







































































































































































































































































































Author: Бязров Л.Г.
Tags: lichenes лишайники экология растений этология растение и окружающая среда метеорология климатология теория вероятностей и математическая статистика угрозы окружающей среде природа охрана окружающей среды ботаника биология экология монография семейство лишайников
ISBN: 5-89176-183-1
Year: 2002




Л.Г. Бязров ЛИШАИНИКИ В ЭКОЛОГИЧЕСКОМ МОНИТОРИН
Л.Г. Бязров
ЛИШАЙНИКИ
В ЭКОЛОГИЧЕСКОМ МОНИТОРИНГЕ
НАУЧНЫЙ МИР
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ИНСТИТУТ ПРОБЛЕМ ЭКОЛОГИИ И ЭВОЛЮЦИИ им. А.Н. СЕВЕРЦОВА
Л.Г. БЯЗРОВ
ЛИШАЙНИКИ В ЭКОЛОГИЧЕСКОМ
МОНИТОРИНГЕ
Ответственный редактор:
член-корреспондент РАН проф. Д.А. Криволуцкий
МОСКВА
НАУЧНЫЙ МИР
2002
УДК 582.29:581.5:551.510.42:519.27:504.3.064.36:502.55
ББК 28.58
Б99
Бязров Л.Г.
Б99 ЛИШАЙНИКИ В ЭКОЛОГИЧЕСКОМ МОНИТОРИНГЕ. - М.: Научный мир,
2002.-336 с.
ISBN 5-89176-183-1
Монография обобщает результаты оригинальных исследований автора, а также обширные
опубликованные материалы о применении лишайников в мониторинге состояния окружающей
среды, главным образом, качества воздушного бассейна. В книге содержатся сведения о совре-
менных трактовках понятия “лишайники”, характеризуются их особенности, позволяющие ис-
пользовать представителей лихенобиоты как индикаторов качества среды, динамики ее свойств
в пространстве и во времени; показаны возможные варианты лихеноиндикации при различном
размещении источников загрязнения; предложен расчет стоимости лихеноиндикационного ис-
следования. Подробно характеризуются признаки представителей лихенобиоты, которые фик-
сируют при проведении мониторинговых исследованиях. Большое внимание уделено методам
учета видового состава лишайников, выявления формируемых ими группировок, транспланта-
ции избранных видов, особенностям сбора материала для определения накопления лишайниками
различных загрязнителей. Отдельный раздел монографии написан по результатам изучения
экологических последствий аварии на Чернобыльской АЭС и испытательных взрывов термо-
ядерного оружия на Семипалатинском ядерном полигоне. Сообщаются также результаты зони-
рования территории Москвы относительно качества воздушного бассейна по особенностям
распространения и развития лишайников.
Монография предназначена для специалистов по экологии урбанизированных и промыш-
ленных территорий, научных сотрудников заповедников и исследовательских лабораторий, пе-
дагогов и студентов.
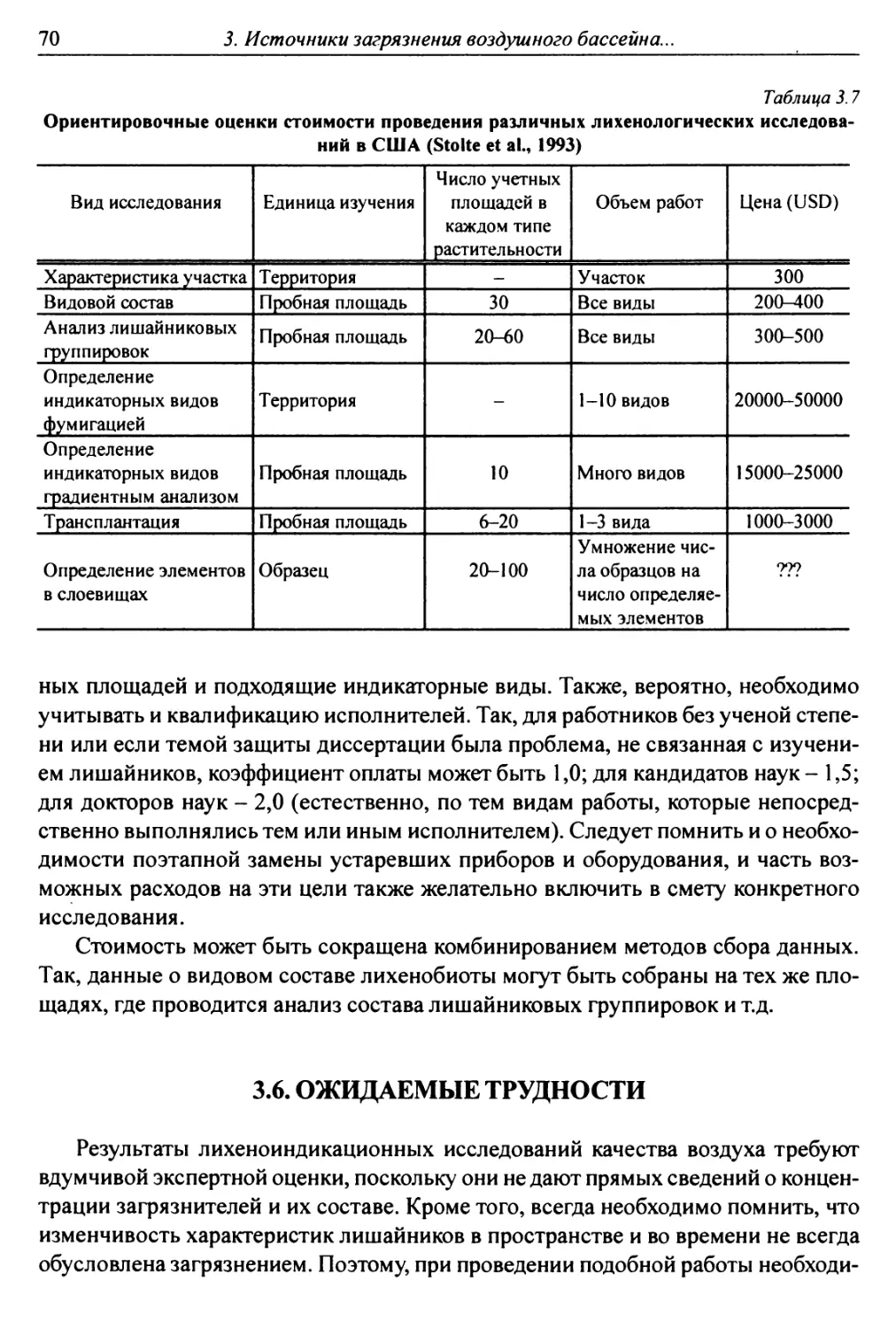
Рис. 22; табл. 40. Библ. 1116 назв.
УДК 582.29:581.5:551.510.42:519.27:504.3.064.36:502.55
ББК 28.58
Публикуется при финансовой поддержке Российского фонда фундаментальных
исследований (проект № 02-04-62003)
ISBN 5-89176-183-1
©Л.Г. Бязров, 2002
© ИПЭЭ РАН, 2002
© Научный мир, 2002
СОДЕРЖАНИЕ
Введение ................................................. 9
1. Биология и экология лишайников ........................ 12
1.1. Микобионт ........................................ 13
1.2. Фотобионт ........................................ 14
1.3. Взаимоотношения микобионта и фотобионта
в слоевище лишайника ................................. 14
1.4. Образование вегетативного тела
(слоевища, или таллома) лишайника .................... 16
1.5. Систематика ...................................... 17
1.6. Типы слоевищ и жизненные формы лишайников ........ 17
1.7. Субстрат ......................................... 19
1.8. Форофит .......................................... 21
1.9. Свет, тепло ...................................... 23
1.10. Водный режим, фотосинтез, дыхание лишайников .... 25
1.11. Географическое положение территории ............ 26
1.12. Конкуренция ..................................... 27
1.13. Влияние деятельности человека.
Использование лишайников .............................. 28
2. Улавливание и поглощение лишайниками минеральных
и органических веществ, их накопление и потеря;
токсичность элементов ..................'................ 30
2.1. Содержание минеральных элементов в лишайниках .... 31
2.2. Потребности в питательных веществах .............. 32
2.3. Источники поступления элементов .................. 33
2.3.1. Атмосферные источники ...................... 33
4
Оглавление
2.3.2. Субстратные источники ........................... 34
2.4. Механизмы накопления .................................. 36
2.4.1. Обмен ионов ..................................... 36
2.4.2. Внутриклеточное поглощение ...................... 37
2.4.3. Улавливание взвешенных в атмосфере частиц ....... 39
2.5. Локализация элементов в слоевище ...................... 39
2.6. Исследования выпадений различных веществ
с помощью лишайников ....................................... 41
2.7. Токсичность металлов .................................. 44
2.8. Толерантность к воздействию металлов .................. 45
2.9. Влияние на гидрологические циклы ...................... 47
2.9.1. Связь минерального питания с водным обменом ..... 48
2.10. Минеральный цикл на уровне биогеоценоза:
перехват сухих и сырых выпадений ........................... 48
2.11. Почвообразование ..................................... 50
3. Источники загрязнения воздушного бассейна,
некоторые примеры их пространственного размещения
и возможные варианты лихеноиндикации ........................... 52
3.1. Состав атмосферы ...................................... 52
3.2. Источники загрязнения атмосферы и их характеристика ... 53
3.3. Приоритетные загрязнители. Оценка уровня загрязнения
атмосферы .................................................. 55
3.4. Некоторые возможные варианты размещения
искусственных источников загрязнения ....................... 57
3.4.1. Изолированный источник загрязнения с постоянным
объемом и составом загрязнителей ....................... 58
3.4.2. Изолированный источник загрязнения с уменьшающимся
объемом выбросов ....................................... 60
3.4.3. Изолированный источник загрязнения
с увеличивающимся объемом выбросов ..................... 61
3.4.4. Маломощный изолированный источник дополнен
новым источником загрязнения ........................... 63
3.4.5. Территория не имеет источников загрязнения,
но загрязняется в результате трансрегионального переноса
загрязнителей воздушными массами ....................... 63
3.4.6. Неравномерное рассредоточение нескольких источников
с перекрывающимся действием загрязнителей .............. 64
3.4.7. Неравномерное рассредоточение нескольких источников
с перекрывающимся воздействием и уменьшающимися
объемом выбросов и составом загрязнителей .............. 65
6
Оглавление
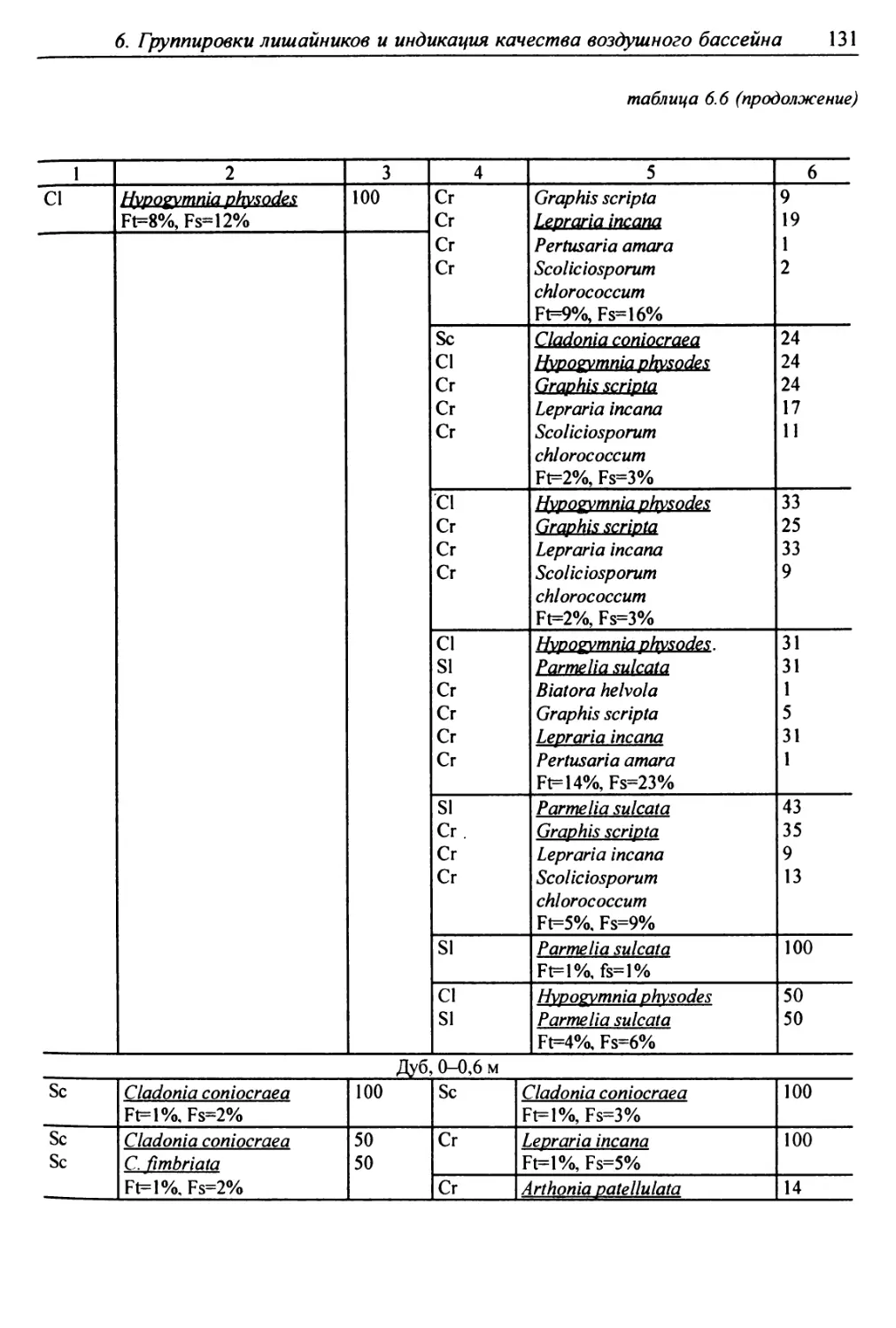
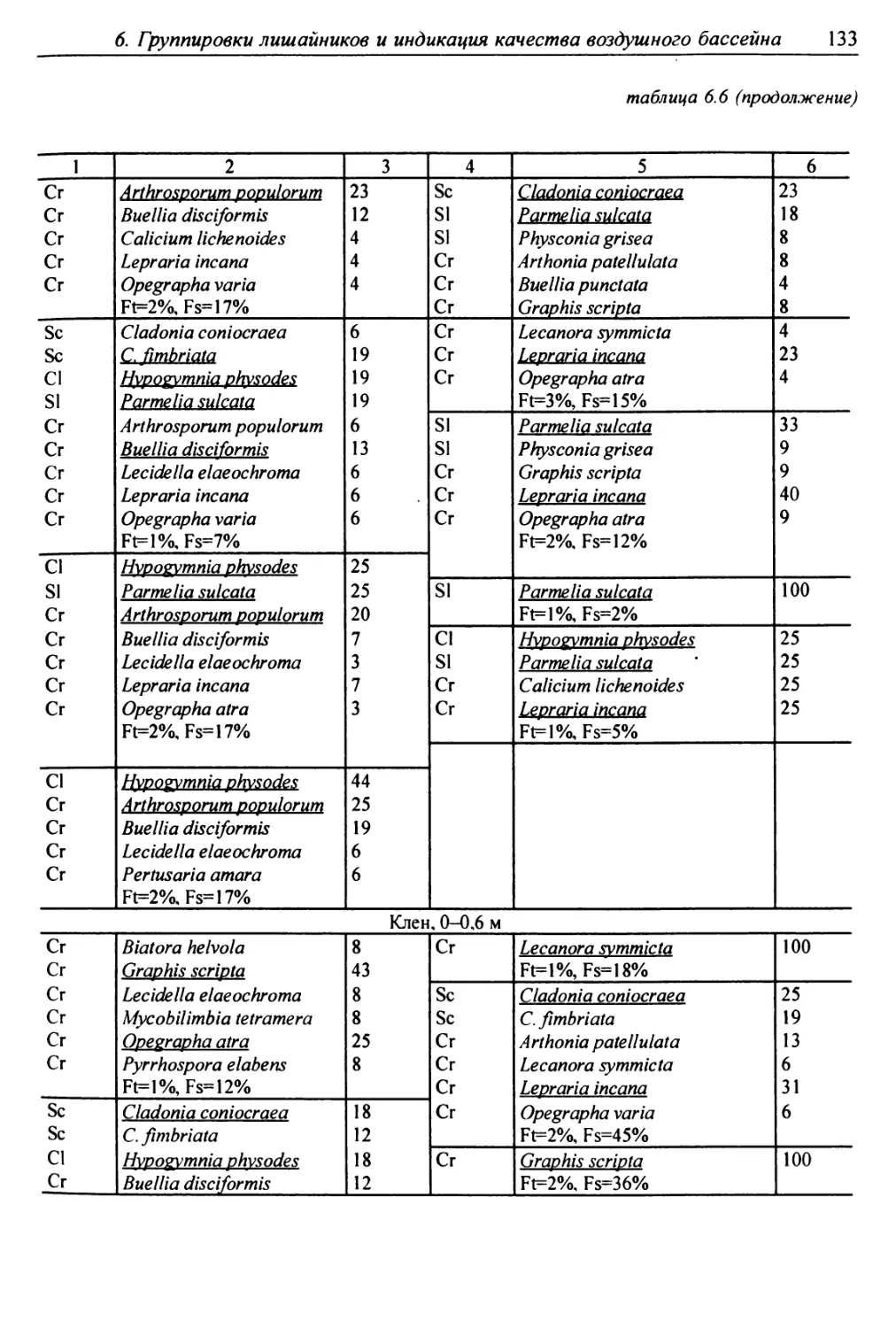
6. Группировки лишайников и индикация качества
воздушного бассейна ........................................ 117
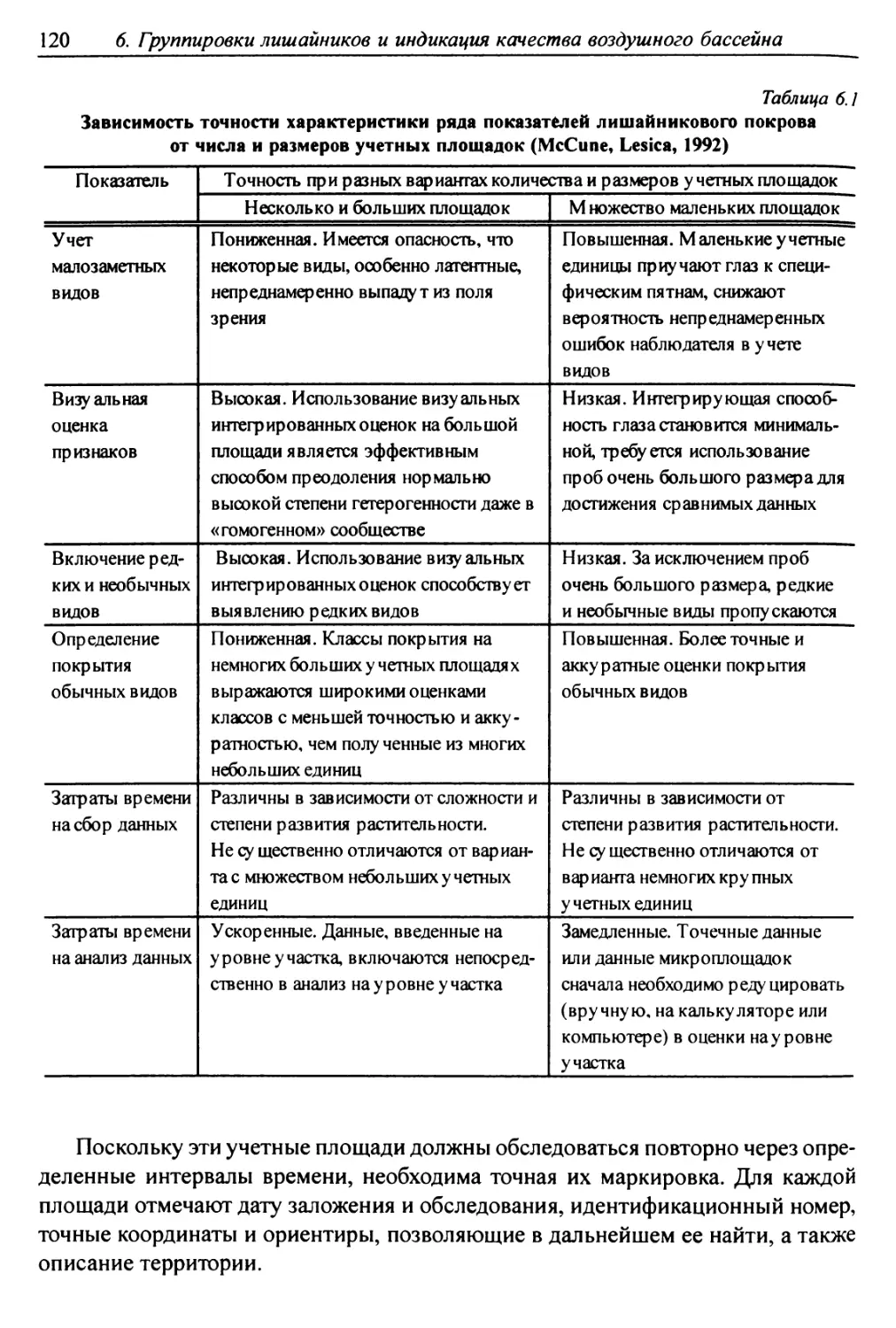
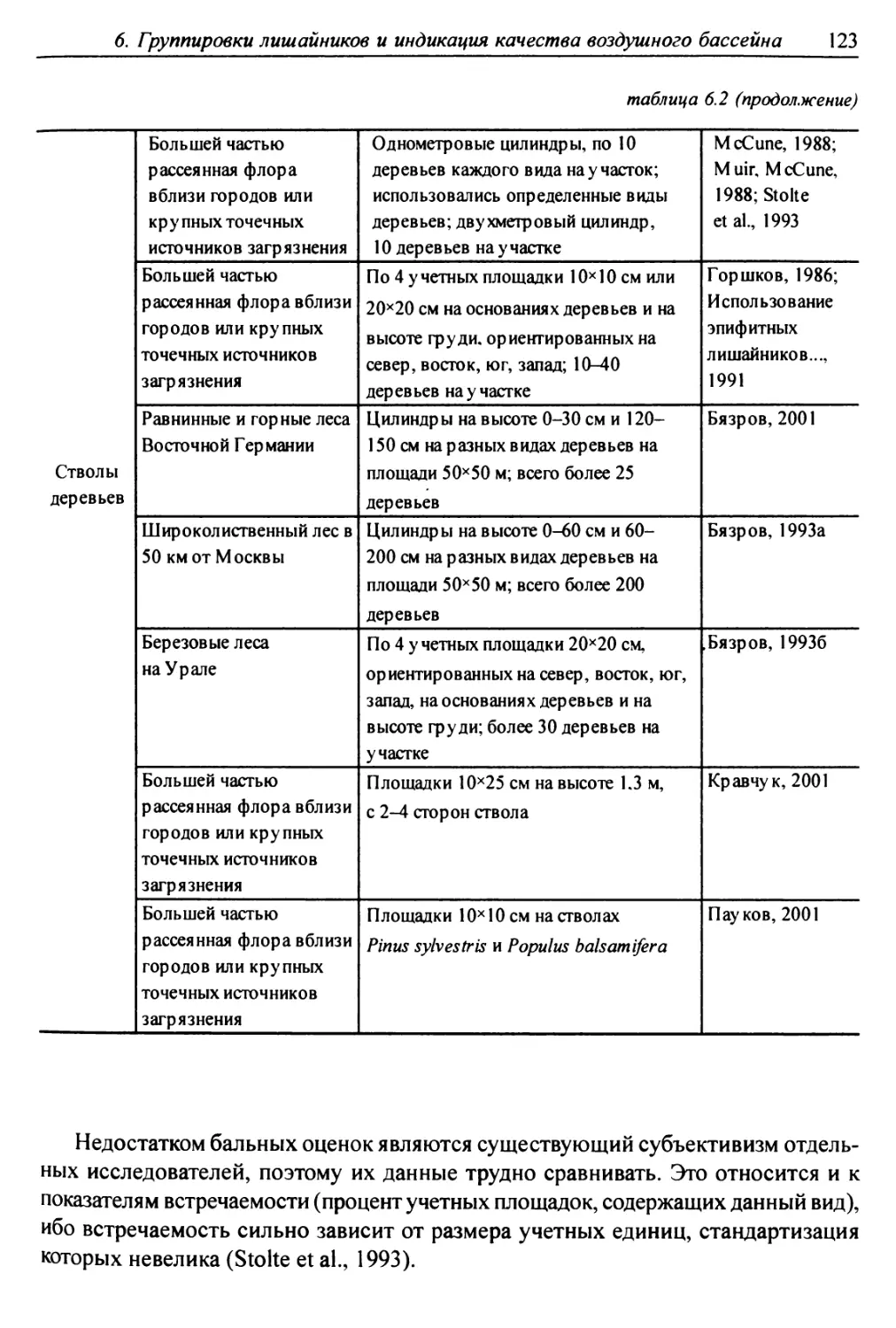
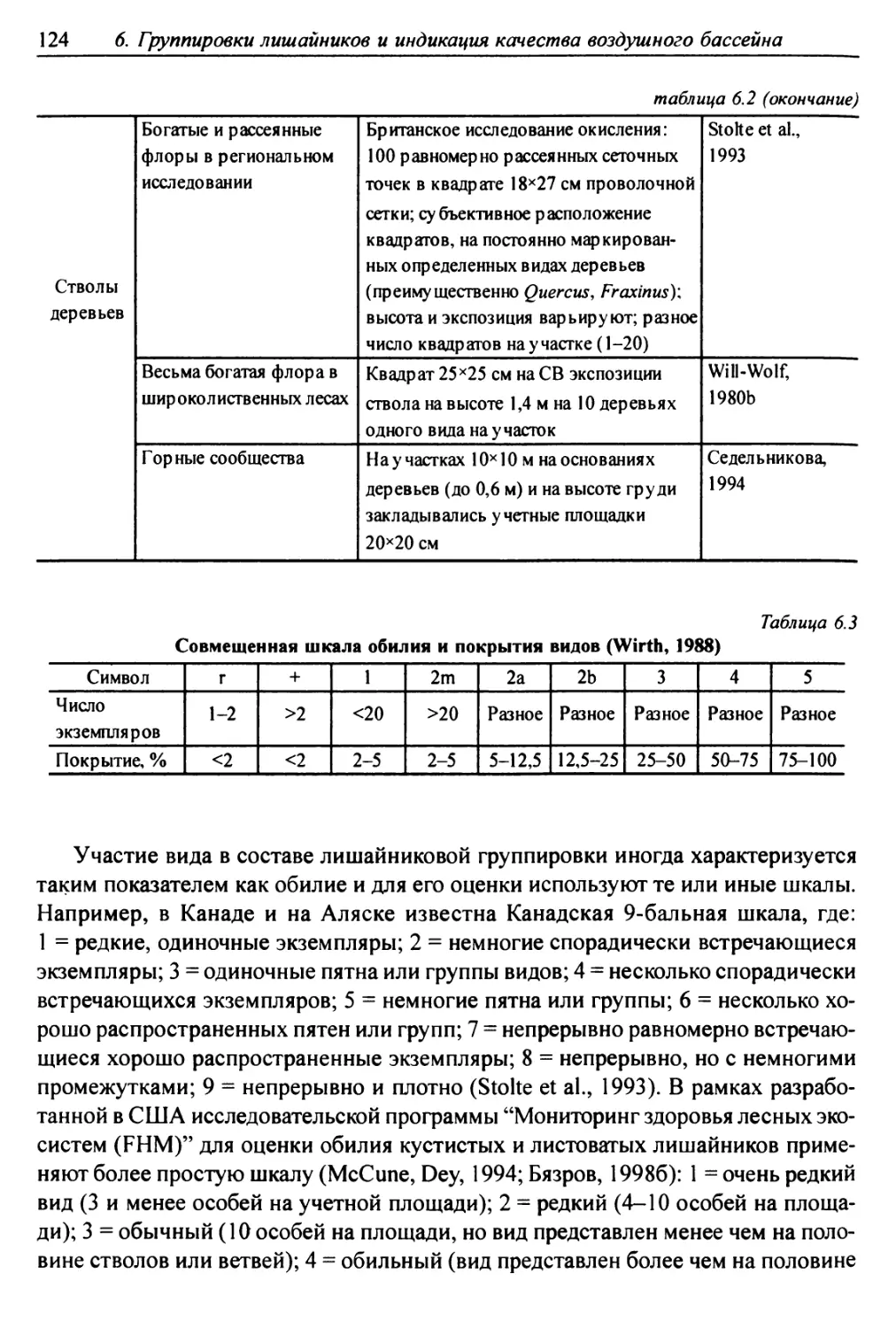
6.1. Методы сбора лихеноценологических данных ........... 118
6.1.1. Выбор участков для характеристики лишайниковых
группировок ......................................... 119
6.1.2. Обследование выбранных участков местности ........ 119
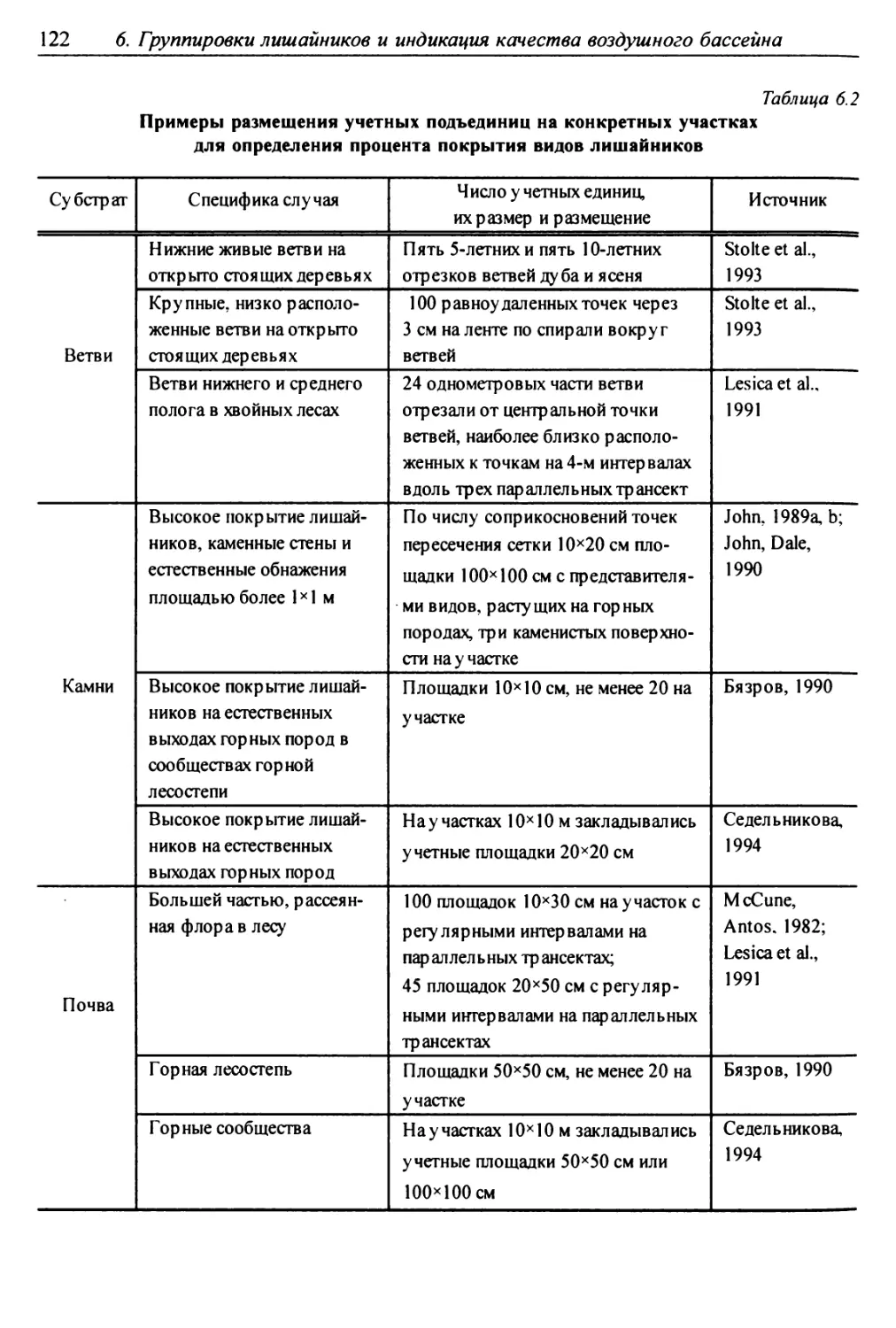
6.1.3. Размещение учетных подъединиц ................ 121
6.1.4. Фиксируемые показатели ....................... 121
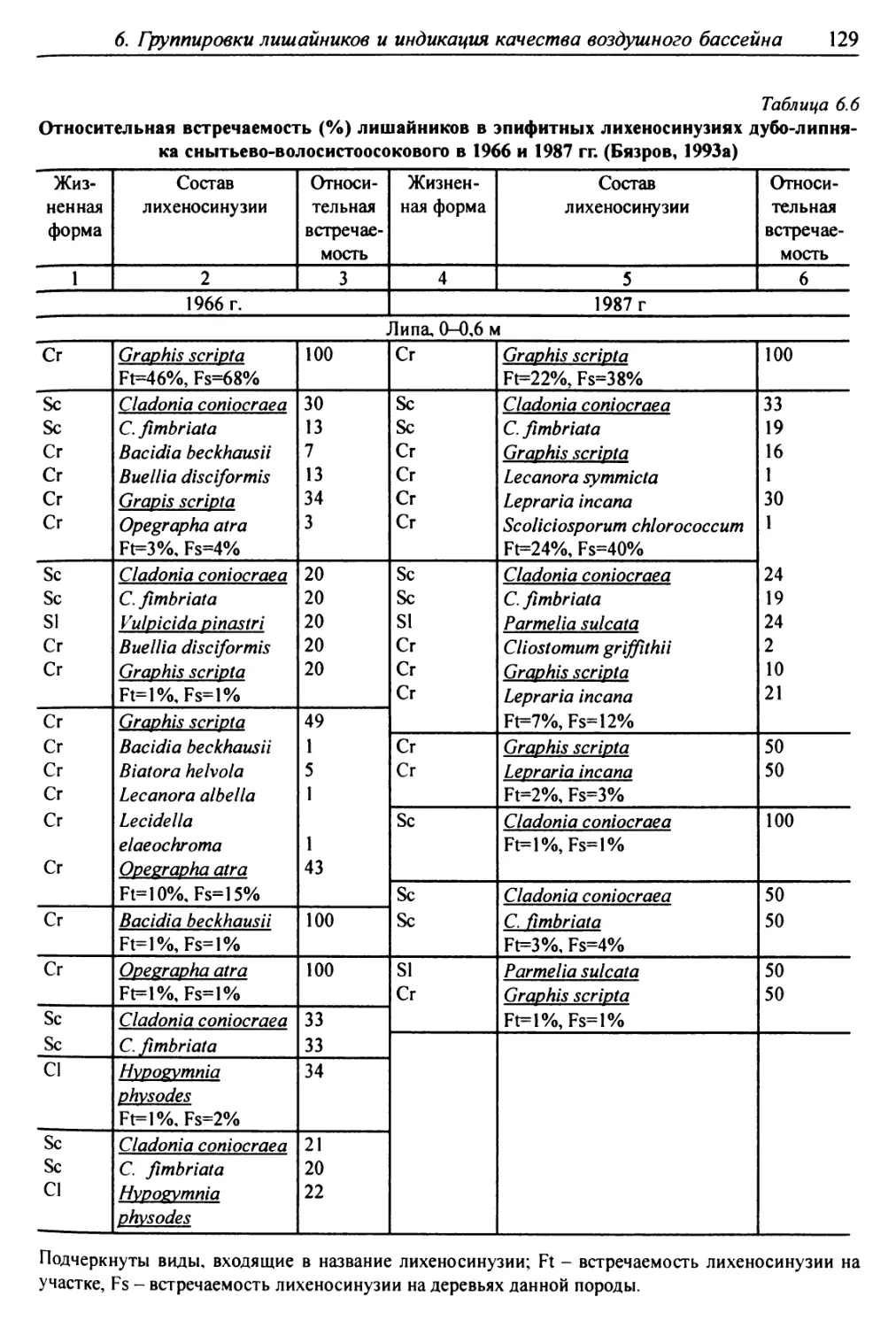
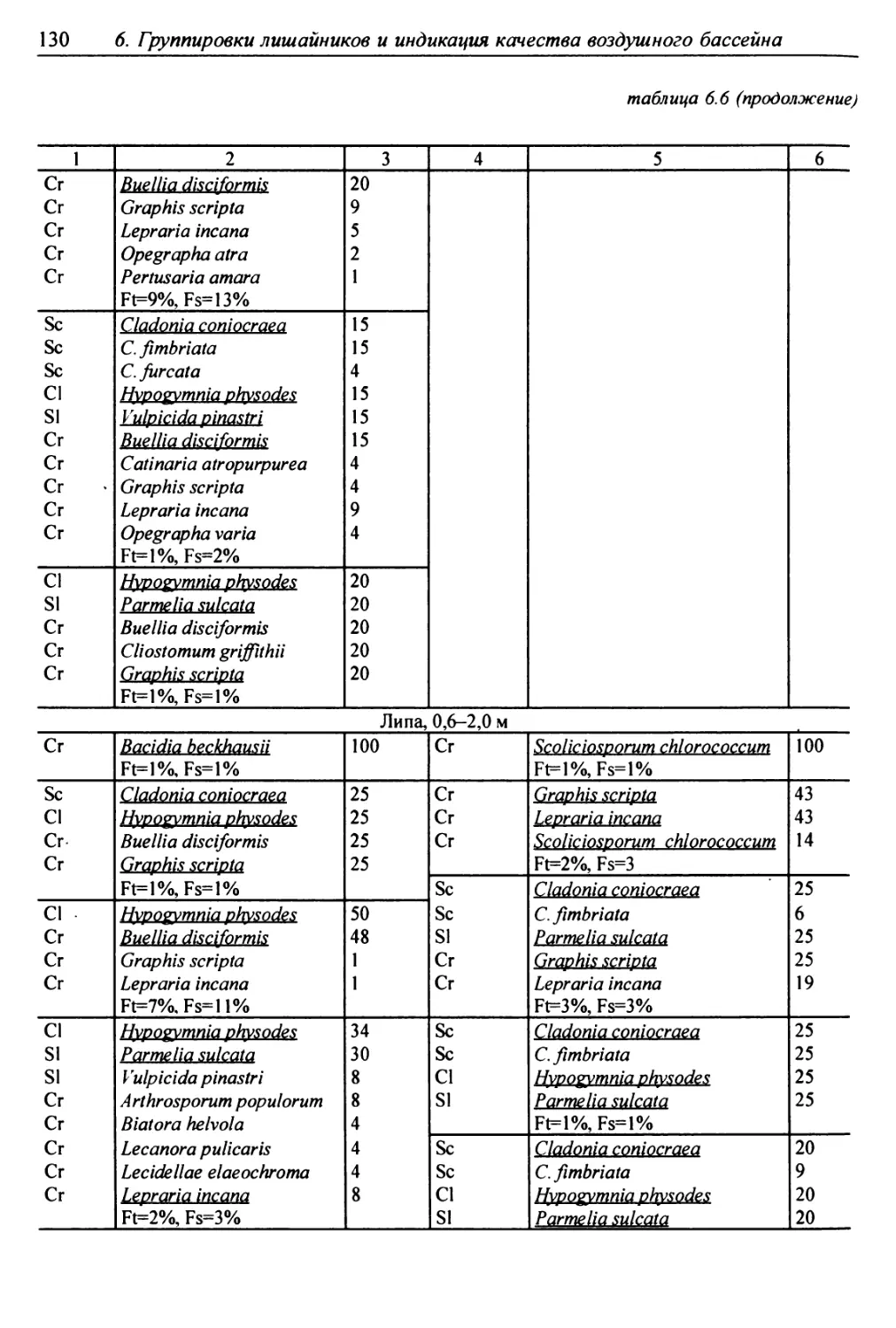
6.2. Выделение типов лишайниковых группировок ........... 125
6.3. Анализ выявленных лишайниковых синузий или сообществ ... 127
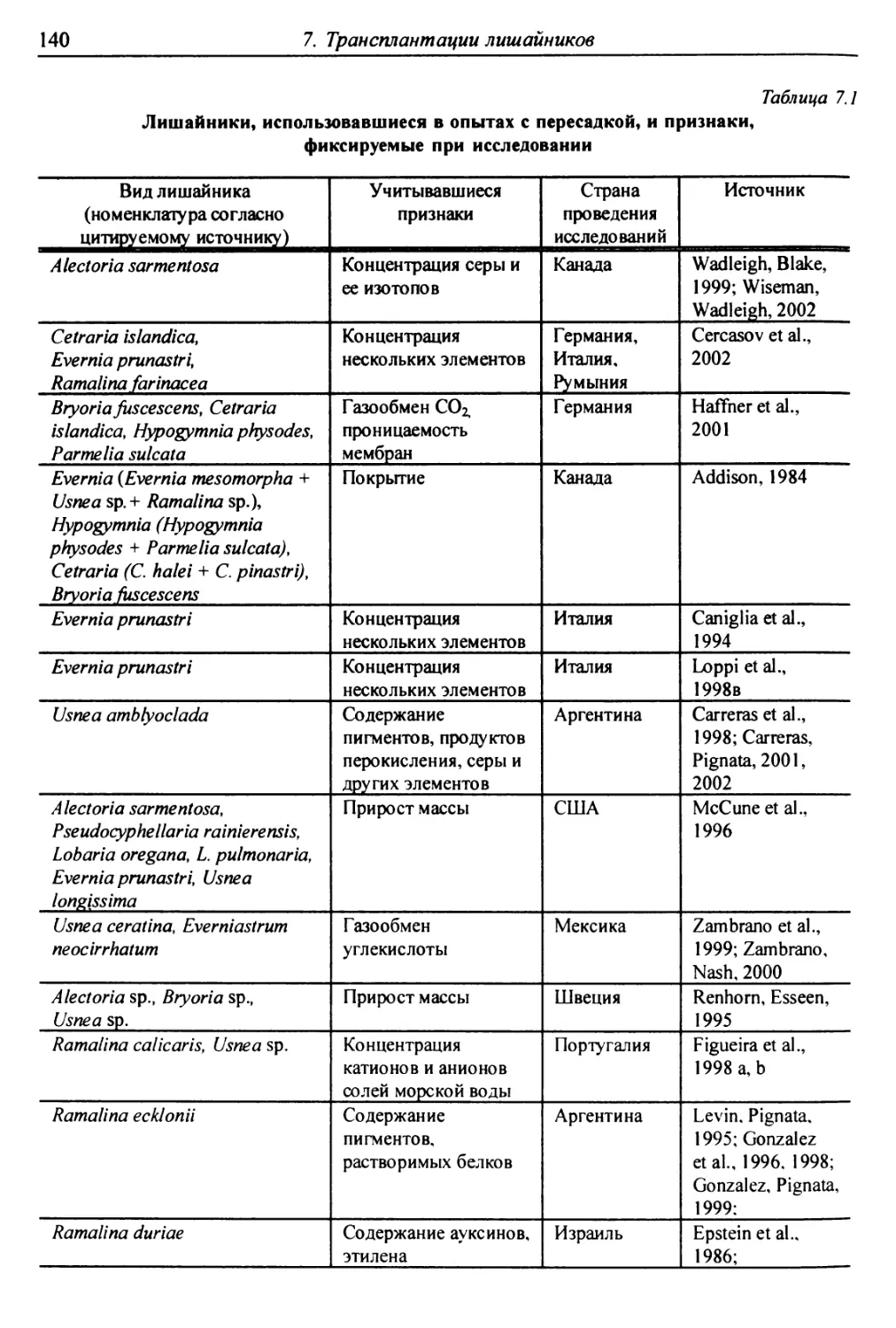
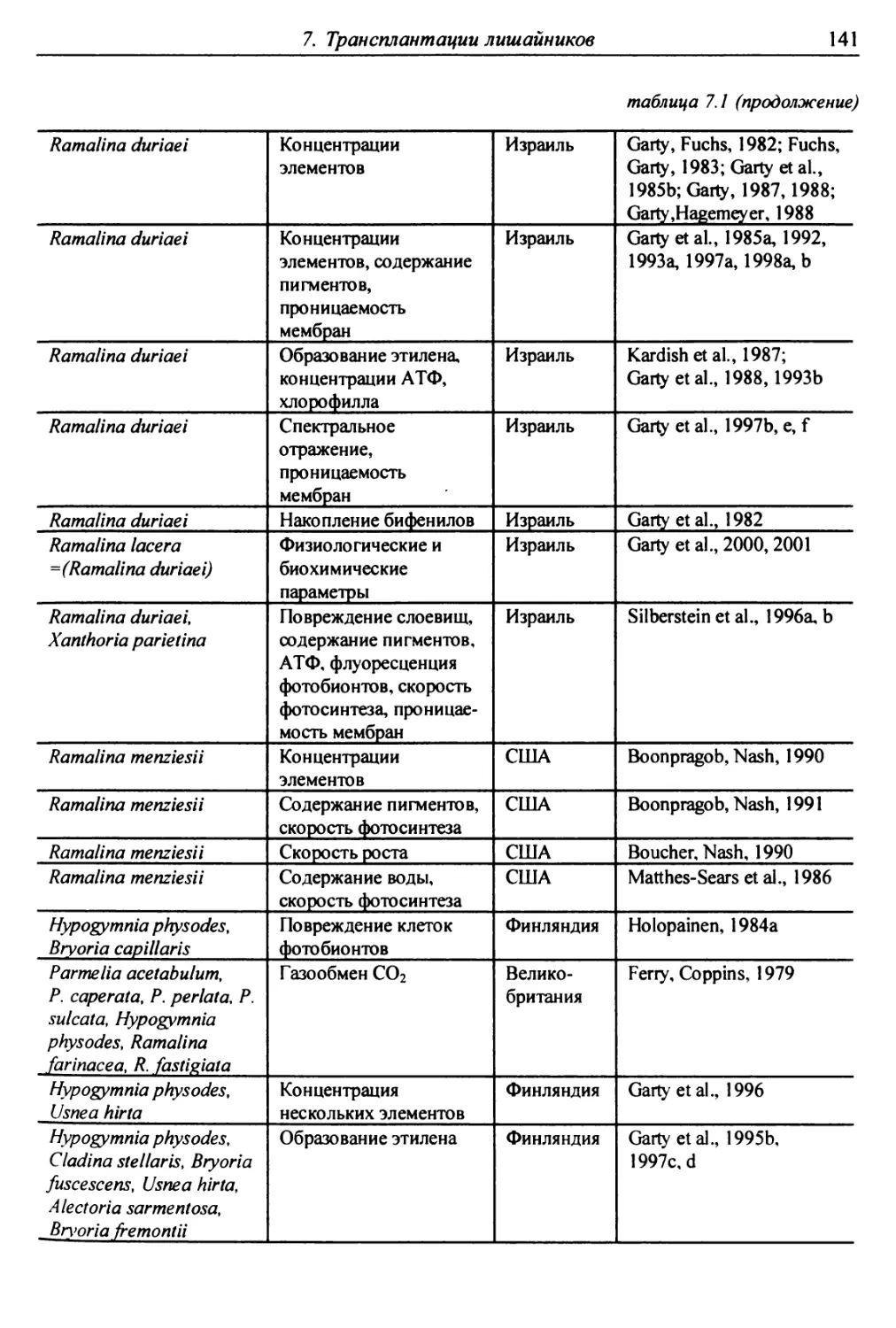
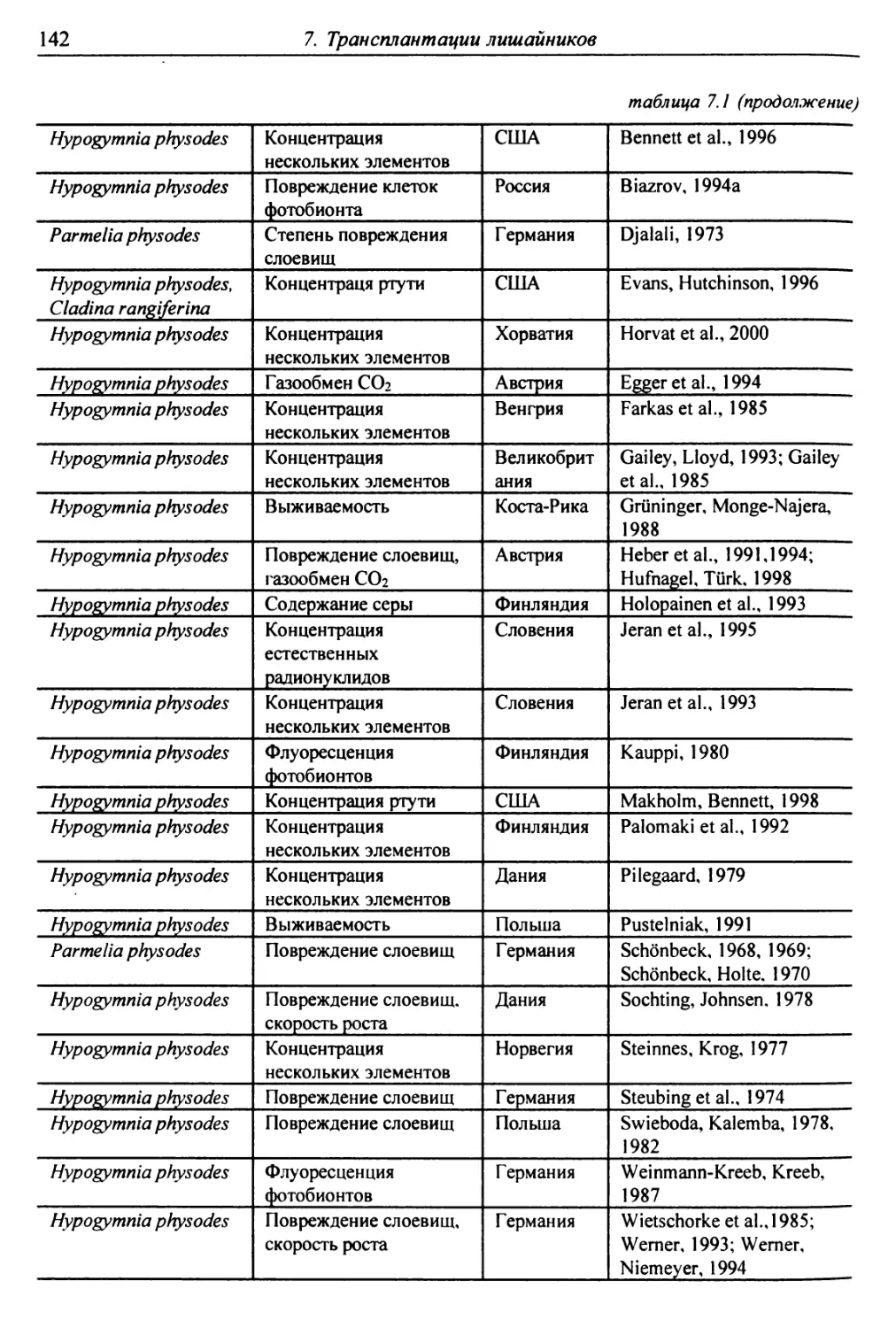
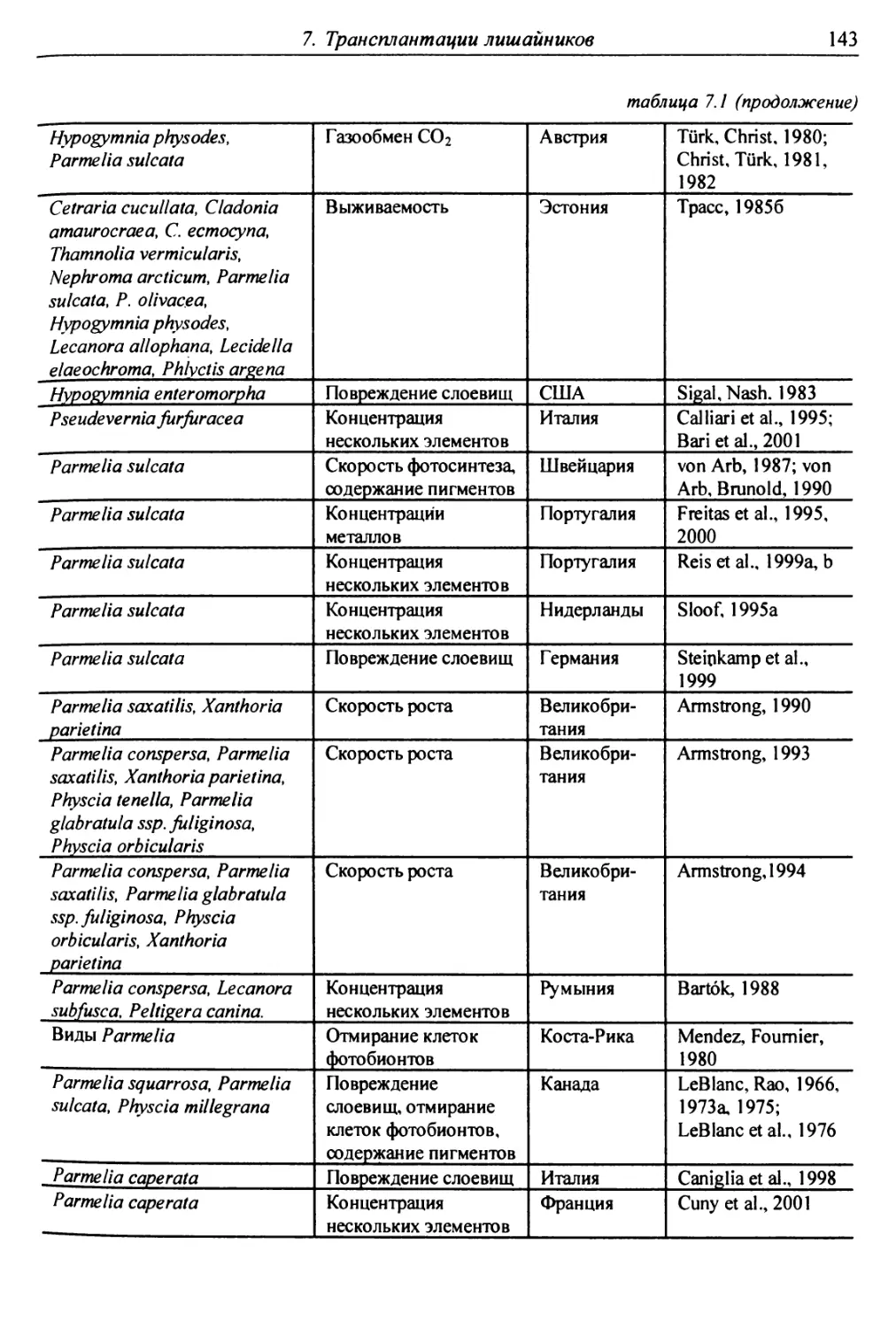
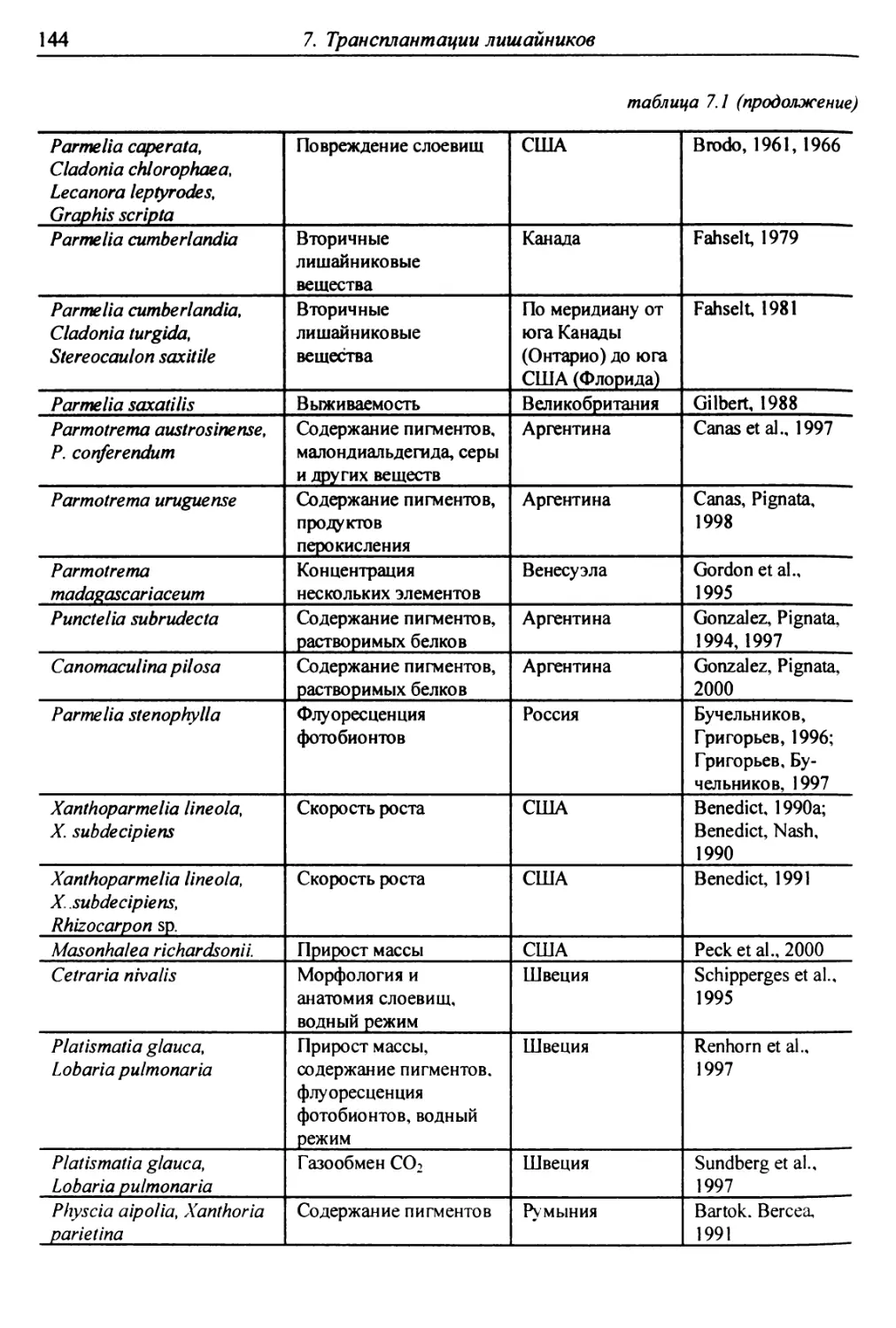
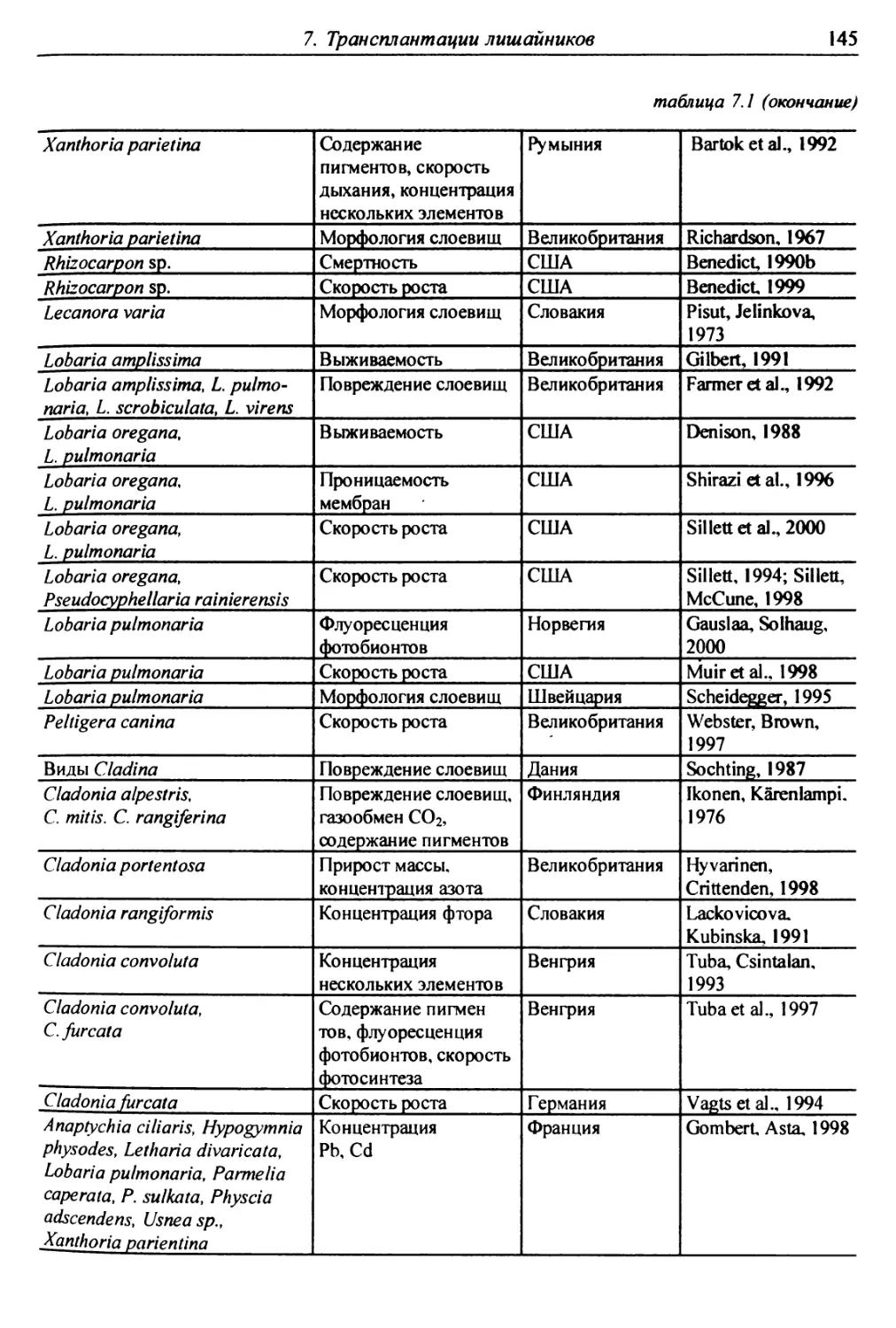
7. Трансплантации лишайников ............................... 137
7.1. Трансплантации ..................................... 137
7.1.1. Отбор слоевищ для пересадки и подготовка
их к экспонированию ................................. 146
7.1.2. Экспонирование ............................... 150
7.2. Оценки воздействия на трансплантированные слоевища . 151
7.2.1. Смертность и изменение внешних признаков слоевищ . 151
7.2.2. Рост слоевищ и прирост биомассы .................. 153
7.2.3. Повреждение клеток бионтов ....................... i 55
7.2.4. Изменение количества пигментов - показателей
фотосинтеза и дыхания ............................... 156
7.2.5. Нарушение проницаемости мембран и других параметров
жизнедеятельности слоевищ ...................... 159
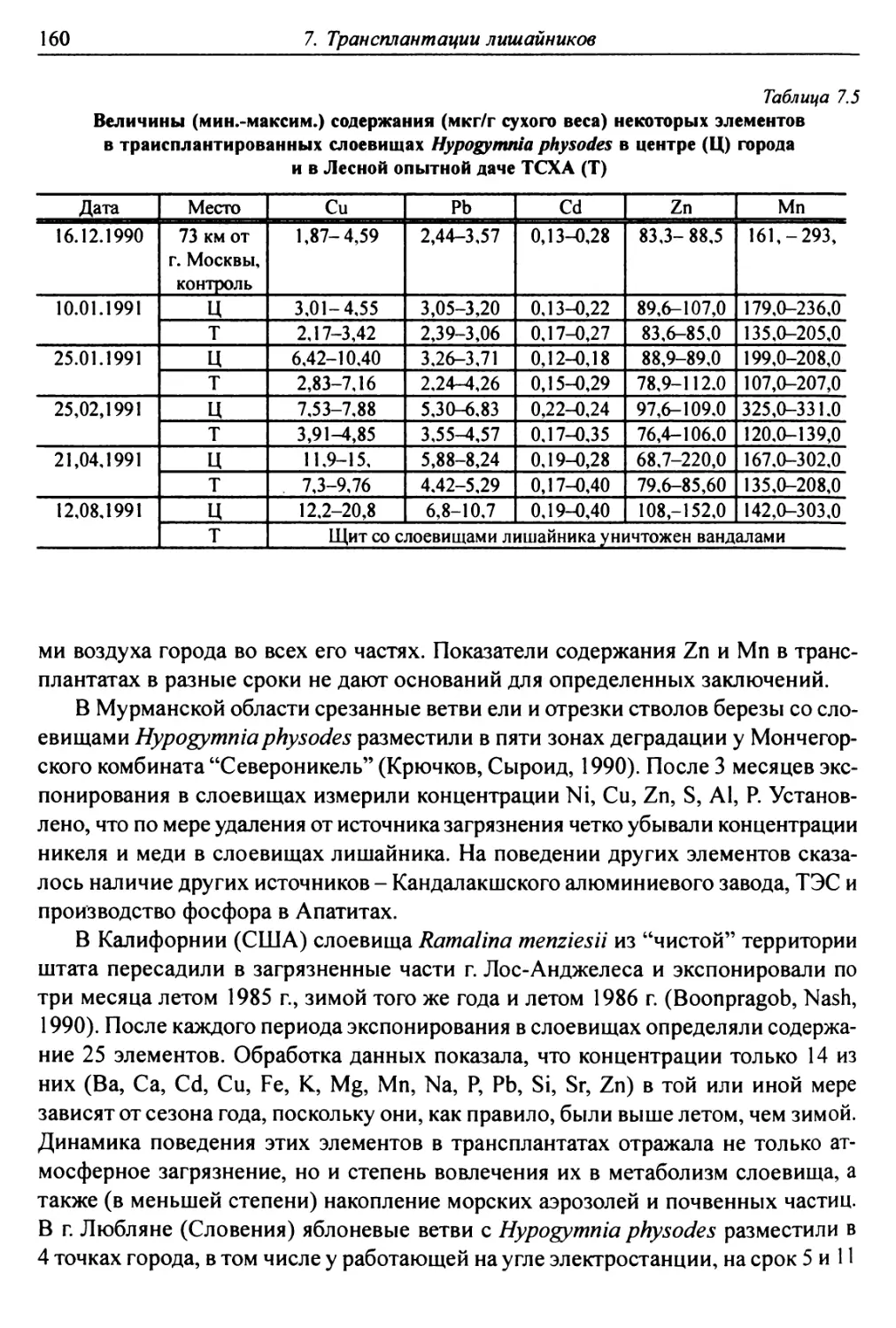
7.2.6. Аккумуляция загрязнителей ........................ 159
8. Лихеноиндикация по данным о накоплении
загрязнителей слоевищами ................................... 164
8.1. План исследования .................................. 165
8.1.1. Выбор химических элементов и соединений для
определения их содержания в слоевищах лишайников ..... 166
8.1.2. Выбор популяции лишайников ................... 166
8.1.3. Расположение учетных точек
на обследуемой местности ............................ 167
8.1.4. Объем проб, размер выборки (повторности)
в одной точке или в один срок ....................... 167
8.1.5. Установление критериев, обеспечивающих качество
результатов и его контроль .......................... 168
8.2. Сбор материала для химического анализа
и его первичная обработка ............................... 169
8.2.1. Точки отбора проб ................................ 170
Оглавление
7
8.2.2. Отбор проб в поле ............................. 170
8.3. Анализ проб .......................................... 173
8.3.1. Подготовка лишайниковых проб для анализа ....... 174
8.3.2. Химический анализ материала .................... 176
8.4. Основные направления лихеноиндикации поданным
об особенностях химического состава лишайников .......... 177
8.4.1. Определение базового уровня концентрации химических
веществ в слоевищах лишайников ....................... 178
8.4.2. Установление трендов содержания загрязнителей
во времени на конкретной территории .................. 180
8.4.3. Выявление пространственного распределения
загрязнителей относительно источников загрязнения 182
8.4.4. Пространственное распределение элементов в слоевищах
лишайников на относительно крупной (область, провинция,
страна) территории ..:................................ 184
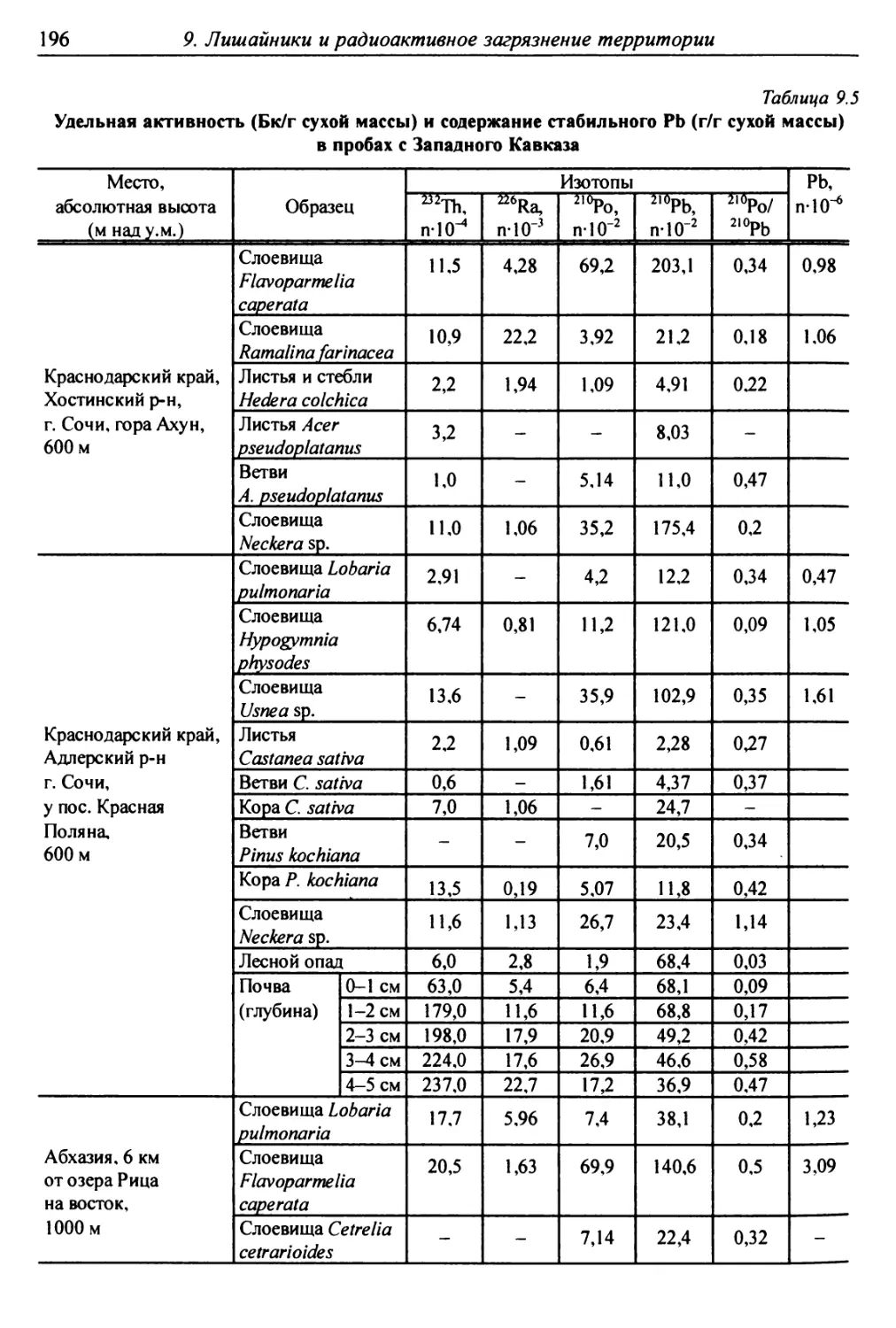
9. Лишайники и радиоактивное загрязнение территории .......... 186
9.1. Устойчивость лишайников к действию ионизирующей
радиации ................................................ 186
9.1.1. Данные экспериментов .......................... 186
9.1.2. Наблюдения на Семипалатинском ядерном полигоне . 187
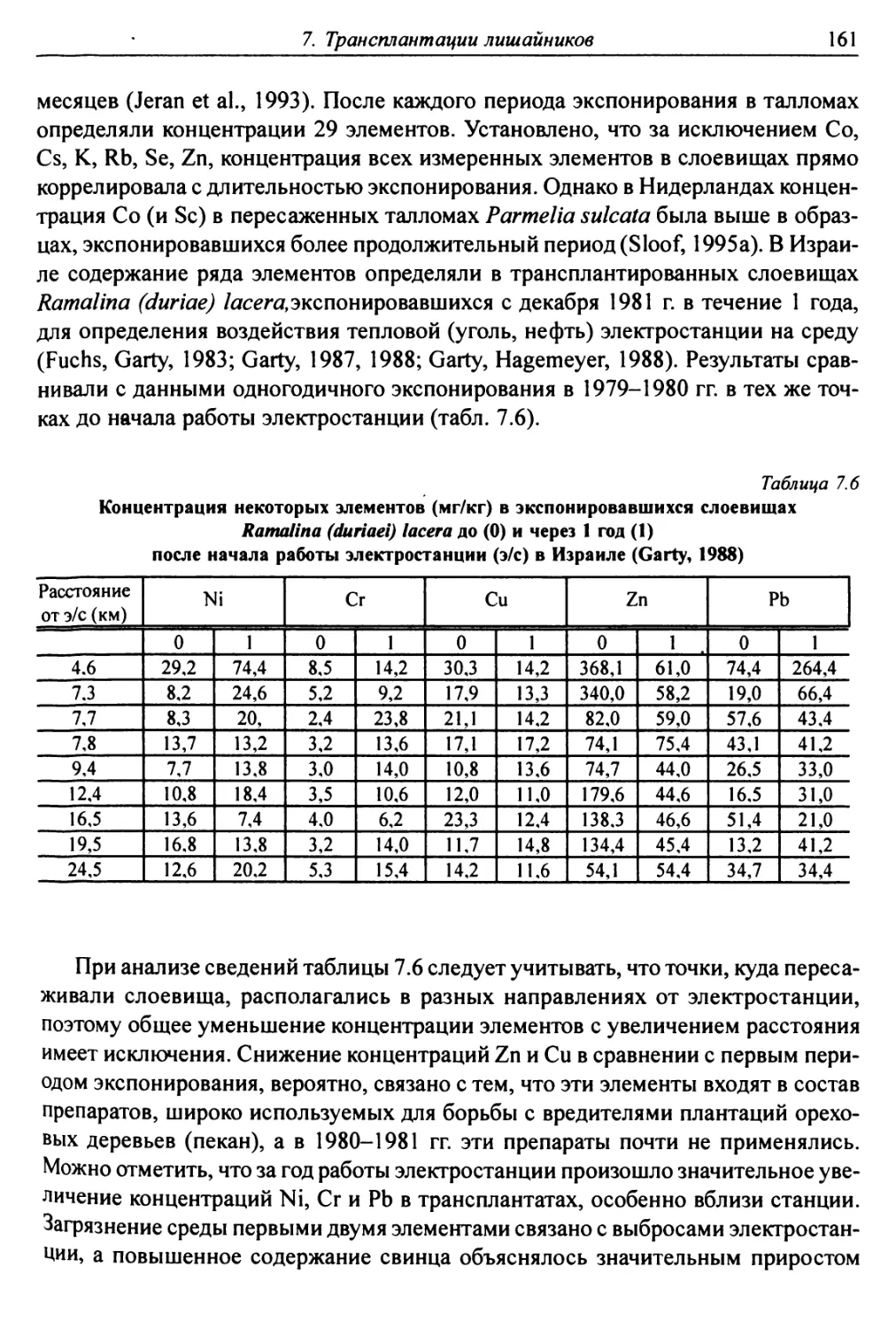
9.2. Накопление радионуклидов слоевищами лишайников ....... 189
9.2.1. Радионуклиды в лишайниках Арктики ............. 189
9.2.2. Радионуклиды в лишайниках после аварии
на Чернобыльской АЭС ................................. 190
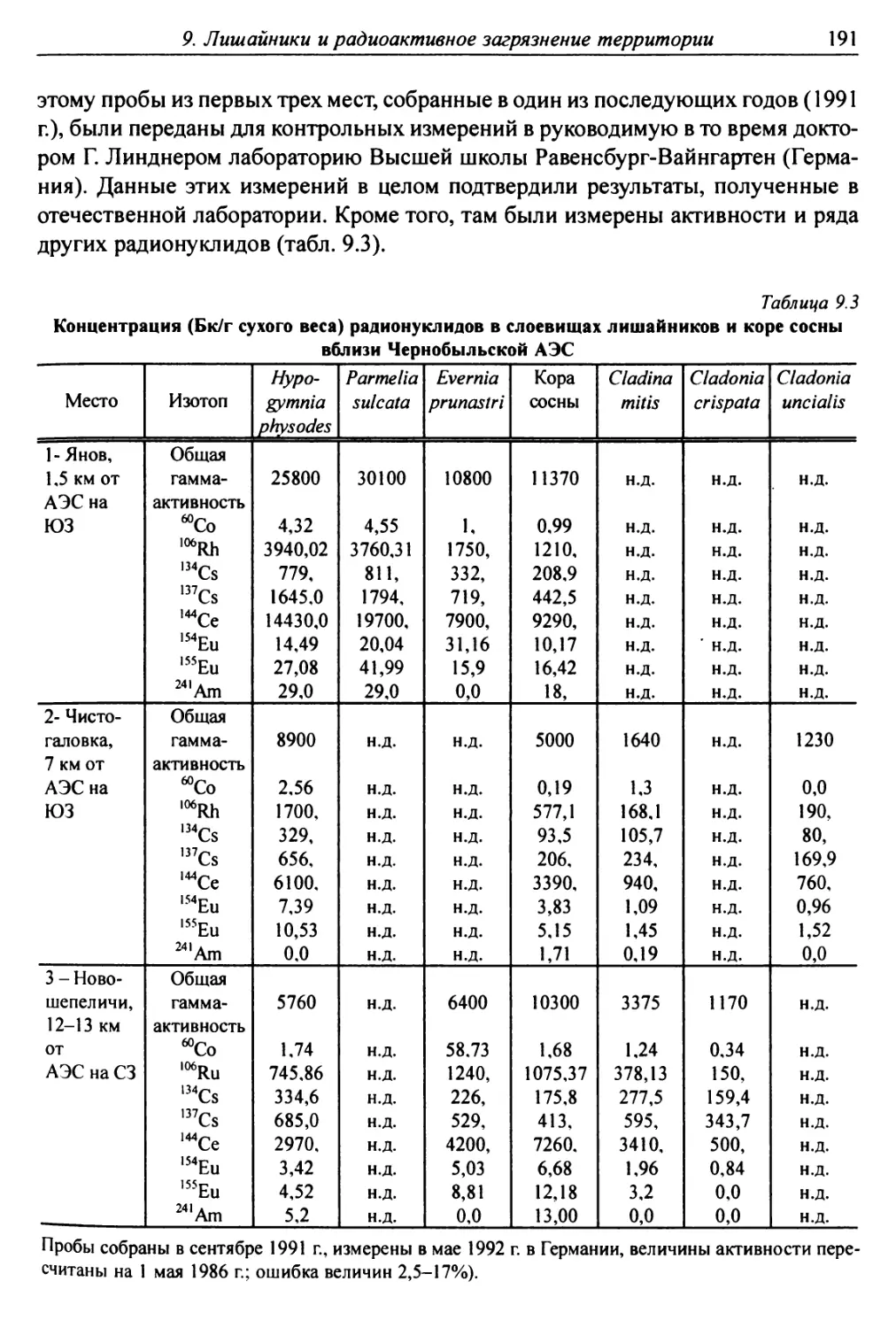
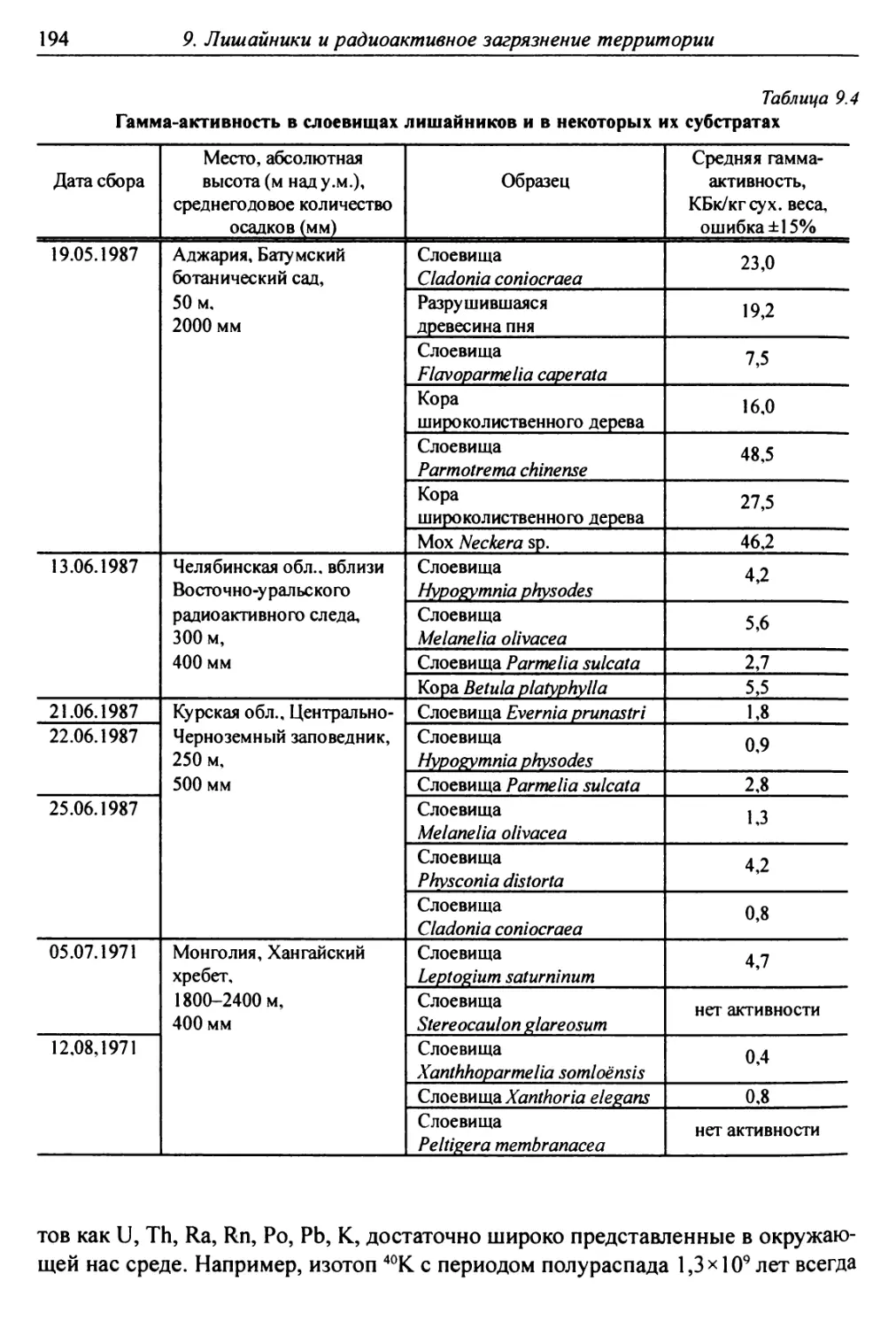
9.2.3. Накопление естественных радионуклидов
слоевищами лишайников ................................ 193
9.3. Распределение радионуклидов в слоевище
и их высвобождение ...................................... 197
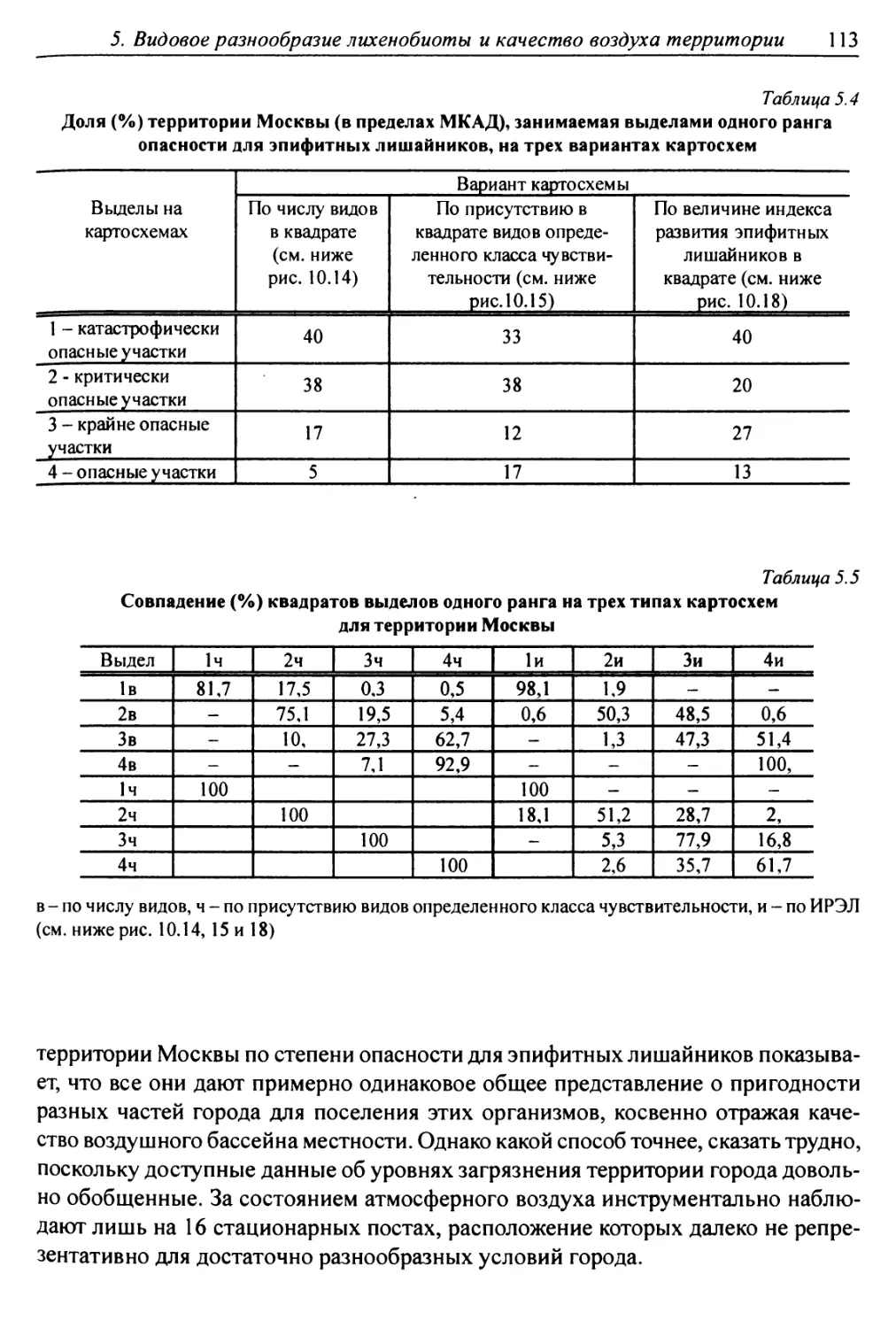
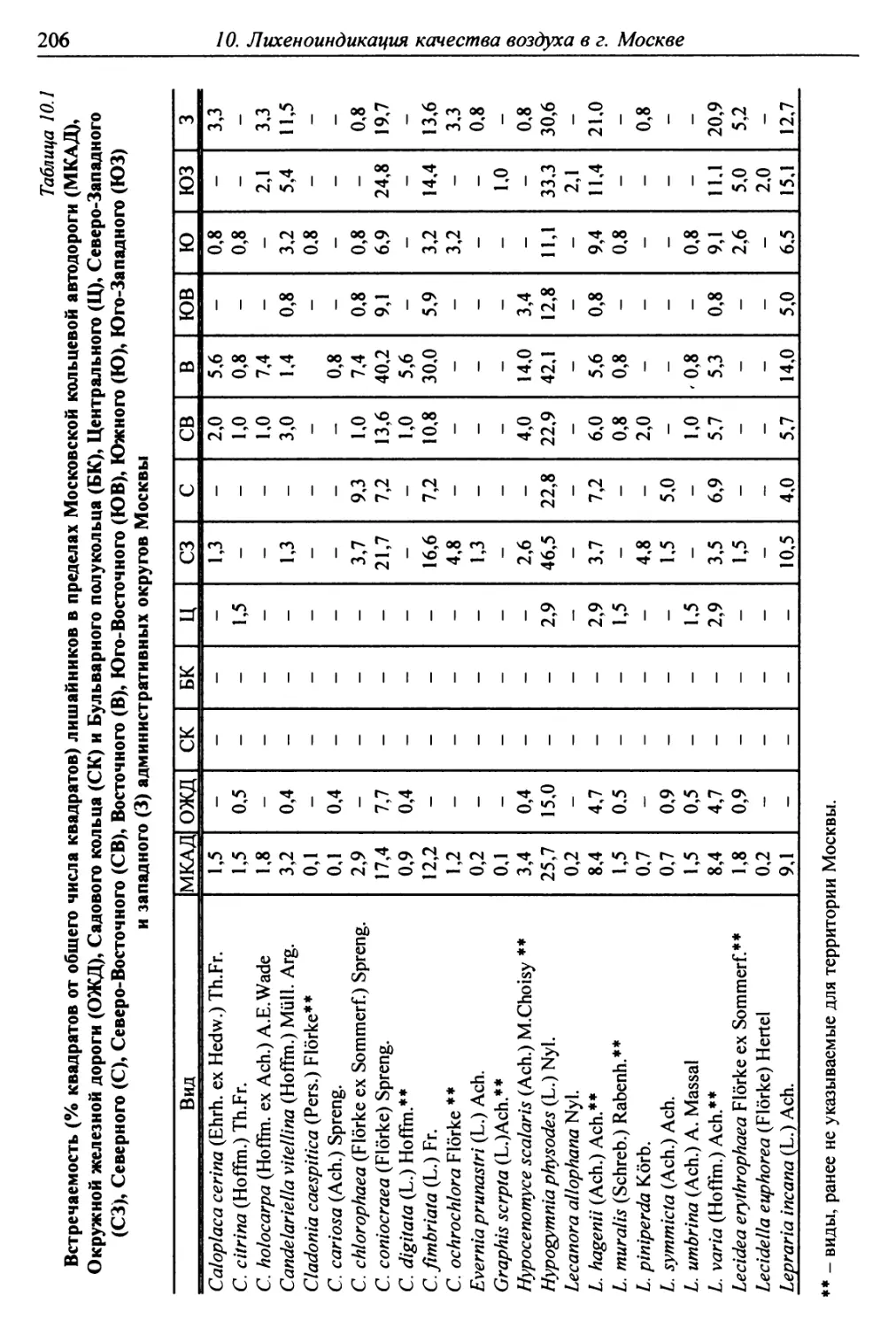
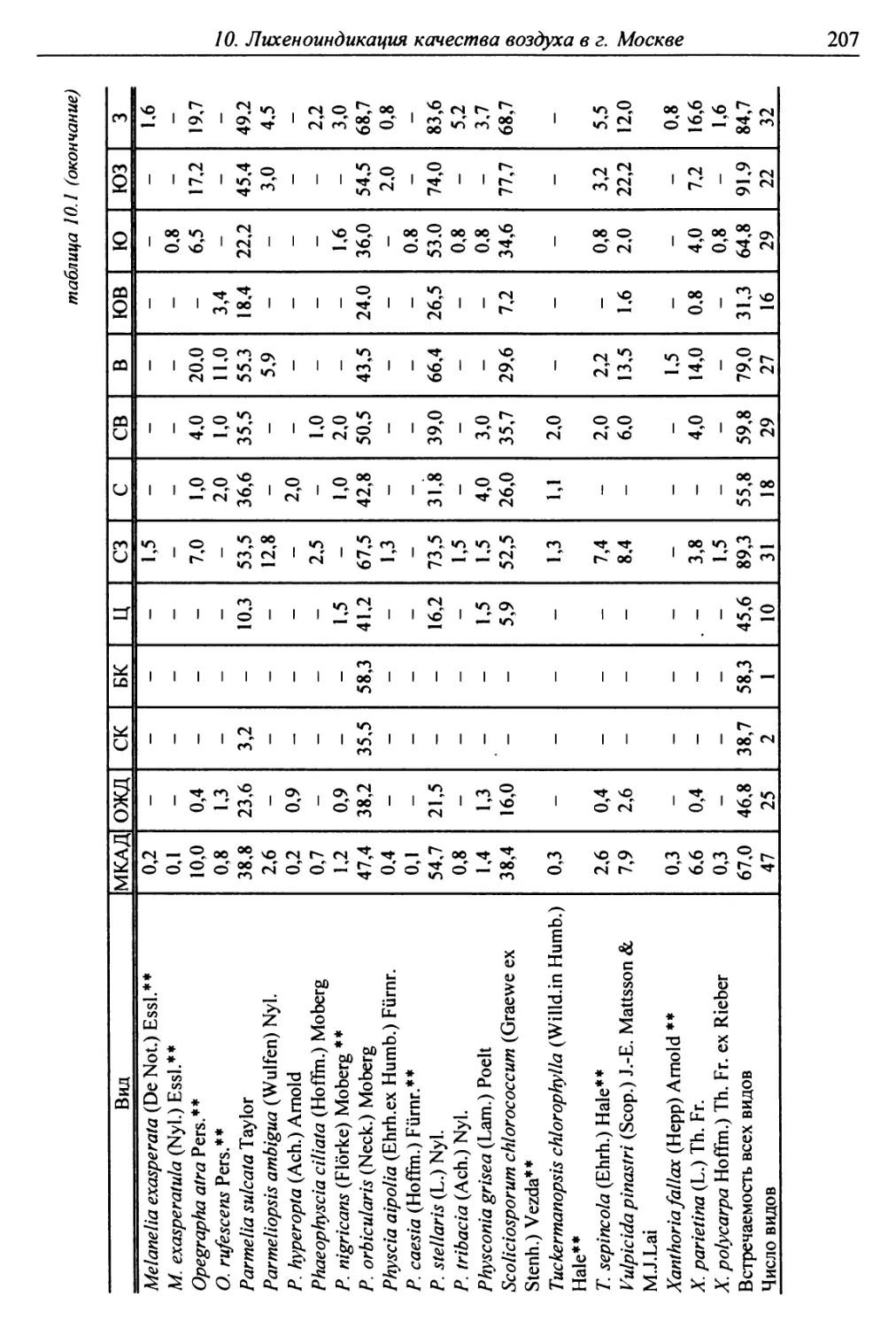
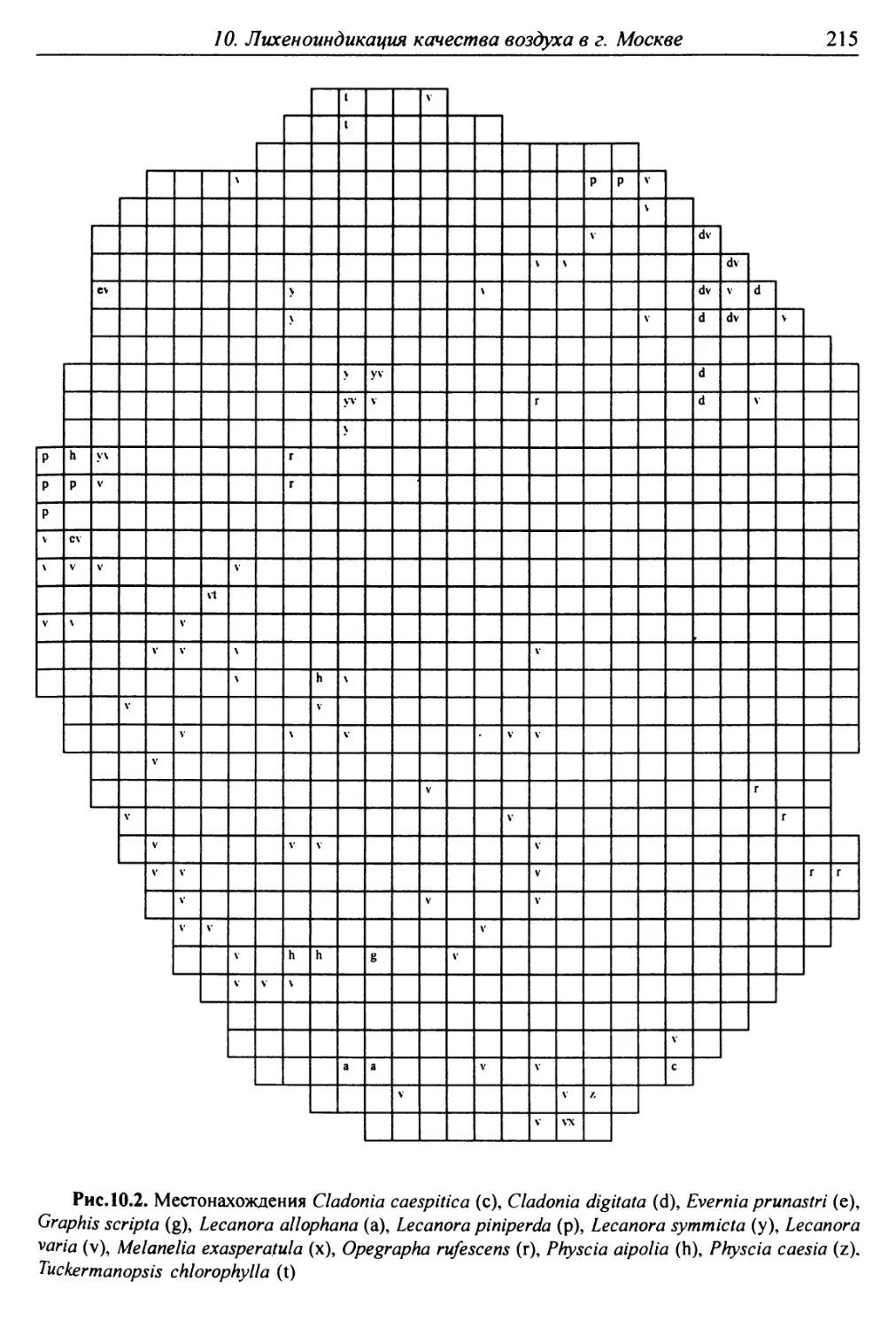
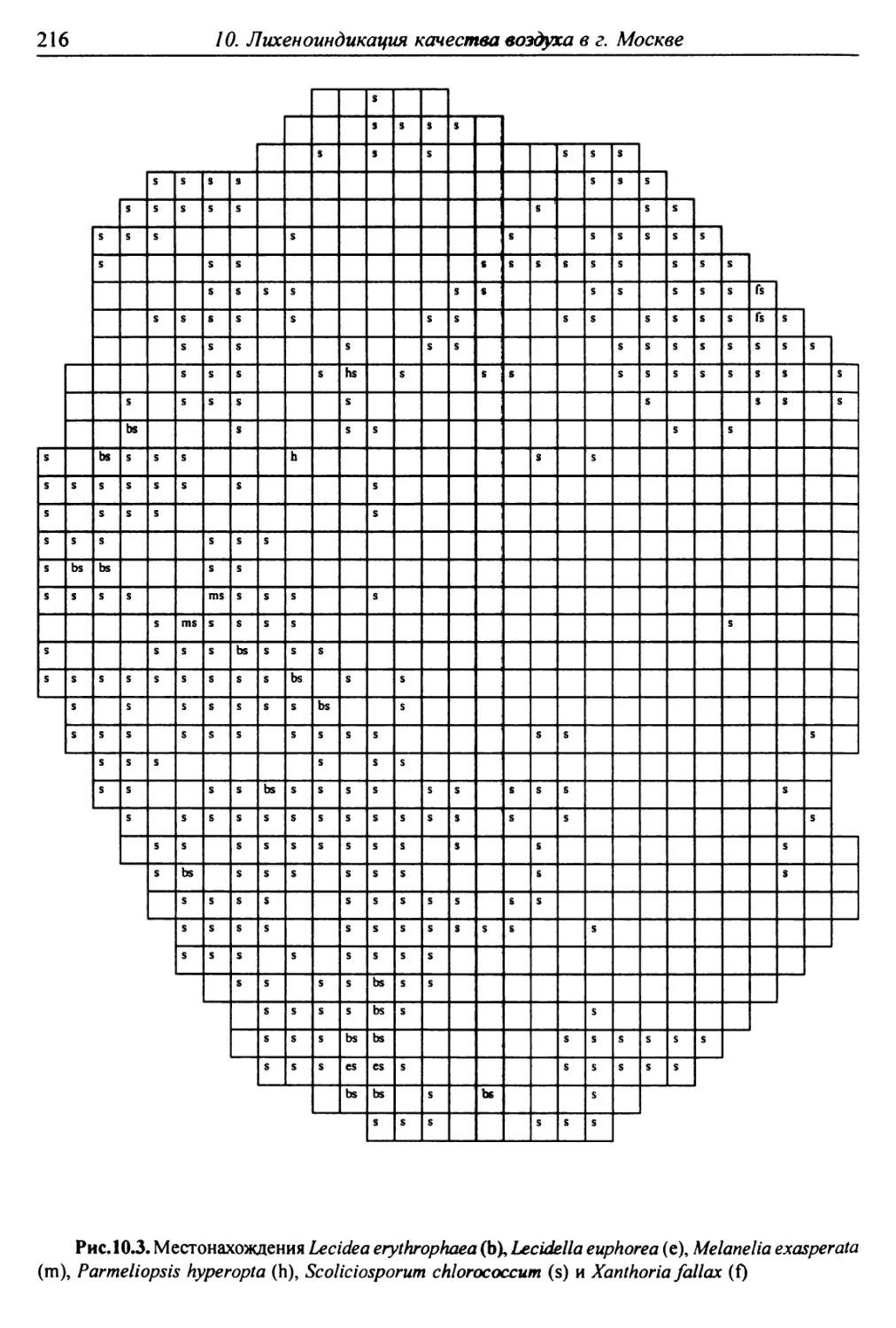
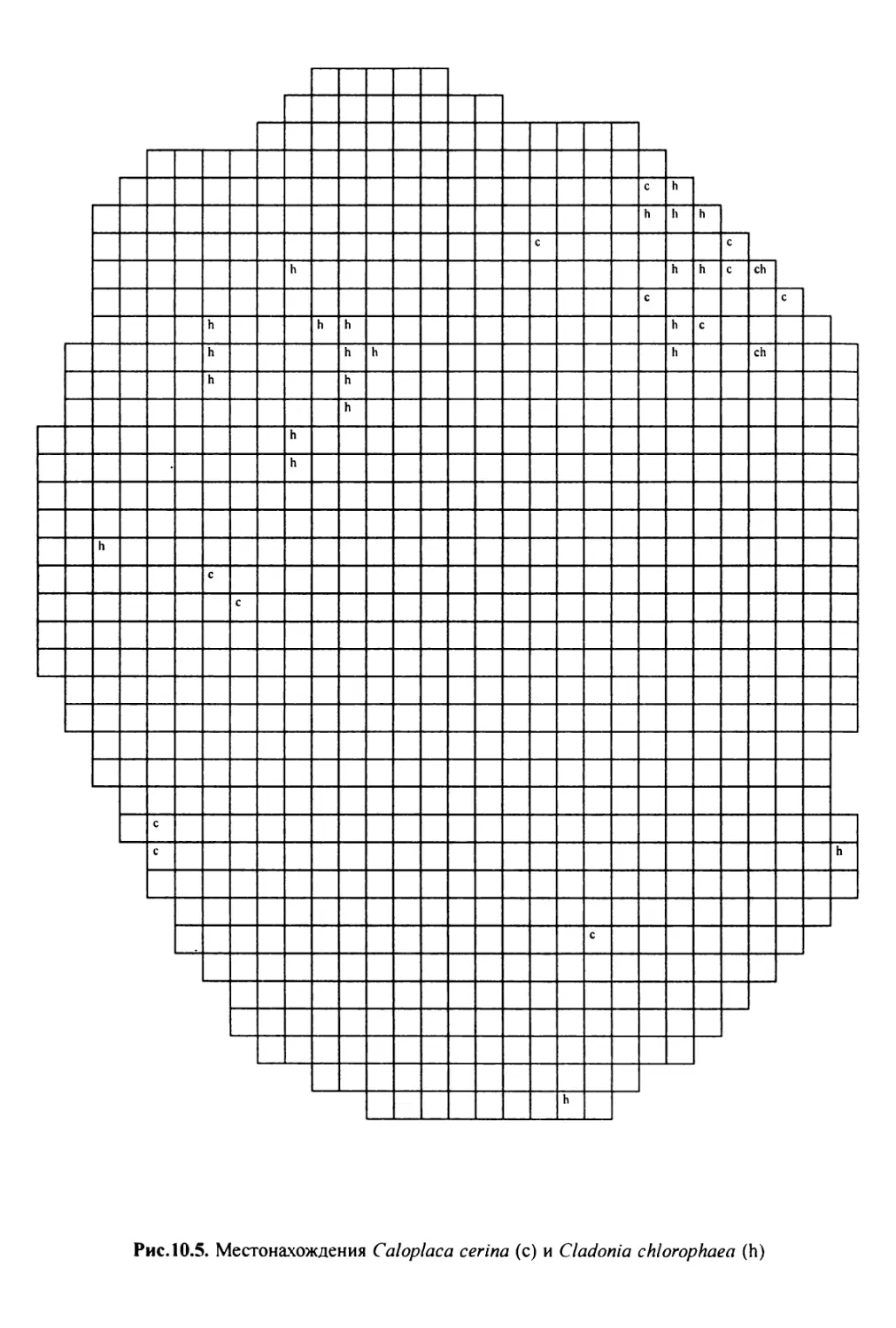
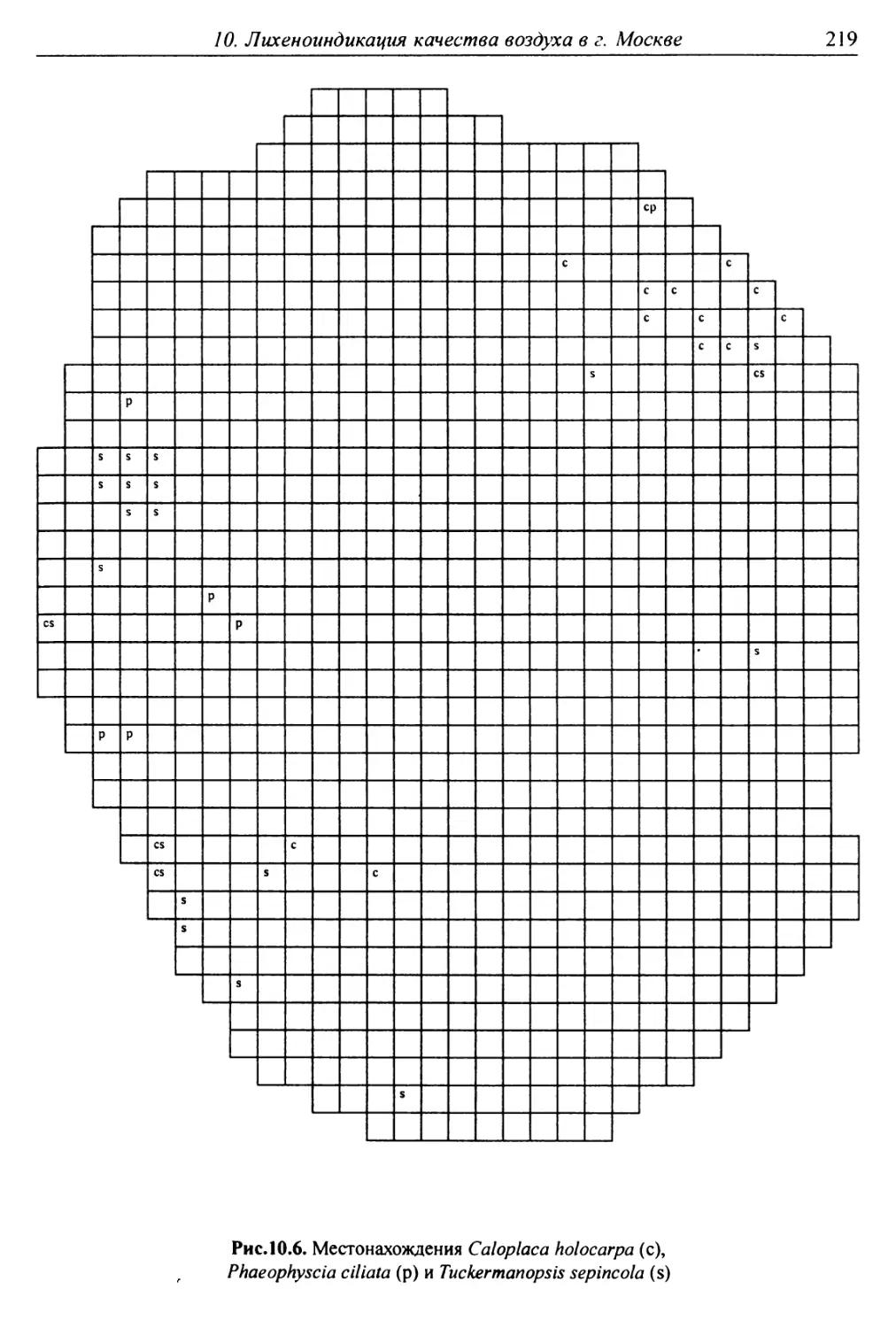
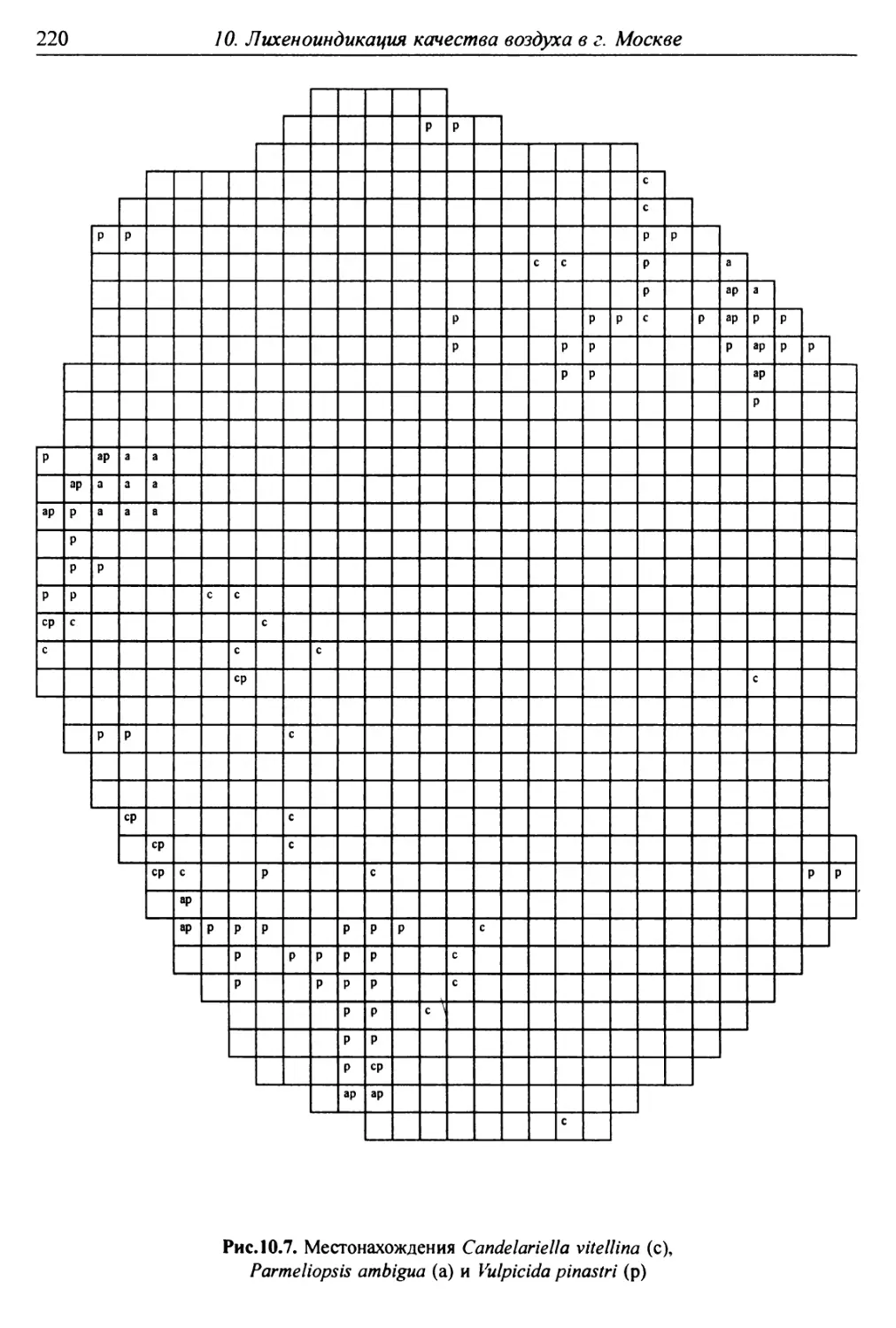
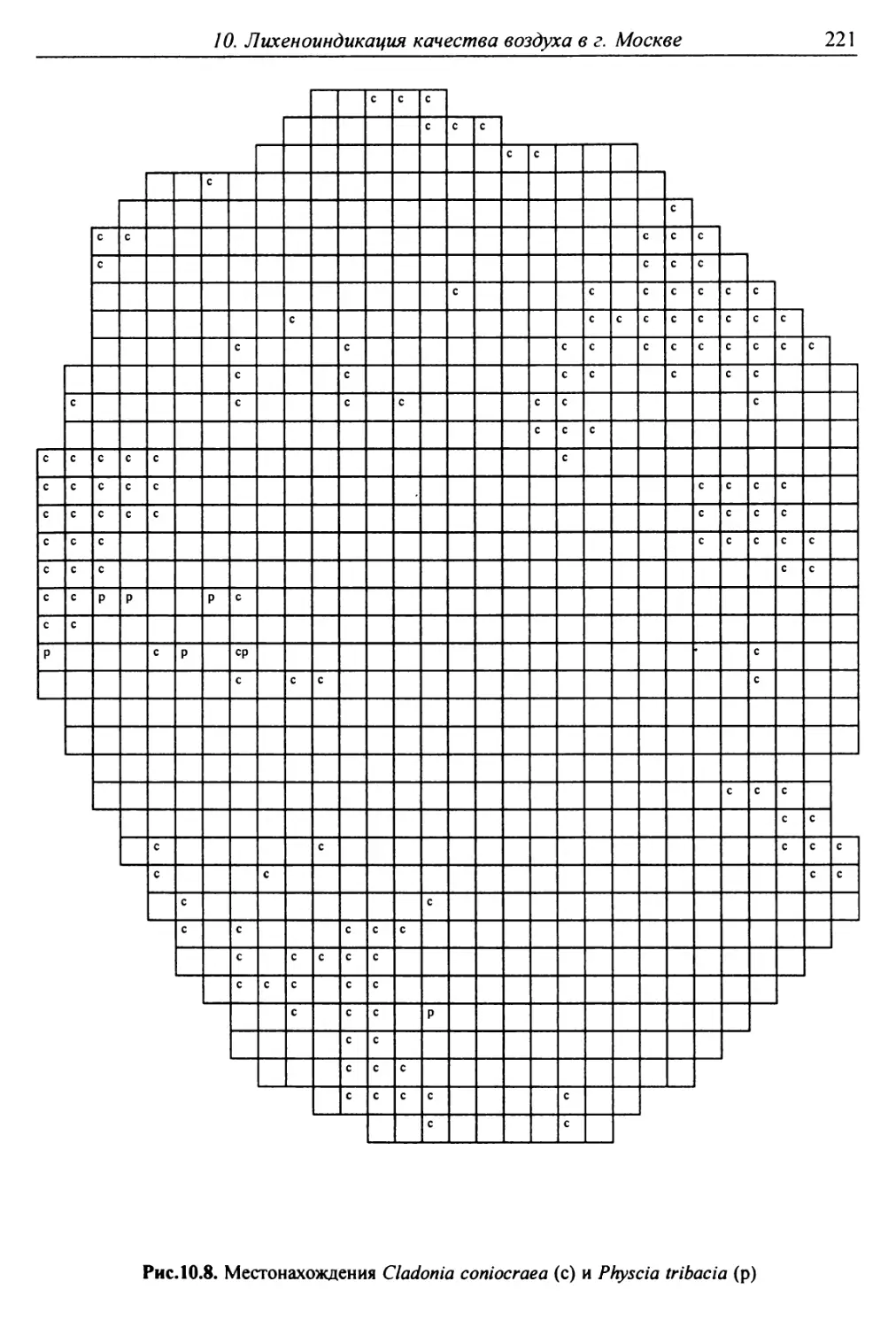
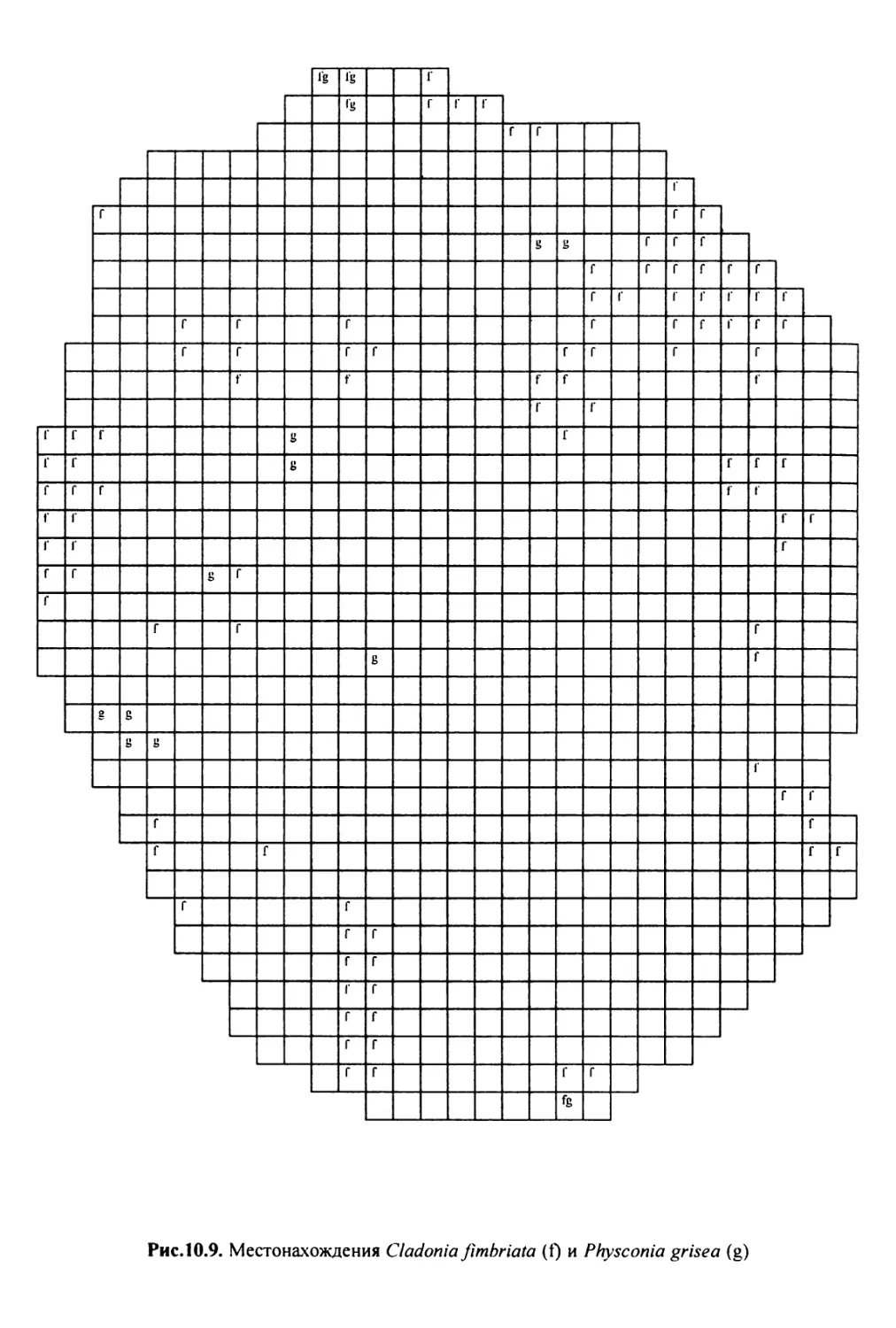
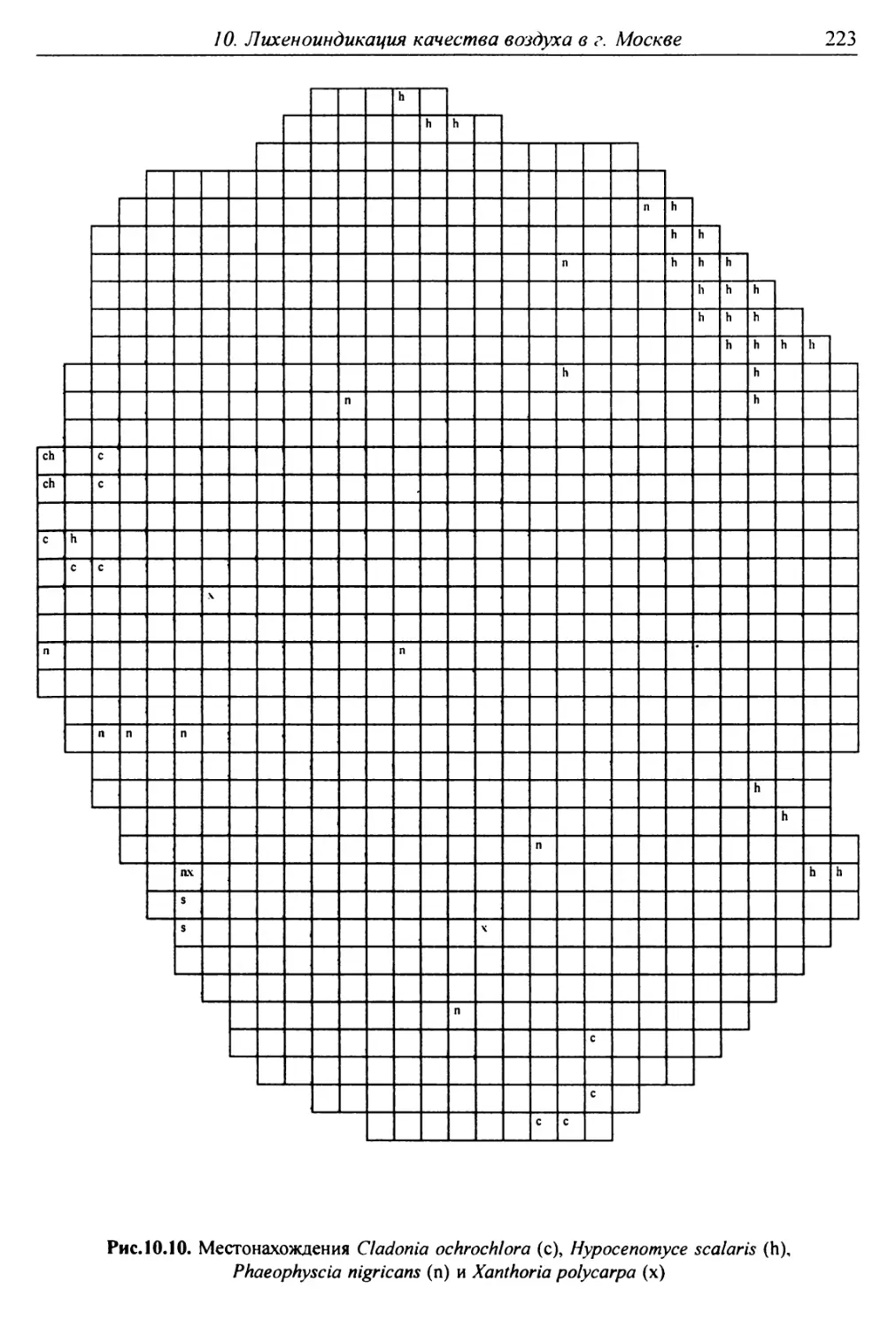
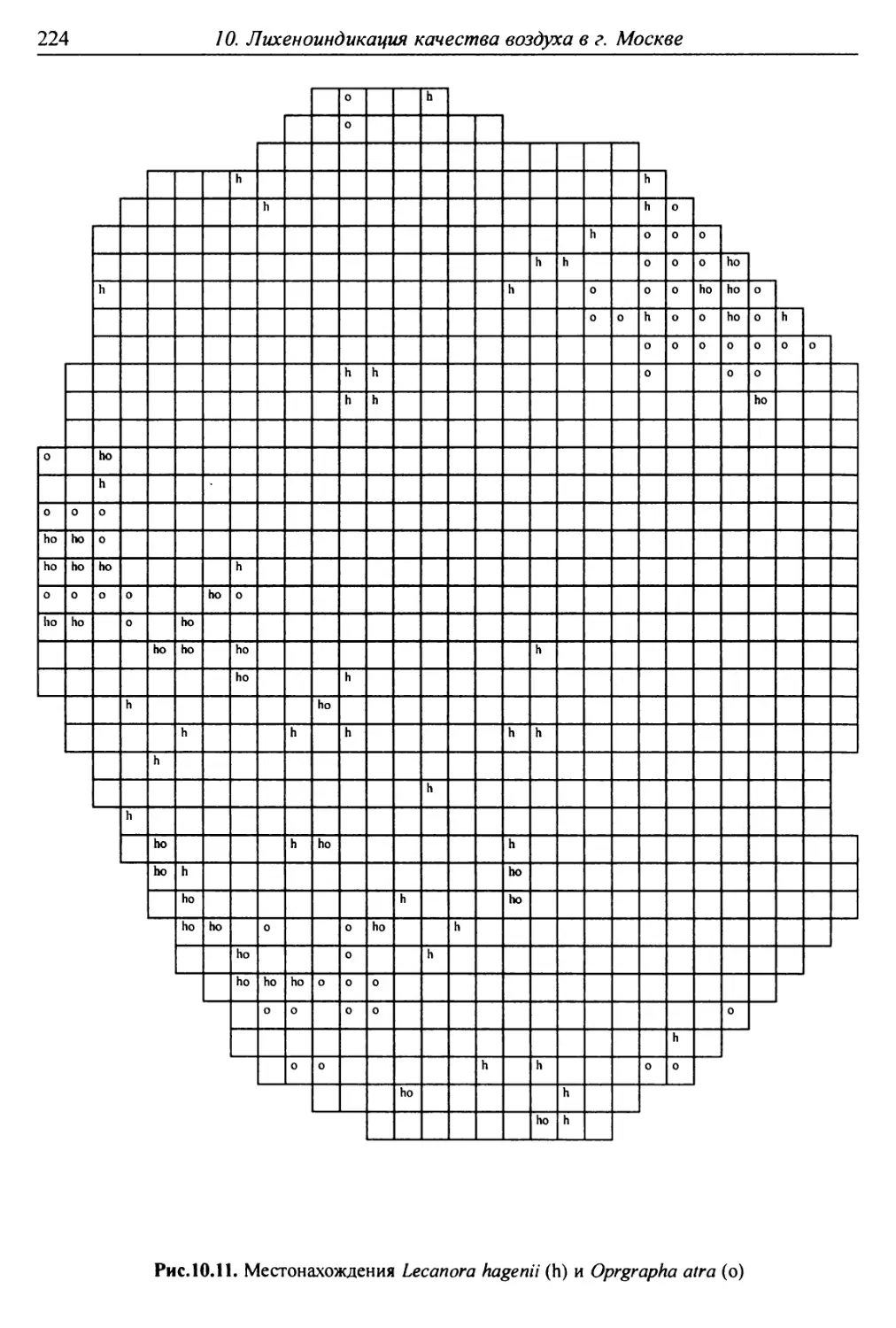
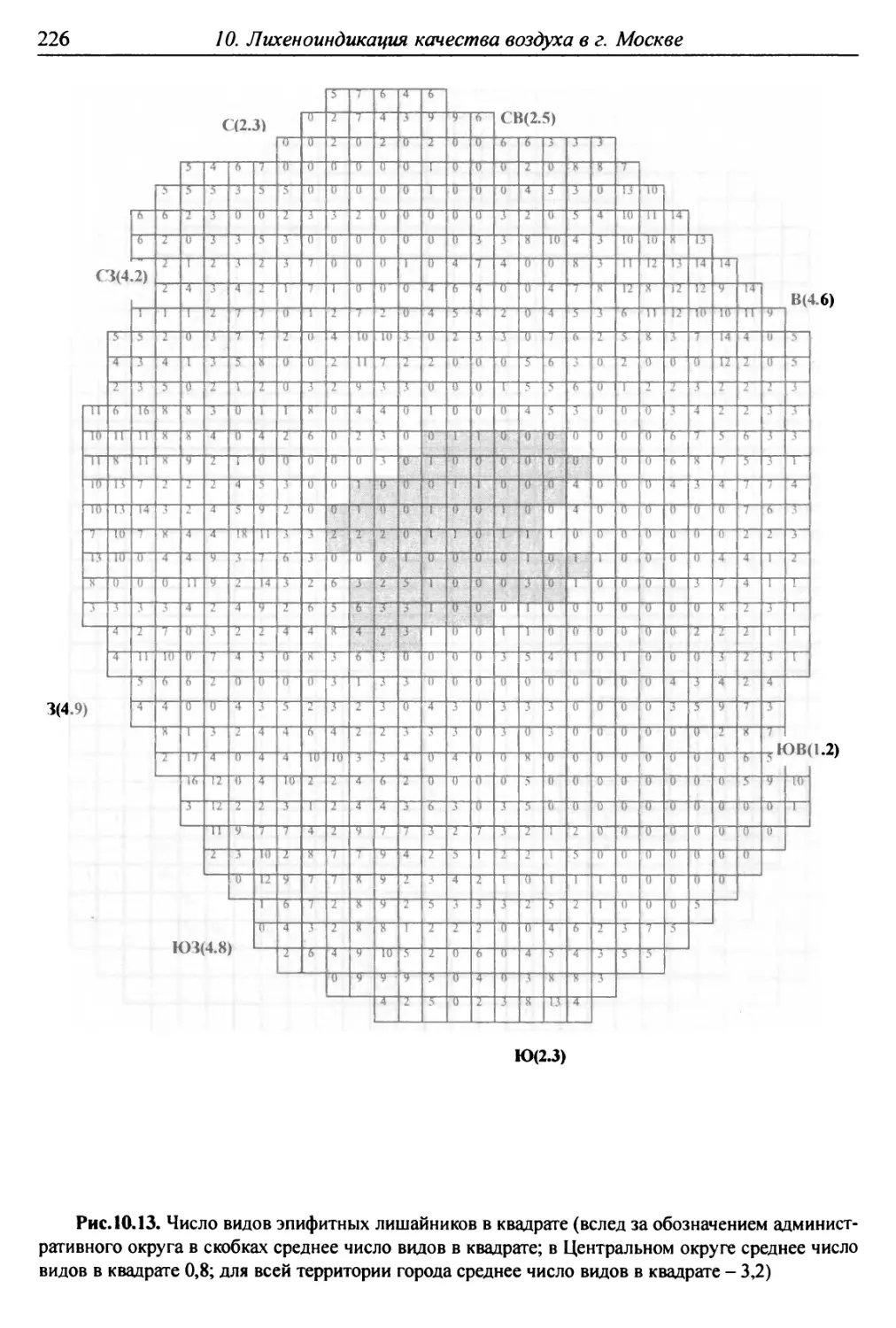
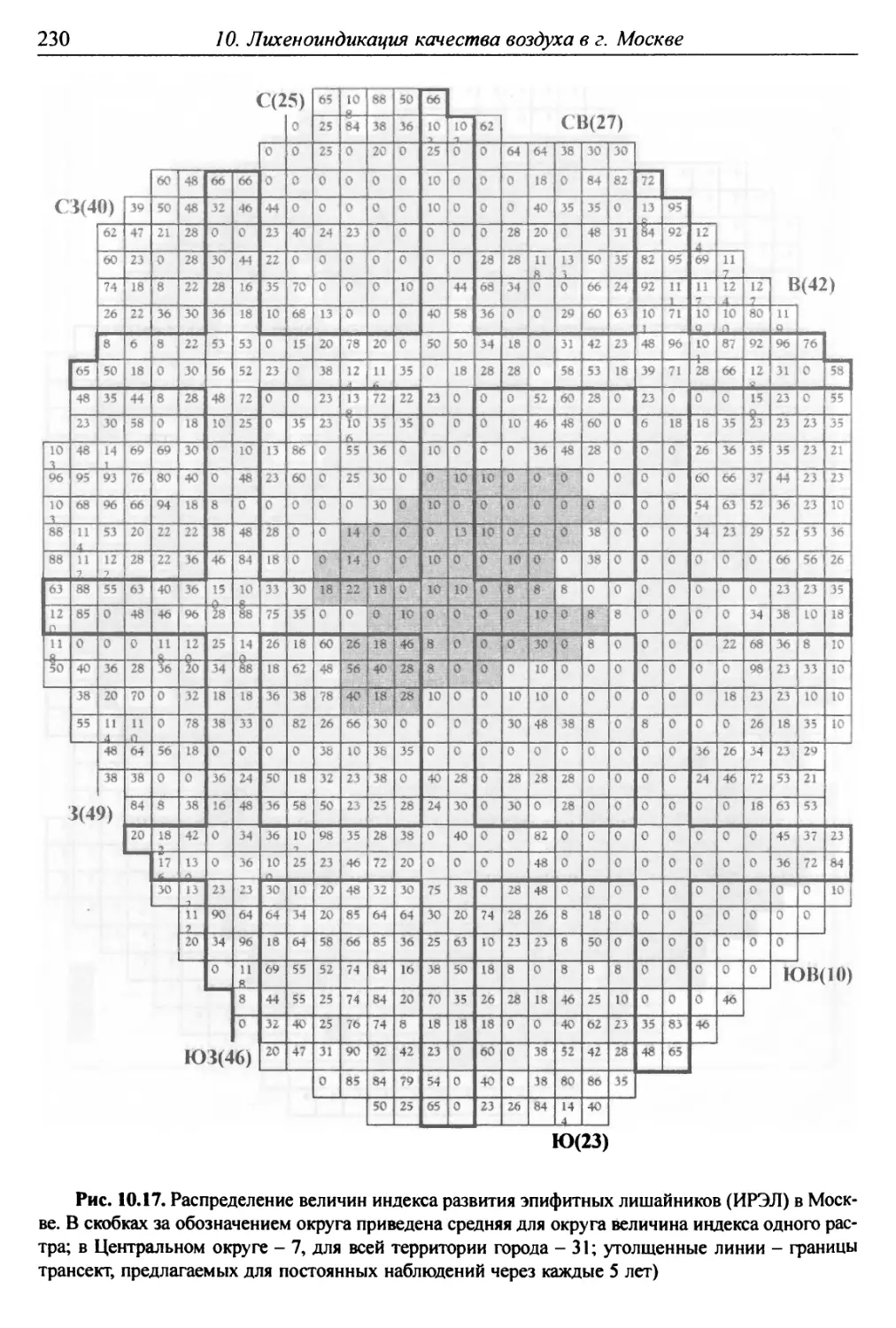
10. Лнхенонндикацня качества воздуха в г. Москве ............. 200
10.1. Методика сбора материала ............................ 200
10.2. Природные условия, структура города и видовой
состав лишайников ....................................... 201
10.3. Особенности современного распространения лишайников
в городе ................................................ 208
10.4. Картосхемы загрязнения воздуха в городе, составленные
на основе особенностей распространения эпифитных
лишайников .............................................. 209
10.5. Результаты наблюдений за трансплантированными
в город слоевищами лишайника ............................ 237
8 Оглавление
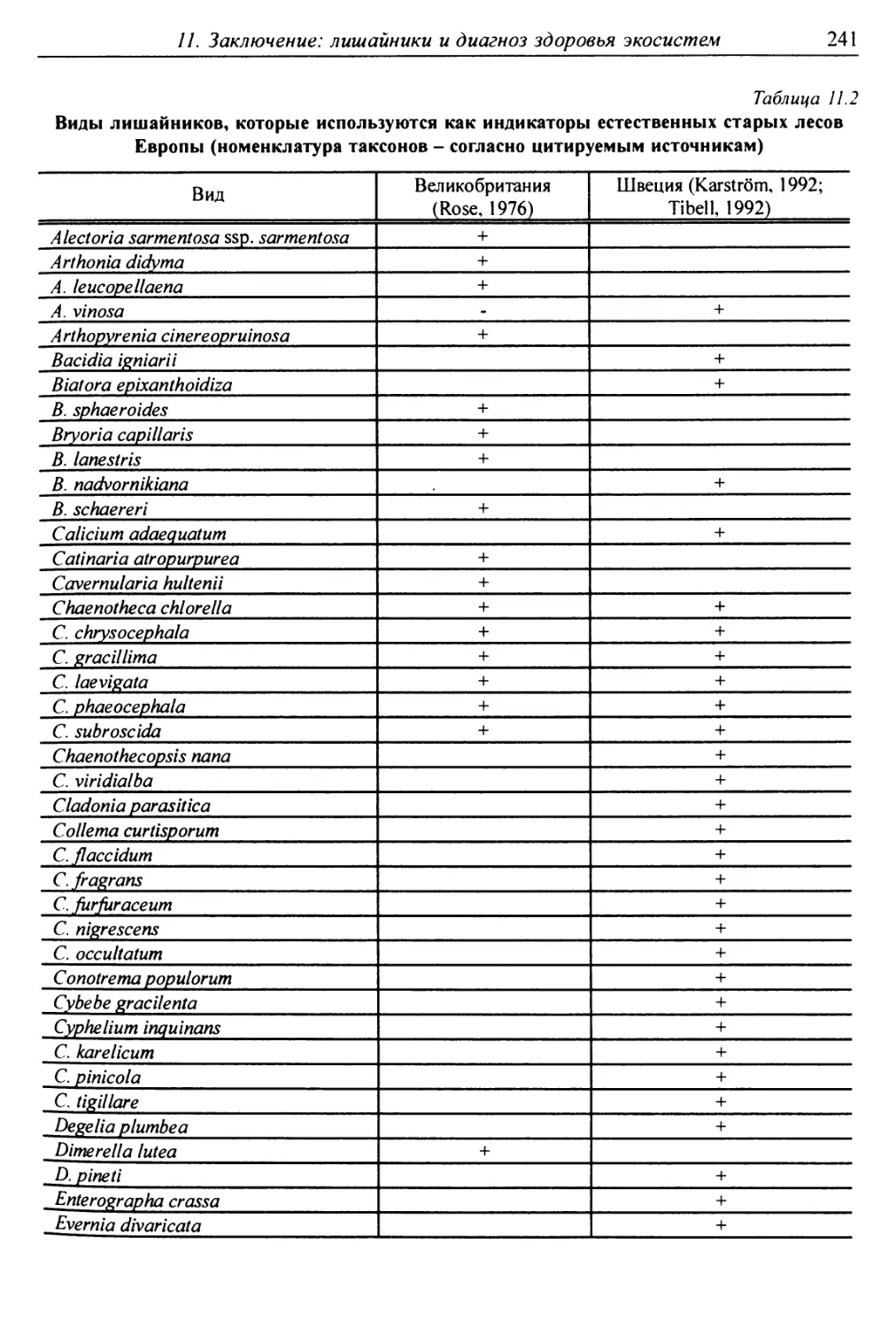
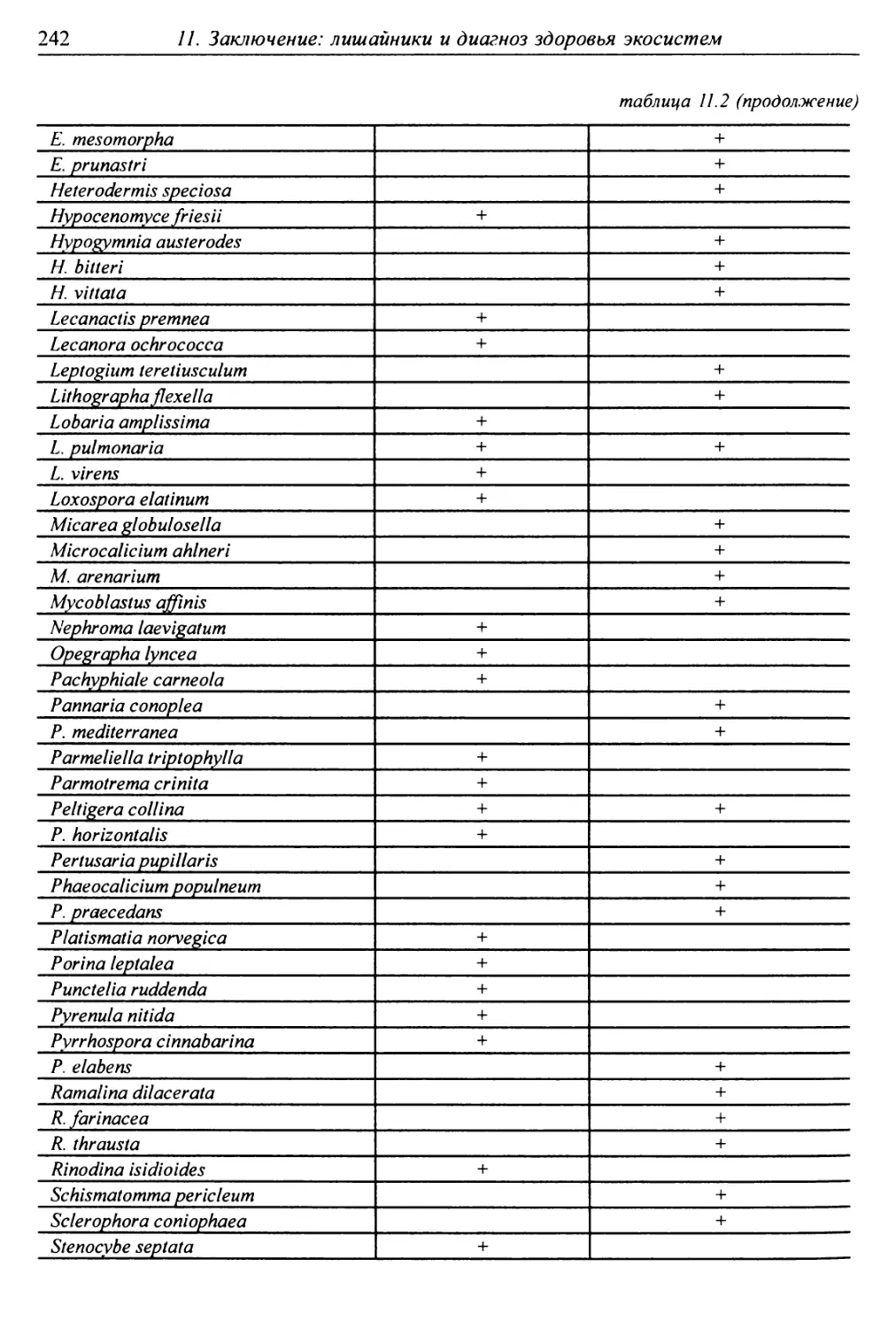
11. Заключение: лишайники и диагноз здоровья экосистем .... 238
11.1. Лишайники-индикаторы ненарушенных лесов ......... 239
11.2. Оценка загрязнения воздушного бассейна .......... 243
11.3. Оценка радиоактивного загрязнения территории .... 243
Литература и авторский индекс ............................. 247
Указатель латинских названий таксонов,
упомянутых в тексте ....................................... 323
ВВЕДЕНИЕ
Закон Российской Федерации “Об охране окружающей среды” от 10 января
2002 г. № 7-ФЗ (Российская газета за 12 января 2002 г., № 6) декларирует, что
и...в соответствии с Конституцией Российской Федерации каждый имеет право
на благоприятную окружающую среду,... на получение достоверной информации
о состоянии окружающей среды”. В этом законе среди компонентов окружаю-
щей среды назван и атмосферный воздух, а одним из способов контроля состоя-
ния среды является мониторинг. Согласно тексту закона, “мониторинг окружаю-
щей среды (экологический мониторинг) - комплексная система наблюдений
за состоянием окружающей среды, оценки и прогноза изменений состояния ок-
ружающей среды под воздействием природных и антропогенных факторов”
(статья 1). Показателями состояния среды и ее компонентов могут быть и пред-
ставители органического мира - растения, животные, грибы, бактерии, комплек-
сные симбиотические ассоциации, к которым принадлежат и лишайники. Видо-
вой состав, особенности распространения лишайников в городах, в зонах воздей-
ствия крупных объектов индустрии, а также на территориях отдельных областей
и даже ряда государств в целом, некоторые показатели жизнедеятельности пред-
ставителей лихенобиоты в последние десятилетия широко используются для суж-
дения о состоянии среды, в особенности воздуха, практически на всех континен-
тах нашей планеты. Это связано с установленной еще в середине XIX века нега-
тивной зависимостью их распространения от загрязнения атмосферы. Результаты
проведенных исследований были широко использованы как для зонирования тер-
риторий по степени загрязнения воздуха или для выявления причин некоторых
заболеваний, например, рака легких (Cislaghi, Nimis, 1997), так и для обоснования
мер, способствующих улучшению качества воздушного бассейна и созданию более
здоровых для человека условий жизни на обследованных территориях. Сведения
о динамике состоянии лишайников позволяют судить об эффективности таких ме-
роприятий. Регистрация показателей представителей лихенобиоты - часть про-
граммы наблюдений многих станций сети глобального мониторинга окружающей
среды (Израэль и др., 1982; Предварительная программа..., 1985; Monitoring...,
2002*). Каждый любознательный человек, в особенности проживающий там,
* Сборник материалов симпозиума в Уэльсе (Великобритания), содержащий 42 статьи, оказался
доступным автору лишь после сдачи рукописи а издательство, поэтому эти публикации, в боль-
10
Введение
где растут деревья, по отсутствию или наличию лишайников, их обилию на
стволах и ветвях может получить общее представление о качестве воздуха в
месте его пребывания: много лишайников - воздух чистый, мало или
их нет - повод для сомнений в его качестве, правда, не во всех случаях оправ-
данный.
Определение свойств среды по особенностям ряда показателей лишайников
называют лихеноиндикацией (от латинского - Lichenes - лишайники). Практи-
чески все живые организмы в той или иной степени пригодны для биоиндикации,
естественно, если в деталях известны особенности их экологии (Покаржевский,
1993; Криволуцкий, 1994). Лишайники, в частности, применялись для выявления
почвенно-геологических и физико-географических параметров территорий (Вик-
торов, 1956, 1960; Джураева, 1978; Викторов, Джураева, 1979; Магомедова, 1980,
1984, 1996; Седельникова, 1982; Таисаев, Константинова, 1988; Rosentreter, 1990;
Урбанавичюс, 2002), динамики поведения горных ледников (Ю. Мартин, 1967,1970;
Турманина, 1971а, б; Савоскул, 1995; и другие), возраста наскальных изображе-
ний (петроглифов), оставленных нашими далекими предками (Седельникова, Че-
ремисин, 2001), степени пастбищной (Бязров, 1976; Андреев, 1980; Магомедова и
др., 1991) и рекреационной (Т. Малышева, 1978; Малышева, Толпышева, 1982)
нагрузок, влияния удобрений (Гордеева, Толпышева, 1978) и других целей. Эколо-
гическая разведка в Вооруженных силах РФ ныне основывается и на показате-
лях свойств лишайников интересующей местности (Бадтиев, Кулемин, 2001). При
изучении последствий применения в Косово (1999 г.) войсками НАТО снарядов
со слабо обогащенным ураном использовали и лишайники (Sansone et al., 2001).
Однако наиболее широко представители лихенобиоты, в особенности эпифиты,
использовались для показа загрязнения воздуха, обусловленного как деятельнос-
тью человека, так и вызванного естественными причинами, например, изверже-
ниями вулканов (Bargagli, Barghigiani, 1991). Еще в середине прошлого века в Южном
Ланкашире (Англия) установлено снижение видового разнообразия лихенофлоры
из-за увеличения загрязнения воздуха (Hawksworth, Rose, 1976), а классик лихе-
нологии - науки о лишайниках - В. Нюландер (Nylander, 1866), швед по происхож-
дению, долгое время работавший во Франции, обратил внимание на постепенное
исчезновение лишайников из Люксембургского сада в Париже из-за использова-
ния новых видов топлива и газа для освещения городских улиц. Это дало ему
основание назвать лишайники “гигиенометрами”, состав и состояние которых в
городе или его частях в определенной степени отражают качество воздуха, ком-
фортность условий проживания населения городов. К началу 1900-х этот “город-
ской эффект” в отношении лишайников, т.е. их исчезновение в городах, был широ-
ко признанным феноменом, и поначалу его связывали с действием угольной сажи.
Позднее основным токсичным для них агентом был признан бесцветный газ дву-
шинстве своем затрагивающие многие проблемы, обсуждаемые в данной монографии, в ней не отра-
жены.
Введение
11
окись серы. Ныне список загрязнителей воздуха, негативно действующих на ли-
хенобиоту, много больше и включает HF, некоторые металлы, кислотные дожди,
окислители и органические соединения. Здесь уместно упомянуть, что существо-
вало и другое объяснение исчезновения лишайников в городах, а именно, более
высокая сухость воздуха в городских поселениях (Rydzak, 1953, 1968), что не
нашло подтверждения в многочисленных исследованиях по этой проблеме
(LeBlanc, Rao, 1973b; Hawksworth, Rose, 1976; Galun et al., 1984; Galun, Ronen,
1988; Nash, 1996).
Различные аспекты лихеноиндикации загрязнения воздуха отражены в мно-
гочисленных публикациях. С 1974 г. журнал “The Lichenologist” регулярно публи-
ковал библиографию “Литература о загрязнении воздуха и лишайники (Literature
on air pollution and lichens)”, 39 выпусков (2000 г.) которой содержали более 3000
названий. Поисковая система базы данных “Recent literature on lichens” (Cullberson
et al., 2002), которая в июле 2002 г. включала 32492 источников, на ключевое слово
“pollution” нашла 1831 название.
Предлагаемая вниманию читателей книга обобщает как оригинальные, так и
доступные опубликованные данные об использовании лишайников в контроле ка-
чества среды, главным образом, состояния воздушного бассейна, в нашей стра-
не и за ее пределами. Особое внимание уделено методическим проблемам. Вме-
сте с тем в книге содержатся и сведения общего характера: различные трактов-
ки лишайникового симбиоза, строение лишайников, их жизненные формы,
особенности физиологии и другое. При подготовке этих разделов были использо-
ваны опубликованные сотрудниками Лаборатории лихенологии и бриологии Бо-
танического института им. В.Л. Комарова обзорные статьи Е.А. Вайнштейн (1972,
1973, 1982), И.А. Шапиро (1986, 1993, 1996), Н.С. Голубковой (1993), а также
книга А.Н. Окснера (1974). Номенклатура лишайников в книге в основном соот-
ветствует названиям, использованным в цитированных в монографии источни-
ках. В некоторых случаях в скобках приведена и современная номенклатура так-
сона. Размерность величин также указана согласно цитированным источникам.
Работа выполнена в рамках Плана НИР Лаборатории биоиндикации ИПЭЭ
РАН на 1996-2000 гг. Считаю своим приятным долгом высказать признатель-
ность за содействие моим усилиям по написанию монографии заведующему
лабораторией члену-корреспонденту РАН Д.А. Криволуцкому, в.н.с. д.б.н.
А.Д. Покаржевскому, а также многочисленным отечественным и зарубежным
коллегам за присланные ими недоступные в Москве публикации. Я благодарю
неизвестных мне экспертов РФФИ, положительно оценивших мою рукопись.
1. БИОЛОГИЯ И ЭКОЛОГИЯ ЛИШАИНИКОВ
Природа лишайникового симбиоза широко обсуждалась в научной и популяр-
ной литературе (Smith, 1976; Hill, 1994), однако еще многое не понято и общепри-
нятой трактовки объекта “лишайники” до сей поры нет. Приведу перевод одного
из известных мне определений: “Лишайник-это ассоциация между грибом, обыч-
но аскомицетом, но в некоторых случаях базидиомицетом или дейтеромицетом, и
одним (или более) фотосинтезирующим партнером - зеленой водорослью или ци-
анобактерией. У всех лишайников гриб образует слоевище (таллом), которое мо-
жет содержать вторичные вещества” (Ahmadjian, 1993). Согласно ему, лишайник
включает микобионт, фотобионт, слоевище и вещества как продукт метаболизма
партнеров. В этой симбиотической ассоциации гриб, видимо, паразитирует либо
на зеленой водоросли, либо на цианобактерии (сине-зеленой водоросли), либо и на
водоросли и на цианобактерии вместе. Эукариотическая водоросль и прокарио-
тическая цианобактерия являются фотосинтезирующими организмами и служат
для гриба источником углеводов. Поток углеводов в лишайнике от фотобионта к
микобионту включает полиолы, в случае зеленой водоросли, и глюкозу, в случае
