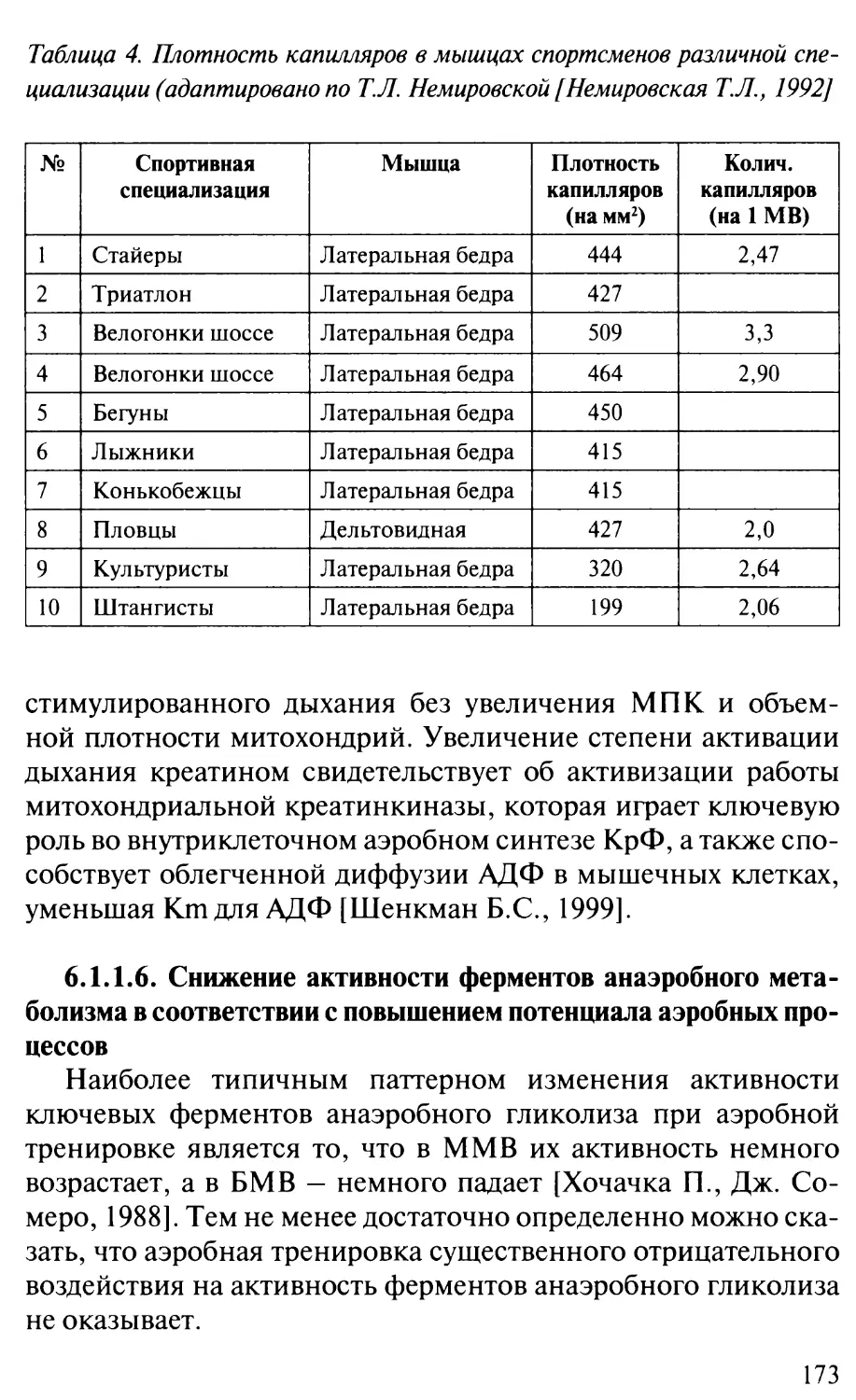
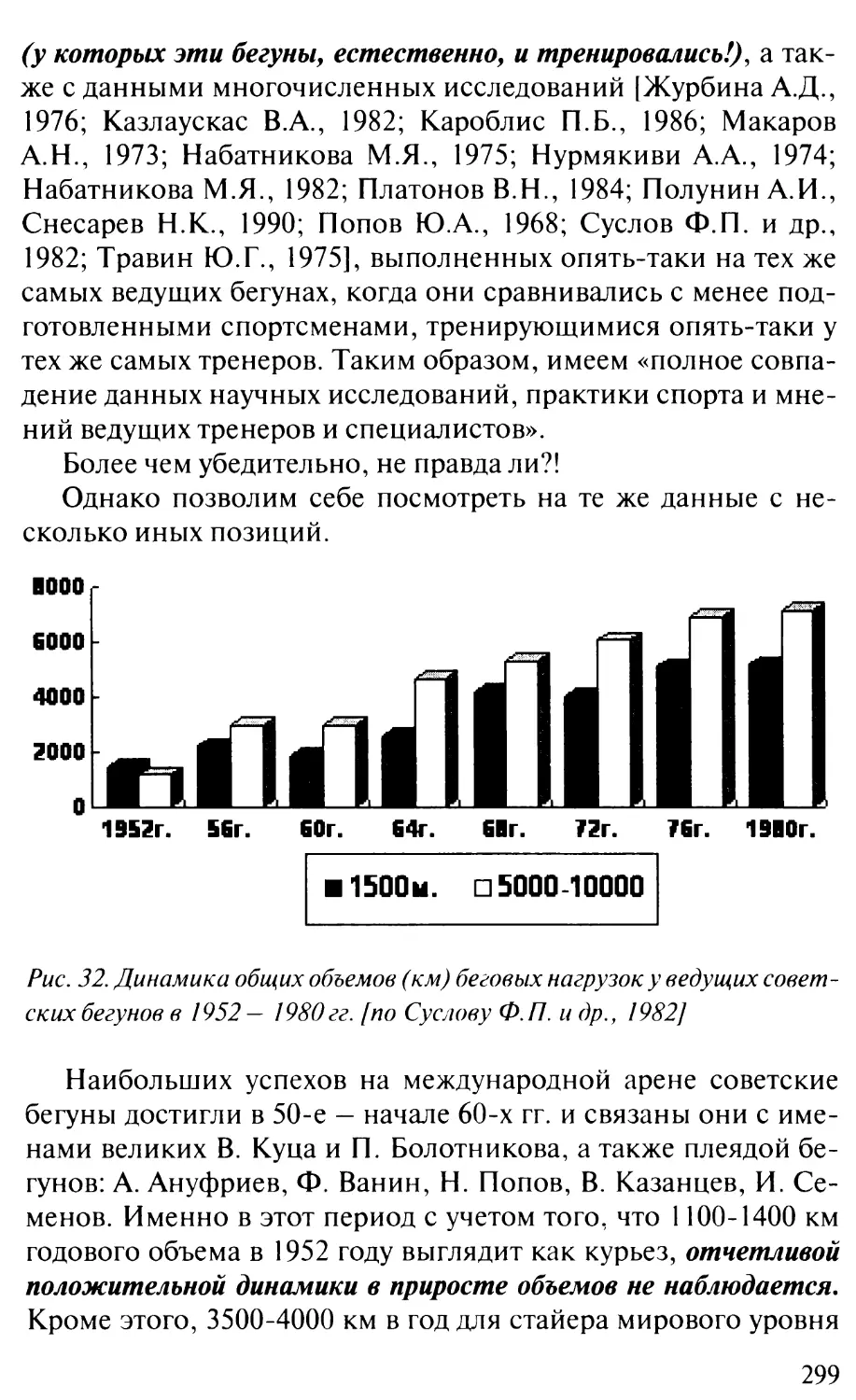
/





































































































































































































































































































































































Author: Селуянов В.Н. Мякинченко Е.Б.
Tags: гимнастика акробатика легкая атлетика спорт развитие выносливости
ISBN: 978-5-98724-010-6
Year: 2009




Text
О! НАУКА-СПОРТУ
Е.Б. Мякинченко, В.Н. Селуянов
РАЗВИТИЕ ЛОКАЛЬНОЙ
МЫШЕЧНОЙ ВЫНОСЛИВОСТИ
В ЦИКЛИЧЕСКИХ ВИДАХ СПОРТА
НАУКА-СПОРТУ
Е.Б. Мякинченко
В.Н.Селуянов
РАЗВИТИЕ
ЛОКАЛЬНОЙ
МЫШЕЧНОЙ
ВЫНОСЛИВОСТИ
В ЦИКЛИЧЕСКИХ
ВИДАХ СПОРТА
□
ТВТ Дивизион
Москва 2009
УДК 796.42
М99
М 99 Мякинченко, Е. Б.
Развитие локальной мышечной выносливости в цикличе-
ских видах спорта [текст] / Е.Б. Мякинченко, В.Н. Селуянов. -
М.: ТВТ Дивизион, 2009. - 360 с.
ISBN 978-5-98724-010-6
В монографии рассмотрены биологические и педагогиче-
ские аспекты повышения производительности основных мы-
шечных групп спортсменов, занимающихся видами спорта на
выносливость. Затронуты вопросы лимитирующих факторов
функциональных возможностей скелетных мышц, средств и
методов воздействия на мышечную систему и их взаимосвязи
с формированием рациональной техники бега, принципов по-
строения одного тренировочного занятия, микро-, мезо- и ма-
кроциклов. Высказаны некоторые гипотезы о совершенство-
вании структуры многолетней подготовки юных спортсменов
в циклических видах спорта.
Для магистрантов, аспирантов, специалистов в области фи-
зической культуры и спорта, спортивных медиков, преподава-
телей ссузов и вузов, тренеров, спортсменов.
УДК 796.42
ISBN 978-5-98724-010-6
© Е.Б. Мякинченко, В.Н. Селуянов, 2005, 2009 г.
© ТВТ Дивизион, 2005, 2009 г.
Содержание
Введение............................................11
Глава 1. Методологические основы исследования
локальной мышечной выносливости .................17
1.1. Эмпирический уровень научного исследования..18
1.2. Теоретический уровень научного исследования.20
1.3. Методология теории и методики
физического воспитания...........................21
1.4. Методология спортивно-педагогической
адаптологии......................................22
1.5. Некоторые проблемы, связанные с различием
в логике эмпирического и теоретического мышления ....22
Глава 2. Основы биологии человека (концептуальные
модели систем и органов человека)...............30
2.1. Биология клетки..........................30
2.2. Нервно-мышечный аппарат..................31
2.3. Биохимия клетки (энергетика).............33
2.4. Модель функционирования нервно-мышечного
аппарата при выполнении циклического упражнения....36
2.5. Биомеханика мышечного сокращения.........39
2.6. Сердце и кровообращение..................41
2.7. Кровеносные сосуды.......................42
2.8. Эндокринная система......................43
2.9. Иммунная система.........................50
2.10. Пищеварение.............................52
2.11. Жировая ткань...........................54
Глава 3. Контроль локальной выносливости........57
3.1. Мощность, эффективность и емкость
механизмов энергообеспечения как критерии оценки
подготовленности спортсменов..................57
5
3.2. Критический анализ интерпретации данных
лабораторного тестирования......................61
3.3. Новые подходы для оценки физической
подготовленности спортсменов....................64
3.4. Определение степени влияния центрального
или периферического лимитирующего фактора.......67
3.5. Метод Conconi .............................71
3.6. Понятие — локальная мышечная
работоспособность...............................72
Глава 4. Локальная выносливость как компонент
физической подготовленности спортсменов
в циклических видах спорта .......................73
4.1. Средства и методы развития силовых
способностей в циклических видах спорта.........76
4.2. Соотношение объемов средств
развития локальной выносливости в ЦВС...........81
4.3. Распределения средств развития
локальной выносливости в рамках одного занятия,
микро-, мезо- и макроциклов и многолетней
подготовки спортсменов..........................83
4.4. Реализация компонентов
локальной выносливости в основном
соревновательном упражнении.....................92
Глава 5. Факторы, лимитирующие локальную выносливость
в циклических видах спорта .......................95
5.1. Схема физиологических и биохимических
процессов, происходящих в мышцах при преодолении
соревновательной дистанции......................106
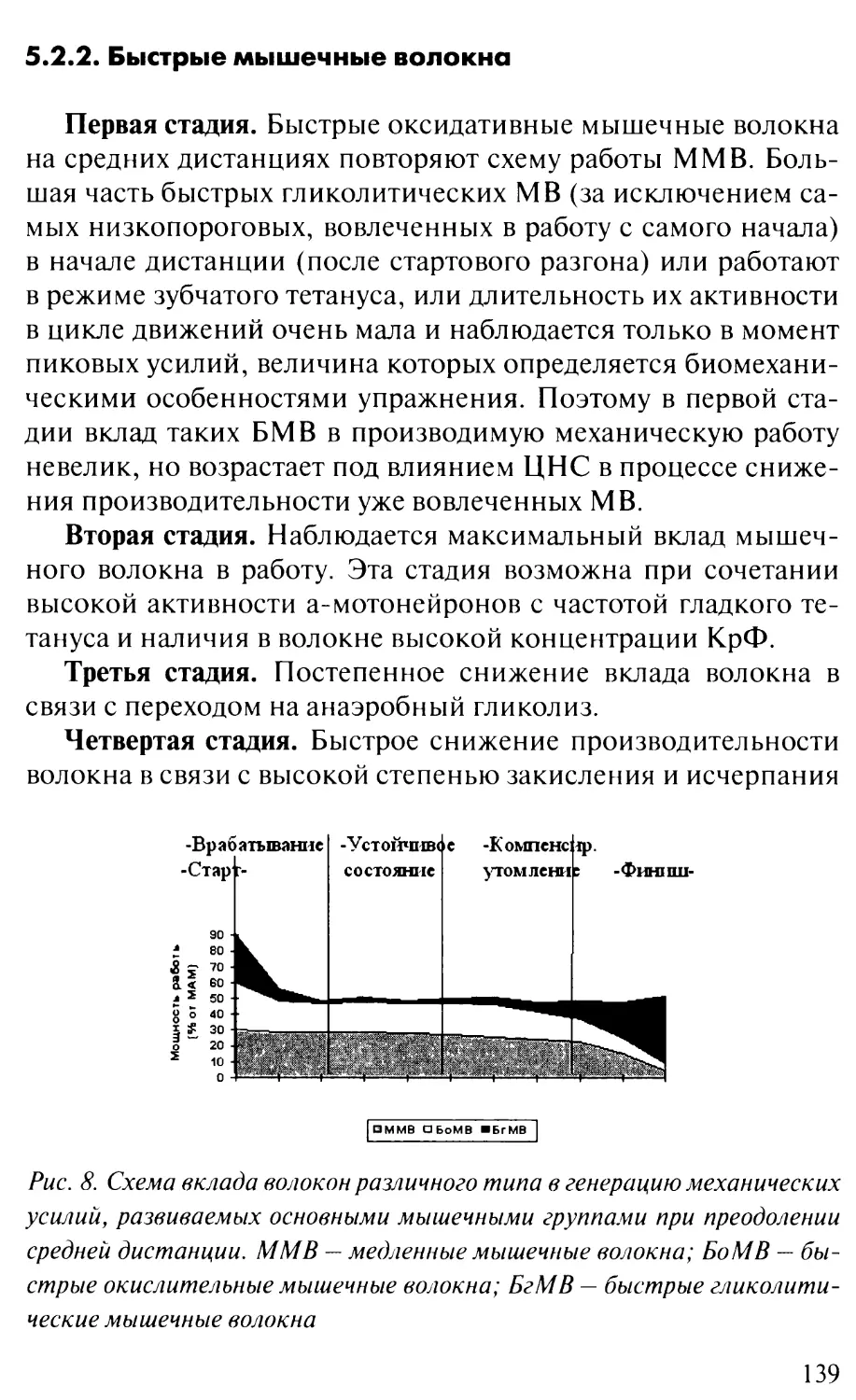
5.2. Схема работы разных типов МВ
при преодолении соревновательной дистанции......137
5.3. Особенности физиологических
и биоэнергетических процессов в мышечном
аппарате при более длинных и более
коротких дистанциях.............................142
5.4. Заключение.................................152
6
Глава 6. Теоретические аспекты выбора средств,
методов и организации тренировочного процесса
в циклических видах спорта с целью улучшения
локальной мышечной выносливости ...................154
6.1. Обоснование выбора средств и методов
тренировки мышечных компонентов, определяющих
выносливость в циклических видах спорта........154
6.2. Тренировочные средства и методы
развития локальной выносливости................180
6.3. Теоретические основы планирования одного
тренировочного занятия, тренировочных микро-,
мезо- и макроциклов.............................196
6.4. Проблема взаимосвязи уровня и особенностей
подготовленности нервно-мышечного аппарата
с техникой и экономичностью локомоций..........224
6.5. Заключение по разделу.....................236
Глава 7. Анализ данных экспериментальных исследований
средств и методов повышения локальной выносливости.244
7.1. Исследование упражнений статодинамического
характера как средства воздействия на медленные
мышечные волокна спортсменов....................244
7.2. Влияние сочетания статодинамической силовой
и аэробной тренировок мышц бедра на аэробный
и анаэробные пороги человека
(лабораторный эксперимент)......................256
7.3. Классификация упражнений бегунов
на средние и длинные дистанции по признаку
их преимущественного воздействия
на морфоструктуры организма....................263
7.4. Критерии обоснованности выводов
по результатам педагогических экспериментов....264
7.5. Исследование влияния акцентированной
силовой и аэробной тренировки
на показатели силы, аэробных способностей
и экономичности техники бега....................268
7.6. Исследование влияния статодинамических
упражнений совместно с традиционными методами
7
подготовки бегунов на показатели силы
и аэробных способностей.......................276
7.7. Исследование эффективности последовательного
применения силовых и аэробных средств подготовки
на показатели физических способностей бегунов.285
7.8. Заключение по главе......................292
Глава 8. Практические аспекты развития
локальной выносливости...........................298
8.1. Возможные варианты коррекции системы
подготовки бегунов на выносливость............302
8.2. Некоторые аспекты построения
многолетней подготовки бегунов................312
8.3. Заключение...............................331
Литература.......................................334
8
Список сокращений
АэП — аэробный порог
АнП - анаэробный порог
БМВ - быстрые мышечные волокна
БгМВ — быстрые гликолитические мышечные волокна
БоМВ — быстрые окислительные мышечные волокна
ГМВ — гликолитические мышечные волокна
КП— количество повторений (например, в одном подходе
или в одной серии)
К++ — ионы калия
КМЭ — коэффициент механической эффективности
Кр— свободный креатин
КрФ— креатинфосфат
Ла - лактат
ЛДГ- лактатдегидрогеназа
ЛВ - локальная (мышечная) выносливость
МАМ - максимальная алактатная мощность
МВ - мышечные волокна скелетных мышц
ММВ - медленные (Тип I) мышечные волокна
МПК - максимальное потребление кислорода
МПС — максимальная произвольная сила
МОК — минутный объем кровотока (сердечный выброс)
МК — молочная кислота
МСП — метаболическая стоимость пути
НМА — нервно-мышечный аппарат
Н+ — ионы водорода (протоны)
ОП — окислительный потенциал
ОДА - опорно-двигательный аппарат
ОМВ - окислительные мышечные волокна
О2 долг — кислородный долг
ОЦМТ — общий центр масс тела
ППС - площадь поперечного сечения
рО2 - парциальное напряжение кислорода
9
Са++ - ионы кальция
СБУ — специальные беговые упражнения легкоатлетов
СТЭОДА - соединительно-тканные элементы опорно-
двигательного аппарата
ССС — сердечно-сосудистая система
ТМГ — трехглавая мышца голени
У О — ударный объем
ЦВС — циклические виды спорта
ЦНС — центральная нервная система
ЧП — число подходов (например, при силовой тренировке)
VO2 — скорость потребления кислорода
10
Введение
Данная книга представляет собой, по существу, второе, пе-
реработанное и исправленное, издание монографии «Локаль-
ная выносливость в беге», вышедшей в 1997 году. Накоплен-
ные за прошедшие годы материалы и их осмысление сделали
возможным распространить выводы и рекомендации на дру-
гие циклические виды спорта, а также на спортивные игры и
единоборства. При этом сохранен основной предмет исследо-
вания — различные стороны повышения производительности
нервно-мышечного аппарата спортсменов. Другими словами,
в книге рассматриваются проблемы улучшения так называе-
мой локальной (мышечной) выносливости. Данная проблема,
на наш взгляд, по-прежнему недостаточно разработана как
в отечественной, так и зарубежной литературе, несмотря на
многочисленные работы, посвященные различным сторонам
тренировки мышц, проводимой в контексте развития силы и
силовой выносливости спортсменов. Это позволяет надеять-
ся, что второе издание также найдет своего читателя.
Переходя к непосредственному описанию содержания
книги, следует рассмотреть вопрос — в каких случаях локаль-
ная выносливость (то есть компонент выносливости, связан-
ный непосредственно с нервно-мышечным аппаратом) будет
существенным или даже решающим фактором повышения
спортивного мастерства и почему эта проблема является акту-
альной?
Непосредственным ограничителем достижения более вы-
сокого результата при преодолении соревновательной дис-
танции является наступающее утомление. Поэтому основное,
что должно быть достигнуто в результате физической подго-
товки, это — отдаление момента утомления или повышение к
нему устойчивости организма. Среди факторов, приводящих к
утомлению при различной длительности физической работы,
выделяют «центральные»:
— утомление корковых центров двигательной зоны ЦНС и
снижение частоты импульсации быстрых ДЕ;
— недостаточную секрецию стресс-гормонов (катехолами-
нов и глюкокортикоидов);
— недостаточную производительность миокарда и систем,
обеспечивающих адекватный региональный и локаль-
ный кровоток, что может приводить к мышечной гипок-
сии;
— изменения в деятельности вегетативной нервной систе-
мы и многих железах внутренней секреции;
— а также «периферические»:
— снижение массы фосфагенов;
— увеличение концентрации ионов водорода и лактата
(молочной кислоты);
— снижение потребления кислорода мышцами;
— снижение концентрации гликогена мышц и др.
Однако при более глубоком рассмотрении обеих групп
факторов, можно выдвинуть гипотезу, что большая мощность
энергетических и сократительных систем, локализованных не-
посредственно в мышцах и определяющих локальную выносли-
вость (ЛВ), позволяет отдалить наступление утомления, а
также снизить нагрузки на «центральные факторы», интен-
сивное функционирование которых также может приводить к
утомлению.
Несмотря на очевидную важность исполнительного звена
двигательной системы (мышц) для спортивной работоспособ-
ности, «центральному фактору», а именно производительно-
сти сердечно-сосудистой системы, «выносливости» централь-
ной нервной и гормональной систем и т.п., длительное время
отводилось решающее значение. В то же время, очевидно, что
существуют спортсмены, для которых периферическое звено
двигательной системы будет являться лимитирующим факто-
ром. Например, на средних и длинных дистанциях к усталости
может приводить локальное утомление из-за накопления мо-
лочной кислоты в мышцах. Это с равной вероятностью может
явиться следствием:
— или недостаточной производительности сердечно-
сосудистой системы и несовершенства региональных и
12
локальных механизмов перераспределения кровотока,
приводящих к тканевой гипоксии;
— или недостаточной аэробной мощности мышц.
Это же справедливо относительно других факторов, кото-
рые можно отнести или к «центральному», или «перифериче-
скому» звену.
Следовательно, даже в том случае, если сформулированная
выше гипотеза окажется неверной, то всегда можно говорить о
наличии двух генеральных совокупностях спортсменов:
— первая, у которых основными лимитирующими фак-
торами будут являться «центральные» (производитель-
ность ССС, утомление нервных центров, ограничения
со стороны гормональной системы и т.п.);
— вторая, у которых лимитирующим звеном являются
периферические факторы, локализованные на уровне
нервно-мышечного аппарата конечностей (алактатная,
гликолитическая, аэробная производительность мышц,
сила мышц и т.п.).
Большинство выводов и рекомендаций этой работы будет
справедливо для тех спортсменов, у которых нет генетически
обусловленных или приобретенных ограничений со сторо-
ны управляющих и обеспечивающих мышечную деятельность
систем организма. Другими словами, мы рассматриваем тот
случай, когда в процессе тренировки производительность и совер-
шенство функционирования «центральных» систем уже обеспе-
чены или повышаются быстрее, чем производительность морфо-
структур, локализованных непосредственно в основных мышцах
спортсменов — т. е. в ситуации, когда в процессе длительной
специализированной тренировки мышцы становятся лимити-
рующим фактором физической работоспособности.
Как определить, являются ли у данного спортсмена «цен-
тральные» системы лимитирующим звеном или нет?
Парадокс заключается в том, что ответа на этот вопрос в на-
стоящее время в литературе нет. В рамках физиологии и био-
химии спорта он должным образом не рассматривался. Это
положение возникло, на наш взгляд, из-за высочайшего автори-
тета таких корифеев российской науки, как И.М. Сеченова, И.П.
Павлова, А.А. Ухтомского, проводивших свои исследования
13
проблем утомления в основном в области физиологии трудовой
деятельности и сформулировавших фундаментальные выводы о
решающей роли центральной нервной системы, главным обра-
зом, для этого вида деятельности. Тем не менее этот вывод был
совершенно необосновано распространен и на спорт (т. е. на экс-
тремальную деятельность), где процессы утомления также чаще
всего рассматривались в этом ключе (Е.Б. Сологуб, 1972; Н.В.
Зимкин, 1975; Н.Н. Яковлев, 1983; А.С. Солодков, 1992).
Другое направление в исследовании утомления в спорте, в
частности в циклических локомоциях (Г.Ф. Фольборт, 1956;
Н.И. Волков, 1969; А.З. Колчинская, 1983; В.Д. Моногаров
1980,1986; Меленберг, 1990, и др.), признавая существенную
роль исполнительного аппарата в развитие утомления, основ-
ным фактором утомления мышц считает тканевую гипоксию,
которая возникает, однако, «по вине» другой «центральной»
системы - сердечно-сосудистой, которая, как предполагает-
ся, не способна снабдить мышцы достаточным количеством
кислорода в соответствии с их запросом во время интенсивной
мышечной работы.
Попытка авторов данной работы найти истоки или основа-
ния для этого широко распространенного мнения не увенча-
лись успехом.
По нашему мнению, в настоящее время есть основания
говорить о наличии т.н. функциональной или относительной
тканевой гипоксии, которая является совершенно необходи-
мым и биологически целесообразным следствием мышечной
работы, так как является одним из «ключей» для запуска и ре-
гулирования системы энергообеспечения мышечных клеток.
Также существуют доводы в пользу того, что гипоксические
условия — необходимый фактор для индукции адаптивного
синтеза белка, приводящего к повышению окислительного
потенциала мышц под воздействием тренировки. Однако и это
не более чем гипотеза, так как против нее свидетельствуют не-
которые экспериментальные данные, представленные в этой
монографии. Поэтому, не отрицая возможности существова-
ния тканевой гипоксии у квалифицированных спортсменов
при выполнении напряженной мышечной работы, мы счита-
ем, что пока нет оснований полагать, что гипоксия является или
14
ограничителем скорости ресинтеза АТФ в процессе мембранного
(дыхательного) фосфорилирования в митохондриях, или причи-
ной утомления мышц.
Другими словами, нет оснований считать, что запрос мышц
в кислороде (но не в энергии!) не удовлетворяется.
Следовательно, мышечная гипоксия не является причиной
явлений, которые связывают с утомлением. Например, таких,
как: продукции и накопления молочной кислоты, повышен-
ной скорости расхода углеводных запасов мышц, рекрутиро-
вания высокопороговых двигательных единиц (ДЕ) и мн. др.
Хотя, наверное, в конце дистанции может возникнуть си-
туация, когда комплекс факторов, связанных с напряженной
мышечной деятельностью, повышением температуры и обе-
звоживанием организма, может ухудшить функциональное
состояние дыхательной системы, миокарда, систем крови, ре-
гуляцию сосудистых реакций и т.п. В этом случае ССС будет
неспособна поставлять кислород к мышцам в прежнем объеме
и теоретически может явиться фактором снижения произво-
дительности мышечной работы. Однако, во-первых, как отме-
чено выше, все эти явления так или иначе связаны с явлениями
утомления в самих мышцах, а во-вторых, будет справедливо
только для случая, когда изменения в мышечных клетках (на-
рушение в деятельности мембран, деградация нуклеотидов,
снижение pH и многое другое) еще в большей степени не ухуд-
шит способность мышц утилизировать кислород.
Таким образом, следует согласиться с общим мнением, что
проблема утомления в спорте, в частности в циклических ло-
комоциях, чрезвычайно сложна и должна решаться биологами
на фундаментальном уровне.
Задачи же авторов при написании этой книги были суще-
ственно скромнее:
— во-первых, определить возможные подходы к такому
построению тренировки, которое способствовало бы
повышению производительности нервно-мышечного
аппарата и тем самым «облегчила бы жизнь» «централь-
ным» системам при преодолении дистанции, если в ходе
дальнейших исследований все же выяснится, что глав-
ный источник утомления находится «в центре»;
15
- во-вторых, представить доказательства и эксперимен-
тальную проверку того, что средства и методы трениров-
ки в циклических локомоциях могут разрабатываться и
обосновываться без привлечения гипотезы о тканевой
гипоксии как центральном факторе утомления мышц.
Не ставя под сомнение важность «центральных механиз-
мов», следует все же констатировать, что многочисленные
научные исследования и методические разработки, направ-
ленные на совершенствование тренировочного процесса в ци-
клических видах спорта, в большинстве случаев проводились
в контексте «первоочередности», «базовости», «решающей
роли» обеспечивающих систем. Проблемы же улучшения ло-
кальной выносливости (ЛВ) изучены существенно хуже и, как
правило, в аспекте тренировки силы или т.н. «силовой вынос-
ливости». Однако проблема Л В существенно шире и для того,
чтобы каждый тренер мог со знанием дела подходить к плани-
рованию тренировочного процесса, в котором существенное
место занимало бы целенаправленное воздействие на мышеч-
ный аппарат, необходимо создать у него целостное представ-
ление (модель организма человека и навыки имитационного
моделирования), на основании которого можно было бы де-
лать следующие обоснованные суждения:
— о значимости мышечных компонентов для выносливости;
— о месте такой тренировки в системе подготовки спор-
тсменов;
— о лимитирующих факторах работоспособности, связан-
ных с мышечной системой;
— об оптимальных средствах и методах тренировочных
воздействий на мышечные компоненты, определяющие
выносливость;
- о вариантах планирования тренировочного занятия, ми-
кро-, мезо-, макроциклов и многолетней подготовки.
Поэтому авторы взяли на себя смелость рассмотреть, по
мере возможности, все из перечисленных аспектов физиче-
ской подготовки и ее взаимосвязь с техникой локомоций, так
как эти две стороны подготовленности не могут рассматри-
ваться изолированно одна от другой.
16
Глава 1
Методологические основы
исследования локальной
мышечной выносливости
В научном познании отражается внешний мир. Познающая
и практически действующая реальность называется субъектом
познания. Это может быть человек, группа людей или обще-
ство в целом. Объект познания — та часть материи, которая
включена в познавательную деятельность субъекта. Материя
же включает два атрибута — вещество и движение, поэтому
предметом исследования могут быть отдельные части объекта
(вещество) или какие-либо свойства, процессы. В этом смыс-
ле у объекта существует множество сторон рассмотрения или,
другими словами, предметов исследования.
Различают эмпирический и теоретический уровни позна-
ния (знания).
1.1. Эмпирический уровень научного
исследования
К эмпирическим наукам могут быть отнесены те науки, ко-
торые находятся еще в первоначальной стадии своего разви-
тия, в которых эмпирические методы исследования еще толь-
ко формируются, а теоретические методы еще не возникли.
Предметом эмпирического исследования, как правило, ста-
новятся явления (свойства, признаки) и зависимости между
ними. Познаются внешние свойства объекта исследования, а
также особенности его взаимодействия со средой.
В эмпирических науках исследование проходит три ста-
дии.
17
Первая стадия — проводятся научные опыты, добываются
знания в виде отдельных данных опыта, составляющих базисное
эмпирическое знание. Ее можно разделить на четыре части:
1. формулировка цели, задач и определение методов иссле-
дования, разработка плана и подготовка опыта (наблю-
дение или эксперимент) на основе каких-то предвари-
тельных данных, иногда просто фантазий;
2. проведение опыта, которое включает получение чув-
ственного отражения, осознание и понятийное выраже-
ние изучаемых явлений;
3. предварительная проверка истинности данных опыта;
4. оценка степени точности отражения изучаемых явлений
в полученных данных опыта.
Для разработки плана и подготовки опыта нужны некото-
рые предварительные знания, которые могут быть научными
или обыденными.
Научное описание данных опыта должно быть таким, что-
бы иметь одно и то же содержание для любых исследователей.
Словесное описание условий получения данных является зна-
ковой формой описания экспериментальных данных. В дан-
ные опыта входит и понятийное содержание, следовательно,
существует возможность неточного оформления чувствен-
ных восприятий. Поэтому понятие факт — вполне оспоримая
вещь, т.к. содержит понятийный компонент, основанный на
каких-то теориях (часто неточных или неверных в эмпириче-
ских науках).
Полностью избежать ошибок в ходе измерений невозмож-
но. Случайные ошибки — ошибки, связанные с функциониро-
ванием прибора, изменением состояния объекта и др. Повто-
рение опыта, сбор статистического материала, усреднение его
(вычисление средней арифметической) позволяют устранить
случайные погрешности. Систематические ошибки и арте-
факты устраняются в ходе логического анализа. Под артефак-
тами понимают грубые ошибки измерения, в частности, нару-
шение однородности выборки данных. Например, измерение
максимальной силы у спортсменов высшей квалификации
должно дать модельную характеристику — среднюю величину
силы. Однако если какой-либо спортсмен болеет или прекра-
18
тил тренироваться, то он выпадает из изучаемой генеральной
совокупности данных. Такие данные должны исключаться из
выборки, эмпирического базиса опыта.
Вторая стадия — выполняется первичная обработка сово-
купности данных опыта, представляющих собой конечное
число наблюдений. Обработка включает логическое и матема-
тическое преобразование исходного материала. В результате
вводятся эмпирические понятия, группируются и классифи-
цируются данные, выявляются корреляции, регрессионные
уравнения, которые называют эмпирическими законами.
Анализ и синтез — основной способ группировки данных
опыта. В одну группу должны включаться данные, соответ-
ствующие существенным связям явлений. Например, данные
могут объединяться по принадлежности к какому-либо одно-
му физическому качеству (сила, быстрота, выносливость, гиб-
кость).
Логика эмпирической научно-исследовательской работы
требует определения:
1. направления;
2. проблемы;
3. темы;
4. объекта;
5. предмета;
6. рабочей гипотезы;
7. методов;
8. плана;
9. организации и проведения:
— эксперимента;
— обработки результатов;
— написания отчета.
Третья стадия — производят обобщение опыта внутри
каждой группы, в процессе которого совершается мыслен-
ный переход от конечного числа членов данной выборки к
бесконечному. Здесь проводится доказательство достовер-
ности переноса выборочных данных на определенную гене-
ральную совокупность изученных объектов исследования.
Эти знания — высшая форма, наиболее сложный вид эмпи-
рического знания.
19
Эмпирические знания, раскрывающие эмпирические за-
кономерности внутри данной генеральной совокупности,
являются предельно широкими. Выход за пределы данной
генеральной совокупности приводит к отрыву от исходного
эмпирического базиса, что в рамках эмпирического исследо-
вания недопустимо.
1.2. Теоретический уровень
научного исследования
Теоретический уровень исследования применяется в разви-
тых науках. Специфика теоретического знания состоит в том,
что оно опирается на свой экспериментальный базис, выходя-
щий за пределы предмета данного исследования или науки.
Предмет теоретического исследования — закономерности,
обуславливающие целостность существования материально-
го образования в заданных условиях, т.е. внутреннее строение
объекта, взаимоотношения между элементами, обуславливаю-
щие его взаимодействие с окружающей средой.
В развитых науках исследование проходит три стадии.
Первая стадия — производится построение модели, на осно-
ве которой можно изучать закономерности, являющиеся пред-
метом данного исследования.
Вторая стадия - заключается в использовании модели для
воспроизведения в идеальном виде явлений объективной ре-
альности с помощью имитационного моделирования. Неиз-
менной составной частью этой стадии является проведение
экспериментальных исследований, подтверждающих коррект-
ность теоретических предсказаний. Результатом второй стадии
является теория изучаемого явления (предмета исследования).
Третья стадия - разработка практических рекомендаций,
новых технологий, обеспечивающих повышение эффективно-
сти практической деятельности человека.
Логика теоретической научно-исследовательской работы
требует определения:
1. направления;
2. проблемы;
20
3. темы;
4. объекта;
5. предмета;
6. модели;
7. рабочей гипотезы на основе имитационного моделиро-
вания;
8. методов;
9. плана;
10. организации и проведения:
— эксперимента;
— обработки результатов;
— написания отчета.
1.3. Методология теории и методики
физического воспитания
Логика построения теоретического исследования в рамках
«Общей теории спорта» (Л.П. Матвеев, 1997) рассматривается
в виде реализации следующих этапов.
Начальный этап — выявление проблемной ситуации как
противоречия между состоянием теоретического знания и
потребностями научного решения практической задачи. В
качестве объекта исследования, например, рассматриваются
«тенденции в развитии спорта», «принципы оптимального по-
строения тренировки». Рассмотрение проблемной ситуации (в
какой логике? — об этом не говорится) создает предпосылки
для поиска решения проблемы, т.е. рабочей гипотезы.
Второй этап — организация и проведение исследования или
экспериментальная проверка следствий теоретического мыш-
ления, рабочей гипотезы.
Завершающий этап теоретического исследования. Фактиче-
ский материал рассматривается, обобщается, систематизирует-
ся, классифицируется, рассчитываются модельные характери-
стики, регрессионные аналитические уравнения, в конечном
итоге выполняется интерпретация материалов на основе зако-
нов формальной логики (диалектическая логика эмпирикам
недоступна), с учетом ранее выполненных исследований.
21
Таким образом, нетрудно заметить, что предлагается ин-
дуктивный метод мышления, свойственный всем эмпири-
ческим исследованиям. Реализуется принцип «от частного к
общему». Установление концептуальных положений — вер-
шина такого подхода, а разработка модельных характеристик
спортсменов, процессов или прогнозирование высших спор-
тивных достижений, количества медалей на олимпийских
играх — вершина математического мышления. Видим, что
математическая статистика предлагается как инструмент по-
знания. Иногда повторяют слова К. Маркса о том, что раз-
витой наука становится тогда, когда в нее приходит матема-
тика. При этом не понимая, что Ф. Энгельс и К. Маркс под
математикой в этом случае понимали дифференциальное
исчисление, которое способно воспроизводить (моделиро-
вать) как строение объекта исследования (часть материи), так
и процессы развития объекта (модели) во времени в задан-
ных условиях окружающей среды. Подход, предложенный
классиками, реализуется только в рамках теоретического на-
правления исследований. В связи с этим перед нами возникла
проблема разработки иного методологического подхода в ис-
следовании спорта.
1.4. Методология спортивно-педагогической
адаптологии
В процессе изучения физических возможностей человека
в логике теоретического направления исследований следует,
как это было показано выше, реализовать три этапа.
Первый этап - моделирование объекта исследования. Объ-
ектом исследования при изучении физических возможностей че-
ловека является сам человек, его системы, органы, ткани, клетки.
Поэтому, с учетом поставленной задачи, должно быть выпол-
нено моделирование человека. Оно может быть мысленным,
как это имеет место в учебниках анатомии, биохимии и физио-
логии, или математическим в виде написания и решения си-
стем дифференциальных уравнений (Н. Амосов с соав., 1978;
В.Н. Селуянов с соав., 1992-2001; В.Л. Уткин, 1995).
22
Предметом теории спорта являются знания о средствах, ме-
тодах и планах построения тренировочного процесса с целью
управления развитием физических возможностей человека.
Поэтому собственно исследование морфофункциональных осо-
бенностей организма человека относится к сфере биологических
наук - биологии человека. А вот разработку адекватных по-
ставленным задачам умозрительных и математических моделей
должны взять на себя «физкультурники», поскольку с помощью
этих моделей они должны изучать и выявлять наиболее эффек-
тивные варианты управления развитием человека.
Таким образом, «физкультурник» (тренер) является как бы
анатомом, биохимиком, физиологом и биомехаником, только
объектом его исследования является модель человека, которая
подвергается воздействию управляющих команд (педагогических
воздействий).
В связи с этим новое направление исследований получило
название спортивно-педагогическая адаптология, а именно
изучение процессов адаптации в спорте в результате педагоги-
ческих воздействий.
Второй этап — имитационное моделирование. После по-
строения модели начинается поиск вариантов решения педа-
гогической задачи. По ходу имитационного моделирования
уточняются параметры модели, доказывается ее адекватность
объективной реальности. Среди множества решений выбира-
ется наиболее рациональный вариант, т.е. адекватный внеш-
ним условиям и индивидуальным особенностям конкретного
спортсмена или группы похожих друг на друга спортсменов
(равной квалификации, одного вида спорта). Выбранный ва-
риант тренировочного воздействия является рабочей гипоте-
зой, справедливость которой следует доказать на следующем
этапе исследования.
Третий этап - экспериментальное обоснование теорети-
ческих положений. В соответствии с рабочей гипотезой про-
водится изучение на реальном объекте (спортсмене) адапта-
ционных процессов с помощью лабораторных (как правило,
биологических, лучше прямых) или педагогических (косвен-
ных) методов исследования адаптационных перестроек в клет-
ках различных органах спортсмена.
23
Четвертый этап - обобщение экспериментальных данных.
Получение частного результата исследования не может иметь
принципиального значения для науки и практики, поскольку
остается вопрос о возможности переноса обнаруженных зако-
номерностей на другие объекты (спортсменов). В эмпирическом
исследовании требуется выполнить переход от выборочных дан-
ных к генеральной совокупности объектов, которые представля-
ла экспериментальная выборка. В теоретическом исследовании
такую генеральную совокупность представляют собой все спор-
тсмены, для которых разработанная модель является адекватной.
Заметим, что для наиболее простых моделей, например биоэнер-
гетики мышечной деятельности, результаты адекватны для всех
практически здоровых людей, поэтому не возникает проблемы с
набором больших выборок, часто бывает достаточно одного или
3-5 испытуемых.
При доказательстве адекватности модели педагогический
эксперимент становится средством обоснования корректно-
сти мышления ученого или тренера.
1.5. Некоторые проблемы, связанные
с различием в логике эмпирического
и теоретического мышления
Обсуждение предыдущей книги в среде ученых и практиков
показало, что существуют трудные моменты, связанные с раз-
ными методологическими установками. Здесь можно коротко
ответить на некоторые наиболее часто задаваемые вопросы и
критические замечания.
Прежде всего обращалось внимание на то, что эксперимен-
тальные результаты, описанные в монографии, получены на
молодых спортсменах, а следовательно, «не могут распростра-
няться на подготовку спортсменов высшей квалификации».
В рамках эмпирического подхода замечание справедливо,
но отражает непонимание сути теоретического подхода, ис-
пользуемого авторами для обоснования своих взглядов.
Этот подход подробнее изложен выше, но считаем необ-
ходимым лишний раз определить его суть, которая сводится
24
к тому, что основные суждения относительно построения тре-
нировочного процесса разрабатываются на основании моделиро-
вания, а затем теоретического анализа (имитационного моде-
лирования) биологических закономерностей функционирования
организма спортсменов, уже экспериментально обоснованных в
многочисленных исследованиях российский и зарубежных ученых,
как правило, биологов. Анализ практики подготовки спортсме-
нов и экспериментальные данные, в том числе и собственные,
привлекались только для того, чтобы продемонстрировать,
что модели и имитационное моделирование (теория) могут
быть подтверждены практикой или экспериментом. Если под-
тверждение модели (теории), пусть даже единичное, существу-
ет, то это означает, что данная модель (точка зрения) коррек-
тна и относится, как правило, ко всем практически здоровым
спортсменам. Поэтому, как минимум, заслуживает внимания
и дальнейшего изучения. Кроме этого отметим, что и сами ав-
торы хорошо осознают «нетрадиционность» некоторых сужде-
ний. Знают, что модели несовершенны (но пока лучше любых
других, применяемых в спорте), поэтому и речи не может быть
о механистическом применении выводов и рекомендаций данной
работы. Более того, за 5 лет накопилось уже достаточное ко-
личество отзывов от тренеров, которые пробовали на прак-
тике применять наши рекомендации. Отзывы, как правило,
положительные. Однако анализ попыток, приведших к отри-
цательным результатам, однозначно позволяет сделать вывод,
что наши уважаемые последователи и оппоненты, чаще всего
не разобравшись в теоретической части «Локальной выносли-
вости в беге» или попросту «опустив» ее, пробовали бессис-
темно, не понимая сути, применять отдельные упражнения,
схемы построения тренировочных циклов и т.п. Поэтому мы
хотели бы подчеркнуть, что основное назначение этой кни-
ги — представить новую целостную теоретическую концепцию
одной из основных сторон подготовки в ЦВС.
Для правильного же применения ее практических выводов
и рекомендаций следует хорошо разобраться (опять-таки!) в
теории:
а) выявления спортсменов, у которых лимитирующим
фактором спортивных результатов является нервно-
25
мышечный аппарат, а не «центральные механизмы», т.е.
тех, кому вообще имеет смысл применять наши реко-
мендации;
б) повышения силы окислительных мышечных волокон
основных для данной локомоции мышечных групп, как
основы работоспособности мышц в ЦВС\
в) повышения окислительного потенциала гликолитиче-
ских (быстрых) мышечных волокон, как важнейшей
стороны подготовленности, о которой повсеместно име-
ется очень смутное представление;
г) так называемой «гликолитической тренировки» и ее от-
рицательных последствий и альтернатив;
д) предлагаемых вариантов построения одного тренировоч-
ного занятия, микро-, мезо- и макроциклов.
Подчеркнем также, что основная «практическая значи-
мость» работы и ее главная цель — предложить новые или на-
помнить «хорошо забытые старые», но не утратившие своей
актуальности средства и методы построения тренировочного
процесса в циклических видах спорта. При этом очевидно, что
определение того, в какой степени и насколько полно пред-
лагаемые идеи и конкретные рекомендации могут быть реа-
лизованы в каждом конкретном случае — это задача тренера и
только его! А задача ученых разрабатывать модели, доказывать
их адекватность и выявлять такие средства и методы, которые
никто и никогда на практике не использовал.
Поэтому такой довод «против» как «это все неверно, пото-
му что так никто не тренируется» мы отвергаем сразу и реши-
тельно.
Во-первых, если так уже тренируются, то нет новизны, нет
науки, а мы стараемся стоять на этих позициях и данная кни-
га — не учебник, а монография, следовательно, в ней должен
представляться новый материал.
Во-вторых — тренируются (и очень многие) наши последо-
ватели.
В-третьих, такие «хорошо теоретически и практически под-
кованные критики» почему-то постоянно забывают, что весь
основной массив научных данных, собранных на «квалифици-
рованных спортсменах», и на основании которых построены
26
«современные» представления о подготовке в ЦВС и обучено
нынешнее поколение тренеров, получены в 70-х и 80-х годах.
А будет ли кто-нибудь спорить, что это период, когда повсе-
местно, бесконтрольно, в немыслимых дозах, бессистемно и,
чаще всего, необоснованно использовались анаболические
стероиды. Этот факт «из скромности» выводился из поля зре-
ния при интерпретации результатов исследований и никогда
не оговаривался даже в тех случаях, когда в разделе «практи-
ческие рекомендации» диссертаций, монографий и учебников
приводились детальные тренировочные планы.
Таким образом, мы можем достаточно уверенно утверж-
дать, что достичь сколько-нибудь заметных спортивных резуль-
татов, используя «традиционные» подходы, можно только при
применении анаболических стероидов.
И напротив, описанные в данной работе рекомендации пред-
полагают подготовку без применения препаратов, искусственно
индуцирующих синтез сократительных белков. При этом авто-
ры прекрасно сознают, что разрешенное фармакологическое
обеспечение тренировочного процесса — необходимая и со-
вершенно неотъемлемая часть современного спорта. Соот-
ветствующие схемы фармакологической поддержки разра-
батываются, в частности, в ПНИЛ РГАФК и могут успешно
«накладываться» на предлагаемые тренировочные программы,
но при этом есть особенности, описание которых выходит за
рамки настоящей работы.
Если же пытаться одновременно применять стероиды по
«традиционной схеме» и наши рекомендации, то результат
заведомо будет отрицательным'.
Существует еще один момент, который следует разъяснить
особо. Одним из ключевых моментов разработанной теории
является утверждение, что «базовое» место в подготовленности
спортсменов в ЦВС — занимает сила мышц (и соответственно,
все разновидности силовой подготовки), а не аэробные способ-
ности (и, следовательно, аэробная подготовка). Вместе с тем
мы утверждаем, что аэробная подготовленность, не являясь
«базовой», является в то же время основной и реализационной,
т.е. той, от которой непосредственно зависит спортивный ре-
зультат. Непонимание этой ключевой идеи вызывает кри-
27
тические замечания, согласно которым нам приписывается
абсолютизация силовой и недооценка аэробной сторон под-
готовленности и подготовки в ЦВС.
Уважаемые критики и наши последователи!
Убедительно просим Вас внимательно разобраться в этом
ключевом положении!
Еще раз повторяем, подобная, приписываемая нам, абсо-
лютизация роли силовой подготовки — это заблуждение.
Доля силовой подготовки в тренировочном процессе может
занимать от 50 до 3% тренировочного времени в зависимости
от множества факторов, и не один нормальный тренер не будет
утверждать, что она не нужна вовсе. Наша задача — дать ключ
к оптимизации этой важнейшей стороны подготовки в ЦВС, в
частности, в аспекте ее сочетания с аэробной подготовкой.
Например, на практике часто путается силовая подготовка
«в интересах» увеличения аэробного потенциала мышц и ско-
ростная, или скоростно-силовая подготовка, направленная на
увеличение силы мышц, укрепление опорно-двигательного
аппарата, повышение скоростных возможностей или совер-
шенствования техники локомоций и мн. др.
Пожалуйста, сначала подумайте, зачем Вы применяете те
или иные схемы силовой подготовки!
Описанные в монографии подходы к организации физиче-
ской подготовки применялись в течение многих лет во многих
видах спорта - циклических, игровых (на уровне команд выс-
шей лиги), единоборствах (на уровне олимпийской команды
России) и т.п. Поэтому, отвечая на часто задаваемый вопрос:
«Насколько универсальны ваши рекомендации?», мы имеем
все основания утверждать, что тренировочные программы,
построенные на теории, приведенной в данной книге, ока-
зываются чрезвычайно эффективными в большинстве видов
спорта, а так же в оздоровительной физической культуре (фит-
несе).
Основной особенностью данной книги является то, что в
отличие от большинства других исследований в области тео-
рии и методики спортивной тренировки, когда выводы де-
28
лаются на основании изучения практического опыта выдаю-
щихся тренеров и спортсменов и по результатам проведения
«естественных» педагогических экспериментов, авторы ста-
рались, по возможности, строго придерживаться логики т.н.
теоретического уровня познания (см. выше), когда гипотезы
относительно возможных путей совершенствования физи-
ческой подготовки спортсменов выдвигаются на основе рас-
смотрения биологических механизмов функционирования
организма человека (другими словами, привлекается весь запас
знаний биологических наук о человеке !), а уже затем проверяют-
ся результатами лабораторных и полевых сравнительных экс-
периментов или опытом тренировки спортсменов. При таком
подходе гипотеза может считаться принятой только в том слу-
чае, если теоретические выкладки нашли свое подтверждение
на практике. И наоборот, никакая, даже самая интересная,
многообещающая и как будто бы оправдавшая себя на практи-
ке тренерская находка, результаты педагогического экспери-
мента или выявленные на основе изучения опыта спортивной
подготовки «общие закономерности тренировочного процес-
са», не могут рассматриваться иначе как околонаучные фан-
тазии, пока им не найдено убедительное естественно-научное
объяснение, подтвержденное данными биологических или
психологических фундаментальных исследований.
29
Глава 2
Основы биологии человека
(концептуальные модели систем
и органов человека)
В учебниках по биологии человека и физиологии и био-
химии спорта излагаются основные сведения о строении тела
человека и функциональных свойствах его систем и органов. В
практической деятельности при решении конкретных задач на
основе принципов природной специфичности, системности,
аналогичности и оптимизации строятся упрощенные модели
человека. Такие модели обладают необходимыми свойствами
и достаточной сложностью для решения поставленных задач.
Описание моделей, необходимых для решения задач физи-
ческой подготовки в спорте и оздоровительной физической
культуры, представлено в этой главе.
2.1. Биология клетки
Клетка — основная структурная единица всех живых орга-
низмов, элементарная живая целостная система, которая об-
ладает рядом свойств: воспроизведение, синтез (анаболизм),
катаболизм, производство энергии, поглощение, выделение,
специфические функции.
Она представляет собой протоплазму (саркоплазму), окру-
женную мембраной. В протоплазме расположено ядро, в ко-
тором содержатся гены (наследственная информация) в виде
молекул ДНК. В протоплазме имеются следующие структур-
ные образования, их еще называют органеллами или органои-
дами:
30
— рибосомы (полирибосомы) — с помощью РНК произво-
дится строительство белка, иными словами, разворачи-
ваются анаболические процессы;
— митохондрии — энергетические станции клетки, в них с
помощью кислорода идет превращение жиров или глю-
козы в углекислый газ (СО2), воду и энергию, заключен-
ную в молекулах АТФ;
— эндоплазматическая сеть или саркоплазматический ре-
тикулум является органеллой, состоящей из мембран и
ферментативных систем, прикрепленных к ней;
— комплекс Гольджи — система мембран, образующих со-
вокупность мешочков и пузырьков, служит для синтеза
и выделения веществ из клетки;
— лизосомы — органеллы в форме пузырьков, содержат
ферменты, разрушающие белки до простейших состав-
ляющих — аминокислот, эти органеллы еще называют
пищеварительным аппаратом клетки;
— глобулы гликогена — внутриклеточные запасы углеводов;
— капельки жира — внутриклеточные запасы жиров;
— специализированные органеллы — структурные компо-
ненты клетки, присущие определенным видам клеток,
например миофибриллы — мышечным волокнам.
2.2. Нервно-мышечный аппарат
Человек выполняет физические упражнения и тратит энер-
гию с помощью нервно-мышечного аппарата.
Нервно-мышечный аппарат — это совокупность двига-
тельных единиц (ДЕ). Каждая ДЕ включает мотонейрон, ак-
сон и совокупность мышечных волокон (МВ). Количество
ДЕ остается неизменным у человека (Физиология челове-
ка, 1995). Количество МВ в мышце возможно и поддается
изменению в ходе тренировки, однако, не более чем на 5%
(Хоппелер, 1987). Поэтому этот фактор роста функциональ-
ных возможностей мышцы не имеет практического значе-
ния. Внутри МВ происходит гиперплазия (рост количества
элементов) многих органелл: миофибрилл, митохондрий,
31
саркоплазматического ретикулума (СПР), глобул глико-
гена, миоглобина, рибосом, ДНК и др. Изменяется также
количество капилляров, обслуживающих МВ (Физиология
мышечной деятельности, 1982).
Миофибрилла является специализированной органеллой
мышечного волокна (клетки). Она у всех животных имеет при-
мерно равное поперечное сечение. Состоит из последователь-
но соединенных саркомеров, каждый из которых включает
нити актина и миозина. Между нитями актина и миозина могут
образовываться мостики и при затрате энергии, заключенной в
АТФ, может происходить поворот мостиков, т.е. сокращение
миофибриллы, сокращение мышечного волокна, сокращение
мышцы. Мостики образуются в присутствии в саркоплазме
ионов кальция и молекул АТФ. Увеличение количества мио-
фибрилл в мышечном волокне приводит к увеличению его
силы, скорости сокращения и размера. Вместе с ростом мио-
фибрилл происходит разрастание и других обслуживающих
миофибриллы органелл, например саркоплазматического ре-
тикулума.
Саркоплазматический ретикулум — это сеть внутренних мем-
бран, которая образует пузырьки, канальцы, цистерны. В МВ
СПР образует цистерны, в этих цистернах скапливаются ионы
кальция (Са++). Предполагается, что к мембранам СПР при-
креплены ферменты гликолиза, поэтому при прекращении
доступа кислорода происходит значительное разбухание кана-
лов. Это явление связано с накоплением ионов водорода (Н),
которые вызывают частичное разрушение (денатурацию) бел-
ковых структур, присоединение воды к радикалам белковых
молекул (МеерсонФ.З., 1978, 1988). Для механизма мышечно-
го сокращения принципиальное значение имеет скорость от-
качивания Са++ из саркоплазмы, поскольку это обеспечивает
процесс расслабления мышцы. В мембраны СПР встроены
натрий-калиевые и кальциевые насосы, поэтому можно пред-
положить, что увеличение поверхности мембран СПР по от-
ношению к массе миофибрилл должно вести к росту скорости
расслабления МВ. Следовательно, увеличение максимального
темпа или скорости расслабления мышцы (интервала времени
от конца электрической активации мышцы до падения меха-
32
нического напряжения в ней до нуля) должно говорить об от-
носительном приросте мембран СПР.
Поддержание максимального темпа обеспечивается за-
пасами в МВ АТФ, КрФ, массой миофибриллярных мито-
хондрий, массой саркоплазматических митохондрий, массой
гликолитических ферментов и буферной емкостью содержи-
мого мышечного волокна и крови. Все эти факторы влияют на
процесс энергообеспечения мышечного сокращения, однако,
способность поддерживать максимальный темп должна за-
висеть преимущественно от митохондрий СПР. Увеличивая
количество окислительных МВ или, другими словами, аэроб-
ных возможностей мышцы, продолжительность упражнения
с максимальной мощностью растет. Обусловлено это тем, что
поддержание концентрации КрФ в ходе гликолиза ведет к за-
кислению МВ, торможению процессов расхода АТФ из-за
конкурирования ионов Н+ с ионами Са++ на активных центрах
головок миозина (Hermansen, 1981). Поэтому процесс поддер-
жания концентрации КрФ при преобладании в мышце аэроб-
ных процессов идет по мере выполнения упражнения все бо-
лее эффективнее. Важно также то, что митохондрии активно
поглощают ионы водорода (Hermansen, 1981; Holloshzy, 1976;
Hoppeler, 1986), поэтому при выполнении кратковременных
предельных упражнений (10-30 с) их роль больше сводится к
буферированию закисления клетки.
Митохондрии располагаются везде, где требуется в боль-
шом количестве энергия АТФ. В мышечных волокнах энер-
гия требуется для сокращения миофибрилл, поэтому вокруг
них образуются миофибриллярные митохондрии (Ленинджер,
1966).
2.3. Биохимия клетки (энергетика)
Процессы мышечного сокращения, передачи нервного им-
пульса, синтеза белка и другие идут с затратами энергии (рис. 1).
В клетках энергия используется только в виде АТФ. Освобож-
дение энергии, заключенной в АТФ, осуществляется благода-
ря ферменту АТФ-азе, который имеется во всех местах клетки,
33
Рис. 1. Простейшая модель энергообеспечения в окислительном мышечном
волокне. При активации мышечного волокна тратятся запасы А ТФ, по-
являются ионы водорода в клетке. КрФресинтезирует АТФ. Молекулы
А ТФ, образующиеся в ходе анаэробного и аэробного гликолиза, идут на
ресинтез КрФ. Цитрат может ингибировать анаэробный гликолиз, а
лактат ключевые ферменты липолиза. Поэтому через 30-60с работы МВ
начинает метаболизироваться только внутрений жир мышечного волок-
на. При поступлении лактата из крови ингибируется окисление жиров
где требуется энергия. По мере освобождения энергии образу-
ются молекулы АДФ, Ф, Н+. Ресинтез АТФ осуществляется в
основном за счет запаса КрФ. Когда КрФ отдает свою энергию
для ресинтеза АТФ, то образуется Кр и Ф. Эти молекулы рас-
пространяются по цитоплазме и активизируют ферментатив-
34
ную активность, связанную с синтезом АТФ. Существуют два
основных пути образования АТФ: анаэробный и аэробный.
Анаэробный путь, или анаэробный гликолиз, связан с фермен-
тативными системами, расположенными на мембране сарко-
плазматического ретикулума и в саркоплазме. При появлении
рядом с этими ферментами Кр и Ф запускается цепь химических
реакций, в ходе которых гликоген или глюкоза распадаются до
пирувата с образованием молекул АТФ. Молекулы АТФ тут же
отдают свою энергию для ресинтеза КрФ, а АДФ и Ф вновь ис-
пользуются в гликолизе для образования новой молекулы АТФ.
Пируват имеет две возможности для преобразования:
1. подойти к митохондриям, превратиться в Ацетил-
коэнзим-А, подвергнуться окислительному фосфори-
лированию до образования углекислого газа, воды и
молекул АТФ. Этот метаболический путь — гликоген-
пируват-митохондрия-углекислый газ и вода — называ-
ют аэробным гликолизом;
2. с помощью фермента ЛДГ-М (лактат-дегидрогеназы
мышечного типа) пируват превращается в лактат. Этот
метаболический путь — гликоген-пируват-лактат — на-
зывается анаэробным гликолизом и сопровождается об-
разованием и накоплением ионов водорода.
Аэробный путь, или окислительное фосфорилирование,
связан с митохондриальной системой. При появлении рядом с
митохондриями Кр и Ф с помощью митохондриальной КФК-
азы выполняется ресинтез КрФ за счет АТФ, образовавшейся
в митохондрии. АДФ и Ф поступают обратно в митохондрию
для образования новой молекулы АТФ. Для синтеза АТФ име-
ется два метаболических пути:
1. аэробный гликолиз;
2. окисление липидов (жиров).
Аэробные процессы связаны с поглощением ионов водо-
рода, а в медленных мышечных волокнах (МВ сердца и диа-
фрагмы) преобладает фермент ЛДГ-Н (лактатдегидрогеназа
сердечного типа), который более интенсивно превращает лак-
тат в пируват. Поэтому при функционировании медленных
мышечных волокон (ММВ) идет быстрое устранение лактата
и ионов водорода.
35
Увеличение в МВ лактата и Н+ приводит к ингибированию
окисления жиров, а интенсивное окисление жиров приводит к на-
коплению в клетке цитрата, а он угнетает ферменты гликолиза.
2 Л. Модель функционирования нервно-
мышечного аппарата при выполнении
циклического упражнения
Простейшая модель нервно-мышечного аппарата пред-
ставлена на рис. 2.
Биохимия и физиология мышечной активности при вы-
полнении физической работы может быть описана следую-
щим образом. Покажем с помощью имитационного модели-
рования как разворачиваются физиологические процессы в
мышце при выполнении ступенчатого теста.
Предположим, что мышца (например, четырехглавая мыш-
ца бедра) имеет ММВ 50%, амплитуда ступеньки — 5% мак-
симальной алактатной мощности, величина которой принята
за 100%, длительность — 1 мин. На первой ступеньке в связи
с малым внешним сопротивлением рекрутируются согласно
«правилу размера» Ханнемана низкопороговые ДЕ (МВ). Они
имеют высокие окислительные возможности, субстратом в
них являются жирные кислоты. Однако первые 10-20 с энерго-
обеспечение идет за счет запасов АТФ и КрФ в активных МВ.
Уже в пределах одной ступеньки (1 мин) имеет место рекрути-
рование новых мышечных волокон, благодаря этому удается
поддерживать заданную мощность на ступеньке. Вызвано это
снижением концентрации фосфогенов в активных МВ, т.е.
силы (мощности) сокращения этих МВ, усилением активиру-
ющего влияния ЦНС, а это приводит к вовлечению новых ДЕ
(МВ). Постепенное ступенчатое увеличение внешней нагрузки
(мощности) сопровождается пропорциональным изменением
некоторых показателей: растет ЧСС, потребление кислорода,
легочная вентиляция, не изменяется концентрация молочной
кислоты и ионов водорода.
При достижении внешней мощности некоторого значения
наступает момент, когда в работу вовлекаются все ММВ и на-
36
Рис. 2. Схема управления мышечным сокращением
чинают рекрутироваться промежуточные мышечные волокна
(ПМВ). Промежуточными мышечными волокнами можно на-
звать те, в которых массы митохондрий недостаточно для обе-
спечения баланса между образованием пирувата и его окисле-
нием в митохондриях. В ПМВ после снижения концентрации
фосфогенов активизируется гликолиз, часть пирувата начина-
ет преобразовываться в молочную кислоту (точнее говоря, в
лактат и ионы водорода), которая выходит в кровь, проникает
в ММВ. Попадание в ММВ (ОМВ) лактата ведет к ингибиро-
ванию окисления жиров, субстратом окисления становится в
большей мере гликоген. Следовательно, признаком рекрути-
рования всех ММВ (ОМВ) является увеличение в крови кон-
центрации лактата и усиление легочной вентиляции. Легочная
вентиляция усиливается в связи с образованием и накоплени-
ем в ПМВ ионов водорода, которые при выходе в кровь взаи-
модействуют с буферными системами крови и вызывают обра-
зование избыточного (неметаболического) углекислого газа.
Повышение концентрации углекислого газа в крови приводит
к активизации дыхания (Физиология человека, 1995).
Таким образом, при выполнении ступенчатого теста имеет
место явление, которое принято называть аэробным порогом
37
(АэП). Появление АэП свидетельствует о рекрутировании всех
ОМВ. По величине внешнего сопротивления можно судить о
силе ОМВ, которую они могут проявить при ресинтезе АТФ
и КрФ за счет окислительного фосфорилирования (Селуянов
В.Н. с соав., 1991).
Дальнейшее увеличение мощности требует рекрутирования
более высокопороговых ДЕ (ГМВ), в которых митохондрий
очень мало. Это усиливает процессы анаэробного гликолиза,
больше выходит лактата и ионов Н в кровь. При попадании
лактата в ОМВ, он превращается обратно в пируват с помо-
щью фермента ЛДГ-Н (Karlsson, 1982). Однако мощность ми-
тохондриальной системы ОМВ имеет предел. Поэтому сна-
чала наступает предельное динамическое равновесие между
образованием лактата и его потреблением вОМВиПМВ, а
затем равновесие нарушается, и некомпенсируемые метабо-
литы — лактат, Н+, СО2 — вызывают резкую интенсификацию
физиологических функций. Дыхание — один из наиболее чув-
ствительных процессов — реагирует очень активно. Кровь при
прохождении легких в зависимости от фаз дыхательного цикла
должна иметь разное парциальное напряжение СО2. «Порция»
артериальной крови с повышенным содержанием СО2 дости-
гает хеморецепторов и непосредственно модулярных хемочув-
ствительных структур ЦНС, что и вызывает интенсификацию
дыхания. В итоге СО2 начинает вымываться из крови так, что
в результате средняя концентрация углекислого газа в крови
начинает снижаться. При достижении мощности, соответ-
ствующей АнП, скорость выхода лактата из работающих гли-
колитических МВ сравнивается со скоростью его окисления в
ОМВ. В этот момент субстратом окисления в ОМВ становятся
практически только углеводы (лактат ингибирует окисление
жиров), часть из них составляет гликоген ММВ, другую часть
— лактат, образовавшийся в гликолитических МВ. Использо-
вание углеводов в качестве субстратов окисления обеспечи-
вает максимальную скорость образования энергии (АТФ) в
митохондриях ОМВ. Следовательно, потребление кислорода
или (и) мощность на анаэробном пороге (АнП) характеризует
максимальный окислительный потенциал (мощность) ОМВ (Се-
луянов В.Н. с соав., 1991).
38
Дальнейший рост внешней мощности делает необходи-
мым вовлечение все более высокопороговых ДЕ, иннерви-
рующих гликолитические МВ. Динамическое равновесие
нарушается, продукция Н+, лактата начинает превышать
скорость их устранения. Это сопровождается дальнейшим
увеличением легочной вентиляции, ЧСС и потребления
кислорода. После АнП потребление кислорода в основном
связано с работой дыхательных мышц и миокарда. При до-
стижении предельных величин легочной вентиляции и ЧСС
или при локальном утомлении мышц потребление кислоро-
да стабилизируется, а затем начинает уменьшаться. В этот
момент фиксируют МПК.
Таким образом, МПК есть сумма величин потребления кис-
лорода окислительными МВ (ММВ), дыхательными мышцами и
миокардом.
2.5. Биомеханика мышечного сокращения
Сила — векторная величина, являющаяся мерой механиче-
ского воздействия на материальную точку или тело со стороны
других тел или полей. Сила полностью задана, если указаны ее
численное значение, направление и точка приложения.
В теории и методике физического воспитания рассматрива-
ют физическое качество силу как способность человека напря-
жением мышц преодолевать механические и биомеханические
силы, препятствующие действию (Л.П. Матвеев, 1991).
Мышцы могут проявлять силу: без изменения своей длины
(изометрический режим); при уменьшении длины (изотони-
ческий режим); при удлинении (эксцентрический режим), при
использовании специальной аппаратуры возможно соблюде-
ние изокинетического режима (в ходе сокращения мышц со-
блюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит:
- от интенсивности активации мотонейронного пула
спинного мозга данной мышцы;
— количества активированных двигательных единиц и мы-
шечных волокон;
39
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от
активности миозиновой АТФ-азы и величины внешнего
сопротивления;
— законов механики мышечного сокращения (сила — дли-
на мышцы, сила — скорость сокращения).
Спортсмен при желании сократить какую-либо мышцу по-
сылает импульсы в спинной мозг к мотонейронному пулу, об-
служивающему данную мышцу. Поскольку в мотонейронном
пуле размеры мотонейронов различаются, то при низкой ча-
стоте импульсации из ЦНС могут активироваться только низ-
копороговые мотонейроны. Каждый мотонейрон иннервиру-
ет свои мышечные волокна. Поэтому активация мотонейрона
приводит к рекрутированию или возбуждению соответствую-
щих мышечных волокон. Каждое активное мышечное волок-
но под влиянием электрических импульсов выпускает из СПР
ионы кальция, которые снимают ингибитор с активных цен-
тров актина. Это обеспечивает образование акгин-миозиновых
мостиков и начало их поворота и мышечного сокращения. На
поворот мостиков и отсоединение актина от миозина тратит-
ся энергия одной молекулы АТФ. Продолжительность работы
мостика составляет 1 мс. Вероятность образования мостиков
зависит от взаимного расположения между собой нитей акти-
на и миозина, отсюда возникает зависимость «сила — длина
активной мышцы», а также от скорости взаимного перемеще-
ния (скольжения) их одной по отношению к другой, соответ-
ственно имеем зависимость «сила—скорость».
Зависимость «сила - длина активного мышечного волок-
на» определяется, как правило, относительным расположени-
ем между собой головок миозина и активных центров актина.
Максимальное количество мостиков возникает при некоторой
средней длине мышцы. Отклонение от этой длины в большую
или меньшую сторону ведет к снижению силовых проявлений
мышечного волокна (мышцы). Однако в случае растяжения
мышцы еще не в активном состоянии у некоторых мышц могут
возникать значительные силы сопротивления растяжению, на-
пример в мышцах — сгибателях голеностопного или лучезапяст-
ного сустава. Эти силы связаны с растяжением соединительных
40
тканей, например перемизиума. В биомеханике в таком случае
говорят о параллельном упругом компоненте мышцы. Упру-
гостью обладают сухожилия, Z-пластинки саркомеров и нити
миозина, к которым прикреплены головки. Такую упругость
называют последовательной упругой компонентой.
Наличие последовательной упругой компоненты в мышечных
волокнах приводит к тому, что с ростом числа рекрутированных
МВ растет жесткость мышцы (коэффициент упругости).
Растягивание активной мышцы приводит не только к на-
коплению энергии упругой деформации в последовательной
упругой компоненте, но и к прекращению работы мостиков,
а именно, они перестают отцепляться за счет энергии молекул
АТФ. Разрыв мостиков происходит благодаря действию внеш-
ней — механической — силы. В итоге отрицательная работа
мышц выполняется с очень высоким коэффициентом полез-
ного действия, с минимальными затратами кислорода.
2.6. Сердце и кровообращение
Деятельность сердца и сосудов обеспечивает кровообра-
щение — непрерывное движение крови в организме. В своем
движении кровь проходит по большому и малому кругам кро-
вообращения. Большой круг начинается от левого желудочка
сердца, включает аорту, отходящие от нее артерии, артериолы,
капилляры, вены и заканчивается полыми венами, впадающи-
ми в правое предсердие. Малый круг кровообращения начи-
нается от правого желудочка, далее — легочная артерия, легоч-
ные артериолы, капилляры, вены, легочная вена, впадающая в
левое предсердие.
Функцией сердца является ритмическое нагнетание в ар-
терии крови. Сокращение мышечных волокон (миокардио-
цитов) стенок предсердий и желудочков называют систолой, а
расслабление — диастолой.
Количество крови, выбрасываемое левым желудочком серд-
ца в мин, называется минутным объемом кровотока (МОК).
В покое он составляет в норме 4-5 л/мин. Разделив МОК на
частоту сердечных сокращений в мин (ЧСС), можно получить
41
ударный объем кровотока или сердца (УОС). В покое он со-
ставляет 60-70 мл крови за удар.
Частота и сила сокращений зависит от нервной, гумораль-
ной (адреналин) регуляции и биомеханических условий рабо-
ты желудочков.
При вертикальном положении тела имеется механический
фактор — сила тяжести крови, затрудняющий работу сердца,
приток венозной крови к правому предсердию. В нижних ко-
нечностях скапливается до 300-800 мл крови.
При мышечной работе минутный объем кровотока растет
за счет увеличения ЧСС и УОС. Заметим, что УОС достигает
максимума при ЧСС 120-150 уд/мин, а максимум ЧСС быва-
ет при 180-200 и более уд/мин. МОК достигает 18-25 л/мин у
нетренированных лиц при достижении максимальной ЧСС
(Физиология мышечной деятельности, 1982). В этот момент
сердце доставляет организму максимум кислорода:
VO2 = МОК х Нв х 0,00134 = 20 х 160 х 0,00134 = 4,288 л/
мин. Здесь Нв — содержание гемоглобина в крови, г/л крови;
0,00134 — кислородная емкость гемоглобина в артериальной
крови.
Если бы мышцы нетренированного человека могли бы пол-
ностью использовать весь приходящий кислород, то этот чело-
век мог бы стать мастером спорта по бегу на длинные дистан-
ции (бегуны мирового класса потребляют кислород на уровне
анаэробного порога 4,0-4,5 л/мин). Однако в мышцах мало
митохондрий, поэтому максимальное потребление кислорода
(МПК) у нетренированного мужчины составляет 3-3,5 л/мин
(45-50 мл/кг/мин), у нетренированной женщины — 2-2,2 л/
мин (40-45 мл/кг/мин). На уровне анаэробного порога потре-
бление кислорода составляет в среднем 60-70% МПК (Аулик
И.В., 1990).
2.7. Кровеносные сосуды
Сердце при сокращении (систоле) выталкивает кровь в
аорту и легочную артерию, растягивая их и создавая давление
крови (Р). Движению крови препятствует сосудистое (пери-
42
ферическое) сопротивление. Максимальное давление назы-
вается систолическим артериальным давлением (САД), мини-
мальное — диастолическим артериальным давлением (ДАД). В
условиях покоя в норме САД = 120 мм рт. ст., ДАД = 80 мм рт.
ст. Между растяжимостью (эластичностью) артерий и давле-
нием крови в сосудах имеется обратная зависимость. Чем рас-
тяжимее артерии, тем больше крови может быть нагнетено без
увеличения артериального давления (АД). При артериоскле-
розе стенка аорты менее эластична, поэтому надо сильнее на-
гнетать кровь (тот же объем крови, как у здорового человека),
чтобы она дальше прошла по сосудам. Сопротивление крово-
току зависит от вязкости крови и, главным образом, от про-
света сосудов. Увеличение напряжения мышц вызывает пере-
крытие сосудов — увеличение сосудистого сопротивления.
Накопление в крови мышц продуктов анаэробных процессов
(pH, рСО2, уменьшение рО2 и др.) приводит к рабочей гипе-
ремии — расширению кровеносных сосудов, т.е. уменьшению
АД (Физиология мышечной деятельности, 1982).
Нервный контроль и гуморальный наиболее важны в управ-
лении функциями сосудистой системы. Симпатические нерв-
ные волокна иннервируют гладкие мышцы в стенках артери-
альных и венозных сосудов, особенно мелких. Кровоток через
капилляры определяется местными факторами.
Сосудосуживающий эффект связан с выделением из окон-
чаний адренэргических симпатических волокон норадренали-
на, который вызывает эффект сокращения гладкомышечных
сосудистых клеток, имеющих альфа-рецепторы на мембране
(почки, печень, желудочно-кишечный тракт, легкие, кожа).
Сосудорасширительный эффект (вазодилятацию) вызывает
действие норадреналина и адреналина на гладкомышечные
клетки, имеющие бета-рецепторы (сосуды скелетных мышц,
сердца, надпочечников) (Физиология человека, 1995).
2.8. Эндокринная система
Межклеточные вещества, передающие информацию, принято
называть информонами (Розен В.Б., 1994). Эти соединения об-
43
ладают следующими свойствами: секретируются во внеклеточное
пространство, не используются в качестве основных источников
пластического и энергетического материала, взаимодействуют с
мембранными белками-рецепторами, обладают специфической
биологической активностью. Выделяют такие типы информонов:
— гистогормоны (амины, простагландины, факторы роста,
регуляторные пептиды и др.);
— нейромедиаторы и нейромодуляторы (ацетилхолин, но-
радреналин, дофамин, гамма-аминомаслянная кислота,
нейропептиды и др.);
— гормоны (инсулин, глюкагон, гормон роста, эстрогены,
андрогены, кортикостероиды и др.);
— антитела (специфические иммуноглобулины).
Межклеточное управление на тканевом уровне осущест-
вляется с помощью гистогормонов — короткоживущих соеди-
нений, действующих в пределах близлежащих клеток.
Специализированный аппарат централизованного управ-
ления жизнедеятельности представлен нервной, эндокринной
и иммунной системами.
ЦНС дистантно передает нервные импульсы, а из синапсов
выделяются порции нейромедиаторов, короткоживущих и бы-
стродействующих соединений.
В организме существуют два типа желез:
— экзокринные, имеют выводные протоки (пищевари-
тельные, потовые, сальные);
— эндокринные, не имеют выводных протоков, выделяют
секрет в кровь, лимфу и т.д.
Эндокринная система представлена совокупностью эндо-
кринных желез, ее называют диффузной системой управле-
ния. Гормоны являются биоорганическими соединениями со
стабильной химической структурой, поэтому они могут осу-
ществлять дистантное воздействие на клетки-мишени.
Иммунная система представлена тимико-лимфоидными
элементами (В- и Т-клетками). Она обеспечивает с помощью
гуморальных и клеточных механизмов защиту организма от
чужеродных белков — антигенов. Антитела (иммуноглобули-
ны) секретируются В-лимфоцитами в кровь в ответ на появле-
ние в организме антигенов.
44
В спортивной практике имеет интерес анализ деятельности
эндокринной и нервной систем, поскольку от их активности
зависят срочные и долговременные процессы в организме
спортсменов.
Гормоны подразделяются на:
— стероидные (кортикостероиды — глюкокортикоиды, ми-
нералкортикоиды; прогестины, андрогены, эстрогены);
— производные полиненасыщенных жирных кислот (про-
стагландины);
— производные аминокислот L-тирозина и L-триптофана
(катехоламины, тиреоидные гормоны, мелатонин);
— белково-пептидные (нейрогипофизарные пептиды,
АКТГ, инсулин, глюкагон, гормоны тимуса и др.).
Биосинтез гормонов может проходить прямым путем и
опосредованным. Прямым путем идет синтез всех основных
гормонов, а опосредованным, или внерибосомальным, — не-
которых стероидных гормонов, рилизинг-факторов и др.
Схема процесса в общем виде выглядит так:
Ген — мРНК — (рибосомы или полирибосомы) — Прегор-
мон — Прогормон — Гормон.
Секреция гормонов протекает спонтанно, обеспечивая
определенный базальный уровень гормонов в циркулирующих
жидкостях. Секреция осуществляется импульсно, дискрет-
ными порциями из клеточных секреторных гранул (секреция
белково-пептидных гормонов и катехоламинов). Тироидные
гормоны освобождаются из белковосвязанной формы. Стеро-
идные гормоны переходят в жидкости путем свободной диф-
фузии, следовательно интенсивность секреторных процессов
определяется уровнем их биосинтеза.
Внутриклеточный транспорт гранул осуществляется при уча-
стии микрофиламентов и микротрубочек. Секрет выбрасывает-
ся через поры в мембранах. Стероидные гормоны содержатся в
составе липидных капель растворимой части цитоплазмы в сво-
бодном виде. Они могут относительно легко диффундировать
через плазматические мембраны в кровь по концентрационно-
му градиенту, не накапливаясь в клетках желез.
Гормоны циркулируют в крови в нескольких физико-
химических формах:
45
— в свободном виде (в виде водного раствора);
— в виде комплексов со специфическими белками плаз-
мы;
— в виде неспецифических комплексов с плазменными
белками;
- в виде неспецифических комплексов с форменными
элементами.
Более 80% концентрации данных гормонов находится в
условиях покоя в виде комплексов со специфическими белка-
ми. Связанные гормоны физиологически неактивны, не под-
вергаются метаболическим превращениям.
В условиях физиологического покоя метаболические про-
цессы катаболизма гормонов находятся в состояния равновесия
с процессами гормональной продукции. В качестве интеграль-
ных показателей интенсивности метаболических процессов ис-
пользуют величину периода полураспада гормонов (Т1/2) и ско-
рость метаболического клиренса (СМК). Период полураспада
гормонов — это время, за которое концентрация введенной в
кровь порции радиоактивного гормона необратимо уменьша-
ется вдвое. Скорость метаболического клиренса гормонов ха-
рактеризует объем крови, полностью и необратимо очищаемый
от гормона за определенный промежуток времени. Большин-
ство гормонов и их метаболитов удаляется из организма почти
полностью через 48-72 часа, причем 80-90% попавшего в кровь
гормона выводится уже в первые сутки.
Основные этапы реализации метаболических ответов на
гормоны можно условно разделить по времени на начальные,
ранние и поздние.
Начальные этапы включают в себя события, развивающие-
ся непосредственно после инициализации гормонального эф-
фекта. Например, аденилатциклазный механизм активации
гликоген-фосфорилазы в клетке под влиянием глюкагона или
адреналина.
Ранние этапы охватывают изменения метаболизма в клетке
через 1-24 часа после начала действия гормона, приводящие
к отставленным конечным эффектам через транскрипцию,
а затем трансляцию. Главная волна усиления синтеза и кон-
центрации различных РНК наблюдается через 2-6 часов после
46
введения гормона. Например, СТГ и инсулин стимулируют
синтез общего белка в соединительной ткани, печени и мыш-
цах, создаются белки-посредники, контролирующие процес-
сы транскрипции и компонентов мембран эндоплазматиче-
ского ретикулума.
Поздние этапы охватывают процессы, длящиеся от 24 до 48
и более часов. Наиболее полно поздние события проявляются
при многократном введение гормональных соединений. Позд-
ние эффекты гормона сводятся к изменению скорости реду-
пликации ДНК и митотического деления клеток-мишений.
