/




















































































































































































































































Text
СВ.Петухов
БИО ЕХАННКА,
БИОНИ
И СИММЕТРИЯ
**
#
Издательство 1а>ка
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ МАШИНОВЕДЕНИЯ им. А. А. 13ЛАГОНРАВОВА
С. В. Петухов
БИОМЕХАНИКА
БИОНИКА
И СИММЕТРИЯ
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА 1981
УДК 57.01+577.4
Петухов С В. Биомеханика, бионика и симметрия.
М.: Наука, 1981. 240 с.
Кпнга посвящена явлениям симметрии в живой
природе и применению математических
методов симметрии для моделирования биологических
объектов и явлений. Особое внимание уделяется
конформным симметриям и числам Фибоначчи в биологических
телах, в частности в кинематической схеме тела
человека и животных. Отмечается, что с точки зрения
реализуемых в телах симметрии неживое вещество
оказывается частным случаем живого вещества.
Книга рассчитана на широкий круг иаучпых
работников, занимающихся исследованиями в области
биомеханики, биофизики, математической и теоретической
биологии, бионики.
Ответственный редактор
член-корреспондент АН СССР
К. В. ФРОЛОВ
п 80501—130
055(02)—81 72°—81« 1Ш- 2- 1502000000
(§) Издательство «Наука», 1981 г.
ВВЕДЕНИЕ
Настоящая книга посвящена главным образом одной
из древних проблем живой природы — проблеме
симметрии в формах биологических тел. Пользуясь
терминологией В. И. Вернадского [42—44], наметившего пути изучения
с позиций математического естествознания этой, по его
мнению, ключевой биологической проблемы, можно
сказать, что в книге изучаются симметрии пространства
живого вещества. В основу работы положена идея о
неевклидовом характере этого пространства, о фундаментальном
значении неевклидовых симметрии в структуризации
живого вещества, о неевклидовом базисе законов
морфогенеза.
Результаты проведенного автором исследования дают
основания использовать группу конформных (круговых)
симметрии в качестве фундаментальной группы
симметрии живого вещества при моделировании рассматриваемых
явлений. По мнению автора, многие загадки как
формирования биологических тел, так и всей биологии кроются в
конформном характере пространства живого вещества.
В этой связи в книге освещены данные о симметриях в
формах биологических тел. Отмечено, что
широкоизвестные в этих телах симметрии евклидового типа —
зеркальные, вращательные, трансляционные — являются лишь
частными случаями конформной симметрии, проявления
которой в формах биологических тел этим далеко не
исчерпываются. Автор полагает, что в живой природе
реализуется весьма общий принцип построения биологических
тел из конформно-симметричных блоков. В книге
представлена проведенная на некотором числе видов
биологических тел проверка этой гипотезы о специфических
кристаллизационных принципах продуцирования живого
вещества.
Излагаемые исследования тесно смыкаются с давно
обсуждаемым в литературе учением об обобщенной кри-
3
сталлографии, способным охватить особенности
Структуризации не только физического, цо и биологического
вещества. Однако осуществленное к настоящему времени
Дж. Берналом и С. Карлайлом [20] развитие этого учения
базируется на идее евклидовости основ обобщенной
кристаллографии. Принципиальным отличием освещаемого в
нашей книге подхода к вопросам структуризации живого
вещества является утверждение о неевклидовом характере
этой структуризации и о необходимости строить
обобщенную кристаллографию на неевклидовой (конформной)
основе. При этом с точки зрения реализуемых симметрии
неживое вещество выступает как частный случай живого
вещества.
Центральным биологическим объектом исследований,
представляемых в нашей книге, является двигательный
аппарат человека и высокоорганизованных животных.
Вопросам организации этого аппарата посвящены
многочисленные труды специалистов самых разных областей
естествознания, поскольку данные вопросы тесно связаны
с целым рядом фундаментальных биологических проблем,
а также прикладных проблем эргономики, протезирования,
спорта и пр. В частности, этот аппарат служит прототипом
многих бионических и робототехнических систем.
Спецификой освещаемого в книге исследования двигательного
аппарата человека и животных является изучение
отношений симметрии между составляющими его блоками.
На этом пути получен ряд новых интересных результатов
относительно закономерностей организации и
функционирования двигательного аппарата. Одним из результатов
является обнаружение конформных симметрии в блочном
строении кинематической схемы тела человека и
широкого класса животных, у которых эти симметрии
существуют наряду с зеркальной симметрией тела.
Книга состоит из пяти глав и ряда приложений. В
главе I представлены известные данные о биологических сим-
метриях. При этом обращается особое внимание на связь
биологических симметрии с рядом чисел Фибоначчи,
Излагаются общие сведения о фундаментальном значении
методов симметрии и инвариантно-групповых концепций в
современном математическом естествознании с целью
точнее определить место исследований по биологическим сим-
метриям в области общего естествознания. Делается
экскурс в известные учения о биологических симметриях и
обобщенной кристаллографии, распространяющих на жи-
4
вое вещество геометрические и кристаллографические
подходы теоретической физики.
Глава Л посвящена исследованию конформных
симметрии в биологии. Приводятся данные о конформных
симметриях в строении различных биологических тел.
Особое место в изложении отводится конформной симметрии в
блочных принципах строения кинематической схемы тела
человека и высокоорганизованных животных. Отмечается,
что эти симметрии, так же как и симметрии евклидового
типа в биологических телах, тесно связаны с рядом чисел
Фибоначчи и золотым сечением, известным в эстетике
пропорций. Представляемые данные говорят о том, что по
своим блочным особенностям кинематическая организация
нашего тела стоит гораздо ближе, чем обычно полагают,
к организации тел растений и даже биологических молекул,
вовсе лишенных кинематической функции. Значит,
кинематическая схема тела человека и животных построена
далеко не во всем по критериям кинематической
оптимальности и ее нельзя рассматривать в качестве
идеальной кинематической схемы, которой надлежит наделять и
роботы. Если уж и смотреть на кинематическую
организацию нашего тела как на патент живой природы, то с
учетом того, что при его развитии требования кинематики и
статики были далеко не единственными определяющими
условиями в отличие от задачи создания робота по
критерию кинематической оптимальности.
В этой же главе особенности конформной геометрии
связываются с вопросами генезиса пространственных
представлений у индивидуума, загадками геометрии
пространства зрительного восприятия, задачами архитектурной
бионики. Разрабатываются подходы к вопросам
организации моторных движений с учетом конформных
симметрии в кинематических блоках тела человека и животных.
Исследование конформных симметрии в биологии
сопровождается краткой сводкой данных о значении конформной
симметрии в физике, в частности данными о конформной
инвариантности уравнений электродинамики Максвелла,
уже на протяжении многих десятилетий вновь и вновь
привлекающей внимание физиков-теоретиков задачей
раскрытия ее физического значения и следствий. Отмечается
перспективность взаимосвязанного изучения проблем
конформной симметрии в физике и биологии.
В главе III приводятся материалы дополнительного
исследования проективных симметрии в биологии, предпри-
5
нятого при поиске фундаментальной группы симметрии
пространства живого вещества. Это исследование
обусловлено во многом тем, что конформные и проективные
симметрии математически взаимосвязаны, а также тем, что
нельзя априорным образом исключить возможность
участия проективных симметрии наряду с конформными в
структуризации биологических явлений.
В главе IV освещается применение
инвариантно-групповых концепций к изучению явлений психофизики. Эта
область биологии ранее других обратилась к высшей
геометрии как источнику идей по осмыслению наблюдаемых
явлений и как богатому арсеналу средств для их
формального моделирования. Основное внимание здесь уделяется
психологическим феноменам константности восприятия и
геометрическому моделированию пространств
восприятия. Анализируются некоторые примеры применения в
бионике результатов исследования биологических
симметрии.
В последней, V главе представлены дополнительные
материалы об использовании методов симметрии в
биомеханике. Речь идет о применении инвариантно-групповой
теории динамического подобия к моделированию
биомеханических систем и о разработанной автором на основе этой
теории масштабной физической модели системы
полукружных каналов вестибулярного аппарата человека.
В заключение хочется отметить, что к изучению
биологических явлений как нельзя более относятся слова
Р. Фейнмана: «Вы не найдете в природе ничего простого,
все в ней перепутано и слито. А наша любознательность
требует найти в этом простоту, требует, чтобы мы ставили
вопросы, пытались ухватить суть вещей и понять их мно-
голикость как возможный итог действия сравнительно
небольшого количества простейших процессов и сил, на
все лады сочетающихся между собой» [241, т. I, с. 37].
Представляется несомненным, что использование методов
симметрии неоценимо для познания биологических
явлений, для нахождения сути и простоты в этом сложнейшем
классе природных явлений. Как писал Г. Вейль,
«симметрия — в широком или узком смысле в зависимости от того,
как вы определите значение этого понятия, — является
той идеей, посредством которой человек на протяжении
веков пытался постичь и создать порядок, красоту и
совершенство» [40, с. 37].
6
Автор считает своим приятным долгом выразить
глубокую благодарность члену-корреспонденту АН СССР
К. В. Фролову за активную поддержку данного
направления исследований и разностороннюю научную и
организационную помощь автору в работе. Автор пользуется
случаем выразить также сердечную признательность
профессору В. С. Гурфинкелю, который на протяжении многих
лет являлся наставником автора в области биомеханики и
внес значительный вклад в настоящую работу. За помощь
в проведении исследований и подготовке книги искренняя
благодарность выражается докторам наук С. В. Мейену,
консультировавшему автора по вопросам морфологии;
Ю. А. Урманцеву, консультировавшему по вопросам
биологической симметрии; Э. М. Чудинову,
консультировавшему по проблемам физического пространства — времени;
кандидатам наук С. А. Владимирову,
консультировавшему по теории групп и ее приложениям; С. Г. Смирнову и
Ф. Р. Черникову, принявшим деятельное участие в
исследованиях и обсуждении результатов. Наконец, автор не
может не отметить с благодарностью большую роль
коллективов кафедры физики живых систем МФТИ и отдела
биомеханики ИМАШ АН СССР в проведении излагаемых
исследований.
Глава I
СИММЕТРИИ В БИОЛОГИ!
1.1. Симметрии в биологических телах
и законы листорасположения
Симметрия в формах биологических тел издавна вызывала
пристальный интерес естествоиспытателей как одно из
наиболее замечательных и загадочных явлений природы.
Ему посвящены труды многих современных ученых, среди
которых надо отметить прежде всего [40, 43, 236, 258, 438
и др.]. Проблемы биологических симметрии на макромоле-
кулярном уровне рассматривались в 1968 г. на
специальном Нобелевском симпозиуме [436]. Уже школьные
программы по биологии включают рассмотрение
многочисленных примеров зеркальных, вращательных, трансляционных
и спиральных симметрии, а также симметрии подобия в
биологических телах: опорно-двигательном аппарате как
простейших, так и высокоорганизованных животных,
цветках и побегах растений, раковинах моллюсков, многих
биологических молекулах и пр. Углубление биологических
знаний сопровождается открытием все новых и новых
фактов подчиненности очень разных биологических тел,
отличающихся масштабом и уровнем организации,
принципам симметрии.
^Надо отметить, что явления симметрии в природе
обычно носят приближенный характер. И подобно тому, как
теоретическая кристаллография базируется на
представлении об идеально симметричных кристаллах, хотя в
природе, как правило, встречаются кристаллы с нарушенной
симметрией, так и теоретическая биология исходит из
представлений об идеальных симметриях, хотя в биосим-
метриях обычно наблюдаются те или иные отклонения от
идеальной симметрии. Например, зеркальная симметрия
правой и левой половины опорно-двигательного аппарата
у человека при внимательном взгляде демонстрирует ряд
отступлений от идеальной зеркальной симметрии. Однако,
как всегда в учении о симметрии природных тел,
отклонения от идеальной симметрии рассматриваются как
вторичные, вызванные действием побочных, дополнительных
8
т "* - >
Рис. 1. Примеры спиральных биосимметрий
а, б — раковины моллюсков; в, г — спиральный цитоскелет клетки из
спиральных микротрубочек (из [94]); д — схематическое изображение побегов
растения с листорасположением 3/1 (слева) и 5/2 (справа)
9
10
факторов. Особенности и причины этих отклонений
составляют особый предмет изучения в биологии (см., например,
учение о диссфакторах Ю. А. Урманцева [237]).
Предпринятые за последние десятилетия попытки осмыслить
явления биологической симметрии с теоретико-групповых
позиций математического естествознания привели к
постановке ряда фундаментальных вопросов о специфике
биологического пространства и времени (В. И. Вернадский
[42, 43 и др.]), а также к рождению новых научных
направлений, например обобщенной кристаллографии
(Дж. Бернал и др. [20]), биосимметрики (Ю. А. Урман-
цев [231, 236 и др.]).
Одним из наиболее распространенных и характерных
являются спиральные биосимметрии. Их широкое
изучение началось с естественнонаучных работ И.-В. Гете,
который говорил об общем стремлении биологических тел к
спиральности. Яркими примерами спиральных биосиммет-
рий являются спирали улитки уха и раковин многих
моллюсков, спиральные расположения листьев на побегах
растений, семян в корзинках подсолнечника, чешуи в
еловой шишке, микротрубочек в цитоскелетах клеток и т. д.
(рис. 1,2).
В биологии давно отмечена загадочная связь этих био-
симметрий с числами ряда Фибоначчи, который при
п = 0, 1, 2, 3, представляет собой рекурентную
последовательность
{Fn+2 = Fn + Fn+1} :0, 1, 1, 2, 3, 5, 8, 13, 21, 34,... (1)
Эта связь представлена в известных законах филлотак-
сиса (т. е. листорасположения), первоначально
сформулированных в ботанике [10, 116, 124, 216, 233, 278, 417, 438
и др.]. Согласно данным законам при спирально
симметричном расположении листоорганов в телах растений
реализуются сразу пары чисел Фибоначчи из последова-
Рис. 2. Примеры спиральных биосимметрий
а, б — сосновая шишка (левая и правая формы) и цветок подсолнечника
(из [40]); в — укладки на побегах пальм (из [317]); г — спиральные
структуры в костной ткани (из [317]); д — срез верхушки человеческого сердца,
справа для сравнения приведена увеличенная раковина фораминифер (из
(317]); е — расположение камер у раковины фораминифер семейства милио-
лид (из [106])
11
тельностеи двух типов
J/V _ Fn+i\. 2 3 5 8 13 21 m_
\Vn Fn f* 1 ' 2 ' ~3~' T' T' 13,--'~*^~~
= ^^- = 1,618... (2)
fn' _ Fn+i\.2 Ъ_ 5 8 13 21 _^ф2_
\Vn ~ ^n Г' 1 ' 1 ' 2'Т'Т"' 8'*'*
=; 3+2^5 =2,618... (3)
Последовательности {(?'п} и {(?"„} называются пара-
стихной и ортостихной последовательностями, а величина
Ф представляет собой иэвестное в эстетике пропорций и
геометрии «золотое сечение», к которому мы вернемся
ниже.
Рассмотрим известные особенности явления филлотак-
сиса подробнее. Для правильного спирального
расположения листоорганов на побеге растения характерно, что
каждый v-й листоорган (по счету на спирали
листорасположения) лежит на одной и той же вертикали побега и
необходимо совершить ц. полных оборотов по спирали, для
того чтобы попасть из точки крепления листооргана Рп в
точку крепления листооргана Pn+v, расположенную по
стеблю прямо над ней. Очевидно, что при этом величина
(ц/v) 360° дает дивергенцию или угол расхождения
одного листооргана от другого. Для разных растений дробь v/u.
имеет разный вид. Например, у липы, вяза, бука, злаков
она равна 2/1; у ольхи, орешника, винограда, осоки — 3/1;
у дуба и вишни — 5/2; у малины, груши, тополя,
барбариса — 8/3; у миндальника, облепихи — 13/5. Для шишек
хвойных встречаются дроби 21/8, 34/13, 55/21. Все эти
дроби являются членами приведенной ортостихной
последовательности (3) из чисел Фибоначчи.
Надо отметить, что обычно в ботанике при изучении
филлотаксиса рассматривается листорасположение в
верхушечной (апикальной) части побега в связи с тем, что в
более старой нижней части растения правильная спираль-
ность листорасположения начинает нарушаться из-за
действия побочных факторов, деформирующих тело
растения и картину правильного листорасположепия па нем.
Следует также добавить, что в литературе часто
приводятся значения филлотаксисных дробей ja/v, а не v/u., как
это из соображений единства изложения материала дано
12
у нас. Шаг спирали из \i оборотов в ботанике называют
генетической спиралью, поскольку расположение листьев
в ней соответствует порядку появления их снизу вверх.
Общее число листоорганов в ней называют листовым
циклом. Вертикальная прямая, соединяющая соответствующие
листоорганы разных листовых циклов, называется орто-
стихом.
Другой случай филлотаксисных спиралей, связанный
с реализацией в биологических телах парастихной
последовательности (2), демонстрируют, например, листья на
поперечном срезе почки, плоды головок подсолнечника и
других сложноцветковых растений, чешуи шишек и т. д.
(рис. 2, а, б). Бросающиеся в глаза лево- и
правозакрученные спирали расположения листоорганов или семян в
подобных укладках называются у ботаников парастихами.
(Они представляют собой пример логарифмических
спиралей — широко распространенных формообразующих
кривых живой природы.) Оказывается, что числа лево- и
правозакрученных парастих для конкретной укладки
являются, как правило, соседними числами ряда
Фибоначчи. Так, у шишек европейской ели отношение чисел этих
парастих бывает равным 5/3, у лиственницы
европейской— 8/5, у шишек сосны—13/8, 21/13, 34/21, у
головок подсолнечника — 55/34, 89/55, 144/89, 233/144 и т. д.
Именно эти дроби составляют последовательность (2).
Надо отметить, что связь с числами Фибоначчи дембвТ-
стрпруют также другие типы биосимметрий, например,
биосимметрия вращений. Так, для цветков с их
вращательной симметрией наиболее распространены числа
лепестков 3 и 5, у сложноцветковых имеются 8 краевых цветков
в одном соцветии далии (георгины), 13 у крестовика, 21 и
34 у календулы и т. д. Однако наиболее изучена и описана
названная связь для спиральных биосимметрий.
Впервые отмеченное в ботапике явление биосимметрии
по принципам фибоиаччиевого филлотаксиса оказалось
гтшпоко оаспростраяенпым пе только у растений, но и
вообще в мире органических тел. Соответственно с этим
надлежит говорить пе просто о ботанических законах
листорасположепия, по об обобщепных законах листорас-
подожештя или (Ьпллотакгттса.Р которые далее й"""будут
йметьттм л ниду. Е|ц7Гвттрошлом веке С. Швенденер писал
о филлотаксисных спиралях чешуи у рыб и
пресмыкающихся [10, с. 81]. 1 «Совершенный филлотакси?»
наблюдается у гидр: возникновение новых почек по стволу
13
тела гидры происходит по спирали с уже знакомыми нам
фибоначчиевыми углами расхождения [302, с. 121—124 J.
Изучение медуз открыло, что и у них органы и зооиды
расположены по принципам филлотаксиса [379]. Ф. Джа-
гер указывает на то, что относительное расположение
отдельных компонентов мышц также имеет
непосредственное отношение к филлотаксису [365х c.__1602JH связи
с явлением филлотаксиса Т. Кук отмечает наличие
характерных спиралей в строении костной ткани и
мышечной ткани сердца (рис. 2, г, д) [317]. Можно добавить,
что современные цитологические исследования выявили
наличие в клетках организма спирализованного
скелета из микротрубочек, / подобного представленному на
рис. 1, в, 2. \4f2jJJ^.j} ;
Г. Итерсон [362] - nf^rogHHli) примеры проявления
филлотаксисных закономерностей у фораминифер —
отряда простейших животных, тело которых покрыто
раковиной. У фораминифер семейства милиолид раковина
спиральна и состоит из последовательности камер. Если, как
прежде в случае ортостихного листорасположения,
описывать расположение камер дробью, в знаменателе
которой стоит число полных оборотов, которые надо
совершить по спирали, для того чтобы попасть из одной камеры
в другую, расположенную точно над ней, а в числителе —
число % камер, которые надо пройти при этом, то
получаемые дроби оказывдются принадлежащими ортостихной|
последовательности 1(3). Так, дробь 2/1 характеризует ра-
кйтгатга фораминифер Pyrgo u Spiroloculina, дробь 3/1 —
Triloculina, 5/2 — Quinqueloculina [106, с. 33]. На рис. 2, е
представлена схема расположения камер у раковины
Quinqueloculina, причем нумерация камер (1—6) дана в
очередности их образования при росте раковины. Сравнивая
рис. 1, д и 2, е, видим что филлотаксисные спирали у
побегов растения и у раковины фораминифер отличаются
лишь порядком образования новых листьев и камер: если
у побега образование новых листьев (зона роста)
находится на внутреннем — ближнем к центру — конце
спирали листорасположения, то у раковин фораминифер
образование новых камер — позиционных аналогов листьев —
происходит на внешнем периферийном конце спирали.
Другими словами, имеет место обращение роста
формообразующей спирали.
Укажем еще на обнаруженные интересные филлотак-
сисноподобные закономерности строения конических ра-
14
Рис. 3. Пример филлотаксисноподобного строения раковины
моллюска Charonia rubicunda Perry
а — вид сбоку; б — вид сверху; в — условная развертка образующей
ленты раковины на плоскость
ковин некоторых моллюсков (опубликованных данных
других авторов по этому вопросу, несмотря на
проведенный специальный поиск их, обнаружить не удалось).
Речь идет о том, что у целого ряда моллюсков с
конической раковиной, например из семейств Gymatidae, Bursi-
dae, Muricidae, на раковине, представляющей собой как
бы скрученную в коническую спираль ленту (рис. 3),
через строго определенные интервалы сформированы
особые швы или наплывы (1—11). Для генетически
детерминированного расположения этих швов характерно, что
каждый v-й шов спирали лежит на одной и той же
образующей боковой линии конуса раковины и необходимо
15
совершить ц. полных оборотов по спирали, для того чтобы
попасть из места нахождения шва Рп в место нахождения
шва Рп+у, расположенного непосредственно под ним.
Оказывается, что в этом случае дробь v/ц принадлежит
обычно парастихной последовательности (2). Рис. 3
иллюстрирует это на примере раковины Charonia rubicunda
Perry, у которой v/u.=3/2. Отметим дополнительно, что
так же, как у фораминифер (и обратно случаю побега
растения), зона роста спиральной раковины моллюсков
находится на внешнем периферическом конце спирали.
Этот список фактических материалов,
свидетельствующих о фундаментальном значении симметрийных
принципов филлотаксиса в продуцировании биологической
ткани, можно продолжать (библиография содержит
богатый перечень литературы по филлотаксису), но обратимся
сейчас к интересной связи филлотаксиса с «золотым
сечением».
С древних времен вопросы эстетики пропорций
связываются архитекторами, скульпторами, мыслителями
различных направлений с пропорцией золотого сечения,
названной так Леонардо да Винчи. Эта Пропорция задолго
до Леонардо да Винчи восхищала своими
геометрическими особенностями древних геометров, изучавших
правильные многоугольники и многогранники и. обнаруживших
ее присутствие в ряде из них. В частности, она
реализуется в пентаграмме — пятиконечной звезде, которая у
древних греков почиталась символом здоровья.
Геометрически золотое сечение определяется как
такое деление отрезка единичной длины АВ на две неравных
части АС—х и СВ=1—х, при котором отношение меньшей
части СВ к большей АС равняется отношению большей
части к целому отрезку АВ:
СВ/АС=АС/АВ или (1-х)/х=х/1,
откуда х2~1—х. Положительным корнем этого уравнения
является величина (—l+V5)/2, так что отношения в
приведенной пропорции равны:
ЛЯ/ЛС=2/(-1+У5) = (1+У5)/2=Ф=1,6180339...
Величина Ф представляет собой численную величину
пропорции золотого сечения и обладает рядом интересных
математических свойств, изложенных в Приложении I.
Золотому сечению в архитектуре, живописи, музыке,
биологии (явлениях роста и пр.) посвящена огромная лите-
16
ратура. Обзор истории вопроса о золотом сечении в
эстетике и биологии также дан в Приложении I.
И. Кеплер первым связал числа Фибоначчи с
особенностями роста биологических тел и указал, что парастих-
ная последовательность (2) из чисел Фибоначчи сходится
именно к величине золотого сечения Ф. Тем самым
известная по отвлеченным геометрическим концепциям
древних величина золотого сечения неожиданно
проявилась в роли объективной характеристики процессов роста
и формообразования в живой природе. —
Позднее было установлено, что не только классический
ряд Фибоначчи (1), но и любой ряд с таким же
рекуррентным свойством {Ln+2=Ln+i+Ln} и другими
начальными членами ряда
{Ln} :а, Ь, а + Ь, и-\- 2Ь, 2а + 36, За + 5Ь, Ъа + 8Ь,...
(4)
порождает последовательность отношений соседних
членов, сходящуюся к величине золотого сечения:
j Ln+i \ . Ь а+ 6 а+ 26 2я +36 За + 56 ^ _
= 1,618... (5)
Ряды (4) естественно считать обобщенными рядамаг
Фибоначчи. В редких случаях у растений наблюдаются
листорасположения, связанные с такими рядами: 2, 1, 3,
4, 7, И, 18, 29,... (ряд Люка [116, с. 244]); 3, 1, 4, 5,
9,.:..; 5, 2, 7, 9, 16,... Это говорит о том, что для явления
филлотаксиса важнее рекуррентное свойство ряда
Фибоначчи, чем значение первых двух членов такого
рекуррентного ряда.
Очень интересны исследования А. Фрей-Висслинга
[242, с. 98—104], обратившего внимание на то, что
филлотаксисные закономерности проявляются не только
на уровне целых организмов и органов, но и на уровне
биологических молекул. Конкретнее, речь идет о том,
что в очень важном классе биомолекулярных
образований — а-спиралях полипептидов — расположение
аминокислотных остатков по ходу спирали определено
величинами v/ju,, равными 11/3, 18/5, 29/8, 47/13 для разных
молекулярных цепей. Эти величины, задающие углы
расхождения аминокислотных остатков, являются членами
17
«ряда а-спирали», вводимого Фрей-Висслингом:
3 /: 7 11 18 29 47 „ , ^_о R1R
Т' Т' Т' ~F'~5~' T' ui----^^n-^ —«5,010...
Числа, стоящие в числителе (знаменателе) каждой
дроби, равны сумме числителей (знаменателей) двух
предыдущих дробей, т. е. здесь имеет место рекуррентное
свойство Фибоначчи, характерное для филлотаксиса,
причем знаменатели образуют ряд Фибоначчи, а
числители — ряд Люка.
Пример рассматриваемых Фрей-Висслингом
молекулярных спиралей с v/u.=18/5 изображен на рис. 4. На
рис. 4, б представлена развертка такой спирали на
плоскую поверхность, где спираль имеет вид отрезков
прямой, наклоненных под углом 25° к горизонтали. 18-й
аминокислотный остаток, отстоящий от нулевого на пять
витков, расположен точно над ним. Пользуясь
терминологией филлотаксиса, можно сказать, что эти
аминокислотные остатки лежат на одной ортостихе. Наклонные
линии, например соединяющие точки 0, 3, 6, 9,... или
2, 6, 10,... , представляют собой парастихи. Фрей-Вис-
слинг отмечает, что такая система с характеристикой 18/5
может легко превратиться в систему с характеристикой
11/3 или даже 7/2.
Нельзя не отметить, что за последние годы в связи с
развитием исследований по теоретической и
математической биологии интерес к математическим
закономерностям филлотаксисного формообразования стремительно
увеличивается, как об этом можно судить, например, по
росту соответствующих публикаций в журналах
«Mathematical Biosciences» и «Theoretical Biology». Имеется
много попыток объяснения закономерностей филлотаксиса
применительно к частным случаям реализации этих
закономерностей в биологических телах, главным
образом в растениях. При этом в основу кладутся
предположения типа: 1) при такой картине листорасположения
взаимодействие растения с окружающей средой
оптимально в смысле облегченного поступления к листьям
солнечного света и воздуха (критику этого положения
см. [438, с. 932—933]); 2) картина филлотаксиса у
растений определяется тем, что каждый листовой бугорок
выделяет ингибитор, который очерчивает зону запрета
для роста других листовых бугорков; 3) картина
филлотаксиса определена механическим давлением между
18
ч
-d П-3,64 It-11,4А
Л
Рис. 4. ос-спираль полипептидной цепи
(из [242])
а — пространственная схема а-спирали; б —
схема а-спирали, развернутой в плоскости
контактирующими листовыми
бугорками и т. д. По нашему мнению,
общим недостатком многих из этих
объяснений является то, что они
кладут в основу объяснения заведомо
узкие посылки, не способные
охватить явление филлотаксиса в очень
разных биологических телах. Здесь
во многом прав Р. Жеан: «Имеется
множество филлотаксисных теорий,
но ни одна из них не дает
удовлетворительной модели филлотак-
19
сиса... Все теории филлотаксиса являются
онтогенетическими; они постулируют запрещенную зону и иногда
могут оставить впечатление игры с примордием» [366,
с. ИЗ]. Особенно неубедительно выглядят попытки
объяснить явления филлотаксиса за счет условий
оптимальности взаимодействия биологических тел с внешней
средой (типа условия хорошего доступа света и воздуха ко
всем листьям побега при таком расположении). Нам
представляется, что разгадка филлотаксиса, этого
фундаментального явления природы, кроется в существовании
общих законов взаимодействия в биологическом
пространстве. Скажем об этом подробнее.
С формальной точки зрения, если пользоваться
физической терминологией, мир биологического тела является
многообразием событий. В этом многообразии событий
существует система отношений воздействия одних
событий на другие, которые определяют его пространственную
структуру. К миру биологического тела вполне
применимо сложившееся в физике понимание пространства и
пространства—времени, сформулированное А. Д.
Александровым так: «Пространство—время есть множество
всяких событий в мире, взятое в отвлечении от всех его
свойств, кроме тех, которые определяются структурой
системы отношений воздействия одних событий на другие»
[8, с. 55]. Видимо, проблема общих законов
взаимодействия в биологическом пространстве (или биологическом
пространстве — времени) близка по духу к аналогичным
физическим проблемам современной квантовой теории
поля, стоящей на пороге установления новых общих
законов взаимодействия (см., например, [8, с. 55]).
Раскрытие биологических общих законов взаимодействия
приведет к знанию биологической <<структуры системы
отношений воздействия одних событий на другие», даст
ключ к биокристаллографической специфике
коллективного поведения элементов биологического тела, в том
числе к филлотаксисным явлениям и к энергетическим
аспектам организации живого вещества, и с
необходимостью выявит новые особенности биологического
пространства. / .
В заключепие_шщчер101ем:^ще раз, что^в
формообразовании биологических тел разных уровней организации
и классов обнаруживаются одни и те же принципы
симметрии, отличающиеся характерной связью с рядом
Фибоначчи. Тело человека является частным представи-
20
телем класса биологических тел и естественно поэтому
исследовать, не имеют ли место родственные принципы
симметрии и филлотаксисноподобные закономерности в
человеческом теде, в частности в кинематической схеме
_тела. / Наши исследования дают положительный ответ на
этот вопрос и указывают на важное значение этих
принципов в ряде фундаментальных особенностей
кинематической организации человеческого тела.
1.2. Изучение биологических симметрии
как научное направление
Творцы научного коммунизма К. Маркс и Ф. Энгельс в
свое время предсказали: «Впоследствии естествознание
будет охватывать науку о человеке, подобно тому как
наука о человеке будет охватывать естествознание, <обе
станут одним), будет одна наука» [142, с. 630].
В плане осуществления этого предсказания
первостепенное значение имеет сближение физики и биологии как
двух наук о взаимосвязанных частях единого мира. Оно
может в огромной мере способствовать обеспечению
власти человеческого интеллекта над природой.
Одним из магистральных путей сближения физики и
биологии, как нам представляется, является изучение
биологических симметрии, внедрение в область биологии
инвариантно-групповых концепций и методов. Такое
заключение базируется на том, что концепция
симметрии является одной из основ современного теоретико-
физического мышления (см. следующий раздел).
Соответственно осмысление биологических явлений с
позиций, родственных теоретико-физическим, предполагает
внедрение в биологию ключевых для физики инвариант-
но-групцовых концепций и методов и уделение
совершенно особого внимания явлениям биосимметрии. В этой
связи все чаще публикации по математической и
теоретической биологии выходят под характерными названиями:
«Концепция группы и теория восприятия» [308],
«Организация сенсомоторной системы и инварианты
проективной геометрии» [177], «Биологическое подобие и теория
групп» [320]. «Исследования по неевклидовой
биомеханике» [181] и др.
По существу речь идет о становлении широкого
научного направления, которое было бы закономерно назвать
теоретико-групповой биологией. Это направление можно
21
кратко именовать также биосимметрикой, как это было
сделано Ю. А. Урманцевым [230, 231]. В становление
исследований биологических симметрии (и их
нарушений) как самостоятельного и важного научного
направления большой вклад внесли многие советские ученые:
В. Н. Беклемишев, Б. К. Вайнштейн, В. И. Вернадский,
Г. Ф. Гаузе, В. G. Готт, Н. П. Депенчук, В. Б. Касинов,
А. А. Любищев, С. В. Мейен, Д. В. Наливкин, Ю. Г.
Сулима, Ю. А. Урманцев, А. В. Хохрин, И. И. Шафрановский
и др. Для более обстоятельной характеристики
специфики этого направления, его концепций, подходов и целей
остановимся на взглядах различных исследователей по
проблеме биосимметрий, прежде всего симметрии форм
биологических тел.
Фундаментально подходил к данной проблеме В. И.
Вернадский, связывавший ее с вопросом о специфике
пространства живого вещества. Он писал: «Натуралист,
исходя из школьной рутины, все время мыслил о
едином пространстве, но не о разных пространствах, но не о
состояниях пространства. Он не сознавал, что
пространство нашей планеты и вообще пространство планет есть
особые пространства, нигде, кроме планет, не
наблюдаемые. В течение тысячелетий, говоря о природных или
естественных телах, он не сознавал и не утверждал, что
каждое природное тело и каждое природное явление
имеет свое естественно-энергетическое специфическое
пространство, которое натуралист изучает, изучая
симметрию» [42, с. 66].
Вернадский утверждал, что между симметрией косных
естественных тел и явлений и симметрией живого
вещества, т. е. живых организмов, существует резкое различие,
без всяких переходов и без исключений [43, с. 72].
Особенность симметрии жизни иллюстрировалась им, в
частности, таким фактом: «Ось симметрии пятого порядка,
неразрывно связанная с «золотым или божественным
сечением», отражающимся в нашем осознании красоты,
занимавшим мысль Леонардо да Винчи, Иоганна Кеплера
и всех других к нему подходивших,— эта ось. играющая
заметную роль в морфологии форм жизни, в
кристаллографии невозможна. И она в ней действительно
отсутствует. А между тем именно эта пятерная симметрия
играет видную роль и в геометрии — еще древней эллинской.
Она определяет один из пяти правильных
многогранников, которым Платон и неопифагорийцы придавали огром-
22
Ное значение в строении мира» [43, с. 46]. Согласно
заключению Вернадского, морфологи-биологи работают над
симметрией вне учения о симметрии, не зная или не
учитывая его. «Здесь быстро создается,— писал ученый в той
же работе,— огромная область разрозненных новых и
давно известных явлений. Эта область учением о симметрии
не охвачена. Необходима обработка учения о симметрии
в тесной связи с морфологией жизни. Это и есть та новая
огромная задача, которая сейчас стала на очереди».
Особую ценность для нас имеют мысли Вернадского о
подчиненности живого вещества неевклидовой геометрии,
установить которую, по его мнению, должно углубленное
изучение симметрии в биологических телах [42, гл. 15, 16].
Мы вернемся к этому в Приложении IV.
Видный советский геолог и палеонтолог Д. В. Налив-
кин [161—163] указал на применимость к описанию форм
ряда биологических тел принципа зеркальной симметрии
обобщенного вида: если обычно зеркальное отображение
производится относительно прямой линии или плоскости,
то Наливкин предложил рассматривать зеркальное
отражение относительно искривленных линии и плоскости.
Симметрию этого вида он назвал криволинейной
симметрией. Искривление линий и поверхностей, относительно
которых имеет место зеркальная симметрия, происходит,
согласно Наливкину, в основном по типу сжатия или
растяжения, изгиба и кручения. Им описано приближенное
проявление криволинейной симметрии в телесных формах
некоторых биологических видов и отмечены определенные
закономерности изменения данных биосимметрий в ходе
биологической эволюции.
А. А. Любищев [77, 136], ставя целью своей жизни
создание естественной системы классификации
организмов, полагал, что путь к этому лежит через
математическое изучение симметрии и характерных кривых в
строении организмов, не имеющих непосредственного
функционального значения.
В связи с аналогичной задачей классификации
организмов В. Н. Беклемишев [15] предпринял широкое
исследование симметрии в телах безпозвоночных. Первый
том своего труда он назвал «Проморфология». Этим
термином Э. Геккель определял учение о симметрии
организмов.
Большое внимание принципам симметрии в биологии
уделяется в работах [300, 399, 438]. Процитируем харак-
23
терное высказывание С. В. Мейена: «Живое вещество мы
обычно мыслим в виде бесформенной амебы, аморфной
протоплазмы. Дескать, только функция и среда наложили
на эту протоплазму определенные формы. Так ли это?
Неужели живая протоплазма не может организовать свое
пространство в нечто более упорядоченное? Иначе говоря,
какова симметрия живого?» [151, с. 30J.
Ю. А. Урманцев формулирует широкую программу
исследований биосимметрий, справедливо рассматривая ее
как программу специального научного направления,
кратко называемого им биосимметрикой. Согласно Урманцеву,
биосимметрика — экспериментальная наука, которая,
опираясь прежде всего на аппарат математической теории
групп и ряд кристаллографических методов, изучает
симметрию живого [231, с. 75]. «Что же касается
пространственных форм и отношений — пространства организмов,—
пишет Урманцев,— то дальнейшее углубление в этом
направлении, по-видимому, пройдет через этап создания
биологической геометрии, о котором еще несколько десятков
лет тому назад писал наш гениальный соотечественник
В. И. Вернадский. ...Причем a priori совершенно не
следует, что живая природа подчиняется одной единственной
геометрии: вполне возможно, что придется развить
несколько геометрий, соответствующих нескольким
геометрическим пространствам» [230, с. 171]. Урманцев
стремится привлечь внимание биологов, физиков, философов,
математиков к проблеме динамической биосимметрии и
биологических законов сохранения. «Ввиду
исключительного значения последних для познания природы жизни,—
пишет он,— необходимы энергичные поисковые работы в
этом направлении. Можно надеяться, что на основе
биологических законов сохранения, разнообразных
инвариантов, симметрии законов живой природы относительно тех
или иных преобразований рано или поздно удастся глубже
проникнуть в сущность живого, объяснить ход эволюции,
ее вершины, тупики, предсказать неизвестные сейчас
ветви, теоретически возможные и действительные числа
типов, классов, семейств... организмов. И вообще нужно
проанализировать вопрос о том, нельзя ли эволюцию
материи в целом и внутри отдельных ее форм представить
как групповые преобразования, найти их инварианты и на
основе последних определить все возможные варианты
эволюции в целом» [ 236, с. 226, 227 ].
24
Развиваемые рядом исследователей идеи обобщенной
кристаллографии нацеливают на трактовку и изучение
живых организмов как объектов, построенных на
геометрических принципах, родственных геометрическим
принципам физических кристаллов. Так, видный советский
кристаллограф Б. К. Вайнштейн и биолог Н. В. Киселев
наблюдали, как белковые молекулы, полученные в
результате «раздробления» вируса, снова собирались вместе при
подходящих условиях и укладывались по правилу
спиральной лестницы. «Этот процесс,— пишут эти
исследователи,— удивительно напоминает кристаллизацию и
действительно имеет с ней чрезвычайно много общего. В обоих
случаях дело сводится к некоторой симметричной укладке
одинаковых строительных элементов» [258, с. 113].
В предыдущем параграфе на примере ос-спиралей
полипептидов уже отмечалось, что такая биокристаллизация
на молекулярном уровне может демонстрировать те же
загадочные связи с числами Фибоначчи, что и
формообразование спиральных симметрии в биологических
макротелах. Это дает дополнительно право рассматривать
формообразование биологических макротел именно как
своеобразную кристаллизацию по аналогии с кристаллизацией
биомолекул. Уместно вспомнить утверждение К. Э.
Циолковского: «Жизнь есть сложная кристаллизация... Все
растения и животные суть только проявления сложной
кристаллизации разнообразных невообразимо сложных
веществ, сопровождаемые механическими, физическими и
химическими процессами» [249/, с. 5, 6]. Здесь
Циолковский выступил одним из предвестников такого
набирающего силу современного научного направления, как
обобщенная кристаллография.
Много сделал для становления этого направления
известный английский ученый Дж. Бернал. Ряд его
публикаций на эту тему переведен на русский язык: «О роли
геометрических факторов в структуре материи» [18],
«Возникновение жизни» [19], «Поля охвата обобщенной
кристаллографии» [20] и др. Говоря об
обобщенно-кристаллографических принципах жизни, Бернал писал:
«По-видимому, все структуры, найденные в клетках, имеют
подобную же кристаллическую природу — будь то
оболочки, трубочки, волокна или трехмерные кристаллы.
Красота жизни, таким образом, есть геометрическая красота,
столь ценимая Платоном» [19, с. 17]. Бернал отмечал:
«В течение многих лет подчеркивалась аналогия между
25
ростом живого организма и ростом кристалла. Сам Пастер
именно с этой точки зрения изучал рост кристаллов»
[20, с. 928].
Надо заметить, что Дж. Бернал и его соавтор С.
Карлайл строят свою концепцию обобщенной
кристаллографии (распространяемой ими, в частности, на филлотаксис-
ные явления) на евклидовой геометрии, беря за основу
понятия математических групп траноляций и вращений,
о которых они пишут: «Все формы обобщенной
кристаллографии вытекают из этих основных понятий в рамках
евклидовой геометрии» [Там же, с. 930].
В отличие от этого положения об евклидовости,
достаточно произвольно перенесенного указанными
исследователями из физической кристаллографии, мы считаем, что
для структуризации живой материи (а потому для
построения действительно обобщенной кристаллографии) важное
значение имеют понятия неевклидовых геометрий, прежде
всего конформной геометрии, биологическому значению
которой посвящена настоящая книга,
1.3. Симметрии и математическое естествознание
Общее значение концепций и методов симметрии для изу
чения природных явлений неоднократно анализировалось
в литературе, например в книгах [40, 48, 141 и др.], из
которых частично заимствованы некоторые материалы
настоящего параграфа.
Широкое внедрение принципов симметрии в
современное естествознание было во многом подготовлено
развитием геометрии, прежде всего успехом знаменитой
Эрлангенской программы 1872 г. Ф. Клейна [167], утвердившей
принципиально новый инвариантно-групповой взгляд на
геометрию. В этой программе геометрия определялась как
наука об инвариантах групп преобразований. Например,
евклидова геометрия — как наука об инвариантах группы
евклидовых преобразований (движений), геометрия
Лобачевского — как наука об инвариантах группы
гиперболических преобразований, проективная геометрия — как
наука об инвариантах группы проективных
преобразований и т> д. С позиций Эрлангенской программы' евклидова
геометрия предстает как всего лишь одна из многих
возможных геометрий, каждая из которых имеет свои
собственные исходные понятия, аксиомы и теоремы. И вся
классификация геометрий строится исходя из групп преоб-
26
разованнй, использование которых не меняет понятий,
аксиом и теорем. На основе такого подхода удалось в
значительной мере восстановить единство геометрий,
утраченное к середине XIX в. в силу того, что многие ее
разделы оказались совершенно не связанными друг с другом.
При этом открылись ценные возможности выяснения
взаимосвязи различных ранее не связанных между собой
геометрий и построения новых.
Революция, произведенная Эрлангенской программой
в геометрии, распространилась затем на область физики
и философских проблем пространства. Были изменены
сами основы представлений человека о пространстве,
которые с той поры связываются с понятием
математических групп преобразований. А. Пуанкаре формулировал
эту связь кратко: пространство есть группа. Язык
пространственных отношений представляет собой язык
симметрии. (Это определяет важность введения в биологию
теории групп преобразований как необходимого условия
включения биологии, изучающей специфически
организованные в пространстве и времени живые системы, в
область развитого математического естествознания с его
формализованными понятиями пространства и
пространственных изменений.)
Видной вехой утверждения принципов симметрии в
фундаментальной физике явилось создание А.
Эйнштейном специальной теории относительности, повлекшее за
собой широкое распространение термина «геометризация
физики». Этот термин возник в результате выяснения
того, что с формальной точки зрения указанная теория
является ни чем иным, как теорией инвариантов
некоторой группы преобразований (группы Пуанкаре), т. е.
некоторой геометрией. Идеи геометризации физики,
построения и изложения ее теорий на языке инвариантов
групп преобразований, т. е. на языке симметрии,
получили развитие и распространение на квантовую механику,
теорию законов сохранения, теорию элементарных частиц
и другие области физики. Инвариантно-групповой подход
стал в физике XX в. одной из наиболее характерных ее
черт. Фундаментальное значение принципов симметрии
(инвариантности, относительности) для современной
физики основано не только на том, что они существенным
образом обогащают математический аппарат физики,
вводя в обиход теоретиков инвариантно-групповые
методы. Положение значительно серьезнее, глубже, содержа-
27
тольнее: концепция симметрии в действительности
является одной из основ современного
теоретико-физического мышления. «В наше время физические теории
понимаются и строятся, прежде всего, как теории инвариантов
некоторых групп преобразований* [48, с. 3].
Исключительную роль концепции симметрии для
современного теоретико-физического мышления
проиллюстрируем словами крупнейших физиковг М. Борн: «Я
убежден, что идея инвариантов является ключом к
рациональному понятию реальности, и не только в физике, но и в
каждом аспекте мира» [28, с. 276]. Г. Вейль: «Насколько
я могу судить, все априорные утверждения физики имеют
своим источником симметрию» [40, с. 144]. П. Дирак:
«Возрастающее применение теории преобразований...
представляет сущность нового метода в теоретической
физике. Дальнейший прогресс состоит в том, чтобы делать
наши уравнения инвариантными относительно все более
широких классов преобразований» [48, с. 3].
Помимо получения ряда замечательных конкретных
результатов в физике, концепция симметрии привела к
перевороту в философских основаниях физики, изменив
представление о том, что следует считать исходными
законами физики. Если ранее — со времен Ньютона — законы
природы формулировались в виде дифференциальных
уравнений, то «сейчас ситуация существенно изменилась.
Для фундаментальной физики основными исходными
законами являются законы симметрии. Это инвариантность
относительно неоднородной группы Лоренца
(релятивистская инвариантность), относительно групп унитарных
преобразований (унитарная симметрия) и других групп»
[51, с. 4].
По поводу указанного переворота в науке советские
теоретики Ю. Б. Румер и А. И. Фет пишут: «Развитие
физики в последние годы обратило, в известном смысле,
соотношение между уравнениями движения и группами
симметрии. Теперь группа симметрии физической
системы выступает на первый план, представления этой группы
и ее подгрупп несут самую фундаментальную
информацию о ней. Таким образом, группы оказываются
первичным наиболее глубоким элементом физического описания
природы» [48, с. 4].
В заключение зададимся вопросом, возможна ли
«геометризация биологии» хотя бы в некоторых областях
знаний о живой природе? Возможно ли, например, по-
28
строение теоретических концепций морфогенеза,
психофизики и пр. с формальной точки зрения теорий инвариантов
определенных групп преобразований? Мы глубоко верим
п это и полагаем, что теоретическая биология будущего
явится во многом теоретико-групповой биологией. При
этом естествознание сделает очередной шаг к тому идеалу,
о котором писал А. Эддипгтон: «Идеал, к которому мы
стремимся, заключается в объединении всех наших
сведений о физическом мире в единую науку, положения
которой могут быть выражены в терминах геометрических
пли квазигеометрических концепций» [158, с. 174].
Глава II
ИССЛЕДОВАНИЕ КОНФОРМНЫХ СИММЕТРИИ
В БИОЛОГИИ
В рамках поиска фундаментальной группы симметрии
живого вещества (или фундаментальной группы
симметрии биологических пространств) рассмотрим данные о
биологической реализации принципов конформной
симметрии. Эти данные представляют собой собственные
результаты исследований автора по симметрии в
биологических телах, дополненные материалами работ Г. Кинла
о частном неевклидовом случае конформной геометрии в
пространстве зрительного восприятия (см. раздел II.9).
Сразу уточним, что под конформными симметриями
будут пониматься симметрии группы конформных
преобразований, состоящие из групп преобразований движения,
подобия и инверсии (преобразований обратных радиусов).
Дополнительная характеристика этой группы и ее
геометрии вынесена в Приложение II.
Наряду с изучением фактических материалов о
конформных биосимметриях проанализируем связи этих
симметрии с низшими — евклидовыми — типами биосиммет-
рий, издавна отмечаемыми в формах биологических тел.
II. 1. Конформные симметрии в физике
и в явлениях роста и формообразования
биологических тел
Прежде чем обратиться непосредственно к биосиммет-
риям, кратко охарактеризуем математические особенности
конформных симметрии и важную роль этих симметрии
в физическом устройстве мира.
Конформная симметрия в физике имеет давнюю
историю. Как известно, специальная теория относительности
Эйнштейна родилась из математического факта
инвариантности уравнений электродинамики Максвелла
относительно группы преобразований Лоренца. В 1909 г., через
30
4 года после создания этой теории, было открыто, что
уравнения Максвелла инвариантны относительно более
широкой группы — группы конформных преобразований,
содержащей группу Лоренца в качестве своего частного
случая. Каково физическое содержание этого загадочного
математического факта конформной симметрии
уравнений электродинамики, какая характерная особенность
нашего мира стоит за этим — неясно и до настоящего
времени, хотя физиками неоднократно предпринимались
попытки осмысления этого и предлагались различные
варианты конформной теории относительности [47].
Развитие физики за прошедшие десятилетия выявило и
другие факты особой физической значимости конформной
симметрии в устройстве физического мира. Так,
математический аппарат квантовой механики оказался тесно
связанным с конформными симметриями, в связи с чем
квантовая механика может быть представлена как теория
нарушенной конформной симметрии (Каструп). К
настоящему времени развито также несколько вариантов
конформной квантовой теории поля (Ингрехэм, Весе,
Каструп и др.), создана кинематическая космология,
опирающаяся на конформную симметрию (Шредингер, Инфельд,
Шильд и др.) и т. д. Создается впечатление, что чем
глубже человек познает устройство природы, тем больше
выявляется важное физическое значение конформной
геометрии. (Более подробное рассмотрение известных данных
о физическом значении конформной симметрии дано в
Приложении II.)
Отличительной математической особенностью
рассматриваемой группы конформных (круговых) преобразований
является то, что ее преобразования всегда сохраняют
углы и переводят сферы снова в сферы. При этом плоскость
интерпретируется как частный случай сферы бесконечно
большого радиуса. Центральным типом преобразований
этой группы является преобразование «инверсии
относительно сферы». Любое преобразование из
рассматриваемой группы можно представить в виде некоторой
последовательности таких преобразований инверсии. Инверсия
относительно сферы радиуса R с центром в точке О
определяется как такое преобразование пространства, которое
произвольную точку Р переводит в точку Р', лежащую на
луче ОР на расстоянии OP'=R2/OP от центра О. Это же
преобразование называется еще преобразованием обратных
радиусов. Оказывается, что так определенное преобразо-
31
Рис. 5. Примеры конформных (круговых) преобразований
а — трансформация конформными преобразованиями окружностей в
окружности или прямые; точки Pt—Р7 инверсионно-симметричны точкам
Р/—Р7'; б — примеры конформно-симметричных четверок точек А, В, С,
D, комплексный вурф Wk которых равен 1,3; в — зеркальное отражение
(слева), вращение (в центре) и параллельный сдвиг (справа) как
преобразовании инверсии относительно окружностей бесконечного радиуса
вание переводит все сферы снова в сферы (или в
плоскости как частный случай сферы). Рис. 5 иллюстрирует
свойства преобразования инверсии для случая двухмерного
пространства.
Отметим, что такие известные из евклидовой
геометрии преобразования, как вращение, параллельный ёдвит,
зеркальное отражение, представляют собой частные
случаи конформных преобразований и выражаются через
преобразования инверсии относительно сферы. Например,
для случая плоскости зеркальное отражение" относительно
прямой представляет собой инверсию относительно этой
прямой как окружности бесконечно большого радиуса.
Преобразование параллельного сдвига может быть
интерпретировано как результат двух последовательных инвер-
32
сии относительно параллельных прямых, а
преобразование вращения — как результат двух последовательных
инверсий относительно прямых, пересекающихся в центре
вращения (рис. 5, в). Рассмотрим сейчас некоторые
особенности конформных симметрии, которые
непосредственно понадобятся при знакомстве с материалами по
биологии, отметив, что более полное освещение положений
конформной геометрии дано в Приложении П.
Прежде всего обратимся к так называемому двойному
отношению или вурфу четырех точек А, В, С, D прямой,
вычисляемому по формуле
W=*(С-A) (D-B)/(C-B) (D-A), (6)
где выражения в круглых скобках являются евклидовыми
длинами отрезков между соответствующими парами точек.
Для нас важно, что если имеются две прямые с такими
четверками конечных точек, А, В, С, D и А', В', С, D'',
что величины их вурфов равны, то зти четверки точек
(или тройки отрезков, определяемых ими) являются
конформно-симметричными, поскольку с помощью
конформных преобразований, как известно в геометрии, эти
прямые могут быть взаимооднозначно отображены друг на
друга с полным совмещением данных четверок точек на
них. (Надо отметить, что аналогичное совмещение может
быть достигнуто и с помощью проективных
преобразований, т. е. названные комплексы могут трактоваться и
как проективно-симметричные.) Об истории термина
«вурф» сказано в Приложении III. На протяжении книги
нами будет использоваться в силу его краткости именно
этот термин, а не более широко употребляемые термины
«сложное» или «двойное» отношение.
А что происходит с четверкой конечных точек прямой,
если некоторым конформным преобразованием эта прямая
трансформируется в окружность? В этом случае взаимное
расположение рассматриваемых четырех точек на
окружности характеризуется той же самой величиной вурфа,
вычисляемого по формуле, несколько модифицированной
по сравнению с (6):
W* = («с — «a) (zd — zB)l{zc — zB) {zD — zA), (7)
где zA=xA+iyA, zB=xB+iyB, zc=xc+iyc, zD=xD+iyD
представляют собой гауссовы комплексные координаты
четырех точек на плоскости, содержащей эти точки
(величина вурфа Wh не зависит от выбора конкретной гауссрвой
2 С. В. Петухов 33
системы координат на этой плоскости [187, с. 701]).
Характерно, что вурф (7) четырех точек плоскости,
лежащих на одной прямой или окружности, всегда имеет
действительную величину; для четверок точек, не
удовлетворяющих этому условию, его величина комплексна. Вурф (7)
является инвариантом рассматриваемых преобразований,
т. е. он не меняется при любом конформном преобразовании
плоскости, которое, вообще говоря, меняет расстояние и
попарные отношения расстояний между точками. При
конформном преобразовании три точки плоскости могут
быть трансформированы в любые три другие ее точки,
но новое положение четвертой точки при этом
определяется однозначно условием неизменности величины вурфа (7)
данной четверки точек. На рис. 5, б приведен пример
конформно-симметричных четверок точек, характеризуемых
одной и той же величиной вурфа; в данном случае она
вещественна и равна 1,3. В общем случае в зависимости от
относительного расположения четырех точек на
окружности величина вурфа может меняться от 1 до °° (при
последовательном расположении точек А, В, С, D на
окружности) .
Нам понадобятся еще сведения о конформных
преобразованиях логарифмической спирали, одной из самых
распространенных формообразующих кривых в
биологических телах. Не рассматривая тривиальные случаи
преобразований подобия и движения, обратимся к
преобразованию инверсии относительно окружности, которое
иллюстрирует рис. 6, а. Из рисунка видно, что при инверсии
относительно окружности спираль трансформируется в
спираль с противоположным направлением закрутки и
обратным порядком точек на ней.
Перейдем, наконец, к биологическим объектам.
Рассмотрим пятилучевой цветок, изображенный на рис. 6, б и
состоящий из двух пятилучевых звезд (называемых
венчиком и чашечкой). Читатель, как и многие
исследователи до него, без труда отметит, что в строении этого цветка с
точки зрения евклидовой геометрии представлены два типа
симметрии: дискретного вращения на пятую часть
окружности и зеркального отражения относительно
некоторых радиальных осей. Форма цветка после этих
преобразований полностью совмещается с формой цветка до
преобразования. Однако в данном цветке наличествует и
симметрия неевклидова типа, характерная для
конформной геометрии; симметрия инверсии относительно ркруж-
34
Рис. 6. Примеры конформных симметрии
а — две логарифмические спирали, инверсионно-симметричные
относительно показанной точечной линией окружности с центром в точке 0;
б, в — цветок (из [32]) н схема, поясняющая инверсионную симметрию;
г — инверсионная симметрия между формообразующими кривыми двух
половин листа
ности конечного радиуса, как это поясняется на рис. 6, в.
При указанной инверсии формообразующая кривая
первой пентазвезды переходит в формообразующую кривую
второй и наоборот, в результате чего очертания
совокупности из обеих пентазвезд не изменяются.
Автор имел возможность убедиться в конформном
согласовании венчика и чашечки простого цветка на
примерах разных видов. В некоторых из них форма венчика
цветка была сложнее и асимметричнее, чем у
рассмотренного выше, но соответственно сложнее и асимметричнее
оказывалась и чашечка, так что конформное соответствие
между формообразующими кривыми лепестков венчика и
чашелистиков цветка в том или ином виде все равно
наблюдалось. Например, у душистого табака форма венчика
цветка представляет собой неправильную звезду, у
которой некоторые лепестки больше других; чашелистики
также оказываются асимметричными в соответствии с
2* 35
принципами инверсионной симметрии относительно
окружности с центром, совпадающим с центром цветка.
Интересно, что конформную тождественность демонстрируют
формы функционально весьма разных частей растения —
лепестков и чашелистиков.
Эти данные иллюстрируют реализацию в живой
природе принципа построения биологических тел из
конформно-симметричных блоков. Приведенный пример
демонстрирует еще одну особенность, которая, как мы полагаем,
характерна вообще для формирования биологических тел;
тело складывается из конформно-симметричных между
собой блоков первой ступени (в данном случае отдельных
лепестков венчика и чашелистиков), объединяемых в
конформно-симметричные друг другу блоки второй ступени
(пятилучевые звезды) и т. д. В следующем параграфе
анализ кинематической схемы человеческого тела
выявит аналогичную особенность: организацию этой схемы из
конформно-симметричных блоков первой ступени (в виде
трехчленных кинематических блоков), собранных в
конформно-симметричные блоки второй ступени (две
половины тела, зеркально симметричные между собой).
Частный пример неевклидовой симметрии дает
иногда встречаемое в природе ассимметричное — с точки
зрения евклидовой геометрии — строение листа растения из
двух половин с нарушенной зеркальной симметрией их
формообразующих кривых, как показано на рис. 6, г.
Нередко такое нарушение зеркальной симметрии
связано с реализацией в листе симметрии инверсии
относительно окружности конечного радиуса, а не инверсии
относительно окружности бесконечного радиуса, каковой
является зеркальное отражение. Однако насколько мы
можем судить, чаще конформная симметрия двух
зеркально-асимметричных половин листьев у растений
строится не только на инверсии относительно окружности
конечного радиуса, но дополнительно к ней еще и на
преобразовании подобия. Добавим еще, что на рис. 6, г дано
условное изображение листа со сглаженными краями.
В природе же обычно края листа расчленены, причем
формообразующая кривая каждой половины листа
состоит как бы из повторяющихся конформно-симметричных
ДРУГ ДРУГУ кусков. Таким образом, и в случае листьев
выполняется принцип объединения конформных блоков
низшей ступени в конформные блоки высшей ступени.