цианобактерии, причем клетки фотобионта более проницаемы для углеводов в
лихенизированном, чем в не лихенизированном состоянии (Smith, Douglas, 1987).
Цианобактерии, кроме того, способны фиксировать молекулярный азот атмосфе-
ры, превращая его в соединения, пригодные для питания гриба. Поток питатель-
ных элементов от микобионта к фотобионту несравнимо меньший. Другим след-
ствием тесной физиологической интеграции является наличие в лишайниках об-
ширного набора вторичных продуктов, так называемых лишайниковых веществ
(Huneck, 2001). Большинство их неизвестны в свободно живущих грибах (или дру-
гих организмах) и потому их наличие - дополнительное свидетельство уникаль-
ности лишайникового симбиоза.
Каждое слоевище лишайника предстает перед нами как отдельный организм,
хотя фактически такой таллом представляет собой морфологическое выражение
взаимодействия двух, а то и трех организмов из разных царств органического
мира (грибов, зеленых растений, цианобактерий). С позиций генетики и эволюции
лишайники, конечно, нельзя рассматривать как индивиды. Вероятно, следует со-
гласиться с мнением, что фактически дискретное слоевище лишайника представ-
ляет собой миниатюрную экосистему (Farrar, 1976а). Фототрофные участники этой
Биология и экология лишайников
13
ассоциации (водоросль, цианобактерия) при высвобождении из слоевища лишай-
ника могут далее развиваться как самостоятельные организмы, тогда как гриб-
ной компонент вне лишайниковой ассоциации живет очень непродолжительное
время. Как отмечала Н.С. Голубкова (1993), с биологической точки зрения ли-
шайник - это трактуемый в широком смысле симбиоз двух или более организ-
мов, генетически обособленных, основанный на метаболических потребностях
одного из них.
Степень лихенизации варьирует в разных группах лишайников в очень широ-
ких пределах - от нескольких клеток фотобионта, которые кажутся почти слу-
чайно ассоциированными с грибами (как некоторые представители Caliciales),
до типично хорошо интегрированного слоевища, в котором отчетливый слой фо-
тобионта находится ниже коровой грибной ткани.
1.1. МИКОБИОНТ
Грибы, формирующие лишайники, не представляют собой естественную так-
сономическую группу. Грибной компонент лишайникового симбиоза, или микоби-
онт у большинства видов (98%) лишайников принадлежит к различным семей-
ствам отдела сумчатых грибов (Ascomycota). у немногих - к базидиомицетам
(Basidiomycota\ у единичных видов - к дейтеромицетам (Deuteromycota) и гри-
боподобным протистам, чьё систематическое положение пока точно не установ-
лено. Все грибы, участвующие в формировании лишайников, называют лихенизи-
рованными, а процесс образования лишайника-лихенизацией. Общее между ними
- пищевая специализация, и их общее обозначение “лишайники” или “лихенизиро-
ванные грибы” имеет такой же смысл, как названия “микоризные грибы”, “фито-
патогенные грибы”. Более 25% известных видов грибов являются лихенизиро-
ванными (Honegger, 2001), причем высказана гипотеза, что именно представите-
ли лихенизированных грибов являются предками большинства филогенетических
линий грибов (Lutzoni et al., 2001). В свободном состоянии, т.е. вне лишайниковой
ассоциации, лихенизированные грибы живут очень непродолжительное время*.
У большинства лишайников практически все слоевище (до 98% от общей массы)
- это гриб, представленный очень тонкими, обычно 3-10 мкм в диаметре, про-
стыми или разветвленными нитями, называемыми гифами. Эти нити разделены
поперечными перегородками на клетки. Микобионт образует множество веществ,
некоторые из которых как, например, пигменты (зеленый, синий, фиолетовый, крас-
ный, коричневый) - видоспецифичны. Их наличие или отсутствие является важ-
ным диагностическим признаком таксонов.
* Лихенизированные грибы не следует отождествлять с лихенофильными, т.е. с грибами, обитаю-
щими на слоевищах лишайников.
14
1. Биология и экология лишайников
1.2. ФОТОБИОНТ
Фотосинтезирующий автотрофный компонент лишайникового симбиоза, или
фотобионт представлен эукариотическими зелеными водорослями и прокариоти-
ческими цианобактериями. Последние по традиции нередко все еще именуют
сине-зелеными водорослями. В научных публикациях до 1950-х годов фотобионт
называли гонидиями, позднее-фикобионтом. В настоящее время известны пред-
ставители 44 родов водорослей и цианобактерий, участвующих в формировании
лишайников (Tschermak-Woess, 1988), причем некоторые из них существуют и в
свободном состоянии, т.е. вне симбиоза с грибом. Примерно 85% лихенизиро-
ванных аскомицетов ассоциированны с одноклеточными или нитчатыми зелены-
ми водорослями, примерно 10% - с цианобактериями и 3-4% - с теми и другими
вместе (Honegger, 2001). Один и тот же вид водоросли и цианбактерии может
участвовать в формировании разных видов лишайников, а один гриб может обра-
зовывать сходные морфотипы, паразитируя на разных видах фотобионтов. На-
пример, зеленая водоросль Trebouxia irregularis как фотобионт была встречена
в слоевищах 30 видов лишайников. Микобионт повсеместно встречающегося на
деревьях, заборах, столбах листоватого лишайника Xanthoria parietina, привле-
кающего внимание ярко-оранжевым цветом слоевища, в разных местообитаниях
паразитирует на разных видах Trebouxia - Т arboricola, Т decolorans, Т. irre-
gularis (Голубкова, 1993). Таким образом, таксономическая принадлежность фо-
тобионта в систематике лишайников формально не учитывается, однако некото-
рые лихенологи все же предлагают считать действительными (валидными) так-
соны лишайников, выделенные и на основе видовых различий фотобионтов
(Jorgensen, 1991).
1.3. ВЗАИМООТНОШЕНИЯ МИКОБИОНТА И ФОТОБИОНТА
В СЛОЕВИЩЕ ЛИШАЙНИКА
Данные современных исследований в основном подтверждают мнение
С. Шведенера, первооткрывателя дуалистической природы лишайников, что в ос-
нове лишайникового симбиоза лежат трофические связи, они антагонистические,
ибо гриб паразитирует на фотобионте, т.е. зеленая водоросль или цианобактерия
являются организмом-хозяином. Он же высказал и гипотезу об особенностях этого
паразитизма, предположив, что водоросль или цианобактерия используется гри-
бом очень медленно и уничтожается постепенно. Поскольку процесс замедлен,
многие клетки фотобионта остаются нетронутыми и даже успевают дать ряд
новых поколений здоровых клеток. Однако эта гипотеза довольно долго имела
ограниченное число сторонников. Значительное число исследователей было со-
гласно с концепцией симбиотических трофических связей между партнерами, но
/. Биология и экология лишайников
15
считали эти связи гармонично взаимовыгодными, мутуалистическими. И хотя
эта взаимная “польза” не имеет строгих научных доказательств (Smith, 2001),
идея гармонии хотя бы в природе была довольно привлекательной, имела и имеет
много приверженцев (Hawksworth, 1988).
Популярной была также и гипотеза “консорция”. Согласно ей лишайник пред-
ставляет собой сожительство - консорций, в котором гриб и водоросль составля-
ют самостоятельный организм, соответствующий целостному растению, а в сис-
теме растений лишайники представляют самостоятельный класс, что косвенно
отражает расположение материала о лишайниках в популярном издании “Жизнь
растений” (1977). Однако современный уровень знаний не дает каких-либо осно-
ваний для утверждения, что взаимоотношения между генетически различными
организмами, какими бы тесными и взаимосвязанными они бы не были, могут
привести к образованию нового самостоятельного организма особой системати-
ческой категории.
Структурную основу лишайникового слоевища составляет преимущественно
микобионт, поскольку именно на него приходится до 98% массы тела у большин-
ства видов лишайника. Известны лишь несколько таксонов, у представителей
которых облик всего таллома определяет фотобионт (роды Coenogonium, Cysto-
coleus, Ephebe, Racodium). Однако развитие характерного лишайникового слое-
вища начинается лишь после установления симбиотических контактов грибного
партнера с зеленой водорослью или цианобактерией, так что с позиций морфоге-
неза строение лишайникового таллома, строго говоря, определяется фотобион-
том и его прямым контактом с микобионтом. В нескольких случаях методами
молекулярной биологии достоверно установлено, что один и тот же микобионт
был способен формировать два очень различно структурированных слоевища,
соответственно, с цианобактерией и зеленой водорослью (Armaleo, Clerc, 1991).
Эти разные морфотипы обозначают как фотосимбиодемы и их наличие среди
лишайников указывает на онтогенетический контроль со стороны фотобионта.
При культивировании в лабораторных условиях грибы, не вступившие в контакт с
водорослью, остаются относительно аморфными, но уже в самом начале контак-
та гриба и фотобионта на месте аморфного скопления грибных гиф начинает фор-
мироваться структурированное слоевище (Ahmadjian, 1993).
Степень облигатности лишайникового симбиоза для участвующих партнеров
варьирует в широких пределах. Опыты по изоляции бионтов из слоевищ показали,
что лихенизированные грибы или умеренно специфичны, т.е. участвуют в симби-
озе с несколькими родственными видами фотобионтов, или высоко специфичны,
т.е. участвуют в симбиозе только с одним видом фотобионта (Honegger, 1998).
Это характерно, в частности, для грибов, у которых фотобионтами являются виды
рода зеленой водоросли Trebouxia. Представители этого рода отмечены у при-
мерно 50% лихенизированных аскомицетов, но их редко находили в свободножи-
вущем состоянии (Friedl, Budel, 1996; Honegger, 2001). Наоборот, другие фотоби-
16
7. Биология и экология лишайников
онтные роды как Gleocapsa, Nostoc, Scytonema, Trentepohlia встречаются обыч-
но как в лихенизированном, так и в свободноживущем состояниях. Наконец, в
некоторых случаях как свободноживущие, так и лихенизированные популяции
встречаются в одних и тех же местообитаниях, такие как свободноживущие
Nostoc, Scytonema в пустынных почвах и их лихенизированные двойники, соот-
ветственно, в напочвенных лишайниках родов Collema, Peltula. Степень встре-
чаемости одних и тех же фотобионтных видов как в свободноживущем, так и в
лихенизированном состояниях точно оценить в настоящее время невозможно,
поскольку относительно мало лишайниковых водорослей были идентифицирова-
ны до уровня вида, да и в целом систематика на уровне видов многих цианобак-
терий и одноклеточных зеленых водорослей изучена еще недостаточно хорошо
(Friedl, Biidel, 1996). И наоборот, систематика микобионтов исследована доволь-
но неплохо. Поскольку изолированные микобионты растут очень медленно, они
не способны к выживанию в свободноживущем состоянии из-за конкуренции с
другими грибами или потребления их другими организмами. Таким образом, боль-
шинство микобионтов лихенизированы облигатно, хотя специфичность микобион-
та для конкретного вида фотобионта может быть не столь велика, как это можно
предполагать.
1.4. ОБРАЗОВАНИЕ ВЕГЕТАТИВНОГО ТЕЛА
(СЛОЕВИЩА, ИЛИ ТАЛЛОМА) ЛИШАЙНИКА
Слоевище, или таллом лишайника может формироваться как из споры, так и
вегетативно. Споры у лишайников большей частью сосредоточены в плодовых
телах, называемых апотециями, если они расположены на поверхности, и пе-
ритециями, если плодовые тела погружены в слоевище. Располагаются плодо-
вые тела либо по всей поверхности слоевища, либо в определенных его частях
(край, центр и другое). Созревшая спора гриба, выброшенная из апотеция или
перитеция, попав в благоприятные условия влажности и тепла, начинает прорас-
тать и через некоторое время образует гифы проталлюса (мицелия), в котором
еще отсутствуют водоросли или цианобактерии. Если их нет на том субстрате,
где разрастается проталлюс, то последний обычно скоро отмирает. Если же ми-
целий гриба встречает водоросль или цианобактерию, то гифа проталлюса просо-
вывается под клетку водоросли или цианобактерии, отделяет ее от субстрата и
обвивает своего будущего хозяина. Затем гифа, вероятно, через выделение гор-
монов, вызывает деление клетки водоросли или цианобактерии. Эти дочерние
клетки также обвиваются гифами гриба, в результате чего образуется зачаток
лишайника, из которого постепенно развивается слоевище лишайника. Различа-
ют несколько стадий формирования таллома в зависимости от морфологического
типа последнего (Окснер, 1974).
1. Биология и экология лишайников
17
Вегетативное образование слоевища происходит как из фрагментов материн-
ского слоевища, так из особых образований таллома, в которых уже имеются и
микобионт, и фотобионт. К числу основных таких образований относятся соредии
и изидии. Соредии представляют собой крохотные шаровидные тельца на слое-
вище лишайника, которые состоят из одной или нескольких клеток фотобионта,
окруженных сплетением короткоклеточных гиф гриба, но лишенные корового слоя.
Располагаются они на поверхности слоевища либо диффузно, либо собраны ком-
пактно в виде соралей разной формы (точковидные, пятнистые, щелевидные, го-
ловчатые, губовидные и другие). Изидии представляют собой маленькие разно-
образной формы (зернистые, коралловидные, цилиндрические, булавовидные и
другие) выросты на верхней поверхности таллома, которые содержат фотобионт
и микобионт. Эти выросты покрыты коровым слоем. Отделившись от субстрата
и попав в благоприятные условия, соредии, изидии или другие фрагменты талло-
ма развиваются в новые слоевища.
1.5. СИСТЕМАТИКА
Согласно Международному коду ботанической номенклатуры, научное название
лишайника дается по его грибному компоненту. Число видов лишайников оцени-
вают от 13500 (Hawksworth, Hill, 1984) до, примерно, 17000 (Hale, 1974) и даже
26000 (Окснер, 1974). С 1931 по 2000 г. в среднем за год описывалось по 175
новых для науки видов (Sipman, Aptroot, 2001). Но в тоже время ревизия ряда
таксономических групп привела к сведению многих видов в синонимы уже опи-
санных, и число признаваемых ныне видов, например, среди калициевых, умень-
шилось на 61% (Sipman, Aptroot, 2001). Поскольку многие регионы мира остают-
ся слабо обследованными, возможно, реально число видов лишайников около 20000.
Наибольшее их количество принадлежит к сумчатым грибам Ascomycota, при-
чем, фактически почти половина описанных аскомицетов лихенизированы (Tehler,
1996). Также имеются несколько представителей лихенизированных базидиаль-
ных грибов Basidiomycota и неопределенных грибов Deuteromycota. Последняя
группа является искусственным классом, в котором представлены стерильные
виды. Если, в конце концов, у них устанавливали плодовые структуры, эти лишай-
ники находили свое место среди аскомицетов или базидиомицетов.
В эволюционном смысле лишайники, используя термины филогении, полифи-
летичны, что подтверждается результатами молекулярно-генетических исследо-
ваний ряда групп лишайников (Иванова, 1998).
1.6. ТИПЫ СЛОЕВИЩ И ЖИЗНЕННЫЕ ФОРМЫ ЛИШАЙНИКОВ
Лишайники чрезвычайно разнообразны по своему внешнему виду. Их талло-
мы бывают самой разнообразной формы, размера, строения, консистенции, ок-
18
I. Биология и экология лишайников
раски. По цвету слоевищ они представляют вариацию оранжевого, желтого, крас-
ного, зеленого, серого, коричневого и черного (Окснер, 1974; Jahns, 1980). Разме-
ры их слоевищ от менее чем 1 мм2 до длинных, простертых форм, которые свиса-
ют более чем на 2 м с ветвей деревьев. Наряду с деревьями и другими растени-
ями, на которых они растут как эпифиты и эндофиты, лишайники часто заселяют
голую почву (эпигеиды) и поверхность горной породы. На камнях большинство
обычных лишайников встречаются эпилитно (на поверхности), но другие встре-
чаются эндолитно, т.е. внутри поверхностного слоя породы. В тропиках и субтро-
пиках некоторые быстро растущие лишайники даже заселяют поверхности лис-
тьев как эпифилы. В умеренных широтах представителей последних иногда мож-
но встретить на старой хвое. Хотя большинство лишайников наземные, некоторые
виды встречаются в водных потоках рек, в озерах, а некоторые были отмечены в
приливно-отливной зоне морей и океанов.
Различно и размещение фотобионтов в талломе. По последнему признаку их
можно разделить на два типа строения слоевища: гетеромерное и гомеомерное.
На вертикальном срезе слоевища гетеромерных лишайников можно наблюдать,
что клетки фотобионта располагаются только в одном слое. У гомеомерных ли-
шайников клетки фотобионта рассредоточены более или менее равномерно по
всему слоевищу. Деление это достаточно условное и грубое, но удобное для иден-
тификации лишайников. По внешней форме слоевища обычно различают три
основных морфологических типа лишайников: 1) наиболее простые накипные
(иногда называемые корковыми), к которым относят и чешуйчатые, 2) листова-
тые, более сложные в анатомо-морфологическом отношении, и 3) кустистые,
с наиболее высоко развитым типом слоевища. Каждая из этих основных групп
также далеко не однородна, и имеются виды с промежуточным типом строения
талломов.
Многообразие форм роста лишайников обусловило создание различных клас-
сификаций жизненных форм лишайников, из которых отечественные лихено-
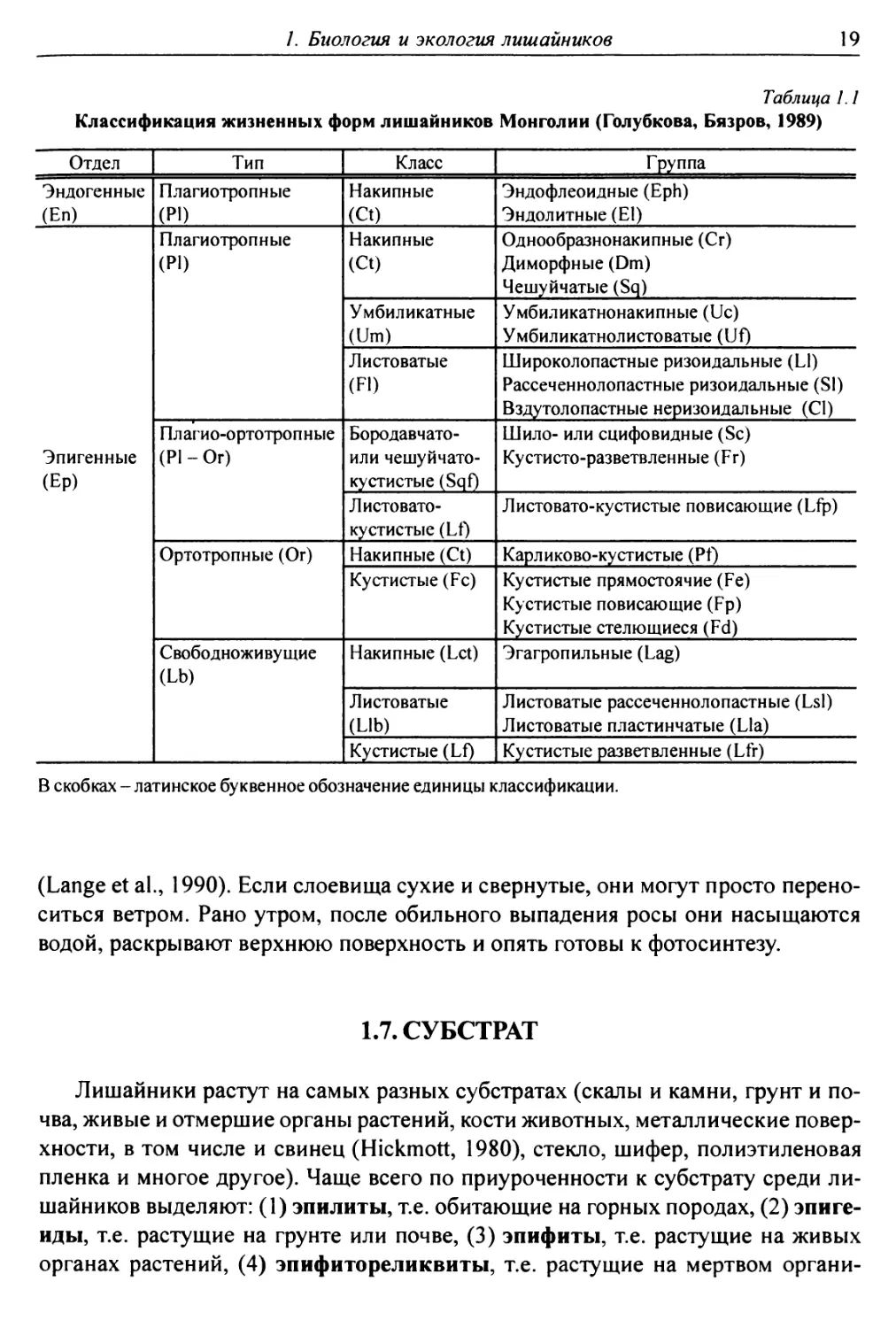
логи (Котлов, 1995; Пристяжнюк, 1996) чаще других применяют предложенную
Н.С. Голубковой (1983). Позднее она была модифицирована при выделении
лихеносинузий в биогеоценозах Монголии (Голубкова, Бязров, 1989; Бязров, 1990)
(табл. 1.1).
Экологически интересной группой лишайников являются кочующие лишайни-
ки, подобные Aspicilia esculenta. Xanthoparmelia camtchadalis =(Parmelia
vagans), Xanthomaculina convoluta, Chondropsis semiviridis, в степях, пустынях
и полупустынях. Части представителей этих лишайников свойственно гигроско-
пическое перемещение (Biidel, Wessels, 1986; Lumbsch, Kothe, 1988). В сухом со-
стоянии их слоевища скручиваются, и открывается нижняя кора талломов. После
смачивания жидкой водой слоевища разворачиваются и открывают верхнюю по-
верхность для солнечного света. У двух последних видов лишайников фотосин-
тез значительно увеличивается после открывания верхней поверхности свету
/. Биология и экология лишайников
19
Таблица 1.1
Классификация жизненных форм лишайников Монголии (Голубкова, Бязров, 1989)
Отдел Тип Класс Группа
Эндогенные (Еп) Плагиотропные (Р1) Накипные (Ct) Эндофлеоидные (Eph) Эндолитные (Е1)
Эпигенные (Ер) Плагиотропные (PI) Накипные (Ct) Однообразнонакипные (Сг) Диморфные (Dm) Чешуйчатые (Sq)
Умбиликатные (Um) Умбиликатнонакипные (Uc) Умбиликатнолистоватые (Uf)
Листоватые (F1) Широколопастные ризоидальные (L1) Рассеченнолопастные ризоидальные (S1) Вздутолопастные неризоидальные (С1)
П л аги о-о ртотро п н ые (PI - Or) Бородавчато- или чешуйчато- кустистые (Sqf) Шило- или сцифовидные (Sc) Кустисто-разветвленные (Fr)
Листовато- кустистые (Lt) Листовато-кустистые повисающие (Lfp)
Ортотропные (Or) Накипные (Ct) Карликово-кустистые (Pf)
Кустистые (Fc) Кустистые прямостоячие (Fe) Кустистые повисающие (Fp) Кустистые стелющиеся (Fd)
Свободноживущие (Lb) Накипные (Let) Эгагропильные (Lag)
Листоватые (Lib) Листоватые рассеченнолопастные (Lsl) Листоватые пластинчатые (Lla)
Кустистые (Lf) Кустистые разветвленные (Lfr)
В скобках - латинское буквенное обозначение единицы классификации.
(Lange et al., 1990). Если слоевища сухие и свернутые, они могут просто перено-
ситься ветром. Рано утром, после обильного выпадения росы они насыщаются
водой, раскрывают верхнюю поверхность и опять готовы к фотосинтезу.
1.7. СУБСТРАТ
Лишайники растут на самых разных субстратах (скалы и камни, грунт и по-
чва, живые и отмершие органы растений, кости животных, металлические повер-
хности, в том числе и свинец (Hickmott, 1980), стекло, шифер, полиэтиленовая
пленка и многое другое). Чаще всего по приуроченности к субстрату среди ли-
шайников выделяют: (1) эпилиты, т.е. обитающие на горных породах, (2) эпиге-
иды, т.е. растущие на грунте или почве, (3) эпифиты, т.е. растущие на живых
органах растений, (4) эпифитореликвиты, т.е. растущие на мертвом органи-
20
/. Биология и экология лишайников
ческом субстрате, (5) эврисубстратные, т.е. обитающие на разных субстратах.
Существуют и более детальные классификации. Например, виды, растущие на
листьях или хвое, называют эпифильными, на древесине - эпилигнофитами,
а лишайники, растущие на мохообразных-эпибриофитами, на других лишайни-
ках - эпилихенофитами. Детальность классификаций зависит от задач, решае-
мых конкретным исследованием, особенностей изучаемого региона или объекта.
При проведении лихеноиндикационых исследований крайне желательно учиты-
вать особенности химического состава субстрата лишайников, его кислотность.
Среди эпилитных видов различают лишайники силикатных и щелочных горных
пород. Кора деревьев также неодинакова по богатству элементами минерального
питания. Выделяют богатую (зольность 5-12%), средне богатую (2-5%) и бед-
ную (0,4-2,7%) кору (Barkman, 1958). Различна и кислотность коры разных дре-
весных пород (табл. 1.2)
Таблица. 1.2
Актуальная кислотность коры (К) и древесины (Д) некоторых древесных пород
(Barkman, 1958)
Древесная порода К Д
Alnus glutinosa 4.2 - 5,0 3,9 - 4,7
A. incana 4.9-6,2 —
Betula verrucosa 3,7 _ 4,4 (4,9) 2,6-5,7
Picea abies 3,8-4,5 (6,0) 3,3-5,6
Pinus sylvestris 3,4-3,8 (4,3) 3,4
Populus tremula 3,9-7,9 —
Quercus robur 3,7-5,0 3,3-5,6
Salix sp. 5,0-5,2 4,1 -6,2
Sorbus aucuparia 4,6-5,2 3,7
Tilia cordata 4,8-6,2 4,2-6,1
Есть данные, что с возрастом дерева кислотность коры увеличивается (Du
Rietz, 1945; Koskinen, 1955). Растет она и по мере распада субстрата. Видимо,
имеется обратная связь между содержанием зольных элементов в субстрате и
его кислотностью, но зависимость эта весьма сложная. Так, наличие танинов в
коре кислотность последней увеличивает, хотя по количеству минеральных ве-
ществ кора будет считаться богатой (Barkman, 1958). Часть видов лишайников
встречается как на бедном, так и богатом субстратах, другие приурочены лишь
к субстрату с определенным количественным и качественным составом мине-
ральных веществ. Неодинаково лишайники относятся и к наличию соединений
азота. Еще в начале XX века было установлено, что 25-дневное опрыскивание
слоевищ раствором куриного помета различно влияло на представителей разных
видов лишайников (Semander, 1912).
1. Биология и экология лишайников
21
Аналогично отношение лишайников к кислотности субстрата. По реакции на
этот показатель их можно разделить на: ацидофилы - рН=4,0, ацидофиты -
рН=4,5-5,5, нейтрофиты - рН=5,5-7,0 и базифиты - pH >7,0 (TrUmpener, 1926;
Du Rietz, 1932; Mattick, 1932; Работнов, 1936; Vezda, 1955; Barkman, 1958). Однако
значительное число видов встречается на субстратах с довольно широкой амп-
литудой величины pH. Лишайники с большой амплитудой pH субстрата были на-
званы эврииониками, а с узкой - стеноиониками, причем число видов после-
дних меньше (Du Rietz, 1932).
