).
На самом деле, бывает подвижность в иных пространствах - например, в
пространстве форм и пространстве функций. А в них подвижность растений
никак не отстает от подвижности животных. Вспомним идею Бергсона о
жизненном порыве (то был способ говорить об активности). По всей види-
мости, орхидеи наделены им весьма щедро, и они потратили его не на из-
быточность размножения (как почти все виды), а именно на поиски.
2) Прогресс часто видят в эволюции зрения от светочувствительного
пятна плоских червей до глаза зверей и осьминогов; он считается понятным,
хотя описано только изменение форм, а химия фоторецепции, оптика хру-
сталика, кибернетика сигналов и психология зрительных образов остались
за кадром. Когда-то Шмальгаузен признал эволюцию глаза «наиболее труд-
ным» и в то же время «вполне понятным» актом эволюции, ибо
“Всегда новые органы возникают путем постепенной дифференцировки в
пределах имеющихся уже органов или частей, обособляются от них и преоб-
разовываются вместе с обособлением и преобразованием их функции”
(Шмальгаузен И. И. Основы сравнительной анатомии позвоночных животных.
М., Учпедгиз, 1938, с. 31).
Ядовитую железу змей он едва упомянул (как модификацию слюнной),
тогда как в функциональном смысле именно появление яда - новшество,
акт прогресса, давший обладателям новую нишу. Для Шмальгаузена, как и
для Дарвина, понять эволюцию органа значило выстроить “непрерывный"
ряд форм, нам же важнее понять, как появилась новая функция.
Она всегда появляется скачком (сальтацией, см. пп. 3-12; 5-1) в том
смысле, что возникает в каком-то поколении сразу: глаз впервые посылает
сигнал, который используется; яд впервые отравляет жертву, не губя обла-
дателя. Непрерывный ряд ничего в этом смысле не разъясняет.
Зато разъясняет физиология: утрата прежней нормы вызывает реак-
цию - генетический поиск, который ведут все, попавшие в эти условия, но
лишь немногие способные приходят к новым приемлемым формам. Боль-
шинство же вымирает. А способными как раз чаще всего и оказываются
“примитивные”. Таков вклад нового ламаркизма.
515
Чернов отметил, что “примитивные” более адаптивны, чем “продвину-
тые” - прежде всего, за счет способности голодать и снижать плодовитость
(ЗЖ, 1999, № 3). В наших терминах это означает, что
в одних таксонах «жизненный порыв» расходуется на усложнение
строения, и потому мы называем эти варианты “продвинутыми”; а в других,
_______в порядке компенсации, усложняются физиология и поведение.__
Если Мейен нашел, что эволюция в Арктике почти не шла на уровне ро-
дов и выше, то Чернов показывает, что на уровне видов она идет, и притом
интенсивно, особенно среди “продвинутых” (Чернов Ю.И. И ЗЖ, 2004, № 5).
Другими словами, "высшие” адаптируются тут, в основном, морфологически
(идет видообразование), а “низшие” - в основном физиологически, в рамках
прежних видов. Нам тут важно, что Арктика - кузница видов, но не родов и
выше. Это понадобится нам при обсуждении проблемы происхождения че-
ловека в п. 9-15.
9-6. Проблема вымирания
Итак, таксоны, состоящие из организмов, “примитивных” своим строе-
нием, живут дольше “продвинутых”. Тут встает проблема вымирания.
Самый, пожалуй, яркий пример являют аммониты: все они, кроме нау-
тилусов, вымерли в конце мела, а род Nautilus, самый примитивный строе-
нием, дожил до наших дней, причем примитивное строение компенсируется
самой развитой физиологией [Несис, с. 183-184].
Из главы 5 мы знаем, что эволюция течет неравномерно. То сто мил-
лионов лет ничего важного не происходит, то за считанные миллионы меня-
ется облик планеты. В главе 8 говорилось о двух таких событиях - быстром
появлении вендской и раннекембрийской фаун.
Следует ли считать биосферные катастрофы самостоятельными явле-
ниями, или же они суть лишь следствия внешних воздействий на биосферу?
Быстрая перемена в биосфере вероятно всегда бывает связана с каким-то
внешним событием - в недрах Земли или в космосе. Поэтому встает во-
прос: является ли такое событие причиной катастрофы или только поводом
к ней? На сегодня лучшей иллюстрацией проблемы служит грандиозное
вымирание, происшедшее на границе мела и палеоцена, т.е. между Мааст-
рихтом и данием (верхним меловым и нижним палеоценовым ярусами). Эту
границу обозначают как М/Д, а саму проблему часто именуют проблемой
вымирания динозавров, хотя они тогда вымерли не все, а одновременно с
ними вымерло множество других животных, морских и наземных. Вряд ли
все они вымерли по одной общей причине [Несис, с. 190, 195].
Ранее (БПС, 2000, № 46, с. 7) я уже замечал, что различие взглядов на
М/Д-проблему вызвано мировоззренческой причиной: разные авторы исхо-
дят из разных ПМ (о них см. п. 5-3). Сейчас добавлю другую компоненту:
многие привыкли искать всему единственную причину, и она всегда видится
516
им в рамках их деятельности. Например, Еськов, ссылаясь на “бритву Окка-
ма’’, предлагает решать проблему вымираний так:
“из нескольких конкурирющих гипотез выбирается наиболее простая”, а все
факты и соображения, в нее не вошедшие, аттестуются как “избыточные сущ-
ности”, которые “можно привлекать лишь после того как исчерпаны все иные...
варианты объяснения” [Еськов, с. 238-239].
Опять презумпция и опять тупик: причин М/Д-вымирания Еськов объяс-
нить так и не смог. Это естественно, ибо он искал чисто биологическую ги-
потезу, не учитывал астероидов, а они падали в это время на самом деле.
Здесь введение новой сущности (падение астероидов) как раз и дает, как
увидим ниже, простую работающую теорию. Другой вопрос: был ли астеро-
ид единственной причиной? Нет, не был.
9-6* Отчего вымерли динозавры?
Этой проблеме посвящена огромная литература. Одни уверены, что на
М/Д-границе произошла глобальная катастрофа, затронувшая основную
массу таксонов (см. схему смены крупных таксонов животных Шиндеволь-
фа, приведенную в гл. 6 на рис. 41), другие видят в то время только мозаи-
ку отдельных перемен, не всегда быстрых. Например, указывают на отсут-
ствие в тот момент катастрофы среди наземных растений и делают вывод,
что в то время не мог сильно меняться климат. Им возражают: это говорит
лишь о краткости катастрофы - после периода похолодания растения могут
заново вырасти из семян, а крупные животные не возродятся. Оппоненты
парируют: несколько родов динозавров вымерло позже, в палеоцене. Ну и
так далее.
Защита своей позиции во что бы то ни стало непродуктивна и вообще
мало интересна. К сожалению, основную часть аргументов всех сторон при-
ходится извлекать именно из актов такой неумеренной защиты. На мой
взгляд, суть дела такова: катастрофа (синхронное вымирание) безусловно
имела место и безусловно сопряжена с космической и геологической ката-
строфами, поскольку М/Д-рубеж прекрасно виден как геологически, так и
биологически, и факт космической катастрофы безупречно зафиксирован
(см. ниже). Другое дело, что в ней было причиной (без чего катастрофа не-
возможна), что - поводом, а что - сопутствующим обстоятельством (слу-
чайным совпадением). А вот это совсем не так ясно.
За 40 лет после Шиндевольфа построено множество диаграмм в его
духе (см. например, рис. 64) и ситуация вполне прояснилась: вымирание в
момент М/Д для большинства вымерших тогда таксонов полным не было (в
этом правы противники идеи глобальной катастрофы), однако факт син-
хронности вымирания огромного числа таксонов налицо, а это прямо гово-
рит о глобальной катастрофе. То есть катастрофа наверняка была, но погу-
била далеко не всё то, что ей приписывают.
517
Рис. 64. М/Д-вымирание планктонных фораминифер (по: Arenillas I. е.а.,
2002, упрощено). Как видим, 70% вымерших видов вымерло точно на М/Д-
границе. Время указано в сотнях тысяч лет
518
М/Д-катастрофа изучена лучше других и доступно описана [Алексеев и
др.], [Хайн, 1994; 2004]. Ее главный момент отмечен иридиевой аномалией:
на границе М/Д геологи нашли по всей Земле тонкий слой с резким (в 10-100
раз) повышением содержания иридия; а этот химический элемент в земной
коре очень редок, зато обычен в метеоритах. К тому же моменту относятся
слои золы - следы множества пожаров.
В 1979 году американские
физики Луис и Уолтер
Альваресы, отец и сын, пред-
ложили гипотезу, катастрофу
вызвал удар астероида. И
привели расчеты: при его
размере в 10 км образовался
кратер диаметром около 200
км и глубиной около 40 км, при
чем сам астероид испарился,
а обогащенная иридием пыль
растеклась по всей атмосфере
планеты.
Такой катастрофы было бы
вполне достаточно, чтобы
вызвать глобальную катастро-
фу: Солнце должно было на
год скрыться из виду, от
похолодания временно
должен был прекратиться
фотосинтез, и все крупные
животные должны были
вымереть. Версия получила в
прессе прозвище «астероид-
ной зимы», а космическое те-
ло - «астероида-убийца».
Рис. 65. Кратеры - следы падения “астероидов-убийц” в конце мела:
а - юкатанский; б - карский. Сплошная окружность - реконструкция внешнего борта кратера;
точечная окружность - центральная впадина большого Карского кратера; -штрих-пунктирная
окружность - реконструкция малого Карского кратера того же возраста; тре-угольники - об-
нажения зювитов (зювит - стекловидная порода с вкраплениями нерасплавившихся облом-
ков), говорящие о космическом (ударном) происхождении кратера
В 1981 году действительно был найден погребенный кратер Чикскулуб
(рис. 65а) на берегу полуострова Юкатан в Мексиканском заливе. Он вызвал
65 млн лет назад грандиозное цунами, следы которого видны поныне.
519
В 1982 году было указано на еще один, уже тогда известный погребен-
ный кратер у берега Карского моря как на второй возможный “астероид-
убийцу” (рис. 656).
К этим сообщениям и мнениям общество тогда не прислушалось, но
через 10 лет детальные исследования привели к ясному выводу: оба крате-
ра возникли в нужное время и обладают нужными размерами: диаметр
Юкатанского 180 км и глубина 35 км, а Карского - около 120 км и 25 км (см.
ВМН, 1991, № 10, с. 77; Бюллетень МОИП, отд. геологич., 1993, № 3, с. 13-
32; Природа, 1995, № 3, с. 120-121; ВМН, 2004, № 3). Найдены и другие
(меньшие) кратеры того времени.
Отрицать эту глобальную катастрофу теперь невозможно, но встает во-
прос, что астероидами-убийцами объяснить можно, а что нет.
Во-первых, почему в то время не вымерли крокодилы? Они входят в тот
же подкласс архозавров, что и динозавры, но все остальные его отряды
вымерли, а отряд крокодилов жив поныне. Во-вторых, почему все морские
рептилии (в том числе и морские крокодилы) тогда вымерли? В-третьих, по-
чему вымирание динозавров началось до бомбардировки астероидами?
Ведь сокращение их разнообразия отмечено на протяжении всего Мааст-
рихта, а он длился 5 млн лет. В-четвертых, почему некоторые динозавры
вымирали заметно позже (в течение всего Дания)?
Словом, вместо вопроса «Почему на М/Д-границе вымерли динозав-
ры?», разумен вопрос: «Почему в окрестности этой границы вымерли очень
многие животные моря и суши, в том числе почти все архозавры?». Ответ
следует искать в общей форме, а не в форме пакета ответов на предыду-
щие частные вопросы; т.е. надо объяснить саму катастрофу (факт синхрон-
ного вымирания наземных и морских фаун), а не судьбу каждого таксона,
как пытаются противники идеи катастрофы.
Мне ближе всего взгляд двух палеоклиматологов:
“Разномасштабные удары, способные глобально нарушить среду, могли сами по
себе возвещать новую геологическую эпоху. Хотя точный механизм вымирания
динозавров и других родов на мел-третичной границе остается неясным, сам гео-
логический переход можно допустить как запущенный (triggered) Чикскулубом”
(Cockell C.S., Lee Р. The biology of impact craters // BR, 2002, № 3, p. 302).
Мое мнение: понять М/Д-вымирание вне связи с другими вымираниями
невозможно. Проблема синхронных вымираний - типично диатропическая,
понимание ее следует начинать с выстраивания ряда вымираний, чтобы за-
тем сопоставить его с другими рядами эволюционных событий.
9-7. К диатропике вымираний
Известно более 20 массовых вымираний (первое: поздний рифей, 850
млн лет назад, последнее идет ныне), но общего в них мало. Принято либо
искать каждому вымиранию свою причину, либо всем вымираниям называть
одну общую причину. Но еще Мейен, не занимаясь ими специально, всё же
520
предложил иной путь: искать общие закономерности вымираний, т.е., в на-
ших терминах, начать с построения рядов вымираний {Мейен С.В. Жизнь в
“безжизненную эру”//Природа, 1973, №4, с. 117).
Мы видели в п. 8-14, что космический фактор может запускать земные
экологические процессы, но не он определяет конкретное их течение.
Пример удачного, на мой взгляд, диатропического подхода дан в АЕ:
Чикскулуб играл, вероятно, ту же роль в М/Д-вымирании, какую в фазовом
переходе играет центр конденсации (АЕ, 161-163; 296-299). Мне остается
добавить, что некоторые склонны видеть в центрах конденсации повод фа-
зовых переходов, а не их причину [Еськов, с. 253]. Эта отговорка вряд ли ин-
тересна, так как именно наличие таких центров определяет реальную дина-
мику любых фазовых переходов.
Если выстроить грубый хронологический ряд вымираний, то бросается
в глаза их периодичность (рис. 66а), а если так, то у них должна быть общая
причина. Ее в настоящее время чаще всего видят в том, что Солнечная сис-
тема каждые 26-30 млн лет проходит через облако космических тел, где
подвергается массовой бомбардировке. Космологические объяснения дан-
ного явления рассматривать не будем, но надо отметить, что на него влияет
много причин, а потому ожидать строгой его периодичности не прихо-
дится. Некоторые максимумы сдвинуты на несколько миллионов лет, а не-
которых максимумов вообще в ожидаемых интервалах нет. К тому же выми-
рания бывают крупные и мелкие, резкие и растянутые. Следовательно, у
вымираний должно быть несколько причин.
Прежде всего, иридиевая аномалия наблюдается во время многих вы-
мираний (в том числе венд-кембрийского и пермо-триасового), но не по всей
Земле. Так, на хорошо изученной границе пермь/гриас найдено лишь одно
(во Франции) место с хорошо выраженной аномалией. Это наводит на
мысль, что космическая причина не служила для этой катастрофы основной
Xeojjoei/vecKoe
Рис. 66. а) периодич-
ность вымираний се-
мейств морских
животных в мезозое и
кайнозое (по: Raup,
Sepkoski, 1984);
Мы не раз обраща-
лись к "гипотезе Геи”,
автор которой Лавлок
уверен в невероятно-
сти случайного совпа-
дения тех парамет-
521
ров биосферы, без которых невозможна животная жизнь, и потому считает
биосферу живым организмом. С этой позиции биосферные катастрофы -
периоды, когда Гея больна. Причина болезни может быть или не быть
внешней, космической, но выход из нее (а он всегда наступал) зависит от
самой Геи и ясно говорит о ее способности к самоорганизации.
б) при более деталь-
ном подсчете такая же
кривая (но за весь
фанерозой) из почти
периодической обра-
щается в почти фрак-
тальную (Gisiger, 2001,
по: Sepkoski, 1993)
Но что же приво-дит
биосферу к неус-
тойчивости? В М/Д-
-500 -400 -зоо -200 -100 о случае ее мог выз-
Geological time (Муг) вать сам космический
удар. В иных случаях
космические следы не так велики, а катастрофы столь же (или еще более)
грандиозны, так что причины надо искать геологические и биологические.
В этом отношении наиболее важно крупнейшее в истории жизни выми-
рание - пермо-триасовое. Если М/Д-вымирание носило чисто таксономиче-
ский характер (одни таксоны вымерли, другие их заместили), то пермо-
триасовое было в большой мере экологическим (например, вымершие ко-
раллы были замещены (совсем иными кораллами) лишь через 10 млн лет
[Алексеев и др., с. 104]). Это наводит на мысль, что причина его была очень
своеобразна. Оно отличается и тем, что морские организмы пострадали на-
много сильнее наземных, чего ни космическая, ни биологическая модели
объяснить не могут — ни порознь, ни вместе.
Зато это могут гипотезы расширяющейся Земли84. Все они утвержда-
ют, что в прошлом были моменты (среди них и пермо-триас), когда земная
кора лопалась (в силу химического и теплового расширения ядра и мантии)
сразу во многих местах. Такие гипотезы не являются целостными теориями,
84 Первым такую гипотезу высказал австралийский геолог Уоррен Кэри (1950-е гг.) в
довольно фантастической форме, однако позже были предложены реальные физичес-
кие и химические механизмы расширения (см., например, [Лайн, 1994]). Ботаник Н.Н.
Цвелёв уверен, что идеи расширения Земли и движущихся литосферных плит допол-
няют друг друга (Цвелёв Н.Н. Проблемы теоретической морфологии и эволюции рас-
тений. М.-СПБ, КМК, 2005, с. 11), а вовсе не противоречат, как считают многие.
522
однако факты в их пользу имеются, и их учёт позволяет многое понять. Так,
растрескиванию коры предшествовало ее напряжение, и спровоцировать ее
синхронное растрескивание коры мог даже некрупный астероид. При этом в
основном трескалась тонкая океаническая кора, и океанские воды отравля-
лись извержениями. С этой точки зрения легко понять почему в пермо-
триасе погибло 9/10 родов морских животных и, в частности, все кораллы.
Далее, прогревом земной коры перед растрескиванием объясняется
наличие ископаемых остатков леса на пермском Полюсе (он находился близ
нынешней Чукотки), не объясненное до сих пор иными гипотезами (в том
числе и идеей переменного наклона земной оси).
Кое-где тогда лопнула и кора континентов, например, в Сибири:
“Величайшее... пермо-триасовое вымирание было сопряжено с излияниями
сибирских траппов (лавы, ныне застывшей - Ю.Ч.\ по многим оценкам -
геологически кратковременным... Ему соответствовало уничтожение барье-
ров между биохориями и разрушение сложившегося за позднюю пермь рас-
пределения растений и животных” (Щербаков Д.Е. И МС, 177).
Поясню: фитохория - область распространения определенной крупной
группы растений; пермо-триасовому вымиранию предшествовал распад
экосистем, приведший к падению прежних барьеров между флорами.
Можно сказать довольно определенно: исходной причиной пермо-
триаса было истончение земной коры, которое поначалу вызвало потепле-
ние (уравняло климат суши, чем сняло барьеры между флорами), а затем -
разрывы коры, ставшие причиной отравления вод океана.
В частности, известно, что падение астероида активизирует вулканиче-
скую деятельность [Хайн, 2004]. Обзор разных гипотез (Berner R.A. Examina-
tion of hypotheses for the Permian-Triassis boundary extinction by carbon cycle
modelling // PNAS, 2002, vol. 99, № 7) наводит на мысль, что ведущей причи-
ной данного вымирания было именно выделение газов из недр Земли в во-
ду и воздух, разрушившее баланс биосферы.
В пермо-триасе, при общем вымирании, росла продуктивность тех зе-
леных водорослей, которые живут в прибрежных водах около вулканов (Ах-
метьев М.А. Причинно-следственные связи глобальных биосферных пере-
строек... // Труды Г14Н, вып. 565, М., 2004, с. 472); активность недр вызвала
отравление тропической зоны океана (а затем атмосферы) сероводородом
[Уорд]. Всё это согласно с идеей, что Земля в тот период расширялась.
В диатропический анализ вымираний следует еще включить длитель-
ные похолодания, именуемые ледниковыми периодами. Они тоже были
причиной вымираний, пусть и не глобальных, и вероятно тоже связаны с
космическими причинами. Первая - это изменение наклона земной оси, из-
за которого сезонность климата исчезает каждые 40 тыс. лет, чем можно
объяснить многие короткие периодичности [Хайн, 1994 с. 170]. Его открыл
сербский астроном Милутин Миланкович в 1920-х годах. Однако этим нель-
зя объяснить более долгие аномалии, например, лес на пермском Полюсе.
523
Вторая причина - это астероиды. «В большинстве случаев области рас-
пространения морен, связанных с покровным оледенением, совмещены с
пло-щадями широкого проявления кольцевых структур» (рис. 67), а они мо-
гут быть ударными кратерами (Нечаева И.А. // Советская геология, 1991, №
1, с. 70).
Оледенения для на-
шей темы важны тем,
что именно в их время
возник человек (см. п. 9-
15), и если бы асте-
роиды вызвали тогда
глобальное вымирание
(как на М/Д-границе),
нас бы просто не было.
Рис. 67. Совпадение
кольцевых структур 2
(ударных кратеров?) с
основной зоной оледене-
ний I эпохи плейстоцена.
Почему одни ливни
вызывали вымирания
без оледенений, а дру-
гие - оледенения без
вымираний? Вопрос
видимо выяснен: в плейстоцене климат планеты был много холоднее преж-
них по географической причине: в кайнозое выросли возвышенности в Азии
и Северной Америке, изменившие атмосферную циркуляцию и тепловой
режим Северного полушария (Раддимен У.Ф., Куцбах Дж.Е. Воздымание
плато и изменение климата // ВМН, 1991, № 5). Ливень астероидов вызыва-
ет пыль в атмосфере и похолодание, а после прояснения неба уже лежит
снежный покров, который в холодном климате поддерживает сам себя, по-
скольку отражает в космос много тепла. Наоборот, на М/Д-границе для оле-
денения не было географических условий, что ясно из высокого в тот мо-
мент уровня океана. И, когда вновь засияло Солнце, снег стаял.
Подводя итог, можно сказать, что единой причины и единого механиз-
ма для всех катастроф искать не стоит (см. [Несис, с. 190, 195]). Общим яв-
ляется лишь то, что вымирание таксонов идет непрерывно, но с из-
бытком возмещается появлением новых таксонов всегда, кроме ко-
ротких периодов катастроф. Каждая катастрофа имеет свой особый
облик, что не должно мешать нам выстраивать их в ряды и искать общие
черты. Это возможно потому, что тут, как и в любой эволюции, огромное
524
разнообразие складывается из комбинации сравнительно небольшого чис-
ла компонент (блочность). Понимание этого важно и для создания верной
картины прошлой эволюции, и для выработки способа существования в
биосфере, на наших глазах входящей в новую катастрофу.
9-7* Статистика вымираний и фракталы
Сплошной массовый анализ данных о вымирании вели в последние 20
лет американские палеонтологи Дэвид Рауп и Джек Сепкоски, а затем и дру-
гие. Как уже сказано, рис. 66а демонстрирует довольно ясную периодич-
ность. Этот факт позволил авторам счесть космос причиной всех массовых
вымираний. Однако затем Сепкоски, охватив более широкий круг данных,
получил более сложную кривую вымираний (рис. 666).
Анализируя ее на периодичность, Гизигер (BR, 2001, № 2) пишет о ее
масштабной инвариантности (отсутствии характерных частот). Другими
словами, можно принять, что процесс вымирания носит характер частот-
ного фрактала. Это неудивительно, если вспомнить об одной фрактальной
характеристике самого процесса вымирания: вымирание классов складыва-
ется из вымираний отрядов, их вымирание - из вымираний семейств и т.д.,
а значит надо ожидать самоподобия.
Близкие результаты получили во Франции уже известный нам палео-
нтолог Жан Шалин и соавторы (АЕ, 12, 304, 329).
Гизигер привел два класса моделей, призванных описать масштабную
инвариантность - критические и некритические. Критические модели ис-
ходят из допущения, что массовое вымирание есть аналог фазового пере-
хода в биосфере, находящейся в критическом, т.е. неустойчивом состоянии
(добавлю: происходит в силу “опрокида” по Жданову, см. п. 6-6). При этом
Гизигер сослался на ряд работ по «самоорганизованной критичности» (одна
из них переведена на русский: Бак П., Чен К. Самоорганизованная критич-
ность // ВМН, 1991, № 3). Но, замечу, сама по себе данная частотная дина-
мика не обязательно говорит о какой-то критичности.
Построены также “некритические” модели массовых вымираний. Они
описывают ситуацию не хуже, если ввести в них случайные внешние собы-
тия, которые Гизигер именует как «стресс» (они могут означать у него и
землетрясения, и вулканизм, и падение астероидов). Основной вывод из
анализа некритических моделей: «Чем дольше время ожидания крупного
стресса, тем больше должно быть последующее вымирание» (Gisiger Т. И
BR, 2001, №2, р. 195).
То есть небольшое событие может запустить огромное вымирание.
Данный тип моделей вполне описывает все те глобальные вымирания, для
которых известен конкретный фактор “стресса”, их запустивший. Такой спо-
соб описания выглядит более удачным, чем принятые ныне чисто биологи-
ческие “модели” вымираний, в которых внешний фактор неизменно упоми-
нается, но никак далее не учитывается.
525
Что касается действующих причин вымираний, то чисто биологические
“модели” относят к ним крушение пищевых пирамид, сужение экониш за
счет специализации и, как обычно, конкуренцию. (Каждая из таких причин
вполне могла иметь место, но ее наличие надо доказывать, а этого никогда
не делается.) Обсуждая одну из таких моделей («гипотезу эволюционного
дисбаланса сообществ», которую в 1978 г. предложил В.В. Жерихин; она
имеет последователей), А.С. Алексеев справедливо заметил: она
“приложима к одному-единственному сообществу, но остается непонятно,
как данный механизм может привести к практически одновременному разру-
шению многих различных сообществ” Земли (Алексеев А. С. Докт. дисс.,
Геол, фак-т МГУ, 1998).
Иными словами, синхронность вымираний (то главное, что надо выяс-
нить), не вытекает из модели, а подразумевается при ее построении.
9-8. Эволюция и сдвиг материков
Выше говорилось, что материки прошлого находились не там, где ныне,
что океаническая кора постоянно задвигается под кору материков. Еще в
1620 году Фрэнсис Бэкон заметил удивительное сходство атлантических
побережий Африки и Южной Америки и счел его неслучайным. Затем дол-
гое время его считали случайным, но в XX веке выяснилось, что стоит мыс-
ленно вложить на карте Бразильский выступ в Гвинейский залив, как кри-
сталлический архейский фундамент обоих материков сольется в единый
массив. В Атлантическом океане в XX веке прослежен срединно-океани-
ческий хребет; так и видишь, как с его подъемом расползались материки.
Взглянем снова на карту флорогенеза (рис. 61): почему тропическая
зона заняла всю Северную Америку и пол-Гренландии, но не попала в Юж-
ную Африку и Южную Америку? Оказывается, подо льдами Гренландии ле-
жат древние тропические флоры и фауны, тогда как в Африке их находят
только близ Средиземного моря. Обнаружив в Гренландии диковинные ис-
копаемые, немецкий геофизик Альфред Вегенер в 1912 году понял, что ма-
терики двигались. Вернувшись, он прочел всё, что до него писали на эту те-
му (см. п. 3-13**), и собрал множество данных о совпадении геологических
толщ, древних климатов, флор и фаун разных материков.
Приведу пример: среди юрских и меловых крокодилов есть виды одного
рода, жившие по разные стороны нынешней Атлантики. Наоборот, опоссу-
мы, возникшие в позднем мелу, в Африке отсутствовали всегда. Естествен-
но было допустить, что на месте Атлантики прежде было лишь внутреннее
море вроде Каспия (рис. 68), раскрывшееся и ставшее океаном в меловом
периоде. Так в науке родился мобилизм.
По Вегенеру, более легкие гранитные материки плавают в более плот-
ной базальтовой мантии, словно айсберги в океане, и расползаются друг от
друга под действием центробежной силы вращения Земли.
526
Рис. 68. Материки в позднем карбоне (а) и в мелу (б), по Вегенеру
Объяснение не
выдерживало крити-
ки, и автора, как во-
дится, не слушали,
отвергнув вместе с
его наивным меха-
низмом и очевид-
ные факты. В 1930
году Вегенер погиб
в Гренландии (его
последний экспеди-
ционный отчет из-
дан уже при фашистах), и о сдвиге материков почти все забыли.
Однако в 1960-е годы мобилизму был предложен реальный механизм:
земная кора бывает двух родов - толстые литосферные плиты, несущие
на себе материки, и тонкая океаническая кора; мантия Земли за счет внут-
реннего жара медленно ворочается, словно закипающая густая каша, и по-
тому плиты медленно и хаотично смещаются по ее верху. Океаническая ко-
ра рождается в разломах дна (срединно-океанических хребтах) и растекает-
ся; происходит спрединг (англ, spreading - растекание), он-то и движет пли-
ты. Избыточная океаническая кора задвигается под плиты у побережий и
островных дуг - происходит субдукция.
Поначалу плит было названо всего шесть, и всё выглядело очень кра-
сиво. Вскоре пришлось вводить добавочные плиты - например, для субкон-
тинента Индостан (он прежде располагался между Африкой, Австралией и
Антарктидой, а затем мигрировал в сторону Азии и в момент М/Д-кризиса
еще был южнее нынешнего экватора, где и треснул, излившись лавой; поз-
же его столкновение с плитой Евразии породило высочайшие в мире горы -
Гималаи). Это тоже изящно, но затем число плит пришлось увеличить до
сотни, и красота мобилизма стала теряться: стройная система плит обрати-
лась в мозаику мелких и порой сомнительных блоков.
Первый серьезный удар по мобилизму нанесла та же Африка, что его
породила: вокруг нее нет ни одной зоны субдукции, а она с трех сторон ок-
ружена зонами спрединга и 30 млн лет неподвижна. Говорят, что Африка от
сдавливания подымается, но как быть с Антарктидой, которая окружена
сплошным кольцом зон спрединга, а подо льдом являет почти сплошную
низменность, во многом ниже уровня океана? Конечно, подняться ей некуда
(давит ледяная шапка), это понятно, но куда девается вещество спрединга?
Отсутствие ясности породило конкурентные взгляды: одна школа заяв-
ляла, что Земля расширяется, объясняя этим расползание материков и
пермо-триасовое вымирание (п. 9-7), другая - что Земля сжимается и этим
527
объясняла столкновение материков и появление складчатых гор, третья -
что Земля пульсирует; четвертая - что континенты проваливаются и проис-
ходит “океанизация Земли”. Единой теории Земли, столь нужной для теории
биологической эволюции, до сих пор нет.
Очевидно лишь, что Земля устроена невообразимо сложно. Укажу на книгу:
Дмитриевский А.Н. и др. Энергоструктура Земли и геодинамика. М, 1993, где
Земля рассматривается как гигантская ДС (с тем же правом здесь можно было бы
говорить о гигантском солитоне). Сверхвысокое давление порождает совсем но-
вую физику и химию. В этих условиях идет очень сложное саморазвитие (вспом-
ним идею Геи). Рождению единой теории гео- и биоэволюции препятствует тот
факт, что оппоненты мало читают друг друга. Доводы против друг друга и подбор
аргументов в свою пользу они воспринимают как доказательство своих теорий. Но
такова, увы, специфика всех форм эволюционизма.
На сегодня можно лишь сказать, что земная кора участвует во многих
одновременных процессах, меняющих облик Земли и жизни на ней.
9-9. Реальные свидетельства факта эволюции
Как бы то ни было, материки в прошлом наверняка двигались, хотя не
всегда ясно, как шло движение конкретно. Организмы эволюировали на ма-
териках сперва общих, затем - раздельных, потом опять сближавшихся. Как
хорошо видно на тех же архозаврах, реальный контакт территорий часто
оказывался важней воображаемого приспособления: пока он был, на них
возникали общие семейства и даже общие роды (иногда на очень разных
широтах, а значит в различных природных условиях), но стоило ему по-
рваться, как общих таксонов архозавров уже не возникало; прежние их об-
щие таксоны, разумеется, продолжали существовать.
Поскольку это найдено до обнаружения сдвига материков, т.е. не могло
быть под него подогнано, можно видеть тут первое реальное свидетель-
ство самого 'факта эволюции. Коснемся этого вопроса подробнее.
Дарвин видел в различии видов вьюрков на соседних островах довод в
пользу эволюции, но теперь его пример выглядит слабым: наличное разно-
образие не есть эволюция и, в частности, верующему говорит только о
щедрости Бога. Важнее ископаемые следы эволюции, показывающие, как
организмы изменялись со временем; но примеры, приводимые в учебниках,
тоже легко трактуются как примеры непрерывного творения: ничто не ме-
шает признать, что Бог сперва сотворил рыб, затем амфибий и т.д. и что он
пользовался при этом некоторыми общими правилами, которые мы воспри-
нимаем как наследование от предка.
Разумеется, Бог мог сотворить всё, если мыслить его всемогущим, но
мы ведь не прибегаем к идее Бога, когда наблюдаем падение камня, варим
кашу или забиваем гвоздь; тут мы пользуемся известными понятными нам
свойствами вещей. Вся наука состоит в раскрытии таких свойств, независи-
мо от того, во что верят ученые, и понять эволюцию - значит свести ее к та-
528
ким свойствам. Мы очень далеки от решения этой задачи и, чтобы подойти к
решению, должны сперва убедиться, что сам феномен эволюции существу-
ет, т.е. что есть естественный процесс, в ходе которого от организмов одних
видов происходят организмы других видов.
Данную малую задачу в наше время можно решить - в том смысле, что
можно увидеть, как идет расселение и при этом вместо одного вида появля-
ется другой, близкий, причем с обрывом контакта между территориями про-
цесс прекращается. Это видно в малом на пермских хвойных (п. 9-4), а в
большом - на меловых позвоночных, живших вокруг Атлантики.
Второе реальное свидетельство эволюции можно видеть в «чернови-
ках Господа бога» (пп. 8-9; 8-12): видно, что эволюция не являла собой пла-
на, заранее обдуманного, что в ней имели место эксперименты и они не все
были успешны. Вопрос о наличии Бога при этом не обсуждается.
Однако это отдельные примеры, а надо показать, что эволюция типична
(имеется тенденция), что наши примеры не извлечены из множества проти-
воположных. Такую работу, насколько мне известно, первым проделал био-
географ Леон Круаза, работавший в Венесуэле. В 1958-1962 годах он издал
несколько огромных книг, в которых скрупулёзно описал трэк (фактический
ход эволюции, сопровождавшейся расселением по суше или по морю) для
многих таксонов. Круаза разъяснял:
«Трэк представляет собой линию на карте, вдоль которой находится область
распространения родственных между собой организмов... трэк является це-
пью центров образования новых форм; каждый центр возник при этом вслед-
ствие разделения ареала, который ранее был заселён формой более низкого
таксономического ранга» (цит. по [Попов, с. 100-101]).
Затем Мейен пунктирно описал подобную эволюцию для целых назем-
ных флор (п. 9-4): у него таксоны тоже не просто сменяют друг друга, и при-
том массово, а еще и расселяются на север и на юг. Это (третье) свиде-
тельство есть общее фактическое доказательство эволюции. Работы
Ахметьева и Мосейчик укрепили этот вывод (хоть и видоизменили).
Круаза установил, что эволюция данного таксона идет сходным обра-
зом в разных частях ареала даже при отсутствии их связи, т.е. фактически
описал ее как движение вдоль рефрена (номогенез).
Конечно, верующий может и это назвать непрерывным творением, но
ему придется признать, что Творец не просто непрерывно творил, но и
имитировал эволюцию - в каждую эпоху соблюдал рамки существовавшей
тогда конкретной биогеографии. А такого “творца” вряд ли примет хоть одна
религия. Проще и естественнее видеть тут совокупность законов природы и
ее эволюции. Разумеется, таким путём не решить вопрос о наличии или от-
сутствии самого Творца (без кавычек), установившего законы эволюции, но
данный вопрос в рамках науки вообще не решается - как всякий вопрос ве-
ры. Зато этими свидетельствами эволюционизм становится в ряд аккурат-
ных естественных наук, чего прежде не было.
529
9-10. Эти живучие крокодилы
Итак, материки двигались, динозавры на них вымирали, а крокодилы
выживали. Многие эволюционисты (например, Брокки, Соболев и Шинде-
вольф) полагали, что всякий таксон, как и индивид, достигает старости и
вымирает. Нельзя ли сказать, что крокодилы были в момент М/Д молоды и
потому выжили? В самом деле, пережили катастрофу только четыре самые
молодые семейства надотряда Crocodylia, известные с позднего мела.
Особо интересно семейство настоящих крокодилов (Crocodylidae), по-
скольку в нем нашелся даже один род, Leidyosuchus, который без измене-
ния перешел М/Д-границу (у остальных трех семейств родовой состав в хо-
де катастрофы сменился полностью) и тем самым показал, что крокодилы
именно пережили катастрофу, а не произошли заново.
К этому семейству принадлежат все нынешние крокодилы. Подробно я
писал об этом ранее (БПС, 2000, № 46) и здесь отмечу немногое.
Взгляните на рис. 69. Кажет-
ся, что изображен нынешний
крокодил вроде индийского
гавиала (узкая морда), однако
на самом деле это рекон-
струкция пермской амфибии
по имени платиопс подкласса
лабиринтодонтов. С ними у
крокодилов хорошо видны
параллельные ряды.
Рис. 69. Амфибия платиопс
(пермь)
Крокодилов отличает ото всех похожих на них амфибий и рептилий на-
личие вторичного нёба (рис. 70): у них воздух идет в глотку, минуя ротовую
полость, что позволяет одновременно хватать плавающую пищу и дышать
воздухом. Среди протозухий, мезозухий и эвзухий есть короткомордые,
длинномордые и узкомордые (пасть, словно пинцет) формы, есть с ластами
и с лапами. Словом, повторили лабиринтодонтов и друг друга в рамках об-
щего рефрена.
Вторичное нёбо - основное свойство, объединяющее настоящих кроко-
дилов - принято считать признаком их родства. Но почему принято?
Если это - приспособление, то как раз его, следуя общепринятой логи-
ке, можно счесть развившимся у разных крокодилов независимо, а их об-
щую тенденцию, “крокодилий” облик - унаследованной от лабиринтодонтов.
Т.е. можно допустить, что все они сами “изобрели” удобную носоглотку, а
форму тела и тенденцию его удлинения унаследовали от общего предка. Но
530
тогда его естественно считать амфибией. А если так, то крокодилы про-
изошли не от наземных рептилий, и значит, их нельзя считать архозаврами.
Рис. 70. Эволюция вторичного нёба надотряда
Crocodylia
а - отсутствие (протозухии, триас - юра),
б - слабое развитие (мезозухии, юра - эоцен),
в - полное развитие (эвзухии, т.е. настоящие
крокодилы, от мела до наших дней); зухос -
египетское название крокодила
Такой взгляд на происхождение крокодилов
прямо противоположен общепринятому, но
согласуется с фактами не хуже. Отстаивать тот
или другой смысла, по-моему, нет. Налицо общий
рефрен, остальное-толкования.
Приспособиться можно было и иначе, чем “кро-
кодильим” обликом: известны две группы репти-
лий - плезиозавры и плакодонты, независимо
освоившие в ту эпоху иную форму водного животного, похожую на черепа-
шью (рис. 71). Медлительные плезиозавры делили эконишу с крокодилами,
а некоторые из плезиозавров (плиозавры) охотились в открытом море и
плавали быстро, деля нишу с ихтиозаврами. Это говорит в пользу первич-
ности (унаследования от предка) именно крокодильего облика, а не носо-
глотки (возникавшей, с этой точки зрения, независимо). Плезиозавры, в от-
личие от крокодилов, явно пришли с суши и видимо имели общих предков с
птицеобразными рептилиями.
Рис. 71. Плакодонт — мезозойская водная
рептилия, чьим основным движителем был не
хвост, а ласты (панцырь был не у всех
плакодонтов; триас — юра)
Насколько крокодилы Маастрихта функ-
ционально превосходили тогдашних ди-
нозавров, можно только гадать, а вот
нынешние крокодилы действительно
высоко организованы: например, единст-
венные из рептилий обладают четырех-
камерным сердцем.
Принцип компенсации позволяет
допустить, что древние крокодилы тоже
имели более развитую физиологию, чем
531
динозавры (имевшие более развитые и сложные формы), отчего и пережи-
ли М/Д-катастрофу.
Кстати, и возврат в воду нельзя целиком свести к приспособлению ко
вновь возникшей среде. Самый простой и ясный пример являют ихтиозавры
с их восьмипалыми конечностями (где восьмой палец представлен одной
косточкой - LEG, 33). Возврат их в воду произошел в триасе, когда все реп-
тилии имели пятипалые конечности. Никакой пользы от усложнения органа
(при упрощении его работы) не было. Очевидно, что в этом ихтиозавры про-
сто повторили элемент прежнего рефрена - воспроизвели общий план
ласта девонских кистеперых рыб, тоже бесполезного.
Кстати, даже дарвинист Г. Рёддер, изучив старые и новые данные о
числе пальцев, пришел в 1999 году к выводу, что «стабилизация пятипалой
конечности вызвана не функциональными причинами, а какими-то внутрен-
ними», в каковых нехотя признал ортогенез [Попов, с. 128]. С позиции ново-
го номогенеза крокодилов и плезиозавров объединяет тенденция к удлине-
нию позвоночника, она свойственна многим рыбам, амфибиям и рептилиям
(п. 6-8). К ней и приспособились, кто как сумел: одни - усилением хвоста,
другие - конечностей-ласт, третьи - того и другого. Видны вариации на ос-
нове общего плана строения. Об этом скажу подробнее.
9-11. Блочность и планы строения
Как не раз говорилось ранее, эволюция носит блочный характер: в ней
постоянно используются комбинации ограниченного числа структурных и
функциональных блоков. План строения - небольшой набор структурных
блоков, задающий общий облик таксона; блоки связаны между собой, но
нежестко. Кроме структурных, со времен Бэра принято включать в план
строения еще и один функциональный блок - путь развития зародыша.
До Дарвина сравнительная анатомия была наукой о планах строения
(см. гл. 2). Это еще помнил морфолог и натурфилософ В.Н. Беклемишев:
“проблема плана строения - важнейший раздел сравнительной анатомии”,
поэтому без него “выпадает организм как целое и вся его эволюция”, хотя
“именно этому вопросу и были посвящены главные усилия эволюционной
морфологии” [Беклемишев, т. 1, с. 5-6].
Главным в плане строения он считал симметрию. Наоборот, новейшее
пособие Дзержинского (см. п. 6-13) ведет изложение в виде перечисления
органов и их свойств, и лишь во введении сказано, что прежние авторы изу-
чали планы строения. Такая анатомия удобна для “доказательства" факта
эволюции (на самом деле лишь иллюстрируются рефрены), но для откры-
тия новых черт эволюции она бесполезна. Поэтому отрадно, что недавно
переведен учебник Альфреда Ромера и Томаса Парсонса, вновь дающий
целостное, симметрийное видение анатомии. Например:
“Главная черта строения тела позвоночных - двусторонняя симметрия... Та-
кой же тип организации характерен для большинства группп беспозвоноч-
532
ных... Резко отличаются от всех этих животных кишечнополостные и иглоко-
жие, обладающие радиальной симметрией... Степень активности животных
коррелирует с типом симметрии... Позвоночные... ведут обычно активный об-
раз жизни. Активность была, по-видимому, одним из ключевых свойств, оп-
ределивших успех позвоночных... У всех животных с двусторонней симмет-
рией тело разделено в продольном направлении на несколько отделов... Тело
позвоночных также делится на четко выраженные отделы... У позвоночных
имеется высокоспециализированная голова, ... здесь находятся все основные
органы чувств, нервные центры, образующие головной мозг, а также рот и
связанные с ним структуры. У позвоночных, как и у всех двусторонне сим-
метричных животных (даже у червей), наблюдается сильная тенденция к це-
фализации, т.е. к концентрации определенных структур и функций на перед-
нем конце тела” [Ромер, Парсонс, с. 9-10].
К сожалению, провести идею симметрии через конкретный материал
непросто. Авторы сами признали, что их схему портит тип иглокожих, кото-
рые радиально симметричны, хотя некоторые из них являются активными
хищниками, и пятилучевая симметрия им очень мешает. Авторы пытались
обойти трудность в рамках, обычных для дарвинизма: «иглокожих нельзя
считать истинно радиально-симметричными; в начале своего онтогенеза
они обладают двусторонней симметрией». (Добавлю, что есть более силь-
ный аргумент: самые древние, кембрийские, иглокожие еще не обладали
пятилучевой симметрией, она известна с ордовика.) Но с позиции приспо-
собления морским звездам надо было попросту потерять неудобную сим-
метрию или вымереть, а они не сделали ни того, ни другого.
Ответ ясен в рамках нового номогенеза: пятилучевая симметрия - тен-
денция всего типа, она плоха для хищничества, но одному классу иглоко-
жих, видимо в силу экологической прочности, удалось выжить и стать хищ-
никами. Ромер и Парсонс сами указывали на тенденции, но хочу отметить:
ни одна тенденция из приспособления не выводится, это отдельный фено-
мен. Всякий план строения можно понимать как набор тенденций.
Самым общим планом строения обладает филум, он же тип (п. 9-2).
Типы группируют в разделы, но лишь по отдельным сходствам. У многокле-
точных животных 3 главных раздела: губки, радиальные и двусторонне
симметричные (билатеральные). Последний делится на 2 подраздела: пер-
вичноротые и вторичноротые (к ним относят типы полухордовых, иглокожих
и хордовых). Допускают, что у них был общий вторичноротый предок (что
иглокожие - наши «колючие родственники» [Ивахненко, Корабельников]), но
у иглокожих план строения радиальный, а у полухордовых и хордовых би-
латеральный. Мы отнюдь не родственники.
Эту трудность иногда обходят просто: включают иглокожих в раздел
билатеральных, на том лишь основании, что у иглокожих ранняя личинка
билатеральна. Можно поступить иначе: счесть, что вторичноротость они
приобрели независимо, что радиальная симметрия - исконное свойство иг-
533
локожих. Ведь у них, как правило, радиальны не только поздняя личинка и
взрослая форма, но и самая ранняя стадия развития - дробление. Однако
включить их в общепризнанный раздел радиальных нельзя, поскольку ра-
диальные двуслойны (образуются из двух зародышевых листков), а иглоко-
жие трехслойны.
Другие обходят эту трудность, ища общего предка хордовых и иглоко-
жих среди полухордовых; но тут приходится либо принять, что билатераль-
ность была утеряна и затем вновь обретена, в иной плоскости (Jefferies
R.P.S. The ancestry of the vertebrates. London, 1986, ch. 2, 3), что, на мой
взгдяд, лишает значения всю идею симметрии; либо искать предка хордо-
вых с кольчатыми червями, но тогда приходится игнорировать их первично-
ротость. К этому надо добавить принципиальное, по Малахову, различие
онтогенезов у хордовых и других вторичноротых (п. 5-17*), что также меша-
ет считать их предками друг друга.
По-моему, лучше вообще не искать предков, а прямо признать
“комбинаторику признаков очень высокого ранга, возникавшую при обособ-
лении планов строения иглокожих и хордовых” (Рожнов С.В. Особенности
эволюции раннепалеозойских иглокожих // Экосистемные перестройки и эво-
люция биосферы. Вып. 3. М., 1998, с. 75).
Картина среди многих других типов (например, одноклеточных - см. гл.
10) примерно та же, и тем самым блочность эволюции типов оказывается
их основным свойством. Сама же она - частный случай диасети: тип сим-
метрии данной стадии развития, первичноротость и т. д. - всё это блоки,
комбинацией которых задаётся диасеть. Блоки так же выстраиваются в ря-
ды, как и все свойства, от белковых цепей до планов строения. Почему так,
мы еще не знаем, но так устроен мир, и не видеть этого - наивно.
9-12. Блочность и родство групп
Блок обычно состоит из блоков поменьше (позвоночник - из позвонков,
путь развития - из дробления, бластуляции, гаструляции и прочих стадий,
ну и т.д.). Но если всё сделано из общих блоков, то как же понимать родст-
во? Геносистематика, на которую ранее было столько надежд, оказалась
малополезна, ибо предок в ней зависит от применяемого метода (см. гл. 10).
Сейчас всё больше вспоминают старые приемы. Сделаем то же и мы.
Вспомним, что организмы обладают свойствами, а составленные из них
таксоны - меронами. Преобразование мерона в череде последовательно
живших видов именуется семофилогенией. Пример - эволюция плавника
рыбы в лапу четвероногого. Если семофилогении всех меронов, опреде-
ляющих таксон А, ведут к таксону Б, то можно достаточно уверенно гово-
рить о филогении (что таксон А - предок таксона Б), но так не бывает: се-
мофилогении идут в разные стороны, это и есть блочность эволюции.
Так что любой систематик, если он строит филогению (термин см.: п. 2-
7), вынужден признать одну или две семофилогении главными, доказываю-
534
щими родство, а прочие - “более поздними приспособлениями”, т.е. считать
основную массу свойств таксона Б возникшими заново. Об этом писал еще
Соболев, см. п. 4-12. Мейен, вслед за Соболевым, отвергал такой подход:
“филогенетические схемы опирались на специально подобранные ряды иско-
паемых растений, родство которых неясно” (Мейен С.В.П ПЖ, 1970, № 4).
«Такого, чтобы показания всех признаков совпали, не случаетсся почти нико-
гда. (Слово “почти” я вставил из осторожности. Думаю, что без него можно и
обойтись.)» [Мейен, 1981, с. 133].
А в конце жизни он пояснил, что реальные отношения таксонов
“редуцируются до схемы, иллюстрирующей мнение филогенетика о некоей
интуитивно суммируемой близости или удаленности таксонов” (Современная
палеонтология. М., Недра, 1988, т. 1, с. 509).
Анализ разнообразия убедил его, что наследуются не сами свойства, а
спектр их возможностей (это - транпол, п. 6-4*), поэтому родство нельзя
установить по одним наличным свойствам, но надо еще наблюдать сам
процесс превращения одного таксона в другой.
До Круаза и Мейена тут был известен лишь один прием - анализ “вверх
по разрезу”, т.е. выявление ископаемых остатков одного крупного таксона в
сплошном непрерывном по времени геологическом разрезе. Соболев в от-
ношении лошадей этот прием осмеял (п. 4-12), но Круаза и затем Мейен по-
казали, как “положить процесс на карту” (указать, в какой части света каж-
дый род впервые появляется в летописи), и Соболев оказался неправ: ро-
ды, которые он счел независимо возникшими, на самом деле пришли в Ев-
ропу из Северной Америки, где род за родом образуют непрерывную во
времени цепь (рисунок, касающийся эволюции лошадей, см. [Кэрролл, т. 3,
с. 104]). Можно ли теперь утверждать, что эволюция понятна? Не вполне.
Поясню это именно в отношении эволюции лошадей.
Во-первых, она выстроена всего по двум семофилогениям - зубов и ко-
пыт. Считается, что именно они определяют облик семейства, но придут
другие авторы, скажут, что главное в каких-то чертах черепа, желудке или
еще в чем-то, и начнут работу почти заново.
Во-вторых, если цепочку родов “рассмотреть в лупу”, она почти нигде
не даст цепочку видов. Сам Кэрролл, уверенный в наличии таких цепочек,
смог привести в качестве примера всего одну мало интересную цепочку -
где вид (предок копытных) «постепенно становился крупнее, пока не изме-
нился настолько, что был признан другим видом», причем укрупнение изме-
рено по ширине одного зуба (там же, с. 145). Даже по двум признакам вме-
сте такое не получается: вдоль цепочки остальные признаки меняются не-
регулярно, и опять вроде бы прав Соболев с монополией скачков.
В-третьих, никто не берется утверждать, что так можно описать всё се-
мейство, т.е. эволюцию каждого рода (уж не говоря о каждом виде).
В-четвертых, этим путем описано появление акциденций (побочных
черт), но не сущности (важного свойства), а Соболев призывал изучать их.
535
Мейен решил поступить иначе: не рассматривать каждый род и вид, а
просмотреть весь доступный корпус фактов по очень большой группе орга-
низмов и построить схему ее эволюции за всё исследуемое время для всей
Земли. Отличие от обычного “вверх по разрезу” то, что судьбу каждого так-
сона было надо рассмотреть по всем разрезам на всех материках. Работа
была намечена колоссальная и должна была позволить увидеть, как факти-
чески таксоны обращались один в другой. Решение задачи удалось наполо-
вину: появилась изящная схема глобального флорогенеза, рассмотренная
нами в п. 9-4, однако она оказалась не вполне верна (п. 9-4*), а судьбу от-
дельных таксонов ниже порядка Мейен выявить вообще не успел. Подробно
о его системе мы будем говорить в главе 10, а сейчас рассмотрим пример.
9-12* Происхождение цветковых и гамогетеротопия
Механизмом, ведущим к блочности, называют горизонтальный перенос
генов (п. 6-9), но Мейен смотрел на этот ход мысли иронически. По его мне-
нию, заполнение рефренной таблицы всегда ведут свои, внутренние меха-
низмы, иначе не было бы транпола. В случае цветковых он приводил еще и
такой аргумент: если горизонтальный перенос «действительно происходил,
то можно ожидать его проявления в становлении не только цветка, но и дру-
гих органов», тогда как на деле все они, кроме цветка, эволюировали посте-
пенно. Шел этот процесс в середине мелового периода.
Механизмом, обеспечившим появление цветка в эволюции, Мейен счел
гомеозис (см. пп. 4-7, 5-15, 5-17). Точнее, мерой «цветок» можно мысленно
получить из мерона «фруктификация голосеменного», допустив гамогете-
ротопию (перенос некоего свойства с одного пола на другой).
Гамогетеротопия хорошо известна у животных - таковы, например, мо-
лочные железы самцов. На нее указывали противники Дарвина и Геккеля
(ибо такие черты не могли быть ни отобраны, ни унаследованы “от общего
предка”) как на свидетельство того, что план строения - самодовлеющая
сущность. Гамогетеротопия бывает и у отдельных особей - тогда она носит
характер патологии (у людей: бородатость женщин, женские груди у мужчин,
гермафродитизм и т.п.). С позиции диатропики такие случаи должны
встраиваться в какие-то более общие ряды изменчивости. Такие ряды су-
ществуют. В п. 4-11 описано правило Кренке (уродство у данного вида часто
является нормой для другого вида), и гамогетеротопия не исключение: на-
пример, у низших животных нормален гермафродитизм, являющийся у
высших патологией. Самый странный случай видовой гамогетеротопии на-
блюдается у пятнистой гиены (п. 5-10*).
Но может ли гамогетеротопия быть фактором эволюции? Как полагает
В.А. Геодакян, может. По его мнению, эволюционная роль самцов состоит в
поиске новых приобретений, а женского - в сохранении уже найденного и
апробированного; передача информации идет через половые хромосомы
(Гэодакян В.А. И Генетика, 1998, № 8). Насчет роли самцов дело обстоит не
536
вполне так: в опытах Шапошникова (ЭнО, 1966, № 1, с. 5-6) и Бородина
(Природа, 2005, № 2, с. 37) поисковая роль самок оказалась выше, чем сам-
цов, - но само указание на гамогетеротопию видится мне важным.
Термина «гамогетеротопия» у Геодакяна нет, его ввел ранее Мейен в
своей гипотезе происхождения цветковых: мужской синангий (сросшийся
спорангий) голосеменного растения мог, по его схеме, мутировать в семяза-
чаток (т.е. в женский орган) простейшего цветкового. Существенно, что это
могло произойти только сразу, скачком.
Было указано и то конкретное семейство (беннетитовые из вы-мершего
класса цикадопсид подтипа голосеменных), чьи синангии расположены при-
мерно так же, как семязачатки самого примитивного цветкового, имевшего
плод в форме листовки (Мейен С.В. //ЖСБ, 1986, № 3).
Недавно палеоботаник А.Б. Герман, ученик Мейена, сделал эту гипоте-
тическую схему достаточно реальной, указав недостававшее прежде звено
- нижнемеловой беннетит Leguminanthus. Он любезно предоставил мне ма-
териалы своего выступления на Чтениях памяти Мейена (ГИН, декабрь 2005
г.), откуда и привожу один из рисунков (рис. 72).
Важно, что даже такой крупный акт прогресса, как рождение нового (по-
следнего в истории жизни) подтипа, ему удалось описать как сборку преж-
них блоков. Это значит, что в древе высших растений (рис. 61) удалось за-
крепить самую мощную ветвь (цветковые), дотоле висевшую отдельно, и
сделано это чисто диатропическим (а не филогенетическим) приемом. Ра-
зумеется, что возникнув, цветок стал новым органом (блоком), а потому, пе-
реходя к анализу цветковых, надо ввести новый мерой (строку рефренной
таблицы), который и будет определяющим для нового таксона.
Рис. 72. Возможное
происхождение цветко-
вых из беннетитов (по
А.Б. Герману)
9-13. Блочность и
эволюция
позвоночных
Истоки типов обычно
теряются в историчес-
кой дали, так что нет
надежных данных о
происхождении типа, к
которому принадлежим
мы - типа хордовых.
537
Первые хордовые документированы в раннем кембрии Китая (п. 8-9) и по-
хожи на нынешнего ланцетника, но уже сложнее его. К тому же ланцетник -
организм нынешний и являет следы прежней эволюции (его хорда состоит
из хрящевой и мышечной тканей, его симметрия изменяется в ходе роста,
напоминая метаморфоз камбал), а в ископаемом виде не встречается. От-
несение его к предкам позвоночных - лишь гипотеза.
Первые позвоночные засвидетельствованы в позднем кембрии - это
класс бесчелюстных. К нему относятся и ныне живущие круглоротые - ми-
ноги и миксины, но они тоже несут следы долгой эволюции: миноги облада-
ют сложным жизненным циклом (мигрируют из моря в реку для нереста), а
миксины - сложным поведением (могут завязываться в узел).
В отличие от нынешних, древние бесчелюстные несли покров из кож-
ных костей, некоторые из которых послужили потомкам основой для фор-
мирования челюстей. Как произошло преобразование челюстей, мы не зна-
ем, но у первых рыб (ранний девон) видим очень сложные челюсти, позже
упростившиеся. У ранних рыб и ранних амфибий нижняя челюсть имеет 9
костей, а у нас с вами - только одну. Опять налицо комбинация блоков и за-
тем их интеграция в новом органе, а вовсе не постепенное усложнение.
Любопытны примеры перекомпоновки блоков. В черепе кистеперых рыб
(ранний девон) и ранних амфибий (вероятно, произошли от кистеперых в
позднем девоне) есть теменное отверстие для иннервации третьего
глаза (у высших тетрапод он редуцирован, и отверстия нет). Можно ли ска-
зать, что отверстие унаследовано? Да как сказать - у рыб оно расположено
между лобными костями, а у амфибий - между теменными. Опять - унасле-
дована идея? (Ср. п. 5-15*.)
После полувекового спора выяснилось, что в эмбриогенезе хвостатых
амфибий фигурируют 3 пары костных зачатков, которые в прошлом, види-
мо, могли соединяться как в пару лобных, так и в пару теменных костей в
различных комбинациях {Дзержинский Ф.Я. Сравнительная анатомия по-
звоночных. М., МГУ, 1998, с. 40-45). В ходе эволюции структурные блоки
(кости) выстроились по-иному и иначе окружили определенный нерв. То
есть унаследован план строения, хотя сами компоненты поменялись.
Тут перестройка блоков напоминает вариацию строительного проекта,
где прежняя конструкция достигается иной укладкой кирпичей: по наследст-
ву передается идея, а ее реализация в материи формируется заново.
Весьма загадочно появление у рыб их кистепёрости, т.е. первое появ-
ление членистых лучей конечностей. У кистепёрых было три пары конечно-
стей. Первой вероятно возникла задняя, расположенная в вертикальной
плоскости (рис. 73А), т.е. ни на что не годная. (Дзержинский нашел здесь
смысл: «действуют... подобно оперению стрелы»; но даже если и допустить,
что “стрела” полезна, то членистое строение ей явно ни к чему.)
В пп. 4-10 и 8-11* мы уже видели, что эти плавники были излишне
сложны и упростились, когда стали использоваться по назначению. Говоря
538
языком нормы (л. 5-13), их строение было стабилизировано давлением но-
вой нормы.
с костными лучами
Небольшой тоз не прикреплен к позвоночнику
----Передняя конечность
Отделение черепа с Авеятипалой стопой
от плечевого пояса
с девятиполой стопой
Более крупный тоз
прикреплен к позвоночнику
Рис. 73. Пример преадапта-
ции: появление хорошо
развитых конечностей, кото-
рыми нельзя было воспользо-
ваться.
А (сверху): кистепёрая рыба
Eusthenopteron (верхний девон
Канады и Европы).
Б (снизу): первая амфибия
Acanthostega (верх, девон
Гренландии, на 20 млн лет
позже). По: КлэкДж., 2006, с
изменениями
Однако обладатели
излишнего блока не исчезли
в борьбе за жизнь, а
распространились по всему океану и тиражировали блок в горизонтальной
плоскости (смена симметрии); грудная и брюшная пары членистых плавни-
ков стали прообразом лап четвероногих. Можно спорить, могли ли кистепе-
рые ползать (ныне ползающие рыбы имеют простые плавники, а не члени-
стые), но несомненно, что пользоваться своими “кистями”, скрытыми в тол-
ще тела плавников, не могли. Это - классический пример преадаптации
(п. 4-10), т.е. приспособления, которое возникает на будущее; мерой «кисть»
реализован впервые не там и не тогда, где мог быть использован, а гораздо
раньше. О преадаптации надо сказать подробнее.
9-13* Преадаптация
Она противоречит не только идее отбора, но и вообще идее приспособ-
ления как процесса, как чего-то текущего во времени. Недаром Зусманов-
ский видит в ней проявление «принципа опережающего отражения действи-
тельности» (п. 6-14). Сам феномен преадаптации отрицать невозможно, и
обычно ее признают без лишних рассуждений [Ивахненко, Корабельников,
с. 151]. Если же сторонники чисто приспособительного понимания эволюции
пытаются ее объяснить, то они вынуждены идти на очередую подмену по-
нятий, вплоть до противоречия самим себе. Так,
“часто можно встретить термин преадаптация. Он обозначает ситуацию, при
которой признак или группа признаков возникли под влиянием отбора, по-
скольку обладали непосредственным адаптивным или селективным преиму-
ществом; впоследствии, когда условия изменятся, эти же самые признаки
539
окажутся еще более благоприятными... Например, легкие... по-видимому,
возникли до того, как эти животные перешли к жизни на суше... они должны
были быть (и были) адаптивными уже в первоначальной водной среде. Их по-
следующее использование животным на суше было в некотором смысле сча-
стливой случайностью - пример, так сказать, прозорливости эволюции” [Ро-
мер, Парсонс, т. 1, с. 25].
Счастливой случайностью авторы сочли и появление лапообразных
плавников кистепёрых (там же, с. 81), но слово «преадаптация» обозначает
совсем не то, что тут сказано. Последние слова цитаты перечеркивают всю
цитату, ибо прозорливость - не итог ни отбора (в любом смысле), ни даже
подбора по Богданову. Если она и итог, то - процесса приспособления к бу-
дущей потребности. Нарушен привычный принцип причинности.
Прекрасный пример преадаптации являет первая амфибия аканто-
стега (поздний девон Вост. Гренландии - рис. 73Б). Она имела лёгкие и
жабры, а также четыре лапы с очень сложным скелетом, но негодные для
движения по суше из-за отсутствия голеностопного сустава.
Английский палеонтолог Дженнифер Клэк пишет: 50 лет назад Ромер
«высказал мысль о том, что рыбы... остававшиеся на мели в периоды засухи,
за счет мускулатуры своих плавников медленно переползали к другим водо-
емам», т.е. что «рыбы вышли на сушу до того, как у них развились конечно-
сти» (КлэкДж. Трудный путь на сушу // ВМН, 2006, № 3, с. 53).
Но данная мысль, вполне в духе Ламарка (п. 2-5), теперь отвергнута:
«было обнаружено много новых окаменелостей, которые... полностью пере-
вернули прежние представления о происхождении и эволюции ранних тетра-
под. .. Акантостега имела конечности, оканчивающиеся пальцами, но эти ноги
не были приспособлены для наземного существования... акантостега была
проамфибией, т.е. лишь “зачатком” четвероногого животного, полностью
адаптирован[ым] к жизни в воде... Всё меньше подтверждений находила ги-
потеза, согласно которой выход на сушу сопровождался формированием ко-
нечностей со ступнями. Напротив... конечности тетрапод сформировались
тогда, когда они обитали в воде» (КлэкДж., с. 53-54).
Итак, первое четвероногое жило в воде и почему-то отрастило себе не-
пригодные для дела ноги. Даже амфибия с гораздо лучше развитыми ко-
нечностями, ихтиостега, которая жила тогда же и там же, еще не могла
выходить на сушу. Она получила шуточное прозвище «рыба на ногах», по-
скольку имеет четыре пятипалых ноги, но рыбьи череп и хвост (рис. 74).
Опять вместо постепенности видим блочность.
Рис. 74. Ихтиостега
(“рыба на ногах”;
поздний девон)
540
Поскольку здесь мы имеем длинный ряд переходных форм от кистепёро-
сти к четвероногости (на сегодня известно 6 стадий), показывающих посте-
пенное развитие лап, всё еще не годных для выхода на сушу, то именно
данный ряд удобен для уяснения преадаптации. Да, нога формировалась
постепенно в течение десятков миллионов лет, но не путем приспособле-
ния, ибо польза сказалась позже; кроме того, эволюция здесь шла не путем
вытеснения лучшими худших, а путем сокращения численности - все 6 ста-
дий представлены единственными экземплярами (находками), несмотря на
сто лет целенаправленных экспедиционных поисков.
Налицо реализация затруднявшей жизнь тенденции, которая состояла
в усложнении трех блоков (плавник-ласт, позвоночник, часть черепа) при
отставании остальных блоков (например, другие плавники, хвост).
Остается напомнить, что преадаптация - одно из основных понятий но-
могенеза еще у Берга (п. 4-10). Новый номогенез добавляет, что она - част-
ное следствие упорядоченности диасети (п. 6-3**): так, все стадии станов-
ления четвероногости суть клетки диасети, существующие независимо от
того, реализованы они материально, или нет (как существуют все элементы
таблицы Менделеева, в том числе еще не синтезированные). Недаром
акантостега и ихтиостега (разные стадии) жили одновременно.
Словом, проблему преадаптации предстоит решать не заменами слов,
а серьезным размышлением о философских основах эволюционизма; наив-
но ожидать незыблемости старого принципа причинности в биологии, если
он подвергнут сомнению и пересмотру в физике. См. Дополнение 7.
Иногда блочность дарит природе то, что никогда не найдет применения,
экстраадаптацию (т.е. нечто вне адаптации). Такова рыба ливониана,
жившая в девоне Вост. Гренландии за 15 млн лет до первых тетрапод: от
нее сохранилась нижняя челюсть, и вместо обычных двух рядов зубов
«у ливонианы зубная формула состояла из семи рядов. Такое расположение
напоминает кукурузный початок, и непонятно, чем нужно было питаться,
чтобы такое строение челюстей было закреплено отбором» (Клэк, с. 57).
На самом деле тут нет оснований полагать не только отбор, но и вооб-
ще приспособление. Вернее допустить, что зубные блоки были собраны в
странную челюсть за счет акта изменчивости (она здесь, как и в концепции
естественного отбора, никуда не направлена), который удобно назвать мак-
ромутацией, и обнаружение одного экземпляра, не ложащегося ни в какую
тенденцию, говорит лишь о том, что обладатель челюсти смог дожить до
взрослого состояния в силу своей экологической прочности.
Вместе они (пре- и экстраадаптация) позволяют понять суть дела: в хо-
де эволюции реализуются самые различные клетки диасети. Какие именно
клетки возможны, ясно только из опыта, но столь же ясно, что сами эти воз-
можности (если не все, то очень многие) существуют до опыта: это видно из
многократного появления одних и тех же блоков в ходе эволюции разных
групп (такова, например, многопальцевая многофаланговая кисть). Вариан-
541
ты, затрудняющие жизнь, крайне малочисленны, но могут служить переход-
ными стадиями к более успешным вариантам.
9-13** Блочность и появление амниот
Следующее важнейшее достижение позвоночных: амнион (оболочка
яйца, предохраняющая его от высыхания). По его наличию позвоночных де-
лят на высших (амниот) и низших. Обычно эту грань проводили между ам-
фибиями и рептилиями, но новейшие данные говорят, что и тут имела ме-
сто блочность: амниотическая организация тела впервые проявилась еще у
стегоцефалов середины карбона (башкирский ярус), а настоящее амниоти-
ческое яйцо известно лишь из нижнего триаса.
И в становлении наземной организации мы снова видим блочность: не-
смотря на черты наземного облика, похожие на рептилий стегоцефалы об-
рели ряд наземных черт (например, барабанную перепонку в ухе) лишь в
конце перми; притом перепонка возникала как минимум трижды (Laurin М.