Образно говоря, речь идет о закономерно организованных
36
друзах кристаллов живого вещества. В Приложении IV
представлены дополнительные результаты исследований
конформных инвариантов в таких друзах, прежде всего,
сохранения углов между разграничивающими
элементами (эти результаты получены после написания основной
части книги).
Конформную биосимметрию выявляет также
рассмотрение раковин моллюсков, подобных изображенной на
рис. 4, которая сформирована из свернутой в конус
ленты, сшитой как бы из лоскутов с характерными
швами на стыках. При развертке этой ленты на плоскость
легко усматривается строгая закономерность в
расположении данных швов, перпендикулярных оси ленты,
заключающаяся в следующем. Каждые три соседние шва,
последовательно расположенные на ленте, связаны друг с
другом условием инверсионной симметрии двух крайних
из них относительно окружности, проходящей через
средний из трех швов и имеющей центр в вершине этой ленты
в форме вытянутого треугольного флажка (см. рис. 3, в).
Другими словами, зная расположение двух соседних
швов и вершины ленты можно, пользуясь только
преобразованием инверсии относительно окружности,
определить местонахождение всех остальных швов на ленте.
Еще один пример конформной биосимметрии дает
анализ особенностей формообразования и роста спиральных
укладок в некоторых биологических телах. В разделе 1.1
уже описывались ростовые особенности спирального фил-
лотаксиса у растений, фораминифер и моллюсков.
Отмечалось, что в данных случаях формообразующие спирали
являются как бы обращенными по отношению друг к
другу: ростовое приращение или движение по филлотаксис-
ной спирали у раковин моллюсков и фораминифер (см.
рис. 2, е, и 3, б) происходит в направлении от центра
спирали, а у побегов растений (см. рис. 1, д) — в
направлении к центру спирали. Если рассматривать эти спирали
как плоские, отвлекаясь от их вытягивания в высоту при
продольном росте, то это взаимоотношение данных
биологических спиралей вполне соответствует рассмотренному
взаимоотношению между двумя
конформно-симметричными спиралями (рис. 6, а). Другими словами, уплощенные
филлотаксисные спирали растительных побегов являются
конформно-симметричными по отношению к уплощенным
филлотаксисным спиралям раковин моллюсков и
фораминифер (с точностью до того или иного вида дробей из
37
чисел Фибоначчи в реализуемых углах расхождения ли-
стоорганов н их позиционных аналогов).
Что касается названного условия уплощения филло-
таксисных спиралей, то, по нашему мнению, его корни
заключаются в том, что продольный рост у рассмотренных
биологических тел является достаточно самостоятельным,
как бы независимым по своей сути процессом,
накладываемым дополнительно на плоскую картину филлотаксис-
ного роста. (Кстати сказать, у рассматриваемого
семейства фораминифер раковина плоская.) Данное положение
об искажении стандартной картины филлотаксиса за счет
большого или меньшего ростового растяжения
биологического тела в высоту неоднократно отмечалось
исследователями филлотаксисного роста (см., например, [438,
с. 918]).
II.2. Конформные симметрии
в кинематической схеме человеческого тела
Исследуем строение кинематической схемы человеческого
тела с позиций принципа построения биологических тел
из конформно-симметричных блоков. С этой целью
изучим совокупные пропорции длин трех звеньев в так
называемых трехчленных кинематических блоках тела.
Пропорции тела человека и высокоорганизованных
животных, в частности длиннотные пропорции звеньев
тела, — классический предмет исследований в биологии.
Ему посвящена огромная литература в связи с большим
теоретическим и прикладным интересом широкого круга
исследователей к закономерностям устройства
биологических тел и прежде всего человеческого тела. Можно
отметить, однако, заметную и неоправданную ограниченность
применявшегося ранее подхода к изучению
геометрических пропорций: изучение строилось только на
рассмотрении аффинных (или простых) отношений между
частями тела. Изучая, например, пропорции длин звеньев тела,
исследователи в основу рассмотрения клали отношения
длин двух звеньев. В связи с такой исключительной акцен-
тацией внимания на аффинных отношениях эти
биологические исследования о пропорциях представляли собой
по существу исследования о пропорциях двухчленных
конструкций.
Но устройство кинематической схемы тела
высокоорганизованных животных и прежде всего человека осно-
38
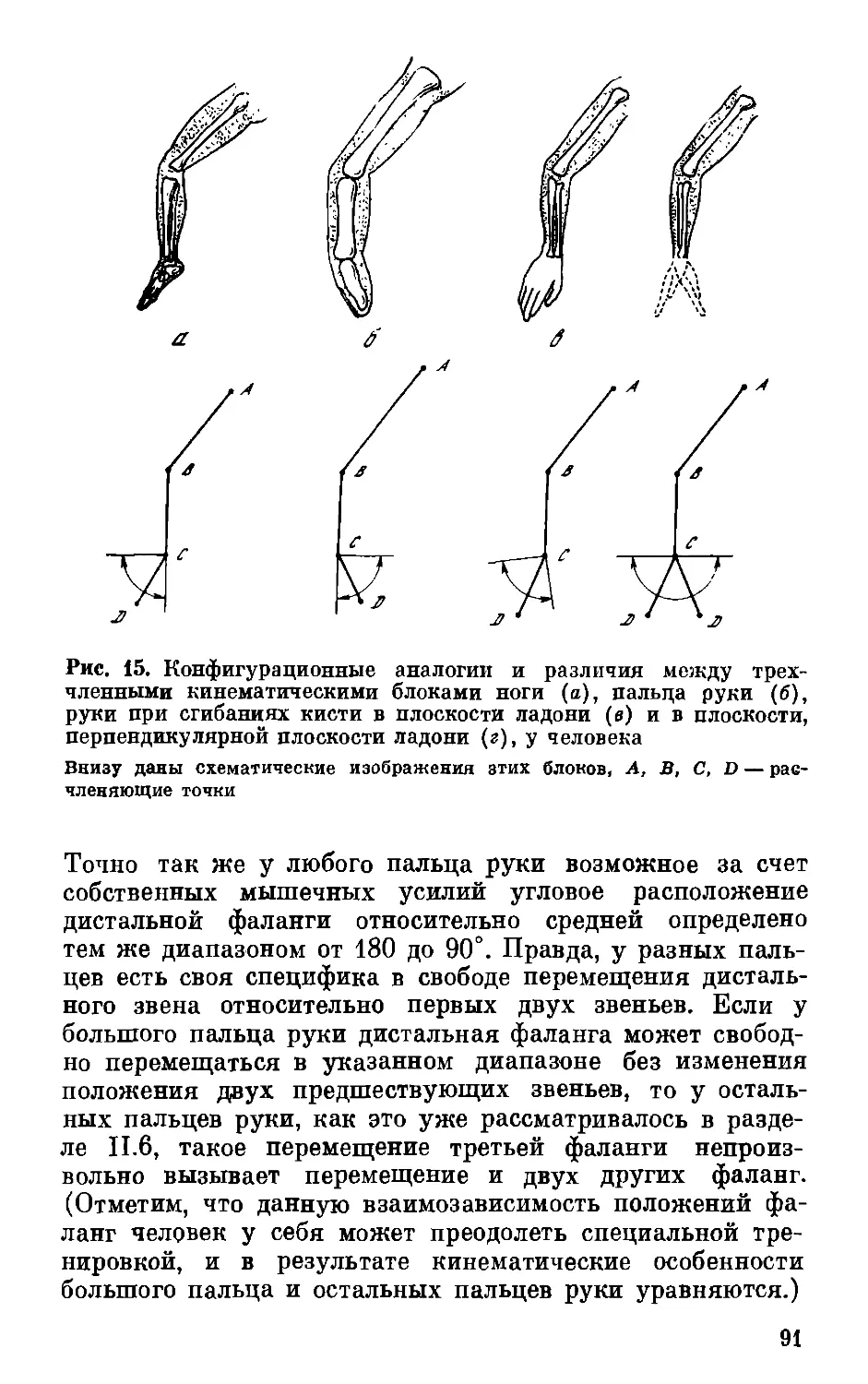
вано во многом на принципе трехчленения. Так, все тело
человека по кинематическим особенностям строения
состоит из следующих трехчленных блоков: трехфаланговые
пальцы рук и ног, трехчленистые руки (плечо —
предплечье—кисть) и ноги (бедро — голень — стопа), трех-
членистое тело (в антропологии естественно различают:
верхний отрезок — от макушки головы до основания шеи;
средний отрезок или туловище — от основания шеи до
тазобедренного сочленения; нижний отрезок — от
тазобедренного сочленения до конца пальцев ног). Для дальнейшего
сразу уточним, что для большого пальца руки (ноги) роль
третьего звена играет I пястная (плюсневая) кость, а под
третьим звеном ноги следует понимать не саму стопу,
а примерно равное ей по длине звено от голеностопного
сочленения до конца пальцев ног. Длины трех отрезков
каждого из перечисленных блоков будем обозначать
следующим образом: длину проксимального отрезка — АВ,
среднего— ВС, дистального — CD. Четыре концевые точки А,
В, С, D трех звеньев каждого блока будем называть
расчленяющими или определяющими.
Так как трехчленные блоки играют важную роль в
кинематическом устройстве нашего тела, то особое внимание
должно быть уделено совокупным пропорциям длин трех
звеньев в каждом блоке. Здесь можно ожидать
интересных морфологических закономерностей пропорционирова-
ния. Мы занялись этим вопросом, т. е. вместо
традиционного для учения о пропорциях исследования связей между
длинами двух звеньев тела обратились к исследованию
связей между длинами трех звеньв в сформированных
самой природой трехчленных блоках тела.
Как можно естественным образом охарактеризовать
совокупную геометрическую пропорцию некоторого
трехчленного блока как единого целого? Имеется ли в
геометрии адекватный инструмент для этого, который мог бы
быть применен для характеристики пропорций
трехчленных блоков так же естественно, как аффинное отношение
для двухчленных конструкций? Да, такой инструмент в
высшей геометрии имеется, и он уже рассмотрен в
предыдущем параграфе в связи с конформной симметрией. Это
пурф (6), представляющий собой обобщение простого
аффинного отношения и играющий роль критерия
конформной (и проективной) симметрии между блоками из трех
последовательных отрезков прямых. Так же, как
геометрические пропорции двухчленных конструкций естественно
39
f,JV
2
/
0,5
выражать простым отношением, так геометрические
пропорции трехчленных конструкций естественно выражать
этим вурфом. Важной особенностью
конформно-симметричных блоков из трех последовательных отрезков прямой
является то, что расстояния или простые отношения
расстояний между расчленяющими точками могут быть у
данных блоков совершенно различны и только величины вур-
фов обязаны быть равными.
Нами были вычислены
по формуле (6) величины
вурфов W для каждого
из перечисленных выше
трехчленных
кинематических блоков, т. е.
определены вурфовые пропорции
этих блоков в
распрямленном состоянии. Прежде
А / / J SC чем приводить результаты
этих вычислений, отметим,
Рис. 7. Сравнение аффинной V и чт0 величина вурфа в за-
вурфовой W пропорций при изме- ,.
нении длины отрезка ВС от 0 висимости от относитель-
до оо ных длин трех отрезков
может, вообще говоря,
меняться от 1 до оо.
На рис. 7 представлено изменение величины W для
отрезков АВ, ВС, CD при изменении ВС от 0 до °° и
АВ=СД=1. Для сравнения дано также поведение
аффинной пропорции V=AB/BC при аналогичном изменении ВС
и АВ—\. В связи с выраженной зависимостью величины И7
от соотношения длин отрезков трудно заранее сказать что-
либо определенное о величинах вурфов трехчленных
блоков тела, тем более, что величины аффинных отношений
длин отдельных звеньев в этих блоках весьма
вариабельны. Так, отношения проксимального звена к среднему в
этих блоках у взрослого человека имеют следующие
примерные значения: у среднего пальца руки отношение дли
ны первой фаланги к длине второй равно 1,6; отношение
верхнего отрезка тела к среднему — 0,5; отношение
плеча к предплечью — 1,3; отношение бедра к голени—1,2.
Кроме того, величины аффинных пропорций, вообще
говоря, зависят от возраста. Например, на протяжении
жизни от рождения у человека верхний отрезок тела
вырастает в 2,4 раза, туловище — в 2,8 раза, а нижний отро-
зок — в 3,8 раза, т. е. изменяются весьма непропорцио-
40
нально с аффинной точки зрения (рис. 8). Для сравнения
на рис. 8, в даны трехотрезочные членения, величина вур-
фов которых равна 1, 3. Эта зависимость аффинных
пропорций длин рассматриваемых звеньев тела от этапа роста
перекликается с общей неустойчивостью аффинных
пропорций различных параметров тела, отраженной в
известном законе аллометрического роста Гексли
Ух = by\t
где yi и уг — сопоставляемые параметры тела; Ь, к —
константы, причем к, вообще говоря, вовсе не равно единице.
Результаты вычислений вурфовых пропорций
трехчленных кинематических блоков тела человека
представленные в табл. 1 и 2, неожиданно обнаруживают
эквивалентность этих блоков и пропорциональность роста их звеньев
с конформной точки зрения. Из таблиц видно, что величины
вурфов W всех трехчленных кинематических блоков в
каждый момент онтогенеза группируются вокруг величины
1, 3, т. е. все эти блоки в распрямленном состоянии
по своим пропорциям почти конформно подобны и
конформно неизменны в онтогенезе. (Это во многом
оказывается справедливым не только для нормально развитых
людей, но и для карликов и гигантов, многократно
отличающихся по росту, а также для широкой совокупности
высокоорганизованных животных, что будет рассмотрено
в разделе П. 7). Значения длин АВ, ВС, CD звеньев
каждого трехчленного блока в разные моменты жизни взяты
из антропометрических работ [33; 206, с. 120—136]. Эти
данные представляют собой результат усреднения
измерений по некоторым выборкам людей одного возраста.
Антропометрическими данными по развитию отдельных
индивидуумов мы не располагаем.
Из таблиц видно, что не все трехчленные блоки
обладают равной онтогенетической устойчивостью. Но во всех
случаях онтогенетические флуктуации величины вурфа
не превышают 1—2%. Особой онтогенетической
стабильностью величины вурфа отличается средний палец руки:
на протяжении жизни она неизменно равна 1,31.
Относительно этого пальца можно отметить, что он является
наиболее выступающим из пальцев кисти (по его концу
определяется длина всей кисти и руки) и наиболее
устойчивым в отношении эволюционного редуцирования числа
пальцев конечности в мире животных. Естественно
средний палец руки считать эталонным трехчленным кинема-
41
Таблица 1. Значения вурфов W конечностей и тела человека в разном возрасте (размеры в сантиметрах
взяты из [33])
Возраст
1
4
6
8
9
Новорожденный
1
4
7
10
13
17
20
о
IT
я^:
2
2,37
4,40
6,17
6,95
8,80
12,5
16,7
20,7
23,2
26,3
30,5
32,3
1 Л
Я acq
3
1,90
3,65
5,05
5,70
7,40
10,4
13,7
16,5
18,4
20,7
23,9
24,5
So
4
1,55
3,23
4,53
5,14
6,30
8,7
11,2
13,1
14,4
16,0
18,3
18,8
£
5
1,33
1,34
1,35
1,35
1,33
1,33
1,33
1,33
1,32
1,32
1,32
1,33
о
а о,
6
Эмбрион
2,76
5,45
7,40
8,40
Человек
10,90
15,8
22,4
28,0
32,1
37,7
44,4
45,4
ень
3S>
7
2,66
5,45
7,60
8,60
9,90
14,4
20,2
24,8
27,8
32,2
37,3
37,5
ei
а «,
8
1,84
4,30
5,80
6,61
7,80
11,5
15,8
18,9
20,7
23,6
26,9
27,0
£
9
1,26
1,27
1,27
1,27
1,30
1,30
1,30
1,30
1,30
1,30
1,29
1,30
S *
S о
ttk
..
—
—
—
10,6
15,6
18,9
21,0
22,5
24,5
26,9
25,3
к
«
—'
—
—
18,3
25,0
31,4
35,3
38,0
42,2
47,8
51,8
s5
я =Л
Я оо
12
-
—
—
28,6
41,7
58,4
71,7
80,6
93,5
108,6
109,9
й£
13
—
—
—
1,29>
1,32.
1,32:
1,33
1,34
1,34
1,33
1,29
1,29±2%=0,98Р±2%
Среднее
арифметическое значение W и
пределы его онтогенетиче- 1,33±1,5%=1,01Р±1,5%
ских флуктуации;
Р=1,309... — золотой
вурф
Примечание. Возраст эмбриона указан в лунных месяцах^ возраст человека — в годах.
1,32±2%=1,01Р±2%.
Таблица 2. Значения вурфов W пальцев у человека в разном
возрасте (размеры фаланг в сантиметрах взяты из [206])
Возраст,
лет
Большой палец
4
6
8
10
\2
14
16
18
21
Среднее
арифметическое
значение W и
пределы его
онтогенетических
флуктуации;
Р=1,309...-
золотой вурф
2,21
2,63
3,08
3,23
3,51
3,83
4,31
4,42
4,65
1,56
1,78
2,00
2,13
2,23
2,48
2,88
2,83
2,99
1,16
1,30
1,45
1,58
1,73
1,84
2,07
2,12
2,17
1,34±1,5% = 1,02Р±1,5%
1,33
1,34
1,34
1,34
1,36
1,35
1,33
1,35
1,34
Возраст,
лет
к
се я
X Ь
о я ,„
х 5оэ
о яд
ое-ч;
Указательный
к я
cj Кг\
иеч
к
я я
в и
и? '•
к
cd ев
Я f-
в Я
О я о
о cd А
ов-ч;
Средний
К cd
к t
X И
(-Я
43 Чгл
а ять
и е-со.
cj cd _,
в ЧО
4
6
8
10
12
14
16
18
21
Среднее
арифметическое зна
чение W и
пределы его
онтогенетических
флуктуации;
Р= 1,309...
-золотой вурф
2,12
2,34
2,65
2,74
2,98
3,18
3,65
3,75
3,93
-
1,15
1,33
1,56
1,59
1,71
1,80
2,04
2,14
2.28
1,37±0,7%
0,80
0,97
1,10
1,18
1,31
1,37.
1,55
1,60
1,65
1,36
1,37 .
1,35 ;
1,37 ,
1,38 .
1,38 :
2,42
2,64
3,00
3,10
3,34
3,56
1,38 4,08
1,37 4,19
1,36 4,41
,
= 1,05Р±0,7% '
1,43
1,65
1,88
1,96
2,13
2,27
2,57
2,65
2,78
0,86
1,02
1,19
1,25
1,37
1,46
1,64
1,69
1,76
1,31=Р
1,31
1,31
1,31
1,31
1.31
1,31
1,31
1,31
1,31
43
Таблица 2 (окончание)
Возраст,
лет
4
6
8
10
12
14
16
18
21
Среднее
арифметическое зна
чение W и
пределы его
онтогенетических
флуктуации;
Р= 1,309... -
золотой вурф
Безымянный
к
cd cd
X fa
П X
ОЯ s
X 4CQ
о cd ~
2,24
2,49
2,82
2,92
3,16
3,37
3,83
3,99
4,11
« cd
ЦП s
ив-Ч
1,35
1,57
1,77
1,88
2,02
2,21
2,45
2,52
2,66
к
cd cd
0,92
1,08
1,23
1,29
1,43
1,56
1,68
1,72
1,82
£
1,34
1,33
1,34
1,33
1,34
1,33
1,33
1,33
1,33
1,33±0,7% = 1,01Р±0,7%
Мизинец
к
cd cd
X fa
П X
X §00
о cd a
1,73
1,95
2,21
2,29
2,46
2,66
3,05
3,11
3,31
С Я
к Ь
X *
W Я со
0,91
1,08
1,22
1,26
1,42
1,55
1,70
1,77
1,83
1,43±0,7%--=
к
в л
Э«в со
0,75
0,92
1,07
1,12
1,26
1,36
1,49
1,55
1,62
и*
1,42
1,42
1,44
1,44
1,42
1,42
1,43
1,42
1,43
1.09Р±0,7%
тическим блоком, а онтогенетически неизменную величину
1,31 его вурфа рассматривать в качестве той эталонной
величины Р, вокруг которой группируются значения вурфов
других блоков. Как видно из табл. 1 и 2, отклонение
средних значений вурфов всех трехчленных блоков от этой
эталонной величины Р составляет не более 2—5% и
только в случае самого малоподвижного и неразвитого
пальца — мизинца — достигает 5%. Маловероятно, что мизинец
устроен существенно по иным законам, чем остальные
пальцы; скорее, это отклонение в 9% для мизинца
говорит о диапазоне допустимых флуктуации в проявлениях
соответствующего морфогенетического закона построения
таких блоков.
Обобщая представленные результаты,
два конформно-геометрических правила
трехчленных кинематических блоков тела.
I. Феномен конформной унификации:
тельный аппарат как набор трехчленных кинематических
блоков отличается конформной унифицированностью этих
сформулируем
или феномена
опорно-двига-
44
Рис. 8. Онтогенетические изменения
человеческого тела (из f195])
а, б — антенатальные и постнатальные стадии;
а — в лунных месяцах; б — в годах, первый
слева — новорожденный; в — трехотрезочные
членения, величина вурфов которых равна 1,3 и 1111.
блоков, заключающейся в том, что величины вурфов всех
блоков в распрямленном состоянии группируются вокруг
значения 1,3.
II. Феномен конформного роста: по крайней мере с
момента рождения индивидуума рост тела происходит с
сохранением величины вурфа каждого трехчленного
кинематического блока.
Из этих правил следует, что, во-первых, ростовые
трансформации кинематических блоков тела (точнее,
четырех расчленяющих точек каждого блока) оказываются
родственными известным в геометрии конформным
преобразованиям; во-вторых, трехчленные блоки ребенка д
взрослого оказываются одинаковыми с точки зрения кон^
формной геометрии с точностью до различий в значениях
вурфов, т. е. кинематическая схема опорно-двигательного
аппарата отличается жестким конформным каркасом, ко-
45
торый может мыслиться как используемый для
формирования устойчивых пространственных представлений и
навыков движений. Полученные результаты порождают
много новых вопросов и среди них вопрос об эталонной
величине 1,31 указанных конформных пропорций,
анализируемый в следующем разделе.
II.3. Ряд Фибоначчи и золотой вурф
Рассмотрим, с чем связана эталонная величина 1,31
вурфов трехчленных кинематических блоков тела. Надо
сказать, что к этому непростому вопросу можно пытаться
подойти с очень разных исходных позиций: например,
можно полагать, что дело здесь в загадочном влиянии
земного тяготения, в условиях действия которого
формируются живые организмы, или что суть в некоторой
оптимальности именно этой вурфовой пропорции в
кинематических задачах тела, и т. п. Однако, по сложившейся у
нас твердой и доказательной точке зрения, истина
заключается в обусловленности этой пропорции филлотаксис-
ными законами морфогенеза, связанными с симметриями
биологических тел и рядом чисел Фибоначчи и
описанными в разделе 1.1.
Поскольку симметрии биологических тел во многих
случаях явно демонстрируют загадочную связь с рядом
Фибоначчи, то естествен вопрос: не имеет ли ряд
Фибоначчи отношения к конформным симметриям
трехчленных блоков тела человека и животных? Мы исследовали
этот вопрос, рассмотрев последовательность не аффинных
отношений, а вурфов соседних членов ряда Фибоначчи.
Три соседних числа F„, Fn+1, Fn+2 ряда Фибоначчи могут
быть интерпретированы как длины трех
последовательных отрезков, заключенных согласно рекуррентному
свойству ряда Фибоначчи "(1) между четырьмя точками
Fn+u Fn+2, Fn±3, Fn+i. Величина вурфа (6) таких трех
отрезков
Wn = (Fn + Fr,+1) (Fn+l + Fn+2)/Fn+1 (Fn + Fn+1 + F„+2). (8)
Величина вурфов всех последовательных троек чисел
Фибоначчи образует нрвую — вурфовую — последователь^
ность
46
Отметим, что если пронормировав длины всех трех
рассматриваемых отрезков (это не меняет величины вур-
фа), приняв длину первого за единицу, то, длины второго
и третьего отрезков будут* равны Fn+i/Fn~Qn' и F„+2/F«=
It / 0
— Qn> Другими словами, вурф Wn строится на Qn и Qn
й вурфовая последовательность Wn опирается на пара- и
ортостихную последовательности, собирает их воедино.
Очевидно, что при п—*-°° длины этих трех отрезков
принимают значения
1, Ф, Ф2. (10)
Соответственно предельное значение вурфовой
последовательности
Р = lim Wn = lim (Fn+2/2Fn) = Ф2/2 = 1 ,309 ... (И)
«-►CD П-»Х
Подобно тому как предел аффинной
последовательности (2) из чисел Фибоначчи называется золотым
сечением, полученную нами предельную величину Р вурфовой
последовательности (9) будем называть золотым вурфом.
Именно это значение имеют вурф тройки
последовательных отрезков (10) прямой, а также вурфы всех троек
отрезков, конформно эквивалентных этой.
Получен интересный результат: эталонная величина
вурфов трехчленных кинематических блоков тела
совпадает с величиной золотого вурфа. Это совпадение
величины не является случайным; аргументируем
дополнительно, что связь между рядами Фибоначчи и строением
тела человека очень многопланова и глубока и что
выдвигаемое положение о золотом вурфе как каноне
кинематической схемы тела человека весьма основательно.
Во-первых, эталонный трехчленный кинематический
блок — средний палец руки, обладающий эталонной
вурфовой пропорцией 1,31 своих фаланг,— демонстрирует
связь с рядом Фибоначчи и в своих аффинных
пропорциях: если длину его дистальной фаланги принять за
единицу, то все три фаланги этого пальца образуют
последовательность трех отрезков с примерными длинами 1, Ф,
Ф2. Таким образом, в зависимости от того, какие
вычисляются пропорции трех фаланг среднего пальца —
эталонного трехчленного кинематического блока тела,
получаются величины предельного значения Ф парастихной
последовательности, предельного значения Ф2 ортостих-
ной последовательности и предельного значения Р вур-
47
фовой последовательности в полном Соответствий с
частным геометрическим представлением (10) пропорции
золотого вурфа как структуры, рожденной рядом
Фибоначчи. (Данные свидетельствуют о том, что остальные
пальцы и вообще трехчленные кинематические блоки
тела, не будучи аффинно подобными среднему пальцу,
являются конформно подобными ему с хорошей степенью
точности, другими словами, по своим пропорциям
представляют собой конформно преобразованный средний
палец.)
Во-вторых, сама мысль о формообразующем
использовании живой природой золотого вурфа и вурфовой
последовательности представляется вполне естественной,
коль скоро вурфовая последовательность строится на
пара- и ортостихной последовательностях, реализация
которых в разнообразных биотелах составляет важнейшую
часть учения о филлотаксисе.
В-третьих, важная черта филлотаксиса у растений и
других изученных филлотаксисных биообъектов
заключается в том, что он происходит с образованием спиралей
вокруг главных осей. Но развитие зародыша у человека и
других позвоночных происходит также со
спиралеобразной закруткой вокруг главной оси, что обусловливает
морфологическую асимметрию тела человека и
животных [72, с. 85].
В-четвертых, все три последовательности — пара-, ор-
тостихная и вурфовая — сходятся к своим пределам осцил-
ляторным образом (из любых двух соседних членов
последовательности один меньше, а другой больше величины
предела). Но и рост длинных костей, определяющих
звенья конечностей, также происходит осцилляторным
образом, что составляет содержание двух «законов
роста»: «...а) длинная кость растет в длину и в толщину
поочередно; перерыв между процессом роста в длину
используется для роста в толщину — и наоборот; б) для
двух длинных костей одной и той же конечности периоды
активного роста и перерывы в росте чередуются.
Например, если плечевая кость находится в состоянии роста
в длину, лучевая и локтевая увеличиваются в это время
в толщину. Это приводит к тому, что пропорции
человеческого тела постоянно варьируют до окончания роста»
[9, с. 18).
Это перечисление аргументов в пользу связи
принципов строения нашего тела с филлотаксисными закономер-
48
ностями морфогенеза можно еще продолжать. Но мы остА*-
новимся на этом, подчеркнув только Следующее. Мысль
о том, что в живой природе, помимо фибоначчиевых пара-
и ортостихной последовательностей, реализуется также
фибоначчиева вурфовая последовательность (9),
просто объясняет наблюдающийся разброс в значениях вур-
фов кинематических блоков тела, сводя его к известным
чертам филлотаксиса: подобно тому, как в объектах
живой природы обычно реализуются начальные члены орто-
и парастихной последовательностей, а не их предельные
значения, прямо выражающие золотое сечение, так и при
реализации вурфовой последовательности природа
использует главным образом ее начальные члены, только
приближенно равные золотому вурфу.
Изложенные материалы свидетельствуют, что по
морфологическим особенностям своей кинематической схемы
наше тело стоит к строению тел растений, низших
животных и даже биологических молекул с их филлотаксис-
ными закономерностями гораздо ближе, чем обычно
полагают, исходя из распространенной мысли о том,,
что формирование кинематической схемы нашего тела
определяется единственно задачами ее кинематического»
функционирования. (Из этого мнения вытекает, что
нельзя и думать о каком-либо подобии кинематической схемы
тела человека со строением тел у растений и других
биообъектов, не способных к движению и не решающих
задач кинематики.) Значит, кинематическая схема тела
человека и животных построена далеко не во всем по
критериям кинематической оптимальности и ее нельзя
рассматривать в качестве идеальной кинематической
схбмы, которой, например, надлежит наделять роботы.
Это полезно учитывать конструкторам роботов, с тем
чтобы при разработке кинематической схемы роботов не быть
скованными несостоятельной идеей о том, что надо
просто копировать тело человека, в котором кинематичность
оптимально решена самой природой в ходе длительной
эволюции. Если уж и смотреть на кинематическую
организацию нашего тела как на патент живой природы,
то с учетом того, что при его развитии требования кине-
матичности были далеко не единственными
определяющими условиями в отличие от задачи создания робота по
критерию кинематической оптимальности.
В заключение этого раздела о золотом вурфе хочется
выразить убеждение, что золотой вурф заслуживает не
49
меньшего внимания, чем аффинное золотое сечение. И чем
дальше, тем все более ярко и неожиданно будет
открываться глубокое биологическое и математическое
значение этой замечательной структуры — золотого вурфа.
Приведем некоторые полученные нами формулы золотого
вурфа Р. Он является большим корнем квадратного
уравнения
АР2 — 6Р+ 1=<0, (12)
3 1
1
Р= | 1 : (13)
2x3-
_3_
2
2x3-
Эта цепная дробь при ее обрывах порождает
последовательность {kn—F2(n+i)/2F2n}, которая представляет собой
последовательность аффинных отношений ряда {Тп=
—F2/2n+i). Этот ряд находится во многом в такой же
связи с золотым вурфом Р, в каком ряд Фибоначчи {Fn} с
золотым сечением Ф. В частности,
Рп=2*ТпР — Тп-1 (14)
в близкой аналогии с известной формулой
($n = FnO + Fn-v (15)
II.4. Золотой вурф как эстетический канон.
Вопросы архитектурной бионики
Ъ свете данных о закономерностях конформных биосим-
метрий рассмотрим вопросы выдвижения и обоснования
новых эстетических систем пропорционирования: системы
золотого вурфа и системы конформных блоков.
В настоящее время в связи с задачами промышленности
и строительства вопросы эстетики, прежде всего
промышленной и архитектурной, приобретают необычно широкое
и качественно новое звучание. Среди этих вопросов можно
выделить такие, как стандартизация продукции,
необходимая в условиях современного производства, по некоторым
■формализованным критериям эстетичного; обеспечение
наилучших условий труда человека-оператора, прежде
всего в особо сложных современных системах человек —
машина, и т. д.
50
При разработке формализованных критериев эстетики
пространственных форм одними из базисных являются
исследования биологических симметрии [2, 3, 128].
Пропорции тела человека с древних времен были
предметом особого внимания со стороны художников,
скульпторов, архитекторов, всех тех, кто решал задачи эстетики
и пытался постичь тайну эстетического предпочтения
одних пропорций другим, кто старался установить, какие
соотношения надо брать в основу при создании «радующих
глаз», «возвышающих душу», «.дарующих отдых и силу»
произведений искусства и по каким архитектурным
канонам возводить дворцы, храмы, жилые дома, чтобы
согласовать их с удивительным чувством прекрасного у человека.
Такое внимание к пропорциям нашего тела связано с
классической концепцией красоты, согласно которой тело
человека является самым прекрасным из творений природы,
истоки красоты пропорций заключены в нем и
благороднейшая задача художника заключается в познании
законов его пропорций. Эта концепция красоты частично
претворяет в пластике лозунг софистов: «человек есть
мера всех вещей». Классик архитектуры Витрувий писал по
этому поводу: «никакой храм без соразмерности и
пропорции не может иметь правильной композиции, если в нем
не будет такого же точного членения, как у хорошо
сложенного человека. Ведь природа сложила человеческое тело
так, что... ступня составляет шестую часть длины тела,
локтевая часть руки — четверть, и грудь — тоже четверть.
У остальных частей есть также своя соразмерность,
которую также принимали в расчет знаменитые древние
живописцы и ваятели, и этим достигли великой и бесконечной
славы» [50, с. 65].
Многие исследователи полагали, что все человеческое
тело как набор определенных пропорций построено на
каком-то едином унифицирующем принципе, раскрыв
который можно познать тайну прекрасного тела. В работе [31]
приведен список из 115 имен известных художников и
мыслителей прошлого (в их числе Аристотель, Микел-
анджело, Леонардо да Винчи, Витрувий, Гельмгольц,
Дюрер, Плиний, Поликлет, Пракситель, Рембрандт,
Рубенс, Тициан, Фехнер, Фидий, Цицерон и др.),
изучавших пропорции человеческого тела с целью установить
этот принцип или «канон» его строения. В частности, в
попытках объяснить известный факт эстетичности
золотого сечения неоднократно высказывались^ предцоложе-
51
пия, что пропорции тела — это пропорции золотого
сечения. Для подтверждения этих идей золотое сечение
различным образом примерялось к человеческому телу,
которое, однако, очень неохотно укладывалось в
прокрустово ложе этих предположений: по длиннотным
пропорциям основное из найденного сводилось к тому, что у
взрослого (но не у ребенка) пупок делит расстояние
между макушкой и ступнями стоящего человека почти по
золотому сечению, а также, что три фаланги среднего
пальца руки попарно дают отношения, близкие к
золотым.
Чем же отличаются наши исследования и
предложения в области эстетики пропорций? Их отличие, прежде
всего в том, что мы вводим принципиально новое
понятие вурфовой пропорции как совокупной пропорции или
инструмента совокупного соразмерения частей
трехчленных ансамблей, тогда как всегда прежде в истории
эстетики речь шла об аффинных пропорциях
двухчленных ансамблей. И даже само слово «пропорция» всегда
понималось в смысле именно аффинной пропорции, что
иллюстрирует, например, определение Витрувия:
«Пропорция есть соответствие... по отношению к части,
принятой за исходную, на чем и основана всякая
соразмерность» [50, с. 65].
Полученные нами результаты по материализации
золотого вурфа в теле, сложенном из трехчленных
кинематических блоков, заставляют считать золотой вурф
трехчленным «каноном» человеческого тела и позволяют
указать новый принцип или систему построения
эстетических пропорций. Эта система, называемая нами
системой золотого вурфа, основывается на утверждении, что
пропорции золотого вурфа обладают особой
эстетичностью в связи с их реализацией в человеческом теле.
Проведенная нами предварительная экспериментальная
проверка этого положения подтверждает его.
По системе золотого вурфа в эстетических целях
рекомендуется проектировать формы всевозможных
машиностроительных изделий, архитектурных сооружений и
пр. с использованием трехчленения в отношении 1 : Ф : Ф2
или любого другого трехчленения, вурфовая пропорция
которого также равна золотому вурфу. Это трехчленное
пропорционирование может применяться как единичным
образом, так и при построении гармонических рядов и
сложных многоступенчатых форм. В силу особенностей
52
Рис. 9. Иллюстрация эстетических закономерностей
а — основное трехчленение модулора Ле Корбюзье (из [126]), величина
вурфовой пропорции трехчленения равна золотому вурфу; б —
исследования Ф. Клейном законов красоты человеческого лица (из [74])
человеческого пространственного восприятия, схожим
образом решающего задачу гармонического пропорциони-
рования и отрезков, и площадей, и объемов [397, с. 395],
система золотого вурфа применима не только к пропор-
ционированию длин, но также площадей и объемов.
Сравним систему золотого вурфа с наиболее
известной системой эстетического пропорционирования — моду-
лором или «соразмерной масштабу человека всеобщей
гармонической системой мер, применимой как в
архитектуре, так и в механике» архитектора Ле Корбюзье [126].
Модулор представляет собой некоторую шкалу размеров,
построенную на алгоритме членения отрезков по
золотому сечению. Ле Корбюзье в основу своего модулора
положил эталонную последовательность из трех отрезков с
отношением сторон именно 1 : Ф : Ф2! «В моей
пропорциональной сетке,— указывал он,— приведены три основных
размера — 113 : 70 : 43; они находятся в отношениях
золотого сечения и отвечают ряду Фибоначчи... Три
размера—113, 183, 226 — определяют размеры пространства,
в которое вписывается фигура человека ростом в 6
футов» [126, с. 60] (рис. 9, а).
Ле Корбюзье не был математиком и не владел
понятием вурфа, но исходя из своего эстетического чувства и
53
своих изысканий эстетических пропорций, он пришел к
заключению строить эстетические пропорции на основе
именно трехотрезочного членения, вурф которого равен
золотому вурфу. Здесь же отметим, что в построенных
им красном и синем рядах модулора любые три отрезка
также дают пропорцию золотого вурфа.
Рассмотрим отличия между модулором и системой
золотого вурфа. Ле Корбюзье исходил из практической
задачи пропорционирования жилых помещений человека с
учетом его роста. Поэтому принцип фибоначчиевого трех-
членения он дополнил привязкой этого трехчленения к
размерам тела взрослого человека с ростом 183 см. Зная,
что пупок делит тело стоящего взрослого человека в
отношении золотого сечения, он связал с этими делениями
два больших отрезка из фибоначчиевой тройки отрезков,
записав за ними размеры в 113 и 70 см. Длина третьего
отрезка через пропорцию золотого сечения с отрезком в
70 см тогда Сказывается равной 43 см. (G заметной
натяжкой в этом расстоянии можно видеть высоту над
головой человека, которую определяет поднятая рука (рис.
9, а)). Так, за каждым отрезком фибоначчиевой тройки
Ле Корбюзье закрепил определенную длину,
удовлетворив при этом исходному условию пропорционирования
43:70:113.
В отличие от Ле Корбюзье мы исходим прежде всего
ие из эстетического чувства, а из вещественных данных
о трехчленных кинематических блоках тела, приведших
к понятию золотого вурфа. И подобно тому, как в теле
человека и животных встречаются очень различные
случаи членения на три отрезка с сохранением пропорции
золотого вурфа, так и в эстетической системе золотого
вурфа разрешаются различные длины и аффинные
пропорции в тройке отрезков, лишь бы их вурфовая
пропорция равнялась золотому вурфу. Тем самым эстетичными
признаются целые множества трехотрезочньгх членений,
включающие фибоначчиеву тройку отрезков Ле
Корбюзье в качестве частного случая. Такая свобода выбора
мер в системе золотого вурфа очень удобна при
изготовлении различных изделий, когда требуется обеспечить
не только эстетичность пропорций, но одновременно и
функциональность изделия, и минимизацию вредных
вибраций в нем, и прочее.
Ле Корбюзье всю жизнь боролся за гармонизацию
пропорций в системах человек — машина -* среда. Будучи
54
архитектором, он выдвигал лозунг: «Дом — это машина1
для жилья!» и считал, что архитектура как искусство
охватывает сооружение не только домов и храмов, но и
конструирование автомобилей, самолетов и пр.
Соответственно его идеи фибоначчиевой гармонизации на основе моду-
лора распространялись на всю проблему организации
пространственных форм в окружающем мире. Система
золотого вурфа, включающая трехотрезочный базис мо-
дулора в качестве частного случая, продолжает эти идеи
и открывает новые плодотворные подходы к решению
этой проблемы.
Скажем еще о связи между золотым вурфом и
золотым сечением. Об эстетичности последнего известный
американский нейрофизиолог, один из основоположников
кибернетики В. Мак-Каллок писал: «Представляется
несомненным, что человек, подобно животным, живет
скорее в мире отношений... Об относительном размере двух
кубов он судит не через измерение длины их ребер или
даже площади их граней. Если он только может
детектировать различие в 1/20 длины, то он может сделать то же
самое по отношению к площадям, а также объемам,
Я затратил два года на измерение способности человека
приводить регулируемый продолговатый предмет к
предпочтительной форме, потому что я не верил, что он
предпочитает золотое сечение или что он может распознать
его. Он предпочитает и он может! В повторных
установлениях наиболее приятной формы он приходит к
предпочтению его и может установить его. Тот, кто способен
обнаружить! различие в двадцатую часть длины, площади или
объема выставляет данное различие на 1 : 1,618, а не на
1: 1,617 или 1 : 1,619. Так, эстетическая оценка прямо
показывает точное знание определенных — могу ли я
сказать привилегированных? — отношений» [397, с. 395].
По самому своему геометрическому определению
аффинная пропорция золотого сечения возникает при
рассмотрении деления отрезка АС на отрезки АВ и ВС (см.
раздел 1.1). И всегда, когда говорят о пропорции золотого
сечения, подразумевают наличие трех отрезков АВ, ВС, АС
с отношением длин 1: Ф : Ф2. Другими словами,
геометрический объект, определяющий пропорции золотого
сечения, содержит в себе три фибоначчиевых отрезка и по
существу представляет собой частный случай трехотре-
зочного объекта с пропорцией золотого вурфа; трехотре-
зочность здесь просто замаскирована полным наложением
55.
двух отрезков на третий. Й нет средств для геометричб-'
ского решения вопроса, вызывает ли у нас эстетическое
чувство объект с золотым сечением в силу наличия в нем
двухчленной пропорции золотого сечения или в силу
наличия в нем же трехчленной пропорции золотого вурфа.
Однако приведенные биологические материалы о золотом
вурфе как каноне тела человека свидетельствуют в
пользу последнего.
Вторым типом эстетической системы, которая может
быть сформулирована в связи с результатами наших
исследований, является система конформных блоков. Коль
скоро биологические тела строятся по принципу
формирования из конформно симметричных блоков, то данный
принцип может быть в эстетических целях положен в основу
построения произведений искусства, например
архитектурных ансамблей. При этом в качестве исходной формы
или блока можно брать едва ли не любую форму и,
конформно преобразовывая ее, получать новые блоки,
играющие роль блоков первой ступени. Эти блоки подлежат
объединению в конформно симметричные блоки второй
ступени и т. д.
Эстетические системы золотого вурфа и конформных
блоков, несомненно, могут быть использованы в теории
искусства, в раскрытии общих законов красоты,
поскольку согласно утверждениям многих авторов законы
пространственной гармонии родственны законам гармонии
цветовой, музыкальной и пр. Большой интерес
представляет развитие соответствующих концепций конформно-
филлотаксисной гармонии цветов, звуков и др. ощущений.
Хотелось бы упомянуть, что в своих попытках
разглядеть законы красоты с позиций высшей геометрии мы
не являемся пионерами. Ф. Клейн, автор знаменитой
Эрлангенской программы, во многом определившей лицо
высшей геометрии и ее физических приложений, пытался
открыть тайны красоты с этих позиций. Он полагал,
что художественная красота лица имеет в основе
определенные математические соотношения, и в связи с этим
исследовал все так называемые параболические кривые
на лице Апполона Бельведерского (см. рис. 9, б), линии
которого представляют высшую степень классической
красоты. Однако это исследование не привело к успеху,
поскольку, во-первых, кривые оказались по своему виду
весьма сложными и, во-вторых, не удалось обнаружить
закона, которому они подчиняются [74, с. 203].
£6
II.5. К проблеме органических форм
Формообразование в живой природе является одной из
наиболее фундаментальных биологических проблем,
имеющей не только теоретическое, но и большое практическое
значение. Достаточно сказать, что эта проблема важна
для принципов биологической систематики, а значит,
для уверенной ориентации исследователя в мире живы^с
организмов, практическая значимость которой в
настоящее время стремительно растет в связи со все более
смелыми и широкими запросами современной медицины,
сельского хозяйства, геологии, промышленного
производства на основе биологической технологии и т. д. В
литературе имеются различные подходы к данной проблеме
(см., например, [184]). Это отражает как ее сложность,
так и далеко не полную ее ясность к настоящему времени.
Рассмотрим некоторые аспекты вопроса в связи с
выявляемыми конформными симметриями в строении
биологических тел.
Существует значительное число тематических
публикаций, авторы которых исходят из того, что строение
каждого органа целиком определено эволюционным
приспособлением к условиям окружающей среды и
непосредственным функциональным нагрузкам этого органа.
При этом явно или неявно предполагается, что
собственных законов морфогенеза, которые могли бы сказаться
на потенциальном многообразии органических форм, либо
нет вовсе, либо они не играют существенной роли. При
таком подходе форма изучается лишь с точки зрения ее
соответствия функции и условиям внешней среды. Другой
подход к проблеме органических форм основан на
признании того, что структуризация биологических тел
далеко не полностью определена условиями окружающей
среды и функциональной оптимизацией. Эта точка зрения
хорошо сформулирована, например, в работах [152, 399]
(первая освещает этот подход в связи с творчеством
А. А. Любищева). Рекомендуем читателю
непосредственное знакомство с данными работами для получения
широкой картины особенностей рассматриваемой проблемы;
здесь же приведем лишь краткие выдержки из этих
публикаций, говорящих о существовании некоторых общих
структурных принципов, которые проявляются в разных
группах органического мира,
Р1
Явление симметрии в формах биологических тел
издавна служило одним из краеугольных камней в
представлениях о существовании специфических законов
морфогенеза, подобных кристаллографическим законам.
В связи с существованием этих законов «структуры лишь
в частных случаях определяются выполняемыми
функциями, а в более общем случае подчиняются некоторым
математическим законам гармонии. В многообразии форм
есть своя, не зависимая от функции упорядоченность,
своя закономерная система, обнаруживаемая, например,
в процессе выявления симметрии на основе строгого
математического описания» [152, с. 119—120]. Авторы
указывают на необходимость изучения симметрии в формах,
безотносительно к исполняемой функции. Данный подход
по существу является продолжением идей И.-В. Гете,
писавшего (см. [258, с. 105]):
«Природным тайнам время умереть,
Отважной мысли нечего скрываться.
Мы организмы нынешние впредь
Заставим кристаллизоваться».
Наличие законов кристаллизации живого вещества
вовсе не отвергает наличия механизмов приспособления
формы к функции и вполне согласуется с
существованием естественного отбора, понимаемого именно как отбор
из тех форм живого вещества, которые вообще могут
быть реализованы. Аналогично этому в физической
кристаллографии наличие законов кристаллизации не
снимает вопроса о том, как растут кристаллы, каково
происхождение определенной модификации в природных
условиях и как изменение условий внешней среды
сказывается на особенностях кристаллизации. Другими словами,
структурный подход в кристаллографии вовсе не
означает отказа от изучения условий, механизмов, причин
процессов кристаллообразования. Но основу кристаллографии
все же составляет учение о форме кристаллов как
таковых, об их пространственных характеристиках. «То же
соотношение подходов,— пишет С. В. Мейен,— оправдано
и в изучении форм организмов, на что неоднократно
указывали многие исследователи» [399, с. 207]. В пользу
этого свидетельствуют и результаты описанного в
предыдущем разделе исследования тела человека:
кинематическая схема человеческого тела оказывается подчиненной
филлотаксисным законам морфогенеза, которые не имеют
58
Рис. 10. Спиральное
строение улитки человеческого
уха и реализация
пропорции золотого вурфа в нем
(из [3171)
а — улитка, свернутая в
спираль; б — развертка улитки
в прямую линию; 1—3 —
соответственно вершинный,
средний и базальный завитки; 4 —
вестибулярный лабиринт ^ А/« fjf
О
непосредственного отношения к задачам кинематики и
определяют пространственные схемы тел, вовсе
лишенных кинематической функции (например, растений).
Значит, критерий кинематической функциональности не
может быть признан единственно определяющим
устройство кинематической схемы тела.
Поясним еще несостоятельность гипотез об устроен-
пости органов единственно по критерию функциональной
приспособленности на примере вопроса о форме улитки
уха. Гипотеза функциональной приспособленности
подсказывает ответ, что существующая форма обусловлена
функциональным предназначением уха воспринимать
акустические сигналы, а потому данная форма может
быть рекомендована акустикам для использования в
технических системах как патент живой природы, как
наилучшая в акустическом смысле форма. Однако к
подобным рекомендациям надо подходить очень осторожно,
поскольку против исходной гипотезы могут быть
выдвинуты серьезные возражения. Так, раковина садовой
улитки имеет ту же форму, что и улитка уха, несмотря
на отличие в исполняемых функциях и условиях
существования. Кроме того, улитка человеческого уха состоит
из трех завитков, длины которых дают пропорцию
золотого вурфа 1,31 (см. рис. 10), уже знакомую нам совсем
по другим творениям морфогенеза, никак не связанным
с восприятием звука.
Видимо, оправданно полагать, что образование
функционально пригодного органа связано с построением этого
органа через использование ограниченного набора
основных морфогенетических возможностей и
предпочтительных образов. И следы функциональной
приспособленности в тесном соседстве со следами унифицированной мор-
59
фологичпости можно увидеть во всяком органе и
организме. Не надо, конечно, думать, что при оформлении
функционально пригодного органа из ограниченного
набора возможностей всегда достигается оптимальное
решение задачи функциональной приспособленности. Можно
лишь говорить о компромиссной оптимизации по
противоречивым критериям функциональности и
унифицированной морфологичности (дополненным еще требованием
общефункциональной согласованности всякого новопри-
обретения организма с остальными частями и
принципами организма как единой системы).
Вернемся, однако, к кинематическому устройству
тела. Результаты наших исследований свидетельствуют,
что в кинематических блоках тела человека и (как мы
ниже покажем) высокоорганизованных животных два
сорта телесных пропорций — вурфовые и аффинные —
по-разному проявляют себя в ходе онто- и
филогенетического развития. Если вурфовые пропорции весьма жестко
сохраняются неизменными и демонстрируют связь с
рядом Фибоначчи, то аффинные пропорции, как правило,
не обнаруживают устойчивой связи с рядом Фибоначчи
и в особенности в филогенезе меняются. Эти изменения
находятся, как давно отмечалось в биологии, в
зависимости от образа жизни, функционального предназначения
конечностей и пр. Таким образом, особенности
кинематической схемы тела определяются параллельным
существованием в ней относительно неизменных вурфовых
пропорций, величина которых определяется филлотак-
сисными законами морфогенеза, и нефиллотаксисных
аффинных пропорций, через изменение которых
организм приспосабливается к специальному образу жизни и
действий.
II.6. Конформная симметрия
в моторике трехфаланговых пальцев
Обратимся к изучению моторики трехфаланговых пальцев
руки человека с позиций конформных симметрии.
Характерной биологической особенностью любого из
трехфаланговых пальцев (за исключением большого, более богатого
по возможностям своих движений) является жесткая
зависимость положения третьей фаланги от положения
двух других фаланг: без специальной тренировки человек
не может изменить положение третьей фаланги, не меняя
60
положения двух других. Иными словами, на их движение
наложена некоторая биологическая связь, в результате
которой положение четвертой расчленяющей точки
пальца определяется положением трех других расчленяющих
точек. Математической природой этой связи мы и
займемся, попутно отметив, что сгибание каждого пальца
происходит в некоторой плоскости — естественной плоскости
пальца.
Было замечено, что эта связь положений трех фаланг
для каждого пальца в широком диапазоне манипуляцион-
ных движений пальцев хорошо описывается уравнением
инвариантности вурфа с той же самой величиной («1,3),
которая выступает в качестве константы роста данного
пальца. Правда, речь здесь идет о более сложном вурфе из
комплексных координат. Замечательно, что при этом
плоскостному характеру сгибаний пальца очень естественно
соответствует то, что данное уравнение характерно для
конформной геометрии плоскости, и, кроме того, это
усложненное уравнение описывает не только моторику
пальца, но и его рост (оно включаете себя уравнение
инвариантности вурфа при росте), а потому претендует на роль
единого уравнения пространственной инвариантности при
онтогенетических трансформациях пальца. Тем самым
моторика и рост пальца как трехчленного
кинематического блока оказываются согласованными и даже с
формальной точки зрения одним и тем же, ибо в пределах
применимости этого общего уравнения они предстают
преобразованиями одной и той же группы: группы конформных
преобразований плоскости. Другими словами, рост и
моторика (в диапазоне манипуляционных движений)
трехфаланговых пальцев представляют собой явления
конформной симметрии.
Обнаружение этого структурного родства и
тождественности говорит о том, что организацию биологической
кинематики, вообще говоря, нельзя рассматривать в
отрыве от принципов морфогенеза и выводить ее
единственно из законов классической механики физических тел,
как это иногда пытаются делать.
Перейдем к самому уравнению инвариантности.
Рассмотрим любой из трехфаланговых пальцев руки,
например средний. Будем формально рассматривать
естественную плоскость этого пальца, в которой происходят его
сгибания, в качестве, гауссовой комплексной плоскости.
При этом четыре расчленяющие точки пальца А, В, С, D
61
получат соответственно комплексные координаты
ZA = xA-\-iyA, гв = хв + 1ув,
zc = xc + iyc, zD = xD + iyD. (16)
Для описания относительного положения этих четырех
точек будем пользоваться формулой комплексного вурфа
WK (7).
При распрямленном состоянии среднего пальца
величина комплексного вурфа равна золотому вурфу. Как
показали наши исследования, для широкого диапазона
движений схватывания (типа взятия в кончики пальцев
швейной иглы) характерна практическая константность
этой величины WK при совершаемых сгибаниях пальца,
т. е. величина WK выступает в роли инварианта моторики
пальцев. Это устанавливается подстановкой в
уравнение (7) координат расчленяющих точек пальца в каждой
фазе сгибания. Следует уточнить, что речь идет не о всем
возможном диапазоне сгибания пальца, а диапазоне
сгибания при рабочих операциях, осуществляемых
кончиками пальцев, типа взятия иголки. Его естественно
назвать диапазоном манипуляционных движений пальцев.
Если же обратиться к ситуациям предельного сгибания
пальцев типа сжатия руки в кулак, то величина вурфа Wh,
оставаясь вещественной, при значительном сгибании
пальца начинает расти и тем сильнее, чем ближе палец
к положению предельного сгиба (рис. 11; ср. с рис. 5, б).
В общем случае для моторики и роста любого трех-
фалангового пальца в диапазоне манипуляционных
движений имеет место следующее уравнение инвариантности
вурфа:
где Pi — средняя онтогенетическая величина вурфа W
г-го пальца из табл. 2. Поскольку (7) представляет собой
инвариант группы конформных преобразований плоскости
(187, с. 700—706], то анализируемые явления роста и
манипуляционной моторики трехфаланговых пальцев
предстают как явления конформной симметрии.
Интересно, что преобразования конформной
геометрии «в малом» представляют собой суперпозицию
преобразований вращения и подобия (на данное обстоятельство
наше внимание обратил С. А. Владимиров). Это, очевидно,
перекликается с тем, что биологические трансформации в
областях суставов, т. е. в малых областях кинематической
62
Рис. 11. Различные фазы сгибательного движения пальца и
соответствующие им величины конформного вурфа Wh
схемы тела, представляют собой именно преобразования
подобия при росте и преобразования вращения при
моторных движениях.
Итак, рост и кинематика выступают здесь как два1
взаимодополняющих аспекта формообразующих
трансформаций в онтогенезе, связанных друг с другом через'
уравнение инвариантности специальной
конформно-геометрической структуры — комплексного вурфа,
инвариантная величина которого, в свою очередь, связана с рядом
чисел Фибоначчи и филлотаксисными законами
морфогенеза. Это говорит, в частности, о том, что не только в
пропорциях звеньев кинематической схемы тела надо
различать приспособительно функциональные и морфогенети-
ческие (филлотаксисноподобные) аспекты, но эти же
соседствующие друг с другом аспекты надо различать в
общем случае и в кинематике.
Как следует объяснить то, что в других трехчленных
кинематических блоках тела человека в обычных условиях
не наблюдаются те жесткие связи филлотаксисного типа в
кинематике их трех звеньев, которые характерны для
трехфаланговых пальцев? Действительно, у других блоков
человек легко может мышечным усилием изменить
положение одного звена блока (например, стопы), не меняя
положения двух других звеньев — бедра и голени. Мы
полагаем, что дело здесь в конкуренции морфогенетиче-
ских и приспособительно-функциональных аспектов
кинематики, в результате которой функциональные аспекты
берут верх и разрывают изначальные морфогенетические
63
связи в кинематике звеньев этих блоков. В пользу того,
что морфогенетические аспекты кинематики могут
преодолеваться функциональными запросами организма,
свидетельствует как то, что при глубоком сгибании пальцев
(при сжатии кисти в кулак) величина вурфа WK
утрачивает свою непосредственную привязку к золотому вурфу
и ряду Фибоначчи, так и то, что при специальной
тренировке человек может научиться разрывать эту морфогене-
тическую связь в кинематике трехфаланговых пальцев и
сгибать одну фалангу пальца без изменения положения
двух других.
Полученные результаты по конформным* симметриям в
организации кинематики интересны не только в
фундаментальных аспектах, но имеют и чисто прикладной
интерес. Так, к настоящему времени в робототехнике
известны попытки построения технических аналогов
пятипалой кисти человека с трехчленными пальцами,
наделенных всеми основными степенями свободы и способных к
таким же, как у кисти, манипуляционным движениям
(см., например, [403]). Для построения подобных
роботов, управляемых с помощью ЭВМ, знание
математических закономерностей морфологии и кинематики пальцев
весьма важно. Надо отметить, что в таком роботе задание
в ЭВМ пространственной связи между четырьмя
расчленяющими точками пальца через вурф WK позволяет
автоматически управлять положением третьей фаланги по
положению двух других фаланг, причем не только в
диапазоне манипуляционных движений, но и в диапазоне
околопредельного сгибания пальца, если только в
уравнении (17) отказаться от константности правой части и
принять ее как величину, нелинейно возрастающую при
приближении пальца к положению предельного сгиба.
Другое приложение полученная «формула пальца»
находит в том, что на основе комплексного золотого вурфа,
видимо, расширяется содержание эстетической системы
золотого вурфа: эта система позволяет гармонично
размещать четверки точек не только на прямых линиях, но и,
учитывая конформные свойства комплексного золотого
вурфа, на любых окружностях. Иными словами, в этом
можно видеть ключ к гармонии членения окружностей.
Третье возможное приложение изложенных в этом
параграфе результатов исследований — разработка новых
адекватных средств анализа локомоций, например
анализа кинематики трехчленных рог при ходьбе, беге (в том
64
числе в условиях измененной гравитации). Речь идет
о том, что формулу комплексного вурфа, имеющую
глубокий геометрический смысл и столь интересно
выявляющую закономерности соотношений между меняющимся
при росте и кинематике расположением расчленяющих
точек у пальцев, целесообразно применять, возможно, в
несколько модифицированном виде и для анализа
особенностей движения других трехчленных кинематических
блоков. Такой анализ естественно назвать блочным
анализом, поскольку оценивается изменение в пространственном
состоянии целого кинематического блока, например
трехчленной ноги. При этом интерес представляет оценка не
только одного блока, но и совокупности блоков (например,
двух ног) в разные фазы локомоции. Необходимость
развития блочного анализа в физиологии движений давно
назрела, на что указывалось в работе [83]. Мы вернемся
к блочному анализу локомоции в разделе II. 11.
II.7. Изучение конформной симметрии
трехчленных кинематических блоков у животных
В разделе 1.2 рассматривались конформные симметрии и
пропорции золотого вурфа в трехчленных кинематических
блоках тела человека. Естественно возникает вопрос о
конформной симметрии аналогичных блоков у животных.
Рассмотрим данный вопрос.
В биологии известно, что пространственные схемы
опорно-двигательных аппаратов разных позвоночных
животных оказываются удивительно схожими. Рис. 12 на
примере скелетов человека и птицы демонстрирует это
сходство, наблюдающееся несмотря на колоссальное
различие в образе жизни и эволюционное расхождение.
