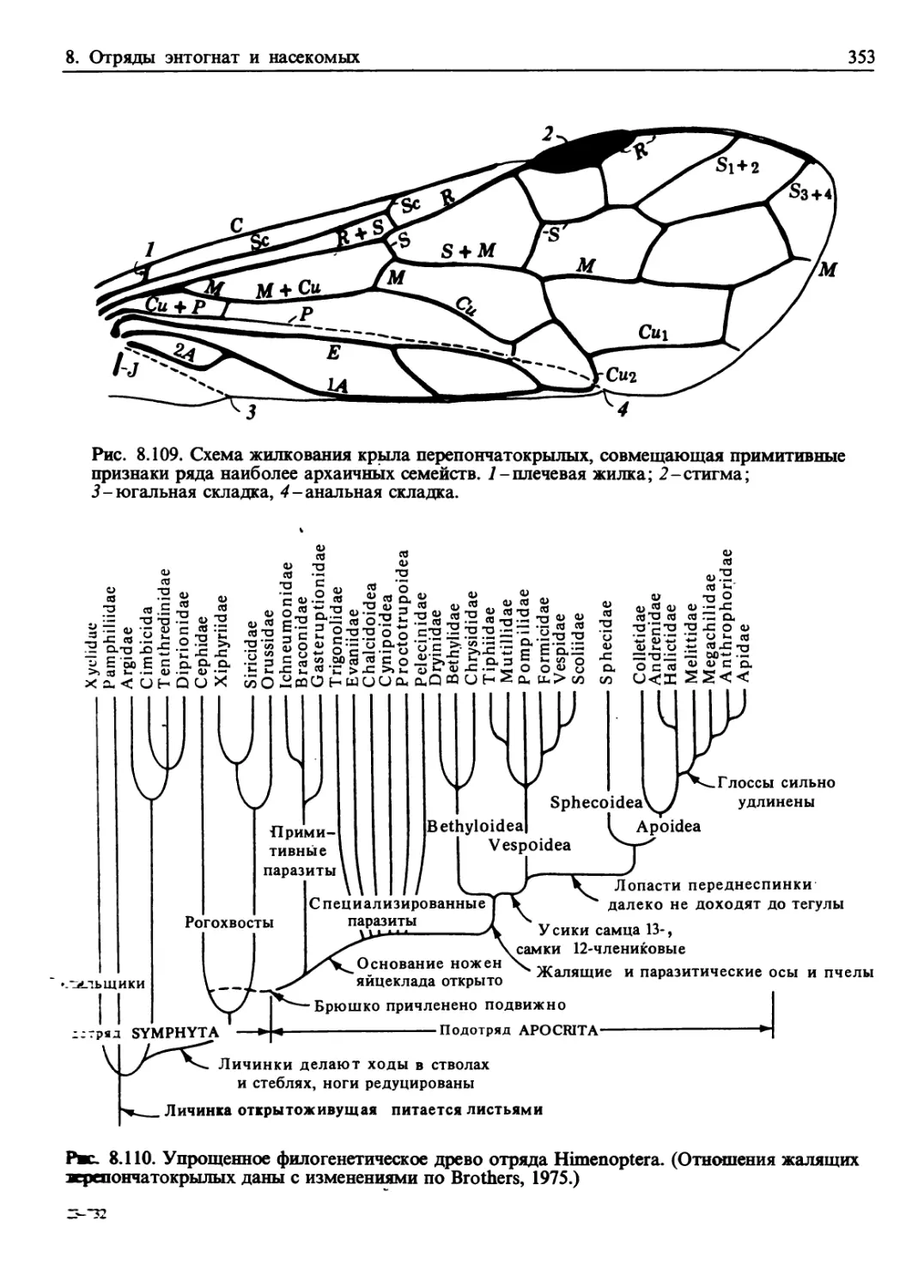
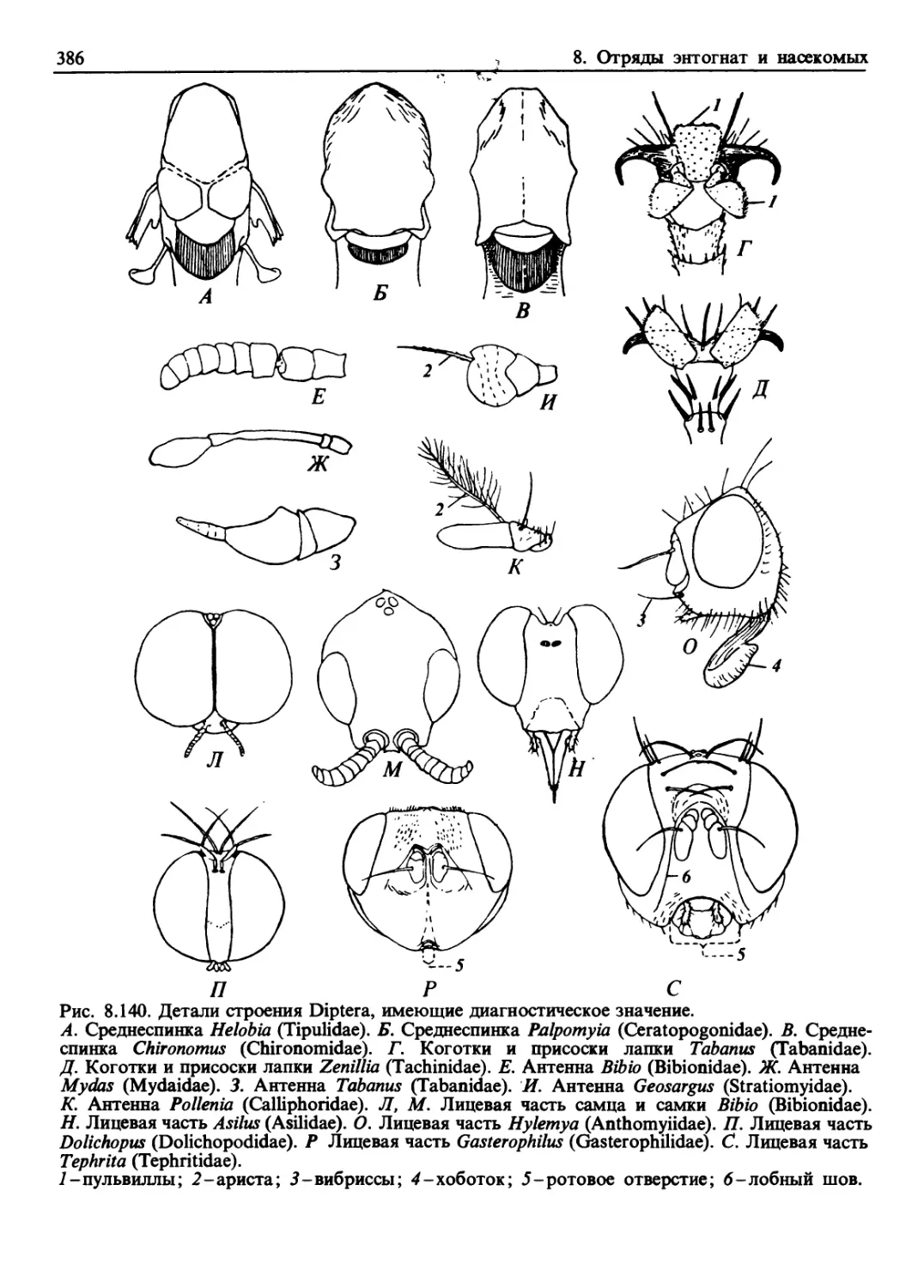
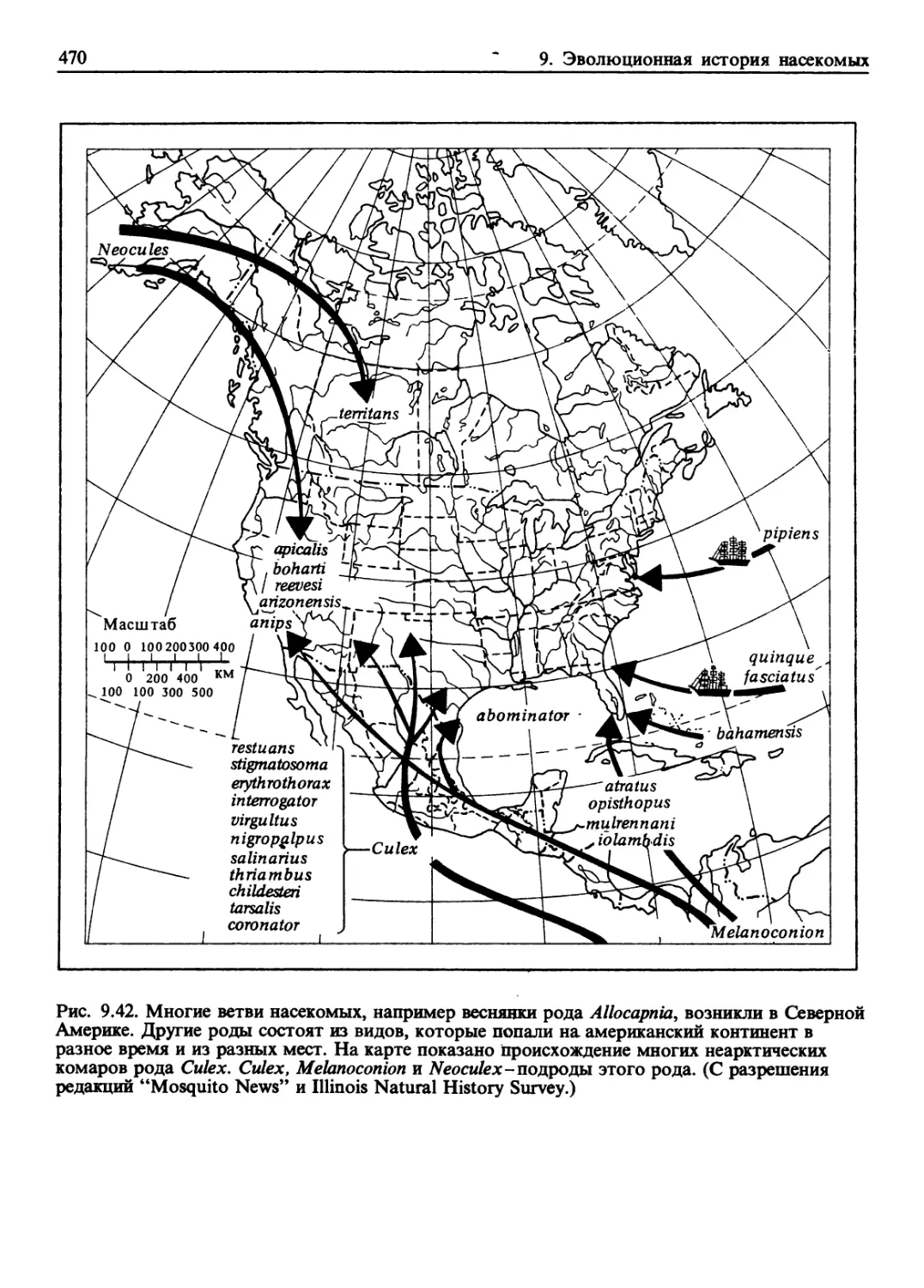
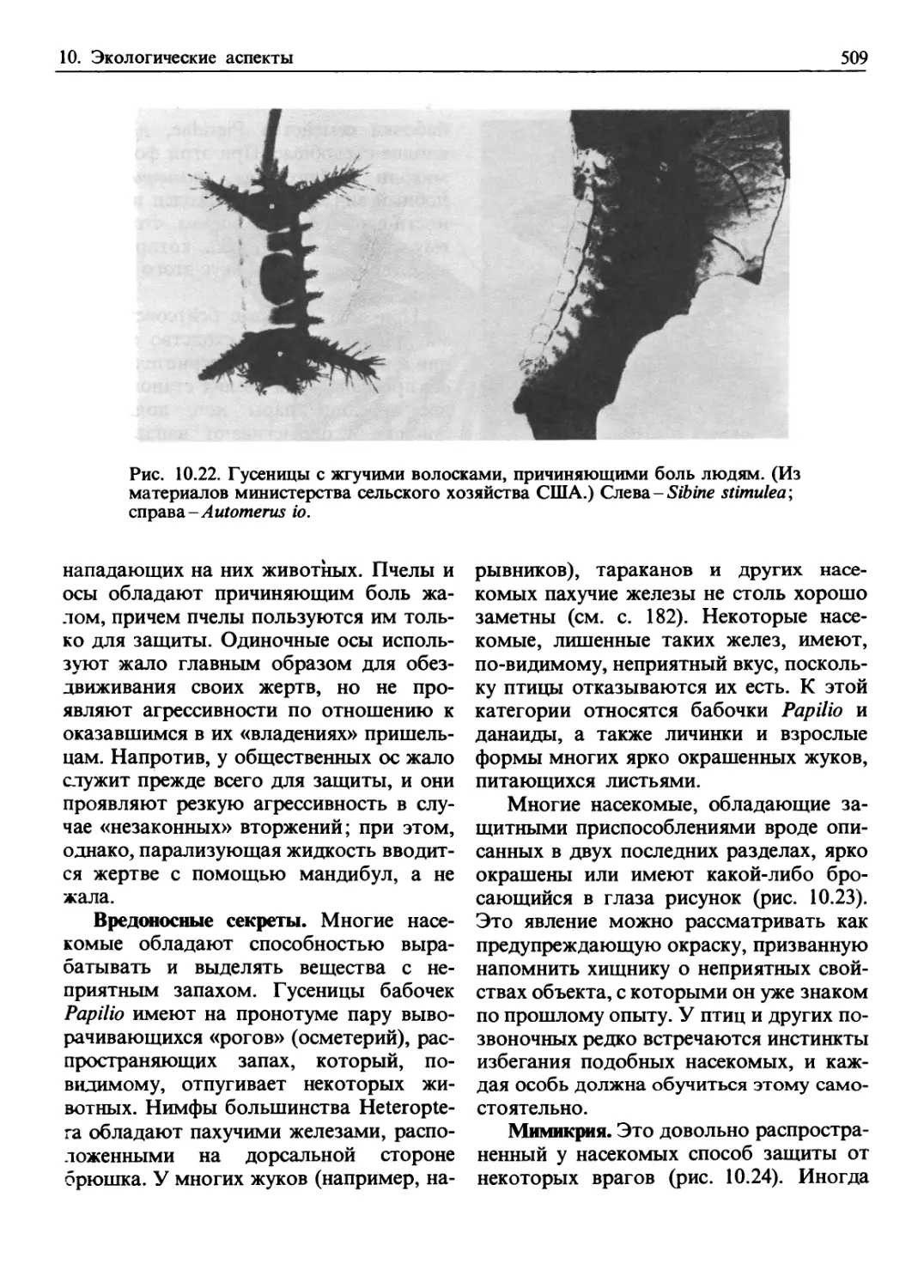
/

























































































































































































































































































































































































































































































































































































Text
^irtWlWJWni
япитвпмш
A Textbook of Entomology
Herbert H. Ross
Late Professor of Entomology
University of Georgia
Charles A. Ross
Western Washington University
June R. P Ross
Western Washington University
Fourth edition
John Wiley & Sons
New York Chichester Brisbane Toronto
Singapore
Г Росс, Ч. Росс, Д.Росс
Энтомология
Перевод с английского
В. В. Белова и
канд. биол. наук Б. Я. Виленкина
под редакцией
проф. Г А. Мазохина-Поршнякова
Москва «Мир» 1985
ББК 28.691.89
Р76
УДК 595.70
Росс Г. и др.
Р76 Энтомология Пер. с англ./Росс Г., Росс Ч.,
Росс Д.-М. Мир, 1985.-576 с., ил.
Пособие по энтомологии американских авторов, выдержавшее в США уже четыре
издания. В книге приведены современные данные по развитию энтомологии в США,
эволюции, морфологии, физиологии и поведению насекомых, рассмотрены жизненные
циклы некоторых особенно интересных видов, рост и размножение, а также вопросы
экологии и борьбы с вредными насекомыми. Книгу можно использовать в качестве
справочника по насекомым.
Предназначена для энтомологов, специалистов сельского хозяйства, для студентов-
биологов.
2005000000-236 t
04Ц01>-81 ’
ББК 28.691.89
592
Редакция литературы по биологии
© 1948, 1956, 1965, 1982, by John Wiley
& Sons, Inc.
© перевод на русский язык, «Мир»,
1985
Предисловие
редактора перевода
Создание руководства по той
или иной научной дисциплине - де-
ло сложное, требующее не только
отличного знания предмета изло-
жения, но и немалой смелости.
Смелость нужна для того, чтобы
решиться на описание в обобщен-
ной форме широкого многообра-
зия явлений. Энтомология как раз
и представляет собой ту науку, в
которой накоплено громадное ко-
личество фактов, обобщить ко-
торые в ограниченном объеме
учебного пособия особенно трудно.
Знакомясь с данной книгой, лег-
ко убедиться, что ее авторы вполне
справились с поставленной зада-
чей. Предлагаемая вниманию со-
ветского читателя книга является
переводом 4-го издания «Энтомо-
логии», написанного в 1948 г. из-
вестным американским энтомоло-
гом проф. Гербертом Россом, ныне
переработанное и дополненное
профессорами Университета шт.
Вашингтон Чарлзом Россом
и Джун Росс. Им удалось в лако-
ничной, четкой форме осветить
множество явлений и понятий, со-
ставляющих содержание современ-
ной энтомологии. Затронуты по су-
ществу все вопросы общей энтомо-
логии. от происхождения, строе-
ния тела и устройства систем жиз-
необеспечения насекомых до обзо-
ра их видового разнообразия,
форм связи с внешней средой, друг
с другом и человеком, кроме того,
изложены основы прикладной эн-
томологии : медико-ветеринарной,
сельскохозяйственной, лесной; дан
обзор полезных и вредных насе-
комых и главных мер борьбы с по-
следними. В описании насекомых
чувствуется сильное влияние зна-
менитого соотечественника авто-
ров морфолога Р. Снодграсса-
большое внимание уделяется эво-
люционной преемственности ча-
стей тела насекомого и их при-
датков. Вместе с тем всегда разъ-
яснено назначение и физиологи-
ческие механизмы действия орга-
нов, и здесь авторы следуют вку-
сам другого корифея энтомоло-
гии-В. Вигглесворса. Иначе гово-
ря, использован комплексный под-
ход, при котором насекомое вос-
принимается как живой организм,
а не просто как объект для сравни-
тельно-морфологических или физи-
ологических изысканий, как это бы-
вает в иных монографиях. Наряду с
общими энтомологическими про-
блемами широко обсуждаются и
региональные, менее известные со-
ветским читателям, но в большин-
стве своем интересные и поучи-
тельные. Приводимых общих све-
дений, пожалуй, ровно столько,
сколько обязан знать энтомолог с
университетским образованием.
Натуралисту же придутся по душе
эмоциональные описания образа
6
Предисловие редактора перевода
жизни примечательных видов.
В охвате материала чувствуется
разумная пропорция, хотя такт не
всегда соблюден раздел иско-
паемых насекомых с подробной си-
стемой вымерших таксонов, палео-
географией да еще с анализом тек-
тоники континентальных плат-
форм - «гипертрофирован». Может
быть поэтому упомянуты в книге
из всех русских и советских энтомо-
логов только палеонтологи
Б. Б. Родендорф и А. Г Шаров?
Помимо традиционных сведе-
ний в книге излагаются новейшие
представления, касающиеся эволю-
ции, систематики, физиологии,
образа жизни и поведения насе-
комых. Описание экологии и пове-
дения сопровождает анализ, воз-
можно чересчур скрупулезный, фи-
зических факторов среды, воздей-
ствующих на насекомых. Интерес-
на своей энциклопедичностью за-
ключительная глава о вредных на-
секомых и борьбе с ними. Текст
подробно рубрицирован, снабжен
большим количеством рисунков,
схем и фотографий, а также списка-
ми (по главам) основных литера-
турных источников. Все это обес-
печивает удобство пользования
книгой-одним из лучших руко-
водств по общей энтомологии.
Можно надеяться, что оно бу-
дет полезно нашим биологам и
в своем прямом назначении и как
современный справочник по эн-
томологии, необходимый не толь-
ко студентам.
Проф. Г. А. Мазохин-Поршняков
Предисловие
Энтомология-непрерывно раз-
вивающаяся наука, постоянно обо-
гащающаяся новыми данными и
новыми идеями и расширяющая
свои перспективы. За 15 лет, про-
шедшие после выхода в свет треть-
его издания настоящего руковод-
ства, энтомология сделала
огромный шаг вперед в понимании
многих аспектов экологии, физио-
логии, поведения и взаимоотноше-
ний насекомых. Новые данные ста-
вят новые, быть может еще более
серьезные, вопросы, требующие
проведения новых исследований,
решения этих вопросов и проверки
возникающих в связи с ними идей.
Таким образом, следует ожидать,
что в недалеком будущем неко-
торые современные представления
и теории претерпят существенные
изменения, и это приблизит успеш-
ное решение практических задач.
Настоящее руководство служит
введением в энтомологию и дает
общее представление об этой науке
и ее основных аспектах. В этом из-
дании мы старались сохранить за-
мысел и структуру предыдущих из-
даний, сделав, однако, более
сильный акцент на вопросах, ка-
сающихся взаимоотношений ме-
жду группами насекомых и между
насекомыми и окружающей сре-
дой. Этот последний вопрос под-
робно рассматривается в главах,
посвященных жизнедеятельности,
поведению, происхождению, эко-
логии, полезным и вредным насе-
комым. Для того чтобы отразить
все то новое, что произошло в эн-
томологии за последние годы, од-
ни главы пришлось написать зано-
во, а другие-переписать почти
полностью. В некоторых главах
также были переписаны значи-
тельные части текста, а глава, ка-
сающаяся отрядов насекомых, бы-
ла перестроена таким образом,
чтобы как можно более точно
представить вероятные эволю-
ционные связи между отдельными
группами.
Эволюционным связям, как и
прежде, уделено основное внима-
ние. Для наглядности при описа-
нии этих связей в книге приведены
филогенетические стволы наиболее
распространенных семейств неко-
торых отрядов. Становится все бо-
лее очевидным, что известные нам
данные, касающиеся сравнитель-
ной биохимии, поведения, анато-
мии и т. д., нельзя полностью
осмыслить до тех пор, пока полу-
ченные факты не найдут своего ме-
ста в эволюционных системах. В
главе о происхождении насекомых,
переработанной в значительной
своей части, дается геологический
фон эволюции насекомых и описы-
ваются факторы, влияющие на нее.
В заключение хотелось бы вы-
разить благодарность всем тем,
8
Предисловие
кто помогал нам в подготовке и на-
писании нескольких изданий этого
руководства. Особенно мы благо-
дарны за поддержку, предложения
и критические замечания, относя-
щиеся к настоящему изданию. Д-ра
Д. Эшдаун (D. Ashdown) из Техно-
логического университета шт Те-
хас, Ф. Карпентер (F. М. Carpen-
ter) из Гарвардского университета,
Р Дик (R. W Dicke) из Вискон-
синского университета, Я. Кукало-
ва-Пек (J Kukalova-Peck) из Уни-
верситета в Карлтоне, Дж. Лэттин
(J Lattin) из Университета шт
Орегон, Ф. Лоусон (F A. Lawson)
из Университета шт Вайоминг,
У Мак-Кафферти (W. Р McCaf-
ferty) из Университета Пардю,
Р Мэттьюз (R. W Matthews) и
Я. Мэттьюз (J. R. Matthews) из
Университета Джорджии, Т. Пе-
терс (Т М. Peters) из Массачусет-
ского университета и Р Торп
(R. W Thorp) из Калифорнийского
университета в Девисе прочитали
полностью или частично рукопись
этого издания, сделав много
ценных и глубоких замечаний и
внеся ряд предложений. Особая
благодарность Д. Крейгу
(D. A. Craig) из Университета
пров. Альберта, А. Дитцу
(A. Dietz) из Университета Джорд-
жии, К. Гамильтону (К. G. A. Ha-
milton) из Agriculture Canada,
П. Хантеру (Р Hunter) из Универ-
ситета Джорджии, А. Ли (A. Lea)
из Университета Джорджии и изда-
телям John Wiley and Sons за их по-
мощь и внимание к нашей работе.
Новый материал для переиздания
был подготовлен с помощью г-жи
Джун Росс. Мы благодарны также
многим организациям и лицам,
давшим разрешение на использо-
вание их материалов в качестве ил-
люстраций в нашей книге; все они
названы в подписях под рисунка-
ми. Мы бесконечно обязаны Па-
триции Комбс, г-же Крис М о де-
ленд и г-же Джоан Роули из Уни-
верситета шт Вашингтон за тща-
тельность, с которой они печатали
главы нашей рукописи.
Чарлз А. Росс
Джун Р. П. Росс
1
Развитие энтомологии
в Северной Америке
Насекомые и близкие к ним беспозвоночные, представляющие собой объект
энтомологии, являются удивительной группой животных. Среди всех живых ор-
ганизмов, населяющих землю, насекомым принадлежит первое место как по числу
видов, так и по численности. Число экологических ниш, занимаемых насекомыми,
огромно: здесь и обитание на других насекомых, и питание на живых или мерт-
вых растениях, и обитание на теле или во внутренних органах высших живот-
ных. Нередко вид имеет свою узкоспецифическую экологическую нишу Строение
тела насекомых, их тканей и органов часто по-своему не менее сложно, чем
у высших позвоночных, хотя размер тела многих взрослых насекомых достигает
всего лишь 1-2 мм. Большинство насекомых обладают огромным репродуктив-
ным потенциалом. Поскольку насекомые используют в пищу те же естествен-
ные продукты, что и люди, и поскольку многие из них разными способами
наносят вред нашему здоровью, их можно считать наиболее серьезными кон-
курентами человека за пищу и промышленное сырье. Порой казалось, что насе-
комые почти одолели человека в этой конкурентной борьбе, однако сейчас
нам все-таки удалось установить сложное равновесие в отношениях с этими кро-
шечными, но многочисленными противниками. В наши дни стоимость поддержа-
ния этого равновесия и цена наших побед над насекомыми постоянно растут.
Человек всегда страдал от насекомых. Первобытных людей беспокоили блохи
и вши, кусали комары, им досаждали мухи. В тот ранний период, когда челове-
ческие популяции были рассеяны и разобщены, борьба за существование состояла
в том, чтобы день за днем находить пищу и избегать нападения хищников. Ко-
нечно, насекомые и вызываемые ими болезни не могли быть тогда столь опас-
ными для людей, как другие, враждебные для них факторы среды. Насекомые
были даже в какой-то степени полезны, поскольку термитов, кузнечиков и личинок
некоторых насекомых можно было сравнительно легко найти и съесть в тех слу-
чаях, когда не было иной пищи.
Начиная с первобытной эпохи, прогресс человеческого общества основы-
вался на изменении условий окружающей среды. Естественно, что человек изме-
нял среду с целью сделать ее более благоприятной для своего собственного выжи-
вания и увеличения численности популяции. Однако любое изменение, полезное
для людей, было полезным и для насекомых-вредителей. Постепенно, по мере
того как наиболее страшные враги первобытного человека, такие, как леопарды
и тигры, становились все менее опасными для него, росло значение насекомых
как одного из факторов внешней среды, мешающих дальнейшему развитию циви-
лизации.
Прежде всего за увеличением численности популяций человека последовало
увеличение численности эктопаразитов - вшей и блох. Это было вызвано появле-
10
1. Развитие энтомологии в Северной Америке
нием большего числа хозяев и созданием благоприятных условий для распростра-
нения и размножения насекомых. Эти же факторы способствовали распростране-
нию эпидемий, в том числе и болезней, переносимых насекомыми. Возникавшие
большие города не раз опустошались страшными эпидемиями, как это случилось,
например, с Римом во II в. н. э. во время эпидемии бубонной чумы.
Насекомые начали угрожать не только здоровью человека. Когда люди стали
делать запасы продуктов, это привлекло множество дотоле безвредных насекомых,
уничтожавших и портивших продукты питания. В наше время, когда создаваемые
запасы чрезвычайно велики, насекомые-вредители ежегодно уничтожают миллионы
тонн продуктов, несмотря на проведение обширных и дорогостоящих истреби-
тельных мероприятий.
Когда численность населения возросла настолько, что люди уже не могли
удовлетворяться природными источниками пищи, началось одомашнивание живот-
ных. Увеличение численности животных привело к увеличению числа эктопарази-
тов и болезней, которые отчасти делали бесполезными попытки увеличить коли-
чество продуктов питания. С появлением и развитием земледелия в корне измени-
лись условия жизни насекомых. Монокультуры способствовали возникновению
огромнейших популяций насекомых-вредителей. Египетский писатель времен Рам-
зеса II (1400 г до н. э.), выражая сочувствие крестьянам, писал, что «черви уничто-
жили половину урожая пшеницы, а гиппопотамы съели оставшуюся; поля кишат
крысами и кузнечиками». Позднее новые высокоурожайные сорта растений оказы-
вались более привлекательными для ряда насекомых, чем дикие растения, что
приводило к увеличению числа видов, наносящих вред культурным растениям.
Ситуация осложнилась с развитием связей между различными частями света.
Насекомые попадали на новые для них континенты, где находили благоприятный
для себя климат и хозяев и где не было их естественных врагов, контролиро-
вавших численность на родине. Иногда результаты принимали характер бедствия,
как, например, в случае с завозом стеблевого мотылька и японского жука
в Северную Америку. Оба этих вида не имели большого хозяйственного значе-
ния в районах своего распространения в Евразии, но в США причиненный ими
ущерб оценивался миллионами долларов в год.
В Северной Америке вред, причиняемый насекомыми, был особенно велик, что
явилось непосредственным результатом возделывания многих завезенных в эту стра-
ну растений, интродукции разнообразных новых вредителей и внедрения ряда агро-
технических приемов, которые оказались благоприятными для многих эндемичных
видов. В 1974 г. вред, причиненный насекомыми продуктам сельского хозяйства, был
оценен более чем в 25 млрд, долларов, и вместе с 5 млрд, долларов, затраченными на
истребительные мероприятия, это составило сумму в 30 млрд, долларов. Сюда не
входят потери от вредителей леса, от болезней и гибели людей от инфекций, переда-
ваемых насекомыми, или от снижения производительности труда, вызванного напа-
дением кровососов, эти потери трудно оценивать, но они составляют около 15 млрд,
долларов. Таким образом, общий годовой счет, который мы можем предъявить насе-
комым, составляет 45 млрд, долларов.
Если трудно зримо представить себе вред от насекомых, выраженный в миллиар-
дах долларов, то можно сделать это иначе. В 1974 г каждый человек в США в сред-
нем «платил» насекомым 225 долларов. Для семьи из 4 человек это составляло 900
1. Развитие энтомологии в Северной Америке
11
долларов. Часть этих денег расходовалась на инсектициды, часть - на возмещение по-
врежденных продуктов, но основная доля затрат была связана с увеличением стоимо-
сти предметов потребления растительного и животного происхождения - строи-
тельных материалов, одежды, продуктов питания.
Перечисление бед, вызываемых насекомыми, может создать весьма неблагоприят-
ное впечатление обо всей этой группе животных, однако поговорка «Нет худа без до-
бра» оправдывается и здесь. Многие насекомые очень полезны. Вредители на самом
деле составляют менее 10% всех известных видов насекомых. Примером чрезвычайно
полезных насекомых могут служить пчелы, не только создающие для нас прекрасный
дорогостоящий продукт, но и опыляющие многие ценные виды растений. Урожай
большей части наших фруктов и овощей зависит от насекомых-опылителей, таких,
как пчелы, бабочки, мухи, жуки. Без них у нас не было бы яблок, груш, гороха, тыквы,
кабачков и плодов многих других растений, опыляемых насекомыми.
Еще одна группа полезных с хозяйственной точки зрения насекомых состоит из
большого числа хищников и паразитов других насекомых. Сюда относятся наездни-
ки, паразитические осы, паразитические мухи и божьи коровки. Взрослые особи или
личинки этих видов являются хищниками или паразитами многих важных вредите-
лей, и в некоторых случаях их успешно используют для борьбы с вредителями. На-
пример, божья коровка родолия используется в качестве наиболее эффективного
средства подавления австралийского желобчатого червеца - вредителя цитрусовых в
Калифорнии.
Интегрированный метод борьбы с вредителями включает уничтожение вредных
видов и улучшение условий для развития полезных. Во многих отношениях эта
область близка к той отрасли медицины, которая занимается борьбой с болезнями и
инфекциями. В Северной Америке на интегрированный метод борьбы тратятся боль-
шие суммы денег Прежде всего тысячи людей заняты изучением насекомых, имею-
щих хозяйственное значение, и разработкой мер борьбы с вредителями. Хотя первей-
шей задачей борьбы является снижение вреда, причиняемого вредителями, давно
известно, что эффективная борьба требует серьезного научного фундамента. В связи с
этим проводятся исследования в самых различных направлениях. Некоторые из них,
казавшиеся маловажными при своем возникновении, в дальнейшем принесли огром-
ную пользу для решения проблем, связанных с контролем численности вредных
насекомых.
ИСТОКИ СОВРЕМЕННОЙ БИОЛОГИИ
Можно было бы предположить, что т<
насекомые, будучи столь многочис- н
ленными, играли важную роль в исследо- к
ваниях, заложивших основы современ- и
ной биологии. Однако против этого сви- ф
детельствуют малые размеры больший- д
ства насекомых. Для изучения анатомии ж
и физиологии насекомых необходимы д
тонкие методы препаровки, а для изуче-
ния систематики почти всех групп насе-
комых-достаточно мощные оптические
приборы. Поэтому объектами ранних
фундаментальных биологических иссле-
дований были, чаще всего, более крупные
животные. Появившись в качестве от-
дельной отрасли биологии, энтомология
12
1. Развитие энтомологии в Северной Америке
использовала вначале методы и прин-
ципы, разработанные в смежных обла-
стях.
Чтобы получить правильное предста-
вление о развитии энтомологии в Новом
Свете, полезно остановиться прежде на
возникновении и развитии науки, кото-
рой энтомология Нового Света обязана
своим рождением,-современной биоло-
гии, возникшей в Старом Свете, а имен-
но в Европе.
Открытие Америки испанцами совпа-
ло с «веком авторитетов» - эпохой, пре-
пятствовавшей прогрессу мировой на-
уки. В научной литературе того времени
велись, например, жаркие споры по та-
кой проблеме, как число зубов у лошади;
при этом широко приводились цитаты
известных «ученых мужей», но никому,
по-видимому, не приходило в голову
просто сосчитать зубы у лошади.
В последующие XVI и XVII вв., когда
шло освоение и колонизация Америки,
началось постепенное развенчивание ав-
торитетов и ученые вернулись к наблю-
дениям и экспериментам. Освоение
страны и научный прогресс уходили
своими корнями в одну почву. В эпоху,
последовавшую за Ренессансом, у людей
вновь появилось желание наблюдать и
осмысливать окружающую действитель-
ность.
В биологии толчком к возобновле-
нию наблюдений послужила работа Ве-
залия (1543) по анатомии человека, а на-
чало экспериментальной биологии поло-
жила работа Гарвея (1628), доказавшего
наличие артериального и венозного то-
ков крови у человека. Наблюдение и экс-
перимент, по словам Лоуси (Locy),
«стоят у истоков биологической науки
после эпохи Ренессанса». С появлением
микроскопа в XVII в. связаны микроана-
томические исследования Мальпиги
(Malpighi) и Сваммердама (Swammer-
dam), а также открытие микроорганиз-
мов Левенгуком (Leeuwenhoek), которое
потрясло научный мир.
Начало развития энтомологии отно-
сится к XVII в. Датой ее рождения могут
считаться 1667 и 1668 гг., поскольку в
1667 г. Реди (Redi) использовал насе-
комых для проверки теории самозаро-
ждения. Он помещал мясо в банки, одну
из которых закрывал пергаментом, дру-
гую - густой металлической сеткой,
а третью не закрывал совсем. Испорчен-
ное мясо привлекало мух, откладывав-
ших на него яйца, из которых вылупля-
лись личинки. В банке, закрытой перга-
ментом, яиц не было, до мяса, лежавше-
го в банке, закрытой металлической сет-
кой, мухи добраться не могли, но их
привлекал запах мяса и они откладывали
яйца на сетку. Вылупившиеся личинки
так и оставались на сетке и их не было на
мясе. Отсюда Реди сделал вывод, что
«черви», которые встречаются на испор-
ченном мясе, на самом деле вылупляют-
ся из яиц, откладываемых насекомыми, а
не самозарождаются, как предполагали
до этого.
В 1668 г Мальпиги опубликовал ре-
зультаты исследования по анатомии
шелкопряда; кроме того, были опубли-
кованы первые энтомологические ра-
боты Сваммердама. Эти два ученых про-
вели тщательные аналитические исследо-
вания, снабдив свои работы подробными
иллюстрациями, демонстрирующими
строение мельчайших структур и органов
(рис. 1.1). Эти работы послужили стиму-
лом для последующих исследований по
анатомии насекомых.
Одновременно с развитием этих на-
правлений биология подходила к новому
важному этапу своей истории. Наблюде-
ния, проводимые в природе, пробудили
интерес к естественной истории, и начали
появляться книги, посвященные свя-
занным с ней проблемам. Ранние трак-
таты Уоттона (Wotton, 1552) и Геснера
1. Развитие энтомологии в Северной Америке
13
Рис. 1.1. Вскрытая нимфа поденки-рисунок, сделанный Сваммердамом.
Одна из самых первых работ, посвященных изучению насекомых,
опубликованная около 1675 г. (Из Essig, 1942. College entomology.)
14
1. Развитие энтомологии в Северной Америке
Рис. 1.2. Знаменитый музей шведа Олауса Ворма. Рисунок сделан его основателем;
опубликован в 1655 г (Репродукция предоставлена Waldo Shumway.)
(Gesner, 1551-1556) содержали лишь на-
чатки биологических знаний, носили об-
щий характер, и в них авторы не пыта-
лись провести границы между различны-
ми группами животных. Даже животные,
персонажи народных сказок, былин,
преданий, рассматривались авторами
как реально существующие. Более позд-
ние работы Рея (Ray), относящиеся к
концу XVII в., имели уже более прочную
основу, и в них было дано ясное предста-
вление о видах, близкое к современному.
Появились музеи естественной истории
(рис. 1.2.), где были выставлены удиви-
тельные и незнакомые животные, приво-
зимые в Европу путешественниками и
мореплавателями. Многие из этих му-
зеев были открыты на средства богатых
меценатов и явились предшественниками
многочисленных частных коллекций, сы-
гравших в дальнейшем важную роль в
развитии систематики.
РАЗВИТИЕ БИОЛОГИИ В XVIII В.
По мнению историков, в XVIII в. до-
стиг своей кульминации бунт против ав-
торитетов, когда деспотизму авторитар-
ному и церковному было противопоста-
влено стремление отдельных ученых
утвердить право на самостоятельное ре-
шение сложных проблем. Историческим
выражением этих настроений явились
Американская и Французская револю-
I. Развитие энтомологии в Северной Америке
15
ции. В США одним из последствий рево-
люции был подъем общественного и
частного образования.
Тенденция к проявлению личной ини-
циативы в XVIII в., несомненно, нашла
свое отражение в новом подъеме в разви-
тии биологических наук в Европе. Осо-
бенно много талантливых исследовате-
лей привлекла энтомология. Голландцу
Лайонету (Lyonet, 1750) принадлежит
весьма тщательное анатомическое иссле-
дование гусеницы зимней пяденицы, эта
работа была первой и лучшей публика-
цией ученого. Примерно в то же время
вышли в свет очень важные с точки зре-
ния интересов общества того периода об-
ширные работы немецкого ученого Росе-
ля (Roesel), французского исследователя
Реомюра (Reaumur) и шведского ученого
де Геера (De Geer). Им принадлежат под-
робные, прекрасно иллюстрированные
наблюдения по развитию, поведению и
отличительным признакам многих насе-
комых.
К середине XVIII в. относится начало
движения, имевшего огромное значение
для всех естественных наук. Мы уже
упоминали о том, что Джон Рей впервые
четко сформулировал понятие вида. Од-
нако названия видов (по-латыни) пред-
ставляли собой, по существу, описания-
длинные (часто в несколько строк), гро-
моздкие и допускающие различные тол-
кования. В большинстве случаев первое
слово было существительным и соответ-
ствовало родовому названию в нашем
понимании, остальная часть наименова-
ния была определением и указывала на
вариации внутри этого родового назва-
ния. Например, во времена Джона Рея
одна из бабочек именовалась Papilio me-
dia, alis pronis, praefertim interioribus, ma-
culis oblongis argenteis perbelle depictis, a
другая - Papilio parva nigra duplici in alis
exterioribus macula alba insignis. Первым
словом в каждом названии было суще-
ствительное Papilio, остальная часть
представляла собой сложное определе-
ние, содержащее развернутое описание
вида. Студенты, изучавшие виды в при-
роде и определявшие по таким описа-
ниям растения и животных, были, безус-
ловно, первыми, кто попытался сокра-
тить эти наименования. Постепенно они
были сокращены до названий Papilio me-
dia и Papilio parva, состоящих из суще-
ствительного и только одного прилага-
тельного, относящегося к какому-то ха-
рактерному признаку данного вида. Та-
кая форма наименований известна как
биноминальная номенклатура.
Этот период ознаменовался появле-
нием первой исчерпывающей классифи-
кации растений и животных, известных к
тому времени, создателем этой класси-
фикации был Линней (Linnaeus,
рис. 1.3). В 1758 г вышло в свет десятое
издание его труда “Systema Naturae”, в
котором впервые биноминальная систе-
ма наименований была использована
единообразно в пределах большой, насы-
щенной материалом книги. Система ока-
залась настолько удобной, что ученые
различных специальностей стали немед-
ленно пользоваться ею. Влияние Линнея
на ученый мир было столь велико, что
выход десятого издания его “Systema Na-
turae” был официально признан от-
правным моментом в деле создания зоо-
логической номенклатуры. Хотя латин-
ский язык не является теперь общепри-
нятым научным языком, как это было до
конца XVII в., латинские названия живых
организмов сохранились и поныне и ис-
пользуются в качестве научных названий
во всем мире.
Сохранение биноминальной номен-
клатуры имело колоссальное преимуще-
ство по двум основным причинам. Во-
первых, такая система давала легкий
способ обозначения видов, не допускаю-
щий двусмысленного толкования, так
16
1. Развитие энтомологии в Северной Америке
Рис. 1.3. Карл Линней (1707-1778) в возрасте
40 лет. (По Schull.)
что исследователям, работающим в раз-
личных областях науки и в разных стра-
нах, стало легче идентифицировать виды,
с которыми работали их коллеги. Это
имело решающее значение для дальней-ч
шего развития сравнительной анатомии,
физиологии и других биологических дис-
циплин. Во-вторых, она открыла неогра-
ниченные возможности для обозначения
простым способом новых родов и видов.
Насколько необходимо было введение
биноминальной номенклатуры для раз-
вития науки, показывают следующие
цифры Линней описал около 4500 видов
животных, включая 2000 видов насе-
комых, на сегодняшний день описано
1 250 000 видов животных, из них пример-
но 900 000-насекомые.
Таким образом, систематикам после-
линнеевского времени предстояло опи-
сать и дать названия множеству видов во
всех частях света. Многие из ранних ра-
бот не отличались глубиной и подверга-
лись критике ряда ученых, однако в ко-
нечном счете они подготовили почву для
создания теории эволюции и появления
таких отраслей биологии, как экология й
лимнология.
Старая система обозначения видов
более всего тормозила развитие таксоно-
мии насекомых. После появления лин-
неевской системы она начала преобра-
зовываться в специальную дисциплину.
Первым выдающимся систематиком на-
секомых был Фабрициус (Fabricius), гол-
ландец по происхождению, ученик Лин-
нея. Его первая работа “Systema Entomo-
logica” вышла в свет в 1775 г., последую-
щие работы были напечатаны
в 1782-1804 гг. Фабрициус в своих трудах
обращался к фауне насекомых всего ми-
ра. В конце его научной деятельности
стало очевидным, что изучение мировой
фауны не под силу одному человеку По-
этому последователи Фабрициуса, рабо-
тавшие в области систематики в первой
половине XIX в., занимались изучением
либо одной группы насекомых, либо
фауны одной страны.
Исследования Реомюра, Линнея, Де
Геера и Фабрициуса послужили стиму-
лом для бурного развития систематики
насекомых в Европе и дали толчок для
развития энтомологии в Северной Аме-
рике.
РАЗВИТИЕ ЭНТОМОЛОГИИ
В СЕВЕРНОЙ АМЕРИКЕ
Рождение американской энтомологи-
ческой науки следует отнести к началу
XIX в. В течение первых двух третей это-
го века она развивалась медленно и ха-
рактеризовалась появлением отдельных
интересных работ, которые постепенно
1. Развитие энтомологии в Северной Америке
17
составляли основу для дальнейшего на-
учного прогресса. После Гражданской
войны, однако, создались условия для
более интенсивного развития энтомоло-
гии в США. Потребность в энтомологи-
ческих исследованиях привлекла талант-
ливых и энергичных ученых, и уже к
концу XIX в. энтомология в Америке
превратилась в стройную науку с боль-
шим теоретическим и практическим ба-
гажом.
Энтомологические исследования
до начала XIX в.
До начала XIX в. о насекомых Север-
ной Америки было известно очень мало.
Впервые изображения' североамерикан-
ских насекомых появились на страницах
книги американского натуралиста Марка
Кейтсби (Catesby, 1679-1749) «Естествен-
ная история Каролины, Флориды и Ба-
гамских островов с рисунками птиц, зве-
рей, рыб, насекомых и растений». Фабри-
циус дал названия некоторым насе-
комым по экземплярам, присланным ему
различными американскими коллекцио-
нерами или имевшимся в частных евро-
пейских коллекциях. Джон Абботт (Ab-
bott), англичанин, живший в штате
Джорджия, собрал в течение 1780 г.
огромный материал для европейских
коллекционеров и сделал много рисунков
насекомых. Экономические потери, свя-
занные с деятельностью насекомых и
особенно вред, причиняемый ими запа-
сам зерна, вызвали в 1782 г. серьезное
беспокойство президента США Томаса
Джефферсона. Он дал ряд рекомендаций
по борьбе с насекомыми-вредителями и
указал на необходимость проведения
специальных исследований. Однако поч-
ти до конца XVIII в. американские спе-
циалисты еще не объединили свои усилия
для изучения фауны насекомых своей
страны.
Подготовительный период
(примерно 1800-1866)
Работы американских ученых, касаю-
щиеся американских насекомых, начали
появляться на рубеже XVIII и XIX вв.
Одними из первых были опубликованы
многочисленные статьи У Д. Пека
(W D. Peck), в которых рассматрива-
лись насекомые-вредители в штатах Но-
вой Англии. Эти статьи появились в
1795-1819 гг в различных сельскохозяй-
ственных журналах. Первой фундамен-
тальной энтомологической работой в Се-
верной Америке был «Каталог насе-
комых Пенсильвании» Ф. В. Мелсхайме-
ра (F V Melsheimer), опубликованный в
1806 г Основная роль этой небольшой в
60 страниц книги состояла в том, что она
послужила своего рода стимулом, вы-
звавшим появление новых энтомологи-
ческих работ Сам автор, его коллекция и
его связь с более поздними исследовате-
лями оказали неоценимую помощь в ста-
новлении и развертывании энтомологи-
ческих исследований. Энтомологическая
коллекция Мелсхаймера была первой
крупной коллекцией в Северной Америке
и много лет спустя ее приобрел Гарвард-
ский музей сравнительной зоологии.
В 1812 г. группа энтузиастов органи-
зовала Академию естественных наук в
Филадельфии. Эта группа сыграла веду-
щую роль в зарождении серьезного опи-
сательного направления во многих обла-
стях биологии в Америке, в том числе
и в энтомологии. Среди членов группы
был выдающийся энтомолог Томас Сей
(Say) (рис. 1.4), которому принадлежит
первый классический труд-иллюстриро-
ванный трехтомник «Американская энто-
мология, или описание насекомых Север-
ной Америки» (1817-1828). Исключи-
тельно высокое качество этой работы, а
также других его статей о насекомых
принесли Сею заслуженный титул «отца
18
1. Развитие энтомологии в Северной Америке
Рис. 1.4. Томас Сей (1787-1834)-отец
американской энтомологии. (По Howard,
1930, с разрешения министерства сельского
хозяйства США.)
американской энтомологии». Сей умер в
Нью-Гермоне, шт. Индиана, в 1834 г.
В 1823г доктор Т У Гаррис
(Т W Harris) (рис. 1.5) из штата Масса-
чусетс опубликовал первую статью из
последовавшей затем серии работ о жиз-
ненных циклах и хозяйственном значении
ряда насекомых. Гаррис окончил Гар-
вардский университет в 1815 г., где учил-
ся у Пека, читавшего курс естественной
истории. Всю жизнь Гаррис собирал и
изучал насекомых, и размах его научных
работ был огромен. Его основной труд
«О вреде, приносимом насекомыми рас-
тениям» появился в 1841 г и был дважды
переиздан (последний раз в 1852 г.). Гар-
рис получил 175 долларов от админи-
страции штата Массачусетс на свои ис-
следования-это была первая энтомоло-
гическая работа в Северной Америке, по-
лучившая финансовую поддержку влас-
тей. Труд Гарриса был также первым
учебником по прикладной энтомологии,
и Гаррис по праву считается основателем
этой отрасли науки в Америке.
Работы Сея и Гарриса немедленно пу-
стили корни. В течение нескольких лет
были опубликованы десятки работ о жиз-
ненных циклах, поведении, хищниках
вредных насекомых и мерах борьбы с
этими насекомыми.
Интересно, какими же средствами
пользовались в те времена для борьбы с
насекомыми. Напомним, что соединения
мышьяка, пиретрум, ДДТ, фосфорорга-
нические соединения, карбаматы и мно-
гие другие эффективные инсектициды
еще не были известны на Западе, хотя со-
единения мышьяка и пиретрум уже в те-
чение многих столетий применялись для
борьбы с насекомыми в некоторых стра-
нах Востока. Как следует из книги Гарри-
са, средства для борьбы с насекомыми в
Америке включали «сбор насекомых ру-
ками, стряхивание их в тазы, опрыскива-
ние растений известковыми белилами и
клеем, обработка серой и шотландским
табаком, окуривание табаком под пере-
носным тентом, поливка раствором мы-
ла из ворвани, мыльной и табачной во-
дой», а также ряд рекомендаций санитар-
но-гигиенического характера. Можно ви-
деть, что среди перечисленных мер
борьбы были многие предшественники
современных методов.
Развитие сельского хозяйства приве-
ло к созданию в 1853 г нового Бюро по
сельскому хозяйству при федеральном
правительстве. Энтомологом и экспер-
том этого бюро был назначен Т Главер
(Т Glover). В его обязанности наряду с
прочими входило изготовление экспона-
тов семян, растений, плодов, а также на-
секомых. Кроме того, он проводил боль-
шую исследовательскую работу, в том
Развитие энтомологии в Северной Америке
19
Рис. 1.5. Таддеус Уильям Гаррис
1 “*95—1856) - основатель прикладной
энтомологии в Америке. (Из Howard, 1930, с
разрешения министерства сельского
хозяйства США.)
числе по насекомым-вредителям апель-
синовых деревьев и хлопчатника. Главер
считал, что изображение насекомого
имеет большую ценность, чем препарат.
Он потратил много времени и сил на из-
готовление гравюр на меди с изображе-
нием насекомых Северной Америки.
Убытки фермерских хозяйств, прино-
симые насекомыми, привлекали все
больше и больше внимания. В 1854 г в
штате Нью-Йорк было ассигновано 1000
долларов для изучения насекомых, осо-
бенно вредителей растений. Эти исследо-
вания с 1854 по 1872 г проводил доктор
Аза Фитч (А. Fitch) (рис. 1.6). Он напи-
сал 14 отличных отчетов, включавших
результаты тщательных оригинальных
наблюдений и касавшихся жизненных ци-
клов многих видов насекомых. После ра-
бот Гарриса эти отчеты послужили
новым стимулом для дальнейшего раз-
вития энтомологических исследований в
США. Фитча обычно называли главным
энтомологом штата, хотя официально он
не имел этого титула. Но его деятель-
ность вполне этому соответствовала, и
он был первым энтомологом такого зна-
чения в США.
В период расцвета деятельности Фит-
ча появился еще ряд энтомологов -
Б. Д. Уолш (В. D. Walsh) и К. В. Райли
(С. V Riley) в Иллинойсе, Е. Т Крессон
(Э. Т Cresson) и А. Р Гроут (A. R.
Grote) в Филадельфии. Основная работа
Уолша, не носившая прикладного харак-
тера, была выполнена в 1860-1864 гг., а
его вклад в прикладную энтомологию
относится к последней трети XIX столе-
тия. Работы Райли, Крессона и Гроута
также относятся к этому более позднему
периоду.
Гражданская война, завершившая
рассматриваемый период, незначительно
повлияла на развитие энтомологии, по-
скольку большинство энтомологических
исследований проводилось севернее
арены военных действий. Ее влияние, од-
нако, проявилось уже в первые после-
военные годы.
Европейская наука в этот период. В то
время когда в Америке работали Сей,
Гаррис, Фитч и Уолш, заложившие ос-
новы американской энтомологии, в Ев-
ропе происходили важные события, ко-
торые хотя и не получили в то время
отклика в Америке, имели очень боль-
шое значение для развития энтомологии
на следующем историческом этапе.
Прежде всего европейские системати-
ки делали определенные успехи в пере-
20
1. Развитие энтомологии в Северной Америке
Рис. 1.6. Аза Фитч (1809-1878)-первый
энтомолог на службе штата. (Из Howard,
1930, с разрешения министерства сельского
хозяйства США.)
смотре таксономических принципов, осо-
бенно в пересмотре семейств и родов,
для того чтобы упорядочить огромное
число видов насекомых, описанных в раз-
ных частях мира. Все крупные отряды на-
секомых были подвергнуты ревизии и
созданы классификации, имевшие боль-
шое практическое значение. Европейские
энтомологи пользовались библиотеками
и коллекциями, равных которым в Аме-
рике тогда не было. Многие определи-
тельные таблицы и иллюстрированные
систематические сводки европейских ав-
торов впоследствии использовались аме-
риканскими систематиками для опреде-
ления американских видов насекомых.
Второе обстоятельство имело отно-
шение ко всем биологическим наукам. В
этот период (1800-1866) получили разви-
тие идеи, революционизировавшие био-
логическое мировоззрение в целом. До
этого времени биология была по преиму-
ществу описательной наукой, не имею-
щей по существу фундаментальной тео-
ретической базы. Теперь биологические
законы открывались один за другим.
Оуэн (Owen) выдвинул идею об аналогии-
и гомологии частей тела, Кювье (Cuvier)
и Ламарк (Lamarck) заложили основы
сравнительной анатомии, Милн Эдвардс
(М. Edwards) выдвинул идею разделения
функций; Мюллер (МйПег) показал взаи-
мосвязь анатомии и физиологии, Шванн
(Schwann) и Шлейден (Schleiden) предло-
жили клеточную теорию, Биша (Bichat)
заложил основы гистологии, а фон Бэр
(Von Baer)-основы современной эм-
бриологии; Шульце (Schultze) дал опре-
деление протоплазмы. Апогеем явилась
теория органической эволюции Дарвина
(Darwin) и Уоллеса (Wallace).
То, что все эти открытия были сде-
ланы в такой короткий срок, неудиви-
тельно. Эти идеи буквально «носились» в
воздухе уже в течение многих лет, и одно
важное открытие служило толчком к по-
явлению следующего, уже отчасти ожи-
даемого. Наряду с генетикой и бактерио-
логией, появившимися позднее, эти от-
крытия наметили в общих чертах все
известные ныне биологические законы.
Естественно, что все это сыграло такую
же важную роль в изучении насекомых,
как и в изучении других групп живых
существ.
Начало изучения энтомологии в амери-
канских колледжах. На ранней стадии
подготовительного периода в развитии
американской энтомологии в различных
колледжах Северной Америки начали чи-
таться курсы естественной истории, но
до середины XIX в. они были нерегут
1. Развитие энтомологии в Северной Америке
21
парными и в основном теоретическими.
Эти курсы давались в форме лекций,
иногда сопровождаемых демонстрация-
ми. но лабораторные или полевые заня-
тия чаще отсутствовали или их было
очень мало. В значительной степени это
относилось также к курсам физики и хи-
мии. Первым преподавателем зоологии,
который ввел лабораторные методы
в преподавание, был Луи Агассиц
iL. Agassiz) из Гарвардского универ-
ситета. Сильнейшим стимулом для вве-
дения лабораторных методов в обучение
послужил бурный всплеск биологических
исследований после опубликования
в 1859 г «Происхождения видов» Дарви-
на. Примерно к этому же времени отно-
сится и усиление преподавания научных
дисциплин в новых университетах - Кор-
неллском и Джона Гопкинса. Это совпа-
ло и с организацией в 1862 г. феде-
ральных колледжей на специально выде-
ленных для них участках в соответствии с
решением конгресса США, известного
под названием Закона Моррилла (Mor-
rill), о подготовке специалистов в обла-
сти сельского хозяйства, техники и есте-
ственных наук. Энтомология была вклю-
чена в курс биологии лишь как одна из ее
частей, но уже закладывались основы для
последующего развития ее как самостоя-
тельной дисциплины.
ПЕРИОД БУРНОГО РАЗВИТИЯ
АМЕРИКИ (ПРИМЕРНО 1867-1900)
В течение двух или трех десятилетий
после Гражданской войны энтомология в
Америке заметно продвинулась вперед.
Этому способствовал ряд событий, на-
иболее важными из которых были сле-
дующие :
1. В результате миграции тысяч лю-
дей на запад после Гражданской войны
на Среднем Западе и Тихоокеанском по-
бережье начало бурно развиваться сель-
ское хозяйство. В связи с периодически-
ми вспышками численности насекомых-
вредителей фермеры нуждались в квали-
фицированной помощи энтомологов.
Это послужило причиной организации в
короткий срок государственных учрежде-
ний и учреждений в отдельных штатах,
занимающихся вопросами прикладной
энтомологии.
2. Постепенное расширение и систе-
матизация энтомологических коллекций
и комплектование библиотек доступной
европейской литературой расширило
возможности и улучшило качество работ
по описанию американских видов насе-
комых. Недавнее открытие основных
биологических законов способствовало
развитию различных направлений
в изучении насекомых.
3. Потребность в квалифициро-
ванных энтомологах привела к созданию
специальных курсов по энтомологии в
колледжах и университетах.
Сельскохозяйственная
энтомология
До начала этого периода
(1867-1900 гг.) шт. Нью-Йорк благодаря
деятельности Азы Фитча был един-
ственным штатом, где проводились ак-
тивные энтомологические исследования.
В 1866 г. учредили должность энтомоло-
га штата в шт Иллинойс (хотя деятель-
ность его началась только в 1867 г.), а в
1868 г.-в шт. Миссури. В Иллинойсе эта
должность была предложена Б. Д. Уол-
шу (W D. Walsh), написавшему несколь-
ко прекрасных статей по систематике и
прикладной энтомологии. Уолш
(рис. 1 7) погиб в результате несчастного
случая в 1869 г., однако до своей трагиче-
ской гибели написал как энтомолог шта-
та три отчета. После него этот пост в те-
чение 5 лет занимал Уильям Ле Барон
(W. LeBaron), затем с 1875 по 1882 г-
22
1. Развитие энтомологии в Северной Америке
Рис. 1.7. Бенджамин Дан Уолш
(1808-1869)-автор интересных работ по
различным проблемам энтомологии, первым
занявший должность энтомолога штата
Иллинойс. (По Forbes.)
Сайрес Томас (С. Thomas) и после него-
С. А. Форбс (S. A. Forbes). В штате
Миссури на эту должность был назначен
К. В. Райли, который работал с 1868 по
1876 г Его годовые отчеты были блестя-
щими как с точки зрения их научного со-
держания, так и иллюстраций, и служили
сильным стимулом для деятельности
других исследователей.
В период с 1874 по 1876 г. в несколь-
ких штатах, где интенсивно возделыва-
лись зерновые культуры, появилась пере-
летная саранча. Райли, который зани-
мался изучением причин этого бедствия,
считал, что против саранчи необходимо
применить меры в масштабе всей
страны. Его усилия по созданию нацио-
нального законодательства убедили кон-
гресс учредить Комиссию Соединенных
Штатов по энтомологии. Впервые при-
кладная энтомология была объявлена во
всеуслышание делом государственной
важности, а ее проблемы - выходящими
за рамки компетенции отдельных шта-
тов. Председателем комиссии стал Рай-
ли, ее членами-А. С. Пакард-младший
(A. S. Packard) и Сайрес Томас. Деятель-
ность комиссии продолжалась всего 3 го-
да, но она проделала большую работу и
опубликовала несколько очень полезных
отчетов и бюллетеней, касающихся не
только перелетной саранчи, но также и
многих других насекомых, имеющих хо-
зяйственное значение.
Очень короткий период времени в
1878 г. Райли (рис. 1.8) был преемником
Главера на посту энтомолога федераль-
ного министерства сельского хозяйст-
ва. В течение двух последующих лет
этот пост занимал Дж. Г. Комсток
(J. Н. Comstock), а затем в течение 15 лет
вновь Райли. При нем проблемами энто-
мологии занимался вновь созданный спе-
циальный отдел министерства, который
вскоре превратился в большую действен-
ную организацию, имевшую в своем рас-
поряжении полевые станции во многих
районах страны.
Примерно в то же время начали по-
являться интересные публикации не-
скольких талантливых канадских энто-
мологов, касающиеся насекомых этой
страны. Наиболее выдающимися из этих
публикаций были работы преподобного
С. Дж. С. Бетьюна (С. J. S. Bethune),
первая из которых относится к 1867 г., и
доктора Уильяма Саундерса (W Saun-
ders), чья первая работа появилась в сле-
дующем году. Усилия этих и других
ученых завершились созданием Энтомо-
логического общества в Онтарио
в 1870 г. и учреждением в 1884 г. должно-
сти почетного энтомолога (неоплачивае-
мой) при министерстве сельского хозяй-
1. Развитие энтомологии в Северной Америке
23
Рис. 1.8. К. В. Райли (1843-1895)-автор
прекрасных изображений насекомых,
основатель федерального энтомологического
бюро. (Из Howard, 1930, с разрешения
министерства сельского хозяйства США.)
ства Канады. Этот пост был предостав-
лен Джеймсу Флетчеру (J. Fletcher), ко-
торый затем в 1887 г. перешел на работу
в Центральную сельскохозяйственную
экспериментальную станцию в качестве
энтомолога и ботаника. У Флетчера бы-
ло мало помощников и огромная терри-
тория, которую он должен был держать
под наблюдением. Его прекрасное долго-
летнее служение энтомологическим нуж-
дам сельского хозяйства Канады гово-
рит о его способностях и трудолю-
бии. Он умер в 1908 г. после опера-
ции.
До 1887 г. организованная работа по
прикладной энтомологии в США прово-
дилась только федеральным правитель-
ством и в шт. Нью-Йорк, Иллинойс и
Миссури. В других штатах от Мэна до
Калифорнии лишь отдельные исследова-
тели выполняли такую работу. Однако в
1887 г в ответ на потребности развиваю-
щегося сельского хозяйства и в соответ-
ствии с Законом Хэтча (Hatch) были ор-
ганизованы экспериментальные сельско-
хозяйственные станции во всех штатах. С
момента их образования изучение
вредных насекомых было интенсифици-
ровано, и потребности в квалифици-
рованных энтомологах скоро дале-
ко превзошли имеющиеся возмож-
ности.
К концу последнего десятилетия
XIX в. прикладная энтомология превра-
тилась в важную отрасль науки, имею-
щую большой выход в практику. Огром-
ную работу проводили сотрудники феде-
рального отдела энтомологии, особенно
Л. О. Говард (L. О. Howard), который в
1894 г. заменил Райли на посту руково-
дителя отдела. Учреждения, находящие-^
ся в ведении штатов, часто были связаны
с экспериментальными сельскохозяй-
ственными станциями, в этих учрежде-
ниях работало несколько блестящих спе-
циалистов. Упомянем некоторых из них:
С. А. Форбс в Иллинойсе, Джон
Б. Смит (J. В. Smith) в Нью-Джерси и
Э. П. Фельт (Е. Р Felt) в Нью-Йорке,
сделавшие много оригинальных работ и
напечатавшие ценнейшие отчеты.
В последнюю треть XIX в. на энтомо-
логическую мысль и энтомологические
исследования оказали большое влияние
два события. Примерно в 1869 г. было
установлено, что парижская зелень
является эффективным инсектицидом, и
успешное применение этого препарата
открыло возможности для использова-
ния в борьбе с насекомыми большого
числа «кишечных ядов». Второе событие
было связано с австралийским желоб-
чатым червецом, завезенным в Калифор-
нию примерно в 1870 г. В 80-е годы он
24
1. Развитие энтомологии в Северной Америке
стал столь массовым и серьезным вреди-
телем цитрусовых деревьев, что это при-
вело к упадку целой отрасли, занимав-
шейся выращиванием цитрусовых в за-
падных штатах. Известные инсектициды
оказались бессильны против этого вреди-
теля. Наконец, из Австралии были вве-
зены естественные враги червеца. Один
из них, божья коровка родолия, уничто-
жал вредителя столь эффективно, что в
течение нескольких лет проблема была
разрешена. Этот удивительный успех по-
казал, сколь большое значение могут
иметь биологические средства борьбы с
вредными насекомыми.
Систематика и морфология
насекомых
Основы классификации насекомых
Северной Америки были заложены в те-
чение последней трети XIX в. трудами
многих ученых, в том числе Дж. Л. Ле
Конта (J. L. LeConte) и Г. Г. Горна
(G. Н. Horn) по Coleoptera; А. С. Па-
карда, Генри Эдвардса (Н. Edwards) и
А. Р. Гроута (A. R. Grote) по Lepi-
doptera; Э. Т. Крессона (Е. Т. Cres-
son), Эдварда Нортона (Е. Norton)
и Л. О. Говарда по Hymenoptera;
С. У Уиллистона (S. W. Williston), Ос-
тена Сакена (О. Sacken) и Д. У. Ко-
кийе (D. W. Coquillet) по Diptera;
С. Г. Скаддера (S. Н. Scudder) по Ог-
thoptera; П. Р. Улера (Р. R. Uhler) и
О. Хейдеманна (О. Heidemann) по Не-
miptera; Дж. Г Комстока (рис. 1.9) по
Coccidae; Герберта Осборна (Н. Osborn)
по эктопаразитам и Homoptera. В Канаде
выдающийся вклад в систематику внес
аббат Л. Прованше (L. Provancher), осо-
бенно своими работами по Hymenoptera.
Это лишь некоторые ученые из
плеяды «старых мастеров», впервые опи-
савших значительную часть североаме-
риканской энтомофауны и создавших
Рис. 1.9. Джон Генри Комсток
(1849-1931)-один из «старых мастеров»
преподавания энтомологии в Америке. (Из
Howard, 1930, с разрешения министерства
сельского хозяйства США.)
первые определительные таблицы. Боль-
шой вклад в изучение насекомых Север-
ной Америки и создание руководств по
систематике внесли некоторые выдаю-
щиеся европейские энтомологи. Очень
важно отметить, что многие крупные
систематики этого и следующего перио-
дов были энтомологами-любителями,
для которых их огромный труд на по-
прище энтомологии носил характер хоб-
би, и они не получали за него никако-
го вознаграждения. Можно упомянуть
только немногих из них: Ле Конт и Горн
были практикующими врачами, Эд-
вардс-актером, Нортон и Крессон
1. Развитие энтомологии в Северной Америке
25
бизнесменами, Уиллистон-геологом и
Прованше - служителем церкви.
После введения линнеевской биноми-
нальной номенклатуры появились раз-
личные точки зрения на многие аспекты
ее приложения. Коль скоро появились
подобные разногласия, преимущества
биноминальной номенклатуры оказа-
лись под угрозой сведения к нулю из-за
отсутствия единообразия в ее примене-
нии. Специалисты, занимавшиеся раз-
личными группами животных, искали
способы унификации методов системати-
ки. Наконец, в 1901 г. на Зоологическом
конгрессе в Берлине были приняты
Международные правила зоологической
номенклатуры и создана Комиссия по
зоологической номенклатуре. Именно на
этом историческом конгрессе десятое из-
дание “Systema Naturae” Линнея было
принято в качестве отправного пункта
для создания зоологических наименова-
ний.
Параллельно с развитием системати-
ки росло число крупных научных коллек-
ций. Почти до 1865 г энтомологические
коллекции в Северной Америке были
сравнительно небольшими, не более чем
из нескольких тысяч экземпляров. При-
кладная и теоретическая систематика ну-
ждалась в более полных коллекциях.
Кроме того, уже давно стало очевидным,
что без точного определения системати-
ческой принадлежности невозможны
серьезные исследования во всех областях
энтомологии и правильные рекоменда-
ции для борьбы с вредными насекомыми.
В США первые серьезные шаги в этом
направлении были предприняты по ини-
циативе Луи Агассица, который поручил
Герману А. Хагену (Н. A. Hagen) со-
брать коллекцию насекомых для Музея
сравнительной зоологии при Гарвард-
ском университете. Впоследствии во
многих институтах, академиях наук, уни-
верситетах и других федеральных учреж-
дениях и учреждениях отдельных штатов
были созданы большие коллекции насе-
комых. Многие ученые имели со-
бственные обширные коллекции.
Преподавание энтомологии
До 1867 г. энтомология была частью
курсов по биологии или естественной ис-
тории, читавшихся во многих американ-
ских колледжах. В 1866 г. Б. Ф. Мадж
(В. F. Mudge) в Сельскохозяйственном
колледже штата Канзас прочел курс,
называвшийся «Насекомые, вредящие
растениям»; в 1867 г. А. Дж. Кук (A. J.
Cook) читал курс энтомологии в Мичи-
ганском сельскохозяйственном коллед-
же; в 1870 г. Хаген (Hagen) читал
факультативные курсы по энтомологии в
Гарварде; в 1872 г. С. Г. Фернальд
(С. Н. Fernaid) начал преподавать энто-
мологию в Колледже шт. Мэн, в 1873 г.-
Комсток в Корнеллском университете, а
в 1879 г.-Герберт Осборн в Сельскохо-
зяйственном колледже шт. Айова. Эти
ученые заложили основы преподавания
энтомологии в США. В их распоряжении
не было отлаженных методов преподава-
ния, не было необходимой литературы, и
они, по словам Осборна, «ощупью отыс-
кивали пути, по которым вели препода-
вание,-это касалось и содержания, и
методов».
Задача, стоявшая перед этими учены-
ми, была огромной: собрать исчерпы-
вающую информацию (путем изучения
литературы или проведения собственных
исследований) о многих сторонах биоло-
гии насекомых, включая жизненные
циклы, морфологию, развитие и класси-
фикацию, и осмыслить эту информацию,
используя новые для того времени кон-
цепции общей физиологии, эмбриологии,
филогении и эволюции. Блестящие и до-
ступные для понимания курсы, со-
зданные в результате переработки всего
26
1. Развитие энтомологии в Северной Америке
Рис. 1.10. А. С. Пакард (1839-1905)-автор
первых учебников по энтомологии. (Из
Howard, 1930, с разрешения министерства
сельского хозяйства США.)
этого материала, были величайшими до-
стижениями этих пионеров в преподава-
нии энтомологии. Специального упоми-
нания заслуживают первые учебники, на-
писанные А. С. Пакардом (рис. 1.10),
который поистине явился первопроход-
цем в деле создания учебной литературы.
Организованные в 1888 г экспери-
ментальные сельскохозяйственные стан-
ции нуждались в квалифицированных
энтомологах, и это еще более стимулиро-
вало преподавание этой науки. К концу
столетия курсы энтомологии читались в
большинстве ведущих университетов и
колледжей с естественнонаучным укло-
ном, в частности в федеральных коллед-
жах, организованных в соответствии с
Законом Моррилла. В тот ранний период
усилиями таких выдающихся ученых, как
С. А. Форбс в Иллинойсе, Г. А. Дин
(G. A. Dean) в Канзасе и М. В. Слингер-
ленд (М. V. Slingerland) в Корнелле бы-
ли созданы первые соответствующие
тогдашнему уровню знаний курсы при-
кладной энтомологии, в которых гармо-
нично сочетались теоретические и прак-
тические аспекты этой науки.
РАЗВИТИЕ ЭНТОМОЛОГИИ В XX В.
Выдающиеся открытия конца XIX и
начала XX вв. позволили понять значе-
ние насекомых как переносчиков болез-
ней человека и животных. В 1879 г Па-
трик Мэнсон (Р. Manson), работавший в
южном Китае, доказал, что комары
являются переносчиками филярий.
В конце 90-х годов Рональд Росс
(R. Ross), работая в Индии, установил,
что заболеваемость малярией связана с
комарами Anopheles. В 1900 г. Уолтер
Рид (W Reed) с сотрудниками выявили
роль комаров Aedes aegypti в переносе
возбудителя желтой лихорадки. Эти от-
крытия разрешили загадки, связанные с
распространением тяжелейших заболева-
ний, и показали, какую серьезную опас-
ность представляют насекомые и другие
членистоногие для здоровья людей. Так
возникла медицинская энтомология. В
дальнейшем было установлено, что насе-
комые и клещи переносят целый ряд за-
болеваний, в том числе бубонную чуму,
лихорадку денге, сыпной тиф, окопную
лихорадку, пятнистую лихорадку Ска-
листых гор, африканскую сонную бо-
лезнь. В большинстве случаев борьба с
этими болезнями проводилась путем
уничтожения переносчиков, поскольку
методы иммунизации были еще не-
известны.
Другая волна исследований была вы-
звана тем, что ряд насекомых, заве-
зенных из других стран и адаптировав-
шихся в США и Канаде, начали при-
1. Развитие энтомологии в Северной Америке
27
носить огромный вред сельскому хозяй-
ству. С 1889 г. непарный шелкопряд стал
угрозой для плодовых и других деревьев
в штатах Новой Англии. К 1900 г. выя-
вился колоссальный урон, приносимый
плодовым деревьям калифорнийской
щитовкой. В период с 1895 по 1920 г.
хлопковый долгоносик поразил все рай-
оны хлопкового пояса США. В начале
30-х годов посевы кукурузы на Среднем
Западе сильно страдали от стеблевого ку-
курузного мотылька. В 30-е годы среди-
земноморская плодовая мушка угрожала
подорвать производство цитрусовых во
Флориде. Еще и сейчас мы встречаемся
со случаями интродукции вредителей, на-
пример люцерновой тли, листоеда-пья-
ницы, капрового жука.
Одним из наиболее драматических со-
бытий в истории борьбы с насекомы-
ми-вредителями следует считать начало
промышленного производства нового
синтетического инсектицида ДДТ
в 1945-1946 гг. Раньше применение ин-
сектицидов было дорогостоящим делом,
поскольку применяемые дозировки были
высокими, а получаемый эффект-низ-
ким. В отличие от всех ранее существо-
вавших инсектицидов ДДТ (относящийся
к группе хлорированных углеводородов)
был очень эффективен при чрезвычайно
низких дозировках, отличался высокой
устойчивостью, в результате чего стои-
мость истребительных мероприятий ре-
зко снизилась. Другие синтетические хло-
рированные углеводороды - гексахлоран
(ГХЦГ), альдрин и дильдрин-также да-
вали хорошие результаты при примене-
нии их в самых различных условиях. Эти
химические соединения, казалось, откры-
ли неограниченные возможности для ши-
рокого использования инсектицидов на
угодьях с низкой урожайностью на еди-
ницу площади или с низкой годовой про-
дуктивностью, например на посевах не-
которых полевых культур, на пастбищах,
лугах и в лесах. Медицина уже почти по-
дошла к реальной возможности значи-
тельного снижения заболеваемости и да-
же искоренения таких инфекций, как ма-
лярия и бубонная чума за счет пода-
вления их переносчиков. Казалось, что
остается совсем немного до того дня,
когда эти «волшебные» инсектициды по-
могут нам полностью уничтожить насе-
комых, конкурирующих с нами за пищу и
промышленное сырье и наносящих вред
нашему здоровью.
Однако уже через несколько лет эти
оптимистические взгляды начали подвер-
гать сомнениям. Появились линии насе-
комых-вредителей, которые приобретали
устойчивость к ДДТ и другим инсектици-
дам. Чтобы преодолеть эту устойчивость
и получить необходимые результаты, на-
чали применять все возрастающие коли-
чества инсектицидов. В то же время было
обнаружено, что инсектициды воздей-
ствуют на многие живые организмы,
уничтожая не только вредителей, но так-
же и их хищников и паразитов. Вредите-
ли, приобретшие устойчивость к ДДТ,
лишились, таким образом, части своих
естественных врагов и получили возмож-
ность увеличивать свою численность.
Многие виды насекомых и клещей, чис-
ленность которых держалась прежде на
низком уровне и которые не могли по-
этому приносить заметного вреда, стали
теперь размножаться более интенсивно и
превратились в новых серьезных вреди-
телей, что в свою очередь потребовало
более частого применения инсектицидов
в еще более высоких дозировках.
Хлорированные углеводороды оказа-
лись чрезвычайно устойчивыми в есте-
ственных условиях; период их полурас-
пада составлял несколько лет. Это вело к
их накоплению в среде и быстрому вклю-
чению в цепи питания, а затем-к нако-
плению в тканях наземных и водных по-
звоночных животных, в том числе и
28
1. Развитие энтомологии в Северной Америке
человека, для которого высокие концен-
трации этих инсектицидов были также
токсичны. Экономическое влияние этих
событий было таково, что, например, к
концу 60-х годов продажа рыбы, выла-
вливаемой в озере Мичиган, была запре-
щена из-за накопления в тканях рыб хло-
рированных углеводородов.
Птицы оказались чрезвычайно чув-
ствительными к токсическому действию
хлорированных углеводородов. У мно-
гих птиц масса съедаемой ежедневно пи-
щи примерно равна их весу, и эта пища у
большого числа птиц состоит из насе-
комых. Поэтому инсектициды, содержа-
щиеся в теле насекомых или на их покро-
вах, очень легко попадали в организм
птиц. Это было причиной огромного
снижения во второй половине 50-х годов
численности птиц, особенно насеко-
моядных. Беспокойство людей, вызван-
ное такой ситуацией, получило отраже-
ние в известной книге Рейчел Карсон
«Безмолвная весна», вышедшей в 1962 г.
Затем последовал ряд законов в масшта-
бе всей страны и отдельных штатов,
предусматривавших контроль, ограниче-
ние и даже запрещение применения ДДТ
и других хлорированных углеводородов.
Перед человеком со всей очевид-
ностью возникла следующая дилемма.
Существовали и существуют множество
насекомых, являющихся серьезными вре-
дителями, с которыми следует вести
борьбу. Вместе с тем показавшиеся пона-
чалу «магическими», хлорированные
углеводороды оказались отнюдь не пана-
цеей, и их неограниченное применение в
течение нескольких лет привело к возник-
новению еще более важных проблем, чем
само существование вредителей. Нужны
были новые подходы к проблемам
борьбы с вредными насекомыми. Задача
состояла в том, чтобы организация ис-
требительных мероприятий не сопрово-
ждалась уничтожением существующих в
природе механизмов, регулирующих
естественную численность вредных насе-
комых. Среди путей решения этой задачи
были следующие:
1. Основанное на исчерпывающем
знании жизненных циклов поддержание
численности популяций вредителей на
экономически приемлемом низком уров-
не с тем, чтобы численность популяций
их хищников не снижалась до критиче-
ского уровня.
2. Поиски и изучение бактериальных,
вирусных и других возбудителей болез-
ней насекомых.
3. Использование половых аттрак-
тантов для привлечения и сбора вредных
насекомых.
4. Использование гормонов с целью
вмешательства в репродуктивный про-
цесс и жизненные циклы вредных насе-
комых.
Помимо этого, получили распростра-
нение новые менее устойчивые инсекти-
циды, причем частота их использования
была доведена до минимума-только
против тех стадий развития насекомых,
которые чувствительны к этим инсекти-
цидам, и только тогда, когда это относи-
тельно безопасно для их естественных
врагов. Против некоторых насекомых
достаточно эффективной мерой борьбы
может быть простое изменение сроков
выращивания культурных растений. Ге-
нетики растений создали ряд сортов
сельскохозяйственных культур, высоко-
устойчивых к вредителям. Каждый из
перечисленных методов следует оцени-
вать с точки зрения конкретной ситуации
и путем сочетания нескольких методов
должна быть создана интегрированная
программа. Термин интегрированный ме-
тод борьбы с вредителями кажется нам
очень точным; он означает комбинацию
всех возможных способов и средств для
решения конкретных проблем борьбы с
вредными насекомыми.
1. Развитие энтомологии в Северной Америке
29
В XX в. многократно наблюдалось
оживление энтомологических исследова-
нии как в связи с необходимостью реше-
ния практических задач, вызванных дея-
тельностью вредных насекомых, так и со
стремлением отдельных исследователей
к углублению знаний о насекомых. Раз-
маху энтомологических работ способ-
ствовало не только усовершенствование
традиционных средств исследования, та-
ких, как световой микроскоп, но и по-
явление новых технических средств, та-
ких, как просвечивающий и сканирую-
щий электронные микроскопы, средства
химического микроанализа, сложные
электронные измерительные приборы,
компьютеры для обработки биологиче-
ской и химической информации. Успехам
систематики и смежных областей, безус-
ловно, способствовали и прокладка от-
личных современных дорог, и развитие
воздушного сообщения, позволившие
проводить сбор насекомых по всей
огромной территории земного шара.
Благодаря всему этому при изучении на-
секомых было получено огромное коли-
чество данных, имеющих общебиологи-
ческий интерес, особенно в области био-
химии, физиологии, поведения, эколо-
гии, биогеографии и эволюционного уче-
ния.
Источники получения энтомологиче-
ской информации. С развитием энтомоло-
гических исследований возрастал
и объем энтомологической лтературы.
Энтомологические статьи печатаются во
множестве различных изданий. Новые
данные обычно печатаются в периодиче-
ских изданиях-журналах, бюллетенях,
материалах симпозиумов, выпускаемых
региональными, национальными и ме-
ждународными энтомологическими ор-
ганизациями. Например, Американское
энтомологическое общество (Entomolo-
gical Society of America) выпускает ряд
журналов, в том числе Bulletin и Annals,
Journal of Economic Entomology и много-
численные книги по различным разделам
энтомологии. Лондонское королевское
энтомологическое общество (Royal Ento-
mological Society of London) также публи-
кует различные серии энтомологических
работ Энтомологические общества
имеются во многих странах1 Некоторые
отрасли энтомологии получили столь
сильное развитие, что появились воз-
можность и необходимость в издании
специальных журналов, например Jour-
nal of Insect Physiology, Journal of Applied
Entomology, Environmental Entomology,
Mosquito News.
Значительное число энтомологиче-
ских изданий выпускается федеральным
правительством и властями отдельных
штатов. Министерства сельского хозяй-
ства США и Канады выпускают множе-
ство публикаций о насекомых, имеющих
значение для всех отраслей сельского хо-
зяйства, а также переводы с других язы-
ков по этим проблемам. Кроме того, ад-
министрация штатов издает многочис-
ленные отчеты об энтомологических ис-
следованиях.
Статьи по энтомологии публикуются
в таких периодических изданиях, как
American Midland Naturalist, Canadian
Journal of Zoology, Hilgardia, Journal of
Animal Ecology, Journal of Experimental
Zoology, Nature, Oecologia, Parasitology,
Science, Scientific American и многих дру-
гих. Кроме того, имеется несколько изда-
ний, печатающих обзоры по энтомологи-
ческим проблемам,-Review of Applied
1 В нашей стране энтомологическое об-
щество, ныне Всесоюзное энтомологическое
общество,-одно из старейших в мире. Оно су-
ществует с 1859 г. и выпускает «Труды Все-
союзного энтомологического общества»
(«Horae Societatis Entomologicae Rossicae») с
1861 г. и «Энтомологическое обозрение»
(«Русское энтомологическое обозрение») с
1901 v.-Прим. ред.
30
1. Развитие энтомологии в Северной Америке
Entomology, Annual Review of Entomo-
logy, Quarterly Review of Biology
Многочисленные музеи, универси-
теты и другие научные учреждения часто
публикуют монографии по специальным
ЛИТЕРАТУРА
Cloudsley-Thompson J L. Insects and history.
St. Martin’s Press, New York, 242 pp., 1976.
Decker G. C. The past is prologue, Ent. Soc.
Amer. Bull., 10 (1), 8-15 (1964).
Dupuis C. Pierre Andre Latreille (1762-1833):
The foremost entomologist of his time, Annu.
Rev. Entomol., 19, 1-14 (1974).
Essig E. O. A history of entomology. Macmil-
lan, New York, 1029 pp., 1931.
Essig E. O. College entomology. Macmillan,
New York, 900 pp., 1942.
Gilbert P. A compendium of the biographic liter-
ature of deceased entomologists, Brit. Mus.
Nat. Hist. Publ. XIV, No. 786, 455 pp.,
1977.
Gill T Systematic zoology: Its progress and pur-
pose, Smithsonian Inst. Rep. for 1907, 449-472
(1908).
Howard L. O. A history of applied entomology
(somewhat anecdotal), Smithsonian Misc. Col-
lections, 84, 1-564 (1930).
Leftwich A. W A dictionary of entomology.
Constable, London, 360 pp., 1976.
проблемам, касающимся насекомых. Би-
блиографическая информация содержит-
ся в таких изданиях, как Biological Abs-
tracts, Abstracts of Entomology, Science
Citation Index, Zoological Record.
Locy W. A. Biology and its makers. Henry Holt
& Co., New York, 477 pp., 1910.
Mallis A. American entomologists. Rutgers
Univ. Press, New Brunswick N. J., 549 pp.,
1971.
Mickel С. E. John Ray Indefatigable student of
nature, Annu. Rev. Entomol., 18, 1-16 (1973).
Osborn H. Fragments of entomological history.
Columbus, Ohio, 1937 (издание автора).
Russell L. M. Leland Ossian Howard. A histori-
cal review, Annu. Rev. Entomol., 23, 1-15
(1978).
Smith E. H. The Comstocks and Cornell: In
the people’s service, Annu. Rev. Entomol., 21,
1-26 (1976).
Smith R. F., Mittler T. E., Smith C. N. (Eds.)
History of entomology. Annual Reviews, Inc.,
Palto Alto, 517 pp., 1973.
Spencer G. J. A century of entomology in
Canada, Can. Entomol., 96, 33-59 (1964).
Weiss H. B. The pioneer century of American
entomology. New Brunswick, N. J., 1936
(издание автдра).
2
Arthropoda: насекомые
и близкие к ним группы
Насекомые относятся к обширному типу членистоногих-Arthropoda. Помимо
насекомых в него входят такие разнообразные животные, как крабы, мокрицы, пауки,
клещи, трилобиты, многоножки и многие другие. Основное, что отличает этих жи-
вотных,-это сегментированное тело с посегментно расположенными и также сегмен-
тированными конечностями. Однако в разных группах Arthropoda встречается мно-
жество отклонений от основного плана строения, особенно у неполовозрелых особей.
ПРЕДКИ ЧЛЕНИСТОНОГИХ
Нет сомнения, что Arthropoda про-
изошли от червеподобного предка, близ-
кого по общему строению к Annelida,
или кольчатым червям. Тело этого пред-
ка (рис. 2.1,Л) состояло из серии одина-
ковых сегментов, каждый из которых
представлял собой замкнутое кольцо.
Голова имела простое строение и, по-
видимому, несла чувствительные щетин-
ки. Ротовое отверстие было расположено
на вентральной стороне тела между го-
ловой и первым сегментом тела. Из-за
своего положения впереди ротового, или
стомодеального, отверстия головной от-
дел этих животных называют простомиу-
мом.
Как отмечал Мэнтон (Manton, 1973),
этот червеподобный предок членистоно-
гих отличался от современных Annelida
по ряду важных черт в строении тела. У
.Annelida каждый сегмент тела имеет
свою полость, или целом, причем це-
ломы разных сегментов разделены пере-
городками-септами. В каждом сегменте
имеются выделительные органы (парные
нефридии), открывающиеся наружу вы-
водными протоками. Ноги, если они
имеются, также расположены посегмент-
но и характеризуются примитивным
строением. У гипотетических членов ря-
да, ведущего к Arthropoda, отсутствова-
ли межсегментные перегородки и по-
лость тела простиралась по всей его
длине. Выделительная система предста-
вляла собой один орган, обслуживавший
весь организм животного. Ноги, если они
имелись, были с более специализирован-
ной, чем у Annelida, мускулатурой. На
рис. 2.1,Л Д показаны ступени эволю-
ционного развития гипотетического
предка от простой преартроподной орга-
низации через генерализованный артрп-
подный тип строения к насекомым.
Первая важная ступень состояла в
образовании на вентральной стороне
каждого сегмента пары конечностей, или
ног, обеспечивавших передвижение жи-
вотного (рис. 2.1,Ь). Очевидно, послед-
ний сегмент, перипрокт, на котором рас-
полагалось анальное отверстие, никогда
не имел конечностей.
Следующим вероятным изменением
было слияние конечностей первого сег-
мента и образование лабрума- вентраль-
ного придатка, функция которого состоя-
ла в направлении пищи к ротовому
32
2. Arthropoda. насекомые и близкие к ним группы
Голова Грудь Брюшко
Органы чувств, Локомоторные ункции переваривания пищи и
захвати функции размножения
проглаты вание
пищи
Рис. 2.1. Схематическое изображение
гипотетических стадий (А- Е) в развитии
различных отделов тела и его придатков - от
червеподобного предка до насекомого. (По
Снодграссу, с изменениями.)
Р-ротовое отверстие; 77-простомиум.
отверстию. Конечности второго сегмен-
та переместились на дорсальную сторону
и превратились в антенны. Конечности
третьего сегмента, имеющиеся на ранних
эмбриональных стадиях насекомых, ис-
чезают как самостоятельные структуры
на более поздних стадиях развития. Один
за другим эти первые три сегмента, нося-
щие соответственно названия ла-
бралъный, антеннальный и интерка-
лярный, сливаются с анцестральным про-
стомиумом, образуя сложную голову,
составленную из исходных шести сегмен-
тов. Ганглии этих сегментов сливаются с
первичным мозгом, или архицеребрумом,
и образуется сложный головной мозг
членистоногих.
Эти изменения в строении головы и
головного мозга сопровождались дру-
гим эволюционным преобразованием,
важным для питания животных. Ротовое
отверстие, расположенное вначале поза-
ди лабрального сегмента, затем переме-
стилось в положение позади антенналь-
ного сегмента и, наконец, оказалось по-
зади интеркалярного сегмента. Несом-
ненно, что на этом доартроподном этапе
эволюции конечности нескольких сле-
дующих сегментов, пока лишь слегка мо-
дифицированные, служили для направле-
ния пищи к ротовому отверстию. Рото-
вое отверстие, смещенное кзади, вместе с
лабрумом, также переместившимся на-
зад, образовало нечто вроде кармана, где
пища могла накапливаться до того, как
животное проглатывало ее. Поскольку
термин «простомиум» не соответствовал
более тому строению, которое приобре-
ла первичная голова после перемещения
ротового отверстия назад, она получила
название акрон.
Перемещение ротового отверстия из
положения между акроном и сегментом 1
в положение между сегментами 3 и 4 тре-
бует некоторого разъяснения. На ранних
стадиях эмбрионального развития струк-
2. Arthropoda: насекомые и близкие к ним группы
33
Рис. 2.2. Древние и современные Onychophora. (По Снодграссу, 1938.)
A. Peripatoides novae-zealandiae. Б. Реконструкция ископаемого Aysheaia pedunculata из
среднего кембрия. В. Предполагаемый представитель онихофор Xenusion auerswaldae из
докембрийского или раннекембрийского кварцита. Г, Д. Передний конец тела Peripatoides
novae-zealandiae, вид с вентральной (Г) и латеральной (Д) сторон. Е. Его правая челюсть с
мышцами: а-d - челюстные мышцы; 1 - антенна; 2 - оральная папилла; 3 - л абрум;
4-челюсть; 5-нога; 6-складка вокруг ротового отверстия; 7-глаз; 8-аподема.
туры вентральной части тела формиро-
вались до того, как сегменты приобрета-
ли свою окончательную форму-форму
колец. Предшественниками будущих
нервных центров, или ганглиев, были рас-
положенные в каждом сегменте парные,
но не связанные между собой лате-
ральные массы нервной ткани, назы-
ваемые нейромерами (нервными телами).
На ранних стадиях эмбриогенеза эти от-
дельные нейромеры, по-видимому, про-
двигались вперед по обеим сторонам от
ротового отверстия и затем сливались с
первичным мозгом.
Примерно на этой стадии эволюции
членистоногих произошло превращение
3-732
34
2. Arthropoda. насекомые и близкие к ним группы
конечностей сегмента 4 в хватательные и
режущие придатки, называемые мандибу-
лами. Однако нейромеры этого сегмента
не переместились вперед и ротовое от-
верстие осталось между сегментами 3
и 4.
Эта стадия эволюции представлена
тремя группами Onychophora, Pentasto-
mida и Tardigrada, особенно четко харак-
теризующие ее черты строения можно
наблюдать у современных представите-
лей Onychophora (Peripatus и близкие к
нему формы, рис. 2.2.). Онихофоры
имеют мягкое тело с посегментно распо-
ложенными парными конечностями, хо-
тя внешние признаки сегментации четко
не выражены. Ноги онихофор, назы-
ваемые лобоподиями, не сегментированы,
но имеют специальные мышцы, прикре-
пленные к неподвижным основаниям
(Manton, 1973). У них также имеется пара
придатков, сходных по своему виду и по-
ложению с мандибулами членистоногих,
однако гомология их пока не доказана.
Даже если это и так, онихофор все-таки
следует рассматривать как отдельную
четкую ветвь этого направления эволю-
ции.
Представители двух других групп,
Tardigrada (рис. 2.3.) и Pentastomida
(рис. 2.4.), претерпели столь сильную ре-
дукцию организации, что их родственные
связи требуют дальнейшего тщательного
изучения. Tardigrada (тихоходки, или во-
Рис. 2.3. Тихоходка. (По материалам
министерства сельского хозяйства США.)
Рис. 2.4. Пятиустка. (По Stiles, Shipley.)
Слева- Рог ocephalus annulatus. А. Голова, вид
с вентральной стороны. Б. Целое животное,
вид с вентральной стороны. Справа-личинка
Porocephalus proboscideus, вид с вентральной
стороны: 7-вбуравливающийся передний
конец; 2-первая пара склеротизированных
отростков, видимая между вилками второй
пары; 3 - вентральный нервный ганглий;
4-пищеварительный канал; 5-рот;
6 и 7-железистые клетки.
дяные медведи; рис. 2.3.)-это кро-
шечные животные, живущие во влажном
мху, а также в пресной и соленой воде.
Их тело, никогда не превышающее в дли-
ну 11 мм, имеет 4 пары бугорковидных
ног, заканчивающихся коготками. На го-
лове нет ни четко различимых ротовых
придатков, ни каких-либо других струк-
тур. Pentastomida (язычковые, или пя-
тиустки; рис. 2.4.) - небольшая группа
животных с червеподобными взрослыми
формами и крошечными четырехногими
неполовозрелыми стадиями, по общему
виду похожими на клещей. Пятиустки
являются внутренними паразитами ряда
позвоночных.
Следующая стадия эволюции той вет-
ви, которая ведет к членистоногим, ха-
рактеризуется некоторыми изменениями
в сторону специализации.
1 Arthropoda: насекомые и близкие к ним группы
35
Редуцированное
брюшко
-Листовидные легкие
иди трахеи
Две пары антенн
Тип
Arthropoda
Т ипы
Onychophora
Т ardigrada
Pentastomida
Развитый грудной отд^я
Половые отверстия
на переднем конце тела
Развитые
сегментированные ноги
__ Развитые антенны и
лопастевидные ноги
?ас. 2.5. Гипотетическое родословное древо членистоногих.
1. Ноги приобрели членистое строе-
ние. что способствовало развитию локо-
моторной функции.
2. Некоторые участки покровов тела
затвердели, и в этих местах образовались
жутренние выросты (так называемые
лодемы и апофизы), служащие для при-
крепления мышц, участвующих в движе-
ниях частей тела и конечностей (см. так-
же рис. 5Л,В,Г).
3. Конечности нескольких посто-
ральных сегментов приобрели функцию
-хотя поначалу и не выраженную струк-
турно) перемещения пищи к ротовому
отверстию.
В результате всех этих преобразова-
ний возник характерный представитель
типа Arthropoda.
Самыми ранними из известных чле-
нистоногих животных были трилобиты -
обитатели морей, составляющие подтип
Trilobita (рис. 2.5, 2.6). Они появились в
кембрийский период примерно 600 млн.
лет назад и исчезли примерно 200 млн.
лет назад. У этих животных была неболь-
шая голова, ориентированная вперед, с
антеннами и лабрумом, а позади головы
несколько коротких ног, возможно уча-
ствовавших в захвате пищи. Ноги и их
основания не были, однако, слиты с
головой.
36
2. Arthropoda. насекомые и близкие к ним группы
Рис. 2.6. Общий вид трилобита Triarthrus becki. (Из Schuchert, по Beecher.)
А. Дорсальная (верхняя) поверхность карапакса (щита), состоящего из трех
долей: плевры (5), осевой доли (рахиса, 4) глабеллы (7) и свободных щек (2),
несущих глаза (3). Б. Вентральная (нижняя) поверхность; видны двуветвистые
конечности (7 и 8), прикрепленные к осевой доле, и верхняя губа, или гипостом
(6), покрывающая ротовое отверстие. Двуветвиртые ноги имеют двойную
функцию; верхняя опушенная ветвь несет жабры для дыхания и служит, кроме
того, гребными лопастями, нижняя голая ветвь служит для ползания. Короткие
передние придатки, возможно, участвуют в процессе питания.
Трилобиты дали начало развитию не-
скольких морфологически различающих-
ся групп. Возможно, что две из них по-
служили исходными для образования
двух крупных групп членистоногих-под-
типов Chelicerata и Mandibulata.
У Chelicerata антенны отсутствуют и
тело состоит из двух частей -головогруди
(цефалоторакса), выполняющей функции
питания и локомоции, и брюшка (абдоме-
на), выполняющего функцию размноже-
ния. К Chelicerata относятся скорпионы,
пауки, клещи и близкие к ним группы.
У Mandibulata (ствол, ведущий к насе-
комым) первые три посторальных сег-
мента и их конечности слились с головой,
которая состоит, таким образом, из пер-
вичного акрона и 6 сегментов тела. Ко-
нечности трех сегментов, присоединив-
шихся к голове, превратились в разно-
образные структуры, предназначенные
для захватывания пищи и помещения ее в
рот. Три сегмента, несущие эти ротовые
придатки, называют гнатальными сег-
1 \r±ropoda: насекомые и близкие к ним группы
37
Такое слияние, объединившее
1 функциональную единицу все на-
т'-т£>е структуры, связанные в конеч-
в.'м счете с питанием, представляет со-
характерную черту всех Mandibulata
Zih <*1ень редкими исключениями.
Древние Mandibulata дали начало
J*v линиям, каждая из которых пред-
г~±злена сегодня большой многообраз-
ней группой; это - Crustacea, или рако-
хразные (крабы, креветки и близкие к
•яем формы), и группа Myriapoda-Insecta
многоножки и насекомые). Ракооб-
гилные - преимущественно обитатели во-
среды, как, возможно, и древние
Mirjdibulata. Исходными же формами
зетзг Myriapoda-Insecta, вполне вероят-
й:. были наземные животные. Они могли
игъ в листовой подстилке по берегам
редких затененных водоемов' и речек
г: годно некоторым современным Isopo-
сд Crustacea). Некоторые зоологи счи-
пют. что ракообразные стоят особня-
среди современных Mandibulata, а
схгдство в строении их ротового аппара-
-1 является результатом конвергентной
ЛЮЦИИ.
В ветви Myriapoda-Insecta произошло
гдгяние третьей пары ротовых конечно-
стей. или придатков (которые у ракооб-
тизных называют вторыми максиллами),
i збразовался длинный поперечный вы-
ууг: - лабиум. Этим завершилось форми-
гс-ванне новой и функционально важной
.тр>гтуры - предротовой, или преораль-
полости, передняя и задняя стенки
i г герой образованы лабрумом и лабиу-
*4 ?м. а боковые - мандибулами и максил-
тдлси. Наличие ротовых придатков и
ттсдротовой полости обеспечивало жи-
летным большие возможности для за-
хватывания пищи. Штёрмер (St0rmer,
* » оценивает это морфологическое
ттаспособление как важнейшую адапта-
к наземной жизни.
Наземный предок многоножек и насе-
комых дал начало трем ветвям, сохра-
нившимся и в наше время: одна предста-
влена классом Chilopoda (губоногие),
другая-тремя классами Symphyla, Pau-
ropoda и Diplopoda (симфилы, пауро-
поды и двупарноногие), и третья-клас-
сом Insecta (насекомые). В первых четы-
рех классах (многоножки) ходильные но-
ги имеются почти на всех туловищных
сегментах.
Насекомые подверглись дальнейшей
специализации. Первые три пары локо-
моторных конечностей увеличились в
размерах, остальные редуцировались, а
затем исчезли совсем или превратились в
специальные образования (рис. 2.1 ,£), не
связанные с локомоцией. Таким обра-
зом, локомоторная функция сосредото-
чилась в трех первых сегментах, располо-
женных позади головы, и сформировался
хорошо выраженный отдел тела, нося-
щий название грудь (торакс). Задняя
часть тела, где расположена большая
часть органов, называется брюшком (аб-
доменом). Задние абдоминальные конеч-
ности превратились в наружные половые
придатки, предназначенные для спарива-
ния и откладки яиц. У некоторых рако-
образных грудь и брюшко также четко
разделены, но при этом грудь обычно со-
стоит примерно из 8 сегментов.
Анализ преобразований, имевших ме-
сто в эволюции членистоногих от их при-
митивного безногого предка, позволяет
предположить что а) неспециализиро-
ванные конечности появились сначала на
всех посторальных сегментах и б) затем
подверглись постепенным изменениям,
приведшим к их морфологической и
функциональной дифференцировке. У на-
секомых это привело к четкому разделе-
нию тела на три отдела: голову, несу-
щую сенсорные придатки и ротовые ча-
сти, грудь, несущую три пары ног, и
брюшко, внутри которого находятся
жизненно важные органы, а снаружи-
38
2. Arthropoda. насекомые и близкие к ним группы
терминальные придатки, выполняющие
функцию наружных половых органов.
Представление о родственных связях
внутри типа членистоногих, принятое в
настоящем руководстве, схематически
изображено на рис. 2.5. Согласно этому
представлению, все гексаподы произош-
ли от одной ветви древних членистоно-
гих и поэтому представляют собой моно-
филетическую группу. Как считает Кри-
стенсен (Kristensen, 1975), предлагаемая
схема происхождения насекомых нуж-
дается в наименьшем числе эволю-
ционных параллелей для своего объясне-
ния. Другие авторы придерживаются
иной точки зрения и считают, что не все
насекомые произошли от одной эволю-
ционной ветви, т. е. их происхождение
носит полифилетический характер (Man-
ton, 1949, 1964, 1977). Обе гипотезы о
происхождении насекомых рассматри-
ваются в книге под редакцией Гупты
(Gupta, 1979).
ТИП ARTHROPODA
ПОДТИП TRILOBITA
Тело было обычно плоское, состоя-
щее из головы, груди и пигидия. Две про-
дольные борозды делили его на три доли
(рис. 2.6.). Голова свободно прикрепля-
лась к телу и состояла из первичной го-
ловы (несущей пару длинных членистых
антенн) и четырех сегментов тела,
каждый с парой двуветвистых конечно-
стей. Голова была покрыта карапаксом
(щитом), похожим на раковину или пан-
цирь. У многих видов имелась пара хоро-
шо развитых глаз. Каждый сегмент тела,
за исключением последнего - тельсона,
нес пару двуветвистых конечностей. Три-
лобиты отличались большим разнообра-
зием форм и размеров тела. У большин-
ства из них длина тела составляла
4-6 см, но были и совсем мелкие формы
(10 мм), и гиганты, достигающие в длину
более 60 см.
В раннем палеозое трилобиты пред-
ставляли собой многочисленную группу
обитателей морских водоемов, но уже к
концу этого геологического периода они
вымерли. По своему поведению они на-
поминали современных изопод-бегаю-
щих по дну или плавающих у дна и пи-
тающихся растительными остатками.
Предполагают, что одни трилобиты бы-
ли плотоядными, другие вели пелагиче-
ский образ жизни, питаясь планктоном,
третьи зарывались в грунт и питались
детритом.
ПОДТИП CHELICERATA
Тело типичного хелицерового живот-
ного разделено на 2 отдела головогрудь
(цефалоторакс) и брюшко (абдомен). У
взрослых особей антенны отсутствуют.
Головогрудь обычно несет 6 пар конеч-
ностей: переднюю пару-хелицеры, рас-
положенные близко к ротовому отвер-
стию, педипальпы, часто с клешнями, и 4
пары ходильных ног Брюшко не несет
членистых придатков, и его собственная
сегментированность внешне может быть
не выражена.
Хелицеровые представляют большой
интерес для энтомологов. Многие из них,
например различные клещи, являются,
подобно насекомым, переносчиками бо-
лезнетворных организмов. Множество
клещей питается культурными растения-
ми, паразитирует на животных и челове-
ке, вызывая большие экономические по-
тери. Обычно такие хелицеровые встре-
чаются вместе с вредными насекомыми и
против них проводятся такие же истреби-
тельные мероприятия. Этим частично
объясняется то обстоятельство, что кле-
щей и пауков нередко рассматривают в
курсах энтомологии.
2. Arthropoda: насекомые и близкие к ним группы
39
Рис. 2.7. Мечехвост Limulus. (Из Wolcott, Animal biology, с разрешения
McGraw-Hill Book Со.)
А. Вид с дорсальной стороны. Б. Вид с вентральной стороны.
7 - головогрудь; 2-глаз, 3- брюшко; 4-листовидные жабры.
Класс MEROSTOMATA
Gigantostraca)
Брюшко несет вентральные парные
конечности - жабры и пластинчатые кры-
лечки и заканчивается шипообразным
хвостом, или тельсоном. Жабры служат
органом дыхания и гребными лопастями
при плавании. Представители вымерше-
го подкласса Eurypterida не имели кара-
пакса и были похожи на скорпионов. У
современных морских организмов, отно-
сящихся к подклассу Xiphosura, голово-
грудь покрыта большим подково-
образным карапаксом, в Северной Аме-
рике встречается только один представи-
тель этого подкласса-мечехвост, отно-
сящийся к роду Limulus (рис. 2.7.).
Класс ARACHNIDA
Брюшко большое без наружных жабр
или локомоторных конечностей. Г олово-
грудь несет пару клешневидных хелицер,
пару педипальп, которые также могут
быть с клешнями, и 4 пары ног. У клещей
педипальпы и хелицеры сильно видоиз-
менены, примыкают к ротовому отвер-
стию, и вся эта область имеет вид само-
стоятельной головки.
Этот класс большой и разнообразный
по своему составу. Большинство его
представителей-наземные животные, но
некоторые клещи-обитатели водоемов.
SCORPIONIDA (скорпионы). ЭТОТ Отряд
представлен небольшой группой видов,
включающей только настоящих скорпио-
40
2. Arthropoda: насекомые и близкие к ним группы
Рис. 2.8. Класс Arachnida. (Л - по Packard, Guide to the study of insects, Henry Holt and
Co; Г-из материалов министерства сельского хозяйства США, Б, В и Д-из
материалов Illinois Natural History Survey.)
А. Скорпион Buthus carolinianus. Б. Паук «черная вдова» Latrodectus mactans.
В. Сенокосец. Г. Восточный собачий клещ Dermacentor variabilis. Д. Лжескорпион
Larca granulata.
2 Arthropoda: насекомые и близкие к ним группы
41
нов. с длиной тела от 13 до 18 мм. На
юнце хвостовой части у них имеется ядо-
витое жало (рис. 2.8,Л.), которым скор-
пион очень ловко манипулирует. Яды
газных видов скорпионов различаются
по силе воздействия. В Северной Амери-
ке скорпионы встречаются в основном в
юго-западных штатах, и лишь один-два
вила доходят на северо-востоке до Илли-
нойса.
C ROPYGIDA (телифоны). Представители
?того отряда встречаются главным обра-
з-ом в тропической и субтропической зо-
нах. Длина тела-от 2 до 65 мм. Крупный
М as tigopr octus giganteus встречается в
южных штатах США от тихоокеанского
по атлантического побережья. Здесь
встречается также и несколько мелких
видов, относящихся к разным родам, эти
виды характеризуются короткой хвосто-
вой частью, и их нередко объединяют в
отдельный отряд Schizomida.
palpigrada (кенении). Другой неболь-
шой отряд мелких хелицеровых с длиной
тела около 2,5 мм. Живут под камнями,
в Северной Америке встречаются только
в юго-западных штатах.
xmblypygida (фрины). Небольшая
тропическая и субтропическая группа,
ранее включаемая в отряд Uropygida.
Длина тела колеблется от 4 до 45 мм. Те-
ло несколько сплющено и напоминает те-
ло пауков. Подобно телифонам, ведут
ночной образ жизни. Предпочитают
влажные местообитания.
PSEUDOSCORPIONIDA (ложноскорпионы).
Ложноскорпионы (рис. 2.8,Д)~много-
численный, широко распространенный
отряд. Это небольшие животные, не пре-
вышающие в длину 7-8 мм. Большин-
ство обитает под корой и гниющими
листьями, а род Chelifer встречается так-
же в домах.
ricinuleida (рицинулеи). Небольшая
труппа редких, в основном тропических
арахнид, похожих на пауков, на переднем
конце тела которых имеется большой
подвижный отросток, расположенный
над хелицерами. Мелкие рицинулеи Cry-
ptocellus dorotheae были обнаружены в
штате Техас, в долине реки Рио-Гранде.
araneae (пауки). Этот отряд насчиты-
вает большое число видов (рис. 2.8,Ь);
только в Северной Америке их несколько
тысяч. Ни ноги, ни педипальпы не имеют
клешней. Брюшко несет лишь следы сег-
ментации и сужено у основания, образуя
тонкую перетяжку, соединяющую его с
головогрудью. Педипальпы самца силь-
но видоизменены и приспособлены для
переноса спермы в половые пути самки.
Модификации эти очень разнообразны и
постоянно используются в систематике.
Пауки охотятся за насекомыми и други-
ми мелкими животными, сильно отли-
чаясь друг от друга по способам охоты.
Некоторые догоняют жертву или пры-
гают на нее, пауки-крабы подстерегают
добычу в цветах или других укрытиях,
пока жертва не окажется в пределах дося-
гаемости; многие пауки ткут сети, в ко-
торые попадает жертва. Семейство The-
raphosidae состоит из крупных хищных
покрытых волосками пауков, назы-
ваемых тарантулами.
acarina (клещи). Длина тела может со-
ставлять от 1 до 15 мм. Тело предста-
вляет собой единое образование из слив-
шихся головогруди и брюшка и лишено
внешних признаков сегментации
(рис. 2.8,Г-2.10). Клещи чрезвычайно
разнообразны по строению и образу жиз-
ни. Хотя число видов клещей меньше,
чем пауков, они являются наиболее важ-
ной с хозяйственной точки зрения груп-
пой в классе Arachnida. Некоторые се-
мейства, в том числе иксодовые клещи
(Ixodidae, рис. 2.8,Г), краснотелки
(Trombiculidae), чесоточные клещи (Sar-
coptidae) и клещи-железницы (Demodici-
dae), являются эктопаразитами насе-
комых, птиц и млекопитающих. Пау-
42
2. Arthropoda. насекомые и близкие к ним группы
Рис. 2.9. Отряд Acarina. (Из материалов министерства сельского хозяйства США.)
А,Б. Паутинный клещ Tetranychus pacificus, вид с латеральной и дорсальной сторон. В,Г
Галловый грушевый клещ Eriophyes pyri, вид с дорсальной и вентральной сторон.
тинные клещи (Tetranychidae, рис.
2.9,А,Б) причиняют вред листьям и
вызывают дефолиацию многих куль-
турных растений. Различные Tyroglyphi-
dae (луковичные клещи) вредят пищевым
запасам и повреждают луковицы и корни
луковичных растений. Представители се-
мейства Eriophyidae (рис. 2.9,В,Г) обра-
зуют наросты и галлы на многих эконо-
мически ценных культурах, особенно на
грушевых деревьях. Помимо названных,
многие другие клещи живут в почве и
подстилке, а также паразитируют на пти-
цах и млекопитающих. Два семейства,
Hydrachnellidae и Unionicolidae, обитают
в пресноводных водоемах, a Halacari-
dae-в морских водоемах.
opilionida (сенокосцы) (рис. 2.8,В). Те-
ло крепкое, круглой или овальной
формы, с широким брюшком. У крупных
видов ноги длинные, часто в 4-5 раз
длиннее тела. Мелкие виды с короткими
ногами, похожи на клещей. Эти жи-
вотные обычны во влажных затененных
лесах, где они живут на листьях, стволах
деревьев, в лесной подстилке, питаясь
мелкими насекомыми и другой расти-
тельной и животной пищей. В Северной
2. Arthropoda: насекомые и близкие к ним группы
43
Рис. 2.10. Клещ на ротовых частях личинки Pericoptus truncatus (Coleoptera); х 3700.
(С разрешения Кафедры энтомологии Университета пров. Альберта и Д. Крейга.)
Америке сенокосцы обычны и предста-
влены примерно 200 видами.
solpugida (сольпуги). Небольшой от-
ряд быстро бегающих, покрытых воло-
сками и щетинками арахнид среднего
размера. Их часто называют уксусными
или солнечными пауками. Педипальпы и
ноги короткие, хелицеры крупные,
мощные. Сольпуги обитают в тропиках,
в Северной Америке встречаются в юго-
западных штатах, а также северо-запад-
нее в штате Канзас.
44
2. Arthropoda. насекомые и близкие к ним группы
Класс PYCNOGONIDA
Морские пауки (рис. 2.1 ^-неболь-
шая группа хищных арахнид, похожих на
пауков, с маленьким палочковидным
брюшком. Питаются тканями ги-
дроидных полипов и морских анемонов,
а иногда-медуз.
Рис. 2.11. Морской паук Nymphon. (Из
материалов министерства сельского
хозяйства США.)
ПОДТИП MANDIBULATA
Взрослые формы Mandibulata отли-
чаются от хелицеровых тем, что у них
голова отделена от груди, несет одну или
две пары антенн и три или четыре пары
ротовых придатков. От трилобитов они
отличаются сложным строением головы,
которая представляет собой продукт
слияния ротовых придатков с просто-
миумом.
Многие представители класса Crusta-
cea (ракообразные), например речные ра-
ки, имеют признаки, общие с трилобита-
ми . тело их разделено на голову, грудь и
брюшко (последнее у трилобитов носит
название пигидий), грудь имеет удлинен-
ную форму, латеральные придатки рас-
положены по крайней мере на грудных
ногах. В нескольких классах Mandibulata
имеются паразитические формы, с боль-
шим трудом поддающиеся диагностике.
Класс CRUSTACEA
К этому классу (см. рис. 2.12) отно-
сится такое большое число разнооб-
разных форм, что очень трудно дать ко-
роткое описание, которое характеризова-
ло бы класс в целом. Большинство форм
отвечает следующей характеристике: те-
ло разделено на голову, грудь и брюшко;
голова и грудь часто плотно соединены,
образуя структуру, называемую цефало-
тораксом; голова несет две пары антенн
и у некоторых групп-4 пары ротовых
придатков. пару мандибул, 2 пары мак-
силл и пару максиллипед; грудь обычно
состоит из 4-20 четко различимых сег-
ментов, каждый из которых несет пару
членистых придатков, в составе брюшка
может быть от одного до многих сегмен-
тов, либо несущих короткие придатки,
либо не несущие их. У представителей не-
скольких групп, ведущих паразитический
или прикрепленный образ жизни, сег-
менты тела и придатки сильно редуциро-
ваны, как, например, у паразитических
Copepoda (рис. 2.12,Ь) и у морских уто-
чек. У'представителей некоторых групп
имеется плотный щит (карапакс), покры-
вающий большую часть тела, как, напри-
мер, у речного рака (рис. 2.12,В); у дру-
гих почти все тело вместе с конечностями
покрыто раковиной, которая выглядит
как двустворчатая. У мокриц, относя-
щихся к отряду Isopoda (рис. 2.12,Д), те-
ло слегка уплощено.
Большинство ракообразных обитают
в воде как пресной, так и соленой. Неко-
торые ведут амфибиотический образ
жизни, например определенные вида
речных раков. Единственными многочис-
ленными наземными ракообразными в
2 Arthropoda. насекомые и близкие к ним группы
45
Рис. 2.12. Класс Crustacea. (Л-из материалов Illinois Natural History Survey;
Д-из материалов министерства сельского хозяйства США, остальные из
Ward и Whipple.)
Все формы пресноводные, кроме мокрицы. А. Бокоплав Gammarus sp. Б. Пара-
зитический веслоногий рачок Ergasilus caerulus. В. Речной рак Cambarus
bartoni. Г Креветка Palaemonetes exilipes. Д. Мокрица Armadillidium vulgarе.
Е. Жаброног Branchinecta paludosa.
46
2. Arthropoda: насекомые и близкие к ним группы
Рис. 2.13. Стадии метаморфоза креветки Penaeus. (Из Storer, General zoology,
McGraw-Hill Book Co., no Muller и Huxley.)
Показаны последовательные изменения в строении тела и конечностей (1-19).
Северной Америке являются некоторые
семейства отряда Isopoda. Виды этих се-
мейств предпочитают в основном
влажные местообитания, например
в гниющей древесине, под камнями или в
почве. Ночью некоторые из них поки-
дают свои убежища и ведут свободный
образ жизни. Мокрицы являются вреди-
телями запасов овощей и обитают также
во влажных, богатых органическими ве-
ществами субстратах, например в почве
цветочных горшков. Как в природе, так и
в закрытых помещениях они могут до-
стигать высокой численности и причи-
нять серьезный вред.
По разнообразию форм рако-
образные соперничают с насекомыми.
Для некоторых групп ракообразных ха-
рактерен метаморфоз (последовательная
смена форм в жизненном цикле)-биоло-
гическая особенность, общая с насекомы-
ми. Типичным примером развития с ме-
таморфозом среди ракообразных служит
развитие креветок (рис. 2.13), у которых
в жизненном цикле имеется 4 неполово-
зрелые стадии, предшествующие появле-
нию взрослых, и, таким образом, всего 5
разных морфологических форм.
Среди ракообразных различают 5
подклассов. Один из них (морские уточ-
ки) представлен исключительно морски-
ми формами, в четырех других наряду с
морскими имеются также наземные и
пресноводные виды.
ГРУППА MYRIAPODA
Представителей этой группы, в кото-
рую входят 4 класса-Diplopoda, Chilo-
poda, Symphyla и Pauropoda, часто назы-
вают многоножками, поскольку почти
все сегменты их тела несут конечности.
Все они имеют четко отграниченную от
туловища голову (состоящую из первич-
ного простомиума, слившегося с не-
сколькими сегментами тела, придатки
которых образуют ротовые части) и про-
долговатое туловище с ходильными но-
гами. У представителей всех четырех
классов имеются антенны, причем в не-
которых случаях они очень хорошо раз-
виты. Несмотря на кажущееся сходство,
классы многоножек различаются между
собой по многим важным признакам.
Класс CHILOPODA (губоногие)
Тело губоногих (рис. 2.14, 2.15) имеет
удлиненную форму и разделено на мно-
жество сегментов, каждый из которых
2 Arthropoda. насекомые и близкие к ним группы
47
несет пару ног, а на предпоследнем сег- дов. Большинство из них охотятся
менте расположено еще и половое отвер-
стие. На голове имеются длинные ан-
тенны и глаза, представляющие собой
либо отдельные простые глазки, либо их
скопления. В состав ротового аппарата
фис. 2.16) входят мандибулы (челюсти)
и две пары максилл. Максиллы первой
пары слились и напоминают лабиум на-
секомых; максиллы второй пары (назы-
ваемые также пальпогнатами) похожи на
ноги, иногда их основания сливаются и
образуют мост под первыми максилла-
ми. Интересным признаком Chilopoda
являются ядоносные коготки
фис. 2.16,В) на конечностях первого ту-
ловищного сегмента. Эти конечности
расположены ниже головы и имеют
внешнее сходство с ротбвыми частями.
По своему образу жизни Chilopoda -
хищники. В США известно около 100 ви-
Рис. 2.14. Скутигера (мухоловка) Scutigera
forceps. (Из материалов Illinois Natural
History Survey.)
ночью, а днем укрываются под опавши-
ми листьями, гниющей древесиной или в
трещинах почвы. Виды, обитающие в
умеренном климате, редко превышают
4 см в длину, однако на крайнем юге
США встречаются несколько тропиче-
Рис. 2.15. Типичная многоножка из класса
Chilopoda, (губоногие). (По Снодграссу.)
Рис. 2.16. Ротовые части и ядоносные
челюсти многоножки геофилы (Geophilus
flavidus). (По Latzel.)
А. Правая мандибула. Б. Две пары максилл.
7-слившиеся основания первой пары
максилл; 2-слившиеся основания второй
пары максилл (пальпогнат). В. Ядоносные
коготки, или токсикогнаты.
48
2. Arthropoda. насекомые и близкие к ним группы
ских видов, достигающих 20-25 см в дли-
ну. Один вид с особенно длинными нога-
ми (рис. 2.14)-обычный обитатель до-
мов.
Класс SYMPHYLA (симфилы)
Длина тела этих животных достигает
5 мм. По внешнему облику они похожи
на мелких сколопендр (Chilopoda,
рис. 2.17). Туловище состоит примерно
из 15 сегментов (при этом слившихся сре-
ди них нет), из которых 11 или 12 сегмен-
тов имеют ходильные ноги. Половое от-
верстие расположено на переднем конце
тела. Голова несет длинные антенны и
ротовые части, состоящие из мандибул,
максилл и лабиума. Многие виды не
имеют широкого распространения,
встречаясь обычно в гумусе; некоторые
виды, питающиеся на корнях растений,
являются вредителями садов и теплиц.
Представителей рода Scolopendrella
в США иногда обнаруживают в пробах
из верхнего слоя почвы. При поверхност-
ном взгляде мелких Symphyla можно спу-
тать с Pauropoda, встречающимися в тех
же местообитаниях.
Класс DIPLOPODA-двупарноногие,
или тысяченожки (рис. 2.18)
За исключением нескольких сегмен-
тов на переднем и заднем концах тела,
туловищные сегменты слиты попарно, и,
таким образом, каждый различаемый
сегмент несет две пары ног Ротовой ап-
парат состоит из пары мандибул и имею-
щего вид пластинки гнатохилария, пред-
ставляющего собой, по-видимому, слив-
шиеся максиллы. Лабиум отсутствует.
Половое отверстие расположено позади
второй пары ног
Двупарноногие живут под опавшими
листьями, в гниющей древесине и других
влажных субстратах. В США встречается
около 100 видов, преимущественно в
лесных местообитаниях. Большинство
видов питаются разлагающимися рас-
тениями, немногие-живыми растения-
ми, и в этом случае они приобретают
практический интерес как вредители.
Класс PAUROPODA-пауроподы
Класс охватывает несколько родов
очень мелких животных. Тело состоит из
11-12 сегментов, дорсальные части ко-
торых слиты попарно. 8 или 9 сегментов
несут каждый по паре ног, располо-
женных на равном расстоянии друг от
друга. Антенны двуветвистые, что не ха-
рактерно для других классов Myriapoda.
Каждый глаз представлен небольшим
'пятнышком. Ротовой аппарат состоит из
пары мандибул и интересного образова-
ния, называемого нижней губой, возмож-
Рис. 2.17. Симфила Scutigerella immaculata. (По Снодграссу,
упрощено.)
2. Arthropoda: насекомые и близкие к ним группы
49
Рис. 2.18. Двупарноногая многоножка (тысяченожка) Parajulus sp. (Оригинал
С. О. Mohr.)
но представляющего собой то же самое,
что и гнатохиларий диплопод. Как и у
Diplopoda, половое отверстие располо-
жено в передней части тела.
В Северной Америке известно около
35 видов с длиной тела от 0,5 до 2 мм.
Они живут на упавших гниющих ветках и
деревьях, особенно много их в местах
скоплений опавшей листвы. Большин-
ство видов, встречающихся в Северной
Америке, имеют нежное белое тело и вы-
глядят как мелкие сколопендры
(рис. 2.19). Представители семейства
Eurypauropodidae характеризуются ко-
ротким и широким телом, несколько на-
поминая мелких мокриц. Представите-
лей этого класса находили лишь в от-
дельных местах на востоке и западе
Североамериканского континента. Об
этих любопытных животных известно
пока очень немного.
НАДКЛАСС HEXAPODA
Все насекомые и энтогнаты (скрытно-
челюстные) относятся к надклассу Неха-
poda. Общим отличительным признаком
этой группы является строение грудного
отдела, состоящего из 3 сегментов,
каждый из которых несет пару ног От-
сюда и название Hexapoda, т. е. шестино-
гие.
Класс ENTOGNATHA
До недавнего времени отряды Collem-
bola, Protura и Diplura объединяли в
группу Apterygota-бескрылых насе-
комых. Однако эти 3 отряда отличаются
от настоящих насекомых энтогнатным
ротовым аппаратом, расположенным
внутри головы в специальной капсуле,
образованной оральными складками
Рис. 2.19. Пауропода Pauropus silvaticus. (По Снодграссу, упрощено.)
4-732
50
2. Arthropoda: насекомые и близкие к ним группы
щек. Современные формы энтогнат опи-
саны в гл. 8.
Класс INSECTA
К этому классу относятся Hexapoda,
имеющие эктогнатный ротовой аппа-
рат, расположенный не внутри, а снару-
жи головы.
Характерные признаки класса. Типич-
ное взрослое насекомое имеет тело, раз-
деленное на три отдела (рис. 2.20).
Передний отдел-это голова, несущая
глаза, антенны и три пары ротовых ча-
стей. Следующий отдел-грудь, состоя-
щая из трех сегментов, каждый из ко-
торых обычно несет по паре ног; во
многих группах насекомых второй и тре-
тий сегменты груди несут также по паре
крыльев. Задний отдел тела-брюшко.
Он состоит из 11 сегментов и не имеет
ног Восьмой, девятый и десятый сег-
менты обычно несут видоизмененные
придатки, приспособленные для спарива-
ния или для откладки яиц. Экзоскелет на-
секомых, как и других членистоногих,
обеспечивает защиту внутренних органов
и сохранение формы тела. К основным
внутренним органам относятся* 1) пище-
варительный тракт, 2) длинное, снабжен-
ное клапанами для насасывания крови
сердце; 3) система трубчатых трахей для
дыхания; 4) парные половые железы, от-
крывающиеся на заднем конце тела, 5)
сложная мышечная система и 6) нервная
система, состоящая из головного мозга,
парных посегментно расположенных ган-
глиев, и коннективов.
За исключением некоторых живоро-
дящих форм, все насекомые отклады-
вают яйца. Неполовозрелые насекомые
линяют несколько раз в течение своего
развития до взрослой стадии и, как пра-
вило, при каждой линьке увеличиваются
размеры тела и появляются специализи-
рованные органы.
Рис. 2.20. Схематическое изображение
типичного крылатого насекомого. (По
Снодграссу, упрощено.)
Неполовозрелые насекомые не имеют
крыльев. Единственным известным ис-
ключением являются представители от-
ряда Ephemeroptera (поденки), у которых
последняя кратковременная неполово-
зрелая стадия уже имеет функцио-
нальные крылья. Неполовозрелые насе-
комые многих групп по своему внешне-
му облику абсолютно непохожи на
взрослых и не имеют ног и других струк-
тур, типичных не только для насекомых,
но и для членистоногих в целом.
Систематическое разнообразие. Среди
насекомых имеется огромное разнообра-
зие форм. Современные представители
класса образуют 27 отрядов, перечис-
ленных в табл. 2.1 и описываемых в гл. 8.
Насекомые - процветающая группа.
Насекомые представляют самую бога-
тую видами группу животных. Число
описанных видов оценивается в 900 ты-
сяч (рис. 2.21), тогда как общее число
описанных видов всех животных - при-
мерно 1,5 млн. Иногда численность насе-
комых бывает столь велика, что они
2. Arthropoda. насекомые и близкие к ним группы
51
Таблица 2.1 Классификация современных
отрядов Hexapoda
Надкласс Hexapoda
Класс Entognatha.
Отряд Diplura-двухвостки (камподеи и
япигиды)
Отряд Protura - бессяжковые
Отряд Collembola - ногохвостки
Класс Insecta.
Подкласс Apterygota-бескрылые насеко-
мые
Отряд Microcoryphia - щетинохвостки
Отряд Thysanura-чешуйницы
Подкласс Pterygota-крылатые насекомые
Ряд Paleoptera-древние крылатые насе-
комые 1
Отряд Ephemeroptera - поденки
Отряд Odonata-стрекозы
Ряд Neoptera-современные или склад-
чатокрылые насекомые
Отряд Dictyoptera - тараканы
Отряд Isoptera-термиты
Отряд Phasmatodea-палочники
Отряд Orthoptera-саранча, кобылки, куз-
нечики
Отряд Dermaptera-уховертки
Отряд Grylloblattodea -гриллоблаттиды
Отряд Embioptera-эмбии
Отряд Plecoptera - веснянки
Отряд Zoraptera - зораптеры
Отряд Psocoptera-сеноеды
Отряд Phthiraptera - вши
Отряд Thysanoptera - трипсы
Отряд Hemiptera-клопы (Heteroptera и
Homoptera)
Отряд Raphidioptera - верблюдки
Отряд Megaloptera - большекрылые
Отряд Neuroptera-сетчатокрылые
Отряд Coleoptera-жуки
Отряд Hymenoptera-пчелы, муравьи,
осы, пилильщики
Отряд Mecoptera-скорпионницы
Отряд Siphonaptera - блохи
Отряд Diptera - двукрылые
Отряд Trichoptera-ручейники
Отряд Lepidoptera - бабочки
образуют целые тучи или привлеченные
светом покрывают улицы города слоем
толщиной в несколько десятков сантиме-
тров.
Как ни одна другая группа животных
Таблица 2.2. Число известных современных
видов мировой фауны, относящихся к
основным группам животных
Группа Число ВИДОВ
Chordata 55000
Echinodermata 5700
Arthropoda (за исключением
насекомых) 80000
Insecta 900000
Mollusca 128000
Annelida 8700
Nematoda 10000
Platyhelminthes 12700
Cnidaria 10000
Porifera 5000
насекомые заселяют любые уголки зем-
ного шара и отсутствуют разве что в
глубинах океана. Они живут в тропиках и
являются одними из немногих по-
стоянных обитателей района Южного
полюса, водные насекомые буквально
устилают собой дно больших рек и озер,
но могут и успешно развиваться в не-
больших лужицах, образовавшихся в ре-
зультате заполнения водой следов от
копыт, саранча, например, может рас-
пространяться на многие километры в
прериях, личинки некоторых мух могут
питаться и достигать зрелости, не поки-
дая ореховой скорлупы, а один из видов
ос прекрасно развивается в крошечных
плодах мелкого растения.
Это лишь несколько примеров, по-
казывающих сколь преуспевающей груп-
пой организмов являются насекомые. В
связи с этим небезынтересно высказать
некоторые соображения относительно
причин, способствовавших достижению
такой огромной численности видов и
особей.
ПРИСПОСОБИТЕЛЬНЫЕ ВОЗМОЖ-
НОСТИ НАСЕКОМЫХ. Насекомые яв-
ляют собой финальную группу эволю-
ционного развития наземных артропод.
Механические преимущества экзоскелета
52
2. Arthropoda: насекомые и близкие к ним группы
PROTOZOA
60000 видов
Примечание:
участок сектора, покрытый точками, соот-
ветствует относительному числу
ископаемых форм в данной
группе
45000
30 000
- 15000
^1°= 3000 видов
Соврем ен
ные
скопаемые! виды
виды-#й|______
Рис. 2.21. Относительное число известных видов животных (современных и
ископаемых) из разных типов. (Из Muller, Campbell, 1954, с разрешения
Systematic Zoology.)
Масштаб
2. Arthropoda. насекомые и близкие к ним группы
53
создали условия для дальнейшей спе-
циализации, обеспечившей им перевес
над своими конкурентами. Благодаря
наличию экзоскелета насекомые полу-
чили: 1) большую поверхность для при-
крепления мускулатуры; 2) прекрасные
возможности для регуляции испарения
воды, что особенно важно для жи-
вотных с малыми размерами и 3) почти
полную защиту жизненно важных внут-
ренних органов от внешних повреждений.
К этим основным факторам следует
добавить другие, как морфологические,
так и физиологические, которые имели
решающее значение для того, чтобы на-
секомые стали такими, какие они есть.
Наиболее важные черты специализации
насекомых описаны ниже.
КРЫЛЬЯ. Способность к полету неиз-
меримо увеличила шансы насекомых на
выживание и распространение по всему
земному шару, за исключением остро-
вов. сильно продуваемых ветром. Увели-
чились также возможности для питания и
размножения, и появились новые спо-
собы для избегания врагов. Расширение
возможностей для добывания пищи от-
крыло путь для использования более спе-
цифических ее источников, особенно в
тех случаях, когда пища или места вы-
плода имелись в ограниченном количе-
стве и были труднодоступны. Например,
только животные, способные летать,
могли адаптироваться к питанию на па-
дали, поскольку нужно было покрывать
большие расстояния в поисках трупов
погибших животных, к тому же годных
для употребления только в течение ко-
роткого времени.
НЕБОЛЬШИЕ РАЗМЕРЫ. Эволюция
насекомых шла в направлении развития
большого числа мелких особей, нежели
небольшого числа крупных. Это позво-
ляло использовать многие новые
необычные виды пищи, встречающиеся в
небольших количествах, а также создава-
ло больше возможностей для укрытия и
избегания врагов. Недостаток малых
размеров состоит в том, что общая по-
верхность тела непропорционально вели-
ка по сравнению с его объемом. Это
сильно повышает испарение и делает на-
земную жизнь практически невозможной
для животных с тонкими покровами те-
ла. Склеротизированный кутикулярный
экзоскелет насекомых обеспечивает регу-
ляцию испарения, и непроницаемость эк-
зоскелета, без сомнения, можно считать
одним из основных факторов, позволив-
ших насекомым сохранить свои малые
размеры.
АДАПТИВНЫЕ СВОЙСТВА МОРФО-
ЛОГИЧЕСКИХ СТРУКТУР. Одна и та же
структура может выполнять различные
функции у насекомых разных групп. На-
пример, передние ноги богомолов и кло-
пов семейства Phymatidae, с помощью
которых насекомые схватывают и дер-
жат жертву, пока она не будет съедена,
служат скорее как дополнительные ро-
товые части, а не ходильные ноги. В дру-
гих случаях существуют модификации
одной функциональной системы, вы-
званные обитанием в разных условиях,
например адаптивные модификации ды-
хательной системы у различных водных
и наземных насекомых.
ПОЛНЫЙ МЕТАМОРФОЗ. Жиз-
ненный цикл насекомых состоит из 4 ста-
дий: 1) яйцо; 2) личинка, или питающая-
ся стадия; 3) куколка, или покоящаяся
переходная стадия и 4) взрослое насеко-
мое, или репродуктивная стадия. Такой
тип жизненного цикла встречается у
групп насекомых с наибольшим числом
видов, в том числе у жуков и мух. Почти
у всех насекомых с таким типом жизнен-
ного цикла рост происходит только за
счет личиночного питания. У взрослого
насекомого поддерживается более или
менее постоянный уровень метаболизма,
и потребляемая пища в основном тратит-
54
2. Arthropoda: насекомые и близкие к ним группы
ся на созревание половых продуктов.
Разные жизненные задачи позволяют ли-
чинкам и взрослым особям жить в совер-
шенно разных местообитаниях с разны-
ми условиями существования, поскольку
условия жизни личинки должны быть
благоприятны для быстрого роста, а ус-
ловия жизни взрослой особи-для спари-
вания, расселения и откладывания яиц.
Полный метаморфоз открыл перед насе-
комыми доступ к огромному числу
разнообразных местообитаний и видов
пищи. Это привело к развитию много-
образных сложных типов поведения. В
некоторых случаях сложился очень ко-
роткий жизненный цикл, основанный на
чрезвычайных способностях личинок к
поглощению больших количеств пищи и
быстрому ее перевариванию. Личинки
мясной мухи, например, могут пол-
ностью развиться в течение 3 дней. Ины-
ми словами, полный метаморфоз позво-
ляет виду использовать преимущества
двух абсолютно разных типов местооби-
тания и в то же время избегать многих их
отрицательных сторон.
УВЕЛИЧЕНИЕ ЧИСЛА ВИДОВ. Фак-
торы, благоприятствующие успешному
развитию популяций насекомых, лежат в
основе процветания видов, но не являют-
ся причиной увеличения их числа. Вместе
с тем большое число видов насекомых-
один из наиболее важных факторов их
процветания как группы. Для объяснения
исключительно большого числа видов
насекомых следует искать другие при-
чины. С точки зрения теории эволюции
особенно важными могут быть следую-
щие:
1. Многие виды насекомых приспосо-
блены к жизни только в узких рамках не-
которых экологических факторов, таких,
как хозяева, температура, влажность.
При относительно небольших, но дли-
тельных изменениях климата, как это
имело место в ледниковый период, ареал
таких видов разбивается на изолиро-
ванные части.
2. Благодаря способности к полету,
крылатые насекомые могут перемещать-
ся на большие расстояния, преодолевая с
воздушными массами водные и другие
преграды. В результате таких перемеще-
ний популяции насекомых могут засе-
лить новые места обитания, географиче-
ски изолированные от мест обитания
основных популяций вида. Такие популя-
ции-колонизаторы могут развиваться в
новые виды, если особи этих популяций
способны адаптироваться к новым усло-
виям существования.
3. Генетическая несовместимость
между изолированными популяциями,
выражающаяся в невозможности спари-
вания и развития потомства, может воз-
никнуть у насекомых очень скоро отча-
сти из-за короткой продолжительности
жизни одного поколения и привести к
ускоренному видообразованию.
Ни один из этих факторов нельзя счи-
тать наиболее важной причиной того,
что насекомые достигли огромного раз-
нообразия и численности. Процесс этот
более сложный, обусловленный раз-
личными комбинациями этих и, конечно
же, других факторов, действующих во
взаимной связи. Необходимо помнить,
что не каждой группе насекомых свой-
ственны все характерные для своего клас-
са особенности. Например, некоторые
отряды целиком состоят из бескрылых
насекомых (пухоеды, вши, блохи), что
связано с ограничением их передвижения
поверхностью тела хозяина и небольшим
пространством около него. Полный ме-
таморфоз отсутствует почти у половины
отрядов, но у них появляются другие
особенности. Следует также помнить,
что описанные адаптации-это лишь не-
многие наиболее важные из огромного
числа адаптаций, получивших развитие у
насекомых.
1 Arthropoda. насекомые и близкие к ним группы
55
Т/ГГЕРАТУРА
t’jrwn D. Т Embryology and phylogeny in
iznelids and arthropods. Pergamon, Oxford,
-*5 pp.. 1973.
E. W., Wharton G. W An introduction
acarology. Macmillan, New York, 465 pp.,
:*52. [Имеется перевод: Бэкер Э., Уар-
?н Г Введение в акарологию-М. ИЛ,
>55.]
R. D. Invertebrate zoology. Saunders
CrlSeee Holt Rinehart & Winston, Philadelp-
—i. 4th ed., 1089 pp., 1980.
J. C. The arachnid order Chelonet-
Stanford Univ. Pubis., Univ.Ser.Biol.
". 1-284 (1931).
h.'^isley-Thompson J. L. Spiders, scorpions,
centipedes and mites. Pergamon, Oxford,
pp.. 1968.
.^siock J. H. Gertsch W J. The spider book.
Comstock, Ithaca, N.Y., 729 pp., 1940.
D. R. Water mite genera and subgenera
Mem. Am. Entomol. Soc., No. 21, 860 pp.
2^—arr E. C. A guide to the identification of the
:.:onon land and freshwater animals of the
Jzzted States (rev. ed.). 119 Prospect St.,
'vrrthhampton, Mass, (издание автора),
55S op.. 1950.
yndson W T., Ward H. B., Whipple G. C.
Sir*. Freshwater biology. Wiley, New York,
ed.. 1248 pp., 1959.
И' J. American spiders. Van Nostrand
Remhold, New York, 2nd ed., 274 pp., 1979.
.4. P. (Ed.). Arthropod phylogeny. Van
Xostrand Reinhold, New York, 762 pp., 1979.
C " С. C. The pseudoscorpions of Illinois, Bull.
~ Nat. Hist. Surv., 24, 413-498 (1949).
< B. J. Spiders of Connecticut, Bull. Conn.
3eol. Nat. Hist. Surv., 70, 1-784 (1948).
B. J. How to know spiders.
W C. Brown, Dubuque, Iowa, 3rd ed., 1978.
<7? P. E. Pycnogonids. Hutchinson, London,
>4 pp., 1973.
G. W A manual of acarology. Oregon
Sure Univ. Bookstores, Inc., 335 pp., 1970.
L-jzensen N. P. The phylogeny of hexapod “or-
ders*’ A critical review of recent accounts, Z.
Zool. Syst. Evolutionsforsch., 13, (1), 1-44
Levi H. W., Levi L. R. A guide to spiders and
their kin. Golden Press, New York, 160 pp.,
1968.
MacSwain J. W., Lanham U. N. New genera
and species of Pauropoda from California,
Pan-Pacific Entomol., 24, 69-84 (1948).
Manton S. M. Studies on the Onychophora. VII.
Phil. Trans. R. Soc. B., 227, 411^464 (1949).
Manton S. M. Mandibular mechanisms and the
evolution of arthropods, Phil. Trans. R. Soc.
B., 247, 1-183 (1964).
Manton S. M. The evolution of arthropodan lo-
comotory mechanisms. Part II. Zool. J. Linn.
Soc., 53, 257-375 (1973).
Manton S. M. The Arthropoda: Habits, functio-
nal morphology and evolution. Oxford Univ.
Press, Oxford, 527 pp. 1977.
McGregor E. A. Mites of the family Tetranychi-
dae, Am.Midl. Naturalist, 44, 257-420 (1950).
Michelbacher A. E. The biology of the garden
centipede, Scutigerella immaculata, Hilgardia,
11, 55-148 (1938).
Pennak R. W Freshwater invertebrates of the
United States. Wiley, New York, 2nd ed.,
1248 pp., 1978.
Ross H. H. A synthesis of evolutionary theory.
Prentice-Hall, Cliffside, N.J., 387 pp., 1962.
Snodgrass R. E. Evolution of the Annelida, Ony-
chophora, and Arthropoda, Smithsonian Misc.
Collections, 97 (6), 1-159 (1938).
Storer T I., Usinger R. L., Stebbins R. C., Ny-
bakken J. W. General zoology. McGraw-Hill,
New York, 6th ed., 902 pp., 1979.
Sttfrmer L. Arthropods from the Lower Devo-
nian (Lower Emsian) of Aiken an der Mosel,
Germany. Part 5: Myriapoda and additional
forms, with general remarks on fauna and pro-
blems regarding invasion of land by arthro-
pods, Senckenb. Lethaea, 57, 87-183 (1977).
Waterhouse D. F. et al. The insects of Australia.
Melbourne Univ. Press, Carleton, Victoria,
1029 pp., 1970; Suppl. 146 pp., 1974.
Wharton G. W. A manual of the chiggers, Ento-
mol. Soc. Wash. Mem., 4, 1-185 (1952).
Williams S. R., Hefner R. A. The millipedes and
centipedes of Ohio, Ohio Biol, Surv, Bull., 18,
4(3), 91-146 (1928).
3
Внешнее строение
В этой главе речь пойдет о наружных частях и отделах тела и его придатках.
Прежде чем приступить к описанию этих частей, необходимо получить некоторые
общие представления о строении стенки тела, которая и образует эти части.
СТЕНКА ТЕЛА И ЭКЗОСКЕЛЕТ
Стенка тела насекомого секретирует
экзоскелет, который служит опорой для
прикрепления внутренней мускулатуры.
Экзоскелет придает жесткость телу насе-
комого и, подобно внутреннему скелету
позвоночных, служит опорой для мышц.
К экзоскелету прикрепляются основные
мышцы, обеспечивающие целостность
тела. Стенка тела может быть достаточ-
но упругой или гибкой, однако растяги-
ваться она способна лишь в течение ко-
роткого времени после линьки.
Поскольку стенка тела служит как для
защиты, так и для прочного прикрепле-
ния мышц, многие ее участки уплотнены
или склеротизированы. Если бы стенка
тела везде была одинаково твердой, то
насекомые не только не могли бы пере-
двигаться, но, например, не могло бы
растягиваться их брюшко, для того
чтобы вмещать потребляемую пищу или
созревающие яйца. Движения становятся
возможными благодаря тому, что
твердые участки тела образуют совокуп-
ность пластин, или склеритов, между ко-
торыми стенка тела остается мягкой и
гибкой, или мембранозной. Такое строе-
ние, с одной стороны, обеспечивает жест-
кость тела и защиту внутренних органов,
а с другой-создает условия для раз-
личных типов движений.
Простой пример того, как работает
эта конструкция, дает брюшко комара
(рис. 3.1). Когда брюшко не наполнено,
его поперечное сечение узкое, эллиптиче-
ское (рис. 3.1,Л), спинной и брюшной
склериты связаны по бокам полосками
тончайшей гофрированной мембраны.
По мере наполнения брюшка кровью эта
мембрана просто расправляется, позво-
ляя спинному и брюшному склеритам
отодвигаться все дальше друг от друга
под действием увеличивающегося объе-
ма пищи, засасываемой в брюшко. В со-
стоянии наибольшего растяжения брюш-
Рис. 3.1. Поперечный срез брюшка комара
(схематизировано) в сокращенном (Л)
и раздутом состояниях (К).
3. Внешнее строение
57
Рис. 3.2. Продольный срединный разрез
телескопических кольцевых сегментов
схематизировано).
4. Втянутое состояние. Б. Вытянутое
состояние. Тонкой линией обозначена
мембрана.
Рис. 3.4. Мембранозные перемычки
в ножном сочленении типа «шара
и впадины».
А. Нога вытянута. Б. Нога согнута,
/-верхняя мембрана, 2-нижняя мембрана.
ко имеет в поперечном сечении почти
круглую форму (рис. 3.1, Б).
Другой широко распространенный
тип мембранного соединения, пока-
занный на рис. 3.2, действует по принци-
пу телескопических колец. Когда тело
подтянуто (рис. 3.2,Л), кольца находят
одно на другое и мембрана втянута
внутрь. Когда же оно вытянуто
«рис. 3.2,Ь), вложенные друг в друга сек-
Рис. 3.3. Сочленение ноги с телом
с помощью мембраны. А. Нога стоит прямо.
Б. Движение ноги вперед.
ции раздвигаются, насколько позволяет
длина связывающих их мембран.
На рис. 3.3 показано, каким образом
гибкая мембранозная полоска обеспечи-
вает движение членистой ноги. Справа
узкий участок мембраны образует шар-
нирное соединение между ногой и скле-
ритом. На рис. 3.3,Л нога стоит прямо;
обратите внимание на то, что левая мем-
брана собрана при этом в складки. В по-
ложении Б нога продвинута вперед; это
движение расправило левую мембрану,
так как основание ноги переместилось
внутрь и назад.
Распространенный тип сочленения ча-
стей ноги взрослых насекомых представ-
лен на рис. 3.4; в этом случае два сегмен-
та имеют соединение в виде шаро-
образного выступа и соответствующей
ему впадины. Когда правый сегмент вы-
тянут, как в положении Л, верхняя мем-
брана (7) сложена, а нижняя мембрана
(2)-растянута. Когда правый сегмент
опущен вниз, как в положении Б, верхняя
58
3. Внешнее строение
мембрана натянута, а нижняя свернута
или собрана в складки.
Во всех четырех приведенных приме-
рах движение возможно благодаря гиб-
кости, но не эластичности. На рисунках
не обозначены мышцы, сокращение ко-
торых на самом деле вызывает движе-
ния, а показаны лишь мембранные соеди-
нения, которые делают эти движения
возможными.
Склериты. Твердые и склеротизиро-
ванные части тела называются склерита-
ми. Главные склериты обычно разделены
участками или полосками мембраны.
Многие крупные склериты могут подраз-
деляться на дополнительные бороздами
или новыми полосками мембраны. Скле-
риты могут также сливаться, причем
обычно в месте слияния остается отчет-
ливая линия, борозда или морщина. В эн-
томологии эти типы границ-полоски
мембраны, борозды, линии слияния-
называют швами. Термин «склерит»
обычно употребляется для обозначения
любого склеротизированного участка
покрова, ограниченного швами любого
типа.
Выросты покровов. Поверхность по-
кровов обычно несет разного рода вы-
росты, в том числе складки, шипы, че-
шуйки, щетинки и волоски. Все эти вы-
росты представляют собой производные
стенки тела. Хотя в плане изучения основ
внешней морфологии они не представ-
ляют особого интереса, однако имеют
огромное функциональное значение, на-
пример при испускании звука или вос-
приятии различных типов раздражите-
лей.
Внутренние выросты. Многочис-
ленные внутренние выросты образованы
впячиванием стенки тела. Они называют-
ся аподемами (см. также рис. 1.5,В). Точ-
ка или линия их впячивания почти всегда
видна снаружи в виде ямки или бороздки.
Эти углубления служат одним из наибо-
лее надежных ориентиров, когда прихо-
дится выяснять, из каких именно склери-
тов образовались аподемы. Аподемы
создают внутри тела поверхности для
прикрепления мускулатуры.
ОТДЕЛЫ ТЕЛА
Тело взрослого насекомого разделено
на три отдела, голову, грудь и брюшко.
Филогенетическое происхождение этих
отделов обсуждается на с. 59. Голова
обычно представляет собой прочную
капсулу, которая утратила видимую сег-
ментацию. Грудь и брюшко сохраняют
явное расчленение и состоят из сегментов
более или менее кольцевидной формы.
У безногих неполовозрелых стадий
некоторых насекомых тело слабо диффе-
ренцировано на отделы. Голова обычно
выражена, а сегменты груди и брюшка
могут быть внешне одинаковыми и вме-
сте составлять единый однородный от-
дел тела-туловище.
Пространственные взаимоотношения.
При описании пространственных взаи-
моотношений различных частей тела на-
секомого, для того чтобы показать на-
правление или расположение, исполь-
зуют ряд специальных терминов. Неко-
торые части тела при этом служат осно-
вой для ориентации, главные из них
следующие
1. Передняя часть-часть тела, несу-
щая голову, или такая часть любого ор-
гана, которая ближе к головному концу
тела.
2. Задняя часть-часть тела, несущая
хвостовые придатки, или «хвостовой ко-
нец» брюшка, или часть любого органа,
расположенная ближе к заднему концу
тела.
3. Дорсальная сторона-верх, или
верхняя сторона тела, или любой из его
частей (органов).
4. Вентральная сторона -низ, или
3. Внешнее строение
59
нижняя сторона тела, или любой из его
частей (органов).
5. Ось-продольная срединная линия
тела, проецируемая либо на дорсальную,
либо на вентральную поверхность, либо
на любую фронтальную плоскость ме-
жду ними.
6. Латеральная часть - боковая часть
тела или любой из его частей (органов).
7 Базальная (основная) и апикальная
(вершинная) части, у придатков или вы-
ростов тела, таких, как усики или ноги,
точка или область прикрепления назы-
вается базальной; концевая или наиболее
удаленная от места прикрепления точка
называется апикальной. Для частей при-
датков, например члеников ноги, исполь-
зуют ту же ориентацию: та часть, кото-
рая причленяется ближе « телу, будет
базальной или проксимальной, а дальняя
от тела часть будет вершинной (апикаль-
ной) или дистальной.
ГОЛОВА
Голова (рис. 3.5) представляет собой
передний отдел тела насекомого. Обыч-
но это капсула со склеротизированной
верхней частью, содержащей мозг, и
мембранозной нижней стенкой, в кото-
рой расположено оральное отверстие,
или рот
Происхождение
Как было показано в гл. 2, голова на-
секомого-сложная структура, состоя-
щая из многих частей. Они срослись друг
с другом так плотно, что лишь немногие
из первоначально составлявших голову
частей могут быть идентифицированы на
неполовозрелых и взрослых стадиях. По-
этому данные, свидетельствующие
о сложной природе головы, мы получаем
только при изучении развития зароды-
шей. Вопрос о том, из каких элементов
сложена голова, был предметом дли-
тельных дискуссий, итоги этих дискуссий
были четко обобщены Ремпелем (Re-
mpel, 1975), взгляды которого мы и при-
нимаем здесь.
Согласно теории Шолля-Ремпеля-
Чёрча, голова состоит из предкового
акрона (называемого простомиумом в
тех группах, где он не слит с сегментами
туловища) и шести туловищных сегмен-
тов (рис. 3.6). У зародыша каждый сег-
мент несет пару вентральных придатков
(рис. 3.7) и аподемы. В поздний период
развития эти структуры сильно видоиз-
меняются. Схема строения головы пред-
ка насекомых, на которой показано пред-
полагаемое расположение сегментов и
соответствующих придатков, изображе-
на на рис. 3.6.
Рис. 3.5. Личинка пилильщика Neodiprion lecontei как пример
насекомого с гипогнатической головой. (По Middleton.)
60
3. Внешнее строение
Рис. 3.6. Строение головы зародыша насекомого; показан предковый
акрон и шесть туловищных сегментов (I-VI). (По Rempel, 1975,
упрощено.)
1 -усик; 2-вставочный сегмент; 3-акрон; 4-аподема верхней губы;
5-вставочный придаток.
Рис. 3.7 Зародыш цикадки Microstelus focifrons, х 75. (С разрешения Dept, of
Entomology, Univ, of Alberta и D. A. Craig.)
3. Внешнее строение
61
Типичная голова насекомого
Ко времени появления насекомых
многочисленные части, входившие в со-
став головы, образовали структуру,
очень сильно отличающуюся от первона-
чальной. В типичной голове насекомого
• рис. 3.8) передняя область, или лицевая
часть, дорсальная и латеральные поверх-
ности образуют непрерывную склероти-
зированную капсулу, открытую снизу
подобно перевернутой чаше. На этой
капсуле располагаются пара сложных
глаз, три глазка и пара усиков. Верхняя
губа свисает вниз с нижнего переднего
края капсулы, образуя козырек перед
ртом. Вентральная часть головы обра-
зует мембранозную стенку позади рта,
эт этой стенки отходит гипофаринкс, на
котором открывается проток слюнных
желез. С каждой стороны этой стенки
свисают вниз три пары придатков,
Счхгтавляющих грызущие органы, или ро-
товые части, представленные верхними и
нижними челюстями и нижней губой.
Они причленяются к вентральному краю
головной капсулы. Задняя часть головы
имеет форму перевернутой подковы, у
которой дорсальная и латеральные части
образованы головной капсулой, а снизу
толкова замыкается нижней губой, от-
крытый центр называется затылочным
’тверстием, через которое проходят пи-
тзевол. нервная цепочка, проток
слюнных желез, аорта, трахеи и свобод-
ная кровь. Внутри головы расположен
комплекс перекладин, называемый Тен-
ториумом (рис. 3.9). В голове находится
также мозг и другие связанные с ним
структуры. Сама капсула несет зри-
тельные структуры двух типов сложные
глаза и простые глазки (рис. 3.10).
СЛОЖНЫЕ ГЛАЗА. Как правило, это
крупные многофасеточные структуры,
расположенные по одному с каждого бо-
ка головы (рис. 3.11). Каждый глаз нахо-
Рис. 3.8. Голова саранчового. (По
Снодграссу.) Показаны придатки и основные
органы.
А. Вид спереди. Б. Вид сбоку, / -щупик;
2-максилла (нижняя челюсть); 3-простые
глазки; 4-сложный глаз; 5-мандибула
(верхняя челюсть), 6-верхняя губа,
7-гипофаринкс; 8-нижняя губа, 9-усик.
дится на глазном склерите или окружен
этим склеритом в виде кольца или усту-
па. У тех групп насекомых, которым
62
3. Внешнее строение
Рис. 3.9. Тенториум перепончатокрылых.
А. Голова Macroxyela, вид сбоку (часть головной капсулы удалена, чтобы показать
тенториум). Б. То же, вид с дорсальной стороны. В. Голова Aleiodes (часть головной
капсулы удалена, чтобы показать тенториум), вид сбоку. Г То же, вид с дорсальной
стороны, / -передние руки; 2-корпоротенториум, 3-верхние руки; 4-дорсальный киль,
простирающийся от тенториума вдоль боков головы; 5-сухожилие.
свойственно полное превращение (голо-
метаболические насекомые), личиночные
глаза, если они имеются, редуцированы
до 1-6-однофасеточных линз-глазков
(рис. 3.12), называемых стеммами.
Группа стемм на каждой стороне головы
называется окулариумом. У взрослых на-
секомых число фасеток может быть
огромно: у комнатной мухи их число в
одном глазу достигает 4000, а у неко-
торых жуков-25 000.
ГЛАЗКИ (ОЦЕЛЛИ). Эти три однофа-
сеточных органа расположены на лице-
вой части, обычно между сложными гла-
зами (рис. 3.10). Два верхних, состав-
ляющих пару и находящихся по обеим
сторонам от средней линии головы, на-
зывают латеральными глазками. Нижний
расположен посередине и называется ме-
диальным глазком. У многих насекомых
глазки отсутствуют или внешне неза-
метны.
НАЛИЧНИК (КЛИПЕУС). Эта область
напоминает губу и лежит между фронто-
клипеальным швом и верхней губой. На-
личник никогда не бывает сочленен со
лбом, а жестко срастается с ним. Верхняя
губа лабрум) свисает и сочленяется с на-
личником посредством мембранозной
перемычки между ними.
ЩЕКИ. Щеки представляют собой
нижнюю часть головы под глазами и по-
зади лба. Иногда впереди на лицевой ча-
сти между лбом и щеками имеется
3. Внешнее строение
63
вечный шов, если же этот шов отсут-
ствует, то определенной границы между
зкками и лбом нет.
ЛОБ. Эта область располагается вни-
зу передней части лица над наличником и
ограничена с боков фронтальными шва-
чи (их иногда называют фронтоге-
нальными), а с вентральной стороны-
гронтоклипеальным швом. Все эти швы
начинаются от передних тенториальных
£мок. Лоб часто отсутствует.
ЗАТЫЛОК, ИЛИ ЗАТЫЛОЧНАЯ ДУ-
ГА. Эта область охватывает большую
Рис. 3.10. Схемы, показывающие основные
швы и области головы. А. Вид спереди
(максиллы и нижняя губа не изображены). Б.
Вид сзади, /-эпикраниальный шов; 2-ветви
эпикраниального шва; 3-темя; 4-простые
глазки; 5-сложный глаз; 6-точка
прикрепления дорсальной руки тенториума,
7-лобный шов; 8-усиковая ямка,
9-щечный шов; /0-щека; //-передняя
тенториальная ямка;
/2-фронтоклипеальный шов, /3-наличник,
14 - верхняя губа, 15 - мандибула,
16-затылочное отверстие; /7-зазатылок;
18-зазатылочный шов; 19-затылочный
мыщелок; 20 - затылочный шов;
21 -корпоротенториум; 22-нижняя губа;
23-защеки; 24-задняя тенториальная ямка;
25-максилла; 26-затылок.
часть задней поверхности головы. Она
отделена от темени и щек затылочным
швом, у многих групп этот шов либо ре-
дуцируется до складки, либо полностью
исчезает, причем в последнем случае за-
тылок можно определить только как
область, сливающуюся впереди с теме-
нем и щеками. Вентральные части всей
затылочной дуги иногда называют заще-
ками.
ЗАЗАТЫЛОК. Это-узкий кольце-
образный склерит, составляющий край
затылочного отверстия. Он отделен от
затылка зазатылочным швом, имею-
щимся практически у всех взрослых насе-
комых. Зазатылок несет затылочные мы-
щелки, с помощью которых голова при-
членяется к цервикальным склеритам
шейной области.
ТЕНТОРИУМ (РИС. 3.9). Голова укре-
плена изнутри комплексом склеротизи-
рованных аподем, или впячиваний стенки
тела, который изначально развился как
более жесткая основа для прикрепления
мышц, связанных с ротовыми частями. У
Collembola, Protura, Diplura и близких к
ним многоножек аподемы более или ме-
нее пластиноподобны или палочковидны
и иногда связаны между собой лигамен-
тами. У предков крылатых насекомых
64
3. Внешнее строение
Рис. 3.11. Голова взрослой самки мошки Simulium, вид сбоку. Видны сложный глаз и
ротовые органы, х 150. (С разрешения Dept, of Entomology, Univ, of Alberta и
D. A. Craig.)
эти структуры увеличиваются, срастают-
ся и превращаются в мощный внутрен-
ний скелет головы, называемый тенто-
риумом. В типичном случае тенториум
(рис. 3.9) состоит из четырех основных
частей: передних рук, задних рук, корпоро-
тенториума, или центрального тела
тенториума, и дорсальных рук. Передние
руки идут от передних тенториальных
ямок, которые обычно хорошо видны в
виде округлых впячиваний с каждой сто-
роны нижнего края лба. Задние руки идут
от задних тенториальных ямок, которые
почти всегда сохраняются в виде за-
метных снаружи щелей на зазатылочном
шве. Корпоротенториум представляет
собой результат продолжения внутрь,
встречи и срастания передних и задних
рук. Дорсальные руки рассматриваются
как вторичные отростки передних рук,
поскольку достаточно больших или по-
стоянных ямок, соответствующих ме-
стам их прикрепления к головной капсу-
ле, обычно находящихся между усиковы-
3. Внешнее строение
65
Рис. 3.12. Голова гусеницы. Видны
личиночные глазки. (По Folsom, Wardle,
1934, Entomology с разрешения The Blakiston
Со.)
ми ямками и латеральными глазками, не
обнаруживается. Форма и относительное
расположение элементов тенториума в
высшей степени различны у разных групп
насекомых.
Коксоподит имел длинный боковой от-
росток, выполнявший, возможно, сразу
две функции-дыхательных жабр и пла-
вательного органа, часто был развит эн-
дит-твердый выступ, направленный к
средней линии тела. У насекомых эти
элементы прототипа видоизменялись
различными путями.
В эволюционном стволе, давшем на-
чало насекомым, боковой отросток ис-
чез. Если бы он сохранился, то мог бы
дать начало некой структуре, напо-
минающей усик насекомого. Верхняя гу-
ба не обнаруживает каких-либо заметных
признаков расчленения. Придатки вста-
вочного сегмента в основном атрофиро-
вались. У посторальных придатков
(верхние и нижние челюсти и нижняя гу-
ба) сильно увеличились в размерах и спе-
циализировались для удержания пищи
коксоподиты и их эндиты, тогда как те-
лоподиты редуцировались и преврати-
лись в органы чувств. Верхние челюсти
полностью утратили телоподиты и пре-
вратились в грызущие, жующие или сосу-
щие придатки.
Придатки головы
Шесть туловищных сегментов, слив-
шихся с предковой головной областью,
или акроном, и вошедших в состав ны-
нешней сложной единой структуры, ко-
торую мы называем головой, несли
парные придатки.
Как и прочие придатки сегментов, от-
носящиеся к остальным отделам тела на-
секомого, придатки головы произошли
из простых ног, типичных для прими-
тивных членистоногих. Таким прототи-
пом, вероятно, стала нога трилобитов
(рис. 3.13). Она состоит из базального
членика, или коксоподита, который ис-
ходно причленялся к боковой стенке тела
посредством жесткой пластинки, и апи-
кальной членистой части -телоподита.
Рис. 3.13. Нога трилобита, а и б-точки
сочленения. (По Снодграссу, с изменениями.)
66
3. Внешнее строение
УСИКИ (АНТЕННЫ)
В типичном случае у взрослых насе-
комых усики представляют собой пару
подвижных членистых придатков, отхо-
дящих от лицевой поверхности, обычно
между глаз. Первый членик называют
скалу сом, второй- педицеллу мом, а все
оставшиеся вместе -флагеллумом. При-
креплен усик в усиковой ямке, иногда
окруженной узким кольцеобразным уси-
ковым склеритом. В периферической ча-
сти ямки имеется небольшой вырост, к
которому причленен сам усик. В эмбрио-
генезе усики развиваются из придатков
2-го головного сегмента. Усики чрезвы-
чайно разнообразны по форме, и неко-
торые наиболее четко выраженные их
типы имеют специальные наименования.
Несколько примеров приведено ниже и
изображено на рис. 3.14:
нитевидный усик
щетинковидный усик
четковидный усик
пиловидный усик
булавовидный усик
головчатый усик
пластинчатый усик
гребенчатый усик
У личинок усики, как правило, сильно ре-
дуцированы как в отношении длины, так
и в отношении числа члеников.
РОТОВЫЕ ЧАСТИ
Тремя важнейшими частями ротово-
го аппарата являются верхние челюсти
(мандибулы), нижние челюсти (мак-
силлы) и нижняя губа (лабиум). Все они
представляют собой результат видоиз-
менения типичных парных конечностей
членистоногих. Форма этих ротовых ча-
стей у насекомых столь сильно отличает-
ся от исходных предковых форм, что для
доказательства их родственных отноше-
ний приходится привлекать данные по
другим группам артропод. Исследование
придатков ископаемых членистоногих и
параллельный анализ данных сравни-
тельной морфологии придатков ныне
живущих форм показывают, что все
имеющиеся придатки тела современных
членистоногих возникли из простого ге-
нерализованного придатка.
Булавовидный
Рис. 3.14. Типы усиков.
3. Внешнее строение
67
Рис. 3.15. Гипотетический придаток
членистоногих, а и б-точки сочленения. (По
Снодграссу, с изменениями.)
Генерализованный придаток членисто-
ногих (рис. 3.15). Базальный членик, или
коксоподит, сидит на боковой стенке те-
ла. Апикальные членики образуют тело-
побит. Каждый членик потенциально
способен давать выросты как на наруж-
ной, так и на внутренней поверхности,
причем наружные отростки называют эк-
зитами, а внутренние -эндитами. При-
мером одной из ранних и примитивных
модификаций этого типа ноги может слу-
жить нога трилобита (рис. 3.13). Обрати-
те внимание на то, что коксоподит снаб-
жен жабровидным экзитом и шипо-
видным эндитом, а телоподит простой и
не несет выростов.