Лишайники сами могут менять кислотность среды. Есть данные, что участ-
ки коры дуба, покрытые лишайниками, имели рН=4,1, а участки коры без лишай-
ников имели рН=4,5 (Barkman, 1958). Было показано, что кора деревьев одной
породы (Pice a abies) в разных условиях имела различную концентрацию элемен-
тов и показатели кислотности, что сказалось на распределении эпифитов (Hauck
et al., 2001а, b).
1.8. ФОРОФИТ
Для целей лихеноиндикации чаще применяют эпифитные лишайники, т.е. ис-
пользующие в качестве субстрата поверхности растений, обычно - деревьев и
кустарников, что дает основание отдельно рассмотреть особенности этого мес-
тообитания. Растение, на котором растут эпифиты, первоначально называли рас-
тением-хозяином (Schimper, 1898). Однако этот термин широко применялся для
растений, на которых живут паразиты, поэтому позднее деревья-носители эпифи-
тов чаще стали называть форофитами (Barkman 1958; Peciar, 1965), хотя были
предложены и другие термины, например, фитофор (LeBlanc, 1963). Эпифитизм
представляет одну из форм взаимоотношений между организмами (Бязров, 19786;
Bronstein, 1994) при неодинаковом значении друг для друга каждого из партнеров
взаимодействия: влияние эпифита на форофит минимально, тогда как последний
определяет среду эпифита, заставляет его приспосабливаться к этой среде и, со-
ответственно, к себе. Поверхность деревьев и кустарников - далеко не идеаль-
ное местообитание для растений, лишайников. Условия жизни здесь экстремаль-
ные ввиду чрезвычайной сухости (Barkman, 1958). Среда на форофите меняется
как во времени, так и в пространстве. В процессе своего роста дерево предо-
ставляет эпифитам новые площади для заселения посредством утолщения, обра-
зования новых ветвей, их удлинения, растрескивания и отслаивания коры. Но од-
новременно дерево механически воздействует на покрывающие ствол слоевища
лишайников, разрывая их и сбрасывая вместе с корой, отмершими сучьями. Ме-
няются структура и текстура коры. Ее неровности создают особый микроклимат
для эпифитов, благоприятствуют аккумуляции пыли, закреплению диаспор. Осо-
бые местообитания с соответствующим видовым составом представляют мес-
22
1. Биология и экология лишайников
та прикрепления ветвей к стволу дерева, вблизи повреждений коры (Klement, 1955;
Barkman, 1958; Бязров, 1970).
Лишайники на форофите должны приспособиться к вертикальному положе-
нию и цилиндрической форме большинства стволов деревьев. Достаточно давно
уже было установлено, что на вертикально расположенном субстрате наблюда-
ется эксцентрический рост слоевищ вследствие неравномерного распределения
влажности, а у талломов Hypogymnia physodes на концах лопастей, обращенных
к поверхности почвы, отмечалось повышенное соредиеобразование (Bitter, 1901).
Поэтому рост слоевищ в этом направлении вообще прекращается, тогда как с
боков и сверху наблюдалось сильное ветвление. В таких условиях значение име-
ет любое отклонение от вертикального положения.
Ввиду сравнительно значительной протяженности дерева по вертикали, раз-
ные его части создают неодинаковые условия для эпифитов. Надземная часть
форофита состоит из ствола и кроны, микроклимат которых различен. На форо-
фите выделяют 4-5 зон поселения эпифитов, соответствующие (1) основанию
ствола, (2) стволу, (3) подкроновой части, (4) кроне, (5) листьям, хвое, шишкам
(Klement, 1955; Barkman, 1958;Peciar, 1965), при этом Я. Баркман (Barkman, 1958)
к кроне относит и ту часть ствола, которая расположена непосредственно в кроне
дерева, а О. Клемент (Klement, 1955) под кроной подразумевает лишь сучья и
ветви. Я полагаю, что зона ствола в области кроны представляет собой особое
местообитание. Границы между названными зонами довольно неопределенные.
Более ясно выражена зона основания ствола, охватывающая граничащую с поч-
вой часть ствола и выступающие из почвы части корней (так называемые корне-
вые лапы). Зона нижней части ствола, расположенная выше основания, выделя-
ется не столь отчетливо, особенно ее верхняя граница, но изучена хорошо. Соб-
ственно, большинство исследований эпифитных лишайников ограниченно именно
этими двумя нижними зонами как наиболее доступными. Было установлено, что
одни виды эпифитных лишайников предпочитают определенные зоны, а другие
встречаются в нескольких зонах поселения на форофите (Hale, 1952, 1955;
Culberson, 1955; Koskinen, 1955; Barkman, 1958; Голубкова, 1959; Бязров, 19696).
Для эпифитов превалирующее значение имеет доступность влаги. Вероятно,
настоящие эпифиты следует считать ксерофитами. Кора деревьев - главный суб-
страт эпифитных лишайников - влаги получает мало: на стволы деревьев попада-
ет незначительная часть дождевой воды вследствие перехватывания ее кроной,
а попавшая на ствол вода быстро стекает и испаряется. Влагоемкость же коры
деревьев разных пород в целом невысокая (Бязров, 1969а; Hauck et al., 2000).
Оценивая форофит как местообитание лишайников, необходимо признать, что
оно динамично во времени и пространстве, выделяется наличием множества
микроместообитаний, вертикальным положением стволов деревьев и крайней
сухостью. Но, возможно, условия на форофите сухие в сравнении с условиями
жизни наземных сосудистых растений, а для слоевищных эпифитов они могут
I. Биология и экология лишайников
23
быть нормальными. При проведении лихеноиндикационных исследований крайне
важно учитывать особенности форофитов в местах наблюдений.
1.9. СВЕТ, ТЕПЛО
В открытых для солнца местообитаниях на экспонированных и затененных
поверхностях однородного субстрата формируются лишайниковые группировки
разного состава. В старых лесных сообществах или в глубоких ущельях, где пре-
обладает рассеянный свет, части стволов деревьев или скальные поверхности
разной экспозиции по видовому составу лишайников практически не отличаются
(Бязров, 1974, 19936). По моим наблюдениям в одном из лесничеств Подмоско-
вья, на стволах осин, растущих в глубине лесного массива, встречены представи-
тели 24 видов лишайников, а на стволах той же породы, растущих на опушке или
вне лесного массива, - представители 39 видов. Соответственно, на стволах бе-
резы бородавчатой были отмечены 12 и 27 видов. В тоже время для дуба череш-
чатого эти показатели 29 и 25, а для сосны обыкновенной - 14 и 11 соответствен-
но. Однако при оценке влияния световых условий на лишайники необходимо учи-
тывать, помимо прямого воздействия на фотосинтез, зависимость температуры
и испарения от освещения. Зачастую трудно определить, является ли лишайник
светолюбивым или термофильным. А. Юницкий (1909) и О. Клемент (Klement,
1951) указывали, что отдельные виды могут служить индикаторами световых
условий в лесу. По их данным, Xanthoria parietina встречается в хорошо осве-
щенных прямыми солнечными лучами местах. Однако я неоднократно встречал
представителей этого вида на стволах деревьев в старых тенистых лесах Голар-
ктики. Только окраска слоевищ не была ярко-оранжевой как в освещенных мес-
тах. Весьма теневыносливыми являются представители рода Lepraria (Barkman,
1958; Jarosh, 1964). Последний автор изучал изменчивость представителей Lepraria
в условиях пещеры, где они проникали до пределов распространения зеленых ра-
стений. Ослабление симбиоза между фотобионтом и микобионтом не наблюда-
лось, однако менялась структура слоевища в благоприятную для водорослевого
компонента сторону. С уменьшением интенсивности освещения микобионт ста-
новился более рыхлым и прозрачным, а клетки водоросли при слабом освещении
располагались в талломе ближе к поверхности. Анатомо-морфологические изме-
нения слоевища под влиянием условий освещения отмечали многие авторы (Bitter
1901; Еленкин 1901; Ertl, 1951; Butin, 1954; Vogel, 1955). Мною в лесах Подмоско-
вья было установлено, что с увеличением высоты поселения на ели, а именно в
этом направлении возрастает интенсивность освещения, наблюдается и рост мас-
сы лишайника Hypogymnia physodes на единицу площади (табл. 1.3). Одновре-
менно возрастает и содержание хлорофилла на единицу площади слоевища, но
24
/. Биология и экология лишайников
Таблица 1.3
Изменение содержания хлорофилла а+b в слоевищах Hypogymniaphysodes в зависимости
от высоты поселения лишайника на ели (в относительных единицах)
Высота поселения, м Масса лишайника с единицы площади Содержание хлорофилла
На единицу площади На единицу сухого веса
20 155 139 76,5
15 132 117 76,5
10 103 112 82,5
1 100 100 100
доля микобионта (если считать содержание хлорофилла показателем доли фото-
бионта в талломе) растет быстрее. Свидетельство тому - уменьшение содержа-
ние хлорофилла на единицу веса лишайника.
Разные виды лишайников, как и растений, требуют для нормального развития
разной интенсивности освещения. Для характеристики их отношения к свету ис-
пользуют термины “фотофильные”, “скиофильные”, “фотофиты”, “гелио-
фиты”, “ангелиофиты”, “скиотолерантные” и другие (Barkman, 1958;
Hultengren et al., 1991). Однако выделение названных групп затруднено, посколь-
ку представители одного вида лишайника в зависимости от конкретных условий
местообитания и времени наблюдения в одних случаях выступают как световые,
а в других - как теневые, или теневыносливые. Возможно, лучше говорить о
теневой или световой структурах слоевищ, признаки которых в основном совпа-
дают с теневой и световой структурами сосудистых растений (Шенников, 1950).
Воздействие тепла на лишайники нередко опосредованно окружающим сооб-
ществом, а в случае эпифитов - форофитом. Также трудно решить, действует ли
тепло на слоевище прямо или косвенно. Лишайники, не выносящие прямой сол-
нечной радиации, иногда называют ангелиофитами, но по отношению к колебани-
ям температуры их можно назвать термофобными. Еще в 1950-х годах было
установлено, что в сухих местообитаниях могут происходить быстрые и значи-
тельные изменения температуры - на 14°С за 2 минуты (Lange, 1954). Тогда же
была зафиксирована температура 69,6°С у слоевища напочвенного лишайника
Cladonia pocillum (Lange, 1953), причем разница с температурой окружающего
воздуха достигала 40°С. Но такую высокую температуру могут выдержать только
сухие слоевища. В сыром состоянии талломы наиболее теплоустойчивого вида
погибали при температуре 50°С через 1 минуту. Было найдено, что лишайники
тенистых и солнечных местообитаний значительно отличаются по теплоустойчи-
вости - у первых интенсивность дыхания уменьшилась наполовину после полу-
часового нагрева при температуре 70°С , а у вторых - при 100°С (Lange, 1953).
О. Ланге (Lange) пришел к выводу, что в некоторых местах температура слоеви-
ща может превысить предел, выдерживаемый отдельными чувствительными к
теплу видами лишайников, и их перегрев может ограничивать распространение
1. Биология и экология лишайников
25
таких видов. Прямого действия температуры на рост лишайников не установлено
(Beschel, 1954), но существует зависимость дыхания и фотосинтеза от тепловых
условий (Lange, 1992).
1.10. ВОДНЫЙ РЕЖИМ, ФОТОСИНТЕЗ,
ДЫХАНИЕ ЛИШАЙНИКОВ
Большинство цветковых и голосеменных растений имеют в своих телах ем-
кости для удержания воды в их листьях или хвое надостаточных и относительно
постоянных уровнях и в определенной степени могут регулировать свой водный
режим. Такие организмы называют гомойгидридными. Лишайники принадле-
жат к пойкилогидридным организмам, которые не имеют таких образований
как эпидермис, устьица, восковая кутикула для регулирования газо- и водообме-
на, и потому их водный режим варьирует пассивно в зависимости от содержания
воды в окружающей среде. К пойкилогидридным организмам также относятся
другие слоевищные - водоросли, мохообразные, а среди сосудистых - некоторые
папоротники и ряд других примитивных растений. Каждый из этих пойкилогид-
ридных организмов относительно быстро высыхает и, как следствие, наличие
доступной влага имеет первостепенное значение для их выживания и объясняет
особенности их встречаемости (Farrar, 1976b, с). По особенностям водного режи-
ма, как и по отношению к другим экологическим факторам, среди лишайников
выделяют группы ксерофитов, мезофитов, гигрофитов и другие (Блюм, 1965).
Можно предполагать, что пойкилогидридные организмы сильно зависимы от
осадков, преимущественно в форме дождя, что подтверждает прямая корреляция
между биомассой лишайников и среднегодовым количеством осадков (Nash,
Moser, 1982). С другой стороны, биомасса лишайников вблизи Тихого океана в
западной части Сонорской пустыни оказалась значительно выше величин, кото-
рые можно было бы ожидать, основываясь только на одних осадках (Nash, Moser,
1982). Это свидетельствует о способность лишайников использовать другие ис-
точники влаги как, например туман, роса, что подтверждают исследования в пу-
стынях Атакама, Намиб (Lange, 1992). Наконец, лишайники способны поглощать
некоторое количество влаги из ненасыщенного воздуха в условиях низких темпе-
ратур и высокой относительной влажности (Бязров, 1970,1978а). Следствием пой-
килогидрии является прекращение в сухом состоянии всех процессов жизнедея-
тельности лишайников. Поэтому довольно продолжительное воздействие на су-
хие слоевища как очень низких (-196°С), так и высоких (+100°С) температур
существенно не влияло на фотосинтез и дыхание талломов при естественных для
их местообитаний температурах после соответствующего увлажнения этих слое-
вищ (Lange, 1992). Аналогичное воздействие на влажные талломы приводило к
быстрой их гибели. Таким образом, тепловые условия Земли практически не ог-
26
1. Биология и экология лишайников
раничивают развитие лишайников на нашей планете. Об этом свидетельствует и
обнаружение в Антарктиде южнее 80° ю.ш. представителей 23 видов лишайни-
ков (Ovstedal, Lewis Smith, 2001), из которых ближе других к Южному полюсу
приблизились представители Carbonea vorticosa, Lecidea canciformis, Sarcogyne
privigna (86°29’ ю.ш.), немного от них отстали Acarospora gwynnii (86°20’ ю.ш.),
Buellia frigida (86°09’ ю.ш.), В. grisea, Rhizoplaca melanophthalma (86°07’ ю.ш.),
причем С. vorticosa, R. melanophthalma, S. privigna входят в состав лихенобиот
регионов Северного полушария, a R. melanophthalma использовали как биомони-
тор при изучении воздействия на среду тепловой электростанции в штате Аризо-
на на юге США (Belnap, Harper, 1990).
В Антарктиде позитивный баланс газообмена в слоевищах лишайника
Neuropogon acromelanus наблюдали при температуре -20°С с оптимальной его
величиной при -4°С, а в Центральной Европе в слоевищах лишайника Cladonia
alcicornis позитивный газообмен был при температуре -15°С с оптимальной его
величиной при +8°С (Lange, 1992). Естественно, газообмен в талломах происхо-
дит лишь при наличии некоторого минимального количества влаги в слоевищах,
которое различно у представителей разных видов. У многих видов положитель-
ный баланс газообмена отмечали при относительной влажности воздуха око-
ло 80%, довольно обычной утром после восхода солнца и нередкой перед его
закатом.
1.11. ГЕОГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ
ТЕРРИТОРИИ
Географическое положение изучаемой территории для лишайников имеет как
экологическое, так и историческое значение. Оно определяет общий характер
климата района, влияющего на микроклиматические особенности конкретных
местообитаний лишайников. Макроклимат местности, естественно, преобразу-
ется ее рельефом и абсолютной высотой. В горах большую роль играет приуро-
ченность участка к склону той или иной экспозиции (Бязров и др., 1989; Седель-
никова, 1994). Большое значение для них имеет и ориентация поверхности суб-
страта (Окснер, 1961; Armstrong, 1974; 1991; Pentecost, 1979; Link, Nash, 1984;
John, Dale, 1990; Седельникова, 1994). Все это сказывается как на характере ра-
стительности в целом, так и на распространении отдельных видов. Важно пред-
ставлять, находится ли вид в данной местности на пределе своего ареала, или
изучаемый район расположен в области его оптимального развития. Наличие или
отсутствие того или иного субстрата также сказывается на особенностях видо-
вого состава лишайников местности. Далеко не всегда совпадают ареалы форо-
фита и связанных с ним эпифитов, что, с одной стороны, определяет особенности
варьирования видового разнообразия эпифитов на форофите в разных частях аре-
1. Биология и экология лишайников
27
ала последнего, с другой - встречаемость данного эпифита на разных видах
форофитов.
1.12. КОНКУРЕНЦИЯ
Лишайники, в отличие от большинства сосудистых растений, не имеют кор-
ней, соответственно, у них отсутствует корневая конкуренция, значение которой
весьма существенно в лесных и травянистых сообществах. У лишайников конку-
ренция сводится к механическому давлению, борьбе за свет, воздействию хими-
ческими выделениями. Во взаимоотношениях между ними есть элементы как
внутривидовой, так и межвидовой борьбы за жизненное пространство (Окснер,
1961; Глуздаков, 1962). А.Н. Окснер установил, что наиболее часто наблюдается
перерастание одного слоевища другим. Большую роль в конкуренции между ви-
дами играет скорость роста слоевищ (Barkman, 1958; Окснер, 1961), которая за-
висит от возраста таллома и особенностей местообитания (Beschel, 1954; Бязров,
19886). Известны многочисленные факты гибели мхов под лишайниками, но не-
редки и обратные случаи, когда лишайники вытеснялись мохообразными. Веро-
ятно, результаты борьбы зависят от особенностей местообитания и положения
вида в пределах его ареала.
Считается, что кустистые лишайники более конкурентноспособны, чем лис-
товатые, а последние побеждают в борьбе с накипными (Barkman, 1958). Изуче-
ние конкурентной способности разных видов лишайников показало, что суборди-
нации среди них по конкурентной силе нет (Мартин, 1967). Разные виды лишайни-
ков ведут себя различно с разными соседями в группировках разной сложности и
происхождения. Взаимодействия между разными видами лишайников и с пред-
ставителями других групп организмов являются одной из причин сукцессий груп-
пировок лишайников. Кроме конкуренции причиной смен бывают и изменения ус-
ловий местообитания. Последние меняются как под влиянием самих лишайни-
ков, так и в результате изменения некоторых экологических факторов. Все это,
вместе с возможными изменениями внешней среды, обуславливает смены ли-
шайниковых группировок. Выделяют нормальные и регрессивные сукцессии
(Barkman, 1958). К последним относят, например, сукцессию: Lobariapulmonaria
—► мохообразные —* водоросли или накипные лишайники. Нормальными счита-
ются обратные сукцессии: водоросли —> накипные лишайники —> листоватые ли-
шайники —► кустистые лишайники и мохообразные. Имеют место и отклонения
от названных видов сукцессий. Климаксом считается зарастание поверхности
субстрата представителями мохообразных, кустистых и листоватых лишайников
(Barkman, 1958). Но, скорее всего, следует согласиться с мнением, что ход смен
лишайниковых группировок зависит от особенностей местообитания (Koskinen,
1955; Бязров, 1972).
28
/. Биология и экология лишайников
Группировки лишайников представляют собой структурные части различных
экосистем, непременным компонентом которых являются животные. В тундро-
вой зоне напочвенные лишайники разных родов - основной корм северных оле-
ней, диких и домашних. Перевыпас ведет к деградации лишайникового покрова
тундры (Щелкунова, 1978; Андреев, 1980; Longton, 1988; Магомедова и др., 1991).
Нередки случаи поедания копытными и эпифитных лишайников. Питаются ли-
шайниками или обитают в их группировках представители многих групп беспоз-
воночных (Бязров, 1988а; Стебаева, Седельникова, 1999; Стебаева и др., 2001).
Погрызы, надкусы вызывают морфологические изменения слоевищ лишайников.
Экскременты животных, особенно в местах гнездовий птиц, способствуют фор-
мированию специфического видового состава поселившихся здесь лишайников
(Котлов, 1993).
1.13. ВЛИЯНИЕ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА.
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ
Прямое воздействие человека связано с его деятельностью в экосистемах с
участием лишайников и использованием их для своих нужд. Например, отдых
большого числа людей в лишайниковых борах приводит к механическому повреж-
дению (вытаптыванию) лишайникового покрова в таких сосновых лесах, посколь-
ку гуляют люди обычно при сухой погоде, когда слоевища легко ломаются (Ма-
лышева, Толпышева, 1982). В тундрах значительные повреждения лишайниково-
му покрову наносят средства передвижения на гусеничном ходу, ибо образующаяся
вслед за таким средством передвижения колея заполняется водой, и бывший здесь
ранее лишайниковый покров не восстанавливается. Вторично по этой колее про-
ехать уже нельзя, и каждый раз такое транспортное средство движется по цели-
не, увеличивая площадь повреждаемых территорий, перспектива восстановления
которых сомнительна. Строительство населенных пунктов, промышленных пред-
приятий, добыча полезных ископаемых, прокладка дорог, трубопроводов, выруб-
ка лесов и многое другое приводят к уничтожению лишайников. Кроме того, часть
видов лишайников заготавливают для использования в парфюмерной, фармацев-
тической, пищевой отраслях промышленности, применяют их и для крашения тек-
стиля, оформления помещений, витрин, композиций из цветов, а иногда и как экзо-
тическую добавку к пище, табаку и т.д. Рассматривают их и как источник пести-
цидов (Dayan, Romagni, 2002). Во всяком случае, лишайниковое сырье является
предметом международной торговли (Kauppi, 1993). Необходимо отметить, что
исчезновению лишайников способствует и коллекционирование их специалиста-
ми-лихенологами и любителями (Nash, Dibben, 1979). В Англии было докумен-
тально восстановлен ход изменения видового разнообразия криптогамов истори-
ческой местности Вилд (Weald) за почти 300 лет - с 1688 по 1984 г. (Pentecost,
1. Биология и экология лишайников
29
Rose, 1985). Из 90 видов встречавшихся здесь лишайников исчезли представите-
ли 18 видов. Среди причин этого явления, прежде всего, названо неумеренное
коллекционирование, а затем загрязнение воздуха, разрушение субстрата, силь-
ное затенение в результате разрастания деревьев и кустарников.
Более значимо косвенное воздействие человека на лишайники. В результате
хозяйственной деятельности воздух загрязняется пылью, выбросами токсичных
газов, других соединений различной природы, губительных для лишайников. Это
приводит либо к полному исчезновению лишайников и образованию так называе-
мых “лишайниковых пустынь” вокруг ряда промышленных предприятий, в горо-
дах с большой концентрацией автомобильного транспорта, либо к значительному
обеднению в таких местах видового состава этих организмов. Подобная реакция
лишайников на загрязнение привела к использованию их в индикации и монито-
ринге загрязнения среды, чему и посвящена данная монография.
С другой стороны, деятельность человека способствует их расселению. На-
пример, перемещение на большие расстояния заготавливаемой необработанной
древесины вместе с сохранившимися на коре бревен эпифитными лишайниками
обогатило видовое разнообразие лишайников Гренландии (Alstrup, 1977). В США
отмечено переселение ряда видов рода Physcia, характерных для восточных ча-
стей страны, в Калифорнию вместе с пересаженными деревьями (Thomson, 1963).
Есть мнение, что слоевища Parmelia sulcata на о. Южная Георгия в Субантарк-
тике завезены китобойными судами (Lindsay, 1973). Кроме того, создавая лесо-
полосы в степях и полупустынях, человек способствует образованию здесь и но-
вого субстрата, пригодного для поселения лишайников (Добровольский, Гаевая,
1985), а введение в культуру древесных пород, не свойственных данной местнос-
ти, разнообразит набор форофитов. Это относится и к рукотворным субстратам
из металла, пластика, других материалов, сопровождающих человека от полюса
до полюса и до самых высоких горных вершин.
2. УЛАВЛИВАНИЕ И ПОГЛОЩЕНИЕ ЛИШАИНИКАМИ
МИНЕРАЛЬНЫХ И ОРГАНИЧЕСКИХ ВЕЩЕСТВ,
ИХ НАКОПЛЕНИЕ И ПОТЕРЯ;
ТОКСИЧНОСТЬ ЭЛЕМЕНТОВ
Доступность как макро-, так и микроэлементов минерального питания, жиз-
ненно важных для осуществления физиологических функций, лимитируют рост и
развитие лишайников также, как и других организмов. У сосудистых растений
поглощение из почвы воды и питательных веществ осуществляет корневая сис-
тема. У лишайников корни отсутствуют, что обусловливает зависимость их снаб-
жения элементами минерального питания от атмосферных источников взамен по-
чвенных запасов, эксплуатируемых сосудистыми растениями (Nieboer et al., 1978).
Кое-что может поступать и из почвы, например, в напочвенные виды powPeltigera
(Goyal, Seaward, 1981а, 1982а, b). Экспериментально было показано, что талломы
лишайников рода Cladina способны поглощать минеральные элементы через мер-
твую часть подециев (Барашкова, 1963; Нифонтова и др., 1979). У родственных
им представителей Cladonia convoluta и С. rangiformis в эксперименте мерт-
вые части подециев показали более высокое накопление РЬ и Си, а живые больше
аккумулировали Zn (Chettri et al., 1997b). Поскольку атмосферные источники ми-
неральных веществ в сравнении с почвой относительно скудны, решающими для
жизни лишайников являются механизмы концентрирования элементов из атмос-
феры. Факт наличия таких механизмов привел в свое время к более широкому
научному интересу к лишайникам как суррогатным рецепторам атмосферных
выпадений, в частности, последствий накопления радионуклидов в телах корен-
ных жителей Арктики в 1950-х и 1960-х годах в результате испытаний ядерного
оружия в атмосфере. Позднее интерес к этому явлению привел к использованию
лишайников для изучения региональных атмосферных выпадений и других ат-
мосферных загрязнителей (Puckett, 1988). При этом были получены свидетель-
ства определенной толерантности ряда видов лишайников к высоким концентра-
циям металлов, что дало основания рекомендовать использование лишайников
в геологических изысканиях (Викторов, 1956; Таисаев, Константинова, 1988;
Chettri et al., 1997а). В дополнение к названным аспектам следует помнить и о
2. Улавливание и поглощение лишайниками веществ...
31
роли лишайников в начальных почвообразовательных процессах путем воздей-
ствия на обнажения горных пород (Полынов, 1945; Ярилова, 1947; Левин, 1949;
Jones, 1988). Такие процессы влияют на минеральный круговорот в шкале геоло-
гического времени. Также было установлено, что лишайники могут играть важ-
ную роль в более быстрых процессах круговорота через синтез чужеродных пи-
тательных элементов, которые не могли бы иным образом попасть в экосистему
(Бязров, 1970; Knopset al., 1996).
2.1. СОДЕРЖАНИЕ МИНЕРАЛЬНЫХ ЭЛЕМЕНТОВ
В ЛИШАЙНИКАХ
Зольность лишайников, как правило, намного ниже, чем других растений (Ра-
ботнов, Говорухин, 1950; Левицкая, 1975), иногда даже менее 1% (Левина, 1960).
Зола содержит как анионы, так и катионы. Отрицательно заряженные анионы
связаны карбоксильными и гидрокарбоксильными участками в структурах поли-
сахаридов клеточных стенок, а положительно заряженные катионы связаны уча-
стками, которые предположительно ответственны за изменение емкости клеточ-
ных стенок лишайников (Nash, 1996). Способность эпилитного лишайника Parmelia
накапливать Р, S, К обнаружил Б.Б. Полынов (1945), что было подтверждено
Е.А. Яриловой (1947), Ф.М. Левиным (1949) и М.А. Бобрицкой (1950). При опре-
делении элементного состава золы лишайников существует опасность появления
ошибок за счет загрязнения таллома частицами минералов субстрата (Бобриц-
кая, 1950; Родин и др., 1968; Markert, 1994b, 1995; Ernst, 1995; и другие). Однако
сведения о накоплении лишайниками ионов в количествах, намного превышаю-
щих их содержание в субстрате, не дают повода для сомнений в возможности
лишайников аккумулировать в своих телах химические вещества. Известен факт,
что концентрация меди в лишайниках на диорите почти в 90 раз превышает этот
показатель в субстрате (Nash, 1996). Методом рентгеновского дифракционного
анализа была подтверждена биогенная природа накопления зольных элементов в
напочвенном лишайнике Parmelia chlorochroa, что связано, прежде всего, с син-
тезом оксалата кальция, а не с загрязнением растения частицами почвы (Erdman
et al., 1977).
Авторы специальных и популярных публикаций об атмосферных выпадениях
и загрязнении среды нередко используют термин “тяжелые металлы** Этимоло-
гически этот термин имеет физический смысл и должен относиться к той группе
металлов, удельный вес которых более 4,5 г/см3. Однако анализ различных ис-
точников показывает, что биологи к тяжелым металлам иногда относят такие
“легкие металлы” как алюминий или полуметаллы как, например, мышьяк (Wittig,
1993). Таким образом, фактически этот термин служит широко распространен-
ным популярным обозначением потенциально токсичных элементов. В силу нео-
32
2. Улавливание и поглощение лишайниками веществ...
днозначности трактовок этого понятия было предложено заменить термин “тя-
желые металлы” химически и биологически значимыми группами, обозначаемы-
ми как элементы класса А (например, Al, Ba, Са, Се, Cs, К, La, Li, Mg, Na, Nd, Rb,
Sc, Sn, Sr, Y), промежуточные (например, As, Cd, Co, Cr, Fe, Ga, Mn, Ni, Sn, Ti, V,
Zn) и элементы класса Б (например, Си, Hg, Pb) (Nieboer, Richardson, 1981; Wittig,
1993; Walker et al., 2001). Это деление основано на их расположении в периоди-
ческой системе элементов Менделеева. Элементы класса А предпочитают
доноров, содержащих кислород (О N S-доноров), тогда как элементы класса
Б - доноров, содержащих серу и азот (S N О-доноров), а элементы промежуточ-
ного класса находятся между ними. При катионном связывании участков ионы
класса Б будут замещать ионы промежуточных элементов, которые, в свою оче-
редь, будут замещать ионы класса А.
Предложена и биологическая система элементов, представляемая в виде кор-
реляционной решетки, построенной на основе таких показателей как место эле-
мента в периодической системе Менделеева, сходство радиусов ионов, физиоло-
гические функции элементов в организмах и других признаках (Markert, 1992,1994а;
Franzle, Markert, 2000).
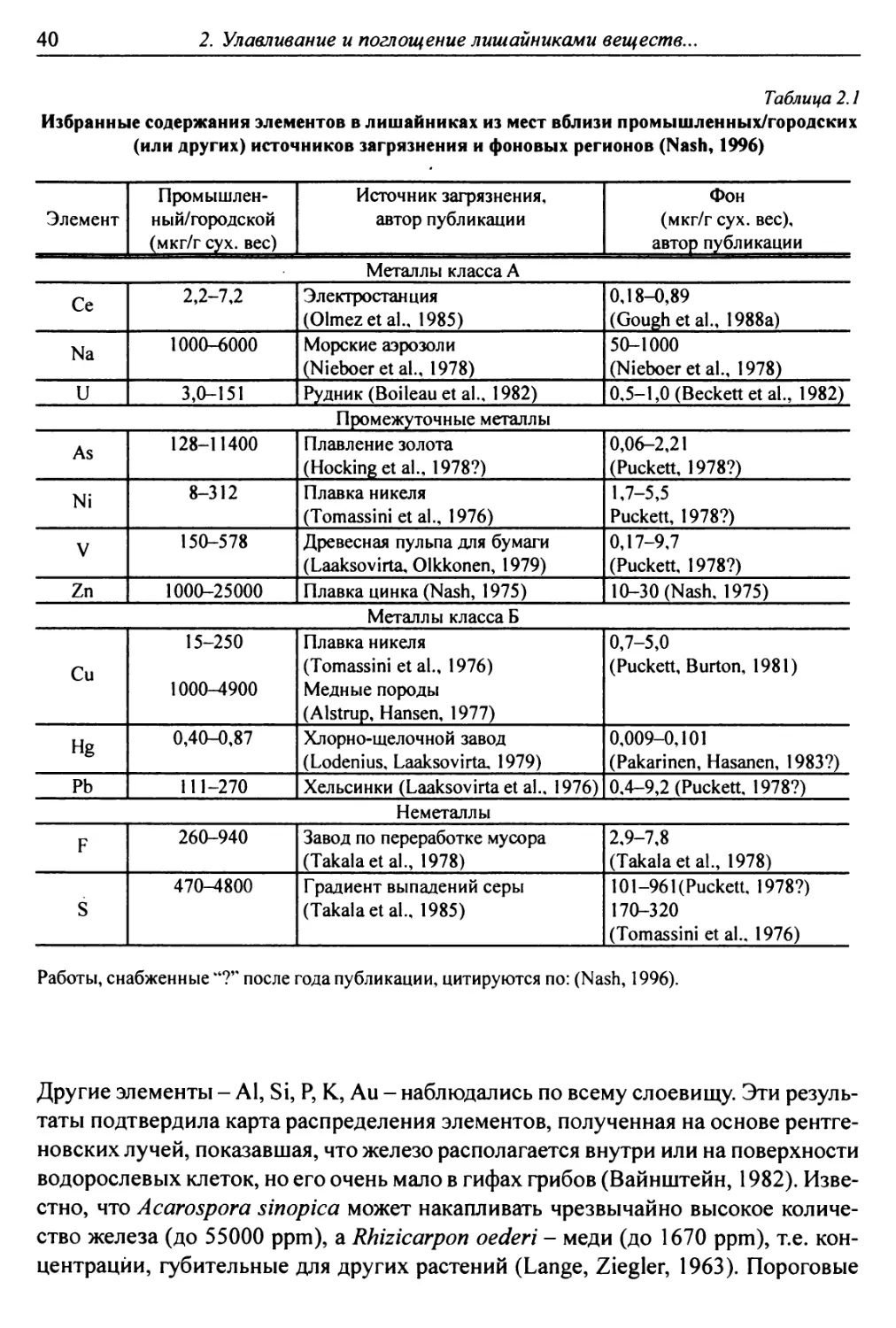
Доступная информация о содержании в лишайниках относится к ограниченно-
му набору элементов. Например, в процессе исследований, посвященных толе-
рантности лишайников к металлам, часто изучались популяции, встречающиеся
на отвалах старых рудников, где большей частью обычны такие металлы как Cd,
Си, Fe, Ni, Zn. В публикациях об атмосферных выпадениях имеются данные об
Al, As, Ba, Be, Са, Се, Со, Cr, Cs, Ga, Hg, К, La, Li, Mg, Mn, Na, Nd, Pb, Rb, Sc, Sn,
Sr, Ti, V, Y. При изучении роли лишайников в биогеоценозах внимание фокусирова-
лось на макроэлементах С, N, Р, S, К, Mg, Мп и других (Garty, 1993).
Содержание большинства микроэлементов в лишайниках большей частью
выше, чем у мохообразных, папоротников, хвойных, кустарников и трав (Lounamaa,
1956). Исключение составляет Мп. Такие элементы как Со, Ni, Mo, Au присут-
ствуют в тех же концентрациях, а содержание Zn, Cd, Sn, Pb намного выше. Од-
нако есть сведения и о низком содержании всех микроэлементов, например, в
слоевищах Cladina stellar is, в северных районах Канады (Doyle et al., 1973) или в
лишайниках Западного Таймыра (Вайнштейн, 1982).
2.2. ПОТРЕБНОСТИ В ПИТАТЕЛЬНЫХ ВЕЩЕСТВАХ
Точные величины концентраций элементов питания, необходимых для нормаль-
ного роста лишайников, остаются пока не установленными (Вайнштейн, 1982).
Имеются лишь косвенные свидетельства, в частности, высокая степень специ-
фичности некоторых видов лишайников относительно субстрата (Brodo, 1973).
Есть данные о питательных веществах, требуемых для роста отдельных симби-
2. Улавливание и поглощение лишайниками веществ...
33
онтов в культурах (Ahmadjian, 1993). Также установлено, что водоросль Trebowcia,
наиболее обычный фотобионт лишайников, является факультативным гетеротро-
фом, и что она, при доступности для нее органических источников питания, неко-
торое время способна расти в темноте. Предполагается, что питательные веще-
ства в слишком высоких концентрациях могут стать причиной нарушения равно-
весия между партнерами симбиоза. Так, выпадения нитратов в Западной Европе
сильно увеличили их поступление в природные экосистемы, что стало причиной
обеднения лихенофлоры ряда регионов, например, в Нидерландах (van Dobben,
1993; van der Eerden et al., 1998).
2.3 ИСТОЧНИКИ ПОСТУПЛЕНИЯ ЭЛЕМЕНТОВ
2.3.1 Атмосферные источники
Атмосферные выпадения доходят до лишайников либо в жидком состоянии:
собственно осадки (дождь, снег) и потенциальные, или скрытые осадки (главным
образом туман, роса), либо в сухом - в результате седиментации аэрозолей, пыли,
давления и поглощения газов (Knops et al., 1991). Как источники питательных
веществ и воды, очень важны для лишайников потенциальные (скрытые) осадки,
в которых концентрации питательных веществ и загрязнителей могут быть суще-
ственно выше, чем в дождевой воде. Также минеральные вещества могут попа-
дать в лишайники в виде пыли, содержащей все важнейшие элементы, в том чис-
ле Р, N, К, Са (Salomon, 1914; Kauppi, 1980). Могут они также использовать неко-
торые соединения азота и аммония (NH4+), находящиеся в воздухе, например,
углекислый аммоний, образующийся при разложении богатых мочевиной продук-
тов животного происхождения, а лишайники с цианобионтом могут фиксировать и
атмосферный азот. Седиментация крупных загрязняющих аэрозолей и их внедре-
ние в лишайниковые слоевища хорошо показаны при сравнении химических про-
филей, полученных по данным преломления рентгеновских лучей в атмосферных
аэрозолях, собранных прямо из воздуха, и в частицах, уловленных лишайником
(Garty et al., 1979). Количественные данные о поглощении газов слоевищами ли-
шайников ограничены, но поглощение SO2 арктическим лишайником Cladina
rangiferina, по меньшей мере, на порядок выше, чем типичными сосудистыми
растениями (Winner et al., 1988).
Поглощение и накопление лишайниками элементов из атмосферы хорошо до-
кументировано (Jenkins, Davies, 1966; Nash, 1996). Показана тесная корреляция
между количеством металлов в золе накипных лишайников и содержанием их в
сухом остатке осадков. С учетом медленного роста и гидролабильности лишай-
ников, полагают, что атмосферные источники играют доминирующую роль в
определении состава микроэлементов таллома (Вайнштейн, 1982).
34
2. Улавливание и поглощение лишайниками веществ...
23.2. Субстратные источники
Многие лишайники встречаются на почве или камнях и потому тесно контак-
тируют с литосферными источниками элементов питания. Содержание элемен-
тов в лишайниках иногда довольно точно отражает их геохимическое распреде-
ление в минералах районов поселения лишайников (Erdman, Gough, 1977), но в
большинстве случаев корреляция между содержанием элементов в лишайниках
и составом субстрата более сложная (Вайнштейн, 1982). Так, напочвенные ли-
шайники рода Cladonia из района угольных разработок в Огайо накапливали Р, К,
Са, Mg, Fe, Си, Zn, Al, Мо в количествах, намного превышающих их концентрации
в субстратах (Lawrey, Rudolph, 1975). Однако лишайники, видимо, каким-то обра-
зом могут регулировать поглощение некоторых элементов, избыточных в суб-
страте, поскольку представители рода Cladina, растущие на сфагновых болотах,
бедных железом, и на почвах, богатых железом, имели одинаковый уровень же-
леза в слоевищах (Lounamaa, 1965). Для лишайников, растущих на известняках,
отмечено пониженное поглощение Са, избыточного в субстрате и доступного для
них (Бобрицкая, 1950). Интересно, что для мхов и лишайников, развивающихся на
бедных микроэлементами субстратах, характерен более высокий коэффициент
биологической аккумуляции микроэлементов и естественных радиоактивных эле-
ментов, чем у тех, которые росли на богатых ими субстратах (Горбанов, Хаджи-
атанасов, 1973). Также отмечено, что в мертвой базальной части лишайников
рода Cladonia содержание Fe, Pb, Си и золы выше, чем в живых верхушках,
причем наибольшие различия у железа - в 2,5 раза (Lounamaa, 1965). У Cladina
stellaris, таллом которой постепенно переходит в органические остатки субстра-
та, это повышенное содержание железа, видимо, связано с особенностями суб-
страта. Кроме того, железо может поступать вниз и из верхушек, поскольку экс-
периментально установлено относительно быстрое передвижение Fe-55 от вер-
хушки к основанию, и это движение однонаправленное (Jaakkola, 1967). Лишайники
могут участвовать в выветривании поверхности субстрата как механически, так
и химически (Syers, Iskandar, 1973), и при этом возможно поглощение лишайника-
ми некоторых растворимых элементов питания. Растворимость многих элемен-
тов зависит от величины pH, поэтому доступность минеральных элементов мо-
жет быть различной на известняковых и кислых субстратах. Как следствие, очень
различный состав лишайниковых сообществ на известняках (Roux, 1981) и на кис-
лых породах (Wirth, 1972). Более того, в гипсе наряду с Са в более высоких кон-
центрациях представлен и сульфат, и известны несколько видов лишайников, при-
уроченных к гипсу, например, Acarospora clauzadeana.
Большинство лишайников, особенно встречающихся на почве и камнях, под-
вержены влиянию переносимой ветром пыли, источником большей части которой
являются незакрепленные поверхности почвы. Эти почвенные частицы могут легко
внедриться в межклеточное пространство и, как результат, - относительно высо-
2. Улавливание и поглощение лишайниками веществ...
35
кая концентрация Al, Fe, Sc, Ti и других элементов литосферного происхождения в
слоевище. Растворимые соединения их являются потенциальным источником
элементов минерального питания, но процесс растворения медленный, и боль-
шинство элементов из этих частиц, вероятно, остаются недоступными. Относи-
тельный вклад почвенных частиц в общее обеспечение лишайников минеральны-
ми элементами может быть оценен сравнением отношений макро- и микроэле-
ментов питания к инертным элементам как S или Ti (Nieboer et al., 1978). Однако
при изучении поглощения из субстрата и накопления лишайниками питательных
веществ четких физиологических, таксономических или морфологических корре-
ляций не выявлено (Вайнштейн, 1982). Имеются лишь косвенные свидетельства
этого, например, избирательность некоторых видов по отношению к субстрату
(Lounamaa, 1965; Salisbury et al., 1968; Локинская, 1970; Brodo, 1973;Purvis, Halls,
1996), толерантность к действию очень высоких концентраций металлов (Lange,
Ziegler, 1963; Brown, Slingsby, 1972; Alstrup, Hansen, 1977; Garty, 1993, 2000a; и
другие), большие различия между вйдами в способности их к накоплению метал-
лов (Бобрицкая, 1950; Локинская, 1970; Tomassini et al., 1976; Garty, 1993).
Для эпифитных лишайников дополнительным источником питания являются
вещества, вымываемые водой из деревьев и кустарников, поскольку большин-
ство элементов способно вымываться дождем из кроны (Smith, 1975). Элементы
типа К довольно быстро вымываются из листвы и, соответственно, могут посту-
пить в тела эпифитов. Двухвалентные катионы (Mn, Zn) вымываются легче, чем
трехвалентные (Fe). Таким образом, вода, стекающая вдоль стволов деревьев
во время дождя, содержит более высокие концентрации минеральных и раство-
ренных органических веществ (включая углеводы), чем дождевая вода, выпада-
ющая из атмосферы. На некоторых деревьях, особенно наклоненных, часто сток
по стволу во время осадков происходит по одной стороне ствола, на которой обра-
зуются своеобразные более темные в сравнении со всей остальной корой поло-
сы. Некоторые виды лишайников приурочены именно к этим высоко обогащен-
ным элементами питания участкам ствола (Barkman, 1958). Вполне возможно,
что эпифитные лишайники удовлетворяют потребности в питательных веществах
в основном за счет дождевой воды, стекающей по стволам деревьев, что было
показано для эпифитов горных лесов в полевых и лабораторных опытах (Lang et
al., 1976). Но получают ли эпифитные лишайники питательные вещества дополни-
тельно непосредственно из богатого органикой субстрата? Анализ содержания
N, Р, Са, К и некоторых других элементов в коре деревьев и слоевищах на этой
коре не обнаружил между ними связи (Kuziel, 1973). Г. Тротет (Trotet, 1968-1969)
изучал возможность поступления фосфора из дерева в растущий на нем лишай-
ник, для чего живые ветки дуба Quercus suber с растущими на них лишайниками
Ramalina calicaris, Parmelia melanothrix были помещены нижним срезанным
концом в раствор радиоактивного фосфата калия. Для сравнения в такой же ра-
створ помещали мертвые ветки, также обросшие лишайниками. Через 4 дня по
36 2. Улавливание и поглощение лишайниками веществ...
всей длине живых веток обнаруживалась сильная радиоактивность, в лишайни-
ках же она не отмечена, в мертвых ветвях она обнаруживалась в 1-2 см от сре-
занного конца, вероятно, за счет капиллярного подъема. Этот опыт позволил сде-
лать вывод, что эпифитные лишайники не поглощают фосфор из тканей дерева-
носителя.
Однако наблюдаются большие различия между составами элементов пита-
ния коры деревьев разных пород. Их доступность, как и на горных породах, зави-
сит от величины pH коры. Результатом этого является различный состав лишай-
никовых сообществ, встречающихся на деревьях с нейтральной корой (Fraxinus,
Tilia) в сравнении с деревьями с кислой корой (Betula, Picea, Pinus). Источники
загрязнения также могут изменять свойства коры, как добавлением элементов,
так и изменением величины pH. Кислотные выпадения часто понижают величи-
ну pH коры, что приводит к изменению лишайниковых сообществ, встречавших-
ся ранее на нейтральном субстрате (Skye, 1968), а выпадения Са, ассоциируемые
с цементными предприятиями, могут увеличить значение pH. Как следствие, про-
исходит смена лишайникового сообщества, свойственного кислому субстрату, на
сообщество, характерное для нейтрального субстрата (Нильсон, Мартин, 1982,
Л. Мартин, 1984).
2.4. МЕХАНИЗМЫ НАКОПЛЕНИЯ
Лишайники, наряду с отсутствием корневой системы, не имеют на поверхно-
сти слоевищ восковой кутикулы, свойственной листьям сосудистых растений, хотя
большинство лишайников имеют верхний коровой слой из довольно плотно пере-
плетенных грибных гиф. Как следствие, обмен элементов у лишайников осуще-
ствляется через всю поверхность их талломов. Ниже рассмотрены основные
механизмы накопления элементов минерального питания.
\ 2.4.1. Обмен ионов
Ионы металлов первично существуют как катионы, хотя анионы некоторых
элементов, таких как, например, U, пока неизвестны. Кинетика и термодинамика
поглощения катионов хорошо изучены (Nieboer, Richardson, 1981, Richardson, 1995).
Изначально поглощение катиона быстрое, пассивное, физико-химическое превра-
щение их в телах лишайников происходит внеклеточно (Nieboer et al., 1978). Уров-
ни насыщения достигаются за минуты (Puckett et al., 1973), и емкости поглоще-
ния для удержания катионов на клеточных стенках оценочно варьируют между 6
и 77 мкмол/г в зависимости от вида (Nash, 1989, 1996). Эти катионы преимуще-
ственно удерживается вне цитоплазмы фотобионта или микобионта на участках
обмена катионов. Экспериментально была установлена конкуренция между об-
2. Улавливание и поглощение лишайниками веществ...
37
менными ионами, которая, в общем, может быть представлена следующей схе-
мой: одновалентный класс А < двухвалентный класс А < двухвалентный проме-
жуточный класс < двухвалентный класс Б (Nieboer, Richardson, 1981).
Поглощение анионов лишайниками изучено в меньшей степени. В целом это
более медленный процесс (часы), чем обмен катионов и, как следствие, мень-
шее общее накопление. Например, анионный ураниловый комплекс (UO2L22 ) на-
капливался гораздо медленнее, чем катионы уранила, и общее поглощение анио-
нов было много меньше, чем катионов (1,6 мкмол/г против 49 мкмол/г). Наобо-
рот, поглощение анионной формы мышьяка (H2AsO4 ) было больше (10 мкмол/г),
что означает активное поглощение (Nieboer et al., 1984; Richardson et al., 1984).
Прежде это было установлено и для фосфатов (Farrar, 1976d). Тем не менее,
в условиях сухих выпадений в сильно загрязненной среде было установлено,
что внешняя анионная нагрузка может быть потенциально высокой (более
200 мкмол/г) (Boonpragob et al., 1989). Таким образом, в клеточных стенках так-
же должны существовать участки обмена анионов, которые пока еще не обнару-
жены.
Активное поглощение оценивается как очень эффективное ввиду высоких ско-
ростей поглощения из относительно концентрированных растворов (10 2 М и выше)
сахаров, аминокислот и фосфатов (Smith, 1962; Farrar 1976d). Эффективное погло-
щение является преимуществом для организмов, живущих в дефиците питатель-
ных веществ. Однако в отличие от эксперимента, в природе лишайники имеют
дело с очень разбавленными растворами. Было показано, что Hypogymnia
physodes, например, эффективно поглощает фосфат и из разбавленных раство-
ров, причем величины поглощения намного выше, чем полученные на отрезках
корня ячменя в сравнимых условиях (Farrar, 1976d). Также обнаружено, что при
быстром активном поглощении из концентрированных растворов последующее
использование поглощенных веществ происходит медленно (Smith, 1962). В слу-
чае разбавленных растворов метаболизм поглощенных веществ может быть
быстрым. Уже через 5 секунд после помещения слоевищ Hypogymnia physodes
на разбавленные растворы КН2Р32О4 радиоактивность обнаружили не менее чем
в 10 органических соединениях, в том числе в полифосфатах и в кислотонера-
створимой фракции.
Следовательно, активное поглощение веществ лишайниками действительно
можно назвать очень эффективным, и, видимо, оно имеет большое значение для
их экологии.
2.4.2. Внутриклеточное поглощение
Процессы внутриклеточного поглощения обычно гораздо более медленные и
включают относительно небольшие потоки ионов в сравнении с внеклеточным
обменов ионов (Beckett, Brown, 1984а, b). Например, после 2,5 часов экспозиции в
38
2. Улавливание и поглощение лишайниками веществ...
растворе Cd внутриклеточное поглощение составляло менее чем 10% от общего
поглощения (Brown, Beckett, 1985). С удлинением периода воздействия (более
часа) увеличивается и внутриклеточное поглощение (Brown, Beckett, 1984). По-
скольку оно увеличивается также в ответ на повышение уровня внешней концен-
трации Cd, поглощение следует рассматривать как процесс переноса ионов с зат-
ратой соответствующего количества энергии. Такая энергетическая зависимость
у лишайников была показана на примере внутриклеточного поглощения фосфатов
(Farrar, 1976d).
У лишайников пассивное поглощение доминирует над активным, о чем свиде-
тельствуют опыты со стронцием (Tuominen, 1967), цезием (Handley, Overstreet,
1968), а также серия работ с тяжелыми металлами (Nieboer et al., 1976а, 1977;
Tomassini et al., 1976; Richardson, 1995). Пассивная фаза поглощения катионов ли-
шайниками, видимо, представляет собой своеобразный катионно-обменный про-
цесс, модифицированный образованием комплексов с металлами (Puckett et al.,
1973). Точками адсорбции являются при этом кислородные и кислородно-азот-
ные доноры. Подобное представление дало основание Е.А. Вайнштейн (1982)
объяснить результаты опытов по определению избирательности поглощения ка-
тионов лишайниками (Brown, Slingsby, 1972; Puckett et al., 1973). По результатам
изучения влияния различных факторов (свет, метаболические ингибиторы, анаэ-
робиоз и другое) на поглощение никеля лишайником Umbilicaria muehlenbergii
был сделан вывод, что это чисто физико-химический процесс, в котором участву-
ют (А) ионообменная поверхность - клеточные стенки гиф, (Б) реакционная зона,
в которой растворены функциональные группы, или точки связывания ионов ме-
таллов, (В) диффузный слой, или пленка, (Г) зона основного раствора (Nieboer et
al., 1976b). По мнению авторов, фиксирующими анионными группами у этого ли-
шайника являются карбоновые или оксикарбоновые кислоты. Вероятно, пассив-
ное поглощение ионов происходит внеклеточно и связано с компонентами клеточ-
ных стенок (Brown, Slingsby, 1972). Возможно, основную роль в связывании кати-
онов играют хитин и боковые группы молекул белков, участвующие в
формировании клеточных стенок микобионта, путем образования комплекса азот-
кислород-катион (металл). Некоторые катионы поглощаются и активным, и пас-
сивным путем, что находит отражение в окончательной клеточной локализации
ионов. Пассивное поглощение путем ионного обмена, или хелатизации имеет ре-
зультатом внеклеточное расположение ионов, тогда как активное поглощение при-
водит к внутриклеточному расположению ионов. Подобное пространственное
разделение наблюдалась у Cladina rangiferina, у которого свинец накапливался
внеклеточно, а калий - в основном внутриклеточно (Brown, Slingsby, 1972). Весь
свинец, накапливающийся в природных условиях у этого лишайника на почвах,
бедных или богатых этим элементом, связан в талломе с нерастворимыми ани-
онными центрами в обменной форме. Калий же связан частично как свинец в
ионной форме, но основная его масса представлена внутриклеточно в свободной
2. Улавливание и поглощение лишайниками веществ...
39
диффузной форме и в норме он удерживается барьерами клеточной проницаемо-
сти. Кроме того, 10-15% всего калия в лишайнике прочно связано с белковопо-
добными веществами. Вероятно, свинец в лишайнике, как и у высших растений,
связан с нерастворимыми анионными группами, расположенными в клеточных
стенках.
2.4.3. Улавливание взвешенных в атмосфере частиц
Талломы лишайников имеют значительное по объему межклеточное простран-
ство. Например, у Xanthoria parietina оно составляет 18% (Collins, Farrar, 1978).
Есть достоверные прямые и косвенные свидетельства попадания частиц разного
происхождения в это пространство (Nieboer et al., 1978). Наличие таких инород-
ных тел в лишайниках вокруг источников загрязнения было установлено также
сканирующей электронной микроскопией (Garty et al., 1979; Jones et al., 1982).
Свободное проникновение в слоевище богатых металлами частиц, несомненно,
является причиной крайне высокого их содержания в лишайниках (табл. 2.1).
Так, авторы публикации (Tomassini et al., 1976) сообщают о концентрации Ni в
пределах (мкг/г) от 220 до 310, Fe - 1700-5200, Си - до 250. Вблизи сталелитей-
ного комплекса в Англии М. Сивард (Seaward, 1973) обнаружил в талломах
Peltigera rufescens концентрации (мкг/г) Fe - 90000, Мп - 5000, Си - 91, Сг - 127,
РЬ - 454, Ni - 38. В г. Лидсе М. Сивард (Seaward, 1988) также установил концен-
трации (мкг/г) Fe - до 35800, Мп - 349, Си - 159, Сг - 97, РЬ - 3124, Ni - 183 в
слоевищах Lecanora muralis.
2.5. ЛОКАЛИЗАЦИЯ ЭЛЕМЕНТОВ В СЛОЕВИЩЕ
Первые исследования по микрохимии лишайников показали, что у разных ви-
дов наблюдается различное распределение Р, Mg, К, NO2, NO3 между мико- и
фотобионтами (Salomon, 1914), при этом значительные количества калия были
обнаружены в плодовых телах, в связи с чем уместно вспомнить, что в культурах
грибов на средах без калия плодовые тела, как правило, не развиваются (Вайн-
штейн, 1982). С помощью рентгеновского микрозонда было показано, что у ли-
шайника Acarospora smaragdula железо концентрируется в налете на верхней
поверхности слоевища, а цинк - внутри самого таллома, прежде всего в нижнем
слое коры (Noske et al., 1970). В корковом налете также были найдены S, Si, Р, О,
что говорит о наличии в нем сульфатов, силикатов и фосфатов. Медь этим мето-
дом обнаружили только в налете (Вайнштейн, 1982). С помощью электронной
сканирующей микроскопии и рентгеноэмиссионного анализа была обнаружена по-
стоянная локализация Na и С1 в микобионте и Fe - в фотобионте (Lawrey, 1977а).
40
2. Улавливание и поглощение лишайниками веществ...
Таблица 2.1
Избранные содержания элементов в лишайниках из мест вблизи промышленных/городских
(или других) источников загрязнения и фоновых регионов (Nash, 1996)
Элемент Промышлен- ный/городской в^мк^с^х^вес)в Источник загрязнения, автор публикации Металлы класса А Фон (мкг/г сух. вес), ^^аВТО^^П^бЛИКаЦИ^^^
Се 2,2-7,2 Электростанция (Olmezetal., 1985) 0,18-0,89 (Gough et al., 1988a)
Na 1000-6000 Морские аэрозоли (Nieboer et al., 1978) 50-1000 (Nieboer et al., 1978)
и 3,0-151 Рудник (Boileau et al., 1982) 0,5-1,0 (Beckett et al., 1982)
Промежуточные металлы
As 128-11400 Плавление золота (Hocking et al., 1978?) 0,06-2,21 (Puckett, 1978?)
Ni 8-312 Плавка никеля (Tomassini et al., 1976) 1,7-5,5 Puckett, 1978?)
V 150-578 Древесная пульпа для бумаги (Laaksovirta, Olkkonen, 1979) 0,17-9,7 (Puckett, 1978?)
Zn 1000-25000 Плавка цинка (Nash, 1975) 10-30 (Nash, 1975)
Металлы класса Б
Си 15-250 1000-4900 Плавка никеля (Tomassini et al., 1976) Медные породы (Alstrup, Hansen, 1977) 0,7-5,0 (Puckett, Burton, 1981)
Hg 0,40-0,87 Хлорно-щелочной завод (Lodenius, Laaksovirta, 1979) 0,009-0,101 (Pakarinen, Hasanen, 1983?)
Pb 111-270 Хельсинки (Laaksovirta et al., 1976) 0,4-9,2 (Puckett, 1978?)
Неметаллы
F 260-940 Завод по переработке мусора (Takala et al., 1978) 2,9-7,8 (Takala et al., 1978)
S 470-4800 Градиент выпадений серы (Takala et al., 1985) 101-961 (Puckett, 1978?) 170-320 (Tomassini et al., 1976)
Работы, снабженные после года публикации, цитируются по: (Nash, 1996).
Другие элементы - Al, Si, Р, К, Au - наблюдались по всему слоевищу. Эти резуль-
таты подтвердила карта распределения элементов, полученная на основе рентге-
новских лучей, показавшая, что железо располагается внутри или на поверхности
водорослевых клеток, но его очень мало в гифах грибов (Вайнштейн, 1982). Изве-
стно, что Acarospora sinopica может накапливать чрезвычайно высокое количе-
ство железа (до 55000 ppm), a Rhizicarpon oederi - меди (до 1670 ppm), т.е. кон-
центрацйи, губительные для других растений (Lange, Ziegler, 1963). Пороговые
2. Улавливание и поглощение лишайниками веществ...
41
концентрации и меди, и железа для грибов и водорослей намного ниже, чем со-
держание их в ряде лишайников.
Анализом последовательно экстрагированных фракций лишайников Д. Броун
и Д. Слингсби (Brown, Slingsby, 1972) оценили распределение катионов (а) в меж-
клеточных и поверхностных фракциях, (аа) во фракции участка обмена ионов,
(ааа) во внутриклеточной фракции и (аааа) в остаточной фракции. Показанное
ими пространственное разделение ионов свидетельствует, что те элементы, ко-
торые располагаются внеклеточно, вне плазмалеммы, непосредственного мета-
болического значения не имеют и в этом случае не обязательно подвергаются
превращениям, а могут связываться иным путем. Например, значительное ко-
личество железа можно наблюдать на клеточных стенках, в межклеточных
пространствах, а порой и на поверхности лишайников Acarospora sinopica,
A. smaragdula, Rhizocarpon oederi, что подтверждается электронно-микроско-
пическим анализом (Noske et al., 1970). Такое отложение железа на поверхности
таллома, возможно, в виде окиси, является одним из путей связывания тяжелых
металлов. Оседание взвешенных в атмосфере частиц на поверхности слоевищ и
их проникновение внутрь талломов, вероятно, является обычным феноменом как
в незагрязненной (атмосферные аэрозоли терригенного происхождения), так и в
загрязненной средах. Как показало изучение слоевищ Caloplaca aurantia, попав-
шие таким образом в таллом частицы осаждаются вне клеток (Garty et al., 1979).
2.6. ИССЛЕДОВАНИЯ ВЫПАДЕНИЙ РАЗЛИЧНЫХ ВЕЩЕСТВ
С ПОМОЩЬЮ ЛИШАЙНИКОВ
Лишайники являются полезными биомониторами в изучении атмосферных
выпадений по нескольким причинам (Puckett, 1988; Richardson, 1988). Многие виды
имеют широкое географическое распространение, что позволяет с их помощью
документировать пространственные особенности выпадений. Кроме того, как
многолетние медленно растущие организмы лишайники долго сохраняют довольно
однообразное строение, и части их не подвергаются сезонной смене, как у сосу-
дистых растений (листья). Да и морфологические изменения самих лишайнико-
вых талломов во времени минимально зависят от параметров выпадений. У та-
ких видов как напочвенный лишайник Cladina stellaris и эпифит Everniaprunastri,
по особенностям ветвления можно определить возраст видимых частей слое-
вищ. Это позволяет до некоторой степени судить о временных трендах поступле-
ния загрязнителей в случаях, если загрязнители конкретного выпадения не явля-
ются подвижными. Вероятно, наиболее важно, что лишайники способны накапли-
вать многие элементы до концентраций, которые намного превышают их
физиологические потребности, что и позволяет по уровням содержания их в сло-
евищах делать выводы об особенностях выпадений и степени загрязнения.
42
2. Улавливание и поглощение лишайниками веществ...
Многочисленны исследования накопления элементов лишайниками вокруг
источников загрязнения (Nash, 1989, 1996). В большинстве случаев четко повы-
шенные концентрации, установленные вблизи источников загрязнения, сравнива-
лись с концентрациями на удаленных от источников территориях. В случае изоли-
рованного источника загрязнения концентрации увеличивались логарифмически с
расстоянием. Фоновыми считались концентрации, сохраняющие с расстоянием
относительное постоянство (Nieboer, Richardson, 1981). Определить фоновые кон-
центрации на деле довольно сложно из-за таких проблем как гетерогенность суб-
стратов, влияние на накопление не загрязняющих источников, и ограничений, обус-
ловленных пределами аналитических измерений используемых приборов и мето-
дов. Кроме того, умеренные широты Северного полушария в настоящее время
имеют высокий уровень индустриализации, откуда в результате движения воз-
душных потоков происходит перенос загрязнителей на большие расстояния в уда-
ленные от промышленных областей регионы, постепенно загрязняя последние
(Carignan, Gariepy, 1995).
Другое направление - оценка элементной загрузки лишайников относительно
вклада в нее земной коры. Фактор обогащения (ФОх) некоего элемента (X) в
лишайнике относительно породы или почвы вычисляется с использованием фор-
мулы:
ФОх = о,/о5,
где Ql - содержание элемента X в лишайнике, Qs - содержание элемента X в
субстрате.
Этот показатель был предложен Б.Б. Полыновым (1945). В западной лихено-
логической литературе его иногда также называют фактором обогащения (Puckett,
Finegan, 1980). А.И. Перельман назвал его коэффициентом биологического погло-
щения (Перельман, Касимов, 1999).
Наиболее подходящими индикаторами процессов поступления элементов из
земной коры являются скандий и титан, так как их биологические функции неиз-
вестны. Полезен для такого использования и алюминий (Перельман, Касимов,
1999). Анализ величин фактора обогащения привел к пониманию причин гетеро-
генности имевшихся абсолютных величин концентрации элементов в слоевищах
лишайников. Например, элементы А1, Сг, Со, Fe, Na, Ti, V имели среднее значе-
ние ФОх менее 5 (хотя абсолютные значения А1 заметно варьировали, отношение
Al/Sc было относительно постоянным), что интерпретировалось как свидетель-
ство преимущественного происхождение этих элементов из земной коры. Наобо-
рот, величины ФОх для других элементов были выше и поднимались до 1958 для
хлора в слоевищах Thamnolia subuliformis.
Элементы со значениями ФОх выше 100 для 14 наиболее обычных видов
лишайников Канады включили Sb, Cl, Pb, S, и эти высокие величины фактора обо-
гащения интерпретировались как свидетельства происхождение этих элементов
не из земной коры (Puckett, Finegan, 1980).
2. Улавливание и поглощение лишайниками веществ...
43
Изучение состава атмосферных выпадений не ограничивалось только метал-
лами, а включало также выпадения анионов, ассоциируемых с кислотными дож-
дями (Бойко и др., 1981; Горбач, Кобзарь, 1982; Киселев и др., 1986), и стойкой
органики, ассоциируемой с сельскохозяйственной и промышленной деятельно-
стью. Например, К. Такала (Takala et al., 1985) в Финляндии и И. Брютег (Bruteig,
1993а) в Норвегии изучили содержания сульфатов в лишайниках вдоль трансекты
север-юг в своих странах. Установлено, что в более южных регионах в районах с
высокой нагрузкой автомобильного транспорта обычны азотные загрязнители
(Campo et al., 1996). В Лос-Анджелесе выпадения соединений азота, среди кото-
рых наибольшая доля, вероятно, приходилась на HNO3, определенные с исполь-
зованием трансплантированных лишайников, составили свыше 170 мкмол/г за 10
недель во время сухого летнего периода (Boonpragob et al., 1989). Длительный
мониторинг выпадений соединений азота с помощью лишайников проводится в
Нидерландах (van Dobben, 1993; van Dobben, ter Braak, 1998).
Стойкие органические загрязнители большей частью естественно не встре-
чаются и потому нет вопросов относительно фоновых уровней их концентраций.
Однако многие из этих опасных ксенобиотиков - различной природы инсектици-
ды, пестициды, полихлорированные и полибпромированные бифенилы, полихло-
рированные дибензофунаны и дибензодиоксины, полициклические ароматиче-
ские углеводороды, органометаллические соединения - могут находиться в ок-
ружающей среде в жидкой или аэрозольной формах (Korte et al., 2000; Walker,
2001; Walker et al., 2001). Поэтому и органические загрязнители легко могут по-
глощаться лишайниками, что, видимо, и стимулировало некоторых исследовате-
лей к определению содержания ряда такого рода веществ в талломах лишайни-
ков, хотя доступных данных все же пока немного. Имеются сведения о содержа-
нии в них нескольких хлорированных углеводородов, родственных им полихлориро-
ванных бифенилов (ПХБ) и гексахлорциклогексановых изомеров по градиенту
север - юг в Канаде (Muir et al., 1993), а также в Норвегии на аналогично ориен-
тированной трансекте изучали содержание в лишайниках хлорированных углево-
дородов, фталатов и полиароматических углеводородов (Carlberg et al., 1983). При-
мерно те же соединения (полиароматические углеводороды, полихлорированные
бифенилы, пестициды, фенол) определили в слоевищах Hypogymnia physodes в
Польше (Migaszewski, 1999). В США (Орегон) экспериментально изучали дей-
ствие на Lobaria pulmonaria широко известного загрязнителя формальдегида
(Muir, Shirazi, 1996). В центральной и западной частях Канадской Арктики изуча-
ли характер накопления устойчивых органических загрязнителей (дуст, гексах-
лорциклогексаны, хлорбензены и другие) в пищевой цепи лишайник - олень - волк
(Kelly, Gobas, 2001).
Хлорированные углеводороды были выявлены даже в слоевищах лишайников
из Антарктиды (Bacci et al., 1986), что указывает на возможность переноса этих
загрязнителей на большие расстояния.
44
2. Улавливание и поглощение лишайниками веществ...
2.7. ТОКСИЧНОСТЬ МЕТАЛЛОВ
Наглядное свидетельство токсичности металлов - распределение лишайни-
ков вокруг цинкового завода в штате Пенсильвания, США (Nash, 1975). На конт-
рольном участке были найдены представители 84 видов лишайников, тогда как в
зоне экстремально высокой нагрузки обнаружены слоевища лишь 9 видов (Nash,
1972). Только представители двух из 9 - Micarea trisepta на камнях и
Scoliciosporum chlorococcum на деревьях - были обильны на загрязненной тер-
ритории, и обилие обоих видов здесь было выше, чем на контрольном участке.
Остальные лишайники, изредка встречавшиеся на загрязненной площади вокруг
завода, принадлежали к следующим видам: Acarospora fuscata, Aspicilia cinerea,
Cladonia sp., Hypocenomyce scalaris, Lepraria (два вида) и Verrucaria nigrescens.
Концентрации цинка, кадмия или двуокиси серы вблизи завода были токсичными
для представителей многих видов лишайников. Конечно, значительная часть цин-
ка и кадмия представлена относительно нерастворимыми оксидами металлов,
однако часть их все же растворялась и участвовала в обменных процессах. Экс-
перименты с представителями 4 видов, найденных на контрольном участке,
но отсутствовавших на загрязненной территории, показали их чувствительность
к растворам Zn и Cd. В частности, у Cladonia uncialis и Lasallia papulosa на-
блюдалась значительная депрессия чистого фотосинтеза при 200 мкмол Zn и Cd
(Nash, 1975). Кроме того, рост культивируемых водорослей, выделенных из
Cladonia cristatella и Flavoparmelia caperata, заметно сдерживался при 50 мкмол
Zn или Cd. Хотя представители нескольких видов, растущих вблизи завода, спе-
циально не изучались, приведенные данные вместе со сведениями о содержании
Zn выше 2,5% и 334 мкг/г Cd в слоевищах Verrucaria nigrescens, и 2,3% Zn и
320 мкг/г Cd в талломах Micarea trisepta на расстоянии до 2 км от завода (Nash,
1975), позволяют сделать вывод, что обнаруженные вблизи завода виды боль-
шей частью толерантны к цинку и кадмию.
Последующее исследование в Англии с 1 ммоль - растворами цинка показа-
ло снижение суммарного фотосинтеза на 5-40% в сравнении с контролем у пред-
ставителей 10 видов лишайников из родов Collema, Lichina, Lobaria, Nephroma,
Peltigera, Sticta с цианобактерией в качестве фотобионта (Brown, Beckett, 1984).
В дополнение, у Peltigera horizontalis фотосинтез снижался менее чем на 50%
при обработке растворами Cd, Си, Zn в концентрации 100 мкмол. Эти результаты
могут помочь в объяснении, почему представители рода Peltigera были найдены
на контрольных участках, но отсутствовали вблизи цинкоплавильного завода в
Пенсильвании (Nash, 1972). В другой работе приводятся результаты сравнитель-
ного изучения популяций Peltigera, собранных в естественных местообитаниях и
со старых отвалов покинутых рудников (Beckett, Brown, 1983). Измерена интен-
сивность фотосинтеза после обработки слоевищ при погружении на 30 минут в
раствор цинка 100 мкмоль. Индекс толерантности цинка (ZTI), который опреде-
2. Улавливание и поглощение лишайниками веществ...
45
лялся по формуле: ZTI = 100х(скорость фотосинтеза после обработки метал-
лом/скорость фотосинтеза после обработки водой), показал, что более устойчи-
выми к цинку были представители популяций со старых отвалов рудников.
Сравнение действия на лишайники токсичных металлов и газов, главным об-
разом, двуокисей серы и азота, свидетельствует, что негативное влияние газооб-
разных загрязнителей на видовое разнообразие эпифитных видов в Нидерландах
намного превышает воздействие металлов; из последних лишь действие сурьмы
было статистически значимым (van Dobben et al., 2001).
2.8. ТОЛЕРАНТНОСТЬ К ВОЗДЕЙСТВИЮ МЕТАЛЛОВ
Лишайники, встречающиеся на обогащенных металлами субстратах, накап-
ливают значительные количества металлов и образуют отчетливые группировки
видов (альянс Acarosporum sinopicae с двумя ассоциациями, называемыми
Acarosporetum sinopicae Hil, Lecanoretum epanorae Wirth, и Lecideion inopis Purvis),
которые отражают вариации содержания металлов в субстрате. Виды, толерант-
ные к высокому содержанию металлов, относятся к родам Acarospora, Aspicilia,
Cladonia, Lecanora, Lecidea, Porpidia, Rhizocarpon, Stereocaulon, Tremolecia.
Многие из этих видов показывают высокую степень специфичности к обогащен-
ным металлами субстратам, и таким образом дизъюнкции их распространения в
пределах общего ареала могут отражать доступность этих субстратов (Purvis,
Halls, 1996).
Естественно встречающиеся повышенные концентрации могут быть доволь-
но поразительными (см. табл. 2.1). В дополнение к приведенным данным можно
указать, что в горах Гарц (Германия) содержание железа в слоевищах лишайни-
ков колебалось в пределах от 0,6 до 5,5% от сухого веса (Lange, Ziegler 1963), а
О. Носке с соавторами (Noske et al., 1970) сообщают о следующих концентраци-
ях (мкг/г): Си - 60-410, РЬ - 125-2750, Zn - 150-436, Sb - 170-270 в талломах
пяти видов лишайников. В Норвегии и Швеции вокруг покинутого медного рудни-
ка в слоевищах двух видов лишайников были выявлены концентрации меди от 5,3
до 5,9% сухого веса (Purvis, 1984). Аналогичным образом, в слоевищах четы-
рех видов в Калифорнии содержание меди составило 0,25-2,3% (Czehura, 1977).
Наконец, в опрыскиваемых купоросом виноградниках Центральной Европы
концентрация меди в слоевищах Lecanora vinetorum составляет до 5000 мкг/г
(Nash, 1996).
Предположительно, механизмы толерантности могут включать: (1) врожден-
ную цитоплазматическую толерантность, (2) цитоплазматическую неподвижность
и химическую детоксикацию ионов и (3) перенос (или удержание) ионов во вне-
шние для плазмалеммы участки и даже на клеточные стенки (Lange, Ziegler, 1963).
Последний, по современным данным, включает в себя нескольких различных
46
2. Улавливание и поглощение лишайниками веществ...
механизмов толерантности (Richardson et al., 1979; Вайнштейн, 1982). Опреде-
ленную роль, видимо, играют обнаруженные в клетках животных, растений, циа-
нобактерий, грибов металлосвязывающие белки, или металлотионеины (Ле-
бедева и др., 1998); однако пока нет надежных свидетельств за или против пер-
вых двух механизмов. Также было показано, что конкуренция среди ионов может
привести к редукции поглощения токсичного иона в цитоплазму (Beckett, Brown,
1984b).
Одним из механизмов избегания токсичности является изоляция металлов
как частей кристаллов на внешних стенках гиф. Например, комбинацией различ-
ных методов было установлено, что в лишайниках Acarospora rugulosa, Lecidea
theiodes с содержанием меди около 5% наибольшая ее часть находилась в фор-
ме кристаллов медного оксалата (Purvis, 1984). Далее было показано, что два
других толерантных к меди лишайника Acarospora smaragdula и Lecidea lactea
способны образовывать комплекс медно-норстиктиковой кислоты, придающий
коровому слою отчетливо зеленый цвет (Purvis et al., 1987). Норстиктиковая кис-
лота является очень обычным вторичным лишайниковым веществом, которое
кристаллизуется на внешних стенках гиф лишайника.
В случае загрязненной среды значительная доля металлов присутствует в
нерастворенной форме (как оксиды металлов), и потому они могут быть физио-
логически неактивными, пока не растворятся. В таких ситуациях концентрации
катионов на местах обмена ионов на клеточных стенках могут быть на 2 или 3
порядка ниже, чем общие концентрации металлов, определенные для всего ли-
шайника. Конкурентные влияния различных ионов при разных концентрациях так-
же могут сказываться на поглощении. Например, было показано, что равномо-
лярные концентрации катионов Со, Mg, Мп, Ni на 40-50%, a Zn - на 32% задержи-
вают внутриклеточное поглощение кадмия (Beckett, Brown, 1984а). Поскольку
такие металлы как цинк и кадмий часто встречаются вместе, конкуренция меж-
ду их ионами может привести к низким внутренним концентрациям. При специ-
альном исследовании отмечено снижение внутриклеточного поглощения кадмия
(Vmax 2,39 мкмоль/г в час против контрольного Vmax3,93 мкмол/г в час) у популя-
ций Peltigera, собранных соответственно, на старых отвалах покинутого рудни-
ка, где почва обогащена цинком и кадмием, и на контрольном участке естествен-
ного местообитания (Beckett, Brown, 1984b). Таким образом, конкурентное ис-
ключение ионов из цитоплазмы является дополнительным механизмом
толерантности.
Металлы в цитоплазме, как известно, являются связующими центрами в
протеинах и ферментах и соответствуют ионам классов А, Б и промежуточного.
Специфичность металлов, занимающих эти центры связывания, определяется их
размерами и геометрией как типа лиганд. Соответственно, механизмы токсично-
сти ионов металлов подразделяют на три категории: (1) блокирующие существен-
ные биологические функции групп протеинов и ферментов, (2) внедряющие суще-
2. Улавливание и поглощение лишайниками веществ...
47
ственные ионы металлов в протеины и ферменты, (3) модифицирующие действу-
ющую структуру протеинов и ферментов (Nash, 1996). Имеются свидетельства,
что представители, по меньшей мере, нескольких видов лишайников более толе-
рантны к высоким внутриклеточным концентрациям ионов металлов в сравнении
с другими видами, но специфичные клеточные механизмы, которые обеспечива-
ют такую толерантность, пока еще неизвестны. Установлено, что стратегия из-
бегания токсичного действия металлов у разных видов лишайников неодинакова.
У представителей гипераккумулятора металлов Diploschistes muscorum и токси-
толерантного вида Xanthoria parietina механизмом защиты клеток от токсично-
го действия РЬ и Zn служит образование комплексов с ионами металлов. Но у
первого вида металлы накапливаются благодаря повышенному синтезу оксала-
та, который осаждает металлы в виде нерастворимых солей, а у второго вида
металлы осаждаются на стенках клеток грибных гиф через образование карбок-
сильных групп. Авторы этого исследования сделали вывод, что гипераккумуля-
ция металлов есть следствие реактивного механизма с органическими кислота-
ми, вырабатываемыми лишайниками, а толерантность к металлам достигается
пассивным образованием комплексов (Sarret et al., 1998а). Вероятно, толерант-
ность к металлам и их гипераккумуляция представляют собой генетически неза-
висимые свойства организмов, что было показано на сосудистых растениях
(MacNair et al., 1999).
2.9. ВЛИЯНИЕ НА ГИДРОЛОГИЧЕСКИЕ ЦИКЛЫ
Пойкилогидридная природа лишайников позволяет им влиять на гидрологи-
ческий цикл даже в экосистемах, где их биомасса мала. В Арктике и Субарктике,
где лишайники сплошным ковром перекрывают почву, они препятствуют испаре-
нию (Kershaw, Rose, 1971). Даже эпифитные лишайники могут изменять водный
баланс экосистемы. Например, в дубовом лесу Калифорнии они перехватили 7,5%
осадков, поступивших за трехлетний период (Nash, 1996). Хотя количественных
данных нет, можно полагать, что перехват воды тумана и росы лишайниками в
прибрежных пустынях может составлять значительную долю в общем водном
балансе этих экосистем, особенно в таких регионах как пустыня Атакама в Юж-
ной Америке, где осадки случаются редко (Lange, 1992).
В континентальных аридных и полуаридных регионах лишайники встреча-
ются обычно как часть корки слоевищных растений (такыры в Средней Азии),
которая особенно широко развита на ненарушенных почвах (Джураева, 1979).
Эти корки не только перехватывают осадки, но также препятствуют инфильтра-
ции воды в почву, которая, при отсутствии криптогамов, часто имеет непрони-
цаемый слой СаСО3 на или вблизи поверхности почвы (Eldridge, 2000; Eldridge et
al., 2000).
48
2. Улавливание и поглощение лишайниками веществ...
2.9.1. Связь минерального питания с водным обменом
Питательные соли попадают в слоевище в растворенной форме вместе с во-
дой, активно поглощаемой и передвигаемой капиллярными силами. Поглощение и
испарение влаги в природных условиях - чисто физические процессы, не контро-
лируемые слоевищами. Однако не исключена возможность, что помимо активно-
го, метаболического, у лишайников существует иной путь регуляции водообмена
- пассивный морфологический контроль (Larson, Kershaw, 1976). Большинство
лишайников в природе испытывает частые и значительные колебания влажности,
которые, возможно, способствуют поддержанию их нормального состояния. Об
этом косвенно свидетельствуют результаты культивирования лишайников в лабо-
ратории, показавшие, что переменные условия влажности необходимы для успе-
ха культивирования (Pearson, 1970; Dibben, 1971; Harris, 1972). Очень важной для
лишайников является фаза реувлажнения, когда к воздушно-сухому лишайнику
добавляют воду (Smith, Molesworth, 1973; Smith, 1975, 1976; Farrar, Smith, 1976;
Farrar, 1976a, b).
Вымывание метаболитов из сухих тканей при погружении в воду - явление,
свойственное не только лишайникам, и обычно его приписывают изменениям при-
роды клеточных мембран при высушивании (Вайнштейн, 1982). Вода является
структурным компонентом мембран, и удаление части ее или всего объема воды
приводит в результате к увеличению проницаемости мембран. Возможны два
следствия увеличения проницаемости: во-первых, растворимые вещества могут
вымываться из таллома через обезвоженные мембраны, и, во-вторых, субстра-
ты могут входить в контакт с теми мембранными системами, от которых они в
норме отделены. Первое следствие было показано в опытах, когда после реувлажне-
ния слоевищ Hypogymnia physodes происходило вымывание соединений, меченных
радиоактивным углеродом или фосфором (Farrar, Smith, 1976). Вымывание раствори-
мых веществ представляется очень важным для физиологии лишайников. В природе
увлажнение сухого таллома первыми каплями дождя может вызывать значительное
локальное вымывание. Однако при дожде таллом последовательно увлажняется пор-
циями дождевых капель в течение нескольких минут. Благодаря быстрому возобнов-
лению процесса поглощения, зона, увлажненная первой, окажется способной реадсор-
бировать растворимые вещества, вымытые из зоны, увлажненной позже, что умень-
шает общую потерю веществ из таллома. Тем не менее, неизбежно происходит
некоторая потеря, а учитывая частоту такого явления в природе, эффект этот может
быть кумулятивным (Вайнштейн, 1982).
2.10. МИНЕРАЛЬНЫЙ ЦИКЛ НА УРОВНЕ БИОГЕОЦЕНОЗА:
ПЕРЕХВАТ СУХИХ И СЫРЫХ ВЫПАДЕНИИ
Минеральный цикл можно разделить на межсистемный и внутрисистемный
(Сукачев, 1964; Дылис, 1973; Waring, Schlesinger, 1985; Перельман, Касимов, 1999).
2. Улавливание и поглощение лишайниками веществ...
49
Межсистемный цикл включает поток элементов питания между биогеоценозами
(экосистемами) и процессы их взаимодействия с глобальными циклами, а внут-
рисистемный - движение питательных веществ внутри биогеоценоза (экосисте-
мы). В экосистемах с доминированием сосудистых растений наибольшее коли-
чество питательных веществ участвует во внутреннем цикле веществ, следую-
щего от почвенных запасов элементов к растениям, и, через вымывание или
разложение, обратно в почву. Внутрисистемный вклад включает как жидкие, так
и сухие выпадения из атмосферных источников, а также выветривание из подсти-
лающей породы. Внутрисистемный выход происходит первично с гидрологиче-
ским движение ионов и частиц через почву, хотя реализация газов также важна в
циклах азота и серы.
Перехват лишайниками взвешенных в атмосфере питательных веществ про-
исходит во многих лесных экосистемах, поскольку первичные источники этих ве-
ществ находятся преимущественно вне конкретной экосистемы. Например, в
Калифорнии в дубовом лесу при сравнении состава стоков с 20 стволов деревьев,
с которых предварительно были содраны слоевища доминирующего эпифитного
лишайника Ramalina menziesii. со стоками с 20 деревьев, где лишайники остава-
лись на стволах, было установлено, что соответственно 2,85 и 0,15 кг/га в год
азота и фосфора захватывалось лишайниками из источников вне экосистемы (Nash,
1996). Хотя значения этих величин явно невелики по отношению к выпадениям
этих элементов в некоторых сельскохозяйственных угодьях, они свидетельству-
ют о внешнем поступлении элементов питания в экосистему. Во всяком случае, в
изученном сообществе перехват лишайниками минеральных веществ втрое пре-
вышал измеренные значения их седиментации.
Перехват эпифитными лишайниками питательных веществ из внешних ис-
точников влияет непосредственно на экосистему через возврат этих веществ в
форме опада слоевищ со стволов и ветвей деревьев на поверхность почвы. На-
пример, в том же самом дубовом лесу в Калифорнии установлено, что эпифитные
лишайники обогащают отложения общего N, NO3", органического N, Са, Mg, Na,
Cl, снижают отложения SO42" и не влияют на отложения NH4+, К и общего Р
(Knops et al., 1996; Nash, 1996). В дополнение, лишайники через их опад и после-
дующее разложение влияют на долговременное отложение питательных веществ
в почвы (Бязров, 1970). Кроме того, слоевища лишайников могут иметь значи-
тельно более высокое количество микроэлементов (например, Fe), чем листья
сосудистых растений, хотя в отношении макроэлементов нередко обратное соот-
ношение (Knopset al., 1991).
Источниками минеральных элементов в лишайниках могут быть как внешние
по отношению к экосистеме, так и внутрисистемные, и нередко бывает относи-
тельно трудно определить происхождение элемента. Например, вещества, вымы-
тые из листьев кроны, могут быть далее поглощены слоевищами эпифитных ли-
шайников, растущих ниже на стволе или ветвях, и потому их происхождение внут-
50
2. Улавливание и поглощение лишайниками веществ...
риэкосистемное. Наоборот, как жидкие, так и сухие выпадения вносят вещества
извне экосистемы (Knops et al., 1991). Жидкие выпадения включают не только
дождь, но и снег, росу и туман. Лишайники особенно эффективны при поглощении
влаги росы и тумана, и эти источники воды, в сравнении с дождевой, часто обога-
щены минеральными элементами. Сухие выпадения образуются при седимента-
ции крупных аэрозолей (крупнее, чем 2-10 мкм в диаметре), касании более мел-
ких аэрозолей и поглощении газов. Результатом седиментации может быть вне-
дрение частиц в лишайниковые слоевища. Касания понимают как столкновение
аэрозолей с поверхностью лишайника. Широко изучалась фиксация атмосферно-
го азота лишайниками (Nash, 1996), но о действии других газообразных соедине-
ний азота (NO, N2O, NOn и т.д.) сведений мало. В случае серных газов, довольно
много известно о поглощение SO2 (Блюм, 1984; Winner et al., 1988), высвобожде-
нии H^S и других газов (Gries et al., 1994, 1997). Скорости поглощения двуокиси
серы лишайниками могут существенно превосходить такой процесс у сосудис-
тых растений, закрытые устьица которых ограничивают поглощение газов.
2.11. ПОЧВООБРАЗОВАНИЕ
Лишайники называются во многих учебниках и популярных публикациях пио-
нерами сукцессий на обнажающихся поверхностях горных пород, где эпилитные
и эндолитные лишайники проникают в поверхностные слои. В целом известны
биологические, химические и физические процессы, определяющие выветрива-
ние (Syers, Iskandar, 1973; Jones, 1988). Высвобождающийся при дыхании СО2в
присутствии воды образует слабую кислоту (Н2СО3), особенно в условиях основ-
ного pH, и эта кислота растворяет некоторые горные минералы. Установлено, что
интенсивность выветривания оливинов и плагиоклазов под лишайниками в 2-18
раз выше, чем при их отсутствии (Brady et al., 1999). На покрытых лишайниками
поверхностях габбро интенсивность выветривания еще выше - в 25-30 раз
(McCarroll, Viles, 1995). Лишайники, растущие на известняках, часто содержат
большое количество кристаллов оксалата кальция и могут играть определенную
роль в отделении кальция от известняка. По другим данным, в целом более зна-
чима роль вторичных лишайниковых веществ как металл-комплексных агентов
(Syers, Iskandar, 1973). Установлено вытравливание поверхности пород под ли-
шайниками и показано сходство частиц, внедрившихся в слоевище эпилитного
лишайника Dimelaena oreina, с подстилающими кварцитными минералами
(Hallbauer, Jahns, 1977). Кроме того, найдена корреляцию между отложением вто-
ричных соединений железа на поверхностях грибных гиф эндолитных лишайников
и выветриванием богатых железом песчаников в Антарктиде (Friedmann, 1982).
Прямое проникновение ризин в случае листоватых лишайников или сердцевинных
гиф у корковых лишайников может физически раздроблять поверхности породы.
2. Улавливание и поглощение лишайниками веществ... 51
При внедрении в породу расширение и сжатие лишайниковых слоевищ с измене-
нием температуры и водного состояния постепенно приводит к поверхностной
дезинтеграции каменистых пород (Полынов, 1945; Ярилова, 1947; Левин, 1949).
В Испании наблюдали, как слоевища Xanthoparmelia (Parmelia) conspersa,
Parmelina (Parmelia) tiliacea, Lasallia pustulata и Ramalina protecta образуют
в местах контакта с каменистым субстратом желеобразное вещество, которое
при высыхании может отделять частички субстрата (Galvan et al., 1981). На воз-
можность подобного способа воздействия на субстрат указывала еще в начале
1920-х годов А. Смит (Smith, 1921).
3. ИСТОЧНИКИ ЗАГРЯЗНЕНИЯ ВОЗДУШНОГО
БАССЕЙНА, НЕКОТОРЫЕ ПРИМЕРЫ
ИХ ПРОСТРАНСТВЕННОГО РАЗМЕЩЕНИЯ
И ВОЗМОЖНЫЕ ВАРИАНТЫ ЛИХЕНОИНДИКАЦИИ
Понятие “загрязнение” в экологической интерпретации имеет санитарно-ги-
гиеническое происхождение, поскольку его употребление, прежде всего, связано
со здоровьем человека. Постепенно оно стало иметь более широкое толкование,
и ныне загрязнением принято считать поступление в природную среду веществ
разного происхождения в количестве, вызывающем неблагоприятное воздействие
на состояние живых организмов разных таксономических групп и биогеоценозов
в целом, а также на материалы, оборудование, здания, сооружения. В цитирован-
ном во Введении Законе об охране окружающей среды загрязнение определяется
как “поступление в окружающую среду вещества и (или) энергии, свойства, мес-
тоположение или количество которых оказывают негативное воздействие на ок-
ружающую среду”, а загрязнителем среды считается “вещество или смесь ве-
ществ, количество и (или) концентрация которых превышают установленные для
химических веществ, в том числе радиоактивных, иных веществ и микроорганиз-
мов нормативы и оказывают негативное воздействие на окружающую среду”
(ст. 1).