La conquete de I’environnement terrestre par les vertebras // LEG, 31-32). Факт
многократности обретений исключает ссылку на редкость нужных мутаций -
здесь тоже налицо номогенез.
Блочность эволюции — причина того, что вопрос о предках приводит к
нескончаемым спорам и переделкам систем. Каждый систематик строит
систему по различиям лишь некоторых органов, и устойчивость системы
достигается там, где удается договориться, какие органы брать (см. гл. 10).
Таковы, например, птицы: общепризнано, что определяющим для всего от-
ряда является наличие перьев. Именно благодаря отпечаткам перьев был
отнесен к птицам знаменитый археоптерикс (верхняя юра):
“в строении скелета археоптерикса так равномерно представлены признаки
архозавров и современных птиц, что его положение в системе оставалось бы
неясным, если бы вместе со скелетом не сохранились бы отпечатки перьев”
[Ромер, Парсонс, т. 1, с. 97].
Но недавно выяснилось, что перья были и у небольших динозавров; у
них «локализация перьев ограничена головой, хвостом или вдоль позвоноч-
ника» (Lange-Barde В. Temoin celebre... // LEG, 26). Т.е. блок “перо” возник не
как адаптация к полету, а, вероятно, как преадаптация к нему:
“Эволюция показывает, что нет корреляции между наличием перьев и поле-
том, перья предшествуют ему на многие десятки миллионов лет. Участие
перьев в полете - итог изменения функции предсуществующей структуры”
(Lange-Barde В., там же).
В китайской провинции Ляонин (к северу от Порт-Артура) открыто более
10 новых видов раннемеловых (на 20 млн лет младше археоптерикса и пре-
орниса) динозавров подотряда теропод с хорошо сохранившимися отпечат-
ками перьев. Sinosauropteryx (“китайский пернатый ящер”) имел перья в ви-
де бесполезных зачатков, они располагались по всей коже и выглядели у
взрослых так же, как выглядят зародыши перьев у птицы — полые конусы. У
542
других ящеров нашли разные формы перьев, соответствующие разным
стадиям развития и типам перьев нынешних птиц. Некоторые даже напоми-
нали маховые перья, служащие для полета, но тоже бесполезные — напри-
мер, на концах лап и хвоста (Прам Р., Браш А. Динозавры или птицы: кто
оперился первым? И ВМН, 2003, N 7). К сожалению, не сказано, как находки
расположены вверх по разрезу, т.е. можно ли уяснить стадии эволюции пе-
ра или перед нами просто спектр возможных вариантов (разнообразие), но
и без того ясно, что налицо повторное рождение пернатости, т.е. номоге-
нез. Если окажется, что вверх по разрезу перо усложняется так же, как в он-
тогенезе птиц, то налицо еще и жоффруизм.
Блочность видна также и в феномене двуступенчатого эмбриогенеза
млекопитающих (п. 8-12*), и его эволюция понятна тоже с позиции жоффру-
изма [Сахарова]. Не будем, однако, думать, что с помощью блочности мож-
но описать всю эволюцию рептилий в теплокровных. Нет, теплокровные
прежде всего характерны тем, что в летописи отсутствует, - ТВ-
иммунитетом. Когда именно его становление происходило, мы не знаем, но
можем кое о чем догадываться (пп. 6-6, 8-12*).
По словам Ромера, систематика млекопитающих - это систематика не
животных, а их зубов. Другими словами, в качестве их филогении тут все
берут одну-единственную семофилогению - эволюцию зубной системы.
Иногда используются две семофилогении. Таков “парадный пример” дарви-
низма, эволюция копытных: сперва появились годные к бегу длинные ноги, а
затем - годные к перетиранию жестких трав зубы. Разное по времени фор-
мирование основных адаптаций известно как мозаичность эволюции; она
замечена английским палеонтологом Дэвидом Уотсоном в 1910-х годах и
является частным случаем блочности эволюции. Мозаичность есть, по всей
видимости, следствие поочередного приобретения возможностей (п. 5-11).
Блочность видна всюду, но в некоторых таксонах видна особенно хо-
рошо и удобна для показа на уроке. Среди млекопитающих это - вымершее
семейство халикотериевых из отряда непарнокопытных. Вот два рода ха-
ликотериев (олигоцен - плиоцен Старого Света): виды одного - с виду поч-
ти гориллы и с ногтями вместо копыт, но с почти лошадиными черепами
(рис. 75 а, б); а другой - с виду почти лошадь, но с почти кошачьими когтями
вместо копыт (рис. 75 в).
Недаром халикотериевые выделены в особый подотряд Ancylopoda
(греч.: непонятноногие). О том, к чему они таким странным образом приспо-
собились, до сих пор идет спор. У всех халикотериев очевидна тенденция -
ослабление зубов, и к ней они, полагаю, приспособились, кто как сумел:
первый объедал высокие растения, второй что-то выкапывал. Источником
блочной изменчивости можно, как обычно, считать генетический поиск: в
данном случае, он, видимо, начался в силу снятия давления нормы при по-
тере тех экологических ниш, которые требовали полноценных зубов.
543
Рис. 75. Халикотериевые (эти животные отнесены к непарнокопытным по строению че-
репа): а— Chalicotherium (миоцен Европы), быть может, ранняя попытка природы создать
нечто вроде человека; б — Borissiakia (олигоцен Казахстана) мог откидывать голову на-
зад; в — Moropus (миоцен Азии и Северной Америки), вероятно, имел втяжные когти
9-14. Блочность и Лазаревы таксоны
Блочность должна вызывать повторение одних и тех же форм. Всем из-
вестно, что кистепёрые рыбы вымерли в мезозое, но один их род - латиме-
рия (3 вида), ныне живет в глубинах Индийского океана; в ископаемом виде
не встречен. Принято писать, что эти кистепёрые сохранились с мезозоя, но
доводов в пользу данного допущения приводить не принято.
Менее известно нечто сходное у яйцекладущих зверей: они живут в Ав-
стралийском регионе, причем семейство утконосовых известно трижды: в
раннем мелу, в среднем миоцене и (нынешний утконос) с плиоцена; осталь-
ные нынешние яйцекладущие (два рода ехидн) известны с плейстоцена
[Кэрролл, т. 3, с. 208-209]. На других материках они известны не позже па-
леоцена; чем заполнить дыру в 40 млн лет, никто не знает. Известна мысль,
что нынешние утконос и ехидна могли заново произойти от сумчатых (у яй-
цекладущих есть сумка для яйца, а затем детеныша, и “сумчатые кости").
Отряд клювоголовых рептилий вымер в мелу, но один вид (гаттерия)
живет ныне (см.: Северцов А.С. И БШ, 1998, № 4, с. 8).
Самый же длинный (среди тканевых) разрыв летописи дают бесчереп-
ные: между Pikaia (кембрий) и нынешним ланцетником.
Подобных примеров десятки (см. обложку, а также: ЧЭ, с. 396), и есте-
ствен вопрос: сохранились ли “живые ископаемые” с прошлых эпох, но до
сих пор их посредники не обнаружены, или возникли заново? Не решая во-
544
прос прямо, палеоботаник В.А. Красилов (ПЖ, 1999, № 3) предложил, одна-
ко, обозначение для повторно появившихся таксонов - Лазаревы группы,
от имени библейского персонажа, воскресшего Лазаря, и привел примеры
из палеоботаники. Признанием таких групп отрицается «закон Долло», и
прежде весьма сомнительный - о нем см. пп. 4-17*, 6-13*.
По сути, лазаревость - частный случай полифилии (независимого появ-
ления сходных форм), а именно, случай, когда точно известен разрыв групп
во времени. Так, появление первоптиц отмечено дважды во времени:
Protoavis (верхний триас, Техас, 3 экземпляра) и Archaeopteryx (верхняя
юра, Бавария, 8 экз.); их относят к разным подклассам. Разрыв в 35 млн лет
позволяет допустить, что перед нами повторное возникновение класса птиц
(независимая реализация близких клеток диасети).
Хотя непонятно, от кого могли произойти нынешние латимерии, но это
еще не основание считать их происшедшими от триасовых кистепёрых. С
диатропической точки зрения скачок в пределах рефренной таблицы понят-
нее, чем скачок через сотни миллионов лет и через материки, не оставив-
ший следов в ископаемых. Встретив выражение «живое ископаемое», все-
гда надо задать себе вопрос: действительно ли данный род пережил многие
миллионы лет без значительных изменений или же возник заново, повторив
своими основными чертами давно вымерших предков?
К первому варианту относятся, например, род гинкго (Ginkgo, голосе-
менные), известный с мезозоя (а возможно даже с перми), найденный во
многих более поздних слоях и представленный одним видом в нынешней
флоре (культурное растение многих теплых стран); и род двоякодышащих
рыб рогозуб (Neoceratodus), до сих пор живущий в двух реках Австралии, в
которой его кости находят также в древних слоях, начиная с верхнего мела.
Но чаще наблюдается второй вариант: “живое ископаемое” в ископае-
мом состоянии неизвестно вообще или обнаружено лишь в молодых слоях
(голоцен), тогда как его ближайшие “родственники” жили на другом матери-
ке и вымерли давным-давно. Этот факт вынуждает задать еще один вопрос:
можно ли уверенно говорить о непрерывной линии предок-потомок в случа-
ях, когда перерывы в летописи невелики - миллион лет и меньше?
Вопрос актуален при изучении эволюции человека: следов культурной
деятельности известно много больше, чем ископаемых костей, поэтому вот
уже второй век возникает, пропадает и вновь возникает вопрос - в самом ли
деле следы культуры оставлены только нашими прямыми предками.
9-15. О происхождении человека
Ныне известно, что человек (род Ното) произошел в Восточной Афри-
ке. Процесс шел на удивленье медленно: 2 млн лет наши предки ходили
прямо, но не оставляли никаких орудий труда, затем еще около миллиона
лет оставляли всего один тип орудий - заостренные камни. Считается, что
этими орудиями никаких изделий не изготовлялось, что они служили только
545
для разбивания крупных костей. Лишь много позже (считается, что 50 тыс.
лет назад) наши предки стали оставлять изделия: наконечники копий, гли-
няные горшки и т.п.; началось то, что принято называть палеолитом (древ-
ним каменным веком). И тут эволюция пошла поразительно быстро.
Появление человека - тема, требующая отдельного разговора, и здесь
мы ограничимся тем, что нужно для завершения нашего экскурса в эволю-
цию. Обычно пишут о форме костей и об одной проблеме - как выстроить
предков человека в единый исторический ряд. Это геккелева традиция.
В силу транпола и блочности любая филогения будет произвольной и
недолговечной. Пока об ископаемых знали мало, их филогении строили лег-
ко (рис. 76а), но к концу XX века от них ничего, кроме слова «филогения», не
СОВРЕМЕННОСТЬ
Гиббоны Шимпанзе
Сиаманги Орангутаны Гориллы
,, Монголоиды
Негроиды
Авсгралэиды Европеоиды
ПЛЕЙСТОЦЕН
200 000 >тет_
ПЛИОЦЕН
450 000 лет
Дриопитек
МИОЦЕН
900 000 лет
Неопигек
Ппиопйтск
Неандерталец ‘у
1 Пекинский человек
I Литекангроа L
t (Кизса)
Австралопитек
(Дарта)
СОВРЕМЕННЫЙ
““ЧЕЛОВЕК
Пилтлаунский
че ювек
ВЕТВЬ НЕБОЛЬШИХ
ПРЯМОХОДЯЩИХ
ПРИМ А ГОЙ
ВЕТВЬ КРУПНЫХ
АНТРОПОИДОВ
---ЧЕЛОВЕЧЕСКАЯ
BFI ВЬ
ОЛИГОЦЕН
1 509 000 лет
ОБЕЗЬЯНЫ
СТАРОГО
СВЕТА
КРУПНЫЕ
-ПРЯМОХОДЯЩИЕ
ПРИМАТЫ
ОБЕЗЬЯНЫ
НОВОГО света
ЭОЦЕН
ОБЩИЙ
СТВОЛ
2 (00 000 лет
Рис. 76. Родословные человека: а) красивое древо (по: А. Кизс, 1931);
осталось, а недавняя (2002 г.) схема эволюции гоминид55 (Senut В.
Philogenie ou philiogenies: exemple des Hominides // LEG, 37-39) являеся на-
бором точек и чёрточек (рис. 766). По-моему, принцип филогении себя ис-
черпал (мы вернемся к этому вопросу в п. 10-2). 8
8э Гоминиды - семейство высших приматов, в котором ныне остался один вид - человек.
546
Ему противостоит (тоже со времен Геккеля) призыв видеть в эволюции
гоминид не древо, а сеть.
Вариант филогении гоминид
1 = Ardipithecus ramidus ramidus
2 Ardipithecus ramidus kodobba
3 — Sahelanthropus tchadensis
A = C>rrorin tugenensis
5 = Praeanthropus africanus
(= Australopithecus anamensis
-+- A. afarensis pro parte)
b — Australopithecus antiquus
(= A. afarensis pro parte}
7 = Australopithecus aethiopicus
8 ™ Australopithecus boisei
9 = Australopithecus robustus
1 О — Australopithecus africanus
1 1 -= Australopithecus bahrelgha-
zali
12 = Australopithecus garhi
13 = Kenyanthropus platyops
14 = Homo rudolfensis
15 = Homo habilis
1 d> = Homo ergaster
17 = Homo erectus
18 = Homo georgicus
19= Homo antecessor
20 = Homo heidelbergensis
21= Неандертальцы
22 = Нынешние люди
Рис. 766: вот что осталось от верхней части древа к нашему времени (Senut В. И LEG,
38). Замечу, что и нижняя его часть выглядит теперь так же мало похожей на древо;
соответствующий рисунок есть в книге: Джохансон Д, Иди М. Люси. Истоки рода че-
ловеческого. М., Мир, 1984; см.также: БПС, 2000, N 32, с. 16
Спор этих традиций верно называют беседой глухих (Clark G.A. Origine
de I’homme: Le dialogue des sourds // LR, 1994, № 263), и вступать в него не
будем; важнее становление механизмов. Даже если вообразить, что древо,
ряд или сеть предков человека выстроены, основная, на мой взгляд, про-
блема даже не будет поставлена: как возникло человеческое мышление?
В эволюции человека явственны два аспекта: происхождение биологи-
ческого вида, которое мы можем пытаться понять в рамках всего, о чем шла
речь ранее, и появление того, чем этот вид в корне ото всех отличается, что
пришло в мир только вместе с ним. Мышление и экологическая универсаль-
ность настолько отличают людей от остальных существ, что человека не
раз выделяли в отдельное царство живой природы. Оставляя этот вопрос
до главы 10, здесь заметим одно: даже в рамках традиционной зоологии
видны две совсем разные стадии очеловечения.
547
Первая стадия шла в тропиках, и возникшее при этом семейство гоми-
нид расселилось далеко на север; согласно концепции биотогенеза (п. 9-4*),
это характерно для становления высших таксонов. Наоборот, вторая ста-
дия - становление вида Homo sapiens, шла в средних широтах под воздей-
ствием локальных катастроф — периодического наступления ледников.
(Замечу, что неандерталец и кроманьонец считаются ныне подвидами од-
ного вида -Homo sapiens.) Сперва поговорим о первой стадии.
Системолог и натурфилософ Ю.А. Урманцев в 1970-х годах показал,
что крупные биологические приобретения связаны с диссимметрией, т.е. с
искажением симметрии (но не с ее уничтожением - не надо путать диссим-
метрию с асимметрией, т.е. с отсутствием симметрии) объектов (Урманцев
Ю.А. Симметрия природы и природа симметрии. М., 1974).
Тогда же историк Б.Ф. Поршнев, к концу жизни ставший антропологом,
верно отметил роль диссимметрии больших полушарий мозга. Она, по его
мнению, играла ведущую роль в антропогенезе. Хотя она известна и у жи-
вотных, но у человека она принципиальна: речевой центр расположен в ле-
вом полушарии. Вопреки царившей тогда догме марксизма, гласившей:
«труд создал человека», - Поршнев считал главной в очеловечении речь.
“Увы, почти всякий современный автор, рассуждая о роли труда в становле-
нии человека, подразумевает именно единичную особь, манипулирующую с
материальными предметами”, а “отдельные упоминания о социальных отно-
шениях остаются чисто словесным придатком”. Речь же социальна по сути
(Поршнев Б.Ф. О начале человеческой истории (проблемы палеопсихологии).
М., Прогресс, 1974, с. 428).
Вскоре химик А.В. Камерницкий высказал общую догадку: жизнь
“использует два языка наследственности. Один, известный нам, основан на
последовательной записи “букв” генетического кода... А в другом, еще со-
вершенно не изученном, используются... скорее принципы пиктографическо-
го письма, где каждый знак выражает сразу целое сложное понятие” (Жвирб-
лис В.Е. Два языка жизни // Техника молодежи, 1990, №11).
Почти о том же шла речь у нас в п. 5-5*. Два “языка” известны и в чело-
веческом сознании, где связаны с диссимметрией мозга: за интуитивное
мышление ответственно, в основном, правое полушарие, а за понятийное -
левое. Роль диссимметрии часто затрагивается в связи с различием муж-
ского и женского (еще Дарвин “выводил” эволюцию интеллекта из предпоч-
тения глупыми самками наиболее умных самцов - см. п. 3-3).
Известно, что женщины часто превосходят мужчин в интуитивном мыш-
лении, а мужчины - в понятийном. Недавно Паршин (о нем см. п. 5-6) вы-
сказал, беседуя со мной, догадку, что обе эти диссимметрии связаны и ле-
жат в основе процедуры познания людьми мира - как внешнего, так и внут-
реннего. По его мнению, мир так устроен, что для познания его требуется
взаимодействие интуитивных и понятийных процедур: правое полушарие
схватывает суть явления или процесса, а левое разворачивает найденное в
548
последовательность слов (в то, что можно сказать и запомнить). Что можно
извлечь из этого для понимания эволюции?
Вероятно, что у человека передача новаций идет не только между по-
лами, но и между полушариями одного мозга. Та идея Паршина, что для
становления понятийного мышления нужны две “матрицы", видится очень
глубокой и далеко идущей. Если интуитивное схватывание сути возможно,
значит мир устроен так, что в нем можно по части восстанавливать целое.
Это свойство мира подмечали многие и давали ему различные назва-
ния. Выше оно названо упорядоченностью диасети (п. 6-3**). Именно упо-
рядоченность диасети делает возможным прогноз. К нему способны и жи-
вотные (опережающее отражение действительности, см. п. 6-14).
Если так, то человек возник в тот момент, когда научился давать опе-
режающему отражению действительности понятийно-речевое выражение.
Как это произошло, мы не знаем, но известно, что для этого понадобилось
не только сформировать речевой центр в левом полушарии, но и разделить
функции полушарий так, что за опережающее отражение ответственно пра-
вое. При этом очеловеченье предстает как изобретение (по Тейяру) той
конструкции мозга, при которой полушария взаимодействуют так, как взаи-
модействуют женская и мужская части вида. В математике существует ап-
парат, теория двойственности, которым, как надеется Паршин, можно бу-
дет все это описать, но это - задача для будущих эволюционистов.
Нам остается снова (как в гл. 6) допустить, что блоки, приобретенные в
ходе становления адаптивного иммунитета, были использованы при созда-
нии памяти, а затем и психики вообще. Об этом еще в 1987 году писал
Эдельман (о его топобиологии мы говорили в п. 5-15), и Стивен Роуз (анг-
лийский нейробиолог) пишет об этом: по Эдельману,
“развитие нервной системы и ее способность изменять свои свойства под
влиянием индивидуального опыта... следует рассматривать как процесс не-
прерывного отбора предсуществующих групп нейронов и их синаптических
связей” (Роуз С. Устройство памяти. М., Мир, 1995, с. 355).
Сам Эдельман назвал это «нервным дарвинизмом», но Роуз протестует
против такой параллели. В самом деле, лучше говорить не про отбор ней-
ронов, а про их активность (о ней см. п. 8-12**), или подбор в смысле п. 6-12,
т.е. про самоорганизацию.
9-15* Шесть тенденций очеловеченья
Как сказано в п. 8-11, очеловеченье приматов было итогом завершения
совместного действия двух общих эволюционных тенденций (террипетность
и цефализация). Теперь надо сказать о других тенденциях. Третьей общей
тенденцией, приведшей к очеловеченью, можно назвать эмбрионизацию
(пп. 4-15* и 5-11 *). Она в чем-то сходна с террипетностью:
“Эмбрионизация является средством перехода из более простых в более
сложные условия существования, она увеличивается при переходе а) из мор-
549
ской воды в пресную; б) из пресной воды в почву; в) из почвы на ее поверх-
ность” (Захваткин А.А. Сб. науч, работ. М., 1953, с. 376).
Напомню, что у человека эмбриогенез идет дольше, чем у других при-
матов, а особенно долго - взросление подростков.
У млекопитающих есть четвертая, более частная, тенденция - “эмбрион
в личинке" (п. 8-12*), которую Сахарова сочла важной и для человека. Воз-
можно, это не тенденция, а общее свойство всех плацентарных.
Пятой можно назвать тенденцию приматов к двуногости (Dambicourt-
Malasse A. Nouveau regard sur I’origine de I’homme // LR, 1996, № 286).
Наконец, шестая тенденция: особая роль половой активности. Прима-
там свойственно размножаться круглый год, это толкуют как особое приспо-
собление, повышающее размножаемость (что не мешает толковать сезон-
ное размножение остальных животных тоже как приспособление), но для
очеловеченья важнее другое - появился избыток активности.
У животных с внесезонным размножением есть
«избыточная энергия, которая может тратиться на исследование обстановки и
ее деталей, на манипулирование, на игровую деятельность... и другие формы
поведения» [Алексеева, с. 105].
Но если у обезьян самки доступны для самцов лишь несколько дней из
каждого полового цикла, то женщины приобрели как бы мужское свойство
постоянной способности к половому акту. В итоге
“сексуальность человека не имеет аналогов в мире животных. Половые сно-
шения у человека стали возможными во все фазы цикла, во время беременно-
сти и лактации, они стали не столько служить своему биологическому назна-
чению, сколько превратились в самоцель, в способ получения удовольствия.
Это весьма типично для человека. Ведь только Человек сделал удовольствие
из утоления голода (гурманство) и жажды..., превратил в источники удоволь-
ствия тяжелые физические действия (туризм, альпинизм, спорт). Только Че-
ловек способен получать удовольствие от труда, от науки, от различных ви-
дов творчества” [Алексеева, с. 176].
Тут верно не всё (многие животные совершают трудоёмкие и опасные
действия ради удовольствия), но основное отмечено: человек покорил мир с
помощью избыточной энергии. Поскольку самцы обезьян в половом отно-
шении сходны с мужчинами, можно полагать, что это сходство обще для
всех гоминид и потому работало в ходе всего антропогенеза. Самки стали
такими намного позже (когда - неизвестно), и до этого самцы вынуждены
были тратить избыток половой энергии на внеполовые цели.
Механизм, запустивший взаимодействие всех шести тенденций, состо-
ял, если следовать Аршавскому, в стрессе, вызванном изменением условий
жизни и заставившем наших предков повысить поведенческую (прежде все-
го двигательную) активность и изменить сам ее характер. Изменения усло-
вий были весомы: похолодание в плейстоцене повлекло сокращение лесов
в тропиках и оледенение в странах, прежде умеренных. К сожалению, неяс-
550
но, что такое стресс в течение сотен поколений, но про снятие давления
нормы говорить можно и тут.
Очеловеченье состояло в появлении никак не прямохождения (оно бы-
ло у других приматов, у ящеров, птиц, и, возможно, у халикотериев), и не из-
готовления орудий (рубила перволюдей гораздо проще плотин бобров или
тлей в рабстве у муравьев), а в появлении культуры и понятийного мышле-
ния. Их надо отличать от социальной организации. Последняя очень высока
у высших обезьян и тоже связана с отношением полов, что показал, напри-
мер, голландский этнолог Франс дэ Вааль {de Waal F.D.M. Chimpanze
politics. Power and sex among apes. Baltimore - London, 1998). Но даже у
шимпанзе самки резко отличны половыми возможностями от женщин, и ес-
тествен вопрос: на какой стадии эволюции (до появления понятийного
мышления, одновременно с ним или позже) самки обратились в женщин?
Никаких данных на сей счет мне не известно.
Хотя очевидно, что человеческое сознание возникло в ходе взаимодей-
ствия указанных шести тенденций (возможно и иных), но остается совер-
шенно неясным, как именно это было. Полагаю, что сознание возникло так
же, как у кистеперой рыбы возникли ее сложные плавники: само по себе, на
будущее. Преадаптациями полна эволюция, и одни видят здесь волю бо-
жью, другие - рациональное поведение Геи, третьи - реализацию объек-
тивно существующих рефренов, четвертые - ограниченность принципа при-
чинности, а пятые (дарвинисты) ничего не видят.
Одно можно сказать определенно: люди возникли из стадных животных
с очень сложной организацией стад, но вне связи с конкуренцией внутри
стад и борьбой самцов за самку5^. «Большинство авторов считают основ-
ной силой, объединяющей приматов в стада, тяготение к себе подобным»
[Алексеева, с. 172]. Это значит, что никакой “проблемы социогенеза” в ходе
антропогенеза нет, а есть противоположная проблема - как в стаде смог
возникнуть мыслящий индивид? (См. далее, п. 9-16**).
Когда и сколько раз? Принято считать, что очеловеченье шло в по-
следние 3 млн лет. Однако есть давняя традиция считать, что что некие су-
щества, обладавшие культурой каменного века, жили в третичном периоде
{Павинский А., 1882 - см. далее, п. 9-15***), даже целых 50 млн лет, т.е. в
течение основной части кайнозоя. Почти все антропологи отказываются да-
же читать о таких данных, заранее утверждая, что они либо ошибочны, либо
подложны - ибо общепринятый каменный век в тысячу раз короче (начался
50 тыс. лет назад), а возраст всего рода Ното оценивают в 3 млн лет. Но
общепринятые взгляды на появление людей слишком часто приходится ме-
нять, а фактов большой их древности набралось слишком много. 86
86 Приматолог Л.В. Алексеева по всей книге подчеркивала, что обостренная борьба за
самку свойственна только видам с сезонным размножением, но не приматам.
551
Некоторые собраны в книге: Кремо М., Томпсон Р. Неизвестная исто-
рия человечества. М., Философская книга, 1999. Ее авторы - религиозные
деятели (кришнаиты)^7, однако многие из собранных ими данных вполне
научны в западном смысле: они взяты из старых научных журналов, где
многие датировки и определения видовой принадлежности ненадежны, но
совершенно ясно, что сами находки в целом никак не могут все быть
вымыслом. Особенно интересен американский ископаемый материал, ибо
принято считать, что люди появились в Америке всего 30 тыс. лет назад.
В указанной книге нет теории, там можно видеть лишь набор загадоч-
ных фактов, который мы сами должны осмыслить. Факты делятся на две
группы: очень много кайнозойских находок, начиная с эоцена, и небольшое
число более древних находок. Первые трудно, но можно пытаться вплести в
ткань привычной науки, так как нигде нет четких доказательств того, что
найден именно Homo sapiens (а значит, можно говорить о Лазаревых таксо-
нах и о независимых актах рождения зачатков культуры). Вторые надо или
начисто отрицать, или искать в нашей науке принципиальные изъяны.
Так, данный на с. 219 той книги снимок отпечатка триасового куска по-
дошвы обуви ставит в тупик: на оригинале в лупу виден след крученой нити
прошивки, уходящей в отверстия. Находку рассматривали многие, и ссы-
латься на подделку несерьезно (хоть и делается некоторыми). В триасе че-
ловек жить мог, но ему не из кого было произойти. Тут как раз можно гово-
рить о визите инопланетян; его принято отрицать, ибо “нет их следов”, одна-
ко именно следов, как видим в книге Кремо и Томпсона, предостаточно.
Этот труд обычная наука почти не заметила. Откликнулся лишь архео-
лог и культуролог Тим Мёррей (Австралия): он признал, что книга не смогла
дать картину становления человека, зато отметил, что она открывает новую
эру в науке: вера и наука как бы меняются местами:
«Кремо и Томпсон делают различие между их истинной и обоснованной ве-
рой и ’ненаучными’ взглядами их эволюционных оппонентов. Для них тако-
вы последователи эволюционной теории в силу их невежества, страха и сле-
пой веры»; «весомая работа Кремо и Томпсона ясно показывает, что теперь в
игру вступают другие» (Murray Т. И The British Journal for the History of
Science, 1995, № 3, p. 378).
Об этих “других” поговорим в главе 12, а сейчас расскажу об одной на-
ходке, которая осталась неизвестна кришнаитам.
Майкл Кремо в устных выступлениях (Москва, апрель 2003 г.) произвел на ученых
самое отрицательное впечатление: упирал на религию, не мог ответить ни на один науч-
ный вопрос, а в ответ на мою просьбу - рассказать о его соавторе Томпсоне, просто сме-
нил тему разговора. В основном высокий критический уровень книги при почти полном
отсутствии религиозной темы склоняет к мысли, что основным ее автором является Ри-
чард Томпсон. О нем ничего определенного мне неизвестно.
552
9-15** Ископаемые мозги
В 1925 году близ Москвы (пос. Одинцово) были найдены два кремневых
образца, которые точно воспроизводили форму человеческих мозгов, что
признали русские и западные анатомы55. Однако тогдашняя наука находки
отвергла. Их сочли “игрой природы”, не убоявшись ни самого этого позорно-
го термина, ни параллели с конфузами прошлого (см. п. 1-5).
Естественные слепки мозгов (эндокраны) известны для человека и мно-
гих животных, но не кремневые, а известковые. Мозги могли окремнеть в
почве (известно окремнение даже столь нежной материи, как медузы). Уче-
ные заявляли, что их отказ признать одинцовские находки за эндокраны вы-
зван именно их необычным материалом, но истинная причина отказа видит-
ся мне в другом - находки противоречили (и противоречат до сих пор) офи-
циальной антропологии, и их проще не замечать, чем обсуждать.
Дело в том, что в горизонте залегания (первое межледниковье плейсто-
цена) нынешнему человеку быть не полагалось: горизонт датировался воз-
растом 500 тыс. лет, а нынешний человек считался вдесятеро моложе.
Мозг “одинцовского человека” внешне неотличим от нынешнего, и если
признать его как факт, то получалось, что обладатель “нашего” мозга 400
тыс. лет не оставлял следов культурной эволюции. К тому же обычная ар-
хеология относит заселение Русской равнины людьми к позднему палеоли-
ту (40 тыс. лет назад). Вот и была принята простейшая (согласно «бритве
Оккама») версия: “мозги” - те же булыжники, что в обилии лежали рядом и
уверенно отнесены к карбону (палеозой). Идея явно нелепа (окатанный
ледником булыжник имеет простую овальную форму), но ее приняли.
Историк науки Г.Г. Кривошеина обратила мое внимание на архивное
дело, где геолог А.П. Павлов (о нем см. п. 4-10), два года уклонявшийся от
“одинцовской” темы, в августе 1927 года высказался о ней, поскольку его
вызвали с докладом в Главнауку. Павлов так резюмировал отношение уче-
ных к находке: хотя детальное сходство с мозгом несомненно,
“Но человек не существовал в каменноугольный период, и следовательно
данные геологии не позволяют признать кремневую массу, найденную в
Одинцове, окаменевшим человеческим мозгом”; впрочем, сходство с мозгом
“делает желательным дальнейшее всестороннее изучение” (Архив РАН, фонд
48, опись 1, дело 48, лист 2, оборот).
Вывод не красит его, и не стоило звать крупнейшего специалиста ради
этой пустой отговорки. Интереснее последний лист дела. Член Комиссии по
изучению находок А.Н. Розанов доложил в декабре 1927 года главное - что
в распиле большого образца нет карбоновой фауны; что поэтому надо
оо
°° Григорович Н.А., Гиндце Б.К., Яковлев С.Я. Окаменевшие мозги людей ледникового
периода. Вологда, Северный печатник, 1927. Об этой находке и этой брошюре подроб-
нее см.: Чайковский Ю.В. Юбилей загадки // БПС, 2000, № 32.
553
подвергнуть “гистологическому изучению те шлифы, в которых были обна-
ружены структуры, напоминающие несколько органическую ткань” и сделать
новые шлифы (там же, л. 24,24 оборот).
Дальнейшее мне неизвестно, но ясно, что признать находки “булыжни-
ком” не получилось, идей не было, ученые не знали, как быть, и охотно всё
забыли. Вот если бы возраст находок был 100 тыс. лет (т.е. ничего особо
интересного), их сочли бы важнейшим открытием и они вошли бы в учебни-
ки. Геохимики нашли бы им механизм окремнения, а о сходстве их с лежав-
шими рядом булыжниками карбона никто бы не завёл и речи - убого.
По-моему, образцы могли быть как естественными, так и искусственн-
ными слепками мозгов тех времен, когда жили их обладатели. Искусствен-
ные слепки могли быть сделаны с целью запечь мозги, чтобы съесть их.
Как говорилось (п. 3-7**), факт вне теории не имеет ни смысла, ни даже
места. Сказанное в предыдущих главах дает основание считать, что непри-
знанные тогда данные достаточно совместимы с нынешней наукой.
Во-первых: нет промежуточных форм к нему от неандертальца? Но по
всей книге мы видели, что существенная эволюция идет только скачками.
Во-вторых, согласно одинцовским находкам, у человека давно не меня-
ется мозг - тот орган, благодаря которому он стал человеком. Если осталь-
ное тело за то же время заметно изменилось, то налицо обычная мозаичная
эволюция: сперва меняется ведущий эволюцию орган, а затем остальные.
Первые 2 млн лет это был опорный скелет (становление двуногости), а мозг
развивался преимущественно в следующий миллион лет.
В-третьих, если эволюция - многократное заполнение клеток одной и
той же диасети, то человек мог возникать не раз. Идея Лазаревых таксонов
допускает, что выявленная кришнаитами «неизвестная история человечест-
ва» - это ряд попыток очеловеченья, точнее, овладения культурой. Попыт-
ки кончились неудачей и, видимо, вымиранием. Если человек - Лазарев род
или даже вид, то люди, которым принадлежали одинцовские мозги, вероят-
но вымерли, и в таком случае мы - не их потомки, а новая попытка природы,
оказавшаяся более результативной (я не говорю “удачной").
В-четвертых, примитивные и продвинутые формы древних людей жили
одновременно (рис. 766), так что мозг “одинцовского человека” не вступает
в конфликт с иными находками. Виды рода Ното употребляли огонь еще
1,4 млн лет назад, а 600 тыс. лет назад уже строили каменные жилища в
Средней Европе (Зубов А.А. //Современная антропология... М., 1995, с. 21).
В-пятых, из Эфиопии известен череп Homo sapiens возрастом в 160
тыс. лет (White T.D. е.а. И Nature, 2003, v. 423, р. 742), а последнее потепле-
ние первого межледниковья оценивают теперь в 350 тыс. лет (Сорохтин
О.Г. // ВРАН, 2006, № 8, с. 703) - разница за 80 лет сократилась почти впя-
теро. Теперь возраст “одинцовского человека” великоват, но не шокирует.
Будучи признан, “одинцовский человек” помогает понять механизм про-
исхождения человека вообще. Скажу об этом чуть подробнее.
554
9-15*** Дитя жары и мороза
Один из первых исследователей антропогенеза Карл Фогт писал в 1863
году, что у человека не было единого предка, что он произошел путем мно-
гих скрещиваний, а видно это из того, что его тело можно мысленно со-
брать, если взять голову ревуна, руку шимпанзе, скелет туловища гиббона,
ногу гориллы и мозгоранга (см. [Чайковский, 1990, с. 181]).
Очевидно, что он писал о блочности, а мы ее теперь умеем объяснять
без скрещивания, через рефрены. О том же в наше время писали не раз.
Так, на рис. 766 под номером 4 упомянута новая (2000 г.) находка, но-
гами похожая на австралопитека, а зубами на человека, но на 1,5 млн лет
более древняя, нежели древнейший австралопитек (Senut В. И LEG, 38).
Фогт преувеличил сходства, но суть дела выразил верно. До сих пор
предка человека ищут на основе тех или иных семофилогений, и каждое
новое открытие приводит к новым недолговечным древам. Налицо одно из
многих свидетельств крушения принципа филогении, о чем см. главу 10.
В 1870-е годы среди антропологов было течение, видевшее в обколо-
тых кремнях третичных (см. сноску к п. 2-10) слоев свидетельство реально-
сти третичного человека (это течение возрождено книгой Кремо и Томпсо-
на). Любопытно, что тогдашняя аргументация затрагивала и то, что мы те-
перь именуем биотогенезом (п. 9-4**): считалось более вероятным, что че-
ловек возник в «третичном раю», когда в Европе были субтропики, чем в
четвертичных оледенениях {Павинский А. Вопрос о существовании челове-
ка в третичной формации // Варшавские Университет, известия, 1882, № 2).
Напомню: в тропиках течет биотогенез, вероятно рождающий высшие
таксоны. Поэтому мысль о происхождении человека в «третичном раю»
весьма актуальна, если признать, что человек - таксон высокого ранга (см.
далее, п. 10-10). А эволюция в рамках рода течет и в холодном климате.
Холодовой стресс рассматривался как ведущий фактор эволюции че-
ловека Аршавским. Поэтому любопытно, в каком климате жил “одинцовский
человек". Между одинцовскими моренами найдены следы как тундры, так и
нашего леса (см. книгу: Московский ледниковый покров Восточной Европы.
М., 1982). Мы не знаем, в тундре ли жил “одинцовский человек" или в при-
вычном нам березняке, но ясно, что он умел жить за три ледниковья до нас
и за три климатических зоны до тропических лесов.
Похоже, что наступая и отступая, ледники способствовали чему-то
вроде “закалки по Аршавскому’’, т.е. последним стадиям антропогенеза.
Что касается мнения об отсутствии следов культурной эволюции в тре-
тичном периоде, то Кремо и Томпсон считают это просто заблуждением:
следов предостаточно, надо только уметь и хотеть их видеть.
9-16. Конец эволюции?
О конце эволюции организмов в целом говорить не приходится: наобо-
рот, с рождением генной инженерии она резко ускорилась и расширила свои
555
рамки. Зато часто можно прочесть, что с появлением цивилизации закончи-
лась биоэволюция человека. Серьезной аргументации при этом не дается, а
вопрос слишком важен, чтобы ограничиваться заявлениями.
Янч (п. 7-6) любил говорить о коэволюции звезд и атомов, молекул и
планет, экосистем и видов, организмов и органов. То есть - что с уменьше-
нием системы увеличивается ее характерный элемент. Мне эта мысль
показалась полезной, и я детализировал ее диаграммой [Чайковский, 1990,
с. 218], которая будет приведена далее, в п. 11-2, на рис. 83. Развитие при-
роды в ней выглядит конусом, острие которого достигнуто в XX веке. Из
диаграммы видно, что с появлением человека и цивилизации макроуровень
слился с макроуровнем.
Можно не верить в то, что эволюция шла “по Тейяру”, т.е. к заранее на-
меченной Богом конечной цели, но надо, следуя Янчу, признать, что______
в одном смысле эволюция своего конца достигла; коэволюция большого с
малым дошла до того предела, когда большое от малого не отличить.
Назовем это тезисом Янча. К нему приводит и анализ фактора време-
ни: время жизни системы и ее элемента сомкнулись. Так, в XX веке за жизнь
одного поколения возникла, расцвела и почти сошла с арены целая исто-
рическая формация - социализм. Это не значит, что эволюция кончилась,
но она приняла форму, не дающую описать ее в прежних терминах.
Новую форму эволюции биолог широкого профиля Р.М. Зелеев (Казан-
ский университет) предложил в виде схемы двух конусов', к вершине преж-
него сужавшегося конуса приставлена вершина нового, расширяющегося во
времени сообразно росту нашего знания о мире. В основном это - «накоп-
ление Культурной информации» (Зелеев Р.М. К опыту геометризации пред-
ставлений о картине мира и путях его познания // ЛЧ, 1999). Однако это не
вполне биология. Что же касается биоэволюции, то она, как отмечено в на-
чале книги, идет и притом весьма бурно. В частности, идет биоэволюция
человека: жители разных широт различны и строением тела, и функцио-
нально, причем нередко тут виден итог приспособления (к жаре, холоду, ги-
поксии, радиации и т.д.), а не просто наличное разнообразие.
Так, хотя некоторые жители Севера и тропиков живут на “первобытном”
(т.е. догосударственном) уровне, но они умеют многое, чего не умеем мы -
например, лучше чувствуют запахи (принцип компенсации). Леви-Строс от-
мечал, что нельзя “архаизм” этих народов считать состоянием, оставшимся
с эпохи палеолита. В наших терминах, речь у него шла о повторной реали-
зации позиций давнего рефрена. Это, однако, обычная эволюция, свойст-
венная многим группам. Что касается особых механизмов эволюции чело-
века, то один из них виден в феномене пассионарности.
«Пассионарный... Одержимо, не признавая никаких преград, стремящийся
к какой-либо цели. Термин образован русским этнологом Л.Н. Гумилевым»
(Словарь-попутчик... М., Изд. МГУ, 1994, с. 129).
556
Пассионарностью людей (прежде всего, вождей) Гумилёв объяснял их
быстрый успех в достижении социальной цели, прежде всего - в создании
новых народов и учений. Ее угасание - причина остановки развития, а затем
и гибели народа, поскольку народ не может существовать, не преодолевая
преград (Гумилев Л.Н. Этногенез и биосфера Земли [1979]. М., ACT, 2003).
Очевидно, что пассионарность - вид жизненного порыва, т.е. активно-
сти, распределенный очень неравномерно как по народам, так и по эпохам
жизни каждого народа (п. 5-2**, Второе правило). Например, пассионарность
финнов, англичан и русских, высокая в прошлом, сейчас угасает.
Поскольку в первые миллионы лет эволюции людей их численность
была ничтожна (считается, что в течение палеолита она выросла со 120
тыс. до трех млн по всей Земле - Козлова М.С. Эволюция человека... М.,
Наука, 2005, с. 53, 72), то их бурную экспансию, от джунглей до тундры,
можно объяснить никак не теснотой, а скорее пассионарностью.
К пассионарности близка особая половая активность людей как вида. В
п. 9-15 избыток активности был увязан с внесезонным размножением и на-
зван одним из факторов антропогенеза. Когда люди стали людьми, этот из-
быток остался одной из важных черт, и его стоит рассмотреть.
Наша наука - детище христианской культуры, а христианство принижа-
ет всё, что связано с полом. (Это видно хотя бы в идее «непорочного зача-
тия» - в иных религиях боги рождаются от соединения полов, и верующие
не видят в этом порока.) Философ и богоискатель В.В. Розанов в 1911 году
изумлялся (опираясь на Ф.М. Достоевского) отношению христиан к полу:
«Только христианство могло отвернуться, или, как теперь очевидно, попыта-
лось отвернуться от этого, и через это потрясти очаги рождения, разрушить
недра мира, как бы проколоть иглой мировой зародыш» (Розанов В.В. В тем-
ных религиозных лучах. М., 1994, с. 286).
Вследствие западного принижения роли пола многие феномены, оче-
видные Востоку, нам увидеть трудно. Так, в христианских книгах обычно
уверение, что те, кто совокупляется не ради продления рода, а ради удо-
вольствия, ведут себя «аки скоты». На самом деле, скорее наоборот: чело-
век отличен от "скотов” именно половой активностью вне размножения,
что, кстати, учитывают новые (фрейдизм) и древние (тантризм в Индии)
учения. В одной из форм тантризма утверждалось даже, что
«блаженство, которое проявляется через сексуальное соединение, есть истин-
ная форма Бхайравы», высшего бога (Пахомов С. В. Представление о высшей
реальности в философии индуистского тантризма И ВФ, 2006, № 5, с. 153).
У нас же бытуют, благодаря дарвинизму, взгляды на происхождение
пола, исходящие из идеи его выгодности, которую, впрочем, доказать ничем
не удалось. Наоборот, раздельнополость уменьшает размножаемость
вдвое. На мой взгляд, без пола (гермафродитно) человек произойти просто
не мог (п. 9-15). И жизненный порыв (по Бергсону), и пассионарность (по Гу-
милёву) людей во многом реализованы через половую энергию.
557
Мужчины и женщины тратят ее различно, и тут интересна структура
людских популяций. Женщины живут дольше, хотя репродуктивную функ-
цию завершают гораздо раньше мужчин. У обезьян жизнь самок, перестав-
ших рожать, гораздо короче - и в годах, и в процентах от видовой длитель-
ности жизни (Cohen А.А. Female post-reproductive lifespan... // BR, 2004, № 4,
c. 737). Про этот эффект бабушки написано много, но понимания нет.
Попытки целевого объяснения (зачем? - чтобы нянчить внуков) терпят
крах, ибо самих бабушек (и дедушек) в первобытных обществах почти не
было: ископаемых костей старше 50 лет практически не находят (а среднее
время жизни было даже ниже 35 лет). Если первобытные бабушки и нянчи-
ли внуков, то, вернее всего, до менопаузы. Попытки причинного объяснения
(почему?) сами требуют объяснения. Такова ссылка на число яйцеклеток,
заложенных еще в зародыш девочки (п. 5-12*): кончились яйцеклетки, и ста-
ла женщина бабушкой. А почему они у людей так рано кончаются?
Как уже не раз сказано, диатропика предлагает начинать с описания
явления (с вопроса: как?). На мой взгляд, лучшим объяснением эффекту
бабушки служит его соответствие особому механизму эволюции людей.
9-16* Двускоростной механизм эволюции людей
У людей очень долгий онтогенез, около 20 лет (как у африканских сло-
нов) и уникально долгое (гораздо дольше, чем у слонов) взросление (отро-
чество и юность), которое занимает более 10% срока нормальной жизни:
физическое и духовное развитие длится много после достижения способно-
сти к размножению. Половое взросление начинается у девочек с 8-10 лет, и
беременность в 11 лет вполне возможна, а в 12 лет - была даже социаль-
ной нормой в течение почти всей истории человечества (кое-где эта норма
бытует по сей день); в мусульманском праве брачный женский возраст ра-
вен всего 9 годам. В то же время медики признают девушку взрослой лишь
в 18-19 лет, а психическая взрослость наступает еще позже.
Поскольку мужчины традиционно вступают в брак много позже женщин,
поколения могли быть по женской линии (и в истории не раз бывали) втрое
короче, чем по мужской, а в отдельных линиях - впятеро (как курьёз - вше-
стеро) короче. Разумеется, речь тут идет лишь о первенцах.
Итак, мы видим двускоростное размножение. Посмотрим, как оно по-
рождает двускоростной механизм эволюции.
Может показаться, что свойства первенцев не могли быть в многодет-
ных обществах определяющими, но это - заблуждение, издержка статисти-
ческой ПМ: на самом деле эволюцию определяет отнюдь не сдвиг средних
значений. У людей, например, гораздо важнее общественные предпочтения.
Особая роль первенцев хорошо видна в Библии.
Руководство Гилберта (мы пользовались им в ч. 2) видит во взрослении
людей второй онтогенез и даже сравнивает его с метаморфозом (т. 3, с.
187). Для эволюции людей это важно тем, что в ее ходе шло регулярное
558
размножение в середине женского взросления. Вспомним, что половые
клетки матери имеют, грубо говоря, тот же возраст, что и сама мать, тогда
как отцовские половые клетки всегда новы (п. 5-12*).
Беременность при «втором онтогенезе» является мощным стрессом.
Все связи у юной матери неустойчивы (за 9 месяцев беременности она
взрослеет по многим параметрам радикально), и можно полагать, что про-
рвать “барьер Вейсмана” у нее легче, чем у взрослой (одна из точек проры-
ва вероятна в бластоцисте, п. 8-12*). А взрослый (и часто немолодой) отец
дает свой набор свойств, апробированный самим фактом его долгой успеш-
ной жизни (высокий статус повышал число жён). Это похоже на “половой от-
бор”, но сходство обманчиво: женщина тут вообще ничего не выбирает, да и
отец не борется за нее - скорее, он ведет себя, как «ветеран тока» (п. 3-3).
Это должно вести к резкому различию генетических вкладов матери и
отца. Подростковая беременность выступает как неотения (термин см. в п.
4-13), а размножение с надставками, обретенными в старшем возрасте -
это геронтогенез. У людей мы видим женскую неотению при мужском
геронтогенезе, т.е. человек, словно два далёких вида, слитые воедино,
реализовал преимущества г- и К-стратегий (см. о них п. 5-10), но без их
недостатков; например, без снижения темпов жизнедеятельности.
Данный механизм как факт достоверен, но его происхождение и роль в
эволюции еще предстоит понять. Смею допустить, что он был ответствен за
важный фактор эволюции людей уже по завершении очеловеченья - за бы-
строе наследование долгого обучения. Подробнее см.: Чайковский Ю.В.
Двускоростной механизм... И Эволюция, 2004, № 2.
Итак, у ранних людей репродуктивный период был резко смещен - у
женщин в сторону молодости, а у мужчин в сторону старости. Отсюда и
“эффект бабушки”: дело не в том, что бабушки распространились в силу
своей полезности для внуков, а в том, что сам механизм эволюции человека
ограничил фертильный возраст рамками молодости. Что же касается функ-
ции вынянчивания внуков, то она наступала в фертильном возрасте, если
женщины в массе становились бабушками в 25-28 лет.
Далее, как мы знаем из п. 5-5*, для быстрого видообразования нужен
тесный инбридинг. У людей его именуют инцестом (т.е. нечистым); в исто-
рическое время он почти неизвестен, зато весьма обычен в мифах.
Когда-то Леви-Строс обронил без пояснений странную фразу: «люди не
в меньшей степени сделали самих себя, чем они создали расы своих до-
машних животных» (Леви-Строс К. Структурная антропология. М., 1985, с.
312). Приведённые им сложные схемы браков у тропических племён ничем
не проясняют данную мысль. Вероятно, он туманно обратил внимание на
то, о чем прямо говорить было в его время неудобно. И одна такая тема
очевидна - инцест. Без него породу не выведешь. См. Дополнение 10.
Учтя идею п. 5-13*, можно полагать, что будущая теория эволюции че-
ловека окажется теорией параллельной смены норм в двух рядах норм -
559
мужском и женском. При этом, вероятно, именно двускоростной механизм
обеспечил эволюцию людей в рамках “двух матриц” Паршина и “двух ролей”
Геодакяна. Если так, то роль двускоростного механизма - не просто ускоре-
ние эволюции, а создание того, чего иначе не создать.
Нужно ли тут искать какие-то ряды в надежде построить рефрен? В це-
лом - не знаю, но отдельные параллели видны. “Эффект бабушки” извес-
тен у млекопитающих (у разных видов китов налицо все варианты, от полно-
го отсутствия самок, окончивших деторождение, до “бабушек”, живущих де-
сятилетиями - Cohen А.А. И BR, 2004, № 4, с. 736). А вот “эффект маленькой
мамы” надо искать много дальше. Массовая активность взрослых самцов в
отношении незрелых самок возможна у головоногих: так, у тихоокеанского
кальмара взрослые самцы «обычно спариваются еще на путях миграций с
далекими от зрелости самками». У аргонавтов самки начинают плодоносить
еще мелкими и продолжают это, «до конца жизни, не прекращая питаться,
расти и надстраивать раковину» [Несис, с. 34,173].
Для эволюции людей была существенна еще и лактационная сте-
рильность (невозможность забеременеть при кормлении грудью), которая в
архаичных обществах сокращает рождаемость почти вдвое (Шорт Р.В.
Грудное вскармливание И ВМН, 1984, № 8). Легко видеть, что это - сильный
фактор выживания: как показал модельный расчёт [Чайковский, 2004, с.
217], пропуск особями половины актов размножения гораздо сильнее по-
вышает вероятность выживания популяции, нежели любое реально дос-
тижимое ею снижение смертности особей. В нашем индустриальном об-
ществе этот механизм тоже почти прекратил свое действие.
Словом, прежние особые факторы биоэволюции человека сходят на
нет. Но эволюция людей продолжается и даже обретает новые формы.
9-16** Нынешняя эволюция человека
В последние полвека много говорят о недопустимом повреждении при-
роды, но мало кто заметил, что пик влияния экологических движений на об-
щество пройден, что после распада СССР западный мир снова провозгла-
сил идеалы потребительского общества. Подробнее см.: Яницкий О.Н. Эко-
логическая парадигма... И Социология, исследования, 2006, № 7, с. 88. На-
лицо исторический откат, начало которому положил провал всемирной
Конференции ООН по окружающей среде (Рио-де-Жанейро, 1992 г.), где
США, Великобритания и Япония заблокировали все реальные действия.
Им помогли 52 нобелевских лауреата, направивших в Рио письмо против «зеленой
истерии». В 1980-х годах немецкий философ и социолог Юрген Хабермас предлагал
альтернативу — сменить общественный идеал с интереса на коммуникацию, т.е. с пре-
следования выгоды на стремление к пониманию других. Тогда много говорили о плю-
рализме в общественных отношениях. Это выглядело как начало реализации линии
Хабермаса и виделось мне как переход к пятой (диатропической) ПМ (см. конец книги
[Чайковский, 1990]), но теперь снова провозглашается преследование интереса. В 1991
560
году нобелевскую премию по экономике неожиданно получил Р. Коуз (о нем см. п. 3-
11) за работу 1959 года, где утверждал, что ущерб природе и выгода от ущерба равно-
ценны для рыночной практики (Нобелевские лауреаты по экономике... М., 1994).
Но молодежь идет еще дальше. Недавно академик Заварзин сделал наблюде-
ние: на совещании по эволюции старики-академики «проявили гораздо больше
скепсиса и стремления к пересмотру существующих концепций, чем их молодые кол-
леги, предпочитавшие оставаться на... основе традиционных подходов» [Заварзин,
2006, с. 168]. Если это верно, то беспрецедентно: самая консервативная часть учёных,
академики, все-таки больше устремлена вперед, чем научная молодёжь.
Вне науки архаизм молодежи виден еще ярче. Она, с электроникой в руках и в ду-
ше, восторженно погрязает в идеалах раннего капитализма (рынок, “разумный эго-
изм” и дарвинизм). И это — стержень исторического отката. Попытки объяснить
молодым, что так уже было и очень плохо кончилось (в дни Дарвина несколько стран,
богатевших за счет остальных, утвердили в школах религию благотворной конкурен-
ции, поделили мир, породили две самых ужасных войны в истории, а принцип конку-
ренции надолго угас), ни к чему не ведут. В чем же дело? Ясно, что все богатыми быть
не могут. Теперь, когда остальные страны захотели жить так же, как богатые, мир ру-
шится. Неужели западные идеологи этого не понимают? Умные - понимают, и их там
достаточно, чтобы не жить самим по тем правилам, какие они предложили нам.
Никакой рыночной экономики на Западе давно, с Великой депрессии 1930 года,
нет, а есть рыночная идеология (играющая примерно ту же роль, что плановая идеоло-
гия в позднем СССР), причем рыночные термины остались в языке от статистической
ПМ (см. петит в параграфах 3-11 и 11-3). Говорить об этом приходится здесь потому,
что ‘'теории” рыночной конкуренции и естественного отбора постоянно заимствуют
аргументацию друг у друга и совершают одну и ту же прискорбную ошибку: частное
наблюдение за низшим уровнем системы (розничная торговля, внутривидовая измен-
чивость) выдают за открытие общих законов развития. Нынче они равным образом
обслуживают идеологию свершающегося отката.
Сам по себе феномен исторического отката хорошо известен. Яркий пример - вто-
ричное закрепощение крестьян Европы в конце Средних веков, перешедшее в России
при реформах Петра I в рабовладение. Таково и само рабовладение, вторично развив-
шееся в России и США, и третичное закрепощение крестьян в колхозах.
Нынешний откат замаскирован, так как протекает на фоне компьютеризации (эта
форма технического прогресса не имет аналога) и потому многим кажется социаль-
ным прогрессом. Он опаснее предыдущих как своей глобальностью (если приведет к
катастрофе, то - весь мир, и спасать будет некому), так и огромной скоростью влияния
на природу (когда экологическое сознание снова овладеет активным меньшинством,
то, в отличие от прежних кризисов, что-то спасать будет, по-видимому, уже поздно).
Нашей темы касается биологическая сторона исторического отката.
Сто лет назад французский социолог Эмиль Дюркгейм высказал пара-
доксальное мнение, что общество произошло не путем соединения мысля-
щих индивидов, а наоборот: мыслящий индивид есть итог осознания особью
561
своей самобытности, вычленения из социума. Добавлю: этим общество лю-
дей отличается от обществ животных и именно поэтому оно сумело создать
культуру. Если Дюркгейм прав, то начавшийся в конце XX века процесс
слияния людей в единый мыслящий субъект противоположен антропогене-
зу и являет собой акт расчеловеченья.
Другой процесс, столь же опасный для человечества - его превраще-
ние в особое царство природы, о котором пойдет речь в конце главы 10.
Оно носит характер фазового перехода, и именно осознание себя новым
царством является, на мой взгляд, основной причиной нынешнего отката.
К этому надо добавить отрицательную размножаемость европейской
расы как целого - эффект, предсказать исход которого для данной расы
легко, а для человечества пока невозможно.
Налицо феномен эволюционный (точнее, эманация, п. 4-17), при кото-
ром грозит распасться культура не только в ее высоком смысле, но и в са-
мом простом - в том, что отличает человека от других животных.
Если так, если эволюция человека действительно пошла назад, то по-
нятно и возвращение отживших познавательных схем. Таковы, в частности,
получившие вторую жизнь идеи рынка и дарвинизма, основанные на треть-
ей (статистической) ПМ. Возвращаются и еще более старые ПМ, казалось
бы навсегда отвергнутые 60 лет назад, при создании ООН: вторая (механи-
ческая) - приоритет силовых действий, и даже нулевая (религиозная).
Продолжает править та самая идеология, какую исповедуют марксизм и
дарвинизм, идеология третьей ПМ. Они всегда черпали силу друг у друга
(не имея фактов, чтобы доказать свои догмы собственными средствами) и
видимо должны рухнуть вместе. Мне уже случалось писать, что с крушени-
ем советской идеологии марксизм никуда не делся ни у нас, ни на Западе
[Чайковский, 1993, с. 107], и теперь добавлю: каждому, кто принимает ре-
шения, важно знать, что за дарвинизмом стоят не факты, а толкования.
Причина долгого господства в науке столь странной доктрины - господ-
ство соответственной идеологии в обществе. Идеологии живут, как извест-
но, тысячелетиями, то увядая, то вновь расцветая. Беда в том, что история
течет нынче слишком быстро, и мир может не дождаться конца отката.
Вряд ли самая лучшая наука способна сама по себе направить мир к
лучшему, но вот что невозможно заведомо, так это улучшить мир, пока в
школах учат, что спасение придет от старых доктрин - тех самых, что при-
вели мир в нынешнее плачевное состояние.
То же, полагаю, относится ко всем «мировым религиям» и к религии не-
давней (рыночная политэкономия плюс дарвинизм). Если религия действи-
тельно нужна, то новая, учащая, как жить в переполненном людьми и
машинами мире. Такая религия неминуемо должна быть основана на науке
(в частности, на экологии в ее биологическом понимании), а не надеяться,
что нас спасут старинные тексты. Они не спасли, увы, ни одну из прежних
культур - при всех религиях творились, в общем, одни и те же мерзости.
562
Часть 4. Приложения и выводы
Гпава 10. Эволюция и систематика
До сих пор нас занимало становление не таксонов, а структур и меха-
низмов, и систематики мы почти не касались. Но от нее никуда не деться,
поскольку без нее нельзя даже назвать те организмы, эволюцию которых
мы обсуждаем, и нельзя всерьез изучать процесс эволюции (хотя и можно
обсуждать ее механизмы). Например, мы пользовались названиями групп
организмов как чем-то заданным, но можно ли так поступать? Ведь сам
факт включения организма в какую-то группу задает возможный круг вопро-
сов. Так, вопрос: «Почему из всех архозавров не вымерли только крокоди-
лы?» -теряет смысл, если крокодилов не считать архозаврами.
Есть и более насущные вопросы. Систематика обслуживает всю биоло-
гию и не только ее, она должна быть удобной и понятной, чего о нынешней
никак не скажешь. Может ли тут помочь новая теория эволюции?
10-1. Система, таксоны и названия
Систематика сложилась в XVI-XVIII веках без всякой связи с идеей эво-
люции: система видов, родов и прочих таксонов мыслилась как нечто за-
данное и постигаемое умом. Именно эту систему дарвинизм и поставил це-
лью рационально истолковать. Недаром Дарвин назвал свою книгу «Проис-
хождение видов», а Коп свою первую эволюционную статью - «Происхож-
дение родов». Ныне ситуация иная: система строится как итог эволюции. В
этом ламаркизм, жоффруизм и дарвинизм едины. Однако есть иная точка
зрения, мало известная систематикам: номогенез видит эволюцию как про-
цесс заполнения объективно заданной таблицы, которая сама от эволюции
не зависит (от эволюции зависит лишь степень заполненности таблицы).
Уточним термины. Согласно полуофициальной (рекомендательной) ра-
боте английского ботаника Чарлза Джеффри, систематика ставит целью
«создание таких систем классификации, которые наилучшим образом отра-
жали бы различную степень общего сходства живых организмов». Она со-
стоит из двух основных областей - классификации и номенклатуры.
Классификация - процесс установления и характеристики систематиче-
ских групп и в то же время результат этого процесса. Номенклатура - рас-
пределение названий между установленными таким путем таксонами и в то
же время результат этой процедуры Щжеффри, с. 14-15]. Тем самым, сис-
тематика не обязательно связана с эволюцией, зато номенклатура строго
следует за классификацией, а это, как увидим, плохо практически.
От Геккеля идет уверенность, что все таксоны возникают в ходе эволю-
ции и притом возникают по единой схеме. Вопрос обоснования этой уверен-
563
ности, в соответствии с тезисом Витгенштейна, не ставится. Более того, за
принятием постулата Геккеля последовало неосознанное принятие еще
одного постулата: что эволюция имеет вполне определенную форму -
форму филогении, т.е. порождает родословное древо.
Кажется, Любищев был первым, кто заметил, что этот постулат ниотку-
да не следует, а в биологию попал из логики иерархической классификации
времен Линнея. От постулата давно отказались все, кроме биологов (из
биологов отказались бактериологи и отчасти палеоботаники).
Не потому ли биологи вынуждены то и дело пересматривать свои сис-
темы, что их материал входит в противоречие с их постулатом? Думаю, да.
Ведь в предыдущих главах мы видели, что эволюция носит блочный харак-
тер, а значит, лучше описывается как заполнение клеток диасети, нежели
как рост древа. Но если так, то система волей-неволей строится произволь-
но, а потому каждым по-своему. Согласие при этом отнюдь не близится и,
видимо, даже не ищется. В итоге нет стабильных названий, и страда-
ют все. Подробнее см. [Чайковский, 1990, с. 45-48].
Поэтому для практических целей нынешняя систематика мало пригод-
на. В самом деле, как признал Джеффри:
“Названия, которые обычно сообщают систематики в ответ на запрос, явля-
ются правильными... по общепринятой в настоящее время классификации
рассматриваемой группы” (с. 58).
Но сама систематика всё время меняется, поэтому такие справки почти
бесполезны. Во-первых, общепринятой системы данный таксон может не
иметь, и специалист обычно дает справку, верную в той системе, какой он
держится сам, а не набор справок, верных для всех конкурирующих систем.
Во-вторых же, даже если общепринятая система и существует, обычно она
принята недавно, а потому не дает понятных указаний тому, кто читает
прежнюю литературу. В итоге, для серьезных исследований требуется лич-
ный контакт с узким специалистом по каждой исследуемой группе (знаю по
собственному опыту), а это мало кому доступно.
Но еще хуже то, что за экспертизой может вовсе не стоять никакой тео-
рии Вот что пишет систематик-зоолог К.Г. Михайлов:
«В случае возникновения противоположных мнений об одном и том же таксоне
спорящие стороны редко прибегают к морфологическим доказательствам.
Нормальному обсуждению подвергаются только таксономические проблемы на
уровне класса и выше... Основной результат работы систематика-практика - это
иллюстрированные определительные ключи, диагнозы и описания новых, а так-
же переописания ранее известных таксонов. Как правило, научные публикации
такого рода не сопровождаются какими-либо обоснованиями выделения того или
иного уровня (ранга) таксона», и «попытки введения прямых доказательств вы-
зывают удивление (“Зачем это, ... мы тебе доверяем”)» [Михайлов, с. 41].
Сами систематики считают (насколько могу судить по нескольким бесе-
дам) такое положение вполне удобным, ибо чувствуют свою незаменимость,
564
но с этим, по-моему, пора кончать - как потому, что тормозятся охрана и ра-
зумное использование видов, так и потому, что в нынешней таксономиче-
ской чехарде нет объективной нужды. Она, по-моему, вызвана лишь жела-
нием филогенетиков без конца “играть в бисер” и нежеланием думать.
«Итак, естествоиспытатель вправе задать вопрос: зачем нужна такая клас-
сификация кому-либо, кроме филогенетиков?» (Заварзин Г.А. Составляет
ли эволюция смысл биологии? И ВРАН, 2006, № 6, с. 526).
Нас интересует не эта систематика для систематиков, а наука, по-
лезная всем. Она не должна быть чисто эволюционной, поскольку главные
вопросы систематики - «кто это?» и «где его место?». Наоборот, вопрос «от
кого?» (и иные исторические вопросы) осмыслен в систематике постольку,
поскольку помогает ответить на главные, а не запутывает их. Тут важен
опыт иных наук: почти все от исторической систематики отказались.
10-1* Стабильность названий. Тезис Линнея
До сих пор у систематиков господствует такое правило:
“Изменение названия, под которым таксон уже известен, разрешается кодек-
сами только в том случае, если необходмо исправить номенклатурную ошиб-
ку, внести изменение в классификацию или исправить допущенное в про-
шлом ошибочное определение” [Джеффри, с. 53].
Подчеркнутые мною слова делают чехарду неизбежной, а ведь само по
себе изменение классификации вовсе не требует изменения названий, если
понимать их просто как адреса («Название - это просто условный символ
или шифр, который дает возможность ссылаться на тот или иной таксон»
[Джеффри, с. 19]). Менять их надо только тогда, когда грозит путаница.
Джеффри сам подчеркнул, что «номенклатура до некоторой степени неза-
висима от классификации» и что та номенклатура, которая изменяется при
изменении классификации, ущербна (с. 15, 34), но практический его вывод
противоположен (по-моему, от нежелания думать).
В принципе задача стабилизации названий не нова, и для ее решения
надо обратиться к истории науки. Подробнее см. [Чайковский, 2006а].
Карл Линней (1707-1778) не знал наших эволюционных догм, зато был
великим систематиком. По Линнею, говоря нашим нынешним языком,______
_________в биологии нужны две системы: искусственная и естественная.
Первая нужна для адресации, вторая - для уяснения “сути вещей”. Ис-
кусственную систему цветковых, основанную на числе тычинок и пестиков,
Линней подробно разработал сам и всячески пропагандировал (ныне от нее
в систематике ничего не осталось). Естественную систему он, наоборот,
лишь бегло наметил в «Философии ботаники» (1751). Фактически, она была
списком типов строения и жизненных форм. Типы строения Линней называл
семействами и в параграфе 78 насчитал их семь:
«РАСТЕНИЯ включают семь семейств (familiae): Грибы, Водоросли, Мхи,
Папоротники, Злаки, Пальмы, [прочие] Растения».
565
Эти семейства он называл «естественными», в противоположность «оби-
ходному» делению растений на деревья, кустарники и травы (там же).
Основу «естественного метода» составил список «фрагментов» (пара-
граф 77). Их у Линнея 68, и каждый «фрагмент» являет собой просто пере-
чень родов растений без каких-либо пояснений, причем последний состав-
лен из «неопределенных» родов, не нашедших места в остальных «фраг-
ментах». Насколько можно признать эти 67 групп естественными, оставляю
судить ботаникам, нам же следует обратить внимание на указания самого
Линнея о характере предложенного им метода.
В отличие от искусственной (половой) системы Линнея, «Естественный
метод старались выработать на основе семядолей, чашечки, пола и проче-
го» (параграф 69), т.е. целостно. Естественные группы
«совпадают по внешнему облику, способу возникновения (при развитии заро-
дыша - Ю.Ч.), качествам, свойствам и применению... Естественный метод
поэтому есть и будет конечной целью ботаники» (параграф 206).
Хотя дал Линней только голые списки родов, но он был уверен, что в
будущем они естественно упорядочатся в компактную плоскую картину:
«Природа не делает скачков», поэтому «Все растения проявляют друг к другу
сродство, как земли на географической карте». «Причиной того, что естест-
венный метод имеет пробелы, является отсутствие пока еще не открытых рас-
тений... ибо природа не делает скачков» (параграф 77).
Если искусственную систему Линней довел до уровня вида и даже до
разновидности, то его «естественный метод» на это в принципе не претен-
дует: рассуждения ведутся лишь о классах и порядках, а списки доведены
до уровня рода. Замечу, что линнеевы роды много крупнее наших.