В частности, общим для всех позвоночных животных
является принцип трехчленного строения конечностей,
который возник в девонском периоде 300 млн. лет назад
примерно в одно время с появлением у животных костного
скелета [105, с. 36]. В соответствии с этим принципом
конечности всех позвоночных являются трехчленными
кинематическими блоками, звенья которых носят
соответственно унифицированные названия: стцлоподий (плечо и
бедро), зейгоподий (предплечье и голень), аутоподпй
(кисть и стопа). В зависимости от вида животного
аффинные соотношения длин этих трех звеньев одной
конечности могут быть весьма различны: у одних животных самое
3 СВ. Петухов
65
Рис. 12. Скелеты человека и птицы (по П. Белону)
длинное звено — стилоподий, а самое короткое — аутопо-
дий; у других наибольшую длину имеет зейгоподий и т. д.
Рис. 13 иллюстрирует (по А. Долландеру и Т. Фенарту)
особенности принципа трехчленения кинематических
блоков на примере различных животных с неокинетическим
атгпаратом движения.
Насколько выполняется условие конформной (и
одновременно проективной) симметрии распрямленных
трехчленных кинематических блоков конечностей у разных
позвоночных животных и прежде всего млекопитающих —
цаиболее родственных человеку животных? Для ответа на
этот неисследовавшийся ранее вопрос нами предпринято
изучение вурфовых пропорций этих блоков. Описанное
ниже зоометрическое исследование проводилось
посредством измерений на экспонатах Зоомузея МГУ,
осуществленных с помощью сотрудника музея В. С.
Никольского. Отличительной особенностью приведенных зоомет-
рических данных является то, что каждая конечность из-
66
Рис. 13. Конечности животных с неокинетическим аппаратом
движения
а — позвоночных, состоящих из стилоподия 1, зейгоподия 2 и аутоподия
3: Т — четвероногого позвоночного, П — птицы, ЛМ — летучей мыши,
С — стегоцефала, Д — дельфина, Л — лошади; б — насекомого: 1 — тазик,
2 — вертлуг, 3 — бедро, 4 — голень, 5 — лапка; АВСД — трехчленный блок
с W=l,3
мерялась как кинематический блок из трех
последовательных звеньев АВ, ВС, CD, концами состыкованных
друг с другом, а потому не учитывались имеющиеся у
некоторых животных заметно выступающие наружу
отрезки локтевой кости (олекронона) и стопы (пяточной
кости), играющие в кинематике вспомогательную роль.
Поскольку не было возможности провести тотальное
Изучение по всему огромному числу видов млекопитаю-
3* 67
Таблица 3. Значения вурфов W передних и задних конечностоп
некоторых млекопитающих. (Размеры звеньев конечностей
даны в миллиметрах. Предплечья и стоны измерялись
без выступающих частей олекронона и пяточной кости.)
Вид
Еж
Крот
Белка
Бобер
Дикобраз
Выхухоль
Нутрия
Выдра
Барсук
Медведь
Макак
Волк
Шакал
Собака
Лисица
Тигр
Пантера
Кот
Рысь
Заяц-русак
Кролик
Лось
Бегемот
Коза
Жираф
Баран
Бизон
Слон
Кулан
Зебра
Летучая
мышь
(кожан)
Звено
о
42,5
14
49
75
94
22
63
66
103
380
124
200
125
190
128
270
205
95
190
110
83
340
360
160
350
190
320
970
220
205
30,5
4)
п г-
Gecq
J3
sq
КО
w
Стопоходящие
39
14
39
91
70
23
65
55
88
300
120
31
15,5
34
60
53
20
49
65
88
240
80
1,30
1,36
1,35
1,22
1,33
1,29
1,27
1,42
1,37
1,33
1,32
Пальцеходящие
210
120
190
128
270
175
100
190
118
79
]
430
270
190
580
240
340
750
200
110
155
102
270
155
75
180
70
64
1,31
1,32
1,29
1,29
1,33
1,34
1,26
1,32
1,22
1,30
Тарнопалые
525
290
245
740
340
380
450
1,32
1,42
1,35
1,27
1,35
1,34
1,27
Непарнопалые
300
250
340
320
1,29
1,34
Рукокрылые
55
116
1,32
Звено
о
staq
(V А
44
18,5
53
104
95
23
77
65
101
430
130
235
145
22
140
340
240
115
235
135
108
380
460
230
410
250
390
1150
275
290
18,5
J3
X
о ^
(hQQ
50
21,5
60
130
95
43
94
76
101
320
126
235
140
23
152
300
215
120
235
150
123
510
340
260
500
315
395
710
295
270
22,5
ев
go
ио
36
16
55
126
82
50
105
94
96
240
110
240
135
19
149
300
200
110
230
132
120
630
350
280
700
395
450
440
380
270
16,5
w
1,24
1,24
1,29
1,28
1,30
1,23
1,31
1,34
1,34
1,33
1,31
1,34
1,33
1,28
1,31
1,36
1,34
1,31
1,33
1,28
1,30
1,30
1,41
1,32
1,36
1,33
1,36
1,31
1,37
1,43
1,24
68
Таблица 4. Вурф W пальцен некоторых антропоидов
(длины фаланг в миллиметрах взяты из работы [85]).
Антропоид
Горилла
Шимпанзе
Орангутанг
Горилла
Шимпанзе
Орангутанг
Квик-коба
(палеоантроп)
Горилла
Шимпанзе
Орангутанг
Горилла
Шимпанзе
Орангутанг
Горилла
Шимпанзе
Орангутанг
Проксимальная фаланга
или I
пястная кость
АН
44,9
46,0
46,1
г>з,з
35,4
18,3
41,7
59,7
58,2
69,3
52,0
56,2
67,8
40,2
41,5
57,9
Средняя
фаланга
ВС
I палец
26,8
29,1
22,4
II палец
Г>0,8
32,4
16,2
III палец
27,5
40,6
48,0
43,0
IV палец
36,4
39,2
V палец
28,1
28,5
34,3
Дистальнам
Фаланга
CD
18,6
17,3
14,2
62.9
342
18,1
20,1
19,7
20,2
20,8
18,9
18,9
19,6
16,0
17,5
17,5
W
1,34(5)
1,30
1,35
1.39(5)
1.37
1.39
1,34
1,24
1,20(5)
1,25
1,25
1,24
1,27
1,29
1,27
щих, прежде всего встал вопрос о нетенденциозной
выборке для исследования достаточно широкой совокупности
их видов. Этот вопрос был решен тем, что за основу
приняли список млекопитающих из работы [123]. Этот
список включает млекопитающих из самых разных
отрядов и семейств с различными типами конечностей:
стопоходящих, пальцеходящих, парнопалых, непарнопалых.
Большинство представителей, фигурирующих в списке,
удалось найти среди экспонатов Зоомузея МГУ в виде
смонтированных скелетов, освобожденных от внешних
покровов и позволяющих удобное измерение звеньев
конечностей. В табл. 3 приве-дены полученные на этих
экспонатах результаты исследований вурфовых
пропорций W конечностей млекопитающих. Из таблицы видно,
что величины данных вурфовых пропорций группируются
69
вокруг значения 1,3, подобно тому как это имеет место
у человека. При этом филогенетические флуктуации W
даже в крайних случаях не превышают 7—У%, а в
подавляющем большинстве случаев находятся в пределах 3%.
Мы полагаем, что, как и у человека, вурфовые
пропорции трехчленных конечностей млекопитающих также
связаны с рядом Фибоначчи и филлотаксисными законами
морфогенеза, а наблюдающийся разброс величины W
объясняется теми же причинами, что и разброс в филло-
таксисных характеристиках у растений: в морфогенезе
реализуются не предельные, а начальные члены парасти-
хной, ортостнхной и вурфовой фибоначчиевых
последовательностей.
В табл. 4 представлены результаты аналогичного
рассмотрения вурфовых пропорций пальцев некоторых
антропоидов. Значения их вурфов также группируются
вокруг величины 1,3.
Мы проводили выборочные исследования вурфовых
пропорций трехчленных конечностей и других классов
позвоночных, помимо млекопитающих. Однако эти
исследования в силу ограниченности наших возможностей
носили несистематический характер, и подробно мы их
рассматривать не будем. Скажем лишь, что, как правило
(исключение из него — класс птиц — мы рассмотрим
ниже), были получены те же результаты группировки
значений вурфов конечностей вокруг величины 1,3.
Другими словами, трехчленные конечности у разных
позвоночных животных оказываются с хорошей степенью
точности подобными не только по морфологическому
сходству строения (эта гомологичность органов известна), но
и по своим вурфовым пропорциям, т. е. оказываются
приблизительно конформно-симметричными.
Особый интерес при проведении этих исследований
вызвал вопрос <» вурфовых пропорциях задних конечностей
птиц. Дело в том, что, как известно в сравнительной
анатомии позвоночных, морфогенез трехчленного
кинематического блока задней конечности птиц протекает
совершенно особо: проксимальные элементы
предплюсны срастаются между собой и затем прирастают к
берцовой кости [266, с. 201]. Другими словами, нога птицы
начинает формироваться по стандартному для позвоночных
животных плану, но затем в некоторый момент
вмешиваются дополнительные механизмы морфогенеза,
корректирующие этот план и вызывающие перераспределение
70
элементен, входящих в среднее и дистальное звенья
трехчленного блока ноги. Можно полагать, что такая
корректировка, обусловливающая удлинение среднего звена —
зейгоподия — трехчленной ноги птиц за счет укорочения
дистального звена — аутоподия, способна изменить
стандарт вурфовой Пропорции ноги птицы, причем в сторону
уменьшения величины вурфа по сравнению с эталонной
величиной золотого вурфа. (Поясним, что в силу свойств
формулы вурфа удлинение в трехотрезочном членении
среднего отрезка за счет крайнего ведет к уменьшению
величины вурфа). Проведенное исследование вурфовых
пропорций трехчленных конечностей птиц подтвердило
это положение. Оно проводилось на случайной выборке
птиц, включающей представителей разных отрядов
классов птиц, из экспонатов (в виде собранных скелетов,
лишенных внешних покровов) Зоомузея МГУ. Его
результаты представлены в табл. 5. Из таблицы видно, что
значения вурфов W ног систематически меньше
величины 1,31, причем зачастую это уменьшение выражено
необычно сильно (в особенности у представителей
отряда голенастых: фламинно — 1,13; аист — 1,18;
колпица — 1,22).
В исследованиях на позвоночных животных была
сделана попытка изучить вурфовые пропорции трехчленного
кинематического блока всего тела у разных организмов.
При этом мы столкнулись с рядом трудностей, например
неясностью во многих случаях, какие точки должны
естественным образом играть роль расчленяющих точек такого
трехчленного блока. Действительно, если у человека
естественной граничной точкой тела со стороны головы
является макушка головы, то строение и позное
расположение головы у многих позвоночных животных, имеющих,
например, выставленную по оси тела длинную морду,
предлагает и другие точки головы в качестве кандидатов
на роль граничной точки трехчленного блока тела. Кроме
того, у многих позвоночных хорошо развит и активно
используется (при лазании по деревьям и пр.) хвост.
Соответственно возникает дилемма, что считать за третье
звено трехчленного блока тела у данных позвоночных — ноги
или хвост. Эти и другие трудности не позволили к
настоящему времени прийти к твердому мнению о закономерном
поведении величин вурфовых пропорций трехчленного
блока тела у позвоночных животных. Однако у нас
сложилось впечатление, что вурфовые пропорции в этом слу-
71
Таблица 5. Значения вурфоп W передних и задних конечностей
птиц (размеры звеньев в миллиметрах)
Птица
Сорока
обыкновенная
Славка
обыкновенная
Славка
Зимородок
Беркут
Орел
Птица-
секретарь
Фламинго
Аист
Колпица
Лебедь
Утка
Пеламедея
Попугай
серый
(жако)
Звено
о
V _
ё<ч
С^
45
12
17
68
195
190
170
195
200
150
283
103
200
55
1. -Э
ЕТ?
8.2 о
С с а.
54
12,5
20
80
225
220
175
210
240
175
272
92
210
65
л
н
gQ
Яо
49
12
19
56
180
165
150
148
190
145
245
104
170
73
W
1,28
1,31
1,29
1,23
1,26
1,25
1,29
1,25
1,25
1,26
1,32
1,39
1,28
1,32
Звено
о
а ~
но
М^
44
13,5
16
40
135
125
105
80
95
87
100
53
105
49
я
V ^
54
СнЧ
70
23,5
28
56
165
165
265
303
240
190
193
85
185
65
d
а ^
2Ъ
оо
80
33,5
37
65
175
190
350
355
295
260
250
104
245
62
w
1.26
1,27
1.26
1.29
1,30
1,30
1,19
1,13
1,18
1,22
1.24
1.27
1,26
1,26
чае отличаются заметно большей вариабельностью
величин, чем для блоков конечностей. Видимо, морфогенез
трехчленного блока всего тела у некоторых позвоночных
протекает с еще большими осложняющими
отступлениями от стандартного плана морфогенеза трехчленных
кинематических блоков, чем это отмечено для морфогенеза
трехчленного блока задних конечностей птиц.
Суставчатой кинематической схемой тела наделены не
только позвоночные животные. В этой связи крайне
интересно определить, является ли принцип трехчленности
конечностей характерным только для позвоночных животных
и если нет, то выполняется ли условие конформной
симметрии между трехчленными блоками конечностей у
позвоночных и безпозвоночных животных, т. е. наблюдается
ли примерное равенство вурфовых пропорций
трехчленных кинематических блоков распрямленных конечностей у
безпозвоночных животных величине 1,3. Для ответа на
эти вопросы мы обратились к классу насекомых, наиболее
72
представительному по числу видов (около 1 млн.) из
существующих классов животных. Насекомые лишены
позвоночника и костного скелета. Строение их конечностей на
первый взгляд существенно отличается от строения
конечностей позвоночных, поскольку они многочленны, и
состоят из коксы (тазика), трохантера (вертлуга), бедра,
голени и лапки, имеющей до 5 члеников. Однако, как
хорошо известно в энтомологии [263, с. 35], эти конечности
по существу также представляют собой трехчленные
кинематические блоки. Дело в том, что в этих конечностях
все сочленения практически только обозначены и не
работают как суставы, кроме двух — вертлужного и коленного,
которые являются действительно высокоподвижными
сочленениями (и по особенностям строения очень
напоминают голеностопное и коленное сочленения у человека).
Кроме того, эти же два сочленения первыми появляются
в эмбриогенезе. Поэтому в трехчленном кинематическом
блоке типичной конечности насекомых следует различать
первое звено, состоящее из коксы, второе звено — из
вертлуга и бедра, третье — из голени и лапки (рис. 13, б).
Универсальность трехчленного строения конечностей
огромного большинства развитых живых организмов
представляет собой поразительный феномен природы. И эту
трехчленность нельзя представить себе просто как
результат последовательного эволюционного перехода от
первоначально двухчленной конечности. Насколько мы можем
судить, позвоночные животные и их предки не обладали
двучленными конечностями. Также и для всего типа
членистоногих, к которому относится и класс насекомых,
между нерасчлененными и многочленными конечностями не
известно переходных состояний. Вот что об этом
говорится в учебнике энтомологии: «Конечности членистоногих
происходят от нерасчлененных параподий аннелид. Между
этими формами нет переходных состояний, ибо у самых
первичных членистоногих — триллобитов — конечности
уже весьма сложны» [262, с. 44]. Подчеркнем, что факт
трехчленности конечностей у позвоночных и насекомых,
не имеющих в эволюции общего предка, который облагал
бы трехчленными конечностями, говорит в пользу особой
морфогенетической фундаментальности принципа трех-
членения.
Что же касается вурфовых пропорций трехчленных
конечностей у насекомых, то мы, конечно, не имели
возможности исследовать пропорции конечностей у всех ви-
73
дов насекомых, однако выборочное знакомство показало,
что обычно вурфовые пропорции и здесь близки к 1,3 и что
формула инвариантности вурфа обладает
предсказательной силой. Например, всегда, когда у некоторого вида
насекомых первое звено конечности оказывается значительно
короче второго звена, то обязательно третье &вено
конечности оказывается значительно (порой в несколько раз!)
длиннее второго звена, как то и должно быть по
отмечаемой тенденции к группировке вурфовых пропорций
конечностей вокруг величины 1,3. Изучение этих пропорций
у насекомых и животных других классов безусловно
должно быть продолжено и поставлено на систематическую
основу, поскольку речь идет о проявлении
общебиологических закономерностей морфологической структуризации.
Резюмируя, можно сказать, что между трехчленными
конечностями различных видов животных усматриваются
отношения конформной симметрии, связанные с филло-
таксисными законами морфогенеза. В книге приведена
только часть имеющихся материалов о конформных сим-
метриях кинематических трехчленных блоков у
животных. По другим материалам хотелось бы дополнительно
отметить, что, как правило, три самых дистальных звена
конечности, подобно трем фалангам пальдев у человека,
также дают вурфовую пропорцию примерно равную 1,3
(например, последние фаланги в многофаланговых
пальцах крокодила или три последних членика лапки
насекомого). Следует, однако, предостеречь от приписывания
излишне универсального характера принципу трехчлене-
ния и пропорции золотого вурфа: живая природа
использует и другие членения, например, у многих рептилий
число фаланг пальцев больше трех. Более интересным и
важным здесь является не вопрос числовых
закономерностей, а организация биологических тел в соответствии с
принципами конформной симметрии. Подчиненность этим
принципам может проявляться и в закономерностях
роста и кинематики многофаланговых пальцев животных.
Надо отметить, что рассмотренные типы животных
относятся, как и человек, к живым организмам с
неокинетическим аппаратом движений. Что вообще известно о
неокинетике?
Древние живые организмы довольствовались
двигательным аппаратом, основанном на работе гладких мышц,
не имеющем суставчатого скелета и проприоцепции
кинетического назначения. На некотором этапе эволюции от-
74
яосителъно внезапно и скачкообразно произошло рождение
(принципиально нового аппарата движения — неокинетиче-
ркого, отличительными чертами которого является то, что
дя построен на основе новых двигательных элементов —
поперечнополосатых мышц, имеет суставчатый
кинематический скелет и новую проприоцептивную систему,
возглавляемую системой полукружных каналов.
Неокинетика обеспечила организму резкое повышение качества
движения и взаимодействия с окружающим миром:
быстроту, мощность, маневренность, управляемость и пр., и в
связи с этим привела к стремительному развитию
нервной деятельности и психических функций.
В современные знания о неокинетике большой вклад
внес своими широкими и глубокими исследованиями
Н. А. Бернштейн. Приведем некоторые его заключения
[24, с. 107—112]. Относительно возникновения
поперечнополосатой мускулатуры он указывал, что
сократительный элемент поперечнополосатой мышцы появляется в
филогенезе с полной внезапностью в том смысле, что
нигде не находится следов каких-либо переходных форм к
нему от гладкой мышечной клетки. И по сию пору
остается загадочным, почему эволюция не пошла по пути
преобразования первичных актиномиозиновых
сократительных элементов, с тем чтобы сделать их
функционально более гибкими и доступными регуляции. Весь ход
эволюционного осваивания неокинетической
мускулатуры,— отмечал Н. А. Бернштейн,— выглядит так, как если
бы эти сократительные элементы были внесены в
структуру организмов откуда-то извне. Начиная с обоих
классов обладателей поперечнополосатой мускулатуры —
членистоногих и позвоночных, возникают жесткие
суставчатые скелеты, каких нет и следа у нижестоящих классов —
обладателей только гладкого типа мускулатуры.
Появляется младшая (опять-таки неокинетическая) форма про-
приоцепторики — геометрической, воспринимающей позы
и скорости и возглавляемой неолабиринтом полукружных
каналов — на фоне древней проприоцепторики тропизмов,
возглавляемой отолитовым аппаратом (палеолабиринтом)
и приспособленной к оценке давлений, ориентировке в
поле тяготения и т. п.
С этими утверждениями Н. А, Бернштейна о
скачкообразном характере возникновения неокинетического
аппарата движения хорошо коррелирует то, что такая
специально исследуемая нами характернейшая черта су-
75
ставчатых скелетов, как трехчленное устройство
конечностей, по данным эволюционной биологии, появилась в
эволюции вместе с появлением самих скелетов, причем
также «с полной внезапностью», без каких-либо
переходных форм (например, двухчленной конечности), как и
поперечно-полосатая мышца.
Говоря о поперечно-полосатых мышцах, Н. А. Берн-
штейн отмечал одну черту (морфологичностъ) их
структуризации в кинематическом устройстве тела, которую
мы считаем частным проявлением весьма общего
положения дел в неокинетике (и не только неокинетике) :
«мышечные массивы, анатомическое членение которых на
отдельные мускулы имеет по большей части чисто
морфологическое основание, без существенной значимости
для биодинамики, облекают эти аксиальные
кинематические цепи снаружи, повинуясь в своем размещении также
преимущественно причинам чисто морфологического
порядка» [24, с. 79]. Эта морфологическая (мы бы сказали,
морфогенетическая), а не
приспособительно-функциональная обусловленность, отмеченная Н. А. Бернштейном для
мышечно-системного компонента неокинетики,
усматривается и в ряде важных черт устройства кинематической
схемы скелета человеческого тела, что уже неоднократно
подчеркивалось в книге.
Мы рассматриваем кинематику как во многом
непосредственное продолжение морфогенеза и, на некотором
этапе, преодоление морфогенеза. При этом мы полагаем,
что симметрийные принципы морфогенеза имеют прямое
отношение и к развитию кинематики. Вообще,
специфической чертой нашего исследования аппарата
неокинетики является то, что большое внимание уделяется мор-
фогенетическим аспектам устройства этого аппарата,
тогда как в подавляющем большинстве работ других
исследователей кинематической организации человека и
животных в центре находятся функционально
обусловленные аспекты (существующие в организме наряду с
морфогенетическими), например следующие:
а) противопоставление большого пальца руки у
человека остальным пальцам как определенное
функциональной задачей оперирования с предметами,
характерной для человека;
б) особенности позы и тонуса мышц у человека в
связи с функциональной задачей преодоления действия
гравитации при прямостоянии и двуногой ходьбе;
76
в) расширение костей скелета у тяжелых животных
как определенное функциональной задачей
выдерживания большего веса и т. д.
I Все такие функциональные аспекты представляют
собой приспособительные изменения уже чего-то
материально существующего, появившегося в эволюции
вовсе не в связи с рассматриваемой функцией и
меняющегося при приспособлении к этой функции лишь в
частных деталях. Например, кисть человека,
приспосабливаясь к функции оперирования с предметами, претерпела
весьма малые изменения типа противопоставления
большого пальца, сохранив свои основные черты: пятипа-
лость, трехчленностъ пальцев и др., существовавшие,
когда не было и намека на появление человека. Или другой
пример: упомянутая выше функция прямостояния и
двуногой ходьбы, наложив некоторый отпечаток на
кинематику человека, не затронула многие основные
черты его кинематической организации, существовавшей
задолго до появления этой функции.
II.8. О возможном участии конформных симметрии
в генезисе пространственных представлений
у индивидуума
Уже давно различными исследователями выдвигались
идеи о важном значении кинематической организации
тела и его движений в генезисе пространственных
представлений у индивидуума. В частности, эти идеи
разрабатывались Г. Гельмгольцем и И. М. Сеченовым. Они же
положены А. Пуанкаре в фундамент его учения о
физиологических основаниях геометрий и о происхождении у
индивидуума пространственных представлений [191—
194]. Изложенные выше данные о конформных симмет-
риях в кинематическом аппарате человека и животных
позволяют дополнить эти идеи положением о
принципиальной важности конформной геометрии для
рассматриваемой биологической проблематики.
Согласно Пуанкаре, само понятие пространства и
геометрии возникает у индивидуума на основе деятельности
кинематической организации тела, включающей
внутреннюю рецепторику для характеризации положения и
движения частей тела относительно друг друга, т. е. в
кинематической организации тела есть нечто,
предшествующее понятию пространства [192, с. 58—59]. При этом
77
особенности всего аппарата кинематической деятельно?
сти организма приведены эволюционным развитием жи^-
ни в соответствие с реалиями окружающего физического
мира. Благодаря этому каждый новорожденный организм
овладевает адекватными пространственными
представлениями не только за счет персонального контакта в х?оде
онтогенеза с предметами окружающего мира, но и за счет
благоприобретений предыдущих поколений,
закрепленных в ходе филогенеза в аппарате движений тела [193,
с. 82-92].
По мнению Пуанкаре, движения нашего тела играют
первенствующую роль в генезисе понятия пространства.
Для существа вполне неподвижного не было бы ни
пространства, ни геометрии [192, с. 57]. «Локализовать
предмет,— писал он,— значит просто представить себе те
движения, которые нужно было бы сделать, чтобы
достигнуть его», т. е. представить себе «те мускульные
ощущения, которыми сопровождаются эти движения и которые
не предполагают предсуществование понятия
пространства» [192, с. 58]. «Если бы у нас не было орудия для
измерения,— подчеркивал Пуанкаре,— мы совершенно не
могли бы построить пространство; но орудие у нас есть, и
к нему мы относим все и пользуемся им инстинктивно; это
наше собственное тело» [193, с. 80].
Свяжем построения Пуанкаре о телесно-двигательных
механизмах генезиса пространственных представлений у
индивидуума с конформно-геометрической организацией
двигательного аппарата тела. Пуанкаре утверждал:
«Система координатных осей, к которым мы, естественно,
относим все внешние предметы,— это система осей,
неизменно связанная с нашим телом, которую мы и носим всюду
с собой» [192, с. 57]. Однако практически он не
затрагивал вопроса, который нами ставится во главу угла: какой
из многих возможных типов систем координатных осей
реализуется в этой координатной системе тела? Известно,
что геометрические системы координат могут вводиться
разнообразными способами, принципиально
отличающимися друг от друга. В частности, конформная геометрия
располагает своими специфическими системами
координат и средствами конформного мероопределения. Наши
данные о конформных симметриях в кинематической
схеме тела заставляют думать, что координатная система
тела строится именно на конформной основе.
Веским свидетельством в пользу этого являются также
78
рапные о важной роли конформных симметрии в зритель-
дом восприятии, излагаемые в следующем разделе. Кроме
этого, нами усматриваются определенные аналогии между
шзштасферической системой координат конформной
геометрии и пятипалой кистью, являющейся основным орудием
осязательного распознавания геометрических форм.
Дополнительным аргументом в пользу того, что организм в
овладении пространственными представлениями
пользуется не евклидовой, а конформной схемой измерения,
служит также следующий факт. Все звенья нашего тела
изменяют свои евклидовы длины с ростом, причем интенсивнее
всего именно в младенчестве, когда происходит основное
знакомство организма с окружающим миром и когда
особенно важно для метрической схемы иметь устойчивые
эталоны длины. Но будучи лишено эталона евклидовой
меры, тело оказывается обеспечено неизменными в его
жизни и в жизни всего рода эталонами конформных вур-
фов, которые являются вполне пригодным инструментом
геометрического сравнения и построения координат.
Хочется также отметить, что, видимо, наше тело несет
в себе не одну, а целую совокупность соседствующих
конформных систем координат, на согласованной работе
которых организм строит свои пространственные
представления и навыки движений. В свете сказанного
представляется целесообразным поиск конформных принципов
функционирования в различных сенсомоторных системах —
мышечной, зрительной, вестибулярной и пр., эволюция
которых проходила в тесной связи с задачами обслуживания
движений тела.
Говоря о конформных биосимметриях кинематических
блоков и учении Пуанкаре, следует также привести
современные данные физиологии о механизмах кинестезии,
которые оказываются тесно связанными с теми самыми
расчленяющими точками тела, в блочном размещении
которых нами обнаруживаются конформные закономерности.
Механизмы кинестезии обеспечивают организм знанием
о положении и движении звеньев тела. Пуанкаре полагал,
что эти представления покоятся на «мышечном чувстве».
А вот что утверждает современный справочник по
физиологии: «В отношении кинестезических ощущений в
последние годы был достигнут значительный прогресс.
Прежде всего, теперь очевидно, что чувство позиции и
движений суставов зависит только от соответствующих
рецепторов в самих суставах. Нет никакой необходимости при-
79
зъгвать таинственное «мышечное» чувство для
объяснения кинестезических ощущений» [418, с. 426].
Другое дополнение современная физиология вносит в
учение Пуанкаре о врожденной взаимосвязи тела и
пространственных представлений, утверждая априорность
представлений индивидуума о своей телесной оболочке. Это
утверждение связано с изучением так называемых
фантомных ощущений у инвалидов: специфического чувства
присутствия утраченной части тела. Сравнительно недавно
установлено [445, 447], что фантомные ощущения имеют
место не только у людей с ампутированными
конечностями, но и у людей с врожденным отсутствием конечностей.
Значит, представление индивидуума о схеме своего тела
вовсе не обусловлено его жизненным опытом пользования
своим телом, а носит врожденный характер.
П.9 Конформная симметрия
и геометрия пространства
зрительного восприятия
Изложенные выше материалы привлекают внимание
к загадочной роли конформной симметрии в явлениях
формообразования и кинематики биологических тел.
Ограничивается ли этим значение конформной симметрии для
живых организмов? Отрицательный ответ на этот вопрос
поддерживают материалы о фундаментальном значении
конформной симметрии для другой, казалось бы, очень
далеко отстоящей области жизнедеятельности организма —
области психофизической организации пространственно^
го восприятия. Рассмотрим известные экспериментальные
данные о геометрии пространства зрительного восприятия,
базирующейся, согласно этим данным, на принципах
конформной симметрии.
Касаясь истории вопроса, обратим внимание на то, что
в психологии прежде всего теории восприятия идеи
неевклидовых геометрий применяются для моделирования и
объяснения биологических явлений уже длительное время
в отличие от других областей биологии. О целесообразности
применения неевклидовой геометрии для описания
особенностей так называемого пространства зрительного
восприятия, учитывая его структурные отличия от окружающего
физического пространства, писали многие авторы (см.
обзор по данному вопросу в работе [375]). Однако для
проверки применимости неевклидовой геометрии не было сде-
80
Аано никаких опытов и вопрос оставался совершенно
открытым вплоть до работ Р. Лунебурга [392—394],
опубликованных в 1947—1950 гг. В этих работах впервые
утверждается мысль о том, что пространство зрительного
восприятия у человека характеризуется геометрией
Лобачевского. Начальной основой для такого утверждения
явились результаты экспериментов, осуществленных еще
в 1913 г. Блюменфельдом. В этих экспериментах
исследуемому наблюдателю, сидящему в кресле в совершенно
темной комнате, предъявлялись две светящиеся в темноте
точки (каждая из них создавалась с помощью маленького
газового пламени), фиксированные на расстоянии 5 м от
наблюдателя симметрично его саггитальной плоскости.
Кроме них, ему предъявлялись еще шесть пар таких же
светящихся точек, которые могли перемещаться
экспериментатором в горизонтальной плоскости по указанию
наблюдателя. От наблюдателя требовалось посредством
указаний экспериментатору о надлежащем размещении этих
дополнительных пар точек установить все точки в две
линии: в одном случае — параллельные прямые, в
другом — эквидистантные (т. е. равноудаленные друг от
руга на всем протяжении). Полученные в результате
гры эквидистантных и параллельных линий («аллеи
шюменфельда») оказались заметно отличающимися
>уг от друга (рис. 14, о), что находится в противоречии
евклидовой геометрией, в которой параллельные пря-
je всегда эквидистантны. Напротив в неевклидовой гео-
"чрии, параллельные прямые вовсе не обязаны быть
видистантными. Учитывая особенности взаимного
расположения параллельных и эквидистантных «аллей Блю-
менфельда», Лунебург пришел к заключению, что
зрительное пространство характеризуется геометрией
Лобачевского. (Дополнительно Лунебург выдвинул гипотезу о том,
что такая специфичность геометрии зрительного
пространства базируется на физиологическом мехапизме
определения равных расстояний через равные углы сходимости
оптических осей двух глаз. Однако эта гипотеза оказалась
несостоятельной, что разобрано, например, в работе [375].
Действительная причина явления, видимо, носит более
фундаментальный характер.) Следует отметить, что
для наблюдения исходных для концепции Лунебурга
зрительных феноменов особо важно отсутствие в поле зрения
наблюдателя каких-либо дополнительных ситуационных
указателей (например, стен, линий их пересечения и пр.,
81
11
w
\)
I
I
/
60 I
0
Рис. 14 Моделирование геометрических особенностей
пространства зрительного восприятия
о — аллеи Блюменфельда; пунктиром показаны эквидистантные линии,
точками — параллельные, слева дано расстояние от наблюдателя в
сантиметрах (из [375]); б — стереографическая проекция, S —центр проекции,
М и М' — соответствующие друг другу точки сферы и плоскости проек~
ции
прямизна или другие геометрические характеристики
которых известны наблюдателю из прошлого опыта),
способных нарушить непосредственную зрительную оценку
пространственной картины, заменив ее на
опосредованную оценку, базирующуюся на опыте обращения с
твердыми телами и потому ведущую к суждениям евклидова
типа.
Исследования Лунебурга нашли широкий отклик и
вызвали десятки публикаций ученых из разных стран,
развивающих и уточняющих идеи неевклидовости
пространства зрительного восприятия [80, 177, 272, 296—299,
373—375 и др.]. Из работ советских авторов,
опирающихся на исследования Лунебурга, можно отметить [159,
197 и др.]. Особо обстоятельную экспериментальную
проверку концепции Лунебурга осуществил Г. Кинл [375],
пришедший при этом к принципиальным выводам о
важном значении конформной симметрии в структуризации
пространства зрительного восприятия.
Основная серия экспериментов Кинла включила
исследование около двухсот наблюдателей, в ходе которого
получено около 1300 зрительных образов разного типа.
Кроме того, было сделано одиннадцать дополнительных серий
опытов, в которых осуществлено свыше 2000 измерений.
Наблюдатели судили о прямизне кривых, равенстве углов,
/
1
!1
\
82
^расстояний, сегментов, площадей; параллельности
прямых линий в пространстве; существовании общего центра
у семейства окружностей, т. е. в этих экспериментах
изучались простые геометрические элементы и соотношения,
известные любому нормально развитому взрослому
человеку. Как и Лупебург, Кинл по существу исследовал
зрительное восприятие объектов на горизонтальной
физической плоскости,
В ходе своих исследований Кинл получил
подтверждение того, что пространство зрительного восприятия
характеризуется геометрией Лобачевского. Он пришел к
мысли, что адекватной моделью этой геометрии в случае
зрительного восприятия является известная конформная
модель Пуанкаре. Напомним существо этой модели.
Если из полной группы конформных (круговых)
преобразований плоскости выделить в особую подгруппу все
преобразования, переводящие в себя некоторый
заданный круг, называемый абсолютом, то эта подгруппа
конформных преобразований оказывается изоморфной
группе гиперболических преобразований плоскости (т. е.
группе преобразований Лобачевского), а геометрия
внутренней области абсолюта — совпадающей с геометрией
Лобачевского. При этом в роли прямых геометрии
Лобачевского выступают дуги окружностей, ортогональных к
абсолюту. Поскольку конформные преобразования
сохраняют углы между пересекающимися прямыми и не
сохраняют расстояния между точками, то в этой указанной
Пуанкаре интерпретации геометрии Лобачевского за угол
принимается обычный евклидов угол, а в качестве
соответствующего данной геометрии расстояния между двумя
точками вводится новый вид расстояния, основанный на
использовании двойного отношения или вурфа четырех
точек, уже многократно фигурировавшего в нашей
работе. В качестве дополнительных двух точек, необходимых
для построения двойного отношения четырех точек, здесь
используются две точки пересечения прямой геометрии
Лобачевского, проходящей через две исходные точки,
с абсолютом (см., например, [102]).
В своих экспериментах Кинл получил целый ряд
.подтверждений адекватности этой конформной модели
геометрии Лобачевского для пространства зрительного
восприятия. Например, оказалось, что «взаимная конфигурация
визуально концентрических окружностей действительно
описывается гиперболическим семейством окружностей в
83
физическом пространстве с поразительной точностью»
[375, с. 398]. Кинл заключает свою работу словами!:
«Модель Пуанкаре гиперболического пространства,
впервые примененная к структуризации зрительного
пространства, показывает убедительное совпадение с
экспериментальными результатами» [Там же, с. 399].
Итак, конформная симметрия, точнее, определенная
ее подгруппа, соответствующая неевклидовой геометрии,
оказывается-основой структуризации пространства
зрительного восприятия, что подтверждает выдвигаемое нами
положение об общей биологической важности
конформной симметрии. Остается открытым вопрос, почему именно
эта подгруппа является определяющей для зрения.
Представляется вполне возможным, что для других видов
пространственного восприятия, а также и для
зрительного восприятия в условиях, отличающихся от
рассмотренных Лунебургом и Киплом (по характеру изучаемой
плоскости, наличию вестибулярного раздражения и т. д.),
важное значение имеют другие подгруппы конформной
симметрии, определяющие иные виды геометрий. Эти
вопросы еще ждут своих исследователей.
Нельзя не отметить в связи с конформными биосим-
метриями, что зрительное восприятие, подобно
рассмотренным выше случаям биологической кинематики,
оказывается структурно родственным морфогенезу и может
трактоваться как структурное продолжение морфогенеза,
а не как нечто совершенно новое, изобретенное живой
природой неясным образом на определенном этапе эволюции.
Можно думать, что в процессах «овладения
пространствам» (по выражению Ле Корбюзье, «овладеть
пространством—таково первое побуждение всего живого» [126,
с 37]) организм во многом использует одни и те же
принципы структуризации, будь то морфогенетическое,
кинематическое или психофизическое овладение
пространством. Такое единство принципов структуризации для
разных систем и уровней организации, подлежащих
взаимосогласованию и в совокупности составляющих
«овладевающий пространством» живой организм, способно
обеспечить живым организмам жизпенно важные
преимущества.
Хотелось бы еще обратить внимание на определенную
аналогию между морфофункционалыюй организацией
человеческого глаза и хорошо известной в конформной
геометрии стереографической проекцией. Эта проекция
84
получается, если имеется некоторая сфера, за центр
проекции принимается одна из точек этой сферы, а за
плоскость проекции — плоскость, касающаяся сферы в
циаметрально противоположной точке, или любая
параллельная ей плоскость, не проходящая через центр
проекции (рис. 14, б). При стереографической проекции
окружности, расположенные на сфере, всегда
трансформируются в окружности плоскости проекции и наоборот.
Сетчатка человеческого глаза представляет собой
сферическую поверхность, и зрение человека строится на
проекционном отображении внешних объектов на данную
сферу сетчатки. При этом центр проекции лежит в
области хрусталика глаза, расположенного вблизи
противоположного полюса данной сферы, хотя и не совсем
совпадающего с ней. В силу такого сходства со стереографической
проекцией при проецировании любых плоских зон
внешнего пространства, расположенных в момент их
разглядывания перпендикулярно оси зрения, окружности из
этих зон проецируются на сетчатку всегда в виде
окружностей, т. е. окружности выступают в роли инвариантных
фигур (с точностью до соответствия устройства глаза
стереографической проекции). Эта гипотеза о
соответствии особенностей устройства глаза и стереографической
проекции позволяет подойти к отмеченной Г. Кинлом
конформной специфике геометрии пространства
зрительного восприятия как обусловленной анатомофизическими
особенностями зрения.
Интересно упомянуть и менее ясные ассоциации
между конформной геометрией и психофизическими
особенностями зрительного восприятия. В работе В. Н.
Прокофьева «О «перцептивной перспективе» и перспективах в
живописи» (см. Приложение к книге [197]) на материале
пространственных построений в картинах ряда
художников (Сезанн, Петров-Водкин и др.) обосновывается
сферичность геометрии пространства зрительного восприятия
и отмечается необходимость развития «теории
сферической перспективы... ближе всего подводящей живопись к
овладению полнотой зрительного восприятия, к освоению
перцептивной перспективы (т. е. геометрии пространства
зрительного восприятия.—С. П.)» [197, с. 183]. Это, по
нашему мнению, может рассматриваться как
дополнительный аргумент в пользу связи геометрии зрительного
восприятия и конформной геометрии, которая недаром
называется ещё и круговой геометрией.
85
11.10. Концепция морфопоз
и организация моторных движений ,
Обратимся к изучению вопроса о взаимоотношении
конформных симметрии, присущих телу развитого живого
организма, и особенностей моторных движений у такого
организма. Изучение симметрии и их нарушений —
традиционная глава в исследованиях, посвященных моторике у
человека и животных. Так, характеризуя аллюры
четвероногих животных, исследователи описывают
сопровождающие эти аллюры нарушения зеркальной симметрии тела
в процессе движения животного, а также особенности
симметрического поведения отдельных пар конечностей
[61, 223 и др.]. Типичным для широкого класса живых
организмов — от простейших до высокоорганизованных —
является то, что моторные движения у них происходят с
нарушением симметрии, присущих телу организма. В
качестве показательного примера можно привести морскую
звезду, пентасимметричная форма тела которой
претерпевает определенную периодическую диссимметризацию
в процессе локомоции звезды.
Б подобных примерах моторные движения тела
предстают как процесс преодоления исходной формы тела и
ее симметрии. Образно говоря, если бы природа решала
задачу обучения пятилучевого цветка движениям типа
локомоторных, то важнейший этап при этом заключался бы
в придании цветку способности преодолевать свою морфо-
генетически определенную пентасимметричную форму.
И подобные задачи применительно к растениям (и не
только к ним) иногда решаются самой природой. Это
отчетливо наблюдается у тех растений, которые по своим
способностям к движению занимают промежуточное
положение между «неподвижными» растениями и животными.
Например, веточка мимозы в ответ на внешнее
раздражение отдельных листочков способна выраженной
двигательной реакцией листочков и черешков изменить свою
исходную форму и симметрические характеристики
расположения этих частей (физиологические механизмы
подобных движений у растений, конечно, имеют свою
специфику по сравнению с животными [11 и др.], но нас в
данном случае интересует лишь общий вопрос о
нарушении исходной или, лучше сказать, канонической формы
биологического тела при моторных движениях). Эта связь
между моторным движением и нарушением определенной
86
симметричной биотелесной формы позволяет трактовать
управление локомоциями и вообще моторными
движениями как диссимметрирующее управление, как управление
по преодолению симметрии исходной формы
биологического тела. Остановимся на этом подробнее.
Все биологические тела по критерию наличия
моторных движений подразделяются на два класса. К первому
классу относятся тела, лишенные способности к моторным
движениям: большинство растений, раковины моллюсков
и т. д. У этих тел каноническая телесная форма,
характерная для каждого этапа онтогенеза, наблюдается
непосредственно, поскольку эта форма претерпевает только
ростовые трансформации, но не изменения моторного типа.
Гак, форма раковины моллюска в любой момент является
канонической формой этой раковины для
соответствующего этапа онтогенеза. Отметим также, что для первого
класса издавна прослеживается, как уже отмечалось в разделе
I. 1, структурное родство между формами различных
биологических тел, которое отражается в связи их
симметрических особенностей с рядом чисел Фибоначчи и говорит
об общих морфогенетических принципах формирования
этих тел. Терминологически эту морфогенетическую
общность можно подчеркнуть, называя форму любого из
данных тел раковиной по ассоциации с раковиной моллюска
(хотя «раковины» различных биологических тел могут,
конечно, по составу вещества, из которого они сложены,
отличаться от раковины моллюска).
Ко второму, более сложному классу относятся
биологические тела, наделенные способностью преодолевать свою
исходную форму за счет моторных движений, которые
сопровождаются изменением взаиморасположения частей,
составляющих эту форму, а именно тела человека и
животных, а также тех растений, которые, подобно мимозе,
обладают моторными реакциями на раздражение. Тот факт,
что в телах второго (так же, как и первого) класса
обнаруживается связь телесных симметрии с рядом Фибоначчи,
свидетельствует о том, что тела обоих классов построены
на родственных морфогенетических принципах
продуцирования живого вещества и формирования биологических
тел. И каноническую форму тел второго класса, в
частности тела человека, также можно назвать раковиной,
подчеркивая родство принципов ее построения с принципами
формирования раковины моллюска. Правда, раковина тел
второго класса имеет существенное отличие, заключаю-
87
щееся в возможности моторного перемещения ее частей
относительно друг друга. Естественно полагать, что в
биологических телах второго класса каноническая форма
(или раковина) тела выступает в роли исходной системы
отсчета для построения движений. Для образности можно
говорить о фиксированной координатной системе тела как
аналоге специфической квазикристаллической решетки,
узловые элементы которой допускают смещение при
действии мышечных усилий, в результате чего и происходят
моторные движения. В телах второго класса примерами
канонической формы являются форма веточки мимозы при
отсутствии внешних раздражений или пентасимметричная
форма морской звезды.
Следует подчеркнуть, что в телах второго класса
каноническая форма зачастую маскируется вследствие работы
аппарата моторных движений, прежде всего мышечных
усилий, в том числе тонического, т. е. продолжительного
непрерывного характера. В этих условиях определение
канонической формы для тела второго класса
представляет собой специальную задачу. Касаясь терминологии,
отметим, что пространственное расположение
совокупности подвижных звеньев суставчатого тела человека обычно
принято называть не формой, а (сегментарной) позой
тела. Соответственно применительно к человеку и
высокоорганизованным животным нами будут использоваться
вместо термина «каноническая форма» его синонимы:
«каноническая поза» или «морфопоза».
Что же можно сказать о морфопозе тела человека и
организации моторных движений как ее преодолении?
Богатство двигательных возможностей человека
обусловливает большое разнообразие его поз, многие из которых
человек способен удерживать длительное время,
приноравливаясь к различным задачам и условиям. Как же
определить среди этого множества поз морфопозу всего тела или
хотя бы канонические позы блоков ноги, руки, пальца? Это
тем более важно, что данные позы, по всей видимости,
определяют собой исходные системы отсчета для
построения моторных движений и относительно них формируется
сложная система мышечных приводов, нарабатываются
двигательные рефлексы, взаимосогласовывается
деятельность анализаторов и т. д. Здесь очевидна определенная
программа исследований аппарата движения у человека и
животных. Мы полагаем, что одним из ключевых пунктов
этой программы является выявление канонических поз,
88
установление присущих им симметрических
особенностей и исследование нарушений этих симметрии при смене
сегментарных поз в процессе разнообразных моторных
движений. В прикладном аспекте изучение морфопоз и их
связи с функционированием различных систем
организма — нервно-мышечной, вестибулярной и пр.— способно
содействовать решению многих задач, связанных с
оптимизацией поз и движений у человека: снижению мышечной
утомляемости при работе человека-оператора;
уменьшению эффектов влияния на аппарат движения различных
неблагоприятных факторов, например вибраций;
повышению вестибулярной устойчивости; выбору оптимальных
для мышечной системы поз при длительной
обездвиженности человека, например в период лечения переломов
конечностей; прогнозированию и объяснению сенсомоторных
реакций человека в непривычных условиях, например в
условиях измененной гравитации в космосе; лучшему
согласованию носильных конструкций — скафандров, экзо-
скелетонов и др.— с особенностями опорно-двигательного
аппарата человека; совершенствованию построений
спортивных движений; улучшению конструкций спортивной
техники и т. д.
Мысль о том, что мышечная организация
подразумевает некоторое отсчетное состояние в блоках кинематической
схемы тела нельзя назвать новой: многие понятия
физиологии движения, такие, как длина ненапряженной мышцы,
явно или неявно подразумевают наличие такого отсчетно-
го состояния. Новым в нашей концепции является
утверждение, что это отсчетное состояние тела или морфопоза
характеризуются определенными — в том числе
неевклидовыми — симметриями и связаны с законами морфогенеза.
Прежде чем продолжить изучение принципов
симметрии в кинематической схеме тела человека, обратим
внимание на интересное явление наложения или
суперпозиции симметрии разного типа в одном биологическом теле.
Так, в корзинке подсолнуха наряду с симметрией
вращения всей корзинки наличествует спиральная симметрия в
отдельных линейных укладках семян. В теле многоножки
зеркальная симметрия существует наряду с
трансляционной симметрией сегментов тела. В геометрически
правильных телах некоторых вирусов, имеющих форму
икосаэдра, симметрия этого многогранника сопровождается
другим видом симметрии (например, сферической) суб-
единиц, из которых сложен этот многогранник. Кинема-
89
тическая схема тела человека дает другие случаи такой
суперпозиции: зеркальная симметрия левой и правой
половин соседствует с конформными симметриями
трехчленных кинематических блоков. Этот биологический принцип
суперпозиции симметрии разного типа может реализовы-
ваться многообразно и, видимо, его важное значение во
многих вопросах биологической организации будет со
временем открываться все более.
Проведем теперь сравнительный анализ
кинематических особенностей таких трехчленных кинематических
блоков, как нога, рука и любой из пальцев руки у
человека. Обращает на себя внимание то интересное
обстоятельство, что два первых (проксимальных) звена у этих
блоков вполне идентичны по особенностям кинематической
связи между ними: их шарнирное соединение имеет одну
степень свободы и, кроме того, диапазон их угловых
перемещений относительно друг друга одинаков и
ограничен 180°. Он простирается от полного разведения обоих
звеньев на 180° (разведение на больший угол
исключается устройством шарнирной связи) до почти полного
смыкания их поверхности при сведении обоих звеньев одно
к другому.
Обратимся к кинематической связи третьего (дисталь-
ного) звена со вторым звеном у разных трехчленных
блоков. Сравним прежде всего кинематические схемы ноги
и пальца руки человека. Рис. 15 иллюстрирует то
отличие между ними, что дистальное звено ноги (стопа)
развернуто в плоскости сгибания трехчленного блока в
противоположную сторону, чем дистальная фаланга. При этом
дистальная фаланга и ступня по своему расположению
относительно оси второго звена (средней фаланги и
голени соответственно) зеркально симметричны друг другу.
Но аналогия в морфокинематическом устройстве
рассматриваемых трехчленных блоков не ограничивается данным
обстоятельством. В этом убеждает анализ диапазонов
угловых перемещений третьих звеньев относительно
вторых при волевых усилиях (без действия внешних сил и
нагрузок, способных несколько видоизменить эти
диапазоны).
Человек может напряжением мышц придать стопе
расположение относительно голени в диапазоне от
прямолинейного развернутого (при этом угол между ними
равен примерно 180°) до взаимоперпендикулярного (угол
равен примерно 90°), как это изображено на рис. 15, а
ЭД
V
с
Рис. 15. Конфигурационные аналогии и различия между
трехчленными кинематическими блоками ноги (а), пальца руки (б),
руки при сгибаниях кисти в плоскости ладони (в) и в плоскости,
перпендикулярной плоскости ладони (г), у человека
Внизу даны схематические изображения этих блоков, А, В, С, D —
расчленяющие точки
Точно так же у любого пальца руки возможное за счет
собственных мышечных усилий угловое расположение
дистальной фаланги относительно средней определено
тем же диапазоном от 180 до 90°. Правда, у разных
пальцев есть своя специфика в свободе перемещения дисталь-
ного звена относительно первых двух звеньев. Если у
большого пальца руки дисталъная фаланга может
свободно перемещаться в указанном диапазоне без изменения
положения двух предшествующих звеньев, то у
остальных пальцев руки, как это уже рассматривалось в
разделе II.6, такое перемещение третьей фаланги
непроизвольно вызывает перемещение и двух других фаланг.
(Отметим, что данную взаимозависимость положений
фаланг человек у себя может преодолеть специальной
тренировкой, и в результате кинематические особенности
большого пальца и остальных пальцев руки уравняются.)
91
Разобранные диапазоны движений третьих звеньев
(от 180 до 90°) занимают зеркальное симметричное
положение у ноги и пальца относительно оси среднего
звена (см. рис. 15). Таким образом, нога в морфокинемати-
ческом отношении устроена во многом подобно пальцам
руки (по свободе перемещения дистального звена она
оказывается столь же «тренированной», как и большой
палец руки), но существенно отличается от пальцев
наложением условия зеркальной симметрии на
расположение дистального звена. Тем самым в организации
трехчленных кинематических блоков, помимо конформной
симметрии этих блоков, наличествует еще и зеркальная
симметрия их отдельных частей, т. е. здесь также имеет
место принцип суперпозиции симметрии, наблюдаемый,
как указывалось выше, во многих биологических телах.
Можно отметить, что трехчленные блоки ноги и пальца
руки в распрямленном состоянии просто конформно
симметричны друг другу, а в таком согнутом состоянии, как
это показано на рис. 15, конформно симметричны друг
другу с точностью до зеркального отображения третьего
звена относительно оси второго звена. Это положение по
требуется нам в следующем параграфе при построении
симметрограмм моторных движений.
В силу ряда соображений наиболее вероятно, что
именно несколько согнутое расположение звеньев
пальца (как и других трехчленных кинематических блоков,
как на рис. 15) выступает в роли морфопозы. Одним из
аргументов в пользу этого является то, что при
значительном мышечном утомлении пальцы теряют
подвижность, способность сгибаться в кулак и разбигаться и
каждый палец принимает как раз такую сегментарную
позу. Это перекликается с тем, как веточка мимозы,
«утомившись» после многократных двигательных
реакций на серию частых раздражений, оказывается в
исходной для нее «позе».
Угловые диапазоны (примерно равные 90°) движений
третьего звена относительно второго у ноги и пальцев
отличаются определенной устойчивостью своей величины.
Это особенно интересно проявляется на голеностопе, для
которого известен следующий феномен из биомеханики
спорта. Если у спортсменов в результате многолетних
тренировок специальных движений увеличивается
максимальная амплитуда допустимых движений в голеностопе
в одном направлении (например, разгибательных движе-
92
ний), то максимальная амплитуда движений в
противоположном направлении (сгибательных) уменьшается по
сравнению с нормой для людей, не занимающихся
спортом. При этом суммарная величина углового диапазона
движений в голеностопе практически не изменяется, т. е.
приспособительная перестройка кинематики идет за счет
разворота относительно голени всего диапазона движений
в голеностопе.
Ф. Л. Доленко, исследовавшая этот феномен у
штангистов, прыгунов на лыжах, конькобежцев и других
спортсменов, писала в своей диссертации: «Сокращение
размаха неупражняемых движений компенсировало увеличение
амплитуды характерных движений так, что их сумма
лишь незначительно отличалась от нормы. Анализ
адаптационных сдвигов у людей с измененной статикой и у
спортсменов высокого класса под влиянием биодинамики
избранного вида спорта показывает, что функциональная
адаптация аппарата движения идет в направлении
преодоления избыточных степеней свободы при сохранении и
увеличении амплитуды движений в требуемом
направлении. По нашему мнению, это является одним из немногих
способов сочетания высокой амплитуды движения с
обеспечением необходимой прочности сочленения» [91, с. 11,
12]. Мы полагаем, что этот феномен устойчивости
величины диапазонов суставных движений имеет морфогенети-
ческую природу.
Сказанное выше о морфокинематическом сходстве
трехчленных блоков ноги и пальцев рук можно также
дополнить следующим указанием: подобно тому, как у
любого трехфалангового пальца имеется отмеченная выше
рефлекторная (по нашему мнению, морфогенетическая по
происхождению) взаимозависимость движений трех
фаланг, так и у ноги имеется сходный рефлекс целостного
поведения всех трех звеньев, выявленный еще Р.
Магнусом. В опытах на кошках он установил существование
целостного рефлекса всей конечности на дорсо-флексию
стопы или кисти. Говоря о последних открытиях Магнуса,
А. Ф. Самойлов пишет о наблюдении, которое было
сделано на животном, лишенном мозжечка. «Если такое
животное,— указывает он,— лежит на боку, например на левой
стороне, то... стоит только произвести сильную
дорсальную флексию стопы или кисти, как конечность сильно
выпрямляется, и лишь с трудом можно заставить ногу
согнуться в ее сочленениях. Конечность становится тогда
93
вытянутой и превращается как бы в несгибаемую палку»
[137, с. 600]. (Далее Самойлов отмечает, что этот рефлекс
целостного поведения трех звеньев конечностей можно
наблюдать не только при удаленном мозжечке, но в
несколько менее резкой форме и на совершенно
нормальных животных, а также на «животных Магнуса».)
Мозжечок, как известно, отвечает за тонкие
дополнительные регулировки позы и движений, накладываемые
на более глубокие и грубые статокинетические
построения. И, по нашему мнению, закономерно, что его
отключение вскрывает существование фундаментального — и мор-
фогенетического по происхождению — рефлекса всей ноги
на изменение положения одной стопы. Существование
двигательного феномена, обнаруженного Магнусом в
эксперименте случайно, могло бы быть предсказано на
основе рассмотрения морфокинематических аналогий между
трехчленными кинематическими блоками.
Если теперь обратиться к руке — наиболее развитому
в кинематическом отношении трехчленному блоку, то в
характеристиках кинематической связи предплечья и
кисти также обнаруживается аналогия с сочленением
второго и третьего звеньев у пальцев и ног. При анализе
кинематики руки надо различать два основных вида
движений сгибания и разгибания кисти относительно
предплечья: 1) в плоскости ладони (рис. 15, в);
2) перпендикулярно плоскости ладони (рис. 15, г).
Для первого вида диапазон угловых движений,
совершаемых волевым образом за счет только собственных
мышечных усилий, составляет, как и
соответствующие диапазоны ног и пальцев, около 90° и лежит
практически с одной стороны от оси второго звена
(предплечья) . Отметим, что этот вид движений следует считать
особо фундаментальным с точки зрения целостного
поведения трехчленного блока руки, поскольку при
автоматизированных локомоторных актах ходьбы и бега сгибания
и разгибания трехчленного блока руки происходят
именно в ладонной плоскости, которая тем самым выступает
в роли естественной плоскости всего данного блока.
Диапазон же движений кисти, перпендикулярных
плоскости ладони, равен примерно 180° и как бы состоит из
двух зеркально симметричных относительно оси второго
звена диапазонов по 90° каждый, характерных для ди-
стальных звеньев трехчленных блоков ноги и пальца,
а также руки при первом виде движений кисти. В этой
94
связи организация данного вида движения
представляется объединением двух стандартных типов движений,
из которых каждый отдельно уже используется в
подобных трехчленных блоках. (Похожая ситуация
усложнения движения за счет суперпозиции стандартных
диапазонов движений выявляется и при рассмотрении
ротаторных движений в трехчленных кинематических блоках.)
Резюмируя сказанное о сходстве кинематических
особенностей трехчленных кинематических блоков рук, ног и
пальцев у человека, можно заключить, что не только
кинематические связи двух проксимальных звеньев этих
блоков идентичны, но и кинематическая связь двух дисталь-
ных звеньев у данных блоков обнаруживает удивительное
родство. По нашему мнению, это свидетельствует о том,
что в морфокинематической организации данных блоков,
столь различных по своему функциональному назначению
и эволюционному развитию, важное значение имеют
общие морфогенетические принципы, в соответствии с
которыми во многом и построены эти блоки. Представленные
данные приводят к выводам широкого плана. Построение
трехчленных кинематических блоков определено не
только критерием оптимального приспособления к конкретным
действиям во внешней среде, как это зачастую полагают,
а некоторыми иными принципами, которые, видимо, мож
но назвать морфогенетическими и выделить в совершенно
особый класс биологических принципов. Эти принципы,
как нам представляется, двояким образом влияют на
формы биологических тел. Во-первых, они ответственны за
продуцирование живого вещества и определяют все
многообразие допустимых форм биологических тел.
(Форма конкретного тела, например человеческого,
устанавливается при активном участии эволюционных принципов
естественного отбора того, что по тем или иным
причинам жизненно выигрышно, производимого на множестве
морфогенетически допустимых форм.) Во-вторых, эти
принципы накладывают свой отпечаток на формы
биологических тел, участвуя в эволюционном естественном
отборе жизненно выигрышных конструкций этих тел по
таким связанным с морфогенезом критериям, как
критерии оптимизации процессов роста и воспроизводства,
развития новых приспособительных органов,
взаимосогласования деятельности составных частей организма и т. д.
Интересно, что особенности кинематических связей
между парами соседних звеньев трехчленных кинематиче-
95
ских блоков у широкого класса позвоночных животных во
многом повторяют рассмотренные выше особенности
данных блоков у человека, несмотря на порой очень разные
условия работы и функциональные нагрузки этих блоков,
например конечностей птиц, четвероногих и т. д. Более
того, поразительное морфокинематическое сходство
отмечается между трехчленными кинематическими блоками
конечностей у позвоночных и насекомых, несмотря на то,
что эволюция аппарата движения у этих организмов шла
совершенно самостоятельными путями и у них не было
общего предка, наделенного трехчленными конечностями,
которые могли быть переданы им по наследству вместе со
своими характерными особенностями. (О строении
конечностей насекомых как трехчленных кинематических
блоков сказано в предыдущем разделе). Указанное сходство
конечностей у насекомых и позвоночных является, в
частности, причиной того, что в энтомологии сочленение
бедра с голенью у насекомых называется коленным. При
этом диапазон движений голени относительно бедра
составляет около 180°, и голень может как смыкаться с
бедром, так и располагаться с ним на одной прямой.