Таким образом, под загрязнением понимают процесс поступления в среду
различных веществ после того, как их содержание достигает некоторой нормиру-
емой критической величины - предельно допустимой концентрации (ПДК), - пре-
вышение которой негативно сказывается на деятельности обитателей Земли.
Нормирование осуществляют государственные органы.
Природная среда - явление многокомпонентное, да и загрязнение ее происхо-
дит из разных источников. Поскольку лишайники в основном используют для ин-
дикации качества атмосферы, уместно кратко охарактеризовать источники ее
загрязнения.
3.1. СОСТАВ АТМОСФЕРЫ
Атмосфера представляет собой сложное природное образование, о составе
которого дает представление таблица 3.1.
3. Источники загрязнения воздушного бассейна...
53
Таблица 3.1
Химический состав сухого природного воздуха (Исаев, 2001; Тищенко, 1991)
Вещество % Вещество %
Азот (N2) 78,084 Гелий (Не) 524x10’5
Кислород (О2) 20,9476 Криптон (Кг) ПЬЮ"*
Пары воды (Н2О) 3-4 Водород (Н2) 50х 10-6
Аргон (Аг) 0,934 Ксенон (Хе) 8,7х 1 О'6
Диоксид углерода (СО2) 3,14хЮ-2 Озон (О3) лето 7х 10-6
Окись углерода (СО) 0-0.01 зима 2х 10-6
Неон (Ne) 1,818x10-’ Двуокись серы (SO2) 0-0,001
Окись азота (NO) 0-0,001 Двуокись азота (NO2) 0-0,0001
3.2. ИСТОЧНИКИ ЗАГРЯЗНЕНИЯ АТМОСФЕРЫ
И ИХ ХАРАКТЕРИСТИКА
Общепринято деление источников загрязнения атмосферы на естественные
(извержения вулканов, пыльные бури, пожары, радиоактивное излучение естествен-
ных источников, брызги морской воды, продукты жизнедеятельности организмов
и другие) и искусственные (деятельность человека). Одна из версий многочис-
ленных классификаций источников загрязнения воздуха показана в таблице 3.2.
Загрязнение от естественных источников признается как слабо воздействую-
щее на свойства атмосферы, а значительные выбросы загрязняющих веществ из
этих источников, как правило, связаны с проявлениями стихийных неуправляе-
мых сил Земли (вулканическая деятельность). Однако лишайники использова-
лись и для индикации последствий вулканической деятельности, влияний геотер-
мальных водных источников (Barghigiani et al., 1990; Bargagli, Barghigiani, 1991;
Loppi, 1993,1996,1997,2001).
Загрязнение, обусловленное хозяйственной деятельностью человека, счита-
ется для организмов наиболее опасным. Промышленные предприятия планеты
ежегодно выбрасывают в атмосферу около 20 млрд, т загрязняющих веществ, из
которых 75% приходится на углекислый газ, 1% - на оксид углерода, более 2,5%
- на углеводороды, 0,6% - на золу и другое (Оценка..., 1996). На территории Рос-
сийской Федерации выбросы загрязняющих веществ в 1998 г. в атмосферу соста-
вили около 33 млн. т (Данилов-Данильян, 1999), в том числе промышленностью-
19 млн. т (58%) и автотранспортом - 12 млн. т (36%). Основные загрязняющие
вещества в промышленности: диоксид серы - 5,7 млн. т, оксид углерода - 4,6,
твердые вещества - 2,9, углеводороды - 3,5, оксиды азота - 1,8 млн. т; на авто-
транспорте: оксиды углерода - 8,8, углеводороды - 1,2, оксиды азота - 1,6 млн. т.
54
3. Источники загрязнения воздушного бассейна...
Классификация основных источников загрязнения воздуха
(Тищенко, 1991, с изменениями и дополнениями)
Таблица 3.2
Естест- венные Внеземные (космическая пыль)
Планетарные Неорганические Выветривание
Вулканы
Естественная радиация
Органические Растения
Животные
Грибы
Другие организмы
Искус- ственные Радиоактивные Руда Добыча
Транспортировка
Переработка
Реакторы
Термоядерные взрывы
Промышленные предприятия Производство энергии
Химическая и нефтеперерабатывающая промышленность
Металлургия, машиностроение
Промышленность стройматериалов
Горнодобывающая промышленность
Строительство
Прочее
Транспорт Автомобильный
Железнодорожный
Водный
Воздушный
Быт и коммунальное хозяйство Индивидуальные печи
Сжигание, гниение бытовых отходов
Сельское и лесное хозяйство Животноводство, птицеводство
Применение продуктов химической промышленности
Предприятия по переработке продукции сельского и лесного хозяйства
(Данилов-Данильян, 1999). Вклад всех видов транспорта в загрязнение атмосфе-
ры населенных пунктов составляет 50-70%, производство пара, тепловой и элек-
трической энергии - 10-15%, промышленных технологий и процессов горения -
15-20%, переработки отходов - 5%, причем около 90% всех выбросов составля-
ют газообразные загрязнители, а 10% - твердые и жидкие вещества (Оценка...,
1996). Представление об основных загрязнителях и их относительной концентра-
ции у различных производств дают таблицы 3.3 и 3.4.
3. Источники загрязнения воздушного бассейна...
55
Таблица 3.3
Основные загрязняющие вещества и их источники (Оценка..., 1996)
Источник Загрязняющие вещества
Аэрозоли, твердые частицы Газы
Котлы и промышленные печи Зола, сажа NO2, SO2, СО, альдегиды (НСНО), органические кислоты, бенз(а)пирен
Автомобильные двигатели Сажа СО, NO2, альдегиды, углеводороды неканцерогенные, бенз(а)пирен
Нефтеперерабатывающая промышленность Пыль, сажа SO2, H2S, NH3, NOX, СО, углеводороды, меркаптаны, кислоты, альдегиды, кетоны, канцерогенные вещества
Химическая промышленность Пыль, сажа В зависимости от процесса (H2S, CS2, СО, NH3, кислоты, органические вещества, растворители, летучие вещества, сульфиды и другое)
Металлургия и коксохимия Пыль, оксиды . железа SO2, СО, NH3, NOX, фтористые соединения, цианистые соединения, органические вещества, бенз(а)пирен
Г орнодобывающая промышленность Пыль, сажа В зависимости от процесса (СОх, фтористые соединения, органические вещества)
Пищевая промышленность Пыль ЫНз, H2S (многокомпонентные смеси органических соединений)
Промышленность стройматериалов Пыль СО, органические соединения
3.3. ПРИОРИТЕТНЫЕ ЗАГРЯЗНИТЕЛИ.
ОЦЕНКА УРОВНЯ ЗАГРЯЗНЕНИЯ АТМОСФЕРЫ
Приоритетность загрязнителей определяется размером фактического или по-
тенциально возможного воздействия на здоровье и благополучие человека, на
климат или экосистемы, степенью содействия физиологической деградации при-
родной среды, накоплением человеком в пищевых цепях и другими факторами
(Исаев, 2001). Эксперты первого межправительственного совещания по монито-
рингу в Найроби (1974) составили список приоритетных примесей-загрязнителей
(табл. 3.5).
Анализ публикаций свидетельствует о том, что лишайники применялись для
индикации загрязнителей всех классов опасности.
Уровень загрязнения атмосферы определяется по нескольким показателям
(Исаев, 2001), из которых наиболее часто используются:
- средняя концентрация примеси в воздухе;
- предельно допустимая концентрация (ПДК) - разовая и среднесуточная;
56
3. Источники загрязнения воздушного бассейна...
Таблица 3.4
Относительная концентрация химических элементов в пылях различных производств
(Сает и др., 1990)
Производство Ряды относительной концентрации
п-1000 n-100 n-10 n-1
Сжигание угля B, Pb, Mo, Ge Be. Li. V, Ni. Си. Zn, Ag. W
Сжигание мазута V, Ni Cr, Zn, Mo, W, Pb Cu, Sn, Ag
Производство алюминия Cd, Zn, Sb, Pb, Zn W, Sn, Cr, Bi, Mo Mn, Ag, B, Co
Производство аккумуляторов Sb, Ag Bi, Pb, Cd, Sn Ni, As, Co, Си, Zn, W, In, Ti
Литье чугуна Zn, W Mn, Sn, Pb, Cu, Cr
Литье стали Sn, Sb, Ni, W Mn, Zn, B, Pb, Mo
Коксохимия Hg Sb, W, Pb Mo, Sn
Механическая обработка черных металлов W, Mo, Си Cr, Ni, Co, Zn Mn, V, Pb, Sn
Механическая обработка цветных металлов Си, Zn, Pb Cr, Ni W, Sn Co, Mo, V
Изготовление масляных красок Hg,Cd Ag, Zn, Sn, Си, Bi, W Sb, Mo Co, Sn, Ni
Обжиг цементной шихты Ag Sb, Zn, Bi, W Sn, Ti, Cu, Mo, Ba
Производство керамзита Pb W, Ag, Bi, Mo Co, Zn, Cu, Li, Ni, V, Cr
Производство огнеупорного кирпича Sb, Pb Bi V, Cu, Cr, Sn, B, Ga, Zn, Nb
Таблица 3.5
Класс приоритетности загрязнителей среды (по уменьшению опасности) (Исаев, 2001)
Класс Загрязнитель Среда
I Двуокись серы плюс взвешенные частицы воздух
Радионуклиды воздух, пища
II ДДТ и другие хлорорганические соединения биота, человек
Кадмий пища, человек, вода
III Нитраты, нитриты вода, пища
Окисли азота воздух
IV Ртуть и ее соединения пища, вода
Свинец воздух, пища
Двуокись углерода воздух
V Окись углерода воздух
VI Фториды вода
VII Асбест, мышьяк воздух
VIII Микробиотоксин ы пища
Реактивные углеводороды воздух
3. Источники загрязнения воздушного бассейна...
57
Таблица 3.6
Предельно допустимые концентрации (мг/м3) ряда загрязнителей
Загрязнители ПДКсут ПДКраз-ч ПДКраз-р ПДКраз-б
Сернистый газ, SO2 0,05 0,5 0,02 0,02
Аммиак, NH3 0,2 0,2 0,05 0,05
Оксид азота, NO 0,06 0,6
Диоксид азота, NO2 0,085 0,085 0,02 0,02
Хлор, С1 0,1 0,025 0,025
Хлорводород, НС1 0,2 0,2
Сероводород, H2S 0,008 0,008 0,02 0,008
Метанол, СНзОН 1,0 0,2 0,2
Этилен, С2Н4 3,0 3,0
Бензол, СбНб 1,5 0,1 0,1
Формальдегид, НСНО 0,035 0,02 0,02
Циклогексан, СбНп 1,4 0,2 0,2
Пары H2SO4 0,3 0,1 0,1
Оксид углерода, СО 3,0 5,0 3,0
Диоксид углерода, СОг 30,0 30,0
Озон, Оз 0,03 0,16
Фторсодержащие загрязнители 0,005 0,02
Пары ртути, Hg 0,0003
ПДКсут - среднесуточная, ПДКраз-ч - максимальная разовая для человека, ПДКраз-р - максималь-
ная разовая для растений, ПДКраз-б - максимальная разовая для биосферы
- повторяемость разовых концентраций примеси в воздухе выше ПДК данной
примеси и другие.
Средняя концентрация - это весовое количество примеси в единице объема.
Предельно допустимая концентрация учитывает воздействие примеси на орга-
низм в зависимости от времени, например, для человека и животных - от продол-
жительности вдыхания (Исаев, 2001). ПДК ориентированы на человека, однако
известны попытки установить ПДК для растений и биосферы в целом. Таблица
3.6, составленная поданным книги А. А. Исаева (2001), содержит величины ПДК
основных загрязнителей атмосферы. Сведения о ПДК других загрязнителей со-
держат различные справочники (Беспамятное, Кротов, 1985).
3.4. НЕКОТОРЫЕ ВОЗМОЖНЫЕ ВАРИАНТЫ РАЗМЕЩЕНИЯ
ИСКУССТВЕННЫХ ИСТОЧНИКОВ ЗАГРЯЗНЕНИЯ
Степень воздействия искусственных источников загрязнения атмосферы за-
висит как от их мощности, так и от концентрации различных производств и их
размещения на изучаемой территории. Опубликованные данные результатов ли-
хеноиндикационных исследований позволяют выделить следующие возможные
58
3. Источники загрязнения воздушного бассейна...
варианты размещения источников загрязнения, воздействие которых изучалось с
помощью лишайников:
1. Территория находится под влиянием изолированного источника загрязне-
ния с постоянным объемом и составом загрязнителей;
2. Территория находится под влиянием изолированного источника загрязне-
ния с уменьшающимся объемом выбросов;
3. Территория находится под влиянием изолированного источника загрязне-
ния с увеличивающимся объемом выбросов;
4. На слабо загрязняемой изолированным источником территории появился
новый изолированный источник загрязнения;
5. Территории не имеет источников загрязнения, но загрязняется в результате
трансрегионального переноса загрязнителей воздушными массами;
6. На территории неравномерно рассредоточено некое число источников за-
грязнения с перекрывающимся действием загрязнителей;
7. На территории неравномерно рассредоточено некое число источников за-
грязнения с перекрывающимся действием и уменьшающимся объемом выбро-
сов и составом загрязнителей;
8. “Чистая” территория находится в сфере воздействия удаленного источника
загрязнения.
Ниже приведены примеры исследований названных вариантов
3.4.1. Изолированный источник загрязнения
с постоянным объемом и составом загрязнителей
В этом случае можно предполагать, что его воздействие на лишайники в наи-
большей степени будет сказываться вблизи источника и выразится в снижении
видового разнообразия за счет гибели представителей чувствительных видов, а
также редукции показателей жизненности сохранившихся видов. Сильное воз-
действие может быть обнаружено и при удалении от источника в направлении
преобладающих в этой местности ветров. Кроме того, вполне ожидаемо сниже-
ние концентрации загрязнителей в слоевищах лишайников по мере удаления от
источника загрязнения.
Примером такого варианта является комплексное исследование территории
вокруг медно-никелевого комбината “Североникель” в Мончегорске (Кольский
полуостров), в том числе и с использованием методов лихеноиндикации (Горш-
ков, 1986,1990,1991; Норин, Ярмишко, 1990). Это предприятие выбрасывало еже-
годно в атмосферу 250-280 тыс. т SO2h полиметаллическую пыль (Ni, Си, Со и
другое). До начала исследования (1982 г.) усредненные значения валовых выбро-
сов в атмосферу здесь составляли (т/сут.): пыли - 30,2, SO2- 730, NOx - 6,9,
H2SO4 - 2,3, СО - 0,22, углеводородов - 0,21 (Норин, Ярмишко, 1990). Эпигейные
и эпифитные лишайники изучались на постоянных пробных площадях, заложен-
3. Источники загрязнения воздушного бассейна...
59
ных для характеристики лесных биогеоценозов на разном расстоянии от комби-
ната, преимущественно к юго-западу и северо-востоку от него (направления пре-
обладающих ветров). Лесные ненарушенные участки располагались на расстоя-
нии 50-80 км, а полностью деградированные - в 8 км от предприятия и ближе.
Наряду с показателями, характеризующими особенности лесного сообщества и
конкретных местообитаний лишайников (субстрат, высота поселения на стволе,
угол наклона ствола и другое), учитывались видовой состав лишайников, покры-
тие видов, изменения ряда морфологических признаков, содержание элементов в
слоевищах и другое. В фоновых районах (50 км и более от комбината) покрытие
напочвенных лишайников в лесах варьировало от 5 до 90%, в среднем 20%; в зоне
поврежденных лесных массивов (30 км от комбината) их покрытие было от 5 до
50%, в среднем 15%, отмечены морфологические изменения слоевищу предста-
вителей родов Cladina и Cladonia\ в зоне разрушения лесных биогеоценозов
(15 км от комбината) - 5-10%, отмечены морфологические изменения слоевищ у
представителей родов Cladina и Cladonia\ в зоне полной деградации лесных био-
геоценозов (3-8 км от комбината) - 0-10%, также отмечены морфологические
изменения слоевищ у представителей рода Cladonia (Горшков, 1986). По мере
приближения к комбинату число видов напочвенных лишайников уменьшалось
(26 видов в фоновых районах, 14 видов на расстоянии 15 км, 10 видов - в 8 км, и
3 вида - в 4 км). На расстоянии 15 км от предприятия из напочвенного лишайни-
кового покрова выпали виды, на долю которых в фоновых районах приходилось
47% покрытия; в 8 км исчезли виды, вклад которых в покрытие участков в фоно-
вых районах составлял 91 %, а в 4 км - 98,5% (Горшков, 1986). В слоевищах Cladina
stellar is, наиболее распространенного эпигейного вида, определяли содержание
ряда элементов. Концентрация Ni в образцах из фоновых районов в 1982 г. соста-
вила 14 мг/кг сухого веса, а на расстоянии 15 км от комбината - 135-255 мг/кг,
Си - 5,3-5,9 и 45-63 мг/кг, Со - 1,0 и 4,7-8,7 мг/кг, Fe - 245-255 и 480-670 мг/кг
соответственно (Норин, Ярмишко, 1990).
В фоновых районах на стволах сосен встречены представители 67 видов; сред-
няя величина покрытия ими стволов деревьев составила 11%, доля деревьев без
лишайников - 13%. В 30 км от комбината на соснах обнаружены представители
24 видов, покрывающих лишь около 2% поверхности стволов, доля деревьев без
лишайников - 54%; в 15 км от источника загрязнения 19 видов лишайников встре-
чены лишь на основаниях стволов деревьев с покрытием 0.1%, доля деревьев
без лишайников - 90%; в 8 км от комбината на соснах обнаружены представите-
ли всего лишь 3 видов лишайников, доля деревьев без лишайников - 99%, а ближе
к предприятию лишайники на соснах отсутствовали (Горшков, 1991). Концент-
рация Ni в слоевищах Hypogymnia physodes в фоновых районах составила
25-30 мг/кг сухого веса, а на расстоянии 15 км от комбината - 200-420 мг/кг,
Си - 5,9-12 и 110-155 мг/кг, Со - 1,5-1,8 и 8,3-14 мг/кг, Fe - 500-775 и 1020-
1820 мг/кг (Норин, Ярмишко, 1990).
60
3. Источники загрязнения воздушного бассейна...
Результаты исследования дали основание выделить по состоянию лишайни-
ков на обследованной территории три зоны: 1) зона нормального развития лишай-
никового покрова (фоновые районы, 60-80 км от источника загрязнения); 2) зона
разрушения лишайникового покрова (30 км от источника на юго-юго-запад);
3) зона полного разрушения лишайникового покрова (15 км и ближе от комбината
на юго-юго-запад) (Норин, Ярмишко, 1990).
Другой пример - изучение влияния предприятия по производству фосфора в
Ньюфаундленде, Канада, на окружающие напочвенные и эпифитные лишайники
(Roberts, Thompson, 1980). Это предприятие работало более 10 лет, и в его выбро-
сах были такие фитотоксиканты как HF, SiF4. Через пять лет после начала рабо-
ты площадь поврежденных вокруг завода сообществ сосудистых растений со-
ставляла 11400 га, причем значительная часть этой нарушенной территории рас-
полагалась северо-восточнее предприятия, по направлению преобладающих в этом
районе ветров. Поврежденная территория по степени загрязнения была разделе-
на на 4 зоны. В каждой из них, а также на контрольных участках, удаленных от
завода на расстояние более 9 км, были обследованы представители обычных
здесь напочвенных лишайников Cladina rangiferina (L.) Harm, и C. stellaris (Opiz)
Brodo. Для них определялись концентрация фтора в слоевищах, а также морфоло-
гические параметры слоевищ. Установлено, что концентрация фтора в сухих сло-
евищах значимо уменьшается по мере удаления от завода (от 2830 ppm в бли-
жайшей к предприятию зоне 1 до 6,4 ppm в 9 км от источника, вне зоны 4). Кон-
центрация фтора в воздухе изменялась от 5,2 мкг/м3 вблизи завода до 0,1 мкг/м3
в зоне 4 (Roberts, Thompson, 1980). Также наблюдалась тесная корреляция между
величиной накопления фтора в тканях лишайников и выраженностью морфологи-
ческих отклонений слоевищ. Все экземпляры лишайников, собранные в зоне 1,
были аморфные, обесцвеченные, тогда как в зоне 4, наиболее удаленной от пред-
приятия, обесцвечены были только 25% образцов, а проявления деградации ана-
томических показателей структуры здесь были незначительными.
Лихеноиндикационное изучение аналогичного воздействия изолированных ис-
точников загрязнения проводилось и в других регионах - на Кавказе (Петрушина,
1987; Залиханов, 1991), Дальнем Востоке (Скирина, Качур, 1991), на Урале (Ми-
хайлова, 1993, 1994, 1996; Михайлова, Воробейчик, 1995, 1999), в Греции (Riga-
Karandinos, Karandinos, 1998), в Финляндии (Takala et al., 1978), в США (Newberry,
1974; Murphy et al., 1999) и многих других регионах Земли.
3.4.2. Изолированный источник загрязнения
с уменьшающимся объемом выбросов
Лишайники могут быть использованы для индикации улучшения качества сре-
ды вследствие снижения эмиссии загрязняющих веществ изолированным источ-
3. Источники загрязнения воздушного бассейна...
61
ником. Уменьшение эмиссий может быть результатом усовершенствования или
изменения технологии очистки выбросов, перехода на более чистое топливо или
снижения активности предприятия. В этих случаях количество выбросов и их со-
став уменьшаются, что предполагает соответствующую реакцию лишайников -
заселение ими зон, где они отсутствовали из-за загрязнения, появление исчезнув-
ших видов, уменьшение числа слоевищ с морфологическими отклонениями, выз-
ванными загрязнением, снижение концентрации загрязнителей в талломах и дру-
гое. Однако подобное исследование требует наличия данных, собранных в этом
месте в момент или до начала снижения количества выбросов. Примером таких
работ является изучение распространения лишайников вокруг работающей на угле
изолированной электростанции Muskingum River в юго-восточном Огайо (США),
начатое в 1973 г. (Showman, 1975). В 1972 г. на электростанции была проведена
замена оборудования, результатом чего стало значительное снижение выбросов
SO2. В 1973 г. вокруг станции было обследовано 128 участков с целью картогра-
фирования распространения Flavoparmelia caperata. Представители этого вида
отсутствовали во многих точках вблизи станции. С 1976 г. обследование этих
точек проводили ежегодно и установили, что слоевища Е caperata постепенно
заселяли участки ближе к станции; там же стали появляться и представители
других видов, ранее отсутствовавшие (Showman, 1981).
3.4.3. Изолированный источник загрязнения
с увеличивающимся объемом выбросов
Появление на некогда “чистой” территории источника загрязнения или увели-
чение эмиссий уже существующего изолированного источника дает основания
ожидать, что имеющиеся на этой территории лишайники отреагируют на измене-
ния условий их существования. Хроническое воздействие загрязнителей может
привести к уменьшению обилия и ухудшению показателей жизненности или ис-
чезновению представителей более чувствительных видов, а также увеличению
покрытия устойчивых видов. Можно ожидать увеличения встречаемости слое-
вищ с проявлениями морфологических отклонений, а также увеличения концент-
рации загрязнителей в тканях лишайников. Как и в предшествующем случае, для
сравнения желательны сведения о лишайниках территории, собранные до изме-
нения масштаба воздействия источника загрязнения на нее. Однако в случае,
если деятельность источника загрязнения непродолжительна, пространственный
градиент можно использовать для сравнения возможных изменений во времени,
как сделала М.А. Поташева (1995) в Карелии. В 1982 г. здесь начал работать
Костомукшский горно-обогатительный комбинат по производству железорудных
окатышей. Ежегодные выбросы этого предприятия составляли более 60 тыс. т,
из которых более 90% приходилось на сернистый газ, а 7% - на обогащенную
62
3. Источники загрязнения воздушного бассейна...
железом пыль (Поташева, 1995). В 1990-1991 гг. здесь проведено изучение ли-
шайников на 10 пробных площадях, расположенных на расстоянии от 1 до 22 км
от предприятия по линии, ориентированной с юго-запада на северо-восток (на-
правление господствующих ветров). Лишайники учитывались на стволах деревь-
ев на высоте 1,5 м с использованием рамки 10x20 см.
Помимо видового состава определялись покрытие видов, а также степень
повреждения слоевищ. Анализ данных не выявил значимых различий видового
состава лишайников и их покрытия между участками, расположенными на раз-
ном удалении от загрязнителя. Это позволило сделать вывод о том, что 9-летняя
работа комбината не привела к изменениям структуры эпифитных лишайниковых
группировок, регистрируемым на уровне видов и синузий (Поташева, 1995). Од-
нако зафиксированы значимые различия признаков, свидетельствующих о повреж-
дениях слоевищ (доля отмерших участков таллома от общей его площади), меж-
ду участками в непосредственной близости от источника загрязнения и удален-
ными от него. Был сделан прогноз дальнейшей активации процесса отмирания,
который приведет к снижению проективного покрытия более чувствительных
видов и в конечном итоге к их выпадению из состава лихенобиоты (Поташе-
ва, 1995).
В Израиле до начала эксплуатации в 1981 г. работающей на угле электростан-
ции Maor David вблизи Тель-Авива в ее окрестностях с февраля 1979 по март
1980 г. в 22 точках экспонировались срезанные ветви с обычным в тех местах
эпифитным лишайником Ramalina lacera (With.) J.R.Laundon = [7?. duriaei (DeNot.)
Bag!.] (Garty, Fuchs, 1982; Fuchs, Garty, 1983). Затем, после начала работы ТЭС, с
июня 1981 по июль 1982 г., в тех же точках вновь экспонировались транспланти-
рованные ветви с тем же лишайником (Garty, 1987). Методами рентгеновской
флуоресценции и атомной абсорбционной спектрометрии в трансплантированных
слоевищах определялись концентрации Ni, Сг, Си, Zn, РЬ, чтобы установить воз-
можное влияние электростанции. Слоевища Ramalina lacera после года воздей-
ствия в 1981-1982 гг. стали бледными, повысилась в них и концентрация метал-
лов. Сравнение данных 1979-1980 гг. и 1981-1982 гг. показало, что концентрации
Pb, Ni, Сг были выше в 1981-1982 гг., a Zn и Си было больше в 1979-1980 гг.
Анализ корреляций между концентрациями металлов показал значимость ассо-
циаций Pb/Ni, Pb/Cr, Pb/Mn, Zn/Mn. Например, отношение Pb/Zn увеличилось
с 0,19 в 1979-1980 гг. до 2,11 в 1981-1982 гг. Повышенное содержание РЬ бы-
ло объяснено увеличением в городе на 16% числа автомобилей, которые все
еще использовали свинцовый бензин, и с увеличением их пробега в 1981-1982 гг.
Уменьшение концентрации Zn объяснялось снижением опрыскивания цинксо-
держащими препаратами расположенных рядом плантаций цитрусовых и на-
саждений пекана, а высокие концентрации Сг и Ni, установленные в тканях ли-
шайников в 1981-1982 гг., были связаны с низким качеством сжигаемого на элек-
тростанции угля.
3. Источники загрязнения воздушного бассейна...
63
3.4.4. Маломощный изолированный источник
дополнен новым источником загрязнения
На территории, на которой функционирует небольшой источник загрязнения,
со временем может начать действовать дополнительный и, возможно, более мощ-
ный изолированный источник. В лихенобиоте этой местности в таком случае мо-
гут произойти определенные перестройки, такие как реакция на изменение эколо-
гических условий - повышение смертности представителей отдельных видов,
снижение покрытия, уменьшение видового разнообразия и другое. Эти признаки
обычно проявляются в пределах 1-5 км от источника, а частота их проявлений
снижается с увеличением расстояния от источника. Пример лихеноиндикации в
подобной ситуации дает исследование местности вокруг нефтеперерабатываю-
щего завода Esso, Fawley, в Гэмпшире, Англия (Morgan-Huws, Haynes, 1973). Здесь
в 1921 г. построен маломощный завод, к которому в 1952 г. добавилось аналогич-
ное, но гораздо более мощное предприятие. Основным загрязнителем был газ
SO2 и сопутствующие вещества, образующиеся при его сгорании на высоте 30-
50 м. Для определения пространственных границ влияния загрязнителей в 1966 г.,
через 45 лет воздействия слабого источника загрязнения и 14 лет работы мощно-
го источника, на старых стволах Quercus robur учитывались эпифитные лишай-
ники до высоты 2 м. Было установлено заметное уменьшение присутствия боль-
шинства видов лишайников вокруг места выбросов. Обнаружены представители
следующих видов (последовательность в порядке убывания их чувствительнос-
ти к загрязнению): Parmeliaperlata, Usnea sp., Ramalina sp., Parmelia subrudecta,
P. glabratula, P caperata, P sulcata, Evernia prunastri, Hypogymnia physodes,
Lecanora conizaeoides (номенклатура (Morgan-Huws, Haynes, 1973)). Особенно
чувствительными оказались первые два вида, и их распространение примерно
соответствует среднегодовой концентрации, не превышающей 40 мкг/м3 SO2.
Только Lecanora conizaeoides, видимо, относительно толерантен к имевшимся
уровням загрязнителей, поскольку представители вида обнаружены во всех учет-
ных точках, даже в непосредственной близости от источника выбросов (Morgan-
Huws, Haynes, 1973).
3.4.5. Территория не имеет источников загрязнения,
но загрязняется в результате трансрегионального переноса
загрязнителей воздушными массами
Удаленные территории с чистым воздухом могут загрязняться и в результате
трансрегионального переноса загрязнителей воздушными массами. Так, на Евро-
пейскую территорию Российской Федерациии, наряду с загрязнителями от соб-
ственных источников, выпадают вещества, принесенные в результате трансгра-
64
3. Источники загрязнения воздушного бассейна...
ничного переноса воздушными массами с Украины, Прибалтики, стран Централь-
ной, Западной и Северной Европы (Данилов-Данильян, 1999). Наиболее показате-
лен пример распространения радиоактивного загрязнения в результате аварии на
Чернобыльской АЭС в 1986 г., следы которого находят практически во всех реги-
онах Северного полушария. Для установления действия такого рода загрязнения
на относительно чистые области и исторического тренда загрязнения территории
также возможно использование лишайников, поскольку многие из них являются
накопителями определенных веществ. Успешные результаты были получены при
определении концентраций свинца и серы в образцах лишайников, собранных мно-
го лет назад и хранившихся как коллекционный материал, и в современных образ-
цах тех же видов, собранных в том же месте. Как пример можно привести иссле-
дование в Западной Вирджинии, США, в Национальном парке Shenandoah в Ап-
палачах (Lawrey, Hale, 1988). Этот парк был создан в 1936 г. для сохранения
вторичных лиственных лесов. Анализ коллекционных материалов позволил выя-
вить на территории парка 11 точек, в которых в 1933-1958 гг. было собрано до-
статочно материала (лишайник Flavoparmelia baltimorensis) для определения в
нем концентраций РЬ и S. В тех же 11 точках в 1984-1986 гг. с той же целью были
собраны образцы того же вида. Установлено, что за период между сборами кон-
центрация свинца в тканях Flavoparmelia baltimorensis уменьшилась во всех
местах отбора проб, вероятно, вследствие изменения региональных эмиссий РЬ,
связанного с запретом использования транспортом бензина с высоким содержа-
нием свинца. Однако концентрация серы в тканях лишайников всюду значимо уве-
личилась, причем в трех точках, располагавшихся в наиболее высоких местах
(выше 1 000 м н.у.м.), был зафиксирован наибольший относительный прирост кон-
центрации серы.
3.4.6. Неравномерное рассредоточение нескольких источников
с перекрывающимся действием загрязнителей
Индустриально развитые территории, как правило, выделяются наличием ком-
плекса неравномерно рассредоточенных разнородных предприятий, выбросы ко-
торых в атмосферу могут быть специфичными, но действие их на природную
среду будет перекрывающимся, так что определить особенности фактического
влияния конкретного источника бывает практически невозможно. Особенности
распределения лишайников на таких территориях нередко позволяют выделить
здесь зоны, различающиеся качеством воздушного бассейна. Кроме того, по-
добному варианту соответствует лихеноиндикационное обследование крупных по
размерам территорий (историческая или административная область, географи-
ческий регион, страна). Примеров подобных исследований множество, и прово-
дились они на уровне стран (van Dobben, 1993), провинций (Nimis et al., 1991;
Kirschbaum, Windisch, 1995), городов (Л. Мартин, Нильсон, 1983; Wonitzki, Coburger,
3. Источники загрязнения воздушного бассейна...
65
1993; Малышева, 1998,1999; Бязров, 1999а), индустриальных бассейнов (Иванов,
1993; Баумгертнер, 1998). Основное требование к методике таких работ - равно-
мерное обследование всей территории. Как правило, результаты представляются
в виде карт разного масштаба, созданных либо на основе вычисленных индексов,
интегрально характеризующих показатели лишайников, либо сведений о распро-
странении отдельных индикаторных видов. Например, при лихеноиндикационном
обследовании провинции Венеция (Италия) учеты эпифитных лишайников сдела-
ны на 2425 стволах липы в 662 точках, распределенных на всей исследуемой
территории (Nimis et al., 1991). Сбор данных проводился как для вычисления ин-
декса атмосферной чистоты (I.A.P.), так и для картографирования частоты встре-
чаемости избранных видов. Их обработка осуществлена с использованием ком-
пьютерной программы. По величинам I.A.P. выделено 7 зон, из которых на са-
мую загрязненную приходится 6,3% всей площади этой административной единицы
Италии, а доля незагрязненной территории составляет 1,8%.
3.4.7. Неравномерное рассредоточение нескольких источников
с перекрывающимся воздействием и уменьшающимся объемом
выбросов и составом загрязнителей
Проблемы качества окружающей среды все более волнуют население, что
отражает принятие в ряде стран законов и других регламентирующих докумен-
тов, направленных на снижение уровня выбросов загрязняющих веществ, в том
числе и в атмосферу. Иногда уменьшение выбросов обеспечивается не только
нормативно-правовыми механизмами, но и является следствием экономического
спада, как это произошло в Российской Федерации в 1990-е годы (Данилов-Дани-
льян, 1999).
Лишайники достаточно чутко реагируют не только на повышенные уровни
загрязнения воздуха, но и на очищение атмосферы. Это проявляется и в заселе-
нии ими тех частей территории, где они прежде отсутствовали из-за загрязнения,
и в увеличении площади покрытия ими субстрата, частоты встречаемости, улуч-
шении показателей жизненности и т.п. Наиболее эффектным лихеноиндикацион-
ное исследование в этом случае выглядит при наличии исторических сведений о
развитии лишайников на изучаемой территории. В 1960-1970-х гг. в Велико-
британии начали действовать строгие законы, направленные на улучшение каче-
ства воздуха. В 1980 г. в 29 точках северо-западной части Лондона (парки, запо-
ведники, сады, кладбища, лесные массивы и другие полуприродные участки) был
проведен учет эпифитных лишайников (Rose, Hawksworth, 1981), а в 1988 г. в тех
же пунктах, дополненных еще 22 точками, провели повторное их обследование
(Hawksworth, McManus, 1989). За этот период среднегодовые концентрации SO2
снизились приблизительно со 130 мкг/м3 до 29-55 мкг/м3. В каждом месте реги-
66
3. Источники загрязнения воздушного бассейна...
стрировались особенности форофитов, встреченные на них виды лишайников, от-
носительная частота встречаемости вида по 5-бальной шкале, диаметр или дли-
на слоевища самого крупного листоватого или кустистого вида в каждом месте
учета. При анализе результатов к материалам, собранным в указанные два срока
обследования, были присоединены данные предшествующей инвентаризации ви-
дового состава лихенобиоты Лондона (Laundon, 1967, 1970), а также привлечены
полученные в гербариях сведения о сборах лишайников в этих местах, что позво-
лило восстановить временной ряд изменения состава видов на обследованной
территории более чем за 200 лет. Встреченные виды лишайников ранжировались
по чувствительности согласно ранее установленным для Англии девяти классам
(Hawksworth, Rose, 1970). Уже в 1980 г. было отмечено увеличение видового раз-
нообразия лихенобиоты в сравнении с данными предшествующего исследования.
При этом прогнозировалось, что если среднегодовые концентрации SO2 снизятся
до 40-50 мкг/м3, то число видов лишайников возрастет (Rose, Hawksworth, 1981).
В 1988 г. здесь встречены представители еще 26 видов, не обнаруженные в 1980
г., 8 из которых на территории Лондона не отмечались последние 200 лет
(Hawksworth, McManus, 1989). Авторы установили, что скорость реколонизации
чувствительных к SO2 видов выше ожидаемой, объяснив это быстрым улучше-
нием качества воздуха в Лондоне. Также был сделан вывод о том, что в крупных
городах такие факторы как температура и влажность гораздо менее значимы
для развития лишайников, чем загрязнение воздуха. Аналогичные исследования
известны для Рурской области (Rabe, Wiegel, 1985), Мюнхена (Kandler, Poelt, 1984;
Kandler, 1988), Парижа (Seaward, Letrouit-Galinou, 1991; Letrouit-Galinou et al., 1992;
Deruelle, Guilloux, 1993) и других мест.
3.4.8. “Чистая” территория находится в сфере воздействия
удаленного источника загрязнения
В связи с освоением удаленных от промышленных центров и ранее не обжи-
тых регионов, территории, считавшиеся “чистыми”, могут попасть в сферу воз-
действия достаточно далеко расположенных, но мощных источников загрязне-
ния. Этому варианту, например, в Российской Федерации соответствуют террито-
рии ряда заповедников, относительно удаленных от крупных предприятий, но в
силу либо особенностей технологии, либо направления преобладающих ветров
подвергающихся их воздействию. Это влияние также может быть обнаружено
методами лихеноиндикации, что подтверждают результаты исследований на хреб-
тах Хамар-Дабан (Пярн и др., 1982; Трасс и др., 1988; Пярн, Трасс, 1990), Сихотэ-
Алинь (Баденкова и др., 1981; Родникова и др., 1998), полуострове Таймыр (От-
нюкова, 1997а, б), в дальнем Подмосковье (Л. Мартин и др., 1978), острове Нью-
фаундленд (Wadleigh, Blake, 1999) и во многих других регионах.
3. Источники загрязнения воздушного бассейна...
67
Примером подобного исследования в Северной Америке является работа
Л. Гейзер с соавторами (Geiser et al., 1994) в Tongass national forest на Тихоокеан-
ском побережье США, на границе с канадской провинцией Британская Колумбия.
Это исследование было предпринято с целью получения исходных данных для
мониторинга состояния среды во времени. Общая площадь обследованной тер-
ритории около 3000 км2. Источниками загрязнения были расположенные непода-
леку два целлюлозных предприятия, а также несколько дизельных электростан-
ций, транспортные средства и другое. Здесь проведена инвентаризация лишайни-
ков, в результате которой выявлены представители 381 вида, типичных для
юго-восточной части Аляски, и собраны 366 образцов четырех обычных здесь
видов (Alectoria sarmentosa, Cladina rangiferina, Hypogymnia enteromorpha,
Lobaria oregana), для которых определен основной уровень концентрации 17 эле-
ментов. Концентрации тех же элементов определены и для аналогичного набора
видов, собранных у источников загрязнения. Уровни S, Р, Na, Cu, Ni, Cd более
высокие вблизи целлюлозных предприятий, а вблизи автомобильных дорог и на-
селенных пунктов отмечено более высокое содержание в слоевищах лишайников
К, Zn, Cr, Al, Fe, В, Pb. Классическое зонированное распределение лишайников
обнаружено лишь вокруг целлюлозных предприятий, по мере приближения к кото-
рым число видов лишайников уменьшается.
3.4.9. Модель лихеноиндикационных исследований
Обобщение сведений из доступных источников, а также собственный опыт
показывают, что лихеноиндикационные исследования в той или иной мере можно
представить в виде следующей схемы (рис. 3.1).
Лихеноиндикационные исследования и мониторинг качества воздуха с помо-
щью лишайников можно проводить разными способами в зависимости от специ-
фики решаемых задач, особенностей территории и возможностей как заказчиков
работы, так и ее исполнителей. Расположение площадей для учета лишайников и
частота этих учетов при длительных наблюдениях зависят как от характера за-
грязнения (одиночный источник или их несколько, объем и состав загрязнителей,
технология и прочее), так и от общегеографических условий (климат, направле-
ние ветров, топография и прочее). Соответственно, для подготовки обоснованно-
го проекта работы необходимо детально изучить особенности подлежащей об-
следованию территории.
Так, при определенном режиме метеорологических показателей выбросы из
высоких дымовых труб могут оседать на поверхность вдали от источника. Неко-
торые предприятия работают эпизодически как, например, “пиковые” электро-
станции, которые действуют главным образом в периоды высокого потребления
энергии. Степень воздействия загрязнителей на лишайники зависит и от особен-
68
3. Источники загрязнения воздушного бассейна...
Рис. 3.1. Схема возможных лихеноиндикационных исследований (Stolte et al., 1993, с изменениями)
ностей рельефа. Загрязненная воздушная масса, пересекая возвышенную мест-
ность, может соприкасаться с компонентами биогеоценозов и загрязнять их. Но-
чью и часто зимой такие массы воздуха могут застаиваться вдоль долин, тем
3. Источники загрязнения воздушного бассейна...
69
самым содействуя накоплению загрязнителей в низинах. При некоторых уровнях
температуры, влажности, освещения может происходить химическое преобразо-
вание загрязнителей как в безвредную, так и в более токсичную формы.
Такие данные позволяют выбрать пригодный метод сбора материала. Весь-
ма существенным моментом исследования является выбор, расположение и ха-
рактеристика участков, на которых проводятся учеты лишайников. Анализ со-
става лихенобиоты и особенностей лишайниковых группировок, а также распре-
деления видов на обследованной территории часто дает необходимую информацию
о качестве воздуха, конечно, если имеются сведения об отношении встреченных
видов к загрязнению. Если таких данных недостаточно, проводится изучение осо-
бенностей накопления элементов в тканях представителей наиболее распростра-
ненных видов лишайников, а также изучаются их физиологические показатели
как реакция на загрязнение. Эта работа требует больше времени, специального
оборудования, особого планирования исследований реакции “доза - эффект”, и,
соответственно, она много дороже по стоимости.
3.5. РАСЧЕТ СТОИМОСТИ ИССЛЕДОВАНИЯ
Лихеноиндикационные исследования нередко пропагандируют как дешевые.
Я полагаю, что их дешевизна относительна и зависит от цены рабочей силы на
рынке труда. В Российской Федерации, как известно, оплата труда научных ра-
ботников очень низкая, поэтому довод о дешевизне лихеноиндикации, а подобную
работу может сделать только квалифицированный научный работник, видимо,
имеет основания. Но какова же реальна стоимость такого исследования? В таб-
лице 3.7 приведены опубликованные ориентировочные сведения о ценах на неко-
торые виды работ, связанных с лихеноиндикацией. В таблице не показаны затра-
ты на проезд к месту проведения работ, проживание, аренду транспорта и некото-
рые другие, которые, несомненно, также должны учитываться при составлении
сметы. Следует признать, что лихеноиндикационные исследования будут никчем-
ными, если они не проведены хорошо, а хорошее исследование обычно стоит
дорого.
Стоимость лихеноиндикационных исследований в Российской Федерации не-
известна. Однажды мне предложили провести подобную работу на территории
одного историко-архитектурного заповедника площадью 10 га. Поскольку речь
зашла об оплате, я предоставил заказчикам свои расчеты (табл. 3.8). Заказчики
сочли мои запросы чрезмерными, хотя я определил минимальную цену своего
труда. Но в конченом итоге не стоимость должна определять программу иссле-
дования, а избранный наилучший метод, адекватный решаемой задаче. Более
того, в смете необходимо предусмотреть расходы на предварительное рекогнос-
цировочное обследование территории, чтобы определить число необходимых учет-
70
3. Источники загрязнения воздушного бассейна...
Таблица 3.7
Ориентировочные оценки стоимости проведения различных лихенологических исследова-
ний в США (Stolte et al., 1993)
Вид исследования Единица изучения Число учетных площадей в каждом типе растительности Объем работ Цена (USD)
Характеристика участка Территория — Участок 300
Видовой состав Пробная площадь 30 Все виды 200-400
Анализ лишайниковых группировок Пробная площадь 20-60 Все виды 300-500
Определение индикаторных видов фумигацией Территория - 1-10 видов 20000-50000
Определение индикаторных видов градиентным анализом Пробная площадь 10 Много видов 15000-25000
Трансплантация Пробная площадь 6-20 1-3 вида 1000-3000
Определение элементов в слоевищах Образец 20-100 Умножение чис- ла образцов на число определяе- мых элементов ???
ных площадей и подходящие индикаторные виды. Также, вероятно, необходимо
учитывать и квалификацию исполнителей. Так, для работников без ученой степе-
ни или если темой защиты диссертации была проблема, не связанная с изучени-
ем лишайников, коэффициент оплаты может быть 1,0; для кандидатов наук -1,5;
для докторов наук - 2,0 (естественно, по тем видам работы, которые непосред-
ственно выполнялись тем или иным исполнителем). Следует помнить и о необхо-
димости поэтапной замены устаревших приборов и оборудования, и часть воз-
можных расходов на эти цели также желательно включить в смету конкретного
исследования.
Стоимость может быть сокращена комбинированием методов сбора данных.
Так, данные о видовом составе лихенобиоты могут быть собраны на тех же пло-
щадях, где проводится анализ состава лишайниковых группировок и т.д.
3.6. ОЖИДАЕМЫЕ ТРУДНОСТИ
Результаты лихеноиндикационных исследований качества воздуха требуют
вдумчивой экспертной оценки, поскольку они не дают прямых сведений о концен-
трации загрязнителей и их составе. Кроме того, всегда необходимо помнить, что
изменчивость характеристик лишайников в пространстве и во времени не всегда
обусловлена загрязнением. Поэтому, при проведении подобной работы необходи-
3. Источники загрязнения воздушного бассейна...
71
Таблица 3.8
Предложенный расчет стоимости лихеноиндикационного исследования на территории
историко-архитектурного заповедника площадью 10 га
Вид работы Стоимость (число минимальных разме- ров оплаты труда)
Обследование территории музея-заповедника (10 га) и сбор образ- цов для выявления видового состава эпифитных лишайников и особенностей их распределения 6
Подготовка образцов для идентификации, их идентификация, этикетирование и т.д. 1 хчисло образцов
Составление списка обнаруженных видов эпифитных лишай- ников, экологический анализ видового состава в форме отчета 15
Описание лишайниковых группировок на отдельных деревьях, за- кладка на них постоянных пробных площадок, зарисовка распре- деления слоевищ на этих площадках и фиксирование их состояния 1,5 х число площадок
Повторное обследование (через каждые 6 месяцев) выбранных постоянных пробных площадок, зарисовка распределения слоевищ и фиксирование их состояния 0.5 х число площадок X число сроков
Анализ динамики состояния лишайниковых группировок на постоянных пробных площадках в течение оговоренного срока (в форме отчета) 15
Отбор здоровых слоевищ лишайника Hypogymnia physodes в «чистой» местности для трансплантации на территорию музея- заповедника (около 150 слоевищ) 8
Определение статистически достоверной средней величины доли отмерших клеток водорослевого компонента отобранных для трансплантации слоевищ лишайника 0,5 х число слоевищ (50)
Индивидуализация отобранных для трансплантации слоевищ лишайника, определение площади проективной поверхности каждого слоевища, прикрепление слоевищ к щитам, установка щитов на территории заповедника 1 х число слоевищ
Определение средней величины доли отмерших клеток водорослевого компонента у периодически снимаемых с щита трансплантированных слоевищ 0,5 х число слоевищ (10) х число сроков
Определение площади проективной поверхности и доли визуально определяемой поврежденной поверхности у периодически снимаемых со щита трансплантированных слоевищ 0,4 х число слоевищ (10) X ЧИСЛО сроков
Анализ данных наблюдений за трансплантированными слоевищами в течение оговоренного срока (в форме отчета) 15
мо обращать внимание на некоторые аспекты, игнорирование которых может при-
вести к неверной интерпретации результатов.
Так, при анализе лишайниковых группировок желательно как можно тщатель-
нее определять покрытие видов, чтобы получить статистически достоверные
результаты. Если планируется долговременный мониторинг, выбор участков на-
72
3. Источники загрязнения воздушного бассейна...
блюдений должен предусматривать возможность будущих повторных исследо-
ваний. Весьма спорной в ряде случаев бывает интерпретация изменчивости мор-
фологических показателей лишайников во времени и пространстве, поскольку не-
редко отсутствуют сведения об естественном варьировании этих признаков.
Аналогичным образом, полученные данные о концентрации загрязнителей в
тканях лишайников нелегко поддаются достоверной интерпретации, поскольку
указываемая величина может включать: (1) загрязнение тканей при сборе образ-
цов, их подготовке к анализу и самом анализе; (2) естественную фоновую концен-
трацию, обусловленную метаболизмом, занесенными ветром пылью, морскими
аэрозолями, частицами субстрата (Getty et al., 1999); (3) загрязнение антропоген-
ного происхождения.
Много проблем возникает при идентификации видов лишайников. Нередко
возникает вопрос, связано ли обнаруженное отклонение признака с изменением
экологических условий или это представитель нового таксона.
Значительная часть доступной информации о реакции видов лишайников на
загрязнение основана на данных о воздействии диоксида серы на немногие обыч-
ные виды (Kiszka, 1977; Kirschbaum, Wirth, 1995; Шапиро, 1996; Geebelen, Hoffmann,
2001; Wiseman, Wadleigh, 2002). Имеются сведения и о восприимчивости лишай-
ников к озону (Sigal, Nash, 1983). Однако представители большинство видов ни-
когда не подвергались строгой проверке в контролируемых условиях опыта с фу-
мигацией (окуриванием) для определения их реакции на различные концентрации
загрязнителей воздуха. Также мало известно о связи между генетическими раз-
личиями представителей внутривидовых таксонов и степенью их чувствительно-
сти к загрязнению.
4. ЧУВСТВИТЕЛЬНОСТЬ ЛИШАЙНИКОВ
К ЗАГРЯЗНЕНИЮ ВОЗДУХА
Использование лишайников для целей биоиндикации и биомониторинга каче-
ства воздушного бассейна основано на существовании среди представителей ли-
хенобиоты видов, различающихся степенью чувствительности к загрязнению воз-
духа. Чувствительность представителей разных видов лишайников к загрязне-
нию устанавливалась эмпирически на основе результатов градиентных
исследований в природе и разных способов фумигации (или окуривания), прове-
денных в лабораторных условиях.
Загрязнители могут оказывать на лишайники и другие организмы как острое,
так и хроническое действия. Высокие объемы атмосферных выпадений и значи-
тельные концентрации загрязнителей воздуха, достаточные для проявления та-
ких острых повреждений, как некрозы, инфекции, гибель, обычно характерны для
местностей вокруг изолированных источников загрязнителей (работающие на ис-
копаемом топливе тепловые электростанции, цементные предприятия, газо- и неф-
теочистительные заводы, сталеплавильные предприятия, заводы по производству
алюминия, химические и целлюлозно-бумажные предприятия). Исторически, наи-
большее число исследований посвящено действию двуокиси серы (SO2) и фтори-
дов (Nash, 1972,1988; Ferry et al., 1973; Gilbert, 1973; Горбач и др., 1982; Griesetal.,
1995; Falla et al., 2000; Conti, Cecchetti, 2001). Эти загрязнители имеют биологи-
ческое значение в той химической форме, в какой они были выброшены в атмос-
феру, и потому их называют первичными. Вторичные загрязнители образуются
как результат химических реакций первичных загрязнителей во время их перено-
са в атмосфере. Примерами вторичных загрязнителей являются озон (О3), пе-
роксиацетилнитрат (PAN) и кислотные дожди. Озон и PAN образуются в фото-
химических реакциях, включающих оксиды азота (NOx) и углеводороды, в боль-
шинстве своем выделяемые транспортом. Кислотный дождь образуют выбросы
как SO2, так и NO2, окисляющиеся до серной (H2SO4) и азотной (HNO3) кислот в
ходе атмосферного переноса. Видимые повреждения, такие как обесцвечивание
и некротические пятна, вызванные острыми и хроническими воздействиями озо-
на, отмечались в районе Лос-Анджелеса (Sigal, Nash, 1983), центральной Сьерра
74
4. Чувствительность лишайников к загрязнению воздуха
Неваде и на востоке США (Stolte et al., 1993). Острые повреждения растительно-
сти вследствие кислотных дождей редки, и случаи их достоверного документи-
рования немногочисленны.
Также опасны, а возможно, гораздо более токсичны, и другие типы загрязни-
телей (промышленные органические соединения, такие как диоксины и дибензо-
фураны, обнаруженные в виде примесей к некоторым гербицидам, пентахлорфе-
нолу, в продуктах сжигания хлорсодержащих полимерных материалов, а также
ряд микроэлементов и металлоидов) (Соколов и др., 1988; Walker, 2001; Walker et
al., 2001), хотя доступная информация о влиянии этих веществ довольно ограни-
чена. Полулетучие и стойкие органические соединения переносятся и осаждают-
ся в ходе атмосферных процессов. Высокие концентрации токсичных органиче-
ских соединений и пестицидов были установлены в тумане и тканях лишайников
(Glotfelty etal., 1987; Belnapetal., 1993).
В воздухе присутствую и другие загрязнители, число которых каждодневно
растет. Однако относительно многих из них информации мало или нет вовсе, а
другие не являются объектами лихеноиндикации, ибо они:
1) широко распространены, но редко вызывают повреждения лишайников, по-
скольку средние концентрации их в среде оказывают слабое воздействие (этилен,
монооксид углерода);
2) редки, и их появление обычно связано с аварийными выбросами (хлор и
производные, аммиак);
3) широко распространены (сероводород, соляная кислота), но обычно не вы-
зывают повреждений, поскольку редко достигают высоких концентраций, губи-
тельных для лишайников.
Хронические повреждения проявляются после долговременного или повтор-
ного воздействия полуострых концентраций многообразных загрязнителей возду-
ха, ассоциированных со стационарными и передвижными источниками загрязне-
ния. Вероятность гибели тканей в результате хронического воздействия невели-
ка, но возможно появление таких симптомов неблагополучия как хлороз (потеря
хлорофилла, пожелтение) и замедление роста. На уровне сообщества чувстви-
тельные виды могут от загрязнения исчезать. По результатам разного типа ис-
следований опубликованы списки видов, ранжирующие лишайники по степени
чувствительности к загрязнителям, в основном, к двуокиси серы (Инсарова, Ин-
саров, 1989; Горшков, 1990; Nash, Gries, 1991; Wirth, 1991; и другие). Некоторые из
них рассматривается ниже.
4.1. ГРАДИЕНТНЫЙ АНАЛИЗ
Метод градиентного анализа предполагает, что фиксируемые параметры ли-
шайников меняются по изучаемому градиенту показателя среды. Последний
4. Чувствительность лишайников к загрязнению воздуха
75
может варьировать как в пространстве, так и во времени. Соответственно, в пер-
вом случае наблюдения за выбранными свойствами лишайников проводят в раз-
ных точках территории, но практически одновременно; во втором - через опре-
деленные интервалы времени.
Виды лучше всего бывают представлены в оптимальных для них условиях
среды и деградируют с приближением к пределам их выносливости. Градиент-
ный анализ изначально использовали и используют и для определения реакции
лишайников на различные свойства среды, включая климатические показатели,
особенности субстрата, а так же факторы нарушенности местообитаний.
В случае изучения влияния загрязнения,исследования обычно проводят вок-
руг существующих или планируемых источников загрязнений (см. гл. 3). Ими могут
быть изолированные источники подобно тепловым электростанциям или пред-
приятиям химической индустрии, или разбросанные источники, подобно террито-
рии крупного города или промышленной агломерации как Кузбасс.
Градиентный анализ удобен для исследований влияний загрязнения воздуха
на лишайники, поскольку нагрузки загрязнителя, за некоторыми исключениями,
часто зависят от удаленности от изолированных источников. Полезен он и при
анализе региональных влияний, например, Кузбасса или Норильска, на соседние
или удаленные территории. В этом случае региональный бассейн должен быть
достаточно крупным и предполагается, что воздействия загрязнителей распро-
страняются на значительные расстояния (Отнюкова, 1997а, б).
Чаще всего фиксируемыми показателями реакции являются видимые повреж-
дения, такие как обесцвечивание или деформации слоевища, изменения в струк-
туре сообщества (богатство видов или покрытие), и физиологические процессы
(фотосинтез, активность нитрогеназы, поглощение элементов, целостность мем-
бран, свойства пигментов, их деградация и флуоресценция) (Belnap et al., 1993).
Однако исследователь должен быть готов к некоторым ограничениям при
планировании использования лишайников в градиентном анализе влияния загряз-
нения воздуха. Они включают:
- трудности, связанные с идентификацией видов, особенно поврежденных сло-
евищ;
- установление наилучших индикаторных видов для специального изучения;
- необходимость обосновать тот факт, что наблюдаемый характер распрост-
ранения или распределение фиксированных показателей отражает стрессы за-
грязнения, а не других биотических и/или абиотических факторов (Johnsen, 1976).
Последняя проблема может быть особенно неприятной, поскольку нагрузка
загрязнения на лишайники бывает второстепенной относительно более прямых
влияний микроклимата, свойств субстратов, вытаптывания или экстремальных
показателей погоды.
Градиентный анализ имеет солидное теоретическое обоснование (ter Braak,
Prentice, 1988; Belnap et al., 1993;). Измерения проводятся на профиле или трансекте,
76
4. Чувствительность лишайников к загрязнению воздуха
ориентированных перпендикулярно изолиниям показателя свойств среды, и пред-
полагается, что фиксируемые различия показателя контролируются различиями
среды между точками. Данные можно оценить несколькими способами, в част-
ности, с помощью регрессионного анализа. Прямой градиентный анализ исполь-
зуют, когда обилие лишайников, степень их видового разнообразия, встречаемость
или морфологические и физиологические показатели описываются как функция
измеренных показателей среды. Каждый встреченный образец можно ассоции-
ровать с положением в рельефе, его размерами, величиной pH субстрата, осо-
бенностями химизма участка или имеющимися данными о долговременном воз-
действии загрязнителей, например, картами качества воздуха. Измеренные по-
казатели лишайников или других криптогамных организмов размещаются вдоль
осей абсолютной высоты, pH почвы или нагрузки загрязнителей.
Сведения о видовом составе лишайниковых группировок сообщества как по-
казателе свойств среды также могут быть подвергнуты градиентному анализу.
Если параметры среды неизвестны, можно использовать информацию с постро-
енных на абстрактных осях кривых распределения видов, которые рассчитыва-
ют для множества, характеризующего варьирование состава видов. Этот прием
называют косвенным градиентным анализом. Он основан на ординации, при кото-
рой учетные единицы располагаются по отношению к одной или более коорди-
натным осям так, что их положение относительно осей и каждой другой учетной
единицы множества предполагает максимальную информацию об их экологичес-
ком сходстве. Косвенный градиентный анализ полезен чаще всего в случаях, ког-
да уже имеются представления о степени чувствительности отдельных видов
лишайников к известным загрязнителям воздуха. Метод может быть не столь
эффективным, когда есть вероятность подавления трудноуловимого действия за-
грязнения влияниями других переменных среды (Belnap et al., 1993).
4.2. ФУМИГАЦИОННЫЕ ИССЛЕДОВАНИЯ
Наличие связи “доза-эффект” достоверно устанавливается при проведении
фумигационных исследований. При этом используют некоторое разнообразие за-
крытых камер, куда помещают слоевища определенных видов лишайников и че-
рез которые пропускается воздух с известными свойствами (состав, концентра-
ция загрязнителей, влажность, температура). Иногда исследования проводят и
без таких камер, когда непосредственно в природных местообитаниях слоевища
лишайников обрабатывают растворами или газами, концентрации которых изве-
стны. Результаты этих исследований были использованы для установления про-
порций между концентрациями загрязнителей воздуха, их составом и реакциями
различных видов организмов, направленных на установление связей “доза-эф-
4. Чувствительность лишайников к загрязнению воздуха
77
фект”. Контролируемые фумигационные исследования в сочетании с градиент-
ными исследованиями дают более достоверную информацию. Для SO2 шкалы
чувствительности были установлены независимо, как по результатам контроли-
руемых лабораторных фумигаций, так и по полевым наблюдениям (Nash, 1988).
Непараметрический корреляционный анализ, значимо согласующий разные шка-
лы, строго подтверждает вывод, что лишайники действительно реагируют на
действие SO2 (Nash, 1988). Для других загрязнителей воздуха имеющихся дан-
ных пока недостаточно для установления точных шкал чувствительности. Одна-
ко измеренных в поле концентраций загрязнителей достаточно, чтобы судить о
неблагоприятном воздействии их на виды, о чем свидетельствует результаты фу-
мигационных экспериментов с такими загрязнителями воздуха как озон, плавико-
вая кислота, цинк (Nash, 1988).
Фумигационные исследования планируются для показа количественно харак-
теризуемых реакций лишайников к загрязнителям воздуха, одиночным и в комби-
нации, при контролируемых условиях в более или менее закрытых системах экс-
понирования слоевищ, как, например, непрерывно продуваемые отсеки реакто-
ров, открытые сверху камеры, отделения камер, миниатюрные кюветные камеры
и другое (Belnap et al., 1993). Такими количественно характеризуемыми реакция-
ми в большинстве случаев являются заранее выбранные физиологические про-
цессы, для измерения которых используется соответствующая аппаратура.
У лишайников такими физиологическими показателями, реагирующими на фуми-
гацию, являются (по степени чувствительности): активность нитрогеназы,
К? истечение/общее, проницаемость электролитов, фотосинтез, состояние дыха-
тельных пигментов (Fields, 1988). Однако существует много проблем в интерпре-
тации полученных результатов, ибо чувствительность зависит от таких факторов
как концентрация и продолжительность экспонирования, условия среды, состоя-
ние слоевища во время экспонирования (Fields, 1988). В дополнение к такому очень
важному фактору, как уровень насыщения слоевища водой, известно также, что
физиологические реакции разных частей слоевища неодинаковы (Karenlampi, 1970;
Moser, Nash, 1978; Nash et al., 1980; Moser et al., 1983). Мало еще сведений о дей-
ствии одних фитотоксикантов при наличии в среде других или о трансформации
токсикантов под воздействием факторов среды. Например, выбрасываемые орга-
нические соединения могут образовать 16-20 продуктов трансформации, сре-
ди которых могут быть более токсичные или опасные, чем исходное вещество
(А. Малышева, 1997).
Проведение долговременных лабораторных фумигационных экспериментов
пока затруднено в силу наличия проблем с сохранением жизнеспособности об-
разцов лишайников в ростовых камерах. Однако имеющиеся результаты кратков-
ременных фумигационных исследований (менее месяца) ценны для объяснения
механизмов реакции лишайников и установления связей “воздействие-ответ”. Тем
не менее, необходимо обращать внимание на состояние образцов и их физиологи-
78
4. Чувствительность лишайников к загрязнению воздуха
ческие параметры в течение всего исследования (Pearson, 1970; Pearson, Benson,
1977; Link, Nash, 1984).
Воздух содержит комплекс многообразных загрязнителей, но в большинстве
фумигационных исследований использовались только одиночные загрязнители при
данном наборе условий. Однако к настоящему времени известны реакции сосу-
дистых растений на смесь двух и более загрязнителей, и надо полагать, что реак-
ции лишайников на действие комплекса загрязнителей не менее сложны, чем вы-
явленные у сосудистых растений.
4.3. ИСКУССТВЕННЫЕ КИСЛОТНЫЕ ДОЖДИ
Загрязненные дождевые осадки воссоздавались разнообразными методами
в поле и лаборатории с использованием простых бидонов для воды, ранцевых
распылителей, и таких сложных систем как ирригационные дождевальные уста-
новки, где интенсивность осадков и состав загрязнителей менялись с помощью
компьютерных программ (Stolte et al., 1993). Эти устройства также использова-
лись в комбинации с фумигационными камерами для изучения сочетанного влия-
ния кислотного дождя и газообразных загрязнителей. Сложные системы для си-
муляции дождей могут воссоздавать физические и химические показатели есте-
ственных осадков, включая:
- скорость выпадения дождя (интенсивность можно регулировать в интерва-
ле 0,5-2,7 см3/час);
- ранг размера капель (0,1-3,2 мм);
- химический состав через регулируемые насосы, обеспечивающие снабже-
ние водой с точным составом ионов, включая струи деионизированной воды (Belnap
et al., 1993; Piervittori et al., 1997).
4.4. ПОКАЗАТЕЛИ, ФИКСИРУЕМЫЕ ДЛЯ ОПРЕДЕЛЕНИЯ
ЧУВСТВИТЕЛЬНОСТИ ЛИШАЙНИКОВ К ЗАГРЯЗНЕНИЮ
Степень чувствительности лишайников к загрязнению воздуха определялась
на основе изучения многих признаков этих организмов. Их выбор зависел от типа
и уровня имеющихся загрязнителей, использованных видов лишайников, особен-
ностей исследования (например, градиентное как альтернатива фумигационному)
и возможностей исследователя.
4.4.1. Характеристики популяции
Многие параметры видов можно наблюдать и фиксировать непосредственно
в природе, причем часто для этого необходимы достаточно простые приборы или
4. Чувствительность лишайников к загрязнению воздуха
19
они вовсе не требуются. Так, наблюдения за свойствами популяций обычно про-
водятся с использованием учетных площадок. На них регистрируются такие по-
казатели как присутствие/отсутствие представителей вида, особенности репро-
дуктивности, скорость роста, смертность, покрытие, биомасса и другое. Эти из-
мерения можно проводить визуально (Karenlampi, 1970, 1971; Armstrong, 1991)
или по фотоснимкам. Фотографии, сделанные традиционными камерами, цифро-
выми или видеокамерами, можно анализировать с помощью компьютерных про-
грамм.
4.4.2. Морфология слоевищ
Заметные морфологические изменения слоевищ нельзя вызвать в лаборатор-
ных условиях при обработке лишайников загрязнителями, поскольку для этого
недостаточно времени. Правда, есть данные, что под воздействием искусствен-
ного кислотного дождя происходила редукция поверхностного аморфного слоя
талломов Pseudoevernia furfuracea (Piervittori et al., 1997). В ходе многочислен-
ных полевых исследований на загрязненных территориях наблюдались различ-
ные морфологические модификации слоевищ, объясняемые действием загрязни-
телей на них (Малышева, 1995, 1996г; Mahaney et al., 1995; Отнюкова, 1997а, б).
Обычная реакция слоевищ - изменение цвета (розоватый или коричневатый) до
полного обесцвечивания (отмирание водорослевых клеток), а также наличие в
популяции более мелких, компактных талломов. Некоторые морфологические
изменения являются видоспецифичными. Например, Л. Сигал и Т. Нэш (Sigal,Nash,
1983) установили, что извилистость лопастей Hypogymnia enteromorpha замет-
но варьирует в горах южной Калифорнии в зависимости от действия окислителей.
У видов Cladonia необычное увеличение высоты и характера ветвления было
вызвано действием озона и кислотных осадков (Scott, Hutchinson, 1990). У видов
Peltigera, растущих на почвах с высоким содержанием металлов, Р. Гойял и
М. Сивард (Goyal, Seaward, 1982а) обнаружили редукцию длины ризин, увеличе-
ние их плотности и ветвления жилок на нижней поверхности и гипертрофию серд-
цевины. Талломы Hypogymnia physodes в городах имели более толстый водо-
рослевой слой и более тонкий слой коры (Holopainen, 1984b). Вокруг мест замеса
бетона в Антарктиде, где загрязнение щелочное, у слоевищ Usnea sphacelata
наблюдали морщинистость и трещиноватые углубления в коровом слое (Adamson,
Seppelt, 1990).
Осмотр слоевища невооруженным глазом или с помощью простой лупы по-
зволяет оценить внешний вид избранных видов и выявить такие свойства лишай-
ников как цвет, размер, особенности репродукции. Преимущество такого способа
в том, что он не требует сложного оборудования и больших средств. Такого типа
наблюдения можно проводить как с фото-, так и с видеокамерами. Выводы дела-
80
4. Чувствительность лишайников к загрязнению воздуха
ются на основе сравнения фиксированных признаков 1) до и после обработки заг-
рязнителем, 2) через определенные периоды времени на тех же местах, 3) и/или
по градиенту расстояния от известного источника загрязнения.
Репродукционная способность является очень чувствительным механизмом,
снижение которой может привести к исчезновению представителей вида с терри-
тории. Изменение репродуктивного потенциала вследствие загрязнения обнару-
живается ранее других визуально отмечаемых повреждений слоевища. Посколь-
ку лишайники сформированы, по меньшей мере, двумя организмами, исследова-
ния их репродуктивности сложны, и его детали пока известны не полностью.
Половая репродукция включает грибные и водорослевые споры и их воссоедине-
ние. Полевые исследования показывают, что образование слоевищами апотеци-
ев значительно снижается в более загрязненных районах (Sigal, Nash, 1983). Здесь
также уменьшается скорость деления клеток фотобионта и неполовая репродук-
ция, хотя по другим данным, SO2 стимулирует образование соредий и изидий,
обычных органов вегетативного размножения всего лишайника (Gries, 1996). На-
блюдения на Среднем Урале в зоне действия сталелитейного предприятия пока-
зали, что в более загрязненных участках снижается уровень выживаемости со-
редий и скорость развития из них слоевищ Hypogymnia physodes (Mikhailova,
Scheidegger, 2001).
Лишайники в целом растут намного медленнее сосудистых растений (Инса-
рова, Инсаров, 1987), поэтому рост как показатель их реакции на загрязнение воз-
духа используется в полевых исследованиях нечасто и практически не применя-
ется в контролируемых лабораторных условиях с обработкой лишайников задан-
ными загрязнителями и их концентрациями. Месячный прирост, выражаемый как
увеличение сухого веса или размера слоевища, можно определить только для
быстро растущих лишайников. Фотографирование и точные способы измерения
площади слоевищ совершенствуют такие измерения и позволяют не разрушать
талломы в ходе таких процедур. Установленные подобным способом скорость
роста и длительность жизни лишайников были заметно снижены в загрязненных
районах (Feige, 1982). Также сравнивались размеры существующих слоевищ од-
ного вида в загрязненных и незагрязненных районах (Feige, 1982). При этом выяв-
лена редукция размеров слоевищ на загрязненных территориях. Этот метод можно
использовать в городах, где трудно отделяемые от субстрата накипные лишайни-
ки встречаются чаще листоватых. Наконец, проводилось микроскопическое изу-
чение ювенильного развития лишайников для выявления действия загрязнения на
первые стадии роста. Полевые и лабораторные опыты с представителями двух
видов показали, что прикрепление и развитие диаспор в первые пять месяцев во
многом происходило независимо от загрязнения (Schuster, 1985). В последующем
образование желатинозного материала вокруг фотобионта снизилось, что сильно
уменьшило запасы влаги. Это привело к иссушающему повреждению во всех
микроместообитаниях, исключая лишь наиболее влажные.
4. Чувствительность лишайников к загрязнению воздуха
81
Изучение морфологии слоевищ направлено на установление изменений фор-
мы или структуры организма из-за загрязнения воздуха. Однако не всегда по-
вреждения слоевищ, обусловленные загрязнением, обнаруживаются визуально,
поэтому желательно визуальное обследование талломов сопровождать изучени-
ем объекта с помощью микроскопа. При анализе повреждений лишайников ис-
пользуют два типа микроскопов: световые микроскопы и трансмиссионные и ска-
нирующие электронные микроскопы. Так, с помощью светового микроскопа была
проведена оценка процента плазмолизированных водорослевых клеток свежих
срезов лишайников, экспонированных в SO2(Eversman, 1978; Will-Wolf, 1980а). Было
установлено, что процент плазмолиза достоверно возрастает с увеличением вре-
мени экспозиции. Для оценки здоровья водорослей в свежих срезах фиксируют
цвет и форму хлоропластов, которые содержат большинство водорослевых кле-
ток. Здоровые клетки зеленые и округлые, плазмолизированные клетки желтова-
тые и сморщенные. Пригодна для определения состояния водорослевых клеток и
флуоресцентная микроскопия (Kauppi, 1980; Вайнштейн, 1996).
Другой метод определения мертвых и плазмолизированных водорослевых
клеток основан на использовании фиксированных и залитых препаратов лишайни-
ков, окрашенных голубым толуидином (Holopainen, Kauppi, 1989). Пустые клетки
считались как мертвые; клетки, содержимое которых сжалось, рассматривались
как плазмолизированные. Но и в этом случае наблюдался рост числа плазмоли-
зированных клеток с увеличением действия SO2.
Некоторые структуры слишком малы для точного наблюдения в световом
микроскопе. В таких случаях полезен трансмиссионный электронный микроскоп,
особенно для изучения воздействия на клетки на уровне физиологических про-
цессов - изменений некоторых параметров хлоропластов и митохондрий в проме-
жуточных стадиях повреждения, признаков, предшествующих коллапсу всей кле-
точной структуры и т.п. (Eversman, Sigal, 1984, 1987; Holopainen, Kauppi, 1989).
С его помощью регистрировались такие специфические изменения как деформа-
ция тилакоидов и пиреноглобул хлоропластов, набухание, вызываемое дегенера-
цией митохондрий, необычное накопление крахмальных зерен и жировых тел, ис-
чезновение ряда типов органелл и мембранных структур, обычно опознаваемых
в здоровых клетках. Изменения на уровне ультраструктуры лишайников были об-
наружены как в образцах, собранных на загрязненных территориях, так и после
обработки SO.,, О3, ПАН (пероксилацетилнитрат) в контролируемых лаборатор-
ных экспериментах (Holopainen, 1981,1982; Eversman, Sigal, 1987; Tarhanen et al.,
1997, 2000; Tarhanen, 1998). Разработана классификация стадий ультраструктур-
ных повреждений по полевым сборам (Holopainen, 1984b). У фотобионта они про-
являются в изменении формы хлоропластов, вздутии митохондрий, появлении элек-
трон-непроницаемых вакуолярных накоплений, увеличении плотности цитоплаз-
мы, снижении размеров цитоплазматических запасных капель и уменьшении
размеров пиреноглобулий. У микобионта вакуоли увеличиваются в размере, и за-
82
4. Чувствительность лишайников к загрязнению воздуха
метны электрон-непроницаемые вакуолярные тела при увеличении загрязнения.
В целом, грибы лишайника, вероятно, более толерантны к загрязнению, чем фо-
тобионт (Holopainen, Karenlampi, 1984). Развитие электрон-непроницаемых ваку-
олярных тел, как в грибных, так и в водорослевых клетках ассоциируется скорее
с наличием азотных загрязнений, чем с SO2. Растягивание окружающего хлороп-
ласт конверта, скручивание тилакодиев, накопление крахмала и раздувание мито-
хондрий отмечается при более высоких концентрациях SO2. Расширение тилако-
идного межпространства, раздувание тилакоидов и появление маленьких плас-
тоглобулий между тилакоидами на ранних стадиях, последующие грануляция и
разрушение тилакоидных мембран являются признаками загрязнения фторида-
ми. На конечных стадиях были различимы неклеточные органеллы; остаются
только крахмальные зерна и цитоплазматические запасы капель. Это подтверж-
дает тот факт, что имеет значение механизм перемещения продуктов фотосинте-
за к грибному компоненту. Поэтому редукция размеров слоевища может быть
объяснена голоданием микобионта (Hutchinson et al., 1987). У цианобактерий Nostoc
из Peltigera canina обработка очень высокой концентрацией SO2 снижала коли-
чество карбоксизомы, увеличивала число и размеры гранул цианофицина и ини-
циировала разделение акинет (Gries, 1996). Разделение акинет обычно не свой-
ственно представителям Nostoc. живущих в симбиозе (Dodds et al., 1995).
Изменения в клетках, вызванные загрязнением, могут быть похожими на обыч-
ные сезонные изменения или нормальное старение, поэтому важно иметь для срав-
нения свежий материал, содержащийся в контролируемых условиях, аналогич-
ных тем, при которых проводился сбор образцов перед фиксацией для микроско-
пии (влажность и световые условия контрольных и обработанных образцов должны
быть идентичными). К сожалению, видимо, все загрязнители вызывают сходные
изменения ультраструктуры водорослевых клеток, и нельзя по наблюдаемым из-
менениям структуры определить конкретный действующий загрязнитель. Воз-
можно, однако, что загрязнители, образовавшиеся в результате окисления как озон
и PAN, вызывают повреждения при меньших концентрациях и за более короткое
время, чем SO2 и кислые загрязнители (Eversman, Sigal, 1987).
4.4.3. Физиологические процессы
Изучение физиологических процессов в лишайниках для определения их чув-
ствительности к загрязнению имеет целью выявить отклонения от нормального
функционирования организма или некоторых его частей под влиянием загрязне-
ния воздуха (Плакунова и др., 1984; Шапиро, 1996; Kong et al., 1999; Haffner et al.,
2001). Существует много способов определения изменений показателей физиоло-
гических процессов, отражающих действие загрязнения воздуха, но все они тре-
буют некоторого уровня оснащения приборами. Кроме того, физиологические про-
4. Чувствительность лишайников к загрязнению воздуха
83
цессы варьируют в зависимости от сезона или географического положения, что
необходимо принимать во внимание при сборе данных и интерпретации результа-
тов. Также известно, что разные части одного слоевища отличаются по ряду
физиологических параметров (фотосинтез, нитрогеназная активность, концентра-
ции хлорофилла). Это необходимо учитывать, если экспериментальный материал
включает не целое слоевище (Karenlampi, 1970; Moser, Nash, 1978; Nash et al.,
1980; Moser et al., 1983).
4.4.3.1. Пигменты
Чаще всего для оценки повреждения слоевищ лишайников загрязнением сре-
ди всех пигментов используется хлорофилл и продукты его распада. Исследова-
ния показали, что уровни хлорофилла в значительной мере подвержены воздей-
ствию загрязнителей. Сообщалось о влияниях на хлорофилл как концентрации
загрязнителей, так и времени их воздействия, в результате чего менялось содер-
жание общего хлорофилла, отношения хлорофиллов а:Ь или отношение хлорофил-
ла к продуктам его распада, снижался процент концентраций хлорофилла (LeBlanc,
Rao, 1973а; Nash, 1973,1976; Moser et al., 1980; Beekley, Hoffman, 1981; Henriksson,
Pearson, 1981; Горбач и др., 1982; Ronen, Galun, 1984;Garty etal., 1985a; Kardishet
al., 1987; Belnap, Harper, 1990; Garty, 2000b).
Одним из видимых сигналов повреждения лишайников загрязнителями явля-
ется обесцвечивание или изменение цвета слоевища, которое вызывается разру-
шением молекул хлорофилла. Для сосудистых растений количественные соотно-
шения между загрязнителями воздуха и специфическими проявления их действия
в виде некрозов (коричневатые пятна мертвых клеток) и/или хлороза (обесцвечи-
вание клеток) известны, но для лишайников специфическая симптомология заг-
рязнителей воздуха в целом не признается. Хотя на лишайниках постоянно обна-
руживаются общие хлорозы и некрозы как реакция к SO2, HF, окислителям и Си
(Mahaney et al., 1995), но специфический характер реакции на действие какого-
либо конкретного загрязнителя еще не установлен.
Начальное разрушение хлорофиллов а и b и соответствующие им феофитины,
не обнаруживаемые визуально, можно определить с помощью экстракции хлоро-
филла и спектрофотометрических измерений. Поскольку в экспериментах прояв-
ления деградации хлорофилла наблюдали лишь при обработке слоевищ высокими
концентрациями SOo, HF, NO2 этот показатель считается менее чувствительной
ответной реакцией на загрязнение, чем фотосинтез (Fields, 1988). Более того, при
низком уровне загрязнения лишайники в полевых условиях могут даже показы-
вать увеличение содержания хлорофилла с увеличением стресса загрязнения (Arb
et al., 1990). Но при более высоких уровнях загрязнения территорий наблюдается
значительное повреждение хлорофилла. Опыты с трансплантацией талломов так-
84
4. Чувствительность лишайников к загрязнению воздуха
же выявили позитивную корреляцию степени деградации хлорофилла с уровнем
загрязнения (Garty et al., 1993а; Garty, 2000b).
Для обнаружения начальных проявлений деградации пигментов М. Каппи
(Kauppi, 1980) использовал метод микроскопического определения флуоресцен-
ции. При голубом и ультрафиолетовом возбужденном свете фотобионт обычно
показывал первичную красную флуоресценцию. Однако после обработки слоеви-
ща SO2 эта флуоресценция переходила сначала к коричневой или оранжевой и,
наконец, к белой. В сравнение с экстракцией пигментов этот способ имеет пре-
имущества в скорости, возможности работы с более мелкими образцами лишай-
ников и в отсутствии необходимости их разрушать (Вайнштейн, 1996).
4.4.3.2. Нитрогеназная активность
Около 10% всех видов лишайников содержат цианобактерии, многие из кото-
рых могут фиксировать атмосферный азот и превращать его в форму, пригодную
для использования (Шапиро, 1986). Эти лишайники могут быть важными вклад-
чиками азота в экосистемы, в которых они встречаются (Denison, 1973; Nash,
1996). Эта способность лишайников чрезвычайно зависима от pH при оптимуме
рН=7 для изолированной Nostoc. В симбиозе этот оптимум может быть снижен к
более кислой рН=5. Дальнейшее окисление из-за загрязнения SO, или NO2 может
сдерживать фиксацию N2. Хорошо известное исчезновение с большинства терри-
торий Европы лишайникового сообщества Lobarion (Wirth, 1988), выделявшегося
наличием в его составе представителей многих крупных листоватых азотфикси-
рующих лишайников, представляет наглядное полевое свидетельство большого
влияния загрязнения воздуха на фиксацию азота. Экспериментальные исследо-
вания подтвердили, что рН= 4 дождевой воды является порогом значимых влия-
ний на фиксацию азота большинством видов (Denison et al., 1977). Некоторые
работы с SO, показали более высокую чувствительность фиксации азота, чем
фотосинтеза (Fields, 1988), и, возможно, фториды даже больше сдерживают фик-
сацию азота, чем SO2. По другим данным, низкие концентрации некоторых ме-
таллов стимулируют нитрогеназную активность, но при более высоких концент-
рациях атмосферных загрязнителей наблюдается ее снижение (Denison et al., 1977;
Hallgren, Nyman, 1977; Henriksson, da Silva, 1978; Sheridan, 1979; Henriksson, Pearson,
1981; Hallbom, Bergman, 1983; Sigal, Johnston, 1986; Belnap, 1990; Шапиро, Нифон-
това, 1991; Шапиро, 1993).
Измерение размеров фиксации азота является трудной и дорогостоящей про-
цедурой. Однако измерение нитрогеназной активности проще и дешевле. В при-
сутствии нитрогеназных ферментов ацетилен превращается в этилен. Соответ-
ственно, уровень нитрогеназной активности отражает количество произведенно-
го этилена. Уровни этилена и ацетилена можно измерить на газовом хроматографе.
4. Чувствительность лишайников к загрязнению воздуха
85
4.4.3.3. Устойчивость мембран
Исследования показали, что диоксид серы, другие загрязнители отрицатель-
но действуют на устойчивость мембран (Pearson, Rogers, 1982; Moser et al., 1983;
Fields, St. Clair, 1984b; Pearson, 1985; Hart et al., 1988; Belnap, Harper 1990; Tarhanen
et al., 1996, 1999). Этот вывод сделан по результатам измерений истечения калия,
общей электролитической проницаемости, а также детального изучения структу-
ры мембран с применением сканирующего электронного микроскопа. Проницае-
мость мембран оценивается при измерении проводимости деионизированной воды
до и после погружения в нее лишайника. Для этого необходим измеритель прово-
димости, который недорог и портативен (Alebic-Juretic, Arko-Pijevac, 1989; Belnap
et al., 1993).
Измерения как проводимости, так и содержания ионов калия в насыщенном
растворе соответствующими электродами является простейшим способом мо-
ниторинга целостности мембран. Такие методы широко применяют в полевом
мониторинге, трансплантантных исследованиях и лабораторных экспериментах
как с обработками слоевищ растворами тяжелых металлов, так и/или SO2. Тип
реакции лишайника варьирует относительно загрязнителя и его концентрации.
Например, элементы Hg, Ag, Си вызывают постепенный ответ с наивысшей по-
терей К+ при обработке ртутью, но иной характер ответа наблюдался после обра-
ботки талломов растворами с Ni, Со, Cd, Pb (Puckett, 1976). При низких началь-
ных концентрациях потери К+ слегка возрастают с увеличением концентрации
металла до достижения критического порога, выше которого дальнейшее увели-
чение концентрации металла приводят к резкому повышению потери К+. Началь-
ная потеря К+ объяснялась перемещением К+ от мест изменения иона, тогда как
последующее резкое увеличение потери иона калия интерпретировалось наруше-
нием целостности мембран. Двуокись серы вызывает третий тип потери К+, оче-
видно, связанный с двумя системами поглощения SO9 . Наблюдения непосред-
ственно в поле позволили установить тесную связь между степенью поврежде-
ния мембран и средней концентрацией SO2 (Garty et al., 1993a).
4.4.3.4. Дыхание, фотосинтез
Процессы фотосинтеза и дыхания регулируются мембранами и потому силь-
но зависят от целостности последних. Энергия для каждого процесса образуется
за счет градиента pH плазмы двух сторон мембраны. Загрязнители воздуха мо-
гут воздействовать на целостность мембраны и pH плазмы. Фотосинтез измеря-
ется главным образом как поглощение СО2 или фиксацией ,4С; дыхание - как
высвобождение СО2. Эти процессы наблюдаются только при определенном во-
донасыщении таллома лишайника. Сухие слоевища практически латентны и спо-
86
4. Чувствительность лишайников к загрязнению воздуха
собны без вреда выдержать действие крайних температур. Фотосинтез, есте-
ственно, свойство фотобионта, тогда как дыхание - преимущественно грибного
компонента, поскольку доля грибной массы в лишайнике гораздо выше. В общем,
фотосинтез более чувствителен к загрязнению в сравнении с дыханием (Fields,
1988); это означает, что фотобионты более чувствительны, чем грибы.
Представители разных видов лишайников обрабатывались SO2, озоном, пла-
виковой кислотой, PAN, опрыскивались кислотным дождем в лаборатории и поле,
погружались в растворы металлов и сульфита (Nash, Gries, 1991). Фотосинтез и
дыхание, измеренные после таких обработок, выявили неодинаковую чувст-
вительность разных видов. У некоторых видов наблюдалась даже стимуляция
этих процессов при низких концентрациях металлов и SO2. Однако при изучении
трансплантатов (Christ, Turk, 1982; Werner, 1993) и при полевых исследованиях
(Arb et al., 1990) часто трудно найти простую корреляцию между средней концен-
трацией загрязнителя и обмена газа СО2 Видимо, при низких концентрациях заг-
рязнителя более высокая скорость фотосинтеза обусловлена увеличением кон-
центрации хлорофилла в поле в сравнении с условиями лаборатории. Но при более
высоких концентрациях загрязнителя интенсивность фотосинтеза обычно резко
снижается.
Особенно много исследований касалось влияния на фотосинтез и дыхание
лишайников двуокиси серы (Вайнштейн, 1972,1973). Большинство лишайников-
сухопутные организмы и потому в опытах обычно обрабатываются газообраз-
ным SO2. Если измерять газообмен СО2 при обработке SO2 высокой концентра-
ции, уже через несколько минут наблюдается значительное снижение скорости
фотосинтеза (Sanz et al., 1992). Соответственно, в природе, вероятно, для выжи-
вания лишайника более важны пиковые концентрации SO2 в сравнении с долговре-
менными средними. Реакция также зависит от длительности экспонирования.
У некоторых видов интенсивность фотосинтеза снижается постепенно, что сви-
детельствует о наличии связи “доза-ответ” (Sanz et al., 1992). Но у представите-
лей других видов наблюдается стремительное падение скорости фотосинтеза при
определенной концентрации SO2 что показывает о существовании отношений “по-
рог-ответ” (Huebert et al., 1985). Кислотность (pH) воды в слоевище также силь-
но влияет на повреждение газом SO2 (Turk, Wirth, 1975; Wirth, 1987). Если SO2
проникает во влажное слоевище, он вступает в реакцию с водой, образуя HoSO3,
HSO3”, SO3~, конкретные формы которых зависят от pH. При низких значениях
рН(3-4) образуется главным образом HSO3", и она выступает как сильный окис-
литель хлорофилла и реагирует с хлорофиллом а, который более чувствителен,
чем хлорофилл Ь. В случае несильных повреждений, как в лабораторных, так и в
полевых условиях наблюдалось некоторое восстановление фотосинтеза после пре-
кращения воздействия SO2 (Gries, 1996)
Влияют на фотосинтез также и некоторые металлы. Низкие концентрации меди,
серебра и ртути всегда вызывают редукцию в фиксации 14С, тогда как Sr, Mg, Са,
4. Чувствительность лишайников к загрязнению воздуха
87
Ni, Zn практически не влияют на это. Использование в экспериментах высоких
концентраций металлов выявило наличие различной чувствительности среди ви-
дов и относительную токсичность металлов для фотосинтеза как Ag, Hg > Со >
Си, Cd > Pb, Ni для кратковременных экспозиций и Ag, Hg, Си > РЬ, Со > Ni для
длительных экспозиций (Gries, 1996). Некоторые виды лишайников оказались спо-
собными выработать толерантность к 'отдельным металлам. Это можно наблю-
дать в природе среди представителей видов, растущих на субстратах с разной
концентрацией металлов, и это нашло подтверждение в экспериментах (Beckett,
Brown, 1983).
Значительное снижение скорости дыхания лишайников, экспонированных при
повышенных уровнях загрязнения, было установлено во многих исследованиях
(Baddeley etal., 1971; Eversman, 1978, 1979; Fields, St.Clair, 1984a, b). В большин-
стве случаев скорость дыхания оценивалась как поглощение О2 кислородными
электродами или измерялось инфракрасным газоанализатором поведение СО2 в
темноте в экспозиционных камерах с лишайниками с заданными уровнями кон-
центрации загрязнителей, обычно SO2.
Все методы, которыми измеряли фотосинтез лишайников, установили сниже-
ние скоростей фотосинтеза от воздействия загрязнителей. Скорость фотосинтеза
также определялась с использованием камеры и газа ,4СО2 (Hallgren, Nyman,
1977; Lechowicz, 1982; Ross,Nash, 1983; Fields, St. Clair, 1984a; Sigal, Johnston, 1986),
погружения в жидкий 14CO2 (Fields, St. Clair, 1984a), измерения скорости поглоще-
ния CO2 или поведения On (Beekley, Hoffman, 1981). Определение чистого газооб-
мена как в продуваемой, так и в закрытой камере связано с наличием газоанали-
затора, тогда как оценка величины фотосинтеза с помощью меченого 14СО2 тре-
бует счета жидкостных сцинтилляций.