Это, в общем, соответствует нынешней точке зрения на классификацию
жизненных форм, которую предложено доводить до уровня рода, поскольку
близкие виды имеют одну и ту же жизненную форму (п. 10-2*).
Насколько понимаю, речь у Линнея шла о том, что естественная
система должна классифицировать сущности, а искусственная -
использовать акциденции (п. 1-6**), удобные для адресации.
Увы, следующие поколения биологов взялись за совсем иную задачу:
захотели переделать заведомо искусственную систему Линнея в естествен-
ную, как они ее понимали. Причем слово “естественная” после Геккеля трак-
туют как “эволюционная”, а “эволюционная” - как “филогенетическая”. Во-
прос о своей правоте почти никем не ставился и не ставится (опять нежела-
ние думать), и ничего путного выйти не могло.
Мне уже случилось писать о стабильности названий [Чайковский, 1990,
с. 19-20, 46-48, 140-141], но единственный известный мне отклик состоял в
том, что лихой критик упрекнул меня в призыве отменить латинские назва-
ния. На самом деле речь у меня шла о другом - о том, что нельзя менять
латинские названия по произволу всякого, кто понял родство данных орга-
низмов по-новому, и что латинские названия надо попытаться сделать
566
столь же устойчивыми, каковы бытовые названия. То есть я предлагал
вновь, согласно тезису Линнея, отделить адресацию и номенклатуру от веч-
ных споров о “естественной системе”. Подробнее см. [Чайковский, 2006].
Если, например, состав рода Canis (собака) меняется от сводки к свод-
ке, это не основание столь же часто и покорно изменять во всех книгах на-
звание Canis vulpes (так названа Линнеем лисица обыкновенная) на Vulpes
vulpes и обратно (как мы видим в нынешних системах). По-моему, будет
лучше, если тот, кто вводит отдельный род Vulpes, будет писать: «вид Canis
vulpes L. из рода Vulpes (точно так же, как сейчас пишут: род Vulpes из се-
мейства Canidae)» [Чайковский, 1990, с. 47]. Примененное мною обращение
к бытовому названию («лисица обыкновенная») с целью прояснить суть де-
ла - довольно обычная в номенклатурной практике процедура. К сожале-
нию, большинство видов не имеет бытовых названий.
Сходную процедуру предлагает для надвидовых таксонов питерский
систематик Н.Ю. Клюге в своей концепции «объемной номенклатуры»:
«Например, “Libellula sensu Linnaeus, 1758”... означает то же, что объемное
название “Odonata”» [Клюге, с. 62].
Предложить столь архаичный термин, как «Libellula в смысле Линнея»,
его вынудил тот печальный факт, что номенклатура отряда «стрекозы»
весьма переменчива и потому запутана - достаточно отметить хотя бы, что
оба приведенные им термина (Libellula и Odonata) введены в XVIII веке, од-
нако я не нашел их в двух обстоятельных энциклопедиях начала XX века, но
нашел в нынешней. (Обычное хождение номенклатуры по кругу.)
Недостатки идеи Клюге состоят в том, что она тоже - запутанная и бо-
лее юридическая, чем биологическая. А меня устроит минимум: пусть сис-
тематики, без конца споря о составе семейства псовых (Canidae) и месте
лисицы в нем, будут помнить, что имеют в виду вид Canis vulpes в смысле
Линнея. При этом можно использовать, например, обозначение: «Canis
vulpes * L», где звездочка указывает на использование старого имени в но-
вой системе. (Обозначать отход от нынешней номенклатуры каким-нибудь
значком предлагает также и Клюге.)
Это и есть путь Линнея: для практики удобна постоянная номенклатура,
что несовместимо со следованием какой-либо моде. Латинские названия
организмов не должны меняться, как не меняются их бытовые названия; как
не меняются имена людей ни с изменением названий улиц, ни с переездом
их в другие дома; как не меняются названия химических элементов с
прогрессом химии и физики. Подробнее см. [Чайковский, 2006а].
А вот “естественная система” меняться вынуждена и, более того, не
может быть единственной, поскольку взгляд на естественность у разных
ученых различен и меняется вместе с эпохой (см. ниже). Но это вовсе не
значит, что она должна меняться часто.
Отсюда следует, что для номенклатурных целей нужна особая клас-
сификация, не связанная с естественной системой жёстко. Зато сама эта
567
искусственная классификация должна быть жёсткой: будучи принята меж-
дународным органом, она не должна меняться ни при каких обстоятельст-
вах, кроме случаев, когда обнаружена ошибка, ведущая к противоречию в
самой номенклатуре - например, к гомонимии (одним словом названо два
таксона и более). Иными словами, принцип приоритета отдельных названий
желательно заменить на принцип постоянства списка в целом, если он уже
вошел в употребление. Надеюсь, что такая практика станет обычной, когда
систематики осознают, что нынешняя чехарда в номенклатуре - тупик Од-
нако в этом теория эволюции им помочь не может.
Зато она нужна там, где идет речь о естественной системе.
10-1** Смысл иерархии. Тезис Короны
Давней беседе с зоологом Н.Н. Воронцовым (около 1965 г.) я обязан
пониманием того факта, что постулат о выразимости процесса эволюции
посредством родословного древа есть всего лишь проекция дворянских ге-
неалогий на мир растений и животных. Вскоре стали появляться статьи Лю-
бищева, где тот напомнил свои сомнения в универсальности иерархической
(древовидной) систематики. Ботаник-структуралист В.В. Корона (Екатерин-
бург) сказал наиболее четко: древо, как форма системы, связано не с идеей
эволюции, а с удобством, с тем, что
“наиболее рациональной формой упорядочения информации является форма
дихотомического ключа”, что “Монофилия и дивергенция - это логическое
следствие принятых постулатов, а не собственная форма системы” [Корона,
1987, с. 32].
Этот тезис Короны можно выразить так:_____________________________
Древо как форма классификации отражает свойства не объекта
______________классификации, а мышления об этом объекте.______________
Принять этот тезис - значит признать, что способ изображать систему
древом искусствен по-существу. Логически этот тезис доказать или опро-
вергнуть невозможно, но с его позиции многое становится понятнее. По
словам Короны, в нем «причина устойчивости филогенетической системати-
ки» как явления науки, хотя она и не решает ни одной из стоящих перед нею
задач (в частности, не дает конкретных устойчивых систем).
Задач этих он видел три: «построить естественную систему органиче-
ского мира, выяснить историю его развития и найти наиболее упорядочен-
ную форму представления существующего многообразия».
Как видим, историчность и естественность у него - разные понятия. Мо-
гу добавить, что удобство рассуждений - тоже понятие историческое: 300
лет назад в виде древ классифицировали всё на свете, и это было удобно,
поскольку в умах царила идея иерархии; а сейчас она доживает свой век.
Век этот будет долог (систематику невозможно сменить декретом, да и не-
ясно пока, на что менять), но на эволюционную мысль идея иерархии таксо-
нов влиять заведомо не должна. Если в процессе эволюции и есть какая-то
568
естественная иерархия, она должна быть выявлена собственно эволюци-
онными средствами, а не извлечена из систематики.
10-2. Филогенетика, кладизм и систематика
Итак, классификация организмов строится как итог реально текшей
эволюции, тогда как сама эволюция мыслится как реализация старинного
классификационного принципа. Иногда сами систематики признают:
«Это порочный круг - что корректная филогения требует установить обстоя-
тельства конвергенции (о ней см. п. 6-2 - Ю. Ч.), тогда как для установления
факта конвергенции надо построить корректную филогению» (Moore J., Will-
mer Р. Convergent evolution in invertebrata // BR, 1997, № 1, p. 48).
Порочного круга можно избежать, если процедура попеременного об-
ращения к зависящим друг от друга массивам данных итеративна, т.е. ес-
ли сами массивы от шага к шагу корректируются практикой. Такую процеду-
ру предлагал Мейен (п. 6-3**), и понятна надежда на ее успех:
«Хочется надеяться, что по мере прохождения такой процедуры систематик
последовательно приближается к истине» [Михайлов, с. 42].
Однако из математики известно, что отнюдь не все итеративные проце-
дуры сходятся; некоторые блуждают вокруг искомой точки, другие удаляют-
ся от нее (расходятся). Приведенные в п. 10-1 примеры (о собаках, стреко-
зах) говорят, что надежны Михайлова сбываются не всегда. Непонимание
этого непрестанно порождает путаницу.
Ее допустил и Джеффри, хотя хотел прекратить путаницу. Целью сис-
тематики он назвал поиск сходств, а не родства, но на той же странице объ-
явил единственной причиной возможности построения систем феномен
родства. Далее (с. 16) единственной формой системы он назвал иерархию
(древо), что не помешало ему в конце работы заметить совсем иное:
“если систематики принимают какую-то иную, не иерархическую... систему
классификации, лежащую в основе существующих кодексов, то они не только
могут, но и должны принять систему номенклатуры..., отличающуюся от тех,
которые установлены кодексами” (с. 85).
Как понимать это важное заявление, не сказано.
Дарвинизм, равнодушный к анализу своих основных понятий, мог тер-
петь эту путаницу (порочный круг), но вне его всегда жила оппозиция ей.
На мой взгляд, надо последовать за теми немногими, кто думал о сути
систематики после Линнея. Далее изложена точка зрения на естественность
системы, описанная в статье [Любищев, 1972], в параграфе 1.3 книги [Коро-
на, 1987] и параграфах 6.1 и 6.2 книги [Чайковский, 1990], где есть ссылки на
соответствующую литературу.
Должна ли естественная система организмов отражать их эволюцию?
Если да, то будет ли она иерархической или какой-то иной формы? Если
нет, то что она должна отражать и какую форму иметь? Только ли эволюци-
онную систему можно считать естественной? Должна ли эволюционная сис-
569
тема строиться на идее полезности свойств? Эти и подобные вопросы почти
не обсуждаются, но каждый систематик имеет на них свой особый ответ, по-
этому систематика вязнет в противоречиях.
Линней понимал, как мы видели, что естественность системы - нечто
более глубокое, нежели удобство адресации, и предложил строить две сис-
темы растений: искусственную (для целей адресации, ее он построил) и ес-
тественную (для уяснения «сродства» видов, которое он понимал как некую
общую близость, но не как родство - ее он едва наметил).
Мишель Адансон, его младший современник, видел естественной ту
систему, в которой рядом стоят таксоны, близкие по максимально возмож-
ному числу признаков. В 1820-х годах Бэр уточнил естественность как сход-
ство общего строения, а не отдельных, пусть и многих, частей или свойств.
Он полагал, что естественную систему следует то и дело пересматривать в
связи с новыми данными, и это мнение бытует до сих пор.
Наоборот, английский логик и историк науки Вильям Уэвелл вскоре
предложил понимать естественность системы как устойчивость - чтобы ее
не требовалось пересматривать при появлении новых классификационных
признаков (при более детальном изучении прежних объектов). Если систему
приходится пересматривать, это значит, что принцип ее не ухвачен (эта
мысль Уэвелла идет от каталепсиса стоиков, п. 1-7*).
Систематику ныне принято строить на “устойчивых” признаках89, а ими
часто оказываются бесполезные. Этот прием противоречит позиции дарви-
низма «всё долговечное полезно», но на деле всеми используется.
Мейен делал иной вывод: “функциональность в принципе непригодна к объяс-
нению всего разнообразия живого”, и ее можно прилагать “лишь к отдельным
особенностям организмов” [Заварзин, 1990, с. 121]. Замечу, что не будет проти-
воречия, если перевернуть установку Мейена, т.е. признать, что как раз отдель-
ные особенности чаще у всего бесполезны, а вот общие свойства обычно нужны
его обладателям. Вместо этих противоположных точек зрения лучше признать,
что систематику нельзя строить априорно (на функциональных свойствах или на
отрицании их значения); систему каждого таксона надо строить на тех свойствах
организмов, которые именно для него нужны при построении.
Для Дарвина и Геккеля естественность системы состояла в общности
предка каждой группы, и это понимание господствует. Утверждение: «сход-
ство свидетельствует о родстве» - игнорирует ту истину, что само понятие
родства почти всегда основано на сходстве и ни на чем другом.
од
Устойчивых в смысле долговечности в эволюции. Вот где видится логический про-
счет: устойчивость во времени может быть принята за основу классификации только
при построении системы по ископаемым данным при наличии непрерывных рядов, что
бывает крайне редко (п. 5-1). На самом деле система обычно строится по ныне живу-
щим, и устойчивым можно счесть любой часто встречающийся признак. Вот почему
Уэвелл брал за основу принцип устойчивости не признаков, а всей системы.
570
Сто лет назад математик Анри Пуанкаре высказал уверенность, что ни-
какая детальная система не может быть выстроена окончательно, и в каче-
стве естественной предложил понимать параметрическую систему, т.е.
систему, в которой положение объекта задается одним (как число протонов
в системе Менделеева), двумя (как координаты на географической карте)
или несколькими параметрами. Пример двух равно естественных систем:
указание химического элемента либо его номером, либо парой координат в
таблице (строка и столбец).
Любищев в 1923 году фактически соединил Пуанкаре и Уэвелла, при-
знав естественной ту систему, «где все признаки объекта определяются по-
ложением его в системе». Образцом он считал систему Менделеева, в ко-
торой номер элемента определяет его свойства.
Вавилов и Кренке видели естественность системы в возможности про-
гнозировать свойства ее элементов (находить новые формы).
В те же годы друг Любищева зоолог Беклемишев, признав негодность
принципа родства для систематики, сохранил, однако, идею иерархии как
единственно годную для построения естественной системы. Эта вроде бы
странная позиция вызвана тем, что саму естественность он понимал отнюдь
не как выяснение какой бы то ни было “природы вещей”, а лишь как краткую
адресацию, и считал идеалом понимание ее по Адансону (Беклемишев В.Н.
Методология систематики (1928). М., 1994, с. 28).
С позиции тезиса Короны, Беклемишев описал идеал вовсе не естест-
венной, а искусственной системы. Но его книга полезна для нашей темы,
показывая невозможность построить работоспособную чисто комбинатив-
ную систему (сеть) - она слишком избыточна и потому громоздка. Но ком-
бинативный принцип может быть применён как составная часть, как один из
приемов, которыми совместно строится система, что мы и увидим.
10-2* Систематика как отражение наличной картины мира
В 1966 году французский культуролог-структуралист Мишель Фуко пи-
сал, что естественность системы определяется ее соответствием тем пред-
ставлениям о структуре мира, которые в данное время господствуют в об-
ществе. И он был прав - в том смысле, что никакое накопление противоре-
чий не заставит систематиков отказать системе в естественности, пока не
сменится картина мира по Хайдеггеру (п. 5-16; вспомним тут заодно и тезис
Витгенштейна, п. 3-7**).
Систематик и ламаркист Е.С. Смирнов в те же годы видел естествен-
ность системы в ее общепризнанности, окончательности и единственности,
«не допускающей вариантов». Эта попытка развить Уэвелла неудачна: до
сих пор всякое “окончательное” решение оказывалось временным. Но пози-
ция Смирнова заслуживает внимания, поскольку всякий систематик, пред-
лагающий новую систему, надеется, что она окончательна. А недавно пред-
ложен совсем оригинальный критерий естественности:
571
“Когда я начинал заниматься функциональной морфологией... я и не помышлял
о построении какой-либо системы”. Она “сложилась сама собой. И это, на мой
взгляд, является главным доказательством ее естественности” [Павлов, с. 6].
Картина мира, усвоенная обществом, порождает в нем новую ПМ. Не
имея здесь возможности сопоставить таксономическую и эволюционную по-
зиции каждой эпохи с ее моделями, скажу только, что эпоха рождения ие-
рархического понимания системы вообще и естественной в частности (XVI-
XVII века) характерна господством знаковой ПМ, в которой мир понимается
как текст или шифр (у Короны - «фигуры подобия»). Эта ПМ обычна для
времени становления научной дисциплины, а затем сменяется моделью,
господствующей в данную эпоху (так, в генетике знаковая ПМ ныне сменя-
ется на системную). Мне ближе всего диатропическая ПМ. Согласно ей, ес-
тественна та система, которая отражает реальную диасеть.
Прежде всего надо вспомнить о квази-гиперболах Виллиса, поскольку
они показывают, что система организмов (по крайней мере, многоклеточ-
ных) имеет общую для всех таксонов структуру, притом фрактальную (п. 6-
11*).Наличие единой структуры дает простое пактическое правило:
если состав таксонов в системе данной группы сильно отклоняется от
_________квази-гиперболы, то система, вернее всего, неестественна.______
Здесь возможен тот же упрек, что и к филогенетике: не являются ли ги-
перболы Виллиса свойством мышления систематиков, а вовсе не свойством
объектов систематики? В прошлом такой упрек звучал многократно, но в
наше время можно уверенно ответить: нет, не являются (п. 6-11*).
Затем, надо определить, что именно подлежит систематизации. Та кар-
тина мира, в которой это были ветви родословного древа, быстро уступает
место картине генетического единства (в силу горизонтального переноса)
всей живой природы, а в этой картине мира любые древа бессмысленны.
Поэтому объектом классификации всё чаще видят жизненную форму (п. 6-
13*). Ей-то и приписывают естественность по Любищеву:
«прикладная ценность понятия жизненной формы состоит в том, что можно
взять малоизученный вид... отнести его по немногим известным параметрам к
той или иной единице системы (иерархии) жизненных форм и предположить по
аналогии..., каковы могут быть основные, пока не исследованные черты биоло-
гии данного вида» (Несис К.Н., Нигматуллин Ч.М. Жизненная форма... // ЖОБ,
2003, № 3). Замечу: иерархией тут названа многоуровневость.
Поскольку, как выше сказано, авторы отказываются доводить систему
до вида, то очевидно, что она не призвана заменить нынешнюю систему ад-
ресации, а лишь дает дополнительный к ней взгляд на разнообразие. Ниже
(п. 10-5 и далее) мы поступим также.
Наконец, надо задать себе вопрос: если не идея общего предка, то что
именно позволяет строить систему в форме иерархии (древа)? Ведь именно
для иерархических систем биологических таксонов обнаружено правило
572
Виллиса. Ответ следует из самого факта общности (выходящего далеко за
пределы биологии) феномена квази-гипербол: они возникают в любой ус-
тойчивой (долговечной) системе, причем система может пониматься не
только в таксономическом, но и в функциональном смысле.
Так, Кудрин постоянно подчеркивает, что любой техноценоз (например,
завод) устойчив только тогда, когда его компоненты (например, электромо-
торы по мощности) распределены квази-гиперболически [Кудрин, с. 26]. Это
значит, что для классификации могут быть выбраны в качестве естествен-
ных очень разные принципы, но отнюдь не всякие, а лишь те, которые удов-
летворяют некоторым соотношениям системности. Их, в основном, еще
предстоит выявить, но один уже известен - квази-гипеболичность распре-
делений подгрупп в группах. Из этого и будем исходить.
10-2** Кладизм
Если мы можем упорядочить мероны, то, проведя сперва разделение
объектов по принципу значений первого мерона, затем - второго и т.д., по-
лучим иерархию. Так, толпу военных легко упорядочить, разделив их по ро-
дам войск (все роды войск суть состояния одного мерона - “род войск”), за-
тем в каждом роде войск разделить людей по званиям, а всех лиц одного
рода и звания выстроив по росту. Подобным образом можно упорядочить
всё что угодно, даже набор случайных чисел: сперва разделить их на чет-
ные и нечетные, затем в каждой из этих двух групп - на кратные трем и все
прочие, затем на кратные пяти и т.д. Это значит, что данный способ клас-
сификации использует конкретные свойства изучаемой структуры не боль-
ше, чем мясорубка использует форму перемалываемых тел.
Такая система (где иерархия получена путем упорядочения меронов),
называется кладистической. Ее можно построить для любого множества
неодинаковых объектов, и она всегда окажется иерархической. Похожим
способом Линней построил свою систему цветковых (см. [Любищев, 1972];
Чайковский Ю.В. Систематика видов и систематика царств И БШ, 1996, №
4), и никто (в том числе и он сам) не полагал ее естественной..
В такой системе самой по себе нет истории. Тем не менее именно кла-
дистические системы распространены в качестве эволюционных. Пока это
противоречие не осознано, нельзя надеяться на построение системы, кото-
рая окажется в каком-то смысле естественной, например, долговечной.
Для примера рассмотрим обширную статью “Филогения змей” (Lee M.S.Y.,
Scanlon J.D. Snake phylogeny based on osteology, soft anatomy and ecology // BR,
2002, № 3). Авторы заявили, что кладистический анализ подтвердил монофиле-
тическую суть подотряда змей, т.е. допущение, что все змеи произошли от еди-
ного предка. Работа проделана огромная — для анализа взято 263 мерона, и зна-
чения их выписаны для всех рассмотренных групп змей.
Однако удивляет полное отсутствие упоминания яда и ядовитых зубов — того,
что для большинства людей считается главным у змей. (Сто лет назад змей клас-
573
сифицировали именно по наличию и форме ядовитых зубов, по их положению
во рту змеи, а в середине XX века было выяснено, что яды бывают двух совсем
разных типов и что это говорит о родстве.)
Оказывается, авторы вообще не анализировали надсемейство ужеобразных
(Colubroidea), в котором состоит 3/4 всех видов змей, в том числе все ядовитые.
Оно взято лишь как единица, равноценная остальным, мелким, причем в харак-
теристику ужеобразных включено только 8 меронов. Столь урезанный анализ
привел к цели, но в весьма странном смысле: от прежних групп пришлось отка-
заться и ввести те, какие диктовал кладистический метод, да и то весь подотряд
охватить не удалось. Мне не удалось сравнить их систему ни со старыми, ни с
новой (в руководстве Кэрролла, которым мы пользовались в гл. 9), настолько
различны названия. Становится досадным правилом - вводить свою филогению-
однодневку, даже не сопоставив ее с известными системами (такова же, напри-
мер, “система” ластоногих И.Я. Павлинова - см. Дополнение 1).
Практически такая филогения бесполезна и явно не будет принята системати-
ками, но она вряд ли нужна и теоретически, ибо не говорит об эволюции змей
почти ничего. В самом деле, ядовитость (одна из главных тенденций эволюции
змей) не рассмотрена; если же ее учесть и включить в анализ хотя бы некоторые
другие из важных меронов надсемейства ужеобразных, его монофилия рухнет и
будет не к чему пристроить остальные группы змей.
Вскоре тот же Майкл Ли опубликовал с соавторами (BJ, 2004, v. 81, № 4)
иную филогению змей, основанную на иных признаках, с иным общим предком,
причем статья в BR даже не помянута. К сожалению, для кладистов это довольно
обычно, а значит, вопроса о долговечности системы они вообще не ставят.
Кладизм является лишь развитием до логического предела (а он на
практике обычно означает абсурд) иерархической идеи, и его обсуждение
не имет смысла, пока не выработано отношение к методу иерархии вообще.
Согласно тезису Короны, данный метод не только приемлем, но и неизбе-
жен в практике, хоть и не связан по сути с идеей общего предка. Другое де-
ло - должна ли иерархия принимать форму кладизма.
Мне ситуация видится прямым следствием утери общебиологической
культуры у предыдущего поколения англоязычных биологов (см. п. 4-17, пе-
тит), повлекшей забывание основной таксономической проблематики. Их
позиция, на мой взгляд, вызвана полным неведением всего, что сделано
вне кладизма. Ныне даже убежденный филогенетик Расницын пишет:
“Кладистика... соответствует западному, прежде всего американскому миро-
ощущению, хотя и там уже возникают вопросы и скепсис” [Расницын, с. 26].
Поэтому представляются интересными слова Зелеева:
“Для создания непротиворечивой системы организмов иерархический подход
неприемлем, так как, наряду с субъективностью выделения ведущих призна-
ков, здесь очевидно смешение степеней родства и уровней организации. В
целом естественная система видится, по меньшей мере, трехмерной, где кро-
574
ме оси времени, отражающей историческую преемственность организмов,
должны быть отражены уровень организации и экологический параметр - ме-
сто организма в экосистеме” (Зелеев Р.М. Подходы к построению системы ор-
ганизмов //ЛЧ, 2000).
Сложность строения иногда бывает отражена и в таксономии: так, в ти-
пе членистоногих состоят организмы, устроенные сложнее, чем в типе коль-
чатых червей, а в классе насекомых - устроенные сложнее, чем в классе
многоножек. Однако в целом надо согласиться с Зелеевым: систематика
строится не на сложности строения (не на целостности), а на совокупности
признаков, и в таксон одного ранга могут входить как организмы близких, так
и очень различных уровней сложности. Некоторые типы (губки, кольчатые
черви, семенные растения) таковы, что каждый из них достаточно одноро-
ден по сложности строения, тогда как другие типы (моллюски, хордовые, зе-
леные водоросли, красные водоросли) состоят из крайне различных по
сложности строения организмов каждый. Так, и ланцетник, и человек - хор-
довые. И наоборот - один уровень сложности можно наблюдать у очень
таксономически далеких организмов, о чем уже шла речь в п. 6-7. Другими
словами, общепринятая систематика никакой картины мира не дает.
Еще до Зелеева высказан аналогичный тезис Кирилла Михайлова:
«Помимо системы собственно таксонов, существуют, по меньшей мере, еще
две - система морфологических уровней и система жизненных форм»
[Михайлов, с. 42]. Далее нам понадобятся все три оси. Тезис перекликается
с тезисом Соболева (п. 4-12), но кладисты не видят этого. Удивительно, что
они уверены в строгой обоснованности своего метода, хотя в центральном
пункте их рассуждений всякий анализ отсутствует. Дело в том, что у них вы-
бор признака, годного для классификации, ведется не из биологических со-
ображений, а из уверенности в ничтожно малой вероятности совпадений:
“Кладистический анализ базируется на предположении о неповторимости и
необратимости эволюционных преобразований (известном, как закон Долло).
А это предположение, в свою очередь, основывается на соображениях веро-
ятности: вероятность того, что одно и то же изменение в генотипе может про-
изойти независимо несколько раз, настолько мала, что ее можно считать рав-
ной нулю. Таким образом, предполагается, что каждая апоморфия (признак,
годный для классификации - Ю. Ч.) возникла один раз и унаследована всеми
потомками” [Клюге, с. 15].
То же в иных словах пишут и другие (например: [Медников]), но важно
понимать, что в основе тут лежит очень частное понимание случайности:
принято, что все варианты равновероятны, а это заведомо не так. Противо-
речиво и понимание филогенетиками задач систематики: целостное по
форме заявлений, оно на деле оперирует отдельными признаками.
Слова Короны, Михайлова и Зелеева наводят на мысль, что если ие-
рархическая система допустима, то не в качестве единственной.
575
Ю-2*** Какую же систему считать эволюционной?
Итак, естественная и эволюционная системы - различные понятия:
“организмы связаны не только генеалогическими, но и многими другими ти-
пами связей со средой и друг с другом, поэтому даже строго и однозначно ус-
тановленная генеалогическая система не более естественна, чем любая дру-
гая, построенная на иных типах связей” [Корона, 1987, с. 32].
Построено много систем, игнорирующих исторический подход к систе-
матике. Из них некоторые интересны и даны у Короны (там же), но беда их в
том, что они пытаются заменить существующую филогенетическую систе-
му, а это вряд ли возможно, поскольку она лежит в основе всех руководств и
сводок, так что заменять ее поздно. Возможно, что она сама собой понемно-
гу будет вытеснена из науки и практики, как в течение XIX века была вытес-
нена половая система цветковых Линнея, но сделать это волевым актом
невозможно. Единственное, что мне видится реальным и разумным, это -
обрисовать систему, которая будет эволюционной и отразит то, чего не мо-
жет филогенетическая. Однако - не взамен ей, а в логическое дополнение.
Поясню, что это может значить. Сам по себе филогенетический метод
ни в какой модификации не дает картины мира, поскольку сам мир не явля-
ется иерархическим (в смысле сноски к п. 6-6). Мир является сложным пе-
реплетением древовидных и сетевидных структур, поэтому выявление в
нем какого бы то ни было древа осмыслено только при условии коррекции
древа соответствующей сетью (и обратно). В частности, кладизм, соеди-
няющий крокодилов с птицами, разрывающий отряд ластоногих (относя их к
совсем несходным отрядам на основе нескольких модных сегодня призна-
ков - см. Дополнение 1) и свершающий много подобных насилий над при-
родой (в наших терминах над диасетью), не годится для построения систе-
мы-древа. Наоборот, устоявшаяся система цветковых годится.
Примечательно, что вне биологической систематики мало кто отобра-
жает историю в форме древ, а если говорить специально о графических
отображениях, то чаще всего используется карта: историю очень удобно
изображать как распространение явления в пространстве, а его возникнов-
ние - как точку в этом пространстве. Именно так Мейен хотел изобразить
флорогенез. Но карта не обязана быть географической: явление может воз-
никать и распространяться в любом абстрактном пространстве.
По-моему, система-карта должна, говоря ботаническим языком, являть
собой план парка, где рассажены кусты и деревья, обозначающие частные
системы. На деле у нас речь пойдет о кустах, ибо нынешние системы непо-
хожи на прежние древа (см. рис. 76а,б). Для пояснения ситуации в ЧЭ на с.
418-419 были также приведены две филогенетические системы млекопи-
тающих - по Геккелю, 1866 год (ветвистое древо), и по новейшей офици-
альной схеме Академии наук (куст без единого достоверного ветвления, где
все немногие гипотетические ветвления отнесены в далекое прошлое).
576
Именно повсеместное вытеснение крепких “древ" робкими “кустами”,
происшедшее в течение XX века, наводит на ту мысль, что сам принцип
филогении исчерпан (ср. п. 9-15), что ныне он неадекватен материалу и по-
тому неинформативен, что вскоре он будет вытеснен более реальным.
Главная причина ущербности всех филогений - объективно сущест-
вующая рефренная структура таксонов и вызванные ею параллелизмы.
“Причину параллелизма С.В. Мейен усматривает в мозаичном типе эволюции
частей целого... В этом принципиальном аспекте позиция С.В. Мейена, осно-
ванная на палеонтологическом материале, близка к позиции... биологов, ис-
пользовавших сравнительный метод изучения многообразия, в том числе и
позиции рецензента” [Заварзин, 1990, с. 120].
Система-парк будет гораздо информативнее. Она описана в п. 10-8.
10-3. Альтернативы кладизму и филогениям вообще
Царящая в биологии филогенетическая систематика лежит вне рамок
данной книги, поскольку диатропический и функциональный подходы вме-
сте противостоят филогенетическому. Здесь, как и в книге [Лима], главная
тема - не происхождение видов и иных таксонов, а происхождение форм и
функций. Поэтому поговорим об альтернативах филогениям.
Как сказано в начале главы 8, один и тот же эволюционный итог может
быть достигнут различными путями. Причин этого ныне известно две: гене-
тическая (горизонтальный перенос) и диатропическая (комбинаторный ха-
рактер разнообразия), причем первая означает, что таксон может иметь
двух неродственных предков. О филогении (родословном древе) нет смыс-
ла говорить в обоих случаях. Если так, то древо - чисто мысленная конст-
рукция, не имеющая эволюционного смысла (тезис Короны).
Но фактически древо может достаточно хорошо описывать систему, ес-
ли разнообразие группы устроено так, что связей между ветвями (кроме то-
чек ветвления) мало и ими можно в первом приближении пренебречь.
Кладизм, филогенетика и систематика - мысленные конструкции, о
соотношении которых можно спорить, тогда как эволюция есть реально
шедший в природе процесс, свойства которого надо не постулировать, а
искать. Что касается кладизма, он не является ни описанием, ни моделью
эволюции ни для одной группы, поскольку исходно задуман как средство
классификации ныне живущих организмов. Но он может быть полезен, ес-
ли поставит свой рабочий прием в один ряд с другими.
Кладизм надо исследовать еще и потому, что он очень распространен
и лежит в основе геносистематики, на которой строятся некоторые но-
вые эволюционные концепции. См. об этом: ЧЭ, с. 423-424. Мы ею пользо-
ваться не будем, так как в ее основе лежит «метод молекулярных часов»
(допущение, что скорость некоторого класса точковых мутаций одинакова
у всех организмов во все эпохи), весьма сомнительный (Гречко В.В. Моле-
кулярные маркеры ДНК... // ЭГ, 1015). Напомню лишь, что сами данные ге-
577
носистематики друг другу противоречат: «Не было построено двух одинаковых
деревьев даже в одной лаборатории» (Карпов С.А. "Рибосомальные" деревья И
Протисты. Ч. 1. СПб., Наука, 2000, с. 126). Это противоречие давно взято на воо-
ружение серьезным научным креационизмом (ЮШ, с. 118-120).
Для англоязычной систематики характерно полное неведение структу-
рализма вообще и гомологических рядов (хотя открыл их Коп) в частности.
Работы первой трети XX века, искавшие закономерность в строении систе-
мы организмов (Виллис, Вавилов и др.), на Западе забыты. Там даже вдум-
чивые авторы, видящие негодность кладизма в качестве эволюционной сис-
тематики, не предлагают ничего взаменю. Поэтому опорой в попытках по-
строения структуралистской систематики в основном оказываются русскоя-
зычные авторы. По Мейену и Шрейдеру, классификация - это
“разбиение любого множества (класса) объектов на подмножества (подклас-
сы) по любым признакам”, а систематика — установление такой классифика-
ции, которая признана привилегированной, “выделенной самой природой”,
т.е. в некоем смысле естественной (Мейен С.В., Шрейдер Ю.А. Методология,
аспекты теории классификации//ВФ, 1976, № 12).
Как видим, систематика названа естественной классификацией; вряд ли
термин удачен, но работать с ним можно. Авторы отметили, что факт суще-
ствования естественной системы неочевиден. Законны вопросы: есть ли
способ достичь естественной системы даже в предположении, что она су-
ществует? Что именно должна упорядочивать систематика?
Есть как системы чисто исторические (всевозможные хронологии), так и
чисто структуралистские, из которых самые известные — грамматика и
система химических элементов. Первая послужила исходным пунктом для
построения диатропики, но во второй мероны и таксоны вводить, по Мейену,
неразумно (Забродин В.Ю. И МС, 113-114).
Все языковые учебники подают грамматику как систему правил, невесть
откуда взявшихся, и только будущим филологам дают знать об их происхо-
ждении. Это правильно: вряд ли многим интересно, что такие русские поня-
тия, как склонение, падеж, спряжение, наклонение и т.п., суть кальки с гре-
ческого. Однако предмет «Историческая грамматика» существует, и никакой
филолог без его усвоения не состоится. (А химикам вообще не рассказыва-
ют об эволюции элементов - вот это вряд ли правильно.) Для биологии эта
аналогия означает, на мой взгляд, что историческая система интересна уз-
кому классу специалистов, занятых именно эволюцией, тогда как остальным
гораздо важнее, чтобы система четко отвечала не на вопрос: «кто от кого?»,
а на вопросы «кто это?» и «где его место?».
Иерархическую систему организмов часто сравнивают с библиотечным
каталогом и с адресной книгой. Но вряд ли найдется библиограф, который
по мере получения новых книг переписывает карточки старых, тасует ста-
рые карточки в ящиках и переставляет сами ящики. Аналогично, имя и ад-
рес жителя не меняются от того, что рядом кто-то поселился или умер. А
578
ведь биологи-систематики заняты как раз подобными изменениями - изучив
одни виды, меняют названия и “адреса” других.
Но каталог и адрес - искусственные системы, от них требуется только
удобство поиска. Никто не ищет в них историю книгопечатания или города.
Для этого есть другие способы записи - книги по истории. И только в био-
систематике царит странное обыкновение извлекать адрес из истории, при-
том не из реально наблюдаемой, а из найденной по тем же адресам.
Каталог иерархичен (шкафы состоят из ящиков, главы - из параграфов
и т.д.) и потому допускает любую степень детальности отдельных тем,
зато крайне ограничен в возможностях учитывать сходства. В сущности,
такая возможность тут одна: помещать сходные объекты в общую группу.
При этом невозможно избежать ситуации, когда две соседние карточки от-
ведены очень различным объектам. Она возникает всякий раз, когда конча-
ется один раздел и начинается другой.
Наоборот, карта (план) обладает тем достоинством, что близкое в при-
роде всегда близко на карте и наоборот', поэтому естественно отобража-
ется феномен общей границы - это каталогу недоступно. Положение объек-
та просто и однозначно задается парой его координат (параметров систе-
мы). Для общего знакомства с городом самая простенькая карта куда по-
лезнее, чем адресная книга. Зато детальность карты жестко задана ее
масштабом, и если детальность надо увеличить, приходится всю карту за-
ново чертить на гораздо большем листе, что лишает ее прежней наглядно-
сти. Словом, каталог и карта друг друга заменить не могут, и нет смыс-
ла спорить, какая из этих систем более естественна.
Линней хотел изобразить (но так и не сделал этого) систему цветковых
в виде карты и полагал как раз такую систему естественной. Наоборот, мно-
гие последующие систематики строили графические системы, в какой-то
мере напоминавшие карту. Например, в 1817 году немецкий зоолог Георг
Август Гольдфус расположил группы животных от простейших (инфузории)
на востоке до человека на западе, причем холодных рептилий поместил на
севере, а теплокровных птиц на юге. Тем самым, в основу классификации
был в вычурной форме положен уровень организации. Его система есть в
книге: Анналы биологии, т. 1. М., МОИП, 1959, с. 297.
Как бы вторя ему, в 1839 году английский хирург и ботаник-любитель
Томас Баскервиль для низших растений избрал южное полушарие: у полю-
са - водоросли, выше них мхи, лишайники и пр.; для высших же отвел се-
верное: у полюса - астровые, т.е. высший порядок двудольных, пониже их
помещены колокольчики и т.д. Осью запад - восток он пренебрег, зато че-
рез карту наискось идет линия раздельнополые - слитнополые, так что по-
следние, как и надо, у цветковых преобладают. Этим он отобразил в своей
системе важную тенденцию. Его книга есть в Библиотеке МОИП.
Да и Геккель не всегда ограничивался родословным принципом. Через
23 года после «Общей морфологии» он предложил другую схему эволюции, в
579
которой признал, что растения и животные независимо достигли тканевого
уровня организации (рис. 77). Здесь в зачаточной форме заметен тезис Ми-
хайлова (п. 10-2**), гораздо лучше видный у Гольдфуса и Баскервиля.
Царство Царство
Рис. 77. Эволюционная система царств по
Геккелю (1889), основанная на параллелизме
тканевых (Histonia) организмов
Из новых систем сошлюсь на систему
голосеменных Мейена, п. 10-6, где на
рисунке соединены исторический
(детально) и географический (в самом
грубом виде) принципы. Наглядность их
несомненна, но возможности детализации
малы. Системы-карты более высоких
таксонов будут рассмотрены в конце
главы. Общее в них то, что они претендуют
не на детальность (доведение до вида), а
на общий абрис множества.
10-3* Экоморфологическая систематика
В биологии царит понимание эволюции как процесса приспособления,
однако систематика избегает использования “адаптивных сходств”, трактуя
их как параллелизмы, которые, как мы знаем, она игнорирует. Отметив этот
факт, И.М. Мирабдуллаев (Ташкент) напомнил о наличии иной систематики,
экоморфологической, и заключил, что, по-видимому,
“филогенетическая и экоморфологическая систематики - законные и взаимо-
дополнительные (в представлении Н. Бора) дисциплины, описывающие две
стороны процесса эволюции — дивергенцию и адаптацию” {Мирабдуллаев И.М.
О происхождении и положении грибов в системе... // УСБ, 1994, № 1, с. 32).
Мысль о двух систематиках очень важна (хотя и неясно, почему парал-
лелизмы надо считать адаптациями), и Мирабдуллаев не одинок:
“таксономия нуждается в создании прогностической системы органического
мира, но в рамках... филогенетического подхода такая система невозможна.
Потенциалом прогностичности обладает экоморфема - система... учитывающая
общие закономерности развития живой природы” [Леонтьев, Акулов, с. 500].
По-моему, прогностичность (возможность предсказания) для система-
тики не так уж важна, но на самом деле авторы имеют в виду практическую
полезность (о ней мы говорили выше). Они справедливо отказывают в ней
нынешней систематике и мечтают о системе, «которую можно не только вы-
учить, но и понять» (с. 502). К сожалению, они, как и Мирабдуллаев, ничего
не сказали о том, какая из двух систем может обеспечить практичную и дол-
говечную адресацию. Зато они развивают идею макросистемы.
580
Макросистемой обычно именуют систему царств живой природы. С
древности организмы делили на растения и животных, и в этом видели раз-
деление их функций: растения производят, а животные потребляют. За по-
следние сто лет выяснилось, что животные отходы становятся пищей для
растений в основном после их минерализации микробами и грибами. Уитте-
кер (п. 8-2) свел новые знания в формальную схему: царство растений -
продуценты, т. е. производят органическое вещество путем фотосинтеза;
царство грибов - редуценты, т. е. разлагают тела и отходы растений и жи-
вотных; а царство животных - промежуточное звено, консументы (потреби-
тели). Подобно трем лепесткам, эти три эвкариотные царства вырастали в
схеме Уиттекера из общего основания - царства протист (простейших), в ко-
тором тенденции всех трех высших царств перемешаны (представлены у
одного и того же организма одновременно). При этом речь шла только о
тенденциях, но не о точных абсолютных законах и не о четких границах. Для
построения макросистемы этого вполне достаточно, поскольку она призвана
выявить именно тенденции.
Итак, макросистематика исходит из того, что царства являются эко-
физиологическими единствами, но отнюдь не монофилетическими, т.е. что
нет оснований искать для каждого из них единого общего предка. Это
очевидно с той точки зрения, что во все эпохи жизнь существовала в форме
экосистем, а потому единый общий предок немыслим. Однако до появления
ГЦК трудно было спорить с утверждением, что первый организм жил в пото-
ке органических веществ и лишь по их исчерпании породил экологическое
разнообразие; с ее появлением стало ясно - Вернадский был прав, говоря,
что жизнь никогда не могла быть представлена одной формой. Еще остава-
лась возможность допустить единого предка всех эвкариот, но и она отпала
по выявлении их разнообразия: слишком велико сходство одноклеточных
эвкариот с соответствующими прокариотами.
До рождения диатропики было эмоционально трудно принять, что столь
сложная и единообразная структура, как эвкариотная клетка, могла поя-
виться несколько раз независимо - отсюда и популярность гипотезы сим-
биогенеза. Однако последняя рухнула в качестве научной (пп. 6-5*, 8-7*) и
бытует, в основном, в учебниках. Возникшая на ее руинах гипотеза сери-
ального эндосимбиоза (п. 8-7*) признаёт многократность актов рождения эв-
кариотной организации.
10-4. Родство и сущность
Казалось бы, удобство построения макросистемы в форме таблицы
очевидно. Однако развитие макросистематики пошло по иному пути: воз-
никла мегатаксономия, т.е. мода строить многоцарственные филогенети-
ческие системы. В каждой вводится около 20 монофилетических царств без
какого-либо (кроме монофилии) принципа, вводится простым перечислени-
ем царств. Из привычных всем царств (бактерии, растения, грибы и живот-
581
ные) обычно сохраняются только грибы, зато появляются мелкие и мель-
чайшие новые «царства» (есть даже из одного вида)90. Не упорядочены и
типы внутри царств. Такую систему можно только заучить, но и это беспо-
лезно, так как через год-два появляется новая, а о старых вскоре уже никто
не вспоминает, даже их авторы.
Словом, системы как таковой не дается. Давно пора спросить: когда это
кончится? Когда появится система царств, о которой можно будет, наконец,
рассказать в школе или хотя бы в вузе? Ответ прост и исторически прове-
рен - тогда, когда авторы “мегатаксономий” прекратят громоздить перечни и
задумаются о сути дела, т.е. займутся теорией. И, прежде всего, спросят
себя: что такое родство групп, существует ли оно объективно и если да, то
как его в принципе можно найти? Чтобы система была хоть сколько-то дол-
говечна, она должна быть лишена грубых внутренних противоречий, харак-
терных для мегатаксономии. Ее позиция прямо противоположна диатропи-
ческой, в основе которой лежит структурализм. Он, как уже говорилось, поч-
ти неизвестен в англоязычных странах. Критику мегатаксономии см. в преж-
них вариантах данной книги (БПС, 2000, № 7, п. 2; ЧЭ, п. 10-4).
При наличии горизонтального переноса идея родства не может быть
использована для классификации в принципе. Но если таксон формулиру-
ется по принципу родства, то вопрос о наличии или отсутствии родства да-
же задать нельзя - всякий таксон состоит из родственников по определе-
нию. Вопрос о том, родственны ли группы, входящие в таксон, осмыслен
лишь тогда, когда каждая группа задается своим определением, не опери-
рующим идеей родства. Именно такую цель ставит себе диатропика и этим
может быть полезна систематике.
До сих пор успех в систематике сопутствовал тем, кто не забывал, что
такое план строения организма. Мегатаксономия даже не упоминает план
строения. Карл Вёзе, главная фигура в мегатаксономии, писал:
“Существуют две группы прокариот, (eu) Bacteria и Archaea, которые имеют
не более отношения друг к другу, чем к Eucaria” (Woese C.R. Microbiology in
transition // PNAS, 1994, vol. 94, № 5, p. 1602).
Начисто игнорирован тот факт, что все бактерии (включая археи) имеют
один план строения, а эвкарии (т.е. эвкариоты) - другой.
К счастью, мегатаксономия клонится к упадку, как и вся молекулярная
филогенетика. Тот же Вёзе теперь пишет, что молекулярная биология с ее
отрицанием эмерджентности и проблемы биологической формы «была ус-
99 Вспоминаются слова юного Фаминцына: “На основании частных признаков невоз-
можно провести рубежа между обоими царствами”, т.е. растений и животных; если же
использовать комбинации признаков, то получим столько царств, сколько захотим
(Фаминцын А.С. Организмы на границе животного и растительного царств // Сборник,
издаваемый студентами Имп. Петерб. университета, вып. 2, СПб., 1860, с. 61-62).
582
таревшей с момента возникновения» и что изображение эволюции древом
непригодно для «додарвинской эры», т.е. до появления механизма дивер-
генции (Woese C.R. к new biology for a new century // Microbiology and Mol.
Biol. Reviews, 2004, vol. 68, N 2, p. 174, 184). Замечу, что момента этого по-
явления Вёзе не назвал, как не может назвать его и никто другой.
Н.Б. Петров и В.В. Алёшин видят в молекулярных методах смысл лишь
там, где те сопоставимы с “классическими”, где систематика строится на ос-
нове планов строения. Тогда молекулярные методы смогут указать на воз-
можные в прошлом пути от одного плана строения к другому (ЭГ, 1043).
Возможна ли система, суть которой можно объяснить, не перечисляя
самих царств? Систематик А.И. Шаталкин напомнил: есть два подхода к
систематике, в равной мере обоснованных - сущностный и филогенети-
ческий. Первый подход он связал с именем Линнея (хотя, замечу, подход
гораздо старше), и состоит он в том, что в один таксон объединяются орга-
низмы, имеющие одинаковое строение. Линней выразил это в своем знаме-
нитом афоризме: «Не признаки задают род, но род задает признаки». Дру-
гой подход формирует таксоны по принципу родства, причем Шаталкин от-
мечает, что само родство нам обычно неизвестно, и мы судим о нем по
общности тех признаков, которые считаем для этого пригодными.
Примирить подходы может, по Шаталкину, такое понимание таксона:
«О таксоне обычно говорят, имея в виду морфологическую общность видов,
обусловленную единством их происхождения» \Шаталкин, с. 13].
Добавлю, что в обоих подходах главное - интуиция: как сущность, так и
родство ухватывается интуитивно, после чего ищутся признаки, формально
выражающие убеждение исследователя. Проще говоря, авторы принимают
в качестве классификационных только те сходства в строении, которые не
противоречат их внутреннему убеждению о родстве, а прочие именуют не-
зависимыми приспособлениями. Шаталкин понимает, что такой прием про-
изволен, и предлагает строить макросистему на сущностной основе, как
Линней. Единственное важное отличие нынешней сущностной систематики
он видит в том, что «суть вещей» трактуют не по Платону, не как что-то ин-
туитивно ухватываемое, а как «главное свойство» каждого таксона. (Добав-
лю: для царств эти свойства выявлены Уиттекером.) Понятие таксона нель-
зя, по Шаталкину, дать вне всей системы. Для макросистемы это означает,
что если мы хотим, чтобы каждое царство было таксоном, надо все царства
определять на основе одной и той же логики. (О ней пойдет речь в п. 8.)
Каждое из привычных нам царств (растения, грибы и животные) Шатал-
кин аттестовал не как таксон, а как «систему жизненных форм», но он уве-
рен, что определить царства как таксоны можно. Этим и займемся.
10-5. Диатропика и систематика
Чтобы быть общей, единой для всех, систематика должна изменяться
очень редко, как можно реже. Но желание строить естественную (в смысле
583
родства) систему ведет к непрестанным перестройкам, ибо то, что один счи-
тает родством, другой - параллелизмом. «Конвергенция и параллелизм... -
бич систематиков, источник ошибок. Сходство не всегда свидетельствует о
родстве» [Медников, с. 9]. Но если так, то не поискать ли иной принцип - та-
кой, который отражает сущность, более реальную, чем предполагаемое
родство? Не попробовать ли построить естественную систематику на зако-
нах разнообразия?
Вавилов, например, полагал, что усвоение биологами идеи гомологиче-
ских рядов открывает перед систематикой новые перспективы:
“Вместо бесконечно путанной номенклатуры, которую не удерживает ни од-
на память, перед систематиком ставится важнейшая задача - создать общую,
выдержанную и однотипную номенклатуру, в которой тождества форм, их
гомология были бы поставлены в основу системы. Этот путь покажется сис-
тематику революционным и его необходимо провести осторожно по отноше-
нию к установленным формам. Гораздо проще, хотя все же сложно, приведе-
ние в общие системы определителей разновидностей и рас. ... Для изучающе-
го многообразия форм вместо бесконечного запоминания бесконечных назва-
ний... открывается возможность легкого и планомерного изучения целой сис-
темы. ... По опыту систематики мы хорошо осознаем всю трудность и огром-
ность задачи, которая открывается перед исследователем. Мы понимаем, что
понадобится, может быть, столетие усилий ботаников и зоологов, чтобы пу-
тем коллективной работы подготовить общую систему организованного мира.
Но это путь неизбежный” [Вавилов, с. 21].
Вопреки ожиданиям Вавилова, за последующие 85 лет движение в ука-
занном им направлении было минимальным, и некоторый успех достигнут
не в самом низу, а на самом верху таксономии: далее будут приведены
макросистема и две системы филумов. Но в целом XX век характерен
ухудшением дела: номенклатура усложняется в прежнем направлении, по-
степенно превращаясь из биологической дисциплины в юри-дическую, но
нормы отнюдь не спасают ее от бесконечной смены бесчисленных назва-
ний. Отмечу, что классификация завела номенклатуру в отмеченный Вави-
ловым тупик во многом из-за господства дарвинизма в его геккелевом вари-
анте: систематики убе>едены, что их цель - в выяснении родства, и не по-
дозревают, что систему можно строить как-то иначе. Номенклатура покорно
следует геккелевой традиции, хотя еще Линней указывал, как мы видели в
п. 10-1*, иной путь, путь двух систематик.
Такой подход в науке известен. Например, первая классификация звезд
(по созвездиям) была произведена, когда небо представляли сферой, вра-
щающейся вокруг неподвижной Земли; система оказалась искусственной,
но ею продолжают пользоваться: так звёзды удобно находить для наблюде-
ния, хотя все понимают, что члены созвездия могут быть очень далеки и в
пространстве, и в отношении своих свойств. Никто не спорит о том, что
лучше - группировать ли звезды «естественно» (например, по спектрам и
584
светимости) или «искусственно» (по созвездиям, т.е. по угловым координа-
там) - оба способа необходимы.
То же в географии: объект удобнее всего указать парой координат (сеть)
или адресом (иерархия), и оба способа искусственны в том смысле, что чу-
жды природе объекта, зато всем понятны и достаточно стабильны. Можно,
наоборот, описать объект «естественно» (например, по его принадлежности
горному хребту, речной системе, времени формирования, структуре), но это
будет сделано по-своему у каждого автора.
К сожалению, это норма: естественная система всегда субъективна,
а потому для практики малопригодна. Наивно думать, что настанет день, ко-
гда большинство станет понимать естественность одинаково и что это про-
длится в следующих поколениях ученых. Наоборот, об искусственной сис-
теме можно договориться и этим пользоваться долго, как мы пользуемся,
например, правилами орфографии.
Величие Линнея видится в том, что он не только различал два типа сис-
тем, но даже искусственную систему строил рационально: клал в ее основу
признаки, отражающие суть строения организмов. Постараемся усвоить
этот урок. Во-первых, признаем, что все нынешние филогении суть искус-
ственные системы (построены по воображаемому критерию родства с
бессознательной целью удовлетворить тезису Короны), а во-вторых, попы-
таемся выстроить естественную (в точно оговоренном смысле) макросисте-
му. Если принять ее за основу, то станет ясно, как строить системы частные.
Одну из частных систем мы сперва и рассмотрим.
10-6. Система голосеменных Мейена. Грубая филогения
Свои принципы классификации Мейен изложил в 1974 году так: хотя «в
признаковом пространстве растений есть свои регулярности», однако они
не выражаются общепринятой систематикой; нужна система, основанная на
трех положениях: (1) признаки не связаны жестко, (2) любой признак может
у какого-то члена таксона отсутствовать и (3) параллелизмы должны быть
учтены системой [Заварзин, 1990, с. 118]. Подробнее о классификационных
взглядах Мейена см. [Чайковский, 2006].
Ими мы воспользуемся в п. 10-7, а пока рассмотрим его конкретную
систему голосеменных. Она построена им в 1984 году, причем на совсем
иных основаниях. Главное заключалось в том, что Мейен отказался брать за
основу существующие варианты систем голосеменных, основанные на дан-
ных о ныне существующих группах, отказался потому, что они не дают об-
щего представления о системе: из 17 порядков голосеменных 11 вымерли и
еще в двух осталось по одному роду. Наоборот, ни один порядок цветковых
не вымер, а из 390 их семейств вымерло одно, так что их систему можно
строить на основе нынешних групп [Мейен, 1992, с. 101].
Сближение таксонов голосеменных Мейен проводил в соответствии с
замечанием Михайлова (п. 10-1), т.е. перечислял семофилогении, которые
585
считал базовыми, не приводя архетипических соображений. И еще в 1976
году Мейен прямо писал об этом:
“для стратиграфических сопоставлений по хвойным надо прилагать недю-
жинные усилия для разыскания их органов размножения и на этом все стро-
ить. Даже если таких находок будет немного, они дадут гораздо больше, чем
сборы вегетативных побегов” (письмо к К.З. Сальменовой // МС).
Через 10 лет он, идя этим путем, пришел к такому выводу: \\
“В целом можно сказать, что в филогении высших растений, если говорить о
происхождении семейств и таксонов более высокого ранга до отдела включи-
тельно, безраздельно господствует монофилия” [Мейен, 1986, с. 50].
Монофилия (п. 6-6) является итогом филогении, а прежде Мейен пола-
гал, что филогению, как правило, построить невозможно (п. 9-12) и что эво-
люция представима в форме не древа, а сети. Что изменилось?
В том же 1986 году он писал: «Лично я считаю, что голосеменные пред-
ставляют собой монофилетический таксон» [Мейен, 1992, с. 106], но факти-
ческих обоснований монофилии не привел. Есть схема флорогенеза (рис.
61), она названа в подписи к рисунку филогенией, но рассмотрев схему,
убеждаемся, что это не филогения: отдел покрытосеменных “повис”, т.е. ни
к какой ветви не отнесен; "повисли” и 6 порядков из отдела голосеменных.
Идее монофилии тут противоречит и тот факт, что толстые ветви часто
(особенно в нижней части схемы) переходят одна в другую без сужения.
Мейен объяснил: это значит, что порядок переходит в порядок, но - «с ши-
рокой трансгрессией признаков» (т.е. тут определяющую роль играет тран-
пол, п. 6-4*). Поэтому нельзя сказать, какое семейство переходит в какое, а
значит, о монофилии на этом уровне говорить нет смысла.
В своей самой последней статье Мейен писал:
“если учесть всю систему ранних хвойных и длительные тенденции в семо-
филогенетических преобразованиях органов, то обнаруживаются более или
менее устойчивые состояния некоторых признаков и соответственно их раз-
ный вес в филогенетических и, следовательно, таксономических построени-
ях”. Поэтому “в эволюции палеозойских хвойных намечается несколько ос-
новных тенденций... Это именно тенденции, а не отчетливые семофилогене-
тические ряды форм... Для растений характерна блочная (“модулярная”)
структура, причем в эволюции отдельные блоки легко меняются местами”.
Итог: “мы видим в эволюции позднепалеозойских хвойных комплекс родов, в
котором прихотливо переплетаются признаки... Как бы мы ни группировали
эти роды, будут получаться и параллелизмы, и реверсии примитивных со-
стояний” {Мейен С.В. // ПЖ, 1990, № 2, с. 6-7).
Как видим, он не нашел не только филогений, но и хороших семофило-
гений. Раннее и позднее его отрицания филогении смыкаются.
Зато вместо привычного понимания филогении как универсального де-
тального процесса эволюции путем дивергенции (расхождения одного так-
сона на два), Мейен дал совсем новое ее понимание: эволюция порождает
586
сеть, отдельные части которой связаны, однако связаны слабо: между по-
рядками и между классами сращений мало, они похожи на ветви, и эволю-
цию в рамках этих областей (таксонов) можно представить древом. То есть
здесь древо - грубая схема, не допускающая детализации: роды и многие
семейства эволюируют «с широкой трансгрессией признаков».
Такое древо отражает не сам процесс эволюции, а способ его описания
(тезис Короны), и его можно назвать грубой филогенией. Тем самым, счи-
тать позднего Мейена филогенетиком можно только в смысле построения
им грубой филогении. То же мы увидим в п. 10-11.
“Филогенетическое древо” он строил на основе одной семофилогении
(например, женской фруктификации), а свой выбор именно данной семофи-
логении в качестве базовой обосновывал “географически” - в том смысле,
что соединял родством роды, сменявшие друг друга в общей географиче-
ской зоне. Вот наиболее ясный пример.
Рис. 78. Семофилогения мужских фруктификации ранних хвойных,
“положенная на карту” (по: Меуеп, 1997. подробности см. в тексте)
На рис. 78 приведена семо-
филогения женских фруктифи-
каций ранних хвойных. Она рас-
падается на три ветви: сверху
экваториальные (N, S, Т, U, V),
посреди северные (L, М, О, Р, Q,
R) и внизу справа южный (К)
роды. Ее в точности повторяет
изображенная на рис. 79
«пробная филогения ранних
хвойных» (Меуеп S.V. И Review
of Palaeobotany and Palynology,
1997, № 96, c. 422, 432). To есть,
“филогения” основана на той
семофилогении, какая “легла на
карту” (сходные формы росли в
одной зоне).
(Ранее у меня ошибочно написано: «все семофилогении сошлись у него
в единую филогению» (МС, 69), за что прошу извинения.)
Слово “филогения” я взял в штриховые кавычки как потому, что форма
древа соблюдена весьма смутно, особенно в отношении семейств, назва-
ния которых помещены в “облаках", так и потому, что нет контроля (а вдруг
иная семофилогения так же хорошо или еще лучше “ляжет на карту”, а
родство укажет иное?). Более четких древ у Мейена мне неизвестно.
587
Рис. 79. “Пробная филогения” ранних
хвойных (Меуеп, 1997) точно повторяет
семофилогению, данную на рис. 78
10-7. Ядро и периферия.
Межтаксон
Карл Бэр в 1825 году выска-
зал простую важную мысль: вся-
кий крупный таксон имеет ядро из
многочисленных характерных для
него форм и периферию, состо-
ящую из сравнительно небольшо-
го числа нечётких форм. Напри-
мер, млекопитающие в основной
своей массе ходят на четырех но-
гах и тем самым являют ядро
класса, тогда как на его перифе-
рии мы видим китов (похожи на
рыб), рукокрылых (похожи на
птиц) и людей (ходят на двух но-
гах). См. [Чайковский, 1990, с. 43].
В силу фрактальности системы таксонов (п. 6-7), в каждом есть свои
ядро и периферия. Так, все кошки, собаки, гиены и медведи относятся к яд-
ру отряда хищных, а большая панда (растительноядный “бамбуковый мед-
ведь”) и калан (морской зверь с задними ногами в форме ласт) - к его пе-
риферии. То же внутри семейств: гепард - периферия семейства кошачьих
(похож на псовых), два вида панд - периферия семейства енотовых.
Вообще, периферии близких таксонов проявляют сходство друг с дру-
гом, что часто отражено в их названии: енотовидная собака, рыбоящер (их-
тиозавр), ракоскорпионы (эвриптериды), лавровишня и т.п.
Именно наличие ядра типичных форм (а вовсе не сведения о родст-
ве) позволяет выявлять таксоны и строить систематику. Ее счи-
тают хорошей, если периферии таксонов малы в сравнении с их ядрами
[Чайковский, 1990, с. 44]. В хорошем выборе типичных форм и состоит ис-
кусство систематика. Свойство природы, благодаря которому обычно это
удается, - упорядоченность диасети (п. 6-3**). Благодаря ей всякая наука
имеет дело с тенденциями (п. 6-2), т.е. с закономерностями, описывающими
ядра таксонов (хотя сам термин «таксон» есть только в систематике)
Ядро одних таксонов является периферией других и обратно. Яркий
пример этого явления мы видели в п. 6-4*: ядро миндалей является пери-
ферией персиков и обратно. Аналогично, ядро физики составляют общие
законы, а анализ разнообразия отодвинут на периферию этой науки; в хи-
588
мии же, наоборот, ядром является анализ разнообразия веществ, тогда как
общие ее законы являются по сути (а часто и по форме) физическими и, по-
моему, могут быть названы периферией химии. Учение о ядре и периферии
являет собой другую ветвь диатропики, которая дополняет ту ветвь, которая
разработана Мейеном (мероно-таксоно-мический анализ). Верно выявить
ядро изучаемого множества объектов - первая задача во всякой науке.
Вот два примера. 1) В физике исходное дело - описать движение. Аристотель
полагал, что действующей на тело силе пропорциональна скорость тела, а Гали-
лей - что ускорение. Первое верно для движения в вязкой жидкости, но применя-
лось ко всем движениям, и физика топталась на месте. Второе верно в вакууме и
верно описывает движения планет, но приближенно - и многие движения на
Земле, что привело к созданию точной науки. То есть у Аристотеля ядро закона
мало, а периферия огромна; у Галилея наоборот. 2) Долгое время считалось, что
генетический код универсален, т.е. един у всех организмов. Это было удобно как
свидетельство в пользу реальности общего предка всех организмов, но мешало
пониманию эволюции кода. Оказалось, что наблюдаемый обычно код - всего
лишь ядро множества кодов, периферия которого существует (минорные осно-
вания и пр., см. п. 6-10), но столь мала, что ее 45 лет не замечали. Однако, как
показали португальские генетики, эволюция кода идет прямо у нас на глазах
(Gomes А.С., Costa Т., Carreto L., Santos M. Молекулярный механизм эволюцион-
ных изменений генетического кода // МБ, 2006, № 4). Царящий в природе код
является нормой, а Мануэль Сантос и его сотрудницы по-видимому описали
снятие давления нормы. Впрочем, механизм этого снятия совсем неясен.
Природа периферий бывает различной: кит - заведомо не рыба (тепло-
кровен), но енотовидную собаку не раз относили как к енотам, так и к соба-
кам. Последний случай нам особенно важен, поскольку тут периферии так-
сонов пересекаются. Таких пересечений множество: гепард между кошачь-
ими и псовыми, ихтиостега между рыбами и амфибиями, эвглена между
растениями и животными и т.д. Так устроена диасеть.
Однако не все периферии пересекаются. Самые диковинные рыбы (лу-
на-рыба, морской клоун, морской конёк и т.д.) своими отличительными час-
тями никаких параллелей ни с кем не обнаруживают. Это — тоже свойство
диасети (точнее, ее краёв, состоящих только из периферий таксонов), но
полезен ли тут диатропический язык? Думаю, что полезен: сегодня парал-
лелей нет, а завтра их могут обнаружить, и где именно, не скажешь. Если их
не выписывать, не группировать в ряды по принципу непонятности, то мно-
гие параллели никогда найдены не будут. Для примера напомню диковин-
ное поведение паразитов (п. 6-15): каждый случай воспринимался как неле-
пая выдумка, пока они не встали в единый ряд.
Наличие периферий у всех крупных таксонов сильно затрудняет систе-
матику и порождает бесконечные споры о принадлежности нечетких
(incertae sedis, т.е. неопределенного положения) групп. Но если филогене-
тик видит здесь изъян исследования и уверен, что все такие группы должны
589
со временем найти четкое место, то с диатропической позиции такие группы
вполне естественны и должны быть особо описаны.
Для этого недавно предложено понятие «интратаксон» [Леонтьев, Аку-
лов, с. 506], означающее периферию таксона или ее часть. Замечу, что пра-
вильнее термин интертаксон, т.е. межтаксон. Надо привыкнуть к этому
понятию, признать, что его промежуточный статус бывает (пусть и не у всех)
межтаксонов определенным (т.е. нет оснований считать, что он изменится
при лучшей изученности), и не пытаться от таких таксонов избавиться. То-
гда соседство ветвей системы станет информативным и приблизит иерар-
хический принцип к удобному (и естественному в смысле долговечности).
Вот межгаксон высокого ранга: класс морские пауки (Pantopoda) имеет
план строения, характерный для типа членистноногих, но без более тесных
сходств с каким-либо из классов этого типа; он обычно описывается как до-
полнение к типу членистоногих. Однако план строения морских пауков име-
ет сходство с планом строения класса морских звезд (Asteroidea): пищева-
рительная и половая системы заходят у обоих классов в конечности.
Налицо периферия типа членистоногих, сходная с ядром класса иного
типа (относящегося даже к иному разделу - к вторичноротым). Для понима-
ния природы этого межтаксона полезно знать, что морские пауки - типичный
Лазарев таксон (п. 9-14): их предки известны лишь в девоне (см. обложку).
Межтаксонами ранга рода можно (среди зверей) признать панду, гепар-
да, енотовидную собаку и т.п.
Что касается адреса межтаксона, то он должен быть четким, и сделать
это просто - приписать его волевым актом. Именно так енотовидная собака
ныне отнесена к семейству псовых. Точнее см. [Чайковский, 1990, с. 48].
Выявление таксона через его ядро играет ту же роль для естественной
системы, что выявление иерархии - для искусственной. Словом,
на смену беспомощному “incertae sedis” должен прийти межтаксон._
Пользуясь понятиями ядра и периферии (а затем межтаксона), можно
решать задачи, для которых в обычной систематике просто нет языка. При-
веду в качестве примера макросистему, соединяющую все три аспекта, ука-
занные Михайловым (п. 10-2). Ее назначение не в том, чтобы заменить
множество нынешних систем, а в том, чтобы дать общий абрис живого ми-
ра, какового ни обычная систематика, ни иные отрасли биологии не дают. В
ее основе лежит известная схема царств Р. Уиттекера, улучшенная Г.А. За-
варзиным. (Недавнюю аналогичную схему см. [Леонтьев, Акулов].)
10-8. Логика макросистемы
Пока биологи видели всего два царства растений и животных, вопрос о
смысле слова «царство» не был ни актуальным, ни сложным: их попросту
задавали перечислением. Но едва возникли предложения о третьем царст-
ве, ситуация изменилась. Почему грибы или одноклеточные надо убрать из
590
царства растений? Пришлось вспомнить, что определения царств не суще-
ствует, что до тех пор всех удовлетворял афоризм Линнея: «растения рас-
тут и живут, животные растут, живут и чувствуют». Афоризм неточен: он по-
зволял лишь отличить животных, имеющих нервную систему, от прочих ор-
ганизмов, а потому излишне расширял царство растений - к ним, по Лин-
нею, относили, например, губок. Выявлению новых царств афоризм помочь
не мог, и они были выделены, исходя из формы организмов (у грибов, на-
пример, главным считали гифы - нитевидные клетки).
Определение царствам дал только в 1947 году Уиттекер; после уточне-
ний (1959 и 1969 гг.) его макросистема приняла следующий вид. Он, как мы
знаем (пп. 8-2, 10-3*), поделил все организмы на продуценты, редуценты и
консументы. Наряду с этой экологической триадой он наметил физиологи-
ческую, а именно три типа питания: растительное (синтез органики из не-
органических веществ путем фотосинтеза), абсорбционное (всасывание ор-
ганики из внешней среды) и животное (активный захват пищи и ее перева-
ривание). Сообразно трем способам питания он отнес все многоклеточные
организмы к трем царствам - растений, грибов и животных, а всех однокле-
точных поместил в одно царство - протисты.
Физиологическая характеристика растений попросту уточнила экологи-
ческую, а вот в отношении двух других многоклеточных царств Уиттекер
подметил нечто новое: оказывается, абсорбционное питание тесно связано
с экологической функцией (редукция органики) и формой организмов (ги-
фы); аналогично, захват пищи и пищеварение тесно связаны с экологиче-
ской функцией потребителей и с их способностью порождать огромное раз-
нообразие весьма сложных структур.