Наиболее эффективно они идут при непрерывном присут-
ствии гормона в клетке на всех этапах его действия.
Эндокринная система состоит из желез внутренней секре-
ции: гипофиза, щитовидной, околощитовидных, поджелудоч-
ной, надпочечников, половых. Эти железы выделяют гормо-
ны — регуляторы обмена веществ, роста и полового развития
организма.
Регуляция выделения гормонов осуществляется нервно-
гуморальным путем. Изменение состояния физиологических
процессов достигается посылкой нервных импульсов из ЦНС
(ядер гипоталамуса) к некоторым железам (гипофизу). Выде-
ляемые передней долей гипофиза гормоны регулируют деятель-
ность других желез — щитовидной, половых, надпочечников.
Для спортивной практики наиболее интересными являют-
ся симпатоадреналовая, гипофизарно-адренокортикальная,
гипофизарно-щитовидная, гипофизарно-половая системы.
Симпатоадреналовая система ответственна за мобилизацию
энергетических ресурсов. Адреналин и норадреналин образу-
ются в мозговом веществе надпочечников и вместе с норадрена-
лином, выделяющимся из нервных окончаний симпатической
нервной системы, действуют через систему «аденилатциклаза
— циклический аденозинмонофосфат (цАМФ)». Для необхо-
димого накопления цАМФ в клетке требуется ингибировать
цАМФ-фосфодиэстеразу — фермент, катализирующий расще-
пление цАМФ. Ингибирование осуществляется глюкокорти-
коидами (инсулин противодействует этому эффекту).
Система «аденилатциклаза — цАМФ» действует следующим
образом. Гормон током крови подходит к клетке, на наружной
47
поверхности клеточной мембраны которой имеются рецепто-
ры. Взаимодействие гормон-рецептор приводит к конформа-
ции рецептора, т.е. активации каталитического компонента
аденилатциклазного комплекса. Далее из АТФ начинает об-
разовываться цАМФ, который участвует в регуляции метабо-
лизма (расщеплении гликогена, активизации фосфофрукто-
киназы в мышцах, липолиза в жировых тканях), клеточной
дифференциации, синтезе белков, мышечного сокращения
(ВируА.А., 1981).
Гипофизарно-адренокортикальная система включает
нервные структуры (гипоталамус, ретикулярную формацию
и миндалевидный комплекс), кровоснабжение и надпочеч-
ники. В состоянии стресса усиливается выход кортиколи-
берина из гипоталамуса в кровоток. Это вызывает усиление
секреции адренокортикотропного гормона (АКТГ), кото-
рый током крови переносится в надпочечники. Нервная
регуляция воздействует на гипофиз и приводит к секреции
либеринов и статинов, а они регулируют секрецию тропных
гормонов аденогипофиза АКТГ.
Механизм действия глюкокортикоидов на синтез фермен-
тов может быть представлен следующим образом (по А. Виру,
1981):
— кортизол, кортикостерон, кортикотропин, кортиколи-
берин проходят через клеточную мембрану (процесс
диффузии).
— В клетке гормон (Г) соединяется со специфическим бел-
ком — рецептором (Р), образуется комплекс Г-Р.
— Комплекс Г-Р перемещается в ядро клетки (через 15
мин) и связывается с хроматиномк(ДНК).
— Стимулируется активность структурного гена, усилива-
ется транскрипция информационной-РНК (и-РНК).
— Образование и-РНК стимулирует синтез других видов
РНК. Непосредственное действие глюкокортикоидов на
аппарат трансляции состоит из двух этапов: 1) освобож-
дения рибосом из эндоплазматической сети и усиления
агрегации рибосом (наступает через 60 мин); 2) транс-
ляции информации, т.е. синтеза ферментов (в печени, в
железах внутренней секреции, скелетных мышцах).
48
После выполнения своей роли в ядре клетки Г отцепляется
от рецептора (время полураспада комплекса — около 13 мин),
выходит из клетки в неизменном виде.
На мембранах органов-мишеней имеются специальные
рецепторы, благодаря которым осуществляется транспорт
гормонов в клетку. Клетки печени имеют особенно много та-
ких рецепторов, поэтому глюкокортикоиды в них интенсивно
накапливаются и метаболизируются. Время полужизни боль-
шинства гормонов составляет 20-200 мин.
Основные функции глюкокортикоидов — синтез фермента-
тивных белков, в частности белков митохондрий, а так же мо-
билизация (т.е. расщепление) структурных белков мышц и лим-
фоидной ткани в условиях длительного и тяжелого стресса.
Гипофизарно-щитовидная система имеет гуморальные и
нервные взаимосвязи. Предполагается ее синхронное функ-
ционирование с гипофизарно-адренокортикальной системой.
Гормоны щитовидной железы (тироксин, трийодтиронин, ти-
ротропонин) положительно сказываются на процессах восста-
новления после выполнения физических упражнений.
Гипофизарно-половая система включает гипофиз, кору над-
почечников, половые железы. Взаимосвязь между ними осу-
ществляется нервным и гуморальным путем. Мужские поло-
вые гормоны — андрогены (стероидные гормоны), женские
- эстрогены. У мужчин биосинтез андрогенов осуществляется
в основном в клетках Лейдига (интерстициальных) семенни-
ков (главным образом тестостерон). В женском организме сте-
роиды образуются в надпочечниках и яичниках, а также коже.
Суточная продукция у мужчин составляет 4-7 мг, у женщин — в
10-30 раз меньше. Органы-мишени андрогенов — предстатель-
ная железа, семенные пузырьки, семенники, придатки, ске-
летные мышцы, миокард и др. Этапы действия тестостерона
на клетки органов-мишеней следующие:
— тестостерон превращается в более активное соединение
5-альфа-дегидротестостерон;
— образуется комплекс Г-Р;
— комплекс активизируется в форму, проникающую в ядро;
— происходит взаимодействие с акцепторными участками
хроматина ядра (ДНК);
49
— усиливается матричная активность ДНК и синтез раз-
личных видов РНК;
— активизируется биогенез рибо- и полисом и синтез бел-
ков, в том числе андрогенозависимых ферментов;
— увеличивается синтез ДНК и активизируется клеточное
деление.
Важно заметить, что для тестостерона участие в синтезе
белка необратимо, гормон полностью метаболизируется.
Основная функция гормонов гипофизарно-половой систе-
мы — синтез структурных белков, в частности белков мышц.
Гормоны, попадающие в кровь, подвергаются катаболизму
(элиминации, разрушению) преимущественно в печени, при-
чем некоторые гормоны при росте мощности интенсивность
метаболизма, в частности глюкокортикоидов, возрастает.
Основой повышения тренированности эндокринной си-
стемы являются структурные приспособительные перестрой-
ки в железах. Известно, что тренировка приводит к росту мас-
сы надпочечников, гипофиза, щитовидной железы, половых
желез (через 125 дней детренировки все возвращается к норме,
Виру А.А., 1977). Отмечено, что увеличение массы надпочеч-
ников сочетается с повышением содержания ДНК, т.е. интен-
сифицируется митоз — растет количество клеток. Изменение
массы железы связано с двумя процессами — синтеза и дегра-
дации. Синтез железы прямо пропорционально зависит от ее
массы и обратно пропорционально — от концентрации гор-
монов в железе. Скорость деградации увеличивается с ростом
массы железы и механической мощности, уменьшается — с по-
вышением концентрации анаболических гормонов в крови.
2.9. Иммунная система
Человек имеет механизмы надзора — иммунную систему. Эта
система защищает его от болезнетворных (патогенных) микро-
организмов (бактерий и вирусов) и от раковых клеток, распозна-
ет и избирательно уничтожает вторгшиеся в организм человека
чужеродные агенты. Различают клеточный и гуморальный вид
ответа. Клеточный иммунный ответ особенно эффективен про-
50
тив грибов, паразитов, внутриклеточных вирусных инфекций,
раковых клеток и чужеродных тканей. Гуморальный иммунный
ответ проявляется преимущественно в период внеклеточной
фазы бактериальных и вирусных инфекций.
Иммунная система — совокупность всех лимфоидных орга-
нов и скоплений лимфоидных клеток: вилочковая железа, се-
лезенка, лимфатические узлы, пейперовы бляшки, стволовые
клетки костного мозга.
Взаимодействие организма с чужеродными размножающи-
мися антигенами академик Р.В. Петров (1987) представляет
четырьмя процессами:
1. Размножение проникших чужеродных клеток. Изме-
нение числа антигенов в организме зависит от темпа их
размножения за данный отрезок времени минус то их
число, которое нейтрализуется за это время существо-
вавшими ранее или появившимися антителами.
2. Иммунная система организма реагирует на антигенное
вторжение накоплением иммунокомпетентных клеток
(антителообразующих). Запускающим реакции субстра-
том является комплекс антигена с рецептором распозна-
ющего Т-лимфоцита. Количество плазмоклеток зависит
от числа активированных В-лимфоцитов и от темпа их
пролиферации минус их убыль за счет старения.
3. Количество антител в данном отрезке времени зависит
от скорости их производства минус то количество, ко-
торое связывается антигеном, и то количество, которое
выводится за счет естественного их катаболизма.
4. Функционирование иммунной системы организма за-
висит от нормальной работы других систем и органов.
Вирус, естественно, поражает какую-то систему (или
орган) не обязательно непосредственно лимфоидную.
Это может быть печень, легкие, железы внутренней се-
креции и др. В любом случае поражение может достигать
такой глубины, которая отразится на обеспечении рабо-
ты иммунной системы.
Простейшая модель иммунологической реакции организма
на вирус является одновременно простейшей моделью инфек-
ционного заболевания. Самый придирчивый критик не смо-
51
жет найти, как пишет Р.В. Петров (1987), здесь неучтенного
процесса, если иметь в виду базовые процессы.
Простейшая математическая модель иммунной системы
была разработана академиком Г.И. Марчуком (1985). Она по-
зволяет имитировать основные закономерности протекания
защитной реакции организма, в модели не различаются кле-
точные и гуморальные компоненты иммунитета. Предполага-
ется, что такие компоненты имеются.
Модель включает элементы: пул антигенов, пул антител,
пул плазмоклеток, орган-мишень.
Имитационное моделирование реакции иммунной системы
введением разного исходного уровня антигенов показало, что
модель демонстрирует хроническую, субклиническую, острую
и летальную форму болезни.
Хроническая форма болезни имеет место в том случае, ког-
да в организм постоянно поступает в небольших дозах инфек-
ция. В этом случае устанавливается динамическое равновесие
между синтезом патогенных микробов и их элиминацией, бла-
годаря адекватному производству антител. Субклиническая,
острая или летальная формы заболевания могут быть вызва-
ны двумя способами: однократным введением возрастающей
дозы антигенов, уменьшением массы органа-мишени.
Кроветворная стволовая клетка костного мозга являет-
ся предшественником различных форм иммунологического
реагирования (Т- и В-систем). По мере старения количество
стволовых клеток уменьшается. В возрасте 65-76 лет иммун-
ная активность антител составляет 20-30% от максимального
уровня (10 лет).
2.10. Пищеварение
К органам пищеварения относятся: полость рта, глотка, пи-
щевод, желудок, тонкая и толстая кишка.
Пищеварение — физиологический процесс, благодаря кото-
рому пища, поступившая в пищеварительный тракт, подверга-
ется физическим и химическим изменениям, а образующиеся
питательные вещества всасываются в кровь и лимфу.
52
Физические изменения пищи связаны с ее механической
обработкой, перемешиванием, растиранием. Химические из-
менения состоят из последовательных этапов гидролитиче-
ского расщепления пищи с помощью ферментов и соляной
кислоты желудка.
В полости рта происходит размельчение, смачивание слю-
ной и формирование пищевого комка. Вкусовые рецепторы
рта возбуждают определенные отделы ЦНС, в результате реф-
лекторно активизируется секреция слюнных, желудочных и
поджелудочных желез, осуществляется двигательный акт гло-
тания и продвижения пищи по пищеводу.
В слюне содержатся ферменты (птиалин, мальтоза) гидро-
литического расщепления углеводов. В желудке действие фер-
ментов слюны прекращается (кислая среда).
В желудке пища находится в течение нескольких часов и по-
степенно переходит в кишечник. Желудочный сок выделяется
железами и содержит соляную кислоту (pH — 0,9-1,5), протеа-
зы - пепсины, желатиназы, химозины (расщепляют белки),
липазы (расщепляют эмульгированные жиры). На мясо выде-
ляется больше соляной кислоты, на хлеб больше выделяется
ферментов, жиры вызывают угнетение желез желудка в тече-
ние нескольких часов, затем наблюдается возбуждение сим-
патической нервной системы. Возбуждение симпатической
нервной системы и появление в крови адреналина оказывает
тормозящее влияние на секрецию желудочных желез. Быстро-
та перехода пищи из желудка в кишку зависит от объема, со-
става и консистенции пищи. Пища находится в желудке 6-8
часов. Углеводистая пища эвакуируется быстрее, чем белко-
вая; жирная пища задерживается на 8-10 часов. Жидкости на-
чинают проходить в кишку почти тотчас после их поступления
в желудок. Содержимое желудка уходит в двенадцатиперстную
кишку, когда его консистенция становится жидкой или полу-
жидкой.
В двенадцатиперстной кишке пища подвергается действию
поджелудочного сока, желчи, выделений бруннеровых и ли-
беркюновых желез. В отсутствии пищеварения среда кишки
имеет слабощелочную реакцию (pH — 7,2-8,0), это связано с
наличием бикарбонатов.
53
Поджелудочный сок богат ферментами, действующими на
белки (трипсин, химотрипсин и др.), углеводы (амилаза, маль-
таза, лактаза и др.), жиры (липаза) и нуклеиновые кислоты
(нуклеазы). Секреция поджелудочного сока начинается через
2-3 мин после приема пищи и продолжается 6-14 часов.
Желчь является продуктом секреторной работы печеноч-
ных клеток. Она активизирует деятельность фермента — ли-
пазы.
А.М. У гол ев (1978) установил, что пористая поверхность
тонкой кишки, адсорбируя ферменты, способствует усилению
энзиматических процессов. Пристеночное пищеварение со-
провождается процессом всасывания элементарных единиц
пищи (мембранное пищеварение).
В толстых кишках находится богатая бактерийная флора,
вызывающая сбраживание углеводов и гниение белков. В ре-
зультате микробного брожения происходит расщепление рас-
тительной клетчатки, освобождение содержимого раститель-
ных клеток и их усвоение с помощью кишечного сока. Кроме
этого, бактерии толстого кишечника являются источником
аминокислот для организма. В толстых кишках происходит
сгущение поступающего содержимого (вода всасывается в
толстом кишечнике), образуется кал.
Всасывание представляет собой сложный физиологический
процесс прохождения веществ через эпителиальную мембрану
кишечной стенки (тонкой или толстой кишок) и поступления
их в кровь или лимфу.
Углеводы активно (с затратой энергии АТФ) всасываются в
кровь в основном в виде глюкозы и галактозы.
Всасывание аминокислот происходит главным образом в
тонком кишечнике и является активным, требующим энергии
АТФ, процессом. Далее они попадают в портальную систему,
следовательно, в печень. Аминокислоты быстро (5 мин) попа-
дают из крови в печень и во все другие органы.
После приема жирной пищи тонкий кишечник содержит
анионы жирных кислот и смесь моно-, ди- и триглицеридов,
хорошо эмульгированных солями желчных кислот и мылами.
Основная часть этой смеси всасывается через стенку тонкого
кишечника. Глицерин водорастворим и вместе с жирными
54
кислотами с короткой цепью уходит в кровь. Жирные кисло-
ты с длинной цепью проникают в лимфатическую систему, где
они обнаруживаются в виде триацилглицеринов в составе хи-
ломикронов (липопротеидов).
2.11. Жировая ткань
Жировая ткань является самостоятельным в отношении ги-
стоэмбриогенеза образованием. Она выполняет три основные
функции:
1. синтез триглицеридов из сывороточных липидов и глю-
козы;
2. сохранение их в жировых депо;
3. освобождение их из жировых депо (липолиз).
Жировая клетка — адипоцит — может увеличиваться в раз-
мере по мере накопления липидов, протоплазма клетки от-
жимается на периферию вместе с ядром, которое постепенно
начинает уплощаться. Механическая деформация ядра ади-
поцита, видимо, мешает ходу обмена веществ, поэтому пере-
полненные жировые клетки плохо метаболизируют глюкозу.
В межклеточном пространстве располагаются кровеносные
капилляры, подходящие к каждой жировой клетке. Здесь же
проходят ретикулярные волокна, выполняющие опорную ме-
ханическую роль. Нервные волокна, иннервирующие жиро-
вые клетки, в основном принадлежат симпатической нервной
системе. Нервные стволы поступают в жировую ткань вместе
с сосудами, далее они постепенно разводокняются, и нервные
волокна охватывают каждую жировую клетку (А.Н. Климов,
Н.Г. Никульчева, 1999).
В жировой ткани происходят как процессы превращения
углеводов в жиры, так и переход жиров в углеводы.
Биосинтез жирных кислот происходит в основном в ци-
топлазме адипоцитов. Сырьем для биосинтеза является
ацетилкоэнзим-А, который образуется из избыточной глюко-
зы или аминокислот.
Липолиз усиливается под действием катехоламинов и глю-
кагона, которые захватываются клетками активизированной
55
жировой ткани. При стрессорных ситуациях увеличивается
скорость высвобождения жирных кислот и глицерина из жи-
ровой ткани.
Жировая ткань может разрастаться как в результате гипер-
трофии, так и гиперплазии адипоцитов.
56
Глава 3
Контроль локальной
выносливости
Физическая подготовка спортсмена направлена на измене-
ние строения клеточных структур в тканях различных органов
под влиянием тренировочного процесса. Физическая подго-
товленность, в частности локальная мышечная выносливость,
требует регулярного контроля для планирования и коррекции
тренировочных нагрузок.
3.1. Мощность, эффективность и емкость
механизмов энергообеспечения как критерии
оценки подготовленности спортсменов
Энергетический обмен в организме человека связан с про-
цессами анаболизма, катаболизма и функциональным мета-
болизмом. Количественно энергетический обмен измеряют в
единицах работы (ккал) и мощности (ккал/час). Используют-
ся также кгм и кгм/мин. Однако в настоящее время принято
пользоваться международной системой единиц (СИ). Здесь
работа измеряется в джоулях (Дж), а мощность в ваттах (Вт) (1
ккал = 4187 Дж, 1 кДж = 0,28 Вт =0,239 ккал/час).
Функциональный метаболизм спортсмена связан с выпол-
нением механической работы и затратами метаболической
энергии. Поэтому при делении внешней механической мощ-
ности на метаболические затраты получается оценка коэффи-
циента полезного действия. При педалировании на велоэрго-
метре коэффициент полезного действия составляет 22-24%, а
при вращении рукоятки — 20- 21%.
Энергообеспечение зависит от мощности (интенсивности)
57
выполняемой работы. Максимальная мощность связана с за-
тратами энергии молекул АТФ и КрФ, и длительность этой
работы не превышает 15-30 с. Если заданная мощность мо-
жет поддерживаться 30-60 с, то говорят о преимущественной
доле анаэробного гликолиза в энергообеспечении мышечной
деятельности. Когда работа продолжается без снижения мощ-
ности более 1 мин, то говорят о преимущественном вкладе в
энергообеспечение аэробного гликолиза или окисления жи-
ров. В связи с этим Н.И. Волков (1990) предложил каждый
механизм энергообеспечения характеризовать мощностью,
эффективностью и емкостью.
Предложенный еще в 1955 г. P-О. Астрандом способ оценки
работоспособности спортсменов и представленный в России
Н.И. Волковым (1969) и И.В. Ауликом(1990) явно устарел, по-
скольку модель, которой они пользовались, была очень про-
стой. Старая модель не учитывает современных достижений
физиологии человека, в частности: строения мышц, правила
рекрутирования мышечных волокон и многого другого.
Устаревший вариант интерпретации метаболических про-
цессов в организме человека представляется следующим обра-
зом. Алактатный механизм оценивается максимальной алак-
татной мощностью (мощность спринта длительностью 3-5 с),
эффективность — коэффициентом полезного действия (КПД),
емкость — запасами АТФ и КрФ. Здесь следует заметить, что
эффективность алактатного механизма энергообеспечения за-
висит от активности работы ферментов — миозиновой АТФ-
азы и КрФ-азы, деятельность которых зависит от температуры,
степени закисления мышечного волокна. КПД зависит также
от техники (Селуянов В.Н., Савельев И.А., 1982), например,
при педалировании с темпом более 150 об/мин у велосипеди-
стов КПД может доходить до 37%, а у спортсменов, которые
подпрыгивают на седле, КПД может снизиться до 10% (поч-
ти вся энергия будет тратиться на подъем туловища). В связи
с этим точно оценить эффективность алактатного механизма
невозможно. Емкость алактатного механизма, как правило,
также оценить невозможно, поскольку все спортсмены до-
стигают максимума мощности к 3-5 с, а затем мощность не-
изменно снижается. Методом биопсии было установлено (см.
58
обзоры Hoppeler Н., 1986; Karlsson J., 1971, 1981, 1982 ), что у
всех людей и спортсменов концентрации АТФ и КрФ пример-
но одинаковые, и только временно можно увеличить запасы
КрФ в мышечных волокнах на 10-30% с помощью приема за
30-40 мин до начала тестирования пищевой добавки — Креа-
гинфосфат моногидрат. Через несколько часов концентрация
КрФ в мышцах нормализуется (Rossiter Н. et al. 1996).
Мощность механизма анаэробного гликолиза предложе-
но оценивать с помощью упражнения, в котором предельная
продолжительность равна 30-60 с. Например, Вингейтский
тест, длительность которого 30 с. В этом случае также можно
дать иную интерпретацию, поскольку в 70-е годы не могли
корректно оценивать вклад анаэробного гликолиза в метабо-
лические затраты испытуемого при выполнении работы с око-
ломаксимальной мощностью. Емкость анаэробного гликолиза
оценивалась по величине кислорода, который был потреблен
после выполнения требуемого тестового задания. Поскольку
потребление кислорода приходило в норму после часа вос-
становления, то все избыточное потребление кислорода отно-
сят к алактатному и анаэробному гликолитическому долгу. В
этом случае лактацидный долг оценивался в величину 16-20 л
запроса кислорода. Эти оценки противоречат величинам кис-
лородного запроса. Например, МАМ = 900 Вт, а мощность в
Вингейтском тесте составляет 80% от МАМ или 750 Вт. Если
КПД=23%, то 75 Вт соответствует 1 л/мин потребления кисло-
рода. Следовательно, за 30 с человек должен был потребить 5 л
кислорода — это кислородный запрос, он значительно меньше
величины потребления кислорода во время восстановления.
Этот факт был обнаружен итальянским ученым Р. Маргариа
еще в 70-е годы. Именно он стал утверждать, что емкость ана-
эробного механизма не может превышать более 4-5 л кисло-
родного эквивалента. В представленном случае кислородный
запрос обеспечивается энергией молекул АТФ и КрФ на 2 л,
потреблением кислорода за время работы 1,8 л, тогда на анаэ-
робный гликолиз остается только 1,2 л. Заметим, что в случае
наличия 100% окислительных мышечных волокон в активных
мышцах анаэробного гликолиза вообще может не наблюдать-
ся. Следовательно, упражнения с предельной продолжитель-
59
ностью 30-60 с позволяют оценить скорее уровень аэробной
подготовленности мышц, поскольку в случае повышения аэ-
робных возможностей мышц они меньше закисляются, при
прочих равных условиях происходит рост средней мощности в
данном задании, за счет поддержания мощности до конца за-
дания (30 или 60 с).
Аэробные возможности оценивают по мощности или ве-
личине максимального потребления кислорода. Этот показа-
тель с 80-х годов подвергается серьезной критике, поскольку
на выборке спортсменов высокой квалификации практически
теряет информативность. Потребление кислорода, мощность
на уровне анаэробного порога являются более надежными и
информативными показателями, поскольку позволяют с вы-
сокой точностью предсказывать спортивные достижения в
циклических видах спорта. Эффективность аэробного меха-
низма или КПД при работе на велоэргометре равен 23-24% и
не меняется, поэтому определение этого показателя такая же
бессмыслица, как и во всех других случаях. Емкость аэробного
механизма связана с запасами в мышцах гликогена и капелек
жира. Запаса этих веществ у обычных людей хватает на 45-
60 мин, а у спортсменов запасов может хватить на 1,5-3 часа
(Физиология мышечной деятельности, 1982). Причем при ре-
гулярном приеме углеводов по ходу выполнения упражнения
продолжительность упражнения многократно возрастает, как,
например, у лыжников или велосипедистов (Алиханова Л.И.,
1983). Следовательно, в спорте определение емкости не име-
ет никакого смысла с точки зрения успешности выступления
спортсмена в соревнованиях, длительность которого не пре-
вышает 30 мин.
Таким образом, определение у спортсмена мощности, эф-
фективности и емкости биоэнергетических механизмов по
методике Н.И. Волкова не учитывает физиологические осо-
бенности реакции организма на выполняемую физическую
работу. Например, невозможность перехода молекул АТФ или
КрФ из одного мышечного волокна в другое, или из одной
мышцы в другую. Поэтому при выполнении упражнений с ис-
пользованием локальных мышечных групп, например руками,
оценки мощности, эффективности и емкости будут иными.
60
Однако проблема локальной работоспособности у биоэнерге-
тиков пока не нашла интереса.
3.2. Критический анализ интерпретации
данных лабораторного тестирования
Тест 1. Проводится педалирование на велоэргометре с це-
лью достижения максимальной работы за 30 с. Далее, в ка-
честве основы для интерпретации, взяты классические пред-
ставления из учебников по физиологии. Там пишется, что
работа предельной интенсивности в 30 с обеспечивается на
90% анаэробными источниками энергообеспечения и пре-
жде всего анаэробном гликолизом. Мощность 30 с предель-
ной работы составляет около 60-80% МАМ. Проверим эти
представления на основе современной модели, имитирую-
щей срочные адаптационные процессы. Предположим, что
мощность составила 500 Вт, это с учетом к.п.д. работы на
велоэргометре 23% будет соответствовать 6,8 л/мин, а за 30
с - 3,4 л кислорода. Известно (Волков Н.И., 1969, 1990), что
на долю алактатного долга приходится около 2,5 л, тогда 0,9
л долга должно распределиться между аэробным гликолизом
и анаэробным гликолизом. Если в мышцах 50% окислитель-
ных мышечных волокон и 50% гликолитических, то аэроб-
ные процессы должны обеспечить 0,45 л кислородного за-
проса и 0,45 л на анаэробный гликолиз. Следовательно, доля
анаэробного гликолиза составит не более чем 13%, а этот тест
характеризует мощность анаэробного алактатного механизма
энергообеспечения, а также аэробного, поскольку чем мень-
ше закисляются мышцы, тем легче поддерживают мощность
и силу сокращения. Известно, что ионы водорода могут при-
соединяться к активным центрам актина, в этом случае они
мешают работе ионов кальция и поперечные мостики между
актином и миозином не образуются, сила тяги начинает па-
дать. Следовательно, чем меньше образуется ионов водорода,
тем легче поддерживать силу сокращения мышц. Такая си-
туация складывается в случае, когда в ходе тренировочного
процесса происходит трансформация гликолитических мы-
61
щечных волокон в окислительные за счет увеличения массы
митохондрий.
Таким образом, когда в публикациях встречается мнение,
что 30 с предельная работа характеризует гликолитическую
анаэробную мощность, следует читать — алактатная и аэробная
гликолитическая мощность активных в упражнении мышц, в
данном случае — ног.
Тест 2. В тесте с выполнением трех одноминутных вело-
эргометрических предельных упражнений с одноминутными
интервалами отдыха суммарную работу решили обозвать по-
казателем анаэробной гликолитической емкости. Очевидно,
что рост этого показателя может происходить только в случае
увеличения алактатной мощности (МАМ) и аэробных воз-
можностей мышц (потребление кислорода на уровне АнП),
поскольку рост мощности анаэробного гликолитического ис-
точника энергообеспечения приводит к увеличению закисле-
ния мышц, а значит к ускорению наступления утомления.
Таким образом, когда пишут о гликолитической анаэроб-
ной емкости надо понимать, что речь идет об алактатной и аэ-
робной мощности энергообеспечения активных в упражнении
мышц, в данном случае — ног.
Тест 3. В случае определения закисления крови после трех
одноминутных велоэргометрических тестов можно определить
показатель как частное отделения суммарной работы за 3 мин на
изменение закисления крови (DpH). Этот показатель обозвали
анаэробной гликолитической эффективностью, однако, смысл
пересчета противоположный. Поскольку увеличение степени за-
кисления приводит к снижению показателя, и наоборот, увеличе-
ние аэробных возможностей — не только к увеличению работоспо-
собности (числителя) и к меньшей степени закисления (DpH).
Таким образом, все три теста характеризуют одно и то же, а
именно уровень развития МАМ и аэробной мощности мышц
ног, другими словами, характеризуют локальную мышечную
работоспособность (выносливость).
Тест 4. С помощью ступенчатого теста и газоанализатора мож-
но определить величину максимального потребления кислорода
или, по данным PWC170+30%, определить мощность, близкую
к мощности МП К. Этот показатель было принято определять
62
как аэробная мощность. Очевидно, что это также ошибка интер-
претации, поскольку величина потребления кислорода на мощ-
ности МПК складывается из нескольких составляющих, а имен-
но: из потребления кислорода активными в данном упражнении
скелетными мышцами, миокардом и дыхательными мышцами,
остальные ткани и органы потребляют кислорода пренебрежимо
мало. В связи с этим информативность показателя МПК исклю-
чительно низкая и не может на относительно однородной выбор-
ке характеризовать аэробную подготовленность спортсменов.
Уже более 15 лет как стало известно, что наиболее информатив-
ным показателем аэробных возможностей спортсменов является
величина потребления кислорода, или скорости, или мощности
на уровне анаэробного порога.
Тест 5. Аэробная емкость определяется как продолжитель-
ность выполнения упражнения критической мощности или
МПК. Очевидно, что это упражнение будет выполнять дольше
гот спортсмен, у которого потребление кислорода на уровне АнП
будет ближе к МПК. Например, стайеры имеют мощность АнП
на уровне 70-90% от мощности МПК. Следовательно, этот пока-
затель характеризует относительную величину мощности АнП.
Тест 6. В ступенчатом тесте можно определить зависимость
между потреблением кислорода и выполняемой мощностью.
Этот показатель называют аэробной эффективностью, однако,
никакую эффективность этот показатель не может продемон-
стрировать, поскольку это отношение есть КПД работы на ве-
лоэргометре. КПД работы на велоэргометре составляет 22-24%
и не зависит ни от спортивной специализации, ни от спортив-
ной квалификации. Поэтому показатель аэробной эффектив-
ности у некоторых «специалистов» меняется только от массы
спортсмена, поскольку КПД делится на вес спортсмена.
Таким образом, все тесты, которые нашли применение в
научных исследованиях и практике получили некорректную
интерпретацию. Главный результат нашей интерпретации
один — все тесты характеризуют или МАМ, или окислительно-
го фосфорилирования мышц ног (потребление кислорода на
уровне АнП), или в каком-то соотношении оба этих механиз-
ма энергообеспечения, другими словами, локальную работо-
способность мышц ног (локальную выносливость).
63
3.3. Новые подходы для оценки физической
подготовленности спортсменов
Лабораторное тестирование
Для оценки работоспособности спортсмена необходимо
выполнить оценку степени влияния центрального фактора
(работоспособность сердечно-сосудистой системы) и перифе-
рического (сила, мощность, потребление кислорода отдель-
ных мышечных групп).
Мышечный аппарат. Физические свойства мышц определя-
ются количеством мышечных волокон, а в каждом мышечном
волокне — количеством миофибрилл, АТФ-азной активно-
стью миозина, массой митохондриальной системы, запасами
гликогена и жира. Запас молекул АТФ и КрФ обусловлен сте-
пенью гипертрофии мышечного волокна.
Метод определения количества миофибрилл. Для опреде-
ления количества миофибрилл в данной мышечной группе
обычно измеряют их максимальную изометрическую силу
или максимальный вес, который они могут преодолеть. Более
удобно оценивать состояние мышц на приборе, который ис-
пользуется и для оценки других характеристик состояния ат-
лета. Таким прибором является велоэргометр.
Максимальная алактатная мощность (МАМ) определяется
на велоэргометре по величине установленного сопротивления
(F, н) и максимальному темпу (R, 1/с), который достигается в
ходе спурта.
МАМ = F х S х R (Вт).
S — расстояние, которое проходит точка на окружности махо-
вика велоэргометра (у Монарха это расстояние составляет 6 м).
Обычно максимальный темп наблюдается на 4-7 с спурта.
Нагрузка подбирается такой, чтобы максимальный темп был
около 120-140 об/мин (0,45 — 0,50 1/с).
Вращение педалей можно выполнять ногами (сидя в седле)
или руками. В первом случае дается оценка по терминологии
ТиМФВ, скоростно-силовым возможностям мышц ног, во вто-
ром — руки туловища. На самом деле тесты следует использовать
для косвенной оценки степени морфологических перестроек в
64
системах и органах тела спортсмена. В данном случае абсолют-
ная величина МАМ определяется количеством миофибрилл и
АТФ-азной активностью активных мышечных волокон. Если
тестирование выполняется повторно, то изменения будут свя-
заны только с ростом количества миофибрилл, поскольку АТФ-
азная активность миозина - наследуемый фактор.
Таким образом, контроль МАМ позволяет косвенно оце-
нивать уровень силовой подготовленности мышц атлета или
количество миофибрилл в активных, в данном тесте, мышцах,
иначе говоря, локальную мышечную мощность.
Метод определения митохондриальной массы. Митохондрии
поглощают кислород, ионы водорода, АДФ, Ф, пируват или
жирные кислоты, выделяют углекислый газ, воду, АТФ. Если
масса митохондрий и фермента лактатдегидрогеназы сердеч-
ного типа (ЛДГ-С) преобладают в мышечном волокне, то та-
кое волокно называют окислительным. В других мышечных
волокнах преобладают ферменты гликолиза и лактатдегидро-
геназы мышечного типа (ЛДГ-М), поэтому при их активации
разворачивается анаэробный гликолиз, а мышечные волокна
классифицируются как гликолитические.
Для определения мощности (массы) митохондриальной
системы активных мышц применяется ступенчатый тест. При
педалировании на велоэргометре с заданным темпом каждые
2-4 мин увеличивают сопротивление. Увеличение сопротив-
ления в заданных условиях означает рекрутирование мышеч-
ных волокон от окислительных к гликолитическим (точнее,
двигательных единиц, иннервирующих мышечные волокна
различного типа). После рекрутирования всех окислитель-
ных МВ начинают функционировать гликолитические МВ, в
крови начинают накапливаться продукты анаэробного глико-
лиза — Н+ и СО2, которые стимулируют деятельность сердца
и органов дыхания, а также лактат. Этот момент определяется
как аэробный порог и по силе сопротивления можно судить о
силовых возможностях окислительных МВ, которые прояв-
ляются в аэробном режиме энергообеспечения. Дальнейшее
увеличение сопротивления (мощности) приводит к моменту
нарушения динамического равновесия, когда количество про-
дуцируемого лактата активными ГМВ становится больше его
65
потребления в ОМВ. Этот момент определяется как анаэроб-
ный порог, он характеризует максимальную мощность мито-
хондриальной системы.
Таким образом, по величине потребления кислорода или
мощности на АнП можно судить о митохондриальной массе
активных мышц, а по сопротивлению на уровне АэП — о силе
окислительных мышечных волокон. При контроле состояния
бегунов и велосипедистов необходимо выполнять ступенча-
тый тест для мышц ног, а для пловцов - мышц пояса верхних
конечностей.
Ударный объем сердца. Сердце выполняет функцию пере-
качивания крови. Его производительность зависит от ударно-
го объема и частоты сокращений. Максимальный минутный
объем сердца наблюдается при работе ногами и достижении
180-190 уд/мин, а ударный объем — при ЧСС 120-150 уд/мин
(Физиология мышечной деятельности, 1982). В ходе трени-
ровочного процесса возможно управление только величиной
ударного объема сердца (УОС), поэтому необходимо регуляр-
но контролировать эту характеристику.
Для выполнения оценки УОС сначала вычисляют мощ-
ность (педалирования на велоэргометре или скорость бега на
тредмилле), соответствующую ЧСС 170 уд/мин по следующей
формуле:
МИО = Ml + (М2-М1) х (170 — ЧСС1)х(ЧСС2-ЧСС1),
где Ml — мощность первой нагрузки;
М2 — мощность второй нагрузки;
ЧСС1 - частота сердечных сокращений на первой нагрузке;
ЧСС2 - частота сердечных сокращений на второй нагрузке.
Коэффициент полезного действия (КПД) при педалирова-
нии с темпом 60-90 об/мин составляет 19-24%, в среднем 23%.
Это дает основание к вычислению потребления кислорода по
мощности, демонстрируемой на велоэргометре:
ПК=М170/ к,
где к = 78 Вт/лО2.
Знание величины потребления кислорода (Аулик И. В., 1990)
позволяет воспользоваться формулой B.Bevegard (1960,1963)
для оценки минутного объема сердца:
МОС = 5,9 х ПК + 4,36 (л/мин).
66
Если потребление кислорода было определено для ЧСС 170
уд/мин, то ударный объем сердца можно вычислить:
УОС = МОС/170.
Было замечено, что ударный объем сердца при работе ру-
ками, как правило, бывает меньше значения, регистрируемого
при тестировании ног.
Состояние эндокринной системы. Смысл физической под-
готовки заключается в изменении строения клеток. Процессом
синтеза управляют в клетках гормоны, путем воздействия на
ДНК. В частности, тестостерон и соматотропин стимулируют
синтез миофибрилл в мышечных волокнах. Тренер не может
прямо контролировать концентрацию гормонов в крови, это
требует специальной аппаратуры и препаратов, однако кон-
троль за состоянием физической подготовленности (МАМ,
мощность АэП) позволяет судить о ходе адаптационных про-
цессов в железах эндокринной системы. Критерии здесь до-
статочно простые — изменения показателей состояния мышц.
Эти критерии особенно хорошо работают, когда ставится чет-
кая задача тренировочного процесса, например увеличение
силы медленных мышечных волокон, тогда должно проис-
ходить увеличение мощности на уровне АэП. Если рост вели-
чин показателей наблюдается, то очевидно, что эндокринная
система справляется с заданными нагрузками. В случае ухуд-
шения показателей и самочувствия при реализации заданной
тренировочной программы, можно сделать вывод об отсут-
ствии адекватного ответа со стороны эндокринной системы.
3.4. Определение степени влияния центрального
или периферического лимитирующего фактора
По результатам тестирования определяли производные по-
казатели, МПК, мощность на ЧСС 170 уд/мин и другие.
Пример экспериментальных данных представлен в табл. 1.
В видах спорта, в которых для выполнения соревнователь-
ной деятельности требуется участие почти всей мышечной
массы (лыжные гонки, плавание, академическая гребля, а так-
же борьба), может возникнуть ситуация, когда центральный
67
Таблица 1. Сравнительная характеристика уровня функциональной под-
готовленности борцов при выполнении тестирования ногами и руками
Показатели Ноги X Ноги S Руки X Руки S
Мощность, АеП, Вт 120 18 54 12
Относительная мощность АеП, Вт/кг 1,69 0,25 0,76 0,17
ЧСС АеП, Уд/мин 135 14 105 11
Мощность АнП, Вт 180 26 65 14
Относительная мощность АнП, Вт/кг 2,53 0,36 0,91 0,20
ЧСС АнП, Уд/мин 155 16 125 12
МАМ, Вт 685 31 584 28
МАМ/масса, Вт/кг 9,64 0,43 8,22 0,39
Мощность ЧСС 170, Вт 230 28 165 23
Относительная мощ- ность ЧСС170, Вт/кг 3,23 0,39 2,32 1,42
Мощность МПК, Вт 260 30 190 24
Относительная мощность МПК, Вт/кг 3,66 0,42 2,67 0,33
фактор, состояние сердечно-сосудистой системы, становится
лимитирующим. Для выявления этой ситуации были проведе-
ны исследования мышц ног и рук у борцов (рис. 3).
На рис. 3 видно, что у борца при работе руками и ногами
ЧСС изменяется на первых ступеньках одинаково, затем кри-
вая «ЧСС-мощность» при работе руками начинает ускоренно
двигаться вверх, одновременно с этим происходит увеличение
скорости легочной вентиляции. По этим отклонениям был об-
наружен аэробный порог для рук. Частота сердечных сокраще-
ний у данного испытуемого изменялась линейно с ростом мощ-
ности упражнения до уровня аэробного порога, а затем ЧСС
стала нарастать более быстро. Легочная вентиляция изменялась
криволинейно с ростом мощности. По первому излому на кри-
вой «легочная вентиляция — мощность» фиксировался аэроб-
68
О 50 100 150 200 250
Рис. 3. Пример изменения ЧСС и легочной вентиляции при выполнении
ступенчатого теста руками и ногами
ный порог, по второму отклонению фиксировался анаэробный
порог. Анаэробный порог определялся также по моменту пере-
сечения кривой «легочная вентиляция — мощность» линии,
изображенной на рис. 3, как прямая, параллельная исходно-
му направлению кривой «легочная вентиляция — мощность».
Очевидно, что первый отрезок кривой «легочная вентиляция —
мощность» характеризует интенсивность дыхания при окисле-
нии преимущественно внутримышечных жиров, при переходе
на окисление углеводов интенсивность дыхания растет, что и
вызывает ускорение легочной вентиляции. Третий участок кри-
вой связан с еще большим ускорением дыхания, что вызывается
началом анаэробного гликолиза в рекрутированных гликолити-
ческих МВ, появлением в крови ионов водорода, освобождени-
ем из крови углекислого газа (эксцесс СО2). Избыточный угле-
кислый газ крови начинает действовать на дыхательный центр,
что и вызывает ускорение легочной вентиляции (Физиология
мышечной деятельности, 1982).
В ходе экспериментов (табл. 1) было показано, что аэроб-
ные возможности мышц пояса верхних конечностей составля-
69
ют 50-60% от аэробных возможностей мышц ног. Примерно
также соотносятся мощности, которые демонстрировали ис-
пытуемые при достижении ЧСС 170 уд/мин в случаях работы
руками и ногами.
ЧСС на уровне АэП и АнП в случае работы руками так-
же оказалась ниже при сравнении с данными, полученными
при работе ногами. Поскольку ЧСС составила 105 — 155 уд/
мин, то можно сделать заключение, что возможности сердечно-
сосудистой системы по доставке кислорода к мышцам выше
аэробных возможностей мышц, участвующих в тесте.
Суммарная мощность мышц рук и ног примерно равна
мощности, которая наблюдалась (или могла наблюдаться) при
достижении ЧСС 190 уд/мин (МПК).
Потенциальную возможность сердечно-сосудистой системы
следует определять по линии, связывающей «ЧСС и мощность».
По этой линии можно определить мощность и потребление
кислорода в момент достижения ЧСС 190 уд/мин. На рис. 3 этот
момент обозначен кружочком на пунктирной линии. Если бы у
спортсмена масса окислительных мышечных волокон была бы
большая, то связь между «ЧСС и мощностью» до ЧСС 190 уд/
мин оставалась бы линейной. В этом случае работоспособность
ССС достигла бы своего максимума. Этот показатель можно
обозвать как потенциально возможное потребление кислорода
мышцами — МПК потенциальное. Столько кислорода может
доставить ССС к мышцам без излишней стимуляции сердца
анаэробными метаболитами (Н+, эксцессом СО2).
Максимальная алактатная мощность при работе руками
была на 10-20% меньше, чем при работе ногами.
Таким образом, по результатам обследования можно сде-
лать следующие выводы:
- аэробные возможности мышц пояса верхних конеч-
ностей составляют 50-60% от аэробных способностей
мышц ног;
— мощность сердечно-сосудистой системы по доставке
кислорода к мышцам выше аэробных возможностей как
мышц пояса верхних конечностей, так и мышц ног.
70
3.5. Метод Conconi
Conconi с соав. предложил в 1982 г. непрямой и неинва-
зивный метод определения анаэробного порога посредством
измерения ЧСС в ступенчатом тесте на стадионе. Было об-
наружено, что у квалифицированных бегунов кривая связи
«ЧСС-скорость бега» имеет сигмовидную форму. Второй пе-
релом на этой кривой совпадал с моментом появления анаэ-
робного порога, т.е. при закислении крови до 4 мМ/л лактата.
В работе Р. Hofmann et al. было обследовано 227 спортсме-
нов мужчин различных видов спорта (возраст — 23, 6=4 г, рост
- 180, 6=6 см, масса — 74, 6=8 кг). Все испытуемые выполнили
ступенчатый тест с амплитудой ступеньки 20 Вт, длительно-
стью по 60 с.
В результате оказалось, что все испытуемые были разделе-
ны на три группы. В первой группе (85,9% испытуемых) ско-
рость прироста ЧСС после АнП уменьшалась, во второй груп-
пе (6,2%) скорость увеличения ЧСС не изменялась, в третьей
группе (7,9%) скорость изменения ЧСС становилась больше.
По ЧСС, мощности, концентрации лактата достоверных раз-
личий между группами не было обнаружено. Однако анализ
представленных в статье данных показал, что АнП, фиксируе-
мый по 4 мМ/л лактата, наблюдался в первой группе на уровне
80% от МПК (отказа от работы), а во второй и третьей группе на
уровне 74% МПК. Следовательно, если после наступления АнП
в мышцах не остается мышечных волокон для рекрутирования,
то мышцы и кровь мало закисляются, нет стимула для ускоре-
ния ЧСС. В том случае, когда в мышце остается еще много не-
рекрутированных гликолитических МВ, то закисление растет,
появляется эксцесс СО2, имеется стимул для роста ЧСС.
Таким образом, эффект Conconi не является обязательным,
а изменение ЧСС связано с наличием рекрутирования глико-
литических мышечных волокон, чем больше их рекрутирует-
ся, тем больше закисление мышц и крови, тем больше нарас-
тает скорость изменения ЧСС.
71
3.6. Понятие - локальная мышечная
работоспособность
Анализ кривых индивидуальных или мировых рекордов
показывает, что во всех соревновательных упражнениях, вы-
полняемых до 1 мин 30 с, т.е. до достижения максимальной ча-
стоты сердечных сокращений, спортивные достижения опре-
деляются работоспособностью активных мышц. Центральное
звено — производительность ССС не имеет принципиального
значения. При выполнении соревновательных упражнений
большей длительности центральное звено может иметь значе-
ние только в случае, когда порог анаэробного порога начинает
приближаться к МПК потенциальному. На уровне высшего
спортивного мастерства такая ситуация встречается часто. В
этом случае спортсмену надо обеспечить делятацию левого
желудочка сердца и после этого лимитирующим звеном будет
уже не «центральный», а «периферический» фактор.
Таким образом, локальная мышечная работоспособность
(локальная выносливость) — это способность человека выпол-
нять предельную мышечную работу при адекватном (избыточ-
ном) снабжении ее кислородом или когда величина потребления
кислорода не имеет существенного значения для обеспечения за-
данной двигательной активности.
Практически во всех случаях лимитирующим звеном в по-
вышении спортивных достижений является локальная мышеч-
ная работоспособность, однако, проблема ее развития остается
вне внимания исследователей. Больше рассуждают об общей
работоспособности, общей алакгатной, гликолитической и
аэробной мощности. Причем все рассуждения строятся в луч-
шем случае на основе простейшей модели организма челове-
ка, которая включает в себя пул молекул АТФ и три-четыре
механизма для их ресинтеза: креатинфосфатный, анаэробный
гликолитический (лакгатный), аэробный гликолитический и
окисления жиров. В такой модели нет конкретных мышц, нет
мышечных волокон, «опущена» физиология с ее законами.
72
Глава 4
Локальная выносливость
как компонент физической
подготовленности спортсменов
в циклических видах спорта
В теории и методике физического воспитания и спорта раз-
личают общую региональную и локальную мышечную вынос-
ливость. Классификация этих видов выносливости выполнена
по величине массы мышц, участвующих в работе. Причем ве-
личину массы мышц никто определять не умеет. Поэтому эту
классификацию нельзя принять и тем более практически ис-
пользовать.
Бесспорно, с локальной выносливостью (ЛВ) можно свя-
зать явления, характеризующие производительность нервно-
мышечного аппарата [Алферова Т.В., 1990; Верхошанский
Ю.В., 1982, 1983; Платонов В.Н., Булатова М.М.,1992; Суслов
Ф.П., Гилязова В.Б., 1990] при физической работе статиче-
ского или динамического характера, когда активно так мало
мышц, что ЧСС практически не изменяется.
Применительно к циклическим локомоциям (при работе с
участием большой мышечной массы) это понятие стали при-
менять сравнительно недавно. Наиболее подробно ЛВ как
один из основных компонентов специальной физической
подготовленности спортсменов, тренирующих выносливость,
впервые была рассмотрена в монографиях Ю.В. Верхошан-
ского, вышедших в 1985 и 1988 гг. В них обобщен материал
многочисленных исследований средств и методов целена-
правленного воздействия на нервно-мышечный аппарат с це-
лью повышения спортивной результативности в ЦВС. Из его
работ следует, что, во-первых, тренировка исполнительного
73
звена имеет большее значение для спортивного результата в
ЦВС, чем тренировка вегетативных обеспечивающих систем
организма, а во-вторых, требует существенно больше време-
ни и сил. Тем не менее можно отметить, что это утверждение
не совсем корректное, поскольку сначала надо выполнить те-
стирование и обосновать, что периферическое звено является
лимитирующим.
Проблема развития локальной выносливости должна рас-
сматриваться с двух взаимосвязанных сторон: а) развития
силовых способностей основных мышечных групп и б) раз-
вития способности к длительному поддержанию высоких или
оптимальных усилий, из чего, собственно, и складывается
спортивный результат на всех дистанциях, на которых суще-
ственное значение имеет такое физическое качество, как вы-
носливость.
В данном аспекте к методике развития локальной выносли-
вости в ЦВС можно отнести применение всех средств и мето-
дов, направленных на улучшение:
1. Силовых возможностей основных мышечных групп
спортсменов в различных вариантах их проявлений, а
именно:
— максимальной силы в статическом или динамиче-
ском режимах;
— взрывной силы и других проявлений скоростно-
силовых возможностей;
— силовой выносливости в динамических циклических
упражнениях, сходных по биомеханическим параме-
трам с соревновательной локомоцией.
2. Выносливости мышц, проявляемой в основной сорев-
новательной локомоции при различной интенсивности
работы (от спринта до умеренной мощности).
Интерес к Л В как компоненту подготовленности спортсме-
нов в ЦВС возник в связи с тем, что в последние десятилетия
стало очевидным исчерпание резервов экстенсивного пути со-
вершенствования подготовленности спортсменов за счет на-
ращивания общего объема нагрузки, что обусловлено ограни-
ченностью «валовых» резервов организма человека, связанных
главным образом с возможностью восполнения энергетиче-
74
ских и пластических ресурсов. Поэтому многие специалисты
сходятся во мнении, что путь дальнейшего повышения спор-
тивных результатов связан с поиском более эффективных, бо-
лее специфичных средств воздействия на физическое состоя-
ние спортсменов. В качестве одного из основных направлений
часто понимается совершенствование методики силовой под-
готовки спортсменов в ЦВС, так как неоднократно и во всех
без исключения ЦВС было показано, что рациональное при-
менение средств акцентированного воздействия на нервно-
мышечный аппарат может приводить к повышению спортив-
ного результата, поэтому правильный выбор средств силовой
подготовки в зависимости от направленности и величины их
тренировочного воздействия, специфики техники движений и
режима работы мышц в данном виде локомоции является ак-
туальной задачей теории и методики подготовки в ЦВС.
В то же время хорошо известно из практики и многочис-
ленных исследований, что сами по себе высокие силовые воз-
можности мышц не связаны, или даже имеют отрицательную
корреляцию, со спортивными результатами в ЦВС, особенно
на длинных дистанциях. Этот результат очевиден, поскольку
увеличение силы гликолитических мышечных волокон, кото-
рые, например, на дистанциях, длящихся более 2 минут, задей-
ствованы очень незначительно, ведет к росту баластной массы
тела. В связи с этим одной из наиболее актуальных является
проблема понимания сути развития силовых возможностей
мышц в основном соревновательном упражнении.
По мнению специалистов, решение связанных с ней задач
подразумевает:
— определение рационального соотношения объемов
средств силовой направленности с другими средствами
подготовки, в частности — аэробной;
— определение оптимального распределения средств си-
ловой направленности в рамках одного занятия, микро-,
мезо- и макроциклов и многолетней подготовки спор-
тсменов и других средств, которые должны способство-
вать реализации силовых способностей;
— опряженное решение задач технической и специальной
силовой подготовки.
75
Мы же считаем, что первоочередной является проблема
понимания физиологических, биохимических и биомехани-
ческих основ силовой подготовки в ЦВС, другими словами, —
теории этого вопроса. Так как без этого, как показывает обще-
ние с тренерами-практиками, делаются грубейшие ошибки в
организации тренировки конкретного спортсмена даже после
самого тщательного и детального объяснения плана силовой
подготовки. Кроме этого, становится практически невозмож-
ным разобраться в сути или различиях всего разнообразия су-
ществующих тренировочных средств и методов, описанных в
последующих разделах.
4.1. Средства и методы развития силовых
способностей в циклических видах спорта
В спортивной подготовке выделяют следующие методы
развития силы.
По характеру работы мышц:
— изометрический характеризуется неизменным расстояни-
ем между точками прикрепления мышцы в процессе на-
пряжения, которое может быть различной величины от-
носительно максимальной произвольной силы (МПС);
- концентрический — мышца укорачивается с различной
скоростью, зависящей от величины сопротивления;
— эксцентрический, в котором максимально активизиро-
ванная мышца насильственно растягивается под воздей-
ствием внешней силы;
— плиометрический (реверсивный) характеризуется быстрой
сменой эксцентрического и концентрического режи-
мов работы мышц (например, отталкивание верх после
спрыгивания с возвышения);
— изокинетический — мышца сокращается с постоянной
скоростью вне зависимости от величины ее напряжения
или силы тяги. Этот метод может быть реализован толь-
ко на специальных тренажерных устройствах;
— метод переменных сопротивлений также предполагает ис-
пользование тренажеров, в которых величина сопротив-
76
ления меняется по определенному закону, зависящему,
как правило, от угла в суставе тренируемой конечности;
— статодинамический характеризуется остановкой в ци-
кле движения, во время которой мышца работает в изо-
метрическом режиме, т.е. представляет собой сочетание
изометрического и концентрического методов;
— изотонический, буквально, предполагает постоянную
степень напряжения мышцы, однако в естественных
условиях такой режим реализован быть не может, поэто-
му правильнее говорить о квазиизотоническом режиме
работы мышц и соответственно методе. При использо-
вании этого метода движения выполняются в медлен-
ном темпе и по возможности плавно, без расслабления
мышц в граничных моментах фаз движения;
— скоростной метод отличается предельной скоростью раз-
гона снаряда, массой тела или преодоления сопротивле-
ния 20-60% от МПС;
— контрастный — разновидность предыдущего, но величи-
на сопротивления меняется по ходу движения;
- метод электростимуляции обычно используется в варианте
сочетания произвольного напряжения мышц и дополнитель-
ного раздражения брюшка или двигательного нерва мышцы.
По построению тренировки:
— повторных усилий — это циклическое выполнение по-
вторных усилий с различным характером работы мышц
и паузами отдыха. Все циклические локомоции, выпол-
няемые в т.н. «утяжеленных условиях», можно отнести к
этому методу;
— максимальных усилий — является разновидностью метода
повторных усилий, предусматривающая упражнение с
предельными весами или степенью напряжения мышц;
— повторно-серийный метод представляет собой сочета-
ние серий подходов с удлиненным интервалом отдыха
между сериями;
- интермедиарный представляет собой упражнение с не-
большими весами, непредельным числом повторений
при статодинамическом характере работы мышц, реко-
мендуется для юных спортсменов;
77
— круговой метод предполагает работу на «станциях», на
которых осуществляется тренировка или различных мы-
шечных групп, или происходит смена режима работы
мышц, т.е. изменение направленности тренировочного
воздействия.
Какие из этих методов наиболее часто используются в ЦВС
и в связи с какими целями силовой подготовки?
Анализ литературы показывает, что все из перечисленных
методов используются или рекомендованы к применению на
основании данных педагогических наблюдений или экспе-
риментальных исследований. Однако основания и цели при-
менения тех или иных методов различаются достаточно суще-
ственно.
В наиболее общем виде, на наш взгляд, основания для при-
менения силовых упражнений в ЦВС определены в работе
Ф.П. Суслова и В.Б. Гилязовой [1990, стр. 3]: «Повышение
силового компонента... ведет к увеличению мощности рабоче-
го усилия, формированию рациональной фазовой структуры
движений, к оптимальному соотношению длины и частоты
шагов.... совершенствуются упругие и реактивные свойства
мышц и их способность к рекуперации (возврату) механиче-
ской энергии..., что повышает экономичность функциониро-
вания мышечной системы.» Аналогичные взгляды высказыва-
ют большинство специалистов.
Считается, что эти положительные сдвиги произойдут, если
в тренировке будет достигнуто улучшение:
— максимальной силы;
- взрывной силы;
— силовой выносливости.
Как должна быть построена тренировка, чтобы примене-
ние перечисленных выше методов способствовало улучшению
компонентов силовой подготовленности?
По этому вопросу имеется обширнейшая литература. Сум-
мируя мнения специалистов и данные исследований, можно
представить следующую обобщенную картину методики при-
менения средств развития силовых способностей в ЦВС.
Максимальная сила наиболее эффективно улучшается при
использовании изометрического, концентрического, эксцен-
78
трического режимов работы мышц, метода электростимуля-
ции, применяемых по методу повторных максимальных уси-
лий. Величина нагрузки (ВН) — 85-130% от МПС, количество
повторений (КП) в подходе 1-5, число подходов (ЧП) — 3-10,
интервал отдыха (ИО) между подходами — 3-5 мин. Если в тре-
нировке стоит задача увеличения не только мышечной силы, но
и мышечной массы (гипертрофии мышечных волокон (МВ)),
то эти методы и режимы мышц дополняются повторным и/или
повторно-серийным методом при уменьшении ВН до 70-85%,
ИО между подходами до 30-120 с и увеличении КП до 8 -12, ИО
между сериями — 5-10 мин. Тренировка может проводиться в
самых различных вариантах и условиях, но в большинстве слу-
чаев используются тяжелые снаряды или специализированные
тренажеры. Перечисленные методы должны способствовать
повышению частоты разрядов а-мотонейронов, совершен-
ствовать способность к синхронизации работы отдельных дви-
гательных единиц (ДЕ) мышцы и произвольной мобилизации
большего их числа, способствовать гипертрофии мышечных
волокон и совершенствовать координацию в работе мышц си-
нергистов и антагонистов.
Специалисты полагают, что взрывная сила будет улуч-
шаться при использовании плиометрического, скоростного,
контрастного и изометрического режимов работы мышц, вы-
полняемых чаще всего по методу максимальных усилий или
повторно-серийным методом. В первом случае: ВН — 85-130%
от МПС, КП в подходе 1-5, ЧП — 3-10, ИО 2-5 мин. Во вто-
ром: ВН — 50-85%, КП — 4-30, ЧП, организованных в серии,
6-12, ИО между сериями 5-10 мин. Наиболее распростране-
ны прыжковые упражнения, отталкивания после прыжка в
глубину, «взрывные» упражнения с отягощениями, с высо-
ким темпом движений и т.п. Предполагается [см., например,
Верхошанский Ю.В., 1983], что в случае использования боль-
ших отягощений совершенствуется взрывная сила, обеспечи-
ваемая всеми двигательными единицами мышц (ДЕ), если же
отягощения небольшие, то происходит совершенствование
способности к взрывным усилиям за счет работы, преимуще-
ственно быстрых ДЕ. Однако существуют данные [Платонов
В.Н., 1986], что порядок рекрутирования ДЕ определяется
79
только силой, но не скоростью сокращения мышц. Считается,
что большая взрывная сила достигается при лучшей синхро-
низации импульсов ДЕ, «спайковой» организации этих им-
пульсов, большей силе мышц, большей прочности и лучших
упругих свойствах соединительно-тканных элементов опорно-
двигательного аппарата (ОДА). Это мысль появляется только в
головах специалистов, не знакомых с физиологией мышечной
деятельности, поскольку синхронизация электрических им-
пульсов бессмысленна. Каждая ДЕ имеет свою собственную
максимальную частоту импульсации, при которой наблюдает-
ся максимальная концентрация кальция в активных мышеч-
ных волокнах, а значит сила сокращения.
Наибольшее внимание в ЦВС уделяется, традиционно,
силовой выносливости мышц, которая развивается при раз-
личных вариантах метода повторных усилий и кругового ме-
тода на тренажерах и в «утяжеленных» условиях выполнения
самой локомоции во всех ЦВС. Силовая выносливость всег-
да рассматривается в связи с производительностью основных
реакций энергообеспечения работы мышц. В зависимости от
длины дистанции речь может идти о преимущественной свя-
зи силы с выносливостью при работе анаэробного, аэробного
или смешанного характера [Платонов В.Н., 1984], поэтому ме-
тодические характеристики тренировочного занятия варьиру-
ются: ВН — 40-70%, длительность работы лежит в пределах от
12 с до 30 мин, ЧП — от 2 до 40, количество серий — от 1 до 12,
паузы отдыха — от 10 с до 10 мин. При занятиях на тренажерах:
30-70% МПС, КП - 30-200, ЧП - 3-10, ИО - 1-4.
Основным методическимтребованиемксовершенствованию
силовой выносливости применительно к коротким дистанциям
является увеличение мощности рабочего усилия в каждом ци-
кле движений за счет такого подбора временных и амплитудных
характеристик, при которых достигается наибольшая мощность
работы сократительного аппарата мышц [Платонов В.Н., 1984].
Это требование реализуется примерно при 40% от максималь-
ной скорости ненагруженного сокращения мышцы, поэтому в
таких ЦВС, как велосипедный, легкоатлетический спринт, пла-
вание, скорость сокращения мышц при выполнении специаль-
ных упражнений ниже соревновательной, а в гребле — выше.
80
Применительно к средним дистанциям, считается, что
надо добиваться наивысшей скорости накопления молочной
кислоты и высоких значений ее концентрации в мышцах. Это
требование может быть реализовано практически при соблю-
дении тех же требований, что и в спринте, однако паузы рас-
слабления мышц делаются короче (для худшего снабжения
мышц кислородом), а продолжительность работы увеличива-
ется до предельной выраженности утомления в мышцах (бо-
левые ощущения, резкое снижение мощности сокращений и
т.п.). В то же время точка зрения, что тренировки, связанные
с предельным накоплением молочной кислоты в мышцах,
полезны для развития выносливости на средних дистанциях,
может быть подвергнута сомнению (избыток и длительное
пребывание в мышечных волокнах ионов водорода ведет к
разрушению органелл).
Применительно к длинным дистанциям требуется макси-
мальная интенсификация дыхательного ресинтеза АТФ в мыш-
цах. Предполагается, что при применении упражнений для
развития силовой выносливости такие условия создаются при
работе в утяжеленных условиях, но только в тех случаях, когда
общая мощность не превышает уровня анаэробного порога.
4.2. Соотношение объемов средств
развития локальной выносливости в ЦВС
Выяснение этого вопроса оказалось наиболее сложным при
анализе как обобщающих работ, так и методических рекомен-
даций в различных видах спорта. Дело в том, что учет нагрузок
в ЦВС традиционно проводится или по километражу, преодо-
ленному спортсменом, или по времени (в часах), которое он за-
тратил на тот или иной вид тренировочной работы. Основные
компоненты локальной выносливости должны оцениваться по
параметрам скоростной, скоростно-силовой, силовой подго-
товки с обязательным учетом того, какие мышцы задействуют-
ся, режима работы мышц, числа подходов, интервалов отдыха
и т.п. (см. выше), с одной стороны, и объему работы в различ-
ных зонах интенсивности — с другой. Однако в каких едини-
81
цах можно сравнить упражнения со штангой, которые широко
применяются в гребле, коньках и велоспорте с занятиями в «за-
лах сухого плавания» у пловцов; спринтерский бег, прыжко-
вые и СБУ у легкоатлетов с занятиями «ОФП» у лыжников или
«спринтом» у пловцов? Единственным критерием, который
может иметь ограниченную информативность для специали-
стов, знающих содержание понятий, которые используются в
разных видах спорта (ОРУ, ОФП, СБУ, «специальная сила»,
«скоростно-силовая подготовка», «силовая подготовка», «си-
ловая выносливость» и мн. др.) в том или ином ЦВС, является
время, затраченное на разные виды подготовки. Такой специа-
лист, проводя мысленную трансформацию термина в конкрет-
ные параметры упражнения, может догадаться на что же имен-
но направленно воздействие данного упражнения в организме
человека, да и то только в том случае, если он обладает необхо-
димыми для этого биологическими знаниями. Однако попытка
раскрыть эти понятия в каждом из видов спорта только на осно-
вании терминов, используемых в методической литературе,
привела нас к выводу, что в настоящее время выполнить квали-
фицированно такой анализ не представляется возможным, так
как повсеместно описание упражнений и их классификация
идет по внешним (не существенным) признакам без раскрытия
внутреннего (существенного) содержания, под которым сле-
дует понимать воздействие на те или иные органеллы клеток,
участвующих в выполнении или обеспечении мышечной дея-
тельности [Селуянов В.Н. и др., 1991]. Однако такая практика,
как правило, отсутствует среди специалистов по теории спор-
та, которые, стараясь использовать понятные широкому кру-
гу тренеров и спортсменов термины, пренебрегают тем самым
научной строгостью. В связи с этим мы с большим сожалением
вынуждены пропустить рассмотрение одного из ключевых во-
просов для теории спортивной подготовки — о соотношении
нагрузок различной направленности. Относительно достовер-
но объемы таких нагрузок определены нами для легкоатлети-
ческого бега и приведены в последующих главах работы. Здесь
приведем лишь наиболее общие цифры, полученные нами при
анализе подготовки в 6 основных циклических видах спорта,
согласно которым на развитие сократительных компонентов
82
мышц, определяющих локальную выносливость (без разделе-
ния на тренировку основных и неосновных мышечных групп
и определения режима работы, в котором эти мышцы трени-
руются), затрачивается от 1,5 до 5 ч в неделю. Больше всего —
в гребле и плавании, меньше всего — в беге. В эти цифры не
входит время, затрачиваемое на развитие силовой выносливо-
сти, под развитием которой понимается очень широкий круг
упражнений — от выпрыгивания со штангой на плечах до бега
на коньках с парашютом и утяжеляющими манжетами на ногах
в легкой атлетике. Распределение нагрузок в циклах подготов-
ки приведено в последующих разделах.
4.3. Распределения средств развития
локальной выносливости в рамках одного
занятия, микро-, мезо- и макроциклов
и многолетней подготовки спортсменов
В подготовке спортсменов выделяют три уровня при по-
строении целостной системы подготовки: микроуровень — по-
строение отдельных тренировочных занятий и микроциклов;
мезоуровень — средние циклы (мезоциклы) и этапы подготов-
ки; макроуровень — большие циклы подготовки и многолет-
нюю подготовку [Матвеев Л.П., 1977].
От рационального распределения основных видов нагруз-
ки, стимулирующих улучшение компонентов локальной вы-
носливости, зависит эффективность накопления двигательно-
го потенциала и степень его реализации в соревновательном
упражнении.
4.3.1. Построение тренировочного занятия
В тренировочном занятии различают вводно-
подготовительную, основную и заключительную части. По
величине нагрузки занятия делятся на основные и дополни-
тельные. По направленности применяемыхсредств и методов —
на занятия избирательной и комплексной направленности
| Матвеев Л.П., 1977].
83
При занятиях избирательной направленности применяют-
ся средства и методы, воздействующие на какую-то одну или
смежные способности спортсмена. Экспериментально по-
казано, что в таких занятиях использование разнообразных
средств одной направленности позволяет выполнить больший
объем нагрузки при сохранении ее качественных параметров
[Стеценко Ю.Н., 1977].
В комплексном занятии основные рассматриваемые виды
нагрузки: 1) силовая, 2) скоростно-силовая, 3) на основные
биоэнергетические компоненты силовой выносливости реко-
мендуется распределять в следующих вариантах [Верхошан-
ский Ю.В., 1988; Озолин Н.Г., 1970; Платонов В.Н.,1984 ]:
1. Алактатная — гликолитическая — аэробная.
2. Скоростно — силовая — развитие выносливости.
3. Скоростная — развитие выносливости.
Такая принципиальная схема построения занятий с не-
большими вариациями вошла в большинство проанализиро-
ванных нами учебников и учебно-методических пособий, поэ-
тому большой неожиданностью явились данные Ф.П. Суслова
и В.Б. Гилязовой, 1990, проведших опрос ведущих тренеров
СССР по 6 основным циклическим видам спорта.
Заметим, характерный случай для эмпирического подхода,
когда мнение тренеров, как правило, плохо подготовленных в
области биохимии, физиологии, биомеханики и теории спорта,
предлагается как научный аргумент в споре и доказательстве
истины.
Было выявлено, что указанной схемы (в той ее части, где
определена последовательность силовой тренировки и тре-
нировки выносливости), придерживаются только в плавании
(сначала занятия в зале «сухого» плавания, затем — в воде),
несмотря на то, что выдающийся тренер пловцов и признан-
ный специалист в этом виде спорта Д. Каунсилмен [1982] ре-
комендовал в начале массированное воздействие на аэробные
функции, а в конце занятия — нагрузки скоростно-силового
или силового характера. Действительно, выносливость — ве-
дущее качество в ЦВС, поэтому, видимо, не лишено целесоо-
бразности развивать его в начале занятия, когда, как счита-
ется, например у культуристов [Вейдер Д., 1992, воздействие
84
наиболее эффективно. Вероятно, поэтому во всех остальных
ЦВС вначале применяются упражнения для развития вы-
носливости, а затем — силы. Видимо, современная практика
внесла коррективы в устоявшиеся представления. Однако в
отношении упражнений для развития скоростно-силовых
качеств большая часть тренеров предпочитает их примене-
ние в первой половине занятия и, как правило, сопряженно с
алактатной или гликолитической тренировкой [Суслов Ф.П.,
Гилязова В.Б., 1990].
4.3.2. Построение микроцикла
Общие правила построения микроциклов, которые в той
или иной степени реализованы в большинстве ЦВС, сформу-
лированы в известной серии работ В.Н. Платонова [1984-1997]
и сводятся к следующему:
1. Очередное занятие с большой нагрузкой должно плани-
роваться на фазу суперкомпенсации от предыдущего.
Заметим, что за понятием суперкомпенсации ничего не
имеется ввиду, в лучшем случае изменение массы глико-
гена в печени и в мышцах, отсюда 2 тренировки с боль-
шой нагрузкой в неделю, а как происходит суперком-
пенсация миофибрилл, митохондрий, капилляров и др.
никто не пишет и не учитывает.
2. На следующий день после применения большой нагруз-
ки следует использовать дополнительное занятие прин-
ципиально другой направленности, что ускоряет восста-
новление.
Заметим, что из-за непонимания сути суперкомпенсации
возникают мифы о влиянии каких-то мероприятий на
ускорение восстановления, тогда как скорость вос-
становления гликогена зависит лишь от нормально-
го питания, а миофибрилл — от достаточного приема
протеинов животного происхождения и отсутствия
«сбивающих факторов».
3. Смежное применение двух занятий со значительной на-
грузкой, но разной направленности несущественно уве-
личивает время восстановления после первого занятия,
85
поэтому можно выполнить больший суммарный объем
работы.
4. Наиболее быстро происходит восстановление после за-
нятий скоростно-силовой и спринтерской направлен-
ности, затем — гликолитической, дольше всего — до 5-7
суток после истощающих занятий — аэробной направ-
ленности. В соответствии с этим следует планировать их
количество и последовательность.
5. Сочетание двух разнонаправленных тренировочных за-
нятий вдень позволяет выполнить больший общий объ-
ем нагрузки, чем сочетание двух однонаправленных,
поэтому первый вариант более целесообразен.
При рассмотрении данных принципов следует учитывать,
что они разработаны на основании результатов, полученных на
пловцах при использовании специфических для них трениро-
вочных средств. В то же время существует достаточно примеров
того, что они или не соблюдаются, или имеют другие времен-
ные рамки в различных видах спорта. Признано, например,
что после занятий силовой направленности восстановление
длится до одной недели [Озолин Н.Г., 1970]. Характер работы
мышц в других ЦВС может существенно отличаться от плава-
ния, это может привести к смене ведущих факторов утомления
и изменить сроки восстановления. Например, очевидно, что в
таких видах спорта, как легкая атлетика, гребля, конькобеж-
ный спорт упражнения скоростно-силовой и скоростной на-
правленности практически всегда предполагают значительную
долю эксцентрического режима работы мышц, который явля-
ется существенным повреждающим фактором в отношении
мышечной ткани [Прилуцкий Б.И., 1989; Newham D.J. и др.,
1987; Paul, G.L., и др., 1989], чего не бывает в тренировке плов-
цов, лыжников и, как правило, велосипедистов, поэтому время
восстановления после такого вида нагрузок может существенно
увеличиться. В беге существует также другая особенность. Вы-
полнение тренировки гликолитического характера предполага-
ет достаточно большой объем бега с высокой интенсивностью
по дорожке. Вероятно, в результате сочетания механического
фактора (ударные нагрузки), химического фактора (накопление
ионов водорода, свободных радикалов) и предельной активиза-
86
ции деятельности симпато-адреналовой и глюкокортикоидной
систем [Виру А.А., Кырге П.К., 1983; Кассиль Г.Н. и др., 1978;
Friden, J. и др., 1983; Пшенникова М.Г.,1986; Eston R.G. и др.,
1994; Moreau D. и др., 1995] такие нагрузки считаются наибо-
лее тяжелыми и их применение не рекомендуется чаще 1 раза в
неделю даже в соревновательном периоде квалифицированных
бегунов на средние дистанции [Легкая атлетика: Учебник для
ИФК, 1989], а в течение этой недели спортсмен практически не
способен выполнять никакую другую нагрузку, кроме аэробно-
го бега по мягкому грунту. И наоборот, современная практика
тренировки в ЦВС хорошо известна огромными объемами на-
грузок, в частности аэробной направленности, которые приме-
няются практически каждый день, включая соревновательный
этап, поэтому приведенная длительность восстановления (5-7
суток) или не соответствует действительности, или такие тре-
нировки никто не использует.
И последнее, самое серьезное замечание: рациональность
построения тренировки по рассматриваемым выше принци-
пам оценивалась по критерию большей или меньшей степени
утомления, большего или меньшего объема тренировочных
работ, однако, очевидно, что единственным критерием в таких
случаях может быть получаемый тренировочный эффект.
Анализ практики планирования микроциклов в различ-
ных ЦВС показал [Озолин Н.Г., 1970], что при двухразовых
тренировках упражнения силовой направленности применя-
ются чаще в первой половине дня, но обоснования для такого
варианта построения тренировочного дня нет. В микроцикле
в различных видах спорта одни и те же компоненты силовых
способностей тренируются от 1 до 7 раз. Наиболее часто — в
коньках, плавании и велоспорте. Наиболее редко (1-2 раза в
неделю) — в беге.
4.3.3. Построение мезоцикла
Мезоцикл представляет собой средний уровень цикловой
структуры построения тренировочного процесса. Его длитель-
ность изменяется в пределах 3-6 недель. Различают втягиваю-
щие, базовые, контрольно-подготовительные, предсоревно-
87
вательные и соревновательные микроциклы [Матвеев Л.П.,
1977; Платонов В.Н., 1984].
В литературе практически отсутствуют сведения, позво-
ляющие выявить специфику организации мезоцикла приме-
нительно к компонентам локальной выносливости. Имеются
только общие рекомендации применительно ко всему трени-
ровочному процессу в целом.
В мезоцикл могут быть включены микроциклы комплекс-
ного или однонаправленного характера, воздействующие со-
ответственно на разные или на какую-то одну сторону подго-
товленности спортсменов.
По величине нагрузки различают мезоциклы, в которых
происходит суммирование (наложение) утомления от микро-
цикла к микроциклу, сопровождающееся снижением рабо-
тоспособности, которая возрастает только после применения
разгрузочного микроцикла. Такие мезоциклы применяются
в подготовке квалифицированных спортсменов и объясня-
ются феноменом запаздывающей трансформации или долго-
временным отставленным тренировочным эффектом (ДОТЭ)
[Верхошанский Ю.В., 1985]. В другом варианте может быть
запланировано постоянное возрастание подготовленности от
микроцикла к микроциклу. Однако наибольшее распростра-
нение получил 4-недельный мезоцикл, в котором в первый
микроцикл запланирована большая нагрузка, во второй — не-
сколько меньшая, в третий — самая большая за мезоцикл, а
четвертый микроцикл является восстановительным.
4.3.4. Построение макроциклов
Основные принципы планирования макроцикла заложены
достаточно давно в трудах наших ведущих специалистов [За-
циорский В.М., 1966; Набатникова М.Я., 1975; Озолин Н.Г.,
1970; Платонов В.Н.,1984] и приняты в теории и методике фи-
зического воспитания и спорта.
Например, в переходном периоде и начале подготовитель-
ного много внимания должно уделяться т.н. средствам ОФП
[Матвеев Л.П., 1977; Платонов В.Н., 1984]. Затем происходит
постепенное увеличение доли более специализированных
88
средств, способствующих становлению спортивной формы к
этапу основных стартов.
Теоретическое обоснование планирования макроцикла
в ЦВС также хорошо устоялось и может быть выражено сле-
дующим образом [Зациорский В.М., 1966, стр. 141-142]: «...
дыхательные возможности являются основой для развития
анаэробных, гликолитические — основой для развития креа-
тинфосфатного механизма... Последовательность воспитания
различных сторон выносливости (в тренировочном цикле)
должна быть такой: сначала дыхательные возможности («общая
выносливость»), затем гликолитические и, наконец, «алактат-
ные» возможности.... Что касается отдельного занятия, то здесь
обычно целесообразной бывает обратная последовательность».
Эта формула обосновывается тем, что при плохо развитых аэ-
робных способностях спортсмен не сможет выполнить боль-
шой объем гликолитической работы из-за медленной оплаты
О2-долга. Точно также при плохоразвитых гликолитических
возможностях скорость восстановления КрФ будет низкой и
спортсмен не сможет полноценно тренироваться.
До 80-х годов данная схема считалась общепризнанной.
Однако позже, в связи с увеличением общих объемов нагрузки
в ЦВС, этап «аэробной подготовки» начал проявлять негатив-
ные стороны, которые можно свести к двум моментам [Пла-
тонов В.Н., 1984]: 1) ухудшение здоровья спортсменов, выра-
жающееся в симптомах ухудшения показателей работы ССС,
почек, печени и иммунной системы [Дембо А.Г., 1980, 1991;
Волков В.Н., 1995; Левин М.Я., Хрущев С.В., 1991]; 2) сниже-
ние спринтерских, скоростно-силовых и силовых способно-
стей к соревновательному этапу [Журбина А.Д., 1976; Зациор-
ский В.М., 1966; Набатникова М.Я., 1975], что стало тормозом
в достижении рекордных результатов, особенно на спринтер-
ских и средних дистанциях. Именно это, на наш взгляд, яви-
лось стимулом к повышению интереса к проблемам локальной
выносливости в последние годы. И, в частности, к вопросам
планирования больших тренировочных циклов с учетом «ин-
тересов» мышечной системы.
Было предложено два основных варианта увеличения доли
упражнений силовой направленности в годичном цикле: 1)
89
распределенный вариант, когда соответствующие средства до-
статочно равномерно используются на протяжении всего года;
2) концентрированный вариант, когда планируются специаль-
ные этапы силовой подготовки, на которых обеспечивается
массированное тренирующее воздействие на организм [Вер-
хошанский Ю.В., 1985; Суслов Ф.П. и др.,1982].
Считается, что распределенный вариант больше подходит
спортсменам низкой и средней квалификации, т.к. «...распы-
ление средств... во времени не обеспечит существенного тре-
нирующего воздействия на тот высокий уровень физической
подготовленности, на котором они находятся» [Верхошанский
Ю.В., 1988, стр. 297].
Концентрированное планирование имеет две основных
схемы. В первой, более распространенной, этап силовой и
скоростно-силовой подготовки планируется на конец под-
готовительного и начало предсоревновательного этапов (при
2-3-цикловом планировании) для ликвидации отрицательно-
го воздействия объемной тренировки на силовые показатели
мышц [Верхошанский Ю.В., 1985; Суслов Ф.П. и др., 1982].
Во второй — на начало подготовительного, чтобы создать «за-
пас» силовых способностей, которые затем можно будет про-
сто поддерживать применением поддерживающей тренировки
[Гетманец В.С., 1987]. Применительно ко второму варианту
существует мнение [Верхошанский Ю.В., 1985|, что долж-
ны четко разграничиваться этапы применения нагрузки для
развития мышечной силы и этап с применением скоростных
упражнений. Это объясняется проявлением долговременного
отставленного эффекта силовой работы, концентрированное
применение которой всегда сопровождается снижением по-
казателей выносливости и скорости, которые повышаются на
этапе «реализации» через 1-2 месяца.
При одноцикловом планировании (в стайерских видах —
при длинном соревновательном периоде) предлагалась схема с
двумя «блокам» силовой нагрузки [Верхошанский Ю.В., 1985].
Первый блок — в начале подготовительного периода, когда
рекомендовано применение силовых упражнений общераз-
вивающего характера и второй блок — в конце подготовитель-
ного периода, в котором следует применять упражнения на
90
«силовую выносливость», скоростно-силовой и спринтерской
направленности. Однако в другой части своей работы [Верхо-
шанский Ю.В., 1988, стр. 298] для бегунов на длинные дистан-
ции, учитывая специфику этих видов легкой атлетики, Ю.В.
Верхошанский рекомендует распределенный вариант органи-
зации СФП в подготовительном периоде.
В то же время считается, что принципиальное решение
проблемы планирования макроцикла лежит в сопряженно-
последовательной организации нагрузок с различной преиму-
щественной направленностью. Такая организация тренировки
по мнению, Ю.В.Верхошанского (1985), реализует принцип
суперпозиции (когда эффект следующего этапа целесообразно
накладывается на эффект предыдущего) и оптимальным обра-
зом учитывает требование преимущественного воздействия на
нервно-мышечный аппарат (т.е. — Л В, прим. наше). Смысл та-
кой организации тренировки заключается в последовательном
«введении в тренировку нагрузок с постепенно повышающей-
ся силой и специфичностью их тренирующего воздействия на
организм» |Верхошанский Ю.В. 1988, стр. 284]. В то же время
этот способ предполагает знание того, какая нагрузка и как
должна накладываться на тот или иной эффект от предыду-
щей работы. По логике цитируемого автора, все последующие
нагрузки должны накладываться на отставленный эффект си-
ловой тренировки, однако никак не интерпретируется явное
противоречие с как будто бы признанным мнением, что «базо-
выми» в ЦВС являются аэробные способности, поэтому осо-
бый интерес представляет изучение того, как на практике про-
исходит планирование макроцикла, в частности в контексте
улучшения компонентов локальной выносливости.
Наиболее представительной по этому вопросу является уже
цитированная работа Ф.П. Суслова и В.Б. Гилязовой [1990].
На основе анкетного опроса ведущих тренеров СССР этими
учеными установлено, что в ЦВС используется как концентри-
рованное, так и распределенное применение средств, направ-
ленных на совершенствование локальной выносливости. В тех
случаях, когда используется концентрированный способ, мак-
симальная сила развивается: в велосипеде, лыжах, коньках — в
начале подготовительного периода; в гребле — на 2-м базовом
91
этапе; в плавании — на 2-м базовом, в предсоревновательный и
соревновательный периоды; в беге — на 2-м базовом этапе и в
предсоревновательный период. Взрывная сила: в велосипеде,
гребле, плавании и беге — в предсоревновательный и соревно-
вательный периоды; в коньках и лыжах — в подготовительный
период. Силовая выносливость — в велосипеде, лыжах, гребле
и плавании — круглогодично с 2-3-месячным перерывом в пе-
реходный период. В коньках — в подготовительном и переход-
ном периодах. В легкоатлетическом беге — на втором базовом
этапе, в предсоревновательный и соревновательный периоды.