Вставочные (ингеркалярные) придатки
(рис. 3.6). Эта пара простых придатков,
появляющихся на 3-м головном сегменте
в ходе эмбрионального развития неко-
торых насекомых, а затем исчезающих.
Вставочный сегмент на ранних стадиях
эмбрионального развития указывает на
местоположение рта.
Верхняя губа. В типичном случае
представляет собой подвижный козырек,
свисающий с края наличника (рис. 3.8,6)
и прикрывающий рот. Его внутренняя
поверхность образует переднюю стенку
предротовой полости и называется эпи-
фаринксом', он обычно несет выступаю-
щие лопасти и чувствительные щетинки
многих типов. Верхняя губа образова-
лась от слияния парных придатков пер-
вого сегмента головы.
Верхние челюсти (мандибулы;
рис. 3.8; 3.14; 3.16). Верхние челюсти-
парные придатки 4-го сегмента головы,
расположенного непосредственно позади
рта; расположены они таким образом,
что кажется, будто они находятся сразу
за верхней губой. В типичном случае
они твердые, склеротизированные и не-
сут зубцы различной конфигурации и ще-
точки. Щупики отсутствуют. Очевидно,
мандибулы являются видоизмененными
коксоподитами и их эндитами, принадле-
жавшими сегменту древних рако-
образных. У представителей энтогнат
имеется всего одна точка сочленения
мандибул с боковым краем головы. У
щетинохвосток и крылатых насекомых
есть и вторая точка сочленения, располо-
Рис. 3.16. Верхняя челюсть (мандибула)
саранчового, а и б-точки сочленения. (По
Снодграссу, с изменениями.)
68
3. Внешнее строение
женная дорсальнее или медиальнее пер-
вой, этим обеспечивается более мощное
разводящее и смыкающее движение этих
придатков. От каждого из этих мест со-
членения внутрь головы отходит мощное
сухожилие, которое служит для прикре-
пления мышц, управляющих движения-
ми мандибул.
Нижние челюсти (максиллы;
рис. 3.17). Нижние челюсти распола-
гаются непосредственно за мандибулами
и являются парными придатками 5-го го-
ловного сегмента. Они имеют лишь бо-
ковое сочленение с основанием головной
капсулы. Коксоподит состоит из двух ча-
стей-кардо и стипеса и несет остальные
элементы максиллы.
В энтомологической литературе ред-
ко прибегают к терминам «коксоподит»,
«телоподит», «эндит» и т. д., с помощью
которых описываются подразделения и
отростки придатков, общие для всех чле-
нистоногих. Мы до сих пор и поль-
зовались ими, чтобы помочь связать
представления о строении насекомых и
Рис. 3.17. Генерализованный тип нижней
челюсти (максиллы) на примере таракана.
(По Снодграссу.)
остальных членистоногих. Однако
в дальнейшем изложении целесообразно
изменить терминологию и употреблять
те наименования, которыми чаще всего
пользуются энтомологи. Термины эти
касаются частей, развитых по большей
части только у насекомых, и, следова-
тельно, необходимы для обозначения
той или иной детали строения или обла-
сти.
Максиллы генерализованного типа
представляют собой жевательные обра-
зования, разделенные на ряд хорошо
обособленных частей (рис. 3.17), ко-
торые перечислены ниже.
КАРДО. Кардо-треугольный основ-
ной склерит, прикрепленный к головной
капсуле и служащий шарниром, на кото-
ром движется остальная часть нижней
челюсти.
СТИПЕС. Центральная часть всей
нижней челюсти называется стипесом и
по форме, как правило, близка к прямо-
угольнику. Стипес размещается за кардо
и является основой для прочих элементов
максиллы.
ГАЛЕА. Галеа-это внешняя (лате-
ральная) лопасть, причлененная к концу
стипеса. Она часто представлена в виде
сенсорной подушечки или несет венчик
сенсорных органов. Галеа и лациния
представляют собой эндиты.
ЛАЦИНИЯ. Внутренняя (медиальная)
лопасть, причлененная к вершине стипе-
са-лациния,-имеет, как правило, манди-
булоподобную общую форму и несет ряд
шипов или зубцов по внутреннему краю.
ЩУПИК. Усикоподобный членистый
придаток, отходящий от бокового края
стипеса и называемый щупиком, состоит
обычно из 5 члеников. Вероятно, он вы-
полняет исключительно сенсорные функ-
ции.
Нижняя губа (рис. 3.14, 3.18). Это
образование, представляющее собой
парные придатки 6-го головного сегмен-
3. Внешнее строение
69
Рис. 3.18. Генерализованный тип нижней
губы на примере таракана. (По Иммсу, с
изменениями.)
та, служит губой, лежащей позади мак-
силл. Оно кажется единым элементом, но
на самом деле состоит из второй пары
максилл, которые срослись вдоль сред-
ней линии и образовали функционально
единую структуру. Части нижней губы
строго соответствуют частям нижних че-
люстей и их гомология была установлена
по данным, полученным при изучении
мускулатуры и точек прикрепления от-
дельных мышц.
Табл. 3.1 иллюстрирует гомологии
соответствующих частей максилл и ниж-
ней губы. При пользовании этой табли-
цей следует обращаться также к рис. 3.17
и 3.18. Далее следует описание этих
частей.
ПОДБОРОДОК (ПОСТМЕНТУМ). Ос-
новная область нижней губы-подборо-
док - подвешена к мембранам головы.
Она часто подразделяется еще на две ча-
сти: базальную-субментум и апикаль-
ную-ментум. Подбородок представляет
собой слившиеся кардо максилл.
ПРЕМЕНТУМ И ДРУГИЕ ЧАСТИ.
.Апикальная область нижней губы содер-
жит ряд лопастей и отростков. Централь-
ная доля, или тело нижней губы, назы-
Таблица 3.1. Таблица гомологий
Максилла Нижняя губа
Кардо Соответ- Подбородку (Субментум
ствуют (постментум) | Ментум
Стипес » Прементуму
Щупики » Щупикам
Лацинии » Г лоссам
Галеа » Параглоссам
вается прементумом и несет пару нижне-
губных щупиков, по одному с каждой
стороны прементума; у генерализо-
ванных форм щупик состоит обычно из
трех члеников.
Апикальная часть нередко образует
подобие язычка и по этой причине назы-
вается лигула. Она бывает весьма разно-
образно устроена, но обычно делится на
две пары долей. 1) глоссы, внутренняя па-
ра лопастей, в большинстве случаев сом-
кнутых друг с другом, и 2) параглоссы,
боковая пара лопастей, обычно парал-
Рис. 3.19. Часть нижней губы
перепончатокрылого Trichiosoma triangulum
как пример срастания глосс.
7-язычок; 2-параглосса; 3-щупик;
4-прементум.
70
3. Внешнее строение
Рис. 3.21. Нижняя губа и гипофаринкс пилильщика Arge pectoralis (справа)
и браконида Aleiodes tenninalis (слева).
/-гипофаринкс; 2-глосса, 3-параглосса; 4-прементум.
дельных глоссам. Во многих группах, на-
пример у Hymenoptera, глоссы срастают-
ся и образуют язычок (alaglossd)
(рис. 3.19). В других случаях глоссы мо-
гут срастаться и с параглоссами в еди-
ную большую общую лопасть, называе-
мую сложным язычком (totoglossa)
(рис. 3.20).
Гипофаринкс. От вентральной мем-
бранозной стенки головы отходит гипо-
фаринкс (рис. 3.21). Он, как правило,
имеет вид выступающей пластинки или
бугорка. У генерализованных насекомых
гипофаринкс столь тесно связан с основа-
нием нижней губы, что может рассматри-
ваться как часть последней. В отличие от
Рис. 3.20. Нижняя губа и сросшиеся
максиллы перепончатокрылого Тгетех
columba как пример полного срастания всех
лопастей нижней губы.
7-максиллярный щупик; 2-сросшиеся галеа
и лациния; 3-нижнегубной щупик;
4 - сложный язычок; 5 - прементум;
6-сросшиеся стипесы; 7-субментум;
8 -кардо.
3. Внешнее строение
71
остальных ротовых частей гипофаринкс
является не придатком, а представляет
собой нерасчлененный вырост стенки те-
ла. На нем находится выводное отвер-
стие протоков слюнных желез.
Суперлингвы. У немногих прими-
тивных насекомых в тесной связи с гипо-
фаринксом имеется пара простых пла-
стинок. В процессе эмбриогенеза они
возникают как пластинки, сопутствую-
щие мандибулам, но в развитом зароды-
ше они становятся теснее связанными с
гипофаринксом. Эти образования встре-
чаются в некоторых отрядах насекомых,
таких, как Thysanura и Ephemeroptera, у
ряда ракообразных и отдельных предста-
вителей Symphyla. Видимо, их наличие-
признак примитивных членистоногих, и у
более специализирдванных насекомых
эти структуры атрофированы.
ОСНОВНЫЕ ТИПЫ
РОТОВОГО АППАРАТА
В различных группах насекомых ро-
товой аппарат модифицировался, чтобы
наилучшим образом и различными спо-
собами поглощать пищу разного рода.
Наиболее специализированные и разно-
образные типы ротового аппарата пере-
числены ниже; они выбраны так, чтобы
продемонстрировать богатство форм,
принимаемых гомологичными частями,
и различные способы их использования.
Существует также множество других ти-
пов. причем они нередко представляют
собой переходные стадии между теми ти-
пами, которые мы здесь рассматриваем.
Грызущий тип. Ротовые придатки это-
го типа ротового аппарата в основном
соответствуют изображенным ранее на
рис. 3.16, 3.17 и 3.18, см. также рис. 3.22
и 3.23. Мандибулы отрезают и пере-
малывают твердую пищу, а максиллы и
нижняя губа проталкивают ее в пищевод.
Типичными примерами могут служить
саранчовые и личинки чешуекрылых-гу-
сеницы. Представляется вполне ве-
роятным, что ротовой аппарат грызуще-
го типа явился исходным для развития
ротовых органов других типов. Эта точ-
ка зрения подкрепляется важными
данными двоякого рода. Во-первых, та-
кой ротовой аппарат наиболее сходен по
строению с ротовым аппаратом много-
ножек и симфил - ближайших родствен-
ников насекомых. Во-вторых, ротовые
органы грызущего типа обнаружены по-
чти во всех генерализованных отрядах
насекомых, таких, как тараканы, пря-
мокрылые и щетинохвостки, а также у
личинок по крайней мере примитивных
семейств нейроптероидных отрядов. Во
многих нейроптероидных группах
взрослые насекомые часто также имеют
грызущий ротовой аппарат, который
мало в чем изменился по сравнению с
примитивным типом, например у Со-
leoptera и большинства Hymenoptera.
Фильтрующий тип. У некоторых вод-
ных личинок, в частности у личинок мно-
гих мошек (Simuliidae), на голове разви-
вается своеобразный веер, предназна-
ченный для фильтрации микроорганиз-
мов (рис. 3.24). Личинки других мошек
могут как фильтровать, так и соскребать
пищу, и веера у них на голове развиты
слабее (рис. 3.25).
Режуще-сосущий тип (рис. 3.26). У
слепней (Tabanidae) и некоторых других
двукрылых мандибулы вытянуты
в острые лезвия, а максиллы - в длинные
колющие стилеты. Обе пары челюстей
прорезают и прокалывают покровы тела
млекопитающих, вызывая кровотечение
из ранки. Эта кровь собирается с по-
мощью напоминающего губку образова-
ния на нижней губе и доставляется к вер-
шине гипофаринкса. Гипофаринкс и эпи-
фаринкс плотно прилегают друг к другу,
образуя трубку, по которой кровь всасы-
вается в пищевод.
72
3. Внешнее строение
Рис. 3.22. А. Головной отдел нимфы поденки Arthroplea, вид с
вентральной стороны. Максиллярные щупики используются в
качестве фильтрующих органов. Б. То же, вид сбоку; х 35. (С
разрешения Dept, of Entomology, Univ, of Alberta и D. A. Craig.)
3. Внешнее строение
73
Рис. 3.23. Грызущий ротовой аппарат личинки первого возраста Microchorista philpoti
(Mecoptera, Nannochoristidae); x 60. (С разрешения Dept, of Entomology, Univ, of
Alberta и D. A. Craig.)
Лижущий тип (рис. 3.27; 3.28). Очень
многие некровососущие мухи, в том чис-
ле комнатная муха, имеют ротовой аппа-
рат подобного типа, предназначенный
для питания только жидкой или легко
растворимой в слюне пищей. Этот тип
наиболее близок к режуще-сосущему, но
мандибулы и максиллы здесь редуциро-
ваны, а прочие части образуют хоботок,
на конце которого расположен губкопо-
добный лабеллум. Он погружается в
жидкую пищу, которая поступает в пи-
щевой канал по тонким капиллярным
трубочкам на поверхности лабеллума.
Пищевой канал в ротовом аппарате это-
го типа также формируется сомкнутыми
краями вытянутых гипофаринкса и эпи-
фаринкса, образующих трубку, которая
ведет к пищеводу. Некоторые виды твер-
дой пищи, например сахар, также
потребляются мухами при помощи этого
хоботка. Осуществляется это следую-
щим образом • вначале муха выделяет ка-
пельку слюны на поверхность пищи, ко-
торая в слюне растворяется, а затем
всасывается в рот уже как жидкость.
Грызуще-лижущий тип. Другой тип
ротового аппарата, приспособленного
для питания жидкой пищей, характерен
для пчел и ос, он будет рассмотрен на
примере медоносной пчелы (рис. 3.29).
Мандибулы и верхняя губа устроены как
в грызущем ротовом аппарате и исполь-
зуются для захватывания добычи, фор-
Рис. 3.24. Головной веер и ротовая область мошки Simulium tahitiensis’
х 90. (С разрешения Dept, of Entomology, Univ, of Alberta и D. A. Craig.)
Рис. 3.25. Голова и ротовой аппарат личинки мошки Crozetia crozetensis;
этот вид питается в основном путем соскабливания субстрата и имеет
редуцированные головные веера, х 150. (С разрешения Dept, of
Entomology, Univ, of Alberta и D. A. Craig.)
Рис. 3.26. Ротовой аппарат мухи Symphoromyia atripes (Rhagionidae) как
пример режуще-сосущего типа. (По Illinois Natural History Survey.)
/-максиллярный щупик; 2-эпифаринкс; 3-мандибула; 4-максилла,
5 - гипофаринкс; 6 -нижняя губа; 7-галеа; 5-стипес; 9-лациния; 10 -кардо;
//-ротовое отверстие; /2-псевдотрахеи, 13-оральный диск, /4-тека.
76
3. Внешнее строение
Рис. 3.27 Общий вид головы плодовой мушки Drosophila melanogaster\ видны детали
строения придатков и ротового аппарата, х 180. (С разрешения Nanometrics, Inc.)
3. Внешнее строение
77
Рис. 3.28. Ротовой аппарат комнатной мухи как пример лижущего типа. (По
Metcalf, Flint, Destructive and useful insects, с разрешения McGraw-Hill Book
Co.)
/-усик, 2-сложный глаз; 3-глазки, 4-углубления над развилками,
5-просветы развилок; 6-щель на поверхности, ведущая в псевдотрахею;
7-хитиновые кольца псевдотрахеи; 5-рострум, 9-максиллярный щупик;
10-верхняя губа и эпифаринкс; 11 -лабеллум; 12-гипофаринкс; 13-нижняя
губа, 14-проток слюнной железы; 15-пищевой канал.
мирования кусочков воска или других
материалов при постройке гнезда. Мак-
силлы и нижняя губа превратились в со-
вокупность уплощенных и вытянутых
элементов, причем слившиеся глоссы
(обычно называемые язычком) образуют
выдвигающийся желобообразный орган.
Этот орган служит для проникновения
глубоко в нектарники цветков.
Остальные створки максилл и нижней
губы прилегают к глоссам и образуют
ряд каналов, по которым вниз поступает
слюна, а вверх-пища. По поводу дета-
лей работы механизма, обеспечивающе-
го продвижение жидкостей, в мнениях ис-
следователей существуют разногласия.
Колюще-сосущий тип (рис. 3.30). Ро-
товые органы многих групп насекомых
приспособлены для прокалывания тка-
ней и высасывания из них соков. К таким
группам относятся тли, цикадовые, чер-
вецы и щитовки и другие насекомые, со-
сущие соки растений, хищнецы, водомер-
ки и многие другие хищные формы,
питающиеся соками насекомых и других
мелких животных; комары, постельные
клопы, вши и блохи, сосущие кровь мле-
копитающих и птиц. У представителей
этих групп верхняя губа, мандибулы и
максиллы (а иногда и гипофаринкс) уд-
78
3. Внешнее строение
Рис. 3.29. Ротовой аппарат медоносной пчелы как пример грызуще-лижущего типа.
(По Metcalf, Flint, Destructive and useful insects, с разрешения McGraw-Hill Book
Co.)
/-простые глазки; 2-сложный глаз; 3-зазатылочный шов; 4-затылок;
5-защеки; 6-кардо; 7-затылочное отверстие; 8-усик; 9-ментум; /0-лорум;
77-осевой стержень язычка; 72-прементум; 73-стипес; 14 - внутренний канал
осевого стержня; /5-наличник; 16-верхняя губа; 77-мандибула, 18-галеа;
19-параглосса; 20-максиллярный щупик; 2/-язычок; 22-нижнегубной щупик;
23 -флабеллум.
линились и, складываясь вместе, обра-
зуют тонкую полую иглу (рис. 3.31,
3.32). Нижняя губа играет роль прочного
футляра, не дающего игле изгибаться.
Вся эта структура носит название хобот-
ка. Чтобы принять пищу, насекомое при-
жимает хоботок к хозяину, затем вводит
иглу в его ткани и всасывает соки хозя-
ина через иглу внутрь пищевода.
Интересный и, по-видимому, генера-
лизованный тип устройства такого рото-
вого аппарата обнаруживается у трипсов
(см. с. 290). Отдельные элементы этого
аппарата стилетоподобны, однако в це-
лом они образуют скорее не хоботок, а
скребущий конус. В высшей степени
сложно построены хоботки вшей (Апо-
plura), обладающих особым втягиваю-
щимся хоботком, в котором сохраняют-
ся лишь немногие из первоначально
имевшихся частей (см. с. 287).
Трубчато-сосущий тип (рис. 3.33).
Взрослые особи Lepidoptera питаются
нектаром и другой жидкой пищей. Она
3. Внешнее строение
79
Рис. 3.30. Ротовой аппарат самки комара как пример колюще-сосущего
типа. Двойная стенка верхней губы позволяет вводить ее в толщу ткани
хозяина.
А. Голова и расправленные ротовые части (по Waldbauer). Б. Поперечный
срез хоботка вблизи середины (по Waldbauer, с изменениями). 1 - верхняя
губа; 2-мандибула; 3-гипофаринкс; 4-лациния; 5-лабеллум; 6-усик;
7-глаз; 8-максиллярный щупик; 9-прементум; 10-верхняя губа с двойной
стенкой; /7-пищевой канал; 72-заполненная гемолимфой полость
прементума.
всасывается с помощью длинного хобот-
ка, образованного только сложенными
вместе галеа каждой из двух максилл. В
результате получается трубка, откры-
вающаяся в пищевод.
ШЕЙНЫЙ ОТДЕЛ
Между головой и туловищем распо-
.тагается мембранозная область, соста-
вляющая шею. Иногда ее рассматривают
как особый сегмент тела,-микроторакс,
злнако данных, подтверждающих такую
точку зрения, довольно мало. Предста-
вляется более вероятным, что шея вклю-
чает как участок нижнегубного сегмента
головы, так и участок переднегрудного
сегмента, образуя между ними гибкую
область.
Шея включает две пары цервикальных
склеритов (рис. 3.34), которые служат
местами сочленения головы с тулови-
щем. С каждой стороны оба склерита
шарнирно соединены друг с другом и
образуют единый комплекс, сочле-
ненный спереди, на зазатылке головы, с
Рис. 3.31. Ротовые органы комара Aedes aegypti’, видна
покрытая сенсиллами нижняя губа и пиловидно зазубренная
лациния; х 300. (С разрешения Dept, of Entomology, Univ, of
Alberta и D. A. Craig.)
Рис. 3.32. Колюще-сосущий ротовой аппарат у личинки первого
возраста цикадки Macrosteles focifrons\ х 115. (С разрешения Dept, of
Entomology, Univ, of Alberta и D. A. Craig.)
3. Внешнее строение
81
Рве. 3.33. Ротовой аппарат чешуекрылых
как пример трубчато-сосущего типа. (По
Folsom, Wardle, 1934. Entomology,
с разрешения The Blakiston Со.)
А. Ночная бабочка. Б. Дневная бабочка.
- пилифер; 2 - мандибула; 3 - верхняя губа,
^-наличник; 5-усик; 6-глаз; 7-хоботок;
.• - нижнегубной щупик.
затылочным мыщелком, а сзади-с пере-
днегрудью. Цервикальные склериты не-
редко срастаются с плеврами передне-
груди.
Рис. 3.34. Схема цервикальных склеритов
насекомого. (По Снодграссу,
с изменениями.)
1 - межсегментная линия; 2 - переднегрудь;
3 - голова, 4 - цервикальные склериты.
РАЗВИТИЕ ГЕНЕРАЛИЗОВАННОГО
СЕГМЕНТА
ТЕЛА НАСЕКОМОГО
Строение современных представите-
лей Chilopoda, Entognatha и прими-
тивных насекомых позволяет предпола-
гать, что сегменты тела произошли из
очень простого предкового типа
(рис. 3.35), состоявшего из следующих
пяти элементов:
1. Тергита, или склеротизированной
дорсальной пластинки, называемой так-
же спинкой, когда речь идет о грудных
сегментах.
2. Стернита, или брюшной склероти-
зированной пластинки.
3. Плевральной области, связываю-
щей тергит со стернитом и полностью
мембранозной.
4. Пары членистых ног; базальный
членик каждой ноги, или коксоподит, ле-
жит на мембране между тергитом и стер-
нитом. Коксоподит подразделяется на
базальную {субкокса) и апикальную {кок-
са, или тазик) части. На рис. 3.35 субкок-
са состоит из трех склеритов.
82
3. Внешнее строение
Рис. 3.35. Схема простого сегмента тела
насекомого. (По Снодграссу.)
/-спинка, 2-дыхальце; 3-плейральная
область; 4 - субкоксальные склериты,
5-тазиковая впадина; 6-стернит.
5. Пары дыхалец, по одному на мем-
бране над каждой ногой.
У немногих архаичных групп, таких,
как Protura и у Chilopoda, обнаруживают-
ся сегменты, построенные по этому типу
(рис. 3.36), представляющему собой про-
стейший тип, или прототип. В этом слу-
чае тергит и стернит не изменены. Суб-
кокса представлена серповидными скле-
ритами - одним медиальнее тазика и дву-
мя-латеральнее. Два последних, види-
мо, являются элементами, находящими-
ся между тазиком ноги и тергитом. Эти
отделенные субкоксальные склериты в
плейральной области-предшественники
плейральных склеритов. Тазик образует
функциональную основу для причлене-
ния ноги.
На следующих этапах эволюционно-
го развития (рис. 3.37) субкоксальные
склериты неподвижно прирастают
к стенке сегмента и образуют прочную
основу, к которой причленяется функцио-
нальная нога. Медиальные субкок-
сальные склериты срастаются со стерни-
том, а латеральные субкоксальные скле-
риты становятся уплощенными приле-
гающими боковыми элементами, в сово-
купности называемыми плейритом. Это
состояние можно рассматривать как ге-
нерализованное, от которого произошли
и специализированные крыловые сег-
менты груди, и упрощенные сегменты
брюшка. Оно встречается у многих ныне
живущих форм, особенно часто в груд-
ном отделе неполовозрелых стадий
столь различных групп, как веснянки, ру-
чейники и сетчатокрылые (рис. 3.37).
ГРУДЬ
Грудью называется отдел тела, лежа-
щий между головой и брюшком. Он со-
стоит из трех последовательно располо-
женных сегментов переднегруди, сред-
негруди и заднегруди.
1
Рис. 3.36. Среднегрудь бессяжкового
Acerentomon. (По Снодграссу.)
1 -спинка, 2-мембранозная плейральная
область, 3-субкоксальные склериты;
4-стернит; 5-тазик; 6-вертлуг; 7-бедро.
3- Внешнее строение
83
Рас 3.37. Переднегрудь личинки веснянки
(По Снодграссу.)
-спинка, 2-субкоксальные склериты
гудущий плейрит); 3-тазик; 4-веотлуг.
В тех отрядах, где крылья никогда не
развиваются, эти три сегмента в главных
чертах устроены примерно одинаково.
Тергит и стернит пластинообразные, а
иеяральные склериты (субкоксальные
д>ти) небольшие или недоразвитые
ж. 3.36).
У представителей крылатых насе-
ьзмыа три грудных сегмента очень силь-
различаются между собой. Передне-
дь обычно сохраняет все те же части,
г? и в исходном состоянии (рис. 3.37),
vrn отдельные склериты могут сливать-
ся зли перегруппировываться до такой
степени, что становится трудно правиль-
тс интерпретировать их происхождение.
Среднегрудь и заднегрудь претерпели
гс длинный морфологический переворот,
связанный с развитием в каждом из этих
сегментов мускулатуры, которая должна
-хеспечивать работу двух механизмов-
тсдета и хождения. Прибавилось много
склеритов, и многие из них пере-
туппиро вались.
Генерализованный крыловой сегмент.
41 этом сегменте, как и на обычном
'«скрылом. можно выделить три ос-
новные области тергит (называемый
спинкой, если термин применен к грудно-
му сегменту), стернит и плейриты. Их
происхождение из примитивного сегмен-
та туловища схематически показано на
рис. 3.38. Хотя каждая область испытала
влияние специализации в различных на-
правлениях, наиболее заметные внешние
изменения, сопровождавшие переход к
крылатому состоянию, произошли
в плейральной области. Генерализо-
ванный крыловой сегмент современных
крылатых насекомых показан на
рис. 3.39.
ПЛЕЙРИТ. Этот склерит значительно
расширился, образовав крупную боко-
вую пластину. Вентрально она несет кок-
сальный отросток, к которому причле-
няется нога, а дорсально-крыловой от-
росток (плейральный столбик), к которо-
му причленяется крыло. Плейрит разде-
ляется на переднюю область, или эпи-
стерн, и заднюю, или эпимер, плей-
ральным швом, простирающимся от кок-
сального до крылового отростка (плей-
рального столбика). Этим швом обозна-
Рис. 3.38. Схема гипотетической стадии
формирования крылового сегмента. (По
Снодграссу.)
/-спинка, 2-субкокса, или плейрит;
3 - тазиковая впадина, 4 - стернит.
84
3. Внешнее строение
Рис. 3.39. Схема типичного крылового
сегмента. (По Снодграссу.)
1 - ал инотум; 2 - постнотум; 3 - фрагма,
4-основание крыла, 5-плейральный
столбик; 6-плейральный шов; 7-эпистерн;
5-трохантин; 9-алярные склериты;
10 - эпимер; 11- коксал ьный отросток;
/2-тазиковая впадина; 73-эустернум;
14 - спинастернум.
чена линия инвагинации внутренней
плейральной аподемы, называемой плей-
ралъным гребнем. Сзади и спереди плей-
рит срастается со стернитом, а участки
срастания образуют мостики впереди и
позади тазиковых впадин.
СПИНКА. Эта область разделена на
два основных склерита, передний -алино-
тум и задний-постнотум. Передний
склерит непосредственно связан с кры-
лом и снабжен передней аподемой, или
фрагмой. Характер подразделения этого
склерита на более мелкие участки весьма
различен у разных групп. Постнотум так-
же несет фрагму, а сбоку связан не с кры-
лом, а с эпимером плейрита, образуя мо-
стик позади места прикрепления крыла.
Часть постнотума, лежащая позади
фрагмы, относится, по сути дела, уже к
лежащему позади следующему сегменту,
ставшему функциональной частью лежа-
щего перед ним. Смещения подобного
рода-частое явление у насекомых.
СТЕРНИТ Эта пластина, объединяясь
с плейритом своими передними и задни-
ми частями, имеющими вид полос, обра-
зует впадину, в которой располагается
тазик. Центральная часть, называемая
эустернумом, имеет борозду, отмечаю-
щую линию впячивания большой вну-
тренней аподемы, именуемой фурка, что
в переводе с латинского означает «вил-
ка», так как эта аподема раздвоена на
вершине (рис. 3.40). Позади эустернума
лежит небольшой склерит - спинастер-
нум, от которого внутрь отходит един-
ственная аподема-спина. Спинастернум
образовался на месте межсегментной
мембраны, но, как правило, он слит с сег-
ментом, лежащим впереди от места его
возникновения.
ВНУТРЕННИЙ СКЕЛЕТ Многочис-
ленные аподемы сегментов часто назы-
вают в совокупности внутренним скеле-
том. Он служит местом прикрепления
многих крупных крыловых и ножных
мышц. Плейральные гребни и фурка в ка-
ждом сегменте тесно соприкасаются,
Рис. 3.40. Схематический поперечный разрез
грудного сегмента, показаны фурка
и плейральные гребни. (По Снодграссу.)
1 - спинка; 2 - плейральный гребень;
3-плейрит; 4-фурка, 5-стернит; 6-тазик.
3 Внешнее строение
85
эбразуя почти непрерывную систему ба-
(рис. 3.40).
СУЩЕСТВУЮЩИЕ ВАРИАНТЫ. В об-
щем очерке строения грудного отдела не-
целесообразно рассматривать вопрос бо-
цее подробно, чем это сделано в данном
разделе, так как вариации в строении гру-
да современных насекомых почти не-
исчерпаемы. Во многих отрядах сохра-
няются отчетливые исходные признаки,
тггда как в крупных отрядах, например
Coleoptera и Diptera, возможны очень
значительные видоизменения внутри са-
мах групп.
Подчас внешнее сходство между су-
ществующей формой и общим планом
строения крайне незначительно. В этих
дтучаях для гомологизации склеритов
сначала необходимо соотнести их с по-
стоянно присутствующими признаками,
или «опорными точками». Наиболее на-
дежными из них являются аподемы и со-
путствующие им наружные швы, а также
места сочленения ног и крыльев.
НОГИ. Типичная грудная нога со-
стоит из шести частей. тазика, вертлуга,
бедра, голени, лапки и предлапки
(рис. 3.41). Тазик-это отдел ноги, сочле-
ненный с телом, он может иметь заднюю
долю, или мерой. Лапка взрослого насе-
комого обычно бывает разделена на 2-5
члеников. Предлапка имеет вид обосо-
бленного маленького концевого членика
ноги у личинок некоторых отрядов и у
Collembola. Почти во всех остальных от-
рядах насекомых предлапка представле-
(задняя нога сверчка); В-хватания (передняя нога богомола); Г-зажимания
(передняя нога клопа); Д-копания (передняя нога медведки); Е-удержания
путем присасывания (передняя нога самца плавунца). (По Folsom, Wardle, 1934,
Entomology, с разрешения The Blakiston Со.) / -бедро; 2-голень; 3-вертлуг;
«/-лапка; 5-тазик.
86
3. Внешнее строение
на в виде комплекса, состоящего из ко-
готков и группы маленьких склеритов на
конце лапки. Collembola и Protura уни-
кальны также в том отношении, что у них
голень и лапка срастаются в единую
голенелапку.
В типичном случае насекомые имеют
простые ноги, предназначенные для бега
или ходьбы (рис. 3.41,Л). Вместе с тем у
многих насекомых ноги претерпели
самые разнообразные модификации, по-
зволяющие выполнять и другие функции.
К числу таких модификаций можно отне-
сти ноги прыгательного типа с сильно
увеличенным бедром, как у саранчовых
(рис. 3,41,Ь); хватательные ноги, воору-
женные острыми смыкающимися шипа-
ми и зубьями (рис. 3.41,В), как у хищно-
го богомола, плавательные ноги
с длинными щетинками или волосками и
уплощенными частями, как у плавунца,
копательные ноги с мощными зазу-
бренными скребущими элементами
(рис. 3.41 ,Г), как, например, у медведки.
Крылья
Крылья насекомых - уникальные
образования, равных которым нет ни в
одной из прочих групп живых существ. У
таких животных, как птицы или летучие
мыши, крылья представляют собой силь-
но видоизмененные передние конечно-
сти. У насекомых дело обстоит иначе. Их
крылья - выросты стенки тела вдоль бо-
кового края дорсального склерита, или
спинки. В отличие от большинства дру-
гих придатков тела насекомых крылья
лишены собственной внутренней муску-
латуры. Крылья, дающие возможность
летать, стали одним из важнейших фак-
торов процветания группы насекомых в
целом. Ни одна другая группа беспозво-
ночных никогда не имела подобных
органов.
Ныне живущие крылатые насекомые,
называемые также Pterygota (от греч.
pteryx-крыло), в типичном случае
имеют две пары крыльев, по одной паре
на средне- и заднегруди. На переднегру-
ди могут быть боковые листовидные вы-
росты, однако подвижно сочлененные
крылья всегда отсутствуют.
Строение. План строения крыла на-
секомого очень прост. Это складкооб-
разное продолжение стенки тела, имею-
щее верхний и нижний мембранные
слои и комплекс придающих жесткость
опорных элементов, называемых жилка-
ми и идущих более или менее вдоль кры-
ла. Продольные жилки связаны рядами
поперечных жилок. Узор из продольных
и поперечных жилок называется жилко-
ванием. Между спинкой груди и крылом
располагается узкая и гибкая мембраноз-
ная полоска, с которой связано несколь-
ко мелких склеритов. Дорсально (рис.
3.42,Л) лежат три базальных склерита
(обозначены на рисунке цифрами 1 2, 3),
из которых по меньшей мере склериты 1
и 3 находятся в тесном контакте со спин-
кой и сочленены с ней, склеритам со-
ответствуют три прилегающие к ним
пластинки (4, 5 и 6), от которых отходят
жилки. Вентрально (рис. 3.39) два
алярных склерита лежат по сторонам
крылового отростка плейрита. Мышцы,
прикрепляющиеся к алярным склеритам,
регулируют некоторые движения крыла.
Варианты жилкования крыльев насе-
комых, имеющего важное значение при
классификации отрядов, семейств и ро-
дов, поистине неисчислимы. Все эти раз-
нообразные типы жилкования, очевидно,
возникли от общего предкового типа,
главнейшие особенности которого могут
быть восстановлены в результате сравне-
ния ископаемых и современных форм.
Эти особенности показаны на рис. 3.43.
Каждая главная продольная жилка
имеет свое название; они перечислены
ниже в том порядке, в каком они распо-
3. Внешнее строение
87
Рис. 3.42. Аксиллярные склериты (/, 2, 3) и связанные с ними пластинки (4, 5, 6)
в основании крыла. (По Hamilton, 1971.)
А. Гипотетический предковый тип. Б. Генерализованный тип складывающегося
крыла. 4-плечевая пластинка; 5-медиальная пластинка; 6-кубитальная
пластинка, 7-тегула, 8-костальная складка. Более подробные объяснения см. в
тексте.
катаются в направлении от переднего к
заднему краю крыла. Для некоторых жи-
хх характерно определенное типичное
ветвление. Для жилок указаны их обще-
принятые сокращенные обозначения
рис. 3.43).
КОСТАЛЬНАЯ ЖИЛКА (С), как прави-
>?. образующая утолщенный передний
грай крыла, не ветвится и отходит от
плечевой пластинки (рис. 3.42).
СУБКОСТАЛЬНАЯ ЖИЛКА (Sc) идет
сразу за костальной, всегда лежит на дне
желоба между костальной и радиальной
жвлками. Она также обычно не ветвится
* отходит от плечевой пластинки.
РАДИАЛЬНАЯ ЖИЛКА (R)-следую-
щая главная жилка. В типичном случае
толстая и связана у основания с ме-
диальной пластинкой. Как правило, не
селится на ветви.
СЕКТОРАЛЬНАЯ ЖИЛКА (S) отходит
от медиальной пластинки и в типичном
случае дихотомически разделяется на
четыре ветви. Первое ветвление дает
жилки, называемые соответственно пере-
дней и задней ветвями сектора (SA и SP).
МЕДИАЛЬНАЯ ЖИЛКА (М) также от-
ходит от медиальной пластинки и дихо-
томически разделяется на переднюю и
заднюю ветви (МА и МР), каждая из ко-
торых в свою очередь делится обычно
еще на две ветви.
КУБИТАЛЬНАЯ ЖИЛКА (Си), отходя-
щая от кубитальной пластинки, обычно
двуветвистая. Почти во всех случаях
это-сильно выступающая жилка, прохо-
дящая вдоль гребня складки.
ПЛИКАЛЬНАЯ ЖИЛКА (Р) отходит
от кубитальной пластинки и не ветвится,
это погруженная жилка, т.е. жилка, про-
88
3. Внешнее строение
Рис. 3.43. Возможное исходное жилкование крыла примитивного
насекомого; поперечные жилки не показаны. (По Hamilton, 1972.)
Объяснение см. в тексте.
ходящая по дну желоба между кубиталь-
ной и эмпузальной жилками.
ЭМПУЗАЛЬНАЯ ЖИЛКА (Е) также от-
ходит от кубитальной пластинки и не ве-
твится. Иногда она выступает, так как
идет по гребню складки.
АНАЛЬНЫЕ ЖИЛКИ (1А, 2А и т.д.)
представляют собой пучок жилок, имею-
щих общее основание, которое непосред-
ственно связано с третьим аксиллярным
склеритом.
Названия жилок. В течение долгого
времени гомологии жилок крыла вызы-
вали противоречивые мнения и дискус-
сии. Итоги этих дискуссий подведены Га-
мильтоном (Hamilton, 1971-1972). При-
меняемая здесь номенклатура жилок в
ряде случаев отличается от обычно ис-
пользуемой многими авторами. Для
сравнения с другой системой обозначе-
ний, эти отличия перечислены ниже.
1. Сектором названа система жилок,
для которой прежде использовали в
большинстве случаев термин радиальный
сектор. Однако это вполне самостоя-
тельная система жилок, а не ответвление
радиальной жилки.
2. Пликальная и эмпузальная жилки
в разных случаях рассматривались как
ветви кубитальной жилки или как
анальные жилки.
3. Пару очень коротких жилкопо-
добных структур на самом основании за-
днего края крыла очень часто называли
югальными жилками. Новые данные сви-
детельствуют о том, что это не настоя-
щие жилки, а новообразования, свя-
занные со складыванием крыла. Пред-
лагается называть их югальными ре-
брами.
ПОПЕРЕЧНЫЕ ЖИЛКИ. В общем по-
перечные жилки принято называть по
тем продольным жилкам, между которы-
ми они лежат. Так, поперечные жилки,
связывающие радиальную жилку с пере-
дней ветвью сектора, получат в этом слу-
чае наименования lr-s, 2r-s и т. д.; связы-
вающие заднюю ветвь медиальной жил-
ки с кубитальной будут обозначены
lm-си, 2т-си и т.д. Поперечные жилки
между ветвями одной и той же продоль-
ной жилки часто имеют особые названия
в разных систематических группах. Су-
ществует и несколько общепринятых ис-
ключений. Поперечная жилка между ко-
стальной и субкостальной жилками в
основании крыла называется плечевой (Л),
а остальные, лежащие между костальной
3. Внешнее строение
89
и следующей за ней жилкой,-просто ко-
стальными поперечными жилками.
Происхождение. Существует несколь-
ко теорий происхождения такого уни-
кального органа, каковым является кры-
ло насекомых. Поскольку оба наиболее
примитивных отряда крылатых насе-
комых, Ephemeroptera и Odonata, а также
другой весьма примитивный отряд, Р1е-
coptera, имеют водных нимф, вначале
предполагали, что крылья возникли как
плавательные лопасти, которые помога-
ли личинкам продвигаться в толще воды.
Однако трудности, возникающие при по-
пытке реконструкции форм, переходных
между плавательной лопастью и воз-
душным крылом, заставляют признать
эту теорию несостоятельной. Согласно
другой теории, предложенной Алексан-
дером и Броуном (Alexander, Brown,
1963), крылья возникли из боковых выро-
стов груди, связанных с половой актив-
ностью. До настоящего времени каких-
либо половых структур, которые могли
бы послужить предшественниками орга-
нов полета, не было обнаружено ни на
груди ископаемых насекомых, ни у совре-
менных представителей. Наиболее до-
стоверной представляется гипотеза, в со-
ответствии с которой крылья берут нача-
ло от боковых лопастевидных выростов
спинки груди; возникли они, возможно, в
качестве защитных образований или в ре-
зультате уплощения тела как адаптации к
проникновению в щели, трещины или
под кору. Если насекомое было чем-то
обеспокоено или встревожено, эти высту-
пающие части помогали ему спланиро-
вать, не потеряв правильной ориентации
тела, чтобы быть готовым к бегству по-
сле падения с высоты вниз. Однако даже
и в этом случае необходимо было особое
стечение обстоятельств. Насекомые дол-
жны были часто посещать поверхности
^например, листья, стволы деревьев, от-
весные скалы), находившиеся на расстоя-
нии нескольких метров от уровня почвы:
в противном случае при падении они бы-
ли бы не в состоянии набрать скорость,
достаточную для того, чтобы боковые
выросты спинки смогли взаимодейство-
вать с воздушным потоком. Флауер (Flo-
wer, 1964) подсчитал, что коротконогие
насекомые с длиной тела немного менее
1 см и лишь со слабой пластинкой вдоль
краев груди вполне могли удовлетворять
этим требованиям и в конце падения до-
стигать устойчивого равномерного
скольжения вниз. Так как насекомыми
питались прежде всего быстро бегающие
хищники, такие, как пауки и многоножки,
скорость и способность маневрировать в
воздухе давали бы им преимущества для
выживания.
Среди современных насекомых по-
добным аэродинамическим требованиям
удовлетворила бы, например, чешуйница
Thermobia. Это и некоторые другие при-
митивные бескрылые насекомые, такие,
как Machilis (рис. 8.7), имеют и иные
приспособления, которые могли бы ока-
заться полезными при попытках подоб-
ного планирующего бегства, грудь рас-
положена вблизи центра тяжести тела, а
три пары больших сильных грудных ног
могут обеспечить сильный толчок для
поспешного спасения. Теперь предста-
вляется вполне вероятным, что способ-
ность к планированию у организмов, на-
поминающих нынешних Thermobia, яви-
лась предпосылкой для развития полета.
Другой факт, который следует при-
нять во внимание при выяснении путей
эволюции полета насекомых,-это нали-
чие на переднеспинке многих древнейших
ископаемых крылатых насекомых пло-
ских боковых выростов, названных пара-
нотальными лопастями и служащих, ве-
роятно, для планирования. Кроме того,
эти лопасти имели набор жилок, по-
видимому гомологичный главным жил-
кам типичного подвижно сочлененного
90
3. Внешнее строение
Рис. 3.44. Гипотетические стадии развития жилкования паранотальных
лопастей (А - Д), приводящие к жилкованию примитивного крыла (Е).
(По Hamilton, 1971.)
функционирующего крыла. Эти допол-
нительные сведения позволяют восстано-
вить ход эволюции крыла насекомых с
высокой степенью достоверности.
Когда было достигнуто устойчивое
скольжение вниз, начало играть свою
роль давление отбора в направлении рас-
ширения спинных выростов и превраще-
ния их в лопасти или плоскости, которые
позволяли бы сократить время до начала
скольжения и увеличить протяженность
самого скольжения. Вначале лопасть по-
ходила, возможно, на изображенную на
рис. 3.44,Л, затем она увеличивалась,
проходя через стадии, показанные на
рис.-3,44,1>, В, Г и Д. Сходные лопасти
изначально имелись, по-видимому, на
всех трех грудных сегментах. По мере
увеличения размеров лопастей в них про-
никали ответвления трахей; у взрослых
особей они превращались в места утол-
щения покровов крыла, т.е. в жилки. На
рис. 3.44,£ изображена наиболее круп-
ная из известных нам лопастей, обнару-
женных на переднегруди. Возможно, что
на этой стадии в основании крыла уже
был сформировавшийся шарнир, и с это-
го времени средне- и заднегрудные кры-
лья становились больше, а лопасти пере-
днегруди уменьшались.
Первые маленькие крылья были, ви-
димо, еще довольно толстыми и неуклю-
жими. По мере увеличения крыла его
жилки становились более мощными, а
крыловая мембрана между ними-более
тонкой и легкой. Наиболее древнее из из-
3. Внешнее строение
91
Рис. 3.45. Форма крыла Paleoptera на
поперечном срезе. (По Hamilton, 1971.)
Сильная гофрировка крыловой пластинки за
счет чередования выступающих
и погруженных жилок придает крылу
дополнительную жесткость.
зестных действующих крыльев было
многократно сложено в продольном на-
правлении подобно вееру, причем жилки
шли по мостам перегибов (рис. 3.45),
громе того, имелась сеть поперечных жи-
лок. связывавших между собой про-
дольные. Такая комбинация вееро-
образной или складчатой структуры и
напоминающего арматурные прутья кар-
каса позволяла прочной, но тонкой мем-
бране быть достаточно жесткой для
полета.
Последующей стадией эволюции
крыла была перегруппировка аксил-
лярных склеритов и пластинок, сделав-
шая возможным диагональное складыва-
ние крыла у его основания (рис. 3.42,1>,
3.46). При такой организации крылья мо-
гли складываться вдоль анальной и ба-
зальной складок назад и плашмя над те-
лом. На том же этапе эволюции большая
часть складок крыла исчезла и область
между R и Си стала плоской.
В разных отрядах насекомых про-
изошли разнообразные изменения в жил-
ковании крыльев. У одних появились до-
бавочные продольные жилки, у других
различные жилки либо сливаются, либо
отсутствуют. То же относится и к попе-
речным жилкам. У крупных насекомых,
характеризующихся быстрым полетом,
обычно имеются длинные и узкие кры-
лья ; порхающий полет, как у дневных ба-
бочек, коррелирует с более короткими и
широкими крыльями. У мелких насе-
комых, например тлей, жилкование мо-
жет быть в значительной степени редуци-
ровано, а полет часто слабый, почти
пассивный. Каждый тип модификации
крыла находится в связи с поведением и
образом жизни взрослого насекомого.
Рис. 3.46. Форма крыла Neoptera на
поперечном срезе. (По Hamilton, 1971.)
Гофрировка крыловой пластинки
изменилась, позволив крылу складываться,
не проигрывая при этом в жесткости.
92
3. Внешнее строение
Во многих группах способность к полету
полностью утрачена, как, например, у
прямокрылых семейства Rhaphidophori-
dae или блох, которые лишены крыльев.
БРЮШКО
Брюшко - третий, самый задний от-
дел тела насекомого. Оно характеризует-
ся довольно простым строением, если
сравнивать его с грудным отделом, и у
взрослых насекомых лишено ходильных
ног. В примитивном состоянии оно со-
стоит из 12 сегментов, но это встречается
только у Protura (рис. 8.5) и у зародышей
некоторых групп. Обычно же брюшко со-
стоит из 10 или 11 сегментов (рис.
3,47,Л). У некоторых форм наблю-
дается и более глубокая редукция, напри-
мер, Collembola имеют лишь 6 брюшных
сегментов (рис. 8.6). У многих групп, та-
ких, как мускоидные двукрылые, не-
сколько последних сегментов брюшка
превратились в копулятивный орган, или
яйцеклад, который в покое втянут внутрь
предшествующих сегментов.
Строение сегмента. Типичный сегмент
взрослого насекомого состоит из 1) тер-
гита, или спинной пластины, 2) стернита,
или брюшной пластины, 3) боковых мем-
бранозных участков, связывающих тер-
гит со стернитом, и 4) дыхалец, располо-
женных по одному на каждой стороне,
обычно на боковой мембране. У неко-
Рис. 3.47. Заднегрудь и брюшко насекомых. (По Снодграссу.)
А. Имаго сверчка Gryllus. Б. Личинка жужелицы Calosoma. 7-тергиты, 2-церк;
3-эпипрокт; 4-анус; 5-парапрокт; 6-дыхальца, 7-стерниты; 8-боковая
мембрана, 9-заднегрудь; 10-дорсальная область; 77-плейральная область;
72-вентральная область; 13-плейральные склериты; 14-боковые участки
тергитов; 75-отросток.
3. Внешнее строение
93
торых личиночных форм (рис. 3.47,Б) и
немногих взрослых насекомых склериты
имеются и на боковой мембране; неко-
торые из этих склеритов, несомненно,
представляют собой остатки субкок-
сальных структур примитивного придат-
ка (см. рис. 3.36).
Придатки. Все придатки брюшка
можно разделить на две группы. придат-
ки. не связанные с размножением, и при-
датки, несущие репродуктивные функ-
ции. такие, как спаривание или отклады-
вание яиц.
ПРИДАТКИ, НЕ СВЯЗАННЫЕ С РАЗ-
МНОЖЕНИЕМ. У большинства взрос-
лых насекомых придатки на брюш-
ных сегментах отсутствуют; исключе-
ние составляют концевые сегменты.
Лишь немногие примитивные формы
сохранили редуцированные ноги, пред-
ставленные в виде грифельков, на-
пример у щетинохвосток (рис. 3.48).
Придатки одиннадцатого сегмента-цер-
ки-имеются у большинства насекомых
рис. 3.47). Они выполняют обычно
функции тактильных органов, а у ряда
групп (например, ручейников) входят в
состав гениталий самцов. Когда один-
надцатый или десятый сегменты брюшка
редуцированы, может создаться впечат-
ление, что церки принадлежат десятому
или даже девятому сегменту. У личинок
и нимф придатки брюшка достигают
большого разнообразия. Широко из-
вестными примерами являются личи-
ночные ноги гусениц (рис. 3.5) и тра-
хейные жабры нимф поденок (рис. 8.10).
ПРИДАТКИ, СВЯЗАННЫЕ С РАЗ-
МНОЖЕНИЕМ. К этой группе относят-
ся в основном придатки восьмого и
девятого сегментов. Многие энтомологи
считают эти придатки гомологами на-
стоящих сегментных придатков, к ка-
ковым принадлежат ротовые органы и
ноги. Другие полагают, что первона-
чальные придатки восьмого и девятого
сегментов брюшка исчезли, а придатки,
связанные с размножением,-это спе-
циальные образования, происшедшие из
выростов, лежащих на тех же сегментах
ближе к средней линии.
САМКА. Яйцеклад (рис. 3.49) со-
стоит главным образом из трех пар ство-
рок (вальв): первой, второй и третьей пар
вальв соответственно. Первая пара вальв
(нижние створки) отходит от пары пла-
стинок - нижних створкодержателей, от-
носящихся к восьмому сегменту. Воз-
можно, створкодержатель и створка со-
Рис. 3.48. Брюшко самки Machilis maritima,
вид с вентральной стороны. (По Oudemans,
из Folsom, Wardle, 1934.) Показаны
рудиментарные конечности (/) на II - IX
сегментах (левый придаток восьмого
сегмента не показан); 2-церки; 3-непарный
каудальный филамент.
94
3, Внешнее строение
Рис. 3.49. Строение яйцеклада крылатых насекомых. (По Снодграссу.)
Л, Б. Отношения элементов яйцеклада к сегментам. В, Г. Вид генитальных
сегментов сбоку и отдельные части яйцеклада. Д-генитальные сегменты и
лопасти яйцеклада личинки Blatta orientalise вид с вентральной стороны. А -Д
схематизировано.
1 -церк; 2- гон опор; 3 -проток сперматеки; 4- проток придаточных желез,
5-верхняя створка, 6-внутренняя створка; 7- нижняя створка, ^ нижний
створкодержатель, 9- верхний створкодержатель, 10-яйцевод; /7-грифелек;
12-верхний створкодержатель и верхняя створка.
ответствуют коксоподиту и телоподиту
генерализованного сегмента членистоно-
гих (рис. 3.15). Верхние створкодержа-
тели несут вентральную пару вальв-
вторые, или внутренние створки, и дор-
сальную ъъру-третьи, или верхние
створки яйцеклада. У большинства насе-
комых с хорошо развитым яйцекладом,
например у пилильщиков (рис. 3.50),
нижние и внутренние створки образуют
режущий или колющий орган с внутрен-
ним каналом, по которому выходят яйца.
Верхняя пара створок становится ножна-
ми или футляром, в который вклады-
вается яйцеклад в покоящемся состоя-
нии. У Orthoptera либо все три пары
створок, складываясь вместе, образуют
функциональный яйцеклад, либо внут-
ренняя дара превращается в короткий ор-
ган, направляющий движение яйца при
откладке.
У многих насекомых створки развиты
слабо или вовсе не развиты, причем в по-
следнем случае концевые сегменты
3. Внешнее строение
95
Рис. 3.50. Яйцеклад пилильщика. (По Снодграссу.)
А. Отношения базальных частей ко всем остальным и к девятому
тергиту. Б. Нижний створкодержатель и нижняя створка. В.
Верхний створкодержатель с внутренней и верхней створками.
1 - анальное отверстие; 2 - церк; 3 - верхний створкодержатель;
4-нижний створкодержатель; 5-верхняя створка, 6-внутренняя
створка и 7-нижняя створка, образующие «пилу» (5).
Рис. 3.51 Примеры «яйцекладов», образованных апикальными
сегментами брюшка. (По Снодграссу.)
Л-бабочка Lymantria monacha (из Eidmann, 1929). К-плодовая мушка
Paracantha culta, В-плодовая мушка Toxotrypania curvicauda
Г-скорпионница Panorpa sp. / -аподема, 2-анальное отверстие.
96
3. Внешнее строение
брюшка обычно формируют выдвижную
трубку, которая функционирует как яйце-
клад. Примером могут служить многие
Lepidoptera и Diptera (рис. 3.51).
САМЕЦ. У представителей этого по-
ла придатки девятого сегмента, комби-
нируясь с элементами самого девятого
сегмента, а иногда также с частями деся-
того сегмента, чаще всего образуют ко-
пулятивный орган. В каждом отряде этот
орган, как правило, характеризуется
особыми, присущими только этому отря-
ду признаками. Гомологизировать от-
дельные части этих копулятивных орга-
нов среди всех отрядов насекомых так,
чтобы с достоверностью выяснить, из ка-
ких простых частей или придатков они
должны были произойти, крайне трудно.
Рис. 3.52. Гениталии самца щетинохвостки
Machilis variabilis, вид с дорсальной стороны.
(По Снодграссу, с изменениями.)
А. Первая пара гоноподов; видны
гонапофизы восьмого сегмента. Б. Вторая
пара гоноподов и непарный копулятивный
орган. 7-восьмой стернит; 2-коксоподит;
3-первая пара гонапофиз; 4-девятый
стернит; 5-вторая пара гонапофиз,
6-эдеагус; 7-грифелек.
Рис. 3.53. Гениталии самца ручейника. (По
Illinois Natural History Survey.)
1 - аподемы класпера; 2 - церк; 3 - эдеагус,
4-класпер.
Структурные различия копулятивных
органов служат великолепными таксоно-
мическими признаками для разделения
многих групп насекомых на семейства,
роды или виды. В каждой конкретной
группе составные части этого органа
обычно бывают хорошо отграничены
друг от друга, и в каждой группе для
образования этих частей существует чет-
кая терминология. Пока не достигнуто
более полное согласие относительно го-
мологии этих структур в разных отрядах,
практичнее употреблять устоявшуюся
терминологию, принятую для каждой
отдельной группы насекомых. Гениталии
простого типа показаны на примере Mic-
rocoryphia (рис. 3.52), а более сложный
тип встречается у ручейников (рис. 3.53).
Эти два примера ни в коем случае не
дают представления о всем разнообра-
зии типов строения гениталий самцов у
насекомых; они приведены здесь лишь
для того, чтобы показать, с какими ва-
риациями могут встретиться исследова-
тели.
3. Внешнее строение
97
ЛИТЕРАТУРА
Alexander R. D., Brown W. L.t Jr., 1963. Mat-
ing behavior and the origin of insect wings,
Occ. Pap. Mus. Zool. Univ. Mich., 682, 1-19.
Chapman R. F., 1969. The insects, structure and
functions, American Elsevier, New York.
Edmunds G. F., Jr., Traver J. R., 1954. The
flight mechanics and evolution of the wings of
Ephemeroptera, with notes on the archetype
msect wing, J. Wash. Acad. Sci., 44,
390-399.
Flower J. W., 1964. On the origin of flight in in-
sects. J. Insect Physiol., 10, 81-88.
F:lsom J. W., Wardle R. H., 1934. Entomology
with special reference to its ecological aspects,
4th ed., Blakiston, Philadelphia.
Hamilton K. G. A., 1971. The insect wing, Part I.
Origin and development of wings from notal
!obes, J. Kans. Entomol. Soc., 44, 421-433.
Hamilton K. G. A., 1972. The insect wing, Part
II. Vein homology and the archetypal insect
wing. J. Kans. Entomol. Soc., 45, 54-58.
Hamilton K. G. A., 1972. The insect wing, Part
III. Venation of the orders, J. Kans. Entomol.
Soc.. 45, 145-162.
Hamilton K. G. A., 1972. The insect wing, Part
IV Venational trends and the phylogeny of the
winged orders, J. Kans. Entomol. Soc., 45,
29M08.
Matsuda R., 1958. On the origin of the external
senitalia of insects, Ann. Entomol. Soc. Am.,
51. 84-94.
Matsuda R., 1963. Some evolutionary aspects of
the insect thorax, Annu. Rev. Entomol., 8,
59-76.
Matsuda R., 1970. Morphology and evolution of
the insect thorax, Mem. Entomol. Soc. Can.,
76, 1-431.
Matsuda R., 1976 Morphology and evolution of
the insect abdomen, Pergamon, Oxford.
Rempel J. G., 1975. The evolution of the insect
head: The endless dispute, Quaest. Entomol.,
11, 7-25.
Richards O. W., Davies R. G., 1977. Imms’
general textbook of entomology, 10th ed.,
Chapman & Hall, London, vols. 1-2.
Schmitt J. B., 1938. The feeding mechanism of
adult Lepidoptera, Smithsonian Misc. Collec-
tions, 97, 1-28.
Scudder G. G. E., 1961. The comparative mor-
phology of the insect ovipositor, Trans. R.
Entomol. Soc. London, 113, 25-40.
Scudder G. G. E., 1971. Comparative morpho-
logy of insect genitalia, Annu. Rev. Entomol.,
16, 379-406.
Shodgrass R. E., 1935. Principles of insect mor-
phology. McGraw-Hill, New York.
Snodgrass R. E., 1944. The feeding apparatus of
biting and sucking insects affecting man and
aminals, Smithsonian Misc. Collections, 104,
1-113.
Snodgrass R. E., 1950. Comparative studies on
the jaws of mandibulate arthropods, Smith-
sonian Misc. Collections, 116, 1-85.
Waterhouse D. F. et al., 1970. The insects of
Australia, Melboum Univ. Press, Carleton.
Weber H., 1952. Morphologic, Histologie und
Entwicklungsgeschichte der Articulaten,
Fortschr. Zool., N.F., 9, 18-231.
Weber H., 1954. Grundriss der Insektenkunde,
3rd rev. ed., Fischer, Stuttgart.
4
Внутреннее строение
Внутренние органы насекомых и родственных им гексапод заключены в экзо-
скелет, защищающий их от внешних воздействий. Любые выросты внутренних
органов, выступающие над поверхностью тела, окружены тонкой кутикулярной
оболочкой и, таким образом, тоже находятся внутри экзоскелета.
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Пищеварительная система представ-
лена кишечным каналом и придаточными
образованиями в виде различных желез,
связанных с ним прямо или косвенно. В
типичном случае они включают слюнные
железы, слепые выросты желудка и маль-
пигиевы сосуды.
Кишечный канал (рис. 4.1). Этот ор-
ган представляет собой трубку, проходя-
щую через центральную часть тела. Его
переднее отверстие, рот, расположено в
основании преоральной полости (про-
странство, ограниченное ротовыми ча-
стями) , заднее отверстие, или анус, нахо-
дится в заднем сегменте тела. Кишечный
канал разделен на три хорошо выра-
женных отдела. переднюю кишку (стомо-
деум), среднюю кишку (мезентерон) и за-
днюю кишку (проктодеум). Обычно ме-
жду передней кишкой и средней располо-
жен стомодеальный, или кардиальный
клапан, а между средней и задней киш-
кой -проктодеальный, или пилорический
клапан. Передняя кишка и задняя кишка
образовались путем впячивания экто-
дермы зародыша; средняя кишка образо-
валась из эндодермы; об этом будет ска-
зано ниже в главе, посвященной зароды-
шевому развитию насекомых.
У некоторых примитивных Hexapoda
все три цтдела кишечного канала имеют
вид простой трубки (рис. 4.1). У боль-
Рис. 4.1. Пищеварительный тракт коллемболы Tomocerus niger, у которой
первичные отделы кишечного канала не имеют вторичных
специализированных образований. (По Снодграссу.)
7-средняя кишка; 2-ротовое отверстие; 3-передняя кишка,
4-стомодеальный, или кардиальный клапан, 5-проктодеальный или
пилорический клапан, 6-задняя кишка; 7-анальное отверстие.
4. Внутреннее строение
99
Рис. 4.2. Схематическое изображение отделов и выростов пищеварительного канала.
(По Снодграссу.)
/-ротовое отверстие; 2-глотка; 3-пищевод; 4-слепой отросток средней кишки;
?-зоб; 6-преджелудок; 7-прямая кишка (ректум); 8-анальное отверстие; 9-тонкая
кишка, 10-мальпигиевы сосуды.
дзянства же насекомых каждый отдел
дифференцирован на специализиро-
ванные подотделы, выполняющие опре-
деленные функции. Типичное строение
такого дифференцированного кишечного
канала показано на рис. 4.2.
ПЕРЕДНЯЯ КИШКА, ИЛИ СТОМО-
ДЕУМ. Этот отдел обычно разделен
3 основные части: 1) переднюю-пище-
(эзофагус), в виде длинной или корот-
кой трубки; 2) следующую за ней расши-
ренную часть - зоб, переходящий в 3)
более узкий мышечный желудок, или
ъ&ейжелудок (провентрикулус), имеющий
»д плотной мускулистой складки и рас-
положенный непосредственно перед
средней кишкой. Часть пищевода, при-
мыкающую к ротовому отверстию, ча-
сто называют глоткой (фаринксом), но ее
трудно обнаружить без точного знания
мускулатуры этой области. Граница ме-
жду пищеводом и зобом часто условна,
как на рис. 4.2; у некоторых же насе-
комых, например бабочек (рис. 4.3,В),
юб имеет вид круглой камеры. Эта мо-
дификация зоба достигла крайнего выра-
жения у многих видов мух (рис. 4.4), у
которых он представляет собой мешок,
соединенный с пищеводом длинной бо-
Передняя кишка Средняя кишка Задняя кишка
В
Рис. 4.3. Изменения в строении
пищеварительного канала на протяжении
жизненного цикла бабочки Malacosoma
americana. (По Снодграссу.)
Пищеварительный канал личинки (А),
куколки (Б) и взрослой особи (В),
/-пищевод; 2-зоб; 3-прямая кишка
(ректум); 4-анальное отверстие;
5-мальпигиевы сосуды; 6-тонкая кишка.
100
4. Внутреннее строение
Рис. 4.4. Пищеварительный канал и слюнные железы яблонной
пестрокрылки Rhagoletis pomonella с зобом и кардиальным клапаном средней
кишки, характерными для многих двукрылых. (По Снодграссу.)
/-слюнные железы; 2-протоки слюнных желез; 3-пищевод;
4-кардиальный клапан; 5-средняя кишка; 6-мальпигиевы сосуды;
7-тонкая кишка; 8-прямая кишка, 9-зоб.
ковой трубкой. В ряде случаев провен-
трикулус устроен как простой клапан, от-
крывающийся в среднюю кишку. У насе-
комых, поедающих твердую пищу, он
снабжен твердыми выростами для пере-
малывания пищи и называется «желудоч-
ной мельницей».
СРЕДНЯЯ КИШКА, ИЛИ МЕЗЕНТЕ-
РОН. В этом отделе кишечного кана-
ла переваривается основная часть пищи.
Его называют желудком или вентрику-
лусом. Обычно он представляет собой
трубку, но иногда разделен на несколько
частей. Это разделение особенно хорошо
выражено у Hemiptera, где таких частей
бывает 3 или 4. Средняя кишка нередко
имеет несколько пальцевидных выро-
стов, называемых слепыми отростками.
Как правило, они расположены на ее
переднем конце (рис. 4.2), но иногда и
ближе к заднему концу.
ЗАДНЯЯ КИШКА, ИЛИ ПРОКТОДЕ-
УМ. Этот задний отдел кишечного
канала имеет неодинаковое строение у
разных насекомых, но в большинстве
случаев подразделяется на трубчатую
переднюю часть -тонкую кишку и рас-
ширенную заднюю часть -прямую киш-
ку, или ректум, на конце которой нахо-
дится анальное отверстие.
Лабиальные, или слюнные, железы. У
большинства насекомых имеется пара
желез, связанных с лабиумом и лежащих
под средней кишкой (рис. 4.4). Каждая
железа снабжена протоком, направ-
ленным вперед. Оба протока соединяют-
ся обычно в области головы, образуя не-
парный проток, открывающийся в пре-
оральную полость между лабиумом и
гипофаринксом. У разных насекомых эти
железы выполняют разные функции, при-
чем в некоторых случаях их функции еще
до конца не выяснены. У большинства
насекомых лабиальные железы секрети-
руют слюну, как, например, у тараканов.
У гусениц бабочек и личинок перепонча-
токрылых эти железы секретируют шелк,
используемый для постройки личи-
ночных ячеек и куколочных коконов. У
кровососущих насекомых лабиальные
Внутреннее строение
101
гдлезы секретируют антикоагулирую-
zee вещество, препятствующее сверты-
поглощенной крови.
ВЫДЕЛИТЕЛЬНАЯ СИСТЕМА
Мальпигиевы сосуды. У всех насе-
rvbix. за небольшим исключением,
смеются пучки длинных тонких трубо-
’ет_ отходящих от кишечного канала
згдгзи места соединения средней и за-
кишок (рис. 4.2). Эти трубочки, на-
изжемые .мальпигиевыми сосудами, вы-
тсд=яют выделительную функцию. Чис-
»_ трубочек колеблется от 1 до 250 или
': ~ее При наличии большого их числа
группируются в пучки одинакового
TLivepa.
КРОВЕНОСНАЯ СИСТЕМА
Кровеносная система включает ткани
:станы. обеспечивающие циркуляцию
гт»: зг по телу, а также саму кровь. У
ч^. тих животных, например у позво-
-г-ных. кровь протекает только по спе-
д*а_тьным сосудам (артериям, венам, ка-
7д_’_тграм). Такая кровеносная система
юзается замкнутой. У насекомых де-
:с<тоит иначе. Кровь свободно течет
i т : гости тела, омывая различные ткани
t станы. Только часть своего пути она
и*2 хс дит через специальный орган-
дзЕной сосуд (сердце), расположен-
з дорсальной части тела. Он ра-
~ тает как насос, накачивая кровь из за-
дне2 части тела и выпуская ее в полость
:дезы. Отсюда кровь снова течет в по-
есть тела, попадает в сердце, вновь
терекдчивается вперед и т. д. Такая кро-
эечтсная система называется незамкну-
или открытой, а полость тела, через
% т ?рую кровь свободно протекает - ге-
4. к кровеносной системе отно-
кроме того, мелкие дополни-
дтьные насасывающие органы, располо-
женные вблизи или внутри конечностей
(по одному или более), а также спе-
циальные клетки - фагоциты, разбро-
санные по всему телу, и объемистое жи-
ровое тело, осуществляющее накопление
и синтез запасных питательных веществ.
Кровь. Жидкость, циркулирующая в
полости тела, называется кровью, или ге-
молимфой. Она состоит из жидкого ин-
гредиента -плазмы- и различных сво-
бодноплавающих клеток, называемых
кровяными тельцами, или гемоцитами.
Изучение состава и функции крови отно-
сится к таким предметам, как гистология
и физиология, которым посвящена сле-
дующая глава.
Спинной сосуд. Спинной сосуд (рис.
4.5,Л) расположен непосредственно под
дорсальной поверхностью тела и прости-
рается от заднего конца брюшка до
переднего конца головы. Он является
главным пульсирующим органом, вызы-
вающим циркуляцию крови.
Спинной сосуд разделен на 2 отдела
задний -сердце и передний -аорту. Серд-
це обеспечивает пульсацию, в результате
чего кровь по аорте течет вперед и выли-
вается в полость головы.
Обычно сердце подразделено на не-
сколько вздутых камер, соответствую-
щих сегментам тела, с сужениями между
ними. В типичном случае камер бывает 9,
по числу первых 9 сегментов брюшка.
Каждая камера имеет пару латерально
расположенных отверстий, или остий,
через которые кровь попадает в камеру.
У некоторых насекомых сердце имеет
строение, сильно отличающееся от опи-
санного. Например, у тараканов и япигид
первые две камеры расположены в сред-
не- и заднегруди. У клопа Nezara сердце
состоит из одной большой камеры с тре-
мя парами остий.
Аорта в типичном случае - простая
трубка, отходящая от сердца. Иногда же,
как у личинок комаров-долгоножек и у
102
4. Внутреннее строение
Рис. 4.5. Спинной кровеносный сосуд, или сердце. (По Снодграссу.)
А. Схематическое изображение аорты и трех камер сердца
с соответствующей частью дорсальной диафрагмы. Вид с дорсальной
стороны, стрелками показано направление тока крови. Б. Вертикальный
разрез через грудь и основание брюшка Sphinx convolvuli, на котором
показана пульсирующая мембрана, расположенная в среднегруди. 1 - сердце,
2-аорта; 3-дорсальная диафрагма; 4-мышцы диафрагмы, 5-остия;
6 - мышца, 7 - среднегрудь, 8 - клапан, 9 - пульсирующая мембрана,
10-заднегрудь; 77-пищеварительный канал; 72-зоб; 13-вентральная
нервная цепочка.
взрослых комаров, аорта также пуль-
сирует и, таким образом, помогает
сердцу поддерживать циркуляцию
крови.
Спинная диафрагма и синус. Снизу к
сердцу прикреплены парные мышечные
пучки, называемые крыловидными, или
алярными, мышцами. Они называются
так, потому что напоминают по форме
веер или крылья. Эти мышцы соединяют
сердце с латеральными частями терги-
тов. В тех случаях, когда они хорошо раз-
виты, они полностью отделяют основ-
ную полость тела от области, прилежа-
щей к сердцу. Перегородка, создаваемая
крыловидными мышцами, называется
дорсальной, диафрагмой, а околосердеч-
ная область -дорсальным синусом. Про-
тяженность диафрагмы и синуса ограни-
чена протяженностью сердца и в область
аорты они не продолжаются.
Дополнительные пульсирующие ор-
ганы. Помимо сердца у насекомых бы-
вают и другие пульсирующие органы,
помогающие циркуляции крови. Чаще
всего встречаются торакальные пульси-
рующие органы и вентральная диафраг-
ма. Другие дополнительные пульси-
рующие органы встречаются очень
редко.
ТОРАКАЛЬНЫЕ ПУЛЬСИРУЮЩИЕ
ОРГАНЫ. У многих насекомых, особенно
4. Внутреннее строение
103
быстро летающих, например у бражни-
ков. имеется специальный пульсирую-
щий орган (рис. 4.5,Б), который протал-
кивает кровь в крылья, откуда она соби-
рается в аорту. Этот пульсирующий ор-
ган представляет собой полость в скутел-
луме. снабженную гибкой или пульси-
рующей мембраной. Кровь попадает в
зрган через трубку, которая называется
аортальным дивертикулом, и ведет не-
посредственно в аорту.
ВЕНТРАЛЬНАЯ ДИАФРАГМА. У мно-
гих Orthoptera, Hymenoptera и Lepi-
doptera имеются пучки мышц, располо-
женных над брюшной нервной цепочкой,
□сдобно крыловидным мышцам, обра-
зующим диафрагму под сердцем; такие
лучки мышц называются вентральной
диафрагмой. Их растяжение и сокра-
щение вызывает токч крови в заднем и
латеральных направлениях.
ТРАХЕЙНАЯ СИСТЕМА
У большинства насекомых имеется
система внутренних трубочек, или тра-
тги. проводящих воздух к клеткам тела.
Эта система трубочек называется тра-
^гйной системой и выполняет функцию
дружного дыхания. Помимо насекомых,
хорошо развитая трахейная система
зстречается в других группах членисто-
ногих . у некоторых Arachnida, Crustacea
> у большинства Chilopoda. Примитив-
ная трахейная система имеется у Опу-
c±ophora и Diplopoda.
Трахейная система устроена весьма
дюжно, поскольку трахеи ветвятся на
многочисленные тонкие трубочки, каж-
дая из которых подходит к небольшой
группе клеток. Сложная сеть трахей у на-
секомых аналогична сети кровеносных
сосудов и капилляров у позвоночных
животных.
Основные компоненты
трахейной системы
Общий тип строения трахейной си-
стемы насекомых показан на рис. 4.6. В
каждом сегменте трахеи собраны в пуч-
ки, и наружный воздух попадает в них че-
рез парные посегментно расположенные
отверстия, называемые дыхальцами (5).
Рис. 4.6. Трахейная система насекомого. (Из
Folsom, по Kolbe.)
7-щупик; 2-антенна, 3-головной мозг;
4-нервная цепочка, 5-дыхальце; 6-нога,
7 - главный трахейный ствол; 8 - ветвь
трахеи, подходящая к дыхальцу;
9-вентральная ветвь трахейного ствола;
10- висцеральная ветвь трахейного ствола.
104
4. Внутреннее строение
Дыхальца на каждой стороне тела более
или менее прямо связаны с одним из двух
основных трахейных стволов (7), прохо-
дящих вдоль всего тела. От основных
стволов в каждом сегменте отходят
многочисленные веточки (всегда парные,
поскольку они отходят от каждого ство-
ла), снабжающие воздухом ткани вну-
тренних органов. Число и расположение
этих веточек варьирует у разных насе-
комых, но обычно в каждом сегменте
имеется по три симметричных больших
ветви: 1) дорсальная ветвь, обслуживаю-
щая спинной сосуд и дорсальные мыш-
цы, 2) вентральная, или висцеральная,
ветвь, обслуживающая пищеваритель-
ную и половую системы, и 3) вентраль-
ная ветвь, обслуживающая вентральные
мышцы и нервную цепочку
Тонкие окончания трахей делятся на
крошечные капиллярные трубочки, или
тракеолы, диаметром не более 1 мкм.
Трахеолы разветвляются между клетка-
ми и оплетают их, являясь функциональ-
ной частью системы, обеспечивающей
диффузию кислорода в клетки тела.
Трахейные стволы. Сегментарное рас-
положение пучков трахей указывает на
то, что первоначально насекомые имели
независимые и не связанные друг с дру-
гом трахейные системы в каждом посто-
ральном сегменте. У современных насе-
комых, за очень немногими исключения-
ми, трахейные системы соседних сегмен-
тов связаны между собой трубочками.
Эти связующие трубочки и образуют
трахейные стволы. У многих насекомых
основные стволы занимают в теле лате-
ральное положение и называются поэто-
му латеральными трахейными стволами.
Часто по обеим сторонам сердца распо-
лагается еще и пара дорсальных тра-
хейных стволов. Эти стволы обычно
меньшего диаметра и являются дополни-
тельными по отношению к латеральным
стволам. Однако у большинства личинок
мух имеет место обратное соотношение:
дорсальные стволы развиты сильнее и
являются основными дыхательными
стволами (рис. 4.7).
Трахейные воздушные мешки. Воз-
душные мешки являются важным при-
способлением у насекомых-они служат
хранилищами воздуха. Часто воздушные
мешки представляют собой раздутые
участки трахейных стволов, как показано
на рис. 4.8 и 4.9. У многих быстро летаю-
щих насекомых, таких, как комнатная
муха или пчелы, мешки заполняют собой
большую часть полости тела. В результа-
те сокращения мышц тела мешки могут
сжиматься и расправляться, подобно
Рис. 4.7. Личинка мухи с крупными дорсальными трахейными стволами. (По
Снодграссу.)
7-ротовые крючья; 2-переднее дыхальце, 3-дорсальный трахейный ствол;
4-заднее дыхальце; 5-латеральный трахейный ствол.
4. Внутреннее строение
105
Рис. 4.8. Строение трахеи (Л) и примеры трахейных воздушных
мешков (Б, В). (По Снодграссу.)
7-эпителий; 2-интима; 3-тенидий, искусственно отделенный от
интимы.
Рис. 4.9. Внутренняя поверхность трахеи насекомого. Изогнутые утолщения (тенидии)
обеспечивают упругость трахей, а тонкая выстилка (интима)-их гибкость; увеличение
примерно 40000. (С разрешения Лаборатории электронной микроскопии Университета
Джорджии; фото К. Депплса и А. Ли.)
106
4. Внутреннее строение
воздуходувным мехам, увеличивая по-
ступление и выход воздуха.
Дыхальца. Функцией дыхалец являет-
ся регуляция поступления воздуха. Они
различаются по размерам, форме
и строению. Функционирующие дыхаль-
ца снабжены своего рода запирательны-
ми устройствами, которые могут быть
наружными (обычно в форме двух распо-
ложенных друг против друга губ) или
внутренними (чаще в виде зажима, сжи-
мающего просвет трахеи).
Открытые трахейные системы
Трахейные системы с открывающи-
мися наружу и функционирующими ды-
хальцами называются открытыми. На-
иболее распространенный тип таких си-
стем имеет 10 пар дыхалец-по паре в
средне- и заднегруди и остальные на каж-
дом из 8 брюшных сегментов. Суще-
ствует много модификаций этого типа,
например, у личинок комаров дыхальца
имеются только на восьмом брюшном
сегменте, у большинства личинок мух
дыхальца расположены на переднегруди
и в последнем (восьмом) брюшном сег-
менте, а у куколок мух осталась только
пара переднегрудных дыхалец; у неко-
торых водных личинок, например обык-
новенной пчеловидки («крыски»), имеет-
ся только задняя пара дыхалец, располо-
женных на конце длинной выдвигающей-
ся дыхательной трубки, выступающей из
воды.
Закрытые трахейные системы
У многих насекомых дыхальца не
функционируют или вообще отсут-
ствуют. Такие трахейные системы назы-
ваются закрытыми. У насекомых с за-
крытыми трахейными системами нет
трахейных стволов с их обширным вну-
тренним ветвлением. Чаще всего в этих
случаях дыхальца заменены сетью тон-
ких трахеол, располагающихся под ко-
жей или в жабрах. Такое строение тра-
хейной системы мы находим у нимф и
личинок многих водных насекомых по-
денок, веснянок, равнокрылых стрекоз,
мошек.
Интересная модификация имеет ме-
сто у нимф стрекоз, на стенках прямой
кишки которых расположены похожие на
жабры лепестки, пронизанные тонкими
трахеями. Периодически насекомые на-
бирают воду в прямую кишку и выталки-
вают ее оттуда, омывая ректальные
жабры и таким образом снабжая трахеи
воздухом.
НЕРВНАЯ СИСТЕМА
Нервная система насекомых хорошо
развита и состоит из центральной и сто-
модеальной нервных систем. Как и у
других животных, нервная система
координирует активность насекомого
с условиями внешней и внутренней
среды.
Центральная нервная система. Цент-
ральная нервная система состоит из сле-
дующих основных компонентов
(рис. 4.10) 1) головного мозга, располо-
женного в голове, и 2) парных нервных
центров, или ганглиев, расположенных
посегментно. Ганглии соединены друг с
другом парными пучками нервных воло-
кон и образуют цепочку, при этом
первый ганглий соединен с головным
мозгом. В тех случаях, когда в эволюции
насекомых имело место слияние сегмен-
тов тела (рис. 2.1), происходило и слия-
ние соответствующих ганглиев. Так, го-
лова насекомого претерпела сильные из-
менения и состоит из компонентов раз-
ного происхождения, первичной головы
(акрона, или простомиума) и шести туло-
вищных сегментов. В соответствии с
этим и нервные центры, расположенные
4. Внутреннее строение
107
Рис. 4.10. Последовательные стадии концентрации центральной
нервной системы Diptera. (Из Folsom, Wardle, 1934, по Brandt.)
А - Chironomus; Б - Empis, В - Tabanus; Г- Sarcophaga.
в голове, неоднородны по своему проис-
хождению и мало похожи на нервные
центры исходного типа.
Головной мозг. Головной мозг
(рис. 4.11) расположен в голове над пи-
щеводом и поэтому его часто называют
надглоточным, или супраэзофагеалъным
ганглием. Он состоит из трех главных от-
делов: 1) протоцеребрума, иннервирую-
щего сложные глаза и простые глазки, 2)
дейтоцеребрума, иннервирующего ан-
тенны, и 3) тритоцеребрума, осущест-
вляющего контроль над симпатической
нервной системой. Все три отдела имеют
четкое парное строение.
В течение длительной эволюции раз-
личные части головы насекомых посте-
пенно изменяли свое первоначальное по-
ложение. Поэтому головной мозг, ко-
торый исходно лежал впереди ротового
отверстия, теперь стал располагаться над
ним или над пищеводом. Поскольку про-
тоцеребрум и дейтоцеребрум располо-
жены над пищеводом, считается, что они
образовались из примитивного просто-
меального мозга аннелид. Тритоце-
ребрум тесно связан с дейтоцеребру-
мом, тогда как обе его части соединены
комиссурой, проходящей под пищево-
дом. На этом основании его считают ган-
глием первого туловищного сегмента,
вошедшего в состав головы.
Подглоточный, или субэзофагеальный,
ганглий. Этот крупный нервный центр
расположен в голове под пищеводом и
соединен с головным мозгом парой
толстых коннективов. Он представляет
собой слившиеся ганглии мандибул, мак-
силл и лабиума. От этого сложного ган-
глия отходят нервные стволы, иннерви-
рующие ротовые органы, а также пара
коннективов, проходящих через шейный
отдел в грудь.
Вентральная (брюшная) нервная цепоч-
ка. В типичном случае в вентральной ча-
сти каждого сегмента груди и брюшка
имеется по одному нервному ганглию.
Ганглии соседних сегментов соединены
между собой парными коннективами и
все вместе образуют цепочку из ганглиев,
протянувшуюся назад от переднегруди
(рис. 4.10); ее и называют вентральной
108
4. Внутреннее строение
Рис. 4.11. Нервная система саранчи, вид сбоку. (По Снодграссу.)
7-оцеллярная ножка; 2-нерв, иннервирующий покровы тела;
3-оптическая доля, 4-аорта; 5-зоб; 6-затылочный ганглий;
7-стомодеальные нервы; 8-прилежащее тело (corpus allatum),
9-подглоточный ганглий, 10-вентральная нервная цепочка;
77-проток слюнной железы, 72-нервы, иннервирующие ротовой
аппарат; 13-окологлоточные коннективы; *14-нерв,
иннервирующий лабрум; 75-глотка; 16-тритоцеребральная
комиссура, 77-фронтальный коннектив; 18 - фронтальный ганглий;
79-возвратный нерв; 20-тритоцеребрум; 27-дейтоцеребрум;
22- протоцеребрум.
нервной цепочкой. Она соединяется с под-
глоточным ганглием посредством кон-
нективов, проходящих через шейный от-
дел. От торакальных ганглиев отходят
нервы, иннервирующие ноги и крылья, а
от абдоминальных ганглиев нервы идут к
абдоминальным мышцам и придаткам.
В исходном типе вентральной нерв-
ной цепочки ганглии были четко отде-
лены один от другого. У различных
групп насекомых некоторые ганглии сли-
лись, образовав меньшее число более
крупных ганглиев. Особенно наглядный
пример такого слияния мы встречаем в
отряде Diptera (рис. 4.10). У прими-
тивных представителей этого отряда не-
рвная цепочка близка к исходному типу,
в более специализированных семействах
4. Внутреннее строение
109
торакальные ганглии слиты в единую не-
рвную массу, а абдоминальные ганглии
становятся маленькими и в конце концов
едва различимыми. Последовательные
стадии этого процесса показаны на рис.
4.10,Л-Г.
Стомодеальная нервная система. У на-
секомых имеется так называемая симпа-
тическая нервная система (рис. 4.12),
осуществляющая регуляцию «непроиз-
вольных» движений передней части пи-
щеварительного тракта и спинного сосу-
да. Однако функции некоторых нервов
этой системы остаются еще до конца не-
понятыми. Более правильно называть
эту систему стомодеальной, поскольку
большинство ее компонентов расположе-
но в верхней части или по сторонам
передней кишки (стомодеума). Цен-
Рис. 4.12. Схема симпатической нервной
системы насекомого. (Скомбинировано из
нескольких источников.)
1 - прилежащее тело (corpus allatum),
2-кардиальное тело (corpus cardiacum)-
3-головной мозг; 4-пищевод;
5-фронтальный ганглий; 6-фронтальный
коннектив; 7- возвратный нерв,
8 - коннективы; 9 - затылочный ганглий;
10-вентрикулярный ганглий.
тральным образованием стомодеальной
системы следует считать фронтальный
ганглий, расположенный впереди голов-
ного мозга и соединяющийся с тритоце-
ре б рум ом посредством парных нервных
тяжей. От фронтального ганглия отхо-
дит непарный возвратный нерв, напра-
вляющийся назад под головной мозг и
затем идущий вдоль верхней части пище-
вода дорсально от него до места, где он
соединяется с системой мелких ганглиев
и нервов, иннервирующих переднюю
кишку, протоки слюнных желез, аорту и,
по-видимому, некоторые мышцы рото-
вого аппарата.
МУСКУЛАТУРА
Тело насекомых снабжено чрезвычай-
но сложной системой мышц. Они управ-
ляют почти всеми движениями тела и ко-
нечностей. У некоторых насекомых
имеется до 2000 пучков мышечных воло-
кон.
При вскрытии тела насекомого мыш-
цы очень хорошо видны. Они не обра-
зуют сплошного слоя, а приурочены к
определенным участкам тела и входят в
состав отдельных органов насекомого. В
зависимости от их расположения мышцы
можно разделить на три категории.
Висцеральные мышцы. Пищевари-
тельный тракт и протоки половых желез
окружены слоем мышц, вызывающих их
перистальтику. Пучки волокон могут
быть круговыми, продольными и косы-
ми; кроме того, могут встречаться раз-
личные комбинации из этих пучков. От-
крывающие и запирающие механизмы
дыхалец, а также ротовой аппарат снаб-
жены специальной мускулатурой. В ра-
боте кровеносной системы также уча-
ствуют пучки мышечных волокон.
Сегментарные мышцы. Сегменты тела
насекомого связаны между собой целой
серией мышечных пучков, обеспечиваю-
Рис. 4.13. Мускулатура таракана у вентральной, дорсальной и латеральной стенок
тела. (Из Folsom, Wardle, 1934, по Miall, Denny.)
7-мышцы головы; 2-аддуктор (приводящая мышца) коксы; 3-абдуктор
(отводящая мышца) коксы; 4-экстензор (разгибатель) бедра; 5-первая
дорсо-вентральная мышца; 6-продольная вентральная мышца, 7-косая
вентральная мышца, 8-латеральная грудная мышца; 9-продольная дорсальная
мышца; 10-косая дорсальная мышца; 77-алярная мышца* 72-дорсо-веигральная
мышца.
Рис. 4.14. Мускулатура средне- и заднегруди гусеницы. (По
Снодграссу.) d- дорсальные пучки v-вентральные пучки.
4. Внутреннее строение
111
Рис. 4.15. Схема мускулатуры нижней
челюсти насекомых. (По Снодграссу.)
А. Аптериготный тип с одним
сочленением -а'. Б. Птериготный тип
с двумя сочленениями-а', с. /-поперечное
сухожилие; 2-гипофарингеальная аподема;
3 - передняя и 4 - задняя дорсальные мышцы
тизануроидной мандибулы, которые
становятся соответственно отводящей
и приводящей в ортоптероидной; 5, 6,
"-поперечные вентральные аддукторы.
щих сохранение формы тела (рис. 4.13).
Тергиты брюшка связаны продольными
дорсальными, а стерниты-продольны-
ми вентральными мышцами. Тергит и
стернит каждого сегмента связаны косы-
ми или поперечными терго-стернальны-
ми (дорсо-вентральными) мышцами. По-
иному выглядит мускулатура груди. Хо-
рошо заметные наиболее крупные пучки
мышечных волокон управляют движе-
нием ног и крыльев, другие мышцы усту-
пают им по размерам и мощности. По-
мимо перечисленных основных групп
мышц имеется много более мелких пуч-
ков, часто очень сложной структуры
(рис. 4.14). Мускулатура груди и брюш-
ка у разных насекомых имеет свои хоро-
шо выраженные особенности.
Мышцы конечностей. Подвижные ко-
нечности насекомых снабжены мускула-
турой самого разного размера и строе-
ния. Мандибулы насекомых с грызущим
ротовым аппаратом имеют несколько
связанных с ними групп мышц, запол-
няющих собой большую часть головной
капсулы, но внутри самих мандибул
мышц нет (рис. 4.15). Вместе с тем дви-
жением сегментированных придатков,
максилл и ног (рис. 4.16), управляют не
только крупные мышцы, расположенные
в теле насекомого, но и межсегмен-
тарные мышцы самих этих придатков.
ПОЛОВАЯ СИСТЕМА
Насекомые являются раздельнополы-
ми животными, у которых в норме от-
дельная особь принадлежит только к ка-
кому-то одному полу. Известно лишь
несколько примеров гермафродитизма
среди насекомых, когда одна особь несет
в себе признаки обоих полов. Наиболее
Рис. 4.16. Схема строения ноги насекомого и ее мускулатура. (Из Berlese,
упрощено.)
112
4. Внутреннее строение
Рис. 4.17. Половая система самки. (По Снодграссу.)
А. Примитивный тип у Heterojapyx gallardi. Б. Схема наиболее
распространенного типа. В. Схема строения яйцевой трубочки. 7-яйцевая
трубочка; 2-яйцевод; 3-чашевидная часть яйцевода (чашечка)’ 4-латеральный
яйцевод; 5-половое отверстие; 6-концевой филамент; 7-железа
семеприемника; 5-семеприемник; 9-придаточная железа; 10-вагина,
77-ножка яйцевой трубочки.
яркий из этих примеров - австралийский
желобчатый червец Icerya purchasi.
Половая система насекомых хорошо
развита и расположена в брюшке. В
строении женской и мужской половой си-
стем наблюдается параллелизм. Боль-
шая часть компонентов обеих систем
характеризуется билатеральной сим-
метрией.
Женская половая система. В состав
женской половой системы входят: 1)
группа овариол, в которых образуются
яйца, 2) сперматека (семеприемник), где
хранится сперма; и 3) система протоков,
через которые проходят яйца перед выхо-
дом во внешнюю среду. Типичная жен-
ская половая система показана на рис.
4.17,1>,В. Парные яичники располагаются
по одному в боковых частях тела. Каж-
дый яичник состоит из овариол, или яйце-
вых трубочек, число которых может коле-
баться от нескольких штук до многих со-
тен и более. Каждая овариола заканчи-
вается прикрепительной нитью, которая
называется терминальным филаментом.
В верхней части яйцевой трубочки нахо-
дятся развивающиеся яйца, а в нижней ее
части-более зрелые яйца. Основание
трубочки образует небольшой проток,
или ножку (pedicellum). Ножки овариол
каждого яичника вместе образуют ка-
лике, который открывается в лате-
ральный яйцевод. Парные яйцеводы со-
единяются в общий яйцевод, открываю-
щийся в яйцевую камеру, или вагину,
которая имеет выход непосредственно в
наружный яйцеклад - приспособление
для откладки яиц.
С дорсальной стенкой яйцевода свя-
заны две структуры. Одна из них-слер-
х. Внутреннее строение
113
матека, непарный, похожий на луковицу
орган, в котором хранится сперма. В
проток сперматеки впадает проток не-
парной железы. Вторая структура пред-
ставляет собой парные придаточные, или
коллатеральные, железы, выделяющие
липкий секрет для склеивания яиц в клад-
ке или для приклеивания яиц к субстрату.
У некоторых более примитивных на-
секомых, например Orthoptera, вагина
имеет вид простого мешковидного впя-
чивания восьмого брюшного стернита.
Существуют самые разнообразные
отклонения от описанного типа женской
половой системы. Эти отклонения ка-
саются числа и формы яйцевых трубочек,
протоков и желез. Во многих группах
особенно изменчиво строение спермате-
ки и вагины, и это имеет большое таксо-
номическое значение.
Примитивное семейство Japygidae ха-
рактеризуется очень интересно устроен-
ной половой системой. Овариолы (см.
4.17,А) расположены посегментно и свя-
заны между собой парой длинных лате-
ральных яйцеводов, сливающихся
и образующих непарный яйцевод недале-
ко от отверстия, через которое яйца вы-
ходят наружу Такое строение половой
системы указывает на то, что у предков
насекомых были самостоятельные яич-
ники в каждом сегменте, а в процессе эво-
люции происходило их смещение и объ-
единение в заднем конце тела, завершив-
шиеся в конце концов образованием ти-
пичной половой системы, примеры кото-
рой показаны на рис. 4. VI,Б,В.
Мужская половая система. По обще-
му принципу строения мужская половая
система сходна с женской. Ее исходными
компонентами являются пара семенни-
ков, связанные с ними протоки и резер-
вуары со спермой и выводящие протоки.
Такой тип строения показан на рис. 4.18.
Каждый семенник состоит из группы
семенных трубочек, или фолликулов, в ко-
Рис. 4.18. Типичная половая система самца.
(По Снодграссу.)
А. Половая система в целом. Б. Строение
семенника. В. Разрез через семенник
и семенной проток. 7-придаточные железы;
2 - семеизвергательный канал, 3 - пенис;
4 - половое отверстие; 5 - семенник;
6-семепровод; 7-семенной пузырек,
8-семенные трубочки, 9-оболочка
семенника, 70-семевыносящий проток,
77-семенная трубочка.
торых образуется сперма. Семенные тру-
бочки открываются в общий проток, се-
мепровод, который в свою очередь от-
крывается в резервуар - семенной пузы-
рек. От каждого семенного пузырька
отходит самостоятельный проток, оба
протока образуют затем общий семе-
извергательный проток. Этот последний
проходит внутри пениса, или фаллуса, на
конце которого имеется выводное отвер-
стие для спермы. Структуры, обычно свя-
занные с пенисом, формируют мужские
гениталии -эдеагус, и включают плот-
ную оболочку вокруг состоящего из
тонкой ткани пениса. Внутри тела к
семеизвергательному каналу примыкают
114
4. Внутреннее строение
придаточные железы, которые могут
быть парными и непарными.
СПЕЦИАЛИЗИРОВАННЫЕ ТКАНИ
Помимо крупных систем, описанных
выше, у насекомых имеются и другие
внутренние образования либо меньшего
размера, либо не столь четко выра-
женные. Наиболее важными из них сле-
дует считать жировое тело и прилежа-
щие тела (corpora allata).
Жировое тело. Это рыхлое клеточное
образование, не имеющее строгой лока-
лизации, расположенное внутри тела и
особенно хорошо выраженное на послед-
ЛИТЕРАТУРА
Chapman R. F. The insects, structure and fun-
ction. American Elsevier, New York, 819 pp.,
1969.
Richards O. W., Davies R. G. Imms’ general
textbook of entomology. Chapman & Hall,
них личиночных и нимфальных возрас-
тах насекомых. Иногда клетки жирового
тела лежат очень плотно и создают впе-
чатление оформленной ткани. Функция
жирового тела состоит в накоплении за-
пасов питательных веществ и в участии в
промежуточном метаболизме.
Прилежащие тела. Это пара ганглио-
подобных образований (рис. 4.11), тесно
связанных со стомодеальной нервной си-
стемой. Они вырабатывают гормоны,
участвующие в регуляции метаморфоза,
а также развития некоторых тканей
взрослых насекомых. Их строение и
функции обсуждаются в связи с други-
ми структурами нервной системы в гл. 6.
London, 10th ed., Vols. 1 and 2, 1977
Wigglesworth V B. The principles of insect phy-
siology. Chapman & Hall, London, 7th ed.,
827 pp., 1972.
См. также литературу к гл. 3, 5, 6 и 7.
5
Жизненные процессы
У’ насекомых, как и у всех живых организмов, непрерывность поколений (другими
словами, существование вида) зависит от успешного роста и размножения. Эти про-
цессы являются основными функциями всего живого. Насекомое должно взять из
зкружающей среды те вещества, которые ему необходимы для роста, созревания
ззрослого организма и в конечном счете для размножения. В этом смысле насекомое,
как и любой другой живой организм, можно рассматривать как комплекс сложных
высоко интегрированных процессов, происходящих в рамках механических, физиче-
ских и химических систем. Эти процессы, которые изучает физиология, являются ре-
зультатом деятельности органов и тканей, описанных в двух предыдущих главах. На-
стоящая глава посвящена описанию функций некоторых наиболее важных систем и
органов и их взаимодействий. В следующей главе отдельно рассматриваются функ-
ции нервной и гормональной систем насекомых-координация движений, поведение и
реакции на воздействие внешних факторов. Глава 7 посвящена рассмотрению жиз-
ненных циклов, роста и размножения насекомых.
К основным физическим и химическим процессам, из которых слагается жизненно
важная активность насекомого, как и любого другого животного, относятся движе-
ние. метаболизм (химические реакции, происходящие в целом организме, его органах
и тканях), дыхание, циркуляция крови, поддержание гомеостаза (постоянства вну-
тренней среды организма), транспорт жидкостей (в том числе и отработанных), адап-
тация и раздражимость (определенные реакции на физические и химические факторы
внешней среды).
СТРУКТУРА И ФУНКЦИЯ СКЕЛЕТА
Отличительной чертой насекомых
является наличие у них хитинового экзо-
скелета, представляющего собой на-
ружный покров тела и конечностей (инте-
гумент) и секретируемого клетками гипо-
дермы (рис. 5.1,Л). Основную часть по-
крова составляет кутикула, но помимо
этого его наружная поверхность несет
множество волосков и сенсорных рецеп-
торов, а внутренняя - специальные вы-
росты (аподемы и апофизы, рис. 5Л,В,Г),
к которым прикреплены мышцы. Инте-
гумент называют экзоскелетом, посколь-
ку он находится снаружи тела и защи-
щает насекомое не только от хищников,
но и от механических повреждений,
охлаждения и высыхания. Он также опре-
деляет форму и очертания тела насеко-
мого. Специальные слои экзоскелета со-
здают барьер для проникновения жидко-
стей, патогенных организмов и инсекти-
цидов. Потеря воды путем испарения
представляет наибольшую опасность для
наземных организмов, а среди насе-
комых большинство ведет наземный или
воздушный образ жизни, по крайней ме-
ре в течение части своего жизненного ци-
кла. Испарение является функцией по-
верхности тела, а не объема, и при
уменьшении размеров тела отношение
поверхности тела к его объему увеличи-
вается. Таким образом, поскольку насе-
116
5. Жизненные процессы
Рис. 5.1. А. Схема строения покровов тела (из Wigglesworth, упрощено). Б. Щетинка с
углублением. (По Снодграссу, упрощено.) В. Впячивание кутикулы, образующее аподему.
Г Впячивание кутикулы, образующее апофиз. (В и Г-из Richards, 1951, с изменениями.)
7-проток, 2-поровый канал, 3-базальная мембрана, 4-железистая клетка,
5-углубление, 6-мембрана щетинки, 7-эпикутикула; 5-тормогенная клетка;
9-кутикула, 10-эпидермальная клетка, 77-трихогенная клетка, 72-гиподерма,
13 -сарколемма; 14 -мышца.
комые-одновременно и мелкие, и на-
земные животные, перед ними стоит
острая проблема защиты от излишнего
испарения того небольшого количества
воды, которое содержится в их теле. Эту
проблему насекомые решают, используя
свойство кутикулы быть непроницаемой
для водяных паров. Без такой серьезной
защиты насекомое, находящееся в полете
даже короткий срок, не смогло бы избе-
жать рокового для него иссушения.
Гиподерма. Это слой, состоящий из
одного ряда эпителиальных клеток, рас-
положен под кутикулой и над базальной
мембраной -аморфным гранулярным
слоем. Большинство эпителиальных кле-
ток выделяет секрет, образующий кути-
кулу, которая представляет собой
инертный неживой наружный компонент
тела насекомого. Специализированные
эпителиальные клетки гиподермы обра-
зуют органы чувств и кожные железы
(рис. 5.1,Л,Б).
Кутикула. Кутикула насекомых вы-
полняет несколько совершенно раз-
личных функций; она служит защитой и
опорой тела насекомого, а также являет-
ся источником пищи во время голодания
Жизненные процессы
117
* таньки. Кроме того, она участвует в
грззовании органов чувств и ротового
х_=арата и выстилает трахеи.
Кутикулярный экзоскелет насекомого
тгевставляет собой тонкий, но очень
гтеткий покров. По своим механическим
сгадтвам кутикула обладает проч-
-<стъю. сравнимой с прочностью метал-
?при сравнении на единицу веса). На-
~<мер. предел прочности кутикулы
<: ло 10 кг/мм 2, а стали -100 кг/мм 2, но
—2_ть в 7 раз тяжелее. Прочность алюми-
почти такая же, как и кутикулы, но он
? 2 раза тяжелее.
Кроме того, кутикула обладает
-тезвычайной твердостью. Некоторые
-„еюмые способны с помощью своих
чхядибул проделать отверстие в метал-
е-жести или меди.
В типичном случае кутикула состоит
~ толстой внутренней ламинарной (слои-
— i кутикулы и тонкой наружной
*^<утикулы.
Ламинарная кутикула состоит из
•- тренней эндокутикулы и внешней эк-
~.тикулы (рис. 5.1,Л). Оба слоя по-
.—роены из гликопротеина, представляю-
щего собой комплекс высокомолекуляр-
т :го полисахарида хитина и белка, при-
мем считают, что микрофибриллы Кити-
ев погружены в белковый матрикс. Экзо-
г. титула отсутствует в сочленениях и
•ноль швов, где происходит отделение
-пгчжного скелета при линьке.
Кутикулярные пластинки (ламеллы)
сразованы чрезвычайно тонкими слоя-
ми чшкрофибрилл, при этом продольные
микрофибрилл, лежащих в разных
^госк остях, повернуты на несколько гра-
сов по отношению друг к другу
рис. 5.2). Такая конструкция, подобно
сапере, характеризуется высокой проч-
- сетью. Наружный слой ламинарной ку-
~<£улы (экзокутикула) химически устой-
-?з и не растворяется в линочной жидко-
гти. В состав экзокутикулы обычно вхо-
дят хиноны (производные фенолов),
образующие поперечные связи между со-
седними белковыми цепями микрофи-
брилл, что делает эти последние прочны-
ми и устойчивыми образованиями.
Ламинарную кутикулу с ее пластинка-
ми пронизывают крошечные поровые ка-
нальцы. Они проходят от гиподермы к
внутренним слоям эпикутикулы и заклю-
чают в себе нитевидные (плазматические)
отростки эпителиальных клеток (рис. 5.1
и 5.3). Поровые канальцы выполняют
функцию протоков для транспортирова-
ния секретов, выделяемых эпите-
лиальными клетками, таких, как воск, ду-
бильные вещества и материал для по-
строения кутикулы. С каждой клеткой
гиподермы может быть связано от 50 до
70 поровых канальцев. У таракана их
плотность достигает 1,2 млн. на 1 мм2.
Эпикутикула. Несмотря на то что эпи-
кутикула очень тонкая, она состоит из 4
слоев, образованных веществами, секре-
тируемыми эпителиальными клетками и
кожными железами (рис. 5.3). Под на-
ружным цементным слоем (секрети-
руемым кожными железами) распола-
гается восковой слой, за которым следует
чрезвычайно тонкий слой плотного ку-
тикулина, а затем более толстый слой го-
могенной кутикулы, прилегающий к на-
ружной части экзокутикулы.
Наружный цементный слой эпикути-
кулы присутствует там, где есть кожные
железы, а его толщина и протяженность
сильно варьируют. В экзоскелете жуков
этот слой наиболее толстый, а у боль-
шинства взрослых насекомых, покрытых
чешуйками, он отсутствует.
Восковой слой построен из чрезвычай-
но длинных углеводородных цепей.
Кутикулиновый слой покрывает все
тело насекомого, за исключением неко-
торых сенсорных образований. Он со-
стоит из двух липидных слоев, которые
на поперечном срезе выглядят как единая
118
5. Жизненные процессы
Рис. 5.2. Экзоскелет в области головы жука Haplorhynchites аепеш-ылпшл слои
кутикулярных пластинок и сенсорная кутикулярная пора; х 7200. (С разрешения
Кафедры. энтомологии Университета пров. Альберта и Д: Крейга.)
5. Жизненные процессы
119
1968.)
тонкая и плотная полоска толщиной от
10 до 20 нм. Этот исключительно важный
слой выполняет несколько функций. Он
проницаем для веществ, стимулирующих
линочные процессы, но непроницаем для
линочной жидкости и не растворяется в
ней. При некоторых условиях он прони-
цаем для воска и воды. Кроме того, ха-
рактер поверхности и скульптурирование
экзоскелета насекомых, а также образо-
вание волосков, чешуек и микротрихий
тесно связаны с кутикулиновым сло-
ем, создающим опору для воскового
слоя.
Плотный гомогенный слой покрывает
большую часть тела насекомого, а также
трахеи (но не трахеолы) и участвует в
образовании выстилки некоторых желез
и органов чувств.
Эпикутикула пронизана продольны-
ми цилиндрическими нитями (филамен-
тами), имеющими 6-13 нм в диаметре
(рис. 5.3). Эти филаменты представляют
собой жидкокристаллические структуры,
состоящие из липидов и воды. Поскольку
они наиболее многочисленны в местах
секреции воска, их первоначально назва-
ли восковыми канальцами. Они, по-види-
мому, служат путями, по которым транс-
портируется воск, откладывающийся на
поверхности кутикулинового слоя. Эпи-
кутикула, таким образом, представляет
собой своего рода проводник, по которо-
му воск направляется к поверхности те-
ла, и барьер, препятствующий потере во-
ды и проникновению ее внутрь. Наруше-
ние воскового слоя у наземных насе-
комых приводит к потере воды.
Специализированные клетки. Часть
клеток гиподермы имеют специальные
120
5. Жизненные процессы
функции-выработки и выделения жид-
ких секретов или образования спе-
циальных структур, например волосков.
Кожные железы. Некоторые гиподер-
мальные клетки участвуют в образова-
нии органов чувств и желез. Кожные
железы обычно состоят из трех клеток,
одна из которых является секреторной, а
две другие образуют проток. Последний
направляется через гиподерму и кутику-
лу к ее поверхности, при формировании
кутикулы по этому протоку секрет, выра-
ботанный в секреторной клетке, изли-
вается на поверхность, образуя це-
ментный слой.
Щетинки (рис. 5.19Б). Большин-
ство гибких волосков, или щетинок, насе-
комых образовано специальными гипо-
дермальными клетками, носящими на-
звание трихогенных клеток. К началу
формирования'волоска сама трихогенная
клетка и ее ядро увеличиваются в разме-
рах и от нее через кутикулу к поверхности
тела появляется проток. С этого момента
начинается построение волоска из мате-
риала, синтезируемого в клетке. С трихо-
генной клеткой тесно связана тормоген-
ная клетка, образующая углубление
(обычно гибкое) вокруг основания воло-
ска. Волосок, или щетинка, сформиро-
ванная таким способом, называется се-
та. После ее формирования родитель-
ские клетки могут дегенерировать.
Таким же образом возникают и спе-
циализированные сеты-чешуйки, ядо-
витые и чувствительные волоски, о ко-
торых пойдет речь в гл. 6.
Окраска. Окраска насекомых связана
с особенностями структуры гиподермы
или экзоскелета и подразделяется на два
типа: пигментную и структурную.
Такие пигменты, как каротиноиды,
склеротин и меланин, откладываются в
кутикуле и создают различную окраску
путем избирательного поглощения све-
товых волн разной длины. Каротиноиды
обычно обусловливают желтый, оран-
жевый или красный цвет. Склеротин и
меланин являются темными пигмента-
ми, во многих случаях определяющими
черную окраску.
Тетрапиррольные пигменты (желчные
пигменты) содержатся в гемолимфе мно-
гих насекомых и имеют голубой цвет.
Комбинация этого голубого пигмента с
каротиноидным пигментом создает зе-
леный цвет, свойственный многим насе-
комым с тонкой прозрачной кутикулой.
Птеридины являются флуоресцирую-
щими пигментами и обнаружены
в крыльях насекомых, глазах и других
тканях. В ряде случаев белая, желтая,
оранжевая и красная окраска насекомых
создается этими пигментами.
О мм охром ы - светозащитные пиг-
менты, локализованные в клетках, приле-
гающих к светочувствительным клеткам
омматидиев сложных глаз насекомых.
Глаза насекомых благодаря наличию ом-
мохромов могут быть красного, желтого
и коричневого цветов. Оммохромы со-
держатся и во многих других тканях на-
секомых, например в гиподерме, а у ба-
бочек они являются Экскретами и одно-
временно пигментами крыльев.
Структурная окраска создается бла-
годаря наличию мельчайших нежных ку-
тикулярных пластинок, которые за счет
рассеивания, отражения и интерференции
разлагают белый свет на лучи с различ-
ной длиной волны. Отдельные пластинки
обычно отделены друг от друга каким-
либо веществом, например у жуков-ска-
рабеид-мочевой кислотой. Объединения
таких пластинок в группы, называемые
ребрами, создают многие металлические
или радужные окраски жуков, ночных и
дневных бабочек. У двух последних
групп крылья покрыты чешуйками
(модифицированными щетинками) с
ребрышками, расположенными по ее
длине. Изучение с помощью электронно-
1
Рис. 5.4. Схема структуры, на которой происходит преломление света, образуемой ребрами чешуйки
крыла бабочки; увеличение примерно 18000. (По Anderson and Richards.)
7-пластинки (ребро), 2-пластинка, 3-поддерживающие структуры (трабекулы), 4-базальная пластинка.
И I ill
122
5. Жизненные процессы
nUHWiiiiiii'iliL i.s ^ihllllliltllicilllitl
ilSII'IIIS
ИидяааеоаооАсерез
!.|l, r litl J ;
Hill llt|inilll*lll||||||||||||IIIIIBIIII llllll'l
— 3
5
зааввввяаиааовнв
[lllllllHIIIIIIinilllllllllllllllllllllllllrillin'HlIlliTlI Illi]
ияавмапдяядакВ|
liillllilltillili;il!^liHiim№!H)l!!ii|
Рис. 5.5. Отслоение старой и образование новой кутикулы (схема линьки). (Из
Hackman, 1971, с изменениями.) А. Экзоскелет и гиподерма перед началом
линочных процессов. Б.Начало отслоения кутикулы: отделение гиподермы от
эндокутикулы и секреция линочной жидкости. В. Растворение старой
эндокутикулы и секреция новой кутикулы. Г Экзоскелет непосредственно перед
линькой-присутствуют одновременно старый и новый покровы тела.
1 - эпикутикула; 2 - экзокутикула; 3 - эндокутикула, 4 - л иночная жидкость;
5-гиподерма; б-линия разрыва кутикулы при линьке; 7-новая эпикутикула и
кутикула.
го микроскопа показало, что каждое ре-
брышко составлено из множества групп
параллельных, чрезвычайно тонких
продырявленных пластинок. Данные, по-
лученные при исследовании тропических
бабочек из рода Mor pho, свидетель-
ствуют о том, что ребрышки с более про-
стой структурой создают неметалличе-
скую окраску, и только при наличии
ребрышек со сложной структурой (рис.
5.4) получается та ослепительная радуж-
ная окраска, которой знамениты эти
бабочки.
Гиподермальный рост: линька. Когда
неполовозрелое насекомое достигает
размера, ограниченного кутикулой, ста-
рая кутикула должна быть заменена но-
вой, более просторной, чтобы создать
возможность для дальнейшего роста
(рис. 5.5). Это периодическое сбрасыва-
ние кутикулы называется линькой. Она
начинается с отслоения старой кутикулы
от лежащей под ней гиподермы (процесс,
называемый аполизисом) и заканчивается
сбрасыванием старой кутикулы (процесс,
называемый экдизисом). Линька вклю-
чает также многочисленные изменения,
происходящие до, между и после аполи-
зиса и экдизиса, и контролируется гормо-
нами. Основными этапами линьки
являются следующие’
1 Эпителиальные клетки гиподермы
либо делятся митотически и образуют
более плотный слой, либо увеличивают-
ся в размерах, после чего кутикула отде-
ляется от гиподермы.
Жизненные процессы
123
2. На поверхности «оголенной» гипо-
лермы образуется новая эпикутикула.
3. В то же время в пространство ме-
хду новой эпикутикулой и старой эндо-
гчтикулой выделяется линочная жид-
кость. находящаяся еще в неактивном
состоянии. Затем эпикутикула становит-
ся непроницаемой, линочная жидкость
переходит в активное состояние, и содер-
жащиеся в ней ферменты хитиназа и про-
теиназа растворяют эндокутикулу ста-
рой кутикулы. Продукты, образующиеся
при растворении старой эндокутикулы,
поглощаются эпителиальными клетками
гиподермы и используются затем для по-
строения новой кутикулы. Таким обра-
зом. утилизируется до 90% старой кути-
кулы.
4. Когда новая эндокутикула наполо-
вину сформирована, кожные железы, уве-
личившиеся в размерах, выделяют се-
крет. который попадает на поверхность
новой кутикулы. Из этого секрета обра-
зуется восковой слой эпикутикулы.
5. Организм с полностью сформиро-
ванной новой кутикулой нуждается в ос-
вобождении от старой кутикулы. Разрыв
старой кутикулы обычно происходит по
средней линии на дорсальной поверхно-
сти груди и головы и вызывается давле-
нием крови. Насекомое сокращает
брюшко, направляя кровь в грудь и за-
ставляя последнюю раздуваться до тех
пор, пока кутикула не лопнет по линии
разрыва. Иногда для усиления этого про-
цесса насекомые заглатывают воздух
(водные насекомые-воду). После разры-
ва старой кутикулы насекомое освобож-
дается из нее. К этому времени линочная
жидкость, как правило, реабсорбируется
тканями тела, и пространство между ста-
рой и новой кутикулой становится сухим.
6. Сразу после линьки новая кутику-
ла, особенно ее несклеротизированные
мембранозные участки, растягивается.
Это происходит в течение короткого пе-
риода и достигается тем, что насекомое
заглатывает воздух или воду, в результа-
те чего увеличивается объем тела; затем
повышается давление крови в разных ча-
стях тела, которые «раздуваются» и рас-
тягивают покров тела. Когда давление
крови снижается, растянутая кутикула не
сокращается до прежней величины, а
образует мелкие складки или склады-
вается в виде гармошки. У личинок, ку-
тикула которых не имеет склеротизиро-
ванных участков, эти складки появляют-
ся на всем теле (рис. 5.6). При наличии
склеротизированных участков складча-
тая кутикула образуется в промежутках
между склеритами. При последующем
росте увеличение кутикулы происходит
благодаря простому расправлению этих
складок (см. рис. 3.1 и 3.2). Когда воз-
можности увеличения поверхности экзо-
скелета таким способом исчерпываются,
возникает необходимость новой линьки.
Процесс сбрасывания старой кутикулы в
одних случаях занимает не более не-
скольких секунд, в других - продолжает-
ся час или более.
7. После линьки кожные железы се-
кретируют тонкий цементный слой, по-
Рис. 5.6. Брюшной сегмент личинки
пилильщика с перепончатыми складками
кутикулы.
А. Сразу после линьки. Б. После роста и
растяжения.
124
5. Жизненные процессы
крывающий кутикулу. Этот слой обеспе-
чивает водонепроницаемость кутикулы и
защищает восковой слой эпикутикулы от
повреждения. Материалом для построе-
ния эндокутикулы служит секрет клеток
эпителия, выделяемый в межлиночный
период.
8. Полностью сформированная новая
кутикула непроницаема для многих жид-
костей, в первую очередь для воды,
имеет склеротизированные участки
и определенную окраску У многих насе-
комых, например саранчовых, затверде-
ние и окрашивание кутикулы происходят
сразу после растяжения, которое следует
за линькой. У других же насекомых
(взрослые Trichoptera, Lepidoptera, мно-
гие Hymenoptera) это происходит до
линьки, когда взрослое насекомое еще
находится в куколочной шкурке. Такое
состояние взрослого насекомого, когда
оно покрыто одновременно старой (со-
стоящей из эпикутикулы и экзокутикулы)
и новой кутикулой, называется фа-
ратным.
ПИТАНИЕ
Как и многие живые существа, насе-
комые нуждаются в воде, большом числе
аминокислот, некоторых нуклеиновых
кислотах, источниках стеролов, в вита-
минах, некоторых жирных кислотах и в
большом числе минеральных и неоргани-
ческих веществ, включая соединения фос-
фора, калия, железа, меди, цинка, ко-
бальта и кальция. Все эти вещества
составляют необходимый пищевой мини-
мум. Многие насекомые в качестве ис-
точников энергии используют такие со-
единения, как углеводы и жиры, однако
они могут удовлетворять свои энергети-
ческие потребности и за счет «сжигания»
других органических соединений.
При наблюдении за насекомыми
остается скрытым от глаз тот факт, что в
конечном счете основой роста и размно-
жения является деление клеток. В свою
очередь деление клетки обусловлено
удвоением количества содержащейся в
ядре дезоксирибонуклеиновой кислоты
(ДНК) и связанного с ней комплекса
ядерных белков. Некоторым клеткам, на-
пример клеткам, образующим щетинки,
требуются большие количества пита-
тельных веществ, чтобы превращать их в
сложные химические соединения для по-
строения клеточных структур. Такие при-
меры помогают объяснить потребности
насекомых в аминокислотах и других ор-
ганических соединениях, необходимых
для синтеза белков и нуклеиновых кислот
(ДНК и РНК).
Насекомые, как и другие животные,
не способны синтезировать некоторые
аминокислоты и многие другие органи-
ческие соединения, в которых они нуж-
даются. Поэтому им необходимо полу-
чать эти соединения с пищей. Эта потреб-
ность удовлетворяется тем, что насе-
комые питаются живыми или мертвыми
организмами, которые либо сами синте-
зируют эти вещества (зеленые растения),
либо получают их непосредственно или
косвенно из зеленых растений (других
животных или растений-паразитов). На-
секомые обладают способностью синте-
зировать нуклеиновые кислоты, однако
скорость этого синтеза часто не удовле-
творяет потребности самих насекомых
(особенно у некоторых видов Diptera и
Coleoptera), поэтому они вынуждены до-
полнительно получать эти вещества с
пищей.
Это не значит, что насекомые исполь-
зуют эти вещества ежедневно - они
нужны им в течение жизни. Запасы пита-
тельных веществ имеют большое значе-
ние для насекомых. Эти запасы, состоя-
щие из разных веществ, накапливаются в
тканях тела насекомых и расходуются в
течение их жизненного цикла. Так, гусе-
5. Жизненные процессы
125
ницы, поедая зеленые растения, создают
запас питательных веществ, достаточный
лля последующего созревания и отклад-
ки яиц взрослыми насекомыми, которые
сами могут питаться лишь углеводами
или вообще не питаться. Однако у других
насекомых, например некоторых кома-
ров. для созревания яиц необходим бе-
лок. который самка получает с кровью
хозяина. У комаров наблюдается еще
одно очень интересное явление, довольно
обычное для насекомых. Личинки пи-
таются в основном микроорганизмами,
взрослые самцы - нектаром, а самки
< большинства видов)-кровью позво-
ночных. Но в ряде случаев питание насе-
комых однообразно в течение всего жиз-
ненного цикла: многие насекомые пи-
таются только 'на зеленых растениях
* листьях, ветвях, корнях или поедают
пыльцу и древесину), другие-на навозе,
гниющей древесине или трупах жи-
вотных, третьи-на живых животных.
Каков бы ни был характер питания, в
каждом конкретном случае пищевари-
тельные системы насекомых приспособ-
лены для максимального извлечения пи-
тательных веществ из получаемой ими
пиши.
Витамины и холестерин. Витаминам
принадлежит важная роль в клеточном
метаболизме. Потребность в витаминах
у разных насекомых неодинакова. Вита-
мины А, С, D, Е и витамины комплекса В
участвуют в ряде метаболических про-
цессов. Насекомые, нуждающиеся в вита-
мине С, обычно питаются на растениях,
но некоторые насекомые (например, та-
раканы рода Blattella) способны синтези-
ровать аскорбиновую кислоту. Для со-
зревания половых продуктов некоторым
насекомым необходим витамин Е.
Большинство насекомых не способны
синтезировать стеролы в достаточных
количествах и поэтому им нужна пища,
содержащая холестерин.
Опыты, проведенные на личинках
мясной мухи, показали, что некоторые из
необходимых витаминов насекомые по-
лучают от симбиотических микроорга-
низмов, восполняя, таким образом, недо-
статок витаминов в поглощаемой ими
пище. В организме насекомого-в пище-
варительном тракте, в специальных
образованиях или внутри клеток-оби-
тают различные симбионты, обеспечи-
вающие насекомое питательными веще-
ствами, в том числе витаминами и ами-
нокислотами, которые хозяин не может
получить с пищей.
Потребность в воде. Для метаболиче-
ских процессов насекомым, как и другим
организмам, необходима вода. Она пред-
ставляет собой важнейший компонент их
пищи. Большинство насекомых полу-
чают достаточное для удовлетворения
своих нужд количество воды с пищей, с
такой, например, как листья растений
или кровь животных. Кроме того, у насе-
комых в процессе эволюции возникли
различные приспособления, как морфо-
логические, так и физиологические, для
сохранения воды. Некоторые насекомые
обладают столь совершенными механиз-
мами сохранения воды, что способны пи-
таться практически сухой пищей. В этих
случаях насекомые используют воду,
образующуюся внутри организма путем
окисления веществ, поступающих
с пищей. Но даже и в этих случаях в пище
должно содержаться небольшое количе-
ство воды в дополнение к воде, образую-
щейся метаболическим путем. Неко-
торые насекомые могут поглощать воду
из влажного воздуха, например мучной
хрущак или большинство жуков, живу-
щих по берегам водоемов.
ПИЩЕВАРЕНИЕ
Пищеварение - это процесс разруше-
ния и изменения пищи с тем, чтобы она
126
5. Жизненные процессы
могла всосаться эпителием кишечника и
попасть в кровь, которая поставляет пи-
тательные вещества тканям и клеткам те-
ла. В соответствии с большим разно-
образием пищи насекомых существует
множество модификаций пищеваритель-
ной системы, каждая из которых приспо-
соблена к определенному роду пищи. Пи-
щеварительная система личинки и взрос-
лого насекомого одного и того же вида
может быть устроена совершенно по-раз-
ному (рис. 4.3), что особенно наглядно
проявляется в отряде Diptera, где разные
стадии развития различаются по способу
питания. Строение пищеварительной си-
стемы может сильно варьировать в пре-
делах одного отряда. Так, в отряде Ну-
menoptera пищеварительная система тра-
воядных личинок пилильщиков отли-
чается от таковой личинок ос, являющих-
ся внутренними паразитами.
Наиболее типичное строение пищева-
рительной системы мы находим у тра-
воядных и всеядных насекомых-тарака-
нов, саранчовых, личинок и взрослых
форм многих жуков. Этот тип строения
будет рассмотрен как основной вместе с
несколькими наиболее яркими его моди-
фикациями. Многочисленные варианты
строения пищеварительной системы,
встречающиеся среди насекомых, пред-
ставляют особый интерес, связанный
именно с каждой отдельной группой
насекомых.
Слюноотделение. У огромного числа
насекомых слюна смешивается с пищей
до начала ее поглощения. У насекомых с
грызущим ротовым аппаратом слюна
выделяется в ротовую полость и там сме-
шивается с пищей. У насекомых с сосу-
щим ротовым аппаратом слюна часто
вводится в жидкую пищу, и смесь затем
всасывается в глотку. Обычно слюна
образуется в лабиальных железах.
Типичная слюнная железа насекомого
имеет вид виноградной грозди, каждая
«ягода» которой представляет собой аль-
веолу, состоящую из секреторных клеток.
Каждая альвеола снабжена собственным
протоком; эти протоки, соединяясь,
образуют общий проток железы. Клетки,
образующие альвеолы, имеют различное
строение. Лабиальные железы в зависи-
мости от характера их секрета делятся на
2 группы.
1. ЖЕЛЕЗЫ, УЧАСТВУЮЩИЕ В ПЕРЕ-
ВАРИВАНИИ ПИЩИ. У многих насе-
комых лабиальные железы являются ос-
новным источником амилазы, расщеп-
ляющей крахмал. Секрет желез обычно
смешивается с пищей до того, как она
«проглатывается», и собственно перева-
ривание пищи происходит в пищевари-
тельном тракте. У взрослых бабочек и
пчел эти железы секретируют инвертазу,
которая выделяется на кончике хоботка и
поступает в желудок вместе с нектаром.
2. ЖЕЛЕЗЫ, ВЫДЕЛЯЮЩИЕ АНТИ-
КОАГУЛИРУЮЩИЕ ВЕЩЕСТВА. Ла-
биальные железы кровососущих насе-
комых секретируют не пищеварительные
ферменты, а антикоагулин, препятствую-
щий свертыванию крови и образованию
тромбов в хоботке или в пищеваритель-
ном тракте.
ВнекиШечное пищеварение. В ряде слу-
чаев насекомые выделяют пищевари-
тельные ферменты непосредственно на
поверхность пищи или внутрь ее, и, та-
ким образом, пища подвергается хотя бы
частичному перевариванию еще до свое-
го поступления в пищеварительный
тракт Такой способ называют внеки-
шечным пищеварением. Так, например,
тли с помощью хоботка вводят слюну,
содержащую амилазу, в ткани растения-
хозяина и таким способом разлагают
крахмал в клетках растения. Многие
хищные жуки, у которых нет слюнных
желез, отрыгивают на жертву пищевари-
тельные ферменты, образующиеся в ки-
шечнике и поступающие в ротовую по-
5. Жизненные процессы
127
л ость. После начала процесса перевари-
вания жидкость, содержащая ферменты,
вновь всасывается насекомыми. Личинки
мясных мух выделяют протеолитические
ферменты из анального отверстия и с их
помощью осуществляют внекишечное
переваривание тканей, в которых они жи-
вут и которые служат им пищей.
Поглощение пищи. В пищевари-
тельный канал пища попадает через ро-
товое отверстие. Насекомые с грызущим
ротовым аппаратом разрезают и разры-
вают пищу с помощью мандибул и мак-
силл. При смыкании этих частей ротово-
го аппарата пища прижимается к задней
стенке ротовой полости (цибариуму),
образуемой основанием гипофаринкса
«рис. 5.7). Гипофаринкс отводится затем
вверх и вперед и проталкивает пищу в
глотку, представляющую собой перед-
ний конец пищевода. Дальнейшее пере-
движение пищи по пищеварительному
тракту происходит благодаря перисталь-
тическим движениям. У насекомых с со-
сущим ротовым аппаратом (рис. 5.8)
глотка образует луковицеобразный на-
сос, который расширяется и сокращается
под действием мышц головы. Этот гло-
точный насос втягивает жидкую пищу че-
рез хоботок в сокращающуюся часть пи-
щеварительного тракта. Пищевари-
тельные ферменты или другие секреты
смешиваются с пищей до того, как она
«проглатывается». В состав слюны и дру-
гих секретов пищеварительного тракта
наиболее часто входят следующие фер-
менты : амилаза, мальтаза, инвертаза,
триптаза, пептидаза и липаза.
У молодых личинок некоторых эндо-
паразитов пища поступает не через рот, а
всасывается всей поверхностью тела из
тканей или крови хозяина.
Передняя кишка, или стомодеум. Из
пищевода пища поступает в переднюю
кишку. Функция передней кишки различ-
на у разных насекомых; она может про-
Рис. 5.7. Голова насекомого с грызущим
типом ротового аппарата в разрезе (схема).
Показаны генерализованные части ротового
аппарата и мышцы, принимающие участие в
проглатывании пищи. (По Снодграссу.)
7-головной мозг; 2-пищевод; 3-пронотум
4 - зоб; 5 - тенториум; 6 - ганглий;
7-слюнной проток; 8-слюнной канал
(саливарий); 9 - лабиум; 10 - гипофаринкс
(подглоточник); 77-лабрум; 72-пищевой
канал; 73-цибариум; 14-ротовое отверстие;
75-клипеус; 16-глотка; 77-мышцы.
сто служить для прохождения пищи в
среднюю кишку, а может, превращаясь в
огромный зоб, служить хранилищем пи-
щи, где последняя частично перевари-
вается. В некоторых случаях, например у
Orthoptera, пищеварительные соки по-
ступают в переднюю кишку из средней.
Передняя кишка в типичном случае
состоит из слоя эпителиальных клеток и
кутикулярной выстилки, образуемой се-
кретом этих клеток (рис. 5.10,Л). Суще-
ствует мнение, что эта кутикулярная вы-
стилка непроницаема для ферментов и
для продуктов переваривания пищи и что
поглощение через нее не происходит во-
обще или происходит в очень слабой
степени. Функция кутикулярной выстил-
ки заключается, вероятно, в предотвра-
щении всасывания частично перева-
128
5. Жизненные процессы
Рис. 5.8. Глоточный и слюнной насосы цикады. (По Снодграссу.)
А. Разрез через голову, показывающий положение глоточного насоса
(цибариума) с мышцей-расширителем, придающей клипеусу выпуклую
форму. Б. Разрез через область рта, показывающий путь, по которому
поступает пища, а также глоточный и слюнной насосы. 7-клипеус;
2-пронотум; 3-глотка; 4-пищевод; 5-слюнные железы, 6-слюнной
проток; 7-лабрум; 5-максиллярная щетинка, 9-мандибулярная
щетинка, 10-глоточный насос; 11 -мышцы глоточного насоса,
72-слюнной насос; 13-гипофаринкс; 14-пищевой канал; 75-хоботок;
16-пищевой канал-функциональный рот, 77-лабиум.
ренных продуктов, что способствует их
дальнейшему полному перевариванию и
усвоению.
Провентрикулус (преджелудок). У Ог-
thoptera и других насекомых, потреб-
ляющих грубую пищу, в провентрикулу-
се имеется ряд острых зубов для измель-
чения поглощенной пищи. Типичный
пример такого строения можно видеть у
таракана (рис. 5.9), у которого провен-
трикулус оснащен шестью крепкими зу-
бами. У блох имеется множество острых
игольчатых зубчиков, направленных на-
зад. Когда кровь в средней кишке про-
талкивается вперед, эти зубчики отводят-
ся назад, так что клеточные элементы
трутся об них, как о терку, и разрушают-
ся, эти движения вызываются сокраще-
нием специальных мышц. У других насе-
комых провентрикулус представляет со-
бой всего лишь суженный конец передней
кишки.
Средняя кишка, или мезеитерон. В
этой части пищеварительного тракта
эпителиальные клетки не защищены ку-
тикулярной выстилкой (рис. 5.10,Б). Од-
ни из этих клеток служат для всасывания
большей части поглощенной пищи, дру-
гие - синтезируют ферменты.
Секреция ферментов эпителиальными
клетками может быть двух типов: 1) го-
локринная секреция, при которой клетки
разрушаются, извергая свое содержимое
в просвет кишечника; 2) мерокринная се-
креция, когда вакуоли с содержащимися
в них ферментами проходят через кле-
5. Жизненные процессы
129
один из белковых компонентов волоса
млекопитающих; осуществляется этот
процесс с помощью обычной протеиназы
насекомых в сочетании с определенным
pH среды в средней кишке. Личинки
большой вощинной моли поедают воск,
Рис 5.9. Преджелудок (провентрикулус)
таракана в разрезе; видны три из шести
зубов для размельчения пищи.
точные мембраны в просвет кишечника.
Первый тип секреции изображен на
рис. 5.10,1>. На этом рисунке показаны
также группы регенеративных клеток,
или крипты (5), которые замещают клет-
ки, разрушенные в процессе голокринной
секреции.
ФЕРМЕНТЫ. В среднюю кишку насе-
комых выделяется большое число фер-
ментов, переваривающих белки, жиры и
углеводы. В основном это такие же фер-
менты, которые синтезируются и у мле-
копитающих. Набор синтезируемых фер-
ментов чаще всего соответствует харак-
теру пищи. У таких всеядных насекомых,
как тараканы, имеется полный набор
ферментов для переваривания всех типов
веществ. У кровососущих насекомых
образуются в основном протеолитиче-
ские ферменты. У некоторых насекомых
образуется целлюлаза для переварива-
ния целлюлозы. Некоторые настоящие
моли способны переваривать кератин,
Рис. 5.10. Клеточное строение трех участков
пищеварительного тракта таракана.
А. Передняя кишка (продольный срез). Б.
Средняя кишка (продольный срез). В. Задняя
кишка (поперечный срез).
7-кутикула; 2-эпителиальная клетка;
3 -мышцы; 4 - разорвавшаяся клетка;
5-группа регенеративных клеток.
130
5. Жизненные процессы
который, очевидно, разлагают в основ-
ном симбиотические бактерии.
ПЕРИТРОФИЧЕСКАЯ МЕМБРАНА.
Поверхность эпителиальных клеток
средней кишки не защищена кутикулой и
очень нежная. Если бы через среднюю
кишку проходили крупные пищевые ком-
ки, то нежная поверхность этих клеток,
несомненно, повредилась бы, в результа-
те чего были бы нарушены процессы се-
креции и всасывания. У позвоночных жи-
вотных специальные железы выделяют
слизь, которая обволакивает пищевые
комки и плотные частицы и делает их
скользкими, что предохраняет эпителий
желудка от повреждения. У насекомых
нет желез, выделяющих слизь, и защита
эпителия осуществляется с помощью пе-
ритрофической мембраны (рис. 5.11). Эта
мембрана покрывает всю пищевую мас-
су Она состоит из хитина и хорошо про-
ницаема для пищеварительных фермен-
тов и продуктов переваривания пищи. О
высокой проницаемости перитрофиче-
ской мембраны свидетельствуют данные,
полученные в опытах с применением
красителей.
Образование перитрофической мем-
браны представляет большой интерес. У
большинства насекомых она выделяется
почти всей поверхностью средней кишки.
Сначала хитиновый секрет образует слой
над выделившими его эпителиальными
клетками, который затем отделяется от
них, создавая подобие трубки вокруг пи-
щевого комка. Обычно эта «трубка» при-
креплена к передней части средней кишки
там, где в нее вдается передняя кишка.
Перитрофическая мембрана не обра-
зуется у насекомых, которые питаются
только жидкой пищей - у Hemiptera, Апо-
plura, взрослых блох и слепней; нет ее и у
некоторых других-у Carabidae, Dytisci-
dae и Formicidae.
Рис. 5.11. Складчатые структуры, участвующие в образовании перитрофической
мембраны. (Из Wigglesworth, с изменениями.)
А. Личинка комара Anopheles. Б. Муха цеце Glossina. В. Уховертка Forficula.
7-пищевод, 2-сфинктер, 3-секреторные клетки, 4-область начала секреции,
5 - склеротизированное кольцо, прессующее и проталкивающее перитрофическую
мембрану, 6-перитрофическая мембрана, 7-стенка средней кишки, 8-кольцо,
9-пищевой комок, 10-кардиальный клапан.
Хлзжнные процессы
131
КАРДИАЛЬНЫЙ ОТДЕЛ КИШЕЧНИ-
* х У некоторых насекомых перитрофи-
мембрана секретируется специа-
тг'5щ>?ванной группой клеток, располо-
вокруг переднего конца средней
гтшшгг Этот секрет спрессовывается в
-*^?эану с помощью наружного давле-
создаваемого либо входной частью
шгешней кишки, растягиваемой поступив-
шей ? нее пищевой массой, либо с по-
тссзъ’-о специального органа, называе-
те- г ълрдиальным клапаном. Это образо-
наиболее развито у Diptera и Der-
ттдгсега (см. рис. 4.4, 5.11). Оно состоит
•• скшеротизированного кольца у входа в
—ешш-ою кишку, которое прижимает к
~г=тдм кишки секрет, выделяемый спе-
пкштьными клетками, находящимися
— впереди этого кольца. Сформиро-
кьшедя мембрана направляется назад по
г$тше*ннку. обволакивая пищу.
кишка, или проктодеум. Основ-
функция этого отдела состоит в уда-
отработанной пищи, хотя наряду с
•“тм здесь осуществляются также, прав-
ка з незначительной степени, всасывание
» искрения. Как и в передней кишке, эпи-
~шш?дльные клетки покрыты секретируе-
ими кутикулярной выстилкой
5.10.2?), однако эта выстилка про-
шгшвема для воды. Задняя часть задней
гг._ти. ректум, обычно снабжена
шштдными мышцами, с помощью ко-
оставшейся непереваренной пи-
ще - зкекрементам - придается форма
щд_г^ков или бляшек до их выведения из
Г’-гнизма. Помимо выведения экскре-
'*ентов задняя кишка выполняет еще две
□ чхГШШ.
ВСАСЫВАНИЕ ВОДЫ. Почти все
щьдекомые вынуждены запасать воду,
основная роль в этом принадле-
жат задней кишке, где происходит всасы-
воды из экскрементов и возвраще-
ние ее в ткани тела. Ректальные эпите-
т*^д>ные клетки мучного хрущака спо-
собны экстрагировать почти всю воду из
экскрементов, оставляя их в виде сухих
комочков. Всасывание воды играет осо-
бенно важную роль в процессах экскре-
ции, на которой мы остановимся более
подробно в соответствующем разделе.
2. ПЕРЕВАРИВАНИЕ ПИЩИ С ПО-
МОЩЬЮ СИМБИОНТОВ. Термиты, неко-
торые лесные тараканы и личинки кома-
ров-долгоножек и жуков-скарабеид, ко-
торые питаются в основном древесиной,
не имеют специальных ферментов для
переваривания целлюлозы. Для этой це-
ли они используют большое число сим-
биотических микроорганизмов, обитаю-
щих в задней кишке насекомых. Эти
симбионты превращают целлюлозу в ук-
сусную кислоту, которая и поглощается
клетками кишечника. Изучение симбион-
тов у других насекомых выявило много
противоречий и неясных вопросов, тре-
бующих более пристального анализа.
Адаптации к жидкой пище. У многих
насекомых, питающихся кровью или со-
ками растений, имеются специальные
приспособления для экстрагирования
большей части воды из пищи до начала
ее переваривания с помощью ферментов.
Такое экстрагирование имеет два пре-
имущества: 1) возможность быстро по-
глощать растворенные сахара и 2) воз-
можность избежать чрезмерного сниже-
ния титра ферментов. К числу
приспособлений, с помощью которых
достигается такая частичная дегидрата-
ция, относятся следующие:
1. У многих взрослых Diptera сред-
няя кишка разделена на несколько отде-
лов, с особым типом эпителия в каждом
из них. Полагают, что в первом из этих
отделов происходит всасывание воды из
поглощенных жидкостей.
2. У некоторых кровососущих Hemip-
tera, например у постельного клопа,
передняя часть средней кишки преобра-
132
5. Жизненные процессы
Рис. 5.12. Фильтрационная камера Homoptera (схема). (Из Snodgrass по
Weber.)
А. Простой тип-два конца средней кишки и передний конец задней
кишки объединены под одной оболочкой. Б. Средняя кишка заключена
внутри фильтрационной камеры, а передняя часть задней кишки выходит
из задней части камеры. В. Фильтрационная камера червеца Lecanium.
7-пищевод, 2-первый отдел средней кишки, 3-второй отдел средней
кишки, 4-третий отдел средней кишки, 5-прямая кишка (ректум),
6-тонкая кишка, 7-мальпигиевы сосуды, 8-фильтрационная камера,
9-средняя кишка, 10-задняя кишка.
зована в большой зоб, куда поступает
кровь. В «зобе» всасывается большая
часть воды и, таким образом, кровь, пре-
жде чем поступить в те отделы, где про-
исходит ее переваривание с помощью
синтезирующихся там ферментов, стано-
вится высококонцентрированной. Отме-
тим, что у двукрылых и у постельного
клопа вода всасывается в средней кишке,
поступает затем в кровяное русло насеко-
мого, а оттуда экскретируется через
мальпигиевы сосуды в заднюю кишку.
3. У червецов и щитовок, цикад и
многих других Homoptera пищевари-
тельный тракт снабжен очень любопыт-
ным образованием, так называемой
фильтрационной камерой (рис. 5.12).
Передняя часть средней кишки распола-
Жизненные процессы
133
зется рядом с участком задней кишки. В
тех ? торых случаях обе эти части объеди-
нены под одной оболочкой, а иногда
^хдняя кишка образует петлю, которая
zrv ходит через впячивание задней киш-
& Поскольку эти насекомые питаются
тсьзлш растений, считали, что фильтра-
т^снная камера создает возможность bo-
te поглощенной порции сока быстро
юс ходить из средней кишки в заднюю,
дтвышая, таким образом, концентрацию
до его переваривания в задней части
тгЕдней кишки. Анализ жидкости, выде-
рем ой через анальное отверстие в виде
“и называемой «медвяной росы», пока-
-кьт. что в этой жидкости содержится
:<_тьшое количество аминокислот и угле-
к:?з. и это ставит под сомнение пра-
»й_д>ность оценки функции фильтра-
з теных камер. Очевидно, что для пони-
^нкя этих функций нужны новые
дънные.
Лншочные приспособления. У личи-
не* высших Hymenoptera и Neuroptera
л5=аружены интересные модификации в
.-“Тоении пищеварительного тракта. Ко-
вед средней кишки у них замкнут и не со-
сытается с задней кишкой, в результате
Hir? в период личиночного развития
дхдняя кишка оказывается сильно рас-
г=>той из-за большого количества фека-
Перед окукливанием оба отдела ки-
еэечника соединяются, и фекальные
члесы. накопившиеся за всю личиночную
ж^знь. выводятся наружу.
Реакция среды в кишечнике. В среднем
* большинства насекомых реакция среды
галечника слабокислая с pH от 6 до 7
Слюна имеет обычно нейтральную реак-
У растительноядных насекомых ре-
х-гтия среды кишечника более щелочная с
гН от 8,4 до 10,3 (у личинки шелкопря-
д* . Всеядные или плотоядные насе-
комые характеризуются обычно смеще-
ежм реакции в более кислую сторону. В
таракана после принятия углевод-
ной пищи pH равнялось 4,8-5,3; такой
уровень кислотности может быть резуль-
татом сбраживания пищи, осуществляе-
мого микроорганизмами, или выделения
кислого секрета некоторыми клетками
слюнных желез. Наиболее кислая реак-
ция (pH 3,0) была зарегистрирована в
одном из участков кишечника личинок
мясной мухи.
Всасывание
Продукты переваривания пищи
всасываются эпителием, выстилающим
пищеварительный тракт, и поступают в
кровь (гемолимфу). Всасывание происхо-
дит в основном в средней кишке, но у не-
которых насекомых ряд веществ всасы-
вается в задней кишке. Углеводы, такие,
например, как моносахарид глюкоза,
всасываются в слепых выростах желудка.
Все эти вещества поступают в кровь и
переносятся к различным тканям, где
превращаются в конечные продукты, не-
обходимые для роста. Скорость поступ-
ления ионов и молекул в гемолимфу за-
висит от движения содержащих их ве-
ществ по направлению к всасывающим
поверхностям кишечника и от скорости
переноса через эпителий кишечника.
Процессы метаболизма у насекомых на-
столько специализированы, что характер
всасывания в пищеварительном тракте
может быть столь же специализиро-
ванным и уникальным для данного вида.
Хранение запасных веществ. Жирово-
му телу насекомых принадлежит особо
важная роль в процессах превращения,
известных под названием промежуточно-
го обмена. В жировом теле происходят
синтез жирных кислот и белков и превра-
щение углеводов (глюкозы в трегалозу).
Помимо этого, в нем накапливаются
энергетические запасы в форме жиров,
гликогена и белка. В соответствии с по-
требностями насекомого эти запасы мо-
134
5. Жизненные процессы
гут быстро метаболизироваться с высво-
бождением энергии.
Местами хранения энергетических за-
пасов могут также быть отделы средней
кишки и другие участки кишечника. У на-
секомых с полным превращением эти за-
пасы обычно наиболее велики непосред-
ственно перед окукливанием, а к концу
метаморфоза оказываются почти цели-
ком использованными.
ЭКСКРЕЦИЯ
И ВОДНО-СОЛЕВОЙ БАЛАНС
В результате метаболических прочее-
сов происходит разрушение органиче-
ских соединений и образование шлаков,
многие из которых токсичны для тканей
и клеток животных. Эти продукты уда-
ляются из организма посредством эк-
скреции. Особенно токсичными являются
продукты азотистого обмена, образую-
щиеся при разрушении белков и нуклеи-
новых кислот. В связи с этим у животных
возникли разнообразные механизмы для
их удаления. Другой токсичный продукт
метаболизма-это двуокись углерода,
образующаяся в процессе клеточного ды-
хания. Хотя по существу ее удаление из
организма представляет собой тоже эк-
скрецию, подробно этот процесс рассма-
тривается в связи с дыханием.
У большинства насекомых, как и у
других животных, экскреторная система
выполняет двоякую функцию: удаление
отходов метаболизма и поддержание на
определенном уровне концентрации рас-
творенных солей и водного баланса в
жидкостях тела. Обе эти задачи решают-
ся путем образования первичной секре-
торной жидкости, последующего всасы-
вания части веществ, содержащихся в
ней, и выведения затем остатков из орга-
низма. В этом циклическом процессе уча-
ствуют вода, ионы (в основном калия, а у
водных насекомых еще и хлора), неорга-
нические и органические молекулы.
Для поддержания определенного со-
става внутренних жидкостей тела как
внутри-, так и внеклеточных, солевой и
водный баланс имеют решающее значе-
ние. Состав внутренних жидкостей дол-
жен поддерживаться в относительно уз-
ких пределах колебаний независимо от
внешних условий. Из-за малых размеров
тела насекомые постоянно подвергаются
риску высыхания или обезвоживания. От
этого их защищает водонепроницаемая
кутикула, специализированная экскре-
торная система, позволяющая сохранять
воду, и система дыхалец, предотвращаю-
щих потери воды через дыхательную
систему.
Экскреторная система
Мальпигиевы сосуды и ректум. У
большинства насекомых основными
компонентами экскреторной системы
являются мальпигиевы сосуды и ректум
(рис. 5.13). Мальпигиевы сосуды имеют-
ся только у насекомых и неизвестны у
других животных1. Обычно они лежат
свободно в полости тела и в большинстве
случаев открываются в пищевари-
тельный тракт в месте соединения сред-
ней кишки с задней. В мальпигиевых со-
судах в результате всасывания раство-
ренных веществ из гемолимфы образует-
ся моча. Это обычно прозрачное, а иног-
да вязкое вещество выводится в заднюю
кишку, оттуда в ректум, где происходит
частичная реабсорбция или дополнение
ее другими веществами перед выведе-
нием наружу.
Число мальпигиевых сосудов варьи-
1 Здесь автор неправ. Мальпигиевы со-
суды имеются у представителей большинства
групп членистоногих, обитающих на суше.-
Прим. ред.
Жизненные процессы
135
Рис. 5.13. Циркуляция растворов солей и воды в выделительной системе насекомого
«схема). (Л-по Stobbart, Shaw, 1974; Ь’-по Wigglesworth.)
Л. Обычный тип; стрелки показывают направление тока воды и движения ионов. Б.
Тип. свойственный мучному хрущаку Tenebrio, характеризующийся тесной связью
ректума с окончаниями мальпигиевых трубочек. 1 -средняя кишка; 2-мальпигиев
сосуд; 3-прямая кишка (ректум); 4-ректальные железы; 5-задняя кишка;
6 - соединительная ткань, 7-гипо- или гиперосмотическая жидкость.
гует у разных видов насекомых-от 2 (у
г аспид) до 200 и более (у некоторых Ort-
roptera и пчел). Мальпигиевы сосуды бы-
эсхют разной длины; они замкнуты на ди-
стальном конце и образованы одним
сдоем эпителиальных клеток. Большое
^зсло трубочек создает обширную по-
верхность для всасывания жидкости из
-емолимфы. У некоторых насекомых, на-
дример у большинства личинок бабочек
г у многих жуков, дистальные концы
мальпигиевых сосудов открываются в
гишечник близ ректума, что повышает
эффективность процесса реутилизации
растворенных солей и воды.
Состав мочи. Состав мочи зависит от
особенностей жизненного цикла и по-
требностей насекомого, а также от харак-
тера пищи. Обычно в ее состав входят во-
да, азотистые соединения, в том числе
птеридины, карбонат кальция (известь),
оксалат кальция и пигменты. В зависи-
мости от характера пищи насекомого в
моче могут также содержаться углеводы,
фосфаты, салициловая кислота и раз-
личные ионы. Из азотистых соединений
наиболее важным является мочевая кис-
лота. Другие азотистые соединения - ам-
миак, мочевина и аминокислоты - обна-
руживаются в моче в малых количествах.
У наземных насекомых 80% экскрети-
руемого азота входит в состав мочевой
136
5. Жизненные процессы
кислоты, которая образуется в тканях,
накапливается в гемолимфе и затем по-
ступает в мальпигиевы сосуды вместе с
аминокислотами, различными ионами и
водой. Поскольку мочевая кислота и ее
аммонийные соли слабо растворимы в
воде, выделяемая моча содержит в ос-
новном твердые компоненты и мало во-
ды. Однако у многих насекомых ос-
новными азотистыми соединениями в
моче являются нерастворимые вещества,
такие, например, как аллантоин и аллан-
тоиновая кислота, образующиеся при
разложении мочевой кислоты. Аллан-
тоин преобладает у Heteroptera и обнару-
жен также у разных видов Orthoptera, Со-
leoptera, Neuroptera, Hymenoptera и Dip-
tera. У изученных в этом отношении
Lepidoptera в выделениях куколок пре-
обладает мочевая кислота, у взрослых и
личинок - мочевая кислота или аллантои-
новая кислота.
Личинки мясной мухи выделяют
большие количества аммиака - очень
токсичного вещества, которое может со-
ставлять до 90% всех экскретируемых
азотистых соединений, а также аллан-
тоин. У многих водных насекомых от 70
до 90% выделяемого азота входит в со-
став аммиака, быстро растворяющегося
в окружающей воде.
У некоторых насекомых в большом
числе образуются гранулы карбоната
кальция и оксалата кальция, процесс
образования которых пока неизвестен.
Растительноядные гусеницы бабочек и
палочники выделяют гранулы и кри-
сталлы мочевой кислоты, а также гра-
нулы солей мочевой кислоты, карбоната
кальция и оксалата кальция. В экскретах
личинок двукрылых содержится большое
количество гранул карбоната кальция.
Дополнительные экскреторные ор-
ганы. Помимо мальпигиевых сосудов и
ректума в экскреции участвуют другие
ткани и органы. Они играют подчинен-
ную, но важную роль особенно в удале-
нии малых молекул и воды. Через прида-
точные железы (utriculi majores) самцов
тараканов выделяются избытки мочевой
кислоты. Лабиальные железы бабочек-
сатурнид, представляющие собой
длинные свернутые трубочки, лежащие в
вентральной части груди, секретируют
большие количества щелочной жидко-
сти-бикарбоната калия. В средней киш-
ке гусениц сатурний образуется экскрет,
содержащий ионы калия. Кишечник, в
основном средняя кишка, играет важную
роль в процессах всасывания ионов у
большинства насекомых, не имеющих
специальных органов, выполняющих эту
функцию.
Удаление веществ с высоким молеку-
лярным весом осуществляют перикар-
диальные клетки, располагающиеся
вдоль сердца и аорты, и нефроциты - спе-
циализированные клетки, разбросанные
по всему телу насекомого. Эти клетки
улавливают из гемолимфы макромоле-
кулы и после их переваривания возвра-
щают малые молекулы в гемолимфу.
Накопление и хранение экскретов. У
некоторых насекомых экскрет в виде мо-
чевой кислоты не выделяется из организ-
ма, а сохраняется в специальных мо-
чевых клетках жирового тела. Такой тип
хранения встречается у тараканов и у на-
секомых, являющихся эндопаразитами
других насекомых, в частности у многих
личинок перепончатокрылых. Накопле-
ние мочевой кислоты имеет большое зна-
чение для насекомых, у которых мальпи-
гиевы сосуды выполняют специальные
функции, например секрецию шелковой
нити. Эти запасы мочевой кислоты в жи-
ровом теле становятся доступными, в
частности, в процессе метаморфоза и ис-
пользуются как источник азота, который
будет выведен позднее.
Пигменты. Небольшие количества из-
быточной мочевой кислоты могут накап-
5 Жизненные процессы
137
лдваться в кутикуле насекомых в виде
>елых кристаллических отложений, при-
дающих кутикуле белый цвет У бабочек
семейства Pieridae небольшие количества
мочевой кислоты и птеринов (лейкопте-
г«н дает белый цвет, ксантоптерин-
желтый) откладываются в чешуйках на
грыльях. У колорадского жука мочевая
г^слота откладывается в склеритах. В
хч?их случаях эти вещества обусловли-
вают характер окраски насекомых.
Насекомые, не имеющие мальпи-
гиевых сосудов. У некоторых насекомых
мальпигиевых сосудов нет, и экскретор-
е~?ю функцию частично выполняют ла-
• ильные железы (головные трубчатые
жглезы) У Thysanura и Collembola ла-
~ иальные железы выделяют красящие ве-
—ества. Мочевая кислота у этих насе-
г?х(ых хранится в специальных мочевых
метках жирового тела.
Регуляция концентрации растворенных
солей и водного баланса. Мальпигиевы
:<хлды представляют собой секреторные
эеразования, в них постоянно происхо-
дит всасывание веществ из гемолимфы и
збразование жидких экскретов. Ак-
тивный транспорт ионов калия через
стенку’ мальпигиевой трубочки играет,
до-видимому, основную роль в образо-
зании мочи внутри нее. Градиент ионов
гдлия вызывает ток мочи в трубочке. У
некоторых водных и наземных насе-
комых жидкость, образующаяся в маль-
пигиевых сосудах, содержит очень высо-
кие концентрации калия-в 6-30 раз вы-
дк. чем в гемолимфе. Напротив, концен-
трация натрия в мальпигиевых трубоч-
ках ниже, чем в гемолимфе. У некоторых
видов, например у кровососущего клопа
Rhodnius, натрий, а не калий играет роль
основного фактора образования мочи.
Скорость образования жидкого экскрета
в мальпигиевых сосудах находится под
контролем нейрогормонов, образую-
щихся в нейросекреторных клетках го-
ловного мозга и в грудном ганглии.
Жидкий экскрет выходит из мальпи-
гиевых сосудов, проходит через заднюю
кишку и попадает в ректум, где подвер-
гается существенному изменению.
У многих наземных насекомых на пути к
ректуму часть воды из этой жидкости
всасывается в переднем отделе задней
кишки - в тонкой кишке (ileum). Однако в
основном всасывание воды из мочи про-
исходит в ректуме, и поэтому у большин-
ства наземных насекомых фекалии отно-
сительно сухие. Эпителиальные клетки
ректума обладают также способностью
переправлять неорганические ионы из ре-
ктума в гемолимфу.
У пресноводных насекомых, окру-
женных массой воды с малым количе-
ством солей или вовсе без солей, поступ-
лению воды через поверхность тела (в
силу разницы осмотических концентра-
ций) препятствует относительно боль-
шой объем ректальной жидкости, содер-
жащей небольшие количества ионов. Ос-
мотическая концентрация ректальной
жидкости у этих насекомых ниже, чем ос-
мотическая концентрация гемолимфы.
Вместе с тем ректальная жидкость насе-
комых, обитающих в солоноватой и со-
леной водах, содержит много ионов, и ее
осмотическая концентрация больше ос-
мотической концентрации гемолимфы. У
некоторых пресноводных личинок насе-
комых в ректуме, по-видимому, происхо-
дит по крайней мере частичное всасыва-
ние воды. У личинок, приспособленных к
солоноватой воде, происходит более ин-
тенсивное всасывание воды в ректуме.
Пищеварительный тракт. У наземных
насекомых вода и растворимые соли из
пищи всасываются в кишечник и попа-
дают затем в полость тела и гемолимфу.
Поскольку часть воды теряется при испа-
рении с поверхности тела насекомого,
объем ректальной жидкости меньше, чем
жидкости, которая всасывается в кишеч-
138
5. Жизненные процессы
Рис. 5.14. Вариации размеров анальных
жабр на заднем конце личинки Culex pipiens
при выращивании их в воде с разной
соленостью. (Из Wigglesworth, 1965,
с изменениями.)
А. Дистиллированная вода. Б. 0,006%
раствор NaCl. В. 0,65% раствор NaCl.
1 - дыхальце; 2 - анальный сифон;
3- анальные жабры.
ся местом, где происходит обмен соеди-
нений фосфора и осмотическое всасыва-
ние воды.
Дыхание
Все животные нуждаются в энергии, и
значительная часть ее образуется в про-
цессе дыхания. Дыхание состоит в погло-
щении, переносе и использовании кисло-
рода тканями и удалении отработанных
веществ, прежде всего углекислого газа.
Процесс газообмена между клеткой и
окружающей ее средой называется на-
ружным дыханием, а процесс газообмена
и утилизация газов внутри клетки -кле-
точным дыханием, или дыхательным
метаболизмом.
нике и попадает в полость тела. Насе-
комые, обитающие в засушливых райо-
нах, например саранча, постоянно нахо-
дятся под угрозой потери воды и повы-
шения концентрации солей, в связи с чем
у них выработались определенные спо-
собы, позволяющие им преодолевать
трудности. У таких насекомых погло-
щенная пища направляется сразу в ре-
ктум, минуя среднюю кишку, и вода
всасывается непосредственно эпителием
ректума.
Дополнительные органы, регулирую-
щие содержание солей и водный баланс. У
многих пресноводных личинок насе-
комых имеются специальные органы,
расположенные на поверхности тела и
осуществляющие контроль за содержа-
нием солей и воды в организме. Наибо-
лее известные органы такого рода-
анальные жабры личинок комаров
(рис. 5.14.) У этих личинок по обе сторо-
ны от анального отверстия расположено
по паре тонкостенных мешков, имеющих
связь с гемоцелем. Эти органы поглоща-
ют ионы натрия, калия и хлора и являют-
Наружное дыхание
У большинства насекомых наружное
дыхание осуществляется с помощью тра-
хейной системы, состоящей из трубочек,
наполненных воздухом и называемых
трахеями, воздушных мешков и трахеол,
которые доставляют воздух непосред-
ственно к клеткам. Трахеи сообщаются с
внешней средой благодаря наличию от-
верстий - дыхалец, снабженных запира-
тельным механизмом, предотвращаю-
щим потери воды за счет испарения.
Трахеи и трахеолы. Трахеи представ-
ляют собой трубчатые впячивания эпику-
тикулы и имеют структурное сходство с
гиподермой (рис. 5.15, 5.16). Стенка тра-
хеи состоит из слоя плоских эпите-
лиальных клеток и кутикулярной выстил-
ки, или интимы, образованной секретом
этих клеток (см. рис. 4.8, 4.9). Внутрен-
няя поверхность интимы имеет спи-
ральные или кольцевые утолщения, на-
зываемые тенидиями (см. рис. 4.8, 4.9),
которые обеспечивают прочность трахеи
и не позволяют ей спадаться, оставаясь
круглой и открытой даже при сгибании и
Жизненные процессы
139
Рис. 5.15. Схематическое изображение открытой трахеи насекомого.
7-наружное отверстие, или дыхальце; 2-стенка тела; 3-эпителиальные
клетки; 4-выстилка трахеи (интима) - 5-трахеолы; 6-главная трахея;
7-трахея; 8-трахейный ствол; 9-трахеобласт (паутиновидная клетка).
мкаяиниж
Рис. 5.16. Подъем и опускание жидкости в трахеолах. (Из
Wigglesworth, Principles of insect physiology, с разрешения
E. P Dutton and Co.)
А. Высокий уровень жидкости при спокойном состоянии мышцы.
Б. Низкий уровень жидкости при утомлении мышцы. Трахеолы,
наполненные жидкостью, изображены точечными линиями,
содержащие воздух-сплошными линиями.
140
5. Жизненные процессы
сдавливании. Трахеи многократно вет-
вятся, становясь все мельче и мельче, в
конце концов каждая заканчивается пуч-
ком крошечных веточек, трахеол, тол-
щиной менее 2 мкм. В структуре трахеол
имеются тенидии, но нет сплошного слоя
эпителиальных клеток. В основании каж-
дого пучка трахеол имеется лучевидная
клетка-трахеальная клетка, или тра-
хеобласт с чрезвычайно тонкими прото-
плазматическими отростками. Оконча-
ния трахеол располагаются вдоль и ме-
жду клетками тканей тела. Газообмен
между трахеолами и тканями происхо-
дит главным образом через окончания
этих трахеол.
Свойства стенок трахей значительно
отличаются от свойств гиподермы. И
трахеи, и трахеолы проницаемы для га-
зов, особенно хорошо, по-видимому,
проходящих через тонкие стенки тра-
хеол. Трахеи непроницаемы для жидко-
стей, дыхальца также чрезвычайно ги-
дрофобны, и вода не может проникнуть
через них. Трахеолы, особенно их окон-
чания, хорошо проницаемы для жидко-
стей.
Воздушные мешки. В некоторых
участках трахеи образуют местные рас-
ширения, которые называются воз-
душными мешками (рис. 5.17). Обычно
воздушные мешки возникают в тех
участках трахеи, где ее стенки тонкие и
гибкие и где очень мало или вообще нет
тенидиев.
Жидкость в трахеолах. Окончания
трахеол могут содержать некоторое ко-
личество жидкости. При контакте с мыш-
цей, находящейся в состоянии покоя (рис.
5.16,Л), жидкость может подниматься
на значительное расстояние в трахеолах,
когда мышца утомлена, жидкость пере-
ходит из трахеол в мышечную ткань
(рис. 5.16,Ь). Это движение жидкости в
мышечную ткань связано с изменением
осмотического давления в тканях, кото-
Рис. 5.17 Трахейная система с воздушными
мешками в брюшке рабочей пчелы.
Дорсальные трахеи и воздушные мешки
удалены. (Из Wigglesworth по Snodgrass,
Principles of insect physiology, с разрешения
E. P Dutton and Co.)
рое втягивает жидкость в результате ме-
таболической активности мышц. Отток
жидкости к тканям засасывает воздух в
трахеолы и вызывает контакт с ним
«уставших» клеток, усиливая, таким
образом, снабжение кислородом тех ра-
ботающих тканей, которые в нем больше
всего нуждаются.
Диффузия. Существует несколько тео-
рий, объясняющих механизм притока
кислорода по трахеям и трахеолам к тка-
ням и оттока углекислого газа в обрат-
ном направлении. Сейчас общепризнано,
что у активных крупных насекомых дви-
жение газов происходит благодаря диф-
фузии (рис. 5.18,Л), а также некоторым
механическим вентиляционным приспо-
Жюненные процессы
141
кислородом ние кислорода
путем вентиляции путем диффу-
зии только на
этом участке
Рис. 5.18. Схема, иллюстрирующая отношение между диффузией и вентиляцией.
А. Система, использующая только диффузию. Б. Система, в которой диффузия
дополняется вентиляцией. 7-стенка тела, 2-дыхальце; 3-трахеи; 4-ткани;
5-воздушный мешок.
^делениям брюшных трахей и воз-
зпных мешков. У различных насе-
г?мых были измерены размеры трахей,
зетребление кислорода и коэффициент
з^ффузии кислорода. Данные, полу-
денные в этих исследованиях, показали,
тто даже у крупных гусениц диффузия са-
ма по себе может обеспечить доста-
точный приток кислорода в окончания
-рахеол, если давление кислорода в них
только на 2 или 3% ниже, чем в атмосфе-
те
Удаление углекислоты из организма
также происходит посредством диффу-
зии. Однако анализ этого процесса пока-
зал. что около V углекислоты, образую-
щейся в теле насекомого, выделяется
через поверхность тела. Это объясняется
тем. что углекислый газ диффундирует
\ерез животные ткани почти в 35 раз бы-
стрее, чем кислород. Поэтому образую-
щийся в процессе метаболизма угле-
кислый газ диффундирует не только в
трахеолы, но и в окружающие ткани во
всех направлениях и, в конце концов,
проходя через стенки тела, выходит нару-
жу*.
Дыхательная функция крови. Кровь у
насекомых не играет, как правило, важ-
ной роли в доставке кислорода к тканям.
Однако, поскольку кровь представляет
собой жидкость с находящимися в ней
живыми клетками, она нуждается в кис-
лороде для своей жизнедеятельности.
Омывая трахеи и трахеолы, кровь насы-
щается кислородом и разносит его по
всей полости тела. Кровь является ос-
новным поставщиком кислорода для
тканей, которые сильно удалены от тра-
хей и поэтому получают от них лишь не-
большие количества кислорода. При не-
полном развитии трахей кислород также
доставляется к тканям с кровью.
Вентиляция трахейной системы.
У многих мелких или малоподвижных
насекомых потребности в кислороде удо-
влетворяются только за счет диффузии
газов; однако этого оказывается недо-
статочно для активных бегающих или ле-
тающих насекомых с высоким уровнем
метаболизма, которым требуются боль-
шие количества энергии. Насекомое, на-
ходящееся в полете, нуждается в значи-
тельно большем притоке кислорода и
142
5. Жизненные процессы
одновременно в более интенсивном уда-
лении СС>2 и воды, нежели насекомое в
состоянии покоя. Трахейная система дол-
жна быть высоко реактивной и достав-
лять огромные количества кислорода к
летательным мышцам. В этой ситуации
диффузия дополняется механической
вентиляцией трахейной системы, что
обеспечивается двумя основными при-
способлениями.
1 Тенидии трахей обеспечивают гиб-
кость трубочек, заполненных воздухом, и
они растягиваются и сокращаются в про-
дольном направлении подобно мехам ак-
кордеона. При сокращении объем их
уменьшается на 30%.
2 . Некоторые участки трахей имеют в
поперечном сечении форму эллипса, а не
окружности и почти лишены тенидий.
Эллиптические участки могут уплощать-
ся при увеличении давления крови вокруг
них или при сгибании трахеи. Часто в от-
дельных участках трахей образуются
воздушные мешки, имеющие вид растя-
нутых больших камер и напоминающие
эллиптические трахейные структуры от-
сутствием или малым числом тенидиев.
Эти мешки легко сжимаются и разду-
ваются (рис. 5.17; 5.18,Б), и работа их
сходна с работой воздуходувных мехов.
Трахеи и воздушные мешки функцио-
нируют подобно легким позвоночных
животных. Дыхательные движения насе-
комого вызывают попеременное напол-
нение и опорожнение этих органов. Ког-
да тело насекомого сжимается, аккордео-
ноподобные участки также сжимаются и
увеличение давления крови приводит к
сжатию эллиптических участков трахей и
воздушных мешков. Эти два процесса
приводят к выходу воздуха, содержащего
углекислый газ, из трахей через дыхальца
наружу. Когда тело расслабляется, воз-
душные камеры расширяются благодаря
их эластичности и наполняются возду-
хом из окружающей среды.
В результате такой вентиляции (рис.
5.18,Л) трахейные стволы и воздушные
мешки наполняются воздухом, сход-
ным по составу с атмосферным. Не-
посредственно к тканям воздух достав-
ляется с помощью диффузии через тра-
хейные веточки, отходящие от воз-
душных мешков или трахейных стволов.
Дыхательные движения. Трахейная
система не имеет собственной мускула-
туры. Вентиляция осуществляется по-
средством открывания и закрывания ды-
халец и сжатия брюшка, которое дости-
гается сокращением дорсо-вентральных
и продольных мышц. Дорсо-вен-
тральные мышцы сближают спинную и
брюшную поверхности тела, а про-
дольные мышцы вызывают телескопиче-
ское сближение сегментов. Обычно эти
процессы происходят одновременно и
сдавливают воздушные мешки.
Регуляция вентиляции. Вентиля-
ционные движения, приводящие к осво-
бождению трахей от воздуха (экспира-
ция) и наполнению трахей воздухом (инс-
пирация), вызываются нервными им-
пульсами. Существуют два типа нервной
регуляции этих движений.