«В этом отношении коленные сочленения насекомых и
человека совершенно сходны» [263, с. 35]. Пользуясь
введенной выше терминологией, можно сказать, что
раковины тел насекомых и позвоночных, как и
диапазоны их кинематического видоизменения при моторных
движениях, во многом стандартизированы и
представляют собой как бы реализации из дискретного и
неширокого набора возможностей, причем важную роль в этом
играют морфогенетические законы.
Это свидетельствует в пользу того, что
филогенетическая эволюция ряда важных особенностей
кинематического аппарата происходит не на основе их постепенного и
последовательного видоизменения, а на основе
скачкообразного изменения реализуемых морфогенетических
возможностей. При этом требование оптимальной
приспособленности к условиям внешней среды и
кинематическим задачам не является единственно определяющим
ввиду наличия еще и морфогенетических требований.
(Данное положение тем более естественно, что
постановки кинематической задачи, вообще говоря, вовсе
недостаточно для определения вида кинематического
механизма, поскольку одна и та же задача может
выполняться механизмами очень разного типа.) Даль-
96
нейшее изучение угловых диапазонов допустимых
моторных движений в кинематических блоках у разных
организмов с суставчатой кинематической схемой тела
способно дать новые интересные материалы к этому
вопросу из области проблем биологической эволюции.
Не следует, конечно, удивляться дискретности
возможных величин этих диапазонов для всей совокупности
указанных организмов; ведь и в других случаях
проявления морфогенетических законов имеется выраженная
дискретность: например, в силу филлотаксисных законов
морфогенеза виды листорасположения у растений
составляют дискретный набор, характеризуемый числами
Фибоначчи.
11.11. Симметрограммы трехчленных
кинематических блоков
при анализе моторных движений
Перейдем теперь от изучения устройства и пропорций
трехчленных кинематических блоков, а также диапазонов
допустимых движений их отдельных звеньев к
исследованию моторных движений как процесса преодоления мор-
фопоз тела, как процесса нарушения симметрии,
присущих телу. С этой целью обратимся к анализу моторных
движений у человека.
Известно, что в процессе таких моторных движений,
как ходьба и бег, у человека нарушается зеркальная
симметрия тела. Можно думать, что при разных видах
моторных движений также нарушается как величина
конформно инвариантной характеристики каждого трехчленного
кинематического блока, так и конформная симметрия
между отдельными блоками. В разделе П.6 уже было
проанализировано, как меняется величина
плоскоконформного вурфа Wk пальца руки при его естественном
волевом сгибании. Эта величина при сгибании пальца из
распрямленного состояния поначалу практически не
меняется (в диапазоне тонких манипуляционных
движений), а затем начинает расти, оставаясь вещественной.
Рис. 16 графически демонстрирует на гауссовой
комплексной плоскости изменение величины данного конформного
инварианта Wh для последовательности отдельных фаз в
процессе моторного сгибания пальца, представленных на
рис. 11. Подобные графики будут в дальнейшем
именоваться симметрограммами трехчленных кинематических
4 С. В. Петухол
97
блоков, и с их помощью будет осуществляться анализ
построения движений. (Данные графики можно было бы
называть и диссимметрограммами, поскольку они
демонстрируют нарушения телесных симметрии, но первое
название выглядит предпочтительнее в силу его
краткости.
Обратимся теперь к симметротраммам трехчленного
блока ноги при локомоциях. Сразу отметим, что
рассмотренное моторное движение пальца резко отличается от мо-
Рис. 16. Симметрограмма
сгибательного движения
среднего пальца руки
человека
Кружками обозначены точки
(1—4), соответствующие
величинам вурфа Wk=Wx+iWyi
в разных фазах движения
торных движений ноги при ходьбе, беге и пр. тем, что при
этом движении деформации трехчленного блока пальца
происходят под действием только собственных мышечных
усилий (весом фаланг можно пренебречь), тогда как при
локомоциях трехчленный блок ноги испытывает еще и
значительные весовые нагрузки от тела, способные внести
заметный вклад в поведение симметрииных характеристик
данного блока. Кроме того, развитие в филогенезе
тонических напряжений мышц ног, связанное с функциональной
задачей стояния и хождения в условиях действия силы
тяжести, способно явиться фактором постоянной
деформации морфопозы трехчленного блока ноги.
В предыдущем параграфе было сформулировано
положение о том, что трехчленные блоки ноги и пальца могут
интерпретироваться как конформно-симметричные друг
другу с точностью до зеркальной симметрии третьих
звеньев относительно оси вторых звеньев (рис. 15). Но,
коль скоро моторику трехчленного блока пальца
естественно характеризовать с помощью конформного
инварианта Wh, то и моторику трехчленного блока ноги
естественно характеризовать с помощью аналогичной величины
W\, отличающейся от Wh учетом дополнительной
зеркальной трансформации третьего звена блока. Очевидно,
формула этой величины W'h (zA, Zb, zc, zd') должна
совпадать с формулой Wh (zA, zB, zc, zD) во всем, кроме того,
что точка ТУ с координатами SD'=#D'4-tyr>' будет зеркаяь-
#,2 -
'0.ZV
98
но симметричной к точке D с координатами zD=zD+iyD
относительно оси, проходящей через точки с координатами
zB=xB+iyB и zc=xc-\-iyc. (Как и в разделе II.6,
координаты расчленяющих точек блоков определяются в системе
гауссовых комплексных координат, вводимой на
плоскости, в которой происходит моторное сгибательное
движение трехчленного блока.)
Компоненты xD' и yD' координат точки D' как функций
координат точек В, С, D нетрудно получить с помошью
аналитической геометрии, решая задачу о координатах
точки, зеркально симметричной данной точке относительно
оси, проходящей через другие две данные точки. Опуская
промежуточные детали, приведем окончательный
результат:
xD = (хс — хв) [{хс — хв) (xD — хв) +
+ (Ус — У в) (yD — Ув)У[(хс — xBf + (ус — у в)2} +
+ (Ус — У в) [(У с — У в) {хв — xD) —
— (хс~ хв){ув— Уо)Шхс— хв? -J- {Ус —У в)2] +хВ)
(18)
VD' = {УС — У в) [(ХС — ХВ) {XD — ХВ) +
+ (Ус — У в) (yD — Ув)У[(хс — хву + (ус — У в?] +
+ (хс — хв) \(хс — хв) (у в — У в) — (у с — У в) X
X (хв — xD)]/[(xc — хв)2 + (ус — ув)2\ + у в- (19)
Исходя из формулы (7) выписываем формулу Wh' для
ноги:
W'k = (zc — zA) (zd- — zB)l(zc — zb) (zd — za). (20)
Для пользования этой формулой при анализе движений
ноги надо взять экспериментальные данные о координатах
четырех расчленяющих точек zA, zb, zc, zd трехчленного
блока ноги в отдельную фазу движения (с помощью,
например, моментальной фотографии движения ноги в
этой фазе), затем по формулам (18) и (19) вычислить
zD' и, наконец, вычислить величину W\. Анализ всего
растянутого во времени движения ноги производится, как
и для движения пальца, путем сравнения поведения
величины W\ в разных фазах движения.
Проанализируем этим способом ходьбу ребенка и
взрослого человека. Для этой цели воспользуемся приве-
4* 92
a
/ J* S S73
e.s
/2 /,£ Л'.
j
г j 4 s e 7 s ?
-ff,z
-0A
f.s
_i i i .—i—
Sfi7
<5
ff,e
ff.t
4»4V^
/ г
4 se 7
-ff,4
Z*
W /
7 0
^**.
Рис. 17. Кинематические схемы и симметрограммы
j_t
> г
ловека
локомоций че-
р,^еРВЫе даИ Х°ДЬбЫ РебеНКа; б~х°Дьба взрослого человека; в - бег
взрослого человека. Обозначения аналогичны принятым на рис. 16
денными на рис. 17, а, 6 данными о положении звеньев ног
ребенка в первые дни самостоятельной ходьбы [99, с 86
рис. 26, опыт 953], взрослого при ходьбе [409 с 197] На
этих же Рисунках представлены вычисленные нами по
формуле (20) величины W\ трехчленных блоков ноги для
каждой из указанных фаз ходьбы. Сравнение полученных
100
симметрограмм ходьбы ребенка и взрослого позволяет
сделать ряд заключений.
Во-первых, обращает на себя внимание, что акт ходьбы
связан с нарушением не только зеркальной симметрии
тела, но и симметрической характеристики — величины
W k — трехчленного блока ноги.
Во-вторых, для всех фаз величина W'h комплексна и
имеет отрицательную мнимую часть. По нашему мнению,
это свидетельствует о том, что трехчленный блок ноги при
ходьбе постоянно находится в деформированном — не
морфопозном — состоянии из-за весовых нагрузок от тела
и непрерывно действующего мышечного тонуса, который
обеспечивает устойчивое хождение на двух ногах.
Отметим, что и у ребенка, и у взрослого фаза, характеризуемая
W\ с минимальной по абсолютной величине мнимой
частью, приходится на момент движения после отрыва носка
и всей ноги от опоры, т. е. на момент разгрузки ноги.
Затем при переносе ноги, совершаемом с учетом
требования не задеть за опору и подготовиться к приему веса
тела, мнимая часть W\ снова возрастает по абсолютной
величине в результате деформации блока уже только
мышечными силами. Все опорные фазы движения ноги,
во время которых на нее действует вес тела, отличаются
величинами W'h со значительной мнимой частью.
Максимальный перепад в значениях мнимой части (от большой
величины к малой) наблюдается в соседних фазах ходьбы
перед отрывом носка от опоры и после его отрыва (фазы
7 is. 8 ъ симметрограмме ходьбы ребенка и фазы 5 и 6 —
взрослого).
В-третьих, области, занимаемые на графиках обеими
симметрограммами ходьбы, практически идентичны и
простираются по оси абсцисс примерно от 1,15 до 1,45
(характеризующая морфопозное состояние трехчленных
кинематических блоков величина, примерно равна 1,3,
находится посередине между этими границами), а по оси
ординат — от —0,05 до —0,3. (Эти области относительно
малы по сравнению с областью, занимаемой
представленной ниже симметрограммой бега.) Значительная аналогия
между обеими симметрограммами говорит о том, что
ходьба как процесс управления трехчленным блоком каждой
ноги оказывается онтогенетически — от самых первых
дней хождения ребенка — очень устойчивым процессом, а
ее симметрограмма практически является
онтогенетическим инвариантом. И это — несмотря на значительные
101
0,7
0,f
ff,J
/7
ж
/л
^
/,2 /J /,f *x
Рис. 18. Кинематическая
схема быстрой рыси
лошади [61, с. 91] и симметро-
грамма движения ее
задних конечностей при этом
Обозначения аналогичны
принятым на рис. 15, 16.
возрастные изменения веса тела, кинематические
изменения относительного движения пар звеньев ноги при
ходьбе и т. д. Можно думать, что отражаемая в симметрограм-
мах устойчивость программы целостного поведения
трехчленного блока ноги служит одним из тех условий, на
которые ориентируется организм при решении
возрастных задач статокинетической регуляции ходьбы.
Обратимся теперь к бегу человека. В качестве
исходных данных для построения симметрограммы возьмем
данные о поведении трехчленного блока ноги при спринте
из работы Н. А. Бернштейна [99, с. 206, рис. 78, а]. Они
представлены на рис. 17, в вместе с построенной намисим-
метрограммой. Судя по симметрограмме, бег имеет
качественные отличия от ходьбы, формально выражающиеся в
наличии фаз движений, при которых мнимая часть W\
является положительной, и резком расширении — именно
в направлении положительных значений оси ординат —
области, занимаемой симметрограммой. Расширение обла-
102
Рис. 19. Кинематическая схема галопа серны ["61] и симметрограм-
ма движения ее задних конечностей при этом
Обозначении аналогичны принятым на рис. 15, 16
сти симметрограммы при беге само по себе представляется
естественным, поскольку оно во многом является
отражением усиления моторной кинематики тела. Усиление же
кинематики моторных движений у биологических тел
обычно связано с увеличенным нарушением симметрии
тела, в частности зеркальной симметрии тела человека
(нарушаемой, например, сильнее при беге, чем при
ходьбе). Как и в симметрограмме ходьбы, так и в симметро-
грамме бега значения W\ с наиболее отрицательными
величинами мнимых частей приходятся на фазы опорного
движения ноги (фазы 4—6). Все Wh, имеющие
положительные значения мнимых частей, приходятся на фазы
безопорного движения ноги, деформируемой в этих фазах
мышечными силами индивидуума и силам л инерции.
Точка абсциссы 1,3 по-прежнему лежит примерно в
середине реализуемого при беге диапазона значений
вещественной части W'h.
103
^fr
0
-A0
-2,0
S>0
-4.0
0,f /,0 * i 2,0
•- р_Л- 1
7 b.
т г
0 2
VJ
£0"x
fb;
ti
,.^o
Рис. 20. Кинемаипеская
схема прыжка кошки [611
и симметрограмма
движения ее задних конечностей
при этом
Обозначения аналогичны
принятым на рис. 16
Несомненный интерес представляет сравнительное
изучение симметрограмм локомоции у человека и различных
животных с суставчатым аппаратом движений. Здесь мы
ограничимся представлением и самым кратким анализом
симметрограмм трехчленных блоков задних конечностей
при некоторых видах локомоции у четвероногих
животных: серны, лошади и кошки. Исходные данные о локо-
моциях взяты из работы [61]. Симметрограммы галопа
серны и рыси лошади (рис. 18 и 19) заметно отличаются
от рассмотренных выше симметрограмм локомоции
человека тем, что они занимают лишь области с
неотрицательными значениями ординат. Наибольшее удаление от оси
абсцисс в обоих случаях имеют точки симметрограмм,
соответствующие фазам движения после отрыва
конечности от опоры и начала подтягивания ее вперед для
очередного шага. Симметрограмма прыжка кошки (рис. 20)
104
имеет свои яркие особенности. Практически она вся
расположена в области отрицательных ординат. При этом
особенно выделяются своей удаленностью от оси абсцисс
точки фаз 3—5, приходящиеся на пружинистое сжатие
задних конечностей перед прыжком.
Приведенное краткое изучение симметрограмм
моторных движений у человека и животных говорит о том, что
организация этих движений обычно происходит с
нарушением не только зеркальной симметрии тела, но и
конформной симметрии трехчленных кинематических
блоков, особенно если речь идет о моторных движениях этих
блоков в условиях внешних силовых воздействий. Данное
изучение заставляет обратить наше внимание на ряд
общих особенностей в организации различных видов ло-
комоций у разных животных. Конечно, представленные
материалы не способны дать полной картины симметрий-
ных особенностей организации локомоций и не позволяют
провести классификацию различных природных типов
моторного преодоления морфопоз у организмов с
суставчатым аппаратом движения. В основном эти материалы
лишь фиксируют интересную область анализа и
обобщений в биомеханике движений, более полно соединяя
последнюю с проблемой биологических симметрии и их
нарушений. Здесь необходимы широкие сравнительные
исследования кинематического построения моторных
движений у различных живых организмов с целью
установления общих закономерностей и дальнейшего развития как
теоретической физиологии движений, так и общей теории
биологических симметрии. Следует отметить, что подобной
же формализации и сравнительному анализу на предмет
выявления общих кинематических закономерностей при
моторных движениях подлежат не только трехчленные
блоки ног и пальцев рук, но и трехчленные блоки рук
человека и передних конечностей животных. Для
построения необходимых симметрограмм в этих случаях также
могут быть установлены соответствующие формулы,
основанные на симметрийных особенностях этих блоков и
позволяющие адекватным образом формализовать
описание их кинематики как целостного биологического
образования.
105
Глава III
ИССЛЕДОВАНИЕ ПРОЕКТИВНЫХ СИММЕТРИИ
В БИОЛОГИИ
Наряду с данными о биологическом значении группы
конформных симметрии имеется ряд данных о возможной
важной роли проективных симметрии в организации
отдельных биологических феноменов. Обратимся к
рассмотрению этих данных.
III.1. О возможном участии принципов
проективной геометрии
» структурировании биологических явлений
В предыдущей главе уже говорилось о симметрии
неевклидова типа между трехчленными кинематическими
блоками тела человека и животных и о том, что эту
симметрию можно интерпретировать и как конформную, и как
проективную. Полагая конформную геометрию наиболее
перспективной для использования в качестве
биологической геометрии (или составной части последней), мы не
можем пройти мимо отдельных данных о возможном
использовании в биологии также и проективной геометрии и
считаем необходимым исследовать наличие проективных
симметрии в биологических объектах и явлениях. Интерес
к возможным биологическим реализациям принципов
проективной геометрии стимулируется также некоторыми
умозрительными рассуждениями, основанными на
важном значении проективной геометрии в логических
основаниях геометрии.
Многие основные положения проективной геометрии
отражены в приемах линейной перспективы,
используемых в живописи. Эти приемы, открытые художниками
Возрождения, явились результатом изучения хода
световых лучей от окружающих предметов к глазу человека и
имеют целью воспроизведение всех следов этих лучей на
рисовальном холсте. В результате их применения на
холсте появляется характерное изображение предметов, в
106
котором, вообще говоря, изменены и расстояния, и углы,
и отношения между парами этих метрических величин.
Но изменяется не все: геометрические характеристики
из особого класса так называемых проективных
инвариантов сохраняются неизменными. К их числу относятся
прежде всего сложные отношения или вурфы четырех
точек прямой, о которых говорилось в предыдущей главе.
Проективная геометрия как раз представляет собой науку
о таких проективных инвариантах, т. е. о структурах,
сохраняющихся при отображениях такого характера.
Эта математическая наука претерпела исключительное
развитие и, далеко уйдя от скромной теории
перспективы в живописи, заняла высокое положение во всей
геометрии. Данный вопрос подробно разбирается в целом
ряде учебников по высшей геометрии [81, 93, 256, 273].
Обратимся к известным данным о реализации
принципов проективной геометрии в структуризации
биологических явлений. Таких данных совсем немного, и они
относятся к области психофизики. Прежде всего здесь
надо сказать о цветовосприятии, о физиологическом
законе смешения цветов, в силу которого для человеческого
восприятия любой цвет может быть представлен в виде
некоторой смеси трех основных цветов, например
красного, зеленого и синего. (На использовании этого закона,
в частности, построено цветное телевидение.) В работе
[267], посвящелной цветовосприятию, подробно
изложены две важные особенности этого закона. Во-первых,
закон смешения цветов не физический, а
физиологический, и его особенности недоступны открытию с
помощью физических приборов. Во-вторых, сущность этого
закона «совпадает в существенном с проективной
геометрией на плоскости цветов» [267, с. 137]. Другими
словами, цветовое многообразие оказывается физиологически
структурированным по принципам проективной
геометрии.
Кроме закона смешения цветов, определенной
близостью к проективной геометрии, видимо, отличаются
структурные закономерности психофизических
феноменов константности восприятия (см. раздел IV.1),
носящие линейно-групповой характер, насколько мы можем
судить об этом по интересным работам Й. Акисиге
[279-282].
Обратимся теперь к упомянутому выше вопросу о
логических основаниях геометрии. Развитие проектив-
107
ной геометрии привело к тому, что ее положения
стали основными в учении о логических основаниях
геометрии, связанном с аксиоматическим построением
геометрии и проблемной задачей о сведении огромного
числа геометрических утверждений к очень небольшому
набору начальных положений — аксиом, на материале
которых можно по правилам логики восстановить все
множество геометрических утверждений. Остановимся на
этом подробнее. Построенная на аксиоматической основе
геометрия предстает в виде некоторой дедуктивной
науки, занимающейся логическими выводами из аксиом.
Так, для построения всей евклидовой геометрии по
Д. Гильберту оказывается достаточным исходить из
аксиом принадлежности, порядка, конгруентности,
параллельности и непрерывности. Исторически проективная
геометрия поначалу развивалась как часть евклидовой
геометрии и соответственно опиралась на все аксиомы
последней. Однако со времени работ X. Штаудта
обнаружилось, что она независима от аксиом конгруентности
и параллельности. Это вызвало смелую мысль строить
метрическую геометрию через проективную. В
результате такого революционного переворота в представлениях
о взаимозависимости метрической и проективной
геометрий выяснилось, что евклидовы и другие геометрии,
основанные на проективных аксиомах и трактуемые как
частные случаи проективной геометрии, приобретают
значительное совершенство, очищаясь от тех
нелогичностей и лишних неопределенных понятий, которые при
классическом построении вносятся в них вместе с
непроективными аксиомами и сильно затемняют существо дела.
Другими словами, выявилась ключевая роль
проективной геометрии в логических основаниях геометрии, что
подчеркнуто словами Кэли: «Проективная геометрия
есть вся геометрия» [167, с. 14].
Но на чем основаны сами аксиомы геометрии?
Принято полагать, что при установлении системы аксиом
интуиция пространства имеет решающее значение.
В этой связи представляет высокую ценность любое
продвижение на пути познания того, как живой
организм обогащается геометрической интуицией и
оказывается способным строить здание геометрической науки
Современные учебники по геометрии, как правило,
удовлетворяются тей, что вслед за Евклидом и Гильбертом
представляют читателю логические выводы из некоторой
108
системы аксиом. Здесь уместно напомнить критическое
эамечание А. Пуанкаре, который, высоко оценивая
книгу Д. Гильберта «Основания геометрии» [73], выявил в
ней один серьезный недостаток: «По-видимому,
профессора Гильберта интересует только логическая сторона
дела. Имея ряд предложений, он находит, что все они
следуют из первого. Его не интересует происхождение
этого первого предложения с психологической точки
зрения... Аксиомы постулируются, мы не знаем их
происхождения; при таком подходе столь же легко
постулировать А равным С... С этой стороны его работа не
совершенна» [200, с. 86]. (Сам Пуанкаре внес значительный
вклад в разработку вопросов происхождения
пространственных представлений и геометрической интуиции у
индивидуума, что кратко изложено в разделе II. 8.)
Но если именно система аксиом проективной
геометрии является наиболее основательной и выигрышной из
всех возможных систем аксиом, то не лежит ли причина
этого (полностью или отчасти) в том, что сама интуиция
пространства, столь важная для установления
геометрической аксиоматики, носит у человека проективно-гео-
метрический характер? Подобное рассуждение,
дополнительно заставляющее со вниманием отнестись к
поднимаемому нами общему вопросу о проективных симметриях
в биологии, не является новым. В свое время ему была
посвящена книга Б. Рассела [420], краткий анализ
некоторых положений которой дан в следующем разделе.
III.2. Концепция Б. Рассела
о врожденности представлений индивидуума
о положениях проективной геометрии
Б. Рассел на основе чисто умозрительных рассуждений
без обращения к каким-либо биологическим данным
выдвинул концепцию о врожденности положений
проективной геометрии для геометрических представлений и
интуиции человека. В своей книге [420] Рассел
анализировал проблему: какое геометрическое знание должно
являться логической отправной точкой для науки о
пространстве и вместе с тем быть логически необходимым для
опыта любой формы внешности? Рассел называл
врожденным или априорным такое знание, которое не зависит от
опыта, хотя, возможно, и выявляется опытом. В свое
время эта проблема была указана еще И. Кантом,В попы.
№
ках решить ее он сформулировал тезис об априорности
евклидовой геометрии. И хотя рождение неевклидовой
геометрии опровергло этот тезис, говоря словами Гильберта,
«наиболее общая основная мысль кантовской теории
познания сохраняет свое значение... Понятие априорности
есть не более и не менее, чем ...выражение некоторых
обязательных предварительных условий мышления и
познания» [200, с. 252].
Рассел рассматривал данную проблему с позиций пос-
лекантовских революционных изменений в области
логических оснований геометрии, обусловленных развитием
проективной геометрии. Как мы уже указывали, это
развитие показало, что целое множество важнейших
геометрий, в том числе Евклида и Лобачевского, к большой
выгоде для ясности логических оснований и взаимосвязей
геометрий может рассматриваться как тот или иной
частный случай проективной геометрии (отметим, что многие
геометрии — Евклида, Лобачевского и др.—могут
интерпретироваться также как частный случай конформной
геометрии). Подобно тому, как копернианская концепция
кардинально прояснила простоту и единообразие строения
солнечной системы за счет постановки Солнца в центре
системы, так и учение о центральной роли проективной
геометрии пролило яркий свет на естественный порядок d
системе геометрий и устранило темные места,
нелогичности и лишние предположения в понимании этого порядка.
Исходя из этих достижений математики, Рассел
полагал, что априорная часть геометрических воззрений у
индивидуума построена на понятиях проективной геометрии.
Как фундаментальный аргумент он использовал мысль о
том, что качественные отношения должны предшествовать
количественным, а потому проективная геометрия,
имеющая дело с дескриптивными или качесвенными
свойствами пространства, логически предшествует метрическим
геометриям, которые занимаются количественными
свойствами пространства. Рассел утверждал, что эта геометрия
полностью априорна и не берет ничего из опыта. Разум
должен знать основные законы проективной геометрии и
потому быть способным получить и организовать
восприятие пространства.
Отдельные места этой работы Рассела можно
трактовать как идеалистические. Но в целом, по нашему мнению,
данный круг вопросов допускает вполне
материалистический подход как представляющий собой часть проблемы о
110
механизмах Моделирования в нервной системе
воспринимаемых пространственных явлений. Именно с
материалистических позиций следует обсуждать концепцию Рассела,
представляющую собой яркую попытку синтезировать
различные аспекты пространственного восприятия на базе!
инвариантно-групповой концепции. При успехе этой
попытки у абстрактных геометрических учений обнажились
бы психологические корни, а сама психология
пространственного восприятия была включена в область развитого
математического естествознания, о чем мечтал Рассел,
(см. его предисловие к [408]). Данная концепция Рассела
служит для нас одним из аргументов в пользу
целесообразности специального поиска и изучения проективных
симметрии в биологии. Вместе с тем мы отмечаем
возможность переноса ее отдельных положений и исходных
проблем, породивших концепцию, на сферу конформной
геометрии, важное биологическое значение которой
продемонстрировано в главе П.
III.3. Использование вурфов для распознавания
трансформируемых геометрических образов
Зрительное восприятие человека строится на
распознавании отображения объекта на сетчатку глаза. (Для
простоты мы будем говорить в этом разделе только о плоских
объектах.) В зависимости от ракурса отображенный на
сетчатку образ может заметно отличаться от образа
исходного объекта. И тем не менее человек, как правило,
распознает объект независимо от ракурса. В этой связи
представляет интерес отметить существование наряду с
широко известной из евклидовой геометрии схемой
описания фигур других геометрических схем, приводящих,
например, к проективно- или конформно-инвариантному
описанию фигур.
Со школьной скамьи каждый человек знает, что для
описания совокупности точек той или иной фигуры можно
использовать понятие расстояния между двумя точками.
Это понятие представляет собой основной инвариант
группы движений или основной инвариант евклидовой
геометрии. Для такого описания достаточно выбрать на
плоскости две точки Л и В (рис. 21, а), положение которых
относительно данной фигуры жестко фиксировано, в
качестве базисных, а затем определять каждую точку фигуры
указанием ее расстояния до этих двух базисных точек,
111
Рис. 21. Способы поточечного описания фигур
с — с помощью расстояния между двумя точками; б — с помощью вурфа
всю Же фигуру описывать как последовательность таким
образом определенных точек. В частности, эллипс
представляет собой фигуру, у которой сумма расстояний
каждой точки до двух специально выбранных базисных точек
(фокусов эллипса) есть величина постоянная. Подобным
образом можно описать любую фигуру как совокупность
точек плоскости если не в виде единого уравнения, как в
случае эллипса, то в виде таблицы соответствующих
расстояний до каждой из последовательных точек фигуры.
Очевидно, что полученное описание произвольной
фигуры является инвариантным относительно
преобразований движения плоскости, поскольку они сохраняют
расстояния и после каждого такого преобразования данной
фигуре с ее базисными точками (А', В') может быть по
тому же алгоритму сопоставлено в точности прежнее
описание. Если же плоскость претерпевает проективное или
конформное преобразование, меняющее расстояния, то
каждая точка F' преобразованной фигуры получает новые
значения координатных расстояний до преобразованных
112
базисных точек и прежнее описание фигуры через задание
координатных расстояний по необходимости меняется;
другими словами, преобразованная фигура оказывается
существенно иной, чем исходная.
Однако этот метод описания можно модифицировать,
используя в качестве инструмента описания не евклидовы
расстояния, а вурфы. Цель настоящего параграфа и
заключается в том, чтобы обратить внимание биологов и всех
тех, кто интересуется рассматриваемой проблематикой, на
существование таких известных из высшей геометрии
способов описания и показать их преимущества. Можно
полагать, что если организм является некоторой
вычислительной машиной, решающей задачи пространственного
восприятия и использующей для распознавания
трансформируемых образов известные геометрические принципы,
то у него есть основания предпочесть использование
конформных или проективных схем описания и
распознавания образов использованию евклидовой схемы. Это вполне
согласуется с существованием целого класса
психофизических феноменов константности восприятия,
демонстрирующих инвариантность распознаваемых организмом
образов (и не только конфигурационных, но и цветовых и
др.) относительно групповых преобразований, более
широких, чем преобразования движений.
Рассмотрим более конкретно, как можно построить
описание фигуры с помощью проективного вурфа (6),
получив при этом значительное преимущество, которое
заключается в том, что данное описание фигуры отличается
устойчивостью уже не только к преобразованиям из
группы движений, но и к любым проективным
преобразованиям.-Читатель, знакомый с идеями проективной геометрии,
без труда увидит, что речь пойдет просто о привязке к
данной фигуре проективной системы координат, что и
определит проективную инвариантность координатного
описания каждой точки, а значит, и всей фигуры. Небольшим
усложнением при данной модификации является то, что
число базисных точек здесь должно быть увеличено на
две точки по сравнению с рассмотренным случаем
евклидовой схемы описания (это отличие в две базисные точки
имеет место для пространства любой размерности, а не
только для рассматриваемого случая плоскости).
Итак, выберем на плоскости четыре базисные точки
А, В, С, D (никакие три из них не должны лежать на
одной прямой), жестко связанные с рассматриваемой фигу-
ИЗ
рой в том смысле, что эта плоскость и расположенная на
ней фигура претерпевают любые преобразования всегда
только вместе. В роли таких базисных точек могут
использоваться и точки самой фигуры, например точки
максимальной кривизны. Положение произвольной точки F
рассматриваемой фигуры (рис. 21, б) тогда может быть
однозначно определено указанием величин двух вурфов: вурфа
четырех точек {С, М, N, F) или равного ему вурфа
четырех прямых ВС, ВЫ, BN, BF, образующих пучок с
центром в точке В, и вурфа четырех точек {В, Q, Р, F) или
равного ему вурфа четырех прямых СВ, CQ, CP, CF пучка
с центром в точке С. Вся рассматриваемая фигура может
быть описана перечислением пар таких координат
последовательности ее точек. Очевидно, что после любого
проективного преобразования плоскости с фигурой (на рис. 21,
б упомянутые точки при этом преобразуются в точки со
штрихами) использование этого способа приведет к тому
же самому описанию фигуры, поскольку вурфы, будучи
проективными инвариантами, в отличие от расстояний не
изменяют своих величин при этих преобразованиях и ни
одна точка фигуры даже «не почувствует», что произошло
какое-то преобразование. Тем самым данный способ
описания в отличие от евклидова, рассмотренного в начале,
дает возможность распознавать проективно
преобразованную фигуру как эквивалентную исходной. Данный способ
может использоваться и в машинном распознавании
образов при установлении идентичности трансформируемых
фигур (конечно, одновременно должны быть решены
своими способами и другие вопросы машинного восприятия,
например необходимое для машинного зрительного
распознавания выделение контура объекта, вычленение
объекта из окружающего фона и т. д.) О важности получения
описаний, устойчивых к преобразованиям, которые
обусловлены изменением расположения разглядываемого
предмета относительно машинного глаза, в монографии Р.
Дуда и П. Харта, посвященной машинному распознаванию,
сказано следующее: «В общем случае мы говорим об
описаниях, которые инвариантны по отношению к некоторым
группам преобразований. Цель всех усилий состоит в том,
чтобы найти описания, которые были бы инвариантными
по отношению к преобразованиям, вызывающим
«несущественные» изменения объектов, и в то же время
чувствительными к преобразованиям, изменяющим объекты
«существенным» образом» [92, с. 366]. При указанном ис-
114
пользовании вурфов как инструмента описания эта цель
достигается в высокой степени.
Конечно, на практике можно ограничиться учетом и
описанием не всех точек фигуры, а некоторого их
конечного числа, допуская, что сохранение или изменение их
относительного положения определяет сохранение или
изменение всей фигуры. Аналогично проективной схеме
строится схема конформно-инвариантного описания
фигур. Для этого на плоскости должны быть выбраны
специальные базисные точки, а в качестве инструмента
описания использован комплексный вурф (7), являющийся
инвариантом конформных преобразований.
В связи с вопросом об использовании организмом
вурфов в распознавании образов можно дополнительно
отметить, что выделение характерных точек фигуры (которые
могут использоваться как базисные точки), прежде всего
точек максимальной кривизны и оконечных точек,
представляет собой, как известно из психологии, первый и
очень важный этап зрительного, а равно и гаптического
ознакомления с геометрическими фигурами.
III.4. Вурф пяти точек плоскости
и пентасимметричные биологические тела
В предыдущей главе при анализе симметрии в
биологических телах было использовано понятие вурфа четырех
точек прямой, являющегося инвариантом проективных
преобразований. Но в проективной геометрии имеется
также родственное понятие вурфа пяти точек плоскости (в
дальнейшем будем для краткости называть его пентавур-
фом в отличие от тетравурфа — вурфа четырех точек
прямой). Оно было использовано нами при изучении
проективных характеристик форм биологических тел и поиске
нетривиальных проективных биосимметрий. Прежде чем
излагать полученные нами результаты, напомним
некоторые положения математической теории пентавурфов,
представленные в работе Н. А. Колмогорова [117], которые в
дальнейшем нам потребуются.
Вурф пяти точек плоскости является непосредственным
обобщением для случая плоскости вурфа четырех точек
прямой линии и определяется следующим образом. Пусть
на плоскости даны пять точек А, В, С, Д, Е, не лежащие
на одной прямой. Тогда вурфом этих пяти точек, обозна-
115
Рис. 22. Иллюстрация к анализу пентавурфных характеристик пя-
тилучевых звезд
а — проективное преобразование звезды (величина пентавурфа ее вершин
инвариантна); б — правильная пятиконечная звезда
чаемым символом (ABCDE), называется выражение
(ABCDE) = (пл. ADB) (пл. ЛЕС) I
/ (пл.АВС) (пл. ЛЕВ). (21)
Из определения пентавурфа видно, что здесь вместо
длин отрезков, фигурирующих в определении тетравурфа,
выступают площади соответствующих треугольников
ADB, AEC, ADC, AEB.
Основным свойством пентавурфа является его
проективная инвариантность: при проективном преобразовании
пяти точек плоскости в другие пять точек (рис. 22, а)
величины всех пентавурфов этих точек сохраняются.
Расположение пяти точек плоскости определяется с точностью
до проективной эквивалентности однозначно, если
известны величины двух пентавурфов этих точек с разными
начальными буквами, например (ABCDE) и (BCDEA).
Величины всех других возможных для этих пяти точек
пентавурфов, символы которых представляют собой
произвольные перестановки букв А, В, С, Д Е, тогда могут быть
вычислены по формулам, приведенным в работе [117].
Поэтому для устаповлепия того, являются ли две пятерки
точек А, В, С, Dt Е, и А', В', С% D', Е' проективно
эквивалентными, надлежит сравнить по крайней мере два
пентавурфа с разной первой буквой, например {ABCDE) и
(BCDEA) одной пятерки точек, с соответствующими пен-
116
шурфами {А' В' С D' Е') и (В' С D' Е' А') другой
пятерки. Если при этом величины соответствующих пен-
тавурфов совпадают, то это означает проективную
эквивалентность (данных пятерок точек плоскости.
(Характеристика с помощью двух вурфов специфична для теории
Пентавурфов: этого нет в теории тетравурфов, где значения
•только одного тетравурфа достаточно для задания четырех
точек прямой с точностью до проективной
эквивалентности.)
Отметим еще следующую связь пентавурфов с тетравур-
фами на примере пентавурфа (ABCDE). Его величина
равна величине тетравурфа (В, К, Г, Е,) четырех точек
Д К, Т, Е, определяемых пересечением лучей АВ, AC, AD,
АЕ со стороной BE (см. рис. 22, б):
(ABCDE) = {BKTE) (22)
Эта связь позволяет вычислять величины пентавурфов
через соответствующие тетравурфы без привлечения
площадей треугольников, фигурирующих в определении
пентавурфа (21).
Перейдем к применению пентавурфа для изучения
проективных характеристик форм биологических тел.
Используем пентавурф в связи с одним из любопытных явлений
живой природы — пентасимметричным устройством многих
биологических тел. Прежде всего скажем немного об этом
явлении, которое представляет интерес в качестве одного
из критериев принципиального отличия между
кристаллическими образованиями неживой природы и растительно-
животными организмами. Дело в том, что запрещенная
принципами физической кристаллографии
пентасимметрия тел в мире растений и простейших животных
встречается исключительно часто (можно сказать, чаще всего).
Так, пентасимметрия весьма обычна для цветов, морских
звезд, морских ежей и многих других развитых
организмов. Биология открывает новые факты пентасимметрич-
ности устройства в мире вирусов и биологических молекул.
О роли пятерной симметрии в живой природе интересно
рассказано в книге [140].
Вот как пишет об этом явлении кристаллограф
А. В. Шубников: «Что касается организмов, то для них мы
не имеем такой теории, которая могла бы ответить на
вопрос, какие виды симметрии совместимы и какие
несовместимы с существованием живого вещества. Но мы не можем
отметить здесь тот в высшей степени замечательный факт,
117
что среди представителей живой природы, пожалуй, чаще
всего встречаются как раз простейшие из невозможных
для затвердевшего, окристаллизованного, «мертвого»
вещества виды симметрии (пятерная симметрия)» [258,
с. 110].
В этой же связи другой советский кристаллограф
Н. В. Белов пишет: «Кристаллический запрет пятерной
оси, как известно, определяется невозможностью
согласовать ее ... с «решетчатым состоянием» кристаллического
вещества. И потому можно думать, что пятерная ось
является у мелких организмов своеобразным инструментом
борьбы за существование, страховкой против окаменения,
против кристаллизации, первым шагом которой была бы
их «поимка» решеткой» [258, с. 110].
Обращаясь к геометрическим свойствам правильной
пятиконечной звезды, надо отметить ее аффинную связь с
золотым сечением Ф, известную, по крайней метзе, со
времен Пифагора. Она заключается в том, что в пентаграмме
стороны (а равно и диагонали) при своем пересечении
делятся в аффинном отношении золотого сечения, например,
на рис. 22 б
ВТ/ТЕ = (1+У5)/2=Ф.
Эта связь рассматривается, в частности, в знаменитом
средневековом трактате Луки Пачиоли «Божественная
пропорция», написанном под влиянием Леонардо да Винчи,
а также в большем числе других работ о золотом сечении.
.Подойдем к анализу устройства пентаграммы
по-новому — с проективной точки зрения — и зададимся вопросом
о проективных характеристиках пентаграммы, прежде
всего о величинах пентавурфов ее пяти вершин (ABCDE) =
= (BCDEA) = (CDEAB) = (DEABC) = (EABCD). Для
установления их значений воспользуемся равенством (22) и
вычислим величину тетравурфа (ВКТЕ). Принимая Длину
В К за единицу, имеем BK=TE=i, КТ=Ф~1. По формуле
тетравурфа (6), учитывая, что Ф2=1+Ф, вычисляем
{ВКТЕ) = -^^щ- = (ф_1)(ф + 1) - Ф.
Полученный результат обнаруживает существование
(наряду с ранее известной аффинной связью) более
глубокой проективной связи пентаграммы с золотым сечением,
т. е. связи, устойчивой при любых проективных
преобразованиях пентаграммы, вообще говоря, рвущих аффинную
118
связь. Полученное означает, что пять вершин пецтаграм-1
мы связаны между собой проективноустойчивым
условием равенства их пентавурфов величине золотого сечения
и что каждая сторона пентаграммы делится точками
пересечения с четырьмя остальными сторонами в вурфовой
пропорции с величиной Ф (то же самое верно и для
членения диагоналей пентаграммы). Этот результат выявляет
новую связь пентасимметричных биологических тел с фи-
боначчиевыми отношениями.
Вурфовый анализ явлений симметрии в живой природе
представляется интересным и перспективным; имеется
большое число биологических объектов, которые могут
быть с его помощью исследованы. Применим его к изучению
морфологических закономерностей тела человека.
III.5. Поиск проективных симметрии
в расположении пяти оконечных точек
человеческого тела
Издавна существовало мнение о том, что биологическая
пентасимметрия, реализуемая в пятилепестковых цветках
растений, телах морских звезд и пр., имеет
непосредственное отношение также к человеческому телу как пятилу-
чевому, где лучами служат голова, две руки и две ноги.
В этой связи на протяжении истории математического
естествознания многими мыслителями в самых разных
целях (эстетических, научных и т. д.) устройство тела
человека приводилось в соответствие с пентаграммой.
Характерный пример попыток уяснить и
продемонстрировать общность планов строения человеческого тела и
пентасимметричных тел низших организмов приведен на
рис. 23, а, где тело человека в распластанной зеркально
симметричной позе весьма произвольным образом
вписано в правильную пентаграмму.
Мы поставили задачу исследовать характер
онтогенетических трансформаций расположения пяти оконечных
точек человеческого тела — макушки головы и концов
рук и ног (обозначенных на рис. 24 буквами Л, В, С, D,
Е) — при различных фиксированных позах тела,
отличающихся распластанностъю и зеркальной симметрией.
Естественным геометрическим инструментом такого
исследования явился пентавурф. При этом исследовании из
возможного набора данных поз была выделена одна поза,
при которой, начиная по крайней мере с тела четырех-
119
Рис. 23. Иллюстрация к представлениям древних мыслителей
о геометрических законах строения тела человека
а _ «пентасимметричный закон» человеческого тела по Агриппе Неттес-
геймскому; б — «человек Витрувия»
Рис. 24. Онтогенетические изменения человеческого тела в позе
прямостояния (а) и в распластанной позе (б—е)
а — слева направо: двухмесячный эмбрион, четырехмесячный эмбрион,
новорожденный, 6-, 25-летний человек (из [195]); б — двухмесячный
эмбрион; в — четырехмесячный эмбрион; г — новорожденный; д — 6-летний
человек; е — 25-летний человек
120
Таблица 6. Величины иентавурфов для оконечных точек
человеческого тела в распластанной позе (рис. 24)
Возраст
а
II
Ы
о
из
3
«О
II
■ч;
Q
О
ч
и
II
05
q
3
тз
II
о
W
Q
й!
II
Q
83
^
W
Эмбрион
2
4
Новорожденный
2
6
12
25
1,3
1,7
1,7
1,8
1,8
1,8
1,7
2,0
1,5
1,6
1,5
1,5
1,5
1,6
1,5
1,6
Человек
1,6
1,7
1,7
1,7
1,6
1,5
1,6
1,6
1,7
1,7
1,7
1,6
2,0
1,5
1,6
1,5
1,5
1,5
1,5
Примечание. Возраст эмбриона дается в лунных месяцах, возраст
человека — в годах.
месячного эмбриона, пентавурф пяти оконечных точек
тела демонстрировал особую онтогенетическую
инвариантность своей величины (насколько позволила судить об
этом точность исходных данных о размерах тела).
Другими словами, расположение оконечных точек тела в
этой позе трансформировалось в ходе онтогенеза
практически проективно-геометрическим образом.
Названная поза человека представляет собой позу
распластанного тела с раздвинутыми на 180° руками и
разведенными на 90° ногами. На рис. 24 представлены
онтогенетические изменения человеческого тела в данной
позе, изображенные на основе известных по литературе
данных о возрастных трансформациях тела [195,
рис. 24, а].
В табл. 6 приведены величины соответствующих пяти
пентавурфов для разных возрастов с разными начальными
буквами в символах, хотя, как отмечалось в предыдущем
разделе, для однозначной проективной фиксации
положения пяти вершин пятиугольника достаточно двух из них
в силу следующих известных [117, с. 163] связей между
пентавурфами а, Ь, с, d, e:aed — ad=e\ bae — be = a;
abc — ac = b; bed—bd=c\ cde — ce = d. Табличные
данные приведены для более наглядного и разностороннего
121
сравнения проективных характеристик пятиугольников
(с вершинами в оконечных точках) данной позы с
пентаграммой, у которой все аналогичные пентавурфы
равны величине Ф=1,618 ...:
a = b = c = d = e= 1,618...
Вследствие некоторого отличия представленных в
таблице величин пентавурфов от величин пентавурфов
пентаграммы пятиугольники с вершинами в оконечных
точках тела при рассматриваемой позе нельзя считать
проективными эквивалентами пентаграммы, но лишь
некоторыми приближениями к этой эквивалентности.
Табличные данные показывают, что (исключая случай
двухмесячного эмбриона) пентавурфные характеристики
человеческого тела в исследуемой позе практически не
зависят от возраста. Является ли эта проективно-геомет-
рическая особенность человеческого тела случайной или
ее наблюдение оповещает о важной морфологической
роли проективной симметрии и о существовании,
возможно, широкого класса других проективных биосимметрий,
покажут будущие исследования.
Отметим ряд частных деталей развития человеческого
тела в связи с рассматриваемой позой. Известно, что в
рамках проективной геометрии пять точек плоскости
однозначно определяют некоторую кривую второго
порядка, проходящую через них. Мы воспользовались этим
свойством и построили для пятерок оконечных точек
тела всех возрастов соответствующие кривые второго
порядка — эллипсы, описывающие человеческое тело (рис. 24).
Поскольку расположение пятерок оконечных точек во
всех изображенных возрастах (за исключением случая
двухмесячного эмбриона) проективно эквивалентно, то
телесные эллипсы также проективно эквивалентны друг
другу с поточечным соответствием точек А, В, С, D, Е у
разных эллипсов. Рассмотрим метрические
характеристики этих эллипсов. Прежде всего оказывается, что для
тела новорожденного такой эллипс представляет собой
правильную окружность с центром в месте пупка. Как тут
не вспомнить древнего зодчего и мыслителя Витрувия с
его попытками продемонстрировать геометрические
принципы построения человеческого тела вписыванием тела в
окружность с центром в точке пупка. Этот «человек
Витрувия» является ныне эмблемой Международной
федерации по автоматическому контролю — ИФАК, а также ряда
122
международных научных конгрессов. Правда, у Битрувия
речь идет о теле взрослого человека, а окружность
описывается только вокруг четырех оконечных тонок рук и
ног (рис. 23, б).
Далее, можно отметить, что центр телесного эллипса в
зрелом возрасте совпадает уже не с пупком, а с половым
органом. Является ли эта геометрическая установка
органов, важных для обеспечения биологического
воспроизводства, в центре телесного эллипса человека
неслучайной? На этот вопрос у нас пока нет ответа, но осветить
его, видимо, могут дальнейшие аналогичные исследования
тел высокоорганизованных животных.
Интересно также, что разведение ног на угол в 90°,
характерное для рассматриваемой позы человека, имеет
место у раннего эмбриона, как это можно видеть у
двухмесячного эмбриона на рис. 24, а. В свете обсуждаемого
эта эмбриональная особенность ног приобретает особый
смысл и связь с другими особенностями тела.
Остановимся еще на том, что во всех возрастах
плечевые суставы практически совпадают с точками
пересечений соответствующих прямых, соединяющих пять
базисных точек Л, В, С, D, Е тела в рассматриваемой позе
(рис. 24). Это привлекает внимание к вопросу о
возможной морфологической значимости точек пересечения
прямых, получаемых с помощью определенных алгоритмов
их проективного построения. Напомним, что в
проективной геометрии хорошо известны сети прямых, образуемые
проведением прямых через несколько базисных точек, в
результате чего создаются новые точки пересечения,
через которые проводятся новые прямые и т. д. Так,
создаваемые совокупности прямых и точек пересечения
называются сетями Мебиуса. Мы полагаем, что определенные
типы сетей Мебиуса (или аналогичных им конформных
сетей) могут участвовать в алгоритмах построения
биотелесных форм и определять их особенности.
123
Глава IV
ПРИМЕНЕНИЕ ТЕОРЕТИКО-ГРУППОВЫХ
КОНЦЕПЦИЙ ДЛЯ МОДЕЛИРОВАНИЯ
ПСИХОЛОГИЧЕСКИХ ЯВЛЕНИЙ
Психология уже имеет значительный опыт
использования высшей геометрии для моделирования наблюдаемых
психологических явлений. На необходимость дальнейшего
углубления знаний о симметриях в этой области биологии
указывали многие исследователи, в частности, в связи с
задачей сближения психологии и геометрической физики.
Например, Б. Рассел писал о проблеме геометризации
психологии: «Надлежит строить мост (между психологией
и геометрической физикой.— С. П.),, начиная сразу с двух
концов: приближать физические гипотезы к
психологическим данным и манипулировать психологическими
данными так, чтобы строить логические конструкции,
максимально соответствующие аксиомам геометрической
физики» [408, с. VIII]. Представляется интересным
познакомиться ближе с историей вопроса о применении методов
симметрии в области психологии и отдельными
результатами этого применения.
IV. 1. Теория групп и психологические феномены
константности восприятия
Еще в 1868 г. Г. Гельмгольц в работе «О фактах,
лежащих в основании геометрии» [66] предпринял попытку
применить определенные математические концепции,
касающиеся понятия группы, к психологическим
проблемам восприятия. При этом, когда Гельмгольц писал
этот очерк, понятие группы еще не было признано
важнейшим и весьма универсальным инструментом
математической мысли, которым, как оказалось впоследствии
оно является. Утверждение теории групп в качестве
организующего и вносящего ясность принципа в самых
различных областях математики было во многом обуслов-
124
лепо прогрессом в геометрии, связанным с открытием
неевклидовых геометрий и разработкой
теоретико-групповых основ построения геометрии.
Стремление осмыслить психологические явления
восприятия с позиций математического естествознания,
точнее, с позиций фундаментального понятия группы
привело другого выдающегося представителя
математического естествознания — А. Пуанкаре к разработке
интересной концепции о генезисе пространственных
представлений у индивидуума, о взаимоотношениях
между физическими, логическими и физиологическими
основаниями геометрии [191—194]. Эту концепцию,
отдельные положения которой уже рассматривались в разделе
II.8, Пуанкаре развивал и утверждал вплоть до конца
своей жизни. Он полагал, что понятие группы должно
быть принято в качестве понятия, ключевого для
научного познания генезиса пространственных представлений
у индивидуума. При этом он приписывал ему
априорность, считая, что у каждого индивидуума общее
понятие группы существует, по крайней мере потенциально, с
самого рождения, еще до приобретения какого-либо
индивидуального жизненного опыта. (В целом концепция
Пуанкаре, как это обычно характерно для широких
концепций, представляет собой по существу
конгломерат отдельных идей разной степени адекватности,
новизны и значимости. Она затрагивает многие глубокие
философские вопросы, анализ которых не входит в
задачу нашей работы.)
С течением времени математическое понятие группы
завоевывало себе все новые позиции в области
психологических исследований, прежде всего исследований так
называемых феномецов константности восприятия.
Анализу некоторых причин этого посвящена публикация
Э. Кассирера «Концепция группы и теория восприятия»
[308]. Часть материалов из нее мы используем в
настоящем параграфе при изучении связей меж!ду теорией
групп преобразований и феноменами константности
восприятия.
В Большой советской энциклопедии дано следующее
определение этой константности: «Константность в
психологии, относительное постоянство воспринимаемых
характеристик объекта (его величины, формы, цвета,
положения и т. д.), вопреки изменению условий его
восприятия (удаленности, ракурса, освещения и т. д.).
125
Проблема константности, рассматривавшаяся уже
Р. Декартом, теоретически разработана еще
недостаточно. Прослежено формирование константности
восприятия у ребенка (Э. Брунсвик, Австрия; Ж. Пиаже,
Швейцария, и др.), намечена связь константности с
предметностью восприятия (Ж. Пиаже и др.)» а также
получены многочисленные частные зависимости,
описывающие отдельные конкретные виды константности
(Й. Акисиге, Япония, и др.)» [1973, т. 13, с. 48].
В качестве примера рассмотрим один из феноменов —
феномен константности зрительного восприятия объекта
по габаритам и форме. Наше зрительное восприятие
любого предмета основано на расшифровке изображения
этого предмета на сетчатке глаза. Естественно, что такое
изображеыие существенно зависит от удаленности и
ракурса предмета, однако организм обладает
замечательной способностью однозначно оценивать предмет по этим,
порой весьма разным его изображениям именно как один
и тот же предмет в отношении его габаритов и формы.
Такая независимость суждения о предметах от условий
их зрительного предъявления и составляет суть
рассматриваемого феномена константности.
Другим примером из многочисленного класса
аналогичных феноменов является феномен константности
восприятия цвета, т. е. относительное постоянство
воспринимаемого нами цвета объекта вопреки изменению его
освещения. Цитируя работы психологов, Э. Кассирер в
этой связи напоминает: «Вообще, когда лист бумаги
является нам в белом цвете при обычном дневном освещении,
мы без колебания признаем его за белый и при тусклом
свете, например, при полнолунии. А кусок бархата,
представляющийся нам черным при облачном небе,
также выглядит для нас черным и в ясную погоду. Тот же
самый лист бумаги выглядит белым и в зеленоватой
тени листвы. Он смотрится таким и в лучах того или
иного обычного источника искусственного света,
обладающего определенной степенью хроматизма. Аналогичные
замечания могут быть сделаны в конечном счете и в
отношении цветных объектов, хотя и в меньшей степени
и менее определенно. Например, лист бумаги, который
днем выглядит голубым, кажется голубым также и при
красновато-желтом освещении газового пламени.
Наблюдения подобного типа показывают, что значительные
изменения интенсивности освещения и, в определенной
126
степени, его цвета не влияют в ощутимых размерах на
ваше обычное восприятие цвета. Этот факт становится
проблемой, когда мы учитываем, что всякое изменение
освещения вызывает изменение в световом потоке,
который внешние объекты отражают в наши глаза, т. е.
каждое изменение освещения сопровождается
модификацией ретинального раздражения» [308, с. 9—10].
Константность восприятия играет жизненно важную
роль, имея решающее значение для познания и
опознания видимых объектов. Если бы этой константности не
было, то живой организм как бы отдавался во власть
каждого изменения внешних условий и не мог отделять
«предметы» и «свойства» от потока изменений. Как
писал еще Э. Геринг, кусок мела в пасмурный день
представлялся бы в таком же цвете, как и кусок угля в
солнечный день. В течение одного дня он являл бы все
возможные промежуточные цвета между черным и белым.
«Белый цветок под сенью зеленой листвы был бы такого
же цвета, как и зеленый лист дерева, находящегося на
открытом воздухе, а моток ниток, белый при дневном
свете, должен иметь цвет апельсина при газовом освещении»
[308, с. 35].
Явления, подобные этим в оптической сфере,
наблюдаются почти во всех других областях перцепции.
Другими словами, имеет место общая проблема организации и
строения мира восприятия. Изучение феноменов
константности восприятия дает современной психологии
основания отвергать существовавшую в свое время сен-
суалистскую догму о строгом тождестве физических
возбуждений и восприятия. Наоборот, именно
преобразованное впечатление, т. е. впечатление, модифицированное
организмом в отношении различных явлений
инвариантности, рассматривается как соответствующее исходному
образу, как «истинное>>, как позволяющее индивидууму
сформировать свои знания реальности.
Надо отметить, что по крайней мере очень широкий
класс организмов обладает константностью восприятия,
поскольку «экспериментально показано, что не только
обезьяны, такие, как шимпанзе, но также и птицы, и
рыбы, и даже такие беспозвоночные, как пчелы, обладают
константностью восприятия» [7, с. 113]. Имеются
данные о врожденном характере этой константности у
ряда животных, однако у человека, согласно работе
Б. Г. Ананьева с сотрудниками [7, с. 114], в первые
W
педели после рождения существование константности
восприятия не отмечается.
В работе Э. Кассирера [308] дан анализ формальной
связи между инвариантно-групповой интерпретацией
геометрии и психологическими феноменами
константности восприятия, т. е. между инвариантами
геометрическими и инвариантами психологическими. Кассирер
пишет: «Мы можем поднять вопрос о том, существуют ли
какие-либо понятия и принципы, которые, хотя и в
различных отношениях и в различных степенях
определенности, являются необходимыми условиями для строения
как мира перцепции, так и мира геометрической мысли.
Мне представляется, что концепция группы и концепция
инвариантности являются такими принципами» [308,
с. 19]. Приведем основные положения этого интересного
анализа, начав с вопросов геометрии.
В Эрлангенской программе Ф. Клейна, указавшей на
инвариантно-групповую природу геометрии, по существу
утверждается, что не всякое понимание и описание
пространственного объекта является его геометрической
характеристикой. Если мы берем объект, принимая во
внимание только его индивидуальность, то он не проявит
своего геометрического характера и значения. Описывая
пространственную форму как таковую в ее особенности и
конкретности, мы в лучшем случае получаем ее
географическую или «топографическую», но не «геометрическую»
характеристику. Чтобы получить последнюю, требуется
совершенно другое направление мышления. Клейн
сформулировал новый принцип следующим образом:
геометрические свойства любых фигур должны быть описаны
с помощью формул, которые сохраняются при изменении
системы координат; наоборот, любая формула, которая в
данном смысле инвариантна относительно группы
имеющихся преобразований координат, представляет
геометрическое свойство. Примерами преобразований такого
типа являются параллельное смещение, вращение,
изменение масштаба, перспективное отображение и др.
В соответствии с этой концепцией свойства, которые
характеризуют объект, не должны определяться
применительно к элементам, образующим его. Это должно
делаться исключительно в плане группы, с которой связан
этот объект. Как только мы заменим одну группу другой,
сразу возникают совершенно другие соотношения. То,
что выступало в качестве выражения «того же» геомет-
128
'ричо^кого попятил, может получить самостоятельное
-значение. То же, что представлялось как что-то сугубо
'отличное, может оказаться идентичным с точки зрения
•новой геометрии. Это сильнее всего проявляется при
переходе от метрической к проективной геометрии. С точки
Прения метрической евклидовой геометрии, например,
.различные конические сечения (окружность, эллипс,
парабола, гипербола) выступают как самостоятельно
существующие, как независимые геометрические образы,
имеющие конкретные и хорошо определенные свойства.
Это различие исчезает при переходе к рассмотрению тех
же конических сечений с позиций проективной
геометрии, поскольку проективные преобразования могут
переводить все названные выше виды конических сечений
друг в друга и соответственно лишают эти виды
независимого существования. Из этого видно, что понятия
современной геометрии обретают присущую им точность и
становятся действительно универсальными лишь в связи
с тем, что подсказываемые интуицией специфические
фигуры рассматриваются не как предопределенные и
жесткие, а скорее как своего рода сырой материал, еще
подлежащий обработке для придания ему
геометрического статуса. Реальная основа математической
определенности покоится уже не на элементах, которые
математики берут за исходные, а на правиле, посредством
которого эти элементы связываются между собой. После
Эрлангенской программы больше не ставится вопрос,
одиняк<твы ли два четырехугольника или равна ли длина
у двух отрезков без отсылки к определенной группе
преобразований, в рамках которой должен решаться данный
вопрос. Другими словами, определенность фигуры
зависит от контекста, в который она интегрирована и
который она представляет как специальный случай.
Обращаясь к психологическим вопросам восприятия,
Кассирер обстоятельно разбирает имеющую место
аналогию с геометрией. Она заключается прежде всего в том,
что для воспринимаемых образов (форм) характерно
сохранение специфических свойств при изменении тех
элементов, па которых они основываются. Так, мелодия
не изменяется значительным образом, когда все ее ноты
подвергаются относительному смещению. То же можно
сказать и в отношении феноменов константности
зрительного восприятия по габаритам, форме, цвету и т. д. «Чем
еще является эта «тождественность» воспринимаемой
5 С. В. Петухов
129
формы, как не тем, что, как мы нашли, со значительно
более высокой степенью точности существует в области
геометрических понятий? — отмечает Кассирер.— То,
что мы находим в обоих случаях, представляет собой
инварианты по отношению к изменениям,
претерпеваемым простыми элементами, из которых создается форма.
Своеобразный тип «тождественности)), приписываемый
явно совершенно различным по составу фигурам из-за их
способности преобразовываться одна в другую с помощью
некоторых операций, определяющих группу, существует,
как видно, таким образом, и в области восприятия.