Примечателен вывод исследования, в котором было отме-
чено, что, по мнению ведущих тренеров, наименьшая ясность
у них имеется именно по вопросу организации силовой подго-
товки, которую они тем не менее считают одним из ключевых
вопросов подготовки в ЦВС.
4.4. Реализация компонентов локальной
выносливости в основном соревновательном
упражнении
В абсолютном большинстве случаев специализированная
тренировка, направленная на совершенствование отдельных
компонентов локальной выносливости, предполагает исполь-
зование упражнений, отличающихся по своей динамической и
кинематической структуре от соревновательного упражнения.
Это формирует двигательный навык, который может отрица-
тельно сказаться на согласованности работы мышц, ухудшив
тем самым экономичность работы в целостной локомоции. В
связи с этим спортивный результат может снизиться даже при
возросшем двигательном потенциале, т.е. ухудшится реализа-
ционная эффективность техники [Биомеханика: Учебник для
ин-ов физ. культ / Под редакцией Д.Д. Донского, В.М. Заци-
орского., 1979; Зациорский В.М., 1982]. Кроме этого, извест-
но, что «техника как костюм, годится лишь тому, на кого он
сшит». Это образное выражение Д.Д. Донского [1968] подчер-
кивает обусловленность техники упражнений индивидуаль-
ными особенностями спортсменов, в частности силой мышц
92
и ее изменением в соответствии с изменениями последней
[Мякинченко Е.Б., 1983]. Однако такая «сонастройка», являю-
щаяся обязательным условием экономичности техники, про-
исходит, во-первых, не автоматически, а во-вторых, требует
определенного времени. Поэтому при тренировке локальной
выносливости, т.е. при целенаправленном изменении состоя-
ния нервно-мышечного аппарата, проблема реализации дви-
гательного потенциала является актуальной.
Целенаправленное изучение литературы по этому вопросу
позволило выявить только два методических подхода для обе-
спечения высокой реализационной эффективности техники:
— принцип сопряженного воздействия, согласующийся с
принципом динамического соответствия. Этот подход
предполагает такой подбор специальных упражнений,
которые были бы по возможности ближе по внутренней
и внешней структуре к соревновательным;
— использование сопряженно-последовательной органи-
зации нагрузок (см. выше) в годичном цикле, которая
предполагает увеличение доли специфических средств
(чаще использование самой локомоции с соревнователь-
ной интенсивностью) по мере приближения к соревно-
вательному этапу. В той или иной форме использование
этого подхода предлагалось всеми ведущими специали-
стами в области спортивной тренировки.
В заключение этой главы хотелось бы подчеркнуть следующее.
Понятие «развитие локальной выносливости» в циклических
видах спорта объединяет весь круг вопросов, связанных с по-
строением тренировочного процесса, направленного на совер-
шенствование компонентов нервно-мышечной системы спор-
тсменов, определяющих результат в циклических видах спорта.
К таким вопросам относят тренировку максимальной
силы мышц, скоростно-силовых способностей, силовой вы-
носливости в связи с различными зонами интенсивности, в
которых лежат соревновательные дистанции; проблемы пла-
нирования тренировочного процесса в различных циклах
подготовки; проблему реализации двигательного потенциа-
ла спортсменов, возрастающего в результате тренировки ло-
кальной выносливости.
93
При анализе научно-методической литературы по обозна-
ченным вопросам обращает на себя внимание прежде всего
противоречие между исключительно большим вниманием,
которое в последние 10-15 лет уделяется тренировке мышц в
ЦВС и крайне незначительным числом обобщающих работ по
этой проблеме. Среди которых можно выделить практически
только две монографии Ю.В. Верхошанского [1985, 1988] и
ряд наших работ [Мякинченко, 1997; Сарсания С.К., Селуя-
нов В.Н., 1991; Селуянов В.Н., 1993, 1995]. В этих исследова-
ниях проблема развития Л В отчетливо поставлена, раскры-
та ее актуальность, проведен анализ медико-биологических
аспектов, связанных с тренировкой мышц, и что самое, на наш
взгляд, существенное, на основе современного понимания
биологических закономерностей функционирования нервно-
мышечного аппарата намечены возможные способы построе-
ния тренировочного процесса с целью улучшения этого ком-
понента выносливости спортсменов.
Следует также отметить, что по отдельным проблемам трени-
ровки мышечной системы человека в последние годы выполнено
очень большое количество биологических (особенно за рубежом)
и педагогических (в основном в России) экспериментальных ис-
следований. Однако их обобщение и реализация в виде относи-
тельно законченной концепции, на основе которой можно было
бы в дальнейшем создавать частные технологии тренировки в
различных ЦВС, как нам представляется, до настоящего време-
ни выполнено не было. В связи с этим в следующих главах пред-
принята попытка обобщить имеющиеся данные и представить их
в виде некоторой системы взглядов, касающихся:
— значимости мышечных компонентов для выносливости в
циклических видах спорта;
— места тренировки Л В в системе подготовки спортсменов;
— лимитирующих факторов работоспособности в ЦВС,
связанных с мышечной системой;
— оптимальных средств и методах тренировочных воздействий
на мышечные компоненты, определяющие выносливость;
— вариант планирования тренировочного занятия, микро-,
мезо-, макроциклов и многолетней подготовки в ЦВС с точки
зрения улучшения Л В.
94
Глава 5
Факторы, лимитирующие
локальную выносливость
в циклических видах спорта
Передвижение спортсмена по дистанции с определенной
скоростью осуществляется за счет сократительной способ-
ности мышц. Передвижение с большей скоростью или более
длительное время с целью достижения лучшего спортивного
результата лимитируется в основном способностью мышц ге-
нерировать необходимое количество энергии. Снижение мощ-
ности сокращения мышц, несмотря на предельные волевые
усилия, происходит, как принято считать |Моногаров В.Д.,
1980, 1986; Ээпик В.А., Виру А.А., 1990; Green HJ.,1987; Devis
J.M., 1995; Enoka R.M., 1992; Orr G.W. и др., 1982] в результате
изменений, происходящих:
— или внутри мышечных волокон, основных для данной
локомоции мышечных групп;
— или в результате неспособности ЦНС обеспечивать
адекватную импульсацию с мотонейронных пулов этих
мышечных групп;
— или нарушения синаптической передачи нервных им-
пульсов на мышечные волокна и распространения воз-
буждения по волокну.
При относительно небольшой длительности циклического
упражнения (20—60 с) одной из основных причин утомления,
связанных с мышцами, считается снижение концентрации
КрФ в БМВ |Greenhalf P.L.,1996].
При работе средней длительности (от 1 до 8-10 мин) таким
изменением является высокая степень накопления ионов во-
дорода в мышцах и крови, приводящая к нарушению взаимо-
95
действия актиновых и миозиновых миофиламентов, деятель-
ности митохондрий, замедлению анаэробного гликолиза.
При длительной работе (10 мин — 3-4 ч), утомление обычно
связывают с понижением или исчерпанием запасов гликоге-
на в мышцах, что приводит к снижению мощности аэробного
ресинтеза АТФ. В результате, для поддержания среднедистан-
ционной скорости спортсмен вынужден проявлять предель-
ные волевые усилия, мобилизуя все ресурсы организма. Для
предотвращения исчерпания жизненно важных веществ ниже
уровня, опасного для жизни, в организме автоматически вклю-
чаются механизмы, ограничивающие дальнейшее выполнение
мышечной работы. Эти механизмы могут быть локализованы:
— в самих мышцах, например — накопление ионов водо-
рода;
— на уровне центральной нервной системы, например —
накопление ГАМК или серотонина [Devis J.M., 1995],
вторично вызывающих нарушение в работе вегетатив-
ных систем обеспечения мышечной деятельности;
— в отделах нейро-гормональной системы, например — ис-
черпание пула катехоламинов;
— в миокарде, например — нарушение ионных процессов
[ВируА.А., 1983] и др.
Другими словами, в каждом конкретном случае лимити-
рующим фактором при преодолении соревновательной дис-
танции могут быть как сами мышцы, так и другие системы ор-
ганизма. Однако принципиальным моментом является то, что
любые формы проявления утомления связаны с недостаточной
способностью основных, для данной локомоции, мышечных
групп выполнять работу требуемой мощности и длительно-
сти, а это, уже вторично, приводит к предельной мобилизации
регуляторных и обеспечивающих систем организма, вызывая
в отдельных случаях отказ от работы даже тогда, когда ресур-
сы мышц еще не истрачены полностью. Это подтверждается,
например, тем известным фактом, что более тренированные
спортсмены способны выполнить ту же работу с меньшими
проявлениями утомления или вообще без них.
Однако гораздо менее очевидным является решение вопро-
са о том, что приводит к указанным изменениям в мышечных
96
клетках. Гипотетически, причиной этого может быть недоста-
точная функциональная мощность процессов, локализован-
ных как в самих мышцах (периферические факторы), так и в
других системах организма (ЦНС, ДС, ССС, гормональной и
др., это — так называемые центральные факторы).
Дискуссия по этой проблеме в научной литературе ведется
уже много десятилетий, однако окончательного решения еще
не получила.
В то же время это — ключевой вопрос, на который должна
ответить спортивная наука и от ответа на него зависит стра-
тегия разработки средств и методов физической подготовки
в спорте.
От ответа на вопрос, что вызывает проявление указанных
признаков утомления, зависит выбор объекта, на который
должно быть направлено основное тренировочное воздей-
ствие, выбор средств и методов тренировки и все построения
целостного тренировочного процесса.
На основе современных данных о физиологических и био-
химических механизмах функционирования организма спор-
тсмена можно сделать следующие выводы относительно отме-
ченных выше изменений в мышечной системе, приводящих к
проявлению признаков утомления:
— скорость исчерпания запасов фосфагенов в относитель-
но кратковременных упражнениях зависит от АТФ-
азной активности миозина [Greenhalf P.L. и др., 1996],
концентрации КрФ внутри мышечных волокон (МВ)
[Яковлев Н.Н., 1983] и мощности других источников
энергии, «экономящих» КрФ при заданной мощности
работы мышц (когда его концентрация оказывается уже
пониженной);
— скорость накопления ионов водорода внутри мышеч-
ной ткани при заданной скорости ресинтеза АТФ в ходе
реакций анаэробного гликолиза и, следовательно, ско-
рость их выхода в кровь (с учетом изменения структу-
ры мембранных белков, тормозящих выход МК при ее
высокой концентрации) обратно пропорционально за-
висит от мощности митохондриального аппарата всех
типов мышечных волокон [Bangsbo J., 1996] и емкости
97
внутриклеточных буферных систем [Хочачка П., Дж.
Сомеро, 1988];
— скорость расхода углеводов мышц и печени при задан-
ной мощности работы мышц в длительных упражнени-
ях (до 2,5 часов) зависит от концентрации субстратов и
мощности митохондриального аппарата мышечных во-
локон [Hargreaves М., 1989].
Таким образом, многие факторы, за счет которых можно
отдалить наступление утомления, локализованы непосред-
ственно в мышечных клетках и могут быть названы мышечны-
ми компонентами, определяющими выносливость.
Однако один вопрос нуждается в специальном рассмотрении.
Дело в том, что в научной и научно-методической литера-
туре имеются многочисленные указания на то, что при напря-
женной мышечной деятельности выше анаэробного порога,
мышцы попадают в гипоксические условия из-за неспособ-
ности сердечно-сосудистой системы поставлять необходимое
количество кислорода, что и является причиной накопления
кислых продуктов метаболизма и снижения эффективности
работы митохондриального аппарата [Волков Н.И., 1969; Кол-
чинская А.З., 1973; Astrand P.-О., Rodahl К., 1970; Wasserman
К. и др., 1985]. Другими словами, если эта гипотеза верна, то
даже при высокой плотности митохондрий их функциональ-
ные возможности при напряженной мышечной работе не будут
использоваться полностью в случае недостаточного снабже-
ния МВ кислородом. В этом случае следует очевидный вывод,
что лимитирующим фактором, по крайней мере на средних
и длинных дистанциях (время работы 2-15 минут), является
центральный фактор — производительность ССС.
В основе этих представлений лежат несколько хорошо из-
вестных фактов.
1. В условиях «in vitro», т.е. — «в пробирке», пируват восста-
навливается до лактата только при отсутствии кислорода
— прямая пастеровская реакция [Мусил Я. и др., 1984].
Исходя из этого сделан вывод, что накопление молочной
кислоты в мышцах обусловлено нехваткой кислорода.
2. Максимальное потребление кислорода человеком растет
параллельно с увеличением мышечной массы, вовлечен-
98
ной в работу. Однако это происходит до определенного
предела, после которого дальнейший прирост МПК пре-
кращается. Этот факт, полученный на нетренированных
людях [Astrand P.-О., Rodahl К., 1970], интерпретирован
таким образом, что существует предел производитель-
ности ССС, ограничивающий МПК. В подтверждение
этого установлено, что способность мышц пропускать
через себя кровь во много раз превосходит способность
сердца перекачивать кровь. Сердце может обеспечить
только 1/3 мышц человека, поэтому при участии в рабо-
те большой мышечной массы часть мышц переводится
на «ограниченный кровоток» путем сокращения сфин-
ктеров артериол [Saltin В., 1985]. О том же говорит факт
большего кровотока через мышцы при работе одной
ноги по сравнению с работой двумя ногами на уровне
МПК [Armstrong R.B. и др., 1987].
3. Даже относительно небольшое снижение парциального
напряжения кислорода в атмосфере, снижающее насыще-
ние артериальной крови кислородом, ведет к уменьшению
показателей М П К и анаэробного порога и ускорению про-
дукции Ла при той же нагрузке [Wasserman К. и др., 1985].
4. Нарушение кровотока и, следовательно, нарушение до-
ставки кислорода к мышцам в результате статического
или изотонического мышечного напряжения, или при
укорочении паузах расслабления в циклической работе,
резко ускоряет накопление Ла.
5. Максимальный кровоток (а значит, и доставка кислорода)
в работающих мышцах конечностей при ступенчатой на-
грузке достигается на уровне анаэробного порога, а затем
— падает. Это может объясняться невозможностью сердца
обеспечить адекватное снабжение мышц кислородом при
сверхпороговых нагрузках [Ю.М. Стойда, 1990].
6. При прочих равных условиях увеличение концентрации
гемоглобина в крови (например, гемотрансфузия кон-
сервированной крови того же спортсмена) или увеличе-
ние скорости кровотока за счет увеличения сердечного
выброса могут приводить к (6-15%) увеличению VO2
АНП и МПК [Freedson P.S., 1981].
99
I 11 юбая врожденная или приобретенная функциональная
недостаточность сердечной мышцы или сосудов сопро-
вождается выраженным снижением аэробной произво-
дительности.
В то же время, в противовес отмеченным фактам известно
следующее:
1. Доступность пирувата для лактатдегидрогеназы в усло-
виях in vivo или in vitro приводит к образованию лактата
в присутствии кислорода [Gaesser GA, Brooks GA., 1984;
Holloszy J.O., 1996]. В мышечных волокнах всегда присут-
ствует мышечная форма ЛД Г, активность которой превы-
шает самую высокую отмеченную скорость производства
лактата [Голник Ф.Д., Германсен Л., 1982], поэтому об-
разование лактата происходит всегда и количество про-
изведенного лактата является разницей между скоростью
продукции пирувата и NADH и скоростью потребления
этих субстратов митохондриями [Bangsbo J., 1996].
2. Квалифицированные спортсмены имеют максималь-
ные значения потребления кислорода в том виде локо-
моции, в котором они тренируются [Fox E.L., Mathews
D.K., 1981; Withers R.T. и др., 1981]. При выполнении
неспецифических для них упражнений МПК ниже даже
при большей мышечной массе, участвующей в работе
(например у велосипедистов в беге). Следовательно, у
тренированных лиц МПК зависит от способности рабо-
тающих мышц потреблять кислород, а не от способности
ССС доставлять его к различным тканям организма.
3. У одних и тех же людей МПК, достигаемое в ступенча-
том тесте, меньше, чем МПК, достигаемое в тесте «удер-
жание критической скорости» [Hill D.W. и др., 1997].
4. Рост спортивных результатов на уровне высшего спор-
тивного мастерства происходит параллельно с ростом
анаэробного порога, значение которого определяется
мышечными факторами, а не ростом МПК, значение
которого связано с производительностью сердца [см.
обзор Верхошанский Ю.В., 1988].
5. Максимальное потребление кислорода мышцей — пропорци-
онально массе митохондрий в этой мышце [Taylor С.Р., 1987].
100
6. Если бы доставка кислорода в ткани являлась лимити-
рующим фактором, то улучшение тренированности со-
провождалось бы повышением потребления кислорода
при нагрузке ниже МПК (но выше анаэробного порога),
однако хорошо установлено, что потребление кислорода
или не меняется, или снижается [Холлоши Дж.О., 1982].
7. Кислород доставляется к мышцам с кровью, если бы
кровоток лимитировал поступление кислорода к рабо-
тающей мышце, то с ростом тренированности кровоток
при немаксимальной нагрузке возрастал, однако уста-
новлено обратное [Холлоши Дж.О., 1982]. При этом
мышцы компенсируют уменьшение кровотока повы-
шенной экстракцией кислорода [Карпман В.Л., Любина
Б.Г., 1982], что напрямую связано с периферическими
факторами.
8. При улучшении выносливости в первую очередь совер-
шенствуются механизмы перераспределения кровото-
ка между активными и неактивными тканями, между
основными и неосновными мышцами [Musch T.J. и др.,
1987].
9. При гипероксии потребление кислорода мышцами не
меняется, но количество крови, омывающее работаю-
щие мышцы, снижается [Ohlsson J., B.Wranne., 1986].
10. МПК отрицательно коррелирует с показателями кис-
лородтранспортной системы крови и положительно — с
тканевыми показателями утилизации кислорода в той же
выборке испытуемых, тренирующих выносливость. Ви-
димо, существует возможность компенсации низких кис-
лородтранспортных показателей ССС путем улучшения
мышечных звеньев транспорта кислорода к митохондри-
ям за счет большей капилляризации и повышения кон-
центрации миоглобина [Немировская Т.Л. и др., 1993].
И. Концентрация миоглобина в мышцах одной ноги по-
вышалась при ее тренировке в гипобарических усло-
виях, но уменьшалась в мышцах другой ноги у тех же
испытуемых, тренирующихся в нормобарических усло-
виях. Это означает, что при работе в нормальных усло-
виях мышцы не испытывают гипоксического состояния
101
[Terrados N. и др., 1990]. Это же подтверждается фактом
повышения концентрации миоглобина в случае анемии
[Немировская Т.Л., 1992].
12. При тренировке квалифицированных лыжников, со-
провождающейся повышением МПК, активность мио-
глобинпероксидазы достоверно снизилась в медленных
мышечных волокнах [Б.С.Шенкман, 1999].
13. Увеличение производительности сердца и связанного
с ней максимального МОК выше некоторого уровня,
определяемого периферическими факторами, бессмыс-
ленно, так как при этом увеличивается скорость крово-
тока через капилляры, т.е. ниже оптимума сокращается
транзитивное время нахождения крови в капиллярах
мышечной ткани [Saltin В., 1985], приводящее к непол-
ной отдаче кислорода мышцам.
14. Квалифицированные спортсмены экстрагируют из кро-
ви гораздо больший процент кислорода, чем новички
[Карпман В.Л., 1994; Карпман В.Л., Любина Б.Г., 1982|.
15. Образование МК в мышцах начинается при относитель-
но высоких значениях парциального напряжения кис-
лорода в оттекающей от мышц крови [Saltin В., 1985].
Это позволяет предполагать, что градиента парциаль-
ного напряжения кислорода в тканях и крови должно
с избытком хватать для адекватного снабжения мышц
кислородом.
16. Об этом же свидетельствуют эксперименты с
b-блокаторами, в которых показано [Карпман В.Л.,
Любина Б.Г., 1982], что даже на уровне МПК имеется
большой резерв увеличения артериовенозной разницы
по кислороду. Следовательно, парциальное напряже-
ние кислорода в тканях и оттекающей крови может быть
уменьшено еще больше без снижения интенсивности
потребления кислорода этими тканями.
17. Показано, что вне зависимости от характера работы:
локальной, региональной или глобальной — потребле-
ние кислорода отдельно взятой мышцей или относи-
тельно небольшой группой мышц растет и после начала
продукции и выхода в кровь МК [Armstrong R.B. и др.,
102
1987]. Если бы потребление кислорода митохондриями
уменьшалось в результате его нехватки, то потребление
кислорода всей мышцей также уменьшалось бы или как
минимум стабилизировалось.
18. При математическом моделировании процессов диффу-
зии кислорода внутрь мышечного волокна показано, что
даже при использовании модели Крога [Krogh А.., 1918]
без учета «облегченного транспорта кислорода» миогло-
бином диффузия кислорода становится лимитирующим
фактором при межкапиллярном расстоянии более 80тм
[Schumacker P.L., R.W.SamseL, 1989], однако в мышцах,
особенно тренированных, такого большого межкапи-
лярного расстояния не бывает [Физиология человека /
Под ред. Р. Шмидта и Г. Тевса. — Т. 3., 1996].
19. В тренированных мышцах митохондрии группируются
ближе к поверхности волокна [Henriksson J. и др., 1986;
Londraville R.L., Siddel B.D., 1990], а транспорт АТФ к со-
кратительным элементам осуществляется за счет «КрФ-
челнока» |Gollnick P.D., 1986].
20. Использование техники быстрого замораживания по-
зволило показать, что даже при максимальном потре-
блении кислорода мышцей, парциальное напряжение
кислорода в любой точке волокна составляет величину
порядка 1,1 - 1,3 Torr, т.е. — значительно выше уровня,
при котором поток кислорода внутрь митохондрий ока-
зывается (в состоянии покоя — 0,4-0,5 Torr) максималь-
ным |Gnaiger Т. и др., 1997] и к тому же не зависит от
площади поперечного сечения волокна. Концентрация
миоглобина также существенно выше критических ве-
личин [Gayeski T.E.J., C.R.Honig, 1986].
Несмотря на полученные данные [Gnaiger Т. и др., 1997],
что при увеличении степени стимуляции дыхания митохон-
дрий со стороны АДФ, напряжение 02, при котором дости-
гается максимальный поток кислорода внутрь митохондрий,
существенно повышается до величины 4-6 Torr, гипотеза о
критичности парциального напряжения 02 в притекающей
крови (т.е. о гипоксическом состоянии мышечных волокон
из-за неспособности ССС поставлять требуемое количество
103
кислорода) не может считаться доказанной, так как последние
данные получены на клетках печени, в которых отсутствует
миоглобин, осуществляющий «облегченную» диффузию кис-
лорода в мышечных клетках. Кроме того, существуют много-
численные данные об улучшении контроля дыхания митохон-
дрий при повышении аэробной мощности мышечных клеток
[Немировская Т.Л., 1992; Хочачка П., Дж. Сомеро, 1988].
Таким образом, все приведенные выше рассуждения и фак-
ты за и против привели нас к выводу, что гипотеза о ведущей
роли периферических факторов (т.е. локализованных в самих
мышцах) в развитии утомления мышц при преодолении дис-
танции является более обоснованной, чем «центральная» (в
частности, связанная с сердечно-сосудистой системой). В по-
следние годы все чаще публикуются работы в поддержку этой
точки зрения [Булатова М.М., Платонов В.Н., 1996; Меерсон
Ф.З., 1978; Graham Т.Е., В.Saltin., 1989; Guezennec C.Y. и др,
1991, Minotti J.D., Jonson Е.С. и др., 1990].
В связи с этим в дальнейшем мы будем исходить из сооб-
ражений, что для снижения вклада, привносимого мышечной
системой в утомлении спортсмена на дистанции, объектом
тренировочных воздействий должны быть структурные и фер-
ментативные белковые комплексы самих мышц, а не морфо-
структуры ССС, дыхательной системы, гормональной систе-
мы, ЦНС и др.
Установив, таким образом, объект исследования — мы-
шечные компоненты, определяющие выносливость, следует
уточнить, что под этим подразумевается. Другими словами,
уточним, какие факторы, связанные с мышечной системой
спортсменов в ЦВС, определяют его двигательный потенциал.
Для этого воспользуемся приемом имитационного моделиро-
вания процессов, происходящих в мышцах при преодолении
соревновательной дистанции.
Для выяснения факторов, которые определяют мышечные
компоненты выносливости спортсменов, рассмотрим процессы,
происходящие в мышцах при преодолении средней дистанции.
Для примера выберем такую интенсивность мышечной ра-
боты, которая имеет место на дистанциях с предельным време-
нем около 3 минут.
104
Этот выбор обусловлен тем, что в литературе достаточно
много экспериментальных данных, полученных при изучении
именно такой длительности работы. Кроме этого, большин-
ство сделанных выводов можно будет экстраполировать и на
более длинные, и на более короткие дистанции.
В качестве объекта наблюдения выберем условную мышцу
конечности, выполняющую основную нагрузку в какой-нибудь
циклической локомоции (например, двуглавая мышца бедра в
беге или четырехглавая — при педалировании). Предположим,
что композиция мышечных волокон в мышце, определенная
по АТФ-азной активности миозина, т.е. обусловленная типом
иннервации волокон, следующая: медленные мышечные во-
локна (ММВ) — 50%, быстрые окислительные МВ (БоМВ) —
25%, быстрые гликолитические (БгМВ) — 25%. Максимальную
алактатную мощность (МАМ) мышцы примем за 100%. Тогда
с высокой долей вероятности можно предполагать, что уровень
анаэробного порога (АнП) у этой мышцы будет около 30% от
МАМ, а критическая (соответствующая МПК для всего орга-
низма) — 40% от МАМ. Следовательно, если МАМ при педа-
лировании на велоэргометре составляет около 700Вт то АнП у
этого спортсмена будет 233Вт, а «критическая» мощность — 280
Вт. Это вполне согласуется с реальными цифрами для трениро-
ванных на выносливость спортсменов-невелосипедистов, по-
лученных в ПН ИЛ РГАФК. Среднедистанционная мощность
работы мышц постоянна (за исключением стартового разгона) и
составляет около 50% от МАМ. Эта цифра, как следует из расче-
тов энергозапроса, выполненных Н.И. Волковым [1969], также
примерно соответствует действительности. Пиковые значения
усилий, которые придется проявлять мышце в цикле движений,
предположим, будут равны 50% от максимальной произвольной
силы (МПС). Эти усилия примерно соответствуют тем, которые
проявляет спортсмен при педалировании на велоэргометре с
частотой 60 об/мин. Или беге, где пиковые значения больше,
но длительность напряжения мышц относительно короче.
Задача спортсмена — показать наивысший результат, поэ-
тому в конце дистанции он будет прикладывать максимальные
волевые усилия, чтобы поддерживать оптимальную скорость
передвижения, несмотря на высокую степень утомления.
105
Среди других упрощений схемы, предположим, что в ре-
синтезе АТФ принимают участие только три реакции: креа-
тинфосфокиназная (КФК-реакция), гликогенолиз и гликолиз
(аэробные и анаэробные), объединенные под одним общим
названием «гликолиз» и дыхательное фосфорилирование. При
этом другие возможные реакции (например, миокиназная),
как имеющие относительно небольшое значение, принимать-
ся во внимание не будут.
При естественной локомоции известно явление периоди-
ческого чередования периодов наибольшей электрической ак-
тивности нескольких мышц-синергистов. Это дало основание
для предположения о таком управлении активностью мышц в
длительном упражнении, при котором периодическое сниже-
ние их активности обеспечивает отдых по мере прохождения
дистанции [Моногаров В.Д., 1980]. В нашем случае мы пред-
полагаем, что синергисты отсутствуют, поэтому механическая
работа выполняется только рассматриваемой мышцей и, сле-
довательно, активность мышцы будет определяться только
логикой поддержания требуемой механической мощности без
периодического выключения мышцы для «отдыха».
Проследим физиологические и биохимические процессы,
происходящие в мышце (локальный уровень) и на уровне все-
го организма (системный уровень) при преодолении соревно-
вательной дистанции.
Для простоты мы абстрагируемся от морфологических,
биомеханических, психологических, тактических и других
факторов, которые, безусловно, могли бы повлиять на резуль-
тативность спортсмена.
5.1. Схема физиологических и биохимических
процессов, происходящих в мышцах
при преодолении соревновательной дистанции
В основу схемы физиологических и биохимических про-
цессов в мышце положены представления о функциональной
специализации различных типов мышечных волокон в ске-
летных мышцах при выполнении физической работы, беру-
106
щих начало с классических исследований Henneman (1965), и
которые могут быть выражены следующим образом:»... как у
рыб, так и у млекопитающих (в том числе и у человека — прим,
наше) красные волокна используются при легкой или умерен-
ной работе, а белые начинают функционировать либо тогда,
когда при тяжелой работе значительно возрастает приток воз-
буждающих импульсов к мотонейронам, либо при утомлении
красных волокон» [Holloszy J.O., Booth F.W., 1976]. Данный
способ функционирования нервно-мышечного аппарата у
млекопитающих, включая человека, считается общеприня-
тым, что подтверждено большим количеством миографиче-
ских исследований [Персон Р.С., 1985; Miyashita Y. и др., 1981;
Shibata М. и др., 1997] и исследованиями с изучением исчер-
пания энергетических субстратов при различном характере и
мощности работы мышц [Gollnick Р и др., 1974; Vollestad NK,
Blom PCS, 1985|.
При разработке схемы мы принимали во внимание также
следующие хорошо установленные факты.
1. При работе спринтерского характера (5-20 с — мощ-
ность, близкая МАМ) полного исчерпания запасов КрФ
в мышце не происходит |Nevill М.Е. и др., 1996], хотя
может снижаться на 75% [Hirvonen J. и др., 1987]. Это
явление наблюдается при работе максимальной интен-
сивности или электростимуляции мышц длительностью
30-40 с [Greenhalf P.L. и др., 1996|.
2. При работе любой интенсивности первые 10-15 с кон-
центрация Ла в крови не возрастает [Голник Ф.Д., Гер-
мансенЛ., 1982].
3. Время достижения потреблением кислорода полови-
ны от своего максимума в среднем — 30 с [Morton R.H.,
G.С.Gass, 1987], однако при интенсивной нагрузке —
VO2 развертывается быстрее [Волков Н.И., 1969].
4. При большей длительности работы до «отказа» и, со-
ответственно, меньшей интенсивности концентрация
КрФ в конце работы оказывается тем выше, чем больше
длительность [Голник Ф.Д., Германсен Л., 1982]. Так,
например, при педалировании до истощения длительно-
стью 77 минут [Sahlin К и др.; 1997] концентрация КрФ
107
составляла 80% от исхода, при этом КрФ был полностью
исчерпан как в части ММВ, так и в БМВ. В то время как в
других волокнах всех типов КрФ был на уровне исхода.
5. При работе:
— 43% от МПК рекрутированы с самого начала 45% во-
локон;
— 61 % от МПК рекрутированы с самого начала 65% во-
локон;
— 90% от МПК рекрутированы с самого начала 95% во-
локон.
Даже в рамках одного типа волокон не все ДЕ участвуют
в работе и разница тем больше, чем выше порог ДЕ
[Vollestad NK, Blom PCS, 1985].
6. При интенсивности работы 43% от МПК и длительности
60 минут гликоген истощается на 50% в ММ В, тогда как
в БМВ никаких изменений нет [Vollestad NK, Blom PCS,
1985].
7. При интенсивности 61% от МПК сначала идет исчер-
пание в ММ В и незначительно в БоМВ, после 30 минут
скорость исчерпания гликогена сравнивается в БМВ и
ММВ. Через 60 минут расход в ММВ — 75%, в БоМВ —
40%, в БгМВ - 22% [Vollestad NK, Blom PCS, 1985].
8. При интенсивности 90% в конце серии 3-4x10 минут
расход гликогена:
Тип волокон ММВ БоМВ БгМВ
Расход % от исхода 74 80 54
9. При физической нагрузке сначала возрастает кровоток
в медленных и быстрых окислительных МВ, а потом — в
БгМВ [Armstrong R.B. и др., 1987].
10. Скорость обновления сократительных белков зависит
от степени активности мышц и характера активности.
Тренировка с 70% от МПК приводит к обновлению тя-
желых цепей миозина во всех типах мышечных волокон.
Кратковременный бег с интенсивностью 190% от МПК
ускоряет обновление ТЦ только в БгМВ и БоМВ [Сээне
Т.П., 1990].
11. Срочный эффект аэробной работы различной мощно-
сти приводит к существенно различным изменениям
108
на уровне ультраструктуры (митохондрий, капилляров,
миофиламентов, СПР, запасов субстратов) мышечных
волокон различного типа [Загоруйко Г.Е., 1990].
Условно разобьем всю дистанцию на 3 части: врабатывание,
квазиустойчивое состояние, финиш.
5.1.1. Врабатывание
Эту фазу условно ограничивают моментами старта и дости-
жения потреблением 02 устойчивого или квазиустойчивого
состояния [КоцЯ.М., 1986].
Сразу со старта спортсмен развивает, как правило, почти
максимальную мощность, но поддерживает ее 2-7 с до дости-
жения среднедистанционной скорости. Пиковые значения
усилий, проявляемые мышцей, также приближаются к макси-
мальным.
Таким образом, в связи с высокой величиной усилий в
стартовом разгоне в работу сначала должны быть вовлечены
большая часть ДЕ, имеющихся в мышце.
Первая цепочка событий, которые произойдут в актирован-
ных МВ (и быстрых, и медленных), это — выход ионов Са++ из
СПР Ю взаимодействие миофиламентов Ю понижение кон-
центрации АТФ. Следствиями этих событий будут являться
многочисленные биохимические процессы. Наиболее значи-
мыми с точки зрения изучаемого объекта будут следующие:
— под влиянием Na+, Са++ и других агентов (адреналин,
ацетилхолин, АДФ, АМФ) в самом начале мышечного
сокращения активируется фосфорилаза, запускающая
первый этап гликолиза и гликогенолиза [Биохимия:
Учебник для институтов физ. культуры, 1986; Голник
Ф.Д., ГермансенЛ., 1982];
— при появлении Са++ и АДФ активируется креатинфос-
фокиназа, запускающая реакцию перефосфорилирова-
ния КрФ и АДФ с образованием АТФ и неорганического
фосфата (Ф);
— появление Ф, Кр и снижение АТФ/АДФ снимают
«блок», a NH4 и/или ц-АМФ активизирует второй клю-
чевой фермент гликогенолиза — фосфофруктокиназу и
109
запускает «КрФ-челнок» [Gollnick P.D., 1986], который
немедленно активизирует дыхание митохондрий, суб-
стратами для которых служит пируват и свободные жир-
ные кислоты.
Наиболее важным выводом из сказанного является то, что
все основные реакции энергообеспечения мышечного сокра-
щения активизируются практически одновременно с первых
с мышечной активности, хотя скорость и вклад этих реакций
будет быстро меняться по ходу работы.
В зависимости от вида локомоции (бег, плавание или ве-
лосипед, гребля) длительность стартового разгона будет изме-
няться от 2-3 с до 5-7 с. Как показывают прямые измерения
[Nevill М.Е. и др., 1996], расход КрФ за это время при макси-
мальной или околомаксимальной работе может составить при-
близительно 15-50% от исхода (в спринте — до 75% [Hirvonen
J. и др., 19871). Это означает наличие и быструю активизацию
в мышечных волокнах всех факторов, активирующих ключе-
вые ферменты основных реакций энергообеспечения и обе-
спечивающих эти реакции субстратами. Однако известно, что
на уровне целостной мышцы основных признаков высокой
активности гликолиза и гликогенолиза (например, накопле-
ние Ла) и окислительного фосфорилирования (например,
уменьшение парциального давления кислорода в оттекаю-
щей от мышц крови) в первые с работы еще не наблюдается.
Основная причина этого [Биохимия: Учебник для институтов
физ. культуры, 1986; Хочачка П., Дж. Сомеро, 1988] — высокая
скорость кратинфосфокиназной реакции, обеспеченной суб-
стратом, которая с высокой эффективностью ресинтезирует
АТФ. Скорость же ресинтеза АТФ в ходе анаэробного глико-
лиза (в БгМВ) и окислительного фосфорилирования (в ММВ
и БоМВ) определяется «успешностью» протекания кратин-
фосфокиназной реакции, а именно, несмотря на то что кон-
центрация АДФ по мере продолжения работы остается насы-
щающей для КФК, скорость этой реакции будет постепенно
снижаться в связи со снижением концентрации КрФ [Хочачка
П., Дж. Сомеро, 1988], при этом внутри мышечного волокна
будут создаваться факторы ускорения других реакций энер-
гообеспечения.
110
После стартового разгона мощность работы мышц и сила
их сокращения снижается, как мы предположили, до 50% со-
ответственно от МАМ и МПС. Это означает снижение акти-
вирующего влияния моторной зоны коры на мотонейронные
пулы работающих мышц. Снижение активирующих стимулов
приводит к выключению наиболее высокопороговых быстрых
ДЕ из работы или как минимум к существенному снижению
частоты импульсации их мотонейронов. Это явление хорошо
показано в миографических исследованиях [Коц Я.М., 1986].
Какие мышечные волокна участвуют в создании тяги мышц
и каков характер их активности в начале дистанции, когда ско-
рость передвижения уже стабилизировалась?
Наиболее вероятной представляется гипотеза, что для ор-
ганизма наиболее выгодно по возможности более полное ис-
пользование малоутомляемых ММВ для генерации механиче-
ской энергии.
Однако можно ли обеспечить 50% от МАМ исключительно
медленными МВ, даже учитывая, что в начале дистанции в них
имеется еще достаточно самого мощного механизма ресинтеза
АТФ — креатинфосфата. Это предположение маловероятно,
так как при 100%-ой силе не может быть обеспечена макси-
мальная механическая мощность волокна, которая опреде-
ляется его АТФ-азной активностью, а не наличием мощного
механизма ресинтеза АТФ.
Обратная ситуация также маловероятна — т.е. когда ско-
рость сокращения мышцы слишком высока для того, чтобы
ММВ могли внести существенный вклад в развиваемое уси-
лие и работа выполнялась бы в основном БоМВ. Так как это
противоречит данным сравнения максимальной скорости
укорочения изолированных ММВ и скорости их укорочения
в реальных локомоциях. Показано, например [Тураев В.Т.,
1996], что даже при максимальном темпе движений или скоро-
сти передвижения ММВ вполне могут вносить существенный
вклад в развитие силы тяги мышц. Тем более это происходит
при передвижении на средних дистанциях.
Таким образом, наиболее вероятной может быть гипотеза,
что с ростом скорости передвижения биомеханика движений,
в частности, соотношение темпа и длины шагов, будет изме-
111
няться таким образом, чтобы скорость сокращения основ-
ных мышц как можно дольше находилась в пределах зоны на
кривой сила-скорость, в которой наблюдается максимальная
мощность сокращения для ММВ. Так как в этой зоне, предпо-
ложительно, обеспечиваются условия для оптимального взаи-
модействия актиновых и миозиновых миофиламентов ММВ,
что выражается не только в их максимальном вкладе в разви-
тие силы тяги мышц, но и в наиболее эффективном использо-
вании их метаболической энергии (наибольшем к.п.д. мышц)
[Платонов В.Н., Булатова М.М., 1992].
Следовательно, можно предположить, что в начале дистан-
ции ММВ вносят максимально возможный вклад в генерацию
усилия мышцы и практически все из них задействованы в ра-
боту, а недостающая мощность и сила тяги целостной мышцы
обеспечивается подключением БоМВ и БгМВ с самого начала
работы [Vollestad NK, Blom PCS, 1985].
Относительно участия БоМВ и БгМВ в работе сделаем еще
одно допущение. В связи с тем что среди промежуточных по раз-
меру мотонейронов, иннервирующих БоМВ, есть мотонейроны
с разным порогом возбуждения и частотой разряда, при которой
достигается гладкий тетанус, разделим БоМВ на две группы: пер-
вая — работает в условиях, предположим, гладкого тетануса и в
МВ этой группы наблюдается максимальный энергозапрос; МВ
второй — работают в условиях зубчатого тетануса и скорость ре-
акций ресинтеза АТФ в них не максимальны. БгМВ задействова-
ны в очень незначительном количестве и существенного вклада
в силу тяги мышц и продукцию лактата в начале дистанции не
вносят. Известно также, что креатинфосфокиназа и митохон-
дриальные ферменты обладают большим химическим сродством
кАДФ, чем ферменты гликолиза [ХочачкаП., Дж. Сомеро, 1988],
следовательно, этот фактор тоже может вносить свой вклад в за-
держку формирования Ла в начале работы.
Рассматривая схему биохимических процессов в мышце
на этой стадии дистанции, можно предположить, что в «от-
ключившихся» после стартового разгона быстрых гликоли-
тических МВ вполне может идти частичное восстановление
запасов КрФ, утилизация небольшого количества образовав-
шегося лактата, удаление ионов водорода. В медленных же МВ
112
(ММВ) и быстрых оксидативных МВ (БоМВ), которые, как от-
мечено, на этом этапе дистанции несут основную нагрузку по
перемещению спортсмена, энергия для актино-миозинового
взаимодействия поставляется вначале в основном за счет рас-
щепления КрФ. Параллельно снижению его концентрации
возрастает скорость его ресинтеза за счет АТФ, ресинтезируе-
мой в митохондриях. Источником кислорода служит внутри-
мышечный оксимиоглобин [Физиология человека / Под ред.
Р. Шмидта и Г. Тевса. — Т.З., 1996], кислород, растворенный
в саркоплазме, и кислород, высвобождаемый из гемоглобина
капиллярной крови. Два отмеченных выше фактора: 1) исполь-
зование запасов КрФ в начале работы мышечного волокна; 2)
наличие резервов кислорода в мышцах и притекающей крови,
являются причиной постепенного (в течении 0,4—1,5 минут в
зависимости от мощности работы) возрастания потребления
кислорода мышцами, что и фиксируется в эксперименте. На
наш взгляд нет оснований категорически утверждать, что в на-
чале физической работы мышцы тренированных спортсменов
(у которых, как можно предположить, имеется «хорошо отла-
женный механизм» поставки кислорода от легких к митохон-
дриям) попадают в гипоксические условия в результате суще-
ствования некого «врожденного дефекта» ССС, называемого
«инерционностью». Хотя известны факты наличия гипокси-
ческого состояния мышц даже в условиях покоя и на любой
стадии «кислородного каскада» [Колчинская А.З., 1973].
Тем не менее интерпретация процессов, происходящих
в мышцах в этой фазе, традиционно осуществляется [Био-
химия: Учебник для институтов физ. культуры, 1986; Волков
Н.И., 1969; Мелихова М.А., 1992; Моногаров В.Д., 1980, 116] в
свете гипотезы о гипоксическом состоянии мышечной ткани,
возникающей из-за «инерционности» системы доставки кис-
лорода (рис. 4) к работающим мышцам, а именно, О2 запрос
с первых с работы оказывается высоким, а системы доставки
кислорода к работающим мышцам интенсифицируют свою
деятельность постепенно, поэтому возникает дефицит кисло-
рода такой высокой степени, что это ограничивает скорость
выработки АТФ на митохондриях. Все дальнейшие процессы,
происходящие в мышцах, трактуются с этой точки зрения.
113
Рис. 4. Динамика возрастания потребления кислорода ( V0) и скорости
КфК-реакции в зависимости от предельного времени упражнения (по
[Волков Н.И., 1964])
Однако, как уже отмечалось выше, анализ большого числа
проведенных отечественных и зарубежных исследований не
позволил обнаружить ни одной работы, в которой путем пря-
мых измерений или путем моделирования было бы показано,
что: 1) парциальное напряжение кислорода внутри мышеч-
ных волокон (рО2) в начальный период работы любой мощ-
ности; 2) рО2 при работе максимальной аэробной мощности;
3) рО2 при работе выше максимальной аэробной мощности у
тренированных спортсменов падает ниже критического для
митохондрий уровня (0,1-1 мм рт. ст. [Физиология человека
/ Под ред. Р. Шмидта и Г. Тевса. — Т.З., 1996]) или, что по-
ток кислорода в митохондрии в состоянии их максимальной
активизации [Gnaiger Т. и др., 1997] ограничивает скорость
выработки энергии при парциальном напряжении О2 в 1-3 мм
рт.ст., которое, как можно предположить, наиболее типично
для максимально активированной мышцы. Это же можно ска-
зать и для степени насыщения миоглобина кислородом, высо-
кое значение которой свидетельствует о доступности кислоро-
да для протекания окислительно-восстановительных реакций
в митохондриях. Так как миоглобин является депо кислорода
114
внутри мышечного волокна, осуществляет «облегченную диф-
фузию кислорода» [Холлоши Дж.О., 1982] и степень насыще-
ния которого не должна снижаться, видимо, ниже 17-19%, что
наблюдается в некоторых участках сокращающегося волокна
при максимальном потреблении кислорода этим волокном
и соответствует напряжению кислорода 1 мм рт.ст. [Gayeski
T.E.J.,C.R. Honig, 1986].
Хотя, разумеется, в связи с ускорением потребления кис-
лорода митохондриями его тканевое напряжение падает ниже
уровня покоя, что является совершенно необходимым при-
способительным механизмом, который обуславливает уве-
личенный градиент по кислороду между саркоплазмой МВ
и капиллярной кровью, т.е. ускоренную доставку кислорода
внутрь МВ при учете роли оксимиоглобина в качестве депо и
переносчика О2 [Физиология человека / Под ред. Р. Шмидта
и Г. Тевса. - Т.З., 1996; Gayeski T.E.J., C.R.Honig, 1986]. Сле-
довательно, есть основания говорить о наличии т.н. функцио-
нальной гипоксии, однако нет никаких прямых или косвен-
ных данных, свидетельствующих, что степень такой тканевой
гипоксии оказывается на столько высокой, что ограничивает
скорость выработки энергии в митохондриях. Это справедли-
во (при нормальном парциальном напряжении О2 в атмосфе-
ре), по меньшей мере, в тех случаях, когда:
— нет нарушений в системе микроциркуляции, как при со-
судистых заболеваниях;
— нет функциональной недостаточности сердечной мыш-
цы, вызванной, например, относительно небольшим (
менее 3-3,5 см2) «устьем» аорты [47];
— нет состояния полной детренированности сердечной
мышцы, которое может наблюдаться в экспериментах с
гипокинезией или после продолжительной болезни;
— отсутствует ишемическая гипоксия, связанная с дли-
тельным напряжением мышц или короткими паузами
расслабления мышц.
Кроме того, гипотеза об «инерционности» ССС не согла-
суется с тем, что при резком психоэмоциональном стрессе
или в начале мышечной работы предельной интенсивности
[Волков Н.И., 1969], все параметры деятельности ССС воз-
115
растают по гораздо более крутому градиенту, чем в рассматри-
ваемом примере. Представления об «инерционности» ССС
также просто противоречат здравому смыслу — зачем орга-
низму в начале работы создавать себе трудности, накапливая
«дефицит» кислорода, а затем «напрягаться» для того, чтобы
его ликвидировать. Тем более, что существует механизм (см.
ниже) который вполне позволяет обойтись без этих проблем.
Правда, чтобы правильно интерпретировать сказанное, надо
ясно понимать разницу между «энергозапросом», т.е. запро-
сом в АТФ и «запросом кислорода», т.е. в количестве кис-
лорода, которое способны использовать митохондрии при
данном уровне стимуляции тканевого дыхания и который,
естественно, различается в разных мышечных волокнах в за-
висимости от их активности, от их метаболического профи-
ля, в зависимости от стадии развертывания внутриклеточных
реакций энергообеспечения и т.д. Нам кажется, что эти два
понятия постоянно перепутываются.
В подтверждение высказанной мысли, что в период враба-
тывания мышечные волокна не испытывают нехватки кисло-
рода для полноценного тканевого дыхания из-за «инерцион-
ности» ССС, можно привести следующие расчеты.
При максимальной мощности работы мышц (например, в
спринтерском беге или тесте на велоэргометре) снижение на-
блюдаемой мощности начинается на 5-6 с работы (рис. 4), од-
нако, приблизительно до 8-10-й с человек еще способен под-
держивать практически максимальную мощность, после чего
начинается ее быстрое снижение [Волков Н.И., 1969; Nevill
М.Е. и др., 1996]. Проведенные исследования со взятием биоп-
сийных проб [Nevill М.Е. и др., 1996] показали, что к этому мо-
менту концентрация КрФ снижается лишь на 50—55% (в мыш-
це с 50% ММВ. В более «быстрой» мышце, вероятно, может
быть обнаружена более низкая концентрация КрФ — до 75%
[Hirvonen J. и др., 1987]), что заведомо выше значений, фикси-
руемых в конце более продолжительной работы [Greenhalf P.L.
и др., 1996; Nevill М.Е. и др., 1996] и выше критической вели-
чины (в среднем по мышце), при которой энергии фосфагенов
начинает не хватать для обеспечения максимальной мощности
и силы взаимодействия миофиламентов [Greenhalf P.L. и др.,
116
1996] . Для того чтобы снять это противоречие примем во вни-
мание следующие четыре факта:
1. скорость расходования КрФ пропорциональна АТФ-
азной активности миозина или креатинфосфокина-
зы [Хочачка П., Дж. Сомеро, 1988; Hirvonen J. и др.,
1987;Greenhalf P.L. и др., 1996], которые максимальны у
быстрых гликолитических мышечных волокон (БгМВ);
2. концентрация КрФ в целом пропорциональна массе
сократительных белков мышц (актина), хотя и может
несколько увеличиваться при тренировке [Биохимия:
Учебник для институтов физ. культуры, 1986; Яковлев
Н.Н., 1983];
3. относительная сила быстрых и медленных МВ в мышце,
или не различается [Биохимия: Учебник для институтов
физ. культуры, 1986], или на 10-30% выше у БМВ. С уче-
том п. 2, это означает, что запасы КрФ в быстрых и мед-
ленных МВ различаются в предельном случае не более
чем на 40% (в среднем 10-30% [Greenhalf P.L. и др., 1996;
Nevill М.Е. и др., 1996]);
4. соотношение АТФ-азной активности миозина у ММВ и
БгМ В определено как 1: 3-4 [Биохимия: Учебник для ин-
ститутов физ. культуры, 1986] (реально скорость расхода
КрФ в ММВ медленнее, чем в БМВ только в 2-3 раза и
это соотношение зависит от времени после начала мак-
симальной работы [Greenhalf P.L. и др., 1996; Nevill М.Е.
и др., 1996]).
Из приведенных данных следует очень существенный для
предмета нашего исследования вывод — время снижения
концентрации КрФ до критического уровня зависит от АТФ-
азной активности миозина и активности КФК, поэтому если
при максимальной мощности волокон запасов этого субстрата
в БгМВ хватает на 5-6 с [Hirvonen J. и др., 1987] (хотя реально
скорость КФК реакции начинает снижаться уже со второй с
максимальных тетанических сокращений [Greenhalf P.L. и др.,
1996]), то в ММВ его запасов должно хватать на 15-20 с. Этот
вывод в целом подтверждается известной схемой Н.И. Волко-
ва [1964] о динамике вклада КФК-реакции в энергообеспече-
ние мышечной работы и данными электростимуляции мышц
117
(когда расход субстрата идет с максимальной скоростью во
всех типах МВ) (рис. 5) [Greenhalf P.L. и др., 1996].
Однако при интерпретации данных рис. 5 следует учи-
тывать различную длительность активных и пассивных фаз
в реальных локомоциях и при электростимуляции. Други-
ми словами, начало существенного снижения максимальной
мощности около 8-12-й с работы связано с исчерпанием КрФ
главным образом в БгМВ, в то время как в ММВ, у которых ак-
тивность АТФ-азы ниже, а запасов КрФ не намного ниже (на
10-30% — см. выше), концентрации КрФ должно вполне хва-
тать для обеспечения достаточно высокой мощности работы
этих волокон в течении 15-20 с, даже без привлечения допол-
нительных источников АТФ, которыми являются гликолиз и
окислительное фосфорилирование.
Обратим внимание, что эти цифры приведены для мышеч-
ной работы «спринтерского» вида. Однако для нашего при-
мера (50% от МАМ) требуется коррекция. По данным И.М.
Козлова [Физиология мышечной деятельности, 1982], дли-
тельность активности основных мышечных групп в сприн-
терском беге (250-280 шаг/мин) составляет 50-80% времени от
длительности бегового цикла. В то время как при сопостави-
мой с нашим примером интенсивности бега (200 шаг/мин) эта
длительность составляет только 30-50% от длительности цик-
ла. Это означает, что время гладкотетанической активности
ф Изометр.сила
(%)
------КрФ БМВ
--ZV--Кр Ф ММВ
--Ф---АТФ БМВ
--ЭК--АТФ ММВ
Время стимуляции (с)
Рис. 5. Изометрическая сила, А ТФ и КРФ в медленных (ММВ) и быстрых
(БМВ) мышечных волокнах в течении 20 с интенсивной электрической
стимуляции (1,6с/ 1,6с; 50Гц) [Greenhalf Р.L. и др., 1996]
118
рекрутированных волокон в беге на средние дистанции будет
меньше на 40-60%, чем в спринте. В других локомоциях, ве-
роятно, сохраняется та же тенденция, хотя могут быть коли-
чественные различия. Следовательно, применительно к ММВ
и БоМВ, рекрутированным с самого начала в нашем примере,
скорость исчерпания КрФ (с учетом нелинейной зависимости
между запасами КрФ и скоростью ресинтеза АТФ) увеличится
на те же 40-60% и составит, например для ММВ, цифру по-
рядка 25-35 с. Следовательно, в течение этого времени ММВ
не будут испытывать дефицита энергии (с допущением о том,
что скорость КФК-реакции достаточна для компенсации раз-
ницы между запросом и ресинтезом АТФ в гликолизе и дыха-
тельном фосфорилировании) для актино-миозинового взаи-
модействия.
Доказательством вышесказанного являются прежде всего
данные Н.И. Волкова (1969) (рис. 6) о зависимости величины
алактатной фракции О2-долга от предельного времени работы.
Хорошо видно, что при длительности работы 23 с (бег на 200
м) алактатный О2-долг не максимален. Однако при длительно-
сти 37 с (бег на 300 м) и выше эта фракция максимальна и не
меняется с увеличением длительности работы. Следователь-
но, можно предположить, что алактатный О2-долг достигает
максимума приблизительно в диапазоне 25-35 с. Так как два
основных компонента алактатного О2-долга — КрФ и оксими-
оглобин — исчерпываются параллельно, а не последовательно,
то указанное время ( в среднем 30 с) — это максимальное время
работы (с предельной интенсивностью), когда в мышцах име-
ются запасы КрФ. Причем, принимая во внимание АТФ-азную
и КФК-активность БМВ и ММВ, логично предположить, что
дольше всего запасы КрФ сохраняются в ММВ ( с учетом по-
стоянного ресинтеза АТФ в ходе окислительного фосфорили-
рования). Это же подтверждается данными рис. 5.
Таким образом, в течение по крайней мере 25-30 с ММВ мог-
ли бы работать вообще без доступа кислорода, используя КФК-
реакцию и оксимиоглобин в качестве источников, обеспечи-
вающих ресинтез АТФ. Конечно, при этом градиент снижения
мощности был бы значительно выше из-за снижения скорости
КфК-реакции в связи со снижением концентрации КрФ.
119
10
02-долг (л)
| —Алактаный О2-долг —Лактатный 02-долг |
Рис. 6. Значения алактатного и лактацидного «кислородного долга» у
одной и той же группы бегунов в зависимости от длительности бега «на
результат» [Волков И.И., 1969]
Если приведенные рассуждения верны, то, во-первых, это
объясняет, почему концентрация КрФ в конце спринтерского
бега (10-20 с) не падает до 10-20%, как при электростимуля-
ции до утомления в течение 30 с [Greenhalf P.L. и др., 1996] или
как при циклической работе длительностью 30 с и более [Nevill
М.Е. и др., 1996], а во-вторых, это является еще одним доводом
в пользу того, что за 30-40 с врабатывания мышцы (по край-
ней мере ММВ, в которых высока концентрация миоглобина
— резерва кислорода, роль которого хорошо продемонстриро-
вана в экспериментах по сокращению мышц в нормальных и
ишемических условиях [Greenhalf P.L. и др., 1996]) не испыты-
вают нехватки энергии из-за «инерционности» ССС. То есть,
по-видимому, не случайно длительность двух процессов: 1)
постепенного снижения КрФ в ММВ и 2) нарастания VO2 до
порогового уровня (рис. 4) приблизительно совпадают и под-
тверждаются расчетами [308, Nevill М.Е. и др., 1996], что при
спринтерском педалировании «во-всю» в течении 30 с 30-40 %
энергии вырабатывается за счет аэробных источников.
Единственными экспериментальными данными, которые
могли бы свидетельствовать в пользу гипотезы о недостаточ-
ном снабжении мышц кислородом в начальный период мы-
шечной работы, могли бы быть следующие. Это:
120
— повышенная концентрация лактата в первые 5-10 мин
работы на уровне или немного выше анаэробного порога
[Holloszy J.О., 1996; Rusko Н. и др., 1986]. Показано, что
по мере продолжения работы, концентрация Ла может
снижаться [Волков Н.И., 1969; Holloszy J.О., 1996];
— существенно более низкие значения КрФ в мышце на
2-й минуте работы с предельной длительностью 20 ми-
нут, по сравнению с 5-й минутой этого же упражнения
[Платонов В.Н., 1997].
Однако, как и в других случаях, без прямых измерений
внутриклеточного напряжения кислорода или концентрации
оксимиоглобина эти примеры не являются достаточно убеди-
тельными.
Так, причиной небольшого повышенного выделения лак-
тата и снижения КрФ может быть все тот же стартовый раз-
гон. Произведем несложные расчеты. Например, используя
данные Н.И. Волкова [Волков Н.И., 1969] по избыточному
энергорасходу в начале бега со скоростью 5,2 м/с, полученные
на основе расчета калорического эквивалента продукции Ла,
можно получить цифру порядка 420-450 «избыточных» кало-
рий для человека массой 70 кг. Используя же формулы элемен-
тарной физики, получаем, что для разгона тела массы 70 кг до
скорости 5,2 м/с необходимо затратить 237 кал. Учитывая, что
механическая эффективность выполнения физической рабо-
ты человеком редко превышает 25%, получаем цифру необ-
ходимых энергозатрат для разгона спортсмена в 948 калорий,
которая существенно превышает полученные в эксперименте
420-450 кал. Вероятно, что недостающие калории «скрыты» в
алактатной и лактацидной фракциях кислородного долга.
Сюда же можно прибавить:
— возможную пониженную эффективность работы мышц
в начале работы, когда их температура не превышает
уровня покоя;
— известный факт, что в метаболизме лактата участвуют
миокард и дыхательные мышцы, которые в первую мин
работы еще не начали потреблять «свою долю» лактата и
поэтому может накапливаться его «излишек» в крови;
— начало работы сопровождается, как правило, повы-
121
шенным выбросом катехоламинов, что вполне может
привести к повышению (относительно последующего
устойчивого состояния) продукции лактата через акти-
визацию аденилатциклазы, вызывающую накопление
ц-АМФ и прямое действие на фосфорилазу и фосфоф-
руктокиназу;
— и, наконец, наиболее существенное — в подробном об-
зоре Holloszy [Holloszy J.О., 1996, с. 6-7] рассматривает-
ся недавно открытое явление «реверсивной активизации
фосфорилазы ионами Са++». Смысл его в том, что в нача-
ле продолжительного упражнения действительно имеет
место активизация фосфорилазы ионами Са++, что, как
известно, является пусковым моментом гликолиза в на-
чале работы. Причем получено, что в этом случае про-
исходит избыточная продукции Ла выше величины, не-
обходимой для удовлетворения реального энергозапроса
МВ. Однако через несколько минут происходит «ревер-
сия активизации», выражающаяся в прекращении акти-
визации фосфорилазы ионами Са++, несмотря на про-
должающуюся сократительную активность и отсутствие
каких бы то ни было признаков утомления. Одним из ре-
зультатов этого, как предполагают авторы, является уве-
личение доли окисляемых жиров (не очевидно, что при
работе на уровне анаэробного порога и выше этот меха-
низм может функционировать — прим, наше), а биоло-
гический смысл — сначала ускоренное обеспечение ми-
тохондрий пируватом, а затем — «экономия углеводов».
Таким образом, кроме формального умозаключения что:
«Если потребление кислорода еще не возросло, а мощность
работы уже высокая, то кислорода должно не хватать,» — ни-
каких других оснований для вывода о дефиците кислорода для
нормального функционирования митохондрий в начале работы
найти не удалось.
В заключение этого отступления отметим, что в настоя-
щее время нельзя точно оценить, какую долю в утомление
спортсменов вносит постепенное ускорение потребления О2
мышцами и вносит ли вообще, однако, достаточно опреде-
ленно можно утверждать, что большая или меньшая скорость
122
нарастания потребления О2 мышцами зависит в основном от
периферических, а не центральных факторов [Физиология че-
ловека / Под ред. Р. Шмидта и Г. Тевса. — Т.З., 1996] и, следо-
вательно, совершенствуется в процессе тренировки совместно
с последними.
В подтверждение этого можно привести следующее: во-
первых, увеличение МОК (доставка О2 кровью) является
функцией периферического сопротивления — он возраста-
ет, когда падает периферическое сопротивление в результате
«опережающего расширения сосудов» под воздействием акти-
визирующейся симпатической системы и местных метаболи-
ческих реакций [Физиология человека/Под ред. Р. Шмидта и
Г. Тевса. — Т.З., 1996; Saltin В., 1985] и выходящего из синапсов
двигательных нервов ацетилхолина; а во-вторых, существует
явление «коиннервации», когда двигательные реакции и соот-
ветствующие им вегетативные изменения «запускаются» ко-
рой одновременно, т. е. они точно «подогнаны» друг другу во
времени. Причем эти сочетанные приспособительные реакции
в начале мышечной работы предшествуют запланированному
действию, т.е. являются «опережающими реакциями» [Физио-
логия человека / Под ред. Р. Шмидта и Г. Тевса. — Т.З., 1996]
Таким образом, имеются основания не привлекать гипотезу
о неадекватности снабжения мышц кислородом в период вра-
батывания (т.е. идеологию, основанную на прямой и обратной
пастеровской реакции) к объяснению процессов, происходя-
щих в мышечных волокнах. Точно также нет никаких осно-
ваний утверждать, что такие явления имеют место и в период
квазиустойчивого состояния, когда, с одной стороны, потре-
бление кислорода близко к максимуму, а с другой — явления
утомления продолжают нарастать (см. ниже).
Продолжим рассмотрение процессов в МВ в период враба-
тывания.
В ММВ и БоМВ появление Na+, Са++в саркоплазме, воз-
растание роли других агентов, активизирующихся по мере
снижения концентрации КрФ, ускоряют гликолиз и глико-
генолиз, которые вММВиБоМВ обеспечивают субстратом
(пируват) окислительное фосфорилирование в митохондриях,
а в БоМВ — еще и анаэробный гликолиз. Причем если КФК
123
является сильным конкурентом для фосфорилазы и фосфоф-
руктозы в отношении саркоплазматических АДФ и АМФ, т. е.
можно ожидать, что до замедления КФК-реакции гликолиз не
будет активизироваться максимально [Хочачка П., Дж. Соме-
ро, 1988] (однако в экспериментах с максимальными вызван-
ными сокращениями мышц показано, что гликолиз активизи-
руется максимально к 3-й с работы [Greenhalf P.L. и др., 1996],
то появление свободного креатина (Кр) немедленно включа-
ет КрФ-челнок, вызывающий появление митохондриальной
АДФ и, следовательно, быструю активизацию мембранного
фосфорилирования. Другими словами, на уровне мышечного
волокна (например, БоМВ) представления о более ранней ак-
тивизации гликолиза, по сравнению с дыхательными процес-
сами, скорее всего не имеет под собой оснований.
В результате доступности пирувата (в силу разницы между
скоростью его формирования и потребления в митохондриях)
для ЛДГ и наличия АДФ в БоМВ начинается продукция и на-
копление ионов Ла и Н+, которые из-за своих небольших раз-
меров легко диффундируют в кровь и соседние ММВ [Волков
Н.И., 1969; Голник Ф.Д., ГермансенЛ., 1982]. Первоначально
это является положительным фактором, так как:
— стимулируется анаэробный гликолиз [Хочачка П., Дж.
Сомеро, 1988];
— обеспечивается торможение окисления жиров в ММВ и
снабжение митохондрий дополнительным субстратом —
Ла, что обеспечивает максимально возможную скорость
ресинтеза АТФ в митохондриях [Хочачка П., Дж. Соме-
ро, 1988];
— повышает протонный потенциал на внутренней мембра-
не митохондрий, и активизируется митохондриальная
пируватдегидрогеназа, что, как предполагается [Албертс
Б. идр., 1994; BangsboJ., 1996], ускоряет окислительный
ресинтез АТФ;
— выход Н+ в капиллярную кровь способствует проявле-
нию «эффекта Бора», увеличивая степень отдачи кисло-
рода притекающей кровью [Карпман В.Л., 1994, Физио-
логия человека / Под ред. Р. Шмидта и Г. Тевса. — Т.З.,
1996];
124
— Н+ взаимодействуют с бикарбонатным буфером, вызы-
вая появление «излишка СО2», который интенсифици-
рует внешнее дыхание;
— эти метаболиты, наряду с К+, воздействуют на сфинкте-
ры артериол, расширяя капиллярное ложе и т.д.
Действие этих факторов вместе с постепенным умеренным
снижением [Голник Ф.Д., ГермансенЛ., 1982] концентрации
КрФ приводят к тому, что к концу первого этапа врабатыва-
ния, когда заканчивается быстрый прирост VO2 (рис. 4), име-
ют место;
— достижение величиной VO2 уровня, примерно соот-
ветствующего анаэробному порогу [Morton, R.H. и др.;
1994];
— максимальная активизация гликолиза и гликогенолиза в
активированных БоМВ;
— несколько повышенная концентрация М К в мышцах и в
меньшей степени в венозной и артериальной крови.
Как предположено выше, механическая мощность сокра-
щения ММВ, определяемая их АТФ-азной активностью, не
должна существенно меняться при смене мощного энергети-
ческого источника (креатинфосфокиназная реакция) на менее
мощный (окислительное фосфорилирование). Следовательно,
эти МВ большую часть дистанции функционируют на полную
мощность, внося максимально возможный вклад в генерируе-
мую мышцей силу.
В БоМВ ситуация может быть иная. В связи тем что мак-
симальная мощность этих МВ в 3-4 раза выше, чем у ММВ,
для тех из БоМВ, которые работают в условиях гладкого те-
тануса, доступность «мощного субстрата» — КрФ — должна
обеспечивать максимально возможный механический вклад
этих МВ в генерируемую мышцей силу, так как имеется воз-
можность одновременного замыкания максимального числа
актино-миозиновых мостиков в единицу времени [Биохимия:
Учебник для институтов физ. культуры, 1986]. Но по мере сни-
жения концентрации КрФ (а длительность снижения запасов
КрФ до критического уровня составляет, вероятно, не более
4-10 с) и перехода на менее мощные источники энергообеспе-
чения, вклад этих волокон будет уменьшаться. Следовательно,
125
в связи с необходимостью поддержания постоянной скорости
передвижения, моторная зона коры головного мозга должна
увеличивать свое активизирующее влияние на альфамотоней-
ронный пул мышцы, что выражается в нормальном случае в
увеличении частоты импульсации а-мотонейроновтех ДЕ, ко-
торые функционировали с немаксимальной интенсивностью
(вторая группа БоМВ, см. выше), а также в подключении до-
полнительных, более высокопороговых ДЕ, что выражается в
увеличении амплитуды интегрированной ЭМГ мышцы, если
не меняется биомеханика локомоции. Таким образом, про-
цесс подключения новых, более высокопороговых, ДЕ и во-
влечение БМВ в работу происходит непрерывно с возможной
цикличностью этого процесса 4-10 с.
То что работа нервно-мышечного аппарата организована
именно таким образом, может следовать из факта возрастания
ИЭМГ работающих мышц и сдвига их частотного спектра в
сторону больших частот [Моногаров В.Д., 1980] по мере про-
должения работы.
Во вновь вовлеченных МВ процессы должны происходить
по тому же сценарию. Сначала расходуется КрФ, обеспечивая
максимальный вклад быстрого МВ в генерируемую силу. По
мере переключения на гликолиз и окислительное фосфорили-
рование, мощность работы волокон падает, а это вызывает не-
обходимость рекрутирования новых ДЕ.
Таким образом, если принять длительность периода враба-
тывания в 1 мин, то за это время может произойти несколько
эпизодов с увеличением частоты импульсации работающих
ДЕ и подключения новых ДЕ, причиной чего будет сниже-
ние скорости креатинфосфокиназной реакции в БоМВ из-за
уменьшения доступности субстрата.
В это время продолжается нарастание потребления кис-
лорода мышцей, что обусловлено развертыванием на полную
мощность процессов окислительного фосфорилирования в
функционирующих и рекрутируемых БоМВ. Период от нача-
ла замедления прироста VO2 до окончания фазы врабатывания
можно назвать вторым периодом врабатывания. К его оконча-
нию потребление кислорода мышцей должно быть практиче-
ски максимальным. Так как известно, что в пока еще неактив-
126
ных БгМВ плотность митохондрий и активность их ферментов
относительно невелика [Хочачка П., Дж. Сомеро, 1988; Верхо-
шанский Ю.В., 1988], поэтому их вовлечение в работу гипоте-
тически не должно привести к существенному приросту VO2
мышцей.
5.1.2. Фаза квазиустойчивого состояния
Если бы мощность работы не превышала порогового уров-
ня (75-90% от МПК для квалифицированных спортсменов), то
после периода врабатывания показатели внешнего дыхания,
потребления кислорода, ЧСС, гемодинамики и др. практи-
чески не изменяли бы своей величины на протяжении доста-
точно длительного периода. Как известно, это состояние на-
зывается устойчивым (steady-stay). Применительно к средним
дистанциям эта фаза называется фазой квазиустойчивого со-
стояния или фазой компенсированного утомления [КоцЯ.М.,
1986; Моногаров В.Д., 1980], так как наблюдается постепенная
интенсификация практически всех вегетативных функций,
обеспечивающих сокращение мышц.
В мышечных волокнах в этот период могут происходить
следующие процессы.
Функционирование БоМ В в условиях гладкого тетануса со-
провождается максимальным энергозапросом этих волокон,
который при пониженной концентрации КрФ обеспечивает-
ся предельной интенсификацией гликолитических реакций
и окислительного фосфорилирования. Емкость буферных
систем МВ конечна, поэтому происходит накопление ионов
водорода. Это постепенно приводит к достижению ими неко-
торого «критического» уровня, который вызывает ряд эффек-
тов, понижающих механическую силу тяги и эффективность
работы мышечных волокон. Это:
— угнетение миозиновой АТФ-азы (фактор менее значи-
мый, чем адекватность энергообеспечения [Nevill М.Е.
и др., 1996]);
— понижение активности ключевых ферментов гликолиза
— фосфорилазы и фосфофруктокиназы, но при pH ниже
6,6 [Bangsbo J., 1996];
127
— нарушается сопряжение реакций окисления с ресинте-
зом АТФ [Биохимия: Учебник для институтов физ. куль-
туры, 1986].
Скорость снижения внутриклеточного pH зависит: от соот-
ношения активности окислительных и анаэробных гликолити-
ческих ферментов; емкости буферных систем (бикарбонатной,
белковой, фосфатной, а также — карнозин, креатин, ансерин
[Хочачка П., Дж. Сомеро, 1988]); и скорости, с которой элими-
нируются лактат и протоновые эквиваленты (Н+) [Westerblad
Н. и др., 1991]. Можно добавить, что активность гликолитиче-
ских ферментов коррелирует с АТФ-азной активностью мио-
зина. Другими словами, также как и в случае со скоростью рас-
ходования КрФ, быстрее всего должны утомляться наиболее
быстрые МВ, что, собственно, хорошо известно [Nevill М.Е. и
др., 1996].
Существуют расчеты, согласно которым наличие АДФ в мы-
шечном волокне (а в БМВ, где высока АТФ-азная активность
во время тетануса при циклических локомоциях АДФ всегда
будет присутствовать) может приводить к полному истощению
запасов КрФ [Хочачка П., Дж. Сомеро, 1988]. Это подтвержде-
но экспериментально [Nevill М.Е. и др., 1996], а снижение ско-
рости ресинтеза АТФ в ходе реакций анаэробного гликолиза и
в митохондриях приводит к очень существенному снижению
механической тяги этих волокон, что стимулирует вовлечение
новых ДЕ в работу.
Предположим, что в течение последующих 20-40 с в работу
оказываются вовлеченными и все БоМВ, и значительная часть
быстрых гликолитических МВ (БгМВ). В этих МВ, также как
и в других МВ, по мере расхода КрФ активизируются глико-
литические и дыхательные процессы. Но концентрация ми-
тохондрий в БгМВ меньше, поэтому прирост потребления 02
относительно небольшой. В то же время высокая активность
АТФ-азы и ключевых ферментов гликолиза приводит к бы-
строму исчерпанию КрФ и «закислению» этих волокон.
ММВ имеют сбалансированное соотношение АТФ-азной
активности миозина и мощности митохондриальной системы
ресинтеза АТФ (см. выше). В фазе квазиустойчивого состояния
при адекватном снабжении кислородом на полную мощность
128
протекают реакции аэробного гликолиза, что обусловлено на-
личием всех факторов, интенсифицирующих соответствую-
щие реакции (прежде всего наличие АДФ между мембранами
митохондрий), включая высокую концентрацию катехола-
минов. Однако в этих МВ, вероятно, может постепенно на-
капливаться Ла в результате его диффузии из соседних БМВ
в количествах, превышающих возможности окисления Ла в
митохондриях. В таких условиях Ла становится предпочти-
тельным субстратом и обуславливает максимальную скорость
выработки энергии дыхательным путем при практически пол-
ном угнетении b-окисления жиров [Хочачка П., Дж. Сомеро,
1988]. Н+ поступают в ММВ также легко, как и Ла. В начале,
как отмечалось выше, это имеет положительное значение —
увеличивается протонный градиент на внутренней мембране
митохондрий. Кроме того, на внешней мембране митохондрий
протекает окисление НАД-Н, не связанное с ресинтезом АТФ,
но имеющее буферирующий эффект, так как устраняет ионы
водорода. Энергия при этом рассеивается в виде тепла [Био-
химия: Учебник для институтов физ. культуры, 1986], а сниже-
ния скорости потребления кислорода не происходит [Албертс
Б. и др., 1994]. Тем не менее накопление Н+ выше некоторого
«критического» уровня, так же, как и в БМВ, приведет в кон-
це концов к снижению мощности аэробной энергопродукции,
глубокому исчерпанию по этой причине КрФ и снижению
вклада этих МВ в механическую работу. Однако следует ожи-
дать, что произойдет это позже, чем в БМВ, включившихся в
работу в то же время. Факт глубокого исчерпания КрФ в конце
утомительной работы следует рассматривать как доказатель-
ство снижения скорости окислительного фосфорилирования
в ММВ в результате их закисления. В противном случае ис-
черпания КрФ не происходило бы (по причине его ресинтеза)
так же, как этого не происходит при работе в устойчивом со-
стоянии, когда концентрация КрФ не понижается ниже 80%
от исхода, несмотря на интенсивное функционирование ми-
тохондрий в ММВ и БоМВ [Rusko Н. и др., 1986].
Что произойдет с волокнами, в которых в силу накопления
высоких концентраций Н+ сильно ограничена энергопродук-
ция и, следовательно, механическая тяга этих МВ?
129
Известно, что в некоторых режимах работы развиваемая
МВ сила начинает существенно снижаться в условиях, когда
эфферентный приток и амплитуда потенциала действия этих
МВ остаются прежними [Физиология человека / Под ред. Р.
Шмидта и Г. Тевса. — Т.З., 1996, Холлоши Дж.О., 1982]. В то же
время показано [Коц Я.М., 1986], что при предельных усилиях
в конце более длинной дистанции амплитуда ИЭМГ оказыва-
ется существенно ниже, чем в конце более короткой, или тем
более при максимальном изометрическом напряжении той же
неутомленной мышцы [Моногаров В.Д., 1980; КоцЯ.М., 1986
]. На основе последнего факта и исследований функциониро-
вания отдельных ДЕ [Персон Р.С., 1985] при изометрическом
режиме выдвинута гипотеза [Holloszy J.O., 1996] о возможности
«выключения» утомленных ДЕ из работы. Эта гипотеза под-
тверждается недавно полученными на людях данными [Sahlin К
и др.; 1997] (см. выше), что в конце истощающей 77-минутной
работы содержание КрФ в мышце имело «мозаичный» вид, т.е.
волокна с истощенными запасами КрФ чередовались с волок-
нами, где КрФ было столько же, сколько в покое. Причем очень
интересен факт, что «истощенные» и «не истощенные» МВ
встречались как среди БМВ, таки среди ММВ.
Тем не менее даже если эта гипотеза справедлива, то ее
принятие не изменит общей схемы процессов работы мышц,
их энергообеспечения и причин утомления, описанных выше.
Следует только учитывать, что факт практически полного ис-
черпания запасов КрФ в мышце в конце предельной работы
длительностью 40-180 с [Коц Я.М., 1986] свидетельствует о
том, что если и происходит выключение МВ из работы, то это
случается не ранее, чем будет полностью исчерпан их метабо-
лический потенциал. Это является еще одним косвенным до-
казательством «периферической гипотезы» утомления на этих
дистанциях.
Существен вопрос: «Как долго могут работать быстрые мы-
шечные волокна различного типа при гладкотетаническом
сокращении в условиях адекватного снабжения кислородом
до момента исчерпания КрФ и понижения pH до уровня, при
котором существенно замедляется гликолиз и окислительное
фосфорилирование?»
130
К сожалению, в литературе мы не нашли прямого ответа на
л от вопрос применительно к циклическим локомоциям. По-
лому прибегнем к косвенным данным. Максимальную ско-
рость закисления мышцы можно было бы измерить в экспери-
менте, в котором давалось бы задание работать «во-всю», когда
спортсмен развил бы максимальную алактатную мощность с
самого начала, а затем старался бы ее удерживать сколько воз-
можно. В этом случае с самого начала были бы рекрутированы
практически все ДЕ. При такой работе через 3-6 с скорость гли-
колиза достигла бы максимума в БгМВ, через 5-10 с — в БоМВ
и с этого момента началось бы накопление МК с максималь-
но возможной для этого человека скоростью. Механическая
мощность, развиваемая работающими мышцами, постепенно
снижалась бы. Методом биопсии в серии таких эксперимен-
тов можно было бы определить, какой величине механической
мощности соответствуют наименьшие значения pH в основ-
ных мышцах. Таким образом, время от начала снижения мощ-
ности (3-5 с) до минимума pH было бы максимальным време-
нем функционирования гликолиза у данного испытуемого.
Однако нам не известны такие эксперименты. Хотя известно,
что у нетренированных людей мощность работы «во-всю» при
беге на пассивном третбане (когда испытуемый сам раскручи-
вает ленту) [Lakomy Н.К.А., 1986] и педалировании на вело-
эргометре [Nevill М.Е. и др., 1996 | к 30-й с падает на 40-50%.
Если принять во внимание соотношение максимальной мощ-
ности КФК-реакции и гликолиза, а также предположение, что
к концу этой работы ММВ обеспечивают около 150 Вт мощно-
сти, то получим, что к этому времени (к 30-й с) гликолиз еще
не отработал до момента своей остановки.
Уточнение можно провести на основе расчетов Н.И. Вол-
кова (1996) (рис. 6). У квалифицированных бегунов минималь-
ное предельное время работы, при котором можно достичь
максимального лактатного О2-долга, — 35 с. Это означает, что
к этому времени гликолиз уже успевает «сработать» с макси-
мальной производительностью во всех МВ.
Запомним эту цифру.
На этой дистанции требуемое условие — рекрутирование
всех ДЕ — практически выполняется. Однако мощность немно-
131
го ниже, чем в спринте. Поэтому можно ожидать, что в БгМВ
интенсивный гликолиз начнется не с 3-6-й с, а чуть позже,
предположим, с 10-й с. Эти МВ обладают максимальной АТФ-
азной, КФК-азной и гликолитической активностью, поэтому
сразу начнется интенсивное накопление Ла и Н+, которые бу-
дут частично буферироваться, а также интенсивно проникать
в соседние МВ. БоМВ, после снижения в них концентрации
КрФ, также начнут продуцировать МК примерно с 15-й с, не-
смотря надостаточно высокий окислительный потенциал. Та-
ким образом, функционирование БгМВ в течении 35 — 10 = 25 с
и БоМВ в условиях интенсивного притока Н+ из БгМВ — в те-
чении 35 — 15 = 20 с приводят к максимальному накоплению
Ла и, как следует ожидать, к снижению pH в мышце в целом до
6,3-6,4, так как даже при pH 6,45 скорость гликолиза (и глико-
генолиза) существенно не снижалась [Spriet L.L. и др., 1987]
при минимальных значениях в БМВ. Это «блокирует» глико-
лиз, однако окислительное фосфорилирование в ММВ, веро-
ятно, в этих условиях не страдает. Так как если учесть, что мак-
симальная скорость выхода лактата из мышцы в кровь — 4,5
ммоль/мин [Jorfeldt L., 1978] и принять грубое допущение, что
скорость диффузии Ла и Н+ через мембраны ММВ одинакова
и происходит с той же интенсивностью, то за 20-25 с концен-
трация Н+в ММВ не может достичь высоких значений.
Если приведенные вычисления в целом верны, то из этого
следует:
1. Анаэробный гликолиз в мышцах, несущих основную на-
грузку, достигает максимальной скорости к 15-й с и под-
держивается на этом уровне только до 35-40-й с, т.е. 20 с.
Это полностью согласуется с имеющимися данными о
максимальной скорости накопления Ла в мышцах [Гол-
ник Ф.Д., Германсен Л., 1982] и результатами экспери-
ментов с электростимуляцией мышц, где показано, что
максимальную скорость гликолиз может поддерживать
только 20 с [Greenhalf P.L. и др., 1996].
2. При большей длительности упражнения скорость гли-
колиза не максимальна.
С одной стороны, это подтверждает положенную в основу
наших рассуждений гипотезу о том, что в циклических локомо-
132
циях соблюдается правило размера Хеннемана [1965], а с другой
— ставит под сомнение роль мощности гликолиза как фактора,
лимитирующего результат при длительности работы более 40 с!
Действительно, если не все МВ задействованы на полную
мощность с самого начала дистанции, то и суммарная энерго-
продукция гликолиза в каждый момент времени будет склады-
ваться только из того количества АТФ, которое вырабатывается
в функционирующих на полную мощность БМВ. Другими сло-
вами, энергопродукция гликолиза распределена во времени ра-
боты и его вклад в общее количество ресинтезируемого АТФ бу-
дет определяться не мощностью, а емкостью этого источника.
В этой связи возникают вопросы, является ли высокая
активность ключевых ферментов гликолиза в БМВ положи-
тельным или отрицательным фактором в отношении общей
энергопродукции этого источника при предельной работе от
40 до 120 с, а также какие внутримышечные или центральные
факторы определяют емкость гликолиза? От решения этого
вопроса зависит выбор средств и методов тренировки с целью
повышения гликолитических возможностей человека.
Действительно, высокая активность гликолиза в БМВ при
гладкотетаническом режиме неминуемо приведет к ускорен-
ной продукции Н+. Однако при этом каждое МВ будет функ-
ционировать с большей мощностью, следовательно, меньшее
число быстрых ДЕ будет рекрутировано и средняя интенсив-
ность гликолиза, необходимая для поддержания дистанцион-
ной скорости (и закисляющая мышцу), останется прежней. В
этом случае следует вывод, что при длительности работы более
40 с высокая активность ферментов гликолиза не нужна.