1. Срединные и поперечные нервы
сегментарных абдоминальных ганглиев
иннервируют инспираторные дорсо-вен-
тральные мышцы. Парные латеральные
нервы брюшка иннервируют дорсо-вен-
тральные экспираторные мышцы и про-
дольные экспираторные и инспира-
торные мышцы.
2. Несколько ганглиев, прежде всего
грудных, а также ганглии 2-го и 3-го
брюшных сегментов, действуют как дат-
чики ритма вентиляции; эти ганглии ре-
гулируют и координируют ритм мы-
шечных сокращений насекомого в целом.
Работа дыхалец и испарение. Помимо
кислорода и углекислого газа через тра-
хейную систему легко диффундируют
также водяные пары. Если дыхальца
?. Жизненные процессы
143
остаются какое-то время открытыми, то
насекомое все это время теряет воду,
представляющую для него большую цен-
ность. Поэтому при освоении насекомы-
ми различных наземных местообитаний
> них появились специальные морфоло-
гические приспособления, снижающие
потери воды через дыхальца, но не пре-
пятствующие дыханию. Такими приспо-
соблениями являются мышечные замы-
кательные лопасти и перфорированные
пластинки, закрывающие дыхальца на
короткие или длинные периоды времени
и исключащие, таким образом, потери
5-оды. когда насекомое не нуждается в по-
ст>тыении кислорода через открытые
дыхальца.
У многих насекомых открывание и за-
крывание дыхалец скоординировано с
вентиляцией, происходящей благодаря
движениям брюшка; в результате, по-
скольку воздух входит через одни ды-
хальца и выходит через другие, создается
лоток воздуха в трахеях.
Регуляция дыхания. Открывание и за-
крывание дыхалец и регуляция дыха-
тельных движений осуществляются нер-
вными центрами, получающими сигналы
об уровне углекислого газа и кислорода в
тканях от специальных сенсорных меха-
низмов. Ниже перечислены наиболее
важные и интересные явления, связанные
с регуляцией дыхания.
1. Непосредственная нервная регуля-
ция дыхательных движений осущест-
вляется сегментарными ганглиями вен-
тральной нервной цепочки. Каждый ган-
глий контролирует движения только
своего собственного сегмента, и, таким
образом, в этой ситуации каждый сег-
мент функционирует как самостоятель-
ная единица.
2. Существует модулирующий, или
координирующий, центр, регулирующий
ритмические движения всех или несколь-
ких сегментов. Механизм действия этого
центра и его ганглионарный состав пока
точно не установлены, хотя известно, что
в нем участвуют грудные и иногда неко-
торые брюшные ганглии.
3. В периоды покоя дыхательные дви-
жения прекращаются совсем, а дыхальца
закрываются. У некоторых насекомых
такая реакция наблюдается и в условиях
избытка кислорода.
4. Практически любое внешнее раз-
дражение нервной системы (зрительное,
тактильное и т. д.) вызывает или усили-
вает дыхательную активность.
5. Дыхание усиливается также под
влиянием различных внутренних химиче-
ских стимулов. У большинства насе-
комых дыхательный нервный центр сти-
мулируется при повышении кислотности
в его рецептивных тканях, что может
быть вызвано высокой концентрацией
либо углекислого газа, либо кислых ме-
таболитов, которые накапливаются в от-
сутствие кислорода. Раздражение дыха-
тельного нервного центра вызывает дви-
жения дыхалец и мышц, контролирую-
щих вентиляционные движения насеко-
мого. У тараканов дыхательная актив-
ность повышается при высокой концен-
трации углекислого газа. У личинок ко-
маров углекислота быстро диффунди-
рует из тела в окружающую воду и
поэтому редко накапливается в больших
количествах; по-видимому, именно
накопление кислых метаболитов заста-
вляет личинок комаров всплывать к по-
верхности воды для захватывания возду-
ха.
6. У многих малоподвижных насе-
комых, или у насекомых, находящихся в
покое, удаление углекислого газа но-
сит характер циклического выброса. Эти
выбросы могут продолжаться от одной
минуты через каждые 2-3 мин до 30 мин
через 24 ч. Механизм и значение таких
выбросов пока неизвестны.
Приспособления к жизни в водной сре-
144
5. Жизненные процессы
де. До сих пор речь шла о типах дыхания,
известных у наземных насекомых. Одна-
ко большое число видов насекомых либо
постоянно живет в воде, либо проводит в
ней значительную часть своей жизни. Су-
ществуют несколько типов адаптаций
для дыхания в водной среде.
1. ЗАХВАТ ВОЗДУХА ПРИ НЫРЯНИИ.
При нырянии определенные насекомые
несут с собой либо воздушную пленку, ко-
торой покрыты те или иные части тела,
либо прикрепленный к телу воздушный
пузырек. У взрослых и нимф водных ко-
риксид (Corixidae) и гладышей (Notonecti-
dae) воздушная пленка находится в склад-
ке на вентральной поверхности тела; эта
пленка сохраняется с помощью гидро-
фобных волосков, препятствующих по-
паданию на нее воды. У взрослых ныря-
ющих жуков (Hydrophilidae, Dytiscidae)
под верхними крыльями, или элитрами,
имеется заполненное воздухом простран-
ство, в которое открываются дыхальца.
Этот запас воздуха служит не только для
снабжения насекомого кислородом, но
также играет роль своего рода легких или
жабр, дополнительно с помощью диффу-
зии получающих кислород из воды и вы-
деляющих в нее двуокись углерода. Это
приспособление не обеспечивает дыхание
насекомого в течение длительного перио-
да, но позволяет ему оставаться все же
довольно долго под водой до поднятия
на поверхность для захвата очередной
порции воздуха.
2. ВОЗДУШНЫЕ ТРУБКИ. Многие
насекомые, живущие все время в воде,
дышат через трубку или пару трубок,
проходящих через толщу воды к поверх-
ности. У таких насекомых функциони-
рует только одна пара дыхалец, связан-
ная с этими трубками, тогда как другие
дыхальца или закрыты, или совсем нера-
звиты. У личинки комара (рис. 8.146)
трубка жесткая; при возникновении по-
требности в кислороде личинка подплы-
вает к поверхности воды и концом труб-
ки протыкает пленку поверхностного на-
тяжения. Личинка обыкновенной пчело-
видки («крыска») (рис. 8.151), живущая в
вязкой или жидкой среде, не подплывает
к поверхности, поскольку ее дыхательная
трубка может удлиняться до 7-10 см и
достигать поверхности. У других во-
дных, полуводных и паразитических на-
секомых также имеются трубки разного
типа.
3. КОЖНОЕ ДЫХАНИЕ. Многие ли-
чинки водных насекомых, а также пара-
зитические насекомые, обитающие в тка-
нях своих хозяев, не имеют специальных
приспособлений для получения воздуха
из атмосферы. У таких насекомых обмен
газов происходит путем диффузии через
покровы тела. Эти насекомые погло-
щают кислород из воды, а избыток угле-
кислого газа выделяют также в воду Су-
ществуют два типа кожного дыхания. В
первом случае (сюда относятся очень
мелкие личинки или личинки первого
возраста) трахейная система отсут-
ствует , газообмен происходит путем
диффузии через ткани, в том числе через
кровь. Во втором случае (куда относится
Рис. 5.19. Схема, иллюстрирующая кожное
дыхание у водных насекомых.
1 -гиподерма; 2-трахеи; 3-веточки трахеи.
5. Жизненные процессы
145
Рис. 5.20. Схематическое изображение
октальных жабр у нимф стрекоз. (Из
Wigglesworth, упрощено.)
. -прямая кишка; 2-жабры, 3-веточки
трахеи; 4 - направление тока воды;
?-анальное отверстие.
большинство крупных личинок, не имею-
щих жабр, такие, например, как личинки
двукрылых последнего возраста и личин-
ки ручейников) трахейная система разви-
та, но вместо дыхалец в гиподерме
имеются пучки тонких трахей (рис. 5.19).
В этом случае сначала имеет место га-
зообмен в гиподерме и только затем-в
тонких периферических трахеях, где про-
цесс диффузии сам по себе не отличается
от такового, происходящего в трахейной
системе с дыхальцами.
К наиболее замечательным
приспособлениям для жизни в воде отно-
сятся листовидные жабры нимф рав-
нокрылых стрекоз (рис. 8.13) и поденок
рис. 8.10). Они являются типичными
для многих обитающих в воде личинок и
нимф, которые дышат жабрами. В этих
жабрах разветвляются трахеи, и газооб-
мен между трахеями и водой происходит
путем диффузии газов через гиподерму.
Необычные приспособления существуют
} нимф стрекоз (рис. 5.20). У этих жи-
вотных ректум сильно увеличен, образуя
мешок, в котором расположены жабры,
пронизанные множеством мелких тра-
хей. Насекомые набирают в этот мешок
воду и затем выпускают ее, а газообмен
происходит через тонкие стенки жабр.
КРОВЬ И ЕЕ ЦИРКУЛЯЦИЯ
Кровь, или гемолимфа, насекомых-
это единственная внеклеточная жид-
кость, циркулирующая по всему телу на-
секомого и омывающая внутренние ор-
ганы. Гемолимфа осуществляет транс-
порт питательных веществ, прежде всего
сахаров и белков, к тканям и забирает от
них отработанные продукты. Вместе с
тем гемолимфа почти не связана с пере-
носом кислорода и двуокиси углерода;
газообмен обеспечивается главным обра-
зом дыхательной системой. Вследствие
этого у насекомых в отличие от других
животных отсутствует тесная связь ме-
жду дыхательной и кровеносной система-
ми. Кровеносная система насекомых
является открытой и только очень корот-
кий отрезок своего пути гемолимфа течет
по закрытым сосудам. В основном же
она протекает по лакунам, располо-
женным между тканями, и, таким обра-
зом, находится в непосредственном кон-
такте с ними. Полость тела, заключаю-
щая в себе открытую кровеносную систе-
му, называется гемоцелем. Кровеносная
система насекомых работает по принци-
пу гидравлической системы, что способ-
ствует движению мягкотелых личинок и
расправлению частей тела насекомых по-
сле линьки.
Кровь. Кровь состоит из жидкости,
или плазмы, и клеток нескольких типов,
в основном бесцветных. Плазма содер-
жит растворенные соли, аминокислоты,
белки, углеводы, мочевую кислоту, ли-
пиды, жирные кислоты, органические ме-
таболиты (цитрат, пируват, сукцинат и
146
5. Жизненные процессы
др.) и органические фосфаты. Ионный со-
став крови различен в разных группах на-
секомых. По сравнению с позвоночными
животными концентрация калия, каль-
ция, магния и фосфора относительно вы-
сока. Концентрация ионов магния в кро-
ви некоторых растительноядных насе-
комых может быть в 50 раз больше, чем у
позвоночных. Столь же высоких концен-
траций может достигать и калий. Боль-
шие различия в соотношении концентра-
ций калия и натрия в крови, которые мы
наблюдаем у растительноядных насе-
комых, являются отражением разно-
образного характера потребляемой ими
пищи. Так, высокие концентрации калия
и низкие концентрации натрия в крови
насекомых обусловлены биохимически-
ми особенностями растительной пищи.
Вообще преобладание ионов калия в кро-
ви можно рассматривать как одну из эво-
люционных адаптаций насекомых к по-
крытосеменным растениям, в тканях ко-
торых калий содержится в больших ко-
личествах. Характерным для насекомых
является присутствие в крови большого
числа аминокислот в высоких концентра-
циях. Присутствие относительно слабо-
реактивного углевода трегалозы в весьма
высоких концентрациях характерно для
насекомых. Этот дисахарид является од-
ним из важных сахаров крови и представ-
ляет собой наиболее доступный источник
энергии у многих видов. Большое число
белков крови насекомых представлены
ферментами.
Свертываемость крови у разных насе-
комых неодинакова. У многих насе-
комых кровь вообще не свертывается, и
раны обычно закрываются пробками, со-
стоящими из специализированных кле-
ток крови (прежде всего плазмоцитов и
подоцитов). У других насекомых кровь
свертывается очень хорошо; этому спо-
собствуют вещества, высвобождаемые
специальными клетками крови.
Кровь насекомых не содержит дыха-
тельных пигментов, в том числе и гемо-
глобина, за исключением Chironomus, у
которого в плазме крови растворено не-
большое количество гемоглобина.
Клетки крови. Клетки крови, или ге-
моциты, содержат ядро и иногда бывают
амебоидными по форме. В плазме они
находятся во взвешенном состоянии или
иногда не прочно прикрепляются к клет-
кам других тканей. У многих групп насе-
комых гемоциты очень разнообразны по
размерам и форме. По внешнему виду
можно выделить две основные категории
гемоцитов-с тонко гранулированной ци-
топлазмой и с цитоплазмой, в которой
хорошо видны отдельные гранулы, число
которых может быть различно. По сово-
купности отдельных признаков разли-
чают 10 типов гемоцитов (Jones, 1962,
1979 и рис. 5.21). Хорошо идентифици-
руемые три типа клеток (прогемоциты,
плазматоциты, гранулоциты) встречают-
ся у большинства насекомых; другие
четыре типа (цистоциты, сферические
клетки, адипогемоциты и эноциты)
встречаются довольно часто, и три высо-
коспециализированных типа (подоциты,
червеобразные клетки и гранулоцитофа-
гоидные клетки) встречаются лишь у не-
многих насекомых. Гемоциты размно-
жаются только митотическим путем.
Функции гемоцитов так же разно-
образны, как и их строение. Они погло-
щают некоторых живых и всех мертвых
бактерий, а также старые или отмираю-
щие ткани. Вместе с тем высвобождаю-
щееся содержимое при разрыве их оболо-
чек служит питательным материалом
для других тканей. Два последних свой-
ства гемоцитов в наибольшей мере про-
являются при метаморфозе. При повреж-
дении покрова насекомых гемоциты ска-
пливаются около ран, образуя пробку,
закрывающую отверстие в покровах; в
других случаях они образуют в полости
5. Жизненные процессы
147
Рис. 5.21. Девять типов гемоцитов насекомых. (По Jones, Chapman.)
А. Прогемоцит. Б. Плазматоцит. В. Гранулоцит. Г. Эноцит. Д. Цистоцит. Е. Сферическая
клетка. Ж. Адипогемоцит. 3. Подоцит И. Червеобразная клетка.
тела своего рода ячейки, изолируя тем
самым паразитов, проникших в полость
тела насекомого.
Функции крови. У насекомых кровь
несет пять функций. Первые четыре,
перечисленные ниже, представляют со-
бой функции крови как живой системы, а
пятая в основном механическая.
1. ТРАНСПОРТ ПИТАТЕЛЬНЫХ ВЕ-
ЩЕСТВ И ОТРАБОТАННЫХ ПРОДУК-
ТОВ. Продукты переваривания пищи по-
падают в кровь из пищеварительной
системы и доставляются к тканям, а от-
работанные продукты переносятся из
тканей к органам выделения. Некоторые
гормоны также переносятся с кровью от
мест их образования к местам их актив-
ности.
2. ДЫХАНИЕ. У большинства насе-
комых по крайней мере к некоторым
клеткам не подходят трахеолы, и поэто-
му здесь невозможен непосредственный
газообмен. По-видимому, такие клетки
получают кислород из крови. Кровь
играет вспомогательную роль в удале-
нии из организма двуокиси углерода, ос-
новная часть которой диффундирует че-
рез ткани и выходит через кутикулу. В
крови личинок некоторых видов Chirono-
mus растворен гемоглобин, участвующий
в процессе переноса кислорода. Связыва-
ние кислорода гемоглобином у Chirono-
mus, конечно же, ни в коей мере не может
сравниться по эффективности с функцией
этого пигмента, содержащегося в эри-
троцитах млекопитающих.
3. ЗАЩИТНАЯ ФУНКЦИЯ. Гемоциты
освобождают организм от некоторых
бактерий и паразитов. В заживлении ран
участвуют кровь или гемоциты крови.
4. МЕТАБОЛИЗМ. Кровь является той
средой, в которой протекают метаболи-
148
5. Жизненные процессы
ческие реакции. При циркуляции крови
по телу насекомого вещества, содержа-
щиеся в ней, подвергаются химическому
превращению. Например, углевод трега-
лоза превращается в глюкозу.
5. ГИДРАВЛИЧЕСКАЯ ФУНКЦИЯ.
Весь объем крови, заключенный внутри
тела насекомого, образует замкнутую ги-
дравлическую систему, способную пере-
давать давление с одной части тела на
другую. Этот в основе своей механиче-
ский процесс используется насекомыми
многообразно. Давление крови регули-
руется сокращениями груди или брюшка,
или той и другой части тела вместе. По-
переменное снижение и повышение да-
вления крови, происходящие при дыха-
тельных и мышечных движениях насеко-
мого, вызывают опорожнение и наполне-
ние трахейных воздушных мешков и
карманов. Давление крови лежит в осно-
ве таких процессов, как растягивание по-
кровов и расправление крыльев после
линьки, а также разрыв оболочки яйца
перед выходом личинки.
Циркуляция крови. Направление тока
крови у насекомых показано на рис. 4.5 и
5.22. Благодаря работе сердечного насо-
са кровь перекачивается из брюшка через
аорту в переднюю часть тела, проходит
через головной мозг и изливается в голо-
ве в том месте, где кончается аорта. Из
головы кровь оттекает назад, омывая
ткани и органы тела, доходит до брюш-
ка, и начинается новый цикл.
Кровь засасывается в сердце через
остии и течет вперед благодаря волнам
продольных перистальтических сокраще-
ний. Перистальтические сокращения со-
здаются чередованием расширения серд-
ца, когда в силу отрицательного давле-
ния кровь засасывается в остии, и систо-
лического сокращения, заставляющего
кровь двигаться вперед. Эти процессы
обеспечиваются эластичностью стенок
сердца и работой крыловидных и других
мышц. Время от времени ток крови ме-
няет направление, и кровь из сердца на-
правляется назад в брюшную полость. У
большинства насекомых сердце не имеет
перегородок на всем своем протяжении.
Редко остии, углубляясь в сердце, обра-
зуют клапанообразные перегородки, ко-
торые делят сердце на камеры.
Помимо сердца, основного кровяного
насоса, существуют еще различные при-
способления для проталкивания крови в
конечности или распределения ее в поло-
сти тела. У некоторых насекомых от
аорты отходят сосуды, несущие кровь в
разных направлениях. У многих насе-
комых антенны и ноги разделены про-
дольными перегородками, или септами,
и кровь входит в конечность с одной сто-
роны, проходит по ней и вытекает по
другой стороне. Поступлению крови в
конечности помогают и дыхательные
движения, и «пульс» в конечностях мо-
жет быть синхронизирован с ними, а не с
биением сердца.
Часто в средне- и заднегруди встре-
чаются дополнительные кровяные на-
сосы, или пульсирующие органы, ко-
торые направляют кровь в крылья. В
этих случаях кровь течет внутри крыльев
по жилкам крыла (рис. 5.23) и возвра-
щается или прямо в аорту, или в полость
тела. Вентральная диафрагма в случае,
если она хорошо развита, также уча-
ствует в регуляции тока крови, при этом
сокращения мышц диафрагмы вызывают
движение крови латерально и назад.
На рис. 5.22 показано, какие органы
или процессы обусловливают движение
крови в том или ином направлении.
У некоторых насекомых сердце ин-
нервируется многочисленными нервами,
принадлежащими как висцеральной нер-
вной системе, так и сегментарным ган-
глиям. Многие исследователи полагают
поэтому, что деятельность сердца регу-
лируется нервными импульсами. Однако
Хкякяные процессы
149
Рис. 5.22. Сердце и связанные с ним органы, обеспечивающие
циркуляцию крови у насекомых. (Из Wigglesworth.)
А. Схематическое изображение хорошо развитой кровеносной
системы. Б. Поперечный разрез через грудь насекомого с хорошо
развитой кровеносной системой. В. Поперечный разрез через брюшко.
Стрелки показывают направление тока крови (в основном по Brocher).
1 -остия; 2-сердце; 3-мышцы диафрагмы, 4-пульсирующие органы;
5 - аорта; 6 - пульсирующий орган; 7 - перегородки; 8 - нервная
цепочка, 9-вентральная диафрагма; 10-дорсальная диафрагма;
11 - перегородка.
Рве. 5.23. Циркуляция крови в переднем крыле веснянки Limnephilus rhombicus. (По
ХгвоИ. 1964.) Стрелками показано направление тока крови.
150
5. Жизненные процессы
до сих пор вопрос о том, обусловлена ли
пульсация сердца нервной стимуляцией
или периодическими сокращениями и
расслаблениями мышц, происходящими
без нервной стимуляции, остается
спорным. Когда насекомое находится в
состоянии стресса, нервная регуляция
сердечных сокращений дополняется гор-
мональной-гормон кардиального тела
(corpus cardiacum) ускоряет пульсацию
сердца.
МЕТАБОЛИЗМ
Метаболизм включает все химиче-
ские процессы, происходящие в живом
организме. Это высокоинтегрированная
последовательность химических реакций,
включающая две фазы: созидательную
(анаболизм) и разрушительную (катабо-
лизм). В клетках и тканях одновременно
протекают и анаболические, и катаболи-
ческие процессы. Все метаболические ре-
акции катализируются специфическими
белками, называемыми ферментами, и
сопровождаются энергетическим обме-
ном.
Катаболическая фаза. В катаболиче-
ской фазе сложные молекулы пита-
тельных веществ (углеводов, жиров и
белков) разрушаются в основном в про-
цессе окисления, превращаясь в более
простые молекулы. Катаболизм сопро-
вождается высвобождением легко до-
ступной энергии, основная часть которой
сохраняется в форме энергии связанного
фосфора аденозинтрифосфата (АТФ).
Энергия, высвобождающаяся в катабо-
лической фазе, используется для биосин-
теза различных веществ, транспорта ио-
нов, проведения нервных импульсов и
сокращения мышц и для свечения (люми-
несценция). Ниже перечислены катаболи-
ческие процессы, приводящие к высво-
бождению химической энергии.
1. Разрушение крупных молекул с
образованием небольших, составляю-
щих их единиц; например, при разруше-
нии белков сначала образуются полипеп-
тидные цепи, которые затем разрушают-
ся до составляющих их аминокислот. В
этих реакциях высвобождается неболь-
шое количество энергии.
2. Окисление углеводов, например
полисахарида гликогена (встречающего-
ся у многих животных) и дисахарида тре-
галозы (характерного для насекомых са-
хара крови), до моносахарида глюкозы.
Наибольшее количество доступной энер-
гии высвобождается именно при окисле-
нии углеводов. Запасы углеводов
являются важным источником энергии,
которую насекомые расходуют во время
полета, в процессе метаморфоза и при
голодании.
При возникновении потребности в
энергии у насекомых трегалоза с по-
мощью фермента трегалазы превращает-
ся в глюкозу. Преимущество накопления
в крови трегалозы, а не глюкозы (обыч-
ного сахара крови) состоит, по-видимо-
му, в том, что молекула трегалозы отно-
сительно слабо реактивна и поэтому
сохраняется в гемолимфе в высоких кон-
центрациях, не вступая в возможные по-
бочные реакции с аминокислотами, ко-
торые также содержатся в крови в боль-
ших количествах.
3. Разрушение липидов (жиров)
с образованием жирных кислот и глице-
рола (глицерина). Эти процессы обеспе-
чивают энергетический резерв, исполь-
зуемый при голодании и во время полета
насекомого.
4. У некоторых насекомых, например
у мухи цеце, небольшой энергетический
резерв создается за счет окисления ами-
нокислот.
Анаболическая фаза. В анаболической
фазе молекулы, представляющие собой
строительные блоки, объединяются в ма-
кромолекулы, такие, как белки, нуклеи-
5 Жизненные процессы
151
sr зые кислоты, липиды и полисахариды,
< здут на построение новых тканей или
охраняются в той или иной форме для
-пользования в дальнейшем. Поскольку
•tv процессы состоят в увеличении раз-
мера и усложнении структуры молекул,
нуждаются в свободной химической
Сергии; эту энергию они получают из
\ТФ. который синтезируется в катаболи-
нотой фазе.
Дыхательный метаболизм. Дыха-
-оьный метаболизм, известный также
таг клеточное дыхание, является источ-
eziom легко доступной энергии для ис-
тсдьзования ее во многих метаболиче-
ztzx процессах. Насекомые, как и боль-
дх?чгтво других организмов, получают
-Сергию, высвобождаемую при «сгора-
пиши, т. е. при окислении органиче-
веществ (углеводов, жиров, белков).
2деление происходит внутри клетки-в
zzr о плазме и митохондриях (органел-
лах. представляющих собой как бы элек-
тсстанции клетки). Основной реакцией,
иге которой у всех аэробных организмов
освобождается энергия, является окис-
теете глюкозы до двуокиси углерода и
зеды. Эта реакция имеет многоступен-
чатый характер и включает три главные
ттатин: гликолиз, цикл Кребса и процесс
тетеноса электронов, что приводит к вы-
тэебождению энергии. Выделение энер-
происходит потому, что молекулы
глюкозы и кислорода заключают в себе
тслыпе энергии, чем молекулы конечных
ттедуктов окисления (двуокиси углерода
♦ воды).
Промежуточный метаболизм. Суще-
ствует много реакций, протекающих в
клетке, при которых энергия высвобо-
ждается не сразу. Все эти реакции состав-
ЛЕот промежуточный метаболизм лю-
тело живого существа, причем многие из
зх протекают в жировом теле. Эти реак-
лей приводят к образованию спе-
дгзльных секретов и синтезу или разру-
шению клеточных компонентов. Ниже
приводятся примеры таких реакций.
1. УГЛЕВОДЫ. Синтез трегалозы (са-
хар крови), хитина (полисахарид TV-аце-
тил-£)-глюкозамин) и гликогена. Глико-
ген накапливается в жировом теле, жуж-
жальцах, летательных мышцах и клетках
средней кишки.
2. ЛИПИДЫ. Синтез жирных кислот,
триглицеридов и восков, являющихся
структурными компонентами жиров.
Жирные кислоты и триглицериды синте-
зируются из аминокислот; эти реакции
происходят как в жировом теле, так и в
других местах.
Липиды составляют основные ре-
зервные вещества в организме насе-
комых. Значительная часть пищи, погло-
щенной насекомым на стадии личинки,
превращается в липиды и запасается в
жировом теле. Некоторые из этих жиров
используются во время окукливания как
источник энергии и химических компо-
нентов для процессов биосинтеза, проис-
ходящих при метаморфозе. Часть липи-
дов представляет собой источник энер-
гии, используемый взрослым насе-
комым. Воска несут защитную функцию
в составе кутикулы, а также входят в со-
став пчелиного воска.
3. АМИНОКИСЛОТЫ И БЕЛКИ. Син-
тез некоторых аминокислот и белков
протекает в жировом теле.
МЫШЕЧНАЯ СИСТЕМА
И ДВИЖЕНИЕ
Способы движения насекомых-пол-
зание, хождение, прыгание, полет-свой-
ственны многим живым организмам и
вызываются действиями пучков сократи-
тельной ткани, называемых мышцами.
Мышцы сгруппированы в координиро-
ванные системы, что повышает эффек-
тивность вызываемого ими действия-
движения. У насекомых мышечные со-
152
5. Жизненные процессы
Рис. 5.24. Мышечные волокна насекомых. (Из Wigglesworth, по Snodgrass.)
А. Личинки медоносной пчелы. Б. Ножных мышц жука-скарабея. В. Ножных мышц
медоносной пчелы. Г. Летательные мышцы непрямого действия медоносной пчелы
(группа саркостилей из фибриллярной мышцы).
1 -саркоплазма; 2-фибриллы, или саркостили; 5-ядра, 4-сарколемма;
5-саркосомы.
кращения вызывают не только движения
различных частей экзоскелета, но также" и
движения внутренних органов.
Мышцы насекомых подобны мыш-
цам других животных; они вытянуты в
продольном направлении и способны со-
кращаться при получении импульса от
нервной клетки. За сокращением следует
расслабление, когда мышечная клетка
возвращается к своему исходному разме-
ру и форме.
Мышцы насекомых состоят только из
поперечнополосатых волокон, хотя в не-
которых висцеральных мышцах эту попе-
речную исчерченность бывает довольно
трудно обнаружить. Каждое поперечно-
полосатое волокно состоит из несколь-
ких, расположенных параллельно друг
другу миофибрилл и окружающей их ци-
топлазмы, называемой саркоплазмой, в
которой содержится много ядер. Кроме
того, в мышечной клетке содержится
большое число других клеточных орга-
нелл -митохондрий. В митохондриях со-
держатся окислительные ферменты, уча-
ствующие в процессе окисления, при
котором высвобождается энергия, ис-
пользуемая для движения йасекомого.
Особенно многочисленны митохондрии
в очень активных мышцах; так, в лета-
тельных мышцах некоторых насекомых
они составляют 40% веса мышцы. Струк-
тура миофибрилл и саркоплазмы варьи-
рует у разных насекомых (рис. 5.24) и в
разных мышцах одного и того же насеко-
мого.
Мышечные сокращения вызывают
движение какой-либо части тела насеко-
мого. За каждым движением следует
контрдвижение, возвращающее эту часть
тела в исходное положение. Контрдвиже-
ние часто обусловлено работой другой
мышцы, как это показано на рис. 5.25;
здесь голень сочленяется с бедром по обе
стороны от точки 3, сокращение под-
нимающей мышцы (5) вызывает подня-
5 Жизненные процессы
153
Рис. 5.25. Движение ноги взрослого насекомого. (По Снодграссу.)
А. Сочленение с помощью одного мыщелка (одна впадина-7). Б, В.
Сочленение с помощью двух мыщелков (две впадины -3 и 4)\ вид сзади и
сбоку. 2 и 5-мышцы, соответственно поднимающие и опускающие ногу.
голени (первое движение), сокраще-
ние мышцы-депрессора (2) вызывает опу-
скание голени (контрдвижение). В других
гггучаях движение осуществляет одна
мышца, и тогда контрдвижение вызы-
вается либо давлением крови, либо натя-
жением эластичных мембран. При контр-
гзажении, вызываемом давлением кро-
ва. это гидравлическое давление просто
зезвращает на место изменившую свое
^сложение часть тела или орган после
тшхлабления мышцы.
Полет. Мышечный механизм полета
насекомого представляет исключи-
тельный интерес, поскольку ничего по-
добного не существует у других жи-
вотных. Полет происходит благодаря
тзижению (взмахам) крыльев, являю-
тзхся выростами латеральных частей
средне- и заднегрудных тергитов. У рав-
нокрылых, стрекоз, златоглазок и других
трнмитивных отрядов насекомых движе-
ния обеих пар крыльев независимы. У
гавнокрылых стрекоз, например, когда
.-дна пара крыльев поднимается, другая
гаускается. У молей и бабочек, пчел и ос
* некоторых других насекомых оба кры-
ла одной стороны удерживаются вместе
или сцеплены различными крючками,
пеганками или складками, так что обе
лары работают синхронно. У большин-
ства жуков и некоторых настоящих кло-
пов передние крылья представляют со-
бой твердые панцирные пластинки, а для
полета используются только задние кры-
лья. У Stylopoidea (веерокрылые) перед-
няя пара крыльев редуцирована, а у
двукрылых (Diptera) редуцирована зад-
няя пара; таким образом, в обеих этих
группах насекомых имеется по одной па-
ре летательных крыльев.
Крылья насекомого функционируют
одновременно и как пропеллеры самоле-
та, и как направляющие поверхности, со-
здающие поток воздуха над крыльями.
Однако крыло не работает точно как
пропеллер, поскольку траектория его
движения представляет собой синусоиду,
и, если насекомое закреплено, вершина
крыла описывает восьмерку. Другими
важными компонентами тела, обеспечи-
вающими полет, являются экзоскелет и
мускулатура. Крепкий, упругий и гибкий
экзоскелет легко поглощает и высвобо-
ждает энергию. Жизненно важная роль
этих механических свойств экзоскелета и
связанной с ним специализированной му-
скулатуры состоит в сохранении кинети-
ческой энергии, образующейся при взма-
хе крыльев, для использования ее при
непроизвольном опускании крыльев. Та-
кой экономный расход энергии обуслов-
лен биохимическими особенностями бел-
ка резилина, синтезирующегося в сочле-
154
5. Жизненные процессы
нениях насекомых, и в частности в осно-
вании крыльев. В силу подвижности
своих аминокислотных цепей этот белок
обладает почти абсолютной эластич-
ностью и способен возвращать 97% энер-
гии, накапливающейся в нем при сжатии
и растяжении.
Движением крыльев насекомого в по-
лете управляют 9 или 10 пар функцио-
нально различных мышечных групп, от-
носящихся к каждому из двух птеротора-
кальных (несущих крылья) сегментов.
Состав этих мышц неодинаков у разных
отрядов насекомых. Мышцы прямого
действия называются так, потому что
они, начинаясь от плейральной и стер-
нальной областей груди, прикрепляются
непосредственно к склеритам крылового
сочленения. Эти мышцы обеспечивают
движение крыльев при так называемом
синхронном полете1. Эти мьшцы со-
стоят из важных базалярной и субалярной
групп и отходят от стернальной или плей-
ральной областей или от кокс, прикреп-
ляясь дорсально к базалярным и суба-
лярным склеритам крылового сочлене-
ния или к верхним склеритам боковой
части груди. В группе неоптероидных на-
секомых мышцы, складывающие крылья,
связывают плейральную область груди
также с третьим аксиллярным склеритом
крылового сочленения. Мышцы непрямо-
го действия, которые называются так по-
тому, что они не прикрепляются непо-
средственно к крыльям, служат основны-
ми эффекторами при движении крыльев
при асинхронном полете. Они идут вер-
тикально и продольно через мезо- и мета-
торакальные сегменты и при своем со-
кращении деформируют грудь, что вы-
зывает движение прикрепленных к ней
1 Они же изменяют наклон крыловой
пластинки в горизонтальной плоскости, обес-
печивая пронацию и супинацию крыла.
крыльев. Дорсо-вентральные мышцы не-
прямого действия поднимают крылья, а
дорсальные продольные мышцы непрямого
действия опускают их.
В разных группах насекомых движе-
ния крыльев при полете вызываются ме-
ханизмами двух типов-синхронным и
асинхронным (рис. 5.26),-которые обес-
печиваются соответственно двумя типа-
ми крыловой мускулатуры. Большин-
ство летающих насекомых имеют синх-
ронные летательные мышцы прямого дей-
ствия, которые сокращаются и расслаб-
ляются непосредственно в ответ на нерв-
ные импульсы (рис. 5.26,Л, Б). Нерв-
ные импульсы непосредственно регули-
руют ритм сокращений этих мышц и ко-
ординируют их движения. Четыре отряда
насекомых-жуки fColeoptera), перепон-
чатокрылые (Hymenoptera), мухи, кома-
ры и близкие к ним формы (Diptera) и не-
которые настоящие клопы и тли (Hemi-
ptera)-имеют асинхронно работающие
летательные мышцы непрямого действия
(рис. 5.26,В,Г). У этих насекомых лета-
тельные мышцы сокращаются и расслаб-
ляются с частотой, намного превышаю-
щей частоту поступления к ним сигналов
от нервной системы. Координированные
взмахи крыльев в этом случае обуслов-
лены механическими свойствами экзоске-
лета и уникальными сократительными
свойствами фибрилл летательных мышц
непрямого действия. На рис. 5.26 меха-
низмы обоих типов полета изображены
схематично; на самом деле расстояния
между различными точками очень малы,
и существует еще сложная дополнитель-
ная мускулатура.
СКОРОСТЬ И НАПРАВЛЕНИЕ ПО-
ЛЕТА. В процессе полета крылья на-
секомого совершают движения трех ти-
пов : 1) взмах и опускание; 2) поворачива-
ние вдоль продольной оси (изменение
угла атаки); 3) перемещение вперед и на-
зад в горизонтальной плоскости. Все три
Жизненные процессы
155
Рис. 5.26. Синхронные и асинхронные летательные мышцы. (По Smith, 1965.)
А, Б. Синхронные летательные мышцы. А. Сокращение внутренней пары мышц в
первой секции груди передает усилие на рычагоподобную часть основания крыла,
что приводит к его подъему. Б. Последующее сокращение более сильной пары
наружных мышц вызывает движение крыльев вниз. В, Г. Асинхронные летательные
мышцы. Наружные мышцы сокращаются в вертикальном направлении,
а внутренние-в горизонтальном. Это происходит поочередно и в колебательном
ритме. Сокращение мышц деформирует грудь, в результате чего крылья
поднимаются (В) или опускаются(Г)-
лпа движений осуществляются одновре-
менно. но превалирование того или дру-
ого зависит от скорости полета и разме-
та крыльев.
Сами по себе взмах и опускание
крыльев обеспечивают только подъем.
Таким образом, бабочки, для которых
характерен в основном этот тип движе-
ния. способны почти исключительно к
порханию и могут продвигаться вперед
тишь с очень незначительной скоростью.
Если при этом крылья поворачиваются
относительно продольной оси, то это ве-
дет к уменьшению давления воздуха на
переднюю поверхность крыла и к увели-
чению давления воздуха сзади, как у про-
пеллера самолета. Давление воздуха сза-
ди и одновременно частичный вакуум
впереди вызывают движение насекомого
вперед. Поэтому изменение угла атаки
крыла является основным условием для
движения вперед. Насекомые с высокора-
звитой способностью к изменению угла
атаки крыла, например стрекозы, могут
длительно сохранять устойчивый полет
вперед.
Частота взмахов крыльев чрезвычай-
но варьирует у разных видов. Наимень-
156
5. Жизненные процессы
шая частота наблюдается у бабочек: так,
капустная белянка делает около 10 взма-
хов в секунду. У пчел частота взмахов
крыльев намного больше -190 в секунду,
у шмелей-240, у комнатной мухи-330.
Наибольшая скорость полета дости-
гается при сочетании следующих факто-
ров . длинное узкое крыло, хорошая спо-
собность к изменению угла его атаки и
достаточно высокая частота взмахов
крыльев. Быстрее всех летают бражники
(Sphingidae) - более 53 км/ч, слепни (Ta-
banus) -более 50 км/ч и стрекозы, ко-
ЛИТЕРАТУРА
Arnold J W Blood circulation in insect wings,
Mem, Ent. Soc. Can., 38, 1-60 (1964).
Barrington E. J. W. Invertebrate structure and
function, 2nd ed., Thomas Nelson, London,
549 pp, 1967.
Chapman R. F. The insects, structure and functi-
on. American Elsevier, New York, 819 pp,
1969.
Eisner T., Wilson E. O. (Eds.) The insects.
W H. Freeman, San Francisco, 334 pp, 1977.
Hackman R. H. Chemistry of the insect cuticle.
In: Morris Rockstein (Ed.). The physiology of
Insecta, 6, 215-270. Academic Press, New
York, 1974.
Hepburn H. R. (Ed.). The insect tegument. El-
sevier, Amsterdam, Oxford, New York, 571
pp, 1976.
House H. L. Nutrition. In: Morris Rockstein
(Ed.). The physiology of Insecta, 5, 1-62.
Academic Press, New York, 1974:
House H. L. Digestion. In: Morris Rockstein
(Ed.). The physiology of Insecta, 5, 63-117.
Academic Press, New York, 1974.
Jones J. C. Current concepts concerning insect
hemocytes, Am. Zool. 2, 209-246 (1962).
Jones J. C. The circulatory system of insects.
Thomas, Springfield, Ill, 255 pp, 1977
торые.обычно летят co скоростью около
16 км/ч, но могут развивать и большую
скорость (почти до 40 км/ч).
При поступательном полете крылья
насекомых выписывают восьмерку с на-
клоном вперед в верхней точке взмаха
крыла. Многие насекомые, например му-
хи-журчалки (Syrphidae), пчелы и многие
посещающие цветки бабочки, могут ле-
тать назад; это достигается тем, что
описываемая крылом восьмерка имеет
наклон назад в верхней точке взмаха
крыла.
Locke М. The structure and function of the
integument in insects. In: Morris Rockstein
(Ed.). The physiology of Insecta, 6, 123-213.
Academic Press, New York, 1974.
Maddrell S. H. P. The mechanisms of insect
excretory systems. In Beament J. W. L, Tre-
herne J. E, Wigglesworth V. B. (Eds.). Ad-
vances in insect physiology, 8, 200-331. Aca-
demic Press, London, New York, 1971.
Pringle J. W. S. Locomotion: Flight. In: Morris
Rockstein (Ed.). The physiology of Insecta, 3,
433-476. Academic Press, New York, 1974.
Smith D. S. The flight muscles of insects, Sci.
Am, 227, 76-84 (1965).
Snodgrass R. E. Principles of insect morphology.
McGraw-Hill,'New York, 667 pp, 1935.
Stobbart R. H.,Shaw J Salt and water balance:
Excretion. In: Morris Rockstein (Ed.). The
physiology of Insecta, 5, 361-446. Academic
Press, New York, 1974.
Waterhouse D. F. Digestion in insects, Annu.
Rev. Entomol, 2, 1-18 (1957).
Wigglesworth V. B. The principles of insect
physiology. Chapman & Hall, London, 7th
ed, 827 pp, 1972.
Wigglesworth V B. Insect physiology. Chap-
man & Hall, London, 7th ed, 166 pp, 1974.
6
Реакции
на внешние воздействия
и поведение
Важнейшим свойством живых организмов является раздражимость, т е. способ-
ность реагировать на различные стимулы. Эта способность лежит в основе реакций
организма на изменения, происходящие в окружающей среде. Реакции организма
представляют собой либо ответы на благоприятные или неблагоприятные условия,
либо просто компоненты его нормального поведения. Раздражимость включает в се-
бя восприятие рецепторами и органами чувств сигналов об изменениях во внешней и
внутренней среде, а также проведение, координацию и интеграцию нервной и эндо-
кринной системами той информации, которая поступает от рецепторов. В свою оче-
редь нервная и эндокринная системы определенным образом реагируют на приходя-
щие к ним стимулы. И наконец, многообразные формы поведения отражают способы
существования и адаптации организма к изменениям внешней среды.
В этой главе рассматриваются рецепторы и органы чувств, нервная и эндокринная
системы, гормональная регуляция и поведение.
ИСТОЧНИКИ ИНФОРМАЦИИ:
РЕЦЕПТОРЫ И ОРГАНЫ ЧУВСТВ
У насекомых, как и у всех многокле-
точных организмов, имеется множество
различных воспринимающих клеток, ре-
цепторов, или сенсилл, чувствительных к
определенным типам стимулов. Эти ре-
цепторные клетки представляют собой
специализированный компонент нервной
системы, воспринимающий стимулы и
передающий их в нервно-мышечный ап-
парат. Некоторые рецепторные клетки
рассеяны в ткани, как, например, так-
тильные рецепторы эпителия. Другие со-
браны в группы, обычно состоящие из
большого числа клеток, и образуют ор-
ганы чувств, такие, как глаза, антенны и
тимпанальные органы.
Рецепторы обычно классифицируют
по характеру воспринимаемых ими сти-
мулов. Рецепторы насекомых очень раз-
нообразны ; к их числу относятся механо-
рецепторы (звуковые рецепторы и про-
приоцепторы), терморецепторы, фоторе-
цепторы, хеморецепторы. С помощью
рецепторов насекомые улавливают меха-
нические вибрации, в том числе большой
диапазон звуков, энергию излучения в
форме тепла и света, механическое давле-
ние, в том числе силу тяжести, количе-
ство воды и летучих веществ в воздухе и
множество других факторов внешней
среды. Кроме того, насекомые обладают
хорошо развитым чувством обоняния и
вкуса.
Механорецепторы
Трихоидные сенсиллы. Простые ор-
ганы чувств -трихоидные сенсиллы
(рис. 6.1)- являются механорецепторами
и обычно состоят из двух частей: 1) спе-
158
6. Реакции на внешние воздействия и поведение
Рис. 6.1 Простая волосковидная сенсилла.
(По Снодграссу.)
1 - соединение чувствительной клетки
с кутикулой; 2-щетинка; 3-кутикула;
4-тормогенная клетка; 5 - трихогенная
клетка; 6-чувствительная клетка; 7-нерв.
диализированного кутикулярного обра-
зования-волоска, пластинки или тонко-
стенного колышка и 2) нервного оконча-
ния, находящегося в тесном контакте
с этим кутикулярным образованием.
Любые изменения в окружающей среде
вызывают изменения в сенсиллах, что в
свою очередь ведет к изменению энерге-
тического потенциала в связанных с ни-
ми нервных окончаниях. Это изменение и
есть стимул. За изменением потенциала
в нервном окончании следует волна воз-
буждения (импульс), проходящего по не-
рвному волокну. После того как импульс
прошел по волокну, нерв на короткое
время становится менее чувствительным,
и ему необходимо вернуться к исходному
Рис. 6.2. Чувствительные клетки
и рецепторы. (По Снодграссу.)
А. Хеморецептор с тонкостенными
выростами и несколькими чувствительными
клетками. Б. Колоколовидный рецептор на
церке таракана.
1 - тонкостенный вырост кутикулы;
2-дистальный отросток нервной клетки;
3 - чувствительные клетки;
4 - проксимальный отросток нервной клетки;
5-пучок нервных волокон;
6 - колоколовидный рецептор; 7 - кутикула,
8-дистальный отросток чувствительной
клетки; 9-трихогенная клетка; 10-оболочка
чувствительной клетки; 11 -нерв.
6. Реакции на внешние воздействия и поведение
159
физиологическому состоянию, прежде
чем он будет готов к проведению сле-
дующего импульса. Таким образом, хотя
нерв может возбуждаться непрерывно,
его ответ на раздражение имеет ди-
скретный характер.
Трихоидные сенсиллы обычно
воспринимают тактильные стимулы. Од-
нако существует огромное разнообразие
таких волосковидных органов чувств, во-
спринимающих вкус, запах и влажность.
Иногда вместо волосков имеются тонко-
стенные колышки, или сеты - щетинко-
видные выросты покровов, обычно со-
стоящие из хитина. Каждое из этих
образований всегда тесно связано с не-
рвным окончанием (рис. 6.2,Л). В неко-
торых случаях в рецепторе имеется груп-
па чувствительных клеток, и соответ-
ственно к нему подходят несколько нерв-
ных окончаний (рис. 6.2,А).
Большинство трихоидных сенсилл
воспринимает раздражение только в мо-
мент изменения положения-при изгибе
или выпрямлении; такой тип возбужде-
ния называется фазической рецепцией. Не-
которые трихоидные сенсиллы восприни-
мают раздражение все время, пока ще-
тинки находятся в несвойственном для
них положении; уто-тоническая рецеп-
ция.
Трихоидные сенсиллы распределены
на поверхности тела насекомого нерав-
номерно. Наиболее многочисленны они
на тех участках ротового аппарата, ан-
тенн (рис. 6.3) и конечностей, которые
соприкасаются с другими поверхностями
или другими частями тела. Некоторые
фазические сенсиллы могут улавливать
малейшие колебания воздуха, произво-
димые находящимся поблизости хищни-
ком, и, таким образом, представляют со-
бой систему предупреждения об опасно-
сти. Группа тонических трихоидных сен-
силл сигнализируют о положении частей
тела относительно друг друга. Напри-
мер, у муравьев сенсорные щетинки, рас-
положенные между головой и грудью,
регистрируют положение головы.
Рецепторы напряжения (проприоцеп-
торы). Куполообразные рецепторы, так
называемые колоколовидные, или кампа-
ниформные сенсиллы (рис. 6.3), встре-
чаются на различных участках покровов,
Вне. 6.3. Чувствительные рецепторы на флагеллуме усика рабочей пчелы Apis mellifercr,
- 3300. (С разрешения А. Дитца, Университет Джорджии; из Dietz, Humphreys.)
Показаны четыре типа сенсилл. 7-базиконическая сенсилла; 2-колоколовидная сенсилла;
3-плакоидная сенсилла; 4-одна из форм трихоидной сенсиллы.
160
6. Реакции на внешние воздействия и поведение
Рис. 6.4. Схематическое изображение поперечного разреза через голову и переднегрудь
муравья Formica polyctena. Видны волосковые пластинки на переднегруди. (По Markl,
1962.)
в том числе и на конечностях. Ди-
стальный отросток такой сенсиллы
(рис. 6.2, Б) упирается в свод (купол).
Изменения натяжения покровов вокруг
сенсиллы вызывают либо уменьшение,
либо увеличение кривизны свода, и это
раздражение передается черву. Эти ор-
ганы контролируют движ 'ни я тела насе-
комого и играют важную роль в геотро-
пической ориентации и в координации
движения конечностей насекомого.
Большое число самых разнообразных
рецепторов, от трихоидных сенсилл и ре-
цепторов натяжения до сложных орга-
нов, включающих много нервных окон-
чаний, снабжает насекомое информацией
о положении тела и его частей. Напри-
мер, у муравья Formica в том месте, где
голова, сочленяясь с переднегрудью,
перекрывает ее, имеются участки, покры-
тые волосками (ролосковые пластинки),
к которым подходят нервные окончания
(рис. 6.4). Когда голова склоняется на-
бок, одна группа пластинок испытывает
большее давление, чем другая. Когда
голова находится в нормальном положе-
нии, давление распределяется равномер-
но между обеими Группами пластинок.
Получая но нервным путям информацию
о распределении давления на эти воло-
сковые пластинки, головной мозг реги-
стрирует положение головы.
Хордотональные сенсиллы воспри-
нимают изменения натяжения и вибра-
ции субстрата и окружающей среды. Не-
которые хордотональные сенсиллы пре-
6. Реакции на внешние воздействия и поведение
161
11-732
Рис. 6.5. Хордотональные органы. (Обе части из Chapman, 1969, с
изменениями.)
А. Схематическое изображение продольного среза через сколопидий из
тимпанального органа саранчи Locusta. Б. Схематическое изображение
среза через джонстонов орган, расположенный в базальной части антенны
самца комара.
7-кутикула тимпанальной мембраны; 2-гиподермальная клетка;
3-клетка, прилежащая к гиподерме, и 4-ядро этой клетки; 5-колпачок
обкладочной клетки; б-дендриты; 7-обкладочная клетка;
5-перикарион; 9-аксон; 10-антенна; 77-нерв чувствительной клетки;
72-волоски; 13-нервные клетки; 14-внутреннее и 75-наружное кольца
сколопидия; 16-нервы; 77-базальная пластинка; 18-второй членик
антенны (ножка); 19-джонстонов орган.
162
6. Реакции на внешние воздействия и поведение
вратились в сложно устроенные слу-
ховые органы. В таких случаях рецеп-
торы, образующие несколько групп, раз-
личающихся по чувствительности,
воспринимают звуковые стимулы благо-
даря вибрации расположенной над ними
тимпанальной мембраны, это позволяет
насекомым улавливать широкий диапа-
зон звуков. Хордотональные органы не-
которых насекомых чрезвычайно чув-
ствительны к ультразвуковым волнам,
производимым летучими мышами, охо-
тящимися за этими насекомыми. Хордо-
тональные органы расположены под ку-
тикулой и обычно не выступают на
поверхности. Каждый орган, или сколо-
пидий, состоит из трех основных частей:
1) нейрона, 2) обкладочной клетки (ско-
лопалия), окутывающей дендриты ней-
рона, и 3) прикрепляющей, или шапочко-
вой, клетки (рис. 6.5,Л). Хордото-
нальные органы могут находиться в раз-
ных участках поверхности тела насеко-
мого, регистрируя движение опреде-
ленных частей тела. У личинки Drosophila
имеется 90 таких органов, каждый из ко-
торых состоит из 1-5 сколопидиев.
Джонстонов орган представляет со-
бой специализированный хордото-
нальный орган (рис. 6.5,Б), располо-
женный на втором членике антенны всех
взрослых насекомых. Этот орган встре-
чается также у многих личинок. У самцов
комаров эти проприоцепторы хорошо
развиты, и второй сегмент антенны в свя-
зи с этим увеличен. Функция джонсто-
нова органа состоит в регистрации дви-
жений флагеллума антенны. Поскольку
эти движения могут быть вызваны раз-
ными причинами, конкретные функции
джонстонова органа разнообразны он
может быть индикатором и контролле-
ром скорости полета, уловителем шума
или вспомогательным органом, уча-
ствующим в ориентации насекомого в
воде или по отношению к направлению
силы тяжести.
У видов Orthoptera, Hemiptera и Lepi-
doptera, где хордотональные органы раз-
вились в специализированные слуховые
органы, называемые тимпанальными ор-
ганами, они расположены на груди,
брюшке или ногах. Они реагируют на по-
явление хищников и воспринимают сиг-
Рис. 6.6. А. Сенсорные структуры (пульвиллы) на последнем членике лапки пчелиной
вши Braula соеса; х 100. Б. Пульвиллы (ножные подушечки) при большем увеличении,
х 500. (С разрешения А. Дитца, Университет Джорджии; из Dietz, Humphreys.)
6. Реакции на внешние воздействия и поведение
163
налы от других особей своего вида.
Жужжальца. У мух механорецепторы
входят также в состав уникального орга-
на равновесия, стабилизирующего полет.
В основании каждого из двух жужжалец,
представляющих собой модифициро-
ванные задние крылья, имеется более 400
механорецепторов, которые контроли-
руют поднятие насекомого в воздух и по-
ложение его в полете. Насекомое в поле-
те должно осуществлять коррекцию по-
ложения тела в трех плоскостях: гори-
зонтальной, продольной и поперечной. В
этом и состоит функция жужжалец.
Пульвиллы. Эти органы чувств
♦рис. 6.6) связаны с добыванием пищи.
Пульвиллы на лапках паразитической
пчелиной вши Braula соеса, по-видимому,
раздражают антенны их хозяев-пчел, что
приводит к раздвиганию мандибул по-
следних и позволяет паразиту получить
пищу из ротовой полости хозяина.
Рецепторы, улавливающие направле-
ве тока воздуха или воды. Ряд структур
на теле насекомых способен ощущать на-
правление потоков воздуха или воды,
проходящих вокруг тела. К таким рецеп-
торам относятся группы коротких воло-
сков на дорсальной поверхности головы
> Orthoptera, джонстонов орган, волоски
на различных участках тела. Воздушные
рецепторы воспринимают скорость и на-
правление потоков воздуха в непосред-
ственной близости от насекомого и регу-
лзгруют скорость полета.
Хеморецепторы
Хеморецепторные сенсиллы пред-
ставлены волосками, колышками, плас-
тинками и ямками. Они в больших ко-
личествах располагаются на разных ча-
стях тела, особенно на антеннах, но
также и на ротовых органах, лапках, яй-
цекладе и церках. Чрезвычайная чувстви-
тельность хеморецепторов позволяет на-
секомым обнаруживать наличие даже
нескольких молекул вещества в окру-
жающей среде. Обонятельные (ольфак-
торные) рецепторы, воспринимающие
запахи, состоят из большого числа сен-
сорных клеток, каждая из которых обыч-
но чувствительна к определенной группе
веществ. Реакция насекомого на присут-
ствие каких-либо веществ (как правило,
газов, но также и жидкостей) выражается
прежде всего в изменении ориентации те-
ла и в движении. Способность насекомых
чувствовать запахи помогает им нахо-
дить пищу и полового партнера. Вку-
совые рецепторы также служат для обна-
ружения пищи, полового партнера или
мест для откладки яиц. Чувство обоня-
ния развито у насекомых значительно
сильнее, чем чувство вкуса.
Рецепторы влажности. Это особая
группа хеморецепторов, чувствительных
к влажности воздуха. Присутствие воды
воспринимается некоторыми насекомы-
ми как химический стимул. Рецепторные
сенсиллы обычно представлены волоска-
ми или колышками на антеннах. У личи-
нок Drosophila эти рецепторы располо-
жены на вентральной поверхности перво-
го сегмента тела. У многих насекомых
волоски реагируют на изменение влаж-
ности воздуха скручиванием или
выпрямлением.
Т ерморецепторы
Насекомые - холоднокровные жи-
вотные, и хотя они реагируют на измене-
ние температуры, терморецепторов у них
обнаружено немного. У некоторых насе-
комых предполагается наличие терморе-
цепторов на антеннах или по всему телу.
По-видимому, терморецепторами явля-
ются волосковидные сенсиллы на антен-
нах клопа Rhodnius, а у Orthoptera-ряд
парных участков на голове, груди и
брюшке, отличающихся по структуре и
164
6. Реакции на внешние воздействия и поведение
виду от окружающей их гиподермы. В
различных группах Orthoptera эти термо-
рецепторные участки расположены на
разных частях тела. У жука Melanophila
(Buprestidae) сенсорные ямки на брюш-
ной стороне среднегруди воспринимают
инфракрасные лучи, что позволяет жу-
кам поселяться на недавно горевших де-
ревьях. Некоторые насекомые, паразити-
рующие на теплокровных животных, об-
наруживают своих хозяев по температур-
ному градиенту, создающемуся вокруг
тела хозяина.
Рецепторы звуков
Насекомые улавливают звуки с по-
мощью разнообразных органов и при-
способлений, расположенных на раз-
личных участках их тела. Все рецепторы
слуха образованы пучком нервных окон-
чаний, имеющих либо непосредствен-
ную, либо косвенную связь с кутикулой,
чаще с утонченным ее участком. Зву-
ковые волны вызывают вибрацию этого
участка кутикулы, которая передается
нервным окончаниям, и этот сигнал по-
ступает в нервную систему. В наиболее
распространенном случае пучок нервных
окончаний подходит к краю барабанной
перепонки (или тимпанальной мем-
браны), воспринимающей колебания воз-
духа (рис. 6.7,Л). Некоторые из тимпа-
нальных органов устроены очень сложно
(рис. 6.7,1), В). Такие сложно устроенные
органы имеются на передних ногах
Рис. 6.7. Звуковые рецепторы. (Michelsen, 1979.)
А. Схематическое изображение поперечного среза через слуховой орган кузнечика,
находящийся непосредственно под его коленом. Б. Схематическое изображение слухового
органа клопа-гладыша; вид снаружи. В. То же в разрезе. Значительная часть
тимпанального органа покрыта основанием кутикулярного образования, имеющего вид
молоточка, выступающего наружу. 7-тимпанальная полость; 2-чувствительные клетки;
3-ветви трахей; 4-голень; 5-тимпанальная мембрана; 6-основание; 7-молоточек;
8-рецепторные клетки; 9-воздушное пространство.
? Реакции на внешние воздействия и поведение
165
сверчков, на среднегруди некоторых кло-
пов. заднегруди ночных бабочек и на
5 рюшке некоторых других насекомых.
В мире животных слух хорошо развит
только у позвоночных и насекомых. Слух
С-Т^ткит для коммуникации особей внутри
слного вида, для информирования о на-
личии представителей других видов, осо-
г<нно хищников или конкурентов. Хотя
производят и воспринимают звуки мно-
гие насекомые, строение и функции слу-
ховых органов изучали в основном на
различных производящих громкие звуки
прямокрылых-кузнечиках и сверчках. У
тих насекомых (рис. 6.8,В) латеральные
~р)лные дыхальца ведут в переднегрудь,
п<е трахея каждой стороны образует
телыпой трахейный мешок и длинную
трубчатую трахею, доходящую до чув-
ствительного органа, расположенного в
плени передней ноги (рис. 6.7,Л).
Каждый трахейный мешок ограничен
тимпанальной мембраной, и оба этих
чешка отделены друг от друга другими
заутренними воздушными мешками (рис.
т ~Л). полностью не изолирующими
зз)к между двумя боковыми частями ор-
ана. Тимпанальная мембрана, большое
рахейное вздутие в переднегруди
и длинные трубчатые трахеи, проходя-
шие в переднюю ногу, играют роль уси-
лителя. или звуковой трубы. Некоторые
насекомые могут закрывать свои
родные дыхальца, у других они всегда
тттрыты, у третьих же обе стороны тра-
хейной системы соединены с помощью
пополнительных трахей. Таким образом,
• некоторых насекомых имеются си-
стемы. напоминающие по форме букву
И. с четырьмя входами для звука (два
пьсхальца и две тимпанальные мем-
'тины) (рис. 6.8,В), как в четырехфониче-
схом звуке. Если колебания волн состав-
ляют меньше 10000 вибраций в секунду,
т? такая система работает как детектор
розницы звуковых давлений, при более
же высоких частотах она воспринимает
само давление.
Существуют и другие структуры,
улавливающие звуки. Наиболее просто
устроены и, по-видимому, наименее чув-
ствительны среди них тонкие волоски на
антеннах (рис. 3.27, 6.8,Л), вибрирую-
щие от движения частиц воздуха, вызы-
ваемого звуковыми волнами. Вибрация
волосков позволяет некоторым гусени-
цам обнаруживать летящую осу, но толь-
ко при интенсивности звука выше 90 де-
цибел, что примерно соответствует уров-
ню звукового давления, создаваемого
летящей осой на расстоянии 1 см от на-
шего уха! У самцов комаров семейств
Chironomidae и Culicidae джонстонов ор-
ган улавливает жужжание летящих самок
своего вида.
Бражники подсемейства Choerocam-
pinae имеют рецептор ультразвука, чув-
ствительный к «радарным» ультразву-
ковым излучениям охотящихся летучих
мышей. Этот рецептор состоит из трех
отдельных образований. лабиальных
щупиков, небольшой лопасти, называе-
мой пилифером, находящейся между ос-
нованиями щупиков и хоботка, и хоботка
(рис. 6.8,Б). Эти три образования (они
искусственно отделены друг от друга в
нижней части рис. 6.8,Б) плотно приле-
гают друг к другу, как показано в верх-
ней части рис. 6.8,5*. Щупики крупные, с
тонкими стенками и находящимися вну-
три большими воздушными мешками,
каждый щупик своей срединной поверх-
ностью примыкает к гладкой, лишенной
волосков латеральной поверхности пили-
фера. Входящие ультразвуковые сигналы
усиливаются щупиками и передаются
при прямом контакте на пилифер, а за-
тем с помощью длинной щетинки на сре-
динной поверхности пилифера-на хобо-
ток (лабрум). Эти импульсы раздражают
лабиальный нерв, посылающий инфор-
мацию в головной мозг.
166
6. Реакции на внешние воздействия и поведение
Рис. 6.8. Звуковые рецепторы. (Из Michelsen, 1979.) А. Тонкая волосковидная
щетинка (ариста) на третьем членике антенны плодовой мушки направлена
латерально от апикального сочленения. Ариста служит приемником звуковых
вибраций, создаваемых призывной песней других особей своего вида. Б.
Схематическое изображение рецептора ультразвука (вид с дорсальной стороны) с
дистальной лопастью специального органа чувств (пилифера) у бражника Celerio
lineata. В. Схематическое изображение среза через передние ноги и переднегрудь
кустарникового сверчка, видна одна пара акустически изолированных слуховых
органов. 7-глаз; 2-ариста; 3-основная часть антенны* 4-щупик (в разрезе);
5-пилифер (орган чувств); б-щупик (отведенный); 7-отверстие слухового органа,
Я-воспринимающие звук чувствительные клетки.
6. Реакции на внешние воздействия и поведение
167
Фоторецепторы
Разнообразные типы фоторецепторов
позволяют насекомым различать форму
предметов, их взаиморасположение и
перемещение, расстояние до них, неко-
торые цвета, интенсивность освещения,
плоскость поляризации света, длину фо-
топериода и отличать свет от темноты.
Фоторецепторы насекомых восприни-
мают свет (одну из форм энергии) с по-
мощью пигмента, поглощающего свет
определенной длины волны, который
возбуждает чувствительные нервные
клетки. Фоторецепторы насекомых пред-
ставлены дермальными светочувстви-
тельными образованиями, латеральны-
ми и дорсальными глазками и сложными
глазами.
Наиболее примитивным типом фото-
рецепции является диффузная кожная
светочувствительность; эта чувствитель-
ность не связана с какими-либо спе-
циальными рецепторами, а реакция воз-
никает просто в результате попадания
света на определенные участки покровов
тела. Полагают, что кожная светочув-
ствительность обусловлена наличием
малых количеств пигмента, близкого к
пигменту фоторецепторов, поскольку
максимальная светочувствительность
тех и других наблюдается при длинах
волн 470-580 мкм (синяя-зеленая-жел-
тая части спектра).
Сложные глаза. Эти крупные органы
зрения, являющиеся основными фоторе-
цепторами у многих насекомых
(рис. 3.27), состоят из тысяч фоторецеп-
торных клеток. У большинства взрослых
насекомых имеется пара глаз, располо-
женных по бокам головы. Каждый глаз
состоит из множества длинных цилин-
дрических светочувствительных единиц-
омматидиев. Сложный глаз стрекозы со-
стоит из 10000-28000 омматидиев, а
комнатной мухи - примерно из 4000.
Каждый омматидий внешне выглядит
как шестигранная кутикулярная фасетка,
что придает сложным глазам их особый
вид.
Для каждого омматидия характерны
два главных компонента (рис. 6.9):
1. Оптический аппарат, собирающий
свет, состоит из наружной прозрачной
двояковыпуклой или плосковыпуклой
линзы (рис. 6.10), так называемого хру-
сталика, или роговицы, окруженного ку-
тикулой. Хрусталик образуется секретом
специальных клеток гиподермы. По-
скольку хрусталик является продолже-
нием кутикулы, он неподвижен. Под хру-
сталиком расположены кристаллический
конус (рис. 6.9., 6.11) и группа первичных
пигментных клеток, которые обычно
окружают кристаллический конус. Одна-
ко у примитивных насекомых, например
у обыкновенной чешуйницы, пигментные
клетки расположены не вокруг кристал-
лического конуса, а под хрусталиком.
2. Сенсорный рецепторный аппарат
находится под оптическим. Он состоит
из чувствительных, или ретинулярных,
клеток, собранных в группы по 6-12, на
центральной поверхности которых
имеется множество тончайших нейрофи-
брилл. Аксон от каждой ретинулярной
клетки проходит через базальную мем-
брану и направляется к головному мозгу.
Дендриты каждой группы ретинулярных
клеток образуют сложное светочувстви-
тельное палочковидное образование -
рабдом. Рабдом содержит зрительные
пигменты -родопсины, конфигурация мо-
лекул которых при попадании на них све-
та подвергается изменениям, приводя-
щим к изменению энергетического со-
стояния светочувствительных клеток.
Информация об этих изменениях пере-
дается через чувствительные нейроны в
мозг. Рабдом и группа ретинулярных
клеток окружены обычно вторичными
пигментными клетками, которые в той
168
6. Реакции на внешние воздействия и поведение
Рис. 6.9. Схематическое изображение сложного глаза и омматидия. (По Снодграссу.)
А. Вертикальный разрез через часть глаза. Б. Типичное строение омматидия.
7-фасетка омматидия; 2-глазная капсула, 3-корнеагенная клетка, 4-базальная
мембрана; 5-хрусталик (роговица); б-кристаллический конус; 7-пигментные клетки
8-рабдом, 9-ретикулярная клетка; 10-нервы.
или иной степени ограничивают попада-
ние света из прилежащих к ним оммати-
диев.
Свет, падающий на омматидий, фоку-
сируется хрусталиком, затем подводится
кристаллическим конусом к раб дому, где
вызывает изменения в зрительных пиг-
ментах. Переданная на нервные оконча-
ния информация об этих изменениях на-
правляется в головной мозг Каждый
омматидий сложного глаза функциони-
рует как одна фоторецепторная единица,
передающая сигнал головному мозгу
Сигналы, воспринятые отдельными ом-
матидиями и переданные в мозг, коорди-
нируются в нем, в результате чего у насе-
комого и создается изображение предме-
та.
Если интенсивность света очень силь-
но изменяется, характер процессов, про-
исходящих в сложном глазу насекомого,
может быть иным. У многих видов насе-
комых на ярком свету вторичный экрани-
рующий пигмент распространяется вниз
между омматидиями, и тогда на раб дом
попадает только свет, прошедший через
линзу своего омматидия. Получаемое в
этом случае изображение носит название
аппозиционного, или фотопического. Оно
обычно для дневных насекомых (рис.
6.12,Л) и эффективно при регистрации
движения, поскольку каждый омматидий
сканирует движущийся объект При сла-
бом свете вторичный экранирующий пиг-
мент сокращается, и проникающие в гла-
за лучи могут раздражать сразу несколь-
ко омматидиев. Такой тип получения
изображения носит название суперпози-
6. Реакции на внешние воздействия и поведение
169
Рис. 6.10 Наружная поверхность нескольких линз сложного глаза комара Aedes
aegypti. Поверхность каждой линзы имеет тончайший регулярный узор, образующий
структуру, не отражающую свет; х4500. (С разрешения кафедры энтомологии
Университета пров. Альберта и Д. Крейга.)
^ионного, или скотопического
рис. 6.12,Б), и встречается у ночных и
сумеречных насекомых. Суперпозицион-
ное зрение дает возможность улавливать
изменения в интенсивности света, но не
позволяет, по-видимому, различать
очертания предметов.
Хотя небольшое число фоторецепто-
ров в сложном глазу насекомого позво-
ляет получить лишь приближенное
изображение предметов, несравнимое,
например, с изображением, получаемым
человеческим глазом, где число фото-
рецепторов достигает 130 млн., насе-
комые обладают исключительной спо-
собностью различать мелькание предме-
тов. Глаз насекомого прекрасно реги-
стрирует изображения, сменяющиеся че-
рез очень короткие промежутки времени,
поскольку рабдомы быстро возвращают-
ся к исходному состоянию после получе-
ния каждого светового импульса. Мухи
170
6. Реакции на внешние воздействия и поведение
Рис. 6.11. Внутренняя часть сложного глаза проволочника (Elateridae); видны
кристаллические конусы, расположенные под хрусталиком; х 2200. (С разрешения
кафедры энтомологии Университета пров. Альберта и Д. Крейга.)
6. Реакции на внешние воздействия и поведение
171
Рис. 6.12. Схема, иллюстрирующая получение изображения в сложных глазах. (Из
Romoser, 1973, с изменениями.)
А. Аппозиционный тип сложного глаза. Б. Суперпозиционный тип; слева-
адаптация глаза к свету, справа -к темноте. 7-хрусталик, 2-кристаллический
конус; 3- пигментная клетка; 4 -раб дом; 5 - ретикулярная клетка.
улавливают до 265 мельканий в секунду,
тогда как человеческий глаз только
45-53, а обычно и того меньше-20-30.
Свет люминесцентной лампы, мигаю-
щий 60 раз в секунду, воспринимается че-
ловеком как непрерывный, а насеко-
мым-как последовательность мелька-
ний. Улавливание мельканий позволяет
насекомым замечать очень слабые дви-
жения в окружающей среде.
РЕЦЕПТОРЫ ЦВЕТА. Многие насе-
комые способны воспринимать зеленый
цвет и ультрафиолет. Вообще насекомые
наиболее чувствительны к ультрафиоле-
товой и сине-зеленой частям спектра. Не-
которые насекомые, например медо-
носные пчелы, способны воспринимать
голубой цвет. Рецепторы зеленого цвета
чаще всего встречаются в вентральной и
фронтальной частях глаза. Способность
улавливать ультрафиолет открывает на-
секомым совершенно иные, отличные от
нашего восприятия, качества окружаю-
щей среды. Поскольку разные цветки по-
разному отражают ультрафиолет, два
цветка, кажущиеся нам одинаково белы-
ми, для насекомого могут быть совсем
непохожими друг на друга. Рецепторы
красного цвета у насекомых обычно
отсутствуют.
ПЛОСКОСТЬ ПОЛЯРИЗАЦИИ. Дру-
гой особенностью сложных глаз насе-
комых является их способность воспри-
нимать и анализировать поляризо-
ванный свет. Электрический вектор в
случае поляризованного света колеблет-
172
6. Реакции на внешние воздействия и поведение
ся только в одной плоскости. Характер
поляризации дневного света изменяется
в зависимости от положения солнца, и
некоторые насекомые используют это
свойство для определения направления
полета. Так, например, медоносные
пчелы используют характер поляриза-
ции света для ориентировки при воз-
вращении в улей после сбора пыль-
цы.
Дорсальные и латеральные глазки.
Кроме сложных глаз у многих взрослых
насекомых имеются также фоторецеп-
торы, состоящие из одного хрусталика
(роговицы) и группы пигментных клеток,
под которыми лежат ретикулярные клет-
ки, образующие несколько раб домов.
Эти фоторецепторы именуются глазками
(ocelli). Многие взрослые насекомые
имеют дорсальные глазки, обычно в коли-
честве трех, расположенных в виде треу-
гольника на передней части головы
(рис. 3.10). Иногда их бывает только
один или два. Функция дорсальных глаз-
Рис. 6.13. Простой глаз гусеницы. (По
Снодграссу.) 7-хрусталик, или роговица,
2-корнеагенная клетка, 3-кутикула,
4 - гиподермальная клетка, 5 - раб дом;
6 - кристаллическое тело, 7 - ретикулярная
клетка.
ков состоит в улавливании изменений ин-
тейсивности света.
У личинок насекомых с полным пре-
вращением сложных глаз нет вообще, а
есть латеральные глазки, или стеммы.
Стеммы бывают самого разного строе-
ния. У гусениц они устроены как оммати-
дий-с хрусталиком, кристаллическим
конусом и группой ретикулярных клеток,
образующих один раб дом (рис. 6.13). С
помощью таких стемм гусеница разли-
чает форму предметов, цвет и движе-
ние. У других насекомых стеммы сход-
ны по строению с дорсальными глаз-
ками.
НЕРВНАЯ КООРДИНАЦИЯ
И ИНТЕГРАЦИЯ
С помощью нервной системы насе-
комые получают многообразную инфор-
мацию об условиях внешней и внутрен-
ней среды. Насекомое координирует по-
токи информации, приходящие из раз-
ных источников, и определенным обра-
зом реагирует на те или иные условия и
ситуации. Нервная система насекомых
включает головной мозг, продольную,
брюшную нервную цепочку и многочис-
ленные нейроны (см. стр. 106). Она под-
разделяется на центральную и стомо-
деальную'(висцеральную), или симпати-
ческую, с чувствительными, двига-
тельными и промежуточными нейрона-
ми, связанными синапсами. У комнатной
мухи более миллиона нейронов, причем
головной мозг и ганглии содержат
огромное число промежуточных нейро-
нов. Органы чувств, например сложные
глаза и механорецепторы, также содер-
жат большое число нейронов.
Нейроны, или нервные клетки. Ней-
рон, или нервная клетка,- это удлиненная
специализированная клетка, способная
получать и передавать стимулы (им-
пульсы). Нервный импульс проходит по
6. Реакции на внешние воздействия и поведение
173
Рис. 6.14. Схематическое изображение нейронов различных типов. (Из Wigglesworth, с
упрощениями.)
А, Б. Чувствительные нейроны. В. Двигательный нейрон. 7-перикарион; 2-ганглий,
3- чувствительный отросток, 4 -аксон; 5-ветвление аксона; б-рецепторная клетка;
7-мышца.
нейрону только в одном направлении.
Каждый нейрон (рис. 6.14) состоит из
трех основных частей: 1) тела клетки,
или перикариона', 2) одного или более чув-
ствительных отростков, или дендритов;
3) проводящего отростка, или аксона,
оканчивающегося разветвлением. По
крайней мере один отросток каждого
нейрона расположен в центральной нерв-
ной системе или в ганглиях (группах ней-
ронов, тесно связанных друг с другом).
Различают два основных типа нейронов.
У нейронов первого типа (рис. 6.14,А, Б)
один или несколько чувствительных от-
ростков отходят непосредственно от тела
клетки. Это чувствительные нейроны, у
которых дендриты связаны с органами
чувств, а аксон идет к центральной нерв-
ной системе и в ней заканчивается. У ней-
ронов другого типа (рис. 6.14,В) чувстви-
тельные отростки представляют собой
ветвь, отходящую от аксона и называе-
мую коллатеральной ветвью. К этому ти-
пу относятся: 1) моторные нейроны, у ко-
торых чувствительные отростки и тела
клеток расположены в нервной системе, а
аксоны формируют нервное волокно, ко-
торое подходит к мышце, и 2) промежу-
точные (вставочные) нейроны, которые
целиком находятся в пределах централь-
ной нервной системы.
Стимулы поступают на чувстви-
тельные отростки и в виде импульсов
следуют до окончаний аксонов. Это на-
правление следования стимулов необра-
тимо. Импульсы переходят от одной не-
174
6. Реакции на внешние воздействия и поведение
Рис. 6.15. Схема простой рефлекторной дуги, включающей чувствительный,
ассоциативный и двигательный нейроны. Направление передачи импульса показано
стрелками. (Из Wigglesworth, с упрощениями.)
7-мышца; 2 -двигательный нейрон; 3 -синапс; 4 - промежуточный нейрон;
5-ганглий; б-гиподерма* 7-чувствительный нейрон; 8-рецепторная клетка.
рвной клетки на другую через синапсы.
Синапсы представляют собой участки,
где происходит контакт между оконча-
ниями аксонов и дендритов двух нейро-
нов, причем контакты эти в большинстве
случаев не сплошные, а в них имеется
щель, называемая синаптической щелью.
Посредниками при передаче импульсов с
нервных окончаний одного нейрона на
нервные окончания другого нейрона че-
рез синаптическую щель служат спе-
циальные химические вещества-нейро-
медиаторы, чаще всего ацетилхолин. В
типичном случае (рис. 6.15) внешний
стимул вызывает определенное измене-
ние в воспринимающем органе, что в
свою очередь стимулирует чувстви-
тельный нейрон, от которого импульс че-
рез синапсы попадает на промежуточный
нейрон и от него на двигательный ней-
рон. По аксону двигательного нейрона
импульс доходит до мышечного волок-
на, которое отвечает на стимул сокраще-
нием.
Координация. В каждом сегменте тела
нервные клетки, группируясь друг с дру-
гом, формируют ганглий, входящий в со-
став центральной нервной системы. Та-
ким образом, по сенсорным нервам раз-
дражение передается к этим нервным
центрам, а ответ на это раздражение идет
к соответствующим эффекторам. Проме-
жуточные нейроны связывают один ган-
глий с другим и с головным мозгом. Они
могут также связывать один двига-
тельный нейрон с несколькими чувстви-
тельными клетками или несколько двига-
тельных нейронов с одной чувствитель-
ной клеткой. При такой системе связи
стимул, возникший в одном месте, мо-
жет вызвать ответ в различных частях те-
ла. Так, прикосновение к одному церку
таракана вызывает возбуждение в по-
следнем брюшном ганглии, которое,
передаваясь в грудной ганглий и затем в
двигательные нейроны ног, стимулирует
таракана к бегству.
6. Реакции на внешние воздействия и поведение
175
ЭНДОКРИННАЯ СИСТЕМА
И ГОРМОНЫ
Физиологические и поведенческие ре-
акции насекомого контролируются нерв-
ной и эндокринной системами. Быстрые
реакции насекомого на конкретные сти-
мулы, например двигательные реакции,
находятся под непосредственным кон-
тролем нервной системы. Напротив, дли-
тельно протекающие изменения, свя-
занные с развитием, ростом, размноже-
нием, метаболизмом, находятся под кон-
тролем эндокринной системы. Наряду с
многими физиологическими процессами,
описанными в гл. 7, эндокринная систе-
ма регулирует и поведенческие реакции.
В свою очередь эндокринная система и
все процессы, связанные с поступлением
информации из внешней среды (через ор-
ганы чувств), контролируются головным
мозгом насекомого. Эндокринная систе-
ма состоит из желез и специальных кле-
ток, секретирующих гормоны. Гормоны -
это химические посредники, действую-
щие в очень слабых концентрациях и
оказывающие влияние на физиологиче-
ские и (или) поведенческие реакции насе-
комого. Гормоны очень устойчивы, а их
транспорт от мест образования к разным
частям тела насекомого осуществляется
или путем диффузии, или с циркулирую-
щими жидкостями тела. По сравнению с
нейронами гормоны лучше приспосо-
блены к выполнению своих функций,
поскольку:
1) мгновенность реакции организма
несущественна;
2) они обеспечивают непрерывное по-
ступление стимула к определенному ме-
сту в течение нескольких часов или дней;
3) многоклеточные ткани, например
гиподерма, более доступны для стиму-
лов. поступающих с циркулирующими
жидкостями, чем по нервным путям.
Источниками основных гормонов на-
секомых являются нейросекреторные
клетки головного мозга, кардиальные те-
ла, нейросекреторные клетки субэзофа-
геального (подглоточного) ганглия, при-
лежащие тела, вентральные железы, пе-
рикардиальные железы, проторакальные
железы, эноциты, нейросекреторные
клетки брюшной нервной цепочки и co-
rpus luteum (рис. 6.16). В головном мозгу
насекомых имеются две парные группы
нейросекреторных клеток (медиальная и
латеральная), а у некоторых насекомых
еще и небольшая третья - задняя группа,
располагающаяся в протоцеребруме.
По характеру влияния на поведение
насекомых гормоны могут быть разде-
лены на три группы: 1) гормоны, обла-
дающие модифицирующим действием
(модификаторы); 2) гормоны, обладаю-
щие «запускающим» действием (рели-
зеры), и 3) гормоны, обладающие и тем
и другим действием одновременно. Мо-
дифицирующий эффект гормона-это
медленный процесс, приводящий в ре-
зультате к изменению характера реакции
нервной системы на какой-то стимул. На
любой стимул насекомое реагирует опре-
деленной поведенческой реакцией, но,
после того как оно подвергнется дей-
ствию модифицирующего гормона, тот
же самый стимул вызывает другую реак-
цию. Запускающий эффект гормона обус-
ловливает непосредственную и быструю
поведенческую реакцию в ответ на его
действие.
В качестве примера модифицирующего
действия гормона, вызывающего измене-
ния в поведении насекомого, можно при-
вести влияние ювенильного гормона на
реакцию самки кобылки Gomphocerus ги-
fus, вызванную ухаживанием самца. Сра-
зу после имагинальной линьки самка от-
вергает приближающегося самца. Воз-
действие ювенильного гормона, обра-
зующегося в прилежащих телах (corpora
altata), приводит к тому, что защитная
176
6. Реакции на внешние воздействия и поведение
Рис. 6.16. Схема, на которой указаны места
выделения основных гормонов у насекомых.
(Из Novak, 1975, с изменениями.)
7-желтое тело (corpus luteum); 2-эноциты;
3-перикардиальные железы; 4-прилежащие
тела (corpora allata); 5 - нейросекреторные
клетки подглоточного (субэзофагеального
ганглия; б-нейросекреторные клетки
головного мозга, 7 - кардиальные тела
(corpora cardiaca), 8 - вентральные железы,
9 - проторакальные железы;
10-нейросекреторные клетки вентральной
нервной цепочки.
Запускающий эффект гормона со-
стоит в стимуляции быстрой специфиче-
ской поведенческой реакции. У шелко-
прядов и многих других насекомых гор-
мон вылупления действует как гормон-
релизер, поскольку он непосредственно
вызывает поведенческие реакции, пред-
шествующие вылуплению. Имагиналь-
ная линька шелкопрядов происходит в
определенное время дня и синхронизиро-
вана с определенным фотопериодом. Фо-
топериод стимулирует секрецию гормо-
на вылупления нейросекреторными клет-
ками головного мозга. Для фаратных
шелкопрядов характерно предлиночное
поведение, контролируемое гормоном
вылупления, которое состоит в много-
кратных стереотипных движениях брюш-
ка, приводящих к постепенному выходу
из кокона.
Другой пример действия запускающе-
го гормона на поведение мы находим у
личинки саранчи Schistocerca gregaria.
Линочный гормон экдизон, стимулирую-
щий линьку и последующее отложение
кутикулы, делает также личинку неактив-
ной в тот период, когда она очень уязви-
ма для хищников.
КОММУНИКАЦИЯ
Коммуникация включает два процес-
са: 1) передачу сигнала особью и 2) при-
нятие сигнала одной или несколькими
особями. Результатом успешной переда-
чи сигнала может быть изменение пове-
дения получивших этот сигнал особей.
Такое изменение иногда нельзя обнару-
жить непосредственно. Ниже мы рассмо-
трим три типа коммуникации зритель-
ную, слуховую и химическую.
реакция самок исчезает. Теперь самка от-
вечает стрекотанием на приближение
самца, позволяет ему делать садку и ко-
пулирует с ним.
Зрительная коммуникация
Сотни лет люди наблюдали удиви-
тельные видимые излучения растений.
6. Реакции на внешние воздействия и поведение
177
животных и микробов. Биолюминесцен-
ция известна в нескольких отрядах насе-
комых, в том числе у Collembola (ного-
хвосток) и Coleoptera (жуков). Наиболь-
шей известностью пользуются жуки се-
мейств Lampyridae (светляки) и Elateridae
(жуки-щелкуны). У светляков, например
у хорошо изученного Photinus pyralis,
обычного в восточной части США, све-
тообразующий орган находится в брюш-
ке, и свечение служит для обнаружения
полового партнера. Частота излучения
света различна у разных видов насе-
комых, и даже в областях, где встречает-
ся несколько видов светляков, характер
излучения видоспецифичен и является
средством коммуникации, позволяющим
особям одного и того «же вида узнавать
друг друга.
Яркие узорчатые крылья некоторых
бабочек используются как зрительные
сигналы при различных формах полово-
го поведения. У бабочки-данаиды Danaus
gilippus Berenice, встречающейся в южной
Флориде, ухаживание начинается с пре-
следования в полете. При этом самцы ру-
ководствуются зрением и привлекаются
комбинацией движений, цветом и очерта-
ниями тела самки.
Зрительная коммуникация играет у
насекомых важную роль в защите от вра-
гов. Характер окраски, цвет и очертания
тела насекомых часто служат предосте-
регающим сигналом для хищников. Спо-
собность насекомых имитировать не
только предметы окружающей среды, но
и других насекомых (известная под на-
званием мимикрия, о которой пойдет
речь в гл. 10) является одним из важных
защитных механизмов. Некоторые севе-
роамериканские кузнечики и бабочки
имитируют форму тела и(или) окраску
общественных ос, а жуки семейства Се-
rambycidae имитируют муравьев, пчел и
ос.
Слуховая коммуникация
Насекомые производят множество
разнообразных звуков и в большинстве
случаев на какой-то стадии развития
именно с помощью звуков осуществляет-
ся связь между особями. У некоторых на-
секомых звук создается в процессе их
нормальной жизнедеятельности без по-
мощи каких-либо специальных звукоиз-
дающих структур. Наиболее известный
пример такого рода - это жужжание летя-
щего или парящего насекомого. Жужжа-
ние создается в результате очень бы-
строй вибрации крыльев и склеритов
груди. У некоторых насекомых, напри-
мер у комаров, звуки, возникающие от
биения крыльев летящей самки, вызы-
вают у самца реакцию спаривания. У
других насекомых такие звуки не служат
средством коммуникации.
Исходя из особенностей человеческо-
го слуха, трудно оценить качество зву-
ков, производимых насекомыми. Про-
должительный звук, например звук от
вибрации крыльев, менее обычен, чем ди-
скретные звуковые пульсы, разделенные
периодами тишины. Насекомые спо-
собны улавливать более широкий диапа-
зон частот, особенно в области высоких
частот, чем человеческое ухо, восприни-
мающее звуки от 60 до 16 000 циклов в
секунду (ц/с). Люди не слышат биения
крыльев бабочек (отряд Lepidoptera), ча-
стота которого около 20 ц/с. Частота
биения крыльев пчелы Apis составляет
около 250 ц/с. Шум летящей саранчи
Schistocerca создается звуком с частотой
от 60 до 6400 ц/с, но в основном в диапа-
зоне 3200-5000 ц/с. Частота звука, со-
здаваемая биением головы термита
(Isoptera) по древесине, около 1000 ц/с. У
различных видов сверчков (Orthoptera) в
результате трения крыльев (элитральная
стридуляция) создаются звуки с боль-
шим диапазоном частот - от
12-732
178
6. Реакции на внешние воздействия и поведение
2000-10000 ц/с до очень высоких, поряд-
ка 50000-100000 ц/с. Стрекотание сверч-
ка имеет характерную для него постоян-
ную частоту и амплитуду и представляет
собой очень чистый звук. Частота звука,
создаваемого тимбалами цикад (Hemip-
tera), равна примерно 4500 ц/с.
Помимо звуков, издаваемых при про-
явлении нормальной жизнедеятельности
насекомого, существуют другие, про-
изводимые иными способами:
1) ударение какой-либо частью тела
по субстрату;
2) фрикционное трение двух частей
тела друг о друга (обычно назы-
вается стридуляцией);
3) вибрация мембран;
4) выталкивание струй воздуха.
Среди множества видов насекомых
встречаются многочисленные вариации
этих способов.
1. При издавании звуков посред-
ством удара о субстрат насекомые поль-
зуются различными частями тела.
Кобылка Oedipoda ударяет по поверхно-
сти субстрата задней голенью. Самец де-
лает примерно 12 ударов в секунду, сам-
ка несколько меньше. Жук Xestobium
стучит по субстрату головой и произво-
дит своеобразную «очередь» хлопков.
Солдаты некоторых термитов совер-
шают колебательные движения, и при
этом голова и кончики мандибул уда-
ряют по древесине так, что получается
серия звуков, которую воспринимают
другие обитатели термитника (рабочие и
личинки). Другие касты термитов также
производят звуки при ударе головой по
древесине, но с более низкой интенсив-
ностью.
2. Звуки, возникающие от трения раз-
личных частей тела друг о друга, из-
вестны у многих насекомых. Этот способ
хорошо развит в отрядах Orthoptera, Не-
teroptera и Coleoptera. У некоторых пря-
мокрылых передний край заднего крыла
Рис. 6.17. Пилочка на внутренней
поверхности бедра задней ноги сверчка. (Из
Comstock, An introduction to entomology; с
разрешения The Comstock Publishing Co.)
А. Заднее бедро Stenobothrus. Б. Пилочка,
сильно увеличено.
ударяет по утолщенным жилкам перед-
него, вызывая вибрацию последнего; в
результате такой вибрации и возникают
трескучие звуки. У некоторых саран-
човых на внутренней поверхности задне-
го бедра имеется ряд маленьких зубчиков
(рис. 6.17), которые трутся о переднее
крыло, производя таким образом звук. У
различных сверчков на одной из элитр
имеется ряд зубчиков (щетка), а на дру-
гой-гребень (скребок). При соприкосно-
вении элитр зубчики трутся о гребень,
элитры вибрируют, и возникает звук
(рис. 6.18). У других насекомых, напри-
мер у жуков, гребень и щетка располо-
жены соответственно на ноге и на теле
(рис. 6.19). Для генерации звуков жуки
используют самые различные части свое-
го склеротизированного экзоскелета, но
особенно элитры.
3. Вибрирующие мембраны, или тим-
бальный механизм, обнаружен у неко-
торых равнокрылых, в частности у ци-
кад, у других полужесткокрылых и у
бабочек. Наиболее интересное приспосо-
бление развилось у цикад (рис. 6.20). У
этих насекомых у основания брюшка
имеются карманы, или полости, в ко-
торых располагается несколько мем-
6. Реакции на внешние воздействия и поведение
179
Рис. 6.18. Переднее крыло сверчка Gryllus с пилочкой и скребком. (Из
Comstock, 1936, An introduction to entomology, с разрешения The Comstock
Publishing Co.)
А. Вид сверху; часть сложенного крыла, прилегающая к брюшку, не видна.
Б. Основание крыла, вид снизу. В. Пилочка, сильно увеличено. 1 - скребок;
2-мембрана; 3-пилочка.
Рзс. 6.19. Стридуляционный орган личинки
?^jja/zz5. (Из Comstock по Sharp.)
. 2-части заднегруди; 3-тазик второй
дсса: -/-пилочка; 5-базальная часть бедра
годней ноги; 6 - волоски с хитиновыми
зьгростами у их основания; 7- уменьшенная
задняя нога, превращенная в скребок. '
бран. Внутренняя поверхность одной из
этих мембран соединена с мышечным во-
локном. Сокращение этого волокна втя-
гивает мембрану внутрь, а расслабление
мышцы позволяет ей вернуться в исход-
ное положение. Эти движения мембраны
происходят с большой скоростью и со-
здают звуковые волны. Остальные мем-
браны функционируют в качестве ре-
флекторов звука.
Рис. 6.20. Музыкальный аппарат цикады.
(По Carlet.)
1 -крышечка (на другой стороне крышечка
удалена); 2 - основание ноги, 3 - складчатая
мембрана, 4 -дыхальце; 5 - вибрирующая
мембрана (тимбал); б-латеральная полость;
7- вентральная полость; 8-зеркальце.
180
6. Реакции на внешние воздействия и поведение
4. Еще одним способом образования
звука у насекомых является проталкива-
ние воздуха через или над маленьким от-
верстием. У бражника Acherontia atropos
звук возникает в результате втягивания и
выталкивания воздуха через хоботок и
создания воздушных струй над эпифа-
ринксом (надглоточником).
Звуки могут служить сигналами, вос-
принимаемыми другими видами (межви-
довые сигналы) или другими особями то-
го же вида (внутривидовые). Неупорядо-
ченные звуки без регулярно повторяю-
щихся пульсов служат, по-видимому, для
защиты или предупреждения и могут
быть как меж-, так и внутривидовыми.
Очевидно, к такому типу звуков относит-
ся стридуляция у самок и самцов неко-
торых Heteroptera, жуков и куколок ба-
бочек. Эти реакции тревоги сопровож-
даются иногда другими защитными ре-
акциями, такими, как шипение, раздаю-
щееся в тот момент, когда бабочка па-
влиноглазка раскрывает крылья, чтобы
продемонстрировать яркие глазчатые
пятна.
Упорядоченные звуки, повторяющие-
ся с определенной частотой, наиболее ха-
рактерны для внутривидовых коммуни-
каций. Как правило, это связано с про-
цессом ухаживания. У Orthoptera разли-
чают пять типов звуковых сигналов,
означающих призыв, ухаживание, копу-
ляцию, нападение и тревогу. Каждый из
этих типов отличается по частоте и ха-
рактеру пульсов. У ряда насекомых, на-
пример цикад, определенные звуки вызы-
вают агрегацию особей. Самцы и самки
одного вида собираются в каком-то
определенном месте, например в кроне
тех или иных деревьев. Пение некоторых
цикад обеспечивает половую изоляцию
видов. В Северной Америке три вида ро-
да Magicicada занимают одно и то же ме-
стообитание. Однако особи каждого ви-
да идентифицируют особей своего и чу-
жого видов по пению. Кроме того, раз-
ные виды поют хором в разное время
дня.
У полу общественных и общественных
насекомых звуки служат средством раз-
личения особей своего вида и своей коло-
нии. Шершни производят в своих гнездах
целый ряд различных звуков-сигналов.
Роение пчел отчасти стимулируется про-
должительным жужжанием, произво-
димым несколькими рабочими особями,
возбужденно шныряющими по улью. У
общественных насекомых звуки служат
также сигналами для поисков пищи и по-
стройки гнезда.
У общественных насекомых звук
является важнейшим средством комму-
никации, однако он не исключает других
средств коммуникации, например хими-
ческих или тактильных. Взаимодействие
и интеграция различных типов коммуни-
кации хорошо проявляются у классиче-
ского общественного насекомого-медо-
носной пчелы Apis mellifera. В танце пчел,
считавшемся когда-то проявлением зри-
тельной коммуникации, обнаружен
целый комплекс сигналов, включающих
и звук, и проприоцепцию. Какой из типов
коммуникации играет ведущую роль в
танце, до сих пор не установлено. Танец
представляет собой ряд специфических
стереотипных движений, совершаемых
пчелой-сборщицей внутри улья на верти-
кальной поверхности сот. Пчела-сборщи-
ца, отыскавшая источник корма, при-
носит в улей пыльцу и нектар и затем
снова вылетает к этому источнику. Воз-
вращаясь второй раз, она совершает та-
нец в месте скопления большого числа
рабочих пчел, причем характер танца ме-
няется в зависимости от расстояния до
источника пищи. Если он находится
близко от улья, то выполняется круговой
танец (рис. 6.21,А). Пчела-сборщица
движется по кругу, меняя направление
движения. Считается, что такой танец оз-
6. Реакции на внешние воздействия и поведение
181
Рис. 6.21. Танцы медоносной пчелы Apis
mellifera. (Данные взяты из von Frisch, 1967.)
А. Круговой танец пчелы-сборщицы,
информирующий о близости источника
корма к улью. Б. «Виляющий» танец
пчелы-сборщицы, информирующий об
удаленности источника корма от улья.
Рис. 6.22. Направление, в котором расположен удаленный источник корма,
чтазывается «виляющим» танцем медоносной пчелы. Угол между направлением
пробега с «вилянием» и вертикалью равен, по-видимому, углу между направлениями к
солнцу и к источнику корма, спроектированному на горизонтальную плоскость. (Из
Brown, 1975.)
182
6. Реакции на внешние воздействия и поведение
начает лишь только то, что источник пи-
щи недалеко, но он не несет информации
о направлении полета и расстоянии до
него. Если источник пищи находится на
среднем удалении от улья, то выполняет-
ся серпообразный танец (открытая «вось-
мерка»), при этом круговой танец посте-
пенно переходит в движение по открытой
«восьмерке». В том случае, если пища да-
леко, выполняется виляющий танец
(рис. 6.21,5). В этом танце пчела, дви-
гаясь по фигуре в виде открытой «вось-
мерки», совершает быстрые виляющие
движения брюшком во время прямой ча-
сти своего пробега. Полагают, что имен-
но эта часть танца дает информацию о
расстоянии до источника корма и напра-
влении к нему. Жужжание, производимое
пчелой-сборщицей во время пробега по
прямой, его ритм и суммарная продол-
жительность звукового сигнала, возмож-
но, используются другими пчелами в
улье для установления расстояния до ис-
точника корма. Вероятно, что эта инфор-
мация дополняется информацией, пред-
ставленной числом виляний брюшка во
время прямого пробега. Угол между на-
правлением прямого пробега пчелы-
сборщицы и проведенной к этой линии
вертикалью соответствует углу между
двумя линиями, проведенными от улья к
солнцу и к источнику корма (рис. 6.22).
Пробег по прямой, направленной верти-
кально вверх, указывает на то, что источ-
ник пищи находится в том же направле-
нии, что и солнце; если прямая направле-
на вертикально вниз, то, следовательно,
источник корма находится в направле-
нии, противоположном солнцу.
Химическая коммуникация
Химические сигналы, включающие
вкусовые раздражения и запахи, являют-
ся важнейшим средством коммуникации
у насекомых. Они охватывают большой
круг химических веществ и широко рас-
пространены на всех стадиях жизненного
цикла у обоих полов. Ни одна другая
группа животных не обладает столь
большим числом и, возможно, столь же
совершенной системой химических сиг-
налов, как насекомые. Вещества, выде-
ляемые насекомыми во внешнюю среду,
образуются в специальных железах, на-
зываемых экзокринными, к которым, на-
пример, относятся мандибулярные или
ядовитые железы. Вещества, образуемые
в экзокринных железах, являются важны-
ми средствами химической коммуника-
ции, особенно у общественных насе-
комых. Химические вещества, обусло-
вливающие те или иные поведенческие
реакции, называются сигнальными веще-
ствами (semiochemicals). Среди них раз-
личают две группы веществ, феромоны,
передающие сигналы особям своего ви-
да, и вещества (allelochemics), служащие
сигналами для особей других видов (ал-
лелохимическое взаимодействие).
ФЕРОМОНЫ
Эти вещества вызывают ответную ре-
акцию у особи того же вида. Они пред-
ставляют собой соединения с низким мо-
лекулярным весом (от 100 до 300). Разли-
чают две функциональные группы феро-
монов-релизеры и праймеры. Феро-
моны-релизеры действуют на нервную
систему реципиента и вызывают немед-
ленную поведенческую реакцию. Они
широко распространены у насекомых и
выполняют роль половых аттрактантов,
следовых сигналов и сигналов опасности.
Они опосредуют также некоторые фор-
мы поведенческих реакций, а именно
половое поведение, образование скопле-
ний, распределение особей в простран-
стве при высокой плотности популяции и
узнавание особей своего вида. Феромоны
вызывают также общие изменения в
6. Реакции на внешние воздействия и поведение
183
ориентации и движении насекомого. У
большинства видов рецептивная самка
продуцирует летучий феромон, стиму-
лирующий самца своего вида к поискам
этой самки. У потревоженной тли из со-
ковых трубочек выделяется феромон, вы-
зывающий реакцию тревоги и спасения у
других членов колонии - они немедленно
высвобождают свой хоботок и падают на
землю. У некоторых видов пилильщиков
потревоженная личинка выделяет лету-
чее вещество, вызывающее синхронные
защитные движения у всех личинок, на-
ходящихся на том же листе.
Феромоны-праймеры вызывают более
медленные реакции, чем феромоны-рели-
зеры, и действуют на эндокринную и ре-
продуктивную системы реципиента, вы-
зывая физиологические изменения, ко-
торые либо тормозят, либо возбуждают
соответствующие поведенческие реак-
ции. Например, самцы перелетной саран-
чи выделяют летучее вещество, повы-
шающее скорость роста и вызывающее
синхронное развитие молодых насе-
комых. Это приводит к синхронной линь-
ке огромного числа особей и образова-
нию обширных стай перелетной саранчи.
Характер феромонных связей. По-
ловые феромоны, стимулирующие поло-
вую активность, охватывают огромное
число химических веществ. Они синтези-
руются и у самок, и у самцов, но выде-
ляют их во внешнюю среду, как правило,
гамки. У некоторых жуков особи обоего
л ода выделяют феромон, который слу-
жит для привлечения и скопления насе-
комых на растении-хозяине, где и проис-
ходит спаривание.
ОБРАЗОВАНИЕ СКОПЛЕНИЙ. Обра-
зование скоплений как реакция на феро-
мон весьма типично для многих насе-
комых-это обусловливает групповую
активность насекомых при добывании
лиши, спаривании, зимовке. За исключе-
нием общественных насекомых, такая
групповая активность носит обычно вре-
менный характер, например образование
роев для спаривания у поденок.
Огромные скопления особей создаются в
колониях общественных насекомых, ког-
да наступает время оплодотворения сам-
ки. Матки в колониях огненных муравьев
и термитов собирают вокруг себя сотни
рабочих особей, привлеченных ее феро-
монами.
У некоторых настоящих обще-
ственных насекомых (эусоциальных) -
муравьев, термитов и некоторых пчел-
рабочие-фуражиры выделяют следовые
феромоны, вероятно, чтобы облегчить
возвращение в свое гнездо. Рабочая
особь, нашедшая богатый источник пи-
щи, возвращается в колонию и мобили-
зует других рабочих с помощью выде-
ляемых ею феромонов, а иногда и других
специальных поведенческих реакций. Мо-
билизация представляет собой одно из
средств коммуникации, с помощью кото-
рого можно собрать рабочих особей для
выполнения определенной работы. Этот
способ коммуникации успешно исполь-
зуют и осы.
ПОДДЕРЖАНИЕ КАСТОВОЙ СТРУК-
ТУРЫ СООБЩЕСТВА. У маток муравьев,
пчел и термитов в мандибулярных желе-
зах образуется так называемое «маточ-
ное вещество», тормозящее развитие
яичников у представителей других каст, а
у медоносных пчел также тормозящее
постройку маточных ячеек. Этот феро-
мон попадает в организм рабочих особей
при слизывании. У пчел, ос и термитов
определенную роль в детерминации каст
играют и другие феромоны.
ПРЕДУПРЕЖДЕНИЕ И ТРЕВОГА. Фе-
ромоны предупреждения и тревоги обра-
зуются в ответ на непосредственную или
потенциальную угрозу Поведенческие
реакции, возникающие при этом, носят
самый разнообразный характер-защи-
та, избегание опасности, возбуждение,
184
6. Реакции на внешние воздействия и поведение
образование скоплений, мобилизация.
Наиболее развиты химические системы
образования феромонов предупреждения
и тревоги у общественных насекомых,
особенно у видов, образующих большие
колонии. У агрессивных насекомых фе-
ромоны тревоги заставляют их собрать-
ся вместе около вторгшегося в колонию
постороннего насекомого и стараться из-
гнать его из колонии. У более спокойных
насекомых в ответ на феромон тревоги
возникает реакция рассеяния, когда
члены колонии покидают гнездо, заби-
рая с собой матку, личинок, куколок и
молодых имаго.
ПРОСТРАНСТВЕННОЕ РАСПРЕДЕ-
ЛЕНИЕ. При высокой плотности популя-
ции как у личинок, так и у взрослых насе-
комых особи в ограниченном простран-
стве распределяются исключительно рав-
номерно. Такое распределение устана-
вливается благодаря секреции спе-
циальных феромонов, которые дей-
ствуют как репелленты.
ИДЕНТИФИКАЦИЯ. Узнавание осо-
бей своего вида необходимо для успеш-
ного спаривания-одного из важных фак-
торов благополучного существования и
распространения вида. Для насекомых,
образующих стаи, а также для обще-
ственных насекомых система безошибоч-
ного узнавания особей своего вида имеет
чрезвычайно важное значение. При этом
химические сигналы (феромоны) воспри-
нимаются обычно обонятельными или
тактильными рецепторами на антенных.
Посторонняя девственная матка, введен-
ная в колонию медоносных пчел с имею-
щейся в ней оплодотворенной яйцекладу-
щей маткой, будет немедленно атакована
и убита членами колонии. Даже соб-
ственные рабочие особи, которые могли
«загрязниться» феромонами этой чужой
самки при контакте с ней, также будут
убиты.
НАПАДЕНИЕ НА ЧУЖИЕ ГНЕЗДА.
Многие муравьи нападают на гнезда
других видов и забирают их потомство.
Когда захваченные особи достигнут зре-
лости в гнезде захватчика, они выпол-
няют функцию собирания пищи для ко-
лонии. Во время нападения на чужую
колонию муравьи-захватчики выделяют
летучий феромон, обезоруживающий
жертвы нападения.
АЛЛЕЛОХИМИЧЕСКИЕ
ВЗАИМОДЕЙСТВИЯ
Аллелохимические взаимодействия
осуществляются с помощью веществ,
действующих на особей других видов.
Среди этих веществ различают алломоны
и кайромоны.
Алломоны. Эти вещества оказывают
вредное действие на особей другого вида
и выполняют прежде всего защитную
функцию. Сюда относятся неприятно
пахнущие выделения преимагинальных и
взрослых клопов-щитников (семейство
Pentatomidae) и других Hemiptera,
взрослых златоглазок и многих других
насекомых. Алломон взрослых жуков-
бомбардиров (Brachinus) выделяется в
форме горячей (с температурой 100°С)
струи хинона. Некоторые алломоны со-
держат цианиды.
Многие насекомые обладают удиви-
тельной способностью направлять свои
защитные выделения на хищника. Обыч-
но это сопровождается или изгибанием
всего тела, или поворачиванием приспо-
собления для выпускания жидкости так,
чтобы выпускаемая струя алломона бы-
ла послана в том направлении, откуда
пришел раздражающий стимул. Сейчас
известно более 500 защитных веществ,
большая часть которых представляет со-
бой органические кислоты, спирты, аль-
дегиды и парахиноны.
Кайромоны. Как и алломоны, эти ве-
щества действуют на особей другого ви-
6. Реакции на внешние воздействия и поведение
185
да, но в отличие от алломонов они по-
лезны для реципиента, а не для донора.
Сюда относятся разнообразные аро-
матные и другие вещества, образующие-
ся у растений, которые привлекают к ним
насекомых-фитофагов. Хищные насе-
комые и насекомые-паразиты других жи-
вотных (в том числе и других насекомых)
благодаря этим веществам не только от-
ыскивают свои жертвы или своих хозяев,
но и приспосабливаются к их жизненным
циклам. Так, многие взрослые насеко-
мые-паразиты находят свою жертву, во-
спринимая издаваемые ею запахи, и за-
тем откладывают яйца на поверхность
или внутрь ее тела.
В ряде случаев одцо и то же вещество
может выполнять двоякую сигнальную
функцию. Например, половой феромон
жука-короеда из рода Ips одновременно
является и кайромоном, привлекающим
других жуков-хищников, охотящихся за
этими короедами. Феромон, регулирую-
щий численность популяции мельничной
огневки Anagasta kueniella, действует
одновременно и как кайромон, облег-
чающий одному из перепончатокрылых
паразитов этой бабочки находить ее.
ПОВЕДЕНЧЕСКИЕ РЕАКЦИИ
И ТИПЫ ПОВЕДЕНИЯ
На протяжении жизненного цикла на-
секомого его поведенческие реакции обы-
чно изменяются, что связано по меньшей
мере с тремя факторами. Некоторые из-
менения вызваны генетическими факто-
рами, обусловливающими развитие и со-
зревание, и называются стереотипными,
или врожденными, поведенческими реак-
циями. Другие изменения в поведении
возникают в результате действия раз-
личных факторов внешней среды, изме-
няющих стереотипный характер поведе-
ния в результате приобретения опреде-
ленного опыта; в этом случае мы имеем
дело с приобретенными поведенческими
реакциями. Очень часто обе эти катего-
рии поведенческих реакций трудно отде-
лить одну от другой, поскольку измене-
ния поведения являются результатом
сложных взаимодействий генетических и
средовых факторов.
Стереотипные
поведенческие реакции
На каждый конкретный стимул насе-
комые обычно отвечают одной или се-
рией характерных специфических реак-
ций. При первой «встрече» с каким-либо
определенным стимулом у насекомого
возникает спонтанный ответ-стереотип-
ная, или врожденная, поведенческая ре-
акция, которая не приобретается, а на-
следуется генетически. Такими стерео-
типными поведенческими реакциями
являются рефлексы, кинезы и таксисы.
Рефлексы. Рефлекс представляет со-
бой самую простую поведенческую реак-
цию организма на тот или иной стимул.
В качестве примера рефлекторной реак-
ции насекомых можно привести реакцию
спасения у сверчка. В ответ на прикосно-
вение к церкам, расположенным на конце
брюшка, сверчок совершает прыжок. В
этой реакции тактильные рецепторы цер-
ков посылают импульс по нервному во-
локну к одному из промежуточных ней-
ронов, расположенному в последнем
брюшном ганглии. Этот промежуточный
нейрон посылает импульс по брюшной
нервной цепочке к грудному ганглию, ко-
торый в свою очередь стимулирует не-
рвы, идущие к ногам, и, таким образом,
вызывает реакцию сверчка, выражаю-
щуюся в прыжке. У многих мух и бабо-
чек при стимуляции передних лапок рас-
твором сахара вытягивается хоботок.
Сама лапка при этом не двигается, но
расположенные на ней хеморецепторы
посылают импульс, проходящий через
186
6. Реакции на внешние воздействия и поведение
нервную клетку хеморецептора к грудно-
му ганглию, который стимулирует про-
межуточный нейрон. От него импульс по
брюшной нервной цепочке достигает
ганглия, который контролирует процесс
питания, в результате чего и раздражает-
ся двигательный нерв, управляющий вы-
тягиванием хоботка.
Путь, который проходит импульс в
простом рефлексе-от момента воздей-
ствия стимула на рецептор до ответной
реакции,-называется рефлекторной ду-
гой (рис. 6.15). Головной мозг не
включается в рефлекторную дугу. Не-
рвные импульсы следуют автоматически
за сигналом, поступающим из органов
чувств. Многие поведенческие реакции
насекомых относятся к рефлексам такого
типа. Рефлексы обычно подразделяются
на две функциональные группы. Фазиче-
ские рефлексы -это быстрые кратковре-
менные реакции, включающие быстрые
движения тела или какой-нибудь его ча-
сти. Примером может служить прыжок
сверчка, описанный выше. Тонические ре-
флексы, напротив, представляют собой
медленные длительные реакции, обеспе-
чивающие сохранение позы или положе-
ния тела насекомого по отношению к
внешним факторам.
Кинезы и таксисы. Непосредственные
реакции на стимулы, приводящие к на-
правленному движению или изменению
положения насекомого, называются ки-
незами и таксисами. Однако взаимодей-
ствие факторов внешней среды, особенно
стрессовых, может изменять эти реакции.
Кинезы, известные у многих насе-
комых,-это случайные движения насеко-
мого, не ориентированные по отноше-
нию к стимулу. Типичные кинезы можно
наблюдать у тараканов. Если нет спе-
циального раздражителя, то их передви-
жения беспорядочны. Повышение интен-
сивности освещения вызывает увеличе-
ние скорости движения тараканов. В
результате они движутся медленнее на
темных участках и быстрее на осве-
щенных, что приводит к их концентрации
на темных участках.
Таксисы-это движения, напра-
вленные в сторону стимула или от него.
Таксис отличается от кинеза тем, что у
животного на определенный стимул воз-
никает определенная стереотипная реак-
ция. Многие бабочки летят на свет-это
положительный фототаксис. Зрелые ли-
чинки многих видов мясных мух движут-
ся от света-это отрицательный фототак-
сис. Таксисы играют особенно важную
роль в правильной ориентации особей во
внешней среде для отыскания, например,
источника пищи или места для окуклива-
ния. Таксисы классифицируют по вызы-
вающим их стимулам: фототаксис-это
реакция на свет, геотаксис-на земное
притяжение, тигмотаксис-реакция на
прикосновение, хемотаксис - на химиче-
ские вещества и т д.
ФОТОТАКСИС-РЕАКЦИЯ НА СВЕТ
У большинства насекомых реакция на
свет хорошо развита, при этом они могут
стремиться как к источнику света, так и
от него. Пчелы и осы летят к свету, т е.
для них характерен положительный фо-
тотаксис. На разных стадиях своего жиз-
ненного цикла насекомые обычно по-раз-
ному реагируют на свет. Личинки ком-
натной мухи обладают отрицательным
фототаксисом и уходят от света,
а взрослые мухи характеризуются поло-
жительным фототаксисом и летят к све-
ту. Во время плавания некоторые водные
насекомые, например нимфы поденок,
сохраняют дорсовентральное положение
тела (правая сторона кверху), ориенти-
руясь на идущий сверху свет.
Некоторые насекомые реагируют не
только на свет вообще, но также и на
определенные длины световых волн. В
большинстве случаев это помогает при
отыскании пищи или (для яйцекладущих
6. Реакции на внешние воздействия и поведение
187
самок) подходящего типа листвы для от-
кладки яиц. Бабочки в поисках пищи ру-
ководствуются чувством цвета, отличая
желтый, красный и синий цвета от зеле-
ного, и подлетают к цветкам растений,
окрашенным в эти цвета, чаще, чем к зе-
леным листьям. Однако те же самые ба-
бочки откладывают яйца только на зеле-
ную поверхность, т е. на здоровые
листья, которыми в природных условиях
питаются их личинки.
ГЕОТАКСИС-РЕАКЦИЯ НА ЗЕМНОЕ
ПРИТЯЖЕНИЕ. Многие насекомые, по-
саженные в вертикально поставленную
пробирку, будут двигаться в ней не бес-
порядочно, а целенаправленно вверх
или вниз. Цикадки семейства Cicadellidae
всегда движутся вверх. Если трубку пере-
вернуть так, что насекомые окажутся на
дне, они снова начнут взбираться по
стенкам вверх. Это - отрицательная реак-
ция на земное притяжение, или отрица-
тельный геотаксис. Другие насекомые
проявляют положительный геотаксис,
двигаясь вниз, к земле. 'Такой тип реак-
ции наблюдается у многих личинок, жи-
вущих в почве, например у проволочни-
ков (личинок жуков-щелкунов). Если они
вылупляются из яиц, отложенных на по-
верхности или вблизи поверхности по-
чвы, то зарываются в землю.
ТИГМОТАКСИС-РЕАКЦИЯ НА КОН-
ТАКТНЫЕ СТИМУЛЫ. Многие насе-
комые, живущие под корой, в почве, в
скрученных листьях, обладают хорошо
развитой осязательной, или тактильной,
реакцией, что позволяет им сохранять
контакт с каким-либо объектом
(рис. 6.23). Такая реакция называется по-
ложительным тигмотаксисом. По-види-
мому, прикосновение к какому-либо
предмету или субстрату тормозит актив-
ность насекомого, вызывая временную
неподвижность.
У всех насекомых, ведущих активный
образ жизни, осязание служит для обна-
Рис. 6.23. Пример положительного
тигмотаксиса. Положение, которое занимает
уховертка Forficula в круглом сосуде. (Из
Wigglesworth.)
ружения врага. Часто на кончике брюш-
ка имеются участки или структуры (на-
пример, церки) с осязательными волоска-
ми, обладающими высокой чувствитель-
ностью, обеспечивающей быструю реак-
цию «спасения».
ХЕМОТАКСИС-РЕАКЦИЯ НА ЗАПА-
ХИ. Число реакций насекомых на раз-
личные запахи бесконечно. Насекомые
используют хемотаксисы главным обра-
зом при поисках пищи-либо для себя,
либо для будущего потомства
(рис. 6.24). Как правило, каждое насеко-
мое реагирует на запахи определенной
пищи, интересующей его в данный мо-
мент Например, самки бабочек рода
Macroglossa откладывают яйца на суб-
страт, имеющий запах растения рода Ga-
lium, на котором питаются их личинки.
Другие запахи не привлекают яйцекладу-
щую самку.
Некоторые насекомые руковод-
ствуются в своем движении запахом,
оставленным их жертвой. Оса Microbra-
соп из семейства Braconidae находит
188
6. Реакции на внешние воздействия и поведение
Рис. 6.24. Рисунок, иллюстрирующий
хемотаксис. (Из Wigglesworth по Fliigge.)
Следы перемещения дрозофил (лишенных
крыльев) под действием: Л-воздушной
струи без запаха и Б- воздушной струи с
запахом груши.
свою жертву, гусеницу мельничной ог-
невки Anagasta, по оставленному ею па-
хучему следу, улавливая запах с по-
мощью антенн, которые она при своем
движении держит близко к земле. Му-
равьи также следуют к гнезду и от гнезда
по путям, отмеченным веществами, се-
кретируемыми и выделяемыми самими
муравьями.
ТЕРМОТАКСИС И ГИДРОТАКСИС.
Насекомые реагируют на различные тем-
пературы и влажность и выбирают усло-
вия, наиболее близкие к оптимальным
для них. Насекомые, питающиеся
кровью теплокровных животных, ис-
пользуют реакцию на температуру при
поисках своих хозяев. Комары и по-
стельные клопы обладают положи-
тельным термотаксисом к температурам
около 37°С, т. е. близким к температуре
тела млекопитающих.
КООРДИНИРОВАННЫЕ ТАКСИСЫ.
Часто активность насекомых основана
на реакциях, включающих два или более
таксиса одновременно. Например, ба-
бочкам Macroglossa для откладки яиц не-
обходимы и зеленый цвет, и запах Ga-
lium. Некоторые свежевылупившиеся гу-
сеницы, питающиеся в кроне деревьев,
обладают одновременно отрицательным
геотаксисом и положительным фототак-
сисом, что позволяет им постоянно дви-
гаться в направлении их естественного
корма-листьев дерева.
Последовательность рефлексов и так-
сисов. Различные проявления активности
насекомых являются, по существу, скор-
релированной последовательностью от-
дельных рефлексов и таксисов, интегри-
рующих чувствительные и двигательные
реакции. Рассмотрим, например, реак-
ции бабочек, связанные с поисками пищи
и питанием. Голодная бабочка обладает
положительным фототаксисом к красно-
желтой области спектра; эти цвета при-
влекают бабочку и в конце концов она са-
дится на цветок. После этого двигатель-
ная реакция выключается, и появляется
собирательный рефлекс-бабочка пере-
дней лапкой дотрагивается до нектара,
затем расправляет хоботок и пробует не-
ктар. Контакт хеморецепторов, находя-
щихся на кончике хоботка, с нектаром
вызывает рефлекс, с которого начинают-
ся всасывающие движения глоточного
насоса и прием пищи.
Такая последовательная серия ре-
флексов и таксисов существенно отли-
чается от простого рефлекса. Будучи за-
пущенной, эта последовательность цели-
6. Реакции на внешние воздействия и поведение
189
ком осуществляется без участия головно-
го мозга. Однако мозг контролирует ее
запуск и окончание. Когда в кишечнике
нет пищи, рецепторы растяжения, нахо-
дящиеся на стенке кишечника, передают
информацию об этом в мозг, который и
инициирует вышеописанную последова-
тельность рефлексов и таксисов. Когда
кишечник полон, в мозг от тех же рецеп-
торов поступает другая информация, и
из мозга идут сигналы, тормозящие про-
цессы, связанные с питанием. Как и боль-
шинство других организмов, насекомые
реагируют не на все Сигналы, поступаю-
щие из окружающей среды, а только на
отдельные, выбираемые ими стимулы.
Спаривание и откладка яиц-два дру-
гих примера коррелированной последо-
вательности рефлексов и таксисов. Пове-
денческий стереотип при спаривании
обычно включает реакции обоих полов.
Сначала рецептивная самка выделяет ле-
тучий половой феромон, который стиму-
лирует самца этого вида искать самку.
Когда самец находит самку, он совер-
шает ряд движений, составляющих ри-
туал ухаживания,- «постукивание» антен-
нами по самке и дрожание крыльев, а
затем самец делает садку. Самка выпу-
скает наружу часть своего генитального
аппарата, самец охватывает его своими
гениталиями, и начинается спаривание.
Последовательность действий, совер-
шаемых самцом, находится под контро-
лем последнего брюшного ганглия, а не
головного мозга. Мозг, однако, либо
тормозит, либо запускает всю эту после-
довательность действий в зависимости
от стимулирующего влияния феромона
самки. У некровососущих комаров и не-
которых других насекомых самец реаги-
рует изначально на звук, возникающий
от движения крыльев летящей самки.
У некоторых мух, дневных и ночных
бабочек самец также выделяет половой
феромон, который необходим, чтобы
самка, издающая призывный сигнал,
приняла самца. У хлопкового долго-
носика самка и самец меняются роля-
ми: самец, выделяя феромон, осуще-
ствляет начальный «зов», по которому
рецептивная самка находит самца.
Поведение насекомых при спарива-
нии очень разнообразно, и каждый его
вариант специфичен для группы насе-
комых или даже отдельного вида. У мно-
гих мух самец предлагает самке пищу
или какое-нибудь вещество, секретируе-
мое в его ротовой полости. У многих
сверчков самцы издают призывные сиг-
налы, на звуки которых привлекается
самка; у жуков-светляков призывными
сигналами самцов служат их световые
вспышки. Все эти поведенческие реакции
находятся, по-видимому, под контролем
соответствующего брюшного ганглия и
грудных ганглиев (непосредственно кон-
тролирующих движения ног или крыль-
ев). Головной мозг осуществляет только
запуск или подавление необходимой по-
следовательности действий.
Научение
Научение можно определить как дли-
тельные адаптивные изменения в поведе-
нии, явившиеся результатом накопленно-
го опыта. Было показано, что насекомые
способны к научению нескольких типов
от самого простого - привыкания, до зна-
чительно более сложных. Необходимой
предпосылкой для научения является
способность к хранению информации,
называемая памятью, т е. ассоциация
стимула с вызываемой им реакцией, воз-
никающая на основе предшествующего
опыта.
Привыкание. Считается, что это на-
иболее простая форма научения, при ко-
торой повторение стимула приводит к
постепенному ослаблению реакции на не-
го до полного ее исчезновения. Это от-
190
6. Реакции на внешние воздействия и поведение
сутствие реакции может сохраняться
длительное время и после прекращения
подачи стимула. Например, насекомое,
подвергаемое действию сильного света,
может в конце концов «отключиться» от
него. Привыканием является также поте-
ря чувствительности к внешнему раздра-
жителю, который не играет существен-
ной роли в нормальной жизни особи.
Вероятно, все насекомые способны к при-
выканию, и оно, по-видимому, предста-
вляет собой важный компонент поведе-
ния у большинства насекомых.
Ассоциативное научение. Известно,
что насекомые могут научиться устана-
вливать связь между стимулом, обычно
не вызывающим ответной реакции, и ка-
ким-нибудь подкрепляющим действием,
например поощрением или наказанием.
Этот процесс называют ассоциативным
научением. Пчелы могут ассоциировать
цвет с источником пищи, и можно на-
учить их ориентироваться при отыскании
пищи на «нужный» цвет, а не на привы-
чные им обонятельные стимулы. В при-
роде они, по-видимому, в основном ре-
агируют на ультрафиолетовый свет, по-
скольку у многих цветков имеется цен-
тральная «мишень» в виде кольца или
пятна, от которой отражаются ультра-
фиолетовые лучи. Медоносные пчелы
способны различать формы углублений
поверхности, и можно научить их связы-
вать это с пищей. Шмели научаются
различать очертания нектароносных рас-
тений и при поисках пищи находят имен-
но те растения, в которых в данное время
образуется нектар.
Латентное или поисковое научение.
Многие виды общественных ос и пчел,
жизнедеятельность которых связана с со-
зданием запасов пищи, умеют обнаружи-
вать свои гнезда, узнавая и запоминая
ориентиры на местности. При первом же
вылете из гнезда или колонии фуражи-
рующие самки или рабочие особи
запоминают ряд ориентиров, которыми
руководствуются при своем возвраще-
нии домой. Если ориентир удалить, то
возвращающееся домой насекомое пы-
тается отыскать его, совершая беспоря-
дочные или круговые поисковые полеты
до тех пор, пока не обнаружит следую-
щий ориентир. Если появляется новый
ориентир, насекомые быстро научаются
узнавать его. При полетах на большие
расстояния (от 1,5 км и более) многие об-
щественные насекомые, очевидно, ориен-
тируются по углу между направлением
полета и положением солнца или пло-
скостью поляризации дневного света.
«Постижение» (insight). Считают, что
насекомые не способны к высшим фор-
мам научения-анализу предшествующе-
го опыта и превращению его в обобщен-
ную информацию, служащую основой
для решения новых задач методом, от-
личным от метода проб и ошибок. Было
предложено несколько примеров, когда
можно было предположить аналитиче-
ские способности у насекомых, однако
они оказывались, как правило, поведен-
ческими реакциями, сопровождавшими-
ся или не сопровождавшимися науче-
нием.
Два примера из жизни общественных
насекомых, которые рассматривались
как рассудочная деятельность, касаются
использования орудий. Муравьи из рода
Oecophylla живут на ветвях тропических
деревьев и строят прочные гнезда из
скрепленных между собой листьев. При
постройке гнезда одни муравьи удержи-
вают края листа вместе, а другие держат
личинку так, что она касается то одного
края листа, то другого, при этом личинка
выделяет шелковую нить, которая скре-
пляет края листа и сшивает их вместе.
Это «сшивание» очень напоминает разу-
мное использование орудий.
Использование такого рода орудий
известно и у других видов муравьев. Не-
6. Реакции на внешние воздействия и поведение
191
сколько видов рода Aphaenogaster ис-
пользуют кусочки листьев для доставки в
гнездо корма, слишком мягкого для
переноса его в мандибулах. Муравьи кла-
дут кусочки листьев на корм, который
прилипает к ним, а через 30-60 мин после
этого несут его к гнезду. Для муравьев
Aphaenogaster с маленьким брюшком бы-
ло высчитано, что количество жидкой
или желатинообразной пищи, которую
они уносят прилипшей к кусочку листа,
равному обычно длине тела муравья,
больше того, что муравей может съесть
за один раз.
ОРГАНИЗАЦИЯ СООБЩЕСТВ
У ОБЩЕСТВЕННЫХ НАСЕКОМЫХ
Среди насекомых самые сложные
формы поведения наблюдаются у обще-
ственных насекомых. Организация сооб-
ществ у них основана на взаимосвязи и
кооперации между ее членами, живущи-
ми одной колонией и занимающей гнез-
до того или иного типа. Истинно обще-
ственные насекомые, так называемые эу-
социальные насекомые, относятся к двум
отрядам. Это-все термиты (Isoptera) и
специализированные представители Hy-
menoptera. все муравьи и некоторые из
наиболее высокоорганизованных ос и
пчел. Эусоциальное поведение характе-
ризуется тремя основными чертами.
1) в колонии данного вида особи объеди-
няются для заботы о потомстве;
2) обязанности в колонии распределены
между специальными группами особей;
3) жизненные циклы особей по крайней
мере двух поколений перекрываются, так
что молодое поколение часть своей жиз-
ни проводит вместе с родительским
поколением.
Для дообщественных (пресоциальных)
уровней организации насекомых харак-
терны только одна или две из указанных
трех черт.
Колонии эу социальных насекомых
характеризуются также высоким уров-
нем репродукции. Численность особей в
колонии колеблется от менее 100 до мно-
гих миллионов у разных видов и групп
насекомых (рис. 6.25, 6.26). В колонии
одного из видов африканских муравьев
может быть до 22 млн. рабочих особей.
У некоторых наиболее высокоорганизо-
ванных муравьев, например Myrmica ru-
bra, в колонии имеется одна матка («ца-
рица»), которая живет несколько лет,
откладывая яйца и обеспечивая потом-
ством всю колонию, а ее снабжают кор-
мом все нерепродуктивные стерильные
самки. Нерепродуктивные особи выпол-
няют в колонии множество функций.
Обычно они представлены различными
морфологическими типами, или кастами
(см. гл. 8* термиты-рис. 8.20, 8.21; му-
равьи-рис. 8.125; медоносные пчелы-
рис. 8.128). Самые мелкие особи-рабо-
чие; более крупные - солдаты или
крупные рабочие. Рабочие занимаются
сбором корма, устройством гнезда, уха-
живанием за потомством. Солдаты за-
щищают гнездо и места хранения жид-
кой пищи. Нерепродуктивные особи жи-
вут относительно недолго, и матка дол-
жна откладывать яйца почти непрерыв-
но, чтобы обеспечить достаточную чис-
ленность колонии к моменту ее созрева-
ния. Помимо этих социальных групп есть
еще одна группа, состоящая из самцов.
Они не выполняют никаких работ, лишь
несут некоторые общественные функции
(например, уход за другими особями) и
находятся в ожидании брачного полета,
чтобы оплодотворить девственных
самок. Девственные самки развиваются
из потомства самки-производительницы.
После брачного полета каждая молодая
самка основывает свою собственную но-
вую колонию, строя гнездо и ухаживая за
потомством. Когда колония становится
зрелой, нерепродуктивные особи берут
192
6. Реакции на внешние воздействия и поведение
Рис. 6.25. Колония примитивного австралийского муравья-бульдога (Myrmecia gulosa),
строящего свое гнездо в земле. (Из Е. О. Wilson, 1971, The insects societies, оригинальный
рисунок С. Лендри; перепечатка с разрешения Harvard University Press.)
А. Матка (царица). Б. Самец. В. Рабочий, предлагающий корм личинке. Г Коконы с
куколками.
на себя заботу о потомстве и другие
функции.
Эволюционными предшественниками
эусоциального уровня организации насе-
комых были две линии развития обще-
ственных отношений -парасоциальная и
субсоциальная (рис. 6.27). При одиноч-
ном (необщественном) образе жизни от-
сутствует забота о потомстве, нет спе-
циальных размножающихся каст
и взаимного перекрывания последова-
тельных поколений. При парасоциальной
организации, характерной для пчел семей-
ства Halictidae, взрослые особи, принад-
лежащие к одному поколению, в той или
иной степени помогают друг другу.
Самый низкий уровень организации это-
го типа называется коммунальным. В со-
обществах, относящихся к этому уров-
ню, взрослые особи объединяются для
постройки гнезда, но потомство во-
спитывают отдельно. На коммунальном
уровне развития находятся несколько ви-
дов пчел из семейства Halictidae. При-
мерно 50 самок одного вида населяют
гнездо, находящееся под землей. Каждая
пчела роет свои боковые туннели или
ячейки, в каждую из которых отклады-
вает одно яйцо, помещает запас корма и
затем запечатывает. На следующем
6. Реакции на внешние воздействия и поведение
193
Рис. 6.26. Гнездо термита Amitermes hastatus. (Из Е. О. Wilson, 1971, The insect societies,
оригинальный рисунок С. Лендри; перепечатка с разрешения Harvard University Press»)
А. Верхняя ячейка с размножающимися нимфами. Б. Средняя ячейка с царицей-
основательницей гнезда, самцом рядом с ней и многочисленными рабочими особями.
В. Нижняя ячейка с солдатами и нимфами, из которых развиваются солдаты.
уровне парасоциальной линии -квазисо-
циалъном- забота о потомстве носит об-
щественный характер, но каждая самка
откладывает яйца в какое-то определен-
ное время. На следующем, полусоциаль-
ном, уровне появляется каста рабочих,
состоящая из неразмножающихся особей
колонии. Следующий уровень организа-
ции-уже эусоциальный, который дости-
гается, когда продолжительность жизни
одного поколения полусоциальной коло-
нии становится столь большой, что два
или более последовательных поколений
живут одновременно и совместно уча-
ствуют в жизни колонии.
Эволюционная последовательность
13-732
субсоциальных состояний, ведущая к фор-
мированию эусоциального уровня орга-
низации насекомых, представлена му-
равьями, термитами, общественными
осами и несколькими группами обще-
ственных пчел. Поскольку все нынеживу-
щие муравьи и термиты находятся на эу-
социальном уровне общественной орга-
низации, последовательное развитие суб-
социальных отношений изучалось на
осах и некоторых пчелах. В этой линии
развития наблюдается усиление связей
между родителями и потомками. При
одиночном образе жизни и на примитив-
ном субсоциалъном уровне самка некото-
рое время ухаживает за отложенной ею
194
6. Реакции на внешние воздействия и поведение
Пара социальная линия
Субсоциальная линия
Кооперация в колонии,
состоящей из взрослых
особей одного
поколения
ЭУСОЦИ-
^Хальнаях
полусоциальна< ПРОМЕЖУТОЧНАЯ СУБСОЦИАЛЬНАЯ н
КВАЗИСОЦИАЛЬНАЯ
ПРОМЕЖУТОЧНАЯ СУБСОЦИАЛЬНАЯ I
КОММУНАЛЬНАЯ\^О;1И(|ОЧ^|>ИМИТИВН'Я «-^СОЦИАЛЬНАЯ
НАЯ
Кооперация в семье,
состоящей из особей
двух и более
поколений
Рис. 6.27. Две линии развития эусоциальной организации у общественных
насекомых - парасоциальной и субсоциальной.
кладкой, но не дожидается вылупления
молоди. На промежуточной первой суб-
социальной стадии самка остается с мо-
лодыми особями до их созревания. На
промежуточной второй субсоциальной
стадии взрослые молодые особи помо-
гают родителям в воспитании нового по-
томства. Кооперация наблюдается ме-
жду матерью и детьми, но не между
дочерними особями. Следующий этап-
появление специальных групп особей,
рабочих, являющихся постоянными по-
мощниками в воспитании нового по-
коления; это -эусоциальный уровень.
Специализация в колониях эусо-
циальных насекомых идет по двум пу-
тям. Первый состоит в увеличении числа
и степени специализаций в кастах рабо-
чих. Источником усиления дифференциа-
ции рабочих каст являются постоянные
морфологические изменения, разные у
разных особей, в результате которых со-
здаются различные морфологические
типы. Например, у хорошо откор-
мленных рабочих муравьев развивается
большая голова и мандибулы, что позво-
ляет им перейти в касту солдат. Вместе с
тем в результате ряда морфологических
изменений у одной особи она может в те-
чение жизни принадлежать более чем к
одной касте. У муравьев Myrmica scabri-
nodis рабочие в первый сезон своей актив-
ности после имагинальной линьки уча-
ствуют в ухаживании за молодью, в
следующий сезон они становятся строи-
телями, еще позднее-фуражирами. Та-
кая смена функций наиболее хорошо раз-
вита у медоносных пчел.
Второй путь специализации у эусо-
циальных насекомых-это развитие ком-
муникации между особями в колонии,
так что активность множества населяю-
щих ее особей становится координиро-
ванной. Коммуникация, как было отме-
чено ранее в этой главе, наиболее развита
у общественных насекомых. У эусо-
циальных насекомых широко развита хи-
мическая коммуникация, включающая
выделение и восприятие химических ве-
ществ. В меньшей степени, но также до-
статочно хорошо развита акустическая
коммуникация, к которой принадлежат
стрекотание * постукивание и другие сиг-
налы. Обмен жидкими феромонами,
подавляющими дифференциацию каст,-
одно из множества удивительных явле-
ний, наблюдающихся в колониях насе-
комых. Многообразию сигналов, соста-
вляющих коммуникационную систему
насекомых, соответствуют различные
поведенческие реакции: тревога, привле-
чение и образование скоплений, поиски
новых источников пищи или мест для
гнезда, ухаживание, трофаллаксис (об-
мен между особями оральными или
анальными жидкими выделениями),
передача другим особям частичек пищи,
групповое взаимодействие, которое либо
усиливает, либо ослабляет индивидуаль-
ную активность, идентификация и узна-
вание партнеров по гнезду и членов своей
касты, детерминация каст, выражающая-
ся либо в торможении, либо в стимуля-
ции их дифференциации.
6. Реакции на внешние воздействия и поведение
195
Пресоциальный уровень
организации насекомых
По ряду черт поведения, связанных с
ухаживанием за самкой, спариванием и
откладкой яиц, эусоциальные насекомые
сходны со своими эволюционными пред-
шественниками -одиночными насекомы-
ми. Но у эусоциальных насекомых по ме-
ре усложнения организации колонии по-
являются и другие поведенческие стерео-
типы. Пресоциальные уровни организа-
ции включают образование скоплений
(агрегацию) особей, выполняющих вме-
сте работу по постройке гнезда, добыва-
нию корма или по ухаживанию за мо-
лодью. Ниже дается несколько приме-
ров пресоциальных уровней организа-
ции.
АГРЕГАЦИЯ ПРИ ПОСТРОЙКЕ ГНЕЗ-
ДА И УХАЖИВАНИИ ЗА МОЛОДЬЮ.
Среди бабочек (Lepidoptera) есть виды, у
которых гусеницы, вылупившиеся одно-
временно из одной кладки, строят гнез-
до, похожее на шелковую паутиновую
сеть и используемое для совместного
жилья. Такое гнездо строится вокруг раз-
вилок мелких или более крупных ветвей
деревьев, и в его постройке принимают
участие все гусеницы. Гусеницы поки-
дают гнездо днем для питания на листь-
ях дерева и возвращаются в него для от-
дыха. Гусеницы коконопрядов Malacoso-
та живут в совместно построенном i нез-
де в течение всего личиночного периода и
покидают его только для окукливания.
Гусеницы бабочек-медведиц (Hyphantria)
тоже строят гнездо, но на последнем гу-
сеничном возрасте покидают его и ведут
одиночный образ жизни.
В отряде эмбий (Embioptera) неко-
торые виды строят в почве, почвенной
подстилке или между корнями растений
гнездо, состоящее из скрепленных между
собой сотканных из шелка туннелей. За-
бота о потомстве у самок развита очень
хорошо и состоит в охране отложенных
яиц и юных нимф. Некоторые колонии,
состоящие из тысяч особей, живут в тол-
стом сплетенном из шелковых нитей ма-
трасе, занимающем площадь во много
квадратных метров. Тем не менее между
особями нет достаточно оформленных
взаимоотношений, и поэтому скопления
этих насекомых может иметь лишь чуть
большее значение в смысле развития со-
циальности, чем скопления тлей или щи-
товок вокруг представителей родитель-
ского или прародительского поко-
лений.
ОБМЕН СИМБИОНТАМИ. С точки
зрения развития социальной организа-
ции интересен один тип колоний, обнару-
женный у тараканов. Древесные тара-
каны питаются древесиной, которую они
переваривают с помощью специальных
протозойных симбионтов, обитающих в
их пищеварительном тракте. У линяю-
щих тараканов пищеварительный тракт
полностью освобождается и после линь-
ки в нем нет симбионтов, поэтому
нимфы голодают, пока симбионты вновь
не попадают в кишечник. Это происхо-
дит при поедании нимфами свежих эк-
скрементов других членов колонии. Та-
кой необходимый обмен протозойными
симбионтами и обусловливает сов-
местное проживание особей в коло-
нии.
ЗАБОТА О ПОТОМСТВЕ. Одной из
предпосылок общественной организации
насекомых является защита и кормление
личинок взрослыми особями. У неко-
торых Hemiptera взрослые защищают
молодых особей, оттаскивая хищников с
помощью направленных вперед отрост-
ков головы или переднегруди. У цикад-
горбаток Umbonia эта реакция стимули-
руется феромоном, выделяемым пора-
ненной нимфой. Самки некоторых ухо-
верток откладывают яйца в укрытия и
охраняют их, отгоняя хищников. После
196
6. Реакции да внешние воздействия и поведение
вылупления личинок они еще некоторое
время продолжают сторожить их, пока
молодые нимфы не покинут выводковую
камеру. На этом моменте материнская
забота оканчивается, и каждая нимфа на-
чинает самостоятельную жизнь.
Сходные данные были получены и для
других насекомых, например для медве-
док Gryllotalpa.
Происхождение эусоциальности
у Hymenoptera
У большинства насекомых из оплодо-
творенных яиц развиваются самки или
самцы, причем каждая особь имеет ди-
плоидный набор хромосом, по одному
набору от каждого из родителей. Среди
эусоциальных насекомых только тер-
миты характеризуются типичной ди-
плоидной генетической системой. В дру-
гой же группе эусоциальных насекомых,
у Hymenoptera, из всех оплодотворенных
яиц развиваются самки с диплоидным
набором хромосом. Из всех неоплодо-
творенных яиц развиваются партеногене-
тические гаплоидные самцы (рис. 6.28).
Рис. 6.28. Схема, иллюстрирующая
генетические отношения между самками и
самцами у эусоциальных перепончатокры-
лых. (Из Crozier, 1979.)
Генетический вклад царицы (матки) ($,) и
самца (ср) в последующие поколения
показаны соответственно штриховкой
и точками.
Это явление носит название гаплодиплои-
дии. Такой необычный тип репродукции в
колониях эусоциальных перепонча-
токрылых, приводящий к развитию осо-
бей с различной генетической конститу-
цией, следует учитывать при ответе на
вопрос, почему общественная органи-
зация не менее 10 раз появилась в от-
ряде Hymenoptera и только 1 раз воз-
никла у других насекомых, у терми-
тов.
Гаплодиплоидия помогает также
объяснить, почему в сообществах пере-
пончатокрылых и по численности, и по
числу выполняемых функций домини-
руют самки, тогда как в сообществе ди-
плоидных термитов самки и самцы делят
функции примерно поровну. В гаплоди-
плоидных колониях перепончатокрылых
сестры генетически более близки между
собой, чем мать и дочери. Диплоидные
сестры генетически ближе друг к другу,
чем к своим гаплоидным братьям. По-
этому самки образуют тесное сообще-
ство, основной задачей которого являет-
ся помощь единственной яйцекладущей
среди них самке-матке, или царице-в
производстве большего числа самок, их
сестер. Это также помогает понять, поче-
му гаплоидные самцы перепончатокры-
лых не выполняют в колонии тех функ-
ций, которые выполняют диплоидные
самцы термитов.
Симбиоз
у общественных насекомых
Гнезда общественных насекомых ис-
пользуются множеством различных ор-
ганизмов. Многие из них-это насекомые
и другие членистоногие, которые живут в
том же гнезде, что и их хозяева, и нахо-
дят здесь пищу для себя. Такое тесное и
разнообразное взаимодействие между
особями разных видов называется сим-
биозом. Симбионты, живущие в гнездах
6. Реакции на внешние воздействия и поведение
197
общественных насекомых, называются
инквилинами (inquilines). Среди инквилин
встречаются крошечные щетинохвостки
(Thysanura), личинки мух-журчалок (Sy-
rphidae), некоторые гусеницы бабочек,
довольно много жуков (особенно из се-
мейств Staphylinidae и Pselaphidae) и не-
сколько видов паразитических муравьев.
Иногда в одном гнезде может встречать-
ся лишь очень немного видов инкви-
лин, однако общее число известных ви-
дов инквилин, населяющих гнезда му-
равьев и термитов, измеряется тыся-
чами.
Некоторые из инквилин малы
и чрезвычайно подвижны. Их выживае-
мость в гнезде обеспечивается, безуслов-
но, только их проворством. Другие-
крупного размера и более медлительные,
часто почти неподвижные. Некоторые
инквилины выделяют секреты, по-
ЛИТЕРАТУРА
Beroza М. (Ed.). Chemicals controlling insect
behavior. Academic Press, New York and Lon-
don, 170 pp., 1970.
Birch M. C. (Ed.). Pheromones. North-Hol-
land, Amsterdam and London, 495 pp., 1974.
Chapman R. F. The insects, structure and func-
tion. American Elsevier, New York, 819 pp.,
1969.
Eisner T., Wilson E. O. (Eds.). The insects.
W. H. Freeman, San Francisco, 334 pp., 1977.
Evans H. E., Eberhard M. J. W The wasps.
Univ, of Michigan Press, Ann Arbor, 265 pp.,
1970.
Fraenkel G. S., Gunn D. L. The orientation of
animals. Dover, New York, 376 pp., 2nd ed.,
1962.
Heinrich B. Bumblebee economics. Harvard
Univ. Press, Cambridge, Mass., 345 pp., 1979.
Hermann H. R. (Ed.). Social insects. Academic
Press, New York, Vol. 1, 438 pp., 1979.
Horridge G. A. (Ed.). The compound eye and
vision of insects. Clarendon Press, Oxford, 595
pp., 1975.
едаемые муравьями-хозяевами гнезда.
У жуков-стафилинид на брюшке имеют-
ся большие участки, где выделяется се-
крет, слизываемый муравьями, которые
в свою очередь отрыгивают пищу для
жуков. Таким образом, между хозяином
гнезда и по меньшей мере частью инкви-
лин существуют дружеские отношения,
поддерживаемые стереотипными (вро-
жденными) поведенческими реакциями.
Пока неизвестно о существовании како-
го-либо другого химического языка по-
мимо обмена пищевыми веществами ме-
жду хозяином гнезда и инквилинами.
Когда колония муравьев перебирается в
другое гнездо, инквилины, как правило,
уходят вместе с хозяевами, следуя за му-
равьями. Это позволяет предполагать,
что инквилины реагируют на какие-то
химические сигналы, оставляемые му-
равьями.
Iwata К. Evolution of instinct: Comparative
ethology of Hymenoptera (Trans, from
Japanese), Smithsonian Inst, and National
Science Foundation, Amerind, New Delhi, 535
pp., 1976.
Jacobson M. Insect sex pheromones. Academic
Press, New York and London, 382 pp., 1972.
[Имеется перевод: Джекобсон M. Половые
феромоны насекомых.-М.: Мир, 1976].
Johnson С. G. Migration and dispersal of insects
by flights. Methuen, London, 763 pp., 1969.
Krishna K., Weesner F. M. (Eds.). Biology of
the termites. Academic Press, New York, Vols.
I and II, 1969, 1970.
Lipke H., Fraenkel G. Insect nutrition, Annu.
Rev. Entomol., 1, 17-44 (1956).
Matthews R. W., Matthews J. R. Insect beha-
vior. Wiley, New York, 507 pp., 1978.
Michener C. D. The social behavior of bees. A
comparative study. Belknap Press of Harvard
Univ. Press, Cambridge, Mass., 404 pp., 1974.
Nachtigall W. Insects in flight. Allen & Unwin,
London, 153 pp., 1974.
198
6. Реакции на внешние воздействия и поведение
Novak V J. A. Insect hormones. Chapman &
Hall, London, 600 pp., 1975.
Rainey R. C. (Ed.). Insect flight, Symp. R.
Entomol. Soc. London, 7, 1-287 (1976).
Romoser W S. The science of entomology. Mac-
millan, New York, 449 pp., 1973.
Saunders D. S. Insect clocks. Pergamon, Oxford,
279 pp., 1976.
Schneirla T. С. (H. R. Topoff, Ed.). Army ants.
W H. Freeman, San Francisco, 349 pp., 1971.
Shorey H. H. Animal communication by phero-
mones. Academic Press, New York, 167 pp.,
1976.
Treherne J. E., Beament J. W L. (Eds.). The
physiology of the insect central nervous system.
Academic Press, London and New York, 277
pp., 1965.
Truman J. W.f Riddiford L. M. Hormonal
mechanisms underlying insect behavior. In.
Treherne J. E., Berridge J. J., Wiggles-
worth V. B. (Eds.). Advances in insect physio-
logy, 10, 297-352. Academic Press, London,
New York, 1974.
Wilson E. O. The insect societies. Belknap Press
of Harvard Univ. Press, Cambridge, Mass.,
548 pp., 1971.
См. также литературу к главам 5, 8 и 10.
7
Жизненные циклы,
рост и размножение
С появлением признаков дифференцировки в яйце начинается сложный и дли-
тельный процесс роста, в результате которого организм становится зрелым и го-
товым к размножению. Последовательность биологических процессов, происходя-
щих при развитии от яйца до взрослого животного, составляет жизненный цикл
особи. Насекомые характеризуются огромным разнообразием жизненных циклов,
различающихся по типу развития и размножения, отношениям между поколениями,
а у некоторых видов даже в пределах одного жизненного цикла происходит изме-
нение способов питания и местообитаний.
РАЗВИТИЕ
Жизненный цикл особи обычно со-
стоит из двух фаз развития (от яйца до
взрослой особи) и зрелости (взрослый
организм). В период развития основные
энергетические ресурсы насекомого за-
трачиваются на рост, при этом укрепле-
ние экзоскелета и двигательная актив-
ность остаются на втором плане. Глав-
ная функция взрослого организма - рас-
пространение и размножение. Поскольку
жизнь большинства насекомых начинает-
ся с яйца, наиболее важным переломным
моментом в развитии организма являет-
ся вылупление из яйца. Развитие орга-
низма внутри яйца называется эмбрио-
нальным, а после выхода из яйца-
постэмбриональным. Морфологические
изменения во время постэмбрионально-
го периода называют метаморфозом.
Эмбриональное развитие
Яйцо. Яйца насекомых могут быть
различной формы (рис. 7.1). Чаще все-
го-это эллипс с гладкой поверхностью,
в других случаях поверхность может
быть ребристой или различным образом
скульптурированной, некоторые яйца
имеют различные выросты, например ла-
теральные выросты (поплавки) у яиц
Anopheles, удерживающие их на воде
(рис. 7.1,^).
Типичное яйцо насекомых (рис. 7.2)
представляет собой билатерально-сим-
метричную клетку с двумя оболочками,
плотной наружной -хорионом и тонкой
внутренней -желточной. В оболочках
имеются крошечные отверстия -микро-
пиле, через которые в яйцо проникает
сперматозоид. Внутренняя часть яйца,
ограниченная оболочками, представлена
крупным ядром и цитоплазмой. Цен-
тральная часть цитоплазмы занята
желтком, представляющим собой запас
питательных веществ, а непосредственно
под желточной оболочкой находится бо-
лее плотный пограничный слой цито-
плазмы - периплазма.
Дробление. В типичном случае у насе-
комых, размножающихся половым пу-
тем, оплодотворение яйца обычно проис-
ходит незадолго до его откладки. Один
или несколько спермиев проникают в яй-
цо в то время, когда оно проходит по яй-
200
7. Жизненные циклы, рост и размножение
Рис. 7.1. Яйца насекомых.
Л-коллемболы Sminthurus viridis\ Ь’-тли Toxoptera graminum; В-коровьей вши
Solenopotes capillatus, прикрепленное к волосу животного; Г-яблоневого клопа-слепняка
Paracalocoris colon в ткани растения; Д-божьей коровки Hyperaspis binotata;
Е- долгоносика Sphenophorus phoeniciensis’, Ж-малярийного комара Anopheles maculipennis\
3-кубышка с яйцами саранчи в почве и отдельное яйцо; Я-наездника Diachasma tryoni;
К-стрекозы Archilestes californica, снятое с водного растения; Л-моли Tineola bisselliella\
М-собачьей блохи Ctenocephalides canis; //-грушевого трипса Taeniothrips inconsequens,
вынутое из ткани растения; О-златоглазки Chrysopa oculata, 77-комнатной мухи Musca
domestica. (По Essig из различных авторов.)
цеводу. После оплодотворения и образо-
вания зиготы начинается процесс деле-
ния ядра, причем сама клетка не делится.
Образовавшиеся дочерние ядра с участ-
ками цитоплазмы вокруг них сначала
беспорядочно располагаются внутри
желтка (рис. 7.2,.Б, В). Этот процесс на-
зывается дроблением. Каждое дочернее
ядро с участком цитоплазмы вокруг него
называют энергидой. Последующие деле-
ния энергид сопровождаются их мигра-
цией в периплазматический слой
(рис. 7.2,7"-£), где вокруг каждой из них
формируется мембрана и, таким обра-
зом, возникают клетки. С этого времени
дробление продолжается только в пери-
плазме, центральная бесклеточная часть
заполнена недробящимся желтком. Та-
кой тип дробления называется поверх-
ностным. На поверхности яйца образует-
ся одноклеточный слой, и эта стадия
развития называется бластодермой. За-
тем в вентральной части яйца появляется
область более крупных клеток, называе-
мая вентральной пластинкой, или зароды-
шевой полоской (рис. 7.3). Это-начало
образования зародыша. Зародышевая
полоска может занимать значительную
часть яйца, а может быть очень неболь-
шой; в последнем случае она имеет вид
тонкой пластинки и называется зароды-
шевым диском (рис. 7.4,1).
7. Жизненные циклы, рост и размножение
201
Рис. 7.2. Строение и ранние стадии дробления типичного яйца насекомого,
показанные на срезах. (По Снодграссу, 1935.) А. Оплодотворенное яйцо; Б.
Яйцо после нескольких делений ядра. В. Яйцо после многих делений ядра
(обратите внимание на миграцию ядер к периферии). Г После окончания
дробления ядра образуют хорошо выраженный периферический слой. Д, Е.
Внутренняя часть и поверхность яйца после того, как энергиды сосредоточились
в периплазме, образовав слой клеток-бластодерму. Обратите внимание на
раннее обособление первичных половых клеток на заднем полюсе яйца.
7-микропиле; 2-хорион; 3-желточная оболочка; #-ядро; 5-периплазма,
6-желток; 7-ранние стадии деления ядер; 3-ядра в периплазме; 9-первичные
половые клетки; 10-клетки бластодермы; 77-бластодерма.
Поверхностное дробление характерно
для большинства насекомых. У энтогнат
(Collembola) в яйцах содержится мало
желтка, и дробление охватывает сразу
все яйцо. Это единственный отряд насе-
комых, у которых наблюдается голобла-
стическое дробление. >
Рост и развитие зародыша. Зародыше-
вая полоска или зародышевый диск рас-
тут благодаря делению и дифференци-
ровке клеток. Вначале рост происходит в
основном за счет увеличения размеров
зародышевого диска, затем по мере
образования сегментов тела и конечно-
202
7. Жизненные циклы, рост и размножение
Рис. 7.3. Образование зародышевой полоски на вентральной стороне
бластодермы. (По Снодграссу.)
А. Поперечный срез через яйцо. Б. Продольный срез через яйцо. 7-желточные
клетки: 2-желток; 3-зародышевая полоска, 4-тонкая дорсальная часть
бластодермы; 5-желточная оболочка.
стей происходит обособление отдельных
частей. Это показано на рис. 7.4, где схе-
матически изображены крупные измене-
ния, приводящие к завершению форми-
рования зародыша и появлению нимфы
первой стадии (рис. 7.5).
Сегментация и образование конечно-
стей. Оба указанных процесса, происхо-
дящие на эмбриональной стадии разви-
тия, в значительной степени отражают
предполагаемый ход эволюции насе-
комых.
Образование сегментов начинается с
появления серии поперечных насечек на
зародышевой полоске (рис. 7.4,1V). Сег-
менты, которые на более поздних ста-
диях развития несут ротовые части и
образуют одно целое с головой, в начале
процесса сегментации не отличаются от
задних сегментов. Слияние мандибуляр-
ного, максиллярного и лабиального сег-
ментов с головой происходит после того,
как конечности уже хорошо развиты
(рис. 7.4,VII).
Развитие конечностей начинается
вскоре после появления видимых призна-
ков сегментации (рис. 7.4,V). В типичном
случае на вентральной стороне каждого
сегмента образуется пара конечностей,
однако большая часть абдоминальных
конечностей лишь слабо развита. У мно-
гих насекомых все абдоминальные конеч-
ности зародыша, исключая церки, не бо-
лее как небольшие рудименты, которые
исчезают еще на одной из ранних стадий
эмбрионального развития. Передние сег-
менты и конечности развиваются бы-
стрее, чем задние, и зародыш на этой ста-
дии выглядит так, как показано на
рис. 7.6.
Форма тела. На ранних стадиях эм-
бриогенеза формируются только конеч-
ности и вентральная часть тела. На этих
стадиях (рис. 7.4,V, VI) у зародыша нет
оформленных дорсальной и боковых ча-
стей, и он представлен только стерналь-
ной областью и конечностями. По суще-
ству тело не закрыто сверху. Позднее
боковые части на переднем и заднем кон-
цах тела начинают расти кверху. На
рис. 7.4,VIII изображена стадия, когда
покровы головы и четырех из пяти зад-
них брюшных сегментов смыкаются на
дорсальной поверхности. Зародыш рас-
положен так, что незакрытая покровами
дорсальная часть промежуточных сег-
Рис. 7.4. Рост зародыша и развитие сегментации тела у веснянки.
(По Miller.)
1 -амниотическая полость; 2-внутренний слой; 3-хвостовая часть
зародыша, 4-серозная кутикула, 5-протоцефалон (простомиум
плюс первый посторальный сегмент); 6-хвостовая часть
зародыша, 7-мандибула (мандибулярный сегмент);
8 - торакальный сегмент; 9 - антенна ,10- церк; 11- задняя кишка,
72-сероза, 73-амнион; 14-желток; 75-дорсальный орган;
16 - анус ',17-лабрум; 18 - яйцевой зуб; 19 - бедро;
20-абдоминальные сегменты.
Рис. 7.5. Последняя стадия эмбрионального развития веснянки.
(По Miller.)
А. Зародыш в яйце. Б. Полностью сформированный зародыш,
покидающий яйцевую оболочку. В. Только что вылупившаяся
нимфа. 21 -максиллярные щупики; остальные обозначения те же,
что на рис. 7.4.
Рис. 7.6. Зародыш на ранней стадии развития конечностей. (Из Snodgrass.)
А. Зародыш с зачатками конечностей на всех сегментах, кроме последнего. Б. Более
поздний зародыш, у которого произошла более четкая дифференцировка головного
и грудного отделов и их придатков. 7-лабрум; 2-антенна, 3-конечность
интеркалярного сегмента, 4-конечности, 5-глаз, 6-ротовое отверстие;
7-мандибула, 3-максилла, 9-лабиум; 10-ноги; 77-церк; 72-анальное
отверстие.
7. Жизненные циклы, рост и размножение
205
ментов тесно прилегает к желтку, запол-
няющему незанятое зародышем внутрен-
нее пространство яйца. На следующих
после этого стадиях латеральные края
«открытых» сегментов начинают расти
вверх, пока не сомкнутся на дорсальной
поверхности зародыша, так что желток
оказывается внутри тела. Этим завер-
шается формирование замкнутого тела.
Зародышевые листки; детерминация
тканей. Сначала зародышевая полоска
состоит из одного слоя клеток, но уже на
ранних стадиях эмбриогенеза образуется
второй слой. Обычно это происходит пу-
тем гаструляции, т. е. путем впячивания
внутрь части зародышевой полоски, что
схематически изображено на рис. 7.7. Га-
струляция обычно начинается с образо-
вания продольного желобка на поверхно-
сти зародышевой полоски (А); наружные
края желобка растут навстречу друг дру-
гу, и будущий второй зародышевый слой
впячивается внутрь зародышевой поло-
ски (Б). Наконец, края желобка смыкают-
ся и образуют наружный слой, или экто-
дерму. Внутренний слой, или мезодерма,
располагается под ним (В). На рис.
7.4,1-V мезодерма изображена в виде
темного дорсального участка. На концах
мезодермального слоя появляются впя-
чивания, состоящие из групп клеток,
образующих эндодерму, или третий за-
родышевый слой.
Группы клеток, из которых на более
Рис. 7.7 Образование мезодермы путем гаструляции. (По Snodgrass.)
А. Поперечный срез яйца, в котором произошла дифференцировка зародышевой полоски с
образованием латеральных пластинок (7) и срединной пластинки (3); 2-дорсальная
бластодерма. Б. Более поздняя стадия, когда срединная пластинка изгибается, образуя
трубковидное впячивание, а края латеральных пластинок начинают расти навстречу друг
другу под впячиванием. В. Еще более поздняя стадия, когда края латеральных пластинок
смыкаются и образуют эктодерму (5), а срединная пластинка разрастается над ней и
образует внутренний слой клеток-мезодерму (4). Г Д. Второй способ образования
мезодермы: срединная пластинка, отделившись от краев латеральных пластинок,
превращается в мезодерму (4), когда латеральные пластинки срастаются под ней; Е. Третий
способ состоит в том, что мезодерма (4) образуется из клеток, мигрировавших от
внутренних концов срединной пластинки.
206
7. Жизненные циклы, рост и размножение
поздних стадиях образуются сегменты,
органы или конечности, детерминируют-
ся в эмбриогенезе очень рано. В тщатель-
но проведенных экспериментах было по-
казано, что эта детерминация завершает-
ся в основном уже к моменту образова-
ния зародышевых слоев. Группа таких
неспециализированных клеток, которые
тем не менее предопределены к развитию
в определенную структуру, называется
зачатком.
Из эктодермы образуются стенка те-
ла, передняя и задняя кишки, нервная и
трахейная системы и многие железы. Из
мезодермы образуются мышечная систе-
ма, гонады, сердце, жировое тело, из эн-
додермы - средняя кишка.
Зародышевые оболочки. В течение по-
чти всего своего развития зародыш ча-
стично или полностью погружен в жел-
ток, что, по-видимому, делает его более
защищенным, вокруг зародыша обра-
зуются две оболочки. Известны два ос-
новных способа погружения зародыша и
образования оболочек (рис. 7.8). Первый
способ состоит в том, что сначала в тол-
щу желтка погружается хвостовой конец
зародыша, увлекая за собой оболочку
(рис. 7.8,Г)- Когда весь зародыш оказы-
вается внутри желтка (рис. 7.8,Д), обо-
лочка, разрастаясь, закрывает полость,
образовавшуюся при впячивании за-
родыша, в результате чего получаются
две оболочки: наружная, сероза, и вну-
тренняя, амнион, замыкающие полость с
вентральной стороны зародыша. Другой
способ состоит в том, что зародыш
целиком погружается в желток
(рис. 7.8,Ж-И). Оболочка нарастает над
его вентральной частью с двух сторон и в
конце концов ее края, смыкаясь, обра-
зуют амнион и серозу. На более поздней
стадии развития зародыш снова изме-
няет положение, разрывает обе оболочки
и поворачивается спинной частью
к желтку, как показано на рис. 7.8,7*.
Развитие пищеварительной системы.
Подробное обсуждение происхождения и
образования различных органов насе-
комых не входит в задачи этой книги. Од-
нако образование пищеварительного
тракта представляет особый интерес, и
краткие сведения о начальных этапах его
развития дают представление о том, как
вообще образуются органы. Последова-
тельные стадии формирования пищева-
рительного тракта схематично изобра-
жены на рис. 7.9. На рис. 7.9,Л показаны
две группы эндодермальных клеток (за-
чатков передней и задней частей средней
кишки), которые растут навстречу друг
другу с переднего и заднего концов за-
родыша. На рис. 7.9,Б можно видеть,
что каждая из этих групп клеток на-
чинает образовывать мешок, еще незамк-
нутый в средней части тела, который по-
степенно охватывает желточную массу. В
это же время эктодерма на обоих концах
яйца впячивается, давая начало передней
и задней частям пищеварительного трак-
та. На рис. 7.9,В видна следующая ста-
дия - продолжение формирования вну-
треннего мешка, заключающего в себя
желток, и рост эктодермальных впячива-
ний. Рис. 7.9,7" показывает уже пол-
ностью сформировавшийся пищевари-
тельный * тракт зародыша, передняя и
задняя части средней кишки соедини-
лись, полностью вобрав в себя весь жел-
ток, образовались отверстия, соединяю-
щие среднюю кишку с передним и зад-
ним впячиваниями эктодермы. Пищева-
рительный тракт, таким образом, со-
стоит теперь из трех четко различимых
отделов: 1) передней кишки, имеющей
эктодермальное происхождение, 2) сред-
ней кишки, имеющей эндодермальное
происхождение, и 3) задней кишки, как и
передняя, также имеющей эктодермаль-
ное происхождение.
Вылупление из яйца. Полностью сфор-
мированный и готовый покинуть яйцо
7. Жизненные циклы, рост и размножение
207
Рис. 7.8. Схематические рисунки, демонстрирующие положение и перемещения зародыша
внутри яйца, происходящие тремя способами (По Снодграссу; Я-по Филипченко, 2>-по
Heymons.) А. Зародыш (7) ногохвостки Isotoma cinerea, погрузившийся путем изгибания
внутрь желтка на нижней стороне яйца. Б, В. Зародыш чешуйницы Lepisma, глубоко
погруженный в желток у заднего конца яйца на ранней стадии (Б); отверстие образовавшейся
при этом полости имеет вид маленькой поры (2); на более поздней стадии (В), обращенный
частично к наружной стороне яйца, в таком положении зародыш остается до конца своего
развития. Г Д, Е. Продольные срезы яйца, на которых видно, как задний конец зародыша
изгибается и погружается сначала в желток (Г), затем зародыш, перевернувшись,
оказывается целиком внутри желтка (Д) и, наконец, вновь выворачивается и занимает
прежнее положение у поверхности яйца (£), оставаясь в этом положении до вылупления. Ж,
3, И. Поперечные срезы яйца, на которых видно, как из складок бластодермы на обоих
концах зародыша начинают образовываться зародышевые оболочки (Ж), постепенно
разрастаясь под зародышем (3); в конце концов эти складки соединяются, образуя две
оболочки (Я), наружную - серозу (5), и внутреннюю-амнион (6).
208
7 Жизненные циклы, рост и размножение
Рис. 7.9. Формирование пищеварительного тракта в эмбриогенезе. (По
Снодграссу.)
7-зачаток передней части средней кишки; 2-зачаток задней части средней
кишки; 3-желток; 4-мезодерма, 5-эктодерма, 6-задняя кишка, 7-передняя
кишка; 8-ротовое отверстие; 9-средняя кишка; 10-анальное отверстие.
(т. е. готовый к вылуплению) зародыш
должен своими собственными силами
разорвать оболочку яйца, или хорион.
Перед вылуплением зародыш заглаты-
вает воздух или амниотическую жид-
кость, чтобы увеличить свой объем. При
выходе из яйца зародыш совершает рит-
мичные мышечные движения, либо нада-
вливая на оболочку яйца, либо ударяясь
в нее головой.
У некоторых насекомых, например
саранчовых, зародыш просто разрывает
оболочку на переднем конце яйца. У дру-
гих, например у многих Hemiptera и не-
которых веснянок (рис. 7.5,Б), на одном
из концов яйца имеется «шапочка», кото-
рую зародыш легко снимает, как кры-
шечку. В третьей группе насекомых на
переднем конце зародыша имеется яйце-
вой зуб типа пилы, иглы или бритвы, с
помощью которого зародыш произво-
дит первичный разрыв хориона.
После разрыва оболочки зародыш
выходит из яйца. Очень часто нимфа по-
крыта эмбриональной, или пронимфаль-
ной, оболочкой, которая сбрасывается,
по мере того как нимфа выходит из яйца.
Сброшенная «шкурка» остается внутри
яйца или торчит из него. Яйцевой зуб
является утолщением этой пронимфаль-
ной оболочки. После выхода из яйца ос-
вобожденный от эмбриональных оболо-
чек зародыш становится нимфой первой
стадии, или личинкой, с которой на-
чинается постэмбриональный период
развития насекомого (рис. 7.5,В).
Полиэмбриония. В яйцах некоторых
паразитических Hymenoptera часто обра-
зуется более чем один зародыш, что
является результатом бесполого размно-
жения. У Platygaster hiemalis, паразита
гессенской мушки, в каждом оплодотво-
ренном яйце образуются два зародыша.
У Macrocentrus gifuensis, относящегося к
семейству Braconidae, в каждом оплодо-
творенном яйце может образоваться не-
сколько зародышей. У некоторых насе-
комых число зародышей в яйце может
достигать 100 или даже 3000, причем
каждый из них становится активной ли-
чинкой. Множественное деление в яйце
происходит до собственно эмбриогенеза.