Красота и глубина этой простой схемы состоит в хорошем (в том смыс-
ле, что периферии малы) соответствии между тремя триадами:
(1) продуценты - редуценты - консументы
(2) фототрофы - осмотрофы - фаготрофы
(3) растения - грибы - животные.
Поскольку соответствие верно не на всех видах (так, цветковое-паразит
заразиха питается осмотрофно, хищное цветковое росянка - фаготрофно),
речь должна идти не о точных определениях царств, а лишь о тенденциях,
господствующих в трех царствах. То есть соответствие триад выявляет лишь
ядра трех многоклеточных царств. Но таково положение во всех науках.
Те же типы питания есть и у одноклеточных, однако Уиттекер предпо-
чел не делить их на царства по данному признаку и имел на то серьезные
основания: микроорганизмы способны переходить от одного способа пита-
ния к другому. Так, эвглена зеленая может захватывать пищу клеточным
ртом. У бактерий собственно фаготрофного питания нет, но переходы в
рамках триады (1) и у них обычны. Поэтому Уиттекер ограничился тем, что
выделил среди одноклеточных те формы, которые не имеют клеточного яд-
ра - прокариоты (Procaryota), а ядерных одноклеточных объединил в царст-
591
во Protista, существовавшее уже более ста лет, но мало кем признанное.
Тем самым, среди одноклеточных деление на два царства было проведено
не по экофизиологическому, а по структурному принципу.
Несмотря на многие возражения, эта схема широко принята, другой
хоть сколько-то признанной, насколько знаю, нет, поэтому любое обсужде-
ние макросистем нужно, на мой взгляд, начинать со схемы Уиттекера.
Хотя биологи еще и не договорились, сколько существует самих царств,
но почти все согласны в том, что их следует делить на два надцарства:
прокариоты (Procaryota), т.е. безъядерные, и эвкариоты (Eucaryota) т.е.
клетки с ядром. Первое надцарство почти у всех систематиков состоит из
единственного царства, которое сейчас принято называть царством бакте-
рий. Второе надцарство составляется очень по-разному, но нам сейчас дос-
таточно вспомнить, как делал это Уиттекер: он делил многоклеточных на
животных, растения и грибы в их традиционном понимании, а одноклеточ-
ных оставил одним царством - протист.
Главное, что это сделано путем рассечения: все организмы Уиттекер
поделил на 2 группы (прокариоты и эвкариоты), все эвкариоты на 4 группы.
Прежде чем делить таким же путем прокариоты, обратим внимание на не-
обычность поцедуры: ведь принято формировать таксоны не рассечением
совокупности организмов, а объединением сходных организмов.
Строить систематику путем объединения проще и естественнее, но
лишь до определенного уровня. Объединяя виды в роды, роды в семейства
и так далее, мы рано или поздно столкнемся с неприятным обстоятельст-
вом: некоторые группы непонятно куда девать. Так, группируя одноклеточ-
ных, мы уверенно отнесем фотосинтетиков к растениям, обладателей кле-
точного рта - к животным, но затруднимся с эвгленой зеленой, имеющей и
то и другое. Или с трипаносомой, не имеющей ни того, ни другого.
Можно пользоваться объединяющей систематикой, пока это ведет к од-
нозначным результатам, и данный процесс удобно, как оказалось, завер-
шить на таксоне тип (филум). (Можно еще объединять близкие типы в раз-
делы, как это сделано, например, в зоологии низших беспозвоночных.) Во-
первых, типов на сегодня около ста (как химических элементов), что близко
к пределу возможностей охвата умом одного человека, и во-вторых, типы,
как правило, можно воспринять интуитивно как некие целостности.
Далее нужен иной способ классификации. Если типы заданы, то макро-
систему надо строить путем последовательных рассечений мно-
жества типов, ибо при рассечении лишних не остается.
Тем самым, макросистематик должен собрать сведения обо всех типах
и найти правила, по которым их можно разделить на царства. Желательно,
чтобы эти правила не рассекали ни один тип, а если это не удалось, то каж-
дый обрывок типа надо назвать новым типом (таким должен быть признан
тип эвгленовых, ныне рассматриваемый зоологами всего лишь как отряд).
Затем типы в царствах надо как-то упорядочить. Это значит, что принципы
592
Уиттекера надо дополнить какими-то еще, в каком-то смысле естественны-
ми. Это и сделал в 1970-х годах бактериолог Заварзин.
Он отметил, что что с тремя эвкариотными царствами проявляют заме-
чательный параллелизм прокариоты: в частности, цианеи (они же цианобак-
терии, или сине-зеленые водоросли) - это как бы прокариотные растения, а
актиномицеты (они же актинобактерии, или лучистые грибки) - прокариот-
ные грибы. То есть можно поделить царство бактерий на три подцарства:
собственно бактерии, цианеи и актиномицеты. Затем Заварзин принял
во внимание давно известное сходство водорослей с грибами и заметил,
что и те и другие не имеют, в отличие от высших растений, настоящего тка-
невого строения. Это не вполне верно, но для первого упорядочения удоб-
но: позволяет, как и при введении надцарств, дополнить экофизиологиче-
ский принцип структурным. Можно строить наглядную схему (рис. 80).
Рис. 80. Симметризованная схема царств
по Уиттекеру — Заварзину
В центре прокариот Заварзин по-
местил собственно бактерии, к кото-
рым слева примкнули прокариоты-
продуценты (цианеи, фотобактерии и
др.), а еще левее следуют эвкари-
оты-продуценты, т.е. растения. С дру-
гой стороны к собственно бактериям
примыкают прокариоты-редуценты
(актиномицеты, гнилостные бактерии
и др.), а к ним - грибы. Естественным
образом собственно бактерии сопоставляются с консументами. Это удобно,
хотя активного захвата пищи бактериальной клеткой и не бывает: никакие
бактерии нельзя отнести к фаготрофам. Удобно потому, что побуждает ис-
кать возможные параллели между бактериями и простейшими животными.
Три полукруга на границе бактерий и простейших символизируют тот факт,
что таких параллелей несколько. Например, паразитическая бактерия
Bdellovibrio имитирует хищничество: может с большой скоростью ударять в
жертву и буквально вгрызаться в нее. Или миксобакгерии: их колония обвола-
кивает жертву, как бы имитируя фаготрофное питание многоклеточных; в то же
время миксобакгерии во многом похожи на миксомицеты, относимые к грибам.
С трех сторон к прокариотам примыкают нетканевые группы эвкариот -
водоросли из царства растений, простейшие из царства животных и грибы,
сами образующие царство (ведь грибов, качественно более сложных, неже-
ли водоросли, не бывает). Тем самым, на схеме учтен, наряду с экофизио-
логическим принципом, и структурный: отмечены три уровня организации -
прокариотный и два эвкариотных (нетканевый и тканевый).
593
Если не считать отсутствия «тканевых» грибов, то схема симметрична,
и это удобно для восприятия. Царства изображены в виде “географической
карты”, где соседство означает существенное сходство Если ни один
“район” не входит сразу в две “области”, то основной принцип систематики
(иерархия) здесь не нарушается. И появилось новое удобство: кроме вклю-
чения меньшей группы в большую (традиционно так отражают родство),
близость свойств можно изображать соседством (так естественно отражать
параллелизм). Подробнее см. [Чайковский, 2006а].
10-8* Устойчивость системы. Межцарства
Схема Уиттекера - Заварзина явила собою огромный прогресс в пони-
мании устройства природы, она проста, наглядна и удобна для преподава-
ния. За полвека она ни разу не опровергалась, а лишь уточнялась и совер-
шенствовалась. Более того, за это время был открыт целый новый мир' -
организмы подводных горячих источников (гидротерм), и схему не пришлось
переделывать: обитатели гидротерм точно так же делятся на продуцентов,
редуцентов и консументов, только продуценты - не фототрофы, а хемоав-
тотрофы (создают органику из неорганики, выносимой из земных недр с го-
рячей водой). А ведь устойчивость к новым фактам - мечта классификато-
ров и одно из определений естественной системы.
Однако она оставила без ответа массу вопросов. Почему эвкариоты по-
делены на царства, а прокариоты - на подцарства? Почему одноклеточные
и многоклеточные водоросли попали в разные царства? Куда отнести насе-
комоядные растения? Грибы во многом похожи на водоросли, т.е. на низ-
шие растения, так почему же им дан тот же ранг, что и всем растениям? По-
чему инфузория и шляпочный гриб отнесены к одному уровню организации?
И так далее. Выходит, что эта схема - не столько макросистема, сколько
очень интересная заявка на макросистему, подход к ней.
Выше было обещано выявлять все царства с помощью единой логики, в
качестве которой были предложены строение клетки (два надцарства) и
триады Уиттекера (три эвкариотных царства). Нет смысла выделять протис-
ты (одноклеточные эвкариоты) в царство, так как им нельзя дать единый
диагноз: их характерные черты носят чисто отрицательный характер, и их
приверженцы считают протисты царством лишь потому, что многие из про-
тист не проявляют четко черт «высших» царств - грибов, растений или жи-
вотных. Однако царство одноклеточных вот уже второй век то исчезает в
одних системах, то появляется в других, а это значит, что в нем есть по-
требность, которую надо учесть - иначе оно будет вновь и вновь предла-
гаться одними и отвергаться другими.
Для этого следует учесть, что протисты - нечеткий объект. Положение
нечеткого объекта можно сделать достаточно четким, если осознать, что
оно состоит из периферий (по Бэру), и изображать, помещая его между чет-
ко классифицируемыми объектами, на границе двух областей. Каждой пе-
594
риферии удобно дать отдельное название и отдельный диагноз, в котором
главное - отразить пограничное положение группы.
Оказывается, такие названия давным-давно существуют: это животно-
растения (Zoophyta) и гриборастения (Mycophyta), только их теперь не
рассматривают как систематические - с тех пор, как систематика стала фи-
логенетической. Поскольку наша макросистема экофизиологична, нам эта
старая традиция очень кстати. Добавим еще название грибоживотные
(Mycozoa) - к нему отнесем, например, хищные грибы, - и дадим каждой из
этих групп общее обозначение: межцарство (Interregnum).
Фактически кое-кто этим понятием пользуется:
«Так, многие одноклеточные водоросли... способны как к фотосинтезу, так и
к осмотрофному, а некоторые даже к фаготрофному питанию, т.е. занимают в
экоморфологической системе промежуточное место. Практически все утра-
тившие фотосинтез водоросли... также питаются осмотрофно. Экоморфоло-
гически такие организмы (“осмофлагелляты” Чайковского [1990]), вероятно
должны рассматриваться как промежуточные между растительной и грибной
жизненными формами» (Мирабдуллаев И.М. О происхождении и положении
грибов в системе органического мира // УСБ, 1994, № 1, с. 34).
Под осмофлагеллятами имеется в виду межцарство «Гриборастения».
Термин «межцарство» использован в работе [Леонтьев, Акулов, с. 512]:
между тремя царствами у них расположены «трофические межцарства».
Межцарства - это межтаксоны, а не таксоны, поскольку не имеют типичных
форм, поэтому их диагнозы не столько указывают свойства их членов,
сколько отмечают отсутствие чего-то (негативные диагнозы). Почти все эв-
кариоты естественным образом распределятся среди трех царств и
трех межцарств, а оставшиеся несколько видов (вроде эвглены зеленой)
окажутся сразу в двух межцарствах, что учесть совсем легко.
Ядерный тип организации либо есть целиком, либо его нет вовсе (в
этом состоит синдром эвкариотности, п. 8-5), так что деление на два над-
царства не только естественно, но и весьма удобно: между ними нет пери-
ферии, а потому нет и межцарств. Все попытки выявить какие-то промежу-
точные формы (мезокариоты) ни к чему не привели, о чем хорошо написано
в статье ботаника А.Б. Шипунова: им нельзя дать конструктивный диагноз, а
без диагноза нет смысла говорить о таксоне [Шипунов].
Так, мезокариотами были объявлены динофлагеллаты (панцырные
жгутиконосцы), но они оказались вполне ядерными организмами с необыч-
ными чертами (п. 8-5*). Строение их отнюдь не примитивно - наоборот, они
во многом сложнее других эвкариот, особенно их хромосомы, образующие
сложные фигуры в пространстве, скрепленные не столько белками, сколько
особыми РНК. Динофлагеллаты довольно молоды (известны лишь с силу-
ра), так что и с филогенетической позиции они - ответвления, а не предки.
Кроме строения клетки, надцарства являют еще одно важное различие:
почти все прокариоты одноклеточны (многоклеточные составляют у них
595
около 1% видов), тогда у эвкариот наоборот: лишь около 1% их видов одно-
клеточны, и разнообразие их видов растет с ростом сложности организации.
В этом состоит реверсия разнообразия (п. 8-13). А вот в межцарствах ее нет
- тут опять преобладают одноклеточные. По этому свойству межцарства
сходны с прокариотами, хотя состоят из эвкариот.
В межцарствах название (адрес) каждой группы следует приписать к
одному из царств произвольно, используя наличные традиции.
10-9. Макросистема эволюционная и нефилогенетическая
Далее предложена макросистема, в которой указание места на схеме
задает уровень организации и основную жизненную форму таксона; внутри
таксона форма системы не предрешается, т.е. может быть древом, сетью и
пр. Исторический аспект отражен дважды: уровни организации возникают
последовательно, а на каждом уровне каждый таксон тоже эволюирует.
Макросистема опубликована мною в 1986 году, и была верно охаракте-
ризована как развитие эко-морфологического подхода Уиттекера - Заварзи-
на (Шафранова Л.М. //ЖОБ, 1990, № 1).
Открытия последующих лет не вызвали потребности существенно из-
менять ее, и в этом видится довод в пользу ее естественности. Она пред-
ставляется мне более эволюционной, нежели чахлое “бескорневое древо
всех организмов" и прочие филогенетические крайности. Ее объяснение,
предназначенное для методологов, см. в книге [Чайковский, 1990], а удоб-
ное для преподавателей см. БШ, 1998, № 6, где даны и две ее цветные схе-
мы: на одной - структура царств, а на другой - уровни организации; см. так-
же БШ, № 4 за 1999 год.
Со времен Линнея до сих пор в биологии царством называется часть
живой природы, занимающая определенное место в «экономии природы».
По Линнею, растения производят, а животные потребляют. К этим двум
царствам в XX веке пришлось добавить (точнее, выделить из царства рас-
тений) еще два - грибы и прокариоты. Оснований для этого много, но я на-
зову одно: кроме производства и потребления органики, ее еще надо вклю-
чать в геохимические круговороты, о которых шла речь в главе 7, - это и
делают представители вновь выявленных царств.
Что касается прокариот, то они могут исполнять функции трех других
царств и к тому же только они могут включать элементы из неорганической
природы в круговороты живого (например, усваивать азот из воздуха). Оче-
видно, что прокариоты - более чем царство: их экологическая роль выше,
чем всех трех эвкариотных царств вместе. Поэтому говорят, что в природе
есть два надцарства - прокариоты и эвкариоты. Надцарство прокариот со-
стоит из одного царства, поскольку разделить уверенно прокариоты на цар-
ства не удается - из-за того, что их свойства свободно комбинируются. Их
проще делить на три нечетких подцарства - бактерии, актиномицеты и
цианеи (т.е. сине-зеленые водоросли).
596
Наоборот, надцарство эвкариот довольно четко делится на 3 царства
по типу питания, причем слова «довольно четко» означают, что в каждом из
них основная часть видов образует чётко определимое ядро. Нечёткие виды
образуют сравнительно небольшую периферию; основную ее часть состав-
ляют виды одноклеточных организмов, поэтому не раз возникала мысль
объединить их в пятое царство (протисты), но оно никогда не бывало обще-
признанным. Причина в том, что данному “царству” не удается дать прием-
лемый диагноз, а без диагноза нет таксона [Шипунов].
Точнее, если задать протист как «одноклеточные эвкариоты», то попа-
дут в разные царства близкие виды (например, водорослей), а то и один
вид, поскольку одинаковые по строению клетки могут как жить порознь, так и
входить в состав примитивных организмов. Если же задать «царство про-
тист» как-то иначе (описывая свойства клеток и их комплексов), то границы,
как их ни проводи, будут устраивать одних систематиков и не устраивать
других, а значит, будут недолговечными. Причина объективна: у однокле-
точных и близких к ним многоклеточных свободно комбинируются их основ-
ные свойства. Подробнее см. [Чайковский, 1990, п. 6.5].
Однако нельзя и попросту пренебречь одноклеточным уровнем при
классификации, поскольку у одноклеточных очень много общего. Например,
у них очень высоко разнообразие митозов и митохондрий (тогда как у много-
клеточных они достаточно однообразны), и в этом можно видеть их главную
тенденцию, а царства, как мы видели в п. 6-2 и позже, определяются имено
через тенденции. Выход видится в том, чтобы признать периферийный ха-
рактер групп многих одноклеточных: они примыкают к соответствующим
группам примитивных многоклеточных, т.е. образуют набор периферий эв-
кариотных царств. И эти периферии пересекаются.
Далее, таксон принято понимать как общность происхождения, а это на
уровне царств совсем бессмысленно: ведь древнейшие организмы тоже об-
разовывали экосистему, т.е. жили одновременно, и нет оснований говорить,
что все растения (все животные) когда-то были одним видом.
Учтя всё это, макросистему будем строить путем рассечения множества
типов, до которых подымается объединяющая систематика (п. 10-8).
Наивысшим таксоном, для которого есть нечто вроде определения, яв-
ляется тип (филум, отдел). Для него можно указать план строения - именно
так понимают тип со времен Кювье (п. 1-15). Каждый биолог знает, что такое
план строения хордовых, моллюсков или цветковых. Царство, наоборот,
единым планом строения не обладает. Другими словами, тип (и более низ-
кий таксон) может быть определен как морфологическое целое, а царство -
только как экологическое и функциональное целое.
10-9* Макросистема как парк
Теперь проясняется форма искомой системы: ее удобно представить
себе в форме “парка”, в середине которого расположено круглое “озеро"
597
(надцарство прокариот), окруженное тремя массивами “деревьев” (тремя
эвкариотными царствами), где каждое “дерево” обозначает тип. Каждый
массив отделен от двух других “полянами” (межцарствами), заполненными
“мелкой растительностью” (типами из сравнительно небольшого числа ви-
дов, в основном одноклеточных). Примером “дерева” может служить систе-
ма голосеменных Мейена, которая, как сказано выше, является филогене-
тическим древом лишь с оговорками.
Изобразить прокариот “озером” надо потому, что среди них вообще нет
групп, представимых в форме ветвящихся систем: систематика бактерий
сводится к перечню родов, очень редко допускающих группировку даже в
семейства. Вызвано это тем, что основные свойства бактерий свободно
комбинируются, а потому система их естественно выражается в форме мно-
гомерной таблицы. В случае N-мерной таблицы каждый род задается пере-
сечением № строк (в случае двумерной таблицы - пересечением строки и
столбца). По любому из свойств можно разделить прокариот на две группы
и назвать их царствами, что не раз делалось (например, предлагалось де-
лить прокариот на царства бактерий и архебактерий - о последних см.
[Медников; Шипунов]).
Такое произвольное членение вряд ли полезно, ибо различий между
членами одного “царства" оказывается больше, чем сходств между членами
разных “царств”. В частности, нет смысла рассматривать архебактерий в
качестве отдельного таксона (равного остальным бактериям) - как потому,
что они имеют общий архетип (основное строение), так и потому, что интен-
сивно обмениваются генами в ходе горизонтального переноса (Aravind L.
е.а. Evidence for massive gene excange between archaeal and bacterial hyper-
thermophiles //Trends in Genetics, 1998, T. 14, p. 442).
В последующие годы подобных сведений накопилось столько, что, на
мой взгляд, вообще стало бессмысленно говорить об этих двух группах, а
можно говорить лишь об архебактериальных свойствах, вперемешку рассы-
паннных по самым разным про- и эвкариотным группам.
Зато полезно выявить среди прокариот те, метаболизм которых ради-
кально отличен от эвкариотного. На рис. 81 эти прокариоты объединены как
идиотрофы (от греч. идиос — особенный, трофос — питание). К ним, в
частности, относятся все литотрофы (пп. 7-4, 7-7). Как видим, идиотрофы
заняли середину надцарства прокариот, и межцарства служат им как бы их
эвкариотными продолжениями. Однако идиотрофы принадлежат ядру про-
кариот, тогда как межцарства образуют периферии эвкариотных царств.
Подробнее см. [Чайковский, 1990, с. 151].
Планировка «парка» должна учесть не только сходства различных «де-
ревьев» (типов), но и различные уровни организации. Напомню, что Завар-
зин ввел всего три уровня организации - прокариотный, простой эвкариот-
ный и сложный эвкариотный (тканевый). Этого явно недостаточно. Ясно
598
Рис. 81. Диатропическая четырехцарственная макросистема живого мира, каким он был до
появления царства человека (по Чайковскому, 1986, с уточнениями). Таксоны обычной сис-
тематики даны курсивом с заглавной буквы (Животные, Хвощи и т.п.), прочие группы -
сплошь курсивными заглавными буквами (ЗООФИТЫ, ХЕМОАВТОТРОФЫ и т.п.); уровни
организации даны прямыми заглавными буквами (НИТЧАТЫЕ, ЦЕНОЦИТНЫЕ и т.п.).
Подробнее см. (Чайковский, 1990, с. 156]. Подробнее о строении “озера” (круга прокариот)
см. там же, с. 150-152. Буквой М обозначены молликуты (микоплазмы) — группа, наиболее
близкая к возможному первоорганизму
фЛГОТРОФЫ
рвотные
fgjioBHbiM мозго*;
вторИчнополос'гнЫЕ
<£» . r.°»HgHOnOJ1OCTWbi.5
° Зу^МЗЕВЫВ, дду
>t5vgj5g4KTbiE пй?5й;
НИТЧАТЫЕ
гГТЗнфуэора
МОЛЛЮСКИ
С ЗЕЛЕНЫМИ
СИМБИОНТАМИ
ПРЕОБЛАДАЕНИЕ
> i ЖГУТИКОВЫХ
ФОРМ
ЗООФИ
к*
М
ПРЕОБЛАДА НИЕ
БЕЗЖГУТИКОВЫХ
МОЛЛЮСКИ с
ХЕМОТРОФНЫМИ
СИМБИОНТАМИ
ИКОН
>«ы
^1
Актиноми-
иеты
М И'ТОз\)„
ХИЩНЫЕ
ГРИБЫ
ЦВЕТКОВЫЕ
ПАРАЗИТЫ
Srtu
ведь, что плесень чем-то существенно ниже шляпочного гриба (но оба За-
варзин отнес к нетканевым), а червь - ниже насекомого (оба отнесены к
тканевым). Для наглядности и цельности надо иметь какой-то четкий прин-
599
цип расположения, подобный столбцам и строкам менделеевской таблицы
в химии. Правда, у нас таблица будет кольцевой.
Обычный иерархический принцип, применяемый в систематике, выяв-
ляет, в качестве высших таксонов, только крупные группы, а не какие-то
уровни организации. Например, губки (состоят из организмов одного уровня
организации) - тип, но и хордовые (состоят из разных уровней) - тоже тип.
Такой тип придется, ради единой логики системы, дробить.
10-9** Уровни развития и макросистема
Для животных базовые уровни известны и общепризнаны: одноклеточ-
ные (Protozoa), пластинчатые (P(acozoa), простые тканевые, т.е. губки
(Spongia), двуслойные, т.е. лучистые (Radiata), простые трехслойные, т.е.
бесполостные (Scolecida), первичнополостные (Pseudocoelomata), вторич-
нополостные (Coelomata). Для цельности макросистемы нужно провести по-
добные уровни через все царства. Примерно ту же роль, какую при опреде-
лении уровня организации животных играет полость (coeloma), у растений
играет побег (cormus); а ту роль, какую у растений и животных играет ткань
(textus), у грибов выполняет плетеная из гиф «ложная ткань», плектенхима
(греч. plectenchyma: слитое путем переплетения).
Это сопоставление позволяет наметить нечто вроде кольцевой коор-
динатной сетки для нашего “парка”. В качестве кольцевой координатной
оси естественно взять границу прокариот с эвкариотами, поскольку это -
главная и единственная четкая граница между таксонами, не осложненная
нечеткими формами. На рисунке она изображена жирной окружностью. Дру-
гой осью координат будет уровень развития, откладываемый внутрь от
этой окружности и наружу от нее (рис. 81).
Кверху от верхней части этой окружности располагаются жгутиковые
одноклеточные организмы, обладающие всеми тремя способами питания,
согласно триаде Уиттекера. К ним относятся упомянутые ранее эвглены.
Обходя данную окружность по часовой стрелке, мы увидим постепенное
исчезновение членов данной комбинации, а затем опять возвращение к ним.
Эта объективно существующая цикличность и потребовала для своего ото-
бражения кольцо: правее эвглен — жгутиковые без фотосинтеза (зоофлагел-
латы), затем - жгутиковые с чисто осмотрофным питанием (осмофлагелла-
ты), еще далее - осмотрофы без жгутиков (споровики и пр.); в нижней части
окружности - одноклеточные водоросли лишайников, затем - безжгутиковые
водоросли (багрянки и др.), а в левой ее части - снова фотосинтетики со жгу-
тиками (фитофлагеллаты), которые в верхней части соседствуют со жгутико-
выми смешанного растительно-животного питания. Круг замкнулся.
Внутри жирной окружности расположится кольцо одноклеточных про-
кариот. Оно будет в нашей системе основным, поскольку служит кольце-
вой осью симметрии. Кроме нее на рисунке видна тройная симметрия, за-
даваемая триадами и определяющая эвкариотные царства.
600
Снаружи от кольца 1п поместим кольцо одноклеточных эвкариот (1э), а
внутри - простые многоклеточные прокариоты (нитевые, плоские и шаровые
структуры) - кольцо 2п. Аналогичное кольцо 2э поместим снаружи кольца
1э. Эти уровни включают и бесклеточность (ею обладают организмы, чьи
тела не поделены на отдельные клетки; таковы ценоцитные водоросли, ми-
целий акгиномицетов и т.п.)- Параллельно с цианеями (прокариотными во-
дорослями) расположатся одноклеточные эвкариотные водоросли, парал-
лельно со жгутиковыми бактериями (псевдомонадами) - жгутиковые эвка-
риоты (их прежде называли монадами) и т.д.
Уровни Зп и Зэ - тканевые: в строгом соответствии с определением
ткани как компактного слоя или полости клеток, объединенных общей функ-
цией, мы можем выявить простые ткани и у грибов, и у некоторых прокари-
от. Тканевые прокариоты - это цианеи в виде сантиметровых кустиков и
плодовые тела миксобактерий в виде ярких милиметровых грибочков. Функ-
ция питания исполняется здесь одной тканью, размножение - другой, а обо-
лочкой органа размножения служит третья. Такие прокариоты естественно
займут небольшой кружок в центре схемы.
Зато у эвкариот соответствующий (тканевый) уровень очень обширен:
это всё то, что мы в обиходе называем водорослями и грибами, а также -
губки, кишечнополостные (медузы, гидры и т.п.) и низшие черви. Правда, у
грибов роль ткани исполняет не слой клеток, а плектенхима.
Симметрия относительно уровня 1п означает, что прокариотные типы
клеточной организации повторены низшими эвкариотами: и тут и там мы
видим клетки круглые, вытянутые и спиральные, с жесткой стенкой и без,
жгутиковые, скользящие и неподвижные; повторены и прокариотные типы
многоклеточности: и тут и там мы видим нитчатые, мицелиальные (ветвя-
щиеся), кустистые, слизевые, корочковые и т. п. Однако ткани у эвкариот
создают основу разнообразия, а у прокариот почти отсутствуют. Этим сим-
метрия между прокариотами и эвкариотами исчерпывается, так как более
сложных форм у прокариот нет. Нет их, кстати, и у грибов (у которых, одна-
ко, разнообразие тканевых форм несравненно выше).
Четвертый уровень (полукольцо) эвкариот тканеорганный: каждый ор-
ган строится из различных тканей. Ствол дерева - из древесины, луба и ко-
ры; пищеварительный тракт даже у кольчатого червя имеет железистые,
соединительные и мускульные ткани и т. д. У высших растений есть много
различных разнотканевых органов, и в этом они проявляют параллелизм с
животными средней сложности (примерно до уровня кольчатых червей).
Этим организация растений и завершилась, тогда как у животных есть
еще и пятый уровень, наиболее богатый видами - централизованный. Ин-
теграция никогда не доходит у растений до централизации: из корня часто
может вырасти новый стебель, отрезанный стебель или ветка часто может
давать новый корень, а разведение культурных пород прививкой общеизве-
стно. В то же время животные выше червей не переносят даже разрезания
601
пополам - настолько их органы интегрированы в единую систему. Поэтому
среди них можно выделить пятый уровень, куда войдут все те животные, ко-
торые имеют головной мозг и сердце.
Более детальное членение для макросистемы не требуется. (Напомню,
что в ней должно быть наглядное место каждому типу, внутри же типа сис-
тематика строится по-другому.) Зато стоит отметить, что каждый уровень,
считая от 1 п, образует следующий путем интеграции, которая наблюдается
относительно уровня 1п как наружу, так и внутрь. Эвкариотная клетка есть
объединение прокариотных механизмов, ткань - объединение клеток для
общей функции и т.д.; объединяться в простую ткань могут как эвкариотные
клетки, так и прокариотные. Разница в том, что эвкариотная организация
влечет реверсию разнообразия: чем выше уровень, тем больше площадь
кольца. Подробнее см. [Чайковский, 2006а].
Эволюция, как ее видят в наше время, шла от простых прокариот к мно-
гоклеточным прокариотам и одновременно к одноклеточным эвкариотам, а
затем - к многоклеточным эвкариотам. Если так, то ее естественно пред-
ставить как постепенное заполнение кольцевой таблицы от кольца 1п
внутрь и наружу. Подробнее см. [Чайковский, 1999]. Тем самым, эволюцию
можно представить в виде процесса заполнения рефренных таблиц на всех
уровнях, но на высшем уровне таблица оказывается кольцевой.
Ю-9*** Единый взгляд на биоразнообразие
Вновь напомню: предложена не замена нынешней систематики и не ее
улучшение (хотя оно необходимо, но это другая задача), а дополнение к
ней, общий абрис разнообразия живого. Он обладает устойчивостью (по
Уэвеллу) и информативностью (по Любищеву), т.е. чертами естественной
системы: свойства каждой группы следуют из ее положения на пересечении
кольца и радиуса; за 20 лет макросистема не потребовала переделок, хотя
мне стало известно многое, что не было учтено при ее публикации в 1986
году (мир гидротерм, архебактерии, идея происхождения эвкариот из проге-
ноты и т.д.). Подробнее см.: Чайковский Ю.В. Естественная система. 30 лет
после Любищева И Любищев и проблемы формы... М., МОИП и ЦСИ, 2003;
то же, но кратко: Чайковский Ю.В. II ЛЧ, 2004.
Имеет смысл сравнить изложенную макросистему с недавно предло-
женной «экоморфемой», ставящей сходную цель - дать общую схему живо-
го мира на основе реальных сходств, а не мысленного родства [Леонтьев,
Акулов]. Ее авторы, харьковские биологи, тоже пишут о двух систематиках,
тоже видят цель в том, чтобы дать способ упорядочения разнооб-разия ор-
ганизмов, но не отделяют ее от цели адресации. Наоборот, они проводят
идею «две таксономии - две номенклатуры», повторяя тем самым идею
книги: Алеев Ю.Г. Экоморфология. Киев, Наукова думка, 1986.
Ее автор счел основными 4 аспекта разнообразия: связь со средой,
уровень функциональной организации, уровень структурной агрегации и
602
способ метаболизма (с. 24). К сожалению, под структурной организацией он
имел в виду отнюдь не морфологию, но всего лишь число клеток (однокле-
точные, многоядерные, колониальные и многоклеточные), а функциналь-
ность понимал исключительно как адаптивность. Еще досаднее то, что свою
неморфологическую систему он пытался строить как методическое повторе-
ние морфологической, т.е. хотел дать иерархию и номенклатуру. Он даже
привел (с. 195) положение воробья в своей системе - громоздкую языковую
конструкцию, доведенную фактически лишь до уровня птицы (т.е. до класса
обычной системы). На мой взгляд, довести такую систему до уровня вида ни-
когда не удастся, поскольку форм гораздо больше, чем функций.
Д.В. Леонтьев и А.Ю. Акулов тоже дают свою громоздкую номенклатуру,
однако две номенклатуры не приживутся, да вряд ли и нужны.
.При построении авторы используют не иерархию, а кольцевую таблицу,
но, к сожалению - без принципа кольцевой симметрии. Впрочем, это и не-
возможно у них, поскольку середина таблицы занята «империей некпеточ-
ных», где собраны не организмы, а наборы веществ, проявляющие матрич-
ную активность (прионы и вирусы).
На с. 512 у них сказано, что кольца символизируют расхождение «из
предкового центра (последнему соответствует условная неспециализиро-
ванная жизненная форма)». Если так, то в центре следовало поместить
прогеноту (она упомянута ими на с. 508), но не прион и не вирус, с которых
жизнь начаться никак не могла (ибо у них нет метаболизма).
За основу авторами взяты «тип питания и сложность организации», но о
литотрофах они не знают, а деление на про- и эвкариоты отвергают как от-
носящееся к филогенетической системе (!). Уровни развития получились в
экоморфеме странные: в одной клетке могут находиться бактерия, зеленая
водоросль и мох [Леонтьев, Акулов, с. 519]. Но это - частности.
А в целом работа пригодна для чтения и разглядывания: во-первых, она
ясно очерчивает обстановку в макросистематике, во-вторых, красива, в-
третьих, дает пищу для размышлений. Например, хотя ни рефрена, ни его
аналога в работе нет, а тема разнообразия подчеркнуто игнорирована (с.
524), но материал сам ведет авторов, и в итоге дано почти такое же напол-
нение таблицы, какое описано мною выше, исходя и совсем иных установок,
- в рамках идеи диасети. Возможно, обе схемы можно объединить.
Особо ценным видится намерение авторов провести процедуру пере-
сечения периферий через все уровни системы (идея интратаксона), хотя на
деле, как и у меня, пересечения даны только для уровня царств.
Построив систему как сеть, авторы дали затем ее иерархическое опи-
сание, что тоже радует: согласно тезису Короны, данное описание остается
удобным, даже если система построена в форме не древа, а сети или еще
чего-то. Точно так же, как адресная книга, т.е. иерархия, удобна для деталь-
ного описания города, хотя на самом деле он являет собой сеть улиц. Ра-
зумеется, названия должны быть в книге и на карте одни и те же.
603
10-10. Царство человека
Какое место в кольцевой макросистеме занимает человек? Если гово-
рить о строении тела, то, как сказано в п. 9-15, удобно считать человека ви-
дом рода Ното в отряде приматов (обезьян и близких к ним групп). Тогда
ему на схеме особого места не требуется. Однако если говорить экологиче-
ски, то человечество определяется не столько свойствами людских тел,
сколько их количеством и тем, что люди делают. Нас так много, что сами
наши тела стали очень заметным фактором в природе. Например, они вы-
рабатывают в год 5 млрд мегавагг-часов энергии - больше, чем наши элек-
тростанции (Иванов К.П. // Успехи физиол. наук, 2004, № 4). А дела наши
меняют облик биосферы настолько, что являют для нее катастрофу, срав-
нимую разве лишь с наступлением эры кислорода в протерозое.
Со времен Линнея человека принято было выделять в особое царство
природы; Бэр, например, писал, что человек биологически относится к жи-
вотным из отряда приматов, а в умственном и духовном плане являет со-
бою отдельное царство. Эта точка зрения не единственна - так, Оуэн в кни-
ге, упомянутой в п. 2-11, классифицируя млекопитающих по их черепам, вы-
делял человека в особый подкласс, состоящий из одного отряда Bimana
(двурукие), т.е. располагал людей как бы между двумя позициями Бэра.
С позиции влияния на другие виды человек тоже является именно цар-
ством, что видно из такого подсчета: площадь возделанных земель состав-
ляет 1,5 млрд гектар, лугов и пастбищ 3,3 млрд, т.е. вместе - более 40%
суши. Еще 0,5 млрд га в принципе можно использовать при орошении; на
большее на Земле не хватит воды (Жученко А.А. Адаптивное растениевод-
ство. Кишинев, Штиинца, 1990, с. 22). Теперь можно уточнить: человек
строением и физиологией относится к подотряду обезьян, а эколо-
гически является особым царством природы. Это, кстати, соответст-
вует общепринятому экологическому пониманию царств.
Человек может выступать как царство и в чисто классификационном
смысле, что не раз отмечали те, кто формулировал отличие людей от жи-
вотных. Эту линию недавно продолжил методолог С.Г Кордонский. У него
особая деятельность человека названа социальной:
«у людей как представителей царства природы есть “животная”, “растите-
льная”, “бактериальная”, “минеральная” и другие “части”, в то время как у...
высших животных есть все “части”, кроме социологической» (Кордонский
С.Г. Циклы деятельности и идеальные объекты. М., Пантори, 2001, с. 127).
Это не вполне верно (социальная структура у высших животных есть, а
у обезьян весьма развита - см. п. 9-15; особую деятельность человека в за-
нимающем Кордонского смысле я бы назвал понятийно-орудийной), но ав-
тор прав в главном: если определять каждое новое царство как появление
нового типа деятельности, то человек - царство. И его деятельность носит,
в частности, глобальноэкологический характер.
604
Экологическая универсальность и генетическое единство (горизонта-
льный перенос) прокариот вынуждает считать их надцарством, состоящим из
одного царства. С этой позиции людей тоже можно считать надцарст-
вом, состоящим из одного царства. В самом деле, люди тоже генетиче-
ски едины. Далее, будучи поначалу всего лишь консументом, человек посте-
пенно овладел всеми трофическими уровнями. Сперва это достигалось толь-
ко посредством разведения нужных человеку организмов, но в последней
четверти XX века стало играть существенную роль прямое их создание, т.е.
генная инженерия. Кроме комбинирования природных генов, в последние го-
ды началось изготовление генов. Теперь можно получить ген, который бу-
дет кодировать никогда не существовавший в природе фермент.
Более того, человек теснит прежние царства - точно так же, как в про-
терозое эвкариотные царства (растения, животные и грибы) начали теснить
прокариот. В Европе большинство первичных (росших до человека) лесов
уничтожено, а в Китае есть обширные территории, относимые не к какой-то
биогеографической единице, но прямо определяемые как «антропогенный
ландшафт». Положение животных еще менее завидно: все виды крупнее
зайца либо уже истреблены, либо оттеснены в малолюдные зоны (где их
численность тоже падает), либо одомашнены. Численность людей и разво-
димых ими животных в тысячи раз первосходит их максимально возможную
численность в диком состоянии. Кроме них процветают, в основном, непро-
мысловые виды животных в почве и океанах.
Инопланетяне, приближаясь к Земле и едва убедившись, что здесь есть
жизнь, увидят, что царят люди, а уж потом смогут выявить, какие еще у нас
есть царства организмов. Ведь люди - не просто геологическая сила (ею
являются все царства), он еще строит объекты, многие из которых видны из
космоса: водохранилища, города и т.п. Можно сказать, что человек в гло-
бальном плане - конструктор, точно также, как грибы - деструкторы. Даже
больше: грибы разлагают лишь органику, а человек конструирует всё.
Это позволяет ввести надцарство (царство) человека формально:
надцарство Humana (человеческие), состоящее из одного царства
Homines (люди) - это группа организмов, единая генетически
и способная выполнять как экологические функции всех
прежних царств, так и новую функцию - конструктора,
т.е. создавать материалы и изделия, отличные качественно от тех, что
_____________существовали в природе до появления людей.____________
При таком подходе на схеме можно дать еще одну короткую дугу - вы-
ше животных, над хордовыми. Однако пятицарственную систему лучше
изображать вообще не так, а иначе, кладя в основу иные, кроме приведен-
ных, рассечения. До ее разработки остается пользоваться схемой рис. 81,
подчеркивая, что она дает абрис живой природы, какой она была до про-
мышленной и информационной революции.
605
Впрочем, царство человека только в XX веке стало доминантой жизни,
а появилось оно раньше (как звери появились в мезозое, а доминантой ста-
ли в кайнозое): первым явлением царства была неолитическая революция
(рождение земледелия), а первым явлением надцарства - утеря естествен-
ного ландшафта значительной частью средневекового Китая.
Прежние царства возникли слишком давно, и мы ничего не знаем о
процессе их формирования. Зато об остальных высших таксонах мы знаем:
вероятно, они возникали в безморозном климате и затем расселялись, эво-
люируя, в более холодные страны (биотогенез). Именно так развивались и
люди. Возникнув в субтропиках, они несут с собой полутропическую среду
обитания (теплое жилье, солнцелюбивые растения); одновидовая культура
влечет в последние годы монотипизацию условий жизни, откуда и «порази-
тельное безразличие подростков», создающее массу новых проблем (Яниц-
кий О.Н. Экологические перспективы города. М., 1987, с. 40, 93). Замечу:
«поразительное безразличие подростков» есть снижение видовой активно-
сти, т.е. основного фактора эволюции, элемент отката (п. 9-16**).
Вообще, снижение разнообразия в рамках царства человека весьма ха-
рактерно, что хорошо видно из упомянутой книги молдавского агробиолога
А.А. Жученко: используемая земля засажена ничтожной долей растений: из
300 тыс. видов цветковых используется 5 тыс., причем широко - всего 30
видов. Из них почти все путем селекции приведены, с целью повышения
продуктивности, в экологически неустойчивое состояние (с. 67, 193). Для из-
бежания катастрофы автор предлагает отказаться от монокультур (от по-
лей, засеянных единственным сортом) и от единообразных мер воздействия
на земли; он даже призывает вводить менее урожайные, но более экологи-
чески самостоятельные растения.
Эти меры можно назвать диатропическими, но главный вопрос диатро-
пического характера таков: может ли вообще одновидовое царство быть ус-
тойчивым и, тем самым, существовать долго? Ответа у меня нет.
С одной стороны, нынешняя неустойчивость биосферы очевидна и явно
связана с царством человека, но с другой - она может оказаться временной.
Ведь человечеству вообще свойственно развиваться циклами. (Есть и вто-
рая причина, очередное глобальное потепление - тоже, возможно, краткая.)
В этом плане интересна книга: Пантин В.И. Циклы и волны глобальной
истории. М., 2003. Тут верно отмечен факт цикличности глобальной истории
цивилизаций: за эпохой объединения следует эпоха распада империй; опи-
саны 3 цикла (античный, средневековый и нынешний), а о более ранних ска-
зано лишь, что они имели место и что длительность каждого «также состав-
ляет около тысячи лет» (с. 51).
На самом деле, как бы ни группировать события, длительность циклов
со временем сокращается. Так, нынешний цикл, начавшийся в XIV веке,
еще не закончен, но уже накладывается на следующий (Европа объединя-
ется, империи распадаются); средневековый составил 1100 лет (III / XIII ве-
606
ка), а античный цикл превысил полторы тысячи лет, и чтобы уместить его в
тысячу лет (-VIII / II века), Пантину пришлось начать его на 500 лет позже
действительности. В самом деле, крушение Микенской цивилизации, став-
шее началом распыления Эгейского мира (и описанное Гомером в -VIII ве-
ке), произошло в -XIII веке. Что касается предыдущего цикла, кульминацией
которого был союз Ассирии и Египта (-XV / -XIV века), то описать его не бе-
русь, но он длился тысячу лет лишь в Месопотамии (-XXII / -XIII века), тогда
как Египет был великой державой в течение большей части Древности, т.е.
около 3 тыс. лет с перерывами.
Чуть более удачной видится идея log-периодичности (логарифмическо-
го роста характерных времен развития по мере углубления в прошлое).
Идея основана на представлении истории мира в форме роста единого
фрактала-древа; она грубо проведена через всю историю мироздания, на-
чиная с Большого взрыва и кончая актами истории XX века (АЕ, 12, 214-216,
314, 318, 335). Словом, процесс эволюции мира убыстряется, и это соответ-
ствует тому, что было сказано в п. 9-16.
10-11. Диатропические системы меньших уровней
Хотя «номотетическую» (диатропическую) систематику аттестуют как
«тупиковую», а ее сторонников — занятыми «лишь общими рассуждения-
ми» [Леонтьев, Акулов, с. 501-502], но такие декларации связаны просто с
незнанием фактов. На самом деле диатропика на исходе XX века доби-
лась важных успехов. О моей попытке судить не мне, однако могу назвать
еще две конкретные системы крупных таксонов.
Пример эволюционной табличной системы с чертами периодичности
привел ботаник А.П. Хохряков. Его система двудольных (подотдел отдела
цветковых) выражена им в форме таблицы из 128 клеток, из коих всего 16
пусто (Хохряков А.П. Табличное изображение системы двудольных И Об-
щебиологические аспекты филогении растений (Совещание в МОИП). М.,
МГУ, 1991). Автор нашел в ней любопытные тенденции, в том числе эво-
люционные; так, он подтвердил идею Мейена о тропическом происхожде-
нии таксонов высокого ранга (Мейен цветковыми не занимался, но А.П.
Хохряков сам говорил мне, что следует его методу).
Одного этого достаточно, чтобы не говорить о тупике, но важнее систе-
ма членистых, которую в 1986-2000 годах построил знаток ракообразных
В.Я. Павлов, прямо назвавший свою таблицу рефренной [Павлов, 2000].
Вообще, его работа представляется мне триумфом диатропического подхо-
да к систематике. Приведу заключительную таблицу Павлова (рис. 82).
Она являет собой часть рефрена «Строение членистых» — пересече-
ние семи его столбцов, задающих зоны обитания, и пяти строк, задающих
форму головы. Исходными являются средний столбец пелагиаль (т.е. тол-
ща вод) и нижняя строка (зачаток головы). Эта строка задает кольчатых
607
Рис. 82. Фрагмент периодической системы членистых, показывающий ве-
роятное происхождение членистоногих из кольчатых червей [Павлов, 2000, с.
173]. 1 - основная личиночная стадия; 2 - педоморфоз; 3 - неотения; 4 - ос-
воение адаптивных зон; ПЗ - переходная зона. Пояснение в тексте
№ СУША ПЗ БЕНТАЛЬ ПЕЛАГИАЛЬ БЕНТАЛЬ ПЗ СУША
IV Eucarida . Eucarida _ Eucarida > - Pterigota
III Peracarida - Syncarida _ Peracarida Copepodoidea Syncarida Peracarida ^3^.. Copepodoidea * Apterigota Myriapoda
II Phyllopoda Ostracoda Phyllocarida Phyllopoda Ostracoda Phyllocarida _ Phyllopoda Ostracoda Chelicerata ~' >- Chelicerata Myriapoda
I ? Pseudocrustacea X Pseudocrustacea Г1 i Pantopoda Trilobita Euthycarcinida Arthopleura?
0 Annelides Annelides __ Annelides Pararthropoda Auscheia Pararthropoda
червей (Annelida), а верхняя строка (сложная головная часть) — высших
раков (Eucarida) и крылатых насекомых (Pterigota).
Кружки означают личинки, а сплошные (наклонные и горизонтальные)
стрелки - эволюцию через изменение личинок (напомню, в этом состоит
жоффруизм). Пунктирные стрелки означают активное приспособление к но-
вым зонам обитания, в том числе посредством смены поведения (ламар-
кизм); варианты переходов заданы рефренной таблицей (номогенез).
Как видим, к каждой клетке указан один-единственный путь, т.е. дано
филогенетическое древо, и это - из опыта дарвинизма. Однако налицо та
же самая грубая филогения, что в п. 10-6: во-первых, она построена на од-
ной семофилогении (эволюция головы), а во-вторых, не допускает детали-
зации. В самом деле, указание происхождения Pterigota из Eucarida не мо-
жет быть уточнено (какой отряд произошел из какого), поскольку именно с
такой точностью само это указание выведено из анализа Павловым планов
строения. Подтверждения дивергентной схемы дарвинизма здесь нет.
Значение этой схемы видится мне в том, что эволюция в ней понята как
заполнение заданной рефренной таблицы, выявленной морфо-эко-
физиологическим анализом. Так что, думаю, автор вполне прав, заявляя,
что его система естественна в различных смыслах (с. 175).
608
Глава 11. Абрис теории
11-1. Исходные положения
Вселенная на всех уровнях, от Метагалактики до субмикроскопических
частиц, обладает целостностью, т.е. является системой, состоящей из
меньших систем. На сегодня видны следующие свойства этой целостности,
общие для всех уровней: упорядоченность разнообразия и активность.
Понятия система и активность не поддаются определению, но смысл
этих слов можно пояснить, а также показать всепроникающую общность
обоих явлений. В отношении активности это проделано в п. 5-2, а в отноше-
нии целостности системы - в п. 6-8. Кроме того, в п. 6-12 было рассказано о
том, что всякая система сама себя регулирует и что в этом состоит богда-
новский подбор (понятие, которое тоже ни из чего не выводится, но обоб-
щает опыт самых разных отраслей знания). Всё это означает новое пони-
мание причинности, близкое к таковому в новой физике (п. 5-2*), но резко
отличное от принятого в биологии его старинного (XVII век) понимания.
А разнообразию можно дать определение: разнообразие множества
есть совокупность отношений сходства и различия между элементами
данного множества (п. 6-3). Это, разумеется, не определение в математи-
ческом смысле. В наше время всерьёз математизирована только теорети-
ческая физика, для чего была когда-то специально разработана соответст-
вующая математика. Многочисленные попытки проделать то же самое в
теоретической биологии, используя ту же математику, не дали ничего срав-
нимого с физикой - ни по красоте, ни по практичности. Очевидно, что если
математизация биологии и возможна, то для этого нужна своя математика.
Не будучи математиком, я смог лишь обозначить проблему в двух пла-
нах. Во-первых, эволюция то и дело ставит вопрос о случайности, и в
прежних работах мне удалось показать, что здесь нужна новая теория, да-
леко выходящая за привычные рамки теории вероятностей (см. п. 5-18 и
книгу [Чайковский, 2004]). Во-вторых, многие процессы эволюции носят
фрактальный характер, и здесь я смог лишь указать на желаемый аппарат
(пп. 6-7, 6-8, Дополнение 4).
Упорядоченность разнообразия выражается в его фрактальности,
рефренной структуре, а также в наличии ядра и периферии.
1) Вселенная на всех уровнях имеет вид сложной совокупности фрак-
талов (п. 6-7): большое подобно малому, а то - еще меньшему, и каждая
структура может иметь дробную размерность. О фрактальности мира см.
[Красный]. Фото фрактала галактик см. Чернин А.Д. И Природа, 2006, № 10.
2) На каждом уровне целостность выглядит во времени как законо-
мерное развитие, а в пространстве - как диатропическая сеть (диасеть).
Диасеть - совокупность многомерных рефренных таблиц, каждый реф-
рен которой являет собой ряд направленных рядов объектов (т.е. мень-
шую таблицу), описывающий преобразование некоторого мерона, т.е.
609
сходных частей этих объектов (п. 6-3). Диасеть удобно мысленно пред-
ставлять себе как многомерную таблицу, часть клеток которой пустует.
3) Ядро множества - совокупность его типичных (хорошо соответству-
ющих диагнозу множества) объектов, а периферия - совокупность его нети-
пичных объектов (п. 10-7). Множество удачно, если ядро намного больше
периферии. Видимо, удачные множества в природе преобладают, в чем и
состоит упорядоченность мира. (Это-довод в пользу шестой ПМ, п. 5-3.)
Каждому масштабному уровню Вселенной соответствуют свои формы
активности. Самые простые и общие примеры форм активности - физиче-
ские (например: гравитация, электрический заряд). Более сложные формы
активности - химические (например, валентность), очень сложные - биоло-
гические, а совсем сложная - сознание (у нее и самый узкий приемлемый
интервал температуры и прочих параметров). Вопрос о том, существуют ли
еще более сложные формы активности, на сегодня науке непосилен.
Самый ясный пример диасети - периодическая система химических
элементов; она состоит из нескольких рефренов (пп. 6-3**, 6-7), и каждый
элемент входит в свои строку и столбец. Два других ясных примера диасети
- грамматика (в ней самыми чёткими рефренами являются парадигмы -
правила склонения существительных и спряжения глаголов) и структура со-
вокупности биологических видов (в ней самыми чёткими рефренами явля-
ются те таблицы гомологических рядов, где видно направление преобра-
зования определенных меронов - см. рис. 35, 36 и 89).
Отдельные примеры рефренов: диаграмма спектр - светимость звезд;
преобразование натурального хозяйства в товарное в разных странах.
В подавляющем большинстве рефрены менее наглядны, чем приве-
денные, из-за многомерности таблиц и наличия в них многих пустых кле-
ток97, но и такие рефрены столь же реальны, как у химических элементов.
Эволюция понимается в книге как единый (целостный или, что то же
самое, системный) процесс преобразования одних объектов в другие.
Данное преобразование идет путем усложнения как самих форм активно-
сти, так и объектов, порождаемых этой активностью. Появление новой
формы активности сопровождается фазовым переходом в смысле п. 5-2,
но сегодня нельзя сказать, что тут является причиной и что следствием.
В силу описанной выше упорядоченности, эволюцию можно предста-
вить как движение вдоль диасети, при котором прежде пустые клетки за-
полняются (прогресс, рост разнообразия), прежде занятые пустеют (выми-
рание), а прежде изолированные занятые клетки окружаются другими за-
нятыми, т.е. заселяются (периферия становится ядром). 91
91 Пустоты традиционно служат душевным препятствием для признания существования
самой диасети (например: [Попов, с. 125]), но это вызвано смешением двух понятий -
удобной классификации (ее диасеть обычно не дает) и реальной структуры разнообразия
объектов - например, организмов. Для понимания эволюции важно только второе.
610
Так, эволюция химических элементов ныне идет (в опытах физиков)
путем синтеза трансурановых, причем заполняются заранее известные
ученым клетки. В иных областях таблицу приходится додумывать.
Эволюция - реально наблюдаемый (на ископаемых и в эксперименте)
процесс, и с этим никто (из понимающих суть дела) не спорит. А признать
или отвергнуть наличие ума, движущего эволюцию, - дело веры.
В природе на каждом уровне видна самоорганизация - появление
элементарных частиц, затем самосборка элементарных частиц в атомы,
атомов в молекулы, молекул в пространственные структуры и т.д. Самоор-
ганизация - аспект активности, обычный для всех ее форм. Она включает
самосборку, но далеко выходит за ее рамки. Она - неотъемлемое свойство
природы, отмеченное многими; в том числе Богдановым - как подбор.
Другим аспектом активности является эмерджентность (создание со-
всем нового, п. 6-7**). Как и самоорганизация, она многоуровнева. В частно-
сти, всякий онтогенез (возникновение звезды из пылинок или организма из
зиготы) - тоже эмерджентный акт, но его мы наблюдаем регулярно, а вот
эволюционная новизна может быть уникальна. Так, всякое возникновение
нового уровня организованности (например: Вселенной, жизни, многокле-
точное™, разума) - очень редкий эмерджентный акт, иногда - уникальный.
Диасетью и фрактальностью обеспечиваются все структуры (в том
числе структурная связь всего со всем, а также рамки онтогенеза), тогда как
активностью - все процессы в этих структурах, в том числе эволюция.
Сказанное приводит к Основному тезису общей теории эволюции:
активность - движущая сила эволюции, разнообразие - ее объект, а
самоорганизация - основной ее регулятор. Разнообразие реализуется в
_________рамках объективно существующей фрактальной диасети.________
Это - философский постулат. Он представляется мне уточнением
«принципа автоэволюции», который призван «осветить процесс... эволю-
ции, внутренне присущей веществу и энергии» [Лима, с. 350]. Постулат
должен явным образом заменить неявные постулаты, которые бытуют в ли-
тературе (и подчас противоречат друг другу даже в одном тексте). Только
такой постулат может стать стержнем теории, а не разговоров о ней.
Уточнение же состоит в том, что 1) «внутренне присущими» считаются
как сама активность, так и усложнение ее форм во времени; 2) в качестве
объекта эволюции рассматривается не столько сама материя, сколько раз-
нообразие ее модификаций, взятое как целое; 3) процесс эволюции высту-
пает как триединство - двигатель, объект, регулятор.
Каждому усложнению структуры соответствует усложнение форм ак-
тивности - в этом состоит прогрессивная эволюция на всех ее уровнях.
11-1* Специфика биологии
Прежде чем строить схему биологической эволюции, надо изложить в
нужных нам терминах основные свойства жизни. Хотя определения жизни
611
дать нельзя (как и никакому из основных понятий), но основные отмеченные
выше свойства мира, важные для эволюции - активность, диасеть и фрак-
тапьность, - можно и нужно описать в биологических терминах. Они прини-
мают у живого следующие существенные для нашей темы формы.
1) Активностью обладают и организмы, и части организма, вплоть до
микроструктур (п. 5-15), и группы организмов (улей, стая, ценоз и т.п.). Наи-
более общими специфически биологическими типами активности, не имею-
щими аналогов в неживой природе, предстают прежде всего четыре фено-
мена - метаболизм, поведение, смена поколений и смена норм.
Метаболизм, т.е. обмен веществ, ведет к тому, что живое существует
только в форме процесса (исключение - замораживание; оно обсуждено в п.
7-10). Поэтому неминуемо действует «метаболическая спираль» (п. 5-11), а
она порождает, в частности, «винт эволюции» (п. 4-13). Можно говорить и
про метаболизм ценоза (например, циано-бактериального мата).
Поведение не надо путать с подвижностью, которая есть и у неживой
материи. Поведение содержит самопроизвольную составляющую, тогда как
подвижность при поведении может быть минимальна (см. фото перед ти-
тульным листом) и выражаться, например, только в виде направления де-
ления клеток (таковы ловчие кольца хищных грибов). Поведение может
быть целесообразным, бесцельным и даже губительным.
Неясно, насколько связано с самопроизвольностью поведение частей
организма (метацеркариев (п. 6-15), иммуноцитов, сперматозоидов и т.п.) и
групп организмов, однако очевидны параллели. Так, поведение отдельных
особей термитов или пчёл сходно с поведением иммуноцитов, а поведение
улья или термитника - с поведением организма.
Тейяр видел в качестве элементарного субъекта поведения клетку:
«Можно сказать, что... сознание тем совершеннее, чем более сложное и луч-
ше организованное материальное строение оно сопровождает». «Обычно пси-
хическую жизнь в мире ведут с первых форм организованной жизни, то есть с
появления клетки. Стало быть, я присоединяюсь к общей точке зрения... ко-
гда решающий шаг в прогрессе сознания на Земле отношу к этой конкретной
стадии эволюции» [Тейяр, с. 58, 79].
Ныне можно утверждать большее: целесообразно поведение на всех
уровнях организации живого, начиная с частей клетки (напомню хотя бы
рост микротрубочки или аксона). Поэтому, если отложить в сторону вопросы
мировоззрения и говорить лишь про описание фактов, то самой научной
представляется та философская позиция, которая признаёт свою долю соз-
нания за каждым живым объектом, начиная с частей клетки. Напомню, что
Геккель шел еще дальше, признавая сознание даже за атомом (п. 3-8).
Смена поколений несет четыре всем известные функции: позволяет
видам образовывать экосистемы, где виды служат друг другу едой и сре-
дой обитания; обеспечивает избыточность размножения, которая позво-
ляет видам занимать новые зоны обитания и экологические ниши; реализу-
612
ет изменчивость в смысле отличия детей от родителей. Итогом последнего
является рост разнообразия в процессе смены поколений.
Смена норм. Норму можно понимать различно (гл. 5 и п. 6-8*), но при
любом понимании она является итогом самоорганизации и ограничивает
изменчивость. Наоборот, снятие давления нормы (п. 5-2*) увеличивает эту
изменчивость в последующих поколениях. Оно выступает как главный био-
логический инструмент запуска акта самоорганизации. Когда найдена новая
норма, наступает завершение акта самоорганизации, а затем следует паде-
ние изменчивости. Таким путем происходит смена нормы.
Единичная смена нормы представляется мне элементарным регуля-
тором эволюции. Как тип активности она стоит особняком, поскольку регу-
лирует остальные типы биоактивности (что будет сказано в п. 11-2).
Это позволяет конкретизировать сформулированный выше Основной
тезис общей теории эволюции так, чтобы получить Первый тезис намечае-
мой теории биологической эволюции:
Эволюция живого являет собой триединство:
__________________активность - диасеть - смена норм._______________
Итоги разных типов активности живого принимают различные формы.
Их как минимум три - материальная (новые формы организмов), энерге-
тическая (см. п. 7-10) и информационная (например, п. 5-5*). Третья наи-
менее понятна, но и в ней все-таки можно вычленить три главных аспекта -
узнавание, изобретение и наследование.
Всепроникающий характер феномена узнавания (его призвана выра-
зить концепция эпигностики), результатами которого являются онтогенез и
иммунитет, отмечен в п. 5-15. Наследование, которое прежде считалось
чем-то вроде копирования, оказалось сложным информационным процес-
сом: например, при создании половой клетки тоже текут процессы узнава-
ния (пп. 8-8, 8-12), а в диплоидной клетке целые хромосомы оказываются
выключенными (генетический импринтинг). Тот факт, что изобретение
(рождение новизны, эмерджентность) обычно выступает на всех уровнях как
итог фрактального роста, рассматривает Дополнение 4.
Для живого характерно, что информационные процессы (узнавание, ко-
пирование, горизонтальный перенос, генетический поиск - пп. 5-13, 5-15, 6-
9) являют собой типы активности, при которых запуск и остановку задают
знаки (маркёры), а их задает, в свою очередь, норма организмов. Знаки
энергетически и информационно ничтожны, но дают значимый (энергетиче-
ски и информационно) эффект. Обратимость действия знаков - характер-
ное свойство живого (пп. 5-5*, 7-10). В расстановке и чтении знаков сущест-
венны тройной текст ДНК (п. 5-5*) и ее резонансные свойства (п. 7-11). Гены
при этом служат знаками, переключающими режимы формообразования (в
том числе фрактального роста), тогда как сами формы в генах записаны
быть не могут (п. 6-7*). Где и как они записаны - предмет споров.
613
2) Диасеть живого характерна полифункциональностью элементов (п.
6-6*): например, белок может выполнять до четырех разных функций, в том
числе - фермента и структуры [Свердлов, с. 711].
Сама по себе диасеть живого выявляется без обращения к идее родст-
ва. Зато, когда она выявлена, можно содержательно ставить вопрос о спо-
собе нахождения родства (см. далее, конец пункта 11-3*).
Фрактальность диасети живой природы установлена на сегодня, на-
сколько знаю, только для одного аспекта ее наблюдения - для общеприня-
той системы таксонов. А именно, описан фрактал-древо системы таксо-
нов и измерена его дробная размерность; она оказалась примерно одинако-
ва у всех главных таксонов (Burlando В. The Fractal dimension of taxonomic
systems //Journ. Theor. Biol., 1990, vol. 146, № 1). Это можно истолковать так:
эволюция жизни (как и эволюция Вселенной) есть итог фрактального роста.
Для других аспектов описания диасети (морфо- и экологических) пока
слишком мало материала, однако можно предполагать, что биосфера - это
фрактал-сеть. Показать фрактальность диасети в целом было бы важно
для понимания связи всего со всем в единую систему (п. 6-8).
3) Фрактальный рост касается не только диасети - это и фракталь-
ный рост особи (п. 6-8). Привычная аналогия живого с неживым («Клетка -
естественная крупинка жизни, как атом - естественная крупинка неоргани-
зованной материи» [Тейяр, с. 73]) довольно поверхностна и уводит от пони-
мания более глубоких свойств живого. Она верна, пока мы понимаем орга-
низм по Окену, как компактный набор “инфузорий”; но для жизни характер-
но глубокое проникание ее частей друг в друга (на что “инфузории” неспо-
собны), и специфически биологический фрактал образует именно оно.
Феномен проникания известен на всех масштабах: в бактериальной
клетке это - пронизывающая ее замкнутая молекула ДНК, в эвкариотной
клетке - растущие сквозь нее микротрубочки, в клеточном ядре - фракталь-
но перемешанные экзоны и интроны, в крупном организме - сеть сосудов и
нервов, в экосистеме - особи разных видов и трофических уровней, живу-
щие вперемешку, и т.п. Мы к ним привыкли и не замечаем скрытых здесь
тайн. К ним относятся и те свойства организма человека, которые позволя-
ют существовать восточной медицине, например, иглоукалыванию (п. 6-8);
они основаны на признании гораздо более глубокого взаимопроникновения
частей, чем принимает европейская наука. Полагаю, что без их раскрытия
эволюцию не понять. Ну хотя бы: единый принцип взаимопроникновения
ясно показывает, что конкуренция единиц отодвинута далеко на задний
план по сравнению с конструктивным взаимодействием.
4) Три типа уровней. Основные уровни биологического взаимодейст-
вия: биосфера, экосистема, популяция, организм (в том числе одноклеточ-
ный), орган или система (например, нервная), ткань, клетка, внутриклеточ-
ная структура, молекулярный механизм, макромолекула. На каждом из этих
уровней работает самоорганизация, которую удобно выразить в форме
614
биополя данного уровня. Оно, как и физическое поле, является теоретиче-
ским конструктом, недоступным прямому наблюдению и вводимым для по-
нимания факта целостности наблюдаемого явления. (Ныне почти всеми за-
быто, что гравитацию тоже отрицали 2 тыс. лет, поскольку ее нельзя пря-
мо наблюдать (пп. 6-12, 7-11), а еще раньше Фалес полагал магнит оду-
шевленным, поскольку не мог вообразить себе магнитное поле.)
Основные уровни биологической классификации, т.е. уровни основных
таксонов', царство, филум (тип, отдел), класс, отряд (порядок), семейство,
род, вид. О том, в каком смысле таксон можно считать целостностью (а с
тем и реальностью), см. п. 8-11.
Основные уровни сложности организма: одноклеточные, колониаль-
ные, пластинчатые и т.д. - до цветковых растений и животных с головным
мозгом (рис. 60 и 81). Указанные три типа уровней требуют трех типов опи-
сания эволюции (см. далее), но все их можно предполагать фрактальными.
5) Системное сходство. Развитое в данной книге представление ис-
ходит из общей точки зрения, согласно которой все формы эволюции (живо-
го и неживого) имеют существенное сходство. В частности, аналогия био- и
социальной эволюций имеет и другую сторону, кроме технической - миро-
воззренческую. Так, японский эволюционист Кунио Кавамура, предпочтя
восточный взгляд на эволюцию западному, пришел недавно (2002 г.) имен-
но к той же системной точке зрения, что и западные философы:
«Отличительная черта концепции Кавамуры - показать единство как можно
большего количества явлений, связанных с развитием биологических объектов -
от происхождения жизни до эволюции человеческого общества... По его мне-
нию, культуры и цивилизации людей формируются в результате “самоорганиза-
ции”, подобной самоорганизации органических молекул, которая постулирова-
лась в гипотезе происхождения жизни... Кавамура утверждает, что и в дебрях
микрокосма мельчайших организмов можно найти те признаки, которые больше
на виду на самых высших уровнях организации..., что и у бактерий есть культу-
ры, аналогичные культурам, возникающим в человеческом обществе. Это любо-
пытное утверждение, к сожалению, практически никак не поясняется. Автор
только призвал исследователей найти у бактерий такие средства хранения ин-
формации, которые лежат вне их генетического материала» [Попов, с. 107-108].
Что хочет найти Кавамура, сказать не берусь, но замечу, что у мель-
чайших бактерий вольбахий (п. 8-4) совсем недавно действительно был
найден целый ряд свойств, которые так и хочется назвать цивилизацион-
ными. В частности, одни вольбахии могут оставлять в геноме хозяев записи
для других вольбахий - см. [Марков, Куликов, 2006].
11-2. Стержень теории биологической эволюции
Дарвинизм не различал движущего (“мотор”) и регулирующего (“руль” и
“тормоз”) принципов, вводя вместо обоих постулат о естественном отборе.
Ламаркизм, пусть и нечётко, различал их (“мотор” - это прогресс, “руль” -
615
приспособление), но пренебрег структурным принципом (таковы “дизайн”
машины, “дорога и знаки” вне ее и т.п.). Наоборот, структурный принцип
служил главным в номогенезе (который зато был равнодушен к двум пер-
вым принципам). Эти частные концепции, можно сказать, не имели стержня
- такого, например, какой в предлагаемой концепции выражен Первым те-
зисом (п. 11-1*). Его надо конкретизировать для нашей цели, для живого.
Излагаемая в данной книге концепция исходит из того положения, что
всё в жизни, в том числе и в эволюции, имеет основой какую-то форму ак-
тивности. Тем самым, процесс эволюции описывается как изменение (сме-
на, приобретение, утрата) и комбинация форм активности живого. Само
возникновение жизни можно рассматривать как появление информационной
формы активности, а также феномена давления нормы. (Хотя есть и другая
точка зрения - что информация пронизывает всю Вселенную, а жизнь воз-
никла в момент появления генетической формы информации, т.е. генетиче-
ского кода [Зусмановский, гл. 3].) Всё сказанное позволяет сформулировать
Второй тезис намечаемой теории эволюции:
Активность выступает в эволюции живого в виде нарастающей
самопроизвольности - как поведения, так и переработки информации.