Однако можно рассуждать по-другому. Например, есть
мнение, что буферная емкость ткани тонко регулируется спо-
собностью этой ткани генерировать протоны [Хочачка П.,
Дж. Сомеро, 1988], т.е. буферная емкость мышцы возрастает
параллельно ее гликолитической способности. В этом случае
при той же средней мощности гликолиза будет ресинтезиро-
вано большее количество АТФ при снижении pH на единицу.
Если буферная емкость действительно тесно коррелирует с ак-
тивностью ферментов гликолиза, то высокая гликолитическая
активность — положительный фактор.
133
Внутримышечная буферная емкость мышцы определяется
бикарбонатным, белковым, фосфатным буферами и гистидин-
содержащими дипептидами и белками. Она возрастает парал-
лельно массе белков органелл клетки [Хочачка П., Дж. Соме-
ро, 1988]. Тогда возникает вопрос: «А возможно ли увеличение
концентрации буферирующих веществ без существенной ги-
пертрофии МВ «саркоплазматического» или «сократитель-
ного» типа?» Считается, что такая возможность существует у
ныряющих млекопитающих [Хочачка П., Дж. Сомеро, 1988].
Кроме того, если бы увеличение буферной емкости целиком
зависело от массы органелл клеток, то в этом случае наиболь-
шую буферную емкость имели спринтеры, которые обладают
большей относительной мышечной массой ( в частности, мас-
сой БМВ), чем бегуны на 400 м и тем более на 800 м. И по-
вышение буферной емкости лучше всего коррелировало бы с
увеличением мышечной силы и содержанием БМВ в мышцах,
поэтому спринтеры были бы способны накапливать самый
большой лактатный О2-долг и достигать наибольших величин
концентрации Ла в крови. Однако такой способностью обла-
дают именно бегуны на 800 м [Волков Н.И., 1969]. От сприн-
теров их отличает лучшая капилляризация мышц, большая
масса митохондрий, меньший процент БМВ. Первый фактор,
предположительно, облегчает выход лактата в кровь. Второй
— прямо увеличивает буферную емкость [Биохимия: Учебник
для институтов физ. культуры, 1986]. Третий фактор должен
приводить к уменьшению скорости накопления лактата. Кро-
ме того, в литературе есть данные об увеличении буферной ем-
кости мышц под воздействием тренировки, близкой по смыс-
лу к тренировке средневиков (наличие «гликолитического» и
«аэробного» компонентов в нагрузке).
К сожалению, нам не известны работы, в которых было бы
прямо показано увеличение концентрации буферирующих
веществ без параллельного увеличения массы или 1) сократи-
тельных белков (и сопутствующего увеличения саркоплазма-
тического ретикулума и некоторых других органелл клеток),
или 2) митохондриального белка и сопутствующего увеличе-
ния капилляризации мышц и улучшения региональных со-
судодвигательных реакций. В связи с этим в качестве рабочей
134
гипотезы мы предполагаем, что увеличение буферной емкости
мышц и, следовательно, увеличение емкости гликолиза обу-
словлено в основном гипертрофией мышечных волокон (как
правило, под воздействием силовой тренировки) и повыше-
нием их окислительного потенциала (под воздействием аэроб-
ной тренировки, способствующей повышению ОП всех типов
мышечных волокон).
Схема продукции и метаболизма лактата и ионов водорода
в БМВ в условиях тетануса может быть представлена на рис. 7.
Данная схема демонстрирует, что накопление (4) лактата и
ионов водорода в мышечной ткани является разницей между
скоростью их продукции, обусловленной массой и степенью ак-
тивизации ключевых ферментов гликолиза (фосфорилаза, фос-
фофруктокиназа), и скоростью удаления, определяемой ско-
Са++, Na+, АДФ/АТФ, АМФ, Ф1
_____________11_______________
Фосфорилаза, фосфофруктокиназа
И
молекулы гликогена
ГЛ 1 ГЛ 1 ГЛ 1 | ГЛ 1 ГЛ 1 ГЛ 1 | ГЛ 1 ГЛ 1 ГЛ 1 | ГЛ 1 ГЛ 1 гл
АТФ Ц АТФ Ц АТФ Ц АТФ
| пируват | | пируват | | пируват ~| | пируват |
к я я к И д
| митохондрия
к
| СО2, Н2О 1 Ла, Н+ 1 1 Ла, Н+ | Ла, Н+ |
Л И К Я
Кровь, со- Буферы | Цитозоль |
седние
Я ММВ к
11
1. ОКИСЛЕ- 2. УДАЛЕ- 3. БУФЕ- ( 4. НАКОП- J
НИЕ НИЕ РИЗАЦИЯ 4 Ч ЛЕНИЕ^Х
Рис. 7. Схема продукции и метаболизма лактата и ионов водорода в бы-
стрых мышечных волокнах в условиях тетануса. Емкость гликолитиче-
ского процесса будет зависеть от соотношения факторов 1+2+3 и 4
135
ростью потребления пирувата митохондриями (1), скоростью
удаления из клетки (2) и степенью буферизации (3). Первые два
фактора повышения емкости гликолиза обусловлены аэробны-
ми способностями мышц, третий — гипертрофией МВ.
Есть еще один аспект, который необходимо принимать во
внимание. Существуют данные, что увеличение нагрузки гли-
колитической направленности [Обухов С.М., 1991] снижает
аэробные способности мышц. Это подтверждается данными
об отрицательном влиянии низких значений pH на массу ми-
тохондрий и активности ферментов окислительного фосфо-
рилирования [Ленинджер Р., 1966]. Следовательно, повышая
гликолитические способности мышц есть вероятность сни-
зить их аэробные способности.
Подводя итоги обсуждения значимости высоких гликоли-
тических способностей для спортивной работоспособности,
можно сделать вывод, что для дистанций с предельной дли-
тельностью работы от 20 до 40 с высокая активность ключевых
ферментов гликолиза всегда является положительным факто-
ром. Для работы длительностью 40-120 мин высокая гликоли-
тическая способность, выражающаяся в большей активности
гликолитических ферментов и наличии каких-то факторов,
увеличивающих емкость гликолиза, также является преиму-
ществом, однако, при строгом соблюдении условия высоких
аэробных возможностей мышц.
Если работа не максимальной мощности и в ней не участву-
ют БгМВ, то длительность функционирования БоМВ будет
пропорциональна их окислительному потенциалу, высокая
величина которого, при прочих равных условиях, будет замед-
лять накопление Н+.
5.1.3. Финишное ускорение
(фаза максимального волевого напряжения)
Опыт подготовки и выступления в соревнованиях квали-
фицированных спортсменов дает им умение преодолевать
дистанцию с очень небольшим снижением скорости вплоть
до финишного ускорения, что считается наиболее рациональ-
ным тактическим вариантом, если ставится задача показать
136
наивысший результат [Волков Н.И., 1969; Уткин В.Л., 1984]. С
точки зрения физиологии и биоэнергетики финишное ускоре-
ние возможно только в том случае, если в работающих мышцах
остались неизрасходованные запасы КрФ и резервы активиза-
ции анаэробного гликолиза. Исходя из описанной схемы, ре-
зервы повышения мощности сокращения мышц могут сохра-
ниться только в БгМВ. Хотя КрФ может находиться в высокой
концентрации и в ММВ, однако мощность их сокращений при
преодолении дистанции близка к максимальной, поэтому они
не могут увеличить свою долю в генерируемой мышцей меха-
нической мощности. Таким образом, БгМВ являются «страте-
гическим резервом» и практически не должны участвовать в
выполнении механической работы на дистанции, за исключе-
нием случаев нерациональной «раскладки сил», которая при-
водит к проявлению некомпенсированного утомления — сни-
жению скорости на «финишной прямой».
5.2. Схема работы разных типов МВ
при преодолении соревновательной дистанции
Представленный анализ работы МВ позволяет предпо-
ложить, что изменение величины вклада каждого отдельного
мышечного волокна в генерацию усилия целостной мышцы
при преодолении соревновательной дистанции будут иметь
свои особенности для ММВ и БМВ (рис. 8).
5.2.1. Медленные мышечные волокна
АТФ-азная активность миозина, зависящая от типа иннер-
вации мышечного волокна, определяет максимальные ско-
рость и мощность его сокращения. Известно, что при работе
в циклических локомоциях ММВ могут практически не утом-
ляться длительное время, несмотря на то, что в период напря-
жения они сокращаются в условиях гладкого тетануса. Други-
ми словами, мощности митохондриального аппарата этих МВ
хватает для поддержания максимальной для данной локомоции
(т.е. при заданном соотношении фаз напряжения и расслабле-
137
ния мышцы) мощности их сокращения в течение длительного
времени. Если бы производительности митохондрий не хвата-
ло для длительного адекватного ресинтеза АТФ, то в процессе
жизнедеятельности и тренировки в ММВ возросла бы актив-
ность гликолитических ферментов. Факт только аэробного пути
энергообеспечения означает, как отмечено выше, что даже при
наличии внутри ММВ высокой концентрации самого мощного
источника АТФ — КрФ (первые 20-25 с активности) вклад этих
волокон гипотетически не может быть выше того, который на-
блюдается в (квази-) устойчивом состоянии (при сопоставимых
биомеханических параметрах локомоции в начале и середине
дистанции). Следовательно, первой стадией в работе этих МВ
будет являться стадия максимальной производительности (рис.
8). Энергообеспечение гипотетически будет осуществляться по
следующей схеме: первые с — КрФ, затем — КрФ и жиры, далее
— вклад КрФ и жиров будет минимизироваться параллельно с
увеличением вклада углеводов, до тех пор, пока углеводы (гли-
коген, глюкоза) и лактат не станут практически единственны-
ми субстратами окислительного фосфорилирования. При этом
концентрация КрФ в среднем по мышце будет сохраняться
на относительно постоянном уровне около 70-80% от исхода
[Rusko Н. и др., 1986; Sahlin К и др., 1997].
Вторая стадия работы ММВ — это стадия снижения вклада
этих волокон в генерацию механического усилия, создаваемо-
го мышцей (рис. 8). При предельной длительности работы до
10-15 мин снижение производительности этих МВ может вы-
зываться их закислением проникающими через сарколемму
Н+. При более длительной работе снижение вклада волокна
вызывается исчерпанием внутренних запасов углеводов. Так
как использование в качестве субстрата жиров снижает ско-
рость выработки АТФ при увеличении потребления кислорода
митохондриями [Хочачка П., Дж. Сомеро, 1988 ].
Третья стадия — быстрое снижение производительности
ММВ в результате их закисления (только при длительности
работы до 10-15 мин), нарушения в работе клеточных мембран
[Пшенникова М.Г., 1986] гипотетически в связи с гипоксией
из-за ухудшения функционального состояния системы транс-
порта кислорода и т.п.
138
5.2.2. Быстрые мышечные волокна
Первая стадия. Быстрые оксидативные мышечные волокна
на средних дистанциях повторяют схему работы ММВ. Боль-
шая часть быстрых гликолитических МВ (за исключением са-
мых низкопороговых, вовлеченных в работу с самого начала)
в начале дистанции (после стартового разгона) или работают
в режиме зубчатого тетануса, или длительность их активности
в цикле движений очень мала и наблюдается только в момент
пиковых усилий, величина которых определяется биомехани-
ческими особенностями упражнения. Поэтому в первой ста-
дии вклад таких БМВ в производимую механическую работу
невелик, но возрастает под влиянием ЦНС в процессе сниже-
ния производительности уже вовлеченных МВ.
Вторая стадия. Наблюдается максимальный вклад мышеч-
ного волокна в работу. Эта стадия возможна при сочетании
высокой активности а-мотонейронов с частотой гладкого те-
тануса и наличия в волокне высокой концентрации КрФ.
Третья стадия. Постепенное снижение вклада волокна в
связи с переходом на анаэробный гликолиз.
Четвертая стадия. Быстрое снижение производительности
волокна в связи с высокой степенью закисления и исчерпания
[ВММВ ОБоМВ ИБгМВ |
Рис. 8. Схема вклада волокон различного типа в генерацию механических
усилий, развиваемых основными мышечными группами при преодолении
средней дистанции. ММВ — медленные мышечные волокна; БоМВ — бы-
стрые окислительные мышечные волокна; БгМВ — быстрые гликолити-
ческие мышечные волокна
139
КрФ. Эта фаза, гипотетически, может быть прервана мотоней-
ронным пулом, выключившим данную ДЕ из работы в связи с
утомлением.
В целом же схема вклада БМВ в генерацию усилий мышцы
представлена на рис. 8.
5.2.3. Парциальный вклад различных типов МВ
в механическую работу при преодолении дистанции
Подробно описанная выше схема представлена на рис. 8.
Стартовый разгон обеспечивается работой всех типов мы-
шечных волокон. Затем участие БгМВ в создании механиче-
ской тяги мышцы сводится к минимуму. И примерно две тре-
ти дистанции преодолевается за счет ММВ и БоМВ. Однако
все ускорения по ходу дистанции обеспечиваются дополни-
тельным рекрутированием БгМВ. По мере накопления Н+
в работающих МВ их вклад снижается, что вынуждает ЦНС
постепенно подключать БгМВ, причем со всевозрастающей
скоростью, увеличивающей долю КФК-реакции и анаэробно-
го гликолиза в ресинтезе АТФ, т.к. как эффективность работы
митохондриального аппарата падает. Если степень утомления
на финише не предельная, то спортсмен способен к финиш-
ному ускорению, которое будет обеспечиваться при преиму-
щественном участии БгМВ.
5.2.4. Схема энергообеспечения работы мышцы
Последовательность развертывания и протекания основ-
ных реакций энергообеспечения представлена на рис. 9.
КФК-реакция на миофиламентах достигает околомакси-
мальной скорости ресинтеза АТФ в стартовом разгоне, пони-
жая в мышце концентрацию КрФ. Затем интенсивность этой
реакции существенно снижается, незначительно превышая
скорость аэробного ресинтеза АТФ, т.к. как субстратом в ней
является КрФ, ресинтезируемый митохондриальной КФК (на
схеме показана только разница между расходом АТФ и ресин-
тезом АТФ, получаемой из КрФ). Снижение концентрации
КрФ в дополнительно рекрутируемых МВ приводит к посте-
140
пенному снижению пула фосфагенов в мышце. На финише
дистанции скорость КФК-реакции на миофиламентах вновь
существенно повышается, приводя к значительному исчерпа-
нию запасов фосфагенов.
Суммарная (по мышце в целом) скорость анаэробного гли-
колиза возрастает по мере расхода запасов КрФ в БоМВ, до-
стигая околомаксимальных значений (т.е. когда одновременно
в большом числе МВ максимально активизированы ключевые
ферменты гликолиза) приблизительно к 25-35-й с. Затем боль-
шую часть дистанции интенсивность гликолиза практически не
меняется, прибавляя свою долю (около 35% в нашем примере)
к АТФ, ресинтезируемой на митохондриях и КФК-реакции. К
концу дистанции в зависимости от степени утомления и ин-
тенсивности «спурта» вклад гликолиза может несколько по-
вышаться. В то же время скорость накопления Ла в мышцах и
крови, достигнув максимума в период 30-40-й с работы, будет
снижаться в связи с активизацией факторов удаления Ла, глав-
ные из которых: раскрытие всех капилляров и достижение мак-
симума МОК к 1,5-2 мин работы, что ускоряет «вымывание»
Ла в кровяное русло, а также максимальная активизация дея-
тельности миокарда, дыхательных мышц и ММВ неосновных
мышц тела, потребляющих Ла из крови. Однако в БгМВ при до-
стижении концентрацией лактата высоких значений (более 15
ммоль/л) скорость выхода Ла в кровь уменьшается.
Скорость ресинтеза АТФ в ходе окислительного (дыхатель-
ного) фосфорилированиячъкже увеличивается по мере снижения
Рис. 9. Схема вклада основных реакций энергообеспечения мышц конеч-
ностей в ресинтез АТФ при преодолении средней дистанции
141
концентрации КрФ в ММВ, практически достигая максимума
к 30-35-й с. Далее идет период максимальной производитель-
ности с постепенным снижением вклада этого источника по
мере снижения pH в ММВ и БоМВ.
5.3. Особенности физиологических
и биоэнергетических процессов
в мышечном аппарате при более длинных
и более коротких дистанциях
Воспользуемся классификацией физических упражнений
Н.И. Волкова [Волков Н.И., 1969] в связи с тем, что она, на
наш взгляд, наиболее точно отражает биохимические и физио-
логические процессы в мышцах с ростом нагрузки. В то время
как другие классификации созданы, как правило [см. напри-
мер, Суслов Ф.П. и др., 1982], в интересах практики управле-
ния тренировочным процессом в ЦВС.
5.3.1. Работа максимальной мощности
Предельная длительность до 15 с (бег на 100-200 м, плава-
ние на 25 м).
После завершения стартового разгона (и смены биомехани-
ческих параметров движений) в мышечных группах, несущих
основную нагрузку, должно быть задействовано максималь-
ное число ДЕ, доступных для произвольного рекрутирования.
Максимальная мощность работы будет пропорциональна
мощности одиночных сокращений мышцы и частоте движе-
ний. Мощность сокращений мышц будет пропорциональна
средней АТФ-азной активности миозина во всех мышечных
волокнах и массе сократительных элементов (миофиламен-
тов) в соответствии с кривой «сила -скорость». Частота движе-
ний, вероятно, обусловлена скоростью расслабления мышц,
которая зависит от массы саркоплазматического ретикулума
и производительности Са++ — насосов, осуществляющих воз-
вращение Са++ в МВ. Основной источник энергообеспечения
— КФК-реакция. Триггерами для ее запуска на максимальную
142
мощность являются, как считается, АДФ и Са++, появление
которых внутри мышечных волокон имеет место с первой с
после начала работы. Однако в БМВ, видимо, с этого же вре-
мени запускается и гликолиз, имеющий похожие триггеры.
Но его вклад несущественен из-за высокой скорости фосфо-
рилирования АДФ КФК-реакцией. Окислительное фосфори-
лирование активизируется также, по всей видимости, с первой
с работы в результате запуска КрФ-челнока и усиливается по
мере снижения концентрации КрФ, т.к. в МВ в этот период
активны все факторы, активизирующие и поддерживающие
дыхательные процессы, включая зарезервированный в окси-
миоглобине кислород.
Насколько хватит этого количества О2 без учета диффузии
из капилляров?
Запасы О2, связанного с миоглобином мышц, составляют
около 10 мл на 1 кг массы мышцы [Волков Н.И., 1969; Nevill
М.Е. и др., 1996]. Если соотношение окислительного потен-
циала БгМВ и ММВ соотносится в среднем как 1 : 5 [Голник
Ф.Д., Германсен Л., 1982|, а концентрация миоглобина про-
порциональна окислительному потенциалу МВ [Хочачка П.,
Дж. Сомеро, 1988], то количество кислорода в ММВ (с учетом
промежуточного положения БоМВ) будет составлять около 15
мл на 1 кг ММВ. Если максимальная скорость потребления
кислорода нетренированной на выносливость мышцей сприн-
тера около 150 мл/(кг. мин) [Верхошанский Ю.В., 1988], то
скорость потребления О2 медленными МВ в этой мышце будет
составлять около 200 мл/(кг. мин), или 3,2 мл/(кг. с). Это озна-
чает, что запасенного в миоглобине кислорода хватит прибли-
зительно на (15 мл/ 3,2 мл/кг/с) 4,5-5 с работы ММВ в глад-
котетаническом режиме. Если это время прибавить к 15-24 с
работы ММВ до момента ожидаемого, как установлено ранее,
исчерпания КрФ, то это будет означать, что ММВ могут рабо-
тать в «спринтерском» режиме без дефицита энергии в услови-
ях полного отсутствия прихода О2 не менее 20-25 с! Что под-
тверждается данными по активизации дыхания (VO2)bo время
и после 10 [Волков Н.И. и др., 1995] и после 22 [Волков Н.И.,
1969] с ного спринта. Через 10 с работы VO2 активизируется
лишь до 30% от максимума, а через 20-25 с после окончания
143
работы — до 45%. Максимум же VO2 после 22 с спринтерской
работы составил 80% от МПК (т.е к концу самого бега — око-
ло 70%). Очевидно, что столь существенный прирост VO2 (с 10
до 22 с работы) обусловлен очень существенной активизаци-
ей дыхания в митохондриях, связанной с достаточно высокой
степенью исчерпания КрФ в ММВ (с15-й по 22-ю с), что пол-
ностью согласуется с расчетами, приведенными ранее в этой
главе. Очевидно также, что зафиксировать эти процессы по
результатам газоанализа внешнего дыхания стало возможно
потому, что за 22 с значительно увеличился МОК, позволив-
ший экстрагировать из альвеолярного воздуха требуемое ко-
личество кислорода.
5.3.2. Работа субмаксимальной мощности
1-й временной диапазон — 15—40с.
При такой мощности работы одновременного рекрутиро-
вания всех ДЕ произойти не может, т.к. как это привело бы к
снижению скорости через 10-15 с в результате исчерпания КрФ
в БгМВ, а спортсмены, как известно, способны равномерно
«распределять силы« по дистанции. Субъективно, это обеспе-
чивается «сбросом напряжения», «расслаблением», включени-
ем «свободного хода» после стартового разгона, осуществляе-
мого, как правило, с максимальными усилиями. Затем часть
БгМВ может или выключаться, или переходить в режим зубча-
того тетануса, или даже одиночных сокращений. В любом слу-
чае, их вклад в генерацию механического усилия снижается,
и работа выполняется ММВ, БоМВ и какой-то частью БгМВ.
По мере снижения мощности КФК-реакции в рекрутирован-
ных МВ спортсмен вынужден увеличивать степень субъектив-
ного напряжения для поддержания постоянной скорости. Как
правило, этот момент приходится на середину дистанции, по-
сле чего степень напряжения непрерывно нарастает.
Энергообеспечение. Часть КрФ в БгМВ расходуется во
время стартового разгона, что включает на полную мощность
анаэробный гликолиз. Ресинтезируемый в гликолизе сарко-
плазматический АТФ, как предполагается, [Уткин В.Л., 1984]
может идти на ресинтез КрФ. Принципиальным моментом
144
является то, что часть КрФ должна быть сохранена в БгМВ по
крайней мере на первой половине дистанции, т.к. именно этот
КрФ будет являться стратегическим запасом, за счет которого
будет компенсироваться падение силы тяги МВ, принадлежа-
щих более низкопороговым ДЕ. На этих дистанциях финиш-
ное ускорение могут себе позволить только самые подготов-
ленные спортсмены. В этом случае ими используется КрФ
БгМВ, т.к. мощности гликолиза может не хватить для увеличе-
ния скорости передвижения при такой высокой средней мощ-
ности работы. В большинстве же случаев на предварительных
кругах соревнования спортсмены стараются «сохранить силы».
Это означает — израсходовать не весь КрФ или даже включить
в работу не все ДЕ. Это является гарантией немаксимальной
степени накопления Н+, высокая концентрация которого (на-
ряду со свободными радикалами [Пшенникова М.Г., 1986] и
механическим повреждением в эксцентрической фазе работы
мышц [Прилуцкий Б.И., 1989] ) считается наиболее деструк-
тивным фактором [Раппопорт Э.А., Казарян В.А., 1996] в от-
ношении морфологических структур МВ, приводящими к
«накоплению утомления» от предварительной до финальной
части соревнований. В этой зоне мощности максимальная
степень исчерпания КрФ во всех типах волокон наблюдается,
видимо, при предельной длительности работы 25-30 с.
2-й временной диапазон — 40— 120 с.
На этих дистанциях схема протекания физиологических и
биохимических процессов аналогична описанной в основной
части главы. С единственным различием — степень вовлече-
ния БМВ будет существенно выше с самого начала дистанции.
Это ускорит развертывание всех энергетических процессов,
включая окислительное фосфорилирование в БоМВ, тогда как
значимых изменений в скорости реакций в медленных МВ на-
блюдаться, как следует из модели, не будет.
Считается, что основной (или одной из основных) при-
чиной утомления на этих дистанциях является накопление
предельной концентрации Н+ в основных мышечных группах,
вызывающих снижение pH до величины порядка 6,3-6,4. Из
этого факта можно сделать простое умозаключение, что имен-
145
но интенсивный гликолиз является лимитирующим фактором
и, следовательно, его участие в энергообеспечении следует
максимально ограничить, или хотя бы не стараться повысить
активность гликолитических ферментов специализированной
«гликолитической» тренировкой. Тем более, что наличие вы-
сокой корреляционной зависимости между способностью на-
капливать большой лактацидный долг и спортивным результа-
том — ровным счетом ничего не доказывает, без рассмотрения
механизмов образования долга и энергообеспечения на дис-
танции с учетом гетерогенности состава мышечных волокон.
Высказать некоторые идеи для преодоления этого противо-
речия позволяет, на наш взгляд, представленная выше схема.
Из схемы следует, что максимальной скорости гликолиз до-
стигает на более коротких дистанциях, а 40 с — это предель-
ное время, когда гликолиз успевает «отработать» на полную
мощность до момента «самоостановки». При этом в мышцах
накапливается предельное количество лактата, который затем
диффундирует в кровь и является маркером интенсивности
гликолиза. Согласно модели, на более длинных дистанциях
мощность гликолиза (как сумма скоростей его продукции во
всех задействованных мышечных волокнах) ниже. Это под-
тверждается расчетами Н.И. Волкова (1969). Заметим, что
концентрация Ла в крови — следствие соотношения процессов
его продукции, диффузии в кровь (с учетом замедленной диф-
фузии при предельной концентрации в отдельных МВ) и эли-
минации (окисление и гликогенез) в мышцах, печени, почках
и т.п. Поэтому те же величины максимальной концентрации
Ла в крови обусловлены тем, что за более длительное время
работы большее количество Ла успевает диффундировать в
кровь. Другими словами, интенсивность продукции Ла ниже,
скорость и время элиминации выше, однако длительность его
продукции существенно выше. Поэтому получаем прибли-
зительно те же цифры концентрации Ла в крови, что и при
максимальной скорости его продукции. Итак, необходимость
высокой максимальной скорости гликолиза (активность клю-
чевых ферментов: фосфорилазы, гексокиназы, фосфофрук-
токиназы и лактатдегидрогеназы) уже может быть поставлена
под сомнение.
146
Рассмотрим, от чего зависит вклад основных источников
энергообеспечения в этой зоне интенсивности.
КФК-реакция. Ее вклад без учета ресинтезируемого КрФ
зависит только от общего содержания КрФ в мышцах, т.е. от
емкости процесса. Чем этот вклад выше, тем лучше. Но значи-
мость этого источника быстро уменьшается с ростом дистан-
ции [Волков Н.И., 1969]. Кроме этого, свободный Кр обладает
буферирующим эффектом [Мелихова М.А., 1992].
Окислительное фосфорилирование. Вопрос также решает-
ся однозначно — чем выше мощность, тем лучше. Значимость
быстро возрастает при увеличении дистанции.
Анаэробный гликолиз. Как установлено выше (схема на
рис. 7), вклад этого источника будет зависеть от соотношения
количества ресинтезируемого АТФ к величине снижения pH в
МВ. Отношение АТФ/pH определяется аэробными способно-
стями мышцы и емкостью буферных систем (бикарбонатной,
белковой, митохондриальной, связанной со дипептидами, со-
держащими гистидин, свободным креатином и др.). Скорость
диффузии МК в кровь, видимо, частично зависит от буфер-
ных систем крови, поддерживающих высокий градиент для
Н+ между цитозолем МВ и кровью, но главным образом — от
плотности капилляров вокруг БМВ.
Подведем итоги. Количество АТФ, которое может быть ре-
синтезировано в этой зоне интенсивности, будет зависеть:
— от количества КрФ (как непосредственного источника
АТФ и донора Кр как компонента буферной системы);
— буферной емкости мышц и крови;
— от окислительного потенциала мышц (это обеспечива-
ет акцепцию пирувата, увеличивает емкость буферной
системы, сопровождается высокой капилляризацией
мышц).
На последнем факторе следует остановиться подробнее,
т.к., на наш взгляд, высокие аэробные способности редко рас-
сматриваются как условие высокой емкости гликолиза. В каче-
стве доказательства значимости гликолитической системы для
спортсменов, соревнующихся в рассматриваемой зоне, обычно
приводят величины максимального лактацидного долга и мак-
симальной концентрации Ла в крови (или минимальное pH) у
147
этих специализаций. Однако, если использовать корреляцион-
ные зависимости в качестве доказательства чего бы то ни было,
необходимо вспомнить, что, например, бегуны на 400 м ( пре-
дельная длительность 43-46 с) существенно превышают сприн-
теров на 100-200 м (10-21 с) по аэробным показателям (МПК и
АнП) [КоцЯ.М., 1986], поэтому если создать выборку спринте-
ров и бегунов на 400 м одной квалификации, то будут получены
достоверные различия между специализациями по показателям
анаэробного гликолиза. Но вероятнее всего, с большим уровнем
значимости будут получены различия также и в их аэробных по-
казателях. Причем эти различия будут иметь физиологическое
обоснование. Например, как уже неоднократно отмечалось
выше, акцепция пирувата, буферирующий эффект митохон-
дрий, плотность капилляров, скорость удаления Ла медленны-
ми МВ, миокардом и дыхательными мышцами будут способ-
ствовать, кроме увеличения аэробной продукции, увеличению
емкости также и гликолитического процесса.
Другим словами, можно предположить, что бегуны на 400 м
— это те же спринтеры, только с более высокими аэробными спо-
собностями, а достичь таких же высоких показателей в спринте
им не позволяет недостаток мышечной силы из-за известного эф-
фекта снижения прироста силы под воздействием одновременно
выполняемой аэробной тренировки [Dudley G.A., R.Djamil, 1985;
Hickson С. и др., 1980; Hunter G. и др., 1987].
Таким образом, в пользу необходимости специализирован-
ной гликолитической тренировки говорит только гипотеза,
что емкость некоторых компонентов буферных систем нельзя
повысить иначе, как применяя тренировочные нагрузки, при-
водящие к росту мощности анаэробного гликолиза.
Работа предельной длительности 2-10 мин попадает в диа-
пазон, подробно описанный выше, поэтому специальных ком-
ментариев мы делать не будем.
5.3.3. Упражнения умеренной мощности
Предельная длительность более 10 мин. Введем два времен-
ных диапазона, установив границу, предположим, в 120 мин.
1-й временной диапазон — 10 —120 мин.
148
Средняя мощность работы находится в диапазоне между
анаэробным порогом и критической мощностью. Это озна-
чает, что в работу вовлечены все ММВ и существенная часть
БоМВ. Степень участия последних определяется величиной
превышения пороговой мощности и утомлением спортсмена.
В БоМВ высока активность митохондриальных ферментов.
Однако активен и гликолиз. Их функционирование будет со-
провождаться выделением Ла и Н+, что, как отмечалось выше,
обеспечивает максимальную скорость ресинтеза АТФ за счет
дыхательного фосфорилирования (до момента существенного
снижения pH). Но в то же время приводит к быстрому исчер-
панию углеводных запасов организма, стимулирует высокую
активность дыхательных мышц для респираторной компен-
сации ацидоза, высокую теплопродукцию. В беге к этому до-
бавляется интенсивная проприорецептивная импульсация и
большие механические воздействия на НМА. Максимальный
02-запрос основными мышечными группами конечностей,
высокий — дыхательной мускулатурой, кожная гиперемия в
результате повышения температуры тела и гемоконцентрация
приводят к высокой нагрузке на сердечную мышцу, а умень-
шение запасов гликогена и связанное с этим психическое на-
пряжение — к высокой активизации симпатоадреналовой си-
стемы. Все эти факторы, как предполагается, могут вызывать
«центральное утомление», что сопровождается накоплением
ГАМК и серотонина в мозге и выражается, в частности, в на-
рушении координации [Солодков А.С., 1992], невозможности
максимально активизировать мотонейронные пулы мышц и
задействовать все мышечные волокна даже при предельном
напряжении в конце дистанции. Несмотря на то что нам не-
известны работы, в которых специально изучалась бы степень
исчерпания КрФ и активизация гликолиза во время макси-
мального финишного ускорения после длительной продолжи-
тельной работы, имеющиеся данные об относительно высокой
концентрации КрФ в момент отказа при «истощающей» дли-
тельной работе [Sahlin К. и др., 1997] поддерживают гипотезу о
«центральном» происхождении утомления на этих дистанци-
ях. Однако еще раз подчеркнем, что пусковым моментом для
всех перечисленных факторов, приводящих к «центральному»
149
утомлению, является продукция Ла в БоМВ из-за недостаточ-
ного окислительного потенциала основных мышечных групп.
2-й временной диапазон — более 120 мин.
Марафонские дистанции. Выделение этой зоны целесоо-
бразно, на наш взгляд, потому, что, во-первых, такая работа
выполняется не выше уровня анаэробного порога [Tanaka К.,
Y.Matsuura., 1984], т.е. в значительной мере за счет функцио-
нирования ММВ, а во-вторых, приблизительно с этой дли-
тельности работы существенную роль в энергообеспечении
начинает играть окисление жиров, особенно во второй поло-
вине дистанции.
Многочисленными исследованиями показано, что аэробная
тренировка приводит к снижению среднего дыхательного ко-
эффициента при длительной стандартной работе, аферменты
b-окисления жирных кислот более чувствительны к физическим
нагрузкам, чем другие митохондриальные ферменты [Kiens
В. и др., 1993; обзор Шенкмана Б.С., 1999]. Это означает, что
большая часть энергии вырабатывается за счет окисления жи-
ров — происходит «жировой сдвиг». При этом хорошо изучены
гормональные и метаболические механизмы, обеспечивающие
повышенную утилизацию жиров [Теппермен Дж., Теппермен
X., 1989]. Кроме того, известно, что запасов углеводов в мышцах
и печени (максимум — 1000 ккал из требуемых 2500-2800 ккал)
практически не хватает для преодоления марафонской дистан-
ции, даже несмотря на углеводное питание во время работы. В
этом случае традиционно считается, что большее использова-
ние жиров позволит «сэкономить» гликоген и, следовательно,
отдалить наступление утомления. Поэтому в дискуссиях по по-
воду подготовки марафонцев много внимания уделяется «тре-
нировке окисления жиров» [Современная система спортивной
подготовки / Подред. Ф.П. Суслова и др., 1995].
Однако почему-то не принимаются во внимание два не ме-
нее хорошо известных факта.
1. Большее использование жиров в качестве топлива в ми-
тохондриях может понизить максимальную скорость
ресинтеза АТФ до 60% [Хочачка П., Дж. Сомеро, 1988].
Другими словами, если анаэробный порог и среднеди-
станционная скорость марафонца при использовании
150
углеводов будут составлять, предположим, 5,8 м/с, то при
окислении жиров эта скорость не может быть выше 4 м/с,
что и наблюдается, например при суточном беге. Следо-
вательно, марафонец должен стремиться к максимально-
му использованию углеводов, а не жиров, чтобы в полной
мере были задействованы его аэробные способности.
2. Нам не известны исследования, в которых было бы по-
казано, что после «тренировки окисления жиров» спор-
тсмены начинали преодолевать соревновательную дис-
танцию с меньшим дыхательным коэффициентом, т.е.
с большим процентом окисления жиров. Наоборот,
хорошо известно, что с уменьшением предельной дли-
тельности работы среднедистанционный дыхательный
коэффициент возрастает. Это означает, что если мара-
фонец улучшит свой спортивный результат, например с
2 ч 20 мин до 2 ч 10 мин, то его средний дыхательный
коэффициент увеличится, предположим, с 0,89 до 0,92
[Scrimgeour A.G. и др., 1986]. Другими словами, доля
окисленных жиров уменьшится. Если же принять во вни-
мание, что метаболическая стоимость пути при аэроб-
ной работе практически не зависит от скорости и состав-
ляет 0,8-0,9 ккал/кг.м [Зациорский В.М. и др., 1982], то
уменьшение времени бега и доли окисляемых жиров бу-
дет означать существенное снижение общего количества
окисленных жиров.
Кроме этого, известно, что не все мышцы одинаково ак-
тивны в процессе работы. Например, в беге масса мышц, вы-
полняющих основную работу по перемещению спортсмена —
не более 10 кг (в спринте — 15 кг [Nevill М.Е. и др., 1996]). Это
означает, что остальная (большая) часть мышц, так же как и
некоторые внутренние органы, работает с меньшей интенсив-
ностью и, следовательно, с большей вероятностью использует
жиры в качестве основного энергетического субстрата, чем эти
10 кг основных мышц, которые, таким образом, работают почти
исключительно «на углеводах». К сожалению, нам не известны
работы, подтверждающие или опровергающие эту гипотезу.
Приведенные рассуждения не позволяют нам рассматри-
вать b-окисление жиров и механизмы мобилизации жировых депо
151
в качестве лимитирующих факторов локальной выносливости.
Имеющиеся данные [см., например, Saltin В., 1985] позволя-
ют предположить, что большая утилизация липидов является
простой функцией от возрастающих в результате тренировки
плотности капилляров и массы митохондрий основных мы-
шечных групп. Другими словами, спортивный результат будет
зависеть в основном от мощности аэробных систем энергоо-
беспечения и углеводных запасов организма, для тренировки
которых существуют другие, не обязательно «дистанцион-
ные», средства и методы тренировки. Поэтому очень широкое
использование больших объемов малоинтенсивной аэробной
работы (на уровне или ниже аэробного порога) — до 50% от об-
щего тренировочного объема [Кулаков В.Н., 1995] — вероятно,
объясняется какими-то другими причинами («тренировкой»
нейроэндокринной системы, терморегуляции, устойчивости
ионных процессов и др.), а не необходимостью повышения
функциональных возможностей мышечных компонентов,
определяющих выносливость.
5.4. Заключение
Проведенный теоретический анализ процессов, проис-
ходящих в работающих мышцах при преодолении соревно-
вательных дистанций, позволяет выделить среди мышечных
компонентов факторы, лимитирующие локальную выносли-
вость в ЦВС.
На спринтерских дистанциях это — масса белков миофи-
ламентов и СПР, АТФ-азная активность миозина и КФК-
азная активность, концентрация КрФ, активность ферментов
гликолиза и все другие элементы внутриклеточной функцио-
нальной системы, обеспечивающей максимальную скорость
сокращения и расслабления мышечных волокон всех типов, а
также поддержание их максимальной сократительной актив-
ности в течение 10-15 с.
На дистанциях «длинного спринта» (15-40 с) существенны-
ми являются все те же самые факторы, однако в максимальной
степени возрастает значимость мощности анаэробного глико-
152
лиза и высокой емкости внутримышечных буферных систем,
а у границы диапазона — и аэробных способностей, главным
образом БМВ.
На «коротких средних» дистанциях (40-120 с) значимость
фосфагенной системы и высокой активности гликолитиче-
ских ферментов остается также высокой, но решающими фак-
торами становятся емкость гликолиза, определяемая емко-
стью буферных систем и аэробными способностями быстрых
мышечных волокон, а также аэробная мощность мышц.
На «длинных и средних» дистанциях (2-10 мин) решающее
значение приобретает аэробная мощность ММВ и БоМВ, со-
храняется значимость фосфагенной и емкости гликолити-
ческой систем и добавляется фактор высокой концентрации
гликогена в основных мышечных группах.
На длинных и марафонских» дистанциях (более 10 мин)
основным фактором является аэробная мощность МВ и угле-
водные запасы всего организма. Эти два фактора являются
компонентами т.н. аэробной емкости спортсмена [Биохимия:
Учебник для институтов физ. культуры, 1986].
Среди всех перечисленных факторов, на наш взгляд, наи-
более слабо разработанными являются теоретические и мето-
дические вопросы, касающиеся повышения аэробной мощно-
сти основных мышечных групп. Потому что, как отмечалось
выше, вопросы аэробной мощности всегда ассоциировались
с производительностью сердечно-сосудистой системы, а мы-
шечным компонентам выносливости отводилась роль «ведо-
мого» фактора.
Проблема повышения аэробной мощности мышц в спор-
те не может рассматриваться изолированно от других сторон
тренировочного процесса. Поэтому в дальнейшем мы будем
рассматривать целостную систему тренировки в ЦВС, но в
контексте интересов повышения локальной мышечной вы-
носливости, среди детерминантов которых аэробные способ-
ности играют решающую роль.
153
Глава 6
Теоретические аспекты выбора
средств, методов и организации
тренировочного процесса
в циклических видах спорта
с целью улучшения локальной
мышечной выносливости
Точно так же, как выявление лимитирующих факторов
физической работоспособности (т.е. определение того, что
тренировать), выбор средств, методов и организации трени-
ровочного процесса может быть корректно произведен только
на основе прогнозирования биологических эффектов тех или
иных способов воздействия на организм. Для того чтобы такой
выбор был обоснованным, нужно, во-первых, знать в каком
направлении мы хотим изменять состояние органов и систем,
а во-вторых, посредством чего и каким оптимальным образом
этого можно добиться.
6.1. Обоснование выбора средств и методов
тренировки мышечных компонентов,
определяющих выносливость
в циклических видах спорта
Тренировочное воздействие направлено на стимулиро-
вание различных морфологических перестроек в организме
спортсмена. Ответственным за развертывание всех процессов
154
адаптации и деадаптации является генетический аппарат кле-
ток. Следовательно, суть управления тренировочным процессом
— это управление функционированием генетического аппарата
клеток и создание условий для облегчения синтеза специфических
белков цитозоля, мембран и органелл клеток в зависимости от
целей тренировки.
Ксожалению, точные молекулярные механизмы стимули-
рования или ингибирования синтеза белков еще не расшиф-
рованы [Биохимия: Учебник для институтов физ. культуры,
1986; Албертс Б. и др., 1994; Vogt М. и др., 1997], поэтому,
обсуждая этот ключевой для теории и методики физическо-
го воспитания момент, обычно говорят о факторах, с кото-
рыми может быть связано управление синтетическими про-
цессами и о том, как посредством физических упражнений
или их сочетаний с сопутствующими внетренировочными
средствами (фармакологическими, физиотерапевтически-
ми, гигиеническими) можно воздействовать на факторы
синтеза белка. Некоторые известные, или гипотетические
факторы, стимулирующие синтез, будут рассмотрены ниже.
В общем же случае можно определить, что выбор средств и
методов физической и любой другой тренировки (вид упраж-
нений, режимы работы мышц и т.п.) должен основываться
на учете факторов стимулирования или ингибирования син-
теза РНК (экспрессирование генов) соответствующих орга-
нелл клеток, а организация процесса подготовки (сочетание
различных средств и методов, планирование одного занятия,
микро- мезо- и макроциклов, использование внетренеровочных
факторов и т.п.) должно проводиться исходя из интересов об-
легчения или угнетения синтеза специфических белков на ри-
босомах. Это — биологическая основа планирования трени-
ровочного процесса.
Однако, прежде чем перейти к решению этого ключевого
вопроса, следует рассмотреть, в каких направлениях необхо-
димо ускорять синтетические процессы.
Наибольшее значение для достижения высокого спор-
тивного результата в ЦВС имеет анаэробная производи-
тельность мышц, определяемая конкретными морфологи-
ческими структурами и ферментативными комплексами
155
клеток исполнительного аппарата (мышц), совершенством
управления деятельностью мышц в целом и каждой ДЕ в от-
дельности, а также производительностью обеспечивающих
систем (ССС, нейро-эндокринной системы и т.д.). Именно
они и являются объектом спортивной тренировки. А выбор
средств и методов тренировки должен проводиться по кри-
терию эффективности воздействия на данные структуры ор-
ганизма.
Рассмотрим более детально, что является ответственным за
высокую аэробную и анаэробную производительность мышц.
Для этого нужно выяснить, что характеризует высокую произ-
водительность мышц (т.е. лимитирующие факторы функцио-
нальной мощности) и что в них изменяется под воздействием
тренировки соответствующей направленности.
6.1.1. Стратегия повышения аэробной
производительности мышц в ЦВС
В литературе обозначены следующие возможные механиз-
мы адаптации с целью увеличения аэробной мощности МВ:
— увеличение размера мышечных волокон (гипертрофия);
— изменение доли красных, белых и промежуточных во-
локон;
— увеличение запасов эндогенных субстратов (триацил-
глицеролов и гликогена);
— повышение содержания ключевых ферментов, участву-
ющих в расщеплении субстратов, используемых в дыха-
тельном фосфорилировании или, что практически тоже
самое, увеличение плотности митохондрий;
— повышение эффективности процессов окислительного
фосфорилирования;
— снижение содержания ферментов анаэробного метабо-
лизма при повышении потенциала аэробных процессов;
— увеличение концентрации миоглобина;
— повышение капилляризации мышц.
Что можно сказать по поводу действительной реализации
или не реализации указанных механизмов?
156
6.1.1.1. Гипертрофия мышечных волокон
Знакомство с литературой по этой проблеме требует пре-
жде всего обратить внимание на следствия, которые вытека-
ют из модели Крога, предложенной в 1918 г, и ее модифика-
ций (модели Тевса, Димера, Грунсвальда) [Колчинская А.З.,
1973], в которой участок ткани, снабжаемый одним капил-
ляром, рассматривается как цилиндр, осью которого служит
этот капилляр. Если принять допущение, что кислород вну-
три ткани перемещается исключительно за счет свободной
диффузии, то расстояние между капилярами становится
критическим для адекватного снабжения всех точек клет-
ки кислородом. Поэтому считается, например, что ткани,
потребляющие в активном состоянии большое количество
кислорода, обладают наиболее развитой капиллярной сетью
(миокард, красные мышцы и др.). Также многократно по-
казана тесная связь между плотностью капилляров и окис-
лительным потенциалом мышц [Немировская Т.Л., 1992],
которые при аэробной тренировке возрастают, как прави-
ло, параллельно. Причем увеличение плотности капилляров
иногда происходит за счет уменьшения площади попереч-
ного сечения мышечных волокон, что интерпретируется как
адаптивная реакция, направленная на улучшение снабже-
ния мышечной ткани кислородом за счет снижения диффу-
зионного расстояния [см. обзор Немировской Т.Л., 1992 ].
Увеличение же размеров мышечных волокон, например при
тренировке культуристов или штангистов, при сохранении
числа капилляров в расчете на мышечное волокно, сопро-
вождается снижением плотности капилляров в расчете на
единицу объема (табл. 4). Это, как предполагается, являет-
ся одним из факторов низкого окислительного потенциала
мышц спортсменов в этих видах спорта.
Однако существуют хорошо известные факты, которые за-
ставляют усомниться в бесспорности приведенных выше сужде-
ний.
1. В модели Крога не принимается во внимание роль мио-
глобина как фактора депонирования и ускоренного пе-
реноса кислорода внутримышечного волокна. Известно,
что оксимиоглобин выполняет две функции — 1) поддер-
157
жание низкого внутриклеточного напряжения О2, для
обеспечения высокого градиента по отношению к ка-
пилярной крови и 2) транспорт кислорода внутри МВ,
в случае возникновения внутриклеточных градиентов
кислорода [Физиология человека / Под ред. Р. Шмид-
та и Г. Тевса. — Т. 3., 1996]. Поэтому наличие оксимио-
глобина уменьшает градиент парциального напряжение
кислорода в разных участках мышечных волокон, лик-
видируя «гипоксические» участки.
2. Вокруг гипертрофированных оксидативных мышечных
волокон капиллярная сеть настолько густая, что средняя
плотность капилляров оказывается столь же высокой,
как в других местах мышцы [Gayeski T.E.J., C.R.Honig,
1986]. Среднее межкапиллярное расстояние при этом
оказывается существенно меньше 80 гпм, которое счи-
тается критическим для адекватного снабжения ткани
кислородом, даже на основании расчетов по модели
Крога (т.е. без учета роли миоглобина).
3. Прямые измерения показали, что напряжение кислоро-
да внутримышечных волокон в состоянии максимальной
респирации митохондрий не зависит от размера волокон
[Gayeski T.E.J., C.R.Honig, 1986].
4. Показано, что в гипертрофированных мышечных волок-
нах митохондрии располагаются по периметру волокна
[Brzank K.D., K.S. Pieper., 1987; Londraville R.L., Siddel
B.D., 1990]. Это уменьшает диффузионное расстояние и
не вызывает необходимости накопления миоглобина.
5. Существуют наблюдения [Немировская Т.Л., 1992] о
реципрокных отношениях между концентрацией мио-
глобина, с одной стороны, плотностью капилляров и
окислительным потенциалом МВ — с другой в процессе
объемной аэробной тренировки, т.е. можно предполо-
жить, что система внутриклеточного транспорта кисло-
рода обладает резервами производительности, делающи-
ми ненужной высокую концентрацию миоглобина при
высокой капилляризации мышц (т.е. при маленьком
диффузионном расстоянии). Это полностью согласует-
ся с принципом симморфоза [Gnaiger Т. и др., 1997], в
158
соответствии с которым организм избавляется от избы-
точных морфологических структур, выходящих за рамки
потребностей данной функциональной системы клетки.
6. Существуют многочисленные наблюдения о высокой
степени гипертрофии мышечных волокон у спортсме-
нов экстракласса, тренирующих выносливость (табл. 2).
7. Аэробная мощность и спортивная результативность в
ЦВС растет параллельно с гипертрофией как медлен-
ных, так и быстрых волокон основных мышечных групп
спортсменов [Шенкман Б.С. и др., 1990].
На основе перечисленных фактов, гипотеза об обязатель-
ности уменьшения размеров мышечных волокон для достиже-
ния ими максимального окислительного потенциала не кажется
столь убедительной.
Тем не менее существуют исследования, в которых показа-
но снижение площади поперечного сечения (ППС) мышеч-
ных волокон под воздействием аэробной тренировки при од-
новременном росте аэробной производительности организма
[Terrados N. и др., 1986]. Выявлена меньшая ППС мышечных
волокон (МВ) у велосипедистов экстра класса, по сравнению
с разрядниками [Coyle E.F. и др., 1991], а также наблюдения,
о меньшей ППС ММВ с ростом спортивной квалификации у
лыжников (от 1-го разряда до КМС) и меньшей ППС ММВ
у бегунов-любителей с большей работоспособностью, опреде-
ленной по PWC170 [Немировская Т.Л., 1992].
В связи с этим стратегия адаптации мышечных волокон к
спортивной тренировке на выносливость требует специально-
го рассмотрения.
Влияние аэробной тренировки на размер МВ многократно
служило предметом исследования.
Всю совокупность данных можно разделить на две группы:
Первая — это исследование влияния аэробной тренировки
как таковой на площадь поперечного сечения МВ (ППС). В
связи с тем что под аэробной тренировкой принято подразуме-
вать работу не выше уровня аэробного порога или интерваль-
ную тренировку со средним уровнем мощности ниже уровня
анаэробного порога, то в большинстве случаев исследовался
именно такой вид воздействия.
159
Вторая группа исследований была направлена на изучение
влияния спортивной тренировки в циклических видах спорта
с целью достижения высокого спортивного результата.
Суммируя результаты первой группы работ, можно сделать
следующие выводы:
1. В большинстве случаев при «обычной» аэробной трени-
ровке ППС МВ или не изменяется, или незначительно
возрастает! Holloszy J. О., Booth F.W., 1976; ШенкманБ.С.,
1999; обзоры Немировская Т.Л., 1992; Saltin В., 1985]. Уве-
личение ППС наблюдается тогда, когда гистохимическо-
му исследованию подвергается тренируемая мышечная
группа, например — широкая мышца бедра при аэробной
тренировке на велоэргометре. В тех же случаях, когда
исследуется неосновная группа мышц (например, та же
широкая мышца, но при тренировке в беге), то прироста
ППС не наблюдается [Шенкман Б.С. и др., 1990].
2. Объемная аэробная тренировка животных или челове-
ка приводит к уменьшению ППС мышечных волокон
[TerradosN. идр., 1986]. В тоже время наблюдается выра-
женный прирост активности окислительных ферментов
и увеличение аэробной производительности организма
[обзор Немировская Т.Л., 1992; Terrados N. и др., 1986].
3. Получено отчетливое явление уменьшения вариации
в размере мышечных волокон около некоторого сред-
него уровня в ответ на выполнение аэробной работы
[Немировская Т.Л., 1992]. Это означает - гипертрофи-
рованные МВ уменьшаются, а гипотрофированные —
увеличивают площадь своего поперечного сечения.
Анализируя результаты второй группы работ, представлен-
ные в табл. 2 и 3, можно сделать следующие выводы:
1. Как уже отмечалось выше, спортивная тренировка в ци-
клических видах, также как и в штанге, бодибилдинге,
спринте и футболе приводит к гипертрофии как медлен-
ных, так и быстрых мышечных волокон. Это следует из
сравнения ППС у неспортсменов и спортсменов всех
представленных специализаций — мужчин и женщин.
Особенно наглядно этот вывод подтверждается данны-
ми рис. 10.
160
(1)
(2)
(3)
(4)
Рис. 10. Величина средней (по данным табл. 2) площади поперечного се-
чения (мкм2) медленных мышечных волокон у мужчин: 1) контроля, 2)
элитных бодибилдеров, 3) спортсменов, тренирующихся на выносли-
вость в мышцах нижних (бег, коньки, гребля, велоспорт) и 4) верхних
(плавание, гребля) конечностей
На рис. 10 представлена ППС медленных МВ у спор-
тсменов различной специализации, полученных про-
стым усреднением данных различных исследователей,
перечисленных в табл. 2. Удивительным является то, что
ППС ММВ у спортсменов, тренирующих выносливость,
не меньше, чему бодибилдеров, чья тренировка направле-
на на гипертрофию МВ как саркоплазматического, так и
«сократительного» типа.
2. К сожалению, имеющиеся в распоряжении результаты
не позволяют сделать однозначного вывода о степени
гипертрофии МВ в зависимости от длительности упраж-
нения. Так как наиболее показательными были бы ре-
зультаты, полученные одной группой исследователей
(предполагаются возможные расхождения в методиках
получения биопатов, окраске волокон и измерении их
ППС), при исследовании спортсменов одинаковой и
высокой квалификации и, самое главное, при исследо-
вании мышц, выполняющих основную нагрузку в дан-
161
ной локомоции. Но таких исследований в нашем рас-
поряжении нет.
Анализ же данных табл. 2 и 3 не подтверждает вы-
вод, сделанный Б.С. Шенкманом [Шенкман Б.С., 1990]
о том, что при увеличении соревновательной дистанции
ППС, в частности ММВ, уменьшается.
С большей долей уверенности можно констатиро-
вать, что имеет место практически одинаковая степень
гипертрофии ММВ, вне зависимости от длительности
соревновательного упражнения.
При интерпретации данных табл. 2 и 3 мы учиты-
вали данные биомеханических, электромиографи-
ческих и наших собственных исследований [Мякин-
ченко Е.Б., 1983] по сравнению изометрической силы
мышц — разгибателей и сгибателей коленного сустава,
в зависимости от квалификации бегунов на средние и
длинные дистанции в диапазоне от 2-го спортивного
разряда до МСМК — членов сборной команды СССР
1982-83 гг. (рис. 11).
Из графиков следует, что в то время как сила разги-
бателей коленного сустава (четырехглавая мышца бедра)
возрастает от 2-го разряда до уровня КМС, а затем сни-
жается, то сила мышц — сгибателей коленного сустава
(задняя поверхность бедра) оказывается наибольшей у
наиболее квалифицированных бегунов. Это отражает
закономерность совершенствования техники бега с ро-
стом квалификации, когда бегуны от «прыжкового» бега
(т.е. использования мышц — разгибателей ног) переходят
к экономичному «катящемуся» бегу (при котором боль-
шую роль играют сгибатели тазобедренного и коленного
суставов) [Nelson R.C., R.J.Gregor., 1976], а также от-
ражает изменение в составе средств подготовки, когда
бегуны от использования широкого круга средств ОФП
переходят к более узконаправленным, преимуществен-
но беговым средствам [Мякинченко Е.Б., 1983]. Кроме
того, из имеющиеся данных биомеханических и миогра-
фических исследований [Тюпа В.В., 1978, Физиология
мышечной деятельности / Под ред. Я.М.Коца., 1982]
162
Рис. 11. Зависимость силы мышц-сгибателей и разгибателей коленного
сустава от спортивной квалификации бегунов на средние дистанции
следует, что до 70% процентов механической работы на
опоре выполняется бегуном за счет работы мышц голе-
ни. В связи с этим к «основным» мышечным группам
бегунов можно отнести двусуставные мышцы задней
поверхности ноги — двуглавую, полусухожильную, по-
луперепончатую, икроножную, а также камбаловидную
мышцы.
Значительная механическая работа [Тюпа В.В., 1978]
выполняется также при «сведении бедер» на опоре, ког-
да наиболее активны мышцы сгибатели тазобедренного
сустава — подвздошнопоясничная, гребешковая и дву-
суставная прямая головка четырехглавой мышцы бедра
[Физиология мышечной деятельности / Под ред. Я.М.
Коца., 1982]. Медиальная и латеральная головки четы-
рехглавой мышцы выполняют уступающую работу прак-
тически только против силы тяжести в фазе амортиза-
ции и в середине опоры. Однако, как известно, с ростом
квалификации у стайеров и особенно длины дистанции
вертикальные перемещения ОЦМТ снижаются [Зациор-
ский В.М. и др., 1982]. Это означает, что снижается доля
работы, выполняемая разгибателями коленного сустава,
в частности широкой мышцей бедра, которая обычно
служит объектом гистохимических исследований. Дру-
гими словами — делать выводы о влиянии тренировочного
процесса на гистохимические и морфометрические пока-
163
затели мышц бегунов по результатам биопатов, получен-
ных из медиальной или латеральной головки четырехгла-
вой мышцы бедра (что обычно и делается) — грубейшая
методологическая ошибка.
Совершенно аналогичная картина наблюдается в
лыжных гонках классическим стилем, где особенности
скользкой опоры не позволяют выполнить сильного от-
талкивания «в догонку», которое могло бы осуществлять-
ся преимущественно мышцами разгибателями коленного
сустава. В связи с этим с ростом спортивного мастерства
спортсмены большую работу выполняют за счет мышц ту-
ловища, разгибателей локтевого сустава, сгибателей и раз-
гибателей тазобедренного сустава, а не широкой мышцей
бедра. Несколько другое положение в «коньковом» ходе.
Однако данные работ Т.Л. Немровской, Б.С. Шенкмана,
приведенные в табл. 2, 3, собирались в 80-х годах, когда
большинство российских лыжников тренировались, ис-
пользуя классический стиль. Кроме того, исследование
проводилось в летний подготовительный период, когда
лыжниками используется в основном высокообъемная
работа в легкоатлетическом беге, приводящая, как сле-
дует из приведенных выше данных, к снижению, или, по
крайней мере, к стабилизации ППС МВ.
Таким образом, использование мышц передней по-
верхности бедра для изучения влияния тренировки и со-
стояния спортсменов в легкоатлетическом беге и лыж-
ных гонках на силу, ППС и окислительный потенциал
МВ некорректно, так как эта мышца выполняет только
небольшую часть работы по перемещению спортсмена
относительно дорожки (лыжни).
Поэтому, например, в табл. 2 имеются данные о чрез-
вычайно высокой степени гипертрофии медленных МВ
икроножной мышцы («основная» мышца) у бегунов-
марафонцев и стайеров экстракласса — 8,34 мкм2, а также
гипертрофии ММВ широкой мышцы бедра («основная»
мышца) у велосипедистов 6,36 мкм2. Обе эти специали-
зации имеют наибольшее время соревновательной дис-
танции.
164
У представителей более коротких дистанций степень
гипертрофии ММВ (в мкм2) лежит в тех же пределах:
— бегуны на средние дистанции (икроножная мышца) —
6,38;
— пловцы (дельтовидная мышца) — 6,90;
— конькобежцы многоборцы (широкая бедра) — 6,70 —
8.97;
— академическая гребля (широкая бедра) — 6,68 — 10,03;
— гребцы на байдарке/каное (дельтовидная мышца) —
4,92 - 6,85.
В то же время ППС в МВ широкой мышцы бедра у
бегунов существенно ниже:
— бегуны-любители — 4,69;
— стайеры — 5,10;
— лыжники — 6,41 — 4,74.
Таким образом, сделанные выше выводы об одина-
ковой степени гипертрофии ММВ у соревнующихся на
разных дистанциях и ее проявлении только в «основных«
мышечных группах подтверждаются данными гистохи-
мического исследования.
3. Об избирательном влиянии тренировки (или отбора) на
ППС БМВ и ММВ можно судить по их соотношению.
Длинные дистанции: икроножная мышца — 0,78; ши-
рокая бедра — 0,97.
Средние дистанции:икроножная мышца — 0,98; ши-
рокая бедра — 1,14.
Из этих данных следует, что хорошо известное явле-
ние снижения поперечника мышц у стайеров [Hakkinen
К., K.L.Keskinen., 1989] по сравнению со средневиками
и спринтерами объясняется снижением степени гипер-
трофии БМВ с ростом соревновательной дистанции. Это
подтверждает многочисленные данные об избирательном
влиянии силовой или скоростно-силовой тренировки у
спортсменов на БМВ. Еще большая степень снижения
всех проявлений мышечной силы у стайеров [Hakkinen
К., K.L.Keskinen., 1989], по сравнению с поперечником
мышц, вероятно, связано как с меньшей гипертрофией
БМВ, так и с нервно-мышечными факторами.
165
4. Чем вызывается гипертрофия МВ у спортсменов, трени-
рующих выносливость, если аэробная тренировка сама
по себе к ней не приводит?
Считается [Шенкман Б.С., 1990], что такими фак-
торами могут быть использование анаболических сте-
роидов, специально организованная силовая трениров-
ка, присутствующая во всех без исключения ЦВС, или
естественный отбор. Значимость последнего фактора не
бесспорна, так как существуют наблюдения, что квали-
фицированные спортсмены (в частности, культуристы и
штангисты [обзор Шенкман Б.С., 1990]) достигают вы-
соких результатов не столько за счет предельной гипер-
трофии МВ, сколько за счет их большего числа.
5. В связи с данными табл. 2 и 3 возникает вопрос, за счет
каких органелл клеток возникает гипертрофия МВ в
ЦВС? Логично было бы предположить, что наибольшее
значение должно иметь увеличение объемной плотно-
сти митохондрий, капилляризация и накопления эндо-
генных энергетических субстратов (гликогена и триа-
цилглицеролов), т.е. ППС МВ в ЦВС должна была бы
увеличиваться по «саркоплазматическому» типу. В част-
ности показано, что в тренированных «на выносливость»
мышцах парциальный объем сократительных элемен-
тов может уменьшаться до 60%, а в тренированных «на
силу» — возрастать до 85% [Хоппелер Г., 1987]. Однако,
по имеющимся данным, объем митохондрий может уве-
личиваться в ММВ с 3-5% у нетренирующихся лиц, до
8-10% у высококвалифицированных спортсменов. Ко-
личество гликогена и доля капилляров в мышцах также
относительно невелики (2-3%) [Хоппелер Г., 1987]. Ги-
потеза о возможной гипертрофии МВ за счет увеличе-
ния массы воды, связанной с гликогеном, не подтверди-
лась экспериментально. Таким образом, столь высокая
степень гипертрофии ММВ (до 100-120% см. табл. 2)
не может объясняться только увеличением объема сар-
коплазмы. Значительную роль играет «миофибрилляр-
ный» тип гипертрофии — т.е. увеличение числа сократи-
тельных элементов мышц. Косвенным подтверждением
166
этому выводу может быть то, что парциальный объем
миофибрилл в ММВ и БгМВ одной и той же мышцы су-
щественно не различается (76-82%) [Хоппелер Г., 1987].
Тот факт, что гипертрофия за счет сократительных эле-
ментов в этом случае не сопровождается адекватным
приростом силы и скоростно-силовых способностей у
спортсменов, тренирующих выносливость, объясняет-
ся, вероятнее всего, нервно-мышечными факторами и
владением техникой скоростно-силовых и силовых дви-
жений. Вероятно, эти способности не являются суще-
ственными в ЦВС.
6. Каков биологический смысл адаптации к спортивной
тренировке в ЦВС через увеличение ППС, в частности,
за счет накопления сократительных элементов мышц?
Можно было бы предположить, что степень гипертро-
фии волокон основных мышц должна коррелировать с
величиной механической нагрузки, которую приходит-
ся преодолевать мышцам при выполнении аэробных
упражнений. Вероятно, некоторые основания у этой ги-
потезы есть, так, например, наибольшие величины ППС
зафиксированы в икроножной мышце бегунов, которая
испытывает очень высокие значения механической на-
грузки в беге. Однако этой гипотезе противоречат дан-
ные, представленные в табл. 2, например в плавании, где
пиковая механическая сила тяги мышц в цикле движения
существенно меньше, чем у представителей «наземных»
локомоций, степень гипертрофии ММВ даже несколько
выше. Этот факт может означать, что гипертрофия воло-
кон необходима не только для противодействия механи-
ческой нагрузке, но и как фактор повышения аэробной
(в случае ММВ) мощности мышц.
Таким образом, можно резюмировать, что для каждого вида
локомоций существует оптимум ППС в каком-то определен-
ном сочетании с морфоструктурами, отвечающими за окисли-
тельный потенциал ОП мышечных волокон данной мышечной
группы. Величина ППС и соответствующий ОП МВ детерми-
нирован величиной механических нагрузок, которые мышцы
испытывают в соревновательном упражнении.
167
При этом избыточная гипертрофия увеличивает инертную
мышечную массу, а ее недостаток снижает анаэробные пороги
(за счет более раннего подключения БгМВ к работе см. гл. 5),
т.е. приводит к более раннему «закислению» на дистанции.
Как тогда добиться оптимума? Этот вопрос решается в каж-
дом конкретном случае. Надо только помнить, что при боль-
ших объемах — снижается ППС. При избытке силовой рабо-
ты — страдают аэробные способности. Следовательно, должно
быть оптимальное сочетание объемных средств (снижающих
ППС) и силовой работы, поддерживающих ППС. В соревно-
вательном же периоде оптимальной будет — соревновательная
интенсивность (в биомеханическом смысле).
6.1.1.2. Изменение доли красных, белых и промежуточных
волокон
Максимальная скорость сокращения волокна, АТФ-азная
и КФК-азная активность, экспрессия различных цепей мио-
зина, гликолитические возможности мышечных волокон и
т.п. определяются типом их иннервации [Whalen, 1981]. т.е.
величиной мотонейрона, его максимальной частотой импуль-
сации и толщиной двигательного нерва. Последние факторы
генетически обусловлены, поэтому изменения соотношения
типов волокон, определяемых по миозиновой АТФ-азе (т.е.
переход ММВ в быстрые и обратно), под воздействием трени-
ровки, как правило, не происходит [обзор П.Хочачка, 1988].
Однако существует такое понятие как «метаболический про-
филь» волокна, определяемый по соотношению активности
окислительных (например, малатдегидрогеназы или цитрат-
синтетазы) и гликолитических ферментов. Под воздействием
тренировки может меняться активность ключевых ферментов
и, следовательно, метаболический профиль волокон. Однако
считается, что при «обычной» аэробной тренировке окисли-
тельный потенциал ММВ и БМВ повышается параллельно. В
то же время имеются данные о переходе быстрых гликолити-
ческих МВ в быстрые оксидативные МВ [см. обзор Шенкман
Б.С., 1990].
168
6.1.1.3. Повышение содержания ключевых ферментов, уча-
ствующих в окислительном расщеплении субстратов
При спортивной тренировке, как обосновано в главе 5,
наиболее важной является мощность метаболических путей
аэробного окисления углеводов. Известно, что повышение
окислительного потенциала мышц всегда сопровождается уве-
личением количества ключевых ферментов гликолиза на всех
его стадиях от расщепления гликогена до челночного перено-
са электронов на внутренней мембране митохондрий [Хочачка
П., Дж. Сомеро, 1988].
6.1.1.4. Увеличение плотности митохондрий
Наиболее очевидным признаком повышения дыхатель-
ных способностей мышц является увеличение объема и числа
митохондрий, которые могут составить до 13% объема ММВ
[Г.Хопеллер, 1987 ]. В мышечном волокне митохондрии рас-
полагаются вокруг миофиламентов, в местах использования
АТФ в саркоплазматическом ретикулуме, но в основном по
периметру волокон в субсарколемальной зоне в непосред-
ственной близости от капилляров, особенно это выражено в
гипертрофированных волокнах [Orenshaw A.G. и др., 1991;
Londraville R.L., Siddel B.D., 1990].
Увеличение числа и объема митохондрий сопровожда-
ется изменением соотношения активности различных
ферментов, выражающимся в повышении эффективности
окислительного метаболизма. Оба этих явления — гипер-
плазия и гипертрофия митохондрий и изменение состава
ферментативных систем приводят к увеличению окисли-
тельного потенциала как ММВ, так и БМВ на 100-200%
[Яковлев Н.Н., 1983].
Принципиальным является вопрос соотношения процес-
сов увеличения плотности митохондрий и увеличения ППС,
в частности за счет сократительных миофиламентов. Точнее,
приводит ли гипертрофия МВ при регулярной аэробной трени-
ровке к автоматическому снижению плотности митохондрий?
Два известных нам исследования, в которых были достигнуты
существенные приросты ППС ММВ у квалифицированных
гребцов [Шенкман Б.С. и др., 1990] и при локальной трени-
169
ровке трехглавой мышцы голени, [Шенкман Б.С., 1999] сви-
детельствуют, что при наличии аэробных нагрузок в структуре
тренировочного процесса, так же как при аэробных условиях
работы мышц, при выполнении локальных силовых упражне-
ний наблюдается тенденция к повышению активности ключе-
вых ферментов окислительного фосфорилирования и увели-
чения плотности митохондрий в субсарколемальной области.
Кроме того, на основе данных Т.Л. Немировской [1992] о
плотности капилляров у элитных спортсменов (табл. 4) и дан-
ных табл. 2 и 3, а также известного факта высокой корреляци-
онной связи между плотностью капилляров и окислительным
потенциалом мышц, можно сделать вывод, что увеличение
объема мышечного волокна может сопровождаться (очевидно,
что только при целенаправленной тренировке) очень высоки-
ми удельными значениями плотности митохондрий. Другими
словами, сила мышечного волокна (содержание в нем миофи-
ламентов) и его окислительный потенциал (детерминантами
которого являются плотность митохондрий, содержание мио-
глобина и капилляризация) у элитных спортсменов могут воз-
растать параллельно, обеспечивая максимальную работоспо-
собность таких мышц.
6.1.1.5. Повышение эффективности процессов окислительно-
го фосфорилирования
Повышение аэробных способностей мышц при трениров-
ке может происходить не только в результате увеличения спо-
собности к потреблению кислорода, но и за счет увеличения
эффективности работы митохондриальной дыхательной цепи
[Шенкман Б.С., 1999]. Это может выражаться в экономизации
использования кислорода за счет снижения базального уровня
дыхания митохондрий и в увеличении синтеза макроэргиче-
ских соединений митохондриальной креатинкиназой без уве-
личения максимальной скорости потребления кислорода. То
что оба этих пути повышения эффективности окислительного
фосфорилирования могут реализовываться при тренировке
квалифицированных спортсменов показано в исследовании
Б.С. Шенкмана [1999]. В частности, было показано увеличе-
ние активности цитохром-С-оксидазы и скорости креатин-
170
Таблица 2. Морфометрические данные мышечных волокон спортсменов — мужчин различной квалификации и специализации
Специализация Пол Мышца Состав МВ ППС, мкм2 ММВ/ БМВ Источник
ММВ БМВ ММВ БМВ
Неспортсмены (контроль) Латеральная бедра Икроножная Латеральная бедра Латеральная бедра Латеральная бедра Задняя дельтовидная 47 57,7 52 51,4 53 42,3 48 48,6 4,72 5,46 5,54 5,27 5,80 4,66 4,71 4,95 6,70 5,57 6,40 6,00 0,99 0,91 1,21 1,06 1,10 1,29 Дж. Уилмор, 1997 Дж. Уилмор, 1997 W.M.Ruzon,1990 Б.С. Шенкман, 1990 Tesh Р.А., 1985 Tesh Р.А., 1985
Штан гисты/пауэрол ифтеры М Латеральная бедра 44 56 5,06 8,91 1,76 Tesh Р.А., 1985
Бодибилдеры
Бегуны на выносливость м Латеральная бедра 5,90 5,80 0,98 Tesh P.A., 1985
Спринтеры (бегуны) м Икроножная 24 76 5,88 6,033 1,03 Дж. Уилмор, 1997
Бегуны низкой квалификации (МПК 60,8 мл/кг мин) м Латеральная бедра 66 34 4,69 5,35 1,14 Т.Л. Немировская, 1992
Стайеры средней квалификации м Латеральная бедра 5,10 5,51 1,08 Б.С. Шенкман, 1990
Марафонцы и бегуны на длинные дистанции экстра-класса м Икроножная 79 21 8,34 6,49 0.78 Дж. Уилмор, 1997
Лыжники 1-й разряд кмс м Икроножная - - 6,41 4,74 6,68 6,50 1,04 1,37 Б.С. Шенкман, 1990
Триатлон м Латеральная бедра 5,36 5,94 1.11
Бегуны на средн, диет. - средн, квалиф. Бегуны на средн, диет. - элита м Латеральная бедра Икроножная 61,8 38,2 6,15 6,38 6,72 6,28 1,09 0,98 Дж. Уилмор, 1997
Плавание м Задняя дельтовидн. 6,91 9,01 1,33 Дж. Уилмор, 1997
Коньки (многоборье) Коньки м м Латеральная бедра Латеральная бедра 6,70 8,97 7,54 10,02 1.14 1,14 Дж. Уилмор, 1997 K.D. Brzank, 1987
Гребля академическая (начало сезона) Начало соревноват. периода м Латеральная бедра Латеральная бедра 6,68 10,03 7,33 11,34 1,10 1.13 Б.С.Шенкман (1990в)
Гребцы на каное Гребля байдарка/каное м м Задняя дельтовидная Задняя дельтовидная 71 29 4.92 6,85 7,04 8,51 1,43 1,25 Tesh Р.А., 1985
Футболисты (элита) м Латеральная бедра 52,9 47,1 6,29 7,70 1,22 W.M. Ruzon (1990)
Таблица 3. Морфометрические данные мышечных волокон спортсменок — женщин различной квалификации и спе-
цианизации
Специализация Пол Мышца Состав МВ % ППС, мкм ММВ/ БМВ Источник
ММВ БМВ ММВ БМВ
Неспортсмены (контроль) Латеральная бедра Латеральная бедра 52 48 3,50 4,38 3,14 3,78 0,90 0,90 Д. Костил, 1997 Б.С. Шенкман, 1990
Спринтеры (бегуны) Ж Икроножная 27 73 3,75 3,93 1.05 Д. Костил,1997
Спортивное ориентирование Ж 4,54 3,95 0,87 Б.С. Шенкман, 1990
Бегуньи на дл. диет. Икроножная 69 31 4,44 4,13 0,93 Д. Костил,1997
Лыжницы 1-й разряд — КМС ж Латеральная бедра 4,69 4,16 0,89 Т.Л. Немировская, 1992
Велосипедисты ж Латеральная бедра 51 49 5,49 5,22 0,95 Д. Костил,1997
Бегуньи на средн, диет - средн, квалиф. ж Латеральная бедра 5,09 5,08 0,99 Б.С. Шенкман, 1990
Коньки (многоборье) ж Латеральная бедра 6,15 5,81 0,93 Б.С. Шенкман, 1990
Гребля академическая ж Латеральная бедра 6,41 6,58 1,07 Б.С. Шенкман, 1990
Таблица 4. Плотность капилляров в мышцах спортсменов различной спе-
циализации (адаптировано по Т.Л. Немировской [Немировская Т.Л., 1992]
№ Спортивная специализация Мышца Плотность капилляров (на мм2) Колич. капилляров (на 1 МВ)
1 Стайеры Латеральная бедра 444 2,47
2 Триатлон Латеральная бедра 427
3 Велогонки шоссе Латеральная бедра 509 3,3
4 Велогонки шоссе Латеральная бедра 464 2,90
5 Бегуны Латеральная бедра 450
6 Лыжники Латеральная бедра 415
7 Конькобежцы Латеральная бедра 415
8 Пловцы Дельтовидная 427 2,0
9 Культуристы Латеральная бедра 320 2,64
10 Штангисты Латеральная бедра 199 2,06
стимулированного дыхания без увеличения МПК и объем-
ной плотности митохондрий. Увеличение степени активации
дыхания креатином свидетельствует об активизации работы
митохондриальной креатинкиназы, которая играет ключевую
роль во внутриклеточном аэробном синтезе КрФ, а также спо-
собствует облегченной диффузии АДФ в мышечных клетках,
уменьшая Кт для АДФ [Шенкман Б.С., 1999].
6.1.1.6. Снижение активности ферментов анаэробного мета-
болизма в соответствии с повышением потенциала аэробных про-
цессов
Наиболее типичным паттерном изменения активности
ключевых ферментов анаэробного гликолиза при аэробной
тренировке является то, что в ММВ их активность немного
возрастает, а в БМВ — немного падает [Хочачка П., Дж. Со-
меро, 1988]. Тем не менее достаточно определенно можно ска-
зать, что аэробная тренировка существенного отрицательного
воздействия на активность ферментов анаэробного гликолиза
не оказывает.
173
6.1.1.7. Увеличение концентрации миоглобина
Стратегия адаптации систем внутриклеточного транспорта
кислорода, как можно было бы предположить, могла бы вклю-
чать в себя:
— увеличение концентрации миоглобина, который уско-
ряет диффузию кислорода внутри волокна;
— снижение ППС вол окна для уменьшения диффузионно-
го расстояния;
— локализацию митохондрий по периметру волокна.
Гипотеза о возможности увеличения концентрации мио-
глобина возникла на основании данных о повышении его кон-
центрации при экспозиции в гипоксических условиях, или
при анемии [Немировская Т.Л., 1992, Terrados N. и др., 1990],
а также из представлений о гипоксическом состоянии мышц
при физической нагрузке.
Однако из перечисленных возможностей реализуется
только периферическая локализация митохондрий. Как ока-
залось, ППС в большинстве случаев возрастает (см. выше).
За исключением случаев объемной истощающей тренировки
или нахождения в высокогорье (т.е. в состоянии компенси-
рованной стресс-реакции [Terrados N. и др., 1986]), а концен-
трация миоглобина у квалифицированных спортсменов или
не отличается от контроля, или даже снижается [Coyle E.F.
и др., 1991], это же наблюдалось под воздействием аэробной
тренировки [Terrados N. и др., 1986, 1990], а увеличение со-
держания миоглобина наблюдалось, напротив, при силовых
и скоростных нагрузках [Верхошанский Ю.В., 1988; Яковлев
Н.Н., 1983], т.е. при увеличении ППС волокон. Поэтому,
учитывая гипертрофию ММВ у элитных спортсменов, мож-
но заключить, что общее содержание миоглобина в основных
мышечных группах увеличивается [Яковлев Н.Н., 1983], но
без увеличения его концентрации, что никак не согласуется
с гипотезой о гипоксическом состоянии мышц при экстре-
мальной аэробной нагрузке.
Тот факт, что при этом концентрация миоглобина не воз-
растает, дает основания предполагать, что внутриклеточный
транспорт кислорода, хотя и является неотъемлемым звеном
системы транспорта и утилизации кислорода, обладает доста-
174
точными адаптационными возможностями для того, чтобы не
лимитировать аэробную производительность мышц. Этот вы-
вод подтверждает и отрицательная корреляция концентрации
гемоглобина в крови (отвечающего за транспорт 02) с пико-
вым кровотоком в конечности и концентрацией миоглобина в
мышцах этой конечности [Немировская Т.Л., 1992].
6.1.1.8. Повышение капилляризации мышц
Этот феномен, также как и увеличение плотности митохон-
дрий, всегда сопровождает регулярную аэробную тренировку
и тесно коррелирует с изменением окислительного потенциа-
ла мышц. Прирост в процессе тренировки связан с исходной
плотностью капилляров. Плотность повышается только до
определенного предела, азатем стабилизируется [Немировская
Т.Л., 1992].
6.1.1.9. Заключение по разделу
Как следует из приведенного анализа, стратегией адапта-
ции в отношении увеличения аэробной производительности
мышц является следующее:
1. Гипертрофия М М В на 50-120% относительно контроля.
Причем существенно, что сама по себе аэробная трени-
ровка не вызывает гипертрофии мышечных волокон.
Увеличение ППС вызывается, вероятнее всего, приме-
нением специальных силовых упражнений, которые яв-
ляются неотъемлемой частью тренировочного процесса
во всех без исключения циклических видах спорта.
2. Накопление эндогенных энергетических субстратов,
среди которых наибольшее значение для аэробного ком-
понента спортивной работоспособности имеет гликоген
мышц.
3. Повышение окислительного потенциала мышц на 100-
200% за счет гипертрофии ММВ, повышения общей
массы и плотности митохондрий, изменения фермента-
тивного состава митохондриальных белков, оптималь-
ной концентрации миоглобина и высокой капилляриза-
ции мышц.
175
6.1.2. Стратегия повышения анаэробной
производительности мышц в ЦВС
Как было определено в предыдущей главе, под существен-
ными для спортивной работоспособности компонентами ана-
эробной производительности, влияющими на выносливость,
следует понимать:
— максимальную мощность и емкость КФК-реакции и
анаэробного гликолиза в упражнениях длительностью
до 40 с;
— максимальную емкость КФК-реакции и анаэробного
гликолиза в упражнениях длительностью от 40 с до 2-3
мин.
Также как и в случае аэробной производительности, име-
ющиеся в литературе данные по поводу стратегии адаптации
к анаэробной работе можно свести к следующим позициям
[Немировская Т.Л., 1992; Сарсания С.К., Селуянов В.Н., 1991;
Хочачка П., Дж. Сомеро, 1988; Шенкман Б.С., 1990]:
— увеличение размера мышечных волокон (гипертрофия);
— изменение доли красных, белых и промежуточных во-
локон;
— повышение запасов эндогенных субстратов (креатин-
фосфата и гликогена);
— повышение содержания ключевых ферментов, участву-
ющих в анаэробном метаболизме и его регуляции;
— изменение изоэнзимов в различных звеньях метаболи-
ческих путей;
— увеличение буферной емкости мышц.
Что можно сказать по поводу действительной реализации
или нереализации указанных механизмов?
6.1.2.1. Гипертрофия мышечных волокон
Факт повышения ППС в ответ на интенсивную анаэроб-
ную работу различного характера за счет накопления главным
образом сократительных элементов мышц хорошо известен.
Также не вызывает сомнений высокая корреляция между
анаэробной производительностью и гипертрофией обоих ви-
дов МВ. Как подробно описано в 5-й главе, это обусловлено
176
тем, что в генерации механического усилия, даже максималь-
ной интенсивности, существенный вклад вносят не только
быстрые, но и медленные мышечные волокна. Тем более это
справедливо в аспекте выносливости, когда требуется не мак-
симальная анаэробная мощность, а высокая емкость процес-
сов энергообеспечения. В этом случае запасы КрФ и аэробная
мощность ММВ служат фактором «экономии» ресурсов БМВ.
Справедливость этих рассуждений подтверждается тем, что
избирательная гипертрофия БМВ наблюдается только у пред-
ставителей скоростно-силовых видов (штангистов, спринте-
ров и т.д. [см. обзор Шенкман Б.С., 1990].
Считается, что возможности переходов ММВ в БМВ, так-
же как и обратно, очень ограничены [Хочачка П., Дж. Сомеро,
1988; Шенкман Б.С., 1990].
6.1.2.2. Повышение запасов эндогенных субстратов (креатин-
фосфата и гликогена)
Насколько можно судить из имеющихся данных, анаэроб-
ная тренировка приводит к увеличению общего количества
КрФ [Хочачка П., Дж. Сомеро, 1988; Яковлев Н.Н., 1983].
Запасы гликогена возрастают при любом виде физической
нагрузки. Однако существует, вероятно, некоторый морфо-
логический или другой предел, выше которого увеличить
запасы гликогена иначе, как путем гипертрофии мышц, не
удается. Этот вывод следует из экспериментов с методом
углеводного насыщения [Алиханова Л.И., 1983]. Кроме того,
при длительности работы, где существенный вклад вносят
анаэробные источники энергии (15 с — 2-3 мин), снижение
запасов гликогена не является фактором снижения скорости
гликолиза [Nevill М.Е. и др., 1996]. Однако известно [Bangsbo
J., 1996], что более высокий исходный уровень гликогена в
мышцах корреляционно связан со скоростью гликолиза. Но
эта корреляция может явиться ложной, поскольку большая
концентрация гликогена может сопровождаться большим
содержанием ключевых ферментов гликолиза, так как при
тестирования разных ног при нормальном и суперкомпен-
сированном гликогене такой зависисмости не подтверждено
[Bangsbo J., 1996].