7. Жизнешнле циклы, рост и размножение
209
Рис. 7.10. Полиэмбриония у хальциды Litomastix. (Из Clausen, 1940, по Sylvestri,
Entomophagous insects, с разрешения McGraw-Hill Book Со.)
Из одного яйца паразита в каждой гусенице развиваются до 2000 личинок.
Ядро исходной зиготы митотически де-
лится на ряд дочерних ядер, и из каждого
затем развивается зародыш. Зародыши
образуют либо неправильные группы,
либо длинные цепи. Благодаря полиэм-
брионии мелкое насекомое-паразит по-
лучает возможность с максимальной
пользой для себя использовать крупного
хозяина-достаточно одного яйца, поме-
щенного в тело хозяина, чтобы создать
большое число потомков, которым пре-
доставляются богатые пищевые ресурсы.
Примером таких насекомых являются
мелкие хальциды Litomastix truncatellus,
паразитирующие на крупных гусеницах
бабочек (рис. 7.10); из небольшого числа
яиц внутри одной гусеницы развивается
более 2000 личинок, и ни одна частичка
хозяина не пропадает для паразита.
Постэмбриональное развитие.
Метаморфоз
Морфологические изменения, проис-
ходящие в процессе развития от личинки
до взрослого насекомого, называются
метаморфозом. В результате этих изме-
нений облик насекомого на более позд-
них стадиях развития становится часто
абсолютно непохожим на его ранние эм-
бриональные формы. Примером такого
резкого изменения формы является пре-
вращение личинки в непитающуюся ста-
дию, куколку, предшествующую появле-
нию взрослого насекомого. Метаморфоз
у других насекомых, например у стрекоз,
хотя и не включает куколочную стадию,
состоит в смене резко отличных друг от
друга форм. У многих насекомых про-
цесс роста включает периодические линь-
ки (см. стр. 122). Большинство насе-
комых линяют по меньшей мере 3^4
раза, тогда как у некоторых бывает до 30
и более линек. В среднем число линек со-
ставляет 5-6.
Процесс линьки носит название экди-
зиса; шкурки, сбрасываемые насекомы-
ми при линьке, называются экзувиями.
Возраст и стадия. За небольшим ис-
ключением, для каждого вида насекомо-
210
7, Жизненные циклы, рост и размножение
го характерны определенное число ли-
нек, определенная продолжительность
времени между ними и определенное уве-
личение размеров тела после каждой
линьки. Период между любыми двумя
линьками называется стадией, а само на*
секомое в период данной стадии -возра-
стом. Так, период между вылуплением
из яйца и первой линькой-это первая
стадия. Любая особь» находящаяся на
этом этапе развития, называется первым
возрастом. Другими словами, мы можем
сказать о каком-либо виде, что первая
стадия продолжается у него 5 дней и что
первый возраст у него мелкий и желтый.
У всех насекомых, кроме примитивных
бескрылых, после достижения взрослой
стадии линьки прекращаются.
Взрослое состояние. Взрослое насеко-
мое, или имаго, имеет полностью раз-
витые и функционирующие органы раз-
множения и связанные с ними наружные
структуры для спаривания и яйцекладки.
У крылатых насекомых на этой стадии
появляются функционирующие крылья.
Единственным исключением являются
поденки (Epbemeroptera), у которых фа-
за, предшествующая крылатой репродук-
тивной фазе, также имеет функциони-
рующие крылья. Этот необычный пре-
имагикальный возраст насекомых,
имеющий крылья, но не достигший зре-
лости, называется субимаго.
Метаморфоз. В отрядах примитивных
бескрылых насекомых (Apterygota) изме-
нения, происходящие у насекомых после
вылупления до достижения взрослого со-
стояния» сводятся к изменению пропор-
ций тела и развитию репродуктивных ор-
ганов. У таких ^метаболических насе-
комых заметных внешних изменений во
время метаморфоза не наблюдается. У
крылатых насекомых (Pterygota) вылу-
пившиеся особи устроены крайне просто,
а у взрослых имеются крылья и свя-
занные с ними склериты, и в целом
взрослые насекомые ничем не похожи на
только что вылупившихся особей. Такая
смена форм на разных стадиях развития
в жизненном цикле насекомого известна
под названием метаморфоза.
ПОСТЕПЕННЫЙ МЕТАМОРФОЗ. У
примитивных крылатых насекомых кры-
лья появляются впервые примерно в
третьем возрасте в виде небольших вы-
ростов в задних частях второго и третье-
го грудных сегментов. Эти зачатки кры-
льев с каждой следующей линькой увели-
чиваются в размерах (рис. 7Д1) и в
последнем преимагинальяом возрасте
они уже могут покрывать несколько сег-
ментов брюшка. В грудных склеритах до
этого момента никаких изменений нс
происходит. У насекомого последнего
преимагикадьного возраста происходит
полное развитие имагинальных струк-
тур, и взрослое насекомое появляется в
результате последней линьки полностью
сформированным: У только что слиняв-
шего взрослого насекомого крылья мяг-
кие и сморщенные, но очень скоро проис-
ходит ях расправление и затвердение, и
они становятся функционально полно-
ценными.
Такое прямое развитие, при котором
крылья развиваются постепенно как на-
ружные выроста, называется посте-
пенным метаморфозом1. Оно называется
также простым или частичным метамор-
фозом и пауро метаболическим разви-
тием. Неполовозрелые стадии обычно
именуются нимфами.
Тараканы, саранчовые, веснянки, ци-
кадки и клопы являются типичными при-
мерами насекомых с постепенным мета-
морфозом, У этих насекомых нимфы по
внешнему виду похожи на взрослых» от-
личаясь от них только отсутствием кры-
льев, репродуктивных органов и свя-
* В русской литературе этот тип превра-
щения называют неполным.- Яром. ред.
7. Жизненные циклы, рост и размножение
211
Рис. 7.11. Постепенный метаморфоз (неполное превращение) у Hemiptera. (Из
материалов министерства сельского хозяйства США, отдел образования и
науки.)
А -Ж*-последовательные возрасты.
занных с ними структур. У бескрылых
представителей этого типа развития
нимфы отличаются от взрослых в основ-
ном по не полностью развитым генита-
лиям. В отрядах Ephemeroptera (поденки)
и Odonata (стрекозы) нимфы ведут во-
дный образ жизни и в связи с этим имеют
много специализированных черт, напри-
мер особое строение ротового аппа-
рата, хорошо развитые латеральные
и анальные жабры, специфические очер-
тания тела. Поэтому в отличие от тара-
канов или веснянок нимфы стрекоз и
поденок непохожи на взрослых и обычно
называются наядами. Такой вариант по-
степенного метаморфоза носит назва-
ние гемиметаболического развития, он
встречается только у водных насекомых,
у которых жабродышащие наяды превра-
щаются во взрослых крылатых насе-
комых, имеющих дыхальца.
ПОЛНЫЙ МЕТАМОРФОЗ. У многих
насекомых зачатки крыльев развиваются
внутри тела вплоть до преимагинального
возраста, когда крылья выворачиваются
наружу в виде больших придатков. Это
непитающаяся стадия, во время которой
из тканей, принадлежащих более ранним
стадиям, формируются имагинальные
структуры. У большинства насекомых
212
7. Жизненные циклы, рост и размножение
эта стадия неподвижна, но Rhaphidiopte-
га, Megaloptera и некоторые прими-
тивные Hymenoptera весьма активны, а
если их в это время потревожить, то и
агрессивны. У этих насекомых к концу
преимагинального роста сразу по-
являются полностью сформированные
крылья. Следовательно, у насекомых с
таким типом развития выделяют три чет-
ко различимые постэмбриональные
фазы: молодая форма, не имеющая кры-
ловых зачатков,-личинка, покоящаяся
форма, имеющая крыловые зачатки,-ку-
колка и, наконец, взрослое насекомое
(рис. 7.12). Такой тип развития назы-
вается полным или непрямым метамор-
фозом и голометаболическим развитием.
К этой группе насекомых относятся ба-
бочки, пчелы, мухи, жуки, трипсы и неко-
торые представители Hemiptera - чер-
вецы и щитовки.
Наиболее простой вариант полного
метаморфоза встречается у трипсов, у
которых личинки очень похожи на
взрослых особей по форме тела, строе-
нию антенн, ног и ротового аппарата. У
трипсов, а также у червецов и щитовок
имеются две куколочные стадии, пред-
ставленные в обоих случаях типичными
покоящимися куколками. Другой, тоже
простой вариант встречается у Megalop-
tera (большекрылые). У них личинки в
большей степени отличаются от
взрослых особей по строению антенн и
ротовых частей, чем у трипсов, но ноги и
многие другие образования почти не от-
личаются от этих же структур
у взрослых. У большекрылых и трипсов,
как и у насекомых с постепенным мета-
морфозом, грудные склериты взрослых
насекомых и преимагинальных стадий
резко различаются.
У других насекомых с полным мета-
морфозом между личинками и взрослы-
ми насекомыми значительно больше
структурных различий. В некоторых
группах, таких, как бабочки и пилильщи-
ки, личинки имеют расчлененные парные
брюшные ноги. У личинок во всех отно-
сящихся к этому типу развития отрядах
антенны, глаза и грудные ноги упрощены
или совсем неразвиты. Может создаться
впечатление, что все эти структуры, как и
крылья, появляются на стадии куколки
уже практически в том виде, в котором
они присутствуют потом у взрослого на-
Молодая
личинка
Зрелая личинка
Куколка
Имаго
Рис. 7.12. Полный метаморфоз (полное превращение) у жука Phyllophaga sp. (Из
материалов министерства сельского хозяйства США.)
7. Жизненные циклы, рост и размножение
213
секомого. В действительности антенны,
ноги, глаза и другие структуры обра-
зуются не путем перестройки редуциро-
ванных личиночных структур, а, подобно
крыльям, развиваются из обособленных,
находящихся в неактивном состоянии
участков эмбриональных тканей, назы-
ваемых зачатками. У личинок эти зачат-
ки, развившиеся до некоторой степени,
имеют вид внутренних рудиментов, на-
зываемых имагиналъными дисками. Пре-
красным примером развития из имаги-
нальных дисков является образование
крыльев.
Имагинальные диски начинают раз-
виваться либо рано у личинки, либо еще
на поздних этапах эмбриогенеза. Ти-
пичные стадии роста крыловых имаги-
нальных дисков схематически изобра-
жены на рис. 7.13, A-Ж. Вначале имаги-
нальный диск представляет собой просто
небольшой участок, состоящий из более
крупных эпителиальных клеток (А). Этот
участок увеличивается в размерах (Б) и
начинает отходить от кутикулы, образуя
своеобразный внутренний карман (В). За-
тем клетки в одном из участков стенки
кармана вновь увеличиваются, впячи-
ваются внутрь кармана (Г), образуя
складку и в конце концов мешок с двой-
ной стенкой (Ди Е). Перед окукливанием
этот мешок, представляющий собой ру-
диментарное крыло, обычно выворачи-
вается наружу, его полость уплощается,
и мешок оказывается лежащим непосред-
ственно под кутикулой (Ж). Когда во
время линьки на куколку кутикула
сбрасывается, крыло становится наруж-
ной структурой. Если отпрепарировать
рудиментарные крылья, находящиеся в
имагинальных дисках, то они выглядят
почти как крыловые зачатки гемиметабо-
лических нимф.
Образование такого большого числа
структур взрослого насекомого из
имагинальных дисков позволяет разви-
ваться у личинок специализированным
личиночным структурам, не переходя-
щим в организм взрослого насекомого.
Эти структуры либо сбрасываются вме-
сте с покровами последнего личиночного
возраста, либо растворяются в результа-
те катаболических процессов, протекаю-
щих у ранней куколки. Эти особенности
голометаболических насекомых предста-
вляют собой тот механизм, благодаря
которому пути развития личинок
и взрослых насекомых одного и того же
вида могут быть абсолютно разными.
Как правило, насекомые полностью ре-
Рис. 7.13. Стадии развития имагинальных дисков крыльев (крыловых зачатков).
7-область будущего имагинального диска, 2-клетки гиподермы; 3-кутикула;
4 - начало образования имагинального диска; 5 - разросшийся имагинальный диск
крыла.
214
7. Жизненные циклы, рост и размножение
ализуют эти возможности. Основная
функция личинок - питание, а взрослых -
распространение и размножение. На-
ибольшего развития эта тенденция до-
стигла у мускоидных мух (Diptera), ли-
чинки которых не имеют не только ног,
но также глаз, антенн, всех частей рото-
вого аппарата и головной капсулы, тогда
как взрослые особи - типичные быстро
летающие насекомые. Хотя личинки этих
мух выглядят как чрезвычайно просто
организованные существа, их ротовой
аппарат, трахейная система, ферментная
система и мускулатура очень высокоспе-
циализированы.
Есть несколько примеров, когда и ли-
чинки, и взрослые являются формами,
высокоспециализированными по спосо-
бам питания. Личинки комаров, живу-
щие в воде, имеют жующий ротовой ап-
парат, и большинство из них питается
микроорганизмами; у взрослых насе-
комых-колюще-сосущий ротовой аппа-
рат, при этом самцы питаются нектаром,
а большинство самок-кровью птиц и
млекопитающих. Отряд Siphonaptera
(блохи) представляет собой другой инте-
ресный пример. Личинки блох питаются
органическими веществами субстра-
та гнезда хозяина, а взрослые-крово-
сосы.
ГИПЕРМЕТАМОРФОЗ. Обычно все
личиночные возрасты у одного вида
имеют одинаковые пищевые предпочте-
ния, сходны по своему облику и разли-
чаются только по размерам. Однако у не-
которых насекомых имеются два или
более четко различимых типа личинок.
Такой тип развития называется гиперме-
таморфозом. Наиболее яркими примера-
ми этого типа являются жуки-нарывники
(семейство Meloidae) и многие из парази-
тических Hymenoptera (рис. 7.14). У этих
насекомых личинка первого возраста
подвижна, тело ее снабжено щетинками,
хвостом и другими отростками, эти ли-
чинки живут в покровах или во внутрен-
них тканях хозяина. Личинки последнего
возраста неподвижны и лишены всех осо-
бенностей личинок первого возраста.
Другой удивительный пример гиперме-
таморфоза-это паразитические насе-
комые из отряда веерокрылых, относя-
щиеся к семейству Stylopidae (рис. 8.107).
Молодые личинки этих насекомых ак-
тивны, имеют ноги, щетинки и хвост, а
следующие личиночные возрасты мало-
подвижны и имеют червеобразное тело.
Такой же тип развития у жуков-нарывни-
ков (Meloidae, рис. 8.98), хотя у них раз-
личия между личиночными возрастами
касаются в основном формы тела и не-
которых морфологических особен-
ностей.
Существуют и промежуточные типы
развития, когда личинки разных возра-
стов значительно различаются по форме
тела и поведению, но более или менее
сходны по морфологии. Например, у не-
которых паразитических жуков-стафили-
нов молодые личинки первого возраста
активные, с тонкими покровами, а после-
дующие похожи на червеобразных гусе-
ниц. В нескольких родах ручейников
(Trichoptera) личинки первого возраста
свободноживущие, с тонкими покрова-
ми, а последующие возрасты имеют тело
с плотными покровами и живут в спе-
циальных сооружаемых ими домиках.
Гистолиз и гистогенез. Во время мета-
морфоза происходит множество процес-
сов, ведущих к перестройке клеток, тка-
ней и органов. Все эти процессы можно
подразделить на две группы: процессы
гистолиза (распада тканей) и гистогенеза
(образования и дифференцировки тка-
ней). Гистолиз и гистогенез обычно на-
чинаются в последнем личиночном воз-
расте и продолжаются в течение всей
куколочной фазы. Источниками энергии,
необходимой для химических реакций,
лежащих в основе этих процессов, служат
7. Жизненные циклы, рост и размножение
215
Рис. 7.14. Гиперметаморфоз у паразитического перепончатокрылого Apanteles
melanoscelus. (Из материалов министерства сельского хозяйства США.)
А. Взрослое насекомое. Б. Яйцо. В. Личинки первого возраста, верхняя еще
прикреплена к желтку. Г Личинка второго возраста. Д. Питающаяся личинка третьего
возраста. Е. Та же личинка перед сплетением кокона (фаза предкуколки). 1 - анальный
сегмент.
запасы жиров, гликогена и других угле-
водов в жировом теле личинки, сахар
крови и мышцы. При гистолизе фаго-
циты и ферменты превращают жировое
тело, большую часть мышечной ткани и,
несомненно также, другие ткани личинки
в питательный субстрат, доставляемый
кровью к растущим тканям. На более по-
здних этапах гистолиза сами фагоциты
усваиваются развивающимися новыми
тканями. Гистолиз и гистогенез проте-
кают одновременно. Наибольшей пере-
216
7 Жизненные циклы, рост и размножение
стройке подвергается мышечная система.
У всех насекомых при этом происходит
сначала дегенерация, а затем регенера-
ция мышечной ткани. У более низкоорга-
низованных насекомых эти изменения
представляют собой только внутрикле-
точную цитоплазматическую перестрой-
ку, тогда как у более высокоорганизо-
ванных насекомых в результате гистоли-
за и гистогенеза происходит полная за-
мена личиночной мускулатуры на имаги-
нальную.
У насекомых с полным превращением
радикальные перестройки (смена личи-
ночных признаков имагинальными) про-
исходят в основном на стадии куколки.
Но эти перестройки не сопровождаются
кардинальными физиологическими изме-
нениями. Развитие имагинальной гипо-
дермы и трахейной системы в ряде слу-
чаев, по-видимому, связано с има-
гинальными дисками. Быстрое увеличе-
ние объема нервной системы происходит
за счет роста составляющих ее компонен-
тов, причем этот процесс иногда сопро-
вождается слиянием некоторых ганглиев.
Рост сердца не сопровождается какими-
либо особенно заметными изменениями.
Изменения в пищеварительном тракте
сводятся к росту или редукции одних ча-
стей и перестройке других (рис. 4.3).
В имагинальной фазе появляется ряд
структур, отсутствовавших у личинок-
крылья, антенны, сложные глаза, органы
размножения. Другие структуры взрос-
лого насекомого резко отличаются от
своих личиночных предшественников по
размерам и строению, например конеч-
ности и мускулатура, особенно мышцы,
управляющие полетом или связанные с
половым поведением насекомого.
Педогенез. Это явление встречается у
небольшого числа насекомых и состоит в
том, что личинки или куколки отклады-
вают яйца или рождают молодь. В жиз-
ненном цикле редкого североамерикан-
ского жука Micromalthus debilis часть
личинок-яйцекладущие или живородя-
щие. Личинки галлиц из родов Miastor и
Oligarces рождают также личинок, но яиц
не откладывают. Среди куколок хироно-
мид из рода Tanytarsus есть как яйцекла-
дущие, так и живородящие. Педогенез-
это аномальный тип метаморфоза и ро-
ста, когда созревание половой системы
опережает созревание других систем
взрослого организма. Часто педогенез
связан с необычным жизненным циклом
(см. обсуждение на стр. 229) и является
результатом нарушения гормонального
баланса.
СОЗРЕВАНИЕ
Половое созревание и спаривание. На-
ступление половой зрелости у взрослых
насекомых редко происходит сразу же
после имагинальной линьки. В большин-
стве случаев самцам требуется для созре-
вания еще несколько дней, а самкам даже
больше. Обычно спаривание происходит
до созревания яиц у самки, и сперма хра-
нится в сперматеке самки до момента
оплодотворения.
У некоторых насекомых с небольшой
продолжительностью жизни, например
поденок, половое созревание самцов и
самок происходит в течение нескольких
часов после последней линьки; спарива-
ние происходит примерно через день,
вскоре после этого самка откладывает
яйца.
Откладка яиц. Большинство насе-
комых откладывают яйца, причем спо-
собы откладки яиц чрезвычайно варьи-
руют у разных насекомых. Палочники
(семейство Phasmidae) бросают яйца по-
одиночке на субстрат, бабочки приклеи-
вают их к листьям, пилильщики и неко-
торые сверчки (рис. 7 15) делают ямку в
листе или стебле для каждого яйца. Яйца
могут откладываться отдельно каждое и
7. Жизненные циклы, рост и размножение
217
Рис. 7.15. Сверчок-трубачик, откладывающий яйца. (Из Metcalf, Flint,
Destuctive and useful insects, с разрешения McGraw-Hill Book Co.)
группами в виде больших скоплений. У
веснянок и поденок яйца накапливаются
на конце тела самки в виде обособленной
массы, которую она откладывает как
единое целое. У тараканов этот способ
получил дальнейшее развитие. Яйца по
мере высвобождения из тела самки
склеиваются между собой слизистым се-
кретом, образующим компактную капсу-
лу, или оотеку, и в таком виде отклады-
ваются самкой. У многих поденок яйца
откладываются единовременно в виде
большой порции. Яйцекладка у постель-
ного клопа идет медленно и может про-
должаться месяцами. Это лишь несколь-
ко примеров. Другие примеры приводят-
ся при описании отрядов насекомых в
следующей главе.
При определении насекомых в по-
левых условиях, когда требуются точные
данные для установления необходимости
и времени проведения мероприятий про-
тив вредителей, форма яиц, места от-
кладки и цвет являются важными систе-
матическими признаками.
Живорождение. Не все насекомые от-
кладывают яйца. В разных группах
имеются живородящие виды, у которых
из тела самки вместо яиц выходят мо-
лодые особи. У живородящих насекомых
яйца остаются в яйцеводах или в матке
по крайней мере до конца эмбрионально-
го развития. Это явление встречается в
разных отрядах насекомых. У взрослых
насекомых известно несколько типов жи-
ворождения, из которых одни предста-
вляют собой лишь небольшие видоизме-
нения яйцекладки, тогда как другие свя-
218
7. Жизненные циклы, рост и размножение
заны с развитием специальных структур.
У мясных мух (семейство Sarcophagi-
dae) личинки вылупляются из яиц очень
рано-еще в яйцевых путях самки: яйца
находятся в матке до созревания, и не-
посредственно перед откладкой происхо-
дит вылупление. Юные личинки прохо-
дят, подобно яйцам, через яйцеклад, и
при выходе из него находятся на ранней
стадии развития, соответствующей вылу-
пляющейся из яйца личинке яйцекладу-
щих насекомых. У жуков из семейства
Stylopidae вылупление происходит в теле
самки, и крошечные личинки выползают
из ее генитального отверстия.
Развитие личинок в матке происходит
у овечьей кровососки Melophagus ovinus и
африканских мух из рода Glossina (отряд
Diptera). У этих насекомых матка пред-
ставляет собой большую камеру
(рис. 7.16), в которую впадают протоки
специальных желез, секретом которых
питается личинка, находящаяся в этой
камере в течение всего периода своего
развития. Окукливается личинка сразу же
после выхода из тела самки.
Классическим примером живорожде-
ния являются партеногенетические поко-
ления тлей. Неоплодотворенные яйца
развиваются в яичниках, и самки ро-
ждают активных нимф, подобных ним-
фам первого возраста, которые вылу-
пляются из яиц яйцекладущих поколений
тлей.
Продолжительность жизни взрослых
насекомых. Продолжительность жизни
взрослых насекомых у разных видов ко-
леблется от нескольких дней до несколь-
ких лет. Существует корреляция между
продолжительностью жизни и плодови-
тостью; смерть обычно наступает после
прекращения половой активности или
откладывания яиц. Так, самки некоторых
видов сеноедов (отряд Psocoptera) живут
около 20 дней и погибают через 5-6 дней
после последней яйцекладки. Неоплодо-
творенные самки этих видов редко от-
кладывают яйца и живут примерно на 20
дней дольше, чем оплодотворенные сам-
Рис. 7.16. Личинка (незаштрихована, за исключением ее дыхальцевой
пластинки) мухи цеце Glossina в материнской матке. (По Снодграссу.)
7-к придаточной железе; 2-к сперматеке; 3-яйцевод; 4-ротовое отверстие;
5-нервная система, 6-личинка; 7-стенка матки самки; 8-задняя кишка,
9-дыхальцевая пластинка личинки; 10-средняя кишка, 77-анальное
отверстие.
7. Жизненные циклы, рост и размножение
219
ки, откладывающие нормальное число
яиц. У многих видов насекомых имаго
способны зимовать, и в этом случае об-
щая продолжительность жизни у них
примерно равна году. Например, у ряда
листоедов половое созревание наступает
в июле, в течение остающегося летнего
времени они питаются и затем уходят на
зимовку; следующей весной они стано-
вятся активными, спариваются, в течение
мая-июня откладывают яйца и вскоре
погибают.
Вольтинизм. Число поколений насе-
комых, развивающихся в течение одного
года, называется вольтинизмом. У моно-
вольтинных насекомых за год развивает-
ся одно поколение, у бивольтинных-два,
у поливольтинных - более двух.
Партеногенез. Развитие особей из не-
оплодотворенных яиц называется парте-
ногенезом. Для некоторых групп насе-
комых, напрцмер для тлей (отряд Hemip-
tera), муравьев, ос и пчел (отряд Hyme-
noptera), партеногенез является обычным
явлением, он встречается в большинстве
других отрядов, за исключением Odona-
ta, Dermaptera, Neuroptera и Siphonapte-
ra. В зависимости от генетического меха-
низма, лежащего в основе этого явления,
различают несколько типов партеногене-
за. Медоносные пчелы представляют со-
бой пример гаплодиплоидии, при кото-
рой из неоплодотворенных яиц (га-
плоидных) развиваются самцы (трутни),
а из оплодотворенных (диплоидных) -
самки (рабочие), эти самки поддержи-
ваются в нерепродуктивном состоянии
благодаря действию специального секре-
та, вырабатываемого репродуктивной
самкой-царицей, или маткой. Такой тип
партеногенеза, когда из неоплодотво-
ренных яиц появляются только самцы,
носит также название арренотокии. Га-
плодиплоидия встречается в четырех
отрядах насекомых - Hymenoptera, Не-
miptera, Thysanoptera и Coleoptera. В
отряде Hymenoptera гаплодиплоидия
распространена очень широко-от при-
митивных пилильщиков до высокоспе-
циализированных паразитических и об-
щественных пчел и ос. Вылупление из
неоплодотворенных яиц только самок
называется телитокией. Такое явление
встречается у пилильщиков Caliroa cerasi
и Endelomyia aethiops, у некоторых мо-
шек (семейство Simuliidae) и у многих
жуков-долгоносиков (семейство Curcu-
lionidae).
Прекращение активности: состояние
покоя и диапауза. В жизненном цикле
многих насекомых есть более или менее
длительный период покоя, когда прекра-
щается видимая активность и тормозят-
ся многие физиологические процессы.
Такие периоды могут наблюдаться на
разных фазах развития: яйца, личинки,
нимфы, куколки или имаго. У неполово-
зрелых фаз это проявляется прежде всего
в прекращении роста, у взрослых-в пре-
кращении активности и полового созре-
вания.
Состояние покоя является непосред-
ственным следствием наступления не-
благоприятных внешних условий (напри-
мер, низкой температуры или летней
засухи); сразу же после восстановления
нормальных условий активность насе-
комых возобновляется. Так, североаме-
риканский пшеничный клоп при благо-
приятной температуре может развивать-
ся непрерывно, но при помещении
взрослых особей в условия низких темпе-
ратур они впадают в состояние покоя.
Диапауза - временное прекращение
развития, наступающее у насекомых на
определенной фазе жизненного цикла.
Это состояние отличается от состояния
покоя двумя основными чертами.
1. Это активно индуцируемое состоя-
ние, при котором наблюдается ослабле-
ние деятельности нейроэндокринной си-
стемы. Обычно такое состояние насту-
220
7. Жизненные циклы, рост и размножение
пает в специфический для данного вида
момент жизненного цикла.
2. Диапауза наступает в ответ на воз-
действие определенных факторов внеш-
ней среды, которые, не являясь сами по
себе неблагоприятными для вида, сигна-
лизируют о скором наступлении небла-
гоприятных условий. К числу наиболее
важных сигнализирующих факторов от-
носятся продолжительность фотоперио-
да (длина дня) и температура.
Диапауза - это генетически обусло-
вленное явление. Она регулируется вну-
тренним механизмом, вызывающим сни-
жение активности насекомого еще до
наступления неблагоприятных условий.
Однако для прекращения диапаузы необ-
ходимо, чтобы организм какое-то время
находился в неблагоприятйых условиях.
Продолжительность диапаузы может
быть равна нескольким неделям, меся-
цам или годам. У некоторых видов диа-
пауза наступает в каждом поколении не-
зависимо от внешних условий. У других
она наступает только в тех поколениях,
когда имеет место воздействие опреде-
ленных условий на какой-либо более ран-
ней стадии жизненного цикла. В качестве
примера можно привести бабочку Нуа-
lophora cecropia. В последнем периоде ро-
ста, обычно в конце лета, гусеница
строит кокон и превращается в куколку,
которая впадает в состояние диапаузы.
Если примерно в течение 6 нед куколку
содержать при температуре 3-5°С, а за-
тем поместить в нормальную комнатную
температуру, то она возобновляет свое
развитие. Если впавшую в диапаузу ку-
колку все время держать при комнатной
температуре, то окончание диапаузы мо-
жет наступить через полгода или даже че-
рез год.
Наступление диапаузы связано также
с сезонными изменениями длины дня,
или фотопериодизмом. Характер этой
зависимости неодинаков у разных насе-
комых. В некоторых случаях наступле-
нию диапаузы на определенной фазе раз-
вития может предшествовать чувстви-
тельный (критический) период на более
ранней фазе. Так, у бабочки Polychrosis
детерминация куколочной диапаузы
обусловлена фотопериодическими усло-
виями, при которых происходит эмбрио-
нальное развитие. Иной характер зависи-
мости диапаузы от фотопериода у жуков
Leptinotarsa' если личинки последнего
возраста подвергаются действию длин-
ного светового дня, то они впадают в
диапаузу сразу же после того, как за-
капываются в землю. У шелкопряда
Bombyx стимулом для наступления диа-
паузы служит увеличение длины светово-
го дня весной.
Механизм, лежащий в основе насту-
пления и прекращения диапаузы, нахо-
дится под контролем гормональной си-
стемы. Изменения в гормональной систе-
ме происходят либо в результате воздей-
ствия внешних условий (фотопериода
или температуры) на нервную систему и
нейросекреторные клетки, либо под
влиянием генетических факторов. В зави-
симости от характера гормональной ре-
гуляции различают три основных типа
диапаузы (Novak, 1974).
1. Личиночная и куколочная диа-
паузы во всех случаях вызываются дефи-
цитом активационного гормона (АГ) и
как следствие этого дефицитом гормона
линьки экдизона (ЛГ). Куколочная диа-
пауза (в период низких зимних темпера-
тур) у американского шелкопряда Нуа-
lophora cecropia связана с низкой концен-
трацией экдизона в организме. После
того как куколка подвергнется действию
критических низких температур и затем
более высоких, концентрация экдизона
повысится, диапауза прекратится и во-
зобновится нормальное развитие.
2. У взрослых насекомых причиной
диапаузы является дефицит АГ и юве-
7. Жизненные циклы, рост и размножение
221
нильного гормона неотенина (ЮГ). У ко-
лорадского жука диапауза наступает при
дефиците ЮГ и может быть прекращена
увеличением концентрации этого гормо-
на.
3. Диапауза, возникающая на ранних
стадиях эмбриогенеза, вызывается дей-
ствием нейросекрета, образующегося в
подглоточном ганглии самок, на разви-
вающиеся яйца. У шелкопряда Bombyx
яйцевая диапауза детерминируется усло-
виями освещения и температуры, при ко-
торых происходил эмбриогенез материн-
ской особи. Воздействие этих условий
также опосредовано нейросекреторным
гормоном.
ТИПЫ ПИТАНИЯ
Пища представляет собой суще-
ственный фактор роста любого организ-
ма и, конечно, имеет большое значение в
жизни насекомых. В качестве пищи насе-
комые используют разнообразный орга-
нический материал как живой, так и
мертвый. В соответствии с характером
используемой пищи насекомых можно
разбить на несколько групп, перечис-
ленных ниже. Для каждой группы приво-
дится один пример.
1. Сапрофаги -питаются мертвым орга-
ническим веществом.
Питающиеся на любом разлагающем-
ся субстрате - Dictyoptera (тараканы).
Питающиеся гумусом (перегноем) -
Collembola (ногохвостки).
Питающиеся навозом - некоторые
Scarabaeidae (жуки-навозники).
Питающиеся только мертвыми тканя-
ми растений-Isoptera (термиты).
Питающиеся только мертвыми тканя-
ми животных - Dermestidae (жуки-
кожееды).
Питающиеся падалью - Calliphoridae
(мясные мухи).
2. Фитофаги-питающиеся на живых
растениях.
Питающиеся листьями - Orthoptera
(саранчовые).
Минеры листьев-Agromyzidae (мини-
рующие мухи).
Стволовые и корневые точильщики-
Cerambycidae (жуки-дровосеки).
Питающиеся корнями растений-не-
которые Scarabaeidae (жуки-хрущи).
Г аллообразователи - Cynipidae (гал-
лообразующие осы).
Питающиеся соками растений-ци-
кадки и тли.
Питающиеся фруктами - Тephritidae
(фруктовые мухи).
Мицетофаги, питающиеся грибами,-
Mycetophagidae (грибные жуки).
3. Зоофаги-питающиеся на живых жи-
вотных.
Паразиты (живущие на других жи-
вотных).
Живущие на теплокровных позвоноч-
ных-Phthirapter а (пухоеды и вши).
Периодические паразиты позвоноч-
ных-кровососущие Culicidae (кома-
ры).
Энтомофаги-паразиты или хищники,
питающиеся другими насекомыми,-
Ichneumonidae (наездники).
Среди этих пищевых специализаций
большой интерес представляют необыч-
ные отношения между насекомыми и хо-
зяином, существующие у галлообразова-
телей и паразитов.
Галлообразователи. Многие насе-
комые вызывают аномальное разраста-
ние тканей или уродства у растений, на-
зываемые галлами (рис. 7.17), и живут
внутри этих образований. Галлы могут
возникать на листьях, почках, стеблях
или корнях. Каждое насекомое индуци-
рует развитие особого свойственного
только ему типа галлов и всегда на опре-
деленной части растения. Пилильщик
Euura salicisnodus вызывает образование
222
7. Жизненные циклы, рост и размножение
Рис. 7.17. Галлы насекомых. (Из Metcafl, Flint.) А Шарообразный галл на золотарнике,
образуемый мухой Eurosta solidaginis. Б. Узелковый галл на ежевике, образованный
орехотворкой Diastrophus nebulosus. В. Волосистый галл на побеге дуба, образованный
орехотворкой Andricus seminator. Г Галл «сосновая шишка» на иве, образованный
галлицей Rhabdophaga strobiloides. Д. Галлы на листе дуба, образованные орехотворкой
Dryophanta lanata. Е. Колючий галл на хамамелисе, образуемый тлей Hamcunelistes
spinosus. Ж. Колючий галл на розе, образуемый орехотворкой Rhodites bicolor. 3. Галл на
листе дуба, вызванный галлицей Cecidomyia poculum; один галл показан в разрезе (7). И.
Колючий галл на дубе, образуемый орехотворкой Philonix prinoides. К. Большое «дубовое
яблоко», образуемое орехотворкой Amphibolips confluens.
галлов на стволе, а другой пилильщик
Еиига hoppingi-на листьях ивы. У неко-
торых видов насекомых, для которых ха-
рактерно чередование поколений, разные
поколения образуют галлы разной
формы.
До сих пор точно не установлен меха-
низм образования галлов, но известно,
Что они представляют собой чрезмерно
разросшиеся ткани растения; такой чрез-
мерный рост обусловлен вредным дей-
ствием одного или нескольких веществ,
7. Жизненные циклы, рост и размножение
223
В
Рис. 7.18. Открытые (А, Б) и закрытый (В)
галлы. (А и 2>-из Wellhouse.)
выделяемых насекомыми. В слюне тлей,
например, содержится вещество, сни-
жающее уровень гормонов роста и уве-
личивающее уровень ингибиторов роста
в растении. По всей вероятности, слюна
насекомых снижает способность расте-
ния к заживлению ран или вовсе лишает
его этой способности.
Галлы бывают двух типов - открытые
и закрытые (рис. 7.18). Открытые галлы
похожи на мешок с наружным отвер-
стием. Образование таких галлов вызы-
вается тлями, причем для каждого вида
тлей характерен только ему одному свой-
ственный тип галлов. Галл образуется
вокруг участка, где питается одна тля.
Ткань вокруг этого участка разрастается
и постепенно свертывается или склады-
вается, образуя мешкообразную по-
лость, внутри которой находится тля.
Края таких галлов обычно плотно приле-
гают друг к другу в течение всего цикла
развития дочернего поколения тлей. По-
сле завершения цикла развития насеко-
мого края галла расходятся, и тли выхо-
дят наружу. Галлы филлоксеры и клещей
относятся к такому же типу.
Закрытые галлы образуются раз-
личными группами перепончатокрылых,
в том числе несколькими родами пилиль-
щиков, некоторыми хальцидами и всеми
орехотворками (семейство Cynipidae).
Самки этих насекомых откладывают яй-
ца по одному под поверхность листа в
его ткань. Вокруг личинки образуется
галл, который личинка никогда не поки-
дает, питаясь его внутренними тканями.
Взрослое насекомое, чтобы покинуть
галл, прокладывает себе путь, выедая
ткани листа. У некоторых пилильщиков
личинка покидает галл и окукливается
вне его. В других случаях личинка оку-
кливается внутри галла, и взрослое насе-
комое «проедает» себе путь наружу.
Паразиты. С зоологической точки
зрения, паразит-это животное, которое
живет на другом животном-хозяине или
внутри его и питается его тканями по
крайней мере на одной из стадий своего
жизненного цикла. Истинные паразиты
известны в нескольких группах насе-
комых. В зависимости от того, на каких
животных-хозяевах паразитирует насеко-
мое, различают две категории парази-
тов . паразиты теплокровных позво-
ночных и паразиты насекомых и других
мелких беспозвоночных, например пау-
ков и червей.
Насекомые, паразитирующие на те-
плокровных позвоночных, не убивают
своих хозяев, и поэтому на одном и том
же животном-хозяине могут жить многие
особи и многие поколения паразита. Вши
(Anoplura) и пухоеды (Mallophaga) слу-
жат примерами наружных паразитов.
Они проводят на хозяине (птице или мле-
копитающем) всю жизнь, часто достигая
на нем огромной численности; в течение
жизни хозяина происходит постоянная
смена поколений паразита. Очень редко
такие паразиты вызывают гибель хозяи-
на, хотя иногда в значительной степени
ухудшают состояние его здоровья и сни-
жают устойчивость к другим болезням.
Личинки оводов-типичные внутренние
паразиты. Эти личинки живут и дости-
224
7. Жизненные циклы, рост и размножение
гают зрелости в носовой полости, в же-
лудке или кожных покровах хозяина и
после окончания личиночной стадии по-
кидают хозяина и окукливаются в почве.
В одном хозяине может жить много осо-
бей паразита, при этом, как и в случае с
наружными паразитами, хозяин не поги-
бает, хотя и получает ущерб, и его могут
использовать многие поколения парази-
та.
Насекомые, паразитирующие на дру-
гих насекомых, отличаются по ряду при-
знаков от паразитов позвоночных.
1. Обычно только юдна особь парази-
та поражает насекомое.
2. Паразит, как правило, убивает хо-
зяина.
3. Как правило, взрослое насекомое
отыскивает хозяина и откладывает яйца
на поверхность или внутрь его тела. Та-
ких паразитов называют паразитоидами.
Как и у внутренних паразитов позво-
ночных, паразитирующей фазой являют-
ся только личинки. Такой тип паразитиз-
ма встречается во многих семействах
отряда Hymenoptera, в ряде семейств
Diptera и в нескольких родах Coleoptera.
Насекомые, паразитирующие на дру-
гих насекомых, обычно заражают хозяи-
на одним из трех способов. Чаще всего
самка откладывает яйцо на хозяина или
прокалывает яйцекладом покров его тела
и откладывает яйцо в ткани хозяина.
Большинство паразитических Hymenop-
tera и Diptera пользуются именно этим
способом. Второй способ состоит в не-
прямом попадании яйца паразита к хо-
зяину. Самка паразита откладывает яйца
на листья растения, на котором питается
насекомое-хозяин. Если хозяин съедает
зараженные яйцами листья, яйца благо-
получно развиваются, и личинки вылу-
пляются в пищеварительном тракте хо-
зяина, а затем проникают в ткани хо-
зяина. Такой способ заражения известен
для семейства перепончатокрылых Trigo-
nalidae и многих видов мух-тахин (семей-
ство Tachinidae). Третий способ состоит
в том, что яйца откладываются в более
или менее случайном месте, и личинки
первого возраста сами отыскивают свое-
го хозяина (Meloidae, см. стр. 340 и Sty-
lopidae, см. стр. 348).
Большинство паразитов насекомых
относится к внутренним паразитам, но у
некоторых паразитических Hymenoptera
личинки питаются на поверхности тела
хозяина. Эти случаи ближе к хищниче-
ству, нежели к паразитизму.
Паразитизм характеризуется чрезвы-
чайно высоким уровнем специализации.
Известны тысячи видов паразитических
Hymenoptera и Diptera, однако каждый
вид паразитирует, как правило, на одном
виде или группе близких видов-хозяев.
Паразиты бывают первичными, если на
них паразитируют другие паразиты, ко-
торых называют вторичными, или гипер-
паразитами', эти последние в свою оче-
редь могут быть хозяевами третичных
паразитов. Обычно вторичные и тре-
тичные паразиты не столь специфичны в
выборе хозяев, как первичные.
Питание и фазы жизненного цикла. Ос-
новной задачей взрослого насекомого
является размножение. Взрослое насеко-
мое питается, чтобы компенсировать ме-
таболические затраты на двигательную
активность и другие процессы жизнедея-
тельности или чтобы обеспечить пита-
тельными веществами развивающиеся в
его теле яйца или сперму. В таких отря-
дах насекомых, как Orthoptera, Hemipte-
ra, Siphonaptera и у кровососущих видов
Diptera, взрослое насекомое нуждается в
больших количествах пищи, чтобы по-
полнить те запасы, которые достаются
ему от нимфы или личинки.
Во многих случаях взрослое насеко-
мое получает от неполовозрелых фаз до-
статочные запасы жиров или других пи-
тательных веществ, что дает ему возмож-
7. Жизненные циклы, рост и размножение
225
ность обходиться малыми количествами
пищи или не питаться совсем. Такая си-
туация наблюдается у ручейников (Tric-
hoptera), многих Hymenoptera, Lepidopte-
га и Diptera. В самых крайних случаях
этого типа, например у поденок (Epheme-
roptera), самка сразу же после линьки, не
питаясь, способна откладывать яйца.
Другую крайность представляют, напри-
мер, мухи-кровососки (семейство Hippo-
boscidae из отряда Diptera). У этих мух
личинки находятся в теле самки до пол-
ного созревания, и самка является един-
ственной активно питающейся фазой на
протяжении жизненного цикла насекомо-
го.
СЕЗОННЫЕ ЦИКЛЫ
I
Если жизненный цикл -это развитие
особи от яйца до яйца, то сезонный цикл -
это общая последовательность жиз-
ненных циклов или поколений у данного
вида в нормальных для него условиях в
течение года, от зимы до зимы.
У многих видов жизненный цикл
укладывается в одно поколение в течение
года. В этом случае жизненный цикл и се-
зонный цикл совпадают.
У таких видов, как комнатная муха, в
течение теплых месяцев развивается не-
сколько поколений, а затем наступает зи-
мовка или диапауза (см. стр. 219). В этом
случае сезонный цикл состоит из не-
скольких жизненных циклов.
Есть виды, у которых жизненный
цикл продолжается больше года, напри-
мер майские жуки, личинкам которых
требуется 2 или 3 года для созревания,
или один из видов цикад с 17-летним
жизненным циклом. У этих насекомых
сезонный цикл составляет только часть
жизненного цикла. Однако в большин-
стве случаев последовательные поколе-
ния перекрывают друг друга, и в резуль-
тате взрослые особи появляются каждый
год. Термин «сезонный цикл» в этом слу-
чае используется для обозначения сезон-
ной активности вида, представленного
разными поколениями в течение года.
Сезонные циклы, включающие более
чем один жизненный цикл, бывают двух
типов с повторяющимися жизненными
циклами и с чередующимися поколения-
ми.
Повторяемость поколений
В этом случае последовательные по-
коления фактически идентичны друг дру-
гу. Например, у комнатной мухи яйца,
отложенные самками одного поколения,
развиваются так же, как и яйца предыду-
щего поколения, и из них вылупляются
мухи с такими же морфологическими
признаками, характером питания и раз-
множения.
Перерывы в развитии некоторых по-
колений, вызванные летним покоем, зи-
мовкой или другими типами диапаузы,
не меняют основного характера жизнен-
ного цикла насекомого. Так, у той же
комнатной мухи в течение лета сменяется
несколько поколений (жизненный цикл
каждого поколения длится 4-5 нед), пока
условия погоды благоприятны для раз-
вития. Взрослые мухи, появляющиеся
осенью, зимой находятся в состоянии по-
коя, а весной снова становятся активны-
ми. Летняя остановка развития не отли-
чается от зимней ничем, кроме временно-
го фактора.
Чередование поколений
Имеется несколько групп насекомых,
последовательные поколения у которых
существенно отличаются друг от друга
по способу размножения, а иногда и по
поведению.
Насекомые, у которых репродуктив-
ной функцией наделены только взрослые
15-732
226
7. Жизненные циклы, рост и размножение
особи. К этой группе относятся расти-
тельноядные насекомые: тли и галло-
образующие осы.
Тли (Aphididae, Hemiptera) характе-
ризуются сложными и варьирующими
сезонными циклами со сменой поколе-
ний, размножающихся половым путем с
откладкой яиц и партеногенетических с
живорождением, крылатых и бескрылых,
а также миграциями с летнего растения-
хозяина на зимнее (табл. 7.1). Наиболее
простой сезонный цикл у капустной тли
Brevicoryne brassicae. Зимующей фазой у
этих тлей являются яйца, отложенные
осенью на стебли крестоцветных расте-
ний. Личинки, вылупившиеся из этих яиц
следующей весной, развиваются
в бескрылых партеногенетических живо-
родящих самок - тлей-основательниц.
Интересно отметить, что все яйца дают
самок-основательниц. От этих особей по-
лучается поколение партеногенетических
живородящих самок, которые могут
быть как крылатыми, так и бескрылыми.
Такие поколения живородящих самок по-
являются одно за другим в течение лета.
Обычно партеногенетические особи жи-
вут около месяца и рождают от 50 до 100
личинок. Осенью, когда дни становятся
короче, живородящие самки дают поко-
ление, состоящее из бескрылых самок и
крылатых самцов, размножающееся по-
ловым путем. После спаривания каждая
самка откладывает от одного до несколь-
ких яиц, которые остаются на зиму.
Более сложные сезонные циклы вклю-
чают еще ряд особенностей. Многие
виды мигрируют на летние растения-
хозяева. У таких тлей зимнее, или пер-
вичное, растение-хозяин обычно много-
Таблица 7.1. Смена поколений у тлей: Л-без смены хозяев (капустная тля) и Б-со сменой
хозяев (сливовая тля)
Сезон А. Немигрирующий вид Б. Типичный мигрирующий вид
все формы на одном хозяине формы, обитающие на первичном хозяине формы, обитающие на вторичном хозяине
Зима Ранняя весна Поздняя весна Яйца Самки-основательницы Бескрылые живородящие самки Крылатые живородящие самки (весенние миг- ранты) Крылатые и бескрылые живородящие самки (мигрирующие с одного на другое растение того же или близкого вида) Половое поколение: самцы и яйцекладущие самки Яйца Яйца Самки-основательницы Бескрылые живородящие самки Крылатые живородящие самки (весенние миг- ранты) Немногочисленные слу- чайные мигранты Осенние мигранты со вто- ричного хозяина Половое поколение: сам- цы и яйцекладущие сам- ки Яйца Весенние мигранты с первичного хозяина
Лето Ранняя осень Поздняя осень Зима Крылатые и бескры- лые живородящие самки Крылатые живородя- щие самки, иногда крылатые самцы (осенние мигранты)
7. Жизненные циклы, рост и размножение
227
летнее. Яйца откладываются на это рас-
тение осенью, личинки выходят из яиц
весной, и из них развиваются бескрылые
тли-основательницы. Эти самки дают на-
чало поколению бескрылых живородя-
щих самок, от которых получается поко-
ление крылатых живородящих самок,
перелетающих на летнее, или вторичное,
растение-хозяина. Поколения живородя-
щих самок, крылатых или бескрылых,
появляются на этом растении вплоть до
осени, когда рождается поколение ми-
грирующих особей, перелетающих на
зимнее растение; здесь появляется моло-
дое поколение, состоящее из самок и сам-
цов. У некоторых видов тлей крылатые
самцы рождаются на летнем растении,
мигрируют на зимнее, где спариваются с
бескрылыми яйцекладущими самками. У
других видов и самки, и самцы - бескры-
лые и происходят от крылатых живоро-
дящих самок, мигрировавших ранее на
зимнее растение-хозяина.
У некоторых видов тлей разные поко-
ления резко отличаются друг от друга по
своему поведению. Например, в роде
Pemphigus самка-основательница делает
галл на листовой пластинке или черешке
листа тополя. Ее потомки мигрируют на
корни сложноцветных и других растений
и дают начало нескольким поколениям,
живущим на корнях и не образующим
галлов.
Филлоксеры (Phylloxeridae, Hemipte-
га) близки к тлям; у них также наблю-
дается чередование партеногенетических
и половых поколений (несколько кры-
латых и несколько бескрылых)
и сложные миграции. От тлей они, одна-
ко. отличаются тем, что все самки яйце-
кладущие. Наиболее типичным приме-
ром является виноградная филлоксера
«рис. 7.19). Ряд поколений партеногене-
тических самок образуют галлы на
листьях растений, а осенью мигрируют
на корни винограда, где следующей вес-
ной новые партеногенетические поколе-
ния вызывают образование галлов на
мелких корнях винограда. Поскольку на-
секомые в этих поколениях происходят
от неоплодотворенных особей, они назы-
ваются агамными. Некоторые особи, жи-
вущие на корнях, дают осенью начало
партеногенетическим агамным кры-
латым мигрантам, выползающим из зе-
мли и перелетающим на виноградную
лозу. Появившееся здесь потомство со-
стоит из бескрылых агамных самцов и
самок. После спаривания каждая самка
откладывает одно яйцо в трещину коры.
Яйца перезимовывают, следующей вес-
ной из них вылупляются личинки, даю-
щие начало серии поколений, живущих
на листьях.
Среди галлообразующих ос (Cynipi-
dae, Hymenoptera) имеется много видов,
которым свойственно чередование по-
ловых и партеногенетических поколений.
У Andricus erinacei, живущей на дубе, зи-
муют яйца, отложенные в листовые или
цветочные почки. Вылупление из яиц
происходит весной, и каждая личинка
оказывается окруженной мягким по-
чечным галлом. Внутренний слой галла
личинка использует в качестве пищи.
Ранним летом по завершении личиноч-
ного развития появляются крылатые
самцы и самки. Самки откладывают яй-
ца на жилки листьев дуба. Вылупившиеся
из яиц личинки вызывают аномальное
разрастание жилок листа с образованием
галлов, которые носят название галлов-
ежиков. Личинки в этих галлах дости-
гают зрелости осенью; из них разви-
ваются самки с короткими крыльями,
которые откладывают партеногенетиче-
ские зимующие яйца в дубовые почки.
Чередование поколений, среди которых
есть партеногенетические, создает воз-
можность для быстрого роста численно-
сти популяции, что обеспечивает успеш-
ное использование короткого периода
228
7. Жизненные циклы, рост и размножение
Рис. 7.19. Виноградная филлоксера; форма, живущая на корнях винограда. (Из Riley.)
а-здоровый корень; б-корень, пораженный насекомыми; видны узелки и вздутия,
вызванные их уколами; в-корень, покинутый насекомыми, с начинающими отмирать
корешками; г-насекомые на крупном корне; Э-агамная женская нимфа, вид с
дорсальной стороны; е-та же нимфа с вентральной стороны; ж-крылатая агамная
самка, вид с дорсальной стороны; з-та же самка с вентральной стороны;
«-увеличенная антенна крылатого насекомого; к-бескрылая агамная самка, кладущая
яйца на корни; л-гниение большого корня в результате проколов насекомых.
7. Жизненные циклы, рост и размножение
229
благоприятных условий. Благодаря пар-
теногенезу отдельные самки могут да-
вать начало новым популяциям, быстро
расселяясь с помощью ветра, или, наобо-
рот, снижать численность при неблаго-
приятных условиях.
Педогенез. Педогенез, т. е. размноже-
ние на личиночной фазе, обычно соче-
тается со сложным и нерегулярным чере-
дованием поколений. Изученные случаи
такого развития показывают, что может
быть несколько последовательных поко-
лений педогенетических личинок, ко-
торые время от времени производят ли-
чинок, нормально созревающих и оку-
кливающихся. Получающиеся в этих слу-
чаях имаго нормально спариваются, и
самки откладывают оплодотворенные
яйца. Сложный цикл развития жука Mic-
romalthus debilis включает педогенетиче-
ские и нормальные поколения. Наиболее
полно изучено в этом ртношении семей-
ство Cecidomyiidae (галлицы), особенно
европейский вид Oligarces paradoxus.
Смена поколений у этого вида выглядит
следующим образом (рис. 7.20) отпедо-
генетической личинки (а) происходит ли-
чинка (б), которая, вырастая, дает один
из 4 вариантов: 1) другую педогенетиче-
скую личинку, подобную личинке а; 2)
личинку в, дающую самку; 3) педогенети-
ческую личинку г, дающую начало ли-
чинкам д, из которых развиваются толь-
ко самцы, или 4) педогенетическую ли-
чинку е, из которой развиваются личинки
д. дающие начало самцам, и другую пе-
догенетическую личинку типа а. Самки и
самцы получаются от соответствующих
личинок путем нормального окуклива-
ния и выхода из куколок. Самки от-
кладывают оплодотворенные яйца, из
которых выходят педогенетические ли-
чинки типа а.
Появление взрослых особей Oligarces
paradoxus связано чаще всего с перенасе-
ленностью колонии или с недостатком
Рис. 7.20. Чередование поколений у галлицы
Oligarces paradoxus. (Из Wigglesworth по
Ulrich.)
а-педогенетическая личинка, рождающая (б)
недетерминированную дочернюю личинку,
которая может развиваться либо снова в
педогенетическую личинку (а), либо
в личинку, которая даст самку (в), либо в
личинку (г), которая сменится личинкой (б),
дающей затем самца, или в личинку (е), от
которой получится недетерминированная
личинка дочернего поколения и личинка,
дающая самца; ж-куколка самки;
з-куколка самца; и- самка; к-самец;
л-сперматозоид; м, н-яжхъ, о -личинка,
вылупившаяся из яйца.
пищи. У педогенетических видов галлиц,
относящихся к роду Mias tor, тип каждого
очередного поколения определяется из-
менениями температуры. Таким обра-
зом, у этих комариков жизненные циклы
с педогенетическими поколениями и се-
230
7. Жизненные циклы, рост и размножение
зонный цикл зависят от конкретных ус-
ловий их существования.
РАЗМНОЖЕНИЕ
Размножение насекомых предста-
вляет собой функцию половой системы.
Обычно для насекомых характерен дву-
полый тип размножения, т. е. яйцо, обра-
зующееся в организме самки, не разви-
вается до тех пор, пока не будет оплодо-
творено сперматозоидами, образующи-
мися у самца. За исключением немногих
видов, одна особь может обладать при-
знаками только одного из двух полов.
Поэтому физиология размножения боль-
шинства насекомых изучает развитие и
созревание сперматозоидов у самцов и
яиц у самок, а также способы встречи по-
лов, необходимой для оплодотворения.
Вопросы, связанные со спариванием, об-
суждались в гл. 6.
Развитие сперматозоидов. Спермато-
зоиды образуются в фолликулах семен-
ников (рис. 7.21). В верхней части фолли-
кулов находятся первичные половые
клетки - сперматогонии. Эти клетки
многократно делятся и образуют цисты,
которые перемещаются к основанию
фолликула под давлением, возникаю-
щим в результате увеличения их соб-
ственных размеров. Попав в основание
фолликула, каждая клетка в цисте снова
многократно делится, так что общее чис-
ло клеток может увеличиться в 5-250 раз.
После этих митотических делений проис-
ходит редукционное деление, при кото-
ром уменьшается число хромосом. За
ним следует период трансформации
Рис. 7.21. Продольный разрез фолликула из
семенника самца саранчи (полусхематично),
а-апикальные клетки, окруженные
сперматогониями; б-зона сперматогониев;
в-зона сперматоцитов; г-цисты с митозами
второго деления созревания; д, е-зона
сперматид; ж-зона сперматозоидов.
7. Жизненные циклы, рост и размножение
231
округлых клеток в тонкие, снабженные
жгутжами сперматозоиды, которые че-
рез семевыносящий проток (vas efferens)
попадают в семепротоки (vas deferens). В
расшнрешюй или извитой части этого
протока, семенном пузырьке, спермато-
зоиды сохраняются до спаривания. При
спаривании сперматозоиды переносятся
в тело самки, где они сохраняются до
оплодотворения.
Сотремте яиц. Яйца развиваются в
овариолах (яйцевых трубочках) яичшиса.
Верхняя часть овариолы, гермарий, со-
держит первичные половые клетки (оого-
нии), из которых в результате дифферен-
цировки образуются яйца (ооциты), пи-
тающие клетки (трофоциты) и клетки
фолликулярного эпителия. Обычно в ова-
риоле одновременно находятся ооциты
на раэшях стадиях развития; они полу-
чают питательные вещества из фоллику-
лярного эпителия и от питающих клеток,
если последние присутствуют в овариоле
(рис. 7.22). Ооцит увеличивается по мере
накопления в нем желтка, состоящего из
белков, липидов, гликогена и многих
Рис. 7.22. Продольные разрезы овариол. (Из Weber.) А. Простая
овариола паноистического типа, содержащая только ооциты
и фолликулярный эпителий. Б. Овариола политрофического типа с
ооцитами, фолликулярным эпителием и питающими клетками. В.
Овариола телотрофического типа с питающими клетками, которые
связаны с ооцитами с помощью питающих тяжей. 7-терминальный
филамент; 2-гермарий; 3-ооциты; 4-фолликулярный эпителий;
5-хорион; 6-яйцо; 7-питающие клетки; 8-питающие тяжи.
232
7. Жизненные циклы, рост и размножение
других соединений. Когда процесс нако-
пления желтка в ооците заканчивается,
фолликулярный эпителий секретирует
хорион (оболочку яйца) и желточную
оболочку, после чего дегенерирует. Пи-
тающие клетки также дегенерируют.
Под ооцитом в конце овариолы нахо-
дится пробка из эпителиальных клеток,
закрывающая проток, соединяющий ова-
риолу с яйцеводом. Когда ооцит пол-
ностью созреет, эта пробка разрушается
и ооцит выходит в яйцевод. Часть ова-
риолы, в которой находилось вышедшее
яйцо, сморщивается, и под следующим
ооцитом образуется новая пробка.
Ко времени выхода в яйцевод яйца
уже окружены хорионом. Эта оболочка
имеет в одном или нескольких местах
крошечные поры, или микропиле. Имен-
но через эти крошечные отверстия спер-
матозоиды попадают внутрь яйца.
Оплодотворение. После прохождения
яиц по яйцеводу (что возможно благода-
ря перистальтическим движениям яйце-
вода) по направлению к влагалищу они
оказываются у отверстия протока спер-
матеки. Из этого протока выходят спер-
матозоиды и проникают в яйцо через ми-
кропиле. После проникновения сперма-
тозоида в яйцо ядро яйца делится
дважды, причем одно из этих делений-
редукционное с образованием женского
пронуклеуса и полярных телец. Сперма-
тозоид теряет жгутик и превращается в
мужской пронуклеус. Сливаясь, женский
и мужской пронуклеусы образуют зиго-
ту.
Существует много вариантов этой
описанной последовательности событий.
Приведем наиболее интересные при-
меры. У постельного клопа Cimex спер-
матозоиды из сперматеки мигрируют в
фолликулярные ткани овариолы и оттуда
в ооцит, находящийся на ранней стадии
развития. Оплодотворение, таким обра-
зом, происходит до образования хорио-
на. У европейского елового пилильщика
Diprion hercyniae, размножающегося пар-
теногенетическим путем, оплодотворе-
ния не происходит, а диплоидный набор
хромосом восстанавливается путем слия-
ния полярного тельца с женским прону-
клеусом.
Перенос спермы и спаривание. При ка-
ждом спаривании самец переносит в сам-
ку большое число сперматозоидов. Эти
сперматозоиды хранятся в сперматеке, и
их выход оттуда находится под строгим
контролем. Лишь небольшое число спер-
матозоидов выходит из сперматеки в тот
момент, когда яйца проходят по яйцево-
ду. Вследствие этого для оплодотворе-
ния каждого яйца не требуется отдельно-
го спаривания, и большинство насе-
комых спаривается в течение жизни толь-
ко один раз, остальные, как правило,
спариваются лишь несколько раз.
Сразу после имагинальной линьки на-
секомые не спариваются, но через не-
сколько часов; а чаще через день-два,
большинство из них становятся спо-
собными воспринимать стимулы, исхо-
дящие от партнера. Эти стимулы бывают
самые разнообразные специфические
движения, например танцы роящихся
самцов поденок, звуки-стрекотание
сверчков и кузнечиков, цвет-у неко-
торых бабочек, но главным образом-
различные запахи. Обо всех этих явле-
ниях речь шла в гл. 6. Гонады, по-види-
мому, не оказывают существенного
влияния на поведенческие реакции, пред-
шествующие спариванию, поскольку
многие виды спариваются еще до того,
как яичники самки достигнут полного
развития, кастрированные самцы также
способны к нормальному спариванию,
при этом лишь отсутствует перенос
спермы.
Способы переноса спермы в тело сам-
ки можно разделить на несколько четко
определимых типов. У многих насе-
7. Жизненные циклы, рост и размножение
233
комых, например у настоящих клопов,
пенис вводится в сперматеку самки, и
сперматозоиды оказываются непосред-
ственно в своей «камере хранения». У
многих бабочек, саранчовых и жуков
сперматозоиды при спаривании вводятся
в совокупительную сумку (bursa copula-
trix) самки, откуда они мигрируют в
сперматеку. О механизме такой мигра-
ции известно мало; полагают, что этот
процесс может стимулироваться проста-
гландинами, синтезируемыми в прида-
точных половых железах самца и вво-
димых самке со сперматозоидами.
У многих насекомых, имеющих сово-
купительную сумку, сперматозоиды
переносятся в тело самки в специальной
капсуле с плотной оболочкой, которая
называется сперматофором и образуется
за счет секрета придаточных половых же-
лез самца. Сперматофор помещается в
совокупительную сумку или во влага-
лище, а затем его содержимое переносит-
ся в сперматеку. После эТого самка ос-
вобождается от пустого сперматофора.
Жизнеспособность сперматозоидов. По
всей вероятности, секреты, образующие-
ся в сперматеке или в связанных с ней же-
лезах, обеспечивают длительное сохране-
ние жизнеспособности сперматозоидов.
У медоносных пчел этот период равен не-
скольким годам, а у муравьев - более чем
16 годам. У бабочек сперматозоиды в
сперматеке остаются жизнеспособными
в течение нескольких месяцев. Лишь у не-
многих насекомых, к которым относится
постельный клоп Cimex, сперматозоиды,
не использованные в течение нескольких
недель, перевариваются и поглощаются
тканями самки, поэтому спаривание для
пополнения запаса сперматозоидов у
этих насекомых происходит периоди-
чески.
РОЛЬ ГОРМОНОВ
Процессы роста, развития и размно-
жения у насекомых регулируются гормо-
нами, образующимися в эндокринной си-
стеме (см. гл. 6). Существуют две ос-
новные группы гормонов, гормоны,
оказывающие свое действие на тот же ор-
ганизм, в котором они вырабатываются,
и называемые эндогенными гормонами, и
гормоны, действующие на другие орга-
низмы и называемые экзогенными гормо-
нами. Один из известных примеров экзо-
генных гормонов связан с биологией
блох. Яйца кроличьей блохи Spilopsyllus
cuniculi не созревают до тех пор, пока
блоха не начнет питаться на беременной
крольчихе, в крови которой в это время
содержатся гормоны, контролирующие
ее собственный репродуктивный цикл.
Именно эти гормоны стимулируют со-
зревание яиц блохи. Возможно, такая за-
висимость представляет собой механизм,
повышающий вероятность того, что сле-
дующее поколение получит достаточное
количество пищи.
Обычно гормоны классифицируют по
месту их образования в организме и пути
попадания в систему циркулирующих
жидкостей тела. Различают три типа
гормонов.
1. Гормоны, образующиеся в железах,
не имеющих протоков. Обычно такие же-
лезы секретируют только или главным
образом гормоны, чаще же один специ-
фический гормон.
2. Тканевые гормоны образуются в
различных нежелезистых тканях, для ко-
торых секреция гормона всегда является
вторичной функцией. Например, гиста-
мин образуется в поврежденных тканях.
3. Нейрогормоны образуются в спе-
циальных клетках центральной нервной
системы, так называемых нейросекре-
торных клетках, и в виде нейросекре-
торных гранул направляются вдоль аксо-
234
7. Жизненные циклы, рост и размножение
нов этих клеток к специальным желе-
зистым органам (например, кар-
диальным телам насекомых). Ош сохра-
няются в этих органах и позже посту-
пают в кровь.
Нейросекрецию обычно рассматри-
вают как третью систему, контролирую-
щую физиологические и поведенческие
процессы и занимающую промежуточ-
ное положение между эндокринной и не-
рвной системами. Эта третья система ин-
дуцирует длительно протекающие гор-
мональные процессы в ответ на корот-
кие, обычно повторяющиеся нервные
стимулы. Например, изменение длины
светового дня вызывает образование в
определенных нейросекреторных клетках
головного мозга активационного гормо-
на, который, воздействуя на протора-
кальные железы и прилежащие тела, сти-
мулирует в них секрецию специальных
гормонов.
Нейрогормоны образуются в раз-
личных частях центральной нервной си-
стемы, в том числе в головном ганглии,
подглоточном ганглии, торакальном и
абдоминальном ганглиях, стомо-
деальных ганглиях и нейрогемальных ор-
ганах. Различные эффекты этих нейро-
гормонов перечислены ниже.
1. Влияние на частоту и амплитуду
спонтанной ритмической активности раз-
личных мышц (например, мышц сердца,
мальпигиевых сосудов, яйцеводов, ки-
шечника и т. д.).
2. Влияние на перемещение пигмен-
тов-их распределение и концентрацию в
гиподерме и хроматофорах, образование
различных пигментов в гиподерме.
3. Влияние на циклическую, сезон-
ную и суточную активность.
4. Регуляция водного баланса насе-
комого.
5. Активация ферментов (например,
протеиназы кишечника).
6. Активация секреции других эндо-
кринных желез.
7 Индукция диапаузы.
8. Влияние на развитие гонад.
9. Влияние на процессы метаболиз-
ма, например на поддержание уровня
трегалозы в крови.
10. Влияние на имагинальную линь-
ку.
11. Контроль за затвердением кути-
кулы у только что перелинявших насе-
комых и за образованием эндокутикулы
после линьки. Нейросекрет, осущест-
вляющий этот контроль, называется бур-
сиконом.
Гормональная регуляция
физиологических процессов
Гормоны эндокринных желез и ней-
рогормоны оказывают огромное влия-
ние на процессы, из которых складывает-
ся жизненный цикл насекомых. Прекрас-
ной иллюстрацией этого является гормо-
нальная регуляция метаморфоза. В на-
стоящее время известны три гормона
метаморфоза', контролирующие постэм-
бриональное развитие: активационный
гормон, гормон линьки, или линочный
гормон, и ювенильный гормон.
1. Актшащюннын гормон (АГ). Этот
стероидный гормон образуется в спе-
циальных нейросекреторных клетках
протоцеребрума гойовного мозга насе-
комых. Он влияет на возобновление ак-
тивности насекомых после каждой линь-
ки и стимулирует образование двух дру-
гих гормонов-линочного и ювенильно-
го. Кроме того, он влияет (опосредован-
но через corpora allata) на развитие яич-
ников у самок и придаточных половых
желез у самцов. Прекращение секреции
этого гормона играет важную роль в ре-
гуляции диапаузы (гл. 7, стр. 220).
2. Гормон линьки (ЛГ; экдазон). Этот
гормон образуется в проторакальных же-
7. Жизненные циклы, рост и размножение
235
лезах или в соответствующих им тканях,
а у некоторых насекомых-в яичниках
самок. Он также является стероидным
гормоном и синтезируется в организме
насекомого из холестерина. Существуют
две формы этого гормона (а и Р), близ-
кие по характеру своего действия. Гор-
мон инициирует процесс линьки и, таким
образом, косвенно регулирует рост и
морфогенез. У личинок мух он вызывает
образование пупария. Этот гормон
оказывает также общее влияние на разви-
тие, стимулируя синтез ДНК, РНК и бел-
ка и активность ферментов.
3. Ювенильный гормон (ЮГ; неоте-
шн). Этот гормон образуется в corpora
allata и выделяется под влиянием нерв-
ных или гормональных стимулов, посту-
пающих из головного мозга. Он имеет
следующие функции: обусловливает рост
личинок у всех метаболических насе-
комых, стимулирует развитие яичников у
самок большинства насекомых, контро-
лирует развитие и активность ряда орга-
нов и процессов - так, например, жировое
тело не растет и не функционирует в от-
сутствие этого гормона. Роль ЮГ можно
проиллюстрировать на следующем при-
мере. При развитии насекомого, когда в
крови юной личинки содержится ли-
ночный гормон, ЮГ обусловливает се-
крецию гиподермой личиночной кути-
кулы. Когда уровень ЮГ понижается,
гиподерма секретирует куколочную ку-
тикулу. В отсутствие ЮГ гиподерма
образует кутикулу взрослого насекомо-
го. Во взрослом организме ЮГ стимули-
рует развитие яиц. ЮГ также оказывает
общее влияние на метаболизм. Напри-
мер, у самца саранчи Schistocerca ЮГ
контролирует изменение окраски при по-
ловом созревании. У нескольких видов
саранчи ЮГ участвует в регуляции про-
цесса перехода одиночной формы в стад-
ную мигрирующую форму. У термитов и
тлей ЮГ играет определенную роль в
дифференциации каст и типов особей.
Развитие солдат у термитов требует
большего количества ЮГ, чем развитие
обычных особей.
ЛИТЕРАТУРА
Agrell I. Р. S., Lundquist А. М. Physiological
and biochemical changes during insect deve-
lopment. In: Morris Rockstein (Ed.). The Phy-
siology of Insecta, 1, 159-247. Academic Press,
New York, 2nd ed., 1973.
Anderson D. T. Embryology and phylogeny in
annelids and arthropods. Pergamon, Oxford,
495 pp., 1973.
Askew R. R. Parasitic insects. American Else-
vier, New York, 316 pp., 1971.
Counce S. J., Waddington С. H. (Eds.). Deve-
lopmental systems: Insects. Academic Press,
London and New York, Vol. 1, 304 pp., Vol. 2,
615 pp., 1971, 1972.
de Wilde J., de Loof A. Reproduction. In.
Morris Rockstein (Ed.). The physiology of In-
secta, 1, 11-95. Academic Press, New York,
2nd ed., 1973.
Dixon A. F. G. Biology of aphids. Edward
Arnold, London, 58 pp., 1973.
Felt E. P. Plant galls and gall makers. Com-
stock, Ithaca, N. Y., 364 pp., 1940.
Hagan H. R. Embryology of the viviparous in-
sects. Ronald Press, New York, 472 pp., 1951.
Johannsen O. A., Butt F. H. Embryology of in-
sects and myriapods. McGraw-Hill, New
York, 462 pp., 1941.
Matsuda R. Morphology and evolution of the in-
sect abdomen. Pergamon, Oxford, 534 pp.,
1976.
Novak V. J. A. Insect hormones. Chapman &
Hall, London, 600 pp., 1975.
Snodgrass R. E. Principles of insect morphology.
McGraw-Hill, New York, 667 pp., 1935.
White M. J. D. Animal cytology and evolution.
Cambridge Univ. Press, Cambridge, 3rd ed.,
961 pp., 1973.
8
Отряды
энтогнат и насекомых
Пожалуй, одно из величайших чудес живой природы-это то, что насекомые и эн-
тогнаты, возникнув из одной-единственной примитивной формы, достигли той ги-
гантской численности и разнообразия, свидетелями которого мы сейчас являемся.
Эволюционная история этих животных представляет собой историю случайного воз-
никновения crpyKTypj наделявших их обладателей все новыми преимуществами в
борьбе за существование. До нашего времени сохранилось достаточно много древних
и примитивных групп насекомых; мы довольно неплохо знаем и ископаемые формы.
Все это позволяет нам в общих чертах воссоздать основные направления эволюции
этих удивительных животных (рис. 8.1).
В целом эволюционные отношения, показанные на рис. 8.1, отражают гипотезы,
предлагавшиеся разными авторами, в том числе Хеннигом (Hennig, 1969), Гамильто-
ном (Hamilton, 1972) и Кристенсеном (Kristensen, 1975). Сходство между различными
шестиногими членистоногими однозначно свидетельствует об общности их происхож-
дения. Хотя очень многие данные в общем согласуются со взглядами Хеннига-
Кристенсена на филогенез насекомых, все же существует значительное разногласие
относительно группировки отрядов в рамках этой системы при классификации. В дан-
ной книге мы придерживаемся разделения шестиногих (рис. 8.1) на два самостоя-
тельных класса: Entognatha и Insecta, изменяя при этом использование упомянутых
названий в работах Хеннига (Hennig, 1969), Туксена (Tuxen, 1970) и Лаутербаха (Lau-
terbach, 1972).
КЛАСС ENTOGNATHA
Первые Hexapoda отличались от
своих подобных многоножкам предков
тем, что имели грудь, состоящую из
трех сегментов, каждый из которых нес
по паре ног, а брюшные ноги были
сильно редуцированы или отсутствовали.
Молодые особи этих древних форм
внешне почти не отличались от взрослых,
крыльев еще не было. Известно шесть
типов таких древних бескрылых существ.
У трех из них-Diplura, Collembola и
Protura (рис. 8.1)-внутреннее строение
головы мало чем отличается от такового
у различных групп многоножек, зато
бока ротовой полости срослись с боками
нижней губы и выдвинулись наружу.
Эта сложная структура образует полость,
окружающую функционирующие рото-
вые части-мандибулы и максиллы. Кро-
ме того, в этих трех группах отсут-
ствуют сложные глаза. На основе этих
признаков три названных отряда объеди-
няются в примитивную ветвь, названную
классом Entognatha, что значит «с на-
ходящимися внутри ротовыми частями».
Наиболее примитивные из энтогнат отно-
сятся к отряду Diplura рис. 8.1). Два
других отряда-Collembola и Protura -
произошли от диплуроподобных пред-
ков, у которых атрофировались брюш-
оо
Класс Entognatha
to
о
Е
<р
о
и
Надкласс Hexapoda
----------------------------------К ласс Insect а
Подкласс
Apterigota
Инфра-
класс
Paleoptera
Ортоптероидные
Подкласс Pterygota
щ О
отряды
Инфракласс Neoptera-
Нейроптероидные
». Отряды энтогнат и насекомых
Рис. 8.1. Схема предполагаемого филогенеза отрядов энтогнат и насекомых.
238
8. Отряды энтогнат и насекомых
ные дыхальца и церки. Это подтвержда-
ется многими фактами. Хотя по многим
признакам оба отряда сходны, в целом
Collembola и Protura совершенно непо-
хожи друг на друга. У Protura нет глаз,
почти исчезли антенны, зато передние
ноги стали несколько похожими на ан-
тенны, у Collembola уменьшилось число
брюшных сегментов, а остатки ног чет-
вертого брюшного сегмента срослись и
образовали прыгательную вилку.
КЛАСС INSECTA
Подкласс Apterygota
Примитивные бескрылые отряды на-
секомых (рис. 8.1,Б) представлены в
современной фауне щетинохвостками
(Microcoryphia) и чешуйницами (Thysa-
nura), которые составляют вместе под-
класс Apterygota. Ротовой аппарат этих
животных находится снаружи головы
(эктогнатый) и в этом отношении они
сходны с остальными насекомыми. В
этих отрядах сложились определенные
пропорции тела и ног, приведшие к
развитию у насекомых полета. Наиболее
важными в этом отношении были удли-
нение и усиление грудных ног, позво-
лявших быстро и искусно бегать и пры-
гать, а также относительное увеличение
размеров груди по сравнению с брюш-
ком, в результате чего центр тяжести
тела сместился вперед. Из современных
представителей Apterygota Microcoryphia
имеют грифельки на 2-9-м сегментах
брюшка и более примитивны, чем Thy-
sanura, которые лишены грифельков на
1-6-м сегментах. Ископаемый отряд Mo-
tt ига напоминал Microcoryphia, но сохра-
нял грифельки на 1-м сегменте брюшка
и был, следовательно, примитивнее
обоих названных ранее. В эволюционной
линии Thysanura тело сделалось более
широким и плоским, что, вероятно, пре-
допределило возможность по меньшей
мере некоторого планирования в воздухе.
Подкласс Pterygota
ИНФРАКЛАСС PALEOPTERA
Возможно, что у каких-то тизануро-
подобных насекомых развилась спо-
собность к более совершенному плани-
рованию и появились связанные с этой
способностью структуры, что в конце
концов привело к возникновению у них
крыльев и полета, именно эти предки
дали начало крылатым насекомым, или
Pterygota (рис. 8.1,В). Первые настоящие
крылья были собраны в глубокие складки
подобно вееру, но не могли склады-
ваться и в покое просто торчали над
спинной стороной тела. Насекомые с
крыльями этого типа принадлежат к
инфраклассу Paleoptera (древнекрылые),
включающему ряд вымерших отрядов
и два современных-Ephemeroptera (по-
денки) и Odonata (стрекозы).
На этом этапе эволюции резкое изме-
нение наступило в жизненном цикле насе-
комых. В подклассе Apterygota особи
продолжали регулярно линять и после
того, как становились функционально
взрослыми. В подклассе Pterygota линьки
прекращались, как только крылья и орга-
ны размножения насекомого достигали
полного развития. Единственным исклю-
чением являются Ephemeroptera, име-
ющие вполне развитые крылья на про-
тяжении двух последних возрастов. Пред-
последний возраст, называемый суби-
маго, в типичном случае оказывается
неспособным к размножению, репродук-
тивная деятельность свойственна только
настоящей взрослой стадии-имаго. Из
этого можно заключить, что первона-
чально крылатые насекомые (о них нам
ничего не известно) не переставали ли-
нять и по достижении зрелости, как
Apterygota, однако впоследствии сильное
давление естественного отбора привело
к возникновению форм, не линяющих
во взрослом состоянии. Должно быть,
8. Отряды энтогнат и насекомых
239
Ephemeroptera отражают как раз по-
следнюю ступень этого эволюционного
перехода. Они также единственные, у
кого радиальная и секторальная жилки
не слились, что делает их самыми при-
митивными из ныне живущих Pterygota.
Odonata и близкие к ним ископаемые
формы дают представление о следующем
этапе (рис. 8.1,Г) эволюции крылатых
насекомых, когда радиальная и секто-
ральная жилки отходят от основания
крыла единым стволом.
ИНФРАКЛАСС NEOPTERA
В некоторых линиях древнекрылых
(рис. 8.1,Д) начали развиваться меха-
низмы, с помощью которых крылья в по-
кое могли складываться и плоско лежать
на спине. Параллельно исчезла в основ-
ном складчатость крыла, в результате че-
го многие части крыла стали плоскими.
Насекомые с крыльями подобного типа
называются новокрылыми, или Neopte-
ra.
Первые многочисленные представи-
тели Neoptera относились к вымершему
подотряду Protoblattoidea и отряду Рго-
torthoptera- разнообразным группам до-
вольно крупных бегающих насекомых с
полным жилкованием крыла, типичным
пиловидным яйцекладом и длинными
церками. От Protorthoptera или близких к
ним древних форм отделились эволю-
ционные стволы, представленные совре-
менными отрядами новокрылых.
Наиболее примитивная ветвь, ото-
шедшая от Protorthoptera, стала ортоп-
тероидной .ветвью, представленной
семью современными отрядами (рис.
8.1,£). У Embioptera сильно редуциро-
вано жилкование крыла, отсутствует хо-
рошо различимый яйцеклад, а передние
ноги несут замечательные шелкоотдели-
тельные органы. И на передней и на за-
дней паре крыльев радиальная и секто-
ральная жилки слились лишь на неболь-
шом протяжении, поэтому считают, что
этот отряд-самый примитивный в ор-
тойтероидной ветви. Второе направление
составляют Dictyoptera (тараканы и бо-
гомолы) и Isoptera (термиты); обе
группы имеют примитивное жилкование
и хорошо развитый у древних иско-
паемых форм пиловидный яйцеклад, ко-
торый у всех ныне живущих представите-
лей в значительной степени редуцирован.
Еще одна ветвь ортоптероидных насе-
комых включает отряды, характеризую-
щиеся сильно модифицированным жил-
кованием • Dermaptera (уховертки), Phas-
matodea (палочники) и Orthoptera (пря-
мокрылые). Orthoptera имеют хорошо
развитый пиловидный яйцеклад; две
остальные группы его утратили, но у них
появились другие характерные признаки.
К той же ортоптероидной ветви отно-
сится и отряд Grylloblattodea (грилло-
блаттиды), все известные представители
которого лишены крыльев. Положение
этого отряда в системе до сих пор не со-
всем ясно. У самок имеется прими-
тивный пиловидный яйцеклад; генита-
лии самцов также характеризуются ря-
дом примитивных черт строения. Неко-
торые авторы рассматривают данную
группу как входящую в состав Orthopte-
ra, другие считают ее наиболее древней
среди всех ортоптероидов вообще, по-
следнюю точку зрения принимаем здесь
и мы.
В этих семи отрядах (а также предпо-
ложительно и в близких к ним иско-
паемых группах) крылья развиваются в
виде наружных крыловых зачатков, а не-
половозрелые особи, если не считать от-
сутствия крыльев и разницы в размерах,
очень похожи на взрослых и, как прави-
ло, живут и питаются вместе с ними. Эти
современные отряды вместе называют
ортоптероидными отрядами.
В другом стволе, обособившемся от
того же неоптероидного основания
240
8. Отряды энтогнат и насекомых
(рис. 8.1,Ж), один из мезоплевральных
склеритов - трохантин - редуцировался
до тонкого тяжа, образовавшего подвиж-
ное сочленение ноги, а некоторые имаги-
нальные признаки на неполовозрелых
стадиях оставались недоразвитыми. Ве-
роятно, отряд Plecoptera (веснянки)
является одним из ранних ответвлений
этого ствола, ибо трохантин здесь уже
стал узким, тогда как подавление разви-
тия имагинальных признаков у личинок
еще не проявилось. И все же еще раньше,
чем веснянки, отделилась форма, у кото-
рой на неполовозрелых стадиях недораз-
вивались дорсальные глазки. Этот пре-
док дал начало двум важнейшим груп-
пам. гемиптероидным и нейроптероид-
ным отрядам (рис. 8.1,3).
У гемиптероидов число члеников лап-
ки у взрослых форм редуцировалось до
трех. Первыми представителями этого
ствола были, возможно, разноядные на-
секомые, сильно напоминавшие сеноедов
(Psocoptera). Наиболее примитивная
ветвь представлена, очевидно, отрядом
Zoraptera (зораптеры) - группой коло-
ниально живущих насекомых, похожих
на сеноедов, но сохранивших хорошо
развитые церки (см. с. 93). У Pso-
coptera лацинии максилл превратились
в длинные долотоподобные придатки,
создав тем самым основу для формиро-
вания колюще-сосущего типа ротового
аппарата двух остальных отрядов.
Первый этап его развития иллюстри-
руется примером отряда Thysanoptera
(трипсы), у которых образовался ком-
плекс режуще-сосущих ротовых частей.
Завершающая стадия имеет место в от-
ряде Hemiptera (полужесткокрылые, или
клопы), где ротовые части образуют вы-
тянутый колюще-сосущий орган. Другое
ответвление гемиптероидного ствола,
очевидно, обособилось непосредственно
от сеноедоподобного предка, становле-
ние этой группы произошло в связи с пи-
танием мертвыми частицами кожного
покрова теплокровных животных и при-
вело к возникновению эктопаразитиче-
ского отряда Phthjraptera (пухоеды и
вши). У личинок большинства гемипте-
роидных крылья развиваются постепен-
но в виде наружных крыловых зачатков
(паурометаболия). У Thysanoptera и двух
групп из отряда Hemiptera (Psyllidae и
Coccoidea) крылья вначале растут внутри
и выворачиваются лишь на поздней ста-
дии (часто на стадии, предшествующей
имаго), которая является покоящейся и,
таким образом, представляет собой ти-
пичную куколку. В этих группах предку-
колочные стадии обычно называют ли-
чинками.
В нейроптероидном стволе (рис.
8.1,Я) тенденция к подавлению разви-
тия имагинальных признаков у ранних
стадий резко усиливается неполовоз-
релые стадии (или личинки) имеют силь-
но редуцированные глаза и антенны,
склеротизация тела у них выражена сла-
бо. Последняя неполовозрелая стадия
превращается в типичную куколку, в ко-
торой различные органы переформи-
ровываются в имагинальные. Имаго
примитивных представителей сохраняют
пятичлениковые лапки.
Первое нейроптероидное насекомое
можно довольно точно реконструиро-
вать, собрав воедино примитивные при-
знаки, сохранившиеся у современных от-
рядов. Оно, видимо, имело средние для
насекомых размеры. У имаго обе пары
крыльев были приблизительно одинако-
вой величины, и каждое крыло имело
весьма богатый набор продольных и по-
перечных жилок, брюшко было сильно
склеротизировано и снабжено пило-
видным яйцекладом, весьма напоминаю-
щим яйцеклад пилильщиков. Личинка
была удлиненной, хищной, быстро бе-
гающей и напоминала современных ли-
чинок верблюдок и некоторых жуков.
8. Отряды энтогнат и насекомых
241
Отношения родства между отрядами
нейроптероидных насекомых были на-
стоящим камнем преткновения, однако в
последние годы дополнительные морфо-
логические данные позволили восстано-
вить с высокой степенью достоверности
основные моменты эволюции важней-
ших ветвей этого комплекса. Первичный
нейроптероидный ствол сперва дал нача-
ло двум ответвлениям - линии Neuropte-
ra-Coleoptera и линии Hymenoptera-Me-
coptera. В линии Neuroptera-Coleoptera
яйцеклад сильно видоизменился в связи с
редукцией нижних створок и исчезнове-
нием внутренних, личиночные лапки со-
хранили примитивный признак-наличие
двух коготков-по крайней мере у наибо-
лее, примитивных семейств внутри каж-
дого отряда. У предка* линии Hymenop-
tera-Mecoptera яйцеклад был примитив-
ного типа и состоял из трех пар створок;
лапки личинок, однако, несли лишь один
коготок.
Линия Neuroptera-Coleoptera дала на-
чало жесткокрылым Coleoptera, передние
крылья которых сделались жесткими и
образовали защитный покров над брюш-
ком, а также комплексу из трех отрядов:
Megaloptera (вислокрылки, большекры-
лые), Neuroptera (сетчатокрылые) и
Rhaphidioptera (верблюдки). Три послед-
ние группы характеризуются рядом
общих второстепенных морфологиче-
ских признаков, но каждая из них тем не
менее в высшей степени обособлена от
остальных. Megaloptera имеют наиболее
примитивное жилкование, у них водные
личинки с длинными посегментно распо-
ложенными брюшными филаментами и
жабрами. У Rhaphidioptera генерализо-
ванные наземные личинки; имаго
с длинным яйцекладом и видоизме-
ненным жилкованием. Neuroptera крайне
разнообразны; у их личинок мандибулы
и максиллы, плотно прилегая друг к дру-
гу, образуют пару структур, служащих
16-732
для прокалывания и высасывания пищи
из тела своей жертвы.
Линия Hymenoptera-Mecoptera (рис.
8.1,А), с одной стороны, дала нача-
ло отряду Hymenoptera (перепонча-
токрылые), в котором жилкование крыла
претерпело существенную модифика-
цию, а яйцеклад сохранил примитивное
пиловидное строение, а с другой - группе
из пяти отрядов, у которых отсутствует
пилообразная часть и ножны яйцеклада,
вследствие чего последний превращается
в простую растяжимую трубку. Предко-
вая линия этой последней группы, види-
мо, разделилась на ствол Trichoptera-Le-
pidoptera, в котором сильно уменьши-
лось число продольных и особенно попе-
речных жилок, и на ствол Mecoptera-Dip-
tera, в котором нижний край передне-
спинки сросся с передним краем мезо-
плевр ниже дыхалец. У Trichoptera (ру-
чейники), отделившихся от ствола Tri-
choptera -Lepidoptera, личинки стали вод-
ными и их дыхальца атрофировались; у
Mecoptera, происшедших от ствола Ме-
coptera-Diptera, не появилось почти ни-
каких изменений; у Diptera (двукрылые)
жилкование в значительной степени ре-
дуцировалось, вторая пара крыльев пре-
вратилась в маленькие головчатые ор-
ганы для поддержания равновесия, назы-
ваемые жужжальцами, а переднеспинка
очень прочно срослась с прилегающими
плейритами среднегруди.
В отряде Siphonaptera (блохи) отсут-
ствуют крылатые формы; блохи на-
столько сильно видоизменились в связи с
эктопаразитическим образом жизни, что
лишь очень немногие черты говорят об
их происхождении. Два признака, при-
обретенные в ходе эволюции, роднят их с
Mecoptera и Diptera. У примитивных
представителей всех трех отрядов манди-
булы стали плоскими и пластинопо-
добными, приспособленными для проре-
зания и прокалывания тканей своих хо-
242
8. Отряды энтогнат и насекомых
зяев, а яйцеклад имеет простую трубча-
тую форму. Эти особенности заставляют
предполагать, что блохи относятся к то-
му же комплексу (рис. 8.1,Л). Некоторые
внутренние структуры провентрикулуса
замечательно схожи с таковыми у Ме-
coptera, но в отличие от Mecoptera пере-
днеспинка блох не срастается с плейрита-
ми среднегруди; вместе с тем эта связь
без труда могла исчезнуть в ходе эволю-
ции бескрылого состояния. Уникальное
строение провентрикулуса позволяет
предположить, что блохи-крайне спе-
циализированная боковая ветвь древней-
ших Mecoptera.
В ходе длительной эволюции насе-
комые достигли большего видового раз-
нообразия, чем какая-либо другая группа
организмов. 27 современных отрядов на-
секомых включают около 1000 семейств
и многие тысячи родов, а также пример-
но 900 000 описанных видов. Здесь не уч-
тено большое число вымерших отрядов,
родов и видов. Все 27 отрядов, свыше 500
семейств, несколько тысяч родов и более
100000 видов были впервые обнаружены
на территории Северной Америки. На де-
ле же число видов значительно больше.
По некоторым оценкам, примерно 25 000
видов еще будут открыты в фауне Север-
ной Америки, а во всем мире-еще не ме-
нее миллиона видов.
НАЗВАНИЯ ОТРЯДОВ И СЕМЕЙСТВ
Во многих случаях для одного и того
же отряда или семейства употребляют не
одно, а несколько названий. Чаще всего
это вызвано разногласиями по поводу
правильности описания или приоритета.
В этой книге сделана попытка во всех
подобных случаях пользоваться наибо-
лее широко употребляемым названием.
Даются также указания относительно си-
нонимических названий, с которыми на-
чинающий студент может столкнуться в
рекомендуемой научной литературе.
ЗООГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
Многие крупные районы земного ша-
ра содержат в своей биоте характерный
комплекс таксономических элементов.
Основываясь на степени сходства и раз-
личия в составе фауны, зоологи выде-
ляют шесть главных царств, или зоогео-
графических областей (рис. 8.2). Эти
области часто упоминаются в таксоно-
мической литературе.
Общая литература
по отрядам насекомых
После характеристики каждого от-
ряда приводится список литературы, ко-
торая поможет получить более глубокие
знания о соответствующем отряде. Пред-
лагаемый ниже список общих работ
посвящен в основном классификации,
эволюции, более редким или экзоти-
ческим семействам насекомых, термино-
логии, а также коллекционированию и
разведению насекомых.
ЛИТЕРАТУРА '
Borror D. J., DeLong D. М., Triplehom С. А.,
1976. An introduction to the study of unsects,
4th ed., Rinehart, New York.
Brues С. T., Melander A. L., Carpenter F. M.,
1954. Classification of insects, Mus. Comp.
Zool. Harvard Coll. Bull., 108, 1-917.
Foote R. H, 1977. Thesaurus of entomology,
Entomological Society of America, College
Park, Md.
Hamilton K. G.A., 1972. The insect wing, Part
IV. Venational trends and the phylogeny of the
winged orders, J. Kans. Entomol. Soc., 45(3),
295-308.
Hennig W., 1969. Die Stammesgeschichte der In-
sekten, W. Kramer, Frankfurt am Main.
Horn D. C., 1976. Biology of insects, Saunders,
Philadelphia.
Insects-the Yearbook of Agriculture, 1952,
USDA, Washington, DC.
Insects of Hawaii, 1948-1981. Вышли тома
1-14. Univ, of Hawaii Press, Honolulu.
Авторы отдельных томов: тт. 1-9-Zimmer-
man Е. С., тт. 10-14-Hardy D. Е., прило-
жение к т. 11-Tenorio J. М.
8. Отряды энтогнат и насекомых
243
Рис. 8.2. Фаунистические царства, или зоогеографические области Земли. (По Sclater,
Wallace.)
Kristensen N. P., 1975. The phylogeny of hexa-
pod “orders” A critical review of recent
accounts, Z. Zool. Syst. Evolutionsforsch., 13,
1-44.
Lauterbach К. -E., 1972. Beschreibung zweier
neuer europaischer Inocelliiden (Insecta,
Raphidioptera), zugleich ein Beitrag zur ver-
gleichenden Morphologic und Phylogenie der
Kamelhalsfliegen, Bonn. Zool. Beitr., 23,
219-252.
Linsenmaier W., 1972. Insects of the world,
McGraw-Hill, New York.
Mackerras I. M., 1970. Evolution and classifica-
tion of the insects. In. The insects of Australia,
D. F. Waterhouse et al. (eds.), Melbourne
Univ. Press, Carlton, Victoria, pp. 152-167;
Suppl. 1974, pp. 26-27
Merritt R. W., Cummins K. W. (eds.), 1978. An
introduction to the aquatic insects of North
America, Kendall/Hunt, Dubuque, Iowa.
Peterson A., 1948, 1951. Larvae of insects, Parts
1-2, J. W. Edwards, Ann Arbor/,
Peterson A., 1953. A manual of entomological
techniques, 7th ed., J. W Edwards, Ann
Arbor.
Richards O. W., Davies R. G., 1977. Imms’
general textbook of entomology, 10th ed.,
Chapman & Hall, London, vols. 1-2.
Romoser W S., 1973. The science of entomo-
logy, Macmillan, New York.
Ross H. H., 1953. How to collect and preserve
insects, 3rd ed., Ill. Nat. Hist. Surv. Circ., 39,
1-59.
Ross H. H., 1974. Biological systematics, Addis-
on-Wesley, Reading, Mass.
Smith R. C. et al., 1944. Insects in Kansas,
Kansas State Board of Agriculture, Topeka.
Tietz H. M., 1963. Illustrated keys to the fami-
lies of North American insects, Burgess,
Minnneapolis.
Torre-Bueno J. R., 1950. A glossary of entomo-
logy, Brooklyn Entomology Society, Brooklyn,
N.Y.
Tuxen S. L. (ed.), 1970. Taxonomist’s glossary of
16*
244
8. Отряды энтогнат и насекомых
genitalia in insects, 2nd ed., rev., Munksgaard,
Copenhagen.
Usinger R. L.(ed.), 1965. Aquatic Insects of Cali-
fornia, with keys to North American genera
and California species, Univ, of California
Press, Berkeley.
Waterhouse D. F. et al., 1970. The insects of
Australia, Melbourne Univ. Press, Carlton,
1974, Suppl.
КЛАСС ENTOGNATHA
Отряд DIPLURA1 (диплуры,
или двухвостки)
Это бескрылые, слепые, мелких раз-
меров энтогнаты с вытянутым телом,
длинными многочлениковыми антенна-
ми, хорошо развитыми ногами и парой
бросающихся в глаза церков, которые мо-
гут быть либо членистыми, либо клеще-
образными и нерасчлененными. Ротовой
аппарат грызущего типа скрыт в кармане
на вентральной стороне головы. Мета-
морфоз не выражен.
Молодые и зрелые особи отличаются
в основном по размерам и состоянию по-
ловой системы. Щеки и нижняя губа обра-
зуют вентральный карман, в котором
1 Некоторые авторы употребляют для
этого отряда название Entotrophi.
располагаются все остальные ротовые ча-
сти. Ноги развиты не столь хорошо, как у
Thysanura, а брюшко не несет ни зача-
точных парных придатков, ни каудально-
го филамента. У представителей семей-
ства Campodeidae (рис. 8.4,Л) и близких к
нему семейств брюшко несет пару много-
членистых церков, у Japygidae
(рис. 8.4,5) церки напоминают клещи.
Виды этого отряда встречаются под
листьями, камнями, бревнами, расти-
тельными остатками или в почве. Их дви-
жения умеренно быстры и они практиче-
ски никогда не выходят на свет. Об
особенностях их жизненного цикла из-
вестно мало, хозяйственного значения не
имеют
ЛИТЕРАТУРА
Smith L. М., 1960. The family Projapygidae and
Anajapygidae (Diplura) in North America.
Ann. Entomol. Soc. Am., 53, 575-583.
Отряд PROTURA
(протуры, или бессяжковые)
Взрослые особи мелкие и вытянутые
(рис. 8.5), длиной от 0,5 до 2 мм. Голова
конической формы; глаз нет; антенны ре-
Рис. 8.3. Строение груди насекомых.
А. Передне- и среднеспинка Hemerobius (Hemerobiidae). Б. Передне- и
среднеспинка Brachypanorpa (Panorpodidae). В. Передняя часть плейральной области
Brachypanorpa (Panorpodidae). 7 - мезоскутеллум; 2-переднеспинка, 3-мезоплевра.
8. Отряды энтогнат и насекомых
245
Рис. 8.4. Diplura. (Из Essig, 1942, College entomology, с Рис. 8.5. Бессяжковое Acerentulus
разрешения Macmillan Со.) A. Campodea folsomi. Б. Japyx barberi. (По Ewing, 1940).
diversiunguis.
.аудированы до крошечных, напоминаю-
щих глазки структур; ротовой аппарат
состоит из стилетоподобных мандибул,
маленьких неспециализированных мак-
силл и слаборазвитой мембранозной
нижней губы, по бокам срастающейся со
щеками. Три пары грудных ног имеют
сходное внешнее строение; первая пара
служит органами осязания. Три первых
сегмента брюшка несут по паре малень-
ких грифельков на вентральной стороне.
Нимфы в общем внешне похожи на
взрослых. Для развития характерен ана-
морфоз, который состоит в увеличении
числа сегментов тела при каждой линьке.
Брюшко нимфы первого возраста, или
протонимфы, имеет 9 сегментов; брюшко
дейтонимфы имеет 10 сегментов, трито-
нимфы-11 сегментов, и, наконец, брюш-
ко взрослой особи содержит 12 сегмен-
246
8. Отряды энтогнат и насекомых
тов. Голова и грудь не затрагиваются
этим процессом. Протуры относительно
редки. Они живут в подстилке и почве,
предпочитая сырые местообитания.
Идеальные условия для многих видов -
старый гумус из опавшей листвы вдоль
лесных опушек. И взрослые особи, и
нимфы питаются разлагающимся орга-
ническим веществом и встречаются вме-
сте в течение большей части года. Бессяж-
ковых можно собрать либо просматривая
гниющие листья, либо высушивая их в
коллекторе Берлезе. Для исследования
материал хранят в 70%-ном этиловом
спирте, а затем приготовляют микроско-
пические препараты.
В свое время отряд Protura считали от-
носящимся к особому классу-Myriento-
mata.
ЛИТЕРАТУРА
Ewing Н. Е., 1940. The Protura of North Amer-
ica, Ann. Entomol. Soc. Am., 33, 495-551.
Tuxen S. L., 1964. The Protura. A revision of
the species of the world with keys for deter-
mination, Hermann and Cie, Paris.
Отряд COLLEMBOLA
(ногохвостки)
В этот отряд входят мелкие и среднего
размера энтогнаты, антенны и ноги хоро-
шо развиты, ротовой аппарат грызущего
типа, но у некоторых форм максиллы и
мандибулы длинные, острые и стилето-
образные; у всех представителей отряда
щеки срастаются с боками нижней губы,
образуя полый конус, в который кажутся
втянутыми остальные ротовые органы.
Брюшко шестичлениковое, часто имеет
вентральный прыгательный орган, или
вилку, и удерживающую ее структуру -
зацепку. Метаморфоз отсутствует, оба
пола обычно схожи.
Длина тела взрослых особей колеб-
лется от 0,2 мм у самых мелких предста-
вителей, относящихся к роду Megalotho-
гах до примерно 10 мм у крупных видов
Poduridae и Entomobryidae. В подотряде
Arthropleona тело длинное и цилиндриче-
ское ; в подотряде Symphypleona брюшко
округленное, более или менее шаровид-
ное (рис. 8.6).
Антенны имеют от четырех до шести
члеников, причем последний членик иног-
да несет многочисленные тонкие коль-
цевые насечки. ГлЬза либо отсутствуют,
либо представлены рядами изо-
лированных омматидиев. У ногохвосток
отсутствуют также мальпигиевы сосуды
и трахейная система. Большинство Co-
llembola имеют вентральный прыга-
тельный орган -прыгательную вилку; она
закреплена в состоянии покоя брюшной
зацепкой. С помощью прыгательной вил-
ки эти мелкие существа могут совершать
прыжки на некоторое расстояние, откуда
и произошло их название-ногохвостки.
Молодые особи сходны со взрослыми как
по внешнему виду, так и по образу жизни,
отличаясь в основном размерами и непо-
ловозрелым состоянием. Многие виды
имеют белую или соломенную окраску,
другие-голубую, серую, желтую, рябую,
или даже развитый рисунок.
Ногохвостки в большом количестве
встречаются в самых разнообразных
влажных местообитаниях, в том числе в
толще листового перегноя, в сырой почве,
гнилой древесине, на берегах стоячих и
проточных водоемов, на снегу и шля-
почных грибах. Немногие виды, особенно
относящиеся к семейству Sminthuridae,
повреждают растения и могут иметь
местное хозяйственное значение. Вид Ас-
horutes armatus (рис. 8.6,Б), имеющий
мелкие размеры и серую окраску, иногда
вредит искусственно разводимым гри-
бам, причиняя тем самым экономический
ущерб. Поведение при откладке яиц из-
вестно лишь у нескольких видов, которые
откладывают яйца поодиночке или кучка-
ми в перегной или почву.
8. Отряды энтогнат и насекомых
247
Рис. 8.6. Collembola.
A. Isotoma andrei. Б. Achorutes armatus подотряд Arthropleona. В. Neosminthurus clavatus
подотряд Symphypleona. (A, В-no Mills, 1934; Б’-из Illinois Natural History Survey.)
Одной из наиболее интересных осо-
бенностей коллембол является их широ-
кое распространение.
ЛИТЕРАТУРА
Christiansen К., 1964. Bionomics of Collembola,
Annu. Rev. Entomol., 9, 147-178.
Joose E. N. G., 1976. Littoral apterygotes (Col-
lembola and Thysanura). In: Marine insects,
L. Cheng (ed.), North-Holland, Amsterdam,
pp. 151-186.
Maynard E. A., 1951. The Collembola of New
York, Comstock, Ithaca.
Mills H. B., 1934. A monograph of the Collem-
bola of Iowa, Collegiate Press, Ames.
Richards W. R., 1968. Generic classification,
evolution and biogeography of the Sminthuri-
dae of the world (Collembola), Mem. Entomol.
Soc. Can., 53, 1-54.
Salmon J. T., 1964-1965. An index of the Col-
lembola. Bull. R. Soc. N.Z., 7, 1-644, 3 vols.
Scott H. G., 1961. Collembola: pictorial keys to
nearctic genera, Ann. Entomol. Soc. Am., 54
(1), 104-113.
КЛАСС INSECTA
ПОДКЛАСС APTERYGOTA
Отряд MICROCORYPHIA
(щетинохвостки)
Эти мелкие или средних размеров
бескрылые насекомые имеют крупные
глаза, длинные многочлениковые ан-
тенны, длинные церки и длинный не-
парный каудальный филамент (рис. 8.7).
Ротовой аппарат грызущего типа, брюш-
ко с парными грифельками на 2-9-м сег-
ментах (рис. 3.48). Они составляют се-
мейство Machilidae.
248
8. Отряды энтогнат и насекомых
Молодые и взрослые особи чрезвы-
чайно похожи друг на друга формой тела
и образом жизни, различаясь в основном
размерами и степенью половой зрелости.
Тело плотное, суживающееся кзади.
Брюшко состоит из 10 полных сегментов,
одиннадцатый сегмент образует кау-
дальный филамент Ноги умеренно
сильные. Все виды очень быстро бегают и
чрезвычайно увертливы, так что поймать
их очень трудно. Представители наиболее
обычного в Америке рода Mesomachilis
питаются перегноем и встречаются среди
опавшей листвы и вокруг камней. Пола-
гают, что они откладывают яйца по одно-
му в трещины и щели. Эти насекомые не
перестают линять в течение всей жизни.
ЛИТЕРАТУРА
См. список на с. 249.
Отряд THYSANURA (чешуйницы)
В целом сходны с представителями
предыдущего отряда, однако отличаются
от них более плоским и широким телом,
грифельковидными придатками только
на 7-9-м сегментах брюшка и маленьки-
ми глазами, последние могут отсутство-
вать (рис. 8.8).
Большинство американских видов
встречаются в домах и относятся к семей-
ству Lepismatidae. Как и Microcoryphia,
они прекрасные бегуны. Яйца отклады-
вают по одному в трещины, щели и дру-
гие скрытые места. Молодые особи рас-
Рис. 8.8. Thysanura. Thermobia domestica, термобия домашняя. (Из Illinois Natural
History Survey.)
8. Отряды энтогнат и насекомых
249
тут медленно, созревая лишь через 3-
24 мес и претерпевая большое и нефикси-
рованное число линек. Линьки продол-
жаются и после того, как наступает
половозрелое состояние. Редко встре-
чающееся семейство Nicoletiidae вклю-
чает несколько мелких овальных форм,
живущих в гнездах муравьев, и несколько
удлиненных почвенных видов.
Хозяйственное значение. Чешуйница
(Lepisma saccharina), термобия домашняя
(Thermobia domestica) и другие живущие в
домах виды обычно питаются крахма-
лом. Они причиняют заметный ущерб
книгам и переплетам, выедая крах-
мальный клейстер, найадают также и на
другие изделия, содержащие клей или
клейстер.
ЛИТЕРАТУРА
Adams J. А., 1933. Biological notes upon the fire-
brat, Thermobia domestica Packard,
J. N. Y Entomol. Soc., 41, 557-562.
Delany M. J., 1957. Life histories of the Thy-
sanura, Acta Zool. Cracov., 2, 61-90.
Remington C. L., 1954. The suprageneric classifi-
cation of the order Thysanura, Ann. Entomol.
Soc. Am., 47, 277-286.
Slabaugh R. E., 1940. A new thysanuran, and a
key to the domestic species of Lepismatidae ...
in the United States, Entomol. News, 51,
95-98.
Smith E. L., 1970. Biology and structure of some
California bristletails and silver fish (Aptery-
gota. Microcoryphia, Thysanura), Pan-Pacific
Entomol., 46, 211-225.
Wygodzinsky P., 1972. A revision of the silver-
fish (Lepismatidae, Thysanura) of the United
States and the Caribbean area, Am. Mus.
Novit., No. 2481.
ПОДКЛАСС PTERYGOTA
ИНФРАКЛАСС PALEOPTERA
Отряд EPHEMEROPTERA1
(поденки)
Эти насекомые характеризуются мяг-
ким телом от мелкого до крупного разме-
ров, вытянутой формой и постепенным
метаморфозом. У имаго две пары крыль-
ев с сетчатым жилкованием, причем у не-
скольких родов задняя пара мала или да-
же полностью атрофирована, ноги обыч-
но хорошо развиты; антенны малоза-
метные, щетинковидные, ротовой аппа-
рат сильно редуцирован; глаза большие,
брюшко с парой церков, у многих видов
также с длинными хвостовыми нитями
(рис. 8.9). Нимфы живут в водоемах,
имеют разнообразный облик и их ос-
1 Иногда для этого отряда употребляют
название Ephemerida.
Рис. 8.9. Ephemeroptera. Hexagenia limbata. (Из Illinois Natural History Survey.)
250
8. Отряды энтогнат и насекомых
Рис. 8.10. Нимфы поденок. (По разным источникам из: Essig, 1942, College
entomology, с разрешения Macmillan Со.) A. Callibaetis fluctuans. Б. Prosopistoma
foliaceum вид со спинной (слева) и брюшной (справа) стороны. В. Ephemerella
grandis. Г. Paraleptophlebia packii. Д. Siphlonurus occidentalis. E. Iron longimanus.
новные структуры сходны co структура-
ми взрослых форм, однако нимфы имеют
хорошо развитый грызущий ротовой ап-
парат и, как правило, ряд трахейных жабр
на брюшке (рис. 8.10).
Метаморфоз поденок характеризует-
ся уникальной среди современных насе-
комых чертой. Нимфы проходят посте-
пенное развитие обычного типа, сопрово-
ждающееся развитием крыльев в виде
наружных зачатков. Когда развитие за-
кончено, они всплывают к поверхности
воды или взбираются на выступающие из
воды предметы, после чего крылатая
особь освобождается из нимфальной
шкурки. Эта крылатая форма способна к
полету и выглядит как взрослая особь, од-
нако у большинства видов не является
еще половозрелой. Эта стадия называет-
ся субимаго. У немногих родов субимаго
стало последней фазой, так как спарива-
ние и яйцекладка происходят на этой ста-
дии. У большинства же поденок субимаго
линяет еще раз, превращаясь в зрелую
особь, или имаго. Имаго не потребляют
никакой твердой пищи, но, вероятно,
пьют воду в течение своей короткой жиз-
ни. У некоторых родов, в частности Не ха-
8. Отряды энтогнат и насекомых
251
genia и Ephemera, иногда наблюдается
массовый вылет имаго, и тогда над озера-
ми или вдоль рек появляются целые тучи
этих насекомых. Спаривание имаго про-
исходит в танцующих роях. У большин-
ства родов самки выдавливают яйцевые
массы из брюшка, стремительно слетают
к воде и оставляют в ней яйца. Самки Ва-
etis забираются в воду и откладывают яй-
ца под камни. Каждая самка может отло-
жить от нескольких сотен до нескольких
тысяч яиц.
Жизнь многих видов поденок в крыла-
том состоянии очень коротка и соста-
вляет от полутора часов до нескольких
дней; спаривание обычно происходит в
тот день, когда оканчивается созревание
особи; немедленно вслед за спариванием
происходит откладка яиц. Развитие яиц
длится несколько недель или месяц. У от-
дельных родов, например у Callibaetis и
Cloeon, взрослые самки живут значитель-
но дольше-две-три недели. У таких
долгоживущих форм яйца после оплодо-
творения остаются в теле самки до стадии
зрелого зародыша. Вылупление нимф из
этих яиц происходит почти сразу же, как
только яйца касаются воды.
Нимфы (рис. 8.10) населяют самые
разнообразные стоячие и проточные во-
доемы. У некоторых видов нимфы могут
достигать окончательных размеров за
шесть недель, тогда как другим требуется
для этого 1,2 или даже 3 года. Пищу их со-
ставляют микроорганизмы и частицы
растительных тканей.
Нимфы семейства Ephemeridae и близ-
ких групп живут в илистом дне, роя там
норки с помощью своих мощных копа-
тельных передних ног Многие нимфы из
других семейств встречаются под камня-
ми и бревнами. Те, что обитают в бы-
стрых горных потоках, часто несут на
всей вентральной стороне присасыва-
тельный диск, который позволяет им
крепко прикрепляться к гладким поверх-
ностям. Нимфы немногих родов насе-
ляют лужи или небольшие озерца и сво-
бодно плавают у дна или на мелководье.
Поденки играют исключительно важ-
ную роль в питании рыб большинства
пресных водоемов Северной Америки.
Они составляют наиболее многочислен-
ную группу насекомых во многих типах
промысловых водоемов. Изучение содер-
жимого желудков рыб показывает, что в
общем поденки и хирономиды являются,
несомненно, двумя самыми важными
группами насекомых с точки зрения пита-
ния рыб.
Хорошо иллюстрированные таблицы
для определения как имаго, так и нимф
различных семейств Ephemeroptera даны
Эдмундсом (Edmunds, in: Merrit, Cum-
mins, 1978).
ЛИТЕРАТУРА
Berner L., 1950. The mayflies of Florida, Univ.
Fla. Publ. Biol. Sci. Ser., 4, 1-267.
Burks B. D., 1953. The mayflies or Ephemerop-
tera of Illinois, Bull. Ill. Nat. Hist. Surv., 26,
1-216.
Day W. C., 1956. Ephemeroptera. In: Aquatic
insects of California, R. L. Usinger(ed.),
Univ, of California Press, Berkeley, pp.
79-105.
Edmunds G. E, Jr., 1972. Biogeography and evo-
lution of the Ephemeroptera, Annu. Rev.
Entomol., 17, 21-42.
Edmunds G. F., Jr., 1978. Ephemeroptera. In:
Introduction to the aquatic insects of North
America, R. W Merritt and K. W. Cummins
(eds.), Kendall/Hunt, Dubuque, Iowa, pp.
57-80.
Edmunds G. F, Jr., Allen R. K., 1957. A check
list of Ephemeroptera of North America north
of Mexico, Ann. Entomol. Soc. Am., 50,
317-324.
Edmunds G. F. Jr., Allen R. K., Peters W. L.,
1963. An annotated key to the nymphs of the
families and subfamilies of mayflies (Epheme-
roptera), Univ. Utah Biol. Ser., 13, 1-49.
Edmunds G. F., Jr., Jensen S. L., Berner L.,
252
8. Отряды энтогнат и насекомых
1976. The mayflies of North and Central
America, Univ, of Minnesota Press, Minnea-
polis.
Edmunds G. F., Jr., Traver J. R., 1959. The clas-
sification of the Ephemeroptera. I. Ephemeroi-
dea: Behningiidae, Ann. Entomol. Soc. Am.,
52, 43-51.
Flannagan J. F., Marshall К. E.(eds.), 1980.
Advances in Ephemeroptera Biology, Plenum,
New York.
Koss R. W., 1968. Morphology and taxonomic
use of Ephemeroptera eggs, Ann. Entomol.
Soc. Am., 61, 696-721.
Needham J. G., Traver J. R., Hsu Yin-chi, 1935.
The biology of mayflies, Comstock, Ithaca,
N. Y.
Отряд ODONATA (стрекозы)
Стрекозы - средних или крупных раз-
меров хищные насекомые, характери-
зующиеся постепенным метаморфозом.
Имаго со стройным или коренастым те-
лом, с двумя парами сходно устроенных
крыльев, имеющих сетчатое жилкование;
антенны щетинковидные; ротовой аппа-
рат грызущего типа с сильными манди-
булами, глаза большие, брюшко без
длинных хвостовых нитей. Нимфы пре-
сноводные; ротовые органы грызущего
типа с сильно вытянутой и коленообраз-
но сгибающейся нижней губой, превра-
щенной в мощный хватательный орган,
способный удерживать жертву; ноги
сильные; у представителей подотряда
Zygoptera имеются три листовидные хво-
стовые жабры.
Взрослые стрекозы питаются насе-
комыми, пойманными на лету. Они по-
едают комаров, слепней и многих дру-
гих, т. е. практически всех насекомых,
которых стрекозы способны поймать и
одолеть. Нимфы ведут водный образ
жизни, обитая главным образом в стоя-
чих водоемах: озерах, прудах и старицах
рек. Они не могут плавать, зато ходят по
дну среди разлагающихся остатков или
растительности. Нимфы также являются
хищниками; они ловят водных насе-
комых, ракообразных и т. п., хватая их
своей выдвижной нижней губой (маской),
снабженной шипами.
Яйца откладываются различными
способами в воду или вблизи воды. Одни
виды погружают их в ткани растений или
гнилую древесину, другие располагают в
виде комков на каких-либо предметах не-
посредственно под поверхностью воды,
третьи откладывают в воду лентами или
кольцами, а иногда помещают в мокрую
грязь близ уреза воды. Самки многих ви-
дов погружаются в воду и смывают яйца
с конца брюшка. Другие ползают под во-
дой, откладывая яйца.
Нимфы более мелких видов разви-
ваются в течение года. У крупных видов
развитие может занимать от 2 до 4 лет.
Зимовка проходит на стадии нимфы. До-
стигнув полного размера, нимфа выпол-
зает из воды и взбирается на палку, сте-
бель или другой предмет, чтобы перели-
нять в последний раз. Покровы только
что вышедших имаго затвердевают и
приобретают окраску сравнительно мед-
ленно многим из них для этого требует-
ся 1 или 2 дня.
Замечательной особенностью отряда
является способ спаривания (рис. 8.11).
Перед спариванием самец подгибает ко-
нец брюшка вперед и переносит сперма-
тозоиды в пузыревидный приемник, рас-
положенный на втором стерпите брюш-
ка. Во время спаривания самец с по-
мощью своих хвостовых гоноподов обх-
ватывает шею самки; после этого самка
подгибает свое брюшко вперед ко второ-
му стерниту самца и в этом положении
совершается собственно передача спер-
матозоидов. Подобная необычная проце-
дура неизвестна больше ни в одном отря-
де насекомых.
К Odonata относятся три типа насе-
комых; они резко различаются как по
8. Отряды энтогнат и насекомых
253
Рис. 8.11. Спаривание и откладка яиц у равнокрылой стрекозы Archilestes californica. (По
Kennedy.) А. Откладка яиц. Б. Рубцы, оставшиеся после яйцекладки спустя 1 год. В. Те же
рубцы спустя 2 года. Г Спаривание. Д. Участок коры удален, видны погруженные в
камбий яйца. Е. Яйцо.
своему облику, так и по поведению, од-
нако число диагностических признаков,
по которым они отличаются друг от дру-
га, невелико. Современные формы одно-
го из подотрядов-Anisozygoptera-весь-
ма редки и распространены лишь в
Юго-Восточной Азии. Остальные два
подотряда (Zygoptera и Anisoptera),
встречаются в Северной Америке.
Хорошо иллюстрированные опреде-
лительные таблицы имаго и нимф Odo-
nata даны Уэстфоллом (Westfall in: Mer-
rit, Cummins, 1978).
ПОДОТРЯД ZYGOPTERA
равнокрылые стрекозы. Равнокрылые
стрекозы - стройные и нежные насекомые
с порхающим полетом, резко контрасти-
рующим с быстрыми и целенаправ-
ленными движениями разнокрылых стре-
коз. Имаго равнокрылых стрекоз отли-
чаются очень своеобразно устроенной
грудью (рис. 8.12): среднегрудь вместе с
заднегрудью имеет вид примерно пря-
моугольной призмы, находящейся при-
близительно под углом 70-80° по отно-
254
8. Отряды энтогнат и насекомых
Рис. 8.12. Равнокрылая стрекоза Archilestes californica. (По Kennedy.)
шению к продольной оси тела. Крылья в
покое направлены все вместе назад и не-
сколько вверх под прямым углом к верх-
ним краям средне- и заднегруди. Из-за
того что последние до известной степени
наклонены, сложенные таким образом
крылья лежат параллельно друг другу и
располагаются прямо над брюшком.
Большинство имаго темноокрашен-
ные, однако некоторые имеют красные
или черные перевязи на крыльях или
металлически-зеленое, или бронзовое те-
ло и крылья.
Нимфы (рис. 8.13) также имеют строй-
ное тело и три крупные хвостовые тра-
хейные жабры. Они предпочитают жить
среди стеблей водных растений, а не не-
посредственно на дне водоемов.
ПОДОТРЯД ANISOPTERA
разнокрылые стрекозы. Имаго ЭТОГО
подотряда имеют более крепкое сложе-
ние (рис. 8.14) и характеризуются
мощным, грациозным и превосходно
управляемым полетом. Грудь не накло-
нена так сильно, как у Zygoptera, и кры-
лья в покое направлены в стороны. Мно-
гие виды ярко окрашены и имеют на
крыльях броский рисунок-пестрый или
8. Отряды энтогнат и насекомых
255
пятнистый. У старых особей на теле и
крыльях часто развивается бледно-голу-
бой восковой налет, который маскирует
первоначальную окраску и рисунок.
Взрослые особи, особенно крупные,
весьма эффектны благодаря своему стре-
мительному и уверенному полету, хотя и
лишены нарядной окраски. Многие Ani-
soptera, особенно самцы, могут устанав-
ливать себе постоянный охотничий уча-
сток, собственную территорию. Они ле-
тают взад-вперед, патрулируя эту терри-
торию через определенные промежутки
времени в надежде встретить жертву-
пролетающее насекомое. Если стрекоза
заметила таковую, она отклоняется от
своего курса и преследует добычу; схва-
тив добычу, стрекоза возвращается назад
на свой участок. Ёсли на территорию
вторгается соперник, то стрекоза демон-
стрирует настоящие чудеса высшего пи-
лотажа под аккомпанемент щелканья
мандибул и шуршания крыльев.
Нимфы (рис. 8.15) также плотно сло-
жены, многие из них живут в иле или ти-
не на дне стоячих водоемов. Наружные
жабры у них отсутствуют, зато имеется
ректальная дыхательная камера (см.
с. 145), в которой и происходит газооб-
мен. Ни в одном другом отряде насе-
комых подобной дыхательной камеры не
обнаружено.
ЛИТЕРАТУРА
Borror D. J., 1945. A key to the New World
genera of Libellulidae, Ann. Entomol. Soc.
Am., 38, 168-194.
Рис. 8.13. Нимфа равнокрылой стрекозы Archilestes califomica. (По Kennedy.)
Отдельно показана маска-нижняя губа; видны хватательные зубцы.
256
8. Отряды энтогнат и насекомые
Рис. 8.14. Разнокрылая стрекоза Macromia magniflca. (По Kennedy.)
Corbet Р. S., 1963. A biology of dragonflies,
Quadrangle Books, Chicago.
Corbet P. S., 1980. Biology of Odonata, Annu.
Rev. Entomol., 25, 189-218.
Corbet P. S., Longfield C., Moore N. W., 1960.
Dragonflies, William Collins, London.
Kormondy E. J., 1961. Territoriality and disper-
sion in dragonflies (Odonata), J. N. Y Ento-
mol. Soc., 69, 42-52.
Needham J. G., Westfall M. J., Jr., 1955.
A manual of the dragonflies of North America
(Anisoptera), Univ, of California Press, Berke-
ley.
Smith R. F., Pritchard A. E., 1956. Odonata. In:
Aquatic insects of California, R. L. Usinger
(ed.), Univ, of California Press, Berkeley.
Walker E. M., 1953. The Odonata of Canada
and Alaska. Part I, General, Part II, The
Zygoptera-damselflies, Univ, of Toronto
Press, Toronto, vol. 1.
Walker E. M., 1958. The Odonata of Canada
8. Отряды энтогнат и насекомых
257
Рис. 8.15. Нимфа разнокрылой стрекозы Macromia magnifica. (По Kennedy.)
258
8. Отряды энтогнат и насекомых
and Alaska. Anisoptera, Univ, of Toronto
Press, Toronto, vol. 2.
Walker E. M., Corbet P S., 1975. The Odonata
of Canada and Alaska. Anisoptera, Libelluli-
dae, Univ, of Toronto Press, Toronto, vol. 3.
Westfall M. J., Jr., 1978. Odonata. In. Intro-
duction to the aquatic insects of North Ameri-
ca, R. W Merritt, K. W Cummins (eds.),
Kendall/Hunt, Dubuque, Iowa.
Wright M., Petersen A., 1944. A key to the
genera of anisopterous dragonfly nymphs of
the United States and Canada (Odonata,
suborder Anisoptera), Ohio J. Sci., 44,
151-166.
Инфракласс Neoptera
Подавляющее большинство совре-
менных крылатых насекомых и близких к
ним групп принадлежит к этому инфра-
классу, куда относятся также многие вы-
мершие отряды. Практически удобно
рассматривать отряды новокрылых по
группам, соответствующим четырем эво-
люционным стволам ортоптероидному,
плекоптероидному, гемиптероидному и
нейроптероидному.
ОРТОПТЕРОИДНЫЕ ОТРЯДЫ
Сюда относится несколько отрядов,
считающихся наиболее примитивными в
составе Neoptera. Для всех этих отрядов
характерна паурометаболия и многочис-
ленные признаки, общие с Paleoptera.
Отряд DICTYOPTERA1
(тараканы и богомолы)
В этот отряд входят разнообразные
группы насекомых, в том числе бескры-
лые, короткокрылые и полнокрылые
формы. У полнокрылых форм две пары
1 Для этого отряда иногда употребляют
названия Cursoria, Oothecaria, Blattiformia,
Blattopteriformia и Blattopteroidea.
крыльев с сетчатым жилкованием перед-
няя пара, называемая надкрыльями,
имеет кожистую или пергаментную тек-
стуру, тогда как задняя пара перепонча-
тая, более широкая и в состоянии покоя
сложена под надкрыльями. Голова несет
антенны, глаза и ротовые органы мало-
специализированного грызущего типа.
Превращение постепенное. Имаго
и нимфы сухопутные. Задние ноги имеют
примерно те же пропорции, что и сред-
ние, приспособленные для ходьбы или
бега. Некоторыми авторами включаются
в Orthoptera.
ПОДОТРЯД BLATTARIA
тараканы. Тараканы-быстро бегаю-
щие насекомые с уплощенным телом,
длинными тонкими антеннами, хорошо
развитыми глазами и грызущим ро-
товым аппаратом с мандибулами, мак-
силлами и нижней губой в типичном слу-
чае очень похожими на изображенные на
рис. 3.16, 3.17 и 3.18 соответственно. У
тех видов, которые имеют хорошо раз-
витые крылья (рис. 8.16), обе пары несут
большое количество продольных и мно-
жество поперечных жилок, передняя па-
ра, более узкая уплотненная кожистая
или напоминающая пергамент, назы-
вается надкрыльями и служит в основном
защитным покровом для крыльев задней
пары, когда насекомое не летит, задняя
пара тонкая, более широкая, несет основ-
ную нагрузку при полете и веерообразно
складывается под надкрыльями в состоя-
нии покоя. У многих видов крылья
имеют вид зачатков или совершенно от-
сутствуют. Переднегрудь большая и за-
крывает большую часть головы. Брюш-
ко широкое, многосегментное и несет
пару апикальных церков.
Живут тараканы в темных и влажных
местах. Обычно это обитатели тропиков,
где встречается наибольшее разнообра-
8. Отряды энтогнат и насекомых
259
Рис. 8.16. Обычные космополитические тараканы. (Из Connecticut Agricultural
Experiment Station.) А. Рыжий таракан, или прусак (Blattella germanica). Б. Черный
таракан (Blatta orientalis), самка. В. Черный таракан, самец. Г Американский таракан
(Periplanet a americana).
зие видов, крупных и мелких. Богатая
местная фауна существует и в южной ча-
сти Соединенных Штатов, особенно во
влажных районах. Небольшое число ви-
дов можно найти в природных условиях
в более северных областях, где они оби-
тают большей частью под корой
мертвых деревьев или лежащих стволов.
Наиболее заметный компонент фауны’
тараканов северных штатов имеет не
местное происхождение, а относится к
группе видов с космополитическим рас-
пространением, являющихся домашни-
ми вредителями. Они находят в жилищах
человека и других отапливаемых поме-
щениях полутропические условия, позво-
ляющие им процветать и размножаться
на протяжении всего года. Тараканы
практически всеядны и употребляют в
пищу самые разнообразные продукты
растительного и животного происхожде-
ния. Нимфы по строению в целом похо-
жи на взрослых особей и, как правило,
встречаются и питаются вместе с ними.
У некоторых видов, не имеющих крыль-
ев на протяжении всей жизни, для того
чтобы отличить взрослую особь от
нимфы, в ряде случаев приходится иссле-
довать гениталии.
Откладка яиц у тараканов происходит
довольно своеобразно. Отдельные яйца
одно за другим поступают из яйцевода и
собираются в яйцевой камере, где склеи-
ваются вместе с помощью специального
секрета, образуя капсулу, или оотеку. У
каждого вида оотека имеет определен-
ную форму и поверхность ее по-разному
скульптурирована. В оотеке обычно бы-
вает от 15 до 40 яиц, уложенных в симме-
тричные сдвоенные ряды (рис. 8.17).
Оотека образуется в течение нескольких
дней. Вылупление из яиц у разных видов
Рис. 8.17. Оотека Blattella
germanica , боковой участок
оболочки удален, видно
расположение отдельных
яиц.
260
8. Отряды энтогнат и насекомых
протекает по-разному. У некоторых ви-
дов оотека откладывается через несколь-
ко дней после ее образования, но задолго
до вылупления нимф (например, у Peri-
planeta, Blatta), у других она выводится
наружу, но остается крепко соединенной
своим основанием с выводковой камерой
(влагалищем) самки почти до момента
вылупления (например, у Blattella), у
третьих оотека сохраняется внутри
самой выводковой камеры и вылупление
может наступать прямо там (например, у
Pycnoscelus, Diplop ter а). В двух последних
случаях яйца получают необходимые ве-
щества из тела матери. Известен ряд пар-
теногенетических видов, таких, как теп-
личный суринамский таракан Pycnoscelus
surinamensis.
Нимфы в высшей степени активны, но
растут относительно медленно. Более
мелкие виды могут достигать зрелости
за несколько месяцев, тогда как крупным
может потребоваться год и более. Мно-
гие виды образуют скопления, и их
нимфы и имаго встречаются вместе.
Необычайно интересен древесный та-
ракан Cryptocercus punctulatus, распро-
страненный в северо-западных при-
брежных и юго-восточных штатах США.
Этот вид живет колониями в гнилых
стволах, питается разлагающейся древе-
синой и выработал чрезвычайно близкий
к настоящему общественному образ жиз-
ни. Семейная ячейка дает начало коло-
нии, в которой совместно обитают особи
нескольких поколений. Начальные ста-
дии переваривания целлюлозы осущест-
вляются определенными простейшими,
которые всегда в изобилии находятся в
кишечной фауне Cryptocercus и подобны
простейшим, живущим у термитов.
Экономическое значение. Тараканы-
одни из самых неприятных обитателей
человеческих жилищ. Они проникают в
самые разнообразные пищевые про-
дукты, частично поедают, портят и за-
грязняют их экскрементами и оставляют
после себя неприятный запах. Вдобавок к
реальному ущербу, который они причи-
няют, эти снующие насекомые вообще
отвратительны и являются показателем
антисанитарных условий. Поэтому на
борьбу с тараканами в хранилищах, мес-
тах общественного питания и в жилых
домах выделяются значительные сред-
ства.
К северу от линии морозов наиболее
многочисленны представители четырех
космополитических синантропных ви-
дов мелкий рыжий таракан (прусак) Blat-
tella germanica, более крупный черный та-
ракан Blatta orientalis, таракан Supella
longipalpa и американский таракан Peri-
planeta americana, который крупнее всех
остальных и иногда достигает 3 или 4 см
в длину. В некоторых местах многочис-
лен и другой космополитический вид то-
го же рода, столь же крупный, как и аме-
риканский таракан,- Periplaneta australa-
siae. К югу от линии морозов жилища
людей наводняют многие другие тропи-
ческие виды; некоторые из них дости-
гают размеров мыши.
ПОДОТРЙД MANTODEA
богомолы. Хищные насекомые сред-
него или крупного размера, с удлиненной
переднегрудью и большими шипастыми
хватательными передними ногами
(рис. 8.18). Средние и задние ноги, как
правило, тонкие. Пронотум не покры-
вает большую часть головы. В осталь-
ном богомолы сходны с тараканами по
общим признакам, строению ротовых
частей и гениталий. В Северной Америке
богомолы представлены единственным
семейством Mantidae, к которому отно-
сится всего несколько десятков местных
видов. Обширна фауна богомолов в тро-
пиках. Два вида были завезены в Север-
ную Америку Mantis religiosa из Европы
8. Отряды энтогнат и насекомых
261
Рис. 8.18. Богомол Stagmomantis Carolina. (Из Illinois Natural History Survey.)
и Paratenodera sinensis из Юго-Восточной
Азии. Возможно, оба этих вида попали в
Соединенные Штаты в виде оотек (паке-
тов с яйцами) на сортовых саженцах или
в упаковках товаров.
Богомолы бывают полнокрылыми,
короткокрылыми или полностью ли-
шенными крыльев. Многие окрашены в
зеленые, бурые или пегие тона; неко-
торые виды расцвечены ярче, иногда
встречается и выраженный рисунок.
Все виды ведут хищный образ жизни,
питаясь другими насекомыми, которых
они ловят при помощи хватательных
передних ног. Широко распространен
каннибализм, для ряда видов совершен-
но нормальное явление-пожирание сам-
кой самца после окончания спаривания.
Яйца богомолы откладывают крупны-
ми массами определенной конфигура-
ции. В этих массах, или оотеках, яйца
располагаются рядами и склеены вместе
секретом; вся оотека приклеивается к
ветке или другому предмету (рис. 8.19).
В северных областях развивается одно
поколение в год, причем зимовка проис-
ходит на стадии яйца. Интересно робрать
оотеки богомолов в природе в конце
зимы или ранней весной, перенести их в
лабораторию и наблюдать за выходом
из яиц молодых особей, наступающим
через некоторое время. Яйца часто бы-
вают заражены перепончатокрылыми па-
разитами, удивительно напоминающими
молодых богомолов, эти паразиты, как
правило, покидают оотеку некоторое
время спустя после вылупления богомо-
лов.
ЛИТЕРАТУРА
Gurney А. В., 1951. Praying mantids of the Uni-
ted States, Smithsonian Inst. Ann. Rep.,
1950, pp. 339-362.
Guthrie D. M., Tindall A. R., 1968. The biology
of the cockroach, St. Martin’s Press, New
York.
Hebard M., 1971. The Blattidae of North Ame-
rica north of the Mexican border, Mem. Am.
Entomol. Soc., 2, 1-284.
McKittrick F. A., 1964, Evolutionary studies of
cockroaches, Cornell Univ. Agric. Exp. Stn.
Mem., 389, 1-197.
Roth L. M., 1970. Evolution and taxonomic
significance of reproduction in Blattaria,
Annu. Rev. Entomol., 15, 75-96.
Roth L. M., Willis E. R., 1960. The biotic asso-
ciations of cockroaches, Smithsonian Misc.
Collections, 141, 1-470.
262
8. Отряды энтогнат и насекомых
Рис. 8.19. Оотеки, или яйцевые капсулы, богомолов. (Из Essig, 1942,. College
entomology, с разрешения Macmillan Со.)
А. Генерализованный тип. Б. Oligonyx mexicanus. В. Внешний вид и /"-поперечный
срез оотеки Paratenodera sinensis. 1-место выхода нимф; 2-пластинка, J-яйцо;
4-оболочка, 5-пластинка, закрывающая выходное отверстие яйцевой камеры.
Отряд ISOPTERA (термиты)
Термиты-насекомые средних разме-
ров с постепенным метаморфозом, живу-
щие большими колониями, сильно на-
поминающими колонии муравьев и диф-
ференцированные на несколько раз-
личных социальных каст. В типичной
колонии, например, три касты сте-
рильные рабочие, стерильные солдаты и
способные к размножению формы обоих
полов (рис. 8.20). Солдаты белого цвета,
иногда прозрачные. У них нет крыльев,
округлая голова, длинные антенны, гры-
зущий ротовой аппарат и маленькие гла-
за; последние могут быть атрофированы.
Ноги хорошо развиты и все три пары
приблизительно равных размеров. Сол-
даты строением тела напоминают рабо-
чих, но имеют увеличенную голову и бо-
лее массивные мандибулы. Репродук-
тивные особи бывают двух типов- одни
белые, бескрылые или лишь с короткими
зачатками крыльев; к другому типу
принадлежат полностью сформиро-
ванные склеротизированные крылатые
самцы и самки. У них круглая голова,
длинные антенны, грызущие ротовые ор-
ч ганы, хорошо развитые глаза и две пары
сходно устроенных прозрачных крыльев.
После расселительного лета и спарива-
ния крылья обламываются и от каждого
остается только короткий обломок или
чешуйка, которая сохраняется до конца
жизни особи. Рабочие термиты отдален-
но напоминают муравьев, однако у них
не выражена тонкая «талия», характер-
ная для последних.
В Северной Америке термиты пи-
таются целлюлозой, которую они почти
во всех случаях извлекают из мертвой
древесины. Колонии-семьи, насчитываю-
8. Отряды энтогнат и насекомых
263
Рис. 8.20. Касты в колонии термитов (отряд Isoptera). (Из Duncan, Pickwell,
A world of insects, с разрешения McGraw-Hill Book Co.)
А. Первичная форма крылатой половой особи. Б. Вторичная половая особь.
В. Солдат. Г. Термит-рабочий. Д. Первичная форма половой особи после
сбрасывания крыльев.
щие порой по нескольку тысяч особей,
располагаются в мертвых деревьях или
бревнах, или же в земле; в последнем
случае гнездо соединяется крытыми гале-
реями с бревном или пнем, служащим ис-
точником пищи. Рабочие, а в ряде групп
также и нимфы, заняты снабжением ко-
лонии пищей, питая как солдат, так и ре-
продуктивных особей. Солдаты обеспе-
чивают защиту от внешних врагов, кон-
тролируя стратегические участки вблизи
места расположения колонии. Только ре-
264
8. Отряды энтогнат и насекомых
продуктивные особи являются фер-
тильными во всей семье и почти бес-
прерывно откладывают яйца. Рабо-
чие ухаживают за яйцами до выхода
нимф.
На протяжении большей части года в
потомстве оказываются исключительно
рабочие особи и солдаты, но раз в году-
весной или осенью - у более северных ви-
дов образуется большое количество кры-
латых самцов и самок. Они представ-
ляют собой полностью готовых к раз-
множению особей, называемых первой
репродуктивной кастой. Они дружно вы-
летают из гнезда, разлетаются, спари-
ваются и основывают новые семьи-коло-
нии.
Новая семья образуется от одной-
единственной пары крылатых особей.
Самец и самка отбрасывают свои крылья
после расселительного полета и совмест-
но выедают (североамериканские виды)
маленькое гнездо в мертвом пне или ле-
жащем стволе. Они питаются как и обыч-
ные особи, а самка откладывает яйца, из
которых развиваются рабочие и сол-
даты. Когда накапливается достаточное
количество таких особей во взрослой фа-
зе, эти стерильные касты берут на себя
функции расширения гнезда и кормления
самки и самца, называемых царицей и ца-
рем колонии. Если кто-либо из них поги-
бает, то его место занимает рабочепо-
добная фертильная особь, эта форма
известна под названием второй репро-
дуктивной касты. Эта каста невелика по
численности в большинстве колоний и,
по-видимому, является резервом для за-
мещения царской пары.
У некоторых родов североамерикан-
ских термитов солдаты отсутствуют, а их
функции выполняет каста носатых осо-
бей (рис. 8.21). У этих особей забавная
голова с вытянутым рылом, они спо-
собны выделять капельки жидкости,
имеющей репеллентные свойства и пред-
Рис. 8.21. Носатый термит. (По Banks,
Snyder, 1920, упрощено.)
назначенной для отпугивания врагов от
колонии.
Неарктическая фауна насчитывает
около 50 видов, относящихся к четырем
семействам. В других районах мира оби-
тает значительно большее число видов,
особенно в тропиках.
Хозяйственное значение. Ежегодно
термиты причиняют огромный ущерб
домам и другим постройкам и сооруже-
ниям. В поисках источников целлюлозы
многие виды термитов нападают на де-
ревянную обвязку у основания зданий и
могут распространяться из этого места
по деревянным элементам постройки до
верхних частей здания. В своем продви-
жении они могут наталкиваться на ка-
менные или металлические конструкции;
термиты преодолевают такие препят-
ствия, сооружая через них крытые гале-
реи из экскрементов, частиц почвы и
пережеванной древесины и тем самым
постоянно поддерживают связь с почвой,
из которой получают необходимую им
для жизнедеятельности влагу. Термиты
могут повреждать книги, мебель, дере-
вянную утварь, если эти вещи в течение
длительного времени оставались на
одном и том же месте и находились в
8. Отряды энтогнат и насекомых
265
контакте с древесиной. В природе тер-
миты питаются отмершей древесиной и
благодаря этому являются важным зве-
ном в цепи процессов, ведущих к возвра-
щению биогенных элементов, заклю-
ченных в мертвых растительных тканях,
в круговорот веществ.
ЛИТЕРАТУРА
Banks N., Snyder Т. Е., 1920. A revision of
Nearctic termites. U. S. Natl. Mus. Bull., 108,
1-226.
Ebling W., 1968. Termites: Identification, bio-
logy, and control of termites attacking build-
ings. Calif. Agric. Exp. Stat. Extension Service
Manual, vol. 38.
Emerson A. E., 1933. A revision of the genera of
fossil and recent Termopsinae (Isoptera), Calif.
Publ. Entomol., 6, 165-196.
Krishna K., Weesner F. M. (eds.), 1969, 1970.
Biology of termites, Academic Press, New
York, vols. 1, 2.
Lee К. E., Wood T. G., 1971. Termites and soil,
Academic Press, New York.
Snyder T. E., 1949. Catalogue of the termites
(Isoptera) of the world, Smithsonian Misc.
Collections, 112, 1-490.
Snyder T E., 1954. Order Isoptera of the United
States and Canada, National Pest Control
Assn., New York.
Weesner F. M., 1965. The termites of the United
States. A handbook, National Pest Control
Assn., Elisabeth, N. J.
Отряд PHASMATODEA (палочники)
Эти крупные медлительные насе-
комые своим внешним видом имити-
руют листья или веточки (рис. 8.22). Все
виды фауны Северной Америки бескры-
лы и напоминают веточки, за исключе-
нием единственного крылатого вида
Aplopus mayeri, обитающего во Флориде.
Все североамериканские виды, кроме
одного, имеют пятичлениковые лапки и
принадлежат к широко распространенно-
му по всему миру семейству Phasmatidae.
У бескрылого эндемичного калифорний-
ского рода Timema лапки трехчлени-
ковые, и он является единственным из-
вестным представителем семейства Ti-
memidae.
Сходство большинства видов
с обломками веточек объясняет название*
отряда-палочники, или по-английски,
“walking sticks”, что означает «бродячие
ветки». У этих насекомых круглая голо-
ва, длинные и тонкие антенны, маленькие
глаза и простые грызущие ротовые ор-
ганы. Туловище и ноги сильно вытя-
нутые, иногда шиповатые или крайне
тонкие. Самые мелкие виды имеют в
длину приблизительно 12мм, самый
крупный из североамериканских видов,
обитающий на юго-востоке США Megap-
hasma dentricus, достигает в длину от 125
Рис. 8.22. Палочник Diapheromera femorata. (Ориг. С. О. Mohr.)
266
8. Отряды энтогнат и насекомых
до 150 мм. Некоторые тропические виды
имеют широкое, листоподобное тело и
листовидные выросты на члениках ног.
Все представители палочников пи-
таются листьями, большинство из них
живет на деревьях. Порой они достаточ-
но многочисленны и могут лишать лист-
вы леса на больших территориях. Сами
насекомые всегда остаются незаметны-
ми. Их стеблевидная внешность и зеле-
ная или бурая окраска предоставляют им
почти идеальное средство защиты, так
как с не очень близкого расстояния они
совершенно незаметны. Передвигаются
они очень медленно и, если их потрево-
жить, притворяются мертвыми.
Яйца откладываются поодиночке и
просто сбрасываются на почву вниз. На
этой стадии насекомые переживают зи-
му, взрослые же особи с наступлением
холодной погоды погибают. В течение
года развивается одно поколение.
ЛИТЕРАТУРА
Bedford G. О., 1978. Biology and ecology of the
Phasmatodea, Annu. Rev. Entomol., 23,
125-149.
Beier M., 1957. Ordnung Cheleutoptera Cramp-
ton 1915 (Phasmida Leach 1815), Bronn’s KI.
Ordn. Tierreichs, (5) (3)6, 305-454.
Beier M., 1968. Phasmida (Staborder Gespens-
theuschrecken), Handb. Zool., 4(2), 1-56.
Key К. H. L., 1970. Phasmatodea. In: The in-
sects of Australia, D. F. Waterhouse et
al.(eds.), Melbourne Univ. Press, Carlton,
pp. 348-359, Suppl. 1974, pp. 48-49.
См. также литературу по отряду Orthoptera.
Отряд ORTHOPTERA
(прямокрылые)
Насекомые от средних до крупных
размеров, как правило, с удлиненными
задними ногами, имеющими мощные,
приспособленные для прыжков бедра
(рис. 8.23). Почти у всех форм большая
переднеспинка, продолженная на боках
вниз, так что образуется широкий ворот-
ничок позади головы. Голова крупная, с
длинными антеннами, хорошо развиты-
ми глазами и грызущим ротовым аппа-
ратом простого типа. У многих видов
крылья большие и функционируют; над-
крылья (уплотненные передние крылья)
всегда кожистые, а задние крылья мем-
бранозные, веерообразные, складываю-
щиеся в покое. У других видов крылья
могут быть укорочены или полностью
отсутствовать. К этому отряду относятся
кобылки, кузнечики, сверчки, медведки и
прыгунчики (тетригиды), в совокупности
Рис. 8.23. Американская саранча Schistocerca americana americana. (Из
Illinois Natural History Survey.)
8. Отряды энтогнат и насекомых
267
представляющие собой группу форм
самых разных размеров, внешнего вида,
окраски и образа жизни. В фауне Север-
ной Америки различают обычно семь
или восемь семейств и много сотен
видов.
Acrididae. К этому семейству отно-
сятся кобылки, или саранчовые1
(рис. 8.23). Антенны короткие, редко до-
стигают половины длины тела, благода-
ря этой особенности данное семейство
часто называют короткоусыми пря-
мокрылыми. Подавляющее большин-
ство представителей саранчовых питают-
ся травянистыми растениями, и лишь
немногие едят листву деревьев. Яйца от-
кладываются группами в поЧву. Самка
вводит конец брюшка в глубь почвы и
формирует там камеру; на дно этой ка-
меры она начинает откладывать яйца и,
постепенно вытаскивая брюшко из ка-
меры, заполняет ее яйцами. Когда каме-
ра заполнена, самка секретирует за-
щитный колпачок, закрывающий вход-
ное отверстие и предохраняющий яйца
от врагов и попадания посторонних час-
тиц.
Многие представители подсемейства
Oedipodinae имеют на задних крыльях
яркие-голубые, красные, розовые или
черные перевязи. В полевых условиях
самцы многих таких видов обращают
на себя внимание своеобразным тре-
ском, который они издают в полете.
К семейству Acrididae принадлежит
интересная и с экономической точки зре-
ния важная группа насекомых, известная
под названием перелетной саранчи. По-
пуляции этих видов периодически выра-
стают до размеров, поражающих во-
ображение. В таких случаях саранча в
короткие сроки полностью опустошает
район, в котором возникла данная попу-
1 У нас к короткоусым принято относить
еще ряд небольших семейств, в том числе тё-
тригид и три дактили л-Прим, перев.
ляция, и по достижении зрелости мигри-
рует гигантскими стаями в другие рай-
оны. Такие стаи могут кочевать на мно-
гие сотни километров, съедая всю листву
и полностью уничтожая все сельскохо-
зяйственные культуры на своем пути. На
каждом континенте обитают свои
особые мигрирующие виды. В Северной
Америке наибольшее значение имеют
виды комплекса Melanoplus mexicanus. В
1873 г стаи одного из них, Melanoplus
spretus (или «кобылка Скалистых гор»),
мигрировали на восток от Скалистых гор
примерно до реки Миссисипи.
Многие другие виды год от года так-
же причиняют большой, хотя и менее
серьезный ущерб. Наиболее постоянны в
этом отношении виды рода Melanoplus, в
том числе femurrubrum, bivittatus и diffe-
rentialis, а также Camnula pellucida. Все
эти виды уничтожают широкий круг
культурных растений, а виды, встречаю-
щиеся в западных штатах, чрезвычайно
вредны на пастбищах, подвергшихся
перевыпасу.
Tetrigidae. Представителей этого се-
мейства называют также прыгунчиками.
С первого взгляда кажется, что они силь-
но напоминают короткоусых саран-
човых, однако резко отличаются от них
сильно редуцированными надкрыльями
и наличием на заднем крае переднеспин-
ки длинного узкого отростка, своей вер-
шиной заходящего за задний конец тела
(рис. 8.24). Североамериканские виды
малочисленны и имеют мелкие размеры,
редко превышающие 15 мм в длину. Они
обитают в разных местах, особенно ча-
сто в сырых, около воды. У отдельных
видов семейства наблюдается крайне вы-
сокая изменчивость в характере окраски.
По этой причине несколько лет тому на-
зад они стали объектом генетических
исследований.
Tettigoniidae. К этому семейству отно-
сятся кузнечики, характеризующиеся
268
8. Отряды энтогнат и насекомых
Рис. 8.24. Прыгунчик Acridium ornatum. (Из Illinois Natural History Survey.)
Рис. 8.25. Самка лугового кузнечика Conocephalus strictus.
(Из Illinois Natural History Survey.)
длинными и тонкими антеннами, по дли-
не равными телу или превышающие его,
и четырехчлениковыми лапками. Это
крупное семейство, куда входят луговые
кузнечики (подсемейство Conocephalinae,
рис. 8.25), различные листовые кузнечи-
ки (подсемейства Tettigoniinae, Phane-
ropterinae и др., рис. 8.26) и другие груп-
пы. Наиболее хорошо изучены листовые
кузнечики; это крупные, как правило,
зеленые или розоватые насекомые с ши-
рокими крыльями. Они издают серии
музыкальных трелей и как насекомые-
музыканты широко известны наравне
со сверчками.
Самый вредный вид кузнечиков - Апа-
brus simplex, крупный бескрылый за-
падный вид, часто встречающийся в
огромных количествах в районе Большо-
го Бассейна в Скалистых горах и нанося-
щий серьезный ущерб природным паст-
бищам, жультурам зерновых и кормовых
трав.
Gryllacrididae. К этому маленькому
семейству принадлежат чрезвычайно ин-
тересные бескрылые длинноусые пря-
мокрылые. Сюда относятся пещерные
формы из рода Ceuthophylus (рис. 8.27) и
родственных групп, а также встречаю-
щиеся на западе виды рода Stenopelmatus.
Gryllidae. Это семейство состоит из
различных сверчков; у них длинные ан-
тенны, как и у Tettigoniidae, но все лапки
трехчлениковые. Многие роды, такие,
как Nemobius (рис. 8.28), живут на откры-
тых пространствах или на лесных поля-
нах. Другие роды населяют кустарники
или деревья. Один из них, Oecanthus,
8. Отряды энтогнат и насекомых
269
Рис. 8.26. Кустарниковый кузнечик Microcentrum rhombifolium. (Из Illinois Natural History
Survey.)
Рис. 8.27. Пещерный лжекузнечик Ceuthophylus maculatus. (Из Illinois Natural History
Survey.)
включающий древесных (стеблевых)
сверчков, характеризуется шиловидным
яйцекладом, с помощью которого эти
насекомые сверлят отверстия в стеблях,
имеющих сердцевину и откладывают в
эти отверстия яйца (рис. 8.29). В неко-
торых местах самки стеблевых сверчков
могут серьезно повреждать таким спосо-
бом побеги малины.
Медведки представлены двумя други-
ми семействами - Gryllotalpidae и Tri-
dactylidae1. Gryllotalpidae (рис. 8.30)
1 Эти последние в отечественной литера-
туре называются триперстами, а название
«медведки» относится лишь к первому семей-
ству.- Прим, перев.
имеют в длину около 25 мм, и их пере-
дние ноги напоминают скребки и исполь-
зуются для рытья ходов в почве. Эти на-
секомые копают норы в достаточно лег-
ких почвах и питаются мелкими корнями
и насекомыми, которые встречаются им
под землей. Взрослые особи редко выхо-
дят из своих ходов и могут быть заме-
чены лишь случайно. Tridactylidae, или
триперсты, гораздо мельче и достигают
в длину самое большее 5 мм (рис. 8.31).
Они встречаются по берегам озер и рек,
где либо роются в песке, либо прыгают
почти у самого уреза воды.
Рис. 8.28. Полевой сверчок (сверкун)
Nemobius fasciatus. (Из Illinois
Natural History Survey.)
Рис. 8.29. Стеблевой сверчок (белоснежный
сверчок-трубачик) Oecanthus niveus. (По Smith из
Essig, 1942, College entomology, с разрешения Mac-
millan Со.) '
А. На побеге малины видны проколы
яйцеклада и отложенные внутрь яйца. Б. Взрос-
лый самец.
Самцы стеблевых сверчков - одни из самых
удивительных «поющих» насекомых.
Рис. 8.30. Медведка Gryllotalpa hexadactyla. (Из Illinois Natural History Survey.)
8. Отряды энтогнат и насекомых
271
Рис. 8.31. Триперст Tridactylus minutus. (Из Illinois Natural History Survey.)
ЛИТЕРАТУРА
Ball E. D., Tinkham E. R., Flock R.r Vor-
hies С. T., 1942. The grasshoppers and other
Orthoptera of Arizona, Univ. Ariz. Tech.
Bull., 93, 257-373.
Blatchley W. S., 1920. Orthoptera of northeas-
tern America, Nature Publishing Co., Indiana-
polis, Ind.
Helfer J. R., 1963. How to know the grasshop-
pers, cockroaches and their allies,
W. C. Brown, Dubuque, Iowa.
Key К. H. L., 1970. Orthoptera. In. Insects of
Australia, D. F. Waterhouse et al. (eds.), Mel-
bourne Univ. Press, Carlton, pp. 323-347;
Suppl. 1974, pp. 45-47.
Rehn J. A. G., Grant H. J., Jr., 1961. A mono-
graph of the Orthoptera of North America
(north of Mexico), Monogr. Acad. Nat. Sci.
Philadelphia. 1(12), 1-225.
Uvarov В. P., 1966. Grasshoppers and locusts. A
handbook of general acridology. Vol. 1. Ana-
tomy, physiology and development, phase
polymorphism, introduction to taxonomy,
Cambridge Univ. Press, Cambridge.
Отряд DERMAPTERA1
(уховертки)
Эти средних размеров, сильно склеро-
тизированные насекомые несут на брюш-
1 Иногда их называют Euplexoptera.
ке пару мощных клешней (у американ-
ских видов), представляющих собой ви-
доизмененные церки (рис. 8.32). Ротовые
органы обычного грызущего типа,
сложные глаза крупные, простые глазки
часто отсутствуют или трудноразли-
чимы ; антенны длинные, тонкие, много-
членистые. Крылья иногда отсутствуют,
если они имеются, то передняя пара
образует короткие, обычно обрубленные
на вершинах и лишенные жилкования
твердые крыловые покрышки (надкры-
лья), а задняя пара веерообразная, с за-
мечательным радиальным жилкованием
(рис. 8.33). В состоянии покоя заднее
крыло сложным образом складывается в
виде компактного пакета, почти по-
лностью прикрываемого надкрыльями.
Превращение постепенное.
Уховертки фауны Северной Америки
варьируют по длине тела от 5 до 15 мм,
но в остальном относительно одно-
образны как по внешнему виду, так и по
образу жизни. Это ночные насекомые,
активно передвигающиеся в темноте и
всеядные в отношении пищевой специа-
лизации. Некоторые виды, видимо, хищ-
ничают; другие питаются в основном
разлагающимися растительными остат-
ками или изредка живой растительной
тканью. В течение дня они прячутся в
самых разнообразных затененных ме-
272
8. Отряды энтогнат и насекомых
стах, под корой и досками, в почве и в
трещинах и щелях разного происхожде-
ния.
В умеренных областях уховертки
дают в год одно поколение. Самка от-
кладывает большую кучку белых
овальных яиц в земляной камере в ка-
ком-нибудь защищенном месте. Она сте-
режет яйца в течение всего периода, необ-
ходимого для их развития, а затем кор-
мит и охраняет вылупившихся молодых
особей, пока они не станут способными
Рис. 8.32. Уховертка Labia minor. (Из Illinois
Natural History Survey.)
Рис. 8.33. Детали строения взрослой уховертки. (Из Essig, 1942, College entomology, с
разрешения Macmillan Со.)
/-место прикрепления крыла, 2-тазик, 3-вертлуг; 4-бедро; 5-голень; 6-лапка,
7-пульвилла, 8-коготок; 9-складки.
8. Отряды энтогнат и насекомых
273
сами заботиться о себе. Нимфы претер-
певают от четырех до шести линек и со-
зревают довольно быстро.
Уховертки в основном тропическая
группа, лишь немногие представители
достигают на севере умеренных обла-
стей. В Америке к северу от Мексики
встречаются менее 20 видов, относящих-
ся к четырем семействам и нескольким
родам. Шире других распространена ма-
ленькая Labia minor, которая, как и боль-
шинство других наших уховерток,-заве-
зенный человеком вид.
Два своеобразнейших семейства,
обычно помещаемые в этот же отряд,
распространены исключительно в Ста-
ром Свете: живородящие Arixeniidae из
Юго-Восточной Азии, связанные с лету-
чими мышами, и африканские Hemimeri-
dae - эктопаразиты грызунов из рода Cri-
cetomys. Семейство Hemimeridae характе-
ризуется одночлениковыми церками, не
имеющими вида клешней, его часто рас-
сматривают как отдельный отряд Diplo-
glossata.
Экономическое значение. В некоторых
районах Северной Америки космополи-
тическая обыкновенная уховертка Forfi-
cula auricularia стала серьезным вредите-
лем. Она особенно многочисленна на
Западном побережье, где повреждает
розы, георгины и другие садовые расте-
ния, подгрызая лепестки у основания и
вызывая тем самым их опадение. Если не
считать этой особенности, обыкновенная
уховертка - всеядный обитатель садов,
огородов и человеческих жилищ.
ЛИТЕРАТУРА
Cantrell I. J., 1968. An annotated list of the Der-
maptera, Dictyoptera, Phasmotoptera and
Orthoptera of Michigan, Mich. Entomol., 1(9),
299-346.
Giles E. T., 1963. The comparative external mor-
phology and affinities of the Dermaptera,
Trans. R. Entomol. Soc. London, 115, 95-164.
Giles E. T., 1970. Dermaptera. In. Insects of
Australia, D. F. Waterhouse et al.(eds.), Mel-
bourne Univ. Press, Carlton, pp. 306-313.
Hebard M., 1917 Notes on earwigs of North
America, north of the Mexican boundary,
Entomol. News, 28, 311-323.
Hebard M., 1934. The Dermaptera and Orthop-
tera of Illinois, Bull. Ill. Nat. Hist. Surv., 20.,
125-279.
Hinks W. D., 1955, 1959. A systematic mono-
graph of the Dermaptera of the world based on
the material in the British Museum (Natural
History). Pt. I. Pygidicranidae, Subfamily Dip-
latynae; Pt. II. Pygidicranidae excluding Dip-
latynae, British Museum, Natural History,
London.
Popham E. J., 1965. A key to the Dermaptera
subfamilies, Entomologist. 98, 126-136.
Отряд GRYLLOBLATTODEA
(грилл облаттиды)
Этот отряд состоит всего из одного
семейства - Grylloblattidae, включающе-
го небольшое число видов, принадлежа-
щих к трем родам. Это бескрылые удли-
ненные насекомые средних размеров
(рис. 8.34), на голове у них расположены
длинные антенны, маленькие глаза и
грызущие ротовые органы генерализо-
ванного типа. Ноги тонкие, но хорошо
развиты и имеют пятичлениковые лапки.
Брюшко самки вооружено на конце
сильным, но примитивно устроенным яй-
цекладом. Оба пола имеют пару восьми-
или девятичлениковых церков.
В Северной Америке гриллоблаттиды
были обнаружены вблизи границы
вечных снегов в горах на западе Канады
и США (в шт. Калифорния, Монтана и
Вашингтон). Они известны также в Япо-
нии и Сибири!. Живут в почве и гнилой
древесине, под камнями и лежащими
стволами, всегда в местах, покрытых сне-
1 Только в Приморье.- Прим, перев.
274
8. Отряды энтогнат и насекомых
Генитальные сегменты брюшка самца
Рис. 8.34. Grylloblatta compodeiformis. (Из Essig, 1942, College entomology, с разрешения
Macmillan Со.)
1 -антенны; 2-максиллярный щупик; 3-эпикраниальный шов, 4-переднегрудь;
5-среднегрудь, 6-заднегрудь; 7-церки; 5-переднеспинка; 9-среднеспинка;
70-мезэпимер; 77-заднеспинка, 72-метэпимер; 13-брюшные дыхальца; 14-яйцеклад;
75-1-е грудное дыхальце; 16-мезэпистерн; 77-2-е грудное дыхальце; 75-метэпистерн,
79-9-й тергит; 29-10-й тергит; 27-базиподит церка; 22-парапрокт;
23-выворачивающийся мешочек; 24-левая лопасть фаллуса; 25-грифельки; 26-левый
коксит; 27-9-й стернит, 28-отросток правого коксита; 29-правая лопасть фаллуса,
30-отросток правой лопасти фаллуса; 31 -правый коксит.
гом на протяжении большей части года;
наиболее активны в самое холодное вре-
мя года, в том числе зимой. Питаются
гриллоблаттиды мертвыми или умираю-
щими насекомыми и прочими органиче-
скими остатками. Самки откладывают
черные яйца в мох или почву.
ЛИТЕРАТУРА
Gurney А. В., 1948. The taxonomy and distribu-
tion of the Grylloblattidae, Proc. Entomol.
Soc. Wash., 50, 86-102.
Gurney A. B., 1961. Further advances in the
taxonomy and distribution of the Grylloblatti-
8. Отряды энтогнат и насекомых
275
dae (Orthoptera), Proc. Biol. Soc. Wash., 74,
67-76.
Key К. H. L., 1970. Grylloblattodea. In: Insects
of Australia, D. F. Waterhouse et al.(eds.),
Melbourne Univ. Press, Carlton, pp. 304-305.
Оград EMBIOPTERA (эмбии)
Эти уплощенные насекомые с сильно
вытянутым телом (рис. 8.35) имеют
своеобразные расширенные передние
лапки, используемые для плетения шел-
ковой паутины, среди которой и живет
насекомое. Ротовой аппарат грызущего
типа, примитивного строения; глаза хо-
рошо развиты, простые глазки отсут-
ствуют;. антенны многочлениковые
и длинные; ноги короткие, но сильные, с
трехчлениковыми лапками; церки одно-
или двухчлениковые. Самки всегда
бескрылые; самцы обычно с двумя пара-
ми длинных перепончатых примерно
одинаково устроенных крыльев с редуци-
рованным жилкованием. Превращение
постепенное.
К этому отряду принадлежит неболь-
шое число замечательных тропических и
полутропических насекомых, живущих в
шелковых туннелях, которые они плетут
среди своего пищевого субстрата. Пи-
Рис. 8.35. Эмбия Oligotoma saundersii. (По Essig, 1942, College entomology, с разрешения
Macmillan Со.)
А. Самка. Б. Самец. 7-гениталии.
276
8. Отряды энтогнат и насекомых
таются эмбии самым разным расти-
тельным материалом, особенно сухими
листьями трав. Их туннели можно найти
под отстающей корой, среди лишайни-
ков или на поверхности почвы. Сетчатые
ходы на почве располагаются часто сре-
ди листьев подстилки, под сухим коровь-
им навозом или камнями. Иногда такие
гнезда сооружаются у основания расте-
ния. В аридных областях эмбии могут
быть активны на поверхности почвы в те-
чение влажного сезона, а в сухое время
года уходят в глубь почвы. Эти насе-
комые весьма подвижны и быстры в дви-
жениях. Крылатые самцы охотно ле-
тают, часто на источник света.
Эмбии живут большими колониями
со множеством сообщающихся туннелей
и склонны к образованию скоплений. У
большинства видов имеются и самцы и
самки, но известен и ряд партеногенети-
ческих форм, у которых самцы не обна-
ружены. Яйца удлиненной формы и от-
носительно крупны. Они откладываются
кучками, которые прикрепляются к стен-
кам туннелей. Самка проявляет боль-
шую заботу о яйцах и только что вылу-
пившихся нимфах, оставаясь поблизости
от них и отгоняя врагов.
Нимфы весьма похожи на взрослых
особей, причем многие из них лишены
крыльев. У видов, самцы которых имеют
крылья, наблюдается интересное явле-
ние. У нимф самцов зачатки крыльев раз-
виваются внутри тела как имагинальные
диски вплоть до предпоследней линьки и
лишь нимфа последнего возраста имеет
типичные крыловые зачатки. Именно так
происходит развитие крыльев у насе-
комых с полным превращением, поэтому
эту последнюю неполовозрелую стадию
эмбий можно было бы назвать куколкой.
В фауне Нового Света насчитывается
около 70 видов этого отряда, относящих-
ся к 17 родам и 6 семействам. Большин-
ство форм встречается в районах с тропи-
ческим климатом, но ареалы 5 видов на
севере достигают южных частей Кали-
форнии, Аризоны, Техаса и Флориды.
Два из них-Oligotoma saundersii и О. ni-
gra-имеют циркумтропическое распро-
странение и развезены с торговыми гру-
зами почти во все экваториальные
страны. Время от времени карантинная
служба обнаруживает в товарах, вво-
зимых в США из других государств, тот
или иной вид эмбий, отсутствующий в
фауне США.
Все виды этого отряда замечательно
однообразны по внешнему облику; И в
самом деле, до настоящего времени на-
йдено очень немного признаков, позво-
ляющих идентифицировать самок эмбий,
и практически вся классификация на
уровне семейств, родов и видов построе-
на на признаках самцов.
ЛИТЕРАТУРА
Davis С., 1940. Family classification of the order
Embioptera, Ann. Entomol. Soc. Am., 33,
677-682.
Ross E. S., 1940. A revision of Embioptera of
North America, Ann. Entomol. Soc. Am., 33,
629-676.
Ross E. S.f 1944. A revision of the Embioptera,
or webspinners, of the New World, Proc. U.S.
Natl'. Mus., 94, 401-504.
Ross E. S., 1970a. Biosystematics of Embiop-
tera, Annu. Rev. Entomol., 15, 157-172.
Ross E. S., 1970b. Embioptera. In. Insects of
Australia, D. F. Waterhouse et al. (eds.), Mel-
bourne Univ. Press, Carlton, pp. 360-366.
ПЛЕКОПТЕРОИДНЫЕ ОТРЯДЫ
В эту группу входит единственный со-
временный отряд Plecoptera, для которо-
го характерно коксальное сочленение
(рис. 3.37) более продвинутого, чем у ор-
топтероидных отрядов, строения. Кроме
того, сюда же относят несколько иско-
паемых отрядов.
8. Отряды энтогнат и насекомых
277
Оград PLECOPTERA
< веснянки)
Насекомые умеренных или крупных
размеров, которые имеют водных нимф
и характеризуются постепенным превра-
щением. Взрослые особи (рис. 8.36) с
грызущим ротовым аппаратом, часто ре-
дуцированным или слабо склеротизиро-
ванным; антенны длинные, состоят из
многих члеников, сложные глаза и про-
стые глазки хорошо выражены, церки
варьируют от коротких, одночлениковых
у одних семейств до длинных и много-
членистых-у других. В типичном случае
имаго имеют две пары длинных крыльев
сходной текстуры; каждое крыло с уме-
ренным количеством продольных жилок
и подчас с многочисленными поперечны-
ми жилками. У многих видов крылья
укорочены или отсутствуют у особей
обоих полов или только одного пола.