Флуоресценция хлорофилла является показателем состояния фотосинтетиче-
ских реакций, происходящих в хлоропластах фотобионтов, и представляет набор
переменных реакции, ассоциированных со световыми реакциями фотосинтеза, в
особенности с фотосистемой II. Это удобный, чувствительный, быстрый и не
повреждающий слоевище способ оценки продуктивности фотосинтеза. Появив-
шиеся новые измерительные системы позволяют использовать флуоресценцию
при мониторинге в полевых условиях. Кроме того, широкие исследования выс-
ших растений предоставили комплекс необходимой информации о флуоресценции
для интерпретации получаемых аналогичных данных при изучении лишайников
(Gries, 1996). Варьирование параметров среды, таких как интенсивность света,
температура, состав газов и другое, могут вызывать специфические изменения в
характере индукции. Для оценки стресса важно поэтому измерять флуоресцен-
цию здорового слоевища и обработанного загрязнителями таллома в одинаковых
условиях. Чаще всего анализируется флуоресцентное поведение фотосинтетиче-
ских систем, которое представляет собой кинетику повышения флуоресценции до
максимальных уровней при насыщенном освещении. Получаемая индуктивная
88
4. Чувствительность лишайников к загрязнению воздуха
(логическая) кривая маркирует изменения после воздействия загрязнителями
воздуха. Флуоресценцию измеряют, используя хлорофильный флуорометр или
модифицированную систему измерения газообмена (Вайнштейн, 1996; Шапи-
ро, 1996).
4.4.3.5. Другие метаболические параметры
Ответная реакция ряда других метаболических процессов в лишайниках изу-
чалась главным образом к воздействию SO2. Двуокись серы вызывала преиму-
щественно снижение активности ключевых ферментов или разрушение молекул.
Например, активность фосфатазы, фермента, который локализуется внеклеточ-
но, что обусловливает его восприимчивость к изменениям непосредственно в
лишайниковой среде, изучалась относительно действия нескольких загрязните-
лей. Редукции активности были выявлены как в полевых исследованиях, так и
при последующих обработках в лаборатории газами и растворами различных не-
органических ионов (Puckett, 1988).
Энергетическое состояние клеток, характеризуемое отношением концентра-
ций АТФ (аденозинтрифосфат), АДФ (аденозиндифосфат) и АМФ (аденозинмо-
нофосфат), также зависит и от загрязнения воздуха (Kock, Schlee, 1981). Более
того, как в лабораторных экспериментах с бисульфитом, так и в трансплантант-
ных исследованиях в поле, концентрация АТФ обратно коррелировала с концент-
рацией загрязнителя (Galun, Ronen, 1988). Наблюдениями в поле и эксперимен-
тально установлено снижение биосинтеза белков и липидов при загрязнении воз-
духа (Malhotra, Khan, 1983; Arb et al., 1990). Липиды являются существенной
составляющей мембран, поэтому сдерживание их синтеза приводит к ослабле-
нию целостности мембран.
Снижение концентрации аминокислоты пролин установлено у водоросли
Trebouxia под воздействием SO2. Индикаторами загрязнения могут быть и такие
показатели как редукция спиртов в слоевище (Roser et al., 1992), продукция этиле-
на (Garty et al., 1993b) и другие метаболические параметры (Шапиро, 1996).
4.5. РАНЖИРОВАНИЕ ВИДОВ ЛИШАЙНИКОВ ПО СТЕПЕНИ
ЧУВСТВИТЕЛЬНОСТИ К ЗАГРЯЗНЕНИЮ ВОЗДУХА
Информация, собранная в ходе градиентных, в том числе и с использованием
трансплантатов (Ferry, Coppins, 1979), фумигационных и общеэкологических ис-
следований, применялась для ранжирования видов лишайников по степени чув-
ствительности их к загрязнению воздуха. Принято считать, что наиболее устой-
чивыми к загрязнению воздуха, главным образом, двуокисью серы, являются
4. Чувствительность лишайников к загрязнению воздуха
89
накипные лишайники, далее следуют листоватые формы и самые чувствитель-
ные - представители кустистых видов. Однако проведенные исследования не
выявили значимых корреляций между жизненной формой вида и реакцией его пред-
ставителей на загрязнение (Wirth, Turk, 1975). Эту реакцию лишайников обозна-
чали разными терминами. Поскольку изначально изучалось действие на лишай-
ники загрязнения городов, или “городской эффект” (Brodo, 1972), то в этих терми-
нах использовали греческое обозначение города “полис”, и способность некоторых
видов лишайников продолжать жить в городской атмосфере была названа “поле-
отолерантностью” (Barkman, 1958; Трасс, 1968), тогда как чувствительность ли-
шайников назвали “полеофобностью” (Barkman, 1958; Галич, 1972). Более широ-
ким обозначением степени чувствительности является термин “токсифобность”
(Инсарова, Инсаров, 1989). Процедура ранжирования лишайников по степени чув-
ствительности к загрязнению заключается в распределении выявленного на тер-
ритории (или изученного экспериментально) множества видов лишайников на то
или иное число классов, различающихся реакциями на загрязнение. Число клас-
сов у разных авторов варьирует от 3 до более 10, обозначают эти классы либо
баллами (1,2,3...), либо качественно (“очень устойчивые виды”, “очень чувстви-
тельные виды”). Балльная оценка “1” у одних авторов может относиться к наи-
более устойчивым видам (Barkman, 1958), ay других - к самым чувствительным
(Wirth, 1991). Иногда виды лишайников располагают в последовательности, пока-
зывающей, что некий вид более/менее чувствителен, чем соседние. Немногие
авторы связывают выделенные группы лишайников с количественными показа-
телями концентраций загрязнителей, в частности, двуокиси серы в атмосфере
(Hawksworth, Rose, 1970), причем в публикациях последних лет утверждает-
ся, что в цитированной работе уровни загрязнения, которые способны перено-
сить разные виды лишайников, сильно завышены (van Dobben, ter Braak, 1999).
И.Д. Инсарова и Г.Э. Инсаров (1989) обобщили доступные им публикации о чув-
ствительности эпифитных лишайников (243 вида) к загрязнению воздуха в регио-
нах Северного полушария и попытались дать сопоставимые оценки относитель-
ной токсифобности видов путем экстраполяции разнообразных шкал в 10-баль-
ную шкалу, в которой балл 10, как и у Barkman (1958), относится к максимально
чувствительным видам. Анализ этого полезного обобщения показывает, что у
ряда исследователей большое значение при оценке токсифобности имеет субъек-
тивный фактор. Как правило, чем чаще вид представлен в публикациях, тем боль-
шее разнообразие оценок его чувствительности. Например, токсифобность
Hypogymnia physodes, самого обычного вида Голарктики, у разных авторов ва-
рьирует от 0,6 до 7,1 (в интерпретации И.Д. Инсаровой и Г.Э. Инсарова); Lecanora
hagenii - от 0,8 до 7,1; Parmelia sulcata - от 1,1 до 8,9. Это подтверждают и
публикации последних лет (Geebelen, Hoffmann, 2001). Реакция на загрязнение
представителей вида в пределах ареала, видимо, зависит от климатических усло-
вий конкретного региона и даже местообитания, особенно от режима и формы
90
4. Чувствительность лишайников к загрязнению воздуха
водоснабжения (Hauck et al., 2000). Поскольку лишайники бывают физиологиче-
ски активны только при определенном минимуме водонасыщения слоевищ, в об-
ластях с влажным климатом талломы активны больший период, чем в аридных
районах. Соответственно, в первом случае при одинаковом уровне загрязнения
симптомы повреждения проявляются быстрее. Эти особенности географических
различий учитывал Х.Х. Трасс (1985а), предложивший разные варианты ранжи-
рования видов лишайников для Прибалтики, Кавказа, Дальнего Востока. Сто во-
семьдесят восемь видов лишайников из Южной Германии были ранжированы на
9 классов токситолерантности (Wirth, 1991), где обозначение “1” имели наиболее
чувствительные к загрязнению виды, а “9” - самые устойчивые, т.е. обратно
шкале в обзоре И.Д. Инсаровой и Г.Э. Инсарова (1989) на русском языке. По-
скольку эти данные не вошли в названный обзор, вероятно, полезно показать их
здесь.
Класс 1 образуют следующие виды (номенклатура по оригиналу): Alectoria
sarmentosa, Bryoria bicolor, Caloplaca herbidella, Collema nigrescens, Cybebe
gracilenta, Evernia divaricata, Gyalecta ulmi, Lecanactis amylacea, Lepraria
lobificans, Leptogium saturninum, Lobaria amplissima, L. pulmonaria,
L. scrobiculata, Loxospora cismonica, Nephroma bellum, N.laevigatum, N. parile,
N, resupinatum, Ochrolechia pallescens, Pachiphyale fagicola, Pannaria conoplea,
Parmelia exasperata, P sinuosa, Parmeliella plumbea, P triptophylla, Peltigera
collina,Rinodina sophodes, Sphaerophorus globosus, Stenocybe major, Sticta
sylvatica. Класс 2 - Anaptychia ciliaris, Arthonia cinnabarina, Bryoria capillaries,
Calicium adpressum, Caloplaca cerina, Cetraria sepincola, Cetrelia cetrarioides,
Chaenotheca brunneola, Collema flaccidum, Enterographa hutchinsiae, Fuscidea
cyathoides, Lecanora impudens, L. sambuci, Loxospora elarina, Menegazzia
terebrata, Parmelia coniocraea, Phlyctis agelaea, Ramalina fastigiata, R. fraxinea,
Rinodina pyrina, Schismatomma pericleum, Sclerophora nivea, Thelotrema
lepadinum, Usnea florida, U. fulvoreagens. Класс 3 - Acrocordia gemmata,
Arthonia leucopellaea, A. vinosa, Bacidia arceutina, B. rubella, B. subincompta,
Bactrospora dryina, Calicium salicinum, Chaenotheca furfuracea, C. phaeo-
cephala, C. trichialis, Hyperphyscia adglutinata, Lecania cyrtella, Lecanora
allophana, L. intumescens, L. varia, Micarea peliocarpa, Ochrolechia arborea,
Parmelia caperata, P. contorta, P. pastilifera, P. subargentifera, Pertusaria
hymenea, P trachythallina, Phaeophyscia endophoenicea, Physconia distorta,
P. enteroxantha, Rinodina colobina, Stenocybe pullatula, Usnea filipendula,
U. subfloridana. Класс 4 - Arthonia byssacea, Arthopyrenia punctiformis, Bacidia
globulosa, Bryoria fuscescens, Buellia disciformis, Calicium viride, Candelaria
concolor, Candelariella reflexa, Cetraria pinastri, Chaenotheca chrysocephala,
Chrysothrix candelaris, Imshaugia aleurites, Lecanora argentata, L. pallida,
L. symmicta, Micarea adnata, M. prasina, Normandina pulchella, Ochrolechia
androgyna, O. turneri, Opegrapha rufescens, O. viridis, Parmelia elegantula,
4. Чувствительность лишайников к загрязнению воздуха
91
Р. mougeotii, Р. revolute, Peltigera praetextata, Pertusaria albescens, P coccodes,
P coronata, P. flavida, P hemisphaerica, P. pertusa, Physcia aipolia, P stellaris,
Physconia perisidiosa, Ramalina pollinaria, Usnea hirta. Класс 5 - Arthonia
radiata, A. spadicea, Buellia griseovirens, Cetraria chlorophylla, Fuscidea viridis,
Graphis scripta, Lecanora carpinea, Mycoblastus sterilis, Parmelia tiliacea,
Pertusaria amara, P leioplaca, Platismatia glauca, Pyrenula nitida, P. nitidella,
Rinodina exigua, Xanthoria candelaria, X. fallax. Класс 6 - Arthothelium ruanum,
Candelariella xanthostigma, Dimerella pineti, Evernia prunastri, Hypocenomyce
caradocensis, Hypogymnia farinacea, H. tubulosa, Lecanora chlarotera, L. pu-
licaris, L. saligna, Lecidella elaeochroma, Opegrapha atra, O. vermicellifera,
Parmelia acetabulum, P exasperatula, P. flaventior, P. glabratula, P. subrudecta,
Phlyctis argena, Ramalina farinacea. Класс 7 - Candelariella efflorescens,
Parmelia saxatilis, Parmeliopsis ambigua, Phaeophyscia orbicularis, Physconia
grisea, Porina aenea, Pseudevernia furfuracea, Xanthoria parietina, X polycarpa.
Класс 8 - Chaenotheca ferruginea, Cladonia digitata, Diploica canescens,
Hypocenomyce scalaris, Hypogymnia physodes, L. hagenii, Micarea denigrata,
Parmelia sulcata, Physcia adscendens, P. tenella, Saccomorpha icmalea,
Scolociosporum chlorococcum, Strangospora pinicola, Trapelia granulosa,
T. pseudogranulosa. Класс 9 - Buellia punctata, Lecanora conizaeoides,
L. expaliens, Lepraria incana. Конечно, В. Вирт (V. Wirth) специалист авторитет-
ный, но результаты моих собственных наблюдений не позволяют безоговорочно
принять его ранжирование. Например, трудно согласиться с тем, что Hypogymnia
physodes, Parmelia sulcata и Scoliciosporum chlorococcum относятся к одному
рангу чувствительности. По моим данным, приведенная последовательность их
расположения соответствует степени возрастания устойчивости этих видов к заг-
рязнению. Я полагаю также, что накопленные данные пока не дают оснований
для выделения среди лишайников большого числа (9-12) классов по степени их
токсифобности/токситолерантности, косвенным подтверждением чего является
и разнообразие оценок этого показателя для наиболее часто изучавшихся видов,
представленное в обзоре И.Д. Инсаровой и Г.Э. Инсарова (1989).
В.В. Горшков (1990) встреченные им в центре Кольского полуострова виды
лишайников по отношению к загрязнению разделил на четыре группы: 1) устой-
чивые (номенклатура по: (Горшков, 1990)) - Cetraria islandica, Cladonia
bacilliformis, C. cenotea, C. coccifera, C. crispata, C. deformis, C. gracilis,
C. merochlorophaea, C. sulphurina, Lecanora fuscescens, Lecidea flexuosa,
Micarea melaena, Parmeliopsis ambigua, P. hyperopta', 2) чувствительные -
Bryoria simplicior, Cladina mitis, C. rangiferina, Cladonia botrytes, C. cornuta,
C. pleurota, Hypocenomyce scalaris, Hypogymnia physodes, Lecanora impudens,
Mycoblastus tornoensis, Ochrolechia androgyna, Parmelia centrifuga,
Parmeliopsis aleurites', 3) очень чувствительные - Bryoria fremontii, B. fus-
cescens, Cetraria chlorophylla, C. cucullata, C. pinastri, C. sepincola, Cladina
92
4. Чувствительность лишайников к загрязнению воздуха
arbuscula, С. stellaris, Cladonia chlorophaea, С. coniocraea, С. cryptochloro-
phaea, C. cyanipes, C. fimbriata, C. rei, Hypocenomyce friesii, Hypogymnia bitteri,
Lecanora chloropolia, L. coilocarpa, Lecidea plebeja, Mycoblastus sanguinarius,
Ochrolechia arborea, Parmelia septentrionalis\ 4) не переносящие загрязне-
ние - Bryoria capillaries, В. furcellata, В. lanestris, В. osteola, В. pseudo-
fuscescens, B. trichodes, Calicium sp., Evernia mesomorpha, Hypogymia
austerodes, H. tublosa, Platismatia glauca, Ramalina dilacerata, Usnea hirta.
По результатам изучения особенностей распределения эпифитных лишайни-
ков на территории города Москвы (около 900 км2) встреченные здесь на деревь-
ях представители 43 видов по их чувствительности к загрязнению были разделе-
ны на четыре класса (см. гл. 10), при этом отмечено, что представители наибо-
лее чувствительных или не переносящих загрязнение видов, которые могли бы
образовать еще один класс, в городе не обнаружены (Бязров, 1994). Таким обра-
зом, я полагаю, что пяти-ранговая шкала чувствительности видов лишайников к
загрязнению может быть относительно приемлемой как с точки зрения имею-
щихся в нашем распоряжении фактов оценки этого показателя, так с практиче-
ской точки зрения, ибо вряд ли серьезный специалист по особенностям распре-
деления лишайников сможет достоверно обосновать выделение на какой-либо
промышленной территории 9-10 зон, различающихся уровнем загрязнения. Если
используется пяти-ранговая шкала, то фактически можно выделить шесть зон,
причем участки территории, отнесенные к “лишайниковой пустыне”, могут быть
далеко не однородными по уровню загрязнения, но зонированию по лишайникам
они не подлежат из-за отсутствия здесь представителей лихенобиоты. Кроме
того, даже для характеристики классов с небольшим числом рангов используют
такие неопределенные понятия как “высокий/низкий”, “средний”, “умеренный” и
т.д., а при характеристике классов с большим числом рангов наряду с теми же
неопределенными эпитетами применяют еще такое обозначение как “промежу-
точный между” (Wetmore, 1989). Это всегда вызывает вопросы у специалистов
органов управления при демонстрации результатов исследования.
5. ВИДОВОЕ РАЗНООБРАЗИЕ ЛИХЕНОБИОТЫ
И КАЧЕСТВО ВОЗДУХА
Любое лихеноиндикационное исследование опирается на сведения о поведе-
нии видов лишайников (одного или многих) в условиях как заданных в опыте, так
и наблюдаемых при обследовании конкретной территории. Наличие списка видов
лишайников как для всей территории; являющейся объектом изучения, так и для
отдельных ее частей, позволяет достаточно надежно оценить состояние воздуш-
ного бассейна района и провести сравнение качества воздуха частей обследо-
ванной территории. Списки видов могут сравниваться как с данными предше-
ствовавших исследований, если они имеются, так и со сведениями о составе ли-
хенобиоты других регионов.
5.1. СОСТАВ ВИДОВ
Один из методов оценки качества воздуха основан на использовании видово-
го состава лихенобиоты изучаемой территории и установлении отсутствия/при-
сутствия чувствительных видов, поскольку их представители достаточно быст-
ро повреждаются или исчезают уже при низких уровнях некоторых загрязнителей
воздуха (Wetmore, 1982). Наблюдаемое обеднение видового состава лихенобио-
ты при неизменности прочих экологических условий обычно объясняют загрязне-
нием воздуха. Загрязнение приводит не только к отмиранию, но и к редукции пло-
доношения и уменьшению покрытия видов (Hawksworth et al., 1973; Deruelle, 1978a)
или исключению молодых стадий роста (Laundon, 1967).
Однако загрязнение воздуха вызывает у представителей некоторых видов и
обратные процессы. Расширение ареалов Lecanora conizaeoides, Scoliciosporum
chlorococcum в 1950-1970-х годах связывалось с увеличившимся загрязнением
(Ahti, Vitikainen, 1974; Hawksworth et al., 1974). Нынешнее отступление Lecanora
conizaeoides в юго-восточной Англии некоторые исследователи объясняют умень-
шением здесь загрязнения воздуха SO2 (Bates et al., 2001).
Расширил свой ареал и Parmeliopsis ambigua (Wetmore, 1988b). Однако все
это может быть связано и с другими факторами, такими как изменение структу-
94
5. Видовое разнообразие лихенобиоты и качество воздуха территории
ры лесопокрытых площадей, вырубка или сельскохозяйственная деятельность
(Wirth, 1988). Подтверждением того, что этот метод действительно отражает
условия загрязнения, является реинвазия видов на территории, где уровни загряз-
нения снизились (Henderson-Sellers, Seaward, 1979; Seaward, 1981,1982; Showman,
1981, 1990; Bates et al., 1990; Letrouit-Galinouetal., 1992).
Оценка качества воздуха по видовому составу лишайников включает:
- коллекционирование всех встреченных видов лишайников во всех типах био-
геоценозов и на всех субстратах обследуемой территории, если заранее не запла-
нированы какие-либо ограничения, например, учет только эпифитов или только
макролишайников;
- фиксирование наличия симптомов повреждения слоевищ загрязнителями:
обесцвечивание частей слоевища и их отмирание, отмирание талломов чувстви-
тельных видов, встречаемость слоевищ без органов плодоношения, ненормаль-
ное развитие слоевищ, отсутствие молодых талломов и другое;
- идентификация собранных представителей лихенобиоты и составление спис-
ка видов с указанием особенностей их распределения по сообществам, субстра-
там и т.д.;
- анализ состава лихенобиоты в отношении наличия чувствительных видов и
особенностей их распространения, выявленных признаков повреждения слоевищ,
изменения репродуктивного потенциала и т.д.;
- сравнение собранных данных с историческими материалами, если они име-
ются, или с составом видов территории с чистым воздухом того же региона для
установления возможных изменений как лихенобиоты в целом, так и показателей
отдельных видов.
Сравнительное изучение крупных по размерам территорий затруднено из-за
отсутствия равноценной информации о составе лихенобиот, поскольку далеко не
во всех регионах были проведены фундаментальные исследования видового со-
става лишайников и особенностей их экологии. Еще меньше возможностей для
сравнения изменений за длительный период. Так, сотрудники Лаборатории мони-
торинга природной среды и климата для целей мониторинга провели учеты эпи-
фитных лишайников практически во всех биосферных заповедниках территории
СССР (Инсаров, Пчелкин, 1988). Однако сведения о повторных наблюдениях в
этих заповедниках они не опубликовали, что не позволяет судить о достоинствах
предложенного ими способа учета (Инсаров, Пчелкин, 1983). Недостаточно изу-
чены лихенобиоты и национальных парков США, для более чем половины кото-
рых известно лишь менее четверти видового состава лишайников (Wetmore, 1981b,
1985, 1988b). Исследования в национальном парке Big Bend (Wetmore, 1981а) по-
казали, что выявленное здесь изменение состава лихенобиоты связано не каче-
ством воздуха, а в парках Indian Dunes (Wetmore, 1988а) и Cuyahoga Valley
(Wetmore, 1989), где установлено уменьшение на 82% числа видов за последние
80 и 90 лет, изменения объяснялись действием загрязнителей. В области Untermain
5. Видовое разнообразие лихенобиоты и качество воздуха территории
95
(Германия) из 315 видов, указываемых для этой территории до 1900 г., в 1980-х не
обнаружены представители 166 видов (Scholler, 1995). В Центральной Италии с
1980 по 1996 гг. увеличилось число более чувствительных к загрязнению видов
лишайников (Loppi, et al., 1998с). На нынешней территории Москвы (в пределах
Московской кольцевой автодороги) также выявлено значительное изменение со-
става лихенобиоты (Бязров, 1996а), но оно связано не только с загрязнением, а и
с другими причинами (уничтожение местообитаний, вытаптывание и другое).
Аналогичные причины изменения видового состава лишайников установлены в
Петербурге и ряде исторических мест Ленинградской области (Малышева, 1992,
1993, 1996а-в, 1998; Малышева, Булыгина, 1996). С 1728 по 1995 гг. на террито-
рии, занимаемой ныне Петербургом, отмечены представители 268 видов лишай-
ников, из которых 139 в последние 60 лет исчезли (Малышева, 19966). В Казани
из 124 видов, встречавшихся с 1883 по 1976 гг., не обнаружены представители
48 видов лишайников (Голубкова, Малышева, 1978).
5.2. РАСПРОСТРАНЕНИЕ ВИДОВ
Изучение распространения видов представляет более сложную процедуру, чем
определение видового состава, поскольку требует точного определения местона-
хождения представителей видов на местности. Картографирование лишайников-
часто используемый способ лихеноиндикации качества воздуха в пространстве и
времени. Оно дает суммарное представление о качестве воздуха как следствие
прошлого воздействия, или, на основе хронологически последовательных серий
исследований, может показывать качественные изменения его состояния. Кар-
тографирование лишайников полезно главным образом на территориях, где инст-
рументальный мониторинг качества воздуха отсутствует, или сеть измеритель-
ных станций редка.
Картографирование может быть основано как на материалах непосредствен-
ного обследования территории, так и коллекциях, опубликованных и архивных
данных. Наряду с регулярными сведениями, полученными с использованием квад-
ратов сетки, материал собирают и случайно из большого числа точек обследуе-
мой территории. Эти сведения затем определенным образом упорядочивают (или
классифицируют) в удобную для картографирования форму и наносят на карту,
“привязываясь” на ней к конкретным точкам или квадратам изучаемой террито-
рии. Точки с одинаковыми показателями лишайников могут быть соединены изо-
линиями или объединены в зоны равного воздействия. Результатом является кар-
та с наглядной информацией обо всей изученной территории.
Первый шаг в проведении исследования по созданию карты распространения
для целей лихеноиндикации - это определение размеров изучаемой площади и
сбор сведений, характеризующих как ее природные условия, так и источники за-
96
5. Видовое разнообразие лихенобиоты и качество воздуха территории
грязнения. В идеале изучаемая площадь должна быть достаточно большой для
того, чтобы воздействие было полностью обнаружено. Другими словами, у гра-
ниц этой площади должна быть в наличии лихенобиота, не подвергавшаяся воз-
действию загрязнителей, т.е. обычная для данного географического региона
(Showman, 1988). После определения размеров и контуров подлежащей изучению
территории следует подобрать детальные топографические карты этой площади,
а также ознакомиться с другими доступными картографическими материалами.
Следующий шаг - рекогносцировочное обследование территории. Рекогнос-
цировка должна выявить самые обычные виды лихенобиоты на изучаемой тер-
ритории и дать общее представление об их распределении и особенностях терри-
тории. Этот этап необходим для того, чтобы обоснованно выбрать детальный
способ обследования для учета лишайников.
Самый обычный способ подобных исследований - картографирование рас-
пространения с использованием квадратов сетки определенного размера. Вели-
чина квадратов зависит от решаемых в данном исследовании задач, размеров
обследуемой территории, расположения на ней источников загрязнения, наличия
средств и времени. Этот метод может быть применен для оценки территории как
вокруг изолированного источника загрязнения, так и крупных областей. Выводы,
сделанные в результате интерпретации таких данных, являются при этом доволь-
но обоснованными, и степень их надежности зависит от однообразия условий гео-
графической среды изученной территории.
Учеты лишайников в этих квадратах местности проводят или сплошь, т.е. по
возможности на всех встреченных деревьях (Бязров, 1994), или выборочно. Так,
Германский союз инженеров (VDI) рекомендует при картографировании в квад-
ратах со стороной до 0,25 км учитывать лишайники на 3 равномерно распреде-
ленных по площади деревьях, максимально обросших лишайниками, при размере
стороны квадрата 1 км рекомендуется выбрать 6 деревьев, а в более крупных по
размеру квадратах на 4 угловых точках на площадях по 1 км2 на каждой обследу-
ют по 6 деревьев (VDI, 1995).
Нередко, однако, картографирование проводят и без использования квадра-
тов сетки, а при рекогносцировке заранее намечают участки или пробные площа-
ди, где будет обследован состав лихенобиоты. Выбираются такие участки, кото-
рые репрезентативны для данного подтипа территории и включают от нескольких
до значительного количества деревьев. Подходящими обычно считаются дере-
вья, растущие на некотором расстоянии друг от друга вдоль дорог со слабым
движением автотранспорта, а также аллеи, кладбища и парки (Showman, 1988).
Деревья, плотно стоящие друг подле друга или находящиеся в закрытом густом
лесу при выборочном обследовании рекомендуется избегать. Пробные площади
следует стандартизировать по размеру, а также по составу древесных пород и
возрасту деревьев. Например, молодые деревья могут иметь иной, в сравнении
со старыми той же породы, состав лишайников (Бязров, 1970).
5. Видовое разнообразие лихенобиоты и качество воздуха территории
97
Когда подходящие участки на местности выделены, можно проводить сбор и
запись данных о лихенобиоте. Данные должны включать в себя следующее: дату,
номер участка, особенности его расположения, абсолютную высоту, характерис-
тику местообитания, состав основных древесных пород, информацию о наличии
видов лишайников. Номер участка должен быть отмечен и на топографической
карте. Сведения о расположении точки также должны включать индексы топог-
рафических карт или иную информацию, позволяющую найти ее при отсутствии
карты. В примечаниях обычно указывают наличие каких-либо особенностей, ко-
торые могут влиять на состав лихенобиоты в данном месте, например, близкое
расположение мест для сжигания мусора, отходов; сильная запыленность доро-
ги. Можно отметить и общие впечатления исполнителя, например, “место слиш-
ком затенено для оптимального развития ряда видов лишайников”.
Часть встреченных видов может быть определена непосредственно при по-
левом обследовании, однако идентификация представителей многих видов лишай-
ников в поле невозможна, что делает необходимым их коллекционирование для
последующего определения в лабораторных условиях. Собранные образцы снаб-
жаются соответствующими этикетками. Одновременно фиксируются сведения
об обилии лишайников, размере слоевищ, наличии органов плодоношения, повреж-
дении талломов и другое.
После идентификации встреченных на каждом обследованном участке тер-
ритории видов приступают к созданию карты. Для этого лучше всего использо-
вать карту-основу подходящего масштаба, которая полностью покрывает изу-
ченную территорию и отображает ее важнейшие объекты (реки, озера, магистра-
ли, застроенные территории и т.д.). При работе с топографической картой в поле
желательно на ней показывать расположение источников загрязнения. На карте-
основе особыми значками отмечаются точки, где обнаружено присутствие пред-
ставителей видов лишайников. Если предварительный анализ карты обнаружива-
ет пробелы в распространении обычных для этой территории видов, желательно
повторно посетить участки для уточнения причин их отсутствия. Высока вероят-
ность того, что такие пробелы в распространении обычных видов на сравнитель-
но однородной территории связаны с воздействием источника загрязнения.
5.2.1. Информация, используемая при создании
лихеноиндикационных карт
5.2.1.1. Степень видового разнообразия лихенобиоты
Степень видового разнообразия лихенобиоты, или ее богатство стали осно-
вой для создания лихеноиндикационных карт, отражающих качественное состоя-
ние воздушного бассейна разных частей обследованных территории России (Бяз-
98
5. Видовое разнообразие лихенобиоты и качество воздуха территории
ров, 19966), США (Showman, 1981), Канады (Rao, LeBlanc, 1967), Италии (Giordani
et al., 2001, 2002; Loppi et al., 2002) и других районов. Она измеряется общим
числом видов на участке. Чем больше видов, тем чище воздух. Соответственно,
участки, где лишайники отсутствуют или они представлены единичными экземп-
лярами одного вида, считают наиболее загрязненными, а участки с наибольшим
числом видов относят к наиболее чистым на обследованной территории. Учет-
ные площади, на основе количества видов, встреченных на участке, группируют
в несколько классов (3-5), качественно различающихся по степени загрязнения
воздуха. Например, для примыкающей к Тирренскому морю части Италии ис-
пользована следующая шкала оценки территории: 0 видов - отклонение от есте-
ственных условий 100%, лишайниковая пустыня; 1-25 видов - отклонение от
естественных условий 75-99%, антропогенная экосистема; 25-50 видов - откло-
нение от естественных условий 50-75%, полуантропогенная экосистема; SO-
75 видов - отклонение от естественных условий 25-50%, полуприродная экосис-
тема; >75 видов - отклонение от естественных условий 0-25%, природная экоси-
стема (Brunialti et al., 2002).
5.2.1.2. Степень чувствительности видов лишайников
Процедура создания лихеноиндикационной карты по степени чувствительно-
сти видов лишайников основана на ранжировании видов, встреченных на каждом
участке, на некоторое число классов по их чувствительности к загрязнению воз-
духа. После этого видовой состав каждой учетной площади анализируется с це-
лью выявления присутствия видов того или иного класса чувствительности. За-
тем из общего числа участков вычленяются участки с наличием видов наиболее
высокого класса чувствительности, которые группируют в территорию с наибо-
лее чистым в обследованном районе воздухом; из оставшихся вычленяют уча-
стки с видами следующего класса чувствительности, которые объединяют в тер-
ритории с более загрязненным воздухом (Бязров и др., 1997; Максимова, Бязров,
2000). В Великобритании также учитывают только эпифитные лишайники, при
этом ориентируются на изолированные и хорошо развитые форофиты со сходны-
ми свойствами коры (Hawksworth, Rose, 1970). Картографирование территории
Англии и Уэльса основано на присутствии/отсутствии видов лишайников, ранжи-
рованных на 9 классов чувствительности к загрязнению. Для каждого класса
чувствительности лишайников и типа коры форофитов приводится средняя вели-
чина концентрации двуокиси серы в воздухе. Например, наличие на коре деревьев
только представителей зеленых водорослей и Lecanora conizaeoides, при отсут-
ствии слоевищ других видов лишайников, свидетельствует о величине концентра-
ции двуокиси серы около 125 мкг на м3 воздуха (0,04 ppm); представители Usnea
ceratina на тех же деревьях могут быть встречены при концентрации двуокиси
5. Видовое разнообразие лихенобиоты и качество воздуха территории
99
серы около 35 мкг в м3, а представители U. florida - при концентрации двуокиси
серы менее 30 мкг на м3. Этот метод был использован во Франции для картогра-
фирования территории департамента Mantes (Deruelle, 1977). Также зонирование
территории можно проводить по данным картографирования распространения
избранных индикаторных видов разной степени чувствительности к загрязнению
воздуха, как это имело место, например, в Гонконге, где при создании картосхе-
мы загрязнения воздуха двуокисью серы ориентировались на распространение в
этом анклаве Pyxine cocoes и желтого налета видов Lepraria, Dirinaria picta,
Parmotrema tinctorum, Usnea bailey (Thrower, 1980).
5.2.1.3. Повреждения лишайников
Картографирование распространения отдельных видов лишайников может
сопровождаться отображением участков, где наблюдались повреждения лишай-
ников (угнетение или повреждение слоевищ). В штате Коннектикут (США) для
этих целей использовали карту-основу, на которой точками были отмечены об-
следованные участки; виды, присутствующие на этих участках, но с выраженны-
ми повреждениями, обозначали пустым кружком, а виды, имеющие здоровые
слоевища - закрашенным кружком (Stolte et al., 1993). Следует,-однако, учиты-
вать, что повреждения могут быть обусловлены и другими факторами, такими
как, например, старение слоевищ или резкое понижение температуры зимой пос-
ле оттепели (Kappen, Lange, 1972; Showman, 1979; Benedict, 1990b).
5.2.1.4. Частота встречаемости
Показателем качества воздуха может быть и частота встречаемости лишай-
ников (Brodo, 1966; Rao, LeBlanc, 1967; Showman, 1988). Частота каждого вида
определяется процентом доли квадратов, в которых они были встречены. Встре-
чаемость можно определять и относительно числа обследованных стволов дере-
вьев. На карту наносятся величины встречаемости лишайников на пробных пло-
щадях, проводится ранжирование этих величин на классы, и участки, объединен-
ные по принадлежности к рангу одного класса встречаемости лишайников, относят
к территории с соответствующим качеством воздушного бассейна. Также точки
на карте с равными величинами встречаемости можно соединить изолиниями.
Частота встречаемости лишайников учитывалась при изучении действия уголь-
ной пыли на экосистемы вокруг тепловой электростанции в аридном районе на
западе США (Spencer, Tinnin, 1997; Spencer, 2001). Показатель встречаемости
лишайников вне дымового шлейфа был намного выше в сравнении с участками,
накрываемыми дымовым облаком.
100 5. Видовое разнообразие лихенобиоты и качество воздуха территории
5.2.1.5. Покрытие
Территории с чистым воздухом, как правило, характеризуются высоким по-
крытием поверхности деревьев слоевищами эпифитных лишайников. Величина
покрытия субстрата может быть определена для каждого из видов или для всех
видов в целом. Ее измеряют или достаточно точно с помощью палетки, или оце-
нивают глазомерно по бальной шкале. Значения покрытия затем наносятся на
карту, точки с равными величинами могут быть соединены изолиниями или объе-
динены в территориальные единицы с соответствующим качеством воздуха. Та-
кая лихеноиндикационная карта по среднему покрытию (в процентах) листова-
тых лишайников на каждом обследованном участке была составлена в США для
территории штата Коннектикут (Showman, 1988). Для территории Германии по-
крытие оценивалось в баллах, причем нередко использовался только один вид,
чаще всего токситолерантная Lecanora conizaeoides (Шуберт, 1988; Ackermann,
1993; Mezger, 1993; Scholz, 1993).
5.3. ЛИХЕНОИНДИКАЦИОННЫЕ ИНДЕКСЫ
По мере совершенствования приемов исследований “лишайники - качество
воздуха” были разработаны эмпирические методы, с помощью которых можно
было установить градиент воздействия загрязнителей по сумме характеристик
лихенобиоты сравниваемых частей изучаемой территории. Их основу составля-
ет определение индексов загрязнения/чистоты и полеофобии. Индекс представ-
ляет собой некое число, результирующее посредством математической формулы
формализованные параметры лишайников (число видов, покрытие, встречаемость,
ассоцированность с другими видами и т.д.) для конкретного места. Абсолютная
величина этого индекса и отражает степень загрязнения/чистоты участка. Зна-
чения индексов можно наносить на карту и проводить деление территории по
степени загрязнения.
Методы определения индексов предъявляют особые требования к отбору
участков и учету лишайников. По возможности, все сравниваемые площади дол-
жны быть сходными по всем экологическим параметрам, кроме уровня загрязне-
ния. Рекомендуется учитывать эпифиты одной древесной породы, хотя допуска-
ются исключения, если показано, что свойства коры растущих вместе разных
видов деревьев имеют одинаковые характеристики (степень бороздчатости, плот-
ность, прочность, буферная емкость, pH, влагоемкость и другое). Деревья долж-
ны быть зрелыми, стоящими в удалении друг от друга, пряморастущими. От-
ступления от этих требований могут привести к определению пространственного
варьирования свойств лишайниковых группировок, а не к выявлению качествен-
ных различий воздушного бассейна. В каждой учетной точке рекомендуется об-
5. Видовое разнообразие лихенобиоты и качество воздуха территории 101
следовать состав эпифитных лишайников и количественно охарактеризовать па-
раметры каждого вида, по меньшей мере, на 10 одинаковых деревьях. Если чис-
ло подходящих деревьев больше, выбирают 10 с наиболее развитым покровом
эпифитов. Сведения об участке и лишайниках желательно заносить на стандарт-
ный полевой бланк. Как и в случае изучения видового состава, необходимо соби-
рать контрольные образцы всех встреченных видов лишайников для гербария.
5.3.1. Индекс полеотолерантности (I.P.)
Индекс полеотолерантности для целей лихеноиндикации предложил Х.Х. Трасс
(1968, 1971, 1985а). Он вычисляется по формуле:
п
1.Р.= £ а, хе,/С,,
1
где а - степень толерантности вида лишайника к городской среде (эмпирическая
величина, определяется экспертными оценками данных о произрастании пред-
ставителей видов в городских районах с известными уровнями загрязнения),
с - ранговая величина покрытия вида (в баллах), С - степень общего покрытия
всех видов (в баллах), п - число видов. Так, территория города Казань была
разделена на квадраты по 1 км2. В этих квадратах лишайники учитывались на
высоте груди на стороне ствола дерева, где их покрытие наибольшее, с помощью
учетной площадки 20x20 см, представляющей сеть квадратов 2x2 см. Покрытие
оценивали в баллах по 10-ранговой шкале: покрытие 1-2% - 1,3-5% - 2,6-10% -
3, 11-20% - 4, 21-30% - 5, 31-40% - 6, 41-50% - 7, 51-65% - 8, 66-80% - 9, 81-
100% - 10 (Голубкова, Малышева, 1978). Величина I.P. может меняться от 0 до
10, на территории г. Казани значения вычисленного индекса составили от 4 до 10,
что дало основание выделить здесь три зоны, различающихся качеством возду-
ха. Однако по результатам обследования в 1993 г., на этой территории по величи-
не индекса полеотолерантности и ряда других показателей выделено 4 зоны (Бай-
баков, 1998).
Автор этого индекса рекомендовал для определения величины I.P. заклады-
вать на каждом дереве по четыре учетных площадки размером 40x40 см (две -
у основания ствола и две - на уровне груди) на противоположных по экспозиции
сторонах ствола (Трасс, 1984). Средние значения I.P. следует вычислять для каж-
дого обследованного ствола дерева, а затем для каждой учетной площади обсле-
дуемой территории. Величины I.P. на территории Эстонии были скоррелированы
со среднегодовыми концентрациями двуокиси серы в воздухе (Трасс, 1968,1984).
Например, значение 2< 1.Р.<5 соответствовало концентрации двуокиси серы 10-
30 мкг/м3 (0,0035-0,0105 ppm), а величина 1.Р.>9 показывала, что концентрация
двуокиси серы варьирует между 100 и 300 мкг/м3 (0,035-0,105 ppm). Кроме Каза-
ни, индекс полеотолерантности использовался при лихеноиндикации городов Эс-
] 02 5. Видовое разнообразие лихенобиоты и качество воздуха территории
тонии (Martin, Martin, 1974; Л ийв, Мартин, 1977; Л. Мартин, 1978; Лийв, 1984,1988),
г. Грозного (Закутнова, 1988), на Кольском полуострове (Аблаева, 1981а), в При-
байкалье (Трасс и др., 1988), на Дальнем Востоке (Родникова и др., 1998) и в ряде
других местностей.
5.3.2. Индекс атмосферной чистоты (I.A.P.)
Индекс атмосферной чистоты впервые предложили Д. Деслувер и Ф. Леб-
ланк (DeSloover, LeBlanc, 1968). Затем он был использован во многих других ис-
следованиях (LeBlanc, DeSloover, 1970; LeBlanc et al., 1972b; Mathis, Tomlinson,
1972; Hoffman 1974; Stringer, Stringer, 1974; Case 1980; Аблаева, 1981a; Кучерявый
и др., 1989; Herzig, Urech, 1991; Кондратюк и др., 1991, 1993а; Nimis et al., 1991;
Малышева, 1998). Для его определения в каждой точке измеряют ряд показате-
лей лишайниковых группировок, которые затем для этой точки преобразуются с
помощью эмпирически выведенной формулы в целое число I.A.P. Значения I.A.P.
можно нанести на карту, а участки с близкими величинами объединить в один
класс территории с определенным качеством воздуха.
Первоначально проводится рекогносцировочное обследование, в ходе которо-
го определяют характер расположения участков для учета лишайников. Отбору
таких точек придается большое значение. Все они по возможности должны быть
сходными. Как указывали авторы индекса I.A.P., для точного картографирования
необходимо, чтобы обследуемая территория была экологически гомогенной, а
выбранных участков для учета лишайников много, чтобы они репрезентативно
представляли всю площадь. Предпочтительно, чтобы эпифиты учитывались на
деревьях одной породы. Однако допускается учет эпифитных лишайников на ство-
лах двух и более видов деревьев, если эти породы имеют примерно одинаковые
характеристики коры (например, буферная емкость, плотность, влагоемкость, проч-
ность и другое). Если этого строго не придерживаться, то в результате будут
измерены естественные экологические вариации распределения эпифитов вме-
сто различий по качеству воздуха (LeBlanc, Rao, 1975). Рекомендуется обследо-
вать в каждой точке, по меньшей мере, 10 одинаковых деревьев с наиболее раз-
витым покровом эпифитов. Стволы этих деревьев тщательно осматривают до
высоты 2 м и фиксируют все обнаруженные виды лишайников. При этом пред-
ставитель каждого вида оценивается в баллах по шкале, характеризующей час-
тоту встречаемости - покрытие (LeBlanc, DeSloover, 1970): 5 - виды с очень
высокой частотой встречаемости и очень высокой степенью покрытия большин-
ства стволов деревьев; 4 - виды с высокой частотой или высокой степенью по-
крытия; 3 - виды, встречающиеся не часто или имеющие небольшую степень
покрытия на некоторых деревьях; 2 - виды, встречающиеся редко или имеющие
низкую степень покрытия; 1 - виды очень редкие и с очень низкой степенью
покрытия.
5. Видовое разнообразие лихенобиоты и качество воздуха территории 103
Значение I.A.P. для каждой точки рассчитывается согласно следующей фор-
муле:
i.a.p. = 1/юЛ 2, хЛ-
/=1
где п - число видов на учетной площади, Qf - экологический показатель вида
/ (среднее число видов, растущих вместе с видом / на учетной площадке),
F - оценка встречаемости - покрытия вида / согласно приведенной выше шкале
(LeBlanc, DeSIoover, 1970). Произведение Q*Ft делится на 10, чтобы получить
более наглядное число.
Классическим считают применение I.A.P. при обследовании территории г.
Монреаля (LeBlanc, DeSIoover, 1970). Сведения об эпифитах были собраны в 349
сходных точках в городе и его окрестностях. Значения I.A.P. нанесены на карту,
и по их величинам, ранжированным на пять классов, выделены изотоксичные зоны.
Установлено, что распространение и показатели плодоношения некоторых видов
тесно коррелируют с этими зонами. Авторы сделали вывод, что карта зон, выде-
ленных по I.A.P., довольно точно отражает характер длительного воздействия
загрязнения воздуха на лишайники и мохообразные. Однако они подчеркивают,
что если локальное изобилие эпифитов, вероятно, всегда является признаком чи-
стоты воздуха, то их бедность не всегда свидетельствует о загрязнении.
В аналогичном исследовании около г. Садбери в Онтарио (Канада) данные об
эпифитах собраны в 31 точке (LeBlanc et al., 1972а). Значения I.A.P. также нане-
сены на карту, в соответствии с чем выделено пять изотоксичных зон, которые
коррелируют с величинами среднегодовой концентрации двуокиси серы в возду-
хе. В другом канадском городе Виннипеге (Манитоба) все 168 обследованных
точек были равномерно распределены по всей территории (Stringer, Srtinger, 1974).
Значения I.A.P. определялись для каждой точки, для чего использовалась немно-
го модифицированная шкала оценки “встречаемости-покрытия”. По величинам
I.A.P. выделено пять изотоксичных зон. Поскольку инструментально замерен-
ные концентрации двуокиси серы были относительно низкими, авторы предполо-
жили, что распределение и развитие эпифитных лишайников определяются заг-
рязнением воздуха как промышленными предприятиями, так и транспортом, а
также особенностями микроклимата разных частей территории. Аналогичный
вывод был сделан и по результатам изучения воздействия целлюлозно-бумажно-
го предприятия на качество воздуха в штате Айдахо (США) с использованием
модифицированного I.A.P., на основе которого было выделено три зоны (Hoffman,
1974). Всесторонняя проверка I.A.P. проведена вокруг комплекса газоперераба-
тывающих предприятий в канадской провинции Альберта у Fox Creek (Case, 1980).
Сведения о частоте встречаемости, покрытии, жизненности лишайников собраны.
на 99 участках. Для каждой такой точки вычислены величины оригинального I.A.P.,
а также нескольких модификаций этого индекса. Кроме того, для каждого участ-
104 5. Видовое разнообразие лихенобиоты и качество воздуха территории
ка определены обычные индексы разнообразия Шенона-Винера. Было установ-
лено, что модифицированные величины I.A.P., вычисленные по данным измере-
ний только нескольких индикаторных видов, сходны с теми, которые вычислены
на основе показателей всех видов лишайников. Эти значения наносились на карту,
и далее была выявлена значимая корреляция как I.A.P., так и индекса Шенона-
Винера с расстоянием до источника загрязнения. Однако эти пространственные
изменения показателей лишайниковых группировок слабо коррелировали с дан-
ными инструментальных измерений качества воздуха, что привело автора иссле-
дования к заключению о том, что наблюдаемые изменения обилия и жизненности
лишайников обусловлены не долговременным ежедневным воздействием суль-
фатов, а в основном иными факторами.
Известны и другие модификации оригинального индекса, часть которых целе-
сообразно воспроизвести:
v-'’ ОхСх/**
I.A.P. = £ехС(1); I.A.P. = £exCxF(2); I.A.P. = „ (3) ’
V X 0
X г-» Х"'' С* х F
С (4); I.A.P. =XCxF <5)’ I A R <6)’
1-а.р. = тI AP-= 1АР- = £ехС(9);
v X 0
I.A.P. = £0(10); I.A.P. = ^Уу (11); I.A.P. = £^(12);
I.A.P. = 2% <13);LAR = £% <14); 1АР- = (15);
' Г X 0
v”» QxF
I.A.P. = 2 Кх5
(16),
где F - показатель частоты встречаемости представителя каждого вида на де-
реве, С - показатель его покрытия, V - оценка его жизненности, S - показатель
его поврежденности, Q - его токситолерантность, определяемая средним чис-
лом сопутствующих ему видов лишайников на всех деревьях обследуемой пло-
щади; чем меньше величина Q, тем более устойчив вид к загрязнению (Herzig,
Urech, 1991). Однако абсолютные значения токситолерантности вида можно ис-
пользовать лишь применительно к изучаемой территории и для сравнения только
с другими встреченными здесь видами, поскольку в другом регионе величина,
характеризующая токситолерантность этого вида, может быть иной. Например,
в Средней Швейцарии значение Q у Lecanora conizaeoides 3,7, а в регионе Биль-
Зееланд в той же стране - 7,9, у Xanthoria parietina - 11,6 и 10,1, у Candelaria
concolor- 12,4 и 10,1, у Anaptychia ciliaris - 14,3 и 12,8, соответственно (Herzig,
Urech, 1991). Однако в обоих сравниваемых регионах самые низкие абсолютные
5. Видовое разнообразие лихенобиоты и качество воздуха территории
105
величины Q были у Lecanora conizaeiedes. После тестирования перечисленных
выше версий I.A.P. для условий изученного региона Швейцарии оптимальной была
признана модификация 8 (Herzig, Urech, 1991). В Греции также использовали ука-
занную модификацию, однако величина F определялась по следующей 5-бальной
шкале: 1 - вид представлен лишь на одном дереве (из 10, обследованных на точ-
ке) и его покрытие менее 5%; 2 - вид представлен на одном дереве или его по-
крытие менее 5%; 3 - вид представлен на более чем половине деревьев; 4 - вид
представлен на более чем половине деревьев или его покрытие на некоторых из
них более 50%; 5 - вид представлен на более чем половине деревьев и покрытие
его на большинстве из них более 50% (Vokou et al., 1999).
Значения I.A.P. всех точек могут наноситься на картосхему обследуемой
территории. Их группируют обычно в классы I.A.P., соответствующие зонам с
различным уровнем загрязнения. В зависимости от степени воздействия, деталь-
ности сбора данных и представлений автора таких зон может быть несколько.
Например, легенда лихеноиндикационной карты провинции Венеция (Италия) со-
держит 11 классов величин I.A.P. с шагом 5 (Nimis et al., 1991), территория Пе-
тербурга на основе величин этого индекса (с шагом 5) была разделена на 5 изо-
токсичных зон (Малышева, 1998), а А.В. Пчелкин (1998) в парке Сокольники
(г. Москва) выделил 6 зон загрязнения с разной шириной шага величины I.A.P.
(0-0,1; 0,1-0,5; 0,5-1,0; 1,0-5,0; 5,0-10,0; >10,0). Таким образом, первый класс ле-
генды итальянских авторов (<5) включает первые четыре класса легенды для
парка Сокольники. В Прибайкалье Х.Х. Трасс, А.Й. Пярн и К.Б. Цобель (1988)
на территории, по размерам не уступающей провинции Венеция, изучая послед-
ствия пятнадцатилетней деятельности Байкальского целлюлозно-бумажного ком-
бината, на основе определения величин I.A.P. выделили четыре изотоксичных
зоны - сильнозагрязненную (I.A.P. = 0-4), среднезагрязненную (I.A.P. = 4-9), сла-
бозагрязненную (I.A.P. = 9-16), незагрязненную (1.А.Р. > 16). Территория г. Льво-
ва (130 км2) была поделена на 1-километровые квадраты (в центре города сторо-
на квадрата 0,5 км), в квадрате эпифитные лишайники учитывались на 20 деревь-
ях каждой лиственной породы (Кучерявый и др., 1989). По вычисленным индексам
1.А.Р. с переменным значением шага I.A.P. выделены четыре изотоксичные зоны:
сильнозагрязненная (1.А.Р. = 0-2), среднезагрязненная (I.A.P. = 2-4), слабоза-
грязненная (I.A.P. = 4-8), и незагрязненная (I.A.P. = 8-24). В статье тех же авто-
ров в другом издании (Кондратюк и др., 1991) указаны несколько иные значения
индекса для выделения тех же изотоксичных зон в г. Львове - 0-0,99, 1-5, 5-10 и
10-18. Эти зоны совпадали с делением территории на основе других показателей
эпифитных лишайников (число видов в квадрате, наличие видов определенного
класса чувствительности к загрязнению).
Зоны изотоксичности иногда коррелируют с имеющимися данными инстру-
ментальных измерений концентраций загрязнителей в воздухе или с другими по-
казателями. Например, в одном из районов Канады были установлены следую-
106 5. Видовое разнообразие лихенобиоты и качество воздуха территории
щие соотношения между значениями I.A.P. и концентрацией двуокиси серы в воз-
духе (табл. 5.1).
> Таблица 5.1
Соотношение между величиной I.A.P. и концентрацией двуокиси серы
в воздухе окрестностей г. Садбери, Канада (LeBlanc et al., 1972а)
I.A.P. 0-9 10-24 25-39 40-54 >55
SO2, ppm 0.03 0,02-0,03 0,01-0,02 0,005-0,01 <0,005
Однако следует иметь в виду, что эти эмпирически установленные зависимо-
сти действительны лишь для местностей, где проводились исследования. Так в
Прибайкалье на величине индекса сказывается высота местности, поскольку
реакция ряда видов лишайников на загрязнение неодинакова на разных абсолют-
ных высотах (Цобель, 1988).
Неоднократно проводилось сравнение индекса полеотолерантности и индек-
са чистоты атмосферы (Martin, Martin, 1974; Deruelle, 1978b; Трасс, 1987, 1988).
Их обычным недостатком является то, что они содержат переменные, определя-
емые субъективно - степень толерантности вида к городской среде у I.P. или
степень встречаемости-покрытия вида в случае I.A.P. Кроме того, величина I.A.P.
зависит от числа видов, но бедный видовой состав лишайников не всегда являет-
ся следствием загрязнения воздуха. Х.Х. Трасс (1987) полагает, что для приме-
нения I.P. необходимо хорошо представлять особенности экологии видов; если
таких сведений у исследователя недостаточно, лучше использовать I.A.P.
5.3.3. Индекс степени качества воздуха (LGW)
В Германии для выделения изотоксичных частей обследуемой территории
используется показатель степени качества воздуха, или LGW (Luftgiitewerte),
который выражается средней величиной сумм встречаемостей лишайников на
картируемых деревьях растрового квадрата и представляет собой статистичес-
кую оценку фактических соотношений между слоевищами на измеренной повер-
хности. Точность оценки зависит от стандартного отклонения результатов и луч-
ше всего описывается данными доверительного интервала. Доверительный ин-
тервал показывает, в пределах каких границ усредненные значения LGW
соответствуют фактическим величинам с заданной статистической вероятнос-
тью (обычно рекомендуется 95%). Для вычисления применяется следующая
формула (VD1,1995):
= средняя сумма встречаемостей в квадрате j = Y Fl njt
5. Видовое разнообразие лихенобиоты и качество воздуха территории 107
где / - номер отдельного дерева на обследованной площади (квадрат),у - номер
обследованной площади (квадрата), F - сумма встречаемостей видов лишайни-
ков на дереве i в квадрате у, п - число обследованных деревьев на площади j.
При этом также используют такие понятия статистики, как стандартное от-
клонение, доверительный интервал, критерий Стьюдента и другие (Kirschbaum,
Windisch, 1995).
Обследование для определения LGW предполагает определенную процедуру.
Прежде всего, интересующая территория на карте делится на равные по площа-
ди участки, размер которых зависит от топографии и величины изучаемой терри-
тории, а также решаемой задачей. Как правило, используют квадраты со сторо-
ной квадрата от 0,25 до 1 км в населенных пунктах и 2-4 км - при обследовании
крупных территорий. При использовании квадратов со стороной до 0,25 км ли-
шайники учитывают не менее чем на трех по возможности равномерно рассре-
доточенных деревьях, при размере стороны квадрата 1 км обследуют 6 деревь-
ев. При более крупном размере квадрата на 4 угловых точках обследуют площа-
ди по 1 кв. км с 6 деревьями на каждой. В местностях с большим высотным
градиентом (например, в Альпах) такие учетные площади рекомендуется закла-
дывать через каждые 50 м высоты (VDI, 1995).
В пределах каждой изучаемой территории, по возможности, лишайники учи-
тывают на деревьях одного вида. Если это невозможно, то лишайники рекомен-
дуется учитывать на древесных породах с примерно одинаковыми свойствами
коры (величина pH, содержание питательных веществ, водоудерживающая спо-
собность). Виды деревьев, которые в условиях Германии возможно обследовать
вместе, показаны в таблице 5.2.
Местности, где обследовались различные по свойствам деревья, не сравни-
мы дуг с другом. Стволы деревьев должны иметь указанные в таблице 5.2 пре-
дельные размеры. Деревья с повреждениями, а также со следами внешнего воз-
действия (обработка известью, обрезание коры, трение скота и другое) обследо-
ванию не подлежат. Не рекомендуют использовать плодовые деревья в местностях,
где большие площади заняты плантациями плодовых культур.
Таблица 5.2
Группы древесных пород, которые в условиях Германии можно обследовать вместе для
вычисления LGW (VDI, 1995) (в скобках-обхват ствола дерева в см на высоте 1,5 м)
Деревья с субнейтральной корой Деревья с умеренно кислой корой Деревья с кислой корой
Acer platanoides (90-280) Acer pseudopl at anus (100-280) Alnus glutinosa (60-280)
Fraxinus excelsior (90-280) Pyrus communis (80-280) Betula pendula (8-280)
Juglans regia (60-280) Robinia pseudacacia (90-280) Prunus avium (60-280)
Malus sp. (60-160) Tilia cor data (100-280) Prunus domestica (60-160)
Populus sp. (80-280) Tilia platyphyllos (100-280) Quercus robur/petraea (90-280)
Ulmus carpinifolia (90-280)
108 5. Видовое разнообразие лихенобиоты и качество воздуха территории
Наклон деревьев допускается не более чем на 5° (отклонение от перпендику-
ляра или отвеса). Учетная площадка на стволе дерева должна располагаться
таким образом, чтобы по возможности обеспечить гомогенность лишайникового
покрова. Подлежащие обследованию деревья должны выбираться так, чтобы
они: а) были представительными для изучаемой территории, б) находились на
каждом участке в сходных условиях.
Учетную площадку 20x50 см, представляющую собой решетку из 10 квадра-
тов величиной 10x10 см, закладывают на наиболее обросшей лишайниками сто-
роне ствола; нижняя, узкая, сторона площадки располагается на высоте 1 м от
поверхности почвы. Поскольку возможно повторное обследование, рекомендует-
ся точная и устойчивая маркировка учетной площади на дереве. Прежде всего,
фиксируется видовой состав всех лишайников, которые представлены на стволе
дерева в пределах учетной площадки (экземпляры, диаметр которых менее 3 мм,
не учитываются, чтобы избежать ошибки определения). Затем отмечается, в
каком числе квадратов представлен каждый вид (встречаемость). Виды, кото-
рые не представлены в пределах 10 квадратов, но находятся в непосредственной
близости к учетной площадке, также фиксируются (их встречаемость принима-
ется за 1).
После определения видов лишайников вычисляется LGW согласно приведен-
ной выше формуле. При расчете LG W принимаются во внимание только те виды,
которые внесены в список. Сначала вычисляют LGW для каждой основной еди-
ницы измерения (квадрата) территории. Значение LGW более крупных квадратов
(2Х2 км, 4x4 км) определяется вычислением арифметического среднего из вели-
чин LGW четырех квадратов в углах крупных квадратов.
В Германии присутствие Lecanora conizaeoides при определении суммы
встречаемостей видов в расчет не принимается. Но реакция представителей этого
вида в пределах экстремально высоко загрязненных территорий четко выражает-
ся ступенчатостью величин степени покрытия. Там, где другие виды представ-
лены слабо (LGW = 0), все же допускается определение зоны загрязнения по
величинам покрытия L. conizaeoides. Степень покрытия вида в пределах учет-
ной решетки определяется с 10% шагом. Процедура вычисления аналогична ме-
тоду сумм встречаемостей. Оценка по L.conizaeoides заканчивается там, где
заметно присутствие представителей других видов (VDI, 1995).
Полученные величины LGV/ группируются в классы качества воздуха (LGK),
которые представляют различные ступени качества воздуха территории. Шири-
на LGK определяется величиной стандартного отклонения полученных значений
и соответствует половине доверительного интервала. Если стандартное отклоне-
ние велико и LGK широкие, то внутри имеющегося градиента значений LGW можно
выделить немного ступеней качества воздуха; при маленьком отклонении можно
получить дифференцированный градиент загрязнения. В Гессене для оценки ка-
чества воздуха по показателям эпифитных лишайников ширина LGK была опре-
5. Видовое разнообразие лихенобиоты и качество воздуха территории 109
делена в 12,5, т.е. 0-12,5-25-37,5-50 (Kirschbaum, Windisch, 1995). Для альпий-
ских районов с особым разнообразием условий рекомендуется другая шкала LGK:
0-30-42,5-55-67,5 (VDI, 1995). В г. Измире (Турция) ширина класса была гораз-
до меньше: 0-7,3-14,6-21,9-29,2-36,5 (Sommerfeldt, John, 2001). Установленные
подобным образом LGK качественным фоном или штриховкой показывают на
карте, причем интенсивность окраски или плотность штриховки уменьшаются с
улучшение качества воздуха. Эти классы получают качественную словесную
характеристику, например, “умеренно до незначительно загрязненного”. От коли-
чественных показателей загрязнения для LGK рекомендуется сознательно отка-
заться, поскольку несоразмерная точность будет вводить в заблуждение (VDI,
1995).
5.3.4. Индекс развития эпифитных лишайников (ИРЭЛ-IDEL)
Вычисление названных выше индексов (I.P., I.A.P., LGW) требует строгого
соблюдения процедур сбора и обработки исходных данных. Однако бывают си-
туации, когда материал, собранный для решения иной задачи, не связанной с ли-
хеноиндикацией, позднее используется для оценки качества воздуха обследован-
ной территории. Например, в начале 1990-х годов я изучал видовой состав и распро-
странение эпифитных лишайников на территории г. Москвы в пределах Московской
кольцевой автодороги (Бязров, 1994). Для выявления особенностей распростра-
нения лишайников в качестве картографической основы был использован план
города масштаба 1:38000, или в 1 дюйме 1 км (Москва..., 1989). Территория горо-
да на этой карте была покрыта сетью с размером ячей 1 х 1 км2, общее число
которых составило 908. В каждом таком квадрате были осмотрены от 100 до 400
стволов деревьев и кустарников, поверхности других субстратов - почва, древе-
сина, мраморные или гранитные монументы, надгробия - во дворах и лесопарках,
вдоль улиц, на территориях кладбищ и предприятий, в садах, парках, на берегах
различных водотоков и водоемов. Деревья обследовались от основания до высо-
ты 2,5 м, как пряморастущие, так искривленные и наклоненные, и наличие лишай-
ников фиксировалось на всей видимой поверхности ствола. Одновременно отме-
чалось покрытие (%) слоевищами каждого вида обследованной поверхности суб-
страта. Средние показатели покрытия и встречаемость определялись
относительно всего числа обследованных в квадрате стволов деревьев. Таким
образом, для каждого квадратного километра территории города имелись сведе-
ния о видовом составе лишайников, встречаемости каждого вида относительно
числа обследованных стволов деревьев, среднем покрытии представителями вида
поверхности стволов. Кроме того, была определена встречаемость вида относи-
тельно общего числа обследованных квадратов на территории города. На основе
этих данных, а также сведений об экологических особенностях конкретного квад-
ПО 5. Видовое разнообразие лихенобиоты и качество воздуха территории
рата был разработан индекс развития эпифитных лишайников, который позволил
преобразовать имеющиеся характеристики лихенобиоты в оценочную картосхе-
му качества воздуха территории города (Бязров, 1998а, в, г, 1999а). Для каждого
квадрата величина индекса определялась на основе следующей формулы:
ИРЭЛ(ЮЕЬ) = К х (F+f+s),
где п - число видов эпифитных лишайников в квадрате; F - оценка степени рас-
пространения вида j на всей обследованной территории;^- оценка степени рас-
пространения вида j в пределах территории конкретного квадрата; s- оценка
степени надежности обнаружения представителей вида на стволах деревьев в
пределах территории того же квадрата; К - коэффициент, показатель экологиче-
ских особенностей территории в пределах квадрата.
Показатели F,/, s оценивались в баллах по пяти-ранговой шкале. Столько же
рангов у коэффициента К. Они определялись на основе признаков, обобщенных в
таблице 5.3. При этом свойства представителей каждого вида оценивались на
двух уровнях - общетерриториальном (показатель F) и узколокальном (показате-
ли/и ^). Поскольку индикаторная ценность представителей более широко рас-
пространенных на обследуемой местности видов ниже в сравнении с редко встре-
чающимися, то для F бальные оценки увеличивались с уменьшением частоты
встречаемости вида в городе (см. ниже табл. 10.1). На узколокальном уровне,
т.е. в пределах каждого конкретного растра, степень развития представителей
каждого вида определяется условиями данного места, и чем более благоприят-
ны они для вида, тем чаще можно встретить здесь его представителей (показа-
тель У), тем более значительную поверхность субстрата занимают их слоевища и
более высока вероятность их обнаружения (показатель 5).Однако распростране-
ние и развитие лишайников даже в городе зависит не только от загрязнения, но и
от наличия соответствующих местообитаний, степени их близости к естествен-
ным. Территория Москвы в этом отношении далеко не однородна. И хотя здесь
отсутствуют экосистемы, обозначенные в п/п 16 и 17 таблицы 5.3, но достаточно
обычны участки, которым отвечают сообщества п/п 18-20 той же таблицы (см.
ниже рис. 10.16). С помощью коэффициента К, имеющего пяти-ранговую лога-
рифмическую шкалу, делается попытка ввести поправку в величину ИРЭЛ в
связи с общей ландшафтно-экологической неравноценностью территории города.
Чем выше значение ИРЭЛ, тем более благоприятна экологическая ситуация на
участке.
Значение ИРЭЛ варьировало в городе от 0 до 182 при средней величине по
отношению ко всей территории 31 (см. ниже рис. 10.17). Для сравнения, прибли-
женное значение ИРЭЛ, определенное по результатам предшествующих иссле-
дований автора (Бязров, 19696, в), на территории в 25-30 км на юг от МКАД
между Калужским и Киевским шоссе составляет 100-300, а для слабо загряз-
ненной или “чистой” части Демидовского района Смоленской области - 250-550.
5. Видовое разнообразие лихенобиоты и качество воздуха территории 111
Таблица 5.3
Характеристики признаков для оценки показателей индекса
развития эпифитных лишайников
№ п/п Признак F f S К
1 Вид встречен более чем в 50% квадратов всей обследованной территории 1
2 Вид встречен в пределах 20.1-50% квадратов территории 2
3 Вид встречен в пределах 5,1-20% квадратов территории 3
4 Вид встречен в пределах 0,5-5.0% квадратов территории 4
5 Вид встречен в пределах менее чем 0,5% квадратов территории 5
6 Вид в пределах квадрата встречен более чем на 50% обследованных деревьев 5
7 Вид в пределах квадрата встречен на 20,1-50% деревьев 4
8 Вид в пределах квадрата встречен на 5,1-20% деревьев 3
9 Вид в пределах квадрата встречен на 0,6-5,0% деревьев 2
10 Вид в пределах квадрата встречен на менее чем 0,5% деревьев 1
11 Надежность обнаружения в пределах квадрата представителей вида на деревьях очень высокая - покрытие стволов слоевищами более 50% 5
12 Надежность обнаружения в пределах квадрата представителей вида на деревьях высокая - покрытие стволов слоевищами 20,1-50% 4
13 Надежность обнаружения в пределах квадрата представителей вида на деревьях средняя - покрытие стволов слоевищами 5,1-20% 3
14 Надежность обнаружения в пределах квадрата представителей вида на деревьях низкая - покрытие стволов слоевищами 0,6-5,0% 2
15 Надежность обнаружения в пределах квадрата представителей вида на деревьях очень низкая - покрытие стволов слоевищами менее 0,5% 1
16 Квадрат полностью или частично является частью естественной экосистемы с малозаметным воздействием деятельности человека 1,0
17 Квадрат включает естественные экосистемы с заметным воздействие деятельности человека при отсутствии сообществ, названных выше в п/п 16 1,25
18 Квадрат включает сообщества лесопарков и лесных культур старше 60 лет, а также сравнительно естественные экосистемы долин рек при отсутствии сообществ, названных выше в п/п 16, 17 1,60
19 Квадрат включает городские парки и сады, территории кладбищ, долины рек в пределах населенных пунктов, водоемы, овраги при отсутствии сообществ, названных выше в п/п 16-18 2,0
20 Квадрат включает только застроенные территории и крупные магистрали, зеленые насаждения представлены скверами, насаждениями вдоль магистралей и внутри дворов при отсутствии сообществ, названных выше в п/п 16-19 2,50
Полученные для Москвы интегрированные данные стали основой для создания
картосхемы экологической ситуации в городе (см. ниже рис. 10.18). Почти 40%
территории города в пределах МКАД приходится на участки с очень высоким
112 5. Видовое разнообразие лихенобиоты и качество воздуха территории
уровнем загрязнения (ИРЭЛ = 0-10). Выдел картосхемы с высоким уровнем заг-
рязнения (ИРЭЛ = 11-30) занимает около 21% территории города. На участки с
умеренно высоким уровнем загрязнения (ИПЭЛ = 31-70) приходится почти 27%
территории города. Участки со средним уровнем загрязнения (ИРЭЛ = 71-150)
занимают в Москве около 13% территории (к этому выделу отнесены и два квад-
рата, включающих Востряковское кладбище, с максимальными на обследован-
ной местности величинами ИРЭЛ, превышающими 150-176 и 182). Районов с
чистым воздухом в Москве нет. Результаты лихеноиндикационных исследований
в Москве детально изложены в отдельном разделе данной монографии (гл. 10).
5.4. ОГРАНИЧЕНИЯ
Анализ многочисленных источников, лишь частично названных в тексте, по-
казывает, что в целях индикации свойств атмосферы используют картографиро-
вание различных показателей жизнедеятельности лишайников. Чаще всего это
картографирование распространения видов и деление территории на изотоксич-
ные зоны на основе вычисленных значений индексов. Каждый метод имеет свои
преимущества и недостатки. Картографирование распространения - более про-
стой метод, но, вероятно, с его помощью невозможно в полной мере выявить
колебания качества воздуха в пространстве. Напротив, картографирование на
основе индексов (I.P., I.A.P., LGW и других) более показательно при оценке про-
странственных изменений качества воздуха, но требует значительных затрат вре-
мени, и точность этого способа также не высока (Case, 1984). Выбор приемлемо-
го метода зависит от географических особенностей изучаемой территории, сте-
пени воздействия загрязнителей и требуемой точности.
Карты распространения обычных в обследуемой местности видов лишайни-
ков, дополненные картой распределения показателей видового разнообразия тер-
ритории, позволяют достаточно надежно провести дифференциацию территории
в отношении качества воздуха. Получаемая при этом информация сравнима с
комплексной количественной процедурой сбора данных. Когда первичные сведе-
ния (LeBlanc, DeSloover, 1970) по определению величин I.A.P. в Монреале (Кана-
да) были подвергнуты компьютерному анализу, то обнаружили, что если бы для
выделения изотоксичных зон воспользовались лишь одним методом - количе-
ством видов в точке, была бы потеряна очень незначительная информация
(LeBlanc, 1971). Это позволило Ф. Лебланку заключить, что информация, полу-
ченная на основе карт распространения видов в Монреале, является столь же
полной и емкой, что и та, которая получена с помощью более сложных и трудоем-
ких методов. Аналогичное заключение можно сделать и на основе сравнения ли-
хеноиндикационных карт (см. ниже рис. 10.14,15 и 18), созданных для территории
Москвы разными методами (табл. 5.4 и 5.5). Сравнение трех вариантов деления
5. Видовое разнообразие лихенобиоты и качество воздуха территории 113
Таблица 5.4
Доля (%) территории Москвы (в пределах МКАД), занимаемая выделами одного ранга
опасности для эпифитных лишайников, на трех вариантах картосхем
Выдели на картосхемах 1 - катастрофически опасные участки Вариант картосхемы
По числу видов в квадрате (см. ниже рис. 10.14) По присутствию в квадрате видов опреде- ленного класса чувстви- тельности (см. ниже рис. 10.15) По величине индекса развития эпифитных лишайников в квадрате (см. ниже рис. 10.18)
40 33 40
2 - критически опасные участки 38 38 20
3 - крайне опасные участки 17 12 27
4 - опасные участки 5 17 13
Таблица 5.5
Совпадение (%) квадратов выделов одного ранга на трех типах картосхем
для территории Москвы
Выдел 1ч 2ч Зч 4ч 1и 2и Зи 4и
1в 81,7 17,5 0,3 0,5 98,1 1,9 — —
2в — 75,1 19,5 5,4 0,6 50,3 48,5 0,6
Зв — 10, 27,3 62,7 — 1,3 47,3 51,4
4в — — 7,1 92,9 — — — 100,
1ч 100 100 — — —
2ч 100 18,1 51,2 28,7 2,
Зч 100 — 5,3 77,9 16,8
4ч 100 2,6 35,7 61,7
в - по числу видов, ч - по присутствию видов определенного класса чувствительности, и - по ИРЭЛ
(см. ниже рис. 10.14, 15 и 18)
территории Москвы по степени опасности для эпифитных лишайников показыва-
ет, что все они дают примерно одинаковое общее представление о пригодности
разных частей города для поселения этих организмов, косвенно отражая каче-
ство воздушного бассейна местности. Однако какой способ точнее, сказать трудно,
поскольку доступные данные об уровнях загрязнения территории города доволь-
но обобщенные. За состоянием атмосферного воздуха инструментально наблю-
дают лишь на 16 стационарных постах, расположение которых далеко не репре-
зентативно для достаточно разнообразных условий города.
114 5. Видовое разнообразие лихенобиоты и качество воздуха территории
Картографирование распространения одного вида-индикатора также счита-
ют ценным способом мониторинга качества воздуха (Showman, 1988). Это про-
стой и быстрый метод индикации воздействия; по крайней мере, в этом случае
имеется один уровень чувствительности рецептора. Некоторые исследователи
предполагают, что мониторинг изменения окружающей среды наилучшим обра-
зом можно проводить с помощью единственного вида (Henderson-Sellers, Seaward,
1979; Seaward, 1980; Pfeifer, Barclay-Estrup, 1992; Bruteig, 1993b; Jordonet et al.,
1995). Однако необходимо провести предварительное исследование для установ-
ления подходящего индикаторного вида. Например, Р. Шоуман (Showman, 1975)
сначала изучил распространение нескольких видов и, установив, что Flavoparmelia
caperata является обычным видом на подлежащей обследованию территории,
да еще и чувствительным к низкому качеству воздуха, использовал его в даль-
нейшей работе как вид-индикатор (Showman, 1981, 1990, 1997). Однако имеются
серьезные аргументы против использования всего лишь одного вида для индика-
ции качества среды (Ausmus, 1984; Cairns, 1984).
На территориях, где явное воздействие загрязнителей на распределение ли-
шайников при рекогносцировочном обследовании не обнаруживается, необходи-
мо использовать картографирование I.A.P. или какого-либо иного комплексного
показателя. Нередко применяются различные модификации индекса чистоты
атмосферы. Один из авторов этого индекса на примере Монреаля показал, что
немного было бы потеряно, если бы для подсчета значений 1.А.Р. использовались
всего 4 вида-индикатора (LeBlanc, 1971). Это подтвердили исследования и в дру-
гих регионах (Mathis, Tomlinson, 1972; Case, 1980; Herzig, Urech, 1991). Лихеноин-
дикация качества воздуха в городах Украины (Кучерявый и др., 1989; Кондратюк
и др., 1991,1993), в Москве (Бязров, 19966,1998а, 1999а; Бязров и др., 1997; Мак-
симова, Бязров, 2000) и в Бельгии (Geebelen, Hoffmann, 2001) показала, что выяв-
ленные различными методами (индексы, показатели видового разнообразия, на-
личие видов определенного класса чувствительности) изотоксичные зоны имеют
довольно сходную конфигурацию.
Трудно или даже невозможно выделить один метод как лучший в той или иной
ситуации. Обычно наилучший метод тот, который наиболее прост и который обес-
печивает требуемую степень точности. При этом желательно помнить, что кар-
ты распространения лишайников или каких-то иных их показателей отображают
пространственные географические особенности учитываемых параметров, сви-
детельствующих о степени благополучия жизни лишайников в тех или иных мес-
тообитаниях. И если исследование должным образом продумано, различия пока-
зателей степени благоприятности существования лишайников можно связать с
различиями в качестве воздуха их местообитаний и, соответственно, различных
частей обследованной территории.
Нередко составленные карты тех или иных показателей лишайников пытают-
ся скоррелировать с данными инструментальных измерений, характеризующих
5. Видовое разнообразие лихенобиоты и качество воздуха территории 115
качество воздуха. Хотя исследования показывают, что карты лишайников в той
или иной степени отражают очертания воздушного загрязнения, корреляция дан-
ных о свойствах лишайников с концентрациями SO2 или других фитотоксикантов
наблюдается далеко не всегда. В некоторых местностях двуокись серы действи-
тельно является главнейшим загрязнителем, однако на некоторых территориях на
развитие лишайников сильно влияют озон, соединения азота, PAN и токсичные
металлы. Трудно установить корреляцию показателей лишайников с одним ток-
сикантом, когда другие не рассматриваются. Снижение давления одного загряз-
нителя может привести к усилению воздействия другого. Например, в Нидерлан-
дах в связи с введением законодательных актов о защите воздуха, с 1970-х годов
происходило уменьшение выбросов двуокиси серы. Наблюдения за лишайниками
показали, что в эпифитном лишайниковом покрове виды кислых субстратов
(Lecanora conizaeoides, Hypogymnia physodes, Evernia prunastri) уступают свои
позиции видам щелочных или нейтральных субстратов (представители родов
Physcia, Xanthoria, Candelariella). Это позволило сделать вывод о том, что в
новых условиях для лишайников более важна кислотность субстрата, чем кон-
центрация аммиака (de Bakker, 1989).
Показатели лишайников обычно сравнивают со среднегодовыми концентра-
циями загрязнителей, так как это наиболее доступная информация. Такое сравне-
ние полезно в регионах с относительно постоянным низким уровнем загрязнения
(Gilbert, 1970; Hawksworth, Rose, 1970). Также показано сходство между разме-
ром и конфигурацией изотоксичных зон, выделенных по значениям I.A.P., и мно-
голетними средними концентрациями двуокиси серы (LeBlanc et al., 1972b; Deruelle,
Guilloux, 1993). Однако статистически такая корреляция не установлена. В Ни-
дерландах провели сравнение показателей обилия лишайников с различными кон-
центрациями двуокиси серы и установили более тесную зависимость развития
лишайников от значений зимних максимумов концентрации, а не от среднегодо-
вых ее величин (de Wit, 1976). В соседней Бельгии также нашли статистически
значимую корреляцию показателей развития лишайников (I.A.P.) с величинами
зимних концентраций SO2 за период 1990-1994 гг., причем она более тесная, если
для определения 1.А.Р. используется ограниченное число (17) видов эпифитных
лишайников (Geebelen, Hoffmann, 2001). В США сравнили величины I.A.P. с пока-
зателями одногодичных средних выпадений сульфатов, взвешенных частиц и пыли
(Mathis, Tomlinson, 1972). Была установлена статистически не достоверная кор-
реляция между величинами I.A.P. и любым из названных показателей загрязне-
ния. В Канаде также обнаружена слабая корреляция между величинами 1.А.Р. и
показателями среднесуточных выпадений сульфатов за год, что позволило сде-
лать заключение о наличии других факторов, влияющих на обилие лишайников и
их жизненность (Case, 1980). Исследователь пришел к выводу, что лишь учет
интенсивности, продолжительности и частоты выбросов загрязняющих веществ
вместе с показателями степени чувствительности лишайников к каждому из них
116 5. Видовое разнообразие лихенобиоты и качество воздуха территории
может дать результаты, тесно коррелирующие с признаками, характеризующими
степень угнетения лишайников.
Одна из причин тщетности попыток установления тесных корреляций между
показателями развития лишайников и количественными характеристиками каче-
ства воздуха, возможно, связана с тем, что, как правило, при этом используются
данные инструментальных измерений концентраций лишь одного года или несколь-
ких лет. Однако в большинстве случаев количественные данные о качестве воз-
духа за один год не сопоставимы с таковыми за несколько предшествовавших
лет. Качество воздуха - всего лишь одно из свойств среды, определяющих бла-
гополучие лишайников в их местообитаниях. Лишайники зависят и от других эко-
логических факторов. Работы по картографированию лишайников в Нидерландах
показали, что высокие концентрации SO2(>80 мкг/м3) - это сильный лимитирую-
щий фактор для их развития, но при более низких концентрациях двуокиси серы
важны и другие свойства среды, которые необходимо учитывать при интерпрета-
ции данных лихеноиндикационного исследования в отношении качества воздуха
(de Witt, 1976).
Материалы картографирования распространения лишайников и признаков их
развития, весьма наглядные для показа дифференциации территории в отношении
воздействия загрязнения воздуха, не коррелируют с концентрациями токсикантов
и, следовательно, не могут быть использованы для определения концентрации
загрязнителя с какой-либо степенью точности на территориях, где не проводи-
лись инструментальные измерения. Таким образом, оправданны сомнения в пер-
спективности использования их для установления средних многолетних стандар-
тов качества воздуха (Sigal, 1988). Тем не менее, картографирование распростра-
нения лишайников и показателей их развития воссоздает общую картину
пространственной дифференциации качества воздуха как результата предшеству-
ющего воздействия источников загрязнения, а хронологически последовательная
серия исследований может показывать динамику качества воздуха и степень
эффективности мероприятий по его улучшению.
6. ГРУППИРОВКИ ЛИШАИНИКОВ И ИНДИКАЦИЯ
КАЧЕСТВА ВОЗДУШНОГО БАССЕЙНА
Лишайники, как и другие живые организмы, в ходе освоения предоставляе-
мого природой пространства, существуют ассоциированно как с представителя-
ми других таксономических групп, так и друг с другом, образуя исторически сфор-
мировавшиеся группировки с определенным составом и соотношениями между
видами на конкретных площадях. Эти комплексы видов отражают не только фи-
зико-географические условия местности, историю расселения представителей
живых организмов, но и экологические особенности их местообитаний, конку-
рентные отношения между организмами. Соответственно, в природе встречают-
ся сходные комбинации видов лишайников, которые можно типизировать, и на
этой основе по признакам сходства или различия создавать иерархические клас-
сификации этих сочетаний. Лишайниковые группировки рассматриваются или
как самостоятельные сообщества, или как структурные части фитоценозов, био-
геоценозов, экосистем. Так, в Западной Европе, где доминируют идеи франко-
швейцарской геоботанической школы Браун-Бланке, ассоциации лишайников груп-
пируют по сходству состава видов в альянсы, альянсы - в порядки, а порядки - в
классы. Для выделения этих синтаксонов используют верные виды, т.е. виды,
встречающиеся, например, в ассоциациях одного альянса, или в альянсах одного
порядка и т.д. Наименование синтаксонов регулируется правилами Кода фитоце-
нологической номенклатуры (Barkman et al., 1986), который представляет собой
модификацию Кода ботанической номенклатуры. Основу названия сообщества
составляет научное название вида, а ранг характеризуется определенным оконча-
нием: у ассоциации окончание -etum; у альянса -ion; у порядка -etalia; у класса -
etea. Также указываются автор, описавший сообщество, и год описания (Klement,
1955; Barkman, 1958).
Несколько по-иному трактовались лишайниковые группировки в Упсальской
геоботанической школе. Здесь основной геоботанической единицей признавалась
не ассоциация, а синузия, структурно, флористически и экологически гомоген-
ная часть растительного сообщества (Трасс, 1966). Соответственно, лишайнико-
вые группировки разного ранга обозначались как социетет, унион, федерион и
формион.
При анализе структуры биогеоценозов (экосистем) оперируют таким терми-
ном как “биогеоценотическая синузия”, которым обозначают разные по объему
118
6. Группировки лишайников и индикация качества воздушного бассейна
структурные части биогеоценоза (Сукачев, 1964). Анализ места формируемых
лишайниками группировок в системе структурных элементов биогеоценоза дал
мне основание предложить следующую иерархическую систему биогеоценоти-
ческих синузий разного ранга: (а) парцелла - элемент горизонтального расчлене-
ния биогеоценоза на всю его вертикальную толщу; (аа) биогеоценотический го-
ризонт-элемент вертикального расчленения парцелл; (ааа) эндостратосинузия -
естественно обособленная и биогеоценотически нерасчленимая элементарная часть
биогеоценотического горизонта (Бязров, 1970). При этом всем названным струк-
турным подразделениям разного ранга придавалось типологическое значение.
Элементарные по объему лишайниковые группировки -лихеносинузии- в моем
понимании соответствуют эндостратосинузиям.
Однако в любом случае по определенной методике возможно сделать описа-
ния конкретных лишайниковых группировок, сравнить их и по принятым крите-
риям сгруппировать сходные описания в некоторые абстрактные типы. Эти абст-
рактные типы (и часто соответствующие им конкретные местообитания тоже) и
называют либо сообществами, либо лихеносинузиями. Для выявления лишайни-
ковых группировок очень полезен геоботанический опыт, который дает большое
преимущество исследователю. Поскольку при лихеноценологических работах
непосредственно в поле делается большое число описаний ассоциирования од-
них видов с другими в определенных экологических условиях, достаточно велика
роль субъективного фактора в таких описаниях. Для уменьшения его влияния на
выделение экологических групп или сообщества используют особые методы сбо-
ра материала, в частности, случайное распределение учетных площадок, закла-
дываемых для описания лишайниковых группировок, а для выделения типов
лишайниковых группировок применяют приемы математической статистики
(Hurka, Winkler, 1973). Таким образом, хотя в процедуре выявления типов лишай-
никовых группировок или сообществ имеется значительная доля субъективизма,
результат может быть проверен объективными способами.
Изменение видового состава лишайников под влиянием загрязнения вызыва-
ет соответствующие изменения и формируемых лишайниками группировок или
сообществ. Такие изменения устанавливают сравнением описаний лишайнико-
вых группировок, сделанных одним методом на одних и тех же участках через те
или иные интервалы времени. Иногда также сравнивают состав лишайниковых
группировок однотипных участков обследуемой местности, различающихся лишь
степенью загрязнения или по иным показателям (Аблаева, 19816; Михайлова,
1993, 1994, 1996; Giordani et al., 2001).
6.1. МЕТОДЫ СБОРА ЛИХЕНОЦЕНОЛОГИЧЕСКИХ ДАННЫХ
Процедура сбора материалов для характеристики лишайниковых группиро-
вок зависит от решаемых задач, заданной точности исследования, особенностей
6. Группировки лишайников и индикация качества воздушного бассейна
119
изучаемой территории, типа загрязнения и размещения его источников, а также
от числа исполнителей, их квалификации, технического оснащения. Она включа-
ет: (а) выбор на местности участков для характеристики лишайниковых группи-
ровок; (б) закладку учетных площадок для описания встреченных комплексов
видов лишайников; (в) анализ сделанных описаний и выделение типов ассоции-
рования лишайников.
6.1.1. Выбор участков для характеристики
лишайниковых группировок
Выбору постоянных участков для наблюдения за лишайниковыми группи-
ровками обычно предшествует рекогносцировочное обследование местности, с
целью получения представления об ее физико-географических особенностях, раз-
нообразии субстратов для заселения лишайниками, их доступности для изучения
и распространении на рассматриваемой территории. Выбираемые участки долж-
ны быть стандартизованы в той мере, насколько это возможно. При этом крайне
желательно сформулировать критерии выбора участков. Число участков зависит
от сложности структуры биогеоценотического покрова территории, ее размеров и
целей исследования. Предварительный сбор данных позволяет с помощью стан-
дартных приемов статистики определить необходимое число таких участков для
достижения требуемой степени точности. На практике, однако, такие возможно-
сти бывают далеко не всегда, и в этих случаях используют эмпирические прави-
ла, согласно которым для каждого важного контролирующего фактора или гради-
ента необходимо 20 дополнительных участков (Stolte et al., 1993). Например, для
определения различий сообществ по градиенту от точечного источника загрязне-
ния на гомогенной равнине достаточно 20 участков вдоль трансекты. Если же
изучаются различия на трех уровнях поселения или трех субстратах, тогда жела-
тельно 60 участков (3 важных фактора х 20 участков/фактор = 60 участков).
6.1.2. Обследование выбранных участков местности
Для изучения особенностей лишайникового покрова выбранных участков ис-
пользуют учетные площади. Их число и размеры зависят от целей работы, осо-
бенностей участков, возможностей исполнителей. Было установлено, что имеет-
ся взаимозависимость между количеством учетных площадок, их размерами и
фиксируемой точностью ряда показателей лишайникового покрова (McCune,
Lesica, 1992; табл. 6.1). Крайностями являются одиночные большие квадраты для
характеристики участка сообщества против множества мелких учетных единиц
(точечный сбор). Также возможны различные промежуточные варианты разме-
ров учетных единиц.
120 6. Группировки лишайников и индикация качества воздушного бассейна
Таблица 6.1
Зависимость точности характеристики ряда показателей лишайникового покрова
от числа и размеров учетных площадок (McCune, Lesica, 1992)
Показатель Точность при разных вариантах количества и размеров учетных площадок
Несколько и больших площадок М ножество маленьких площадок
Учет малозаметных видов Пониженная. Имеется опасность, что некоторые виды, особенно латентные, непреднамеренно выпадут из поля зрения Повышенная. Маленькие учетные единицы приучают глаз к специ- фическим пятнам, снижают вероятность непреднамеренных ошибок наблюдателя в учете видов
Визу альная оценка признаков Высокая. Использование визуальных интегрированных оценок на большой площади является эффективным способом преодоления нормально высокой степени гетерогенности даже в «гомогенном» сообществе Низкая. Интегрирующая способ- ность глаза становится минималь- ной, требуется использование проб очень большого размера для достижения сравнимых данных
Включение ред- ких и необычных видов Высокая. Использование визуальных интегрированных оценок способствует выявлению редких видов Низкая. За исключением проб очень большого размера, редкие и необычные виды пропускаются
Определение покрытия обычных видов Пониженная. Классы покрытия на немногих больших у четных площадях выражаются широкими оценками классов с меньшей точностью и акку- ратностью, чем полу ченные из многих небольших единиц Повышенная. Более точные и аккуратные оценки покрытия обычных видов
Затраты времени на сбор данных Различны в зависимости от сложности и степени развития растительности. Не су щественно отличаются от вариан- та с множеством небольших у четных единиц Различны в зависимости от степени развития растительности. Не су щественно отличаются от варианта немногих крупных у четных единиц
Затраты времени на анализ данных Ускоренные. Данные, введенные на уровне участка, включаются непосред- ственно в анализ на у ровне участка Замедленные. Точечные данные или данные микроплощадок сначала необходимо реду цировать (вручную, на калькуляторе или компьютере) в оценки на у ровне участка
Поскольку эти учетные площади должны обследоваться повторно через опре-
деленные интервалы времени, необходима точная их маркировка. Для каждой
площади отмечают дату заложения и обследования, идентификационный номер,
точные координаты и ориентиры, позволяющие в дальнейшем ее найти, а также
описание территории.