177
6.1.2.3. Повышение содержания ключевых ферментов, уча-
ствующих в анаэробном метаболизме и его регуляции
Количество АТФ-азы и КФК-азы, связанных с типом ин-
нервации волокон, вероятно, под воздействием тренировки
существенно изменяться не должно. Однако содержание ми-
окиназы и ферментов анаэробного ресинтеза АТФ (фосфо-
рилазы, фосфофруктокиназы, лактатдегидрогеназы), может
повышаться на 100-150% [Хочачка П., Дж. Сомеро, 1988] за
счет простой гипертрофии мышц и за счет изменения отно-
сительного содержания ферментов. Под воздействием трени-
ровки может изменяться и изоинзимный состав анаэробных
ферментов. Например, это хорошо показано для изоэнзимов
лактатдегидрогеназы [Хочачка П., Дж. Сомеро, 1988].
6.1.2.4. Увеличение буферной емкости мышц
Система буферирования протонов в мышечных волокнах
подробно описана в главе 5 и включает бикарбонатный, бел-
ковый, дипептидовый (при содержании имидазольных групп
гистидина), креатиновый, митохондриальный компоненты и
систему удаления ионов из мышечного волокна.
К сожалению, нам не известны механизмы увеличения
концентрации буферирующих веществ или, даже, какой точно
характер нагрузки способствует их преимущественному нако-
плению.
Имеются данные, что у спортсменов, тренированных в
«гликолитической» зоне, повышена способность накапливать
Ла в крови [Волков Н.И., 1969] и увеличения отношения ЭЛа/
DpH [Sahlin К., 1986], что косвенно может свидетельствовать
о приросте емкости их буферных систем, т.к., вероятно, ми-
нимальное pH, при которых происходит резкое замедление
процессов энергопродукции в МВ, не отличается у трениро-
ванных и нетренированных, поэтому если значительная часть
Н+ буферируются, то больше Ла может накопиться в мышцах,
а затем выйти в кровь. В то же время большая концентрация Ла
может явиться следствием его облегченной диффузии в кровь
(см. ниже) в результате большей капилляризации мышц (их
большего окислительного потенциала) у средневика, по срав-
нению со спринтерами.
178
В литературе имеются также указания, что концентрация
гистидина (одного из компонентов белкового и дипептидного
буфера) растет параллельно активности гликолитических фер-
ментов [Хочачка П., Дж. Сомеро, 1988].
Свободный креатин выполняет функцию буферирования
ионов водорода. Увеличение емкости этого звена буферной
системы происходит параллельно с увеличением пула Кр и
КрФ [Greenhalf P.L. и др., 1996].
Диффузия лактата и ионов водорода в другие (ММВ) мы-
шечные волокна и в кровь снижают скорость накопления Н+.
Известно, что скорость прохода этих молекул через мембрану
ограничена [Jorfeldt L. и др. 1978], особенно при их большой
концентрации, но нет сведений о возможности ускорения их
диффузии в результате тренировки. Можно только высказать
предположение, что ускорению элиминации молекул молоч-
ной кислоты будет способствовать гипертрофия ММВ и уве-
личение капилляризации мышц, которые, предположительно,
могли бы способствовать увеличению мембранного градиента
для Ла и Н+ и тем самым скорости их выхода из БМВ.
Увеличение окислительного потенциала МВ, в частности —
объема митохондрий. Это снижает доступность пирувата для
М-лактатдегидрогеназы, митохондрии потребляют протоны, а
в ММВ, предположительно, ускоряется окисление Ла.
6.1.2.5. Заключение по разделу
Как следует из приведенного анализа, стратегией адапта-
ции в отношении увеличения анаэробной производительно-
сти мышц является:
1. Гипертрофия МВ на 50-100% относительно контроля.
Гипертрофии волокон способствует сам анаэробный ха-
рактер физической нагрузки.
2. Накопление эндогенных энергетических субстратов,
среди которых наибольшее значение имеют запасы
КрФ. Высокая концентрация гликогена на дистанциях,
где существенна анаэробная мощность и емкость (15 с —
2-Змин), как представляется, значимой роли не играет.
3. Накопление гликолитических ферментов и изменение
их изоэнзимного состава.
179
4. Небольшое повышение содержания АТФ-азы и КФК-
азы и существенное повышение содержания ферментов
гликолиза (20-50%), независимо от гипертрофии воло-
кон.
5. Повышение буферной емкости мышц за счет накопле-
ния буферирующих веществ, капилляризации мышц,
гипертрофии МВ и повышения окислительного потен-
циала всех типов волокон.
6.2. Тренировочные средства и методы
развития локальной выносливости
Простая логика и данные по гипертрофии МВ (табл. 2 и 3)
указывают на то, что для квалифицированных спортсменов
наибольшее место в тренировочном процессе важно уделять
воздействию на т.н. основные мышечные группы, участвую-
щие в продвижении спортсмена вперед с учетом биомеханиче-
ских особенностей локомоции и внешней среды.
Насколько нам известно, этот тезис не подвергался сомне-
нию ни практикой спорта, ни научными исследованиями.
В то же время это утверждение не противоречит требова-
нию о безусловном соблюдении целесообразной гармонично-
сти развития различных мышечных групп. В этом смысле де-
ление на «основные» и «второстепенные» мышечные группы
условно. Тем более что существует мнение, что в некоторых
локомоциях (например, беге) длительная тренировка сама
оптимизирует технику движений [Cavanagh P.R. и др., 1985;
Nelson R.C., R.J.Gregor., 1976] и «формирует» под эту технику
«топографию» производительности мышечных групп. В то же
время показано, что правомочно и обратное явление — имею-
щееся соотношение мышечных групп откладывает серьезный
отпечаток на основные биомеханические параметры локомо-
ций [Мякинченко Е.Б., 1983]. Однако в свете того что гипер-
трофия МВ — неотъемлемая часть адаптации к спортивной
деятельности (см. выше), а наиболее действенное средство
гипертрофии МВ — силовая тренировка, вопрос выявления
«основных» мышечных групп для целенаправленного воз-
180
действия на них посредством силовых упражнений важен и
актуален. Этот вопрос рассмотрен в конце главы. Ниже, где
речь будет идти о средствах и методах силовой подготовки,
мы подразумеваем, что основным объектом воздействия яв-
ляются именно «основные» мышечные группы.
6.2.1. Средства и методы тренировочного
воздействия на ММВ
ММВ играют наиболее существенную роль практически на
всех дистанциях в циклических видах спорта и их работоспо-
собность является решающей для обеспечения высокой аэроб-
ной производительности спортсмена в ЦВС.
Как установлено в предыдущих разделах, высокая работо-
способность ММВ обеспечивается гипертрофией МВ за счет
накопления главным образом сократительных элементов и со-
путствующих им органелл, одновременным повышением их
окислительного потенциала за счет гиперплазии и гипертро-
фии митохондрий, капилляризации и накоплением запасов
гликогена.
Напомним, что предмет нашего исследования — локальная
выносливость мышц, т.е. внутримышечные факторы, приво-
дящие к увеличению спортивной работоспособности. Поэто-
му, говоря о средствах и методах силовой тренировки, мы чаще
будем говорить, например о гипертрофии мышечных воло-
кон, а не об увеличении мышечной силы, так как улучшению
локальной выносливости в большей мере будет способствовать
тренировка, приводящая именно к увеличению ППС (в том числе
и за счет несократительных элементов мышц), по сравнению с
тренировкой, приводящей только к увеличению мышечной силы,
что происходит, например, при применении упражнений
скоростно-силового типа.
6.2.1.1. Средства и методы, направленные на гипертрофию
(увеличение силы) ММВ
Точные механизмы и стимулы индукции синтеза сократи-
тельного белка неизвестны. Среди факторов, приводящих к
гипертрофии МВ, выделяют как минимум следующие:
181
1. Основное условие — наличие импульсной активности
мотонейрона как фактор нейротрофического контроля
[Валиулин В.В. и др., 1996].
На стадии синтеза РНК:
2. наличие свободного креатина, аденозинмонофосфор-
ной и инозиновой кислот [Walker J.В. 1980, Н.И. Волков,
В.И. Олейников, 2001];
3. наличие ионов Н+ (способствует либерализации мем-
бран, увеличению пор и раскручиванию структур бел-
ков) [Панин Л.Е., 1983].
На стадии синтеза белка:
4. присутствие необходимых стероидных гормонов [Био-
химия: Учебник для институтов физ. культуры, 1986;
Виру А.А., Кырге П.К., 1983];
5. наличие свободных аминокислот (как составных частей
сократительных белков — строительный материал) и
пептидов [Биохимия: Учебник для институтов физ. куль-
туры, 1986; Виру А.А.,Кырге П.К., 1983; Н.И.Волков,
В.И.Олейников, 20011;
Наиболее очевидный и, видимо, эффективный способ обе-
спечения этих условий и, следовательно, гипертрофии ММВ
— это силовая тренировка.
Силовая тренировка.
Высокие концентрации свободного креатина и Н+ в
мышце в целом, а также повышение концентрации ана-
болических гормонов (соматотропный гормон, инсулин,
тестостерон [Виру А.А. Кырге П.К., 1983; Ahtiainen. J. и
др., 2001]) возникают при высокоинтенсивных упражне-
ниях. Однако известно, что гипертрофия ММВ при таком
характере тренировки выражена относительно не сильно,
видимо, из-за краткосрочности действия стимула, а про-
являемая гипертрофия БМВ часто является негативным
фактором в видах на выносливость, т.к. увеличивает мы-
шечную массу без увеличения окислительного потенциала
мышц. Поэтому наиболее приемлемой кажется гипотеза
[Арнис В.Р., 1994; Платонов В.Н., Булатова М.М., 1992;
Сарсания С.К., Селуянов В.Н., 1991; Селуянов В.Н., 1992],
что гипертрофии ММВ будут способствовать изотониче-
182
ские и статодинамические упражнения, выполняемые при
строгом соблюдении следующих правил:
— медленный, плавный характер движений;
— относительно небольшая величина преодолеваемой
силы или степени напряжения мышц (40-70% от МПС);
— отсутствие расслабления мышц в течение всего подхода;
— выполнение подхода до «отказа».
— проведение тренировки, как правило с применением су-
персетов [Вейдер Д.,1992 ], на все основные мышечные
группы;
— достаточно большая длительность всей тренировки (не
менее 1 часа) (более подробно см. Е.Б. Мякинченко,
В.Н. Селуянов, 2001).
Такой характер упражнений приводит к следующим целе-
сообразным явлениям:
— первоначально, и что наиболее важно, будут рекрутиро-
ваться ММВ;
— затрудняется доступ кислорода в ММВ и тем самым
ускоряется снижение концентрации КрФ и накопление
Н+ именно в этих волокнах;
— достаточно большая длительность подходов (60-90 с) и
большое число подходов (4-10) обеспечивает длительное
действие указанных стимулов в ММВ;
— есть основания предполагать, что из-за длительности
подхода, даже при максимальных волевых усилиях в
конце подхода, степень вовлечения БМВ в работу и,
следовательно, их гипертрофия будет относительно
небольшой.
В пользу сделанного предположения, что подобная тре-
нировка будет способствовать гипертрофии ММВ, свиде-
тельствует опыт подготовки бодибилдеров [Вейдер Д., 1992],
у которых ММВ гипертрофированны в отсутствие специали-
зированной тренировки на выносливость (табл. 2), особенно
если применяются подходы с большим числом повторений
[Платонов В.Н. Булатова М.М., 1992]. Их тренировка отлича-
ется от предложенной выше только большими весами (70-85%
от МПС), а также исследованиями, проведенными в Проблем-
ной лаборатории РГАФК (см. выше).
183
В то же время возможно отрицательное влияние подобного
вида силовой тренировки на окислительный потенциал ММВ,
т.к. известно, что высокая степень и длительность закисления
мышц приводит к деструкции митохондрий [Дин Р., 1981; Ле-
нинджер Р., 1966; Лузиков В.Н., 1980].
При рассматриваемом варианте тренировки этот эффект
снижается гипотетически за счет:
— локального характера упражнений, который исключа-
ет существенное снижение pH крови и, следовательно,
обеспечивает высокий градиент между саркоплазмой и
кровью для Н+, облегчающий выход последних в кровь;
— невысокой средней мощности упражнения и небольшо-
го задействование БМВ, что замедляет скорость приро-
ста концентрации Н+ (см. ниже);
— возможности использования аэробных упражнений не-
большой длительности (2-6 минут) между подходами и
сериями для ускоренной элиминации МК.
Основываясь на практике спортивной подготовки и дан-
ных исследований, можно отметить еще два гипотетических
пути гипертрофии ММВ:
1. использование т.н. аэробно-силовой тренировки [Вер-
хошанский Ю.В., 1988 ];
2. использование обычной аэробной тренировки, но при
гормональном статусе организма, благоприятном для
развертывания анаболических процессов.
Аэробно-силовой метод [Верхошанский Ю.В., 1988].
Под этим обобщенным названием подразумеваются хоро-
шо известные в ЦВС варианты тренировки в «утяжеленных»
условиях [Нурмякиви А.А., 1974; Современная система спор-
тивной подготовки / Под ред. Ф.П. Суслова и др., 1995; Фоми-
ченко Т.Г., 1999].
Биологическим обоснованием этого метода являются пря-
мые данные, свидетельствующие о высокой степени гипертро-
фии мышечных волокон в тех мышцах, которые испытывают
наибольшую резистивную нагрузку в данном виде локомоции
([Уилмор Дж.Х., Костил Д.Л., 1997] — у бегунов), [Шенкман
Б.С. идр., 1990] — у гребцов), а также то, что, несмотря на уста-
новленный факт преимущественного увеличения окислитель-
184
ного потенциала в ММВ при аэробной тренировке [Holloszy
J.O., and Coyle E.F., 1984], увеличение интенсивности нагру-
зок или добавление механического отягощения способствует
повышению ОП в БМВ [Dudley и др., 1982; Ишкоидр., 1991].
В легкой атлетике этот метод реализуется путем бега по пе-
ску, в гору, с сопротивлением; в велоспорте — во время «гор-
ной» подготовки или езды с большой передачей; в плавании
- при плавании «на привязи», с тормозом или лопатками; в
лыжных гонках — при тренировке на сильно пересеченной
местности, растягивании резиновых амортизаторов или спе-
циальных блочных устройств и др.
Смысл всех этих вариантов заключается в создании больше-
го относительно обычной локомоции механического усилия,
проявляемого основными мышцами в рабочей фазе движения
при соблюдении в целом аэробного или смешанного характера
энергообеспечения.
К сожалению, нам не известны работы, в которых целена-
правленно исследовалось бы воздействие аэробно-силовых
упражнений на ППС ММВ. Однако в исследовании [Уилмор
Дж.Х., Костил Д.Л., 1997] показана очень высокая степень
гипертрофии ММВ икроножной мышцы у элитных стайеров
и марафонцев (см. табл. 2). Нельзя утверждать, что эти бегу-
ны применяли аэробно-силовой метод, однако известно, что
трехглавая мышца голени испытывает самые высокие механи-
ческие нагрузки в беге (см. выше). То есть, предполагаем, что
сочетание высоких нагрузок и аэробного характера тренировки
может обеспечивать гипертрофию ММВ. Кроме этого, хоро-
шо установлен факт увеличения мышечной силы и спортив-
ного результата под воздействием аэробно-силового метода во
всех ЦВС.
Тренировка в аэробном режиме.
В разделе 7.1. описаны случаи увеличения ППС у людей
под воздействием самой аэробной тренировки в хорошо
контролируемых условиях. Это же получено при умеренных
беговых нагрузках у крыс, причем прирост ППС и скоро-
сти дыхания митохондрий не сопровождались увеличением
плотности капилляров [Немировская Т.Л., 1992]. Стимулы,
которые могут приводить к гипертрофии МВ в этих условиях,
185
не известны, кроме гипотетического «фактора-регулятора»
[Меерсон Ф.З., 1978], действия лизосомальных ферментов
на геном ядра [Панин Л.Е., 1983] при повреждении органелл
клеток, «стимула нагрузкой» [Шенкман Б.С., 1990], фактора
роста фибробластов [Панин Л.Е., 1983]. Есть данные [Watt Р.
и др., 1982], свидетельствующие в пользу того, что гипертро-
фия ММВ реализуется через замедление деградации белков,
тогда как гипертрофия БМВ — через ускорение их синтеза. О
меньшей интенсивности процессов деградации в МВ после
аэробной тренировки свидетельствует также факт повыше-
ния активности дегидрооротатдегидрогеназы, являющейся
индуктором синтетических процессов (в БМВ больше, чем
в ММВ) и небольшое снижение активности глютаматдеги-
дрогеназы, что говорит о снижении активности окисления
аминокислот после аэробной тренировки [Немировская
Т.Л., 1992].
Однако точно установлено действие эндогенных или экзо-
генных анаболических гормонов, приводящих к гипертрофии
волокон при сочетании их повышенного уровня в крови и фи-
зической нагрузки практически любого вида [Виру А.А., Кыр-
ге П.К., 1983]. При этом установлено, что анаболические гор-
моны в большей мере воздействуют на БМВ, ускоряя синтез
быстрого миозина, а глюкокортикоиды, экскреция которых
особенно выражена в конце длительной утомительной работы
[Виру А.А., Кырге П.К., 1983], в большей мере ответственны
за экспрессию генов медленного миозина в ММВ [Валиулин
В.В., 1996 (2)].
Из всего сказанного можно сделать вывод, что, вероятно, воз-
можен путь гипертрофии ММВ просто за счет рационального
тренировочного процесса, стимулирующего, а не угнетающего
гормональную систему спортсменов. Правда, примечательным
является факт, что наиболее выраженным стимулирующим
эффектом на анаболический пул гормонов обладает силовая
тренировка в оптимальном объеме. Таким образом, наиболее
надежным путем [см., например, Шенкман Б.С. и др., 1990]
повышения работоспособности ММВ через их гипертрофию
представляется включение специально организованной сило-
вой тренировки в подготовку спортсменов в ЦВС.
186
6.2.1.2. Средства и методы, направленные на повышение
окислительного потенциала ММВ
В многочисленных исследованиях показана большая плот-
ность и размер митохондрий, объединенных в митохондри-
альную сеть, связанная с ними масса ферментовдыхательного
фосфорилирования, изменение относительной активности
дыхательных ферментов [Немировская Т.Л., 1992; Шенкман
Б.С., 1990; Henriksson J., J.S.Reitman., 1977] у спортсменов,
по сравнению с нетренированными лицами, а также увеличе-
ние окислительного потенциала (ОП) ММВ при начале регу-
лярной тренировки, или увеличение ее объема [Немировская
Т.Л., 1992].
О механизмах и факторах индукции синтеза окислительных
ферментов известно еще меньше, чем о механизмах гипертро-
фии МВ. Длительно бытовавшая гипотеза о гипоксии мышц
как основном стимуле для увеличения их ОП не подтвердилась
в экспериментах с различными моделями гипоксии и ишемии
мышц [Немировская Т.Л., 1992; Daub W.P. и др., 1982], в ко-
торых показано, что при пассивном нахождении в гипоксиче-
ских условиях окислительный потенциал, размеры волокон и
капилляризация падает. В то же время умеренная тренировка в
среднегорье, или периодическая экспозиция в гипоксических
условиях в сочетании с тренировкой оказываются более силь-
ным стимулом в отношении ОП мышц [Булатова М.М., Плато-
нов В.Н., 1996]. Было выяснено, что для роста ОП необходима
аэробная нагрузка, а увеличение активности митохондриаль-
ных ферментов связано с синтезом митохондриальной ДНК и
ускорением синтеза и-РНК в ядре. В то же время показано, что
аэробная работа ниже уровня аэробного порога является очень
слабым стимулом для роста ОП мышц и процессы экспрессии
генов при такой нагрузке активизируются только в конце дли-
тельной работы в фазе утомления [Некрасов А.Н., 1982]. Равно-
мерная тренировка на уровне анаэробного порога, повторная
тренировка с интенсивностью между анаэробным порогом и
МПК, а также интервальная тренировка со средней интенсив-
ностью в зоне мощности аэробного-анаэробного порогов об-
ладают высокой и примерно одинаковой эффективностью, за-
висящей от объема тренировки и исходной подготовленности
187
испытуемых. Обнаружено, что при тренировке наиболее ла-
бильной (т.е. тренируемой) является плотность поверхностных
субсарколемальных митохондрий [Henriksson J., J.S.Reitman.,
1977]. Синтез митохондриальной РНК ускоряется уже через 1 -2
часа после начала аэробной работы [Некрасов А.Н., 1982]. Од-
нако в первые 10-12 дней тренировки ОП мышц не меняется,
но выявлены адаптационные сдвиги в дыхательном контроле
внутри митохондрий, в пиковом кровотоке, углеводном и ли-
пидном обмене внутри мышечных волокон, чувствительности
на Са++стимул [Green H.J., 1996; Williams И.З.идр, 1986].
В большинстве исследований найдено, что активность окис-
лительных ферментов интенсивно повышается в течение пер-
вых 1 -3 месяцев, а потом происходит стабилизация этого пока-
зателя, если тренировочный стимул не изменяется [Биохимия:
Учебник для институтов физ. культуры, 1986; Немировская
Т.Л., 1992; Henriksson J., J.S.Reitman., 1977; Williams R.S. и др.,
1986]. Прекращение тренировки сопровождается быстрым (в
течение 2 недель) возвращением активности ферментов к ис-
ходному уровню [Henriksson J., J.S.Reitman., 1977].
6.2.2. Средства и методы тренировочного
воздействия на БМВ
БМВ вносят существенный вклад на относительно корот-
ких дистанциях (до 3 минут). Наиболее важными аспектами,
касающимися их работоспособности, являются: гипертрофия
(увеличение силы), окислительный потенциал (ОП), буфер-
ная емкость, активность ферментов фосфагенной и гликоли-
тической систем.
6.2.2.1. Средства и методы, направленные на гипертрофию БМВ
Факторы, стимулирующие увеличение ППС БМВ, те же,
что и у ММВ. Создание необходимых условий в БМВ суще-
ственно облегчается высокой активностью КФК-азы и фер-
ментов анаэробного гликолиза, поэтому гипертрофия БМВ
— наиболее типичное следствие любой физической трени-
ровки, включая аэробную [Сээне, 1990; Шенкман Б.С., 1990;
Henriksson J. и др., 1986; Test A., Karlsson J., 1985].
188
Эффективные варианты тренировки, направленной на уве-
личение силы — гипертрофию БМВ, описаны в 4-й главе.
Под воздействием силовой тренировки можно добиться
очень высокой ППС и силы БМВ, однако в ЦВС гипертрофия
быстрых волокон важна только как условие высокой мощно-
сти и емкости процессов энергообеспечения.
Другим традиционным путем повышения силы мышц яв-
ляются прыжковые упражнения различного вида. Известно,
что такие упражнения способствуют приросту силы мышц
[Верхошанский Ю.В., 1988], не приводя, как правило, к зна-
чительной гипертрофии МВ. Поэтому считается, что причи-
ной повышения силы в этом случае является совершенство-
вание нервно-мышечных механизмов управления мышечным
сокращением, например повышение частоты импульсации и
перестройка паттерна активности мотонейронов, приводящие
к улучшению способности к наращиванию усилия [Персон
Р.С., 1985], повышение доли рекрутированных ДЕ, синхрони-
зация импульсов разных ДЕ, улучшение техники движений,
психические факторы и т.п.
Положительное влияние улучшения таких способностей на
спортивный результат в спринте очевидно и многократно под-
тверждено экспериментально и на практике. Однако в связи с
тем что совершенствование этих механизмов не оказывает пря-
мого влияния на мощность и емкость основных источников
энергообеспечения на более длинных дистанциях, т.е. они не
улучшают локальную выносливость, их значимость для спор-
тивного результата на более длинных дистанциях является дис-
куссионной, за исключением случаев, когда такие упражнения
используются с целью укрепления соединительно-тканных
элементов (СТЭ) опорно-двигательного аппарата (ОДА) (чаще
в переходном и подготовительном периоде). Другой случай —
когда они используются как предпосылка повышения эффек-
тивности силовой тренировки, направленной на гипертрофию
мышц. В этом случае применение такого вида упражнений,
как средства «стимуляции центральных компонентов НМА»,
целесообразно в начале тренировочного занятия, а в макроци-
кле — в конце переходного периода, перед началом этапа ак-
центированной силовой тренировки (подробнее см. ниже).
189
6.2.2.2. Средства и методы, направленные на повышение
окислительного потенциала БМВ
Такая постановка проблемы в значительной степени не ти-
пична для спортивных исследований в области ЦВС. Тради-
ционно основным достоинством БМВ считается их высокая
анаэробная (алактатная и гликолитическая) производитель-
ность. Это совершенно справедливо, особенно для дистанций
длительностью 15-180 с. Однако анализ метаболических пу-
тей расщепления КрФ и углеводов не выявляет какого бы то
ни было антагонизма между высокой активностью КФК-азы,
ЛДГ и митохондриальными ферментами. То есть вполне воз-
можно сочетание высоких окислительных и гликолитических
возможностей мышц. Это убедительно доказывает существо-
вание быстрых окислительных МВ, в которых плотность ми-
тохондрий может практически не уступать таковой в ММВ.
Однако существуют исследования, в которых показано, что
высокая степень закисления мышц при гликолитической тре-
нировке может негативно сказываться на митохондриальном
аппарате [ДинР., 1981; Ленинджер Р., 1966; Лузиков В.Н., 1980],
а при аэробной тренировке могут немного понижаться гликоли-
тические возможности [Хочачка П., Дж. Сомеро, 1988]. Однако
в этих исследованиях не рассматривались варианты параллель-
ного воздействия на ферментативную активность гликолитиче-
ский и аэробных реакций. Но гипотетически такое сочетание
вполне может обеспечить прирост обоих показателей.
Точные механизмы и факторы повышения ОП БМВ, так-
же как и ММВ, не известны. Однако показано, что их окисли-
тельный потенциал растет при относительно интенсивных ин-
тервальных или повторных нагрузках, причем даже в большей
степени, чем в ММВ. Это следует из прироста массы фермен-
тов [Daub W.P. и др., 1982; Dudley G.A., R.Djamil, 1985; Ingier
F., 1979] и повышения скорости синтеза РНК [Некрасов А.Н.,
1982; Kavashina К., J-Zawa М., 1982].
Таким образом, анализируя все доступные данные относи-
тельно механизмов стимулирования экспрессии митохондри-
альных генов как в медленных, так и БМВ, можно заключить,
что все сводится к соблюдению двух достаточно простых усло-
вий [Селуянов В.Н., 1993]: 1) интенсивное функционирование
190
митохондрий (т.е. активное состояние данного мышечного
волокна); 2) относительно невысокая степень закисления ци-
тозоля мышечных волокон, в которых митохондрии функцио-
нируют. Вероятно, некоторое положительное влияние может
оказывать снижение степени оксигенации гемоглобина, что
происходит в среднегорье или барокамере [Булатова М.М.,
Платонов В.Н., 1996; Современная система спортивной под-
готовки / Под ред. Ф.П. Суслова и др., 1995]. Хотя и в нормо-
барических условиях достигается высочайшая степень окис-
лительного потенциала и уровня выносливости спортсменов.
Эффект тренировки будет определяться только временем
работы, т.е. длительностью активного состояния мышечных
волокон. Длительность работы, в свою очередь, может огра-
ничиваться скоростью «закисления» мышцы, исчерпанием
запасов углеводов, «центральным» утомлением, механической
перегрузкой ОДА и т.д., а эффективность повышения ОП во-
локон — еще и состоянием организма в период отдыха, точнее
— состоянием нервной, гормональной и иммунной систем ор-
ганизма [Виру А.А., 1981; Суркина И.Д., Готовцева Е.П., 1991;
ШубикВ.М., Левин М.Я., 1985]
В случае ММВ проблема обеспечения указанных двух усло-
вий решается просто — возможно большим поддержанием ин-
тенсивности нагрузки не выше анаэробного порога.
Как добиться таких условий функционирования для БМВ?
Для обеспечения рекрутирования быстрых ДЕ мощность
механической работы в активной (для данной мышцы) фазе
движений должна быть выше значений, которые могут быть
обеспечены ММВ (см. п. 5.1.1).
Для предотвращения их закисления есть четыре способа:
1. Средняя метаболическая мощность работы мышцы
должна быть не выше анаэробного порога. Это означает,
что в таком упражнении значительная сила одиночного
сокращения (это необходимое условие задействования
БМВ в работу, см. выше) должна сочетаться с низким
темпом движений [Верхошанский Ю.В., 1988]. Тогда об-
разующаяся МК успевает частично окисляться в ММВ,
а частично уходить в кровь и окисляться в миокарде и
ММВ менее активных скелетных мышц во время отно-
191
сительно длительной фазы расслабления. Следователь-
но, упражнение может выполняться достаточно долго
без выраженного ацидоза в мышцах и крови.
Такие условия могут быть созданы, например, при
медленном беге, или прыжках широкими шагами в гору,
или с сопротивлением, плавании «широкими шагами»
на привязи, езде с пониженной передачей на велосипе-
де и т.д. Во всех этих случаях мы получаем средство, уже
описанное в аэробно-силовом методе тренировки (см. п.
6.2.1.1)
Для предотвращения случайного понижения pH мышц
ниже оптимального уровня упражнение может выпол-
няться интервально с длительностью рабочего периода
1,5-3 мин и отдыхом 2-3 мин [Мякинченко Е.Б., 1983].
2. Следующий метод также описан в литературе — это «ве-
тровой спринт» [Гилмор Г., Снелл П., 1972], «миоглоби-
новая» тренировка [Биохимия: Учебник для институтов
физ. культуры, 1986]. Его суть — использование много-
численных, но относительно коротких (5-15 с) ускоре-
ний во время работы в зоне аэробного — анаэробного по-
рогов. Интервал между ускорениями 1,5-2,5 мин. Таким
образом, это — интервальная спринтерская тренировка,
выполняемая переменным методом.
Смысл ускорения — вовлечение в работу практически
всех мышечных волокон и существенное понижение в
них концентрации фосфагенов (на 30-50%), это обеспе-
чит развертывание всех процессов энергообеспечения в
активных МВ на полную мощность (см. п. 5.1.1), вклю-
чая окислительное фосфорилирование. Однако короткий
период работы предохраняет от существенного снижения
pH. Важно, что период активного функционирования
митохондрий в БМВ гипотетически будет продолжаться
и после завершения ускорения, когда идет активный ре-
синтез КрФ и окисление лактата. Эффективность такой
тренировки, кроме теоретического обоснования, имеет
и экспериментальное подтверждение [Верхошанский
Ю.В., 1988; Обухов С.М., 1991; Both F.W., 1981].
Назовем этот метод «интервальный спринт».
192
3. Традиционная интервальная тренировка с длительно-
стью рабочего периода 30-60 с и интервалом отдыха 2-4
мин. В таком варианте гипотетически также задейство-
ваны БМВ, но, вероятнее всего, только часть БоМВ, так
как невысокое снижение pH крови свидетельствует о не-
высокой интенсивности анаэробного гликолиза, несмо-
тря на достаточно продолжительный рабочий период
(т.е. о невовлечении БгМВ).
4. Как следует из схемы развертывания процессов энер-
гообеспечения, описанной в первой и пятой главах, на
первой половине дистанции, преодолеваемой с любой ин-
тенсивностью, включая спринт, соблюдаются условия,
необходимые для повышения ОП мышц, а именно — до-
ступность кислорода, наличие всех факторов, активизи-
рующих окислительное фосфорилирование, невысокая
степень накопления Н+. Следовательно, повторная или
интервальная работа с соревновательной скоростью на
отрезках, не превышающих 1/2 или даже 2/3 от длины
соревновательной дистанции (т.е. до начала или середи-
ны зоны компенсированного утомления), и интервалом
отдыха 3-5 мин (или восстановления ЧСС до 120-130 уд/
мин) является прежде всего средством аэробной подготов-
ки, а не «смешанной», гликолитической и т.п. В таком ка-
честве ее и стоит рассматривать. При этом можно ожидать,
что эффект тренировки будет выражен в большей степени
в отношении БМВ, так как для ММВ длительность дей-
ствия стимула, с учетом плотности митохондрий в них,
будет недостаточной для заметного прироста их ОП.
6. 2.2.3. Средства и методы, направленные на повышение бу-
ферной емкости мышц и массы ферментов анаэробного гликолиза
Как было установлено в 5-й главе, максимальная гликолитиче-
ская мощность требуется только в спринте — при работе до 40 с.
К сожалению, нам ничего не известно о механизмах и фак-
торах повышения массы (или плотности) ферментов гликоли-
за в цитозоле. Можно только предположить, что:
— количество ферментов гликолиза будет расти парал-
лельно с гипертрофией и повышением массы сократи-
193
тельного белка в БМВ. Этого можно добиться методами
силовой подготовки;
— стимулом к ускоренному синтезу и-РНК (активация
цепочки: медиаторы, гормоны Ю ц-АМФ, ц-ГМФ Ю
ферменты Ю генетический аппарат) могут являться
факторы, стимулирующие высокую скорость протека-
ния гликолитических реакций, т.е. высокий градиент
накопления и высокая концентрация Са++, Кр, АДФ.
Это достигается посредством спринтерских ускорений
максимальной интенсивности при длительности работы
не менее 8-10 с (для существенного понижения КрФ в
БМВ);
— будет наблюдаться высокая концентрация ионов водо-
рода в цитозоле волокна.
В отношении последнего предположения можно заметить,
что, с одной стороны, этот фактор является индуктором синте-
за белков, а с другой— именно снижение pH ограничивает ско-
рость протекания гликолитических реакций, поэтому кажется
маловероятным, что фактор-ограничитель скорости реакции
может одновременно являться фактором-индуктором повы-
шения скорости той же реакции!
К сожалению, это все — предположения, для которых нуж-
ны серьезные основания.
Классическим вариантом «гликолитической» тренировки
является повторное преодоление 3-4 отрезков длительностью
40-90 с с околомаксимальной интенсивностью и сокращаю-
щимся интервалом отдыха (5-2 мин). В этом случае удается
добиться максимальных значений концентрации Ла в крови
и, видимо, минимума pH. Предполагается, что это является
признаком высокого тренировочного воздействия на глико-
литические системы. Однако исследование двух упражнений
длительностью 30 с максимальной интенсивности с интер-
валом отдыха 4 мин [Nevill М.Е. и др., 1996] с взятием биоп-
сийных проб показало, что перед второй попыткой в мышце
были понижены концентрации гликогена, КрФ, АТФ и pH.
В этих условиях при повторной попытке повышенными ока-
зались только показатели аэробной мощности (!) (на 18%).
Выход АТФ из фосфагенов и анаэробного гликолиза сократи-
194
лись на 25 и 43%. Это означает, что активность алактатных
и гликолитических анаэробных процессов энгергообеспечения при
таком варианте тренировки снижается. Таким образом, если
серия повторных интенсивные нагрузок более эффективна в
отношении гликолиза, чем однократные предельные нагруз-
ки (хотя нам не известны эксперименты, где такое сравнение
проводилось бы), то фактором воздействия на гликолитиче-
скую производительность является не столько интенсивность
функционирования этого механизма или глубина снижения
pH, сколько длительное поддержания низкого pH и понижен-
ного содержания субстратов в мышце.
Если предположить, что накоплению буферирующих ве-
ществ будет способствовать низкая pH, поддерживаемая до-
статочно длительное время, то максимальной буферной ем-
костью обладали бы культуристы, для которых длительное
поддержание низких значений pH в мышце, при выполнении,
например суперсетов, — одно из условий эффективности тре-
нировки. Однако нам не известны сравнительные данные о
концентрации буферирующих веществ (например, бикарбо-
натов или накоплении гистидина) в мышцах этих спортсменов
в сравнении с другими специализациями.
Таки м образом, проведенный анализ с достаточно большой
вероятностью позволяет сделать вывод, что росту «гликоли-
тических возможностей» (повышению массы или плотности
ключевых ферментов гликолиза и «емкости» гликолиза) бу-
дут способствовать спринтерские упражнения (10-20 с), все
виды силовой тренировки быстрых мышечных волокон, все
виды «аэробной тренировки быстрых мышечных волокон».
С меньшей вероятностью таким эффектом будет обладать
«гликолитическая тренировка», а известный из практики
высокий стимулирующий эффект таких упражнений в отно-
шении спортивного результата может объясняться другими
причинами. Например, психологическим эффектом (воле-
вой подготовкой — «умением терпеть»), совершенствованием
техники, «стимуляцией» нейро-эндокринной системы, уве-
личением силы основных мышц спортсмена, при некоторых
условиях — стимуляцией максимальной аэробной произво-
дительности и т.д.
195
6.3. Теоретические основы планирования
одного тренировочного занятия, тренировочных
микро-, мезо- и макроциклов
Исходным пунктом для составления тренировочного плана
является ожидаемый тренировочный эффект (ТЭ). В нашем
случае — улучшение показателей локальной выносливости
спорстменов в ЦВС.
Улучшения локальной выносливости (также как и других
компонентов физической подготовленности) можно добить-
ся, решив три задачи:
1. целесообразно подобрав средства и методы воздействия
на генетический аппарат соответствующих морфострук-
тур организма (т.е. стимулировав синтез определенного
вида и-РНК);
2. обеспечив оптимальные условия для протекания про-
цессов синтеза органелл клеток, подвергшихся трениро-
вочному воздействию во время восстановления;
3. обеспечив оптимальные последовательность и уровень
развития мышечных компонентов, определяющих ло-
кальную выносливость применительно к выбранной со-
ревновательной дистанции.
Первая задача решается путем планирования тренировоч-
ного занятия, вторая — организацией отдыха и планированием
микро- и мезоциклов, третья — макроциклов.
6.3.1. Теоретические основания
для планирования одного
тренировочного занятия
Как определено ранее, основными морфоструктурами, ко-
торые должны подвергаться воздействию с целью развития ло-
кальной выносливости, являются:
— структурные и сократительные элементы мышц;
— капилляры и митохондриальный аппарат;
— ферментативные комплексы КФК-реакции и гликолиза.
По каким правилам должно строиться тренировочное за-
нятие (далее, для краткости — тренировка) для обеспечения
196
высокого тренировочного эффекта, направленного на синтез
указанных органелл мышечных клеток и капилляров мышц?
В наиболее общем виде основным правилом является: обе-
спечение максимальных стимулов для синтеза и-РНК соответ-
ствующего типа при минимальном повреждении структурных
и сократительных элементов и исчерпании углеводных запасов
мышц, а именно создание оптимальных условий для развертыва-
ния анаболических (ассимиляционных) процессов при минимиза-
ции катаболического эффекта.
Ускоренный синтез и-РНК обеспечивается адекватны-
ми стимулами, воздействующими на генетический аппарат.
Можно ожидать, что количество молекул и-РНК будет про-
порционально длительности действия стимула. В нашем слу-
чае — длительности выполнения упражнений определенной
направленности. Ограничителями длительности тренировки
являются: повреждения элементов мышц за счет действия ме-
ханических [Friden J. и др., 1983; Armstrong, 1991; Eston R.G. и
др., 1994] или химических [Пшенникова М.Г. 1986; Moreau D.
и др., 1995] факторов; исчерпание запасов гликогена в мышцах
[Westerblad Н. и др., 1991; Kiens В. и др., 1993]; «центральное»
утомление, связанное с ЦНС, ССС или нейроэндокринной
системой (см. главу 5) и др. По мере продолжения тренировки
степень проявления «повреждающих» факторов увеличива-
ется, достигая максимума в конце занятия в связи с повреж-
дением клеточных мембран со стороны свободных радикалов
[Пшенникова М.Г., 1986], деградации адениннуклеотидов,
снижением запасов гликогена, «накоплением» повреждений
в структурных и сократительных белках, утомлением нервной
системы и ухудшением координации и др.
Существуют представления, что, для того чтобы получить
высокий анаболический эффект (т.е. сильно стимулировать
синтез и-РНК и белков), необходимо довести спортсмена до
высокой степени утомления. В общем случае это верно, так
как утомление часто связано с разрушением клеточных струк-
тур, что сильно интенсифицирует синтетические процессы.
Особенно выраженный эффект (пролиферация и гипертрофия
ядер МВ, содержание в них ДНК) удается получить, например,
при высокоинтенсивной электростимуляционной тренировке
197
[Cabric M. и др., 1988], приводящей к интенсивным после-
тренировочным мышечным болям. Однако разрушение — не
единственный стимул синтетических процессов, которые в
этом случае носят компенсаторный характер. Вторым и наи-
более важным для спортивной тренировки стимулом является
создание таких условий в мышечных клетках, которые уско-
ряли бы приспособительные (адаптационные) синтетические
процессы, лежащие в основе развития физических способно-
стей. Причем создание таких условий совершенно необяза-
тельно должно сопровождаться выраженным повреждением
морфоструктур мышц.
Таким образом, при выборе средств и методов тренировки
очень важно четко представлять, какие из упражнений несут
в себе высокий катаболический потенциал (они будут стимули-
ровать синтез, но компенсаторный), а какие не только разру-
шают, но и способствуют развитию мышц, то есть реализуют
цель физической тренировки.
Этот вопрос для теории и методики спортивной трениров-
ки является наиважнейшим, и в тоже время он практически не
разработан.
В настоящее время есть основания предполагать, что наи-
более высоким катаболическим эффектом, при минимальном
анаболическом, будут обладать скоростно-силовые упражне-
ния, в которых в явном виде присутствует уступающий режим
работы мышц, т.е. когда мышцы насильно растягиваются с
высокой скоростью. «Классический» пример — спрыгивания
«в глубину» и бег под гору, а также прыжки и быстрые при-
седания со штангой или другими отягощениями, резкие махо-
вые движения при «развитии гибкости», любые прыжковые и
спринтерские упражнения или даже просто беговые нагрузки
в большом объеме, если проводятся спортсменами, у которых
специальной предварительной тренировкой ОДА не подготов-
лен к такому характеру работы. Повреждение мышц легко диа-
гностируется по появлению послетренировочных мышечных
болей и снижению силы этих мышц.
Второй вид таких упражнений — это те, при которых до-
стигается глубокое локальное и «глобальное» «закисление»
организма, сопровождающееся значительным понижением и
198
долгим удерживанием низких значений pH крови. При такой
тренировке в максимальной степени активизируется симпа-
тоадреналовая и глюкокортикоидные системы, что сопрово-
ждается выбросом «стресс-гормонов», максимально мобили-
зующих не только энергетические, но и пластические ресурсы
организма, а также воздействуют на иммунную систему. Кроме
того, высокая концентрация ионов водорода в мышцах и мед-
ленная их элиминация из-за низкого градиента между сарко-
плазмой и кровью является одним из основных «химических»
факторов повреждения органелл и мембран клеток.
Применение таких упражнений, предположительно, будет
отвлекать часть пластических ресурсов для «компенсаторно-
го», а не «развивающего» синтеза белков, так как пластиче-
ский и гормональный пул организма ограничен [Виру А.А.,
Кырге П.К., 1983].
Сказанное не означает, что мы расцениваем такие упраж-
нения как безусловно вредные. Их применение в нужное вре-
мя и в нужном месте может быть необходимо (см. ниже в этой
главе и часть 7-й главы). Однако всегда надо помнить об их
особенностях, и соответственно строить тренировку, напри-
мер, в те периоды, когда необходимо добиться максимального
«анаболического» эффекта — быстрого прироста тренируемой
способности.
Об упражнениях, которые обладают, на наш взгляд, мак-
симальным «анаболическим» потенциалом, уже говорилось
выше.
Таким образом, первым принципом построения трениро-
вочного занятия, основной целью которого является улучше-
ние компонентов Л В мышц, можно считать «принцип учета
анаболического/катаболического потенциала упражнения».
В связи с тем что скорость накопления повреждений уве-
личивается к концу занятия, а это увеличивает время и стои-
мость восстановления, то вторым принципом будет «принцип
укороченных, более частых занятий», который предполагает,
что две коротких тренировки дадут больший прирост спо-
собности, чем одна длинная такого же объема. В то же время
«укороченные» тренировки предполагают экономию време-
ни и ресурсов организма только за счет низкоэффективных
199
средств подготовки, но не за счет эффективных средств, под
которыми мы понимаем такие, которые с предельной интен-
сивностью воздействуют на развиваемую способность. Такие
средства улучшения основных компонентов Л В выносливости
описаны выше.
В то же время для недопущения ошибок в построении ре-
ального тренировочного процесса еще раз необходимо подчер-
кнуть, что данные принципы относятся к построению занятий
«в интересах» развития ЛВ. Однако существуют задачи разви-
тия других компонентов выносливости, например, накопле-
ние гликогена в мышцах, повышение устойчивости деятель-
ности нейро-эндокринной системы, системы терморегуляции
и т.п., которые могут требовать для своего решения использова-
ния других принципов построения занятия.
Как уже отмечалось, Л В зависит от степени развития в
основном трех мышечных компонентов:
— сократительных элементов мышц;
— митохондриального аппарата и системы микроциркуляции;
— ферментативных комплексов, обеспечивающих высо-
кое содержание субстратов, высокую емкость и скорость
КФК и гликолитической реакции.
Для обеспечения прироста функциональных возможно-
стей указанных структур требуются в большинстве случаев
совершенно различные внутриклеточные условия и характер
сокращения мышц. Для гипертрофии волокон — анаэробные
условия; для митохондриального аппарата — аэробные; для
ферментативных комплексов, определяющих алактатные и
гликолитические характеристики мышц, предположительно
— высокая скорость и степень исчерпания субстратов и т.п. В
процессе выполнения какого-то упражнения в ядрах МВ про-
исходят совершенно конкретные изменения, ускоряется син-
тез строго определенных молекул и-РНК и т.д. В реализации
этих изменений участвуют специфические энергетические,
пластические и гормональные процессы. Таким образом, су-
ществует очень мало доводов в пользу так называемых «ком-
плексных» занятий, когда в одной тренировке ставится задача
развития сразу нескольких способностей, связанных с уско-
рением синтетических процессов различных морфоструктур
200
внутри мышц. Поэтому целесообразно считать третьим прин-
ципом построения тренировочного занятия — «принцип однона-
правленного воздействия» [Верхошанский Ю.В., 1985], в основе
которого лежит предположение, что серия разного вида (на-
пример, «на силу» и «на выносливость») однонаправленных
занятий будет более эффективной в отношении развиваемых
способностей, чем серия комплексных занятий. Этот прин-
цип не отменяет имеющиеся рекомендации [Платонов В.Н.,
1984] использовать в одном занятии весь спектр эффективных
средств и методов для развития, например, аэробных способ-
ностей. Или совмещения, например, технической подготовки
с любым из средств улучшения локальной выносливости и т.п.
В то же время он отражает современные тенденции в органи-
зации подготовки высококвалифицированных спортсменов
| Верхошанский Ю.В., 1985; Платонов В.Н., 1984].
Но в реальном тренировочном процессе часто становится
невозможным использование только однонаправленных за-
нятий, поэтому возникает проблема оптимального комплек-
сирования упражнений различной направленности в одном
занятии.
Наиболее часто используемые виды тренировки — это «на
силу» и «выносливость», и от их рационального сочетания в
большинстве случаев зависит эффективность тренировки.
Увеличение мышечной силы подразумевает 1) улучшение
«нервного» компонента силовых способностей и 2) гипертро-
фию мышечных волокон.
Вероятно, особого значения не имеет, в какой части трени-
ровки применять упражнения, улучшающие «нервный» ком-
понент силы (это упражнения с околопредельными весами и
с максимальной мощностью — «взрывные» упражнения), так
как ЛВ в большей мере зависит от внутримышечных факто-
ров. Хотя считается, что выполнение таких упражнений в на-
чале занятия, когда спортсмен «свежий», дает лучший эффект.
Объем таких упражнений зависит от механической прочности
мышц и при передозировке они начнут болеть. Поэтому лучше
за один раз применять меньший объем таких упражнений. Бо-
лее рациональный вариант — понемногу, но чаще, например
на каждой тренировке.
201
В то же время планирование тренировки с целью гипертро-
фии мышечных волокон и одновременного повышения содер-
жания «аэробных» энзимов — существенный вопрос.
Для того чтобы синтез сократительных белков мышц (ги-
пертрофия мышц) происходил успешно, нужно (кроме самого
тренировочного воздействия) обеспечить наличие достаточ-
ных запасов углеводов в мышцах (как источника энергии), в
крови должно быть достаточное количество аминокислот —
строительного материала для мышц и (наиболее существенно)
повышенное содержание в крови так называемых анаболи-
ческих гормонов (например, тестостерона и соматотропина)
[Теппермен Дж., Теппермен X., 1989; Galbo Н., 1981].
Ускоренный синтез же аэробных ферментов обеспечивает-
ся другими условиями. Основными характеристиками которых
является высокая энергетическая стоимость и выброс другой
группы гормонов (называемых стресс-гормонами), главные
среди которых глюкокортикоиды [Теппермен Дж., Теппер-
мен X., 1989]. «Анаболическая» функция глюкокортикоидов —
ускорение синтеза митохондриальных компонентов. Однако
во время занятия эти гормоны выполняют мобилизацию (т.е.
расщепление) белковых ресурсов организма. Проще говоря,
они вызывают расщепление мышечных белков (а задача — их
накапливать) и белков лимфатической системы, которые вы-
полняют иммунные (защитные) функции в нашем организме.
Именно поэтому при перетренировке увеличивается частота
заболеваний [Fitzgerald L., 1988], а слишком «объемная» аэроб-
ная тренировка приводит к уменьшению толщины мышечных
волокон, потере миоглобина [Terrados N. и др., 1986]. Кроме
того, высокая энергетическая стоимость такой тренировки
приводит к утилизации (сжиганию) не только углеводов, но и
гормонов, аминокислот, т.е. тех веществ, которые нужны для
«строительства» мышц.
Силовая тренировка может «вредить» выносливости еще и
потому, что при тренировке, направленной на гипертрофию
мышц, в мышце накапливаются в высокой концентрации и
удерживаются длительное время ионы водорода, «закисляю-
щие» внутреннюю среду мышечных клеток, что ускоряет раз-
рушение митохондриальных компонентов, поэтому аэробная
202
выносливость мышц может ухудшаться [Luthi J.M. и др., 1986;
MacDougal J.D. и др., 1979].
Какое сочетание рациональнее — сначала тренировка силы,
а потом выносливости, или наоборот?
Нам известно только одно исследование, в котором этот
вопрос изучался в контролируемых условиях [Jackson С. и др.,
1990]. Показано, что если сначала применять силовую, азатем
аэробную нагрузку, то объем МВ уменьшается, если наоборот
— увеличивается.
Эти данные подтверждаются и объясняются результатами
имитационного компьютерного моделирования, выполнен-
ного в ПНИЛ РГАФК.
Суть полученных результатов следующая.
Как отмечалось, нагрузки на выносливость приводят к рас-
щеплению белков до аминокислот и их использованию в ка-
честве топлива. То же происходит и с гормонами. Таким обра-
зом, если сначала выполнить силовую тренировку, то в самих
мышцах повышается концентрация метаболитов, а в крови
- концентрация аминокислот и гормонов, необходимых для
начальной стадии синтеза белков. Если после нее следует аэ-
робная, то происходит элиминация этих веществ и, таким об-
разом, эффективность силовой тренировки будет очень низкой.
Если же сначала выполнить аэробную нагрузку, а потом
силовую, то выделившиеся полезные для ускоренного синтеза
белка вещества будут в восстановительный период беспрепят-
ственно выполнять свою функцию ускорения синтеза белков.
Кроме этого, есть предположение, что создаваемый в
результате силовой тренировки «анаболический фон» в ор-
ганизме, будет положительно влиять и на синтез аэробных
ферментов, особенно если между аэробной и силовой тре-
нировкой сделать 15-20-минутный перерыв и выпить слад-
кий напиток.
Если стоит задача сочетания скоростно-силовой и силовой
нагрузки, то оптимальным будет такое же сочетание — сначала
интенсивные нагрузки, пока не ухудшена координация, пока
ЦНС способна к обеспечению спринтерских и скоростно-
силовых упражнений, пока не созданы предпосылки для
микротравмирования мышц во время силовых упражнений.
203
Также можно предположить, что интенсивные средства, при-
мененные в начале, будут иметь стимулирующий эффект в от-
ношении повышения эффективности «анаболической» сило-
вой тренировки.
На тех же основаниях можно предположить, что при со-
четании скоростно-силовой и аэробной тренировки первая
должна выполняться в начале.
При трехкомпонентном занятии рациональным будет со-
четание: спринт + скоростно-силовые упражнения, далее —
аэробная нагрузка, затем — силовая.
Но эффективность таких занятий очень сомнительна и их
применение целесообразно только в тренировке юношей в
переходный период в «разгрузочных» микроциклах для под-
держания способностей.
В обобщенном виде материалы этого раздела представлены
в табл. 5 и на рис. 12.
Эффективность тренировочного занятия (%)
Рис. 12. Схема планирования одного тренировочного занятия: наиболее
эффективно однонаправленное занятие; в случае комбинирования — сна-
чала выполняется аэробная нагрузка, затем — силовая
204
Таблица 5. Рациональные варианты построения тренировочных занятий
Варианты занятий Примечания
1. Однонаправленное занятие любого вида и небольшой длительности Будет обладать максималь- ной эффективностью
2. Аэробная нагрузка — силовая на- грузка Будут обладать средней
3. Скоростно-силовая - силовая эффективностью
4. Скоростно-силовая - аэробная
5. Скоростно-силовая - аэробная- силовая Будет обладать минималь- ной эффективностью
6. «Гликолитическая» Такая нагрузка всегда должна применяться на отдельном занятии
В приведенных выше рассуждениях не затрагивалась про-
блема технической подготовки. Она подробно будет рассмо-
трена ниже.
6.3.2. Теоретические основания для планирование
микроциклов
При планировании микроциклов, согласно известной кон-
цепции Г.В. Фольборта, можно ориентироваться на правило,
что следующая нагрузка развивающего характера должна при-
ходиться на фазу суперкомпенсации.
Когда наступает фаза суперкомпенсации после различных
упражнений?
Считается, что это зависит от вида упражнений, величины
нагрузки и варианта чередования занятий различной направ-
ленности [Платонов В.Н., 1984].
Однако более целесообразно ориентироваться не на внеш-
ние, а на внутренние факторы. Например, мышечные фермен-
тативные комплексы, обеспечивающие выносливость, более
лабильны, т.е. они быстрее, чем сократительные белки синте-
зируются при тренировке, но и быстрее расщепляются при ее
прекращении (период «полужизни» первых — 6,5, вторых — 11
205
суток) [Меерсон Ф.З., 1978; Fleck S.J., Kraemer W.J., 1987]. Это
отражается в известном положении о гетерохронности восста-
новления, поэтому, в частности после силовой тренировки на
крупные мышечные группы, фаза суперкомпенсации наступа-
ет не ранее чем через 3-4 дня [Платонов В.Н., 1984]. Следова-
тельно, в микроцикле может применяться не более 2 силовых
тренировок на одни и те же мышечные группы при условии,
что величина нагрузки была значительной, но не чрезмерной,
после которой фазы суперкомпенсации может вообще не на-
ступить.
В то же время есть основания предполагать, что механиз-
мы ускорения синтеза белков митохондриального аппарата
и пролиферации капилляров обладают высокой мощностью
и способны обеспечивать прирост плотности митохондрий,
даже при ежедневных многочасовых тренировках. Например,
показано, что если синтез миофибриллярных белков сразу по-
сле истощающего бега оказывается угнетен и возрастает толь-
ко через сутки, то синтез саркоплазматических белков ускорен
сразу же после нагрузки [Некрасов А.Н., 1982; Сээне Т.П.,
1987]. Поэтому после аэробных занятий восстановление зави-
сит в основном от скорости восполнения запасов гликогена в
мышцах, которое может затягиваться на 2-3 суток [Платонов
В.Н., 1984]. Но у квалифицированных спортсменов механиз-
мы ресинтеза гликогена имеют высокую мощность. Кроме
этого, если не использовать истощающих занятий, а ту же на-
грузку распределить на 2-3 тренировки в день (при этом вос-
становление завершается через 10-12 часов) и использовать
дробное углеводное питание, то одних суток после тренировки
выносливости хватит, чтобы синтезировать некоторое допол-
нительное количество митохондрий, увеличить капиллярную
сеть около мышечных волокон, улучшить работоспособность
сердца и в тоже время восстановить запасы гликогена в мыш-
цах и печени. На практике же обычно используют тренировоч-
ные занятия с большой нагрузкой в два соседних дня, после
которых идут 1-2 дня отдыха для восстановления указанных
ресурсов мышц.
В то же время нельзя забывать, что в некоторых локомоци-
ях (например, бег) структурные и сократительные белки мышц
206
могут сильно страдать в силу биомеханических причин из-за
наличия «отрицательной фазы работы мышц» в период аморти-
зации, когда опорно-двигательный аппарат работает в режиме
«гашения» ударных механических нагрузок. Этот фактор мо-
жет существенно замедлять процессы восстановления и должен
всегда учитываться при планировании нагрузки. Например,
темповой бег по шоссе в большом объеме чаще всего должен
приравниваться к силовой нагрузке, т.е. для восстановления до
фазы суперкомпенсации требуется не менее 3 суток.
После силовых тренировок концентрация гормонов в крови
повышена достаточно длительное время — до 1-2 суток [Виру
А.А.,Кырге П.К., 1983]. Весь этот период протекают процессы
синтеза сократительных белков. Поэтому если на следующий
день после силовой тренировки провести нагрузочную аэроб-
ную, то это остановит синтетические процессы белков миофи-
брилл [Сээне Т.П., 1987] и эффект силовой тренировки будет
в значительной степени ликвидирован. Чтобы этого не проис-
ходило, после силовой тренировки в течение 36-48 часов дол-
жен планироваться отдых.
Таким образом, получаем «элементарный» микроцикл, со-
стоящий из 4 дней: 1 -й день — две аэробных тренировки утром
и вечером; 2-й день — аэробная тренировка вечером; 3-й день
— силовая тренировка вечером; 4-й день — отдых.
Если это подготовительный период и этап развития си-
ловых способностей (гипертрофии ММВ), то для того чтобы
«привязаться» к недельному циклу, следующий микроцикл
может состоять из 3 дней: 1-й день — «аэробный»; 2-й — «сило-
вой», 3-й — отдых.
При таком недельном микроцикле может быть обеспечен
максимальный эффект развития силы спортсмена при поддер-
жании или даже улучшении аэробных способностей. Это мно-
гократно подтверждено в исследовании наших сотрудников и
аспирантов кафедры легкой атлетики и ПНИЛ РГАФК, про-
веденных на бегунах [Масленников А.В., 1986; Обухов С.М.,
1991; Паленый В.И., 1991; Чесноков Н.Н., 1991]. Однако, кро-
ме улучшения ЛВ, необходимо решать и другие задачи, поэто-
му на практике «день отдыха» чаще бывает днем «активного
отдыха», когда проводится скоростно-силовая, техническая
207
или восстановительная тренировка, но в объеме не более 1/2
от обычного развивающего. Это вполне допустимо или даже
может иметь положительные стороны, т,к. показано [Плато-
нов В.Н., 1986], что дополнительное занятие принципиально
другой направленности ускоряет восстановление и наступле-
ние фазы суперкомпенсации.
На этапе развития аэробных способностей происходит
лишь одна замена — вместо второй силовой тренировки про-
водится еще одна аэробная с использованием «эффективных
средств аэробной подготовки» (см. выше) и т.д.
В то же время известно, что в тех ЦВС, где мышцы не ис-
пытывают больших механических воздействий (плавание,
велосипед — шоссе) удается увеличить тренировочное время
до 5-7 часов в день за счет планирования двух-, трехднев-
ных тренировочных занятий. Может создаться впечатление
о расхождении наших выводов с выводами, например, та-
кого признанного авторитета, как В.Н. Платонов [Платнов
В.Н., 1980, 1984; Полунин А.И., Снесарев Н.К., 1990]. Од-
нако следует отметить, что расхождение только кажущееся
из-за различий в использовании терминов. Например, такое
средство, как интервальный спринт мы относим исключи-
тельно к средству аэробной подготовки (см. раздел 6.2.2.2),
а не скоростной или «параллельной» [Платонов В.Н., 1984,
с. 201]. Хотя бы уже на том основании, что скоростные воз-
можности мышц генетически предопределены [Гурфинкель
В.С., Левик Ю.С., 1985] и «тренировать» их, по крайней
мере у взрослых людей, бессмысленно. Поэтому, призна-
вая большую целесообразность использования вариативных
средств как способа увеличения тренировочного объема,
мы предпочитаем все же классифицировать нагрузки по их
физиологической и биохимической направленности, а не
по внешним, часто не существенным, признакам. В связи с
этим, когда мы говорим о дне «аэробной» нагрузки и воз-
можности ее разделения на три тренировочных занятия, то
это может выглядеть в том же плавании так:
1-е занятие (дополнительное, утром). Отработка техники
на 50-100-метровых отрезках с соревновательной скоростью
на 400 м, с ИО — 3-5 мин. Нагрузка — средняя (1 час).
208
2-е (основное) занятие. Первая половина — плавание с до-
полнительным сопротивлением, интервально — 3 мин через 3
мин длительность — 50-60 мин). Вторая половина (через 15-20
мин отдыха и приема сладкого напитка) — интервальная тре-
нировка на 50-метровых отрезках с соревновательной скоро-
стью на 400 м, ИО — 2-3 мин ( длительность 30-40 мин). На-
грузка - большая (2 часа).
3-е занятие (дополнительное, вечером). Первая половина
— занятие в зале «сухого плавания» — интервальная работа на
тренажере для «специальной силовой подготовки пловцов»:
подход 30-40 с, ИО — 2-3 мин. Сопротивление подбирается
так, чтобы локальное утомление мышц рук ощущалось, но не
накапливалось на протяжении всего занятия длительностью
30-40 мин. Вторая половина (через 15-20 мин и приема слад-
кого напитка) — интервальный спринт: ускорение 5-8 с с мак-
симальной скоростью, ИО - 2-2,5 мин покойного плавания
(длительность 30-40 мин).
По нашей классификации, все используемые в течение дня
средства будут способствовать развитию аэробных способно-
стей основных мышц пловца, а не «смешанных», «гликолити-
ческих», «скоростных» способностей или «силовой выносли-
вости», «скоростной выносливости» и т.п. Хотя в тренировках
применялись достаточно вариативные средства.
6.3.3. Теоретические основания для планирования
мезоциклов
Период полужизни большинства морфологических струк-
тур ОДА, за исключением костей и коллагеновых волокон, со-
ставляет 1 -2 недели. Это означает, что в течение этого времени
при правильно организованной тренировке можно добиться
положительных сдвигов в развиваемой способности. Об этом
свидетельствуют результаты большого числа лабораторных и
естественных экспериментов (см., например, [Воробьев А.Н.,
1977]). Правда, в тех случаях, когда происходит относительно
резкая смена средств и методов тренировки, 1-2 недели за-
нимает период адаптации к ним, во время которого работо-
способность может понижаться в силу различных причин как
209
«центрального», так и «локального» уровня. Например, извест-
но, что в начале тренировки мышечная сила растет в большей
мере за счет нейрогенных, а не миогенных (мышечных) факто-
ров [Современная система спортивной подготовки / Под ред.
Ф.П. Суслова и др., 1995; Хоппелер Г., 1987]. Однако даже с
учетом этой оговорки в течение обычного мезоцикла (3-6 не-
дель) должны быть достигнуты положительные изменения в
показателях тренируемой функции. Если этого не происходит,
то это означает лишь одно — неверное планирование тренировоч-
ного процесса. Мы полагаем, что это должно быть общим пра-
вилом для большинства видов спорта. В схематическом виде
такой мезоцикл представлен на рис. 13.
Исключение может представлять тренировка прыгунов, у
которых специфика соревновательного упражнения вынуж-
дает применять высокоударные прыжковые упражнения для
адаптации соединительно-тканных и сократительных элемен-
тов ОДА к высоким механическим нагрузкам. В связи с тем что
период полужизни белков коллагеновых волокон относитель-
но большой, а при массированном применении такого вида
нагрузок сократительные элементы мышц находятся в «по-
луразрушенном» состоянии, то продолжительность снижения
показателей может продолжаться 1-2 месяца [Верхошанский
Ю.В., 1982]. В то же время следует отметить, что сформули-
рованный принцип не входит в противоречие с выявленными
С.М. Вайцеховским [1985] тремя фазами реализации (напри-
мер, силовых качеств) в процессе тренировки, которые близки
по смыслу представлениям Ю.В. Верхошанского (1985) об от-
ставленном тренировочном эффекте, когда (1-я фаза) в тече-
ние 4-6 недель после начала интенсивной силовой подготов-
ки степень реализации силовых качеств в соревновательном
упражнении и соответственно спортивный результат ухудша-
ются, в течение следующих 3-4 недель (2-я фаза) происходит
постепенное повышение реализации возросших силовых
способностей и только после этого (3-я фаза) увеличение си-
ловых возможностей будет идти параллельно техническому
совершенствованию, в процессе этого будет наблюдаться вы-
раженный прирост спортивных результатов. В нашем случае
речь идет о приросте функциональных возможностей мышц,
210
@ Величина нагрузки (%)
1
тестир.
2
тестир.
*****•* Показатель 1 Показатель 2
Рис. 13. Принципиальная схема планирования мезоцикла (4 недели) и
оптимальная динамика показателя тренируемой способности (1) и под-
держиваемой на достигнутом уровне (2)
которые могут и не находить выражение в улучшении результа-
тов в соревновательной локомоции, но могут быть выявлены в
специфических тестах, наилучшими из которых будут являть-
ся такие, в которых характер проявления усилий мышц будет
наиболее близок к характеру работы мышц, в котором проис-
ходит тренировка. Только в этом случае может быть произве-
дена адекватная оценка происходящих изменений в состоянии
мышц, а также ликвидирована почва для различных спекуля-
ций на тему «о возможности накопления двигательного потен-
циала в течение 2-6 месяцев (когда способности понижены) с
последующей реализацией...». Когда такие спекуляции имеют
место, то в большинстве случаев тренируется одно, измеряется
второе, а говориться о третьем.
211
6.3.4. Планирование макроциклов
Какие теоретические основания можно положить в осно-
ву разработки принципиальной схемы планирования ма-
кроцикла?
На наш взгляд, таковыми могут явиться следующие:
1. Аэробная тренировка с интенсивностью ниже анаэроб-
ного порога у тренированных спортсменов обладает
очень низкой эффективностью в отношении показате-
лей всех мышечных компонентов системы транспорта
и утилизации кислорода (активность ферментов окис-
лительного фосфорилирования, плотность капилляров,
концентрация миоглобина). Это означает, что их про-
изводительность адекватна имеющейся массе сокра-
тительного белка в ММВ, и для дальнейшего прироста
аэробной мощности ММВ (основных в ЦВС) требуется
увеличение количества (массы) сократительных белков,
т.е. гипертрофия ММВ. Это должно найти отражение в
увеличении силы ММВ. Этот вывод особенно справед-
лив для регулярно тренирующихся спортсменов.
2. Белковые структуры различных тканей организма чело-
века, как уже отмечалось, обладают различной лабиль-
ностью, т.е. средним периодом полураспада (от несколь-
ких часов у кишечного эпителия до нескольких месяцев
в костной ткани) [Биохимия: Учебник для институтов
физ. культуры, 1986; Меерсон Ф.З., 1978; Сээнэ Т.П. и
др., 1990; Яковлев Н.Н., 1955; Fleck S.J., Kraemer W.J.,
1987]. Это означает, что и реактивность на воздействие
факторов индукции соответствующих белков также раз-
личается.
Лабильность белков основных структур мышц может быть
проранжирована следующим образом:
— наиболее долгоживущими белками являются структуры
соединительно-тканных элементов мышц и сухожилий
(коллаген и эластин);
— далее идут сократительные белки мышц;
— затем комплекс белков, обеспечивающий транспорт и
утилизацию кислорода, включающий структурные бел-
212
ки капилляров, митохондрий и ферментативные ком-
плексы митохондрий и миоглобин;
— более лабильными по сравнению с окислительным ком-
плексом являются гликолитические ферменты, локали-
зованные в цитозоле, а также фосфагеновая система;
— миокард, как наиболее жизненно важный орган, занимает
особое положение в этом ряду. Известно, что при искус-
ственной ишемии масса миокарда удваивается за три не-
дели [Меерсон Ф.З., 1978], исследования на уровне экс-
прессии генов [Некрасов А.Н., 1982] показали, что синтез
РНК и белков в миокарде начинается уже через несколько
минут после начала нагрузки, в то время как в скелетных
мышцах — только через несколько часов после окончания
работы. Показано, что достоверные изменения в сердеч-
ной деятельности (например, увеличение ударного объема
в покое и при стандартной нагрузке [Saltin В., 1985]) на-
блюдаются уже через несколько дней после начала трени-
ровки. Тогда как на уровне митохондриального аппарата
скелетных мышц в этот период еще никаких изменений
не обнаруживается [Williams R.S., 1986]. Это означает, что
реактивность генетического аппарата миокардиоцитов
очень велика, миокард обладает очень высокими адапта-
ционными возможностями и способен реагировать на из-
меняющуюся ситуацию существенно быстрее, чем даже
ферментативные комплексы скелетных мышц.
Подтверждением представленному ранжированному
ряду могут служить данные многочисленных исследова-
ний, в которых изучалась скорость прироста и снижения
показателей соответствующих способностей человека.
3. К моменту основных стартов спортсмен должен обладать
максимальной работоспособностью и согласованностью
в деятельности основных биомеханических, физиологи-
ческих и биохимических систем организма, от которых
зависит спортивный результат. Среди мышечных ком-
понентов таковыми являются:
— аэробные способности ММВ и БМВ;
— высокая активность ферментов анаэробного гликолиза
(для спринта);
213
— высокая буферная емкость мышц (для средних дистан-
ций);
— максимальное содержание энергетических субстратов —
КрФ и гликогена.
Этими факторами определяется специальная работо-
способность спортсменов, от которой зависит спортивный
результат.
4. У квалифицированных спортсменов большая аэробная
мощность ММВ, большее содержание КрФ, большие за-
пасы гликогена в мышцах и большая буферная емкость
прямо или косвенно связаны с гипертрофией мышечных
волокон, увеличивающей «морфологическое простран-
ство» для накопления ферментативных белков [Виру
А.А., 1981], от которых непосредственно зависят ЛВ и
связанный с ней спортивный результат. Поэтому можно
считать, что высокое содержание сократительных эле-
ментов и связанных с ними органелл клеток являются
структурной основой или главным условием повышения
производительности других систем мышц (и всего орга-
низма в целом). Следовательно, высокие сократительные
возможности мышц являются биологической «базой»
специальной работоспособности, а специализированная
тренировка (главным образом силовая в той или иной
форме), направленная на улучшение сократительных
свойств мышц (силы, скоростно-силовых качеств, ло-
кальной выносливости и т.п.), должна занимать «базовое»
положение в системе подготовки спортсменов. Другими
словами — сначала увеличиваем мышечную силу и связан-
ные с ней способности, а затем повышаем способности,
определяемые ферментативными системами (гликолити-
ческими и окислительными) и совершенствуем технику
локомоции, которая должна соответствовать новому со-
стоянию мышц и обеспечивающих систем.
В некоторых случаях возможно параллельное развитие этих
способностей.
Однако, исходя из ограниченных гормональных, пласти-
ческих и энергетических ресурсов организма, необходимых
для обеспечения адаптации, последовательное распределение
214
средств воспитания «базовых» и «реализационных» способно-
стей может быть более целесообразным.
Определяя сократительные свойства мышц как базовые,
следует отметить, что в таких видах локомоций, как легкоатле-
тический бег, в силу биомеханических особенностей взаимо-
действия с опорой важное значение имеет профилактика трав-
матизма и перенапряжения опорно-двигательного аппарата
(ОДА), что подразумевает, в частности, повышение прочности
соединительно-тканных элементов ОДА путем специальной
тренировки (например, прыжковых упражнений).
Таким образом, ставя во главу угла необходимость дости-
жения максимальной специальной работоспособности, мож-
но представить описанные закономерности в виде «пирамиды
специальной работоспособности» (рис. 14).
«Пирамида» состоит из основания и вершины. В основании
— соединительно-тканные и сократительные элементы (фи-
бриллярные белки) мышц, с которыми корреляционно связа-
но содержание КрФ и гликогена. Вершина: 1) масса митохон-
дрий и плотность капиллярной сети, определяющие вместе с
запасами гликогена аэробные способности спортсменов; 2)
активность ферментативных комплексов анаэробного гли-
колиза и креатинфосфокиназной реакции, а также элементы
мышц и крови, повышающие буферную емкость в отношении
кислых продуктов метаболизма, которые вместе определяют
анаэробную производительность спортсмена.
Высота каждого из блоков пропорциональна производи-
тельности данной системы.
Высота пирамиды (уровень специальной работоспособно-
сти) складывается из высоты отдельных блоков.
Ширина блоков пропорциональна лабильности белков —
времени, необходимого для того, чтобы произошли значимые
изменения в содержании специфических белков. Она наи-
большая — у основания (соединительно-тканные элементы)
и минимальна — у цитоплазматических ферментативных ком-
плексов (КФК-аза, ферменты анаэробного гликолиза, белки
митохондрий и капилляров).
Если пирамиду «положить на бок», то получим принципи-
альную схему (модель) планирования многолетней подготов-
215
Факторы, определяющие
специальную физическую
работоспособность (СФР) в ЦВС
Рис. 14. «Пирамида специальной работоспособности» с точки зрения
локальных компонентов выносливости, схематически изображаю-
щая иерархию мышечных компонентов выносливости. Обозначения:
СФР — специальная физическая работоспособность, которая является
функцией биохимических, физиологических, биомеханических и психо-
логических систем организма; ЦВС — циклические виды спорта; ОП —
окислительный потенциал; АнГ — анаэробный гликолиз; БЕ — буферная
емкость; КФ К — ферменты креатинфосфокиназной реакции; СТЭ —
соединительно-тканные элементы ОДА. На схеме указаны только мы-
шечные компоненты, определяющие выносливость
ки спортсменов в ЦВС, схему планирования 4-летнего олим-
пийского цикла, годичного или полугодичного макроцикла в
контексте воспитания локальной мышечной выносливости.
В основе этой модели лежат три положения:
1. Максимальная производительность главных фермента-
тивных систем энергообеспечения должна достигаться
к моменту основных стартов (достижение состояния
«спортивной формы»).
2. Соединительно-тканные и сократительные элементы
мышц занимают «базовое» положение в структуре спе-
циальной подготовленности в ЦВС.
216
Модель макроцикла и многолетней
подготовки в ЦВС
Рис. 15. Модель макроцикла и многолетней подготовки в ЦВС. Обозна-
чения: СТА ОДА — акцент на укрепление соединительно-тканных эле-
ментов опорно-двигательного аппарата; сила — акцент на развитие
силовых способностей; аэроб. — акцент на развитие аэробных способно-
стей; соревн. — предсоревновательная (интегрирующая, реализующая)
подготовка; ПП — переходный период; Предс. — предсоревновательный;
Сил. вын. — динамика объемов средств развития силовой выносливости;
Гликол. — средства «гликолитической» подготовки; скор. сил. — сред-
ства скоростно-силовой подготовки; силов. — средства силовой подго-
товки
3. Длительность этапа акцентированного воздействия на
те или иные морфоструктуры определяется временем
достижения «плато» в приросте их показателей при
данном (оптимальном) объеме тренировочных средств
(объем определяется значительным числом факторов).
При этом надо учитывать, что время достижения «пла-
то» может варьироваться в очень широких пределах (2-7
месяцев), которые определены в значительной мере ин-
дивидуальными особенностями спортсменов [Бондар-
чук А. П., 1989]. Кроме этого, достижение «плато» можно
отодвинуть, достигнув тем самым более высокого при-
217
роста способности, если использовать варьирование ви-
дами средств и объемами нагрузки (той же направлен-
ности) в занятиях, микроциклах и мезоциклах [Воробьев
А.Н., 1977], или последовательно добавляя все более эф-
фективные средства [Бондарчук А. П., 1986].
Таким образом, получаем, что в начале макроцикла (в пере-
ходный период) или многолетней подготовки основной зада-
чей является укрепление соединительно-тканных элементов
опорно-двигательного аппарата, а в тех видах спорта, где про-
блема травматизма ОДА не актуальна (как, например, в плава-
нии, велоспорте) — начинать можно сразу с увеличения силы
ММВ или силы ММВ и БМВ (в зависимости от соревнователь-
ной дистанции). Остальные средства подготовки обязательно
присутствуют, но в «поддерживающем» объеме.
Следующий этап посвящается преимущественно аэробной
подготовке, но не только ММВ (непрерывный и переменный
метод на скорости анаэробного порога), но и БМВ (средства:
интервальный спринт, аэробно-силовой метод, интервальная
тренировка и др. (см. п. 6.2.2.2). Несмотря на внешнюю вы-
сокую интенсивность упражнений (необходимо вовлечение
в работу БМВ), основным методическим требованием явля-
ется отсутствие существенного закисления мышц. Таким об-
разом, тренировка должна носить истинно аэробную направ-
ленность. При нарушении этого ключевого правила получим
«классический» вариант форсирования спортивной формы со
всеми вытекающими отсюда последствиями и дискредитаци-
ей всей идеи.
Этап имеет два подэтапа. На первом допускается использо-
вание средств, отличающихся по биомеханической структуре от
основного соревновательного упражнения (бег в гору, плавание
с тормозом и т.п.). Эти средства на практике называются сред-
ствами для развития «силовой выносливости». На втором этапе
используются в большей мере соревновательные упражнения
(например, многочисленные пробежки или заплывы с соревно-
вательной скоростью повторно или интервально, интервальный
спринт, темповой бег на уровне АнП). На длинных дистанциях
этот подэтап постепенно переходит в предсоревновательный
период, на котором осуществляется так называемая интегри-
218
рующая тренировка [Н.Г. Озолин, 1070], в процессе которой
происходит сонастройка различных систем организма.
Третий этап присутствует только в спринте и на средних
дистанциях. Его цель — повышение гликолитических способ-
ностей (активности ферментов гликолиза и буферной емко-
сти). В связи с тем что:
— ферменты гликолиза очень мобильны;
— гликолитическая тренировка носит явно выраженный
стрессовый характер, оказывающий очень глубокое
воздействие на пластические и энергетические ресурсы
организма, на нейроэндокринную и опосредованно на
иммунную систему [Сашенков С.Л. и др., 1995];
— гликолитическая тренировка может негативно сказаться
на накопленном аэробном потенциале, так как ацидоз
стимулирует протеолиз [Раппопорт Э.А., Казарян В.А.,
1996], в частности митохондриальных белков [Лузиков
В.Н., 1980];
— длительность третьего этапа должна быть небольшой (на-
пример, месяц), во время которого может быть проведено
не более 4-5 специализированных «гликолитических» тре-
нировок. Дальнейшая «доводка» гликолитического потен-
циала осуществляется во время проведения «прикидок»
или выступления в предварительных соревнованиях.
Отклонения от этой принципиальной схемы могут быть
следующие.
1. Как уже отмечалось, в видах спорта, где проблема травма-
тизма ОДА не актуальна, первый этап может быть опущен.
2. На уровне высшего спортивного мастерства вполне мо-
жет возникнуть ситуация, когда уровень силы мышц (ги-
пертрофии МВ) достаточен для того, чтобы:
— этап силовой тренировки был сокращен до минимума;
— силовая тренировка проводилась не концентрированно, а
распределенно — в течение всего подготовительного периода;
— в качестве средства силовой тренировки использовался
аэробно-силовой метод (см. п. 6.2.2).
3. При использовании анаболических стероидов этап си-
ловой тренировки также может быть сокращен или во-
обще не использоваться.
219
4. В тренировке стайеров и марафонцев этап «гликолити-
ческой» подготовки опускается.
В аспекте многолетней подготовки обозначенная теорети-
ческая модель получила экспериментальное подтверждение
в работах ученых института медико-биологических наук (см.,
например, докт. дисс. Б.С. Шенкмана, 1999), которыми с ис-
пользованием прямых методов измерения размера мышечных
волокон и эффективности дыхания митохондрий на самых
различных экспериментальных моделях тренировки и де-
тренировки показано, что стратегия повышения и снижения
окислительного потенциала мышечных волокон принципи-
ально проходит три стадии (рис. 15-1).
1. В начале эффективность энергообеспечения
мышц повышается без заметных структурно-
метаболических изменений внутри МВ (в нашем
случае — «втягивающая» тренировка или начальная
аэробная тренировка).
2. На второй стадии прирост окислительного потенциала
обусловлен параллельными процессами гипертрофии
МВ и увеличением массы окислительных ферментов,
капилляров и миоглобина. Эта стадия дает основную
величину прироста окислительного потенциала мышц
(т.е. аэробной производительности). При этом степень
гипертрофии мышечных волокон пропорциональна
величине резистивных нагрузок (степени проявления
силы мышц в соревновательном упражнении, величине
«ударных нагрузок» при беге и т.п.), которые мышца ис-
пытывает в процессе тренировки.
3. На высшей стадии аэробной тренированности (в нашем
случае — этап высшего спортивного мастерства) окисли-
тельный потенциал растет за счет повышения эффектив-
ности работы митохондрий без существенных морфоло-
гических изменений внутри волокна.
Необходимо сделать еще некоторые уточнения к представ-
ленным выше схемам построения макроцикла.
1. В подготовке квалифицированных спортсменов все
средства силовой, аэробной и гликолитической под-
готовки должны быть предельно «специализированы».
220
В плане повышения силовых способностей (гипертро-
фии МВ) основное внимание должно уделяться воздей-
ствию на ММВ, а для спринтеров и, в значительно мень-
шей степени, средневиков, так же и на БМВ, но только
основных мышечных групп. И по возможности при со-
блюдении принципа сопряженного воздействия [Дьяч-
ков В.М., 1972].
2. Средства и методы гипертрофии ММВ очень существен-
но отличаются (см. п. 6.2.1.1) от традиционных видов
силовой и скоростно-силовой подготовки, под которыми
часто понимают упражнения со штангой, на тренаже-
рах, спринтерские и прыжковые упражнения примерно
в таком же варианте, как это используют представители
скоростно-силовых и силовых видов спорта. Это, по на-
шему мнению, является грубейшей ошибкой, так как при
этом отсутствует воздействие на ММВ (наиболее «рабо-
тающая» часть мышц в ЦВС), спортсмен приобретает
ненужную мышечную массу. Такая тренировка может
очень негативно сказаться на экономичности техники,
резко повышается риск травматизма.
3. Глубоко бытующее убеждение, что главное в высокой
аэробной производительности — это производитель-
ность ССС, привело на практике к «игнорированию
интересов» исполнительного аппарата — мышц. В ре-
зультате этап «накопления аэробного фундамента» в
целом описывается формулой:»... большие объемы в
умеренном темпе» [Суслов Ф.П. и др., 1982, стр. 28].
Длительность такого этапа в различных ЦВС может
быть 3-6 и более месяцев [Макаров А.Н., 1973]. При
этом необходимость поддержания высокого силового
потенциала на практике упускается, так как этап по-
вышения силовых способностей спланирован на конец
подготовительного периода [Набатникова М.Я., 1975;
Суслов Ф.П. и др., 1982]. В результате получается хоро-
шо прогнозируемый эффект:
— на этапе аэробной подготовки снижается сила мышц
[Суслов Ф.П. и др., 1982], в особенности — БМВ (ре-
зультаты в спринте и прыжковых тестах падают);
221
— в предельной степени повышаются окислительные
способности ММВ (при низкой мощности (аэроб-
ный порог и ниже) активны в основном ММВ, см.
главу 4).