Все нимфы веснянок ведут водный
образ жизни. У них длинные антенны и
пара длинных многочлениковых церков,
грызущие ротовые органы, хорошо раз-
витые глаза и простые глазки и сходные
со взрослыми пропорции тела.
Рис. 8.36. Веснянка Isoperla confusa.
А. Нимфа. Б. Имаго.
278
8. Отряды энтогнат и насекомых
Неарктическая фауна веснянок на-
считывает приблизительно 300 видов,
принадлежащих к шести семействам. Их
генерализованный ротовой аппарат; ан-
тенны и крылья наряду с простым мета-
морфозом указывают на то, что данный
отряд представляет собой примитивную
ветвь, отошедшую от предкового ортоп-
тероидного ствола.
Нимфы веснянок - один из многочис-
ленных компонентов речных сообществ
организмов. Длина тела их составляет от
5 до 20 и более миллиметров, внешний
вид довольно разнообразен: встречают-
ся как простые, тускло окрашенные
формы, так и пятнистые, или имеющие
рисунок из желтых, бурых или черных
полос. Многие из них дышат при помо-
щи наружных пальцевидных жабр. У од-
них жабры одиночные, у других могут
быть собраны в пучки. Некоторые
нимфы лишены наружных жабр и ис-
пользуют в качестве органов дыхания по-
кровы тела или зачатки крыльев. Как
правило, нимфы живут в холодных и не-
загрязненных проточных водоемах, не-
многие обитают также вдоль омываемой
волнами прибрежной части некоторых
наиболее холодных озер. Нимфы насе-
ляют местообитания с различными усло-
виями, часто строго специфичными для
определенных видов. Их можно обнару-
жить под камнями, в трещинах упавших
в воду стволов, в кучах листьев, накапли-
вающихся около камней или вокруг спу-
скающихся в воду ветвей, а также в орга-
ническом детрите. Большинство нимф
питаются мертвыми органическими ча-
стицами, предварительно заселенными
водорослями и диатомеями. Есть
и хищные формы, поедающие более мел-
ких насекомых и других водных беспо-
звоночных.
Самки откладывают от нескольких
сотен до нескольких тысяч яиц, выбрасы-
вая их массами в воду. Вскоре из яиц вы-
лупляются личинки. Мелкие виды и
часть крупных заканчивают развитие за
год, тогда как другим крупным видам
требуется для достижения зрелого со-
стояния два года. Достигнув окончатель-
ного размера, нимфы выползают из во-
ды и занимают неподвижное положение
на камне, палке, стволе дерева или на
ином предмете в ожидании последней
линьки. При линьке нимфальная шкурка
лопается вдоль спины и взрослое насеко-
мое выходит из нее примерно за минуту
или еще скорее. В течение нескольких по-
следующих минут расправляются и твер-
деют крылья, становясь пригодными для
полета. Имаго могут жить в течение не-
скольких недель.
Один бескрылый вид веснянок - Сар-
nia lacustra-интересен в том отношении,
что это единственное насекомое, все ста-
дии которого ведут исключительно вод-
ный образ жизни. Имаго никогда не вы-
ходят на поверхность и живут, спари-
ваются и размножаются на глубинах от
30 до 120 м в озере Тахо на границе шта-
тов Калифорния и Невада. И нимфы и
половозрелые особи дышат поверх-
ностью тела.
Среди некоторых групп веснянок на-
блюдается замечательная особенность,
редко встречающаяся у насекомых. Для
большинства насекомых наступление
зимы означает окончание сезона актив-
ности и наступление периода покоя. У
многих веснянок все происходит наобо-
рот. Нимфы начальных возрастов, по-
видимому, заторможены в развитии в те-
чение более теплых месяцев года. При
приближении зимы темпы развития
нимф ускоряются и имаго выходят в
самые холодные месяцы года начиная с
конца ноября или начала декабря и кон-
чая мартом. Имаго активны в более
теплые дни зимы, можно видеть, как они
взбираются на камни и стволы деревьев,
спариваются и поедают зеленые водо-
8. Отряды энтогнат и насекомых
279
росли. Они явно предпочитают железо-
бетонные мосты и их можно собирать
там в большом количестве. Эта группа
называется осенне-зимними веснянками
и включает в общем представителей се-
мейств Capniidae, Leuctridae, Nemouridae
и Taeniopterygidae. В трех последних се-
мействах есть виды, появляющиеся в бо-
лее поздние сроки; их период лёта пере-
крывается с периодом лёта весенних и
летних видов.
Описанный своеобразный сдвиг раз-
вития осенне-зимних видов веснянок го-
ворит о совершенно иных физиологиче-
ских адаптациях к теплым и холодным
сезонам, чем у большинства прочих насе-
комых. Раскрытие механизмов этих
адаптаций и их регуляции обещает много
интересного.
Хорошо иллюстрированные опреде-
лительные таблицы семейств веснянок по
признакам нимф и имаго составлены
Харпером (Harper in. Merritt, Cummins,
1978).
ЛИТЕРАТУРА
Bauman R. W. et al., 1977. The stoneflies
(Plecoptera) of the Rocky Mountains, Mem.
Am. Entomol. Soc., 31, 1-208.
Claassen P. W., 1931. Plecoptera nymphs of
America north of Mexico, Thomas Say Foun-
dation, Entomol. Soc. Am., 3, 1-199.
Prison T H., 1935. The stoneflies, or Plecoptera,
of Illinois, Bull. Ill. Nat. Hist. Surv., 20 (4),
281-471.
Prison T H., 1942. Studies of North American
Plecoptera, Bull. Ill. Nat. Hist. Surv., 22 (2),
235-355.
Harper P. P., 1978. Plecoptera. In. Introduction
to the aquatic insects of North America,
R. W. Merritt, K. W. Cummins (eds.), Ken-
dall/Hunt, Dubuque^ pp. 105-118.
Hitchcock S. W., 1974. The Plecoptera or
stoneflies of Connecticut, Bull. Conn. State
Geol. Nat. Hist. Surv., 107, 1-262.
Hynes H. B., 1976. Biology of Plecoptera, Annu.
Rev. Entomol., 21, 135-153.
lilies J., 1965. Phylogeny and zoogeography of
the Plecoptera, Annu. Rev. Entomol., 10,
117-140.
Jewett S. G., Jr., 1956. Plecoptera. In. Aquatic
Insects of California, R. L. Usinger (ed.),
Univ. of California Press, Berkeley,
pp. 155-181.
Needham J. H., Claassen P W., 1925. A mono-
graph of the Plecoptera or stoneflies of Ameri-
ca north of Mexico, Thomas Say Foundation,
Entomol. Soc. Am., 2, 1-397
Ricker W E., 1943. Stoneflies of southwestern
British Columbia, Ind. Univ. Studies, Sci. Ser.,
12, 1-145.
Ricker W E., 1943. Systematic studies in Ple-
coptera, Ind. Univ. Studies, Sci. Ser., 18,
1-200.
Riek E. E, 1970. Plecoptera. In. Insects of Aus-
tralia, D. F. Waterhouse et al. (eds.), Mel-
bourne Univ. Press, Carlton, pp. 314-322.
Ross H. H., Ricker W E., 1971. The classifica-
tion, evolution and dispersion of the winter
stonefly genus Allocapnia, III. Biol. Monogr.,
43, 1-240.
Zwick R., 1973. Insecta. Plecoptera. Phylogene-
tisches System und Katalog, Tierreich, 94,
1-465.
ГЕМИПТЕРОИДНЫЕ ОТРЯДЫ
Пять отрядов, составляющих этот
комплекс, характеризуются в основном
развитием хоботка и сосущего ротового
аппарата, начиная от простого удлине-
ния некоторых придатков, связанных с
питанием. Кроме того, у неполово-
зрелых стадий происходило исчезнове-
ние некоторых имагинальных органов
(структур), например простых глазков.
Отряд ZORAPTERA (зораптеры)
У этих крошечных насекомых длиной
от 1,5 до 2,5 мм существуют как кры-
латые, так и бескрылые взрослые формы
(рис. 8.37). И у тех и у других голова
овальная и четко выражена, ротовые ор-
ганы грызущего типа, длинные девяти-
280
8. Отряды энтогнат и насекомых
Рис. 8.37. Формы зораптеры Zorotypus hubbardi. (По Claudell.)
А. Крылатая взрослая самка. Б. Взрослая самка, сбросившая крылья. В.
Нимфа крылатой формы. Г Бескрылая взрослая самка. Д. Антенна
бескрылой взрослой особи Zorotypus snyderi.
8. Отряды энтогнат и насекомых
281
члениковые антенны и одночлениковые
церки. Бескрылые формы лишены также
и глаз, и только в отдельных случаях
имеют рудиментарные глаза или про-
стые глазки; крылатые формы имеют
сложные глаза и хорошо развитые глаз-
ки. У них две пары нежных перепончатых
крыльев лишь с одной-двумя продольны-
ми жилками каждое, жилки могут вет-
виться. Взрослые особи сбрасывают эти
крылья почти так же, как это делают тер-
миты, оставляя лишь маленькие облом-
ки в местах прикрепления крыльев к телу.
Превращение постепенное.
В отряд входит одно Семейство - Zoro-
typidae-c единственным родом Zoroty-
pus. Во всем мире известно не более 20 ви-
дов, причем большая часть-из тропи-
ческих областей. Два вида встречаются
в южных районах Северной Америки;
это Z. snyderi, описанный в Вест-Индии и
на юго-востоке Северной Америки, и Z.
hubbardi, обнаруженный во многих точ-
ках южных штатов и доходящий на
север до Вашингтона (федеральный ок-
руг Колумбия) и центрального Илли-
нойса.
Зораптеры обитают в гнилой древеси-
не или под отмершей корой и большей
частью держатся колониями, насчиты-
вающими от нескольких до сотни особей.
Их пищу, насколько известно, состав-
ляют мелкие членистоногие, в основном
клещи и мелкие насекомые. До сих пор не
установлено, питаются ли они мертвыми
особями или хищничают, однако наблю-
дения, сделанные на лабораторной куль-
туре, говорят в пользу первого типа
питания.
Бескрылые и крылатые особи имеют
сходные гениталии и характеризуются
сходным типом репродуктивного поведе-
ния. Кладку яиц наблюдали только у Z.
hubbardi; яйца располагались без опреде-
ленного порядка или прикрепления' пря-
мо на тропах в колонии. Нимфы выходят
из этих кремовых овальных яиц пример-
но через 3 нед. Многолетние наблюдения
путем выборок указывают, что для до-
стижения зрелости нимфам нужно не-
сколько месяцев.
Хотя само наличие совместно встре-
чающихся крылатых и бескрылых форм
могло бы свидетельствовать о переходе к
системе каст, никаких доказательств об-
щественного поведения в отряде Zorapte-
га пока нет. Разделение труда, так же как
и забота о потомстве или социальные
взаимоотношения между особями, по-
видимому, отсутствуют. Колонии-ско-
пления очень напоминают скопления, на-
блюдающиеся у многих видов Psocopte-
га.
ЛИТЕРАТУРА
Gurney Л. В., 1938. A synopsis of the order Zor-
aptera, with notes on the biology of Zorotypus
hubbardi Caudell, Proc. Entomol. Soc. Wash.,
40 (3), 57-87.
Smithers C. N., 1970. Zoraptera. In. Insects of
Australia, D. F. Waterhouse et al. (eds.), Mel-
bourne Univ. Press, Carlton, pp. 302-303.
Отряд PSOCOPTERA1
(сеноеды)
Сеноеды-мелкие насекомые, длина
тела которых составляет от 1,5 до 5 мм, с
грызущими ротовыми органами,
длинными 13-50-члениковыми усиками,
маленькой переднегрудью и без высту-
пающих церков (рис. 8.38). У некоторых
форм две пары хорошо развитых крыль-
ев, причем передняя пара больше задней
пары, но обе пары одинаковой текстуры
и с редуцированным простым жилкова-
нием. У других форм крылья малень-
кие, имеют вид чешуек или совсем
1 Для этого отряда иногда употребляют
названия Corrodentia или Copeognatha.
282
8. Отряды энтогнат и насекомых
Рис. 8.38. Крылатый сеноед Ectopsocus pumilis и стадии его жизненного
цикла. (По Sommerman.)
А. Яйца. Б. Зрелое яйцо. В. Нимфа первого возраста. Г. Разрыватель
яйцевой оболочки; увеличено. Д. Нимфа третьего возраста. Е. Нимфа
шестого возраста. Ж. Взрослая самка.
8. Отряды энтогнат и насекомых
283
отсутствуют. Превращение постепен-
ное.
Большинство представителей этого
отряда окрашены в неяркие тона или
имеют явно защитную окраску. По этой
причине они редко попадаются в сборах
начинающих студентов, хотя в изобилии
встречаются во многих местах. Их пища
состоит в основном из мицелия грибов,
лишайников, мертвых растительных тка-
ней и мертвых насекомых, даже своего
собственного вида. Сеноеды живут в раз-
нообразных природных местообитаниях,
в кучах сухих листьев, сухой стоящей тра-
ве, мертвых или отмирающих листьях
зерновых растений, на коре древесных
стволов, в листовом опаде на поверхно-
сти почвы, на затененных обнажениях
скальных пород, под столбами оград и в
гнездах птиц и грызунов. Многие виды
встречаются на полностью или частично
заплесневелых пищевых продуктах,
книжных переплетах и практически по-
всюду, где имеется доступный крахмал
или грибной мицелий.
Некоторые виды имеют коренастое
тело и двигаются медленно, даже если их
потревожить. Многие более стройны, а
отдельные виды довольно сильно упло-
щены. Последние, как правило, движутся
со значительной скоростью, и некоторые
из них относятся к самым быстрым бегу-
нам среди всех насекомых. Все прове-
денные исследования свидетельствуют о
том, что полный срок жизни особи от яй-
ла до гибели имаго занимает от 30 до 60
сут, из которых примерно половина при-
ходится на взрослую стадию. Яйца от-
кладываются на поверхность листа или
другого посещаемого самкой места. В за-
висимости от вида яйца располагаются
поодиночке или группами, содержащими
примерно до 10 штук. После откладки
яиц самка сплетает над ними шелковую
сеточку и прикрепляет их к поверхности
субстрата. У одних видов поверх яиц кла-
дется лишь несколько нитей, другие за-
крывают каждую группу яиц густой се-
точкой. Через несколько дней из яиц
вылупляются нимфы, которые проходят
шесть нимфальных возрастов и стано-
вятся половозрелыми насекомыми через
3 или 4 нед.
В северных штатах одни виды пере-
живают зиму на стадии яйца, а другие-
на стадии нимфы или имаго. Виды, оби-
тающие в отапливаемых помещениях, не
прекращают размножаться в течение все-
го года.
На территории Северной Америки из-
вестно около 150 видов сеноедов, относя-
щихся примерно к 12 семействам и мно-
гим родам. Отряд имеет повсеместное
распространение и насчитывает прибли-
зительно 900 видов.
Хозяйственное значение. Некоторые
виды сеноедов причиняют значительный
ущерб, портя продовольственные запасы
и книги в библиотеках. Они потребляют
лишь небольшие количества пищевых
продуктов, так как питаются в основном
плесенью. Временами, однако, сеноеды
могут чрезвычайно сильно размножить-
ся и распространяются тогда по всему
зданию, проникая во всевозможные
укромные места. Таким образом, они за-
ражают грибами остальные вполне при-
годные продукты до такой степени, что
их приходится выбрасывать в больших
количествах. Библиотекам они причи-
няют более прямой вред: выедают крах-
мальный клейстер из переплетов книг и
вдоль краев страниц, уродуя внешний
вид книг и делая необходимым замену
переплета. Два наиболее обычных вида-
это книжная вошь Liposcelis divinatorius,
крошечный бескрылый сеноед (рис. 8.39)
и пыльная вошь Trogium pulsatorium,
другой мелкий вид, крылья которо-
го редуцированы до небольших че-
шуек.
284
8. Отряды энтогнат и насекомых
Рис. 8.39. Бескрылый сеноед-книжная вошь
Liposcelis divinatorius. (По материалам
министерства сельского хозяйства США.)
ЛИТЕРАТУРА
Chapman Р. J., 1930. Corrodentia of the United
States of America. I. Suborder Isotecnomera,
J.N.Y. Entomol. Soc., 38, 219-290.
Gurney A. B., 1950. Corrodentia. In. Pest Con-
trol Technology, Entomological Section,
National Pest Control Assn., New York,
pp. 129-163.
Mockford E. L., 1951. The Psocoptera of In-
diana, Proc. Ind. Acad. Sci., 60, 192-204.
Mockford E. L., 1963. The species of Embiodop-
socinae of the United States (Psocoptera:
Liposcelidae), Ann. Entomol. Soc. Am., 56,
25-37.
Mockford E. L., Gurney A. B., 1956. A review of
the psocids, or book-lice and bark-lice of Texas
(Psocoptera), J. Wash. Acad. Sci., 46,
353-368.
Smithers C. N., 1970. Psocoptera. In. Insects of
Australia, D. F. Waterhouse et al. (eds.), Mel-
bourne Univ. Press, Carlton, pp. 367-375.
Sommerman К. M., 1944. Bionomics of Amap-
socus amabilis (Walsh), Ann. Entomol. Soc.
Am., 37, 359-364.
Отряд PHTHIRAPTERA1
(пухоеды и вши)
Эти мелкие или средних размеров
бескрылые насекомые, как правило, не-
сколько уплощены (рис. 8.40-8.45) и ве-
дут эктопаразитический образ жизни на
птицах и млекопитающих. Их ротовой
аппарат имеет различное строение, ан-
тенны всегда короткие, трех-, пятичлени-
ковые, иногда убирающиеся в спе-
циальные выемки на голове; сложные
глаза частично или полностью редуциро-
ваны, простых глазков нет. Грудь не-
большая, иногда без заметной сегмента-
ции , ноги короткие, но сильные; брюшко
имеет от пяти до восьми хорошо выра-
женных сегментов. Крылья отсутствуют.
Метаморфоз постепенный, включает
только три стадии.
Phthiraptera делятся на три четких
подотряда с интересной эволюционной
историей. Представители примитивного
подотряда Mallophaga питаются слущи-
вающимся эпителием, засохшей кровью
около ранок и другим органическим ма-
териалом, который они обнаружат на те-
ле хозяина. Для этого подотряда харак-
терен редуцированный ротовой аппарат
простого грызущего типа. Одна из линий
Mallophaga, по видимому, стала прогры-
зать кожу хозяина и питаться выступаю-
щей кровью. В соответствии с этим ро-
товые органы редуцировались, и от них
сохранились только нижняя губа и ман-
дибулы, и развился глоточный насос, ис-
пользуемый для всасывания этой пищи.
Единственный известный представитель,
сохранивший такое состояние,-слоновая
вошь Haematomyzus. От примитивного
насекомого этой эволюционной ветви
произошла группа, в пределах которой
совершенно новый комплекс колюще-
1 Некоторые авторы делят его на два от-
ряда. Mallophaga (пухоеды) и Anoplura (вши).
8. Отряды энтогнат и насекомых
285
сосущих стилетов (рис. 8.42) сформиро-
вался параллельно с кровососущим обра-
зом жизни. Отсюда возник подотряд
Anoplura, или вши.
ПОДОТРЯД MALLOPHAGA1
(ПУХОЕДЫ)
пухоеды. Эти насекомые достигают в
среднем длины 3 мм, немногие-10 мм.
Виды существенно различаются между
собой по внешнему виду и образу жизни
одни узкие и удлиненные, другие-корот-
1 Некоторые авторы делят этот подотряд
на два самостоятельных подотряда. Amblyce-
га, имеющих максиллярные щупики, и Ischno-
сега, у которых эти щупики отсутствуют.
кие и широкие; одни активны и быстро
передвигаются, другие малоподвижны и
медлительны. Корреляции между сте-
пенью подвижности и формой тела не об-
наружено. Ротовые органы пухоедов
грызущего типа, однако сильно редуци-
рованы, поэтому без тщательного иссле-
дования с трудом поддаются интерпре-
тации.
В фауне Северной Америки насчиты-
вают много сотен видов Mallophaga,
принадлежащих примерно к шести се-
мействам и множеству родов. Каждый
вид встречается только на одном-един-
ственном виде-хозяине или на несколь-
ких близкородственных видах. Напри-
мер, Virgula meleagridis живет только на
индюках, тогда как крупный пухоед до-
Рис. 8.40. Пухоед Cuclotogaster heterographus, обитающий на голове кур.
А. Взрослая особь. Б. Яйца на пере. (Л-по Illinois Natural History Survey; К-по
материалам министерства сельского хозяйства США.)
286
8. Отряды энтогнат и насекомых
Рис. 8.41. Пухоед Menacanthus stramineus, живущий на теле кур. (По материалам
министерства сельского хозяйства США.)
А. Взрослая особь. Б. Яйца на пере.
машней птицы встречается на широком
круге домашних птиц: курах, индюках,
павлинах, цесарках и голубях. Неболь-
шое семейство Trichodectidae обитает ис-
ключительно на млекопитающих, а об-
ширное семейство Menoponidae-только
на птицах.
Цикл развития всех Mallophaga цели-
ком протекает на теле хозяина, они бес-
прерывно дают все новые и новые пере-
крывающиеся между собой поколения на
протяжении всего года. Питаются эти на-
секомые слущивающимся эпителием, ку-
сочками перьев, волос, свернувшейся
кровью и частицами мусора, находящи-
мися на коже. Яйца приклеиваются к во-
лосу или перу хозяина и, таким образом,
содержатся в условиях, напоминающих
инкубатор. Яйца разных видов имеют
разную форму, некоторые длинные, про-
стые, как на рис. 8.40,2>; у других видов
яйца несут пучки ресничек или волосков,
как показано на рис. 8.41,К
Хозяйственное значение. Многие виды
пухоедов заражают домашних животных
и птиц и причиняют этим значительный
ущерб животноводству и птицеводству.
Сильнее всего страдает от пухоедов до-
машняя птица. Заражение Мепороп gal-
linae и Menacanthus stramineus (рис. 8.41)
приводит к потере веса и снижению яйце-
носкости у кур, индеек и другой птицы. У
Cuclotogaster heterographus (рис. 8.40)
иногда наблюдаются резкие вспышки
8. Отряды энтогнат и насекомых
287
численности, в результате чего погибают
целые выводки цыплят. Домашних птиц
поражают и другие виды пухоедов, но
упомянутые выше наиболее обычны и
вредны из-за своей высокой репродук-
тивной способности.
Домашние животные подвергаются
нападению различных видов рода Tricho-
dectes. Собаки, кошки, лошади, крупный
рогатый скот, овцы и козы могут испы-
тывать значительное ухудшение состоя-
ния при сильной степени заражения эти-
ми насекомыми. Показано, что живущий
на овцах T.ovis повреждает основания во-
лос, снижая тем самым ценность про-
изводимой шерсти за счет уменьшения
длины волос.
ПОДОТРЯД ANOPLURA
вши. В Северной Америке обитает
приблизительно 100 видов этих насе-
комых, принадлежащих к 20 родам; дли-
на тела вшей составляет от 2 до 5 мм. Все
они обычно живут на млекопитающих и
питаются кровью, которую всасывают
через трубку, образованную выдвижным
комплексом тонких стилетов (рис. 8.42).
Иногда вши поражают домашнюю пти-
цу, но во всех зарегистрированных слу-
чаях это были обычные виды вшей мле-
копитающих, случайно перешедшие на
птиц. Цикл развития полностью проте-
кает на теле хозяина. Яйца вши приклеи-
вают к волосам, и вскоре из яиц вылу-
пляются нимфы, очень сходные со
взрослыми особями как по внешнему ви-
Рис. 8.42. Голова и ротовые органы вши Pediculus humanus. (По Metcalf,
Flint, Destructive and useful insects, с разрешения McGraw-Hill Book Co.)
1 - мышцы-протракторы ротового аппарата; 2 - глотка;
3-мышцы-расширители глотки; 4-мозг; 5-проток слюнной железы;
б-пищевод; 7-глоточная сосущая трубка; 8- ротовая воронка;
9-ротовые крючья; 10-трубка мешка; 11 -мешок со стилетами;
12-вентральный стилет; 13-левые отвилки стилетов; 14-мышцы,
втягивающие мешок со стилетами; 15-дорсальный стилет и лежащий
под ним проток слюнной железы.
288
8. Отряды энтогнат и насекомых
ду, так и по поведению. Размножаются
вши беспрерывно в течение всего года.
Хозяйственное значение. Вши имеют
важное значение в двух отношениях. во-
первых, они причиняют вред домашним
животным и, во-вторых, они опасны для
человека.
ВРЕД, ПРИЧИНЯЕМЫЙ ДОМАШНИМ ЖИ-
ВОТНЫМ. Лошади, крупный рогатый
скот, овцы, козы, собаки и кошки подвер-
гаются нападению различных видов
вшей. Вред обусловлен двумя причина-
ми : с одной стороны, вши докучают жи-
вотным, а с другой - вызывают потери
крови, что приводит к ухудшению со-
стояния животных и снижению в боль-
шинстве случаев прироста в весе. Часто
зараженные вшами козы и овцы чешутся
об изгороди и деревья, что сильно ухуд-
шает качество шерсти. В общем наиболее
сильно поражаются те животные, ко-
торые содержатся в худших условиях, но
это далеко не обязательно. Если вспыш-
ка размножения не была вовремя по-
давлена, паразиты обычно распростра-
няются по всему стаду. Большинство
вшей, нападающих на домашних жи-
вотных, относятся к семейству Haemato-
pinidae, типичным представителем кото-
рого служит, например, лошадиная вошь
Haematopinus asini (рис. 8.43).
опасность для человека. Два вида
Anoplura являются эктопаразитами чело-
века. Они широко распространяются и
достигают высокой численности при
большой скученности населения и в анти-
санитарных условиях. Вши передаются
от человека к человеку при тесном кон-
такте, через одежду или постель.
ЛОБКОВАЯ ВОШЬ. Phthirus pubis
(рис. 8.44)-очень мелкий, несколько на-
поминающий краба вид, заражающий
обволосненные части тела, в особенности
область лобка. На голове встречается
редко. Этот вид наносит болезненные
укусы и вызывает сильное раздражение.
Рис. 8.43. Лошадиная вошь Haematopinus
asini. (По Illinois Natural History Survey.)
Перенос лобковой вошью каких-либо за-
болеваний не отмечался.
ГОЛОВНАЯ, ИЛИ ПЛАТЯНАЯ ВОШЬ.
Pediculus humanus (рис. 8.45)-более
крупная вошь, около 4 или 5 мм длиной,
живет на обволосненных частях тела. У
этой вши известны две формы головная,
встречающаяся в основном на голове и
приклеивающая яйца к волосам головы,
и платяная, встречающаяся главным
образом на одежде и переходящая для
Рис. 8.44. Лобковая вошь Phthirus pubis. (По
Ferris.)
8. Отряды энтогнат и насекомых
289
Рис. 8.45. Платяная вошь Pediculus humanus.
(По Ferris.)
питания на близлежащие участки тела.
Платяная вошь приклеивает яйца к ни-
тям одежды. При регулярном мытье го-
ловы и смене одежды эти вши редко мо-
гут вызвать ощутимое заражение. В ан-
тисанитарных условиях они могут раз-
множиться в невероятных количествах и
очень докучать человеку.
Вши могут соперничать с малярийны-
ми комарами в смысле влияния на ход
мировой истории. Головная и платяная
вши переносят сыпной тиф и траншей-
ную лихорадку, которые до последнего
времени были бичом северных армий, в
особенности зимой. В антисанитарных
условиях лагерной скученности или в
траншеях тепло одетые солдаты предста-
вляли собой идеальных хозяев для вшей.
Сыпной тиф и траншейная лихорадка
свирепствовали в форме эпидемических
вспышек и приводили к печальным для
европейских армий последствиям. Вой-
ска Наполеона в России несли такие же
огромные потери от передаваемых вша-
ми болезней, как и от голода и недоста-
точно теплого обмундирования. Против-
ник-русская армия-точно так же жесто-
ко страдал от тифа, как и армия францу-
зов. Многие говорили, что кампанию
выиграли вши, нанеся поражение обеим
сторонам.
Во время второй мировой войны борь-
ба с вшивостью посредством обработки
населения целых городов (главным обра-
зом ДДТ) позволила подавить вспышки
тифа, которые уже принимали размеры
эпидемий. Особенно впечатляющие ре-
зультаты были получены в 1944 г в Не-
аполе, что знаменовало одно из самых
значительных достижений нашего вре-
мени в области профилактического здра-
воохранения.
ЛИТЕРАТУРА
Clay Т., 1970. The Amblycera (Phthiraptera: In-
secta), Bull. Brit. Mus. Nat. Hist. Entomol.,
25, 73-98.
Emerson К. C., 1964. Checklist of the Mallo-
phaga of North America (North of Mexico),
Proving Ground, Dugway, Utah.
Ewing H. E., 1929. A manual of external para-
sites, Thomas, Baltimore, pp. 127-157.
Ferris G. E, 1934. A summary of the sucking lice
(Anoplura), Entomol. News, 45, 70-74; 85-88.
Ferris G. F., 1951. The sucking lice, Pacif. Coast
Entomol. Soc., Mem., 1, 1-320.
Hopkins G. H. E., Clay T., 1952. A list of Mal-
lophaga taken from birds and mammals of
North America, Proc. U.S. Natl. Mus., 22,
39-100.
Kim К. C., Ludwig H. W., 1978. The family
classification of the Anoplura, Syst. Entomol.,
3, 249-284.
Riley W. A., Johannsen O. A., 1938. Medical
entomology, 2nd ed., McGraw-Hill, New
York.
290
8. Отряды энтогнат и насекомых
Отряд THYSANOPTERA 1
(трипсы, или бахромчатокрылые)
Мелкие вытянутые насекомые
(рис. 8.47) длиной по большей части от 2
до 3 мм, с шести-, девятичлениковыми
антеннами, крупными сложными глаза-
ми и компактным ротовым аппаратом,
образующим колюще-сосущий конус.
Многие формы бескрылы, у других кры-
лья могут быть короткими или пол-
ностью развитыми. В последнем случае
их две пары, обе очень узкие и длинные, с
одной или двумя продольными жилками
или вовсе без них; передняя пара часто
крупнее, обе пары несут по заднему краю
или по краям всего крыла длинную бах-
рому из тонких волосков. Ноги сильные,
лапки заканчиваются тупым наконечни-
ком, содержащим пару выворачиваю-
щихся пузырьков. Превращение полное,
т. е. имеется явная куколочная стадия.
Трипсы обычно встречаются на цветках;
их можно обнаружить, раскрыв практи-
чески любой цветок и посмотрев вокруг
оснований тычинок и пестиков. Большое
количество различных форм можно со-
брать путем кошения сачком по цвету-
щим травам или посевам. Многие виды
вредят различным растениям, обитая на
листьях пораженных кормовых растений.
Значительное число видов, хищничаю-
щих на клещах и мелких насекомых,
встречается под корой мертвых деревьев
и в подстилке.
Ротовые органы трипсов устроены
необычно (рис. 8.46). Все части подог-
наны друг к другу таким образом, что
вместе образуют конус; некоторые из
них представляют собой игловидные сти-
леты, прокалывающие и разрывающие
ткани хозяина, добытые таким образом
1 В европейской литературе для этого от-
ряда обычно употребляется название Physo-
poda.
15'
Рис. 8.46. Ротовые органы цветочного
трипса Frankliniella tritici. (По Metcalf, Flint,
Destructive and useful insects, с разрешения
McGraw-Hill Book Co.)
7-антенна; 2-сложный глаз, 3-хитиновый
тяж, проходящий под лбом;
4 - максиллярный склерит; 5 - левая
мандибула; 6-максиллярный стилет;
7-максиллярный щупик; 8-нижняя губа;
9-верхняя губа; 10-мышцы гипофаринкса;
77 - переднегрудь; 72-наличник; 13-правая
максилл а; 14 -гипофаринкс;
15 - нижнегубной щупик.
соки всасываются в желудок с помощью
насоса, находящегося в головной капсуле
и качающего жидкую пищу через конус,
образованный ротовыми органами.
Метаморфоз трипсов столь же своео-
бразен, как и их морфологическое строе-
ние. Ранние стадии развития по строе-
нию ног, ротового аппарата и общему
виду сходны со взрослыми формами,
способ их питания также не отличается
от способа питания зрелых особей. Эти
признаки характерны для насекомых,
8. Отряды энтогнат и насекомых
291
Рис. 8.47. Трипс Heliothrips rubrocinctus. (По материалам министерства
сельского хозяйства США.)
А. Имаго. Б. Нимфа, или личинка. В. Предкуколка. Г. Куколка.
имеющих постепенное превращение.
Первые два возраста не имеют крыловых
зачатков (рис. 8.47,5); зачатки внезапно
появляются в третьем возрасте уже в ви-
де довольно крупных структур
•рис. 8.47,5), в четвертом (последнем)
возрасте крыловые зачатки сильно уве-
личиваются (рис. 8.47,Г)- Особи четвер-
того возраста значительно отличаются
от других возрастов своим поведением:
они не питаются и полностью непо-
движны, а их усики загнуты назад и ле-
292
8. Отряды энтогнат и насекомых
Рис. 8.48. Детали строения Thysanoptera. (По разным источникам.)
А. Апикальная часть брюшка Anaphothrips (Thripidae). Б. Основание
антенны Heterothrips (Heterothripidae). В. Основание антенны
Aeolothrips (Aeolothripidae). Г Антенна Anaphothrips (Thripidae). Д. Вид
с дорсальной стороны апикальной части брюшка Oxythrips (Thripidae).
Е. Вид с дорсальной стороны апикальной части брюшка. Allothrips
(Phlaeothripidae). У-яйцеклад, 2-апикальное кольцо.
жат на спинной стороне головы и пере-
днеспинки. У некоторых трипсов суще-
ствует дополнительный, пятый возраст.
Виды, у которых он имеется, на стадии
четвертого возраста уходят в почву и
устраивают кокон, в котором и проте-
кает покоящаяся пятая стадия. Это на-
столько напоминает развитие с полным
превращением, что покоящуюся стадию
называют куколкой, а две первые - личин-
ками. Форма третьего возраста - актив-
ная стадия с зачатками крыльев - получи-
ла название предкуколки. В некоторых
группах эта форма не развивается, и ли-
чинка превращается непосредственно в
покоящуюся куколку.
Отряд насчитывает множество се-
мейств, представленных в фауне Север-
ной Америки приблизительно 500 вида-
ми , некоторые из них достигают в длину
5-6 мм. Многие виды встречаются толь-
ко на одном виде кормового растения,
в. Отряды энтогнат и насекомых
293
однако немногие обычные виды, как, на-
пример, Frankliniella tritici, питаются на
широком круге растений-хозяев, поэтому
встречаются на цветках почти всех видов.
Некоторые виды-хищники питаются пау-
тинными клещами и другими видами
клещей, а также мельчайшими насекомы-
ми.
Хозяйственное значение. Многие пред-
ставители этого отряда причиняют суще-
ственный ущерб сельскохозяйственным
культурам. Перечисленные ниже виды,
относящиеся к семейству Thripidae,
иляются одними из наиболее вредных
трипсов на территории США в целом.
Табачный трипс Thrips tabaci- широ-
ко распространенный вид, окраска кото-
рого варьирует от лимонно-желтой до
темно-бурой; особенно сильно повре-
ждает лук, фасоль и табак.
Тепличный трипс Heliothrips haemorr-
•*>idalis- темный вид, тело которого по-
крыто морщинками, придающими его
поверхности исчерченный или сетчатый
вид. Этот вид имеет космополитическое
распространение. В умеренных зонах он
вредит в основном в закрытом грунте,
поражая многие тепличные культуры.
Грушевый трипс Taeniothrips incon-
Kquens имеет бурую окраску и серые
крылья. Он нападает на груши, сливы и
эекоторые другие растения, вызывая
своеобразный серебристый блеск повре-
жденных им листьев.
ЛИТЕРАТУРА
Antmthakrishnan Т. N., 1979. Biosystematics of
Thysanoptera, Annu. Rev. Entomol., 24,
159-184.
Lewis T., 1973. Thrips, their biology, ecology
and economic importance, Academic Press,
New York.
Mound L. Л., 1977. Species diversity and the sys-
tematics of some New World leaf-litter Thysa-
noptera, Syst. Entomol., 2, 225-244.
Preisner H., 1949. Genera Thysanopterorum,
Soc. Fouad 1-er Entomol. Bull., 33, 31-157
Stannard L. J., 1957. The phylogeny and classifi-
cation of the North American genera of the su-
border Tubulifera, Ill. Biol. Monogr., 25,
1-200.
Stannard L. J., 1968. The thrips or Thysanoptera
of Illinois, Bull. Ill. Nat. Hist. Surv., 29(4),
215-552.
Watson J. R., 1923. Synopsis and catalogue of
the Thysanoptera of North America, Univ.
Fla. Agr. Exp. Stn. Bull., 168, 1-100.
Отряд HEMIPTERA
(равнокрылые и клопы)
Будучи представлен очень разнооб-
разными насекомыми, этот отряд ха-
рактеризуется главным образом 1) ко-
люще-сосущим ротовым аппаратом,
образующим хоботок, 2) постепенным
превращением и 3) наличием крыльев в
большинстве случаев (рис. 8.49). За ред-
кими исключениями, эти насекомые
имеют большие сложные глаза, антенны,
содержащие от 4 до 10 члеников, причем
сами членики часто бывают длинные, и
две пары крыльев с относительно про-
стым и редуцированным жилкованием;
церки на брюшке отсутствуют. Во мно-
гих семействах самки имеют на брюшке
хорошо развитые ножны и пиловидный
яйцеклад такого типа, как показан на
рис. 3.49.
Название отряда - «полукрылые» -
объясняется особенностью строения
передних крыльев у многих семейств
(рис. 8.49): базальная часть крыла, или
кориум, - жесткая и утолщенная, а вер-
шина (перепоночка) - тонкая и прозрач-
ная. Кориум по текстуре приближается к
жестким надкрыльям, или элитрам, жу-
ков; отсюда название «полунадкрылье»,
часто применяемое в отношении полу-
жесткого - полумягкого типа крыла.
Все виды Hemiptera, кроме очень не-
многих, можно разделить на две боль-
294
8. Отряды энтогнат и насекомых
Рис. 8.49. Типичный клоп-щитник - пример характерного строения хоботка
(7) и крыльев. (По Illinois Natural History Survey.)
шие группы (рис. 8.50). Для первой из
этих групп, подотряда Homoptera, в ти-
пичном случае характерна полностью
прозрачная мембрана передних крыльев
(морщинистая в некоторых случаях), хо-
боток, отходящий от задней части го-
ловы и типичный тенториум. В другой
группе-в подотряде Heteroptera- осно-
вания передних крыльев утолщены, а
вершинные участки мембранозные; хо-
боток в типичном случае отходит от
передней части головы, щеки срастаются
позади хоботка и образуют горло, а тен-
ториум в голове отсутствует. Исключе-
ние составляет третья группа, содержа-
щая лишь один-единственный циркум-
атлантический род Peloridium с разор-
ванным ареалом и представленная мел-
кими формами, которые встречаются на
коре Notophagus. Во многих отношениях
род Peloridium занимает промежуточное
положение между Homoptera и Heterop-
tera у него отсутствует горло, что харак-
терно для Homoptera, а остальные его
признаки соответствуют Heteroptera. Не-
которые авторы помещают этот род в
качестве аберрантной группы в семей-
ство Peloridiidae, другие рассматривают
его в составе особого подотряда-Со-
leorhyncha.
Сравнивая эти подотряды с прочими
родственными группами насекомых,
можно прийти к наиболее вероятному
выводу, что предковая форма этого отря-
да в целом была растительноядным насе-
комым, морфологически сходным в об-
щих чертах с некоторыми из совре-
менных равнокрылых (Homoptera),
в особенности с мелкими представителя-
ми семейства Cicadidae, характеризую-
щимися богатым жилкованием крыла и
примитивным пиловидным яйцекладом.
Эволюция ствола Homoptera, рано
отделившегося от таких древнейших He-
miptera, пошла, по-видимому, по пути
развития тонкого флагеллума антенн и
органов, издающих звук. У рано отде-
лившейся от этого ствола равнокрылых
ветви, представленной семейством Ful-
goridae, антенны стали располагаться
8. Отряды энтогнат и насекомых
295
л
л
л
св --
о is
:z: «
щ
8
Q
се
св
тз
‘Е _ _
О > X S X
л
л
U О £
Sg
св
SS
8'5 8 Л
UZOZffl
g 8 8
•”2 "§ § -п'
h б < S Z
Nepomorpha
И: антенны pentatomorpha
_ . (очень разно* cu I короткие
И заЯ°нтиеГтРази обР“ные’ -3
ки короткие, наземные)
вращающиеся.
увеличены
Dipsocoromorpha
uuSu
HETEROPTERA Переднеспинка длинная
Мельчайшие
обитающие в почве
клопы
Настоящие клопы
Антенны 4-чле—
никовы е; тело
уплощено
HOMOPTERA*
Предковая форма
HEMIPTERA
Н: 1-й членик
лапки длинный
Н: отверстия дорсальных
абдоминальных желез
Антенны прикреплены
ниже глаз
Яйцеклад
атрофирован
Флаге л лум
более тонкий
Переднеспинка
короткая
Антенны прикреплены
между глазами
Рис. 8.50. Упрощенная схема филогенеза отряда Hemiptera. Я-имаго; Я-нимфы.
зол глазами, а не между ними. В даль-
нейшем ствол равнокрылых разделился
яве на две основные ветви. В одной ветви
зереднеспинка стала большой и в значи-
тельной степени закрыла среднеспинку;
тта ветвь включает певчих цикад (Cicadi-
daek пенниц (Cercopidae), горбаток
(Membracidae) и цикадок (Cicadellidae). В
другой ветви переднеспинка осталась не-
большой, но флагеллум утолщился и хо-
боток приобрел перегиб, так что стал ка-
заться растущим между передними нога-
ми, наиболее примитивные представите-
ли этой ветви-семейство Psylidae-
296
8. Отряды энтогнат и насекомых
Рис. 8.51. Особенности строения некоторых Hemiptera. А. Контур тела Lygus
(Miridae). Б. Контур тела Solubia (Pentatomidae). В. Контур тела Stiretrus
(Pentatomidae). Г Контур тела Corimelaena (Pentatomidae). Д. Общий вид
Gelastocoris (Gelastocoridae). Е. Задняя голень и лапка Pangaeus (Pentatomidae). Ж.
Задняя голень и лапка Thyanta (Pentatomidae). 3. Задняя голень и лапка Paraulacizes
(Cicadellidae). И. Задняя голень и лапка Aphrophora (Cercopidae). К. Задняя голень и
лапка Stenocranus (Fulgoridae). Л. Переднее бедро Pacarina (Cicadidae). М. Г олова
Gyrona (Cicadellidae). Н. Голова Poblicia (Fulgoridae) сбоку и спереди. О. Крыло
Psylla (Psillidae). 77. Крыло Aphis (Aphididae). 7-переднеспинка; 2-щиток.
8. Отряды энтогнат и насекомых
297
имеют типичный пиловидный яйцеклад,
но у более специализированных групп
этой ветви, а именно у тлей (Aphidoidea),
белокрылок (Aleurodidae), червецов и
щитовок (Coccoidea), эта структура от-
сутствует.
Все виды подотряда Homoptera-ра-
стительноядны. Из-за такого однообра-
зия способов питания и отсутствия мор-
фологической специализации подотряд
Homoptera в целом может рассматри-
ваться как малоспециализированный.
Однако у тлей, белокрылок и кокцид вы-
работалась глубокая биологическая спе-
циализация, включающая развитие с пол-
ным превращением, сложные жиз-
ненные циклы, содержащие до семи раз-
личных фаз или морф в течение годово-
го цикла одного вида, а также сезон-
ную смену хозяев. В другой крупной
ветви равнокрылых - Cicadellidae - обра-
зовалось такое гигантское количество
видов, что лишь очень немногие се-
мейства могут соперничать в разнообра-
зии с этим семейством.
В стволе Heteroptera хоботок занял
переднее положение и щеки срослись по-
зади него, образовав чрезвычайно
мощный мостик, или горло, поперек зад-
ней части головы под затылочным от-
верстием. Это обстоятельство, видимо,
следует рассматривать как эволюцион-
ное изменение, сопряженное с измене-
нием способа питания в связи с перехо-
дом к хищничеству. Основания передних
крыльев стали толще, а усики редуциро-
вались до четырех члеников. Предковая
форма с такими признаками дала начало
большому числу семейств Heteroptera,
многие из этих семейств сильно видоиз-
менились в связи с переходом к водно-
му или амфибиотическому образу жиз-
ни, поэтому в настоящее время они
могут быть помещены на филогенети-
ческом древе лишь весьма приблизи-
тельно.
Эволюционные изменения, происхо-
дившие в подотряде Heteroptera, с боль-
шим трудом поддаются расшифровке.
Разнообразные семейства были сгруппи-
рованы в ряд довольно четких эволю-
ционных линий, но каким образом эти
отдельные линии соединяются воедино и
образуют филогенетическое древо се-
мейств всего подотряда, остается до сих
пор непонятным, хотя мы располагаем
множеством фактов, касающихся их эм-
бриологии и морфологии и выявленных в
тщательных исследованиях Коббена (Со-
bben, 1968,1978) и других авторов. В фау-
не Северной Америки представлены семь
основных групп; название каждой
группы складывается из основы названия
семейства, к которой добавляется окон-
чание -omorpha. Например, группа водо-
мерок, включающая семейство Gerridae
и родственные группы, получила назва-
ние Gerromorpha.
Видимо, наиболее примитивные
из существующих клопов - мельчайшие
формы Enicocephalomorpha и Dipsoco-
romorpha. Эти прыгающие почвенные
клопы населяют лесную подстилку из
опавшей листвы. В дальнейшем изложе-
нии мы не будем их касаться.
Водомерки (Gerromorpha) имеют спе-
циализированные волоски на лапках и
вращающиеся задние тазики; обе эти
особенности позволяют им быстро
скользить по поверхности воды, на кото-
рой они живут. В Северной Америке
встречаются пять семейств этой группы,
есть и морские представители. Наиболее
многочисленно семейство Gerridae.
Четыре оставшиеся крупные группы
имеют так много разнообразных общих
между собой предковых черт, что род-
ство их друг с другом очевидно, однако
до сих пор неизвестно, каким же именно
образом соотносятся в филогенезе
данные группы и имеет ли каждая из них
монофилетическое происхождение. Cimi-
298
8. Отряды энтогнат и насекомых
Рис. 8.52. Особенности строения некоторых Hemiptera. А, Б. Задние и средние ноги
Corixa (Corixidae). В. Передняя и задняя лапки Belostoma (Belostomatidae). Г
Передняя и задняя лапки Nepa (Nepidae). Д, Е. Передняя лапка Gerris (Gerridae)
сбоку и со спинной стороны. Ж. Голова Corixa (Corixidae). 3. Голова Nabis
(Nabidae). И. Голова Lygaeus (Lygaeidae). К. Голова Myodocha (Lygaeidae).
Л. Передняя нога Phymata (Phymatidae), отдельно показано строение лапки.
М. Голова Alydus (Coreidae). Н, О. Голова и антенна Jalysus (Neididae). П. Антенна
Myodocha (Lygaeidae). Р. Антенна Lyctocoris (Anthocoridae). 7-лапка, 2-голень;
3-коготки; 4-бедро; 5-верхняя губа; 6-горло; 7-простой глазок.
8. Отряды энтогнат и насекомых
299
Рис. 8.53. Надкрылья, или передние крылья Hemiptera. A. Triphelps (Antocoridae).
Б. Lygus (Miridae). В. Pselliopus (Reduviidae). Г Saida (Saldidae). Д. Gargaphia (Tingidae).
E. Blissus (Lygaeidae). Ж. Myodocha (Lygaeidae). 3. Alydus (Coreidae). И. Nabis (Nabidae).
K. Euryophthalmus (Pyrrhocoridae). Л. Piesma (Piesmidae).
300
8. Отряды энтогнат и насекомых
comorpha - богатая группа наземных и
эктопаразитических форм с сильно реду-
цированным жилкованием передней
пары крыльев. Leptopodomorpha - обита-
тели побережий с очень характерным
жилкованием в основании крыла
(рис. 8.53). Короткоусые клопы (Nepo-
morpha) имеют крайне укороченные ан-
тенны, в покое почти полностью спря-
танные в боковые углубления головы.
Все представители, кроме живущих по
берегам Gelastocoridae, обитают в воде.
Типичные Pentatomorpha имеют ряд ха-
рактерных признаков на нимфальной
стадии. Первый членик лапок длинный.
Брюшко несет на боках ряд своеоб-
разных небольших щетинок, назы-
ваемых трихоботриями, а выводные от-
верстия (устья) дорсальных пахучих же-
лез брюшка сильно склеротизированы.
Почти у всех наших видов взрослые насе-
комые имеют четырех-, а не пятичлени-
ковые усики. К этой группе, возможно,
принадлежат Reduviidae и родственные
им семейства, имеющие массивные устья
дорсальных пахучих желез брюшка.
Более подробная характеристика ти-
пичных семейств четырех наиболее обы-
чных групп полужесткокрылых дана ни-
же.
ПОДОТРЯД HOMOPTERA
равнокрылые. Этот подотряд вклю-
чает певчих цикад, цикадок, тлей, черве-
цов и щитовок и родственные им группы;
все они без исключения являются фито-
фагами. Фауна Северной Америки на-
считывает приблизительно двенадцать
семейств или надсемейств.
Цикадовые. У этих равнокрылых
(рис. 8.50) флагеллум антенн имеет игло-
видную форму, а лапки почти неизменно
трехчлениковые. Сюда относятся многие
формы с причудливым внешним видом.
Некоторые Membracidae, или горбатки
(рис. 8.54), имеют сильно увеличенную
переднеспинку, украшенную гребнями,
рогами и зубцами. Обширное семейство
Fulgoridae внешне сильно напоминает
цикадок (рис. 8.55). У некоторых фульго-
рид имеются широкие листовидные кры-
Рис. 8.54. Горбатки (Membracidae).
A. Campylenchia latipes. Б. Ceresa bubalus. В. Enchenopa binotata. (А, В-по материалам
Kansas State University; В-по материалам министерства сельского хозяйства США.)
8. Отряды энтогнат и насекомых
301
Рис. 8.55. Фульгорида Peregrinus maidis. (По Thomas.)
лья, а у других, например у североамери-
канской Scolops и южноамериканской
фонарницы Lanternaria phosphorea, голо-
ва несет странные выросты. Необычен и
образ жизни пенниц, относящихся к се-
мейству Cercopidae их нимфы выделяют
белое пенистое вещество и живут, скры-
тые под этой пеной. Широко известны
также обширные семейства Cicadidae
(певчие цикады) и Cicadellidae (цикад-
ки).
CICADIDAE (ПЕВЧИЕ ЦИКАДЫ). Это
крупные насекомые, у многих представи-
телей фауны Северной Америки длина
тела составляет до 5 см и более. Цикад
легко отличить от близких семейств, так
как дорсальная поверхность головы не-
сет у них три хорошо развитых простых
глазка. У самцов очень мощные звуко-
вые органы, и они издают пронзитель-
ные трели в теплые летние дни и ве-
чера. У нимф расширенные передние но-
ги, предназначенные для рытья; живут
они под землей, питаясь соками из кор-
ней листопадных деревьев.
У большинства видов нимфальная
стадия развивается на протяжении 2-5
лет. Семнадцатилетняя цикада Magicica-
da septendecim и ее ближайшие родствен-
ники (рис. 8.56) составляют группу
южных видов, у которых нимфы живут
13 лет, и группу более северных видов,
нимфы которых развиваются 17 лет. Эти
виды привлекали к себе внимание ученых
периодической природой своего жизнен-
ного цикла. В некоторых местностях по-
пуляции состоят из потомства единствен-
ной самки, поэтому там имаго по-
являются только раз в 13 или 17 лет. В
таких ситуациях они обычно сразу выхо-
дят огромными стаями и самки при от-
кладке яиц могут причинять серьезный
вред побегам и ветвям плодовых деревь-
ев и твердолиственных пород.
CICADELLIDAE (цикадки). Это самое
обширное семейство во всем отряде He-
miptera. В Северной Америке оно пред-
ставлено более чем 2500 видами. Цикад-
ки не только разнообразны в видовом
отношении, но и крайне многочисленны
по количеству особей. При общем коше-
нии сачком они, пожалуй, попадаются
чаще любых других насекомых. Большая
часть цикадок имеет менее 10 мм в длину
и обладает длинными задними голенями,
несущими продольные ряды шипов, но
лишенными как крупных вершинных
шпор, так и венчика шипов на конце го-
лени. Хотя изредка попадаются виды с
широким или угловатым телом, боль-
шинство цикадок имеют стройное тело с
почти параллельными боками
(рис. 8.57). У самок цикадок сильные яй-
цеклады, которыми они пользуются, вы-
резая в листьях или стеблях растений
(преимущественно травянистых) ячейки
для своих яиц. Многие цикадки вредят
сельскохозяйственным культурам не
только непосредственно, питаясь на рас-
тениях, но и путем переноса многих бо-
302
8. Отряды энтогнат и насекомых
Рис. 8.56. Семнадцатилетняя цикада Magicicada septendecim. (По материалам
министерства сельского хозяйства США.)
А, Г. Имаго. Б. Нимфа. В. Сброшенная нимфой шкурка.
Рис. 8.57. Цикада Draeculacephala mollipes. (По материалам министерства сельского
хозяйства США.)
А. Имаго. Б-Г. Нимфы. Д. Яйца.
8. Отряды энтогнат и насекомых
303
Рис. 8.58. Грушевая медяница Psylla pyricola. (По материалам
Connecticut Agricultural Experiment Station.)
А. Имаг'о. Б. Нимфа. В. Яйцо.
лезней растений. Цикадка Circulifer tenel-
lus переносит вирус, вызывающий курча-
вость верхушек свеклы-наиболее опас-
ное заболевание в культуре сахарной
свеклы. Другой вид, Macropsis trimacula-
ta, переносит еще один опасный вирус,
вызывающий пожелтение у персиков.
Листоблошки и тли. Антенны либо ко-
роткие и толстые, либо длинные и ните-
видные (рис. 8.60) по крайней мере на не-
которых стадиях жизненного цикла.
Жилкование крыла сильно редуцировано
и лапки состоят всего из одного-двух чле-
ников. Так как в цикле развития одного и
того же вида встречаются особи с раз-
личным строением тела, трудно охарак-
теризовать отдельные семейства в крат-
ком описании.
Два семейства-листоблошки, или
Psyllidae (рис. 8.58), и белокрылки, или
Aleurodidae (рис. 8.59), имеют простой
цикл развития, при котором имаго обоих
полов крылаты и имеют сходный внеш-
ний вид. У многих листоблошек и бе-
локрылок особи последнего нимфально-
го возраста имеют плоское тело, неак-
тивны или медлительны и напоминают
чешуйки. Представители обоих семейств
характеризуются мелкими размерами.
Все остальные семейства псиллидно-
афидной ветви распадаются на две ос-
новные группы. 1) тли и близкие группы,
образующие надсемейство Aphidoidea, и
2) червецы и щитовки, составляющие
надсемейство Coccoidea. Каждая из этих
групп содержит много семейств, разли-
чающихся в основном по биологическим
особенностям и содержащих большое
число видов, имеющих важное экономи-
ческое значение.
НАДСЕМЕЙСТВО APHIDOIDEA. Это над-
семейство (рис. 8.60) характеризуется 1)
наличием у крылатых форм нескольких
жилок в области стигмы переднего кры-
г
Рис. 8.59. Белокрылка Trialeurodes vaporariorwn. (По Morrill.)
А. Яйцо. Б. Личинка. В. Пупарий, вид с дорсальной стороны. Г. Пупарий,
вид сбоку. Д. Имаго.
Рис. 8.60. Зеленая яблоневая тля Aphis ponii. (По материалам министерства
сельского хозяйства США.)
А. Крылатая живородящая самка. Б. Бескрылая живородящая самка. В.
Нимфа крылатой особи. Г Яйцекладущая самка. Д. Самец.
8. Отряды энтогнат и насекомых
305
ла, 2) двухчлениковыми лапками у боль-
шинства видов и 3) сложной системой
смены поколений, включающей кры-
латых, бескрылых, партеногенетических
и половых особей в ходе жизненного ци-
кла одного вида. Эти особенности под-
робнее обсуждаются в гл. 7 Aphididae,
или тли, представляют собой наиболее
важное и крупное семейство этой группы.
К этому семейству относятся многие
виды первостепенного хозяйственного
значения, например бахчевая тля Aphis
gossypii-vpejwmnb бахчевых культур и
хлопчатника, а также персиковая (оран-
20-732
Рис. 8.61. Калифорнийская щитовка Quadraspidiotus pemiciosus, поражающая
яблоню. (По материалам министерства сельского хозяйства США.)
А. Щиток взрослой самки. Б. Щиток самца. В. Молодая особь первого возраста.
Г То же, увеличено. Д. Приподнятый щиток и находящаяся под ним самка.
Е. Самка. Ж. Взрослый самец.
306
8. Отряды энтогнат и насекомых
жерейная, или табачная) тля Myzus persi-
сае- вредитель многих культур
и переносчик многих болезней растений.
НАДСЕМЕЙСТВО COCCOIDEA (рис. 8.61)
во многих отношениях существенно от-
личается от тлей. Во-первых, самки кок-
цид всегда бескрылы, крайне медли-
тельны в движениях или даже неподвиж-
но прикреплены к субстрату и покрыты
восковыми выделениями или плотной че-
шуйкой (щитком), либо имеют твердые
покровы, как, например, в семействе Со-
ccidae, во-вторых, самцы мелкие, с неж-
ным телом и одной парой крыльев,
имеющих лишь одну-две простые жилки,
в-третьих, цикл развития этих насекомых
относительно простой.
В надсемействе червецов и щитовок
одним из важнейших семейств являются
Diaspididae (рис. 8.61). Самки непо-
движны и ведут прикрепленный образ
жизни, они мелкие, напоминают чешуйки
или подушечки и обитают на многих дре-
весных породах. Само тело насекомого
нежное, овальное, скрыто под щитком,
представляющим собой защитный по-
кров. Придатки очень сильно редуциро-
ваны, и тело взрослой самки больше все-
го напоминает мешок с яйцами. По мере
постепенного освобождения от яиц тело
спадается, так что вся масса отложенных
яиц размещается под защитным покро-
вом щитка. Нимфы первого возраста
имеют ничтожные размеры и крайне ак-
тивны. Они быстро разбегаются во всех
направлениях, осуществляя тем самым
широкое расселение своего вида. После
первой линьки нимфы становятся при-
крепленными и каждая образует щиток.
Некоторые виды этого семейства при-
надлежат к числу наиболее вредных для
сельского хозяйства насекомых. Кали-
форнийская щитовка Quadraspidiotus рег-
niciosus (рис. 8.61)-постоянный вреди-
тель плодовых деревьев и многих декора-
тивных растений; до введения метода
опрыскивания маслом этот вид угрожал
полным уничтожением некоторым пло-
довым культурам во многих районах
США. Распространенная по всему миру
запятовидная щитовка Lepidosaphes ulmi
(рис. 8.62)-обычнейший вредитель
практически всех лиственных деревь-
ев и кустарников в Соединенных Шта-
тах.
Другой важной группой являются
войлочники - семейство Eriococcidae
(рис. 8.63). Они повреждают широкий
круг кормовых растений, особенно те-
пличные и комнатные растения в более
северных районах. Войлочники не обра-
зуют щитков, а выделяют восковые ни-
ти, особенно заметные по периферии
тела.
Рис. 8.62. Запятовидная щитовка
Lepidosaphes ulmi. (По Blackman, Ellis.)
А. Щиток самки. Б. Щиток самца. В. Нимфа
первого возраста. Г Яйца.
8. Отряды энтогнат и насекомых
307
Рис. 8.63. Мучнистый червец. (Ориг.
С О. Mohr.) Внизу справа показано
насекомое в натуральную величину.
ПОДОТРЯД HETEROPTERA
клопы. Этот подотряд включает до-
вольно разнообразные формы, длина ко-
торых составляет от нескольких милли-
метров до нескольких сантиметров, а
образ жизни может быть как наземным,
так и водным или полуводным. Антенны
состоят из 4 или 5 члеников, глаза хоро-
шо развиты, за исключением эктопарази-
гического семейства Polyctenidae.
Нимфы всех форм общим обликом на-
поминают взрослых особей, но в типич-
зом случае отличаются тем, что имеют
за брюшке дорсальные пахучие железы.
В этом отношении они отличаются также
а от нимф равнокрылых.
Яйца откладываются поодиночке или
группами, приклеиваются к стеблям или
листьям, иногда бывают погружены
внутрь стебля растений или в редких слу-
чаях в сырой песок.
В этом подотряде есть и хищные, и
растительноядные виды, часто оба этих
типа встречаются в пределах одного се-
мейства. Хищные виды питаются в ос-
новном мелкими насекомыми. Неко-
торые виды слепняков, Miridae, ведут
исключительно хищный образ жизни, у
других хищничество лишь факультатив-
ное кровь насекомых служит дополни-
тельной пищей, тогда как основную пи-
щу составляют соки растений. Подобное
смешение типов питания в ряде семейств
привело к странным аномалиям. В се-
мействе щитников клоп-арлекин (Ми-
rgantia histrionica) является серьезным
вредителем капусты, а хищный вид Peril-
lus bioculatus - один из наиболее эффек-
тивных естественных врагов колорадско-
го жука.
Водомерки (Gerromorpha)
GERRIDAE, ВОДОМЕРКИ. Это стройные
клопы с длинными ногами (рис. 8.64).
Они живут на поверхности воды, населяя
в основном стоячие водоемы, при-
брежные участки озер, наиболее мед-
ленные участки (омуты) и старицы рек и
ручьев. Лапки снабжены пучками несма-
чивающихся волосков, позволяющих
клопам с большой скоростью скользить
и без всякого труда стоять на поверхно-
сти воды. Все виды-хищники или па-
дальщики, питающиеся главным обра-
зом другими насекомыми, попадающи-
ми на поверхность воды. Яйца водомер-
ки откладывают кучками, прикрепляя их
к водной растительности, или погру-
жают их в подводные части побегов. Ряд
других близкородственных семейств так-
же скользят и бегают по поверхности во-
308
8. Отряды энтогнат и насекомых
Рис. 8.64. Водомерки. (Из Hemiptera of
Connecticut.) A. Gerris rufomaculata (Gerridae).
Б, B. Hydrometra (Hydrometridae).
ды, Gerridae отличаются от этих се-
мейств очень длинными задними бедра-
ми, которые заходят далеко назад за
апекс брюшка.
ч
Постельные клопы
и близкие группы (Cimicomorpha)
tingidae, кружевницы. Это мелкие и
нежные растительноядные насекомые
(рис. 8.65,Л,Б), встречающиеся, как пра-
вило, большими скоплениями. Передне-
спинка и полунадкрылья широкие, сет-
чатые, напоминают кружево и прости-
раются далеко в стороны и назад за
пределы тела; у некоторых родов пере-
днеспийка имеет крупную пузыревидную
среднюю долю, нависающую спереди
над головой (рис. 8.65,Л). Антенны и хо-
боток четырехчлениковые, глазки отсут-
ствуют, а лапки двухчлениковые. Нимфы
сильно отличаются от половозрелых
особей своим внешним видом, у одних
видов они гладкие и напоминают щиток,
у других вооружены большим количе-
ством длинных шипов. Кружевницы от-
кладывают яйца на листья кормового
8. Отряды энтогнат и насекомых
309
Рис. 8.65. Клопы-кружевницы. (По
Heidemann.)
4. Atheas exiguus. Б. Corythuca floridanus.
растения или в ткани листа. В фауне Се-
верной Америки это семейство предста-
влено более чем 200 видами, подавляю-
щее большинство которых связано с
единственным родом или видом кормо-
вого растения. Колония клопов-кружев-
ниц придает листу характерную белопят-
нистую окраску, что сразу выдает ее
присутствие. При осмотре дубов, ольх,
платанов, яблонь, боярышника, берез и
других пород можно собрать разно-
образные виды кружевниц. Кустарники и
травы также населяют многочисленные
виды кружевниц.
miridae, слепняки. Это весьма об-
ширное семейство и на территории Се-
верной Америки оно насчитывает около
2000 видов, т. е. примерно треть всех из-
вестных североамериканских Heteroptera.
Клопы-слепняки (рис. 8.66) принадлежат
к группе семейств, имеющих четырехчле-
никовый хоботок и лишенных глазков. За
немногими исключениями, они обла-
дают хорошо развитыми крыльями, по-
лунадкрылья обыкновенно с хорошо раз-
личимым склеритом -кунеусом- в скле-
ротизированной части и одной или двумя
Рис. 8.66. Слепняки Miridae. (По Illinois Natural History Survey.)
A. Lygus lineolaris. Б. Halticus bracteatus, самец и самка (В).
310
8. Отряды энтогнат и насекомых
простыми ячейками на перепоночке
(рис. 8.53).
Большинство видов растительноядны,
многие развиваются только на одном ви-
де кормового растения или на крайне
ограниченном круге видов. Слепняки вы-
зывают обесцвечивание (этиоляцию)
листьев или скручивание цветков своего
растения-хозяина, что часто приводит к
значительным убыткам при разведении
некоторых сельскохозяйственных куль-
тур. Одними из наиболее вредных с эко-
номической точки зрения видов являют-
ся Pseudatomoscelis seriatus, Halticus bra-
cteatus, повреждающие люцерну, клевер
и огородные культуры, например фа-
соль, и Lygus lineolaris- многоядный вид,
вредящий в разных районах различным
культурам. Представители ряда родов, в
том числе формы, поразительно имити-
рующие муравьев, хищничают, поедая
тлей и других насекомых.
Самки клопов-слепняков отклады-
вают яйца в ткани растений. Большин-
ство видов имеют в году одно поколе-
ние; зимующей стадией является, как
правило, яйцо. Некоторые виды, в том
числе Lygus lineolaris, зимуют во взрос-
лом состоянии и откладывают яйца ран-
ней весной.
CIMICIDAE, ПОСТЕЛЬНЫЕ КЛОПЫ. Это
семейство включает всего несколько ви-
дов широких и плоских насекомых, пи-
тающихся кровью птиц и млекопитаю-
щих. Передние крылья, или полунад-
крылья, сохраняются лишь в виде корот-
ких чешуйковидных зачатков, задняя па-
ра крыльев атрофирована полностью.
Cimicidae обитают в гнездах птиц и мле-
копитающих, а также в жилищах челове-
ка. На людей нападают постельный клоп
Cimex lectulaarius (рис. 8.67) и С. hemipte-
rus, которые могут становиться важней-
шими паразитами в условиях жилых по-
мещений любого типа. В дневное время
постельные клопы прячутся в щелях и
трещинах стен, пола, мебели и т. п., а
также в мусоре, выползая в ночное время
на поиски хозяев для кровососания. Сам-
ка откладывает до 200 цилиндрических
беловатых яиц, размещая их в трещинах.
Короткоусые клопы (Nepomorpha)
Представители этой группы имеют
короткие усики, как правило уложенные
на нижней поверхности головы и неви-
димые при осмотре сверху; эти клопы
обитают в воде или на берегах водоемов.
Из девяти семейств, представленных в
фауне Северной Америки, наиболее обы-
чны Corixidae, Notonectidae и Belostoma-
tidae.
CORIXIDAE, ГРЕБЛЯКИ. Для ЭТИХ КЛО-
ПОВ (рис. 8.68) характерна короткая и
мощная нижняя губа, которая выглядит
скорее как нижний склерит головы, чем
как хоботок (рис. 8.52,3). Передние ноги
короткие, уплощенные или ковше-
образные, задние ноги длинные, упло-
щенные и усажены гребнями щетинок. И
нимфы, ц имаго - исключительно водные
обитатели, хорошо плавающие в воде и
способные лишь неуклюже барахтаться
на суше. Несущие щетинки задние ноги
используются для плавания, плавают
гребляки спинной стороной вверх. Имаго
покидают воду для расселительного лёта,
и часто можно наблюдать большие стаи
этих насекомых над водоемами. Иногда
такие стаи привлекаются источниками
света. Яйца прикрепляются к твердому
субстрату, каковым могут служить кам-
ни, палки и раковины моллюсков, нахо-
дящиеся в воде. Некоторые виды, в част-
ности Ramphocorixa, чаще всего отклады-
вают яйца на тело или придатки речных
раков из рода Camb ar us, причем в неко-
торых местностях последние могут быть
буквально залеплены яйцами гребляков.
Клопы-гребляки по типу питания от-
личаются от всех прочих Hemiptera: они
Рис. 8.67. Обыкновенный постельный клоп Cimex
lectularius. (С разрешения British Museum of Natural
History.)
Рис. 8.68. A. Arctocorixa (Corixidae), вид co спинной стороны. Б. Nepa
(Nepidae), вид co спинной стороны. (Л-no Hungerford, 1959,
схематизировано; 2>-по Hemiptera of Connecticut.)
312
8. Отряды энтогнат и насекомых
Рис. 8.69. Гигантский водяной клоп Lethocerus (Belostomatidae). (Ориг
С. О. Mohr.)
питаются в толще ила на дне водоема,
причем стилеты их ротового аппарата
движутся взад и вперед все вместе подоб-
но языку змеи. Эти стилеты доставляют
в глотку различных диатомей и других
водорослей, а также мельчайших жи-
вотных организмов, которые и соста-
вляют пищу этих клопов.
NOTONECTIDAE, ГЛАДЫШИ. Формой
тела эти водные клопы с первого взгляда
похожи на греб л яков, в частности несу-
щими длинные щетинки веслообразными
задними ногами, служащими для плава-
ния. Однако во многих отношениях их
различия очень велики. Наиболее бро-
сается в глаза их привычка плавать толь-
ко на спине. В связи с таким необычным
способом плаванья окраска гладышей
изменена соответствующим образом.
Вентральная сторона, оказавшаяся ввер-
ху, стала темно-бурой под цвет дна реки
или пруда. Дорсальная сторона, невиди-
мая сверху, когда насекомое плывет,
имеет, как правило, беловатую, кремо-
вую или слегка рябую расцветку. Хобо-
ток Notonectidae сильный и острый, он
служит для высасывания содержимого
тела мелких водных животных, таких,
как ракообразные и небольшие насе-
комые. Многие гладыши откладывают
яйца на поверхности находящихся в воде
предметов, другие погружают их в сте-
бли водных растений.
belostomatidae. Виды этого се-
мейства имеют широкое и плотное
тело, хватательные передние ноги и
приспособленные для ползания и пла-
ванья средние и задние ноги (рис. 8.69).
Сюда относятся одни из наиболее
крупных 'североамериканских Hemiptera,
например Lethocerus americanus, длина
которого достигает 8-10 см. Представи-
тели этого семейства имеют сильный хо-
боток. Они ведут хищный образ жизни,
питаясь насекомыми, моллюсками, голо-
вастиками и мелкими рыбками. Они ча-
сто прилетают на свет, обращая на себя
внимание неуклюжими движениями и
большими размерами.
Щитники и родственные им группы
(Pentatomorpha)
reduviidae, хищнецы. Семейство Re-
duviidae лишь предварительно помещено
в группу Pentatomorpha по причинам, из-
ложенным на с. 300. Это медлительные
хищные клопы, как правило, средних или
крупных размеров, питающиеся другими
насекомыми. У большинства североаме-
риканских видов полностью развиты
крылья (рис. 8.70) и у многих видов ноги
8. Отряды энтогнат и насекомых
313
несут по краям листовидные волоски.
Нимфы некоторых видов выделяют на
спинной стороне особое клейкое веще-
ство, на которое налипают кусочки
листьев и мусора, обеспечивая особи ве-
ликолепную маскировку. Яйца отклады-
ваются поодиночке или группами и при-
клеиваются к растениям или иному
субстрату. Клопы-хищнецы, в том числе
Reduviuspersonatus, иногда нападают и на
человека, оставляя болезненные жгучие
укусы. Все Reduviidae ведут наземный
образ жизни.
COREIDAE, КРАЕВИКИ. По основным
признакам Coreidae напоминают Lygaei-
dae, отличаясь главным образом нали-
чием многочисленных жилок на перепо-
ночках полунадкрыльев. Многие краеви-
кн по внешнему облику напоминают
Lygaeidae. Другие, например Acanthocep-
hala, чрезвычайно похожи на Reduviidae.
Клопы-краевики растительноядны. Наи-
более известный вцд-Anasa tristis
(рис. 8.71,В), нападающий на тыкву,
огурцы и другие бахчевые культуры.
Этот клоп откладывает яйца группами
на листья и стебли своих кормовых рас-
тений. В году этот вид дает несколько по-
колений, а зимует в стадии имаго.
aradidae, подкорники. В это семей-
ство входит группа клопов средней вели-
чины, являющихся наиболее уплощенны-
ми представителями подотряда Heterop-
tera. Они обитают под корой мертвых
деревьев и, по всей вероятности, питают-
ся грибами. У них двучлениковые лапки
и четырехчлениковые антенны и хобо-
ток, простые глазки отсутствуют. Кры-
лья сильно редуцированы в размере и в
сложенном состоянии занимают на спин-
ной стороне лишь незначительное место
(рис. 8.72,Л,Б).
lygaeidae. Большинство представи-
телей североамериканской фауны этого
семейства-довольно мелкие, темноокра-
шенные или бледные формы. Немногие
Рис. 8.70. Три представителя семейства Reduviidae. (Рисунки присланы
R. С. Froeschner.)
A. Pselliopus barberi. Б. Melanolestes picipes. В. Triatoma sanguisuga.
314
8. Отряды энтогнат и насекомых
Рис. 8.71. А. Молочайный клоп Oncopeltus fasciatus. Б. Blissus leucopterus leucopterus
(Lygaeidae). В. Anasa tristis (Coreidae).
(Л-по Froeschner; Б-по Illinois Natural History Survey; В-по материалам
министерства сельского хозяйства США.)
роды, в том числе молочайный клоп On-
copeltus fasciatus (рис. 8.71,Л), имеют
броскую черную с красным окраску.
Диагностическими признаками данного
Рис. 8.72. Подкорники Aradidae. (По
Froeschner.) Aradus acutus (Л) и Neuroctenus
simplex (Б)-два обычных вида восточной и
центральной частей Северной Америки.
семейства являются четырехчлениковый
хоботок й немногочисленные неровные
жилки, пересекающие перепоночку
(рис. 8.53,Б). Простые глазки у большин-
ства видов хорошо выражены. Важней-
ший представитель семейства в Северной
Америке-клоп Blissus leucopterus leucop-
terus (рис. 8.71,Б), один из основных вре-
дителей кукурузы и прочих зерновых
культур в штатах зерносеющей зоны.
Этот клоп перезимовывает под расти-
тельными остатками на почве в стадии
имаго. Ранней весной имаго начинают
питаться на злаках и зерновых и от-
кладывать яйца на корни и верхушки
своих растений-хозяев. Через 2 нед из яиц
вылупляются нимфы, которые питаются
на том же самом растении и достигают
зрелого состояния за 6 нед. К тому вре-
мени, когда это поколение завершает
нимфальное развитие, культура кормо-
вого растения почти всегда либо созре-
вает и начинает высыхать, либо в случае
перенаселения уже не предоставляет не-
8. Отряды энтогнат и насекомых
315
обходимых количеств пищи для вредите-
ля. В последнем случае вся популяция
мигрирует с данного участка в поисках
более сочной пищи. Эта миграция при-
обретает характер массового переселе-
ния, но не путем перелета, а «пешим»
способом. Старшие нимфы и только что
перелинявшие имаго образуют спеша-
щую массу насекомых на марше. Поиски
новых пищевых ресурсов обычно закан-
чиваются на кукурузном поле, где к это-
му поколению. Как мигрирующее первое
поколение, так и питающееся на месте до
зрелого состояния второе поколение при-
чиняют огромный ущерб культуре куку-
рузы. Достигнув взрослого состояния,
второе поколение уходит на зимовку до
следующей весны.
PENTATOMIDAE, ЩИТНИКИ. К ЭТОМУ Се-
мейству (рис. %.51,Б-Г, и 8.73) относятся
многочисленные крупные и средних раз-
меров клопы, большей частью с широ-
ким телом, часто покрытым зелеными,
серыми или бурыми пятнами. Есть и яр-
му времени всходы уже хорошо при-
нялись и пребывают в начальной стадии
роста. Особи, достигшие кукурузного по-
Рис. 8.73. Щитники Pentatomidae. (По материалам министерства сельского хозяйства
США.)
Клоп-арлекин Murgantia histrionica. А. Взрослое насекомое. Б. Кладка яиц. В. Нимфа
первого возраста. Г Нимфа второго возраста. Д. Нимфа третьего возраста. Е. Нимфа
четвертого возраста. Ж. Нимфа пятого возраста.
316
8. Отряды энтогнат и насекомые
коокрашенные виды. Некоторые виды-
хищники, питающиеся разнообразными
насекомыми. Другие - исключительно
растительноядны, примером может слу-
жить обычный клоп-арлекин Murgantia
histrionica (рис. 8.73). Этот клоп питается
на крестоцветных и часто причиняет
серьезный вред культуре капусты. В не-
которых работах три подсемейства-Scu-
tellerinae, Cydninae и Thyreocorinae -
рассматриваются в качестве самостоя-
тельных семейств. Отличительные при-
знаки этих подсемейств, четко выра-
женные в рамках неарктической фауны,
оказываются непригодными при изуче-
нии щитников в объеме мировой фауны.
ЛИТЕРАТУРА
Blatchley W S., 1926. Heteroptera or true bugs
of eastern North America, Nature Publishing
Co., Indianapolis, Ind.
Britton W. E., 1923. The Hemiptera or sucking
insects of Connecticut, Bull. Conn. State Geol.
Nat. Hist. Surv., 34, 1-807
Brooks A. F., Kelton L. A., 1967 Aquatic and
semi-aquatic Heteroptera of Alberta, Saskat-
chewan and Manitoba (Hemiptera), Mem.
Entomol. Soc. Can., 51, 1-92.
China W E., Miller N. С. E., 1959. Checklist
and keys to the families and subfamilies of the
Hemiptera-Heteroptera, Bull. Brit. Mus. Nat.
Hist. Entomol., 8, 1-45.
Cobben R. H., 1968. Evolutionary trends in
Heteroptera. Part I: Eggs, architecture of the
shell, gross embryology and eclosion, Centre
for Agricultural Publishing and Documen-
tation, Wageningen.
Cobben R. H, 1978. Evolutionary trends in
Heteroptera, Part II. Mouthpart structures
and feeding strategies, Entomologie, Landbou-
whogesch. Wageningen Meded., 289, 1-407.
DeCoursey R. M., 1971. Keys to the families and
subfamilies of the nymphs of North American
Hemiptera-Heteroptera, Proc. Entomol. Soc.
Wash., 73, 413-428.
DeLong D. M., 1971. The bionomics of leafhop-
pers, Annu. Rev. Entomol., 16, 179-210.
Dixon A. F. G., 1973. Biology of aphids, Edward
Arnold, London.
Evans J. W., 1963. The phylogeny of the
Homoptera, Annu. Rev. Entomol., 8, 77-94.
Froeschner R. C., 1941-1944. Contributions to a
synopsis of the Hemiptera of Missouri, Pts.
1-4, Am. Midi. Naturalist, 26, 122-146; 27.
591-609- 31, 636-683, 42, 123-188.
Herring J. L., Ashlock P. D., 1971. The key to
the nymphs of the families of Hemiptera
(Heteroptera) of America north of Mexico,
Fla. Entomol., 54, 207-213.
Hungerford H. D., 1959. Hemiptera. In. Fresh-
water biology, W T. Edmundson (ed.), Wiley,
New York, pp. 958-972.
Kennedy J. S., Stroyan H. L. G., 1959. Biology
of aphids, Annu. Rev. Entomol., 4, 139-160.
McKenzie H. L., 1967 Mealybugs of California,
Univ, of California Press, Berkeley and Los
Angeles.
Menke A. S., 1979. The semiaquatic and aquatic
Hemiptera of California (Heteroptera. Hemip-
tera), Bull. Calif. Insect Surv., 21, 1-166.
Miller N. С. E., 1956. Biology of the Heterop-
tera, Leonard Hill, London.
Milne L. J., Milne M., 1978. Insects of the water
surface, Sci. Am., 238, 134-143.
Mound L. A., Halsey S. H, 1978. Whitefly of
the world: A systematic catalogue of the Aleyr-
odidae (Honjoptera) with host plant and
natural enemy data, Wiley, New York.
Slater J. A., Baranowski R. M., 1978. How to
know the true bugs (Hemiptera-Heteroptera),
W C. Brown, Dubuque, Iowa.
Smith С. F, 1972. Bibliography oh Aphididae of
the world, N. C. Agric. Exp. Stn. Tech. Bull.,
216, 1-177.
Torre-Bueno J. R. de la, 1939. The synopsis of
the Hemiptera-Heteroptera, Entomol. Am.,
19, 141-310.
Usinger R. L., 1966. Monograph of Cimicidae
(Hemiptera-Heteroptera), Thomas Say Foun-
dation, Entomol. Soc. Am., 7, 1-585.
Usinger R. L., Matsuda R., 1959. Classification
of the Aradidae (Hemiptera-Heteroptera), Bri-
tish Museum, Natural History, London.
Van Duzee E. P., 1917. Catalogue of the Hemip-
tera of America north of Mexico, Univ. Calif.
Tech. Bull., 2, 1-902.
Woodward T E., Evans J. W., Eastop V. F,
8. Отряды энтогнат и насекомых
317
1970. Hemiptera. In. Insects of Australia,
D. F. Waterhouse et al. (eds.), Melbourne
Univ. Press, Carlton, pp. 387-464; Suppl.
1974, pp. 52-57.
И vgodzinsky P., 1966. A monograph of the
Emesinae (Reduviidae, Hemiptera), Bull. Am.
Mus. Nat. Hist., 133, 1-614.
НЕЙРОПТЕРОИДНЫЕ ОТРЯДЫ
Все 10 нейроптероидных отрядов ха-
рактеризуются полным превращением, и
по этой причине их часто называют Holo-
metabola. Современные представители
образуют две крупные группировки:
отвол Neuroptera-Coleoptera, у личинок
которых сохраняются многие прими-
тивные признаки и возникает ряд инте-
р?сных изменений яйцеклада у самок, и
ствол Mecoptera-Hymenoptera, у личинок
юторых имеется лишь один коготок на
сапке.
С твоя Neuroptera-Coleoptera
Четыре отряда, составляющие этот
ствол,- Raphidioptera, Megaloptera, Ne-
uroptera и Coleoptera-имеют личинок с
двумя коготками на каждой лапке (при-
митивный признак) и яйцеклад с особым
образом сросшимися створками.
Отряд RAPHIDIOPTERA 1
(верблюдки)
Крупные насекомые, по многим при-
знакам сходные с Megaloptera, но отли-
чающиеся от последних длинной змеепо-
добной шеей (рис. 8.74). У верблюдок
длинные антенны, грызущие ротовые ор-
ганы, большие глаза и две пары про-
зрачных примерно одинаково
устроенных крыльев с сетчатым жилко-
ванием. У самок хорошо заметный яйце-
клад на конце брюшка. Личинки на-
земные с членистыми антеннами, фасети-
рованными глазами, хорошо развитыми
грудными ногами, но без каких-либо вы-
ростов или придатков на брюшке. В ми-
ровой фауне известно всего пять родов,
насчитывающих приблизительно сотню
1 Иногда употребляется название Raphi-
diodea.
Рис. 8.74. Европейская верблюдка Raphidia ratzeburgi. (По Essig, 1942, College
entomology, с разрешения Macmillan Со.)
318
8. Отряды энтогнат и насекомых
видов, которые группируются в два се-
мейства - Raphidiidae (имеющие простые
глазки) и Inocellidae (у которых глазки
отсутствуют). В Северной Америке жи-
вут представители двух родов, Aguila
(Raphidiidae) и Inocellia (Inocellidae), при-
чем все они встречаются лишь в запад-
ной части материка. Личинки обитают
под отстающей корой хвойных деревьев
и поедают других насекомых. Закончив
личиночное развитие, личинки не прядут
кокона, а строят овальную колыбельку, в
которой находятся в защищенном со-
стоянии в течение всей стадии куколки.
Взрослые насекомые также хищничают.
Эти странного вида насекомые иногда
случайно попадают в сачок при отряхи-
вании листвы.
ЛИТЕРАТУРА
Carpenter F. М., 1936. Revision of the nearctic
Raphidiodea (recent and fossil), Am. Acad.
Arts. Sci. Proc., 71, 89-157.
Woglum R. S., McGregor F. A., 1958. Observa-
tions on the life history and morphology of
Aguila bractea Carpenter, Ann. Entomol. Soc.
Am., 51:129-141.
Woglum R. S., McGregor F. A., 1959. Observa-
tions on the life history and morphology of
Aguila astuta (Banks) (Neuroptera. Raphidio-
dea. Raphidiidae), Ann. Entomol. Soc. Am.,
52, 489-502.
Отряд MEGALOPTERA1
(большекрылые)
У этих крупных насекомых водные
личинки и наземные имаго и куколки.
Взрослые особи (рис. 8.75) с длинными
антеннами, грызущим ротовым аппара-
том, большими глазами и двумя парами
крыльев. Крылья обеих пар сходны меж-
ду собой по текстуре и жилкованию,
имеют полный набор продольных жилок
1 Иногда этот отряд называют Sialoidea.
и несколько дополнительных тер-
минальных ветвей, а также множество
поперечных жилок. Переднеспинка боль-
шая и широкая, брюшко без выступаю-
щих церков. Личинки (рис. 8.75) имеют
мощные ротовые органы, приспособ-
ленные для кусания, хорошо выраженные
членистые антенны, большие глазные
пятна, состоящие из группы примерно в
шесть фасеток каждое (стеммы), хорошо
развитые грудные ноги и парные брю-
шные выросты, или жабры. Вершина
брюшка несет длинный непарный вырост
у представителей семейства Sialidae (вис-
локрылки) и пару мощных крючко-
видных урогомф у Corydalidae.
Оба этих семейства представлены в
Северной Америке лишь немногими ро-
дами и менее чем 50 видами. Окраска
взрослых особей варьирует от черной до
пятнистой или желтой; у одного рода
(Nigronia) на крыльях имеются черные и
белые перевязи.
Личинйи ведут водный образ жизни,
встречаясь как в проточных, так и в стоя-
чих водоемах. Они поедают мелких вод-
ных животных. Взрослые личинки Согу-
dalus могут достигать в длину 80 мм. Эти
страшные на вид личинки очень охотно
используются рыболовами в качестве на-
живки. Более мелкие виды этого отряда
достигают зрелости в течение года и
имеют одногодичный цикл развития. Не-
которым Corydalidae для завершения
жизненного цикла необходимо 2 или 3
года.
По окончании развития личинки по-
кидают водоем и делают куколочную
колыбельку в сырой почве иди гнилой
древесине вблизи от него. Здесь личинки
превращаются в куколок. Куколки Mega-
loptera активно реагируют на внешние
раздражения и способны к передвиже-
нию. Стадия куколки занимает обычно
около двух недель.
Имаго хорошо летают, но по сравне-
8. Отряды энтогнат и насекомых
319
Рис. 8.75. Вислокрылка Sialis sp. (По Illinois Natural History
Survey.)
А. Личинка. Б. Имаго.
нню с двукрылыми и бабочками их полет
мало лабилен. Взрослые особи неко-
торых Corydalus могут достигать в раз-
махе крыльев 13 см и относятся к числу
наиболее крупных насекомых Северной
.Америки. Самки откладывают яйца
большими кучками по нескольку сотен
штук в каждой на камни и другие высту-
пающие из воды предметы. Вскоре после
откладки яиц из них вылупляются мель-
чайшие личинки, которые падают или со-
скальзывают в воду.
ЛИТЕРАТУРА
Parfin S., 1952. The Megaloptera and Neuropte-
ra of Minnesota, Am. Midi. Naturalist, 47,
421-434.
Ross H. H., 1937 Nearctic alderflies of the ge-
nus Sialis, Bull. Ill. Nat. Hist. Surv., 21, 57-78.
Weele W H. van der, 1910. Megaloptera, Coll.
Zool. Selys Longchamps, Brussels, fasc., 5,
1-93.
Отряд NEUROPTERA
(сетчатокрылые)
Взрослые насекомые от очень мелких
до крупных размеров, обычно с двумя
320
8. Отряды энтогнат и насекомых
Рис. 8.76. Neuroptera. А, Б. Взрослое насекомое и личинка златоглазки Chrysopa*sp.
В. Личинка Sisyra sp., питающаяся пресноводными губками. (А, Б-по Illinois Natural
History Survey; В-no Townsend.)
8. Отряды энтогнат и насекомых
321
парами прозрачных крыльев, имеющих
ушожество продольных и поперечных
жилок, ротовые органы грызущего типа,
антенны длинные и многочлениковые,
глаза большие (рис. 8.76,Л). Личинки
чрезвычайно разнообразны, большин-
ство из них ведут наземный образ жизни
и являются хищниками; одно семейство
(Sisyridae) представлено водными форма-
ми, личинки которых питаются пресно-
водными губками. У всех личинок есть
грудные ноги, но нет брюшных, голова
хорошо развита, ротовой аппарат грызу-
щего типа с сильно увеличенными ман-
дибулами.
виды, питающиеся губками. Все
эти виды относятся к одному небольшо-
му семейству Sisyridae. Имаго-мелкие,
типичного облика сетчатокрылые, но ли-
чинки коренасты, обитают в пресно-
водных губках и йитаются ими. Их ро-
товые органы образовали длинный силь-
но выдающийся вперед хоботок
(рис. 8.76,В). В неарктической области
отмечено лишь несколько видов этого
семейства.
хищники-засадники. Мантиспы
<рис. 8.78) образуют другое небольшое
семейство-Mantispidae. Их имаго пора-
зительным образом напоминают бого-
молов. Сильно увеличенные передние но-
ги приспособлены для захватывания до-
бычи (насекомых) и прикреплены к пере-
днему краю сильно удлиненной пере-
днее пинки. Личинки питаются яйцевыми
коконами пауков или содержимым
осиных гнезд. Личинки первого возраста
активны, имеют стройное тело и разы-
Рнс. 8.77. Передние крылья сетчатокрылых
Neuroptera. A. Nallachius (Dilaridae). Б.
Semidalis (Coniopterygidae). В. Climacia
Sisyridae). Г Hemerobius (Hemerobiidae). Д.
Chrysopa (Chrysopidae). E. Lomamyia
Berothidae). Ж. Polystoechotes
Polystoechotidae). 1 -базальная жилка;
2-ступенчатая жилка.
322
8. Отряды энтогнат и насекомых
Рис. 8.78. Мантиспа Mantispa brunnea. (По Essig, 1942, College entomology, с
разрешения Macmillan Со.)
/-тазик; 2-кардо, 3-базигалеа, 4-стипес; 5-максиллярный щупик; б-галеа;
7-лациния; Я-лигула (язычок); 9-нижнегубной щупик; 10-субментум.
скивают подходящий источник пищи.
Как только таковой обнаружен, личинка
переходит в паразитоидную стадию, и
следующие возраста характеризуются
неподвижностью и червеобразным обли-
ком с дегенерировавшими ногами.
активные хищники. Личинки всех
остальных семейств-активные хищники.
Имаго имеют прозрачные крылья
с очень богатым жилкованием, откуда и
пошло их название «сетчатокрылые»
(рис. 8.76,Л). Для большинства этих насе-
комых характерен относительно мед-
ленный полет. Самка либо прикрепляет
яйца непосредственно к листьям, либо
яйца располагаются на конце длинного
волосковидного стебелька, прикреплен-
ного к листу (рис. 7.1,0). Последний спо-
соб используют только Chrysopidae. Ли-
чинки активны, хотя и медлительны,
имеют мягкое тело и часто покрыты бо-
родавками, бугорками и длинными воло-
сками. Ротовые органы видоизменены
для высасывания соков из тела жертвы.
Мандибулы и апикальные части максилл
длинные, пластинчатые и серповидные;
i. Отряды энтогнат и насекомых
323
чаксиллярная пластинка плотно приле-
гает к нижней поверхности каждой ман-
дибулы, причем в каждой из этих сопри-
касающихся частей имеется желобок, так
что между сложенными вместе мандибу-
лой и максиллой образуется канал при-
мерно от вершины мандибулы, откры-
вающийся в рот. Оба мандибулярно-мак-
лллярных комплекса погружаются в те-
ло жертвы с противоположных сторон, и
соки тела жертвы высасываются через
?тот канал.
Личинки представителей семейств
Chrysopidae (рис. 8.76,Б) и Hemerobiidae
свободно ползают по растениям и пи-
таются тлями и другими мелкими насе-
комыми и их яйцами. Поскольку эти ли-
ванки постоянно нападают на колонии
глей. этих насекомых назвали «тлёвыми
львами». Достигнув полного роста, ли-
ванка плетет овальный покрытый Воло-
жами кокон под листом или в каком-
=гбудь ином укромном месте и окукли-
злется. Личинки Myrmeleontidae живут
si песчаной почве и роют воронко-
збразные ямки, в которые попадают му-
тазьи и другие насекомые. Эти личинки,
отзываемые «муравьиными львами», от-
зчаются от тлёвых львов только более
I тоенастым телосложением. Муравь-
лев роет свою воронку, выбрасывая
гегок из ее центра посредством резких
гмжений головы вверх, причем манди-
: г ды выполняют здесь роль совков. Лов-
оы ямка может доходить в глубину до 2
i-тз 3 см и имеет такую крутизну стенок,
сткую только позволяет сыпучесть песка
} санном месте. Муравьиные львы сидят,
мывшись в песок, на дне воронки, по-
~гянно ожидая неосторожную жертву.
^*эершив развитие, личинка строит ко-
£ в почве и окукливается.
ЛИТЕРАТУРА
Carpenter F. М., 1940. A revision of nearctic
Hemerobiidae, Berothidae, Sisyridae, Polys-
toechotidae, and Dilaridae, Proc. Am. Acad.
Arts Sci., 74(7), 193-280.
Froeschner R. C., 1947. Notes and keys to the
Neuroptera of Missouri, Ann. Entomol. Soc.
Am., 40(1), 123-136.
Gurney A. B., Parfin S., 1959. Neuroptera. In.
Freshwater biology, W T. Edmundson (ed.),
Wiley, New York, pp. 973-980.
MacLeod E. G., Adams P. A., 1967. A review of
the taxonomy and morphology of the Berothi-
dae with a description of a new subfamily from
Chile (Neuroptera), Psyche, 74, 237-265.
New T. R., 1975. The biology of Chrysopidae
and Hemerobiidae (Neuroptera), with referen-
ce to their usage as biocontrol agents: A re-
view, Trans. R. Soc. Entomol. London, 127,
115-140.
Parfin S., 1952. The Megaloptera and Neuropte-
ra of Minnesota, Am. Midi. Naturalist, 47,
421-434.
Riek E. E, 1970. Neuroptera. In: Insects of Aus-
tralia, D. F. Waterhouse et al. (eds.), Melbour-
ne Univ. Press, Carlton, pp. 472-494.
Smith C., 1934. Notes on the Neuroptera and
Mecoptera of Kansas, with keys for the identi-
fication of species, J. Kans. Entomol. Soc.,
7(4), 120-145.
Tauber C. A., 1969. Taxonomy and biology of
the lacewing genus Meleoma (Neuroptera:
Chrysopidae), Univ. Calif. Publ. Entomol., 58,
1-94.
Toschi G. A., 1965. Taxonomy, life history and
mating behavior of green lacewings of Straw-
berry Canyon (Neuroptera. Chrysopidae), Hil-
gardia, 36, 391^131.
Withycombe C. L., 1925. Some aspects of the
biology and morphology of the Neuroptera
with special reference to the immature stages
and their possible phylogenetic significance,
Trans. R. Entomol. Soc. London, 1924, pp.
303^111.
324
8. Отряды энтогнат и насекомых
Отряд COLEOPTERA
(жесткокрылые, или жуки)
Взрослые насекомые обычно имеют
две пары крыльев • первая лишена жилок,
плотная, напоминает щитки и склады-
вается вместе над брюшком, образуя
твердый покров для защиты крыльев,
вторая пара, используемая для полета,-
перепончатая, обычно имеет жилки и в
состоянии покоя убирается под надкры-
лья, или элитры (рис. 8.79). В типичном
случае тело твердое и компактное. Ро-
товые органы грызущего типа, антенны
хорошо развиты, обычно состоят из
10—14 члеников; сложные глаза обычно
хорошо выражены; ноги сильно склеро-
тизированы. Личинки, как правило, с чет-
ко отграниченной головной капсулой,
грызущим ротовым аппаратом, антенна-
ми и грудными ногами, но без брюшных
конечностей. Куколки (рис. 8.96) с при-
жатыми к телу имагинальными придат-
ками, не срастающимися, однако, с те-
лом. Имаго разнообразного облика, раз-
меры варьируют от менее миллиметра
до 100 мм. Столь же значительно варьи-
руют в пределах отряда окраска и форма
,тела; в целом наибольшее количество яр-
коокрашенных, причудливых и гигант-
ских видов встречается в тропических
районах земного шара.
Большая часть жуков либо раститель-
ноядны, либо питаются другими насе-
комыми. Обычно и личинки, и имаго
одного и того же вида питаются одина-
ковой пищей, т. е. обе фазы развития ли-
бо фитофаги, либо хищники. Июньские
хрущи, будучи фитофагами, в течение
своего жизненного цикла используют в
пищу различные части растений. Имаго
питаются листьями деревьев, тогда как
личинки едят корни деревьев, кустарни-
ков, злаков и других травянистых расте-
ний. Хищные жуки активно охотятся,
подкрадываясь к своей добыче. В неко-
торых группах способы питания более
специализированы. Ряд видов - эндопа-
разиты других насекомых или питаются
яйцами насекомых.
В целом отряд жуков можно назвать
сухопутным. Однако некоторые семей-
ства обитают в воде как на стадии личин-
ки, так и во взрослой фазе. Для окуклива-
ния личинки обычно выходят из воды и
сооружают земляную колыбельку близ
водоема. Как и наземные виды, водные
Рис. 8.79. Навозник Copris minutus. (Из Illinois Natural History
Survey.)
Отряды энтогнат и насекомых
325
гермы могут быть как раститель-
ноядными, так и хищными, причем по-
-тедние преобладают.
У подавляющего большинства жест-
»гсрылых в году бывает одно поколение,
ггеме того, для них характерен простой
развития, подобный описанному
-<ie Овальные или сферические яйца
отгладываются весной или в начале лета,
* нерез 1-2 нед из них выходят личинки.
Личинки проходят три личиночных воз-
г*хста. они усиленно питаются, обычно
гсетигая полных размеров ко второй по-
гозине лета, и окукливаются в почве. Че-
тез несколько недель появляются
простые насекомые, которые питаются
* окончательно созревают за оставшую-
часть лета и осени. С наступлением хо-
сдной погоды они уходят на зимовку.
Следующей весной перезимовавшие жу-
v* зткладывают яйца, и весь цикл повто-
ряется. Старые имаго обычно умирают
кз ере после завершения откладки яиц.
Существует множество отклонений
:<ой биологической схемы. Например,
хлъи коровки дают непрерывный ряд
тер?крывающихся поколений на протя-
к г-оии всего теплого времени года. В дру-
дл группах довольно многочисленны
ь* ты. личинки которых живут в почве
• д гниющей древесине и зимовка проис-
'' Д5гг на стадии личинки, а имаго встре-
=а_<тся лишь в течение короткого перио-
эесной или осенью.
1а более чем 300 млн. лет, прошедших
- того времени, когда жесткокрылые от-
гедстись от своего нейроптероидного
отхдка. эти насекомые превратились в
о^ыгоолее многочисленную по числу ви-
зги группу животных. Возможно, до на-
зяшего времени дожило около 200 000
iiz_zc3i из этого числа видов примерно
25 АЮ. принадлежащих приблизительно
* .50 семействам, встречаются в Север-
z Америке. Собрав воедино прими-
диные признаки современных жест-
кокрылых (рис. 8.80), можно выдвинуть
предположение, что все жуки произошли
от предковой формы, имевшей во взрос-
лом состоянии свободные и раздельные
стерниты брюшка и личиночные ноги с
тарзальным сегментом. Этот предок дал
начало двум линиям. В первой из них со-
хранились личиночные лапки, но три
первых стернита брюшка имаго срослись
и задние тазики неподвижно прикрепи-
лись к первому стерниту и разделили его
(рис. 8.81). Эта линия привела к образо-
ванию подотряда Adephaga, включаю-
щего такое примитивное семейство, как
Gyrinidae (представители которого
имеют пиловидный яйцеклад), неболь-
шое семейство Rhysodidae (представите-
ли которого живут в гниющих стволах),
наземных хищников из семейств жуже-
лиц (Carabidae) и близких им групп и
хищных водных жуков (Dytiscidae и дру-
гие).
В другой линии, идущей от древней-
ших форм жуков, сегменты брюшка
остались свободными, однако ис.чезли
лапки у личинок. Подотряд Archostema-
ta, по-видимому, является наиболее при-
митивным из всех сохранившихся групп
этой линии, так как у его представителей
тарзальный сегмент на ногах личинок
первых возрастов сохраняется и пропа-
дает лишь у более поздних возрастов.
Этот подотряд включает очень неболь-
шое число бедных видами и редко встре-
чающихся семейств, в том числе Cupedi-
dae и Micromalthidae. В линии, берущей
начало от предка с таким строением ли-
чиночных лапок, эти лапки полностью
утратились. Эта линия развилась в под-
отряд Polyphaga, составляющий основ-
ную массу современных жесткокрылых.
В подотряде Polyphaga отдельные се-
мейства стали в высшей степени специа-
лизированными и морфологически силь-
но отличающимися от более прими-
тивных представителей того же подотря-
326
8. Отряды энтогнат и насекомых
Scarab eidae
Histeridae \
Hydrophilidae
Staphylinidae
Silphidae
Мальпигиевых
сосудов
Staphylinoidea
Heteroceridae
Psephenidae
Elmidae
Dryopidae
Buprestidae
Elateridae
Cantharidae
Lampyridae
\ ।
i Lampiroidea
9 1
7 видимых
стернитов
Bostrichidae
Anobiidae
Dermestidae
Bostrichoidea
Scolytidae
Curculionidae
Cerambycidae
Chrysomelidae
Bruchidae
Meloidae
Mordellidae
Ten ebrio n id ae
Languriidae
Coccinellidae
Cucujidae
Nitidulidae
/
_ Cucuioidea
Функциональное
8-е дыхальце Stylopoidea
Надкрылья
редуцированны
Менее 7 Нефункциональное 8-е
видимых дыхальце
стернитов
неподвижные
Урогомфы
сочлененные
Урогомфы
Cicindelidae
Carabidae
Gyrinidae
Dytiscidae
Haliplidae
Водные формы
ADEPHAGA
POLYPHAGA
MYXOPHAGA
Личинки старших
возрастов лишены лапок
Имеется продолговатая ячейка
[ выражен пронотоплевральный шов
Эти структуры отсутствуют
Все Личиночные стадии
лишены лапок
ARCHOSTEMATA
У имаго I-III
стерниты брюшка
срастаются Предковая форма жуков
Стерниты свободные,
личинки с лапкой
Рис. 8.80. Упрощенная схема филогенеза отряда Coleoptera. (Составлено в основном по
Crowson, 1955.)
Рве. 8.81. А. Жужелица Harpalus sp. (Carabidae), вид с вентральной стороны. Ноги с левой
стироны удалены. Б. Хрущ Phyllophaga sp. (Scarabaeidae), вид с вентральной стороны. Ноги с
травой стороны удалены. В. Часть груди мягкотелки Chauliognathus pennsylvanicus (Cantharidae),
жд с вентральной стороны. (По Matheson, Entomology for introductory courses,
с разрешения Comstock Publishing Co.)
'-антенны; 2-нижнегубной щупик; 5-максиллярный щупик; 4-мандибула; 5-нижняя губа,
f-горло; 7-сложный глаз; 8-горловой шов; 9-шов, отделяющий переднеспинку от
зпистерна; 7О1-7О3-эпистерны передне-, средне- и заднегруди, 771-773-стерниты передне-,
средне- и заднегруди; 12х-12ъ-передние, средние и задние тазики; 75 j - 753-передние,
средние и задние тазиковые впадины; 74-переднеспинка; 15j-153-эпимеры передне-, средне-
заднегруди; 16-эпиплевры надкрыльев; 77-поперечный шов; 75-предтазиковая часть
заднегруди; 79-бедро; 20-вертлуг; 27-надкрылья, 22-голень; 2323«-членики лапки;
2А-верхняя губа; 25-восьмой тергит брюшка; 26-дыхальца; 27-мембрана, 28-крылья.
328
8. Отряды энтогнат и насекомых
Рис. 8.82. Отличительные признаки некоторых Coleoptera. А. Антенна Nicrophorus (Silphidae);
отдельно показаны вогнутые членики булавы и один из них со стороны вершины усика.
Б. Антенна Scolytus (Scolytidae). В. Антенна Thy се (Scarabaeidae). Г Голень и лапка
Heterocerus (Heteroceridae). Д. Антенна Silpha (Silphidae). Е. Антенна Attagenus (Dermestidae).
Ж. Антенна Languria (Languriidae). 3. Антенна Stegobium (Anobiidae). И. Антенна Anthrenus
(Dermestidae). К. Антенна Melanotus (Elateridae). Л. Голова Dineutes (Gyrinidae). M. Его же
ноги. Н. Общий вид сбоку Mordellistena (Mordellidae). О, П. Голова и переднеспинка
Dendroctonus (Scolytidae), вид снизу и сбоку. Р. Голова короткохоботного долгоносика
(Curculionidae). С. Голова Tropisternus (Hydrophilidae). 7-усик, с -тазик; 3-бедро;
4-максиллярный щупик, 5-лапка; 6-голень.
8. Отряды энтогнат и насекомых
329
да. Имаго некоторых мелких грибных
жуков, например, полностью утратили
жилкование на задних крыльях, боль-
шую часть члеников лапок, претерпели
сильнейшие изменения склеритов тела,
связанные с уменьшением размеров. Та-
ким образом, исходя из современного со-
стояния наших знаний, при описании раз-
личных линий жесткокрылых часто воз-
можно лишь указать признаки прими-
тивных представителей каждого таксона.
Наиболее примитивными представи-
телями той линии, которая дала начало
Polyphaga, являются несколько редких,
очень мелких жуков (Hydroscaphidae и
Sphaeriidae), имеющих поперечную жил-
ку. ограничивающую на заднем крыле
лро долговатую ячейку, а также шов меж-
ду переднеспинкой и проплеврами. Эти
эормы рассматривают как отдельный
зодотряд Myxophaga. Продолговатая
ячейка и пронотоплевральный шов исчез-
ли в той ветви, которая развилась в под-
стряд Polyphaga. Предок этого подотря-
да дал начало двум стволам. В более
тримитивном (стафилиноидном) уро-
гомфы (органы, подобные церкам) личи-
нок остались сочлененными, у остальных
групп урогомфы неподвижно срослись с
несущим их сегментом.
Стафилиноидный ствол разделился
за две главные линии • одна имела четыре
мальпигиевых сосуда и включала, в част-
ности, коротконадкрылых жуков (Staphy-
hnidae), мертвоедов (Silphidae) и род-
ственные им группы; другая линия имела
зюсть мальпигиевых сосудов, и к ней от-
носились карапузики (Histeridae) и водо-
любы (Hydrophilidae). Связи пластинча-
тоусых (Scarabaeidae) не поддаются уста-
новлению; высказывалось предположе-
ние. что они представляют собой побоч-
ную ветвь, уклонившуюся от стафили-
ноидной группы.
Другая линия типичных Polyphaga в
свою очередь разделилась на две следую-
Рис. 8.83. Лапки Coleoptera. (По Matheson,
Entpmology for introductory courses,
с разрешения Comstock Publishing Co.) A.
Harpalus (Carabidae). Б. Leptinotarsa
(Chrysomelidae). B. Chelymorpha
(Chrysomelidae). Г Epilachna (Coccinellidae).
Д. Коготок с зубцом в основании.
7-5-членики лапки.
щие ветви. В лампироидной ветви тело
взрослых насекомых самых прими-
тивных семейств сохраняет не менее семи
видимых стернитов брюшка. Эта ветвь
развилась в богатую семействами груп-
пу, включающую мягкотелок (Canthari-
dae) и светляков (Lampyridae), щелкунов
(Elateridae) и златок (Buprestidae), ряд
водных жуков (Psephenidae и близкие се-
мейства) и многих других.
В ветви, сестринской по отношению к
лампироидной, брюшко имеет не более
шести видимых стернитов. От этой ЛИ-
НИИ произошли бострихоиды, у которых
все дыхальца тела имаго сохраняют
функционирующее состояние, а также ку-
куйоиды, на восьмом брюшном сегменте
которых дыхальце перестало функциони-
ровать. Бострихоиды дали кожеедов, то-
330
8. Отряды энтогнат и насекомых
Рис. 8.84. Отличительные признаки некоторых Coleoptera.
А. Заднегрудь и брюшко Haliplus (Haliplidae), вид с вентральной стороны. Б. Средне- и
заднегрудь Dytiscus (Dytiscidae), вид с вентральной стороны. В. Пример переднегруди с
открытыми тазиковыми впадинами, схематизировано. Г Пример переднегруди
с замкнутыми тазиковыми впадинами, схематизировано. Д. Брюшко Leptura (Cerambycidae),
вид сбоку. Е. Брюшко Bruchus (Bruchidae). Ж. Брюшко Crioceris (Chrysomelidae), вид сбоку.
3. Голова и переднегрудь Cryptocephalus (Chrysomelidae), вид сбоку. И. Голова и переднегрудь
Acanthoscelides (Bruchidae). К. Голова и переднегрудь Helichus (Dryopidae). Л. Голова Saperda
(Cerambycidae). М. Заднее бедро Neoclytus (Cerambycidae). Н. Голова Cicindela (Cicindelidae).
О. Голова представителя Carabidae. 77. Задняя нога Amphicerus (Bostrichidae). Р. Задняя нога
Amblycerus (Bruchidae). С. Задняя лапка Psephenus (Psephenidae). 1 -стернит заднегруди;
2-тазиковые покрышки; 3-тазик; 4-брюшко; 5-бедро; 6-вертлуг; 7-лоб; 8-верхняя
губа; 9-мандибула, 10-наличник.
чилыциков и капюшонников (Dermesti-
dae, Anobiidae и Bostrichidae) и еще ряд
родственных им групп. Кукуйоиды диф-
ференцировались в большое число се-
мейств с самым различным образом жиз-
ни и внешним обликом. Наиболее широ-
ко известны божьи коровки (Coccinelli-
dae); чернотелки (Tenebrionidae) и ос-
новные группы растительноядных жест-
кокрылых (Chrysomelidae, Scolytidae,
Curculionidae и близкие группы).
Строение веерокрылых (надсемейство
8. Отряды энтогнат и насекомых
331
Stylopoidea) подверглось столь значи-
тельной модификации, что истинные
родственные отношения этой группы не-
ясны. У имаго на восьмом сегменте
брюшка дыхальца не функционируют.
Этот факт в сочетании с некоторыми
другими косвенными данными подтвер-
ждает гипотезу о возможном происхо-
ждении веерокрылых либо в качестве се-
стринской группы кукуйоидов, либо от
какого-то предка из числа кукуйоидов.
ПОДОТРЯД ADEPHAGA
Этот подотряд содержит восемь се-
мейств, в большинстве своем ведущих
хищный образ жизни и питающихся насе-
комыми. Два широко распространенных
семейства -Cicindelidae (скакуны) и Сага-
bidae (жужелицы)-наземные, ряд дру-
гих -Gyrinidae, Haliplidae и Dytiscidae-
эодные, причем в воде живут как имаго,
так и личинки.
CARABIDAE, ЖУЖЕЛИЦЫ. ВИДЫ ЭТОГО
семейства-активные наземные жуки с
длинными тонкими антеннами равно-
мерной толщины, длинными надкрылья-
ми и длинными ногами, приспосо-
бленными к беганию (рис. 8.85). Раз-
меры жужелиц варьируют от 1 мм в
длину до 35 мм у некоторых крупных, ме-
таллически-зеленых видов L Ротовые ор-
ганы хорошо развиты, мандибулы
длинные, мощные и острые. Имаго мо-
гут вести либо дневной, либо ночной
образ жизни, питаясь самыми разно-
образными животными, в том числе
улитками, червями, неполовозрелыми и
взрослыми стадиями различных насе-
комых. Личинки с сильно вытянутым те-
лом и мощным ротовым аппаратом, хо-
рошо развитыми ногами и парой церко-
подобных органов-урогомф- на конце
брюшка; они ведут хищный образ жизни,
питаясь другими насекомыми. Жуже-
1 Известны виды, достигающие в длину
90 мм - Прим, перев.
Рис. 8.85. Жуки. (Из Kansas State University.)
А. Жужелица Pterostichus substriatus. Б. Мертвоед-могильщик Nicrophorus
marginatus.
332
8. Отряды энтогнат и насекомых
лицы встречаются обычно на почве или
под слоем подстилки, где охотятся на
свою добычу. Очень богатое видами се-
мейство, насчитывающее более 2000 не-
арктических видов.
gyrinidae, вертячки. Водные жуки;
имаго твердые, выпуклые, блестящие
и темные. Средние и задние ноги широ-
кие, усажены бахромой щетинок и ис-
пользуются для плавания. Голова имеет
две важные особенности. 1) короткие ан-
тенны с уховидным выростом на третьем
членике и 2) глаза, полностью разде-
ленные на верхнюю и нижнюю поло-
вины, так что кажется, будто у жуков две
пары сложных глаз (рис. 8.82,Л). Имаго
весьма обычны в стоячих и проточных
водоемах. Они собираются тысячными
стаями и кружатся по поверхности, дви-
гаясь с большой скоростью и оставляя за
собой серебристые пересекающиеся вол-
ны-привычное зрелище для любого на-
туралиста. Во время этого кружения по
воде разделенные надвое глаза дают жу-
кам возможность видеть происходящее в
воздухе с помощью дорсальных частей
Рис. 8.86. Личинка
жука-вертячки. (По
Boving, Craighead.)
глаз и в воде-с помощью вентральных
частей. Личинки (рис. 8.86) тонкие, удли-
ненные, с белым телом. Каждый сегмент
брюшка несет пару длинных трахейных
жабр, а конец брюшка-пару крючьев. И
имаго, и личинки-хищники, причем има-
го поедают в основном мелких жи-
вотных, падающих в воду или всплываю-
щих к поверхности, а личинки-мелких
животных, которых они находят в скры-
тых местах на дне водоема.
ПОДОТРЯДЫ
ARCHOSTEMATA И MYXOPHAGA
В Северной Америке Archostemata на-
считывают два небольших семейства Си-
pedidae и Micromalthidae, представители
которых населяют гнилые стволы. По-
следнее включает единственный мелкий
и редкий вид (имаго имеют в длину от 1,8
до 2,5 мм), замечательный своими педо-
генетическими личинками (с. 216).
Из трех известных семейств подотря-
да Myxophaga два встречаются на терри-
тории Северной Америки. Крошечные
(1,3 мм) представители Hydroscaphidae
обитают в воде, а более крупные (2-7 мм)
представители Scaphidiidae живут в под-
стилке и гнилой древесине.
ПОДОТРЯД POLYPHAGA
Сюда относится подавляющее боль-
шинство жуков около 130 семейств, этот
подотряд охватывает формы с разноо-
бразным обликом и различными цикла-
ми развития. Одни семейства, как, напри-
мер, Chrysomelidae и Coccinellidae, бо-
гаты видами и содержат ряд важных в
экономическом отношении насекомых,
другие семейства, однако, редки или
малоизвестны, так как требуют спе-
циальных методов сбора и не имеют, на-
сколько это известно, хозяйственного
значения. Упоминаемые ниже семейства
8. Отряды энтогнат и насекомых
333
составляют лишь малую часть всех се-
мейств жесткокрылых, включаемых в
подотряд Polyphaga. Подробную и об-
ширную информацию о жизненных ци-
клах и диагностике тех семейств, ко-
торые не обсуждаются в нашей книге,
студенты найдут в книгах, список ко-
торых приведен в конце раздела о Со-
leoptera.
СТАФИЛИНОИДЫ
STAPHYLINIDAE, СТАФИЛИНИДЫ, ИЛИ
коротконадкрылые жуки. Удли-
ненные жуки с узким телом (рис.
S.87, А), короткими обрубленными на
вершинах надкрыльями, под которыми
складываются задние летательные кры-
лья. Антенны довольно длинные, ните-
видные или слегка утолщенные на вер-
шинах. Личинки также вытянутые и силь-
но напоминают мелких личинок жуже-
лиц. Взрослые насекомые и личинки пи-
таются разлагающимися веществами
или хищничают, за исключением немно-
гочисленных видов, личинки которых ве-
дут паразитический образ жизни.
Взрослые стафилиниды встречаются на
цветках, в подстилке, под корой, в разла-
гающихся органических веществах,
в гнездах муравьев и термитов и многих
других местах. Личинки более скрытны и
встречаются в основном в сырых местах.
Виды, ведущие хищный образ жизни, по-
едают клещей, мелких насекомых, яйца
насекомых и клещей, особенно мелких
личинок двукрылых. Ряд видов играют
значительную роль в естественной регу-
ляции численности вредителей. Интере-
сен род Baryodma, личинки которого па-
разитируют на куколке капустной мухи.
В первом возрасте личинка представляет
собой стройную свободнобегающую
форму с относительно широкими терги-
тами. Она разыскивает в земле пупарий
мухи и залезает в него, проделывая от-
А
Рис. 8.87. Стафилиниды.
А. Имаго Stenus. Б. Личинка первого
возраста Aleochara cur tula до начала питания
(слева) и после окончания питания (справа).
(Л-по Sanderson; К-по Kemner,
с изменениями.)
верстие. Внутри пупария личинка жука
поедает куколку мухи и стремительно
растет, становясь после линьки сильно
раздутой и червеобразной. Все после-
дующие личиночные возраста также чер-
веобразные (эруковидные). Подобный
путь развития свойствен и некоторым ви-
дам Aleochara (рис. 8.87, Б).
SCARABAEIDAE, ПЛАСТИНЧАТОУСЫЕ,
ИЛИ НАВОЗНИКИ И ХРУЩИ. Это ОДНО ИЗ
крупнейших семейств жуков, к числу ха-
рактерных признаков которого относит-
ся строение антенн с их пластинчатой бу-
лавой, т. е. с листовидно расширенными
и в покое прилегающими друг к другу по-
следними члениками. Пластинчатоусые
очень разнообразны по размерам и фор-
ме, большинство видов имеет коренастое
тело с очень плотными покровами. Ли-
334
8. Отряды энтогнат и насекомых
Рис. 8.88. Японский жук Popillia japonica. (По материалам Connecticut
Agricultural Experiment Station.)
А. Имаго. Б. Личинка. В. Яйцо.
чинки малоподвижные, толстые, обычно
белые и характерной С-образной формы
(рис. 8.88). Важную группу пластинча-
тоусых составляют навозники-потреби-
тели разлагающихся веществ, в том чис-
ле навоза, гниющих шкур или грибов.
Крайне интересны Canthon и некоторые
другие роды; имаго делает из навоза
шар, укатывает его в сторону и закапы-
вает На этот шар откладываются яйца, и
вышедшие из них личинки используют
его в качестве пищи. Остальные группы
пластинчатоусых, кроме навозников,
Рис. 8.89. Dynastes tityus-самый крупный жук штата Иллинойс и бурозубка Sorex
longirostris - самое маленькое млекопитающее Иллинойса. (Рисунок, присланный
С. О. Mohr.)
т. е. хрущи,-растительноядны, и многие
из них имеют огромное экономическое
значение как вредители. Наибольшей из-
вестностью среди хрущей пользуется
японский жук Popillia japonica (рис. 8.88),
его личинки питаются корнями трав и
8. Отряды энтогнат и насекомых
335
особенно вредны на газонах и площадках
для гольфа, а имаго объедают листья
плодовых и декоративных деревьев. Дру-
гой группой видов, приносящих опреде-
ленный вред, являются хрущи из рода
Phyllophaga (рис. 7.12); их взрослые осо-
би объедают листву листопадных пород
деревьев, а личинки питаются корнями
различных сельскохозяйственных куль-
тур, в том числе кукурузы и прочих зер-
новых, а также пастбищных трав.
В тропиках многочисленны виды
крупных, часто очень ярко окрашенных
пластинчатоусых, вооруженных различ-
ного рода рогами и выростами на голове
и переднеспинке. Наиболее крупный вид
фауны Соединенных Штатов - Dynastes
tytius, жук-геркулес, размерами превы-
шающий землеройку (рис. 8.89). Личин-
ка его живет в гнилой древесине.
ЛАМПИРОИДЫ
LAMPYRIDAE, СВЕТЛЯКИ. Это МЯГКО-
телые жуки средних размеров, имеющие
пильчатые антенны и уплощенные края
переднеспинки, выступающие в виде на-
веса или полей шляпы и отчасти закры-
вающие голову. Надкрылья относитель-
но мягкие. На востоке наиболее обычен
вид Photurus pennsylvanicus (рис. 8.90).
Имаго встречаются летом, активно ле-
тают теплыми ночами и практически не-
изменно совершают ныряющие движе-
ния вверх и вниз примерно в 1-1,5 м над
землей. В начале каждого подъема особь
вспыхивает ярким светом, что является
сигналом к спариванию. Когда имаго ле-
тают в больших количествах, вся окрест-
ность озяряется этими мельчайшими го-
рящими точками. От этого и пошло
название «светляки». Самки в некоторых
родах бескрылые и червеобразные; они и
дичинки также способны светиться. И ли-
чинки, и имаго-хищники, хотя неко-
торые виды во взрослой фазе могут ча-
Рис. 8.90. Светляк Photurus pennsylvanicus.
(По Hess, Matheson.)
А. Взрослый самец. Б. Зрелая личинка.
В. Левая мандибула личинки; показан
мандибулярный канал. 1 - наружное
отверстие канала; 2-зубец мандибулы;
3-отверстие канала, открывающееся в рот;
4-мыщелок. Г. Начальная стадия
сооружения кукол очной колыбельки.
Д. Оконченная куколочная колыбелька.
Е. Куколка в колыбельке.
стично поедать и растительные вещества
или не питаться вовсе.
ELATERIDAE, ЩЕЛКУНЫ (ПРОВОЛОЧНИ-
КИ). У имаго гладкое, стройное и твердое
тело (рис. 8.91), пятичлениковые лапки,
большая переднеспинка с острыми за-
дними углами, пильчатые антенны и
длинный сильный отросток, напра-
вленный от переднегруди назад. Оказав-
шись лежаЩИМИ На ииинс, эти жуки спо-
собны подпрыгивать на несколько санти-
метров, издавая в момент прыжка гром-
кий щелчок и, как правило, приземляясь
правильной стороной вверх. Этот пры-
жок осуществляется с помощью вен-
трального отростка переднегруди за счет
336
8. Отряды энтогнат и насекомых
Рис. 8.91. Elateridae. (По материалам министерства сельского хозяйства
США.)
А, Б. Сухолюбивый вид Ludius inflatus; взрослое насекомое (Л) и личинка-
проволочник (Б). В. Обитающий в песках Horistonotus uhlerii, взрослое
насекомое и его личинка-проволочник (Г). '
сил, возникающих при освобождении
упругой энергии во время растяжения те-
ла. Имаго чаще всего встречаются вес-
ной и летом. Эти жуки и их «акробатиче-
ские» прыжки хорошо знакомы детворе в
сельских местностях.
Личинки щелкунов, называемые про-
волочниками (рис. 8.91), напоминают
червей, имеют твердые покровы и оби-
тают в почве или гнилой древесине. Ли-
чинки, обитающие в почве, питаются
корнями трявянистых и иных растений и
в высшей степени вредны на полях боль-
шинства зерновых культур. Особенно
значительный ущерб причиняют виды
родов Melanotus, Agriotes и Monocrepi-
dius. В Северной Америке известно не-
сколько сотен видов. Собирателю личи-
нок насекомых будут необычайно инте-
ресны странные червеобразные личинки
группы, к которой принадлежит род Ho-
ristonotus (рис. 8.91, Г).
BUPRESTIDAE, ЗЛАТКИ. Эти Жуки
(рис. 8.92) в общем напоминают щелку-
нов тем, что их имаго также имеют пя-
тичлениковые лапки, пильчатые антенны
и твердые покровы, однако отличаются
от последних сросшимися двумя первы-
ми стернитами брюшка и медной или
блестящей металлической окраской тела
* Отряды энтогнат и насекомых
337
Рис. 8.92. Buprestidae. (По материалам министерства сельского хозяйства
США.)
А. Плоскоголовая яблоневая златка Chrysobothris femorata. Б. Тихоокеанская
плоскоголовая златка С. mali. В. Личинка в ходе.
* надкрыльев. Они обычно несколько ко-
ренастее и имеют более короткую пере-
днеспинку. Личинки крупных видов
Г2С. 8.92) прокладывают ходы в древе-
_сче. поселяясь на живых, ослабленных
дд? отмерших деревьях, и питаются либо
корой, либо в толще твердой древе-
сины. Эти личинки удлинены и лишены
с расширенной и уплощенной
~7>лью. Развиваются они на различных
древесных породах, как лиственных, так
двойных. В плодовых садах деревьям
=дсто сильно вредит златка Chrysobothris
z~orata. Небольшое число родов златок
* личиночной стадии минирует листья,
Д'-дшки минеров имеют более цилин-
дрическое тело, чем личинки ксилофагов,
1 также крошечные ноги.
DRYOPIDAE И БЛИЗКИЕ ГРУППЫ, ПРИ-
ЛЛЫШИ. И личинки, и взрослые особи
-той водной группы обитают в воде. И те
f другие медлительны, ползают по кам-
дкм иди затопленным стволам деревьев,
даггаясь поверхностными обрастаниями.
7±дс взрослого жука покрыто тонкими
~стыми волосками, удерживающими
д.енку из воздуха, когда насекомое на-
ходится под водой. Жук использует эту
пленку как средство для газообмена с
окружающей водой. Личинки дышат по-
средством трахейных жабр, расположен-
ных на различных частях тела, в том
числе групп жабр, втягивающихся в вен-
тральные карманы. Как и многие другие
водные жесткокрылые, личинки прице-
пышей, закончив развитие, покидают во-
ду и окукливаются во влажной почве.
Большинство прицепышей обитают в хо-
лодных быстрых потоках и часто в зна-
чительных количествах попадаются при
просеивании гальки на мелководье. Има-
го периодически выходят из водоемов для
расселительного лёта и спаривания.
БОСТРИХОИДЫ
DERMESTIDAE, КОЖЕЕДЫ. Выпуклые
овальные жуки (рис. 8.93, 8.94) с корот-
кими булавовидными антеннами, пятич-
лениковыми лапками и брюшком, имею-
щим лишь пять склеротизированных
стернитов. Тело личинки удлиненное или
овальное, покрытое крупными пучками
или перевязями из длинных перистых во-
338
8. Отряды энтогнат и насекомых
Рис. 8.93. Ковровый кожеед Attagenus
megatoma. (По материалам Connecticut
Agricultural Experiment Station.)
А. Взрослое насекомое. Б. Личинка.
лосков. Они питаются сухими продукта-
ми животного происхождения, в том чис-
ле мехом, шерстью, кожей и сухими
мясными продуктами. Последние осо-
бенно охотно поедаются видами рода
Dermestes, прежде всего D. lardarius, или
ветчинным кожеедом. Некоторые более
мелкие виды, в частности Anthrenus scrop-
hulariae (рис. 8.94) и кожеед ковровый,
Attagenus megatoma (рис. 8.93), нападают
на меха, ковры, обивку мебели - словом,
на любые изделия из шерсти животных.
Эти вредители распространены столь
широко, что являются постоянной угро-
зой для материальных ценностей и мно-
гих хранящихся материалов. В природе
кожееды питаются мертвыми насекомы-
ми или останками животных. Имаго по-
едают ту же пищу, что и личинки, однако
во время расселительного лёта они пи-
таются пыльцой, и в этот период их час-
то можно встретить на цветах в саду.
КУКУЙОИДЫ
COCCINELLIDAE, БОЖЬИ КОРОВКИ.
Округлые, выпуклые, блестящие жучки
немного меньше средних размеров
(рис. 8.95), часто с рисунком из желтых,
красных, черных или синих пятен. Ан-
тенны короткие и булавовидные, лапки
четырехчлениковые, но кажутся трехчле-
никовыми, так как третий членик крайне
мал и расположен между подушко-
видным вторым члеником и большим
концевым члеником, несущим коготки.
8. Отряды энтогнат и насекомых
339
Рис. 8.94. Кожеед Anthrenus scrophulariae. (По материалам Connecticut Agricultural
Experiment Station.)
А. Личинка. Б. Взрослое насекомое.
Личинки покрыты либо бородавками,
либо восковыми выделениями и имеют
очень короткие усики, но длинные ноги.
Хозяйственное значение имеют две кате-
гории божьих коровок. Виды первой ка-
тегории питаются тлями и кокцидами и
могут служить эффективным средством
естественного ограничения численности
некоторых вредителей из этих групп. Из
таких хищников наиболее известна родо-
лия (Rodolia cardinalis) - вид астралийско-
го происхождения, интродуцированный в
Рис. 8.95. Божья коровка Hippodamia
convergens. (По материалам министерства
сельского хозяйства США.)
А. Взрослое насекомое. Б. Куколка. В.
Личинка.
340
8. Отряды энтогнат и насекомых
Рис. 8.96. Мексиканская фасолевая коровка Epilachna varivestis. (По
материалам министерства сельского хозяйства США.)
А. Личинка. Б. Взрослое насекомое. В. Куколка. Г Яйца.
Калифорнию для борьбы с желобчатым
червецом (на цитрусовых). Родолия ока-
залась очень эффективной в уничтожении
червеца. Обычнейший вид местного про-
исхождения - Hippodam ia con ver gens
(рис. 8.95). Вторая важная категория
божьих коровок включает раститель-
ноядные виды. В Северной Америке од-
ним из наиболее вредных дефолиаторов
фасоли во многих центральных и южных
районах является коровка Epilachna va-
rivestis (рис. 8.96). Листвой питаются как
личинки, так и имаго.
MELOIDAE, НАРЫВНИКИ. Имаго
(рис. 8.97)-жуки средней величины с от-
носительно мягким телом и надкрылья-
ми, длинными простыми антеннами и
выступающей круглой или овальной го-
ловой, резко отделенной от груди. Лапки
передних и средних ног пятичлениковые,
а задних ног-четырехчлениковые. Это
характерная черта группы, включающей
приблизительно 20 семейств жуков и
иногда называемой Heteromera. В теле
нарывников содержится маслянистое ве-
щество кантаридин, которое вызывает
сильные раздражения кожи и появление
больших водяных волдырей на коже че-
ловека. Достаточно просто дотронуться
до живого взрослого жука-нарывника,
Рис. 8.97. Нарывник-шпанка Epicauta vittata. (Ориг С. О. Mohr.)
S. Отряды энтогнат и насекомых
341
чтобы полученное при этом количество
кантаридина вызвало серьезное воспале-
ние.
В большинстве подсемейств личинки
питаются личинками пчел или в качестве
инквилинов запасенной в гнездах пчел
провизией. Личинки одного из подсе-
мейств, к которому относятся и широко
распространенные обычные роды Epicau-
Z2 и Henous, питаются кубышками (яйца-
ми) саранчовых. У всех нарывников
сложный цикл развития. Самки отклады-
вают яйца группами от 50 до 300 штук в
почву на глубину около 2-3 см. Первый
зозраст представляет собой активную,
> длиненную, сильно склеротизирован-
=ую форму, именуемую триунгулином.
У видов, живущих в пчелиных гнездах в
качестве инквилинов, триунгулины заби-
раются на цветки, прицепляются к пче-
лам и, таким образом, переносятся в их
гнезда, где покидают тело пчелы и на-
чинают активно питаться. У видов, ко-
торые питаются яйцами саранчовых,
-рнунгулины просто разыскивают клад-
ки яиц саранчовых-кубышки.
Развитие личинок замечательно во
многих отношениях. Оно может быть хо-
з?шо проиллюстрировано примером
Epicauta pennsylvanica (рис. 8.98). Триун-
у. лнн находит кубышку с яйцами саран-
чового, проникает внутрь, прокалывает
с дно яйцо и поедает его содержимое. Че-
ргз несколько дней, когда триунгулин
раздувается и полностью наедается, на-
ступает первая линька. Червеобразная
ти чинка второго возраста имеет тонкие
покровы, относительно малоподвижна и
продолжает питаться кладкой яиц кобы-
лки.
Третий, четвертый и пятый личи-
ночные возраста быстро следуют один за
другим и сходны со вторым. Достигнув
полного размера, личинка пятого возра-
ста покидает кубышку, закапывается еще
на несколько сантиметров глубже в поч-
ву и делает земляную колыбельку, в ко-
торой затем линяет и превращается в ли-
чинку шестого возраста. Эта стадия,
известная под названием ложнокуколки-
уникальное явление в отряде жесткокры-
лых. Она не питается, имеет сильно скле-
ротизированное и несгибающееся тело
овальной формы. Ноги присутствуют
лишь в виде бугорков. Зимовка обычно
происходит именно на этой стадии. На
следующее лето ложнокуколка линяет,
превращаясь в личинку седьмого возра-
ста, сильно напоминающего пятый, из
этой личинки развивается настоящая ку-
колка, из которой через несколько недель
выходит имаго. Ложнокуколка в высшей
степени устойчива к высыханию, что
обеспечивает виду переживание засуш-
ливых лет, так как если в летний сезон,
наступивший после зимовки, стоит чере-
счур сухая погода, то ложнокуколка «от-
кладывает» свою линьку еще на один год
или даже на 2 года, когда это необходи-
мо, чтобы дождаться наступления благо-
приятных условий для завершения нор-
мального жизненного цикла.
Взрослые особи нарывников-ли-
стогрызущие насекомые; они могут на-
носить заметный ущерб культурам кар-
тофеля, томатов, кабачков, бобовых и
другим растениям. Шпанка Epicauta vitta-
ta (рис. 8.97)-вид с яркой окраской, ча-
сто вредящий сельскохозяйственным
культурам. Один из видов, часто питаю-
щихся на картофеле,- Meloe angusticollis-
необычен среди жуков тем, что его над-
крылья налегают друг на друга при
основании. Другой коротконадкрылый
нарывник-вредящий бахчевым культу-
рам Henous confertus.
TENEBRIONIDAE, ЧЕРНОТЕЛКИ. Эти жу-
ки обыкновенно имеют твердый панцирь
и темную окраску; тело овальное или уд-
линенное с параллельными боковыми
сторонами (рис. 8.99), лапки передней и
средней пар ног пятичлениковые, а за-
342
8. Отряды энтогнат и насекомых
Рис. 8.98. Преимагинальные стадии развития черного нарывника Epicauta pennsylvanica. (По
Horsfall.)
А. Непитавшаяся личинка первого возраста, х 17. К. Полностью напитавшаяся личинка
первого возраста; х 17 В-Д. Личинки второго, третьего и четвертого возрастов; х 17. Е.
Только что перелинявшая личинка пятого возраста, х 9. Ж. Наевшаяся личинка пятого
возраста, х 9. 3. Личинка шестого возраста, х 5. И. Личинка седьмого возраста х 5. К.
Куколка, х 5.
дних ног-четырехчлениковые; антенны
умеренной длины, обычно нитевидные
или булавовидные. Личинки питаются
главным образом мертвыми расти-
тельными остатками и грибами, особен-
но трутовиками, либо мицелием в гни-
лой древесине; немногие хищничают,
некоторые вредят запасам зерна в храни-
лищах. Личинки удлиненные, цилиндри-
ческие, с сильными ногами. Личинки рас-
пространенного на западе Америки рода
Eleodes питаются корнями растений и на-
зываются ложнопроволочниками из-за
сильного сходства с настоящими прово-
лочниками-личинками Elateridae. Лож-
нопроволочники другого западного ви-
S. Отряды энтогнат и насекомых
343
Рее. 8.99. А. Малый булавоусый хрущак
Tribolium castaneum. Взрослое насекомое и
ето личинка (Б). В. Малый мучной хрущак
Tribolium confusum. (По материалам
министерства сельского хозяйства США.)
да - Embaphion muricatum - вредят пшени-
це. Некоторые виды, являющиеся вреди-
телями запасов зерна и готовых пищевых
продуктов, широко распространены и
причиняют серьезный экономический
ущерб. К этой группе принадлежат, в
частности, большие мучные хрущаки ро-
та Tenebrio, имеющие относительно
крупные размеры (имаго достигают в
длину 15 мм, а личинки-25-30 мм), а
также малый мучной хрущак Tribolium
:onfusum и несколько близких к нему ви-
дов Tribolium, размеры которых очень не-
велики: имаго не превышают в длину
S-4 мм.
chrysomelidae, листоеды. Они со-
ставляют обширное семейство, виды
которого имеют мелкие или средние раз-
меры, обычно овальное коренастое тело
и достаточно длинные нитевидные ан-
тенны (рис. 8.100). Самый замеча-
тельный признак семейства- строение
лапок (рис. 8.83,Б, В) они кажутся четы-
рехчлениковыми; третий членик расши-
рен и образует широкую почковидную
подушечку; последний членик, являю-
щийся на самом деле пятым, длинный и
тонкий, кажется прикрепленным в сре-
динной выемке третьего членика, в дей-
ствительности же четвертый членик
представлен крайне редуцированным
кольцом, расположенным в основании
пятого, но он так мал, что его можно раз-
личить только в том случае, если сделать
специальный препарат ноги. Личинки ли-
стоедов разнообразны, но большинство
из них-толстые и короткие, черве-
образные, с короткими ногами и антен-
нами; многие несут на теле шипы и вы-
росты. Некоторые виды, минирующие
листья, длинные и плоские. Яйца ли-
стоеды откладывают в почву, под кору
деревьев или располагают на стеблях и
листьях.
За редкими исключениями, взрослые
листоеды питаются листвой растений, а
их личинки - корнями или листьями.
Многие вредят сельскохозяйственным
культурам, и в состав данного семейства
входит значительное число экономически
Рис. 8.100. Колорадский картофельный жук
Leptinotarsa decemlineata.
А. Взрослое насекомое. Б. Личинка. В.
Куколка.
344
8. Отряды энтогнат и насекомых
Рис. 8.101. Полосатый огуречный листоед
Acalymma vittata. (По материалам
министерства сельского хозяйства США.)
А. Взрослое насекомое. Б. Личинка.
важных видов. Колорадский жук Leptino-
tarsa decemlineata (рис. 8.100)-один из
наиболее вредных видов, поражающих
картофель, как личинки, так и имаго по-
едают листья. Спаржевая трещалка Crio-
ceris asparagi-ярко окрашенный вид,
обычный повсюду, где выращивают
спаржу; и личинки, и взрослые особи пи-
таются на листьях, яйца имеют черную
окраску и воткнуты одним концом в по-
являющиеся побеги растений. Личинки
многих видов, например Acalymma vittata
(рис. 8.101), известны как вредители кор-
ней. Имаго многих мелких видов пры-
гают, подобно блохам, за что и получили
название «земляные блохи». Из этой
группы роды Phyllotreta и Epitrix содер-
жат довольно много видов, взрослые
особи которых проедают дыры в листь-
ях, а вытянутой формы личинки грызут
корни капусты, репы, картофеля, огурцов
и других растений. Большое хозяйствен-
ное значение имеет табачная блошка Epi-
trix hirtipennis. Не у всех земляных
блох личинки питаются корнями расте-
ний, так, например, личинки видов ро-
да Haltica, как и имаго, питаются
листьями.
bruchidae, зерновки. Это наиболее
интересное семейство, близкородствен-
ное Chrysomelidae. Взрослые насекомые
короткие, коренастые; личинки эруко-
видные и почти безногие, обитают вну-
три семян бобовых (и некоторых других
растений). Ряд видов-весьма важные
вредители различных зернобобовых (го-
роха, фасоли и др.) в хранилищах, обыч-
нейшим примером может служить горо-
ховая зерновка Bruchus pisorum
(рис. 8.102).
CERAMBYCIDAE, УСАЧИ, ИЛИ ДРОВОСЕ-
КИ. Жуки с вытянутым телом, многие
красиво окрашены, имеют длинные ноги
и необычайно длинные антенны
(рис. 8.103). В остальном признаки дро-
восеков почти полностью совпадают с
признаками листоедов, в том числе и в
отношении строения лапок, имеющих
также расширенный третий членик и ре-
дуцированный четвертый. Личинки уса-
чей вытянутые, цилиндрические, с кру-
глой головой и либо вовсе без ног, либо с
крошечными' ногами. Они большей
частью прогрызают ходы в слое камбия
или в ядровой древесине деревьев; не-
многие обитают внутри корней и низких
Рис. 8.102. Гороховая зерновка Bruchus
pisorum. (По материалам министерства
сельского хозяйства США.)
А. Взрослое насекомое. Б. Личинка.
В. Куколка.
8. Отряды энтогнат и насекомых
345
Рис. 8.103. Круглоголовый яблоневый усач Saperda Candida.
Личинки, взрослые насекомые и выходные отверстия на
стволе в натуральную величину. (По Rumsey, Brooks.)
побегах суккулентов с травянистыми сте-
блями, таких, как молочаи и амброзия,
или в ветвях кустарников, в частности ив
а малины. Некоторые виды имеют боль-
шое экономическое значение.
Яблоневый усач Saperda Candida
рис. 8.103)-вид, раскрашенный корич-
невыми и белыми полосками и в неко-
торых местностях являющийся серь-
езным вредителем яблонь, его личинки
внедряются в ствол и прокладывают в
нем длинные ходы. Взрослые усачи Ме-
zacyllene robiniae имеют красивый геоме-
трический рисунок из желтых пятен на
черном фоне; его личинки проделывают
ходы в молодых деревцах акации белой
Robinia pseudoacacid) и настолько иста-
чивают их, что они легко переламывают-
ся ветром, многие посадки этого дерева,
предназначавшиеся для защиты почв от
эрозии, полностью уничтожались таким
образом.
Большинство жуков-усачей являются
вредителями лесных пород, нападающи-
ми как на лиственные, так и на хвойные
деревья. В условиях плохого санитарного
состояния или бесхозяйственного засоре-
ния лесов валежником и сучьями, а также
при необычных погодных условиях мно-
гие виды усачей могут размножиться до
такой степени, что станут причинять за-
метный ущерб лесным массивам, имею-
щим хозяйственное значение.
CURCULIONIDAE, ДОЛГОНОСИКИ, ИЛИ
слоники. Голова несет хорошо выра-
женную головотрубку, иногда вытяну-
тую и изогнутую (рис. 8.104), антенны
обычно коленчатые и булавовидные; те-
ло, надкрылья и ноги очень плотные, что
придает слоникам вид прочных, надежно
346
8. Отряды энтогнат и насекомых
Рис. 8.104. Хлопковый долгоносик Anthonomus grandis.
Справа-растение хлопчатника, подвергшееся нападению долгоносика. А. Висящая
сухая поврежденная коробочка. Б. Обнаженная коробочка с отверстиями,
проделанными долгоносиком. В. Разрезанная коробочка; показаны нападающий жук и
личинка в своей колыбельке. Г. Взрослая самка с расправленными как во время полета
крыльями. Д. Взрослое насекомое сбоку. Е. Куколка с вентральной стороны.
Ж. Личинка.
защищенных жуков. Личинки безногие,
эруковидные, обычно имеющие темную
головную капсулу и белое тело.
Curculionidae-одно из обширнейших
семейств насекомых, содержащее более
75000 видов, многие из которых - серьез-
нейшие вредители. Личинки питаются
самой разнообразной растительной
пищей: одни питаются корнями, листья-
ми или плодами, например лесными оре-
хами, желудями, вишнями и сливами;
другие прокладывают ходы внутри стеб-
8. Отряды энтогнат и насекомых
347
лей; остальные развиваются в гнилой
древесине или хранящихся продуктах.
Взрослые жуки обычно питаются листья-
ми или плодами тех видов растений, ко-
торые являются хозяевами для личи-
нок.
Хлопковый долгоносик Anthonomus
grandis (рис. 8.104)-один из опаснейших
вредителей хлопчатника в США и зани-
мает первое место в перечне видов, при-
чиняющих громадные убытки. Имаго
долгоносика заселяют растения в начале
вегетационного периода, питаясь листья-
ми. цветками и молодыми коробочками.
Самка откладывает яйца в отверстия,
прогрызенные ею в коробочках,-по
одному яйцу в каждое. Личинки разви-
ваются внутри коробочек, а затем де-
лают колыбельку, в которой окукли-
ваются. Через несколькд недель по-
оляются жуки, наносящие дополни-
тельный вред, питаясь на тех же расте-
ниях; перед наступлением зимы у этих
хуков наступает диапауза. Anthonomus
rrandis происходит из тропических обла-
стей Америки и проник в Техас из Мекси-
ки около 1890 г С этого времени он ус-
зел постепенно распространиться прак-
тически по всем хлопкосеющим районам
США.
Долгоносик Conotrachelus nenuphar
повреждает сливы, вишни и близкие к
ним плодовые; личинки питаются мя-
котью плода. Этот вид часто становится
серьезным вредителем.
Особо опасными вредителями запа-
сов зерна являются амбарный долгоно-
сик Sitophilus granarius и рисовый долго-
носик Sitophilus oryzae\ личинки обоих
видов живут и питаются внутри зерен, а
взрослые особи-либо в старых камерах
личинок, либо беспорядочно грызут зер-
на снаружи.
SCOLYTIDAE, КОРОЕДЫ. ВИДЫ ЭТОГО Се-
мейства развиваются главным образом
на деревьях, как на живых, так и на
мертвых. У жуков лишь слаборазвитая
головотрубка и тело почти цилиндриче-
ской формы (рис. 8.105). Личинки безно-
гие, эруковидные и имеют все типичные
черты, присущие долгоносикам. Как
взрослые жуки, так и личинки питаются,
пробуравливая ходы (рис. 8.106). У од-
них видов эти ходы прокладываются в
слое камбия (луба), что при большом
числе вредителей приводит к полному
Рис. 8.105. Короед Phloeotribus limnaris. (По материалам министерства сельского
хозяйства США.)
348
8. Отряды энтогнат и насекомых
Рис. 8.106. Маточный ход и ходы личинок
короеда Phloeotribus limnaris. (По
материалам министерства сельского
хозяйства США.)
Справа показан участок маточного хода с
яйцевыми нишами и отходящими от них
ходами молодых личинок.
опоясыванию дерева. Многие виды этого
семейства вредят породам, имеющим
значительную ценность, особенно сосне,
и порой вызывают преждевременную ги-
бель обширных посадок этих пород.
СТИЛОПОИДЫ (ВЕЕРОКРЫЛЫЕ)
Стилопоиды, или веерокрылые
(рис. 8.107), составляют надсемейство
Stylopoidea (иногда рассматриваемое в
качестве отряда или подотряда Strepsip-
terat) и представляют собой замечатель-
ную группу в отряде Coleoptera. Вее-
рокрылые делятся на два семейства, Sty-
lopidae и Mengeidae, представители
обеих групп редко встречаются в коллек-
циях.
stylopidae. Самец крылатый, сво-
бодно летающий, напоминает скорее му-
ху, чем жука, его надкрылья редуциро-
ваны до скрученных пальцеобразных ор-
ганов, а задняя пара крыльев крупная и
веерообразная; заднегрудь сильно увели-
чена. Антенны короткие, и по крайней ме-
ре некоторые из их члеников листовидно
расширены; поверхность сложных глаз
8. Отряды энтогнат и насекомых
349
напоминает плод малины • каждая фасет-
ка явственно выступает и отделена от
других. Самка крайне дегенерировавшая,
представляет собой не более чем мешок в
оболочке из последней личиночной
шкурки, остающийся погруженным в те-
ло хозяина, только передний отдел тела,
называемый головогрудью, выдается за
пределы покровов хозяина.
Самка живородяща; личинки первого
возраста развиваются внутри ее тела и
выползают наружу, пробираясь через
щель в высовывающейся головогруди
самки. Эти личинки, называемые из-за
сходства с личинками нарывников также
триунгулинами, имеют три пары ног, хо-
рошо развитые склериты, редуциро-
ванные ротовые органы и простые глаз-
ки. несколько хорошо развитых фасеток
сложного глаза и одну или две пары
спинных хвостовых нитей. Личинки спо-
собны очень активно бегать и подпрыги-
вать. Каждая из них прикрепляется к хо-
зяину и внедряется в его организм, а
затем тотчас линяет и превращается в
сезногую эруковидную личинку. Эта ли-
чинка неподвижно лежит в теле хозяина и
поглощает пищу путем диффузии пита-
тельных веществ из кровяного русла хо-
зяина. Таким образом, в данном случае
ес происходит настоящего разрушения
ттаней хозяина, причиняемого типичны-
ми паразитическими личинками насе-
комых, измельчающими и поглощающи-
ми свою пищу при помощи рта. Достиг-
ну окончательных размеров, личинка
зеерокрылого просовывает передний ко-
нец тела между стернитами брюшка хо-
зяина, так что головной и грудной от-
делы находятся снаружи, образуя окру-
.тую или уплощенную структуру. Если
данная личинка мужского пола, то она
превращается внутри личиночной шкур-
ки в типичную куколку жука; став
зрелым, имаго разрывает высунутый, ко-
нец личиночной шкурки и выбирается на-
ружу. Если из личинки должна вылупить-
ся самка, то настоящей куколочной ста-
дии не наступает, а она линяет, превра-
щаясь непосредственно в мешковидную
взрослую самку, которая остается в ли-
чиночной шкурке. Развитие яиц и вылу-
пление происходит внутри тела матери,
поскольку она всего лишь мешок с яйца-
ми или молодыми личинками. Потом-
ство одной самки огромно: подсчеты по-
казали, что на одну самку приходится от
2500 до 7000 личинок.
Североамериканские виды Stylopidae
заражают в основном взрослых особей
различных пчел и ос. В других районах
мира различные группы стилопид разви-
ваются в теле Hemiptera (Pentatomidae,
Cicadellidae, Fulgoridae и некоторых
близких к ним семейств) и богомолов
(Mantidae).
Близкородственное семейство Меп-
geidae во многих отношениях сходно со
Stylopidae, но самки этого семейства на-
поминают личинок и ведут свободный
образ жизни. Хозяева единственного не-
арктического рода неизвестны, европей-
ские виды паразитируют на чешуйницах.
Многие авторы относят Stylopidae и
близкие к ним группы к особому отряду-
Strepsiptera. Тем не менее большинство
специалистов придерживаются мнения,
что данная группа обособилась внутри
комплекса жесткокрылых. Триунгулино-
подобная личинка, передние крылья типа
элитр, редуцированное жилкование зад-
них крыльев и паразитический образ жиз-
ни заставляют предполагать близкое
родство веерокрылых с нарывниками.
ЛИТЕРАТУРА
Arnett R. Н., Jr., 1960. The beetles of the United
States. (A manual for identification), Catholic
Univ. Press, Washington, D.C.
Arnett R. H., Jr., 1967. Present and future syste-
matics of the Coleoptera in North America,
Ann. Rev. Entomol., 60, 162-170.
350
8. Отряды энтогнат и насекомых
8. Отряды энтогнат и насекомых
351
Рис. 8.107 Stylopoidea, основные черты внешнего строения и цикл развития. (По Essig, 1942,
College entomology, с разрешения Macmillan Со.)
А. Взрослая особь Stylops medionitans Pierce. Б. Половозрелая самка. В. Нога. Г. Xenos
pallidus Brues (по Bohart). Д. Голова Eoxenos laboulbenei Peyer (no Parker, Smith), вид с
вентральной стороны. Е. То же, вид сбоку. Ж. Взрослая самка и ее нога (3). И. Stylops
pacifica Bohart (по Bohart). 7-максилла; 2-антенна; 3-сложный глаз; 4-надкрылье;
5 -голова; 6 - переднегрудь; 7-среднегрудь; 5-скутум; 9-скутеллум, 70-постлюмбиум;
27-постскутеллум; 72-брюшко; 13-гипоэпимер; 14-заднее крыло; 75-грудь; 16-рот;
Iголовогрудь; 18 - мандибула, 79-выходное отверстие для потомства; 20 -дыхальце;
21 - трахея;
22-несброшенная личиночная шкурка; 25-туловище; 24-половое отверстие; 25-анальное
отверстие; 26 - эдеагус; 27-тазик; 28 - бедро; 29 - голень; 30 - лапка; 57 - пульвилла;
32-задняя лапка, 55-щупик; 54-стилопсы. К. Стилопизированная пчела-андренида. Л. Жиз-
ненный цикл веерокрылого: 7-взрослый самец вылетает и спаривается с самкой;
77-молодые особи развиваются в теле самки и покидают его в виде активных триунгулинов;
777-триунгулины переносятся пчелами в свое гнездо; 7К-триунгулины сами прикрепляются
к личинкам пчел; К-триунгулины внедряются в тело личинок пчел и развиваются внутри них,
17-головогрудной отдел развивающихся веерокрылых высовывается наружу из тела
хозяина; VII-передние части тела почти полн<
высовываются наружу из брюшец хозяев.
Blatchley W. S., 1910. The Coleoptera or beetles
known to occur in Indiana, Nature Publishing
Co., Indianapolis, Ind.
Blatchley W S., Leng C. W., 1916. Rhynco-
phora or weevils of northeastern Ameri-
ca, Nature Publishing Co., Indianapolis,
Indiana.
Bohart R. M., 1941. A revision of Strepsiptera,
Univ. Calif. Publ. Entomol., 7, 91-160.
Bradley J. C., 1930. Manual of the genera of bee-
tles of America, north of Mexico, Daw, Illston
& Co., Ithaca, New York.
Britton E. B., 1970. Coleoptera (beetles). In. The
insects of Australia, D. F. Waterhouse et al.
teds.), Melbourne Univ. Press, Carlton, Vic-
toria, pp. 495-621, Suppl. 1974, pp. 62-89.
Clausen С. P., 1940. Entomophagous insects:
Coleoptera, McGraw-Hill, New York,
pp. 522-584.
Crowson R. Л., 1955. The natural classification
of the families of Coleoptera, Nathaniel Lloyd,
London.
Clawson R. Л., 1960. The phylogeny of the Cole-
optera, Annu. Rev. Entomol., 5, 111-134.
C.llon E. S., Dillon L. S., 1961. A manual of
common beetles of Eastern North America,
Row, Peterson, Evanston, Ill.
Eawards J. G., 1949. Coleoptera or beetles east
of the Great Plains, Edwards Bros., Ann
Arbor, Mich.
Hatch M. H., 1957-1973. The beetles of ? the
сформировавшихся самцов и самок
Pacific Northwest, Univ. Wash. Publ. Biol.,
Univ, of Washington Press, 5 vols.
Hinton H. E., 1955. On respiratory adaptation,
biology and taxonomy of the Psephenidae with
notes on related families (Coleoptera), Proc.
Zool. Soc. London, 125, 543-568.
Leech H. B., Chandler H. D., 1956. Aquatic
Coleoptera. In. Aquatic insects of California,
R. L. Usinger (ed.), Univ, of California Press,
Berkeley, pp. 293-371.
Leech H. B., Sanderson H. W., 1959. Coleop-
tera. In: Freshwater biology, W T. Edmond-
son (ed.), Wiley, New York, pp. 981-1023.
Thiele H.-U., 1977. Carabid beetles in their
environments, Springer-Verlag, Berlin.
Ствол Hymenoptera-Mecoptera
Этот ствол может быть подразделен
на две ветви 1) Hymenoptera, у которых
сохранился почти древнейший тип пла-
стинчатого или пиловидного яйцеклада,
но развились в высшей степени специали-
зированные крылья, в которых произош-
ли многочисленные слияния продольных
жилок, а поперечные не сливались, и 2)
Mecoptera и близкие к ним группы, в ко-
торых пиловидный яйцеклад полностью
утратился, зато жилкование у более при-
352
8. Отряды энтогнат и насекомых
митивных представителей мало измени-
лось по сравнению с нейроптероидным
архетипом. Mecoptera и четыре род-
ственных им отряда в дальнейшем разде-
ляются еще на две группы: Mecoptera-
Siphonaptera-Diptera, мандибулы ко-
торых сделались острыми и саблевидны-
ми, и Trichoptera-Lepidoptera, у которых
крылья практически лишились попе-
речных жилок.
Отряд IIYMENOPTERA
(перепончатокрылые)
Этот большой отряд включает насе-
комых с различной формой тела и разме-
рами, колеблющимися от 0,1 мм у мель-
чайших паразитических форм до 50 мм и
более у некоторых ос (рис. 8.108). По-
кровы сильно склеротизированы и плей-
ральные склериты срастаются очень
прочно. Ротовые органы грызущего типа
у многих форм преобразованы для лиза-
ния или сосания. Крылья могут быть хо-
рошо развитыми, редуцированными или
отсутствовать совсем; если они развиты
хорошо, то они прозрачны, обе пары
одинаковой текстуры и лишены чешуек;
жилкование весьма разнообразно. У при-
митивных форм на ногах имеется отде-
ленный от основания бедра как бы лиш-
ний вертлугоподобный сегмент. Генера-
лизованные формы (рис. 8.109) характе-
ризуются значительной редукцией
и слиянием жилок, однако число попе-
речных жилок умеренное. Антенны раз-
нообразной формы; число члеников
варьирует от 3 до примерно 60. Личинки
напоминают гусениц или эруковидные,
всегда с отчетливо различимой головой и
грызущим ротовым аппаратом; они
имеют грудные или брюшные ноги, либо
одновременно те и другие, или же вовсе
лишены ног
Рис. 8.108. Складчатокрылая оса Vespula (Dolichovespuld)
maculata- перепончатокрылое значительных размеров. (По материалам
Illinois Natural History Survey.)
8. Отряды энтогнат и насекомых
353
Рис. 8.109. Схема жилкования крыла перепончатокрылых, совмещающая примитивные
признаки ряда наиболее архаичных семейств, / -плечевая жилка; 2-стигма;
3-югальная складка, 4-анальная складка.
Прими-
тивные
паразиты
Специализированные
паразиты
Bethyloidea
Vespoidea
».Т>С1ЬЩИКИ
Рогохвосты
ряд SYMPHYTA
Основание ножен
яйцеклада открыто
Брюшко причленено подвижно
-------------Подотряд APOCRITA
Личинки делают ходы в стволах
и стеблях, ноги редуцированы
Глоссы сильно
удлинены
Apoidea
Лопасти переднеспинки
далеко не доходят до тегулы
У сики самца 13-,
самки 12-члениковые
Жалящие и паразитические осы и пчелы
Личинка открытомивущая питается листьями
Рве. 8.110. Упрощенное филогенетическое древо отряда Himenoptera. (Отношения жалящих
жрепончатокрылых даны с изменениями по Brothers, 1975.)
354
8. Отряды энтогнат и насекомых
По числу известных видов перепонча-
токрылые - один из крупнейших отрядов
насекомых. В него входят также такие
группы, как муравьи, осы, пчелы, ко-
торые чрезвычайно многочисленны в
природе и вследствие этого предста-
вляют очень заметный элемент фауны.
Эти формы, однако, считаются продви-
нутыми представителями отряда.
Наиболее примитивными из нынежи-
вущих Hymenoptera являются раститель-
ноядные пилильщики из относительно
редких семейств Xyelidae и Pamphiliidae
(рис. 8.110). Их личинки имеют антенны,
сохраняющие до семи члеников, и хоро-
шо развитые ноги на каждом из сегмен-
тов груди. От этой линии рано отдели-
лась ветвь, давшая начало типичным
пилильщикам, исключительно листогры-
зущим формам, в том числе семействам
Argidae, Tenthredinidae, Cimbicidae и Di-
prionidae. У всех них, а также у Xyelidae
личинки имеют по паре ног на каждом из
нескольких сегментов брюшка, по этой
причине их часто ошибочно принимают
за гусениц Lepidoptera. У этих групп сам-
ки надрезают своим яйцекладом листья и
откладывают в каждый разрез по яйцу.
Разрез зарастает, и яйца, таким образом,
оказываются погруженными в ткани рас-
тения.
Другая ранняя линия, возникшая
предположительно от прексиелидо-
образного предка, дала начало группе
перепончатокрылых, живущих внутри
побегов, в том числе и так называемых
рогохвостов. У этих насекомых пило-
видный яйцеклад стал узким, приспосо-
бленным для внедрения в более плотные
растительные ткани, а ноги у личинок в
значительной степени редуцировались.
Самым примитивным производным дан-
ной линии является, видимо, семейство
Cephidae (стеблевые пилильщики), дру-
гие ее представители - Siricidae и Xiphy-
driidae, имеющие твердые покровы тела,
а также интересное редкое семейство
Orussidae, личинки которого паразити-
руют на личинках жуков-ксилофагов.
Пилильщики и рогохвосты состав-
ляют подотряд сидячебрюхих, или Sy-
mphyta, у представителей которого
брюшко прикрепляется к груди широким
основанием (рис. 8.111,Л, Ё). У другой,
более обширной части отряда Hymenop-
tera, называемой подотрядом стебельча-
тобрюхих, или Apocrita, первый сегмент
брюшка накрепко срастается с задне-
грудью и появляется сочленение шарнир-
ного типа между первым и вторым сег-
ментами брюшка (рис. 8.111,2Э, причем
остальная часть брюшка становится
единым подвижным образованием, иног-
да называемым гастером1. Личинки
Apocrita являются первичными парази-
тами или хищниками других неполово-
зрелых насекомых и лишены ног Яйце-
клад взрослой самки округлый в сечении
и тонкий, предназначенный для протыка-
ния субстратов, в которых обитают ли-
чинки хозяина, проникновения в тело
самого хозяина или используемый в ка-
честве жала.
ТГока еще далеко не ясно, от какой
именно группы Symphyta произошли
Apocrita. Очень возможно, что стебель-
чатобрюхие выделились из основания
рогохвостов. Представляется вполне ве-
роятным, что при своем продвижении че-
рез ткани кормового растения сте-
блегрызущая личинка пилильщика се-
мейства Cephidae или рогохвоста могла
натолкнуться на личинку близкого вида и
съесть ее, затем подобные отношения
стали регулярными, и, наконец, они уста-
новились в качестве нормальных пи-
щевых взаимоотношений данного вида с
другими. Таким путем, возможно, раз-
вился паразитический образ жизни. Тот
1 В отечественной литературе термин не
употребляется.- Прим, перев.
8. Отряды энтогнат и насекомых
355
Рис. 8.111. Диагностические признаки некоторых Hymenoptera.
А. Грудь Arge (Argidae). Б. Грудь Janus (Cephidae). В. Вентральная сторона брюшка
Chrysis (Chrysididae). Г Грудь Eremotylus (Ichneumonidae). Д. Голова и грудь Chaicis
(Chalcididae). Е. Грудь Proctotrupes (Proctotrupidae). Ж. Грудь Ancistrocerus (Vespidae).
3. Грудь Sceliphron (Sphecidae).
7-переднеспинка, 2-мезоплевра, 3-ценхрус; 4-метэпимер; 5-метэпистерн;
6-дыхальце; 7-метаплевра; 8-базальное сочленение брюшка; 9-тегула; 10-лопасть
переднеспинки; III-заднеспинка, 1, 2, 3-сегменты брюшка.
□акт, что подобный переход происходил
знутри группы рогохвостов, приводя к
рормированию паразитических Orussi-
dae. подкрепляет данное предположение
г в отношении Apocrita. Стебельчатое
трюшко Apocrita представляет собой
тишь небольшую модификацию состоя-
ния, присущего Cephidae; это позволяет
предположить, что Apocrita берут свое
начало от предкового ствола Cephidae.
Наиболее древние из известных Apoc-
rita в настоящее время представлены
примитивными паразитами, к числу ко-
торых относятся редкие Gasteruptionidae
и многочисленные Braconidae и Ichneu-
monidae. У этих групп основание яйце-
клада и его ножны находятся снаружи,
почти как у пилильщиков. От линии, к
356
8. Отряды энтогнат и насекомых
которой принадлежал некоторый древ-
ний предок этих групп, произошла фор-
ма, у которой седьмой стернит разросся
вперед, скрыл основание пиловидного
яйцеклада и образовал для него вен-
тральный желоб или направляющую
трубку. Эта форма дала начало обшир-
ной группе специализированных парази-
тов, а именно Cynipoidea, Chalcidoidea,
Proctotrupoidea и нескольким самостоя-
тельным более мелким семействам, та-
ким, как Pelecinidae, Evaniidae и Trigona-
lidae. Любопытным обстоятельством
является независимое воссоединение ба-
зального сегмента бедра с остальной его
частью и уменьшение числа члеников ан-
тенны, происходившие в различных ли-
ниях. Как внутри Cynipoidea, так и среди
Chalcidoidea возникли растительноядные
формы, остальные представители-ти-
пичные паразиты, личинки которых уби-
вают своих хозяев чрезвычайно просто,
а именно постепенно поедая его тка-
ни. У всех паразитов самка не парализует
свою добычу. Их часто называют Теге-
brantia.
У некоторых специализированных па-
разитов развился новый тип паразитиз-
ма. Самки используют яйцеклад, чтобы
уколоть и обездвижить или парализовать
личинку хозяина, а затем откладывают
на нее яйцо. Тем самым добыча остается
живой к тому времени, когда личинка вы-
ходит из яйца и начинает питаться, кро-
ме того, обездвиженная добыча не может
принять иного положения, а следова-
тельно, и раздавить личинку паразита.
Большинство таких ос питается личинка-
ми насекомых, живущих в скрытых ме-
стах, например обитающими в почве ли-
чинками жуков.
Наиболее примитивные представите-
Рис. 8.112. Антенны Hymenoptera.
Л. Самка Sofus (Argidae). Б. Самец Sphacophilus (Argidae). В. Trichiosoma
(Cimbicidae). Г Самка Augomonoctenus (Diprionidae). Д. Pleroneura (Xyelidae).
E. Самец Cladius (Tenthredinidae). Ж. Tenthredo (Tenthredinidae).
3. Pseudodineura (Tenthredinidae). И. Самец Monoctenus (Diprionidae.).
8. Отряды энтогнат и насекомых
357
Рис. 8.113. Крылья Hymenoptera.
A. Ichneumonidae. Б. Vespa (Vespidae). В. Myzine (Tiphiidae). Г Tetrastichus (Chalcididae).
J, E. Два типа Cynipidae.
ли этих жалящих форм (Aculeata) отно-
сятся к группе из нескольких семейств,
составляющих надсемейство Bethyloidea,
куда входит семейство Bethylidae и се-
мейство ос-блестянок (Chrysididae). От
бетилоидного предка возникла линия, в
которой число члеников антенн стабили-
зировалось на уровне 13 у самцов и 12 у
самок. От этой линии ведут начало все
оставшиеся Hymenoptera, а именно над-
семейства Vespoidea, Sphecoidea и Apoi-
dea.
Vespoidea развились в необычайно
разнообразный комплекс семейств,
включающий ос-немок (Mutillidae), бо-
лее типичных одиночных ос Tiphiidae и
Scoliidae, муравьев (Formicidae) и склад-
чатокрылых ос (Vespidae), к которым от-
носится ряд подсемейств общественных
насекомых.
358
8. Отряды энтогнат и насекомых
Рис. 8.114. Детали строения ос и пчел.
А. Мезоплевра Ceropales (Pompilidae). Б. Среднегрудь Му zine (Tiphiidae) с вентральной
стороны. В. Общий вид сбоку Pelecinus (Pelecinidae). Г. Стерниты груди Scolia (Scoliidae).
Д. Нижняя губа Colletes (Colletidae). Е, Нижняя губа Anthidium (Megachilidae). Ж. Голова
Halictus (Halictidae). 3. Голова Andrena (Andrenidae).