6. Группировки лишайников и индикация качества воздушного бассейна
121
таблица 6. / (окончание)
Анализ выборок Изменчивость внутри участка не оценивается. Оценивается варьирование внутри участка
Рекомендации Крайний случай - одна большая площадь - в большинстве случаев полезен при условии тщательной инвентаризации видов Крайний случай - точечный сбор данных - полезен, когда малознат чимы редкие и необычные виды и желательны точные оценки для обычных видов. В большинстве случаев лучшим компромиссом является использование неболь- шого числа крупных у четных единиц
6.1.3. Размещение учетных подъединиц
В большинстве руководств по статистике рекомендуется случайное или регу-
лярно-случайное размещение точек для описания особенностей ассоциирования
представителей организмов и их признаков или отбора проб. Однако планируе-
мое при мониторинге во времени повторное обследование предполагает обследо-
вание тех же точек, и таким образом, при последующем сборе данных происхо-
дит переход к регулярному размещению точек учета. В большинстве случаев при-
емлемым является пространственно равномерное расположение учетных единиц,
однако конкретное их размещение и размеры зависят от субстрата (табл. 6.2).
6.1.4. Фиксируемые показатели
На учетных площадках фиксируется наличие представителей видов лишай-
ников, а также в большинстве случаев процент покрытия, определяемый с помо-
щью квадрат-сетки или визуально. Часто покрытие вида выражают не в процен-
тах, а в баллах, соответствующих установленным классам покрытия. Большин-
ство применяемых шкал покрытия обычно имеют узкий шаг в начале и в конце и
широкий - в середине, например, 0, 1,5, 25, 50, 75, 95, 99, поскольку числовые
значения (баллы) этих классов являются приближенными арксинусами квадрат-
ного корня из величины процента покрытия (McCune 1988; Muir, McCune, 1988;
Stolte et al., 1993). Нами применялась приближенная логарифмическая шкала: 0,
0.5, 5, 20, 50 (см. табл. 5.3). Известна Скандинавская шкала Хульта-Сернандера
1,56, 3,13, 6,25, 12,5, 25, 50, 75... (Oksanen, 1976). Х.Х. Трасс (1968) предложил
Ю-бальную шкалу со следующими классами покрытия (%): 1-3, 3-5, 5-10, 10-
20, 20-30, 30—40, 40-50, 50-65, 65-80, 80-100. Имеются шкалы, совмещающие
показатели обилия и покрытия видов (Wirth, 1988) (табл. 6.3).
122 6. Группировки лишайников и индикация качества воздушного бассейна
Таблица 6.2
Примеры размещения учетных подъединиц на конкретных участках
для определения процента покрытия видов лишайников
Су бстрат Специфика случая Число у четных единиц, их размер и размещение Источник
Ветви Нижние живые ветви на открыто стоящих деревьях Пять 5-летних и пять 10-летних отрезков ветвей дуба и ясеня Stolte et al., 1993
Крупные, низко располо- женные ветви на открыто стоящих деревьях 100 равноудаленных точек через 3 см налейте по спирали вокруг ветвей Stolte et al., 1993
Ветви нижнего и среднего полога в хвойных лесах 24 однометровых части ветви отрезали от центральной точки ветвей, наиболее близко располо- женных к точкам на4-м интервалах вдоль трех параллельных трансект Lesica et al., 1991
Камни Высокое покрытие лишай- ников, каменные стены и естественные обнажения площадью более 1х 1 м По числу соприкосновений точек пересечения сетки 10х20 см пло- щадки 100х 100 см с представителя- ми видов, растущих на горных породах, три каменистых поверхно- сти на у частке John, 1989a, b; John, Dale, 1990
Высокое покрытие лишай- ников на естественных выходах горных пород в сообществах горной лесостепи Площадки 10х 10 см, не менее 20 на участке Бязров, 1990
Высокое покрытие лишай- ников на естественных выходах горных пород На участках 10х 10 м закладывались учетные площадки 20х20 см Седел ь никова, 1994
Почва Большей частью, рассеян- ная флора в лесу 100 площадок 10х30 см на участок с регулярными интервалами на пар ал лел ь ных тр ансектах; 45 площадок 20х50 см с регуляр- ными интервалами на параллельных тр ансектах McCune, Antos. 1982; Lesica et al., 1991
Горная лесостепь Площадки 50х50 см, не менее 20 на участке Бязров, 1990
Г ор ные сообщества На участках 10х 10 м закладывались учетные площадки 50х50 см или 100х100 см Седел ь никова, 1994
6. Группировки лишайников и индикация качества воздушного бассейна
123
таблица 6.2 (продолжение)
Стволы деревьев Большей частью рассеянная флора вблизи городов или крупных точечных источников загрязнения Однометровые цилиндры, по 10 деревьев каждого вида на участок; использовались определенные виды деревьев; двухметровый цилиндр, 10 деревьев на участке McCune, 1988; Muir, McCune, 1988; Stolte et al., 1993
Большей частью рассеянная флора вблизи городов или крупных точечных источников загрязнения По 4 у четных площадки 10х 10 см или 20х20 см на основаниях деревьев и на высоте груди, ориентированных на север, восток, юг, запад; 10-40 деревьев на у частке Горшков, 1986; Использование эпифитных лишайников..., 1991
Равнинные и горные леса Восточной Германии Цилиндры на высоте 0-30 см и 120— 150 см на разных видах деревьев на площади 50х50 м; всего более 25 деревьев Бязров, 2001
Широколиственный лес в 50 км от Москвы Цилиндры на высоте 0-60 см и 60- 200 см на разных видах деревьев на площади 50х50 м; всего более 200 деревьев Бязров, 1993а
Березовые леса на Урале По 4 у четных площадки 20х20 см, ориентированных на север, восток, юг, запад, на основаниях деревьев и на высоте груди; более 30 деревьев на участке .Бязров, 19936
Большей частью рассеянная флора вблизи городов или крупных точечных источников загрязнения Площадки 10х25 см на высоте 1.3 м, с 2-4 сторон ствола Кравчук, 2001
Большей частью рассеянная флора вблизи городов или крупных точечных источников загрязнения Площадки 10х 10 см на стволах Pinus sylvestris и Populus balsam iferа Пауков, 2001
Недостатком бальных оценок являются существующий субъективизм отдель-
ных исследователей, поэтому их данные трудно сравнивать. Это относится и к
показателям встречаемости (процент учетных площадок, содержащих данный вид),
ибо встречаемость сильно зависит от размера учетных единиц, стандартизация
которых невелика (Stolte et al., 1993).
124 6. Группировки лишайников и индикация качества воздушного бассейна
таблица 6.2 (окончание)
Стволы деревьев Богатые и рассеянные флоры в региональном исследовании Британское исследование окисления: 100 равномерно рассеянных сеточных точек в квадрате 18Х27 см проволочной сетки; су бъектив ное расположение квадратов, на постоянно маркирован- ных определенных видах деревьев (преимущественно Quercus, Fraxinus)\ высота и экспозиция варьируют; разное число квадратов на участке (1-20) Stolte et al., 1993
Весьма богатая флора в широколиственных лесах Квадрат 25х25 см на СВ экспозиции ствола на высоте 1,4 м на 10 деревьях одного вида на участок Will-Wolf, 1980b
Г ор ные сообщества На участках 10х 10 м на основаниях деревьев (до 0,6 м) и на высоте груди закладывались учетные площадки 20х20 см Седельникова, 1994
Таблица 6.3
Совмещенная шкала обилия и покрытия видов (Wirth, 1988)
Символ г 4- 1 2m 2а 2Ь 3 4 5
Число экземпляров 1-2 >2 <20 >20 Разное Разное Разное Разное Разное
Покрытие, % <2 <2 2-5 2-5 5-12,5 12,5-25 25-50 50-75 75-100
Участие вида в составе лишайниковой группировки иногда характеризуется
таким показателем как обилие и для его оценки используют те или иные шкалы.
Например, в Канаде и на Аляске известна Канадская 9-бальная шкала, где:
1 = редкие, одиночные экземпляры; 2 = немногие спорадически встречающиеся
экземпляры; 3 = одиночные пятна или группы видов; 4 = несколько спорадически
встречающихся экземпляров; 5 = немногие пятна или группы; 6 = несколько хо-
рошо распространенных пятен или групп; 7 = непрерывно равномерно встречаю-
щиеся хорошо распространенные экземпляры; 8 = непрерывно, но с немногими
промежутками; 9 - непрерывно и плотно (Stolte et al., 1993). В рамках разрабо-
танной в США исследовательской программы “Мониторинг здоровья лесных эко-
систем (FHM)” для оценки обилия кустистых и листоватых лишайников приме-
няют более простую шкалу (McCune, Dey, 1994; Бязров, 19986): 1 = очень редкий
вид (3 и менее особей на учетной площади); 2 - редкий (4-10 особей на площа-
ди); 3 = обычный (10 особей на площади, но вид представлен менее чем на поло-
вине стволов или ветвей); 4 = обильный (вид представлен более чем на половине
6. Группировки лишайников и индикация качества воздушного бассейна
125
стволов или ветвей). Эти шкалы, вероятно, большей частью более приемлемы для
обширных региональных исследований, использующих как учетные единицы
одиночные большие пробные площади без мелких учетных площадок на этих
больших площадях.
6.2. ВЫДЕЛЕНИЕ ТИПОВ ЛИШАЙНИКОВЫХ ГРУППИРОВОК
Основу последующей работы по выявлению единств, называемых согласно
принципам академической школы, исповедуемой исследователем, либо синузия-
ми, либо сообществами, составляют описания конкретных лишайниковых груп-
пировок. Так, последователи школы Браун-Бланке выделяют ассоциации лишай-
ников по верным видам, устанавливаемым анализом большого числа описаний
лишайниковых группировок (Wirth, 1988). Биогеоценотические лихеносинузии
ранга эндостратосинузии выделяются по сходству видового состава и жизнен-
ных форм лишайников, развивающихся на одном субстрате и в одном биогеоце-
нотическом горизонте (Бязров, 1990, 1993а, б). Имеются и иные трактовки вари-
антов ассоциирования видов лишайников (Трасс, 1966; Седельникова, Седель-
ников, 1979).
Примером выделения эпифитныхбиогеоценотическихлихеносинузий может
быть исследование в ельнике Piceetum vaccinosum, занимающем средние части
сравнительно пологих склонов Рудных гор (Восточная Германия) (Бязров, 2001).
В древостое представлена только ель (Picea abies}, возраст деревьев 120-160 лет.
В нижнем ярусе преобладают многочисленные и обширные куртины Vaccinium
myrtyllus. Для учета лишайников обследованы 25 стволов ели, в том числе с диа-
метром 24 и 60 см - по 1 дереву; 28, 32, 44 и 48 см - по 3; 36 см - 2; 40 см - 5;
52 см - 4 дерева. На каждом обследованном стволе дерева выделялось два уров-
ня поселения лишайников: основание ствола, или комель - от поверхности почвы
до высоты 0,3 м, и стволовой - на высоте груди - от 1,2 до 1,5 м. Лишайники
на этих уровнях учитывались по всей поверхности ствола, то есть фиксиро-
вались видовая принадлежность встреченных слоевищ и покрытие (%) пред-
ставителями каждого вида поверхности обследуемого обрубка ствола. Описание
каждой учетной площадки на дереве рассматривалось как характеристика кон-
кретной лишайниковой группировки, являющейся частью той или иной лихено-
синузии. Для выделения последних из общего числа описаний учетных площа-
док, сделанных на участках типа биогеоценоза, последовательно вычленялись:
1) приуроченные к одному уровню поселения, то есть к основанию ствола или к
стволу; 2) сходные по видовому составу и жизненным формам лишайников (на
уровне групп).
Выявленные виды принадлежат к следующим группам жизненных форм
лишайников (Голубкова, Бязров, 1989): однообразнонакипные (Сг), накипные
126 6. Группировки лишайников и индикация качества воздушного бассейна
чешуйчатые (Sq), листоватые рассеченнолопастные ризоидальные (S1), листова-
тые вздутолопастные неризоидальные (С1), бородавчато- или чешуйчато-кустис-
тые шило- или сцифовидные (Sc), листовато-кустистые повисающие (Lfp).
Фактический материал представлен в таблице 6.4. Помимо видового состава,
в ней показаны число видов в лихеносинузии, величина среднего покрытия их
представителями стволов деревьев, а также встречаемость (%) лихеносинузии на
участках, т.е. доля деревьев, на которых обнаружены фрагменты лихеносинузии,
от общего числа обследованных на участках типа стволов деревьев. Следуя ука-
занной выше процедуре, на основаниях стволов ели в Piceetum vaccinosum выде-
лены следующие лихеносинузии (см. табл. 6.4): (i)Cladonia foliacea-Lecanora
conizaeoides, формируемая представителями чешуйчато-кустистых и однообраз-
нонакипных лишайников. Спектр жизненных форм (СЖФ), то есть доля (%) ви-
дов каждой жизненной формы в составе лихеносинузии, - 63Sc 37Cr; (ii) Cladonia
foliacea-Hypogymnia physodes-Hypocenomyce scalaris-Lecanora conizaeoides, СЖФ
- 25Sc 25CI 12Sq 38Cr; (iii) Cladonia foliacea-Hypogymnia physodes-Lecanora
conizaeoides, СЖФ - 50Sc 10C140Cr; (iv) Cladonia foliacea-Hypogymnia physodes-
Parmeliopsis ambigua-Lecanora conizaeoides, СЖФ-498с 17C117S117Cr. На высо-
те груди на стволах ели лишайники формируют следующие синузии (см. табл.
6.4): (j) Lecanora conizaeoides, СЖФ - ЮОСг; (jj) Lecanora conizaeoides-Lepraria
Таблица 6.4
Покрытие (%) лишайников в лихеносинузиях на елях
в Piceetum vaccinosum (Рудные горы)
Лишайник Жизненная форма Лихеносинузия
На комлях На стволах
i ii iii iv j ii iii
Lecanora conizaeoides Сг 3 2 6 5 4 12 14
Cladonia coniocraea Sc 1 - 0,6 - - - -
C. digitata Sc 1 - 2 - - - -
C. foliacea Sc 11 3 10 15 - - -
C. pleurota Sc 1 2 0,3 4 - - -
C. pyxidata Sc 0,2 - 0,1 1 - - -
Lepraria incan a Сг 0,5 1 1 - - i 0,4
Lecidea sphaerella Сг 0,1 0,1 0,1 - - - -
Hypocenomyce scalar is Sq - 0,1 - - - - -
Hypogymnia physodes Cl - 0,5 0,5 2 - - 1
Chaenotheca chrysocephala Сг - - 0,1 - - - -
Parmeliopsis ambigua SI - - - 0,1 - - -
Число видов 12 8 7 10 6 1 2 3
Покрытие (%) 18 9 20 27 4 13 15
Встречаемость (%) 40 4 52 4 16 20 64
Объяснение в тексте.
6. Группировки лишайников и индикация качества воздушного бассейна
127
incana, СЖФ - ЮОСг; (jjj) Hypogymnia physodes-Lecanora conizaeoides, СЖФ -
33C1 67Cr.
63. АНАЛИЗ ВЫЯВЛЕННЫХ ЛИШАЙНИКОВЫХ СИНУЗИЙ
ИЛИ СООБЩЕСТВ
Суждения относительно особенностей среды по лишайниковым синузиям или
сообществам базируются на анализе их состава, параметрах их развития, изме-
нении их показателей в пространстве или во времени. Как и для видов лишайни-
ков, имеется ранжирование лишайниковых сообществ по степени чувствитель-
ности их к кислотному загрязнению воздуха (табл. 6.5). Эту шкалу можно приме-
нять для грубых оценок состояния воздуха, в особенности наличия в нем кислотных
загрязнителей.
Более точные оценки можно дать на основе таких показателей как богатство
видового состава, степень развития видов, наличие видов того или иного ранга
чувствительности. Так, выявленный состав лихеносинузий и видов лишайников
Таблица 6.5
Фитоценологическая шкала для оценки относительного (кислого) загрязнения воздуха
в Южной Германии (Wirth, 1988)
Оценка устойчивости Лишайниковое сообщество
1 1 2 3 3-4 4 5 5-6 6 6 7 8 9 9-10 10 11 12 13 14 Lobarietum pulmonariae subass. Lobaria amplissima Nephrometum laevigati Gyalectum ulmi Usneetum florida-neglectae Ramalina fasti gitae Parmelietum acetabuli c Anaptychia ciliaris Usneetum filipendulae Physcietum adscendentis c Physconia distorta, Physcia stellar is, Ph. aipolia Bacidia rubella - Aleurodiscus - ass Leprarietum candelaris Pertusarietum hemisphaericae Parmelietum caperatae (Flavoparmelia caperata, если имеется) Opegraphetum vermicelli ferae Porinetum aeneae Hypogymnia physodes - Parmelia sulcata - сообщество Chaenothecetum ferruginae Buellietum punctatea Lecanoretum conizaeodes Pleurococcetum vulgaris
1 - устойчивость лишайникового сообщества низкая; 14 - устойчивость высокая.
128 6. Группировки лишайников и индикация качества воздушного бассейна
(см. табл. 6.4) дают основания признать изученную территорию Рудных гор до-
вольно сильно загрязненной. Это заметно и по состоянию деревьев, у части кото-
рых отмечена пожелтевшая хвоя, много сухостоя. На участках обследованного
типа леса отсутствуют представители таких чувствительных к загрязнению воз-
духа родов как Bryoria, Usnea, Даже нет представителей такого обычного вида
как Parmelia sulcata, О загрязнении свидетельствуют и высокие показатели встре-
чаемости и покрытия токситолерантного вида Lecanora conizaeiodes при общей
бедности видового состава лишайников в изученном типе ельников.
Анализ данных, полученных в одном месте, но в разные годы, ставит перед
исследователем дополнительную задачу, которая заключается в необходимости
выделить из общей суммы выявленных изменений те, которые связаны с дей-
ствием загрязнителей. Как известно, в природных сообществах имеет место не-
прерывный процесс их развития, который определяется как взаимоотношениями
организмов друг с другом и с изменяемой ими средой, что обычно называют сук-
цессией, так и воздействием внешних по отношению к сообществу факторов, в
том числе и деятельности человека, называемых экзогенными сменами (Работ-
нов, 1983). Установленные изменения видового состава лишайников и комплекса
формируемых ими лихеносинузий, произошедшие за известный промежуток вре-
мени, несомненно, являются следствием того и другого. И надо четко осознавать,
что наблюдения за лишайниковыми группировками при мониторинге среды обес-
печивают скорее корреляции, чем доказательства причинных связей, за исключе-
нием случаев, когда эти наблюдения являются частью контролируемых воспро-
изводимых экспериментов, при которых воссоздаются те или иные параметры
качества воздуха. Но опытов таких немного, и мы вынуждены делать выводы,
основываясь на опубликованных сведениях о реакции разных видов лишайников
на загрязнители воздуха. Эти выводы должны помочь изолировать из общей сум-
мы выявленных изменений те, которые связаны с действием загрязнителей. Ар-
сенал способов такой изоляции невелик, и они основаны главным образом на
сведениях об экологии видов лишайников. Так, известно, что со временем проис-
ходит изменение свойств субстрата, например, коры липы, в молодом возрасте
гладкой и бороздчатой - в зрелом. Соответственно, со временем меняется и со-
став лишайников, обитающих на этой породе, что и было выявлено при сравни-
тельном анализе данных наблюдений за составом эпифитных лихеносинузий в
дубо-липняке снытьево-волосистоосоковом, проведенных в 1966 и 1987 гг. в Под-
московье (Бязров, 1993а). Видовое разнообразие лишайников (с учетом номенк-
латурных новаций) на стволах деревьев до высоты 2 м за прошедшие годы прак-
тически не изменилось - 28 видов в 1966 г. и 27 - в 1987 г., однако таксонов,
зафиксированных на участке в оба срока, всего 14. Изменился и комплекс лихе-
носинузий древесных пород (табл. 6.6).
К 1987 г. со стволов деревьев исчезли представители 14 видов, но появились
талломы 13 видов, ранее здесь не встречавшихся. Выпадение из состава группи-
6. Группировки лишайников и индикация качества воздушного бассейна
129
Таблица 6.6
Относительная встречаемость (%) лишайников в эпифитных лихеносинузиях дубо-ливня-
ка снытьево-волосистоосокового в 1966 и 1987 гг. (Бязров, 1993а)
Жиз- ненная форма Состав лихеносинузии Относи- тельная встречае- мость Жизнен- ная форма Состав лихеносинузии Относи- тельная встречае- мость
1 2 3 4 5 6
1966 г. 1987 г
Липа, 0-0,6 м
Сг Gran his scripta 100 Cr Graphis scripta Ft=22%, Fs=38% 100
Ft=46%, Fs=68%
Sc Cladonia coniocraea 30 Sc Cladonia coniocraea 33
Sc Сг C. fimbriata Bacidia beckhausii 13 7 Sc Cr C. fimbriata Graphis scripta 19 16
Сг Сг Buellia disciformis Grapis scripta 13 34 Cr Cr Lecanora symmicta Lepraria incana 1 30
Сг Opegrapha atra Ft=3%, Fs=4% 3 Cr Scoliciosporum chlorococcum Ft=24%, Fs=40% 1
Sc Cladonia coniocraea 20 Sc Cladonia coniocraea 24
Sc SI C. fimbriata Vulpicida pinastri 20 20 Sc SI C. fimbriata Parmelia sulcata 19 24
Сг Cr Buellia disciformis Graphis scripta 20 20 Cr Cr CUostomum grijfithii Graphis scripta 2 10
Ft=l%, Fs=l% Cr Lepraria incana 21
Сг Graphis scripta 49 Ft=7%, Fs=12%
Cr Bacidia beckhausii 1 Cr Graphis scripta 50
Cr Biatora helvola 5 Cr Lepraria incana 50
Сг Lecanora albella 1 Ft=2%, Fs=3%
Сг Lecidella Sc Cladonia coniocraea 100
Сг elaeochroma Opegrapha atra 1 43 Ft=l%, Fs=l%
Ft=10%, Fs=15% Sc Cladonia coniocraea 50
Сг Bacidia beckhausii 100 Sc C. fimbriata 50
Ft=l%, Fs=l% Ft=3%, Fs=4%
Сг Opegrapha atra 100 SI Parmelia sulcata 50
Ft=l%, Fs=l% Cr Graphis scripta 50
Sc Cladonia coniocraea 33 Ft=l%, Fs=l%
Sc C. fimbriata 33
Cl Hypogymnia 34
physodes
Ft=l%, Fs=2%
Sc Cladonia coniocraea 21
Sc Cl C. fimbriata Hypogymnia 20 22
physodes
Подчеркнуты виды, входящие в название лихеносинузии; Ft - встречаемость лихеносинузии на
участке, Fs - встречаемость лихеносинузии на деревьях данной породы.
130 6. Группировки лишайников и индикация качества воздушного бассейна
таблица 6.6 (продолжение)
1 2 3 4 5
Сг Buellia disciformis 20
Сг Graphis scripta 9
Сг Lepraria incana 5
Сг 0pegrap ha atra 2
Сг Pertusaria amara Ft=9%, Fs=13% 1
Sc Cladonia coniocraea 15
Sc C. fimbriata 15
Sc C. furcata 4
Cl Hypogymnia physodes 15
SI Vulpicida pinastri 15
Cr Buellia disciformis 15
Cr Catinaria atropurpurea 4
Cr Graphis scripta 4
Cr Lepraria incana 9
Cr Opegrapha varia Ft=l%, Fs=2% 4
Cl Hypogymnia phvsodes 20
SI Parmelia sulcata 20
Cr Buellia disciformis 20
Cr Cliostomum griffithii 20
Cr Graphis scripta Ft=l%, Fs=1 % 20
Липа, 0,6-2,0 м
Cr Bacidia beckhausii 100 Cr Scoliciosporum chlorococcum 100
Ft=l%, Fs=l% Ft=l%, Fs=l%
Sc Cladonia coniocraea 25 Cr Graphis scripta 43
Cl Hypogymnia physodes 25 Cr Lepraria incana 43
Cr Buellia disciformis 25 Cr Scoliciosporum chlorococcum 14
Cr Graphis scripta 25 Ft=2%, Fs=3
Ft=l%, Fs=l% Sc Cladonia coniocraea 25
Cl Hypogymnia physodes 50 Sc C. fimbriata 6
Cr Buellia disciformis 48 SI Parmelia sulcata 25
Cr Graphis scripta 1 Cr Graphis scripta 25
Cr Lepraria incana 1 Cr Lepraria incana 19
Ft=7%, Fs=l 1% Ft=3%, Fs=3%
Cl Hypogymnia physodes 34 Sc Cladonia coniocraea 25
SI Parmelia sulcata 30 Sc C. fimbriata 25
SI Vulpicida pinastri 8 Cl Hypogymnia physodes 25
Cr Arthrosporum populorum 8 SI Parmelia sulcata 25
Cr Biatora helvola 4 Ft=l%, Fs=l%
Cr Lecanora pulicaris 4 Sc Cladonia coniocraea 20
Cr Lecidellae elaeochroma 4 Sc C. fimbriata 9
Cr Lepraria incana 8 Cl Hypogymnia physodes 20
Ft=2%, Fs=3% SI Parmelia sulcata 20
6. Группировки лишайников и индикация качества воздушного бассейна 131
таблица 6.6 (продолжение)
1 2 3 4 5 6
С1 Hypogymnia physodes 100 Сг Graphis scripta 9
Ft=8%, Fs=12% Сг Lepraria incana 19
Сг Pertusaria amara 1
Сг Scoliciosporum chlorococcum Ft=9%, Fs=16% 2
Sc Cladonia coniocraea 24
Cl Hypogymnia physodes 24
Сг Graphis scripta 24
Сг Lepraria incana 17
Сг Scoliciosporum chlorococcum Ft=2%, Fs=3% 11
С1 Hypogymnia physodes 33
Сг Graphis scripta 25
Сг Lepraria incana 33
Сг Scoliciosporum chlorococcum Ft=2%, Fs=3% 9
CI Hypogymnia physodes. 31
S1 Parmelia sulcata 31
Сг Biatora helvola 1
Сг Graphis scripta 5
Сг Lepraria incana 31
Сг Pertusaria amara Ft=l 4%, Fs=23% 1
SI Parmelia sulcata 43
Сг Graphis scripta 35
Сг Lepraria incana 9
Сг Scoliciosporum chlorococcum Ft=5%, Fs=9% 13
SI Parmelia sulcata 100
Ft=l%, fs=l%
С1 Hypogymnia physodes 50
S1 Parmelia sulcata Ft=4%, Fs=6% 50
Дуб, 0-0,6 м
Sc Cladonia coniocraea 100 Sc Cladonia coniocraea 100
Ft=l%, Fs=2% Ft=l%, Fs=3%
Sc Sc Cladonia coniocraea 50 50 Cr Lepraria incana 100
C. fimbriata Ft=l%, Fs=5%
Ft=l%, Fs=2% Cr Arthonia patellulata 14
132 6. Группировки лишайников и индикация качества воздушного бассейна
таблица 6.6 (продолжение)
1 2 3 4 5 6
Sc Cladonia coniocraea 21 Cr Buellia punctata Lecanora pulicaris 14
Sc C. fimbriata 17 Cr 14
Сг Arthrosporum populorum 16 Cr Lepraria incana 44
Сг Biatora helvola 1 Cr Opegrapha atra 14
Сг Buellia disciformis 7 Ft=l 6%, Fs=82%
Сг Cliostomum griffithii 9 Sc Cladonia coniocraea 25
Сг Graphis scripta 4 Cl Hypogymnia physodes 25
Сг Lecanora symmicta 1 Cr Calicium lichenoides 25
Сг Lecidella elaeochroma 1 Cr Lepraria incana 25
Сг Lepraria incana 9 Ft=l%, Fs=3%
Сг Opegrapha atra 0. varia 4 Sc Cladonia coniocraea 34
Сг 10 Sc C. fimbriata 5
Ft=l%, Fs=2% Sc C. ochrochlora 1
Sc Cladonia coniocraea 24 Cr Arthonia patellulata 10
Sc C. fimbriata 38 Cr Biatora helvola 1
CI Hypogymnia physodes 38 Cr Calicium lichenoides 1
Ft=l%, Fs=l% Cr Chaenotheca furfuracea 1
Sc Cladonia coniocraea 11 Cr Graphis scriptum 3
Sc C. fimbriata 15 Cr Lecanora symmicta 2
Cl Hypogymnia physodes 15 Cr Lepraria incana 35
Cr Arthrosporum populorum 11 Cr Opegrapha atra Pertusaria amara 5
Cr Buellia disciformis 4 Cr 1
Cr Cliostomum griffithii 4 Cr P. coccodes 1
Cr Graphis scripta 4 Ft=17%, Fs=28%
Cr Lecanora symmicta 4
Cr Lepraria incana 8
Cr Opegrapha atra 8
Cr 0. varia 8
Cr Pertusaria albescens 4
Cr P. leioplaca Ft=l%, Fs=2% 4
Дуб, 0,6-2,0 м
Cr Arthrosporum populorum 33 Cr Lepraria incana 100
Cr Graphis scripta 34 Ft=4%, Fs=20%
Cr Lepraria incana 33 Cr Arthonia patellulata 36
Ft=l%, Fs=3% Cr Calicium lichenoides 5
Cl Hypogymnia physodes 100 Cr Graphis scripta 9
Ft=l%, Fs=8% Cr Lepraria incana 45
Sc Cladonia coniocraea 50 Cr Opegrapha atra 5
Cl Hypogymnia physodes 50 Ft=5%, Fs=25%
Ft=l%, Fs=2% Sc Cladonia coniocraea 37
Sc Cladonia coniocraea 8 Cr Arthonia patellulata 21
Sc C. fimbriata 19 Cr Lecanora symmicta 5
Cl Hypogymnia physodes 26 Cr Lepraria incana 37
Ft=4%, Fs=17%
6. Группировки лишайников и индикация качества воздушного бассейна
133
таблица 6.6 (продолжение)
1 2 3 4 5 6
Сг Arthrosporum populorum 23 Sc Cladonia coniocraea 23
Сг Buellia disciformis 12 SI Parmelia sulcata 18
Сг Calicium lichenoides 4 SI Physconia grisea 8
Сг Lepraria incana 4 Cr Arthonia patellulata 8
Сг Opegrapha varia 4 Cr Buellia punctata 4
Ft=2%, Fs=17% Cr Graphis scripta 8
Sc Cladonia coniocraea 6 Cr Lecanora symmicta 4
Sc C, fimbriata 19 Cr Lepraria incana 23
Cl Hypogymnia physodes 19 Cr Opegrapha atra 4
SI Parmelia sulcata 19 Ft=3%, Fs=15%
Сг Arthrosporum populorum 6 SI Parmelia sulcata 33
Сг Buellia disciformis 13 SI Physconia grisea 9
Сг Lecidella elaeochroma 6 Cr Graphis scripta 9
Сг Lepraria incana 6 Cr Lepraria incana 40
Сг Opegrapha varia 6 Cr Opegrapha atra 9
Ft=l%, Fs=7% Ft=2%, Fs=12%
С1 Hypogymnia physodes 25
S1 Parmelia sulcata 25 SI Parmelia sulcata 100
Сг Arthrosporum populorum 20 Ft=l%, Fs=2%
Сг Buellia disciformis 7 Cl Hypogymnia physodes 25
Сг Lecidella elaeochroma 3 SI Parmelia sulcata 25
Сг Lepraria incana 7 Cr Calicium lichenoides 25
Сг Opegrapha atra 3 Cr Lepraria incana 25
Ft=2%, Fs=17% Ft=l%, Fs=5%
С1 Hypogymnia physodes 44
Сг Arthrosporum populorum 25
Сг Buellia disciformis 19
Сг Lecidella elaeochroma 6
Сг Pertusaria amara 6
Ft=2%, Fs=17%
Клен, 0-0,6 м
Cr Biatora helvola 8 Cr Lecanora symmicta 100
Cr Graphis scripta 43 Ft=l%, Fs=18%
Cr Lecidella elaeochroma 8 Sc Cladonia coniocraea 25
Cr Mycobilimbia tetramera 8 Sc C. fimbriata 19
Cr Opegrapha atra 25 Cr Arthonia patellulata 13
Cr Pyrrhospora elabens 8 Cr Lecanora symmicta 6
Ft=l%, Fs=12% Cr Lepraria incana 31
Sc Cladonia coniocraea 18 Cr Opegrapha varia 6
Sc C. fimbriata 12 Ft=2%, Fs=45%
Cl Hypogymnia physodes 18 Cr Graphis scripta 100
Cr Buellia disciformis 12 Ft=2%, Fs=36%
134 6. Группировки лишайников и индикация качества воздушного бассейна
таблица 6.6 (окончание)
1 2 3 4 5 6
Cr Graphis scripta 12
Cr Lecanora allophana 5
Cr Lepraria incana 5
Cr Opegrapha atra Ft=l%, Fs=27% 18
Клен, 0,6-2,0 м
Cr Graphis scripta 100 SI Parmelia sulcata 100
Ft=l%, Fs=9% Ft=l%, Fs=9%
Cl Hypogymnia physodes 30 Sc Cladonia coniocraea 21
Cr Arthrosporum populorum 10 SI Parmelia sulcata 16
Cr Buellia disciformis 30 SI Physconia grisea 16
Cr Graphis scripta 10 SI P. distorta 5
Cr Lecanora allophana 10 Cr Biatora helyola 5
Cr Lecidella elaeochroma 10 Cr Buellia alboatra 5
Ft=l%, Fs=27% Cr Graphis scripta 5
Cr Lepraria incana 22
Cr Pertusaria coccodes Ft=2%, Fs=36% 5
Sc Cladonia coniocraea 34
Cr Graphis scripta 33
Cr Lepraria incana Ft=l%, Fs=9% 33
ровок на липах и кленах таких видов как Bacidia beckhausii, Catinaria atropurpurea,
Lecanora albella, Lecanora allophana, Mycobilimbia tetramera, Pyrrhospora elabens
и появление Arthoniapatellulata, Buellia alboatra, Pertusaria coccodes можно свя-
зать с изменением физических свойств коры деревьев (Бязров, 1993а). Далее я
обратил внимание, что во второй срок наблюдений (1987 г.) на стволах деревьях
отмечена высокая встречаемость погибших слоевищ Parmelia sulcata. Часто 50-
70% обследованной поверхности стволов липы покрывали мертвые слоевища
пармелии. Нередки были и талломы Hypogymniaphysodes с белыми некротичес-
кими пятнами. О значительном количестве отмирающих лишайников свидетель-
ствовало и увеличение вдвое показателя встречаемости на почве опавших с дере-
вьев слоевищ лишайников (25% против 12% в 1966 г.). На ухудшение условий
для ряда видов показывало и значительное снижение средней поверхности, зани-
маемой слоевищами гипогимнии и пармелии на одном дереве, - с 1680 см2 на
липе в 1966 г. до 328 см2 в 1987 г. у Hypogymniaphysodes (Бязров, 1993а). Видимо,
по той же причине значительно реже стали встречаться Cliostomum griffithii,
Opegrapha atra, Vulpicidapinastri. исчезли npewTbbmennArthrosporumpopulorum,
Buellia disciformis, B. geophila, Lecidella elaeochroma, Opegrapha pulicaris,
6. Группировки лишайников и индикация качества воздушного бассейна
135
Pertusaria albescens, Р. leioplaca, но широкое развитие получил лепрозный налет
(Lepraria incana).
Исчезнувшие и вновь появившиеся в дубо-липняке виды эпифитных лишай-
ников были распределены по классам полеотолерантности (Трасс, 1985а). Среди
первых в основном оказались виды, принадлежащие к II-VI классам, т.е. приуро-
ченные к местообитаниям от антропогенно слабо измененных до умеренно изме-
ненных, тогда как вторые принадлежали к IV-X классам, т.е. от слабо и умеренно
измененных до очень сильно антропогенно измененных местообитаний. Поскольку
в местности, где проводилось исследование, антропогенное воздействие в основ-
ном происходило через общее загрязнение атмосферы, можно предположить, что
оно оказало влияние и на развитие эпифитных лишайников. Вероятно, это было
действие кислотных дождей, на что указывают и погибшие слоевища пармелии,
и характерные красные пятна на еще функционировавших ее талломах, и много-
численные повреждения слоевищ гипогимнии. На повышение уровня кислотно-
сти местообитаний лишайников указывает и более частая встречаемость пред-
ставителей Calicium lichenoides, Cladonia coniocraea, Scoliciosporum chlorococcum
и появление здесь Chaenotheca chrysocephala, C. furfuracea. Судя по размерам
калициевых лишайников, отмерших талломов Parmelia sulcata, воздействие кис-
лотных осадков началось здесь, по меньшей мере, за 10—12 лет до проведения
повторных наблюдений. Еще раньше (около 15 лет) началось загрязнение пылью,
о котором свидетельствуют сравнительно нередкие на дубах и кленах относи-
тельно крупные слоевища представителей рода фискония (см. табл. 6.6), ранее на
участке не встречавшиеся. Изменения видового состава и характера распределе-
ния представителей отдельных таксонов вызвали перестройку комплексов лихе-
носинузий, свойственных древесным породам участка. Однако изменения на уров-
не лихеносинузий, причиной которых были бы процессы, связанные с сукцесси-
ей, не столь выражены, как изменения, обусловленные воздействием внешних
факторов. Сукцессии привели к выпадению ряда лихеносинузий грегационного
типа (одновидовых), например, синузии Bacidia beckhausii на липах и кленах,
исчезновение из состава лихеносинузий одних накипных видов и появление в
них представителей других при сохранении прежнего состава жизненных форм.
Более заметны изменения, вызванные внешними факторами. Это отражает раз-
растание кладоний, лепрарии, сколициоспорум, выпадение из состава или значи-
тельное снижение встречаемости на комлях деревьев гипогимнии, появление на
стволах деревьев представителей рода фискония, а на березах (в табл. 6.6 не пока-
заны) чешуйчатого вида Hypocenomyce scalaris. Такое соотношение внутренних
и внешних причин, вызвавших перестройку комплексов лихеносинузий древес-
ных пород, можно объяснить тем, что в 1966 г. древостой эдификаторного яруса
уже был достаточно зрелым (120 лет), и формирование лихеносинузий на ство-
лах деревьев, видимо, практически завершилось. Относительно быстрые сукцес-
сии лишайниковых группировок можно было ожидать на молодых гладкокорых
136 6. Группировки лишайников и индикация качества воздушного бассейна
липах и кленах, у которых в ходе роста происходило существенное изменение
физических свойств коры - субстрата лишайников. Но к таким гладкокорым де-
ревьям тогда в основном были приурочены лихеносинузии грегационного типа,
сформированные отдельными видами накипных лишайников. Некоторые из них
и не были обнаружены в 1987 г. Здесь уместно отметить, что разрастание кладо-
ний на участке связано не только с увеличением кислотности и запыленности
местообитаний, но и с тем, что огрубевшая кора некогда молодых лип и кленов
стала более благоприятным для заселения субстратом.
Таким образом, результаты последовательного изучения в разные сроки на
одном участке видового состава лишайников и комплексов формируемых ими
синузий (сообществ) можно использовать для определения тенденции загрязне-
ния воздушного бассейна обследованной территории. Опыт работы показывает,
что наибольшую информацию дает анализ изменений видового состава и коли-
чественных показателей особенностей распределения видов лишайников в био-
геоценозе. Результаты сравнительного изучения комплексов лихеносинузий без
привлечения сведений об изменении видового состава интерпретировать гораздо
труднее. При анализе выявленных изменений, прежде всего, желательно отде-
лить то, что явно не связано с загрязнением. Затем необходимо попытаться уста-
новить главный тренд изменений лишайниковых группировок. Далее привлека-
ются сведения о реакции встреченных видов лишайников на загрязнение. Есте-
ственно, крайне желательно сопоставление данных об изменениях лишайниковых
группировок во времени и пространстве с информацией о динамике загрязнения
местности, полученной прямыми инструментальными измерениями. Однако та-
кая информация, как правило, отсутствует. Поэтому достоверность лихеноинди-
кационных интерпретаций чаще всего зависит от опыта исследователя и его зна-
ний экологии лишайников.
7. ТРАНСПЛАНТАЦИИ ЛИШАЙНИКОВ
Пересадки, или трансплантации лишайников, отобранных в местностях, где
условия считаются “фоновыми”, на обследуемые территории с целью изучения
их реакции на новые, возможно, экстремальные для них условия, используются в
лихеноиндикационных исследованиях нередко. Иногда такой способ лихеноин-
дикации называют “активным мониторингом” (Pearson, 1993), считая, что на-
блюдение за состоянием представителей естественно произрастающих лишайни-
ков представляет собой “пассивный мониторинг”. Также к методам активного
мониторинга относят и контролируемую фумигацию (окуривание) лишайников,
и культивирование слоевищ лишайников. Трансплантации включают перенос
организмов с территорий, где они встречаются в природе, в места, где они необ-
ходимы для мониторинга загрязнения воздуха или других показателей среды
(Brodo, 1961, 1966; Hawksworth, Rose, 1976; Skye, 1979; Трасс, 19856). Контроли-
руемая фумигация заключается в создании точечного источника загрязнения и
возможности имитации влияний специфических загрязнителей на лишайники,
которые встречаются на данной территории (Pearson, Rogers, 1982). Культивиро-
вание лишайников предназначено для подтверждения или опровержения возмож-
ности роста лишайников на изучаемых территориях, а также включает методы
выращивания лишайников в “лихенологических садах”, фермах или в лабора-
торных ростовых камерах при контролируемых условиях (Pearson, 1993). К ак-
тивному мониторингу относят и долговременные наблюдения лишайников на
территориях, где они встречаются в природе, после начала работы здесь предпри-
ятия или какого-либо другого источника загрязнения (Skye, 1958; Will-Wolf, 1980а,
b, 1988; Kandler, 1988).
7.1. ТРАНСПЛАНТАЦИИ
Основными достоинствами использования трансплантатов для мониторинга
качества воздуха считаются (Pearson, 1993):
138
7. Трансплантации лишайников
- возможность размещения слоевищ лишайников во всех местах, где желате-
лен мониторинг качества воздуха, в том числе и там, где лишайники по тем или
иным причинам отсутствуют;
- в каждой местности всегда можно найти достаточное количество образцов
массовых видов лишайников, чтобы обеспечить требуемую точность исследова-
ния и полноту охвата территории;
- во всех тестируемых точках размещается сравнительно однородный матери-
ал, собранный в месте с известными условиями существования, что позволяет
более обоснованно интерпретировать результаты;
- исследователь имеет возможность выбора лучшего варианта эксперимента
для изучения конкретных местообитаний и реакции представителей тех или иных
видов лишайников;
- возможность установления темпов увеличения уровней загрязнения, вызы-
вающих повреждения лишайников.
Трансплантация слоевищ лишайников позволяет планировать изучение вли-
яния разных условий среды на эти организмы, когда образцы соответствующим
образом размещены исследователем или относительно источника загрязнения,
вызывающем наблюдаемые повреждения, или для более точной оценки простран-
ственной протяженности воздействия данного источника загрязнения. Например,
трансплантированные образцы можно разместить на одном расстоянии от источ-
ника загрязнения, но на открытом и закрытом местах, различно ориентировать
их относительно источника и т.п. Метод будет более эффективен, если трансплан-
тацию применять в комплексе с фумигацией известным загрязнителем. Напри-
мер, четыре горелки с SO2 разместили на участках с густым, расположенным в
низине, лесом (1), густым лесом на плато (2), в низине с рассеянными деревьями
(3) и на открытом сухом лугу (4). Требуемые уровни SO2 были получены при
горении серы в дни, когда скорость ветра и его направление были благоприятны-
ми. Контрольными служили участки со стороны, откуда дул ветер (Pearson, Rogers,
1982).
Трансплантации также дают возможность для сокращения периода проявле-
ния реакции лишайников на загрязнение или другие изменения среды. Когда в
изучаемых точках территории размещено достаточно большое количество пере-
саженного материала, наблюдения можно проводить с требуемыми интервалами
(еженедельно, через две недели, ежемесячно). Некоторые методы оценки влияния
загрязнения достаточно чувствительны, чтобы обнаружить повреждения лишай-
ников через неделю воздействия даже при низких уровнях загрязнения, и в тече-
ние нескольких часов - при экстремально высоких уровнях (Pearson, 1993). Недо-
статками пересадки являются, с одной стороны, вандализм отдельных предста-
вителей человеческого общества, нередко уничтожающих трансплантаты, с другой
- перемещение организмов из привычной для них среды в новые условия, вслед-
ствие чего реакция пересаженных сюда слоевищ не всегда адекватна ответу по-
7. Трансплантации лишайников
139
стоянно растущих здесь особей того же вида (von Arb, Brunold, 1990); однако име-
ются сведения о наличии тесной корреляции между данными, полученными в
опытах с пересадкой лишайников, и результатами картографирования распростране-
ния лишайников в зоне воздействия источника загрязнения (Ferry, Coppins, 1979).
Первые опыты по пересадке лишайников были проведены, вероятно, в конце
XIX века в г. Мюнхене, где несколько эпигейных видов из нетронутых человеком
ландшафтов переместили в город, где все они вскоре погибли (Трасс, 19856).
В августе 1959 г. в Ботанический сад Тартуского университета (Эстония) из Хи-
бинского горного массива (Мурманская область) пересадили представителей пяти
видов напочвенных лишайников - Cladonia amaurocraea, С. естосупа,
Flavocetraria (Cetraria) cucullata, Nephroma arcticum и Thamnolia vermicularis.
Уже через месяц после пересадки были зафиксированы признаки значительных
повреждений слоевищ, а к середине июля 1960 г. все лишайники погибли (Трасс,
19856). Аналогичной, но, пожалуй, более “контрастной” пересадкой была транс-
плантация в марте 1987 г. десяти слоевищ Hypogymniaphysodes из Западной Гер-
мании в Коста-Рику, где среднегодовая температура более 20°С, а уровень осад-
ков 1900 мм/год (Griininger, Monge-Najera, 1988). Все пересаженные талломы по-
гибли через 3,5-10 месяцев.
Для изучения экологии и физиологии лишайников лишайниковые трансплан-
таты начали применять в 1950-1960 годах (Hale, 1950, 1954, 1959; Lindahl, 1953;
Smith, 1960а-с). Вероятно, первые методически корректные лихеноиндикацион-
ные исследования с использованием трансплантатов были проведены в США
(Brodo, 1961) и в Германии (Schonbeck, 1968, 1969). Э. Бродо (Brodo, 1961) пере-
садил слоевища эпифитных лишайников Flavoparmelia (Parmelia) caperata и
Lecanora leptyrodes с частью их субстрата. Диски коры диаметром около 3,5 см
(1 и 3/8 дюйма) и толщиной около 1,5 см отбирали с деревьев в чистой местности
с помощью пробоотборника и переносились в отверстия того же диаметра, подго-
товленные на деревьях той же породы, растущих на участках, удаленных на раз-
ное расстояние от Бруклина, одного из районов г. Нью-Йорка. После определен-
ного периода экспонирования фиксировалось состояние пересаженных слоевищ
лишайников: изменение цвета, ослабление прочности прикрепления к субстрату,
изменение структуры соредий и ризин, плазмолиз водорослевых клеток относи-
тельно периода экспонирования и удаления от источника загрязнения. Кроме того,
диски с Cladonia chlorophaea были перемещены с оснований деревьев у Брукхе-
венской лаборатории подле г. Нью-Йорка на более высокие части тех же стволов
для определения воздействия высоты поселения на форофите на состояние ли-
шайников. X. Щёнбек (Schonbeck, 1968,1969) незначительно модифицировал этот
метод, прикрепив диски с лишайниками к деревянным щитам, что позволило
трансплантировать лишайники в места, где деревьев было мало. Доступные мне
публикации о применении трансплантатов для целей лихеноиндикации обобще-
ны в таблице 7.1.
140
7. Трансплантации лишайников
Таблица 7.1
Лишайники, использовавшиеся в опытах с пересадкой, и признаки,
фиксируемые при исследовании
Вид лишайника (номенклатура согласно цитируемому источнику) Учитывавшиеся признаки Страна проведения исследований Источник
Alectoria sarmentosa Концентрация серы и ее изотопов Канада Wadleigh, Blake, 1999; Wiseman, Wadleigh, 2002
Cetraria islandica, Evernia prunastri, Ramalina farinacea Концентрация нескольких элементов Германия, Италия, Румыния Cercasov et al., 2002
Bryoria fuscescens, Cetraria islandica, Hypogymnia physodes, Parmelia sulcata Газообмен СОг, проницаемость мембран Германия Haffner et al., 2001
Evernia {Evernia mesomorpha + Usnea sp.+ Ramalina sp.), Hypogymnia (Hypogymnia physodes + Parmelia sulcata), Cetraria (C. halei + C. pinastri), Bryoria fuscescens Покрытие Канада Addison, 1984
Evernia prunastri Концентрация нескольких элементов Италия Caniglia et al., 1994
Evernia prunastri Концентрация нескольких элементов Италия Loppi et al., 1998b
Usnea amblyoclada Содержание пигментов, продуктов перокисления, серы и других элементов Аргентина Carreras et al., 1998; Carreras, Pignata, 2001, 2002
Alectoria sarmentosa, Pseudocyphellaria rainierensis, Lobaria oregana, L. pulmonaria, Evernia prunastri, Usnea longissima Прирост массы США McCune et al., 1996
Usnea ceratina, Everniastrum neocirrhatum Газообмен углекислоты Мексика Zambrano et al., 1999; Zambrano, Nash, 2000
Alectoria sp., Bryoria sp., Usnea sp. Прирост массы Швеция Renhorn, Esseen, 1995
Ramalina calicaris, Usnea sp. Концентрация катионов и анионов солей морской воды Португалия Figueira et al., 1998 a, b
Ramalina ecklonii Содержание пигментов, растворимых белков Аргентина Levin, Pignata, 1995; Gonzalez etal., 1996, 1998; Gonzalez, Pignata, 1999:
Ramalina duriae Содержание ауксинов, этилена Израиль Epstein et al., 1986;
7. Трансплантации лишайников
141
таблица 7.1 (продолжение)
Ramalina duriaei Концентрации элементов Израиль Garty, Fuchs, 1982; Fuchs, Garty, 1983; Garty et al., 1985b; Garty, 1987,1988; Garty ,Hagemey er, 1988
Ramalina duriaei Концентрации элементов, содержание пигментов, проницаемость мембран Израиль Garty et al., 1985a, 1992, 1993a, 1997a, 1998a, b
Ramalina duriaei Образование этилена, концентрации АТФ, хлорофилла Израиль Kardish et al., 1987; Garty et al., 1988,1993b
Ramalina duriaei Спектральное отражение, проницаемость мембран Израиль Garty et al., 1997b, e, f
Ramalina duriaei Накопление бифенилов Израиль Garty et al., 1982
Ramalina lacera =(Ramalina duriaei) Физиологические и биохимические параметры Израиль Garty et al., 2000,2001
Ramalina duriaei, Xanthoria parietina Повреждение слоевищ, содержание пигментов, АТФ, флуоресценция фотобионтов, скорость фотосинтеза, проницае- мость мембран Израиль Silberstein et al., 1996a, b
Ramalina menziesii Концентрации элементов США Boonpragob, Nash, 1990
Ramalina menziesii Содержание пигментов, скорость фотосинтеза США Boonpragob, Nash, 1991
Ramalina menziesii Скорость роста США Boucher, Nash, 1990
Ramalina menziesii Содержание воды, скорость фотосинтеза США Matthes-Sears et al., 1986
Hypogymnia physodes, Bryoria capillaris Повреждение клеток фотобионтов Финляндия Holopainen, 1984a
Parmelia acetabulum, P. caperata, P. perlata, P. sulcata, Hypogymnia physodes, Ramalina farinacea, R. fastigiata Газообмен СО2 Велико- британия Ferry, Coppins, 1979
Hypogymnia physodes, Usnea hirta Концентрация нескольких элементов Финляндия Garty et al., 1996
Hypogymnia physodes, Cladina stellar is, Bryoria fuscescens, Usnea hirta, Alectoria sarmentosa, Brvoria fremontii Образование этилена Финляндия Garty etal., 1995b, 1997c, d
142
7. Трансплантации лишайников
таблица 7.1 (продолжение)
Hypogymnia physodes Концентрация нескольких элементов США Bennett et al., 1996
Hypogymnia physodes Повреждение клеток фотобионта Россия Biazrov, 1994a
Parmelia physodes Степень повреждения слоевищ Германия Djalali, 1973
Hypogymnia physodes, Cladina rangiferina Концентраця ртути США Evans, Hutchinson, 1996
Hypogymnia physodes Концентрация нескольких элементов Хорватия Horvat et al., 2000
Hypogymnia physodes Газообмен ССЬ Австрия Egger et al., 1994
Hypogymnia physodes Концентрация нескольких элементов Венгрия Farkas et al., 1985
Hypogymnia physodes Концентрация нескольких элементов Великобрит алия Gailey, Lloyd, 1993; Gailey et al., 1985
Hypogymnia physodes Выживаемость Коста-Рика Griininger, Monge-Najera, 1988
Hypogymnia physodes Повреждение слоевищ, газообмен СОа Австрия Heber et al., 1991,1994; Hufnagel, Turk, 1998
Hypogymnia physodes Содержание серы Финляндия Holopainen et al., 1993
Hypogymnia physodes Концентрация естественных радионуклидов Словения Jeran et al., 1995
Hypogymnia physodes Концентрация нескольких элементов Словения Jeran et al., 1993
Hypogymnia physodes Флуоресценция фотобионтов Финляндия Kauppi, 1980
Hypogymnia physodes Концентрация ртути США Makholm, Bennett, 1998
Hypogymnia physodes Концентрация нескольких элементов Финляндия Palomaki et al., 1992
Hypogymnia physodes Концентрация нескольких элементов Дания Pilegaard, 1979
Hypogymnia physodes Выживаемость Польша Pustelniak, 1991
Parmelia physodes Повреждение слоевищ Германия Schonbeck, 1968, 1969; Schonbeck, Holte. 1970
Hypogymnia physodes Повреждение слоевищ, скорость роста Дания Sochting, Johnsen. 1978
Hypogymnia physodes Концентрация нескольких элементов Норвегия Steinnes, Krog, 1977
Hypogymnia physodes Повреждение слоевищ Германия Steubing et al., 1974
Hypogymnia physodes Повреждение слоевищ Польша Swieboda, Kalemba, 1978, 1982
Hypogymnia physodes Флуоресценция фотобионтов Германия Weinmann-Kreeb, Kreeb, 1987
Hypogymnia physodes Повреждение слоевищ, скорость роста Германия Wietschorke et al., 1985; Werner, 1993; Werner, Niemeyer, 1994
7. Трансплантации лишайников
143
таблица 7.1 (продолжение)
Hypogymnia physodes, Parmelia sulcata Газообмен CO2 Австрия Turk, Christ, 1980; Christ, Turk, 1981, 1982
Cetraria cucullata, Cladonia amaurocraea, C. ecmocyna, Thamnolia vermicularis, Nephroma arcticum, Parmelia sulcata, P. olivacea, Hypogymnia physodes, Lecanora allophana, Lecidella elaeochroma, Phlyctis argena Выживаемость Эстония Трасс, 19856
Hypogymnia enteromorpha Повреждение слоевищ США Sigal, Nash. 1983
Pseudevernia furfuracea Концентрация нескольких элементов Италия Calliari et al., 1995; Bari et al., 2001
Parmelia sulcata Скорость фотосинтеза, содержание пигментов Швейцария von Arb, 1987; von Arb, Brunold, 1990
Parmelia sulcata Концентрации металлов Португалия Freitas et al., 1995, 2000
Parmelia sulcata Концентрация нескольких элементов Португалия Reis et al., 1999a, b
Parmelia sulcata Концентрация нескольких элементов Нидерланды Sloof, 1995a
Parmelia sulcata Повреждение слоевищ Германия Steipkamp et al., 1999
Parmelia saxatilis, Xanthoria parietina Скорость роста Великобри- тания Armstrong, 1990
Parmelia conspersa, Parmelia saxatilis, Xanthoria parietina, Physcia tenella, Parmelia glabratula ssp. fuliginosa, Physcia orbicularis Скорость роста Великобри- тания Armstrong, 1993
Parmelia conspersa, Parmelia saxatilis, Parmelia glabratula ssp. fuliginosa, Physcia orbicularis, Xanthoria parietina Скорость роста Великобри- тания Armstrong, 1994
Parmelia conspersa, Lecanora subfusca, Peltigera canina. Концентрация нескольких элементов Румыния Bartok, 1988
Виды Parmelia Отмирание клеток фотобионтов Коста-Рика Mendez, Fournier, 1980
Parmelia squarrosa, Parmelia sulcata, Physcia millegrana Повреждение слоевищ, отмирание клеток фотобионтов, содержание пигментов Канада LeBlanc, Rao, 1966, 1973a, 1975; LeBlanc et al., 1976
Parmelia caperata Повреждение слоевищ Италия Canigliaet al., 1998
Parmelia caperata Концентрация нескольких элементов Франция Cuny et al., 2001
144
7. Трансплантации лишайников
таблица 7.1 (продолжение)
Parmelia caperata, Cladonia chlorophaea, Lecanora leptyrodes, Graphis scripta Повреждение слоевищ США Brodo, 1961,1966
Parmelia cumberlandia Вторичные лишайниковые вещества Канада Fahselt, 1979
Parmelia cumberlandia, Cladonia turgida, Stereocaulon saxitile Вторичные лишайниковые вещества По меридиану от юга Канады (Онтарио) до юга США (Флорида) Fahselt, 1981
Parmelia saxatilis Выживаемость Великобритания Gilbert, 1988
Parmotrema austrosinense, P. conferendum Содержание пигментов, малондиальдегида, серы и других веществ Аргентина Canas et al., 1997
Parmotrema uruguense Содержание пигментов, продуктов перокисления Аргентина Canas, Pignata, 1998
Parmotrema madagascariaceum Концентрация нескольких элементов Венесуэла Gordon et al., 1995
Punctelia subrudecta Содержание пигментов, растворимых белков Аргентина Gonzalez, Pignata, 1994, 1997
Canomaculina pilosa Содержание пигментов, растворимых белков Аргентина Gonzalez, Pignata, 2000
Parmelia stenophylla Флуоресценция фотобионтов Россия Бучельников, Григорьев, 1996; Григорьев, Бу- чельников, 1997
Xanthoparmelia lineola, X. subdecipiens Скорость роста США Benedict, 1990а; Benedict, Nash, 1990
Xanthoparmelia lineola, X subdecipiens, Rhizocarpon sp. Скорость роста США Benedict, 1991
Masonhalea richardsonii. Прирост массы США Peck et al., 2000
Cetraria nivalis Морфология и анатомия слоевищ, водный режим Швеция Schipperges et al., 1995
Platismatia glauca, Lobaria pulmonaria Прирост массы, содержание пигментов, флуоресценция фотобионтов, водный режим Швеция Renhorn et al., 1997
Platismatia glauca, Lobaria pulmonaria Газообмен СО2 Швеция Sundberg et al., 1997
Physcia aipolia, Xanthoria parietina Содержание пигментов Румыния Bartok. Bercea, 1991
7. Трансплантации лишайников
145
таблица 7.1 (окончание)
Xanthoria parietina Содержание пигментов, скорость дыхания, концентрация нескольких элементов Румыния Bartok et al., 1992
Xanthoria parietina Морфология слоевищ Великобритания Richardson, 1967
Rhizocarpon sp. Смертность США Benedict, 1990b
Rhizocarpon sp. Скорость роста США Benedict 1999
Lecanora varia Морфология слоевищ Словакия Pisut, Jelinkova, 1973
Lobaria amplissima Выживаемость Великобритания Gilbert, 1991
Lobaria amplissima, L. pulmo- naria, L. scrobiculata, L. virens Повреждение слоевищ Великобритания Farmer et al., 1992
Lobaria oregana, L. pulmonaria Выживаемость США Denison, 1988
Lobaria oregana, L. pulmonaria Проницаемость мембран США Shirazi etal., 1996
Lobaria oregana, L. pulmonaria Скорость роста США Sillett et al., 2000
Lobaria oregana, Pseudocyphellaria rainierensis Скорость роста США Sillett, 1994; Sillett, McCune, 1998
Lobaria pulmonaria Флуоресценция фотобионтов Норвегия Gauslaa, Solhaug, 2000
Lobaria pulmonaria Скорость роста США Muir et al., 1998
Lobaria pulmonaria Морфология слоевищ Швейцария Scheidegger, 1995
Peltigera canina Скорость роста Великобритания Webster, Brown, 1997
Виды Cladina Повреждение слоевищ Дания Sochting, 1987
Cladonia alpestris, C. mitis. C. rangiferina Повреждение слоевищ, газообмен СО2, содержание пигментов Финляндия Ikonen, Karenlampi. 1976
Cladonia port ent osa Прирост массы, концентрация азота Великобритания Hyvarinen, Crittenden, 1998
Cladonia rangiformis Концентрация фтора Словакия Lacko vicova. Kubinska, 1991
Cladonia convoluta Концентрация нескольких элементов Венгрия Tuba, Csintalan, 1993
Cladonia convoluta, C. furcata Содержание пигмеи тов, флуоресценция фотобионтов, скорость фотосинтеза Венгрия Tubaet al., 1997
Cladonia furcata Скорость роста Германия Vagts et al., 1994
Anaptychia ci Haris, Hypogymnia physodes, Letharia divaricata, Lobaria pulmonaria, Parmelia caperata, P. sulkata, Physcia adscendens, Usnea sp., Xanthoria parientina Концентрация Pb, Cd Франция Gombert Asta, 1998
146
7. Трансплантации лишайников
7.1.1. Отбор слоевищ лишайников для пересадки
и подготовка их к экспонированию
Для трансплантации чаще всего используются эпифитные лишайники, слое-
вища которых специальными бурами вместе с субстратом отделяют от деревьев, а
затем эти диски тем или иным способ размещают на обследуемой территории.
Если исследование проводится для оценки поглощения тяжелых металлов или
других элементов-загрязнителей, при отборе образцов для пересаживания и их
подготовке к трансплантации не рекомендуют пользоваться металлическими
предметами (Pearson, 1993). Выбирают виды, талломы которых легко собрать,
трансплантировать и за которым просто наблюдать. Перечень видов, слоевища
которых использовали в экспериментах с пересадкой, довольно велик (см. табл.
7.1). В основном это эпифиты, но также в нем представлены эпигейные и эпилит-
ные виды. В Европе обычно используют слоевища эпифитного лишайника
Hypogymniaphysodes, однако пересаживаются и талломы других видов - Bryoria
capillaris (Holopainen, 1984b), Cladonia convoluta (Farkas et al., 1985), Cladonia
furcata (Vagts et al., 1994), Everniaprunastri, Parmelina (Parmelia) quercina (Silva-
Pando, Ascaso, 1982), Flavocetraria (Cetraria) nivalis (Schipperges et al., 1995),
Flavoparmelia (Parmelia) caperata (Caniglia et al., 1998), Parmelia sulcata (Sloof,
1995a), Xanthoparmelia (Parmelia) conspersa, Lecanora subfusca, Peltigera canina
(Bartok, 1988), Xanthoriaparietina, Physcia aipolia (Bartok, Osvath, 1990). Уместно
отметить, что не все эксперименты с трансплантатами лишайников решали зада-
чи индикации загрязнений. Так, опыты с видами Lobaria осуществлялись с це-
лью восстановления их представителей в биоте территорий, откуда они исчезли
(Gilbert, 1991; Scheidegger, 1995); пересаживались лишайники и для установле-
ния воздействия различных условий на скорость роста (Benedict, Nash, 1990;
Benedict, 1991; Sillett, 1994; Muir et al., 1998; Sillett, McCune, 1998) и другие пока-
затели их жизнедеятельности (Brodo, 1961; Nash et al., 1990; Renhom, Esseen, 1995;
Schippegers et al., 1995; Shirazi et al., 1996; Renhom et al., 1997). Этим методом
изучались и адаптации лишайников к непривычным для них условиям, переса-
див представителей обычного в умеренных широтах вида Hypogymniaphysodes в
тропики Центральной Америки (Griininger, Monge-Najera, 1988). В Германии Союз
инженеров (VDI) разработал процедуру отбора и подготовки образцов чаще дру-
гих используемого для пересадки вида Hypogymniaphysodes, рекомендуемого для
получения сравнимых результатов (VDI, 1991). Как критерий действия загрязни-
теля здесь предлагается доля (%) поврежденной части экспонированного слоеви-
ща от его общей поверхности за период наблюдения. Отмершие части лишайника
лишены зеленых и сине-зеленых пигментов; соответственно, они отличаются сво-
ей характерной белой и серой окраской. Измерения проводят на фотосним-
ках слоевищ, сделанных до и после экспонирования.
7. Трансплантации лишайников
147
Лишайники для экспонирования отбирают вместе с их субстратом стандарт-
ным ручным буром (диаметр 40 мм) с упавших старых деревьев одной породы
(такие деревья, как правило, ищут зимой). Вырезанные шайбы должны быть, по
меньшей мере, на 2/3 покрыты хорошо развитыми экземплярами лишайника (по
возможности одно слоевище на куске коры). Только в исключительных случаях
могут отбираться лишайники с растущих деревьев; возникающие при этом раны
должны быть залечены. В каждом случае для отбора лишайников необходимо
получить разрешение владельца леса. В лесной службе можно получить сведения
о местах наличия ветровала и планируемых рубках.
Для экспонирования используются только безупречные, по возможности, ме-
стные индивиды. Они, например, не должны иметь темных пигментированных
поверхностей, вызванных паразитическими грибами.
Лишайники из различных местообитаний могут неодинаково реагировать на
новые для них условия среды. Это требует, чтобы, по меньшей мере, в рамках
одной серии измерений экспонируемые лишайники были из одного строго огра-
ниченного района и росли там в возможно сходных микроклиматических усло-
виях (например, на определенном отрезке ствола дерева). Остающиеся различия
выравнивают до нормальной биологической вариации акклиматизацией слоевищ
перед экспонированием в течение определенного периода в условиях местного
микроклимата. Продолжительность акклиматизации может достигать 4-5 меся-
цев. Такая процедура обязательна, если место отбора образцов и область экспо-
нирования климатически сильно различаются. Проводится она в стандартных
условиях. Для этого слоевища лишайника приклеивают древесной замазкой на
стеллажи из необработанной древесины, где все экземпляры имеют равные усло-
вия освещения и вентилирования (рис. 7.1). Воздух места акклиматизации дол-
Рис. 7.1. Устройство для акклиматизации слоевищ (VDI, 1991)
148
7. Трансплантации лишайников
жен быть чистым, а его климатические показатели сходны с таковыми места бу-
дущего экспонирования.
Для экспонирования выбирают неповрежденные экземпляры лишайников, по
возможности, одинаковые по размерам и состоянию развития. Независимо оттого,
используют ли акклиматизированные или не акклиматизированные слоевища,
перед экспонированием необходимо провести строгий контроль их качества. Для
этого все имеющиеся в наличии лишайники располагают рядом и увлажняют,
затем эти сырые слоевища внимательно просматривают и отбраковывают талло-
мы с дефектами, т.е. такие, цвет которых в водонасыщенном состоянии не зеле-
ный. При одинаковом качестве предпочтение отдается более крупным экземпля-
рам.
Чаще всего для экспонирования лишайников используют специальные дере-
вянные щиты. Их размер может быть различным. В Германии, например, в экспе-
риментах с пересаженными лишайниками рекомендуются щиты 290 мм*
170 ммх25 мм, в которых высверлены 10 цилиндрических отверстий глубиной от
15 до 20 мм и диаметром 42 мм (VDI, 1991). Кусочки коры с лишайниками зак-
репляют в углублениях так, чтобы кора субстрата находилась на одном уровне с
древесиной щита; отверстия затем замазывают древесной замазкой или воском.
Поэтому древесина и кора должны быть сухими. Каждое трансплантированное
слоевище получает индивидуальный номер. Щиты прикрепляют к вертикальным
стойкам высотой 1,5 м и размещают в намеченных точках обследуемой террито-
рии. Чтобы защитить экспонируемое устройство от птиц, на верхнюю сторону
щита рекомендуется вбивать острые гвозди (рис. 7.2).
Рис. 7.2. Щит для экспонирования трансплантированных слоевищ (VDI, 1991)
7. Трансплантации лишайников
149
Экспонирование пересаженных слоевищ осуществляют и с помощью специ-
ального устройства, называемого “лишайниковой каруселью” (VDI, 1991), в ко-
тором трансплантаты вращаются ветром вокруг вертикальной оси. Шесть талло-
мов с корой субстрата закрепляют тем же способом, что и на щитах, на шести
звездообразно расположенных вокруг центральной оси деревянных носителях.
Они подвешены к спицам, вставленным в шарообразную ступицу, вращающую-
ся на вертикальной стойке-трубе (рис. 7.3). Плоскости отдельных носителей рас-
полагаются под одним углом к длинной оси трубы-носителя. Импульс для враще-
ния вокруг оси создается ветром с помощью чашеобразных деталей анемометра,
прикрепленных к той же шарообразной ступице над носителями трансплантатов.
Некоторые исследователи экспонировали ветви с растущими на них лишай-
никами (Addison, 1984; Garty et al., 1997e, f, 1998b). Есть примеры транспланта-
ции кусочков коры, валунов или обломков горных пород с растущими на них
представителями лихенобиоты. Их прикрепляют к деревьям, столбам, щитам,
устанавливают в садах и т.д. (Pearson, 1993). Х.Х. Трасс (19856) предлагал пере-
возить куски черепицы с лишайниками из сельской местности или небольших
городов в более крупные города. Оригинальный метод экспонирования слоевищ
разных жизненных форм разработан в Университете шт. Орегон, США (McCune
et al., 1996). Талломы лишайников без субстрата закреплялись специальным кле-
ем в петлях тонких нейлоновых нитей, которые развешивались в намеченных
местах. Такой способ позволил достаточно точно определять влияние условий
экспонирования на рост лишайников и изменение их массы. Не менее своеобра-
зен способ трансплантации лишайников, использованный в г. Красноярске (Бу-
Рис. 7.3. “Карусель” для экспонирования трансплантированных слоевищ (VD1, 1991)
150
7. Трансплантации лишайников
мельников, Григорьев, 1996; Григорьев, Бучельников, 1997). Фрагменты слоеви-
ща эпилитного m\mMmK?iXanthoparmeliasomloensis (=Parmeliastenophylla) сма-
чивали водой и укладывали в стеклянные банки емкостью 1 л на поверхность
предварительно засыпанной в них увлажненной гранитной крошки. Банки зак-
рывали полиэтиленовыми крышками с отверстиями и прикрывали бумажным
экраном для защиты от прямого солнечного света. Такое устройство боксов обес-
печивало поддержание трансплантатов в активном состоянии в течении трехне-
дельного срока их экспонирования в разных по уровню загрязнения районах Крас-
ноярска.
Особые деревянные боксы использовались для экспонирования транспланти-
рованных представителей напочвенных видов, слоевища которых помещались в
такие боксы вместе с субстратом (Tuba, Csintalan, 1993).
При анализе результатов исследований с использованием трансплантатов не-
обходимо обращать внимание на способы экспонирования, поскольку сравни-
вать можно данные, полученные одинаковыми методами. Поэтому эксперименты
с пересадкой лишайников имеют локальное значение и их результаты можно эк-
страполировать лишь на условия района, где они проводились.
7.1.2. Экспонирование
Согласно рекомендациям VDI (1991), приспособления для экспонирования
прикрепляются к стойке, чтобы трансплантаты располагались на высоте 1,5 м
над поверхностью почвы. Деревья, здания и другие предметы могут препятство-
вать движению воздуха, затенять экспонируемые лишайники, поэтому устрой-
ства для экспонирования желательно располагать от них на расстоянии не менее
15 м. В городах или в лесах такое требование не всегда выполнимо. Здесь воз-
можно их размещение в открытых дворах, садах, полянах или прогалинах. При-
способления для экспонирования трансплантатов все же нельзя располагать на
крышах, рядом с деревьями, вблизи строек или на улицах и открытых площадях
с сильным загрязнением почвы. В случае применения экспозиционных щитов
весь период наблюдений по всей изучаемой территории они должны быть одина-
ково ориентированы относительно сторон горизонта, обычно на север.
Расположение точек размещения трансплантатов, а также число пересажен-
ных в каждую такую точку слоевищ зависят от решаемых задач. Чаще, однако,
оно определяется конкретными условиями местности. Так, пересаженные слое-
вища Flavoparmelia (Parmelia) caperata были размещены на расстоянии 6, 12, 23,
34 и 59 км на восток от Бруклина в г. Нью-Йорк, США (Brodo, 1966). Также для
контроля рекомендуется помещать устройство с трансплантатами в точке отбора
образцов лишайников для пересаживания (VDI, 1991).
Продолжительность экспонирования и частота наблюдений определяются
решаемыми задачами исследования и свойствами фиксируемых признаков.
7. Трансплантации лишайников
151
В идеале трансплантированные слоевища желательно измерять и фотографиро-
вать ежемесячно. По ходу наблюдений можно вносить коррективы, если получа-
емые данные свидетельствуют о приемлемости более короткого или более дли-
тельного периодов между документированием наблюдаемых показателей. Если
наблюдают за внешними проявлениями повреждений слоевищ, их фотографиру-
ют непосредственно перед экспонированием, а также через установленные сроки
для получения черно-белых или цветных изображений. Фотоаппарат устанавли-
вается на переносной штатив, а для равномерного освещения используют вспышку.
Чтобы иметь сравнимый материал, в течение всего периода наблюдений необхо-
димо пользоваться одним и тем же оборудованием и фотопленкой. Перед каждой
съемкой лишайники приводят в состояния максимального водонасыщения (око-
ло 5 мин. равномерно опрыскивают дистиллированной водой и ждут, пока капли
воды исчезнут со слоевища). Хорошие результаты были получены с использова-
нием инфракрасной техники фотографирования, поскольку получаемые снимки
наглядно показывали жизненное состояние организма (Plummer, 1968; Kauppi,
Kauppi, 1978; Gouaux, Vincent, 1990; Pearson, 1993).
Помимо внешних признаков нередко фиксируют динамику изменения и дру-
гих показателей (отмирание водорослевых клеток, накопление элементов, физио-
логические параметры и другое), что требует изъятия слоевищ в намеченные сро-
ки. Это приводит к необходимости экспонирования большого числа слоевищ,
поскольку в каждый срок изымается несколько слоевищ, чтобы получить ста-
тистически достоверные результаты. Также надо помнить, что некоторые транс-
плантаты могут погибнуть из-за механических повреждений, оторваться от суб-
страта, быть поврежденными или съеденными птицами или другими живот-
ными, погибнуть от вандализма. Учет таких потенциальных потерь и заданная
точность исследования определяют необходимое количество трансплантируемо-
го материала.
Изменения оценивают сравнением показателей слоевищ перед началом экс-
понирования с параметрами, зафиксированными в сроки наблюдений. При ин-
терпретации данных вносят поправку на возможные изменения характеристик
лишайниковых слоевищ, трансплантированных в месте отбора образцов.
7.2. ОЦЕНКИ ВОЗДЕЙСТВИЯ НА ТРАНСПЛАНТИРОВАННЫЕ
СЛОЕВИЩА
7.2.1. Смертность и изменение внешних признаков слоевищ
Простейшим способом оценки действия новых условий на пересаженные сло-
евища является определение смертности пересаженных слоевищ, выражаемой как
процент погибших за период наблюдения слоевищ от общего числа транспланта-
152
7. Трансплантации лишайников
тов данного вида. К нему относятся и наблюдения за морфологическими измене-
ниями талломов: цвета, толщины слоевища, внешнего вида апотециев, перитеци-
ев, толщины лопастей и т. д. Для характеристики цвета можно пользоваться спе-
циальными таблицами, предназначенными для стандартизации этого показателя
в научных исследованиях, полиграфии, других производствах. Изменение вне-
шних признаков пересаженных слоевищ за период экспонирования оценивают
визуально в баллах или измеряют палеткой долю поврежденных частей талло-
мов. Так, Э. Бродо (Brodo, 1961, 1966) использовал 4-х бальную шкалу оценок
(3, 2, 1,0), где баллом “3” оценивалось здоровое слоевище, а баллом “0” - полно-
стью погибшее. Результаты его наблюдений четко показали негативное воздей-
ствие города Нью-Йорка и продолжительности экспонирования на пересажен-
ные слоевища Flavoparmelia (Parmelia) caperata (табл. 7.2). Те из них, которые
оказались дальше всего от г. Нью-Йорка, претерпевали небольшие изменения, в
то время как ближайшие к городу погибали. Эти результаты дали основания сде-
лать вывод, что воздействие крупного города на загрязнение воздуха распростра-
няется далеко за пределы его границ (Brodo, 1966).
Таблица 7.2
Состояние пересаженных талломов Flavoparmelia (Parmelia) caperata на разном расстоянии
от г. Нью-Йорка через 4 и 12 месяцев после трансплантации (Brodo, 1966).
Расстояние (км) от Бруклина, г. Нью-Йорк Номе р транспланти эованного слоевища
11 1 2 | |з 4 1 5 || 1 2 1 1 3 1 1 4 | Н
Через 4 месяца Через 12 месяцев
59 3 2 3 2 2 3 3 3 2 2
34 2 3 3 2 3 0 1 1 0 1
23 2 1 2 2 2 0 0 1 0 0
12 2 1 2 1 0 0 0 0 0 0
6 1 1 2 1 1 0 0 0 0 0
0 - полностью погибшее слоевище, 1 - слоевище с сильными повреждениями. 2 - слоевище с
незначительными повреждениями, 3 - здоровое слоевище.
В г. Копенгагене (Дания) состояние пересаженных из “чистого” района стра-
ны слоевищ Hypogymnia physodes на дисках коры диаметром 6 см оценивали с
помощью фотоснимков по следующей шкале: 1 - обесцвечено 0% поверхности
таллома, 2 - обесцвечено 10% поверхности, 2-4 - обесцвечено 25% поверхности,
5 - обесцвечено 50% и более поверхности таллома (Sochting, Johnsen, 1978). Уста-
новлена высокая степень корреляции между видимыми повреждениями переса-
женных слоевищ и уровнями концентрации SO2 в различных районах города.
В Копенгагене для Hypogymnia physodes лимитирующим был уровень SO2 в
100 мкг/м3 (Johnsen, Sochting, 1973), тогда как в Англии и Уэльсе - 60-70 мкг/м3.
В г. Будапеште (Венгрия) трансплантированные слоевища Hypogymnia physodes,
судя по приложенным к статье цветным фотографиям, показали значительные
7. Трансплантации лишайников
153
Таблица 7.3
Изменение показателей жизнестойкости эпифитных лишайников
(средние показатели для 15 деревьев) (Трасс, 19856)
Вид 15.06.1982 15.12.1982 15.06.1983 15.06.1984
Parmelia sulcata 1 2 3 4
Melanelia (Parmelia) olivacea 1 3 4 5
Hypogymnia physodes 1 1 2 2
Ramalina farinacea 1 3 4 5
Lecanora allophana 1 4 5 5
Lecidella elaeochroma 1 2 4 4
Phlyctis argena 1 4 5 5
изменения внешних признаков талломов в результате воздействия химического
завода, автотранспорта и других источников загрязнения (Farkas et al., 1985).
В г. Бремене (Германия) цветные фотоизображения пересаженных слоевищ
Parmelia sulcata анализировали с помощью компьютерной программы (Steinkamp
und and., 1999). Изменение окраски совпадало с параметрами “нарушение среды
местности”, которые соответствовали интенсивности движения городского транс-
порта и степени промышленного загрязнения воздуха. Заметные разрушения сло-
евищ пересаженных представителей Hypogymnia enteromorpha наблюдали через
1 год в условиях кислотного загрязнения в горах Сан Бернардино в Калифорнии
(Sigal, Nash, 1983). В Германии и Австрии установлена прямая корреляция меж-
ду степенью загрязнения и процентом отмерших поверхностей пересаженных
слоевищ Hypogymnia physodes (Werner, 1993; Werner, Niemeyer, 1994; Hufna-
gel, Turk, 1998). Оригинальное исследование было проведено в Тарту, где весной
1982 г. при реставрации парка в центре города были высажены завезенные из
центральной части Эстонии 15-30-летние липы, на которых росли и лишайники.
Местные специалисты регулярно фиксировали их состояние (Трасс, 19856). Ре-
зультаты их наблюдений показаны в таблице 7.3, в которой состояние слоевищ
оценено по 5-бальной шкале: 1 - слоевище нормально развитое; 2 - на слоевищах
единичные повреждения; 3 - повреждено до половины слоевища; 4 - остались
лишь небольшие участки слоевища; 5 - слоевище погибло. Полученные данные
дали основание для ранжирования обследованных видов в виде ряда от более
чувствительных к загрязнению к выносливым: Lecanora allophana, Phlyctis argena,
Melanelia olivacea, Ramalina farinacea, Lecidella elaeochroma, Parmelia sulcata,
Hypogymnia physodes (Трасс, 19856).
7.2.2. Рост слоевищ и прирост биомассы
Наблюдения за скоростью роста и изменением биомассы трансплантирован-
ных слоевищ проводились в основном для изучения влияния условий, не связан-
154
7. Трансплантации лишайников
ных с прямым действием загрязнения воздуха. Так, исследовалось влияние свойств
субстрата на рост пересаженных Xanthoparmelia (Parmelia) conspersa, Р saxatilis,
Melanelia (Parmelia) glabratula ssp. fuliginosa, Physcia tenella, Phaeophyscia
(Physcia) orbicularis, Xanthoria parietina (Armstrong, 1990, 1993), снежного по-
крова на рост Xanthoparmelia lineoloa и X. decipiens (Benedict, 1991), сезонов года
на прирост биомассы Lobariapulmonaria (Muir et al., 1998), местонахождения на
опушке или в глубине леса на рост и биомассу алекториевых лишайников,
Platismatiaglauca и Lobariapulmonaria (Renhom, Esseen, 1995; Renhom et al., 1997),
произрастания в естественном или посаженном лесу на рост и биомассу Lobaria
oregana, Pseudocyphellaria rainierensis (Sillet, 1994; Sillett, McCune, 1998). В пос-
ледних случаях был использован оригинальный метод пересаживания, позволя-
ющий без повреждений слоевищ проводить измерения их массы и других показа-
телей (McCune et al., 1996). Однако можно привести примеры исследований воз-
действия и загрязнителей на рост слоевищ. На юго-западе Финляндии изучалось
воздействие целлюлозно-бумажной фабрики на рост трансплантированных эпи-
гейных лишайников Cladina stellaris (^Cladonia alpestris), C. mitis, C. rangiferina
(Ikonen, Karenlampi, 1976). Прирост слоевищ на расстоянии 0,5 км от предприя-
тия был на 50% меньше, чем на расстоянии 5 км. В Германии, под Бременом, в
сухих сообществах на песках изучалось действие различных удобрений на реге-
нерацию пересаженных подециев Cladonia furcata (Vagts et al., 1994). Этот пока-
затель у контрольных образцов через год экспонирования был оценен как “силь-
ная” и “средняя”. Образцы, обработанные фекалиями кролика, показали “сред-
нюю” регенерацию, азотом - “сильную”, СаСО3 и S - “слабую”, CaCN2 - не
регенерировали. В г. Копенгагене (Дания) для оценки роста пересаженных из “чи-
стого” района слоевищ Hypogymnia physodes применили трехбалльную шкалу:
0 - рост не наблюдается, 1 - слабый рост, 2 - заметный рост (Sochting, Johnsen,
1978). В контроле (место отбора слоевищ для пересадки) рост всех слоевищ был
оценен баллом 2, в менее загрязненных частях г. Копенгагена - 1, а в сильно
загрязненных - 0. В г. Венеции (Италия) слоевища Flavoparmelia (Parmelia)
caperata пересадили в промышленную и сельскую зоны, а также на урбанизиро-
ванную территорию, где наблюдали за изменением поверхности трансплантиро-
ванных талломов, сравнивая этот показатель с варьированием поверхности слое-
вищ в месте отбора образцов для пересадки (Caniglia et al., 1998). Измерения по-
верхности проводили по фотоснимкам. Период экспонирования составил 257 дней
с сентября 1994 по июнь 1995 г. В контрольной точке происходило медленное, но
незначительное увеличение поверхности слоевищ - с 21,22 до 23,15 см2. В про-
мышленной зоне во всех 4 точках поверхность слоевищ уменьшилась (с 5,16 до
1,40 см2, с 17,58 до 14,77 см2, с 10,32 до 7,27 см2, с 14,88 до 13,41 см2). У транс-
плантатов на сельской территории в 3 точках происходило увеличение поверхно-
сти слоевищ (с 16,84 до 19,76 см2, с 11,85 до 13,23 см2, с 13,00 до 15,61 см2), а в
одной точке поверхность уменьшилась (с 9,15 до 6,93 см2). У слоевищ, Переса-
7. Трансплантации лишайников
155
женных на урбанизированную территорию, значимых различий величин поверх-
ности за период экспонирования не обнаружено. В этой работе исследователи
столкнулись с рядом проблем: за период экспонирования кора под транспланти-
рованными слоевищами деформировалась, что вызвало изменение изображения
слоевищ на фотографиях. Кроме того, перекрывание лопастей в ходе роста слое-
вищ приводило к неверным оценкам изменения поверхности.
7.2.3. Повреждение клеток бионтов
Визуально наблюдаемые повреждения слоевищ пересаженных лишайников
являются следствием изменений клеток бионтов лишайника. В г. Оулу (Финлян-
дия) слоевища Hypogymniaphysodes. собранные в 50 км от города, пересадили в
незагрязненную, слабо загрязненную местности и вблизи предприятия со значи-
тельными выбросами загрязнителей. Реакцию пересаженных лишайников на
новые условия изучали на срезах с краев лопастей с помощью флуоресцентного
микроскопа (Kauppi, 1980). В незагрязненной местности показатели флуоресцен-
ции пересаженных лишайников практически не отличались от показателей конт-
роля в месте отбора образцов. В слабо загрязненной местности флуоресценция
срезов лишайников значительно увеличивалась при длине волны 656 нм, тогда
как вблизи предприятия это увеличение наблюдалось при 589 нм. Исследователи
из Красноярска измеряли замедленную флуоресценцию экспонировавшихся три
недели пересаженных cjxQtbH^Xanthoparmelia somloensis (= Parmeliastenophylla).
Величина этого показателя зависит от количества работающих реакционных цен-
тров, активности электротранспортной цепи фотосинтеза и значения градиентов
протонов на мембране тилакоидов. В слабо загрязненной части города показа-
тель замедленной флуоресценции талломов составлял 80-93% от контроля, в сред-
не загрязненной - 50-60%, в сильно загрязненной - 32-40% и в критически заг-
рязненной - 2-13% (Григорьев, Бучельников, 1997). В г. Куопио (Финляндия)
изучали воздействие выбросов предприятий по производству удобрений и бума-
ги (двуокись серы, двуокись азота, аммиак, фториды, пыль) на клетки бионтов
пересаженных слоевищ Hypogymnia physodes и Bryoria capillaries (Holopainen,
1984a). Трансплантаты поместили на расстоянии 500 и 3000 м от предприятий и
периодически с помощью светового микроскопа определяли соотношение мерт-
вых и здоровых водорослевых клеток. Было выделено 6 ступеней повреждений в
зависимости от доли мертвых водорослевых клеток. Уже через неделю после пе-
ресадки в наиболее загрязненных точках наблюдались структурные изменения,
которые приводили к быстрой гибели водорослевых клеток: скручивание и деге-
нерация митохондрий, скручивание тилакоидного пространства и сближение ти-
лакоидов, деформация пиреноглобулей и дегенерация пиреноидного матрикса,
гранулирование и разрыв тилакоидов, появление жироподобных образований
156
7. Трансплантации лишайников
между тилакоидами, заполнение бесформенной массой клеточного пространства.
У представителей В. capillaries эти повреждения происходили быстрее и чаще,
чем у слоевищ Н. physodes. Изменения происходили и в клетках микобионта, но
они были не столь очевидными (Holopainen, 1984а, Ь). Разные изменения ультра-
структуры наблюдались и у пересаженных в различные по уровню загрязнения
районы г. Мадрида (Испания) слоевищ Evernia prunsatri и Parmelina (Parmelia)
quercina, поскольку фотобионтом первого вида является представитель Protococcus,
а второго - Trebouxia (Silva-Pando, Ascaso, 1982). В г. Сан-Хосе (Коста-Рика) так-
же определяли долю поврежденных водорослевых клеток у пересаженных слое-
вищ рода Parmelia (Mendez, Fournier, 1980). Здесь установлена корреляция между
числом мертвых клеток фотобионтов и уровнем загрязнения места, где экспони-
ровали слоевища. Все трансплантаты погибли через 8 месяцев.
В декабре 1990 г. я разместил 10 щитов со слоевищами Hypogymniaphysodes,
собранными в лесу в 1 км от платформы 73 км Рижского направления железной
дороги, в разных по уровню загрязнения частях г. Москвы. К сожалению, ванда-
лы довольно быстро уничтожили большинство щитов, в том числе и в месте отбо-
ра образцов. Дольше всего (до августа 1991 г.) сохранялся щит, установленный в
центре города на Боровицком холме, напротив Государственной библиотеки им.
В.И. Ленина, во дворе здания, в котором тогда располагался музей М.И. Калини-
на. С этих щитов я периодически снимал по 10 слоевищ и на срезах (10 на каж-
дом слоевище, т.е. 100 срезов в каждый срок) на расстоянии 0,5 см от краев лопа-
стей при помощи микроскопа МБИ-4 подсчитывал общее число водорослевых
клеток и число мертвых клеток. Результаты показаны в таблице 7.4.
Таблица 7.4
Доля (%) мертвых водорослевых клеток в пересаженных на Боровицкий холм в г. Москве
слоевищах Hypogymnia physodes
Дата 16.12.90 10.01.91 25.01.91 25.02.91 21.04.91 12.08.91
% мертвых клеток 4,0±2,0 14,4±1,0 10,5±0,9 11,2±1.1 17.0±2.0 34,0±4.0
Как видим, в первые недели наблюдался стресс, вызванный пересадкой, за-
тем в зимние месяцы сохранялось относительно стабильное состояние, но с на-
ступлением теплого сезона отмирание водорослевых клеток ускорилось.