На первый взгляд, цель — высокая аэробная мощность (на-
пример, высокий анаэробный порог и высокое относительное
МПК) — достигнута. Однако на следующем этапе, когда пла-
нируется увеличение силы мышц, повышение интенсивности
основного упражнения и воспитание гликолитических спо-
собностей, происходит следующее:
— если оказывается успешным силовое воздействие, то
происходит гипертрофия мышечных волокон, при-
водящая к неминуемому одновременному понижению
удельного окислительного потенциала всех МВ, где ги-
пертрофия произошла. Кроме того, силовые упраж-
нения сопровождаются существенным понижением
pH мышцы, требуют больших пластических ресурсов
и дадут эффект только при снижении объемов других
видов подготовки. Таким образом, имеются альтер-
нативы:
— или силовая тренировка будет не успешной, или, в ре-
зультате понижения окислительного потенциала мышц
(белки митохондрий относятся к лабильным белкам
— см. выше), будет ликвидирован единственный положи-
тельный эффект предыдущего этапа — высокая аэробная
производительность;
— если процесс увеличения силы оказывается не успеш-
ным, то емкость фосфагенного и гликолитического источ-
ников энергообеспечения к этапу соревнований окажется
существенно меньше возможного;
— изменение силового потенциала мышц неминуемо от-
рицательно скажется на экономичности техники [Мя-
кинченко Е.Б., 1983], а времени по «настройке» техники
под новое состояние мышц может не хватить для успеш-
ного выступления в соревнованиях.
Таким образом, или рассматриваемый этап предсоревно-
вательной подготовки оказывается неэффективным и спор-
тсмен остается со «слабыми» мышцами, пониженной ем-
222
костью креатинфосфатного источника и гликолиза на весь
сезон соревнований, или понижаются аэробные способности
[Набатникова М.Я., 1975].
4. Использование более интенсивных режимов трениров-
ки в конце подготовительного и в предсоревновательный
период среди возможных положительных эффектов («на-
стройка» техники, отработка тактики, психологическая под-
готовка, увеличение силы и скорости) может способствовать
повышению окислительного потенциала БМВ. Однако пред-
ставляется нелогичным разнесение во времени воздействий
на окислительный потенциал ММВ и БМВ. При оптималь-
ном планировании повышение окислительного потенциала
всех мышечных волокон должно производится одновременно
и не совмещаться с параллельным повышением силовых
возможностей гликолитического потенциала и отработкой
техники движений. То есть на этапе аэробной тренировки
в обязательном порядке должно производится воздействие
на ОП БМВ и ММВ (даже в большей мере — БМВ, т.к. у
квалифицированных спортсменов ОП ММВ, вероятно, на-
ходится на околопредельном уровне), а принцип постепен-
ности должен реализовываться не (!!!) путем от «медленной
скорости к быстрой» или — «от ММВ к БМВ» (т.е. сменой
объектов воздействия !!!), а регулированием объема нагрузки
при оптимальной ее структуре.
5. У средневиков этап тренировки гликолитических воз-
можностей должен быть минимизирован как по объему на-
грузки, так и по длительности (не более 1 месяца), а в случае
если планируется длительный соревновательный сезон и боль-
шое число предварительных стартов, не проводиться вовсе.
Тезис, что гликолитические способности являются ведущими
на средних дистанциях, справедлив (см. главу 5), но не должен
пониматься таким образом, что повышению гликолитических
возможностей следует уделять основное место в тренировоч-
ном процессе. Как многократно отмечалось выше, решающий
вклад в проявление гликолитических способностей вносят аэ-
робные и силовые способности спортсменов, улучшению кото-
рых уделяется основное внимание в подготовительный период
макроцикла.
223
6.4. Проблема взаимосвязи уровня
и особенностей подготовленности нервно-
мышечного аппарата с техникой
и экономичностью локомоций
Основные закономерности взаимосвязи техники локомо-
ций с уровнем и особенностями подготовленности нервно-
мышечного аппарата общие для большинства локомоций.
Поэтому мы рассмотрим только бег как наиболее хорошо из-
ученный.
Изменение функциональных показателей нервно-
мышечного аппарата всегда закономерно изменяет биомеха-
нические параметры спортивных упражнений [Бальсевич В. К.,
1965; Дьячков В.М., 1972; Суслов Ф.П. и др., 1982; Мякинчен-
ко Е.Б., 1983]. В то же время, рациональная биомеханическая
структура движений (техника) является условием достижения
высоких результатов. Таким образом, возникают вопросы: как
изменение в показателях НМА спортсменов будет отражаться
на биомеханических характеристиках локомоции и какие из
этих изменений можно считать положительными, а какие нет;
какие показатели изменяются в связи с изменением физиче-
ского состояния, а какие под воздействием обучения? Так как
основным показателем совершенства техники в ЦВС является
ее экономичность, то вопрос можно конкретизировать — как
изменения в НМА будут влиять на экономичность, основной
характеристикой которой является метаболическая стоимость
пути? В то же время не решена проблема: насколько критерии
экономичности на низкой скорости, на которой возможно
точное определение энергорасхода, информативны для сорев-
новательной скорости?
Чтобы ответить на эти и другие, возникающие по ходу рабо-
ты вопросы, было проведено исследование [Мякинченко Е.Б.,
1983] на примере легкоатлетического бега, в котором изучалась
взаимосвязь с показателями техники следующих параметров:
— квалификации бегунов;
— различий в силе мышц разгибателей суставов ног;
— различий в соотношении силы мышц сгибателей колен-
ного сустава (задняя поверхность бедра) к силе мышц
224
разгибателей коленного сустава (передняя поверхность
бедра);
— пороговой скорости бега;
— специализации (стайеры/средневики);
— метаболической стоимости пути (МСП) — отношение
скорость бега к потреблению кислорода в устойчивом
состоянии при беге по дорожке (VO2);
— коэффициента механической эффективности бега
(КМЭ) — отношение механической работы по переме-
щению общего центра масс тела (ОЦМТ) к МСП.
Кроме этого, проводилось сравнение техники бега в фазе
компенсированного и некомпенсированного утомления и у
одних и тех же спортсменов в подготовительном и соревнова-
тельном периодах.
При изучении литературы по обозначенным вопросам было
выявлено следующее (подробнее см. Мякинченко Е.Б., 1983).
1. Квалифицированные бегуны совершают меньшую меха-
ническую работу и тратят меньше метаболической энер-
гии при преодолении единицы расстояния, чем бегуны
низкой квалификации. В тоже время есть исследования
[Williams R.S. 1986], где не обнаружено никакой связи
между МСП на скорости 3,57 м/с и спортивным резуль-
татом на 10000 м. У более квалифицированных, как пра-
вило, наблюдается меньший период опоры и больший
— полета.
2. Критериями экономичности бега считаются меньшие
вертикальные колебания и меньшие потери горизон-
тальной скорости движений ОЦМТ.
3. При постановке ноги на опору критерием техничности
считается активность всех движений. Нога должна ста-
виться «загребающим» движением, мягко, с передней
части стопы.
4. Показателями технически правильного выполнения
движений в фазе амортизации являются: укорочение ее
длительности относительно времени опоры; уменьше-
ние амплитуды амортизационного сгибания опорной
ноги; большее ускорение ОЦМ в фазе амортизации, т.е.
большая средняя сила воздействия на опору. Для дости-
225
жения этого требуются активные маховые движения и
активное «сведение бедер» на опоре.
5. В фазе отталкивания должно наблюдаться полное разги-
бание ноги в тазобедренном и голеностопном суставах,
обеспечивая «острый» угол отталкивания.
6. Рациональному соотношению длины и частоты шагов
придается очень большое значение. Было показано;
— длина шагов возрастает до уровня КМС, но у МСМК
длина шагов меньше.
— с ростом квалификации конкретного спортсмена (т.е.
скорости преодоления соревновательной дистанции),
вероятно, во всех циклических видах спорта просле-
живается тенденция увеличения скорости за счет дли-
ны шагов при стабилизации частоты около некоторого
«оптимального» для данного спортсмена уровня, кото-
рый зависит от антропометричесих признаков, мышеч-
ной композиции, особенностей подготовки и т.п. [Вер-
хошанский Ю.В., 1985; Попов Ю.А., 1968; Brandon L.J.,
R.A. Boileau, 1987];
— тем не менее в одном лонгитудинальном исследовании
было показано [Nelson R.C., R.J. Gregor., 1976], что за 4
года тренировок спортсмены уменьшили длину шагов
(рассчитываемую как среднюю для 3 пробежек со ско-
ростью от минимальной до максимальной) на 7 см. Тем
не менее почему-то никак не комментировался факт,
приведенный в том же исследовании, где показано, что
лучший бегун мирового уровня, достигший наибольшего
прогресса, за 4 года наблюдений увеличил длину шагов на
15 см. Интересно, что здесь же показано, что бегуны,
достигшие наибольших приростов в результате, увели-
чивали скорость бега до 6-7 м/с (т.е. до значения сорев-
новательной скорости) в большей мере за счет прироста
длины шагов, тогда как спортсмены, имевшие наимень-
ший прирост или не имевшие его, увеличивали скорость
бега за счет прироста частоты шагов;
— в той же группе (10 человек) квалифицированных бе-
гунов [Nelson R.C., R.J. Gregor., 1976], несмотря на
снижение длины шагов, уменьшился период опоры
226
(на 0,026с) и увеличился период полета (на 0,013с),
т.е. увеличился ритмовой коэффициент (соотношение
периода полета и периода опоры). Это означает, что
спортсмены стали развивать на опоре большую мощ-
ность [Мякинченко Е.Б., 1983], что не согласуется с
мнением, что основным признаком повышения эко-
номичности бега является снижение механических
энергозатрат, в частности за счет уменьшения верти-
кальных колебаний ОЦМ;
— было показано [Cavanagh P.R. и др., 1977], что элитные
спортсмены на стандартной скорости бега имеют мень-
шую длину и большую частоту шагов;
— для каждого спортсмена имеет место некоторое опти-
мальное соотношение длины и частоты шагов, при кото-
ром наблюдается минимум метаболической стоимости
пути [Уткин В.Л., 1984; Cavanagh P.R. и др., 1977];
— среди «общепринятых» факторов, влияющих на длину
и частоту шагов, выделенных таким признанным авто-
ритетом по биомеханике бега как Р. Cavanagh (1989), а
именно: скорость; рельеф местности; обувь; поверх-
ность; антропометрические показатели; биологический
возраст; мышечная композиция; степень утомления;
предшествующие травмы и др., тем не менее отсутствует
наиболее логичный — уровень физической подготовленно-
сти, в частности — сила мышц ног.
7. Показатели физических качеств связаны с техникой бега
следующим образом:
— ритмовой коэффициент (время полета/время опоры)
связан с силой мышц подошвенных сгибателей голено-
стопного сустава;
— спортсмены с более сильными ногами имеют более ши-
рокий шаг;
— повышение силовых показателей у одного и того же
спортсмена сопровождается увеличением длины шагов
при сохранении оптимальной частоты;
— величина амортизационного сгибания в коленном су-
ставе опорной ноги меньше у более физических сильных
спортсменов;
227
— сильные спортсмены имеют возможность использовать
«низкий» бег, характеризующийся острым углом оттал-
кивания.
Тем не менее по значительному числу обозначенных вопро-
сов данные были противоречивы или отсутствовали. В связи с
этим было предпринято самостоятельное исследование.
Метод. В сравнительных и лонгитудинальных экспери-
ментах принимали участие взрослые спортсмены-бегуны на
средние и длинные дистанции от П-го разряда до МСМК —
членов сборной СССР 1982-1983 гг. 34 механических и энер-
гетических характеристик бега со скоростью 7 м/с, 6,2 м/с и
4,7 м/с рассчитывались по опорным реакциям естественного
бега по дорожке манежа с наступанием на тензоплатформы
одной и второй ногой. Бег проводился с равномерной ско-
ростью по кругу 200 м под звуколидер. Регистрация опорных
реакций проводилась через 150 и 350 метров после старта (7
м/с), 350 и 950 м (6,2 м/с) и через 350-750 м (4,7 м/с). Всего
регистрировалось не менее 4 опор. Данные усреднялись. Так-
же определялась метаболическая стоимость пути при беге на
дорожке (4,7 м/с), анаэробный порог, показатели относитель-
ной изометрической силы и градиента силы мышц сгибателей
и разгибателей суставов ног, максимальная скорость бега и
результат прыжка в длину с места, антропометрические при-
знаки спортсменов, биомеханические свойства мышц голени.
Всего статистической обработке подвергалось 57 показателей,
характеризующих спортсменов и технику их бега.
Исследование выявило следующее:
Установлено, что квалифицированные бегуны (КМС-МС)
на средние дистанции по сравнению с менее подготовленны-
ми (II-I-й разряды) имеют следующие статистически досто-
верные отличия по показателям, имеющим отношение к ло-
кальной выносливости:
— по антропометрическим признакам особенностями вы-
борки было то, что более квалифицированные бегуны
имели достоверно меньшую массу тела и индекс «длин-
ноногое™».
Среди физических показателей:
— большую максимальную и пороговую скорость бега;
228
— лучше результат прыжка в длину с места;
— большую силу мышц разгибателей ног;
— лучше показатели скоростных свойств мышц ног (отно-
сительный градиент силы);
— меньшую жесткость трехглавой мышцы голени.
Среди биомеханических показателей на стандартной ско-
рости 7 м/с+0,05:
— большую длину шагов;
— меньший период опоры;
— большее ускорение ОЦМТ на опоре (т.е. большую меха-
ническую мощность работы мышц);
— большее вертикальное ускорение ОЦМТ относительно
горизонтального;
— большие вертикальные перемещения ОЦМТ (р<0,07);
— меньшую «потерю» скорости в фазе амортизации;
— меньшую длительность фазы амортизации относитель-
но длительности фазы опоры и др.
Среди энергетических характеристик:
— меньшую метаболическую стоимость пути (экономич-
ность бега). Примечательно, что МСП снижалась за счет
увеличения коэффициента механической эффектив-
ности (АУмеханическая/АУметаболическая), так как ве-
личина механической работы по перемещению ОЦМ у
спортсменов разной квалификации была одинаковой.
При изучении техники бега на скорости 4,70+0,05 м/с у спор-
тсменов, существенно (р<0,001) различающихся по метаболиче-
ской стоимости пути (т.е. по экономичности бега), но с одина-
ковыми антропометрическими показателями и квалификацией,
снова были получены результаты, противоположные ранее полу-
ченным данным (например, [Miura М. и др., 1973, Williams R.S. и
др., 1986]). В частности, более экономичные бегуны имели боль-
шие вертикальные перемещения ОЦМ (р<0,02), больший коэф-
фициент «активности доталкивания» (определяемый работой
трехглавой мышцы голени) в конце опоры (р<0,03) и на уровне
тенденции большим отношением механической работы, совер-
шаемой в вертикальном направлении по сравнению с продоль-
ным направлением (р<0,07). У экономичных бегунов был более
длительный опорный период (р<0,10).
229
Таким образом, принимая допущение, что «экономичные»
на скорости 4,7 м/с бегуны не могли стать «не экономичны-
ми» на соревновательной скорости, а также проведенный ана-
лиз тенденций изменений биомеханических характеристик с
ростом скорости бега у одних и тех же спортсменов [Мякин-
ченко Е.Б., 1983], приходим к достаточно неожиданному, ис-
ходя из сложившихся представлений, выводу — «прыжковый»,
«упругий», «активный» бег оказывается (на соревновательной
скорости!) более экономичным, чем бег с низкими вертикальны-
ми колебаниями ОЦМТ, меньшими ускорениями, меньшей фазой
полета и т.д. Подчеркнем, что при этом более подготовленные
бегуны имели одновременно существенно большие силовые и аэ-
робные показатели мышц. Такие особенности НМА позволяют
им отталкиваться от опоры с большими усилиями без риска
накопления в мышцах ионов водорода (закисления) и лактата
или проявления других признаков локального утомления.
Однако остается вопрос: почему на стандартной скорости
квалифицированные бегуны предпочитают путь более полно-
го использования двигательного потенциала НМА, а не путь
«экономизации» за счет уменьшения, например, внешней ме-
ханической работы по перемещению ОЦМТ?
Анализируя литературные данные и собственные результа-
ты, мы пришли к выводу, что такой путь более рационален как
минимум по двум причинам:
1. Увеличение длины шагов снижает так называемую вну-
треннюю механическую работу по перемещению конеч-
ностей относительно ОЦМТ [Зациорский В.М. и др.,
1982], «стоимость» которой на скорости 5-6 м/с начина-
ет превышать стоимость внешней механической работы
по перемещению ОЦМТ, т.е. ее вклад в общие энерго-
затраты на соревновательной скорости выше, чем «по-
лезная работа» по перемещению спортсмена вперед от-
носительно дорожки.
2. ОДА обладает способностью к рекуперации (повторному
использованию) энергии, запасаемой в его упругих эле-
ментах, а количество рекуперируемой энергии возрастает
как раз при «активном», «упругом беге». При этом возрас-
тает коэффициент механической эффективности и спор-
230
темен при той же внешней механической работе (напри-
мер, вертикальных перемещениях ОЦМТ) оказывается
способным снижать метаболическую стоимость пути.
Эти особенности в полной мере проявились при сравни-
тельном исследовании бегунов различной квалификации
и различной физической подготовленности. При этом
путь «экономизации» используется только в отношении
механической работы, затрачиваемой на торможение и
разгон ОЦМТ в продольном направлении.
Описываемый «активно-экономичный» вариант техники
бега характеризуется активной постановкой ноги на опору,
«под себя», так, чтобы всем телом как бы «накатываться» на
ногу. Кроме того, «активно» выполняется встречный мах ру-
ками и сводятся бедра. Все это создает большее напряжение
мышц — разгибателей опорной ноги и мышц задней поверх-
ности бедра, что позволяет дополнительно их активировать и
накопить в них энергию упругой деформации, которая затем
реализуется в отталкивании. Во время отталкивания квали-
фицированные бегуны в среднем больше усилий направляют
вверх, однако акцент делается на завершающей стадии оттал-
кивания, когда создаются условия для направления усилий
больше вперед, а не вверх. На тензограмме это выражается в
сдвиге максимума силы в продольном направлении ближе к
окончанию опоры и увеличении активности завершающего
разгибания ноги в голеностопном суставе.
Таковы основные черты техники квалифицированных бе-
гунов на средние дистанции, которые, предположительно, об-
условлены лучшей локальной работоспособностью основных
мышечных групп.
Следующими вопросами явились:
1. Какие мышечные группы ответственны за те или иные
особенности техники?
2. Как изменяется техника при увеличении длины дистан-
ции?
3. Как сформировать экономичную технику?
4. Чем отличается бег в утомленном состоянии?
Для выяснения этих вопросов было проведено дальнейшее из-
учение взаимосвязи техники и физических способностей бегунов.
231
Была изучена техника бега (на скорости 6,96+0,06 м/с)
спортсменов одинаковой квалификации, но существенно раз-
личающихся по пороговой скорости бега (Р<0,001). Можно
предположить, что среди бегунов, специализирующихся на
средние дистанции и имеющих относительно более высокую
пороговую скорость, преимущество будут иметь спортсмены,
которые идут к результату «от выносливости» и имеют более
«дистанционную» направленность тренировочного процес-
са. По данным тестирования физических способностей, это
— слабые, медленные и легкие спортсмены. Их технику бега
характеризует хорошее «доталкивание», т.е. они лучше ис-
пользуют трехглавую мышцу голени, а сдвиг максимума уси-
лий в продольном направлении ближе к концу опоры свиде-
тельствует о том, что ближе к завершению отталкивания они
больше усилий направляют вперед, а не вверх. При этом у них
меньшие потери горизонтальной скорости в фазе амортиза-
ции. Однако снова обнаружена положительная взаимосвязь
хорошей аэробной подготовленности с вертикальными коле-
баниями ОЦМ тела.
Считается, что «экономичным» стилем бега обладают стай-
еры относительно средневиков. Поэтому мы провели сравне-
ние бегунов на 800-1500 и 3000-10000 м по всем исследуемым
показателям на одинаковой скорости бега (6,89 м/с). Было
выяснено, что стайеров характеризует, как и ожидалось, боль-
шая пороговая и меньшая абсолютная скорость бега и большее
отношение силы мышц — сгибателей к силе мышц — разгиба-
телей коленного сустава. При этом их технику характеризуют
существенно меньшие потери горизонтальной скорости в фазе
амортизации (0,281 — 0,313 м/с, р<0,05) и большее отноше-
ние механической работы в вертикальном и продольном на-
правлениях (0,546 — 0,488 м/с, р<0,05), т.е. они большую, чем
средневики, часть энергии мышц направляют вверх, экономя
в то же время энергию, затрачиваемую на торможение и разгон
ОЦМТ в продольном направлении. Вертикальная скорость
вылета, перемещение ОЦМТ по вертикали и отношение пери-
ода полета к периоду опоры (т.е. все традиционные показатели
«экономичности») оказались ниже только на уровне слабой
тенденции.
232
Таблица 6. Биомеханические показатели бега у испытуемых с различной си-
лой мышц — разгибателей коленного сустава на скорости 7м/с и 4,7м/с
№ Показатель Труп- па I Труп- ппа II t-кри- терий Труп- ппа I Труп- ппа II t-кри терий
1 Скорость бега (м/с) 6,98 6,92 0,6 4,76 4,81 0,40
2 Сила мышц - разгибателей ног 26,6 20,4 3,83 26,6 20,4 3,83
3 Длина шагов/ длина ног 2,22 2,12 4,41 1,81 1,73 3,62
4 Ускорение центра масс тела (м/с2) 31,5 28,4 3,12 24,3 22,1 2,09
5 Фаза полета/ фаза опоры 1,092 1,011 3,03 0,802 0,783 2,64
6 Верт./горизонталь- ная механич. работа 0,53 0,45 3,08 1,06 0,91 2,18
Нам удалось получить только однажды «классическое»
[Miura М. и др., 1973] проявление «экономизации» бега (сниже-
ние вертикальных перемещений ОЦМ) — когда мы сравнивали
технику бега в начале дистанции и в фазах компенсированного
(бег выполнялся под звуколидер, поэтому обязательное требо-
вание равенства скоростей было соблюдено) и некомпенсиро-
ванного утомления (когда скорость немного снижалась). Бег
в состоянии утомления по сравнению с нормальным действи-
тельно характеризуется снижением времени полета, механиче-
ской работы и ускорения ОЦМТ в вертикальном направлении
и соответственно вертикальных перемещений ОЦМТ, при этом
ухудшились все остальные показатели активности и механи-
ческой эффективности бега, которые, как показано выше, по-
ложительно связаны с экономичностью на соревновательной
скорости бега. Следовательно, описанные особенности «утом-
ленного» бега не могут рассматриваться в качестве «эталона»
экономичности и тренировка бегунов должна быть направлена
как раз на предотвращение таких изменений техники, а не на их
«культивирование». Так как, не изменяясь по существу, они за-
метно усиливаются в фазе некомпенсированного утомления.
233
Таблица 7. Биомеханические показатели бега у испытуемых с различ-
ным соотношением силы мышц — сгибателей и разгибателей коленного
сустава (п=10)
№ Показатель Группа I Группа II t-критерий
1 Скорость бега (м/с) 7,08 7,02 0,31
2 Сила мышц сгибателей/ разгибателей кол. сустава 0,670 0,380 9,45
3 Частота шагов 3,514 3,465 0,80
3 Верт, скор вылета ОЦМТ (м/с) 0,607 0,489 2,29
4 Верт, ускор. ОЦМТ/ верт. ус-кор. ОЦМТ(м/с2) 0,249 0,281 2,27
5 Коэффициент активности «до-тал кивания» в конце опоры. 66 48 2,39
6 Верт, перемещ. ОЦМТ (м) 0,057 0,043 2,30
7 Потеря продольн. скоро- сти в фазе амортизации (м/с) 0,280 0,298 2,30
Следующие данные позволяют сделать вывод о значимо-
сти силовых показателей мышц. В табл. 6 приведены данные
бегунов одинаковой квалификации и антропометрических
признаков, но с разной силой мышц — разгибателей ног (опре-
деляемой в тесте, когда из положения сидя надавливают на
тензометрическую педаль).
Таким образом, нет оснований сомневаться, что большая
длина шагов и некоторые другие показатели во всем диапазо-
не соревновательного бега от средних дистанций до марафона
связаны с силой мышц — разгибателей ног.
Для выяснения влияния фактора соотношения силы мышц —
сгибателей и разгибателей коленного сустава сформированы две
группы бегунов, различающиеся по этому признаку (табл. 7).
Представленные данные свидетельствуют, что соотноше-
ние силы мышц задней и передней поверхности бедра также
являются фактором, связанным с техническими параметрами
234
Таблица 8. Показатели физических способностей и техники одних и тех
же бегунов в середине подготовительного периода и на этапе основных
соревнований
№ Показатель Подг. период Соревно- ват. период t-критерий
1 Скорость бега (м/с) 6,94 7,04 0,39
2 Сила мышц — сгибателей коленного сустава 4,33 5,13 2,23
3 Сила мышц — сгибателей/ разгибателей коленного сустава 0,487 0,573 2,19
4 Сила мышц — разгибате- лей ног 9,27 9,28 0,04
5 Пороговая скорость бега 4,81 4.91 1,05
6 Частота шагов 3,56 3,60 0,80
7 Верт, скор вылета ОЦМТ (м/с) 0,562 0,661 3,20
8 Верт, ускор. ОЦМТ/ верт. ускор. ОЦМТ(м/с2) 0,249 0,281 2,27
9 Верт, перемещ. ОЦМТ (м) 0,051 0,061 2,83
10 Время опоры / время по- лета 0,975 1,124 3,65
и Потеря продольн. скоро- сти в фазе амортизации (м/с) 0,302 0,281 2,28
12 Положение макс, усилий по продольной состав- ляющей 0,47 0,49 2,04
13 Коэффициент активности «доталкивания» в конце опоры (вертикальн. составляющая) 81 88 2,15
235
бега. И на уровне тенденции могут способствовать относи-
тельному увеличению частоты шагов.
Данные табл. 7 получены при межгрупповом сравнении,
однако особую ценность имеют данные лонгитудинальных ис-
следований на одном и том же контингенте испытуемых. Мы
провели сравнение показателей одних и тех же бегунов в раз-
ные периоды макроцикла — подготовительный и соревнова-
тельный (табл. 8).
В качестве основного средства «сопряженного» воздей-
ствия на технику бега и показатели локальной выносливости
мышц использовался бег в утяжеленных условиях, где в каче-
стве тормоза использовалась волокуша. При тренировке спор-
тсменам давалась установка «держаться прямо, вперед не на-
клоняться» и «хорошо доталкиваться стопой». Было замечено,
что уже примерно через месяц техника бега менялась в нужном
направлении, а тестирование выявило достоверное изменение
силы мышц задней поверхности бедра.
Из данных табл. 8 следует, что закономерное изменение
силы мышц-сгибателей от подготовительного к соревнователь-
ному периоду вызвало практически аналогичные изменения в
технике, что и при межгрупповом сравнении. Кроме того, нет
никаких оснований предполагать, что в соревновательном пе-
риоде спортсмены стали менее «экономичными», однако все
основные показатели «активности» бега достоверно возросли.
6.5. Заключение по разделу
Таким образом, на основе учета схемы функционирования
нервно-мышечного аппарата при преодолении соревнова-
тельной дистанции, представленной в 5-й главе, определении
стратегии адаптации и рассмотрения средств, методов и теоре-
тических предпосылок планирования макроцикла подготовки
(6-я глава) можно сформулировать следующие теоретически
обоснованные положения улучшения локальной мышечной
выносливости:
1. Морфологической основой высокой работоспособности
мышц является масса сократительных белков мышц, ко-
236
торые создают «морфологическое пространство» для на-
копления энергетических субстратов (КрФ и гликоген),
белков, определяющих буферную емкость мышц, а так-
же ферментативных комплексов всех основных реакций
энергообеспечения (фосфокиназной реакции, анаэроб-
ного гликолиза и мембранного фосфорилирования), от
которых непосредственно зависит спортивный резуль-
тат. Кроме этого, высокая активная мышечная масса
создает дефицит энергии внутри мышечных волокон,
создавая тем самым стимулы для запуска адаптивного
синтеза ферментативных комплексов. Поэтому базо-
вое положение среди факторов, определяющих высо-
кую специальную работоспособность, занимает силовая
подготовленность мышц. Специализированная силовая
подготовка имеет смысл только в отношении мышечных
групп, выполняющих основную нагрузку по перемеще-
нию спортсмена по дистанции. От уровня и соотноше-
ния силового потенциала этих мышц в стратегическом
плане зависит специальная работоспособность и эконо-
мичность техники спортивной локомоции.
2. По мере возрастания дистанции значимость силового
потенциала медленных мышечных волокон (как условия
высоких аэробных способностей мышц) относительно
быстрых мышечных волокон также возрастает. Поэтому
на всех дистанциях, где решающую роль играют аэроб-
ные способности, должны использоваться главным об-
разом средства силовой подготовки, способствующие
увеличению силы ММВ. Такими средствами являют-
ся статодинамические (квазиизотонические) силовые
упражнения, а также длительное и регулярное примене-
ние аэробных упражнений, выполняемых в утяжелен-
ных условиях (с высокой степенью напряжения мышц).
3. В связи с тем что производительность сердечно-
сосудистой системы и окислительный потенциал ММВ
под воздействием целенаправленной аэробной трени-
ровки повышается достаточно быстро, то стратегией по-
вышения функциональных возможностей ММВ должна
являться такая, при которой сначала выполняется ак-
237
центированное тренировочное воздействие с целью ги-
пертрофии ММВ, а затем доведение их окислительного
потенциала до максимальных значений к соревнователь-
ному периоду. Степень гипертрофии ММВ у спортсменов
в различных видах спорта не обнаружила связи с величи-
ной механической нагрузки, испытываемой ведущими
мышцами в рабочей фазе локомоции. Поэтому данный
принцип остается справедливым для всех дистанций и для
всех циклических локомоций. Однако в связи с необходи-
мостью параллельного улучшения других способностей,
будут изменяться средства и методы воздействия на сило-
вой и окислительный потенциал ММВ. Здесь будет про-
являться следующая общая тенденция: на более коротких
дистанциях — преимущественно используются силовая
тренировка и аэробно-силовой метод; на более длин-
ных — аэробно-силовой метод и длительная тренировка
в самой локомоции с использованием непрерывного или
переменного метода, но при условии поддержания хоро-
шего состояния гормональной системы педагогическими
и/или фармакологическими средствами.
4. Методика увеличения ППС ММВ существенно отличает-
ся от традиционных видов силовой и скоростно-силовой
тренировки. Поэтому использование упражнений со
штангой, на тренажерах при величине сопротивления
более 80%, а также всевозможные прыжковые, спринтер-
ские и интенсивные изокинетический и изодинамические
упражнения не могут рассматриваться в качестве средств
воздействия на ММВ, т.е., по существу, не относятся к
средствам специальной силовой подготовки в ЦВС.
Эффективным средством воздействия на М М В могут
являться статодинамические упражнения, выполняе-
мые при соблюдении следующих правил: медленный,
плавный характер движений; относительно небольшая
величина преодолеваемой силы или степени напряже-
ния мышц (40-60% от МПС); отсутствие расслабления
мышц в течение всего подхода; выполнение подхода до
«отказа»; проведение тренировки, как правило, с приме-
нением суперсетов на все основные мышечные группы;
238
паузы между сериями могут заполняться легкой аэроб-
ной работой длительностью 3-10 мин; достаточно боль-
шая длительности всей тренировки (не менее 1 часа).
Гипертрофия БМВ происходит при использовании
большинства средств и методов спортивной тренировки.
В то же время повышение окислительного потенциала
этих волокон является сложной задачей, требующей хо-
рошо спланированной целенаправленной тренировки,
которая может быть решена на этапе акцентированной
аэробной подготовки путем использования интерваль-
ного спринта, интервальной тренировки на более длин-
ных отрезках, аэробно-силового метода.
5. Эффективность силовой подготовки в отношении как
быстрых, так и медленных мышечных волокон при вы-
полнении традиционно рекомендуемых объемов средств
силовой подготовки зависит, предположительно, не от
увеличения объемов последней, а от уменьшения объемов
неэффективных средств аэробной подготовки.
6. Аэробный потенциал М М В у квалифицированных спор-
тсменов достаточно высок в результате общего большого
объема тренировочной работы, поэтому использование
низкоинтенсивных упражнений для повышения аэроб-
ного потенциала мышц в большинстве случаев бессмыс-
ленно. В связи с этим стратегическим путем повышения
аэробных способностей мышц спортсменов является,
кроме гипертрофии ММВ, воздействие на окисли-
тельный потенциал БМВ (Тип Па и, особенно, ПЬ). К
средствам воздействия на ОП БМВ относятся два вида
упражнений: а) такие, при которых БМВ рекрутирова-
ны, но они функционируют в аэробных условиях. Это
— интервальный спринт, аэробные упражнения в утяже-
ленных условиях, интервальная тренировка или трени-
ровка на длинных напряженных отрезках; б) длительные
равномерные упражнения на мощности анаэробного
порога. В этом случае воздействие на ОП БМВ будет
происходить на второй половине дистанции, после су-
щественного понижения запасов гликогена в медленных
и промежуточных МВ.
239
7. Силовые способности являются «базовыми» для аэроб-
ных; ферментативные белки более лабильны, чем сокра-
тительные или соединительно-тканные; силовая тре-
нировка требует снижения объемов аэробной и в ряде
случаев может способствовать снижению ОП мышц;
аэробные способности — основные на большинстве со-
ревновательных дистанций. В связи с перечисленными
закономерностями этап акцентированного воздействия
на аэробные способности мышц должен планироваться
после силового и как можно ближе к соревновательному
периоду в тех случаях, когда в рамках макроцикла стоит
задача повышения и силовых, и аэробных способностей.
Однако в тренировке спортсменов высшей квалифика-
ции задачи повышения силовых способностей мышц
может не стоять. В таком случае для поддержания сило-
вых способностей допустимо равномерное распределе-
ние объемов эффективных тренировочных средств (типа
статодинамических комплексов) в макроцикле (в объеме
60-90 мин в неделю чистого тренировочного времени).
8. Ферментативные комплексы креатинфосфокиназной
реакции и анаэробного гликолиза относятся к наиболее
лабильным белкам, а использование скоростно-силовых
и, особенно, гликолитических упражнений в большом
объеме может негативно сказаться на силовых и аэроб-
ных способностях мышц. Поэтому скоростно-силовая и
гликолитическая подготовка в тех ЦВС, где есть необхо-
димость использования этих средств, должна планиро-
ваться как можно ближе к этапу основных стартов.
При условии хорошей предварительной силовой и
аэробной подготовки повышение гликолитических и
алактатных способностей может быть реализовано за
достаточно непродолжительный период (3-5 недель,
включая этап предварительных стартов). Эта задача ре-
шается применением алактатных нагрузок (спринт с
околомаксимальной скоростью в течении 10-15 с) и «за-
кисляющих» гликолитических нагрузок в виде повтор-
ных отрезков «на результат» длительностью 90-120 с, при
которых в максимальной степени понижается pH мышц
240
или серий из 3-4 повторений длительностью по 40-70 с
с околомаксимальной интенсивностью и коротким ин-
тервалом отдыха (2-3 мин), при которых в максимальной
степени понижается pH крови.
9. В связи с тем что в беге стоит проблема травмирования
ОДА в результате механических перегрузок, в годичном
цикле специально должен планироваться этап укрепле-
ния соединительно-тканных элементов ОДА. Такой этап
целесообразно проводить в начале макроцикла, в рамках
которого может параллельно решаться задача повыше-
ния мощности моторных отделов ЦНС.
Суммируя все вышесказанное, принципиальной схе-
мой планирования макроцикла, олимпийского четырех-
летия или многолетней подготовки является такая, при
которой происходит одновременное воздействие на все
факторы, определяющие спортивный результат, но с рас-
становкой следующих акцентов от начала к завершению
цикла: укрепление и повышение эластичности опорно-
двигательного аппарата (существенно для ЦВС с высоки-
ми механическими нагрузками) => тренировка моторных
центров ЦНС средствами скоростно-силовой и сприн-
терской подготовки => увеличение силовых возможно-
стей основных мышечных групп средствами повышения
силы медленных мышечных волокон => массированная
аэробная подготовка с использованием средств воздей-
ствия на все типы мышечных волокон => интегрирующая
подготовка (допускается использование «анаэробно-
гликолитической тренировки») => участие в соревнова-
ниях. Выраженность акцентов снижается по мере повы-
шения мастерства и стажа занятий спортсменов.
10. Исследование технических параметров бегунов и их вза-
имосвязи с показателями физических способностей вы-
явили, что особенности техники находятся под сильным
контролем силовых показателей мышц и закономерно
изменяются при изменении всех других показателей ло-
кальной выносливости. Наиболее существенными мы-
шечными группами, которые не только определяют ра-
циональность техники бега, но и выполняют основную
241
работу по перемещению спортсмена относительно до-
рожки, можно считать мышцы голени, задней поверхно-
сти бедра и ягодичных мышц, сгибатели бедра. Повыше-
ние функционального состояния этих мышечных групп
«сдвигает» технику бега в сторону ее «экономизации»,
особенно по показателям отталкивания от опоры. Для
бегунов на средние дистанции некоторое значение име-
ет силовая подготовка мышц — разгибателей ног. В то же
время рациональная техника постановки ноги на опору
формируется путем целенаправленного обучения.
Гипотеза, что бег в «утомленном состоянии» являет-
ся средством его «экономизации» — не нашла эксперимен-
тального подтверждения.
Есть все основания предполагать, что выявленная
закономерность — высокая степень согласованности
параметров техники с характеристиками мышечных
компонентов, определяющих локальную выносливость,
справедлива для большинства циклических локомоций.
11. На основании данных экспериментальных и теорети-
ческих исследований, изложенных в этой главе, можно
утверждать, что имеющиеся в литературе указания, что
при развитии «силовой выносливости» (которая в боль-
шинстве случаев может рассматриваться как синоним
локальной выносливости) нужно стремиться к тому, что-
бы характеристики движений как можно больше соот-
ветствовали соревновательному упражнению, представ-
ляются недопустимо грубым упрощением ситуации. На
наш взгляд, здесь происходит смешивание двух задач — 1)
повышения функциональной мощности мышечных
структур и 2) его реализации в соревновательном упраж-
нении, что при определенных условиях может привести
к отрицательному результату.
Например, при тождественности временных и амплитуд-
ных характеристик, но больших силовых (тренировка силовой
выносливости предполагает именно этот вариант), т.е. при
применении аэробно-силового метода — получим простое по-
вышение мощности работы мышц и, следовательно, интенси-
фикацию накопления молочной кислоты. Это приведет к сни-
242
жению или даже блокированию эффекта повышения аэробной
мощности мышц, который в большинстве случаев и является
целью тренировки. В связи с этим при выборе упражнений
для тренировки «силовой выносливости» нужно хорошо раз-
личать:
— какие средства и методы способствуют повышению силы
мышц;
— какие средства и методы способствуют повышению аэ-
робной мощности мышц;
— как при этом может быть реализован принцип сопря-
женности технической и физической подготовки;
— как будет в дальнейшем (или параллельно) решаться за-
дача реализации двигательного потенциала;
— как добиться положительного и не допустить отрицатель-
ного эффекта при решении поставленных выше задач.
Некоторые подходы к решению этих ключевых для улучше-
ния локальной выносливости задач изложены в предыдущих
разделах и будут уточнены в следующих главах работы.
243
Глава 7
Анализ данных
экспериментальных
исследований средств
и методов повышения
локальной выносливости
7.1. Исследование упражнений
статодинамического характера
как средства воздействия на медленные
мышечные волокна спортсменов
В шестой главе описаны теоретически разработанные ха-
рактеристики упражнений силового характера, которые ги-
потетически могли бы способствовать гипертрофии ММВскс-
летных мышц, т.е. увеличивали бы «... «рабочую площадь» для
выполнения клеточных функций и тем самым создавали бы
основу для повышения функциональной мощности клеточ-
ных структур...» [ЭэпикВ.А., ВируА.А., 1990].
Это:
— тренировка только основных мышц для данной локомо-
ции;
— медленный, плавный характер движений по полной или
по «рабочей» амплитуде;
— относительно небольшая величина преодолеваемой
силы или степени напряжения мышц (30-60% от МПС);
— отсутствие расслабления мышц в течение всего подхода;
— выполнение подхода до «отказа» (длительность работы
до «отказа» должна быть 45-70 с);
244
— проведение тренировки, как правило, с применением
суперсетов на все основные мышечные группы (в этом
случае интервал между подходами 30 с, до «отказа» вы-
полняется только последний подход в серии);
— заполнение паузы отдыха (длительностью 5-8 мин) лег-
кой аэробной работой «общего « характера;
— достаточно большая длительность всей тренировки (не
менее 1 часа);
— техника каждого движения отличается от техники сило-
вых упражнений (даже с малыми весами), принятых в
бодибилдинге, тем, что в них запрещается «быстрое нача-
ло», а так же не только разрешается, но и приветствуется
пауза (пусть даже в долю с) между уступающей и преодо-
левающей фазой работы — именно в этом смысл и основ-
ное отличие техники статодинамических упражнений.
Однако теоретические предложения должны быть подкре-
плены экспериментальными данными.
Действительно ли описанный вид упражнений силового
характера (условно названный статодинамическими упражне-
ниями) воздействует в большей мере на ММВ? Каковы физио-
логические, биохимические и педагогические характеристики
такого рода упражнений?
Для выяснения поставленных вопросов был проведен ряд
экспериментов.
Эксперимент 1.
С участием 7 добровольцев (длина тела = 177,3±11,8 см, мас-
са тела = 71,7±9,7 кг, возраст = 25,0±4,8 лет) исследовалась тре-
нировка, состоящая из приседаний с неполным вставанием с
массой штанги 50-60% от МПС. Было выполнено 4 подхода с
интервалом 8 мин. Испытуемые сразу после подхода ходили 2-3
мин., затем применялся 5-минутный пассивный отдых. Пред-
варительно определялись аэробный и анаэробный пороги в
упражнениях, в которых участвовали тренируемые мышцы.
Для регистрации ЧСС применяли спорттестер РЕ3000, ре-
гистрация проводилась в течение всей тренировки каждые 5 с.
После тренировки информация со спорттестера через специ-
альный интерфейс передавалась на компьютер, где обрабаты-
245
велась и выводилась на монитор и на принтер в цифровом и
графическом виде.
Электрическая активность мышц (медиальная и латераль-
ная головки четырехглавой мышцы бедра) измерялась элек-
тромиографом ЕМГСТ-01 и подвергалась интегрированию
за 500 мс по оригинальной программе. Во время тренировки
брались пробы крови из пальца: перед разминкой, непосред-
ственно перед каждой попыткой, сразу после попытки и через
3 мин. Биохимические анализы проводились в отделе биохи-
мии ЛПСР ГЦОЛИФК.
Анализ пульсовой кривой показал (рис. 16), что ЧСС во вре-
мя выполнения отдельных подходов, длящихся 40-60 с, под-
нималась до уровня ЧСС АэП или незначительно превышала
этот уровень. После окончания упражнения пульс поддержи-
вался 10-30 с, затем ЧСС начинала резко снижаться до уровня
100-110 уд/мин. Однако перед очередным подходом ЧСС была
несколько выше, чем перед предыдущей попыткой.
Во время силовой тренировки с четырьмя подходами на-
блюдаются следующие изменения в ИЭМГ (рис. 17):
— амплитуда ИЭМГ в преодолевающей и уступающей фа-
зах в течение каждого подхода находится приблизитель-
но на одном уровне;
— амплитуда ИЭМГ в преодолевающей фазе от подхода к
подходу снижается (с 813±56 усл. ед. в первом подходе до
544±62 усл. ед. в четвертом подходе р<0,001);
— амплитуда ИЭМГ в уступающей фазе во время выполне-
ния упражнения составляет 25+3% от максимальных зна-
чений, причем достоверно отличается от нуля (р<0,05). К
четвертому подходу величина ИЭМГ в этот момент вы-
полнения упражнения также снижается (с 200± 16 усл. ед. в
первом подходе до 140± 17 усл. ед. в четвертом (р<0,05));
— в ходе каждого подхода этот показатель практически не
изменяется.
В табл. 9 представлены данные об изменении биохимиче-
ских показателей крови.
Показатели, отражающие состояние буферных систем, от
попытки к попытке снижались: буферные основания с +0,7
до -7,5 м-экв/л, показатель бикарбонатной буферной систе-
246
мы с 25,6 до 16,8 м-экв/л и общее количество СО с 27,9 до 17,7
м-экв/л.
Во время силовой тренировки с четырьмя подходами на-
блюдаются следующие изменения в ИЭМГ (рис. 20):
Рис. 16. Изменение ЧСС, BE и pH при выполнении 4 статодинамических
подходов
pH за время тренировки изменялся незначительно — в пре-
делах 7,36-7,41.
10 20 30 40 50 мин.
Рис. 1. Примерный вид графика ИЭМГ при выполнении 4 статодина-
мических подходов до «отказа» с интервалом 8 мин. В конце каждого
подхода испытуемых просили попытаться прыгнуть, чтобы получить
максимальное значение ИЭМГутомленных мышц
Таким образом, невысокие максимальные показатели ЧСС
и быстрое восстановление практически до исходных величин
247
после подхода свидетельствуют о невысоком напряжении ССС
во время тренировки такого вида.
Таблица 9. Изменение показателей кислотно-щелочного состояния кро-
ви и ЧСС во время силовой тренировки
Время измерения Показатели
BE pH ЧСС
Исход 0,7 7,39 75
П./д. 1-м подходом 1,0 7,40 80
После 1-го подхода 3,8 7,41 145
Ч/з 3 мин 5,5 7,37 100
П/д 2-м подходом 3,5 7,36 80
После 2-го подхода 3,2 7,41 151
Ч/з 3 мин 6,0 7,32 121
П/д 3-м подходом 4,5 7,38 81
После 3-го подхода 6,6 7,36 152
Ч/з 3 мин 6,0 7,37 125
П/д 4-м подходом 5,1 7,38 98
После 4-го подхода 6,1 7,37 157
Ч/з 3 мин 7,5 7,38 102
Отмеченные изменения показателей буферных систем в со-
четании с незначительными изменениями кислотно-щелочного
равновесия (pH) говорят о том, что выполненная тренировка
проходила в условиях компенсированного ацидоза.
Отказ от выполнения упражнения статодинамического ха-
рактера наступает в результате появления болевых (жжение)
ощущений в мышцах, а не из-за рекрутирования всех ДЕ, по-
скольку при необходимости испытуемый даже в момент отказа
может прыгнуть со штангой.
Эти наблюдения подтверждает динамика ИЭМГ, по кото-
рой видно, что амплитуда ИЭМГ по ходу выполнения упраж-
нения не увеличивается. Следовательно, можно утверждать,
что при выполнении упражнений в статодинамическом ре-
248
жиме действительно задействованы не все ДЕ, активность МВ
не обеспечивается кровотоком, отказ наступает в момент на-
копления в активных МВ высоких концентраций ионов Н+ и
Ла. Поэтому статодинамические упражнения создают в ММВ
условия, которые, как предполагается, интенсифицируют об-
разование и-РНК (свободный Кр, ионы Н+, ц-АМФ). В то же
время можно предположить, что высокие концентрации ио-
нов водорода могут оказать негативное влияние на митохон-
дриальную систему. Этот вопрос был подвергнут эксперимен-
тальной проверке и будет описан далее.
Эксперимент 2.
Ключевым для повышения силовых возможностей в спор-
те и оздоровительной тренировке является вопрос о создании
т.н. анаболического эффекта и минимизации катаболиче-
ского эффекта при применении тех или иных упражнений.
Другими словами, оптимальной будет такая тренировка, при
которой в максимальной степени активируется синтез спец-
ифических белков (мышц) во время отдыха, но при этом их
разрушение в процессе собственно выполнения упражнений
оказывается небольшим.
Для оценки «анаболического потенциала» статодинамиче-
ских упражнений было проведено исследование, в котором
оценивалось влияние на мышечную силу (как характеристику
анаболического потенциала) и проявление мышечных болей
(как характеристику степени катаболизма) одного статодина-
мического подхода и одного тренировочного занятия с приме-
нением статодинамических упражнений.
В двух сериях экспериментов воздействию подвергались
мышцы задней поверхности бедра правой и левой ноги испы-
туемых. Выбор режима работы мышц осуществлялся случай-
ным образом.
Измерения изометрической силы (ИС) проводились на спе-
циальном измерительном стенде при углах в тазобедренном и
коленном суставах 90 %, а нагрузка задавалась на тренажере,
позволяющем нагружать заднюю поверхность бедра правой и
левой ног в положении лежа на животе путем сгибания ног в
коленном суставе.
249
В первой серии экспериментов изучался эффект острого
тренировочного воздействия в виде одного эксцентрического и
одного статодинамического подхода, выполняемого «до отказа».
В первый приход испытуемых в лабораторию воздействию
подвергалась одна нога, вторая служила контролем. Во второй
приход (через 2-4 недели) воздействию подвергалась другая
нога. Виды воздействия чередовались случайным образом.
При статодинамическом режиме испытуемый в медленном
темпе поднимал и опускал груз, равный 50% от максимальной
силы, одной ногой. На протяжении всего подхода мышцы не
расслаблялись.
Для создания эксцентрического режима на тренажере уста-
навливался вес в 130... 150% от максимального. Подъем груза
осуществлялся партнером, затем испытуемый старался пре-
пятствовать опусканию груза за счет усилия мышц задней по-
верхности бедра.
После разминки у 5 испытуемых измерялась исходная сила
мышц — сгибателей коленного сустава. Затем выполнялся ста-
тодинамический или эксцентрический подход, после которого
ежеминутно в течение 8 мин измерялась ИС для оценки дина-
мики восстановления мышечной силы.
Усредненные данные по 5 испытуемым представлены на
рисунке.
250
Из этих данных следует, что степень снижения мышечной
силы после эксцентрического больше, чем после статодина-
мического режима.
Оценка степени проявления МБ в течение 6 дней восстано-
вительного периода выявила (рисунок), что после эксцентри-
ческого режима МБ выражены существенно больше и восста-
новление протекает медленнее.
Динамика мышечных болей (МБ) после острого
тренировочного воздействия эксцентрического
Во второй серии изучалась реакция НМА на одно трениро-
вочное занятие у 2 испытуемых, состоящее из 7 подходов с раз-
личным режимом работы мышц.
На рисунке представлена динамика ИС после серии подхо-
дов эксцентрического или статодинамического характера.
На следующем рисунке представлена динамика МБ после
этого же тренировочного занятия.
3
i
©
£
Z
Я
О
Динамика мышечных болей (МБ) после острого
тренировочного воздействия эксцентрического
или статодинамического характера
3,6
3
2,5
2
1,5
1
0,5
0
1 2 3 4 5
Время восстановления,дней
Как указывают данные этого эксперимента, также как и в пред-
ыдущих случаях, статодинамический режим существенно отлича-
251
ется по своему воздействию на НМА от эксцентрического режима.
Во-первых, после статодинамического занятия несущественное
снижение ИМС наблюдается только на второй день, а затем, на-
чиная с третьего дня, наблюдается явно выраженная фаза супер-
компенсации с превышением исходного уровня в среднем на 16%.
К этому времени МБ полностью прекращаются.
При этом противоположная картина наблюдается в состоя-
нии другой ноги (после эксцентрического режима). Макси-
мально выраженные и существенно большие по интенсивно-
сти МБ наблюдаются на второй день восстановления. В этот
же день снижение ИМС достигает в среднем 22% и не возвра-
щается к исходному уровню на протяжении 6 дней. Весь этот
период испытуемые отмечали явно ощущаемые боли в мыш-
цах, подвергшихся эксцентрическому воздействию.
Эксперимент 3.
Отставленный эффект статодинамических упражнений
был изучен в ПНИЛ РГАФК И.В. Еркомайшвили (1990). Ис-
следовалась динамика мочевины в крови утром натощак после
тренировки различной направленности у женщин — конько-
бежек, не употреблявших анаболические стероиды.
1-й вид тренировки — гликолитическая, педалирование 3
серии 3x60 с с интервалом 2 мин;
2-й вид тренировки — 10 подходов по 20 прыжков вверх из
глубокого приседа («лягушкой»);
3-й вид тренировки — аэробно-силовая, педалирование с
пульсом 150, 3x10 мин, темп 40 об/мин;
4-й вид тренировки — 8 подходов по 8-10 раз медленных
приседаний со штангой 70-80% от МПС по методике выпол-
нения статодинамических упражнений;
5-й вид — 8 подходов (длительность каждого около 60 с) ста-
тодинамических упражнений со штангой 40-50% от МПС.
Результаты представлены на рис. 18.
По значениям мочевины в крови в 1-2-й день после трени-
ровки можно судить о степени «катаболичности — анаболич-
ности» тренировки. Если величина мочевины повышена, то
баланс смещен в сторону катаболических процессов, если по-
нижена — преобладают анаболические процессы.
252
Рис. 18. Динамика мочевины в крови после различных по характеру при-
меняемых упражнений тренировок силовой направленности
1-й вид тренировки — гликолитическая, обладает наиболь-
шим катаболическим эффектом, не переходящим в анаболи-
ческую фазу даже к 4-му дню после тренировки.
2-й вид тренировки — прыжковая с явно выраженным по-
вреждающим эффектом в отношении мышц ног (мышечные
боли через 24-48 часов). Также катаболическая, но немного
«мягче».
3-й вид тренировки — аэробно-силовая, на следующий день
— небольшое повышение мочевины в крови, однако потом на-
ступает анаболическая фаза.
4-й вид тренировки — силовая статодинамическая, анабо-
лическая фаза наиболее длительна и выражена.
5-й вид — статодинамические упражнения, явно выражен-
ный, но непродолжительный анаболический эффект.
253
В данном эксперименте подтверждены результаты преды-
дущих, а именно:
1 . Упражнения бывают «анаболическими» и «катаболиче-
скими». Гликолитическая работа и тяжелые скоростно-
силовые упражнения, споровождающися появлением
мышечных болей имеют явно выраженный «катаболи-
ческий» эффект, который может переходить, а может и
не переходить в «анаболическую» фазу.
2 . Статодинамические силовые упражнения с тяжелой
штангой (70-80% от МПС) имеют выраженный и дли-
тельный анаболический эффект.
3 . Статодинамические силовые упражнения с легкой на-
грузкой (40-50% от МПС), которые, как мы предпола-
гаем, в наибольшей степени задействуют ММВ, имеют
также анаболический, но менее выраженный и непро-
должительный эффект. Причин могжет быть две. Первая
— для квалифицированных спорстменов, привыкших
к интенсивной силовой работе (напомним, речь идет
о конькобежцах-спринтерах и многоборцах), нагрузка
40-50% от МПС может быть недостаточно стрессовой.
Вторая — каждый подход выполнялся «по времени», а не
«до отказа», как рекомендуется, и не использовались су-
персерии (например, 3x60 с с интервалом 30 с. и паузой
между сериями 6-8 мин), которые, как предполагается,
более эффективно загружают мышцы при таком виде
тренировки.
Эксперимент 4.
Отставленные эффекты упражнений, которые похожи на
статодинамические по характеру работы мышц, описаны в ра-
боте Б.С. Шенкмана [1999].
9 нетренированных молодых мужчин (18-25 лет) 8 недель
выполняли подошвенные сгибания стопы с частотой 1 раз в с
у и 30% от МПС 4 раза в неделю в положении стоя в течение 45
минут. Тренировке подвергалась левая нога. Правая служила
контролем. Биопаты методом игольчатой биопсии забирались
до и после тренировки из m.soleus обеих ног. Результаты пред-
ставлены в таблице:
254
Ко- нечн. ммв% ППС цм2 Кап/ вол Плоти, кап-ов
ММВ БМВ
Рабоч. До 76 7900 9400 2,72 285
После 85 9100 8700 3,53 339
Контр. До 80 8600 9600 2,82 304
После 86 9000 8100 3,1 294
Результаты.
ППС ММВ увеличились на 13,2%, но недостоверно. Коли-
чество капилляров на волокно достоверно возросло на 21,4%,
плотность — достоверно на 19%. Достоверных изменений ак-
тивности окислительных ферментов и миоглобинпероксида-
зы за время тренировок не обнаружено в обоих типах волокон.
При этом активность цитоплазматической лактатдегидрогена-
зы и фосфофруктокиназы значительно снизилась в ММВ на 35
и 24% и в БМВ — на 46 и 26% соответственно. Объемная плот-
ность митохондрий недостоверно увеличилась с 7,9 до 8,1%,
субсарколемальных — с 5,2 до 5,4%. Липидных включений — с
0,43 до 0,70%. миофибрилл и саркоплазмы не изменился. Вы-
явлена тенденция к увеличению числа митохондрий как на пе-
риферии, так и в центральной части мышечного волокна, при
одновременном уменьшении периметра и средней плотности
митохондрий.
Таким образом, тренировка описанного типа привела к не-
которому увеличению площади поперечного сечения (ППС)
медленных МВ, некоторому снижению ППС быстрых мы-
шечных волокон, достоверному и значительному увеличению
плотности капиллярной сети.
Для объяснения механизма роста капилляров при трени-
ровке рассматривают по крайней мере две гипотезы: «гипок-
сическую», связывающую рост капилляров с пониженным
напряжением кислорода в ткани при мышечной работе и
«гиперемическую», предполагающую зависимость роста ка-
пилляров (или лежащую в его основе пролиферацию эндоте-
лиоцитов) от механических эффектов повышенного послера-
бочего кровотока [см., например, обзор Б.С. Шенкмана, 1999].
При изотонической тренировке имеет место значительное на-
255
пряжение в мышце и большое накопление метаболитов. Это, в
свою очередь, может способствовать накоплению «эндотели-
альных факторов роста» и, кроме того, при такой тренировке
практически до максимальной степени увеличен послерабо-
чий кровоток, вероятно, именно эти факторы привели к до-
стоверному приросту плотности капилляров.
Незначительный прирост ППС, как мы предполагаем, мо-
жет быть связан с недостаточно высокой нагрузкой (30% от
МПС) и с тем, что упражнения в большинстве подходов вы-
полнялись не «до отказа».
7.2. Влияние сочетания статодинамической
силовой и аэробной тренировок мышц бедра
на аэробный и анаэробные пороги человека
(лабораторный эксперимент)
Выводы результатов лабораторных, хорошо контролируе-
мых экспериментов, в которых изучались эффекты сочетания
силовой и аэробной тренировки [Шенкман Б.С. и др., 1990;
Dudley G.A., R.Djamil, 1985; Hickson С. и др., 1980; Hunter G. и
др., 1987] можно суммировать следующим образом:
1. Аэробная тренировка на уровне аэробного порога (АэП),
т.е. с умеренной интенсивностью, имеет ограниченную
эффективность.
2. Аэробная тренировка всегда снижает эффект силовой
[Dudley G.A., R.Djamil, 1985].
3. Силовая тренировка не снижает эффекта аэробной
[Шенкман Б.С., 1990; Dudley G.A., R.Djamil, 1985;
Hickson С. и др., 1980].
4. При использовании одного и того же объема аэробной
тренировки через 1-2 месяца начинают проявляться яв-
ления «насыщения» (т.е. показатели выходят на плато),
однако при сочетании силовой и аэробной тренировок
скорость прироста аэробных показателей возрастает
[Dudley G.A., R.Djamil, 1985].
Выводы советских и российских естественных педагогиче-
ских исследований, а также вся практика спортивной подготов-
256
ки в ЦВС однозначно свидетельствует, что силовая подготовка
является неотъемлемой частью тренировочного процесса на
всех дистанциях от спринта до марафона. Дискуссия ведется
лишь около средств, методов, объемов и места силовой подго-
товки в рамках микро-, мезо-, и макроциклов, т.к. при неко-
торых условиях применение силовых упражнений и соответ-
ствующий прирост силы сопровождается снижением аэробных
показателей [Журбина А.Д., 1976; Набатникова М.Я., 1975].
Таким образом, считая вопрос о целесообразности сило-
вых упражнений в тренировке бегунов решенным, описан-
ный ниже эксперимент посвящен изучению разработанной в
ПНИЛ РГАФК методике силовых упражнений, которые, как
предполагается, на основе теоретического анализа и представ-
ленных выше данных, должны в большей мере, чем какие бы
то ни были другие упражнения, способствовать гипертрофии
ММВ.
При организации лабораторного эксперимента мы предпо-
лагали, что статодинамическая силовая тренировка в сочетании
с аэробной тренировкой тех же мышечных групп будет более
эффективным путем повышения аэробного (АэП) и анаэроб-
ного порогов (АнП) мышц, чем только аэробная тренировка.
В основании этого предположения лежат представления,
что индукция синтеза не только митохондриальных белков,
которая имеет место при аэробной тренировке, но и сократи-
тельных элементов ММВ, суммарно в большей степени увели-
чит окислительный потенциал мышечных волокон этого типа
(т.е. массу митохондрий), чем просто аэробная тренировка
ограниченного объема.
Две группы физически активных испытуемых неспортсме-
нов (п: 10 и 7; возраст: 23,1±6 и 25,6±5 лет; масса тела: 69,5±9 и
69,4±12 кг) дважды в неделю тренировались на велоэргометре
по 45-50 мин на уровне ЧСС, соответствующему вентилятор-
ному аэробному порогу (АэП), который определялся путем
ступенчатого теста. Контроль интенсивности осуществлялся
по ЧСС с использованием спорттестера РЕ-3000.
Вторая группа дополнительно дважды в неделю выполняла
медленные приседания со штангой (масса 60-70% от ПМС) с
неполным вставанием, ограничиваемым специальным устрой-
257
ством. Рабочий угол в коленных суставах изменялся в пределах
90-140°. То есть работа выполнялась без расслабления мышц и
«до отказа» в каждом подходе. Следовательно, можно предпо-
ложить, что тренировка проходила в практически анаэробных
условиях и сопровождалась значительным понижением вну-
тримышечного pH.
В первую тренировку испытуемые выполняли 4 подхода с
интервалом отдыха 8 мин. Характер отдыха — активный (ходь-
ба в среднем и медленном темпе). Во вторую тренировку при-
менялось 9 подходов в виде суперсерий, т.е. три подхода «до
отказа» с коротким 30-секундным интервалом отдыха состав-
ляли 1 серию. Таких серий выполнялось три. Интервал актив-
ного отдыха между сериями — 12 мин.
Сила мышц — разгибателей ног оценивалась до и после тре-
нировки путем выполнения теста с приседанием штанги до
угла в коленных суставах 90° и полным вставанием. Вес штан-
ги подбирался таковым, чтобы испытуемый мог выполнить
не более 5-8 приседаний. Максимальная произвольная сила
(МПС) мышц — разгибателей ног рассчитывалась по формуле,
выведенной на основании экспериментальной зависимости:
масса штанги — число повторений, по данным Н.Н. Кулика
(1967):
У = 0,122Х2 - 3,871 Х-ь 100; МПС = (масса штанги в тесте) * 100 / У,
где У — величина поднимаемого веса в процентах от макси-
мальной силы, X — число подъемов штанги.
Вентиляторные анаэробные пороги (методика Мякинчен-
ко Е.Б., 1997) испытуемых определялись до, после и каждые
две недели эксперимента (всего 4 среза) в велоэргометриче-
ском тесте со ступенчато-возрастающей нагрузкой. Частота
педалирования у мужчин — 80 об/мин, у женщин — 70 об/мин.
Начальная нагрузка у мужчин — 32 Вт, у женщин — 14 Вт вы-
полнялась в течение двух минут, затем каждую мин мощность
нагрузки увеличивалась на 16 Втумужчинина 14Втуженщин.
Тест выполнялся до явных признаков утомления (одышка,
пот, появление лишних движений), но не «до отказа». Фикси-
ровались мощность выполняемой нагрузки, ЧСС при помощи
спорттестера РЕ-3000 и легочная вентиляция с использовани-
ем портативного вентилометра, соединенного с лицевой ма-
258
ской и имеющего малое «мертвое пространство». Дальнейшая
обработка результатов тестирования проводилась с помощью
компьютера, где аэробный и анаэробный пороги определялись
графическим методом по графикам зависимостей VE-4CC,
VE-мощность, ЧСС-мощность. Локализация пороговых точек
определялась двумя независимыми экспертами, не имевших
информации о принадлежности графиков. Велоэргометри-
ческий тест перед началом и в конце эксперимента для по-
вышения надежности проводился трижды. Значения порогов
определялось по средней арифметической оценок экспертов.
Рассчитывались следующие показатели: мощность аэробного
порога (АэП); мощность анаэробного порога (АнП); ЧСС на
стандартной немаксимальной мощности (100 Вт); ЧСС АэП;
ЧСС АнП.
Общая длительность эксперимента — 6 недель.
Через 6 недель эксперимента зарегистрированы следующие
изменения в показателях (рис. 19-22): за период исследования в
экспериментальной группе достоверно повысилась сила мышц
—разгибателей коленного сустава: с 866±276 Н в начале (рис. 19)
до 1058±320 Н в конце эксперимента (р < 0,001) против недо-
стоверных изменений этого показателя в контрольной группе:
с 705+169 Н до 737±200 Н (р > 0,1). Различия между группами в
величине приростов достоверны (р < 0,05).
В экспериментальной группе (рис. 20) мощность АэП в на-
чале эксперимента была 125,4±31,1 Вт, через 6 недель стала
155,3±42,9 Вт (р < 0,01, достоверное повышение), мощность
АнП соответственно 162,7±36,1 Вт и 189,0±45,1 Вт (р < 0,01). За
это время в контрольной группе повышение было недостовер-
ным: мощность АэП повысилась с 129,2±33,1 Вт до 135,±28,9
Вт (р > 0,1) и мощность АнП повысилась с 158,8±39,0 Вт до
167,3±34,1 Вт (р > 0,1). Различия в величинах приростов этих
показателей между экспериментальной и контрольной груп-
пами достоверны (р < 0,01).
В обеих группах (рис. 21) достоверно снизилась ЧСС на
стандартной немаксимальной нагрузке (100 Вт): в контрольной
группе на 15,1±4,7 уд/мин (р < 0,001), в экспериментальной на
14,2± 11,9 уд/мин р < 0,05). Различия между группами до начала
и после окончания эксперимента недостоверны (р >0,1).
259
Однако в то время как в контрольной группе очевидно про-
явление эффекта «насыщения», в экспериментальной группе
ЧСС монотонно снижалась весь период.
Величина ЧСС на аэробном и анаэробном порогах в обеих груп-
пах изменились не достоверно, однако обращает на себя внимание
разнонаправленность динамики пороговых ЧСС в контрольной и
экспериментальной группах. Что подтверждает возможную тен-
денцию более высокого тренирующего воздействия сочетания си-
ловой и аэробной тренировки на сердечно-сосудистую систему по
сравнению с чисто аэробной тренировкой.
Обсуждение результатов
Обнаружен интересный факт существенного повышения
эффективности аэробной тренировки, если ее сочетать с си-
ловой (анаэробной) тренировкой тех же мышечных групп, при
условии, что два вида нагрузки применяются в различные дни.
Полученные результаты можно интерпретировать именно та-
ким образом в связи с тем, что многократно показывалось, что
применяемые раздельно чисто силовая тренировка и аэробная
тренировка на уровне АэП не приводят к повышению аэробных
способностей физически активных лиц.
Таким образом, исследование свидетельствует в пользу пред-
ложенной нами гипотезы, что целенаправленное воздействие
(как предполагается) на ММВ с целью их гипертрофии создает
предпосылки для роста аэробных способностей путем:
— увеличения «рабочей площади» для увеличения массы
митохондриальных белков;
— и/или создавая дополнительный «гипоксический» сти-
мул для прироста митохондрий за счет увеличения мас-
сы наиболее энергопотребляющих органелл клеток, ка-
ковыми являются миофиламенты;
— и/или пролиферации (разрастания) капиллярной сети;
— и/или улучшая гормональный фон, способствующий
анаболическом эффекту не только в отношении мио-
филаментов и нервных механизмов управления работой
мышц, но и в отношении митохондриального аппарата.
Разумеется, справедливость этих предположений нуждает-
ся в дополнительных исследованиях.
260
Динамика силы разгибателей ног в контрольной (контр.) и
экспериментальной (эксп.) группах
1200
600 -I--------------1--------------н
Исход 2 недели 4 недели 6 недель
Сила (и) Эксп.
—Сила (н) Контр.
Рис. 19
Динамика аэробного (АэП) и анаэробного (АнП) порогов в контрольной
(контр.) и экспериментальной (эксп.) группах
АэП Эксп.
АэП Контр.
—*— АнП Эксп.
— х— АнП Контр.
Рис. 20
Динамика ЧСС на стандартной мощности 100 Wb контрольной (контр.)
и экспериментальной (эксп.) группах
Рис. 21
261
Диамиса ЧСС на уровне аэробного (АэГ) и анаэробного (МЦ
порогов в контрольной (контр.) и экспериментальной (эксп)
группах
Рис. 22
Тем не менее получены три принципиальных факта, суще-
ственных для теории и практики спорта и оздоровительной
тренировки.
1. Тренировка с использованием статодинамических
упражнений дважды в неделю не ухудшает, а значитель-
но увеличивает аэробный потенциал мышц.
2. Именно сочетание специально разработанных силовых
упражнений с аэробной тренировкой является стимулом
для повышения аэробных способностей мышц.
3. Снижение ЧСС на стандартной мощности было одина-
ковым в двух группах при более благоприятной динами-
ке в экспериментальной группе, что можно интерпрети-
ровать как доказательства относительной независимости
адаптационных процессов, происходящих в мышцах и
ССС при тренировочных воздействиях.
Это полностью согласуется с данными экспериментов, в
которых показана разная динамика изменений, происходящих
в показателях деятельности ССС и адаптации митохондриаль-
ного аппарата мышц.
Однако эксперимент поднимает несколько вопросов, нуж-
дающихся в более углубленном изучении.
1. Действительное ли обнаруженное повышение мощности
на уровне вентиляторных порогов отражает повышение
окислительных способностей мышц, в частности — мед-
ленных мышечных волокон, которые, как считается,
262
выполняют основную работу при мощности ниже поро-
говой? Так как повышение пороговой мощности может
отражать простое увеличение силы мышц.
2. Значительное повышение силы мышц, вероятнее всего,
отражает факт повышения физиологического попереч-
ника мышечных волокон, в том числе — медленных, что
многократно показано в экспериментах (см. главу 6).
Действительно ли данные эксперимента означают, что
повышение поперечника волокна не является препят-
ствием для улучшения оксидативных свойств волокна?
3. Если повышение оксидативных свойств мышц произо-
шло, то каковы механизмы воздействия силовых упраж-
нений на индукцию синтеза митохондриальных белков?
7.3. Классификация упражнений бегунов
на средние и длинные дистанции по признаку
их преимущественного воздействия на
морфоструктуры организма
Материалы, представленные в предыдущих разделах, по-
зволили выявить ключевые аспекты развития ЛВ, которые
должны были пройти свое практическое опробование в серии
педагогических экспериментов. Они и были выполнены в пе-
риод 1982 — 1993 гг. на кафедре легкой атлетики и проблемной
научно-исследовательской лаборатории ГЦОЛИФК (ныне
РГАФК) студентами, аспирантами и сотрудниками этих под-
разделений, а именно: В.Н. Селуяновым, Е.Б. Мякинченко,
А.В. Маслениковым, Д.Г.Холодняком, В.И. Паленым, Н.Н.
Чесноковым, Н.В. Каташинским, С.М. Обуховым, В.Н. Май-
оровым, В.Т. Тураевым.
Основными аспектами повышения Л В, которые необходимо
было проверить в условиях педагогических экспериментов, были:
1. Средства и методы гипертрофии ММВ.
2. Средства и методы повышения аэробных свойств БМВ.
3. Совместимость и взаимовлияние указанных трениро-
вочных средств между собой и с другими тренировочны-
ми средствами.
263
4. Схема планирования макроцикла в последовательности
акцентов: сила ММВ => аэробные способности БМВ =>
гликолитические способности => интегральная подго-
товка => участие в соревнованиях.
5. Схема планирования микроцикла (для бегунов на сред-
ние дистанции): соединительно-тканные элементы ОДА
=> сила БМВ => аэробные способности БМВ + сила ММВ
=> интегральная подготовка => участие в соревнованиях.
Все исследования были проведены на материале бега на вы-
носливость (средние и длинные дистанции).
Для краткости описания таблиц и рисунков мы использо-
вали условные названия тренировочных средств и методов по
их преимущественной морфологической направленности. В
табл. 10 приведена расшифровка названий, используемых да-
лее в тексте.
Бег без «закисления» — концентрация Ла в крови не должна
превышать 7-8 мМ/л, субъективно — бег продолжается до по-
явления явно выраженной «одышки» или локального утомле-
ния мышц.
7.4. Критерии обоснованности выводов
по результатам педагогических экспериментов
Известно, что в педагогических экспериментах, также как
во всех других, где объектом исследования является биологи-
ческий объект, выводы корректно распространять только на ту
генеральную совокупность, к которой принадлежит исследуе-
мая выборка.
Педагогические эксперименты, описанные ниже, выпол-
нены на спортсменах не высшего уровня, кроме того, боль-
шинство из них проведено на молодых и юных спортсменах.
Закономерно возникает вопрос, в какой степени сделанные
выводы могут быть экстраполированы хотя бы на бегунов на-
ционального уровня, не говоря уже о других видах спорта?
Первым и основным аргументом, который мы можем при-
вести в защиту правомерности экстраполяции результатов ис-
следования является то, что на основе педагогического иссле-
264
дования мы не делаем новых выводов, а лишь подтверждаем,
уточняем или опровергаем те, которые в гипотетической фор-
ме были сформулированы ранее на основе моделирования по-
ведения объекта исследования — спортсменов, тренирующих
выносливость. Сама же умозрительная модель была создана
на основе биологических и педагогических исследований,
имеющих высокий уровень достоверности, подтвержденный
различными специфическими для каждой науки методами.
Таким образом, если какой-то результат исследования был
сначала предсказан теоретически (на основе достоверных дан-
ных), а затем был получен фактически, то это — веский довод
в пользу того, что модель построена правильно и сделанные
на ее основе предсказания будут верны не только в этом, но и
во всех других случаях. Следовательно, цель описанных ниже
экспериментов — это не получение новых данных и выводов,
которые конечно же могли бы быть в этом случае распростра-
нены только на генеральную совокупность исследуемых объ-
ектов, а — доказательство адекватности модели. Тогда поло-
жительным итогом исследования и критерием достоверности
является лишь сам факт совпадения предсказанных и полу-
ченных результатов.
Вторым доводом может быть то, что все наши испытуемые
достигли возраста половой зрелости, т.е. минули многочис-
ленные особенности организма, свойственные детям до и по-
сле пубертатного периода, и фактически, с точки зрения био-
логии, мало чем отличались от взрослых людей.
Третий довод тот, что большая часть наших молодых испы-
туемых были учениками ШИСП № 9 г. Москвы практически
профессиональными спортсменами со стажем занятий легкой
атлетикой 3-7 лет.
Профессиональный спортсмен высшей квалификации от-
личается от спортсмена средней квалификации в случае оди-
накового стажа и методики тренировки только количественно,
но не качественно (это и не возможно с точки зрения одинако-
вости хромосомного набора человека). Поэтому первые могут
отличаться от вторых величиной адаптационных перестроек,
глубиной переносимых неблагоприятных сдвигов в организ-
ме, скоростью восстановительных процессов и т.п., но прак-
265
Таблица 10. Классификация упражнений, традиционно используемых в
подготовке бегунов на средние и длинные дистанции по признаку их пре-
имущественного воздействия на морфоструктуры организма
Морфоструктура Наименование упражнения
1 2
1. Соединительно-тканные эле- менты ОДА (СТЭОДА) 1. Спринтерский бег во всех вариантах, выполняемый повторно, переменно, под гору, в процессе аэробного кросса или спортивных игр. 2. Прыжковые упражнения во всех вариантах, выполняемые с различной интенсивностью твердому и мягкому грунту, а также с отягощениями. 3. Стретч («упражнения на гибкость»)
2. Миофиламенты БоМВ и БгМВ (сила БМВ) 1. Упражнения со штангой, на трена- жерах или с отягощениями с сопротив- лением 80-85% ПМ и выше. 2. Спринтерский бег и максималь- ные прыжки, спринт и максимальные прыжки в гору и с сопротивлением на отрезках 80-200 м. 3. СБУ с высокой интенсивностью в гору или на равнине на отрезках 50- 150м
3. Миофиламенты ММВ, БоМВ (сила ММВ) 1. Локальные силовые упражнения на тренажерах, с отягощениями, со- противлением, собственным весом, напряжение 30-60% ПМ, упражне- ния выполняются медленно в стато- динамическом режиме, без рассла- бления мышц «до отказа». Условное название—«статодинамический ком- плекс»
4. Двигательная зона коры и мо- тонейронный пул. Координация в работе двигательных единиц и мышц синергистов 1. Спринтерский бег со старта на 10-30 м на время. 2. Максимальные прыжки на резуль- тат, 1-10 отталкиваний. 3. Упражнения со штангой и на трена- жерах с предельным весом
5. Митохондрии и капилляры ММВ 1. Равномерный бег со скоростью ниже или на уровне аэробного порога
266
1 2
6. То же для ММВ и БоМВ 1. Темповой бег со скоростью анаэроб- ного порога. 2. Переменный кросс с ускорениями 200-600 м. 3. Повторный или интервальный бег на средних и длинных отрезках 600- 2000 м без «закисления». 4. Повторный или переменный бег по дорожке с соревновательной скоростью на дистанции 30-50% от соревнователь- ной. Интервал отдыха — 3-4 мин
7. То же для ММВ, БоМВ, БгМВ (аэробные способности БМВ) 1. «Ветровой» спринт (50/50, 50/100). 2. Интервальный спринт — кросс с многочисленными быстрыми ускоре- ниями по 20-40 м. 3. Интервальный бег с быстрыми 100- 200 м отрезками. 4. Повторный или переменный бег по дорожке с соревновательной скоростью на дистанции 50-60% от соревнователь- ной. Интервал отдыха — 4-6 мин. 5. Интервальный бег в холм, с сопро- тивлением или по сыпучему грунту на отрезках 400-800 м. Отдых трусцой или многоскоками 2-4 мин. 6. Интервальный бег прыжками или СБУ на отрезках 200-800 м. 7. Кросс по сильно пересеченной мест- ности и сыпучему грунту. 8. «Беговые» спортивные и подвижные игры
8. Мощность ферментов гликоли- за, буферная емкость 1. «Длинный» спринт 200-400м с мак- симальной скоростью. 2. Специальная гликолитическая тре- нировка (например, быстро 3-4x300- 500 м, через 2-4мин). 3. Соревнования на дистанциях 400- 3000м
9. Соревнова тельный бег 1. Повторный или переменный бег по дорожке с соревновательной скоро- стью на дистанции 30-60% от соревно- вательной. Интервал отдыха — 3-6 мин
Примечание. ММВ — медленные мышечные волокна,
БоМВ — быстрые оксидативные мышечные волокна,
БгМВ — быстрые гликолитические мышечные волокна,
267
тически никогда — качественными изменениями в каких-то
органах и тканях, психофизиологическом или психическом
статусе. Поэтому, на наш взгляд, принципиальных различий в
реакции на нагрузку у спортсменов более или менее высокой
квалификации быть не может, а следовательно, экстраполя-
ция выводов допустима.
7.5. Исследование влияния акцентированной
силовой и аэробной тренировки на показатели
силы, аэробных способностей и экономичности
техники бега
Цель эксперимента — выявить влияние традиционной си-
ловой, скоростно-силовой (прыжки, спринт, упражнения со
штангой и на тренажере «Геркулес») и аэробной (аэробно-
силовым методом) тренировки на показатели силы, аэробных
способностей и экономичности техники бега при следующем ва-
рианте распределения акцентов в макроцикле: соединительно-
тканные элементы ОДА => сила БМВ => аэробные способности
БМВ + сила ММВ => интегрирующая подготовка участие в со-
ревнованиях [Мякинченко Е.Б., 1983] (рис. 15).
Методика.
В эксперименте приняли участие 10 бегунов на средние
дистанции (Ip-КМС), разделенные на 2 группы.
Контрольная (8 человек, 22,8±2 лет, 174,8±4 см, 67,3±3 кг) трени-
ровалась у своих тренеров по традиционной схеме (см. рис. 33):
— ОФП в переходный период (сентябрь);
— объемная аэробная тренировка переменным или непре-
рывным методом умеренной интенсивности (октябрь —
середина декабря);
— этап скоростно-силовой и гликолитической подготовки
(середина декабря — середина января);
— участие в соревнованиях (конец января, февраль).
Экспериментальная (2 человека, 24/23 лет, 174/181см,
69/71кг) тренировалась один полугодичный макроцикл строго
по программе, составленной нами с соблюдением следующих,
явно выраженных акцентов (рис. 23):
268
— соединительно-тканные элементы ОДА в переходный
период (сентябрь) в виде многочисленных прыжков,
стретча, спорт, игр для подготовки ОДА к последующим
высокоинтенсивным нагрузкам;
— сила БМВ (октябрь — середина ноября) в виде: всевоз-
можных прыжков по равнине, в гору и по песку с макси-
мальной интенсивностью; спринт в гору и с сопротивле-
нием; штанга; тренажер «Геркулес»; продолжительность
подхода — до явного утомления мышц;
— аэробные способности БМВ + сила ММВ (середина
ноября, декабрь) в виде: бега в утяжеленных условиях (в
гору, по песку, снегу, с сопротивлением в виде волоку-
ши); интервально 2-3 мин через 1,5-2 мин трусцы; про-
должительность работы 50-70 мин без пауз;
— интегрирующая подготовка (конец декабря, январь) в
виде: интервального и переменного бега с соревнова-
тельной скоростью по дорожке манежа; длительность
отрезка — до наступления фазы компенсированного
утомления (300-500 м); объем — 2-4 км за тренировку;
— участие в соревнованиях.
В экспериментальной группе:
— Во время «активного» отрезка при выполнении бега в
утяжеленных условиях делался акцент на: 1) продвиже-
ние «вперед, а не вверх; хорошее проталкивание», «шаги
должны быть широкие, но редкие»; 2) непременное со-
блюдение аэробного характера нагрузки, т.е. недопуще-
ние каких бы то ни было явлений «закисления», несмо-
тря на длительный рабочий период (2-3 мин) и короткий
период отдыха (1,5-2 мин). ЧСС — не выше 170 уд/мин,
восстановление — до 120-130 уд/мин.
— Аэробный бег в виде длительного кросса или фартлека
(50-60 мин) со средней скоростью анаэробного порога,
разминочного и заминочного бега применялся в одина-
ковом объеме во все этапы макроцикла (условно предпо-
лагаем, что это — воздействие на аэробные способности
ММВ и БоМВ) в объеме — 24±6 км/неделю.
— Бег с соревновательной скоростью в виде отрезков 200-
600 м для поддержания технических навыков выполнял-
269
ся на каждой тренировке до и после основной работы в
объеме 0,8-1,2 км.
— Тренировки гликолитической направленности были ис-
ключены полностью.
— Еженедельно проводилось не более 4-5 тренировочных
занятий, длительностью не более 1,5 часов, включая раз-
минку и заминку. Каждый этап состоял из 2 трехнедель-
ных мезоциклов (всего — 6 недель): два нагрузочных +
один разгрузочный микроцикл.
В контрольной группе число тренировочных занятий со-
ставило в среднем 8,2 при наибольшей плотности в ноябре —
декабре (9,8).
Сравнение нагрузок в экспериментальной и контрольной
группе (рис. 23) показало, что объемы силовой работы были
приблизительно одинаковыми, но распределены по-разному.
В экспериментальной выполнялась в начале, а в контроль-
ной — в конце подготовительного периода. Тренировки типа
«Аэро-БМВ», к которой в контрольной группе мы отнесли и
интервальную тренировку на отрезках 200-400 м была суще-
ственно больше в экспериментальной группе. При том же рас-
пределении, что и в контрольной. Обращает на себя внимание
5-6-кратное преобладание бега в аэробной зоне энергообеспе-
чения в контрольной группе при традиционном распределе-
нии с пиком в середине подготовительного периода (октябрь
— ноябрь).