1 -переднеспинка, 2-тегула; 3-основание крыла; 4-мезоплевральный шов;
52 -5з - тазики, 62 - - стерниты; 7- лопастевидный отросток среднегруди; 8 - основание
брюшка; 9-глоссы (лигула); 10-щупик; 77-прементум; 72-ментум; 73-субментум;
14-субантеннальные швы.
S. Отряды энтогнат и насекомых
359
От общего предка с Vespoidea отдели-
лась линия, в которой у переднеспинки
развилась задняя доля, отделенная от те-
плы широким промежутком
«рис. 8.111,3). Эта линия дала начало
роющим осам (Sphecoidea) и пчелиным
। Apoidea). У примитивных Sphecoidea,
как и у некоторых Vespidae, самки внача-
ле строят гнездо (вероятно, роя норку в
земле), затем ловят и жалят добычу, не-
сут ее в гнездо и откладывают на ней яй-
цо. У более высокоорганизованных Sphe-
cidae заготовка пищи для личинок обыч-
но происходит до откладки яиц.
Ответвление от типичной сфекоидной
линии дало начало пчелам. Основными
эволюционными изменениями в этой ве-
тви был переход к питанию пыльцой, что
исключало конкуренцию за источники
пищи с другими насекомыми и пауками,
а также развитие на теле и придатках ве-
твистых волосков в связи со сбором
пыльцы. Хотя большинство пчел ведут
одиночный образ жизни, во многих груп-
пах развился и общественный или по-
л> общественный образ жизни, в том чис-
ле у шмелей и медоносных пчел (Apidae).
Таким образом, эволюция высших
перепончатокрылых (ос и пчел) предста-
вляется связанной прежде всего с измене-
ниями особенностей поведения, не сопро-
вождавшимися значительными переме-
нами во внешнем строении.
ПОДОТРЯД SYMPHYTA
«СИДЯЧЕБРЮХИЕ)
ПИЛИЛЬЩИКИ И РОГОХВОСТЫ. Sy-
mphyta, за исключением небольшого па-
разитического семейства Orussidae, пред-
ставляют собой растительноядную груп-
пу. Личинки либо питаются снаружи на
листьях, либо минируют листовые пла-
стинки, черешки или стебли. Взрослые
насекомые во многих случаях питаются
опушением кормового растения, состри-
гая волоски с помощью своих серпо-
видных мандибул, подобно пасущимся
на траве коровам; представители других
групп могут поедать мелких насекомых
или питаться нектаром или пыльцой. Эта
группа весьма обширна, на территории
Северной Америки она представлена 12
семействами и связана с самыми разно-
образными группами растений.
Личинки пилильщиков характери-
зуются следующими отличительными
признаками: четко отделенной головой с
простыми грызущими ротовыми органа-
ми; тонкими или пластинчатыми антен-
нами из 1-7 члеников; глазами с един-
ственной линзой; брюшными ногами,
если они имеются, без больших и малых
крючков на концах.
Имаго большинства пилильщиков
имеют компактное, довольно коренастое
тело. Из листогрызущих семейств наибо-
лее крупный обычный вид - Cimbex ameri-
cana (рис. 8.115) с булавовидными антен-
нами; самки и самцы этого вида имеют
различную окраску. Самки, за редкими
исключениями, с хорошо развитым пи-
ловидным яйцекладом, используемым
для надрезания листьев или черешков
при откладке яиц.
Самое большое семейство - Tenthredi-
nidae-характеризуется в основном про-
стыми 9-16-члениковыми усиками. Боль-
шинство видов питается снаружи на
листьях, среди них есть и имеющие важ-
ное экономическое значение, например
Endelomyia ае thiops, завезенный желтый
крыжовниковый, или ягодный, пилиль-
щик Nematus ribesi (рис. 8.116) и пих-
товый пилильщик Эрихсона, Pristiphora
erichsoni. Отдельные виды минируют тка-
ни листа, например Heterarthrus петога-
ta-ONim из минеров листьев березы.
Виды ряда других родов, в том числе
Еиига, образуют галлы.
Diprionidae - другое экономически
важное семейство. У представителей это-
360
8. Отряды энтогнат и насекомых
Рис. 8.115. Крупный пилильщик Cimbex americana. (По Riley.)
А. Листья ивы с расположенными на них яйцами. Б. Побег с надрезами,
сделанными взрослой самкой. В. Яйцо. Г. Только что вылупившаяся из яйца
личинка. Д. Зрелая личинка. Е. Кокон. Ж. Вскрытый кокон, видна куколка. 3. Ку-
колка, вид сбоку. И. Взрослое насекомое. К, Л. Пиловидный яйцеклад самки и
его вершина.
8. Отряды энтогнат и насекомых
361
Рис. 8.116. Личинка питающейся стадии
завезенного ягодного пилильщика Nematus
ribesi. (По материалам Connecticut
Agricultural Experiment Station.)
го семейства антенны состоят не менее
чем из 13 члеников, причем у самки они
пильчатые, а у самца-гребенчатые
(рис. 8.117). Все виды плотно сложены и
более или менее пестро окрашены. Ли-
чинки похожи на гусениц и открыто пи-
таются хвоей. Многие виды относятся к
числу важнейших дефолиаторов еловых
и сосновых лесов. Особого упоминания
заслуживают опустошения, произво-
димые в ельниках северо-востока Амери-
ки завезенным из Европы еловым пи-
лильщиком Diprion hercyniae. Красного-
ловый сосновый пилильщик Neodiprion
lecontei (рис. 8.117) особенно вредит мо-
лодым соснам.
К семейству Siricidae относятся на-
иболее крупные представители всего под-
отряда. У этих вытянутых насекомых
длина тела может достигать 40 мм. Ци-
линдрические личинки почти лишены ног
и делают ходы в стволах деревьев. И
взрослые особи, и личинки на заднем
конце тела несут твердый шиповидный
вырост; благодаря этой особенности их и
назвали рогохвостами. Примером широ-
ко распространенного вида служит Тге-
mex columba (рис. 8.118), нападающий на
клен, ильм, бук, дуб и ряд других лис-
твенных древесных пород.
ПОДОТРЯД APOCRITA
(СТЕБЕЛЬЧАТОБРЮХИЕ)
МУРАВЬИ, ПЧЕЛЫ, ОСЫ И НАЕЗДНИКИ.
В общем стебельчатобрюхие более изящ-
ны, проворны, активны и быстры в дви-
жениях, чем сидячебрюхие. Личинки, как
правило, являются внутренними или на-
ружными паразитами, либо выкармли-
ваются взрослыми особями, либо вызы-
Рис. 8.117. Красноголовый сосновый, пилильщик Neodiprion lecontei. (По материалам
министерства сельского хозяйства США.)
А. Самец. Б. Самка. к
362
8. Отряды энтогнат и насекомых
Рис. 8.118. Рогохвост Tremex columba. (По Riley.)
А. Личинка. К. Голова личинки, вид с вентральной стороны. В. Куколка самки. Г Куколка
самца. Д. Взрослое насекомое. Обратите внимание на маленькую личинку паразита,
сидящую на личинке рогохвоста.
вают образование галлов на растениях.
У них нет ног, хорошо заметна головная
капсула, несущая сильно редуциро-
ванные антенны и ротовые части, а раз-
витие часто сопровождается гипермета-
морфозом (с. 214).
В подотряде Apocrita первый сегмент,
относящийся к брюшку, неподвижно сра-
стается с грудью. Поэтому отдел, кажу-
щийся грудью, на самом деле содержит
один брюшной сегмент, а то, что кажется
брюшком, лишено своего переднего сег-
мента. Соответственно эти отделы тела
получили названия мезосома и метасо-
ма. В старой литературе последняя име-
новалась «гастер».
Примитивные паразиты
Ichneumonidae, ихневмониды. Это
обычно стройные наездники с длинными
многочлениковыми антеннами
(рис. 8.119) и субкостальной жилкой,
слитой на переднем крыле с радиальным
стволом. Все представители этого семей-
ства-паразиты насекомых или пауков.
Обычно хозяевами им служат гусеницы
Lepidoptera, например Glypta rufiscutella-
ris паразитирует на восточной плодожор-
ке. Кроме того, многие ихневмониды по-
ражают личинок Coleoptera, Hymenopte-
ra, Diptera и некоторых других насе-
комых. Взрослые самки наездников-
ихневмонид откладывают яйца на по-
верхность или внутрь тела хозяина. Если
яйца откладываются на эпидермис хозя-
ина, вышедшие из них личинки могут
внедриться в его тело. Личинки имеют
вид безногих червей, которые либо при-
креплены снаружи к хозяину, либо живут
внутри его тела. Окончив развитие, ли-
чинки могут либо окуклиться в теле хозя-
8. Отряды энтогнат и насекомых
363
Рис. 8.119. Наездник Glypta ruflscutellaria. (По материалам Agricultural Experiment
Station.)
А. Самец. Б. Самка. В. Вершина яйцеклада.
ина, либо покинуть его, чтобы сплести
кокон. Размеры тела ихневмонид очень
различны. Многие мелкие формы, дли-
ной всего несколько миллиметров, зара-
жают мелких гусениц чешуекрылых.
Braconidae. Это крупное семейство на-
ходится в близком родстве с ихневмони-
дами. В среднем эти наездники мельче
ихневмонид, и у многих из них редуциро-
ванное жилкование крыла. Целый ряд ви-
дов имеет важное значение, являясь пара-
зитами вредителей сельского хозяйства.
Один из них, Apanteles melanoscelus
(рис. 7.14), был специально завезен для
биологического подавления гусениц не-
парного щелкопряда. Для этого мелкого
паразита характерен гиперметаморфоз,
преобладающий во всех паразитических
семействах перепончатокрылых. На
рис. 7.14 изображены различные формы
364
8. Отряды энтогнат и насекомых
личинок на разных стадиях развития,
чтобы определить задний конец тела ли-
чинки, можно воспользоваться положе-
нием анального пузырька (7).
Специализированные паразиты
Надсемейство Chalcidoidea, хальциды.
Эти мелкие наездники часто имеют ме-
нее миллиметра в длину, жилкование
крыльев у них очень сильно редуцирова-
но (рис. 8.120), а антенны, как правило,
коленчатые. Большинство этих наездни-
ков являются эндопаразитами главным
образом гусениц чешуекрылых и личинок
других паразитических перепончатокры-
лых, на которых они нападают внутри
тела их первичного хозяина. Такие пара-
зиты паразитов называются гиперпарази-
тами, или вторичными паразитами. Не-
многочисленные непаразитические виды
хальцид развиваются в различных семе-
нах или в стеблях растений, в особенно-
сти злаков. К этой непаразитической
группе принадлежат клеверный семеед
Bruchophagus platyptera1, личинки кото-
1 У нас его называют Bruchophagus gib-
bus-Прим. перев.
рого развиваются внутри семян клевера
и люцерны, Harmolita tritici, личинка ко-
торого прокладывает ходы в стеблях
пшеницы, и Н. grandis (рис. 8.120). Ло-
кально и в отдельные годы Н. tritici
причиняет серьезный ущерб посевам
пшеницы.
Большой интерес представляет уни-
кальный и специализированный жиз-
ненный цикл некоторых крошечных халь-
цид, принадлежащих к семейству Agaoni-
dae. Эти хальциды развиваются в цико-
ниях инжира. Самцы бескрылы и прово-
дят всю жизнь внутри цикония, в кото-
ром происходило их развитие, там они и
оплодотворяют самок еще до того, как
последние покидают зараженные хальци-
дами семязачатки. Самки крылаты и
перелетают с цветка на цветок в поисках
пригодных для откладки яиц цикониев.
перенося на своем теле пыльцу и опыляя
каждый из посещаемых цветков. Это
единственный способ, посредством кото-
рого происходит опыление инжира. Мно-
гие культивируемые сорта инжира не
требуют опыления для образования пло-
дов, однако плоды отборного измирско-
го инжира не развиваются без опыления.
Чтобы выращивать этот сорт в Северной
Рис. 8.120. А. Пшеничная галловая изозома Harmolita tritici. Бескрылая (в центре) и кры-
латая (Б) формы большой узловой толстоножки Harmolita grandis ( = Philachyra aptera).
(По материалам министерства сельского хозяйства США.)
8. Отряды энтогнат и насекомых
365
Рис. 8.121. Орехотворка и ее галл. (По Essig,
1942, College entomology.)
А. Взрослое насекомое. Б. Незрелый галл.
В. Зрелый галл в разрезе, видны камеры и
выход наружу.
Америке, пришлось интродуцировать и
его опылителя Blastophaga psenes.
Надсемейство Cynipoidea, орехотвор-
ки. Эти мелкие перепончатокрылые в
большинстве своем характеризуются на-
личием крупной треугольной ячейки
вблизи апикальной части переднего кры-
ла (рис. 8.113,Д) и высоким, сильно
сжатым с боков брюшком. Многие
группы этого надсемейства паразити-
руют на личинках двукрылых, тлях и
прочих насекомых, однако наиболее ши-
роко известны представители, образую-
щие галлы на растениях. Взрослых оре-
хотворок можно видеть крайне редко, но
любому натуралисту хорошо знакомы
разнообразные галлы, образование ко-
торых инициируют личинки этих насе-
комых на листьях, стеблях или корнях
дубов, роз и иных растений. Один из та-
ких галлов показан на рис. 8.121. Суще-
ствуют сотни видов орехотворок, и поч-
ти каждый из них образует галлы особо-
го типа. Для некоторых живущих на
дубах видов характерно чередование по-
колений, когда одно поколение вызывает
образование галлов на корнях, а следую-
щее за ним-на листьях или веточках.
Жалящие осы
В линии Terebrantia, давшей начало
жалящим перепончатокрылым, яйцеклад
превратился в жало, а его придаточные
железы стали вырабатывать яд, выде-
ляющийся в момент укола. У таких вра-
гов, как млекопитающие, этот яд вызы-
вает неприятные болезненные ощущения
и в некоторых случаях его попадание в
организм животного может привести к
летальному исходу. Будучи введен в
тело добычи (как правило, в насекомое
или паука), этот секрет обычно вызывает
полный двигательный паралич, не приво-
дя к смертельному исходу. Таким спосо-
бом парализуют добычу осы, отклады-
вающие на нее яйца или запасающие в
гнезде живой корм. Парализация имеет
тройную выгоду она позволяет добыче
оставаться съедобной до вылупления ли-
чинки и начала питания, делает невоз-
можным бегство хозяина от безногой ли-
чинки паразита и исключает привлечение
различных сапрофагов на запах мертвых
насекомых.
Жалящие перепончатокрылые, обык-
новенно называемые Aculeata, представ-
ляют собой комплекс весьма разно-
образных форм, объединяемых в четыре
следующих надсемейства.
Надсемейство Bethyloidea. Сюда отно-
сится группа примитивных форм, вклю-
366
8. Отряды энтогнат и насекомых
Рис. 8.122. Оса-сколия Scolia dubia. (По материалам
министерства сельского хозяйства США.)
А. Взрослая самка. Б. Антенна самца. В. Кокон с выходным
отверстием.
чающая семейство Bethylidae, представи-
тели которого внешне отдаленно
напоминают муравьев и паразитируют
на самых разных насекомых, а также се-
мейство Chrysididae (осы-блестянки) ме-
таллически-зеленой или синей окраски,
представители которого являются на-
ружными паразитами.
Надсемейство Vespoidea. У данной и
всех оставшихся групп Hymenoptera чис-
ло антеннальных члеников стало пос-
тоянным: 12 у самки и 13 у самца. В двух
ветвях независимо возник общественный
образ жизни. Примером типичных гене-
рализованных представителей Vespoidea
является семейство Scoliidae.
SCOLIIDAE, осы-сколии. У этих до-
вольно крупных насекомых крылья раз-
виты у обоих полов и большинство видов
черные либо в пятнах и перевязях черно-
го и желтого цвета, как, например, Scolia
dubia (рис. 8.122). Самка осы роет почву
в поисках жертвы-личинок жуков семей-
ства Scarabaeidae. Когда самка находит
подходящую личинку хозяина, она жалит
ее, парализуя тем самым, а затем вы-
капывает вокруг нее ячейку, откладывает
на обреченную личинку яйцо и улетает на
поиски новой жертвы. Вскоре из яйца вы-
лупляется безногая эруковидная личин-
ка, которая прикрепляется к парализо-
ванной личинке жука и начинает поедать
ее. Приблизительно за две недели личин-
ка осы съедает всего хозяина и достигает
окончательных размеров. Затем она пле-
тет кокон внутри земляной ячейки и
обыкновенно в таком состоянии прово-
дит зиму. На следующую весну или ле-
то личинка окукливается, а затем поя-
вившееся имаго прогрызает себе вы-
ход из кокона и прокапывает ход к по-
верхности.
mutillidae и tiphiidae. К Scoliidae
весьма близки многие семейства, веду-
щие примерно такой же образ жизни.
Виды семейства ос-немок, или Mutillidae,
являются паразитами ос и пчел. У мно-
гих немок самки лишены крыльев и очень
8. Отряды энтогнат и насекомых
367
Рис. 8.123. Самка осы-немки Dasymutilla
bioculata. (По Washburn.)
inae, крылья в состоянии покоя сложены
в продольном направлении наподобие
веера. Для Masarinae характерны булаво-
видные антенны. Большинство предста-
вителей Vespidae ведет одиночный образ
жизни. Взрослые особи сооружают норку
в древесине или почве или строят для
пребывания личинки вместилище в виде
горшочка. Имаго, как правило, запасают
в гнезде парализованных гусениц или
пыльцу и мед. У ряда складчатокрылых
ос выработался общественный образ
жизни. Пережевывая волокна древесины
с особым секретом полости рта, они про-
изводят «бумагу», из которой изготавли-
вают блюдцеобразное или чашеобразное
гнездо. Наиболее часто встречаются
блюдцеобразные гнезда Polistes
(рис. 8.124), сложенные всего одним го-
похожи на муравьев. Однако у самок Ми-
tillidae отсутствует чешуйка или «узелок»
на стебельке брюшка (рис. 8.123), и кро-
ме того, они покрыты густыми барха-
тистыми или шелковистыми волосками.
Последняя особенность и послужила
причиной того, что это семейство стали
называть «бархатные муравьи» (velvet
ants). У ос-немок сильное жало, и они
легко пускают его в ход, когда их
беспокоят.
Многие виды рода Tiphia, относяще-
гося к родственному семейству Tiphiidae,
были завезены в США из юго-восточной
Азии и разводились для борьбы с личин-
ками японского жука, причем некоторые
виды подают определенные надежды на
успешное применение против этого вре-
дителя.
VESPIDAE, СКЛАДЧАТОКРЫЛЫЕ ОСЫ. К
этому семейству относятся виды, длина
тела которых варьирует от 10 до пример-
но 30 мм, а окраска в большинстве слу-
чаев состоит из желто-черного или бело-
черного рисунка (рис. 8-Ю8). Во всех
подсемействах, за исключением Masar-
Рис. 8.124. Взрослая самка и гнездо Polistes.
(По материалам министерства сельского
хозяйства США.)
368
8. Отряды энтогнат и насекомых
ризонтальным слоем личиночных ячеек.
Такое подвешенное гнездо легко обнару-
жить под карнизами зданий и в других
подобных укромных местах. Колонии
Polistes редко насчитывают больше не-
скольких десятков особей. Самые
крупные колонии складчатокрылых ос,
зарегистрированные в Северной Амери-
ке, устраивает Vespula (Dolichovespula)
maculata. Эти колониальные гнезда
овальной формы и с входным отвер-
стием на дне чаще всего висят на ветвях
деревьев. В каждом содержится несколь-
ко рядов личиночных ячеек, или сотов,
расположенных один над другим. Рабо-
чие особи колонии приносят в гнездо до-
бытых насекомых, в частности мух и гу-
сениц. Осы перемалывают и разминают
свои жертвы, а затем скармливают их ли-
чинкам, находящимся в ячейках гнезда.
В умеренных областях Северной Аме-
рики в конце осени колонии погибают. В
конце лета появляется молодое поколе-
ние самцов и самок. С наступлением хо-
лодной погоды рабочие особи и самцы
гибнут, а оплодотворенный к этому
Рис. 8.125. Маленький черный муравей Monomorium minimum; показаны различные
касты и стадии развития, а также его характерная деятельность. (По Marlatt и
материалам министерства сельского хозяйства США.)
8. Отряды энтогнат и насекомых
369
Рис. 8.126. Вид сбоку муравьев, у которых хорошо заметна спинная чешуйка на
стебельке. Обратите внимание, что на Б и В эта чешуйка маленькая, однако
отчетливо выражена. A. Tapinoma sessile. Б. Lasius, группа niger. В. Paratrechina
longicomis.
времени осенний выводок самок перези-
мовывает в гнилых стволах или пнях.
Следующей весной эти самки вылетают
и основывают новые колонии.
formicidae, муравьи. У этих насе-
комых первый (морфологически второй!)
сегмент брюшка образует стебелек, или
петиоль, и несет дорсальный вырост, или
чешуйку (рис. 8.125; 8.126). Эта структу-
ра отличает муравьев от прочих муравье-
подобных ос. Помимо нормальных сам-
цов и самок подавляющее большинство
видов муравьев имеет и третью форму-
неспособных к размножению рабочих,
которые всегда бескрылы. Рабочие особи
и есть те самые муравьи, которых мы
обычно видим снующими вокруг. Они
выполняют большую часть всех работ в
колонии: сооружают гнездо, роют под-
земные помещения и собирают пищу для
семьи.
В типичном случае семья образуется
после брачного лёта крылатых самцов и
самок, способных размножаться. Через
определенные интервалы (чаще всего раз
в году) в зрелой семье появляется боль-
шое количество крылатых самцов
и самок. Когда погода благоприятствует,
эти половые особи роем покидают гнез-
до, устремляются в брачный полет и спа-
риваются на лету. Самцы погибают че-
рез несколько дней после спаривания.
Оплодотворенная самка подыскивает
пригодное для гнезда место на почве, в
старом бревне и т. п., затем откусывает
крылья и устраивает маленькую замкну-
тую камеру, которая становится потом
первой камерой гнезда. Самка остается в
этой камере в течение нескольких недель,
за это время она откладывает яйца и вы-
кармливает вылупившихся из них эруко-
видных личинок до достижения ими зре-
24-732
370
8. Отряды энтогнат и насекомых
лости. По-видимому, пища образуется в
результате лизиса крыловой мускула-
туры и жирового тела самки. Она вы да-,
вливается изо рта самки в виде секрета.
Старшие личинки окукливаются и вскоре
превращаются в мелких рабочих. Они
разбегаются из гнездовой камеры, нахо-
дят пищу и с этих пор начинают сами
кормить самку, или царицу, а также снаб-
жать пропитанием следующие группы
личинок. Следующие партии потомства
приходят им на помощь, чтобы семья
всегда была обеспечена провизией. Сам-
ка продолжает откладывать яйца без до-
полнительного оплодотворения в тече-
ние нескольких лет.
Семьи многих видов состоят лишь из
нескольких десятков или нескольких со-
тен особей, тогда как у других видов на-
селение семьи может доходить до многих
тысяч. Маленькие семьи обитают под
камнями, в пнях, стволах или в ходах под
землей. Обширные семьи, как правило,
сооружают большие холмики из почвы,
стеблей, веточек и различного мусора,
внутри содержащие сложную систему
переходов и камер.
В целом муравьи всеядны, питаясь
живыми и мертвыми продуктами живот-
ного происхождения (в основном други-
ми насекомыми), растениями, например
грибами, а также эксудатами или выделе-
ниями растений, такими, как нектар, вы-
текающий из ран сок и железистые про-
дукты. Некоторые насекомые, например
тли и ряд кокцид, выделяют «медвяную
росу» или другие секреты, муравьи тща-
тельно «ухаживают» за такими насе-
комыми и собирают «урожай» в виде вы-
ступающих капелек секрета. «Медвяная
роса» насекомых является идеальным ис-
точником пищи, содержащим сахара,
аминокислоты и множество других ве-
ществ, необходимых муравьям.
Некоторые виды муравьев поселяют-
ся в домах и на складах, попадая в число
наиболее постоянных вредителей в поме-
щениях. В северных штатах Solenopsis
molesta, фараонов муравей (Monomorium
pharaonis) и Tapinoma sessile весьма обыч-
ны в домах. В южных штатах в высшей
степени обычным домашним вредителем
является интродуцированный аргентин-
ский муравей Iridomyrmex humilis. Также
завезенный муравей Solenopsis invicta, жа-
лящий и наносящий болезненные укусы,
вредит на пастбищах, однако в то же вре-
мя является важным энтомофагом хлоп-
кового долгоносика Anthonomus grandis.
Он пожирает также муравьев других ви-
дов и в некоторых местностях уже совер-
шенно вытеснил местные популяции му-
равьев. Большой интерес представляют и
другие южные муравьи, в том числе му-
равьи-листорезы (например, Atta texana),
которые разрезают листья деревьев на
мелкие кусочки и относят их в гнездо, где
устраивают «грибные сады». Сами му-
равьи питаются грибами.
Некоторые виды муравьев питаются
главным образом семенами растений. Из
этой группы особенно сильно размножи-
лись в США различные виды рода Pogo-
потугтех, известные также под назва-
нием «полевые муравьи» и вредящие
зерновым и кормовым злакам в рай-
оне Великих равнин и к западу от
них.
прочие группы vespoidea. Суще-
ствует довольно много мелких семейств
ос, родственных Scoliidae и Vespidae.
Это, как правило, редко встречающиеся
семейства, или же редко попадающиеся в
сборах по причине малых размеров и
скрытого образа жизни их представите-
лей. Часто попадаются на глаза зеленые
с металлическим блеском осы-блестянки,
или Chrysididae, которые откладывают
яйца в норки других ос и личинки ко-
торых питаются запасами пищи, храня-
щимися в гнезде осы-хозяина.
8. Отряды энтогнат и насекомых
371
Одиночные осы и пчелы
У этих ос и пчел задние углы пере-
днеспинки заканчиваются округлыми ло-
пастями, которые располагаются ниже
уровня тегул и не касаются их.
Надсемейство Sphecoidea, роющие
осы. У этих насекомых в отличие от пчел
волоски простые и неразделенные, а за-
дние голени и лапки не модифицированы
для собирания пыльцы. Это обширная
группа, включающая ос самых разных
размеров, внешнего вида и окраски. Как
считают в настоящее время, все они от-
носятся к одному большому и гетероген-
ному семейству Sphecidae. Образ жизни
практически всех Sphecidae в общих чер-
тах одинаков. Самка осы строит гнездо
из грязи или выгрызает его в сердце-
вине стебля, древесине, почве, а затем
снабжает это гцездо запасом пищи -
определенным типом парализованных
жертв. В каждую заполненную пищей
ячейку гнезда самка откладывает одно
яйцо, из которого вылупляется без-
ногая личинка, питающаяся запасами,
заготовленными для нее родительской
особью. В умеренном климате личинка
зимует в своем коконе, окукливаясь на
следующий год в начале лета.
Различные группы роющих ос обыч-
но строго специфичны в отношении до-
бычи, которой они снабжают личи-
ночные ячейки. Pemphredoninae (часто
имеющие в длину лишь 2 или 3 мм) ло-
вят тлей, Sphecinae обычно охотятся на
гусениц, Trypoxylinae используют пауков
и т. д. Один из наиболее интересных и
эффектных видов в центральных и во-
сточных штатах -Sphecius speciosus, уби-
вающий цикад. Это крупное черно-жел-
тое насекомое, достигающее в длину 40
или 50 мм. Оно обычно ловит и парали-
зует обыкновенную цикаду Tibicen linnei,
а затем относит ее в земляную норку, по-
мещая в каждую норку по одной цикаде.
Рис. 8.127.Черно-желтая оса-штукатур
Sceliphron servillei, строящая ряды ячеек из
грязи на камнях и стенах и складывающая в
них запас пищи-пауков. (По Essig, Insects of
Western North America.)
Когда оса нападает, цикада издает гром-
кий пронзительный звук, но он длится
очень недолго, так как ужаленная цикада
мгновенно парализуется.
Из одиночных роющих ос наиболее
обычны осы-штукатуры с тончайшим
длинным стебельком брюшка, особенно
виды рода Sceliphron (рис. 8.127). Эти
«штукатуры» лепят свои гнезда из грязи;
в каждом гнезде имеется несколько
ячеек. Гнезда располагаются обычно под
мостами, карнизами домов или в других
скрытых местах. Осы собирают грязь у
края или вблизи луж и прудов, летая взад
и вперед от уреза воды к гнезду с порция-
ми «цемента» во рту. В качестве пищи в
ячейках запасаются пауки.
Надсемейство Apoidea, пчелы. К дан-
ной группе относятся все дикие и домаш-
ние пчелы (рис. 8.128), разделяемые на
шесть самостоятельных семейств. Мор-
фологически они очень близки к роющим
осам, отличаясь наличием ветвистых во-
372
8. Отряды энтогнат и насекомъп
Рис. 8.128. Медоносная пчела. (По материалам министерства сельского хозяйства
США.)
А. Рабочая особь. Б. Матка. В. Трутень.
лосков на теле (рис. 8.129), придающих
пчелам пушистый или бархатистый вид.
В типичном случае на задаем паре ног
или на вентральной стороне брюшка у
них имеются участки, покрытые доволь-
но длшшыми волосками, используемы-
ми при сборе пыльцы; у самцов в неко-
торых родах и у особей обоих полов в
паразитических родах эти структуры от-
сутствуют. Большая часть из 3000 наших
видов ведет одиночное существование,
сооружая ячейки в норках или пустотах,
как и многие одиночные осы. В качестве
провизии пчелы снабжают ячейки медом
Рис. 8.129. Волоски пчел. (По J. В. Smith.)
8. Отряды энтогнат и насекомых
373
и пыльцой, которые и составляют пищу
личинок. Среди пчел есть и «паразитиче-
ские» роды, представители которых от-
кладывают яйца в ячейки других пчел.
Личинка-пришелец развивается быстрее
личинки-хозяина и съедает весь запас пи-
щи. Такими «паразитами» являются, в
частности, так называемые шмели-
кукушки.
Ряд родов североамериканской
фауны, в том числе шмели (род Bombus,
семейство Apidae), перешел к обществен-
ному образу жизни. У шмелей оплодо-
творенные самки перезимовывают
в трещинах стволов или в подстилке. Они
вылетают весной, отыскивают безопас-
ное место, например ямку в земле или
брошенную мышиную нору, и основы-
вают семью. Начинается развитие яиц и
молоди, материнская особь-матка-вы-
кармливает потомство медом. Первый
выводок состоит из стерильных самок,
или рабочих, выполняющих задачу сбора
пищевых запасов для последующих по-
колений личинок. Ближе к осени рабочие
особи больше не рождаются, вместо них
появляется поколение самцов и спо-
собных к размножению самок. Самцы
вскоре после спаривания погибают; при
приближении зимы погибают также и ра-
бочие. Оплодотворенные молодые матки
рахтетаются на зимовку, и, таким обра-
зом, семья распадается.
Образ жизни домашней медоносной
пчелы Apis mellifera специализирован
значительно сильнее, чем у шмелей.
Прежде всего семьи не погибают на вре-
мя зимы; члены семьи переживают этот
период за счет запасов меда, нако-
пленных в течение лета. Матки утратили
способность собирать себе пищу; следо-
вательно, они не могут в одиночку осно-
вать новую семью и для этого их обяза-
тельно должны сопровождать рабочие из
родительской семьи. Явление «роения»
представляет собой расселительный лёт,
при котором старая матка покидает гнез-
до, чтобы основать новое.
Медоносная пчела имеет важное зна-
чение, так как поставляет человеку
чрезвычайно важный продукт-мед. Кро-
ме того, необычайно важна роль пчел-и
диких видов, и медоносной пчелы-как
опылителей самых различных дикорасту-
щих и культурных растений, в том числе
большинства плодово-ягодных и бо-
бовых культур.
ЛИТЕРАТУРА
Crozier R. Н., 1977. Evolutionary gentics of the
Hymenoptera, Annu. Rev. Entomol., 22,
263-288.
Evans H. E., Eberhard M. J. W., 1970. The
wasps, Univ, of Michigan Press, Ann Arbor.
Krombein К. V., Hurd P. D., Jr., Smith D. R.,
Burks B. D., 1979. Catalog of Hymenoptera
in America north of Mexico, Smithsonian
Inst. Press, Washington, D.C., 3 vis.
Malyshev S. L, 1968. Genetics of the Hyme-
noptera and phases of their evolution,
Methuen, London.
Matthews R. W., 1974. Biology of Braconidae,
Annu. Rev. Entomol., 19, 15-32.
Michener C. D., 1944. Comparative external
morhlogy, phylogeny and a classification of
the bees, Bull. Am. Mus. Nat. Hist., 82,
151-326.
Michener C. D., 1953. Comparative morpholo-
gical and systematic studies of bee larvae
with a key to the families of hymenopterous
larvae, Univ. Kans. Sci. Bull., 35, 987-1102.
Michener C. D., 1974. The social behavior of the
bees. A comparative study, Harverd Univ.
Press, Cambridge, Mass.
Michener C. D, Fraser A., 1978. A comparative
anotomical study of mandibular structure in
bees, Univ. Kans, Sci. Bull., 51, 463-482.
Mitchell T W., 1960-1962. Bees of eastern
United States, N.C. Agric Exp. Stn. Tech.
Bull., 141, 1-538; 152, 1-557.
Muesebeck C. F. W et al., 1951. Hymenoptera
of America north of Mexico, synoptic cata-
logue, USDA, Agric. Monogr, vol. 2.
Richards O. W., 1977. Hymenoptera. Intro-
374
8. Отряды энтогнат и насекомых
duction and key to families, 2nd ed., Hand-
book for the identification of British insects.
Royal Entomological Society, London, vol. 6,
No. 1.
Ross H. H., 1937. A generic classification of
Nearctic sawflies, Ill. Biol. Monogr., 15(2),
1-173.
Spradberry J. P., 1973. Wasps. An account of
the biology and natural history of social
and solitary wasps, Univ, of Washington
Press, Seattle.
Townes H., 1969 The genera of Ichneumoni-
dae, Mem. Am. Entomol. Inst., 11, 1-300;
12, 1-537, 13, 1-307.
Viereck et al., 1916. The Hymenoptera of
Connecticut, Bull. Conn. State Geol. Nat.
Hist. Surv., 22, 1-824.
Отряд MECOPTERA
(скорпионницы)
Взрослые особи от мелких до сред-
них размеров имеют либо две пары
больших крыльев с сетчатым жилкова-
нием (рис. 8.130), либо короткие или пол-
ностью редуцированные крылья. Ан-
тенны длинные, сложные глаза большие,
ноги тонкие, в некоторых семействах
длинные и сильно вытянутые. Ротовые
органы грызущего типа часто располо-
жены на конце клювовидного выроста
головы. Личинки эруковидные или гусе-
ницеобразные, всегда с грудными нога-
ми, а у ряда групп также и с брюшными
личиночными ногами (рис. 8.131).
Имаго всеядны, питаются в основном
мелкими насекомыми, но разнообразят
рацион нектаром, пыльцой, лепестками,
плодами и мхами. Крылатые формы хо-
рошо летают. Самцы в семействе Рапог-
pidae (рис. 8.130) имеют крупную луко-
вицеобразную генитальную капсулу, не-
сколько напоминающую концевую часть
брюшка скорпионов, откуда и произош-
ло название всего отряда «скорпион-
ницы».
Яйца овальной формы, откладывают-
ся в почву или на ее поверхности, пооди-
ночке или группами до 100 и более штук
в группе. Личинки обитают во мху,
гниющей древесине, или в богатой орга-
ническими веществами грязи и перегное
вокруг сильно увлажненных мест (вблизи
родников и т. п.), в густых лесах. Пищу
их составляют органические вещества
различного происхождения. Окуклива-
Рис. 8.130. Самец скорпионницы Panorpa chelata. (Из Illinois
Natural History Survey.)
8. Отряды энтогнат и насекомых
375
Рис. 8.131. Личинка Apterobittacus apterus. (Из Applegarth.)
ние происходит в почве. В году разви-
вается только одно поколение.
У мелких видов, относящихся к роду
Boreus (семейство Boreidae), имаго снаб-
жены маленькими короткими крыльями,
и так как они достигают зрелости зимой
или ранней весной, их часто можно ви-
деть бегающими по снегу. Толстые эру-
ковидные личинки представителей этого
рода развиваются в гниющих основаниях
моховых подушек. Поскольку как личин-
ки, так и имаго по многим признакам от-
личаются от всех остальных Mecoptera,
некоторые авторы относят семейство Bo-
reidae к самостоятельному отряду Ne-
omecoptera. Однако строение ротовых
частей указывает на то, что Boreidae
представляют собой специализирован-
ное производное линии, давшей начало
семействам Panorpodidae и Panorpidae, а
не составляет отдельного отряда.
ЛИТЕРАТУРА
Byers G. W., 1954. Notes on North American
Mecoptera, Ann. Entomol. Soc. Am., 47,
481-510.
Byers G. W., 1965. Families and genera of Me-
coptera, Proc. 12th Int. Congr. Entomol.,
1964, 123.
Byers G. W., 1971. Ecological distribution and
structural adaptation in the classification of
Mecoptera, Proc. 13th Int. Congr. Entomol.,
1968, 486.
Carpenter F. M., 1931. A revision of the nearctic
Mecoptera, Mus, Comp. Zool. Harvard Coll.
Bull., 76, 206-277.
Hinton H. E., 1958. The phylogeny of the pa-
norpoid orders, Annu. Rev. Entomol., 3,
181-206.
Penny N. D., 1975. Evolution of extant Me-
coptera, J. Kans. Entomol. Soc., 48, 331-350.
Отряд SIPHONAPTERA (блохи)
Взрослые насекомые всегда бескрылы
(рис. 8.132), имеют сильно сжатое с бо-
ков тело, сильные длинные ноги с много-
численными шипами и короткие булаво-
видные антенны, спрятанные в покое
внутри углублений по бокам головы. Ро-
товые органы приспособлены для про-
калывания кожи и кровососания и со-
стоят из хоботка, пары щупиков и пары
коротких пластинчатых максилл. Блохи-
мелкие насекомые, длина их тела в боль-
шинстве случаев составляет от 2 до 4 мм,
однако немногие виды достигают
6-8 мм длины. Личинки с тонким чер-
веобразным телом (рис. 8.133). У них
округлая голова, ноги отсутствуют, на
каждом сегменте тела растут длинные
волоски. Сегменты груди и брюшка
внешне устроены одинаково. Ротовые
органы крошечные, малозаметные, гры-
зущего типа. Куколки образуются вну-
три кокона, формой напоминающего пи-
рожок и устроенного из частиц почвы
или травы; у куколок имеются руди-
менты крыльев.
376
8. Отряда энтогнат и насекомых
Рис. 8.132. Два обычных вида блох. (Из Illinois Natural History Survey.)
А. Человеческая блоха Pulex irritans. Б. Крысиная блоха Nosopsyllus fasciatus.
Блохи питаются кровью млекопитаю-
щих или птиц и обитают на теле хозяина
или в его гнездах, ходах или на тропах.
Яйца очень мелкие, беловатые,
овальные. Они просто сбрасываются
самкой либо на хозяина, либо в его гнез-
до. Так как яйца не клейкие, они, будучи
отложены на теле хозяина, скатываются
по его волосяному покрову и падают в
подстилку гнезда или убежища, где из
них вскоре вылупляются личинки. Ли-
чинки развиваются в почве или в мусоре
в гнезде и питаются анальными выделе-
ниями взрослых особей. Достигнув окон-
чательных размеров, личинки плетут ко-
кон неправильной формы в гнезде хозяи-
на и там окукливаются. Яйца, личинок
Рис. 8.133. Личинка блохи. (Из Illinois
Natural History Survey.)
и куколок блох почти невозможно уви-
деть без специального тщательного об-
следования.
Взрослые особи большинства видов
крайне активны. Они очень ловко бегают
между волосами и перьями; кроме того,
у многих видов тело несет гребни щети-
нок и шипов, которые еще более способ-
ствуют такому передвижению. У неко-
торых видов имаго почти постоянно
находятся на теле хозяина, у других има-
го находятся в гнезде и нападают на хо-
зяина лишь на время питания. По этой
причине, чтобы быть уверенным, что об-
наружены все виды блох, связанные с
хозяином, необходимо собирать их
как с тела хозяина, так и из его
гнезда.
Отряд Siphonaptera невелик по объе-
му. Фауна Северной Америки насчиты-
вает около 60 родов и свыше 200 видов.
Большая часть этих видов встречается на
диких млекопитающих, особенно на раз-
личных видах мышей, землероек, сусли-
ков, белок, гоферов и кроликов. Медве-
ди, бобры, койоты и многие другие
животные также имеют свою фауну блох.
Немногие блохи питаются на птицах и
8. Отряды энтогнат и насекомых
377
иногда случайно вредят домашней птице.
pulicidae. Многие виды этого семей-
ства имеют особенно важное значение
для человека. Собачья и кошачья блохи,
Ctenocephalides canis и С. felis, а также че-
ловеческая блоха Pulex irritans нападают
на людей и поселяются в жилых помеще-
ниях, создавая большие неудобства и да-
же делая их непригодными для жилья.
Укусы, наносимые блохами, болезненны,
обычно вызывают образование твердых
зудящих бугорков и могут стать причи-
ной вторичной инфекции в результате
расчесывания. Наиболее важны среди
блох некоторые крысиные блохи, прежде
всего Xenopsylla cheopis (и Nosopsyllus fas-
ciatus из семейства Ceratophyllidae), ко-
торые переносят бубонную чуму от крыс
человеку. Участвуя в распространении
этой болезни, которая много раз, подоб-
но пожару, охватывала густонаселенные
города в различных районах мира, блохи
несут ответственность за гибель миллио-
нов людей. В наше время системы обще-
ственного здравоохранения во всем мире
ведут постоянюе наблюдение за при-
родными очагами чумы, с тем чтобы
предотвратить новые вспышки этого за-
болевания путем контроля популяций
крыс и блох.
tungid ае. Два вида этого семейства-
Echidnophaga gallinacea и Tunga penetrans -
характеризуются сильно отличным от
других блох образом жизни. Самки обо-
их видов имеют очень мелкие размеры и
неподвижно прикрепляются к хозяину,
питаясь более или менее непрерывно.
Блоха Echidnophaga gallinacea (рис. 8.134)
нападает на домашнюю птицу, впиваясь
в кожу лицевой части головы и гребень.
Блоха Tunga penetrans нападает на лю-
дей, особенно на ноги. Она вгрызается
под кожу и образует болезненный буго-
рок, из которого высовывается наружу
лишь конец тела блохи. Оба вида сбра-
сывают яйца на почву и характеризу-
ются типичным для блох превраще-
нием.
Рис. 8.134. Блоха домашней птицы Echidnophaga gallinacea. (По материалам
министерства сельского хозяйства США.)
А. Пораженная блохами голова петуха (темные пятна-скопления блох).
Б. Взрослая самка блохи.
378
8. Отряды энтогнат и насекомых
ЛИТЕРАТУРА
Dunnet G. М., 1970. Siphonaptera. In: The in-
sects of Australia, D. F. Waterhouse et
al.(eds.), Melbourn Univ, Press, Carlton,
pp. 647-655.
Dunnet G. M., Mardon D. K.f 1974. A mono-
graph of Australian fleas (Siphonaptera),
Aust. J. Zool. Suppl. Ser., 30, 1-273.
Ewing H. E., Fox I., 1943. The fleas of North
America. USDA Misc. Publ. 500.
Fox I., 1940. Fleas of eastern United States,
Collegiate Press, Ames, Iowa.
George R. S. (ed.), 1974. Provisional atlas of
insects of the British Isles, Pt.4. Siphonapte-
ra, Inst. Terrestrial Ecol., Monks Wood Exp.
Stn., Abbots Ripton, U.K.
Haddow J., Traub R., Rothshild M., 1975. The
distribution of the Ceratophyllidae with an
illustrated key to the genera, Cambridge Univ.
Press, Cambridge.
Holland G. P., 1949. The Siphonaptera of
Canada, Can. Dept. Agric. Tech. Bull., 70,
1-306.
Holland G. P., 1964. Evolution, classification,
and host relationships of Siphonaptera, Annu,
Rev. Entomol., 9, 123-146.
Hopkins G. H. E., Rothshild M. (eds.), 1953—
1971. An illustrated catalogue of the Roth-
schild collection of fleas in the British Mu-
seum (Natural History), British Museum, Na-
tural History, London, vols. I-V
Hubbard C. A., 1947. Fleas of western North
America, Iowa State College Press, Ames,
Iowa.
Layne J. N., 1971 Fleas (Siphonaptera) of Flo-
rida, Entomologist, 54, 35-51.
Rothschild M., 1965. Fleas, Sci. Am., 213(6),
44-53.
Rothschild M., 1975. Recent advances in our
knowledge of the order Siphonaptera, Annu.
Rev. Entomol., 20, 241-259.
Rothschild M., Traub R., 1971. A revised glossa-
ry of terms in the taxonomy and morpho-
logy of fleas. In. illustrated catalogue of the
Rothschild collection of fleas in the British
Museum (Natural History), G. H. E. Hop-
kins, M. Rothschild (eds.), British Museum,
Natural history, London, vol.5, pp. 8-85.
Отряд DIPTERA (двукрылые)
Типичные взрослые формы
(рис. 8.135) имеют единственную (перед-
нюю) пару мембранозных крыльев, ред-
ко редуцированных до чешуек. На кры-
льях имеется несколько поперечных жи-
лок и умеренное число продольных. Зад-
ние крылья представлены лишь парой
тонких рукояткообразных качающихся
органов, называемых жужжальцами.
Ротовой аппарат разнообразного строе-
ния; в одних группах он приспособлен
Рис. 8.135. Типичные мухи. (Из Illinois Natural History Survey.)
A. Meromyza americana. Б. Eurosta solidaginis. h- жужжальце.
8. Отряды энтогнат и насекомых
379
для прокалывания и сосания, в других -
для соскребания или слизывания. Форма
тела также различна. В отдельных груп-
пах имаго полностью лишены крыльев.
Сложные глаза обычно большие, число
члеников антенн варьирует от 3 до 40.
Двукрылые-насекомые с полным пре-
вращением, их личинки безногие и обыч-
но либо с явственно выраженной голо-
вой, несущей хорошо развитые манди-
булы (рис. 8.146), либо с внутренним
склеротизированным скелетом, свя-
занным с парой крючковидных манди-
бул. Куколка либо свободная, либо обра-
зуется внутри шкурки личинки третьего
возраста.
Diptera-крупный от)эяд, насчиты-
вающий свыше 15 000 видов только в
фауне Северной Америки. Питание и
образ жизни взрослых насекомых, как
правило, очень сильно отличаются от пи-
тания и образа жизни их личинок.
Имаго многих семейств питаются в
основном нектаром и соками растений
или жидкостями, образующимися при
гниении органических остатков. Неко-
торые группы, например многие комары
и слепни, питаются кровью животных,
ротовые органы у представителей этих
групп сильнейшим образом видоизме-
нены в связи с прокалыванием и соса-
нием. В немногих группах, например у
оводов, ротовой аппарат сильно недораз-
вит, так что представляется сомнитель-
ной сама возможность того, что эти има-
го потребляют какую-либо пищу.
Личинки всей группы двукрылых в
общем влаголюбивы, и подавляющее
большинство их обитает в воде, гнию-
щем мясе, внутри тела других животных,
в разлагающихся плодах или других
влажных органических остатках, или в
тканях живых растений. Немногие личи-
ночные формы живут в относительно су-
хой почве или передвигаются по откры-
тому воздуху, однако подобные случаи-
скорее исключение, чем правило.
По большей части яйца двукрылых
простые, овальной или удлиненной
формы и обычно откладываются пооди-
ночке в пищу, предназначающуюся ли-
чинке, или на нее, или вблизи от этой пи-
щи. Яйца ряда форм, например у Drosop-
hila, снабжены боковыми или вер-
хушечными плавательными пузырьками,
предохраняющими их от погружения в
полужидкую пищу и потопления. Яйца
некоторых комаров достаточно надежно
защищены от чередующихся на протяже-
нии ряда месяцев противоположных воз-
действий : высыхания, увлажнения
и переохлаждения. В некоторых группах,
таких, как мясные мухи и некоторые па-
разитические мухи, личинки могут вылу-
пляться из яиц в теле самки, т. е. от-
кладываются уже крошечные личинки.
Эта особенность образа жизни достигает
наибольшего развития у группы Glossi-
noidea, в которой личинки выходят из
яиц и вырастают до окончательных раз-
меров внутри тела матери. Крайне инте-
ресны примеры педогенеза в этом отря-
де, наблюдающиеся у некоторых видов
комариков (см. с. 229).
Филогения. Для нынеживущих
двукрылых характерно чрезвычайно ши-
рокое разнообразие в строении головы
личинок и ее частей, антенн взрослых на-
секомых, в жилковании крыла, в строе-
нии ротового аппарата и других структур
(рис. 8.136). В той группе семейств, куда
входят грибные комарики и долгоножки,
состояние этих структур приближается к
таковому у более примитивных насе-
комых, таких, как Mecoptera. Названные
семейства комаров считают поэтому
примитивным концом «эволюционного»
ряда Diptera и выделяют в подотряд
Nematocera.
Собирая воедино эти примитивные
состояния различных органов, мы полу-
чаем следующую частичную реконструк-
0)
а «
-s —: о
Е схЗ
” ГЧ —
се
се
О
co
о
се
•б
о
О
о
2
о
cd
* 6
о
cd
S ffi
ex
я я e .а
х з.Е-с
О О о» о
се
JD
CO
с Е
.2 °
оо’С
cd cd
js £
at с/з
<u
co
<L>
CO
0)
cd
<D
cd
се
<D
cd
Q
cd
0)
cd o>
<1>
тЕ "’Р Е
сл о х:
ТаЬвпО” ||||
moidea
Ё О
щ Q
О
e
о
Si
e
о
N я
Е cu Е S
S о о Я
оЕ S-gS
Эмподий
Подотряд
Asilomoidea
Отдел Orthorrhapha
К» Дыха ль
7ца на бокал
видный Подотряд/ца на бо.ках
Brachycera
o> a, tx
Подотдел
Aschiza
Ос/з
cd
2
си •—
пз ~
o>:s и х
(V
о
cd
се
о
•W ~ *w М4 VMM > - *w *w W w
jUXh Q< иш ии<^Ои^ОЬО1
?
СЛ
- iMoscoidea>
Glossinoidea
Личинки:
развиваются
в матке
Секция Секция
Acalyptratae Calyptratae
Подотдел "‘’—Чешуйки увеличёйы'
____________Schizophora
Отдел —Возникает лобный пузырь
Cyclorrhapha
Возникает пупарий
Редуцируется жилкование крыла
Редуцируются антенны
* Дыхальца расположены дорсально или на конце брюшка
Л: мандибулы занимают вертикальное положение, голова редуцируется
И: максиллярные щупики 1-члениковые
Задние дыхальца расположены на пластинке
Задние дыхальца крупные
Л: голова хорошо выражена, мандибулы боковые , дыхальца одинаковые
И:Максиллярные щупики 3-члениковые, мандибулы горизонтальные
Рис. 8.136. Упрощенное
филогенетическое древо
отряда Diptera.
Я-имаго; Л-личинка.
8. Отряды энтогнат и насекомых
381
цию предковой формы двукрылых. У ли-
чинки голова образовывала хорошо вы-
раженную типичную склеротизирован-
ную капсулу, мандибулы двигались в
стороны, как у саранчовых, а брюшко нес-
ло маленькие боковые функционирую-
щие дыхальца на первых восьми сегмен-
тах. У взрослого насекомого усики были
длинными и состояли из многих свобод-
но сочлененных члеников, а продольная
жилка Е простиралась как свободная
жилка до края крыла.
Все Nematocera имеют капсулообраз-
ную личиночную голову, раздвигающие-
ся в стороны личиночные мандибулы и
примитивные имагинальные антенны.
Лишь представители семейств Cecidomy-
iidae (галлицы) , и Mycetophilidae
(грибные комарики) имеют практически
одинаково устроенные брюшные ды-
хальца, что указывает на наибольшую
древность этих линий в пределах всего
отряда. У личинок Bibionidae (толсто-
ножки), Psychodidae (бабочницы) и дру-
гих групп в тех же эволюционных линиях
задняя пара брюшных дыхалец увеличи-
лась, а остальные уменьшились, так что
респираторный газообмен стал осущест-
вляться личинкой главным образом или
полностью через дыхальца последней
пары. Вероятно, от основания психодид-
ной линии отошли ветви Tipulidae (ко-
мары-долгоножки) и Culicidae (кровосо-
сущие комары), у которых передние
брюшные дыхальца личинок либо силь-
но уменьшились в размерах и перестали
функционировать, либо атрофировались,
и все дыхательные функции осуществля-
лись исключительно за счет задних дыха-
лец.
Эти адаптации дыхальцевого аппара-
та были связаны главным образом с оби-
танием личинок в небольших водоемах, в
иле или очень сильно увлажненных орга-
нических средах. Эти личинки питались в
толще органического вещества, но при
этом держали задние дыхальца над гра-
ницей раздела водной и воздушной сред.
Такая особенность в питании личинок
явилась основой для практически всех
последующих направлений эволюцион-
ного развития Diptera. У комаров личин-
ки стали водными, способными суще-
ствовать на некоторой глубине. У выс-
ших двукрылых личинки многих се-
мейств перешли к жизни в иных, также в
основном жидких средах, в том числе в
тканях других насекомых, экскрементах,
полужидких органических остатках, та-
ких, как разлагающиеся плодовые тела
грибов и гниющее мясо.
В двух обычных семействах подотря-
да Nematocera -Simuliidae (мошки) и Chi-
ronomidae (звонцы)-личинки стали пол-
ностью водными и утратили задние
дыхальца. Подлинные родственные свя-
зи этих семейств пока неясны. В приводи-
мом здесь филогенетическом древе
данные семейства помещены предвари-
тельно главным образом на основе их
сходства в жилковании крыла с другими
семействами.
От основания либо психодидной, ли-
бо типулидной линий возникло некое
двукрылое насекомое, сильно изменив-
шееся во многих отношениях. Манди-
булы личинок стали крючковидными и
движущимися вверх-вниз и внутрь-нару-
жу, совершая разрывающие движения;
личиночная голова утратила большую
часть своей склеротизации; число члени-
ков максиллярных щупиков имаго реду-
цировалось с трех до одного; на крыльях
жилки Е и Си? в типичном случае сопри-
касались на краю крыла. Эти преобразо-
вания знаменовали начало эволюции
подотряда Brachycera (короткоусые
двукрылые). В одной из его прими-
тивных линий, включающей Asilidae
(ктырей) и близкие к ним группы, личи-
ночные дыхальца последней пары нахо-
дятся на боках сегмента, примерно как у
382
8. Отряды энтогнат и насекомых
длинноусых семейства Bibionidae. Ли-
чинки азилоидной ветви хищничают или
паразитируют на других насекомых. В
соседней линии задние дыхальца личинки
сблизились друг с другом и расположи-
лись в спинном или хвостовом углуб-
лении или полости, а находившиеся кау-
дальнее них сегменты брюшка редуциро-
вались до маленьких вентральных лопа-
стей, расположенных под восьмым сег-
ментом. Этот предок дал начало двум
ветвям: ветви Tabanidae (слепни), в кото-
рой эмподии имагинальных лапок стали
пульвиллообразными, и ветви Empididae
(толкунчики), в которой эмподии оста-
лись тонкими, но редуцировалось жилко-
вание крыла. В ходе эволюции Brachycera
антенны взрослых насекомых делались
все более и более мощными и ком-
пактными и прогрессивно редуцирова-
лись. Со временем возникла эмпидидная
линия, у представителей которой ан-
тенны состояли лишь из трех члеников,
причем последний из них нес длинную
палочку, или аристу.
В тех семействах, которые обсужда-
лись нами до сих пор, куколка была, как
правило, свободной, хотя в нескольких
группах, занимающих изолированное по-
ложение в системе (например, Cecidomy-
iidae и Stratiomyidae), куколка формиро-
валась внутри старой личиночной шкур-
ки, которая отвердевала и образовывала
защитную оболочку, или пупарий. Вылу-
пляющееся из пупария взрослое двукры-
лое делало на нем сверху продольную Т-
образную щель, через которую оно и
вылезало наружу. Эти семейства соста-
вляют отдел Orthorrhapha (прямо-
шовные).
От основания эмпидидного ствола
произошла форма, у которой образова-
ние пупария закрепилось и взрослая
особь появлялась, вытолкнув переднюю
стенку пупария. Эта линия образовала от-
дел круглошовных-Cyclorrhapha. Наи-
более примитивные представители это-
го отдела составляют подотдел Aschiza,
куда входят Syrphidae (журчалки) и род-
ственные им группы.
Во второй линии круглошовных раз-
вился выворачивающийся мешок, или
лобный пузырь (ptillinum), который выдав-
ливается через расходящиеся швы над и
сбоку от антенн и помогает вытолкнуть
передний конец пупария. Эта линия-по-
дотдел Schizophora-отличается наибо-
льшим разнообразием среди двукрылых.
Schizophora включает две обширные
секции. Наиболее примитивная из них-
Acalyptratae - представлена Otitidae и
Drosophilidae (плодовые мушки). В дру-
гой линии - Calyptratae - связующие пере-
понки у основания крыла сильно расши-
ряются, образуя отчетливые лопасти,
или чешуйки. Ранние этапы развития
этих образований представлены у Ant-
homyiidae (цветочные мухи); более про-
двинутое состояние наблюдается у Tachi-
nidae и Sarcophagidae.
В одной из ветвей Cyclorrhapha,
включающей надсемейство Glossinoidea,
личинки развиваются в особом кармане -
матке, внутри тела самки. Происхожде-
ние Glossinoidea точно не известно, но,
возможно, они выделились как раннее
ответвление ствола Calyptratae.
Одна из деталей строения предста-
вляет значительный эволюционный инте-
рес; это имагинальные мандибулы.
Представители семейств Simuliidae, Psyc-
hodidae, Ceratopogonidae, Culicidae, Ta-
banidae и Rhagionidae имеют мандибулы
во взрослом состоянии. Эти мандибулы
пластинчатые, с типичным мандибу-
лярным сочленением и мускулатурой, а
по форме напоминают верхние челюсти
имаго Siphonaptera и некоторых Mecop-
tera. Значительное число семейств, у ко-
торых мандибулы сохраняются во взрос-
лом состоянии, определенно указывает
на то, что у древнейших двукрылых эти
Рис. 8.137. Крылья Diptera.
A. Psychoda (Psychodidae). Б. Aedes (Culicidae). В. Chironomus (Chironomidae). Г. Simuliwn
(Simuliidae). Д. Bibio (Bibionidae). E. Rhagio (Rhagionidae). Ж. Stratiomys (Stratiomyidae).
3. Tabanus (Tabanidae). И. Thereva (Therevidae). K. Asilus (Asilidae.). Л. Anthrax (BombyHidae).
M. Scaeva (Syrphidae).
/-сверхкомплектная жилка. Другие ..обозначения жилок см. в гл. 3.
8. Отряды энтогнат и насекомых
384
т
S1 + 2
Рис. 8.138. Детали строения Diptera, имеющие диагностическое значение.
А. Крыло Megaselia (Phoridae). Б. Крыло Hylemya (Anthomyiidae). В. Крыло Empis
(Emptdidae). Г Крыло Sarcophaga (Sarcophagidae). Д. Костальная область крыла Rhagoletis
(Tephritidae). Е. Щиток ZenilHa (Tachinidae). Ж. Закрыловые чешуйки Anthomyia
(Anthomyiidae). 3. Закрыловые чешуйки Musca (Muscidae). И. Грудной отдел высших
двукрылых (схематизировано). 1 -щиток; 2-постскутеллум; 3-заднеспинка, 4-грудная
чешуйка, 5-крыловая чешуйка; 6-прескутум; 7-плечевой бугорок; 8-проплевра,
Р-скутум; 10-нотоплевра; У/-мезоплевра; 72-птероплевра, 13-тазик; 74-стерноплевра;
75-брюшко; 76-жужжальце; 77-гипоплевра.
Отряды энтогнат и насекомых
385
8.139. Детали строения Diptera (3, И по Curran).
4 Закрыловые чешуйки Scopeuma (Scopeumatidae). Б. Закрыловые чешуйки Pegomyia
Anthomyiidae). В, Г Закрыловые чешуйки Fannia (Fanniidae). Д. Закрыловые чешуйки Musca
Muscidae). Е. Голова Agromyza (Agromyzidae). Ж. Схема верхней части головы; показано
расположение простых глазков и окружающих щетинок. 3. Голова Euthycera (Sciomyzidae).
i? Голова Nemopoda (Sepsidae).
-грудная чешуйка; 2-крыловая чешуйка; 3-ариста, 4-глазковый треугольник;
5-теменные щетинки; 6-затеменные щетинки; 7-глазковые щетинки.
зрганы имелись и что во многих ветвях
гтряда они полностью атрофировались.
В той линии, которая берет начало в ос-
новании табанидной ветви и затем при-
водит к формированию круглошовных,
мандибулы отсутствуют.
ПОДОТРЯД NEMATOCERA
Фауна Северной Америки насчиты-
вает представителей примерно 20 се-
мейств Nematocera (длинноусые, или ко-
мары). Звонцы, долгоножки, кровососу-
щие комары и мошки-вот примеры
наиболее обширных, многочисленных и
часто попадающихся на глаза семейств
длинноусых. Личинки во всех семействах
с хорошо развитой склеротизированной
головой, втянутой в грудь только у
Tipulidae.
CECIDOMYIIDAE (ITONIDIDAE), ГАЛ-
лицы. Имаго галлиц (рис. 8.141)-мало-
заметные нежные комарики, с сильно ре-
дуцированным жилкованием крыльев и
удлиненными четковидными антеннами.
Взрослые особи питаются исключитель-
386
8. Отряды энтогнат и насекомых
П Р С
Рис. 8.140. Детали строения Diptera, имеющие диагностическое значение.
А. Среднеспинка Helobia (Tipulidae). Б. Среднеспинка Palpomyia (Ceratopogonidae). В. Средне-
спинка Chironomus (Chironomidae). Г. Коготки и присоски лапки Tabanus (Tabanidae).
Д. Коготки и присоски лапки Zenillia (Tachinidae). Е. Антенна Bibio (Bibionidae). Ж. Антенна
Mydas (Mydaidae). 3. Антенна Tabanus (Tabanidae). И. Антенна Geosargus (Stratiomyidae).
К. Антенна Pollenia (Calliphoridae). Л, М. Лицевая часть самца и самки Bibio (Bibionidae).
Н. Лицевая часть Asilus (Asilidae). О. Лицевая часть Hylemya (Anthomyiidae). П. Лицевая часть
Dolichopus (Dolichopodidae). Р Лицевая часть Gasterophilus (Gasterophilidae). С. Лицевая часть
Tephrita (Tephritidae).
7-пульвиллы; 2-ариста; 3-вибриссы; 4-хоботок; 5-ротовое отверстие; 6-лобный шов.
8. Отряды энтогнат и насекомых
387
Рис. 8.141. Гессенская муха Mayetiola destructor. (По материалам министерства
сельского хозяйства США.)
А. Взрослая самка. Б. Яйца, одно с выходящей личинкой. В. Личинка. Г Голова и
лопаточка. Д. Куколка. Е. Пупарий. Ж. Пораженный стебель пшеницы с выходящими
куколками и имаго. 3. Антенны самца и самки.
но жидкой пищей, например сахаристы-
ми выделениями растений; большая
часть пищи поглощается в течение личи-
ночного периода развития. Многие роды
этого семейства-галлообразователи, и
именно благодаря галлам, особенно на
ивах, других лиственных деревьях и мно-
гих травах, это семейство широко извест-
но натуралистам. Внутри семейства рас-
пространены и многие другие типы пита-
ния. Некоторые личинки хищничают, по-
едая клещей и мелких насекомых, другие
питаются на развивающихся семенах
растений, третьи-разлагающимися ор-
ганическими веществами, остальные
едят ткани листьев или стеблей растений.
К последней категории принадлежит на-
иболее важный с экономической точки
зрения вид во всем семействе-гессенская
муха Mayetiola destructor (рис. 8.141), ли-
чинки которой питаются у основания
стеблей злаков и особенно сильно вредят
пшенице и ячменю.
simuliidae, мошки. У представите-
лей этого семейства личинки и кукол-
ки живут в воде, а имаго сосут кровь по-
добно Culicidae. Мошки имеют короткое,
обрубленное и горбатое тело с коротки-
ми ногами и компактными, как правило,
11-члениковыми антеннами (рис. 8.142).
Как и у кровососущих комаров, кусаются
только самки. Они нападают на лоша-
388
8. Отряды энтогнат и насекомых
Рис. 8.142. Имаго мошки Simulium vittatum.
(По Knowlton, Rowe.)
дей, коров, уток и многих диких жи-
вотных. Некоторые виды нападают и на
человека, собираясь вокруг глаз, ушей и
открытых частей лица, рук и лодыжек.
Они высасывают большое количество
крови и оставляют жгучие ранки на коже
жертвы. Эти укусы могут чесаться и бо-
леть неделю и более.
Яйца откладываются кучками близ
уреза воды или в воду. Личинки прикре-
пленные, удлиненные и по форме не-
сколько напоминают кувшин
(рис. 8.143), встречаются только в про-
точной воде; задний конец тела заякорен
шипастым присасывательным диском на
каком-либо твердом субстрате, напри-
мер камне, бревне или погруженных в по-
ток листьях, голова несет пару гребенок
из перистых волосков, предназначенных
для отцеживания частиц пищи из теку-
щей воды, прежде всего микроорганиз-
мов и органических остатков. Куколки
также водные, образуются в коконах, по-
хожих формой на комнатную туфлю
(рис. 8.143). На переднегруди куколка не-
сет пару длинных дыхательных отрост-
ков, обычно разветвленных на множе-
ство тонких нитей.
TIPULIDAE, КОМАРЫ-ДОЛГОНОЖКИ.
Взрослые насекомые имеют длинные но-
В
Рис. 8.143. А. Личинка мошки Simulium
johannseni. Б. Ее же куколка. В. Кукол очные
чехлики S. venustum. (По Illinois Natural
History Survey.)
8. Отряды энтогнат и насекомых
389
Рис. 8.144. Взрослая самка
комара-долгоножки Epiphragma fascipennis.
ги и узкие крылья (рис. 8.144), ните-
видные антенны из многих хорошо выра-
женных члеников и V-образный попе-
речный желобок или шов на среднеспин-
ке. Имаго крайне многочисленны в сы-
рых лесах и укромных лощинах, а также
вдоль лесистых речных отмелей. Личин-
ки вытянутые и червеобразные
(рис. 8.145); многие из них водные, оби-
тают преимущественно на дне в кучах
гниющей листвы; большинство питают-
ся плесенью на мертвых листьях, неболь-
шое число форм поедают корни живых
растений или минируют листья. Они
сильно варьируют по размерам и внеш-
нему виду, однако в большинстве случаев
имеют мощную голову, которая частич-
но втянута в грудь (у ряда видов она мо-
жет втягиваться еще сильнее и пол-
ностью прятаться внутри груди, когда
насекомое потревожено), а также манди-
булы с сильными зубцами, движущиеся
из стороны в сторону.
CULICIDAE, КРОВОСОСУЩИЕ КОМАРЫ,
или настоящие комары. Стройные на-
секомые с длинными ногами (рис. 8.146),
четковидными антеннами (у самцов ан-
тенны перистые, как и в семействе Chiro-
nomidae) и многочисленными про-
дольными жилками на крыльях. Ротовые
придатки образуют хоботок, состоящий
из сильно видоизмененного комплекса
Рис. 8.145. Epiphragma. (По Needham.)
А. Личинка. Б. Задний конец тела личинки
сверху. В. Куколка.
390
8. Отряды энтогнат и насекомых
Рис. 8.146. Желтый малярийный комар Aedes aegypti. (По материалам
министерства сельского хозяйства США.)
А. Взрослое насекомое. Б. Личинка. В. Куколка.
колюще-сосущих органов (см. рис. 3.30,
3.31).
В некоторых родах яйца отклады-
ваются на воду, либо по одному, либо
склеенными между собой в плотики; ли-
чинки вылупляются из яиц через несколь-
ко дней. В других родах комары от-
кладывают яйца на перегной или прямо
над урезом воды на стенки резервуаров,
когда уровень воды поднимается и яйца
оказываются под водой, из них вылу-
пляются личинки. Личинки водные, жи-
8. Отряды энтогнат и насекомых
391
вут в большинстве случаев в стоячей во-
де, но немногие виды обитают и в
слабопроточных водоемах. В некоторых
районах земного шара обнаружены
виды, развивающиеся и в быстрых пото-
ках, что совершенно не свойственно
образу жизни неарктических видов. У ли-
чинок крупная голова и довольно
длинные антенны, широкая раздутая
грудь и цилиндрическое брюшко. У всех
настоящих комаров и представителей
близких групп брюшко несет спинную
дыхательную трубку или площадку на
восьмом сегменте. Личинки комаров пи-
таются в основном микроорганизмами и
органическими остатками на поверхно-
сти или в толще воды. Немногие группы
хищничают и поедают исключительно
личинок других видов комаров. Куколки
также водные, свободноживущие и ак-
тивные. Их дыхательные трубки распо-
ложены на груди.
Взрослые самки некоторых видов и
самцы всех видов Culicidae питаются
только нектаром или пьют воду. К сожа-
лению, самки подавляющего большин-
ства видов сосут кровь, которая необхо-
дима им для размножения в природных
условиях.
Кровососущие комары имеют огром-
ное экономическое значение для челове-
ка. Некоторые виды, в основном относя-
щиеся к родам Anopheles и Aedes, перено-
сят возбудителей заболеваний человека,
список которых выглядит весьма внуши-
тельно: малярия, лихорадка денге, жел-
тая лихорадка и филяриоз. По этой при-
чине кровососущие комары являются
объектами первостепенного значения с
точки зрения медицинской энтомологии.
Они имеют хозяйственное значение и как
фактор, наносящий непосредственный
ущерб- обилие комаров снижает цен-
ность некоторых мест отдыха, особенно
в курортных зонах, и, несомненно,
оказывает влияние на освоение террито-
рий Крайнего Севера, где они особенно
многочисленны.
CH1RONOMIDAE (TENDIPEDIDAE),
звонцы. Этих нежных насекомых
(рис. 8.147) часто ошибочно принимают
за кровососущих комаров, однако они не
кусаются и имеют ряд характерных отли-
чий, позволяющих распознать их. Усики
самцов перистые, с венчиками длинных
шелковистых волосков. Личинки, за ис-
ключением очень немногих видов, живут
в воде, причем одни-свободноживущие,
а другие плетут широкую камеру из
твердых частиц со дна и шелковых ни-
тей; личинки представителей некоторых
родов сооружают настоящий домик. Те-
ло их тонкое, червеобразное, с неболь-
шой, но хорошо склеротизированной го-
ловой и 12-сегментным туловищем. В
ряде групп переднегрудь, или последний
сегмент брюшка, или оба этих сегмента
могут нести по паре несочлененных ного-
подобных выростов, или ложных ножек.
Личинки питаются органическими остат-
ками на дне водоемов и населяют реки,
озера, канавы и пруды. Куколки также
водные, некоторые из них свободноживу-
щие, но большинство находится внутри
сети или домика, построенного личин-
кой. Во многих водоемах эти личинки
звонцов невероятно обильны и состав-
ляют один из важнейших компонентов
корма рыб. Когда появляются крылатые
особи, они вылетают ночью целыми ту-
чами и своей жужжащей массой завола-
кивают окрестные источники света. Сам-
ки откладывают яйца в воду, где из них
вскоре вылупляются личинки.
ПОДОТРЯД BRACHYCERA
Как правило, взрослые насекомые
этого подотряда (короткоусые) крупнее и
имеют более плотное сложение, чем
длинноусые, а также отличаются более
совершенным полетом.
392
8. Отряды энтогнат и насекомых
Рис. 8.147. Стадии жизненного цикла звонцов (Chironomidae). (Из Illinois Natural
History Survey.)
А. Личинка Chironomus tentans. Б. Куколка Cricotopus trifasciatus. В. Взрослый
самец Chironomus ferrugineovittatus.
ОТДЕЛ ORTHORRHAPHA (ПРЯМОШОВНЫЕ)
Личинки имеют крючковидные или
пластинчатые мандибулы, движущиеся в
вертикальной плоскости, а не из стороны
в сторону, голова часто сильно втянута
внутрь груди (рис. 8.148) и может иметь
мощный внутренний каркас, идущий да-
леко в глубь туловища. В фауне Северной
Америки представлено около 15 се-
мейств, некоторые из них очень обычны,
а в одном семействе-Tabanidae, или
слепни,-имаго являются прожорливыми
кровососами.
Другие семейства Orthorrhapha вклю-
чают самые разнообразные формы как в
отношении особенностей строения, так и
по образу жизни. Большинство из них,
это в особенности касается семейств Dol-
ichopodidae (зеленушки) и Empididae
(толкунчики),-хищники как во взрос-
лой, так и в личиночной стадии, на-
падающие на других насекомых и
имеющие свободную куколку; личи-
нки большинства видов обитают в гни-
8. Отряды энтогнат и насекомых
393
дых стволах или в почве и там же
окукливаются. Личинки Conopidae
(болыпеголовки) - внутренние паразиты
других насекомых.
asilidae, ктыри. Это также крупные
мухи (рис. 8.148), обычно с сильно выпу-
клой, напоминающей горб, грудью и вы-
тянутым брюшком; у представителей ря-
да родов, однако, тело может быть
коренастым, сильно опушенным и ярко-
окрашенным, напоминающим шмеля.
Нижняя часть лицевой области во мно-
гих родах несет также напоминающую
бороду щетку из волосков, называемую
«лицевой бородой». Ктыри охотятся на
Рис. 8.148. Ктырь Promachus vertebratus. (Из
Illinois Natural History Survey.)
А. Взрослое насекомЛ. Б. Безногая личинка,
напавшая на личинку хруща.
других насекомых, ловят и поедают шме-
лей и стрекоз, а не только более мелкие
формы. Взрослые мухи не пугливы, их
легко заметить и нетрудно также пой-
мать сачком. Личинки встречаются в ос-
новном в почве и гнилой древесине, где
поедают обитающих там насекомых. Ли-
чинки некоторых ктырей-важные есте-
ственные враги личинок хрущей и дру-
гих почвообитающих видов, повреждаю-
щих сельскохозяйственные культуры
(рис. 8.148).
mydidae. Это небольшое семейство,
однако к нему принадлежат одни из
самых крупных и наиболее эффектных
видов. Усики у них булавовидные, а кры-
лья с видоизмененным жилкованием.
концы многих жилок загибаются вперед,
по направлению к переднему краю кры-
ла. Обычный восточный (для США) вид
Му das clavatus-4t^bin с оранжевой
перевязью и достигает в размахе крыльев
55 мм. Личинки, похожие на крупных ли-
чинок ктырей, живут в гнилой древесине
и ведут хищный образ жизни.
TABANIDAE, СЛЕПНИ (ПАУТЫ).
Взрослые насекомые (рис. 8.149) имеют
большую голову и плотное тело, часто
достигают в длину 25 или 30 мм; их
мощные крылья характеризуются весьма
полным жилкованием и нередко с ярким
рисунком. В жилковании характерно на-
личие широкой V-образной ячейки S3,
захватывающей вершину крыла. Самки-
кровососы; их ротовые части предназна-
чены для прорезания кожи и сосания вы-
текающей из ранки крови. Самцы пи-
таются нектаром. Яйца откладываются
кучками на побеги или другие нависаю-
щие над водой предметы; вылупившиеся
личинки сползают или падают в то место
где они будут развиваться. Личинки оби-
тают в болотах или слабопроточных во-
доемах, сидят в топком дне и часто
многочисленны в почве сырых пастбищ.
Они хищны и питаются улитками, насе-
394
8. Отряды энтогнат и насекомых
Рис. 8.149. Слепень Tabanus Uneolus. (По
материалам министерства сельского
хозяйства США.)
комыми и другими водными организма-
ми. Тело личинки жесткое, кожистое,
обычно белое или с перевязями, голова
способна полностью втягиваться в грудь.
Куколки цилиндрические, бурого цвета;
окукливание, как правило, происходит в
толще торфа или мертвой растительно-
сти, которые увлажнены, но находятся
выше уровня свободной воды.
Имаго слепней часто досаждают до-
машним животным, в особенности круп-
ному рогатому скоту и лошадям. В от-
дельных местностях эти вредители
играют подчас существенную роль в
ухудшении общего состояния животных.
Многие виды слепней изобилуют в боло-
тистых районах северных и влажных
горных областей на континенте, доса-
ждая отдыхающим и местным жителям.
Ближе всего к Tabanidae стоит семей-
ство Rhagionidae (бекасницы), которые
хищничают и в личиночной, и во взрос-
лой фазе. Некоторые виды рода Sympho-
romyia (рис. 3.26) на стадии имаго крово-
сосущи и нападают на людей и других
позвоночных. В более отдаленном род-
стве со слепнями находятся Stratiomyidae
(львинки), личинки которых либо хищни-
ки, либо сапрофаги, а взрослые мухи ве-
дут хищный образ жизни.
ОТДЕЛ CYCLORRHAPHA
(КРУГЛОШОВНЫЕ)
У этих мух куколка всегда находится
внутри пупария, из которого взрослое
насекомое выходит через круглое отвер-
стие, выталкивая переднюю стенку. У бо-
лее специализированных представителей
подотдела Schizophora это осуществляет-
ся путем выпячивания лобного пузыря,
высовывающегося наружу через лобные
швы в момент выхода из пупария, а за-
тем полностью втягивающегося обрат-
но. У нескольких примитивных семейств,
составляющих подотдел Aschiza, лобные
швы не соприкасаются на дорсальной
стороне и лобный пузырь отсутствует,
ч
ПОДОТДЕЛ ASCHIZA
SYRPHIDAE, ЖУРЧАЛКИ. Мухи ОТ мел-
ких до крупных размеров (рис. 8.150), ха-
рактеризующиеся обращенными вперед
окончаниями некоторых продольных жи-
лок крыла, а также обычно наличием
жилкообразного утолщения в виде сверх-
комплектной продольной жилки, лежа-
щей перед М Эти мухи питаются почти
исключительно на цветках и отличаются
замечательной способностью зависать в
воздухе без всякого заметного движения.
На теле многих видов имеются яркие по-
лосы или пятна желтого, белого, красно-
го и черного цветов, в семействе журча-
лок сильно развито подражание облику
различных ос, шмелей и других пчел.
Личинки крайне разнообразны. Мно-
гие питаются тлями, переползая от коло-
нии к колонии и пожирая огромное коли-
8. Отряды энтогнат и насекомых
395
Рис. 8.150. Муха-журчалка Didea fasciata. (По Metcalf.)
А. Взрослое насекомое. Б. Личинка. В. Пупарий. /-переднее дыхальце; 2-задние
дыхальца.
чество особей. Другие виды развиваются
в разлагающихся жидких органических
остатках, например личинки мух из
рода Eristalis, называемые «крысками»
(рис. 8.151). У крысок задние дыхальца
расположены на конце вытягивающегося
«хвоста»-дыхательной трубки, которая
высовывается над поверхностью пищево-
го субстрата и, таким образом, обеспечи-
вает контакт с воздухом.
Кроме того, личинки питаются в хо-
дах насекомых-ксилофагов, листьями
злаков и их влагалищами, мусором в
гнездах шмелей, муравьев и ос. Личинки
двух видов серьезно повреждают луко-
вицы культурных тюльпанов и других
луковичных растений, выедая их- сере-
дину.
К мухам-журчалкам близки Phoridae
(горбатки)-мелкие формы с разно-
образным образом жизни, но, как прави-
ло, сапрофагами на стадии личинки, а
также Pipunculidae (Dorilaidae)-неболь-
шие мухи, личинки которых паразити-
руют на цикадах (Hemiptera).
ПОДОТДЕЛ SCHIZOPHORA
Schizophora составляют одну из наи-
более богатых эволюционных ветвей
двукрылых и подразделяются на две
крупные секции: Acalyptratae, у которых
мембраны в основании крыла не продол-
жены в виде складок или лопастей, и
Calyptratae, у которых эти мембраны
образуют три лопасти. Две из этих трех
396
8. Отряды энтогнат и насекомых
Рис. 8.151. Личинки-крыски. (По Metcalf.)
А. Пупарий Eristalis aeneus. Б. Личинка Eristalis tenax. /-переднее дыхальце; 2-задние
дыхальца на длинном «крысином хвосте».
лопастей лежат одна над другой при ос-
новании крыла и называются закрыловы-
ми чешуйками-крыловой и грудной.
Третья лопасть лежит в стороне от них
вдоль основания заднего края крыла и
называется крылышком. У большинства
Calyptratae, например у Muscidae, закры-
ловые чешуйки настолько велики, что за-
крывают жужжальца.
Секция Acalyptratae. К данной секции
принадлежит множество семейств. К чис-
лу примитивных для этой группы се-
мейств относятся Otitidae, Lauxaniidae,
Coelopidae и Heleomyzidae, личинки ко-
торых питаются различными, разлагаю-
щимися органическими остатками, Sepsi-
dae, личинки которых питаются навозом,
и Sciomyzidae, личинки которых живут в
воде и поедают главным образом вод-
ных и амфибиотических улиток. К числу
более специализированных групп отно-
сятся маленькие навозные мушки ВогЬо-
ridae, листовые минеры Agromyzidae,
преимущественно сапрофаги Chloropidae
(злаковые мухи) и Ephydridae (берего-
вушки), личинки которых могут обитать
в соленой воде. Ниже приводятся харак-
теристики наиболее часто встречающих-
ся семейств.
sciomyzidae. Это распространенное
по всему земному шару семейство вклю-
чает мух среднего размера, паразити-
рующих на наземных и пресноводных
брюхоногих моллюсках и способных на-
падать и убивать улиток, во много раз
более крупных, чем сама муха. Чаще все-
го их свободноживущих личинок и пла-
вающие пупарии можно видеть в прудах
и водохранилищах, где водится много
улиток. Так как брюхоногие моллюски
нередко служат промежуточными хозяе-
вами некоторых возбудителей заболева-
ний человека и домашних животных, в
особенности в субтропическом и тропи-
8. Отряды энтогнат и насекомых
397
песком поясе, Sciomyzidae рассматри-
ваются сейчас как потенциально эффек-
тивное средство биологической борьбы с
улитками.
lauxaniidae. Это другая широко рас-
пространенная и часто встречающаяся
группа. Взрослые Lauxaniidae встречают-
ся в широком диапазоне условий среды-
от заболоченных побережий, лугов и ле-
сов всех типов до горных вершин. О
личинках известно довольно мало; тем
не менее большая часть изученных видов
оказалась сапрофагами.
heleomyzidae. Представители этого
семейства населяют в основном леса уме-
ренного пояса, где они достаточно обык-
новенны и встречаются в разнообразных
местообитаниях. У одних ёидов личинки
живут в грибах, однако у большинства
личинки обитают в ходах других насе-
комых, в частности в ходах личинок жу-
ков-ксилофагов, питаясь мусором. Близ-
кородственное семейство Coelopidae в
личиночной фазе специализировано к
обитанию в гниющих бурых водорослях
и выброшенных на берег Ruppia maritima.
TEPHRITIDAE (TRYPETIDAE, TURPANEI-
dae), пестрокрылки. Сюда относятся
многие ярко окрашенные виды с крап-
чатыми или поперечнополосатыми
крыльями. Личинки некоторых видов
живут в семенных коробочках или со-
чных плодах растений. Экономическое
значение имеют вишневая муха Rhagole-
tis cingulata (рис. 8.152), личинки которой
прокладывают ходы в плодах вишни, и
пестрокрылка яблонная Rhagoletis рото-
nella, личинки которой развиваются в
яблоках. Средиземноморская плодовая
муха Ceratitis capitata была обнаружена
во Флориде в 1929 г. и, подобно восточ-
ной фруктовой мухе Dacus dorsalis,
является широко распространенным тро-
пическим видом, личинки питаются мя-
котью сочных плодов.
DROSOPHILIDAE, ПЛОДОВЫЕ МУШКИ. К
этому семейству принадлежат мелкие
мушки (рис. 8.153), время от времени во
Рис. 8.152. Вишневая муха Rhagoletis cingulata. (По материалам
министерства сельского хозяйства США.)
А. Взрослое насекомое. Б. Личинка. В. Передние дыхальца личинки.
Г Пупарий. Д. Задняя дыхальцевая пластинка куколки.
398
8. Отряды энтогнат и насекомых
Рис. 8.153. Плодовая мушка Drosophila melanogaster. (По материалам
министерства сельского хозяйства США.)
А. Взрослое насекомое. Б. Антенна. В. Основание голени и первый членик лапки
имаго. Г Пупарий, вид сбоку. Д. Пупарий, вид сверху. Е. Задние дыхальца зрелой
личинки Ж. Зрелая личинка.
множестве появляющиеся в большинстве
жилищ и складских помещений. Личинки
их питаются гниющими фруктами и дру-
гими органическими материалами. Dro-
sophila melanogaster используется как ла-
бораторный объект в генетических ис-
следованиях и, что касается картиро-
вания хромосом и генов, без сомне-
ния, является наиболее изученным ви-
дом.
AGROMYZIDAE, МИНИРУЮЩИЕ МУХИ.
К этому семейству относятся мухи не-
больших или совсем мелких размеров.
Личинки являются листовыми или сте-
блевыми минерами, иногда галлообразо-
вателями и изредка обитателями плодов.
Во многих районах мира эти мухи нано-
сят серьезный вред культурным расте-
ниям.
ПРОЧИЕ СЕМЕЙСТВА ACALYPTRATAE.
Borboridae - небольшие навозные муш-
ки-широко распространенные мелкие
или очень мелкие формы, которые даже в
состоянии покоя постоянно покачивают
крыльями. Обычно они связаны с поме-
том животных или разлагающимся орга-
ническим материалом, где развиваются
их личинки. Береговушки Ephydridae жи-
вут в основном вблизи воды и личинки их
по большей части либо водные, либо жи-
вут внутри стеблей влаголюбивых расте-
ний, хотя некоторые виды встречаются и
вдали от водоемов. Chloropidae, или зла-
ковые мушки, приспособились к самым
разнообразным местообитаниям. У не-
которых видов личинки питаются побе-
гами молодых растений, а по меньшей
мере у одного вида личинки развиваются
в подкожном слое у лягушек и питаются
кровью. Ряд видов хищничает, поедая
яйцевые коконы других насекомых и
пауков.
BRAULIDAE, ПЧЕЛИНЫЕ ВШИ. Кро-
шечный паразит медоносной пчелы Вгаи-
la caeca является единственным предста-
вителем этого семейства. Он умеет от-
влечь пчелу и слизнуть отрыгиваемую
пчелой капельку меда прямо с ее ротовых
органов.
1 Отряды энтогнат и насекомых
399
Cesom Calyptratae. Представители
секции характеризуются развитием
_• имаго закрыловых чешуек и увеличе-
нием задних дыхалец у личинок. К этой
□алии относится около десятка се-
мейств, группирующихся в два надсемей-
гтжа-Muscoidea и Glossinoidea-и встре-
чающихся в самом широком диапазоне
местообитаний.
НАДСЕМЕЙСТВО MUSCOIDEA. На-
иболее примитивными представителями
утого надсемейства являются Anthomyii-
дае и Muscidae, более специализиро-
ванные семейства Sarcophagidae и Callip-
leridae, а также еще ряд упоминаемых
ниже семейств.
ANTHOMYIIDAE, ЦВЕТОЧНЫЕ МУХИ.
Мут, принадлежащие к этому семей-
ству. своим обликом сильно напоминают
гэмнатную муху. Хозяйственно важные
зады данной группы-это прежде всего
туковая муха Hylemya antiqua
рис. 8.154) и капустная муха Hylemya
-rassicae, питающиеся в личиночной ста-
дии корнями соответствующих растений.
MUSCIDAE, НАСТОЯЩИЕ мухи. Это се-
мейство включает, вероятно, самое
обычное и повсеместно встречающееся
Рис. 8.154. Стадии жизненного цикла
луковой мухи Hylemya antiqua. (Из Illinois
Natural History Survey.) А. Взрослое
насекомое. Б. Личинка. В. Пупарий.
насекомое-комнатную муху Musca do-
mestica (рис. 8.155). У комнатной мухи
белые личинки, развивающиеся в разно-
го рода разлагающихся остатках.
Взрослые мухи переносят возбудителей
многих опасных и широко распростра-
ненных заболеваний, в том числе брюш-
ной тиф, многие формы дизентерии, хо-
Рис. 8.155. Стадии развития комнатной мухи Musca domestica.
(По материалам министерству сельского хозяйства США.)
А. Пупарий. Б. Взрослое насекомое. В. Личинка и некоторые
детали строения.
400
8. Отряды энтогнат и насекомых
леру и трахому. Другой хорошо из-
вестный представитель этого же семей-
ства-жигалка осенняя Stomoxys calci-
trans, взрослые особи которой наносят
болезненные укусы и нападают на чело-
века и домашних животных. Личинки жи-
галки развиваются в гниющих органиче-
ских остатках, наиболее благоприятны
для их размножения гниющие кучи све-
жей травы и перегной (компост).
GASTEROPHILIDAE, ЖЕЛУДОЧНЫЕ
оводы. Эти средних размеров (примерно
с рабочую пчелу) покрытые волосками
мухи имеют на теле черные, желтые или
красные перевязи (рис. 8.156). Личинки
этих оводов-внутренние паразиты ло-
шадей, мулов и некоторых крупных ди-
ких млекопитающих. В Северной Амери-
ке представлен лишь один род - Gasterop-
hilus, четыре вида которого заражают
лошадей. Овод-крючок, или лошадиный
овод (G. intestinalis), откладывает яйца на
волосы, растущие на ногах и на лопатках
лошади. Когда лошадь слизнет эти яйца,
из них сразу же вылупляются личинки,
которые через рот, ткани глотки и пище-
вода проникают в желудок лошади.
Здесь молодые особи овода прикре-
пляются к эпителию и начинают питать-
ся, вырастая в толстых шипастых червео-
бразных личинок длиной до 15 мм и
более. Лето и зиму они проводят в теле
лошади, а весной отрываются от места
прикрепления, выходят из организма хо-
зяина и окукливаются в почве. Через не-
сколько недель появляются взрослые
особи.
CALLIPHORIDAE, ПАДАЛЬНЫЕ МУХИ.
Эти большие, крепко сложенные мухи
имеют всесветное распространение и во
множестве летают днем, громко жужжа.
Взрослых насекомых привлекает влаж-
ность, видимо, потому, что они питают-
ся в основном разжиженным гниющим
материалом или нектаром и другими са-
харистыми жидкостями. Личинки неко-
Рис. 8.156. Желудочные оводы. (Из Canadian
Department of Agriculture.)
A. Gasterophilus intestinalis. Б. G. haemor-
rhoidalis.
торых видов живут на мясе животных,
часть видов паразитирует на дождевых
червях, другие обитают в колониях тер-
митов или муравьев. Одна из групп-па-
разиты наземных улиток. Несмотря на
высокую численность, падальные мухи в
меньшей степени передают инфек-
ционные заболевания и менее важны с
экономической точки зрения, чем
8. Отряды энтогнат и насекомых
401
остальные близкие к ним семейства мух.
Немногие виды каллифорид отклады-
вают яйца в ранки на уже пораженной ко-
же овец, и личинки начинают питаться
омертвевшими тканями, вызывая
кожные миазы у овец. Такие зараженные
животные привлекают другие виды мух,
которые не нападают на здоровых овец.
SARCOPHAGIDAE, СЕРЫЕ МЯСНЫЕ МУ-
ХИ. Это также крупное, всесветно распро-
страненное семейство, близкородствен-
ное падальным мухам; однако большин-
ство его видов живородящи. Некоторые
личинки паразитируют на пище, запасен-
ной в гнездах ос, но, как правило, эти му-
хи развиваются на падали или разлагаю-
щихся органических остатках. Ряд видов
имеет важное значение, так как поражает
кожные покровы живых животных и лю-
дей. В одном роде - Blaesoxiphia- личин-
ки полностью перешли к эндопаразити-
ческому существованию на саранчовых
семейства Acrididae.
OESTRIDAE, НОСОГЛОТОЧНЫЕ И ПОД-
КОЖНЫЕ ОВОДЫ. Эти мухи (рис. 8.157)
составляют небольшую группу быстро
26-732
Рис. 8.157. Имаго обычного подкожного овода крупного рогатого
скота -Hypoderma lineatwn. (Из Canadian Department of Agriculture.)
402
8. Отряды энтогнат и насекомых
Рис. 8.158. Мухи-тахины. (По материалам министерства сельского хозяйства США.)
Слева. Trichopoda pennipes, паразитирующая на Hemiptera. Справа. Winthemia
quadripustulata- паразит гусениц чешуекрылых.
летающих пушистых шмелеподобных
двукрылых - паразитов млекопитающих;
некоторые из них имеют столь же спе-
циализированный жизненный цикл, как и
желудочные оводы. У многих видов жиз-
ненный цикл простой. Например, овечий
овод Oestrus ovis откладывает молодых
личинок в ноздри овец; личинки мигри-
руют в лобные пазухи и полости у осно-
вания рогов, где достигают зрелости. За-
кончившая развитие личинка вылезает
наружу через ноздри овцы, попадает в
почву и там окукливается. Намного
сложнее цикл развития бычьего овода
Hypoderma bovis, поражающего крупный
рогатый скот Самка откладывает яйца
на волоски задних ног или на бока жи-
вотного. Вскоре из яиц выходят личинки,
заползают под волосяной покров, вбура-
вливаются под кожу, медленно продви-
гаясь через соединительную ткань к пи-
щеводу ; это «путешествие» занимает
около 4 мес. После трехмесячного разви-
тия в пищеводе личинки (все еще остаю-
щиеся сравнительно мелкими) вновь ми-
грируют через соединительную ткань и
заканчивают эту миграцию под кожей
поясничного отдела. Здесь личинки дора-
стают до своих окончательных размеров,
причем каждая вызывает образование
желвака, открывающегося свищом. Че-
рез этот свищ личинка получает доступ к
воздуху во время созревания, а затем вы-
ходит через него и окукливается в почве.
TACHINIDAE (LARVAEVORIDAE), ТАХИ-
НЫ, ИЛИ ЕЖЕМУХИ. Это одно из об-
ширнейших семейств в отряде Diptera.
Все его представители (рис. 8.158) пара-
зитируют на насекомых и нападают на
личинок или взрослых особей многих от-
рядов, особенно на гусениц чешуекры-
лых. Тахин многих видов искусственно
8. Отряды энтогнат и насекомых
403
размножали и интродуцировали в раз-
личные районы земного шара с целью
контроля численности некоторых видов
насекомых-вредителей. Взрослые самки
многих видов откладывают яйца непос-
редственно на хозяина, вылупившиеся
личинки внедряются в тело хозяина и на-
чинают питаться его тканями. Другие
виды откладывают яйца на листву; если
эта листва будет съедена личинкой вида-
хозяина, то личинка паразита вылупится
из яйца прямо в пищеварительном трак-
те хозяина, а отсюда молодые личинки
внедрятся в полость тела, где протекает
их развитие. Достигнув зрелости, личин-
ки паразита окукливаются либо внутри
тела хозяина, либо оставляют его и ухо-
дят на окукливание в почву
НАДСЕМЕЙСТВО GLOSSINOIDEA
Сюда относятся четыре небольших
семейства, представители которых пи-
таются кровью млекопитающих и птиц
или живут на их теле как эктопаразиты.
Насколько известно, большинство пред-
ставителей Glossinoidea имеют особен-
ность, очень необычную для насекомых ,-
внутриутробное развитие молоди. Един-
ственная в каждый момент личинка со-
держится в теле матери, в специальном
кармане-матке, где выкармливается вы-
делениями особых желез.
GLOSSINIDAE, МУХИ ЦЕЦЕ. Это семей-
ство включает только один род-Gloss-
ina-с приблизительно 20 видами, оби-
тающими в Африке. Мухи цеце, немного
более крупные, чем комнатные мухи,
переносят ряд тяжелых заболеваний-
трипанозомозов, таких, как африканская
сонная болезнь человека и болезнь-нага-
на, поражающая крупный рогатый скот.
Мухи цеце - облигатные кровососы и за
день обычно высасывают количество
крови, во много раз превышающее вес их
собственного тела. Единственная моло-
дая личинка выходит из яйца прямо в те-
ле матери и на протяжении примерно 10
дней находится там, питаясь секретами
сильно видоизмененных желез, располо-
женных на стенке маточного кармана
самки. Напитавшаяся личинка последне-
го возраста весит уже столько же, сколь-
ко ее мать. Через час или два после того,
как личинка покидает тело матери, она
зарывается в почву и окукливается. Спу-
стя примерно 30 дней появляется взрос-
лое насекомое.
HIPPOBOSCIDAE, кровососки. Это се-
мейство включает сильно видоизме-
ненных мух, которые сосут кровь и ведут
эктопаразитический образ жизни в воло-
сяном покрове млекопитающих или
перьях птиц. У кровососок уплощенное
тело и сильные ноги; по общему облику
они напоминают скорее вошь или клеща.
Большинство видов имеют хорошо раз-
витые крылья с мощными передними
жилками. Ряд форм сбрасывает крылья,
лишь только попадет на поверхность те-
ла хозяина; у некоторых крылья сильно
редуцированы или могут вовсе отсут-
ствовать, как, например, у Melophagus
(рис. 8.159). Зрелые личинки обычно не
откладываются на хозяина.
Рис. 8.159. Овечий рунец Melophagus ovinus.
(Из Kentucky Agricultural Experiment Station.)
404
8. Отряды энтогнат и насекомых
Наиболее известный вид фауны Се-
верной Америки - овечий рунец Melopha-
gus ovinus (рис. 8.159). Это бескрылое,
похожее на клеща насекомое - обычный
паразит домашних овец во многих райо-
нах мира, в особенности в умеренных
широтах. Когда самка «рождает» зрелых
личинок, они приклеиваются к шерсти
хозяина, где сразу же окукливаются. Две
трети видов данного семейства встре-
чается исключительно на птицах.
прочие glossinoidea. Представите-
ли семейства Streblidae и близкого к нему
Nycteribiidae - кровососущие эндопара-
зиты летучих мышей. Часть из этих спе-
циализированных видов, видимо, стали
строго специфичны в отношении своего
хозяина. Большинство видов обоих се-
мейств распространены в тропиках и
субтропиках.
ЛИТЕРАТУРА
Berg С. О., Knutson L., 1978. Biology and syste-
matics of the Sciomyzidae, Annu. Rev. Ento-
mol., 23, 239-258.
Cole F. P., 1969. The flies of western North
America, Univ, of California Press, Berkeley
and Los Angeles.
Crampton G. C. et al., 1942. The Diptera or true
flies of Connecticut, Conn. State Geol. Nat.
Hist. Surv., 64, 1-509.
Curran С. H, 1934. The families and genera of
North American Diptera, Privately published,
The Ballou Press, New York.
Demerec M. (ed.), 1950. Biology of Drosophila,
Wiley, New York.
Downes J. A., 1971. The ecology of blood-suc-
king Diptera: An evolutionary perspective. In:
Ecology and physiology of parasites,
A. M. Fallis (ed.), Univ, of Toronto Press, p.
232-258.
Frost S. W., 1924. A study of the leaf-mining
Diptera of North America, Cornell Univ. Agr.
Exp. Stn. Mem., 78, 1-228.
Gillett J. D., 1971. Mosquitoes, Wiedenfield &
Nicholson, London.
Herring E. M., 1951. Biology of leaf miners,
Junk, The Hague.
Hull F. M., 1962. Robberflies of the world. The
genera of the family Asilidae, U. S. Natl. Mus.
Bull., 224, 1-907 (2 vols.).
Knight K. L., Stone A., 1977. A catalog of the
mosquitoes of the world (Diptera: Culicidae).
Thomas Say Foundation, Entomol. Soc. Am..
6, 1-612.
Oldroyd H, 1964. The natural history of flies.
Weidenfield & Nicholson, London.
Oliver D. R., 19171. Life history of the Chirono-
midae, Annu. Rev. Entomol., 16, 211-230.
Roback S. S., 1951. A classification of the mus-
coid calyptrate Diptera, Ann. Entomol. Soc.
Am., 44, 327-361.
Seguy E., 1951. Ordre des Dipteres (Diptera
Linne, 1758), Traite de Zool., 10, 449-744.
Stone A., Sabrosky C. W., Wirth W W., Foo-
te R. H, Coulson J. R. (eds.), 1965. A catalo-
gue of the Diptera of America north of Mex-
ico, U. S. Govt. Printing Office, Washington
D.C.
Wirth W W., Stone A., 1956. Aquatic Diptera.
In. Aquatic insects of California, R. L. Us-
inger (ed.), Univ, of California Press, Berkeley,
pp. 372-482.
Отряд TRICHOPTERA
(ручейники)
Ручейники - похожие на ночных бабо-
чек насекомые, личинки и куколки ко-
торых живут в воде. Имаго ручейников
(рис. 8.160) достигают в длину
1,5-40 мм. Для них характерен ротовой
аппарат, грызущего типа с сильно реду-
цированными или почти атрофиро-
ванными частями, кроме двух пар щупи-
ков ; длинные и многочлениковые ан-
тенны, большие сложные глаза
и длинные ноги. За исключением бескры-
лых или короткокрылых самок несколь-
ких видов, взрослые особи имеют две
пары больших перепончатых крыльев, с
почти полным набором продольных жи-
лок, но с сокращенным числом попе-
речных. У большинства видов тело и
крылья покрыты волосками, а не чешуй-
ками ; у немногих видов на антеннах, щу-
пиках, ногах и крыльях могут иметься
♦ Отряды энтогнат и насекомых
405
Рис. 8.160. Ручейники. (Из Illinois Natural History Survey.) A. Rhyacophila fenestra.
Б. Eubasilissa pardalis.
группы чешуек или отдельные чешуйки,
жоеянные среди более длинных воло-
сков. Личинки (рис. 8.161-8.164) разно-
образны по внешнему облику и образу
жизни. Каждый глаз представлен един-
ственной фасеткой, антенны маленькие,
ссшочлениковые, ротовые органы грызу-
сзего типа, хорошо развиты. На груди
три пары сильных ног, на брюшке также
имеется пара сильных ног, несущих
крючья и часто комплект пальцео-
бразных жабр.
Большинство имаго темноокра-
шенные или коричневатые, однако у не-
многих форм на крыльях имеется рису-
нок из желтых или оранжевых пятен или
серебристых штрихов.
Ручейники подразделяются на два
406
8. Отряды энтогнат и насекомые
Рис. 8.161. Личинка и куколка ручейника Limnephilus submonilifer,
сооружающего домики. (По Illinois Natural History Survey.)
7-переднеспинка, 2-среднеспинка, 3-заднеспинка; 4-боковой бугорок;
5-спинной бугорок, 6-жабры; 7-глаз, 8-усик, 9-тазик, 10-вертлуг;
77-бедро; 72-голень, 13-лапка; 14-коготок; 75-анальная ножка,
16-площадки, несущие крючки; 17-передние площадки пятого тергита,
18-задние площадки пятого тергита, 79-передние площадки шестого
тергита, 20-анальные выросты.
подотряда. Annulipalpia и Integripalpia
(рис. 8.165). К Annulipalpia относится
лишь одно надсемейство - Hydropsychoi-
dea (рис. 8.165,2), личинки которого
строят ловчие сети или простые убежи-
ща. Подотряд Integripalpia включает
два надсемейства Limnephiloidea (рис.
8.165,5), личинки которых сооружают
трубчатые домики (рис. 8.164), и
Rhyacophiloidea (рис.8.165,5-7). К по-
следнему надсемейству принадлежат три
семейства с относительно примитивны-
ми личиночными признаками: у Rhya-
cophilidae свободноживущие личинки, у
Glossosomatidae они сооружают седло-
видные домики-пещерки, из которых вы-
совываются наружу и голова, и брюшко,
у Hydroptilidae личиночные домики ме-
шочковидные (рис. 8.163). Лимнофи-
лоидные ручейники - наиболее разно-
образная группа, к которой относятся
приблизительно два десятка семейств
8. Отряды энтогнат и насекомых
407
Рис. 8.162. Свободноживущая личинка
ручейника Rhyacophila fenestra. (Из Illinois
Natural History Survey.)
(рис. 8.165,5-75). Всего в мире известно
около 30 семейств, несколько сотен ро-
дов и свыше 7000 видов ручейников.
Большинство семейств имеют всесвет-
ное распространение, однако некоторые
ограничены в своем распространении ли-
бо Северным полушарием, либо от-
дельными районами Южного полуша-
рия. Неарктическая фауна ручейников
насчитывает более 800 видов, принадле-
жащих к 17 семействам.
У личинок большинства семейств
этих насекомых высокой степени разви-
тия достигла строительная деятельность.
Трубочки, представляющие собой пере-
носные «дома», сооруженные личинками,
восхищали исследователей жизни пре-
сноводных насекомых. Домики
(рис. 8.163, 8.164) личинок ручейников
имеют различную форму-от прямой
трубочки до сложного спирального жи-
лища Helicopsyche, напоминающего ра-
ковину улитки. При постройке этих до-
миков используются различные виды ма-
териалов мелкие камешки, частицы пе-
ска, кусочки листьев, прутики, иглы
хвойных деревьев, а подчас и маленькие
раковины улиток. В большинстве случаев
особи данного рода или вида строят до-
мики определенной формы, но предста-
вители родов, относящихся к разным се-
мействам, часто делают очень похожие
домики.
Самки ручейников откладывают от
300 до 1000 яиц. У некоторых видов яйца
выходят из тела самки в виде нитки бус
Рис. 8.163. Мешочковидный домик
и личинка ручейника Ochrotrichia unio. (Из
Illinois Natural History Survey.)
Рис. 8.164. Личинка ручейника Limnephilus
rhombicus в цилиндрическом домике. (По
Illinois Natural History Survey.)
<Л
Личиночные жабры
1-члениковые
гоноподы
ХРлвиЙЙл*
ЗС об *J cujE-
Усики личинок
длинные
Бородавки линейны
Бородавки овальные
'Мезофурка
клювовидная
Мезофурка обычная
Щиток короткий
Щупик самца
4-члениковый
Роговидный вырост
переднегруди /Лимнефили
Имагинальные мандибулы ная^етвь^Лептоцеридная ветвь
'—Щиток длинный
Простые глазки отсутствуют;
'•супратенторий редуцирован;
М« сохраняется; переднеспинка
личинки без борозды
Простые глазки и супра-
тенторий нормально раз-
виты; на переднем крыле
самца М< отсутствует;
переднеспинка личинки
с предвершинной
бороздой
Ю-й тергит самки с
дополнительными
бугорками
Сочленения ноги ли-
чинки редуцированы
Простые глазки и 1
ный шов отсутствует -
Голова личинки-— А*® Строят трубчатые домики
вытянута ANNULIPALPIA Строят мешочковидные домики
Строят ловчие ^-^^^TEGRlPALPIA^l^Z^
Вершинный членик---- Д домики пещерки
щупиков кольчатый; | \ Сдичинка свободноживущая
супратенторий 1 V ~ .
отсутствует ^9-и тергит личинки склеротизирован
ваны Имагинальные мандиоулы
V А У .1 » 1 1 . редуциро-
•тв —\^1 Rhyaao^hiloidea MM^i^mnSphiloidea
Рис. 8.165. Филогенетическая схема семейств Trichoptera. (С разрешения Annual Reviews, Inc.
из Annual Reveiews of Entomology, vol. 12, 1967.)
8. Отряды энтогнат и насекомых
409
Рис. 8.166. Яйца ручейников. (Из Illinois
Natural History survey.)
A. Triaenodes tarda. Б. Cyrnellus marginalis.
(рис. 8.166,Б); самка заходит в воду и от-
кладывает яйца на камни и другие пред-
меты, укладывая этот шнур в виде непра-
вильной массы, содержащей док 800 яиц.
Самки из других групп выдавливают яй-
ца и формируют из них большой комок,
висящий вплоть до откладки на конце
брюшка (рис. 8.166,Л). Яйца заключены
в желеобразное вещество, которое набу-
хает при поглощении воды. Такие комки
прикрепляются к побегам или камням,
находящимся под водой, вблизи воды
или нависающими над ней.
В семействе Limnephilidae самки ча-
сто откладывают яйца массами на ветви,
нависающие над водой. Во многих слу-
чаях наблюдали, что эти яйцевые массы
набухают и разжижаются, а личинки вы-
лупляются во время дождя. Образую-
щиеся в результате этого процесса капли
желеобразного вещества стекают по ве-
точкам и падают в воду, унося с собой
личинок.
Все личинки активно двигаются, пи-
таясь при этом мелкими водными жи-
вотными или микроорганизмами, ко-
торыми обрастают отложения органиче-
ских остатков в воде. Немногие роды, а
именно Rhyacophila (Rhyacophilidae) и
Oecetis (Leptoceridae), представлены по
преимуществу хищными формами, по-
едающими мелких личинок насекомых.
Личинки Hydropsychoidea плетут стацио-
нарную ловчую сеть и укрытие; сеть ис-
пользуется для ловли мелких водных ор-
ганизмов, которыми питается личинка.
Почти во всех остальных семействах ли-
чинки сооружают переносные домики
различных типов. Эти домики исполь-
зуются личинками для защиты большей
части тела, имеющей нежные покровы.
Когда личинка активно передвигается, из
домика высовываются только силь-
но склеротизированная голова, ноги и
передняя часть груди (рис. 8.163,
8.164).
Перед тем как окуклиться, личинки
всех ручейников плетут кокон. Личинки,
строящие домики, делают его просто пу-
тем укладывания внутри домика выстил-
ки из шелковой нити и закупоривания
концов трубки решетчатой или снабжен-
ной щелями мембраной. Свободноживу-
щие и сооружающие ловчие сети личинки
плетут овальный кокон из шелка и песка,
камешков или кусочков мусора и прикре-
пляют этот кокон к камню, бревну или
иному твердому субстрату. Куколки раз-
виваются в коконе до тех пор, пока пол-
ностью не сформируются и не достиг-
нут окончательной склеротизации имаги-
нальные структуры (за исключением
крыльев). Куколки (рис. 8.161) необычны
тем, что имеют мощные мандибулы. С
их помощью созревшая куколка прогры-
зает себе выход из кокона. Затем она
всплывает к поверхности, выползает на
бревно или камень и превращается во
взрослое насекомое.
Как правило, для полного жизненно-
го цикла требуется год, причем большая
часть этого периода приходится на личи-
ночную стадию. Стадия яйца продол-
жается лишь короткое время; куколоч-
ная стадия занимает от 2 до 3 нед,
410
8. Отряды энтогнат и насекомые
взрослые ручейники живут около месяца.
Значительный интерес представляет
образ жизни «микро»-ручейников из се-
мейства Hydroptilidae. В это семейство
входят только мелкие формы, длина ко-
торых составляет от 1,5 до 6 мм. Личин-
ки первых возрастов - мельчайшие сво-
бодноживущие насекомые с маленьким
брюшком; личинки последнего возраста
строят переносной домик и брюшко их
раздувается. Существуют данные лишь о
двух родах североамериканской фауны-
Mayatrichia и Ochrotrichia. У представи-
телей последнего наблюдаются различия
в строении коготков между свободножи-
вущими возрастами и возрастами, строя-
щими домики. Описанный диморфизм во
многих отношениях сходен с типичными
случаями гиперметаморфоза.
Личинки ручейников живут как в
стоячих, так и в проточных водоемах, яв-
но предпочитая холодную и незагрязнен-
ную воду. Видовой состав ручейников-
хороший индикатор уровня загрязненно-
сти реки, а для отдельных видов характе-
рен относительно узкий диапазон усло-
вий обитания. Как группа в целом, они
имеют более широкую экологическую
пластичность, однако более чувстви-
тельны к загрязнению, чем звонцы (Chi-
ronomidae). Ручейники играют важную
роль в речных экосистемах, так как со-
ставляют крупное звено в цепи питания и
являются первичным источником пищи
для многих видов рыб.
Хорошо иллюстрированные таблицы
для определения до семейств личинок,
куколок и имаго североамериканских
Trichoptera даны Уиггинсом (Wiggins in:
Merritt, Cummins, 1978).
ЛИТЕРАТУРА
Anderson N. H., 1976. The distribution and bio-
logy of the Oregon Trichoptera, Oreg. Agric.
Exp. Stn. Tech. Bull., 131, 1-152.
Betten C., 1934. The caddisflies or Trichoptera
of New York State, N. Y State Mus. Bull.
292, 1-576.
Crichton M. I. (ed.), 1978. Proceedings of the
2nd International Symposium on Trichoptera.
Junk, The Hague.
Denning D. G., 1956. Trichoptera. In. Aquatic
insects of California, R. L. Usinger (ed*.
Univ, of California Press, Berkeley, pp
237-270.
Nielson A., 1948. Postembryonic developmen:
and biology of the Hydroptilidae, Kgl. Danske
Videnskab., Biol. Skr., 5, 1-200.
Ross H. H., 1944. The caddisflies or Trichoptera
of Illinois, Bull. Ill. Nat. Hist. Surv., 23, 1-326.
Ross H. H., 1056. Evolution and classification of
the mountain caddisflies, Univ, of Illinois
Press, Urbana.
Ross H. H., 1964. Evolution of caddiswonr
cases and nets, Am. Zool., 4, 209-220
Ross H. H., 1967. The evolution and past disper-
sal of Trichoptera, Annu. Rev. Entomol.. 12.
169-206.
Wiggins G. B., 1977. Larvae of the North Ameri-
can caddisfly genera (Trichoptera), Toronto
Univ. Press, Toronto.
Wiggins G. B., 1978. Trichoptera. In. An intro-
duction to aquatic insects, R. W Merritt.
K. W Cummins (eds.), Kendall/Hunt, Dubu-
que, pp. 147-185.
Отряд LEPIDOPTERA (бабочки, или
чешуекрылые)
У этих насекомых две пары крыльев
(рис. 8.167), если не считать немногие
виды, имеющие бескрылых самок. Тело,
крылья и прочие придатки покрыты че-
шуйками, которые часто ярко окрашены
и образуют красивый рисунок. Ротовые
органы взрослых особей сильно редуци-
рованы, у большинства форм хорошо
развиты только мак силлы. Они срослись
и вытянулись, образовав спирально свер-
нутую трубку для всасывания жидкой пи-
щи. Крупные сложные глаза, длинные
антенны и ноги хорошо развиты. Пре-
вращение у бабочек полное. Личинки-
знакомые всем гусеницы - имеют цилин
8. Отряды энтогнат и насекомых
411
Рис. 8.167. Типичная ночная бабочка. Видны
чешуйки на крыльях и теле, а также
сосательный хоботок, который в покое
спирально закручивается под головой. (По
Illinois Natural History Survey.)
дрическое тело, по большей части хоро-
шо выраженную голову, грудные ноги и
пять пар личиночных ног на брюшке
(включая и концевую пару) h Куколки
обычно твердые, коричневые, придат-
ки выглядят у них приклеенными к
телу.
Это крупнейший отряд, включающий
около 10000 североамериканских видов.
Взрослые формы очень сильно варьи-
руют в размерах, от крошечных, дости-
гающих лишь 1-2 мм в длину, до
огромных видов, с размахом крыльев
15 см и более. Крылья характерно рас-
крашены, и отряд в целом представляет
собой притягательное собрание красок и
изящных рисунков.
Гусеницы Lepidoptera раститель-
ноядны, за исключением разбросанных
по разным группам видов, ведущих
хищный образ жизни или являющихся
потребителями органических остатков
1 Гусеницы пядениц (Geometridae) имеют
только 2 пары брюшных ног.
или запасов различных продуктов и изде-
лий. Огромная масса видов питается от-
крыто на листьях, большое число мелких
видов минирует листовые пластинки или
черешки, гусеницы еще одной обширной
группы, включающей как крупные, так и
мелкие виды, прокладывают ходы в сте-
блях, стволах или корнях. Многие из
этих видов нападают на сельскохозяй-
ственные растения, ежегодно причиняя
значительный ущерб урожаям и запасам
продовольствия. По этой причине отряд
чешуекрылых имеет важное хозяйствен-
ное значение.
Помимо вредящих видов отряд Lepi-
doptera содержит один вид, играющий
существенную роль в экономике,-это ту-
товый шелкопряд, или шелковичный
червь Bombyx mori, являющийся главней-
шим источником натурального шелка.
Разведение этого вида и сбор шелка, из-
вестные под названием шелководство,-
важная отрасль народного хозяйства во
многих районах земного шара, каждый
год страны восточной Азии производят
шелка на многие миллионы долларов.
На рубеже столетий производство нату-
рального шелка было организовано в
Калифорнии, однако это предприятие
потерпело неудачу из-за высокой стои-
мости рабочей силы. В 1945 г новую по-
пытку предприняли в Техасе, где боль-
шая часть операций была механизирова-
на.
Филогения. В трех семействах Lepi-
doptera (рис. 8.168) передние и задние
крылья примерно одинаковы по разме-
рам и жилкованию, а передние крылья
снабжены лопастевидным оценочным ап-
паратом, называемым югум, или
югальным выступом переднего крыла
(рис. 8.169, А,2), и расположенным у ос-
нования заднего края. Эти особенности
напоминают состояние, характерное для
Trichoptera, и свидетельствуют о том, что
данные три семейства, относимые к под-
412
8. Отряды энтогнт и насекомых
Микро
о
се
тз
*БЬ
св
св
се
се
О
О
се
се
X Щ
JUGATAE
FRENATAE
Макро
Е32
О м
0'S-
S-5S
се
CU Z<JZJ слООсл
нтенны
булавовидные
Антенны с
загнутой вершиной
Усики на
RHOPALOCERA вершине
расширены
На заднем крыле
Р редуцирована
Л: преддыхальцевая бородавка с 2 щетинками
К максиллярные щупики неразличимы
преддыхальцевая бородавка с 3 щетинками
максиллярные щупики хорошо выражены
Л :
К
Югум * Самка с 2 генитальными отверстиями
отсутствует
И: мандибулы редуцированы
Л: брюшных ног меньше
Л 8 пар брюшных ног
Рис. 8.168. Упрощенное филогенетическое древо отряда Lepidoptera. (В основном по Forbes,
1923.)
отряду Jugatae, являются наиболее при-
митивной группой среди чешуекрылых. В
семействе Micropterygidae гусеницы
имеют ноги на восьми сегментах брюш-
ка (и в этом отношении напоминают ли-
чинок Mecoptera), а у имаго имеются
функционирующие мандибулы. Исходя
из этого, можно считать, что Microptery-
gidae берут начало от наиболее архаич-
ной из известных ветвей подотряда, а
следовательно, и всего отряда. В другой
линии подотряда Jugatae мандибулы
имаго подверглись редукции, а затем и
исчезли, брюшко гусениц утратило по
меньшей мере одну пару ног Данная
ветвь дала начало двум типичным для
Jugatae семействам: Hepialidae (тонко-
пряды) и Eriocraniidae, содержащему
мелких ночных бабочек.
От основания линии Eriocraniidae от-
делилась ветвь, в которой югальный вы-
ступ исчез и основной структурой, сце-
пляющей крылья, стал френулум
(рис. 8.169,Д,7)- длинная щетинка в
Рис. 8.169. Крылья Lepidoptera. (Прислано д-ром Kathryn М. Sommerman.)
А, Б. Переднее и заднее крылья Mnemonica (Eriocraniidae.) В. Переднее крыло
Achroia (Pyralidae). Г, Д. Переднее и заднее крылья Archips (Tortricidae).
1 - френулум; 2 - югум.
Рис. 8.170. Детали строения Lepidoptera, имеющие диагностическое значение.
А. Переднее крыло Pamphila (Hesperiidae). Б. Переднее крыло Pieris (Pieridae).
В. Переднее крыло Papilio (Papilionidae). Г Передняя (маленькая)
и средняя ноги Brenthis (Nymphalidae). Д. Вершинная часть антенны Brenthis.
Е. Вершинная часть антенны Thanaos (Hesperiidae).
Рис. 8.171. Крылья Lepidoptera.
Л, Б. Переднее и заднее крылья Acidalia (Geometridae). В, Г Переднее и
заднее крылья Malacosoma (Lasiocampidae). Д. Заднее крыло Halisidota
(Arctiidae). Е. Переднее крыло Protoparce (Sphingidae). Ж. Заднее крыло
Hyperaeschra (Notodontidae). 3. Заднее крыло Citheronia (Citheroniidae).
И. Заднее крыло Sarnia (Saturniidae).
8. Отряды энтогнат и насекомых
415
основании заднего крыла. В этом ство-
ле появились различия в форме перед-
них и задних крыльев, а также в их жил-
ковании. Так произошел подотряд Frena-
tae. Наши единственные современные
представители ранних Frenatae-группа
редко попадающихся в сборах кро-
шечных молей, составляющих надсемей-
ство Incurvarioidea.
От древнего предкового представите-
ля Frenatae возникла форма, имевшая
два генитальных отверстия у самки: одно
для осеменения, а второе для откладыва-
ния яиц. Эта форма дала начало двум
крупным ветвям. Одна мало изменив-
шаяся ветвь дала начало большому чис-
лу семейств мелких и очень мелких бабо-
чек, таких, как Tineidae (настоящие моли)
и Tortricidae (листовертки), а также ряду
в высшей степени своеобразных се-
мейств, например Psychidae (мешоч-
ницы, или чехлоноски) и Aegeriidae (сте-
клянницы).
В другой ветви атрофировались неко-
торые щетинки на теле гусениц и максил-
лярные щупики куколки сделались нераз-
личимыми. Наиболее примитивными
представителями данной ветви являют-
ся, возможно, Pyralidae (огневки) и более
редкие Pyromorphidae, у которых хоро-
шо сохранилась пликальная жилка (Р) за-
днего крыла. Специализированными
представителями, по-видимому, следует
считать более крупные формы ночных
бабочек, у которых Р на заднем крыле ре-
дуцировалась и стала неразличимой. От
ствола крупных ночных бабочек про-
изошли крупные семейства Noctuidae,
Arctiidae, Saturniidae и многие другие.
От основания последней ветви отде-
лилась форма, для имаго которой был
характерен дневной образ жизни и рас-
ширенные на вершине антенны. От этой
линии берет начало подотряд Rhopaloce-
га. включающий толстоголовок (Hespe-
riidae), у которых антенны взрослых осо-
бей на вершине крючковидно загнуты, а
такясе многочисленных дневных була-
воусых бабочек (Papilionidae и род-
ственные им группы), имеющих булаво-
видные усики.
ПОДОТРЯД JUGATAE
Jugatae - наиболее примитивные из
всех современных Lepidoptera. Помимо
югума, у взрослых насекомых этого под-
отряда имеются примерно одинаковые
по форме и жилкованию передние и за-
дние крылья. В одной из групп для куко-
лок характерны длинные сильные манди-
булы, используемые для прогрызания
выхода из кокона; ротовые части имаго
включают хорошо развитые мандибулы
и лопастевидные максиллы и нижнюю
губу, которые не вытянуты и не обра-
зуют обычной для бабочек сосательной
трубки. Эти признаки весьма интересны,
так как имеются также у ручейников и
показывают тесные родственные связи
между примитивными представителями
Trichoptera и Lepidoptera.
Jugatae представлены в фауне Север-
ной Америки лишь немногими видами. В
семействе Micropterygidae имаго неболь-
ших размеров и имеют мандибулы, гусе-
ницы питаются мхами. Eriocraniidae так-
же включают мелкие формы, но их
мандибулы рудиментарны и развит сосу-
щий хоботок; личинки-листовые ми-
неры. К Hepialidae относятся несколько
более крупных (до 5 см в размахе крыль-
ев) видов, называемых тонкопрядами
(по-английски их называют за быстрый
полет «стрижами»-swifts), их личинки
живут в древесине или питаются корня-
ми.
ПОДОТРЯД FRENATAE
Сюда относится значительно боль-
шее число чешуекрылых, весьма разно-
416
8. Отряды энтогнат и насекомых
Рис. 8.172. Ковровая моль Trichophaga
tapetzella-oj\m из видов Microlepidoptera.
(По материалам министерства сельского
хозяйства США.)
образных по внешнему виду, размерам и
образу жизни. Кроме отсутствия югума,
взрослое насекомое характеризуется тем,
что его задние крылья отличаются от
передних (рис. 8.172), а число секто-
ральных жилок обычно бывает редуци-
ровано до одной, тогда как на передних
крыльях секторальная жилка, как прави-
ло, имеет три или четыре ветви. Куколка
покрытая, т. е. придатки кажутся вда-
вленными в тело куколки, и она не спо-
собна двигаться.
В Северной Америке представлено 50
семейств этого подотряда, причем мно-
гие из них содержат виды большого эко-
номического значения. Диагнозы и иден-
тификация этих семейств затрудни-
тельны и требуют изготовления спе-
циальных препаратов крыльев после уда-
ления с них чешуек - процесса, известно-
го под названием «оголение».
Microfrenatae. Многие более прими-
тивные семейства Frenatae включают
мелких стройных бабочек, на заднем
крыле которых либо присутствует жилка
Р, либо имеется бахрома из длинных во-
лосков по заднему краю, либо задние
крылья очень узкие и заостренные на вер-
шине. Эту группу семейств часто назы-
вают Microlepidoptera или Microfrenatae.
Некоторые представители Microfrenatae
довольно крупные, так что размеры
являются лишь в среднем характерными
для этой группы. Приводить здесь диаг-
ностические признаки семейств бессмыс-
ленно, но все же ряд семейств, содержа-
щих обычные и важные виды, мы рассмо-
трим ниже.
tineidae, настоящие моли. Гусе-
ницы питаются в основном грибами, тка-
нями или органическими остатками. К
Tineidae относятся наши обычные пла-
тяные моли (рис. 8.173). У платяной мо-
ли Tineola bisselliella гусеница плетет пау-
тину неопределенной конфигурации; по-
мимо тканей этот вид очень охотно
потребляет старые перья. Гусеница шуб-
ной моли Tinea pellionella изготавливает
переносной домик из шелка и обрывков
ткани. Оба вида - обычнейшие разруши-
тели тканей животного происхож-
дения (таких, как шерсть), мехов и
пера.
PSYCHIDAE, МЕШОЧНИЦЫ, ИЛИ ЧЕХЛО-
НОСКИ. В это семейство входят относи-
тельно крупные виды; размах крыльев
некоторых достигает 30 мм. Личинки
строят домикЦ или мешочки из шелка и
кусочков листьев или коры (рис. 8.174).
Из домика наружу высовываются лишь
сильно склеротизированная голова
и грудь. Часто можно видеть, как такие
домики свисают с веток и листьев раз-
личных хвойных и лиственных пород де-
ревьев. Обычной мешочницей в во-
сточных штатах является Thyridopteryx
ephemeraeformis (рис. 8.175). У самцов
пушистое темноокрашенное тело, гре-
бенчатые усики и обычно прозрачные
крылья. Самки похожи на личинок и по-
чти лишены способности к передвиже-
нию. Цикл развития имеет ряд инте-
ресных особенностей. К концу лета гусе-
ница достигает окончательных размеров,
после чего она прикручивает свой домик
к веточке и окукливается внутри домика.
Созревшая куколка самца частично вы-
лезает из домика наружу и из нее выхо-
Рис. 8.173. Шубная моль Tinea pellionella. (По Снодграссу.) А. Вскрытый кокон,
показана находящаяся в нем полностью сформированная куколка. Б. Пустая
кукол очная шкурка, торчащая из кокона после вылета имаго. В. Взрослая
бабочка. Г. Гусеница, повреждающая одежду.
Рис. 8.174. Стадии жизненного цикла чехлоноски Thyridopteryx
ephemeraeformis. (По материалам министерства сельского хозяйства США.)
А. Закончившая развитие гусеница. Б. Ее голова. В. Куколка самца.
Г. Куколка самки. Д. Взрослая самка. Е. Взрослый самец.
418
8. Отряды энтогнат и насекомых
Рис. 8.175. А-В. Последовательные стадии
развития гусеницы чехлоноски. Г. Чехлик
самца. Д. Чехлик самки. (По материалам
министерства сельского хозяйства США.)
дат взрослый самец. Куколка самки
остается внутри домика, взрослая самка
сама прокладывает себе путь наружу из
куколочной шкурки и ожидает оплодо-
творения. Самцы летают вокруг, высма-
тривая чехлики, содержащие зрелых
самок, и спариваются с ними посред-
ством вытянутого выдвижного копуля-
тивного аппарата, который может быть
просунут глубоко внутрь домика, где на-
ходится самка. Вскоре после оплодотво-
рения самка откладывает яйца, попросту
давая им падать в свою старую куколоч-
ную шкурку, которая делается наполови-
ну заполненной ими. Закончив откладку
яиц, истощенная самка выползает из до-
мика, падает на землю и погибает. Яйца
сохраняются в течение зимы в покоящем-
ся состоянии, и личинки из них вылу-
пляются следующей весной. Только что
вылупившиеся гусеницы расползаются
из старого домика в разные стороны и
начинают питаться.
GELECHIIDAE, ВЫЕМЧАТОКРЫЛЫЕ МО-
ЛИ. Для гусениц этого семейства харак-
терно широкое разнообразие типов пита-
ния и кормовых растений. Многие во-
сточные виды рода Gnorimoschema де-
лают крупные галлы на стеблях золотар-
ника и близких группах сложноцветных.
гусеницы питаются внутри галлов и, до-
стигнув полного размера, там же окукли-
ваются. Розовый коробочный червь
хлопчатника Pectinophora gossypiella про-
грызает ходы в развивающейся коробоч-
ке хлопчатника и питается семенами.
Этот вид азиатского происхождения рас-
пространился по всем хлопкосеющим
районам мира и стал одним из опасней-
ших вредителей этой культуры. К этому
же семейству относятся виды, которые
минируют иглы сосен и других хвойных
пород и повреждают картофель
(рис. 8.176) и томаты, а также ряд видов,
развивающихся в цобегах или плодах не-
которых древесных пород. Один вид-
зерновая моль Sitotroga cerealella- вреди-
тель запасов зерна, встречается по всему
миру.
AEGERIIDAE, СТЕКЛЯННИЦЫ. Это уз-
кокрылые формы умеренных размеров,
их передние крылья связаны с задними
посредством рядов зацепляющихся друг
за друга шипов, расположенных пример-
но посередине на краях крыльев. У боль-
шинства видов на крыльях имеются чет-
ко ограниченные свободные от чешуек
участки - «окошки». Отсюда и название
бабочек-стеклянницы. Имаго ведут
дневной образ жизни и характеризуются
чрезвычайно быстрым полетом. Многие
имеют на теле и крыльях перевязи пур-
пурного (фиолетового), красного и жел-
того цветов, явно подражая некоторым
видам обычных ос. Личинки пробуравли-
вают стебли, нападая на травянистые
растения, кустарники и деревья. Серьез-
8. Отряды энтогнат и насекомых
419
Рис. 8.176. Представители семейства Gelechiidae. (По материалам министерства
сельского хозяйства США.)
А. Картофельная моль Phthorimaea operculella. Б. Комарик баклажанный Keiferia
inconspicuella.
ное экономическое значение имеют два
вида, персиковая стеклянница Sanninoi-
dea exitiosa (рис. 8.177) и тыквенная сте-
клянница Melittia cucurbitae.
tortricidae, листовертки. Крупней-
шее семейство мелких и средних разме-
ров бабочек, имеющих широкие крылья,
передняя пара которых с прямо обре-
занным или даже вогнутым или выре-
занным наружным краем. Гусеницы пи-
таются орехами и другими плодами,
листьями и стеблями. У многих видов
личинки сооружают гнезда путем скручи-
вания и связывания листьев, отчего они и
получили свое обиходное название - «ли-
стовертки». К этой группе относится
Argyrotaenia velutinana (рис. 8.178), пи-
тающаяся на многих дикорастущих и
культурных деревьях и кустарниках, а
также некоторые другие виды, имеющие
хозяйственное значение* восточная пло-
дожорка Grapholita molesta, питающаяся
на веточках и плодах персиков, слив и
близких к ним плодовых деревьев, побе-
говьюны-ряд видов рода Rhyacionia, по-
вреждающие верхушечные почки сосен, а
Рис. 8.177. Стеклянница персиковая
Sanninoidea exitiosa. (По материалам
министерства сельского хозяйства США.)
А. Самец. Б. Самка.
420
8. Отряды энтогнат и насекомых
Рис. 8.178. Листовертка Argyrotaenia
velutinana. (По материалам министерства
сельского хозяйства США.)
Л, Б. Взрослое насекомое. В. Гусеница. Г
Куколка. Д. Конец брюшка куколки.
также виноградная листовертка Polychro-
sis viteana, которая питается листьями и
плодами винограда.
PYRALIDAE, ОГНЕВКИ. Это обширное И
экономически очень важное семейство
отряда бабочек. В систематическом от-
ношении огневки занимают промежуточ-
ное положение между Microfrenatae и
Macrofrenatae. Сюда относятся бабочки
различных форм и размеров, но большей
частью нежные, нарядные, нередко
имеющие дробный рисунок и окра-
шенные в мягкие тона. Способы питания
гусениц довольно разнообразны • они
развиваются на листьях, плодах и цвет-
ках, прогрызают ходы в стеблях и ветках,
некоторые являются сапрофагами, а ряд
видов-хищниками. Многие плетут во-
круг себя пространную сетку, куда вхо-
дит также их пищевой субстрат и другие
элементы среды обитания, эти сети назы-
вают паутинными гнездами. Примером
подобного вида может служить мотылек
садовый Loxostege rantalis (рис. 8.179). К
семейству огневок относятся некоторые
из наиболее неприятных вредителей сель-
скохозяйственных культур. Кукурузный
мотылек Ostrinia nubilalis и кукурузная
юго-западная огневка Diatraea grandiosel-
la-серьезные вредители кукурузы, ог-
невка Udea rubigalis вредит хризантемам
и прочим тепличным культурам, боль-
шая пчелиная огневка, или вощинная
моль Galleria mellonella, часто наносит
значительный ущерб пчеловодству. Не-
которые виды, в том числе южная амбар-
ная огневка Plodia interpunctella, напа-
дают на запасы зерна и готовые пищевые
продукты, a Ephestia elutella развивается
Рис. 8.179. Мотылек садовый Loxostege rantalis. (По материалам министерства
сельского хозяйства США.)
А. Взрослое насекомое Б. Гусеница.
• Отряды энтогнат и насекомых
421
Рис. 8.180. Стадии жизненного цикла кольчатого коконопряда Malacosoma
americanum. (По материалам министерства сельского хозяйства США.)
А. Кладка яиц. Б. Гусеница. В. Куколка. Г Кокон. Д. Взрослая самка. Е.
Взрослый самец.
j хранящемся табаке и других сушеных
зродуктах растительного происхожде-
гзя. Многие другие виды повреждают
зьмые разнообразные культуры.
Macrofrenatae. В подотряде Frenatae
fv-iee специализированные семейства, к
ггторым относятся в среднем и более
грудные виды, такие, как разноусые ба-
гочки (Heterocera), различные американ-
шяе шелкопряды, а также бражники,
эбычно обозначаются как Macrofrenatae.
них характерны широкие задние
трылья, короткая бахрома и отсутствие
жилки Е. Как и Microfrenatae, комплекс
«крофренат содержит множество се-
ъетств. определение которых для неспе-
циалистов весьма затруднительно.
LASIOCAMPIDAE, КОКОНОПРЯДЫ. К
т-тому небольшому семейству принадле-
жат немногие виды средних размеров,
имеющие мохнатых гусениц и барха-
“астых имаго с толстым телом. Наибо-
лее широкой известностью пользуются
коконопряды из рода Malacosoma
(рис. 8.180). Их гусеницы развиваются на
различных лиственных древесных поро-
дах, в том числе на плодовых, и периоди-
чески появляются в огромных количе-
ствах, объедая листву на тысячах гекта-
ров древесных насаждений.
Представители рода Malacosoma про-
водят зиму на стадии яйца. Гусеницы вы-
ходят из яиц в конце весны; молодые гу-
сеницы, вылупившиеся из одной кладки,
ткут общее паутинное гнездо, обычно во-
круг развилки ветви дерева. Гусеницы
выползают из гнезда, когда отправляют-
ся питаться листьями, причем каждая
особь возвращается назад по шелковой
нити, оставляемой за собой. Затем гусе-
ницы покидают гнездо и окукливаются
поодиночке в защищенном месте под ко-
рой или мусором, где они сооружают ко-
коны. Взрослые насекомые появляются в
конце лета, и после спаривания самки от-
кладывают яйца поясками вокруг тонких
422
8. Отряды энтогнат и насекомых
веточек. Каждая кладка содержит не-
сколько сотен яиц и заключена в оболоч-
ку из отвердевшего секрета, защищаю-
щую их от внешних воздействий.
В годы массового размножения ба-
бочки могут в огромных количествах
прилетать на огни городского освеще-
ния. В одну из летних ночей 1925 г в Эд-
монтоне (пров. Альберта) колоссальное
количество этих бабочек покрыло всю
деловую часть города слоем около 15 см
толщиной; скользкие тела насекомых
полностью прервали движение трамваев
и автомашин и затрудняли ходьбу пеше-
ходов. Под каждым уличным фонарем и
витриной магазина образовались горы
бабочек, высотой достигавшие 60-90 см.
Подавляющее большинство насекомых
погибло от удушения грудами тел своих
товарищей по несчастью. На некоторое
время улицы города наполнились
сильным запахом гниющих бабочек.
sphingidae, бражники. Все бражни-
ки-крупные бабочки, нередко достигаю-
щие в размахе крыльев более 65 мм. Те-
ло у них коренастое и веретеновидное,
часто сходящее на нет к заднему концу.
Когда крылья находятся в расправлен-
ном положении, задний край второй
пары крыльев редко заходит дальше се-
редины длины брюшка, передние крылья
длинные и относительно узкие. Антенны
длинные и просто устроенные, часто
слегка утолщены к вершине. Бабочки
очень быстро летают и питаются некта-
ром. Гусеницы поедают листья; у боль-
шинства форм они несут на восьмом сег-
менте брюшка острый шип и обычно
называются по-английски hornworms,
т. е. «рогатыми червями». Американские
виды питаются главным образом раз-
нообразными травянистыми растения-
ми, листьями винограда и различных де-
ревьев. Немногие имеют и хозяйственное
значение, в частности бражники Manduca
quinquemaculata и Manduca sexta, гусе-
ницы которых развиваются на томатах и
табаке (рис. 8.181).
GEOMETRIDAE, ПЯДЕНИЦЫ. Это ИЗЯ-
щные бабочки с тонкими или гребен-
чатыми антеннами, широкими нежными
крыльями и стройным телом
(рис. 8.182). Гусеницы их широко из-
вестны благодаря замечательной манере
передвижения, состоящей из серии дви-
жений, когда тело гусеницы то образует
замкнутую петлю, то распрямляется. У
гусениц длинное тонкое тело либо с дву-
мя, либо с тремя парами брюшных ног.
расположенных ближе к заднему концу
тела. Когда гусеница ползет, она изги-
бает тело так, что образует высокую уз-
кую петлю, вследствие чего брюшные
ноги оказываются рядом с передними,
затем она отцепляет грудные ноги от
субстрата, выбрасывает тело вперед и
опускается на субстрат во всю длину
своего тела. Далее она вновь выгибает
тело дугой, подтягивая задние ноги к пе-
редним, и вся процедура повторяется.
Пяденицы образуют обширное семейст-
во; их кормовые растения включают
представителей многих семейств и родов.
Некоторые виды относятся к числу обыч-
ных дефолиаторов лиственных и хвой-
ных деревьев. Некоторые виды пядениц
(рис. 8.182) в отдельных местностях
очень сильно повреждают плоды фрук-
товых и декоративных деревьев, у этих
видов нормальные крылатые самцы, но
самки полностью лишены крыльев.
SATURNIIDAE, ПАВЛИНОГЛАЗКИ. К ЭТО-
му семейству принадлежат самые
крупные из североамериканских бабочек
и гусениц. Имаго их бархатистые или
мохнатые, с широкими нарядно окра-
шенными крыльями. Задние крылья без
каких-либо следов френулума, но базаль-
ная часть переднего края расширена и
прилегает снизу к переднему крылу во
время полета, синхронизируя, таким
образом, работу обеих пар. Антенны у
8. Отряды энтогнат и насекомых
423
Рис. 8.181. Бражник Manduca sexta, повреждающий томаты и табак. (По
материалам министерства сельского хозяйства США.) А. Взрослое насекомое.
Б. Гусеница. В. Куколка.
Рис. 8.182. Пяденица Paleacrita vemata. (Из Wellhouse, How insects live.)
А. Самец. Б. Самка. В. Яйца. Г Гусеница. Д. Куколка.
Рис. 8.184. Вредитель клена Anisota rubicunda. (По материалам министерства
сельского хозяйства США.)
А. Взрослая самка и антенна самца. Б. Яйцо с зародышем внутри. В. Кладка
яиц. Г, Д. Гусеницы. Е. Куколка.
S. Отряды энтогнат и насекомых
425
самцов и часто у самок - перистые. Гусе-
ницы питаются листьями; более крупные
из них чрезвычайно прожорливы и унич-
тожают целиком большой лист с неве-
роятной быстротой. Тело гусениц тол-
стое, несет шипастые бугорки или пучки
жестких волосков и может достигать в
длину 100 мм. Полностью выросшие гу-
сеницы плетут большие бурые коконы на
ветвях или побегах низко над уровнем
почвы, окукливаются и в таком состоя-
нии проводят зиму. Обычным на востоке
США видом является сатурния Прометея
Callosamia promethean рис. 8.183), имаго
которой красновато-бурой окраски, а гу-
сеница зеленая, в шипастых бугорках
желтого, красного и синего цветов.
Близкое к предыдущему семейство
Citheroniidae включает, в частности, ба-
бочек Anisota rubicunda, широко распро-
страненных к востоку от Скалистых гор и
имеющих полосатых зеленых гусениц
рис. 8.184), повреждающих клены.
liparidae, волнянки. Это также не-
большое семейство, включающее только
виды средних размеров. В некоторых ро-
дах самки не способны летать и имеют
лишь маленькие зачаточные крылья. К
этому семейству относится несколько ви-
дов, имеющих громадное хозяйственное
значение, в том числе непарный шелко-
пряд Porthetria dispar и златогузка Nyg-
mia phaeorrhoea (рис. 8.185). Как тот, так
и другой виды завезены в США из
Европы и являются опаснейшими вреди-
телями декоративных деревьев на северо-
востоке Соединенных Штатов. Еще один
вид семейства волнянок, развивающийся
на разнообразных лиственных породах,-
Hemerocampa leucostigma (рис. 8.186). Гу-
сеница этой бабочки несет пучки
длинных жгучих волосков на обоих кон-
цах тела и «карандашики» из белых воло-
сков на некоторых срединных сегментах.
Она строит кокон на коре деревьев. Сам-
ка червеобразная; она откладывает боль-
Рис. 8.185. Златогузка Nygmia phaeorrhoea. (По материалам министерства
сельского хозяйства США.)
А. Самка. Б. Самец. В. Гусеница. Г. Она же, увеличено.
426
8. Отряды энтогнат и насекомых
Рис. 8.186. Волнянка Hemerocampa leucostigma. (По материалам
министерства сельского хозяйства США.)
А. Гусеница. Б. Куколка самки. В. Куколка самца. Г, Д. Взрослый
самец. Е. Взрослая самка. Ж. Она же в процессе яйцекладки. 3. Кладка
яиц. И. Коконы самцов. К. Коконы самок с откладывающими яйца
бабочками.
шую кучку яиц, заключенную в пенистую
белую оболочку, составляющую их за-
щитный покров.
NOCTUIDAE (PHALAENIDAE), СОВКИ. Это
самое большое семейство чешуекрылых.
Имаго имеют самые различные раз-
меры, весьма разнообразны также осо-
бенности строения, так что представите-
лей этого семейства можно отличить от
близких групп только по сочетанию не-
скольких критических признаков. Гусе-
ницы питаются в основном листьями или
прокладывают ходы в стеблях или кор-
нях ; в большинстве случаев тело их не не-
сет шипов или заметных выростов. Се-
мейство совок имеет важное экономиче-
8. Отряды энтогнат и насекомых
427
ское значение. Оно содержит множество
видов, гусеницы которых, называемые
подгрызающими совками, нападают на
различные зернобобовые, овощные и
технические культуры. Другими эконо-
мически значимыми видами являются
«походные черви» Pseudaletia unipuncta и
Spodoptera frugiperda (рис. 8.187), вредя-
щие пастбищным злакам, кукурузе и дру-
гим зерновым; хлопковая совка, или ко-
робочный червь Helio this zea, повре-
ждающий хлопчатник, кукурузу, томаты
и другие культуры; совка «ни» Trichoplu-
sia ni, развивающаяся на крестоцветных и
шагающая «петлей», подобно гусеницам
пядениц, но при этом имеющая лишь две
пары брюшных ног на средних сегмен-
тах. Гусеницы всех перечисленных видов
являются серьезными вредителями.
arctiidae, медведицы. Это семей-
ство близкородственно Noctuidae, отли-
чаясь от него главным образом тем, что
взрослые бабочки-медведицы окрашены
обычно в белые или желтые тона, либо
имеют сложный яркий или желтый рису-
нок. У гусениц толстые пучки волосков,
из-за чего их и назвали медведицами.
Эти гусеницы часто попадаются на глаза
в конце лета, когда они быстро ползут
по земле в поисках укромного места
для постройки кокона и окуклива-
ния.
прочие семейства. Существует
большое число других ночных бабочек,
часть из которых имеют броскую внеш-
ность, а другие по внешнему виду
напоминают представителей немногих
перечисленных выше семейств. Чтобы
определить эти группы, студент может
воспользоваться литературой, список ко-
торой приведен в конце раздела по
Lepidoptera.
Рис. 8.187. «Походный червь”-совка Spodoptera frugiperda. (По материалам
министерства сельского хозяйства США.)
А. Кладка яиц. Б. Яйца. В. Взрослое насекомое Г Гусеница. Д. Куколка.
428
8. Отряды энтогнат и насекомых
ПОДОТРЯД RHOPALOCERA
булавоусые и толстоголовки. У пред-
ставителей этого подотряда задние кры-
лья лишены френулума и медиальный
ствол как на передних, так и на задних
крыльях утратился, в результате чего
возникла крупная срединная дискоидаль-
ная ячейка. Большинство видов ярко
окрашены. Имаго ведут дневной образ
жизни и предпочитают солнечную пого-
ду в противоположность сумеречному
или ночному образу жизни большинства
представителей предыдущего подотряда.
В состоянии покоя Rhopalocera держат
крылья прямо вверх над спинной сторо-
ной тела, а не плоско складывают их по-
верх тела, как это делают ночные бабоч-
ки. Подотряд подразделяется на две
четко ограниченные группы: толстого-
ловки и булавоусые бабочки.
Толстоголовки. Это очень быстрые в по-
лете бабочки, способные лететь строго
по прямой подобно осам или бражникам.
Большая часть североамериканских ви-
дов принадлежит к семейству Hesperii-
dae, они окрашены в тусклые тона, при-
чем преобладают желтый и бурый, и
имеют менее 30 мм от вершины одного
крыла до вершины другого. За исключе-
нием немногих видов, гусеницы имеют
большую голову, что подчеркивается и
маленькой шеевидной переднегрудью.
Большинство видов развивается в гнез-
дах, которые гусеницы делают, сшивая
друг с другом несколько листьев кор-
мового растения. Обычнейший на
востоке США вид-толстоголовка Ераг-
gyreus tityrus, питающаяся на различ-
ных бобовых, предпочтительно на
Wisteria.
Булавоусые бабочки. Порхающий полет-
характерная особенность большинства
представителей этой группы. У них очень
низкая частота взмахов крыльев, и по-
этому их полет состоит из чередующихся
подъемов и спусков, что приводит к блу-
ждающей, зигзагообразной траектории.
Группа булавоусых чешуекрылых пред-
ставлена в фауне Северной Америки
семью семействами, содержащими по
большей части хорошо знакомых всем
натуралистам бабочек.
PAPILIONIDAE, ПАРУСНИКИ. У представи-
телей этого семейства (рис. 8.188) край
заднего крыла обычно с рядом выемок, а
жилка заканчивается пальцевидным
отростком, или хвостом. Парусники-
крупные бабочки, многие из них имеют
нарядный рисунок из пятен или разно-
цветных полос; в окраске ряда видов пре-
обладают желтые тона. Гусеницы пи-
таются листьями и могут быть окра-
шены столь же ярко, как и имаго. Их
уникальной чертой является выпячиваю-
щаяся вилкообразная пахучая железа,
или осметерий (рис. 8.188, Б), на спинке
переднегруди. Она выворачивается, ког-
да гусеница потревожена, сама железа
обыкновенно оранжевого цвета и издает
резкий запах.
pieridae, белянки. Это в подавляющем
большинстве случаев белые, желтые или
оранжевые бабочки, причем у некоторых
хорошо развит черный рисунок. Гусе-
ницы густо покрыты жесткими волоска-
ми и выглядят щетинистыми. Завезенная
в Америку репница Pieris гарае
(рис. 8.189), имаго которой белые
с черными отметинами, часто встречает-
ся в центральных и северных штатах. Гу-
сеницы зеленого цвета вредят капусте и
родственным ей растениям. Этот вид
происходит из Европы. Местный вид то-
го же рода, Р protodice или «южная ка-
пустница», является вредителем кресто-
цветных культур в южных штатах. Дру-
гой вид белянок, часто вредящий в от-
дельных местностях,-желтушка (Colias
eury theme), встречающаяся на большей
части Американского континента
и имеющая сильно варьирующую окра-
8. Отряды энтогнат и насекомых
429
Рис. 8.188. Парусник Поликсен Papilio polyxenes. (По материалам министерства сельского
хозяйства США.)
А. Гусеница сбоку. Б. Гусеница, демонстрирующая голову с пахучей железой-вилочкой.
В. Взрослый самец. Г Очертания яйца. Д. Молодая гусеница. Е. Куколка.
Рис. 8.189. Белянка репная Pieris гарае. (По материалам министерства сельского
хозяйства США.)
А. Взрослая самка. Б. Яйцо (вверху-вид сверху; внизу-вид сбоку). В. Гусеница в
естественном положении на листе капусты. Г Подвешенная куколка.
430
8. Отряды энтогнат и насекомых
ску. Гусеницы питаются на люцерне, кле-
вере и некоторых других бобовых.
ПРОЧИЕ БАБОЧКИ. В Северной Амери-
ке живут и многие другие виды була-
воусых, помимо немногих упомянутых
выше. Любое место на континенте обла-
дает своим набором форм, от ярких до
темноокрашенных, многие из которых
локально очень обильны. Более под-
робные сведения, касающиеся их диагно-
стических признаков, кормовых растений
и ареалов, студенты могут получить,
обратившись к перечисленным ниже ра-
ботам.
ЛИТЕРАТУРА
Brock J. R., 1971. A contribution towards an
understanding of the morfology and phylogeny
of the Ditrysian Lepidoptera, J. Nat. Hist., 5,
29-102.
Common I. F.B., 1975. Evolution and classifica-
tion of the Lepidoptera, Annu. Rev. Entomol.,
20, 183-202.
Dominick R. B. (ed.), 1971-to date. Moths of
America north of Mexico, E. W Classey &
R. B. D. Publishers, London. (Выходит от-
дельными выпусками по каждому семейст-
ву.)
Ehrlich Р. R., 1958. The comparative morpho-
logy, phylogeny and higher classification of
butterflies (Lepidoptera. Papilionoidea), Univ.
Kans. Sci. Bull., 39, 305-370.
Ehrlich P. R., Ehrlich A. H., 1961. How to
know the butterflies. W C. Brown, Dubuque.
Iowa.
Holland W J., 1913. The moth book, Double-
day, New York.
Holland W J., 1931. The butterfly book, Doub-
leday, New York.
Howe W H., 1975. The butterflies of North
America, Doubleday, Garden City.
Klots A. W., 1951. A field guide to the butter-
flies, Houghton-Mifflin, Boston.
McDunnough J., 1938-1939. Check list of the
Lepidoptera of Canada and the United States
of America, Pts. I—II, Mem. So. Calif. Acad.
Sci., 1, 1-272, 2, 1-171.
9
ЭВОЛЮЦИОННАЯ ИСТОРИЯ НАСЕКОМЫХ
Насекомые прошли долгий путь эволюции, прежде чем стали такими, какими мы
их видим сегодня. Примитивные предки насекомых существовали уже 350 млн. лет
назад, в девонском периоде, и, вероятно, еще много раньше (рис. 9.1). Эволюционная
история насекомых была тесно связана с историей других животных и растений, по-
степенно осваивавших сушу и открывших для насекомых возможности приспособле-
ния к наземным местообитаниям. В новых экологических условиях у насекомых
мало-помалу выработались морфологические, физиологические и поведенческие
Адаптации, которые позволили им - пожалуй, успешнее, чем любой другой группе на-
земных животных,-занять множество небольших, весьма специализированных эко-
логических ниш. Филогенез современных отрядов насекомых и родственные отноше-
ния между ними сложны и отражают ряд важных и интересных этапов
эволюции. Об этом и пойдет речь в настоящей главе.
СВИДЕТЕЛЬСТВА ЭВОЛЮЦИИ НАСЕКОМЫХ
Существует несколько источников
данных, помогающих нам расшифровать
эволюционную историю насекомых.
Окаменелости. Они дают нам наибо-
лее прямые сведения о прошлой эволю-
ции. Ископаемые находки, хотя и далеко
не полные, доставляют очень ценную ин-
формацию о том, что происходило в
элределенные геологические периоды, и
гем самым помогают раскрыть ряд
важных этапов в филогенезе совре-
менных групп насекомых.
Удивительно, что ископаемые насе-
комые. несмотря на их относительно
малые размеры и хрупкость, обнаружены
в очень многих местах. Чаще всего они
встречаются там, где есть залежи угля
или мелкозернистые осадочные породы,
эбразовавшиеся в спокойных анаэ-
тчэбных (без доступа кислорода) усло-
виях. Вокруг погребенного под осадками
насекомого могут формироваться кон-
креции (рис. 9.2), предохраняющие его
т разрушения. Ископаемых насекомых
находят в наземных и озерных отложе-
ниях сланцев, в слоях вулканического пе-
пла, торфа и других материалов. В боль-
шинстве случаев встречаются не целые
насекомые, а их отдельные части. Наибо-
лее часты находки крыльев, которые слу-
жат основой для идентификации боль-
шинства ископаемых форм. Остатки на-
секомых уже давно находили в пенсиль-
ванских и нижнепермских отложениях
центральных районов США, хорошо из-
вестны также ископаемые фауны насе-
комых из четвертичных озерных отложе-
ний района Скалистых гор.
В ископаемых углях, особенно отно-
сящихся к меловому периоду (120 млн.
лет назад) или более ранних, часто встре-
чаются желваки янтаря (образованные из
затвердевшей смолы хвойных растений),
в которых могут содержаться хорошо со-
хранившиеся целые насекомые (рис. 9.3).
Существует много местонахождений ян-
таря ; наиболее известен раннетретичный
балтийский янтарь из северных районов
КЛАСС ENTOGNATHA
НАДКЛАСС HEXAPODA
КЛАСС INSECTA
ПОДКЛАСС APTERYGOTA
Инфракласс
** Paleoptera"*
Подкласс PTERYGOTA
___________________________________И н ф р а к л ас с_
Плекоптероидные Гемиптеро- Neoptera
Ортоптероидные
отряды
8
КАЙНОЗОЙ Четв. пер.
Трет. пер.
-65 млн.——----—
CQ
о
С
Мел
-135 млн._
Юра
-195 млн.-
___ Триас
—220 млн—-------
Пермь
-280 млн._
Пенсильваний
-320 млн.-
Миссисипий
’345 млн.-
Девон
г400 млн.—
Силур
отряды
of
e §
идные
'отряды
2 8
g о 3
i-Нейроптероидные—►
в отряды св
S св св Я* _ Н св «
s
— co
о
у
Личинка и куколка
Появление \ Складывание крыльев
крыльев
gee al
S3
8
§
е с
тз Si 2
fsi
Рис. 9.1. Филогенетические отношения между отрядами насекомых и находки их
ископаемых форм. Сплошными линиями представлены различные ветви в период после
их первого появления в палеонтологической летописи; прерывистыми
линиями-предполагаемые продолжения этих ветвей в прошлом (по данным
сравнительной морфологии и другим косвенными данным). В таблицу не включен ряд
предварительно описанных, но недостаточно изученных вымерших отрядов.
Млн-миллионов лет назад.
9. Эволюционная история насекомых
433
Рис. 9.2. А. Фотография железорудной почки с отпечатком крыла древней поденки
Lithoneura mirifica (Mazon Creek в шт. Иллинойс). Б. Рисунок жилкования крыла. (По
Carpenter.)
Европы. Другие местонахождения были
найдены в штате Чьяпас на юге Мексики,
в Манитобе и западной Канаде, на Аля-
ске, в Бирме и Сибири. Находки насе-
комых в янтаре известны начиная с ран-
него мела.
Филогенетические отношения. Вто-
рым, не столь прямым источником
данных служит анализ морфологических
сходств и различий современных насе-
комых, позволяющий установить фило-
генетические отношения, которые под-
робно рассматривались в гл. 8. При
таком анализе ищут примитивные черты
434
9. Эволюционная история насекомых
Рис. 9.3. Насекомые из верхнемелового канадского янтаря. (Из McAlpine, Martin, 1969, The
Canadian Entomologist.) А. Муха из сем. Sciadoceridae, которое представлено в наше время
только двумя видами, обитающими в южном Чили, юго-восточной Австралии и Новой
Зеландии. Б. Комар из сем. Chironomidae.
в строении насекомых и пытаются вос-
создать ту логическую последователь-
ность, в которой появлялись новые при-
способления в различных группах насе-
комых. Этот важный подход может силь-
но облегчить интерпретацию иско-
паемых находок.
Географическое распространение. Тре-
тий источник данных-это изучение со-
временного географического распростра-
нения групп насекомых и анализ их
филогенетических взаимоотношений
в связи с их расселением в прошлые эпо-
хи. Для понимания распространения на-
секомых наиболее важны три биогеогра-
фические концепции. Согласно одной из
них, некоторые насекомые расселялись
на большие расстояния, минуя террито-
рии, к условиям которых они не были
приспособлены, и таким образом созда-
вался ряд почти изолированных ареалов.
Крайний случай такого распростране-
ния-фауна островов Тихого океана. Со-
гласно второй концепции, географиче-
ское распространение многих совре-
менных насекомых-это результат разде-
ления некогда обширных ареалов на
более мелкие. Причиной дробления пер-
воначальных континентов и передвиже-
ния их частей в мезозойское и кайнозой-
ское время обычно считают перемещение
литосферных плит и раздвигание дна
океанов. Третья концепция объясняет
распространение большинства совре-
9. Эволюционная история насекомых
435
менных насекомых влиянием экологиче-
ских факторов, таких, как, например, из-
менение климата в четвертичный период,
которое привело к перемещению и изоля-
ции отдельных частей прежних ареалов.
Данные палеогеографии. Изучение
географических изменений в сопоставле-
нии с данными об изменявшемся распро-
странении насекомых в прошлом-еще
один важный источник сведений. Приме-
ром может служить изучение насекомых
пенсильванского периода, проведенное
Дёрденом (Durden, 1974), который обна-
ружил выраженный широтный градиент
разнообразия насекомых по обе стороны
от палеоэкватора той эпохи. Подобные
исследования могут существенно помочь
в понимании происхождения различных
групп насекомых.
ПАЛЕОГЕОГРАФИЯ
В 60-е годы наши представления о
географическом прошлом Земли ко-
ренным образом изменились. Геологи
узнали много нового о структуре и воз-
расте океанов, и в связи с этим появились
две теории, получившие в настоящее вре-
мя широкое признание,-теория раздви-
гания морского дна (спрединга) и теория
тектоники плит. Эти теории позволили
отказаться от прежних представлений о
незыблемости географического положе-
ния земных континентов. До 1965 г. в
большинстве гипотез, касающихся фило-
генетических отношений и расселения на-
секомых, предполагалось, что на протя-
жении большей части геологического
времени относительное расположение
континентов оставалось неизменным.
Новые геологические теории существен-
но изменили эти представления; они
вновь пробудили интерес к старой гипо-
тезе о движении материков, выдвинутой
Вегенером в 1912 г. Новые факты, добы-
тые наукой, дали возможность по-ново-
му использовать географические и био-
логические данные, тщательно со-
бранные Вегенером и другими уче-
ными.
Раздвигание морского диа (спрединг).
Теория спрединга утверждает, что океа-
ническая кора вдоль срединно-океаниче-
ских хребтов Мирового океана очень мо-
лода и формируется путем интрузии
базальтовых магм (рис. 9.4). Возраст по-
Зона спрединга
Базальтовая кора срединно-
Континентальная кора
Уд.в. 2.7
Рис. 9.4. Структура срединно-океанических хребтов в зоне спрединга и их отношение к зонам
субдукции и некоторым типам трансформных разломов. Уд.в.-удельный вес.
436
9. Эволюционная история насекомых
род, слагающих кору, увеличивается в
обе стороны от этих хребтов по мере уда-
ления от них. Наиболее древняя океани-
ческая кора в Атлантическом океане от-
носится к средней юре (150 млн. лет
назад); самая старая океаническая кора
на всем земном шаре находится в запад-
ной части Тихого океана восточнее Фи-
липпин и относится к позднему три-
асу или ранней юре (195-180 млн. лет
назад).
Тектоника плит. Теория тектоники
плит объясняет, как происходит спре-
динг Если возраст океанов не старше
раннего или среднего мезозоя, то что же
произошло со старыми горными порода-
ми, из которых была сложена ранняя
океаническая кора? Согласно теории тек-
тоники плит, кора с верхней частью ман-
тии состоит из 10 больших жестких ли-
тосферных плит и нескольких остатков
других плит меньшего размера
(рис. 9.5). Крупные плиты примыкают
одной из своих сторон к срединно-океа-
ническим хребтам, которые и предста-
вляют собой зоны спрединга. Другие
края литосферных плит находятся или в
зонах субдукции, или у больших транс-
формных разломов. Зона субдукции-это
место, где одна плита надвигается на
другую. Как правило, та плита, в кото-
рой кора состоит из менее плотных по-
род, надвигается сверху на более плот-
ную (базальтовую) океаническую кору
другой плиты. Там, где обе плиты со-
стоят из океанической коры, любая из
Рис. 9.5. Современное расположение основных тектонических плит и типы границ между
ними. У краев плит происходят землетрясения разного характера, в тех местах, где края
плит находятся в области континента, как правило, имеет место активное горообразование.
Показаны зоны субдукции (зубцы обращены в сторону надвигающейся плиты),
срединно-океанические хребты, зоны спрединга и трансформные разломы. (По разным
источникам.)
9. Эволюционная история насекомых
437
Рве. 9.6. Реконструкция географии земли в эпоху палеоцена (60 млн. лет назад). (Из Smith,
Briden, 1977, с изменениями.)
dl может оказаться поддвинутой под
.фугую; на этой стороне в зоне субдук-
жи образуется глубокая впадина океани-
ческого дна. Такие впадины хорошо вы-
ражены, например, в окраинных частях
Тихого океана (рис. 9.5). Там, где стал-
киваются две литосферные плиты, со-
стоящие из континентальной коры, про-
исходит активное горообразование
с поднятием обширных плато, как, на-
пример, в Гималаях и других частях юж-
ной Азии и южной Европы (рис. 9.5).
Палеогеографические реконструкции.
На рис. 9.6-9.8 представлены рекон-
струкции континентов в палеоцене
(60 млн. лет назад), раннем мелу
(120 млн. лет назад) и раннем триасе
(220 млн. лет назад). На реконструкциях
438
9. Эволюционная история насекомых
позднего Пенсильвания1 или ранней пер-
ми (280 млн. лет назад) (рис. 9.9.) можно
видеть, что континенты были тогда по-
вернуты так, что палеоэкватор проходил
от южной Калифорнии через северную
Испанию. В позднем девоне (360 млн.
лет назад) Северная Америка и северо-
западный угол Африки начали двигаться
1 Пенсильваний приблизительно эквива-
лентен верхнему карбону по европейской но-
менклатуре.- Прим. ред.
навстречу друг другу (рис. 9.10) и пол-
ностью соединились в период позднего
Пенсильвания и ранней перми, образовав
единый материк Пангею, включивший
Еврамерику и Гондвану.
ВЫМЕРШИЕ ОТРЯДЫ
НАСЕКОМЫХ
В настоящее время полагают, что в
дополнение к многочисленным совре-
Рис. 9.7. Реконструкция географии земли раннего мела (120 млн. лет назад). (Из Smith,
Briden, 1977, с изменениями.)
9. Эволюционная история насекомых
439
Рис. 9.8. Реконструкция географии земли во время раннего триаса (220 млн. лет назад.) (Из
Smith, Briden, 1977, с изменениями.)
менным отрядам насекомых прежде, су-
дя по ископаемым остаткам, существова-
ло еще 11 или даже больше отрядов
(рис. 9.1). Большинство исчезнувших от-
рядов впервые появились в отложениях,
относящихся к Пенсильванию или ранней
перми, и в основном вымерли уже к кон-
цу пермского периода. Среди этих исчез-
нувших форм были и ранние предки не-
которых современных отрядов насе-
комых. Большинство этих ископаемых
предков морфологически были более
примитивны и менее специализированы,
чем их потомки; поэтому в позднем па-
леозое различия между группами насе-
комых еще не были такими четкими, как
440
9. Эволюционная история насекомых
Рис. 9.9. Реконструкция географии земли в позднепенсильванское и раннепермское время
(280 млн. лет назад). (По Ross, Ross.)
Рис. 9.10. Реконструкция географии земли в раннемиссисипское время (330 млн. лет
назад). (По Ross, Ross.)
Э. Эволюционная история насекомых
441
между соответствующими современны-
ми отрядами. С другой стороны, изуче-
ние поздних палеозойских насекомых яс-
но показывает, что к концу Пенсильвания
ллювные этапы эволюции насекомых
были уже завершены. Это относится к
развитию крыльев, их складыванию,
дифференциации способов питания и вы-
работке жизненных циклов с полным
превращением. Ниже даются краткие
описания тех вымерших отрядов насе-
1омых, которые уже достаточно изучены.
Нх отношение к современным отрядам
показано на рис. 9.1. Мы не включили
ряд других исчезнувших отрядов, ко-
торые были предположительно описаны
тишь по находкам неполных экземпля-
ров.
Подкласс Apterygota-
ермчнобескрылые
Отряд MONURA - однохвостки
Примитивные бескрылые насекомые,
дохранившие тергальные рудименты в
тдбиально-мандибулярной серии го-
ловных придатков (рис. 9.11). Подобно
МюгосогурЫа (или Archeognatha), одно-
хэостки имели 10 полных брюшных сег-
ментов с парными придатками, но у них
гтсутствовали или были лишь рудимен-
тарными церки на 11-м сегменте брюш-
ка- Отряд описан по роду Dasyleptus,
представители которого известны из
верхнего карбона (Франция, штаты Нью-
Мексико и Иллинойс в США) и нижней
перми (СССР, Чехословакия, штаты
Канзас и Оклахома).
Подкласс Pterygota-
крылатые насекомые
ИНФРАКЛАСС PALEOPTERA-
ДРЕВНЕКРЫЛЫЕ
Отряд PALAEODICTYOPTERA
Это большая и весьма разнообразная
группа, широко представленная в Пен-
сильвании и перми. К этому отряду отно-
сятся некоторые из самых крупных иско-
паемых насекомых. Это были массивные
(рис. 9.2,9.12) формы с размахом крыль-
ев нередко до 20 см, а иногда и более
50 см. Голова маленькая с тонкими ан-
теннами, состоящими из многочис-
ленных мелких члеников. Глаза большие
и выпуклые. Ротовые части крупные, ко-
лющего типа-хоботок, приспосо-
бленный для высасывания жидкостей, ве-
роятно, из растений.