7.2.4. Изменение количества пигментов -
показателей фотосинтеза и дыхания
Появление на пересаженных в загрязненные места слоевищах лишайников
визуально обнаруживаемых повреждений требует длительного экспонирования.
7. Трансплантации лишайников
157
Измерения же газообмена С02 у таких талломов показали, что редукция этого
жизненно важного физиологического процесса начинается задолго до появления
визуально наблюдаемых повреждений слоевищ, как, например, у транспланти-
рованных в г. Зальцбурге (Австрия) представителей Hypogymnia physodes и
Parmelia sulcata (Turk, Christ, 1980; Christ, Tiirk, 1981, 1982). Там же позднее у
талломов Hypogymniaphysodes, пересаженных в места, где загрязнение было ком-
плексным, ежемесячно в течение 8 месяцев определяли несколько физиологиче-
ских показателей (Egger et al., 1994). Была установлена положительная корреля-
ция между уровнем озона и содержанием хлорофилла, малон-диальдегида, а так-
же с активностью супероксид-дисмутазы. Уровень двуокиси серы имел отрица-
тельную корреляцию с показателями нетто-фотосинтеза и положительную - с
содержанием малон-диальдегида и кислой фосфатазой. В г. Линце (Австрия) на-
ряду с визуальной оценкой степени повреждения измеряли наблюдаемый и сум-
марный фотосинтез слоевищ Hypogymnia physodes^ пересаженных в 9 точек тер-
ритории города и его окрестностей, различающихся по степени загрязнения воз-
духа (Hufnagel, Tiirk, 1998). Было установлено, что наиболее поврежденные
талломы имели наиболее низкие показатели фотосинтетической активности.
Измерение физиологических показателей трансплантированных в Рудных
горах и ряде других мест Германии, различающихся уровнем SO2 в атмосфере,
слоевищ Bryoria fuscescens, Cetraria islandica, Hypogymnia physodes, Parmelia
sulcata показало снижение измеряемых параметров газообмена СО2 с увеличени-
ем загрязнения, причем у представителей В. fuscescens повреждение фотосинте-
тического аппарата наступало через 4-8 недель экспонирования, у С. islandica и
Н physodes - через 8-10 недель, а у Р sulcata - после 3-4 месяцев экспонирова-
ния фотосинтез снизился только на 30% в сравнении с контролем (Haffner et
al., 2001).
На юго-западе Финляндии на расстоянии 0,5 км от целлюлозно-бумажной
фабрики газообмен СО2 трансплантированных эпигейных лишайников Cladina
stellaris (=Cladonia alpestris), C. mitis, C. rangiferina составил около 50% от вели-
чины их газообмена на расстоянии 5 км от предприятия при не изменившемся
содержании хлорофилла (Ikonen, Karenlampi, 1976).
Иногда наблюдались неожиданные явления. Так, в г. Лос-Анджелесе (США)
у трансплантатов кустистого вида Ramalina menziesii обнаружили, что летом тал-
ломы этого вида, экспонированные в “грязном” районе города, в сравнении с
контролем значимо снижали показатели нетто-фотосинтеза, количества хлоро-
филла и увеличивали долю феофитина (Boonpragob, Nash, 1991). Однако зимой
содержание хлорофилла в слоевищах из “грязного” места было выше, чем в тал-
ломах из “чистого” района, также не наблюдалось у них и большой разницы па-
раметров нетто-фотосинтеза. Авторы исследования предположили, что зимой в
загрязненных местах происходит стимулирование синтеза хлорофилла, чему бла-
гоприятствует присутствие среди поллютантов соединений азота. Несколько иные
158
7. Трансплантации лишайников
результаты были получены в г. Кордове (Аргентина). Здесь слоевища листоватых
видов Parmotrema austrosinense и Р. conferendum поместили в мешочки из нейло-
новой сетки, которые экспонировались с августа 1994 г. (конец зимы) по январь
1995 г. (середина лета) в разных по степени загрязнения местах города (Canas et
al., 1997). У обоих видов концентрация хлорофилла “а” снизилась как в загряз-
ненном, так и в “чистом” районах уже через 1 месяц (август-сентябрь) экспони-
рования, причем это уменьшение в загрязненном месте было статистически зна-
чимым. Также снизились концентрации хлорофилла “Ь” и общего хлорофилла.
Затем с ноября в слоевищах представителей обоих видов произошло увеличение
содержания хлорофилла “а”, общего хлорофилла и каротиноидов. Ранее (май
1992 г.) в том же городе было проведено изучение воздействия разных факторов
(интенсивность движения автотранспорта, промышленное загрязнение, характе-
ристики древесных насаждений, высота зданий, удаление от реки и другое) на
трансплантаты кустистого лишайника Ramalina ecklonii, слоевища которого раз-
местили на деревьях одной породы в мешочках из нейлоновой сетки в 28 точках
обследуемой территории (Gonzalez et al., 1996, 1998). После 6 недель экспониро-
вания в талломах определяли концентрации хлорофилла “а” и “Ь”, общего хлоро-
филла, феофитина “а”, малон-диальдегида. Установлено, что концентрации хло-
рофилла “а” и “Ь” имеют отрицательную корреляцию с показателями интенсив-
ности движения транспорта и в меньшей степени - с наличием древесных
насаждений. Величина феофитина “а” положительно коррелировала с видом про-
мышленного загрязнения, а отношение концентраций феофитин “а’Ухлорофилл
“а” имело высокую степень корреляции с показателем интенсивности движения
автотранспорта, и менее значимую - с наличием древесных насаждений и уда-
ленностью от реки. Содержание малон-диальдегида в талломах за этот короткий
период экспонирования не показало какой-либо четкой связи с параметрами сре-
ды. Сходные результаты были получены здесь и при использовании в качестве
тест-объекта слоевищ Punctelia subrudecta, пересаженных в 22 точки вокруг элек-
тростанции (Gonzalez, Pignata, 1997). Деградацию содержания хлорофилла в транс-
плантированных слоевищах и снижение показателей газообмена СО2 с увеличе-
нием степени загрязнения воздуха наблюдали неоднократно (Galun, Ronen, 1988;
Garty et al., 1993a; Gries, 1996). Изменение пигментного состава талломов вызы-
вает изменение спектральных свойств трансплантированных слоевищ (Garty, et
al., 1997а, b, е, f, 1998b). Однако необходимо учитывать, что реакция пересажен-
ных слоевищ лишайников в новых условиях отличается от реакции представите-
лей того же вида, постоянно растущих в местах, куда были перемещены транс-
плантаты.
В Швейцарии сравнили физиологические показатели трансплантированных
и аборигенных представителей Parmelia sulcata в разных по уровню загрязнения
зонах территории и выявили значимые различия физиологических реакций мес-
тных и пересаженных слоевищ (von Arb, Brunold, 1990).
7. Трансплантации лишайников
159
7.2.5. Нарушение проницаемости мембран
и других параметров жизнедеятельности слоевищ
Показателями воздействия загрязнителей на трансплантированные слоевища
разных видов лишайников служат и такие параметры как целостность мембран
(Kauppi 1976; Pearson, Rogers, 1982; Garty et al., 1993a, b, 1997a, b, e, 1998a, b),
содержание растворимых белков (Levin, Pignata, 1995; Gonzalez et al., 1996, 1998),
концентрация аденозинтрифосфата (АТФ) (Kardish et al., 1987; Garty et al., 1988),
образование гормонов и этилена (Epstein et al., 1986; Garty et al., 1993a, b, 1997f).
Практически все исследования показали, что с увеличением степени загрязнения
увеличивается и проницаемость мембран, что отражается в более высокой элект-
ролитической проводимости водных вытяжек и оттоке ионов калия. Концентра-
ция растворимых белков под влиянием загрязнения у слоевищ Ramalina ecklonii
уменьшалась. Концентрация АТФ в загрязненных местах значительно снижалась,
причем это уменьшение коррелировало с увеличением содержания в слоевищах
Pb, Zn, Си, и не было связано с концентрацией S в талломах Ramalina (duriaei)
lacera (Garty et al., 1988). Образование этилена в загрязненных местностях у пе-
ресаженных слоевищ Ramalina (duriaei) lacera увеличивалось, причем довольно
быстро, а уровень одного из ауксиновых гормонов - индол ил у ксу ной кислоты -
через год экспонирования в загрязненном месте составил 0,3 мкг/г, тогда как в
контроле - 2-5 мкг/г (Epstein et al., 1986).
7.2.6. Аккумуляция загрязнителей
Накопление веществ-загрязнителей трансплантированными слоевищами - это,
пожалуй, чаще других используемый показатель при опытах с пересадкой ли-
шайников, за исключением визуально обнаруживаемых признаков. Для экспери-
ментов выбирали представителей разных видов всех трех основных форм роста
лишайников-кустистые, листоватые и корковые. Значителен и перечень веществ,
которые измеряли в пересаженных талломах. Нередко определение особенностей
аккумуляции загрязнителей слоевищами сочеталось с наблюдениями за другими
признаками, часть которых уже была названа. Например, измерения степени от-
мирания водорослевых клеток в г. Москве (см. табл. 7.4) я дополнил результа-
тами определения концентрации ряда элементов в слоевищах трансплантатов
(табл. 7.5).
В сравнении с контролем концентрация Cu, Pb, Cd в талломах со временем
возрастает как в центре города, так и в лесопарке примерно в 12 км к северу от
Боровицкого холма, где экспонировались слоевища. Однако в центре рост кон-
центрации этих элементов более значителен, чем в парке ТСХА. Можно сделать
вывод, что названные три элемента в той или иной мере являются загрязнителя-
160
7. Трансплантации лишайников
Таблица 7.5
Величины (мин.-максим.) содержания (мкг/г сухого веса) некоторых элементов
в трансплантированных слоевищах Hypogymnia physodes в центре (Ц) города
и в Лесной опытной даче ТСХА (Т)
Дата 16.12.1990 Место 73 км от г. Москвы, контроль Си 1,87-4,59 РЬ 2,44-3,57 Cd 0,13-0,28 Zn 83,3- 88,5 Мп 161,-293,
10.01.1991 ц 3,01-4,55 3,05-3,20 0,13-0,22 89,6-107,0 179,0-236,0
т 2,17-3,42 2,39-3,06 0,17-0,27 83,6-85,0 135,0-205,0
25.01.1991 ц 6,42-10,40 3,26-3,71 0,12-0,18 88,9-89,0 199,0-208,0
т 2,83-7,16 2,24-4,26 0,15-0,29 78,9-112.0 107,0-207,0
25,02,1991 ц 7,53-7,88 5,30-6,83 0,22-0,24 97,6-109.0 325,0-331.0
т 3,91-4,85 3,55-4,57 0.17-0.35 76,4-106.0 120.0-139,0
21,04,1991 ц 11,9-15, 5,88-8,24 0,19-0,28 68,7-220,0 167.0-302,0
т . 7,3-9,76 4.42-5,29 0,17-0,40 79.6-85,60 135,0-208,0
12,08.1991 ц 12,2-20,8 6,8-10.7 0,19-0,40 108,-152,0 142,0-303,0
т Щит со слоевищами лишайника уничтожен вандалами
ми воздуха города во всех его частях. Показатели содержания Zn и Мп в транс-
плантатах в разные сроки не дают оснований для определенных заключений.
В Мурманской области срезанные ветви ели и отрезки стволов березы со сло-
евищами Hypogymniaphysodes разместили в пяти зонах деградации у Мончегор-
ского комбината “Североникель” (Крючков, Сыроид, 1990). После 3 месяцев экс-
понирования в слоевищах измерили концентрации Ni, Си, Zn, S, Al, Р. Установ-
лено, что по мере удаления от источника загрязнения четко убывали концентрации
никеля и меди в слоевищах лишайника. На поведении других элементов сказа-
лось наличие других источников - Кандалакшского алюминиевого завода, ТЭС и
производство фосфора в Апатитах.
В Калифорнии (США) слоевища Ramalina menziesii из “чистой” территории
штата пересадили в загрязненные части г. Лос-Анджелеса и экспонировали по
три месяца летом 1985 г., зимой того же года и летом 1986 г. (Boonpragob, Nash,
1990). После каждого периода экспонирования в слоевищах определяли содержа-
ние 25 элементов. Обработка данных показала, что концентрации только 14 из
них (Ba, Са, Cd, Си, Fe, К, Mg, Мп, Na, Р, Pb, Si, Sr, Zn) в той или иной мере
зависят от сезона года, поскольку они, как правило, были выше летом, чем зимой.
Динамика поведения этих элементов в трансплантатах отражала не только ат-
мосферное загрязнение, но и степень вовлечения их в метаболизм слоевища, а
также (в меньшей степени) накопление морских аэрозолей и почвенных частиц.
В г. Любляне (Словения) яблоневые ветви с Hypogymnia physodes разместили в
4 точках города, в том числе у работающей на угле электростанции, на срок 5 и 11
7. Трансплантации лишайников
161
месяцев (Jeran et al., 1993). После каждого периода экспонирования в талломах
определяли концентрации 29 элементов. Установлено, что за исключением Со,
Cs, К, Rb, Se, Zn, концентрация всех измеренных элементов в слоевищах прямо
коррелировала с длительностью экспонирования. Однако в Нидерландах концен-
трация Со (и Sc) в пересаженных талломах Parmelia sulcata была выше в образ-
цах, экспонировавшихся более продолжительный период (Sloof, 1995а). В Израи-
ле содержание ряда элементов определяли в трансплантированных слоевищах
Ramalina (duriae) lacera,экспонировавшихся с декабря 1981 г. в течение 1 года,
для определения воздействия тепловой (уголь, нефть) электростанции на среду
(Fuchs, Garty, 1983; Garty, 1987, 1988; Garty, Hagemeyer, 1988). Результаты срав-
нивали с данными одногодичного экспонирования в 1979-1980 гг. в тех же точ-
ках до начала работы электростанции (табл. 7.6).
Таблица 7.6
Концентрация некоторых элементов (мг/кг) в экспонировавшихся слоевищах
Ramalina (duriaei) lacera до (0) и через 1 год (1)
после начала работы электростанции (э/с) в Израиле (Garty, 1988)
Расстояние от э/с (км) Ni Сг Си Zn Pb
0 1 0 1 0 1 0 1 0 1
4.6 29,2 74,4 8,5 14,2 30,3 14,2 368,1 61,0 74,4 264,4
7.3 8,2 24,6 5,2 9,2 17,9 13,3 340,0 58,2 19,0 66,4
7,7 8,3 20, 2,4 23,8 21,1 14.2 82,0 59,0 57,6 43,4
7,8 13,7 13,2 3,2 13,6 17,1 17,2 74,1 75,4 43,1 41,2
9,4 7,7 13,8 3,0 14,0 10,8 13,6 74,7 44,0 26,5 33,0
12,4 10,8 18,4 3,5 10,6 12,0 11,0 179,6 44.6 16,5 31,0
16,5 13,6 7,4 4.0 6,2 23,3 12.4 138,3 46,6 51,4 21,0
19,5 16,8 13,8 3,2 14,0 11,7 14,8 134,4 45,4 13,2 41,2
24,5 12,6 20,2 5.3 15,4 14,2 11,6 54,1 54,4 34,7 34,4
При анализе сведений таблицы 7.6 следует учитывать, что точки, куда переса-
живали слоевища, располагались в разных направлениях от электростанции,
поэтому общее уменьшение концентрации элементов с увеличением расстояния
имеет исключения. Снижение концентраций Zn и Си в сравнении с первым пери-
одом экспонирования, вероятно, связано с тем, что эти элементы входят в состав
препаратов, широко используемых для борьбы с вредителями плантаций орехо-
вых деревьев (пекан), а в 1980-1981 гг. эти препараты почти не применялись.
Можно отметить, что за год работы электростанции произошло значительное уве-
личение концентраций Ni, Сг и Pb в трансплантатах, особенно вблизи станции.
Загрязнение среды первыми двумя элементами связано с выбросами электростан-
ции, а повышенное содержание свинца объяснялось значительным приростом
162
7. Трансплантации лишайников
числа автомобилей в изученной местности - с 524950 в 1979 г. до 661671 в
1982 г. (Garty, 1988). Однако число автомобилей не всегда определяет уровень
загрязнения местности. Имеет значение и скоростной режим автомобильных
трасс, что показали результаты исследования, проведенного в г. Оулу, Финляндия
(Garty et al., 1997с). Здесь сосновые ветви с бородовидными слоевищами Usnea
hirta, собранными в лесу в 15 км от города, разместили в пригороде у скоростной
автомобильной трассы с нагрузкой около 25000 автомобилей в сутки, а также на
двух улицах города со значительно меньшей нагрузкой (около 14000 автомобилей
в сутки) и где скорость движения была ограничена. Продолжительность экспони-
рования составила 45 дней (с 7 августа по 23 сентября 1993 г.). Наряду с другими
параметрами в слоевищах определены и концентрации 11 элементов (табл. 7.7).
Оказалось, что в слоевищах, экспонировавшихся у магистралей с относительно
невысокой нагрузкой автотранспорта, концентрации ряда элементов были выше,
чем на трассе, где за сутки проходило почти вдвое большее число автомобилей.
Этот феномен авторы связали с более высокой степенью продуваемости высоко-
скоростной трассы, поскольку она располагалась вне городской черты (Garty et.
al., 1996, 1997с). Воздействие автодороги на загрязнение среды также зависит от
направления преобладающих ветров и особенностей загрязнителя. В 27 км юж-
нее Будапешта, где преобладают ветры северо-западного направления, на разном
расстоянии от шоссе (1,2,5,5,10,30,60,90 м) с интенсивностью движения 10000-
12000 автомобилей/день в течение трех месяцев экспонировались слоевища
Cladonia convoluta (Tuba, Csintalan, 1993). Установлено, что наивысшая концент-
рация всех элементов, исключая ванадий, была в наиболее близких к дороге об-
разцах лишайника. На расстоянии 1 м от полотна дороги уровни загрязнения
Таблица 7.7
Концентрация элементов (мкг/г сухого веса) в трансплантированных слоевищах Usnea
hula, экспонировавшихся 45 дней (август-сентябрь 1993 г.) в г. Оулу (Garty et. al., 1997с)
Эле- мент Место экспонирования
Контроль (лес) Скоростная трасса (25000 ам/сут)* Низкоскоростная улица I (14000 ам/сут) Низкоскоростная улица 2 (14000 ам/сут)
К 3735 4285 4708 4233
Са 899 869 3615 2223
Na 963 1050 1271 750
Mg 555 590 697 668
Fe 523 538 691 487
Zn 114 124 231 114
Mn 45 61 68 53
Pb II 23 13 29
Ni 6,2 11,9 14,7 10,5
Cu 6,2 9,8 13,6 9,2
Cd 2,1 2,9 7,1 3,7
* - автомобилей/сутки
7. Трансплантации лишайников 163
кадмием, медью, никелем и цинком были выше в 2-3 раза, чем в контроле, алю-
минием и свинцом - в 3-4 раза, а хромом и железом - в 5 раз. С увеличением
расстояния от шоссе концентрации металлов снижались. Повышенные, в сравне-
нии с контролем, уровни кадмия, свинца и цинка отмечались на расстояние до
30 м на запад от дороги и до 60 м - на восток, хрома -до 2,5 и до 5 м соответствен-
но, меди, железа и алюминия - до 5 и до 10 м, никеля - до 5 м с обеих сторон
дороги. Не найдена зависимость концентрации ванадия от расстояния от дороги,
хотя уровни содержания этого элемента в трансплантантах у шоссе были выше,
чем в контроле.
8. ЛИХЕНОИНДИКАЦИЯ ПО ДАННЫМ
О НАКОПЛЕНИИ ЗАГРЯЗНИТЕЛЕЙ СЛОЕВИЩАМИ
Особенности биологии и физиологии лишайников как пойкилогидридных
организмов, не имеющих приспособлений, препятствующих проникновению в
их тела различных загрязнителей, - медленный и длительный рост талломов,
невыразительность реакций на сезонные явления, географически обширные аре-
алы многих видов - увеличили частоту использования их как биомониторов
загрязнения среды не только газами, но и такими загрязнителями как токсичные
металлы и неметаллы и органические соединения, поскольку динамику химиче-
ского состава талломов в пространстве и во времени связывали с изменени-
ем свойств среды их обитания (Ю. Мартин, 1985, 1987). Если в обзоре начала
1980-х годов цитировалось 26 публикаций, посвященных мониторингу металлов
в среде с помощью лишайников (Martin, Coughtrey, 1982), через год - 41 (Инсаро-
ва, 1983), то в начале 1990-х анализ данных о содержании только свинца в ли-
шайниках содержал ссылки на 19 работ (Garty, 1993). И число подобных исследо-
ваний непрерывно растет (Falla et al., 2000; Garty, 2000a, 2000b; Conti, Cecchetti,
2001). Выявляется содержание в лишайниках все большего количества элементов
и соединений. Кроме того, растет список видов лишайников, для которых имеют-
ся данные о химическом составе их слоевищ; расширяется перечень регионов,
где проводились такие работы. Детальный анализ опубликованных по этой про-
блеме обширных количественных материалов может составить основу отдельной
монографии большого объема, поэтому в данном разделе книги конкретные ис-
следования не рассматриваются, и главное внимание будет уделено лишь некото-
рым принципиальным аспектам этого направления лихеноиндикации. Некото-
рые фактические данные о накоплении различных элементов лишайниками были
приведены в конце предшествующей главы (7.2) и в главе 2.
Исследования, базирующиеся на способности лишайников накапливать в сво-
их телах различные химические вещества, в своих выводах опираются на сведе-
ния о химическом составе лишайников и динамику концентрации элементов в
слоевищах отдельных видов во времени и/или пространстве. Среди них можно
выделить два основных типа: (1) устанавливающие фоновый уровень или “точку
8. Лихеноиндиксщия по данным о накоплении загрязнителей слоевищами 165
отсчета” концентрации избранного ряда элементов или соединений в талломах
ограниченного числа видов лишайников для определения долговременного варь-
ирования содержания этих веществ на избранной территории; это предполагает
повторное измерение химического состава лишайников через некоторые проме-
жутки времени; при этом изменчивость химического содержания лишайников в
пространстве обычно составляет сопутствующую проблему таких исследований;
(2) устанавливающие особенности только пространственной вариабельности хи-
мического состава лишайников для определения влияния изолированных или
конгломерата источников специфических загрязнителей и разделения загрязни-
телей природного и техногенного происхождения; при этом изменчивость хими-
ческого состава лишайников во времени может быть сопутствующей проблемой
таких исследований. На практике для достоверного суждения о свойствах среды
по имеющимся сведениям о показателях лишайников важно знать варьирование
содержания загрязнителей в их телах как во времени, так и в пространстве.
►8.1. ПЛАН ИССЛЕДОВАНИЯ
Исследование, опирающееся на данные о химическом составе лишайников,
как и любое другое индикационное исследование, зависит от специфики изучае-
мых объектов, особенностей экосистемы, бюджета проекта, доступного лабора-
торного оборудования, квалификации персонала, а также некоторых других фак-
торов. Лихеноиндикационные исследования, опирающиеся на аккумулирующие
способности лишайников, требуют значительных финансовых затрат, что долж-
но стимулировать тщательное планирование таких работ. Это планирование, в
зависимости от задач исследования, включает:
- выбор химических элементов и соединений, концентрации которых будут
измеряться в слоевищах лишайников;
- выбор популяции лишайников, у представителей которой будут измеряться
концентрации веществ;
- получение сведений об особенностях экологии выбранного вида;
- установление мест отбора проб для анализов или интервалов времени, че-
рез которые будут отбираться пробы;
- установление объема отбираемой пробы, размера выборки (повторности) в
одном месте или в один срок, обеспечивающей статистически приемлемые ре-
зультаты;
- установление критериев, обеспечивающих качество результатов и контроль
качества;
- обзор доступных материалов для определения параметров возможного ва-
рьирования содержания определяемых элементов в слоевищах выбранного вида;
- проведение исследования в соответствии с намеченной методикой;
166 8. Лихен виндикация по данным о накоплении загрязнителей слоевищами
- оценку результатов химических анализов и установление соответствующих
трендов в пространстве и/или во времени.
8.1.1. Выбор химических элементов и соединений
для определения их содержания в слоевищах лишайников
Перечень химических элементов и соединений, намечаемых для измерения
их содержания в телах лишайников, определяется задачами исследования, харак-
теристиками источников загрязнения, воздействующих на изучаемую террито-
рию, наличием соответствующей измерительной техники и финансовым обеспе-
чением, поскольку затраты на определение концентраций разных химических
веществ в биологических объектах далеко не одинаковы. Предварительно жела-
тельно собрать сведения о свойствах интересующих веществах, особенностях их
миграции в разных типах биогеоценозов и специфике накопления у представите-
лей разных таксономических групп организмов. Например, диоксины аккумули-
руются главным образом в жировых тканях. Для целей биоиндикации в лишай-
никах целесообразно определение элементов, переносимых воздушными потока-
ми в форме газов, пыли, аэрозолей, а также с осадками.
8.1.2. Выбор популяции лишайников
Обычно в лихеноиндикационном исследовании для измерения содержания
химических веществ выбирают вид, чувствительный к интересующим загрязни-
телям и представленный стабильной и здоровой популяцией на всей изучаемой
территории на одном субстрате. Желательно, чтобы этот субстрат имел низкие
концентрации веществ, которые будут измеряться в слоевищах лишайников. Раз-
мер популяции должен быть достаточным для обеспечения сбора необходимого
для проведения анализов объема проб в нужной повторности и чтобы обор проб
не стал причиной гибели популяции. Также важно, чтобы вид можно было про-
сто идентифицировать в поле, без труда отделить от субстрата и очистить от
примесей. Естественно, такая избирательность базируется на сведениях об осо-
бенностях экологии видов лишайников, свойственных обследуемой местности.
Наличие среди лишайников толерантных к металлам видов и видов-гиперакку-
муляторов, что, вероятно, связано с генетическими различиями между организ-
мами (MacNair et al., 1999), также необходимо учитывать при выборе популяции.
Несмотря на эти ограничения, перечень видов, в которых определялось содержа-
ние различных элементов и соединений для целей лихеноиндикации, обширен.
Среди них есть представители кустистых, листоватых, корковых видов, расту-
щих на почве, деревьях, камнях. Собраны они на всех континентах, включая
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 167
Антарктиду. Часть их называлась в предшествующей главе об экспериментах с
пересадкой лишайников (см. табл. 7.1). Вероятно, чаще других использовались
слоевища Hypogymniaphysodes (Garty, 2000а). Желательно при обследовании кон-
кретной территории или зоны воздействия источника загрязнения использовать
один вид лишайника, поскольку есть данные о наличии различий в улавливании
веществ из атмосферы между представителями разных таксонов, таких, как
Parmelia и Usnea (Bosserman, Hagner, 1981), или Parmelia, Physcia, Xanthoria (Nimis
et al., 1993, 2001), или Cetraria islandica, Evernia prunastri, Ramalina farinacea
(Cercasov et al., 2002). Есть и противоположные сведения - измерение концентра-
ций 9 элементов (Hg, Pb, Zn, Си, Сг, Мп, Fe, Se и Al) в слоевищах трех видов
пармелий {Flavoparmelia (Parmelia) caperata, Parmelia saxatilis и P. sulcata) по-
зволило сделать вывод о накоплении ими примерно одинакового количества этих
веществ (Bargagli et al., 1987b).
Необходимо сохранять эталонные образцы видов лишайников, в которых оп-
ределялись концентрации веществ, а в публикации (отчете) указывать, кто опре-
делил образцы, а также место хранения эталонных экземпляров.
8.1.3. Расположение учетных точек на обследуемой местности
Размещение точек, на которых проводят сбор слоевищ для химических анали-
зов, как и в других исследованиях, может быть либо случайным, либо регуляр-
ным, либо комбинацией первого и второго. Их недостатки и преимущества полно
описаны в соответствующих руководствах по количественным методам биологи-
ческих исследований (Грейг-Смит, 1967; Василевич, 1969; и другие). Если пред-
полагается изучение динамики поведения загрязнителей во времени, то предпоч-
тительнее регулярное расположение точек сбора, поскольку этим будет обеспечен
отбор проб с одних и тех же мест.
8.1.4. Объем проб, размер выборки (повторности)
в одной точке или в один срок
Объем пробы, необходимой для анализа, прежде всего зависит от характери-
стик аппаратуры, с помощью которой предполагается проводить измерение со-
держания элементов и соединений в тканях лишайников, способа подготовки проб
для анализов, методики непосредственного измерения, концентрации определяе-
мых веществ в тканях. Для ряда приборов достаточно долей грамма биологичес-
кого материала; в то же время во многих отечественных лабораториях все еще
используют приборы и методики, требующие килограммы сухих слоевищ, по-
скольку зольность большинства лишайников низкая. Значительный объем исход-
168 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
ного материала также необходим для идентификации веществ, концентрация ко-
торых в тканях чрезвычайно мала.
Получение статистически достоверных данных требует обора в одном месте
нескольких проб, поскольку выявлена высокая степень варьирования величин
концентрации элементов в пределах гомогенной популяции лишайника из одного
места (Garty, Ammann, 1987). Размер выборки зависит от заданной точности ис-
следования. Процедура ее установления описана во многих пособиях по статис-
тике (Финни, 1970; Зайцев, 1973). Общее число отбираемых проб определяется
уже на стадии планирования, поскольку от этого в значительной мере зависят
финансовые затраты на исследование.
8.1.5. Установление критериев,
обеспечивающих качество результатов и его контроль
Технология любого исследования, в особенности связанного с измерением тех
или иных величин, характеризующих изучаемый объект, требует гарантии каче-
ства работы (quality assurance) и его контроля (quality control). Гарантия качества
- это система действий, обеспечивающих наличие документации, показывающей,
что работа проведена согласно оговоренным с заказчиком стандартам данного
типа исследований, и измерения получены с известной степенью точности
(Shampine, 1993; Stolte et al., 1993; Wagner, 1995). Это предполагает последова-
тельное документирование процесса планирования исследования, полевых и ла-
бораторных методов, процедуры анализа данных. Если исследование проводится
в связи с экспертизой какого-либо проекта, реализация которого может отрица-
тельно сказаться на состоянии здоровья населения или стать причиной необрати-
мых изменений природной среды, необходимо предусмотреть проведение анали-
зов в разных лабораториях или внешнюю проверку данных анализов и эталон-
ных образцов лишайников.
Контроль качества определяется как процедура устанавливающая, что поле-
вой или лабораторный методы являются статистически контролируемыми в пре-
делах известных уровней вероятности и обеспечивают декларируемую точность,
приемлемую для данного периода времени (Stolte et al., 1993). Контроль качества
предполагает возможность проверки полевых коллекций, конкретных данных
лабораторных анализов, результатов анализов возможных дублирующих учетов
в поле другим коллектором, анализ повторных и стандартных эталонных матери-
алов, экспертную оценку сделанных по полученным результатам выводов. Га-
рантия качества обеспечивается также уровнем квалификации исполнителей ра-
боты, проведением химических анализов в лабораториях, имеющих сертификат
на определение интересующих веществ в биологических объектах. Такую серти-
фикацию лабораторий проводят Европейская Комиссия и Международное агент-
ство по атомной энергии (Quevauviller et al., 1993; 1996; Freitas et al., 1995; Stone
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 169
Таблица 8.1
Содержание элементов (мкг/г)
в сертифицированном (IAEA-336) лишайниковом материале (Rossbach et al., 1999)
Элемент Концентрация Элемент Концентрация
Li 0,473± 0,001 Rb I,39± 0,01
Na 271,0 ±9,8 Sr 8,54 ±0,17
Mg 654,0± 12,4 Cd 0,100± 0,009
р 551,0± 18,2 Sn 0,124± 0,004
s 641,0 ±11 Cs 0,096 ±0,004
к 1800,0 ±36 Ba 5,86 ±0,11
Са 2440,0 ±22 La 0,550± 0,007
Ti 20,2 ±2,1 Ce l,09± 0,028
V 1,53± 0,04 Nd 0,520± 0,014
Сг 1,87 ±0,32 Sm 0,090 ±0,004
Мп 65,5 ±0,46 Hg 0,193 ±0,003
Fe 433,0± 10,4 TI 0,009± 0,001
Со 0,295± 0,013 Pb 4,80± 0,042
Си 3,53 ±0,007 Th 0,119 ±0,001
Zn 29,0 ±0,14 U 0,038 ±0,001
As 0,671 ±0,012
et al., 1995; Schmeling et al., 1997). Уже имеются стандартные эталонные образцы
ряда видов (Everniaprunastri, Pseudevemia furfuracea), используемые при серти-
фикации лабораторий и для сравнения полученных данных (табл.8.1).
8.2. СБОР МАТЕРИАЛА ДЛЯ ХИМИЧЕСКОГО АНАЛИЗА
И ЕГО ПЕРВИЧНАЯ ОБРАБОТКА
Сбор материала для химического анализа имеет целью получение репрезен-
тативных проб, соответствующих решаемым в ходе исследования задачам, кото-
рые были бы минимально загрязнены частицами субстрата, как при отборе, так и
при транспортировке в лабораторию, и которые собирались бы одинаково в тече-
ние всего периода на всей обследуемой территории. Процедура отбора проб ли-
шайников для химических анализов освещена в многочисленных специальных
публикациях (Jackson et al., 1993; Markert, 1993,1994b, 1995; Pirintsos et al., 1993;
Ernst, 1995; Wolterbeek, Bode, 1995; и другие), которые полезно изучить при пла-
нировании такого рода исследований.
План учета должен четко обозначить количество проб, достаточное для полу-
чения статистически достоверных данных и для возможного переанализа при
проверке сомнительных величин. Иногда пробы с соседних точек объединяют.
Такая процедура должна быть единообразной, ее следует документировать и от-
170 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
ражать весовую долю каждой точки в новой пробе. Иногда, наряду с лишайника-
ми в каждой точке отдельно отбирают пробы субстрата. Это необходимо при изу-
чении взаимовлияний лишайников и субстрата и для получения сведений о гео-
химических особенностях последнего.
8.2.1. Точки отбора проб
Точки отбора проб намечаются на этапе планирования работ, чему обычно
предшествует рекогносцировочное обследование изучаемой местности. Прежде
всего, в этих точках в достаточном количестве должны встречаться представите-
ли того вида лишайников, слоевища которого будут подвергнуты химическому
анализу. Также желательно соблюдать правило “при прочих равных условиях”,
кроме нагрузки по изучаемому загрязнителю. При долговременных исследовани-
ях, предполагающих неоднократный отбор проб в одном месте, необходимо про-
думать способы маркировки точек, чтобы облегчить их обнаружение при повтор-
ных посещениях и при этом не загрязнить будущие пробы.
8.2.2. Отбор проб в поле
При сборе слоевищ лишайников крайне желательно, чтобы отбирались тал-
ломы (или их части) примерно одинакового возраста, поскольку молодые слоеви-
ща в сравнении с более старыми находились под воздействием загрязнителей
меньший период, и, соответственно, концентрация попавших в них загрязните-
лей будет меньше. Установлено, что в центральных, более старых, частях слое-
вищ Flavoparmelia (Parmelia) caperata, Parmelia saxatilis и P. sulcata концентра-
ция ряда металлов более высокая, чем в молодых краевых частях талломов (Bargagli
et al., 1987b), что подтвердило и последующее определение концентраций Al, As,
Cd, Со, Cr, Си, Fe, Hg, Мп, Mo, Ni, Pb, Sb, Zn в центральных и краевых частях
талломов Flavoparmelia (Parmelia) caperata (Loppi et al., 1997c). Аналогичные
результаты были получены и при измерении концентраций 16 элементов в цент-
ральных и периферийных частях слоевищ Flavoparmelia (Parmelia) caperata и
Xanthoriaparietina (Nimis et al., 2001). Более двадцати лет назад уже сообщалось
о подобном распределении Сг в талломах первого вида (Schutte, 1977). У кустис-
того напочвенного лишайника Cladoniafurcata концентрация большинства изме-
ренных элементов (13), кроме калия, в верхних, более молодых, частях подециев
была значительно меньшей, чем в нижних отрезках подециев (Tuba et al., 1994).
Это необходимо учитывать и при определении периода полувыведения загрязни-
телей из слоевищ после аварийного разового выброса загрязнителей. Расчеты,
сделанные по результатам наблюдений вблизи Чернобыльской АЭС, показывают,
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 171
Таблица 8.2
Изменение прироста поверхности (мм2) листоватых или корковых слоевищ лишайников,
имеющих различный первоначальный диаметр (прирост по радиусу 1 мм/год)
Показатель 1986 г. 1987 г. 1988 г. 1989 г. 1990 г. 1991 г.
Д = 30 мм 706,9 804,2 907,9 1017,9 1134,1 1256,6
Прирост (мм2) + 97,3 201,0 зн, 427,2 549,7
% к 1986 г. — 13,8 28,4 44,0 60,4 77,8
Д = 40 мм 1256,6 1385,4 1520,5 1661,9 1809,6 1963,5
Прирост (мм2) + 128,8 263,9 405,3 533,0 706,9
% к 1986 г. — 10,2 21,0 32,2 44,0 56,2
Д = 50 мм 1963,5 2123,7 2290,2 2463,0 2642,1 2827,4
Прирост (мм2) + 160,2 326,7 499,5 678,6 863,9
% к 1986 г. - 8,1 16,6 25,4 34,6 44,0
что доля прирастающей поверхности листоватых или корковых талломов увели-
чивается с большей скоростью у слоевищ, имеющих меньший первоначальный
диаметр (табл. 8.2) (Бязров, Штанчаева, 1999). Это дает основание предположить,
что загрязнители (в данном случае - радионуклиды), попавшие в тела лишайни-
ков в результате аварийного выброса, фактически не выводятся из слоевищ, а
растворяются в нарастающей массе.
Возраст слоевищ определяется по соотношению среднегодового линейного
прироста таллома и размера последнего. Известны также попытки измерения
возраста лишайника Flavoparmelia (Pseudoparmelia) baltimorensis из штата Мэ-
риленд (США) по отношению активностей изотопов 210Pb/210Po (Schwartzman et
al., 1987, 1991). 2|0РЬ является продуктом распада естественно встречающегося
радона, который проникает в атмосферу из почвы, и скорость поступления после-
днего известна для каждого региона. В свою очередь, 210Ро является дочерним
продуктом 210РЬ. Таким образом, вычислив теоретически возможную концентра-
цию 2|0РЬ в слоевище и измерив реальные активности изотопов в талломе, можно
приближенно судить как о возрасте слоевища, так и о способности его поглощать
свинец из атмосферы. Однако при расчетах следует исходить из концентраций
изотопов на единицу поверхности таллома, поскольку с возрастом его толщина
меняется.
Имеет значение и расположение слоевищ относительно источников загрязне-
ния. Так, величина концентрации в талломах эпифитов может зависеть от высо-
ты поселения лишайника на дереве, причем более высокие значения концентра-
ции элементов отмечены в слоевищах, растущих ниже, поскольку здесь дополни-
тельным источником поступления элементов могут быть частицы почвы или
грунта, заносимые ветром или беспозвоночными (Solberg, 1979). Загрязнение
слоевищ терригенным материалом рассматривается как важный источник оши-
бок биогеохимической лихеноиндикации (Olmez et al., 1985; Bargagli, 1990a).
172 8. Лихеноиндиксщия по данным о накоплении загрязнителей слоевищами
Поэтому для получения более объективной информации о загрязнении воздуха
величины концентрации в лишайниках нормализуют относительно содержания
элементов, роль которых в метаболизме лишайников крайне мала, например,
Al, Sc, Ti.
Желательно пробы отбирать с одного типа субстрата, ибо, по некоторым дан-
ным, представители одного вида, собранные с разных субстратов, отличаются по
величинам концентрации элементов. Например, вблизи г. Вашингтона (США) по
результатам измерений 10 элементов в слоевищах близких видов Flavoparmelia
baltimorensis и Е caperata. собранных с деревьев и камней, установлены значи-
тельные различия величин концентрации фосфора, калия, кальция и цинка в тал-
ломах первого вида, растущих на гранитных валунах и на коре соседних деревь-
ев двух видов Quercus (Prussia, Killingbeck, 1991). Так, содержание Са в талломах
эпифитной популяции Е baltimorensis было 0,08%, а в слоевищах эпилитов -
0,73%. Различны были концентрации N, К, Са, Си, Fe, Zn в слоевищах обоих
видов лишайников, собранных с коры Quercus borealis и Q. alba. По другим эле-
ментам таких различий не выявлено. Есть сведения и о сезонном варьировании
величин концентрации растворимых в воде веществ (Boonpragob, Nash, 1990;
Prussia, Killingbeck, 1991).
Совершенствование измерительной аппаратуры, позволяющей определять
концентрации веществ с высокой степенью точности, предъявляет особые требо-
вания к тщательности отбора проб в поле, их транспортировке, подготовке к ана-
лизам. Собранные образцы могут быть загрязнены инструментами, используе-
мыми при отборе, емкостями, куда помещают собранные слоевища, газами или
пылью при транспортировке или хранении и т.д., что может исказить выводы,
сделанные на основе результатов химических анализов. Поэтому при сборе мате-
риала для анализов желательно выбирать инструменты, в зависимости от инте-
ресующих веществ, которые бы не загрязняли пробы. Например, образцы лишай-
ников для анализа в них концентраций металлов-микроэлементов рекомендуют
отбирать инструментами из тефлона, пластика, металла с тефлоновым покрыти-
ем или нержавеющей стали (в порядке их предпочтения) для минимизации заг-
рязнения металлами (Stolte et al., 1993). Пластиковые инструменты могут быть
неподходящими при изучении органических загрязнителей. Масло или смазоч-
ные вещества на инструменте также могут быть источником загрязнения пробы.
Потенциальными источниками загрязнения также являются химически обрабо-
танная древесина, в которой могут быть высокие уровни концентрации As, Сг,
Си, личные аксессуары - перчатки или аэрозоль от насекомых, табачный дым.
При сборе образцов для определения органических загрязнителей нежелательно
применять репелленты или другие подобные средства. Следует помнить, что та-
бачный дым содержит изотопы РЬ и Ро.
Пробы с низкими концентрациями интересующих элементов могут загряз-
ниться и в лаборатории, расположенной в местности с высоким фоновым уров-
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 173
нем этих веществ. Это, прежде всего, касается таких летучих веществ как ртуть
или таких органических соединений как полихлорированные бифенилы, гексах-
лорбензен и другие.
Пробы обычно помещают в бумажные или полиэтиленовые пакеты, мешки из
ткани или пластиката, в бутылки из стекла или пластиката. Эти емкости для проб
также могут быть источником их загрязнения, например, серой - от крафт-бума-
ги или органическими веществами - от пластикатов. При перевозке проб в матер-
чатых мешках в кузове открытого грузовика они могут аккумулировать вещества,
выбрасываемые с выхлопными газами автомобилей, на них может попасть до-
рожная пыль.
Пробы требуют аккуратного обращения с ними на всех этапах соприкоснове-
ния с ними. Крайне нежелательно перекрестное их загрязнение между собой.
Перевозка и хранение проб не должны ухудшить их качество (деформацию, за-
гнивание, улетучивание или преобразование интересующих химических веществ).
Поэтому рекомендуется помещать образцы в те или иные емкости в воздушно-
сухом состоянии В ряде случаев (длительное хранение, возможность трансфор-
мации веществ в собранном материале, улетучивание органических загрязните-
лей или элементов типа ртути) может потребоваться замораживание проб. Эта
процедура с использованием жидкого азота осуществляется сразу после отбора
образцов. Обычно для минимизации потерь летучих углеводородов необходима
температура -20°С и ниже, а длительное хранение порядка 1 года осуществляют
при -80°С (Jackson et al., 1993).
8.3. АНАЛИЗ ПРОБ
Анализ проб обычно осуществляется согласно процедурам, принятым в ана-
литической лаборатории. Официальные документы, устанавливающие стандар-
ты химических анализов биологического материала, в том числе и лишайников,
отсутствуют. В СССР для лишайников, как для лекарственно-технического сы-
рья, отвечающего требованиям технологии ряда отраслей промышленности, су-
ществовал стандарт N OCT 8236/НКВТ-454 (1948) (Лекарственно-техническое
сырье, 1958). Существуют нормативы содержания некоторых веществ в органах
ряда растений и грибов, используемых для приготовления продуктов питания.
По грубым оценкам, в странах Европейского Сообщества в середине 1990-х еже-
годно проводили более 5000 анализов лишайниковых проб (Quevauviller et al.,
1996). Однако методы сбора, подготовки проб к анализам, измерительная аппа-
ратура были неодинаковы, что давало основания для сомнений при сравнении
результатов разных исследований и снижало возможности их интерпретации для
Целей мониторинга в пространстве и во времени. В конечно счете это приводит к
значительным экономическим потерям, поскольку стоимость аналитических ра-
174 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
бот высокая. Последнее стимулировало Европейскую Комиссию заняться про-
блемой стандартизации методов анализов лишайникового материала, начать сер-
тификацию лабораторий, работающих с лишайниками. Лабораториям, претен-
дующим на получение такого сертификата, предлагается для анализа сертифици-
рованный эталонный материал (CRM 482), представляющий гомогенизированные
ткани лишайника Pseudeverniafurfuracea, слоевища которого в начале 1990-х были
собраны в одной из местностей Швейцарии и анализировались в 32 лаборатори-
ях из 11 стран (Quevauviller et al., 1996).
Анализ проб состоит из двух этапов: подготовки проб и измерения. И тот, и
другой варьируют в зависимости от намеченых для измерения элементов или
соединений, а также от имеющейся в наличии измерительной техники.
8.3.1 .Подготовка лишайниковых проб для анализа
Подготовка проб включает очистку образцов, их высушивание, измельчение,
гомогенизацию и деструкцию органической ткани. На стадии очистки из проб
удаляют чужеродный материал, не являющийся частью тела лишайника. Обычно
это ручная разборка под микроскопом, во время которой удаляют частицы при-
крепившегося к слоевищам субстрата, почвы, хвои и листьев, отмерших тканей.
На этом этапе спорным является применение обмывания образцов водой для уда-
ления осевшей на поверхности талломов пыли, золы, частиц автомобильного топ-
лива, других переносимых ветром частиц. Считается, что в ходе этой процедуры
происходит вымывание из слоевищ ряда веществ и элементов, в частности, ка-
лия. Выщелачивание металлов и серы из лишайников наблюдалось во многих
исследованиях (Goyal, Seaward, 1981b; Lawrey, Hale, 1981; Jackson et al., .1985),
поэтому специалисты Европейской Комиссии, занимавшиеся стандартизацией
анализов, не рекомендуют обмывать образцы (Quevauviller et al., 1996). Такой
процедуре подготовки образцов отдали предпочтение авторы ряда публикаций
(Pakarinen et al., 1978;Pilegaardetal., 1979; Folkeson, 1981; Bargagli, 1989;Gasparo
et al., 1989; Nimis et al., 1993, 2001). Другие исследователи обмывали талломы
лишайников водой перед анализом материала (Nieboer et al., 1972; Garty et al.,
1977). Однако достоверные сведения о том, что более агрессивный процесс очи-
стки (с дополнительным циклом смыва или более интенсивным обмыванием) при-
водит к уменьшению концентрации элементов, отсутствуют. Кроме того, совер-
шенствование измерительной аппаратуры ныне позволяет проводить определе-
ния концентраций элементов в незначительных по объему образцах, поэтому по-
верхностное загрязнение слоевищ животными, человеком, другими агентами,
внешне незаметное, при высокой степени точности измерительной аппаратуры
может дать искаженное представление о содержании элементов или соединений
в анализировавшихся тканях лишайника. Есть и другая проблема - продолжи-
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 175
тельность периода между датой последнего дождя и датой отбора проб. При ко-
ротком промежутке между ними поверхностных отложений на слоевищах будет
меньше, чем при продолжительном, что может сказаться на результатах анализа,
если пробы не обмыты.
Я полагаю, что при таких работах следует отдавать себе отчет, воспринимаем
ли мы таллом лишайника как физическую поверхность, на которой происходит
осаждение различных веществ, или слоевище - живое тело, накапливающее в
себе вещества из окружающей его среды. В первом случае обмывать образцы не
следует, во втором - необходимо. И всегда желательно продумать, имеется ли по-
верхностное загрязнение и каков его объем; является ли концентрация интересу-
ющих элементов или веществ в воздухе значимой в сравнении с их концентраци-
ей в телах лишайников; удалит ли смыв значительную часть поверхностного заг-
рязнения; произойдет ли при смыве выщелачивание значительного количества
интересующих компонентов или веществ из самого лишайника (Jackson et al.,
1993).
Пробы лишайников перед анализом сушат в основном для обеспечения их
весового однообразия, поскольку концентрация элементов определяется относи-
тельно единицы веса тела. Кроме того, сушка призвана минимизировать измене-
ния в тканях пробы. Так, активность ферментов в растениях прекращается при
температуре выше 60°С, но при температурах выше 100°С уже происходит поте-
ря массы из-за улетучивания ряда веществ (Jackson et al., 1993), поэтому специа-
листы Европейской Комиссии, занимавшиеся стандартизацией анализов, не ре-
комендуют высокотемпературную сушку лишайниковых проб (Quevauviller et al.,
1996). Предпочтительнее сушить образцы в десиккаторе или замораживанием,
но этот способ не практичен при большом объеме пробы или значительном числе
образцов. Альтернативным методом является сушка проб в сушильном шкафу
при 40°С в течение 48 часов (Gough et al., 1988а, b, 1994). Сушка при низкой
температуре (менее 35°С) наиболее подходит для определения таких летучих ве-
ществ как углеводороды или ртуть. В некоторых случаях (высокая вероятность
потери летучих веществ) измерения можно проводить и не в сухом материале,
тогда они сопровождаются определением влажности в частях образцов, отделен-
ных из основной пробы.
Измельчение лишайниковых образцов проводят для получения гомогенной
пробы с одинаковым размером частиц. На этой стадии подготовки возможно за-
грязнение материала и потеря его части, поскольку для измельчения используют
различные способы и приспособления, включая мельницы разного типа, ступки
и другое. Выбор способа определяется количеством материала, методом измере-
ния веществ, требуемого для анализа объема проб, конкретного элемента или со-
единения. При выборе способа измельчения также ориентируются на морфологи-
ческие особенности лишайникового слоевища и устойчивость его тканей, посколь-
ку от этого зависит размер получаемых измельченных частиц (Gough et al., 1988а).
176 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
Частицы требуемой величины получают просеиванием через сито с соответству-
ющим диаметром отверстий.
Измельчение до слишком маленьких размеров частиц может привести к до-
полнительному обогащению металлом. При механическом измельчении в мель-
ницах пробы могут загрязниться такими элементами как Al, Со, Си, Cr, Fe, Мо,
Ni, Si, W, Zn. Использование агатовой шаровой мельницы или ступок снижает
загрязнение металлами и позволяет получить частицы подходящего размера. Од-
нако при чрезмерном измельчении происходит разогрев материала и улетучива-
ние некоторых органических соединений.
Промежуток времени между измельчением и анализами для большинства не-
органических элементов значения не имеет. Однако некоторые органические ве-
щества могут быть утеряны или трансформироваться из-за действия ферментов
после разрушения клеток при измельчения (Jackson et al., 1993), поэтому экст-
рагирование органических веществ следует проводить сразу после измельчения.
Для оценки точности анализов измельченную пробу в лаборатории перед анали-
зом нередко делят на несколько. Для этого рекомендуют способ дробления и чет-
вертования. Содержимое целой пробы рассыпают на чистом листе бумаги, и оно
смешивается встряхиванием и подергиванием за уголки листа. После этого из
этой массы формируют конусовидную фигуру, которую уплощают и делят на чет-
верти. Большую часть массы противоположных четвертей объединяют и помеща-
ют в 2 пакета. Эту процедуру повторяют с остатком образца, пока вся проба не
будет поделена на две части (Jackson et al., 1993).
Многие методы измерения неорганических веществ в биологических объек-
тах предусматривают разрушение органической части пробы. Это достигается
либо мокрым (посредством действия смеси кислот как HNO3, НС1О4, или H2SO4),
либо сухим (сжиганием в высокотемпературных печах) озолением. Выбор спосо-
ба озоления зависит от метода анализа и степени летучести определяемых хими-
ческих веществ. Например, при действии кислотами улетучивается В, тогда как
при сухом озолении могут быть потеряны As, Hg, Se, S, и некоторые другие эле-
менты (Jackson et al., 1993). Необходимо учитывать, что внутренние поверхности
муфельных печей, в которых обычно озоляют материал, а также тигли, куда поме-
щают пробы для сжигания, могут быть источниками частичного загрязнения
материала.
8.3.2. Химический анализ материала
Химический анализ лишайникового материала проводят в специализирован-
ных лабораториях. Разработаны многочисленные инструментальные аналитичес-
кие методы для определения содержания большинства металлов в лишайниках.
Однако довольно ограничен выбор методов для определения Hg, полуметаллов -
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 177
As, Se, неметаллов - В, S и галогенов, а также многих органических веществ.
При выборе метода необходимо учитывать многие факторы, в частности, требуе-
мый для анализа объем пробы (обычно 1-10 г или более), особенности подготов-
ки проб, измерение ограниченного перечня элементов или определение всего воз-
можного, ранги определяемых концентраций и пределы определения, точность
метода, число одновременно определяемых прибором элементов, доступность
аппаратуры, сроки проведения анализов, стоимость работы.
Ныне для определения содержания элементов и соединений в биологических
объектах в основном используют разные виды атомной спектрометрии, нейтрон-
ной активации, электронный парамагнитный резонанс, радиохимические мето-
ды, масс-спектрометрию, методы хроматографии, рентгеноструктурный анализ,
колориметрию, титрование, электрохимические методы и другое (Tomassini et al.,
1976; Lawrey, 1977b; Flora, Nieboer, 1980; Hrynkiewicz et al., 1980; Jackson et al.,
1985, 1993; Казачевский и др., 1987; Connor et al., 1991; Dempsey et al., 1992;
Маркина и др., 1995; Nazarov et al., 1995; Quevauviller, 1995; Quevauviller et al.,
1996; Williams et al., 1996; van Vaeck et al., 1997; Nejedly et al., 1998; Sarret et al.,
1998b; Jezierski et al., 1999; 2002; Pyatt et al., 1999; Coccaro et al., 2000; Narewski,
2000; Senhou et al., 2001). Каждый из них имеет свои достоинства и недостатки.
Например, для определения органических веществ пока пригодны только масс-
спектрометрия и методы хроматографии. Нейтронная активация дает более точ-
ные результаты при определении Mg, Al, Si, Cl, J, Ca, Ti, Fe и менее точна для
определения содержания Zn, Rb, Sr, Ва и La (Senhou etal., 2001). При планирова-
нии исследования желательно подробно познакомиться с возможностями анали-
тической лаборатории, где намечено проводить анализы, и согласовать с ее пер-
соналом требования к размеру проб, их подготовке и т.д.
8.4. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЛИХЕНОИНДИКАЦИИ
ПО ДАННЫМ ОБ ОСОБЕННОСТЯХ ХИМИЧЕСКОГО СОСТАВА
ЛИШАЙНИКОВ
Полученные в лабораториях количественные сведения о содержании хими-
ческих веществ в лишайниках прежде всего обрабатываются методами статисти -
ки, чтобы оценить достоверность данных и степень точности возможных эколо-
гических интерпретаций. Опубликовано несколько обзоров исследований, посвя-
щенных лихеноиндикации по данным о накоплении лишайниками различных
химических веществ (Инсарова, 1982, 1983; Инсаров, Инсарова, 1986; Puckett,
1988; Tyler, 1989; Tyler et al., 1989; Garty, 1993, 2000a, b; Richardson, 1995; Purvis,
Halls, 1996; Sarret et al., 1998a; Jacquiot, Dai I Ian t, 2000). Число публикаций о
содержании химических веществ в лишайниках ныне, вероятно, составляет око-
ло 3000 (Quevauviller et al., 1996). Большинство их содержат данные о неоргани-
178 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
ческих загрязнителях, но есть исследования по аккумулировании в телах лишай-
ников и органических ксенобиотиков (Gaskell et aL, 1973; Слепян и др., 1981;
Carlberg et al., 1983; Thomas et aL, 1984; Villeneuve, Holm, 1984; Villeneuve
et al., 1985, 1988; Bacci et al., 1986; Focardi et aL, 1991; Morosini et aL, 1993; Muir
et aL, 1993; Elkin, Bethke, 1995; Muir, Shirazi, 1996; Migaszewski, 1999; Kelly,
Gobas, 2001).
Анализ доступных источников позволяет условно выделить следующие ос-
новные направления биогеохимической лихеноиндикации: определение базового
уровня (baseline) концентраций химических веществ в слоевищах лишайников(1),
установление трендов содержания интересующих загрязнителей во времени на
конкретной территории (2), выявление пространственного распределения ксено-
биотиков относительно источников загрязнения (3), пространственное распреде-
ление элементов на относительно крупной (область, провинция, страна) террито-
рии (4). Естественно, я осознаю спорность такого деления, поскольку нередко
одно и то же исследование можно отнести к любому из этих направлений или ко
всем сразу. Однако такая условная классификация несколько облегчает дальней-
ший анализ многочисленных публикаций по этой проблеме.
8.4.1. Определение базового уровня концентрации
химических веществ в слоевищах лишайников
Значительная доля лихеноиндикационных исследований, включающих изме-
рение концентраций химических веществ в талломах лишайниках, посвящена
определению фонового или базового уровней концентраций в тканях этих орга-
низмов. Рекомендуется различать понятия “фоновый” (background) и “базовый”
(baseline) уровни концентрации элемента (Stolte et aL, 1993). Это соответствует
рекомендациям Всемирной метеорологической организации выделять фоновое
планетарное и фоновое региональное загрязнения (Исаев, 2001). Фон подразуме-
вает представление об естественной или природной концентрации, которая в иде-
але исключает влияние человека. Геохимики считают, что фоновые величины от-
ражают отсутствие аномалии, поэтому они не всегда эквивалентны низким зна-
чениям концентрации элемента или вещества (Matschullat et aL, 2000). Сведения
о фоне желательны для сравнения с ними получаемых в ходе исследования дан-
ных. Базовый уровень концентрации элемента представляет собой совокупность
рассеянных примесей в данном районе, для которых нет возможности разделить
отдельные источники загрязнения. Обычно базовый уровень не соответствует
фоновому. Фон отражает идеализированную ситуацию, и его определение более
проблематично в сравнении с измерением базового уровня концентрации эле-
мента. Для установления фона необходим анализ образцов доиндустриального
периода развития человеческого общества, поскольку вещества, связанные с дея-
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 179
тельностью человека, обнаружены в лишайниках даже в Антарктиде, континенте
без постоянного населения, где лишь несколько десятков человек последние не-
сколько десятков лет проводят только научные исследования (Bacci et al., 1986;
Bargagli et al., 1993, 1999). Предложен ряд способов вычисления фоновых кон-
центраций элементов земной коры (Matschullat et al., 2000).
Базовый уровень концентрации определить проще. Обычно за него принима-
ют среднюю величину выборки, статистическая вероятность которой 95% (Jackson
et al., 1993). При этом следует обращать внимание на соотношение варьирования
измеренных величин в выборке и точности измерительной аппаратуры или лабо-
раторной ошибки, поскольку нередки случаи, когда величины концентрации на-
ходятся в пределах лабораторной ошибки. В этом случае базовый уровень не ус-
танавливают, как это имело место при определении базовых уровней концентра-
ции элементов в слоевищах Parmelia sulcata в Национальном парке Т. Рузвельта
и в талломах Hypogymnia enteromorpha и Usnea sp. в Национальном парке Ред-
вуд, США (Gough et al., 1988а, b). Заповедники, национальные парки и другие
“чистые” территории обычно являются теми местами, где собирают лишайники
для подобных измерений (Локинская, 1970; Горбанов, Хаджиатанасов, 1973;
Erdman, Gough, 1977; Erdman et al., 1977; Глазовский и др., 1978; Скрипниченко и
др., 1978; Солдатенкова, 1978; Лапицкая и др., 1979; Золотарева и др., 1981; Тур-
манина, Баденкова, 1982; Казачевский и др., 1987; Sarkela, Nuorteva, 1987; Gough
et al., 1988a, b; Бязров, Адамова, 1990; Frenzel et al., 1990; Адамова, Бязров, 1991;
Skacel, Pekarek, 1992; Wiersma et al., 1992; Biazrov, Adamova, 1993; Geiser et al.,
1994; Bennett, 1995; Второва, Маркерт, 1995; Маркерт, Второва, 1995; Bennett,
Wetmore, 1997,2000; Calvelo et al., 1998; Bargagli et al., 1999; Аржанова, Скирина,
2000; и другие). Как базовый уровень, вероятно, можно использовать данные,
полученные при изучении участия лишайников в круговороте веществ в биогео-
ценозах (Полынов, 1945; Ярилова, 1947; Бобрицкая, 1950; Левина, 1960; Родин,
Базилевич, 1965; Манаков, 1972; Doyle et al., 1973; Левицкая, 1975; Garty et al.,
1995a; Migaszewski et al., 2001) или при определении их свойств как источника
пищи для животных (Работнов, Говорухин, 1950) и сырья для промышленности
(Курсанов, Дьячков, 1945; Моисеева, 1961; Сафонова и др., 1999), естественно,
если известны места отбора проб для анализов и приемлема точность измерений.
Из доступных мне материалов эталоном можно назвать публикацию Линды Гей-
зер с соавторами (Geiser et al., 1994), в которой наряду с детальной характеристи-
кой видового состава лишайников и величин базового уровня концентраций при-
ведены данные о концентрации S, Р, К, Са, Mg, Al, Fe, Na, Mn, Zn, Си, В, Pb, Ni,
Cr, Cd в каждом из почти 300 образцов Alectoria sarmentosa, Cladina rangiferina,
Hypogymnia enteromorpha, Lobaria oregona из лесного массива на юго-востоке
Аляски (США). Сведения о содержании веществ в лишайниках Арктики и Ан-
тарктиды, где концентрации ряда элементов на уровне нанограммов, вероятно,
важны для суждения о степени загрязнения экосистем других, более освоенных
180 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
территорий (Jozwik, 1990;Olech, 1991; Crete et al., 1992; Bargagli etal., 1993,1999;
Nash, Gries, 1995a, b; France, Coquery, 1996; Chiarenzelli et al., 1997; Poblet
et al., 1997; Riget et al., 2000; Upreti, Pandev, 2000). Сведения о базовом уровне
элементов содержат и материалы достаточно экстравагантного исследования ин-
тернационального коллектива (Rossbach et al., 1999), целью которого было срав-
нение концентрации элементов в слоевищах Usnea sp., собранных в чистых рай-
онах Баварии, Канады, Сибири, Шри-Ланка, с их концентрациями в сертифици-
рованном материале лишайников и в пыли, собранной в отелях Hilton в городах
Антананариву (Мадагаскар), Дрезден, Дюссельдорф, Мюнхен (Германия), а так-
же в атомном институте в Вене (Австрия).
8.4.2. Установление трендов содержания загрязнителей
во времени на конкретной территории
Конечная цель определения базового уровня концентраций веществ - ус-
тановление изменений во времени показателей загрязнения среды обследуемой
территории путем сравнения величин базового уровня с данными повторных из-
мерений через некоторые промежутки времени. Интервалы между такими после-
довательными измерениями могут быть и год, и десятки лет. В одном из промыш-
ленных анклавов штата Луизиана (США), сравнив концентрации ряда элементов
(Al, Fe, Си, Pb, Cd, Hg, Cr, Zn) в слоевищах Parmotremapraesorediosum и Ramalina
stenospora, собранных в 1983 и 1988 гг., установили значительное снижение со-
держания большинства элементов в талломах за пять лет, что объяснялось про-
изошедшими изменениями технологических циклов основных предприятий
(Walther et al., 1990b). В одном из национальных парков штата Мичиган (США)
сравнили концентрации 16 элементов в слоевищах лишайников Evemia meso-
morpha и Hypogymnia physodes, собранных в разных частях этой обширной тер-
ритории в 1983, 1984 и 1992 гг. (Bennett, 1995). Величины концентраций элемен-
тов соотносились с их содержанием в земной коре, географическим положением
мест отбора проб; также учитывалась токсичность элементов. Установлено, что
концентрации таких элементов как S, Pb, Zn, Se в слоевищах лишайников более
высокие, чем можно было ожидать из величин их содержания в земной коре, что
указывает на атмосферный перенос этих элементов на территорию парка; вели-
чины концентраций тяжелых металлов в лишайниках увеличились больше, чем
нетяжелых металлов, что также свидетельствует об атмосферном загрязнении;
повышенные уровни загрязнителей отмечены в точках с большей абсолютной
высотой, перехватывающих воздушные массы; концентрации Al, Са, Na, Fe, Mg,
К за 8-9 лет увеличились на 53-62%, Р, Мп, S - на 17-24%, Cr, Ni, Zn, В, Си,
Со - на 111-252%, Pb, As, Mo, Cd, Se уменьшились на 51-74%; концентрации
токсичных элементов за 8-9 лет увеличились на 62-123%, не токсичных - на 45-
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 181
59%. На севере штата Миннесота (США) определение содержания ряда элемен-
тов (А1, В, Cr, Fe, Ni, S, Са, Cd, Mg и Mn) в слоевищах Cladina rangiferina, Evernia
mesomorpha, Hypogymnia physodes, Parmelia sulcata были проведены с интерва-
лом в 11 лет (Bennett, Wetmore, 1999а). За этот период концентрации А1, В, Cr, Fe,
Ni, S, наличие которых авторы связывают с деятельностью человека, снизились,
а концентрации остальных элементов, участвующих в циклах минерального пи-
тания лишайников, увеличились. В парке Т. Рузвельта (штат Сев. Дакота, США)
те «же авторы провели повторные измерения 22 элементов в слоевищах эпифита
Parmelia sulcata и эпигеида Xanthoparmelia chlorochroa с интервалом в 16 лет -
в 1982 и 1998 гг. (Bennet, Wetmore, 2000). Содержание Са, Fe, В, Cr, Al, Mn, Mg, Р,
Cu, Ni в талломах эпифита увеличилось от 94% у кальция до 6% - у меди и
никеля, тогда как концентрации К, Zn, S, Ti, Pb, V, Mo, Co, Hg, Cd, Na, As и
зольность снизились от 1 % у калия до 51 % - у мышьяка. В слоевищах напочвен-
ного вида увеличились концентрации Mo, Fe, Mg, В, Cr, Mn, Al, Ni, Cu, As, Na,
Ti, Zn, P, S, Co, К от 174% у первого до 7% - у последнего, а зольность и содержа-
ние Са, V, Hg, Cd, Pb снизились от 3% у кальция до 56% - у свинца. Такие разли-
чия между напочвенным и эпифитным видами авторы объяснили возможной за-
грязненностью слоевищ эпигейного вида почвенными частицами и не рекомен-
дуют широкое использование напочвенных лишайников в биомониторинговых
исследованиях, базирующихся на определении содержания химических веществ
в слоевищах (Bennett, Wetmore, 2000). С таким же 16-летним интервалом (1980 и
1996 гг.) были проведены измерения концентраций в талломах Flavoparmelia
(Parmelia) caperata в Центральной Италии (Loppi et al., 1998с). Содержание боль-
шинства элементов, кроме серы, уменьшилось, а увеличившуюся концентрацию
S авторы связали с широким применением в этот период пестицидов для обра-
ботки виноградников. В другой местности Италии с 1978 по 1993 гг. видовой
состав лишайников и содержание элементов в слоевищах Flavoparmelia (Parmelia)
caperata практически не изменились (Loppi, De Dominicis, 1996). Так, концентра-
ция Pb в 1978 г. составила 31,13 мкг/г, а в 1993 г. - 32,68 мкг/г. В западной части
Финляндии данные измерений содержания элементов в слоевищах Hypogymnia
physodes, собранных в 1989 г., сравнили с аналогичными данными, полученны-
ми в 1974-1975 и 1982-1983 гг. (Halonen etal., 1993). Отмечено снижение концен-
трации РЬ, что объясняется запретом использования автотранспортом горючего
невысокого качества.
Тренды содержания ряда элементов во времени устанавливались и сравнени-
ем их концентраций в современных и исторических образцах из одного места.
В качестве последних использовали коллекционные материалы, хранившиеся в
различных гербариях. Таким способом пытались изучить динамику хрома в шта-
тах Огайо и Западная Виржиния, США (Schutte, 1977), свинца и серы - в штатах
Мэриленд и Виржиния (Lawrey, Hale, 1981,1988; Lawrey, 1993). Например, уста-
новлено, что содержание Сг в слоевищах Flavoparmelia (Parmelia) caperata и
182 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
Punctelia (Parmelia) rudecta, собранных в 1975 и 1976 гг. в Огайо, было значи-
тельно выше, чем в гербарных образцах из тех же мест, собранных в 1893 г. (Schutte,
1977). Концентрация S в слоевищах Flavoparmelia baltimorensis за 30-50 лет в
разных местах парка Shenandoah увеличилось на 3-66%, а концентрация РЬ сни-
зилась на 17-366% (Lawrey, Hale, 1988). Аналогичная идеология была в основе
исследования, проведенного и на Украине, сначала в окрестностях г. Киева при-
менительно к динамике РЬ (Блюм, Тютюнник, 1985; Тютюнник, 1987), а затем
оно охватило несколько природных зон от Киевской области до Крымского полу-
острова. Анализировались слоевища 13 видов, в которых определялись концент-
рации Fe, Мп, Zn, Са, Ti, РЬ, Мо (Блюм, Тютюнник, 1989). Образцы, собранные в
1983-1986 гг., сравнивались с образцами, собранными в тех же местах с 1838-
1888 по 1979 гг. Авторы установили, что ожидаемое увеличение концентрации
металлов в слоевищах современных образцов в сравнении с собранными десятки
и сотню лет назад наблюдалось не всегда. В такого рода исследованиях необходи-
мо учитывать и место отбора проб, и видовую принадлежность лишайника, и
длительность интервала между сравниваемыми датами отбора образцов. От себя
добавлю, что не обсуждена проблема возможного загрязнения исторических проб
при их сборе и хранении, поскольку коллекторы явно не предполагали, что гер-
барные образцы будут использованы для химических анализов.
8.4.3. Выявление пространственного распределения загрязнителей
относительно источников загрязнения
Исследования особенностей пространственного распределения загрязнителей
по данным о содержании их в слоевищах лишайников составляют наибольшую
долю среди работ, касающихся биогеохимических аспектов лихеноиндикации.
В значительной их части анализируется распределение единичных или несколь-
ких элементов относительно источника загрязнения. В качестве последних рас-
сматривают как естественные (вулканы, геотермальные образования, вода морей
или океанов и другое), так и связанные с деятельностью человека. Естественно, в
Европе воздействие вулканов изучалось в основном в Италии, где время от вре-
мени происходят извержения Этны, Стромболи и других (Barghigiani et al., 1988а,
1988b, 1990; Davies, Notcutt, 1988, 1989; Bargagli et al., 1989, 1991; Bargagli,
Barghigiani, 1991; Dongarra, Varrica, 1998; Grasso et al., 1999; Monna et al., 1999;
Tretiach, Ganis, 1999; Varrica et al., 2000). Проводились такие исследования и на
Азорских (Notcutt, Davies, 1999) и Гавайских (Notcutt, Davies, 1993) островах, на
западе США (Moser et al., 1983; Harris et al., 1987), в Японии (Glime, Iwatsuki,
1990; Fahselt, 1995) и в других регионах. Интересно, что тепловое воздействие
вулканической деятельности может привести и к изменению состава вторичных
лишайниковых веществ в талломах из вулканических областей, что, в свою оче-
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 183
редь, может стать причиной таксономических ошибок при идентификации по
химическим признакам, как это имело место с образцами Hypotrachynapartita из
вулканически активного района Коста-Рики (Culberson et al., 1977).
Знаменитый своими гейзерами Иеллустонский национальный парк в Скали-
стых горах (США), на обширной территории которого известно около 10000 гео-
термальных образований, также стал объектом биогеохимической лихеноинди-
кации, где в слоевищах Bryoria fremontii и Letharia vulpina, собранных в разных
точках парка, определялись концентрации 22 элементов (Bennett, Wetmore, 1999b).
Распределение значений концентраций элементов в талломах обоих видов было
сходным (коэффициент корреляции 0,88). Из металлов только ртуть выделялась
высокими значениями концентраций, что объясняется наличием значительного
количества этого элемента в извержениях геотермальных источников. То же от-
носится и к сере. Средние различия величин концентраций между видами были
статистически значимыми для 13 элементов (А1, В, Са, Cd, Со, Fe, Mg, Мп, Mo, Р,
Ti, V, Zn), а различия между точками отбора образцов значимыми были для
14 элементов (As, Cd, Со, Си, Fe, Hg, К, Na, Р, Pb, S, Ti, V, Zn).
Высокое содержание ртути в слоевищах Flavoparmelia (Parmelia) caperata,
собранных в местах с геотермальными проявлениями, обнаружено и в Италии,
где помимо природных источников изучалось и воздействие функционирующих
геотермальных электростанций (Ferrara et al., 1988; Bargagli, Barghigiani, 1991;
Loppi et al., 1991a, b, 1997a, 1998a, b, 1999; Loppi, 1993, 1996, 1997; Loppi, Bargagli,
1996; Bargagli et al., 1997; Loppi, Nascimbene, 1998; Loppi, Bonini, 2000). Также в обла-
стях с геотермальными образованиями определялось накопление лишайниками есте-
ственных и искусственных радионуклидов (Matthews, 1981; Loppi et al., 1997b).
В прибрежных зонах морей и океанов среди прочих факторов на распределе-
ние лишайников влияет и чувствительность их к растворенным в морской воде
солям, переносимых ветрами на сушу (Nash, Lange, 1988). Эти соли вместе с час-
тицами морской воды поступают в тела лишайников и накапливаются в их тка-
нях, причем место накопления (поверхность слоевища, межклеточное простран-
ство, стенки клеток) зависит от иона.
Большая часть Na+ в прибрежной полосе Португалии обнаружена на поверх-
ности и в межклеточных фракциях лишайников, тогда как Mg++ в основном на-
капливался в клеточных стенках (Figueira et al., 1998а, b). Концентрация катио-
нов (Na, Mg, К) и анионов (С1) морских солей в слоевищах представителей рода
Ramalina снижалась с удалением от береговой линии (Nash et al., 1990; Figueira et
al., 1999a-e).
На острове Ньюфаундленд установили, что состав элементов в слоевищах
Alectoria sarmentosa в большей степени определяется удаленностью от океана,
чем переносом загрязнителей с континента (Wadleigh, Blake, 1999). Полагаю, что
фактор близости моря необходимо учитывать при интерпретации результатов
биогеохимической лихеноиндикации в приморских местностях.
184 8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами
Большинство исследований, связанных с определением накопления химиче-
ских веществ лишайниками, направлены на установление пространственного рас-
пределения загрязнителей в сфере действия конкретного источника загрязняю-
щих веществ. К ним относятся работы по изучению воздействия металлургиче-
ских производств на Дальнем Востоке страны (Баденкова и др., 1978, 1981; Ски-
рина, Качур, 1991; Аржанова, Скирина, 2000), на Кавказе (Петрушина, 1987), на
Кольском полуострове (Норин, Ярмишко, 1990; Yevseev, Krasovskaya, 1994), в Фин-
ляндии (Kansanen, Venetvaara, 1991; Manninen et al., 1991; Mukherjee, Nuorteva,
1994), Великобритании (Perkins, 1992), Дании (Vestergaard, 1986), Югославии
(Jovanovic et al., 1995), Канаде (Nieboer et al., 1972); рудников по добыче ртути в
Италии (Bargagli et al., 1987a; Bargagli, 1990b), Словении (Lupsina et al., 1992) и
урана в Канаде (Nieboer et al.,1982; Looney et al., 1986; Fahselt et al., 1995), меди -
в Италии (Bargagli et al., 2002), Португалии (Branquinho et al., 1999); предприятий
по производству удобрений в Финляндии (Tynnyrinen et al., 1992); электростан-
ций в США (Nash, Sommerfeld, 1981; Olmez et al., 1985), в Канаде (Juinchang et al.,
1995), в Португалии (Freitas, 1994, 1995; Freiras et al., 1997), в Италии (Zanini et
al., 1992; Nimis et al., 1993), в Финляндии (Nygard, Harju, 1983; Kauppi, Halonen,
1992; Haapala et al., 1996); цементного завода в Финляндии (Kortesharju et al.,
1990); фосфорообогатительного предприятия в США (Dillman, 1996), целлюлоз-
но-бумажных заводов в Румынии (Bartok, Osvath, 1990), Финляндии (Kytomaa et
al., 1995; Lodenius, Laaksovirta, 1979); предприятий по переработке мусора в Ита-
лии (Gasparo et al., 1989; Loppi et al., 2000), низкорадиоакгивных материалов - в
России (Biazrov, Mineyeva, 1994); многочисленных источников загрязнения в го-
родах (Saeki et al., 1975, 1977; Takala, Olkkonen, 1976, 1981; Киселев и др., 1983,
1986; Jaakkola et al., 1983; Lodenius, Kumpulainen, 1983; Бойко и др., 1984, 1988;
Nyangababo, 1987; Rope, Perason, 1990; Glenn et al., 1991; Pfeiffer, Barclay-Estrup,
1992); автотранспорта (Laaksovirta et al., 1976; Laaksovirta, Olkkonen, 1979;
Hrynkiewicz et al., 1980; Orsi, Glenn, 1991; Nimis et al., 1993; Glenn et al., 1995;
Bargagli et al., 1997; Steubing, Vogel, 1998; Monaci et al., 2000; Cuny et al., 2001;
Owczarek et al., 2001).
8.4.4. Пространственное распределение элементов
в слоевищах лишайников на относительно крупной
(область, провинция, страна) территории
Анализ содержания элементов в талломах лишайников использовался для
оценки распространения загрязняющих веществ на территориях, занимаемых
крупными административно-территориальными единицами или природными
областями (Pakarinen et al., 1978; Puckett, Finegan, 1980; Pakarinen, 1981a; Zakshek
et al., 1986; Kral et al., 1989; Nieminen, Rantataro, 1990; Biazrov, Adamova, 1993;
8. Лихеноиндикация по данным о накоплении загрязнителей слоевищами 185
Nimis et al., 1993, 2000; Dongarra et al., 1994; Scerbo et al., 1999, 2002; Walther et
al., 1990a; Jalkanen et al., 2000; Salemaa et al., 2001) или отдельными государства-
ми, такими как, например, Нидерланды (Sloof, Wolterbeek, 1991а, b, 1993а, Ь;
Sloof, 1995b), Норвегия (Bruteig, 1993а), Португалия (Freitas et al., 1997, 1999,
2000), Финляндия (Lodenius, 1981; Pakarinen, 1981b; Takala, Olkkonen, 1985; Takala
et al., 1985, 1990, 1991, 1994, 1998; Kubin, 1990; Lippo et al., 1995; Kubin et al.,
1997). В Финляндии имеется депозитарий биологических проб из разных райо-
нов страны, в том числе и лишайников, в которых измерялось содержание эле-
ментов (Kubin et al., 1997). Обычно при подобном обследовании обширных пло-
щадей в слоевищах представителей одного вида измерялись концентрации всего
одного-двух элементов - ртути (Lodenius, 1981), серы (Pakarinen, 1981b; Takala et
al., 1985,1991), хлора (Takala et al., 1990), железа (Takala et al., 1994), цинка (Takala
et al., 1998), титана (Takala, Olkkonen, 1985), однако в публикациях последних
лет, затрагивающих эту проблему, количество определяемых в талломах элемен-
тов, как правило, более десяти. В Нидерландах, где использовали эпифит Parmelia
sulcata (Sloof, Wolterbeek, 1991a), географическое распределение величин концен-
траций в лишайнике совпало с результатами моделирования рассеивания загряз-
нителей и данными инструментальных измерений концентрации их в воздухе.
9. ЛИШАИНИКИ И РАДИОАКТИВНОЕ ЗАГРЯЗНЕНИЕ
ТЕРРИТОРИИ
Использование ядерной энергии в военных и мирных целях значительно уве-
личило риск получения организмами повышенных доз ионизирующего облуче-
ния. Особый интерес к этой проблеме проявился после трагической аварии в ап-
реле 1986 г. на Чернобыльской АЭС в Украинском Полесье. Можно вспомнить и
о других крупных ядерных катастрофах - взрыв хранилища отходов ядерного
производства на Южном Урале в нашей стране, аварии на атомных станциях
Три-Майл-Айленд (США) и Уиндскейл (Великобритания). Однако еще в 1950—
1960 гг., когда испытания ядерного оружия проводили в атмосфере, уже имел
место бум радиоэкологических исследований. Поскольку лишайники в припо-
лярных регионах Северного полушария находятся в начале пищевой цепи: “ли-
шайник-олень-человек” (Palmer et al., 1963), они стали объектом широкого ра-
диоэкологического изучения в отношении накопления ими радионуклидов и транс-
формации аккумулированных продуктов выпадений испытательных ядерных
взрывов, наибольшее число которых было проведено в атмосфере в 1950-х годах
именно в Арктике.
9.1. УСТОЙЧИВОСТЬ ЛИШАЙНИКОВ
К ДЕЙСТВИЮ ИОНИЗИРУЮЩЕЙ РАДИАЦИИ
9.1.1. Данные экспериментов
Многочисленные эксперименты показали, что параметры жизненности ли-
шайников остаются нормальными даже при воздействии достаточно высоких доз
внешнего облучения. В штате Нью-Йорк (США) в течение 32 месяцев наблюдали
за состоянием живых организмов на участке облучаемого дубово-соснового леса
(Quercus alba, Q. coccinea, Pinus rigida). На расстоянии 15 м от гамма-источника
(,37Cs) сосна погибла при дозе 16—40 P/сутки, все местные сосудистые растения -
при дозе 300 P/сутки. Из 47 видов лишайников представители 12 видов погибли
при дозе 1250 P/сутки, а при дозе 2250 P/сутки еще были живы представители 12
9. Лишайники и радиоактивное загрязнение территории
187
видов лишайников (Woodwell, Ganjnutz, 1967; Woodwell, Whittaker, 1968). Наблю-
дения за состоянием и скоростью роста представителей 19 видов лишайников
при экспозиционной дозе до 1000 P/день показали, что через 9 месяцев после
облучения лишь у 20,7% изученных слоевищ были обнаружены повреждения
(обычно в форме изменения цвета), причем половина их приходилась на предста-
вителей рода Cladonia (Brodo, 1964). Через 22 месяца общее число поврежденных
слоевищ уменьшилось и даже появились зачатки новых талломов. Скорость рос-
та представителей листоватого лишайника Parmelia sulcata на расстоянии 18 и
34 м от источника гамма-облучения (доза от 230 до 62 P/день) даже увеличилась
в сравнении с контролем, что дало основание сделать вывод о большей зависимо-
сти лишайников от микроклимата, чем от гамма-радиации (Brodo, 1964). Анало-
гичные комплексные исследования были проведены и на лесном эксперименталь-
ном участке в штате Висконсин (Erbisch, 1974,1977). Изучение слоевищ Melanelia
subaurifera, собранных на различном расстоянии от гамма-источника (,37Cs)
и получивших в результате хронического облучения суммарные дозы 122272 Р,
78797 Р, 66570 Р, 11412 Р и 1956 Р, выявило разную степень повреждений тканей
растений, причем у талломов, получивших суммарную дозу 66570 Р и менее, по-
вреждений не обнаружили (Erbisch, 1978).
Имевшие место крупные ядерные аварии, сопровождавшиеся выбросом в среду
значительного количества радионуклидов, не вызвали повреждений лишайников
ни на Урале (Бязров, 19936), ни в непосредственной близости от взорвавшегося
реактора Чернобыльской АЭС (Biazrov, 1994с, d).
Эксперименты в лаборатории позволяли изучить реакцию лишайников на
гораздо более высокие дозы облучения. У подеций Cladonia cervicornis ssp.
verticillata, облученных источником 60Со и получивших дозу 10000 Р, через 5 не-
дель после облучения обнаружили видимые повреждения водорослевых клеток,
тогда как у получившие дозу 1000000 Р уже через неделю после облучения были
заметны повреждения, а через 4 недели повреждения были выявлены у 20-30%
водорослевых клеток (Pullum, Erbish, 1972). Облучение в 10000 Р стимулировало
поглощение подециями фосфора, а облучение в 1000000 Р заметно снижало по-
глощение этого элемента. Измерения силы растяжения подециев представителей
рода Cladina показали, что подвергнувшиеся облучению подеции имеют мень-
шую в сравнении с необлученными силу растяжения, что сказывается на степени
ломкости подециев - слоевища облученных лишайников более ломкие (Erbisch et
al., 1977).
9.1.2. Наблюдения на Семипалатинском ядерном полигоне
Изучение видового состава лишайников, проведенное мною в 1992 году на
Семипалатинском ядерном полигоне в Казахстане (Бязров, 19996), через 30 лет
188
9. Лишайники и радиоактивное загрязнение территории
Таблица 9.1
Встречаемость видов лишайников (доля учетных площадок с лишайниками от общего
числа площадок, в %) в эпицентре наземного ядерного взрыва
Название вида Субстрат
Почва Растительные остатки Древесина Железобетон
Acarospora impressula — — — +
Aspic ilia desertorum — — — +
Buellia punctata — 2 + —
Caloplaca cerina 5 5 + —
C. geophila 3 9 + —
C. vitellina — — — +
Candelariella vitellina 5 12 + +
Endocarpon subfoliaceum 1 1 - -
Lecanora hagenii — — + +
Peltula impressula 1 1 — -
Xanthoria elegans — — — +
Число видов на субстрате 5 6 5 6
Тире - отсутствие представителей вида на субстрате, + - наличие представителей вида на субстра-
тах, на которых учетные площадки не закладывались
после последнего в СССР наземного (надповерхностного) испытания ядерного
устройства, выявили на оплавленной стекловидной поверхности почвы, а также
на остатках железобетонных сооружений в условиях высоких уровней радиации
представителей 11 видов лишайников (табл. 9.1). Для их учета были заложены
100 учетных площадок по 1 м2, тщательно осмотрены поверхности железобетон-
ных блоков и плит инженерных сооружений, обломки обработанной древесины,
некогда оставленные здесь персоналом полигона. Состав видов и жизненных форм
напоминает начальные стадии формирования лишайниковых группировок, уста-
новленные в степях Монголии (Бязров, 1988в). Однако наиболее близким анало-
гом видовому составу лишайников на стекловидной корке и почве, а также на
железобетонных блоках и плитах обследованного участка является установлен-
ная на Ближнем Востоке (Carmel Mountains) начальная стадия послепожарной
сукцессии лишайников на каменистом субстрате, где на первом этапе восстанов-
ления наиболее заметными были представители рода Caloplaca (Garty, 1990).
Сопоставление размеров слоевищ с установленными ранее показателями скорос-
ти роста лишайников в степях Монголии (Бязров, 19886), а также особенности их
видового состава дали основания для предположения, что формирование напоч-
венных лишайниковых группировок здесь началось за 5-7 лет до начала наблю-
дений, а на железобетонных плитах - за 10-15 лет, и летом 1992 г. были зафикси-
рованы лишь начальные стадии сукцессии лишайниковых группировок, анало-
гичные восстановлению лишайникового покрова после пожара.
9. Лишайники и радиоактивное загрязнение территории
189
Измерения активности радионуклидов в лишайниках, собранных на Семипа-
латинском ядерном полигоне, не проводились, однако, судя по наблюдениям на
аналогичном полигоне в Южной Австралии, где в 1950-1960 гг. испытывали ядер-
ное оружие, в лишайниках рода Dermatocarpon на полигоне обнаружили в не-
сколько раз больше плутония, чем в почве под ним и на окружающих участках
(Ferris et al., 1995). Авторы сделали вывод, что лишайники уменьшают рассеива-
ние частиц РиО2 в воздухе путем включения их в ткань и удержания в гифах,
а также препятствуя эрозии загрязненной почвы. Вероятно, и на Семипалатин-
ском полигоне появившийся после испытаний лишайниковый покров играет сход-
ную роль.
9.2. НАКОПЛЕНИЕ РАДИОНУКЛИДОВ
СЛОЕВИЩАМИ ЛИШАЙНИКОВ
Содержащиеся в среде естественные и искусственные радионуклиды различ-
ного происхождения в той или иной мере поступают в живые организмы. Эти
изотопы, поступившие в тело растения, гриба или животного, с одной стороны,
являются источником внутреннего облучения конкретного живого организма, с
другой - они источник облучения для других организмов, непосредственно взаи-
модействующих с этим конкретным индивидом. Лишайники считаются концент-
раторами радионуклидов из среды (Gorham, 1959; Kwapulinski et al., 1985; Eckl et
al., 1986; Seaward, 1992), что подтвердили и результаты наших исследований (Ада-
мова, Бязров, 1991; Biazrov, 1994с).
9.2.1. Радионуклиды в лишайниках Арктики
Измерения содержания радионуклидов в лишайниках тундры, проведенные в
1960-1970-х годах, показали значительное варьирование этого показателя как в
пространстве, так и во времени. Например, содержание основного долгоживуще-
го изотопа l37Cs в те годы колебалось от 400 до 3200 Бк/кг сухой массы кустистых
напочвенных лишайников, 134Cs - от 1 до 8, |55Еи - от 35 до 550, |44Се - от 150 до
7500, ,06Ru-ot50 до 1860 (Hanson, 1967; Mattsson, 1975). Эти уровни обеспечили
удвоение, в сравнении с естественными радионуклидами и космическим излуче-
нием, поглощенной лишайниками дозы облучения за период 1961-1975 гг., при-
чем 40% этой дозы приходится на ,44Се-|44Рг (Mattsson, 1975). После аварии на
Чернобыльской АЭС суммарное содержание радиоактивных изотопов цезия в
разных видах лишайников тундр Норвегии составляло от 7600 до 20800 Бк/кг
(Gaare, 1987). В тундрах Финляндии этот показатель варьировал от 1000 до
12100 Бк/кг (Rissanen, Rah о la, 1990).
190
9. Лишайники и радиоактивное загрязнение территории
9.2.2. Радионуклиды в лишайниках после аварии
на Чернобыльской АЭС
В 1987-1991 гг. я изучал экологические последствия аварии на ЧАЭС непос-
редственно в 30-километровой зоне отселения вокруг станции. Рекогносцировоч-
ное обследование территории после аварии не выявило видимых повреждений
слоевищ лишайников как вблизи взорвавшегося 4-го блока станции, так и в уда-
ленных от него местах. Поэтому для суждения о степени загрязнения местности в
сосновых лесах на разном удалении от станции в разные годы были собраны
слоевища разных видов лишайников, а также субстрат (кора сосны) для измере-
ния активности аккумулированных ими радионуклидов (табл. 9.2). Эти измере-
ния были проведены в лаборатории Опытной научно-исследовательской станции
химкомбината “Маяк” в Челябинской области.