Динамика показателей в контрольной группе свидетель-
ствует о крайней неэффективности тренировочного процесса.
К середине подготовительного периода все показатели снизи-
лись. Небольшой прирост силовых показателей наблюдался
в декабре, когда выполнялась соответствующая работа в уве-
личенном объеме. В соревновательном периоде заметно улуч-
шилась только экономичность бега. Спортивные результаты в
этом сезоне у спортсменов контрольной группе в среднем так-
же остались без изменений.
Результаты.
Результаты исследования приведены на рис. 24, на котором
представлена динамика показателей, отражающих основные
270
K\\\\SSKJ Сила I |Дарл-КМ1 —-Cop, бег «“АэроММВ
ESSSSSSCiim I ) Аэро-БЛ® —Сори бег ™*™ Аэро МЛ®
Рис. 23. Принципиальная схема распределения недельных (в минутах
тренировочного времени, включая интервалы отдыха) объемов трени-
ровочных средств различной направленности в макроцикле эксперимен-
тальной и контрольной групп. Значения объемов средств подготовки
умножены на коэффициенты для лучшей обозримости предельной ин-
тенсивности показатели физических качеств снижаются, что полно-
стью согласуется с данными Ю.В. Верхошанского (1992), и только после
этого начинается «реализация»
стороны подготовленности бегунов: силу, скорость, аэробные
способности и экономичность бега.
В экспериментальной группе акцентированное исполь-
зование интенсивных силовых и спринтерских упражнений
в сочетании с аэробной подготовкой в небольшом объеме, к
удивлению, резко понизило все без исключения показатели
(октябрь — ноябрь), включая силовые и скоростно-силовые.
271
Сила сгибателей колонн, сует Лео
Скорость АнП (м£)
Контр ,|
Рис. 24. Динамика показателей, отражающих основные стороны подго-
товленности бегунов: силу, скорость, аэробные способности и экономич-
ность бега за время педагогического эксперимента
Последующее снижение объема силовой нагрузки и ак-
цент на аэробные способности БМВ при строгом сохранении
объема аэробного бега и пробежек с соревновательной скоро-
стью обозначили прирост в силовых показателях при продол-
жающемся ухудшении экономичности бега. Вероятно, поэто-
му скорость на уровне АнП не возрастала. Завершение этапа
акцентированного использования аэробно-силового метода и
переход к интегрирующей тренировке привел к резкому улуч-
шению всех без исключения показателей в январе. Участие в
272
соревнованиях в январе — феврале не снизило положительной
динамики всех показателей способностей бегунов.
Один участник экспериментальной группы, начав макро-
цикл с норматива второго разряда на 1500 м (4.07.5), пробежал
в марте 1500 м за 3.53.0, выполнив норматив КМС. Второй
участник эксперимента также существенно улучшил результат
с 1,57,5 до 1,55,0 на 800 м, что особенно весомо с учетом «за-
стоя» в результатах на протяжении предыдущих 4 лет.
Обсуждение и выводы.
1. Эксперимент выявил, что в течение 6 недель акценти-
рованной силовой и скоростно-силовой подготовки
(дважды в неделю тренировка была посвящена развитию
только этих способностей) в форме упражнений
Этот факт свидетельствует, что такая тренировка соз-
дает преобладание «катаболических» процессов, пред-
положительно, за счет повреждения сократительных
элементов мышц в «эксцентрической» фазе движений с
последующей элиминацией деструктурированных бел-
ков лизосомальным аппаратом [Хоппелер Г., 1987]. При
условии, что мощности гормональной системы и пула
аминокислот не хватает для адекватного обеспечения
синтеза белков в результате присутствия в тренировке
аэробных упражнений, имеющих высокую энергетиче-
скую стоимость, что способствует ускоренному сжига-
нию аминокислот, гормонов, нейромедиаторов и других
белковых веществ. Несмотря на то что в последующем
наступает фаза «реализации», во время которой быстро
повышаются силовые показатели, признать такую орга-
низацию тренировочного процесса рациональной, когда
почти на 2 месяца в работающих мышцах создается «ка-
таболический фон», видимо, нельзя. Кроме этого, в со-
ответствии с нашей классификацией, основное воздей-
ствие в процессе такой тренировки осуществляется на
сократительный аппарат БМВ, что может иметь какое-то
значение для средних дистанций, но минимальное — для
стайеров. Помимо прочего, такие упражнения травмоо-
пасны. Об этом свидетельствует «эпидемия массового
273
травматизма» (1982-1984 гг.) в сборной СССР на сред-
ние дистанции, со спортсменами которой мы работали
в то время, когда «вошла в моду силовая подготовка». На
основании этих данных был сделан вывод, что нежела-
тельными характеристиками тренировочного процесса
бегунов являются:
— использование в больших объемах интенсивных си-
ловых и скоростно-силовых упражнений, имеющих
эксцентрическую фазу; в то же время был сделан вы-
вод о возможной большей рациональности статоди-
намических упражнений;
— длительное (более 2-3 недель) «западение» показа-
телей физических способностей под воздействием
каких бы то ни было тренировочных средств. Опти-
мальнее суметь обеспечить непрерывную положи-
тельную динамику показателей сразу после окон-
чания 1-2 недельного «острого» адаптационного
периода. Это особенно важно в тренировках юных и
молодых спортсменов, у которых состояние «придав-
ленности нагрузкой» может самым негативным об-
разом сказаться на иммунной системе [Шубик В.М.,
Левин М.Я., 1982, 1985].
Поэтому этот принцип неуклонно соблюдался во
всех последующих педагогических экспериментах. Тем
не менее нельзя отрицать, что этап силовой подготовки
не имел эффекта «тренировки тренируемости», точнее —
повышение ресурса эндокринных желез, обеспечиваю-
щих анаболические процессы в организме при помощи
ряда гормонов (соматотропин, тестостерон, андроген-
ные гормоны, инсулин и др.) [Теппермен Дж., Теппер-
мен X., 1989]
2. Можно утверждать, что о тренировке с бегом в утяже-
ленных условиях следует говорить как об эффективной
разновидности аэробной тренировки, а не «смешан-
ной» или тем более «гликолитической», несмотря на то,
что, действительно, в энергообеспечении принимают
участие и гликолитические реакции. Здесь уместно на-
помнить, что даже в покое 20-50% энергии выделяется
274
в ходе гликолитических реакций, а КрФ задействован
при любом сокращении любого мышечного волокна,
даже гладкого, поэтому классификации нагрузок и их
«тренировочных эффектов», в которых используются
слова «смешанные», аэробно- анаэробные, анаэробно-
алактатные и т.п. дают слишком упрощенную интер-
претацию реальных явлений и поэтому вносят слишком
много путаницы в сознание тренеров-практиков.
Аэробный характер бега в утяжеленных условиях,
если он выполняется правильно, должен носить, прежде
всего, «антигликолитическую» направленность [Верхо-
шанский Ю.В., 1988]. Так как, повышая окислительный
потенциал мышц и, вероятно, увеличивая силу ММВ, он
уменьшает тем самым вовлечение БМВ в работу при той
же мощности и, следовательно, снижает интенсивность
анаэробного гликолиза. Кроме того, при таком упражне-
нии отсутствует ишемия мышц (нарушение кровотока)
или гипобарическая гипоксия (если тренировка прово-
дится на равнине), следовательно, мышцы не испыты-
вают иной гипоксии кроме «функциональной», которая
является нормальным явлением и имеет место и в покое,
и при любой физической нагрузке. Критерием того что
тренировка проводится правильно, является способ-
ность спортсменов непрерывно поддерживать такой ре-
жим работы в течение часа и более, что будет совершен-
но невозможно при активизации некомпенсированного
анаэробного гликолиза.
В нашем эксперименте в период акцентированного
воздействия проводилось три таких тренировки в неде-
лю. В течение первого мезоцикла никаких положитель-
ных изменений обнаружено не было, что согласуется с
данными [Green H.J., 1996], где показано, что заметные
изменения в митохондриальном аппарате происходят,
начиная с 8-12-го дня регулярной тренировки на вы-
носливость. В нашем эксперименте резкий и хорошо
ощущаемый прирост всех показателей, включая сило-
вые, начался со второго мезоцикла. Отметим, что ни-
чего подобного не наблюдалось в контрольной группе,
275
несмотря на сопоставимые объемы силовой и аэробной
тренировки БМВ, из чего остается сделать вывод, что
большие объемы аэробного бега полностью «блокирова-
ли» эффект силовой тренировки (как, впрочем, и аэроб-
ной), что хорошо согласуется с данными других авторов
[Dudley G.A., R.Djamil, 1985]. При этом эффекта «тре-
нировки тренируемости» не наблюдалось, о чем говорит
отсутствие прироста показателей после снижения объе-
мов аэробной нагрузки.
3. При рациональном распределении и объемах нагруз-
ки возможен одновременный и существенный прирост
силовых и аэробных показателей с градиентом и до зна-
чений намного выше «среднего уровня» для данной ква-
лификационной группы, что также хорошо согласуется с
данными других авторов [Шенкман Б.С., 1990].
4. Сознательное использование минимальных объемов
«традиционных «средств аэробной подготовки (35-60%
от «обычных» [Травин Ю.Г., 1975]) нисколько не снизи-
ло ее эффективности.
7.6. Исследование влияния статодинамических
упражнений совместно с традиционными
методами подготовки бегунов на показатели
силы и аэробных способностей
Данные предыдущего исследования поставили вопрос о по-
иске более рациональных средств силовой подготовки бегунов,
чем интенсивные силовые и скоростно-силовые упражнения.
В соответствии с теоретическими положениями и данными
лабораторных исследований таковыми могли стать статодина-
мические упражнения, воздействующие преимущественно на
ММВ и создающие «анаболический» эффект в организме.
Для практической реализации принципов тренировки, на-
правленной на гипертрофию ММВ посредством статодина-
мических упражнений, был разработан комплекс, который с
небольшими модификациями использовался в течении 6 лет
(1985-1991 гг.) в серии педагогических экспериментов.
276
Комплекс включал:
1. Для мышц голени: медленные вставания на носке одной
ноги и опускания ниже горизонтали (нога стояла на воз-
вышении). Как правило, упражнение длилось 50-70 с и
состояло из 20-25 подъемов (т.е. один цикл занимал 2-3
с). Для подготовленных использовался вариант с пар-
тнером на плечах.
2. Для мышц — разгибателей ног: «пистолетик» — медлен-
ные приседания на одной ноге при неполном вставании
(10-15 повторений). Для слабоподготовленных бегунов
использовался вариант медленных приседаний на двух
ногах с неполным вставанием или опускания глубоко
вниз и поднимания с помощью рук, стоя одной ногой на
перекладине шведской стенки.
3. Для мышц задней поверхности бедра, ягодиц и, в неко-
торой степени, мышц спины: в положении упор сзади на
руках, с опорой пяткой одной ноги на возвышение 20-30
см — медленные поднимания и опускания таза и свобод-
ной ноги за счет полного разгибания тазобедренного и
небольшого сгибания коленного сустава (8-15 повторе-
ний). Для подготовленных иногда использовалось упраж-
нение «Себастьяна Коэ» — из положения стоя на коленях
пятки удерживает партнер — опускание в положение лежа
и поднимание (при необходимости — с помощью рук) за
счет усилий мышц задней поверхности бедра.
4. Для сгибателей тазобедренного сустава: 1) стоя сбоку от
барьера высотой от 91 до 106,7 см в зависимости от роста
и подготовленности — медленный пронос выпрямлен-
ной ноги над барьером справа налево и слева направо без
опускания ноги до вертикали; 2) медленное поднимание
прямых (иногда согнутых) ног к перекладине в висе на
шведской стенке или перекладине.
5. Для мышц спины, задней поверхности бедра и ягодич-
ных мышц. Лежа на верхней трети бедер на козле, коне
или тренажере «экстензор» медленные опускания туло-
вища вниз (до конца) и поднимания с прогибом в пояс-
нице (если И.П. — горизонтально) или без прогиба — до
выпрямления туловища (если И.П. — под углом 45° к
277
горизонтали). В последней трети траектории в фазе раз-
гибания — акцентировано напрягались мышцы задней
поверхности бедра для осуществления небольшого сги-
бания в коленных суставах.
6. Для мышц брюшного пресса — стандартное упражнение.
Из положения лежа на спине с ногами, согнутыми до 90°
в коленях — различные модификации подъема тулови-
ща, плеч и головы от пола.
При выполнении каждого упражнения, как и в других экс-
периментах, строго требовалось соблюдение методического
принципа: »Движения медленные, мышцы не расслабляются
весь подход, работаем «до боли». Указанные упражнения объ-
единялись в серию и выполнялись круговым методом с интер-
валом между упражнениями — 30-60 с. Отдых между сериями
— 3-5 мин, заполняемый, как правило, бегом трусцой и/или
«расслаблением» мышц в положении лежа на спине. Число се-
рий от 2 до 8 в зависимости от этапа («поддержание» или «раз-
витие»). Число занятий в неделю 2-3: на этапе «развития» — 2,
на этапе «поддержания» 1-3 укороченных, часто — в разминке
или заключительной части занятия.
Первый педагогический эксперимент был проведен С.М.
Обуховым (1991).
Цель.
Исследование одновременного применения статодинами-
ческих упражнений и аэробной тренировки для повышения
аэробных способностей быстрых мышечных волокон (БМВ)
в условиях реального педагогического процесса и изучения
влияния такой тренировки на АнП в беге.
Методика.
Из 35 обследованных спортсменов были сформированы 2
группы по 7 человек в каждой (5 девушек и 2 юношей). Кон-
трольная: 15,2±0,7 лет; 167,5±6,4 см; 54,2±6,1 кг; 2-3-й разря-
ды. Экспериментальная: 15,5±0,9лет; 165,9±5,9см; 53,6±5,1 кг;
2-3-й разряды. По антропометрическим показателям и пока-
зателям физической подготовленности группы статистически
не различались (при уровне значимости 0,05).
Распределение нагрузок в тренировке контрольной и экс-
периментальной групп представлено в табл. 11 и на рис. 25.
278
Таблица 11. Объемы основных тренировочных средств у эксперимен-
тальной и контрольной групп за время педагогического эксперимента
Акцент/объем Месяцы эксперимента
Янв. Февр. Март Апр. Май Июнь
Экспе- римен- тальная группа Сила ММВ (мин) 10 44 26 95 20 52
Аэро-БМВ (мин) 50 160 100 279 180 136
Аэро ММВ (км) 30 22 28 37 29 24
Гликол. бег (км) 0,9 1,1 1,2 1,4 1,7 3,0
Кон- троль- ная группа Ан-алакт. (км) 0,42 0,40 0,16 0,24 0,46 0,46
Смеш. (км) 6,2 3,8 6,3 7,0 4,8 4,6
Аэробн.бег(км) 49 45 82 84 52 57
Гликол. бег (км) 1,45 1,3 0,3 0,6 1,7 1,4
Так же, как и в предыдущем эксперименте, бегуны кон-
трольной группы тренировались по «традиционной» схеме с
использованием обычных средств. Наибольшее количество
гликолитической и алактатной работы планировалось на зим-
ний и летний соревновательные периоды. Наибольший объем
беговых средств в аэробной и смешанной зонах — на середину
подготовительного периода.
В экспериментальной группе в соответствии с целями экс-
перимента в середине подготовительного периода было запла-
нировано совместное существенное увеличение объемов ста-
тодинамических упражнений и средств повышения аэробных
способностей БМВ в виде интервального спринта (см. главу
2), а также в относительно небольшом объеме бега в гору и по
песку. Объемы аэробного бега и других средств подготовки в
течении весеннего подготовительного периода не изменялись.
В конце эксперимента, в соревновательном периоде, по на-
стоянию тренера было проведено несколько тренировок гли-
колитической направленности.
Типичный мезоцикл состоял из трех недельных микроци-
клов. Ниже приведено содержание основной части занятия по
дням мезоцикла.
279
Контрольная группа
Яна Февр Март Апр Май Июнь
kvMMimnd Ан - а л . х 10
ь j С м еш. х 10)
Аэ рОб
^^—Глик х 40
Экспериментальная группа
Рис. 25. Распределение объемов основных тренировочных средств у экс-
периментальной и контрольной групп за время педагогического экспери-
мента. Объемы тренировочных средств представлены в километрах и
умножены на коэффициенты для лучшей обозримости графиков. Объе-
мы тренировочных средств в экспериментальной группе указаны в мин
х тренировочного времени. К гликолитическим тренировкам отнесены
соревнования и «прикидки»
1-й день. Бег с соревновательной скоростью на отрезках
3-5 х 500 м.
2-й день. Спринтерский бег 5-10 х 150 м или интерваль-
ный спринт 15-20 раз (в течении 30-40 мин аэробного
бега).
3-й день. Статодинамический комплекс 4-6 серий по 4-6
упражнений по 40-60 с.
4-й день. Отдых.
5-й день. Бег в утяжеленных условиях 15-20 мин.
6-й день. Статодинамический комплекс 4-6 серий по 4-6
упражнений по 40-60 с.
7-й день. Отдых.
8-й день. Интервальный спринт 15-20 раз (в течение 30-40
мин аэробного бега).
280
Таблица 12. Изменение показателей бегунов контрольной группы за 6
месяцев эксперимента
Показатели Янв. Июнь Р
Макс. скор, (м/с) 7,79 7,83 >0,1
V АнП (м/с) 3,79 3,89 >0,1
ЧСС (3м/с) (уд/мин) 156 152 >0,1
Дл. шаг. АнП/дл. ноги 153 157 >0,1
N ср. з.п. (у.е.) 8,6 8,3 >0,1
Скор. 1500 м (м/с) 5,17 5,32 >0,1
Примечание. VАнП (м/с) — скорость на анаэробном пороге, ЧСС (3м/с)
(уд/мин) — ЧСС на стандартной скорости 3 м/с, Дл. шаг. АнП/дл. ноги
— относительная длина шага на пороговой скорости, N ср. з.п. (у.е.) —
средняя мощность мышц задней поверхности бедра (см. гл. 3), Скор. 1500
м (м/с) — скорость бега на 1500 м на контрольных тренировках и на со-
ревнованиях
9-й день. Аэробный бег 40-70 мин.
10-й день. Статодинамический комплекс 4-6 серий по 4-6
упражнений по 40-60 с.
11-й день. Отдых.
12-й день. Бег с соревновательной скоростью на отрезках
3-5 х 500 м.
13-й день. Аэробный бег 40-70 мин, фартлек или бег в утя-
желенных условиях 35-40 мин.
14-й день. Отдых.
15-21 -й — повторение дней 1 -7, в уменьшенном объеме.
В один из дней этого микроцикла проводилось тестирова-
ние по программе, описанной [Мякинченко, 1997].
Результаты.
Динамика показателей физических способностей бегунов экс-
периментальной группы представлена в табл. 18 и на рис. 26, 27.
Применение рекомендованных средств в эксперименталь-
ной группе началось с февраля. За это время результат вырос
с 4,99±0,37м/с до 5,29±0,18 м/с, т.е. на 6,0%, тогда как в кон-
281
трольной группе — на 2,9%. Различия между группами в вели-
чине приростов за это время достоверны (р < 0,05).
В экспериментальной группе с января по май — июнь про-
изошли достоверные изменения во всех показателях, кроме
средней мощности мышц задней поверхности бедра. В кон-
трольной группе изменения были недостоверны.
Особо обращает на себя внимание резкий прирост поро-
говых показателей (АнП, дл./ш. на АнП) и снижение ЧСС на
стандартной скорости между тестированиями в апреле и мае.
Напомним, что именно с конца марта по май было проведено
2 трехнедельных мезоцикла (см. выше) (один из них — полно-
стью на учебно-тренировочном сборе), где строго выполня-
Рис. 26. Изменение показателей, регистрируемых в рэмп-тесте и изме-
нение скорости бега на 1500 м по результатам ежемесячных контроль-
ных тренировок. Обозначения см. в табл. 12
282
лась спланированная нами схема подготовки, включающая
статодинамический комплекс и специальные беговые сред-
ства, условно обозначаемые как «тренировка аэробных спо-
собностей БМВ».
Частота шагов на скорости АнП изменялась недостоверно
и не имела определенной тенденции, т.е. прирост пороговой
скорости происходил исключительно за счет роста длины ша-
гов (предполагаем — за счет увеличения силы ММВ). Это под-
тверждается тем, что длина шага на стандартной скорости (3
м/с) изменилась достоверно с 116,6 см в январе до 119,5 см в
июне (р < 0,05).
Прирост аэробных способностей закономерно отразился в
самом большом приросте спортивного результата.
В мае применение статодинамического комплекса было
почти полностью приостановлено (до проведения майского
тестирования, показавшего его эффективность) по настоянию
тренера как «неспецифическое средство», которое не должно
применяться в соревновательном периоде. Вероятно, это одна
из причин снижения аэробных способностей в июне.
Вторая наиболее вероятная причина резкого ухудшения аэ-
робных показателей и состояния ССС — это применение в мае
— начале июня серии гликолитических тренировок, включая
участие в соревнованиях.
Показатели локальной (как мы предполагаем, — гликолити-
ческой) работоспособности мышц голени и задней поверхно-
сти бедра повышались экспоненциально с выходом на «плато»
к летнему соревновательному периоду. Некоторый ускорен-
ный рост показателей обнаружен после зимнего соревнова-
тельного периода (февраль), после завершения которого был
проведен «пробный блок» с применением статодинамического
комплекса. Применение гликолитических тренировок в июне
не отразилось положительно на этих показателях. Наоборот,
их значения стабилизировались.
Выводы.
1. Апробование разработанного статодинамического ком-
плекса выявило его полную «совместимость» с трениро-
вочным процессом бегунов на выносливость и психоло-
гическую приемлемость для спортсменов.
283
N средняя ТМГ
N средняя задней поверие бедра
Рис. 27. Изменение показателей, отражающих алактатные способно-
сти бегунов (максимальная скорость бега) и локальную работоспособ-
ность мышц голени (ТМГ) и задней поверхности бедра, регистрируемых
в тесте на тензоплатформе. Обозначения см. в табл. 13
2. Одновременное применение статодинамического ком-
плекса и средств «повышения аэробных способностей
БМВ» при минимальных объемах остальных средств аэ-
робной подготовки привело к резкому приросту аэроб-
ных показателей и улучшению состояния ССС.
3. Применение на предсоревновательном этапе нагрузок
гликолитической направленности в объёме 1,0-1,5 км
в неделю, вызывающих значительное и длительное по
времени закисление мышц, замедляет темпы прироста
аэробных возможностей бегунов. А дальнейшее увели-
чение объема упражнений анаэробно-гликолитической
направленности с 1,5 км/нед. до 3,0 км/нед. вызывает
снижение этих возможностей.
284
7.7. Исследование эффективности
последовательного применения силовых
и аэробных средств подготовки на показатели
физических способностей бегунов
В этом разделе будут описаны три из серии экспериментов,
проведенных в этом направлении, которые были выполнены в
1987-1990 гг. В.И. Паленым [1991].
Целью первого эксперимента было исследование влияния
сочетания этапа (7 недель) совместного применения средств
силовой (включая статодинамических комплекс) и аэробной
направленности с этапом (2 недели) концентрированного
применения больших объемов аэробного бега и эффективных
средств аэробной подготовки (бег в гору, темповой и интер-
вальный бег на местности и дорожке).
Методика.
В эксперименте приняли участие 8 квалифицированных
молодых бегуна на 800—5000 м (16,7±1,1 года; 61,7±4,3 кг;
174,6±3,3 см; 2-1-й сп. разряды, стаж не менее 3 лет). Длитель-
ность эксперимента составила 16 недель. Средние недельные
объемы по 3 этапам подготовки приведены в табл. 13 и на рис.
28. Спортсмены тренировались у своего тренера (В. Виногра-
дов) по плану, составленному совместно с нами.
Тестирование по стандартной программе проводилось в
конце 1 -го этапа (7-я неделя) и через три дня после окончания
3-го (17-я неделя).
Результаты.
Результаты представлены в табл. 14. Между исходным и заклю-
чительным тестированием получены достоверные сдвиги в показа-
телях физических способностей бегунов и спортивном результате,
кроме силы мышц сгибателей коленного сустава, где были получе-
ны также положительные изменения, но на уровне тенденции.
Обсуждение и выводы.
Первое тестирование было проведено после 7-недельного
втягивающего этапа, где использовались те же самые трени-
285
ровочные средства, реакция на изменение объемов которых
и служила предметом исследования. Следовательно, бегуны
были обучены и адаптированы к нагрузке.
Таблица 13. Объемы тренировочных средств в 3 этапах осенне-зимнего
подготовительного и предсоревновательного периода
Виды нагрузки Этапы
1-7-я неделя 8-14-я неделя 15-16-я неделя
Аэробн. бег (км) 58,2 64 134
Аэро-БМВ (км) 16,1 12,2 19,1
Скор-сил. (км) 1,1 3,8 1,2
Ст.-дин. (час) 0,95 1,38 0,53
Примечание. Аэробн. Бег (км) — все виды беговой работы со скоростью
до АэП. Аэро-БМВ (км) — эффективные средства аэробной подготовки
(бег в холм, с сопротивлением, темповой и интервальный бег, аэробный
спринт). Скор-сил. (км) — интенсивные прыжки, спринт, С БУ. Ст.-
дин. (мин) — статодинамические (квазиизотонические) упражнения и
близкие к ним силовые упражнения.
Таблица 14. Изменения показателей физических способностей в ходе 16-
недельного педагогического эксперимента
Тестиро- вание УАнП, м/с Ьш.АнП, м ЧСС(3,5 м/с), уд/мин Умакс., м/с Гсгиб/ вес Рез. 1000 м, мин, с
6 нед. 4,42 1,68 153 8,62 0,54 2,39,5
17 нед. 4,6 1,74 148 8,77 0,56 2,36,5
Р <0,01 <0,01 <0,01 <0,05 <0,1 <0,05
Примечание. Умакс — максимальная скорость бега; Тсгиб/вес — относи-
тельная сила мышц — сгибателей коленного сустава; Остальные обозна-
чения см. в табл. 13.
Дальнейший эксперимент позволил получить ожидаемый
результат — у бегунов одновременно возросли показатели
силы и скоростно-силовых способностей, а в результате всего
286
1н 2н Зн 4н 5н бн 7н 8н Эм Юн 11н 12н 13н 14н 15м 16н
| .... АэробН. 6«Г(КМ ) — А эро- БМ В (км ) X 3
Рис. 28. Динамика объемов тренировочных средств на 3 этапах осенне-
зимнего подготовительного и предсоревновательного периодов
двухнедельной акцентированной аэробной подготовки полу-
чено резкое улучшение АнП и ЧСС на стандартной скорости.
То есть — положительные сдвиги в оксидативных свойствах
мышц и состоянии ССС. Напомним, что это — наилучший
тип адаптации к тренировочной нагрузке в ЦВС, особенно на
фоне улучшения силовых показателей.
Данные этого эксперимента, проведенного на «втором
эшелоне» группы, позволили убедить тренера (В. Виноградо-
ва), воспитанного на «классической» концепции, в возмож-
ности применения предложенной нами схемы распределения
нагрузок для планирования всего годичного макроцикла и для
всей группы спортсменов (15 человек), большая часть которых
входила в то время в сборную Москвы и в резерв сборной Рос-
сии среди школьников и юниоров.
Второй эксперимент оказался направленным (вопреки наше-
му желанию!) на изучение реакции спортсменов на рекомендо-
ванное нами распределение нагрузки, но при объемах аэробных
средств подготовки (в особенности аэробного бега), превышаю-
щих не только разумные пределы, но и имеющиеся рекомендации
[Травин Ю.Г., 1975] для спортсменов этой возрастной категории,
287
которые были сделаны в рамках «классической» концепции под-
готовки бегунов [Набатникова М.Я., 1982].
Методика.
В эксперименте приняли участие 15 квалифицирован-
ных молодых бегунов на средние дистанции (17,2±1,2 года;
62,6±2,9 кг; 175,1±4,1см; 2-1 сп. разряды). Эксперимент охва-
тил годичный макроцикл с сентября по июнь и был разбит на
две части. В первой части спортсмены применяли нагрузки в
соответствии с нашим распределением, но в объемах, уста-
новленных тренером группы, придерживающегося правила,
что «путь к результату лежит через объемы». Во второй ча-
сти эксперимента нам удалось убедить тренера существенно
уменьшить объемы беговых нагрузок и подойти дифферен-
цировано к подготовке бегунов в соответствии с выявленны-
ми в ходе первой части эксперимента индивидуальными осо-
бенностями. Средства, применяемые бегунами, были те же,
что и в предыдущих экспериментах: к скоростно-силовым
упражнениям относили спринт, различные прыжки до 15-20
отталкиваний подряд, СБУ (главным образом — бег «коле-
со»), выполняемые активно, с акцентом на большую ампли-
туду движений; к силовым — статодинамический комплекс
(см. выше); к эффективным аэробным — бег в холм, с сопро-
тивлением, темповой и интервальный бег, аэробный спринт,
длительные прыжки (200-400 м).
Объемы и динамика нагрузок различной направленности
представлены на рис. 29.
Из представленных материалов следует, что в эксперименте
строго соблюдено требуемое распределение средств подготов-
ки. В осенне-зимнем и весенне-летнем макроциклах вначале
акцент был на силовую, а затем — на аэробную подготовку.
Результаты.
В первой части эксперимента, где использовались большие
беговые нагрузки, был получен ожидаемый результат: только
часть бегунов «выжила» и только у них наблюдалась положи-
тельная динамика состояния. Вторая часть бегунов или не из-
менила, или ухудшила показатели. В связи с этим вся группа
была разделена на две части: 1) успешная и 2) неуспешная. На
рис. 30 приведены изменения показателей в 2 группах. Обна-
288
Скор-ОИЛ^КМ) ———« Ст .-ДИН. (ЧЗО) |
|имшмшт Аэробн. бег(км) ——— Азро- БМВ (км) |
Рис. 29. Распределение среднемесячных объемов тренировочных средств
различной направленности в течение 10 месяцев 1-й и 2-й частей педа-
гогического эксперимента
ружено, что в 1-ю группу («успешных») вошли спортсмены,
которых характеризует два момента:
1. изначально это были относительно «медленные» спор-
тсмены, как следует из тестирования максимальной ско-
рости бега.
2. в то время как после этапа силовой подготовки аэробные
показатели выросли в обеих группах, только в «успеш-
ной» был прогресс в силовых показателях. Это следует из
динамики силы мышц — сгибателей к.с. (рис. 30) (кроме
этого теста в эксперименте измерялись не приводимые
здесь сила мышц — разгибателей к.с. и прыжок в длину с
места), а также роста скорости АнП за счет длины шагов,
при уменьшении их частоты.
289
Рис. 30. Динамика показателей физических способностей у бегунов
«успешной» (п=7) и «неуспешной» (п=8) групп в 1-й части эксперимента
После этапа объемной аэробной подготовки у бегунов
«успешной» группы обнаружен дальнейший прогресс почти
всех способностей, а «неуспешная» группа ухудшила все по-
казатели, включая спортивный результат к зимнему соревно-
вательному периоду (февраль).
Данные результаты убедили тренера скорректировать под-
готовку в соответствии с нашими рекомендациями в весенне-
летнем макроцикле.
Значения и динамика показателей в этот период представ-
лены на рис. 31.
290
Рис. 31. Динамика показателей физических способностей у бегунов всей
группы (п = 15) во 2-й части эксперимента
Как видно, в период с марта по июнь получена постоянная
положительная динамика всех показателей. Приросты до-
стоверны. Все без исключения участники эксперимента за 10
месяцев эксперимента улучшили личные рекорды на различ-
ных дистанциях от 800 до 5000 м на величину большую, чем в
среднем показывали спортсмены их возраста, учащиеся той же
школы-интерната спортивного профиля 9 г. Москвы (средние
значения приростов результатов выявлялись в специальном
лонгитудинальном исследовании в течении трех лет без наше-
го вмешательства в тренировочный процесс).
Обсуждение и выводы.
Так же, как и в предыдущем эксперименте, выявлена вы-
сокая эффективность такого распределения нагрузки, при
291
котором силовая тренировка основных мышц бегуна предше-
ствовала массированному аэробному воздействию. При этом
объем так называемых средств ОФП поддерживался на одном
уровне в течение всего макроцикла в объеме около 1 часа в не-
делю и на динамику силовых показателей не влиял. В соответ-
ствии с предположениями, сделанными в теоретической части
исследования, и результатами экспериментов, полученными
другими авторами, большие объемы аэробного бега могут бло-
кировать эффект силовой подготовки. Но этот эффект наблю-
дался не у всех спортсменов. 7 из них удалось выдержать вы-
сокие нагрузки. Их основной характеристикой были меньшие
скоростные показатели до эксперимента и больший прирост
силовых показателей по данным второго тестирования после
этапа силовой подготовки. Из этого можно сделать два основ-
ных вывода, согласующихся с данными ранее проведенных
исследований [Раппопорт Э.А., Казарян В.А., 1996], что высо-
кие беговые нагрузки и связанные с ними ацидотические яв-
ления в БМВ могут привести к преобладанию катаболических
реакций в мышечной системе, приводящих к снижению мы-
шечной силы и вместе с ней окислительного потенциала МВ.
Другим возможным фактором мог бы быть различный психо-
логический статус «быстрых» и «медленных» бегунов. Первые
хуже переносят монотонные нагрузки [Казлаускас В.А., 1982].
Однако фактом остается то, что при существенном сниже-
нии объемов низкоэффективных аэробных нагрузок во второй
части эксперимента при сохранении объемов статодинамиче-
ских упражнений был получен выраженный прирост как сило-
вых, так и аэробных показателей.
7.8. Заключение по главе
Проведенная серия педагогических экспериментов позво-
ляет сделать следующие выводы.
1. Во всех экспериментах обнаружено существенное по-
вышение эффективности силовой подготовки. Одна-
ко принципиальным моментом является то, что ни в
одном случае прирост не был обусловлен повышением
292
объема нагрузки. Количество «чистого» тренировочного
времени, включая отдых между подходами на этапе ак-
центированного воздействия на силовые способности,
никогда не превышало 80-100 мин в неделю, или 5-6 ч
в месяц. На этапах поддержания способности — 30-50
мин. Если даже к этому прибавить время, направленное
на спринтерскую беговую подготовку (спринт 30-150 м),
т.е. еще около 40-60 мин в неделю, то даже в этом случае
затраты времени будут меньше (с учетом разведения ак-
центов этих средств в различные этапы макроцикла) или
на уровне, рекомендуемом в литературе [Макаров А.Н.,
1973; Травин Ю.Г., 1975] бегунам на длинные или тем
более на средние дистанции. Повышение эффективно-
сти достигнуто, как мы предполагаем, за счет сознатель-
ного соблюдения следующих методических принципов
организации силовой подготовки:
Выполнение подхода «до отказа», т.е. до невозможности
продолжать упражнение, сильного локального утом-
ления или заметного снижения мощности работы. Это
соблюдалось как при применении скоростно-силовых
(прыжки, спринт в гору и т.п.), так и статодинамических
упражнений. При формулировании этого принципа мы
ориентировались на хорошо известное методическое
требование, что для эффективной силовой тренировки
надо: «Выполнить подход «до отказа», а затем,... еще два
раза!» [Ягодин В.М., лекция для студентов ИФК (кон-
спект автора)], а также на известные представления о
том, что в основе повышения силы мышц могут лежать
или совершенствование нервно-мышечных механизмов
(повышение частоты импульсации а-мотонейронов,
синхронизация ДЕ, улучшение техники), или морфо-
логические перестройки мышц (гиперплазия органелл),
или, чаще, оба механизма. Для совершенствования пер-
вого механизма требуется высокая интенсивность или
околопредельная степень напряжения мышц, что, види-
мо, имеет меньшую значимость в ЦВС. Для накопления
миофиламентов и обеспечивающих их органелл МВ (т.е.
гипертрофии мышц, что и является «морфологической
293
основой выносливости»), как установлено ранее (глава
5), необходимо: активное состояние мышцы, «дефицит
энергии» — исчерпание фосфагенов, накопление ме-
таболитов. Эти условия достигаются в мышце только
перед «отказом». Поэтому: «... подход не доведенный
до «отказа» — впустую затраченное время и силы!». Эта
мысль всячески внушалась тренерам и спортсменам, и
если для силовых видов спорта это является аксиомой,
то в циклических видах спорта это часто было «полным
откровением».
— На этапе повышения силовых способностей — недопу-
щение «комплексных занятий». То есть использование
в занятии нагрузки только одной направленности — или
«на силу», или «на выносливость». Это ни в коем случае
не предполагало использование длительных, монотон-
ных тренировок, особенно у молодых спортсменов. На
этапах поддержания силовых способностей — одна тре-
нировка также была только силовой направленности, но
допускалось применение 1 -3 статодинамических серий в
разминке или, чаще, в заключительной части занятия.
— После силовой тренировки планировался день отды-
ха для полноценного протекания процессов белкового
синтеза. На этапе поддержания допускалось занятие лю-
бой направленности, но только низкой энергетической
стоимости.
— Сочетание силовых тренировок и тренировок с исполь-
зованием бега в утяжеленных условиях и интервального
спринта.
— Основным условием, на наш взгляд, является суще-
ственное сокращение объемов неэффективного «аэроб-
ного бега» на 30-50% от рекомендуемых объемов.
2. Использование в течение 6-8 недель эффективных
средств аэробной подготовки во второй половине под-
готовительного периода неизменно приводило к бы-
строму приросту аэробных показателей с их выходом на
«плато». Величина прироста, как правило, существенно
превышала эффективность «объемного периода аэроб-
ной подготовки» в контрольных группах при меньшем
294
времени, затрачиваемом на совершенствование этой
способности. При этом строго отслеживалось соблюде-
ние принципа, что «эффективные средства — это не то
же самое, что интенсивные средства!». Физиологически
говорить об интенсификация тренировочного процесса
можно тогда, когда имеет место большое напряжение
симпато-адреналовой системы, одновременно достига-
ется высокая концентрация «стресс-гормонов» в кро-
ви (катехоламинов и глюкокортикоидов), исчерпание
углеводных запасов мышц и печени, мобилизация пла-
стических ресурсов организма, повреждение органелл
мышечных волокон механическими и химическими
факторами, которые всегда присутствуют в тренировке
бегунов и т.п. Такого состояния с равной вероятностью
можно достичь при длительном беге относительно уме-
ренной интенсивности (например, при преодолении
марафона) или короткой «гликолитической трениров-
кой». Но ни в первом, ни во втором случае достижение
состояния стресса совсем не означает, что будет получен
высокий тренировочный эффект: в первом случае — в
отношении аэробных способностей, а во втором — гли-
колитических. Другими словами, оба варианта «плохие»
и не отвечают целям спортивной тренировки, особенно
на подготовительном и предсоревновательной этапах.
3. Использование гликолитических тренировок всегда
приводило к снижению аэробных показателей, но да-
вало прирост в спортивном результате в беге на средние
дистанции. Проследить, насколько длительным и устой-
чивым был это прирост, нам не удалось. В то же время
показано, что использование гликолитической подго-
товки только в форме участия в серии соревнований (на
средних дистанциях), при сохранении аэробно-силовой
направленности подготовки в соревновательном перио-
де, обеспечивает длительное (2-3 месяца) сохранение
«спортивной формы».
4. Распределение акцентов основных средств подготовки в
последовательности: «соединительно-тканные элементы
ОДА» => сила мышц => аэробная подготовка => интегри-
295
рующая подготовка (совмещенная или не совмещенная
с гликолитической) => этап соревнований, как правило,
— приводило к существенно большему приросту резуль-
татов в годичном цикле, чем «традиционная» при усло-
вии «успешности» этапа силовой подготовки. Если этап
силовой подготовки по каким-то причинам оказывался
неудачным для спортсмена (отсутствовал прирост сило-
вых показателей), то компенсировать этот пробел в под-
готовке другими средствами уже не удавалось и прироста
спортивного результата в этом макроцикле не наблюда-
лось.
5. Одним из основных доводов в пользу объемной аэробной
подготовки в начале подготовительного периода счита-
ется то, что при таких нагрузках улучшаются локальные и
региональные сосудистые реакции, приводящие, в част-
ности, к уменьшению периферического сопротивления
[Musch T.J. и др., 1987] и обеспечивающие этим рацио-
нальную адаптацию миокарда к более интенсивной по-
следующей работе. И наоборот, «жесткая» интенсивная
тренировка на выносливость приводит к увеличению
периферического сосудистого сопротивления, стимули-
руя нерациональный тип гипертрофии миокарда [Дембо
А.Г., 1991]. Поэтому мы считаем очень важным наблю-
дение, которое было сделано в процессе проведения пе-
дагогических экспериментов, согласно которому дости-
жение приростов показателей аэробного и анаэробного
порогов в результате акцентированной аэробной под-
готовки или ее сочетания с применением статодинами-
ческого комплекса всегда сопровождалось существенным
понижением значений ЧССна стандартной скорости, что
обеспечивается, как показано в экспериментах с трени-
ровкой 1 ноги [Saltin В., 1985], в значительной степени
периферическими факторами. Это означает, что такой
вид тренировки, наряду с улучшением показателей ло-
кальной выносливости, обеспечивает положительные
изменения на всех уровнях системы транспорта кисло-
рода к мышцам. Например, одновременно мог возрасти
парасимпатический драйв и снизиться хронотропное
296
влияние симпатической системы, произойти делятация
полостей сердца, улучшиться сократимость миокарда,
региональные и локальные сосудистые реакции, про-
изойти пролиферация капилляров. Другими словами,
можно утверждать, что при реализации разработанных
нами принципов построения тренировки не проис-
ходит излишней интенсификации или форсирования
подготовки. Кроме того, видимо, такое построение тре-
нировочного процесса позволяет достичь именно тех
эффектов, которые, как считается, достигаются только
в результате применения этапа объемной аэробной под-
готовки с применением «низкоинтенсивных» беговых
средств (как аэробной базы для применения более «ин-
тенсивных» средств). Однако традиционный «аэробный
этап» приводит к потере силы мышц, тогда как мы неиз-
менно получали одновременный прирост силовых и аэроб-
ных показателей.
297
Глава 8
Практические аспекты развития
локальной выносливости
Концепции построения тренировочного процесса форми-
руются, как правило, на основе изучения опыта подготовки
выдающихся спортсменов и методических взглядов их трене-
ров. На этой основе выявляются некоторые закономерности,
которые в дальнейшем проверяются в ходе педагогических
экспериментов, и только после этого рекомендуются для вне-
дрения в практику подготовки спортсменов. Таким образом,
собирается эмпирический материал, на основе которого могут
быть сформулированы общие принципы тренировки и опре-
делены способы их реализации применительно к конкретным
видам спорта, т.е. определяется ответ на вопрос: «Кактрениро-
вать?». Однако остается без ответа вопрос: «Что тренировать?
И почему это надо тренировать?»
Описанный процесс хорошо изучен в логике и методоло-
гии научного исследования (см. гл. 1) и характерен для так на-
зываемого эмпирического этапа развития науки, который хотя
и является необходимым, но тем не менее остается начальным
уровнем с присущими ему ошибками и неоптимальными пу-
тями решения практических задач.
Продемонстрировать сказанное можно следующим приме-
ром. С 1952 по 1980 гг. рост среднегодовых объемов беговой
работы на всех дистанциях от 800 м до марафона у сильнейших
советских бегунов положительно коррелировал с приростами
результатов на уровне 0,83-0,99. Эти данные представлены в
книге группы наиболее авторитетных в СССР специалистов
по бегу на выносливость [Суслов Ф.П. и др., 1982]. Кроме
того, выявленная таким образом закономерность полностью
совпадает с многолетним опытом ведущих советских тренеров
298
(у которых эти бегуны, естественно, и тренировались!), а так-
же с данными многочисленных исследований [Журбина А.Д.,
1976; Казлаускас В.А., 1982; Кароблис П.Б., 1986; Макаров
А.Н., 1973; Набатникова М.Я., 1975; Нурмякиви А.А., 1974;
Набатникова М.Я., 1982; Платонов В.Н., 1984; Полунин А.И.,
Снесарев Н.К., 1990; Попов Ю.А., 1968; Суслов Ф.П. и др.,
1982; Травин Ю.Г., 1975], выполненных опять-таки на тех же
самых ведущих бегунах, когда они сравнивались с менее под-
готовленными спортсменами, тренирующимися опять-таки у
тех же самых тренеров. Таким образом, имеем «полное совпа-
дение данных научных исследований, практики спорта и мне-
ний ведущих тренеров и специалистов».
Более чем убедительно, не правда ли?!
Однако позволим себе посмотреть на те же данные с не-
сколько иных позиций.
1500м. □5000-10000
Рис. 32. Динамика общих объемов (км) беговых нагрузок у ведущих совет-
ских бегунов в 1952— 1980гг. [по Суслову Ф.П. и др., 1982]
Наибольших успехов на международной арене советские
бегуны достигли в 50-е — начале 60-х гг. и связаны они с име-
нами великих В. Куца и П. Болотникова, а также плеядой бе-
гунов: А. Ануфриев, Ф. Ванин, Н. Попов, В. Казанцев, И. Се-
менов. Именно в этот период с учетом того, что 1100-1400 км
годового объема в 1952 году выглядит как курьез, отчетливой
положительной динамики в приросте объемов не наблюдается.
Кроме этого, 3500-4000 км в год для стайера мирового уровня
299
— весьма умеренны по сравнению с объемами в 4000-6500 км в
год, на который вышли выдающиеся бегуны еще в 20-40-х го-
дах (П. Нурми, Г. Хег, Э. Затопек) [Кайтамазова Е.Н., Теннов
В.П., 1974; Полунин А.И., Снесарев Н.К., 1990 ].
Таким образом, успехи советских бегунов в исторической
перспективе связаны с чем угодно, но только не с наращивани-
ем объемов бега!
В то же время, начиная с 1964 по 1980 гг. и тем более по на-
стоящее время признаки «заката» советской школы бега более
чем очевидны, несмотря на все увеличивающиеся объемы бега
(рис. 32). Таким образом, объединять эти два противополож-
ные по своей сути исторических периода в российском беге —
не корректно. Для большей доказательности этой мысли мож-
но было бы привести, например, данные корреляционного
анализа между величиной отставания 10 лучших советских бе-
гунов от 10 лучших в мире в период с 1956г. по настоящий день
и годовыми объемами бега наших лучших бегунов. Даже если
исключить из этой выборки чернокожих бегунов, то очевидно,
что коэффициент корреляции будет достоверным и очень вы-
соким. То есть — чем больше годовые объемы у российских бегу-
нов, тем выше их отставание на мировой арене!!!
Вывод — надо немедленно снижать объемы бега!
Заметьте, полностью противоположный вывод был сделан
из анализа тех же самых данных только потому, что нам захо-
телось посмотреть на них с другой позиции. Естественно, что
можно найти и другие точки зрения с получением выводов, от-
личных от первых двух!
К сожалению, мы не имеем данных для статистического
подтверждения нашего вывода, да и нет никакого смысла тра-
тить время на поиск таких данных, т.к. доказательность такой
корреляции будет не выше, чем в критикуемом нами примере.
Тем не менее приведем, с любезного разрешения В.Н. Ку-
лакова, его уникальные данные о годовых объемах лучших со-
ветских бегунов в период 1970-1991гг. Однако вместе с коэф-
фициентами корреляции между парциальными объемами бега
в различных зонах и спортивными результатами, рассчитан-
ными по средней скорости соревновательного бега на 5000 и
10 000 м (табл. 15).
300
Таблица 15. Объемы бега в различных зонах интенсивности, спортивные
результаты лучших стайеров СССР и коэффициенты корреляции меж-
ду средней скоростью соревновательного бега и парциальными объемами
нагрузки [Кулаков В.Н., 1995]
Годы, когда обрабатывались данные
Нагрузка R 70-73гг. 75-76гг. 77-79гг. 82-85гг. 88-91ГГ.
Общ. объем бега 0,71 6121 6657 6947 6512 6382
1-я зона 0,16 1230 1691 1723 1823 2259
2-я зона 0,45 3673 3801 3828 3575 3453
3-я зона 0,28 942 852 972 697 364
4-5-я зоны 0,91 276 313 424 417 306
Результат
5000 м 13,38,6 13,31,5 13,29,1 13,23,8 13,38,0
10000 м 28,22 28,15 28,00 28,16 28,39
Средняя скорость 5,98 6,03 6,07 6,06 5,96
Как следует из таблицы, единственный достоверный ко-
эффициент корреляции (п=5) отражает взаимосвязь между
спортивным результатом и объемами бега в 4-5-й зонах, т.е., по
существу, с объемом спринтерской и скоростно-силовой подго-
товки, дефицит которой вместе с неверно построенной силовой
подготовкой и лежит, как можно предположить из приведенных
данных, в основе неудач советских и российских бегунов.
Таким образом, чтобы не следовать традиционной ошибке
— использовать коэффициенты корреляции в качестве пред-
посылки для выдвижения гипотезы или доказательства ее
обоснованности, при планировании исследования мы много
внимания уделяли теоретическому обоснованию выдвинутых
гипотез о средствах, методах и построении тренировочного
процесса с целью улучшения локальной выносливости (главы
5, 6). Затем сделанные предположения были подтверждены в
серии лабораторных и естественно-педагогических экспери-
ментов, выполненных нами, нашими сотрудниками и аспи-
рантами, а также путем привлечения данных экспериментов
других авторов (глава 7).
301
В ходе этого исследования было выявлено, что при фор-
мальном совпадении основной концепции подготовки спор-
тсменов, выражающейся, например, в известных положе-
ниях, что общая физическая подготовка является базой для
совершенствования специальных способностей [Матвеев
Л.П., 1977], аэробная подготовка является основой для дости-
жения высокого спортивного результата, а «специальная под-
готовка» (под которой часто подразумевают гликолитические
способности на средних дистанциях) должна проводиться по-
сле достижения максимально высокого уровня аэробной про-
изводительности [Волков Н.И., 1969, Зациорский В.М., 1966]
и т.п., были обнаружены явные расхождения в том, каким
способом реализовать отмеченные выше фундаментальные
положения теории спорта, в частности в беговых дисципли-
нах. Например, построение макроцикла в экспериментальных
группах, чья подготовка планировалась на основе теоретиче-
ских положений, значительно отличалась от планирования в
контрольных группах. Столь явное расхождение между «тео-
рией» и предыдущей «теорией», подкрепленной к тому же
многолетней «практикой», вызвало необходимость проведе-
ния еще одного исследования, направленного на выяснение,
действительно ли планирование тренировочного процесса в
контрольных группах отражает «традиционный» вариант под-
готовки бегунов.
8.1. Возможные варианты коррекции системы
подготовки бегунов на выносливость
Для построения схемы «традиционного» макроцикла
(рис. 36) были использованы материалы всех монографий и
учебников для вузов, изданных в СССР за период 1976-1990
гг., посвященные бегу на средние и длинные дистанции,
опубликованные материалы подготовки сборной коман-
ды СССР; конкретные тренировочные планы ведущих со-
ветских бегунов по материалам журнала «Легкая атлетика»
и других изданий за последние 10 лет; данные нескольких
диссертационных работ и т.д.
302
При изучении этого вопроса по литературным источникам
мы столкнулись с серьезными трудностями терминологическо-
го характера. Наиболее часто используемыми терминами при-
менительно к тренировке бегунов являются: специальная вы-
носливость, общая выносливость, скоростная выносливость,
упражнения на скорость, специальная скорость, скоростно-
силовые упражнения, силовые упражнения, специальная
сила, силовая выносливость, локальная выносливость, об-
щая физическая подготовка (подготовленность), специаль-
ная физическая подготовка (подготовленность) и т.д. Имеет-
ся множество и классификаций одних и тех же упражнений.
К сожалению, в доступных нам учебных пособиях по теории
спортивной тренировки не содержится четкого определения
объема и содержания каждого из этих понятий. Поэтому в тех
случаях, когда авторы четко не описывали конкретные упраж-
нения и их дозировку, было чрезвычайно сложно разобраться,
что подразумевается под отдельными словами, и тем более по-
нять, какой физиологический фактор определяет спортивный
результат, является объектом тренировки в том или ином слу-
чае. Предположим, например, что бегун на 1500 м тренирует-
ся на отрезках 300-400 м по дорожке стадиона со скоростью,
равной соревновательной. В первом случае бег выполняется
повторно с интервалом отдыха до полного восстановления, во
втором случае — переменно с интервалами 3-4 мин, в третьем
случае — переменно с интервалами 1,5-2 мин. Соответственно
в первом случае можно предположить, что тренировка будет
иметь скорее технический характер, во втором будут интен-
сивно функционировать все аэробные функции организма, но
скорость анаэробного гликолиза будет низкой, а в третьем —
тренировка будет иметь явно выраженную гликолитическую
направленность. Следует ли относить все три вида тренировок
к разряду специальной физической подготовки, если ведущая
для бегунов функция организма — аэробная — будет разви-
ваться только во втором случае? Или следует ли относить бег в
холм к разряду общей или специальной подготовки, если при
этом развивается ведущая функция бегуна — аэробная, причем
очень эффективно, но бег выполняется на сильнопересечен-
ной местности? Точно так же неясны основания объединения
303
силовой тренировки и медленного продолжительного бега в
группу средств ОФП, если в первом случае накапливаются со-
кратительные белки мышц, а во втором — белки митохондрий.
И те, и другие совершенно необходимы для достижения ре-
зультата международного уровня, если, конечно, тренировка
выполняется в развивающем режиме. Если же эти средства ис-
пользуются для активного отдыха или тренируются неоснов-
ные мышцы, тогда это должно было бы быть специально ого-
ворено и т.д.
Для избежания разночтений и терминологической пута-
ницы в изложении последующего материала мы пытались
раскрывать контекст высказываний авторов (когда это было
возможно) в соответствии с классификацией упражнений,
предложенной нами ранее, где в основу классификации упраж-
нений положены конкретные морфологические белковые суб-
станции организма, существенные для спортивного результата
в беге на выносливость и имеющие отношение к развитию ло-
кальной выносливости, а именно:
1. Соединительно-тканные образования ОДА.
2. Сократительные элементы (миофиламенты) ММВ и
БоМВ, и обслуживающие их органеллы.
3. То же, но для БМВ.
4. Двигательная зона коры больших полушарий и мото-
нейронный пул спинного мозга.
5. Митохондрии и капилляры (окислительный потенциал
ОП) ММВ.
6. То же для БоМВ.
7. То же для БгМВ.
8. Ферменты гликолиза и буферная емкость.
Примерное соответствие между нашей классификацией и
традиционным описанием тренировочных средств и методов в
легкой атлетике представлено в табл. 10 (глава 7).
При этом учитывалось время воздействия на данную мор-
фоструктуру, которое и является мерой тренирующего потен-
циала данного упражнения.
Эта классификация позволила нам привести к единому зна-
менателю самые разнообразные средства и методы и сравнить
имеющийся в литературе материал с нашей схемой планиро-
304
вания макроцикла. За основу сравнения был взят полугодич-
ный осенне-зимний макроцикл с выступлениями на зимних
соревнованиях в феврале — марте.
Полученная описанным выше способном схема для бегу-
нов, предположим, на 1500-5000 м, представлена на рис. 33
вместе со схемой, теоретически разработанной нами, про-
веренной в серии педагогических экспериментов и адапти-
рованная под высококвалифицированных спортсменов (т.к.
часть экспериментов была проведена нами с участием юных и
молодых спортсменов).
Данные рис. 31, на котором представлены значения трени-
ровочных нагрузок в соответствии с рекомендациями совет-
ской и российской учебно-методической литературы, позво-
ляют констатировать, что обнаруженное нами распределение
тренировочных средств в контрольных группах педагогических
экспериментов (глава 7) соответствуют в общих чертах обще-
принятым рекомендациям. Таким образом, эксперименталь-
ное сравнение двух схем, рекомендуемой нами и «традицион-
ной», можно считать корректным, по крайней мере в рамках
исследуемой выборки испытуемых.
Основными чертами «традиционной» схемы в сравнении с
рекомендуемой являются:
1. 1,5-2-кратное превалирование дистанционных средств,
воздействующих главным образом на окислительный
потенциал ММВ и при большой длительности трени-
ровочного занятия на БоМВ, а также повышающие
степень утилизации жиров мышцами (на допороговой
скорости). В силу высокой энергетической стоимости и
слабого задействования БМВ такие средства или не спо-
собствуют или даже снижают силу всех типов МВ вместе
с силовыми показателями.
2. Пик объемов беговых нагрузок аэробной направленно-
сти наблюдается в ноябре, т.е. за 3 месяца до периода со-
ревнований. В нашей схеме максимальные объемы «эф-
фективных» средств (так как о других мы не говорим)
практически примыкают к соревновательному периоду.
3. При традиционном планировании в конце подготови-
тельного и на предсоревновательном этапе существенно
305
Рекомендуется схема
“Традиционная” схема (по
материалам учебно-мет одической
литературы).
Рис. 33. Рекомендуемая и «традиционная» схемы распределения нагрузок
различной направленности для квалифицированных бегунов на 1500-5000
м. Обозначения см. в табл. 10.
возрастает скорость кроссового бега, бега по дорожке,
«гликолитического» бега и объем бега в утяжеленных
условиях. По этому показателю две схемы похожи.
4. Объем средств укрепления соединительно-тканных
элементов ОДА (прыжки, СБУ, спринтерский бег),
применяемые для профилактики травматизма, в «тра-
диционной» схеме оказывается наибольшим в предсо-
ревновательном и соревновательном периодах. Отме-
тим, что при неаккуратном применении этих средств,
тем более в сочетании с большими объемами «жестко-
го» аэробного и гликолитического бега, повышается
риск травматизма. В нашем случае прыжковые и другие
скоростно-силовые упражнения, вместе со спортивны-
306
ми играми и стретчем планируются в переходный пери-
од, а спринт (для средневиков) — в предсоревнователь-
ный и соревновательный, но в ограниченном объеме.
5. Силовая подготовка в первом случае планируется в до-
статочно большом объеме с двумя пиками — в начале
макроцикла и в предсоревновательный период. В нашей
схеме — на конец переходного и начало подготовитель-
ного.
6. «Интересы» технической подготовки соблюдаются кру-
глогодично. Мы рассматриваем два аспекта: 1) научение,
посредством которого вырабатываются навыки правиль-
ной постановки ноги на дорожку, работа рук и положе-
ние туловища; 2) формирование, посредством которого
обеспечивается правильная работа мышц ног при оттал-
кивании и «складывании» ноги в фазе переноса. Второй
аспект реализуется методом сопряженного воздействия.
Мы рекомендуем использовать бег с соревновательной
скоростью по дорожке в туфлях с шипами не реже 1 раза
в неделю (объем 3-5 км в зависимости от дистанции) на
протяжении всего макроцикла, начиная с переходного
периода.
При разработке «рекомендуемой» схемы во главу угла ста-
вилось развитие локальной выносливости и неразрывно с ней
связанное формирование рациональной техники бега, точнее —
биологические закономерности, на которые следовало бы опи-
раться при построении тренировочного процесса такой направ-
ленности. При этом мы сознательно опускали «интересы» так
называемых «центральных» систем организма и другие аспекты
подготовки (психологический, тактический, внетренировоч-
ных факторов, фармакологической поддержки, питания и т.п.).
В настоящее время мы не можем сказать, насколько такой под-
ход правомерен и универсален, какие он имеет ограничения и
в какую сторону его следует развивать в дальнейшем примени-
тельно к построению целостной системы подготовки бегунов.
Тем не менее попробуем приблизить описанные выше схемы к
реальной практике тренировки бегунов.
Нижеописанные рекомендации в своей сути справедливы
для всех дистанций от 400 м до марафона, однако при прак-
307
тическом применении должна осуществляться коррекция на
спортивную специализацию, уровень подготовленности и стаж
занятий, возрастные, психические, психо-физиологические,
анатомо-морфологические особенности отдельных спортсме-
нов или их групп.
Основными правилами при планировании физической
подготовки в годичном цикле, как предположено ранее, долж-
ны быть следующие:
1. вначале целесообразно накапливать морфоструктуры,
которые занимают «базовое» положение, а уже на этой
основе те, которые непосредственно обеспечивают мак-
симально возможный для данного уровня «базы» спор-
тивный результат;
2. акцентированное воздействие на один вид морфострук-
тур на определенном этапе обязательно подразумевает
поддержание на достигнутом уровне количества дру-
гих. Так как поддержание требует значительно меньше
пластических и энергетических ресурсов, чем процесс
«потеря-приобретение» [Меерсон Ф.З., 1978; Пшенни-
кова М.Г., 1986; обзор Платонов В.Н., 1984];
3. интегрирующая тренировка проводится в предсорев-
новательный период, основное средство этого раздела
подготовки — бег с соревновательной скоростью по до-
рожке.
Логика «рекомендуемой» схемы, представленной на рис. 36.
1. Переходный период (условно - сентябрь).
Цель — укрепление ОДА. Средства: прыжки во всех ва-
риантах, спринт, спортивные игры, упражнения по системе
стретчинга (эта система отличается от традиционной системы
упражнений для развития гибкости). Прыжковые и спринтер-
ские упражнения к концу периода в случае если тщательно
проводится профилактика травматизма, могут выполняться с
предельной интенсивностью, что, предположительно, повы-
шает мощность моторных центров головного и спинного моз-
га, что положительно скажется на эффективности последую-
щей силовой подготовки. Низкая энергетическая стоимость
должна способствовать восстановлению психики и ресурсов
гормональной системы после предыдущего соревнователь-
308
ного периода. Аэробные способности мышц поддерживают-
ся небольшими объемами фартлека и кроссов, проводимых в
лесу, по сильнопересеченной местности. Эти же средства ис-
пользуются с целью отдыха, расслабления.
2. 1-й этап подготовительного периода (конец сентября,
октябрь, первая половина ноября).
Цель — силовая подготовка, т.е. накопление сократитель-
ных элементов ММВ, БоМВ и БгМВ в соотношении, диктуе-
мом специализацией: все мышечные волокна — 400 и 800 м,
ММВ и БоМВ — 1500-10 000 м, только ММВ — марафон.
Средства: сила БоМВ и БгМВ повышается при любом виде
традиционной силовой подготовки и при спринтерском беге
в гору. Сила ММВ — только при статодинамических упражне-
ниях, а также беге и прыжках в гору, с сопротивлением, по сы-
пучему грунту. Силовая подготовка бегунов на выносливость
имеет смысл только в отношении основных мышечных групп:
мышц голени, задней поверхности бедра, мышц — сгибате-
лей тазобедренного сустава, ягодичных мышц. Поддержание
силы остальных мышечных групп, разумеется, необходимо, но
это относится к средствам ОФП и здесь не рассматривается.
Также важно отметить, что применение любых видов силовых
упражнений связано с накоплением ионов водорода в мыш-
цах в высоких концентрациях, а это приводит к деструкции
(гибели) митохондрий, особенно в БоМВ и БгМВ. Если это
не учитывать, то на восстановление аэробного потенциала по-
надобиться несколько месяцев, за которые будет потеряна на-
копленная сила мышц. Поэтому поддерживающую аэробную
тренировку следует проводить с использованием упражнений,
повышающих ОП всех мышечных волокон, а не только ММВ.
На этом этапе целесообразны прыжки или беговые упражне-
ния, выполняемые на отрезках 300-800 м в холм достаточной
крутизны (8-15°) или в любых других утяжеленных условиях.
Могут использоваться интервальный спринт или переменный
бег на сильнопересеченной местности. Непременное требова-
ние к этому виду тренировки — аэробные условия выполнения
упражнений, т.е. отсутствие накопления лактата в крови. Объ-
ем беговой нагрузки — не более 4 часов в неделю чистого бего-
вого времени.
309
Примерный недельный микроцикл (рис. 34) может содер-
жать в себе: две чисто силовые тренировки на основные мы-
шечные группы бегуна. Это — основной вид подготовки на 2-м
этапе, поэтому полноценному проведению этого вида работы
подчиняется вся организация микроцикла. Одна трениров-
ка должна быть чисто силовой (например, по круговому ме-
тоду — статодинамический комплекс). Длительность — 50-70
мин основной работы. Вторая может быть комбинированной
— первая половина спринтерско-прыжковая, вторая — ста-
тодинамическая. После силовых тренировок в течении 24-36
часов никакой нагрузочной аэробной работы не допускается.
Поддержанию аэробных способностей может быть отведено
3-4 тренировки, включая «техническую тренировку» с бегом
по дорожке и длинный кросс по песку и сильнопересеченной
местности. Средства: бег в утяжеленных условиях (непрерыв-
но — 40-50 мин); интервальный спринт( 20-30 ускорений); бег
на уровне анаэробного порога или немного выше (8-15 км);
интервальный бег на средних отрезках (400-800 м); повторный
или переменный бег на длинных отрезках (600 -2000 м). Объем
бега на «быстрых участках» за тренировку 5-8 км. Одна трени-
ровка отводится длительному восстановительному кроссу (15-
25 км) в аэробной зоне. Один день (желательно после силовой
тренировки) — полный отдых. Из 2-3 дополнительных занятий
одно посвящается бегу на технику с соревновательной скоро-
стью в шиповках по дорожке (3-5 км), одно или два — сприн-
ту на отрезках 80-200 м или ритмовым пробежкам на технику
(для стайеров) и упражнениям ОФП на неосновные мышеч-
ные группы. Недельный объем всех видов бега и прыжков в
нагрузочном микроцикле для средневиков — 40-60 км, стайе-
ров — 50-70 км, т.е. общая энергетическая «стоимость» этапа
невысокая. Это необходимо, чтобы не «блокировать» эффект
силовой тренировки.
3. 2-й этап подготовительного периода (вторая половина ноя-
бря, декабрь).
Цель — на базе повышенной силовой подготовленности до-
биться максимально высокого ОП ММВ, БоМВ и БгМВ. Этап
условно делится на две части. Первая часть (4-6 недель) посвя-
щена массированному воздействию на все типы мышечных
310
волокон. В это же время происходит эффективное воздействие
на ССС и дыхательные мышцы. Вторая часть 3-го этапа имеет
основной задачей достижение максимума аэробной мощности
мышц, но с соблюдением интересов технической подготовки.
Это достигается применением предельнопереносимых объе-
мов бега во всех режимах (за исключением восстановительно-
го). Непременное условие — поддержание уровня силы и ре-
шение задач технической подготовки. Длительность 2-й части
этапа не более 3-4 недель — этого достаточно, чтобы достичь
максимума аэробной мощности, но не «истощить» ресурсы
гормональной системы.
На этом этапе используется 4-5 аэробных тренировки в раз-
вивающем режиме (они могут быть разделены пополам и про-
водиться в виде 2 тренировочных занятий в день). Это означает,
что вид упражнения и его интенсивность должна обеспечивать
вовлечение всех мышечных волокон в работу, а длительность
— максимальная и ограничивается только запасами углевод-
ных ресурсов организма и мышечными болями. Очень важно
не превысить этой оптимальной длительности тренировки и не
войти в «катаболическую» фазу, в которой продолжение тре-
нировки будет сопровождаться интенсивным катаболизмом
белков мышц и лимфатической системы. Для бегунов средней
подготовленности это — 45-50 мин, высококвалифицирован-
ных спортсменов — 50-70 мин, после чего работоспособность
должна резко падать. Это означает, что спортсмен приблизился
к «катаболической» фазе. Если спортсмен способен продолжать
тренировку с прежней интенсивностью и после этого времени,
то это означает что интенсивность была занижена.
Одна тренировка полностью посвящается поддержанию
силы посредством скоростно-силовых (в небольшом объеме)
и статодинамических упражнений.
Как минимум одна дополнительная тренировка (объем 3-6
км) полностью выполняется на дорожке в шиповках в виде интер-
вального или повторного бега с соревновательной скоростью.
4. Предсоревновательный период (конец декабря, январь).
Этот период можно назвать периодом интегрирующей
тренировки. Его цель — обеспечение «сонастройки» техники
бега и обеспечивающих вегетативных систем с возможностя-
311
ми нервно-мышечного аппарата. Основные средства: бег с
околосоревновательной скоростью во всех вариантах. Объем
максимальный (ограничивается прочностью ОДА). Основная
сложность — соблюсти баланс между максимальным объемом
и хорошим состоянием мышц ног (в первую очередь — голе-
ни). Бег вне дорожки используется для поддержания аэроб-
ных способностей (1-2 тренировки в объеме 1/2 — 2/3)) и вос-
становления психики (восстановительный кросс — 10-15 км).
Обычная силовая тренировка может быть разделена пополам и
проводиться на вечернем дополнительном занятии или перед
днем относительного отдыха.
5. Соревновательный период.
Цель физической подготовки — как можно дольшее сохра-
нение работоспособного состояния гормональной системы, от
чего зависит ОП и сила мышц и в конечном итоге — длитель-
ность сохранения «спортивной формы», несмотря на вынуж-
денное применение работы гликолитической направленности
(для достижения максимального спортивного результата у
средневиков) или «жесткой» работы на длинных отрезках, или
в виде темпового бега (у стайеров) и участие в серии соревно-
ваний. Оптимальный вариант для этого — пилообразный вид
кривой условной нагрузки — когда занятия с высокой нагруз-
кой чередуются с отдыхом.
Возможные варианты построения микроциклов для реа-
лизации описанных выше практических рекомендаций пред-
ставлены на рис. 34.
8.2. Некоторые аспекты построения
многолетней подготовки бегунов
Общеизвестным является факт, что основа высоких резуль-
татов закладывается в детском и юношеском возрасте, поэто-
му организации многолетней подготовки и изучению основа-
ний, на которых она должна базироваться, уделяется большое
внимание.
При организации многолетней подготовки считается целе-
сообразным ориентироваться на так называемые сенситивные
312
Пере- ход- ный пери- од 2-3 недели
СТЭ СИЛА ОФП СТЭ оп БМВ СТЭ техн.
стэ оп МВ ОП ММВ СИЛА ОФП ОП БМВ
отдых
1-й базо- вый этап 3-4 недели техн. СТЭ СТЭ
СТЭ ОП БМВ СИЛА ОП БМВ СИЛ А
ОФП оп БМВ ОФП ОП ММВ
отдых
3-4 недели техн. СТЭ техн. СИЛА
ОП БМВ ОП БМВ техн.
ОП БМВ ОП ММВ
ОФП СИЛА ОФП СТЭ
2-й базо- вый этап отдых
3-4 недели техн. СТЭ ОП БМВ техн.
ОП СИЛА
ОП СТЭ БМВ ОП ММВ
БМВ СИЛА
отдых
3-4 недели техн. ОП БМВ техн ОП БМВ ОП ММВ техн ОП БМВ
ОП ММВ
СИЛА
отдых
Предсо- ревнов. период 4-6 недель техн. + Глико ЛИЗ СТЭ техн техн
ОП БМВ ОП СТЭ
ОП БМВ БМВ СИЛА ОП ММВ
Дни недели 1 2 3 4 5 6 7
Рис. 34. Гипотетическая схема планирования микроциклов бегунов на
выносливость на разных этапах макроцикла. Обозначения см. в табл.
10-11. Высота столбца обозначает величину нагрузки
313
периоды, в которые происходит ускоренное естественное раз-
витие организма [Набатникова М.Я., 1982; Травин Ю.Г., 1975;
Набатникова М.Я., 1982; Шварц В.Б., Хрущев С.В., 1984]. Уста-
новлено, что системы организма, определяющие координаци-
онные способности, интенсивно развиваются до 11-12 лет — до
момента созревания двигательного анализатора. Общая вынос-
ливость естественным образом наиболее быстро улучшается в
период 11-13 и 15-17 лет [Травин Ю.Г., 1975], а мышечная сила
— в период полового созревания — в 12-14 лет [Казарян Ф.Г.
1975]. В соответствии с этими данными дистанционные сред-
ства, развивающие, как считается, общую выносливость, реко-
мендуется применять в период 11-13 лет, а целенаправленное
развитие силовых способностей проводить позже — в 12-14 лет.
К специальной тренировке бегунов на выносливость следует
приступатьс 14-15 лет [Травин Ю.Г., 1975].
При всей логичности и «общепринятости» этих рекоменда-
ций существуют как минимум несколько моментов, вызываю-
щих необходимость их переосмысления и уточнения.
Во-первых, если принять за аксиому, что подготовка
бегунов экстракласса — процесс многолетний и его успеш-
ность закладывается на ранних этапах формирования
спортсмена, то отсутствие даже признаков возрождения
российской «школы бега» следует искать в несовершенстве
теоретико-методических концепций подготовки прежде все-
го юных бегунов.
Во-вторых, несмотря на бесспорные представления, что
подготовка юных и молодых бегунов не должна копировать
подготовку квалифицированных спортсменов, проведенные
исследования [Чесноков Н.Н., 1991] показали, что трениров-
ка юных и молодых российских бегунов (начиная с 12-13 лет)
практически полностью повторяет «в миниатюре» тренировку
взрослых спортсменов, распределение нагрузок в которой пред-
ставлено на рисунке.
В-третьих, длительные исследования, выполненные на ка-
федре легкой атлетики и ПН ИЛ РГАФК (ГЦОЛИФК) и под-
робно описанные выше, позволили обосновать и опробовать
на практике иные акценты в схеме построения многолетней
подготовки бегунов.
314
По нашему мнению, акценты в многолетней подготовке бе-
гунов могут быть следующими.
В современных урбанизированных условиях регулярные за-
нятия спортом должны быть обязательной составляющей жизни
детей, начиная с рождения. Однако до 11-12-летнего возраста
проводится действительно разносторонняя подготовка на мате-
риале возможно большего числа видов спорта или видов легкой
атлетики. Цель — создание «базы движений», развитие коорди-
национных способностей и укрепление опорно-двигательного
аппарата средствами спортивных игр и скоростно-силовых
упражнений, развитие гибкости. В этот период проводятся
спортивная ориентация и отбор, формируется устойчивый ин-
терес и любовь к занятиям спортом в широком смысле.
Главное, чтобы до этого возраста дети не знали о том, что им
уготовлена карьера бегунов. И уж тем более допуская «... воз-
можность использования больших объемов аэробных средств
подготовки уже в 10-11-летнем возрасте ...» [Набатникова
М.Я., 1982] (действительно, детский организм обладает вы-
сокими адаптационными способностями и может выдержать
многие «издевательства»!) — мы категорически не согласны с
целесообразностью этого, по крайней мере в беге.
Следующий этап (начальной подготовки) приходится на
12-14 лет. С этого возраста дети, отобранные как будущие бе-
гуны, уже могут чаще участвовать в соревнованиях по бегу на
выносливость после непродолжительной специальной (под-
водящей) тренировки (подробнее см. ниже). Однако ведущим
остается требование разносторонности подготовки. Признаки
специализированное™ в этот период могут просматриваться
лишь в одном: этот этап должен быть посвящен преимуще-
ственному развитию силовых способностей основных мышечных
групп бегунов, дальнейшему укреплению опорно-двигательного
аппарата нижних конечностей (средствами прыжков и сприн-
та), формированию техники бега «на выносливость».
И только начиная с 15-16-летнего возраста, на этапе на-
чальной специализации, допускается, на наш взгляд, некото-
рое изменение структуры планирования в сторону увеличения
объемов средств аэробной и других специальных средств под-
готовки бегунов.
315
Однако если во главу угла ставятся интересы достиже-
ния наивысших результатов, то рекомендуемая в научно-
методической литературе современная структура нагрузок как
крайне специализированная может применяться не ранее 17-
18 лет после полного завершения созревания организма бегуна
в биологическом и психофизиологическом смысле.
Основания этой гипотезы подробно описаны нами выше.
Например, высокие результаты бегунов определяются локаль-
ной выносливостью итехникой бега (соответствующейуровню
и особенностям подготовленности опорно-двигательного ап-
парата). А производительность управляющих и обеспечиваю-
щих систем (нервной, гормональной и сердечно-сосудистой)
или генетически предопределены, или легко «подстраивают-
ся» под исполнительный аппарат. Следовательно, форми-
рованию опорно-двигательного аппарата должно уделяться
преимущественное внимание. Но такой вывод даже на уров-
не теории входит в противоречие с бытующими представле-
ниями о структуре и распределении объемов основных тре-
нировочных средств в макроцикле и особенно многолетнем
аспекте.
Согласно нашей концепции, в макроцикле и в многолетней
подготовке бегунов «базовой» является не аэробная, а силовая,
скоростно-силовая, координационная подготовка и развитие
гибкости, а «специальной» — аэробная и гликолитическая во
всех видах независимо от интенсивности работы. Крайне оши-
бочными являются представления, что низкоинтенсивный
продолжительный бег является средством «общей», а не спе-
циальной подготовки юных бегунов. Такой бег является одним
из наиболее «специальных» средств для юных спортсменов,
которое, как хорошо известно, быстро приводит к улучшению
выносливости. Однако увлечение им придает структуре подго-
товки признаки чрезмерной специализированное™, которая
форсирует спортивный результат, но тормозит нормальное
физическое развитие подростков. Таким образом, формиру-
ются «плоды ранней специализации», не имеющие перспек-
тив в разряде взрослых спортсменов.
Именно это, на наш взгляд, является одним из факторов,
лишающих «российскую школу бега» шансов на возрождение.
316
Ссылки на опыт других циклических видов спорта (плава-
ние, велоспорт, лыжные гонки) является не корректным, т.к.
бег имеет важное биомеханическое отличие от других локомо-
ций — ударные взаимодействия с опорой, которые предопре-
деляют иные закономерности формирования исполнительно-
го звена двигательной системы спортсменов в онтогенезе.
Таким образом, главное, что хотелось бы подчеркнуть в
связи с вышесказанным — это требование проводить развитие
аэробных способностей в сенситивный период 11-12 лет, су-
щественно не изменяя структуры тренировочных объемов от-
носительно предыдущего этапа, т.е. не используя относитель-
но большие объемы аэробных средств.
В этой связи можно напомнить, что при акцентированном
воздействии показатели аэробных способностей выходят «на
плато» через 2-3 месяца в любом возрасте и дальнейший про-
гресс возможен, видимо, только после существенных морфо-
функциональных перестроек в организме.
8.2.1. Принципы подготовки
юных бегунов
Для обозначения путей реализации предлагаемого под-
хода мы разработали «принципы тренировки юных бегунов»
для критического возраста 12-15 лет, когда делаются, на наш
взгляд, наибольшие ошибки в их подготовке.
Несмотря на то что многие из «принципов» согласуются с
хорошо известными из литературы, акцентирование на них
внимания является важным, т.к. в практической работе они не
всегда соблюдаются. Причина этого кроется не только в под-
спудном стремлении форсировать подготовку, но и часто — в
неверных представлениях о том, что является «базовой», а что
является «специальной» подготовкой (см. выше).
I принцип. Приоритетность и предшествование развития
скоростно-силовых и силовых способностей относительно аэ-
робных.
Это наиболее важный принцип, соблюдение которого со-
вершенно необходимо при планировании макроцикла и мно-
голетней подготовки юных бегунов.
317
Формулировка этого принципа основана на следующих те-
зисах:
1. В макроцикле и тем более в многолетнем аспекте внача-
ле необходимо сформировать морфологическую основу
выносливости — соединительно-тканные образования
опорно-двигательного аппарата и сократительные эле-
менты мышц (это обеспечивается средствами скоростно-
силовой и силовой подготовки), а затем уже накапливать
на этой основе митохондрии и увеличивать капилляриза-
цию мышц — факторы, непосредственно обеспечиваю-
щие аэробную производительность. Обратная последова-
тельность развития качеств не рациональна.
2. Уровень функциональной подготовленности ССС, опре-
деляемый производительностью сердечной мышцы, со-
судистым руслом и характеризуемый величиной МПК,
должен соответствовать аэробной мощности скелетных
мышц. Однако возраст 13-16 лет, на который чаще всего
приходится этап начальной специализации в беге, наи-
менее благоприятен для интенсивного тренировочного
воздействия на миокард (из-за интенсивных перестроек
в организме в пубертатный период). Поэтому целесоо-
бразно отодвинуть время тренировки ССС (в том числе
и большими объемами) на более поздний возраст — 17-18
лет. Это не отразится на наивысших спортивных резуль-
татах данного бегуна, так как хорошо известно, что от-
носительный МПК достигает максимума в течение 1,5-2
лет регулярной тренировки независимо от возраста. По-
сле этого происходит стабилизация или даже снижение
этого показателя, несмотря на постоянное повышение
результатов в беге.
Взаимосвязь МПК со спортивным результатом у ква-
лифицированных спортсменов достаточно слаба, хотя
ССС может накладывать генетически обусловленные
ограничения на величину МПК. Поэтому если после 1-2
лет регулярной тренировки в беге (независимо от возрас-
та) МПК не достигло 65-70 мл/кг, то этому бегуну лучше
сменить специализацию или вообще не думать о карьере
спортсмена.
318
3. Объемные и интенсивные тренировки на выносливость
оказывают чрезвычайно мощное воздействие на гормо-
нальную систему. Ресурсы ее в этом случае тратятся на
восстановление энергетических субстратов, снижая тем
самым гормональное обеспечение синтеза структурных
и сократительных белков. В частности, известно, что
после нагрузки на выносливость, в крови наблюдается
существенное и длительное (до нескольких дней) по-
нижение уровня тестостерона и других гормонов, уча-
ствующих в анаболических процессах. Регулярное по-
вторение таких воздействий может привести к раннему
окостенению скелета, хроническому отставанию в фор-
мировании соединительных и сократительных элемен-
тов опорно-двигательного аппарата, что уже невоспол-
нимо у взрослых спортсменов. Кроме того, интенсивное
истощающее воздействие на гормональную систему в
период ее перестроек в пубертатный период вполне мо-
жет отрицательно сказаться на ее формировании. Это, в
частности, отражается на иммунологической реактивно-
сти юных спортсменов [Левин М.Я., Хрущев С.В., 1991;
Шубик В.М., Левин М.Я., 1982, 1985] и подтверждается,
например, экспериментом на крысах [Bloor С.М., 1968],
у которых регулярные тренировки в продолжительном
беге с раннего периода жизни, действительно, увеличи-
ли число кардиомиоцитов (мышечных клеток сердца),
но уменьшили число клеток печени, почек и надпочеч-
ников. Как можно судить по имеющимся данным, при
тренировке юных спортсменов вполне могут происхо-
дить похожие явления.
Упражнения же скоростно-силового характера, даже
в значительных объемах, напротив, положительно ска-
зываются на состоянии гормональной системы.
4. Фактом является то, что многие бегуны, достигшие вы-
дающихся и стабильно высоких результатов и показы-
вающие их в течение длительного времени, начали регу-
лярные занятия в беге не раньше 15-17-летнего возраста.
А до этого времени они или вообще не тренировались,
или получили хорошую физическую подготовку, зани-
319
маясь другими видами спорта. Высокие результаты этих
атлетов объясняются, вероятно, тем, что они не подвер-
гались в подростковом возрасте отрицательному воздей-
ствию факторов, описанных в пунктах 2 и 3.
5. Широкое применение средств скоростно-силовой под-
готовки позволяет организовать тренировочный про-
цесс на высоком эмоциональном фоне, что чрезвычайно
важно для формирования устойчивого интереса к заня-
тиям спортом.
II принцип. Непрерывная тренировка скоростно-силовых и
силовых способностей в годичном цикле и акцентированное воз-
действие на аэробные способности мышц только в конце подгото-
вительного и в предсоревновательном периодах.
Формулирование этого принципа основано на следующих
данных:
1. Большей лабильностью (т.е. подвижностью, подвер-
женностью влияниям) обладают белки митохондрий и
капилляров по сравнению с сократительными белками
и особенно соединительно-тканными белками опорно-
двигательного аппарата. Следовательно, скоростно-
силовые способности требуют большего времени для
получения эквивалентного тренировочного эффекта.
2. Не рекомендуется прекращение силовой и особенно
скоростно-силовой тренировки ни на одном этапе, т.к.
возобновление этого вида упражнений после перерыва
вызывает мышечные боли и снижает силу мышц. Это
негативно сказывается на состоянии спортсменов, уве-
личивает вероятность травматизма, отрицательно ска-
зывается на технике и экономичности бега и т.п.