Ноги у этих насекомых были коротки-
ми и крепкими, напоминающими ноги
Ephemeroptera. Переднегрудь несла пару
лопастей, чаще сердцевидных, присоеди-
ненных к переднеспинке. Эти лопасти бы-
ли сильно склеротизированы, у многих
форм они были крупными и заходили
9.11. Однохвостка Dasyleptus brongniarti из нижнепермских отложений Кузнецкого
бассейна в СССР. (По Шарову, из Родендорфа, 1962.)
442
9. Эволюционная история насекомых
Рис. 9.12. Stenodictya-представитель отряда Paleodictyoptera из позднепенсильванских
отложений Франции. (Из.Кика1оуа, 1969-1970, Psyche.)
сверху за основания передних крыльев.
Лопасти имели систему жилок, гомоло-
гичную жилкованию крыльев.
Крылья палеодиктиоптер были очень
разнообразны по размерам и форме
(рис. 9.12). Базальные пластинки крыль-
ев такого же типа, как у Protodonata и
Odonata. Крылья обычно с цветными
окантовками и круглыми пятнами, часто
покрытые волосками, особенно вдоль
жилок у основания крыла, по его краю и
на крыловой мембране.
Строение относительно короткого
брюшка этих насекомых такое же, как и у
нимф Ephemeroptera. Церки, состоящие
из большого числа сегментов, примерно
вдвое длиннее брюшка. У самок-длин-
ный крепкий яйцеклад, у самцов-ко-
роткие членистые кластеры и парный
эдеагус. Представители отряда обитали
главным образом в низких влажных тро-
пических и субтропических лесах и боло-
тах и достигали высокой численности.
Отряд MEGASECOPTERA
Отряд этот четко выделяется среди
отрядов насекомых, относящихся к ран-
нему Пенсильванию, особой системой
жилкования и специфической серповид-
ной формой крыльев (рис. 9.13). Размеры
тела от средних до крупных (до 12 см),
тело тонкое с длинными церками, рото-
вой аппарат сосательного типа имел вид
вытянутого хоботка. От Megasecoptera,
вероятно, произошел вымерший отряд
Diaphanopterodea. Megasecoptera и Ра-
laeodictyoptera занимали сходные ме-
стообитания. Находки представителей
отряда позже триаса неизвестны.
9. Эволюционная история насекомых
443
Рис. 9.13. Отряд Megasecoptera. Переднее (А) и заднее (Б) крылья Aspidothorax
triangularis из позднего карбона Европы. (По Carpenter.) В. Pseudohymen angustipennis
из нижнепермских отложений Урала. (Из Родендорфа и др.)
Отряд DIAPHANOPTERODEA
Этот отряд известен только из пен-
сильванских и пермских отложений. Ро-
товой аппарат этих насекомых предста-
вляет собой сосущий хоботок, состоящий
из сильно измененных ротовых частей.
Хотя по жилкованию крыльев Diapha-
□opterodea были в основных чертах
сходны с Palaeodictyoptera и Megasecop-
зета, они могли складывать крылья над
телом. Как они это делали, не вполне яс-
но, но во всяком случае не так, как Neop-
зега. У пермских представителей жилко-
вание было сильно редуцировано.
Отряд PROTODONATA
(или MEGANISOPTERA) - гигантские
стрекозы
Представители этого отряда, жившие
с Пенсильвания по триас, были крупны-
ми, иногда гигантскими насекомыми с
размахом крыльев от 12 до 75 см
(рис. 9.14). Это самые крупные из из-
вестных насекомых. По общему облику
они были очень похожи на стрекоз-с
большими глазами, длинным тонким те-
лом и хорошо развитыми мандибулами.
Жилкование крыльев у них было прими-
тивное - имелись все крупные жилки,
444
9. Эволюционная история насекомых
Рис. 9.14. Представители отряда Protodonata. (Из Родендорфа и др., Л-по
Handlirsch, 1906-1908; К-по Мартыновой.)
A. Meganeura moyni из верхнепенсильванских отложений Франции. Б. Arctotypus
sinuatus из верхнепермских отложений СССР.
кроме субнодальней, а жилки Ри Е были
разделены у оснований. Стрекозы про-
изошли от Protodonata в Пенсильвании.
По всей вероятности, Protodonata были
быстрыми и агрессивными хищниками.
ИНФРАКЛАСС NEOPTERA-
НОВОКРЫЛЫЕ
ОРТОПТЕРОИДНЫЕ ОТРЯДЫ
Из вымерших позднепалеозойских
ортоптероидных насекомых хорошо из-
вестны четыре отряда. Protorthoptera,
Miomoptera, Caloneurodea и Protelytrop-
tera.
Отряд PROTORTHOPTERA
Эти насекомые жили в период от ран-
него Пенсильвания до перми и относятся
к наиболее обычным среди встречаю-
щихся в ископаемых находках. Отряд со-
держит много разнообразных групп, он
самый ранний из известных ископаемых
Neoptera. У Protorthoptera были ко-
жистые передние крылья и увеличенная
анальная область на задних крыльях
(рис. 9.15,Л), а также хорошо развитый
жующий ротовой аппарат
Представители одной из больших
групп - Protoblattoidea - внешне похожи
на тараканов, хотя отличаются от них ря-
дом признаков, в том числе жилкованием
крыльев (рис. 9.15,Ь). Еще одна группа,
жившая в раннем Пенсильвании, сохрани-
ла примитивный архедиктион с семью
анальными жилками на обеих парах
крыльев.
Отряд MIOMOPTERA
Отряд включает мелких и очень мел-
ких насекомых с относительно длинными
крыльями (рис. 9.16). Они еще редки в
раннем Пенсильвании, но обычны в отло-
жениях пермского периода. По жилкова-
нию крыльев сходны с Psocoptera; имели
жующий ротовой аппарат (с мандибула-
ми) и короткие церки.
9. Эволюционная история насекомых
445
Рис. 9.15. Отряд Protorthoptera. ("Из Родендорфа и др.; Л-по Carpenter; К-по
Handlirsch, 1906-1908.)
A. Liomoptenan omatum (подотр. Paraplecoptera) из нижней перми Северной Америки.
Б. Крыло Stenoneura fdyoli (подотр. Protoblattodea) из пенсильванских отложений
западной Европы.
Рис. 9.16. Передние крылья представителей отряда Miomoptera. (По Tillyard, из
Родендорфа и др.)
A. Delopterum incertum из верхнепермских отложений Урала, СССР. Б. Permembria
delicatula из нижнепермских отложений Северной Америки.
Отряд CALONEURODEA
Эти насекомые жили в пенсильван-
ско-пермское время и имели длинные,
почти одинаковые передние и задние
крылья (рис. 9.17), у которых базальные
отрезки жилок М и Си слились, как у не-
которых Protorthoptera. Ротовой аппа-
рат был мандибулярного типа, церки-
чрезвычайно короткие.
Отряд PROTELYTROPTERA
(или PROTOCOLEOPTERA)
Еще один отряд мелких насекомых,
известный из пермских отложений. У ти-
446
9. Эволюционная история насекомых
Рис. 9.17. Отряд Caloneurodea.
A. Paleuthygramma tenuicorne из нижнепермских отложений Урала, СССР. Б. Переднее
крыло Euthygramma parallelum из пермских отложений Архангельской области, СССР.
(Из Родендорфа и др.)
Рис. 9.18. Отряд Protelytroptera. крылья Protelytron permianum из нижнеперских отложений
Северной Америки.
А. Переднее крыло (из Carpenter). Б. Заднее крыло (из Tillyard).
9. Эволюционная история насекомых
447
пичных представителей передние крылья
превращены в хорошо развитые элитры;
задние крылья широкие, с продольными
и поперечными складками в расширен-
ной анальной области (рис. 9.18). У этих
насекомых были короткие толстые ан-
тенны, широкое туловище, сильные ноги
и короткие церки. Судя по жилкованию
крыльев, они, вероятно, были предками
Dermaptera, а образование элитр у них
считают независимым от развития элитр
> Coleoptera.
ПЛЕКОПТЕРОИДНЫЕ ОТРЯДЫ
Отряд PROTOPERLARIA
Этот отряд, также известный только
из пермских отложений, очень близок к
Plecoptera (рис. 9.19). Передние крылья
длиннее задних, у которых пять
анальных жилок располагаются так, как
Як. 9.19. Отряд Protoperlaria. Lemmatophora
гура из нижнепермских отложений Канзаса,
•ЛПА. (Из Riek, 1970, с изменениями; по
carpenter.)
у современных Plecoptera. У некоторых
Protoperlaria была пара рудиментарных
плоских лопастей, отходящих в горизон-
тальном направлении от переднегруди.
Церки длинные; задние ноги толстые,
длиннее передних и средних. По-видимо-
му, от Protoperlaria произошли в доперм-
ское время Plecoptera.
НЕЙРОПТЕРОИДНЫЕ ОТРЯДЫ
Отряд GLOSSELYTRODEA
Эта группа, единственная, вымершая
среди эндоптериготных Neoptera, отно-
сится к пермско-юрскому времени. Жил-
кование крыльев (рис. 9.20) более спе-
циализированное, чем у Neuroptera или
Mecoptera, а передние крылья тоже пре-
вращены в элитры. По многим призна-
кам крылья, голова и грудь у Glosselytro-
dea были сходны с таковыми Neuroptera.
Возможно, что оба этих отряда имели
близкого общего предка в позднем кар-
боне или ранней перми.
СВЯЗИ ИСКОПАЕМЫХ
НАСЕКОМЫХ С СОВРЕМЕННЫМИ
Paleoptera
Находки ископаемых насекомых,
принадлежащих четырем вымершим и
двум современным отрядам Paleoptera,
свидетельствуют о том, что два выжив-
ших отряда-Ephemeroptera (поденки;
рис. 9.21, А и Б) и Odonata (стрекозы)-
это всего лишь остатки древней и очень
разнообразной фауны крылатых насеко-
мых Пенсильвания. Современные отряды
представляют собой две разошедшиеся
ветви Paleoptera. У Ephemeroptera спо-
собность к полету как на преимаги-
нальной, так и имагинальной стадии раз-
вития, а также редукция ротового аппа-
рата у этих стадий, произошедшая в
448
9. Эволюционная история насекомых
Рис. 9.20. Отряд Glosselytrodea.
А. Переднее крыло Archoglossopterum shoricum из пермских слоев Кузнецкого бассейна,
СССР Б. Переднее крыло Jurina marginata из пермских отложений Архангельской области,
СССР (Из Родендорфа и др.)
Рис. 9.21. Ранние представители отряда Ephemeroptera.
A. Triplosoba pulchella из верхнепенсильванских отложений Европы (из Riek, 1970, с
изменениями; по Edmonds, Traver). Б. Protereisma permianum из нижней перми
Северной Америки (из Родендорфа и др., по Tillyard).
послепалеозойское время, говорят об их
давнем отделении от других Paleoptera.
Odonata мало изменились со времени
своего появления, и, возможно, их
нимфы и взрослые особи всегда были
хищниками. По строению ротового ап-
парата Protodonata сходны с активными,
быстрыми стрекозами, но тело у них бы-
ло значительно более массивное. Судя по
облику ископаемых нимф Palaeodictyop-
tera и Megasecoptera, можно предполо-
жить, что они были наземными насе-
9. Эволюционная история насекомых
449
юмыми, а их взрослые формы-расти-
тельноядными. Значение Paleoptera со-
стоит в том, что они стали предками
одной, а возможно, и двух групп насе-
комых, обладающих способностью
складывать крылья над брюшком. Это
обеспечивало крыльям защиту, когда
они не использовались для полета, и зна-
чительно увеличивало подвижность осо-
бей.
\eoptera
Наиболее важной группой, происхо-
дящей от ранних палеоптер, является
группа Neoptera. Крылья этих насекомых
благодаря сложной системе крыловых
сочленованных склеритов могут склады-
ваться на спине вдоль тела
• см. рис. 3.42, 3.46). Совершенно иначе
устроены крыловые сочленения у двух
современных отрядов Paleoptera (см.
рис. 3.45).
ОРТОПТЕРОИДНЫЕ ОТРЯДЫ
Среди ортоптероидных отрядов наи-
более примитивными считаются Dictyop-
iera (тараканы) и Isoptera (термиты), и
жилкование крыльев у этих отрядов в об-
щих чертах сходно. Первый из них был
хорошо представлен в Пенсильвании на-
стоящими тараканами (подотряд Blatta-
па). тогда как самые ранние находки тер-
митов относятся к мелу. В крыльях
Protorthoptera из пенсильванских и перм-
ских отложений обнаружены вариации в
числе анальных жилок, различия между
передними и задними крыльями, разные
способы складывания крыльев. Наиболее
примитивные Protorthoptera из раннего
Пенсильвания являются самыми древни-
ми из известных крылатых насекомых,
жилкование их крыльев представляло со-
бой типичный архедиктион. Protelytrop-
:ега. по-видимому, имели общего предка
с Dermaptera (уховертками), последние
известны со средней юры.
Orthoptera (саранчовые) и, возможно,
также Phasmatodea (палочники), Mio-
moptera и Caloneurodea представляют со-
бой ветви, рано отделившиеся от одной
или нескольких подгрупп внутри Protort-
hoptera. Phasmatodea известны из триа-
совых отложений Австралии. Представи-
тели Miomoptera и Caloneurodea, первые
находки которых относятся к пенсиль-
ванскому периоду, были уже настолько
специализированы, что их филогенетиче-
ские связи трудно с точностью устано-
вить. Строение тела и крыльев сближает
их с ортоптероидными отрядами. Orth-
optera впервые появились в отложениях
пенсильванского периода, и с тех пор
жилкование крыльев у них изменилось
лишь незначительно.
Еще один ортоптероидный отряд,
представленный в палеонтологической
летописи, это Embioptera (эмбии). Один
род эмбий был найден в пермских отло-
жениях восточной Европы (рис. 9.22).
Остальные относятся к кайнозойской эре
и в основном были найдены в балтий-
ском янтаре; эти формы в большинстве
своем сходны с современными предста-
вителями отряда. Ископаемые формы
Grylloblattodea - весьма специализиро-
ванного отряда-неизвестны.
ПЛЕКОПТЕРОИДНЫЕ ОТРЯДЫ
Есть весьма достоверные находки
Protoperlaria и Plecoptera (веснянки) из
отложений пермского периода. Эти два
отряда насекомых, нимфы которых жили
в воде, представляют собой небольшую
четкую ветвь, отделившуюся от Protort-
hoptera. Веснянки известны из пермских,
юрских, меловых и кайнозойских отло-
жений.
450
9. Эволюционная история насекомых
Рис. 9.22. Отряд Embioptera. Sheimia sojanensis из пермских слоев Архангельской
области, СССР. (По Родендорфу и др.)
ГЕМИПТЕРОИДНЫЕ ОТРЯДЫ
Все ископаемые гемиптероиды
принадлежат к современным отрядам,
среди которых только у Zoroptera и
Phthiraptera (вши) неизвестны иско-
паемые формы. Hemiptera (клопы) и Pso-
coptera (сеноеды) хорошо представлены в
пермских отложениях, а появление Thy-
sanoptera (трипсов) можно с достовер-
ностью отнести к юрскому периоду. Все
пермские Hemiptera (рис. 9.23) принадле-
жали к Homoptera (равнокрылым) и име-
ли хоботок сосущего типа, характерный
для этого подотряда. Вымершие семей-
ства пермских Hemiptera весьма разно-
образны, иногда с длинными яйцеклада-
ми, что указывает на их родство с
некоторыми древними прямокрылыми.
Они отлично приспособились к жидкой
пище и, видимо, постепенно вытеснили
палеодиктиоптер из экологических ниш с
пищей такого рода.
У раннепермских Psocoptera, относя-
4 Эволюционная история насекомых
451
Рве. 9.23. Отряд Hemiptera: Permocicada Integra из пермских отложений Архангельской
□спасти. СССР. (По Родендорфу и др.)
924. Представители отряда Psocoptera.
£ 1>агЬмотит tinctum (подотр. Permopsocida) из нижнепермских отложений Канзаса, США
ж: Carpenter). Б. Zoropsocus tomiensis (подотр. Parapsocida) из пермских отложений
% лти пгой области, СССР. В. Lophioneurodes sarbalensis (подотр. Parapsocida) из нижней
тг тип Кузнецкий бассейн, СССР (Б и В по Родендорфу и др.).
452
9. Эволюционная история насекомых
щихся к подотряду Permopsocida
(рис. 9.24, А), обе пары крыльев были по-
чти одинаковыми, с простым жилкова-
нием. К концу пермского периода по
меньшей мере у двух семейств отряда
Psocoptera уже были редуцированные за-
дние крылья и видоизменено жилкование
передних крыльев (рис. 9.24, Б), эти
формы были удивительно похожи на со-
временных сеноедов. Ископаемых Thysa-
noptera немного, наиболее известны на-
ходки из юрских отложений Средней
Азии. Известны также находки насе-
комых, похожих на трипсов, из верхне-
пермских отложений в Европейской ча-
сти СССР
НЕЙРОПТЕРОИДНЫЕ ОТРЯДЫ
Эти отряды очень хорошо предста-
влены ископаемыми формами. Обе
главные ветви нейроптероидных отрядов
впервые появились в пенсильванском пе-
риоде и были многочисленны в перми.
Примитивные нейроптероиды. Из при-
митивных нейроптероидов (см. рис. 9.1)
отряды Megaloptera (большекрылые),
Neuroptera (сетчатокрылые) (рис. 9.25),
Glosselytrodea и Coleoptera (жуки) появи-
лись в пермском периоде. Появление
Raphidioptera (верблюдки) (рис. 9.26)
можно с достоверностью отнести к юр-
скому периоду. Обилие и видовое раз-
нообразие представителей этих отрядов
были значительно больше в пермском,
триасовом и юрском периодах, чем сей-
час.
Большинство семейств сформирова-
лось еще до окончания юрского периода.
Жуки-единственный отряд из прими-
тивных нейроптероидов, освоивший
множество наземных местообитаний, и в
настоящее время они составляют почти
40% всех известных видов насекомых. У
пермских ископаемых Coleoptera, отно-
сившихся к подотряду Archostemmata,
уже появились признаки жилкования, ха-
рактерные для элитр. Archostemmata до-
минировали среди Coleoptera до юрского
периода, когда появилось большое число
современных подотрядов и семейств.
Высшие нейроптероиды. Сюда отно-
сятся Hymenoptera (осы и пчелы), обра-
зующие независимую ветвь, Mecoptera
(скорпионовые мухи), Siphonaptera (бло-
хи) и Diptera (двукрылые), образующие
вторую ветвь, Trichoptera (ручейники) и
Lepidoptera (бабочки), составляющие
Рис. 9.25. Отряд Neuroptera. Mesypochrysa latipennis из юрских отложений Казахстана,
СССР (Из Родендорфа и др.)
’ Эволюционная история насекомых
453
9.26. Отряд Raphidioptera. Mesoraphidia
ncistigmalis из юрских отложений
<±здхстана, СССР. (Из Родендорфа и др.,
Handlirsch 1906-1908.)
менные семейства. Две группы Eumecop-
tera, составлявшие две четкие самостоя-
тельные ветви на протяжении всего мезо-
зойского периода, дали начало двум
современным ветвям 1) семейству Bitta-
cidae и 2) семействам Panorpidae и Panor-
podidae. Мезозойский подотряд Paratric-
hop tera существовал в триасе, но в кайно-
зое уже не встречался. Пермские и мезо-
зойские Mecoptera были намного более
разнообразны, чем современные Mecop-
tera, и от них произошли два специализи-
рованных отряда - Siphonaptera и Dipte-
ra.
Ископаемых остатков Siphonaptera
(блохи) мало, сюда относятся находки из
нижнемеловых отложений Австралии и
раннечетвертичного янтаря (рис. 9.28) из
Прибалтики. Отсутствие крыльев и вы-
работка адаптаций к паразитическому
образу жизни и кровососанию не дают
возможности четко установить происхо-
ждение блох в пределах группы Mecopte-
ra.
~т^тъю ветвь. Формы, близкие к Trichop-
m и представители Mecoptera известны
ез зермских отложений, что указывает на
те более раннее отделение этих ветвей,
мгжет быть в Пенсильвании. Hymenopte-
-1 впервые появились в триасовых слоях.
В пермском периоде Mecoptera были
*же достаточно дифференцированным
тт*ядом и включали не менее трех под-
г тралов. У одного из них, Protomecopte-
жилкование крыльев было сходно с
жглкованием у ископаемых насекомых,
техожих на Megaloptera и живших при-
мерно в то же время, и у их потомков-
тредставителей семейства Meropeidae,
£ъсае реликтовой ветви. Подотряд Eume-
гсггега представлен в пермских отложе-
stu восемью или более семействами
рас. 9.27), большинство из которых вы-
мерли к концу мезозоя или, претерпев
ильные изменения, образовали совре-
Рис. 9.27. Agetochorista tillyardi-представи-
тель пермских Mecoptera (подотр. Parame-
coptera) с Урала, СССР. (Из Родендорфа
и др., по Handlirsch, 1906-1908.)
454
9. Эволюционная история насекомых
Рис. 9.28. Блоха Palaeopsylla clebsiana из
позднеэоценового балтийского янтаря. (Из
Родендорфа и др.)
В отряде двукрылых больше иско-
паемых форм (рис. 9.29). Крылья,
сходные с крыльями Diptera, находили в
пермских отложениях Австралии и, воз-
можно, в триасовых отложениях СССР
Двукрылые, очень близкие к совре-
менным, известны из ранней юры, где
были широко представлены Nematocera
и ряд семейств Brachycera. В эоцене и в
более ранних слоях встречались предста-
вители большинства семейств двукры-
лых. Представители семейств высших
мух из группы Schizophora появляются в
средне- и верхнекайнозойских отложе-
ниях.
Hymenoptera (пилильщики, муравьи,
пчелы, осы) очень часто встречаются сре-
ди ископаемых насекомых мезозоя и кай-
нозоя. Наиболее древние из них, прими-
тивные пилильщики из семейства Xyeli-
dae (подотряд Symphyta), известны из
нижнего триаса Центральной Азии, верх-
него триаса Австралии и юрских отложе-
ний в Азии (рис. 9.30, А). Представители
подотряда Apocrita впервые обнаружены
в юре, а паразитические Ichneumonoidea
Рис. 9.29. Двукрылое Macrochile spectrum из позднеэоценового балтийского янтаря.
(Из Родендорфа и др.)
т Эволюционная история насекомых
455
Рис. 9.30. Отряд Hymenoptera. A. Anaxyela gracilis (подотр. Symphyta из юрских
отложений в Казахстане, СССР). Б. Microtypus longicornis (подотр. Apocrita, надсем.
Ichneumonida) из эоценового балтийского янтаря; В. Cryptoserphus pinorus (сем.
Proctotrupidae) из эоценового балтийского янтаря. (Из Родендорфа и др.)
>рис. 9.30, Б) и Chalcidoidea хорошо
представлены в мелу и более поздних
сдоях (рис. 9.30, В). Sphecoidea известны
яз нижнемеловых отложений Австралии.
Fonnicidae (муравьи) (рис. 9.31) и Vespi-
dae (осы) были многочисленны в начале
кайнозоя, a Apoidea (пчелы)-в олигоце-
не. По-видимому, последовательность
появления различных ископаемых форм
Hymenoptera отражает некоторые этапы
эволюции этого отряда.
Trichoptera (ручейники) и Lepidoptera
< бабочки) представляют собой еще одну
сильно продвинувшуюся в своей эволю-
ции группу нейроптероидных насекомых.
Trichoptera или их непосредственные
предшественники известны из ранне-
пермских отложений Северной Америки
и из позднепермских отложений Австра-
лии. По характеру жилкования крыльев
они были сходны с древними Mecoptera,
что указывает на их общее происхожде-
ние. Пермские ручейники были промежу-
точным этапом на пути к мезозойским
представителям этого отряда, у них были
редуцированы поперечные жилки
и терминальные разветвления. В верхнем
триасе Австралии были найдены пред-
ставители семейства Mesopsychidae, а в
верхнем триасе и нижней юре северо-
западной Европы-представители семей-
ства Necrotaulidae. В этих семействах до
456
9. Эволюционная история насекомых
Рис. 9.31. Древнейший из известных муравьев Sphecomyrma freyi из верхнемеловых
отложений Нью-Джерси, США. (Из Wilson, Carpenter, Brown, фото F. М. Carpenter.)
сих пор сохранились самостоятельные
основания у жилок М и Си^ У более по-
здних представителей отряда основания
этих двух жилок слиты. Большая часть
других ископаемых находок ручейников
относится к меловому периоду и олиго-
цену. В ископаемых находках Trichoptera
довольно часто сохраняются домики ли-
чинок, конструкция которых сложилась,
по-видймому, уже к триасовому периоду.
Lepidoptera-отряд, хорошо всем из-
вестный из-за огромного разнообразия
крупных, ярко окрашенных форм,-эво-
люционно очень молод и относительно
скудно представлен в ископаемых наход-
ках. Известно несколько примитивных
семейств из меловых отложений, однако
большинство ископаемых бабочек-из
эоцена и олигоцена северной Европы
(балтийский янтарь), а несколько-из
олигоценовых отложений Северной Аме-
рики. Подотряд Jugatae (рис. 9.32, А)
был уже хорошо представлен к началу
кайнозоя. По жилкованию крыльев его
представители сходны с некоторыми ру-
чейниками и, вероятно, произошли от ру-
чейников в среднемезозойское время. У
более высокоразвитых Frenatae
(рис. 9.32, Б), известных из балтийского
янтаря, передние и задние крылья уже не
были одинаковыми. Самый прогрес-
сивный подотряд Rhopalocera вообще не
имеет ископаемых форм.
9. Эволюционная история насекомых
457
Рис. 9.32. Отряд Lepidoptera.
A. Electrocrania immensipalpa (подотр. Jugatae) Б. Glendotricha olgae (подотр. Frenatae);
оба экземпляра из эоценового балтийского янтаря. (Из Родендорфа и др.)
НАСЕКОМЫЕ И ИСТОРИЯ
ЖИЗНИ НА ЗЕМЛЕ
Pf пп жизнь и освоение наземных
честообнтаний
Наземные растения и животные впер-
вые появились в геологической летописи
в середине палеозойской эры, всего лишь
около 400 млн. лет назад. До этого в
течение долгого времени (3,5-4 млрд,
лет) живые организмы обитали в воде
рее. 9.33). Появление всех основных
стволов животных относится к кембрию
а ордовику, в частности, в эти ранне-
залеозойские периоды возникли и многие
группы морских животных, сохранив-
шиеся до сих пор. Относительно назем-
эой жизни в эти периоды палеонтоло-
гические данные чрезвычайно скудны.
В позднем силуре (410 млн. лет назад)
уже существовали наземные растения.
Еще до окончания этого периода по-
явились Bryophyta (мхи), а в начале девон-
ского периода мы уже находим достаточ-
ное обилие и многообразие болотных
псилофитов. Помимо этих древних рас-
тений известны нижнедевонские находки
(из риниевых сланцев Шотландии) мел-
ких ракообразных, паукообразных и но-
гохвосток (Rhyniella praecursor, рис. 9.34).
В нижнедевонских слоях близ Аль-
кена в долине реки Мозель (ФРГ)
обнаружен ряд представителей фауны и
флоры пресных, солоноватых и морских
вод, отложившихся на дне лагуны.
К среднедевонскому времени (375 млн.
лет назад) многие влажные болота на
низменностях уже заросли древовидны-
ми папоротниками. Это были деревья
от средних до крупных размеров, не-
которые имели ствол толщиной до по-
луметра. Хотя находки ископаемых насе-
комых и не связаны непосредственно с
этими среднедевонскими лесами, однако
высота и густота последних указывает на
458
9. Эволюционная история насекомых
Кайнозой
Мезозой
Палеозой
2 _ s s
Hi
Дышащие автотрофы
н Фото
§ синтез
II Покрытосеменные
-/Л (“цветущие" растения)
* Цикадофиты, хвойные,
Ч гинкговые
Папоротники, семенные
папоротники, кордаиты и
родственные группы
Псилопсиды, сфенопсиды
Позвоночные ликопсиды
Растения
В атмосфере начинает
накапливаться О2
Полосчатые
железорудные
формации
с
Рис. 9.33. Эволюция жизни на земле и ее влияние на состав земной атмосферы.
Рис. 9.34. Rhyniella praecursor- девонское
ископаемое, относящееся к отряду
Collembola. (По Aubert.)
то, что в это время для обитания насе-
комых уже быстро создавались сложные
многоярусные стации, при освоении ко-
торых способность к полету давала боль-
шие преимущества.
Древние крылатые насекомые
Миссисипский и пенсильванский пе-
риоды1. В верхнедевонских и миссисип-
ских отложениях ископаемых насекомых
до сих пор найдено не было. Однако бы-
страя эволюция амфибий с разделением
1 Примерно соответствуют нижнему и
верхнему карбону по европейской номенкла-
туре.- Прим. ред.
9. Эволюционная история насекомых
459
Рис. 9.35. Угольное болото пенсильванского периода. (Из R. Zallinger, репродукция
1975 г с фрески «Век рептилий», Peabody Museum of Natural History, Йельский
университет.)
их на четыре отряда в течение миссисип-
ского периода могла быть связана с ис-
пользованием в качестве пищи пресно-
водных членистоногих, в особенности
.личинок насекомых.
Мелководные континентальные
шельфы были покрыты широкими реч-
ными дельтами, где росли густые леса
рис. 9.35), состоявшие из псилофитов,
древовидных плаунов, хвощей и род-
ственных форм, семенных папоротников
и ранних форм хвойных (кордаитов и
древних Conifer ales). Эти леса были ис-
точником образования крупнейших зале-
жей каменного угля, а также многих от-
ложений, богатых ископаемыми остатка-
ми насекомых. После середины пенсиль-
ванского периода в бассейнах горных
озер возникло много болотистых лесов.
Фауна насекомых в пенсильванский
период состояла не менее чем из 12 отря-
дов (см. рис. 9.1); судя по отдельным
фрагментарным находкам, были еще и
другие отряды. Известно около 35 значи-
тельных мест нахождения ископаемых, и
собранный там материал показывает,
что Paleoptera, в особенности представи-
тели отрядов Paleodictyoptera, Megase-
coptera и Diaphanoptera, достигали высо-
кой численности и разнообразия. Весьма
многообразны были и Neoptera, особен-
но многочислен был отряд Protorthopte-
га. Отряды Miomoptera, Caloneurodea и
Orthoptera тоже отличались большим
разнообразием форм. Другую большую
группу пенсильванских насекомых, вклю-
чавшую более двух десятков семейств,
составляли тараканоподобные насе-
комые, в том числе подотряд Protoblat-
toidea (Protorthoptera). Из-за обилия на-
ходок этих форм пенсильванский период
называют «веком тараканов». С точки
зрения палеогеографии представляет ин-
терес постепенное продвижение мест на-
ходок пенсильванских насекомых к севе-
ру и югу от первоначального их распо-
ложения в виде узкого экваториального
пояса.
Пермский период. Суперконтинент
Пангея простирался от северных по-
лярных областей через умеренные, суб-
тропические и тропические зоны до юж-
ной полярной области. Пермские болота
оставили после себя много угольных
пластов и не менее 75 мест круп-
ных находок ископаемых насекомых.
460
9. Эволюционная история насекомых
Рис. 9.36. Фотография ископаемого насекомого Dunbaria fasciipennis из нижнепермских
отложений Канзаса, США. (Из Mavor, General Biology, с разрешения Macmillan Со.)
Почти все пенсильванские отряды на-
секомых перешли в пермские слои, и раз-
нообразие их форм продолжало возра-
стать (рис. 9.36); кроме того, появилось
не менее 10 новых отрядов, которые
стали доминировать в находках (см.
рис. 9.1). Были хорошо представлены
Psocoptera (сеноеды), Hemiptera (клопы),
Mecoptera (скорпионовые мухи) и Со-
leoptera (жуки). Впервые во время перм-
ского периода появляются настоящие
Odonata (стрекозы), Protelytroptera, Pro-
toperlaria, Plecoptera (веснянки), Ne-
uroptera (сетчатокрылые), Glosselytrodea,
Trichoptera (ручейники) и, возможно, Me-
galoptera (большекрылые). Многие из
этих древних отрядов к концу периода
вымерли, но 10 или большее число отря-
дов образовали исходные ветви, от ко-
торых произошли современные отряды.
В пермском периоде быстро развива-
лись также две другие группы наземных
организмов-рептилии и высшие расте-
ния. Рептилии, прявившиеся лишь в кон-
це пенсильванского периода, в перми бы-
стро достигли расцвета. Среди них были
травоядные, хищные и много насеко-
моядных форм. Увеличивалось и разно-
образие растений. Во влажных стациях
доминировали споровые и семенные па-
поротники, однако многие древние кони-
ферофиты заселяли все более и более
сухие местообитания.
Увеличение разнообразия
наземных сообществ
Триасовый период. В триасе происхо-
дили большие изменения в условиях жиз-
ни на суше, и это отражалось в эволю-
ционной истории насекомых. Большие
области суперконтинента Пангеи стано-
вились теплыми и сухими и покрывались
красными аллювиальными равнинами.
Приспосабливаясь к этим новым усло-
виям, быстро изменялась раститель-
ность. Кониферофиты были представ-
A
Б
Рис. 9.37. Личиночные ходы: Л-жука из семейства Buprestidae
(златки); К-жука-короеда в окаменевших деревьях триасового
периода в Petrified Forest National Monument, шт. Аризона. (По
Walker.)
462
9. Эволюционная история насекомых
лены многими формами Cordaitales, Co-
niferales и Taxales. В теплых умеренных
зонах широко распространялись Gink-
goales и, возможно, Gnetales, а в тропи-
ческих и субтропических областях - Суса-
deoidales и цветущие Cycadales. Все эти
изменения в условиях жизни привели к
важным эволюционным изменениям и у
насекомых.
Мест, где в триасовых отложениях
были найдены ископаемые насекомые,
значительно меньше, чем мест с находка-
ми из более ранних или более поздних
отложений. Известно около десятка та-
ких мест, в основном в Австралии и Цен-
тральной Азии, но только в некоторых из
них оказалось много хорошо сохранив-
шихся насекомых. По меньшей мере де-
вять отрядов, появившихся в пермский
период, не пережили триас. В триасовых
отложениях впервые появились прими-
тивные Hymenoptera (пилильщики),
Phasmatodea (палочники) и Diptera
(двукрылые), однако высокая степень
дифференциации этих групп, вероятно,
указывает на более раннее возникнове-
ние. Личиночные ходы, столь типичные
для современных жуков из семейств Ви-
prestidae и Scolytidae (рис. 9.37), до сих
пор сохранились в окаменевших деревьях
из триасовых слоев.
Обновление фауны насекомых
Юрский период. В юре Пангея остава-
лась почти без изменений. Вдоль запад-
ного края Северной Америки-от Мекси-
ки до Аляски-протянулась в основном в
меридиональном направлении горная си-
стема. Наносы и ветры, которые дули с
этих гор, а также трансгрессии и регрес-
сии внутренних морей сильно влияли
на климат Северной Америки.
В юрский период быстрыми темпами
продолжалась эволюция высших расте-
ний. В отложениях, относящихся к концу
этого периода, встречаются пыльцевые
зерна, сходные с пыльцой двудольных
покрытосеменных. Семенные папоротни-
ки были в это время редкими и не пере-
жили юру. Кониферофиты, особенно
Ginkgoales и Cycadales, были широко
распространены.
Насекомые юрского периода из-
вестны из нескольких хорошо изученных
источников, к которым относятся ниж-
неюрские отложения северо-западной
Европы, несколько мест со среднеюрски-
ми отложениями вблизи гор Каратау в
Азии и несколько в Иркутской области в
Азиатской части СССР. В этих юрских
фаунах появилось много «современных»
новых подотрядов и надсемейств в отря-
дах Neuroptera, Mecoptera и Diptera. Раз-
нообразная флора предоставляла насе-
комым обильную пищу. У некоторых
растений уже были хорошо заметные
цветки, и вполне возможно, что насе-
комые посещали и опыляли их. Впервые
в юре появились Dermaptera (уховертки)
и настоящие Thysanoptera (трипсы), и в
целом фауна насекомых этого периода
выглядела вполне современной. То, что в
юрском периоде уже были млекопитаю-
щие, летающие рептилии и птицы, позво-
ляет предполагать, что и Phthiraptera
(вши), и Siphonaptera (блохи) уже могли в
то время паразитировать на соответ-
ствующих хозяевах.
Современная эволюция насекомых
и цветковых растений
Меловой период. Продолжительность
мелового периода составляет 60-70 млн.
лет. В течение этого времени бассейн Се-
веро-Атлантического океана расширял-
ся, постепенно отделяя южные районы
Европы от Северной Америки и обе эти
области от северной Африки и северной
части Южной Америки (см. рис. 9.7.).
Бассейн Южно-Атлантического океана
9. Эволюционная история насекомых
463
Рис. 9.38. Ландшафт мелового периода. (Из R. Zallinger, репродукция 1975 г. с фрески
«Век рептилий», Peabody Museum of Natural History, Йельский университет.)
стал в это время расширяться, так
что Африка и Южная Америка начали
отделяться друг от друга примерно с се-
тклинь! мелового периода. На протяже-
нии всего периода между Северной Аме-
рикой и Европой существовала матери-
ковая связь через северную Европу,
гпельф Баренцева моря и Гренландию.
Высокие горные цепи Кордильер по-
прежнему оказывали сильное влияние на
климат прилежащих областей.
В этот период (рис. 9.38) быстро эво-
люционировали покрытосеменные расте-
ния. особенно двудольные. К концу пе-
риода мы находим уже почти все совре-
менные семейства двудольных. По-
являются осоки и злаки. Это обогащение
фауны цветковых растений, многие из ко-
торых опылялись насекомыми, сопро-
вождалось увеличением числа групп на-
секомых, использующих эти растения в
качестве источника пищи; возник, в част-
ности, ряд новых семейств бабочек, пи-
тающихся пыльцой и нектаром. Иско-
паемых насекомых из меловых отложе-
ний (рис. 9.39) известно не так много, но
зсе же такие находки обнаружены в ряде
мест Аляски, Канады, Сибири и Средне-
го Востока. Фауна насекомых была до-
вольно разнообразна и включала весня-
нок, стрекоз, тараканов, мелких двукры-
лых, тлей, ручейников, паразитических
перепончатокрылых, термитов и низшие
семейства бабочек. Многие из них были
либо близкими родственниками совре-
менных родов, либо представителями
этих родов; другие принадлежали к бо-
лее примитивным родам. Особенный ин-
терес представляют те семейства юрских
насекомых, которые были промежу-
точными звеньями между современными
семействами. В качестве примера можно
привести ископаемое семейство Jascopi-
dae, известное по одной нимфе из канад-
ского мелового янтаря; оно заполняет
промежуток между семействами Cercopi-
dae и Cicadellidae (из Homoptera), пред-
ставляя собой раннюю стадию в эволю-
ции Cicadellidae от церкопидоподобного
предка.
Становление современных
фаунистических областей
Кайнозойская эра. Несмотря на не-
большую продолжительность кайнозой-
464
? 9. Эволюционная история насекомых
Рис. 9.39. Ископаемые насекомые из мелового янтаря (озеро Сидар, пров. Манитоба,
Канада). (Из McAlpine, Martin, 1969, The Beaver; рисунки К. A. Hamilton, Agriculture
Canada.)
Л.Хальцида (сем. Mymaridae, отр. Hymenoptera). Б. Мелкий хищный клоп (сем.
Anthocoridae, отр. Hemiptera). В. Жук (сем. Scydmaenidae, отр. Coleoptera). Г Личинка
жука-хищника (сем. Staphylinidae).
ской эры (60-70 млн. лет), ископаемые
свидетельства об энтомофауне этого
времени наиболее полны.
Изменения климата. Современная
физическая картина земной поверхности
сформировалась в основном в кайнозой-
скую эру. Происшедшее в кайнозое похо-
лодание и сильные колебания темпера-
туры, последовавшие в четвертичном пе-
риоде, оказали большое влияние и на
современное распространение насе-
комых. Эти события приводили к часто-
му и значительному расширению и суже-
нию областей с определенными условия-
ми среды, к смещению климатических
поясов к экватору или от него; в мире на-
секомых они вызывали многократное
образование изолированных и релик-
товых ареалов. Хотя подобные экстре-
мальные климатические изменения мо-
гли происходить и раньше, в пенсильван-
ское время и в начале пермского периода,
в мезозое столь сильных колебаний кли-
мата не было.
Географические изменения. В кайно-
зойскую эру продолжалось также отда-
ление друг от друга и усиление изоляции
большинства континентов. Северная
Америка и северная часть Европы все
еще раздвигались, и к середине эоцена су-
хопутная связь между ними была прерва-
на (см. рис. 9.6). В позднеэоценовое и
олигоценовое время по мере продвиже-
ния на север континентального блока,
9. Эволюционная история насекомых
465
включающего Африку, Индию и Сред-
ний Восток, теплые прибрежные равнины
по южным окраинам Центральной
Европы, Центральной Азии и Китая под-
вергались деформации. Ко времени мио-
цена Гималайская горная система про-
стиралась почти без разрывов от Европы
до южной Азии, и к плиоцену эти горы
достигли огромной высоты. С северной
стороны этой горной цепи образовался
ряд обширных впадин, и на месте многих
из них возникли озера неморского проис-
хождения (остатками этих озер являются
Черное, Каспийское и Азовское моря).
В наше время Гималаи представляют
собой климатический и физический
барьер между Палеарктикой и Восточ-
ной зоогеографической областью (см.
рис. 8.2).
Австралия и Новая Гвинея тоже дви-
гались на север, заняв положение, близ-
кое к границе с Восточной зоогеографи-
ческой областью. Резкие контрасты кли-
матических и природных условий в соче-
тании с относительно недавним передви-
жением материков ограничивали взаимо-
проникновение животных и растений Во-
сточной и Австралийской областей.
С юрского периода вплоть до начала
плиоцена Северная и Южная Америки
были разделены. От Африки и Антарк-
тиды Южная Америка отделилась в кон-
це мелового периода. Панамский пере-
шеек образовался, по-видимому, в сере-
дине плиоцена, т. е. 4-6 млн. лет назад.
Хотя обмен животными и растениями
через этот узкий перешеек шел весьма
интенсивно, оба американских континен-
та сохранили ряд четких биотических
особенностей. Наибольшие изменения
произошли с млекопитающими, северо-
американские плацентарные формы в
значительной мере вытеснили сумчатых
и плацентарных млекопитающих Южной
Америки.
Северная Америка и северные районы
30-732
Азии на протяжении почти всей кайно-
зойской эры неоднократно имели связь в
области Берингова пролива, и в резуль-
тате фауны неарктической и палеарктиче-
ской частей Голарктики стали очень
сходны между собой (рис. 8.2). В Север-
ной Америке значительное увеличение
высоты Кордильер и Скалистых гор,
происходившее в миоцене, плиоцене и че-
твертичном периоде, привело к разделе-
нию Провинции бассейнов и хребтов (в
юго-западной части США) на мелкие,
экологически обособленные участки.
Провинция бассейнов и хребтов, Ска-
листые горы и Великие Равнины были
подняты на 2-4 км в виде широкого ре-
гионального свода, а последующая эро-
зия придала этому горному ландшафту
современный облик. Это поднятие созда-
ло «дождевую тень», что привело к рас-
пространению луговых сообществ на Ве-
ликие Равнины. Горы стали холоднее, и
на них стало выпадать больше атмос-
ферных осадков.
Среди ископаемых насекомых из
верхнеэоценового и нижнеолигоценового
балтийского янтаря есть представители
нескольких отрядов и подотрядов, ко-
торые не встречались в более ранних от-
ложениях. Мantodea, Embioptera и спе-
циализированные паразитические жуки
Strepsiptera (веерокрылые). Давно из-
вестны находки олигоценовых насе-
комых из северной Европы (Германия и
Франция), Северной Америки, Централь-
ной Азии и с Кавказа, они послужили ос-
новой для ряда классических исследова-
ний. Миоценовые находки известны из
северной Европы и с Северного Кавказа,
а также из пустыни Мохаве в Калифор-
нии. Плиоценовых находок значительно
меньше.
Большинство ископаемых насекомых,
найденных в балтийском янтаре, принад-
лежат к современным родам. Некоторые
семейства высших Hymenoptera и Lepi-
466
9. Эволюционная история насекомых
Рис. 9.40. Сетчатокрылое Lithosmylus columbianus (сем. Osmylidae, отр. Neuroptera) из
олигоценовых глинистых сланцев, Флориссант, шт. Колорадо. (Фото предоставлено
F. М. Carpenter.)
doptera в коллекциях насекомых из бал-
тийского янтаря отсутствуют и, вероят-
но, появились в эоцене. Недалеко от
Флориссанта (штат Колорадо) в тонких
слоях вулканического пепла, осевшего в
водоемах у подножия старых Скалистых
гор, сохранилось много ископаемых на-
секомых из олигоцена. Эти насекомые
жили в самых разнообразных экологиче-
ских условиях, от тропических до умерен-
но-холодных, и их фауна не уступала по
многообразию родов современной фау-
не. Некоторые из этих родов отсут-
ствуют теперь в Северной Америке, но
встречаются в других местах - например,
мухи цеце (род Glossina) известны сейчас
только в Экваториальной Африке. Среди
олигоценовых находок из Флориссанта
есть интересные златоглазки из семей-
ства Osmylidae (рис. 9.40), а современные
представители этого семейства живут
только в тропических областях.
Ледниковые эпохи четвертичного пе-
риода (плейстоцен). После плиоцена во
всем мире более прохладные эпохи неод-
нократно чередовались с чуть более
теплыми. Во многих районах, особенно в
Антарктике и в высоких северных широ-
тах Америки и Евразии, росли и распро-
странялись льды, которые затем таяли. С
севера льды продвигались на юг и по-
крывали обширные территории
(рис. 9.41). Известно четыре главных
оледенения, и после каждого из них на-
9. Эволюционная история насекомых
467
ступал долгий период с климатом более
теплым, чем в настоящее время. Послед-
ние два из этих оледенений, а может быть
и предыдущие, включали несколько ци-
клов с чередованием ледниковых и меж-
ледниковых периодов. Сейчас мы нахо-
димся в регрессивной фазе последнего
ледникового периода, и лишь остатки се-
верной ледяной шапки сохраняются ныне
в виде изолированных ледников или
островных масс, как на Гренландии.
Расселение, дизъюнктивные ареалы и
видообразование. Изменения климата во
время смены ледниковых и межледни-
ковых периодов оказали огромное влия-
ние на живую природу Северной Амери-
ки, особенно в северных районах, отли-
чавшихся умеренным климатом. В то
время как обмен между фаунами Север-
Рис. 9.41. Общие очертания максимального оледенения в четвертичный период на
территории Северной Америки. (Из Illinois Natural History Survey.)
Белой оставлена зона последнего крупного оледенения (Висконсинская ледниковая
:-~оха)1 горизонтальной штриховкой показана дополнительная область, покрытая
льдами во время предшествующих оледенений (Иллинойсской, Канзасской
и Небрасской ледниковых эпох).
468
9. Эволюционная история насекомых
ной Америки и западной Европы отсут-
ствовал, северо-западная часть Северной
Америки и северо-восточная часть Азии
были связаны по суше Беринговым мо-
стом, который мог служить путем для
расселения животных. В результате, ког-
да условия на севере стали такими же,
как в наше время, или даже более теплы-
ми, многие виды насекомых, приспосо-
бленные к умеренно-холодному климату,
расселились по всей Голарктике, распро-
страняясь из Северной Америки в Азию
или в обратном направлении. Во время
следующего похолодания, при новом
увеличении площади ледников ареалы
этих видов расширялись к югу, так как
южные окраины их ареалов становились
холоднее и лучше соответствовали по-
требностям этих насекомых. Когда се-
верные границы ареалов смещались к
югу от Берингова моста, ареал каждого
вида распадался на две части, между ко-
торыми не было генетического обмена.
Если такое разобщение было достаточно
долгим, популяции внутри каждой из
этих частей могли стать самостоя-
тельными видами. Когда ледники таяли
и наступал очередной теплый межледни-
ковый период, ареалы видов перемеща-
лись к северу. При этом вновь происхо-
дило смешивание азиатских и североаме-
риканских видов, многие из которых
были близкородственными, но уже гене-
тически изолированными; в результате
формировалась новая группа голаркти-
ческих видов, а затем весь этот дли-
тельный процесс образования новых ви-
дов под влиянием климатических факто-
ров повторялся снова.
Такой же механизм лежал в основе
видообразования в горных районах в
пределах одного континента. Например,
холодолюбивый род веснянок Allocapnia,
распространение которого ограничено
холодными ручьями восточной части Се-
верной Америки, дал начало большой
группе видов (около 38), этому способ-
ствовали прежде всего волны рассеяния,
связанные с флуктуациями климата, а за-
тем изоляция географически обособив-
шихся популяций родительских видов в
нескольких горных районах-на севере и
юге Аппалачских гор, на плато Камбер-
ленд и в горах Озарк-Уошито (штаты
Миссури, Арканзас и Оклахома). Судя по
имеющимся данным, все или почти все
эти виды возникли, вероятно, в плейсто-
цене, т. е. в последние 3 млн. лет.
Ареал северного холодолюбивого ви-
да ручейников Glossosoma intermedium в
целом сейчас простирается от западной
Европы через Азию на восток до Минне-
соты и Миссури. На протяжении всего
огромного ареала вид поразительно
однороден, и это указывает на то, что он
распространился по всей Голарктике во
время последнего оледенения - возмож-
но, всего лишь 12-30 тыс. лет назад. Од-
нако ареал этого вида разделен на ряд из-
олированных популяций, что создает ус-
ловия для образования новых видов в
будущем.
В юго-западных районах США благо-
даря климатическим изменениям в че-
твертичном периоде возник еще один ме-
ханизм видообразования. Ледниковые
периоды были холодными и дождливы-
ми, а межледниковые-жаркими и сухи-
ми. Целый ряд насекомых в этих райо-
нах, в том числе многие жуки, осы и
пчелы, приспособлен к условиям пусты-
ни. В дождливые эпохи их ареалы были
ограничены небольшими изолирован-
ными слегка приподнятыми пустынны-
ми или полупустынными районами,
расположенными между холодными,
влажными горными хребтами. В это
время такие изолированные, приспосо-
бленные к сухому климату популяции
становились видами. В следующий меж-
ледниковый период сухие жаркие районы
расширялись за счет прохладных
9. Эволюционная история насекомых
469
и влажных, в результате возникавших
контактов происходило расселение и
смешение видов.
Благодаря находкам в торфе и бога-
том органическим веществом иле по-
является все больше данных об иско-
паемых насекомых четвертичного перио-
да. У некоторых насекомых, например у
жуков, части экзоскелета настолько про-
чны, что их можно вымывать из рыхлых
отложений, не повреждая. Недавно были
изучены находки Coleoptera из поздне-
плейстоценовых (висконсинских) отложе-
ний. Большинство этих ископаемых жу-
ков принадлежит к современным видам
жуков из семейств, приспособленных к
умеренному климату и, судя по полу-
ченным данным, эволюционирующих
сравнительно медленно. Перемещения,
сужения и расширения климатических
поясов в четвертичном периоде имели
для насекомых многосторонние послед-
ствия, важнейшими из которых были из-
менения их ареалов и характера расселе-
ния; при этом темпы эволюции не были
во всех группах одинаковыми.
Смешение фаун
Многие представители современной
северо-американской фауны насекомых
возникли, по-видимому, не здесь, а попа-
ли сюда (сами или их предки) из других
мест Превосходный пример - комары
рода Culex (рис. 9.42). Как показано на
рисунке, человек случайно или намерен-
но завез с других континентов большое
число видов этого рода. Такие расселив-
шиеся или завезенные виды вместе с воз-
никшими в самой Северной Америке и
составляют ту смешанную фауну, кото-
рая характерна для современных эколо-
гических сообществ.
Древние реликтовые фауны
Среди современных насекомых, живу-
щих в областях, входивших в древнюю
Гондвану, огромное число форм принад-
лежит к семействам, известным только
из Австралии, Новой Зеландии и Южной
Америки. Некоторые другие встречают-
ся, помимо указанных районов, также в
Южной Африке. Большинство этих насе-
комых приспособлены к южной умерен-
ной зоне, ведут оседлый образ жизни и не
имеют средств для преодоления океани-
ческих пространств. К таким насекомым
относятся, например, нелетающие расти-
тельноядные клопы из семейства Pelori-
diidae (Hemiptera), долгоносики из семей-
ства Belidae, нелетающие сверчки из се-
мейства Cylindrachetidae, три семейства и
одно подсемейство веснянок (Plecoptera).
В этот южный фаунистический комплекс
входят также примитивные скорпио-
новые мухи из семейства Nannochoristi-
dae-единственные дожившие до наших
дней представители отряда Mecoptera,
имеющие холодолюбивых водных личи-
нок. Это семейство известно и из перм-
ских отложений Австралии. Из двукры-
лых представителями этой фауны
являются род Pelecorhynchus, род-
ственный толстоножкам (сем. Bibionidae)
и слепням (сем. Tabanidae), род Tric-
hophthalma (сем. Nemestrinidae), подсе-
мейство Ceratomerinae мух-толкунчиков
(сем. Empididae) и примитивное семей-
ство комаров Blephariceridae. Есть и дру-
гие примеры, указывающие на общность
происхождения этой фауны. Большин-
ство таких видов-дожившие до наших
дней представители древних прими-
тивных ветвей, и это заставляет предпо-
лагать, что когда-то до раскола Гонд-
ваны у них были сплошные обширные
ареалы. В каком-то смысле эти насе-
комые-реликтовые, эндемичные и дизъ-
юнктивные в одно и то же время!
470
9. Эволюционная история насекомых
Рис. 9.42. Многие ветви насекомых, например веснянки рода Allocapnia, возникли в Северной
Америке. Другие роды состоят из видов, которые попали на американский континент в
разное время и из разных мест. На карте показано происхождение многих неарктических
комаров рода Culex. Culex, Melanoconion и Neoculex- подроды этого рода. (С разрешения
редакций “Mosquito News” и Illinois Natural History Survey.)
9. Эволюционная история насекомых
471
ЛИТЕРАТУРА
CarpenterF. М. Adaptations among Paleozoic
insects, Proc. N. Am. Paleont. Conv., 1969, Pt.
I, p. 1236-1251, 1971.
Carpenter F. M. Geological history and evolu-
tion of the insects, Proc. 15th Cong. Entomol.,
Washington, D.C., p. 63-70, 1976.
Coope G. R. Interpretations of Quaternary insect
fossils, Annu. Rev. Entomol., 15, 97-120
(1970).
Сох С. B., Healey I. N., Moore P. D. Biogeo-
graphy • An ecological and evolutionary
approach, Halsted (Wiley), New York, 2nd
ed., 194 pp., 1973.
Dillon L. S. Wisconsin climate and life zones in
North America, Science, 123, 167-176 (1956).
Durden C. J. Pennsylvanian correlations using
blattoid insects, Can. J. Earth Sci., 6,
1159-1177 (1969).
Durden C. J. Biomerization. An ecologic theory
of provincial differentiation. In: Ross C. A.
(Ed.). Paleogeographic provinces and provin-
ciality, Soc. Econ. Paleon. Mineral., Spec.
Pub., 21, 18-54 (1974).
Emerson A. E. Tertiary fossil species of the
Rhinotermitidae (Isoptera), phylogeny of
genera, and reciprocal phylogeny of associated
Flagellata (Protozoa) and the Staphylinidae
(Coleoptera), Bull. Am. Mus. Nat. Hist., 143,
(3), 247-303 (1971).
Handlirsch A. Die Fossilen Insekten und die
Phylogenie der rezenten Formen. W. Engel-
mann, Leipzig, 1430 pp., 1906-1908.
Kukalova J. Revisional study of the Order
Palaeodictyoptera in the Upper Carboniferous
shales of Commentry, France, Pts. I—III,
Psyche, 76, 163-215; 76, 439-486; 77, 1^44
(1969-1970).
Larson S. G. Baltc amber- A palaeobiological
study, Entomonograph. Scand. Sci. Press,
Klampenborg, Vol. 1, p. 1-192, 1978.
Love A., Love D. (Eds.). North Atlantic biota
and their history. Pergamon, Oxford, 430 pp.,
1963.
McAlpine D. K. Insects and continental drift,
Aust. Nat. Mus., 17 (8), 274-278 (1972).
McAlpine J. E, Martin J. E. H. Canadian
amber. Hudson Bay Co., The Beaver, Summer,
p. 28-37, 1969.
Riek E. F. Fossil history. In: Waterhouse D. F.
et al. (Eds.). The insects of Australia. Mel-
bourne Univ. Press, Carlton, Victoria, p.
168-186, 1970, Suppl., 28-29, 1974.
Родендорф Б. Б. (ped.) Членистоногие-тра-
хейные и хелицеровые. Основы палеонтоло-
гии.-М. Изд. АН СССР, 560 с., 1962.
Родендорф Б. Б. (ред.). Юрские насекомые
Кара-Тау.-М. Наука, 252 с., 1968.
Родендорф Б. Б. Историческое развитие дву-
крылых насекомых.-Тр. Палеонтологии,
ин-та, 100,-(1964).
Ross С. A. (Ed.). Paleogeographic provinces and
provinciality, Soc. Econ. Paleont. Mineral.,
Spec. Pub., 21, 1-233 (1974).
Ross C. A. (Ed.). Paleobiogeography. Bench-
mark Papers in Geology 131. Dowden, Hut-
chinson & Ross, Stroudsburg, 429 pp., 1976.
Ross H. H. On the origin and composition of the
nearctic insect fauna, Evolution, 7, 145-158
(1953).
Ross H. H. The evolution and classification of
the mountain caddisflies. Univ, of Illinois
Press, Urbana, 213 pp., 1956.
Scudder S. H. The Tertiary insects of North
America, Rep. U.S. Geol. Surv. of the Terri-
tories (Hayden), 13, 103-116, 624 (1890).
Шаров А. Г. Филогения ортоптероидных
насекомых-Тр. Палеонтологии, ин-та, 118,
1-221 (1968).
Smith A. G., Briden J. С. Mesozoic and Ceno-
zoic paleocontinental maps. Cambridge Univ.
Press, Cambridge and New York, 63 pp., 1977.
Wolfe J. A. A paleobotanical interpretation of
Tertiary climates in the Northern Hemisphere,
Am. Scientist, 66, 694-703 (1978).
Wootton R. J. The evolution of insects in fresh-
water ecosystems. In: Clark R. B., Woot-
ton R. J. (Eds.). Essays in hydrobiology.
Univ, of Exeter Press, Exeter, 69-82, 1072.
Wootton R. J. The fossil record and insect flight.
In. Rainey R. C. (Ed.). Insect flight, Symp. R.
Entomol. Soc London, 7, 236-254 (1976).
10
Экологические аспекты
Высокая численность насекомых, их широкое распространение, необычайное оби-
лие видов, среди которых многие имеют для нас практическое значение,-все это де-
лает очень важным изучение их образа жизни, приспособления к среде, биологии от-
дельных популяций и взаимоотношений между разными популяциями. Эти вопросы
вводят нас в область экологии-науки очень сложной из-за большого числа причинно-
следственных связей, нередко с дополнительными длинными цепями побочных
эффектов.
Экологию можно грубо подразделить на три уровня, каждый из которых имеет
свой собственный круг проблем, задач и трудностей. Первый уровень-это экология
отдельных особей. Сюда входит изучение образа жизни, питания, поведения. В общих
чертах многие из этих вопросов обсуждались в предыдущих главах-в гл. 5 (о жиз-
ненных процессах), в гл. 6 (реакции и поведение) и гл. 7 (жизненные циклы, рост и
размножение).
Второй уровень-изучение экологии популяций, включающее два направления,
очень важных для энтомологов. Одно из них изучает динамику популяций, другое -
взаимодействие популяций разных видов. Оба направления имеют дело со структу-
рой, численностью и относительной стабильностью популяций. Именно на этом
уровне были впервые детально исследованы нарушение равновесия между конкури-
рующими видами насекомых и нарушение равновесия в системе хищник-жертва,
приводящее к катастрофическим вспышкам численности вредных насекомых.
Третий, наиболее интегральный уровень-это изучение сообществ, биомов и их
экосистем. Сообщество включает все организмы, обитающие на одной территории.
Биом объединяет несколько близких сообществ вместе с физическими условиями их
обитания. Если мы будем рассматривать сообщество или биом с точки зрения дина-
мики вещества и энергии (в качественном и количественном аспектах), то речь будет
идти о соответствующей экосистеме.
БИОМЫ И СООБЩЕСТВА
Жизнь в природе распределена между
различными типами взаимосвязанных
ландшафтов, каковы, например, прерии,
леса, пустыни, озера, реки. Тип расти-
тельного покрова-то, что создает
пустыню, прерию, лес и т. д.,-обусло-
влен климатическими факторами темпе-
ратурой, осадками, интенсивностью ис-
парения (рис. 10.1). Леса обычно приуро-
чены к областям с большим количеством
осадков, прерии-к областям с меньшим
их количеством, а пустыни-это террито-
рии, где осадки очень скудны, а испаре-
ние велико. Пресноводные водоемы раз-
личаются по наклону рельефа, количе-
ству осадков и множеству локальных
факторов, таких, как кислотность и соле-
ность почвы, характер фильтрации и сто-
ка, температура. Растительный покров
отражает климат местности, поэтому да-
10. Экологические аспекты
473
°C
1,5
3
6
12
24
Широтные зоны
Полярная
Субполярная
Бореальная
— Холодная
умеренная
Теплая
умеренная
Линия
морозов
Субтропичес
— кая
Т ропичес
кая
500
1000
о
о
125
0,5мм
250
0,2 5 мм
2000
8000
/Сухой лес \
Тундра
(влажная)^(мокрая)
(сухая) (дождевая)
4000
Рис. 10.1. Связь между климатическими факторами (температура, осадки и др.) и типами
растительности. (По данным из Pianka, 1978; Holdridge, 1967.)
же удаленные друг от друга территории с
одинаковым климатом сходны по своему
ландшафту. Основные типы ландшаф-
тов-это наиболее крупные экологиче-
ские подразделения, называемые биома-
ми (рис. 10.2). Каждый биом подразде-
ляется на меньшие участки. Например,
лес имеет краевую зону (опушка) и в нем
могут быть открытые пространства (по-
ляны); в одних местах лес может быть
хорошо дренируемым и высоким, с пре-
обладанием дубов, гикори или сосен, в
других-болотистым и низкорослым, с
ильмами, эвкалиптами, кипарисами и
другими деревьями, нехарактерными для
хорошо дренируемых почв. Каждую из
таких относительно однородных терри-
торий эколог рассматривает как природ-
ную биологическую единицу, которую он
называет сообществом. Каждое сообще-
ство имеет определенный набор видов
животных, сохраняющий свое постоян-
ство из года в год лишь с незначительны-
ми отклонениями. В одинаковых сообще-
ствах обитают, как правило, одни и те же
виды животных. Так, например, дубово-
гикориевые леса в штатах Висконсин,
Индиана, Миссури и Оклахома очень
сходны по видовому составу своей
фауны.
Хотя в сообществах ильмовых или эв-
калиптовых лесов есть довольно много
видов животных, встречающихся в дубо-
во-гикориевых сообществах, наряду с
этим есть много видов, специфичных для
каждого из этих сообществ. Если мы
обратимся к прерии, то увидим, что по
фауне она уже очень сильно отличается
от леса, а лес и прерия уже не имеют
почти ничего общего с водными био-
мами.
При более детальном изучении сооб-
ществ мы замечаем, что виды животных
распределены в них по ярусам. В на-
земных сообществах, например в лесу,
часть животных обитает в почве, часть-
на травянистых растениях и часть - на де-
ревьях. Между различными организма-
474
10. Экологические аспекты
| | Тундра или льды
М Сырой хвойный лес
I . | Бореальный хвойный лес
lllllllllll Лиственный лес умеренной зоны
Горные хвойные биомы
Е I Чапараль и засухоустойчивые леса
I | Луговые биомы
Полупустынные биомы
»,*.| Пустыня
Субтропические эверглейды
Рис. 10.2. Основные области наземных биологических зон (биомов) на территории
Северной Америки. (Из Illinois Natural History Survey.)
ми одного сообщества существует жиз-
ненно необходимая связь, как, например,
между растительноядными животными и
растениями, которые служат им пищей,
или между хищниками и их жертвами.
Все эти упорядоченные связи создают си-
стему зависимостей, объединяющую все
многообразие особей сообщества в еди-
ное биологическое целое.
ЭКОСИСТЕМЫ
Экологи, изучающие экосистемы,
стремятся выяснить, как в результате
10. Экологические аспекты
475
взаимодействия живых компонентов
(т. е. всех организмов) между собой и с
факторами внешней среды создается
биолого-физическая система. В такой си-
стеме рассматриваются потоки пита-
тельных веществ и энергии, проходящие
по пищевым цепям (трофическим уров-
ням), биологическое разнообразие и об-
мен веществами между организмами и
физической средой. Для экосистемы нет
ограничений в размерах (самая крупная
экосистема-это вся биосфера), но для
изучения более удобны экосистемы мень-
шей величины, например участок леса
площадью 1 км2, участок луга площадью
1 м2, какой-нибудь пруд или даже куча
навоза. Экосистемы сильно различаются
по географической протяженности, зани-
маемой площади и внутренней сложно-
сти. Кроме того, границы соседних эко-
систем перекрываются: одна система
переходит в другую, образуя экотоны
(переходные зоны), такие, как берег пру-
да или заливаемая пойма реки.
Биологические компоненты
экосистемы
Во всех экосистемах имеются два
крупных биологических компонента
(рис. 10.3).
1. Автотрофные организмы находят
все необходимое для своей жизнедея-
тельности в окружающей их неживой
природе. В наземных экосистемах это в
основном зеленые растения, использую-
щие с помощью фотосинтеза атмосфер-
ную углекислоту и воду для создания
сложных органических веществ.
2. Гетеротрофные организмы ис-
пользуют источники питания, содержа-
щие сложные органические вещества, и
создают из этих веществ другие сложные
соединения. Гетеротрофов подразде-
ляют на первичных консументов - жи-
вотных, использующих автотрофные ор-
ганизмы непосредственно (травоядные),
и вторичных консументов -животных,
которые охотятся на первичных консу-
ментов или других вторичных консумен-
тов и используют, таким образом, в пи-
щу автотрофные организмы опосредо-
ванно (плотоядные и насекомоядные).
Обычно в экосистеме существует не-
сколько уровней вторичных консумен-
тов.
Деструкторы. Помимо описанных вы-
ше весьма заметных компонентов суще-
ствует еще третий важный биологиче-
ский компонент-деструкторы (рис. 10.3).
Это в основном микроорганизмы-гри-
бы, бактерии, простейшие и мелкие
беспозвоночные (в том числе и многие
насекомые), разлагающие сложные ор-
ганические соединения; продукты их
жизнедеятельности усваиваются расте-
ниями, а также в меньшей степени
гетеротрофами.
Физические компоненты
экосистем
Имеются три типа физических компо-
нентов :
1. Климат. Климатические факторы
- это температура, осадки, смена времен
года и некоторые другие (рис. 10.1).
Взаимодействие различных аспектов
климатического компонента в значитель-
ной мере определяет интенсивность кру-
говорота биогенных элементов (как
функцию температуры) и количество (как
функцию количества осадков) раство-
римых питательных веществ в экосисте-
ме.
2. Неорганические вещества. Это хи-
мические соединения, включенные
в сложные процессы круговорота ве-
ществ (таков, например, круговорот во-
ды; рис. 10.4).
3. Органические соединения. Эти ве-
щества одновременно принадлежат хи-
Энергия солнца
Рис. 10.3. Поток вещества и энергии через пищевые цепи экосистемы.
Рис. 10.4. В круговорот воды (гидрологический цикл) ежегодно вовлечено около 1000 км3
воды. (Из Ehrlich, Ehrlich, Holden, 1977 и Budyko, 1974 с изменениями.)
10. Экологические аспекты
477
мическому и биологическому компонен-
там экосистемы. Их цикл регулируется
климатическими факторами, например
температурой и доступностью воды.
Процессы
Различные компоненты экосистемы
взаимодействуют между собой в ряде
процессов, к которым относятся, поток
энергии, круговорот воды и мине-
ральных (питательных) веществ, пи-
щевые цепи, изменения видового разно-
образия, эволюционные изменения, ин-
дивидуальное развитие, регуляторные
взаимодействия между видами.
Поток энергии. Источником энергии
служит солнечное излучение, лишь не-
большая часть которого (до 5%) исполь-
зуется для связывания углерода в органи-
ческие соединения автотрофными орга-
низмами. Большая часть энергии сол-
нечных лучей (90-95%) расходуется на
испарение воды и поддержание климати-
ческой зональности. Первичные консу-
менты, например гусеницы, поедающие
листья, могут использовать только
10-20% биохимической энергии, запа-
сенной листьями автотрофов. Примерно
такова же (10-20%) эффективность вто-
ричных консументов каждого из после-
дующих уровней. Эти взаимоотношения
можно представить в виде пирамиды
трофических уровней (рис. 10.3).
Круговороты воды и минеральных ве-
ществ. Эти процессы, в которых тоже ис-
пользуется энергия солнечного излуче-
ния, играют важную роль в круговороте
воды на Земле (рис. 10.4) и в создании
многих физико-химических условий
окружающей среды. Кроме того, они в
очень сильной степени влияют на раство-
рение, перенос и отложение двуокиси
углерода и карбонатов, а также на обмен
СО2 между атмосферой и водой.
Пищевые цепи. Трофические уровни в
экосистеме создаются на основе пи-
щевых цепей, при этом насекомые
являются первичными и вторичными
консументами и деструкторами (табл.
10.1). Несколько позже, в разделе,
посвященном аутэкологии, речь пойдет о
том, что насекомым более, чем другим
животным, свойственна чрезвычайно вы-
сокая специализация по отношению к ис-
точникам пищи. Нередко один вид (пер-
вичный консумент) питается на одном
предпочитаемом растении и не может
столь же успешно использовать близко-
родственные виды растений. Более того,
многие представители некоторых групп
цикадок питаются на одном и том же де-
реве, но адаптированы к различным его
частям. Такое тонкое разделение авто-
трофных источников пищи редко встре-
чается у других животных, но чрезвычай-
но широко распространено среди насе-
комых. Отчасти именно поэтому число
видов насекомых так велико. Подобным
же образом, насекомые, являющиеся
вторичными консументами, могут охо-
титься только на одного или нескольких
представителей того же класса. Для пи-
щевых цепей насекомых характерна так-
же смена источника пищи по мере разви-
тия и созревания особи (табл. 10.2). У
многих насекомых личиночные
и взрослые формы используют разные
источники пищи. В некоторых случаях
необходимым условием выживания слу-
жит использование специфических видов
пищи. Взрослым комарам и многим му-
хам для завершения жизненного цикла
необходимо питание кровью позво-
ночных. В свою очередь насекомые со-
ставляют пищу для множества других
животных, в том числе для многих птиц,
полевок, рыб, амфибий, рептилий и мле-
копитающих.
Эволюция. Эволюция - это важный
для экосистемы долговременный про-
цесс, так как на протяжении длительных
478
10. Экологические аспекты
Таблица 10.1 Общая трофическая классификация отрядов насекомых
Первичные консументы (травоядные) Вторичные консументы п (насекомоядные и хищники) Деструкторы
Phasmatodea Orthoptera Grylloblattodea Embioptera Thysanoptera Hemiptera (Homoptera)2 Hemiptera (Heteroptera) з Coleoptera1 2 Ephemeroptera (личинки) Ephemeroptera (личинки) Odonata Isoptera з Dictyoptera (Mantodea) Plecoptera (некоторые) Plecoptera (некоторые) Phthiraptera Psocoptera Hemiptera (Heteroptera)2 Coleoptera1 2 Coleoptera1 2 Hymenoptera (паразитические и запасающие осы)
Hymenoptera (пилильщики и рогохвосты)2 Hymenoptera (запасающие пчелы) Mecoptera1 Lepidoptera1 Diptera1 2 Neuroptera Megaloptera Rhaphidioptera Siphonaptera Diptera1 2 Diptera1 2 Trichoptera
1 Во многих отрядах имеются семейства, использующие пищу определенного типа; в других
семействах личинки и взрослые различаются по характеру питания.
2 На разных стадиях развития потребляют разную пищу. 1
з Характер питания определяется специализацией на низших таксономических уровнях.
Таблица 10.2. Примеры смены источников пищи при переходе от личинки к взрослой
особи у голометаболических насекомых
Пища личинок Пища взрослых
Diptera Комары Водные водоросли и микро- Кровь позвоночных и бес- организмы позвоночных животных
Hymenoptera Наездники Насекомые (паразитирова- Цветочный нектар
Lepidoptera Бабочки ние) Зеленые растения Цветочный нектар
периодов отношения между видами по- относительное постоянство, в рамках ко-
степенно изменяются. У одних видов мо- торого виды претерпевают постепенную
гут выработаться более эффективные совместную эволюцию, однако при этом
способы защиты от врагов, другие могут сохраняется возможность (хотя и ограни-
стать жертвами каких-то иных хищни- ченная) резких изменений. Природные
ков. Большинство экосистем сохраняет экологические сукцессии, каждая фаза ко-
10. Экологические аспекты
479
торых характеризуется определенным
набором животных и растений, в значи-
тельной степени ускоряют восстановле-
ние экосистем после разрушений, вы-
званных пожаром или другими бедствия-
ми.
Видовое разнообразие. Видовое раз-
нообразие экосистем обычно определяет-
ся двумя факторами - временем и геогра-
фическим положением (см. рис. 10.1); эти
факторы в известной степени взаимосвя-
заны. Большое разнообразие видов ха-
рактерно для тропических экосистем,
тогда как в субарктических экосистемах
оно очень невелико. Частично это обус-
ловлено годовыми колебаниями клима-
тических условий и соответственно чис-
лом видов, приспособившихся к смене
резко различающихся сезонов. Кроме то-
го. это еще и эволюционный результат
дивергенции видов, происходившей в те-
чение относительно долгой и непрерыв-
ной геологической истории тропических
экосистем в отличие от короткой и пре-
рывистой истории арктических биомов.
ПОПУЛЯЦИИ
Динамика популяций
Энтомологи установили, что популя-
ции насекомых можно разделить на две
четко различимые группы:
1) многие популяции имеют чрезвы-
чайно стабильные размеры, сохраняю-
щиеся из поколения в поколение,
2) численность других популяций мо-
хет резко колебаться - нарастать за
: дно-два поколения до чрезвычайно вы-
сокого уровня, а затем столь же быстро
додать до очень малой величины.
Стабильность многих популяций бы-
щ хорошо документирована путем от-
>оза насекомых в световые ловушки,
доводившегося в Англии много лет
тсдряд. Число особей одного вида в этих
сборах было удивительно постоянным.
Численность популяции тропической ба-
бочки Heliconius ethilla, учеты которой
проводили в Тринидаде на протяжении
по меньшей мере 26 поколений, оказа-
лась очень стабильной.
В отличие от этого многие другие на-
секомые дают такие вспышки численно-
сти, что становятся серьезнейшими вре-
дителями. В качестве примера часто
упоминают непредсказуемые вспышки
колоссального размножения саранчи и
кольчатого коконопряда, происходив-
шие в отдельные годы.
Эти два типа динамики популяций по-
служили основой для двух почти диаме-
трально противоположных представле-
ний о причинах наблюдаемых колебаний.
Согласно первому из них, по мере роста
популяции возникает все больше эколо-
гических факторов, которые, действуя со-
вместно, снижают интенсивность раз-
множения, доводя ее до уровня, поддер-
живающего стабильную численность ви-
да. Эти процессы называются зависимы-
ми от плотности популяции. Привер-
женцы другой точки зрения утверждают,
что размеры популяций колеблются в
широких пределах, ограниченных прежде
всего изменениями внешних условий (на-
пример, продолжительностью вегета-
ционного периода, сезонными и геогра-
фическими колебаниями количества
осадков) и лишь в конечном счете разме-
рами пищевых ресурсов (рис. 10.5).
Чтобы оценить обе эти точки зрения, сле-
дует рассмотреть структуру и взаимодей-
ствие популяций.
Структура популяций. При изучении
структуры какой-либо популяции насе-
комых полезно прежде всего составить
таблицу выживания для данного вида,
показывающую число особей в каждой
фазе жизненного цикла (см., например,
табл. 10.3). Из приведенной таблицы
можно видеть, что у еловой листовертки-
480
10. Экологические аспекты
Скорость роста популяции
почкоеда изначально имеются огромные
возможности для роста популяции - одна
кладка ее содержит 200 яиц. Это отра-
жает идеальная (intrinsic) скорость роста
популяции. Если бы из всех этих яиц вы-
лупились личинки, достигли зрелости, а
затем каждая взрослая особь в свою оче-
редь отложила такое же число яиц, из ко-
торых тоже вылупились бы личинки и до-
Рис. 10.5. Модели трех различных типов
роста численности и плотности популяций.
Прогиб кривой в том месте, где г-отбор и
К-отбор действуют примерно в равной
степени, является результатом
взаимодействия хищник-жертва. (Из
Southwood, 1977, с изменениями.)
стигли зрелого состояния, то через два
поколения популяция от одной особи-
родоначальницы выросла бы до 40 тыс.
особей. К счастью, существует ряд фак-
торов, не позволяющих популяции ли-
стовертки-почкоеда расти столь быстро.
К числу таких факторов относятся пара-
зиты, болезни, хищники, расселение и
воздействие климатических условий. В
результате обычно лишь 2-3 особи из
кладки достигают зрелости, т. е. уровень
смертности очень высок.
Популяции еловой листовертки-поч-
коеда, как видно из табл. 10.3, характе-
ризуются определенной возрастной
структурой. У многих насекомых личин-
ки не вылупляются точно в одно и то же
время, у других в течение года бывает не-
Таблица 10.3. Выживание еловой листовертки-почкоеда, Грин-Ривер, Нью-Брансуик, Канада1
(Morris, 1957, с изменениями)
Стадия жизненного цикла Число особей Число особей, Причина гибели Гибель на каж- дой стадии, %
в начале каждой стадии погибших до окончания стадии
Яйцо 200 10 20 Паразиты Другие причины 15
Ранняя личинка 170 136 Расселение 80
Поздняя личинка 34 13,6 6,8 10,2 Паразиты Болезни Другие причины 90
Куколка 3,4 0,3 0,5 Паразиты Другие причины 25
Имаго 2,5 0,5 Разные причины 20
1 Выживаемость на протяжении одного поколения-2 особи (1%); смертность на протяжении одного
поколения-198 особей (99%). Соотношение полов 1:1 (равное число особей каждого пола).
10. Экологические аспекты
481
сколько поколений. Популяции таких на-
секомых в сезон активности обычно со-
стоят одновременно из яиц, личинок
разных возрастов, куколок и взрослых
особей.
Важную роль играет также соотноше-
ние полов. У многих насекомых число
самок и самцов примерно одинаково,
тогда как у множества других самки зна-
чительно превосходят самцов по числен-
ности. В ряде случаев, например у обще-
ственных насекомых, самцы составляют
лишь малую долю популяции. У неко-
торых долгоносиков, таких, как люцер-
новый долгоносик и представители рода
Graphognathus, самцы вообще не-
известны.
Среди других характеристик популя-
ций следует отметить их' генетическую
неоднородность. В основе ее лежат раз-
личия в хромосомах, связанные с поли-
морфизмом определенных локусов, с
перестройками хромосом и, что, возмож-
но, важнее всего, со смешением или ре-
комбинацией множества сочетаний раз-
личных генов в последовательных поко-
лениях. Генетические вариации обусло-
вливают определенный размах морфоло-
гической (фенотипическое разнообразие)
и физиологической изменчивости попу-
ляции, что исключительно важно для
естественного отбора, определяющего
эволюционное развитие популяции и все-
го вида в целом.
Изменения внутри популяций. Суще-
ствование резко различающихся типов
динамики популяций, о которых говори-
лось выше, можно объяснить как резуль-
тат различия в регулирующих процессах
(табл. 10.4). Стабильная популяция со-
здается тогда, когда основным факто-
ром, поддерживающим смертность в по-
пуляции на высоком уровне, служит воз-
действие одного или нескольких хищни-
ков, популяции которых тоже весьма
стабильны. Этому способствует также
Таблица 10.4. Некоторые характеристики г- и К-отбора у насекомых на основании
экосистемных и популяционных признаков (по Pianka, 1978, с изменениями)
г-Отбор К-отбор
Климат Изменчивый, непредсказуемый Весьма постоянный, предсказуемый
Жизненный цикл Короткий - несколько недель Быстрое развитие и быстрый рост популяции Раннее размножение, один репро- дуктивный цикл Небольшие размеры тела Потенциально большое число по- томков Смертность катастрофическая, обычно независящая от плот- ности популяции Более длинный - обычно несколько месяцев Более медленное развитие, медлен- ный рост популяции Поздно наступающий или растяну- тый репродуктивный период Более крупные размеры тела Меньшее число более крупных по- томков Смертность более равномерно рас- пределенная, зависящая от плотности
Величина попу- Изменчивая во времени, неравно- Весьма постоянная, равновесная,
ляции весная, обычно значительно ни- же предельной емкости среды, периодические повторные засе- ления близкая к предельной емкости среды, нет необходимости в повторных заселениях
Конкуренция Изменчивая, обычно незначитель- ная Обычно сильно выраженная
4-732
482
10. Экологические аспекты
то, что условия, в которых живет популя-
ция, предсказуемо стабильны. Числен-
ность менее стабильной популяции тоже
регулируется хищниками и отчасти
внешними условиями, но последние ме-
нее предсказуемы и менее стабильны. В
таких менее стабильных популяциях вы-
живаемость более высока и взрослые
формы достигают численности, лимити-
руемой в основном конкуренцией за пи-
щу между отдельными особями. Хотя
численность таких популяций колеблется
в значительных пределах, она в при-
родных условиях редко достигает «эпи-
демического» уровня. Крайне неста-
бильные популяции с огромными коле-
баниями численности нередко избегают
(по крайней мере на некоторое время)
влияния паразитов и ограничивающих
факторов среды и потребляют пищи
больше, чем позволяют обычно исполь-
зуемые пищевые ресурсы. Часто при до-
стижении высокой численности насе-
комые начинают потреблять пищу, кото-
рую они в нормальных условиях не
используют.
Регулирование численности популяций
или сохранение равновесия экосистем (их
кибернетика) является функцией самих
экосистем и связано с взаимодействием
происходящих в них процессов и соста-
вляющих их компонентов. Обычно регу-
ляторами численности служат меха-
низмы обратной связи, которые описы-
ваются в виде математических моделей в
особом разделе экологии - системной
экологии. Простой пример математиче-
ской модели-это взаимодействие между
плотностью популяций хищника
и жертвы. При увеличении плотности по-
пуляции жертвы с некоторым запозда-
нием растет и популяция хищника. Рост
популяции жертвы делает ее более до-
ступной для хищника, и это создает усло-
вия для постепенного роста популяции
хищника. В конце концов достигается та-
кая плотность популяции хищника, что
им уничтожается значительная часть по-
пуляции жертвы. В результате популяция
жертвы быстро сокращается, а это ведет
к большему давлению на нее со стороны
хищника, что вызывает дальнейшее
уменьшение популяции жертвы. Когда
она достигает очень низкого уровня,
хищнику становится трудно находить
жертву, и тогда популяция хищника
резко уменьшается. Популяция жерт-
вы снова получает возможность рас-
ти, и весь цикл повторяется вновь. Таким
образом, численность жертвы регули-
руется ее естественными врагами.
Вспышки численности происходят
тогда, когда популяция какого-либо вида
на короткое время оказывается вне влия-
ния цикла хищник-жертва. В этих случаях
популяция жертвы увеличивается бы-
стрее, чем популяция хищника, и дости-
гает того порога, за которым популяция
хищника уже не может угнаться за нею.
Происходит вспышка численности, или
«эпидемия», вида-жертвы, особи которо-
го уничтожают почти все доступные ис-
точники пищи (рис. 10.5). Ограничиваю-
щим фактором при этом служит продук-
тивная емкость источника корма для
популяции жертвы. Как только потреб-
ности размножающейся популяции пре-
высят эту емкость, достаточное количе-
ство пи!ци смогут раздобыть лишь не-
многие особи, и плотность популяции
начнет быстро снижаться. Такого рода
подъемы численности лимитируются пи-
щевыми ресурсами.
Большую роль в жизни и адаптациях
насекомых играет стабильность внешней
среды. Все животные (и в особенности на-
секомые), населяющие местообитания с
кратковременной стабильностью (напри-
мер, из-за резких сезонных изменений
или непредсказуемой погоды), обычно
имеют небольшие размеры тела, очень
подвижны, обладают высоким репродук-
10. Экологические аспекты
483
тивным потенциалом и коротким време-
нем генерации. Такой тип адаптации на-
зывается r-стратегией. С другой сто-
роны, животные, в том числе и насе-
комые, живущие в очень стабильных
условиях, чаще имеют большие размеры
тела, в большей степени проявляют тер-
риториальное поведение, обладают бо-
лее низким репродуктивным потенциа-
лом и большим временем генерации.
Такой тип адаптации носит название К-
стратегии. Между животными, исполь-
зующими одну из этих стратегий в чи-
стом виде, находится целый спектр про-
межуточных форм-так называемый г-
К-континуум. Можно сравнить плот-
ность различных популяций, входящих в
г-К-континуум (см. рис. 10.5), и их взаи-
модействие в модели хищник-жертва.
Пустынная саранча {Schistocerca gregaria)
и комнатная муха {Musca domesticd) от-
носятся к насекомым с r-стратегией, так
как условия их существования крайне не-
постоянны, у них мало естественных вра-
гов, и распространяются они широко и
быстро. На другом конце спектра нахо-
дится яблонная плодожорка {Cydia рото-
nella) она обычно обитает в стабильных
условиях, развивается медленно, и плот-
ность ее популяций довольно постоянна,
т. е. это определенно организм с К-стра-
тегией. Большинство насекомых распо-
лагаются между этими крайними типа-
ми. Те из них, которые находятся ближе к
типу г (например, тли), легко могут
ускользать из-под регулирующего влия-
ния естественных врагов и размножаться
в катастрофических масштабах.
Во многих случаях на территориях с
большим разнообразием местообитаний
создается большое разнообразие видов
насекомых. Если на такой территории
обитает и много видов-хищников, то воз-
никают сложные взаимоотношения хищ-
ник-жертва. Фауна цикадок (Hemiptera)
на лугах с кислыми почвами близ Силву-
да (Беркшир, южная Англия) представле-
на большим числом видов. При сравни-
тельном изучении энтомофауны двух
местообитаний в этом районе было обна-
ружено, что в одном из них, более разно-
образном по своим условиям, встре-
чаются 63 вида цикадок, в том числе 30
из сем. Cicadellidae, 10 из сем. Delphaci-
dae и 2 из сем. Cercopidae размножаются
здесь постоянно. В другом, более одно-
родном местообитании постоянная фау-
на цикадок включала 9 видов из сем. Ci-
cadellidae и 1 из сем. Delphacidae.
В более разнородном местообитании
численность популяций цикадок находи-
лась под контролем многих видов пара-
зитических ос-дриинид (Dryinidae, Hyme-
noptera) и мух из сем. Pipunculidae
(Diptera) (рис. 10.6, £). В более однород-
ном местообитании паразитических ви-
дов было значительно меньше. Отноше-
ния между несколькими обычными вида-
ми цикадок и паразитирующими на них
видами показаны на рис. 10.6. Явления
паразитизма носили, по-видимому, ло-
кальный характер. В популяциях, обитав-
ших в одном и том же месте на протяже-
нии ряда поколений, отношения хозяин-
паразит или хищник-жертва оставались
довольно постоянными. Они сохраня-
лись даже тогда, когда в период между
двумя поколениями вида-жертвы проис-
ходили изменения в видовом составе па-
разитов и их встречаемости (рис. 10.6,
А-Д). В случаях, представленных на
рис. 10.6 A-В, осы-дрииниды, паразити-
ровавшие на цикадках первого поколе-
ния, принадлежали не к тем видам, ко-
торые паразитировали на цикадках вто-
рого поколения. Кроме того, сама сте-
пень поражения в первом поколении
была больше, чем во втором. Поскольку
некоторые паразиты способны заражать
и уничтожать представителей нескольких
видов хозяев (т. е. являются полифага-
ми), численность популяций нескольких
484
10. Экологические аспекты
Июнь Июль Авг. Сент. Окг. Июнь Июль Авг. Сент. Окт.
1969
1970
10. Экологические аспекты
485
Рис. 10.6. Паразитизм мелких ос-дриинид (Hymenoptera, Dryinidae) и мух (Diptera,
Pipunculidae) на луговых цикадках (Hemiptera, Cicadellidae) близ Силвуда (Беркшир, южная
Англия). (По данным из Waloff, 1975, 1980.)
A-В. Степень заражения (%) осами-дриинидами трех наиболее многочисленных видов
цикадок в более разнородном местообитании с высоким уровнем паразитизма.
A. Psammotettix confinis. Б. Arthaldeus pascuellus. В. Jassargus pseudocellaris. Г P. confinis.
Д. Errastunus ocellaris. А-Д\ 7-первая генерация; 2-вторая генерация. Е. Число взрослых
цикадок Р. confinis по данным еженедельных сборов в более разнородном местообитании за
1967-1970 гг. Кружки-общее число взрослых особей Р. confinis; зачерненные участки
соответствуют доле Р. confinis, зараженных осами-дриинидами; участки, обозначенные
пунктиром,-доле цикадок, зараженных мухами.
видов хозяев и паразитов обычно взаи-
мозависима. У одного вида из сем. Cica-
dellidae в начальный период освоения им
более однородного местообитания зара-
женность паразитами колебалась в более
широких пределах, чем в популяциях то-
го же вида, обитавших в давно ос-
военных, хотя и более разнородных
местообитаниях (рис. 10.6, Г, Д).
В сильно измененных экосистемах, на-
пример в посевах сельскохозяйственных
культур, действие многих естественных
регуляторов численности ограничено. В
результате наблюдаются частые вспыш-
ки вредителей, уничтожающих многие
культуры. Такие вспышки могут иметь
катастрофические последствия, и о них
известно со времен появления письмен-
ности. По мере роста численности насе-
ления земного шара и площади пахотных
земель люди стали изыскивать эффек-
тивные методы борьбы с вредителями,
способные заменить действие есте-
ственных регуляторов, утраченных в ре-
зультате развития сельского хозяйства. В
настоящее время примерно один человек
из шести страдает серьезным заболева-
нием, переносчиками которого служат
насекомые, и один из пяти недоедает в
результате ущерба, наносимого насе-
комыми сельскохозяйственным культу-
рам. Экологический подход к трудным
проблемам восстановления сбалансиро-
ванной динамики популяций насекомых,
нарушенной вмешательством человека,
открывает большие возможности для их
решения.
ЭКОЛОГИЯ ОСОБЕЙ
Каждый вид насекомых приспособлен
к жизни в определенной нише внутри со-
общества. Эта ниша ограничена раз-
личными факторами среды, в частности
погодой и источниками пищи. Предель-
ные значения этих факторов, при ко-
торых вид еще может существовать, ха-
рактеризуют его экологическую толе-
рантность. Последняя неодинакова у
разных видов. Особи любого вида стре-
мятся занять места, где имеются опти-
мальные условия для их жизни. Посколь-
ку экологическая толерантность - это ви-
довой признак, важно иметь дело с точно
определенными видами.
При экологическом исследовании от-
дельных видов насекомых в большинстве
случаев изучают
1) лимитирующие факторы среды,
которые определяют, где может жить
данный вид;
2) взаимоотношения вида с его кон-
курентами или врагами;
3) влияния обоих указанных аспектов
на распределение и численность вида.
Факторы внешней среды
Наиболее важные факторы среды,
влияющие на распространение и числен-
486
10. Экологические аспекты
ность насекомых,-это погода, физиче-
ские и химические условия, пища, враги и
конкуренция.
ПОГОДА
Погода, как одеяло, окутывает все
биологическое сообщество и прямо или
косвенно влияет на все его элементы. По-
года-сложный фактор, и экологически
наиболее важными его компонентами
являются свет, температура, относитель-
ная влажность, осадки и ветер. На со-
стояние популяции влияют не среднего-
довые величины этих компонентов (т. е.
климат), а условия каждого дня. Один
ночной заморозок может, например,
уничтожить значительную часть популя-
ции субтропического вида, даже если
средние температуры данного года до-
статочно высоки. На близких участках
условия могут резко различаться, и это
приведет к большим различиям между
местными популяциями. В холмистом
районе единственный ночной заморозок
может сильно сказаться на обитателях
долин и никак не повлиять на популяцию
насекомых, живущих на гребнях холмов.
Свет. Свет играет чрезвычайно важ-
ную роль в поведении насекомых, и его
значение обсуждалось в разделах, посвя-
щенных таксисам и фотопериодизму.
Многих дневных насекомых удавалось
успешно разводить в течение многих по-
колений либо при искусственном свете,
лишенном многих частей спектра, либо в
полной темноте. Следовательно, свет
влияет на большинство насекомых не
прямо, а опосредованно, через качество
растительной пищи, зависящее от реак-
ции растений на свет
Температура. Для насекомых темпе-
ратура является одним из решающих
факторов. Насекомые - холоднокровные
животные, и, когда они находятся в по-
кое, температура их тела почти не отли-
чается от температуры окружающей
среды. Большинство насекомых неспо-
собно регулировать температуру среды:
вместо этого у каждого вида есть особые
физиологические механизмы, благодаря
которым насекомые могут переносить
крайние температуры, обычные для их
экологической ниши.
Из тех насекомых, которые способны
регулировать температуру своего ме-
стообитания, лучше всего изучена медо-
носная пчела. Пчелы сами устанавли-
вают температуру в своем улье. Летом
они поддерживают ее на уровне около
35°С. Если температура поднимается вы-
ше этого уровня, пчелы создают ток воз-
духа у входа в улей, махая крыльями, как
веерами, и проветривают его, тогда как
другие особи приносят воду и разбрызги-
вают ее на сотах, испарение воды охла-
ждает улей. Зимой необходимая для жиз-
ни температура в улье поддерживается за
счет тепла, выделяющегося при окисле-
нии пищи в организме пчел. Другие виды
общественных пчел и муравьи тоже в из-
вестной степени регулируют температу-
ру своих гнезд.
Воздействие температуры проявляет-
ся двояким образом она влияет на ско-
рость развития и на смертность.
Влияние температуры на развитие.
Температура тела у насекомых непо-
стоянна. При повышении температуры
автоматически возрастает и скорость ме-
таболических реакций, протекающих в
организме, и это заметно влияет на раз-
витие насекомых и на их активность. Но
не все химические реакции, протекающие
в организме, одинаково реагируют на по-
вышение температуры, а некоторые фи-
зические факторы, как, например, рас-
творимость газов в жидкостях, изме-
няются так, что при повышении темпера-
туры создают неблагоприятные условия
для процессов метаболизма. Поэтому
влияние температуры на развитие насе-
10. Экологические аспекты
487
комых неоднозначно. При некоторой
низкой температуре, которую называют
пороговой развитие останавливается; она
может быть на 5-ЗО°С выше нижней ле-
тальной температуры. Так же точно раз-
витие прекращается и при определенной
высокой температуре, которая, однако,
обычно очень близка к летальному уров-
ню.
Между этими двумя точками ско-
рость развития изменяется с изменением
температуры. Однако у разных насе-
комых эта зависимость может быть раз-
личной. Каждому виду свойственна опре-
деленная скорость развития. На рис. 10.7
показано, насколько различаются скоро-
сти развития у четырех видов кобылок.
Кроме того, скорость развития у одного
и того же вида может изменяться при
переходе от одной стадии к другой
(рис. 10.8) при одинаковых температу-
рах яйца и куколки жука Popillia japonica
развиваются быстрее, чем личинки.
Интересный пример неодинаковой
скорости развития представляют яйца и
нимфы красноногой кобылки (рис. 10.7).
Рис. 10.7. Скорость развития четырех видов
кобылок при постоянных температурах в
диапазоне 22-37°С. (По Chapman, 1931,
Animal ecology.)
C-Camnula\ M - Melanoplus.
Рис. 10.8 Сравнение скорости развития
разных фаз жизненного цикла жука Popillia
japonica в зависимости от температуры. (По
Ludwig.)
Самая верхняя кривая-яйцо; следующая
кривая - куколка; 1-3- личинки трех
возрастов.
Скорость развития нимф возрастает с по-
вышением температуры до точки, близ-
кой к верхней летальной температуре.
Скорость развития яиц сначала тоже воз-
растает, но затем, после некоторой точ-
ки, начинает при дальнейшем повыше-
нии температуры уменьшаться; эта тем-
пературная точка лежит намного ниже
летальной. Приведенные примеры по-
казывают, что для получения точных све-
дений о влиянии температуры на разви-
тие того или иного вида необходимо
изучать все стадии этого процесса.
СЕЗОННАЯ КООРДИНАЦИЯ. Скорость
роста растительноядных насекомых или
видов, питающихся кровью холодно-
кровных животных, тесно скоординиро-
вана со скоростью роста хозяев. В ре-
зультате к тому времени, когда состоя-
ние хозяина становится наиболее благо-
приятным для нападения паразита, по-
следний тоже оказывается на подходя-
щей для этого стадии. Посмотрим, как
выглядят отношения хозяина и паразита,
488
10. Экологические аспекты
на примере двух видов перепончатокры-
лых-одного пилильщика (сем.) Tenthre-
dinidae и паразитирующего на нем наезд-
ника (сем. Ichneumodidae). Взрослые пи-
лильщики появляются ранней весной и
откладывают яйца на молодые листья
растения-хозяина. Вылупление личинок
происходит через 1-2 нед., когда расте-
ние находится в фазе активного роста, и
личинки, таким образом, получают бо-
гатый источник пищи. Развитие наездни-
ка протекает медленнее или начинается
при более высокой температуре,
и взрослые особи выводятся через 3-4
нед., после появления взрослых пилиль-
щиков. К этому времени личинки нового
поколения пилильщиков уже заканчи-
вают свой рост и находятся именно на
той стадии, когда наездникам удобно от-
ложить на них свои яйца.
Другим примером могут служить тли
(Aphididae), питающиеся весной на ябло-
не. Скорость развития их перезимовав-
ших яиц такова, что молодые тли выво-
дятся как раз в то время, когда ябло-
невые почки весной начинают раскры-
ваться. Тли тут же начинают питаться на
крошечных листочках, появляющихся из
лопнувших почек.
Это совпадение определенных этапов
в развитии насекомых с определенными
событиями в жизни растений (которые
легко наблюдать) позволяет использо-
вать эти события как временной ориен-
тир при проведении многих мероприятий
по борьбе с вредителями. Таково, напри-
мер, опрыскивание почек для раннего
уничтожения тлей или опрыскивание при
опадении лепестков или появлениии за-
вязей; фазы развития растения исполь-
зуются здесь как критерии развития
насекомого.
Влияние температуры на смерт-
ность. Диапазон температур, в котором
насекомые могут существовать, сильно
варьирует от вида к виду. Наиболее те-
плоустойчивые виды погибают при
47-52°С, для большинства насекомых
верхние летальные температуры соста-
вляют 38-44°С. Виды, обитающие в про-
хладных местах, менее устойчивы к высо-
ким температурам. Так, у обитающего в
горах Grylloblatta (Grylloblattodea) темпе-
ратурный оптимум находится около 3°С,
нормальная активность поддерживается
при температурах от —2 до 16°С, а
тепловое оцепенение наступает около
28°С.
Нижние летальные температуры
столь же различны, как и верхние. Тропи-
ческие насекомые обычно погибают при
температуре, близкой к нулю (рис. 10.9).
Малый мучной хрущак, например, поги-
бает при содержании в течение несколь-
ких недель при 7°С. Многие насекомые
гибнут только тогда, когда температура
опускается на несколько градусов ниже
нуля. Зимующие стадии большинства се-
верных насекомых чрезвычайно устой-
чивы к холоду. Например, зимующая ку-
колка сатурнии Прометея выдерживает
длительное пребывание при — 35°С, а не-
которые насекомые выживают даже при
— 50°С.
Насекомые, обитающие в районах с
морозной зимой, почти всегда обладают
различной температурной устойчи-
востью на разных стадиях развития (это
не относится только к паразитам тепло-
кровных животных). В жизненном цикле
этих насекомых есть по крайней мере од-
на стадия, устойчивая к низким темпера-
турам, и именно на этой стадии насе-
комые переживают зиму. Устойчивой к
холоду фазой может быть яйцо, нимфа,
личинка, куколка или взрослое насеко-
мое. В большинстве случаев только одна
стадия бывает холодоустойчивой, и к на-
ступлению зимы все остальные стадии
погибают. Например, у клопа-черепашки
холодоустойчивы взрослые клопы, и при
сильных зимних морозах только они и
10. Экологическе аспекты
489
Рис. 10.9. Длительность воздействия различных температур от —10 до +5°С,
необходимая для полной гибели яиц, личинок, куколок и имаго табачного жука.
(По Swingle.)
выживают, а все сохранившиеся к этому
времени нимфы гибнут.
Насекомые хорошо приспособлены к
температурам, преобладающим в местах
их обитания. Температура действует как
фактор, ограничивающий активность на-
секомых в необычно жаркую или холод-
ную (для данного места или сезона) пого-
ду. Как правило, необычные темпера-
туры контролируют или изменяют гра-
ницу распространения вида. Обитающий
на юге комар Culex quinquefasciatus в
годы с мягкой зимой может расширять
свой ареал к северу, но при суровых зи-
мах граница его распространения вновь
отступает к югу.
Необычные для данного сезона тем-
пературы, например слишком ранние
или слишком поздние морозы, могут
оказывать такое же влияние, как и экс-
тремальные температуры, поскольку в
этом случае неблагоприятные условия
наступают либо до появления стадии,
устойчивой к этим условиям, либо после
того, как эта стадия перешла в другую,
чувствительную стадию. Например, в се-
верных и центральных штатах США зи-
мующие клопы-черепашки не выживают
при многократном чередовании мороза и
оттепелей. Зима с необычно теплыми пе-
риодами оттепелей, за которыми сле-
дуют периоды с температурой — 20°С
490
10. Экологические аспекты
Декабрь
Декабрь
Рис. 10.10. Температура воздуха и почвы на разной глубине при открытой (Л) и
покрытой снегом (Б) поверхности земли в декабре в штате Монтана. (По Mail.)
или ниже, чрезвычайно губительна для
популяций этих клопов. Воздействию не-
обычных для данного сезона температур
насекомые чаще подвергаются на грани-
цах своего ареала, что отчасти и опреде-
ляет положение этих границ. Время от
времени такие отклонения температуры
могут случаться и в других, внутренних
частях ареала, и тогда они оказывают
влияние на численность вида в этих
районах.
Осадки. Осадки обычно имеют для
насекомых значение не сами по себе, а
лишь как фактор, влияющий на влаж-
ность воздуха и почвы и на растительные
пищевые ресурсы. От снежного покрова
зависит также температура почвы. При
отсутствии снега изменения температуры
воздуха сказываются на глубине до
0,6 м, а снежный покров защищает от ко-
лебаний температуры воздуха даже по-
верхностный слой почвы (рис. 10.10). Та-
ким образом, снег оказывает большое
влияние как на крайние, так и на средние
температурные условия существования
насекомых в почве.
Однако бывают случаи, когда осадки
влияют на насекомых и непосредственно.
10. Экологические аспекты
491
Обильные дожди могут причинять насе-
комым большой физический вред. Выпа-
дение 20 мм осадков в виде тихого и про-
должительного дождя скорее всего не
нанесет никакого ущерба, а внезапный
шквальный ливень может посбивать на
землю и уничтожить большую часть
тлей или молодых нимф клопа-черепаш-
ки. Такую же опасность представляет для
насекомых и град.
Влажность н испарение. Влияние этих
факторов на насекомых трудно оценить
раздельно как в эксперименте, так и в
природе. Влажность - это количество
влаги в воздухе, а испарение - это факти-
ческие потери воды с какой-либо поверх-
ности. Если насекомое поместить в усло-
вия с низкой влажностью, испарение
воды с поверхности его тела увеличится.
Из-за малых размеров тела насекомого
усиленное испарение быстро приводит к
обезвоживанию организма, и насекомо-
му приходится восполнять потерю воды
путем поглощения ее с1 пищей. До того
как насекомое получит такую возмож-
ность, потеря воды определяется влаж-
ностью среды, в которой оно находится.
На рис. 10.11 показана зависимость ис-
парения от влажности у зеленополосой
кобылки в условиях голодания.
Судя по данным, которыми мы рас-
полагаем, влажность обычно имеет важ-
ное, но не столь решающее значение, как
температура, при этом для каждого вида
существует свой оптимум влажности, ко-
торый может быть различным для раз-
ных фаз жизненного цикла. Личинки фа-
солевой зерновки быстрее развиваются
при более высокой влажности, а яйца и
куколки-при более низкой. Во многих
случаях, однако, скорость развития
остается почти постоянной в широком
диапазоне влажности.
Влажность влияет также на уровень
смертности насекомых. У Drosophila низ-
кая влажность повышает смертность, а у
Рис. 10.11. Потеря веса Chortophaga
viridifasciata при различной относительной
влажности (указанной в конце каждой
кривой). (Из Wigglesworth, по Ludwig.) Вес
выражен в процентах к исходному.
некоторых видов тлей высокая влаж-
ность препятствует вылуплению личинок
и линькам. Высокая влажность, по-види-
мому, снижает устойчивость некоторых
насекомых к заражению грибами.
Влажность и испарение принадлежат
к главным факторам, ограничивающим
распространение многих насекомых. В
восточных районах Северной Америки
есть немало видов, ареалы которых про-
стираются к западу примерно до реки
Миссисипи. По-видимому, уменьшение
влажности не позволяет этим видам рас-
селяться в западном направлении
(рис. 10.12). С другой стороны, виды,
обитающие в районе Великих Равнин, не
распространяются далеко на восток, так
как этому, вероятно, препятствует слиш-
ком высокая для них влажность.
Температура и влажность. Сочетание
этих двух факторов оказывает суще-
492
10. Экологические аспекты
Рис. 10.12. Зоны различной влажности на территории США. (Упрощенный вариант по
материалам министерства сельского хозяйства США.)
Средняя влажность,%
Средняя температура,°F
Рис. 10.13. Зоны максимальной
продолжительности жизни взрослых
жуков-усачей Hoplocerambyx spinicomis при
различных комбинациях температуры
и относительной влажности. (Из Linsley, по
Beeson, Bhatia.)
ственное влияние как на развитие, так и
на распространение насекомых. Тот или
другой из этих факторов часто приобре-
тает решающее значение на разных ста-
диях развития и в разное время года
(рис. 10.13). Например, критически низ-
кие температуры зимой могут отрица-
тельно влиять на зимующих зрелых ли-
чинок, а неблагоприятные условия влаж-
ности летом-затруднять развитие яиц
или активно питающихся личинок.
Исследования на мучных хрущаках
рода Tribolium показали, что для выжива-
ния отдельных видов в условиях конку-
ренции с другими видами этого же рода
очень важны определенные комбинации
температуры и влажности. Таким обра-
зом, различия в численности или в разме-
рах ареалов видов-конкурентов могут
определяться не различиями в реакциях
каждого из этих видов непосредственно
10. Экологические аспекты
493
на данные факторы, а результатом трой-
ного взаимодействия-каждый вид взаи-
модействует с двумя названными по-
годными факторами и с видами-конку-
рентами.
Суточный ритм. Чередование дня и
ночи сопровождается суточным ритмом
температуры и влажности, характерным
для каждого района. Если не считать пе-
риода диапаузы, жизнедеятельность
большинства насекомых четко опреде-
ляется этим ритмом. Наиболее яркие
примеры мы встречаем в районах с высо-
кой дневной температурой. В жаркий
летний день, когда влажность низка,
многие насекомые сравнительно мало
активны и занимают более прохладные и
влажные ниши. К концу дня, когда тем-
пература падает, происходит быстрое по-
вышение влажности. В это время множе-
ство насекомых покидает свои дневные
>бежища и появляется в массе на земле, в
листве и в воздухе.
Движение воздуха. Движение воздуха
само по себе мало влияет на жизнедея-
тельность насекомых, но оно тесно связа-
но с влажностью и испарением. Усилен-
ное испарение, вызываемое движением
воздуха, способствует снижению темпе-
ратуры тела. Ветры и термические пото-
ки играют важную роль в расселении на-
секомых. Утренние и вечерние восходя-
щие конвекционные токи воздуха подни-
мают на высоту в несколько сотен мет-
ров самых разнообразных насекомых,
среди них не только множество кры-
латых видов, но и мелкие бескрылые
формы, такие, как ногохвостки (Collem-
bola). Эти поднятые в воздух насекомые
составляют основной корм стрижей и
козодоев.
Известно, что некоторых хорошо ле-
тающих насекомых, например бабочек
рода Erebus, сильные ветры часто уносят
на несколько тысяч километров к северу
от их тропических мест обитания. Живых
особей Erebus изредка находили даже в
Канаде, куда они попадали из Мексики,
Вест-Индии или еще более южных обла-
стей. Перенос многочисленных бабочек
ветром на более короткие расстояния-
весьма обычное явление.
Периодические переселения. Ряд ши-
роко распространенных в северных шта-
тах видов, достигающих каждый год вы-
сокой численности, зимой либо выми-
рает в северной части своего ареала, либо
сохраняется там в очень небольшом ко-
личестве. В самых южных штатах попу-
ляции благополучно перезимовывают,
после чего весной вид снова расселяется в
северные штаты. Таким образом ведут
себя совка Pseudaletia unipuncta, астровая
цикадка Macrosteles fascifrons, картофель-
ная цикадка Empoasca fabae и картофель-
Рис. 10.14. Распространение листоблошки
Коккереля, Paratrioza cockerelli, в США. (По
Wallis.)
Перекрестной штриховкой обозначен район
наибольшего вреда, простой
штриховкой - примерные районы зимовки,
жирными линиями - восточный и западный
пределы распространения вида в летнее
время.
494
10. Экологические аспекты
ная листоблошка Paratrioza cockerelli
(рис. 10.14). Имеющиеся данные указы-
вают на то, что весеннее расселение насе-
комых к северу осуществляется с по-
мощью ветра. Вероятно, ветер периоди-
чески переносит и многие другие виды
насекомых помимо тех, о которых это
уже известно.
ФИЗИЧЕСКИЕ И ХИМИЧЕСКИЕ
ФАКТОРЫ СРЕДЫ
В местообитаниях насекомых опреде-
ленные условия среды иногда могут быть
смягчены, а иногда, наоборот, выражены
более резко. Существуют три основных
типа местообитаний-наземные, поч-
венные и водные.
Наземные местообитания. Наземные
местообитания-это поверхность земли и
все, что над нею. Сюда относятся воз-
душное пространство и древесная расти-
тельность, между которыми перемещает-
ся множество насекомых. Условия в на-
земных местообитаниях отличаются от
условий в толще воздуха благодаря на-
личию растительного покрова. На от-
крытых местах-на верхушках деревьев,
на верхней поверхности листьев пустын-
ных и луговых растений, на голой незате-
ненной земле и камнях-температура вы-
ше, чем в тени (где обычно принято
определять температуру воздуха). Насе-
комые, живущие под листьями или дру-
гими укрытиями, а также под пологом
леса, меньше подвержены воздействию
крайних температур. Суточные и се-
зонные колебания температуры и влаж-
ности, наиболее сильные на открытых
местах, постепенно уменьшаются в зате-
ненных или укрытых участках. На по-
верхности земли и около нее температу-
ра повышается за счет тепла, выделяю-
щегося при гниении органических мате-
риалов. В грибах, растительных галлах,
тканях листьев и внутри древесных ство-
лов, где живут многие насекомые, влаж-
ность достаточно высока, что создает ус-
ловия, близкие к условиям водных ме-
стообитаний. В гниющей древесине бла-
годаря хорошей изоляции колебания
температуры смягчаются, поэтому усло-
вия здесь близки к существующим в
почве.
Почва. Условия, создающиеся в поч-
ве, отражают общий климат местности,
но его крайние проявления смягчены. В
то же время почва обладает рядом
важных специфических особенностей.
Смягчение климатических условий. В
зависимости от обстоятельств почва мо-
жет выполнять роль поглотителя, изоля-
тора и излучателя. Почва сохраняет в се-
бе дождевую воду и медленно отдает ее,
поэтому колебания влажности в почве
значительно меньше, чем в воздухе. По-
верхностный слой почвы поглощает те-
пло и изолирует нижележащие слои, это
поглощенное тепло тоже расходуется
медленно, поэтому амплитуда суточных
колебаний температуры в почве значи-
тельно меньше, чем в наземных место-
обитаниях.
Свойства почвы. Свойства многих
почв не есть результат непосредственно-
го воздействия существующего климата.
Эти свойства (структура, влажность, дре-
наж, химический состав, физико-геогра-
фические особенности) являются резуль-
татом длительных процессов, происхо-
дивших при различных климатических
условиях.
Урбанизация и возделывание почвы
больше, чем какие-либо другие экологи-
ческие факторы, нарушили ее естествен-
ное состояние. Не только сам факт выра-
щивания культурных растений, но также
вспашка и другие виды обработки почвы
изменяют ее исходное состояние; осуше-
ние или ирригационные мероприятия
уменьшают или же увеличивают содер-
жание влаги в почве; методы ведения хо-
10. Экологические аспекты
495
зяйства могут сказываться на химиче-
ском составе почвы, на содержании в ней
органического вещества. Все это сильно
влияет на структуру почвы. Изменения,
губительные для многих видов насе-
комых, могут способствовать размноже-
нию других видов, которые нередко ста-
новятся серьезными вредителями.
По своей структуре почвы образуют
непрерывный ряд от плотных глин до
рыхлых песков. Лишь немногие насе-
комые способны продвигаться или рыть
ходы в плотной глинистой почве. По-
видимому, для насекомых, роющих ходы
и норки, наиболее подходящи сугли-
нистые почвы. Эти почвы благоприятны
для насекомых и в других отношениях-
по содержанию влаги, органических ве-
ществ, дренажу.
Влияние структуры почвы на числен-
ность насекомых было продемонстриро-
вано на примере прямоугольной совки
Agro t is orthogonia). Эта* бабочка, оби-
тающая в северной зоне прерий, была
редким экспонатом в ранних коллекциях
североамериканских насекомых. Разру-
шение дернового слоя в результате воз-
делывания земель на обширных про-
странствах Канады и севера США приве-
ло к резкому увеличению численности
этой совки, и ее гусеницы стали чрезвы-
чайно активными вредителями зерновых
культур. Оказалось, что они живут толь-
ко в рыхлых почвах, где перемещаются в
соответствии с суточными и сезонными
колебаниями температуры и влажности.
В нетронутых северных прериях эти насе-
комые встречались только на песчаных
)настках с рыхлой почвой, но их не было
з верхнем плотном дерновом слое. В ре-
зультате распашки прерий дерновый
слой превратился в почву со структурой,
оптимальной для гусениц этого вида, и в
гезультате для них возникли тысячи ква-
дратных километров новых местообита-
ний.
Содержание влаги в почве в сильной
степени зависит от дренажа. Такие не-
проницаемые слои, как глина или ка-
мень, могут задерживать естественный
дренаж, что ведет к постоянной или вре-
менной заболоченности или созданию
влажных почв. В таких условиях могут
жить лишь те насекомые, которые хотя
бы частично приспособлены к жизни в
водной среде, например многие личинки
двукрылых. В подпочвенных слоях с бо-
лее рыхлой структурой-в песке, гравии,
сланцах-создаются условия для более
свободного дренажа, лучшей аэрации,
более быстрого восстановления нор-
мальной влажности после дождей. Насе-
комые, обитающие в хорошо аэрируемой
почве, не нуждаются в приспособлениях
к водной или полуволной жизни.
Интересный пример влияния почвен-
ной влаги на насекомых представляет
жук Diabrotica virgifera. До проведения
обширных ирригационных мероприятий
этот вид встречался довольно редко,
только в аридных районах штатов Нью-
Мексико, Колорадо и Небраска. Личин-
ки его питаются корнями кукурузы и, ес-
ли численность их высока, могут разру-
шать корневую систему и тем самым
сильно снижать урожай. Примерно с
1890 г этот вид стал постоянным вреди-
телем в южных и центральных районах
штата Небраска, но в засушливые 30-е
годы практически исчез. Позднее он при-
обрел большое значение как вредитель
на орошаемых землях долины реки
Платт, где в результате широких иррига-
ционных мероприятий влажность почвы
повысилась.
Жуки-проволочники в северо-
западных штатах Тихоокеанского побе-
режья-еще один яркий пример измене-
ний видового состава и численности,
вызванных изменениями влажности поч-
вы. Четыре вида этих жуков, Limonius са-
lifomicus, L. infuscatus, L. canus и L. subau-
496
10. Экологические аспекты
Рис. 10.15. Распространение влаголюбивых
проволочников в северной части
Тихоокеанского побережья США. (По
материалам министерства сельского
хозяйства США.)
Зачерненные участки - районы проведения
ирригационных мероприятий, где один или
более видов проволочников ежегодно
причиняют серьезный вред
сельскохозяйственным культурам.
Кружочки - обозначены места, где
проволочники обитают в естественных
условиях вне связи с ирригацией и где они
лишь изредка причиняют вред.
ratus, являются вредителями на сильно
увлажненных землях, обычно на заболо-
ченных участках и в речных долинах.
Когда началось орошение засушливых
земель и использование их для нужд
сельского хозяйства, проволочники ста-
ли экономически важными вредителями
картофеля, кукурузы, салата, лука и мно-
гих других орошаемых культур. Высокая
влажность почвы, поддерживаемая ирри-
гацией, позволяет проволочникам раз-
множаться в больших количествах
(рис. 10.15).
Дренаж и структура почвы в суще-
ственной мере определяют также распро-
странение насекомых. Например, ареал
весьма вредного техасского муравья-
листореза Atta texana достиг сейчас Луи-
зианы, где он строит гнезда только в тон-
ких песчанистых суглинках со светлым
подпочвенным слоем и хорошим дрена-
жем.
Физико-географические особенности
местности сами по себе мало влияют на
распространение насекомых, однако они
могут в значительной мере определять
некоторые свойства почвы. Равнинные
земли отличаются замедленным стоком
дождевой воды, и для поддержания хоро-
шей аэрации почвы здесь нужен доста-
точный дренаж. Земли, расположенные
на склонах холмов и гор, характеризуют-
ся быстрым стоком дождевой воды, что
обеспечивает хорошую аэрацию почвы.
Кроме того, в умеренной зоне Северного
полушария температура почвы на участ-
ках южной, восточной и западной экспо-
зиции выше, а испарение больше, чем на
участках северной экспозиции, и это
влияет на флору и фауну.
Химический состав. Химический со-
став почвы может определять естествен-
ную флору данной местности и, таким
образом, влиять на распространение
многих растительноядных животных,
нуждающихся в специфических кор-
мовых растениях. Дефицит минеральных
солей в почве влечет за собой их дефицит
и у растений, и это тормозит развитие,
снижает численность и ограничивает рас-
пространение некоторых насекомых, но
не оказывает заметного влияния на дру-
гих. Нехватка азота подавляет размно-
жение определенных видов насекомых,
но, по-видимому, способствует вспыш-
кам численности других видов.
Водные местообитания. Жизнь насе-
комых в водной среде связана с трудно-
стями осуществления дыхательной функ-
ции, и многие особенности водных ме-
стообитаний имеют в связи с этим боль-
шое значение. Для многих водных насе-
комых очень важна диффузия избытка
СС>2 из воды в воздушную среду; еще бо-
лее важным моментом является диффу-
зия или растворение кислорода в воде.
10. Экологические аспекты
497
Кислород поступает в воду в основном
из воздуха, поэтому любое перемешива-
ние воды, приводящее больший ее объем
в соприкосновение с воздушной средой,
способствует обогащению воды кислоро-
дом. В прудах и озерах основным факто-
ром перемешивания служит ветер, в теку-
щих водах-водопады, стремнины, водо-
вороты.
Аэрация зависит также от темпера-
туры-чем холоднее вода, тем большие
количества газов, в том числе и кислоро-
да, могут растворяться в данном ее объе-
ме. Высокие температуры значительно
уменьшают растворимость газов в воде.
Зависимость водных насекомых от
аэрации воды неодинакова у разных ви-
дов. Многие насекомые, например ли-
чинки комаров, слепней и некоторые во-
дяные клопы, либо имеют растяжимые
дыхательные трубки, достигающие по-
верхности воды, либо периодически под-
нимаются к поверхности для дыхания.
Другие, например клопы-гребляки
и взрослые жуки-плавунцы, уносят воз-
дух в виде пузырька или пленки с собой
под воду и время от времени возвра-
щаются к поверхности, чтобы возобно-
вить его запас (см. с. 143-144). Эти насе-
комые почти не зависят от аэрации воды,
и многие из них живут в воде, почти не
содержащей кислорода.
Водные насекомые, не имеющие при-
способлений для прямого контакта с воз-
душной средой, нуждаются для дыхания
в кислороде, растворенном в воде. По-
требность различных насекомых в кисло-
роде, как и в других экологических фак-
торах, неодинакова и может ограничи-
вать их распространение. Нимфы неко-
торых стрекоз и личинки многих мелких
двукрылых могут жить в очень плохо
аэрируемых стоячих водоемах. С другой
стороны, личинкам двукрылых из сем.
Blepharoceratidae свойственна чрезвычай-
но большая потребность в кислороде, и
они встречаются только в быстрых
горных ручьях.
Температура. Температура воды из-
меняется не в столь широких пределах,
как температура воздуха. Обычно вод-
ные насекомые приспособлены к весьма
узкому диапазону температур. Для мно-
гих насекомых это, безусловно, связано
со степенью аэрации воды, но для дру-
гих, по-видимому, важна сама по себе
температура. Личинки некоторых кома-
ров живут и нормально развиваются в
воде при 18°С, но погибают во время
линьки, если она происходит при 27°С.
Поскольку комары не зависят от содер-
жания кислорода в воде (см. выше), мож-
но думать, что здесь имеет значение сама
температура.
Известно несколько видов насе-
комых, обитающих в горячих источниках
с температурой воды 43-49°С. В основ-
ном это водные жуки и личинки двукры-
лых, набирающие кислород из атмосфер-
ного воздуха. Как правило, скорость
развития и активность водных насе-
комых возрастают с повышением темпе-
ратуры воды, и это приводит к появле-
нию огромного числа взрослых особей
поздней весной и летом.
На веснянок температура воды
влияет противоположным образом.
Нимфы этих насекомых обитают в про-
точных водоемах северных, северо-
восточных и восточных штатов США.
Летом развитие нимф замедляется, но с
наступлением прохладной осенней по-
годы вновь активируется. В результате
взрослые насекомые появляются зимой,
с конца ноября до февраля или марта
(рис. 10.16).
Глубина. В прудах и озерах количе-
ство кислорода, температура и освещен-
ность меняются с глубиной. В проточных
водах влияние глубины не столь сильно,
так как течение постоянно перемешивает
слои воды.
498
10. Экологические аспекты
Виды
Янв.]Февр.-МартрАпр
Дек.
Л31
Май-Июнь Июль Авг.
M*.lb'jo|l-iyib-3l
Ноя.
|и;.ц>-зо||-охз<
’AUocopnia mystica
Alldcapnia recta
Allocapnia vivipara
Рис. 10.16. Сезонная последовательность появления взрослых особей 15 видов веснянок в
Оквуде (шт. Иллинойс). (Из Illinois Natural History Survey.)
Самая широкая часть «веретена» соответствует периоду максимального обилия взрослых
особей каждого вида.
Perlinella drumo
Isoperlo bilineata
AUocapnia granulate
Strophoptergx fasciata
Perlinella drumo
boperla bilineata
Neophasganophora capital a
Atqperla ephure
Perlesta placida
Acroneuna oridq
Acrpneuria interndta
Neoperla clqmene
Acroneuria obnormis
Вода поглощает тепло и свет, причем
свет тоже преобразуется в тепло. В верх-
них слоях воды поглощаются тепловые
лучи и красная часть видимого спектра, а
с увеличением глубины постепенно по-
глощаются световые лучи с другими дли-
нами волн, пока, наконец, не будет задер-
жан весь видимый свет (даже в очень
прозрачной озерной воде это происходит
на глубине около 33 м). Это имеет
огромное значение для водных расте-
ний-практически ни одно из них не
встречается на глубине более 20 м, а ос-
новная масса растений обитает на глуби-
не 0-3 м, где имеется достаточно света
для осуществления фотосинтеза. Тем
самым создается зональность пищевых
ресурсов, лимитирующая в свою очередь
распределение многих насекомых.
В глубоких озерах (глубиной 30 м и
более) наблюдается еще одно интересное
явление, имеющее большое биологиче-
ское значение,-термоклин. Летом по-
верхностный слой воды в большом озере
значительно теплее, чем придонный, где
температура остается близкой к точке на-
ибольшей плотности воды-около 4—5 °C
(рис. 10.17). Этот придонный слой самый
тяжелый, и более теплые воды верхних
слоев лежат поверх него, не смешиваясь с
ним. Между ними располагается относи-
тельно тонкий слой, называемый термо-
клином; где создаются условия, средние
между весьма однородными верхним и
нижним слоями. Верхний слой - эпилим-
нион - перемешивается под действием ве-
тра, поэтому он почти однороден по тем-
пературе и хорошо аэрирован. При-
Рис. 10.17. Схема, иллюстрирующая явление
термоклина. (Из Ward, Whipple, по Birge.)
Показана циркуляция воды в озере
с одинаковой температурой воды на разных
глубинах (А) и в озере с неодинаковой
температурой (Б). Ткл-термоклин;
Эм-эпилимнион; Гип - гиполимнион.
10. Экологические аспекты
499
донный слой - гиполимнион - неподвижен,
и содержащийся в нем кислород посте-
пенно расходуется на окисление органи-
ческих веществ. Почти вся жизнь в глубо-
ких озерах сосредоточена в эпилимнионе,
тогда как гиполимнион-это по существу
биологическая пустыня.
Мутность. Крошечные частички
глины и ила, микроскопические водорос-
ли и другие организмы обычно делают
воду в той или иной степени мутной. Эта
мутность косвенно влияет на насекомых,
так как уменьшает проникновение света в
глубину, а тем самым и рост придонных
растений. В условиях постоянно высокой
мутности воды взвешенные частицы не-
прерывно оседают на дно, изменяя как
характер дна, так и придонную фауну.
Субстрат. Многие водные насе-
комые живут на донном субстрате или
внутри него, и при этом большинство ви-
дов обитают лишь там, где есть субстрат
определенного типа. Чаще всего для
классификации субстратов используют
размеры составляющих его частиц-на-
пример, глинистый, илистый, песчаный,
каменистый субстрат Органическим ве-
ществом, служащим пищей животным,
богаче всего субстраты, состоящие из
глины и ила, а беднее всего песчаные и
каменистые субстраты. Глинисто-
илистые субстраты в проточных водое-
мах обычно сочетаются с медленным те-
чением, низким содержанием кислорода
в воде и высокими температурами. Пе-
счаные субстраты не отличаются ста-
бильностью, и их фауна относительно
бедна. Каменистые субстраты обеспечи-
вают наиболее прочную опору и служат
излюбленным местообитанием для мно-
гих групп насекомых.
Растительность. Для одних насе-
комых водная растительность - это в ос-
новном пища, для других-убежище.
Подводные растения служат защитой и
опорой, особенно для насекомых, являю-
щихся объектом охоты других жи-
вотных, или не имеющих приспособле-
ний для плавания. Водная раститель-
ность особенно обильна в озерах, где
значение ее для насекомых очень велико.
Большинство насекомых, живущих в озе-
рах, редко появляются в свободной тол-
ще воды-в основном они проводят вре-
мя на дне и в придонной растительности.
Те насекомые, которые часто плавают в
открытом пространстве, обычно делают
это в сумерках или ночью, а днем прячут-
ся на дне.
ПИЩА
Пища и распространение насекомых.
Наличие достаточного количества до-
ступной пищи-один из самых важных
факторов, определяющих распростране-
ние и численность насекомых. Многие
насекомые обычно всегда встречаются
там, где есть подходящая для них пища,
и именно этот фактор ограничивает их
распространение. Пищевые ресурсы мно-
гих насекомых коренным образом из-
менились в результате человече-
ской деятельности - сельскохозяйствен-
ного производства, перевозки грузов, пу-
тешествий. Например, многие виды тлей
прекрасно приспособились к обитанию
на сельскохозяйственных культурах дале-
ко за пределами ареалов их прежних
кормовых растений. С другой стороны,
ареал растения-хозяина данного вида на-
секомых может быть значительно боль-
ше ареала этого вида; очевидно, его рас-
пространение ограничивает какой-то
другой фактор, например температура
или почвенные условия.
Многоядные насекомые. Многие насе-
комые используют в качестве пищи ши-
рокий круг растений или животных либо
питаются мертвым органическим мате-
риалом, широко представленным в боль-
шинстве биологических сообществ. Для
500
10. Экологические аспекты
таких насекомых пища редко бывает фак-
тором, ограничивающим распростране-
ние, а если где-то их нет, то это обусло-
влено обычно не отсутствием пищи, а
какими-то другими факторами, неприе-
млемыми для этих видов, например тем-
пературой или влажностью.
Насекомые с узкими пищевыми требо-
ваниями. Огромное множество насе-
комых-в основном хищников, парази-
тов и растительноядных видов - питается
либо на очень небольшом числе видов-
хозяев, либо связано с группой близко-
родственных видов хозяев, а иногда даже
с одним видом. Чем больше избиратель-
ность в отношении пищи, тем в большей
степени пища является фактором, огра-
ничивающим распространение. Ареал
пилильщика Pseudodineura parvula вклю-
чает все северные штаты средней и во-
сточной части США, однако в средней ча-
сти этот вид встречается только в от-
дельных местах, где растет печеночница
(Hepatica), на которой он питается. Пи-
лильщик Nematus tibialis раньше был рас-
пространен лишь в ограниченном ареале
своего кормового растения - ложной ака-
ции Robinia pseudoacacia; однако в тех
районах, куда ложная акация была ин-
тродуцирована, в конце концов обнару-
жили и пилильщика, т. е. экологические
возможности насекомого оказались в
данном случае шире естественного ареа-
ла растения-хозяина.
Смена кормовых растений. Насе-
комые могут лишить определенный вид
кормовых растений листьев, после чего
они либо пытаются приспособиться к пи-
танию на другом растении, либо умень-
шают свою численность до уровня, дик-
туемого продуктивной емкостью перво-
го растения. Некоторые виды насекомых,
например гусеницы лесного кольчатого
коконопряда, без видимого ущерба для
себя переходят на близкородственные
растения. Клоп-черепашка может пи-
таться на овсе или пшенице почти до
своего полного созревания, а затем легко
переходить на кукурузу, и при этом не на-
блюдается повышенной смертности из-
за перемены пищевого режима.
Другие насекомые совершают пере-
ход со своего кормового растения на
родственный вид с большими трудностя-
ми: либо после длительного голодания
(молодые личинки), либо в случае край-
ней необходимости (яйцекладущие сам-
ки). Личинки старших возрастов, как
правило, не способны переходить на дру-
гое растение, при смене пищи у них по-
являются симптомы кишечного рас-
стройства (диаррея) и они гибнут. Если
личинок первого возраста поместить на
новое растение, они питаются на нем, но
процент гибели их очень высок. На по-
следующих стадиях роста смертность ли-
чинок снижается, однако у куколок она
снова возрастает. Из тысяч исходных ли-
чинок первого возраста лишь несколько
достигает взрослого состояния. Эти
взрослые насекомые предпочитают от-
кладывать яйца на новое кормовое расте-
ние, и вылупившиеся личинки охотно пи-
таются на нем без каких-либо патологи-
ческих явлений.
Интересным примером может слу-
жить ивовая волнянка (Stilpnotia salicis).
Этот европейский вредитель ломбард-
ского тополя примерно в 1922 г. был за-
везен на Тихоокеанское побережье США
и через несколько лет достиг высокой
численности, вызывая везде, куда он про-
никал, дефолиацию ломбардского и дру-
гих интродуцированных видов тополя. В
Британской Колумбии (Канада) к концу
20-х годов он распространился в долине
реки Фрейзер. Здесь возникла огромная
популяция, которая практически пол-
ностью уничтожила ломбардский то-
поль. Лишившись своего прежнего кор-
мового растения, насекомые стали от-
кладывать яйца на местный вид тополя
10. Экологические аспекты
501
Populus trichocarpa hastata, широко рас-
пространенный в этом районе. Сначала
на этих тополях развивались лишь от-
дельные колонии насекомых, но через не-
сколько лет ивовая волнянка стала здесь
наиболее опасным вредителем тополей.
Опыты, проведенные в лаборатории, по-
казали, что на первых этапах перехода
ивовой волнянки с одного растения на
другое имеет место очень высокая смерт-
ность.
Непарный шелкопряд (Porthetria dis-
par) дает нам пример совсем иного рода.
Его растительноядные личинки питают-
ся на более чем 450 видах растений; одна-
ко для нормального развития необходи-
мо, чтобы личинки первых двух возра-
стов питались на одном из 42 наиболее
подходящих видов, к которым относятся
ивы, березы и дубы. Личинки старших
возрастов могут без какого-либо вреда
для себя переходить на другие растения
(один из предпочитаемых видов-горная
сосна) и развиваться* на них до зрелого
состояния.
Смена кормовых растений оказывает
огромное влияние на распространение
насекомых и их численность и предста-
вляет собой одно из важных проявлений
экологической толерантности. При вве-
дении новой сельскохозяйственной куль-
туры всегда есть опасность перехода на
нее насекомых, питавшихся ранее на дру-
гих растениях. Так произошло, напри-
мер, с соей. Когда она была интродуци-
рована в центральные штаты США, неко-
торое время местные насекомые ее не
трогали. Однако постепенно они стали
использовать ее в качестве кормового
растения, и теперь соя в этих районах
подвержена атакам ряда серьезных вре-
дителей, в число которых входят кобыл-
ки. личинки капустных мух и хрущи.
Влияние пищи на численность и раз-
меры тела насекомых. На популяцию
вида в определенном сообществе сильно
влияет количество пищи. Нередко вид ис-
пользует все имеющиеся пищевые ре-
сурсы, что ведет затем к резкому сниже-
нию его численности.
Хотя размеры особей одного вида
могут значительно варьировать, для нор-
мального развития особи необходимо
какое-то минимальное количество пищи.
Например, если личинок комнатной му-
хи последнего возраста перестать кор-
мить, они погибнут при окукливании.
Личинки пилильщиков, лишенные всего
лишь двух или трех последних приемов
пищи, необходимых им для полного раз-
вития, в большинстве своем погибают.
Поэтому при избыточном числе особей и
ограниченном количестве пищи особи,
начавшие питаться первыми, успевают
получить свою пищевую норму и созре-
вают, а многим другим пищи не хватает,
и они гибнут.
Интересное исключение составляют
некоторые паразитические насекомые с
широким кругом хозяев. Размеры особей
у них определяются величиной особи хо-
зяина. Превосходным примером может
служить паразитическая оса Dasymutilla
bioculata (Mutillidae). Если ее личинки пи-
таются на хозяине малых размеров, то из
них развиваются мелкие осы, тогда как
на крупном хозяине они дают крупных ос
(рис. 10.18).
Развитие сельского хозяйства вносит
ряд изменений в естественные отношения
между насекомыми и их кормовыми рас-
тениями. 1) создаются новые пищевые
ресурсы в то время и в тех местах, где их
не было в естественных условиях, 2)
иногда насекомым предоставляется луч-
шая пища, 3) создаются просто более об-
ширные пищевые ресурсы и изменяются
условия внешней среды.
Увеличение пищевых ресурсов. В конце
30-х-начале 40-х годов во многих шта-
тах США, особенно в кукурузном поясе,
широко распространился, достиг высо-
502
10. Экологические аспекты
Рис. 10.18. Корреляция размеров Dasymutilla bioculata и ее хозяев - Microbembix
monodonata (слева) и Bembix pruinosa (справа). В каждом вертикальном ряду
сверху показаны самец и самка Dasymutilla, а внизу - оса-хозяин. (Материал
предоставлен С. Е. Mickel, фото W Е. Clark.)
кой численности и стал важным вредите-
лем запасов зерна жук Cynaeus angustus
(сем. Tenebrionidae). Впервые этот жук
был обнаружен и описан в 1852 г в штате
Калифорния, и до 1938 г. он оставался
коллекционной редкостью, встречаясь
только на юкке. В 1938 г. было неожидан-
но обнаружено, что он вредит зерновым
запасам в штатах Вашингтон, Канзас и
Айова. К 1941 г. он уже широко распро-
странился по всему кукурузному поясу. В
условиях, созданных человеком, популя-
ции этого некогда редкого вида достигли
огромных размеров.
Лучшее качество пищи. Интродукция
новых сельскохозяйственных культур
иногда приводила к повышению плодо-
витости и тем самым к увеличению чис-
ленности местных видов насекомых.
Плодовитость двух видов кобылок, Ме-
lanoplus differentialis и М. bivittatus, в экс-
перименте заметно увеличилась при пи-
тании соответственно на сое и на люцер-
не. Это согласуется с полевыми наблюде-
ниями, согласно которым при посеве
этих культур на больших площадях чис-
ленность обоих насекомых возрастает.
Дополнительные кормовые ресурсы.
10. Экологические аспекты
503
Почти каждая сельскохозяйственная
культура становится дополнительным
кормовым растением по меньшей мере
для нескольких видов насекомых. Коло-
радский картофельный жук, кукурузная
тля и другие вредители картофеля и куку-
рузы процветают на тысячах гектаров,
занятых этими культурами. Они обра-
зуют огромные популяции, несравнимые
с разрозненными небольшими колония-
ми, существовавшими во времена, когда
их кормовые растения по своей численно-
сти и питательной ценности значительно
уступали современным сельскохозяй-
ственным культурам.
ВРАГИ
Известно множество разнообразных
паразитов и иных врагов насекомых. Не-
которые паразитические организмы, на-
пример возбудители малярии Plasmodium
sp., по-видимому, не причиняют насе-
комым вреда, но большинство все-таки
оказывает на хозяина вредное воздей-
ствие. Подобные организмы предста-
вляют собой внешний фактор, влияющий
на численность, а иногда и на распро-
странение вида-хозяина. У каждой ста-
дии развития хозяина могут быть свои
враги, отличные от врагов другой ста-
дии, в других же случаях несколько ста-
дий развития хозяина подвержены напа-
дению одного и того же врага. Обычно
насекомоядные животные и паразитиче-
ские растения, например грибы, менее
специфичны в выборе своих жертв, и от
них страдают разные стадии, тогда как
внутренние паразиты поражают опреде-
ленную фазу развития насекомого.
Внутренние паразиты. Наибольшее
значение среди внутренних паразитов на-
секомых имеют представители неко-
торых отрядов насекомых, паразитиче-
ские черви, бактерии и грибы.
Насекомые. Многие семейства пере-
пончатокрылых (Ichneumonidae, Chalci-
didae, Scelionidae и другие) и несколько
семейств двукрылых (Pyrgotidae, Tachini-
dae) полностью состоят из видов, личин-
ки которых являются внутренними пара-
зитами насекомых или других род-
ственных членистоногих. Эндопаразиты
есть также среди личинок некоторых ба-
бочек и жуков, в том числе из группы Sty-
lopoidea. В Северной Америке сейчас из-
вестно более 11 тыс. видов паразитиче-
ских насекомых. Большинство из них
довольно специфичны в отношении
группы насекомых-хозяев. Некоторые
паразитируют на гусеницах разно-
образных бабочек, другие поражают
лишь определенных первичных парази-
тов этих гусениц (см. с. 364). Более под-
робно эти вопросы обсуждаются в гл. 11.
Другие животные. Ряд паразитиче-
ских простейших и других беспозво-
ночных проходят в насекомом одну из
стадий своего жизненного цикла. Приме-
рами протозойных паразитов являются
малярийные плазмодии и виды Тгурапо-
soma - возбудители сонной болезни. Из
паразитических червей часть своей жизни
в теле насекомых проводят некоторые
сосальщики, нематоды {Filaria и др.) и
Acanthocephala (например, свиной скре-
бень Macracanthorhynchus hirudinaceus). В
каждом из этих случаев только одна из
ранних стадий протекает в насекомом-
промежуточном хозяине. Эти паразиты,
по-видимому, не причиняют насекомым
вреда и, во всяком случае, не вызывают
их гибели, как паразиты-насекомые. С
другой стороны, некоторые простейшие,
особенно микроспоридии, паразитируют
только в насекомых и часто убивают
своего хозяина.
Грибы, бактерии, вирусы. Представи-
тели этих групп поражают насекомых на
разных стадиях жизненного цикла и мо-
гут вызывать гибель своих хозяев. При
разведении в лаборатории насекомые
504
10. Экологические аспекты
Рис. 10.19. Empusa muscae-гриб, поражающий мух. (Из Folsom,
Wardle, 1934, Entomology.)
А. Комнатная муха (Musca domestica), окруженная спорами гриба
(конидиями). Б. Группа конидий на разных стадиях развития.
В. Спороносен, несущий конидию, перед раскрытием.
легко заражаются грибами и бактерия-
ми, так как в культурах нередко создают-
ся условия повышенной температуры и
влажности, очень благоприятные для
этих микроорганизмов.
Один из самых известных возбудите-
лей грибковых заболеваний насекомых-
Empusa muscae, поражающая комнатную
муху (рис. 10.19). Представители того же
рода поражают множество других насе-
комых, в том числе кобылок, тлей и кло-
па-черепашку. К числу известных возбу-
дителей микозов относится и Beauveria
globulifera, часто называемая также Spo-
rotrichum globulifera-«белый грибок»
клопа-черепашки. В теплые и влажные
периоды года (поздняя осень и ранняя
весна) этот гриб уничтожает огромное
число клопов-черепашек и других насе-
комых, доводя в отдельных местах попу-
ляции клопов до практически полного
вымирания. В условиях искусственного
разведения насекомых главными возбу-
дителями микозов служат грибы-энто-
мофаги рода Isaria.
Особый интерес представляют грибы
семейства Laboulbeniaceae. Большинство
видов этого семейства-энтомофаги; они
окутывают тело насекомого снаружи
своими длинными плодовыми телами.
Один из представителей этого семейства,
Cordyceps ravenelii, иногда встречается в
восточных штатах США на личинках
хрущей (рис. 10.20).
Бактериальные болезни насекомых не
так многочисленны, как микозы, но вре-
менами могут быть опустошительными.
Флашерия, инфекционная и высоколе-
тальная болезнь шелкопрядов, вызы-
вается бактерией. Кобылок и клопов-
черепашек тоже поражают бактерии, и
были сделаны попытки разводить их и
вносить в популяции этих вредных насе-
комых для борьбы с ними. Бактерию Ва-
10. Экологические аспекты
505
Рис. 10.20. Плодовые структуры гриба Cordyceps ravenelii, выходящие из
личинки хруща Phyllophaga. (По Riley.)
cillus рорИНае, вызывающую «молочную
болезнь», успешно применяли для
борьбы с личинками жука Popillia japoni-
са. Споры бактерий, смешанные
с инертным порошком, наносили на по-
верхность почвы на участках, зара-
женных личинками. После дождя споры
попадали в землю, где приходили в со-
прикосновение с личинками жуков.
Вирусные заболевания также чрезвы-
чайно сильно поражают восприимчивых
к ним насекомых. Вирусы полиэдроза
оказались очень эффективным средством
борьбы с некоторыми пилильщиками и
гусеницами бабочек.
Наружные паразиты. Некоторые экто-
паразитические насекомые (вши, блохи)
обитают на теле хозяина либо во взрос-
лой фазе, либо в течение всего жизненно-
го цикла. На различных насекомых пара-
зитируют некоторые клещи. Необычный
случай эктопаразитизма представляет
пчелиная вошь Braula caeca-крошечная
муха, которая во взрослой фазе парази-
тирует на медоносных пчелах.
В некоторых семействах Hymenoptera,
где личинки, в основном эндопаразиты,
например в семействе Braconidae, встре-
чаются роды, у которых личинки прикре-
пляются к поверхности тела личинок-
хозяев. Отношения этих эктопаразитиче-
ских личинок с их хозяевами такие же,
как и у их родственников-эндопаразитов;
так, обычно на одном хозяине живет
только одна особь паразита, и хозяин
почти всегда погибает ко времени созре-
вания паразита или раньше.
Хищники. И здесь, так же как в случае
эндопаразитизма, самых опасных врагов
мы находим среди самих насекомых. Две
очень обширные группы жуков, питаю-
щихся и во взрослой, и в личиночной фа-
зе,-Staphylinidae и часть семейства Cara-
bidae - состоят почти исключительно из
хищников, охотящихся на других насе-
комых. Хищниками являются представи-
тели многих семейств ос, личинки Taba-
nidae, Dolichopodidae и некоторых дру-
гих крупных семейств двукрылых, а так-
же Mecoptera. У стрекоз и нимфы, и
взрослые-хищники. То же относится и к
некоторым семействам Hemiptera, та-
ким, как Pentatomidae (щитники), Re-
duviidae (клопы-хищнецы) и Phymatidae.
В других семействах Hemiptera (напри-
мер, Miridae) большинство родов расти-
тельноядные, но некоторые - хищники.
Есть также большое число мелких групп
хищных насекомых, в том числе многие
водные Hemiptera.
Кроме того, на насекомых охотятся
представители нескольких больших
групп других животных. Главным обра-
зом насекомыми питаются пауки; в Се-
верной Америке известно около 3000 ви-
дов пауков, и каждый имеет своих жертв
среди насекомых. Губоногие (Chilopoda)
тоже в основном питаются насекомыми.
Много групп насекомоядных жи-
вотных и среди позвоночных. Немалую
долю составляют насекомые в рационе
некоторых рыб-окуней и ряда других
видов. Рептилии и амфибии в основном
насекомоядные, так же как летучие мы-
506
10. Экологические аспекты
ши и кроты. Много насекомых уничто-
жают и другие млекопитающие (мыши,
скунсы, землеройки, еноты).
Прожорливыми врагами насекомых
являются птицы. Стрижи, козодои, мухо-
ловки питаются только насекомыми, ко-
торых они ловят в полете. Дрозды, кра-
пивники, гаички, кукушки, перепела,
степные тетерева предпочитают питаться
насекомыми, если тех достаточно много.
В периоды вспышек численности насе-
комых многие всеядные птицы становят-
ся насекомоядными. Например, очень
охотно питаются саранчовыми в пе-
риоды их высокой численности такие
птицы, как вороны, дрозды, чайки, совы
и мелкие хищные птицы.
Все эти позвоночные животные, буду-
чи сравнительно крупными и многочис-
ленными, потребляют соответственно
большие количества насекомых. Таким
образом, это постоянный экологический
фактор, направленный против роста по-
пуляций насекомых.
Насекомоядные растения. Список вра-
гов насекомых будет неполным, если не
упомянуть о тех интересных растениях,
которые улавливают насекомых и пере-
варивают их. Большинство этих расте-
ний относятся к порядкам Nepenthales и
Sarraceniales и обычно растут на почвах,
бедных питательными веществами.
Пузырчатки (Utricularia)-водные расте-
ния, которые ловят мелкие организмы с
помощью «карманов», имеющих вид
пузырьков. Росянки (Drosera) - болотные
растения, имеющие листья с липкими
чувствительными волосками для схваты-
вания жертвы. У саррацении (Sarracenia)
листья имеют вид кувшина, частично за-
полненного водой, с жесткими волоска-
ми, направленными к воде: эти волоски
позволяют насекомому добраться до дна
кувшина, но не выпускают его обратно.
По-видимому, азот, используемый после
переваривания насекомых и других мел-
ких беспозвоночных, способствует росту
этих растений. Интересно, что у ряда на-
секомых, например у бабочки Ехуга, ли-
чинок некоторых мух (Sarcophaga) и дру-
гих выработался «иммунитет»- у них
есть фермент, препятствующий перева-
риванию их растением, в результате они
могут находиться на стенках «кувшина»
и охотиться за другими насекомыми, по-
падающими в эту ловушку. Ни одно из
хищных растений не имеет экологическо-
го значения как фактор, способный сни-
жать численность насекомых, так как их
собственная численность слишком мала
для этого.
ЗАЩИТА ОТ ВРАГОВ
Маскировка. Каждый из нас был ког-
да-нибудь поражен, видя, как вдруг «су-
чок» приходит в движение или кусочек
коры на стволе дерева обретает крылья и
летит прочь. Насекомые широко исполь-
зуют такого рода маскировку, позволяю-
щую им слиться с окружающим их фо-
ном и не быть замеченными хищником.
Палочники (рис. 10.21) похожи на веточ-
ки деревьев, на которых они живут, при-
чем весной молодые насекомые окра-
шены в зеленый цвет наподобие весенних
побегов, а взрослые осенью меняют
окраску на коричневую. Многие бабочки,
сидя на дереве, сливаются с его корой.
Крупная бабочка-ленточница (Catocala)
незаметна на коре дерева, когда ее кры-
лья сложены, но в полете сразу же бро-
сается в глаза из-за ярко окрашенных за-
дних крыльев (рис. 10.21). Прямокрылые
в зависимости от видовой принадлежно-
сти и потребляемой пищи могут быть не-
заметны на фоне лишайника, сухих
листьев, травы или почвы того или иного
типа. Сеноеды тоже используют защит-
ную окраску, особенно те виды, которые
питаются водорослями или лишайника-
ми, растущими на коре деревьев или на
Экологические аспекты
507
Рис. 10.21. Защитная мимикрия и окраска. (Из Folsom, Wardle, 1934,
Entomology.)
Слева-палочник на ветке; справа-ленточница (Catocala) с расправленными
крыльями (А) и в покое на коре дерева (Ь).
лилах. Некоторые личинки в покое сво-
рачиваются, напоминая свежий птичий
л?мет. например личинки пилильщика
Uegaxyela aviingrata. Как правило, боль-
шинство листогрызущих личинок или зе-
леного цвета, или имеют на теле рисунок,
позволяющий им слиться с листвой, на
юторой они кормятся. Восточно-афри-
гднские цикады в большом числе отды-
\дют на стеблях растений, располагаясь
чтобы напоминать цветки люпина.
По крайней мере у одного вида этих ци-
гдд самки зеленые, а самцы-желтые;
тогда они собираются группами, самки
едятся на растение выше самцов, и по-
учается картина, похожая на соцветие, в
котором нижние цветки раскрываются
раньше, чем верхние.
Защитная окраска и подражание
(рис. 10.21,Л, Б) очень важны для вы-
живания насекомых. Это хорошо иллю-
стрирует следующий известный пример.
Еще в первой половине прошлого века в
Средней Англии проводились обширные
сборы насекомых для коллекций. В сере-
дине века пяденица Biston betularia часто
встречалась на светло-серых стволах бе-
рез. Обычно это была белая или светло-
серая бабочка с пятнами и узкими поло-
сками черного цвета. Встречалась также,
но очень редко, необычная форма черно-
го цвета. С ростом индустриализации в
508
10. Экологические аспекты
этой части Англии, особенно усилив-
шимся в конце XIX века, деревья стали
темными от копоти и грязи. К 1895 г
доля черной (меланистической) формы в
местных популяциях пяденицы вблизи
главных промышленных городов состав-
ляла почти 95%. Врагам (в основном пти-
цам) было легче обнаруживать на
темных стволах деревьев светлых бабо-
чек, а не черных. В районах, удаленных
от промышленных центров, где стволы
деревьев оставались светлыми, темные
бабочки были хорошо видны на них, и
популяции состояли здесь главным обра-
зом из светлых особей. Эту тенденцию к
избирательному сохранению темных
особей благодаря их лучшей приспособ-
ленности к условиям промышленных
районов часто называют инду-
стриальным меланизмом.
Другая известная форма мимикрии
менее пассивна. Богомол Hymenopus со-
ronatus имеет такую же красную окраску,
как и цветки некоторых орхидей, среди
которых он живет в Малайзии. Он на-
столько сходен с этими цветками, что
другие насекомые, посещающие орхидеи
в поисках нектара, обследуют своим хо-
ботком и богомолов, немедленно стано-
вясь их жертвами.
Приспособления для отпугивания вра-
гов. Некоторые насекомые пытаются ис-
пугать нападающего на них хищника, на-
пример у многих бабочек на задних
крыльях есть «глазки», которые не
видны, пока бабочка сидит неподвижно.
В случае нападения птицы бабочка отво-
дит передние крылья, внезапно открывая
«глазки», которые пугают птицу. У неко-
торых гусениц тоже бывают большие,
резко выделяющиеся глазчатые пятна;
при отпугивании хищника гусеница при-
нимает такую позу, что хищник видит два
огромных глаза, смотрящих назад.
Растения, имитирующие насекомых.
Сами насекомые тоже бывают введены в
заблуждение мимикрией. Наиболее изо-
щрены в этом некоторые орхидные, осо-
бенно из рода Ophrys. Часть цветка у
этих растений похожа на самку неко-
торых видов пчел и ос. К тому же эти
цветки издают запах, имитирующий за-
пах полового аттрактанта самок тех ви-
дов, которых они имитируют внешне. В
некоторых случаях запах цветка даже
сильнее привлекает самцов, чем настоя-
щая самка. В результате самцы часто
впадают в ошибку, пытаются копулиро-
вать с подобием самки и в результате по-
крываются пыльцой, которую затем
переносят на другие цветки.
Защитные структуры. Некоторые ли-
чинки строят специальные футляры, до-
мики или навесы, которые обеспечивают
их обладателю некоторую физическую
защиту; кроме того, они могут быть по-
хожи на какие-то объекты окружающей
среды и тогда служат для целей маски-
ровки. Известные примеры - гусеницы
бабочек-мешочниц (рис. 8.175) и многие
личинки ручейников (рис. 8.163). Фут-
ляры ружейников иногда очень трудно
обнаружить до тех пор, пока насекомое
не начнет двигаться. Личинки некоторых
листогрызущих жуков строят урно-
образные домики, которые трудно раз-
личить среди их естественного окруже-
ния.
Яды, укусы, жала. Некоторые насе-
комые защищаются, причиняя боль то-
му, кто к ним прикоснется. Многие гусе-
ницы снабжены острыми волосками, в
которых содержится ядовитая жидкость,
вызывающая мгновенную и резкую боль.
Эти волоски так легко обламываются,
что даже легкого прикосновения к ним
достаточно, чтобы ощутить их сильное
обжигающее действие. Таким способом
защиты обладают, например, гусеницы
бабочек-волнянок и бабочки Sibine stimu-
lea (сем. Limacodidae, рис. 10.22). Другие
насекомые, например муравьи, кусают
10. Экологические аспекты
509
Рис. 10.22. Гусеницы с жгучими волосками, причиняющими боль людям. (Из
материалов министерства сельского хозяйства США.) Слева -Sibine stimulea\
справа-Automerus io.
нападающих на них животных. Пчелы и
осы обладают причиняющим боль жа-
лом, причем пчелы пользуются им толь-
ко для защиты. Одиночные осы исполь-
зуют жало главным образом для обез-
движивания своих жертв, но не про-
являют агрессивности по отношению к
оказавшимся в их «владениях» пришель-
цам. Напротив, у общественных ос жало
служит прежде всего для защиты, и они
проявляют резкую агрессивность в слу-
чае «незаконных» вторжений; при этом,
однако, парализующая жидкость вводит-
ся жертве с помощью мандибул, а не
жала.
Вредоносные секреты. Многие насе-
комые обладают способностью выра-
батывать и выделять вещества с не-
приятным запахом. Гусеницы бабочек
Papilio имеют на пронотуме пару выво-
рачивающихся «рогов» (осметерий), рас-
пространяющих запах, который, по-
видимому, отпугивает некоторых жи-
вотных. Нимфы большинства Heteropte-
ra обладают пахучими железами, распо-
ложенными на дорсальной стороне
брюшка. У многих жуков (например, на-
рывников), тараканов и других насе-
комых пахучие железы не столь хорошо
заметны (см. с. 182). Некоторые насе-
комые, лишенные таких желез, имеют,
по-видимому, неприятный вкус, посколь-
ку птицы отказываются их есть. К этой
категории относятся бабочки Papilio и
данаиды, а также личинки и взрослые
формы многих ярко окрашенных жуков,
питающихся листьями.
Многие насекомые, обладающие за-
щитными приспособлениями вроде опи-
санных в двух последних разделах, ярко
окрашены или имеют какой-либо бро-
сающийся в глаза рисунок (рис. 10.23).
Это явление можно рассматривать как
предупреждающую окраску, призванную
напомнить хищнику о неприятных свой-
ствах объекта, с которыми он уже знаком
по прошлому опыту. У птиц и других по-
звоночных редко встречаются инстинкты
избегания подобных насекомых, и каж-
дая особь должна обучиться этому само-
стоятельно.
Мимикрия. Это довольно распростра-
ненный у насекомых способ защиты от
некоторых врагов (рис. 10.24). Иногда
510
10. Экологические аспекты
Рис. 10.23. Ярко окрашенная бабочка
с неприятным вкусом Papilio ajax. (Из
Folsom, Wardle, 1934, Entomology.)
два, три или более видов, обычно не на-
ходящихся в близком родстве, бывают
внешне очень сходны, что обеспечивает
какому-то (или каким-то) из них опреде-
ленное преимущество. Известно несколь-
ко типов мимикрии. Одну из первых
форм мимикрии открыл в 1862 г Бейтс
(Н. W Bates), изучавший тропических
бабочек Южной Америки. Бабочки се-
мейства Heliconiidae яркие, хорошо за-
метные и очень неприятные на вкус для
птиц. На них очень похожи по окраске
Рис. 10.24. Мимикрия у пчел и ос. (Из
Folsom, Wardle, 1934, Entomology.)
А. Трутень медоносной пчелы, Apis mellifera.
Б. Пчеловидка Eristalis tenax.
бабочки семейства Pieridae, для птиц
вполне съедобные. При этой форме ми-
микрии (бейтсовская мимикрия) съе-
добный вид насекомых сходен по внеш-
ности с несъедобным видом, что служит
ему защитой от хищника, которому уже
известен неприятный вкус этого второго
вида.
При другой форме бейтсовской ми-
микрии используется сходство в строе-
нии и поведении. Жуки-чернотелки Eleo-
des при нападении на них становятся на
две передние пары ног, поднимают
брюшко и опрыскивают нападающего
противника неприятной жидкостью. Жу-
ки рода Megasida очень похожи на Eleo-
des и при нападении на них принимают
точно такую же позу, хотя у них нет аб-
доминальных желез, выделяющих за-
щитную жидкость.
Мюллеровская мимикрия состоит во
внешнем сходстве нескольких далеких
друг от друга видов, которые все несъе-
добны. Такие группы видов называют
миметическими кольцами. Это явление,
тоже впервые обнаруженное у тропиче-
ских бабочек, было истолковано Фрицем
Мюллером в 1878 г. как важное приспо-
собление, благодаря которому хищнику
достаточно познакомиться всего лишь с
одним насекомым из данного кольца,
чтобы научиться избегать всех сходно
окрашенных насекомых; таким образом,
безопасность для всех видов кольца до-
стигается более дешевой ценой.
Некоторые особи могут иногда полу-
чать преимущество от имитации других
особей того же вида. Эта так называемая
аутомимикрия проявляется в различных
формах. У самцов многих пчел и ос нет
жала, и поэтому они беззащитны, тогда
как самки хорошо вооружены. Самцы
этих видов очень похожи на самок, и это
обеспечивает им защиту от хищников.
Другая форма аутомимикрии встречает-
10. Экологические аспекты
511
Рис. 10.25. Бабочка-нимфалида (внизу) и бабочка-данаида (вверху). (Из
Folsom, Wardle, 1934, Entomology.)
ся у бабочек-данаид (Danaus plexippus и
D. gilippus', рис. 10.25). Гусеницы этих ба-
бочек питаются на различных растениях,
но основным кормовым растением для
них служит ваточник (Asclepias), содер-
жащий сердечные гликозиды или карде-
нолиды. Эти вещества очень токсичны
для позвоночных, например птиц, и в не-
больших дозах (ниже летальных) вызы-
вают у животных рвоту. Гусеницы бабо-
чек нечувствительны к этим веществам и
сохраняют в своем организме большие
их количества, переходящие затем в ор-
ганизм взрослых бабочек и обеспечиваю-
щие им защиту от хищников. Гусеницы
тех же видов, могут питаться и на других
растениях, но эти особи тоже защищены
от врагов, так как внешне ничем не отли-
512
10. Экологические аспекты
чаются от особей, питавшихся на ваточ-
нике. Ленточник (Limenitis archippus), на-
ходящийся лишь в отдаленном родстве с
данаидами, внешне сходен с ними и по-
этому также защищен от хищников
(бейтсовская мимикрия).
Существуют и более сложные формы
мимикрии, причем иногда в миметиче-
ских кольцах встречаются сочетания ее
различных типов. Одно из самых инте-
ресных колец такого рода включает жу-
ков-мягкотелок из подсемейства Lycinae,
близких к светлякам. В юго-западной ча-
сти Северной Америки жуки-лициды Li-
eus loripes и L. fernandezi обитают вместе
с подражающими им усачами Elytroleptus
ignitus и Е. apicalis. Жуки L. loripes и Е. ig-
nitus окрашены в ярко-оранжевый цвет, а
L. fernandezi и Е. apicalis-ярко-оран-
жевые с коричневыми верхушками элитр.
Лициды несъедобны для большинства
хищников, и их численность в сотню раз
больше численности усачей. Наблюдения
показали, что усачи не только служат
примером бейтсовской мимикрии, на-
поминая лицид по окраске, но иногда они
еще и охотятся на лицид, и в этом смысле
являются агрессивными подражателями.
Поглощая жидкости тела лицид, усачи
получают те ядовитые вещества, ко-
торых птицы научились избегать, таким
образом, усачи используют, хотя и вре-
менно, еще одну форму защиты.
Сходные миметические группы, состоя-
щие из представителей лицид и усачей,
известны в других районах Северной
Америки, в Южной Америке, на Борнео
и Кубе, что говорит о большой древно-
сти этих миметических отношений. В
большинстве таких колец участвуют так-
же и другие насекомые, например бабоч-
ки, полужесткокрылые (некоторые из них
тоже несъедобны), а иногда даже и осы,
имеющие свое собственное защитное
оружие в виде жала. Известно одно коль-
цо на Кубе, в котором не менее 15 видов
связаны между собой очень сложными
отношениями.
Мимикрия у насекомых требует даль-
нейшего изучения. Например, множество
насекомых (и пауков) похожи на муравь-
ев. Смысл такой мимикрии не всегда по-
нятен, так как многие взаимоотношения
нельзя объяснить, пользуясь только опи-
санными выше моделями.
КОНКУРЕНЦИЯ
Предметом конкуренции между насе-
комыми чаще всего служит пища. Конку-
ренция возможна между особями одного
вида и между особями разных видов.
Острая конкуренция обычно не вызывает
каких-то специальных реакций, а ведет к
голоданию всех особей. Если, например,
личинки пилильщика превысят по чис-
ленности популяцию хозяев, они будут
питаться до тех пор, пока есть особи хо-
зяина, а после этого все начнут до изне-
можения передвигаться в поисках друго-
го источника пищи, и если не найдут его,
то погибнут. В случае острой конкурен-
ции за пищу между двумя и более видами
разница в их потребностях может ока-
заться полезной для одного из них. Инте-
ресным примером могут служить два ро-
да перепончатокрылых, Opius и Tetrastich-
us, паразитирующие на личинках среди-
земноморской плодовой мухи на Гавай-
ях. В одной личинке мухи может разви-
ваться только одна личинка Opius (сем.
Braconidae), тогда как крошечные личин-
ки Tetrastichus (сем. Chalcidae) могут раз-
виваться в одном хозяине в числе 10-30
особей (рис. 10.26). Если оба паразита
откладывают яйца в одну и ту же личин-
ку плодовой мухи, то личинка Opius уби-
вает большинство личинок Tetrastichus, и
лишь немногие из них выживают. Зато
они развиваются быстрее, чем личинка
Opius, достигают зрелости и оставляют
после себя так мало пищи, что крупная
10. Экологические аспекты
513
Рис. 10.26. Личинки наездника Tetrastichus
giffar dianus в пупарии плодовой мухи. (По
Pemberton, Willard, 1918.)
Рис. 10.27. Личинка первого
возраста Opius humilis.
Обратите внимание на
острые, сильно
склеротизированные
мандибулы Си)- (Из
Pemberton, Willard, 1918.)
личинка Opius неизбежно погибает.
Часто конкуренция за пищу носит ак-
тивный и даже агрессивный характер. На
Гавайях три вида наездников-браконид
(Opius tryoni, О. fullawayi и О. humilis) па-
разитируют на личинках плодовой мухи.
Самки наездников откладывают яйца в
личинок мухи, при этом в одну и ту же
личинку могут отложить яйца несколько
особей всех трех видов. Выживает только
одна личинка паразита - победитель в
борьбе с другими вылупившимися ли-
чинками. У личинок первого возраста
всех трех видов Opius относительно боль-
шая голова с парой длинных острых ман-
дибул, которые раздвигаются и смы-
каются быстро и с большой силой
(рис. 10.27). Эти личинки очень драч-
ливы и при нападении на своих соседей в
той же личинке хозяина прокалывают их
тело и разрывают его с помощью этих
мандибул. Независимо от того, идет ли
борьба между личинками одного вида
или разных видов, в результате ее остает-
ся в живых только одна личинка. В борь-
бе личинок трех видов Opius почти всегда
побеждает О, tryoni благодаря своему
проворству, быстроте реакций, силе ман-
дибул и другим преимуществам.
Во многих группах насекомых встре-
чается склонность к каннибализму, и она
всегда усиливается при увеличении плот-
ности. Печально известный мучной хру-
щак живет в запасах муки или круп, ко-
торыми и питается. Новые поколения
появляются непрерывно, поэтому одно-
временно в одном и том же месте встре-
чаются взрослые особи яйца и личинки
всех возрастов. Крупные личинки
и взрослые жуки могут поедать яйца и
мелких личинок собственного вида, но,
по-видимому, никогда не пытаются изго-
нять их. Когда численность жуков неве-
лика по отношению к объему имеющейся
пищи, старшие особи не так часто встре-
чаются с молодыми. При большей зара-
женности пищевых запасов такие встречи
учащаются и каннибализм усиливается.
Благодаря такому механизму потери от
каннибализма всегда соответствуют
интенсивности размножения, и это пре-
дотвращает переуплотнение популя-
ции.
33-732
514
10. Экологические аспекты
ЛИТЕРАТУРА
Allee W. С., Emerson А. Е., Park О., Park Т.,
Schmidt К. Р. Principles of animal ecology.
Saunders, Philadelphia, 837 pp., 1949.
Allee W. C., Schmidt К. P. Ecological animal
geography. Wiley, New York, 2nd ed., 597 pp.,
1951.
Anderson R. M., Turner B. D., Taylor L. R.
(Eds.). Population dynamics. 20th Symposium
British Ecological Society. Blackwell, Oxford,
434 pp., 1979.
Andrewartha H. G., Birch L. C. The distribution
and abundance of animals. Univ, of Chicago
Press, Chicago, 782 pp., 1954.
Breymeyer A. I., Dyne G. M. Grasslands, sys-
tems analysis and man. Cambridge Univ.
Press, New York, 950 pp., 1980.
Chapman R. N. Animal ecology. McGraw-Hill,
New York, 464 pp., 1931.
Clark L. R., Geier P. W., Hughes R. D., Mor-
ris R. E. The ecology of insect populations in
theory and practice. Methuen, London, 232
pp., 1967.
Clements F. E., Shelf ord V E. Bio-Ecology. Wi-
ley, New York, 425 pp., 1939.
Elton C. Animal ecology. Macmillan, New
York, 207 pp., 1927.
Folsom J. W., Wardle R. H. Entomology with
special reference to its ecological aspects.
P.Blakiston’s Son & Co., Philadelphia, 4th ed.,
605 pp., 1934.