К хаосу это не ведет потому, что объект активности - упорядоченная
диасеть (п. 11-1). Эволюция любых объектов имеет дело с нею, но в теории
биоэволюции она служит основным предметом внимания потому, что имеет
место транзитивный полиморфизм (транпол). Благодаря нему разнообра-
зие свойств вида может наследоваться через единственную особь (п. 6-4*),
а разнообразие свойств высшего таксона может наследоваться через
единственный низший таксон (п. 10-6). Такое наследование возможно
потому, что все варианты (блоки) черпаются из общего набора
возгЛЗэеггиннйвенно трем типам уровней (взаимодействия, классификации
и сложности - п. 11-1*, рубрика 4) имеем три аспекта эволюции - функцио-
нальный (эко-физиологический), структурный (морфо-таксономический) и
сложностный (согласно тезису Михайлова, п. 10-2**). Первые два взаимодо-
полнительны92, а третий тип стоит особняком - это предмет учения о про-
грессивной эволюции (пп. 5-14, 6-8); он связан с эмерджентностью (см. п.
.11-1, а также далее, п. 11-2*) и потому относится к малой части актов эво-
люции. Акты эволюции будут описаны в конце главы.
Все три типа (аспекта) эволюции реализуются через процедуру онто-
генеза, которая до сих пор в основе своей непонятна (п. 5-15). Однако во-
просы эволюции, с нею связанные, мы формулировать умеем:
1) Как происходит макромутация (п. 5-5*), т.е. переход в новую клетку
диасети (каким образом изменение наследственной информации связано с
изменением онтогенеза, вызывающим изменение организма)?
92 Ламаркизм занят в основном функциональным, а номогенез - структурным аспектом.
616
2) Откуда берутся преадаптации (пп. 4-10, 9-13, 9-15*), т.е. каким обра-
зом заполняется та клетка диасети, в которой организм получает приспо-
собление к будущей среде обитания?
3) Как происходит акт прогресса (каким образом в ходе онтогенеза за-
полняется строка таблицы диасети, прежде пустовавшая)?
Любая теория эволюции будет должна ответить на эти вопросы. В их
постановке видится едва ли не основной смысл нового номогенеза, по-
скольку прежние учения даже не имели языка для их формулировки.
Тем самым, имеет смысл Третий тезис намечаемой теории:
три явления, вместе описывающие эволюцию организмов, -
макромутация, преадаптация и прогресс - вытекают из свойств диасети и
_________будут поняты не раньше, чем появится теория онтогенеза____
Благожелательный рецензент счел главным изъяном ЧЭ мою неспособность
«объяснить, что стоит за диатропикой с таблицами рефренов и самим феноменом
“блочной эволюции”» {Миркин Б.М. // УСБ, 2005, № 2, с. 223; рецензия перепеча-
тана в ЛЧ, 2005). Я благодарен Б.М. Миркину за эту рецензию, побудившую ме-
ня более внятно изложить в главе 6 положения диатропики. Относительно дар-
винизма он заключил, что «заменить его пока нечем» (с. 224).
Это неверно: даже нынче, в зачаточном виде, диатропика «заменяет дарви-
низм» в том смысле, что выполняет основную функцию теории - ставит новые
вопросы. (Напомню, что дарвинизм, объявив отбор удачных мутаций причиной
изменений организмов, откровенно упустил всю проблематику. Ни блочная эво-
люция, ни сами ряды дарвинизмом не только не объяснены, но даже им не упо-
минаются — для этого в нем нет языка.) Диатропика фактически описывает за-
кономерности там, где дарвинизм постулирует их отсутствие.
Далее, биосфера одна, поэтому ее эволюция состоит только в ее изме-
нении. Остальные эволюирующие объекты как изменяются сами, так и сме-
няют друг друга. Чем мельче экосистемы и таксоны, тем больше роль их
смен в их эволюции. Онтогенез экосистемы именуется сукцессией (п. 8-13*).
Особи (и только они) сменяют друг друга регулярно, но это не акт эво-
люции и не его причина, а его непременное условие (вообще, элементар-
ные условия и причины эволюции именуются ее факторами - см. далее).
Основной экологический механизм эволюции организмов - смена
норм. Она запускается снятием давления нормы, частным случаем которо-
го является стресс. Механизм действует тем эффективнее, чем выше эко-
логическая прочность вида (п. 6-2*). Это противоположно принципу есте-
ственного отбора, зато совместимо с идеей богдановского подбора.
Основной физиологический механизм эволюции организмов - гене-
тический поиск. Суть его в следующем. В дискомфортных условиях (а ут-
рата нормы обычно влечет дискомфорт) большинство особей вида попада-
ют в состояние стресса и либо гибнет, либо претерпевает модификации.
Некоторые из них адаптивны (эволюция по Аршавскому, п. 6-10*), и их-то
617
носители ведут генетический поиск. Он состоит в том, что доминанта (п. 5-
11) направляет всю активность организма (поведенческую, физиологиче-
скую и генетическую) в одном направлении, а это помогает найти нужное
сложное изменение из наличного (расширенного снятием давления нормы)
спектра возможностей, т.е. из числа доступных клеток диасети.
Основной морфологический механизм эволюции многоклеточных со-
стоит в перестройках онтогенеза, суммируемых схемой «винт эволюции»
(пп. 4-13, 5-15). «Шаг винта» (то, что можно заметить как элементарное из-
менение) различен - от нуля (некоторые виды не изменяются миллионы
лет) до величин, заметных в немногих поколениях (сальтационная эво-
люция). Причины изменения «шага винта» лежат в физиологии (эволюция
снизу - см. далее, п. 11 -3*) и в экологии (эволюция сверху - п. 11 -3**).
Эти три механизма обычно рассматриваются как преобразующие на-
личное разнообразие организмов, т.е. как обеспечивающие движение в
рамках уже реализованной части диасети. Преобразование разнообразия
является набором тенденций (п. 6-2), т.е. преимущественно наблюдаемых
путей заполнения диасети. Наличие тенденций является формой активно-
сти. Эволюция, понимаемая как движение по диасети, неминуемо носит ха-
рактер тенденции, поскольку параллельные ряды в рамках рефрена никогда
не бывают тождественны. Однако нужен еще механизм новаций.
Как возникает новое, как заполняется первая клетка в той строке диасе-
ти, которая прежде была пуста (никогда прежде не заполнялась)?
11-2* Возникновение нового
Почти вся эволюционная литература занята исследованием частных
свойств - число и форма органов, типы питания, окраска, теплоустойчи-
вость, детали поведения и т.п., но этим не продвигается решение главной
проблемы - откуда и как взялись сама жизнь и главные свойства живого
(дыхание, питание, фотосинтез, размножение, ценозы и т.д.), и как появи-
лось мышление. Здесь успех минимален, а без него нет теории.
В ходе эволюции несомненно появление новых сущностей, т.е. совсем
нового, того, чему прежде и зачатков не было. Может ли теория это опи-
сать? Луэс, введя понятие «эмердженты» (п. 3-10), как бы ответил положи-
тельно, но за истекшие 130 лет сути явления раскрыть не удалось.
Почти все эволюционисты просто не замечают проблемы эмерджент-
ности (п. 10-4), простодушно полагая, что новое возникает тем же способом,
каким улучшается старое. Поэтому они избегают и главного вопроса: как
произошла жизнь? Этим вопросом (биопоэзом) занимается узкий круг уче-
ных, почти или вовсе равнодушный к остальным проблемам эволюции.
Данная книга написана с иной позиции (см. начало гл. 7). А именно:
биопоэз есть главный эмержёнтный акт эволюции, и от того, как его
понимает данная теория, зависит ее содержательность в отношении
описания последующих актов эволюции.
618
В частности, приняв, как Воейков, что в биопоэзе первым источником
энергии были АФК (но не АТФ), мы задаём понимание эволюции энергетики;
приняв концепцию прогеноты Дарнела и Дулитла, мы отказываемся искать
организм-предок и признаём правоту Вернадского; приняв, с геологами, что
жизнь возникла практически вместе с условиями для нее, мы отказываемся
от принципа ожидания редких событий; а приняв, вслед за Скулачевым, что
фотосинтез возник еще до аппарата наследственности и был ультрафиоле-
товым, но позже, в бактериях, стал работать от видимого света, мы прини-
маем принцип реализации прежней идеи на новом материальном носителе
и тем самым допускаем платонизм как способ рассуждения.
Наоборот, если принять модель Опарина (в «первичном бульоне» само
собою накапливалось всё, что нужно для возникновения жизни - вот она и
возникла), то такой “принцип слонопотама” (пп. 7-2*, 7-7*) неминуемо перей-
дет в остальную теорию эволюции - это и будет дарвинизм. Кстати, хотя
дарвинисты избегают говорить про биопоэз, однако если говорят, то всегда
выплывает нечто мутное, в стиле опаринского «бульона».
Обычная уловка, высказанная еще Луэсом (новое всегда составлено из
элементов старого, например, из тех же атомов), очень слаба: элементов
слишком много, сборка идет слишком регулярно, а значит, для объяснения
эмерджентности нужен особый принцип. На него указывали многие, в том
числе Поппер: будучи рациональным мыслителем, он всё же отметил, что
«иррационалисты, которые говорили о “креативной” или об “эмерджентной”
эволюции, смогли разглядеть нечто важное» (Поппер К. За пределами поиска
инвариантов // ВИЕТ, 2002, № 4, с. 681).
Ближе всех к сути этой проблемы подходили те, кто видел в эволюции
таксонов хологению (или трансиндивидуальный онтогенез, п. 4-13): ведь он-
тогенез тоже создает новую сущность (живую особь с новой душой).
Новая эволюционная сущность мыслится при этом как принципиальное
изменение онтогенеза: новизна в эволюции следует из новизны очередных
особей, но не при их рождении, а при их зародышевом развитии. Это есте-
ственно: в силу общности диасети оказываются общими и эмерджентные
акты - онтогенеза и эволюции. Поясню: в ходе онтогенеза происходит дви-
жение зародыша вдоль диасети, причем реализацию новой строки рефрен-
ной таблицы (например, закладку сердца) следует считать эмерджентным
актом данного онтогенеза; но сходная строка когда-то появилась впервые
(появление организмов с сердцем) - то был эмерджентный акт эволюции.
Можно ли эту параллель признать решением проблемы? Стороники хо-
логенеза отвечали положительно. Ныне можно добавить: эмерджентность,
как сказано в п. 5-2*, - это смена формы активности за счет фазового пере-
хода, а ее сохранение в одной эволюционной линии при различии ее от ли-
нии к линии - это принцип компенсации (п. 8-10): в одной линии жизненный
порыв тратится, в основном, на увеличение численности, в другой - на ус-
ложнение прежних конструкций, в третьей - на создание новизны.
619
В качестве абстрактного механизма новизны в пп. 6-7** и 6-8 были
предложены фрактальная турбулентность и блочный системогенез. Хотя
конкретное описание на этом языке - дело будущего, но уже сейчас можно
(вспомнив Второй тезис), высказать Четвертый тезис будущей теории:
Новое рождается в каждом онтогенезе, но без свободы выбора. Акт эволю-
ционной новизны состоит в изменении онтогенеза, причем выбор свободен.
Вот, вроде бы, и стержень теории. И если мы не удивляемся, когда в
голову приходит совсем новая мысль, то, согласно этой идеологии, не надо
удивляться и тому, что в эволюции появляются совсем новые формы.
Но тут читатель-скептик, думаю, почувствует обман: вместо понимания
эволюции дается ссылка на непонятость мышления. Да, это так, но такова
сама проблема, и нет оснований ожидать, что мышление и эмерджентность
могут быть поняты по отдельности. См. Дополнение 9.
Тот факт, что сходные уровни развития, в том числе и сходная принци-
пиальная новизна, достижимы в разных линиях эволюции, также говорит о
единой диасети живого. Вместе с формами активности эта фрактальная
диасеть образует пространство жизни. В данном пространстве (а не в на-
блюдаемом трехмерном) и течет эволюция (по Янчу, п. 6-12).
11-3. Эволюция снизу и сверху
Изменение организмов в ряду поколений - это их эволюция. Описание
ее как эволюции самих организмов и их частей - это описание снизу, а че-
рез эволюцию биосферы и экосистем - это описание сверху (п. 6-14).
Не раз говорилось, притом многими, что мир организован в единое це-
лое сверху (п. 8-1). Вот совсем новое на сей счет высказывание:
«Понятие целесообразности как условия существования объекта снова появляется
в поле зрения биологов. Что же задаёт цель, предписывает свойства “сортов” ор-
ганизмов..? Ответ лежит на поверхности: система высшего ранга» (Заварзин Г.А.
Составляет ли эволюция смысл биологии? // ВРАН, 2006, № 6, с. 533).
Как видно из заглавия статьи Заварзина, автор склонен вообще
отказаться от эволюционного понимания мира, и это не редкость -
эволюцию сверху многие видят как творение. Приведу любопытный пример.
Зоолог А.С. Раутиан вспоминает беседу с палеоэкологом В.В. Жерихиным:
«в 1972 г. я услышал от Владимира Васильевича такую сентенцию. “Ты зна-
ешь, я похоже сам изобрел Господа Бога. В самом деле, направлением фило-
генеза таксонов “заведует” сообщество... Эволюцией сообществ, наверное -
биосфера. А Кто направляет эволюцию биосферы? Не космическая же пусто-
та, в самом деле!”. Это типичная ошибка, вызванная поиском объемлющей
управляющей системы: целого, определяющего судьбу своих частей... и не-
дооценкой возможностей саморегуляции через систему внутренних обратных
связей (как учит нас “Кибернетика” Н. Винера)» [Жерихин, с. 24].
В самом деле, ошибка типична: Раутиан полагает (в отличие от покой-
ного Жерихина), что кибернетика что-то говорит о системогенезе. Но нет,
620
она всего лишь описывает работу существующих систем, в которых все свя-
зи, прямые и обратные, в наличии. Жерихина же, насколько понимаю, вол-
новал системогенез - появление и самих объектов, и связей между ними.
Если допустить, вместе с Раутианом, что система связей задана, то мы
придем к эволюции по Григорию Нисскому, от чего сам Раутиан далёк.
Появление новой системы прямых и обратных связей (системогенез)
до сих пор совсем выпадало из внимания большинства ученых именно в си-
лу господства необдуманной веры в возможность эволюции как «саморегу-
ляции через систему внутренних обратных связей». Регулятор бессмыслен,
пока нечего регулировать, пока нету самой системы - объекта регуляции.
Суть дела можно пояснить на языке ПМ (п. 5-3). Статистическая ПМ видит в
мире лишь балансы, не видя целостных систем, т.е. не видя прямых связей, регу-
лируемых обратными связями. В биологии это ведёт к ссылкам на отбор мута-
ций, а в общественной жизни - к рыночной модели: она тоже не видит прямых
связей, уповая на обратные - конкуренцию, налоги, благотворительность и суд.
Двигатели (формы активности, которые создают нечто новое, они всегда вне рын-
ка — на заводе, в поле, в лаборатории, в кабинете учёного или чиновника) выпада-
ют из анализа и сознательного реформирования. Из-за этого на деле прямые связи
ведут себя непредсказуемо, часто - подминая регуляторы. Устойчивые системы
(организм, экосистема, государство) на деле описываются системной ПМ, однако в
языке описания сохраняются статистические термины. Так, до сих пор говорят:
«баланс властей», хотя имеют в виду многослойную регуляцию (п. 6-8*).
Иногда пишут, что эволюция сверху есть примат функции над формой
(экосистема задает требования к организму, каковые форма и выполняет).
Это неверно: достаточно взглянуть без презумпций на разнообразие форм и
функций, чтобы увидеть, что они друг друга не задают (п. 6-13**).
Согласно тезису Янча (п. 9-16), обе эти формы эволюции сопряжены
(кроме прочего) в том смысле, что чем мельче система, тем крупнее ее ха-
рактерный компонент. Данный факт отображен на рис. 83, где t0 - момент
разделения вещества и энергии, интервал tn - ts означает время становле-
ния жизни (около 1 млрд лет), момент tp - появление тканевой организации,
tm - момент появления человека. Под каждой системой указан ее характер-
ный элемент (под звездой - атом, под планетой - молекула и т.д.).
Как видим, с появлением цивилизации макроуровень слился с микро-
уровнем. В частности, человек выступил и как вид, и как царство (п. 10-10).
11-3* Эволюция снизу
1) Активность особи. Из нее вытекают предпосылки всех теорий -
ламарково упражнение, сент-илеровы ответы зародыша на среду, дарвино-
ва избыточность размножения, кропоткинская взаимопомощь, заполнение
мейенских таблиц, ламарково стремление к прогрессу, бергсоновы тенден-
ции и т.д. Словом, ни один автор без идеи активности не обходился.
621
макроуровень
макроуровень
Охлаждение Вселенной
Стабилизация размеров небесных тел
Синтез тяжелых ядер
Планетная хемодинамика
Геодинамика
Анаболизм (5) ♦ рекомбинация
Фотосинтез
Самовоспроизведение
Размножение
механизмов
Обособление
envmpu клетки
разнообразия
Половое размножение
Морфологический прогресс
Обособление полов
Язык сигналов
Автономизация
от среды
Реверсия
(11
Характерные процессы
Протон иейтрок ядро молегула
Язык
х 5-с понятий
5 сл.
к
Рис. 83. Схема коэволюции уровней. Подробнее см. [Чайковский, 1990, с. 219]
622
2) Размножение. Оно служит базовым феноменом эволюции, но глав-
ное - не его избыточность, как думал Дарвин (п. 2-13), а смена поколений. В
норме избыточность невелика и важна для экологии (для расселения), но не
для эволюции: огромно лишь число гамет и семян, гораздо меньше - число
мальков и заростков; а число взрослых особей столь близко к числу оста-
вивших потомство, что конкуренция вариантов не может быть эффективна
для выявления различий их качеств. О конкуренции см. далее, п. 11-4*.
3) Норма (рамки в пространстве форм и функций, обеспечивающие
жизнь и размножение) существует для любого организма (и любого его ор-
гана). Активность организма ограничена рамками нормы, и снятие рамок
(снятие давления нормы) ведет к росту изменчивости, в том числе наслед-
ственной. Примеры: стресс, иммунная реакция, доместикация.
Стресс может менять спектры изменчивости (и тем способствовать
эволюции), притом при низкой размножаемости. Эвстресс и дистресс ведут
к одним и тем же изменениям форм и функций (поскольку оба вызваны сня-
тием давления нормы и порождают движение по одной и той же диасети) -
только первые ненаследственны, а вторые, являясь итогом прорыва "барь-
ера Вейсмана” (пп. 3-9, 5-11**), могут быть наследственными; при этом
“мутанты” обычно мало жизнеспособны, но и они могут выживать, если ис-
ходный вид обладал запасом экологической прочности (п. 6-2*).
4) Организмы имеют форму (упорядоченность, не сводимую целиком к
их функциям); формы проявляют сходства, и эти сходства упорядочены
диасетью (п. 6-13**). Ее рефрены не ограничены никакими рамками (сход-
ства наблюдаются между видами очень далеких таксонов), но чем ближе (в
пространстве признаков) таксоны, тем больше рефренов их связывает. Это
принято считать (без должных оснований) свидетельством родства. Вероят-
но, функции упорядочены своей диасетью, но об этом известно мало.
В любом рефрене к одному мерону могут принадлежать органы с раз-
ными функциями (гомология). Орган с данной функцией может входить в
разные рефрены (аналогия). Число различных форм ограничено (обозри-
мо), и формы дискретны. Форма зависит от функции, но обилие форм го-
раздо выше обилия функций, особенно у насекомых и цветковых растений.
5) Каждый таксон задан набором меронов (или, более детально: мно-
гомерной таблицей возможных значений своих меронов). Эта таблица яв-
ляет собой архетип таксона по Мейену. Соединение меронов в единую
систему (задаваемую рисунком) являет собой архетип таксона по Оуэну.
По аналогии с техникой: если задан класс изделий, то архетип по Мейе-
ну - это инвентарный список всех деталей данного класса изделий (сгруп-
пированный по классам деталей), а архетип по Оуэну - чертеж типового из-
делия, характеризующего данный класс. Эти два понимания архетипа взаи-
модополнительны по Бору, чем и дается нужный инструмент анализа.
Архетип по Мейену обнаруживает рефренную структуру, сходную для
разных таксонов (диасеть). Клетки диасети заполнены неравномерно, но не
623
хаотично - видны тенденции (по Бергсону). Так, для эвкариот обычно при-
близительное распределение Виллиса видов по родам, родов по семейст-
вам и т.д. (в этом - одно из проявлений фрактальности Вселенной).
6) Таксономический взгляд на эволюцию: она состоит в преобразова-
нии одних таксонов в другие; диатропический взгляд, эволюция состоит в
преобразовании наборов меронов. Таксоны регулярно появляются и исче-
зают, а мероны появляются редко, и в появлении новых меронов (новый
строк рефренной таблицы) состоит прогрессивная эволюция (п. 11-1).
В остальном же и в основном эволюция - изменение состояний налич-
ных меронов, т.е. движение в пределах одних и тех же строк таблицы. Дви-
жением здесь именуется заполнение одних клеток таблицы и опустошение
других (смежных). Такую эволюцию иногда именуют плоской (в отличие от
прогрессивной). Шмальгаузен называл каждый ее акт алломорфозом.
Таксон состоит из одного или более подтаксонов (таксонов предыдуще-
го ранга). Таксоном самого низкого ранга у нас служит вид - генетически
единое множество организмов с определенным устойчивым свойством осо-
бей, т.е. со своей особой нормой.
Это целостное определение представляется более пригодным практи-
чески, чем прежние - репродуктивное (нескрещиваемость с иными видами)
или экологическое, при котором вид задается своей эконишей. Последнее
неудачно потому, что вид может существовать в различных нишах9-7. При-
мер: у нас в России яснотка (глухая крапива) - обычный член ценозов,
включая лесные, тогда как в Англии - типичный маргинал, растет по обочи-
нам вне леса [Элтон, с. 154].
Таксон появляется (исчезает), если все его особи получили (утратили)
некое устойчивое свойство; и он исчезает, если в нем не осталось ни одной
пары особей. Снятие давления нормы приводит к росту как числа подтаксо-
нов (в том числе видов), так и изменчивости особей одного вида.
7) Приспособленность - факт очевидный, но процесс приспосабли-
вания - отнюдь (п. 4-17*). В силу феномена экологической прочности нельзя
измерять приспосабливание через рост выживаемости, а в силу феноменов
неприспособительной эволюции (например, ранних стадий прогресса) нель-
зя рассматривать эволюцию как череду приспособлений. Дать определение
приспособлению пока не удалось (см.: Мейен С.В. // Системность и эволю-
ция. М., Наука, 1984), но всякий имеет о нем интуитивное представление.
Для эволюции важна не столько смертность плохо приспособленных,
сколько их дискомфорт, порождающий стресс.
83 Видя это, В.В. Жерихин экологически определил вид как набор всех возможных для
него ниш, а экосистему - как «совокупность реализованных ниш» [Жерихин, с. 412-413].
Это вряд ли поможет делу: исчезло наглядно очевидное понимание экосистемы через на-
бор реальных видов, которые можно задать их названиями, а сами виды расплылись, по-
теряв формы. Включать ли бутылочное молоко (Дополнение 7) в определение синиц?
624
8) Часто наблюдается преадаптация - приспособленность к тому, что
еще не возникло (пп. 4-10, 9-13). Она никак не может быть итогом процесса
приспособления в его обычном понимании, и Янч (п. 6-12) предлагал изме-
нить это понимание; в наших терминах оно означает признание объективно
существующей диасети. Не исключено, что понимание этого факта потре-
бует переосмысления категории причинности (пп. 3-14, 5-2*, 6-14, 7-11).
9) Онтогенез начинается с одной клетки (у одноклеточного он — рост
самой клетки), и это наблюдение привело к идее, что эволюция началась с
одноклеточных. Однако вернее, что эволюция началась с объектов, позже
распавшихся на клетки, и первый организм вернее мог быть парой клеток,
составлявших симбиоз (п. 7-4). В онтогенезе организм образуется путем
двух типов активности - роста и усложнения, но не путем сборки (тезис Ик-
скюля, п. 5-15). В отличие от онтогенеза, в эволюции происходит сборка но-
вого организма из прежних блоков (блочность эволюции)', сходством блоков
в разных эволюционных линиях как раз и образуется диасеть.
Поскольку онтогенез начинается с одной клетки, то уровень развития
(см. ниже) многоклеточного каждый раз строится заново. Онтогенез контро-
лируется генами, но до сих пор непонятно, как информация, записанная в
гене, преобразуется в трехмерную форму; эту непонятность выражает кон-
цепция биополя (п. 5-14*). В норме онтогенез клетки в многоклеточный орга-
низм идет единственным путем (давление нормы), но и в опыте, и в патоло-
гии он может идти разными путями и, тем не менее, достигать одной цели -
это эквифинальность (п. 3-14). Ее нарушение приводит в данном поколении
к уродству, а в ряду поколений - к эволюции (п. 9-12*).
10) Причиной изменения онтогенеза, вызывающей появление новых
видов и новых уровней развития, предположительно можно назвать насле-
дуемое изменение эмбрионального поля при переключении гомеозисных
генов. Эта форма биополя является динамической (п. 5-15). Движение от
единственной клетки к многоклеточному организму происходит и в онтоге-
незе, и в эволюции в рамках одной диасети, что создает впечатление как
«повторения филогенеза в онтогенезе» (Мюллер, Геккель), так и наоборот,
«пророческих фаз» (Агассиц, Бэр, А П. Павлов).
11) Достижение последних 30 лет - понимание того, что центральным
событием онтогенеза является акт узнавания (п. 5-15). Всякое существен-
ное изменение онтогенеза вероятно обязано изменению типа иммунного
надзора - видимо, это общий молекулярный механизм эволюции.
12) Уровни развития (сложности организма) как правило вовсе не
связаны с выживаемостью; при своем появлении организмы нового уровня
обычно долгое время бывают малочисленны (п. 9-13*), а это значит, что
прогрессивная эволюция идет в таких случаях не путем вытеснения высши-
ми низших, а путем автономной реализации новой тенденции с возможным
длительным падением приспособленности и даже численности, т.е. за счет
исчерпания запаса экологической прочности.
625
Повышение уровня развития в ходе эволюции - одна из форм активно-
сти живого. Как оно происходит, неизвестно (и это служит козырем креацио-
нистов; здесь их позиции сильнее, чем в отрицании факта эволюции), но всё
же здесь очевидны: действие принципа компенсации и аналогия с самоус-
ложнением (эмерджентностью) математических фракталов (п. 6-7).
13) Поставщиком первичных наследственных изменений служат вовсе
не мутации, а более сложные информационные акты — прежде всего,
транспозиции мобильных элементов (пп. 5-6, 5-13*, 6-10*). Их появление в
данной точке случайно, однако их действие всегда направленно, поскольку
каждый транспозон является регуляторной системой.
14) Другой молекулярный механизм эволюции - усложнение полово-
го процесса (см. рубрику 1) пункта 11-1*). Если у бактерий он сравнитель-
но прост (обмен генетическим материалом кольцевой “хромосомы” с эписо-
мами, включение в нее новых транспозонов и их перемещение), то у эвка-
риот он гораздо сложнее (п. 8-8), особенно у млекопитающих (п. 8-12**).
Информационный аспект полового процесса состоит не только в реком-
бинации генетического материала, но и в «обнулении счетчиков» (п. 8-6).
Недавно стало возможно говорить и о более общем процессе - обнулении
эпигенетических программ в ходе мейоза [Чуриков], т.е. о стирании записей,
произведенных на «втором языке» (п. 5-5*). По всей видимости, именно на-
рушения данного процесса приводят к изменениям хода онтогенеза.
15) У животных есть свой особый стержень эволюции: усложнение
нервной системы — основы поведения особей. Однако поведение особи
возможно и при отсутствии нервной системы (п. 6-15). Наиболее сложным
(из известных достоверно) информационным процессом является мышле-
ние. Оно, в частности, преобразует лик планеты (п. 10-10).
16) Родство можно выявить, насколько знаю, тремя способами.
1. Прямым наблюдением. За 400 лет (от Шпренгера до Стегния) их из-
вестно всего несколько, все они являют примеры небольших скачков, но
мало говорят о процессе эволюции в целом, ибо являются акциденциями.
2. Указанием совместного изменения (от предка к потомку) всех суще-
ственных признаков, т.е. соединение семофилогений в филогению (п. 9-12).
Мне не известно примеров такого соединения, и, даже если они найдутся,
они ничего не скажут об эволюции в целом - в силу своей единичности.
3. Наложением ископаемых данных на палеогеографическую карту
(третье свидетельство факта эволюции, п. 9-9). Оно показало, что среди
семофилогений можно выделить ведущую и назвать именно ее филогенией
данного таксона. При этом эволюция остальных свойств таксона будет яв-
лять собой транпол (п. 6-4**), один из основных феноменов эволюции.
17) Организм должен быть способен к существованию и размножению
всегда, в каждом поколении, и в каждом он создается заново (тезис Икскю-
ля). Эволюция снизу состоит в том, что дети создаются не вполне так, как
родители. Это при половом размножении требует одновременного измене-
626
ния наследственности самки и самца, потому необходимым этапом эволю-
ции является либо самоопыление, либо инбридинг (п. 5-9). Механизмом од-
новременного появления самцов и самок с одинаковой новацией может
служить, например, миграция транспозона в гаметах одной самки (п. 5-6).
11-3** Эволюция сверху
Жизнь существует в форме биосферы, т.е. совокупности организмов,
связанных между собой функционально и пространственно. Эта совокуп-
ность являет как минимум три формы упорядоченности: 1) состоит из час-
тично проникающих друг в друга экосистем; 2) образована таксонами, ко-
торые образуют систему в том смысле, что каждый состоит из меньших так-
сонов (членение учеными множества организмов на таксоны не вполне про-
извольно: например, таксоны распределены по Виллису), а таксон наимень-
шего ранга (у нас это, как выше сказано, вид) состоит из организмов (осо-
бей); 3) организмы принадлежат разным уровням сложности, не совпада-
ющим с их положением ни в экосистемах, ни в таксонах (тезис Михайлова).
Эволюция описана в предыдущих главах как единый процесс преобра-
зования разнообразия. Иными словами, она выступает не как набор разроз-
ненных актов, каждый из которых надо изучать отдельно (как приспособле-
ние таксона к конкретным условиям), а как единый системогенез, как пере-
ход от одного упорядоченного множества к другому, столь же (или более)
упорядоченному. Поэтому процесс эволюции неравномерен: долгие перио-
ды взаимной упорядоченности прерываются короткими периодами неустой-
чивости и поиска новой упорядоченности (пунктуализм, п. 9-1).
По Жерихину, экосистема задает сама себе программу, которая запи-
сана в ее строении. Точнее, существование экосистемы есть сукцессия, она
подобна индивидуальному развитию организма. Он полагал, что
«роль детерминирующей индивидуальное развитие программы в сукцессион-
ной системе исполняет сам набор реализованных ниш популяций, населяю-
щих данную территорию», что «сукцессионная система, подобно вирусу, це-
ликом состоит из своего “генетического материала”» [Жерихин, с. 415].
Если принять, как делают ныне, что биосфера - экосистема наивысше-
го порядка, то вот ответ на вопрос Жерихина в начале параграфа (впрочем,
известный еще с древности как вопрос: «Кто будет сторожить сторожей?»):
биосфера управляет собой сама. Это - идея богдановского подбора (пп.
6-12, 11-1), т.е. философский постулат, который нельзя доказать, но мож-
но пояснять, что и делал Жерихин до конца (2001 г.) своей, увы, слишком
короткой жизни. Мне остаётся отметить, что его концепция описывает пере-
стройки, но совсем не касается проблем прогресса и эмерджентности.
Самый важный для биосферы акт эволюции может быть для системы
таксонов всего лишь взаимозаменой ядра и периферии (п. 10-7) - так, М/Д-
вымирание обратило таксон ящеров в периферию биосферы, а прежде пе-
риферический таксон зверей сделало, наоборот, частью ее ядра.
627
Неживая среда как независимый фон выступает в отношении немногих,
но очень важных обстоятельств (космические и геологические факторы).
Они могут вызывать глобальные катастрофы, если резко воздействуют на
биосферу во время ее неустойчивости. Главным неорганическим фактором
в течение последних 2 млрд лет служит излучение Солнца как источник
энергии жизни, а до этого, вероятно, служили потоки из недр.
Эволюция таксонов носит совместный характер, т.е. является процес-
сом коэволюции таксонов данной экосистемы и в то же время процессом
приспособления всей экосистемы к условиям неживой природы. В долго-
срочном плане сама неживая природа предстает как итог коэволюции био-
сферы и геосферы. Но жёсткой связи тут нет: так, даже великие вымирания
животных не всегда влекут ощутимую смену таксонов растений.
При распаде экосистемы возникает дискомфорт особей. Если он пора-
жает сразу многие виды, то говорят об экологическом кризисе. Уцелевшие
выходят из него новыми видами, но не любыми, а лишь теми, которые до-
пустимы диасетью и экологической прочностью.
(В этом многие видят естественный отбор, но налицо отбор в понима-
нии прежних натурфилософов, начиная с Де Малье: неспособное жить не
живёт; с этим никто не спорит, но тут нет отбора как движущего фактора.)
Фрактальность разнообразия ведет к тому, что аналогичные по форме
процессы могут происходить как очень быстро (десятки лет), так и очень
медленно (многие миллионы лет). Быстрая эволюция вида возможна и без
экологического кризиса: она может быть вызвана, например, инфекцией,
вызвавшей массовый горизонтальный перенос генов.
Экосистема предстает как фрактал типа «сеть», слабо связанный (но
связанный!) с другими системами, а эволюция экосистемы - как распад
«сети» и соткание новой, обычно - заплетенной еще сложнее. Если эволю-
ция таксонов может быть и постепенной, и скачкообразной, то эволюция
экосистем, вероятно, - только скачкообразной', экосистема не может не
быть сетью. Об элементарном акте эволюции экосистем см. п. 11-5.
Механизм перехода от нормы к норме видится для экосистем в рамках
идеи подвижного равновесия (по Еленкину и Соболеву), а для биосферы -
в рамках концепции Гей (по Лавлоку). Все эти взгляды нашли фундамент
(правда, пока еще весьма абстрактный) в понятиях новой термодинамики
(п. 5-14) и теории систем. Система устойчива постольку, поскольку ее ре-
гуляция многослойна (п. 6-8*). С такой позиции легко понять, почему в
бедной связями экосистеме изменчивость растет (хотя жить там бывает
легче - п. 5-13*) и почему тут возможна эволюция: «Если... контроль орга-
низации становится односторонним, не исключено, что популяция сможет
войти в процесс инадаптивного преобразования» [Расницын, с. 20].
Неокатастрофизм, идущий от работ Соболева и поздних работ Шинде-
вольфа, дополненный пунктуализмом, задает картину темпов эволюции:
длительное постоянство (когда идут подстройки видов к их экосистемам),
628
перемежается краткими периодами быстрых перестроек экосистем и, как
следствие, - входящих в них таксонов (катастрофизм).
Ход эволюции можно видеть единым процессом развития биосферы, в
чем-то похожим на развитие организма: как появление у зародыша отдель-
ных типов клеток подчинено задаче формирования цельного организма, так
и эволюция природы как целого подчиняет себе эволюцию каждого таксона
и экосистемы. Единый сверх-организм биосферы (Гея) являет собой как фи-
зиологическую и генетическую, так и морфологическую целостность. Одно
неясно: есть ли он единое целое в смысле поведения (свободы выбора).
Добавлю, что Жерихин видел аналог организма во всякой экосистеме.
Всё сказанное относится к прошлому, тогда как в настоящее время
складывается новый процесс эволюции, в котором главную роль играет
царство человека (п. 10-10). Становлению этого царства сопутствовали осо-
бые механизмы эволюции людей, еще никем подробно не описанные и
лишь слегка намеченные в параграфах 9-15 и 9-16.
11-4. О факторах эволюции
Фактор эволюции - это элементарные (неразложимые на части в по-
нятиях биологии) причина или условие эволюции [Чайковский, 1990, с.
168]. Само это различение позволяет избежать некоторых грубых ошибок.
Так, естественный отбор (как его видят все формы дарвинизма) яв-
ляется причиной вымирания и условием выживания, т.е. одним словом име-
нуют два разных фактора: он реально действовал как причина на всех тех,
кого теперь нет, и не действовал прямо ни на тех, кто жив, ни на их предков.
Тем, кто живет сейчас, отбор лишь создавал некоторые условия для жизни,
убирая некоторые другие организмы. Тогда для всех выживших и их пред-
ков причиной их эволюции остается лишь их собственная изменчивость, а
она в дарвинизме мыслится как не имеющая собственного направления.
Тем самым, по логике дарвинизма, если рассуждать честно, без диа-
лектики94, каждый из нас - итог цепи случайностей и только, а все слова о
“творческой роли отбора” - просто итог смешения понятий (причины и усло-
вия). В п. 4-6 мы видели, что дарвинизм смешивает также понятия новизны
и повтора, а в п. 1-6** - что он смешивает понятия сути и акциденции.
Поэтому, увы, всю процедуру выявления факторов эволюции надо про-
вести заново. К счастью, анализ факторов облегчается тем, что естествен-
ного отбора как фактора эволюции (как следствия избирательной смертно-
сти и избирательной размножаемости) в природе вообще не обнаружено, а
94 Без той диалектики, которая усвоена Дарвином и Марксом из средневековой схоласти-
ки (а ею - от школы Платона); она состоит в подборе доводов в пользу своей уверенно-
сти и применяет подмену понятий в ходе рассуждения (п. 3-7). Наоборот, вполне прием-
лема диалектика Гегеля - набор приемов, организующих мышление (например: «тезис,
антитезис, синтез»), но не природу (как думают марксисты, а за ими и дарвинисты).
629
все “доказательства роли отбора" - всего лишь толкование фактов приспо-
собленности, допускающих и иные толкования (пп. 2-1, 4-6, 4-20, 5-1, 5-8* и
6-15*). Следовательно, из гибели одних не следует изменения других.
Зато можно говорить о подборе?5 по Богданову (п. 6-12), т.е. о причин-
ном факторе самоорганизации систем, которая не вызвана ни избиратель-
ной смертностью, ни избирательным размножением, но сама определяет их
(лучший пример: половое поколение термитов, п. 4-20). Можно сказать, что
дарвинизм также путает понятия расплода (п. 5-8*) и подбора.
Подбор, согласно принятой в книге точке зрения, является основным
регулирующим фактором эволюции. Механизмы, посредством которых он
действует, еще мало известны; возможно, что их раскрытие потребует но-
вой картины мира и, прежде всего - нового понимания причинности.
Однако на уровне организма един из этих механизмов недавно раскрыт
- это иммунитет. Его основное назначение - не борьба с заразой, а узна-
вание, опознавание (такова концепция эпигностики, п. 8-12).
Врожденный иммунитет есть у всех организмов, начиная с прокариот, а
у высших тканевых к нему добавлен иммунитет адаптивный, представлен-
ный у теплокровных (звери и птицы) изощренной ТВ-системой. Элементы
адаптивного иммунитета есть у многих организмов, включая растения.
Однако регулятор сам по себе ничего двигать не может, и встает вопрос
о движущих факторах эволюции. Все они являются причинными, но некото-
рые могут быть также и условными.
Самым общим движущим фактором является время: поскольку оно те-
чет, то что-нибудь обязательно изменяется (п. 8-6). К той позиции Спенсера
и Криштофовича, что время есть первый фактор эволюции (п. 9-1*), недавно
присоединилась Е.Е. Коваленко, утверждая «невозможность неопределенно
долго воспроизводить себе подобное» без изменений (цит. по [Попов, с.
122]). С этой позиции время является и условием (что всем очевидно), и
причиной (что ученым еще предстоит обосновать) изменений.
Далее, общую группу движущих факторов являет активность, причем
для каждого круга эволюционных явлений есть своя форма активности. Ти-
пичной формой является биологическое поле; обычно мы связываем эво-
люционный акт не с самим биополем, а с изменением биополя.
В эмбриогенезе (пп. 4-16, 5-14*) это легче всего увидеть: поле выступа-
ет как причинный фактор развития особи, а изменение поля от поколения к
поколению - как причинный фактор эволюции. Однако поле может оказать-
ся ведущим (как причинным, так и условным) фактором и на иных уровнях,
от молекулярного до биосферного, о чём сказано ниже.
9э Далее до конца параграфа каждый фактор эволюции при первом упоминании выделен
полужирным курсивом, а группа факторов - прямым полужирным.
630
Конкретные факторы эволюции мы разделим на факторы, изменяющие
сам организм, и факторы, изменяющие отношения между организмами.
(Чёткой границы между ними нет, поскольку можно, например, считать орга-
низмом и пчелу, и улей.) В обеих группах всякий причинный фактор будем
рассматривать как итог действия некоторой активности, а всякий условный -
либо как снятие прежней активности, либо как проявление некоторых огра-
ничений пространства возможностей (физических или диатропических), ли-
бо как то и другое вместе. Под ограничениями диатропическими имеются в
виду прежде всего диасеть и тенденции.
Основным путём изменения и смены объектов является системогенез
(п. 6-8), а основным его причинным (точнее, запускающим его) фактором -
снятие давления нормы (п. 5-13*)
11-4* Факторы, изменяющие организм
Основные причинные факторы эволюции организмов удобно разделять
на две группы, смена активности особи (смена поведения, смена фи-
зиологических функций, изменение способа развития особи, моди-
фикация) и смена наследственных свойств особи (текстов на разных ге-
нетических языках — п. 5-5*), в том числе унаследование модификаций.
1) Если смена активности вызывает смену свойств, то налицо НПС -
классический фактор эволюции в неоламаркизме. Хотя в ряде случаев о
самом факте наследования можно говорить уверенно, однако роль его как
фактора эволюции, по-моему, сильно преувеличена (п. 5-12). Ведь главное
- понять, как нечто появляется, а затем уж встанет вопрос о его передаче.
Даже когда НПС нет, смена активности всё равно - фактор эволюции,
если она позволяет особям выжить (повышает экологическую прочность ви-
да) и тем самым принять участие в дальнейшем преобразовании особей.
Важным примером физиологической смены активности является запуск
и остановка размножения. Из них напомню расплод - процедуру избира-
тельного массового размножения (п. 5-8*), а также лактационную сте-
рильность и пропуск актов размножения (п. 9-16*).
Изменение способа развития особи часто трактуют как изменение по-
ля. В эмбриогенезе действуют динамические поля (п. 5-15), и их изменение
от поколения к поколению можно рассматривать как движущий фактор. Это,
в сущности, идеология жоффруизма. Для понимания эволюции предстоит
выяснить, как такие изменения поля устроены. Поскольку морфогенез явля-
ет собой фрактальный рост (п. 6-7), то и поле, вероятно, фрактально. В та-
ком случае любое изменение структуры фрактала поля можно счесть
причинным фактором эволюции.
Кроме фрактальности, поле характерно дальнодействием. Нейроны и
сосуды растут через весь организм, и пространственно далекое оказывает-
ся близким функционально. (На этом построено, в частности, иглоукалыва-
ние, п. 6-8.) Можно допустить, что изменение дальнодействия поля слу-
631
жит причинным фактором эволюции. Оба типа изменений могут быть итогом
мутаций или иных изменений наследственных свойств.
2) Основным путём смены наследственных свойств особей является,
как уже сказано, системогенез, а основным ее средством - снятие давления
нормы. В отношении наследования есть свой особый механизм: снятием
давления нормы запускаются все формы генетического поиска (п. 5-13).
Результатом поиска является смена функциональных блоков, т.е ра-
ботающих частей систем. Из конкретных механизмов поиска известны ре-
дактирование РНК (п. 5-6) и ее последующая обратная транскрипция, пе-
ремещение транспозонов (п. 5-6), инерционный мутагенез (п. 5-13), а
также эпигенетические механизмы (метилирование ДНК и прочие, п. 5-5).
Вероятно, что их список увеличится.
Законы создания новых (а не взаимозамены) блоков выступают как
причинные факторы прогрессивной эволюции организмов. Они слабо из-
вестны. Как пример абстрактного фактора прогрессивной эволюции можно
назвать образование ДС (п. 5-14), а как пример конкретного - самосборку
структур (п. 7-9). Оба - примеры активности, которую должен запустить
какой-нибудь иной (например, генетический или внешний) фактор.
Основными условными факторами эволюции, задающими характер
изменений организма, как уже сказано, являются снятие давления нормы и
диатропические факторы (диасеть и тенденции). Первое часто порождает
дискомфорт, а с тем и стресс.
Если эволюция есть процесс преобразования разнообразия, которое
описывается рефренами, то различные таксоны должны обладать сходны-
ми блоками (в этом и состоит феномен диасети). Откуда берутся рефрены,
мы пока знаем мало (см. п. 6-7), зато примерно знаем, как заполняется в хо-
де эволюции рефренная таблица: чаще заполняются клетки, близкие к уже
занятым, т.е. значительные изменения происходят редко. Новая клетка таб-
лицы заполняется с помощью тех особей, которые успешно пережили сня-
тие давления нормы, например, стресс; стрессом, в свою очередь, запуска-
ется генетический поиск.
Что касается конкуренции, то, согласно географу А.Д. Арманду,
“палеонтологическая летопись свидетельствует скорее не о вытеснении возник-
шими ранее таксонами более древних вследствие меньшей приспособленности
последних к своей среде. Более вероятен другой ход событий, когда сначала раз-
рушаются структуры, контролирующие видообразование, происходит расчистка
экологического пространства, которое затем заполняется возникающими парал-
лельно новообразованиями” (Арманд А.Д. // Анатомия кризисов. М, 2000, с. 50).
Подсчет числа ископаемых таксонов по геологическим ярусам привел к
отрицанию всякой роли конкуренции в появлении таксонов: свободную нишу
новый таксон заполняет с той же скоростью, что и занятую конкурентами:
“после массовых вымираний освободившиеся ниши не заполнялись со стре-
мительностью, пропорциональной степени опустошения. Темпы диверсифи-
632
кации (роста разнообразия - Ю.Ч ) в такие времена ничуть не отличались от
фоновых” (Журавлев А.Ю. // Природа, 2002, № 9, с. 55).
Словом, был прав Брони: конкуренция ничего нового не создает, но
может вытеснять старое. То есть она если и служит в эволюции, то
только условным фактором.Примерно то же видим в экономике и культуре.
Больше из принципа конкуренции, вероятнее всего, ничего не извлечь.
В этом нет нового (то же самое писали многие в 1920-х годах [Назаров,
2005, с. 260]), однако это следует напоминать вновь и вновь, поскольку кон-
курентные схемы эволюции (всегда чисто умозрительные) строятся до сих
пор. Причинным фактором конкуренция служит в экологии: есть множество
примеров вытеснения одним видом другого или даже нескольких [Элтон],
но это вовсе не близкие расы (как думали Уоллес и Дарвин), а достаточно
далекие виды. По-видимому, “принцип конкурентного исключения" (п. 3-2*,
сноска) имеет место только на бумаге: хотя близкие виды обычно чем-то
экологически различаются, но ни разу тут не показана роль конкуренции.
11-4** Факторы, изменяющие отношения меиеду организмами
Первым и главным таким фактором является подбор по Богданову,
обеспечивающий коэволюцию внутри вида (например, у общественных жи-
вотных) и между видами - такова любая экосистема. Особенно это хорошо
видно там, где один вид явственно приспособлен для поедания другими
(почти все растения и их семена, почти все личинки, а среди взрослых жи-
вотных - половое поколение подземных термитов). Однако подбор не явля-
ется фактором, специфическим для изменения сообществ.
Вопрос о собственных факторах эволюции экосистем разработан сла-
бо. Нам достаточно иметь в виду, что у эволюции экосистем есть собст-
венные факторы. Таковы все причины, ведущие к распаду экосистемы, ли-
бо к изменению хода ее сукцессии (п. 11-2). Например, похолодание, распа-
хивание земель и т.п., т.е. климатический и человеческий факторы.
Есть и чисто биологические факторы. Ранее [Чайковский, 1990, с. 177] я
смог назвать всего два таких фактора - неполную замкнутость биосферных
круговоротов (т.е. они на самом деле не круги, а спирали) и «неустойчивость
селективных процедур». Второй из них был изучен Кауфманом (п. 5-18*),
работы которого были мне тогда не известны.
Теперь можно добавить изменение биосферного поля. Этот фактор по-
ка остается гипотетическим, но отрицать его опасно. Поэтому лучше дер-
жать его в уме до лучших (в смысле понимания биосферы) времен.
Еще меньше можно сегодня сказать про факторы эволюции иных со-
обществ. Многие сомневаются, что про них вообще стоит говорить. Однако
есть примеры, когда некое множество организмов, не являющееся ни ви-
дом, ни экосистемой, эволюирует как некое единство (п. 6-13*).
Согласно броккизму (пп. 1-15*, 4-12*), всякий таксон стареет и умирает.
Всё, что можно сказать про причины этого - что они относятся к еще мало
633
понятному феномену системности (например, свёртка информации - см. п.
6-16), т.е. могут быть частными случаями богдановского подбора.
Эволюция экосистемы есть смена одного сукцессионного ряда на дру-
гой; например, образование леса - на обезлесение или опустынивание. Ес-
ли основной фактор сукцессии - время, то основные факторы смены сук-
цессионных рядов - смена климата и деятельность людей.
Как связаны эволюция снизу и сверху? Жерихин был уверен, что первая
целиком управляется второй, и приводил, например такие доводы: опыты
Шапошникова (и им подобные) показывают, что видообразование насеко-
мых возможно в миллион раз быстрее, чем оно идет на самом деле; он объ-
яснял это «снятием ценотического контроля» [Жерихин, с. 71-72]. Такой кон-
троль вполне возможен, однако его надо бы доказать. Есть совсем иные
данные, например, вымирание старых таксонов (броккизм), где вовсе не
видно контроля со стороны экосистемы. Вопрос остается открытым.
11-5. Акты эволюции
На основе всего сказанного можно принять следующую (заведомо не
единственную) модель акта эволюции.
Когда эволюирующий объект утрачивает прежнее состояние нормы, он
повышает свою наследственную изменчивость и начинает поиск новой нор-
мы среди доступных для него состояний (близких клеток диасети). Числен-
ность особей при этом надолго снижается. Если ему удалось приемлемую
норму найти, то изменчивость вновь падает, теряет направленность и на
долгое время перестает быть фактором эволюции. Акт эволюции объекта
завершен - впредь до следующего момента утраты им состояния нормы.
Падение численности тем глубже и длительнее, чем сложнее пере-
стройка рассматриваемого объекта. Это прямо противоположно той мысли,
что эволюцию движет лучшая размножаемость. Вытеснение лучшими худ-
ших возможно лишь по завершении акта эволюции - это расплод (п. 5-8*).
Объектом эволюции может быть любая целостность - таксон, экоси-
стема, группа близких (в смысле места или источника питания) видов, попу-
ляция организмов или клеток (например, в иммуногенезе). Его норма есть
множество норм входящих в него единиц - норм организмов и групповых
норм. Пример групповой нормы: концентрация нужных веществ среды.
Утерю нормы легче всего понимать как событие экологическое (любое
существенное изменение среды обитания), но причиной может быть и сиг-
нал (например, изменение какого либо поля), и просто наступление опреде-
ленного момента времени (ведь время - тоже фактор эволюции), когда
объект в некотором смысле созрел для акта эволюции.
Поиск может быть поведенческим, физиологическим, модификацион-
ным (как у кроликов Аршавского), эпигенетическим и генетическим. Генети-
ческий поиск представляется здесь завершающим компонентом, поскольку
именно он делает итоги всех форм поиска материалом для эволюции.
634
Для акта эволюции нужен запас экологической прочности (п. 6-2*). Чем
он выше, тем перспективнее данный вид, однако сам по себе он к эволюции
не ведет (например, у реликтов). Нужно еще снятие давления нормы.
Утеря нормы может носить как стрессовый (дискомфортный) характер,
так и повышать комфорт жизни, но в любом случае рост изменчивости сни-
жает жизнеспособность (за счет роста доли дефектных особей). Если поиск
связан с катастрофическим падением численности, он должен завершаться
быстро. Это вполне возможно, так как выбор вариантов идет не из бессмы-
сленных сочетаний “букв”, а из “осмысленных текстов” - клеток диасети.
Элементарный акт эволюции организмов - появление популяции с
новой стойко наследуемой нормой. Для наблюдателя он выступает как мак-
ромутация. Лучшим примером такого акта может служить опыт Агаева (п. 5-
9), затем опыты Шапошникова и Стегния. См. также Дополнение 5.
Примером более сложного акта эволюции служит еще не оконченная
серия опытов Бородина (пп. 8-8, 8-12), где показана роль эпигностики.
Еще сложнее акт появления новых структур и симметрий. Его лучшим
палеонтологическим примером может служить становление четвероногое™,
шедшее путем преадаптации за счет запаса экологической прочности (п. 9-
13*). Из косвенных данных наиболее ясны описания недавно возникших Ла-
заревых таксонов - например, латимерий, у которых, по всей видимости,
заново возник бесполезный вид симметрии плавников (п. 9-14).
Массовые акты (процессы) эволюции выявили Круаза и Мейен (п. 9-9).
В массовых процессах огромны различия скоростей. Например, таксоны
крупных малочисленных животных эволюируют быстрее прочих (п. 9-1), но
для растений это не так. Еще меньше сходств актов эволюции у таксона и у
фауны (т.е. всех животных компактной ограниченной территории) или фло-
ры: малые фауны и флоры (на малых удаленных островах в океане) по-
видимому эволюируют слабо и тем слабее, чем меньше остров [Элтон].
Акт прогрессивной эволюции - появление нового стойкого более
сложно устроенного объекта: например, нового отношения симметрии, или
нового органа, или экосистемы с новым трофическим уровнем, или способа
усвоения пищи, прежде недоступной. Изредка такой акт бывает элементар-
ным, а потому легко наблюдаемым - например, нынешнее освоение микро-
бами или птицами новых источников питания.
Элементарным актом эволюции “сверху” можно считать появление
экосистемы со своей особой нормой, т.е. со своими сукцессионным рядом и
климаксом (см. пп. 5-13*, 6-16**). Вероятно, лучшим примером будет, после
описания ее климакса, становление городской фауны птиц (п. 6-14).
В силу фрактального устройства природы наблюдаются сходные акты
в разных масштабах. Ярким примером служит эволюция семейства гавай-
ских птиц Drepaniidae: все его 43 вида произошли от одного вида, случайно
попавшего на острова, и дали более десяти экологически различных форм,
столь далеко разошедшихся, что «на материке подобные различия разви-
635
лись бы между отдельными отрядами, а не родами птиц» [Элтон, с. 107].
Акт эволюции экосистем может быть и совсем иным: например, вторже-
ние плацентарных в биоценоз Австралии повлекло его перестройку.
Различные участники акта эволюции проявляют согласованную актив-
ность (в опыте Агаева огромное большинство особей, нашедших выход из
кризиса, нашли его в 7-м поколении). Особенно согласованность удиви-
тельна там, где само системное единство объекта только через нее и видно.
Таковы, например, рост изменчивости перед вымиранием у крупных соста-
рившихся таксонов, приводящий к массовому появлению «несуразных» ор-
ганизмов [Попов, с. 141], или 12 рядов климений (п. 4-12).
Гпава 12. Практические рекомендации
Возможность давать практические рекомендации - главный критерий
работоспособности теории (п. 5-1). Она должна дать спектр возможных ва-
риантов событий, что и позволит строить рекомендации. Насколько знаю, ни
одна из прежних теорий эволюции не претендовала на рекомендации.
Стоит напомнить, что охрана природы строится на пресечении борьбы
за существование (на нее, наоборот, часто уповают те, кто губит природу.)
Самая простая и общая рекомендация - не путать эволюцию ни с от-
бором (п. 5-13*), ни с филогенией (гл. 10). Ни то, ни другое ничего не говорят
о том, как появились и были приноровлены друг к другу конкретные меха-
низмы, а это - главное, чем опыт эволюции может быть полезен.. Сейчас в
связи с перспективами биотехнологий много говорят о необходимости
«брать уроки у природы», но учебники эволюции дают лишь старинную
сказку, будто гибель одних есть причина эволюции других. Тут брать нечего.
Дарвинизм, будучи единственным преподаваемым учением, настолько
сужает кругозор исследователей, что они то и дело проходят мимо главного,
словно лошадь в шорах. И предлагают тупики вместо выходов. Итог хорошо
виден: публика, разочарованная в дарвинизме, возвращается к идее творе-
ния в ее самой примитивной донаучной форме. Как быть?
12-1. Шоры дарвинизма и давление нормы
Чтобы понять эволюцию (как и любую проблему), надо иметь какой-то
набор альтернатив, дающий возможность задавать себе вопрос: в ту ли
сторону я двигаюсь, куда мне нужно, или просто плыву по течению в потоке
общепринятого? А набор альтернатив задается мировоззрением.
Забавно и грустно читать, как тот или иной автор, даже знающий и
вдумчивый, не может в понимании эволюции сдвинуться с места просто по-
тому, что не знает, куда двигаться - дарвинизм навесил на его глаза шоры.
Так было с адаптивной окраской (п. 3-7), гомологическими рядами (п. 4-9),
устойчивостью бактерий к вирусам (пп. 4-14** и 5-1***), доместикационной
изменчивостью (п. 4-19), иммуногенезом (пп. 5-3*, 5-18) , «мусорной ДНК»
(пп. 5-5, 5-15), тесным инбридингом (п. 5-9), утратой глаз (п. 5-13*), тождест-
636
вом гомеобоксов при различии онтогенезов (п. 5-15**), ролью теплокровно-
сти (п. 6-5**), неуниверсальностью кода (п. 6-10), гнездовым паразитизмом
(п. 6-11), разнообразием митозов (п. 8-5*), сложностью орхидных (п. 9-5), ос-
новными фактами очеловеченья (п. 9-15) и со многим другим.
Во всех названных случаях факт, никак не совместимый с дарвинизмом,
вызывал оторопь и ожидание, что теоретики что-то изменят в теории. Одна-
ко дарвинисты либо говорили про какую-то новую форму отбора, либо, ко-
гда сказать было совсем нечего (пророческие фазы, приобретенная устой-
чивость бактерий, необычный код), ничего не говорили. Проходили годы,
неприятные факты забывались, вновь открывались и вновь забывались.
Даже единственное явное достижение дарвинизма - включение слу-
чайности в число причин изменчивости - не повлекло анализа феномена
случайности дарвинистами. (Мою попытку анализа см. в п. 5-18 и указанной
там литературе.) А вот совсем новый пример отсутствия анализа.
«Творец не требуется» - пишет восторженный рецензент (New Scientist,
vol. 188, 10 Dec. 2005, с. 57) книги «Правдоподобие жизни: решение дилем-
мы Дарвина» {Kirschner М., Gerhart J. The plausibility of life: resolving Darwin’s
dilemma. Yale Univ., 2005). Насколько можно судить по рецензии, “решение”
двух американских авторов таково: полезная случайная мутация (например,
удлинение шеи жирафа) может быть поддержана естественным отбором,
ибо все органы, клетки и даже микротрубочки «методом проб и ошибок»
подстраиваются к ней, причем все неудачные вариации тут же отмирают
(shrink) «в силу строгого отбора». Вопрос, как это проверить, не ставится.
Словом, вполне оправдан давний вывод: «Отбором и случайностью
“объясняют” любую проблему, не имеющую объяснения» [Лима, с. 11].
Препона, как мы знаем из п. 5-2**, в давлении нормы: и в природе, и в
науке норма (нормальная наука по Куну) подавляет изменчивость. Тем са-
мым, отказ от дарвинизма - это снятие давления нормы, это фактор науч-
ной эволюции, которая, если еще не поздно, может спасти от вымирания.
Барьер мышлению (шоры) в том, что дарвинизм во всех формах осно-
ван на идее обратной связи (именуя ее естественным отбором), и начисто
отрицает наличие прямых связей, а они-то и движут любой процесс (пп. 4-
20, 11-3). Прямой связью является, в терминах Богданова, формирующий
принцип, а обратной - регулирующий (п. 6-12). Любой разговор об альтер-
нативах дарвинизму есть разговор о запретных в нем прямых связях.
И, если отбросить анекдот про вечные повторы проб и ошибок, то легко
видеть, что упомянутые американские авторы приблизились, пусть и бес-
сознательно, именно к богдановскому подбору (он не требует смены поко-
лений). Вот и рекомендация: помня “скверные" факты и не ожидая падения
догмы, искать прямые связи. Поясню это тремя вопросами из иммунологии,
которые не решаются именно в силу господства дарвинизма.
1) Развитие генетики родило гипотезу, по которой все гены, кодирую-
щие изменчивую часть антител - итог случайных мутаций. Ее разбивает
637
простой расчет: во-первых, нужные антитела формируются при жизни особи
миллионами вариантов, а это значит, что неудачных вариантов при случай-
ном мутировании должно быть много триллионов, чего не наблюдается; во-
вторых, для этого нужна такая частота мутаций, какая разрушила бы всю
наследственность; а в-третьих, если бы ход болезни зависел от поиска ред-
кого варианта гена, темп выздоровления различался бы в сотни раз (кому
как повезёт), а на деле различие скоростей несущественно.
Слова «Дарвинов микрокосм», невзначай оброненные Тонегавой, дез-
ориентировали иммунологов на четверть века. Как мы видели в п. 5-3*, ме-
ханизм поиска конструкции антитела к данному антигену остался совсем
неясным. Однако учебники и руководства дружно подают всю тему как за-
вершенную. По аналогии с дарвинизмом поиск был назван случайным, но,
как и в дарвинизме, анализа этой “случайности” проведено не было.
И тут, и там для практики важно знать, из какого набора альтернатив
производится “случайный" выбор. При этом в иммунологии всё можно (при
желании) посчитать, и первый же результат оказался поразителен - “слу-
чайный поиск” не требует времени: иммунный ответ В-клеток занимает при-
мерно столько времени, сколько растет их клон (п. 5-18). Так где же время
на ожидание нужной “случайной” комбинации мутаций? И откуда сама идея
случайности поиска конструкции антитела? Из учебника дарвинизма.
2) В п. 5-18 мы также видели, что В-клетки делятся впятеро медленнее,
чем те бактерии, с которыми они должны бороться. В-клетка созревает за
двое суток, после чего начинает массовый синтез своего антитела и больше
уже не делится (Игнатов П.Е. Иммунитет и инфекция. Возможности управ-
ления. М., Время, 2002, с. 167). За это время она делится 10 раз (по 5 часов
на деление), т.е. ее клон составит лишь около тысячи клеток.
Однако на деле нужный тип В-клеток присутствует во многих миллионах
экземпляров. Откуда они берутся? Специалисты, которых я спрашивал,
ссылались на отбор и меняли тему разговора. Ясно, что «клонально-селек-
ционная» модель динамику не отражает, что клоны образуются впараллель,
что расплоду подвергается не уникальный клон (как пишут руководства), а
массовый синхронный материал (как у пшеницы в опытах Агаева, п. 5-9), но
его появление никого не интересует. Почему? Из-за господства дарвинизма.
3) Еще важнее вопросы иммунитета, связанные с эпидемиями. Процес-
сы здесь тоже очень быстры, но и в них иммунологи видят отбор:
«считается, что изменчивость микроорганизма обусловлена адаптационными
процессами, сходными с процессами эволюции... При направленном отборе пре-
имущество в популяции получают те микроорганизмы, которые наиболее удачно
приспособились к условиям изменившейся среды... В дальнейшем вступает в си-
лу стабилизирующий отбор, который в наибольшей степени благоприятствует
оптимальным признакам. Варианты, несущие другие признаки, чаще всего устра-
няются. .. Изменение состояния коллективного иммунитета неизменно включает
механизмы отбора и приводит к изменениям в популяции паразитов. И наоборот,
638
изменение свойств паразитов оказывает влияние на популяцию их хозяев. Все это
последовательно происходит в течение всего эпидемического процесса» {Игна-
тов П.Е. Иммунитет и инфекция. Возможности управления. М., 2002, с. 298-299).
Интересно, как автор представляет себе «последовательный» (т.е. мно-
гоступенчатый) отбор, который многократно успешно действует за время
одной эпидемии? И, тем более, обеспечивает изменчивое «состояние кол-
лективного иммунитета»? Разумеется, данных никаких не приведено. Вновь
мы видим попытку выразить в терминах третьей (статистической) ПМ то, что
явно требует новых ПМ - системной, диатропической и активностной.
Затронутый П.Е. Игнатовым процесс не только очень важен, но и в са-
мом деле интересен как эволюционный: бактерии регулярно обманывают
иммунитет хозяев, причем могут делать это быстро и синхронно, что исклю-
чает всякую возможность отбора редких вариантов (см. п. 8-12).
4) К тому же кругу относится вопрос об удивительных возможностях
бактерии Wolbachia (п. 8-4). Он грозит полностью разрушить наши знания о
работе генов, и, чтобы не думать об этом, генетики опять ссылаются на
“спасительный” (а вернее, губящий всякую мысль) отбор:
«Столь незаурядные свойства вольбахии, должно быть, возникли под дейст-
вием естественного отбора: они помогают бактерии выжить, оставив невре-
димым хозяина...» {Петров П.Н. Бактерия Wolbachia помогает дрозофилам и
вредит генетикам // Природа, 2006, № 5, с. 83).
Можно ли сказать, что здесь идет богдановский подбор? По-моему, да.
Искомой прямой связью здесь всюду является механизм самоорганизации,
которую, увы, никто конкретно не изучает. Теории данных процессов нет и
до отказа от дарвинизма (шире-до смены ПМ и картины мира) не будет.
12-1* Евгеника и эволюция
Евгенические ужасы фашистской Германии хорошо известны, но мало
кто знает, что и либеральный Запад мыслил в те годы вполне сходно. В не-
скольких государствах практиковалась принудительная стерилизация [Евге-
ника..., с. 225], а 16 сентября 1939 года, как раз когда Германия и СССР де-
лили Польшу, журнал «Nature» поместил текст, известный как «Манифест
евгеники». Он был подписан видными англо-американскими биологами и до
сих пор подается как образец «справедливости и гуманизма» [Глэд, с. 137].
За извиняющимися фразами о добровольности и общем благе в манифесте
следует примерно та же, что у фашистов, программа: пусть
“лица с более совершенным генетическим багажом произведут на свет боль-
ше потомков, чем остальные, - в результате сознательного выбора или как
следствие их образа жизни”. “Это в свою очередь предполагает государствен-
ную организацию отбора” (там же, с. 141).
Что это за стерильный «образ жизни», не сказано, но легко догадаться
(в Германии уже было тогда стерилизовано более 300 тыс. человек). Глав-
ное же в том, что «Манифест» признаёт необходимость «более обширного и
639
глубокого изучения наследственности», но отнюдь не изучения самого фе-
номена отбора. А ведь загвоздка была именно в нем: как известно, селекция
эффективна только при полном отстранении от размножения всех нежела-
тельных и принудительном спаривании желательных.
Лет через 40 иммунолог Бернет предлагал применять медицинскую ор-
ганизацию, вооруженную генетикой, для формирования высшей и низшей
репродуктивных каст с возможным в будущем разделением людей на два
вида (LR, 1981, № 119). И в наши дни вновь слышен призыв:
«можно идентифицировать гетерозиготных индивидов и, проводя соответст-
вующий отбор, удалить вредный ген из популяции за одно поколение... А это
фактически означает, что если, например, те индивиды (ничтожная часть в
процентном отношении от всей популяции), которые являются носителями
дефектных генов, воздержатся от участия в размножении в одном поколении,
то этот ген исчезнет, если не принимать во внимание новые мутации» (Бак-
санский О.Е. Блеск и нищета евгеники в XXI веке // [Евгеника..., с. 198]).
Опустим ошибочное «за одно поколение», не в нем суть. Суть в том, что
практически каждый человек (а вовсе не «ничтожная часть») несет в гетеро-
зиготе какой-нибудь ген тяжелой наследственной патологии, но это не при-
нимается во внимание, ибо тогда евгенике нечего будет делать.
На практике приходится иметь дело не с генами, а с людьми, фактиче-
ски проявляющими патологию, в том числе - преступное поведение. На по-
следних и обрушивается гнев сторонников евгеники. Однако связь огромно-
го роста российской преступности с развалом быта и ростом безработицы
ясно говорит: генетика тут не главное и репрессии не помогут. К тому же
«это мы уже проходили» при военном коммунизме и при фашизме.
Самый чистый эксперимент поставили германские фашисты: уничтожи-
ли целые категории душевнобольных, однако прежняя психическая заболе-
ваемость восстановилась в Германии через два поколения [Евгеника..., с.
205]. Столь быстрый процесс никак не мог идти за счет мутаций, и неясно,
можно ли его вывести из одного процесса выщепления гомозигот. А значит,
надо не призывать повторить побоище, а изучить суть дела.