3. Аэробные упражнения в больших объемах в силу их высо-
кой энергетической стоимости и глубокого воздействия
на гормональную систему замедляют или даже блокиру-
ют синтез сократительных белков даже при наличии си-
ловых упражнений. Поэтому целесообразно разводить
во времени периоды акцентированного воздействия на
сократительные и митохондриальные белковые ком-
плексы. Однако мощность аэробных систем мышц явля-
ется основным фактором, определяющим спортивный
320
результат. Следовательно, тренировочные средства, по-
вышающие окислительный потенциал мышц, необходи-
мо планировать ближе к этапу основных соревнований,
а не на подготовительный период.
Реализация этого принципа позволяет решить важную за-
дачу юношеского спорта — достижение высоких результатов в
соревновательном периоде без излишней специализированности
на других этапах макроцикла.
III принцип. Предшествование спринтерских, скоростно-
силовых и собственно-силовых упражнений беговым средствам
подготовки в рамках годичного цикла.
Скоростно-силовые упражнения (прыжки, многоскоки), так
же как и упражнения на гибкость, способствуют укреплению
соединительно-тканных элементов опорно-двигательного аппа-
рата. Это один из факторов профилактики травматизма. Поэтому
такие средства должны применяться в начале макроцикла.
Собственно-силовые упражнения направлены на реше-
ние основной «базовой» задачи тренировки — создание мор-
фологической основы выносливости, активизации «спящих»
аа-мотонейронов спинного мозга, поэтому их применение
в большем или меньшем объеме необходимо на всех этапах
спортивного совершенствования. Но такие упражнения не-
посредственно не сопровождаются повышением окислитель-
ного потенциала мышц или ростом максимальной скорости
бега, кроме этого, их применение чаще всего отрицательно
сказывается на экономичности бега, поэтому акцентирован-
ная силовая тренировка должна планироваться до повышения
объемов спринтерской и аэробной подготовки.
Необходимость обязательного присутствия собственно-
силовых упражнений определяется также тем, что они позво-
ляют увеличивать силу и быстрых мышечных волокон (МВ),
и медленных МВ, в то время как спринтерские и скоростно-
силовые упражнения повышают силу только быстрых МВ.
IV принцип. Преимущественное применение средств, спо-
собствующих повышению окислительного потенциала (аэробной
мощности) всех типов мышечных волокон основных мышечных
групп бегунов по отношению к средствам, избирательно воздей-
ствующим на медленные мышечные волокна.
321
Известно, что мышцы человека неоднородны по своему
составу. Они состоят из медленных (окислительных) мышеч-
ных волокон (ММВ), имеющих высокую аэробную мощность,
но низкую скорость сокращения, быстрых окислительных
(БоМВ) — в равной степени и выносливых, и быстрых, а также
быстрых гликолитических (БгМВ) — быстрых, но не выносли-
вых. Состав волокон в той или иной мышце определяется ти-
пом иннервации этой мышцы и генетически предопределен.
Для бега на выносливость желательно иметь больше ММВ. Но
абсолютное преобладание ММВ в основных мышечных груп-
пах встречается крайне редко и имеет безусловную ценность
только для марафонцев.
Очевидно, что для достижения рекордного для себя ре-
зультата бегун должен иметь максимальный окислительный
потенциал всех мышечных волокон. Однако существует за-
кономерность, что при низкой мощности работы (например,
при длительном аэробном беге) в работу вовлечены, а следо-
вательно, и тренируются в большей мере ММВ, в то же время
более интенсивные средства, такие, как: бег в утяжеленных
условиях, «интервальный спринт», спринтерские ускорения в
процессе аэробного бега, фартлек и т.п. — повышают окисли-
тельный потенциал как быстрых, так и медленных мышечных
волокон (МВ). Другим словами, они позволяют «прорабаты-
вать» всю мышцу.
Справедливости ради надо сказать, что длительный аэроб-
ный бег и особенно длительный темповой бег так же может
привести к существенному росту аэробной мощности мышц и
всего организма. Но для этого требуются большие объемы на-
грузки как в одной тренировке, так и в макроцикле в целом.
Однако при этом всегда страдает силовая подготовленность
спортсменов и повышается вероятность негативных явле-
ний относительно общего развития организма, описанных в
предыдущем номере журнала. В связи с этим преимуществен-
ное использование бега в утяжеленных условиях, перемен-
ного метода, темповых пробежек на дорожке и спринтерских
упражнений и т.п. с целью аэробной подготовки позволит до-
биться высокого тренировочного эффекта при относительно
меньших затратах времени и энергетических ресурсов. Це-
322
лесообразность сокращения объемов медленного или темпо-
вого продолжительного бега определяется также и тем, что в
тренировке юных спортсменов менее актуальными являются
вопросы накопления запасов гликогена в организме, повы-
шение устойчивости гормональных механизмов, регулирую-
щих водно-солевой обмен и терморегуляцию, повышающих
устойчивость эритроцитов крови и регуляторных механизмов
ионных процессов в миокарде и т.д., т.е. всех тех специфиче-
ских задач, которые решаются путем применения длительного
непрерывного бега.
Реализация этого принципа позволит уменьшить общий
объем беговой работы при сохранении ее эффективности.
В то же время, если придерживаться указанного принци-
па, то существует строгое правило, которое следует соблюдать
в обязательном порядке, — упражнение должно выполняться
без так называемого «закисления» мышц и крови. В противном
случае, избавившись от избыточных общих объемов, мы полу-
чим чрезмерную «интенсификацию» тренировочного процес-
са с еще худшими последствиями. Для предотвращения этого
нужно так организовать тренировку, чтобы средняя мощность
(интенсивность) работы была ниже т.н. аэробного порога, не-
смотря на то, что во время ускорений может достигаться вы-
сокая или даже предельная интенсивность. Это может быть
обеспечено двумя способами: 1) низким темпом движений
при относительно большой силе отталкивания (например,
медленный бег широким шагом в гору) или 2) достаточными
для отдыха интервалами отдыха.
Требуемый для реализации этого принципа эффект дости-
гается при использовании следующих средств.
В подготовительном периоде:
1. Интервальный бег с сопротивлением (волокушей). Шаг
широкий, спина прямая (без наклона вперед), акцент
на проталкивание вперед и вынос колена маховой ноги
«вверх». Ускорение — 60-90 с, отдых — 2,5-3 мин в виде бега
трусцой или многоскоков «на стопе». Длительность упраж-
нения в одной тренировке 35-45 мин (9-12 ускорений).
2. Интервальный бег широким шагом в гору. Методиче-
ские указания и дозировка — та же.
323
3. Интервальный бег широким шагом по песку. Методиче-
ские указания и дозировка — та же.
4. СБУ «колесо». Ускорение — 45-70 с, отдых 3-3,5 мин.
Длительность — та же.
5. Спринтерские ускорения в процессе аэробного бега.
Ускорение 3-5 с с интенсивностью 80-90% от макси-
мальной скорости. Отдых в виде бега трусцой 2-2,5 мин.
Продолжительность одной серии 10-15 мин, число се-
рий — 2-3. Отдых между сериями (в виде релаксации в
тепле) — 5-7 мин.
6. Фартлек с короткими ускорениями (30-150 м) по силь-
нопересеченной местности (40-60 мин).
7. Кросс по сильнопересеченной местности и сыпучему
грунту (40-60 мин).
На предсоревновательном этапе:
1. Интервальный бег по дорожке. Ускорения 150-200 м.
Отдых в виде бега трусцой 3-4 мин.
2. Интервальные пробежки «на технику» с соревнователь-
ной скоростью. Длина отрезков ускорений — 30-40% от
длины соревновательной дистанции.
3. Спринтерские ускорения в процессе аэробного бега.
Акцентированное использование интенсивных средств аэ-
робной подготовки рекомендуется на 2-м базовом этапе под-
готовительного периода после этапа силовой подготовки (1-го
базового) (см. схему макроцикла в предыдущем номере журна-
ла). Ориентировочная схема макроцикла дана на рис. 35.
V принцип. Силовая тренировка бегунов должна быть спец-
ифична.
Это означает, во-первых, что основное внимание должно
уделяться повышению силы тех мышечных групп, которые
несут основную нагрузку в беге. Это мышцы, выполняющие
разгибание ноги в голеностопном суставе, мышцы задней по-
верхности бедра и мышцы — сгибатели тазобедренного сустава.
Все остальные мышечные группы выполняют вспомогатель-
ную функцию. Во-вторых, результат в беге на выносливость
определяется в основном производительностью медленных
мышечных волокон (МВ) работающих мышц. Поэтому при
организации силовой тренировки необходимо отдавать пред-
324
Техн. Аэро(И) СТЭ Аэро(И) Аэро(И) Отдых
Техн. СИЛА ОФП Аэроб.
СТЭ СИЛА
Пн. Вт. Ср. Чт. Пт. Сб. Вс.
35. Схема микроцикла и направленность тренировочного воздействия по
дня м на 2-м базовом этапе подготовительного периода у юных бегунов
13-14 лет
Обозначения:
техн. — пробежки на «технику» (150-300 м с соревновательной
скоростью); СТЭ — соединительно-тканных элементы опорно-
двигательного аппарата (многоскоки, прыжки, спринт, «прыжковые»
спортивные игры, С БУ, стретчинг); Аэро(И) — интенсивные сред-
ства аэробной подготовки (фартлек, интервальный бег, «аэробный»
спринт, бег в утяжеленных условиях, «беговые» спорт, игры); СИЛА
— силовая тренировка основных мышечных групп бегуна; ОФП —
«общая» силовая тренировка, «силовые» спортивные и подвижные
игры; Аэроб. — бег в аэробном режиме.
Пн. — вначале темповые пробежки на технику, затем — бег в гору
или с волокушей, или спринтеровские ускорения в процессе
кросса на местности, или СБУ, или сочетание перечисленного.
В конце — 3-4 пробежки по 250-400 м на «технику». Нагрузка
большая (максимальная высота столбика — на рис. 39).
Вт. — вначале — спринт, прыжки, многоскоки, затем — кросс по
сильнопересеченной местности или фартлек.
Ср. — день относительного отдыха с небольшим объемом много-
скоков, упражнений на гибкость и 2-3 сериями специальных
силовых упражнений бегуна (см. V принцип).
Чт. — бег в гору, с волокушей или фартлек. Нагрузка большая.
Пт. — вначале темповые пробежки на технику интервально, затем
силовая тренировка половинного объема. В конце — круговая
тренировка ОФП.
Сб. — аэробный кросс.
Вс. — отдых.
325
почтение таким упражнениям, которые позволяют увели-
чивать силу и гипертрофировать не только быстрые МВ (эти
МВ тренируются при любой силовой или скоростно-силовой
тренировке), но и медленные МВ. Эффективным в этом отно-
шении видом силовой тренировки можно считать применение
изотонических и статодинамических упражнений.
Эти упражнения выполняются сериями или круговым ме-
тодом на все основные (см. выше) мышечные группы. Типич-
ная серия (круг) может выглядеть так:
1-е упр. — на мышцы голени. Стоя на возвышении на носке
одной ноги — пятка «на весу» — сгибание и разгибание
ноги в голеностопном суставе с полной амплитудой в
темпе 1- 0,5 движения в с у. На правой и левой ногах.
2-е упр. — на мышцы — разгибатели ног. Ноги на ширине
плеч или шире, носки разведены в стороны, руки за го-
ловой, локти максимально разведены спина строго пря-
мая, взгляд вперед-вверх. Медленные приседания с не-
полным вставанием.
3-е упр. — на мышцы задней поверхности бедра. Упор лежа
сзади, пятка одной ноги на возвышении 20-30 см, вторая
— выпрямлена «на весу». Поднимание и опускание таза и
свободной ноги без касания земли ягодицами.
4-е упр. — на мышцы — сгибатели тазобедренного сустава.
Стоя на одной ноге рядом с барьером перпендикулярно
ему, перенос выпрямленной в коленном суставе ноги че-
рез барьер справа и слева от него без маха.
5-е упр. — на мышцы спины и задней поверхности бедра.
Лежа на верхней трети бедер на «коне» или «козле» ли-
цом вниз, ноги закреплены, гуловище и голова — на весу,
руки за головой, сгибание и разгибание туловища, мак-
симально «включая» мышцы задней поверхности бедра.
6-е упр. — на мышцы — сгибатели тазобедренного сустава и
брюшного пресса. В висе на перекладине или шведской
стенке — поднимание и опускание выпрямленных ног к
перекладине без маха.
Виды упражнений, разумеется, могут быть самыми разноо-
бразными. Однако необходимый эффект увеличения силового
потенциала как предпосылки улучшения выносливости бегуна
326
будет получен только при самом строгом соблюдении следую-
щих методических рекомендаций:
а) на протяжении всего подхода движения выполняются
медленно (темп 1-0,5 движений в с у), плавно, без рас-
слабления упражняемых мышц ни в какой фазе движе-
ния, с полной амплитудой;
б) величина напряжения мышцы (массы отягощения) 40-60%
от максимального. Ориентиром правильно подобранной
нагрузки является длительность подхода — «отказ» дол-
жен наступать через 45-70 с после начала подхода;
в) каждый подход выполняется «до отказа» — до сильного
утомления мышц. Упражнение, прекращенное раньше
— выполнено «в пустую»!
г) интервал отдыха между подходами — 30-60 с. Отдых пас-
сивный. Пауза отдыха между сериями (кругами) — 3-4
мин, заполняемых бегом трусцой;
д) число серий — 2-7 в зависимости от адаптированное™
мышц, этапа макроцикла и дня микроцикла;
е) число занятий силовой направленности в микроцикле на
развивающем этапе — 2-2,5. Наэтапе поддержания силы —
0,5-1;
ж) после силовых упражнений не допускается никакой бе-
говой работы, на следующий день — желателен день от-
дыха или скоростно-силовая/спринтерская тренировка
половинного объема.
Этап акцентированной силовой подготовки начинается
сразу после переходного периода (этапа «мощностной» тре-
нировки — см. предыдущий номер, схема на рис. 1). На этом
этапе достаточно, как правило, 8-10 тренировочных занятий
в течение 1,5 месяцев, чтобы существенно улучшить силовую
подготовленность бегунов.
Ориентировочная схема «силового» микроцикла (1-й базо-
вый этап) приведена на рис. 36.
VI принцип. Минимизация средств «гликолитической на-
правленности» в соревновательном периоде и их практическое
исключение на других этапах макроцикла.
Под «гликолитической» работой может пониматься, напри-
мер, бег на 600-1000 м на результат или — интервальная работа
327
СТЭ техн. ОФП Аэро(И) СИЛА Отдых Техн. Аэро(И) СТЭ СИЛА Отдых
Пн. Вт. Ср_ Чт. Пт. Сб. Вс.
Рис. 36. Схема микроцикла и направленность тренировочного воздей-
ствия по дням на 1-ом базовом этапе подготовительного периода у юных
бегунов 13-14 лет Обозначения см. рис. 35
Пн. — вначале спринт, прыжки, затем — темповые пробежки на тех-
нику, в конце тренировки — ОФП или «силовая» игра. Нагруз-
ка полная.
Вт. — кросс по сильнопересеченной местности, фартлек по холмам,
бег в гору. Нагрузка средняя.
Ср. — силовая тренировка. Нагрузка полная.
Чт. — отдых.
Пт. — темповые пробежки на технику. Затем — спринт в процессе
аэробного бега, интервальная тренировка на местности, бег в
утяжеленных условиях. Нагрузка средняя.
Сб. — вначале многоскоки, прыжки, пробежки, затем — силовая
тренировка с полной нагрузкой.
Вс. — отдых.
3x300-400 м в полную силу с интервалом отдыха 2-3 мин и т.п.,
которые сопровождаются предельным снижением pH крови
и/или мышц (их «закислением»).
Применение «гликолитической» тренировки имеет целый ряд
отрицательных последствий, основными из которых являются:
1. Деструктивные изменения в ферментативных системах,
определяющих окислительный потенциал мышц.
2. Сильнейшее стрессовое воздействие на гормональную
систему и психику, а также при некоторых условиях,
глубокое исчерпание углеводных запасов организма и
деструктивное воздействие на сократительные белки.
Это на длительный период (7-21 дней) может понизить
работоспособность спортсменов без возвращения к ис-
ходному уровню.
328
К положительным эффектам гликолитической тренировки
относят: повышение мышечной силы, увеличение буферной
емкости мышц, улучшение психологической устойчивости.
Однако следует учитывать, что:
— существуют более эффективные методы силовой подготовки;
— буферная емкость мышц определяется в основном аэ-
робными способностями (в частности, быстрых МВ)
и степенью гипертрофии мышц, поэтому применение
гликолитической тренировки для повышения буферной
емкости мышц не эффективно;
— в психологическом плане умение «терпеть» на дистан-
ции в значительной степени определяется врожденны-
ми чертами личности, а польза от столь сильного воздей-
ствия на психику подростков не является очевидной.
Если бег выполняется с «гликолитической» скоростью, но
не продолжается до предельной степени утомления, то такая
нагрузка будет иметь «аэробный» или «технический» эффект,
но не «гликолитический»! Это простое и хорошо известное
правило часто игнорируется при исследовании объемов тре-
нировочной нагрузки. В результате в данных и выводах таких
«научных» исследований реальные объемы работы гликоли-
тической направленности оказываются завышенными в не-
сколько раз. На этом, на наш взгляд, «сгорело» много поколе-
ний российских бегунов, чьи тренеры некритически отнеслись
к таким рекомендациям.
На практике действительно «гликолитических» трениро-
вок (см. выше) может быть 2-3 за весь макроцикл с интерва-
лом 7-10 дней на этапе, непосредственно предшествующем
основным соревнованиям, да и то только в том случае, если по
каким-то причинам нет возможности выступить в неосновных
соревнованиях или провести 2-3 «прикидки».
VII принцип. Обязательное планирование разгрузочных ми-
кроциклов, заполняемых спринтерскими, скоростно-силовыми
и силовыми видами тренировки высокой интенсивности, но не-
большого объема.
Соблюдение этого принципа является профилактической
мерой против возможного исчерпания ресурсов гормональной
системы под воздействием объемной тренировки.
329
В тренировке юных бегунов целесообразно каждый третий
микроцикл делать разгрузочным. В рамках этого микроцикла
практически полный отдых (по крайней мере, психологиче-
ский) должен длиться не менее 5 дней.
Особенно это важно на этапе «объемной» тренировки.
На практике же в погоне за «объемом» именно в этот пери-
од рассматриваемый принцип чаще всего нарушается.
VIII принцип. Преобладание на базовых этапах подготовки
чередующихся узконаправленных тренировочных занятий в рам-
ках микроциклов.
Однонаправленные занятия с целью улучшения физиче-
ской подготовленности более эффективны по сравнению с
комплексными занятиями.
В рамках микроцикла необходимо как можно дальше раз-
водить во времени тренировочные занятия аэробного и сило-
вого характера (если в них планируется развивающая величи-
на нагрузки) (см. рис. 35, 36). Это требование объясняется тем,
что синтез сократительного белка после силовой тренировки
требует функционирования определенного гормонального
«ансамбля» и высокой концентрации свободных аминокислот
в крови. Однако эти условия нарушаются, если одновременно
выполняется объемная аэробная тренировка. Кроме того, си-
ловая или скоростно-силовая тренировка (так же, как объем-
ная аэробная) сопровождается выраженным катаболическим
эффектом. Проведение на следующий день объемной аэроб-
ной тренировки «блокирует» процессы восстановительного
синтеза сократительных элементов мышц.
В результате получаем хорошо прогнозируемый эффект —
«ни выносливости, ни силы».
Комплексные, или «вариативные», тренировки наиболее
полезны для целей психологического «переключения», совер-
шенствования техники, поддержания, но не развития физиче-
ских кондиций.
Это замечание не относится к случаям, когда используются ва-
риативные средства одной физиологической направленности, на-
пример аэробной или скоростно-силовой [Платонов В.Н., 1984].
Так если в одной тренировке использовать любое сочетание
средств 1-7, описанных в IV принципе, то это — однонаправ-
330
ленная тренировка, несмотря на то, что в ней могут соседство-
вать бег с соревновательной скоростью по дорожке в шипов-
ках, интервальный спринт и кросс по сильнопересеченной
местности.
Изложенные «принципы» прошли апробацию на практи-
ке. Кроме того, они имеют под собой физиологические и био-
химические основания. Поэтому в своем большинстве могут
быть применимы и в других циклических видах спорта.
8.3. Заключение
Анализ литературных источников позволяет выделить три
подхода, которые могут быть положены в основу разработки
принципов планирования тренировочных программ спор-
тсменов.
1. Тщательное изучение тренировки выдающихся спор-
тсменов и взглядов известных тренеров с целью выявле-
ния общих закономерностей спортивной тренировки.
Как нам кажется, этот подход используется наиболее часто
и во многих случаях себя оправдал. Однако универсальным его
назвать нельзя, так как иногда чрезвычайно сложно выявить
общие тенденции в подготовке выдающихся спортсменов, по-
казывающих одинаковые результаты. Так, например, в беге на
выносливость почти в одно и то же время (60-е годы) показыва-
ли высокие результаты представители новозеландской «шко-
лы», характеризуемой небольшими объемами, очень длинным
соревновательным периодом, высокой долей силовой подго-
товки в «специфических» упражнениях и объемами бега как с
соревновательной скоростью по дорожке, так и по сильнопе-
ресеченной местности; представители австралийской «шко-
лы», характеризуемой средними или высокими объемами бега,
широким применением «небеговых» упражнений со штангой,
отсутствием малоинтенсивного бега, предпочтением трени-
ровки на местности тренировке на дорожке; американской
«школы», характеризуемой высокой долей специфической
работы на дорожке, спринтерской подготовкой, отсутствием
силовых упражнений; английской «школы», характеризуемой
331
традиционно большими объемами медленного бега на местно-
сти, низкой долей спринтерской и силовой подготовки.
Та же тенденция может быть прослежена и в настоящее
время. Например, достаточно большие различия могут быть
обнаружены в подготовке африканских бегунов (Эфиопия,
Кения, Марокко), португальских и итальянских, английских
и финских. Более того, даже в рамках одной «системы» может
существовать значительная разница во взглядах (например, в
России). В связи с этим данный подход не может быть признан
основным при определении принципов планирования подго-
товки бегунов.
2. Исследование эффективности различных вариантов
тренировочных программ путем проведения сравни-
тельных педагогических экспериментов и выявление на
этой основе общих принципов подготовки. Однако, как
давно было показано, число переменных, влияние ко-
торых необходимо изучить, при организации естествен-
ного тренировочного процесса может быть очень боль-
шим. В связи с этим число вариантов сочетаний разных
значений переменных стремится к бесконечности. Сле-
довательно, этот путь также не может быть основным,
так как он заведомо ведет в тупик из-за необозримого
объема экспериментальной работы.
3. Принципы планирования тренировки могут быть опре-
делены, исходя из известных биологических законо-
мерностей функционирования определенных белковых
структур, определяющих рамки жизнедеятельности ор-
ганизма человека.
Используя этот подход, можно определить последователь-
ность в развитии различных физических качеств в многолет-
нем аспекте и в рамках различных циклов подготовки, необ-
ходимую частоту повторяемости тренировочных воздействий
на ту или иную морфоструктуру, длительность применения
одних и тех же способов воздействия, сопутствующие условия
тренировки (питание, восстановительные мероприятия и т.д.).
Следовательно, этот подход дает необходимые основания для
планирования тренировки. Особенно ценным является то, что
на его основе возможно создание имитационных математиче-
332
ских моделей, реализуемых на ЭВМ для быстрой проверки всех
мыслимых способов применения тренировочных воздействий
без проведения трудоемких педагогических экспериментов
[Сарсания С.К., Селуянов В.Н., 1991; Селуянов В.Н., 1992].
Однако этот подход имеет существенные ограничения. Во-
первых, при определении конкретных объемов тренировоч-
ных нагрузок, во-вторых, на его основе сложно «просчитать»
такие моменты, как необходимость сочетания физической,
технической, тактической и психологической подготовки.
В-третьих, в частности при спортивной подготовке детей, су-
ществует много особенностей детского организма, значитель-
но усложняющих «модель», кроме того, важно учитывать, что
ведущим моментом в подготовке юных спортсменов является
не столько физическая подготовка с целью достижения наи-
высших результатов, сколько педагогические, психологиче-
ские и медицинские аспекты формирования юного бегуна и
т.п. (однако этот подход, как нам представляется, наиболее
продуктивен при исследовании проблем локальной выносли-
вости, когда число факторов, которые должны быть внесены в
модель, существенно уменьшается).
В связи с этим при разработке принципов построения го-
дичного цикла подготовки в ЦВС, определении конкретных
парциальных объемов и разработке тренировочных микро-
циклов мы использовали достоинства различных подходов,
применимых, по нашему мнению, в каждом отдельном случае.
В результате такой организации исследования стало возмож-
ным подойти к вопросу воспитания локальной выносливости
с различных позиций и сделать суждения относительно основ-
ных компонентов целостного тренировочного процесса, учет
которых, на наш взгляд, может помочь в организации нового
этапа исследований, а тренерам-практикам дать некоторые
ориентиры в совершенствовании индивидуальной подготовки
их учеников.
333
Литература
1. Алиханова Л.И. Связь между углеводными ресурсами ор-
ганизма (мышечным гликогеном) и физической аэробной
работоспособностью: Автореф. дис... канд. биол. наук. —
М., 1983.- 22 с.
2. Алферова Т.В. Утомление и восстановление при локаль-
ной работе мышц. — Омск: СГИФК, 1990. — 16 с.
3. Арнис В.Р. Развитие мощности работы у человека при тре-
нировке силы // Физиология человека. — 1994. — № 2. — С.
80-87.
4. Амосов Н. Теоретические исследования физиологических
систем // Н. А. Амосов, Б.Л. Палец, Б.Т. Агапов и др. —
Киев: Наукова думка, 1977. — 324 с.
5. Борисов Е.П. Экспериментальное обоснование регулиро-
вания тренировочной нагрузки бегунов на средние дис-
танции в процессе многолетней тренировки: Автореф.
дис... канд. пед. наук. - М., 1979. -24 с.
6. Бондарчук А. П. Тренировка легкоатлета. — Киев: Здоров'я,
1986.- 160 с.
7. Бондарчук А.П. Объем тренировочных нагрузок и дли-
тельность цикла развития спортивной формы // Теория и
практика физической культуры. - 1989. - №8. - С. 18-20.
8. Бравая Д.Ю. Физиологический анализ разных методов
и режимов тренировки мышечной силы: Автореф. дис...
канд. биол. наук. — М., 1985. — 22 с.
9. Биомеханика: Учебник для ин-ов физ. культ / Под ред.
Д.Д. Донского, В.М. Зациорского. — М.: Физкультура и
спорт, 1979.- С.264 с.
10. Валиулин В.В., Дзамуков Р.А. Пластичность скелетной
мышцы / Прикладные аспекты исследований склетных,
сердечных и гладких мышц, 7-11 октября 1996 г. — М.: Пу-
тинский научный центр РАН, 1996. — 54.
334
11. Валиулин В.В., Станишевская О.Ю., Дзамуков Р.А. Воз-
можная роль импульсной активности мотонейрона в
регляции качественного состава миозинов скелетной
мышцы / Прикладные аспекты исследований склетных,
сердечных и гладких мышц, 7-11 октября 1996 г. — Пу-
тинский научный центр РАН, 1996. — С. 55.
12. Вайцеховский С.М.. Система спортивной подготовки
пловцов к Олимпийским играм: Автореф. дис... д-ра. пед.
наук. - М., 1985. - 52 с.
13. Вейдер Д. Строительство тела по системе Джо Вейдера. —
М.: Физкультура и спорт, 1992. — 112 с.
14. Верхошанский Ю.В. Основы специальной силовой подго-
товки в спорте. — М.: Физкультура и спорт, 1977. — 215 с.
15. Верхошанский Ю.В., Мироненко И.Н., Антонова Т.М.
Модель динамики состояния спортсмена в годичном ци-
кле и ее роль в управлении тренировочным процессом //
Теория и практика физической культуры. — 1982. — №1.
-С.14-19.
16. Верхошанский Ю.В., Сиренко В.А. Силовая подготовка
бегунов на средние дистанции // Легкая атлетика. — 1983.
-№12.-С. 9-10.
17. Верхошанский Ю.В., Чарыева А.А. Проблемы развития
выносливости // Научно-спортивный вестник. — 1984. —
№3. — С. 14-20
18. Верхошанский Ю.В. Программирование и организация
тренировочного процесса. - М.: Физкультура и спорт,
1985.- 176 с.
19. Верхошанский Ю.В. Основы специальной физической
подготовки спортсменов. — М.: Физкультура и спорт,
1988.- 331 с.
20. Верхошанский Ю.В., Залеев Е.Н. Система тренировки в
беге на средние дистанции в годичном цикле // Спортив-
ный вестник. - 1989. - № 6. - С. 3-8.
21. Верхошанский Ю.В. Принципы организации тренировки
спортсменов высокого класса в годичном цикле. // Теория
и практика физической культуры. - 1991. - №2. - С. 6-16.
22. Виру А.А. Функции коры надпочечников при мышечной
деятельности. — М.: Медицина, 1977. — 175 с.
335
23. Виру А.А. Гормональные механизмы адаптации к трени-
ровке. - Л.: Наука, 1981. - 155 с.
24. Виру А.А., Кырге П.К. Гормоны и спортивная работоспо-
собность. — М.: Физкультура и спорт, 1983. — 159 с.
25. Влияние изотонической силовой и аэробной тренировки
на аэробный и анаэробный пороги человека / Мякинчен-
ко Е.Б., Обухов С.М., Тураев В.Т., Селуянов В.Н. / При-
кладные аспекты исследований скелетных, сердечных и
гладких мышц, 7-11 октября 1996 г. — Пущино: Пущин-
скийНЦРАН, 1996.-С. 67.
26. Васильева В.В. Сосудистые реакции у спортсменов. — М.:
Физкультура и спорт, 1971. - 145 с.
27. Васильковский Б.М. Контроль за уровнем специальной
выносливости и нормирование тренировочных нагру-
зок в подготовке конькобежцев-многоборцев высокой
квалификации: Автореф. дис... канд. пед. наук. — М.,
1983.- 18 с.
28. Волунгявичюс Г., Беляев С., Селуянов В. Влияние
скоростно-силовой подготовленности на выносливость
велосипедистов. // Тематический сборник научных тру-
дов: Средства и методы спортивной тренировки. — Виль-
нюс: СМ В и ССО ЛССР, 1984. - С. 52-56.
29. Волков В.Н., Исаев А.П., Баженова С.В., Гавриш Т.В. Им-
мунология спорта // Теория и практика физической куль-
туры. - 1995. - №10. - С. 12-14.
30. Волков Н.И. Энергетический обмен и работоспособ-
ность человека в условиях напряженной мышечной дея-
тельности: Автореф. дис... канд. биол. наук. — М., 1969.
-21с.
31. Волков Н.И. Биохимия: Учебник для ин-тов физкультуры
/ Под ред. В.В.Меньшикова, Н.И. Волкова. - М.: Физ-
культура и спорт, 1986. - 384 с.
32. Волков Н.И. Физиологические основы современных ме-
тодов развития выносливости. - М., 1961.
33. Волков Н.И. Биоэнергетика напряженной мышечной дея-
тельности человека и способы повышения работоспособ-
ности спортсменов: Автореф. дис.... док. биол. Наук. — М.
НИИНФ, 1990.- 101 с.
336
34. Волков Н.И., Карасев А.В., Хосни М. Теория и практика
интервальной тренировки в спорте. — М.: Воен. Академия
им. Дзержинского, 1995. — 196 с.
35. Волков Н.И., Олейничев В.И. Биологически активные
пищевые добавки в специализированном питании спор-
тсменов. - М.: СпортАкадемПресс, 2001 - 80 с.
36. Воробьев А.Н. Тяжелоатлетический спорт / Очерки по
физиологии спортивной тренировки. — М.: Физкультура
и спорт, 1977. - 255 с.
37. Воробьев А.Н. Тренировка, работоспособность, реабили-
тация. — М.: Физкультура и спорт, 1989. — 272 с.
38. Гетманец В.С. Специальная силовая подготовка бегунов
на длинные дистанции в годичном цикле: Автореф. дис...
канд. пед. наук. - М., 1985. - 25 с.
39. Гетманец В.С., Травин Ю.Г. Нужна ли стайеру сила //
Легкая атлетика. - 1987. - № 11. - С. 4-5.
40. Гилмор Г., Снелл П. Без труб, без барабанов. — М.: Физ-
культура и спорт, 1972. - 225 с.
41. Голник Ф.Д., Германсен Л. Биохимическая адаптация к
упражнениям: анаэробный метаболизм./ В кн. Наука и
спорт. Пер с англ. М.: Прогресс, 1982, С. 14-59.
42. Грузных Г.М. Экспериментальное исследование вынос-
ливости и методов ее совершенствования у спортсменов
с различными морфологическими особенностями: Авто-
реф. дис... канд. пед. наук. — М., 1971. — 23 с.
43. Гурфинкель В.С., Левик Ю.С. Скелетная мышца: структу-
ра и функции. — М.: Наука, 1985. — 233 с.
44. Гуморально-гормональные механизмы регуляции функ-
ций при спортивной деятельности / Кассиль Г.Н., И.Л.
Вайсфельд, Э.Ш. Матлина, Г.Л. Шрейберг. — М.: Наука,
1978.- 304 с.
45. Дембо А.Г. Актуальные проблемы современной спортив-
ной медицины. М.: Физкультура и спорт, 1980. — 295 с.
46. Дембо А.Г. Заболевания и повреждения при занятиях
спортом. - Л.: Медицина, 1991. - 336 с.
47. Дин Р. Процессы распада в клетке. — М.: Мир, 1981. — 154 с.
48. Донской Д.Д. Законы движений в спорте. - М.: Физкуль-
тура и спорт, 1968. - 176 с.
337
49. Дьячков В.М. Совершенствование технического ма-
стерства спортсменов. — М.: Физкультура и спорт, 1972.
-231 с.
50. Еркомайшвили И.В. Планирование физической подго-
товки конькобежек групп спортивного совершенствова-
ния: Автореф. дис... канд. пед. наук. - М., 1991. - 23 с.
51. Журбина А.Д. Экспериментальное исследование особен-
ностей силовой подготовки женщин, специализирующих-
ся в беге на средние дистанции: Автореф. дис... канд. пед.
наук - М., 1976. - 24 с.
52. Захарченко С.А. Методика воспитания силовой выносли-
вости бегунов на длинные дистанции 15-18 лет в годичном
цикле тренировки: Автореф. дис... канд. пед. наук. — М.,
1986.- 23 с.
53. Зациорский В.М. Физические качества спортсмена — М.:
Физкультура и спорт, 1966. — 250 с.
54. Зациорский В.М., Алешинский С.Ю., Якунин Н.А. Био-
механические основы выносливости. — М,: Физкультура и
спорт, 1982. - 207 с.
55. Казарян Ф.Г. Особенности возрастной динамики мышеч-
ной силы и проблема рационализации силовой подготов-
ки в школьном возрасте: Автореф. дис... докт. пед. наук —
М., 1975.-47 с.
56. Казлаускас В.А. Индивидуальная подготовка марафонцев.
— В.: Минтис, 1982. — 136 с.
57. Кайтамазова Е.Н., Теннов В.П. Легкая атлетика за рубе-
жом. — М.: Физкультура и спорт, 1974. — 320 с.
58. Кароблис П.Б. Основы системы управления тренировоч-
ным процессом бегунов высокой квалификации на сред-
ние, длинные дистанции и 3000 м с препятствиями: Авто-
реф. дис... докт. пед. наук — М., 1986. — 41 с.
59. Карпман В.Л. Сердечно-сосудистая система и транс-
порт кислорода при мышечной работе / Клинико-
физиологические характеристики сердечно-сосудистой
системы у спортсменов. — М.: РГАФК, 1994. — С. 11-41.
60. Карпман В.Л., Белоцерковский З.Б., Гудков И.А. Иссле-
дование физической работоспособности у спортсменов. —
М.: Физкультура и спорт, 1974. — 95 с.
338
61. Карпман В.Л., Любина Б.Г. Динамика кровообращения у
спортсменов. — М.: Физкультура и спорт, 1982. — 135 с.
62. Каунсилмен Д.. Спортивное плавание // М.: Физкультура
и спорт, 1982. - 208 с.
63. Классификация физических нагрузок в теории физиче-
ской подготовки / Селуянов В.Н., Сарсания С.К., Конрад
А.Н., Мякинченко Е.Б. // Теория и практика физической
культуры. - 1991. - № 12. - С. 2-8.
64. Колчинская А.З. Кислородные режимы организма ребен-
ка и подростка. — Киев: Наукова думка, 1973. — 320 с.
65. Кондратов Н.Н. Скоростно-силовая подготовка
лыжников-гонщиков : Автореф. дис... канд. пед. наук. —
М., 1987.- 23 с.
66. Коптелов А. Ю. Программирование и методика совершен-
ствования специальной силовой подготовки конькобеж-
цев в группах спортивного совершенствования: Автореф.
дис... канд. пед. наук. - М., 1991. - 22 с.
67. Коробов А.Н. Исследование эффективности применения
изометрического метода в системе тренировки юных бегу-
нов на средние дистанции: Автореф. дис... канд. пед. наук.
-М., 1977.- 28 с.
68. Коробов А.Н., Р.К. Минкявичус. Соотношение трениро-
вочных средств в годичной подготовке квалифицирован-
ных гребчих // Факторы, лимитирующие повышение ра-
ботоспособности у спортсменов высокой квалификации /
Под. ред. М.А. Годика и А.Н. Коробова. - М.: РИО ГЦО-
ЛИФК, 1985.-С. 64-73
69. Коробов А., Селуянов В., Волков Н. Бег на средние дис-
танции //Легкая атлетика. - 1985. - № 12. - С. 6-9.
70. Коробова А.А. Влияние различных сроков неактивности на
сохранение физических качеств у спортсменов // Теория и
практика физической культуры. — 1972. — №1. — С. 10-14.
71. КоэП. Знать об ученике все//Легкая атлетика, 1989. — №
8. - С. 44-45
72. Кузнецов В. В. Специальная силовая подготовка спор-
тсмена. — М.: Советская Россия, 1975. — 208 с.
73. КоцЯ.М. Спортивная физиология: Учебникдля институтов фи-
зической культуры. — М.: Физкультура и спорт. — 1986. — 240 с.
339
74. Кулаков В.Н. Программирование тренировочного про-
цесса высококвалифицированных бегунов на средние,
длинные и сверхдлинные дистанции: Автореф. дис... док.
пед. наук. - М., 1995. - 48 с.
75. Кулик Н.Н. Совершенствование путей повышения рабо-
тоспособности борцов-самбистов: Автореф. дис... канд.
пед. наук. - М., 1967. — 30 с.
76. Левин М.Я., Хрущев С.В. Предпатологические и патоло-
гические изменения неспецифической и специфической
реактивности при нерациональной организации спортив-
ных занятий //Детская спортивная медицина. — М.: Ме-
дицина, 1991.-С. 463-473.
77. Легкая атлетика: Учебник для ИФК / Под ред. Н.Г. Озоли-
на и др. - М.: Физкультура и спорт, 1989. — 671 с.
78. Легкая атлетика за рубежом / Под ред. Е.Н. Кайтмазовой.
— М.: Физкультура и спорт, 1974. — 430 с.
79. Ленинджер Р. Митохондрия. — М.: Мир, 1966. — 316 с.
80. Линец М.М. Воспитание выносливости в циклических ло-
комоциях субмаксимальной мощности в процессе много-
летней тренировки: Автореф. дис... канд. пед. наук. — М.,
1979.- 23 с.
81. Лузиков В.Н. Регулирование формирования митохон-
дрий: молекулярный аспект. — М.: Наука, 1980. — 316 с.
82. Майсурадзе М.И. Влияние силовых упражнений на разви-
тие выносливости (на работе большой мощности): Авто-
реф. дис... канд. пед. наук. — М., 1962. — 14 с.
83. Макаров А.Н. Бег на средние и длинные дистанции: тех-
ника, тактика, тренировка. — М.: Физкультура и спорт,
1973.-240 с.
84. Масленников А.В., Мякинченко Е.Б., Козьмин Р.К., Па-
леный В.И. Оценка тренировочной нагрузки бегунов на
средние дистанции / Совершенствование системы подго-
товки легкоатлетов. - М.: ГЦОЛИФК, 1986. - С. 43-51.
85. Масленников А.В. Система этапного педагогического кон-
троля за физической подготовкой бегунов на средние дистан-
ции: Автореф. дис... канд. пед. наук. — Минск, 1986. — 160 с.
86. Марчук Г.И. Математические модели в иммунологии. —
М.: Наука, 1985.-240 с.
340
87. Матвеев Л.П. Основы спортивной тренировки. — М.: Физ-
культура и спорт, 1977. — 271 с.
88. Меерсон Ф.З. Адаптация, деадаптация и недостаточность
сердца. - М.: Медицина, 1978. - 339 с.
89. Меерсон Ф.З., Пшеничникова М.Г. Адаптация к стрессо-
вым ситуациям и физическим нагрузкам. — М.: Медици-
на, 1988.- 256 с.
90. Мелихова М.А. Динамика биохимических процессов при
мышечной деятельности: Учебное пособие. — М.: ГЦО-
ЛИФК, 1992.-60 с.
91. Молекулярная биология клетки / Албертс Б., Брей Д.,
Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. В Зт. 2-е изд.
Т.1.: Пер. с англ. — М.: Мир, 1994. — 517 с.
92. Моногаров В.Д. Преодолеваемое утомление у высококва-
лифицированных спортсменов-велосипедистов при рабо-
те различной мощности / Медико-биологические основы
оптимизации тренировочного процесса в циклических
видах спорта: Сборник науч. тр. КГИФК. — Киев, 1980. —
С.5-28.
93. Моногаров В.Д. Утомление в спорте. - К.: Здоров’я, 1986.
- 120 с.
94. Мусил Я., Новакова О., Кунц К. Современная биохимия в
схемах. — М.: Мир, 1984. — 216 с.
95. Мякинченко Е.Б. Техника бега на средние дистанции и
ее взаимосвязь с физической подготовленностью: Дис....
канд. пед. наук. — М., 1983. — 202 с.
96. Мякинченко Е.Б., Масленников А.В. Взаимосвязь силовых
показателей бегунов на выносливость с ритмовыми харак-
теристиками бега / Актуальные вопросы подготовки спор-
тсменов высокой квалификации. — Омск, 1986. — С. 33.
97. Мякинченко Е.Б., Милехин С.А., Паленый В.И. Влияние
длины шагов и темпа бега на аэробный и анаэробный по-
роги // Факторы, лимитирующие повышение спортивной
работоспособности у высококвалифицированных спор-
тсменов / Под ред. В.Н. Селуянова. — М.: ГЦОЛИФК,
1990.-С. 14-24.
98. Мякинченко Е.Б., Паленый В.И., Карпова О.И., Вино-
градов В.В. Динамика ЧСС и показателей газообмена при
341
беге на тредбане со ступенчато повышающейся скоростью
// Факторы, лимитирующие повышение спортивной ра-
ботоспособности у высококвалифицированных спортсме-
нов / Под ред. В.Н. Селуянова. — М.: ГЦОЛИФК, 1990. -
С.29-32.
99. Мякинченко Е.Б., Обухов С.А., Паленый В.И. Динамика
и вариативность легочной вентиляции и ЧСС в ступен-
чатых тестах с различной крутизной увеличения мощно-
сти // Факторы, лимитирующие повышение спортивной
работоспособности у высококвалифицированных спор-
тсменов / Под ред. В.Н. Селуянова. — М.: ГЦОЛИФК,
1990.-С.29-34.
100. Мякинчнко Е.Б. Локальная выносливость в беге. — М.:
Физкультура, образование, наука, — 1997. — 312 с.
101. Набатникова М.Я. Проблема совершенствования специ-
альной выносливости спортсмена при циклической рабо-
те субмаксимальной и большой мощности: Автореф. дис...
докт. пед наук — М., 1975.- 48 с.
102. Нарский Г.И.. Структура тренировочных нагрузок
скоростно-силовой направленности бегунов на средние
дистанции в годичном цикле тренировки // Автореф.
дис... канд. пед. наук. — Минск., 1988. — 24с.
103. Некрасов А.Н. Реакция систем синтеза РНК в сердце и
скелетной мышце на физическую нагрузку (гистохимиче-
ское исследование): Автореф. дис. канд. биол. наук — М.,
1982.- 23 с.
104. Немировская Т.Л., Шенкман Б.С., Некрасов А.Н., Вино-
градова О. Л., Тхоревский В.И., Белицкая Л.А Взаимосвязь
гематологических и мышечных показателей кислород-
транспортной системы с уровнем работоспособности у
людей, тренирующих выносливость. // Физиология чело-
века. - 1993. -Т. 19. - №1. - С. 27-33.
105. Немировская Т.Л. Влияние аэробной тренировки на си-
стему доставки кислорода и энергетический метаболизм
мышц человека: Автореф. дис.... канд. биол. наук. — М.,
1992.-23 с.
106. Немировская Т.Л., Б.С.Шенкман, А.Н.Некрасов, А.В.
Кузнецов, В.А.Сакс. Эффект физической тренировки на
342
структуру и метаболизм скелетных мышц у спортсменов //
Биохимия. - 1993. - Т. 58. - С.471-479.
107. Нурмякиви А.А. О применении продолжительного бега
и бега в гору в тренировке бегунов на средние и длинные
дистанции в подготовительном периоде: Автореф. дис...
канд. пед. наук. - Тарту, 1974. — 33 с.
108. Обухов С.М. Методика развития локальной мышечной
выносливости у бегунов на средние дистанции 13-17 лет:
Автореф. дис... канд. пед. наук. — М., 1991. — 23 с.
109. Озолин Н.Г. Современная система спортивной трениров-
ки. — М.: Физкультура и спорт, 1970. — 279 с.
ПО. Основны управления подготовкой юных спортсменов /
Под ред. М.Я. Набатниковой. — М.: Физкультура и спорт,
1982.- 237 с.
111. Паленый В., Мякинченко Е., Обухов С., Чесноков Н., Се-
луянов В. Как избежать перетренировки // Легкая атлети-
ка. - 1991. - №10. - С.19-21.
112. Паленый В.И. Динамика показателей специальной фи-
зической подготовки как критерий управления трениро-
вочным процессом в беге на выносливость: Автореф. дис...
канд. пед. наук. — Омск, 1991. — 18 с.
113. Панин Л.Е. Биохимические механизмы стресса. —
Новосибирск: Наука, 1983. - 234 с.
114. Петров Р.В. Иммунология. — М.: Медицина, 1987. — 416
с.
115. Персон Р.С. Спинальные механизмы управления мышеч-
ным сокращением. — М.: Наука, 1985. — 184 с.
116. Питание при занятиях оздоровительной тренировкой:
Учебн. пособ. для студ. ИФК, трен, инстр. ОФК / Селуя-
нов В.Н., Мякинченко Е.Б., Антохина Е.Г., Тураев В.Т. —
М.: РИО РГАФК, 1995. - 68 с.
117. Платонов В.Н. Теория и методика спортивной трениров-
ки. — Киев: Вища школа, 1984. — 352 с.
118. Платонов В.Н. Адаптация в спорте. — Киев: Здоровья,
1986.-286 с.
119. Платонов В.Н. Булатова М.М. Силовая подготовка спор-
тсмена : учебно-метод. пособие. Часть 1. — Киев.: КГИФК,
1992.- 52 с.
343
120. Платонов В.Н. Булатова М.М.. Силовая подготовка спор-
тсмена: учебно-метод пособие.Часть 3. -Киев.: КГИФК,
1992.-42 с.
121. Платонов В.Н. Общая теория подготовки спортсменов в
олимпийском спорте. — Киев: Олимпийская литература,
1997.- 585 с.
122. Полунин А.И., Снесарев Н.К. Методические особенности
подготовки высококвалифицированных бегунов на длин-
ные дистанции (ретроспективный анализ). — М.: Совет-
ский спорт, 1990. — 45 с.
123. Попов Ю.А. Исследование средств силовой подготовки в
связи с развитием специальной выносливости: Автореф.
дис... канд. пед. наук. — М., 1968. — 22 с.
124. Прилуцкий Б.И. Мышечные боли, вызванные непривыч-
ными физическими упражнениями // Теория и практика
физической культуры. — 1989. — №2. — С. 16-22.
125. Пшенникова М.Г. Адаптация к физической нагрузке /
Физиология адаптационных процессов. — М.: Наука, 1986.
-С. 124-224.
126. Раппопорт Э.А., Казарян В.А. Адаптационно-
патологические изменения скелетных мышц при атрофи-
ческих реакциях и их аппаратурная и фармакологическая
коррекция / Прикладные аспекты исследований склет-
ных, сердечных и гладких мышц, 7-11 октября 1996 г. —
Путинский научный центр РАН, 1996. — С.71-72.
127. Розен В.Б. Основы эндокринологии. — М.: МГУ, 1994. — 384 с.
128. Сарсания С.К., Селуянов В.Н.. Физическая подготовка в
спортивных играх (хоккей на траве, футбол, хоккей с шай-
бой): Учебное пособие. — М.: НПЦ «Антей», 1991. С.84 с.
129. Сашенков С.Л., Исаев А.П., Волчегорский И.А., Волков
В.Н., Ариунцэцэг Н., Юсупов Х.М. Проблемы и критерии
адаптации спортсменов к экстремальным физическим
нагрузкам в динамике тренировочно-соревновательного
цикла подготовки // Теория и практика физической куль-
туры. - 1995. - №10. - С. 14-17.
130. Селуянов В.Н., Савельев И.А., Внутренняя механическая
работа при педалировании на велоэргометре // Физиоло-
гия человека, 1982. — 8. — 2. — С. 235 240.
344
131. Селуянов В.Н., Еркомайшвили И.В. Адаптация скелет-
ных мышц и теория физической подготовки // Научно-
спортивный вестник. — 1990. — С. 3-8.
132. Селуянов В.Н. Научно-методические основы подготовки
бегунов на средние дистанции высшей квалификации: Ме-
тодические рекомендации. — М.:ГЦОЛИФК, 1983. - 24 с.
133. Селуянов В.Н., Мякинченко Е.Б., Бикбаев И.З., Козьмин
Р.К., Прилуцкий Б.И., Цирков В.Н. Пути повышения эко-
номичности бега на средние дистанции // Теория и прак-
тика физической культуры. — 1989. — №11. — С.47- 49.
134. Селуянов В.Н., Мякинченко Е.Б., Холодняк Д.Г., Обухов
С.М. Физиологические механизмы и методы определения
аэробного и анаэробного порогов // Теория и практика
физической культуры. — 1991. — №10. — С. 10-18.
135. Селуянов В.Н. Методы построения физической подготов-
ки спортсменов высокой квалификации на основе имита-
ционного моделирования: Автореф. дис... докт. пед. наук
- 1992.-48 с.
136. Селуянов В.Н., Мякинченко Е.Б., Тураев В.Т. Биологи-
ческие закономерности в планировании физической под-
готовки спортсменов // Теория и практика физической
культуры. — 1993. — №7. — С. 29-34.
137. Селуянов В.Н. Мякинченко Е.Б. Основы теории оздоро-
вительной физической культуры: Учебное пособие для
инструкторов ОФК. — М.: «ИЗОТОН», 1995. — 35 с.
138. Селуянов В.Н., Зубкова А.В., Обухов С.М., Мякинчен-
ко Е.Б., Тураев В.Т. Имитационное моделирование фи-
зической подготовки бегунов на средние дистанции //
Организация автоматического тестового контроля зна-
ний студентов, г. Петропавловск, 26-27 ноября 1991 г. —
Петропавловск-Казахский: Петроп. пед. институт, 1995.
-С. 32-33.
139. Селуянов В.Н., Мякинченко Е.Б., Тураев В.Т., Обухов
С.М., Космина И.П., Зубкова А.В., Безденежных А.И.
Теория и практика применения дидактики развивающе-
го обучения в подготовке специалистов по физическому
воспитанию // Труды сотрудников ПНИЛ / Под рук. В.Н.
Селуянова. - М: ФОН, 1996. -106 с.
345
140. Современная система спортивной подготовки / Под ред.
Ф.П.Суслова, В.Л.Сыча, Б.Н.Шустина. - М.: СААМ,
1995.-448 с.
141. Солодков А.С. Проблемы утомления и восстановления в
спорте: Лекция. - С.-Пб.:ГДОИФК им. Лесгафта, 1992. —
34 с.
142. Стеценко Ю.Н. Экспериментальное обоснование основ-
ных направлений интенсификации тренировочного про-
цесса гребцов-байдарочников высших разрядов: Автореф.
дис... канд. пед. наук. — М., 1977. — 32 с.
143. Суркина И.Д., Готовцева Е.П. Роль иммунной системы в
процессах адаптации у спортсменов // Теория и практика
физической культуры — 1991. — №8. — С. 27-38.
144. Суслов Ф.П., Гилязова В. Б. Методика силовой подготовки
в циклических видах спорта, требующих преимуществен-
ного проявления выносливости: Методические рекомен-
дации. - ГКФКТ, ВНИИФК, ЦНИС, 1990. - 32 с.
145. Суслов Ф.П., Попов Ю.А., Кулаков В.Н., Тихонов С.А.
Бег на средние и длинные дистанции. — М.: Физкультура
и спорт, 1982. — 176 с.
146. Сээне Т.П. Адаптация мышечных клеток с высоким окис-
лительным потенциалом к тренировке на выносливость. —
Тарту: Тарт. гос. университет, 1987. — 51с.
147. Сээне Т.П., К.П Алев, М.М.Умнова, А.Я. Пехме. Измене-
ния скорости обновления сократительных белков в раз-
личных типах скелетных мышц в зависимости от характе-
ра тренировки. Структурно-энергетическое обеспечение
механической работы мышц: Тез. всесоюз. симпоз. (Мо-
сква, 23-25 марта 1990 г.) с. 64-65
148. Теппермен Дж., Теппермен X. Физиология обмена ве-
ществ и эндокринной системы: Вводный курс. — М.: Мир,
1989.-656 с.
149. Травин Ю.Г. Исследование закономерностей возрастных
изменений выносливости и построения многолетней тре-
нировки юных бегунов на средние дистанции: Автореф.
дис.... докт. пед. наук. — М., 1975. — 48 с.
150. Тураев В.Т., Мякинченко Е.Б., Селуянов В.Н. Планиро-
вание физической подготовки легкоатлетов-спринтеров
346
на основе имитационного моделирования / Труды ученых
ГЦОЛИФК. - М.: ГЦОЛИФК, 1993. - С. 188-193.
151. Тураев В.Т. Специальная силовая подготовка бегунов на
короткие дистанции: Автореф. дис... канд. пед. наук. — М.,
1996.-23 с.
152. Тюпа В.В. Исследование внутрицикловых биомеханиче-
ских характеристик спринтерского бега: Автореф. дис....
канд. пед. наук. — М., 1978. — 24 с.
153. Уголев А.М., Тимофеева Н.М., Груздов А.А.. Адаптация
пищеварительной системы / Физиология адаптационных
процессов. — М.: Наука, 1986. — С. 371-481.
154. Уголев А.М., Иезуитова Н.Н., Тимофеева Н.М. Энзима-
тический барьер тонкой кишки. // Физиоологический
журнал, 1992. — № 8. - С. 1-10.
155. Уилмор Дж.Х., Костил Д.Л. Физиология спорта и двига-
тельной активности. — Киев: Олимпийская литература,
1997.- 504 с.
156. Уткин В.Л. Биомеханические аспекты спортивной так-
тики / Под ред. В.М. Зациорского. — М.: Физкультура и
спорт, 1984. — 128 с.
157. Уткин В.Л. Энергетически обеспечение и оптимальные
режимы циклической мышечной работы.: Автореф. дис....
док. биол. наук. — М., МГУ, 1985. — 46с.
158. Физиология мышечной деятельности: Учебник для ин-тов
физ. Культуры / Под ред. Я.М. Коца. — М.: Физкультура и
спорт, 1982. - 500 с.
159. Физиология человека / Под ред. Р.Шмидта и Г.Тевса. —
Т.З.- М.: Мир, 1996.- 198 с.
160. Филин В.П., Фомин И.А. Основы юношеского спорта. —
М.: Физкультура и спорт, 1980. — 255 с.
161. Фомиченко Т.Г. Возрастные закономерности развития и
проявления силовых способностей в циклических водных
видах спорта. Автореф. докт. дис., 1999 — 54 с.
162. Функциональные системы организма: Руководство/ Под
ред. К.В. Судакова, — М.: Медицина 1987. — 432 с.
163. Холлоши Дж.О. Биохимическая адаптация к физической
нагрузке: аэробный метаболизм // Наука — спорту. Пер. с
англ. — М.: Прогресс, 1982. — С. 60-83.
347
164. Хоппелер Г. Ультраструктурные изменения в скелетной
мышце под воздействием. — М.: ЦООНТИ, Физкультура
и спорт, 1987. — Выпуск 6. — С. 3-48.
165. Хочачка П., Дж.Сомеро. Биохимическая адатация. — М.:
Мир, 1988.- 568 с.
166. Чебыкин Г.П. Методтка воспитания силовой выносливо-
сти высококвалифицированных гребцов: Автореф. дис...
канд. пед. наук. — М., 1991. — 23 с.
167. Чесноков Н.Н. Планирование скоростно-силовой и бе-
говой подготовки у юных бегунов на средние и длинные
дистанции в макроцикле: Автореф. дис... канд. пед. наук.
-М., 1991.- 23 с.
168. Шварц В.Б., Хрущев С.В. Медико-биологические аспек-
ты спортивной ориентации и отбора. — М.: Физкультура и
спорт, 1984. — 151 с.
169. Шенкман Б.С. Влияние тренировки на композицию мышц,
размеры и окислительный потнециал мышечных волокон у
человека: Автореф. дис... канд. биол. наук. — М., 1990. — 23 с.
170. 170. Шенкман Б.С., Некрасов А.Н., Головачев А.И. Раз-
меры мышечных волокон и аэробные способности спор-
тсменов, тренирующих выносливость / Структурно-
энергетическое обеспечение механической работы мышц.
-М., 1990.-С.71-72.
171. Шенкман Б.С., Ширковец Е.А., Кузнецов С.Л. и др.. Раз-
меры и окислительный потенциал мышечных волокон у
гребцов // Научно-спортивный вестник. — 1990. — № 6. —
С.33-35.
172. Шенкман Б.С. Влияние тренировки на композицию
мышц, размеры и относительный потенциал мышечных
волокон у человека: Автореф. дис.... канд. биол. наук. —
М., 1990.-23 с.
173. Шубик В.М., Левин М.Я. Иммунологическая реактив-
ность юных спортсменов. — М.: Физкультура и спорт,
1982.- 136 с.
174. Шубик В.М., Левин М.Я. Иммунитет и здоровье спор-
тсменов. — М.: Физкультура и спорт, 1985. — 171 с.
175. Ээпик В.А., Виру А.А.Специфика адаптивного протеино-
синтеза при выполнении упражнений на выносливость //
348
Теория и практика физической культуры. — 1990. — №5.
- С.24-27.
176. Яковлев Н.Н. Очерки по биохимии спорта. — М., 1955. —
311с.
177. Яковлев Н.Н. Химия движений: молекулярные основы
мышечной деятельности. — Л.: Наука, 1983. — 191 с.
178. Abernethy P.J., Thayer R., Taylor A.W. Acute and chronic
responces of skeletal muscle to endurance and sprint exercise: a
review. //Sports Medicine/ -1990. -V.10. — P. 365-389.
179. An approach to biomedical profiling of elite runners / Cavanagh
P.R., Andrew G.C., Kram R., Rodgers M.M., Sanderson D.J.,
Hennig E.M. // Int. J. Sports Biomech. — 1985. — N1. — P. 36-62.
180. Astrand P.-О., Rodahl K. Textbook of Warie Physiology. //
New York: McGraw. — Hill Book Co., 1970. — 669 p.
181. Bangsbo J. Regulation of muscle glycogenolysis and glycolysis
during intense exercise: in vivo studies using repeated intense
exercise // In: Biochemistry of Exercise IX /R.J. Maughan, S.M.
Shirreffs eds. — Human Kinetics Publ., 1996. — P. 261-275.
182. BellG.J., H. A. Wenger. The effect of one-legged sprint training
on intramuscular pH and nonbicarbonate buffering capacity //
Eur. J. Appl. Physiol. - 58. - 1988. - P. 158-164.
183. Bloor C.M., A.S. Leon, S. Pasyk. The effects of exercise on
organ and cellular development in rats // Lab. Invest. — 9.- N6.
- 1968.-P. 675-680.
184. Both F.W. Enzymatic capacities of skeletal muscle: effects of
different type of training// Med. Sport. — 13.— 1981. —P. 154-
164.
185. Bradley J.L., N.C. Spurway and S.Hood. Preliminary
investigation into the effect of exercising small and lerge muscle
groups on the blood flow and extracellular lactate concentrations
of the forearm // Journal of Sports Sciences. - 12. — N2. —
1994.-P.129-130.
186. Brandon L.J., R.A.Boileau.The contrebution of selected
variables to middle and long distance run performance //
J.Sports Med. - 27. - 1987. - P.157-164.
187. Brzank K.D. and K.S. Pieper. Characteristics of muscle-cellular
adaptation to intense loads. // Biomed. Biochem. Acta. — 45. —
N1-2-P.- 1986.-P. 107-110.
349
188. Brzank K.D. and K.S. Pieper. Mitichondrial content and
diffusion distances in hypertrophic skeletal muscle of well
trained man. // Act. Steriol. - 6/Ш. — 1987. — P. 503-
507.
189. Cabric M., Appell H.-J., Resic A. Fine structural changes in
electrostimulated human skeletal muscle // Eur J Appl Physiol.
-V.57. - 1988. -P.1-5.
190. Cavanagh P.R.,M.L.Pollock, J.Landa. A biomechanical
comparison of elite and good distance runners. // Ann. New
YorkAcad. Sci. - V.301. - 1977. - P. 328-345.
191. Cellular mechanisms of fatigue in skeletal muscle /
Westerblad H., J.A. Lee, J.Lannergren, and D.G. Allen. //
Am.J. Physiol. V.261 (cell Phisiol.30). - 1991. - P.C195-
C209.
192. Conconi F., Ferrary M., Ziglio P.G., Droghetti P., Codeca L.
Determination of the anaerobic threshold by a noninvasive field
test in runners // J. Appl. Physiol.:Respirat. Environ. Exercise
Physiol. - V. 52. - N4.-1982. - P. 869-873.
193. Costill D.L., Finck B., Pollock M.. Muscular fibers composition
and enzimatic activity in elite long distance runners // Medicine
and Science in Sports. — 1976. — N2. — P. 121-130.
194. Coyle E.F., M.E. Feltner, S.A. Kantz. Physiological and
biomechanical factors associated with elite endurance cycling
performance // Med. Sci. Sports. Exercise. — V.23. — Nl.
-1991.-P. 93-107.
195. Daub W. P., H. J. Green, Houston M. E.Cross-adaptive responses
to different forms of leg training: skeletal muscle biochemistry
and histocemistry // Can. J. Physiol. Pharmacol. — V. 60. — N5.
-1982.-P. 628-633.
196. Decreasing in skeletal muscle myoglobin with intensive training
in man / Terrados N., Melichna J., Sylven C., Jansson J. //
Acta Physiol Scand. - V.128. - 1986. - P.651-652.
197. Devis J.M. Central and peripheral factors in fatigue. // Journal
of Sport Sciences. -V. 13. - 1995. - P. S49-S53.
198. Distribution of blood flow in muscles of miniature swine during
exercise / Armstrong R.B. Delp M.D. Goljan E.F. Laughlin
M.N.// J Appl Physiol: Respirat Environ Exercise Physiol. —
62. - 1987.-P. 1285-1298.
350
199. Docherty D., Wenger H.A., Collis N.L. The effect of resistance
trainng on aerobic and anaerobic power of young boys // Med.
Sci. Sports. Exerc. - V.19.- 1987. - P. 389-392.
200. Duchateau J., S.L. Bozec, K.Hainaut. Contribution of slow
and fast muscles of triceps surae to a cyclic movement // Eur. J.
Appl. Physiol. -V.55. - 1986. -P.476-481.
201. Dudley G.A., R.Djamil. Incompatibility of endurance and
strength-training modes of exercise // J.Appl.Physiol. — 59. —
N5.-985.-P.1446-1451.
202. Effect of endurance training on possible determinants of Vo2
during heavy exercise / Casabuti R., Storer T.W., Bendov I.,
Wasserman K. // J.Appl. Physiol. — V.62. — 1987. — Nl. —
P. 199-207.
203. Enoka R.M., Stuart D.G.. Neurobiology of muscle fatigue //
J.Appl.Phy. - 72. - 1992. - P.1631-1648.
204. Enzymatic adaptations consequent to long-term strength
training / Tesh P.A.,Komi P.V.,and Haakinen K. // Int. J.
Sports Med. - V.8.- 1987. - P. 66-69.
205. Eston R.G., N. Critchly and V. Baltzopoulos. Delayed-onset
muscle sorenes, strength loss characteristics and creatine kinaze
activity following uphill and downhill running // Journal of
Sports Scienses. - 1994. - 12. - N2. - P.134-135.
206. Exercise induced morphological and biochemical changes in seletal
muscle of the rat / Watt P., Kelly F.G., Goldspink D.F., Goldspink
G. //J.Appl. Physiol. - V.53. - 1982. - N5. - P. 1144-1151.
207. Extrime endurance training: evidance of capillary and
mitochondria compartmentalization in human skeletal muscle
/ Orenshaw A.G., Friden J., Thornell L., Hargens A. // Eur. J.
Appl. Physiol. -V.63.- 1991. - N3. - P. 173-178.
208. Fitzgerald L. Exercise and immune system. // Immunology
today. - 1988. - 9. - Nl 1. - P. 337-340.
209. FleckS.J., KraemerWJ. Designing resistance training programs
// Champaign, IL, Human Kinetics, 1987. — 264 p.
210. Fox E.L., Mathews D.K.. The physiological basis of physical
education and athletics // 3d ed. Philadelphia: CBS College
Publ., 1981.-677 p.
211. Friden, j., J.Seger, M. Sjostrom and B. Ekblom. Adaptive
response in human muscles subjected to prolonged eccentric
351
training // International Journal of Sports Medicine. — 1983.
-4.-P. 177-183.
212. Freedson P.S. The influence of hemoglobin concentration on
exercise cardiac output. — Int. J. sports med., 1981. — v. 2. — N
2.-p. 81-86.
213. Gaesser GA, Brooks GA. Metabolic basis of excess post exercise
oxigen consumption: a revew // Med Sci Sports Exerc. — 1984.-
N16.-P.29-43.
214. Galbo, H.. Endocrinology and metabolism in exercise //Intern.
J. of Sports Med. - 1981. - 2. - P. 203-211.
215. Gayeski T.E.J., C.R.Honig. 02 gradients from sarcolemma to
cell interior in red muscle at maximal VO2 // Am.J.Physiol. 251
(Heart Circ. Physiol.20). - 1986.- P. H789-H799.
216. Gollnick P., Piehl K., Saltin B. Selective glicogen depletion
pattern in human muscle fibers after exercise of varying intensity
and at various pedalling rates // J. Physiol. London. — 1974. —
V. 25. - P.45-57.
217. Gollnick P.D.. Metabolic regulation in skeletal muscle:
influence of endurance training as exerted by mitochondrial
protein cocentration //Acta Physiol Scand. — 128 (Suppl 556).
- 1986.-P.53-66.
218. Graham T.E. and B.Saltin. Estimation of the mitochondrial
redox state in human skeletal muscle during exercise // J.Appl.
Physiol. - 1989. - 66. - N2. - P.561-566.
219. Green H.J. Neuromuscular aspects of fatigue // Canadian J.
Sport. Sci.- 1987.- 12.-P.7-19.
220. Green H.J.. What is the physiological significance of training-
induced adaptations in muscle mitohondrial capacity // In:
Biochemistry of Exercise IX /R.J.Maughan, S.M.Shirreffs eds.
— Human Kinetics Publ., 1996. — P. 345-360.
221. Greenhalf P.L.,K.Bodin, A.Casey et.al.. Dietary Creatine
supplementation and fatigue during high-intensity exercise in
humans // In: Biochemistry of Exercise IX /R.J.Maughan,
S.M.Shirreffs eds. — Human Kinetics Publ., 1996.- P. 219-
243.
222. Guezennec C.Y., Lienhard F., Donisy F.. In situ NADH laser
fluorimetry during muscle contraction in humans // Eur.J.Appl.
Physiol. - 1991. - 63. - Nl. - P. 36-42.
352
223. Hakkinen К., H.Kauhanen and P.V.Komi. Aerobic, anaerobic,
assistant exercise and weightlifting performance capacities in elite
weightlifters//J.Sports Med. - 1987. - 27. - P. 240-246.
224. Hakkinen K., K.L.Keskinen. Muscle cross-sectional area and
volantory force production characteristics in elite strength-
and endurance-trained athletes and sprinters // Eur. J. Appl.
Physiol. - 1989. - 59. - P. 215-220.
225. Hargreaves M.. Exercise related muscle metabolism //
Proceedings of the Australian Physiol, and Pharm. Society. —
1989. — 20. — Nl. — P.26A.
226. Henneman, E. Somjen, G. and Carpenter, D.O. Ecxcitability
and inhibitability of motoneurones of different sizes // Journal
of Neurophysiology. — 1965. — 28. — P. 599-620.
227. Henriksson J., J.S.Reitman. Time course of changes in human
skeletal muscle succinate dehydrogenase activities and maximal
oxigen uptake with physical activity and inactivity // Acta
physiol. Scand. - 1977. - 99. - P. 91-97.
228. Hickson C., R.C.Rosenkoctter, M.M.Brown. Strength training
effects on aerobic power and sport-term endurance // Med.Sci.
Sports. - 1980. - 12. - P. 336-339.
229. Hill F.V., Long C.N.H., Lupton H. Muscular exercise, lactic
acide and the supply and utilization of oxygen. IV. The oxigen
debt at the end of exercise // Proc. R. Soc. London. Ser. —
V.B97. - 1924.-P. 127-137.
230. Hill D.W.; Williams C.S.; Burt S.E. Responses to exercise at 92%
and 100% of the velocity associated with V0(2)max // International
journal of sports medicine. — 1997. — 18. — N5. — P. 325-329.
231. Hirvonen J., Rehunen S., Rusko H., Harkonen M. Breakdown
of high-energy phosphate compounds and lactate accumulation
during short supramaximal exercise // Eur.J.Appl.Physiol. —
1987.-56.-P.253-259.
232. Holloszy J.O. Regulation of carbohydrate metabolism during
exercise: new insights and remaining puzzles//In: Biochemistry
of Exercise IX /R.J.Maughan, S.M.Shirreffs eds. — Human
Kinetics Publ., 1996. - P.3-12.
233. Holloszy J.O., Booth F.W. Biochemical adaptations to
endurance exercise in muscle // Ann. Rev. Physiol. — 1976. —
38.-P. 273-291.
353
234. Holloszy J.О., and Coyle E.F. Adaptations of skeletal muscle to
endurance exercise and their metabolic consequence // J.Appl.
Physiol. - 1984. - 56. - P. 831-838.
235. Houston M., Bentzen H., Larsen H. Interrelationships between
skeletal muscle adaptations and performance as studied by
detraining and retraining // Acta Phisiol. Scand. — V. 105. —
1979.-P. 163-170.
236. Hughes EF, Turner SL, Brooks GR. Effect of glycogen depletion
and pedalling speed on ’’anaerobic threshold” // J Appl Physiol:
Respirat Environ Exercise Physiol. — 1982. — 52. — P. 1598-1607.
237. Human muscle metabolism during sprint running / Cheetham
M.E., Boobis L.H., Brooks S., Williams C. // J.Appl.Physiol.
-V.61.-1986.-NL-P.54-60.
238. Hunter G., R.Demment and D.Miller. Developing of strength
and maximum oxygen uptake during simultaneous training for
strength and endurance // J. Sports Med. and Phys. Fitness. —
1987.-27.-N3.-P. 269-275.
239. Ingier F.. Effects of endurance training on muscle fibre ATP-
ase activity, capillary supply and mitochondrial content in man
//J.Physiol (Lond.). - 1979. - 294. - P. 419-432.
240. Immediate-early-gene expression in human muscle fibers after
a single exercise bout / Puntschart A., Vogt M., Wey E., Jostamt
K., Hoppeler H., Billeter R. // First Ann. Congr. Frontiers in
Sport Sci. The Europe Perspective. — University of Nice, 1996.
- P. 208-209.
241. Is hypoxia a stimulus of oxidative enzymes and myoglobin? /
TerradosN., Sylven C., Jansson J., Kaijser L. //J Appl Physiol.
-V.68.- 1990.-P. 2369-2372.
242. Jackson C., G.Ratzin, A.L. Dickinson, S.P. Ringel. Skeletal
muscle fiber area alterations in two opposing modes of resistance
exercise training in the same individual // Eur. J. Appl. Physiol.
— 1990. — 61. - Nl. — P. 37-41.
243. Jorfeldt L.,Juhlin-Dannffelt A., Karlsson J. Lactate realease
in relation to tissue lactate in huiman skeletal muscle during
exercise // J. Appl. Physiol. Resp. Envir. Ex. Physiol. -V. 44.-
1978.-N3.-P.350-352.
244. Kavashina K., J-Zawa M. Stimulation of RNA synthesis in
nucleol bu inorganic phosphate in vitro // Biochem. and
354
Biophys. Res. Commun. - V.104. - 1982. - N4. - P. 1167-
1174.
245. Kiens B., Essen-Gustavsson B., Christensen N.J., Saltin B..
Skeletal muscle substrate utilization during submaximal exercise
in man: effect of endurance training // Journal of Physiology. —
1993.-469.-P. 459-478.
246. Krogh A.. The number and distribution of cappillaries in muscle
with calculations of the oxygen pressure head necessary for
supplying the tissue // J. Physiol. (London). - 1918. - 52. -
P.409-415.
247. Lakomy H.K.A.. The use of non-motorized tredmill for
analysing sprint performance // Ergonomics. — 1986. — 30. —
N4. - P. 627-637.
248. Lactate, pyruvate, lactate-to-pyruvate ratio during exercise and
recovery / Wasserman K., Beaver W.L., Devis J.A., Jung-Zong
Pu, Heber D., and Whipp B.J. // J.Appl.Physiol. — V.59. —
1985.-N3.-P. 935-940.
249. Londraville R.L., Siddel B.D.. Maximal diffusion distance
witthin skeletal muscle can be estimated from the motochondrial
distribution// Resp. Physiol. — 1990. — N3. - P.291-301.
250. Lukaszewska,-J.-H; Wojcieszak,-!; Wojczuk,-J;
Wysokinska,-B; Krogulski,-A; Sienicka,-M; Zalewska,-M;
Rucinski,-A. Aerobic capacity and release of hormones in
exercise // Biology-of-sport. - 1989. - 6(Suppl. 3). - P.82-92.
251. Luthi,J.M.,Howald,H.,Claassen,H.,Rosler,K.,Vock,P.,and
Hoppeler,H. Structural changes in skeletal muscles tissue with
heave-resistance exercise // Int.J.Sports Med. — 1986. — 7. —
P.123-127.
252. MacDougal,J.D., Sale,D.G., Moroz,J.R., Elder,C.B.,
Sutton,J.R., Howald,H. Mitochondrial volume density in
human skeletal muscle following heavy resistance training //
Med.Sci.Sports. - 1979. - 11. - P. 164-166.
253. Medbo J.L, I.Tabata. Relative importance of aerobic and
anaerobic energy release during short-lasting exhausting bicycle
exercise//J.Appl.Physiol. - 1989.-67. - P. 1881-1886.
254. Minotti J.D., Jonson E.C., Hydson T.L., et. al.. Training indused
skeletal muscle adaptations are independent of systemic adaptations
//J.Appl. Physiol. - 1990. - 68. - N1. - P. 289-294.
355
255. Mitochondrial respiratory control by oxygen in passiv and active
states. / Gnaiger T., Lassing B., Kuznetsov A., Margraiter R. /
Proceed, of Second. Ann. Congr. of ECSS (Aug 20-23), 1997.
— Denmark, Copenhgagen, 1997. — C. 504-505
256. Miura M, K.Kobagashi, M.Migashita, H.Matsui. Experimental
studies on biomechanics in long distance running // —
Department of Phis.Education, Univer. of Nagoya, Japan,
1973.-P. 46-56.
257. Miyashita Y, Kanehisa H, Nemoto J. EMG related to anaerobic
threshold // Quarterly review. — 1981. — 21. — P. 209-216.
258. Molecular events in human skeletal muskle adaptation /
Vogt M., Puntschart A., Widmer H.R., Wey E., Jostamdt K.,
Hoppeler H., Billet R. // Second Ann. Congr. of the ECSS, 20-
23 Aug 1997. Book of Abstracts I. — University of Copenhagen,
1997.-P. 378-379.
259. Moreau D.,P.Dubots, V.Boggio, J.C.Guilland and G.Cometti.
Effects of electromyostimulation and strength training on
muscle soreness, muscle damage and sympathetic activation //
Journal of Sports Sciences. — 1995. — 13. — N2. — P. 95-100.
260. Morton R.H., G.C.Gass. A sistems model approach to the
ventilatory anaerobic threshold // Eur.J.Appl.Physiol. — 1987.
- 56. - - P. 367-373.
261. Muscle metabolism, blood lactate and oxygen uptake in steady
state exercise at aerobic and anaerobic threshold / Rusko H.,
Luhtanen P., Rahkila P., Viitasalo J., Rehunen S., Harkonen
M. U Eur. J. Appl. Physiol. - V.55.- 1986. - P. 181-186.
262. Musch TJ, Friedman DB, Pitetti KH, Haidet GC, Stray-
Gundersen J, Mitchell JH, Ordway GA. Regional distribution
of blood flow of dogs during graded dynamic exercise // J. Appl.
Physiol: Respirat Environ Exercise Physiol. — 1987. — 63. — P.
2269-2277.
263. Nelson R.C., R.J.Gregor. Biomechanics of distance running. A
longitudinal study // Res. Quart. — 1976. — 47. — N3. — P.417-
428.
264. Nevill M.E., G.C.Bogdanis, L.H.Boobis at. al.. Muscle
metabolism and performance During Sprinting // In:
Biochemistry of Exercise IX /R.J.Maughan, S.M.Shirreffs eds.
— Human Kinetics Publ., 1996. — P. 243-259.
356
265. Newham D.J., Janes D.A., Clarkson J. Repeated high force
eccentric exercise: effects on muscle pain and damage // J.
Appl. Physiol. - V.63. - 1987. - N4. - P. 1381-1386.
266. Ohlsson J., B.Wranne. Non invasive assesment of cardiac output
and stroke volume in patients during exercise. Evaluation of
a CO2-rebreathing method // Eur.J.Appl.Physiol. - V.55. —
1986.-P. 538-544.
267. Orr GW, Green HJ, Hughson RL, Bennet GW. A computer
linear regression model to determine ventilatory anaerobic
threshold//J Appl Physiol. - V.52.- 1982. - P. 1349-1352.
268. Phosphocreatine content in single fibers of human muscle after
submaximal exercise / Sahlin, K; Soderlund, K; Tonkonogi, M;
Hirakoba, K. // Am. J. Physiol. - Cell Phys. - V.42. - 1997. -
Nl. — P. C172-C178.
269. Rossiter H. et al, 1996
270. Sahlin K.Muscle Fatigue and Lactic Acid Accumulation //Acta
Phisiol Scand. - V.108(Suppl 556).- 1986. - P. 83-91.
271. Saltin B. Physiological adaptation to physical conditioning. Old
problems revisited // Acta Med Scand Suppl. - V.711.- 1985.
-P. 11-24.
272. Schumacker P.L., R.W.Samsel. Analysis of oxygen delivery and
uptake relationships in the Krog tissue model //J.Appl. Physiol.
- V.67. - 1989. - N3. - P. 1234-1244.
273. Schwarz, L; Kindermann, W. Beta-endorphin, catecholamines,
and cortisol during exhaustive endurance exercise // Intern. J.
of Sports Medicine. -V. 10.- 1989. - N5. - P.324-328.
274. Seruum and urinary markers of skeletal muscles tissue damage
after weight lifting exercise / Paul, G.L., J.P. De Lany, J.T.
Sneek, J.G.Seifert and T.E. Kirby //Eur. J. Appl. Phisiol. —
V.58.- 1989. - P. 786-790.
275. Shibata, M; Oda, S; Moritani, T. The relationships between
movement-related cortical potentials and motor unit activity
during muscle contraction // J. Electromiogr. and kinez. -
V.7.- 1997.-N2.-P. 79-85.
276. Skeletal muscle glycogenolisis, glycolisis, and pH during
electrical stimulation in man / Spriet L.L., Soderlund K.,
Bergstrom M., Hultman E. // J.Appl.Physiol. - V.62. — 1987.
-N2.-P. 616-621.
357
Til. Specifity of the anaerobic threshold in endurance trained
cyclists and runners / Withers R.T., Sherman V.M., Miller
J.M., Costill D.L. // Eur.J.Appl.Physiol. - V.47. - 1981. - Nl.
-P. 93-104.
278. Tanaka K., Y.Matsuura. Marathon performance, anaerobic
threshold and onset of blood lactate accumulation // J. Appl.
Physiol.: Respirat. Environ. Exercise Physiol. — V.57. — 1984.
-N3.-P. 640-643.
279. Taylor C.P. Structural and functional limits to oxidative
metabolism: insight from scaling //Ann. Rev. Physiol. — V.49.-
1987.-P. 135-146.
280. Tesch,A., Karlsson J. Muscle fiber types and size in trained and
untrained muscles of elite athletes // J. Appl. Physiol. — 59. —
1985.-N6.-P. 1716-1720.
281. The influence of weekly training distance on fractional utilization
of maximum aerobic capacity in marathon and ultramarafon
runners / Scrimgeour A.G., Noakes T.D., Adams B., Myburgh
K. // Eur J.Appl. Physiol. - V.55.- 1986. - P. 202-209.
282. Three myosine heave-chain isozymes appear sequentially in rat
muscle development/Whalen R.G., Sell S.M., Butter-Browne
G.S., Shwartz K., Bouweret P., Pinset-Harstrom I. // Nature.
— V.292. — 1981. -P.805-809.
283. Vollestad NK, Blom PCS. Effect of varying exercise intensity
on glycogen depletion in human muscle fibres // Acta Physiol
Scand. - V.125.- 1985. - P. 395-405.
284. Williams R.K., P.R.Cavanagh relationship between distamce
running mechanics, running economy and performance // J.
Appl. Physiol. -V.63. - 1987. -N3. - P. 1236-1245.
285. Williams R.S., S.Salmons, E.A.Newsholme. Regulation of
nuclear and mitochondrial gene expression by contractile
activity in sceletal muscle // J.Biol.Chem. — V.261. — 1986. —
Nl.-P. 376-380.
358
Наука — спорту
Мякинченко Евгений Борисович
Виктор Николаевич Селуянов
РАЗВИТИЕ ЛОКАЛЬНОЙ
МЫШЕЧНОЙ ВЫНОСЛИВОСТИ
В ЦИКЛИЧЕСКИХ ВИДАХ СПОРТА
Подписано в печать 14.10.2009. Формат 60x84 !/|6.
Гарнитура «Newton». Усл. печ. л. 40. Бумага офсетная.
Печать офсетная. Тираж 1 000 экз.
ООО «ТВТ Дивизион»
По всем вопросам обращаться: sportbooks@mail.ru
Заказ 1299.
Отпечатано в ООО «Типография «САРМА»,
г. Подольск, ул. Правды, д.ЗО
НАУКА-СПОРТУ
Е.Б. Мякинченко, В.Н. Селуянов
РАЗВИТИЕ ЛОКАЛЬНОЙ
МЫШЕЧНОЙ ВЫНОСЛИВОСТИ
В ЦИКЛИЧЕСКИХ ВИДАХ СПОРТА