Эта идентичность позволяет нам не только выделять
элементы, но и осознавать «структуры в восприятии» [308,
с. 25]. Восприятие вовсе не является процессом
непосредственного отражения, а представляет собой процесс
объективизации, характерная природа и тенденция
которой находит выражение в образовании инвариантов.
При этом, согласно Кассиреру, совсем не случайно то,
что понятия преобразования и инвариантности,
относящиеся к теории групп, выступают на сцену при самом
толковании психологических феноменов.
Конечно, аналогия между образованием инвариантов в
восприятии и геометрии не должна быть основанием для
того, чтобы из виду упускались важные различия.
Так, при акте восприятия достижимы лишь
приблизительные, но не абсолютные определения. Это характерно,
и для константности воприятия, которая никогда не
бывает идеально полной, а всегда остается ограниченное
определенными рамками. Фиксирование этих рамок
представляет еобой одну из задач психологического
исследования. За пределами этих рамок не существует
дальнейшего «преобразования». Например, относительное
постоянство цветового тона нарушается, когда цветовое
освещение становится слишком интенсивным. Не
существует полной • константности восприятия цвета и
никакой идеальной инвариантности, есть только тенденция
в этом направлении.
Напротив, математические понятия не зависят ни от
каких пределов, которые могут быть навязаны восприя^.
тию-. Геометрическое понятие включает и охватывает все
разнообразие изменений, которые претерпевает
пространственная фигура, когда она подвергается определенным
преобразованиям. После того как задана группа
преобразований, все возможные в отнош&нии нее изменения могут
130
»^ыть точно определены. Инвариантно-групповые
построения в геометрии представляют собой как бы идеальный
обраэ, с дефектами воспроизводимый в области психологи-
• чеек«х явлений.
Рассмотренная выше аналогия-легла в основу целого
ряда работ по математической психологии. Особенно
обстоятельное экспериментальное исследование феноменов
константности восприятия (перцепции)' и их
теоретическое моделирование с инвариантно-групповых позиций
осуществлено японской школой психологов во главе с
•Й. Акисиге в Институте психологии при Университете
острова Кюсю [279—282]. В ходе проводившихся на-
протяжении нескольких десятилетий экспериментальных
исследований, направленных на выяснение закономерностей
преобразования явлений фивического мира в явления мира
перцептивного, школой Акисиге была '-показана
возможность установления единого уравнения перцептивного
преобразования, применимого ко всем беэ исключения
константам -восприятия. Изучению подверглись константы
восприятия величиныЗоложения, формы, глубины,
скорости, цвета, яркости, громкости, акустического расстояния
и т. д. Всего в исследованиях обнаружено около 60
основных функций^ представляющих различные константы вос-
npHHTHHi При этом 53 из них состоят из двух переменных,
остальные являются функциями многих переменных;
Инварианты, связанные с этими функциями, помяению Аки-
GHre, есть и сущность и характеристика константности
восприятия.- В целом полученные результаты
подтверждают формулируемое Акисигой положение о том, что
закономерности преобразований различных констант
восприятия носят инвариантно-групповой характер нна этой
основе может проводиться- геометрическое изучение
-пространства восприятия. Развернутый обзор и анализ, этих
исследований школы Акисиге дан в работе Б.Г.Ананьева
с соавторами, [7, с. 111—132]. В атой же
работе.содержатся, богатые материалы об онтогенетических,
физиологических и других аспектах явления константности,
восприятия.
IV.2. Применение высшей геометрии
при моделирований пространств восприятия
В разделе II. 8 нами рассмотрены работы Р. Лунебурга,
Г. Кинла и др. по анализу аналогий между геометрией
Лобачевского и геометрией пространства зрительного вос-
5* 131
приятия. В настоящее время область изучения
пространств восприятия с помощью концепции и аппарата
высшей геометрии привлекает внимание многих
исследователей п интенсивно развивается. Задачей данного параграфа
является анализ оощего состояния работ в этой области
и краткий обзор наиболее характерных из них для
читателя, желающего ближе познакомиться с конкретными
исследованиями.
Еще М. Планк говорил: «Основной чертой каждой
возникшей в науке новой идеи является то, что она
связывает определенным образом два различных ряда фактов»
1258, с. 7J. Сильной стороной исследований
рассматриваемого нами научного направления является то, что они
ставят в параллель ряду психологических феноменов
фундаментальный для всего математического естествознания
ряд основных геометрических конструкций и на этой
способной генерировать крупные идеи основе пытаются
осмыслить существо исходных феноменов. При этом,
поскольку данный ряд математических конструкций высшей
геометрии уже имеет параллели в других областях
естествознания, психологические феномены оказываются
структурно ассоциируемыми с весьма подчас
неожиданными классами природных явлений. Так, в работе [425]
по изучению психологических особенностей восприятия
привлекаются идеи и формализмы из специальной теории
относительности, обслуживающей совсем другую область
природных явлений.
Нельзя не отметить, что практически все имеющиеся
к настоящему времени исследования по моделированию
пространств восприятия средствами высшей геометрии
(за исключением рассмотренных в предыдущем
параграфе феноменов константности восприятия,
представляющих собой один из аспектов восприятия) касаются
только зрительного восприятия. Это обусловлено рядом
причин, в том числе относительной простотой и развитостью
экспериментальных психологических исследований
зрения. Исторически именно психофизическое изучение
зрительного восприятия цветов впервые поставило вопрос
о специфичности геометрии пространств восприятия и,
кроме того, во многом способствовало осознанию того,
что геометрия есть наука о структурных соотношениях
на множестве элементов совершенно независимо от их
природы: эти элементы могут быть точками, цветами
и т.д.
132
За последние десятилетия проблемы структуризации
^етовоснриятия породили целый ряд интересных
исследований по психологическим приложениям геометрии.
Среди пих можно указать следующие. Классик физики
£. Шредингер, много занимавшийся вопросами зрения,
^освятил обстоятельный труд [276] анализу аналогий и
различий между физиологическими законами смешения
Цветов и проективной геометрией плоскости цветов.
Именно на пути геометрического исследования он считал
возможным решение многих психологических проблем
цветовосприятия. Г. Шеллинг [421, 422] на основе своих
изысканий в области неевклидовой геометрии и
психологии восприятия ввел для описания цветовосприятия
неевклидову метрику и получил ряд прекрасных
соответствий между выводами построенной им теоретической
модели и экспериментальными данными для случая
«теплых» цветов, т. е. для диапазона цветов с длинами волн
выше 495 m\i. Г. Резников [414] интересным образом
применил дифференциальную геометрию для
углубленного изучения и моделирования цветовосприятия;
в этой же работе содержится исторический экскурс в
исследования по вопросам приложения геометрии к
проблемам цветовосприятия.
Исследования по структуризации цветового
восприятия, как и других типов восприятия, имеют не только
теоретический интерес, но и важное прикладное
значение. На знании математических принципов
структуризации цветового восприятия непосредственно основаны
методические подходы и решения ряда задач
промышленности и торговли по оптимизации цветовой раскраски
рабочих помещений, коммерческих товаров и т. д. [89].
В области геометрии зрительного восприятия, однако,
особенно большое число работ посвящено зрительному
восприятию геометрических форм (прямых линий и
плоскостей, окружностей, углов и т. д.), в том числе
анализу иллюзий зрительного восприятия. Обращаясь к
истории, можно вспомнить, что еще Птолемей привлекал
геометрию к объяснению зрительной иллюзии видимых
размеров Луны, заключающейся в том, что Луна
кажется больше, когда стоит над горизонтом [304, с. 267]. Из
современных исследований наиболее характерными,
наряду с рассмотренными в разделе II. 8 исследованиями
Р. Лунебурга и Г. Кинла, являются работы Г. Шеллинга
и В. Гоффмана, на которых остановимся подробнее.
133
В основу псйхолого-гсометрйческих работ Г.
Шеллинга [421—425] положен разработанный им специальный
способ неевклидовой метризации, имеющий общие черты
с известной схемой проективного мероопределения
Кели — Клейна [348, 423]. По мысли Шеллинга, его
случай неевклидовой метризации имеет непосредственное
отношение к пространству зрительного восприятия.
Попытка обосновать это и предпринимается в названных
работах, причем в последней из них [425]' автор выдви^
гает «перцептуальную теорию относительности>>,
объединяющую воспринимаемое пространство и время в единое
целое по аналогии с физическим пространством —
временем из специальной теории относительности. При этом
рассматриваются пространственные и временные иллюзии
восприятия, анализируются правила пространственных
построений в живописи, вводится ряд новых понятий,
например перцептуальный конус Пассивного прошлого
и активного будущего по аналогии с конусом "Мйн-
ковского — Вейля в специальной теорий относительности
н т. д. В целом эти работы Шеллинга по математической
психологии отличаются исключительной смелостью и
новаторством, однако принять безоговорочно его
утверждения, по нашему мнению, не представляется возможным.
Излагаемое в его публикациях на материале
Психологических данных обоснование структурного' родства между
феноменами зрительного восприятия И геометрией с
метрикой Шеллинга является во многом очень
дискуссионным, что снижает убедительность концепций Шеллинга
о ключевом для психологии значении вводимых им
геометрических конструкций. Однако независимо от этого
думается^ что исследования Шеллинга составляют видную
веху в истории математизации психологии восприятия и
многие поднятые им вопросы; несомненно, лягут в
основу дальнейших исследований в этой области.
Большой интерес представляют работы В. Гоффмана
[352—356] по применению групп Ли для моделирования
зрительного восприятия. В своих биогеометрйческих
исследованиях зрения Гоффман исходит из двухмерного
характера ретинального образа и визуального
многообразия, представляющего поле зрения в зрительном центре
коры головного мозга. При этом инварианты 'зрительного
восприятия связываются с группами Ли преобразований
на визуальном многообразии; аргументируется
возможность подхода к зрительному восприятию как последова-
134
дельности продолженных производных Ли, взятых- от
соответствующих дифференциальных форм.. Широкие
модельные и эвристические возможности данного
психологического применения теории групп Ли демонстрируются в
работах Гоффмана на примерах объяснения им ряда
зрительных феноменов (ортогональных послеобразов,
иллюзий восприятия фигур, послеэффектов видимого
движения и др.) и новой интересной интерпретации многих
фундаментальных вопросов. Так, перцептивное и
моторное обучение трактуется Гоффманом как образование
градуированной псевдогруппы преобразований,
элементами которой являются расширенные псевдогруппы,
генерируемые путем продолжения производных Ли,
которые сформированы в предыдущем познавательном опыте.
.Идеология групп Ли распространяется Гоффманом и на
функциональную организацию нейронных сетей, причем
отдельный нейрон интерпретируется как •
физиологический эквивалент инфинитезимальных преобразований
группы Ли.
Также с теоретико-групповых позиций подошел к
зрительному восприятию A. G. Блайвас [26, 294, 295],
полагающий, что в настоящее время теория представлений
групп Ли является наиболее адекватным математическим
аппаратом для анализа зрительного восприятия. На
основе теории лредставлений группы вращений SO (3) им
разработана модель зрительной системы млекопитающих,
описывающая физиологию зрения на клеточном уровне
и позволяющая строить прогнозы о функциях зрительных
элементов, а также о связях и взаимодействиях между
ними. Группа SO (3) принята за основную в связи с
отмечаемым Блайвасом исходно упрощенным подходом к мор-
фофункциональным особенностям глаз"а. В качестве
основного инструмента анализа зрительного восприятия" в
этом исследовании применено разложение зрительных
образов в ряды яо базисным функциям, связанным с
указанной группой.
Названные работы, как и целый ряд других
исследований^ по существу реализуют в отношении психологии
восприятия программу «геометризации биологии»,
утверждают широкие перспективы применения здесь
концепций, и методов симметрии, прокладывают пути для
включения этой части психологии в область развитого
математического естествознания.
135
IV.3. Некоторые бионические проблемы
машиноведения
Через всю историю эволюции техники красной нитью
проходит стремление человека наделить машину
возможностями, которыми природа изначально одарила только
живые организмы, воссоздать из груды мертвого
материала технический организм, способный действовать
подобно живому. Это стремление, являющееся одним из
мощных катализаторов прогресса в мире машинной техники,
основано на естественном желании человека получит!
себе помощника, способного заменить человека в работе
и расширить свои собственные возможности, например
научиться летать, как птица. Оно основано также на
неумирающей жажде человека утвердить свое человеческое
«я», доказать, что человеческому разуму нет преград и
ему под силу воспроизвести созданное самой природой.
Наше время — время научно-технической
революции — кардинально расширило применение
биологических принципов в технике, различные области которой
бурно насыщаются достижениями кибернетики, бионики,
биомеханики и при этом не только резко — количественно
и качественно — развиваются сами, но и отпочковывают
все новые и новые ветви технического прогресса. Одной
из них является область создания антропоморфных
машин — важного класса робототехнических машин
будущего, наделенных операциональными способностями,
которые копируют операциональные способности человека.
К настоящему времени эта область уже весьма развита
и сложно дифференцирована.
В данном параграфе мы хотели бы сосредоточить
внимание на тех проблемах антропоморфной техники, где
видную роль игоает или может сыгпать геометрическое
моделирование функциональных особенностей живых
организмов и познание биосимметрийных закономерностей.
Начнем с вопросов психологии.
Задачей ключевого значения для дальнейшего поог-
ресса антропоморфных машин является решение
проблемы наделения машины представлением о пространстве,
способностью ориентироваться в нем, различать
пространственные образы и пр. с теми же особенностями
восприятия и различения, что и v человека. При этом речь
идет пе только о машинпом восприятии телесных
образов, по и о машинном восприятии цветов, звуков и т. д.
Ш
'л той же структуризацией воспринимаемого, которой
отличается восприятие форм, цветов, звуков и пр. у
человека.
Конечно, для решения подобной задачи должны
привлекаться данные из области психофизики и пространст-
,венной перцепции человека и животных. Эта область
фюлогии интенсивно развивается. Многие ее достижения
уделяются результатом применения в исследовании
психологических феноменов концепций и аппарата таких
фундаментальных математических дисциплин, как теория
групп, функциональный анализ и т. д. Введение в область
психологии семантики из сложившихся областей
математики, осмысление удивительных биологических явлений
в математических терминах и понятиях — одно из
мощных средств развития психофизики. Естественно, что в
математизации науки о пространственном восприятии
ведущая роль отводится инвариантно-групповым
дисциплинам, обобщившим классическое геометрическое учение
о фигурах и пространственных отношениях. Получаемые
на этом пути в математической биологии результаты
имеют важное значение не только в рамках самой биологии,
но и в других областях естествознания от философии до
техники.
Отметим конкретнее ту пользу, которая может быть
получена при перенесении уже рассмотренных нами в
разделе IV. 1 психофизических феноменов константности
восприятия с их яркой геометрической спецификой в
организацию машинного восприятия. Укажем на один из
многих подобных феноменов — феномен константности
зрительного восприятия по габаритам и форме, т. е.
относительного постоянства воспринимаемых габаритов и
формы рассматриваемого предмета вопреки изменению
условий его предъявления — ракурса и удаленности от
глаз. Придание аналогичной способности
искусственному зрению робота обеспечило бы важное преимущество
распознавания им окружающих телесных форм во многом
независимо от расположения предметов или от
характера движения робота относительно них.
Другой пример касается феномена константности
зрительного восприятия цвета. Предположим, что мы
находимся в освещенном дневным светом помещении и видим
желтый диван, зеленые занавески, синюю тетрадь на
столе и т. д. В этих условиях обычно используемые в
технике хроматографы — приборы для определения цвета
137
по максимуму длины волпы отраженного от предмета
света ~ также покажут, что, перечисленные предметы
характеризуются названными цветами, поскольку максимум
отраженного света будет цриходиться соответственно на
длины волн желтого, зеленого, синего цветов, и т. д.
Налицо совпадение оценок цветов глазом и прибором. Но
осветим это же помещение иным, например красным,
цветом. Оказывается, что в широком диапазоне таких
возможных изменений освещенности наш глаз, улавливая
изменение общего освещения, отличается способностью
устойчивой оценки цвета предметов: человек в новых
условиях опять говорит, что диван желтый, занавески
зеленые, тетрадь синяя и т. д. А что покажет прибор?
Согласно его показаниям, все цвета изменились, став при
красном освещении красными, поскольку максимум
отраженного света при новом освещении естественно
приходится на красный цвет. Теперь представим, что мы
использовали такой обычный прибор различения цветов в зрении
робота. В результате этот робот будет воспринимать мир
по его цветовым характеристикам иначе, чем человек,
и какой-либо диалог между ними о цветах предметов будет
невозможен или очень затруднен, поскольку суждение
робота о цвете предмета окажется зависимым от характера
освещения. Посланный на иную планету, такой робот
будет, с точки зрения человека, постоянно ошибаться в
решении цветовых задач и не сможет надежно и
независимо от освещения выполнять даже простые команды
«нажми красную кнопку» или «собери зеленые камни для
доставки на Землю»,
Естественно, что задачи воспроизведения
психологических феноменов константности восприятия в технике
все более привлекают внимание конструкторов.
Возможные подходы к решению некоторых из них на основе
инвариантно-групповых концепций даны в работах [92,
245 и др.]. В этой же связи, в частности, в книге [7,
с. 99] указывается, что многие специалисты
«рассматривают изучение факторов и диапазонов константности как
первоочередное и обязательное условие успешного
моделирования перцептивных актов, так как построение лучших
механических моделей самоорганизующихся систем —
перцептронов — возможно лишь при условии
проникновения в суть этого биологического процесса,
обеспечивающего приспособление живых систем к существованию в
постоянно изменяющейся среде окружения (Веккер, 1964;
138
юнблатт, ,1965; и др.)». И далее в этой книге
рассматривая использование Ф. Розенблаттом в его обширном
(де 1965 г. теории групп для воспроизведения психоло-
{еских феноменов в искусственных нервных сетях [7,
В качестве интересного примера применения теории
рш в бионических вопросах перцепции,следует назвать
ее машинное распознавание музыкальных произведе-
и их синтез. Ряд исследований этого направления
эдставлен в монографии Р. X. Зарипова [96], где среди
*чего содержится обзор тематических работ зарубеж-
авторов-М. Бэббита (I960)-, М. Касслера (1963)т
[;.,Падберга (1965) и др. В монографии отмечается, что
шменение теории групп здесь связано с тем, что музы-
шьная тема и ее преобразования образуют группу и
сазания для сочинения, например, фуги свободного стиля
>лжны подчиняться различным выводам теории групп
)6, с. 59]. Отмечается также аналогия двенадцатитоно-
[X (додекафонных) систем и теории конечных групп,
^оторая была использована М. Касслером в работе по рас-
>знаванию шёнберговых композиций [96, с. 64].
В заключение раздела нам хотелось бы вернуться к
{ному интересному для машиноведов вопросу, уже
затрахавшемуся в разделах П. 3 и П. 5. Это вопрос о том,
сколько особенности выполнения живым организмом той
ги иной функциональной задачи могут априори прини-
1аться конструктором машин за образец для подражания:
|асколько можно доверять весьма распространенному
гждению, что биологические объекты выполняют те или
[ые функциональные задачи оптимальным образом, ибо
>ни имели миллионы лет эволюции и функционально опти-
шзировались благодаря механизмам естественного отбора.
Тзучение закономерностей симметрии в биологических
^елах вскрывает тот факт, что далеко не все в строении
юлогических органов определено критерием оптималь-
юй приспособленности к выполнению непосредственных
функций, но что важное значение имеют принципы
кристаллизации живого вещества, физические законы морфо-
'енеза. В связи с этим очевидно, что конструктор машин
[ри решении конструкторских задач находится в лучшем
положении по сравнению с живой природой, поскольку он
Может исходить целиком из критерия функциональной
оптимальности и не быть связанным необходимостью
соблюдать дополнительные условия, проистекающие из
139
закономерностей чисто биологического характера.
Конечно, понимая эти преимущества положения конструктора,
не надо вовсе обесценивать — применительно к
машиноведению — достижения живой природы в тех или иных
функциональных задачах. Как-никак, живая природа
первой решила многие задачи, которые сегодня только встают
перед конструкторами техники. И пускай эти решения
сплошь и рядом не оптимальны в обычном смысле, но они
имеют огромную ценность как привлекающие внимание
к существованию интереснейших задач и
демонстрирующие возможность их решения, как стимулятор фантазии
человека и средство борьбы с инерцией его мышления, как
апробированный всей историей человечества катализатор
технического прогресса.
Глава V.
ИССЛЕДОВАНИЕ
ВЕСТИБУЛЯРНОГО АППАРАТА
С ПОМОЩЬЮ МЕТОДОВ СИММЕТРИИ
В предыдущих главах методы симметрии
использовались для выявления закономерностей структуризации
биологических тел и явлений. Однако полезные применения
данных методов в биомеханике и бионике этим не
исчерпываются. Они играют важную роль при многих
исследованиях, связанных с обеспечением необходимых условий
модельного воспроизведения изучаемых биомеханических
процессов. Рассмотрим использование в биологии инва-
рианто-групповой теории динамического подобия и
применение ее для изучения одпой из наиболее важных
механорецепторпых систем организма —вестибулярного
аппарата. Исследования проводились на масштабной
физической модели системы полукружных каналов человека.
Подчеркнем, что применение теории динамического
подобия носило здесь принципиальный характер.
Теория динамического подобия базируется на
использовании свойства инвариантности дифференциальных
уравнений, описывающих динамические процессы,
относительно некоторой группы масштабных преобразований
динамических переменных (время, расстояние, скорость
и т. д.) и коэффициентов уравнений. Упомянутая группа
масштабных преобразований является подгруппой группы
произвольных масштабных преобразований физических
параметров, определяющих динамику процессов, и
выделяется из даппой группы требованием инвариантности
критериев подобия.
V.I. Теория динамического подобия
в биомеханике
Теория динамического подобия, представляющая собой
простой и эффективный аппарат исследования и
моделирования, широко используется в различных областях физтт-
Ш
ки [209]. С давнего времени она применяется и в
биомеханике. Примером использования ее положений в
биомеханике являются применения безразмерных - критериев
подобия типа числа Рейнольдса при решении широкого
класса гидродинамических задач биомеханики1.
Так, многие теоретические и -экспериментальные
подходы к известному . биомеханическому . п-арадок-су Грея
скоростного плавания дельфина опираются на применение
числа Рейнольдса для оценки степени ламинарности
движения воды вокруг тела дельфина [4,76]. Дельфины —
крупные, быстро плавающие животные, и поэтому числа
Рейнольдса достигают у них больших значений. При
коротких бросках дельфин с длиной тела в 190 см может
развивать скорость до 830 см/с, что дает число
Рейнольдса 1,6-107. Коэффициенты лобового сопротивления для
обтекаемых тел при таких числах Рейнольдса близки к
0,055, что дает величину лобового сопротивления для
дельфина с лобовой площадью 1100 см2, равную 2,0 • 107 дин.
При оценке удельной мощности мышц дельфина
возникает парадокс, заключающийся в том, что для плавания
с наблюдаемой у дельфинов скоростью мощность мышц
должна почти на порядок превышать их действительную
мощность. Этот парадокс ведет к предположению о
наличии у дельфинов специальных механизмов снижения гид
родинамического сопротивления движению, что' породило
целый ряд интересных исследований о таких механизмах.
Многие другие вопросы об особенностях плавания рыб
решаются посредством помещения моделей их тел в
аэродинамические трубы. При этом для сохранения чисел
Рейнольдса осуществляется масштабирование величин
относительных скоростей с учетом различия в
кинематической вязкости воздуха и воды. Так, При исследовании
Харрисом устойчивости небольшой акулы при скоростном
плавании слепок тела акулы длиной в 90 см помещали в
аэродинамическую трубу и прикрепляли к
аэродинамическим весам/ Скорость воздушного потока составляла
18 м/с. С учетом того, что кинематическая вязкость
воздуха в 14 раз больше, чем у воды, число Рейнольдса при
этом было таким, как при плавании акулы со скоростью
130 см/с. Эта модель позволила получить ряд
результатов по устойчивости к движениям по рысканию [4],
В книге [76] приведен ряд других примеров
использования теории подобия в биомеханике, например к
задачам прыжка кенгуру, теплообмена тела и др.
142
ш*--- Известны применения теории подобия к некоторым
Кэопросам- исследования вестибулярного аппарата, которо-
h|ty будут посвящены следующие разделы книги. Речь
|здег о работе Джонса и Спеллса [196], которые на основе
|втой теории сумели предсказать ряд особенностей строе-
LflMft системы полукружных каналов у различных живот-
[вых, в том числе слабую степенную зависимость внутрен-
[tfero радиуса и радиуса кривизны полукружных каналов
Бот массы тела животного. В результате проведенных ими
^проверочных измерений полукружных каналов 87 видов
мкивотных, включающих 46 млекопитающих, 17 птиц,
117-рыб и 7 рептилий, эти предсказания были
подтверждены. Измерения показали, что радиус кривизны и квад-
|ват величины внутреннего радиуса одинаковым образом
медленно возрастают с ростом массы животного: так, 10.00-
■кратное возрастание массы тела у млекопитающих (ска-
Вкем, от 0,1 до 100 кг) соответствует росту внутреннего
шрадиуса полукружных каналов в 1,6 раза, а радиуса кри-
ьвизны — в 2,2 раза. Разброс средних величин этих радиу-
■сов существенно уменьшается, если при подсчете
исключались рыбы, размеры каналов которых значительно
большое размеров каналов млекопитающих той же массы.
I V.2. Физическое моделирование системы
Ш полукружных каналов вестибулярного аппарата
В формировании пространственных представлений,
поддержании позы и построении координированных движений
непосредственно участвуют многие системы организма.
Функциональная настройка едва ли не всех из них
происходит с]учетом показаний вестибулярного аппарата,
который инпюрмирует организм о действии инерционных и
[гравитарионных сил и является одним из основных эле-
!ментов1в общей системе статокинетической регуляции
организм!. В силу широких функциональных связей
вестибулярного аппарата вестибулярные расстройства
сопровождают^ самыми разнообразными нарушениями: общей
пространственной дезориентацией, многочисленными сен-
сомоторными иллюзиями, вегетативными нарушениями
и пр.
Достигнутый за последние десятилетия прогресс в
изучении этой системы во многом обязан уточнению
наших знаний о функционировании ее рецепторных отделов.
Исходя из представлений о динамике процессов в
вестибулярном лабиринте, удается объяснить многие особен-
143
пости вестибулярных реакций (длительность послевраща-
тельного нистагма глаз, возникновение калорического
нистагма и пр.) и разработать методики вестибулярных
исследований (купулометрия, калорическое тестирование,
вестибулярная стимуляция в условиях действия кориоли-
совых сил и пр.).
Однако проблема получения точных знаний о динамике
лабиринтных процессов и установления соответствий
между этими процессами и вестибулярными реакциями
организма решена далеко не полностью.
В целях изучения этого вопроса была создана
масштабная физическая модель системы полукружных каналов
человека, воспроизводящая динамику натуральных
лабиринтных процессов в удобных для ее изучения условиях
физического эксперимента. Разработка таких масштабных
моделей должна основываться на критериях подобия
процессов в вестибулярном лабиринте, выводимых с
помощью теории динамического подобия.
Некоторые сведения о системе полукружных каналов.
Все позвоночные животные имеют принципиально
одинаковое устройство вестибулярного лабиринта. В
лабиринтах позвоночных, расположенных зеркально-симметрично
относительно сагиттальной плоскости в левой и правой
височных костях черепа, различают две вестибулярные
подсистемы: систему полукружных каналов и систему
отолитов.
Система полукружных каналов в схеме статокинетиче-
ской регуляции организма является датчиком угловых
параметров движения головы. В каждом из двух
симметричных вестибулярных лабиринтов она представляет собой
трехсвязную перепончатую оболочку довольно сложной
формы (рис. 25, 26), заполненную эндолимфой.
Перепончатая оболочка фиксирована в черепе (точнее, в перилттм-
фатическом пространстве костного лабиринта)
посредством множества фиброзных перемычек. В диапазоне
действия инерционных сил, возникающих при физиологически
нормальных ускорениях головы, эта оболочка согласно
бытующим представлениям может рассматриваться как
замкнутый резервуар с твердыми стенками.
Каждый из трех каналов (их названия:
горизонтальный, передний вертикальный и задний вертикальный)
данного резервуара выходит из общей полости — утрику-
люса и, описав дугу, приходит опять тс ней с другой
стороны. Характерно, что оба вертикальных канала задолго
144
Рис. 25. Вестибулярный лабиринт человека (из [288])
1, 2, 3 — горизонтальный, передний и задний полукружные каналы, 4 —
ампулы каналов, S — утрикулюс, в — саккулюс, 7 — стремечко, * —
молоточек, 9 — наковальня, 10 — слуховая труба, 11 — мозг, 12 — эндолимфа-
тический мешок
Рис. 26. Расположение полукружных каналов у человека (из
[341])
1—3 — передний, задний и горизонтальный полукружные каналы; 4—
ампулы; 5 — утрикулюс; в ~ саккулюс
145
до впадения в утрикулюс сливаются в один, образуя
общую ножку. Каждый канал имеет по одной рецепторной
зоне, расположенной вблизи конца канала в специальном
расширении — ампуле. Эта зона содержит сенсорный
гребешок (кристу) с нейроэпителием, состоящим из
эпителиальных чувствительных клеток, волосковидные отростки
которых пронизывают студневидную купулу. Купула
представляет собой взвешенную в эндолимфе перегородку,
перекрывающую все сечение ампулы и имеющую
возможность смещаться вдоль ампулы. В первом приближении это
образование подобно подвижной пробке в канале. Купула
имеет в "ампуле определенное положение равновесия, при
отклонении от которого со стороны пронизывающих
купулу волосков возникают упругие силы, стремящиеся
вернуть купулу в равновесное положение.
При ускоренном вращении головы под действием сил
инерции вся внутриканальная среда вместе е купудами
смещается относительно стенок каналов, купулы
деформируют чувствительные волоски и по отходящим от крист
ампулярным нервам в центральную нервную систему
поступает информация о характере вращения. В покое
при равновесном положении кулул от кулулярных
рецепторов отводится стабильная частота импульсации покоя
(у обезьян она примерно равна 100 Гц) [326]. При утри-
кулопетальном смещении купул горизонтальных каналов
частота нервной импульсации увеличивается, а при утри-
кулофугальном смещении — уменьшается. Для
вертикальных полукружных каналов частота импульсации
возрастает при отклонении купулы от утрикулюса и
уменьшается при смещении в противоположную сторону. Для всех
каналов частота импульсации ампулярного нерва
изменяется пропорционально отклонению купулы от положения
равновесия, если это отклонение не слишком велико.
Поэтому по характеру движения купул можно судить о том,
что подается на вход нервной системы от полукружных
каналов.
Следует, однако, отметить, что поведение небольшой
части волокон ампулярных нервов не вписывается в
указанную пропорциональность [332]. Кроме того,
необходимо иметь в виду, что частота вестибулярной импульсации
может регулироваться эфферентными командами из
центральной нервной системы.
Для установления функциональных соответствий
между разными уровнями вестибулярного анализатора и,
146
в частности, для интерпретации особенностей
вестибулярных реакций как возможных следствий специфики
процессов в вестибулярном лабиринте важно найти способ
изучения динамики движения купул в полукружных каналах
при всевозможных адекватных раздражениях
вестибулярного аппарата. Трудность заключается в том, что
исследовать эту динамику на натуральной системе чрезвычайно
слбжно в силу следующих причин. Вестибулярный anna-
pat заключен в труднодоступной части черепа и легко
повреждается "При попытках обнажения лабиринта;
лабиринт имеет мизерные размеры (его поперечник у человека
равен 8—10 мм) и его оболочка непрозрачна; исследовать
движение купул в вестибулярном аппарате человека
можно только на трупах или при фенестрации лабиринта у
больных со всеми сопутствующими трудностями и
ограничениями. В этой ситуации естественно обращение к фи-
рическому моделированию системы полукружных
каналов. /
О физическом моделировании пАлукружных каналов.
С первых же шагов в области исследования
вестибулярного аппарата развитие представлена о его
функциональных свойствах сопровождалось попытками моделирования
процессов в полукружных канал/х. В ряде работ было
высказано предположение, что зЬ возбуждение нервных
окончаний в ампулах полукружных каналов ответственно
инерционное движение эндолимфы, которое возникает при
ускоренном вращении головы [175, с. 18]. Это
представление, получившее с течением времени широкое признание,
породило с самого начала целый ряд чисто физических
допросов о динамике движения эндолимфы в полукружных
каналах: ламинарно ли течение эндолимфы; как зависит
гок эндолимфы от радиуса вращения головы, долго ли
цлится течение эндолимфы после остановки вращения
головы и пр.
В поисках ответа на многие из этих вопросов, которые
Практически невозможно изучить на натуральном лаби-
)инте, исследователи неоднократно обращались к
построению физических моделей полукружных каналов. Как
1равило, эти модели представляли собой стеклянные
Сольцеобразные трубки различной формы и размеров,
to многих случаях имевшие расширения для имитации
1мпул и утрикулюса. Стеклянная оболочка заполнялась
Жидкостью (обычно водой или спиртом) с взвесью види-
1ых частиц. Движение этих частиц после остановки вра-
147
тения модели подвергалось изучению невооруженным
глазом или под микроскопом.
В частности, с помощью таких моделей рядом авторов
была продемонстрирована такая важная функциональная
особенность системы полукружных каналов, как
независимость тока эндолимфы от радиуса вращения [175,
с. 19]. Это значит, что организм не может определить
расстояние от центра произвольного вращения головы
непосредственно за счет системы полукружных каналов,
поскольку ее показания не зависят от радиуса вращения.
Другая важная особенность, выявленная посредством
названных моделей, заключается в том, что в кольцевых
каналах с размерами натуральных полукружных каналов
ток жидкости после остановки вращения продолжается
доли секунды. Поэтому значительная продолжительность
рефлекторных реакций, вызванных адекватным
раздражением ампулярных рецепторов вестибулярного лабиринта,
не может быть объяснена продолжительным инерционным
движением эндолимфы.
Однако возможности физического моделирования как
метода изучения процессов в полукружных каналах
использовались далеко не полностью из-за следующих
непреодоленных в прошлом трудностей.
Создание модельной оболочки с размерами и формой
натуральной системы, даже если это технологически
выполнимо, не обеспечивает точного и удобного изучения
процессов вестибулярного лабиринта, так как в такой
миниатюрной оболочке трудно установить модельные купу-
льт, свойства которых соответствовали бы динамическим
свойствам натуральных купул. а наблюдать течение
жидкости в каналах можно лишь под микроскопом. При этом
исследователь должен уловить и описать характер
движения жидкости, продолжающегося менее 1 с. на участке
модели, появляющемся под микроскопом в момент
остановки.
Можно облегчить установку модельных купул и
наблюдение процессов в системе, изготовив увеличенную
оболочку, геометрически подобную оболочке натуральных
полукружных каналов. Но увеличение размеров оболочки по
понятным причинам чревато опасностью получения на
модели динамики процессов, количественно и качественно
отличающейся от дипамики патуральпых процессов.
Основным педостатком известных ранее крупногабаритных
моделей полукружных каналов является то, что они соз-
148
давались без учета требований теории динамического
подобия и их авторы не устанавливали, в каком отношении
подобия находятся процессы, происходящие в моделях, к
процессам в оболочке с размерами натурального
лабиринта. Отметим также, что применявшееся авторами
визуальное изучение происходящего в оболочке затруднено во
время вращения модели, а поэтому им приходилось
ограничиваться изучением лишь стадии движения жидкости после
прекращения вращения.
В настоящей работе указанные трудности преодолены.
Создана масштабная физическая модель системы
полукружных каналов человека, воспроизводящая динамику
натуральных процессов в удобных для их изучения
условиях физического эксперимента. Для разработки этой
модели потребовалось учесть требования теоршт
динамического подобия, которые изложены в следукицем разделе.
Критерии подобия процессов в системе/полукружных
каналов. Общепринято считать оболочку м/мбранного
лабиринта гладкой изнутри и достаточно тве/дой, чтобы при
обычных вращениях головы не возника/и деформации.
При этом динамика процессов в оболочке системы
полукружных каналов определяется шестью физическими
параметрами: габаритами оболочки, которые при
фиксированной форме задаются любым характерным линейным
размером I оболочки; временем t течения нестационарных
процессов в системе; вязкостью л и плотностью р внутри-
канальной среды; упругостью а купул; угловым
ускорением у оболочки системы.
Возникает вопрос: можно ли изменить произвольным
образом величины отдельных параметров, но в то же время
сохранить динамику натуральных процессов в системе
полукружных каналов неизменной? Ответ, полученный с
помощью методов теории динамического подобия (см.
например, [209]), заключается в следующем: можно изменить
произвольным образом три параметра из шести указанных,
но сохранить динамику натуральных процессов в
оболочке, геометрически подобной оболочке системы
полукружных каналов, за счет соответствующего изменения
остальных трех параметров. Таким образом, имеется возможность
вместо изучения процессов в натуральной оболочке малых
размеров, заполненной физиологической жидкостью,
изучать тс жо процессы в оболочке с произвольно большими
габаритами и внутриканальной жидкостью, выбранной из
соображений экспериментальных удобств.
149
Докажем сделанное, утверждение. Если обозначить
размерности длины — L, массы — М, времени — Т, то
размерности, плотности, вязкости, -упругости и углового
ускорения, соответственно имеют вид [p]=M/L3,[r\]=M/L-T,
[<х]=М/Т2, [у]=1/Т2 (угол считается безразмерной
величиной). Как и. любую другую характеристику протекания
процессов в системе полукружных каналов, особо
интересующее нас угловое отклонение 0 купул можно
Записать в виде функции от шести определяющих
параметров:
6=6 (Z, *, тьр, а, ?). (23)
Из этих шести параметров всего три параметра
независимы по размерностям, т. е. их размерности нельзя
представить в виде степенного одночлена из формул
размерности для других параметров. В качестве
.независимых по размерности выберем Z, t, r| (можно взять и
другие тройки параметров, но при данном выборе
окончательный результат имеет более обозримый вид).
Размерности остальных величин можно выразить через
выбранные:
1Р] = Ы [t]/[l]\ [а]=[ц] Ш/Ш, MHL/U]1. (24)
Если единицы измерения величин I, t, r\ изменить
соответственно в р\, (5<, (5„ раз, т/е. перейти к Z'=(5,Z," £'=(}<£,
■п'=р\,ть то в новой системе единиц измерения изменятся
и величины р, а, у:
р=1Гр, «=тГ«> ?=■£•
Соотношение (23) выражает некоторую физическую
закономерность и потому не зависит от выбора системы
единиц измерения. В новой системе единиц
6 = 6 (pV, PV, to. -^р, 1ф-а, -П . (25)
\ Pt Pt Pt I
Масштабы (5(, (5Л, (5< произвольны. Выбором этих
масштабов можно сократить число аргументов у функции.
Положим (5,=1/Z, (5<=1/£, рл=1/т1, т. е. выберем систему
единиц измерения таким образом, чтобы значения первых
трех аргументов равнялись единице. Тогда
6 = 6(1,1,1-^-, ^, Y««). (26)
Нетрудно видеть, что аргументы данной функции
150
безразмерны. Таким образом, от соотношения (23)
шестью размерными аргументами мы пришли к
эквивалентному соотношению (26) с тремя безразмерными
аргументами. Эти три безразмерных отношения
l2p/r\t, ta/r\l, ft2 (27)
являются критериями подобия процессов в системе
полукружных каналов человека и всех позвоночных
животных. Из (26) видно, что именно сохранение величины
критериев подобия гарантирует воспроизведение
динамики натуральных процессов в оболочке, геометрически^
подобной оболочке исходной системы, с той точностью,/;
какой процессы в системе определяются принятыми в/
внимание шестью физическими величинами. Можно изменить
значения до трех размерных параметров (например, I, ц,
р), но, сохранить динамику натуральных процессов за
счет компенсирующего изменения остальных параметров.
Эта возможность и была использована автором для
построения масштабной физической модели системы
полукружных каналов человека. Проведенное доказательство
представляет собой частное приложение П-теоремы теории
динамического подобия [209] к случаю процессов в
полукружных каналах.
Масштабная физическая модель системы
полукружных каналов человека. Перейдем к описанию
построенной масштабной модели системы полукружных каналов
человека (рис. 27, 28). Данные о размерах оболочки этой
системы взяты в основном из работ [290, 360, 452, 283
и др.]. Построенная модель [1] воспроизводит в стекле
форму правой половины системы полукружных; каналов
человека с линейными размерами, увеличенными из
соображений экспериментальных удобств в 49 раз по
сравнению с натуральными. Цифрами на рис. 27 обозначены:
1 — полукружные каналы, 2 — ампулы, 3 — утрикулюс,
4, 5 — отверстия с заглушкой для заполнения модели
жидкостью. В качестве такой жидкости выбран глицерин,
что обусловливает изменение плотности и вязкости
внутриканальной среды от р=1,0 г/см3 и п=0,85-
•10~z г/(см-с) при 35° С в натуральном лабиринте [431]
к их модельным значениям р=1,26 г/см3, ii=14,9 г/(см-с)
при температуре проведения экспериментов 20° С.
Задавшись модельными значениями I, г\, р, из требования
сохранить величины критериев подобия (27) получим
соотношения между модельными (т) и натуральными (п)
151
Рис. 27. Конструкция масштабной физической модели системы
полукружных каналов человека
значениями упругости а купул, углового ускорения y и
времени t течения процессов:
ат ^4,9-104, Y«-3vm, tm«l,75*n. (28)
Таким образом, для воспроизведения динамики
натуральных процессов модель следует вращать с ускорением,
втрое меньшим, чем то, действие которого на
вестибулярный аппарат нас интересует, а процессы в ней буду г
растянуты во времени в 1,75 раза.
Для сравнения укажем, что если бы в качестве
модельной жидкости использовать воду, то соотношения
между натуральными и модельными величинами имели
бы гораздо менее удобный для экспериментирования вид:
а™ « 3 • 1<ГЧ; 2,3.107Ym = уп; tm = 2064*п.
Тогда пришлось бы создавать модельную упругость по-
152
* "" - ''
i
i
J
A
"V
i
Рис. 28. Внешний вид масштабной физической модели системы
полукружных каналов человека
рядка сотых долей дин/см, а для вращения модели на
разных режимах ускорений конструировать установку,
позволяющую работать на ускорениях, в 2,3-107 раз меньших,
чем соответствующие ускорения головы человека.
Эксперимент по вращению модели, аналогичный десяти-
секундному эксперименту на человеке, продолжался бы
около шести часов. Выбор глицерина в качестве модельной
жидкости не только позволяет изменить величину а, у,
t в удобное число раз, но имеет и ряд других преимуществ,
о которых будет сказано далее.
Поскольку возможность некоторой деформации
студневидного купулярного колпачка при движении, а также
изменение купулой фронта движущейся жидкости нельзя
считать заметно влияющими на величину продольного
смещения внутрик анальной среды, то при моделировании
купул было сделано естественное предположение: каждую
купулу можно считать твердой подвижной пробкой,
взвешенной в окружающей жидкости и перекрывающей все
речение канала, влияние которой па величину продольного
153
смещения внутриканальной среды определяется целиком
за счет упругих сил, стремящихся вернуть отклоненную
купулу в положение покой. (Относительно малой
значимости роли купулы как экрана, деформирующего ток э/ндо-
лимфы, в работе [254] отмечено: «Экранирующее действие
ампулярной перегородки на выход струй жидкости из
канала в ампулу или очень мало, или исключается»).
В той степени, в которой сформулированное
предположение справедливо, не важно, где установить модельную
купулу — в ампуле или в самом полукружном канале,
лишь бы было сохранено влияние упругих сил нужной
величины на процессы в системе. (Расположение
натуральных купул в ампулах объясняется, вероятно, тем, что
диапазон линейного функционирования купулярных
рецепторов ограничен, и для предотвращения выхода за
пределы этого диапазона купулы установлены в
расширенных ампулах, где при той же величине вращательногс
стимула смещение среды меньше, чем смещение среды в
самом канале.) Естественно, что коэффициент упругости
купулы при ее расположении в канале должен быть
уменьшен во столько раз, во сколько площадь сечения
канала меньше площади сечения ампулы. Именно в такой
степени смещение среды в канале больше, чем в ампуле,
и, таким образом, нужная величина упругих сил, равная
произведению коэффициента упругости на смещение
купулы, будет сохранена.
Все это было принято во внимание, и из соображений
технологической простоты купулы устанавливались не в
ампулах, а внутри однородных по сечению каналов на
небольших прямолинейных участках. Модельная купула
(рис. 27) представляет собой взвешенную в окружающей
Жидкости й хорошо подогнанную под размер канала
перегородку из двух дисков 6, средняя плотность которой
равна плотности глицерина, что предохраняет сразу от
двух нежелательных эффектов: смещения модельной
купулы под действием собственной тяжести и
возникновения сухого трения из-за давления купулы на стенки
канала при движении купулы.
Стабилизирующие упругие силы придаются купуле за
счет её растяжения на тонких упругих нитях #, концы
которых прикреплены к игольчатым воротам 9. Эти
вороты Дозволяют регулировать упругие силы купулы
посредством наматывания на них нитей 8. Вороты 9 размещены
в Полых стеклянных отростках 10, Наличие в модели
154
заполненных неподвижной жидкостью отростков 10 никак
не сказывается на динамике течения жидкости в системе,
но позволяет взять упругую пить 8 увеличенной
начальной длины для расширения диапазона регулирования
упругих сил купулы и, кроме того, разместить вороты 9
в них, а не в самих каналах, где вороты мешали бы
течению жидкости.
Из литературы известно, что возвращение отклоненной
купулы в положение равновесия под действием только
собственных упругих сил приближенно описывается экспо-
нентой с показателем (—pi), где р примерно равно 0,1 с-1
[340, 396 и др.].'На построенной модели упругость купул
подбиралась так, что время возвращения отклоненных
модельных купул в положение равновесия равнялось
определяемому экспонентой е ~0,п времени натуральной
системы, умноженному на коэффициент подобия 1,7
(соотношение (28)).
Система регистрации смещений модельных купул
(т. е. того самого параметра, который определяет, что
подается от вестибулярного аппарата на вход нервной
системы) использует источник света 11 и фоторезистор 12,
укрепленные над купулой на внешней стороне канала
напротив друг друга. Количество света, падающего от
источника света 11 на фоторезистор, определяется
положением в канале светораспределительного экрана 7,
прикрепленного к дискам купулы. В целях
предотвращения нежелательных поворотов вокруг своей центральной
оси, которые могут внести искажения при данной схеме
регистрации искомых продольных перемещений купул,
диски установлены на направляющий рельс 13. Для
проведения экспериментов модель помещают на
вращательную установку, способную обеспечить нужный режим
вращения. При действии вращений, характеристики
которых соответствуют физиологическим, перемещения
каждой из купул происходят в пределах выпрямленного
участка канала (1/20 часть окружности канала).
Смещающийся с каждой купулой светораспределительный экран
7 изменяет количество света, падающего на фоторезистор
12, пропорционально смещению купулы. По величине
сопротивления фоторезистора на ленте осциллографа
регистрируются искомые перемещения купул системы
полукружных каналов. Для улучшения качества
регистрации перемещений каждой купулы использовались не
один, а два фоторезистора, включенных в мостовую схему,
,155
что позволяло легко обеспечить линейную зависимость
между смещением светораспределительного экрана и
током, идущим через шлейф осциллографа. Для
исключении воздействии на фоторезисторы посторонних
источников света участки каналов с купулами накрываются
светозащитными чехлами.
Осуществление данного способа моделирования купул
и регистрации их перемещений возможно при заполнении
оболочки далеко не любой жидкостью. Но
использование глицерина позволяет это. Во-первых, глицерин не
агрессивен по отношению к резине. Было проверено, что
упругость резиновой нити не меняется при
многомесячном пребывании ее в глицерине. Во-вторых, глицерин
имеет одинаковый удельный вес со многими видами
пластмасс, из которых можно сделать модельные купулы,
что сразу решает проблему их взвешивания в
окружающей среде. В-третьих, глицерин прозрачен, что позволяет
использовать простую схему регистрации на
фоторезисторах.
При отладке модели была проведена специальная
проверка соответствия некоторых функциональных
особенностей модели особенностям натуральной системы
полукружных каналов. Было проверено, что каждая
модельная купула достаточно полно перекрывает сечение своего
канала: тушь, вспрыснутая в модельный канал с одной
стороны от купулы, практически не проникала по другую
сторону от купулы при смещениях внутриканальной
среды, как это имеет место и в натуральных
полукружных каналах [432]. Кроме того, установлено, что
изменения положения модели относительно вектора гравитации
не меняют положения купул в каналах, т. е. модельные
купулы нечувствительны к действию силы тяжести, как
нечувствительны к ней и купулы натурального лабиринта
(см. обзор [196]). Конечно, было бы крайне желательно
сравнить хотя бы в некоторых случаях ускоренного
вращения характеристики смещений модельных купул с
соответствующими характеристиками натуральной
системы полукружных каналов. В частности, подобное
сравнение могло бы показать, насколько полно динамика
процессов в этой системе определяется физическими
параметрами, учтенными при выводе критериев подобия
процессов в полукружных каналах. К сожалению, в
литературе, насколько известно автору, не имеется каких-либо
количественных дашшх, полученных в исследованиях
156
непосредственно на натуральном лабиринте, о
характеристиках движения купул при вращении головы.
Следует особо отметить, что хотя изготовлена модель
только правой половины системы полукружных каналов
человека, данной модели вполне достаточно для изучения
процессов, протекающих в обоих вестибулярных
лабиринтах при вращении головы. Это утверждение вытекает и.з
того, что оба лабиринта являются зеркальным
отражением друг друга относительно сагиттальной плоскости.
Введем декартову систему координат, связанную с черепом:
пусть плоскость yz совпадает с сагиттальной плоскостью,
а по оси х происходит зеркальное отражение лабиринтов
(см. рис. 26).
Задача заключается в следующем. Если имеется
модель одного, например правого, лабиринта, то можно ли
на ней получить процессы, протекающие в левой системе
полукружных каналов при движении головы с угловым
ускорением
Y(Y» V», V,)? (29)
Ответ, очевидно, будет утвердительным, если удастся
найти такие условия вращения модели, при которых бы
силы, стимулирующие движение каждого объема
жидкости в правой модели, были сагиттально-зеркальным
отражением сил, действующих на соответствующие объемы
жидкости левой системы полукружных каналов при
интересующем нас вращении (29), т. е. преобразование сил
F (Fx, Fy, F2), действующих на любой объем жидкости,
должно иметь вид
Fx->-Fx, FV->FV, FZ^FZ. (30)
Такой же вид имеет преобразование отражения
координат радиус-вектора г (гх, гу, гг) от центра вращения до
выделенного объема:
г*-> — гх, Гу-*гу, rz-+rz, (31)
Теперь обратимся к закону преобразования
отражения координат углового ускорения. Силы инерции,
порожденные ускоренным вращением, представляются в виде
F = i»lr.v], (32)
где т — масса выделенного объема жидкости. В записи
157
по координатам уравнение (32) имеет виД.
,£х = т [гууг .~г г2уу], Fv = m [rzyx — rxyz\,
fi = m[rxyv^ryyx\. (33)
Легко проверить, что для выполнения, условия. (30)
перехода, от. левого.лабиринта к..его зеркальному
отражению, достаточно потребовать, чтобы
'Ь -** "К If» -*" "7v. Tf. -^ ~Ъ- (34)
Таким образом, искомые характеристики, процессрв,
протекающих в правой и левой системе полукружных
каналов при движении головы с угловым ускорением f (^x,
tvi X») можно получить, если. соответственно. провести на
модели правого лабиринта, два отдельных, эксперимента:
1).до вращению модели. ,с ускорением ^ (ух, ч^ч*);
2). до вращению модели с ускорением f (fx, —t», —%*)•
Переходя к описанию экспериментов по вращению
модели, необходимо отметить, что на построенной модели
могут быть не только изучены отличия между динамикой
процессов в изолированном торе и в реальном
полукружном канале, что составляет задачу Настоящей работы, но и
решены многие другие задачи, такие, как получение
характеристик смещения купул при действии кориолисовых
сил, исследование движения эндолимфы в области отоли-
товых макул и многие другие.
V.3. Эксперименты
на масштабной физической модели
системы полукружных каналов человека
Масштабная модель системы полукружных каналов
человека была разработана для уточнения уже имеющихся
знаний о динамике лабиринтных процессов»
До настоящего времени повсеместно распространенной
функциональной моделью полукружного канала является
математическая модель Штейнхаузена [434], основанная
на физическом представлении о каждом полукружном
канале как об изолированном торе, заполненном вязкой
жидкостью и имеющем внутри упругую перегородку.
Эксперименты на описанной масштабной модели
системы полукружных каналов позволили показать
существование двух важных особенностей динамики процессов ъ
полукружных каналах, неучитываемых -в модели
Штейнхаузена: наличия некоторого гидромеханического взаимо-
158
действия между полукружными каналами и асимметрии
условий для движения эндолимфы в противоположных
направлениях обхода каждого канала. Прежде чем
останавливаться на этих результатах, обратимся к
представлениям Штейнхаузена.
О модели Штейнхаузена. Движение эндолимфы и ку-
пулы в полукружном канале, интерпретируемом как
изолированный тор, описывается Штейнхаузеном по
аналогии с движением сверхдемпфированного крутильного
маятника линейным дифференциальным уравнением второго
порядка
1в4-Св + КЭ = 1у, (35)
где в, в, в — соответственно угловые смещения, скорость
и ускорение эндолимфы на оси канала относительно его
стенок; / — момент инерции кольца эндолимфы;
С—момент трения на единицу угловой скорости; А' —момент
упругих сил на единицу углового отклонения; if —
составляющая углового ускорения черепа, относительно инер-
циального пространства, перпендикулярная к плоскости
полукружного канала.
Дополнительное обоснование применимости уравнения
(35) проведено Р. Стиром [430], который проанализировал
динамику движения эндолимфы в жестком торе с
размерами полукружного канала, исходя из предположений о
нулевой скорости тока эндолимфы по стенке канала и
отсутствии купулы. Он показал, что в случае тора решение
уравнения Стокса для жидкостного потока, вызванного
внезапным угловым ускорением, выражается через
функции Весселя и хорошо аппроксимируется решением
уравнения, подобного уравнению Штейнхаузена, но без
члена упругости.
В многочисленных работах [322, 323, 338—340, 347 и
Лр.] подтверждена полезность модели Штейнхаузена для
интерпретации некоторых данных физиологического
эксперимента, таких, как продолжительность постнистагма
или иллюзии противовращения в купулометрических
тестах.
Однако класс вестибулярных реакций, особенности
которых могут быть объяснены спецификой лабиринтных
процессов, представляемых по Штейнхаузену,
оказывается заведомо обедненным, поскольку эта модель заметно
упрощает картину протекания процессов в системе,
полукружных каналов. При описании движения эндолимфы в
159
полукружном капало посредством уравнения Штсйн-
хаузена (35) предполагается выполнение следующих
условий: 1) полукружный канал — изолированное
тороидальное кольцо с твердыми стенками, целиком
заполненное однородной вязкой несжимаемой жидкостью;
2) поперечное сечение канала полностью перекрывается
взвешенной в эндолимфе купулой, при отклонении
которой возникают упругие силы, стремящиеся вернуть купу-
лу в положение равновесия, причем величина этих сил не
зависит от направления отклонения купулы.
Реально же полукружные каналы не являются
изолированными торами. В частности, наличие у полукружных
каналов общих полостей — утрикулюса, общей ножки
вертикальных каналов — заставляет предположить
существование гидромеханического взаимодействия между
каналами, т. е. влияния течения эндолимфы по кольцу одного
канала на течение эндолимфы в других каналах.
Имеются отдельные экспериментальные работы, указывающие
на возможность такого взаимодействия.
Так, Ловенштейн и Санд (390) получили в опытах на
хрящевых рыбах, что горизонтальные каналы реагируют
на вращение вокруг вертикальной оси, не возбуждаясь
при поворотах вокруг двух горизонтальных осей, в то
время как вертикальные каналы отвечают на вращение
вокруг всех трех осей. Данные результаты могут быть
объяснены гидромеханическим взаимодействием полукружных
каналов [107]. Подобная специфика реакций
полукружных каналов, вероятно, имеет определенное
физиологическое значение: например, Ловенштейн [389] указывал,
что стимуляция вертикальных каналов при
горизонтальном вращении ведет к одновременной активации верхних
и нижних прямых и косых мышц глазодвигательного
аппарата, зарегистрированной Лоренте де Но [387], который
приписывал этой активизации роль создания устойчивой
оси для движения глаза в плоскости вращения.
На наличие гидромеханического межканального
взаимодействия при грубых механических воздействиях на
перепончатый лабиринт в условиях фенестрации
костного лабиринта указывал Сентаготаи [211].
Сложность связи между реакцией каждого
полукружного канала и плоскостью вращения отмечена в работе
[254]. Автор в опытах на стеклянной модели,
воспроизводящей форму системы полукружных каналов с
десятикратно увеличенными размерами, установил: при любом
160
положении модели относительно плоскости вращения
купулометрическии стоп-стимул вызывает токи жидкости
во всех каналах. Однако, как показывает анализ [173] на
основе теории динамического подобия, результаты С. Г. Че-
банова [254] соответствуют случаю сверхсильного
стимула типа резкого удара, и серьезно апеллировать к ним в
вопросах о движении эндолимфы в нормально
стимулированных полукружных каналах нельзя.
В. А. Кисляков и И. В. Орлов [109] подчеркивают воз-
[можность существования гидромеханического межкаиаль-
:ного взаимодействия. В их работе осуществлялся нагрев
отдельных точек на поверхности частично обнаженного
вестибулярного лабиринта лягушки. Установлено, что
подобная калорическая стимуляция, например, латерального
канала при определенной величине температуры нагрева
ведет к заметному изменению импульсной активности
ампулярных нервов и латерального и переднего верти^-
кального каналов (активность нерва заднего
вертикального канала не контролировалась). Авторы работы
трактуют полученные данные как результат влияния
течения эндолимфы в одном канале на течение В' другом.
Необходимо, однако, отметить, что использование
результатов электрофизиологических исследований
ампулярных нервов на предмет выявления гидромеханического
межканального взаимодействия затруднено из-за
возможности прямых нервных связей между ампулярными
рецепторами разных каналов [331].
Следует также обратить внимание, что представление
о полукружных каналах как совокупности изолированных
торов особо не соответствует действительности при
рассмотрении лабиринтов хорошо летающих птиц, имеющих
наиболее развитый вестибулярный аппарат. Лабиринт
этих птиц много сложнее по форме, чем у человека и
других позвоночных, причем эта сравнительная сложность
связана как с сильным выгибанием каналов из своих
плоскостей, так и с образованием у птиц дополнительных
общих полостей между полукружными каналами [130,
226, 344], что увеличивает предпосылки для
взаимодействия токов эндолимфы из разных каналов.
Таким образом, имеющиеся работы указывают на
возможность ограниченности модели Штейнхаузена в
отношении межканального взаимодействия, однако наличие
этого гидромеханического взаимодействия до сих пор не
доказано.
6 С. В Петухои
161
Другое упрощение, вносимое моделью Штейнхаузена,
связано с тем, что при применении уравнения
Штейнхаузена предполагается одинаковость условий движений
эндолимфы по каналу в обоих направлениях его обхода.
Однако каждый из полукружных каналов вовсе не
является тором, а имеет весьма неоднородную по сечению
форму, что определяет движение эндолимфы по каналу
как процесс преодоления последовательности различных
гидродинамических сопротивлений отдельных участков
канала. Поскольку величина гидромеханического
сопротивления участка канала с переменным сечением
зависит, вообще говоря, от направления прохождения
жидкостью этого участка, то полное гидродинамическое
сопротивление, которое приходится преодолевать эндолим-
фе при движении в полукружном канале, зависит от
направления движения эндолимфы по каналу.
Формально это следует из того, что уравнение Навье-
'Стокса, описывающее течение вязкой несжимаемой
жидкости, наряду с членами, линейными по скорости,
содержит квадратичный по скорости конвективный член. Из-за
квадратичного члена при изменении знака скорости на
противоположный мы приходим к уравнению, не
эквивалентному исходному, так как все члены скорости
поменяют знак, кроме конвективного. Отсюда следует, что и силы
гидродинамического сопротивления, которые
определяются на основе решения уравнения Навье-Стокса,
вообще говоря, зависят от знака скорости жидкости, т. е.
от направления обхода канала. Эта асимметрия
направлений недействительна лишь в частном классе
физических ситуаций, например в случае течения жидкости по
симметричной оболочке типа тора.