Тогда, в годы фашизма, она была плохо известна, но теперь и такого
оправдания нет. Напомню то, что ныне известно: попытки ликвидировать
какие-то качества популяции путем отстранения носителей этих качеств от
размножения не могут дать ничего в силу транпола (пл. 6-4*; 9-12) - разно-
образие быстро восстанавливается. В той мере, в какой феномен транпола
универсален (он еще мало изучен), он и определяет быстрое восстановле-
ние нарушенного разнообразия. Пример с персиками (восстановление раз-
нообразия транзитом через одну-единственную особь) наводит на мысль,
что дело отнюдь не только в выщеплении гомозигот.
Единственный известный нынче процесс, способный ломать структуру
разнообразия - расплод (п. 5-8*), в данном случае искусственный отбор, ко-
гда в течение многих поколений принудительно скрещиваются избранные с
640
избранными, а остальные (в том числе и потомство избранных, оказавшее-
ся некондиционным) не размножаются вообще. И евгеника вновь и вновь
предлагает этим инструментом пользоваться, напрочь игнорируя тот факт,
что создать новую породу путем расплода можно, а вот избавить ее особей
от уродств и прочих индивидуальных патологий - нет.
Вот что верно отмечено в евгенике, так это угрожающая роль людской
активности, не нашедшей применения. Нынешние индейцы США
«тяжко страдают от избытка агрессивных побуждений, реализовать которые
они лишены возможности в условиях индейской резервации». Они «в течение
нескольких столетий вели дикую жизнь, складывавшуюся из войн и грабежей.
Очевидно, не мог не происходить отбор, усиливавший их агрессивность...
Этому не стоит удивляться, поскольку при жестком отборе породы домашних
животных меняются так же быстро» (Корочкин Л. И. О месте евгеники в гене-
тике//Евгеника..., с. 163]).
Но и тут - шоры дарвинизма. Очевидно как раз иное: искусственного
отбора на агрессивность никто пока ни у одного народа не описал. Высокую
агрессивность надо изучать всерьез, а не выводить из идеи отбора.
Единственная мера, предлагаемая против агрессивности Корочкиным
(спорт), полезна, но ничтожно слаба. Ее не раз применяли в очень широких
размерах. Вспомним древних греков, тоже страдавших от избытка агрессив-
ности: они даже устраивали всеобщий мир на время Олимпийских игр, но в
остальное время воевали столь же яро. Их цивилизация процветала, пока
расширяла свою территорию, а затем быстро распалась и растворилась в
римской (которая проделала тот же путь, хотя в ней междоусобные войны
бывали очень редко). Исследовать надо саму активность.
Есть еще «новая евгеника», апеллирующая не к селекции, а к генной и
прочей инженерии. О ней см. п. 12-5.
В остальном евгеника ныне предлагает то же, что плачевный «Мани-
фест евгеники» (1939 г.). Мы возвращаемся к Платону и прочим чудовищ-
ным утопиям. Как говорил философ-богоискатель Н.А. Бердяев, самое
страшное в утопиях то, что они реализуются. Добавлю: в социальной сфере
обычно, что опыт прошлого, даже самый кошмарный, ничему не учит.
Прошу понять меня правильно: евгеника представляется мне антинауч-
ной не потому, что ее методы мне или кому-то отвратительны (разработка
оружия мне столь же отвратительна, но это - наука), а потому, что она игно-
рирует всю науку и исторический опыт - например, вырождение чистокров-
ных пород животных, а также граждан Спарты. Если среди евгеников есть
ученые, они должны приступить к исследованию самой возможности устой-
чиво улучшить население, которая далеко не очевидна.
Единственным бесспорно полезным мне представляется генетическое
консультирование, позволяющее предотвратить рождение генетически
ущербных детей. Но это - почти не евгеника и совсем не эволюция.
641
12-1** Поиски остатков внеземной жизни
Столь же важным в смысле снятия шор дарвинизма видится мне рас-
смотрение диасети. Вот пример. В последнее время много пишут о возмож-
ном происхождении жизни вне Земли. Исследование метеоритов ясно гово-
рит о том, что многие из них образовались в недрах большой планеты (дли-
тельные сверхвысокие давления), а это значит, что она раскололась. Нали-
чие в таких метеоритах микроструктур, похожих по форме на бактерии, слу-
жит основным материалом для рассуждений о внеземном начале жизни, в
том числе и о занесении ее на Землю.
Не вступая в спор о природе микроструктур, замечу, что занести на
Землю жизнь через метеориты вряд ли возможно, поскольку заносить надо
не бактерию, а самодостаточный фрагмент биосферы. Крупные метеориты
испаряются при ударе о Землю, и мне не известно ни одной модели “мягкой
посадки”. К тому же, как сказано в п. 7-2, проблема биопоэза как такового не
продвигается допущением космической гипотезы ни на шаг. Однако убе-
диться в наличии жизни вне Земли было бы важно именно для решения
проблем биопоэза: ведь если удастся обнаружить остатки других жизней, да
еще очень древних, модели биопоэза обретут почву: станут возможны
сравнения, построение параллельных рядов и выявление рефренов.
В Солнечной системе мы можем надеяться увидеть остатки сразу двух
древних жизней: Марса (с которого, как полагают, давняя катастрофа со-
рвала атмосферу и океан, погубив этим жизнь) и погибшей планеты Фа-
этон. Они могут раскрыть тайны многих стадий биопоэза.
Метеориты явно - с большой планеты и содержат следы микробов. Ве-
роятно, эта планета (Фаэтон), обращавшаяся между Марсом и Юпитером, 4
млрд лет назад погибла (то ли от взрыва недр, то ли от столкновения с иной
планетой), причем осколки в основном ушли в космос или упали на Солнце,
а остатки образовали пояс астероидов. Если будет доказано, что на ней
была жизнь, это подтвердит ту изложенную в главе 7 идею, что жизнь воз-
никла на Земле быстро, в первые 500 млн лет Солнечной системы.
Был ли на Марсе и Фаэтоне океан? Принятая ныне концепция биопоэза
исходит из предположения ведущей роли тепла недр, а не Солнца. Если
так, то водный океан мог быть и на далекой от Солнца планете. (Есть кос-
венные соображения и о возможной подлёдной жизни на Европе, спутнике
Юпитера - LEG, 13-14). При гибели от столкновения или взрыва океан дол-
жен был выплеснуться в космос большими фрагментами и замерзнуть, об-
ратившись в рой комет. Кометы состоят из льда и камней, а беспорядоч-
ность плоскостей их траекторий (в отличие от упорядоченности орбит ос-
тальных тел Солнечной системы - они движутся вблизи плоскости солнеч-
ного экватора) говорят в пользу их происхождения от удара или взрыва.
Креационисты видят в кометах довод в пользу краткости истории Сол-
нечной системы: кометы не могут иметь ее возраст (около 4,6 млрд лет), по-
642
скольку испаряют лед при каждом приближении к Солнцу, и могут жить
лишь миллионы лет (ЮШ, 162). Довод серьезный, но мы наблюдаем лишь
небольшую их часть - ту, которая при гибели океана оказалась в сотни раз
крупнее средней величины, а потому во много тысяч раз долговечнее.
Толща земного океана почти безжизненна, поэтому, по аналогии, важ-
ных находок можно ожидать в камнях каменно-ледяных комет: камни могут
быть кусками дна или берега, и это надо иметь в виду, планируя исследова-
ние ядер комет. Интересно было бы обнаружить остатки жизни во фрагмен-
тах первичных гидротерм Фаэтона, но еще интереснее найти что-то совсем
неведомое. Основана ли внеземная жизнь на знакомой нам биохимии?
Можно ли понять это, исследуя “бактерии” метеоритов? Принадлежат ли
кометы и метеориты одному и тому же погибшему небесному телу?
Хотя за миллиард лет жизнеспособность организмов должна утратить-
ся, однако космический холод при небольших давлениях и хорошей изоля-
ции от излучений оставляют поле для неожиданностей. Поэтому анализ
недр комет требует всех возможных предосторожностей. Особенно с пози-
ции номогенеза: если законы форм всеобщи, то можно заразить Землю, к
примеру, опасными прионами (о них см. п. 5-7, 6-10, 7-9).
Эти и многие другие вопросы надо исследовать совместно, сразу вы-
страивая ряды и, если удастся, рефрены по трем планетам - иначе мы по-
грязнем в спорах о каждой находке и каждой идее.
12-2. Загадочный карбон и умеренные креационисты
Пафос завершаемой книги опирается на уверенность в том, что после-
довательность событий эволюции нам в основном известна. Это основано
на солидном авторитете стратиграфии (науки о земных слоях). В целом
она действительно дает надежную (в том смысле, что не видно существен-
ных противоречий) цепь событий, однако есть заметные изъяны, затруд-
няющие построение картины биоэволюции и дающие пищу креационистам.
Такова их идея, согласно которой каменный уголь возник вовсе не за мил-
лионы лет в карбоновых болотах, а в огромных свалках, образовавшихся за
малое время из пластов деревьев, брошенных друг на друга мощным пото-
ком. Вывод авторам ясен - это след Всемирного потопа (ЮШ, 178).
Вывод получен креационистами на материалах нескольких каменно-
угольных зон Германии, где ясно видна аллохтонность древней расти-
тельности (она захоронена не там, где росла). Авторы не знают новых ра-
бот, показавших автохтонность большинства угольных пластов (захоро-
ненных там же, где росли), например, в Северной Америке96, так что о Все-
мирном потопе говорить вряд ли стоит. Однако сами эти новые эволюцион-
96 См. Gastaldo R.A.Debates on autochtonous and allochtonous origin of coal... // The Paleon-
tological Society Papers, vol. 5, Octob. 1999. Я признателен палеоботаникам И.А. Иг-
натьеву и Ю.В. Мосейчик за разъяснение мне ситуации с захоронениями карбона.
643
ные работы были вызваны к жизни креационистами, за что им спасибо.
В частности, аллохтонные пласты в карбоне тоже известны. Есть угле-
носные пласты (polystrate fossils), которые разделены несколькими слоями
песка и глины, и всё это вместе пронизано многолетними стволами деревь-
ев (см. цветное фото: ЮШ, 176). Чередование пластов наводит на мысль,
что катастроф было несколько подряд. Пласты являются объектом много-
летних споров, и участники больше упирают на просчеты противников, не-
жели строят свои собственные согласованные объяснения феномена. «По-
гребение этих деревьев быстрым накоплением осадков не оспаривается
геологическим сообществом» (Gastaldo R.A., с. 155), но, увы, и не побуждает
его что-то менять в общепринятой концепции карбона. Исключение состав-
ляют немногие катастрофисты.
Катастрофизм, сраженный когда-то актуализмом Лайеля (который ви-
дел в прошлом Земли только медленное накопление осадков), вновь заявил
о себе в 1976 году на съезде в память столетия смерти Лайеля. Вскоре он
стал привычной частью геологической науки. Главное, что он внес в геоло-
гию, это критическое отношение к геохронологии: последовательность сло-
ев может быть итогом как долгого накопления осадков, так и катастрофиче-
ского потока в течение суток.
Это отнюдь не значит, что данным стратиграфии нельзя верить; нет,
они вполне надежны, когда дают картину, согласованную во времени и про-
странстве многими методами сразу по всей суше (кембрий, он и в Африке
кембрий, хотя описан в Англии, близ Кембриджа). Но отдельные массивы
могли отложиться быстро, даже мгновенно, и это легко обнаружить, когда
разные методы дают различную датировку.
В 1979 году геолог Стивен Остин (США) защитил диссертацию, где, в
частности, доказывал, что вертикальные бревна, пронизывающие толщи
карбона, говорят о быстром захоронении леса в ходе катастрофы. Это ис-
пользовали креационисты.
Какая катастрофа могла переместить массивы деревьев и поставить их
стоймя? Это могли быть наводнения, вызванные самыми различными есте-
ственными причинами. Есть креационисты, которые смыкаются с катастро-
фистами: они согласны признать, что впечатление Всемирного потопа мог-
ли создать цунами, вызванные падением астероидов в океан в разные эпо-
хи и что это вполне согласуется с Библией.
Но на вопрос, когда это было, дает ответ палеоботаника: каждой ката-
строфе свойственны свои растения. Тут-то и проходит грань между катаст-
рофистами и креационистами - вторые не могут объяснить, почему в водах
“потопа” погибли растения разных геологических эпох, причем в нижних
слоях карбона нет голосеменных и ни в каких его слоях нет цветковых. Сло-
вом, эволюционная наука может за себя постоять.
Зато умеренные креационисты (см. п. 9-18) задают эволюционистам
действительно трудные вопросы: почему некоторые массивы следов живот-
644
ных карбона таковы, будто все животные ползли по мокрому склону вверх?
Почему среди них нет отпечатков растений? Быть может, поток воды рас-
сортировал животных и растения по разным слоям? (Рос А. В начале... . За-
окский, 2002, с. 219, 243).
Кстати, если ссылаться на потоп, то надо искать останки погибших при
потопе людей. Оказывается, их ищут (только другие креационисты - не хри-
стиане, а кришнаиты). Они друг друга не читают (ибо веры их противопо-
ложны: у первых мир сотворен 7 тыс. лет назад, а у вторых ему миллиарды
лет, и люди жили в нем всегда), но ничто не мешает нам сделать это за них.
Будем, как Любищев, «беспринципны в выборе цитат» (п. 4-17*).
Кришнаиты Кремо и Томпсон (о них см. п. 9-15) составили весьма инте-
ресную таблицу «аномальных свидетельств о существовании человека в
глубокой древности». Из шести страниц две занимают молодые находки (из
плейстоцена), три с лишним - из третичного периода (начиная с эоцена), и
меньше одной - из более ранней истории Земли. Так вот, эти ранние наход-
ки по периодам фанерозоя разложились любопытно: на 11 периодов (от
венда до палеоцена включительно) всего пришлось 16 находок, из которых
10 - на карбон: 8 позиций, 10 находок (9 в США и одна в Англии). Угольные
разработки, разумеется, обследованы лучше других горизонтов, но ведь
пермские угли не дали ни одной находки.
Принято возражать: почти все находки “людей” в карбоне сделаны до
рождения изотопной хронологии и поэтому, вернее всего, датированы оши-
бочно. Да, это так. Однако, во-первых, креационисты резонно укажут, что
сам изотопный анализ часто дает нелепые результаты97 А во-вторых, как
понять пик находок в карбоне США? Почему пик именно в карбоне? Как сю-
да попало больше находок, чем в остальные 10 периодов вместе взятые?
Кришнаитов карбон сам по себе не занимает, они даже не заметили пи-
ка, а значит, эти данные ими явно не подобраны. Мое мнение: не зря креа-
ционисты прицепились к карбону. Пуст даже их собственное построение ни-
куда не годится, но они могут быть правы в том, что указывают на факты,
неверно описанные другими.
Что именно неверно, не мне судить, но напомню, что одинцовские иско-
паемые мозги (п. 9-15**) тоже описаны как карбоновые кремни (и поэтому
отвергнуты наукой), хотя карбоновой фауны в них не было найдено. Они,
полагаю, не карбоновые, а плейстоценовые, но ведь отвергнуты именно как
карбоновые. Не тут ли путь к разгадке? Иными словами, не описан ли, хотя
бы в редких случаях, в качестве аллохтонного карбона совсем иной мир,
третичный или четвертичный? Для идеи человека как Лазарева таксона это
очень важно.
Q7
Таков “древний” возраст лавы, реально излившейся всего сто лет назад - Рос А., с.
280; ЮШ, 158). Эволюционисты их не оспаривают, но просто молчат о них.
645
Подлинный карбон достоверно отличают от мнимого (созданного геоло-
гически недавней катастрофой) по ископаемой фауне; а ее в распиле не
было. Если образцы сохранились, надо бы продолжить их анализ.
12-3. Анализ рефренов и диатропический прогноз
Много раз мы видели примеры того, что разнообразие организовано в
диасеть (имеет рефренную структуру) всюду - от макромолекул и клеток до
таксонов и (отчасти) сообществ. Выявление диасети как целого видится мне
первой задачей для построения приемлемой систематики и работоспособ-
ной теории эволюции. В частности, для диатропического прогноза.
Под диатроп и чески м прогнозом понимается выявление (путем ана-
лиза рефренов) спектра возможных блоков - таких, что всё огромное разно-
образие реальных особей есть набор комбинаций не очень большого числа
блоков. В качестве примера напомню историю с “особаченными” лисятами
(пп. 4-19, 5-13*): селекционеры были в недоумении и, как водится, успокоили
себя, введя новую форму отбора. Но тогда же теоретик-животновод Бого-
любский писал об этом эффекте иначе, без апелляции к отбору, на основе
знания свойств соответствующих собак. Он предсказывал спектр новых
свойств лисиц, каковой и был вскоре выявлен.
Таким же прогнозом можно считать мысль, что безногий “млекопитаю-
щий угорь” возможен, а крылатый “ангел” - нет [Чайковский, 1990, с. 54, 247-
248]. Есть иная точка зрения - что “ангела" можно создать тем же путем,
каким природа создавала новые пары конечностей у насекомых. Изложение
ее см.: [Гоодницкий, с. 98-99], а также ЧЭ, с. 449. Кто прав?
С первого взгляда идея “ангела” выглядит просто перепевом Григория
Нисского: все гены уже существуют, и эволюция состоит в их поочередном
включении. Однако на самом деле вопрос глубже. Поскольку гомеобокс, за-
пускающий онтогенез плечевого пояса, не формирует новую пару конечно-
стей, а лишь маркирует место, где должен быть включен нужный процесс
формирования (задаваемый другими, свойственными именно данному ор-
ганизму, генами), то вполне возможно, что его встраивание в новое место
генома привело когда-то к появлению нового органа. Но это именно встраи-
вание, а не экспрессия: если бы ген там уже был, “ангелы” иногда возникали
бы за счет мутаций. Ожидать “ангела” нет оснований.
Диатропический прогноз характерен указанием на сами альтернативы,
а не на их комбинации и не на их вероятности. Особенно это важно при про-
гнозировании и рекомендациях в экологии. Вот два примера.
В ближайшие 50 лет доля СО2 в атмосфере удвоится, и это может из-
менить тип растительности. Неясно, будет от этого вред или польза, однако
“в любом случае скорости адаптивных реакций растительного покрова Земли
и миграция растительных форм должны ограничивать скорость антропоген-
ных воздействий” (Чмора С.Н., Мокроносов А.Т. Глобальное повышение СО2
в атмосфере и адаптивная стратегия растений// ФР, 1994, № 5, с. 776).
646
Здесь вместо предсказания конкретного варианта хода событий, даны
возможный спектр вариантов и рекомендация на худший случай.
Почти то же видим при обсуждении возможной катастрофы на атомной
электростанции: пусть вероятность ничтожна, но исходить надо не из нее, а
из размеров ущерба, если она произойдет (LEG, 45).
12-4. Тенденции эволюции и спасение природы
До сих пор вся охрана природы строилась на желании сохранить то, что
имеется, но ведь природа эволюирует, и нельзя этому мешать. (См. об
этом: Геодакян В.А. Об эволюционной близорукости экологических концеп-
ций // [Кудрин].) Из этого прямо следует несколько рекомендаций.
Спасая вид от исчезновения, надо знать, что мы спасаем его от истреб-
ления, а не от естественного вымирания. К сожалению, ныне тратятся силы
и средства на безнадежное спасение вымирающих и даже вымерших.
Так, стерх и большая панда явно вымирают (за счет сокращения рож-
даемости), а дикий зубр вымер. Скудные средства охраны природы имеет
смысл тратить на спасение тех, кто не проявляет склонности к вымиранию
(очевидной у названных видов). Столь же очевидна она у стеллеровой
(морской) коровы. Бытующее в литературе мнение, что стеллерову корову
истребили европейские моряки, сомнительно: их побывало там очень мало,
и сколько кто убил, в общем известно. Более реальна версия, что морская
корова к моменту ее описания Степлером уже почти вымерла, а ее послед-
нее (командорское) стадо пало от эпизоотии, точнее от глистов.
Воронцов мечтал когда-то, что удастся вывести животное, способное
заполнить богатые морские пастбища, где прежде она паслась. Но кем? Ее
ныне живые родственники (дюгонь и ламантин) сами в Красной книге. Нет
смысла пытаться акклиматизировать их на Командорах, не поняв, в чем со-
стоит тенденция к вымиранию (ей подвержен весь отряд) а этого нельзя
сделать, пока само наличие тенденций отрицается. Приняв тенденции как
реальность, можно хотя бы планировать эксперименты.
Если тенденция к вымиранию необратима, не стоит иметь дело с ее но-
сителями. Однако возможно, что тенденции имеют генетическую природу, и
их можно будет преодолевать, как хотят преодолеть генетические болезни.
В таком случае морскую корову стоит попробовать заместить новым и при-
том жизнеспособным видом; но это дело будущего.
Зато в примере с зубром вопрос о заполнении опустевшей экониши ус-
пешно решается, и пример поучителен. Как и предсказывал Усов (п. 2-12),
единственным способом сохранить эконишу занятой оказалось скрещение
вымиравшего зубра с бизоном, данной склонности не проявлявшим. К на-
стоящему времени этим путем выведен новый подвид зубра, горный зубр,
довольно успешно занимающий эконишу вымершего кавказского зубра
[Трепет]. Налицо успешный шаг управляемой эволюции. Это и есть под-
линный акт спасения участка природы.
647
Однако, к сожалению, основная масса прикосновенных к зубру лиц за-
интересована не в заполнении экониш, а в скорейшем получении благ и по-
тому настаивает на отстреле всего стада горных зубров как гибрида, не
представляющего интереса для науки. Вместо них предлагается поддержи-
вать инвалидные стада “чистокровных” зубров, неспособных вписаться в
экосистему (на что средств нужно гораздо больше; естественно, их нет и не
предвидится). Увы в этом евгеническом деянии чиновников и браконьеров
поддерживают биологи-дарвинисты, отрицающие как феномен тенденции,
так и всю экологию, кроме «борьбы за существование», которую они пони-
мают как массовый отстрел всех, кого хотят заказчики [Трепет].
Спасти зубра можно - но, на мой взгляд, только сменой общей ситуации
в науке и обществе, что нереально. А стерха - нельзя и вряд ли стоит пы-
таться, ибо его и так замещают другие птицы. Экониша пустовать не будет.
Далее, спасение природы принято понимать как воссоединение с нею,
как отказ от наиболее опасных нововведений. Мне и самому случалось пи-
сать в таком духе [Чайковский, 1990, с. 244]. Но это - благие пожелания.
Увы, более реалистичным оказалось совсем другое: вот
“парадоксальное обстоятельство, в корне противоречащее расхожим сентен-
циям экологов: ...разрешение кризисов в социоприродных отношениях дости-
галось не приближением к природе, а напротив, очередным удалением обще-
ства и его природной среды от естественного (дикого) состояния”. Так, “охота
и собирательство естественнее земледелия и скотоводства” (Назаретян А.П.
Антропогенные кризисы: гипотеза техно-гуманитарного баланса // ВРАН,
2004, № 4, с. 329).
Иными словами, нужно не фантазировать, а разрабатывать теорию
эволюции (но не инволюции — их различие см. п. 4-17*).
12-5. Как жить в новой биосфере?
Итак, прежняя эволюция закончилась, но жизнь, а значит и ее эволю-
ция, пока продолжаются. Эволюция стала, впервые в истории Земли, зави-
сеть от судьбы одного вида, оказавшегося целым царством в экологическом
смысле слова. Человек встал в один ряд с бактериями и растениями, став
главным экологическим фактором эволюции биосферы - таково царст-
во человека. Человечество эволюирует как царство путем усиления своих
органов (орудий) и изменения круговоротов, зато ослабевает как вид (см.
ниже). Обычно пишут про вред биосфере, но люди могут попросту не ус-
петь нанести серьезный вред ей. Не потому, что вымрут от ядерной войны
(это тоже экологическая катастрофа), а в силу законов эволюции людей.
ВО-ПЕРВЫХ, есть много работ о том, что
«варварское отношение к природе является неотъемлемой чертой Ното
sapiens sapiens, которая становится заметной уже с самого начала экспансии
неоантропа». «По-видимому, для того, чтобы стало возможным реальное ос-
лабление истребляющего воздействия человечества на дикие биосистемы, не-
648
обходима коренная перестройка стереотипа поведения и всей структуры на-
шего мышления. Фактически, если это произойдет, мы станем другими суще-
ствами (придется менять видовое название)» (Марков А.В., Наймарк Е.Б. Ко-
личественные закономерности макроэволюции. М., 1998, с. 273-274).
Если так, то выжить можем не мы, а другие, которых еще нет. Появле-
ние их потребовало бы появления новой мировой религии - экологической.
Она явно нарождается, но займется ли она, придя к власти, спасением при-
роды или истреблением неугодных (как делали прежние религии), неясно.
ВО-ВТОРЫХ, в царстве человека на первый план вышел рост мысли-
тельных возможностей за счет вычислительной техники. Фантасты долго
изощрялись в описаниях сцен порабощения цивилизации людей цивилиза-
цией роботов, упирая на акт выхода роботов из повиновения, а беда при-
шла с неожиданной стороны - человечество уже сейчас неспособно отклю-
чить сеть компьютеров, распоряжающихся его благополучием, хотя компью-
теры вовсе не бунтуют, а покорно исполняют волю людей. Сама эта воля
неуклонно ведет человечество к собственному порабощению.
Происходит то же, что с отравлением среды обитания: формально мы
можем его ограничить, но фактически не можем. Стремление к безопасно-
сти всегда отступает перед стремлением к комфорту.
Начало века знаменуется вторжением техники прямо в сферу мысли:
если прежде мы имели дело с компьютерами и их сетями, от нас отграни-
ченными, то теперь речь пошла о сочетании компьютера с живым организ-
мом. Нейрокомпьютеры обещают новый тип вычислений, который
“представляет исключительную важность не только для прогресса науки и
техники, но и для развития самого человека как интеллектуального члена об-
щества, поскольку откроется возможность поиска путей повышения эффек-
тивности человеческого мышления” (Микаэлян А.Л. Ассоциативная обработ-
ка информации и проблема памяти // ВРАН, 2002, № 2, с. 119).
Фактически это — новая евгеника, и ее глашатаи говорят прямо то же:
«Поражает мысль о скором достижении более высокого, даже неограничен-
ного интеллекта» [Гпэд, с. 132]. Потребности в таком повышении я не вижу.
Есть иная потребность: надо, чтобы сообщество научилось принимать
решения, очевидные с позиций общих долговременных интересов, но иду-
щие вразрез с сиюминутными желаниями масс (комфорт) и их элит
(власть). Вернее всего, нейрокомпьютер ведет в прямо противоположную
сторону - к расширению власти роботов за счет человека, а новая евгеника
— к расширению власти своекорыстной элиты. Вообще,
«Если идти по пути евгенизма, то судьи (эксперты -Ю.Ч.) будут назначаться
диктаторами, криминальными кланами и очень богатыми организациями. И за
этих судей будет идти ожесточенная борьба» (Олескин А.В. Генетическое раз-
нообразие человечества и евгеника // [Евгеника..., с. 191]).
Нынешние евгеники почти не ссылаются на отбор, уповая не на него, а
прямо на генную инженерию. Когда-то Кольцов предостерегал: проще всего
649
добиться всеобщего счастья, отобрав людей с той душевной аномалией, ко-
торая позволяет им считать себя счастливыми в любых несчастьях, но это
был бы, по Кольцову, самообман, ибо мнимое счастье гасит всякую актив-
ность, а она необходима для развития общества:
«элемент недовольства настоящим и искание лучшего необходимы для про-
гресса» (Кольцов Н.К. Улучшение человеческой породы // [Евгеника, с. 296]).
Прошло 80 лет, и вот евгеники всерьез предлагают осчастливить лю-
дей, сделав путем генной инженерии их вечно радостными. Биополитик А.В.
Олескин верно заметил по этому поводу:
«На примере уже предпринятых попыток улучшения настроения, снятия де-
прессии лекарственными средствами мы видели, что тоска, тревога, дурное
настроение снимаются ценой той или иной роботизации» (Олескин А. В. И [Ев-
геника..., с. 190]).
Если прежняя евгеника ничего не добилась, ибо исходила из фантазий
дарвинизма, то новая пугает своей возможной эффективностью. Пугает и
потому, что среди перечня ее целей культура даже не упоминается (упоми-
наются здоровье, ум индивида, экология и демография) [Гпэб].
Однако нельзя отрицать, что в этом — очередной акт эволюции чело-
века, являющийся также актом эволюции природы. Тем самым, налицо
мощный психо-технический фактор эволюции.
В-ТРЕТЬИХ, царство человека (и роботов в нем) ставит вопрос: сможет
ли новая биосфера (с новым царством, царством человека) сколько-то дол-
го жить или же она окажется неустойчивой и рухнет? Хотя никаких надежд
на рождение дальновидной политики (“ноосферы" Тейяра и Вернадского, п.
6-1) в обозримом будущем нет, однако в одном смысле спасения можно
ожидать как раз со стороны Геи (биосферы в понимании Лавлока) - экспан-
сия человечества проявляет тенденцию к исчерпанию и, возможно,
как раз в силу противодействия Геи. Вот мои доводы.
Народы европейской цивилизации более других ответственны за явле-
ние нового царства, и их рождаемость то тут, то там падает ниже смертно-
сти. Рождаемость вероятно будет падать и впредь. Глядя, как вымирают,
несмотря на все усилия людей, зубр, панда и стерх, легко видеть, что чело-
век встал на тот же путь Другими словами, падает экологическая прочность
человека как вида. Долговечна ли эта тенденция человека как вида? Думаю,
да. Повлечет ли она вымирание человека? Думаю, нет.
Поначалу причиной падения рождаемости служили рост комфорта, рас-
крепощение женщин и снижение роли детей в благополучии родителей, от-
чего у женщин возобладало обыкновение ограничиваться единственным
ребенком. Однако устойчивое падение числа людей в нынешней России на-
водит на мысль, что есть и иные причины. Если обычно на катастрофу на-
род отвечает ростом рождаемости, то теперь наоборот. (В истории так бы-
вало - например в поздней Античности.) Как зубр и стерх, человек всё
меньше способен к размножению. Всё больше больных детей (в России -
650
больше половины), всё ниже доля способных к труду, а способность людей
к воспроизводству падает не только в социальном (воздержание от деторо-
ждения, обилие одиноких женщин), но и в чисто медицинском смысле (рост
числа неспособных к воспроизводству; подробнее см. ЧЭ, с. 452).
Это может быть следствием как исчерпания (такого же, как у зубра и
стерха) видом Homo sapiens своей эволюционной потенции (жизненного по-
рыва), так и реакции Геи на безумие человеческой стратегии. В обоих слу-
чаях люди захотят преодолеть тенденцию техническими средствами. Если
размножение станет (сперва в видах комфорта, а затем по необходимости)
искусственной процедурой, то можно думать, что люди не вымрут, власть
роботов усилится, а государство (всё более и более компьютерное) получит
эффективный способ регуляции численности и покорности людей.
В-ЧЕТВЕРТЫХ, при этом для государств откроются новые способы де-
лить людей на высших и низших, о чем мы уже говорили. Возможно, что
расплод людей плюс генная инженерия откроют путь (ныне не просматри-
ваемый) перелома эволюционных тенденций. Такая евгеника породит иных
людей, неспособных жить вне искусственных условий. Их эволюция будет
очень быстрой и жуткой (безумной в смысле п. 6-6, как бы по Ломброзо). Но
быстрое не бывает долгим - либо люди с новыми тенденциями вымрут, ли-
бо перейдут в совсем иной этап своей истории, о котором можно лишь фан-
тазировать. Если же вымрут5®, то предсказывать варианты дальнейшей
судьбы биосферы легко, но неинтересно.
Так как же нам жить в новой биосфере? Прежде всего, надо узнать ее
законы и стараться соблюдать их, а не желаемые нами (в частности, не пы-
таться успокаивать себя уверениями, что нам предстоит жить по законам
вымышленной нами же «ноосферы»). Из этих реальных законов напомню
биотогенез Мейена (п. 9-4**). Сам он писал:
«Если она (идея биотогенеза - Ю. Ч.) подтвердится, это будет иметь немалый
смысл в стратегии охраны природы в масштабе планеты. Защита экватори-
альной биоты станет необходимой не только для сбережения всего того, что
там живет сейчас, но и для сохранения главных генераторов будущего разно-
образия» (Мейен С.В. // Природа, 1986, № 11, с. 57).
Затем, надо определить желаемую политику ограничения людской рож-
даемости - вместо нынешней, когда носители господствующей культуры
начали вымирание, а народы, их теснящие, не несут никакой внятной (в
смысле спасения природы и культуры) альтернативы.
Наконец, надо понять, куда направлена эволюция человека (и как вида,
и как царства) на самом деле, а не в наших желаниях. Запад, проведя пол-
века без крупных войн и поправив отчасти свою природу (путем антирыноч-
98 Разрозненные племена в тундре, джунглях и иных диких местах могут уцелеть
и, быть может, повторят историю человечества со всеми ее ужасами.
651
ных мер, формулируемых в рыночных терминах), склонен видеть будущее
светлым, однако остальной мир, получив выброшенные с Запада грязные
(во всех смыслах) технологии, быстро деградирует. Тем самым, эманация
человека (п. 9-16**) идет во всем мире, но по разным параметрам. Если эту
тенденцию можно переломить, то только радикальным перераспределени-
ем средств и благ с богатого Запада в остальные регионы, что и было бы
началом “ноосферы”. Но к этому ныне никаких возможностей не видно.
12-6. Заключение
В книге я старался показать, что основы для теории эволюции имеются
и ло частям известны многим. Эта теория не призвана отвечать на вопрос о
том, идет ли эволюция сама как естественный процесс, являет ли собой
арену борьбы высших сил или управляется единым Богом, зато может дру-
гое: объяснить многие давние загадки и быть полезной для практических
действий, чего о дарвинизме не может сказать даже его ярый защитник.
Самым важным мне представляется показ реальности факта эволюции
как таковой (п. 9-9), и пусть креационисты попробуют выйти на спор
не с устаревшим учебником, а с новой наукой. Замечу, что ни дарвини-
сты, ни креационисты никаких рекомендаций по спасению природы не дают,
вполне удовлетворяясь упреками друг другу.
Поскольку спор с ученых с богословами так же бессмыслен, как состя-
зание шахматиста с бегуном, в книге мной была занята иная позиция. По-
зволю себе выразить ее в форме тезиса:
Для спасения природы (включая людей) надо знать законы ее развития -
независимо от того, идет ли развитие автономно или по высшей воле.
Для атеистов такая позиция выглядит почти религиозной, но это не
вполне так. По всей книге мы видели обширные параллели эволюции и он-
тогенеза, однако онтогенез идет по заданной программе, почти без всего,
что можно назвать свободой воли. Если параллели законны, то Богу (как
субъекту со свободной волей) остается место только в эмерджентных актах
(п. 11-2*), но и их (акты) мы видим во фракталах (см. Дополнение 4).
Факт отсутствия в учебных программах едва ли не всех государств дос-
тижений эволюционизма последнего полувека (кроме пунктуализма - очень
частной концепции, которой тоже более 30 лет) ясно говорит, что дело не в
чьей-то злой воле, а в ориентации общества на устаревшие цели.
Насколько они устарели, видно из обсуждения книги [Назаров, 2005] и
из факта его публикации официальным педагогическим журналом под заго-
ловком «Существует ли естественный отбор?». Никто, даже ярые дарвини-
сты, не сказал там ни слова в защиту отбора, что позволило редакции под-
вести осторожный итог: «Выживание приспособленных в истории биологи-
ческой жизни, как оказывается, отнюдь не очевидная реальность» [В по-
мощь аспиранту]. Там же Г.А. Заварзин отметил социально-религиозную, а
не биологическую, основу исконного дарвинизма.
652
Хотя смена установок уже началась (успех всяческих учений вроде ев-
геники и забытых философий, возрождение роли религий, движение “зе-
леных” как нарождающаяся религия, смена конфликта Запад-Восток на
конфликт Север-Юг), но идет она совсем не в желаемую учеными сторону.
Наоборот, во всем мире наука сдает позиции: почти никто уже не ждет
(как ждали почти все в XX веке) спасения от успехов науки. Не желая обо-
готворять науку, замечу всё же, что ее место (в смысле рекомендаций)
может быть занято только другой наукой и ничем иным. А ее не видно.
Поэтому в Заключении книги ЧЭ главной мыслью была уверенность,
что человечество идет к гибели (по социальным, а затем уж по более от-
даленным экологическим причинам), что пути выхода мне не видно, но ес-
ли он найдется, то обязательно потребует новой теории эволюции. И хотя
благосклонные читатели (особенно Н.А. Малыгина) говорили мне, что та-
кой конец никуда негоден, что пока и поскольку наука жива, нужна какая-то
программа научных действий, однако добавить мне было нечего.
Новая книга была уже сдана издателю, когда мне в руки попала статья
[Свердлов]. Автор очень ярко обрисовал тупик нынешней молекулярной
биологии, в котором по его (и моему) мнению повинен редукционизм (тер-
мин см. в п. 8-1) - намерение понять свойства жизни снизу, исходя из
свойств ее элементов. Ему противостоит холизм (см. там же). Вот и тема
для Заключения: что может дать холизм пониманию эволюции?
Замечу, что почти все конкретные знания естественных наук получены
в рамках редукционизма, и недаром генетик Е.Д. Свердлов уверен, что
«редукционистский подход еще долго будет основным в изучении клетки».
Однако он понимает, что засилье редукционизма всегда ведет в тупик, из
которого возможен лишь холистический выход, что смена подхода в какие-
то моменты неизбежна (научная революция по Куну) и что такой момент
наступает. К сожалению, статья закончена тем вопросом, которому ее, на
мой взгляд, надо бы, в основном, посвятить - что будет дальше?
По Свердлову, эволюционный подход - главный для биологии, но он
им, как обычно, фактически не пользуется. По мнению Свердлова, генети-
ческому редукционизму можно (но не нужно) противопоставить редукцио-
низм биохимический, а нужно - изучение клетки как целого. Он прав в том
смысле, что целостность клетки (например, механизм ее онтогенеза, п. 6-
10*) даже не осознана как проблема, но ведь он предлагает тоже редук-
ционизм - клеточный. На самом деле ни становление, ни жизнь многокле-
точного из знаний о его клетках не выводятся.
Приходится обратиться к теории редукционизма. Известно, что отказ
от редукционизма достижим лишь тогда, когда никакой из уровней органи-
зации и рассмотрения не избирается в качестве основного, когда уровень
задается задачей (Мейен С.В. Проблема редукционизма в биологии И
Диалектика развития в природе и научном познании. М., ИНИОН, 1978).
Лишь с такой позиции удается понять, почему клетка, в том числе челове-
653
ческая, может как быть организмом (в культуре ткани), так и не быть им.
Это значит, что редукционизм уйти из практики не может, поскольку
почти всякий ученый видит только свой уровень исследования, а потому
неизбежно обходится с ним как с основным. Но редукционизм никогда не
должен целиком господствовать, поскольку этим порождаются тупики.
Иными словами, немногочисленные холисты нужны всегда (чтобы не пре-
секалась традиция), ибо наступают моменты, когда выслушать их просто
необходимо. По-моему, такой момент как раз и настал сейчас.
Чисто холистическим вопросом оказался следующий: «как геномная
информация превращается в фенотипическую?» [Свердлов, с. 715]. Ины-
ми словами, автор статьи напомнил о проблеме осуществления (п. 5-15) и
признал, что в рамках редукционизма ее решения не видно.
Мы знаем, что без этой проблемы не решить и проблем эволюции (п.
11-2, Третий тезис), но теперь еще надо признать, что вывести понимание
эволюции из будущего понимания онтогенеза тоже вряд ли реально. В са-
мом деле, это означало бы редукцию эволюции к онтогенезу, но ведь сам
онтогенез сложился в ходе эволюции. Остается надежда понять эволюцию
и онтогенез вместе. Иными словами, эволюции и онтогенезу (а заодно и
экогенезу), следует искать общее обоснование. Его, как говорилось в гла-
ве 11, нет надежды найти в рамках принятой ныне картины мира (п. 5-16),
по существу статичной. В этом, по-моему, и есть пафос статьи Свердлова
- поиск новой картины мира, динамической по существу.
Вот и программа для будущих эволюционистов: построить картину
мира, эволюционную на самом деле. О ней мечтал еще Янч, когда писал,
что движение проще и естественнее рассматривать как первичное явле-
ние, чем как последовательность состояний. Замечательна его метафора:
динамическую картину мира будет легче понять и использовать, чем ны-
нешнюю, статическую, как «велосипед легче удержать в равновесии, когда
он едет» (см. [Чайковский, 1990, с. 202]).
В этой картине мира могут стать первичными понятиями активность и
диасеть, в ней процессы самоорганизации (в том числе онтогенез, экоге-
нез и эволюция) будут первичными явлениями, а состояния (в том числе
упорядоченность и приспособленность) - вторичными.
Как мало мы знаем о природе жизни и как много нам предстоит узнать в
ближайшие годы, ясно хотя бы из того, сколько поставленных в книге вопро-
сов осталось без ответа. Из того, что пишут генетики на темы, близкие к
«внутриклеточному мышлению» и о различных языках наследования (пп. 5-
6, 7-8*), физиологи - об «автодиагностике» (п. 6-8), а этологи - об устройст-
ве сообществ (пп. 6-5, 6-15). Если наука выживет, нас ждут самые невероят-
ные открытия. Но для этого людям надо не только выжить самим, а еще и
не впасть в беспамятство, то есть сохранить науку.
654
Дополнения
Дополнение 1. Юридические презумпции - не пример для учёных
К пп. 3-9, 3-13. Презумпции увлекают многих ученых, но ведь наука
ищет истину в качестве самостоятельной цели, тогда как для юстиции поиск
истины - всего лишь средство к достижению справедливости и обществен-
ного спокойствия. Когда подсудимый умер или «дело утратило обществен-
ную опасность», закон требует прекратить поиск истины. И, как известно,
«сомнение толкуется в пользу подсудимого» - фактически теория права
признаёт, что лучше оправдать виновного, чем осудить невиновного. Всё
это значит, что справедливость здесь ставится выше истины, а это нравст-
венное положение никак не связано с идеей научного поиска.
Наука строится иначе, но, к сожалению, именно аналогия с правом слу-
жит опорой сторонникам презумпций в биологии. Вот их недавнее кредо:
«в основе всякого научного исследования лежит нечто вроде “презумпции ис-
тинности”, также заимствованной из юриспруденции. Она обязывет доверять
мнению предшественников, а не начинать собственные исследования каждый
раз “с нуля” (что отражает преемственность научного знания и, между про-
чим, экономит массу времени и сил). В этом смысле названная презумпция
допускает формулировку, аналогичную презумпции невиновности: автор не-
которого суждения не может быть обвинен в лживости в отсутствие доста-
точных аргументов против него» {Павлинов И.Я. О значении презумпций в
филогенетике... // ЖОБ, 2005, № 5, с. 437).
Неужели это сказано всерьез? Ведь никакой «презумпции истинности» в
области права нет, а есть противоположное требование - доказывать каж-
дый факт документами и проверять, не подложны ли сами документы. По-
дозревать ложь в показаниях - норма у юристов. Итальянский писатель-
историк Умберто Эко, вечно ищущий в настоящем прошлое, пишет:
«“Так вот учти, что первейший долг порядочного следователя - подозревать
именно тех, кто кажется честным”. “Какая гадость работа следователя”, ска-
зал я» {Эко У. Имя розы. Роман. СПб., Symposium, 2003, с. 203).
Доверять приходится эксперту, но и о его лжи предусмотрена статья в
кодексе. И в науке «доверять мнению предшественников» вовсе не принято,
а принято иное - либо следовать общему мнению, либо, если такового не
выработано, выискивать у предшественников мнения, близкие своим собст-
венным, и им «доверять»; остальные принято игнорировать. Отказ от сис-
тем предшественников есть основной вид интеллектуальной игры по имени
«Филогенетика», за которую дают степени и гранты.
Вот иллюстрация: сам И.Я. Павлинов в ключевых местах своих книг не
упоминает никаких теорий, кроме собственной (вторя традиции избегания
предтеч, п. 1-17). Издав “учебное пособие”, из которого исключен отряд
655
ластоногих, он даже не упомянул, что этот отряд вообще существует. Из-
лагая предысторию своей концепции, которую назвал «естественной сис-
темой» (параграф «В поисках Естественной системы» книги: Павлинов
И.Я. Введение в современную филогенетику. М., КМК, 2005), он вовсе не
упомянул всю огромную традицию тех пониманий естественной системы, с
которыми фактически спорит. Поиска истины не видно.
Тот, кто видит в словах: «не может быть обвинен в лживости» аналог
презумпции невиновности, допускает простую логическую ошибку: отказ от
обвинения в лживости вовсе не означает веру в правдивость. В этом незна-
нии формальной логики - беда сторонников презумпций. На самом деле от-
каз от презумпции это не противоположная презумпция, а отсутствие вся-
кой оценки. Кстати, как раз юристы знают, что отказ от презумпции невинов-
ности вовсе не есть презумпция виновности.
Защищая метод презумпций, Павлинов в качестве безукоризненной
презумпции приводит идею эволюции - на том лишь основании, что некото-
рые эту идею отрицают. Опять логическая ошибка: эволюционист, даже са-
мый слабый и доверчивый, принимает идею эволюции на основе известных
ему свидетельств биологии, которые считает фактами, а не презумпциями.
С такой “логикой” никакое движение вперед невозможно. Его и нет -
филогенетика вот уже второй век ходит кругами, о чем сказано в главе 10.
По-моему, это должно бы насторожить сторонников презумпций.
Далее, презумпция невиновности - принцип формальных юридических
действий (следствие, суд, кассация, надзор), а не оперативной розыскной
работы: нелепо думать, что оперативник, преследуя преступника и рискуя
при этом жизнью, может всерьез держать в голове идею невиновности пре-
следуемого - настолько, чтобы ею руководствоваться. Различение процес-
суальных и оперативных действий - азы правовой науки, и использование
оперативных сведений в качестве доказательств (запрещенное в позднем
СССР, но, увы, разрешенное в России ныне) безграмотно. А ведь сторонни-
ки презумпций в науке даже не задают себе вопроса о различии процедур
добывания фактов и построения умозаключений: факты, не ложащиеся в
презумпцию, выпадают у них из анализа, а то и просто отрицаются; пре-
зумпция в науке служит тайным цензором.
В сталинскую эпоху презумпция невиновности неофициально отверга-
лась как элемент буржуазного права, неприемлемый при социализме (где
при Сталине каждый человек был потенциальным подозреваемым), но фак-
тически безраздельно господствовала в отношении к работникам органов
госбезопасности. Позже, при Хрущеве, она была отвергнута формально и,
более того, заменена противоположной презумпцией (таковы дела о “нетру-
довых доходах”, где обвиняемый был обязан доказывать свою невинов-
ность, причем участие адвоката запрещалось, а все доходы, кроме государ-
ственной зарплаты, считались нетрудовыми) и с большим трудом возвра-
щена в юридическую практику при Брежневе.
656
Естественно, что большинство тогда сочло ее основой права, и это бы-
ло естественной реакцией на нелепость хрущевской юстиции. Но тогда же
(около 1982 г.) у нас стали говорить (А.П. Расницын и др.) о пользе метода
презумпций в эволюционизме, а вот это уже оснований не имело.
Наконец, презумпция невиновности ведет к беззаконию, если трактует-
ся хоть немного расширительно (вспомним презумпцию времен сталинского
террора: «органы безопасности не ошибаются»: никто не собирал сведений
о виновности работника органов, пока не поступал приказ собрать; а когда
поступал, их фабриковали, в силу той же презумпции). Эта презумпция до-
пустима только в отношении слабого, и практика показывает: если юрист от-
казывает в возбуждении дела против сильного (юриста или начальника) на
основании «недостаточности улик» (они фактически могут быть собраны
только в ходе следствия, а признаны достаточными - только в суде), право
рушится. А в науке презумпции всегда трактуются очень широко.
Общественные нормы не являются презумпциями, поскольку их истинность
вообще не обсуждается, однако из них вытекают презумпции. Например, из со-
вокупности норм “невиновный не может быть наказан” и “виновный должен
быть наказан” вытекает, в качестве одной из возможностей, презумпция неви-
новности. Ее принимали и принимают отнюдь не все общества. Так, в древних
обществах, где наказание выступало как жертва богам, часто преобладало убеж-
дение, что кто-то должен быть наказан за преступление обязательно.
В неявной форме оно и ныне обычно - в частности, у юристов, которые борют-
ся за “высокую раскрываемость преступлений”. Борьба ведется в России с по-
мощью нескольких презумпций, передаваемых среди юристов устно. Например:
«убийство совершили либо свои, либо чужие. Чужих искать сложно и мало пер-
спективно, поэтому их не следует искать до того, как исчерпаны все возможно-
сти привлечь своих». В итоге опять, как при Сталине (но не при Брежневе), ца-
рит “чистосердечное признание” (теперь - родных и друзей погибшего). Наивно
думать, что эту презумпцию проверяет суд: здесь царят свои презумпции (право-
ты следователя, лживости обвиняемого и др.), и они делают поиск истины редко-
стью. Итог чудовищен: оправдательных приговоров - всего полпроцента (хотя
низкое качество следствия всем известно и широко обсуждается), причем часто
оправдывают как раз, насколько знаю, виновных.
Зарубежная юстиция известна мне мало, но и там могу указать вопиющую
практику презумпций. Не привлечен к суду ни один (кроме пары рядовых) воен-
ный преступник стран-победительниц недавних войн, даже когда сам факт пре-
ступления признан властями этих стран. Хотя древний постулат «Победителей
не судят» в наше время формально отвергнут (заявлено, что военные преступле-
ния должны быть наказуемы), на деле его сменила презумпция: поскольку наши
действия в целом хороши (постулат), постольку расследование действий наших
агентов принесет больше вреда, чем пользы (пока не доказано противное).
Словом, как раз аналогия с правом вынуждает воздержаться от выдви-
жения презумпций в естественных науках.
657
Дополнение 2. Лучи Гурвича и биополе
К пп. 4-16, 5-5*, 5-14*, 6-7, 7-10*. Александр Гаврилович Гурвич (1874-
1954), биолог-универсал и философ науки, открыл эти лучи в 1923 году.
То, что Гурвича до сих пор мало кто понимает, связано, по-моему, с тремя об-
стоятельствами: во-первых, он очень мутно выражался, и читать его крайне труд-
но; во-вторых, под одним названием «биополе» он выдал две плохо совместимые
концепции; а в-третьих и в-главных, вся идея биополя противоречит царящей в
биологии идее механического контакта, от которой физика с трудом избавилась
300 лет назад (п. 5-2). И всё же читать Гурвича надо.
Роль работ Гурвича освещена во многих книгах, в том числе в книге:
Будаговский А.В. Дистанционное межклеточное взаимодействие. М., Науч-
но-производств. центр «Техника», 2004. В ней же изложены работы Поппа, в
частности, те, где показана лазерная природа лучей Гурвича.
Сам эффект сверхслабого излучения оказался имеющим всеобщее
распространение и нужным для всех жизненных процессов. Тот странный
факт, что живое вещество излучает в ультрафиолете, словно Солнце, вы-
зван накоплением энергии многими молекулами (поляритон) с последую-
щим испусканием единого высокоэнергетического кванта. «Живое тело не
холоднее Солнца» [Воейков, 1998, с. 291]. Конечно, такой квант может излу-
чаться лишь изредка. Вклад этого излучения в энергетику жизни пренебре-
жимо мал, зато велик информационный вклад. Так, без ультрафиолетового
сигнала деление клетки не может начаться.
Основную массу высокоэнергетических квантов, нужную для жизни,
организм черпает из природного радиоактивного фона (без которого, ви-
димо, жизнь невозможна), а друг от друга клетки получают специфические
кванты-сигналы, обеспечивающие целостность процессов, которую описы-
вают со времен Гурвича как поле. По Кузину (см. п. 7-10*), эти излучения
“создают в структурно-метаболической системе биологическое информаци-
онное поле, которое и образует по еще неясным нам механизмам то единое
целое, которое мы воспринимаем как живое” {Кузин А.М., 2002, с. 62).
Вопрос о «неясных механизмах» является здесь главным.
В п. 5-5* говорилось о предложении Гаряева понимать онтогенез как
считывание голограммы. Рассмотрев эту идею и ей подобные, А.В. Буда-
говский, симпатизируя им, пришел однако к выводу (если я его верно по-
нял), что это скорее заявка на модель механизма онтогенеза, нежели сама
модель. В самом деле, если понимать онтогенез, как ныне принято, в тер-
минах смены активностей генов, то искомый механизм должен работать с
миллиардами бит информации в секунду. Автор указывает на возможность
достаточной мощности канала передачи такой информации в клетке и меж-
ду клетками, но ни слова не говорит о том, как такую огромную информацию
реализовать в материальных структурах, притом многоклеточных {Будагов-
ский А.В., с. 63). Наоборот, в другом месте (с. 72) он прямо указывает, что
658
мощность биополя мала: создающее его излучение сохраняет свою струк-
туру всего на нескольких милиметрах ткани.
Это значит, что нынешнее “лазерное” понимание поля может быть при-
влечено для понимания и моделирования взаимного расположения близких
клеток, но не для построения макроорганизма. Конечно, это лучше, чем
«каскады генов» (п. 5-15*), однако мы знаем массу примеров, когда микро- и
макрообъекты строятся по единому правилу (таковы рост микротрубочки и
аксона), поэтому и механизм следует искать общий. По-моему, он должен
иметь основой фрактальный рост.
Иными словами, не видно оснований для понимания поля (если искать
ему общебиологический механизм) по позднему Гурвичу, но видно - по
раннему, с тем уточнением, что поле должно быть фрактально: на сегодня
только фрактал известен как структура, порождающая информацию. Приве-
ду два примера, указывающие на вероятную фрактальность биополя.
Есть лекарства, дающие противораковый эффект как в обычных дозах
(когда они очень ядовиты), так и в малых, и затем снова - в сверхмалых, ко-
гда они совсем безвредны (Бурлакова Е.Б., Конрадов А.А., Мальцева ЕЛ.
Сверхслабые воздействия... // Биофизика, 2004, № 3).
Замечу, что традиционная химиотерапия при наличии описанных в этой статье
средств выглядит ныне такой же тупой жестокостью, как прежде - отказ принять
асептику Земмельвейса (п. 5-3*). Но уменьшить дозу в миллион раз - значит да-
вать лекарство бесплатно, что при рыночной медицине немыслимо:
«На сегодня рыночно несостоятельны любые дешевые методы... Дешевое
лекарство невыгодно ни производить, ни продавать» (Окушко В.Р. Бремя
биологического безвластия // Химия и жизнь, 1993, № 7, с. 26).
Добавлю: дешевое лекарство никто никогда не подделывает.
Независимость от масштаба - элемент фрактальной структуры. Как она
достигается? Проблема известна для лекарств давно - как гомеопатия.
Единый механизм таких реакций Любищев видел как раз в изменении био-
полей: химикат создает в растворе поле, которое действует даже в той час-
ти раствора, в которой нет ни одной молекулы химиката. В его время ничего
конкретнее сказать было невозможно, а ныне можно назвать, например, из-
менение фрактальной структуры воды.
В пользу довода Любищева могу добавить: запах тоже есть эффект
восприятия считанных молекул пахучего вещества, и принято считать, что
считанные молекулы химически действуют на обонятельные рецепторы; но
в некоторых случаях это сомнительно. Звери всегда грызут банки и коробки
с нужными им продуктами, хотя вся тара захватана и пахнет снаружи иными
запахами. Особенно загадочны “стойкие запахи”: в отличие от “стойких кра-
сок”, они должны (по химической теории запахов) непрерывно излучать па-
хучие молекулы (причем излучать многие годы с чисто вымытых поверхно-
стей!). В чем же стойкость? Думаю, в том, что крепко связанные с субстра-
том немногие молекулы создают поле.
659
Что же касается лучей Гурвича, то они представляются мне не основой
и не причиной биополя, а его следствием, (почти) не несущим онтогенети-
ческой информации. Ситуация сходна с избирательной размножаемостью: в
ней видели причину создания эволюционной информации, а она - лишь
следствие эволюции, не создающее никакой новой информации. Она помо-
гает успешным расселяться, а синхронное излучение помогает организмам
и их частям действовать совместно. Онтогенез же требует более детально
действующего механизма. Его надо искать, и, по всей вероятности, - вне
нынешней картины мира (см. Дополнение 9).
Дополнение 3. Аспекты эпигностики
К п. 5-15. Эпигностика, под именем «интегративная функция иммунной
системы», недавно фактически заявлена группой армянских иммунологов
(Давтян Т.К., Геворкян Г.А., Погосян Д.А. II УСБ, 2005, № 1); ими дано 2
примера этой функции: участие иммунных белков в формообразовании (у
дрозофилы) и возможная роль индивидуальной несовместимости в предот-
вращении инбридинга у видов-гермафродитов.
В концепции «естественной (биосферной) иммунизации макроорганиз-
ма» Л.А. Пирузяна тоже очевиден эпигностический аспект:
“Иммунная система организма рассматривается как биологический маркер
генетически детерминированных метаболических процессов”, когда выявля-
ются “изменения метаболического статуса организма человека и его иммун-
ного статуса под влиянием сапротрофных бактерий” (Пирузян Л.А., Михай-
ловский Е.М. Сапротрофные бактерии как возможное средство управления
иммунным статусом организма... // Физиология человека, 2005, № 1).
Поясню: по иммунной реакции можно узнать о состоянии обмена ве-
ществ. К эпигностике надо отнести и проблемы медицины, связанные с на-
рушением работы молекулярных рецепторов.
Но как же зародыш узнаёт, как ему развиваться, какие гены включать?
В геометрических терминах - как он узнаёт, куда ему из каждой данной точ-
ки расти? Вопрос эпигностический, и у Черданцева есть намек на занимаю-
щую нас проблему:
«Если описывать структуризацию с помощью дифференциальных уравнений,
то ее математический “механизм” совершенно ясен... Система должна быть
далека от термодинамического равновесия, в... уравнении должен быть авто-
катализ “активатора”, активатор должен производить свой собственный ин-
гибитор, который должен диффундировать быстрее, чем активатор. Тогда
возможно пространственное расслоение..., то есть возникновение макроско-
пической доменной структуры [...] То, что последовательность выделения
сегментов доменной структуры может описываться гармоническими волно-
выми функциями, так чтобы внутри каждого сегмента укладывалось конеч-
ное число полуволн (Kauffman, 1977; Goodwin, 1994), является одной из са-
мых остроумных идей теоретической эмбриологии... С ней очень тесно связа-
660
но представление о “комбинаторном” (на самом деле просто бинарном) коде
пространственной дифференцировки (Kauffman, 1977). Очень важным досто-
инством модели является ее независимость (относительная, конечно) от мате-
риальной природы переменных» (Черданцев ЯЛ.Морфогенез и эволюция. М.,
2003, с. 181-183).
Если я верно понимаю это место, смысл его в том, что рамки онтогенезу за-
дает сама четырехмерность пространства-времени.
Поясню аналогией: теория квантов (в той форме, какая вытекала из
уравнения Шрёдингера) являет собой модификацию теории колебаний:
квантовые числа вычислены, в частности, как точки покоя при колебании
шарового слоя (собственные точки полиномов Лежандра). Теория Зоммер-
фельда хорошо описала спектр излучения водорода, хотя общим в обоих
явлениях (спектр излучения и механическое колебание) является только
трехмерность пространства. Пусть за сто лет квантовая теория не смогла
приемлемо описать химию (не говоря уж о макрофизике), но в науке твердо
установилось квантовое мировоззрение, и оно оказалось полезно.
Полагаю, что если элементарный акт морфогенеза будет понят и при-
знан на языке доменов, то родится теория морфогенеза, мало что описы-
вающая всерьез, но пригодная для понимания механизма эволюции по
Жоффруа. Точно так же, как термостатика XIX века оказалась полезна для
дарвинизма, а новая термодинамика (теория ДС) - для понимания номоге-
неза и экологического эволюционизма.
Эпигностический элемент этой будущей квази-теории видится в том,
что каждый этап морфогенеза будет, вероятно, толковаться как активация
гена, компетентного к той структуре, которая в данный момент в данном
месте должна быть сформирована (по Любищеву, она существует в потен-
ции) и уже формировалась в прежних онтогенезах, а акт эволюции - как уз-
навание (или появление) гена, компетентного к новой структуре, дотоле ни
разу не формировавшейся.
Дополнение 4. Фрактал как пример рефрена и рождения новизны
К пп. 5-15* и 6-7*. Понятие фрактала ввел в 1975 году математик Бенуа
Мандельброт. Поскольку феномен рефрена принято считать непонятным,
выскажу свое мнение: он вполне понятен как следствие фрактальной струк-
туры мира. Причиной того, что на всех уровнях, от галактик до атомов, на-
блюдается фрактапьность, служит природа самих чисел.
На любом отрезке вещественной прямой, независимо от его длины,
умещается континуум точек, т.е. часть устроена точно так же, как целое, и
так до бесконечности. Это еще не фрактал, ибо всякий отрезок имеет еди-
ничную размерность, а фрактал - дробную, но на отрезке легко получить
(выбрасывая из него малые отрезки по очень простому закону - например,
выбрасывая его среднюю треть, из оставшихся двух - среднюю треть каж-
дого и т.д.) одномерный фрактал. Столь же легко, выбрасывая квадратики
661
из квадрата, получить двумерный фрактал. Иными словами, даже очень
простая функция вещественного числа может быть устроена фрактально.
Если такие функции описывают что-то в природе, то мир будет устроен во-
все не так, как рисует его гладкая математика Ньютона - Лейбница. Под-
робнее см. [Чайковский, 2004, гл. 6].
Поэтому всё, что нелинейно, что содержит компонент самоподобия и
притом подчиняется арифметике, должно быть фрактально. А ей в природе
подчиняется всё, а в науке, насколько знаю, - всё, кроме разве лишь диа-
лектики (в смысле п. 3-7) и ее следствий - дарвинизма и марксизма.
Рассмотрим простой пример получения плоских фракталов. Пусть зада-
но очень простое преобразование одного числа в другое:
zn+1 = (zn)2 + с, (1)
где переменная z = х + yi и постоянная c = a + bi комплексны.
На вещественной прямой (т.е. при у = 0, b = 0) это преобразование не-
интересно: его свойства сводятся к тому, что точки хп приближаются к нулю
при малых значениях начального х0 и постоянной величины а, при больших
значениях этих величин уходят в бесконечность, а в двух пограничных точ-
ках остаются на месте. Зато в комплексной области творятся чудеса: по-
скольку квадрат мнимой единицы есть единица вещественная, то преобра-
зование (1) состоит из сплошных разрывов, а потому zn ведут себя фрак-
тально и невообразимо сложно. Всё это подробно описано в книге: Пайтген
X-О., Рихтер П.Х. Красота фракталов. М., Мир, 1993.
Пограничная область (из которой переменные z„ не уходят ни внутрь, ни
в бесконечность) оказывается вовсе не окружностью, как можно ожидать по
аналогии с вещественным преобразованием, а замкнутой фрактальной кри-
вой. Наиболее общий случай показан на рис. 84: внутри области располо-
жена единственная неподвижная точка, к которой сходятся все точки zn, ес-
ли процесс начат с точки zb внутри области, ограниченной фрактальной
кривой. Множество J точек на плоскости (х, у), ограниченное этой кривой,
называется множеством Жюлиа (в честь учителя Мандельброта).
Если же точка zb взята вне множества J, то вне его окажутся и все по-
следующие точки zn - они с ростом п уйдут в бесконечность.
Рис. 84. Обычное множество Жюлиа на плос-
кости (х, у) для случая, когда начальная точка
z0 процесса (1) взята из глубины центральной
полости множества Мандельброта (той по-
лости, что ограничена кардиоидой на рис. 85а
и 86). Жирной показана неподвижная точка
(z„ = z0) данного процесса
662
Каждому значению параметра с - а + Ы соответствует свое множество J
(с), что можно изобразить на плоскости (а, Ь): тут каждому J соответст-
вует своя точка. Эти точки образуют множество Мандельброта М (показа-
но целиком на рис. 85а и его половина крупным планом - на рис. 86); его
граница тоже фрактапьна.
Можно сказать и иначе: если в уравнении (1) зафиксировать параметр с,
то получим ломаную линию от начальной точки z0 к неподвижной точке. Ес-
ли изменять z0, то получится семейство ломаных траекторий, заполняющих
свое множество J =J (с) на плоскости (х, у), а если зафиксировать z0 = 0 и
изменять с, то получится множество М = М (z0 = 0) на плоскости (а, Ь).
Множество М имеет ось симметрии Ь = 0, главную полость, ограничен-
ную кардиоидой (известная кривая 4-го порядка), и бесконечное множество
почек. Почка первого порядка - окружность на оси b = 0, три почки второго
порядка - окружности поменьше (одна на той же оси и две по бокам кардио-
иды) и так до бесконечности; причем в почке л-го порядка каждое J имеет п
+ 1 неподвижных точек. Кроме почек, у множества М есть дендриты, не
имеющие полостей; они видны на рис. 86.
Рис. 85. Несколько представителей семейства «мандельбротовых» множеств
а - множество Мандельброта М; b - f- более сложные множества (описаны в
книге Пайтгена и Рихтера). Прошу обратить внимание: множества b и f содер-
жат фигуру, похожую на множество М, однако это другие множества - почки
второго порядка в них иные, а главная полость множества f - окружность
Рис. 86. Фрагмент множества Мандельброта с указанием некоторых
пограничных множеств Жюлиа
663
664
На рис. 86 изображен фрагмент множества М с указанием положения на
нем избранных характерных точек (от 1 до 12), причем для каждой показано
соответствующее множество Жюлиа. Множество 1 является внутренним, а
не пограничным, оно лежит внутри кардиоиды и потому имеет одну точку
покоя (как на рис. 84), но оно близко к границе множества М и потому похо-
же на сложно устроенное пограничное множество 12, расположенное ря-
дом. Множество 3 тоже соответствует внутренней точке, но - точке из круж-
ка второго порядка; поэтому оно имеет три точки покоя.
Двигаясь вдоль границы множества М (плавно изменяя параметры в
уравнении (1)), мы непременно будем пересекать почки и дендриты, т.е. ни-
чтожное изменение параметров будет качественно менять получаемые кар-
тины множеств J. Иными словами, получена модель эмерджентности.
Наконец, видно геометрическое сходство соседних картин (1 и 12; 2, 3 и
5), несмотря на их качественное (топологическое) различие - например,
различное количество неподвижных точек, наличие или отсутствие полос-
тей. Сходства на разной топологической основе возникают в ходе измене-
ний параметров уравнения, т.е. являются рефренами.
Не должен пугать и факт преобразования на комплексной плоскости:
напомню, что введение комплексных чисел в математику рассматривалось
в XVI и даже в XVII веке как нелепая игра ума, а теперь ими описывают са-
мые что ни на есть вещественные процессы - например, электрический ток
в цепях с ёмкостью и индуктивностью.
Хотя данный фрактал и носит чисто математический характер, он нам
полезен тем, что наглядно показывает процедуру рождения новизны (новой
информации) - она рождается фракталом. Очевидно, что и более сложные
фракталообразующие правила биологии тоже должны порождать новизну.
Особенно интересна точка 6. Ее множество Жюлиа само по себе краси-
во, но если немного сдвинуться от нее (в какую сторону и насколько, в книге
Пайтгена и Рихтера не сказано - видимо, секрет фирмы), то попадешь в
«долину морских коньков», где множества Жюлиа прямо-таки поражают
изысканной красотой. Одно из них показано на рис. 87.
Трудно поверить, но и этот фрактал порожден уравнением (1). Он обра-
зован одной-единственной линией бесконечной длины (причем бесконечной
на любом сколь угодно малом квадратике) и имеет иррациональную раз-
мерность между единицей и двойкой. Если хоть чуть-чуть изменить хотя бы
один из четырех его параметров (х0, у0, а, Ь) в уравнении (1), то фрактал ли-
бо обретет полость (ненулевую площадь), либо, наоборот, распадется на
бесконечное множество отдельных “морских коньков”. Здесь мы тоже видим
рефрен: явное сходство форм (“морских коньков”) при различии топологиче-
ской основы (при нарушении размерности или связности) получено путем
преобразования согласно уравнению. Такие фракталы проясняют ту мысль
многих учёных, что системность и целесообразность могут носить не функ-
циональный, а чисто эстетический смысл.
665
Более сложные фракталы обладают более сложными свойствами. Так,
Бульенков (п. 7-9) показал, что во фракталах структурированной воды кле-
ток идет «метрический отбор» допустимых органических форм {Бульенков
Н.А. И Биофизика, 2005, № 5, п. 4), т.е. вариант Богдановского подбора.