Harcourt D. G. The development and use of life
tables in the study of natural insect popula-
tions, Annu. Rev. Entomol., 14, 175-196
(1969).
Harper J. Population biology of plants. Acade-
mic Press, London, 892 pp., 1977.
Hassell M. P The dynamics of arthropod pred-
ator-prey systems. Princeton Univ. Press, Prin-
ceton, 328 pp., 1978.
May R. M. (Ed.). Theoretical ecology. Saun-
ders, Philadelphia and Toronto, 317 pp., 1976.
Mound L. A., Waloff N. (Eds.) Diversity of In-
sect Faunas. Symposia Royal Entomological
Society. Blackwell, Oxford, 204 pp., 1978.
Parker J. R., Newton R. C., Shotwell R. L. Ob-
servations on mass flights and other activities
of the migratory grasshopper, USDA Tech.
Bull., 1109, 1-46 (1955).
Pianka E. R. Evolutionary ecology. Harper &
Row, New York, 2nd ed., 397 pp., 1978.
[Имеется перевод: Пианка Э. Эволюцион-
ная экология.-М.: Мир, 1981.]
Price Р. W Insect ecology. Wiley, New York,
514 pp., 1975.
Roughgarden J. Theory of population genetics
and evolutionary ecology An introduction.
Macmillan, New York, 634 pp., 1979.
Southwood T R.E. (Ed.). Insect abundance. Sy-
mposia Royal Entomological Society. Bla-
ckwell, Oxford, 160 pp., 1968.
Southwood T R.E. Entomology and mankind,
Am. Scientist, 65, 30-39 (1977).
Spurr S. H.t Barnes В. V Forest ecology. Wi-
ley, New York, 3rd ed., 688 pp., 1980.
Waloff N. Studies on grassland leafhoppers
(Auchenorrhyncha, Homoptera) and their na-
tural enemies, Adv. Ecol. Res., 11, 82-215
(1980).
Wickler W Mimicry in plants and animals
(Trans. R. D. Martin). McGraw-Hill, New
York, 255 pp., 1968.
11
Полезные насекомые
Число видов насекомых столь велико, а их адаптации к многочисленным место-
обитаниям столь разнообразны, что по отношению к человеку они могут являться как
помощниками, так и врагами. Насекомые часто напоминают о себе своей назойли-
востью и тем ущербом, который они причиняют нашему здоровью, продуктам пита-
ния и промышленному сырью, тогда как полезная деятельность бывает в значитель-
ной степени скрыта от наших глаз. Именно этой полезной деятельности и посвящена
настоящая глава, причем рассматриваются в основном насекомые Северной Амери-
ки. Наиболее важными аспектами полезной деятельности насекомых можно считать
следующие: 1) поддержание сбалансированного уровня популяций вредных насе-
комых посредством паразитизма и хищничества, 2) паразитирование на сорняках, в
результате чего снижается способность последних к распространению и сопротивле-
нию болезням, 3) опыление цветущих растений, в том числе многих растений, ис-
пользуемых человеком в пищу (особенно фруктовых и ягодных); 4) создание по-
лезных продуктов, включая мед, пчелиный воск, шелк, шеллак, красители и
лекарства. Кроме того, некоторые насекомые используются человеком в качестве пи-
щи или в качестве лабораторных животных, а коллекционирование насекомых и на-
блюдения за ними доставляют немало приятных минут натуралистам-любителям.
Еще один аспект полезной деятельности насекомых, который, как правило, не при-
нимается во внимание,-это их роль как деструкторов в круговороте многих химиче-
ских веществ в экосистеме.
ХИЩНЫЕ И ПАРАЗИТИЧЕСКИЕ НАСЕКОМЫЕ
В предыдущей главе было отмечено,
что многие насекомые-хищники способ-
ствуют поддержанию численности
и плотности популяций других насе-
комых в некоторых разумных пределах.
Более половины всех известных видов
насекомых питаются другими насекомы-
ми, большинство из которых в свою оче-
редь использует в пищу материалы, не
имеющие непосредственного хозяйствен-
ного значения, например ядовитую ли-
ству токсикодендрона или гниющую дре-
весину на поверхности почвы. Некоторые
из паразитических и хищных насекомых
питаются насекомыми, которые конку-
рируют за пищу и различное сырье с че-
ловеком, приносят вред его здоровью
или просто докучают ему. Такие хищни-
ки и паразиты становятся нашими по-
мощниками, и, используя их, мы можем
более эффективно регулировать числен-
ность вредных и назойливых насекомых.
По ряду причин отношения хищниче-
ства и паразитизма среди насекомых
приобретают все большее значение как
для самих людей, так и для их пищевых
ресурсов. Во-первых, стало ясно, что ис-
пользование химических средств борьбы
с вредными насекомыми влечет за собой
ряд побочных эффектов. Применение пе-
стицидов таит в себе опасность для лю-
дей и животных. Некоторые насекомые,
особенно двукрылые, проявляют все воз-
растающую устойчивость к этим соеди-
516
11. Полезные насекомые
нениям, и поэтому для достижения тре-
буемого эффекта необходимо использо-
вать все более высокие дозы препаратов.
Кроме того, помимо связей, в которых
непосредственно участвуют насекомые-
вредители, применение инсектицидов
обычно ведет к затрагиванию и наруше-
нию многих других взаимоотношений в
популяциях. Многие химические соеди-
нения, в том числе ДДТ, весьма устой-
чивы во внешней среде и с дождевыми
стоками легко попадают в ручьи, озера,
реки и океаны, где обычно накапливают-
ся разными организмами. Во-вторых, ис-
пользование пестицидов для борьбы с
вредными насекомыми - это в лучшем
случае лишь временное и дорогостоящее
решение более сложной проблемы, ка-
сающейся взаимоотношений насекомых
и растений. В рамках решения этой про-
блемы огромный интерес вызывают на-
секомые-хищники и паразиты, ставшие
объектом очень важного направления эн-
томологии, известного как биологическая
борьба (см. также гл. 12, посвященную
вредным насекомым).
Насекомые-хищники имеют несколь-
ко важных отличий от паразитических
насекомых. Хищники, как правило, ак-
тивно отыскивают добычу и на протяже-
нии жизни поедают множество особей.
Насекомые-паразиты, напротив, прово-
дят жизнь, прикрепившись к телу (или
внутри) одной особи хозяина, и обычно
это не ведет к гибели хозяина. При более
крайних формах паразитизма, весьма
распространенных среди насекомых,
особь паразита, прикрепленная к телу хо-
зяина или живущая в нем, растет какое-
то время вместе с хозяином, но в конце
концов убивает его. Такие отношения но-
сят название паразитоидных, и почти
всегда паразитоидом является личинка,
тогда, как взрослая особь ведет сво-
бодный образ жизни, взрослая самка
обычно отыскивает хозяина и отклады-
Кры ТО
Рис. 11.1. Наездник Megarhyssa lunator,
пробуравливающий мертвую древесину
в поисках личиночного хода своей жертвы.
(Из The common insects of North America,
Lester A. Swan a. Charles S. Papp: рис. 1155,
c. 527.)
вает яйца на яйца или личинок хозяина,
проделывая иногда сложную работу. На-
пример, п]3едставитель ихневмонид Me-
garhyssa lunator (рис. 11.1) обладает
длинным яйцекладом, с помощью кото-
рого самка буквально просверливает
древесину, чтобы отложить яйца в личи-
ночный ход своего хозяина - рогохвоста
Tremex columba (рис. 8.118).
НАСЕКОМЫЕ-ХИЩНИКИ
Coleoptera
Семейство Coccinellidae. Семейство Сос-
cinellidae (божьи коровки; рис. 8.95)
включает многих полезных хищников,
уничтожающих других насекомых. Пред-
ставитель одной из групп божьих коро-
вок, Rodolia cardinalis (рис. 11.2, А), пи-
тается на червецах и щитовках и
завершает летом свой жизненный цикл
менее чем за 2 нед. Личинки питаются
И. Полезные насекомые
517
Рис. 11.2. Хищные божьи коровки: Л-родолия (Rodolia cardinalis), хищник австралийского
желобчатого червеца, вредителя'цитрусовых; Я-калифорнийская божья коровка Coccinella
califomica, активный хищник, уничтожающий тлей; B-Cryptolaemus montrouzieri,
уничтожающая червецов, Г-жужелица Calosoma sycophanta. (Из The common insects of North
America, Lester A. Swan a. Charles S. Papp: 4-рис. 805, c. 407; K-рис. 826, c. 412;
В-рис. 804, c. 407; F-рис. 542, c. 337.)
яйцами и нимфами своей жертвы, а
имаго-особями всех стадий развития.
Coccinella californica (рис. 11.2, К) охот-
но поедает тлей и может съесть до
500 тлей за время развития до взрослого
состояния. Взрослые насекомые продол-
жают поедать тлей-по 30-40 особей в
день. Другие представители данного се-
мейства также обладают огромным ап-
петитом. Так, в Калифорнии один из ви-
дов божьих коровок (Hippodamia conver-
ges) используется обычно в борьбе с
тлями, поскольку многочисленные попу-
ляции этого хищника перезимовывают в
лесной подстилке, в пнях и под камнями.
Их легко собрать и держать в холодиль-
нике до транспортировки. В литровом
сосуде может содержаться более 30 ты-
сяч особей этой божьей коровки. На
практике, однако, выпуск большого чис-
ла особей Н. converges ранней весной не
дает полного эффекта, поскольку, чтобы
удовлетворить полностью их пищевые
потребности, численность популяции
тлей должна достичь определенного
уровня.
Одна из ранних успешных попыток
использования насекомых в биологиче-
ской борьбе с вредителями связана с
представителем кокцинеллид Rodolia
cardinalis, которая была завезена в США
из Австралии в 1888 г. с целью попытать-
ся использовать ее для борьбы с вредите-
лем цитрусовых - австралийским желоб-
чатым червецом (Icerya purchasi, семей-
ство Coccidae). Этот червец, завезенный в
518
11. Полезные насекомые
Калифорнию, по-видимому, до 1872 г., в
течение следующих 15 лет почти пол-
ностью уничтожил цитрусовые рощи на
территории штата. До этого австралий-
ский желобчатый червец был известен
только в Австралии, где он не причинял
серьезного вреда, поскольку его числен-
ность контролировалась несколькими
местными хищниками, в том числе мел-
ким двукрылым насекомым Cryptochae-
tum iceryne и божьей коровкой Л. cardina-
lis. Оба хищника были интродуцированы
в Калифорнию, где божья коровка очень
скоро прекрасно адаптировалась. При-
мерно 500 завезенных особей этого хищ-
ника дали начало калифорнийским попу-
ляциям, которые вскоре возникли во всех
районах возделывания цитрусовых
в штате. Всего лишь через два года жуки
полностью взяли под свой контроль по-
пуляции австралийского желобчатого
червеца. После второй мировой войны в
Калифорнии широко применялись хло-
рированные углеводороды и фосфорор-
ганические соединения. Эти инсекти-
циды, особенно ДДТ, сохранялись на
листьях растений в течение месяцев и,
уничтожая вредителей цитрусовых, одно-
временно уничтожили в ряде мест и
божью коровку R. cardinalis. Австралий-
ский желобчатый червец вскоре вновь до-
стиг огромной численности, и после того
как токсичность отложений инсектици-
дов упала до низкого уровня, пришлось
заново восстанавливать популяции R.
cardinalis.
Другая божья коровка-Cryptolaemus
montrouzieri (рис. 11.2, В) - является хищ-
ником цитрусового червеца Pseudococcus
citri. Cryptolaemus montrouzieri была заве-
зена в Калифорнию в 1891 г., прекрасно
там адаптировалась и стала чрезвычайно
эффективным хищником. Она хорошо
переживает зиму во внутренних районах
Калифорнии, и с 1917 г разводится в
промышленных масштабах для распро-
странения в нуждающихся в ней цитру-
совых хозяйствах.
Семейство жуков-кокцинеллид вклю-
чает также ряд других видов, являющих-
ся активными хищниками тлей и черве-
цов. Для борьбы с другим вредителем
цитрусовых (масличной ложнощитовкой
Saissetia oleae) были ввезены черный жук-
коровка Rhizobius ventralis и близкие к не-
му виды R. debilis и Lindorus lophantae.
Местами эти жуки весьма эффективны и
часто с успехом включают в число своих
жертв другие виды щитовок и червецов.
С десяток других видов божьих коровок,
завезенных для борьбы с различными
вредителями, хорошо акклиматизирова-
лись в условиях Калифорнии. В странах
Карибского бассейна и в Мексике числен-
ность цитрусовой мошки азиатского про-
исхождения контролируется божьей ко-
ровкой Catana clausensi и паразитическим
перепончатокрылым Eretmocerus serius,
завезенными в 1930 г. из Малайи. Благо-
даря этим двум хищникам и главным
образом благодаря эффективной дея-
тельности паразита, цитрусовая мошка
не является ныне серьезным вреди-
телем.
Семейство Carabidae. Это большое се-
мейство жужелиц (рис. 8.85) состоит пре-
имущественно из хищников, охотящихся
за насекомыми, и по эффективности оно
близко к кокцинеллидам. Личинки жуже-
лиц, как правило, поедают насекомых. В
Северной Америке насчитывается около
2000 видов жуков этого семейства. Они
питаются личинками и куколками бабо-
чек и взрослыми особями большинства
других отрядов насекомых. Пахучий кра-
сотел (Calosoma sycophanta, рис. 11.2, Г)
был интродуцирован в Новую Англию в
1905 г для борьбы с непарным шелко-
прядом (Porthetria dispar), который, буду-
чи завезен сюда раньше, стал к этому
времени опасным вредителем. Личинка
пахучего красотела съедает за свою двух-
11. Полезные насекомые
519
Рис. 11.3. А. Жук-стафилин Aleochara bimaculata. Б. Enoclerus sphegeus (Cleridae) - хищник
короедов рода Dendroctonus. В. Chauliognathus permsylvanicus (Cantharidae) Г Взрослый жук
и личинка калифорнийского черного скакуна Omus califomicus. (Из Lester A. Swan а.
Charles S. Papp: The common insects of North America; Л-рис. 653, c. 364; К-рис. 691,
c. 375; В и Г-рис. 524, с. 332.)
недельную жизнь более 50 гусениц непар-
ного шелкопряда. Взрослые особи С. sy-
cophanta живут несколько лет и съедают
за это время несколько сотен гусениц
вредителя. Прожорливые жужелицы ус-
пешно контролируют численность непар-
ного шелкопряда.
Семейство Staphylinidae. Представите-
ли этого семейства также в основном
хищники насекомых, в США и прилежа-
щих районах встречаются более 1000 ви-
дов Staphylinidae. У многих видов агрес-
сивной стадией являются личинки, ко-
торым свойственна избирательность по
отношению к жертве. У одного из таких
видов, мелкого жука Aleochara bimaculata
(рис. 11.3, А), личинки поедают един-
ственную жертву - личинок капустной
мухи Hylemya brassicae. Специфичность
стафилин по отношению к своим жер-
твам обеспечивает снижение численности
определенных видов вредителей до безо-
пасного уровня.
Семейство Cleridae. Семейство Cleri-
dae (пестряки) состоит в основном из
хищников, питающихся личинками и в
некоторых случаях взрослыми особями
жуков-короедов и древогрызов. Короеды
520
11. Полезные насекомые
в лесах западных штатов служат жертвой
пестряка Enoclerus sphegeus (рис. 11.3, Б),
а жукй-древогрызы семейства Lyctidae
(Lyctus и Xylobiops)-жертвой другого
представителя пестряков Tarostenus
univittatus. Представители родов Thanasi-
mus, Tillus и Thaneroclerus имеют боль-
шое значение как хищники жуков, про-
грызающих ходы в деревьях.
Другие хищные жуки. Некоторые дру-
гие группы жуков представлены преиму-
щественно хищниками. Многие виды жу-
ков-светляков (Lampyridae) являются
хищниками не только насекомых, но и
слизней и улиток и используются для
борьбы с этими садовыми вредителями.
Мягкотелки (Cantharidae; рис. 11.3, В)
питаются преимущественно яйцами и ли-
чинками других насекомых. Некоторые
виды мягкотелок, относящиеся к родам
Podabrus и Canthris, питаются тлями; из-
любленная пища других жуков этого се-
мейства-яйца саранчовых, а также гусе-
ницы бабочек и личинки различных жу-
ков. По-видимому, хищниками насе-
комых являются все жуки-скакуны (Cicin-
delidae), причем у них отсутствует выра-
женная избирательность по отношению к
жертве. Численность природных популя-
ций скакунов обычно не бывает высокой,
и возможности использования их как
агентов биологической борьбы пока не
изучены. Для жуков-нарывников (Meloi-
dae) характерен смешанный тип питания.
Их личинки - хищники или паразиты, а
взрослые особи - растительноядны, и
среди них известен ряд важных вреди-
телей сельскохозяйственных культур,
особенно пасленовых.
В других семействах жуков некоторые
роды и виды представлены хищниками,
тогда как большинство близкород-
ственных таксонов таковыми не являют-
ся. В отрядах жуков, питающихся па-
далью (Histeridae, Silphidae, Nitidulidae,
Tenebrionidae), имеются виды со сме-
шанным питанием, а в семействе Elateri-
dae есть по крайней мере несколько ви-
дов, поедающих личинок пластинча-
тоусых жуков (Scarabaeidae). Веерокры-
лые (Stylopidae) успешно используются
во многих странах для борьбы с фонар-
ницами (Fulgoridae) и цикадками (Cica-
dellidae). Большинство северо-американ-
ских видов веерокрылых нападают на
различные виды Hymenoptera.
Odonata
Эти древние хищники отличаются вы-
сокой скоростью полета, большой силой
и прожорливостью. Личинки и имаго
стрекоз способствуют снижению числен-
ности личинок и взрослых особей
двукрылых, особенно комаров, в озерах
и ручьях, где они вообще охотятся за все-
ми мелкими животными. В условиях экс-
перимента личинка стрекозы на протяже-
нии довольно большого срока демон-
стрирует способность съедать каждую
минуту по 6 комариных личинок. Хотя
трудно количественно оценить хищниче-
скую деятельность представителей этой
группы, очевидно, что они имеют боль-
шое значение в ограничении численно-
сти популяций многих водных насе-
комых.
Dictyoptera
Богомолы (Mantodea) представляют
другую группу древних хищников. Ис-
кусные и очень агрессивные хищники на
всех стадиях своего жизненного цикла,
они способствуют снижению численно-
сти нежелательных вредных насекомых.
Они обладают огромной, не имеющей
себе равной прожорливостью, но при
этом могут уничтожать заодно и по-
лезных насекомых.
11. Полезные насекомые
521
Raphidioptera, Megaloptera,
Neuroptera
Почти все личинки этих насекомых, в
том числе личинки вислокрыл ок,-хищ-
ники, поедающие других насекомых.
Сетчатокрылые охотятся в основном на
тлей, трипсов, червецов и щитовок, кле-
щей и молодые стадии саранчовых. Ли-
чинки муравьиного льва (в соответствии
со своим названием) ловят в свои ямки
главным образом муравьев. Яйца и ли-
чинки Neuroptera менее чувствительны к
инсектицидам, чем некоторые вредные
насекомые, и поэтому в садах, где прово-
дятся обработки инсектицидами, целе-
сообразно разбрасывать яйца златогла-
зок, например яйца Chrysopa califomica,
личинки которых поедают яйца вино-
градного червеца Pseudococcus maritimus,
заменяя уничтоженных инсектицидами
его естественных врагов. Это-одна из
форм интегрированного управления чис-
ленностью вредителей, которая наряду с
другими его формами успешно применя-
лась на протяжении многих лет в ряде
фруктовых садов.
Hemiptera
Большинство видов Hemiptera
являются вредителями растений. Однако
в составе этого многочисленного и раз-
нообразного отряда имеется несколько
семейств, которые либо целиком состоят
из хищников, либо имеют в своем соста-
ве несколько родов хищников. Клопы-фи-
матиды (Phymatidae; рис. 11.4, А)-хищ-
Рис. 11.4. Хищные клопы: Л-клоп-фиматид Phymata erosa, хищник насекомых,
посещающих цветы; Ь’-клоп-хищнец, или поцелуйный клоп Reduvius personatus,
хищник постельных клопов и других насекомых, может кусать людей. (Из
Thecommon insects of North America, Lester A. Swan a. Charles S. Papp:
Л-рис. 103, c. 120; Б-рис. 105, c. 122.)
522
11. Полезные насекомые
ники, ведущие скрытый образ жизни; они
прячутся среди цветов и листьев расте-
ний и ловят оказавшихся поблизости на-
секомых. Такие семейства клопов, как
Reduviidae (рис. 11.4, Б), Nabidae и Ant-
hocoridae, также почти целиком состоят
из хищников насекомых. В семействах
Pentatomidae (щитники) и Miridae хищ-
никами являются представители не-
скольких родов. Как правило, хищные
клопы поедают других клопов, гусениц, а
также личинок жуков, но некоторые, по-
видимому, более приспособлены к поеда-
нию тлей, муравьев, постельных клопов
и насекомых из других групп.
Diptera
Многие двукрылые ведут хищный
образ жизни, что особенно характерно
для их личинок. Полезными хищниками
являются личинки Syrphidae (журчалок;
рис. 11.5, А), многие из которых по-
едают различных вредителей растений,
относящихся к Homoptera. Личиночная
стадия журчалок длится примерно 2-3
нед, так что во время вегетационного се-
зона численность их популяций быстро
увеличивается. В семействе Asilidae (кты-
ри, рис. 11.5, Б) хищничают и личинки, и
взрослые особи. Личинки ктырей оби-
Рис. 11.5. Хищные двукрылые: Л-цветочная муха Scaeva pyrastri (Syrphidae), хищник,
уничтожающий тлей; Б-ктырь Efferia interrupta (Asilidae), его личинки охотятся на
личинок, а взрослые особи-на взрослых слепней; В-муха-жужжало Bombylius major,
хищник личинок нескольких видов одиночных пчел; Г- муха-толкунчик Ramphomyia
sp., хищник мелких насекомых. (Из The common insects of North America, Lester
A. Swan a. Charles S. Papp: Л-рис. 1325, c. 617; Б-рис. 1305, В, с. 612; Б-рис. 1312,
с. 614; Г-рис. 1321, с. 617.)
11. Полезные насекомые
523
тают в почве и питаются личинками жу-
ков, взрослые ктыри ловят других насе-
комых на лету. В нескольких других
семействах двукрылых также известны
хищники, имеющие важное значение, на-
пример в семействе Bombyliidae (жуж-
жалы; рис. 11.5, В); хищные представи-
тели этого семейства интенсивно по-
едают яйца прямокрылых. У некоторых
видов в семействах Itonididae (галлицы) и
Culicidae (комары) хищничают личинки,
и известно несколько локальных попы-
ток использовать их в качестве агентов
борьбы с вредителями. В семействах
Rhagionidae (бекасницы) и Empididae
(толкунчики, рис. 11.5, Г) ряд видов ве-
дут хищный образ жизни на личиночной
и взрослой стадиях развития.
Hymenoptera
Хотя большинство видов этого се-
мейства являются паразитами, в не-
скольких семействах имеются и хищные
виды. Осы (Vespidae) кормят своих личи-
нок главным образом гусеницами бабо-
чек. Оса Polistes exclamans оказалась хо-
рошим регулятором численности браж-
ника Protopar се sexta, поддерживая ее на
приемлемом уровне. Поскольку Polistes
(рис. 8.124) живут в маленьких гнездах и
сами являются объектом хищничества,
целесообразно строить небольшие укры-
тия для их защиты. Настоящим осам
свойствен сходный характер питания, од-
нако он изучен хуже. Роющие осы также
кормят своих личинок насекомыми, и,
по-видимому, каждый вид имеет опреде-
ленные предпочтения в отношении
объекта хищничества. Один из видов
роющих ос, Bembix hinei, нападает в ос-
новном на слепней, и в южных штатах
этих ос часто можно встретить около до-
машнего скота. Другой вид, Sphecius spe-
ciosus (рис. 11.6), специализированный
хищник, нападающий на цикад. Несколь-
Рис. 11.6. Гигантское перепончатокрылое
Sphecius speciosus, хищник цикад. (Из The
common insects of North America, Lester A.
Swan a. Charles S. Papp: рис: 1228, c. 566.)
ко видов настоящих ос поедают двух вре-
дителей леса-еловую листовертку-поч-
коеда (Choristoneura fumiferana) и сосно-
вого пилильщика (Neodiprion swainei); эти
осы представляют определенную цен-
ность как возможные агенты борьбы с
этими вредителями.
Среди муравьев (Formicidae) также
имеется ряд групп, уничтожающих
вредных насекомых, в том числе и других
муравьев. Большинство хищных муравь-
ев являются, по-видимому, сохранивши-
мися представителями примитивных ве-
твей эволюции, они широко распростра-
нены в тропиках и умеренных зонах и
играют важную роль в регуляции числен-
ности хлопкового долгоносика, терми-
тов и многих других растительноядных
насекомых. Некоторые виды муравьев
приносят вред, так как они питаются вы-
делениями червецов, щитовок, тлей и
других растительноядных насекомых и
фактически защищают их, распугивая па-
разитов и хищников.
Mecoptera
Сюда относятся настоящие скорпион-
ницы (Panorpidae) и биттаки (Bittacidae),
524
11. Полезные насекомые
Рис. 11.7. Биттак Hylobittacus apicalis,
хищник летающих насекомых. (Из Thornhill,
1980.)
А. Вид с дорсальной стороны. Б. В висячем
состоянии. В. Хватательная лапка при
большом увеличении.
среди которых имеется ряд хищных ви-
дов, регулярно или эпизодически охотя-
щихся за другими насекомыми. Биттаки
(рис. 11.7) ловят мух, свешиваясь с рас-
тений, за которые они держатся передни-
ми ногами, и захватывая добычу двумя
другими парами ног
НАСЕКОМЫЕ-ПАРАЗИТЫ
Отношения паразит-хозяин у насе-
комых обычно носят вполне опреде-
ленный характер и отличаются специ-
фичностью. На преимагинальных ста-
диях паразит, как правило, живет за счет
одного хозяина и часто делит его с одним
или несколькими другими паразитами.
Паразитоиды дают возможность хозяи-
ну питаться и расти до определенного
момента, тогда они убивают его и после
этого окукливаются. Эти отношения ос-
нованы на строгой синхронизации жиз-
ненных циклов хозяина и паразитоида.
Паразитические
перепончатокрылые
Две основные ветви Hymenoptera -
примитивные паразиты, к которым отно-
сятся Ichneumonidae, Braconidae и Gaste-
ruptionidae, и специализированные пара-
зиты (рис. 8.111)-имеют большое хо-
зяйственное значение. У самок паразити-
ческих перепончатокрылых яйцеклад
приспособлен для прокалывания, про-
сверливания и введения жидкости и яиц;
кроме того, он служит органом чувств
для обнаружения и «опробывания» добы-
чи.
Мелкие бракониды паразитируют на
представителях Coleoptera, Lepidoptera,
Diptera и Hemiptera (Homoptera), а от-
дельные виды-на представителях других
групп. Часть из них-наружные пара-
зиты, другие - внутренние. Гусеница
бражника Protoparce quinquemaculata слу-
11. Полезные насекомые
525
жит хозяином бракониды Apanteles co-
ngregates (рис. 11.8, Б), а гусеница южной
капустной белянки Pieris protodice -хо-
зяином бракониды Apanteles glomeratus.
Паразитизм браконид приводит обычно
к тому, что гусеницы их хозяев становят-
ся неактивными или неспособными к за-
вершению жизненного цикла.
Браконидам принадлежит также важ-
ная роль в борьбе с некоторыми вредны-
ми насекомыми, естественные хищники
или паразиты которых не были интроду-
цированы вслед за ними в Северную
Америку. К числу таких ввезенных вре-
дителей относятся Stilpnotia salicis (иво-
вая волнянка), Nygmia phaeorrhoea (зла-
тогузка, рис. 8.185) и Porthetria dispar
(непарный шелкопряд). Эти бабочки бы-
ли завезены в Америку из Европы, и в це-
лях снижения их высокой численности
пришлось интродуцировать и их раз-
личных паразитов. С помощью интроду-
цированных браконид Apanteles solitarius,
A. lacteicolor, A. melanoscelus (рис. 11.8,
А) и Meteorus versicolor, паразитирующих
на одном или нескольких видах на-
званных бабочек, удавалось поддержи-
вать размеры популяций вредителей на
уровне, не представляющем опасности,
еще до начала широкого применения
ДДТ в конце 40-х-начале 50-х годов. По-
сле отказа от использования ДДТ для
борьбы с этими вредителями популяции
браконид были восстановлены. Серь-
езным вредителем яблонь считается
яблонная плодожорка Cydia pomonella\ в
ряде случаев сокращение численности ее
популяций достигается с помощью ее па-
разита-бракониды Ascogaster quadriden-
tata carpocapsae.
Помимо интродуцированных брако-
нид, которые используются для борьбы
Рис. 11.8. Бракониды (Hymenoptera): A-Apanteles melanoscelus; Б-Apanteles
congregates ;В-наезддик Horogenes punctorius, паразит стеблевого кукурузного мотылька.
(Из The common insects of North America, Lester A. Swan a. Charles S. Papp: Л-рис. 1150,
c. 524; Б-рис. 1151, c. 524; В-рис. 1161, c. 531.)
526
11. Полезные насекомые
со случайно завезенными вредителями,
численность большого числа потен-
циальных вредителей местной североа-
мериканской энтомофауны регулируется
местными браконидами. Американский
хлебный пилильщик Cephus cinctus, ко-
торый расширил свой ареал, перейдя с
диких злаков на зерновые поля канадских
прерий, является весьма серьезным вре-
дителем. Большинство видов перепонча-
токрылых, регулирующих численность
пилильщика на естественных лугах, не в
состоянии эффективно влиять на него на
обширных полях пшеницы. В этих усло-
виях только Bracon cephi оказался эффек-
тивным паразитом.
Ввезенная в Северную Америку во-
сточная персиковая листовертка Grapho-
litha molesta стала использоваться в каче-
стве хозяина несколькими североамери-
канскими паразитами, однако в данном
случае хозяино-паразитные отношения
не сделались специфическими или по-
стоянными. Поэтому ни один из этих па-
разитов не стал эффективным агентом
биологической борьбы, хотя многочис-
ленные паразитические виды, нападаю-
щие на листовертку, играют существен-
ную роль в снижении ее численности.
Североамериканский наездник Масгосеп-
trus ancylivorus (Braconidae) также неред-
ко использует восточную персиковую ли-
стовертку в качестве хозяина. Другие
насекомые, живущие в растительности
вблизи фруктовых садов, служат есте-
ственным резервуаром для паразита. В
ЗО-е годы удалось распространить М.
ancylivorus на большей части территории
персиковых садов в восточных штатах.
После того как в середине 40-х годов во-
сточная персиковая листовертка была
обнаружена в Калифорнии, там началось
массовое культивирование М. ancylivo-
rus, причем в качестве дополнительного
хозяина были использованы личинки вы-
емчатокрылой моли Gnorimoschema орег-
culella, размножившейся в результате
массового культивирования божьих ко-
ровок. Введение в практику борьбы в
конце 40-х годов. ДДТ и других род-
ственных пестицидов уменьшило зависи-
мость садоводов от разводимых парази-
тов, и их культивирование было сокра-
щено.
Ихневмониды, подобно браконидам,
также оказались чрезвычайно эффек-
тивными паразитами широкого круга
вредных насекомых. Эта группа перепон-
чатокрылых почти целиком состоит из
паразитов, поражающих в основном Hy-
menoptera и личинок Lepidoptera. Наезд-
ников использовали для регулирования
численности елового пилильщика Diprion
hercyniae, ставшего крайне серьезным
вредителем лесов в восточных областях
Канады и в северо-восточной части Но-
вой Англии в 30-х и начале 40-х годов.
Наездники Aptesis basizonia, Exenterus
abruptorius, E. amictorius, E. tricolor E. cla-
ripennis и E. cpnfusus были интродуциро-
ваны в Северную Америку, где хорошо
приспособились к паразитированию на
этом пилильщике. До 1942 г сотни мил-
лионов этих паразитов были разведены
или завезены для использования в широ-
ко проводимой программе борьбы с
еловым пилильщиком. Впоследствии пи-
лильщики были поражены смертельным
для них вирусом, и после 1942 г числен-
ность этого вида сохраняется на весьма
низком уровне.
Dahlbominus fuscipennis - другой пред-
ставитель наездников, которого с успе-
хом использовали для борьбы против ли-
ственничного пилильщика в северо-
восточных штатах США. Зимующий по-
беговьюн Rhyacionia bouliana поражается
несколькими видами интродуциро-
ванных паразитических перепончатокры-
лых, но лишь с ограниченным успехом.
Кукурузного мотылька впервые обна-
ружили в Северной Америке приблизи-
11. Полезные насекомые
527
тельно в 1917 г., и в борьбе с ним широко
использовались паразиты. Было ввезено
более 22 видов паразитов кукурузного
мотылька, 6 из них хорошо акклимати-
зировались, в том числе 2 вида ихневмо-
нид-Horogenespunctorius (рис. 11.8, В) и
Phaeogenes nigridens. Степень поражения
ими личинок кукурузного мотылька до-
стигает 50-60%.
В группу хальцид входят несколько
семейств специализированных парази-
тов. Большинство представителей этих
семейств имеют очень мелкие размеры
(1-2 мм) и являются эффективными вну-
тренними паразитами Coleoptera, Lepi-
doptera, Hemiptera (Homoptera) и неко-
торых Diptera. Наряду с другими пере-
пончатокрылыми паразитами некоторые
виды хальцид также были интродуциро-
ваны для подавления численности вве-
зенных вредных насекомых в связи с тем,
что аборигенные хищники и паразиты не
справлялись с этой задачей. Хальцида
Aphytis chrysomphali паразитирует в крас-
ной померанцевой щитовке Aonidiella аи-
rantii; по-видимому, и паразит, и хозяин
были завезены примерно в одно и то же
время. Интродуцированные виды халь-
цид Aphytis lingnamensis и A. melinus вме-
сте с A. chrysomphali используют для
борьбы с красной померанцевой щитов-
кой в большинстве прибрежных цитру-
совых рощ Калифорнии. Другой также
интродуцированный вид Comperiella bi-
Рис. 11.9. Хальциды: А - Trichogramma minutum, паразит яиц нескольких
сотен видов насекомых; Б-Comperiella bifasciata, паразит личинок желтой и
нескольких красных померанцевых щитовок; В-золотая хальцида Aphytis
chrysomphali, паразит многих видов щитовок; Г- Aphelinus mali, паразит
многих вредоносных тлей. (Из The common insects of North America, Lester
A. Swan a. Charles S. Papp: Л-рис. 1165, c. 532, 1>-рис. 1172, c. 536;
В-рис. 1167, c. 534; Г-рис. 1169, c. 534.)
528
11. Полезные насекомые
fasciata паразитирует на желтой апельси-
новой щитовке Aonidiella citrina и контро-
лирует численность этой щитовки на
большей части ее ареала. С помощью
ввезенных хальцид рода Aphelinus уда-
лось снизить численность масличной
ложнощитовки Saissetia oleae в южных
районах прибрежной Калифорнии, хотя
в ряде мест необходимы дополнитель-
ные меры борьбы. Яблонная кровяная
тля Eriosoma lanigerum, обитающая на
северо-востоке Северной Америки, ли-
шилась своего постоянного паразита Ap-
helinus mali, когда была неумышленно
перенесена на западное побережье; ин-
тродуцированный позднее A. mali смог
обосноваться в штатах Вашингтон и
Орегон, но дальше к югу не прижился.
Среди прочих в группу хальцид
(рис. 11.9) входят два семейства, пред-
ставленные очень мелкими особями, ко-
торые приспособились паразитировать в
яйцах других насекомых. Это семейства
Mymaridae и Trichogrammatidae, в по-
следнем есть вид, особи которого не до-
стигают в длину и четверти миллиметра.
Эти хальциды успешно контролируют
численность многих вредных насекомых.
В штате Юта удалось сократить числен-
ность люцернового долгоносика Нурега
postica, когда туда был ввезен из южной
Европы Mymar pratensis (Mymaridae).
Представители семейства Trichogramma-
tidae являются паразитами яиц многих
насекомых, в частности они оказались
эффективными регуляторами численно-
сти совки Heliothis zea и еловой листо-
вертки-почкоеда Choristoneura fumiferana.
В состав семейств Encyrtidae и Eupelmi-
dae также входят виды, паразитирующие
в яйцах насекомых и имеющие большое
хозяйственное значение.
Жалящим перепончатокрылым также
принадлежит важная роль в регуляции
численности многих видов насекомых.
Некоторые из них проявляют избира-
тельность в выборе своих жертв, напри-
мер представители семейства Scoliidae (с.
366); однако другим, например Vespi-
dae, такая избирательность не свойствен-
на.
Diptera
Tachinidae, одно из многих полезных
для человека семейств двукрылых, пред-
ставлено крепко сложенными и быстро
летающими насекомыми - паразитами
ряда уже упоминавшихся выше вредных
насекомых.
Обычно тахины поражают личи-
ночные стадии своих хозяев, чаще всего
жуков и бабочек. Ряд видов тахин был
использован в качестве агентов биологи-
ческой борьбы в тропиках; например,
тахина Microceromasia sphenophori была
интродуцирована на Гавайские острова
для борьбы с долгоносиком Rhabdoscelus
obscura. 'Североамериканская тахина
Winthemia quadripustulata (рис. 11.10, А)
паразитирует на многих гусеницах,
вредящих различным сельскохозяйствен-
ным культурам, Lydella thompsoni (рис.
11.10, Б) была ввезена в восточные
Рис. 11.10. Мухи-тахины: Л-тахина
Winthemia quadripustulata, паразит гусениц
совок и других насекомых, Б-Lydella
thompsoni, паразит стеблевого кукурузного
мотылька. (Из The common insects of North
America, Lester A. Swan a. Charles S. Papp:
Л-рис. 1395, с. 648;Ь*-рис. 1396, c. 648.)
11. Полезные насекомые
529
районы Северной Америки из Европы
для использования в борьбе с куку-
рузным мотыльком. Несколько видов та-
хин паразитируют на завезенных из
Европы златогузке и непарном шелко-
пряде. Помимо их значения как специфи-
ческих паразитов вредных насекомых му-
хи-тахины, имеющие широкий круг хо-
зяев, играют также важную роль в под-
держании «равновесия в природе».
БОРЬБА С СОРНЯКАМИ
Одну из наиболее полезных групп со-
ставляют насекомые, питающиеся на
растениях-сорняках, которые, с нашей
точки зрения, крайне нежелательны. На-
ибольший практический интерес предста-
вляют не обычные для фермерских и са-
довых участков виды Ъорняков, с которы-
ми можно бороться и другими способа-
ми, а те, которые вытесняют другую
растительность на больших террито-
риях, делая их непригодными для ис-
пользования под сельскохозяйственные
культуры. Классическими примерами та-
ких сорняков являются: 1) американский
кактус-опунция (Opuntia sp.), завезенный
в Австралию, где он широко распростра-
нился и разрушил миллионы гектаров
пастбищ для скота, 2) европейский
многолетний зверобой {Hypericum perfo-
ratum), завезенный на аридные пастбища
Австралии и Калифорнии, где он вытес-
нил и заменил собой пастбищные травы
на территориях в тысячи квадратных ки-
лометров, и 3) тропическое американ-
ское растение памакани Eupatorium ade-
nophorum, завезенное на Гавайи, где оно
разрослось и образовало плотный непро-
ницаемый полог до 3 м высотой, что
привело к гибели травяного покрова и к
уничтожению пастбищ на огромных пло-
щадях. В этих, а также многих других
подобных случаях огромные территории,
имевшие большое хозяйственное значе-
ние, стало невозможно использовать в
качестве сельскохозяйственных угодий.
Описанные события навели на мысль
о том, что для снижения численности
этих растений можно использовать их
насекомых-вредителей. Сначала эта идея
была встречена без особого энтузиазма.
Однако широкое распространение опун-
ции в Австралии к 1925 г стимулировало
поиски питающихся этим растением на-
секомых, которые могли бы сократить
его численность. Высокоэффективной в
этом отношении оказалась южно-афри-
канская бабочка Cactoblastis cactorum, ее
интродукция в Австралию привела к со-
кращению площадей, занятых опунцией,
почти на 99%, благодаря чему эти терри-
тории восстановили свое значение как
пастбища для скота. Позже с помощью
жука-листоеда Chrysolina gemellata уда-
лось существенно снизить численность
зверобоя как в Австралии, так и в Север-
ной Америке. Плодовая мушка Proceci-
dochares utilis помогла в борьбе с памака-
ни на Гавайских островах. В настоящее
время в разных местах земного шара с
помощью насекомых контролируется
численность примерно десятка видов
растений-сорняков. Использование насе-
комых в борьбе с сорняками стало благо-
даря достигнутым успехам одним из
важнейших направлений исследователь-
ских работ.
Этот метод стал в последнее время
применяться в отношении водных расте-
ний, «засоряющих» речки, пруды и озера
и создающих препятствия для плавания,
рыболовства и купания. Одним из таких
растений является альтернантера Alter-
nanthera philoxeroides, завезенная сначала
из Южной Америки во Флориду, став-
шая теперь обычным видом в юго-
восточных штатах, и на запад до Кали-
форнии. Несколько видов насекомых
(рис. 11.11), питающихся на этом расте-
34-732
530
11. Полезные насекомые
Рис. 11.11. Насекомые, питающиеся сорным растением альтернантерой. (По материалам
министерства сельского хозяйства США, Tech. Bull. 1547.)
А. Бабочка Vogtia malloi хорошо замаскирована на мертвом стебле. Б. Южноамериканский
листоед Agasicles hygrophilia на альтернантере, своем естественном хозяине. В. Гусеница V
malloi в стебле альтернантеры.
нии в Южной Америке, были ввезены в
южные штаты США, и можно надеяться,
что один из этих видов, листоед из рода
Agasicles, окажется эффективным аген-
том регулирования численности альтер-
нантеры.
При интродукции насекомых для
борьбы с сорняками следует соблюдать
чрезвычайную осторожность, чтобы слу-
чайно не ввезти такое насекомое, которое
может сменить растение-хозяина, перей-
дя на полезный для человека вид. В связи
с этим любой интродукции насекомого в
новую для него страну предшествует
большой объем специальных исследова-
ний, направленных на изучение всего кру-
га его растений-хозяев и его климатиче-
ских адаптаций.
НАСЕКОМЫЕ-ОПЫЛИТЕЛИ
Не будь растений, опыляемых насе-
комыми, пища человека была бы значи-
тельно беднее, чем она есть теперь. Насе-
комые опыляют все фруктовые растения,
относящиеся к семейству розоцветных
(Rosaceae), в том числе розу, клубнику,
яблоню и вишню, а также дыню и тыкву
(Cucurbitaceae) и богатые белком горох,
бобы и родственные им растения (Legu-
minosae). Хотя у многих растений мы
употребляем в пищу вегетативные части
или структуры, образующиеся до опыле-
ния, например морковь (Umbelliferae) и
капусту (Cruciferae), нам все равно необ-
ходимо получать семена этих растений,
чтобы вырастить новый урожай. Семена
.. Полезные насекомые
531
многих растений используются также как
источник масла или в качестве приправ,
52лример семена перца или сельдерея.
О -нлко лишь небольшое число семейств
Гостений. опыляемых насекомыми, ис-
пользуется человеком в качестве продук-
т ?з литания, приправ и напитков. В крат-
ким. но исчерпывающем и весьма инфор-
мативном руководстве Мак-Грегора
McGregor, 1976) перечисляется более 200
возделываемых сельскохозяй-
ственных культур, относящихся почти к
5* семействам, встречающихся на терри-
тссии США, включая Гавайи; все эти
г?ль туры опыляются главным образом
=зсекомыми. Многие из этих растений, в
т см числе и те, у которых нет ярких цве-
т:в. опыляются мухами, бабочками, жу-
и другими насекомыми, обитающи-
ми на растениях.
Насекомые опыляют и другую боль-
~и разнообразную группу сельскохо-
тЕйственных растений - подавляющее
сельшинство садовых цветов. Сюда от-
бесятся все семейства растений, перечис-
ленные выше, и еще множество видов.
Несколько меньшее число растений вы-
тягивается для приготовления из них ле-
ятсг венных препаратов (например, ма-
£ < или инсектицидов (например, ромаш-
гя из которой готовят пиретрум).
Насекомые опыляют небольшое чис-
- ? зилов деревьев, выращиваемых на ле-
;.^термах США. Исключение составляют
?йлы. выращиваемые в маточных питом-
американские виды кизила, виш-
г-о. б\зину, тюльпанный тополь и магно-
тс-п?. а также ряд интродуцированных
‘ПОВ.
Опыление растений, имеющих яркие
лзетки. осуществляется в основном ме-
л-косными пчелами. Если садовод не
тьстюлагает достаточным числом пчел в
тзоем хозяйстве, чтобы обеспечить пол-
-оленное опыление, то он арендует нуж-
еге число ульев у пчеловода и, таким
образом, обеспечивает опыление своего
сада. Во многих районах страны аренда
пчел прекрасно организована и соста-
вляет важный компонент сельскохозяй-
ственной экономики.
Многие дикие пчелы при достаточной
численности также являются прекрасны-
ми опылителями сельскохозяйственных
культур. Поискам способов повышения
численности популяций диких пчел-опы-
лителей было посвящено множество ис-
следований. К настоящему времени по-
лучены прекрасные результаты в отно-
шении солончаковой пчелы Nomia соске-
relli в ряде западных и центральных
штатов и пчелы-листореза Megachile ра-
cifica. В тропических районах безжалые
пчелы, относящиеся к родам Melipona и
Trigona, служат прекрасными опылителя-
ми и довольно легко «одомашнивают-
ся», однако они не выносят морозов, и
поэтому их нельзя использовать в райо-
нах с умеренным климатом. Хотя попыт-
ки использования других американских
пчел в качестве опылителей имели лишь
ограниченный успех, эти виды, часто
весьма многочисленные, могут быть хо-
рошими опылителями на небольших тер-
риториях.
НАСЕКОМЫЕ - ПРОИЗВОДИТЕЛИ
ПОЛЕЗНЫХ ПРОДУКТОВ
На протяжении истории своего разви-
тия человек пользовался рядом продук-
тов, которые создавали насекомые и ко-
торые сохранили свое значение и поныне.
Мед и пчелиный воск. В Северной
Америке эти продукты создают медо-
носные пчелы Apis mellifera. Мед, по-
видимому, издревле служил лакомством
человека и это в свою очередь привело к
открытию воска, который имел разноо-
бразное применение. Разведение пчел с
целью получения этих продуктов стало в
наши дни доходным занятием, однако
532
11. Полезные насекомые
пчелиный воск уступает современным
синтетическим соединениям, сходным с
ним по свойствам, но более дешевым.
Шелк. В течение столетий шелк, выра-
батываемый гусеницами шелкопряда,
оставался тончайшим по своей фактуре
материалом, используемым для про-
изводства чулок и различных тканей. За
последние несколько десятилетий в Се-
верной Америке и Европе нейлон и дру-
гие прочные синтетические материалы в
значительной степени вытеснили про-
изводство шелка, оставив в качестве его
потребителей лишь людей с аллергией к
синтетическим тканям. Однако во всем
мире шелк сохраняет еще свое значение -
ежегодно его производится около
32 млн. кг
Шеллак. Laccifer lacca (Coccidae), оби-
тающий в Азии, выделяет секрет, покры-
вающий плотной коркой веточки расте-
ния, на котором живет это насекомое.
Этот секрет отделяют от растения, рас-
творяя его, и получают из него шеллак. В
настоящее время шеллак во многих слу-
чаях заменяется синтетическими мате-
риалами, но в азиатских странах при из-
готовлении лакированных изделий до
сих пор используется большое количе-
ство шеллака.
Другие продукты. В прошлом неко-
торые виды насекомых служили источни-
ком для приготовления красок и лекар-
ственных препаратов, но теперь синтети-
ческие продукты почти полностью вытес-
нили их. Юго-западная кокцида Dactylo-
pius coccus остается источником темно-
красной краски кошенили. Хорошо из-
вестное лекарство кантаридин получают
из высушенных жуков-нарывников Lytta
vesicatoria, которых обычно называют
шпанской мушкой.
ДРУГИЕ СПОСОБЫ
ИСПОЛЬЗОВАНИЯ НАСЕКОМЫХ
Насекомые в пище человека
В доисторические эпохи насекомые
составляли обычный компонент пищи че-
ловека, однако сейчас их потребление но-
сит очень ограниченный характер. Самы-
ми распространенными из насекомых,
употребляемых в пищу, являются саран-
човые, поедаемые в тушеном или жаре-
ном виде. В Мексике жареные насе-
комые, в том числе высокоценимые гусе-
ницы, живущие в агавах и называемые
по-испански gusanos de maguey, считают-
ся деликатесом; из них готовятся кон-
сервы, идущие на экспорт Япония экс-
портирует консервы, приготовленные из
личинок ручейников и веснянок и куко-
лок шелкопрядов.
Высокое содержание жиров и белка в
тканях насекомых делает последних
идеальным дополнением к углеводной
пище. В Африке с помощью белков насе-
комых удается предотвратить заболева-
ние детей-квашиоркор, вызываемое не-
достатком белков в пище.
Насекомые
как лабораторные животные
Исследования, проводимые на насе-
комых, внесли и продолжают вносить
огромный вклад во многие отрасли на-
уки. Так, в XV и XVI вв. был внесен
большой вклад в зародившуюся тогда
сравнительную анатомию. В начале на-
шего века плодовая мушка Drosophila по-
служила важным объектом классических
исследований по генетике. На основе
этих исследований позднее были созданы
некоторые из наиболее ярких концепций,
касающихся видообразования и эволю-
ции. И сегодня Drosophila остается
важным объектом генетических исследо-
11. Полезные насекомые
533
ваний и вносит свой вклад в дальнейшее
развитие эволюционной теории. Многие
группы насекомых дают материал для
более углубленного понимания основ фи-
логении, экологии, динамики популяций
и биогеографии.
Объясняется это 1) наличием огром-
ного числа видов насекомых как иско-
паемых, так и современных, и наличием
большого числа современных насе-
комых, мало изменившихся по сравне-
нию с древними исходными формами и
являющихся в сущности «живыми иско-
паемыми»; 2) коротким жизненным ци-
клом и высоким репродуктивным потен-
циалом многих насекомых по сравнению
с такими экологически сходными форма-
ми, как птицы или млекопитающие.
Поскольку насекомые имеют неболь-
шие размеры, их мало использовали в
ранних физиологических и биохимиче-
ских исследованиях. Однако с внедре-
нием ультрамикротехнических методов
химического анализа и методов радио-
изотопного мечения насекомые приобре-
ли важную роль как объекты физиологи-
ческих и биохимических исследований.
Это особенно справедливо в отношении
таких областей науки, как эндокриноло-
гия и поведение животных, поскольку на
насекомых не только можно проводить
чрезвычайно сложные хирургические
операции, но и сопоставлять данные о
протекающих в организме биохимиче-
ских процессах с физиологическими и по-
веденческими реакциями подопытных
животных. По этим же причинам насе-
комые являются идеальными объектами
при решении различных экологических
проблем.
Рекреационное
эстетическое значение насекомых
Во всем мире наблюдается сейчас по-
вышенное стремление человека к обще-
нию с природой. Формы такого общения
могут быть весьма различными: от спор-
тивного рыболовства и охоты до прогу-
лок и от разных видов активного отдыха
до простого наблюдения и созерцания. В
США в последние годы наблюдается все
больший приток отдыхающих в нацио-
нальные парки и другие места, подходя-
щие для устройства лагеря, проведения
пикника или установки трайлера.
Чувствам радости и умиротворенно-
сти, которые охватывают нас на лоне
природы, мы в значительной степени
обязаны насекомым, хотя осознаем это
не сразу. Почти все растения с яркими
цветами-от маленьких звездчаток до
маргариток и остролистов, от кизила до
тюльпанного дерева и магнолий-суще-
ствуют благодаря огромной армии насе-
комых-опылителей. Целые семейства
птиц-стрижи, ласточки, дятлы, славки,
крапивники, мухоловки, тираны-пи-
таются только насекомыми, а у многих
других птиц насекомые составляют зна-
чительную часть диеты. Даже питающие-
ся семенами кардиналы своих птенцов
выкармливают насекомыми. Змеи, яще-
рицы, лягушки и жабы, летучие мыши,
землеройки и некоторые другие млекопи-
тающие питаются в основном насекомы-
ми. В отсутствие другой пищи лисы,
скунсы, еноты-полоскуны и медведи так-
же находят и поедают насекомых. Таким
образом, удивительно большое число
живых существ в той или иной степени
зависит от насекомых. Вместе с тем и са-
ми насекомые представляют собой пред-
мет эстетического наслаждения, о чем
свидетельствует большое число людей,
для которых изучение насекомых соста-
вляет одно из приятнейших занятий.
Удовольствие, которое получают лю-
бители отдыхать у воды или любители
водных видов спорта, также во многом
связано с насекомыми. Большинство
рыб, являющихся объектом рыбной л о-
534
11. Полезные насекомые
вли, питаются насекомыми либо более
мелкими рыбами, которые в свою оче-
редь питаются насекомыми. Иногда ры-
бья «трапеза» представляет собой за-
хватывающее зрелище. Например, когда
в реке Ганнисон происходит вылупление
личинок веснянок, известие об этом со-
бытии передается по радио и телевиде-
нию, и рыбаки из Колорадо и соседних
штатов собираются для увлекательной
ловли форели. Подобные зрелища из-
вестны и во многих других частях света,
например массовое вылупление белых
ручейников в форелевых ручьях Тасма-
нии. Нет более яркой иллюстрации роли
насекомых в спортивном рыболовстве,
чем сотни приманок для рыбы, имити-
рующих различных насекомых.
Есть еще один аспект, касающийся
пресноводных водоемов, где насекомые
также играют важную роль. Многим, ес-
ли не всем пресноводным водоемам
угрожает загрязнение различными хими-
ческими веществами, которые оказы-
вают губительное действие на обитаю-
щие в них живые организмы. Выясни-
лось, что водные насекомые чрезвычайно
чувствительны к изменениям, происходя-
щим в экосистемах пресноводных водое-
мов, и поэтому сейчас они все более ши-
роко используются в программах мони-
торинга, предназначенных для устано-
вления степени загрязнения водоемов.
НАСЕКОМЫЕ-ДЕСТРУКТОРЫ
И КРУГОВОРОТ ПИТАТЕЛЬНЫХ
ВЕЩЕСТВ
Насекомые играют важную роль в го-
довом цикле основных питательных ве-
ществ в экосистеме, о чем уже говори-
лось в предыдущей главе. Каждый год
зеленые растения посредством фотосин-
теза и других химических процессов, в
которых используется вода и двуокись
углерода, создают углеводы, витамины и
другие вещества, необходимые для их ро-
ста. Но растения не способны синтезиро-
вать ряд важных для их жизнедеятельно-
сти веществ, особенно веществ, содержа-
щих азот, калий, фосфор, серу и многие
другие элементы. Они должны получать
эти вещества из внешней среды. По-
скольку вещества, о которых идет речь,
находятся в почве в растворенном со-
стоянии, они постепенно вымываются, и
их количество может дойти до критичес-
ки низкого уровня. На этой стадии ос-
новным источником питательных ве-
ществ становятся различные части расте-
ния, падающие на землю и образующие
слой подстилки и гумуса на поверхности
почвы. Этот слой может состоять из
старых листьев, мертвых веток или
травы, целых кустов или деревьев, погиб-
ших и упавших на землю или в ручьи.
Этот слой мертвых растительных остат-
ков включает также тела мертвых жи-
вотных и фекалии живых.
В результате деятельности различных
микроорганизмов слой детрита разла-
гается на простые составные части, и та-
ким образом появляются элементы, не-
обходимые для роста нового зеленого
растения. Однако в ряде тщательных ис-
следований было показано, что насе-
комые и некоторые близкие к ним
формы, например клещи и многоножки,
питающиеся детритом, ускоряют этот
процесс химического превращения, меха-
нически размельчая растительные ткани
и способствуя, следовательно, более быс-
трому проникновению микроорганизмов
в мертвый растительный материал. Этот
материал, или гумус, сначала подвер-
гается воздействию коллембол, протур,
диплур, сапрофитных клещей, многоно-
жек и многих сапрофитных личинок насе-
комых. Мертвые пни и ветви заселяются
термитами, личинками жуков, прогры-
зающих ходы в древесине, и многи-
ми другими насекомыми, многонож-
IE Полезные насекомые
535
ками и близкими к ним беспозвоноч-
ными.
Насекомые часто могут принимать и
более прямое участие в круговороте пи-
тательных веществ. Многие насекомые
питаются живыми листьями, веточками
а стеблями растений, и их фекалии попа-
дают в слой детрита-размельченного
растительного материала, который под-
вергается обработке беспозвоночными
2лн микроорганизмами, использующи-
ми его в пищу.
Помимо этого, насекомым принадле-
жит очень важная роль в процессах раз-
рушения и усвоения растительного мате-
риала. падающего в воду у истоков рек.
Многие крупные личинки веснянок и ру-
жейников измельчают и перерабатывают
растительный материал с тщатель-
ностью, сравнимой с наземными потре-
бителями листвы. Фекалии этих насе-
комых включаются в поток веществ,
направляющихся вниз по течению. Мно-
гие личинки поденок и ручейников имеют
приспособления для вылавливания мел-
ких частиц из текущей воды, используя
затем их или связанных с этими частица-
ми микроорганизмов в пищу.
Суммарное воздействие этих процес-
сов как в наземных, так и в водных био-
ценозах сводится к максимальному ис-
пользованию питательных веществ, уча-
ствующих в круговороте, непосредствен-
но в месте их возникновения до того, как
они будут смыты в реку, а затем в океан.
О роли насекомых как деструкторов
Рис. 11.12. Жуки-навозники семейства Scarabaeidae (подсемейство
(Scarabaeinae). (Из Waterhouse, 1974.)
A. Garreta nitens, делающий навозные шары. Б, В. Onthophagus gazella и
Euoniticellus intermedium оба относящиеся к роющим видам. Г Heliocopris
gigas, крупный вид, способный отражать нападение мелких позвоночных,
например жаб.
536
11. Полезные насекомые
Рис. 11.13. Разложение навоза жуками-навозниками. (Из Waterhouse, 1974.)
А. Жуки, такие, например, как Garreta nit ens, делающие нцвозные шары. Б. Жуки,
такие, например, как Onthophagus gazella, роющие норки.
знали еще многие тысячи лет назад древ-
ние египтяне, которые считали навозного
жука Scar abacus sacer священным. Этот
вид, так же как и многие другие Scara-
baeidae (рис. 11.12), поглощает ежеднев-
но огромное количество навоза, превы-
шающее в 10-12 раз вес их тела. Самка
этого жука заготавливает навоз, с тем
чтобы вылупившиеся личинки могли сра-
зу же питаться им. Обычно скарабей при-
катывает навозный шар.в определенное
место, где и зарывает его (рис. 11.13).
Эти жуки мастерски очищают пастбища
от навоза и препятствуют, таким обра-
зом, развитию в нем мух и других
вредных насекомых.
В тех местах, где либо нет жуков-
навозников, либо имеющиеся виды рабо-
тают недостаточно эффективно, разло-
жение навоза происходит медленно и
образуются его скопления. В Австралии,
где сельское хозяйство имеет в основном
скотоводческое направление, утилизация
навоза, образующегося при содержании
импортированных из Европы пород ско-
та, представляет серьезную проблему,
так как австралийские жуки-навозники не
способны к его быстрому разложению.
Навоз от каждого животного в стаде еже-
годно губит почти десятую часть гектара
полезной площади. Для разложения на-
воза требуется 2-3 года. Много неприят-
ностей доставляют также мухи и другие
вредные насекомые, живущие в навозе.
Для того чтобы решить эту проблему,
австралийское правительство начиная с
1967 г. приступило к ввозу нескольких
видов жуков-навозников. За 6 лет с нача-
11. Полезные насекомые
537
ла осуществления этого проекта один из
видов навозников, Onthophagus gazella,
акклиматизировался в северной части
Австралии и в восточных прибрежных
районах вплоть до Ньюкасла на юге.
Особенно эффективен этот жук во
влажные сезоны года. Были акклимати-
зированы также еще несколько видов жу-
ков-навозников. Все эти мероприятия
привели к увеличению полезных площа-
дей пастбищ, улучшению круговорота
ЛИТЕРАТУРА
Apple J. L., Smith R. F. (Eds.). Integrated pest
management. Plenum, New York, 214 pp.,
1976.
Arnaud P. H., Jr. A host-parasite catalog of
North American Tachinidae (Diptera), USDA
Misc. Publ. 1319, 860 pp., 1978.
Bohart G. E. Management of wild bees for the
pollination of crops, Annu. Rev. Entomol., 17,
287-312 (1972).
Chapman R. F., Ветауs E. H. (Eds.). Insect
and host plant. Nederlandse Entomologische
Vereniging, Amsterdam, pp. 201-766, 1978.
Daly H. K, Doyen J. T., Ehrlich P. R. Intro-
duction to insect biology and diversity.
McGraw-Hill, New York, 564 pp., 1978.
DeBach P. Biological control by natural ene-
mies. Cambridge Univ. Press, London and
New York, 323 pp., 1974.
Free J. B. Insect pollination of crops. Academic
Press, London and New York, 544 pp., 1970.
Heinrich B. Bumblebee economics. Harvard
азота, повышению качества кормов. Ав-
стралийцы заняты сейчас поисками хищ-
ника для личинок мух, обитающих в на-
возе; эта проблема остается пока нере-
шенной.
В Северной Америке обычными пред-
ставителями жуков-навозников являются
Canthon pilularis, Aphodius fimetarius и
Geotrupes splendidus. Эти виды наряду с
другими прекрасно выполняют роль де-
структоров навоза.
Univ. Press, Cambridge, Mass., 246 pp., 1979.
Heinrich B., Bartholomew G. A. The ecology of
the African dung beetle, Sci. Am., 241, 146-156
(1979).
Huffaker С. B., Messenger P. S. (Eds.). Theory
and practice of biological control. Academic
Press, New York, 788 pp., 1976.
McGregory S. E. Insect pollination of cultivated
crop plants. USDA Agric. Handbook 496, 411
pp., 1976.
Ridgeway R. L., Vinson S. B. (Eds.). Biological
control by augmentation of natural enemies.
Plenum, New York, 480 pp., 1977.
Swan L. A. Beneficial insects. Harper & Row,
New York, 429 pp., 1964.
Swan L. A., Papp C. S. The common insects of
North America. Harper & Row, New York,
750 pp., 1972.
Waterhouse D. F. The biological control of
dung, Sci. Am., 236, 100-107 (1974).
12
Вредные насекомые
и борьба с ними
Многие тысячи видов насекомых и паукообразных Северной Америки тем или
иным способом вредят человеку. Огромное число разных видов питается на расти-
тельности, другие проникают в места хранения сельскохозяйственной продукции,
третьи являются паразитами человека и домашних животных. Подавляющее боль-
шинство этих вредителей встречаются лишь местами и спорадически, как, например,
один из видов пилильщиков, поражающий листопадные азалии, или редкие виды ко-
маров, достигающие высокой численности только при необычных погодных усло-
виях. Лишь около 1000 видов могут быть отнесены к постоянным вредителям, нано-
сящим основную часть ущерба, связанного с членистоногими.
Проявления этого ущерба могут быть самыми разнообразными. Наиболее замет-
на дефолиация растительности, вызываемая насекомыми с грызущим ротовым аппа-
ратом ; такие насекомые лишают растения значительной части листвы или побегов,
столь же тяжелые последствия может иметь потеря растением корней или повреж-
дение любых его частей. Насекомые с сосущим ротовым аппаратом потребляют соки
растений или содержимое растительных клеток; некоторые насекомые выделяют ток-
сины, отравляющие растение, или переносят различные патогенные организмы от
одного растения к другому. В некоторых случаев вред приносит откладка насекомы-
ми яиц (рис. 12.1). Человеку и домашним животным основной ущерб приносят
«укусы» членистоногих, обладающих колюще-сосущим ротовым аппаратом, в част-
ности «укусы» мух, блох, вшей и клещей. Многие из этих членистоногих переносят
микроорганизмы, вызывающие опаснейшие заболевания человека, такие, как малярия
или бубонная чума.
Одной из первейших задач прикладной энтомологии всегда было и остается до сих
пор сокращение численности вредных видов членистоногих до уровня, при котором
устраняются или предотвращаются материальные потери. Как мы увидим далее в
этой главе, достигать этого становится все труднее и труднее. Цена поддержания чис-
ленности вредителей на каком-то приемлемом уровне часто рассматривается как про-
блема сравнения стоимости различных мероприятий в системе отношений расходы/
прибыль. Ниже представлен краткий обзор видов и групп членистоногих, в наи-
большей степени вредящих сельскохозяйственным культурам или продуктам или
причиняющих какой-либо другой ущерб.
НАСЕКОМЫЕ-ВРЕДИТЕЛИ РАСТЕНИЙ
В эту группу входит наибольшее чис-
ло видов насекомых-вредителей. Причи-
няемый ими ущерб может быть самым
разнообразным, а сами виды обладают
самыми разными биологическими харак-
теристиками.
Насекомые - вредители сельскохозяй-
ственных полевых культур. Все основные
полевые культуры в той или иной сте-
пени подвержены нападению со стороны
насекомых. Серьезными вредителями
хлопчатника (рис. 12.2) являются хлоп-
12. Вредные насекомые и борьба с ними
539
Рис. 12.1. Повреждения на растениях, связанные с откладкой насекомыми
яиц. (По Metcalf a. Flint, Destructive and useful insects, McGraw-Hill Book
Co.)
А. Расщепление ветки 17-летней цикадой. Б. Лунки на ветке малины,
сделанные сверчком-трубачиком. В. Разрезы на коре яблони, сделанные
цикадой-горбаткой. Г Ветка ореха пекана, почти перерезанная надвое
жуком Oncideres cingulata. Д. Плод вишни с двумя проколами,
сделанными жуком-долгоносиком при откладке яиц.
ковый долгоносик и хлопковая совка, пи-
тающиеся внутри семенной коробочки и
разрушающие развивающееся волокно;
хлопковая американская совка, гусеницы
которой поедают листву, и хлопковая
тля, высасывающая сок из стеблей и
листьев. Посевы кукурузы могут быть
почти полностью уничтожены саран-
човыми, поедающими листву, или кло-
пом-черепашкой, питающимся соками
растения. Ежегодно урожай кукурузы
значительно снижается из-за нападения
на нее нескольких видов точильщиков,
включая стеблевого кукурузного моты-
лька и юго-западную кукурузную огнев-
ку. Пшеница и другие злаки подвергают-
ся нападению совок, проволочников,
тлей и саранчовых, набор видов-вредите-
лей определяется районом возделывания
и климатическими условиями. Личинки
гессенской мушки повреждают стебли и
листья зерновых культур; этот вид
является самым страшным вредителем
пшеницы. Вредители табака, этой высо-
кодоходной культуры, питаются
главным образом его листьями; к числу
540
12. Вредные насекомые и борьба с ними
Рис. 12.2. Распространение хлопкового долгоносика на территории США до 1933 г.
После 1933 г граница распространения вида со стороны побережья Мексиканского
залива оставалась более или менее постоянной. (Из USDA Handbook 515, 1978.)
таких вредителей относятся прежде всего
табачные бражники и имаго жуков-бло-
шек; совки и табачные листовертки так-
же причиняют серьезный вред этой куль-
туре. Листьями картофеля питается ко-
лорадский жук, а различные цикадки
высасывают его соки. Клубни картофеля
повреждают живущие в почве личинки
проволочников и жуков-блошек.
Насекомые, вредящие садовым и ого-
родным культурам. Каждый вид расте-
ний, выращиваемых в огороде или в са-
ду, является объектом нападения одного
или нескольких насекомых, различных по
пищевым предпочтениям. Среди этих на-
секомых можно назвать совку «ни», ка-
пустную белянку и капустную тлю, пи-
тающихся на белокочанной и цветной
капусте, а также на других кресто-
цветных; морковную муху-специфично-
го вредителя моркови; дынных бабочек,
спаржевых листоедов и мексиканскую
зерновку. Помимо вредителей, специ-
фичных для той или иной культуры,
имеется множество неспециализиро-
ванных насекомых, которые могут пи-
таться практически на любой культуре.
Чаще других могут достигать высокой
численности и наносить серьезный ущерб
луговой садовый мотылек, различные са-
ранчовые, имаго жуков-нарывников,
подгрызающие совки и травяная совка.
Насекомые - вредители тепличных
растений. Поскольку в теплицах на про-
12. Вредные насекомые и борьба с ними
541
тяжении всей зимы поддерживается
теплый и влажный микроклимат, там
можно встретить множество видов насе-
комых, свойственных тропикам и субтро-
пикам. Из них наибольший вред причи-
няют несколько видов трипсов, червецы,
белокрылки и щитовки. Кроме того, не-
которые виды, обитающие обычно в лет-
ние месяцы вне теплиц, проникают туда
перед наступлением холодов и остаются
активными всю зиму, не впадая в спячку.
Примерами такого рода могут служить
бахчевая тля, персиковая зеленая тля и
бабочка-огневка.
Множество самых разных растений
выращивают в теплицах и лишь очень не-
многие из них не подвергаются нападе-
нию либо всеядных насекомых типа гусе-
ниц огневок, либо специфических парази-
тов вроде хризантемцвой галлицы, чьи
личинки образуют галлы на листьях и
стеблях (рис. 12.3). Обычно зимой в те-
плицах насчитывается от 40 до 50 видов
насекомых, являющихся потенциальны-
ми вредителями. Учитывая разнообразие
растений-хозяев, произрастающих в те-
плице, и тот факт, что все они сконцен-
трированы на ограниченной площади,
можно представить, насколько серьезной
проблемой является борьба с вредителя-
ми в теплицах, требующая постоянной
бдительности со стороны энтомологов.
Насекомые-вредители фруктовых де-
ревьев. Все фруктовые деревья, дающие
мелкие и крупные плоды, вечнозеленые
цитрусовые и культуры, сбрасывающие
листву на зиму, подвергаются нападе-
нию вредителей, и для каждой группы
растений имеются свои главные вредите-
ли.
Цитрусовые деревья в наибольшей
степени страдают от червецов и щито-
вок, белокрылок, трипсов и раститель-
ноядных клещей. Среди них наибольший
вред причиняют апельсиновая щитовка,
красная померанцевая щитовка и мас-
личная ложнощитовка, повреждающие и
деревья, и плоды или образующие медвя-
ную росу, на которой развивается гриб-
возбудитель сажистой пятнистости,
«пачкающий» созревающие плоды. Мно-
гие щитовки имеют широкий круг расте-
ний-хозяев, но будучи обитателями суб-
тропиков, они не выходят за пределы
цитрусового пояса США, включающего
Флориду, южный Техас и южную часть
Калифорнии.
Фруктовые деревья, сбрасывающие
листву на зиму,-яблоня, груша, вишня,
персик, слива и близкие к ним формы-
подвергаются нападению многих вреди-
телей. Плоды яблони страдают главным
образом от гусениц яблонной плодожор-
ки. При составлении плана мероприятий
по защите яблони от вредителей этот вид
учитывается в первую очередь. Личинки
сливового жука-долгоносика, проникаю-
щие в персики, вишни и другие мягкие
плоды, поражают также и яблоки. У
многих фруктовых деревьев ветви и ли-
ства страдают от калифорнийской щи-
товки, восточной персиковой листоверт-
ки, тлей, паутинных клещей, от множе-
ства других листоядных насекомых и от
таких, которые сверлят ходы в стволе
дерева или повреждают фрукты.
Фруктовые деревья, дающие мелкие
плоды, являются сложной в таксономи-
ческом отношении группой и имеют бо-
лее специализированных вредителей. На
виноград нападают виноградная листо-
вертка, много видов тлей и цикадок, ли-
стогрызущие жуки, повреждающие так-
же и корни. Многочисленные тли, гусе-
ницы-листоеды и стеблевые точильщики
повреждают смородину, малину и клуб-
нику.
Насекомые-вредители насаждений и
лесов. Деревья дают приют и пищу тыся-
чам видов насекомых, которые могут ли-
шить дерево листвы, «окольцевать» его,
проникнуть в глубь ствола или высосать
542
12. Вредные насекомые и борьба с ними
Рис. 12.3. Повреждения, вызванные хризантемовой
галлицей Diarthronomyia hypogaea. Примерно
в натуральную величину. (Из Metcalf, Flint, Destructive
and useful insects.)
соки дерева. Многие виды насекомых по-
вреждают деревья лишь незначительно,
но существуют виды, наносящие дереву
серьезный ущерб, приводящий иногда к
его гибели. В результате деревья в лесах
и искусственных насаждениях ежегодно
несут огромные потери от вредителей.
Лесонасаждения в северо-восточных
штатах США особенно страдают от не-
парного шелкопряда и златогузки. На-
ибольший ущерб вязам причиняет вя-
зовый листоед, а также вязовый заболон-
ник, переносящий от дерева к дереву
возбудителя голландской болезни иль-
мовых (графиоза). Прямой вред, нано-
симый короедами и древоточцами, осла-
бляет и убивает деревья многих видов.
Деревья в лесах периодически стра-
дают от вспышек численности различных
насекомых, в результате чего лес гибнет
12. Вредные насекомые и борьба с ними
543
Рис. 12.4. Распространение елового общественного пилильщика в Северной
Америке в период вспышки его размножения в 1938 г (По Balch, 1952, с
изменениями.)
на огромных пространствах. В прежние
годы на такие естественные потери обра-
щалось мало внимания, однако теперь,
когда площадь, занятая лесами, ката-
строфически уменьшается, предприни-
маются все возрастающие усилия в по-
исках средств для снижения этих потерь.
Гусеницы кольчатых коконопрядов, не-
парного шелкопряда и златогузки, гем-
локовой пяденицы, листоверток являют-
ся дефолиаторами как вечнозеленых, так
и сбрасывающих ежегодно листву де-
ревьев. Короеды-это злейшие враги
хвойных пород. Пилильщики питаются
на хвойных деревьях нерегулярно, но во
время вспышек численности они могут
причинить огромный ущерб. Причиной
одной из сильнейших вспышек был завоз
из Европы елового общественного пи-
лильщика, который уничтожил в 1938 г.
еловый строевой лес примерно на 12
тыс. кв. миль главным образом в вос-
точных провинциях Канады (рис. 12.4).
Насекомые как переносчики возбуди-
телей болезней растений. Причиной серь-
езных поражений растений являются па-
тогенные организмы, распространяемые
насекомыми. Как правило, болезнь
имеет значительно более разруши-
тельный характер для растения, чем пи-
тание на нем тех или иных насекомых.
Решение проблемы борьбы с болезнями
может быть достигнуто лишь путем пол-
ного подавления насекомых-переносчи-
ков, поскольку даже небольшое число
особей переносчика могут оказаться кос-
венной причиной огромных потерь.
Некоторые патогены растений, ко-
торые обычно не переносятся насекомы-
ми, проникают в растение через повреж-
дения, наносимые ему насекомыми при
питании, или через проколы, произво-
димые насекомыми при откладывании
яиц. Возбудитель бурой гнили персиков
попадает через проколы, образуемые
имаго сливового жука-долгоносика при
544
12. Вредные насекомые и борьба с ними
питании, а бактерии, вызывающие гниль
хлопчатника, проникают через проколы,
наносимые разными насекомыми как при
питании, так и при яйцекладке.
Насекомые распространяют неко-
торые болезни растений, перенося возбу-
дителей этих болезней на теле или в пи-
щеварительном тракте. Возбудитель бак-
териального ожога переносится на ногах
и теле пчел, жуков и других насекомых (а
также птицами и другими позвоночны-
ми). Споры некоторых патогенных гри-
бов, вызывающих, например, рак ябло-
ни, попадают к насекомым при питании
и в жизнеспособном состоянии проходят
через пищеварительный тракт насе-
комых, в подобных случаях насекомые
представляют лишь один из многих пу-
тей распространения инфекции.
Более важны те случаи, когда насе-
комые являются основным или един-
ственным переносчиком болезнетворно-
го организма от одного растения к дру-
гому. Насекомые получают патоген,
обычно бактериальной или вирусной
природы, при питании на зараженном
растении; затем этот патоген вводится
либо механически, либо со слюной в тка-
ни следующего растения, на котором пи-
тается насекомое. Различные виды цика-
док участвуют в передаче вирусной жел-
тухи астр, а один из видов переносит
вирус курчавости верхушки сахарного
тростника. Множество других насе-
комых также принимают участие в пере-
даче вирусных болезней растений. Насе-
комые переносят также патогенных бак-
терий, например возбудителя вилта тык-
вы. Этот возбудитель проводит зиму в
пищеварительном тракте зимующих ли-
стоедов, которые со следующей весны
начинают передавать инфекцию. Многие
бактериальные и вирусные болезни нано-
сят растениям огромный ущерб.
Насекомые-вредители пищевых запа-
сов. Зерно и мясо, мука и крупы и другие
продукты питания с высоким содержа-
нием белка используют в пищу многие
насекомые. Хранение этих продуктов со-
провождается огромными потерями от
опустошительных нападений насекомых,
поэтому необходимо проводить по-
стоянные защитные мероприятия, чтобы
свести эти потери к минимуму.
Главными вредителями хранящегося
зерна и зерновых продуктов в Северной
Америке являются имаго и личинки су-
ринамского мукоеда, малого мучного
хрущака, мавританской козявки, боль-
шого мучного хрущака и рисового и ам-
барного долгоносиков; сюда следует до-
бавить также гусениц индийской мучной
моли и мельничной огневки. Хранящиеся
горох и бобы служат пищей различным
гороховым зерновкам (Bruchidae). Мясо
и сыры привлекают ветчинных кожеедов
и личинок сырной мухи.
Продукты в хранилищах сначала, как
правило, подвергаются нападению пере-
численных 'выше насекомых. Нанеся
лишь небольшой ущерб на первом этапе,
насекомые способствуют появлению
плесени, вслед за которой вскоре по-
является множество других насекомых-
вредителей ; очень скоро от первоначаль-
ного количества может остаться лишь
небольшая доля неповрежденных про-
дуктов.
НАСЕКОМЫЕ - ПАРАЗИТЫ
ЧЕЛОВЕКА И ЖИВОТНЫХ
Насекомые, поселяющиеся в жилище
человека. Некоторые виды насекомых
стали почти «домашними», особенно к
северу от линии морозов, где эти виды
встречаются почти исключительно
в жилище человека. В случае эктопарази-
тических видов эти отношения возникли
еще до развития цивилизации и обусло-
влены необходимостью сосуществования
паразита с теплокровным хозяином. У
12. Вредные насекомые и борьба с ними
545
других видов эта связь возникла позже и
была обусловлена относительно высоки-
ми температурами, поддерживаемыми в
домах и других строениях в суровые зим-
ние месяцы. Таким образом, некоторые
виды, полутропические по своему проис-
хождению, встречаются теперь значи-
тельно севернее и способны существо-
вать в жилищах человека.
По степени приносимого вреда пер-
вое место в жилищах человека занимают
эктопаразиты и вредители пищевых запа-
сов. Кроме того, личинки настоящих мо-
лей и шубных кожеедов поедают вещи,
сделанные из материалов животного
происхождения - шерстяную одежду,
обивку, ковры. Обыкновенная чешуйни-
ца и тараканы, являющиеся всеядными
насекомыми, поедают книжные пере-
плеты, содержащие крахмал, а также
просто вызывают у человека брезгливое
чувство. Тараканы, беспорядочно пачкая
все вокруг своими экскрементами, за-
грязняют и портят пищу и жилище; если
численность их высока, они наполняют
дом, склад или ресторан неприятным и
резким запахом. Часто в дома прони-
кают муравьи, и иногда они могут стать
серьезной помехой в кухнях и кладо-
вых.
Термиты обладают наибольшей раз-
рушительной силой, поедая древесину в
фундаментах, стенах и полах жилищ, что
требует проведения постоянных ремон-
тов. В деревянных частях жилых домов
живут и другие насекомые, например жу-
ки семейства Lyctidae, а пенсильванские
муравьи-древоточцы могут выедать в де-
ревянных частях зданий обширные гале-
реи, используемые как гнезда.
Насекомые, паразитирующие на чело-
веке и домашних животных. Насекомые
наносят человеку и домашним животным
самый различный ущерб-от простого
беспокойства до заражения опасными
болезнями. Некоторые насекомые, на-
35-732
пример Anoplura, нападают лишь на
один или два близкородственных вида
животных, другие, например комары, пи-
таются обычно на широком круге тепло-
кровных позвоночных.
Домашняя птица подвергается в ос-
новном нападениям Mallophaga (пухое-
дов) и мелких клещей, причем некоторые
из них постоянно живут на курах, утках,
индейках и гусях. Заражение пухоедами
часто приводит к гибели молодых птиц;
у взрослых птиц пухоеды ухудшают то-
варный вид и снижают яйценоскость.
Иногда массовое нападение клещей мо-
жет отразиться на состоянии всей птичь-
ей стаи. Возбудитель по меньшей мере
одной болезни уток, схожей во многих
отношениях с малярией, переносится
мошками. Куры страдают от нападения
ряда специфических для них блох, из ко-
торых наиболее постоянной считается
присасывающаяся блоха.
У домашних животных и человека
имеется множество специфических пара-
зитов: Anoplura (вши), блохи, по-
стельные клопы, несколько видов пухое-
дов и разные группы клещей. К послед-
ним относятся чесоточные клещи, крас-
нотелки и иксодовые клещи. У овец
имеются свои «овечьи клещи»-необыч-
ные бескрылые мухи семейства Hippo-
boscidae. Нападение эктопаразитов вы-
зывает раздражение кожи, ухудшает об-
щее состояние, но достаточно редко при-
водит к гибели, если кровососание не
связано с переносом возбудителей болез-
ней.
Личинки оводов паразитируют в ор-
ганизме ряда позвоночных. У лошадей,
например, эти личинки повреждают раз-
личные участки желудочно-кишечного
тракта, что приводит к серьезным поте-
рям в весе и ухудшению общего состоя-
ния животных. Некоторые из этих личи-
нок развиваются в лобных пазухах овец,
другие - непосредственно под кожей на
546
12. Вредные насекомые и борьба с ними
спине крупного рогатого скота, где обра-
зуют желваки. Личинки мясных мух про-
никают в ранки на теле животных, пи-
таются их мышечной тканью и ежегодно
вызывают огромные потери в разных
группах сельскохозяйственных жи-
вотных.
Помимо этих и других специфических
паразитов имеется огромное число кро-
вососущих двукрылых, нападающих на
теплокровных животных,-комары, слеп-
ни, мошки, мухи рода Symphoromyia и
различные жигалки. Неспецифическими
кровососами являются также некоторые
блохи, различные крупные и мелкие кле-
щи и постельные клопы. Часто они вызы-
вают очень сильное раздражение. Осо-
бенно серьезный ущерб причиняют мош-
ки; в отдельных районах они периодиче-
ски приводят к падежу большого числа
лошадей и мулов. Комары, мошки и
слепни, достигая временами огромной
численности, могут «очистить» от тури-
стов живописные районы, послужить
причиной падения цены на землю в при-
городных районах, задержать прокладку
дорог, как это было в крайних северных
областях Канады и на Аляске. Массовое
нападение кровососов может вызывать
такое же или даже большее ухудшение
состояния здоровья животных, чем спе-
цифические паразиты.
Жалящие и другие насекомые. У насе-
комых имеются различные защитные
приспособления, с помощью которых
они могут наносить человеку поврежде-
ния, вызывать аллергические реакции
или сильное раздражение кожи, к числу
таких насекомых относятся, например,
пчелы, осы, муравьи и некоторые гусе-
ницы, имеющие жгучие или ядовитые во-
лоски. Однако все это, несмотря на не-
приятные ощущения и болезненность,
лишь ничтожная часть вреда, причиняе-
мого насекомыми. Некоторые люди ис-
пытывают сильный страх перед насе-
комыми, которые могут причинить им
боль, другие обладают повышенной ал-
лергической реакцией к яду жалящих
насекомых.
Насекомые - переносчики возбудите-
лей болезней животных. Насекомые
являются переносчиками возбудителей
некоторых'из наиболее опасных болезней
человека и других позвоночных. Как и в
случае растений, нередко передача возбу-
носчик
Бешенство
Энцефалит
Мышиный тиф
Р) Пятнистая
лихорадка
Туляремия
Чума (нелегоч-
ные формы )
Случай-
ный
хозяин
Случай - Пара-Пере-Паразит Хозяин
ный хозяин зит носчик
Рис. 12.5. А. Схема передачи инфекционного заболевания при наличии трех основных
биотических факторов (хозяин, паразит и переносчик), где человек - главный или
единственный хозяин. Б. Схема передачи инфекционного заболевания при наличии трех
основных хозяев, где человек-случайный хозяин. (Из материалов министерства
здравоохранения, образования и социального обеспечения США, Служба общественного
здравоохранения.)
Вредные насекомые и борьба с ними
547
Зимовка -
ДИКИЙ ГЫЗУН
Прямой контакт
Может
передавать
Бубонная чума
Вторичная
чумная
пневмония
4 Хчума ЖИВОТНЫХ
ЗАРАЗНАЯХ >
БЛОХА* / •
Прямой контакт
лЗНАЯ
Зимовка
Прямой
контакт
ЧУМА
Эпидемия
легочной чумы
ЧЕЛОВЕКА
СЕЛЬСКАЯ ЧУМА ’YhoXA*
“УТИ — обычный
. £?ЕДАЧИ — случайный
..редкий или теоретически
возможный
Загрязненная
почва Прямой
вконтакт
ДОМАШНИЙ
ГРЫЗУН
ГОРОДСКАЯ ЧУМА
ЗАРАЗНАЯ
БЛОХА
ДОМАШНИЙ
ГРЫЗУН
Прямой контакт
* Механизм сохранения в зимний период
Рве. 12.6. Эпидемиологический цикл при чуме. (Из материалов министерства
здравоохранения, образования и социального обеспечения США, Служба общественного
здравоохранения.)
дителя насекомыми - это только один из
нескольких путей его распространения
среди животных, иногда насекомое-пере-
носчик является единственным из-
вестным агентом, с помощью которого
дронсходит перенос патогена от одного
организма-хозяина к другому (рис. 12.5).
К первой категории относятся брюш-
ной тиф, летняя диаррея и некоторые
формы дизентерии, вызываемые бакте-
риями из рода Bacillus. Комнатные мухи,
контактируя со сточными водами, мо-
кротой и другим заразным материалом,
разносят бактерии на своих лапках или
на ротовых частях и загрязняют пищу и
другие предметы, на которые садятся.
Эти патогены передаются и другими пу-
тями, но при определенных условиях му-
хи могут оказаться наиболее эффек-
тивными их распространителями.
Бубонная чума (черная смерть), вызы-
ваемая Bacillus pestis- другое контагиоз-
ное заболевание, относящееся к этой же
категории. Крысы и мелкие млекопитаю-
щие служат естественным резервуаром
Bacillus pestis, а крысиные блохи перено-
сят бактерии от крысы к крысе или от
крысы к человеку (рис. 12.6).
Существует несколько опасных болез-
ней человека, единственными переносчи-
ками которых являются насекомые. Ма-
лярия, вызываемая простейшими из рода
548
12. Вредные насекомые и борьба с ними
Plasmodium, распространяется от челове-
ка к человеку несколькими видами кома-
ров рода Anopheles. Желтая лихорадка и
лихорадка денге вызываются вирусами,
которых переносят несколько видов ко-
маров, комар Aedes aegypti- основной
переносчик этих вирусов на территории
Северной Америки. Мухи из рода Gloss-
ina переносят простейших рода Trypano-
soma, вызывающих африканскую сонную
болезнь, слоновая болезнь (филяриоз)
вызывается нематодами из рода Filaria,
переносимыми комарами. Во всех на-
званных случаях комар или муха, питаю-
щиеся на человеке, несущем в себе возбу-
дителя, поглощают вместе с кровью
некоторое число патогенных организ-
мов , через некоторое время при питании
на здоровом человеке насекомые вносят
возбудителя в его ткани. Именно таким
образом и происходит заражение здо-
ровых людей.
Сыпной тиф вызывается ультрами-
кроскопическими организмами, назы-
ваемыми Rickettsia и переносимыми вша-
ми. Вши получают патогенный микроор-
ганизм при питании кровью человека и
позднее выделяют его с экскрементами.
Расчесывание мест укусов создает усло-
вия для проникновения патогена в кожу
человека, что и приводит к заражению.
Клещи известны как единственные
переносчики нескольких серьезных забо-
леваний, из которых о трех следует упо-
мянуть особо. Техасская лихорадка-ле-
тальная болезнь скота, вызываемая спо-
ровиком Babesia bigemina. Возбудитель
передается скоту иксодовым клещом
Margaropus annulatus. Пятнистая лихо-
радка Скалистых гор, частота которой за
последнее время резко увеличилась и ко-
торая в большом проценте случаев за-
канчивается смертельным исходом, вы-
зывается риккетсией, которая сохраняет-
ся в организме мелких диких грызунов,
несколько видов клещей рода Dermacen-
tor осуществляют передачу возбудителя,
питаясь на стадии нимфы на зараженных
грызунах, а на стадии имаго-на челове-
ке. Третья болезнь - японская речная ли-
хорадка, или цуцугамуши, возбудителем
которой служит другой вид риккетсии,
который передается человеку от диких
грызунов личинками ранних возрастов
клещей-краснотелок (Trombidiidae). Эта
болезнь представляла серьезную опас-
ность во время второй мировой войны в
Бирме и на тихоокеанском театре во-
енных действий.
В большинстве случаев борьба с эти-
ми болезнями может быть осуществлена
путем борьбы с их переносчиками. В
частности, такой подход был очень эф-
фективен в случае техасской лихорадки,
борьба с клещом рогатого скота привела
к фактическому искоренению этой болез-
ни на территории США. Вспышки сыпно-
го тифа исключительно эффективно по-
давлялись путем организации борьбы со
вшивостью. Значительно более трудна
борьба с комарами, мухами или клеща-
ми Dermacentor; эти переносчики распро-
страняют возбудителей на обширных
пространствах, и меры борьбы с ними
далеко не всегда приводят к столь же за-
мечательным результатам, какие были
достигнуты в случае техасской лихорад-
ки или сыпного тифа.
БОРЬБА С ВРЕДНЫМИ
НАСЕКОМЫМИ
Когда вредный вид достигает доста-
точно высокой плотности, начинает ощу-
щаться наносимый им ущерб и естествен-
но возникает желание сократить числен-
ность данного вида. Если это касается
видов, наносящих ущерб исключительно
своей деятельностью (например, в случае
гусениц, питающихся на капусте, или та-
раканов, поедающих пищевые запасы),
то плотность вредителя должна, очевид-
12. Вредные насекомые и борьба с ними
549
но. достичь весьма высокого уровня,
прежде чем станет экономически оправ-
дано проведение борьбы в той или иной
форме. Если же это касается видов-пере-
носчиков возбудителей различных болез-
нен животных и человека, то даже одна
особь переносчика может послужить
причиной серьезных бедствий; в таких
случаях плотность переносчика должна
постоянно поддерживаться на макси-
мально низком уровне.
В большинстве случаев проведение
борьбы с насекомыми основывается на
произвольном решении вопроса о допу-
стимом ущербе. Принятые стандарты на
фрукты, не имеющие ни малейшего
пятнышка, или на овощи без малейших
следов порчи предусматривают практи-
чески полное освобождение продукции
от вредителей, что почти недостижимо и
крайне дорогостояще. Если бы мы пере-
стали придерживаться критериев столь
высокой «чистоты», то многие насе-
комые, которые конкурируют с нами за
пишу, перестали бы считаться «вредите-
лями», и в результате можно было бы со-
кратить или ликвидировать вовсе многие
из проводимых сейчас мероприятий.
В некоторых случаях неблаго-
приятные факторы окружающей среды
могут привести к резкому сокращению
численности популяции вредного вида.
Хорошо известным примером может
служить вспышка численности кобылки
Anabrus simplex, которая угрожала по-
лностью погубить урожай зерновых в
окрестностях Солт-Лейк-Сити (шт. Юта)
в период освоения этих территорий.
Вспышка была ликвидирована огромной
стаей морских чаек. В 1935 г клоп-чере-
пашка был сведен до статуса «редкого»
зила во многих штатах кукурузного по-
ка в результате неблагоприятных зим-
зих условий в сочетании с какой-то гриб-
ковой болезнью насекомых. Несколько
менее впечатляющий пример касается
растительноядной яблонной листоверт-
ки, плотность которой на яблонях была
доведена до экономически незначимого
уровня за счет активности местных хищ-
ников и паразитов.
Биологическая борьба
Существующие способы биологиче-
ской борьбы можно разделить на три ка-
тегории : использование паразитов, пара-
зитоидов или хищников насекомых (см.
также гл. 11), использование патогенных
организмов; генетическая борьба. Обыч-
но термйн «биологическая борьба» ис-
пользуется в более узком смысле и под-
разумевает только использование пара-
зитов, паразитоидов, хищников и патоге-
нов.
Паразиты, паразитоиды и хищники на-
секомых. Возможность разведения и рас-
пространения естественных врагов для
борьбы с вредными насекомыми волно-
вала воображение энтомологов на протя-
жении многих десятилетий. Было выяс-
нено, однако, что в отношении вредных
видов, эндемичных для территории
США, такие возможности крайне ограни-
чены. Усилить естественно существую-
щий уровень биологической регуляции
численности вредного вида крайне слож-
но, и в тех случаях, когда нам не грозят
значительные потери урожая, это не-
оправданно дорого. Следовательно, ме-
тоды биологической борьбы носят
главным образом превентивный, а не
корректирующий характер.
Как было показано в предыдущей
главе, совершенно иная ситуация наблю-
дается при интродукции вредителей.
Каждый вид, вероятно, имеет множество
паразитов и хищников, сдерживающих
его численность на его родине. Когда
происходит случайный завоз вредителя в
другую страну, он попадает туда, как
правило, без своих паразитов. Освобо-
550
12. Вредные насекомые и борьба с ними
давшись от своих врагов, вредитель спо-
собен беспрепятственно размножаться на
новом месте.
Для того чтобы успешно бороться с
таким завезенным вредителем, следует
установить, какие враги могут эффектив-
но сокращать его численность до мини-
мальных размеров. Наличие таких вра-
гов могло бы обеспечить постоянный
контроль численности вредного насеко-
мого и избавить нас от необходимости
ежегодного проведения более дорого-
стоящих мероприятий.
Столь идеальный результат дости-
гается весьма редко. Наиболее выдаю-
щимся достижением подобного рода сле-
дует считать проведение борьбы с заве-
зенным в США австралийским желоб-
чатым червецом путем ввоза и акклима-
тизации его паразита-австралийской
божьей коровки родолии. На территории
Калифорнии эта божья коровка и ее ли-
чинки столь эффективно контролируют
численность червеца, что вредитель ста-
новится экономически значимым лишь
изредка и на ограниченных участках. Ряд
паразитов завезенных ранее в США
вредных видов, в частности паразитов
японского жука, непарного шелкопряда и
стеблевого кукурузного мотылька, ин-
тродуцируются министерством сельско-
го хозяйства США и выпускаются в ме-
стах обитания их хозяев. Однако многие
интродуцируемые паразиты не могут су-
ществовать в природных условиях США,
поскольку они либо не приспособлены к
местным климатическим условиям, либо
не находят в нужное время подходящего
хозяина. Лишь немногие виды успешно
акклиматизируются на новой для них
территории и становятся действенными
агентами в борьбе с вредными видами.
Можно надеяться, что в конечном счете
удастся создать дееспособные популяции
паразитов, которые снизят численность
популяций многих вредных видов ниже
нынешнего опасного уровня. В неко-
торых районах уже удалось достичь та-
кого результата, например, в штате Ва-
шингтон и провинции Британская Ко-
лумбия (Канада) численность ивовой во-
лнянки успешно регулируется завезенны-
ми туда паразитическими перепонча-
токрылыми. Разведение и последующее
распространение возбудителей бакте-
риальных болезней обещает быть весьма
эффективным средством борьбы с япон-
ским жуком. Значительный успех биоло-
гической борьбы достигается на остро-
вах, так, на Гавайских островах исполь-
зование комплекса паразитических орга-
низмов позволило регулировать числен-
ность многих насекомых, вредящих сель-
скохозяйственным культурам.
Результаты многочисленных исследо-
ваний показывают, что на успех или не-
удачу биологических методов может
влиять большое число факторов. К числу
таких факторов относятся экологические
требования паразитов, их влияние друг
на друга (с. 512), специфичность в отно-
шении хозяина, скорость роста популя-
ции, характер распределения и многие
другие. Для достижения высокой эффек-
тивности мероприятий необходимы так-
же хорошо обученный персонал и боль-
шое количество специального оборудо-
вания, а также сбор паразитов или хищ-
ников за рубежом и доставка их в США в
целости и сохранности.
С учетом всех этих обстоятельств раз-
работка биологических методов борьбы
с вредными насекомыми проводится в
США главным образом под эгидой феде-
рального правительства, исключение
составляет штат Калифорния, где интен-
сивные разработки выполняются
штатными организациями. Завершаю-
щий этап разработок, связанный с выпу-
ском паразитов, часто осуществляется
совместно с научными работниками, со-
стоящими на государственной службе, и
12. Вредные насекомые и борьба с ними
551
заинтересованными штатными учрежде-
ниями. Канадское правительство также
проводит обширные работы по исполь-
зованию биологических методов борьбы
с вредителями.
Патогенные организмы. Наблю-
даемые в природных условиях сильные
вспышки грибковых и бактериальных
болезней клопа-черепашки и других
вредных насекомых позволяют энтомо-
логам надеяться на возможность борьбы
с этими насекомыми путем распростра-
нения в их популяциях патогенных орга-
низмов. Первые попытки в этом направ-
лении были неудачными, поскольку
годы, неблагоприятные для естественно-
го развития болезней, оказались экологи-
чески неблагоприятными и для искус-
ственного распространения их возбуди-
телей. Начиная примерно с 1930 г было
обнаружено несколько групп организ-
мов, пригодных дЛя практического ис-
пользования, в их число входят вирусы,
бактерии, простейшие, грибы и риккет-
сии. Молочная болезнь японского жука
оказалась губительной для этого вида в
некоторых частях его ареала на террито-
рии Америки. Некоторые разновидности
вирусов полиэдроза успешно регулируют
численность ряда пилильщиков - вреди-
телей хвойных пород. В Калифорнии ви-
рус полиэдроза и Bacillus thuringiensis
оказались высокоэффективными в борь-
бе с гусеницами на люцерне. Эти успехи
стимулировали открытие более чем 1000
подобных болезней насекомых.
Генетическая борьба. В 50-е годы эн-
томологами министерства сельского хо-
зяйства США был предложен совершен-
но новый вариант биологической
эорьбы. Объектом была выбрана мясная
муха, нападающая на теплокровных мле-
копитающих, в том числе и на домашних
тавотных, и приносящая огромный
•тперб животноводству США. Новый ме-
тод включал в себя последовательное
осуществление двух мероприятий: 1)
облучение куколок мух гамма-лучами, с
тем чтобы получить стерильных, но ак-
тивных и сексуально агрессивных сам-
цов, и 2) массовое разведение и выпуск
таких самцов в природные популяции
мухи. При спаривании с такими сте-
рильными самцами самки получали ано-
мальные сперматозоиды и в результате
откладывали нежизнеспособные яйца.
При осуществлении этой программы вы-
пуск стерильных самцов продолжается
до тех пор, пока вид полностью не исчез-
нет на данной территории. В 1955 г. мяс-
ная муха была полностью уничтожена на
площади 440 кв. км на острове Кюра-
сао, а в 1958 г.-во Флориде и других
юго-восточных штатах США, и ее значе-
ние как экономически важного вредителя
крупного рогатого скота существенно
снизилось. Косвенным результатом про-
ведения этой программы явилось замет-
ное увеличение поголовья оленей в юго-
западных штатах США.
Другой подход к генетической борьбе
связан с воздействием на механизм опре-
деления пола у данного вида, так что в
результате получается очень мало самок.
Генетические линии, в которых был до-
стигнут такой результат, выделены у ко-
маров и описаны у некоторых бабочек.
Проблема эффективного использования
этих линий и сохранения их в процессе
проведения полевых мероприятий оказа-
лась чрезвычайно сложной.
Организационные мероприятия и ме-
тоды возделывания. Численность неко-
торых насекомых, вредящих сельскохо-
зяйственным культурам или лесным на-
саждениям, может поддерживаться на
уровне, не представляющем опасности,
за счет проведения ряда организа-
ционных мероприятий и применения
определенных методов возделывания.
Общим важным условием при этом
является поддержание растений в здоро-
552
12. Вредные насекомые и борьба с ними
вом состоянии посредством использова-
ния соответствующих удобрений, прове-
дения дренажных или ирригационных
мероприятий, обработки почвы, а также
путем выращивания культур, хорошо
приспособленных к данным климатиче-
ским и почвенным условиям.
Против ряда вредителей могут ока-
заться полезными специальные методы
возделывания. очистка почвы парова-
нием, чередование культур, определен-
ное время посева или сбора урожая, а
также использование резистентных или
толерантных к нападению насекомых
сортов растений.
ОЧИСТКА ПОЧВЫ ПАРОВАНИЕМ.
Этим путем достигается уничтожение
сорняков, которые могут служить хозяе-
вами насекомым, вредящим сельскохо-
зяйственным культурам. Так, цикада-
горбатка буйвол размножается на мно-
гих травянистых сорняках, взрослые ци-
кады летят на соседние фруктовые де-
ревья, делают надрезы на их ветках и
откладывают в эти надрезы яйца. Очист-
ка сада предотвращает наносимый этими
насекомыми ущерб, так как уничтожает-
ся первичный хозяин вредителя. Сорняки
и почвенный мусор служат также местом
зимовки и окукливания для большого
числа вредных насекомых, и периодиче-
ская очистка почвы парованием напра-
влена на предотвращение закрепления
популяций этих видов в том или ином
районе.
ЧЕРЕДОВАНИЕ КУЛЬТУР Установле-
но, что чередование выращиваемых куль-
тур особенно эффективно в отношении
насекомых, личинки которых питаются
на корневых частях растений. Таким пу-
тем можно бороться, например, с некото-
рыми видами жуков-блошек рода Diabro-
tica. Личинки этих жуков питаются
главным образом на корнях кукурузы.
Если кукуруза выращивается более трех
лет на одном и том же поле, где суще-
ствуют благоприятные условия для этих
насекомых, последние могут достичь вы-
сокой численности и принести серьезный
экономический ущерб. Если, однако, ку-
курузу заменить всего на один год пше-
ницей или бобовыми, то жуки погибнут
от голода. Поэтому замена кукурузы
пшеницей или другой культурой, прово-
димая так, чтобы кукуруза не оставалась
на одном и том же поле больше 2-3
лет, почти полностью ликвидирует
ущерб, причиняемый этими вредителя-
ми.
СРОКИ ПОСЕВА. Этот метод может
быть полезен в борьбе с некоторыми на-
секомыми. Численность гессенской муш-
ки, серьезного вредителя пшеницы, чьи
личинки питаются внутри листовой об-
вертки, может частично контролировать-
ся выбором сроков посева озимой пше-
ницы. Все осеннее поколение взрослых
насекомых, как правило, появляется на
протяжении короткого отрезка времени
сразу после позднелетних дождей. Имаго
живут всего J-4 дня, и за это время они
должны отложить яйца в углубление
пшеничного листа. Если высеять озимую
пшеницу после того, как прошло осеннее
поколение гессенской мушки, то яйца не
попадут на растения, и, следовательно,
до весны никто не причинит им вреда.
Чтобы в полной мере использовать эти
условия, энтомологи в районах выращи-
вания пшеницы ежегодно устанавливают
сроки ее посева, эти сроки должны, с
одной стороны, позволить растениям
расти при хорошей погоде, чтобы до-
стичь до зимы нужного состояния, а
с другой - избавить их от массового
заражения яйцами гессенской мушки
(рис. 12.7). Весеннее поколение гессен-
ской мушки нападает только на поздние
побеги озимой пшеницы, причиняя лишь
небольшой вред. Ранний или поздний
посев кукурузы и других культур может
иногда способствовать снижению ущер-
12. Вредные насекомые и борьба с ними
553
Рис. 12.7. Образец карты, на которой показаны рекомендуемые сроки посева пшеницы
в раде восточных и центральных штатов США, позволяющие избежать ущерба,
причиняемого гессенской мушкой. (Из материалов министерства сельского хозяйства
США.)
ба, причиняемого им жуками-блошками
и кукурузным мотыльком.
ЛЕСОПОЛЬЗОВАНИЕ. Практика лесо-
пользования направлена на сокращение
обилия вредных насекомых, что может
быть достигнуто изменением экологиче-
ской структуры лесного сообщества. На-
пример, вспышки численности неко-
торых из наиболее вредоносных видов
жуков-короедов происходят в пере-
зрелых сосновых насаждениях. Вырубка
деревьев для хозяйственных нужд до то-
го, как они состарятся, предотвращает
увеличение численности популяций ко-
роедов, а молодые деревья оказываются
в более благоприятных для роста усло-
виях. Селективным прореживанием леса
можно добиться оптимального разно-
образия как в отношении видов деревьев,
так и их возрастных групп. Известно, что
чем больше разнообразие, тем меньше
шансов на то, что произойдет вспышка
размножения насекомых.
Подбор деревьев используется как
мера борьбы с непарным шелкопрядом.
Для быстрого успешного развития шел-
копряда нужен подходящий хозяин. Для
личинок ранних возрастов таким хозяи-
ном служат ива, береза или дуб, личинки
более поздних возрастов переходят на
сосну и другие менее подходящие виды
деревьев, подвергая их также дефолиа-
ции. Чтобы снизить поражение леса этим
вредителем, наиболее подходящие для
него виды деревьев-хозяев оставляют в
минимальном количестве, а на их место
высаживают менее подходящие для не-
парного шелкопряда виды.
Карантин. Наиболее очевидным спо-
собом избежать ущерба от вредителя, от-
сутствующего пока в той или иной стра-
не, представляется недопущение его в эту
554
12. Вредные насекомые и борьба с ними
страну. В разных частях света суще-
ствуют сотни насекомых, которые, как
мы полагаем, могут стать опаснейшими
вредителями в случае укоренения в Се-
верной Америке. Для предотвращения их
ввоза правительство США осуществляет
инспекцию всего, что ввозится в страну,
прежде всего живых растений и жи-
вотных, а также упаковочного материа-
ла, который часто может скрывать вре-
дителей и служить для них своего рода
«переносчиком». Большая часть или да-
же весь упаковочный материал подвер-
гается фумигации, прежде чем будет да-
но разрешение на его ввоз в страну.
Кроме того, отдельные штаты могут
устанавливать собственные ограничения
на перевоз каких-то материалов в преде-
лах штата или на ввоз их на территорию
штата. Канадское правительство имеет
аналогичную службу.
Практически невозможно полностью
предотвратить завоз в страну всех потен-
циальных вредителей. Тем не менее от-
четы карантинных служб говорят о том,
что ежегодно на пути сотен и тысяч
новых «иммигрантов» ставится заслон.
Невозможно также подсчитать, сколько
мы выигрываем на этом. Однако пе-
чальный опыт, полученный от таких
опасных «иммигрантов», как хлопковый
долгоносик (рис. 12.2), кукурузный
мотылек и японский жук, не оставляет
сомнений в том, что нельзя позволять
любому виду насекомых свободно про-
никать через границу страны.
Устойчивость (резистентность)
растений
У некоторых видов сельскохозяй-
ственных растений один сорт может
быть высоко чувствителен к нападению
какого-либо вредителя, тогда как другой
может обладать высокой устойчивостью
к тому же вредителю. Устойчивость рас-
тения определяется различными факто-
рами, причем некоторые из этих факто-
ров являются результатом прямого взаи-
модействия растения и насекомого-вре-
дителя, а другие, косвенные, или экологи-
ческие, весьма вариабельны и связаны с
временем года, колебаниями климатиче-
ских факторов и естественными врагами.
Естественная устойчивость. Обычно
устойчивость в наибольшей степени кон-
тролируется генотипом растения. Суще-
ствование различных сортов одного и то-
го же вида является выражением внутри-
видовой генетической изменчивости.
Местные популяции многих видов насе-
комых-вредителей, называемые расами,
или биотипами, также различаются гене-
тически. Так, биотипы отдельных линий
гороховой тли из двух разных районов,
например из Канады и штата Канзас,
различаются по интенсивности нападе-
ния на горох.
Различный уровень устойчивости
сельскохозяйственных культур и его
влияние на насекомых-вредителей можно
продемонстрировать на примере горохо-
вой тли {Acythosiphon pisum), питающейся
на люцерне {Medicago sativa). Молодая
тля, питающаяся на обычной чувстви-
тельной разновидности люцерны, может
давать около 60 потомков за 10 дней, но
та же самая тля, попавшая на устойчи-
вую разновидность люцерны, либо со-
всем не размножается, либо за такой же
период дает не более одного потомка.
Сходным образом, клоп-черепашка {Blis-
sus leucopterus) откладывает в течение
жизни около 100 яиц, питаясь на чувстви-
тельной линии сорго {Sorghum vulgare),
тогда как при питании на устойчивой ли-
нии этой культуры средняя яйцепродук-
ция насекомого падает до уровня менее
одного яйца на самку.
Устойчивость некоторых сортов сое-
вых бобов к цикадкам обусловлена, по-
видимому, сильной опушенностью
12. Вредные насекомые и борьба с ними
555
листьев у резистентных линий в отличие
от почти голых листьев чувствительных
штаммов. В других случаях некоторые
особенности строения растения могут ча-
стично нейтрализовать вред, наносимый
ему питающимся на нем вредным насе-
комым. Так, у некоторых сортов куку-
рузы укороченные и утолщенные стебли
увеличивают «силу» растений, уменьшая
возможность поломки их ветром, а сле-
довательно, и снижая потери урожая от
деятельности насекомых-точильщиков.
Устойчивость кукурузы к кукурузно-
му мотыльку может служить примером
более сложного взаимодействия расте-
ния с вредителем. У резистентных сортов
уменьшается содержание аскорбиновой
кислоты (витамина С) в тканях, и, кроме
того, в определенные периоды роста в
различных тканях синтезируются ток-
сичные вещества. Реакция растения, вы-
ражающаяся в образовании токсинов,
носит название антибиоза.
Экологические адаптации. Косвенным
фактором устойчивости растений могут
быть экологические адаптации. Напри-
мер, различные сорта некоторых видов
сельскохозяйственных культур созре-
вают в разное время. Следовательно, ка-
кие-то сорта созревают до того времени,
как насекомое-вредитель достигнет вы-
сокой численности, и именно эти сорта
обеспечивают хороший урожай.
Резистентность за счет селекционных
мероприятий. Иногда контролировать
численность вредителя можно путем ин-
тродукции устойчивого сорта данного
вида растения или выведения новых сор-
тов, в которых свойства резистентного
сорта сочетаются с высокой урожай-
ностью чувствительного. Этим способом
генетические факторы резистентности
подвергаются отбору и комбинируются с
факторами, ответственными за коммер-
ческие свойства сельскохозяйственной
культуры. Этот подход является одним
из наиболее многообещающих в отноше-
нии контроля численности насекомых. К
настоящему времени удалось вывести
сорта пшеницы, устойчивые к гессенской
мушке и американскому хлебному пи-
лильщику, сорта люцерны, устойчивые к
голубой и пятнистой люцерновым тлям,
и ряд других. Для многих других культур
удалось вывести частично резистентные
сорта.
При создании резистентных сортов
могут возникать другие проблемы.
Устойчивость растения к другим
вредным организмам обычно связана с
высоким значением отношения угле-
воды :азот; при снижении этого отноше-
ния, а следовательно, и устойчивости
растение становится в большей степени
подвержено грибковым и бактериаль-
ным болезням. В ряде случаев устойчи-
вость растения может быть только вре-
менной, так как насекомое способно вы-
рабатывать иммунитет или находить дру-
гие способы ее преодоления. Примером
такого «прорыва» резистентности может
служить «биотип Е» злаковой тли, ко-
торый ныне охотно питается на сортах
сорго, ранее резистентных к нему.
Физические меры борьбы
Для уничтожения насекомых нередко
используются «прямые» методы борьбы.
Простейшими из них являются физиче-
ские меры, в том числе удаление насе-
комых с растений вручную или использо-
вание механических приспособлений для
поимки или уничтожения насекомых.
Вручную собирали крупных гусениц, вро-
де гусениц табачного или томатного (пя-
титочечного) бражников. Проведение
подобных мероприятий возможно в свя-
зи с невысокой, как правило, числен-
ностью гусениц и их крупными размера-
ми, облегчающими поиск. Гнезда личи-
нок на деревьях могут быть срезаны и
556
12. Вредные насекомые и борьба с ними
уничтожены. Против отдельных видов
вредителей эффективно используется ряд
механических приспособлений. Одно из
наиболее обычных приспособлений тако-
го рода - завешивание дверей и окон сет-
кой, чтобы насекомые не могли проник-
нуть в помещение. Для отлова мух при-
меняются разные типы ловушек с «лаби-
ринтами». Вокруг стволов фруктовых
деревьев привязывают пояса из ткани
или бумаги, создавая места зимовки или
окукливания для яблонной плодожорки;
эти пояса регулярно осматривают и
спрятавшихся там насекомых уничто-
жают Чтобы помешать заползанию на
деревья бескрылых самок бабочек или
личинок, стволы обвязывают поясами из
Рис. 12.8. Ловчие пояса, позволяющие
предотвратить подъем по стволу личинок и
бескрылых самок бабочек. (По материалам
министерства сельского хозяйства США.)
сетки, газа или клейких материалов
(рис. 12.8). В прежние годы против ми-
грирующих бескрылых насекомых, вроде
кобылок или клопов-наземников, прино-
сящих ущерб полевым сельскозохяй-
ственным культурам, широко использо-
вались различные механические препят-
ствия, например борозды в почве или
барьеры из дерева или бумаги.
Диапазон устойчивости насекомых к
высоким и низким температурам и дру-
гим физическим факторам крайне огра-
ничен. Это также используется для
борьбы с вредителями. Естественно, что
такими физическими методами трудно
контролировать численность насекомых
на больших территориях, поэтому их
применяют главным образом в зданиях
или на небольших огороженных про-
странствах. Облучение инфракрасным
светом применяется на мельницах и зер-
новых элеваторах. В период жаркой лет-
ней погоды температуру в помещениях
поднимают на несколько часов почти до
60°С, что убивает всех насекомых внутри
здания. Тепловая энергия инфракрасного
излучения используется для борьбы с на-
секомыми в различных хранилищах.
Как средство борьбы с насекомыми
на складах широко используется охлаж-
дение. Меха, гобелены и другие ценные
товары животного происхождения хра-
нятся в закрытых помещениях при темпе-
ратуре ниже 5°С. Эта температура не
убивает насекомых, но насекомые при
ней неактивны и не приносят ущерба.
Для уничтожения насекомых исполь-
зуются иногда и электрические
приспособления. Экраны и лампы могут
быть снабжены электрически заряженны-
ми решетками, которые убивают насе-
комых, попадающих между элементами.
Для привлечения и уничтожения насе-
комых, особенно бабочек, пользуются
световыми ловушками, хотя эффектив-
ность этого метода очень ограничена.
12. Вредные насекомые и борьба с ними
557
Химические средства борьбы
После второй мировой войны нача-
лось широкое использование синтетиче-
ских инсектицидов для снижения ущерба
причиняемого насекомыми сельскохо-
зяйственным культурам и животным и
для предотвращения эпидемий болезней,
передаваемых насекомыми. Потеря при-
мерно половины мировых продуктовых
запасов за счет вредителей интенсифици-
ровала поиски таких инсектицидов, ко-
торые при высокой эффективности не
обладали бы неблагоприятными по-
бочными действиями.
Инсектицидные препараты. Инсекти-
цид состоит из токсиканта (ядовитого ве-
щества) и одного или более инертных не-
ядовитых продуктов, которые необхо-
димы для растворения яда или выпол-
няют функции носителя, эмульгатора,
дисперсанта или прилипателя. Таким
образом, для целей практического при-
менения химическому веществу придает-
ся особая форма, а продукт, поступаю-
щий в продажу, носит название инсекти-
цидного препарата. Тип и качество пре-
парата в значительной мере влияют на
его эффективность как инсектицида. Наи-
более известны следующие формы инсек-
тицидов. дусты, гранулы, смеси удоб-
рений с инсектицидами, смачивающиеся
порошки, эмульгирующиеся концен-
траты, растекатели, растворы, раство-
римые порошки, загущенные суспензии,
аэрозоли, фумиганты и т д.
Факторы, влияющие на эффективность
инсектицидов. Одни инсектициды прони-
кают внутрь тела насекомого легче, дру-
гие-труднее. Если инсектициды легко
проникают в трахеи или через кутикулу,
их действие проявляется быстро. Напри-
мер, яды-фумиганты попадают в трахеи
в форме газа. Органические растворите-
ли, например масла, легко проникают че-
рез наружный покров, поэтому присут-
ствие масла в смеси для опрыскивания
обычно увеличивает токсичность послед-
ней. Ранние фазы развития насекомых
наиболее чувствительны к действию ин-
сектицидов; яйца очень чувствительны
непосредственно перед вылуплением из
них личинок, а личинки и нимфы более
чувствительны, чем куколки и имаго.
Смачивающие агенты используются в
препаратах инсектицидов для того,
чтобы улучшать проникновение послед-
них через густую листву или их растека-
ние по неровным поверхностям.
Условия окружающей среды также
влияют на эффективность инсектицидов.
Для большинства инсектицидов предпоч-
тительна смена высокой температуры
воздуха, которая способствует быстрому
их проникновению в организм насекомо-
го, на низкую, • замедляющую скорость
разложения токсиканта. Однако такие
инсектициды, как ДДТ и метоксихлор,
наиболее эффективны при низкой темпе-
ратуре.
Повышение эффективности примене-
ния инсектицидов связано со многими
проблемами. Во-первых, инсектицид не-
обходимо применять строго в определен-
ное время, иногда (как при борьбе с ли-
чинками комаров) в течение всего не-
скольких дней. Во-вторых, важны по-
годные условия в момент обработки, так
как опрыскивание и опыливание не могут
проводиться в дождь и при сильном ве-
тре, иногда обработка не может выпол-
няться при высоких температуре и влаж-
ности, так как при этих условиях воз-
можны ожоги некоторых культур инсек-
тицидами.
Для опрыскивания, опыливания, при-
менения аэрозолей (рис. 12.9), для купа-
ния скота или для применения фумиган-
тов необходимо специальное оборудова-
ние. В каждом отдельном случае следует
составлять наиболее подходящую про-
грамму борбы с вредителями, учитывая
558
12. Вредные насекомые и борьба с ними
Рис. 12.9. Момент обработки посевов инсектицидом; во многих случаях
использование авиационной техники оказывается настоятельно необходимым. (С
разрешения Piper Aircraft Corp.)
топографию местности, высоту и густоту
растительности, условия работы.
Среди перспективных средств механи-
зации следует назвать аппаратуру для
ультрамалообъемных обработок, с по-
мощью которой при определенных усло-
виях можно добиться такого же сниже-
ния численности вредителя, как и с тра-
диционным оборудованием, но расходуя
на 10-25% инсектицида меньше. Эффек-
тивность ультрамалообъемного опрыс-
кивания (УМО) связана с исключительно
тонким распылением жидкости-на ка-
пли диаметром 10-60 мкм, которые, раз-
носясь вихревыми потоками воздуха, до-
стигают всех частей зараженного расте-
ния.
Следует с осторожностью подходить
к совместному применению различных
инсектицидов на одной и той же культу-
ре. Два инсектицида при совместном
применении могут взаимодействовать
друг с другом, образуя неэффективные
соединения. Смесь инсектицидов может
нанести растению вред, даже если каждое
соединение само по себе безвредно. Не-
которые инсектициды при совместном
применении способны изменять свои фи-
зические свойства, так что одно из них
может потерять стабильность или даже
стать опасным. Отдельные инсектициды
резко увеличивают токсичность других
инсектицидов, в результате чего эффект
их совместного применения будет значи-
тельно выше суммы результатов при их
применении по отдельности. Это так на-
зываемое синергическое действие, или си-
нергизм. В качестве примера синергистов
можно назвать бутоксид, сезамекс, MGK
264, пиперонил и сульфоксид.
Предосторожности при применении ин-
сектицидов. Некоторые инсектициды вы-
сокотоксичны для человека, другие мо-
гут представлять потенциальную опас-
ность, особенно для лиц, занятых их
применением, что связано с возмож-
ностью попадания больших количеств
инсектицида на кожу. Следует в принци-
пе избегать попадания инсектицидов на
кожу и одежду. Инсектициды должны
применяться в открытом и хорошо про-
ветриваемом помещении, следует кате-
горически избегать вдыхания инсекти-
цидных паров. Ежегодно регистрируется
примерно 200 смертельных случаев от не-
правильного применения инсектицидов и
значительно большее число случаев не-
смертельных отравлений. Каждый, кто
намерен воспользоваться тем или иным
инсектицидом, должен внимательно про-
читать инструкцию, касающуюся данно-
го инсектицида, и тщательно ей следо-
вать. С точки зрения безопасности чело-
века надо прилагать максимум усилий не
12. Вредные насекомые и борьба с ними
559
к увеличению, а к уменьшению масштабов
и интенсивности применения инсектици-
дов.
Инсектициды следует применять с
осторожностью также и потому, что они
могут причинять вред не только насеко-
мому, но и его хозяину, а накапливание
инсектицидов в почве, воде и различных
продуктах растительного и животного
происхождения может привести к серь-
езным последствиям для человека и жи-
вотных. Некоторые растения высокочув-
ствительны только к какому-то одному
инсектициду. Сведения обо всех этих
«можно» и «нельзя» должны содержать-
ся на этикетке, сопровождающей упаков-
ку инсектицида, и должны быть легко
доступны в местной сельскохозяйствен-
ной организации. Для каждого инсекти-
цида, разрешенного к применению,
имеются строго установленные предель-
ные количества, содержание которых до-
пустимо в продуктах, поступающих на
рынок. Эти количества kдолжны прини-
маться в расчет при планировании про-
грамм борьбы с вредителями той или
иной культуры.
Важной характеристикой инсектици-
дов служит продолжительность сохране-
ния ими токсичности после применения.
Многие яды сохраняют токсичность в те-
чение длительного времени; про них го-
ворят, что они обладают высокой оста-
точной активностью. Другие инсекти-
циды практически не обладают оста-
точным действием, теряя токсичность
почти сразу же после применения. Неко-
торые синтетические яды контактного
действия, например ДДТ, характери-
зуются длительным остаточным дейст-
вием, тогда как остаточное действие дру-
гих, например некоторых фосфороргани-
ческих соединений, коротко. Продолжи-
тельность сохранения остаточной актив-
ности в значительной степени определяет
необходимость повторных обработок.
Механизм действия инсектицидов.
Каждый инсектицид вызывает гибель на-
секомого одним из четырех перечис-
ленных ниже способов.
1. Вещества с физическим механиз-
мом действия. Вещества этой группы
убивают насекомых в результате какого-
то физического эффекта; например, ми-
неральные масла закрывают доступ воз-
духа в организм насекомого, а дуст
силикааэрогеля приводит к потере воды
за счет повреждения кутикулы.
2. Протоплазматические яды. Эти ве-
щества убивают насекомых, вызывая
свертывание белка в их тканях. Сюда от-
носятся соединения мышьяка.
3. Ингибиторы метаболизма. В эту
группу входит большое число соедине-
ний, вызывающих угнетение метаболиче-
ских процессов у насекомых. Например,
дыхательные яды типа циан-водорода,
динитрофенола и ротенона угнетают ды-
хательные ферменты; фторацетат натрия
подавляет метаболизм углеводов.
4. Яды, действующие на нервную си-
стему. В эту группу входит множество
инсектицидов, применяемых в настоящее
время. Они делятся на три категории.
а. Ингибиторы холинэстеразы-фос-
форорганические соединения и кар-
баматы, эти вещества вызывают
чрезвычайно сильное возбуждение
нервной системы.
б. Вещества, влияющие на ионную
проницаемость мембран нервных
клеток,-ДДТ, хлордан, пире-
трины.
в. Соединения, действующие непос-
редственно на нервные клетки или
на ганглии, например никотин.
Еще одна группа органических ве-
ществ, применение которых в конечном
итоге вызывает гибель вредителя,- хемо-
стерилянты. Эти вещества вызывают у
насекомых ту или иную степень стериль-
ности. Будучи применены в полевых ус-
560
12. Вредные насекомые и борьба с ними
ловиях, хемостерилянты вызывают мас-
совую стерилизацию самцов, так что
число стерильных самцов на большой
территории начинает превышать число
нормальных самцов во много раз.
Классификация инсектицидов. По хи-
мическому строению инсектициды делят-
ся на неорганические и органические. Ор-
ганические инсектициды в свою очередь
можно подразделить на масла, яды рас-
тительного происхождения и синтетиче-
ские соединения.
Неорганические инсектициды, ныне в
значительной мере замененные органиче-
скими, включают* 1) арсенат свинца, ис-
пользуемый для борьбы с насекомыми,
обладающими грызущим ротовым аппа-
ратом и вредящими кустарникам, а так-
же для защиты фруктовых деревьев и де-
ревьев в лесах и зеленых насаждениях, 2)
фторсиликат натрия, применяемый в ка-
честве приманки для муравьев, тарака-
нов и саранчовых; 3) серу, которая пере-
малывается в тончайший порошок для
применения в борьбе с растительноядны-
ми клещами и некоторыми грибковыми
болезнями растений.
Масла представляют собой продукты
перегонки нефти. Минеральные масла
используются для защиты фруктовых де-
ревьев, а также в качестве растворителей
и носителей в инсектицидных препара-
тах. Дизельное топливо служит носите-
лем инсектицидов при обработках с воз-
духа и при обработках водоемов для
борьбы с личинками комаров.
Масла могут применяться и как само-
стоятельные инсектициды. Летние масла
являются высокоочищенными и не столь
токсичными для растений; они приме-
няются для обработки листвы цитру-
совых деревьев против клещей, а также
щитовок и червецов. Зимние масла отно-
сительно плохо очищены и применяются
для обработки деревьев при отсутствии
на них листвы.
Среди веществ растительного проис-
хождения, используемых в борьбе с насе-
комыми, можно назвать гераниол и эуге-
нол, привлекающие насекомых; цитро-
неллу и кедровое масло, отпугивающие
насекомых, а также хлопковое масло,
служащее растворителем и пролонгато-
ром. Однако большинство органических
веществ этой группы токсичны для насе-
комых, наибольшее значение среди них
имеют никотин, пиретрум и ротенон.
Синтетические органические инсекти-
циды. Эта группа веществ исключитель-
но многочисленна.
1. ХЛОРИРОВАННЫЕ УГЛЕВОДО-
РОДЫ. Эта группа веществ, известных
как хлорорганические соединения, вклю-
чает ДДТ (дихлордифенилтрихлорэтан) -
высокоэффективное соединение, исполь-
зуемое в борьбе с переносчиками болез-
ней человека и многими вредителями
сельскохозяйственных культур. ДДТ -
высокоустойчивое соединение, способное
длительное время сохраняться в окру-
жающей среде; его вымывание из почвы
происходит с большим трудом. ДДТ на-
капливается в жировых тканях жи-
вотных, где может долго сохраняться. Из
других соединений этой группы следует
назвать бензолгексахлорид (ГХЦГ), ме-
токсихлор, пертан, токсафен и циклодие-
новые производные-альдрин, дильдрин,
хлордан, эндосульфан, эндрин и гепта-
хлор. Метоксихлор сходен по строению с
ДДТ, но значительно менее токсичен для
млекопитающих. Он значительно безо-
паснее ДДТ при обработке растений и не
накапливается с такой же легкостью в
жировых тканях животных. Токсафен на-
ходит широкое применение в борьбе со
многими видами вредных насекомых,
однако он значительно токсичнее для
млекопитающих, чем ДДТ, и очень ток-
сичен для рыб. Вместе с тем он не накап-
ливается в жировых тканях с такой же
легкостью, как ДДТ Токсафен исполь-
12. Вредные насекомые и борьба с ними
561
зуется для борьбы с саранчовыми, совка-
ми, рядом гусениц, вредителями хлоп-
чатника, бобовых и паразитами скота.
Циклодиеновые производные эффек-
тивны против широкого круга вредных
насекомых, но по сравнению с ДДТ они
более токсичны для млекопитающих.
Применение этих соединений сильно
ограничено из-за накопления их в почве.
2. ФОСФОРОРГАНИЧЕСКИЕ ИНСЕК-
ТИЦИДЫ. В эту группу входит наиболь-
шее число синтетических инсектицидов.
Некоторые из них поглощаются расте-
ниями, в результате чего соки растений
становятся токсичными для насекомых,
или животными, чья кровь становится
токсичной для других организмов. Такие
вещества называются системными инсек-
тицидами. К инсектицидам системного
действия на растения относятся деметон,
дикротофос, диметоат, дисульфотон, ме-
винфос, монокротофос, форат и фосфа-
мидон. Вещества, являющиеся системны-
ми ядами для животных, включают ку-
мафос, круфомат, роннел, фамфур, фос-
мет и трихлорфон. Вещества, не обла-
дающие системным действием,-диази-
нон, малатион, паратион и тепп. Пара-
тион черезвычайно токсичен для млеко-
питающих, но, несмотря на это, широко
применяется для защиты фруктов, ово-
щей, декоративных растений и полевых
культур. Из-за его высокой токсичности
следует соблюдать тщательные меры
предосторожности в отношении людей и
скота. Паратион применяется в форме
капсул. Значительно менее опасен метил-
паратион, который используется для
борьбы с вредителями хлопчатника и как
заменитель ДДТ Различные модифика-
ции паратиона используются для борьбы
с вредителями сельскохозяйственных
культур незадолго до сбора урожая и для
борьбы с насекомыми, вредящими здо-
ровью человека (комары, мухи). Одно-
кратное применение карбобентиона мо-
36-732
жет заменить несколько обработок дру-
гими инсектицидами в борьбе с сос-
новым шелкопрядом.
3. КАРБАМАТНЫЕ ИНСЕКТИЦИДЫ.
Эти соединения быстро разлагаются и не
оставляют токсичных остатков. Токсич-
ность их для млекопитающих варьирует
от низкой до высокой. Наиболее из-
вестным представителем этой группы
является карбарил, обладающий низкой
токсичностью для млекопитающих и ис-
пользуемый в садах и на полях в борьбе с
жуками-нарывниками, японским жуком,
саранчовыми, щитовками и червецами,
колорадским жуком и мексиканской бо-
бовой зерновкой. Из других карбаматов
можно назвать карбофуран, металкамат,
метомил и пропоксур.
4. СИНТЕТИЧЕСКИЕ ПИРЕТРОИДЫ.
Эти соединения, родственные при-
родным пиретринам, синтезируются из
производных нефти. Аллетрин исполь-
зуется для установления факта зараже-
ния помещений тараканами и определе-
ния степени этого заражения.
5. ГОРМОНЫ НАСЕКОМЫХ. Установ-
ление инсектицидной природы химиче-
ских веществ, сходных с ювенильным
гормоном насекомых, привело к синтезу
нескольких сотен соединений, биологиче-
ская активность которых была сравнима
с активностью гормона прилежащих тел.
Эти токсичные или слаботоксичные сое-
динения прерывают ход морфогенеза и
предотвращают, таким образом, нор-
мальное развитие, вызывая в конечном
итоге гибель насекомого. Многие из син-
тезированных веществ характеризуются
избирательностью, действуя только на
какую-то одну группу насекомых, что
является многообещающим фактором
для применения этих веществ в самых
разнообразных программах борьбы с
вредителями; кроме того, эти вещества
безопасны или обладают слабой токсич-
ностью для полезных насекомых, челове-
562
12. Вредные насекомые и борьба с ними
ка и других животных, и они в исключи-
тельно низких дозах эффективно влияют
на насекомых, служащих объектами
борьбы. Эта новая группа веществ была
разрешена к широкому применению
только в 1975 г Поскольку эти вещества
влияют на нормальный ход процессов
роста насекомого и косвенно приводят
его к гибели, их назвали регуляторами
роста насекомых (РРН). Один из та-
ких регуляторов - метопрен - применя-
ется для борьбы с личинками комаров.
До сих пор не удалось создать РРН,
который бы избирательно воздействовал
на ход различных процессов, связанных с
активностью линочных гормонов насе-
комых-экдизона и 20-гидроксиэкдизона.
Был синтезирован дифлубензурон,
влияющий на линьку путем ингибирова-
ния синтеза хитина. Основным препят-
ствием к получению эффективного ли-
ночного РРН служит высокая стоимость
препаратов, получаемых из природных
материалов, и слабая проницаемость
синтезируемых соединений через кутику-
лу. Существует несколько родов расте-
ний, содержащих экдистероиды - соеди-
нения с активностью линочного гормона
насекомых. Экдистероиды чаще всего
встречаются у папоротников, а также у
представителей одного семейства голосе-
менных и одного семейства покрытосе-
менных. Их наличие в папоротниках
объясняет почти полное отсутствие насе-
комых на этих растениях.
6. РЕПЕЛЛЕНТЫ НАСЕКОМЫХ. Ве-
щества этой группы используются
главным образом для того, чтобы отпу-
гивать насекомых от человека и жи-
вотных. Непременное требование к ре-
пеллентам - отсутствие токсичности. На
протяжении многих веков для защиты от
насекомых хранящейся одежды исполь-
зовали нафталин и камфару. Древесину
от термитов защищают, пропитывая ее
креозотом.
Среди множества соединений, испы-
танных в качестве репеллентов против
комаров и других кровососущих двукры-
лых, иксодовых клещей, блох и личинок
красноте л ок, наиболее эффективными
признаны диметилфталат, индалон
и ДЭТА (диэтилтолуамид). Каждое из
этих соединений эффективно только в от-
ношении некоторого числа видов, но раз-
личные смеси, наносимые на одежду и на
кожу, обеспечивают хорошую защиту в
течение короткого отрезка времени. Раз-
личных клещей особенно эффективно от-
пугивает бензилбензоат. Для отпугива-
ния кровососущих насекомых применяет-
ся также пиретрум. С этой же целью
использовался диметилфталат, обеспечи-
вавший хорошую защиту от малярийных
комаров, иксодовых клещей и личинок
краснотелок.
7 АТТРАКТАНТЫ НАСЕКОМЫХ. К
этой группе относятся некоторые при-
родные вещества, например сахар, ме-
ласса, дрожжевой экстракт, жирные кис-
лоты, гераниол и эугенол. Среди синте-
тических соединений можно назвать фе-
нетилпропионат и эугенол, смесь ко-
торых известна как очень эффективный
аттрактант. Медлур и тримедлур-два
других синтетических аттрактанта, они
используются для привлечения средизем-
номорской плодовой мушки. Аттрак-
танты применяются в различных типах
ловушек.
В конце 50-х годов были начаты ши-
рокие исследования, направленные на
выяснение возможностей использования
феромонов как средства борьбы
с вредными видами насекомых, в первую
очередь внимание было обращено на
виды, существование которых тесно свя-
зано с феромонной коммуникацией.
Борьба, основанная на использовании
феромонов, может быть направлена на
изменение способности к образованию
скоплений (особенно в местах размноже-
12. Вредные насекомые и борьба с ними
563
ния на нарушение полового и пищевого
поведения. В качестве примеров успеш-
ной борьбы можно назвать использова-
ние синтетических половых аттрактантов
луплюра (на полях капусты) и госсиплю-
ра < на хлопчатнике), эти аттрактанты на-
рушали поведенческие стереотипы сам-
цов соответственно капустной совки и
розового коробочного червя хлопчатни-
ка, в результате чего большое число
самок оставалось неоплодотворенными.
Феромоны использовали также в ка-
честве приманки для привлечения
вредных насекомых в ловушки с инсекти-
цидом или на клейкие настилы. Самки
некоторых видов вредных бабочек выде-
ляют феромоны, которые сильно привле-
кают самцов с окружающей территории.
Таких самок помещают в ловушки, рас-
ставляемые в фруктовых садах; в резуль-
тате одна неоплодотворенная самка мо-
жет привлечь в ловушку тысячи самцов.
Обычно для привлечения самцов раз-
личных бабочек используются синтетиче-
ские феромоны, например диспарлюр-
для привлечения непарного шелкопряда,
кодлюр-для привлечения яблонной пло-
дожорки.
8. МИКРОБНЫЕ ИНСЕКТИЦИДЫ.
Некоторые патогены насекомых послу-
жили основой для создания инсекти-
цидных препаратов, к числу таких пато-
генов относятся вирусы, бактерии, грибы
и простейшие. Эти средства борьбы безо-
пасны и эффективны, потому что они спе-
цифичны для целевого объекта, не ток-
сичны и не патогенны для человека, не
наносят вреда хищникам, паразитам и
окружающей среде в целом. В качестве
примера можно назвать бактериальный
инсектицид на основе Bacillus popilliae,
вызывающий молочную болезнь у личи-
нок японского жука.
Первым вирусом, зарегистриро-
ванным в 1975 г в качестве пестицида,
был встречающийся в природе вирус
ядерного полиэдроза. Он используется
против двух опасных вредителей хлоп-
чатника: коробочного червя и табачной
листовертки-почкоеда. Другой вирус
ядерного полиэдроза помогает контро-
лировать вспышки размножения одной
из волнянок.
9. ФУМИГАНТЫ. Фумигантами назы-
вают летучие вещества, токсичные и ре-
пеллентные для насекомых. Фумиганты,
как правило, используются для борьбы с
насекомыми, нематодами и грибами в
почве, для борьбы с насекомыми в до-
мах, складах, хранилищах, зернохрани-
лищах, теплицах, на мельницах и в дру-
гих строениях, а также на плантациях
цитрусовых, т. е. они обычно приме-
няются в каком-то закрытом простран-
стве ящике, бункере, товарном вагоне, в
постройках или под тентом. Фумиганты
следует применять с осторожностью, а
помещение после обработки должно
быть тщательно проветрено. В качестве
фумигантов используют цианистый во-
дород, парадихлорбензол, дисульфоугле-
род, дихлорэтан, тетрахлористый угле-
род, метилбромид, нафталин и дихлор-
пропан. При использовании фумигантов
следует помнить о двух опасностях. спо-
собности некоторых газов к самовозго-
ранию и их токсичности для человека.
Цианистый водород смертелен для чело-
века и особенно опасен потому, что
почти не имеет запаха.
РЕГУЛЯЦИЯ ЧИСЛЕННОСТИ
НАСЕКОМЫХ-ВРЕДИТЕЛЕЙ
Внедрение в практику синтетических
инсектицидов в годы после второй миро-
вой войны казалось предвестником эры
безраздельного контроля над числен-
ностью насекомых. Первые синтетиче-
ские инсектициды были дешевы и эффек-
тивны, что позволяло получать прибыль,
во много раз превышающую стоимость
564
12. Вредные насекомые и борьба с ними
инсектицида и расходов на его примене-
ние. Однако уже в самые первые годы од-
на за другой стали возникать проблемы,
вызвавшие необходимость коренного
пересмотра программ по борьбе с насе-
комыми. Среди этих проблем было три
основных. у вредителей развивалась
устойчивость к инсектицидам; остатки
инсектицидов обладали высокой перси-
стентностью, стоимость применяемых
инсектицидов возрастала в связи с необ-
ходимостью увеличивать дозировки и ча-
стоту обработок.
Устойчивость к инсектицидам. Уже че-
рез 1-2 года широкого применения ДДТ
для борьбы с комнатной мухой в Среди-
земноморье появились расы этого насе-
комого, устойчивые к ДДТ. В дальней-
шем у комнатной мухи появились расы,
устойчивые почти к любому из приме-
нявшихся инсектицидов. То же самое на-
блюдалось у таракана-прусака. У огром-
ного числа других насекомых вредителей
также возникли расы, устойчивые к ин-
сектицидам, применявшимся в борьбе с
ними. Среди этих насекомых можно наз-
вать многие виды малярийных комаров,
некоторых жуков-блошек (Diabroticd), не-
скольких видов растительноядных кле-
щей, хлопкового долгоносика-цветоеда и
ряд видов бабочек. В любой популяции
насекомых, подвергающейся воздей-
ствию инсектицида, имеются отдельные
особи, которые оказываются более
устойчивыми к действию последнего и
могут выжить, дело в том, что гены,
обусловливающие устойчивость к данно-
му инсектициду, имеются в популяции и
до начала его применения, но частота
этих генов чрезвычайно низка. Большин-
ство особей не обладает этими генами и
погибает. В следующем поколении боль-
шая доля особей будет обладать геном
устойчивости, и если популяция вновь
подвергнется действию инсектицида, то
процесс отбора будет повторяться, при-
водя ко все возрастающему уровню
устойчивости в последующих поколе-
ниях. При достаточно длительном пов-
торении обработок возникнет популя-
ция, почти полностью устойчивая к ин-
сектициду. До последнего времени вся
практика обработок чрезвычайно спо-
собствовала быстрому развитию устой-
чивости у вредных насекомых на основе
указанной модели. Согласно существо-
вавшим схемам, инсектицид применялся
через регулярные промежутки времени
независимо от того, каким было обилие
вредителя - низким или высоким. Эта ре-
гулярность из года в год поддерживала
давление отбора в отношении особей, не-
сущих ген устойчивости. Если в популя-
ции имеются хотя бы «следы» генов
устойчивости, то такая профилактиче-
ская схема обработок оказывается в ко-
нечном счете саморазрушительной с точ-
ки зрения борьбы с видом-вредителем.
Остаточные количества инсектицидов
и загрязнение окружающей среды. Вскоре
после начала щирокого применения пер-
систентных синтетических инсектицидов
возникла другая проблема. Эти инсекти-
циды накапливались в продуктах пита-
ния человека и в окружающей среде, где
они попадали в пищу беспозвоночным
животным: в почве - дождевым червям, в
воде прудов и озер - личинкам комаров.
Инсектицид накапливался в тканях тела
этих животных, которые затем станови-
лись добычей более крупных живых су-
ществ, например дроздов или мелких
рыб. На каждом последующем этапе
концентрация инсектицида в тканях тела
очередного хищника возрастала до тех
пор, пока она не становилась смертель-
ной для крупных животных, находящих-
ся на высших ступенях пищевой цепи.
Широко известным примером такого ро-
да была попытка борьбы с некровососу-
щим комаром Chaoborus astictopus в озе-
ре Клэр (шт. Калифорния), где этот
12. Вредные насекомые и борьба с ними
565
комар, развиваясь в огромных количе-
ствах. чрезвычайно досаждал местным
жителям и отдыхающим. При периоди-
ческом применении ДДД в концентрации
20 частей на миллиард частей воды уда-
валось поддерживать популяцию личи-
нок этого вида на очень низком уровне. В
нишевой цепи, идущей от личинок кома-
ров к мелким, а затем к более крупным
рыбам, и, наконец, к западной поганке,
ДДД накапливался на каждом последую-
жм уровне, достигнув 2000 частей на
миллион у птиц, которые и погибали от
Агтрого отравления. В другом случае
сток из лесов, обрабатывавшихся ДДТ,
вызывал гибель водных насекомых во
многих проточных водоемах, значитель-
но снизив и обилие рыбы, населявшей
эти водоемы и питавшейся преимуще-
ственно водными насекомыми. Разложе-
ние многих инсектицидов происходит
столь медленно, что их токсическое дей-
ствие может сохраняться в течение ряда
лет уже после того, как применение ин-
сектицида прекращено.
Действие инсектицидов на паразитов и
хжппков. Почти все используемые ин-
сектициды имеют широкий спектр дей-
ствия. а многие из них являются к тому
же персистентными соединениями, так
тто в результате обработок погибают не
только особи вредного вида, но также и
нецелевые объекты. Наиболее достойны-
ми внимания среди жертв применения
инсектицидов, конечно же, следует счи-
тать паразитов и хищников, которые в
естественном состоянии способны огра-
ничивать численность вредного или по-
тенциально вредных видов на уровне, не
представляющем опасности. Когда гиб-
нет паразиты и хищники, то их хозяева и
жертвы, являвшиеся потенциальными
зредителями, становятся действенными
< их называют вторичными вредителями),
что требует проведения дополнительных
мероприятий. Так, использование ДДТ
для борьбы с вредителями цитрусовых в
Калифорнии сопровождалось резким
возрастанием численности австралийско-
го желобчатого червеца, которая до это-
го поддерживалась на низком уровне
хищным жуком родолией, эта вспышка
была вызвана значительным снижением
численности родолии в результате при-
менения ДДТ. Известно множество дру-
гих примеров подобного рода.
Возрастающая стоимость мероприя-
тий. Переход на инсектициды с непродол-
жительным остаточным действием при-
вел к значительному увеличению стоимо-
сти мероприятий по борьбе с насекомы-
ми-вредителями сельскохозяйственных
культур и переносчиками болезней чело-
века. Произошло это прежде всего из-за
увеличения числа обработок, требую-
щихся на протяжении года. На полях
хлопчатника, где также возникают про-
блемы, связанные с развитием устойчи-
вости у вредителей, приходится прово-
дить 10-20, а обычно даже большее число
обработок, что ведет к резкому увеличе-
нию себестоимости производимой про-
дукции.
Новый подход к борьбе с насекомыми.
В свете изложенных обстоятельств была
очевидна необходимость принципиально
иного подхода к борьбе с вредными насе-
комыми. Продолжающееся использова-
ние некоторых инсектицидов с дли-
тельным остаточным действием вело бы
к глобальному загрязнению окружаю-
щей среды, что представляло бы серьез-
ную опасность для всей жизни на земле.
Поэтому в ряде стран было полностью
запрещено применение ДДТ, и почти во
всех странах были наложены строгие
ограничения на применение ДДТ и неко-
торых других персистентных инсектици-
дов.
Это изменение образа мышления на-
чалось в конце 50-х годов с предложения
программ интегрированной борьбы, осно-
566
12. Вредные насекомые и борьба с ними
ванных на комбинированном использо-
вании паразитов и инсектицидов. В нача-
ле 60-х годов были составлены про-
граммы борьбы с учетом всех суще-
ственных биологических особенностей
конкретных видов-вредителей. Этим из-
менением в подходе упор был перенесен с
«борьбы с насекомыми», как это было
раньше, на «управление численностью
вредного вида» или точнее на «интегри-
рованное управление» в его более широ-
ком значении.
ИНТЕГРИРОВАННОЕ УПРАВЛЕНИЕ
ЧИСЛЕННОСТЬЮ ВРЕДИТЕЛЕЙ
Современный подход к управлению
численностью вредителей является
в своей основе экологическим. При этом
используется все многообразие суще-
ствующих методов борьбы, с тем чтобы
для каждого конкретного случая можно
было использовать некую систему
средств и методов, дополняющих друг
друга. Именно поэтому такой подход на-
зывают обычно интегрированным управ-
лением численностью вредителя. Ос-
новные цели интегрированного подхода
следующие 1) до конца понять все
многообразие взаимодействий организ-
мов с окружающей их средой в таких эко-
системах, как пастбище, возделываемое
поле или фруктовый сад, 2) определить
или уровень экономической вредоносно-
сти, или экономический порог, при кото-
ром будет требоваться проведение истре-
бительных мероприятий, 3) составить
такую программу мероприятий, которая
не нарушает иные в высшей степени же-
лательные взаимоотношения между дру-
гими живыми организмами в данной
экосистеме.
Поскольку большинство экосистем, в
которых осуществляется управление чис-
ленностью вредителя,- это сельскохозяй-
ственные экосистемы, их иногда относят
к агроэкосистемам. Они представляют
собой высокоспециализированные экоси-
стемы с одним или ограниченным набо-
ром видов растений или домашних жи-
вотных и обычно только с несколькими
видами насекомых или других организ-
мов, которые могут исключительно бы-
стро достигать огромной численности и
причинять серьезный вред. Большее чис-
ло других видов является потенциальны-
ми вредителями, но они характеризуют-
ся относительно невысоким обилием.
Только случайно и при необычных об-
стоятельствах эти редкие виды дости-
гают такого уровня численности, при ко-
тором их можно расценивать как вреди-
телей.
Для каждой культуры и ее вредителей
должен быть определен уровень ущерба,
который экономически вполне терпим.
Этот уровень может варьировать для
разных культур и в разных местностях;
он известен как экономический уровень
вредоносности. Во многих случаях воз-
можно установить экономический порог
вредоносности, показывающий, что
плотность популяции потенциального
вредителя достигла уровня, когда необ-
ходимо предпринимать какие-то меры.
Эти меры должны быть направлены на
предотвращение дальнейшего увеличе-
ния плотности вредителя до того уровня,
который нанесет урожаю ущерб, превы-
шающий экономический уровень вредо-
носности.
Многие культуры могут сильно по-
вреждаться насекомыми, причем уро-
жайность этих культур не снижается или
снижается очень незначительно. Обычно
экономический порог вредного вида пре-
вышается только в каких-то отдельных
популяциях. Это означает, что необходи-
мость применения инсектицидов суще-
ствует только в тех районах, где и когда
эти популяции достигли критических
уровней.
12. Вредные насекомые и борьба с ними
567
Со словом «когда» связаны два
важных момента. Может случиться так,
что популяция, имеющая низкую плот-
ность в первом поколении, достигнет вы-
сокой плотности в следующем поколе-
нии этого же года. В данном случае
потребуется постоянный контроль. Дру-
гой момент касается точных сроков про-
ведения истребительных мероприятий,
совпадающих с такими стадиями в жиз-
ненном цикле вредителя, когда возмож-
но наиболее эффективное управление.
Так. например, в Судане хлопковая совка
Heliothis armigera откладывает яйца в те-
чение 10 дней синхронно на всем обшир-
ном пространстве хлопкосеющего райо-
на. Установив начало этого периода,
можно приурочить обработки так, чтобы
уничтожить появляющихся гусениц,
обеспечивая тем самым наибольший эф-
фект истребительных мероприятий. Регу-
лярно собирая пробы и наблюдая за со-
стоянием популяцци вредного вида, мы
можем предсказать тенденции, намечаю-
щиеся в этой популяции до, во время и
после сезона вегетации.
Программы интегрированного управ-
ления предусматривают использование
наилучших селективных методов, позво-
ляющих в наименьшей степени нарушать
-покой» экосистемы. Эти программы
могут меняться в зависимости от вре-
мени года, от относительного обилия
других насекомых и от многих других
факторов. В рамках программы возмож-
но проведение селективной обработки в
некий момент времени, когда вредитель
наиболее уязвим для нее, в то же время
такая обработка не будет сокращать по-
пуляции хищников и паразитов, которые
счгтанутся в состоянии контролировать
остающуюся небольшую часть популя-
ции вредителя. Некоторые программы, в
юторых сочетается использование
устойчивых к вредителям сортов сель-
скохозяйственных культур с деятель-
ностью хищников и паразитоидов, ис-
ключают необходимость применения ин-
сектицидов или снижают ее до миниму-
ма. В других программах сочетается
проведение некоторых агрокультурных
мероприятий с деятельностью хищников
и паразитоидов (которые могут
представлять собой естественные
местные природные популяции или быть
«усилены» выпуском дополнительного
числа особей) и применением мини-
мальных количеств избирательно дей-
ствующих инсектицидов. Принимаются
любые меры, чтобы минимизировать
внесение инсектицидов, особенно перси-
стентных, в экосистему.
Среди других методов, включаемых в
программы интегрированного управле-
ния, назовем смену пастбищ, изменения
сроков посева, использование местных и
интродуцированных паразитов и хищни-
ков, применение растений-ловушек, ис-
пользование устойчивых сортов и т. д.
При осуществлении программ инте-
грированного управления обращается
также внимание на более качественное
использование оборудования и проведе-
ние обработок; имеется в виду достиже-
ние максимальной эффективности на
единицу распыленного инсектицида.
Операциональные программы. После
того как необходимость интегрирован-
ного подхода к управлению числен-
ностью вредителей была осознана, по-
требовалось составить, испытать, одоб-
рить и внедрить в практику конкретные
программы. Работа в этом направлении
проводится многими федеральными,
штатными, университетскими и про-
мышленными лабораториями. Мно-
гие из этих лабораторий уже имеют боль-
шой опыт исследований по проблемам,
связанным с сельскохозяйственными и
другими вредителями во многих ча-
стях Северной Америки.
Исследования сконцентрированы на
568
12. Вредные насекомые и борьба с ними
самых неотложных проблемах, свя-
занных с защитой основных сельскохо-
зяйственных культур, возделываемых на
обширных территориях и гарантирую-
щих высокие урожаи. Перемещение ак-
цента с уничтожения вредителя на защи-
ту возделываемой культуры предпола-
гает, что системы управления числен-
ностью-это только часть общей слож-
ной системы сельскохозяйственного про-
изводства. Разработка операциональных
программ проводится при эффективном
разделении труда между различными
подразделениями исследовательской
группы, постоянном обмене информа-
цией между ними и хорошо спланирован-
ной приоритетности исследований. На-
личие в таких подразделениях исследова-
телей с широкой, но разнообразной под-
готовкой является предпосылкой быст-
рой разработки эффективных программ.
Для упрощения планирования и более
быстрого получения рекомендаций ши-
роко используются модели, позволяю-
щие представить сложные процессы
взаимодействия в экосистемах в более
обобщенном виде. В разных районах воз-
делывания одна и та же культура часто
страдает от совершенно разных вредите-
лей ; поэтому невозможно применять од-
ну и ту же модель к любой местности.
Большую помощь в поисках информа-
ции и анализе моделей могут оказать
компьютеры. Следует, однако, помнить,
что моделирование связано с упроще-
нием реальной ситуации и при попытке
использования модельных разработок в
реальных полевых условиях могут воз-
никнуть определенные трудности.
Для многих сфер сельскохозяйствен-
ного производства такие программы по-
ка что существуют лишь в перспективе,
применение их пока ограничивается не-
большими фермерскими участками, при-
усадебными садами, а также хозяйства-
ми с интенсивным производством.
Пройдет еще немало лет, прежде чем ре-
зультаты разработок интегрированных
программ коснутся всех сфер сельскохо-
зяйственного производства и войдут в
повседневную практику.
Доведение до каждого фермера ин-
формации, касающейся управления попу-
ляциями вредных видов, осуществляется
главным образом через обширную сеть
федеральных и штатных служб. Они же
распространяют информацию, связан-
ную с применением удобрений, борьбой
с сорняками, возделыванием земли и
другими вопросами ведения хозяйства.
Конечно, весь этот набор информации
чересчур велик и сложен, чтобы фермер
мог его использовать без квалифициро-
ванной помощи. Поэтому была создана
служба различных сельскохозяйственных
консультантов, которые приглашаются
для решения различных вопросов, касаю-
щихся борьбы с вредителями и сорняка-
ми, ирригационных работ, применения
удобрений и т. д. Специалисты в области
управления численностью вредителей
обычно разрабатывают или анализи-
руют имеющиеся модели для вредных
насекомых, приносящих ущерб в данном
районе, в предписанные сроки проводят
контрольные наблюдения, а также вы-
полняют другие обусловленные догово-
ром работы.
ПЕРСПЕКТИВЫ НА БЛИЖАЙШЕЕ
БУДУЩЕЕ
Человек в самом прямом смысле кон-
курирует с насекомыми за все доступные
источники пищи на Земле. Всемирная ор-
ганизация здравоохранения ООН под-
считала, что около 80% людей во всем
мире голодает или серьезно недоедает. И
хотя предпринимаются отчаянные
попытки увеличить- производство про-
дуктов питания путем развития сельско-
хозяйственного производства, регулиро-
12. Вредные насекомые и борьба с ними
569
вания численности популяций вредите-
лей и улучшения сортов сельскохозяй-
ственных культур, численность населе-
ния увеличивается значительно быстрее,
что в свою очередь приводит к быстрому
увеличению числа голодающих и остро
недоедающих. Для того чтобы эффек-
тивное управление популяциями вредных
насекомых смогло ощутимо сказаться на
состоянии сельскохозяйственного про-
изводства и здравоохранения и помочь
подавляющему большинству людей, жи-
вущих на нашей планете, необходимо
предпринимать определенные меры по
поддержанию численности населения
Земли в пределах ее продуктивной емко-
сти.
ЛИТЕРАТУРА
Annu. Rev. Entomol., vols. 1-25 (1956-1980).
Статьи по многим из затронутых в данной
главе вопросов.
Balch R. Е. The spruce budworm and aerial for-
est spraying, Can. Geog. J., 45, 201-209 (1952).
Berozoa M. (Ed.). Pest management with insect
sex attractants and other behavior-controlling
chemicals, Am. Chem. Soc. Symp., Ser. 23, 192
pp., 1976.
Blum M. 5., Blum N. A. (Eds.). Sexual selec-
tion and reproductive competition in insects.
Academic Press, 'New York, 464 pp., 1979.
Carter W Insects in relation to plant disease.
Wiley, New York, 2nd ed., 759 pp., 1973.
Caswell R. L. (Ed.). Pesticide handbook-En-
toma. 1977-1978. Entomological Society of
America, College Park, Md., 27th ed., 248 pp.,
1977.
Cherrett J. M., Sagar G. R. (Eds.). Origins of
pest, parasite, disease, and weed problems.
Blackwell, Oxford, 413 pp., 1977.
Chiang H. C. Pest management of corn, Annu.
Rev. Entomol., 23, 101-123 (1978).
Cox G. W., Atkins M. D. Agricultural ecology.
W H. Freeman, San Francisco, 722 pp., 1979.
DeBach P. Biological control by natural ene-
mies. Cambridge Univ. Press, Cambridge, 323
pp., 1974.
Ebeling W Subtropical fruit pests. Univ, of Cali-
fornia, Div. Agr. Sci. California, Berkeley, 436
pp., 1959.
Geier P. W., Clark L. R., Anderson D. J.,
Nix H. A. (Eds.). Insects: Studies in popula-
tion management. Ecological Society of Aus-
tralia Memoirs, Canberra, vol. 1, 295 pp.,
1973.
Gibbs A. J. (Ed.). Viruses and invertebrates.
North-Holland, Amsterdam, 673 pp., 1973.
Harborne J. B. (Ed.). Biochemical aspects of
plant and animal coevolution, Academic Press,
New York, 436 pp., 1978.
Harris K. F, Maramorosch K. (Eds.). Aphids
as virus vectors. Academic Press, New York,
560 pp., 1977.
Harrison G. Mosquitoes, malaria and man.
E. P. Dutton, New York, 314 pp., 1978.
Headley J. C. Economics of agricultural pest
control, Annu. Rev. Entomol., 17, 273-286
(1972).
Herms W B. Medical entomology. Macmillan,
New York, 4th ed., 643 pp., 1950.
Horsfall W R. Medical entomology. Ronald
Press, New York, 467 pp., 1962.
Huffaker С. B. (Ed.). New technology of pest
control. Wiley, New York, 500 pp., 1980.
Huffaker С. B., Messenger P S. (Eds.). Theory
and practice of biological control. Academic
Press, New York, 788 pp., 1976.
Jones D. P., Solomon M. E. (Eds.). Biology in
pest and disease control. 13th Symposium Bri-
tish Ecological Society. Wiley, New York, 398
pp., 1974.
Maramorosch K., Harris K. F. Leafhopper vec-
tors and plant disease agents. Academic Press,
New York, 654 pp., 1979.
McEven F. L., Stephenson G. R. The use and sig-
nificance of pesticides in the environment.
Wiley, New York, 538 pp., 1979.
Metcalf C. L., Flint W P., Metcalf R. L. Des-
tructive and useful insects, their habits and
control. McGraw-Hill, New York, 4th ed.,
1087 pp., 1962.
570
12. Вредные насекомые и борьба с ними
Metcalf R. L., Luckmann W. Н. (Eds.). Intro-
duction to insect pest management. Wiley,
New York, 587 pp., 1975.
Mitchell E. R. Disruption of pheromonal com-
munication among coexistent pest insects with
multichemical formulations, BioScience, 25,
493-499 (1975).
Narahashi T (Ed.) Neurotoxicology of insects
and pheromones. Plenum, New York, 308 pp.,
1979.
Neal J. W., Jr. (Ed.). Guidelines for the control
of insect and mite pests of foods, fibers, feeds,
ornamentals, livestock, forests, and forest pro-
ducts, USDA Agric. Handbook 554, 822 pp.,
1979.
Pal R., Whitten M. J. The use of genetics in in-
sect control. North-Holland, Amsterdam, 241
pp., 1974.
Peairs L. M., Davidson R. H. Insect pests of
farm, garden, and orchard. Wiley, New York,
5th ed., 661 pp., 1956.
Pfadt R. E. (Ed.). Fundamentals of applied
entomology. Macmillan, New York, 3rd ed.,
798 pp., 1978.
Smith K. G. V (Ed.). Insects and other arthro-
pods of medical importance. Wiley, New York,
600 pp., 1978.
Stairs G. R. Pathogenic microorganisms in the
regulation of forest insect populations, Annu.
Rev. Entomol., 17, 355-372 (1972).
Theiler M., Downs W G. The arthropod-borne
viruses of vertebrates. Yale Univ. Press, New
Haven, 578 pp., 1973.
Tinsley T W The potential of insect pathogenic
viruses as pesticidal agents, Annu. Rev. Ento-
mol., 24, 63-88 (1979).
Wallace J. W., Mansell R. L. (Eds.). Biochemi-
cal interaction between plants and insects.
Plenum, New York, 436 pp., 1976.
Watson M. A., Plumb R. T Transmission of
plant-pathogenic viruses by aphids, Annu.
Rev. Entomol., 17, 425-Л52 (1972).
Whitten M. J., Foster G. G. Genetical methods
of pest control, Annu. Rev. Entomol., 20,
461-476 (1975).
См. также списки литературы к главам 1, 5, 6,
10, 11.
Оглавление
Предисловие редактора перевода 5 Экскреция и водно-солевой баланс 134
Дыхание 138
Предисловие 7 Кровь и ее циркуляция 145
Метаболизм 150
1. РАЗВИТИЕ ЭНТОМОЛОГИИ В Мышечная система и движение 151
СЕВЕРНОЙ АМЕРИКЕ 9
Истоки современной биологии 11 6. РЕАКЦИИ НА ВНЕШНИЕ ВОЗ-
Развитие биологии в XVIII в. 14 ДЕЙСТВИЯ И ПОВЕДЕНИЕ 157
Развитие энтомологии в Северной Источники информации: рецепторы и
Америке 16 органы чувств -
Период бурного развития Америки Нервная координация и интеграция 172
(примерно 1867-1900) 21 Эндокринная система и гормоны 175
Развитие энтомологии в XX в. 2. ARTHROPODA: НАСЕКОМЫЕ И 26 Коммуникация Поведенческие реакции и типы пове- ГТАиТЛ СТ 176 185
БЛИЗКИЕ К НИМ ГРУППЫ 31 деним Организация сообществ у обществен-
Предки членистоногих — ных насекомых 191
Тип Arthropoda 38
Надкласс Hexapoda 49 7. ЖИЗНЕННЫЕ ЦИКЛЫ, РОСТ
3. ВНЕШНЕЕ СТРОЕНИЕ 56 И РАЗМНОЖЕНИЕ 199
Стенка тела и экзоскелет Развитие -
Отделы тела 58 Созревание 216
Г олова 59 Типы питания 221
Шейный отдел Развитие’ генерализованного сегмента 79 Сезонные циклы Размножение 225 230
тела насекомого 81 Роль гормонов 233
Грудь 82 8. ОТРЯДЫ ЭНТОГНАТ И НАСЕ-
Брюшко 92 КОМЫХ 236
4. ВНУТРЕННЕЕ СТРОЕНИЕ 98 Класс Entognatha -
Пищеварительная система Выделительная система 101 Класс Insecta Класс Entognatha 238 244
Кровеносная система Трахейная система 103 Отряд Diplura (диплуры, или дву- хвостки) -
Нервная система Мускулатура Половая система 106 109 111 Отряд Protura (протуры, или бес- сяжковые) Отряд Collembola (ногохвостки) 246
Класс Insecta 247
5. ЖИЗНЕННЫЕ ПРОЦЕССЫ 115 Отряд Microcoryphia (щетинохвост-
Структура и функция скелета - ки) -
Питание 124 Отряд Thysanura (чешуйницы) 248
Пищеварение 125 Отряд Ephemeroptera (поденки) 249
572
Оглавление
Отряд Odonata (стрекозы) 252
Отряд Dictioptera (тараканы и бо-
гомолы) 258
Отряд Isoptera (термиты) 262
Отряд Phasmatodea (палочники) 265
Отряд Orthoptera (прямокрылые) 266
Отряд Dermaptera (уховертки) 271
Отряд Grylloblattodea (гриллоблат-
тиды) 273
Отряд Embioptera (эмбии) 275
Отряд Plecoptera (веснянки) 277
Отряд Zoraptera (зораптеры) 279
Отряд Psocoptera (сеноеды) 281
Отряд Phthiraptera (пухоеды и вши) 284
Отряд Thysanoptera (трипсы, или
бахромчатокрылые) 290
Отряд Hemiptera (равнокрылые и
клопы) 293
Отряд Raphidioptera (верблюдки) 317
Отряд Megaloptera (большекрылые) 318
Отряд Neuroptera (сетчатокрылые) 319
Отряд Coleoptera (жесткокрылые,
или жуки) 324
Отряд Hymenoptera (перепончато-
крылые) 352
Отряд Mecoptera (скорпионницы) 374
Отряд Siphonaptera (блохи) 375
Отряд Diptera (двукрылые) 378
Отряд Trichoptera (ручейники) 404
Отряд Lepidoptera (бабочки, или че-
шуекрылые) 410
9. ЭВОЛЮЦИОННАЯ ИСТОРИЯ
НАСЕКОМЫХ 431
Свидетельства эволюции насекомых
Палеогеография 435
Вымершие отряды насекомых 438
Связи ископаемых насекомых с со-
временными 447
Насекомые и история жизни на Земле 457
10. ЭКОЛОГИЧЕСКИЕ АСПЕКТЫ 472
Биомы и сообщества
Экосистемы 474
Популяции 479
Экология особей 485
11. ПОЛЕЗНЫЕ НАСЕКОМЫЕ 515
Хищные и паразитические насекомые
Насекомые-хищники 516
Насекомые-паразиты 524
Борьба с сорняками 529
Насекомые-опылители 530
Насекомые - производители полезных
продуктов 531
Другие способы использования насе-
комых 532
Насекомые-деструкторы и круговорот
питательных веществ 534
12. ВРЕДНЫЕ НАСЕКОМЫЕ И
БОРЬБА С НИМИ 538
Насекомые - вредители растений
Насекомые-паразиты человека и жи-
вотных 544
Борьба с вредными насекомыми 548
Регуляция численности насекомых-вре-
дителей 563
Интегрированное управление числен-
ностью вредителей 566
Перспективы на ближайшее будущее 568
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании кни-
ги, ее оформлении, качестве перевода и
другие просим присылать по адресу
129820, Москва, И-110, ГСП
1-й Рижский пер., д. 2,
издательство «Мир».
Герберт Росс, Чарлз Росс, Джун Росс
ЭНТОМОЛОГИЯ
Научный редактор М. Б. Николаева
Мл. научн. редактор О. А. Горгун
Художник В. А. Захаров
Художественный редактор А. Я. Мусин
Технический редактор И. И. Володина
Корректор С. А. Денисова
ИБ № 5137
Сдано в набор 31.07.84. Подписано к печати 12.08.85.
Формат 70x90I/i6. Бумага офсетная № 1. Печать
офсетная. Гарнитура тайме. Объем 18 бум. л.
Усл. печ. л.42,12. Усл. кр.-отт. 81,90. Уч. изд. л. 43,37.
Изд. № 4/3398. Тираж 10000 экз. Зак. 732.
Цена 3 р. 50 коп.
ИЗДАТЕЛЬСТВО «МИР»
129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
Можайский полиграфкомбинат Союзполиграфпро-
ма при Государственном комитете СССР по делам
издательств, полиграфии и книжной торговли.
143200, Можайск, ул. Мира, 93.