На то, что должно существовать физическое различие
между течением эндолимфы в противоположных
направлениях обхода полукружного канала, обращал внимание
еще Крум Браун [175, с. 31]. В качестве облегченного
направления движения эндолимфы разными авторами
указывалось и ампулопетальное и ампулофугальное
направления. Так, если Н. Ф. Тюмянцев [227, 228] на
основании умозрительных рассуждений утверждал, что
ампулопетальное движение жидкости должно быть слабее
ампулофугального, то С. Г. Чебанов [254] считал, что
ампулопетальное движение эндолимфы сильнее
ампулофугального.
Указанная асимметрия может иметь особое значение
162
При угловых осцйлляЦйях головы. Из линейного уравнб-
вия Штейнхаузена следует, что при этих движениях голо-
1Ы смещение купулы каждого стимулированного канала
должно происходить но синусоидальному закону. Но если
рядолимфе легче течь в одну сторону по каналу, чем в
другую, то очевидно, что синусоидальное качание
полукружного канала должно сопровождаться несинусоидаль-
Ным движением купулы в канале: уже после первого
периода качаний купула не вернется в исходное положение
покоя, а окажется несколько смещенной в сторону
облегченного движения эндолимфы, и это смещение будет
Накапливаться с каждым периодом. Таким образом,
различие условий для движения эндолимфы в
противоположных направлениях должно вести к накоплению
постоянных составляющих смещения купул при синусоидальных
Качаниях головы, что может обусловить дополнительные
вестибулярные реакции.
Перейдем, наконец, к описанию экспериментов на
Масштабной физической модели системы полукружных
|каналов человека, которые позволили уточнить, чем
отличается динамика движения жидкости в изолированном
pope от динамики процессов в такой сложной оболочке,
какую имеет система полукружных каналов.
Результаты. Для вращения масштабной модели
системы полукружных каналов человека использовалась
специализированная вращательная установка ВУ-4,
предназначенная для проведения вестибулярных тестов на
Людях и животных. Она представляет собой несколько
Осложненную модификацию вращательной установки ВУ-2
175]. Размещение на ВУ-4 модели полукружных
каналов потребовало изготовления особой платформы,
позволяющей надежную фиксацию модели в разнообразных
Положениях относительно оси вращения, что давало
возможность подбирать нужную плоскость вращения
модели.
\; Для решения вопроса о гидродинамическом межка-
кальном взаимодействии модель из состояния покоя
щриводилась во вращение в наиболее характерных пло-
Еостях с разными значениями постоянного углового уско-
ния от '6 до 110 град/с2. В связи с тем, что при
интернировании данных модельного эксперимента
применительно к натуральной системе полукружных каналов
человека величины угловых ускорений должны
утраиваться (соотношение (28)), использование этих ускорений со-
6* W
Рис. 29. Пример осцилло-
графической записи
смещения модельных купул
прп вращении
1—скорость иращепин кресла,
2 — 4 — смещении купул
соответственно переднего,
заднего и горизонтального
каналов
ответствует вращению человека с ускорениями от 9 до
330 град/с2, что перекрывает диапазон угловых
ускорений, применяемых при обычных вестибулярных
исследованиях. О степени стимуляции каждого канала судили
по относительной величине смещений модельных купул
к моменту достижения моделью некоторой скорости
вращения (200 град/с). Регистрация перемещений
модельных купул производилась на осциллографе Н-135,
лентопротяжный механизм которого включался с пульта
управления ВУ-4 непосредственно перед началом
вращения модели. Кроме перемещений купул,
регистрировался также сигнал с дистанционного пульта ВУ-4,
управляющий вращением установки. Пример осциллографиче-
ской записи приведен на рис. 29.
Результаты вращения модели в некоторых плоскостях
приведены на рис. 30—32. По оси абсцисс отложена
величина углового ускорения модели, умноженная на три,
для соответствия результатов случаю вращения человека;
по оси ординат — относительные смещения купул в долях
от принимаемого за единицу смещения купулы канала,
наиболее стимулированного в данной ситуации. П —
передний, 3 — задний, Г — горизонтальный полукружные
каналы. Знаком «+» обозначаются отклонения купул,
приводящие в натуральном лабиринте к увеличению частоты
ампулярной импульсации, знаком «—»
—противоположные отклонения. При вращении системы полукружных
каналов в направлении, обратном указанному в подписях
к рисункам, знаки отклонения купул полукружных
каналов меняются на противоположные, а величина их
отклонения изменяется не более чем на 5—7%.
164
' —Г
]0 - " Рис. 30. Относительное
смещение эндолимфы в
трех полукружных
каналах правого
лабиринта при вращении
головы в плоскости горизон-
ЦВУ тальных каналов налево
ь
*-3
60 120 -180 290 300 360
град/ег
4,0
+/7
Рис. 31. Относительное
смещение эндолимфы в
^BY трех полукружных ка
палах правого
лабиринта при вращении головы
,аб\- в плоскости переднего
правого канала «вперед-
направо»
Ц2 {-***- I I—i—*—*—I /I I I I+J
±L-J^-L J T 1 T J T T T~r,
60 120 ISO 240 300 360
град/с
» I Рис. 32. Относительное
смещение эндолимфы в
трех полукружных
каналах правого
лабиринта при вращении
головы в плоскости заднего
4s V правого канала «назад-
направо»
*** i 5 i 5 1 i *—* f-^+/7
0,2
И Т т у т i т ] т =j=
60 120 180 240 300 360
+ Г
-i
36L
град/с1
Доказательство существования гидромеханического
взаимодействия дает, например, вращение модели в
плоскости горизонтальных каналов, при котором, как
показали эксперименты, стимулируется не только
горизонтальный канал, но и два вертикальных канала,
перпендикулярное расположение которых в плоскости вращения
исключает их самостоятельную стимуляцию в отсутствие
связи с горизонтальным каналом. На рис. 30 представлен
график, демонстрирующий, что отклонение внутриканаль-
ной среды, например, передних вертикальных каналов
при вращениях модели в плоскости горизонтальных
каналов достигает 15% от отклонения внутриканальной
среды горизонтальных каналов. Для проверки того, что
вертикальные каналы стимулируются в данном случае
действительно из-за наличия движения жидкости в
горизонтальном канале, была проведена закупорка
горизонтального канала, исключающая движение в нем
глицерина. В этих условиях при рассматриваемом вращении
перестали стимулироваться и вертикальные каналы.
В экспериментах на модели установлено, что при
вращении в плоскостях вертикальных каналов
горизонтальный канал почти не стимулируется (см. рис. 31, 32), что
перекликается с результатами электрофизиологических
исследований [391].
В целом полученные на модели данные говорят о том,
что рассматривать систему полукружных каналов как
совокупность изолированных колец можно лишь весьма
приближенно.
Следует отметить, что нами не обнаружено реверсии
в направлении течения жидкости в период вращения
модели с постоянной скоростью, что расходится с
результатами экспериментов [254] на модели, созданной без
учета требований теории динамического подобия.
Разборная оболочка масштабной физической модели
системы полукружных каналов позволяет сменную
установку различных частей оболочки для исследования
влияния на динамику процессов в системе формы и размеров
ее отдельных частей. Например, известно, что сечение
общей ножки вертикальных каналов сильно отличается
для разных видов животных. С целью проверки, какое
влияние на процессы в системе оказывает изменение
сечения общей ножки, на модели была установлена
специально изготовленная общая ножка с поперечным сечением,
втрое меньшим нормального. При таком изменении обо-
166
^M/WVWWWWWVWVl/V'
—чшшшш
Рис. 33. Движение купул при синусоидальных угловых качаниях
модели системы полукружных каналов
Смещение квазисинусоид вверх (вниз) соответствует смещению
соответствующей купулы в сторону увеличения (уменьшения) частоты ампуляр-
ной импульсации, 1—3 — передний, задний и горизонтальный каналы
соответственно
лочки системы для вращения в большинстве плоскостей
характерно усиление гидромеханического взаимодействия
между вертикальными каналами и его ослабление в
отношении влияния горизонтальных каналов на вертикальные.
Обратимся теперь к функционированию вестибулярного
аппарата при угловых осцилляциях головы. Автором было
изучено поведение купул в масштабной физической
модели системы полукружных каналов человека при
синусоидальных угловых качаниях модели на вращающейся
установке ВУ-4. Крепление модели на установке и
регистрация движения модельных купул осуществлялись так же,
как и в описанных экспериментах. На рис. 33
представлено поведение трех купул одного лабиринта при угловых
осцилляциях модели. В первом приближении отличие
между любой из представленных кривых и синусоидой,
по которой, согласно уравнению Штейнхаузена, должна
колебаться купула, заключается в накоплении
постоянной составляющей, стремящейся с течением времени к
некоторому предельному значению. Это значение тем
больше, чем больше частота и амплитуда угловых
осцилляции головы.
Накапливающуюся постоянную составляющую
отклонения купулы будем называть ее дифференциальным
отклонением. Это название предлагается в силу следующих
ассоциаций: данное накопление осуществляется малыми
167
порциями (дифференциал в математике означает малое
приращение) и его причина заключается в разнице
условий движения эндолимфы в противоположных
направлениях. Условимся считать дифференциальное отклонение
положительным, если куиула смещается в сторону
увеличения частоты ампулярной импульсации, и
отрицательным при смещении купулы в другую сторону.
Накопление дифференциальных отклонений купул при
угловых осцилляциях головы происходит, вероятно, в
вестибулярных лабиринтах всех животных, но характер
этой асимметрии должен устанавливаться в каждом
случае отдельным исследованием. Дифференциальные
отклонения купул у человека, как показали исследования на
нашей физической модели системы полукружных
каналов, отрицательны у передних вертикальных каналов и
положительны у горизонтальных и задних вертикальных
каналов. «
Отметим, что именно опыты на масштабной
физической модели полукружных каналов выявили накопление
постоянных составляющих отклонения купул при
угловых осцилляциях полукружных каналов и обратили наше
внимание на существование асимметрии условий для
движения жидкости по каналу в противоположных
направлениях обхода. В литературе не удалось найти работ, в
которых показывалось бы или предполагалось накопление
постоянных составляющих отклонения купул при
осцилляциях головы.
Косвенное подтверждение дифференциального
отклонения натуральных купул, выявленного в опытах на
нашей модели системы полукружных каналов человека,
можно видеть в работе [326]. В ней демонстрируется, что
при угловых синусоидальных качаниях натуральной
системы полукружных каналов обезьяны активность купу-
лярных афферентов, модулированная по искаженной
синусоиде, с каждым периодом последовательно сдвигается
относительно уровня активности в покое, как это и
должно быть при накоплении дифференциального отклонения
купул (рис. 34). Авторы указывают, что такая
квазисинусоидальность не может быть объяснена на основе
модели Штейнхаузсна, и относят это несоответствие на счет
динамических особенностей афферентов, иннсрвирующих
полукружные каналы. Представляется, что постепенное
смещение постоянной составляющей ампулярной
импульсации проще объяснить накоплением дифференциальных
168
, спайки/с
tdO>
■160 ■
140
120
100
80
ВО
40
20
-астдгтчнащ' активность' • .".... ..-■■•'у ^-~
I i I ''''' ■ ■ I l_
О 19.84 3968 5952 7335 99.20 119.04 138.88 158.72 178.5В J98.40t,i
Рис. 34. Ампулярная импульсация переднего полукружного
канала обезьяны при угловых синусоидальных качаниях головы с
частотой 0,05 Гц.
По оси абсцисс отложено время в секундах, по оси ординат — частота
ампулярной импульсации; нижняя и верхняя горизонтальные линии —
уровни импульсации в покое до и после качаний соответственно
(Фернандес, Голдберг [326))
отклонений купул, возможность которого авторы
упускают из вида.
Можно ли скорректировать уравнение Штейнхаузена
так, чтобы оно отражало различие в гидродинамических
сопротивлениях при движении эндолимфы в разных
направлениях обхода полукружного канала? Для этого надо
величину члена вязкости в уравнении Штейнхаузена
сделать зависимой от направления движения эндолимфы,
модифицировав тем самым линейное уравнение в
нелинейное:
ie + C(signe)B-\rKe=I'y, (36)
где
С при 6>0
С" при в<0.
С (sign В) =
Здесь С и С" — моменты силы трения на единицу
угловой скорости при движении эндолимфы в
противоположных направлениях. Остальные обозначения введены в
уравнении (35).
Исследование уравнения (36) на аналоговой машине
при синусоидальном характере углового ускорения пока-
.169
зало хорошую качественную пригодность этого уравнения
для описания движения эндолимфы при угловых осцил-
ляциях полукружного канала. В частности, посредством
сравнения с результатами угловых осцилляции
масштабной физической модели полукружных каналов в
физиологически нормальном диапазоне получено, что значение
коэффициентов Р=С1С" для любого из каналов
заключено в интервале 9/10<Р<1.
В зависимости от параметров осцилляции возможны
два основных случая. Случаем дифференциального
суперотклонения купулы назовем ту ситуацию, когда
накопление дифференциального отклонения купулы столь
значительно, что, начиная с некоторого момента, ее
колебания происходят полностью с одной стороны от положения
купулы в покое (рис. 35). Этот случай характерен для
больших частот качаний (^0,8 Гц). Ситуация
дифференциального суботклонения купулы наблюдается тогда,
когда накопление постоянной составляющей смещения
купулы в одну из сторон от положения равновесия
невелико и часть каждого периода качания полукружного
канала купула проводит и по другую сторону. Этот случай
характерен для небольших частот качания. Два
указанных случая могут отличаться некоторыми особенностями
вестибулярных реакций, как это будет рассмотрено далее.
На основании изложенных модельных результатов
удалось предсказать и объяснить целую серию
вестибулярных реакций, описанию которых посвящен следующий
раздел.
V.4. Физиологические эксперименты
для проверки результатов
модельных исследований
Результаты, которые дают модельные исследования,
должны быть проверены на живом организме. Что
касается моделей вестибулярного лабиринта, то такая
проверка на интактном организме возможна лишь косвенными
методами. Для этой цели обычно используются
особенности протекания тех или иных вестибулярных реакций,
зависящие от динамики процессов в вестибулярном
лабиринте. Например, широко применяемым способом
тестирования вестибулярной рецепции на интактных
организмах является изучение вестибуло-окуломоторных
реакций. Среди этих реакций наиболее изученным является
170
n
Рис. 35. Дифференциальное суперотклоненио купулы
По оси абсцисс отложено время, по оси ординат — отклонение купулы от
положения равновесия 0. Стрелкой обозначено начало синусоидальных
угловых качаний канала
вестибулярный нистагм, проявляющийся при стимуляции
полукружных каналов.
Многими исследователями были предложены
разнообразные, порой очень сложные методики искусственного
стимулирования ампулярных рецепторов:
трапецеидального ускоренного вращения, искусственного
гидродинамического давления в полукружных каналах, калорической
стимуляции, электростимулирования и пр. (обзор этих
методик содержится в работе [НО]).
С помощью указанных методик было установлено,
отклонение купул каких каналов и в каком направлении
вызывает нистагм того или иного типа. Подобные
результаты позволяют судить по регистрируемому нистагму глаз
О процессах в вестибулярном лабиринте [340, 433 и др.].
Эта возможность косвенного суждения о процессах в
вестибулярном лабиринте использована и в настоящей
работе.
Надо сказать, что представление о каждом
полукружном канале как изолированном торе хорошо объясняет
многие черты вестибулярных реакций. К их числу,
например, относится проявление вестибулярного нистагма глаз
в плоскости качаний при угловых осцилляциях головы на
частотах до 0,2 Гц [453 и др.]. Однако, как изложено в
предыдущем разделе, в функционировании реальных
полукружных каналов имеются особенности, не
учитываемые представлением о полукружных каналах как
совокупности изолированных торов, а потому следует ожидать
существования вестибулярных реакций, обязанных своим
появлением стимуляции полукружных каналов, но при
этом не объяснимых на основе упрощенной модели
Штейнхаузена.
171
Обратимся к вопросу о том, какие вестибулярные
реакции могут быть предсказаны, исходя из выявленного в
модельных экспериментах накопления
дифференциальных отклонений капул при угловых осцилляциях головы.
Назовем условно эти вестибулярные реакции
дифференциальными, чтобы формально отличать их от реакций,
которые могут быть объяснены на основе идеализированных
представлений о полукружном канале как о торе.
Хорошо известно, что можно разными искусственными
способами вызывать смещение купул и изменение
частоты амггулярной импульсации (калорическая стимуляция,
раздражение электрическим током и т. д.), но
вестибулярные реакции при этом будут таковы, как если бы на
полукружные каналы действовало угловое ускорение,
приводящее к аналогичному изменению частоты ампулярной
импульсации. Иначе говоря, смещение купул любого
происхождения организм воспринимает как результат
действия некоторого углового ускорения.
Поэтому естественно сформулировать следующий
исходный тезис для предсказания дифференциальных
вестибулярных реакций: накапливающиеся при
синусоидальных угловых качаниях головы постоянные составляющие
отклонения купул в одну из сторон организм
интерпретирует как результат действия, помимо осцилляторных,
еще и постоянных угловых ускорений, которых на самом
деле нет. Учет организмом при угловых осцилляциях
головы несуществующих постоянных угловых ускорений
должен вести к появлению дополнительных сенсомотор-
ных реакций, не соответствующих реальной обстановке.
Ниже приводятся экспериментальные данные об одном из
подклассов всего множества вестибулярных реакций —
подкласса вестибуло-окуломоторных реакций, собранные
с целью проверки истинности сформулированного тезиса.
Эта проверка осуществлялась посредством изучения
литературного материала и проведения собственных
направленных экспериментов с участием испытуемых на предмет
обнаружения вестибуло-окуломоторных реакций,
совпадающих с прогнозируемыми дифференциальными вести-
було-окуломоторными реакциями.
Дифференциальный нистагм. Рассмотрим возможные
следствия дифференциального отклонения купул шести
полукружных каналов двух зеркально-симметричных
лабиринтов при продолжительных угловых осцилляциях
головы в сагиттальной и перпендикулярных к ней плос-
172
костях, К последним относятся, в частности, фронтальная
и горизонтальная плоскости. При указанных качаниях
характер накопления дифференциальных отклонений ку-
пул в двух лабиринтах зеркально симметричен, причем
в вертикальных каналах накапливающееся отклонение
купул происходит в ту же сторону (у задних каналов —
ампулофугально, у передних — ампулопетально), что и
отклонение этих купул при ускоренном вращении головы
назад. Если такие дифференциальные отклонения купул
организм действительно интерпретирует как результат
действия несуществующего углового ускорения головы
назад, то должны проявляться вестибулярные реакции,
аналогичные тем, которые имеют место при ускоренном
вращении головы назад. В частности, должен проявляться
бьющий вверх вертикальный нистагм, который не може1
быть объяснен на основе представления о полукружном
канале, как о торе, а потому будет именоваться
дифференциальным вертикальным нистагмом.
Проявление такого нистагма непосредственно во
время качаний следует ожидать лишь на достаточно больших
частотах качаний (/>'0,7-^0,8 Гц), поскольку известно,
что нистагм как результат отклонения купул проявляется
только при длительном (т>2 с) пребывании купул с
одной стороны от положения покоя. Такое условие
длительного одностороннего нахождения купул во время угловых
качаний головы соблюдается, как показали наши
модельные исследования, лишь при частотах качаний порядка
1 Гц, когда накопление дифференциальных отклонений
каждой купулы столь велико, что ее колебания
происходят целиком по одну сторону от положения покоя.
Подтверждение того, что во время угловых качаний
в сагиттально-симметричных плоскостях на больших
частотах должен появляться дифференциальный
вертикальный нистагм глаз, не наблюдаемый на более низких
частотах, имеется в экспериментальных данных [442].
Подвергая испытуемых угловым осцилляииям с амплитудой
25° и частотами 0,07; 0,12; 0,21; 0,31; 0,50; 0,91 Гц в
плоскости горизонтальных каналов, эти авторы
зарегистрировали, что па частоте 0,01 Гц на обычные противовраща-
тельпые движения глаз в плоскости качания
накладывается бьющий вверх вертикальпый пистагм, не имеющий
места на меньших частотах из указанной серии (рис. 36).
Эти дапные одновременно служат косвенным
подтверждением наличия гидромеханического межканального Аза-
173
Рис. 36. Электронистагмо-
граммы при
синусоидальных качаниях (для
различных частот) с амплитудой
25° человека на
вращающемся кресло в плоскости
горизонтальных каналов
1 — горизонтальные движения
глаз; 2 — вертикальные
движения глаз; 3 — синусоида
качаний. Видно появление с
увеличением частоты
вертикального нистагма (Тринкер
с соавтр. [442])
наших ранее описанных
модельных экспериментах. Действительно, накопление
дифференциальных отклонений купул вертикальных
каналов, позволяющее объяснить возникновение
вертикального нистагма при качаниях в плоскости горизонтальных
каналов, возможно только в том случае, если за счет
гидромеханического взаимодействия приходит в движение и
эндолимфа вертикальных каналов.
Случай малых частот (менее 0,8 Гц) угловых
осцилляции головы отличается тем, что дифференциальные
отклонения купул невелики, а потому раскачиваемая купула,
несмотря на накопление постоянной составляющей своего
смещения в одну из сторон, часть каждого периода
проводит и по другую сторону своего равновесного положения.
В этих условиях дифференциальный вертикальный
нистагм во время качаний проявиться не может. Однако
маловероятно, чтобы существование постоянных
составляющих отклонения купул при продолжительных осцил-
ляциях головы прошло бесследно. Более вероятно, что
проявление воздействия этих дифференциальных
отклонений купул на глазодвигательный аппарат в
рассматриваемой ситуации окажется отставленным во времени на
период после качаний. Возможность окуломоторных
реакций быть отставленным во времени и длительно сущест-
0,073 Гц
0,ШГц
о,г1Гц
,„,• JVUAAaaaa/\/;
Z
05оГц AAv^W\^wWV^aa/^
0,3/Гц
имодеиствия, установленного в
174
tfcfrfiaTb после окончания вестибулярного или оптокинбти-
Ьского раздражения была ранее показана [108] и др.
F Нами проведено несколько серий экспериментов дли
явления дифференциального вертикального нистагма,
идаемого после угловых осцилляции головы в сагит-
Сально-симметричных плоскостях на частотах меньше
),8 Гц. Во всех опытах, описанных в данном разделе,
испытуемыми были здоровые люди в возрасте от 18 до 25
кет, никогда ранее не жаловавшиеся на вестибулярные
касстройства и практически не испытывавшие укачива-
шя на городском транспорте. На глаза испытуемым
назвалась черная повязка и давалось задание глаза под
■связкой держать открытыми, чтобы не провоцировать
шонтанного нистагма. Исследовался как вертикальный,
рак и горизонтальный нистагм глаз; их одновременная
Ьегистрация проводилась по обычной методике электро-
Ёистагмографии с использованием усилителей
биопотенциалов УБП-1 и шлейфового осциллографа Н-135.
[ В первой серии экспериментов испытуемый
усаживался на стул, его туловище фиксировалось к спинке
стука, на голову надевалось матерчатое кольцо, соединенное
р одной стороны с закрепленной на стене пружиной, а с
кругой — с эксцентриком мотора. При вращении
эксцентрика голова совершала синусоидальные движения,
имеющие угловую составляющую. Этим методом, варьируя
гочки приложения сил, вызывающих осцилляцию головы,
можно подвергать голову пассивным осцилляциям как в
сагиттальной, так и в перпендикулярных к ней плоскостях
1рис 37). Частота колебаний / изменялась от 0,3 до 0,7 Гц,
ггловая амплитуда — от 10 до 40°, продолжительность
Осцилляции 5-j-lO мин. Изучались осцилляции в
сагиттальной, фронтальной и горизонтальной плоскостях го-
ковы.
Было обследовано семь испытуемых. С каждым из них
рпыты по угловому качанию ставились дважды с
перерывом между опытами в 10-Н5 дней. В этих опытах у
четырех испытуемых был зарегистрирован ярко
выраженный бьющий вверх вертикальный нистагм (рис. 38).
возникающий через некоторое латентное время после
прекращения осцилляции головы. Продолжительность
основной серии этого постосцилляторного нистагма равнялась
0,5-^1,5 мин, но отдельные движения глаз нистагменного
Типа появлялись и по истечении длительного времени
После эксперимента. У двух других испытуемых этот
175
Рис. 37. Качание головы человека, туловище которого
фиксировано к неподвижному стулу, под действием синусоидальной силы
и возвратной пружины в сагиттальной плоскости (а) и во
фронтальной (б)
-«м
\cman
jl_a-_J-X—Л Лл Kj^JK A^ja
_a-ju>—AA^
Л_
-Л~^-Л—-^-
20
L
1с
Рис. 38. Пример нистагма глаз, регистрируемого после угловых
осцилляции испытуемого в сагиттальной или перпендикулярных
к ней плоскостях
А, Б, В — последовательные части одной записи, верхняя кривая —
движение глаз в вертикальной плоскости (отклонение кривой вверх
соответствует движению глаз вверх); нижняя кривая — движение глаз в
горизонтальной плоскости
нистагм был выражен слабее и прекращался за более
короткий срок после остановки качаний. Еще у одного
испытуемого даже осцилляции в течение 10 мин с
амплитудой в 40° и частотой 0,7 Гц вертикального нистагма
практически не вызывали. Горизонтальный нистагм не
наблюдался ни у одного из испытуемых. Условия качания,
латентный период после остановки осцилляции головы до
появления вертикального нистагма, а также длительность
176
Таблица 7. Постосцилляторный дифференциальный нистагм
глаз как следствие пассивных качаний головы
Испытуемый
1
1
2
3
4
5
6
7
Номер опыта
1
2
1
2
1
2
1
2
1
2
1
2
1
2
Плоскость
0СЦИ.1ЛЛЦИИ
Сагиттальная
Фронтальная
Сагиттальная
»
Фронтальная
Сагиттальная
Фронтальная
Горизонтальная
»
»
Сагиттальная
Горизонтальная
»
Сагиттальная
Амплитуда
осцилляции,
град.
20
20
30
30
30
30
10
10
20
20
10
20
40
40
Частота
осцилляции,
| Гц
0,3
0,3
0,7
0,3
0,5
0,5
0,7
0,3
0,5
0,7
0,7
0,5
0,7
0,7
Продолжитель
ность
осцилляции, мин
5
5
10
10
5
5
10
10
5
5
5
5
10
10
Латентный
период, с
20
26
3
8
24
17
15
18
17
24
5
12
—
48
Продолжительность
вертикального
нистагма, с
7
9
90
44
9
10
52
31
35
47
78
55
—
4
основной серии этого постосцилляторного нистагма
приведены в табл. 7.
Вторая серия экспериментов — те же качания головы,
но производимые не под влиянием внешней силы, а
активно за счет усилий самого испытуемого. Точнее,
испытуемый получал задание активно качать головой,
выдерживая, по возможности, определенную плоскость,
амплитуду и частоту качания. В результате экспериментов
выявилось, что, как и пассивные осцилляции, активные
качания головой в сагиттальной и перпендикулярных
к ней плоскостях также приводят в большинстве случаев
к выраженному постосцилляторному вертикальному
нистагму, бьющему вверх. Горизонтальный нистагм не
проявлялся ни у одного испытуемого. Данные этих
экспериментов приведены в табл. 8. Отметим, что если частично
разгрузить шейные мышцы от веса головы за счет
подтягивания с помощью петли Глиссона головы вверх, то
постосцилляторный нистагм заметно усиливается так же,
как и ощущения укачивания, сопровождающие у
большинства испытуемых процесс пассивного или активного
качания головы.
177
Таблица 8. Постосцилляторный дифференциальный нистагм
глаз как следствие активных качаний головы
Испытуемый
Плоскость
осцилляции
Амплитуда
осцилляции, град.
Латентный
период, с
Продолжительность
вертикального
нистагма, с
1
2
3
4
5
Сагиттальная
»
Фронтальная
Горизонтальная
»
20
20
10
20
20
22
16
11
20
17
27
60
22
4
19
Примечание. Частота осцилляции 0,5 Гц, продолжительность осцилляции
7 мин.
Для проверки того, что постосцилляторный
дифференциальный нистагм определяется вовсе не активацией
шейных мышц при осцилляциях головы относительно
неподвижного туловища, испытуемые подверглись также
качаниям на вращающемся в горизонтальной плоскости
кресле. Кресло имело приспособление для фиксации
испытуемого. Опыты ставились при двух положениях
головы: голова выпрямлена и голова наклонена вперед на 45°.
В обоих случаях при угловых качаниях на кресле
продолжительностью 3—10 мин с амплитудой 20—100° и
частотами 0,3—0,7 Гц характер изучаемой вестибулярной
реакции оставался тем же, что и в предыдущих опытах:
у большинства испытуемых регистрировался
постосцилляторный вертикальный нистагм, бьющий практически
всегда вверх. Горизонтальный нистагм практически
отсутствовал. Данные этих опытов приведены в табл. 9.
Предсказания дифференциальных вестибулярных
реакций возможны при продолжительных угловых качаниях
головы не только в сагиттально-симметричных
плоскостях. Так, можно ожидать, что в результате угловых
осцилляции головы в плоскости левого заднего и правого
переднего полукружных каналов (расположенных
примерно под углом в 45° к сагиттальной плоскости)
возникнет дифференциальный нистагм глаз того же вида, что
и нистагм, наблюдающийся при простом ускоренном
вращении головы назад — влево в плоскости левого заднего
и правого переднего каналов. Действительно, эти
осцилляции, почти не затрагивая остальных каналов,
максимально стимулируют каналы, находящиеся в плоскости
качаний, и обусловливают накопление в них дифферен-
178
Таблица 9. Постосцилляторный дифференциальный нистагм
глаз как следствие качаний на вращающемся кресле
Испытуемый
Наклон
головы
вперед,
град
Амплитуда
осцилляции,
град
Частота
осцилляции, Гц
Продол-
>\{итель-
ность
осцилляции, мин
Латентный
период, с
жительность
вертикального
нистагма, с
1
2
3
4
5
6
0
30
45
0
0
45
20
50
50
100
40
40
0,5
0,5
0,5
0,3
0,7
0,5
5
5
10
3
5
5
6
25
15
13
20
11
8
23
35
18
6
28
циальных отклонений купул того же знака, что и
отклонение купул при простом ускоренном вращении головы
назад — влево. Отметим, что сформулированное
предсказание базируется на учете расположения полукружных
каналов; оно не могло бы быть сделано при расположении
вертикальных каналов, например, в сагиттальной и
фронтальной плоскостях, так как в этом случае
дифференциальные отклонения купул были бы сагиттально
симметричны при осцилляциях головы в любой плоскости.
В проведенных нами экспериментах испытуемые
усаживались во вращающемся крвгле так, чтобы левый
задний канал расположился в горизонтальной плоскости,
в которой и совершались осцилляции кресла. Эти
осцилляции происходили с амплитудой 20—60°, частотой 0,3—
0,7 Гц и продолжительностью 5—10 мин. В результате
экспериментов на пяти испытуемых выявлено, что у
четырех из них после этих качаний появляется нистагм того
же типа, что и нистагм, сопровождающий простое
ускорение вращения головы назад — влево в плоскости левого
заднего канала (рис. 39). У пятого испытуемого были
зарегистрированы лишь редкие одиночные удары
нистагма хаотического типа.
Следует отметить, что во всех наших экспериментах
по дифференциальному нистагму, отставленному во
времени на период после качания, никогда не
регистрировался сколь-либо значительный реверсивный нистагм,
т. е. направление нистагма не менялось. Однако
нижеописанные данные по окулогиральным иллюзиям свиде-
179
тельствуют, что реверсия движения глаз все же иногда
проявляется.
Окулогиральные иллюзии как результат угловых
осцилляции. Согласно работам [335, 337] стимуляция ампуляр-
ных рецепторов влияет на зрительное восприятие. При
ярком освещении и зрительно сложной обстановке для
появления визуальных иллюзий требуются сильные
раздражения лабиринта. Однако при слабом освещении и
восприятии простой зрительной картины весьма слабая
стимуляция полукружных каналов может вызывать
сильную иллюзию движения наблюдаемых объектов, в
действительности неподвижных относительно наблюдателя.
Причем кажущееся движение сохраняется даже и тогда,
когда уже исчезает ощущение вращения. Эта иллюзия,
названная окулогиральной, изучалась авторами в опытах
со стимуляцией полукружных каналов посредством
вращения испытуемых в режиме «трапеции»: разгон с
небольшим угловым ускорением, вращение с постоянной
угловой скоростью, затем прекращение вращения в
результате интенсивного торможения. В этом режиме купула
дважды оказывается отклоненной из своего положения
равновесия на время до 30 с.
Указанные авторы описали как иллюзии,
появляющиеся во время вращения, так и иллюзии после вращения.
В начале вращения налево появляется иллюзия движения
налево светящейся в темноте цели, неподвижно
закрепленной относительно испытуемого. Примерно через 30 с эта
иллюзия полностью затухает, но у многих испытуемых ей
на смену приходит кажущееся движение цели направо.
Это движение уже не так выражено, как в первом случае,
и заканчивается быстрее.
После резкой остановки вращения начинается после-
вращательная иллюзия, имеющая три стадии. В первой
стадии, продолжающейся около 30 с, наблюдалось
кажущееся движение цели направо. После прекращения этой
иллюзии через латентный промежуток времени в
несколько секунд начиналось кажущееся движение цели налево,
менее интенсивное, но более длительное, чем в первой
стадии. Третья стадия, заключающаяся в кажущемся
движении цели снова направо, наступала после короткого
латентного периода вслед за второй стадией, по эта
иллюзия была уже кратковременной и очень ослабленной.
Естественно предположить, что возникновение этих
иллюзий обусловлено хотя бы частично нистагмом глаз.
180
Применение электропистагмографического метода для
записи нистагма глаз во время иллюзий не дало полного
подтверждения этого предположения, поскольку данный
метод позволял регистрацию лишь достаточно сильного
начального нистагма, который мог быть связан с первой
стадией окулогиральной иллюзии. Однако в
дополнительных опытах -с использованием феномена остаточного
изображения на сетчатке яркой цели указанные авторы
получили данные, свидетельствующие в пользу того, что
именно движения глаз соответствующих направлений
ответственны за возникновение во всяком случае первой
и второй стадий окулогиральной иллюзии.
-\ ■ i .а>»<-^-^. К, ■ ■ / .—iJJjr*J^- A - fy^~A>uJv-—A
«А-^у
Рис. 39. Нистагм, регистрируемый в плоскости левого заднего
полукружного канала после угловых осцилляции испытуемого
Верхняя запись — движение глаз в сагиттальной плоскости (отклонение
кривой вверх соответствует движению глаз вверх); нижняя — в
горизонтальной плоскости (отклонение кривой вниз соответствует движению глаз
влево)
Основываясь на этих данных, можно полагать:
поскольку в результате угловых осцилляции в
сагиттально-симметричных плоскостях ожидаются, как это разобрано
ранее, дифференциальные вестибулярные реакции, схожие
с реакциями при простом ускоренном вращении головы
назад, то в результате этих осцилляции должна
возникнуть дифференциальная окулогиральная иллюзия
движения светящейся в темноте неподвижной цели вверх,
наблюдаемая во всяком случае до тех пор, пока существует
бьющий вверх дифференциальный нистагм,
зарегистрированный в описанных выше опытах.
Для проверки этого предположения были поставлены
эксперименты по качанию восьми испытуемых на
вращающемся кресле с амплитудой 40°, частотой 0,5 Гц и
продолжительностью угловых осцилляции 5—10 мин.
Туловище и голова испытуемого фиксировались к креслу при-
тяжными ремнями, причем голова была наклонена в
сагиттальной плоскости вперед на 30°, чтобы осцилляции
происходили в плоскости горизонтальных каналов. Такое
181
положение головы было выбрано потому, что, кроме
иллюзии движения цели вверх, ожидалось появление еще одной
иллюзии, обусловленной стимуляцией именно
горизонтальных каналов и описанной в следующем разделе.
С целью облегчить проявление окулогиральной
иллюзии на лицо испытуемого надевался черный
полупрозрачный экран (засвеченная широкоформатная фотопленка,
вставленная в маску аквалангиста), через который из
окружающей обстановки нельзя было видеть ничего,
кроме ярко светящейся цели. Эта цель представляла собой
сложную по форме трехмерную нить накала лампочки.
Отчетливая трехмерность цели необходима для того, чтобы
исключить наложение на окулогиральные иллюзии
автокинетических иллюзий, проявляющихся в хаотических
подергиваниях и перемещениях цели [335].
Во время укзанных качаний иллюзий вертикального
движения цели не наблюдалось, но после качаний эти
иллюзии более или менее выраженно проявились у всех
испытуемых. Для отметки характера и продолжительности
иллюзий испытуемые пользовались трехпозиционным
переключателем, задающим положение светового зайчика
шлейфового осциллографа. Три возможных положения
светового зайчика соответствовали направлению
кажущегося движения цели вверх, вниз, а также случаю
отсутствия иллюзии.
Результаты опытов по вызову окулогиральной иллюзии
движения цели в сагиттальной плоскости приведены в
табл. 10, содержащей ряды чисел, которые характеризуют
продолжительность в секундах очередных стадий иллюзии
и латентных промежутков между стадиями. Числа со
знаком плюс (минус) означают время, в течение которого
испытуемый отмечал иллюзии движения цели вверх
(вниз); числа без знака — время, в течение которого
иллюзии не наблюдались. Первое из чисел в ряду для
каждого испытуемого представляет собой латентный период
от конца осцилляции до начала иллюзии. У четырех
испытуемых иллюзия движения цели сменилась иллюзией
раздвоения, описанной в следующем разделе.
Как уже упоминалось, в работе [335] использован
феномен остаточного изображения и получены данные,
свидетельствующие в пользу того, что за возникновение
подобных окулогиральных иллюзий ответственны нистаг-
менные движения глаз даже в тех случаях, когда нистагм
зарегистрировать электронистагмографически не удается.
182
(лица 10. Иостосцилляторные окулогиральныо иллюзии
юпытуе-
1й
Продолжительность движения цели, с
1 60, +40, 16, +50, 3, +32, 7, +2, 24, +13, 4, -2, 7, +3,
2, 1, +18, 78, +20, 80, +6, +11, +60, 10, +9, 1, -4,
1, -2, 1, +5, 10, +11
2 54, +44, 6, -20, 22, +28, 44, +5, -18, 6, -20, 22, +28,
1, -44, 5, -18, 6, +118, 78, +82, 10, раздвоение
3 40, +50, 7, -10, 5, +32, 18, -12, 25, +48, 24, +70, 15,
раздвоение
4 65, +20, 10, +30, 8, -12, 20, +38, 3, -7, 5, +28, 22,
+15, 5, -18, 14, +35, 4, +11, 10, -16, 9, -25, 30, +40
5 38, +12, 8, +40, 11, -14, 14, +25, 8, -6, 14, +32, 2,
+12, 5, -13, 23, +4, 5, +19, 2, +42, 9, -18, 4, +3, 2,
2, 4, +15, 5, +30
6 62, +80, 7, +23, 10, +19, 18, -15, 3, +42, -8, 10, +32,
14, -33, 29, +5, 35, -3, 15, +27, 40, +53, 15,
раздвоение
7 45, +68, 9, +27, 12, +15, 11, -13, 4, +39, 15, +22, -8,
13, +3, 2, +2, 4, -3, 3, +21, 8, +30, раздвоение
8 58, +39, 12, +23, 11, +14, 4, -7, 3, +19, 5, -3, 8, +23,
14, +2, 10, +7, 5, +18, 12, +16
Наличие среди стадий иллюзий, приведенных в табл. 10,
таких, во время которых цель движется вниз,
свидетельствует в пользу того, что после осцилляции имеется
многократная реверсия нистагма, хотя электронистагмогра-
фически это не удалось зарегистрировать.
Раздвоение объектов в поле зрения. Обратимся теперь
к горизонтальным полукружным каналам, отклонения
купул которых обычно имеют противоположные знаки:
если частота ампулярной импульсации одного канала
увеличивается, то другого при этом уменьшается. Это
выполняется как при любых естественных однократных
поворотах головы, к восприятию которых и предназначена
система полукружных каналов, так и при продолжительных
ускоренных однонаправленных вращениях, используемых
в вестибулярных исследованиях. Но при стимуляции
горизонтальных каналов синусоидальным угловым ускорением
головы имеет место новая ситуация: знаки
дифференциальных отклонений купул этих каналов одинаковы.
Поскольку при продолжительном качании головы величины
этих дифференциальных отклонений одинаковы, то
дифференциального нистагма в плоскости горизонтальных
183
каналов это не вызовет (в случае
зеркально-симметричного строения лабиринтов и их проводящих путей нет
преимущества правого перед левым). Однако возможен
другой эффект.
Между полукружными каналами и глазными мышцами
прослеживаются отчетливые нервные связи — трехнейрон-
ные дуги, по которым осуществляются основные
рефлекторные реакции вестибулоглазодвигательного аппарата.
Стимуляция горизонтальных полукружных каналов
избирательно влияет на активность тех глазодвигательных
мышц, которые ответственны за движения глаз
параллельно плоскости горизонтальных каналов. Каждый
горизонтальный полукружный канал связан с внутренней
прлмой мышцей глаза той же стороны и наружной
прямой мышцей глаза другой стороны. Эти мышцы
активируются под действием ампулопетальпого (плюсового)
смещения купулы соответствующего горизонтального канала.
Противоположное смещение купулы способно не более
как тормозить те же мышцы [211].
Возможна ли хотя бы частичная активация при
перераспределении тонуса всех четырех прямых мышц
глазодвигательного аппарата в результате одновременного
длительно существующего ампулопетального отклонения
купул обоих горизонтальных каналов? Если эти эффекты
имеют место, то в зависимости от того, окажется ли
сильнее действие наружных прямых или внутренних прямых
мышц, следует ожидать конвергенции или дивергенции
глаз и сопутствующего двоения объектов в поле зрения
в результате продолжительных угловых осцилляции
головы, сопровождающихся ампулопетальными
дифференциальными отклонениями купул обоих горизонтальных
каналов.
В пользу возможности подобного нарушения биноку-
лярности зрения за счет вестибулярных влияний говорит
следующий известный факт: вестибулярные раздражения
при болезни Меньера зачастую сопровождаются спазмом
конвергенции глаз и двоением изображений [250]. Вместе
с тем в работе [315] в опытах на спинальных кошках,
подвергнутых местной анестезии, не обнаружено
активации прямых мышц глазодвигательного аппарата
непосредственно во время одинаковой электрической
стимуляции ампулярных нервов обоих горизонтальных каналов.
Проверка возможности конвергенции или дивергенции
глаз в результате осцилляторной стимуляции горизонталь-
184
ных каналов осложнена тем, что проявление
вестибулярных влияний на глазодвигательный аппарат сильно
ослабляется при ярком освещении, и изучение ожидаемого
феномена следует проводить в темноте. Поэтому отпадает
такой способ определения относительного положения двух
глаз, как обычная киносъемка. Также пришлось
отказаться и от способа регистрации положения глазных яблок в
глазных впадинах посредством отведения окулоретиналь-
ных потенциалов, требующего в данном случае
высококачественных усилителей постоянного тока, которыми мы
не располагали. Поэтому прежде всего надежды были
возложены на проявление окулогиральной иллюзии
раздвоения объектов в поле зрения.
Опыты по выявлению этой специфической иллюзии
проводились одновременно с описанными в предыдущем
разделе опытами по выявлению обычных окулогиральных
иллюзий движения светящейся в темноте неподвижной
цели. Восемь испытуемых в течение 5—10 мин
подвергались на вращающемся кресле угловым осцилляциям в
плоскости горизонтальных каналов. Параметры осцилляции:
амплитуда 40°, частота 0,5 Гц. Испытуемому надевался на
лицо темный полупрозрачный экран, через который из
окружающей обстановки нельзя было видеть ничего, кроме
нити накала светящейся лампочки, находящейся от него
на расстоянии в 1 м. Во всех случаях до начала опытов
испытуемые докладывали, что видят одну-единственную
нить накала. Во время указанных осцилляции двоения
цели не наблюдалось. По окончании осцилляции все
испытуемые доложили о появлении окулогиральных
иллюзий движения цели в сагиттальной плоскости. У
четырех из них эта иллюзия постепенно исчезала, и после этого
цель воспринималась, как и до опыта: в виде относительно
неподвижной одиночной нити накала сложной формы.
Однако у четырех из восьми испытуемых на смену
иллюзии движения цели пришла иллюзия нового вида:
устойчивое и выраженное раздвоение цели в плоскости
горизонтальных каналов (рис. 40). При повторном
наблюдении такой иллюзии в новых опытах, по заявлениям
испытуемых, время процесса раздвоения уменьшается. Пои
наклоне головы набок липия раздвоения также
воспринималась наклоненной, оставаясь в плоскости
горизонтальных каналов (рис. 40, в). У трех испытуемых это
раздвоение прошло через несколько минут после своего
возникновения, но у одного оно, временами пропадая, держалось
185
Рис. 40. Раздвоение нити накала лампочки в опыте на окулоги-
ральные иллюзии, возникающие в результате угловых осцилляции
головы в горизонтальной плоскости
А — нормальное восприятие человеком нити накала одной лампочки; Б —
раздвоение нити накала параллельно плоскости горизонтальных каналов;
В — наклон оси раздвоения нити накала при наклоне головы; Р ~
плоскость горизонтальных каналов
Много дней: достаточно было ему надеть на глаза
указанный полупрозрачный экран и посмотреть на лампочку,
как раздвоение нити накала происходило вновь. Величина
этого раздвоения могла быть меньше, чем при первом
наблюдении, но продолжала оставаться значительной. Ни у
одного испытуемого раздвоения объектов в поле зрения не
происходило при нормальных условиях освещенности
окружающей обстановки.
Следует отметить, что при раздвоении цели у трех
испытуемых закрывание одного глаза приводило к
устранению иллюзорной цели той же стороны, но сохраняло
вторую цель на стороне открытого глаза. Исходя из
закона смещения изображения цели на сетчатке глаза, следует
предположить, что у этих испытуемых происходит
конвергенция глаз. Ниже описанные исследования на фотоска-
нирующем пупиллографе подтвердили это. У четвертого
испытуемого наблюдалась дивергенция глаз, что
обусловливало при закрывании любого из глаз устранение
цели противоположной стороны.
Тот факт, что у остальных четырех из восьми
испытуемых не происходило раздвоения в поле зрения, может
быть объяснен взаимной нейтрализацией противоположно
направленных действий внутренних и внешних прямых
мышц глазодвигательного аппарата, а также
недостаточной продолжительностью осцилляции в наших
эксперименте
тах. Не установлены причины, по которым предсказанная,
исходя из особенностей лабиринтных процессов, иллюзия
раздвоения (в случаях ее проявления) оказывается
отставленной на значительный промежуток времени по
окончании осцилляции и следует лишь после иллюзии
вертикального движения цели.
Для измерения величины обнаруживаемого
раздвоения был предложен следующий метод. Испытуемым
предлагалось смотреть на нити накала сразу двух лампочек,
выставленных в плоскости горизонтальных каналов. В
результате осцилляции головы иллюзия раздвоения
возникала для обеих лампочек с одинаковой интенсивностью,
причем иллюзия раздвоения двух нитей накала в четыре имеет
ту же выраженность, что и раздвоение одной нити накала
в две. Очевидно, что в этих условиях можно подобрать
расстояние между двумя лампочками так, чтобы правое
иллюзорное изображение одной нити накала совместилось
с левым иллюзорным изображением другой; данное
расстояние и будет характеризовать величину раздвоения.
Этим методом было получено, что при удалении лампочки
от испытуемых на 1 м раздвоенное изображение нити
накала (ее длина равнялась 0,5 см) расходилось на
расстояние до 5 см. Величина расхождения сложным образом
зависит от удаленности лампочки от наблюдателя, имея
максимум на расстоянии в 0,5—2 м.
Проявление окулогиральной иллюзии раздвоения
служит, конечно, лишь косвенным аргументом в пользу
соображений об изменении угла между осями глаз. То, что этот
угол действительно меняется, удалось доказать с помощью
фотосканирующего пупиллографа, позволяющего
регистрировать на кинопленку положение зрачка при фиксации
взгляда на светящейся в темноте цели. Этот прибор имеет
электронно-оптический преобразователь и дает
возможность производить киносъемку в инфракрасном свете,
т. е. в условиях, когда вестибулярные влияния на
глазодвигательный аппарат не ослаблены ярким освещением.
Для участия в экспериментах с пупиллографиче-
ской регистрацией были приглашены испытуемые, у
которых в описанных выше опытах проявилась окулогиральная
иллюзия раздвоения цели. Киносъемка производилась в
темном помещении, и поэтому испытуемые не нуждались в
темном экране, надеваемом на лицо в прежних
экспериментах. К сожалению, имевшийся в нашем распоряжении
пупиллограф позволял регистрацию положения лишь од-
187
Рис. 41. Пупиллографическая запись конвергенционного смещения
правого глаза, происходящего при наступлении иллюзии двоения
светящейся в темноте цели
Белое пятно на каждом кадре — зайчик, отражающийся от хрусталика
глаза, смещение зайчика вверх соответствует смещению глаза влево
ного глаза. Поэтому приходилось производить поочередную
киносъемку положений сначала правого, а потом левого
глаза при фиксации взгляда на светящейся цели.
Результаты экспериментов показывают, что одновременно с
появлением иллюзии раздвоения цели у трех испытуемых
глаза конвергировали, а у одного — дивергировали. На рис. 41
представлена пупиллографическая запись отклонения
зрачка при конвергенции глаз одного из испытуемых. При
ярком освещении иллюзия раздвоения сразу пропадала и
одновременно глаза возвращались в исходное положение.
Полученные данные свидетельствуют в пользу
утверждения о возможности изменения конвергенции или
дивергенции глаз из-за специфики вестибулярных влияний
в результате угловых осцилляции головы. Однако это
утверждение нуждается в дополнительной проверке,
поскольку у испытуемых не проводилось предварительного
детального обследования зрительной системы на
предмет устойчивости бинокулярного зрения.
Феномен предпочтительного направления вращения
при осциллировании. До сих пор мы рассматривали
функционирование систем полукружных каналов, у которых
правая и левая половины зеркально симметричны.
При продолжительном осциллировании головы в плоско-
188
сти, например, горизонтальных каналов купулы обоих
горизонтальных каналов накапливают одинаковые по
знаку и величине дифференциальные отклонения, а
потому оснований для предпочтения в вестибулярных
реакциях правого направления левому не имеется.
Однако далеко не всегда зеркальная симметрия двух
лабиринтов выполняется хотя бы приблизительно: она
может быть нарушена в результате действия определенных
наследственных факторов, известкования каналов и пр.
При зеркально асимметричном строении двух
лабиринтов условия для накопления дифференциальных
отклонений купул, например, в правом и левом горизонтальных
каналах будут, вообще говоря, неодинаковы. При
неодинаковых по величине дифференциальных отклонениях
купул горизонтальных каналов предпочтение в
формировании дифференциальных вестибулярных реакций уже
может быть отдано показаниям рецепторов того
горизонтального канала, в котором дифференциальное
отклонение купулы больше. Как это следует из материалов
предыдущего раздела, с увеличением частоты осцилляции
головы эта разница в величине дифференциальных
отклонений купул правого и левого горизонтальных каналов
растет, т. е. увеличиваются и предпосылки для
проявления в вестибулярных реакциях право-левой
асимметрии, «ч
В частности, при осцилляциях испытуемого в
отсутствие зрительных ориентиров проявление данной
асимметрии может ожидаться в субъективном восприятии
синусоидального углового качания как
квазисинусоидального качания с преимущественным проворачиванием в
одну из сторон. Причем эта ошибка в восприятии должна
расти с увеличением частоты осцилляции.
Среди кандидатов в испытуемые для опытов по осцил-
лированию у трех людей был обнаружен феномен
предпочтительного направления вращения. Эти испытуемые
не участвовали ни в одном из опытов, описанных в
предыдущих разделах, но на них исследовался указанный
феномен. Перед началом качаний фиксированный во
вращательном кресле испытуемый, наклоняющий голову
вперед на 30°, запоминал положение, в котором он сидел,
относительно окружающей обстановки. После этого ему
на глаза надевалась черная повязка, и кресло
подвергалось синусоидальному угловому качанию с амплитудой
в 40°. Плоскость качаний совпадала с плоскостью гори-
189
зонтальпых полукружных каналов. При осцилляциях на
малых частотах испытуемые верно оценивали свое
движение, докладывая, что они поворачиваются из стороны в
сторону, причем среднее положение совпадает с их
исходным положением перед опытом. Однако начиная с частот
осцилляции около 0,2 Гц, у испытуемых возникала очень
убедительная для них иллюзия постепенного
проворачивания с каждым периодом в одну из сторон. Причем
скорость проворачивания увеличивалась с ростом частоты
осцилляции. Через некоторое число периодов
испытуемые, например, докладывали, что они провернулись на
180° и в среднем смотрят в направлении, прямо
противоположном направлению взгляда при исходном
положении. После прекращения осцилляции испытуемые
докладывали, что они остались в субъективно достигнутом
положении. Эта иллюзия нового положения
относительно окружающей обстановки была настолько убедительна,
что даже ориентирующие звуки, исходящие от хорошо
знакомых по расположению объектов (шум работающих
приборов, стук двери и пр.), не вызывали у испытуемых
переоценки своего положения. Наоборот, все испытуемые
заявляли, что они воспринимают объекты, от которых
исходят звуки, как переместившиеся в пространстве в
соответствии с иллюзией достигнутого положения у
испытуемого. Так, если до качаний перед испытуемым работал
генератор звука, то при достижении в результате
качаний иллюзии изменения своего положения в пространстве
на любое число градусов испытуемый был уверен, что и
генератор переместился в пространстве так, что
оказался снова перед испытуемым.
Снятие повязки с глаз вызывало мгновенную
переоценку испытуемым своего положения и исчезновение
иллюзии измененного положения.
Несколько измененный вариант опыта заключался в
том, что на подлокотнике кресла устанавливался
потенциометр с ручкой в виде стрелы. Во время качаний
испытуемый должен был поворачивать ручку потенциометра
так, чтобы стрела была направлена в среднем в том
направлении, которое испытуемым воспринималось как
исходное. Снимаемое с потенциометра напряжение
подавалось на вход шлейфового осциллографа, что позволяло
регистрировать иллюзию изменения положения из-за
феномена предпочтительного направления вращения при
осциллировании. На рис. 42 представлены результаты
190
град
Рис. 42. Иллюзия поворота
у трех испытуемых (1, 2,
3) при синусоидальных уг- ^60
ловых качаниях
По оси ординат отложены
углы поворота стрелки потен- ™"
циометра; по оси абсцисс —
время сип
О J
/ Z 3 У 5 6 7
мим
обработки оциллографических записей качания трех
испытуемых в течение 5 мин с частотой 0,4 Гц, а затем в
течение 2 мин с частотой 0,6 Гц. Из рисунка видно, как с
увеличением частоты происходит ускорение иллюзорного
проворота испытуемых. У испытуемых 1 и 2 наблюдалась
иллюзия проворачивания против часовой стрелки, у
испытуемого 3 — по часовой стрелке.
Дополнительное исследование постосцилляторных
вестибулярных реакций: дифференциального нистагма и
окулогиральной иллюзии, описанных в предыдущих
разделах, показало, что и в этих реакциях у данных
испытуемых наблюдается элемент лево-правой асимметрии.
Дифференциальный нистагм, возникающий в результате
осцилляции в плоскостях, перпендикулярных
сагиттальной, не был чисто вертикальным, а кажущееся движение
светящейся в темноте цели, наблюдаемое после этих
осцилляции, происходило обычно под углом к сагиттальной
плоскости.
Конечно, нельзя однозначно связывать асимметрию
вестибулярных реакций при осциллировании с
накоплением неодинаковых дифференциальных отклонений ку-
пул двух лабиринтов. Однако эта разница в
дифференциальных отклонениях может давать свой вклад в
указанную асимметрию.
Заключение по вестибулярным исследованиям. Были
установлены соответствия между результатами
экспериментов на масштабной физической модели системы полу-
кружных каналов человека, построенной с учетом
требований теории динамического подобия, и результатами фи-
т
апологических экспериментов с участием испытуемых.
Установление этих соответствий означает углубление
наших знаний о функционировании вестибулярной
системы, под воздействием которой находится вся статокинс-
тическая регуляция организма. Каковы же основные
приложения этой работы применительно к человеку как
целостному организму?
Выявление дифференциальных отклонений купул при
угловых осцилляциях головы позволило сделать ряд
предсказаний о новом классе вестибулярных реакций —
дифференциальных вестибулоокуломоторных реакциях.
То, что предсказанные реакции были обнаружены в
направленных экспериментах с участием испытуемых,
подтверждает исходный тезис: накапливающиеся
дифференциальные отклонения купул при угловых осцилляциях
головы организм интерпретирует как результат действия,
помимо осцилляторных, еще и постоянных угловых
ускорений, которых в действительности не существует.
Это ведет к появлению сенЬомоторных реакций, не
соответствующих реальной обстановке. Например,
вестибулярный нистагм согласно бытующим представлениям о
его функциональном предназначении служит для
фиксации объектов зрения на сетчатке глаза при вращательных
движениях головы относительно окружающей
обстановки. Поэтому в норме нистагм глаз и происходит в
плоскости вращения головы. Но накопление
дифференциальных отклонений купул при осцилляциях головы вызывает
дополнительный дифференциальный нистагм,
направление которого не согласуется с действительным
движением головы и который не позволяет стабилизировать
изображение на сетчатке.
Отметим, что в силу асимметричной формы отолито-
вых мембран и эндолимфатических полостей у отолитов
также следует предположить накопление постоянных
составляющих отклонения от положения покоя при
осцилляциях головы. Такое отклонение может обусловливать
многие дополнительные особенности отолитовых реакций
при этом типе движений.
Полученные результаты, по нашему мнению, имеют
непосредственное отношение к вопросу об укачивании.
В литературе имеется много данных, свидетельствующих
о тесной связи функционирования вестибулярного
аппарата с развитием укачивания. Например, закупорка
шести полукружных каналов полностью устраняет развитие
192
болезни движения у собак при качании на двухполюсных
качелях [401]. Многие непривычные условия
функционирования вестибулярного аппарата (длительная
невесомость, действие кориолисовых сил и пр.) обусловливают
развитие болезни движения. В этих условиях хорошо
знакомые организму движения головы вызывают
необычные показания вестибулярного аппарата, не
согласующиеся с опытом движений организма и показаниями
других анализаторов, что согласно современным
представлениям и ведет к болезни движения (обзоры [52, 119, 443,
450]).
Но и осцилляторные как угловые, так и линейные
движения головы в обычных земных условиях вызывают
укачивание, т. е. также ведут к рассогласованию
показаний вестибулярного аппарата с имеющимся у организма
опытом движений и информацией со стороны проприоцеп-
ции, зрения н пр. В настоящей работе как раз выявлен
один из возможных механизмов рассогласования
анализаторов при осцилляциях головы — накопление
дифференциальных отклонений чувствительных элементов
вестибулярного аппарата, ведущее к дезинформации организма
о характере действующих ускорений, а значит,
являющееся возможной причиной укачивания при таком типе
движений. Выявление данного механизма ставит укачивание
при осцилляциях головы в один ряд с болезнью
движения, развивающейся в невесомости и при действии
кориолисовых сил, в том отношении, что во всех случаях
источником искажения вестибулярной информации является
сам вестибулярный лабиринт. Это, в частности,
оправдывает использование осцилляции головы как теста на
укачивание при разных типах непривычной стимуляции
вестибулярного аппарата.
Вопрос об относительной ценности качаний человека
как теста для профотбора вестибулоустойчивых людей
изучался в работе [351]. Используя в качестве
испытуемых летчиков и курсантов летных училищ, автор пришел
к следующим выводам: тест с качаниями имеет
определенные преимущества перед другими тестами на болезнь
движения, так как позволяет работать со строго
стандартизированным стимулом и производить вестибулярный
профотбор с высокой надежностью; укачиваемость при
осцилляторном тестировании хорошо коррелирует с ука-
чиваемостыо в полетах; вместе с тем этот тест весьма
жесткий, поскольку зачастую вызывает болезнь во
7 С. В. Петухов
193
время полетов. Существенным недостатком этого теста
является его длительность, так как при нем даже для
людей, склонных к болезни движения, обычно требуется
длительное время (20—30 мин), чтобы получить ярко
выраженное укачивание, регистрируемое по побледнению
кожных покровов, появлению холодного пота,
тошноте и пр.