Рис. 87. Наиболее эффектное множество Жюлиа из «поля морских коньков»:
А - общий вид
Может показаться странным, что объяснение феномену рефрена дано
не биологическое, а математическое. Однако иначе и быть не может: ведь
рефрен не является не только собственно биологическим явлением, но и не
обязан даже иметь материальную природу - вспомним хотя бы грамматиче-
ские рефрены. Если же феномен вездесущ, то и носители его должны быть
вездесущи, а таковыми прежде всего являются числа. Словом
«Бенуа Мандельброт создал неевклидову геометрию негладких, шероховатых,
зазубренных, изъеденных ходами, порами, трещинами и отверстиями, извили-
стых и т.п. объектов... по молчаливому уговору ранее исключавшихся из рас-
смотрения... Между тем именно такие неправильные объекты составляют по-
давляющее большинство объектов природы» {Данилов Ю.А. Лекции по нели-
нейной динамике. Элементарное введение. М., УРСС, 2006, с. 91-92).
В мире этих объектов наблюдаются и могут быть эффективно описаны
параллелизм и эмерджентность. Можно надеяться, что математика фракта-
лов ляжет в основу будущей теории эволюции, в том числе биологической.
666
Б — его фрагмент (это может быть любой “хвост морского конька”); если увеличить
часть данного фрагмента, помещенную в рамку, до размеров всего фрагмента, то
получится точно такой же рисунок, и так до бесконечности
Дополнение 5. Свойства рефренов и диасети
К пп. 6-3**, 11-5. Первый известный мне ясный пример рефрена нахо-
дим у Владимира Ковалевского (1873 год): настоящие рога (из рогового ве-
щества) полорогих (овцы, козы и др.) и костяные “рога” оленьих образуют в
миоценовых отложениях два ряда направленных рядов.
667
У тех и у других в различных группах после завершения формирования
характерных для них скелетов начинается образование рогов. Сперва это
простые шишки на лбу, затем «простые, булавовидные, затем вилковид-
ные... Эти выросты становились все больше и разнообразнее по мере
дальнейшего развития группы». Ковалевский назвал данные ряды рогов
«признаками роскоши» (цит. по Давиташвили Л.Ш., 1948, с. 112-113).
Коп придал рядам рядов форму таблиц и пытался нащупать диасеть.
Ограничусь уточнением примера из пункта 3-10*. В 1887 году Коп писал:
«Среди лягушек мы видим четыре подразделения. Одно имеет эмбриональ-
ный (слабо развитый) рот и эмбриональную грудину; другое - эмбриональ-
ный рот и вполне развитую грудину; третье - эмбриональную грудину, но
вполне развитый рот; а четвертое - вполне развитые рот и грудину. Первое
подразделение есть низшее и, вероятно, древнейшее... Второе, третье и чет-
вертое подразделения имеют соответствующие друг другу (сходные поэле-
ментно в разных рядах - Ю. Ч.) ряды родов различного строения, приспособ-
ленные к тому или иному образу жизни. Одни имеют “лопаты” для рытья,
другие - перепонки для плаванья, а третьи - диски для приставанья к ветвям и
листьям деревьев» (цит. по Давиташвили Л.Ш., 1948, с. 176-177).
Переходы от «эмбриональности» к полному развитию Коп относил к ро-
довым изменениям, а приспособления - к видовым. Элемент диасети со-
стоит здесь в том, что видовые и родовые признаки образуют две таблицы,
реализуемые почти или вовсе независимо. Причину упорядоченности дар-
винист Ковалевский полагал загадочной и лишь заметил, что «признаки
роскоши» неизбежно возникают потому, что «в организме всегда имеются
скрытые силы и возможности». Ламаркист Коп прямо объяснял все приоб-
ретения действием «особого вида энергии» (там же, с. 185). Как видим, оба
ссылались на то, что мы именуем активностью.
Однако Коп сделал еще и очень важное добавление: ввел понятие го-
мологических рядов, которые полагал сущностно сходными с гомологиче-
скими рядами в химии (п. 3-10*). Иными словами, он полагал феномен диа-
сети общеприродным явлением. Как мы теперь знаем, общность еще более
широка: диасеть выявляется и в грамматике, и в обществе.
Вскоре после появления понятия рефрена Корочкин, сославшись на
Штейнмана (о нем см. п. 3-10), выявил нечто вроде диасети. Отметив:
«То, что параллелизмы буквально пронизывают все уровни организации жи-
вого, включая генетический материал, противоречит представлениям о после-
довательном и постепенном расхождении признаков», -
Корочкин высказал «гипотезу программированной трансформации»:
«Предполагается, что исходная совокупность живых форм представлена на-
бором архетипов (по Оуэну - Ю.Ч\ каждый из которых содержит генетиче-
скую программу, обеспечивающую осуществление определенного направле-
ния онтогенеза». Макромутации «вызывают к жизни программированный, но
не проявленный до того морфогенез, ибо тот или иной морфогенез может
668
быть не осуществлен не по тому, что эволюционно не сформировалась его ге-
нетическая программа, а потому что в генетическом материале содержатся
элементы, блокирующие проявление этой программы» (Корочкин Л.И. Па-
раллелизмы в молекулярной организации генома... // Молекулярные меха-
низмы генетических процессов. М., Наука, 1985, с. 140, 142-143).
Отличие от диатропики здесь в том, что объективно существующей с
самого начала эволюции постулирована не структура разнообразия (идея),
а конкретный генетический текст. Это - направляемая Богом эволюция по
Григорию Нисскому и Августину, что и сам Корочкин признал (в послесловии
к книге [Лима]), едва то позволила политическая обстановка.
Перечислить даже главные работы по рефренам нет возможности.
Скажу только, что самую широкую панораму рефренов - от макромолекул
до исторических рядов организмов - можно извлечь из работы Э.И. Воробь-
евой (ЖСБ, 1987, № 4), хотя вместо слова «рефрен» она применила выра-
жение «устойчивость структурного уровня». Недавно Е.Е. Коваленко (см. п.
5-2) выявила рефренную структуру скелета амфибий и его эволюцию как
движение вдоль рефренов при снятии давления нормы (хотя самих этих по-
нятий у нее нет). В совокупности все эти работы делают понятие диасети
широко принятой практической реальностью.
Обычно довольно полные ряды выявляются у достаточно близких групп,
однако сам феномен параллелизма (а потому формируемая параллелиз-
мамими диасеть) не имеет таксономических границ. Прекрасный пример
обширного параллелизма между филумами (отделами) В.В. Корона привел
сходство листьев цветковых и “листьев" папоротников (рис. 88; лат. назва-
ния по моей просьбе любезно выверены Д.Д. Соколовым, Биофак МГУ).
Тип листа цветковых
1 - почковидный
2 - ланцетный
3 - парноперистосложный
4 - непарноперистосложный
5 - пальчатосложный
6 - пальчатолопастный
7 - дваждысложный пильчатый
8 - сердцевидный
9 - яйцевидный
10 - веерный + дваждысложный
пальчатый
11 - пальчатораздельный
12 - ромбо-яйцевидный
13 - перистолопастной (нижние) +
перисторассеченный (верхние)
14 - стрело-копьевидный
Пример сходного папоротника
Pterozonium reniforme
Pterozonium scopulinum
Hemionitis subcordata
Hemionitis tomentosa
Tectaria incisa
Doryopteris rediviva (взросл.)
Adiantum anceps
Lindsaea sagittata
Elaphoglossum crinitum
Anogramma leptophylla
Hemionitis pinnatifida
Doryopteris rediviva (ювенильн.)
Tectaria coriandrifolia
Doryopteris sagittifolia
669
Замечу, что этот пример является иллюстрацией правила Кренке (нор-
ма в одном таксоне бывает редкостью в другом) и что Корона дал здесь не
рефрен, а лишь материал для него: еще предстоит выяснить, есть ли у па-
поротников и цветковых общее правило изменения листовых пластинок.
формулами. Лучший пример рефрена-формулы
Рис. 88. Вайи папоротников,
внешне сходные с листьями
цветковых (Корона В. В.
Основы структурного
анализа в морфологии
растений. Свердловск, СГУ,
1987).
Общее правило пре-
образования в рамках
одного параллелизма
(простого или множе-
ственного) - вот что
Мейен хотел видеть в
понятии рефрена.
Таких правил в био-
логии много (напри-
мер, усложнение впра-
во и вниз на таблицах
рис. 35 и 36), но они
задаются словами, а не
- спиральное листораспо-
ложение: если листья расположены на стебле очерёдно (а не кустами), то
их основания образуют вдоль стебля спираль, причем число листьев на шаг
спирали постоянно для данного растения и эти числа образуют ряд:
ai=2/1, а2=3/1, а3 = 5/2, а4 = 8/3, а5= 13/5, а6= 21/8, (2)
где числители и знаменатели образованы числами Фибоначчи (каждое чис-
ло - сумма двух предыдущих; а4 = 8/3 означает, что на 3 оборота спирали
приходится 8 листьев). Замечательно, что а, постоянно для всех особей
данного вида растений. Подробнее см. [Чайковский, 1990, с. 51].
Факт этого постоянства установлен твердо и является примером закона
формы, не сводимого ни к какой функции. В самом деле, если даже и пове-
рить, что какая-то форма (выражаемая числом а,) вызвана наличием у дан-
ного растенифя некой функции, то всё равно различие а,- у разных видов
растений и их принадлежность единой формуле останутся без объяснения.
Но что касается формулировки рефрена (т.е. самой этой формулы, ис-
черпывающей все возможности), то встает одна трудность: черешки листь-
670
ев - не точки, а кружочки, так что уже значения а6= 2,625 и а7= 2,615 факти-
чески на стеблях различить невозможно (хотя можно на чешуе шишек).
Но не является ли ряд Фибоначчи выдумкой, основанной лишь на не-
точностях измерений? Может быть, все растения попросту имеют от 1,5 до
2,63 листьев на один оборот спирали? Для отрицательного ответа на второй
вопрос надо бы провести сплошной промер всех видов растений со спи-
ральным листорасположением и убедиться, что в интервале от 1,5 до 2,6
никаких значений, кроме задаваемых набором (2), не имеется.
Есть ли смысл говорить о единой природе рефренов или сходство их
только словесное? Новый номогенез исходит из их единой природы (см. До-
полнение 4). В пользу единства говорит всеобщность феномена паралле-
лизма, причем он отнюдь не всегда связан с общностью материальных
структур, проявляющих параллелизм. Примеры мы видели: сходство форм
талломов водорослей с листьями растений (и даже с морозными узорами),
сходство и даже идентичность гомеобоксов при различии онтогенезов и на-
оборот - различие типов дробления при сходстве последующих онтогене-
зов. Однако рефреном явится не всякий параллелизм, а только направлен-
ный, притом с определенным правилом, пусть и словесным.
На сегодня наиболее общим доводом в пользу единства служит наблю-
даемая фрактальность разнообразия, поскольку о математических фракта-
лах (чисто математических структурах) известно, что они могут образовы-
вать диасеть (Дополнение 4). Однако феномен диасети наблюдается сам по
себе, на разнообразии живых объектов, и для чуждого математики читателя
это служит более сильным доводом. Рассмотрим один пример.
В п. 6-15* мы касались работы зоолога Е.А. Артемьевой, ученицы Р.В.
Наумова (Ульяновск), где показана модульная природа крылового рисунка
бабочек. Позже Артемьева на этом материале показала номогенетический
характер эволюции в семействе голубянок - точнее, вида голубянка икар
(Polyommatus icarus) и близких к нему (образующих с ним, по Артемьевой,
один надвид). Проведя кропотливый анализ по 29 признакам рисунка в об-
ширных коллекцях Р. icarus, собранных за 116 лет, она пришла к выводу:
«Морфологическая структура крылового рисунка в целом в популяциях Р.
icarus остается неизменной на протяжении 116 лет. Однако пространственные
соотношения признаков значительно изменяются во времени» {Артемьева
Е.А. Крыловой рисунок голубянки Polyommatus icarus Rott. ... как модель эво-
люционных исследований //ЛЧ, 2005, с. 128).
Это значит, что эволюция текла не путем появления или утраты каких-
либо качественных признаков, а посредством изменения их количественных
параметров - как в картине крыла одной особи, так и в частотах признаков в
каждой популяции. В частности, картины менялись в западной группе попу-
ляций не так, как в восточной. Наложив данный материал на географиче-
скую карту, автор получила наглядную схему эволюции:
671
«фенотипическое разнообразие крылового рисунка Р. icarus выше в популя-
циях, обитающих на периферии ареала» {Артемьева Е.А. Эколого-генети-
ческий и морфологический анализ крылового рисунка... // Автореф. доктор,
дисс.; Биофак Ниже город, университета, Ниж. Новгород, 2006, с. 16).
А именно, если исходная (европейская) группа популяций почти цели-
ком состоит из обычных (как 116 лет назад) особей, то восточная (азиат-
ская) - в основном из «девиантных» (уклонившихся по рисунку крыла) осо-
бей (там же, карта на с. 18). Иными словами, обращу внимание, что хорошо
видно, как краевые популяции ведут генетический поиск (за счет снятия
давления нормы исходного состояния) в рамках неизменной диасети. Итоги
поиска распространяются в популяциях очень быстро (почти как в опыте
Шапошникова): за 10-15 лет на сотни километров (там же, с. 29), при актив-
ности особи менее, чем на 2 км {Артемьева Е.А. // ЛЧ, 2005, с. 123).
Если всё это верно, то придется вспомнить о других сверхбыстрых про-
цессах - такова «мода на мутации», которую еще в 1930-х годах описала
Р.Л. Берг (лит-ру см. [Назаров, 2005, с. 410]), опыты Шапошникова и многое
другое. То есть, добавлю, ни о каком “отборе случайных вариаций” речи
быть не может - идет направленный процесс образования нового вида.
Дополнение 6. Более тонкие свойства рефренов
К пп. 6-3***, 6-4*. При первой публикации (БПС, 1998, № 28) редактор И.Г.
Мещерский сделал к тексту о лапках сиренов и хиротов примечание:
«в семействе сцинков подотряда ящериц имеется значительное количество рас-
пространенных в разных частях света форм с недоразвитыми или отсутствующи-
ми конечностями - передними, задними, а также теми и другими одновременно».
Это важное примечание побудило меня обратиться к разнообразию
сцинков - самого обширного (1200 видов) семейства в подотряде ящериц
(включающем всего 4 тыс. видов). Выяснилось, что разнообразие сцинков,
для класса рептилий огромное, включает в себя черты разнообразий других
семейств, в том числе и хиротов, на что и обратил внимание Мещерский.
Видовое обилие сцинков достаточно велико, чтобы можно было рассмот-
реть его диасеть. Например, разнообразие их конечностей выстраивается в
ряд - от полного развития до полного отсутствия:
“У различных сцинковых в гораздо большей мере, чем у других яшериц, вы-
ражена тенденция к змееобразному удлинению туловища, сопровождающе-
муся одновременным уменьшением размеров или даже полным исчезновени-
ем конечностей. Этот своеобразный процесс, характерный для многих пред-
ставителей семейства, лучше всего проследить на примере африкано-
мадагаскарского рода Scelotes. У распространенного на острове Маврикий
Scelotes bojeri имеются еще вполне развитые обе пары пятипалых конечно-
стей, причем, вытянутые навстречу друг другу с одной стороны тела, они со-
прикасаются или отчасти даже перекрываются друг другом” {Доревский И.С.
/ГЖмзнь животных. Т. 4, ч. 2. М., 1969, с. 261).
672
То есть, добавлю, конечности не используются.
//'vS' Рис. 89- Три стадии редукции задних
/дД конечностей у роющих сцинков рода
RxJ Н~\/ [ЗЧ Scelotes (по: Даревский ИС. //Жизнь
Peg гСу рД животных. 1969)
YgA ид г2д Прерву цитату, чтобы отме-
уа титы речь идет о тенденции, веду-
уж: pg Щей от ящериц к змеям, и «этот
wV своеобразный процесс» своеобра-
зен лишь тем, что не встречается в
других семействах ныне живущих
ящериц. (С позиции диатропики -
потому, что в них мало видов и редкие формы не смогли выявиться.) Зато в
семействе сцинков он реализован, пусть и не столь полно, еще как минимум
трижды, чем и высвечена с иной стороны та же диасеть. После описания
стадий редукции лапок в роде Scelotes (три формы даны на рис. 89, но есть
еще вид вообще без лапок), сказано:
“В той или иной мере редукция ног выражена также у роющих видов в родах
Anguincephalus, Nessia, Rodona и др. Исчезновение конечностей и вытягивание
тела почти всегда сопровождается сильным удлинением хвоста, на долю кото-
рого приходится иногда почти две трети общей длины животного” (там же).
Тенденция очевидна, и она в целом явно неадаптивна (встречно на-
правленные лапы, излишне длинный хвост и прочее). Поэтому нет осно-
ваний верить в то обиходное утверждение, что редукция лап является на-
бором приспособлений к частным условиям среды. Вернее будет сказать,
что редукция конечностей - тенденция, проявляющаяся во многих таксо-
нах всех классов позвоночных. Те, кто мог к ней приспособиться, уйдя в
подходящие зоны обитания, живут, а кто не мог, тех мы не наблюдаем -
например, птиц без задних лап. (Если угодно, можно назвать это естест-
венным отбором, но вот и вся его роль.) Вообще, редукция активно рабо-
тающих органов вполне обычна и никак не объяснена.
Любищев, отвергая наивное объяснение редукции крыльев пустынных
насекомых (как предотвращения перегрева), резюмировал (1972 г.): «ре-
дукция крыльев крылатых насекомых - явление чрезвычайно распростра-
ненное и причины такой редукции большей частью неясны» (ТПЭ-3, с. 8).
Одно можно сказать определенно - каждая редукция укладывается в ка-
кую-то тенденцию.
На рис. 89 три стадии редукции пятипалой лапы сцинков в рамках рода
Scelotes (четырехпалая, двупалая и беспалая) даны как S. limpopoensis, S.
bipes и S. gronovii; к ним можно добавить безногую форму S. inornatiis). Это
похоже на четыре падежа одного существительного или прилагательного,
причем “биологическая грамматика” предлагает (как мне представляется)
673
называть их как единый подрод или единый вид. Если все они живут на реке
Лимпопо и различаются только числом пальцев, то их можно обозначить как
формы вида S. limpopoensis. Еще удобнее сравнить эти четыре формы со
степенями выраженности одного прилагательного - например, с рядом
полулёгкий, лёгкий, легчайший, наилегчайший.
Если одновременно наблюдается упорядоченное изменение не по од-
ному, а по двум параметрам, то вместо ряда надо для описания подрода
ввести прямоугольную таблицу вроде тех в грамматике, где строки означа-
ют падежи прилагательных, а столбцы - их степени выраженности.
Если упорядочение наблюдается по трем параметрам, то плоские таб-
лицы сложатся в трехмерную стопку; для прилагательных: в стопке 3 слоя (3
рода: мужской, женский и средний). Разумеется, реальные биологические
таблицы не двумерны, а многомерны.
Хороший пример необходимости введения таблиц для понимания раз-
нообразия видов дают ящерицы горные агамы (род Laudakia семейства
Agamidae). Сравнение новейшей сводки (Панов Е.Н., Зыкова Л.Ю. Горные
агамы Евразии. М., Лазурь, 2003) с прежней литературой открывает вековое
хождение по кругу (такое же, о каком в связи с собаками (род Canis) говори-
лось в книге [Чайковский, 1990]). Например, античное название Stellio
(звёздчатая ящерица) то появляется в обозначении отдельного рода, то эта
группа снижается до подрода или вовсе исчезает из классификации, то сно-
ва появляется. Ныне Stellio означает «нейтральный термин “комплекс ви-
дов”» (Панов Е.Н., Зыкова П.Ю., с. 21), объединяющий 8 из 16-ти видов ро-
да Laudakia. Эта «нейтральность» характерна: интуитивно близость всех
Stellio очевидна, но формализовать ее не удается.
Авторы полагают, что неустойчивость системы агам вызвана недостат-
ком изученности, но их материал говорит совсем иное: как и всюду, добав-
ление новых признаков (ультраструктурных и молекулярных) не только не
прекращает хождение по кругу, но делает надежду на прекращение тщет-
ной. Так, иммунный метод дает не то, что сравнение митохондриальных
ДНК, а они сами - не то, что морфологические методы (с. 23).
Авторы пишут, что «придерживаются филогенетической концепции», но
это не вполне так: в ней нет места «нейтральным комплексам», да и сами
авторы признали, что группируют не столько по родству (никому не извест-
ному), сколько по распространению - от Средиземноморья на восток (с. 21-
22). Даже если допустить невероятное - что для какой-то группы видов рода
Stellio филогенетическая система будет найдена и окажется устойчивой
(долговечной), всё равно ее будут портить гибриды: авторы сами пишут, что
«ограниченно гибридизирующие виды могут, вероятно, даже не состоять в
близком родстве» (с. 16).
Зато надежду внушает идея биологической грамматики, т.е. структу-
ралистской систематики, поскольку она апеллирует не к родству, а к
674
свойствам. Любые промежуточные формы (в том числе гибриды), находят в
ней свое место (подробнее см. гл. 10), как они находят его в грамматике.
В языковой грамматике никого не удивляет включение одного слова в
разные группы. Например, в выражении «он выглядит франтом» третье
слово можно счесть и дополнением (кем?), и обстоятельством (как?). Его
можно однозначно описать как словарное слово «франт» плюс окончание,
но нельзя однозначно характеризовать как член фразы.
В биологии аналогом словарного слова служит архетип по Оуэну (схема
строения, рисунок), а аналогом множества словоформ (форм этого слова) -
архетип по Мейену (таблица). Фактически Панов и Зыкова задали класси-
фикацию именно таблицами, тогда как «филогению», данную рисунком (с.
24), по сути даже не обсуждают, хотя она отлична от их системы.
Наконец, рефрены видны в идее «смены адаптивных норм» Шмальгау-
зена (п. 4-14). Высказана она лишь однажды и нарочито туманно - основные
ее части сами себе противоречат или тут же опровергнуты, так что на вся-
кий упрёк можно ответить: «Я говорил не это». Главное ее положение: не-
наследственные изменения «рано или поздно становятся наследственны-
ми», взятое из ламаркистского труда Копа, - не только не объяснено, но и
ничем не подтверждено. Она никогда более не излагалась и, если понимать
идею буквально, то обсуждать ее невозможно. Однако, как уже сказано в п.
4-14, она была иносказанием, не рассчитанным на буквальное понимание.
Главное то, что у Шмальгаузена утверждается (вслед за Болдуином и
другими сторонниками органического отбора) всепроникающий паралле-
лизм процедур преобразования прижизненной изменчивости в наследст-
венную. А параллелизм перобразований и есть рефрен по определению.
Никакой попытки объяснить данный параллелизм ни у автора, ни у его
коллег не было, но это, замечу, для нашей темы (диатропики) нормально -
сперва надо найти закономерность, затем уложить ее в ряд сходных зако-
номерностей, а потом уж только стоит искать механизм.
Новым было желание объяснить этим рефреном смену целостностей:
«наличие ненаследственной пластичности приобретает решающее значение.
Возможность скачкообразного перехода, т.е. быстрой и гармонической пере-
стройки всей организации, предполагает широкую способность к индивиду-
альному приспособлению», а затем «все такие преобразования рано или позд-
но становятся наследственными» (Шмалъгаузен И. И. Изменчивость и смена
адаптивных норм в процессе эволюции // ЖОБ, 1940, № 4, с. 513).
Как видим, сама смена целостностей мыслится происходящей у инди-
вида (т.е. вне естественного отбора). Однако формируется ли движущий
смену рефрен средой или же он задан природой организма? Шмальгаузен
чётко (в отличие от остальных положений данной концепции) встал на сто-
рону определяющего влияния среды:
«В процессе смены среды обитания одна адаптивная норма может быть заме-
нена другой, если факторы новой среды уже встречались в прежней в виде
675
случайных, периодических или локальных уклонений, послуживших уже ос-
нованием для выработки соответствующей адаптивной нормы» (с. 524).
Никакой эволюции тут нет, есть лишь “повторение пройденного” под
влиянием повторяющих воздействий среды. Будучи положена в основу уче-
ния, эта позиция привела к смягченному креационизму, царящему в СТЭ
поныне (п. 5-3*). Нас же интересует эволюция. Чтобы выйти из бесконечного
блуждания по кругам “повторения пройденного” и уйти от испуганных реве-
рансов ламаркизму, понадобилось переосмыслить понятие отбора в поня-
тие подбора (п. 6-12*) и понять, что норма не всегда адаптивна - может, на-
пример, даже утрачиваться при улучшений условий жизни (5-13*).
Дополнение 7. Формирующая причинность
К п. 6-15**, 7-11 и 9-13. Первая книга Шелдрейка называлась «Новая
наука о жизни. Гипотеза формирующей причинности» (Sheldrake R. A new
science of life. The hypothesis of formative causation. London, 1981). Форми-
рующая (по смыслу можно перевести и как созидательная) причинность -
это новый для науки принцип связи причин со следствиями, но он не вполне
нов для философии. Вспомним, что Аристотель различал 4 типа причин -
материальную, формальную, действующую и целевую, причем для живых
объектов он по сути объединял три последних в едином понятии энтелехии
(п. 1-6). Такое объединение лишь поначалу кажется упрощением, на деле
же очень затрудняет анализ причин, и одной из заслуг Шелдрейка является
вычленение такого аналога формальной причины, какой можно приложить к
живому. Это и есть его понятие «formative causation» (formative можно пере-
вести многими способами, в том числе и как относящийся к формирова-
нию). Вспомним также formative aptitudes Эразма Дарвина и концептуаль-
ную активность Бёрджерса.
Кроме примера со строительством дома (п. 1-6), приведу более новый
пример четырех причин. При игре в биллиард материальную причину каж-
дого шага игры составляют стол с лузами, шары и кий; формальную - пра-
вила игры и законы механики; действующую - удар кием по шару, а целе-
вую - замысел игрока. Тут формальная причина не является сущностью (ни
образом, ни планом), она только задает необходимые рамки процесса, не
всем заметные.
Шелдрейк вспомнил энтелехию:
“В системе Дриша, которая явным образом основана на системе Аристотеля,
особенные формы живых организмов были обусловлены неэнергетическим
фактором, энтелехией. Морфогенетические поля... играют сходную роль в
проведении морфогенетических процесов к их особенным конечным фор-
мам. Но их природа остается столь же неясной”.
Далее он энтелехией не пользуется, предлагая вместо нее аналогию с
химией, поскольку ничего похожего в физике не нашел (Sheldrake R., с. 63).
Это может показаться странным - разве химия не есть физика межатомных
676
связей? Нет, физику в основном интересуют процессы, а не разнообразие
получаемых веществ, интересующее химию. В терминах диатропики ядро
химии является периферией физики и обратно (п. 10-7). В этом разнообра-
зии путей, ведущих к энергетически сходным веществам, Шелдрейк и ищет
(как мне видится) ключ к проблеме.
Как физика построена на идее взаимодействия частиц посредством фи-
зических полей, так в химии и биологии надо выяснить, с какими полями они
фактически имеют дело. Шелдрейк, напомнив о поле Гурвича, считает, что
нечто подобное необходимо ввести для биологии в целом:
“Эта причинная обусловленность формы морфогенетическими полями имену-
ется формирующей причинностью, чтобы отличить ее от энергетического типа
причинности, который так тщательно исследует физика”. И далее: “Это разли-
чение между формирующей и энергетической причинностями напоминает ари-
стотелево различение между формальной и действующей причинами. Однако
гипотеза формирующей причинности... радикально отличается от теории Ари-
стотеля, предполагающей навечно заданные формы” (Sheldrake R., с. 75,78).
Как и Лавлок, он полагает, что природа как целое обладает памятью,
поэтому любое новшество становится как бы общим достоянием. Примеры,
которые Шелдрейк приводит в подтверждение, таковы.
Первый (химический); если вещество с данной формулой может обра-
зовывать несколько форм кристаллов, то при первых попытках его получе-
ния образуется смесь разных форм, но вскоре - во всех лабораториях, где
его получают, независимо от расстояния между ними, устанавливается еди-
ная форма - даже в лабораториях, осуществляющих кристаллизацию впер-
вые. (Обычное объяснение - распространение зародышей кристаллизации
через пыль на бородах и одежде заезжих химиков - не кажется Шелдрейку
серьезным.)
Второй (морфологический): развитие зародыша объясняют через по-
следовательную смену морфогенетических полей, но ведь они еще сами не
объяснены. Конечно, эти поля можно описать в терминах физики (излуче-
ний, концентраций веществ), но это - не объяснение. (Напомню, что на эти
два типа объяснений указал еще Сократ, п. 1-6.) Шелдрейк объясняет мор-
фогенез иначе, как реализацию памяти вида.
Третий (этологический): если животные чему-то научились, то научение
быстро передается необученным даже при полном отсутствии контактов.
Так, известно, что в 1920-30 годах синицы-лазоревки в Англии (а затем в
других странах Западной Европы) научились открывать молочные бутылки.
Синица живет 3 года и перемещается за свою жизнь на участке с радиусом
всего 3 мили, однако семилетний перерыв производства (во время войны)
бутылочного молока не помешал всей голландской популяции этих птиц
вновь овладеть этим умением всего за год (добавлю: довод со сроками
слаб, но сам феномен отрицать рискованно). Столь же необычные скорости
распространения видны в данных Артемьевой (Дополнение 5).
677
Четвертый (социально-психологический): феномен «коллективного бес-
сознательного» (так швейцарский психолог Карл Юнг назвал полвека назад
господствующие в обществе неосознанные «архетипы», т.е. образцы мыш-
ления) является памятью общества не в переносном, а в прямом смысле.
Ярким образцом социального архетипа служит для Шелдрейка дарвинизм.
Действительно, он распространился в европейском сознании почти мгно-
венно и стал восприниматься почти всеми на уровне бессознательного. Это
современный миф, живущий по законам мифа (ср. п. 6-11).
Для него ДНК - резонатор внешних сигналов, генерируемых морфиче-
ским полем (как и Гаряев, п. 5-5*), которое автор не считает энергетическим
и сравнивает с миром идей Платона, улучшенным аристотелевой схемой
становления. Последнюю он склонен понимать как энтелехию - как реали-
зацию возможности (п. 1-6) и как идею целостности по Дришу (п. 3-14).
В советские времена такие слова звучали страшно, но не следует их пу-
гаться - с идеями, существующими независимо от их материальных носи-
телей, мы встречаемся постоянно: никого не смущает, что теорема Пифаго-
ра была верна до того, как кто-то стал чертить и измерять треугольники.
Разнообразие конечностей позвоночных материально, но и тут нет основа-
ний считать, что рефрен мерона «конечности позвоночных» (гл. 6), описы-
вающий это разнообразие, появился вместе с последней из включенных в
него позицией (рука человека). А если так, то вернее будет сказать, что его
клетки существовали до того, как были заполнены.
Противопоставление идеализма материализму, идущее из Античности,
многие, в том числе Шелдрейк, считают устаревшим. Добавлю, что морфи-
ческое поле не выдумано Шелдрейком, а в некоем смысле взято у нынеш-
них физиков: у них складывается убеждение, что мир - не столько набор
тел, сколько взаимодействие полей. Шелдрейк не одинок, когда пишет:
«Поля более фундаментальны, чем материя», они - «области влияния в про-
странстве и времени. Их природа более напоминает модификации в про-
странстве, чем что-либо, истекающее из материи или творимое ею» (Sheldrake
R. Habits of Nature // One Earth. 1988, № 3, c. 30).
Понять эти слова сначала трудно, но они проясняются после ознаком-
ления с его понятием «морфического резонанса»: как телевизор может при-
нять любую из станций за счет резонанса (совпадения несущей частоты
станции с собственной частотой входного контура телевизора, задаваемой
пользователем), так и в природе Шелдрейк видит своеобразную настройку.
Например, вещество кристаллизируется так, а не иначе, совсем не потому,
что какое-то «морфическое поле» управляет процессом кристаллизации
(этот процесс задается физическими законами), а потому, что это поле ука-
зывает данному образцу вещества один из физически возможных путей, т.е.
“настраивает” образец на данный путь.
Для Шелдрейка хромосомная ДНК - набор резонаторов, способных вы-
бирать из возможных путей онтогенеза тот, который соответствует данному
678
виду. В роли морфического поля особи в данном случае будет обычное
морфогенетическое поле Гурвича. Шелдрейк уподобил молекулярного био-
лога дикарю, заглянувшему внутрь телевизора: действуя на детали, он вы-
зывает на экране искажения и полагает, что нашел источник изображения в
самом телевизоре, в то время как на самом деле сигнал исходит из теле-
центра, о наличии поля которого дикарь не знает.
Химический пример морфического резонанса стоит в биологической
концепции Шелдрейка как бы особняком, но на деле автор (как Янч до него)
ищет то общее, что объединяет формирование атомов, молекул, кристал-
лов, клеток, тканей, органов, организмов и сообществ. Поиск аналогий живо-
го с неживым характерен для Янча и Шелдрейка и сближает их концепции с
диатропикой. Похоже, что Шелдрейк фактически рассмотрел диасеть, и
морфическое поле является тем инструментом, которым природа выбирает
конкретный вариант развития (рефренной строки). Настоящая новизна по-
является в эволюции крайне редко - с появлением новой рефренной таб-
лицы (по Мейену) или нового поля (по Шелдрейку). В биопоэзе так выглядит
появление субвитальных областей (по Берналу), которые могут быть сопос-
тавлены с «областями влияния» Шелдрейка.
Едва упомянув Янча и совсем не ссылаясь на Лавлока, близких ему по
духу, Шелдрейк, однако, часто цитирует Девиса (п. 7-1), оспаривая антроп-
ный принцип. Он уверен в том, что нет смысла говорить о законах физики,
существовавших до тех пор, пока не было самих физических объектов, и
вместо антропного принципа принимает как аксиому, что законы физики (как
и биологии) - итог эволюции природы. Он допускает, что его «морфическое
поле» столь же старо и универсально, как физические поля. Из этого следу-
ет, что «созидательная причинность» могла быть ответственной и за едино-
образие космической эволюции:
«Возможна вселенская сеть морфического резонанса среди галактик, звезд и
атомов. Если так, то естественно допустить, что есть вселенская сеть резонанса
между молекулами, кристаллами и формами жизни» (Sheldrake R. The presence
of the past. Morphic resonance and the habits of nature. London, 1994, c. 302).
Добавим, что в этом случае перестают удивлять быстрые темпы биопо-
эза на Земле - если этот процесс шел параллельно в разных местах Все-
ленной. Фактически здесь воспроизведены как древняя идея мировой души,
от которой черпает свою активность всякая индивидуальная душа (пп. 1-5,
8-16), так и идея диасети (рефрена в пространстве-времени).
Существование вселенской, да и земных резонансных сетей пока всего
лишь допущение. Отказ видеть физические законы до физических объектов
противоречит платонизму (в иных отношениях признаваемому самим Шел-
дрейком); здесь он ближе к линии Аристотеля.
Словом, перед нами не теория, а лишь общая философская заявка на
нее. Главное в ней не результаты, а постановка вопросов, новых для науки.
Увы, сам Шелдрейке 1988 года не продвинулся по сути ни на шаг.
679
Дополнение 8. Активность, компенсация и номогенез
К п. 8-10. Классическим образцом компенсации служат сверхклеточные
(п. 8-8*). Особенно поражают инфузории. Трудно поверить, что изображен-
ный на рис. 90 хищник с разверстой пастью и складчатой “кожей" - однокле-
точное Onychodromus quadricornutus. Этот гигант (0,3 мм) пожирает мелкого
собрата, поедавшего бактерий - вид образует, при недостатке пищи, два
звена цепи питания (что обычно у некоторых высших, например, у окуней).
Рис. 90. Гигант-
ский экземпляр
четырехрогой ин-
фузории пожира-
ет своего карли-
кового собрата
(по Wicklow, 1988)
Как самый
загадочный
пример прин-
ципа компен-
сации можно привести глаза кубомедуз. Этот небольшой отряд мелких тро-
пических медуз, печально известных тем, что ожог некоторых видов бывает
для людей смертельным, удивителен в нескольких отношениях.
«Кубомедузы отличаются от остальных книдарий активным рыбоподоб-
ным поведением и развитым сенсорным аппаратом. Каждая из четырех сто-
рон животного несет ... ропалий, который развился в изумительный пучок
различных глаз. Два из глаз каждого ропалия давно известны как напоми-
нающие глаза высших животных, но функция и назначение этих глаз остались
неизвестны». В частности, непонятным образом «положение их сетчатки не
совпадает с нужным для чёткого изображения» {Nilsson D.-E. е.а., Advanced
optics in a yellyfish eye П Nature, 2005, vol. 435, No 7039, p. 201).
Добавлю, что каждый глазик имеет размер всего около 0,1 мм, однако
снабжен всеми частями глаза (роговица, радужка, хрусталик и сетчатка).
Нет только мышц, растягивающих хрусталик (что и ни к чему, сетчатка рас-
положена много ближе фокуса хрусталика, и фокусирование невозможно).
Глазной нерв в данной статье не упомянут, хотя он есть: его еще в XIX
веке описал петербургский зоолог В.Т. Шевяков, в то время работавший в
Германии (Schewiakoff W.T. Beitrage zur Kenntnis der Acalephenauges // Mor-
phol. Jahrbuch, Bd 15. Leipzig, 1889). Нерв сложен из отростков светочувст-
вительных клеток. Обычно зрительный нерв идет от сетчатки в мозг, но у
медуз мозга нет. Есть только нервное кольцо и на нем ганглий (скопление
нервных клеток) около каждого ропалия (скопления органов чувств). Зачем
нужны такие глаза?
680
«Несмотря на сложность строения, глаза [медуз — Ю.Ч], по мнению большин-
ства авторов, служат лишь для различения света и тьмы» (Догель В.А. Зооло-
гия беспозвоночных. Учебник. М., Высшая школа, 1981, с. 131).
Если верить написанному буквально, то дарвинисты признали номо-
генез. глаза развились безотносительно к пользе (ведь свет от тьмы разли-
чают и простые зрительные пятна, обычные у медуз). Это, разумеется, не
так - «большинство авторов» (т.е. дарвинисты) просто об этом не думают.
Наоборот, группа шведских ученых (специалист по эволюции зрения
Дан-Э. Нильсон и др.), пытаясь объяснить феномен в рамках полезности,
полагает, что глаза медуз освещают раннюю стадию эволюции зрения, что
расфокусировка выгодна (чтобы не разглядывать ближайший планктон, а
видеть, например, рачков-копепод, которыми этот вид медуз питается) и что
каждый тип глаз настроен на свою задачу (на какую, не сказано). Это был
бы обычный дарвинизм, если бы не отсутствие ссылок на отбор и не заяв-
ление в конце статьи, что эволюция зрения блочна и дискретна.
Иначе думает швейцарский физиолог Рюдигер Вэнер. В том же номере
«Nature» он поместил заметку «Безмозглые глаза», где пишет:
«Нильсон и коллеги дали нам один из самых ярких примеров того, что
зрение имеет не [один только] общепринятый смысл. Кубомедуза очевидно ...
отдала те задачи, которые обычно осуществляются нервными процессами, оп-
тике глаз», т.е. проводит анализ изображения на уровне сетчатки и передает
результат «прямо на водитель ритма плавательного движения». Она «в ходе
эволюции явно не имела нужды передавать информацию от ... глаз в цен-
тральный процессор, т.е. в мозг» (Wehner R. // Nature, 2005, vol. 435, с. 159).
Тем самым, он видит здесь не раннюю стадию эволюции, а ее итог. Мо-
гу лишь добавить, что этим он признаёт возможность обработки сложной
информации вне нервов (о чем и мы говорили в п. 6-15) и что феномен об-
работки информации в глазу хорошо известен для позвоночных.
На мой взгляд, глаза кубомедуз - пример компенсации (активность уш-
ла не на усложнение тканей, а на создание нового органа), приспособитель-
ный смысл которой можно признать только при допущении нового принципа
работы («безмозглые глаза» по Вэнеру). Сложный же глаз, не осуществ-
ляющий фокусировки, возможен у медуз, полагаю, в силу номогенеза: он
создан по общему образцу и укладывается в рефрен «линзовый глаз». Дан-
ный образец реализуется как при необходимости в фокусированном изобра-
жении, так и без оной. Реализуется просто потому, что таков данный рефрен:
«аналогичные ряды развития представляют головные глаза полихет и моллю-
сков (брюхоногих и головоногих) и многие неголовные глаза сидячих поли-
хет, пластинчатожаберных и брюхоногих» [Беклемишев, т. 2, с. 149].
Другой пример активной компенсации в рамках общего рефрена - об-
щественные насекомые: это целиком отряд термитов и два семейства в
отряде перепончатокрылых (муравьи и некоторые пчёлы). Термиты счита-
ются самыми примитивными по строению тел, зато их социальная органи-
681
зация наиболее развита. Сто лет назад сходство общественной организа-
ции пчёл, муравьёв и термитов настолько поражало учёных, что этих насе-
комых пытались даже объединить в общий отряд, хотя термиты принадле-
жат к низшим насекомым (с неполным превращением), а пчёлы и муравьи -
к высшим. 80 лет назад Метерлинк писал о термите:
«Он способен выжить только лишь в экваториальных районах и, - фатальное
противоречие! - погибает, как только попадает под солнечные лучи. Он сильно
нуждается во влаге и почти всегда вынужден жить в странах, где семь-восемь
месяцев с неба не падает ни капли... Однако он может так же хорошо, как чело-
век, а иногда и лучше него, извлекать пользу из единственного преимущества:
... той маленькой невидимой силы, которую мы называем инстинктом, а у са-
мих себя, неизвестно почему - разумом» [Метерлинк, с. 278-279].
В самом деле, термиты поразительно неадаптивны и в то же время по-
разительно многочисленны, являя собой основу многих тропических экоси-
стем (п. 6-14**). Свою телесную беспомощность они компенсировали не
только огромной плодовитостью (что делают многие, но что в их условиях
не дало бы само по себе успеха), но и удивительной активностью, которая
выражается, в частности, удивительной социальной организацией.
Важно, что эта социальная организация в основных своих чертах по-
вторена у высших насекомых, перепончатокрылых - у муравьёв и, отчасти,
у пчёл (а также, во многом, у одного вида млекопитающих - см. п. 6-5).
В отличие от термитов, перепончатокрылые хорошо адаптированы те-
лесно. Однако они нигде не стали основой экосистем. Условия жизни многих
термитов (высокая влажность, поддерживаемая в гнёздах для предотвра-
щения высыхания их мягких, а иногда и прозрачных тел; питание древеси-
ной) резко отличают их от пчёл и муравьёв, так что обычное бездумное
объяснение сходств их социальных организаций «сходными условиями су-
ществования» ничем не обосновано и, полагаю, просто повторяет идеоло-
гию Геккеля (п. 3-9). Подлинная причина сходств организмов при резко не-
сходных условиях жизни видится мне в том, что активность всего множества
организмов реализуется в рамках достаточно ограниченной диасети.
Удивительно и достойно изучения то, что феномен диасети просматри-
вается не только в формах, но и в поведениях: социальная организация то-
же образует рефрены (например, общественные насекомые). То есть речь
идет о сходстве не условий, а способов существования, задаваемых сход-
ством реализованных клеток диасети. Мы это видели в п. 6-15.
Дополнение 9. Параллель мышления и эволюции. Новизна
К пп. 5-3 и 11-2*. Пока не будет дано содержательное описание эмерд-
жентности, позиция креационистов останется по сути более крепкой и по-
следовательной, чем эволюционная. Наиболее сильна позиция умеренных
креационистов, признающих эволюцию вне рамок эмерджентности, а появ-
ление принципиальных новшеств понимающих как акты творения. Что на
682
это принято возражать? Эмерджентность в эволюции редко становится те-
мой отдельных работ (литературу см. [Чайковский, 1990, с. 227-228]), да и
те обычно не идут далее утверждений вроде такого:
«наделение природы способностью к изобретательской деятельности... не ведет к
необходимости признания существования “жизненных сил” или чего-то подобно-
го, вдруг проявляющего свою активность при переходе одного уровня организа-
ции материи в другой» (Денбиг К. Дж. Случайность и возникновение нового при
естественных процессах//Журнал Всесоюз. химич. общ-ва, 1980, № 3, с. 320).
Почему же английский химик уверен, что «не ведет»? А вот почему:
«Чтобы в природе возникали действительно новые сущности, вполне доста-
точно, чтобы частота случайных событий была минимальной» (там же).
(«Минимальной» - значит: хоть чуть выше нуля.) Обсуждать эту умст-
венную беспомощность нет смысла (тот, кто ее не видит, вряд ли станет чи-
тать данное Дополнение), но за четверть века ситуация не изменилась
(Денбиги господствуют), и от нее приходится отталкиваться, как первый ас-
троном отталкивался от уверений толпы, что звёзды - осколки прежней Лу-
ны, погибшей в новолуние (мягче: что движение светил непознаваемо).
Даже стоя на той (неверной) позиции, что каждое редкое событие когда-
то наверняка произойдет, надо понимать, что в эволюции мы видим боль-
шее: редкие эмерджентные события повторяются и, более того, протекают
сходными путями (что и привело к понятию диасети). Следовательно, у
эмерджентности есть механизм, и искать надо его, а не отговорки.
Ошибки Кеннета Денбига очевидны: он 1) наделил изобретение часто-
той, т.е. уравнял его со случайным событием теории вероятностей. На са-
мом деле изобретение есть акт свободной воли, который (даже если счесть
его случайным) частотой не обладает [Чайковский, 2004, с. 158]; и 2) счел
активность фактором, допускаемым к рассмотрению только в моменты пе-
рехода к высшему уровню, а потому и не увидал его общности.
В рамках статистической ПМ эмерджентная задача не может быть даже
поставлена. Послушаем более сведущих. Композитор и методолог А.А. Коб-
ляков уверен, что прежних ПМ в принципе недостаточно для новой картины
мира; нужна новая ПМ, описывающая акт творчества в научных понятиях:
«Отсутствие ясной и прозрачной модели творчества порождает мифы и спе-
куляции, группирующиеся вокруг Сциллы позитивизма и Харибды креацио-
низма... Построение рациональной модели творческого процесса как новой
познавательной модели является насущной общенаучной и мировоззренче-
ской проблемой» (Кобляков А.А. Синергетика и творчество // Синергетическая
парадигма. М., Прогресс-традиция, 2000, с. 312).
Познавательную модель он понимает вне всякой связи с социальной
проблематикой и для ее построения использует модальную логику (с кван-
тором уместности, отражающим контекст высказывания). То, что он
предложил, можно рассматривать не как ПМ, а лишь как заявку на будущую
ПМ, в которой нам интересны как отдельные соображения:
683
«У творчества и синергетики есть общая основа - гармония как взаимодопол-
нительность конкурирующих процессов»; «взаимодополнительностъ процес-
сов-антагонистов есть главный объединяющий принцип в поведении сложных
систем»; «в неустойчивых нелинейных средах на уровне тонкой структуры
фазовых переходов природа демонстрирует свою, если угодно, психическую
компоненту, способную самоорганизовываться»,
так и общие выводы:
1) «вся история последовательного расширения (обогащения) имеющихся ма-
тематических представлений... может рассматриваться не только гносеологи-
чески (как акт познания), но и онтологически, как акт творения, как акт твор-
ческого созидания нового» (Кобляков А.А., с. 315).
2) «признание творчества в качестве новой парадигмы и модели устранения
противоречий в качестве новой познавательной модели неизбежно влечет за
собой следующую переформулировку закона-вектора творческого процесса, а
именно: любая эволюция направлена в сторону больших размерностей,
больших степеней свободы. Структурное усложнение материи в процессе раз-
вития — результат действия этого закона» (с. 319).
Проще говоря, эволюция понимается Кобляковым как усложнение форм
активности и рост их разнообразия. Как видим, он надеется понять эмерд-
жентность не на пути открытия каких-то новых молекулярных механизмов
(как наивно надеется большинство генетиков), а через переосмысление са-
мой картины мира (о ней см. пп. 5-16, 7-10*). Без этого (добавлю) мы будем
множить «каскады генов» (о которых шла речь в пп. 5-6, 5-15*, 6-10*), не
приближаясь к пониманию самого их появления ни на шаг.
В космологии проблема эмерджентности заставила ввести антропный
принцип (п. 7-1), и нам надо, как всегда в диатропике, провести параллели.
Для этого удобен философский сборник: Причинность и телеономизм в
современной естественнонаучной парадигме. М., Наука, 2002.
В нем философ физики Е.А. Мамчур напомнила о синхронизме истори-
ческих явлений вроде сходных культурных феноменов в удаленных друг от
друга регионах (наиболее известно почти одновременное появление фило-
софии в Греции, Индии и Китае в начале -VI века):
“Юнг предположил, что в данном случае речь должна идти о некоем третьем
типе связи, которая, не будучи каузальной (причинной — Ю. Ч.), тем не менее
не является и случайной, а представляет собой... полную смысла и значения
событийную связь. [...] Какой бы необычный характер эта связь ни имела, она
представляет собой род регулярности” (с. 14).
Этот отмеченный ею род регулярности именуется в диатропике рефре-
ном, и выявление его в контексте связи космологии, квантовой физики, пси-
хологии и социальной истории очень важно как акт признания общности
эволюционных проблем. (Замечу, что непричинная связь легче всего видна
в синхронных явлениях, но непричинность не обязана быть синхронной —•
возможен временной рефрен, см. рис. 58 в п. 8-11**). Эту общность обозна-
684
чил и В.В. Казютинский, добавив в тот же эмерджентный ряд еще биопоэз и
антропогенез. Об антропном принципе он пишет (с. 66):
«принцип несомненно содержит и научное содержание, утверждая, что приро-
да, Вселенная “запрограммирована” на возникновение жизни, человека, наблю-
дателя. Но суть проблемы в том, какова причина этой запрограммированности».
Он назвал три возможности: телеология (у эволюции есть цель), кото-
рая ему неприемлема эмоционально; случайность, которую он не отличает
от вероятности и потому не исследует; и универсальный эволюционизм,
гласящий, что между свойствами Вселенной и появлением человека «есть
глубинные пока не до конца понятые связи». Последний и относится к на-
шей теме. Его смысла Казютинский не раскрыл, но это постарался сделать
в том же сборнике А.Ю. Севальников.
Он обратил внимание на то, что сам по себе антропный принцип не да-
ет понимания биопоэза (если даже принять, что мировые константы подоб-
раны удачно, всё равно биопоэз требует нереального совпадения редких
случайностей — ср. п. 7-12) и что принятие цели тоже ничего не дает: цель
при этом предопределена, тогда как (добавлю) реальная эволюция демон-
стрирует многовариантность. Вывод Севальникова (с. 86):
“Либо... должен меняться тип рациональности... либо необходимым образом
неполна сама наука, поскольку внутри самой себя она не может формализо-
вать, описать некоторые основные, необходимые ей принципы”.
Этих принципов мне видно два: общий - признание активности мате-
рии (о чем у космологов нет, насколько знаю, речи) и частный - признание
того, что эмерджентность есть одна из высших форм активности".
Замечу в связи с этим, что Янч (о нем см. пп. 6-12, 7-6) назвал ту актив-
ность, которая имеет направленность, но не имеет предварительно задан-
ной цели, открытым обучением. Оно ведет к некоторой цели, но к какой,
станет ясно только по ее достижении. Это выше, чем телеология, однако
реальнее, ибо наблюдается при любом творчестве.
Эволюции как творчества сборник не затронул, но творчество там крат-
ко рассмотрено в параграфе моей статьи (Чайковский Ю.В. Случайность,
сложность и творчество, с. 128-131), отчасти проясняющий эмерджентность.
Взято простейшее фракталообразующее правило из книги Пайтгена и Рих-
тера (п. 6-7) и исследована эволюция фрактальных узоров при изменении
параметров этого правила. Оказывается, эволюция тут бывает двух сортов:
чаще всего малое изменение параметра дает малое и предсказуемое изме-
нение узора (это как бы результант по Луэсу), но при выходе параметра за
пределы этой области узор качественно меняет характер (это как бы
" Высшей формой активности можно счесть мышление; оно требует свободы выбора.
Некоторые полагают, развивая Бергсона, что эмерджентность — одна из форм косми-
ческого мышления. Однако неясно, обладает ли она свободой выбора.
685
эмерджент), причем может стать фантастически сложным и столь же кра-
сивым. (Мы это видели в Дополнении 4.) Творчество налицо, но кто его ав-
тор? Ответ оставлен читателю.
Словом, фракталообразующее правило может содержать бесконечное
множество «упакованных смыслов» (с. 128), причем малое изменение пра-
вила (точнее, малое изменение параметра в нем) может приводить к реали-
зации совсем нового смысла; показано (с. 130), чем это полезно для пони-
мания эволюции как хологении (о ней см. п. 4-13).
А именно, поскольку биоэволюция есть последовательность фракталь-
ных ростов (п. 6-7), постольку источник эмерджентов вероятно таит-
ся в смене параметров фракталов, и встает вопрос: каким образом
эмерджентность вписывается в жизнеспособность? То есть: каким образом
принципиально новое оказывается сразу настолько годным для существо-
вания, что может начаться процесс приспосабливания?
Вопрос отчасти проясняет статья биофизика Говарда Патти (США). Он
не касается фракталов, зато отмечает, что онтогенез состоит из двух со-
пряженных процессов регуляции - восходящего и нисходящего (и этим, за-
мечу, идет далее тезиса Икскюля, п. 5-15). Первый это регуляция активно-
сти генов, второй - активности формирующегося организма. Их взаимодей-
ствие он назвал семантическим замыканием, имея в виду тот факт, что ге-
ном сам по себе ничего не строит, что построение организма ведёт «дина-
мический домен конструкций и функций» (с. 147), орудующий веществами,
силами и энергией, тогда как гены орудуют знаками (информацией). Их кон-
такт происходит не на уровне сил и не на уровне знаков, а на уровне смысла
(семантики) получающейся конструкции. Замечу, что речь здесь никак не
может идти о “генах домашнего хозяйства” (т.е. активных во всех клетках) и
что Патти видимо имеет в виду гены вроде гомеозисных.
Эволюция как хологения состоит при этом, на мой взгляд, в последова-
тельных малых изменениях параметров фрактального роста (дающих кон-
струкции эмерджентные и гармоничные, но не приспособленные), корректи-
руемых в сторону приспособления семантическим замыканием в согласии с
тезисом Любищева (п. 4-17*), тезисом Аршавского (п. 6-10*) и в рамках эко-
логической прочности (п. 6-2*). Поскольку возникновение нового управля-
ется не началом процесса, а его концом (п. 9-13), то Бёрджерс предлагал
дополнить принцип причинности идеей концептуальной активности (п. 5-2).
Теперь добавим точку зрения Патти на причинность. Она оригинальна и не-
ожиданна - оказывается, в физике причинность не нужна-.
“Отсутствие... полезности концепции причинности на уровне физических зако-
нов привело к вопросу о том, на каком уровне сложности причинность стано-
вится полезной”. Она полезна “только тогда, когда она ассоциируется с силой и
контролем”; “организмы — тот первый естественный уровень организации, где
понятия функционального контроля и пригнанности к окружению действитель-
но имеют смысл и необходимы для моделирования” (с. 144-145).
686
И, добавлю, как раз на этом уровне причина вовсе не всегда предшест-
вует следствию, ибо часто является целевой причиной по Аристотелю. Это
странно, но - только в рамках физики XIX века и более ранней, которой
привыкли пользоваться биологи. См. Дополнение 6.
Словом, проблема эмерджентности требует нового взгляда на мир, а
он, в свою очередь, должен будет породить новую ПМ. Не имея оснований
для четкой ее формулировки, ограничусь предположением, что она, на мой
взгляд, представит мир и его эволюцию в виде всё более усложняющихся
форм активности.
Дополнение 10. Мифы, Библия и эволюция человека
К п. 9-16*. Леви-Строс утверждал, что инцест у людей полностью за-
прещен во всех первобытных племенах. Возможно, что нынче это и так (хо-
тя неясно, на какой материал он опирался), но его слова разительно проти-
воречат мифам: в тех, что записаны в древности, инцест прямо-таки бьет в
глаза. В мифах боги вступают в него регулярно, а люди редко и в порядке
греха. Нужды в этом у них нет, по крайней мере в тех мифах, где они созда-
ны сразу в виде многочисленной популяции. А такие мифы (впараллель с
мифами, где боги творят одну пару людей), известны для Египта, Вавило-
нии, Финикии, Греции и других стран.
В Библии не так: Бог един и полом не обладает (только носит мужское
имя), а человека он создает «по образу и подобию» себя самого. Что значат
эти слова? Никаких слов о том, что Бог имел тело, сходное с человеческим,
в Ветхом завете нет. Нет, насколько знаю, и в апокрифах (священных тек-
стах, не вошедших в канонический свод Библии).
В выражении «по образу и подобию» принято видеть лишь духовную
сторону, хотя Бог произносит эти слова перед творением тела Адама. О
теле ветхозаветного Бога неизвестно решительно ничего, и остаются толко-
вания. Здесь можно видеть также и намерение создать человечество не как
животных (популяцией), а так же, как созданы языческие боги - от одной па-
ры путем инцеста. В него вступают Адам с Евой, своей собственной частью;
затем - дети, внуки и правнуки Адама и Евы, т.е. и тогда, когда необходимо-
сти в этом уже не было (см. апокриф «Книга юбилеев»); затем - внуки Ноя,
а еще позже - праведник Лот с дочерьми. В каноническом тексте тема инце-
ста донельзя сокращена древними цензорами, но в апокрифах сохранилась
подробно (с женскими именами). Историками прослежено, что эти темы взя-
ты Библией из более древних религий (Гоейвс Р., Патай Р. Иудейские ми-
фы. Книга Бытия. М., БСГ Пресс, 2002).
Вопрос о том, какой из текстов принят впоследствии той или иной рели-
гией за истинный, нашей темы не касается - нам важно лишь, что древних
людей эти темы занимали. Все тексты согласны в одном: потомство Адама
нарушило чистоту породы тем, что женщины стали в массе рождать от «сы-
687
нов Божьих»700. И хотя об их потомстве сказано: «Это сильные, издревле
славные люди» (Бытие, 6, 4), Бог уничтожил всю их цивилизацию путем По-
топа, причем единственным конкретным обвинением была именно порча
породы. Спасение получил Ной, чья порода оставалась чистой, и с его ро-
дом вновь был повторен инцест: все три сына Ноя, давшие три ветви чело-
вечества, имели по одной жене (Бытие, 7, 13) и разошлись в разные страны,
так что их сыновья могли брать в жёны только родных сестёр.
Мне видится правдоподобным допущение, что растление современни-
ков Ноя состояло, по мысли авторов книги Бытия, в связях с преадамитами
или же с потомками Каина (Регельсон П.П., Хварцкия И.И., с. 130-135). От-
мечу, что основная идея данной книги (будто Адам и Ной - абхазы) слаба,
однако книга проясняет избегаемые богословами вопросы о людях до Ада-
ма, об Адаме как итоге божьего внимания (селекции), о городе, основанном
Каином, о причине божьего гнева, вызвавшего Потоп, и другие.
Богословие обходит интересующую нас тему утверждением (идущим от
Филона Александрийского), что Адам - собирательное имя, обозначающее
раннее человечество. Александр Мень, соглашаясь с этим, всё же указал,
что Библия 4 раза упоминает Адама как конкретное лицо (Мень А.В. Исто-
рия религии в семи томах. Т. 1, М., Слово, 1991, с. 148). В апокрифах таких
упоминаний много больше. Именно в них идет речь об Адаме как единст-
венном мужчине, и потому возможность увязать их с идеей Филона видится
мне только в допущении, что этот Адам был вождём, вступавшим, как и его
прямые потомки, в священный инцест (возможно, запрещенный остальным
людям). Тогда яснее становится и история с Лотом.
Ни в каноне, ни в ранних апокрифах Бог не осуждает Лота и дочерей, а
сама Палтит (старшая дочь Лота, устроившая соитие с отцом) произносит
странную фразу: «отец наш стар; и нет человека на земле, который вошел
бы к нам по обычаю всей земли» (Бытие, 19, 31).
Разумеется, в соседнем Сигоре, откуда они ушли, мужчины были707, так
что речь Палтит надо понимать в том смысле, что эти мужчины не соответ- * 101
1,10 Одни богословы утверждают, что то были ангелы, другие это отрицают. Апокриф
«Книга юбилеев» именует их «стражами». В каноне и апокрифах есть несколько ссылок
еще и на каких-то иных людей. Высказывалось обоснованное предположение, что в ран-
них текстах, легших позже в основу Книги Бытия, их творили иные боги. Они тоже упо-
мянуты - в одном апокрифе Бог воюет с ними весь второй день творения (вот почему и в
каноне Бог почти ничего не творит в этот день - цензор вычеркивал, но не вставлял).
Любопытна концепция преадамитов, т.е. допущение наличия людей, сотворенных Бо-
гом до Адама (Регельсон Л.Л., Хварцкия И.И. Земля Адама. Сухум, 1997).
101 Впрочем, есть мнение, что сюжет со спасением семьи Лота — след совсем иного мифа,
мифа об уничтожении всех людей огнем (Хук С. Мифология Ближнего Востока. М., Центр-
полиграф, 2005, с. 130). Тогда инцест Лота с дочерьми прямо задан Богом, а истребление
жены Лота понятно как намерение Бога создать два новых народа только от Лота.
688
ствуют «обычаю всей земли», а отец (патриарх Лот) соответствует, как со-
ответствовал патриарх Адам , но стар для поиска невесты.
Этот-то обычай нам и важен. По всей видимости, он был доиудейской
практикой, возможно, пришедшей в Палестину из Двуречья. Выявлять его
следы здесь нет места (нужен долгий экскурс в сравнительную мифологию),
и укажу лишь отправную и конечную точки.
Священный характер брака с дочерьми заявлен в самой древней из-
вестной письменности - у шумеров. В их очень раннем мифе бог Энки, вла-
дыка земли и вод, вступает в брак с Нинхурсаг, мировой матерью,
«благодаря чему рождается три последовательно производящих друг друга
поколений богинь, каждая из которых зачинается Энки и... основательницей
рода выступает Н.» (Мифологич. Словарь. М.. Сов. Энц., 1990, с. 393).
Любопытно, что третий акт этого троекратного инцеста подан в древ-
нейшем мифе именно как брак (свадебные подарки - Хук С. Мифология
Ближнего Востока. М., 2005, с. 29), а не как соблазнение или изнасилование
(что видим в более поздних мифах иных стран). Тема брака бога с дочерью
на Ближнем Востоке была весьма обычна (там же, с. 30).
Если говорить о людях, то брак с сестрами и (реже) с дочерьми извес-
тен у древних царей (Египет, Персия), где носил священный характер.
След того, что брак с дочерьми допускали и предки евреев, в Ветхом
завете чудом сохранился: Пятикнижие трижды перечисляет всех родствен-
ниц и свойственниц, с которыми запрещено соитие (Левит, 18, 7-19; Левит,
20, 11-21; Второзаконие, 27, 10-23), и дочь во всех трех списках отсутствует.
Отсутствие “в силу очевидности” исключено, поскольку упомянуты мать, се-
стра и внучка. На дремучую древность списков указывает отсутствие поня-
тия «родственница». Позже, когда понятие появилось (Левит, 18, 6), все три
старых списка оказались излишни, но сохранились как священные.
Мое мнение таково: люди очень рано (не позже неолита) заметили осо-
бую роль инцеста - рождение самых различных по своим качествам детей.
(Мифы рисуют как их особые достоинства, так и уродства.) Это наблюдение
было использовано - наверняка при селекции домашних животных и, пола-
гаю, при создании человеческих элит.
Если для нас повышение шанса рождения ребенка с тяжелым изъяном
делает идею инцеста неприемлемой, то в древности сама идея шанса от-
сутствовала, как отсутствовало и стремление спасти каждого младенца. Бо-
лее того, высокая смертность младенцев считалась естественной.
Поэтому стоит отметить, что при детской смертности намного выше
50% (а при инцесте было заведомо так) процент выдающихся личностей
среди выживших должен быть в таких линиях заметно выше, чем у всего
населения. На успешные итоги долгого инцеста (таковы царицы Хатшепсут
и Клеопатра, царевна Арсиноя) указывают нынешние учебники генетики.
Разумеется, древние племена не вели подсчетов, однако был один фе-
номен, бьющий при инцесте в глаза - дети элит. Мифолог Роберт Грейвс,
689
искавший в мифах разных стран древнейшие пласты, полагал, что брак во-
ждя с дочерью мог стать регулярным тогда, когда брак с новой царицей (а
ею чаще всего становилась дочь прежней) стал давать ему возможность из-
бежать казни, прежде неизбежной (Грейвс Р. Мифы древней Греции. М.,
Прогресс, 1992, с. 11). Если это в самом деле было так, то итог регулярного
инцеста, каков бы он ни был, становился виден всем.
Построение Грейвса может быть ошибочно, но какие-то обычаи, свя-
занные с инцестом, явно были у многих народов - это отразилось в мифах.
На мой взгляд, инцест был компонентом двускоростного механизма эволю-
ции, но не всех людей, а элит. Могло ли это повлиять на эволюцию вида
людей в целом? Да, если элиты человеческих обществ играли тогда роль,
близкую к роли элитных пород в нынешней селекции. Тогда становятся по-
нятны как весьма странные брачные обычаи некоторых племён (о них см.:
Чайковский Ю.В. Двускоростной механизм... И Эволюция, 2004, № 2), так и
загадочные фразы у Платона и у Леви-Строса.
Почему обычай рано исчез у всех или почти всех племен, непонятно, но
удивительно, что запрета на инцест строго держатся даже те племена, где
роль отца в зачатии отрицается702. Очевидно, причина запрета была, если
говорить о сознании самих племён, религиозно-этической. Если же говорить
о нашем ее понимании, то, вернее всего, имел место акт системогенеза,
прекративший двускоростную эволюцию людей, которая, вернее всего, без
инцеста вообще не идёт (как не идет никакая селекция). Само двускорост-
ное размножение продолжалось и впоследствии, но уже не было эволюци-
онным феноменом. В наше время и оно почти закончилось, однако в неко-
торых малых обществах его еще, полагаю, можно изучать.
Так или иначе, идея происхождения людей от одной особи-основателя
сыграла решающую роль в становлении некоторых биологических принци-
пов, до сих пор определяя, например, облик дарвинизма и систематики (п.
10-1**). Поэтому полезно помнить, что лежащая в основе филогенетики ре-
лигиозная идея (единый предок) не только не имеет биологической основы
(ведь люди не произошли ни от одной особи, ни от одной пары), но и в са-
мой религии по-видимому исходит из давно забытой идеи селекции элит.
102 Воззрение выглядит для нас нелепо, однако туземными мыслителями оно вполне
разработано логически и для критики со стороны европейцев совершенно беспробойно.
Оно соответствует и уверенности мыслителей этих племён (замечу: фактически такой
же, как и у мыслителей-правоведов развитых обществ прошлого), что задача женщин -
размножение, а мужчин - всё остальное; и практике племён, где девочки почти пого-
ловно вступают в половую жизнь до начала месячных, а потому не беременеют лишь
“дефектные” особи (Малиновский Бр. Избранное. Пер. с англ.; М., Росспэн, 2004). До-
воды туземных мыслителей стоит почитать - они удивительно напоминают доводы
наших дарвинистов в пользу роли отбора, и для каждого исключения придумывается,
как и в дарвинизме, особое объяснение, иногда остроумное.
690
Рекомендуемая литература
ОБЩАЯ
Александров В.Я. Поведение клеток и внутриклеточ. структур. М., Знание, 1975.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. Том 1. Про-
морфология; том 2. Органология. М., Наука, 1964.
БергЛ.С.. Труды по теории эволюции (1922-1930). Л., Наука, 1977.
Бергсон /(.Творческая эволюция (1907). М., Канон Пресс, 1998.
Вавилов Н.И. Закон гомологических рядов... Л., Наука, 1987.
Воейков В.Л. Научные основы новой биологической парадигмы // От эффекта
Кирлиан к биоэлектрографии. СПб., 1998.
Галимов Э.М. Феномен жизни: между равновесием и нелинейностью. Происхож-
дение и принципы эволюции. М., УРСС, 2001.
Гринченко С.Н. Системная память живого. М., ИЛИ РАН, 2004.
Гродницкий Д.Л. Две теории биологической эволюции. Красноярск, 2000.
Зусмановский АГ. Биоинформация и эволюция. Правы и Ламарк, и Дарвин. Уль-
яновск, 2003.
Кудрин Б.И. (ред.) Теория эволюции: наука или идеология? М. - Абакан, ЦСИ,
1998. [Любищевские чтения в Москве].
Лима-де-Фариа А. Эволюция без отбора. Автоэволюция формы и функции. М.,
Мир, 1991.
Любищев А.А. О природе наследственных факторов. Критич. исследование [1925]
/ Примечания и послесловие Ю.В. Чайковского; послесловие М.Д. Голубовского.
Ульяновск, УГПУ, 2004.
Любищев А.А. Проблемы формы, систематики и эволюции организмов. М., Наука,
1982.
Марков А.В., Куликов А.М. Историческое развитие систем распознавания “своего”
и “чужого”... // УСБ, 2006, № 2. [Новая эволюционная иммунология.]