Выявление в настоящей работе дифференциальных
вестибулярных реакций открывает пути к разработке
методов вестибулярного профотбора и тренировок на основе
синусоидальных качаний. Поскольку развитие болезни
движения при осцилляциях головы сопровождается
допускающими четкую регистрацию вестибулярными
эффектами, то есть надежда, что по выраженности этих
эффектов удастся определять степень устойчивости человека к
болезни движения более надежно и за более короткое
время. Такими новыми тестами на вестнбулоустойчивость
могут быть: тест на дифференциальный нистагм (по
нашим предварительным данным, чем стабильнее и
продолжительнее дифференциальный нистагм, наблюдающийся
после угловых осцилляции головы, тем выше
вестнбулоустойчивость испытуемого); тест на раздвоение объектов
в поле зрения в результате осцилляции головы; тест на
способность испытуемого сохранять равновесие при
стоянии на синусоидально движущейся площадке (для
оценки этой способности хорошо приспособлена методика ста-
билографии) [82]. Разработка перечисленных тестов тем
более ценна, что они дают количественные показатели,
допускающие объективное сравнение вестибулярных
реакций у разных людей.
Другой актуальной проблемой, привлекающей
внимание к вопросам функционирования вестибулярного
аппарата, является проблема ориентации в пространстве.
Изучение особенностей пространственной дезориентации и
поиски методов борьбы с нею в лабораторных условиях
требуют прежде всего разработки моделей
пространственных иллюзий. Эти модельные иллюзии должны по
возможности обладать достаточной продолжительностью и
выраженностью для удобства работы с ними. Выявленные
в работе такие особенности вестибулярных иллюзий,
возникающих в результате осцилляции, как простота их
создания, продолжительность, выраженность и
убедительность иллюзий для испытуемых, позволяют рекомендовать
эти иллюзии (постосцилляторное кажущееся движение
194
цели, раздвоение объектов в поле зрения и пр.) в
качестве моделей для изучения вопросов пространственной
дезориентации.
Развитие методик синусоидальных качаний полезно
также для улучшения отбора людей с повышенной
устойчивостью к пространственной дезориентации. Если у
человека синусоидальные качания, зеркально
симметричные относительно сагиттальной плоскости, вызывают
вестибулярные реакции, имеющие элементы лево-правой
асимметрии, то это свидетельствует о функциональной
асимметрии вестибулярного анализатора. В силу этой
асимметрии у человека имеются предпосылки
воспринимать изотропное физическое пространство как
анизотропное. Естественно, что допуск таких людей к работе,
связанной с ориентацией в пространстве, должен быть
ограничен.
Отметим, что методика синусоидальных угловых
качаний для выявления сильно выраженных (обычно
патологического характера) вестибулярных асимметрий
применялась ранее рядом авторов [378, 453 и др.]. Они
исследовали феномен предпочтительного направления
нистагма при осцилляциях испытуемых на вращательном
кресле в горизонтальной плоскости. В качестве меры
асимметрии служила разница в числе ударов нистагма вправо и
влево в плоскости качаний за некоторое число периодов.
Однако данный метод применим только на малых частотах
качаний (до 0,2 Гц), поскольку на больших частотах в
плоскости качаний регистрируется лишь обычное проти-
вовращательное движение глаз, а нистагм отсутствует.
Область же изучения вестибулярных реакций при
частотах осциллирования, больших 0,2 Гц, остается до сих
пор практически не изученной в плане выявления
функциональных асимметрий вестибулярного анализатора.
Описанные в настоящей работе дифференциальные
вестибулярные реакции, возникающие в результате
осциллирования головы и увеличивающие свою интенсивность с
ростом частоты осциллирования, являются основой для
разработки новых, возможно более чувствительных тестов
по выявлению вестибулярных асимметрий. Например,
если в результате синусоидальных угловых качаний в
плоскостях, перпендикулярных к сагиттальной, проявится
окулогиральная иллюзия движения светящейся в темноте
цели не в сагиттальной плоскости, а под углом к пей,
или будет зарегистрирован не чисто сагиттальный диф-
195
ференциальный нистагм, то это может служить
свидетельством функциональной лево-правой асимметрии.
Перечислим результаты и выводы, полученные нами
при исследовании вестибулярного аппарата посредством
использования инвариантно-групповых подходов к
моделированию физических процессов.
1. Па основе теории динамического подобия выведены
критерии подобия процессов в вестибулярном лабиринте.
С учетом этих критериев разработана масштабная
физическая модель системы полукружных каналов человека,
воспроизводящая динамику натуральных процессов в
удобных для их изучения условиях.
2. Исследования модели на стенде ВУ-4 обнаружили
наличие гидромеханического взаимодействия между
полукружными каналами и выявили асимметрию условий для
движения эндолимфы по каждому каналу в
противоположных направлениях, ведущую при угловых осцилляциях
головы к постепенному ампулопетальному смещению ку-
пул горизонтальных и передних вертикальных каналов и
ампулофугальному смещению у задних вертикальных
каналов.
3. Предложена нелинейная модификация уравнений
Штейнгаузена, которая позволяет лучше описать
поведение купул при угловых осцилляциях головы.
4. Накапливающиеся при угловых осцилляциях головы
постоянные составляющие отклонения купул организм
интерпретирует как результат действия постоянных угловых
ускорений, в действительности не существующих. Это
ведет к сенсомоторным реакциям, не соответствующим
реальной обстановке. На данной основе возможно
предсказание выявленных в экспериментах на испытуемых
следующих вестибулярных реакций:
а) дифференциального вертикального нистагма как
результата угловых осцилляции головы в сагиттально-
симметричных плоскостях, например фронтальной и
горизонтальной;
б) дифференциальной окулогиральной иллюзии
движения в сагиттальной плоскости светящейся в темноте цели
как результата угловых осцилляции головы в
сагиттально-симметричных плоскостях;
в) дифференциального сагиттально-асимметричного
нистагма как результата угловых осцилляции головы в
плоскостях, зеркально асимметричных относительно
сагиттальной плоскости;
196
г) конвергенции или дивергенции глаз и
сопутствующей иллюзии раздвоения объектов в поле зрения
параллельно плоскости горизонтальных каналов как результата
угловых осцилляции головы.
5. При асимметричном строении левого и правого
вестибулярных лабиринтов различие в накоплении
дифференциальных отклонений купул двух лабиринтов может
являться одной из причин асимметрии вестибулярных
реакций, вызываемых угловыми осцилляциями головы.
6. Представления об асимметрии динамики процессов
в вестибулярном аппарате позволяют указать на
накопление дифференциальных отклонений чувствительных
элементов вестибулярного аппарата как на дополнительную
причину укачивания у людей и животных. Данные
представления могут оказаться полезными при моделировании
нарушений восприятия пространства и для разработки
методик вестибулярных тренировок и профотбора.
Таким образом, результаты работы уточняют наши
знания о динамике процессов в вестибулярном аппарате и о
функционировании вестибулярной системы,
устанавливают существование нового класса дифференциальных
вестибулярных реакций, проясняют причины развития
некоторых форм укачивания, обосновывают новые
возможности для моделирования нарушений восприятия
пространства и движения, а также открывают новые пути
для разработки методик вестибулярного профотбора.
Эти результаты имеют важное значение в решении
многочисленных проблем, возникающих при работе
человека-оператора как эвена управления в системах человек-
машина в условиях воздействия механических колебаний.
Приложение I
ДОПОЛНИТЕЛЬНЫЕ СВЕДЕНИЯ
О ЧИСЛАХ ФИБОНАЧЧИ
1. Числа Фибоначчи и золотое сечение в эстетике пропорций.
С древних времен вопросы эстетики пропорций, как уже
отмечалось, связываются архитекторами, скульпторами, мыслителями
различных направлений с пропорцией золотого сечения,
названной так Леонардо да Винчи. Этому посвящено много работ, что
частично отражено в прилагаемой библиографии; отдельные
материалы из данной литературы используются нами в настоящем
обзорном разделе.
Геометрическая интерпретация пропорции золотого сечения
представляет собой деление одного отрезка на два таким образом,
что меньший из них относится к большему так же, как
больший — к целому отрезку. Эта пропорция была известна задолго
до Леонардо да Винчи. Некоторые археологи, основываясь на
находках в районе палеолитических стоянок человека, относят
начало практического использования человеком золотого сечения к
древнейшему периоду, отстоящему от нашего времени на 20—
25 тыс. лет. Данная пропорции восхищала своими
геометрическими особенностями древних геометров, изучавших правильные
многоугольники и многогранники и обнаруживших ее
присутствие в ряде из них. В частности, эта пропорция реализуется в
разных формах в пентаграмме — пятиконечной звезде, которая у
древних греков почиталась символом здоровья.
При содействии Леонардо да Винчи Лука Пачиоли в 1509 г.
написал известную книгу «О божественной пропорции», где в
восторженных выражениях описал геометрические свойства
золотого сечения в связи с правильными геометрическими фигурами.
Геометрические идеи и достижения служили опорой многим
художникам в их попытках осмыслить основы «прекрасных форм»:
Браманте, Рафаэль, Микеланджело, Виньола и многие другие
считали, что совершенное знание геометрии и непрестанное
размышление о законах «науки пространства)» необходимы каждому, кто
призван создавать или закреплять формы при помощи кисти,
резца или шнура. Общепринятым обозначением для золотого
сечения взято Ф — первая буква имени Фидия, греческого
скульптора, применявшего золотое сечение.
198
Пропорции золотого сечения применялись Микеланджело,
Палладио и другими для модуляции фасадов здание
Древнерусскими зодчими, как показывают исследования А. Пилецкого [183],
также широко применялись золотое сечение и числа..Фибоначчи.
Пытаясь решить загадку о том, как древнерусские архитекторы
могли возводить сложнейшие здания без чертежей, Пилецкии
исследовал большое число древнерусских сооружений и обнаружил,
что, несмотря на чрезвычайное разнообразие и неповторимость
форм, в них много одинаковых размеров. Совпадающие размеры
встречаются в самых различных зданиях и объектах вне
зависимости от их назначения и времени постройки. По данным
Пилецкого, употреблявшиеся размеры в целом слагали единую стройную
систему величин, воспроизводящую многие свойства ряда
Фибоначчи и включающую его в качестве своего частного случая.
Эта система теснейшим образом связана с абсолютными велич*-
нами различных типов древнерусских саженей: казенных,
народных, малых, греческих, церковных, простых, великих и царских,
которые, таким образом, оказываются неявно родственными
числам Фибоначчи.
Современная архитектура, в которой оформилось
специальное научное направление — архитектурная бионика [128, 129],
проявляет совершенно особый интерес к симметрии в живой
природе и фибоначчиевым отношениям в пропорциях тел.
Активными проводниками идеи золотого, сечения в современной
архитектуре были видные советские архитекторы А. К. Буров, И. В.
Жолтовский [36, 146—148]. Они называли: золотое сечеиие пропорцией
органического роста д утверждали значение принципа
органического роста в качестве- первого и основного признака нодлявно
живого воплощения художественного образа в архитектуре.
В настоящее время в исследовательских центрах архитектурной
бионики проводятся углубленные исследования закономерностей
золотого сечения в живой, природе в связи с задачами эстетики
пропорций.
Знаменитые итальянские мастера музыкальных инструментов
Маджйни, Амати, Гварнери и Страдивари сознательно создавали
пропорция своих скрипок цо закону золотого сечения [70, с. 212].
Многие авторы связывала эстетику музыкальных произведений
с принципом золотого сечения. Так, Л. Сабанеев утверждал, что
золотое сечение эстетически, значимо не только в
пространственных формах и искусствах, но к во временных, и что временное
протяжение музыкального нроивведения делится вехами,
важными для воспринимающего, но принципу золотого сечеиия [208].
Р. X. Зарипов также отмечает, что кульминация музыкальной
мелодии часто приходится на точку золотого сечения [96, с. 28].
199
Советский композитор М. А. Марутаев не только проанализ!»
ровал реализацию пропорции золотого сечения в композиции
большого числа музыкальных произведений, но и предпринял попытку
связать величину золотого сечения с фундаментальными
физическими константами за счет введения определенных правил
соответствия между числами [190, с. 363—394; 221]. А. А. Соколов
и Я. А. Соколов полагают, что по принципам золотого сечения
организованы электрические колебания мозга [219]. По их
мнению, эта особенность мозговой деятельности является причиной
эстетического предпочтения пропорций золотого сечения.
Интересно применение К. С. Симоняном эстетического принципа
золотого сечения в качестве принципа диагностических критериев
течения болезней типа перитонита [215]. Уже не раз различными
авторами ставился вопрос о введении принципов золотого
сечения в промышленные стандарты предпочтительных чисел [3, 185,
189].
То, что пропорции золотого сечения многими
исследователями отмечаются в композиции произведений и архитектуры, и
музыки, и поэзии перекликается с широко распространенным
мнением об универсальном характере законов гармонии. Здесь
уместно привести слова композитора Р. Шумана: «Эстетика одного
искусства есть эстетика и другого; только материал различен»
[3, с. 13].
Основоположник психофизики Г. Фехнер произвел опыты по
эстетическому предпочтению человеком пропорций золотого
сечения [325]. Он показывал ряду испытуемых лиц 10
прямоугольников с определенным отношением сторон и заставлял их выбирать
такой прямоугольник, который больше всего их удовлетворял.
Результаты опыта, воспроизведенные на рис. 43, оказались чрез-
вычайно благоприятны для золотого сечения. Оно действительно
является общей эстетической нормой для восприятия. Фехнер
поясняет, что только в очень немногих случаях во время опыта
было отказано в суждении, но также только в очень немногих
случаях, хотя таковые все же имелись, суждение было
определенное и решительное. Большей частью имело место длительное
колебание, и иногда, остановившись уже иа каком-нибудь
прямоугольнике, при том же самом опыте испытуемые, поправляй себя,
отдавали предпочтение другому или же продолжали до конца
колебаться между двуми, тремя или даже четырьми
прямоугольниками. Когда опыт производился с теми же лицами в другое
время, после того как впечатление прежнего уже изгладилось,
нередко вместо прямоугольника, выбранного при прежнем опыте,
отдавали предпочтение другому, соседнему с ним. Основные
результаты этих опытов Фехнера, впоследствии не раз повторенных
200
Jff
Zff
/0 V
/J
A
«^
■"CtS,
:-'&-
и
U*<
\
К
V
:ч
/••/ ff:ff 4-f J:t Z0Z9 ZJ Zf-Л /J:ZJ /-Z Z:f
Рис. 43. Результаты психологических опытов Г. Фехнера по
эстетическому предпочтению золотого сечения (из Г345])
По оси ординат — процент случаев эстетического предпочтения,
пунктирная кривая — женщины, штрихпунктирная — мужчины
другими исследователями, так или иначе связываются многими
авторами с тем, что отношения размеров огромного множества
окружающих нас предметов обнаруживают отношения золотого
сечения. Форматам книг, картинам, входным билетам,
бумажникам, сундукам, ящикам, шкатулкам и всевозможным другим
предметам, частью сознательно, частью бессознательно придается
форма с пропорциями золотого сечения.
В качестве одного из примеров многочисленных
искусствоведческих работ на тему золотого сечения подробно рассмотрим
публикацию кинорежиссера С. Эйзенштейна [270], которая
посвящена ключевой роли принципа золотого сечения в
композиционном строе его знаменитого фильма «Броненосец «Потемкин».
Эйзенштейн проанализировал средства, которыми достигнута
исключительная «композиционная органичность», отличающая, по
общему мнению, этот фильм. По его утверждению, органичность
произведения, как и ощущение органичности, получаемое от
произведения, должна возникнуть в том случае, если закон
построения этого произведения будет отвечать законам строя
органических ивлений природы. При выполнении этого условия
произведение имеет совершенно особое воздействие на воспринимающего
не только потому, что оно возвышается до одного уровня с
явлениями природы, но и потому, что закон строения его есть
одновременно и закон, управляющий теми, кто воспринимает
произведение, поскольку и они являются частью органической
природы. Согласно Эйзенштейну, воспринимающий чувствует
себя органически связанным, слитым, соединенным с произпеде-
201
ннем такого типа. «В большей или меньшей степени это
ощущение неизбежно в каждом из нас, и секрет состоит в том, что
п этом случае и нами и произведением управляет одна и та же
закономерность» [270, с. 10].
В данной связи Эйзенштейн особое внимание обращал на
соответствие композиционных пропорций своего фильма
фундаментальным структурным закономерностям живой природы. Эти
закономерности он определял на основном феномене, отличающем
жпвую органическую природу от иных явлений. «Феномен этот —
рост, и вокруг этой формулы роста как основного признака
органического явления мы и сосредоточим наши попеки»,— писал
Эйзенштейн. Определяя формулу роста как первичного типичного
признака органической природы, он называл такой формулой
пропорцию золотого сечения [270, с. 12].
Согласно Эйзенштейну^ строй вещей, скомпанованных по
пропорциям золотого сечения, обладает в искусстве совершенно
особой силой воздействия, ибо создает ощущение предельной
органичности. Режиссер подчеркивает, что органичность строя
«Броненосца «Потемкин» достигнута именно благодаря тому, что эта
картина построена целиком по закону золотого сечения. «В
„Потемкине",— писал Эйзенштейн,— не только каждая отдельная часть
его, но весь фильм в целом... самым строгим образом следует
закону золотого сечения — закону строя органических явлений
природы. В этом секрет органичности его композиция» [270, с. 13].
«Совершенно очевидно, что ничего «мистического» в основе его
(золотого сечения.— С. П.) особой и исключительной воздейст-
венности нет. Мы достаточно обстоятельно старались показать,
почему этот эффект столь сугубо органичен и почему именно эта
закономерность дает наибольший отклик внутри нас самих»
[270, с. 12].
Развернутый- анализ своего фильма Эйзенштейн дополнил
примерами широкого использования принципа золотого сечеиии
в живописи и скульптуре, музыке и поэзия. Он отмечал, что
вообще в области пластических искусств золотое сечение и
композиционное использование его более чем популярны. Отдельно»
приложение к своей статье Эйзенштейн посвятил принципу
золотого сечения в поэзии Пушкина. Указывая па использование
данного принципа в произведениях крупнейших представителей
реалистического направления в искусстве, Эйзенштейн
подчеркнул, что это «может служить укором тем предрассудкам, согласно
которым для реализма будто бы достаточно одной бытовой
правды, строгость же* композиционного письма отнюдь не важна и
даже чуть ли не вредна Г» [270, <*.. 18].
202
2. Некоторые математические свойства чисел Фибоначчи.
В сиоей книге, вышедшей в 1228 г., знаменитый итальянский
математик Леонардо из Пизы, более известный по прозвищу
Фибоначчи, рассмотрел ряд чисел (1). Задача, в которой впервые
возник этот ряд чисел, была по содержанию биологической. В ней
шла речь о размножении пары кроликов при условии, что через
месяц пара кроликов производит па свет другую пару, а рождают
кролики со второго месяца со дпя своего рождения. Так как
первая пара в первом же месяце дает потомство, то в этом месяце
окажутся 2 пары; из них первая пара рождает и в следующем
месяце, так что во втором месяце оказывается 3 пары; из них
в следующем месяце 2 пары будут давать потомство, так что в
третьем месяце родятся еще 2 пары кроликов и число пар
кроликов в этом месяце достигнет 5. Нетрудно видеть, что в
последующие месяцы число пар кроликов соответственно будет
равняться 8, 13, 21, 34, 55 и т. д.
Возникшая в этой задаче рекуррентная: последовательность
чисел, в которой каждый член равен сумме двух предыдущих,
обладает, как выяснилось впоследствии, многими интересными
математическими свойствами. Очень вероятно, что открытие со
временем природы проявления чисел Фибоначчи в биологических
явлениях наполнит некоторые из этих абстрактных
математических свойств биологическим и физическим содержанием. Нам
хотелось бы проиллюстрировать эти свойства рядом красивых, на
наш взгляд, примеров, взятых из работ [39, 55, 70, 116, 284, 358,
359, 407, 427], не ставя данные примеры в прямую связь с
реализацией чисел Фибоначчи в биологических явлениях.
Через несколько столетий после выхода в свет книги
Фибоначчи И. Кеплер отметил связь (2) последовательности чисел
Фибоначчи с золотым сече-нием Ф. Укажем и другие известные
формы взаимосвязи чисел Фибоначчи с золотым сечением:
фо=о+1 = 1+0,
ф!=0+Ф=1+1/Ф,
ф2=1+ф=2+1/ф>
фз=1+2Ф=3+2/Ф,
Ф4=2+ЗФ=5+3/Ф, (37)
ф5=3+5Ф=8+5/Ф,
фв=5+8Ф=13+8/Ф,
Ф~=8+13Ф=21+13/Ф,
В равенствах (37) имеются четыре параллельно развернутых
вертикальных ряда чисел Фибоначчи. Соответственно
ф-1=Ф—1=~1+Ф,
ф-2=1_ф-1=2-Ф,
203
ф-з=ф-1-ф-*=-3+2Ф,
ф-4 = ф-2_ф-Э = 5-ЗФ,
ф-5 = ф-3_ф-4 = _8+5Ф,
ф-в=ф-1_ф-5=13_8ф,
ф-7= ф-5_ф-6=_21 + 13Ф,
(38)
ф1_ф-1=1,
ф2 + ф-2=3>
фЭ_ф-Э=4,
ф4+ф-4=7,
ф5_ф-5 = Ц?
ф6+ф-6 = 18,
ф7_ф-7=29,
(39)
В равенствах (39) получаем справа ряд Люка, у которого,
как и у ряда Фибоначчи, каждый член ряда равен сумме двух
предыдущих членов и который также наблюдается в филлотакси-
сных явлениях биологического морфогенеза (см. разд. 1.1).
ф*_ф-2=1У5,
фэ+ф-»=2У5,
ф4_ф-4 = 3У5,
ф5+ф-5=5У5,
фв_ф-в=8У5,
ф7+ф-7=1зУ5,
(40)
В равенствах (40) справа образовывается ряд чисел
Фибоначчи, умноженных на У 5.
Обычно п-и член ряда Фибоначчи вычисляется по формуле
Бине
Fn = [(1 + VW - (1 - /5)4/2" /5. (41)
Приведем формулу, по которой, зная величину п-то члена Fn
ряда Фибоначчи и величину золотого сечения Ф, можно сразу
вычислить величину (гс+1)-го члена ряда Фибоначчи
Детерминанты, образованные из последовательных чисел
Фибоначчи, равны 0:
3 5 8
13 21 34
55 89 144
= 0,
1
8
55
377
2
13
89
G10
3
21
144
987
5
34
233
1597
= 0.
(43)
204
Множеством оригинальных математических свойств обладает
золотое сечение Ф:
ф = |/ 1 + 1Л +j/i+ Vi +
(44)
Ф = 1+ J (/(5)
1+ «
1 +
1 +
1 +
Ф = 1/Ф-т-1/Ф?+1/Ф' + +1/Фп+..., (*G)
фп= 1-фп =
= 1.фп-1-|-1.фГ1-2=,
= 1 .фп"2 -j- 2. ф71"3 -j- 1 • Фп~4 =
= 1. фп-з _(_ з- фп"4 -+- З.фп_5 -f 1.фп~9 =
= 1. фп-4 + 4 • Фп_5 + 6-Фп_6 + 4. Фп"7 + 1. Фп~8 =
= 1. фп~5 _|_ 5. Фп"6 + 10« Фп~7 +J0- Фп_а + 5. фп-9 +
+ i. фп"10 = 1. фп-« + 6 • Ф4-7 -f 15. фп"8-20. Фп_9 +
-|- 15-фп~10 -f- 6- Фп~и + 1 -Фп~12 =
(47)
В равенствах (47) коэффициенты образуют так называемый
треугольник Паскаля (в свою очередь имеющий ряд интересных
математических приложений)
1—
1—1 —
1—2—1—
1—3—3—1—
1—4—6—4—1—
1—5—10—10—5—1—
1—6—15—20—15—6—1—
Многие приложения чисел Фибоначчи и золотого сечения
связаны с геометрией, теорией поиска, решением экстремальных
задач и др. Математические свойства и возможности практических
приложений золотого вурфа Р, введенного в разделе П.З в связи
с биологическими феноменами и неевклидовыми инвариантами,
еще подлежат исследованию.
205
Приложение II
ДОПОЛНИТЕЛЬНЫЕ СВЕДЕНИЯ
О КОНФОРМНОЙ ГЕОМЕТРИИ
Некоторые данные о конформной геометрии уже приводились в
разделе II.1. Дополним их известпыми математическими,
физическими и историческими сведениями.
По определению, конформная геометрия — это геометрия
евклидового пространства, дополненного одной бесконечно
удаленной (несобственной) точкой, с фундаментальной группой
точечных преобразований, переводящих сферы в сферы (группа
конформных преобразований). При размерности пространства nS*3
преобразования, переводящие сферы в сферы, исчерпывают все
преобразования, сохраняющие углы. При гс=2 группа
преобразований, сохраняющих углы, шире, однако и здесь название
конформной геометрии сохраняется за геометрией с фундаментальной
группой точечных преобразований, переводящих круги в круги.
Конформная геометрия, как ни одна другая геометрия,
богата различными вариантами своих названий. Бе называют еще
круговой, аналагматической, геометрией обратных радиусов,
геометрией Мебиуса (по имени геометра, изучившего многие
свойства конформных преобразований), геометрией Лиувилля (впервые
рассмотревшего конформные преобразования трехмерного
пространства и обнаружившего существенное различие между
плоскими и пространственными конформными преобразованиями),
геометрией Кельвина (одним из первых применивших
конформные преобразования в физике). Формально права самостоятельной
геометрии конформная геометрия обрела в 1872 г., когда Клейн
в своей Эрлангенской программе ввел понятия «конформного
пространства», конформной группы, которую он называл «группой
обратных радиусов», разработал проективную интерпретацию
конформной геометрии, развил представление о конформной
геометрии как особой геометрии, которая имела бы основной группой
совокупность всех преобразований, получаемых соединением
группы преобразований евклидовой геометрии с преобразованиями
посредством обратных радиусов.
Любое конформное преобразование можно получить с
помощью конечного числа инверсий (см. раздел II.1). При инверсии
относительно сферы (окружности) с радиусом R число R2
называется коэффициентом инверсии. Евклидовы преобразования вра-
206
щення и параллельного сдвига, представляющие собой частные
случаи конформных преобразований, могут интерпретироваться
с помощью инверсий с бесконечным коэффициентом инверсии.
Всякое конформное преобразование сводится либо к
преобразованию подобия, либо к последовательно выполненным одному
преобразованию инверсии и одному преобразованию подобия (теи
рема Лиувилля). Целая совокупность метрических геометрий, в
том числе Евклида н Лобачевского, могут быть интерпретированы
как частные случаи конформной геометрии на основе принципов
конформной метризации, развитых Пуанкаре [201, с. 416—428].
Математические свойства конформной геометрии подробно
излагаются в работах [37, 102, 201, 202 и др.].
Конформные преобразования имеют богатые приложения в
физике. Еще в середине прошлого века В. Томпсон (лорд Кельвин)
разработал метод решения электростатического уравнения Лапласа
с помощью преобразования инверсии. Этот метод известен как
метод «электрических» или «зеркальных изображений» и широко
используется в математической физике. В 90-х годах прошлого века
было получено обобщение зтого метода для решения п-мерного
уравнения Лапласа.
В XX в. конформные симметрии заняли видное место в
области фундаментальных физических проблем. Этому посвящен ряд
специальных публикаций [47, 309-311, 330, 371, 372 и др.]. Краткий
обзор проблемы конформной симметрии в физике проведем,
используя выдержки из работы [47].
Хорошо известно, к каким последствиям привело открытие
того факта, что волновое уравнение электродинамики (и
уравнения Максвелла) инвариантно относительно некоторой группы
преобразований пространственно-временных переменных, которая
впоследствии стала называться неоднородной группой Лоренца или
группой Пуанкаре. Закономерно было возникновение вопроса: не
существует ли еще каких-нибудь групп инвариантности основных
дифференциальных уравнений физики и, если они существуют,
нельзя ли возвести их в ранг фундаментальных групп симметрии
и связать с некоторыми новыми, более широкими принципами
относительности, чем эйнштейновские? На этом пути английские
теоретики Г. Бэйтман и Э. Кашшгхэм открыли в 1909 г.
конформную инвариантность уравнений Максвелла и предприняли
попытку связать найденную ими симметрию с расширением
специального принципа относительности. Однако ни в этой попытке
построения конформной теории относительности, ни в ряде
последующих попыток других авторов конформная теория
относительности построена не была. Основная причина этого заключалась в
том, что такая теория, обобщающая теорию относительности Эйи-
207
штейна, должна была по идее охватить всю физику, тогда как
конформная симметрия справедлива лишь в области
электродинамики и не справедлива в области классической механики. Другой
причиной явилось то, что конформную группу не удалось
обосновать (подобно группе Пуанкаре) путем анализа измерительных
процедур с помощью масштабов и часов и тем самым возвести в
ранг универсальной физической симметрии.
Были и другие попытки наполнить загадочную конформную
симметрию определенным физическим смыслом. При этом, в
частности, привлекался вывод на основе теорем Нётер законов
сохранения, которые следуют из инвариантности
электродинамического действия, приводящего к уравнениям Максвелла,
относительно 15-параметрической конформной группы. Здесь должно
быть пять дополнительных законов сохранения, помимо десяти
известных, связанных с группой Пуанкаре. Эта задача была
решена Бессель-Хагеном в 1921 г. Вывод конформных законов
сохранения имел принципиальное значение, поскольку он связал
загадочную симметрию с физически интерпретируемыми величинами
и тем самым указал путь, на котором могла быть вскрыта
физическая сущность конформной симметрии. Впоследствии любая
попытка использования конформной симметрии в физике, отмечает
В. П. Визгин [47, с. 210], оказывалась связанной с введением
конформных законов сохранения.
Заметный вклад в разработку проблемы конформной
симметрии в физике внесли работы Г. Вейля [47] по единой теории
гравитации и электромагнетизма и К. Каратеодори [47] по
обоснованию специальной теории относительности на основе лишь
процедур со световыми сигналами без измерения длин и без
предварительного введения понятия скорости.
Рассмотренные работы относятся к первой четверти нашего
столетия. Характерной особенностью этого периода было то, что
чуть ли не каждый существенный прогресс в теоретической
физике давал повод к возобновлению попыток введения конформной
симметрии в физику как фундаментальной группы, подобной
группе Пуанкаре. Эта особенность характеризует и последующее
развитие конформной симметрии вплоть до настоящего времени.
Согласно обзору В. П. Визгина [47, с. 217—218] в 30-40-х годах
конформная группа привлекает внимание теоретиков в связи с
развитием квантовой теории поля, в частности, устанавливается
конформная инвариантность уравнения Дирака для частиц
нулевой массы покоя (Дирак, Баба, Паули). Разработка классификации
илементарных частиц по неприводимым представлениям группы
Пуанкаре и невозможность в рамках этой группы описать
дискретный спектр масс возродили в конце 40-х — начале 50-х годов
208
надежду на то, что это удастся на основе неприводимых
представлений конформной группы (Бопп, Мураи, Ингрэхем и др.).
Разработка единой нелинейной спинорной теории поля Гейзенберга в
середине и конце 50-х годов открыла еще одну возможность для
реабилитации конформной симметрии, поскольку некоторые
варианты основного уравнения единого поля оказались конформно-
инвариантными (Гейзенберг, Дюрр, Гюрши и др.). Примерно к
этому же времени относятся попытки построения квантовой
теории поля на основе более широких групп, чем группа Пуанкаре,
с целью обойти некоторые трудности этой теории и устранить ее
разрыв с теорией гравитации. В частности, разрабатываются
варианты конформной квантовой теории поля (Ингрэхем, Весе, Ка-
струп и др.). В начале и середине 60-х годов появляется еще
несколько направлений, в которых конформная симметрия
выдвигается на первый план: 1) применение конформных констант
движения для анализа взаимодействия частиц, поскольку в физике
сверхвысоких энергий можно пренебречь массами покоя частиц
(Каструп); 2) попытки использования конформной группы как
группы динамической симметрии элементарных частиц (Тодоров,
Салам, Бом, Вижье и др.); 3) конформная группа как группа
динамической симметрии атома водорода (И. А. Малкин и В. И. Мань-
ко, Р. Мурто и др.).
В связи с явно усилившимся интересом к конформной
симметрии вновь возродилась проблема ее физического истолкования,
а также проблема согласования этой симметрии с принципом
причинности (Фултон, Рорлич, Виттен, Каструп, Кастелл, Розен и др.).
Наконец, отметим очень интересную идею Каструпа об
интерпретации квантовой механики как нарушенной конформной
симметрии (1968).
В свете изложенного представляется несомненным, что
объединение знаний о конформных симметриях в физике и биологии
окажется взаимно полезным для этих наук.
0 С.Ц. Петухов
209
Приложение III
ДОПОЛНИТЕЛЬНЫЕ СВЕДЕНИЯ
О ПРОЕКТИВНОЙ ГЕОМЕТРИИ
Некоторые начальные сведения о проективной геометрии уже
были даны в разделе III.1. Дополним их некоторыми
математическими и историческими материалами, полезными для лучшего
понимания вопросов, излагаемых в книге. Более обстоятельное
знакомство с проективной геометрией может быть осуществлено
читателем по работам [54, 81, 93, 187, 256, 273].
Проективными преобразованиями (для простоты будем
говорить о случае плоскости) называются преобразования
(проективной) плоскости, переводящие каждую прямую снова в
прямую с сохранением отношений принадлежности между
прямыми. Для того чтобы при проективном отображении каждая
точка прямой и каждый луч имели свои образы, нужно
формально ввести несобственные (бесконечно удаленные) точки. Эти
точки следует понимать как точки пересечения параллельных, так
что несобственная точка соответствует направлению прямой.
Совокупность всех несобственных точек обладает проективными
свойствами прямой и называется несобственной прямой.
Плоскость, дополненная такой несобственной прямой, называется
проективной плоскостью.
Множество проективных преобразований образует группу
проективных преобразований, что устанавливается проверкой
выполнимости трех общих условий группы: 1) множество
преобразований содержит тождественное преобразование; 2) множество
преобразований содержит для каждого преобразования обратное ему
преобразование; 3) наряду с каждыми двумя преобразованиями
множество преобразований содержит произведение этих
преобразований. Существование группы проективных преобразований
влечет за собой согласно Эрлангенской программе Ф. Клейна
существование проективной геометрии — науки, изучающей
свойства фигур, сохраняющиеся при преобразованиях данной группы.
Исторически для возникновения и развития проективной
геометрии большое значение имели практические задачи
изображения окружающих предметов на каких-либо плоскостях. Первые ее
зачатки и идеи относятся к глубокой древности. Постепенно
накапливались знания о соответствии точек изображаемого
предмета с точками его изображения на плоскости. Подмеченные правила
210
'"г<?'Я1
послужили началом Для образования метода проекций, следы
которого можно видеть у греческих математиков.
Настоящий расцвет учения о перспективе относится к эпохе
Ренессанса. А. Дюрер, Альберти, Леонардо да Винчи не только
использовали и творчески развили приемы перспективы в своих
художественных произведениях, но и написали вашные трактаты
о законах перспективы. Эти законы широко используют в своих
картинах Рафаэль, Микеланджело, Тициан, Веронезе и другие
великие мастера итальянской живописи. Блестящее развитие
живописи и архитектуры в эпоху Ренессанса вызвало громадное
внимание к вопросам перспективы, которые послужили
источником замечательных геометрических обобщений.
Научные основы современной проективной геометрии были
заложены работами французского архитектора и инженера
Жирара Дезарга (1593—1662). Интересно отметить, что при введении
многих понятий этой геометрии Дезарг, видимо, пользовался
образами живой природы. Об этом говорит использованная им
биологическая терминология (кстати, сильно затруднившая
восприятие современниками математических идей Дезарга). Так,
введенный им один из основных терминов — инволюция, взят им из
ботанического словаря и означает скрученное состояние молодых
листьев. Кроме того, Дезарг использует в своей геометрии
термины: древо, пень, нарост, ветви, стебли и т. д. Не предчувствовал
ли он глубокого родства исследуемых им проективных структур
с устройством тел живой природы? К сожалению, о творчестве и
мировоззрении Дезарга осталось весьма немного свидетельств и
нет возможности оценить отношение Дезарга к биологии и его
представления о живой природе.
Несколькими годами позже выхода в свет сочинений Дезарга
другой выдающийся французский ученый, Б. Паскаль, которому
в то время было всего 16 лет, установил замечательную теорему,
играющую важную роль в теории конических сечений. Эта
теорема, вскрывшая проективную связь шести точек конического
сечения, ныне носит имя Паскаля.
Через полтораста лет после смерти Дезарга и Паскаля
наступил новый творческий период в области проективной геометрии,
приведший к признанию проективной геометрии как
самостоятельной научной дисциплины. Это произошло в результате трудов
замечательных геометров Понселе, Жергонна, Брианшонна, Мон-
жа, Мебиуса, Штейнера, Шаля и др. Работами нашего великого
соотечественника Н. И. Лобачевского, впервые построившего
геометрию, логически столь же безупречную, как и геометрия
Евклида, но существенно отличную от нее, была разрушена наивная
вера в евклидову геометрию как единственно возможную, незави-
Ь* 211
сймую от действительного мира и его опытного познания,
априорную геометрическую систему. После открытия Лобачевским
неевклидовой геометрии на повестку дня встало изучение и других
возможных геометрических систем (абстрактных геометрических
«пространств») и соответствующих им свойств или явлений
действительного мира.
Первоначально проективная геометрия возникла как один из
разделов евклидовой геометрии. В частности, характерное для
проективной геометрии двойное отношение (6) вычислялось
только через длины соответствующих отрезков, т. е. основывалось
на евклидовом понятии длины отрезка. Однако с общим
развитием геометрии все острее стало проявляться стремление к
построению проективной геометрии как совершенно не зависимой
от евклидовой геометрии, т. е. свободной от метрических понятий
в ее основных принципах и положениях. Реализация такого
построения была осуществлена во многом X. Штаудтом. Он
использовал возможность чисто проективного построения двойного
отношения четырех точек прямой, по величине равного —1. Четверки
точек с таким двойным отношением получили название
гармонических четверок. На основе гармонических четверок Штаудт
определил соответствие проективных рядов и создал своеобразное
«проективное исчисление», в котором роль чисел играют четверки
точек прямой, называемые им вурфами (что в переводе с
немецкого означает «бросок»).
Успехи в утверждении независимости проективной геометрии
от евклидовой привели к замечательной мысли интерпретировать
евклидову геометрию как частный случай проективной. Полное
решение этой задачи было дано Кэли и Клейном, которыми было
построено так называемое проективное мероопределение. Суть
его заключается в том, что метрические свойства фигур можно
рассматривать как проективные отношения их к особым
геометрическим образам, называемым абсолютами. Эта новая концепция
оказалась настолько общей, что она охватила, кроме евклидовой,
и целый ряд неевклидовых геометрических систем.
212
Приложение IV
О ПРИНЦИПЕ КОНФОРМНЫХ БЛОКОВ
И ОБОБЩЕННОЙ КРИСТАЛЛОГРАФИИ
Приведем некоторые дополнительные данные о явлении
формирования биологических тел из конформно-симметричных блоков.
Неевклидовы конформные симметрии реализуются не только в
телах относительно сложных организмов, но и в простейших
биологических телах. В пользу этого свидетельствуют, например,
особенности строения вируса табачной
мозаики, состоящего из спирально
уложенных в цилиндр белковых
субъединиц. На рис. 44 дан вид
этого цилиндра сверху. Видна
симметрия инверсии относительно
окружности, обозначенной точечной
линией, между неравными
половинками белковых субъединиц.
Интересно, что это не просто
воображаемая окружность, но именно в этом
месте расположена нить РНК
вируса. (На рис. 44, взятом из работы
[242], величина внутреннего
радиуса указана равной 18 А, а не 20 А
как приближенно представлено в
исходной работе; основание для
такой поправки дают результаты рент-
геноструктурного анализа,
приведенные на рис. 86 той же
работы.)
К явлениям конформной симметрии, по нашему мнению,
можно отнести также особенности формирования клеток, известные
под названием явлений пропорциональности размеров клеточных
классов и заключающиеся в закономерном квантовании размеров
реализуемых в живых организмах клеточных ядер [41]. Добавим,
что некоторые черты процесса деления клеток позволяют думать,
что это деление является в живой природе не чем-то сугубо
специфическим, а лишь одним из проявлений загадочных
механизмов роста и размножения живого вещества по правилу
конформно-подобных блоков.
Рис. 44. Вирус табачной
мозаики (вид сверху)
J — белковые субъединицы;
2 — спираль РНК с тремя
нуклеотидами на каждую
субъединицу
213
Явление формирования биологических Гел из
конформно-симметричных блоков имеет то отличие от кристаллизации неживого
вещества, базирующейся на евклидовых симметриях (т. е. на
группе движений), что при нем реализуются симметрии инверсии
относительно сфер конечных и бесконечного радиусов, тогда как
при формировании кристаллов неживого вещества имеют место
только симметрии инверсии относительно сфер бесконечно
большого радиуса. Хотелось бы еще дополнительно подчеркнуть
характерную особенность формообразования при кристаллизации
живого вещества, которую Л. В. Белоусов называет
нестатистическим упорядочением [399, с. 235]. Речь идет о материализации
упорядоченной формы целого органа из неупорядоченных частей;
например, у некоторых соцветий возникает идеальный
параболический контур из цветков с самыми разными пропорциями в
длине цветоложа и венчика. В этом и подобном случаях ясно видна
как бы независимость целого от своих составных элементов.
Отметим особо одну связь между закономерностями
формообразования физических кристаллов и биологических тел.
Основным законом геометрической кристаллографии является открытый
Н. Стеноном закон постоянства углов на кристаллах: углы между
соответственными гранями и ребрами во всех кристаллах одного
и того же вещества одинаковой полиморфной модификации
постоянны, в то время как число и длина сторон кристалла
варьируют. Основное направление развития кристаллографии в
прошлом, вплоть до начала нынешнего столетия, чуть ли не всецело
базировалось на этом законе [261, с. 68]. Тем более интересно,
что закономерность постоянства углов имеет место и в
формировании живого вещества: при наблюдении многих биологических
тел, для очертаний которых характерен повторяющийся в
пространстве мотив, обращает на себя внимание постоянство величины
соответственных углов в повторяющихся частях очертаний
конкретного вида тел. Число же и протяженность данных
повторяющихся частей могут при этом заметно изменяться.
Поясним сказанное на примере листьев клена. Очертания
каждого листа представляют собой набор повторяющихся зубцов,
между вершинами которых край листа имеет криволинейную
форму. Углы при вершинах всех зубцов одного листа, а также разных
листьев равновелики, насколько об этом можно судить при
затрудняющем измерения обстоятельстве: искривленности сторон зубцов.
(Для облегчения гониометрических исследований нами
использовалось масштабное увеличение очертаний листа посредством их
оптического проецирования на экран.) Протяженность же
криволинейных отрезков между зубцами, а также число зубцов у
кленовых листьев не являются постоянными. В общем контексте
214
книги биологическая закономерность постоянства углов
представляется особенно интересной, поскольку именно величины углов
являются характерными инвариантами конформных
преобразований.
Проявление принципа конформной симметрии в очертаниях
рассматриваемых листьев не ограничивается инвариантностью
величин углов при вершинах зубцов: насколько мы можем судить,
криволинейные отрезки между соседними вершинами листа
также конформно-симметричны друг другу, т. е. равновеликость
соблюдается и между угловыми характеристиками в
соответственных точках соседних криволинейных отрезков. Тенденция
постоянства углов наблюдается и в местах стыкования прожилок
листа. Например, в нормально развитых листьях клена углы
стыковки боковых прожилок со срединными прожилками всегда
примерно равны 70°. У листьев других растений эти углы могут иметь
иные величины, причем иногда у одного и того же листа с
сетчатым жилкованием данные углы могут принимать несколько
значений, но постоянно повторяющихся и связанных друг с другом
по определенным правилам. Здесь напрашивается некоторая
аналогия с кристаллами косного вещества, у которого для
монокристаллов конкретного вещества определенной полиморфной
модификации есть строго фиксированный набор реализуемых межгранных
углов.
Листья растений являются только одним из примеров того,
что биологические тела зачастую представляют собой
закономерно организованные друзы кристаллов живого вещества,
подчиненные отношениям конформной симметрии и чаще всего носящие
плоскостной характер, т. е. имеющие вид листа или ленты,
нередко изогнутых спиральным образом в пространстве. При этом
в целом ряде своих особенностей эти объекты напоминают
мезоморфные кристаллы.
В живой природе существует множество других интересных
примеров, когда при наличии необходимых условий наблюдается
закономерность постоянства углов (или тенденция к такому
постоянству) в формах биологических тел: крепление листьев и
иголок к побегу растений, стыкование веток с сучьями деревьев,
соединение частей в древовидных разветвляющихся системах в
организме человека и животных и т. д. При этом разграничиваю
щие углы конкретных объектов нередко имеют не одно
фиксированное значение, а несколько, по постоянно повторяющихся в
данном объекте и взаимосвязанных.
Все это говорит о тесной близости структурных
закономерностей кристаллизации живого и неживого вещества и
перспективности изучения пррблем формирования биологических тел с
215
позиций обобщенной кристаллографии. В частности, необходимы
широкие и детальные гониометрические исследования в области
морфогенеза биологических объектов по аналогии с
гониометрическими исследованиями в классической кристаллографии.
Особого внимания заслуживают систематизация величин углов,
реализуемых в биологических объектах в норме и при уродствах,
изучение влияния условий внешней среды на угловые
характеристики биологических объектов, исследование возможностей
полиморфного по данным характеристикам состояния живого
вещества. Получаемые здесь результаты могут оказаться полезными для
разработки естественной классификации живых организмов. На
данном пути возможна также разработка теорий онто- и
филогенеза как регулируемой кристаллизации (понимаемой в обобщенном
смысле) живого вещества, протекающей с соблюдением законов
обобщенной кристаллографии.
В связи с прилегающими сюда вопросами генетического
кодирования кристаллизации форм живого вещества заметим, что
построение тел из пространственно однотипных в некотором смысле
блоков, видимо, способно обеспечивать известные жизненные
выгоды для организмов в аспекте оптимизации генетических
механизмов, поскольку отпадает необходимость детерминировать план
строения каждой части тела как во всем индивидуальной. Одни
и те же генетические программы могут использоваться
организмом разнопланово и многократно и отпределять геометрически
родственное строение разных частей тела.
Не распространяются ли принципы обобщенной
кристаллографии на другие явления биологического формообразования,
например на явления моторных движений (особенности почерка,
рисунка локомоций и пр.) или на другие биологические явления,
где речь в том или ином смысле идет о «кристаллизации формы»,
которую можно разложить на составные блоки с последующим
изучением геометрических взаимоотношений между данными
блоками в разных ситуациях образования форм? Это покажут
будущие исследования, в ходе которых можно ожидать обнаружения
интересных правил биологической организации данных явлений.
Идеи о подчиненности живого вещества неевклидовой
геометрии являются во многом продолжением идей В. И. Вернадского,
подробно изложенных им в двух главах его последней книги
[42, гл. 15, 16], в которой он подвел итоги своей научной
деятельности. Нельзя не порекомендовать нашему читателю внимательное
знакомство с этой исторической работой, в которой рассмотрен ряд
ключевых задач исследования живого вещества. Ставя вопрос о
причинах симметрии в формах биологических тел, Вернадский
решал его на рснове глубокого положения П. Кюри о симметрии
216
Как следствий й Характеристике состояния пространства.
Пространство может иметь различные состояния и отсюда многообразие
проявлений симметрии в природных телах. При этом Вернадский
дришел к заключению, что в живых веществах состояние
пространства не отвечает евклидовой геометрии и является совершенно
иным, чем для случая косных тол, соответствующего евклидовой
геометрии. Он считал программной задачей, одной из основных
.В биологии, установить на базе изучения биологических
симметрии, какой же из многих возможных типов неевклидовых
геометрий характеризует пространство живого вещества, но обеспечить
решение этой задачи не успел.
Вернадский видел в явлениях жизни проявления некоторого
реального пространства-времени, отличающегося от пространства-
времени специальной теории относительности. В развитие этих
идей можно, по нашему мнению, выдвинуть гипотезу о
конформной геометрии биологического пространства-времени. При этом
оправданность обращения именно к конформной геометрии
усматривается в фактах пространственной конформной симметрии в
формах биологических тел, а также в том, что
конформно-геометрическое состояние пространства-времени, в случае его
обнаружения для живого вещества, не было бы беспрецедентным и
вовсе оторванным от известных физических явлений.
Действительно, оно может ставиться в параллель с конформной
пространственно-временной инвариантностью уравнений Максвелла, если
трактовать эту инвариантность уравнений электродинамики как
следствие реальных конформно-геометрических состояний
пространства-времени, т. е. идти по пути идей П. Кюри и В. И.
Вернадского. (Подобное сопоставление применимо и к другим фактам
конформной инвариантности в физике, отмеченным в предыдущих
разделах книги.) В этой связи несомненно интересно попытаться
связать формальные следствия из конформной инвариантности
уравнений Максвелла с биологическими
пространственно-временными явлениями, приложить уже разработанные разными
авторами варианты конформной теории относительности к феноменам
живого вещества.
В литературе давно дискутируется вопрос о том, приведут ли
биологические исследования к открытию неизвестных физических
законов или удастся описать все особенности биологических тел
На основе законов, уже известных в физике. Интересное
обсуждение этого вопроса имеется, например, в работе В. Гейзенберга
(62, с. 75]. Не представляют ли описанные в книге закономерности
Конформной структуризации биологических объектов основу
фундаментальных физических законов живого вещества, не
встречавшихся ранее в физике вещества и не сводимых к уже известным
законам физики?
ЛИТЕРАТУРА
1. А. с. 390555 (СССР). Устройство для моделирования процессов
в системе полукружных каналов вестибулярного аппарата /
С. В. Петухов. Заявл. 20.07.71, № 1684783/31—16; Опубл. в
Б. И., 1973, № 30, МКИ Аб1в 5/10.
2. Азгалъдов Г. Г. Численная мера и проблема красоты в
архитектуре. М.: Стройиздат, 1978. 88 с.
3. Азгалъдов Г. Г., Повилейко Р. П. О возможности оценки
красоты в технике. М.: Изд-во стандартов, 1977. 120 с.
4. Александер Р. Биомеханика. М.: Мир, 1970. 339 с.
5. Алякринский Б. С, Степанова С. И. О механизмах
пространственной ориентировки в полете и некоторых причинах ее
нарушения.— Космическая биология и медицина, 1968, т. 2,
№ 1, с. 63—68.
6. Ананьев Б. Г., Веккер Л. М., Ломов Б. Ф., Ярмоленко А. В.
Осязание в процессах познания и труда. М.: АПН РСФСР,
1959.263 с.
7. Ананьев Б. Г., Дворяшина М. Д., Кудрявцева Н. А.
Индивидуальное развитие человека и константность восприятия. М.:
Просвещение, 1968. 334 с.
8. Андреев Э. Н. Пространство микромира. М.: Наука, 1969. 80 с.
9. Андронеску А. Анатомия ребенка. Бухарест: Меридиан, 1970.
363 с.
10. Араратян А. Г. Листорасположение и числа Фибоначчи.—
Биологический журнал Армении, 1967, т. 20, N° И, с. 69—84.
И. Араратян А. Г. О паратонических
чувствительно-двигательных системах высших растений.— Биологический журнал
Армении, 1967, т. 20, № 12, с. 85—94.
12. Араратян А. Г. Криогенные нарушения в
листорасположении.— Биологический журнал Армении, 1970, т. 23, № 9, с. 23—
29.
13. Афанасьев Я. Н. Анализ пропорций храма Покрова на Нер-
ли.— В кн.: Искусство и точпые науки. М.: Наука, 1979,
с. 126—141.
14. Банк Ч. Кристаллы, их роль в природе и науке. М.: Мир,
1970. 312 с.
15. Беклемишев В. Н. Основы сравнительной анатомии
беспозвоночных. М.: Сов. наука, 1952. 699 с.
16. Белоусов Л. В. Онтогенетическая модель рядов Фибоначчи в
апикальных меристемах растений.— Журн. общ. биол., 1976.
т. 37, № 6, с. 900-911.
218
17. Бердышев А. Н. О некоторых математических
закономерностях биологических процессов.— Журнал общей биологии,
1972, т. 33, № 5, с. 631—637.
18. Бернал Дж. О роли геометрических факторов в структуре
материи.— Кристаллография, 1962, т. 7, № 4, с. 507—519.
19. Бернал Дж. Возникновение жизни. М.: Мир, 1969. 392 с.
20. Бернал Дж., Карлайл С. Поля охвата обобщенной
кристаллографии.— Кристаллография, 1968, т. 13, вып. 5, с. 927—951.
21. Бернштейн Н. А. Общая биомеханика. М.: РИО ВЦСПС, 1926.
416 с.
22. Бернштейн Н. А. Клинические пути современной
биомеханики. Казань: Главнаука, 1929, с. 249—270. (Труды Казанского
государственного ин-та усовершенствования врачей; Т. 1).
23. Бернштейн Н. А. Очередные проблемы физиологии
активности.— Проблемы кибернетики, 1961, т. 6, с. 101—160.
24. Бернштейн Н. А. Очерки по физиологии движений и
физиологии активности. М.: Медицина, 1966. 349 с.
25. Биркгофф Г. Гидродинамика. Методы. Факты. Подобие. М.:
Изд-во иностр. лит., 1963. 244 с.
26. Блайвас А. С. Зрительный анализ в неспециализированных
рецептивных полях как разложение по ортогональным
базисным функциям.— Нейрофизиология, 1974, т. 6, № 2, с. 211—
218.
27. Бом Д. Физика и восприятие.— В кн.: Специальная теория
относительности. М.: Мир, 1967, с. 223—283.
28. Борн М. Физика в жизни моего поколения. М.: Изд-во иностр.
лит., 1963. 535 с.
29. Борщевский И. Я., Корешков А. А. Маркарян С. С. и др.
Влияние на организм человека вибраций некоторых типов
современных вертолетов и самолетов.— Воен.-мед. журн., 1958, № 4,
с. 51—55.
30. Брандт А. Ф. Десноручие, шуеручие и перекрестная
асимметрия конечностей.— Рус. антропол. журн., 1927, т. 15, № 3—4,
с. 7-28.
31. Брейтман М. Я. Введение в учение о пропорциях и
конструкциях человеческого тела. Л.: Ленингр. педол. ин-т, 1924. 110 с.
32. Врунов И. И. Пропорции античной и средневековой
архитектуры. М.: Изд-во Всесоюз. акад. архитект., 1935. 108 с.
33. Бунак В. В. Изменение относительной длины сегментов
скелета конечностей человека в период роста.— Изв. АПН РСФСР,
1957, вып. 84, с. 13—31.
34. Бунак В. В. Об изменении продольных размеров сегментов
скелета конечностей у некоторых групп млекопитающих в
период роста.— Изв. АПН РСФСР, 1957, вып. 84, с. 33—45.
35. Бурбаки Н. Очерки по истории математики. М.: Изд-во иностр.
лит., 1963. 292 с.
36. Буров А. К. Вторая производная золотого сечения.—
Архитектура СССР, 1935, № 3, с. 15—24.
37. Бушманова Г. В., Норден А. П. Элементы конформной
геометрии. Казань: Изд-во Казан, ун-та, 1972. 177 с.
38. Ван дер Варден Б. Л. Пробуждающаяся наука. М.: Физматгиз,
1959. 310 с.
39. Васильев Ф. П. Лекции по методам решения экстремальных
задач. М.: МГУ, 1974. 310 с.
40. Вейль Г. Симметрия. М.: Наука, 1968. 191 с.
219
41. Вермелъ Е. М. Исследование о клеточных размерах.— Учен,
зап. МГПИ, 1940, т. 25, вып. 1.
42. Вернадский В. И. Химическое строение биосферы и ее
окружения. М.: Наука, 1965. 205 с.
43. Вернадский В. И. Размышления натуралиста: Пространство
и время в неживой и живой природе. М.: Наука, 1975. 220 с.
44. Вернадский В. И. Живое вещество. М.: Наука, 1978. 220 с.
45. Вигнер Е. Этюды о симметрии. М.: Мир, 1971. 318 с.
46. Визгин В. П. Развитие взаимосвязи принципов
инвариантности с законами сохранения в классической физике. М.:
Наука, 1972. 236 с.
47. Визгин В. П. Из истории конформной симметрии в физике.
О некоторых особенностях взаимосвязи физики и
математики в XX веке: Историко-математическио исследования. М.:
Наука, 1974, вып. 19, с. 188—219.
48. Визгин В. П. Эрлангенская программа и физика. М.: Наука,
1975. 110 с.
49. Визгин В. П. Проблемы взаимосвязи математики и физики.—
Историко-математические исследования, 1975, вып. 20, с. 28—
50.
50. Витрувий. Десять книг об архитектуре. М.: Изд-во Всесоюз.
акад. архитект., 1936, т. 1. 331 с.
51. Владимиров С. А. Группы симметрии дифференциальных
уравнений и релятивистские поля. М.: Атомиздат, 1979. 167 с.
52. Вожжова А. И., Окунев Р. А. Укачивание и борьба с ним. Л.:
Медицина, 1964. 168 с.
53. Волохонский А. Г. О формальной структуре генетического
кода.— Цитология и генетика, ;1972, т. 6, № 6, с. 32—40.
54. Вольберг О. А. Основные идеи проективной геометрии. М.; Л.:
ОНТИ, 1935. 125 с.
55. Воробьев Н. Н. Числа Фибоначчи. М.: Наука, 1978. 141 с.
56. Восприятие пространства и времени. Л.: Наука, 1969. 135 с.
57. Вульф Г. В. Избранные работы по кристаллофизике и
кристаллографии. М.: Физматгиз, 1952. 343 с.
58. Галактионов С. Г. Асимметрия биологических молекул. Минск:
Высш. школа, 1978. 172 с.
59. Гальперин П. Я. Опыт изучения формирования умственных
действий.— В кн.: Докл. на Всесоюз. совещ. по вопросам
психологии. М.: АПН РСФСР, 1954, с. 60—66.
60. Галярский 3. И., Заморзаев А. М. О группах симметрии и
антисимметрии подобия,— Кристаллография, 1963, т. 8, вып. 5,
с. 691—698.
61. Гамбарян П. П. Бег млекопитающих. Л.: Наука, 1972. 334 с.
62. Гейзенберг В. Философия и физика. М.: Изд-во иностр. лит.,
1963. 202 с.
63. Геккель Э. Красота форм в природе. СПб.: Просвещение, 1907.
246 с.
64. Гельмголъц Г. Счет и измерение. Казань: Изд-во Казан, ун-та,
1893. 23 с.
65. Гельмголъц Г. О происхождении и значении геометрических
аксиом. СПб., 1895. 58 с.
66. Гельмголъц Г. О фактах, лежащих в основании геометрии.—
В кн.: Об основаниях геометрии. М.: Высш. школа, 1956,
с. 366—382.
220
67. Гератеволъ 3. Психология человека и самолете. М.: Изд-во
иностр. лит., 1956. 357 с.
68. Герке П. Я. Частная эмбриология человека. Рига: Изд-во АН
ЛатвССР, 1957. 248 с.
69. Гете И.-В. Избранные сочинения по естествознанию. М.:
Изд-во АН СССР, 1957, 553 с.
70. Гика М. Эстетика пропорций в природе и искусстве. М.:
Изд-во Всесоюз. акад. архитект., 1936. 320 с.
71. Гинзбург В. В. Об асимметрии конечностей человека.—
Природа, 1947, № 8, с. 42—46.
72. Гинзбург В. В. Элементы антропологии для медиков. Л.:
Медицина, 1963. 210 с.
73. Гильберт Д. Основания геометрии. М.; Л.: ГИТТЛ, 1948. 491 с.
74. Гильберт Д., Кон-фоссен С. Наглядная геометрия М.; Л.:
ГИТТЛ, 1951. 352 с.
75. Гиппенрейтер Ю. Б., Романов В. Я., Смирнов С. Д.
Соотношение глазодвигательных и ручных операций в процессе счета
элементов тест-объекта.— В кн.: XVIII Междунар. психол.
конгресс. 30-й симпозиум «Восприятие и действие». М.:
Наука, 1966, с. 110—113.
76. Глазер Р. Биология в новом свете. М.: Мир, 1978. 172 с.
77. Гранин Д. А. Эта странная жизнь. М.: Сов. Россия, 1974,110 с.
78. Гранит Р. Основы регуляции движений. / Под ред. В. С. Гур-
финкеля. М.: Мир, 1973, 367 с.
79. Григорьев А. Т., Розенфельд Б. А. Теория винтов и
неевклидова механика.— В кн.: История механики с конца 18-го до
середины 20-го века. М.: Наука, 1972, с. 337—350.
80. Грюнбаум А. Философские проблемы пространства и
времени/Под общ. ред. Э. М. Чудинова. М.: Прогресс, 1969. 568 с.
81. Гуревич Г. Б. Проективная геометрия. М.: Физматгиз, 1960.
320 с.
82. Гурфинкель В. С, Кои Я. М., Шик М. Л. Регуляция позы
человека. М.: Наука, 1965. 256 с.
83. Гурфинкель В. С., Левик Ю. С. Сенсорные комплексы и сен-
сомоторная интеграция.— Физиология человека, 1979, № 3,
с. 6—20.
84. Дайсон Ф. Математика в физических науках.— В кн.:
Математика в современном мире: Сб. стат. М.: Мир, 1967, с. 111—
128.
85. Данилова Е. И. Эволюция руки в связи с вопросами
антропогенеза. Киев: Наук, думка, 1965. 198 с.
86. Дарбу Г. Принципы аналитической геометрии. М.; Л.: ОНТИ,
1938. 375 с.
87. Декарт Р. Избранные произведения. М.: Изд-во иностр. лит.,
1959. 720 с.
88. Депенчук И. II. Симметрия и асимметрия в живой природе.
Киев: Изд-во АН УССР, 1963. 176 с.
89. Джадд Д., Вышецки Г. Цвет в науке и технике. М.: Мир,
1978. 592 с.
90. Дмитриев И. С. Симметрия в мире молекул. Л.: Химия, 1976.
124 с.
91. Доленко Ф. Л. Исследование подвижности в голеностопном
суставе человека и обоснование средств и методов ее развития:
Автореф. дис. ...канд. мед. наук. М.: Го,С, ЦЦ-т физ..
культуры, 1969. т
221
92. Дуда Р., Харт П. Распознавание образов и анализ сцен. М.:
Мир, 1976. 511 с.
93. Ефимов Н. В. Высшая геометрия. М.: Наука, 1971. 576 с.
94. Жемайтис С, Миронов В. Нужна ли жизни форма? —
Знание — сила, 1979, № 12, с. 23—26.
95. Жуковский П. М. Ботаника. М.: Высш. школа, 1964. 667 с.
96. Зарипов Р. X. Кибернетика и музыка. М.: Наука, 1971. 235 с.
97. Зрительное опознание и его нейрофизиологические
механизмы/Под ред. В. Д. Глезера. Л.: Наука, 1975. 271 с.
98. Игнациус Г. И. Проективная геометрия. М.: Знание, 1966. 70 с.
99. Исследования по биодинамике ходьбы, бега, прыжка / Под
ред. Н. А. Бернштейна, М.: Физкультура и спорт, 1940. 390 с.
100. Каган В. Ф. Очерки по геометрии. М.: Физматгиз, 1964. 490 с.
101. Канаве И. И. Гете как естествоиспытатель. Л.: Наука, 1970.
467 с.
102. Каратеодори К. Конформное отображение. М.; Л.: ГТТИ, 1934.
180 с.
103. Касинов В. Б. О симметрии в биологии. Л.: Наука, 1971. 46 с.
104. Кафанов А. И. Об интерпретации логарифмической спирали
в связи с анализом изменчивости и роста двухстворчатых
моллюсков.— Зоол. журн. 1975, т. 54, вып. 10, с. 1457—1467.
105. Кацитадзе 3. И. Эволюция вертикальной ходьбы. М.: Наука,
1968. 220 с.
106. Кешмэн Д. Фораминиферы. М.; Л.: ОНТИ, 1933. 260 с.
107. Кисляков В. А. Периферические и центральные механизмы
вестибулярных реакций: Автореф. дис. ... д-ра биол. наук. Л.:
Ин-т физиол. им. И. П. Павлова АН ССС,Р, 1971.
108. Кисляков В. А., Неверов В. П. О следовых явлениях в .цент-
ральной нервной системе после длительного действия
"оптокинетических раздражителей.— В кн.: Авиационная и
космическая медицина: Матер, конф. М.: Всесоюз. физиол. о-во,
1963, с. 241—242.
109. Кисляков В. А., Орлов И. В. О гидродинамических влияниях
между отделами вестибулярного аппарата,— ДАН СССР, 1971,
т. 198, № 2, с. 479—482.
110. Кисляков В. А., Левашов М. М., Орлов И. В. Вестибулярная
система.— В кн.: Физиология сенсорных систем. Л.: Наука,
1972, ч. II, с. 57—129.
111. Клейн Ф. О геометрических основаниях Лоренцевой группы.—
В кн.: Новые идеи в математике.— Образование. (СПб.), 1914,
вып. 5, с. 170—185.
112. Клейн Ф. Элементарная геометрия с точки зрения высшей.
М.; Л.: ОНТИ, 1933, т. 1; 1934, т. 2. 580 с.
113. Клейн Ф. Неевклидова геометрия. М; Л.: ОНТИ, 1936. 355 с.
114. Клейн Ф. Лекции о развитии математики в XIX столетии.
М.: ОНТИ, 1937.
115. Клейн Ф. Высшая геометрия. М.; Л.: ОНТИ, 1939. 410 с.
116. Кокстер Г. С. М. Введение в геометрию (Гл. XI: Золотое
сечение и филотаксис). М.: Наука, 1966. 648 с.
117. Колмогоров Н. А. Теория ангармонического отношения пяти
точек плоскости.— Тр. / Кировский эоотехн.-ветер. ин-т, 1938,
т. 3, вып. 2—3, с. 161—168.
118. Комаров В. Е. Проблема неевклидовости пространственной
организации головного мозга человека.— В кн.: Философские
222
вопросы медицины и биологии: Респ. межвед. сб. Киев: НауК.
думка, 1977, с. 84—90.
119. Комендантов Г Л. Воздушная болезнь. М.: ЦИУВ МЗ СССР,
1965. 88 с.
120. Компанеец А. С. Симметрия в микро- и макромире. М.: Наука,
1978. 207 с.
121. Короленок К. X. О клиническом и профессиональном
значении иллюзий ориентации в пространстве.— Вопр.
психопатологии и психотерапии. М.: Медицина, 1963, с. 55—64.
122. Котова Е. Глаз и законы красоты.— Искусство, 1966, № 12,
с. 8—14.
123. Коток В. С, Манзий С. Ф. К функциональному анализу
относительной длины звеньев тазовой конечности некоторых
млекопитающих.— Вестн. зоол., 1973, N° 4, с. 43—48.
124. Котурницкий Н. В. Квинкункс и его применения в
естествознании и ткацком деле.— Изв. Технол. ин-та, 1893, т. 8, с. 45—
240.
125. Криндач В. П. Принципы симметрии и некоторые другие
вопросы теории симметрии в физике: Автореф. дис. ... канд. физ.-
мат. наук. М.: МГУ, 1972.
126. Ле Корбюзье Ш.-Э. Модулор: Опыт соразмерной масштабу
человека всеобщей гармонической системы мер, применимой
как в архитектуре, так и в механике. М.: Стройиздат, 1976.
237 с.
127. Ле Корбюзье Ш.-Э. Архитектура XX века. М.: Прогресс, 1977.
240 с.
128. Лебедев Ю. С. Архитектура и бионика. М.: Стройиздат, 1977.
211 с.
129. Лебедев Ю. С, Бялъский И. И. Гармония форм в живой
природе и архитектуре. М.: ЦНТИ по гражданскому
строительству и архитектуре, 1976. 43 с.
130. Левин Н. А. Зависимость анатомического строения костного
лабиринта птиц от образа их жизни.— Зоол. журн., 1955, т. 34,
вып. 3, с. 601—604.
131. Левитин К. Геометрическая рапсодия. М.: Знание, 1976. 120 с.
132. Леонардо да Винни. Анатомия, записки и рисунки. М.: Наука,
1965. 586 с.
133. Листинг И. Б. Предварительные исследования по топологии.
М.: ГТТИ, 1932. 110 с.
134. Лобачевский Н. И. Поли. собр. соч.: В 5-ти т. М.; Л.: Гостех-
. издат, 1946.
135. Ломаное М. Элементы симметрии в музыке.— В кн.:
Музыкальное искусство и наука. М.: Музыка, 1970, вып. 3, с. 136—
165.
136. Любищев А. А. Систематика и эволюция.— В кн.:
Внутривидовая изменчивость наземных позвоночных животных и
микроэволюция. Свердловск: Просвещение, 1966, с. 45—57.
137. Магнус Р. Установка тела. М.; Л.: Изд-во АН СССР, 1962.
624 с.
138. Магомедмирзаев М. М. Анализ структуры изменчивости
морфологических признаков высших растений и его
использование в решении общих и прикладных задач популяционной
биологии: Автореф. дис. ... д-ра биол. наук. Л.: ЛГУ, 1977.
139. Майкова-Строганова В. С, Рохлин Д. Г. Кости и суставы в
рентгеновском изображении. Л.: Медгиз, 1957. 483 с.
223
140. Малахов А. Л. £5-симметрия жизни. Свердловск: Сред.-Урал.
кн. изд-во, 1965. 60 с.
141. Манин Ю. U. Математика и физика. М.: Знание, 1979. 63 с.
142. Маркс К., Энгельс Ф. Соч. в 30-ти т. Т. 3. Исследования, статьи
1844—1845 гг. М.; Л.: Госиздат, 1929, 709 с.
143. Маркс К., Энгельс Ф., Ленин В. И. О биологии. М.: Партиздат.,
1933, 238 с.
144. Маркушевич А. И. Комплексные числа и конформные
отображения. М.: Физматгиз, 1960. 55 с.
145. Марусяк В. И. Закон сохранения симметрии и динамика:
Препринт Ин-та теор. физ. АН УССР, № 76—33 р. Киев, 1976. 15 с.
146. Мастера архитектуры об архитектуре. М.: Искусство, 1972.
390 с.
147. Мастера советской архитектуры об архитектуре. Киев: Изд-во
Акад. архитект. УССР, 1953. 108 с.
148. Мастера советской архитектуры об архитектуре. М.:
Искусство, 1975. 345 о.
149. Межжерин В. А. Метрика, кванты, время и пространство
биологических систем.— В кн.: Проблемы эволюционной и попу-
ляционной генетики: Материалы выездной сессии Секции
эволюционной и популяционной генетики Научного совета по
проблемам генетики и селекции АН СССР (Сб. стат.).
Махачкала, 1978, с. 145—154.
150. Мейен С. В. Путь к новому синтезу, или куда ведут
гомологические ряды.— Знание — сила, 1972, № 8, с. 20—22.
151. Мейен С. В. Дискуссия к статье А. Д. Любищева «Поли- и
моно».— Знание — сила, 1977, № 7, с. 29—31.
152. Мейен С. В., Соколов Б. С, Шрейдер Ю. А. Классическая и
неклассическая биология: Феномен Любищева.— Вестн. АН
СССР, 1977, № 10, с. 112—124.
153. Мессель Э. Пропорции в античности и в средние века. М.:
Изд-во Всесоюз. акад. архитект., 1936. 160 с.
154. Михайленко В. Е., Ковалев С. Н. Конструирование форм
современных сооружений. Киев: Будивельник, 1978. 111 с.
155. Миракян А. И. Математический и психологический аспекты
изучения константности восприятия величины.— В кн.:
XV Междунар. психол. конгресс: Тез. докл. М.: Наука, 1972,
с. 128—129.
156. Мороз О. В. В поисках гармоники. М.: Атомиздат, 1978. 207 с.
157. Мостепаненко А. М. Проблема универсальности основных
свойств пространства и времени. Л.: Наука, 1969. 180 с.
158. Мостепаненко А. М. Пространство — время и физическое
познание. М.: Атомиздат, 1975. 215 с.
159. Мусатов В. И. Зрительное восприятие пространственных
отношений: Автореф. канд. дис. ... канд. психол. наук. М.: НИИ
общ. и пед. психологии АПН СССР, 1977.
160. Назайкинский Е. О константности в восприятии музыки.—
В кн.: Музыкальное искусство и наука. М.: Музыка, 1970,
вып. 2, с. 59—98.
161. Наливкин Д. В. Элементы симметрии органического мира.—
Изв. Биол. НИИ при Пермском ун-те, 1925. с. 291.
162. Наливкин Д. В. Криволинейная симметрия.— В кн.:
Кристаллография. М.: Металлургиздат, 1951, с. 15—23.
224
1бЗ. Наливкин Д. В. Симметрия форм органического мира
(изогнутый шар и его разновидности).— Тр. /Ленингр. о-во
естествоиспытателей, 1965, т. 75, № 1, с. 27—33.
164 На пути к теоретической биологии. / Под ред. Б. Л. Астауро-
ва. М.: Мир, 1970. 186 с.
165. Наттал Дж., Сенфорд У. Пространственная дезориентация в
полете.— В кн.: Медицинские проблемы безопасности полетов.
М.: Воениздат, 1962, с. 47—74.
166. Неванлинна Р. Пространство, время и относительность. М.:
Мир, 1966. 230 с.
167 Об основаниях геометрии / Под ред. А. П. Нордена. М.: ГИТТЛ,
1956. 680 с.
168. Овсянников Л. В. Групповой анализ дифференциальных
уравнений. М.: Наука, 1978. 399 с.
169. Овчинников Н. Ф. Принципы сохранения. М.: Наука, 1966.
331 с.
170. Патури Ф. Растения — гениальные инженеры природы. М.:
Прогресс, 1979. 312 с.
171. Паули В. Физические очерки. М.: Наука, 1975. 256 с.
172. Переверзев-Орлов В. С. Модели и методы автоматического
чтения. М.: Наука, 1976. 215 с.
173. Петухов С. В. О динамике процессов в вестибулярном
аппарате: Автореф. дис. ... канд. биол. наук. М.: Ин-т проблем
передачи информации АН СССР, 1973.
174. Петухов С. В. О развитии опорного аппарата движений.— В
кн.: Тр. МФТИ. Сер. «Общая и молекулярная физика». М.:
МФТИ, 1975, № 6, с. 57-63.
175. Петухов С. В. Результаты физического моделирования
системы полукружных каналов человека. Функционирование
вестибулярного аппарата при осцилляциях головы.— В кн.:
Проблемы космической биологии: Сб. стат. М.: Наука, 1975, т. 31,
с. 14—64.
176. Петухов С. В. К вопросу о механизмах пространственного
восприятия.— В кн.: Влияние вибраций на организм человека. М.:
Наука, 1977, с. 149—152.
177. Петухов С. В. Организация сенсо-моторной системы и
инварианты проективной геометрии.— В кн.: Тр. XI чтений,
посвященных разработке научного наследия и развитию идей
К. Э. Циолковского. Секц. «Проблемы космической биологии
и медицины», 1976, М.: ИИЕТ АН СССР, 1978, с. 71—80.
178. К вопросу о применении проективной геометрии в
биомеханике / С. В. Петухов, М., 1978. Рукопись депонирована в ВИНИТИ
10.07.79, № 2304—78 Деп.
179. Петухов С. В. Симметрия и биологическая эволюция. Вурф-
ный анализ биотел и эстетическая система золотого вурфа.—*
В кн.: Тр. XIV чтений, посвященных разработке научного
наследия и развитию идей К. Э. Циолковского. Секц. «Проблемы
космической биологии и медицины», 1978. М.: ИИЕТ АН СССР,
1981, с. 42—48.
180. Петухов С. В. Некоторые характеристики развития опорно-
двигательного аппарата в связи с организацией рабочего
места оператора.— В кн.: VIII Гагаринские чтения: Тез. докл.
М.: ИИЕТ, 1979, с. 53—56.
225
181. Петухов J?. В. Исследования по неевклидовой биомеханике.—
В кн.: Кпомехапика систем «человек — машина». М.: Наука,
1981, с. 35—80.
182. Пидоу Д. Геометрия и искусство. М.: Мир, 1979. 332 с.
183. Пилецкий А. Модулор древнерусских зодчих.— Наука и жизнь,
1979, № 9, с. 66—69.
184. По поводу теории эволюции Л. С. Берга (Обсуждение с
участием С. В. Мейена, Ю. А. Урманцева, В. П. Алексеева,
А. К. Скворцова).—Природа, 1979, № 9, с. 113—126.
185. Повилейко Р. П. Симметрия в технике. Новосибирск: Новосиб.
электротехн. ин-т, 1970. 129 с.
186. Погребысский И. Б. Геометризация механики.— В кн.: От Лаг-
ранжа к Эйнштейну. М.: Наука, 1966, с. 251—271.
187. Постников М. М. Аналитическая геометрия. М.; Наука, 1973.
751 с.
188. Поттер Э. Патологическая анатомия плодов, новорожденных
и детей раннего возраста. М.: Медицина, 1971. 343 с.
189. Предпочтительные числа и ряды предпочтительных чисел.
ГОСТ 8032—56. М.: Изд-во стандартов, 1977. 10 с.
190. Принцип симметрии: Историко-методологические проблемы /
Под ред. Б. М. Кедрова, Н. Ф. Овчинникова, М.: Наука, 1978.
397 с.
191. Пуанкаре Л. Наука и гипотеза. М.: Мамонтовское изд-во,
1904. 268 с.
192. Пуанкаре А. Ценность науки. М.: Творческая мысль, 1906.
194 с.
193. Пуанкаре А. Наука и метод. СПб.: Изд-во Карбасникова, 1910.
238 с.
194. Пуанкаре А. Последние мысли. СПб.: ОГИЗ, 1923. 134 с.
195. Пэттен Б. М. Эмбриология человека. М.: Медгиз, 1959. 768 с.
196. Разумеев А. Н., Шипов А. А. Нервные механизмы
вестибулярных реакций.— В кн.: Проблемы космической биологии. М.:
Наука, 1969, т. 10. 341 с.
197. Раушенбах Б. В. Пространственные построения в
древнерусской живописи. М.: Наука, 1975. 184 с.
198. Раушенбах Б. В. Восприятие и перспективные изображения
пространства.— В кн.: Искусство и точные науки. М.: Наука,
1979, с. 142—184.
499. Винников Я. А., Газенко О. Г., Титова Л. К. и др. Рецептор
гравитации.— В кн.: Проблемы космической биологии. Л.:
Наука, 1971, т. 12. 523 с.
200. Рид К. Гильберт. М.: Наука, 1977. 366 с.
201. Розенфельд Б. А. Неевклидовы геометрии. М.: ГИТТЛ, 1955.
744 с.
202. Розенфельд Б. А. Неевклидовы пространства. М.: Наука, 1969.
547 с.
203. Розенфельд Б. А. История неевклидовой геометрии. М.:
Наука, 1976. 413 с.
204. Рорер Ф. О вестибулярной функции.— В кн.: Руководство по
нормальной и патологической физиологии. М.: Медгиз, 1926,
т. И, с. 985—1001.
205. Рохлин Д. Г. Костная система при эндокринных и
конституционных аномалиях. Л.; М.: ОГИЗ, 1931. 188 с.
206. Рохлин Д. Г. Рентгенооствология и рентгеноантропология. Л.;
М.: Биомедгиз, 1936, ч. 1, с. 310.
226
207. Руководство по зоологии / Под ред. Л. А. Зенкевича. В 6-ти т.
М.: Виомедгиз, 1937—1951.
208. Сабанеев Л. Этюды Шопена в освещении закона золотого
сечения.— Искусство, 1925, № 2, с. 20—30; 1927, № 2/3, с. 15—25.
209. Седов Л. И. Методы подобия и размерности в механике. М.:
Наука, 1977. 438 с.
210. Семенов В. М., Туник А. А. Инвариантность, устойчивость,
чувствительность.— Автоматика, 1977, т. 2, с. 82—88.
211. Сентаготаи Я. Роль отдельных лабиринтных рецепторов при
ориентации глаз и головы в пространстве. Л.: Наука, 1967.
140 с.
212. Сеченов И. М. Избранные философские и психологические
произведения. М.: ОГИЗ, 1947. 740 с.
213. Сеченов И. М. Избранные произведения. М.: Изд-во АН СССР,
1952, с. 270—426.
214. Симметрия в природе. М.: ИИЕТ АН СССР, 1971. 380 с.
215. Симонян К. С. Перитонит. М.: Медицина, 1971. 305 с.
216. Синнот Э. Морфогенез растений. М.: Изд-во иностр. лит., 1963.
480 с.
217. Сойер У. У. Прелюдия к математике. М.: Просвещение, 1972.
192 с.
218. Сойер У. У. Путь в современную математику. М.: Мир, 1972.
258 с.
219. Соколов А. А., Соколов Я. А. Математические закономерности
электрических колебаний мозга. М.: Наука, 1976. 110 с.
220. Соловьев И. М. К психологии осязательного восприятия.— В
кн.: Совещание по вопросам психологии познания: Тез. докл.
М.: Моск. рабочий, 1957, с. 12—13.
221. Соркин Э. Поверить алгеброй гармонию? — Техника и наука,
1977, № 9, с. 34—36.
222. Стулов Н. Н., Шафрановский И. И. В. И. Вернадский о
симметрии природы.— Зап. Всесоюз. минерал, о-ва, 1963, вып. 5,
с. 583—600.
223. Суханов В. Б. Общая система симметрической локомоции
наземных позвоночных и особенности передвижения низших
тетрапод. Л.: Наука, 1968. 440 с.
224. Сулима Ю. Г. Биосимметрические и биоритмические явления
и признаки у сельскохозяйственных растений. Кишинев:
Изд-во АН МССР, 1970. 148 с.
225. Тимердинг Г. Е. Золотое сечение. Петроград: Науч. книго-
изд-во, 1924, НО с.
226. Туркевич Б. Г. Некоторые данные зависимости анатомического
строения костного лабиринта от положения тела и степени
подвижности птиц.— Рус. физиол. журн. им. Сеченова, 1931,
т. 14, вып. 1, с. 71—75.
227. Тюмянцев II. Ф. Несколько точек опоры для суждения о
вестибулярном нистагме.— Иркут. мед. журн., 1925, т. 3, № 1/2,
с. 79—91.
228. Тюмянцев Н. Ф. О механизме вращательного нистагма.— Рус.
отоларингология, 1926, № 3, с. 171—173.
229. Урманцев Ю.'А. О проявлениях и значении правизны и
левизны у растений: Автореф. дис. ... канд. биол. наук. М.: Ии-т
физиологии растений им. К. А. Тимирязева, 1963.
230. Урманцев Ю. А. О философском и естественнонаучном
значении некоторых проявлений правизны и левизны в живой при-
227
роде.— В кн.: О сущности жизни: Сб. стат. М.: Наука, 1964,
с. 170—191.
231. Урманцев Ю. А. Биосимметрика. Симметрия и диссиметрия
цветков растений.—Изв. АН СССР. Сер. биол., 1965, № 1,
с. 75—87.
232. Урманцев Ю. А. О свойствах D и L модификаций
биологических объектов.— Усп. соврем, биол., 1966, т. 61, № 3, с. 374—
389.
233. Урманцев Ю. А. Золотое сечение.—Природа, 1968, № 11, с. 33—
40.
234. Урманцев Ю. А. Поли- и изоморфизм в живой и неживой
природе,— Вопр. филос, 1968, № 12, с. 77—88.
235. Урманцев Ю. А. Специфика пространственных и временных
отношений в живой природе.— В кн.: Пространство, время,
движение: Сб. стат. М.: Наука, 1971, с. 215—241.
236. Урманцев Ю. А. Симметрия природы и природа симметрии.
М.: Мысль, 1974. 229 с.
237. Урманцев Ю. А. Об определении знаков энантиоморфизма
некоторых нехимических (биологических) диссизомеров
посредством химических.— Журн. общ. биол., 1979, т. XL, № 3,
с. 351-367.
238. Урманцев Ю. А., Каден Н. Я. Изомерия в живой природе.—
Бот. журн., 1971, т. 56, № 2, с. 161—174; № 8, с. 1060—1067.
239. Урманцев Ю. А., Смирнов А. М. Правизна и левизна
изолированных корней в стерильной культуре.— Физиол. раст.,
1973, т. 20, вып. 4, с. 677—687.
240. Ухтомский А. А. Собр. соч.: В 6-ти т. JL,- ЛГУ, 1945.
241. Фейнман Р., Лейтон Р., Сэндс М. Фейнмановские лекции по
физике: В 10-ти т. М.: Мир, 1965.
242. Фрей-Висслинг А., Мюлеталер К. Ультраструктура
растительной клетки. М.: Мир, 1968. 310 с.
243. Хамермеш М. Теория групп и ее приложения к физическим
проблемам. М.: Мир, 1966. 290 с.
244. Хайнд Р. Поведение животных. М.: Мир, 1975. 855 с.
245. Харичев В. В., Шмидт А. А., Якубович В. А. Об одной новой
задаче распознавания образов.— АиТ, 1973, № 1, с. 109—122.
246. Хохрин А. В. Внутривидовая дисимметрическая изменчивость
древесных растений в связи с их экологией: Автореф. дис....
... д-ра биол. наук. Свердловск: Урал. науч. центр. Ин-т
экологии растений и животных АН СССР, 1977.
247. Хэмбидж Д. Динамическая симметрия в архитектуре. М.:
Изд-во Всесоюз. акад. архитект., 1936. 202 с.
248. Цейзинг А., Виппер Ю. Ф. Золотое сечение как основной
морфологический закон в природе и искусстве. М., 1876. 60 с.
249. Циолковский К. Э. Механика в биологии: Рукопись.— Архив
АН СССР. Моск. отд-ние, ф. 555, оп. 1, № 297, с. 1—120.
250. Циммерман Г. С. Ухо и мозг. М.: Медицина, 1967. 403 с.
251. Цион И. Ф. Об отправлениях полукружных каналов и об их
роли при образовании наших представлений о пространстве.—
Военно-мед. журн., 1879, т. 134, № 6, с. 65—70.
252. Циркунов В. Ю. Об эстетической природе зодчества. М.:
Стройиздат, 1970. 220 с.
253. Цыганкова Э. Г. У истоков дизайна. М.: Наука, 1977. 111 с.
254. Чебанов С. Г. Материалы к физиологии полукружных
каналов: О значении гидравлических явлений в эндолимфе для
228
физиологии полукружных каналов (ампулярпыл рецепторов):
Автореф. дис. ...д-ра биол. наук. Л.: Военно-мед. акад. им.
Кирова, 1956.
255. Чеботарев П. Г. Теория групп Ли. М.; Л.: ГИТТЛ, 1940. 395 с.
256. Четверухин Н. Ф. Проективная геометрия. М.: Просвещение,
1969. 367 с.
257. Чудинов Э. М. А. Эйнштейн об отношении геометрии к
реальности.— В кн.: Эйнштейновский сб. 1971 г. М.: Наука, 1972,
с. 302—316.
258. Шафрановский И. И. Симметрия в природе. Л.: Наука, 1968.
184 с.
259. Шафрановский И, И. Лекции по кристалломорфологии. М.:
Высш. школа, 1968. 173 с.
260. Шафрановский И. И. Кристаллографические представления
И. Кеплера и его трактат «О шестиугольном снеге». М.:
Наука, 1971, 21 с.
261. Шафрановский И. И. История кристаллографии. Л.: Наука,
1978. 295 с.
262. Шванвич Б. Н. Курс общей энтомологии. М.; Л.: Сов. наука,
1949. 900 с.
263. Шванвич Б. Я. Введение в энтомологию. Л.: ЛГУ, 1959. 342 с
264. Шевелев И. Ш. Геометрическая гармония. Опыт исследования
пропорциональности в архитектуре. Кострома: Костромское
кн. изд-во, 1963. 107 с.
265. Шевелев И. Ш. Логика архитектурной гармонии. М.: Стройиз-
дат, 1973. 179 с.
266. Шмалъгаузен И. И. Основы сравнительной анатомии
позвоночных животных. М.: Наркомпрос РСФСР, 1938. 490 с.
267. Шредингер Э. Основные принципы теории метрики цветов в
дневном свете.— В кн.: Новые пути в физике. М.: Наука, 1971,
с. 135—192.
268. Шубников А. В. О работах Пьера Кюри в области физики.—
Усп. физ. наук, 1956, т. 59, № 4, с. 591—602.
269. Шубников А. В., Копцик В. А. Симметрия в науке и
искусстве. М.: Наука, 1972. 339 с.
270. Эйзенштейн С. М. О строении Вещей.— Искусство кино, 1939,
№ 6, с. 7—20.
271. Эйнштейн А. Собр. науч. трудов: В 4-х т. М.: Наука, 1965.
272. Эндрюс Д. П. Механизмы коррекции ошибок в восприятии.—
В кн.: Инженерная психология за рубежом. М.: Прогресс,
1967, с. 222—243.
273. Юнг Дж. В. Проективная геометрия. М.: ГИМЛ, 1949. 183 с.
274. Якимов В. П. Онтогенетическое формирование пропорции
передних конечностей у человека и млекопитающих.— Зоол.
журн. 1946, т. 25, вып. 2, с. 167—170.
275. Якимов В. П. Развитие скелета передних конечностей
человека и некоторых млекопитающих.— Зоол. журн., 1947, т. 26,
вып. 4, с. 351—356.
276. Якимов В. П. О противопоставлении большого пальца руки.—
Журн. общ. биол., 1949, т. 10, с. 3—9.
277. Янг Л. Р. Модель вестибулярной системы управления.— В кн.:
Тр. Междунар. симпоз. по техническим и биологическим
проблемам управления, Ереван, 1968. М.: Наука, 1970, с. 93—119.
278. Adler I. A model of contact pressure in phyllotaxis.— J. Theor.
Biol, 1974, v. 45, p. 1—79.
229
079 Akishige Y. Studies on constancy problems in Japan.— Psycho-
' logia, 1958, v. 1, p. 143—157.
280. Akishige Y. Perceptual space and the law of conservation of
perceptual information.— In: XVIII Intern. Congr. Psychol. Symp.
19: Perception of space and time. Moscow, 1966, p. 42—49.
281 Akishige Y. Perceptual space and the law of conservation of
perceptual information.—Ztschr. Psychol., 1967, v. 173(3—4),
p. 153—158.
282 Akishige Y. Studies on constancy problem in Japan: II B.—
Psychologia, 1968, v. 11, N 1/2, p. 43—55; N 3/4, p. 127—138.
283. Anson B. J., Harper D. C, Winch T. R. The vestibular and cob-
lear aqueducts: developmental and adult anatomy of their
contents and parietes.— In: Third symp. on the role of the
vestibular organs in space exploration. Pensacola (Fla.), 1967, p. 125—
148.
284. Archibald R. C. Golden section.—Amer. Math. Mon., 1918, v. 25,
p. 232—238.
285. Aspects of form: A symposium on form in nature and art / Ed.
by L. L. White. L.: Humphries, 1968. 254 p.
286. Baravalle H. Die Geometrie des Pentagramms und der Goldene
Schnitt. Stuttgart, 1950. 30 S.
287. Barut A. 0., Haugen R. B. Theory of the conformally invariant
mass.— Ann. Phys., 1972, v. 71, N 2, p. 519—541.
288. Bast Th. H., Anson B. J. The temporal bone and the ear. N. Y.:
Springfeld, 1949. 350 p.
289. Beard R. S. Patterns in space. Palo Alto (Calif.), 1973. 237 p.
290. Beck C, Bader J. Ein Beitrag zur feineren Anatomie des men-
schlichen Innenohres. Zusammengefabt dargestellt in einem
neuen Labyrinthmodel.— Arch. Ohren-, Nasen- und Kehlkop-
fheilk., 1963, Bd. 181, H. 3, S. 245-267.
291. Berrill N. J. Growth, development and pattern. San Francisco:
Freeman, 1961. 555 p.
292. Birkhoff C. D. A mathematical theory of aesthetics and its
applications to poetry and music.— Rice Inst. Pamphlet, 1932, v. 19,
p. 189—342.
293. Birkhoff C. D. Aesthetic measure. Cambrige (Mass.): Harvard
Univ. Press, 1933. 225 p.
294. Blaivas A. S. Visual analysis: theory of Lie group
representations.— Math. Biosci., 1976, v. 28, N 1/2, p. 45—67.
295. Blaivas A. S. Visual analysis: theory of Lie-group
representations. II. Computations.—Math. Biosci., 1977, v. 35, N 1/2, p. 101—
105.
296. Blank A. A. The non-euclidean geometry of binocular visual
space.— Bull. Amer. Math. Soc, 1954, v. LX, p. 376.
297. Blank A. A. The geometry of vision.—Brit. J. Physiol. Opt., 1957,
v. XIV, p. 154—165; 222—235.
298. Blank A. A. Analysis of experiments in binocular space
perception.— J. Opt. Soc. Amer., 1958, v. XLVIII, p. 911—925.
299. Blank A. A. Curvature of binocular visual space. An
experiment— J. Opt. Soc. Amer., 1961, v. 51, N 3, p. 335—339.
300. Blum H. Biological shape and visual science.—J. Theor Biol
1973, v. 38, p. 205—287.
301. Bolle G. Theorie des Blaltstcllung. В., 1939. 320 S.
302. Bonner J. T. Morphogenesis. N. Y.: Athencum, 1963. 450 p.
230
303. Bookstlen F. L. The study of shage transformation after d'Arcy
Thompson.— Math. Biosci., 1977, v. 34, N 3/4, p. 177—219.
304. Boring E. G. Sensation and perception in the nistory of
experimental psychology. N. Y.: Appleton-Century Co, 1942. 644 p.
305. Borissavlievitch M. The golden number and the scientific
aesthetics of architecture. L.: Tiranti, 1958. 91 p.
306. Borissavlievitch M. Le nombre d'or et I'esthetique scientifique
de l'architecture. P.: Libr-Blanchard, 1963. 88 p.
307. Braus H. Anatomie des Menschen. В.: Springer-Verl., 1960.
Bd. 1—3. Bd. 1. Bewegungsapparat, 1954, 789 S.; Bd. 2. Linge-
weide, 1956. 686 S.
308. Cassirer E. The concept of group and the theory of perception.—
Phil, and Phenomenol. Res., 1944, N 5, N 1, p. 1—36.
309. Castell L. Analysis of space — time structure in elementary
particle physics.— Nuovo cim., 1966, v. 46, N 1, p. 1—38.
310. Castell L. A new quantum number derived from conformal inva-
riance.— Nucl. Phys., 1969, N 5, p. 231—236.
311. Castell L. Representations of conformal Symmetry by para-bose
operators and supersymmetry.— Nuovo cim., 1975, v. 29A, N 4,
p. 445—451.
312. Church A. H. The relation of phyllotaxis to mechanical laws. L.:
Williams and Norgate, 1904. 353 p.
313. Church A. H. On the interpretation of phenomena of phyllotaxis.
Oxford, 1920. 58 p. (Bot. mem. Oxford Univ. V. 6).
314. Cohen В., Suzuki J. Т., Shanzer S., Bonder M. B. Semicircular
canal control of eye movements.— In: The oculomotor system /
Ed. M. B. Bender. N. Y.; L.: 1964, p. 163—192.
315. Cohen В., Suzuki J. Т., Bender M. B. Eye movements from
semicircular canal nerve stimulation in the cat.— Ann. Otol., Rhinol.
and Laryngol, 1964, v. 73, N 1, p. 153—164.
316. Colman S. Nature's harmonic unity: A treatise on its relation to
proportional form. N. Y.: Blom, 1971. 327 p.
317. Cook T. A. The curves of life. L.: Constable and Co, 1914. 490 p.
318. Coxeter G. The golden section, phyllotaxis and Wythoff's game.—
Scr. math., 1953, v. 19, p. 135—143.
319. Coxeter H. S. M. The role of intermediate convergents in Tait's
explanation for phyllotaxis.— J. Algebra, 1972, v. 20, p. 167—175.
320. Derome J. R. Biological similarity and group theory.—J. Theor.
Biol., 1977, v. 65, N 2, p. 366—378.
321. Desargues G. Oeuvres de Desargues / par M. Poudra. P., 1878,
t. 1, 2.
322. Egmond A. A. J. van, Groen J. J., Jongkees L. B. W. The
mechanics of the semicircular canal.— J. Physiol., 1949, v. 110, N 1/2,
o. 1-17.
323. Egmond A. A. J. van, Groen J. J., Jongkees L. B. W. The function
of the vestibular organ.— Pract. oto-rhino-laryngol., 1952, suppl. 2,
p. 40.
324. Essays on growth and form. Oxford: Clarendon Press, 1945.
408 p.
325. Fechner G. Th. Vorschule der Asthetick. Leipzig: Breitkopf und
Hartel, 1897. 264 S.
326. Fernandes G., Goldberg J. M. Physiology of peripheral neurons
innervating semicircular canals of the squirrel monkey: The
response to sinusoidal stimulation and dynamics of peripheral
vestibular system.— J. Neurophysiol., 1971, v. 34, N 4, p. 661—674.
231
327. Fischer R. Out on a (phantom) limb. Variation on the theme:
stability of body image and the golden section.— Perspect. Biol,
and Med., 1969, v. 12, p. 259—273.
328. Foley J. M. Desarguesian property in visual space.— J. Opt. Soc.
Amer., 1964, v. 54, N 5, p. 684—692.
329. Franguin P. Geometrie de la plante.— Rev. bio-math., 1974, v. 46,
p. 43—54.
330. Fulton Т., Rohrlich F., Witten L. Conformal invariance in
physics,— Rev. Mod. Phys., 1962, v. 34, N 3, p. 442—457.
331. Gaston J. Mise en evidence de relations directs entre certains
recepteure vestibulaires.—J. Physiol. (France), 1972, v. 65, N 2,
p. 210A.
332. Gernandt B. Response of mammalian vestibular neurons to
horizontal rotation and caloric stimulation.— J. Neurophysiol., 1949,
v. 12, N 2, p. 173—184.
333. Gibson J. J. Observations on active touch.— Psychol. Rev., 1962,
v. 69, N 6, p. 477—490.
334. Graybiel A. Oculogravic illusion.— Arch. Ophthalmol., 1952, v. 48,
N 5, p. 605—615.
335. Graybiel A., Hupp D. J. Oculogyral illusion; from of apparent
motion which may be observed following stimulation of
semicircular canals.— J. Aviat. Med., 1946, v. 17, p. 3—6.
336. Graf H. Bibliographie zur Problem der Proportionen: Literatur
iiber Proportionen, Mass und Zahl in Architektur, bildender
Kunst und Natur. Wien: 1958. 120 S.
337. Graybiel A., Clark B. Duration of oculogyral illusions as a
function of the interval between angular acceleration and
deceleration; its significance in terms of dynamics of semicircular
canals in man.— J. Appl. Physiol, 1952, v. 5, p. 147—152.
338. Groen J. J. The semicircular canal systems of organs of
equilibrium. I,— Phys. Med. and Biol., 1956, v. 1, N 2, p. 103—117.
339. Groen J. J. The semicircular canal systems of organs of
equilibrium. II.— Phys. Med. and Biol, 1957, v. 1, N 3, p. 225—242.
340. Groen J. J., Lowenstein O., Vendrik A. J. H. The mechanical
analysis of the responses from the end-organs of the horizontal
semicircular canal in the isolated elasmobranch labyrinth.— J.
Physiol, 1952, v. 117, N 3, p. 329—346.
341. Guedry F. E. Vestibular system.— In: Bioastronautics data book.
Wash.: NASA SP 3006, 1964, p. 363—381.
342. Haase R. Geschichte des harmonikalen Pythagoreismus. Wien:
Verl, 1969. 308 S.
343. Habenicht B. Beitrage zur mathematischen Begriindung einer
Morphologie der Blatter. В.: О. Salle, 1955. 200 S.
344. Hadziselimovic H., Savkovic L. J. Appearance of semicircular
canals in birds in relation to mode of life.— Acta anat. (Basel),
1964, v. 57, N 4, p. 306—315.
345. Hagenmaier O. Der golden Schnitt. В., 1958. 60 S.
346. Handbuch der Zoologie / Gegriindet von W. K. Pukenthal. В.: De
Grugter, 1923—1962. Bd. 1—8.
347. Hallpike C. S., Hood J. D. The speed of the slow component of
ocular nystagmus induced by angular acceleration of the head:
its experimental determination and application to the physical
theory of the cupulae mechanism.— Proc. Roy. Soc. B, 1952,
v. 141, p. 216-230.
232
348. Heerden P. J. van. Concept of distance in affinc geometry.— J.
Opt. Soc. Amer., 1956, v. 46, N 11, p. 1000.
349. Held R. Plasticity in sensory-motor systems.— Sci. Amer., 1965,
v. 213, p. 84—94.
350. Held R., Rekosh J. Motor-sensory feedback and the geometry of
visual space.— Science, 1965, v. 141, p. 722—723.
351. Hemingway A. Selection of men for aeronautical training based
on sesceptibility to motion sickness.— J. Aviat. Med., 1946, v. 17,
N 2, p. 153—163.
352. Hoffman W. С The Lie algebra of visual perception.— J. Math.
Psychol., 1966, v. 3, p. 65—98.
353. Hoffman W. C. Comments on «optical art».— Appl. Opt., 1966,
v. 5, p. 873—874.
354. Hoffman W. С Neuron as a Lie group germ and a Lie product.—
Quart. Appl. Math., 1968, v. 25, p. 423—440.
355. Hoffman W. C. Higher visual perception as prolangation of the
basic Lie transformation group.— Math. Biosci., 1970, v. 6,
p. 437-471.
356. Hoffman W. C. Visual illusions of angle as an application of
Lie transformation groups.—ST AM Rev., 1971, v. 13(2), p. 169—
184.
357. Hoffman W. С Memory grows.— Kybernetik, 1971, Bd. 8(4),
S. 151—157.
358. Holland J. N. Studies in structure. L.: Macmillan Press, 1972.
233 p.
359. Huntley H. E. The divine proportions, a study in mathematical
beauty. N. Y.: Dover, 1970. 341 p.
360. Igarashi M. Dimensional study of the vestibular end organ
apparatus.— In: Second symposium on the role of vestibular
organs in space exploration: Papers Calif. Pensacola (Fla.), 1966,
p. 47—54.
361. Ingraham R. L. Conformal geometry and elementary particles.—
Nuovo cim., 1954, v. 12, N 6, p. 825—851.
362. Iterson G. van. Mathematische und mikroskopishanatomische
Studien fiber Blattstellungen nebst Betrachtungen fiber den
Schalenbur der Miliolinen. Amsterdam, 1907. 220 S.
363. Iterson G. van. Nieuwe studien over bladstanden. Amsterdam:
Noord-Hollandsche uitg. moatschappij., 1964. 151 s.
364. Ivins W. M. Art and geometry: A study in space intuitions.
Cambridge, 1946. 264 p.
365. Jaeger F. M. Lectures on the principle of symmetry. Amsterdam,
1920. 282 p.
366. Jean R. V. La G-entropie en phyllotaxie.— Rev. bio-math., 1976,
N 5, p. 111—130.
367. Johnson W. H., Stubbs R. A., Kelk F. G., Franks W. R. Stimulus
required to produce motion sickness.— J. Aviat. Med., 1951, v. 22,
p. 365—380.
368. Johnson W. H. Some experiments on the relative effectiveness
of various types of accelerations on motion sickness.— J. Aviat.
Med., 1961, v. 32, p. 205—215.
369. Johnson W. H., Mayne J. W. Stimulus required to produce
motion sickness; restriction of head movement as a preventive of
airsickness field studies on airborn Iroops.— J. Aviat. Med., 1953,
V. 24, p. 400—407,
233
370 Kampfe L., Kiltel R.t Klapperatilck J. Leilfaden dcr Anatomie dor
Wirbeltiere. Wien, 1966. 680 S.
371. Kastrup H. A. Conformal group in space-time.—Phys. Rev., 1966
v. 142, N 4, p. 1060—1071.
372. Kastrup H. A. Position operators, gauge transformations and the
conformal group.— Phys. Rev., 1966, v. 143, N 4, p. 1021—1027.
373. Kienle G. Parallelenaxiom und Sehraum.— Naturwissenschaften
1962, Bd. 49, S. 359.
374. Kienle G. Geometrische Axiome, nicht-euklidische Abbildungs-
modelle und Sehraum.— Optik, 1963, Bd. 20, S. 333—372.
375. Kienle G. Experiments concerning the non-euclidian structure of
the visual space.—In: Bioastronautics. N. Y.; L., 1964, p. 386—
400.
376. Kienle G. Die optischen Wahrnehmungsstorungen und die nich-
teuklidische Struktur des Sehraumes. Stuttgart: Thieme, 1968.
172 S.
377. Kilmer W L. On growing pine cones and other Fibonacci fruits
McCulloch localized algorithm.— Math. Biosci., 1971, v. 11, p. 53—
57.
378. Kley W., Reinecken R. Drehpendelpriifung.— Ztschr. Laryngol.
Stuttgart, 1961, Bd. 40, S. 68—76.
379. Komai T. Phyllotaxis like arrangement of organs and zooids in
some medusae.— Amer. Natur., 1951, v. 85, p. 75—76.
380. Kuroda T. Distance constancy: functional relationships between
apparent distance and physical distance.— Psychol. Forsch.
Ztschr. Psychol., Berlin, 1971, Bd. 34, S. 199—219.
381. Larson P. The golden section in the earliest notated western
music— Fibonacci Quart., 1978, v. 16, N 6, p. 513—515.
382. Leigh E. G. The golden section and spiral leaf-arrangement.—
Trans. Conn. Arts and Sci., 1972, v. 44, p. 165—176.
383. Leppik E. E. Evolutionary differentiation of the flower head of
the composital II.—Ann. bot. fenn., 1970, t. 7, s. 325—352.
384. Lewin К Principles of topological psychology. N. Y.; L., 1936.
280 p.
385. Linn С F. The golden mean. N. Y.: 1974. 200 p.
386. Lioseau J. E. La phyllotaxie. P.: Masson et Cie, 1969. 195 p.
387. Lorente de No. Ausgewahlte Kapitel aus der vergleichenden Phy-
siologie des Labyrinthes.—Ergeb. Physiol, 1931, Bd. 32, S. 73—
242.
388. Lorente de No. The regulation of eye positions and movements
induced by the labyrinth.— Laryngoscope, 1932, v. 43, N 4,
p. 233—330.
389. Lowenstein O. Labyrinth and equilibrium.— In: Symp. Soc. Exp.
Biol., 1950, v. 4, p. 60—82.
390. Lowenstien O., Sand A. The individual and integrated activity
of the semicircular canals of the Elasmobranch labyrinth.— J.
Physiol., 1940, v. 99, N 1, p. 89—101.
391. Lowenstein 0-, Sand A. The mechanism of the semicircular
canal: A study of the responses of single-fibre preparations to
angular accelerations and to rotation at constant speed,— Proc.
Roy. Soc. B, 1940, v. 129, p. 265—280.
392. Luneburg R. K. Mathematical analysis of binocular vision.
Princeton: Univ. Press, 1947, p. 104.
334
393. Luneburg R. К. Metric methods in binocular visual perception.—
In: Studies and essays. N. Y.: Intersci. Publ., 1948, Courant an-
niv. vol., p. 215—239.
394. Luneburg R. K. The metric of binocular visual space.— J. Opt.
Soc. Amer., 1950, v. 40, N 10, p. 627—642.
395. Mathai A. M., Davis T. A. Constructing the sunflower head.—
Math. Biosci., 1974, v. 20, p. 117—133.
396. Mayne R. The dynamic characteristics of the semicircular
canals.— J. Сотр. Psychol., 1950, v. 43, N 4, p. 309—319.
397. McCulloch W. S. Embodiments of mind. Cambridge (Mass.): MIT
Press, 1965. 438 p.
398. McNabb M. D. Phyllotaxis.—Fibonacci Quart, 1963, v. 1, N 4,
p. 57—60.
399. Meyen S. V. Plant morphology in its homogenetical a.spects.—
Bot. Rev., 1973, v. 39, N 3, p. 205—260.
400. Miller E. F., Graybiel A. The semicircular canals as a primary
etiological factor in motion sickness.— In: Fourth symposium
on the role of the vestibular organs in space exploration. Pensa-
cola (Fla.), 1968, p. 69—82.
401. Money К. Е., Friedberg I. The role of the semicircular canals in
causation of the motion sickness and nystagm in the dog.— Ca-
nad. J. Physiol, and Pharmacol., 1964, v. 42, p. 793—802.
402. Money K. E., Bonen L., Beatty J. D. et al. Physical properties
fluids and structures of vestibular apparatus of the pigeon.—
Amer. J. Physiol., 1971, v. 220, N 1, p. 140—147.
403. Morecki A., Ekiel J., Fidelus K. Bionika ruchu. Warszawa: 1971.
310 p.
404. Miiller A. H. Einiges iiber Spirale und Schraubenformige Struktu-
ren bei fossilen Tieren unter besonderer Berucksichtigung taxo-
nomischer und phylogenetischer Zusammenhange. Teil 1—4.—
Monatsber. Dtsch. Akad. Wiss. Berlin, 1971, Bd. 13, N 4—5,
S. 345—359, S. 369—382; Bd. 13, N 6, S. 463—478; Bd. 13, N 8,
s. 613—624.
405. Muybridge E. Animals in motion. N. Y.: 1957. 220 p.
406. Needham C. The growth process in animals. N. Y., 1964. 400 p.
407. Neuman A. L'humanisation de l'espace. P.: 1956. 49 p.
408. Nicod I. La geometrie dans le monde sensible / Pref*. de B.
Russell. P.: Press. Univ. France, 1962. 160 p.
409. Peizer E., Wright D. W. Human locomotion.— In: Human
locomotion engineering. L.: Inst. Mech. Eng., 1974, p. 196—214.
410. Petukhov S. V. Investigations on non-euclidean biomechanics.
Projective geometry, Fibonacci numbers and the kinematic
scheme of human body.— In: Biomechanics VII. Warszawa, 1980,
p. 603—607.
411. Pfeifer F. Der goldene Schnitt und dessen Erscheinungsformen in
Mathematik, Natur und Kunst. Augsburg, 1885. 240 S.
412. Pipping N. Drei geometrische Miniaturen.— Acta Acad, aboen.,
1952, Bd. 18, N 10, S. 5—8.
413. Poincare H. The foundations of sciences: Science and
hypothesis, the value of science, science and method. Lancaster: Sci.
Press, 1964. 553 p.
414. Resnikoff H. L. Differential geometry and color perception.— J.
Math. Biol., 1974, v. 1, N 2, p. 97—131.
235
415. Richards E. J. The geometry of phyllotaxis and its origin.— in:
Symposium of the Society of experimental biology. Cambridge
Univ. Press, 1948, p. 216—280.
416. Richards F. J. Phyllotaxis: its quantitative expression and
relation to growth in the apex.— Phil. Trans. Roy. Soc. London B,
1951, v. 235, p. 509—564.
417. Richards T. J., Schwabe W. W. Analysis of growth; behavior of
plants and their organs.— In: Plant physiology. N. Y.: Academic,
1969, v. VA, chap. 2, p. 79—116.
418. Rose J. E., Mountcastle V. B. Touch and kinesthesis.— In:
Handbook of physiology. Wash.: Amer. Physiol. Soc, 1959, v. 1,
p. 387—429.
419. Rosen R. The generation and recognition of patterns in
biological systems.— Lect. Notes Biomath., 1977, N 5, p. 222—341.
420. Russell B. A. W. An essay on the foundations of geometry. N. Y.:
Dover, 1956. 201 p.
421. Schelling H. von. Advanced color geometry.— J. Opt. Soc. Amer.,
1955, v. 45, N 12, p. 1072—1079.
422. Schelling H. von. Concept of distance in affine geometry and its
application in theories of vision.— J. Opt. Soc. Amer., 1956, v. 46,
p. 309—315.
423. Schelling H. von. Reply to P. J. van Heerden.— J. Opt. Soc. Amer.,
1956, v. 46, N 11, p. 1000—1001.
424. Schelling H. von. Die Geometrie des beideaugigen Sehens.—
Optik, 1960, Bd. 17, H. 7, S. 345—364.
425. Schelling H. von. Experienced space and time.— In: Bioastro-
nautics. N. Y.; L., 1964, p. 361—385.
426. Scholfield P. H. The theory of proportion in architecture.
Cambridge, 1958. 160 p.
427. Schooling W. The Ф-progression.— In: Cook T. A. The curves of
life. L.: Constable and Co, 1914, p. 441—464.
428. Schwabe W. W. Chemical modification of phyllotaxis and its
applications.— In: Control mechanisms of growth and
differentiation: Symp. 25 of the Society for experimental biology.
Cambridge, 1971, p. 301—322.
429. Schweitzer B. von. Sinn der Perspektive.— In: Schumann F. K.,
Troll W., Wolf K. L. Die Gestalt. Tubingen, 1953, S. 40—66.
430. Steer R. W. The influence of angular and linear acceleration and
thermal stimulation on the human semicircular canal: Sci. D.
Thes. (Mass.): MIT, 1967. 250 p.
431. Steer R. W., Li Т., Young L. R., Meiry J. L. Physical properties
of the labyrinthine fluids and quantification of the phenomenon
of caloric stimulation.— In: Third symposium on the role
of the vestibular organs in space exploration. Pensacola (Fla.),
1967, p. 409—420.
432. Steinhausen W. Uber den Nachweis der Bewegung der Cupula
in der intakten Bogengansampulle des Labyrinthes bei der na-
turlichen rotatorischen und kalorischen Reizung.— Pflug. Arch.
ges. Physiol., 1931, Bd. 228, S. 322—340.
433. Steinhausen W. Uber die Beobachtung der Cupula in der Bogen-
gansampullen des Labyrinthes des lebanden Hechts.— Pfliig.
Arch. ges. Physiol. 1933, Bd. 232, S. 506—510.
434. Steinhausen W. Uber die Cupula.— Ztschr. Hals-, Nasen- und
Ohrenheilk., 1935, Bd. 39, S. 19—62.
236
435. Suzuki J. I., Cohen B. Head, eye, body and limb movements from
semicircular canal nerves.— Exp. Neurol., 1964, v. 10, N 5,
p. 393—405.
436. Symmetry and function of biological systems at the macromo-
lecular level.— In: Proc. Eleventh Nobel Symp. held Aug. 26—
29, 1968 / Ed. by A. Engstrom, B. Strandberg. Stockholm: Almg-
vist and Wiksell, 1969. 436 p.
437. Szentagothai J. The elementary vestibuloocular reflex arc.— J.
Neurophysiol., 1950, v. 13, N 6, p. 395—407.
438. Thompson D'Arcy W On growth and form. Cambridge Univ.
Press, 1942. 1116 p.
439. Thornley J. H. Phyllotaxis I, II.— Ann. Bot., 1975, v. 39, p. 491—
507, 511—526.
440; Thornley J. H. A model of apical bifucartion applicable to trees
and other organisms.— J. Theor. Biol, 1977, v. 64, p. 165—176.
441. Traite de zoologie: Anatomie, systematique, biologie / Public
sons la direction de Pierre-P. Grasse. P.: Masson, 1948—1970.
442. Trincker D., Silber J., Bartual J. Schwingungsanalyse der
vestibular, optokinetisch und durch elektrischc Reizung ausgelasten
Augenbewegungen beim Menschen.— Kybernetik, 1961, Bd. 1,
N 1, S. 21—28.
443. Tyler D. В., Bard P. Motion sickness.— Physiol. Rev., 1949, v. 29,
p. 311—369.
444. Vita J. de. Fibonacci, insects and flowers.— Fibonacci Quart.,
1978, v. 16, N 4, p. 315—317.
445. Vetter R. J., Weinstein S. The history of the phantom in
congenially absent limbs.—Neuropsychologia, 1967, N 5, p. 335—338.
446. Wardlaw C. W. Morphogenesis in plants. L.: Methuen, 1968.
240 p.
447. Weinstein S., Sprsen E. A. Phantoms in cases of congenital
absence of limbs.— Neurology, 1961, N 11, p. 905—911.
448. Wess J. The conformal invariance in quantum field theory.—
Nuovo cim., ЦЙ60, v. 18, N 6, p. 1086—1107.
449. Williams R. !F. The shoot apex and leaf growth: a study in
quantitative bfology. Cambridge Univ. Press, 1975. 265 p.
450. Wit G. de.; Seasickness (motion sickness), a labyrinthological
study,— Acta oto-laryngol., 1953, bd. 108, suppl., s. 1—56.
451. Wolf K., Kuhn D. Gestalt und Symmetrie. Tubingen: Wiemeyer,
1952. 64 S.
452. Wulf B. Uber die Dimensionen des Bogengangsystems bei den
Wirbelthieren.— Arch. anat. und Entwicklungsgesch., 1901, N 1,
S. 57—74.
453. Wust #. Untersuchungen des Vestibularapparates mit neuen
Drehschwachreizmethoden.— Arch. Ohr.— USW. Heilk. 7. Hal-
susw. Heilk., 1956, Bd. 168, S. 414—428.
454 Young D. A. On the diffusion theory of phyllotaxis.— J. Theor.
Biol., 1978, v. 71, N 3, p. 421—432.
ОГЛАВЛЕНИЕ
Введение 3
Глава I. Симметрии в биологии 8
1.1. Симметрии в биологических телах и законы
листорасположения 8
1.2. Изучение биологических симметрии как научное
направление 21
1.3. Симметрии и математическое естествознание 26
Глава II. Исследование конформных симметрии в биологии 30
11.1. Конформные симметрии в физике и в явлениях роста
и формообразования биологических тел . 20
11.2. Конформные симметрии в кинематической схеме
человеческого тела . 38
П.З. Ряд Фибоначчи и золотой вурф . 46
11.4. Золотой вурф как эстетический канон. Вопросы
архитектурной бионики . 50
11.5. К проблеме органических форм . 57
11.6. Конформная симметрия в моторике трехфаланговых
пальцев" . 60
11.7. Изучение конформной симметрии трехчленных
кинематических блоков у животных . 65
11.8. О возможном участии конформных симметрии в ге~
незисе пространственных представлений у
индивидуума . 77
11.9. Конформная симметрия и геометрия пространства
зрительного восприятия . 80
11.10. Концепция морфопоз и организация моторных
движений . . 86
11.11. Симметрограммы трехчленных кинематических
блоков при анализе моторных движений 97
Глава III. Исследование проективных симметрии в биологии 106
Ш.1. О возможном участии принципов проективной
геометрии в структурировании биологических явлений 106
Ш.2. Концепция Б. Рассела о врожденности
представлений индивидуума о положениях проективной
геометрии 1С9
Ш.З. Использование вурфов для распознавания
трансформируемых геометрических образов 111
238
III.4. Вурф пяти точек плоскости и пентасимметричные
биологические тела 115
II 1.5. Поиск проективных симметрии в расположении пяти
оконечных точек человеческого тела 119
Глава IV. Применение теоретико-групповых концепций для
моделирования психологических явлений 124
IV.1. Теория групп и психологические феномены
константности восприятия 124
IV.2. Применение высшей геометрии при моделировании
пространств восприятия 131
IV.3. Некоторые бионические проблемы машиноведения 136
Глава V. Исследование вестибулярного аппарата с помощью
методов симметрии . 141
V.I. Теория динамического подобия в биомеханике 141
V.2. Физическое моделирование системы полукружных
каналов вестибулярного аппарата 143
V.3. Эксперименты на масштабной физической модели
системы полукружных каналов человека 158
V.4. Физиологические эксперименты для проверки
результатов модельных исследований 170
Приложение I. Дополнительные сведения о числах
Фибоначчи . 198
1. Числа Фибоначчи и золотое сечение в эстетике
пропорции .- . 198
2. Некоторые математические свойства чисел Фибоначчи 203
Приложение II. Дополнительные сведения о конформной
геометрии 206
Приложение III. Дополнительные сведения о проективной
геометрии
210
Приложение IV. О принципе конформных блоков и
обобщенной кристаллографии . 213
Литература 218
Сергей Валентинович Петухов
БИОМЕХАНИКА,
БИОНИКА
И СИММЕТРИЯ
Утверждено к печати #-
Институтом машиноведения им. А. А. Благонрапова
Редактор Н. Н. Лебедева
Редактор издательства Ю. А. Юдина
Художник Е. Н. Волков
Художественный редактор Н. Н. Власик
Технический редактор Р. Г. Грузинова
Корректоры Д. Ф. Арапова, В. А. Березина
И Б № 21128
Сдано в набор С8.01.81
Подписано к печати 19.03.81.
Т-03291. Формат 84x108Vи
Бумага типографская Jsft l
Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 12,6. Уч.-изд. л. 13,9
Тираж 1850 экз. Тип. зак. 88
Цена 2 р. 20 к.
Ивдательство «Наука»
117864 ГСП-7, Москва, В-485, Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-69, Шубинский пер., 1Q