Материалы симпозиума, посвященного памяти С.В. Мейена. М., ГЕОС, 2001.
Метерлинк М. Тайная жизнь термитов [1926]. М., ЭКСМО-Пресс, 2002.
Назаров В.И. Эволюция не по Дарвину. М., УРСС, 2005.
Несис КН. Головоногие: умные и стремительные. М., Октопус, 2005.
Попов И.Ю. Ортогенез против дарвинизма. Историко-научный анализ концепций
направленной эволюции. СПб., Изд. СПбГУ, 2005.
Расницын А.П. Процесс эволюции и методология систематики. СПб., 2002 (Труды
Русского энтомологии, общества, т. 73).
Чайковский Ю.В. Элементы эволюционной диатропики. М., Наука, 1990.
Чайковский Ю.В. К общей теории эволюции // Путь. Философ, ж-л, 1993, № 4.
Чайковский Ю. Ламаркизм умер. Да здравствует ламаркизм И ВИЕТ, 2002, № 3.
Чайковский Ю.В. О природе случайности. 2-е изд. М., ЦСИ, 2004.
Эволюционная биология: история и теория / ред. Э.И. Колчинский. СПб., 1999.
Юнкер Р., Шерер 3. История происхождения и развития жизни. Осн. положения и
понятия для уроков биологии. СПб., Кайрос, 1997. [Умеренный креационизм.]
691
К ГЛАВАМ 1 и 2
Амлинский И..Е. Жоффруа Сент-Ил ер и его борьба против Кювье. М., Изд. АН
СССР, 1955.
Баранов П.А. История эмбриологии растений. М.-Л., Изд-во АН СССР, 1955.
Канаев И.И. Очерки из истории сравнительной биологии до Дарвина. М.-Л., Изд.
АН СССР, 1963.
Лебедев А..В. (составитель и переводчик) Фрагменты ранних греческих филосо-
фов. М., Наука, 1989.
Лункевич В.В. От Гераклита до Дарвина. Очерки по истории биологии. М., Учпед-
гиз, 1960, тт. 1, 2.
Нидхэм Дж. История эмбриологии. М., Изд. иностр, лит., 1947.
Райков Б.Е. Русские биологи-эволюционисты до Дарвина. Тт. 1-4. М.-Л., Изд. АН
СССР, 1952-1959.
Райков Б.Е. Германские биологи-эволюционисты до Дарвина. Л., Наука, 1969.
Рассел Б. История западной философии. М., ИЛ, 1959 [есть новые издания].
Столяров А.А. (составитель и переводчик) Фрагменты ранних стоиков. Т. 1. М.,
Греко-латинский кабинет Ю.А. Шичалина, 1998.
Чайковский Ю.В. Первые шаги дарвинизма в России // Историко-биологич. ис-
следования, вып. 10. М., Наука, 1989.
Чайковский Ю.В. Избегание предтеч // ВФ, 2000, № 10.
К ГЛАВАМ 3 и 4
БабковВ.В. Московская школа эволюционной генетики. М., Наука, 1985.
Воронцов Н.Н. Развитие эволюционных идей в биологии. М., МГУ, 1999.
Гиляров А.М. Виды сосуществуют в одной экологической нише // Природа, 2002,
№ 11.
Канаев И.И. Очерки из истории проблемы морфологического типа от Дарвина до
наших дней. Л., Наука, 1966.
Колчинский Э.И Неокатастрофизм и селекционизм: вечная дилемма или возмож-
ность синтеза? СПб., Наука, 2002.
Любищев А.А. К классификации эволюционных теорий // Проблемы эволюции.
Том 4. Новосибирск, Наука, 1975.
Любищев А.А. О монополии Т.Д. Лысенко в биологии. / Предисл. А.Н. Марасова.
М., Памятники исторической мысли, 2006.
Любищев — Гурвич. Диалог о биополе. Ульяновск, 1998. [Содержание книжки
много шире заглавия, данного составителями].
Мейен С.В. Принцип сочувствия И Пути в незнаемое. № 13. М., Совет, писатель, 1977.
Мейен С.В., Соколов Б.С., Шрейдер Ю.А. Классическая и неклассическая биоло-
гия. Феномен Любищева // Вестник АН СССР, 1977, № 10.
Назаров В.И. Эволюционная теория во Франции после Дарвина. М., Наука, 1974.
Панов Е.Н. Бегство от одиночества. М., Лазурь, 2001.
Системность и эволюция. М., Наука, 1984. [Материалы Любиш. чтений в МОИП].
Титов Ю.В. Эффект группы у растений. Л., Наука, 1978.
692
Филипченко Ю.А. Эволюционная идея в биологии [1927]. М., Наука, 1977.
Чайковский Ю.В. О формировании концепции Ч. Дарвина (опыт междисциплин,
подхода) // Науки в их взаимосвязи. История, теория, практика. М., Наука, 1988.
К ГЛАВАМ 5 и 6
Агаев М.Г. Экспериментальная эволюция. Л., Изд. ЛГУ, 1978.
Албертс Б., БрейД., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекуляр-
ная биология клетки. В трех томах. М., Мир, 1994.
Аршавский И. А. Физиологические механизмы и закономерности индивидуального
развития. М., Наука, 1982.
Белоусов Л.В. Основы общей эмбриологии. М., Изд. МГУ, 2005.
Геном, клонирование... / ред. Л.И. Корочкин. Фрязино (Моск, обл.), 2004.
Гилберт С. Биология развития. В трех томах. М., Мир, 1995.
Гилберт С.Ф., Опиц Д.М., Рэф Р.А. Новый синтез эволюционной биологии и био-
логии развития // Онтогенез, 1997, № 5.
Голдбергер Э.Л., Ригни ДР., Уэст Б.Дж. Хаос и фракталы в физиологии челове-
ка // ВМН, 1990, № 4.
Катастрофы и история Земли. М., Мир, 1986.
Кауфман С.А. Антихаос и приспособление // ВМН, 1991, № 10.
Корочкин Л.И. Онтогенез, эволюция и гены // Природа, 2002, № 7. [О том же см.:
Корочкин Л.И. // ЭГ, 727.]
Лебедев К.А., Понякина ИД. Новый этап развития иммунологии // Природа, 2006,
№4.
Маленков Г.Г, Чайковский Ю.В. Выживание мутантного клона. 3. Катастрофиче-
ский отбор //Генетика, 1979, № 10.
Маресин В.М. Пространственная организация эмбриогенеза. М., Наука, 1990.
Мейен С.В. Основные аспекты типологии организмов //ЖОБ, 1978, № 4.
О’Нилл Л. Врожденный иммунитет // ВМН, 2005, № 4.
Проблемы и перспективы современ. иммунологии. Новосибирск, Наука, 1988.
Происхождение и развитие половых клеток... Пер. с франц. Л., Медицина, 1968.
Пустилъник С.Н. Принцип подбора как основа тектологии А. Богданова // ВФ,
1995, №8.
Румянцев С.Н. Конституциональный иммунитет и его молекулярно-экологичес-
кие основы. Л., Наука, 1983.
Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция. М., Мир, 1986.
Стил Э., Линдли Р., Бландэн Р. Что, если Ламарк прав? М., Мир, 2002.
Том Р. Структурная устойчивость и морфогенез [1972]. М., Логос, 2002.
Чайковский Ю.В. Проблема наследования и генетический поиск // Теоретич. и
экспериментальная, биофизика. Вып. 6. Калининград, 1976.
Чайковский Ю.В. Почему лиса рыжая // Пути в незнаемое. Сб. 17. М., Советский
писатель, 1983.
Чайковский Ю.В. Междисциплинарность современного эволюционизма // Кон-
цепция самоорганизации в исторической ретроспективе. М., Наука, 1994.
693
Чайковский Ю.В. Иммуногенетика и эволюция: не впасть бы в другую крайность
// ВРАН, 2003, № 3. [О книге: Стил Э. и др. Что, если Ламарк прав?]
Чайковский Ю.В. Диатропика Мейена: сегодняшний взгляд // ВФ, 2006, № 5.
К ГЛАВЕ 7
Бернал Дж. Проблема стадий в биопоэзе // Возникновение жизни на Земле. Тру-
ды междунар. симпозиума. М., Изд. АН СССР, 1959, с. 42-58.
Воейков В.Л. Вода с активным кислородом - вода жизни // Дельфис (журнал, Мо-
сква), 2005, № № 1,2.
Галимов Э.М. Предпосылки и условия возникновения жизни И Геохимия, 2005, N 5.
Кузин А.М. Роль природного радиоактивного фона и вторичного биогенного из-
лучения в явлении жизни. М., Наука, 2002.
ЛобъеЛ. Оазисы на дне океана. Л., Гидрометео, 1990.
Пармон В.Н. Пребиотическая фаза зарождения жизни // ВРАН, 2002, №11.
Резанов ИА. Жизнь и космические катастрофы. М., Агар, 2003. [Также см.:
ВРАН, 2001, № 4; ВРАН, 2002, № 7.]
Руттен М. Происхождение жизни (естественным путем). М., Мир, 1973.
Самуилов В.Д. Проблемы энергетики в эволюции живого // Биохимия, 2005, № 2.
Спирин А. С. Биосинтез белков, мир РНК и происхождение жизни // ВРАН, 2001, № 4.
Спирин А. С. Рибонуклеиновые кислоты как центральное звено живой материи //
ВРАН, 2003, № 2.
Спирин А.С. Происхождение, возможные формы существования и размеры перво-
зданных особей // ПЖ, 2005, № 4.
К ГЛАВАМ 8 и 9
Алексеев А.С, Дмитриев В.Ю., Пономаренко А.Г. Эволюция таксономического
разнообразия // Экосистемные перестройки и эволюция биосферы. Вып. 5. М.,
ГЕОС, 2001.
Алексеева Л.В. Полицикличность размножения у приматов и антропогенез. М.,
МОИП, Наука, 1977.
Бородин П.М. и различ. соавторы [Статьи по эволюционной цитогенетике грызу-
нов] // Природа, 2001, № 1; 2002, № 9; 2005, № 2.
Еськов К.Ю. История Земли и жизни на ней. Учебное пособие для старших клас-
сов. М., Мирос-МАИК-Наука, 2000.
Заварзин Г.А. Эволюция биосферы с точки зрения геологов и биологов // ВРАН,
2006, № 2.
Ивахненко М.Ф., КорабельниковВ.А. Живое прошлое Земли. М., Просвещение, 1987.
Кондратьев К.Я., Федченко П.П. Влияние спектра солнечной радиации на эволю-
цию биосферы // ВРАН, 2005, № 6.
Кэрролл Р. Палеонтология и эволюция позвоночных. М., Мир, 1993, тт. 1-3.
Марков А.В., Захаров И.А. Половое размножение насекомых регулируется пара-
зитическими бактериями // Онтогенез, 2005, № 4.
Марков А. В., Куликов А.М. Происхождение эвкариот... //ПЖ, 2005, № 4.
694
Мейен С. В. Следы трав индейских. М., Мысль, 1981.
Мейен С.В. Флорогенез и эволюция растений И Природа, 1986, №11.
Пономаренко А.Г. Основные события в эволюции биосферы // Проблемы доан-
тропогенной эволюции биосферы. М., Наука, 1993.
Прозоров А.А. Горизонтальный перенос генов у бактерий // УСБ, 2000, № 6.
Райков И.Б. Пути эволюции митотического аппарата... // Труды Зоология, инсти-
тута, т. 144. Л., Наука, 1986, с. 26-56.
Ромер А., Парсонс Т. Анатомия позвоночных. М., Мир, 1992, тт. 1-2.
Сахарова Н.Ю. Млекопитающие: эмбрион в личинке //Природа, 2004, № 5.
Тейяр де Шарден 77. Феномен человека. М., Наука, 1987.
Уголев А.М. Эволюция пищеварения и принципы эволюции функций. Л., Наука, 1985.
Уорд П. Океан - убийца // ВМН, 2006, № 12.
Хайн В.Е. Основные проблемы современной геологии. М., Наука, 1994.
Хайн В.Е. О причинах великих вымираний... // Природа, 2004, № 6.
Эволюция флор в палеозое. М., ГЕОС, 2003 (Труды ГИН, вып. 556). [С разделом
об истории рождения эволюционной диатропики.]
К ГЛАВЕ 10
Джеффри Ч. Биологическая номенклатура. М., Мир, 1980.
Заварзин ГА. Рецензия на неизданную книгу [Мейена] // Природа, 1990, № 4.
Клюге Н.Ю. Современная систематика насекомых... СПб., “Лань”, 2000.
Корона В.В. Основы структурного анализа в морфологии растений. Свердловск,
СГУ, 1987.
Леонтьев Д.В., Акулов А.Ю. Экоморфема органического мира//ЖОБ, 2004, № 6.
Любищев А. А. К логике систематики // Проблемы эволюции. Т. 2. Новосибирск, 1972.
Медников Б.М. Геносистематика: проблемы и трудности // БШ, 2000, № 1.
Мейен С.В. Эволюция и систематика высших растений по данным палеоботаники.
/ Отв. ред. А.В. Гоманьков. М., Наука, 1992.
Михайлов КГ. Типология и практическая систематика // Теоретич. проблемы эко-
логии и эволюции (Вторые Любищевские чтения). Тольятти, 1995, с. 40-46.
Павлов В.Я. Периодическая система членистых. М., ВНИРО, 2000.
Чайковский Ю.В. Что такое царства живого мира? // БШ, 1998, № 4.
Чайковский Ю.В. Макросистема и эволюция // БШ, 1999, № 4.
Чайковский Ю.В. Диатропика Мейена: сегодняшний взгляд // ВФ, 2006, № 5.
Чайковский Ю.В. 1. Из истории систематики. 2. Сколько царств в живой природе
// Биология для школьников, 2006а, № 2.
Шаталкин А.И. Категория "царства" в системе организмов // УСБ, 1996, № 1.
Шипунов А.Б. Сколько царств живых организмов? // БШ, 1997, № 4.
К ГЛАВАМ 11 и 12
В помощь аспиранту. Существует ли естественный отбор? (Материалы круглого
стола) // Высшее образование в России, 2006, №№ 7,8.
ГлэдДж. Будущая эволюция человека. Евгеника XXI века. М., Захаров, 2005.
695
Евгеника в дискурсе глобальных проблем современности. М., Канон+, 2005.
Жерихин В.В. Избранные труды по палеоэкологии и филоценогенетике. М.. Това-
рищество КМК, 2003.
Красный Л.И. Разномасштабная делимость // ВРАН, 2002, № 6.
Свердлов Е.Д. Биологический редукционизм уходит? Что дальше? // ВРАН, 2006, N 8.
Трепет С.А. Горный зубр // Природа, 2005, № 7.
Чуриков Н.А. Молекулярные механизмы эпигенетики // Биохимия, 2005, № 4.
Элтон Ч. Экология нашествий животных и растений. М., Изд. ин. лит., 1960.
Сокращения
АФК - активные формы кислорода; БЖ - Ботанич. журнал; БПС - Биология
(еженед. газета издательства “Первое сентября” для учителей); БШ - Биология в
школе (журнал); ВИЕТ - Вопросы истории естествознания и техники (журнал);
ВМН - В мире науки (русский вариант журнала “Scientific American”); ВРАН -
Вестник Российской Академии наук (журнал); ВФ - Вопросы философии (журнал);
ГИН - Геологич. институт РАН (Москва); ГЦК - гель-ценотическая концепция
происхождения жизни; диасеть - диатропическая сеть (рефренная структура разно-
образия); ДС - диссипативная структура; ЖОБ - Журнал общей биологии; ЗЖ -
Зоологич. журнал; ЛЧ - Любищевские чтения (ежегодник, Ульяновск); МБ - Мо-
лекулярная биология (журнал); М/Д - маастрихт/даний (грань мезозойской и кай-
нозойской эр); МОИП - Московское общество испытателей природы; МС - Мате-
риалы симпозиума в ГИН (сборник, см. общую рекомендуемую литературу); НПС
- наследование приобретенных свойств; ПЖ - Палеонтологии, журнал; ПМ - по-
знавательная модель; ППК - первичная половая клетка; СК - стволовая клетка;
СТЭ - синтетич. теория эволюции (самоназвание популяционного неодарвинизма с
1940-х гг.); ТПЭ-3 - Теоретич. проблемы экологии и эволюции (Третьи Любищев-
ские чтения). Тольятти, 2000; транпол - транзитивный полиморфизм; УСБ - Успе-
хи современной биологии (журнал); ФР - Физиология растений (журнал); ЦСИ -
Центр системных исследований (издательское бюро Б.И. Кудрина, Москва); ЧЭ -
Чайковский Ю.В. Эволюция. Книга для изучающих и преподающих биологию. М.,
ЦСИ., 2003; ЭБ - Эволюционная биология (сборник, см. общую рекомендуемую
литературу); ЭГ - Эволюционная геномика (два спец, выпуска журнала “Генетика”:
2002, № 6, с. 709-864 и № 8, с. 1013-1168); ЭнО - Энтомологии, обозрение (жур-
нал); ЮШ - Юнкер, Шерер (учебник, см. общую рекомендуемую литературу).
АЕ - Nott ale L., Chaline J., Grou P. Les arbres de revolution. Paris, Hachette, 2000;
BJ - Biological Journal of the Linnean Society. A Journal of Evolution (Англия); BR -
Biological Reviews. Cambridge Philosophical Society (журнал, Англия); CR - Comptes
Rendus (первые слова заглавий журналов Парижской Академии наук); LEG -
L’Evolution // Geochronique, 2002, № 84 (специальный выпуск журнала, Франция);
LR - La Recherche (журнал, Франция); PNAS - Proceedings of the National Academy
of Sciences (журнал, США); ТВ-система - приобретенный специфический иммуни-
тет теплокровных, осуществляемый Т-клетками (от лат. timus) и В-клетками (от лат.
bursa Fabriz.il).
696
Указатель имён
Соавторы указаны лишь при необходимости
Абеляр (Abaillard Р.) 40
Августин 39,44,206,216,441,668
Агаев М.Г. 254,256,634-635,637
Агассиц (Agassiz J.-L.) 93,317,319,624
Агафонов Б.П. 255
Адам (миф.) 22,46,686,688
Адансон (Adanson М.) 569-570
Акифьев А.П. 241
Албертс (Alberts В.) 286,288,330
Алеев Ю.Г. 601
Александров ВЛ. 226,244,288,330,387
Алексеев А.С. 518.521,525
Алексеева Л.В. 549-550
Алексий II17
Алешин В.В. 375,582
Алкмеон 28,29
Аллен (Allen J.) 400,411
Алтухов Ю.П. 325-326
Альберт Великий 40,42,43.46. 54,234
Альбрехт-Бюлер (Albrecht-Bueler G.) 388
Альварес (Alvarez L.) 518
Альдрованди (Aldrovandi U.) 41
Альт (Alt F.M.) 247-248,251
Альтштейн А. Д. 423
Амалицкий В.П. 145,146,176
Амлинский И.Е. 79-82,
Анаксагор 25,29,30,35,37,38
Анаксимандр 25-27,29, 34, 36,50,56,75,
294, 396
Андрианов Б.В. 245,276
Анохин И. А. 386
Анохин П.К. 237,380
Аристотель 32-36,38,39,45,50,69,78,
118.122.143,147,169,200,207,224,
316-317,329,588,676
- компенсации пр-п 80,446,475
- о побочном и существенном 34
- о причинах 32,228,397,675,686
Арманд А. Д. 631
Аронова Е.А. 236.268
Арсиноя 688
Артемьева Е.А. 390,670-671,676
Артур (Arthur W.B.) 136
Аршавский И.А. 106.148,226,262-266,
273-275,283-285,293,298,357-359,370,
377,382-383,502,549.554,616,633
- доминанта и стресс 263.283
АстауровБЛ. 159,161
Ахметъев М.А. 508,522,528
Бабков В.В. 151-155,157-158.161-163,367
Бабынин Э.В. 275
Базыкин Г.А. 12
Бак (Back Р.) 524
Балкашина Е.И. 159-161,165,264,285,381
Банковский Б.Е. 324
Балтимор (Baltimore D.) 247-248,251
Баранов О.К. 335
Баранов П.А. 28,73, 85
Баранцев Р.Г. 207,229
Баскервиль (Baskerville Т.) 578
Баулер (Bowler Р.) 60,64,65
Бауэр Э.С. (Bauer Е.) 165,199,275,340,
378,427
БедаН.В. 370
Беклемишев В.Н. 101, 126,198,299,306,
387.460,531,570,680
Беликов В.В. 85
Белобородов Г.С. 266
Белон (Belon Р.) 50-51,57
Белоусов Л.В. 284,297-299
Беляев ДК. 210
Беляев Н.К. 162
Беннет (Bennett А.) 114,129,150
БергЛ.С. 38,169-173,176,204,219,227,
229,242,334,360,391.438,505,512,540
Берг Р. Л. 149,189,671
697
Бергсон (Bergson Н.) 145,227,229,291,308,
337,349,359,392,427,475,512-514,557,684
Бёрджерс (Burgers J.M.) 228,368,381,
675,685
Бердников В.А. 210,219,279
Бердяев Н.А. 640
Бернал (Bernal J.D.) 202,406-408,410,
419,430,436
Бернар (Bernard С.) 331
Бернет (Burnet F.M.) 332,341,639
Бернет (Burnet Т.) 50,411
Берталанфи (Bertalanffy L.) 296,352,367
Берцова Ю.В. 331
Бибиков П. А. 101
Блэнвиль (Blainville H.-M.D.) 78
Блюменбах (Blumenbach J.) 67,68,144
Бовери (Boveri Т.) 197
Бог (Демиург, Творец и т.п.) 19,21,24, 26,
31,39,49,68,441,472,527-528,686-687
Богданов А.А. 365-370, 381, 389,391,
394,397,426,440,539,610,629,632,
636,650
Боги греч. старшие 20,23 (также Гея)
Боголюбский С.Н. 210,279,645
Болдуин (Baldwin J.M.) 186,187,233,
240,674
Бор (Bohr N.) 56,579,622
Бородин П.М. 469.485, 536.634
Бородина Н.А. 324
Брайен (Brian M.V.) 384
Брежнев Л.И. 136,655-656
Бриттен (Britten R.I.) 298
Брокки (Brocchi G.) 59, 89,378,461,529
Брони (Bronn H.G.) 90-93,96,98,103,105,
108,110, 124,125,308,438,489,632
Брюкке (Brucke Е.) 143
Буавэн (Boivin А.) 190,236
Будаговский А.В. 430,657
Бульенков Н.А. 370,425-426,432,665
Бурзин М.Б. 491,500
Буридан (Buridan J.) 218
Бурлакова Е.Б. 658
Бутео (Buteo J.) 42,43,66
Бучаченко АЛ. 432
Бэйтс (Bates H.W.) 119-120
Бэкон (Bacon F.) 43,46,525
Бэкон (Bacon R.) 40,56
Бэр К.М. (Baer К.Е.) 72,88,115,116,125,
141,170,203,229,481,569
- о зародышевом сходстве 75,182,317,
531,624
- о прогрессе 90,438
- прочности вида 306,310-311,513
- о царстве человека 603
- о ядре и периферии 587,593
Бэтсон (Bateson W.) 160,171,204,285,
310,312
Бюргин (Burgin Т.) 309
Бюффон (Buffon G.L.) 44,56,64,68,292,
372,476
Бяллович Ю.П. 192
Вавилов Н.П. 167- 169, 173.190.210.
220,313,324,361,376,431
- о мимикрии 390
- о систематике 570,577, 583
Валентайн (Valentine J.W.) 501
Вальт (Реммель) (Valt М.)
Ванини (Vanini L.) 43,45
Ванюшин Б.В. 420
Вахрушев А.А. 380
Вегенер (Wegener А.) 525-526
Вёзе (WoeseC.R.) 581
Вейсман (Weismann А.) 84,129-132,137,
141,186,266-267,270,558,622
Вентребер (WintrebertP.) 184, 196,235-
239,242,312,341,357,394
Вересаев В. 20
Вермель Е.М. 73,352
Вернадский В.Н. 71,72, 180, 181,307,
360,406,408-409,460-461,490,492,502,
580,618,649
Вертьянов С.Ю. 17
Веселова Т.В. 394,502
Виганд (Wigand А.) 108,119,142,143,253
698
Вигзел (Wigzell Н.) 251-252, 302
Виллис (Willis J.C.) 159,325,363-364,382,
461,496,571-572,576,623,626
Виллис (Willis Т.) 52,77
Вильберфорс (Wilberforce S.) Ill
Винер (Wiener N.) 619
Винни-Пух (лит.) 402
Витгенштейн (Wittgenstein L.) 123,129,
216,563,570
Вичлер (Wichler G.) 42-44
Воейков ВЛ. 101, 198,199,225,330,
338,396,403-405,417,427-430,432,434-
435,494,618,657
Вольф (Wolf C.F.) 67,73, 161,268
Вольф (Wolf E.) 270
Воробьева Э.И. 315.480,668
Воронцов Н.Н. 140, 190, 567,646
By(WuT.T.) 258
Высоцкий В.С. 221
Вэнер (Wehner R.) 680
Гаврилов Л. А. 462
Галилей 588
Галимов Э.М. 241,280,282,345,398,
417-418,434,446
Гальтон (Gallon F.) 115,155
Гаркави Л.Х, 264
Гаряев П.П, 241,434,657,677
ГаузеГ.Ф. 107
Гвоздев В.А. 340,352
Гёбель (Goebel К.) 126
Гегель (Hegel G.W.F.) 74,119,201,395,
472,628
Гёдель (Goedel К.) 216, 396
Геер (Heer О.) 92,93,177,440,497
Геккель (Haeckel Е.) 124-127, 129,134,
157,161.163, 165,170,181,182,253,
329.341,355,438,562-563,569,576,611
- система царств 579
-триада 125
Гексли (Huxley Т.) 96,109,
Гельфанд М.С. 102
Геодакян В.А. 535,559,646
Гептадор 25,36
Гераклит 28,215
Гердер (Herder J.) 68,70-72,74,100,319,380
Герман А.Б. 536
Герман (Hermann J.R.) 145
Гершель (Herschel F.W.) 293-294
Гершензон С,М. 14,158, 164,223,236,
267
Гесиод 20,23,26,29,227,231
Гёте (Goethe J.W.) 52,57,68,124,202,
319,360-361,372
Гёттон (Hutton J.) 306
Гея (миф.) 20, 306,389, 399,401,409-
410,521,527,550,627.649
Гиббс (Gibbs W.W.) 333
Гизигер (GisigerT.) 347, 364. 380,521,
524
Гийено(Оиуепо1Е.) 171,173
Гилберт (Gilbert S.F.) 222,269.270-272,
285,287,289,291,299-300,334, 354. 380-
381,558
Гиляров А.М. 107
Гиляров М.С. 206,384,504
Г иппократ 35
Гис (His W.) 355
Глазко В.И. 370
Глэд (Glad. J.) 638,648-649
Голдбергер (Goldberger A.I.) 348,380
Голубовский М.Д. 130,138,223-224,
239,244,276,356,358,383,498
Гольдфус (Goldluss G.A.) 578
Гольдшмидт (Goldschmidt R.) 154,161,
184.227,243,244,255,312, 325
Гоманьков А.В. 16,23,319
Гомер 26,606
Горовиц (Horowitz N.H.) 401,406,427
ГорькийА.М. 155
Грассэ (Grasse Р.-Р.) 191,238-239, 357
Грейвс (Graves R.) 21,22,686,688-689
Гречко В.В. 576
Григорий Нисский 38.39, 53,160,292,
358, 619,645, 668
699
Григорович Н.А. 552
Гринченко С.Н. 240,252,275,312,349-350
Гриффит (Griffith F.) 164,267,275
Гродницкий Д.Л. 66,140.215,291,357,
381,645
Гужов А.В. 12
Гук (Hooke R.) 48,50,306
Гукер (Hooker J.D.) 105
Гулд (Gould S.J.) 177,496
Гумбольдт (Humboldt А.) 145,360
Гумилев Л.Н. 556-557
Гурвич А.Г. 197-199,283-284,297,340,
354, 381,430,657-659,676,678
Гуркин В.А. 204
Гэлбрейт (Galbraith J.K.) 136
Д’Арси Томпсон (D’Arcy Tompson W.)
292-293
Д’Орбиньи (D’Orbigny А.) 59,
Давиташвили Л.Ш. 93,133, 134,667
Давтян Т.К. 659
Даль В.Н. 211,253
Дальк (Dalcq А.) 184,243
Дана (Dana J.) 479,505,512
Данилевский И.Я. 66,119, 130,139, 140,
142, 152,200,252-253
Данилов Ю.А. 665
Дарвин Ф. (Darwin F.) 106
Дарвин Ч. (Darwin Ch.) по всей книге, в
там числе'. 14,47,54,70, 81, 84, 97,101-
102. всяглаваЗ,218,252,438,480
- и естественное богословие 48
- и пангенез 106
- и экстремальный опыт 256
- изменение взглядов на эволюцию ЮЗ-
105, 498
- основные заслуги и просчеты 13,104-
113, 118-120,256,628,632
Дарвин Э. (Darwin Е.) 59-65,70,84, 87,
226,246,437-438,675
- как основатель ламаркизма 63
Даревский И.С. 671-672
Дарнел (Darnell J.E.) 419,618
Де Вааль (De Waal F.) 550
Де Малье (De Maillet В.) 56,57,60,203,627
Девис (Davis Р.) 396-397,678
Дейчман А.М. 245-247,423
Декарт (Descartes R.) 43-45,225,289
Дельбрюк (Delbriick М.) 189-190,222
Демокрит 24.33,207
Денбиг (Denbigh K.G.) 682
Де-Фриз (De-Vries Н.) 137- 139,148.149.
161,325,354,364
Дженкин (Jenkin F.) 111-114,149-152,
154,158,325
Джеффри (Jeffrey Ch.) 562-564,568
Джилас (Djilas М.) 157
ДжирландаС. 110
Джохансон (Johanson D.C.) 546
Дзержинский Ф.Я. 375,531, 537
Дикерсон (Dickerson R.E.) 401
Диодор Сицилийский 25
Дмитриевский А.Н. 527
Довлатов С.Д 487
Догель В.А. 449,680
Долло (Dollo L.) 203, 375,544,574
Дорн (Dohm А.) 115-117,132,258, 334,496
Доси (Dosi G.) 136
Достоевский Ф.М. 556
Доуи (Dawe G.S.) 273
Дриш (Driesch Н.) 146,147, 184,299,
380, 675,677
Дулитл (Doolittle W.F.) 419,618
Дымшиц Г.М. 308
Дьяков Ю.Т. 321-322,339,454
Дэвис (Davis D.M.) 337
Дюркгейм (Durkheim Е.) 560-561
Ева (.миф.) 22,686
Ежиков ИИ. 196
Еленкин А.А. 179.180, 193,196,627
Ерне (Jerne N.K.) 334-336.345. 349,388
Еськов К.Ю. 216-218.472.474.478.516.520
Жакоб (Jacob Е) 237,267,273
Жвирблис В.Е. 547
Жданов В.С. 337,524
700
Жерихин В.В. 376,493, 525,619,623,
626,628.633
Жорж (Georges М.) 242
Жоффруа Сент-Илер (Jeoffroy St-Hilaire
Е.) 76-84,86,88.124,126,130,171,183,
187,236,246.283.298,328,372, 381,
497,660
Журавлев А.Ю. 495,512,632
Жученко А.А. 276,468.603,605
Жюлиа (Julia G.) 661.663-665
Забродин В.Ю. 577
Завадовский Б.М. 157
Завадовский М.М. 279
Заварзин А.А. 449,462
Заварзин Г.А. 373,408,414-415,418,
425,437,442,446,490,559,560,564,
569,576,584,592-593, 598, 619-620,651
Зайцев Г.Н. 279
Зайцева О.В. 479
Захаров И.А. 156,444,447
Захваткин А.А. 196,265,299, 549
Захваткин Ю.А. 189
Зевс {миф.) 23
Зелеев Р.М. 555, 573-574
ЗеликманА.Л. 189
Земмельвейс (Semmelveis I.) 234,658
Зенон Китийский 37
Зенон Элейский 37
Зубов А.А. 553
Зусмановский А.Г. 140,223,226,227,
242,255,260,271,276,298, 328,372-373,
377,381,435,500,502,615
Иванов А.В. 469-470
Иванов К.П. 331,336,603
Иванова-Казас О.М. 393,486
Ивановский А.Б. 378
Ивахненко М.Ф. 437,532, 538
Игнатов П.Е. 637-638
Игнатьев И.А. 505, 509-510,642
Икскюль (Uexkull J.V.) 242,287,347,
624-625,685
Инге-Вечтомов С.Г. 248,426
Иноземцев А. А. 276
Инь и Ян (миф.) 20
Иоганнсен (Johannsen W.L.) 159
Иост (Yost Н.) 456
Йоллос (Jollos W.) 222,267,358
Кабанис (Cabanis P.-J.-L.) 62,63,65,75
Кавалье-Смит (Cavalier-Smith Т.) 452,
457-458,466
Кавамура (Kawamura К.) 614
Казютинский В.В. 683-684
Каин {миф} 687
Каландадзе Н.Н. 512
Каллиникова В.Д. 330
Камерницкий А.В. 547
Кампер (Camper Р.) 57
Камшилов М.М. 408
Канаев И.И. 70, 82, 126
Кант (Kant I.) 69,70.127,200,458
Капи (СаруР.) 315
Карпов С. А. 577
Карус (Cams C.-J.) 58.82
Кауфман (Kauffman S.) 303-304.632,
659-660
Квитко О.В. 281
Кейзерлинг А.А. 96,224
Кеплер (Kepler J.) 96,306
Керкис Ю.Я. 164,275
Кизс (Keith А.) 545
Кильмейер (Kielmeyer С.Е) 69,70,127,
161,203
Клеанф 25,37,253
Кленов М.С. 340
Клеопатра 688
Клиге Р.К. 444
Климения {миф.) 178
Клэк (Clack J.А.) 538-540
Клюге Н.Ю. 566, 574
Кобляков А.А. 682-683
Ковалев О.В. 385
Ковалевский В.О. 177,666-667
Коваленко Е.Е. 11,280,629,668
Козлов В.А. 335,337
701
Козлова М.С. 556
Кокшайский Н.В. 117
Колмогоров А.Н. 300
Колумб (Colombo Ch.) 42
Колчанов Н.А. 351
Колчинский Э.И. 102,124,140, 176,180,190
Кольман (Kollmann J.) 183
Кольцов Н.К. 155,162-165, 187,648-649
Коммонер (Commoner В.) 249
Компаниченко В.Н. 412
Кондратьев К.Я. 494
Коп (Соре E.D.) 132-134,153,165,168,
173,203,220,313,325,361,378,382,
431,479.496,562,577,667,674
Коперник (Copernicus N.) 14,22,96
Кордонский С.Г. 603
Кордюм В.А. 353, 355,393
Коржинский С.И. 139,324
Корона В.В. 315,360-361,441,567-568,
570-571,573-576,602,668-669
Корочкин Л.И. 197,241,284-285,290,
292,383,640,667-668
Костандов Э.А. 263
Костецкий Э.Я. 400
Котельникова Е. А. 102
Котс А.Ф. 173,174,317
Котт (Cott Н.) 122,374
Кох (Koch W.) 279
Коуз (Coase R.) 136,559
Коэн (Cohen S.N.) 299,353
Красилов В.А. 543
Красный Л.И. 608
Кребс (Krebs Н.А.) 464
Кремо (Cremo М.А.) 551, 554,644
Кренке Н.П. 55,173,313,317,326.462,
535,570,669
Кривошеина Г.Г. 552
Крик (Crick F.) 247, 395,418
Криштофович А.Н. 498-499,629
Кропоткин П.А. 135-137,192,193,227
Круаза (Croizat L.C.M.) 528,534,634
Крумбигель (Krumbiegel G.) 2
Крюкова Е.П. 12
Крюссман (Krussmann G.) 488
Ксенофан 26,27,118,224
Ксенофонт 30
Кудрин Б.И. 123, 189,207,362, 370,439,572
Кузин А.М. 429-430,435,657
Кузин Б.С. 200,229
Кун (Kuhn Т.) 66,67,123,142,166,234,
636,652
Кундт (Kundt W.) 491
Курильский (Courilsky Р.) 245
Куэйрос-Конде (Queiros-Conde D.) 347-348
Кэйрнс (Cairns J.) 222,223,255,269,275
Кэно (QuenotL.) 122,170
Кэри (Carey S.W.) 521
Кэрролл (Carroll R.) 310,437,486,512,
534,543,573
Кювье Клементина (Cuvier С.) 79
Кювье (Cuvier G.-C.-L.-D.) 53,58,70,76-
79,90,93,122,124, 132,203,217,296,596
Лавлок (Lovelock J.E.) 306,338,399,409.
412,520,627,649,676,678
Лавренова Г.В. 513
Лазарь (миф.) 544
Лайель (Lyell Ch.) 85,203,217,498,643
Ламарк (Lamarck J.-B.) 27,37,62-66,71,
74-77,83,84,116,123,132,209,220,240,
246,367,463
- и Э. Дарвин 62
- об эвол-и высших и низших 65,307
- о прогрессе 62,374,438,450
- человек как геологич. сила 71,490
Ландштейнер (Landsteiner К.) 233
Лебедев А.В. 24,25,28
Лебедев К.А. 332,335-336,338-339,341
Леви-Строс (Levy-Ctrauss С.) 360-362,
555,558,686,689
Левит Г.С. 307
Лежандр (Legendre А.М.) 660
Лейбниц (Leibniz G.W.) 52-54,68,69,90,
343,661
Лем (Lem S.) 409
702
Ленин В.Н. 190
Ленинджер (Lehninger A.L.) 355
Леонтьев Д.В. 579, 589, 594.601-602,606
Ли (Lee M.S.) 572-573
Ли (Li А.) 352
Лима-де-Фариа (Lima-de-Faria А.) 361,
394,438,576,610,636,668
Линг (Ling G.) 403
Линней (Linnaeus С.) 34, 54, 57,122,360,
442, 506,563-566,568-569, 590.595.603
- о двух системах 564, 578, 582-584
Линник Ю.В. 110,196,392, 513
Ло (Lau N.C.). 340
Лобье (LaubierL.) 413-414
Ломброзо (Lombroso С.) 337-338,650
ЛопашовГ.В. 197,284
Лот (.миф.) 686-687
Лукреций 38,47, 175
Лункевич В.В. 28,30,40,63, 73, 75, 82
Лупандин И. В. 40
Лурия (Luria S.E.) 189-190,222,233,267
Луэс (Lewes G.H.) 117,133,618,684
Лысенко Т.Д. 156,157, 187,190,191,
209,256
Льюис К. (Lewis К.) 102,258
Любищев А.А. 76, 109-111,122,132,
156,172,188,198-208,216,220,289,292,
304,313, 346, 356. 377,394,401,440,504
- о гомеопатии 658
- о приспособлении 202.368
- о роли ест. отбора 204.213,218
- о наивности селекц. объяс-й 313,672
- о проблеме осуществления 287-288,
354,660
- о сути и форме системы 441. 563, 567,
570-571
- свое понимание эволюции 206
- эстетич. начало в природе 110
Любовь (.миф.) 29, 35
Магометов А. А. 324
Мазохин-Поршняков Г.А. 388
Майварт (Mivart St-G.) 115,150,496
Майоров Г.Г. 38,39
Майр (Маут Е.) 186,188, 393
Мак-Клинток (McClintock В.) 244
Макрусова В.А. 202
Максвелл (Maxwell J. Оегс) 147,219
Максимович М.А. 167
Малахов В.В. 297, 533
Малахов М.П. 301
Маленков Г.Г. 259,261
Малиновский (Malinowski В.) 689
Малыгин А.Г. 317
Малыгина Н.А. 12, 501,652
Мальтус (Malthus T.R.) 47, 86. 87,101,
135,194,337,366
Мамонтов С.Г. 17
Мамчур Е.А. 683
Мандельброт (Mandelbrot В.В.) 660-663,665
Марасов А.Н. 207,440
Маргелис (Margulis L.) 465-467
Маресин В.М. 287,293,301
Марков А.В. 234,290,333-334, 340, 349.
382-383,444,446,451,458,465,468.475-
477.484,614,648
Маркс (Marx К.) 157,628
Марфенин Н.Н. 361
Маршалл (Marshall А.) 135
Маслова Н.П. 319
Масюк Н.П. 466
Маттик (Mattick J.) 249
Медавар (Medawar Р.) 233,268
Медников Б.М. 574, 583, 597
Мейен С.В. 11,123,147,191,200,201,
206,209,218,220,295, 329, 346, 361,
372,377,380,384,437, 576, 623, 650,652
- грубая филогения 586
- о естествен, системе 568-569, 577,579
- основатель диатропики 313-319,439,
462, 520,534,669, 674
- происхождение цветковых 536
- система голосеменных 584-586
- флорогенез 440,474, 506-512, 515, 528,
535,606, 634
703
Мейнард-Смит (Maynard-Smyth J.) 247
Мёллер (Muller Hl.) 164
Мельников В.П. 345
Менделеев Д.И. 167,322,346,371, 570
Мендель (Mendel GJ.) 17,137,138,239,
354,431,512
Мензбир М.А. 139-141,149,174,190,201
Мень А.В. 687
Мережковский К.С. 143, 144,180
Мёррей (Murray Т.) 551
Метерлинк (Maeterlinck М.) 120,154,
211,385, 389,392,681
Метлина А.Л. 446
Мещерский И. Г. 671
Микаэлян А.Л. 648
Микрюков К.А. 456
Миланкович (Milankovic М.) 522
Мирабдуллаев И.М. 579, 594
Миркин Б.М. 105,616
Михайлов К.Г. 386. 563. 568, 574. 584.
589.615.626
Мичурин И.В. 191.209
Момыналиев К.Т. 388,448
Мопертюи (Maupertuis P.L.) 54-57,60,
67, 68,73,177,178,219,224,227,397
Морган (Morgan Т.Н.) 17,222,358
Мосейчик Ю.В. 510-511,528,642
Мудрость (миф.) 22,24,32
Мужч инкин В.Ф. 315
Мусина Р.А. 277
Мэтью (Mattew Р.) 86
Мюллер (Muller Е.) 454
Мюллер (Muller F.) 181-182,299,624
Назаретян А.П. 647
Назаров В.Н. 67,122, 132,140,170,171,
185,215,223,263,267,269,276, 325,
370,441,461,496,632,651,671
Наймарк Е.Б. 475-477, 648
Наливкин Д.В. 473
Наполеон Бонапарт 76, 88
Наточин Ю.В. 443,458
Наумов Р.В. 494
Нашимото (Nashimoto М) 423
Немец М.Г. 226
Несис К.Н. 110,327,331,371, 373,387,
391,515,523,559, 571
Нечаева И.А. 523
Нигматуллин Ч.М. 373,571
Нидхэм (Needham J.) 69,76
Никишин В.П. 450
Николай! 88
Нильсон (Nilsson D.-E.) 679-680
Нинхурсаг (миф.) 688
Новосельцев В.Н. 350-351,368
Ной (миф.) 686-687
Нортон (Norton H.T.J.) 150,193,213,
254-255,259
Нострадамус (Nostradamus М.) 215
Нуссбаум (Nussbaum М.) 269
Ньютон (Newton I.) 56.95.96.225.227.
289,431.661
Нэгели (Nageli К.) 83,84,103, 176,181,235
Огурцов А.П. 230
Одинцова М.С. 330
Океан (миф.) 20,178
Окен (Oken L.) 72-75, 82,90,613
Оккам (Okham, Occam W.) 205,218-220,
244,260,314-315,516,552
Окушко В.Р. 658
Олескин А.В. 648-649
Оловников А.М. 243,288,421,459
О’Нилл (O’Nell L.A.J.O.) 332,338
Опарин А.И. 17, 399-401,403,412,618
Остер (Oster G.) 432
Остин (Austin S.A.) 643
Оуэн (Owen R.) 82,83,95-97,112,125,
203,298-299,329,603,622,667,674
Павинский (Pawinski А.) 550,554
Павлинов И.Я. 573,654-655
Павлов А.П. 170, 178,299,480, 552, 624
Павлов В.Я. 311, 571,606-607
Пайтген (Peitgen Н.-О.) 661-662,664,684
Палисси (Palissi В.) 42,43,48
Палтит (миф) 687
704
Пальцев М.А. 274
Панов Е.Н. НО, 137, 192,311,673-674
Пантин В.И. 605-606
Парменид {лит.) 32
Пармон В.Н. 403
Парсонс (Parsons Р. А.) Zlb-ТП
Паршин А.Н. 246, 547-548,559
Пастер (Pasteur L.) 14,15
Патти (Pattee Н.Н.) 685
Пелыунов А.Н. 497
Пахомов С.В. 556
Петр 1 157, 560
Петров Н.Б. 375, 582
Петров П.Н. 638
Петрова Н.В. 243
Петухов С.В. 282
Пилипенко А.П. 163
Пиневич А.В. 294.466
Пири (Pirie N.W.) 17,404
Пирузян Л.А. 659
Пифагор 25,27,28,207,346.371,677
Плате (Plate L.) 124
Платон 24,25,30-32,36,38,51,64.118.
122, 155,200,289,299,305,313,329,
346,362, 390,392,582,628,640,677, 689
Побежимова Т.П. 465
Поллак (Pollack G.H.) 404
Поляков В.Ю. 243
Пономаренко А.Г. 490-492,503-504
Попов И.Ю. 169, 170, 179,222,375, 513,
528,609,614,629, 635
Попп (Popp F.-A.) 284,430,657
Поппер (Popper К.) 216,268,362,509,618
Поршнев Б.Ф. 547
Прам (Prum R.O.) 542
Пригожин (Prigogine 1.) 282,368,459
Прозоров А. А. 448,450,484
Пропп В.Я. 360
Пуанкаре (Poincare Н.) 570
Пустильник С.Н. 366-367,370
Пушкин А.С. 132
Пэйли (Paley W.) 48,49
Раддимен (Ruddiman W.F.) 523
Радль (Radi Е.) 125
Райков Б.Е. 74,79,94, 96
Райков Н.Б. 452-455
Райордэн (Riordan М.) 226
Расницын А.П. 218-220,497,573,627.656
Рассел (Russell В.) 40,118,218
Ратке (Rathke Н.) 74
Рауп (RaupD.M.)461,520,524
Раутиан А.С. 294,380,393, 512,619
Рачинский С.А. 100,365
Регельсон Л.Л. 687
Рёддер (Rodder G.) 531
Резанов Н.А. 443
Реммель (Вальт) (Remmel М.) 137
Рерих Н.К. 196
Робертсон (Robertson М.Р.) 422
Рожнов С.В. 473, 533
Розанов А.Н. 552
Розанов А.Ю. 421
Розанов В.В. 556
Ройт (Roitt I.) 483,485
РокицкийП.Ф. 158-160
Роли, Рэли (Raleigh W.) 43,44,66
Ромер (Romer A.Sh.) 483,531-532, 539,
541-542
Ронов А.Б. 399
Рос (Roth А.) 16,23,644
Роуз (Rose S.) 548
Ру (Roux W.) 355.365,370
Руденко А.П. 402-403
Рузвельт (Roosevelt Т.) 119,152
РульеК.Ф. 88-91.93,94,99,100,141
Румфорд (Томсон) (Rumford В.) 211
Румянцев С.Н. 332
Руттен (Rutten M.G.) 402,406-407,410
Рэй (Ray J.) 48,56
Рэф (RaffR.A.) 285,292,355.380,480
Салихов М.В. 200
Сальменова К.3.585
Салямон Л.С. 234-235
Самуилов В.Д. 408,415,433
705
Сапожников Ф.В. 470
Сапольски (Sapolsky R.) 388-389
Сапунов В.Б. 257
Сахарова Н.Ю. 196,487-488.542.549
Свердлов Е.Д. 613,652-653
Светлов П.Г. 183,197,198.284
Севальников А.Ю. 684
Северино (Severino А.) 51-52
Северцов А.Н. 181-184,293
Северцов А.С. 299,543
Северцов Н.А. 94, 181
Селье (Selye Н.) 164,237,264,266,345
Семенов Д.В. 328
Сен-Жермен (St-Germain В.) 53
Сепкоски (Sepkosky J.) 520-521,524
Серавин Л.Н. 465,471
Сергеев В.Н. 414
Серов О.Л. 267-267
Серебровская К.Б. 403
Сидорова Е.В. 303
Силлен (Sillen L.G.) 402,406
Симонов П.В. 372
Симпсон (Simpson G.G.) 169,512
Сингер (Singer М.). 419-420
Скёддер (Scudder S.) 120-122
Сковрон (Skowron S.) 18
Скулачев В.П. 260,330,417-418,465,617
Смирнов Е.С. 198,200,256, 570
Соболев Д.Н. 175-181,262,306, 315,360,
378,440,462,529, 534,574,627
Соболь С.Л. 86
Сократ 30,32, 200,365, 396-397,676
Соколов Д.Д. 668
СокулерЗ.А. 123
Солбриг (Solbrig О.Т.) 17
Соловьев Ю.И. 431
Сорохтин О.Г. 553
Спенсер (Spencer Н.) 19,93,133,142,
163,180,221,237,313,629
Спиноза (Spinoza В.) 40, 165
Спирин А.С. 419,423,444
Сталин И.В. 190-191,655-656
Старобогатов Я.И. 378,504
Стеббинс {Stebbins G.L.) 363
Стегний В.Н. 185,243,326,378,382-383,
453,625,634
Стент (Stent G.S.) 190,223
Стил (Steele E.J.) 132,148,250-251,266,301
Столяров А.А. 25,27,37
Страйер (Stryer L.) 464
Страхов Н.Н. 141,142, 144,146,152,
171,235,291,317, 358
Струнников В.А. 300
Суворов А.И. 146
Сукачев В.Н. 191-193
Суриков И.М. 333
Сэлт (Salthe S.) 362
Тай Тэ By (Т.Т. Wu)258
Тарчевский И.А. 340
Таусон Е.Л. 292
Тейз (Theise N.D.) 273
Тейяр де Шарден (Teilhard de Chardin Р.)
18,221,306-307,372,374,440-441,476,
490,548,555,611,613,649
Теофраст 36,37,43,129,176
Тидеман (TidemannF.) 69
Тимей (тип.) 31-32,118,207
Тимирязев К.А. 149
Тимонин А. К. 503
Тимофеев-Ресовский Н.В. 163,219,222,
327,394
Тинберген (Tinbergen N.) 121,122
Титов Ю.В. 192,194,195
Тиходеев О.Н. 253,315
Тихомирова А.Л. 308
Ткемаладзе Дж. В. 457
Том (Thom R.) 287-290, 337, 348,365, 377
Томпсон (Thompson R.) 551,554,644
Тонегава (Tonegawa S.) 251-253, 301-
303, 356,637
Торканов А.М. 311
Траутшольд (Trautschold Н.) 98,482
Трепет С. А. 646-647
Тролль (Troll W.) 126
706
Трошин А.С. 403
ТрутЛ.Н. 210
Уголев А.М. 462-464,491,502
Уитмен (Whitman С.О.) 355
Уиттекер (Whittaker R.) 376,442,493,
580, 582,590-593
Ульянкина Т.И. 232
Уоддингтон (Waddington С.Н.) 169,186
Уоллес (Wallace A.R.) 94-96,103,108,
109,113,119-121, 129,137,153,159,390,
505,632
Уорд (Ward P.D.) 522
Уотсон (Watson D.M.S.) 542
Уотсон (Watson H.Ch.) 96
Уотсон (Watson J.D.) 395
Урманцев Ю.А. 387, 547
Усов С.А. 99,100,110,371,646
Ухтомский А.А. 262-263
Уэвелл (Whewell W.) 569,601
Фалес 25,294, 614
ФаминцынА.С. 142, 143, 180. 581
Федон (лит.) 30
Федонкин М.А. 421
ФеогнидЗб, 155
Фибоначчи (Fibonacci L.) 321,377,669-670
Филипченко Ю.А. 125, 126,129,132,
169,172,174,186,187,317, 367
Филон 23,27,42, 52,687
Финкельштейн А.В. 425
Фиц-Рой (Fitz Roy R.) 84,96,99
Фишер (Fisher R.A.) 259-260
Фогт (Vogt К.) 554
Фокс (Fox S.W.) 399-400,410,426,444
Франклин (Franklin R.E.) 395
Фрей-Вислинг (Frey-Wyssling A.) 248
Фуко (Foucault M.P.) 570
Фюрон (Furon R.) 184
Хабермас (Habermas J.) 559
Хайн В.Е. 473,518,521-523
Хайтун С.Д. 440
Хаксли (Huxley J.) 140,185,186,201
Хартли (Hartley В.) 257
Хартман Ф. 202
Хаттман (Hattman S.) 471
Хатшепсут 688
Хварцкия И.И. 687
Хлебович В.В. 233-234
Холл (Hall В.G.) 223
Холодковский Н.А. 79
Хомский (Chomsky N.) 336
Хоорст (Hohorst W.) 389
Хоутон (Haughton S.) 95
Хохряков А.П. 606
Хрущев НС. 136,157,654
Хук (Hooke S.H.) 687-688
Хэйл (Hale М.) 44-52,55,56,62,87, 254
Хэнтер (Hunter J.) 69
Цвелёв Н.Н. 521
Цезальпин(Се5а1рто А.) 226
Цельсий (Celsius А.) 217
Циммерман (Zimmermann W.) 185
Циолковский К.Э. 280,282,367
Чайковский Ю.В. 14, 16,22,25, 82,99,
105,118,136,137,189,201,259,330,
334, 363, 374, 386, 388, 391,451,467,
558-561,563-566,632,689
- об активности и фазовых переходах 224
- о генетич. поиске 275
- о методологии 218,224,295
- о макросистеме и ядре 587, 594, 597-598
- о познават. моделях 230,431
- о диатропике и рефренах 314,322, 669
- о случайности в эволюции 301-305,
348,608.682,684
- об экологии, прочности вида 311
- схема эволюции 621
Чайлд (Child Ch.) 283
Челомбитько В. А. 185,239
Чемберлин (Chamberlin Т.С.) 201
Чемберс (Chambers R.) 87,103
Черданцев В.Г. 283,288,659-660
Черешнев В. А. 356
Чернавский Д.С. 349,394, 502
Чернин А.Д. 297,608
707
Чернов В.М. 375
Чернов Ю.И. 502, 511,515
Черняк Б.В. 330
Чесноков В.А. 163
Четвериков С.С. 151-155.158-160,162,
174,190, 325, 394
Четверин419
Чжи-Чэн Сяо (Xiao Z.-Ch.) 273
Чижевский А.Л. 494
Чмора С.Н. 645
Чуриков Н.А. 625
Шалин (Chaline J.) 220,298, 300,448,
461,473,480-481,486,524
Шамрай С.Н. 501
Шапошников Г.Х. 208-211,254,256,
263,275-276,278, 347,359,382, 385,536,
633-634,671
Шаталкин А.И. 582
Шафранова Л. М. 595
Шевяков В.Т. 679
Шварц С.С. 278
Шелдрейк (Sheldrake R.) 434,675-678
Шиндевольф (Schindewolf О.Н.) 244,
378,440,476-477,516,529,627
Шипунов А.Б. 594, 596-597
Шишкин М.А. 265,369
Шлегель (Schlegel H.G.) 463
Шлиман (Schliemann Н.) 132
Шмальгаузен И. И. 181,187-189,201,
265, 311, 357-358, 369, 384,514,623,674
ШнольС.Э. 163,331
Шовен (Chauvin R.) 191,195
Шойхцер (Scheuchzer J.) 41
Шопенгауэр (Shopenhauer А.) 224
Шорт (Short R.V.) 559
Шпеман (SpemannH.) 147.184,196,197
Шпренгель (Sprengel Ch.) 48
UInpeHrep(SprengerPh.S.) 83,139.176.
382,625
Шрёдингер (Schrodinger Е.) 660
Шреадер Ю.А. 207,218,260,346, 577
Штейнман (Steinmann G.) 133,667
Шубинский Г.З. 335
Щербаков Д.Е. 522
Эдельман (Edelman G.M.) 285-286,288,
334,548
Эйген (Eigen М.) 403
Эймер (Eimer Т.) 169
Эйнштейн (Einstein А.) 228
Эйринг (Eyring Н.) 431
Эко (Есо U.) 654
Эплюи (Hellui S.) 386
Элтон (Elton Ch.S.) 623,632,634-635
Эмпедокл 28,29,35,47,74,253,465
Энгельс (Engels F.) 17
Энки (миф} 688
Эрида (миф.) 231
Эрлих (Erlich Р.) 232-233
Эрмит (Hermite Ch.) 289
Эрос (миф.) 20,224
Эфроимсон В.П. 162,163,394
Юл (Yule G.U.) 364
Юнг (Jung J.) 52
Юнг (Jung K.G.) 677,683
Юнкер (Junker R) 16
Яковенко Л.В. 426,443-444
Яковлев Г.П. 185
Яницкий О.Н. 559,605
Янч (Jantsch Е.) 368,412,432,555,620,
624,653,678,684
Ярвис (Jarvis J.U.M.) 329
AlivesJ.M. 391
Aravind L. 597
Arenillas I. 517
Bauhin C. 139
Berner R. A 522
Bininda-Edmonds O.R.P. 316
Brooks J.L. 94
BurlandoB. 613
ChabraeoD. 139
Chowdhury D. 341
Clark G.A. 546
Cockell C.S. 519
708
Cohen А.А.557,559 Moore J. 568
Dambicourt-Malase А. 549 Norris K.S. 349
Doherty J.K. 241 Ohno S. 259
Du Pasquier L. 485 Pickett-Heaps J.D. 455
Galleron C. 240 Rassoulzadegan M. 271
Garcia-Belido A. 287 Santos M. 588
Gastaldo R.A. 642-643 Selosse M.-A. 467
GhiselinM.T. 97 Senut B. 545-546, 554
Gilbert W. 241 Skogsmir I. 110, 333
Gomes A.C. 277,588 Soler J J. 362
Goodwin B.C. 659 Summit M. 489
Gordon S. 338 Thierry J. 481
Gregory T.R. 356 ThuillerP. 14
Jefferies R.P.S. 533 TodesD.P. 101
Kirschner M. 636 von Dohlen C.D. 467
Kordis D. 352 White T.D. 553
Lange-Barde B. 502,541 WiaS. 257
Laurin M. 480,541 Wicklow B.J. 679
Martin W. 442,467 Wilkens H. 279
Meizel S. 489 Wither N.W. 466
Указатель основных терминов
Указаны места, в которых термин введен или обсуждается
адаптационизм 48 адгезия избирательная 285,484 активность 25,33,55,60,70, 85,129, 147,219,224,609,629 -генома 247,250 - концептуальная 228 - принципы 229,612 актуализм 217,498 акциденция (побочное св-во) 34 аналогия и гомология 82, 622 антропный пр-п 31,396,401,678,683 апоптоз 340,433 архаллаксис 182 архетип, план строения 42,126,531 - по Мейену 622 - по Оуэну 126,622 бабушки эффект 557 биогенетич. закон 299 биоплёнка 258,470 биополе 430 биопоэз 395 биосферы различные 489 биотогенез 512 блочность эв-и 315,462, 504,531 вид, определение 623 видообразование 382,485 витализм 146 время 458,498, 629 выдвиженцы 156. 162 вымирание 379,516 - раннего человека 553 - склонность видов к в. 100,649 гамогетеротопия 535 гармония 146,203
709
генетический код 201,242,277,588
генетич. поиск 238,275
геномные цензоры 340,421,424
геносистематика 292,576
геохронология 437
гибридный орган 330
гигантизм 309,472
гидротерма413
гиперболич. распред-я 363
гомеобокс 291, 328, 636
гомеозис 160, 535
гомология - см. аналогия
горизонтальный перенос генов 352
- и рекомбинация 468
группы эффект 191
ГЦК405
деизм и теизм 39
диалектика, за и против 118,201
диасеть 319
- полифункциональность ее
элементов 340,609,612
диатропика, ее состав 314
диатропич. прогноз 645
диссипативная структура 282,411
длительная модиф-я 35,267,358
доместикация 210
доминанта 263
дробление 182
евгеника 31,155,327, 638-640,648-650
естествен, богословие 30,48,253
жизненный порыв 227
затравка 283
идиотрофы 597
иерархия, определение 335
изменчивость направленна 328
иммунитет и эв-я 249,332
- врожденный 338
- растений 333,340
инволюция 205,292
индустриальный меланизм 254
интерференция РНК 340
интроны и экзоны 240
картина мира 296,380,430, 570
катастроф теория 347
катастрофы 58, 77,217,275,378
квази-гиперболы 363-364
код неуниверсален 355
компенсации пр-п 80,446,474,679
компромисс 349
конкордизм 23
конкуренция 86. 100, 107, 135,193,
231,253,455,477
- и “рыночная экономика” 136
коэволюция 391. 555
креационизм 16,23,47, 66,234,239,
258.348,396
- смягченный 234,643.675,681
круговороты 398, см. циклы
Лазаревы таксоны 2, 543, 551, 553, 589
литотрофы 408
макросистема 580
межтаксон 589,594
меланизм индустриал. 254
метаболии, спираль 262
метилирование ДНК 240
мерой 317
мимикрия 389
митогенетич. лучи 198, 284
митозы, диатропика 453
мичуринская биология 172
мифы космогонические 19-25
- в эволюционных теориях 118,362,677
- исторические 21,28,36,685
многослойная регуляция 350
молликуты (микоплазмы) 444
монофилия, см. филогения
морфология 52, 55,124
мутация, мутационизм 61, 138,140,
148,151,154,158, 161,164,190,212,
222,244.251,258,267,274,291, 325,
356. 363,387,394
надцарство 442
нанобакгерии 444
неотения 558
710
ниша (в разных пониманиях) 372
ноосфера 307,461,490
норма (биологии.) 277, 612, 622
- ее давление 229,280,383
НПС 35, 60,130,269
онтогенез клетки 358
- неклеточный 286
опережающее отражение 369, 380.549
осуществления проблема 288,354.653
отбор 36
-естественный 86,107, 154,
- - как фактор эволюции 628
- - отсутствие в природе 212,222,651
- искусственный 252
- как итог толкований 212
- как способ обойти непонятное 418
- катастрофический 259
- органический 186, 674
- половой 108,558
- стабилизирующий 189
- химический 401
пангенез 106
- новый 269,272
параллелизм множественный 93
- параллелей 387
пассионарность 555
первичные половые клетки 269
план строения, см. архетип
подбор по Богданову 366, 608,629,665
подмена основания 105
- понятий 252
познавательные модели 230
поле 225,283
потребности 372
преадаптация 170,381.396,538,550,
616,624
презумпции 129, 178,203,216,295
- непригодность 217,328, 516,654
-отказот315, 620
-смена 168,247,466
презумпция рационального 220,397
приспособление 202,365-370,623
причинность 30,32,228,675,683-685
прогенота419,450
прогресс 62,90,448, 610
продуценты вторичные 416,442
- первичные 414,442
проморфология 125
пророческие фазы 170,299,624
пространство жизни 297,319,619
разнообразие 305,314
- реверсия 491,495,595,601
- упорядоченность 608
расплод 253,296
редактирование РНК 245
редукционизм 441
рефрен, определения 319,324
- поведенческие 681
- примеры 320-323, 367,462,531,609
- рефренов 503
рибозим 419
риторика 29. 118, 121
рутинизация 204,265,386,391,470
рыночная идеология 230,279,315,
560,620,651,658
ряды 93,133,165, 172, 316.509
самоорганизация 136, 143,296.610
самоподобие 345
самосборка 425,610
сверхмалые дозы 430, 659
семофилогения 533
сигналы 428
симбиогенез 142,457,465
симметрия 126, 531,537, 547, 599
- параметров 350
синергетика 282,682-683
система естественная 564, 584
- и эволюция 441
- удобная 578
системогенез 349,619
случайность 35,45,297,608
солитон 225,282,514,385
софисты 29-31,118, 313,390
сплайсинг 240
711
-альтернативный 245
сравнительный метод 293
- не дает истории 295
сродство 224,433,
стабильность названий 564
старение организма 281
- таксона (броккизм), см. эвопюц.
концепции
стволовые клетки 272
стоики ранние 37, 118.569
стресс и эволюция 164.223.229,237,
250,256,262
- - человека 554
структурализм 360
сходство системное 614
тезис Аршавского 357,372,685
-Бэтсона 171,310
- Вентребера 236
- Витгенштейна 123
- Воейкова 405
- Икскюпя 287, 624, 685
- Канта - Кил ьмейера 69
- Короны 567, 584,586,602
- ламаркизма 238
- Линнея 564
- Любищева 204,685
- Михайлова 574,615
- Соболева 179
- центральный 249
- Чайковского 652
- Шрейдера 218
- Янча 555
тезисы Мейена 318-319
- Окена 73
- теории эволюции 611-619
тенденции 307,474,481,548.627
теплокровные 331
теория, ее цель 214.635
термодинамика и термостатика 280
тип (филум, отдел) 499,591,596
термоплазма 445
террипетность 92,478,483, 548
транзитивный полиморфизм 326
транспозоны 244
узел сходств 182,299
упорядоченность 37,174,229,321
уровни развития 341,471,500,503-
505,599,624
фазовый переход 226,297,404, 561, 618
факторы эволюции 218,222,224,226,
239,245,255,261,267,276,628
филогения 127
- грубая 584,607
фитоспрединг 506
флорогенез - см. эволюц концепции
форма и функция 203, 503,514,622
фракталы 342,524,660
хологения 185.618
царства природы 441.475.499,581,595
царство человека 546,561,604
ценозы 405,409,413,415,435,611
«центральная догма» 247
цефализация 479
циклы 429,459-461,464
- разномасштабные 352
цитоид 286,449,471
эвкариоты 223,421,450
эволюционные концепции 221,226
- броккизм 58
- дарвинизм 2, 14,95-140,214
- - генетический 148
- - шоры 636
- жоффруизм 79,181
- историч. биогенетика 175
- ламаркизм 37,63,117,208
- - молекулярный 235
- мутационизм 138
- неодарвинизм, СТЭ 66,131, 186,219
- номогенез 165-175,209
- - новый 363,390,445,616
- ортогенез 169,206,221,239,375,476,
513.531
- пунктуализм 496. 626
- тератологии, эволюционизм 171
712
- термодинамич. эволюционизм 282
- физиологии, эволюционизм 61
- флорогенез 506-512
- экологии, эволюционизм 63
эволюция, понятие 39,205,226,609
- биосферы 626
- предначертанная 39,221
- прогрессивная 624
- реальные свидетельства 527
- скачкообразная 627
- скорости парадоксальные 497
- факторы, см. факторы эволюции
- экосистем 493
- экспериментальная 249-267,385
эквифинальность 147,299
экологии, прочность 310
эманация 205,561
эмбрионизация 548
эмерджентность 133,228,610,617
энергия структурная 199
энтелехия 147,675
эпигенетич. наследств-сть 240
эпигностика 286,337,356,485,659
эргодическая теория 296
ядро и периферия 315,587, 609
С-парадокс 355
formative aptitudes 61, 675
г- и А-стратегии 260, 558
ТВ- иммунитет 250, 332, 336,483
tektological selection 366
Wolbachia 444,446,614,638
Научное издание
ЧАЙКОВСКИЙ Юрий Викторович
НАУКА О РАЗВИТИИ ЖИЗНИ.
Опыт теории эволюции
Москва: Т-во научных изданий КМК. 2006. 712 с.
Книга свёрстана автором и печатается
в авторской редакции
Для заявок:
123100 Москва а/я 16
или:
kmk2000@online. ги
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН.
121099 Москва, Шубинский пер., 6.
Подписано в печать 18.12.2006. Заказ №5083
Формат 60x90/16. Объём 45 печ.л. Бум. офсетная. Тираж 1000 экз.
Обыкновенная сосна в зоне ураганных ветров вросла в грунт
срединными ветвями. Пример поведенческой активности
растения (по Б.П. Агафонову, 2005)
Книга рассказывает о появлении и развитии жизни, о развитии
взглядов учёных на этот процесс. В отличие от других книг на
данную тему, здесь история науки вплетена в рассказ, она призвана
показать, что наши новые теории коренятся в прежнем знании, в
традициях и мировоззрении. При необходимости даются параллели
с идеями эволюции в других науках (космологии, геологии,
лингвистике, экономике и др.) и в различных религиях.
Описаны реальные акты эволюции.
Новая наука видит эволюцию не как “происхождение видов” и иных
таксонов, а как активный процесс становления систем всех уровней
(от биосферы до макромолекулы) в едином пространстве жизни.
Рассмотрены экологические, организменные, тканевые, клеточные и
молекулярные механизмы эволюции. Сделан акцент на нерешенных
проблемах и на загадках, не попадающих в учебники. Но главное —
показано, что начавшийся отказ от дарвинизма есть закономерный
результат смены картины мира, происходящий у нас на глазах.
Автор исходит из той идеи, что новая теория может быть полезна
для практики не объяснениями происшедшего и не предсказанием
будущего хода эволюции (это мало кому важно), а конкретными
рекомендациями — например, по охране природы и культуры, по
медицине и систематике.