Величины активности радионуклидов в лишайниках вблизи АЭС были столь
велики, что у меня возникли сомнения в верности проведенных измерений. По-
Таблица 9.2
Концентрация (Бк/г сухого веса) радионуклидов в слоевищах лишайников и коре сосны
вблизи Чернобыльской АЭС (пробы собраны в июле 1988 г., измерены в январе 1989 г.)
Место Изотоп Hypogymnia physodes Parmelia sulcata Кора сосны Cladina mitis Cladonia crispata Cladonia gracilis
1-Янов, 1,5 l06Ru 1700 3940 335 111 1260 593
км от АЭС на l34Cs 1140 2840 258 561 н.д. 1230
ЮЗ l37Cs 5840 14600 1260 2700 н.д. 5960
|44Се 2400 6480 588 662 1830 981
2 - Чисто- ,06Ru 1130 Н.Д. 355 747 10500 н.д.
галовка, ,34Cs 730 Н.Д. 257 570 н.д. н.д.
7 км от АЭС l37Cs 3600 Н.Д. 1210 2850 н.д. н.д.
на ЮЗ l44Ce 1160 н.д. 684 988 2690 н.д.
3 - Новоше- l06Ru 183 н.д. 23 394 17 н.д.
пеличи, ,34Cs 92 н.д. 24 263 н.д. н.д.
12-13 км от ,37Cs 539 н.д. 107 1230 н.д. н.д.
АЭС на СЗ l44Ce 184 н.д. 34 385 184 н.д.
4 - Черевач, ,06Ru 49 н.д. н.д. 43 н.д. н.д.
19 км от l34Cs 41 н.д. н.д. 26 н.д. н.д.
АЭС на Ю ,37Cs 200 Н.Д. н.д. 118 н.д. н.д.
,44Ce 46 Н.Д. н.д. 35 н.д. н.д.
5* - Ораное, l06Ru 34-337 Н.Д. 86 19-20 16-39 н.д.
37 км от l34Cs 12-150 Н.Д. 12 10-12 н.д. н.д.
АЭС на Ю ,37Cs 71-792 Н.Д. 57 44-58 н.д. н.д.
l44Ce 20-223 Н.Д. 22 12-20 23-44 н.д.
* - приведены интервалы активности для трех точек с данного места отбора проб; н.д. - нет дан-
ных.
9. Лишайники и радиоактивное загрязнение территории
191
этому пробы из первых трех мест, собранные в один из последующих годов (1991
г.), были переданы для контрольных измерений в руководимую в то время докто-
ром Г. Линднером лабораторию Высшей школы Равенсбург-Вайнгартен (Герма-
ния). Данные этих измерений в целом подтвердили результаты, полученные в
отечественной лаборатории. Кроме того, там были измерены активности и ряда
других радионуклидов (табл. 9.3).
Таблица 9.3
Концентрация (Бк/г сухого веса) радионуклидов в слоевищах лишайников и коре сосны
вблизи Чернобыльской АЭС
Место Изотоп Hypo- gymnia physodes Parmelia sulcata Evernia prunastri Кора сосны Cladina mitis Cladonia crispata Cladonia uncialis
1- Янов, 1,5 км от Общая гамма- 25800 30100 10800 11370 Н.Д. Н.Д. Н.Д.
АЭС на ЮЗ активность “Со 4,32 4,55 1. 0,99 Н.Д. Н.Д. Н.Д.
l06Rh 3940,02 3760,31 1750, 1210, Н.Д. Н.Д. Н.Д.
,34Cs 779, 811, 332, 208,9 Н.Д. Н.Д. Н.Д.
l37Cs 1645,0 1794, 719, 442,5 Н.Д. Н.Д. Н.Д.
,44Се 14430,0 19700, 7900, 9290, Н.Д. Н.Д. Н.Д.
,54Eu 14,49 20,04 31,16 10,17 Н.Д. ' Н.Д. Н.Д.
i55Eu 27,08 41,99 15,9 16,42 Н.Д. Н.Д. Н.Д.
241 Am 29,0 29,0 0,0 18, Н.Д. Н.Д. Н.Д.
2- Чисто- галовка, Общая гамма- 8900 Н.Д. Н.Д. 5000 1640 Н.Д. 1230
7 км от АЭС на активность “Co 2,56 Н.Д. Н.Д. 0,19 1,3 Н.Д. 0,0
ЮЗ ,06Rh 1700, Н.Д. Н.Д. 577,1 168,1 Н.Д. 190,
l34Cs 329, Н.Д. Н.Д. 93,5 105,7 Н.Д. 80,
,37Cs 656, Н.Д. Н.Д. 206, 234, Н.Д. 169,9
l44Ce 6100, Н.Д. Н.Д. 3390, 940, Н.Д. 760,
l54Eu 7,39 Н.Д. Н.Д. 3,83 1,09 Н.Д. 0,96
155Eu 10,53 Н.Д. Н.Д. 5,15 1,45 Н.Д. 1,52
24'Am 0,0 Н.Д. Н.Д. 1,71 0,19 Н.Д. 0,0
3 - Ново- шепеличи, Общая гамма- 5760 Н.Д. 6400 10300 3375 1170 Н.Д.
12-13 км от активность “Co 1,74 Н.Д. 58,73 1,68 L24 0,34 Н.Д.
АЭС наСЗ ,06Ru 745,86 Н.Д. 1240, 1075,37 378,13 150, Н.Д.
134Cs 334,6 Н.Д. 226, 175,8 277,5 159,4 Н.Д.
137Cs 685,0 Н.Д. 529, 413, 595, 343,7 Н.Д.
144Ce 2970, Н.Д. 4200, 7260, 3410, 500, Н.Д.
,54Eu 3,42 Н.Д. 5,03 6,68 1,96 0,84 Н.Д.
155Eu 4,52 Н.Д. 8,81 12,18 3,2 0,0 Н.Д.
241 Am 5,2 Н.Д. 0,0 13,00 0,0 0,0 Н.Д.
Пробы собраны в сентябре 1991 г., измерены в мае 1992 г. в Германии, величины активности пере-
считаны на 1 мая 1986 г.; ошибка величин 2,5-17%).
192
9. Лишайники и радиоактивное загрязнение территории
Если сравнить данные таблиц 9.2 и 9.3 с приведенными в тексте количествен-
ными показателями о содержании некоторых радионуклидов в слоевищах лишай-
ников в тундрах Аляски и Лапландии в период максимума испытаний ядерного
оружия в атмосфере и после аварии на Чернобыльской АЭС, то можно сделать
заключение о масштабе загрязнения территории - величины активности радио-
нуклидов вблизи Чернобыльской АЭС, как правило, на 2-4 порядка выше. Соот-
ветственно, поглощенная лишайниками доза облучения здесь несравненно выше.
Вероятно, следует ожидать значительного и продолжительного воздействия слое-
вищ, подвергнувшихся воздействию радиоактивных выбросов из взорвавшегося
реактора, на взаимодействующие с ними организмы, как в качестве источника
внешнего облучения, так и внутреннего для питающихся отмершими или живы-
ми частями лишайников.
Уместно указать, что и слоевища, появившиеся на территории, подвергнув-
шейся сильному радиоактивному загрязнению, после аварийного выброса радио-
нуклидов, но растущие в условиях повышенного радиационного фона, также об-
наруживают высокий уровень активности радиоизотопов, что было выявлено нами
на территории Восточно-уральского радиоактивного следа (Biazrov, 1994d).
Полученные материалы (см. табл. 9.2 и 9.3) показали, что состав радионукли-
дов в слоевищах лишайников и их соотношение соответствуют составу топлива
взорвавшегося реактора АЭС (Богатов и др., 1990; Levi, 1991; Боровой, Гагарин-
ский, 2001). Это дало основание назвать лишайники летописцами ядерных ава-
рий (Feige et al., 1990; Seaward, 1992), поскольку по результатам определений
радионуклидов в долгоживущих слоевищах лишайников можно судить об осо-
бенностях происходивших ранее процессов, связанных с деятельностью челове-
ка. Однако желательно, чтобы исследователь, решающий подобные задачи, хоро-
шо ориентировался не только в видах лишайников, но и в свойствах радионукли-
дов. В этой связи странным представляется вывод в публикации С.Я. Кондратюка
и др. (19936), что в 1991 г. в Украинском Полесье количество l34Cs в сравнении с
предшествовавшим годом увеличилось, а количество l37Cs осталось на прежнем
уровне. И если относительно второго изотопа такое заключение в пределах ошиб-
ки, поскольку период его полураспада около 30 лет, то данные по l34Cs явно оши-
бочные, ибо период полураспада этого изотопа гораздо короче (2,1 года), поэтому
его накопления не происходит, если только не было других выбросов. Но второй
изотоп отдельно от первого в среду попасть не может, так что в случае повторных
выбросов увеличилось бы и содержание ,37Cs. Эти изотопы образуются при деле-
нии урана и плутония, и l37Cs считается ведущим с точки зрения формирования
тканевых доз и радиобиологических последствий. Известно, что l37Cs лишайни-
ки в больших количествах накапливают из глобальных выпадений. В Арктике в
слоевищах кладин в годы, когда ядерное оружие испытывалось в атмосфере, его
концентрация составляла около 2,6 Бк/г (Алексахин, Равикович, 1969). В слоеви-
щах Hypogymnia physodes в районе Белоярской АЭС (Средний Урал) в начале
9. Лишайники и радиоактивное загрязнение территории
193
1980-х активность l37Cs составляла 0,3-0,75 Бк/г, а в коре сосны 0,1-0,25 Бк/г
(Нифонтова и др., 1988). В Австрии в 1980 г. после проведенных в Китае испыта-
ний ядерного оружия в атмосфере активность l37Cs в слоевищах Cladina rangiferina
и Cladonia furcata составляла 0,25 и 0,3 Бк/г, соответственно, а в слоевищах
Hypogymniaphysodes - 0,1-1,3 Бк/г (Eckl et al., 1986). В момент аварии на ЧАЭС
значение ,34Cs/l37Cs было около 0,5 (см. табл. 9.3), а через 1000 дней - 0,2 (см.
табл. 9.2), причем значения этих отношений (в отличие от значений отношений
между активностями других изотопов) близки в слоевищах всех обследованных
видов лишайников и в коре сосны (Biazrov, 1994d). Концентрация этих изотопов
в слоевищах и коре дерева последовательно снижается с удалением от эпицентра
аварии. Как правило, в эпифитах она выше, чем в представителях напочвенных
видов и в коре сосны практически во всех обследованных пунктах, что соответ-
ствует данным, полученным в лесах Австрии (Eckl et al., 1986). Порядок величин
концентрации l37Cs в слоевищах Hypogymniaphysodes вне 30-километровой зоны
отселения (Ораное) через 1000 дней после аварии такой же, как и в лесах Австрии в
августе 1986 г., где активность l37Cs в талломах этого вида составляла 33,5 Бк/г, а
в стволах ели - 3,5 Бк/г (Hofmann und and., 1988). До аварии на ЧАЭС концентра-
ция l37Cs в слоевищах Н. physodes в тех же австрийских лесах составляла
0,07 Бк/г. Это свидетельствует о воздействии аварии на Чернобыльской АЭС на
значительные территории нашей планеты и неравномерности радиоактивных вы-
падений, что подтверждается многочисленными публикациями (Ветров и др., 1991;
Levi, 1991; Медведев, 1991; Квасникова и др., 1999; Исаев, 2001 и другие), а так-
же результаты измерений общей гамма-акгивности в пробах лишайников, соб-
ранных мною в разных регионах Евразии в 1987 г. (табл. 9.4). Для сравнения в
таблицу включены данные о гамма-активности проб лишайников, собранных в
1970-х в Монголии.
Данные таблицы 9.4 свидетельствуют о том, что, по уровню активности ра-
дионуклидов в лишайниках выделяется Черноморское побережье Кавказа, Аджа-
рия. Это связано с тем, что один из маршрутов Чернобыльского радиоактивного
облака с 1 по 3 мая 1986 г. проходил через Южную Украину, далее над Черным
морем к побережью Кавказа и Турции (Борзилов и др., 1989). Здесь в это время
шли дожди, и радионуклиды вместе с осадками попали в экосистемы этих райо-
нов, что и нашло отражение в повышенной их активности в собранных там через
год слоевищах лишайников
9.2.3. Накопление естественных радионуклидов
слоевищами лишайников
Талломы лишайников накапливают не только искусственные, но и естествен-
ные радионуклиды. Природную радиоактивность создают изотопы таких элемен-
194
9. Лишайники и радиоактивное загрязнение территории
Таблица 9.4
Гамма-активность в слоевищах лишайников и в некоторых их субстратах
Дата сбора Место, абсолютная высота (м наду.м.), среднегодовое количество осадков Образец Средняя гамма- активность, КБк/кгсух. веса, ошибка ±15%
19.05.1987 Аджария, Балумский ботанический сад, 50 м, 2000 мм Слоевища Cladonia coniocraea 23,0
Разрушившаяся древесина пня 19,2
Слоевища Flavoparmelia caperata 7,5
Кора широколиственного дерева 16,0
Слоевища Parmotrema chinense 48,5
Кора широколиственного дерева 27,5
Мох Flecker a sp. 46,2
13.06.1987 Челябинская обл.. вблизи Восточно-уральского радиоактивного следа, 300 м, 400 мм Слоевища Hypogymnia physodes 4,2
Слоевища Melanelia olivacea 5,6
Слоевища Parmelia sulcata 2,7
Кора Betula platyphylla 5,5
21.06.1987 Курская обл.. Центрально- Черноземный заповедник, 250 м, 500 мм Слоевища Evernia prunastri 1,8
22.06.1987 Слоевища Hypogymnia physodes 0,9
Слоевища Parmelia sulcata 2,8
25.06.1987 Слоевища Melanelia olivacea 1,3
Слоевища Physconia distorta 4,2
Слоевища Cladonia coniocraea 0,8
05.07.1971 Монголия, Хан гайский хребет, 1800-2400 м, 400 мм Слоевища Leptogium saturninum 4,7
Слоевища Stereocaulon glareosum нет активности
12.08,1971 Слоевища Xanthhoparmelia somloensis 0,4
Слоевища Xanthoria elegans 0,8
Слоевища Peltigera membranacea нет активности
тов как U, Th, Ra, Rn, Ро, Pb, К, достаточно широко представленные в окружаю-
щей нас среде. Например, изотоп 40К с периодом полураспада 1,3><109 лет всегда
9. Лишайники и радиоактивное загрязнение территории
195
содержится в организмах и является основным бета-излучающим естественным
радионуклидом в биосфере (Эйзенбад, 1967; Алексахин, Равикович, 1969). Газ
радон (Rn) можно обнаружить практически в каждом здании (Исаев, 2001). Све-
дения о величинах активности естественных радионуклидов в представителях
биоты полезны для определения естественного фона радиоактивности. Осенью
1987 г. на Западном Кавказе были собраны образцы лишайников, растений и
почвы для измерения в них концентраций ряда естественных изотопов и стабиль-
ного свинца. Аналитическая работа проведена в Институте биологии Коми НЦ
УрО Академии наук (Адамова, Бязров, 1991). Результаты измерений показаны в
таблице 9.5.
Период полураспада 232Th равен 1,4* 10'°лет, что дает основание считать этот
радионуклид одним из первоисточников радиоактивности компонентов среды.
В колебаниях его содержания в слоевищах лишайников разных видов и жизнен-
ных форм обследованного региона трудно установить какую-либо закономерность.
Удельная активность этого изотопа здесь практически во всех случаях ниже ак-
тивности 226Ra, что не противоречит общему мнению о неперспективности Th
для изучения радиоактивности среды (Эйзенбад, 1967). Немного в лишайниках и
226Ra, а в некоторых случаях использованный метод не позволил достоверно об-
наружить в пробах этот изотоп. Довольно значительная доля активности в иссле-
дованном материале приходится на изотопы полония и свинца. Период полурас-
пада 210Ро составляет 138 дней, а 210РЬ - 22 года, причем первый является дочер-
ним продуктом второго, а 210РЬ, в свою очередь, образуется при распаде радона
(Ермолаева-Маковская, Литвер, 1978). Этот радиоактивный урановый ряд закан-
чивается стабильным 206РЬ. Однако равновесие между 2|0Ро и его материнским
нуклидом 210РЬ, а также между стабильным свинцом и названными изотопами,
как это было установлено для планктона (Черри, 1968), у нас не наблюдается.
Аналогичный результат был получен и в финской части Лапландии, где величи-
на отношения между этими изотопами варьировала от 0,92 до 0,28 (Kauranen,
Miettinen, 1967; Persson, 1970). Вероятно, отсутствие такого равновесия между
2|0Ро и материнским 2|0РЬ можно объяснить смывом изотопов полония дождями.
Я не решаюсь устанавливать какие-либо закономерности распределения 2,0Ро, 210РЬ,
206РЬ в слоевищах лишайников в зависимости от местообитания, жизненной фор-
мы или вида. Но совершенно определенно можно утверждать, что в сравнении с
сосудистыми растениями лишайники, как правило, выступают как концентрато-
ры тяжелых радионуклидов, уступая почве (см. табл. 9.5). Лишь слоевища мха
Neckera sp. показывают сходное с лишайниками накопление анализируемых тя-
желых радионуклидов. Если сравнить полученные данные с результатами анало-
гичных исследований в других местах, например, в Альпах (Eckl et al., 1986), то
надо признать, что в 1987 г. на Западном Кавказе в целом лишайники содержали
меньше изотопов тяжелых радионуклидов, чем в Австрийских Альпах в 1980-
1982 гг. Там активность 210РЬ в слоевищах Hypogymnia physodes варьировала от
196
9. Лишайники и радиоактивное загрязнение территории
Таблица 9.5
Удельная активность (Бк/г сухой массы) и содержание стабильного РЬ (г/г сухой массы)
в пробах с Западного Кавказа
Место, абсолютная высота (м над у.м.) Образец Слоевища Flavoparmelia caperata Изотопы Pb, n-10-6 0,98
232Th, nlO^* 11,5 226Ra, n-10~3 4,28 2,0Po, n-10~2 2l0Pb, nl(T2 2l0Po/ 2l0Pb
Краснодарский край, Хостинский р-н, г. Сочи, гора Ахун, 600 м 69,2 203,1 0,34
Слоевища Ramalina farinacea 10,9 22,2 3,92 21,2 0,18 1,06
Листья и стебли Hedera colchica 2,2 1,94 1,09 4,91 0,22
Листья Acer pseudoplatanus 3,2 - - 8,03 -
Ветви A. pseudoplatanus 1,0 - 5,14 11,0 0,47
Слоевища Meekera sp. 11,0 1,06 35,2 175,4 0,2
Краснодарский край, Адлерский р-н г. Сочи, у пос. Красная Поляна, 600 м Слоевища Lobaria pulmonaria 2,91 - 4,2 12,2 0,34 0,47
Слоевища Hypogymnia physodes 6,74 0,81 11,2 121,0 0,09 1,05
Слоевища Usnea sp. 13,6 - 35,9 102,9 0,35 1,61
Листья Castanea sativa 22 1,09 0,61 2,28 0,27
Ветви C. sativa 0,6 — 1,61 4,37 0,37
Кора C. sativa 7,0 1,06 — 24,7 —
Ветви Pinus kochiana - - 7,0 20,5 0,34
Кора P. kochiana 13,5 0,19 5,07 11,8 0,42
Слоевища Neckera sp. 11,6 1,13 26,7 23,4 1,14
Лесной опад 6,0 2,8 1,9 68,4 0,03
Почва (глубина) 0-1 см 63,0 5,4 6,4 68,1 0,09
1-2 cm 179,0 11,6 11,6 68,8 0,17
2-3 cm 198,0 17,9 20,9 49,2 0,42
3-4 cm 224,0 17,6 26,9 46,6 0,58
4-5 cm 237,0 22,7 17,2 36,9 0,47
Абхазия, 6 км от озера Рица на восток, 1000 м Слоевища Lobaria pulmonaria 17,7 5,96 7,4 38,1 0,2 1,23
Слоевища Flavoparmelia caperata 20,5 1,63 69,9 140,6 0,5 3,09
Слоевища Cetrelia cetrarioides - - 7,14 22,4 0,32 -
9. Лишайники и радиоактивное загрязнение территории
197
таблица 9.5 (окончание)
Абхазия, 6 км от озера Рица на восток. 1000 м Слоевища Usnea spp. 16,4 3,1 14,9 60,7 0,25 0,87
Хвоя Taxus baccata 1,2 — 0,7 3,45 0,2
Ветви Т baccta — — 1,06 5, 0,21
Хвоя Abies nordmaniana 2,3 0,32 7,3 27, 0,27
Ветви A. nordmaniana - - 9,8 12,4 0,79
Тире - концентрация ниже предела измерений: пустая клетка - не определяли.
74x10-2 до 666х 10 2 Бк/г. Концентрация того же изотопа в лишайниках и мхах,
собранных в 1988, 1996, 1998 гг. на Южных Шетландских островах и Антаркти-
ческом полуострове, изменялась от <0,2x10’2 до 12,5х Ю 2 Бк/г (Mietelski et al.,
2000).
Поведение тяжелых радионуклидов в талломах лишайников анализируется
во многих публикациях. Большинством авторов изучалось воздействие на ли-
шайники рудников по добыче урана (Beckett et al., 1982; Boileauetal., 1982;Nieboer
et al., 1982; Looney et al., 1985) или в районах с высоким уровнем естественной
радиоактивности (Mahon, 1982; Sheard, 1986а, b; McLean et al., 1998). В других
публикациях приводятся результаты экспериментальных исследований (Boileau
et al., 1985а, b; Richardson et al., 1985; Haas et al., 1998). Например, было установ-
лено, что наибольшее поглощение урана из раствора слоевищами Peltigera
membranacea происходило при pH = 4,5 за 24 часа (Haas et al., 1998). Показано,
что концентрация урана в лишайниках отражает его содержание в субстрате
(Sheppard, Thibault, 1984). Использовались лишайники {Melanelia exasperatula),
вместе с такими индикаторами как вода, почва, кора деревьев, и для выявления
последствий применения войсками НАТО в Косово снарядов со слабо обогащен-
ным ураном. Следы этого урана в повышенных концентрациях были обнаруже-
ны во всех средах, кроме воды, но лучшими индикаторами были признаны ли-
шайники и кора деревьев (Sansone et al., 2001).
9.3. РАСПРЕДЕЛЕНИЕ РАДИОНУКЛИДОВ
В СЛОЕВИЩЕ И ИХ ВЫСВОБОЖДЕНИЕ
Сведения о распределении аккумулированных радионуклидов в слоевищах
лишайников достаточно противоречивы. Этот показатель зависит как от свойств
радиоизотопов (формы выпадения, количества, растворимости в воде, подвиж-
ности в среде), так и особенностей лишайника (жизненная форма, размеры расте-
ния) и местообитания (субстрат, экспозиция, количество осадков). У кустистых
198
9. Лишайники и радиоактивное загрязнение территории
напочвенных лишайников радионуклиды из глобальных выпадений концентри-
руются в верхних более молодых частях слоевищ (Mattsson, 1975; Rissanen, Rahola,
1990). Наблюдаемые количественные различия в распределении отдельных ра-
дионуклидов, например l37Cs, связывают с морфологией конкретных особей -
чем плотнее, компактнее верхушечная часть растений, тем меньше l37Cs в нижних
частях куртин кладоний (Kreuzer, Schauer, 1972). Такое объяснение создает впе-
чатление, что плотная компактная верхушечная часть механически препятствует
проникновению радионуклидов в нижележащие слои лишайникового покрова.
Не следует забывать, что верхушечные части кладоний, как и других кустистых
лишайников, представляют собой наиболее молодые и тонкие, иногда волосовид-
ные, веточкообразные выросты слоевища. Поверхность их намного больше в срав-
нении с более старыми частями растения, а слоевище лишайника взаимодейству-
ет со средой как физическое тело, подобно губке впитывая из воздуха, осадков
все, что способно поглотить, в том числе и радиоизотопы (Kershaw, 1985; Ellis,
Smith, 1987). Поэтому концентрирование радионуклидов в верхушечных частях
слоевищ можно связать и с более значительной поглощающей поверхностью это-
го слоя. Детальная картина подвижности нуклидов внутри таллома была получе-
на с использованием радиоактивных изотопов цезия и стронция (Nevstrueva et
al., 1967). При погружении верхушек таллома Cladina rangiferina в водные ра-
створы l37Cs и 90Sr распределение этих радионуклидов различалось: если строн-
ций распределялся равномерно, то цезий концентрировался преимущественно в
той части таллома, которая контактировала с раствором. В случае погружения
всего слоевища в раствор, оба изотопа распределялись равномерно в талломе,
несколько больше накапливаясь в его старых частях. Таким образом, оба радио-
нуклида могут свободно передвигаться от верхушки к основанию и наоборот.
Накопление цезия в верхушках и равномерное распределение стронция наблюда-
лось у других видов лишайников (Троицкая и др., 1971; Kreuzer, Schauer, 1972;
Нифонтова, 1973, 1976; Нифонтова, Куликов, 1977).
В опытах по изучению поглощения слоевищами Peltigera membranacea уран
неравномерно распределялся в талломе, главным образов в верхнем коровом слое,
причем было установлено, что поглощение урана строго коррелирует к местам,
где в слоевище много фосфора (Haas et al., 1998). Вероятно, фосфатные лиганды
и функциональные группы корового слоя включены в процесс поглощения ура-
на. Изучение распределения урана в слоевищах Trapelia involuta показало, что
этот элемент накапливается во внешних стенках плодовых тел (McLean et al.,
1998)
Аккумулированные в слоевищах лишайников радионуклиды со временем
высвобождаются с той или иной скоростью. По аналогии с физической величи-
ной периода полураспада радиоизотопа в отношении живых организмов суще-
ствует понятие “период полувыведения”, т.е. период времени, за который из орга-
низма выводится половина содержащегося в нем радионуклида. Для больший-
9. Лишайники и радиоактивное загрязнение территории
199
ства радиоизотопов период полувыведения короче периода полураспада, поскольку
к последнему добавляются вымывание, поедание животными, удаление с отло-
мившимися частями слоевища. Однако концентрация ряда радиоизотопов, про-
изводных от распада других, например, содержание24’Ат в среде, с течением
времени до определенного момента будет увеличиваться, и обнаружение в талло-
ме изотопов америция свидетельствует о наличии в среде высокотоксичных и
опасных для животных изотопов плутония, поскольку америций образуется в
результате распада плутония. И все чаще этот опасный радионуклид находят в
тканях живых организмов и в почве (Иванов и др., 1994; Якушев и др., 2000).
Обнаружен америций, как и изотопы плутония, и в лишайниках Антарктики
(Mietelski et al., 2000). Правда, там его концентрация на 2 порядка ниже, чем
возле Чернобыльской АЭС.
Период полу выведения из лишайников 137Cs, одного из основных загрязните-
лей, по некоторым данным, составляет от 17 лет (Liden, Gustafsson, 1967) до 3,4-
3,7 лет (Guillitte et al., 1990). Полагаю, что последние числа сильно занижены.
Скорее всего, не учтено, что лишайники, хотя и медленно, но постоянно растут.
Соответственно, непрерывно увеличивается доля слоевища, не подвергнувшего-
ся воздействию радиоактивного выброса. Если представить идеализированный
вариант роста листоватого или коркового лишайника после аварии на Чернобыль-
ской АЭС в 1986 г. (прирост по радиусу - 1 мм/год), то доля прирастающей по-
верхности таллома увеличивалась с большей скоростью у слоевищ, имеющих
меньший первоначальный диаметр (см. табл. 8.2). Мне представляется, что это
обстоятельство необходимо учитывать при изучении последствий воздействия
одноразового выброса на слоевищные растения и формируемые ими сообщества.
Фактически мы наблюдаем не выведение аккумулированных загрязнителей из
организма, а растворение их в нарастающей массе растения. Элиминации попав-
ших на слоевище радионуклидов, особенно вместе с топливными частицами,
практически нет. Наблюдается процесс естественного физического распада изо-
топов с незначительным вымыванием некоторых легкорастворимых соединений.
Если принять во внимание, что представители отдельных видов лишайников живут
очень долго (сотни и тысячи лет), то их слоевища могут содержать следы многих
выбросов загрязнителей в атмосферу.
10. ЛИХЕНОИНДИКАЦИЯ КАЧЕСТВА ВОЗДУХА
В Г. МОСКВЕ
В начале 1990-х годов автор изучал особенности распространения лишайни-
ков на территории г. Москвы в пределах Московской кольцевой автомобильной
дороги (МКАД). Собранные материалы (Бязров, 1994,1996а) в дальнейшем были
использованы для зонирования территории города относительно качества воз-
душного бассейна (Бязров, 19966,1998а, в, г, 1999а; Бязров и др., 1997; Максимо-
ва, Бязров, 2000).
10.1. МЕТОДИКА СБОРА МАТЕРИАЛА
Подробно методика сбора данных изложена ранее (п. 5.3.4). Идентификация
лишайников проведена традиционными для лихенологии методами с использо-
ванием соответствующего руководства (Голубкова, 1966). Для суждения о видо-
вом составе лишайников, собранных когда-либо на обследованной территории,
изучены коллекции гербария слоевищных растений Биологического факультета
Московского государственного университета им. М.В. Ломоносова и гербария
лишайников Ботанического института им. В.Л. Комарова РАН (С.-Петербург).
Помимо гербариев использованы и различные источники, содержащие сведения
о лишайниках Москвы (Бязров, 1996а). Номенклатура лишайников постоянно
меняется, и в данной главе их названия соответствуют доступным опубликован-
ным обобщениям (Santesson, 1993; Esslinger, Egan, 1995).
Помимо изучения видового состава лишайников и картографирования их рас-
пространения, для выявления воздействия городских условий был использован и
метод трансплантации слоевищ из относительно чистого района Подмосковья на
территорию города. Для этого в окрестностях платформы 73 км Рижской желез-
ной дороги в лесу со стволов березы вместе с субстратом были отобраны талломы
Hypogymnia physodes диаметром около 5 см. Они были прикреплены на специ-
ально изготовленные фанерные щиты и размещены в 10 различных точках горо-
да, а также в лесу в месте отбора образцов для трансплантации. Предполагалось
через определенные промежутки времени снимать с этих щитов по 5 слоевищ и
подсчитывать на 5 поперечных срезах каждого таллома с помощью микроскопа
10. Лихеноиндикация качества воздуха в г. Москве
201
долю поврежденных водорослевых клеток, а также определять содержание не-
скольких элементов в каждом слоевище. К сожалению, из-за вандализма отдель-
ных представителей населения города и области большинство щитов с транс-
плантатами было довольно быстро уничтожено. Дольше других (около 8 меся-
цев) экспонировались слоевища в центре города, во дворе здания бывшего музея
М.И. Калинина между Библиотекой им. В.И. Ленина и Боровицкой башней Крем-
ля. Более 4 месяцев экспонировались талломы в массиве Лесной опытной дачи
ТСХА примерно в 9 км на СЗ от Кремля (см. гл. 7).
10.2. ПРИРОДНЫЕ УСЛОВИЯ, СТРУКТУРА ГОРОДА
И ВИДОВОЙ СОСТАВ ЛИШАЙНИКОВ
Столица Российской Федерации г. Москва в пределах МКАД занимает
886,5 км2. Эта величина более чем в четыре раза превышает ее площадь в начале
века (История Москвы, 1978). Москва-река разделяет территорию города на мень-
шую юго-западную и большую северо-восточную части. Кроме нее в пределах
города протекают реки Яуза, Сетунь, Сходня, а также расположены части гидро-
системы, соединяющей рр. Волга и Москва.
Город находится на стыке трех крупных геоморфологических образований
разного происхождения: Клинско-Дмитровской моренной возвышенности в севе-
ро-восточной его части, Мещерской песчаной низменности в восточной и юго-
восточной частях и Теплостанской возвышенности на юго-западе, которая, обры-
ваясь к р. Москва, образует Воробьевы (Ленинские) горы.
Зима здесь умеренно-холодная, а лето - умеренно-теплое, преобладают запад-
ные ветры атлантического происхождения, и количество осадков довольно значи-
тельно (Климат Москвы..., 1989). Климат города существенно отличается от кли-
мата Подмосковья. Это как бы “остров тепла” среди моря более холодного возду-
ха. Максимальная разница между значениями температуры воздуха в городе и
его окрестностях зимой при ясной погоде достигает 14,5°С, а в среднем за год
составляет 2-4,5° С (Безуглая и др., 1991).
Территория Москвы расположена в подзоне широколиственно-хвойных ле-
сов (Алехин, 1947), и здесь еще сохранились остатки естественных лесных мас-
сивов, превращенные в лесопарки - Лосиный остров, Серебряный бор, Измай-
ловский, Битцевский, Кузьминки и другие. Всего площадь зеленых насаждений
общего пользования (лесопарки, скверы) составляет в пределах МКАД более
160 км2, или около 18% от площади города. Кроме того, существуют внутридво-
ровые скверы, насаждения вдоль улиц и дорог, кладбища, долины ручьев, овраги
и т.п. В озеленении Москвы наиболее часто используется около 50 видов деревь-
ев, среди которых, прежде всего, следует назвать липу, тополя, клены, березы,
ивы, вязы и ряд других. В сохранившихся лесных массивах представлены дуб,
202
10. Лихеноиндикация качества воздуха в г. Москве
липа, клен, сосна, ель, осина, ольха, береза, ивы, лещина и другие древесные
породы и кустарники.
Население Москвы составляет около 9 млн. человек, т.е. на 1 жителя города
приходится около 0,01 га территории, около 18 м2 зеленых насаждений. В атмос-
феру в городе ежегодно выбрасывается около 1100 тыс. т вредных веществ (Безуг-
лая и др., 1991), т.е. более 1 ц на 1 жителя за год. Больше всего поступает окиси
углерода (60% от суммарных выбросов), окислов азота (14%), углеводородов (13%),
двуокиси серы (9%).
Москва, как и другие крупные города, исторически представляет собой неус-
тойчивую конечную стадию развития населенного пункта; промежуточными эта-
пами ее развития были поселение или укрепление, мелкий и средний город
(История Москвы, 1978). Ошибочно было бы рассматривать современную терри-
торию Москвы как единую экосистему - фактически это мозаика различных био-
топов, экологическое своеобразие которых зависит, прежде всего, от использова-
ния их человеком в ходе исторического развития города. Вслед за Б. Клауснитце-
ром (1990), городские местообитания лишайников можно разделить на две большие
группы: “внешняя оболочка строений” и “отдельные зеленые насаждения и озе-
лененные городские территории”.
Строения (стены и крыши зданий, поверхности заборов, монументов) - наи-
более характерные для города местообитания, в современной Москве лишайни-
ками практически не заселены, исключая остатки некогда сельских поселений в
районах новой застройки, заборы из неокрашенной древесины вблизи больших
пустырей, а также монументы в старых парках и на ряде кладбищ.
Отдельные зеленые насаждения - это одиночные деревья, посадки вдоль
транспортных магистралей, улиц и внутри дворов, аллеи и живые изгороди. Они
встречаются во всех частях города, от центра, района старой застройки, до окра-
инных новостроек, сформированы довольно разнообразным по составу набором
древесных и кустарниковых пород, иногда экзотических, посаженных организо-
ванно или стихийно, при преобладании липы и тополей. Лишайники на них встре-
чаются, но далеко не повсеместно, и видовой состав их беден.
Озелененные площади представлены в городе парками, ботаническими са-
дами, садовыми участками, кладбищами, насаждениями на территориях круп-
ных спорткомплексов и ряда больниц, пустырями, а также остатками негород-
ских экосистем (леса и лесопосадки, рощи, берега рек, прудов и другое). Город-
ские леса довольно разнообразны по составу древесных и кустарниковых пород,
обогащенному либо не свойственными естественным лесам видами, например,
кленом ясенелистным, либо нитрофильными растениями, например, бузиной чер-
ной. Этим местообитаниям свойственно наибольшее разнообразие видов лишай-
ников в городе, они же могут рассматриваться как рефугиумы для расселения
лишайников в другие городские территории при создании на них благоприятных
для лишайников условий.
10. Лихеноиндикация качества воздуха в г. Москве
203
Список лишайников, найденных на нынешней территории Москвы к началу
данного исследования, составленный по опубликованным данным, а также по
результатам изучения коллекций гербариев Московского университета и Ботани-
ческого института РАН, включал 72 вида (звездочкой отмечены виды, представи-
тели которых нами в Москве не встречены; синонимы указаны для видов, приве-
денных в книге Н.С. Голубковой (1966) и последующих публикациях других ав-
торов под иными названиями; символы из 2-3 латинских букв, находящиеся за
названием вида обозначают жизненную форму лишайника, их объяснение дано
далее в тексте; также см. табл. 1.1.):
★Anaptychia ciliaris (L.) Korb. -Lfp-Богородское, Кунцево, Сокольники, Цари-
цыно; *Bryoria implexa (Hoffin.) Brodo and D. Hawksw. = {Alectoria implexa (Hoffin.)
Nyl.) -Fp-Сокольники; * Buellia punctata {Woffm.) A.Masal. -Сг-Петровско-Разумов-
ское; Caloplacacerina(Ehrh.ex Hedw.) Th.Fr. -Cr-Лосиный остров; C. citrina (Hoffin.)
Th.Fr. -Cr-Петровско-Разумовское; C. holocarpa (Hoffm.ex Ach.) A.E.Wade =
(C. pyracea (Ach.) Th.Fr.) -Сг-Сокольники, Царицыно; ^Candelaria concolor (Dicks.)
Stein. -SI-Петровско-Разумовское, Сокольники; Candelariella vitellina (Hoffin.) Miill.
Arg. -Cr-Петровско-Разумовское; Сокольники; *C. xanthostigma (Ach.) Lettau =
(C. vitellina (Hoffin.) Miill.Arg. var. xanthostigma) -Cr-Петровско-Разумовское, Со-
кольники; *Cetraria islandica (L.) Ach. -Fe-Богородское, Ходынское поле, Хоро-
шево; *Cetrelia olivetorum (Nyl.) W.L.Culb.and C.F.Culb. = {Parmelia cetrarioides
(Duby) Nyl.) -Sl-Царицыно; *Cladonia arbuscula (Wallr.) Flot.= {Cladonia silvatica
auct.) -Fe-Кунцево, Кусково, Сокольники, Царицыно; C. cariosa (Ach.) Spreng.
-Sc-Екатериновка; C. chlorophaea (Florke ex Sommerf.) Spreng. -Sc-Лосиный ост-
ров, ТСХА, Ясенево; C. coniocraea (Florke) Spreng. -Sc-Кусково, Лосиный остров,
Останкино; C. fimbriata (L.) Fr. -Sc-Богородское, Кунцево, Кусково, Останкино,
Сокольники, Царицыно; *С. macilentaiAoffm. ssp. macilenta = (С. bacillaris(Leight.)
Arnold) -Sc-Лосиный остров; *C. parasitica (Hoffm.) Hoffm. -Sc-TCXA;
*C. portentosa (Dufour) Coem.= {Cladonia impexa Harm. ssp. spumosa Florke)
-Fe-Москва?; *C. pyxidata (L.) Hoffin.-Sc - Воробьевы горы; *C.ramulosa (With.)
J.R.Laundon = {C. pityrea (Florke) Fr.) -Sc-Лосиный остров; *C. rangiferina (L.)
Weber ex F.H. Wigg. -Fe-Богородское; *C.squamosa Hoffm.-Fr-Богородское; *C.
subulata (L.) Weber ex F.H.Wigg.=(C. cornutoradiata (Vain.) Zopf f.cornutoradiata)
-Sc-Коптево; *Cturgida Hoffin.-Fr-Кунцево; *Dibaes baeomyces (L.fil.) Rambold and
Hertel = {Baeomyces roseus Pers.) -Sc-Кунцево, Ходынское поле; Everniaprunastri
(L.) Ach. -Fp-Богородское,Кунцево, Сокольники, Царицыно; *Flavoparmelia
caperata (L.) Hale = {Parmelia caperata (L.)Ach.) -Sl-Кунцево, Царицыно;
Hypogymnia physodes (L.) Nyl. -Cl-Богородское, Кунцево, Лосиный остров, Со-
кольники, ТСХА, Царицыно; *Lecanora albella (Pers.) Ach. = {L. pallida (Schreb.)
Rabenh.var. cinerella) -Cr-Востря kobo; L. allophanaNy\. -Cr-Богородское, Востря-
ково, Петровский парк, Сокольники; *L. carpinea (L.) Vain. -Cr-Кунцево, Соколь-
ники,Петровско=Разумовское; *Z. intumescens (Rebent.) Rabenh. -Сг-Востряково;
204
10. Лихеноиндикация качества воздуха в г. Москве
L. piniperda Korb. -Сг-Сокольники; *L. rugosella Zahlbr. -Сг-Сокольники;
L. symmicta (Ach.) Ach. = (Lecidea symmicta (Ach.) Ach =Biatora symmicta Ach.) -
Cr-Петровско-Разумовское, Сокольники; L umbrina (Ach.) A. Massal. -Сг-Соколь-
ники; *Lecania dubitans (Nyl.) A.L.Sm.= (L. dimera (Nyl.) Th. Fr.) -Сг-Кунцево;
Lecidella euphorea (Florke) Hertel = (Lecidea glomerulosa (DC.) Steud.) -Сг-Соколь-
ники; Lepraria incana (L.) Ach.= (L. aeruginosa auct.) -Cr-Лосиный остров;
*Melanelia olivacea (L.) Essl.= (Parmelia olivacea (L.) Ach.) -Sl-Богородское, Вос-
тряково, Кунцево, Царицыно; *Opegrapha varia Pers.= (O. diaphora Ach.)
-Сг-Москва?; *Parmelia saxatilis (L.) Ach. -Sl-Богородское; P. sulcata Taylor
-SI-Кунцево,Сокольники, Царицыно; *Parmelina tiliacea (Hoffm.) Hale = (Parmelia
scortea (Ach.) Ach.,7? tiliacea Hoffm.) Ach.) -SI-Петровско-Разумовское, парк TCXA;
Parmeliopsis ambigua (Wulfen) Nyl. -Sl-Петровский парк; P. hyperopta (Ach.) Arnold
-SI-с.Всехсвятское, Сокольники; *Peltigera canina(L.) Willd. -Ll-Богородское, Во-
робьевы горы; *P. didactyla (With.) J.R.Laundon = (P. spuria (Ach.) DC. fspuria)
-Ll-Воробьевы горы; *P. leucophlebia (Nyl.) Gyeln.= (P. aphthosa (L.) Willd. f.
variolosa A. Massal.)-Ll-Кунцево; *P. polydactyla (Neck.) Hoffm. -Ы-Богородское;
*P.praetextata (Florke ex Sommerf.) Zopf = (P. canina (L.) f. subcanina (Gyeln.) Oxn.)
-LI- Москва?; *P. rufescens (Weiss) Humb.-Ll- Кунцево; *P. venosa (L.) Hoffm.
-Ll-Москва; ^Pertusaria albescens (Huds.) M.Choisy and Werner = (P. globulifera
(Turner) A.Massal.) -Сг-Кунцево; Phaeophyscia ciliata (Hoffm.) Moberg = (Physcia
ciliata (Hoffm.) Du Rietz) -SI-Востряково;^ orbicularis (Neck.)Moberg - (Physcia
virella (Ach.) Flagey) -SI-Петровско-Разумовское; Physcia aipolia (Ehrh.ex Humb.)
Fiimr. -SI-Востряково, Лосиный остров; *P. dubia (Hoffm.) Lettau = (Pteretiuscula
(Ach.) Lynge) -SI-Петровско-Разумовское; P. stellaris (L.) Nyl. -Sl-Богородское;
*P. tenella (Scop.) DC. in Lam. and DC.= (P.hispida auct.) -SI-Петровско- Разумов-
ское;Р in'Z>ac«7(Ach.)Nyl. -Sl-Сокольники; *Physconiadistorta (With.) J.R.Laundon
= (Physcia pulverulenta (Hoffm.) Fiimr.) -Sl-Воробьевы горы, Востряково, Кусково,
пос. Мещерских, Петровско-Разумовское, Сокольники, Царицыно; P.grisea (Lam.)
Poelt = (Physcia grisea (Lam.) Zahlbr.) -Sl-Воробьевы горы; *Ramalina pollinaria
(Westr.) Ach.-Fp- с.Всехсвятское, Кунцево, Петровско-Разумовское; *Stereocaulon
tomentosum Fr. -Fe-Богородское, Кунцево; *Strangospora moriformis (Ach.) Stein.=
(Biatorella moriformis (Ach.) Th. Fr.) -Cr-Петровский парк, Петровско-Разумов-
ское; * Usnea hirta (L.) Weber ex F.H.Wigg. -Fp-Богородское, Царицыно; Vulpicida
pinastri (Scop.) J.-E. Mattsson and M.J.Lai = (Cetraria pinastri (Scop.) Gray)
-Sl-Богородское, Сокольники; *Xanthoria Candelaria (L.)Th. Fr.= (X. lychnea(Ach.)
Th.Fr.) -Sl-Сокольники; X. parietina (L.) Th.Fr. -Sl-Богородское, Кунцево, Нескуч-
ный сад, Сокольники, Царицыно; X. polycarpa ( Hoffm.) Th.Fr. ex Rieber -Sl-Во-
робьевы горы, с.Всехсвятское, Петровско-Разумовское, Ходынское поле.
В пределах МКАД мною было обследовано 908 квадратов. На всех субстра-
тах (кора и древесина, почва, камень) обнаружены представители 47 видов ли-
шайников (табл. 10.1), причем 43 из них встречены на коре деревьев и древесине
10. Лихеноиндикация качества воздуха в г. Москве
205
(Бязров, 1994). Один вид {Cladonia cariosd) обнаружен на нарушенной поверхно-
сти почвы, а три вида {Caloplaca citrina, Lecanora muralis, L. umbrind) - на над-
гробных плитах старых монастырских кладбищ и на памятниках дворцовых пар-
ков. Двадцать из этих видов ранее для территории Москвы не указывались.
Я не нашел представителей 44 из 72 видов, перечисленных выше, т.е.61%. Не
обнаружены совсем лишайники таких жизненных форм как листоватые широко-
лопастные ризоидальные (L1), бородавчато- или чешуйчато-кустистые кустисто-
разветвленные (Fr), листовато-кустистые повисающие (Lfp), кустистые
прямостоячие(Ее). Исчезло три четверти видов кустистых повисающих лишай-
ников. Если слоевища ряда видов, возможно, были мною пропущены {Cladonia
macilenta, С. pyxidata, С. subulatd), а некоторые, скорее всего, указаны ошибочно
из-за неверного определения - Cladoniaparasitica (Буткевич, 1985), С. ramulosa
(Слука, Абрамова, 1984), то отсутствие остальных видов представляется законо-
мерным. Практически все сборы лишайников на обследованной территории Мос-
квы, исключая лишь те, которые указаны в только что цитированных публикаци-
ях, относятся к 19 и началу 20 столетий. Местности, где коллекционировались
лишайники (Кунцево, Петровско-Разумовское, Троицкое, Царицыно, Петровский
парк, Сокольники и другие), в то время в черту города не входили или были его
самыми дальними окраинами. Здесь еще можно было нередко встретить старые
не вытоптанные естественные леса. Например, судя по этикеткам одного из ос-
новных коллекторов московских лишайников И.П. Петрова (значительнейшая
часть его сборов хранится в гербарии МГУ), в 1910 г. у Ходынского поля нахо-
дился сосновый бор, в напочвенном покрове которого была обильно представле-
на Cetraria isian die а, можно было встретить Dibaes baeomyces = (Baeomyces rose us).
Сейчас эта территория занята аэродромом и предприятиями. Население Москвы
и пригородов было намного меньше, индустриализация территории была не столь
высока, практически отсутствовал автомобильный транспорт и, соответственно,
все с ним связанное (густая сеть дорог, мобильность населения, губительные вых-
лопы двигателей, нефтеперерабатывающий завод и многое другое). Иной был образ
жизни, иным было отношение к природе. Поэтому было бы удивительно в наши
дни встретить даже в сохранившихся лесных массивах представителей таких за-
метных напочвенных макролишайников как виды родов Cladina, Peltigera или
виды Cetraria islandica, Stereocaulon tomentosum, поскольку в этих лесных участ-
ках, видимо, нет квадратного дециметра, по которому бы не ступала нога челове-
ка, а вытаптывание весьма негативно действует на лишайники (Малышева, Тол-
пышева, 1982). Отсутствие эпифитных лишайников родов Bryoria, Pertusaria,
Usnea и ряда других связано с сильным загрязнением воздушного бассейна сто-
лицы, поскольку представители этих родов отрицательно реагируют даже на не-
значительную примесь в воздухе таких загрязнителей как двуокись серы, окислы
азота, углерода и другие. И, конечно же, как причину исчезновения многих видов
лишайников следует назвать уничтожение их местообитаний - застройку обшир-
Таблица 10.1
Встречаемость (% квадратов от общего числа квадратов) лишайников в пределах Московской кольцевой автодороги (МКАД),
Окружной железной дороги (ОЖД), Садового кольца (СК) и Бульварного полукольца (БК), Центрального (Ц), Северо-Западного
(СЗ), Северного (С), Северо-Восточного (СВ), Восточного (В), Юго-Восточного (ЮВ), Южного (Ю), Юго-Западного (ЮЗ)
и западного (3) административных округов Москвы
Вид МКАД ОЖД СК БК ц СЗ С СВ В ЮВ Ю ЮЗ 3
Caloplaca cerina (Ehrh. ex Hedw.) Th.Fr. 1,5 — — — 1,3 — 2,0 5,6 — 0,8 — 3,3
C. citrina (Hoffm.) Th.Fr. 1,5 0,5 — — 1,5 — — 1,0 0,8 — 0,8 — —
C. holocarpa (Hoffm. ex Ach.) A.E.Wade 1,8 — — — — — — 1,0 7,4 — — 2,1 3,3
Candelariella vitellina (Hoffm.) Miill. Arg. 3,2 0,4 — — — 1,3 — 3,0 1,4 0,8 3,2 5,4 11,5
Cladonia caespitica (Pers.) Florke** 0,1 — - — — — — - — 0,8 — -
C. cariosa (Ach.) Spreng. 0,1 0,4 — — — — — — 0,8 — — — —
C. chlorophaea (Florke ex Sommerf.) Spreng. 2,9 — — — — 3,7 9,3 1,0 7,4 0,8 0,8 — 0,8
C. coniocraea (Florke) Spreng. 17,4 7,7 — — — 21,7 7,2 13,6 40,2 9,1 6,9 24,8 19,7
C. digitata (L.) Hoffm.** 0,9 0,4 — — — — — 1,0 5,6 — — — —
C. fimbriata (L.) Fr. 12,2 — — — — 16,6 7,2 10,8 30,0 5,9 3,2 14,4 13,6
C. ochrochlora Florke ** 1,2 — — — — 4,8 — — — — 3,2 — 3,3
Evernia prunastri (L.) Ach. 0,2 — — — — 1,3 — — — — — — 0,8
Graphis scrpta (L.)Ach.** 0,1 — — — — — — — — — — 1,0 —
Hypocenomyce scalaris (Ach.) M.Choisy ** 3,4 0,4 — — — 2,6 — 4,0 14,0 3,4 — — 0,8
Hypogymnia physodes (L.) NyL 25,7 15,0 — — 2,9 46,5 22,8 22,9 42,1 12,8 11,1 33,3 30,6
Lecanora allophana Nyl. 0,2 — - — — — — — - — — 2,1 —
L. hagenii (Ach.) Ach.** 8,4 4,7 — — 2,9 3,7 7,2 6,0 5,6 0,8 9,4 11,4 21,0
L. muralis (Schreb.) Rabenh.** 1,5 0,5 — — 1,5 — — 0,8 0,8 — 0,8 — —
L. piniperda Korb. 0,7 — - — — 4,8 — 2,0 — — — — 0,8
L. symmicta (Ach.) Ach. 0,7 0,9 — — — 1,5 5,0 — - — — — —
L. umbrina (Ach.) A. Massal 1,5 0,5 — — 1,5 — — 1,0 *0,8 — 0,8 — —
L. varia (Hoffm.) Ach.** 8,4 4,7 — — 2,9 3,5 6,9 5,7 5,3 0,8 9,1 11,1 20,9
Lecidea erythrophaea Florke ex Sommerf.** 1,8 0,9 - - — 1,5 — — — — 2,6 5,0 5,2
Lecidella euphorea (Florke) Hertel 0,2 — — — — — — — — — — 2,0 —
Lepraria incana (L.) Ach. 9,1 — — — — 10,5 4,0 5,7 14,0 5,0 6,5 15,1 12,7
** - виды, ранее не указываемые для территории Москвы.
206 10. Лихеноиндикация качества воздуха в г. Москве
таблица 10.1 (окончание)
Вид МКАД ожд СК БК ц СЗ С СВ В ЮВ ю ЮЗ 3
Melanelia exasperata (De Not.) Essl.** 0,2 - - - - 1,5 - - - - - - 1,6
M. exasperatula (Nyl.) Essl.** 0,1 — — — — — — — — — 0,8 — —
Opegrapha atra Pers.** 10,0 0,4 — — — 7,0 1,0 4,0 20,0 — 6,5 17,2 19,7
O. rufescens Pers.** 0,8 1,3 — — — - 2,0 1,0 11,0 3,4 — — —
Parmelia sulcata Taylor 38,8 23,6 3,2 — 10,3 53,5 36,6 35,5 55,3 18,4 22,2 45,4 49,2
Parmeliopsis ambigua (Wulfen) Nyl. 2,6 — — — — 12,8 — — 5,9 — — 3,0 4,5
P. hyperopta (Ach.) Arnold 0,2 0,9 — — — — 2,0 — — — - - —
Phaeophyscia ciliata (Hoffin.) Moberg 0,7 — — — — 2,5 — 1,0 — — — — 2,2
P. nigricans (Florke) Moberg ** 1,2 0,9 — — 1,5 — 1,0 2,0 — — 1,6 — 3,0
P. orbicularis (Neck.) Moberg 47,4 38,2 35,5 58,3 41,2 67,5 42,8 50,5 43,5 24,0 36,0 54,5 68,7
Physcia aipolia (Ehrh.ex Humb.) Fiirnr. 0,4 — — — - 1,3 - — - — — 2,0 0,8
P. caesia (Hoffin.) Fiirnr.** 0,1 — — — — — — — — — 0,8 — —
P. stellaris (L.) Nyl. 54,7 21,5 — — 16,2 73,5 31,8 39,0 66,4 26,5 53.0 74,0 83,6
P. tribacia (Ach.) Nyl. 0,8 — — — — 1,5 — — — — 0,8 — 5,2
Physconia grisea (Lam.) Poelt 1,4 1,3 — — 1,5 1,5 4,0 3,0 — — 0,8 — 3,7
Scoliciosporum chlorococcum (Graewe ex 38,4 16,0 — — 5,9 52,5 26,0 35,7 29,6 7,2 34,6 77,7 68,7
Stenh.) Vezda** Tuckermanopsis chlorophylla (Willd.in Humb.) 0,3 — — — — 1,3 1,1 2,0 — — — — —
Hale** T. sepincola (Ehrh.) Hale** 2,6 0,4 — — — 7,4 — 2,0 2,2 — 0,8 3,2 5,5
Vulpicida pinastri (Scop.) J.-E. Mattsson & 7,9 2,6 — — — 8,4 — 6,0 13,5 1,6 2,0 22,2 12,0
MJ.Lai Xanthoria fallax (Hepp) Arnold ** 0,3 — — — — — — — 1,5 — — — 0,8
X. parietina (L.) Th. Fr. 6,6 0,4 — - . - 3,8 - 4,0 14,0 0,8 4,0 7,2 16,6
X. polycarpa Hoffm.) Th. Fr. ex Rieber 0,3 — — — — 1,5 — — — — 0,8 — 1,6
Встречаемость всех видов 67,0 46,8 38,7 58,3 45,6 89,3 55,8 59,8 79,0 31,3 64,8 91,9 84,7
Число видов 47 25 2 1 10 31 18 29 27 16 29 22 32
10. Лихеноиндикация качества воздуха в г. Москве
208
10. Лихеноиндикация качества воздуха в г. Москве
ных площадей жилыми домами, предприятиями, прокладку дорог и другое. Про-
цесс этот продолжается и ныне.1
Обнаружение представителей 20 видов, ранее для этой территории не указы-
ваемых, скорее всего в отношении большинства из них связано с тем, что в отли-
чие от предшественников, я провел специальное изучение распространения ли-
шайников в городе, использовав хорошо зарекомендовавший себя метод растро-
вого картографирования с последовательным обследованием каждого квадрата и
всех пригодных для лишайников местообитаний. Это минимизировало вероят-
ность пропуска растений на местности, хотя полностью его не исключило, по-
скольку внимание исследователя варьирует в зависимости от его физического и
эмоционального состояния в каждый конкретный момент времени. С учетом ска-
занного в настоящее время флору лишайников территории Москвы в пределах
МКАД, видимо, составляют представители не более 55 видов.
Характеристику произошедших за почти 150 лет изменений видового состава
лишайников на нынешней территории Москвы, описанную выше, можно назвать
мониторингом. Но и полученные в ходе исследования данные по распростране-
нию лишайников в городе также являются определенной точкой отсчета для пос-
ледующего мониторинга. При этом они гораздо более точны в сравнении с пре-
дыдущими, поскольку они собраны за довольно короткий промежуток времени
на всей территории и распространение видов зафиксировано на карте.
10.3. ОСОБЕННОСТИ СОВРЕМЕННОГО РАСПРОСТРАНЕНИЯ
ЛИШАЙНИКОВ В ГОРОДЕ
В пределах МКАД лишайники были найдены в 609 квадратах, что составляет
примерно 67% от их общего числа (см. табл. 10.1). Встречаемость представите-
лей конкретных видов гораздо меньше: Physcia stellaris зафиксирована в 497 квад-
ратах, a Cladonia cariosa (на почве), Everniaprunastri, Graphis scripta и ряд дру-
гих на деревьях и кустарниках - всего лишь в 1-2 квадратах (рис. 10.1- 10.12).
Среднее число видов лишайников в квадрате несколько более трех при варьиро-
вании этого показателя от полного их отсутствия до 18 видов на деревьях Филев-
ского парка на склоне правого берега Москвы-реки, однако треть территории го-
рода приходится на квадраты с 1-3 видами (рис. 10.13).
По мере удаления от центра города видовое разнообразие лишайников увели-
чивается. Так, в пределах Бульварного полукольца встречен представитель одно-
1 В Красную книгу города Москвы (2001) включены виды Bryoria fuscescens, Cladonia arbuscula,
C. botrytes, C. cariosa, Cfurcata, C. macilenta, C. stellaris, C. turgida, Hypogymnia tubulosa, Lecanora
muralis, Cetraria islandica, Evernia mesomorpha, E. prunastri, Parmeliopsis ambigua, Usnea hirta,
Peltigera canina, P rufescens, Ramalina farinacea. Представителей ряда видов из этого списка я на
территории Москвы в начале 1990-х не встретил.
10. Лихеноиндикация качества воздуха в г. Москве
209
го вида (Phaeophyscia orbicularis), в пределах Садового кольца - 2 вида (плюс
Parmelia sulcata), а в пределах Московской окружной железной дороги - 25 видов
(см. табл. 10.1). В то же время на некоторых окраинах города (Капотня, Братеево,
Дегунино) лишайники не обнаружены. В северной и южной половинах города
отмечено по 40 видов лишайников, встреченных в 68 и 66% квадратов соответ-
ственно. Более заметны различия между западной и восточной частями города -
42 вида в 78% квадратов в западной и 33 вида в 56% квадратов в восточной.
В северо-западном секторе города обнаружено 34 вида, встречаемость 70%, в юго-
западном секторе - 32 вида, встречаемость 86%; по 29 видов в северо-восточном
и юго-восточном секторах, встречаемость 65% и 49%, соответственно. Среди ад-
министративных округов города наименьшее число видов лишайников встрече-
но в Центральном (10 видов), Юго-Восточном (16) и Северном (18), тогда как в
Западном (32) и Северо-Западном (31) число видов в 2-3 раза выше (см. табл.
10.1). При этом следует отметить, что показатель видового разнообразия Цент-
рального округа “улучшен” территорией Лужников. Таким образом, по состоя-
нию на начало 1990-х годов, юго-восточная часть города (от шоссе Энтузиастов
до Варшавского), а также территория в пределах Садового кольца и примыкаю-
щих к нему кварталов наименее благоприятны для поселения лишайников.
Наиболее пригодные для лишайников в городе территории - сохранившиеся
крупные лесные массивы. Серебряный бор с Филевским парком, Лосиный ост-
ров, Битцевский лесопарк выделяются относительно большим видовым разнооб-
разие растущих на деревьях лишайников (Бязров, 1994). Эти естественные лес-
ные массивы можно назвать своеобразными рефугиумами эпифитной лихеноби-
оты, где еще в особых микроместообитаниях существуют условия для сохранения
в черте города представителей ряда видов лишайников: Graphis scripta, Evernia
prunastri, Melanelia exasperata и некоторых других (см. рис. 10.2-10.3). Отсюда
же, в случае улучшения состояния воздушного бассейна в городе, лишайники могут
продвинуться в районы, расположенные ближе к центру города. Такого рода под-
вижки были зафиксированы в Лондоне и Париже уже через несколько лет после
введение в действие законов, направленных на улучшение состояния воздушного
бассейна (Hawksworth, McManus, 1989; Letrouit-Galinou etal., 1992).
10.4. КАРТОСХЕМЫ ЗАГРЯЗНЕНИЯ ВОЗДУХА В ГОРОДЕ,
СОСТАВЛЕННЫЕ НА ОСНОВЕ ОСОБЕННОСТЕЙ
РАСПРОСТРАНЕНИЯ ЭПИФИТНЫХ ЛИШАЙНИКОВ
Результаты картографирования распространения эпифитных лишайников по
квадратам 1 х 1 км стали основой для составления карт, отражающих современ-
ную экологическую ситуацию на территории города, главным образом - качество
воздушного бассейна. Территория Москвы в этом отношении может анализиро-
210
10. Лихеноиндикация качества воздуха в г. Москве
ваться как на региональном, так и на локальном уровнях. В первом случае Моск-
ва, с одной стороны, выступает как единый крупнейший источник загрязнения
для прилегающих к ней районов, воздействие которого простирается на несколь-
ко десятков километров от города в толще воздуха до 200 м от поверхности (Обу-
хов, 1982 ), с другой - ее обширная площадь подвержена влиянию трансгранич-
ного переноса, главным образом, с запада и северо-запада. В локальном аспекте
на территории Москвы неравномерно рассредоточено множество источников заг-
рязнения в виде промышленных объектов различного назначения, автотранспор-
та и другого, с той или иной степенью нагрузки на окружающую их городскую
территорию. Поскольку расстояния между этими источниками часто невелико,
во многих районах города наблюдается перекрестное воздействие нескольких
источников. Представленные здесь материалы касаются локального уровня про-
блемы. При этом считаю необходимым оговорить, что целями проведенного рас-
трового картографирования распространения лишайников были изучение совре-
менного состава лихенофлоры и особенностей ее размещения на обследованной
территории, а не составление карты загрязнения воздуха по признакам лихено-
биоты. В последнем случае и методы сбора материала были бы ориентированы
на решение этой задачи. Идея преобразовать полученные данные в карту, отража-
ющую состояние воздуха, появилась позже, в ходе обсуждения результатов рас-
трового картографирования на нескольких научных форумах.
Первым вариантом такого преобразования стала схема (см. рис. 10.14), со-
ставленная на основе сведений о числе видов эпифитных лишайников в квадра-
те-растре (Бязров, 1996а). Основанием для такой модификации были многочис-
ленные свидетельства о прямой связи снижения видового разнообразия эпифит-
ных лишайников с увеличением уровня загрязнения воздуха (Wetmore, 1988b).
Схема показывает, что “лишайниковая пустыня” простирается почти сплошь с
северо-северо-запада через центр, где она заметно расширена, до крайнего юго-
востока города. На нее приходится более 40% территории города. Чуть более 38%
площади занято участками с крайне критическими условиями для лишайников,
17% - с критическими и лишь 4,5% приходится на участки с относительно уме-
ренными условиями для лишайников, где число видов в квадрате 11-18. После-
дние сосредоточены главным образом на северо-востоке (Лосиный остров), запа-
де (Серебряный бор, Фили) и юго-западе города.
Следующим шагом преобразования результатов растрового картографирова-
ния распространения эпифитных лишайников стала схема, составленная с уче-
том отношения отдельных видов лишайников к загрязнению (Бязров и др., 1997;
Максимова, Бязров, 2000). Известно, что реакция представителей разных видов
на такое воздействие далеко не одинакова. Для некоторых регионов разработаны
шкалы с разным числом классов, ранжирующих лишайники по чувствительнос-
ти к загрязнению среды (см. гл. 4). Имеются обобщения опубликованных сведе-
ний по этой проблеме (Инсарова, Инсаров, 1989; Wirth, 1991). Как правило, юз-
10. Лихеноиндикация качества воздуха в г. Москве
211
личественные сведения о концентрации загрязнителей в среде, характеризующие
шаги предлагаемых шкал, не указываются. Видимо, разработать шкалу чувстви-
тельности лишайников, ориентируясь на конкретные величины концентрации
разных загрязнителей в среде, пока практически невозможно. С одной стороны,
действие загрязнителя на организм зависит от других свойств среды, прежде все-
го влажности и температуры. С другой - загрязнителей чрезвычайно много, и
наличие в среде других токсикантов может как усиливать негативное воздействие
данного загрязнителя, так и ослаблять его. Поэтому при анализе видового соста-
ва встреченных на территории Москвы лишайников в отношении их чувстви-
тельности к загрязнению я ориентировался на качественную пяти-ранговую шка-
лу, базируясь как на материалах уже цитированных обобщений, так и опыте мно-
голетней работы автора в разных регионах. Первый класс этой шкалы составляют
виды, толерантные к высокому уровню загрязнения. Это Lecanora hagenii, Lepraria
incana, Phaeophyscia orbicularis, Physcia stellaris, Scoliciosporum chlorococcum,
Xanthoriaparietina. Представители этих видов в Москве встречаются относительно
часто (см. табл. 10.1) и во всех ее частях (см. рис. 10.1, 10.3, 10.11, 10.12). Ко
второму классу принадлежат виды, выдерживающие довольно высокий уровень
загрязнения. Это Caloplaca cerina, Candelariella vitellina, Lecanora varia, Parmelia
sulcata, Phaeophyscia nigricans, Physcia tribacia, Physconia grisea, Xanthoria
polycarpa. Они встречаются в городе реже (табл. 10.1), но также во всех его час-
тях (рис. 10.4, 10.5, 10.7-10.10). Третий класс шкалы образуют виды, выдержи-
вающие средний, достаточно ощутимый уровень загрязнения. Это Caloplaca
holocarpa, Cladonia coniocraea, C. fimbriata, Hypocenomyce scalaris, Hypogymnia
physodes, Lecanora allophana, L. piniperda, Lecidea erythrophaea, Lecidella euphorea,
Melanelia exasperatula, Phaeophyscia ciliata, Physcia aipolia, P. caesia, Xanthoria
fallax. Их представители встречаются в Москве редко (см. табл. 10.1) и главным
образом в лесопарках (см. рис. 10.2-10.4, 10.6, 10.8-10.10). Четвертый класс со-
ставляют виды, выдерживающие умеренное загрязнение. Это Cladonia caespitica,
С. chlorophaea, С. digitata, С. ochrochlora, Evernia prunastri, Graphis scripta,
Lecanora symmicta, Melanelia exasperata, Opegrapha atra, O. rufescens, Parmeliopsis
ambigua, P. hyperopia, Tuckermanopsis chlorophylla, T. sepincola, Vulpicidapinastri.
Встречаются они в городе, за некоторым исключением, единично (см. табл. 10.1)
и главным образом в лесопарках (см. рис. 10.2, 10.3, 10.5-10.7, 10.10, 10.11).
Лишайники, представляющие пятый класс наиболее чувствительных к загряз-
нению видов, на обследованной территории города в конце 1980-х - начале
1990-х гг. автором не обнаружены.
Схема деления города по степени загрязнения была составлена на основе при-
сутствия в квадрате лишайников определенного класса чувствительности, или
толерантности (рис. 10.15). “Лишайниковая пустыня”, т. е. участки, где лишай-
ники отсутствуют, отражает районы города с опасно высоким уровнем загрязне-
ния. Это почти 33% площади Москвы. Участки, где представлены исключитель-
212
10. Лихеноиндикация качества воздуха в г. Москве
но лишайники первого класса, т.е. наиболее толерантные к загрязнению виды,
отражают очень высокий уровень загрязнения. Они занимают около 25% терри-
тории города, среднее число видов в квадрате 2.5. Участки, где наряду с предста-
вителями первого класса отмечены лишайники также и второго класса чувстви-
тельности, но отсутствуют виды третьего и четвертого, отражают районы с высо-
ким уровнем загрязнения. Это почти 13% площади города, среднее число видов в
квадрате здесь около 4. Следующий выдел на схеме включает квадраты, в кото-
рых отмечены виды третьего класса чувствительности вместе с представителями
первого и второго, но отсутствуют лишайники четвертого класса. Он отражает
участки со средним уровнем загрязнения. На них приходится также около 13%
площади при среднем числе видов в квадрате 5.4. Последний выдел включает
квадраты с представителями наиболее чувствительных к загрязнению среди об-
наруженных в городе видов лишайников. Он отражает участки с умеренным заг-
рязнением. На них приходится более 16% территории города, среднее число ви-
дов в квадрате 9. Поскольку на обследованной территории не обнаружено наибо-
лее чувствительных к загрязнению видов лишайников, можно достаточно уверенно
констатировать, что в городе нет участков с нормальным чистым воздухом, по-
этому при его характеристике следует искать эпитеты не для степени чистоты, а
для степени загрязнения. Этот вывод подтверждают и тревожные статистические
данные о повышенном уровне заболевания у населения города органов дыхания
и чувств, а также стабильно высокая заболеваемость раком, что связывают с силь-
ным загрязнением среды (Аксенова и др., 1997).
Составленные и показанные выше оценочные картосхемы состояния воздуха
основываются лишь на наличии либо оговоренного числа видов, либо присут-
ствия одного или более представителей видов определенного класса толерантно-
сти и не учитывают степень их развития, особенности распространения и эколо-
гическое своеобразие территории. Для устранения названных недостатков автор
(Бязров, 1998а, 1999а) предложил преобразование растровых карт распростране-
ния эпифитных лишайников на конкретной территории на основе индекса разви-
тия эпифитных лишайников (ИРЭЛ-IDEL), определяемого для каждого квадрата
(см. п. 5.3.4). Чем выше значение ИРЭЛ, тем более благоприятна экологическая
ситуация на участке. Этот индекс учитывает экологическую неравноценность тер-
ритории (рис. 10.16).
Значение ИРЭЛ растра варьирует в городе от 0 до 182 при средней величине
по отношению ко всей территории 31 (рис. 10.17). Для сравнения, приближенное
значение ИРЭЛ, определенное по результатам предшествующих исследований
автора (Бязров, 19696, в), на территории в 25-30 км на юг от МКАД между Ка-
лужским и Киевским шоссе, составляет 100-300, а для слабо загрязненной или
“чистой” части Демидовского района Смоленской области - 250-550. Получен-
ные для Москвы интегрированные данные стали основой для создания картосхе-
мы экологической ситуации в городе (рис. 10.18). Почти 40% территории города
10. Лихеноиндикация качества воздуха в г. Москве
213
в пределах МКАД приходится на участки с очень высоким уровнем загрязнения
(ИРЭЛ = 0-10). Эпифитные лишайники на большей части этого выдела не обна-
ружены или же встречаются слаборазвитые одиночные экземпляры Phaeophyscia
orbicularis или Physcia stellaris. Фактически это “лишайниковая пустыня” в горо-
де, которая широкой полосой простирается с северо-запада на юго-восток с неко-
торым расширением в центре города. Образована она главным образом террито-
риями, занимаемыми крупными промзонами или находящимися под влиянием
последних. Многие из них расположены в долине р. Москвы от Бережковской
набережной до МКАД, причем непосредственно на берегах реки чаще размеще-
ны наиболее крупные и экологически опасные объекты, такие как “Павелецкий
вокзал”, “ЗИЛ”, “Южный порт”, “Курьяново”, “Люблино-Перерва”, “Верхние
Котлы”, “Чагино-Капотня” и другие, что создает напряженную экологическую
ситуацию на обоих берегах р. Москвы и дает основание задать вопрос городским
архитекторам и властям, доколе при проектировании городской застройки будут
игнорироваться эстетические преимущества имеющихся в городе водных магис-
тралей - рр. Москва, Яуза, Сетунь и других, ныне фактически превращенных в
клоаки.
Выдел “лишайниковой пустыни” разделяется на две неравные части - северо-
северо-запад и центр с юго-востоком. В северной части “лишайниковую пусты-
ню” формируют выбросы промзон “Коровино”, “Вагоноремонтная”, “Алтуфье-
во”, “Владыкино”, “Лихоборы-Дегунино”, “Бескудниково” и других с ТЭЦ-21,
рыбоперерабатывающим комплексом, чугуно-литейным заводом им. Войкова,
стройкомбинатом. Разовые замеры показали здесь превышение в воздухе ПДК в
5 и более раз по СО, ЬЮ2, аммиаку и другим загрязнителям. В центре города
формирование “лишайниковой пустыни” определяется как наличием промзон
“Огородный проезд”, “ул. Магистральная”, “Силикатный проезд”, “Боткинский
проезд”, “ул. Правды”, “ул. Грузинский вал” , “Павелецкий вокзал”, товарными
станциями и другими, так и значительной нагрузкой автомобильного транспор-
та. В центре - более 80%, а за пределами Садового кольца - 47% магистралей
исчерпали свою пропускную способность. Практически исчерпана пропускная
способность Садового кольца. Коэффициент загрузки транспортом таких круп-
ных узлов центра Москвы как Кудринская, Смоленская, Таганские площади, пло-
щади Белорусского и Рижского вокзалов превосходит в 1,2-1,3 раза допустимую
норму. Снижение скорости движения автомобилей приводит к резкому увеличе-
нию расхода топлива и, соответственно, увеличению объемов выбросов. В Цент-
ральном административном округе концентрации оксидов азота составляют в
районе трех вокзалов (Комсомольская площадь) 10-20 ПДК (Беккер, 1989), а на
Садовом кольце в районе Крымского вала - 7 ПДК. Юго-восточная часть выдела
простирается широкой полосой от Садового кольца до МКАД. Эта территория
находится под влиянием промзон “ЗИЛ”, “Волгоградский проспект”, “Южный
порт”, “Карачарово”, “Грайвороново”, “Люблино-Перерва”, “Верхние Котлы”,
214
10. Лихеноиндикация качества воздуха в г. Москве
0 0 OS
0 0 OS OS OS
S S s OS os OS OS OS
OS OS OS OS 0 OS OS OS os
OS OS OS S OS о 0 OS OS OS OS
s OS OS 0 OS s OS OS OS OS OS OS
s OS OS OS OS OS OS OS OS OS OS OS OS OS
OS OS S OS os s OS OS о OS OS OS S OS S S S
OS OS OS OS OS s о OS о OS OS OS s OS OS s s OS
S s s S s s о о OS 0 OS OS s OS S OS s S
OS OS S OS OS OS о OS OS OS OS OS OS s OS OS OS OS OS
s OS s s OS о OS OS OS 0 OS OS OS s OS s OS
OS OS OS OS 0 s о о 0 OS OS 0 s OS OS s s OS OS OS OS OS OS
OS OS OS OS 0 OS OS os OS 0 OS OS OS s OS OS OS OS OS
S OS OS OS OS 0 OS О s s OS 0 о OS OS OS OS OS OS
OS OS OS OS OS OS 0 OS 0 OS OS OS OS OS OS
OS OS OS OS OS OS os OS OS 0 о 0 OS OS OS OS OS OS
OS s OS OS OS OS OS OS OS о 0 0 s OS OS OS
OS OS OS OS OS OS OS OS OS OS os s 0 о о 0 о OS OS OS
OS OS OS OS OS OS OS OS OS о о о OS OS s OS
OS OS OS S OS OS S OS OS S OS о о о OS OS OS s s
OS OS OS OS OS s OS s OS OS OS о OS о о OS OS s s
s OS OS os S s OS OS OS OS OS OS о о О OS OS OS s s
OS OS OS os OS OS OS s OS OS О os OS s о OS OS OS s
OS OS S OS OS s OS OS OS OS OS OS OS
OS OS OS OS OS s OS S OS s OS OS OS OS OS OS OS OS s
OS s s s OS OS OS OS s s OS OS OS OS 0 s
OS OS os OS OS OS OS OS OS OS OS OS OS s
OS OS OS OS s OS OS OS s s S S s
s s S S OS S OS OS OS OS OS OS OS s s
OS OS OS OS OS OS OS s s OS OS s s
s OS s OS OS OS s OS s OS OS s OS
OS OS о OS s OS OS OS s s s S
S OS OS s s OS OS OS OS OS OS s S 0
OS OS s s OS OS OS OS s S OS OS OS
S OS S s s OS s OS S S
s OS OS OS OS OS OS
OS s OS OS os OS s OS
Рис. 10.1. Местонахождения Phaeophyscia orbicularis (о) и Physcia stellaris (s)
Сторона квадрата на рис. 10.1-10.18 -1 км
10. Лихеноиндикация качества воздуха в г. Москве
215
Рис. 10.2. Местонахождения Cladonia caespitica (с), Cladonia digitata (d), Evernia prunastri (e),
Graphis scripta (g), Lecanora allophana (a), Lecanora piniperda (p), Lecanora symmicta (y), Lecanora
varia (v), Melanelia exasperatula (x), Opegrapha rufescens (r), Physcia aipolia (h), Physcia caesia (z).
Tuckermanopsis chlorophylla (t)
216
10. Лихеноиндикация качества воздуха в г. Москве
s
s s s s
s s s s s s
s s s s s S s
s s s s s s s s
s s s s s s s s s s
s s s « j s s s s s s s s
s s s s s « s s s s s fs
s s s s s s s s s s s s s fs s
s s s s s s s s s s s s s s
s s s s hs s s s s s s s s s s s
s s s s s s s s s
bs s s s s s
s bs s s s h s s
s s s s s s s s
s s s s s
s s s s s s
s bs bs s s
s s s s ms s s s s
s ms s s s s s
s s s s bs s s s
s s s s s s s s s bs s s
s s s s s s s bs s
s s s s s s s s s s s s s
s s s s s s
s s s s bs s s s s s s s s s s
s s s s s s s s s s s s s s s
s s s s s s s s s s s s
s bs s s s s s s s s
s s s s s s s s s s s
s s s s s s s s s s s s
s s s s s s s s
s s s s bs s s
s s s s bs s s
s s s bs bs s s s s s s
s s s es es s s s s s s
bs bs s be s
s s s s s s
Рис. 10.3. Местонахождения Lecidea erythrophaea (b), Lecidella euphorea (e), Melanelia exasperata
(m), Parmeliopsis hyperopia (h), Scoliciosporum chlorococcum (s) и Xanthoria fallax (f)
10. Лихеноиндикация качества воздуха в г. Москве
217
hp hp hp hp hp
P hp P hp hp hp
hp hp
hp P hp hp hp hp P
hp hp hp P hp hp P P P hp hp
hp hp P P P P P hp hp hp
hp P hp P P P hp hp hp hp
hp hp P P hp hp hp P hp hp hp hp hp
P P P hp P hp hp hp hp hp hp hp hp hp hp hp
hp hp P hp P P hp hp hp hp P P hp hp hp hp hp hp
hp hp P hp hp P P hp hp hp hp P hp hp hp P hp
hp P P hp hp hp hp hp hp P hp hp
h h hp P hp P hp hp hp Р
hp hp hp hp hp P hp hp hp P P hp hp P P
hp hp hp hp hp hp P h hp P hp hp P hp P P
h hp hp h hp hp hp hp hp P
h hp hp P h hp P h P hp hp hp
hp hp hp P hp hp hp hp hp hp P
h hp hp hp hp hp hp hp P
hp hp P P hp P P P P hp
P hp hp hp P P P hp P hp hp
h P hp P P P hp hp
P hp P P hp
P hp hp P P P P P
P hp P hp P hp P
P P P P P hp hp P P
P P P P P hp hp
hp P P P hp hp P hp hp hp
hp hp P hp hp hp h
hp hp P P
hp hp hp hp hp hp hp hp
P hp hp hp hp hp P P
hp hp hp hp hp hp
hp hp hp hp P P hp hp
P hp hp P hp hp hp
hp P hp hp hp P P P P hp P hp hp
hp hp hp P P hp hp P
P P P hp P
Рис. 10.4. Местонахождения Hypogymnia physodes (h) и Parmelia sulcata (p)
Рис. 10.5. Местонахождения Caloplaca cerina (с) и Cladonia chlorophaea (h)
10. Лихеноиндикация качества воздуха в г. Москве
219
Рис. 10.6. Местонахождения Caloplaca holocarpa (с),
Phaeophyscia ciliata (р) и Tuckermanopsis sepincola (s)
220
10. Лихеноиндикация качества воздуха в г. Москве
Рис. 10.7. Местонахождения Candelariella vitellina (с),
Parmeliopsis ambigua (а) и Vulpicida pinastri (р)
10. Лихеноиндикация качества воздуха в г. Москве
221
Рис. 10.8. Местонахождения Cladonia coniocraea (с) и Physcia tribacia (р)
Рис.10.9. Местонахождения Cladonia fimbriata (f) и Physconia grisea (g)
10. Лихеноиндикация качества воздуха в г. Москве
223
Рис. 10.10. Местонахождения Cladonia ochrochlora (с), Нуросепотусе scalaris (h),
Phaeophyscia nigricans (n) и Xanthoria polycarpa (x)
224
10. Лихеноиндикация качества воздуха в г. Москве
0 h
0
h h
h h 0
h о 0 О
h h 0 0 О ho
h h 0 0 0 ho ho о
0 0 h о о ho о h
о О о о о О о
h h 0 о о
h h ho
о ho
h
О о 0
ho ho 0
ho ho ho h
о о о 0 ho 0
ho ho 0 ho
ho ho ho h
ho h
h ho
h h h h h
h
h
h
ho h ho h
ho h ho
ho h ho
ho ho 0 0 ho h
ho 0 h
ho ho ho о 0 0
о 0 0 0 о
h
0 0 h h 0 О
ho h
ho h
Рис.10.11. Местонахождения Lecanora hagenii (h) и Oprgrapha atra (о)
10. Лихеноиндикация качества воздуха в г. Москве
225
Рис.10.12. Местонахождения Lepraria incana (г) и Xanthoria parietina (х)
226
10. Лихеноиндикация качества воздуха в г. Москве
Рис. 10.13. Число видов эпифитных лишайников в квадрате (вслед за обозначением админист-
ративного округа в скобках среднее число видов в квадрате; в Центральном округе среднее число
видов в квадрате 0,8; для всей территории города среднее число видов в квадрате - 3,2)
10. Лихеноиндикация качества воздуха в г. Москве
227
Рис.10.14. Схема деления территории Москвы по числу видов эпифитных лишайников на
км2: 1 - “лишайниковая пустыня” (0-1 видов), катастрофически опасные участки; 2 - критиче-
и опасные участки (2-5 видов); 3 - крайне опасные участки (6-10 видов); 4 - опасные участки
10 видов)
228
10. Лихеноиндикация качества воздуха в г. Москве
Рис.10.15. Схема деления территории Москвы по присутствию в квадрате-растре лишайников
определенного класса чувствительности: 1 - “лишайниковая пустыня”, катастрофически опасные
участки; 2 - критически опасные участки; 3 - крайне опасные участки; 4 - опасные участки
10. Лихеноиндикация качества воздуха в г. Москве
229
Рис. 10.16. Типы экосистем Москвы: 3 - квадрат включает сообщества лесопарков и лесных
культур старше 60 лет, сравнительно естественные экосистемы долин рек; 4 - квадрат включает
городские парки и сады, территории кладбищ, водоемы, овраги при отсутствии названных выше
сообществ; 5 - квадрат включает только застроенные территории и крупные магистрали, зеленые
насаждения представлены скверами, насаждениями вдоль магистралей и внутри дворов при отсут-
ствии названных выше сообществ
230
10. Лихеноиндикация качества воздуха в г. Москве
С(25) 65 10 я. 88 50 66
0 25 84 38 36 10 10 62 СВ(27)
0 0 25 0 20 0 25 0 0 64 64 38 30 30
60 48 66 66 0 0 0 0 0 0 10 0 0 0 18 0 84 82 72
€3(40) 39 50 48 32 46 44 0 0 0 0 0 10 0 0 0 40 35 35 0 13 н 95
62 47 21 28 0 0 23 40 24 23 0 0 0 0 0 28 20 0 48 31 84 92 12 4
60 23 0 28 30 44 22 0 0 0 0 0 0 0 28 28 11 8 13 50 35 82 95 69 11 7
74 18 8 22 28 16 35 70 0 0 0 10 0 44 68 34 0 0 66 24 92 11 11 12 4 12 7 В(42)
26 22 36 30 36 18 10 68 13 0 0 0 40 58 36 0 0 29 60 63 10 _1 71 10 9 10 Q 80 11 9
8 6 8 22 53 53 0 15 20 78 20 0 50 50 34 18 0 31 42 23 48 96 10 87 92 96 76
65 50 18 0 30 56 52 23 0 38 12 11 35 0 18 28 28 0 58 53 18 39 71 28 66 12 * 31 0 58
48 35 44 8 28 48 72 0 0 23 13 12 22 23 0 0 0 52 60 28 0 23 0 0 0 15 0 23 0 55
23 30 58 0 18 10 25 0 35 23 10 6 35 35 0 0 0 10 46 48 60 0 6 18 18 35 23 23 23 35
10 1 48 14 1 69 69 30 0 10 13 86 0 55 36 0 10 0 0 0 36 48 28 0 0 0 26 36 35 35 23 21
96 95 93 76 80 40 0 48 23 60 0 25 30 0 0 10 10 0 0 0 0 0 0 0 60 66 37 44 23 23
10 з 68 96 66 94 18 8 0 0 0 0 0 30 0 10 0 0 0 0 0 0 0 0 0 54 63 52 36 23 10
88 11 4 53 20 22 22 38 48 28 0 0 14 0 0 0 13 10 0 0 0 38 0 0 0 34 23 29 52 53 36
88 11 12 28 22 36 46 84 18 0 0 14 0 0 10 0 0 10 0 0 38 0 0 0 0 0 0 66 56 26
63 88 55 63 40 36 15 о 10 8 33 30 18 22 18 V 10 10 0 8 8 8 0 0 0 0 0 0 0 23 23 35
12 85 0 48 46 96 28 88 75 35 0 0 0 10 0 0 0 0 10 0 8 8 0 0 0 0 34 38 10 18
11 0 0 0 11 в 12 25 14 26 18 60 26 18 46 8 0 0 0 30 0 8 0 0 0 0 22 68 36 8 10
50 40 36 28 36 20 34 88 18 62 48 56 40 28 8 0 0 0 10 0 0 0 0 0 0 0 98 23 33 10
38 20 70 0 32 18 18 36 38 78 40 18 28 10 0 0 10 10 0 0 0 0 0 0 18 23 23 10 10
55 И 4 11 п 0 78 38 33 0 82 26 66 30 0 0 0 0 30 48 38 8 0 8 0 0 0 26 18 35 10
48 64 56 18 0 0 0 0 38 10 38 35 0 0 0 0 0 0 0 0 0 0 36 26 34 23 29
38 38 0 0 36 24 50 18 32 23 38 0 40 28 0 28 28 28 0 0 0 0 24 46 72 53 21
3(49) 84 8 38 16 48 36 58 50 23 28 24 30 0 30 0 28 0 0 0 0 С 0 18 63 53
20 18 42 0 34 36 10 98 35 28 38 0 40 0 0 82 0 0 0 0 0 0 0 0 45 37 23
17 13 0 36 10 25 23 46 72 20 0 0 0 0 48 0 0 0 0 0 0 0 0 36 72 84
30 13 23 23 30 10 20 48 32 30 75 38 0 28 48 0 0 0 0 0 0 0 0 0 0 10
11 7 90 64 64 34 20 85 64 64 30 20 74 28 26 8 18 0 0 0 0 0 0 0 0
20 34 96 18 64 58 66 85 36 25 63 10 23 23 8 50 0 0 0 0 0 0 0
0 11 R 69 55 52 74 84 16 38 50 18 8 0 8 8 8 0 0 0 0 0 ЮВ(10)
8 44 55 25 74 84 20 70 35 26 28 18 46 25 10 0 0 0 46
0 32 40 25 76 74 8 18 18 18 0 0 40 62 23 35 83 46
Ю3(46) 20 47 31 90 92 42 23 0 60 0 38 52 42 28 48 65
0 85 84 79 54 0 40 0 38 80 86 35
50 25 65 0 23 26 84 14 4 40
Ю(23)
Рис. 10.17. Распределение величин индекса развития эпифитных лишайников (ИРЭЛ) в Моск-
ве. В скобках за обозначением округа приведена средняя для округа величина индекса одного рас-
тра; в Центральном округе - 7, для всей территории города - 31; утолщенные линии - границы
трансект, предлагаемых для постоянных наблюдений через каждые 5 лет)
10. Лихеноиндикация качества воздуха в г. Москве
231
Рис. 10.18. Схема деления территории Москвы по величине индекса развития эпифитных
лишайников (ИРЭЛ) в квадрате: 1 - катастрофически опасные для лишайников участки (ИРЭЛ =
0-10); 2 - критически опасные участки (ИРЭЛ = 11-30); 3 - крайне опасные участки (ИРЭЛ =
31-70); 4- опасные участки (ИРЭЛ = 71-150)
232
10. Лихеноиндикация качества воздуха в г. Москве
“Серп и Молот”, “Варшавское шоссе”, “Бирюлево”, “Чертаново”, “Коломенское”,
“Ленино”, “Борисово”, “Перово”, “Соколиная гора”, “Прожектор”, в том числе
Люблинской и Курьяновской станций аэрации, Московского нефтеперерабатыва-
ющего и Московского коксогазового заводов. Здесь наблюдаются высокие (>5 ПДК)
разовые уровни загрязнения по NO2, аммиаку, фенолу, СО. Загрязнение снега и
почв пылью и тяжелыми металлами характеризуется как “максимальное”, а эко-
логическая ситуация по загрязнению почвы для ряда участков оценена как “эко-
логическое бедствие”(Соколов, Астрахан, 1993). В районе ЗИЛ - Волгоградский
проспект загрязнение воздуха пылью превышает ПДК более чем в два раза (Бек-
кер, 1989). Накопление вредных примесей в воздухе усугубляется наличием здесь
пониженной широкой поймы р. Москвы, что способствует возникновению зас-
тойного режима атмосферы.
Формирование более мелких фрагментов “лишайниковой пустыни” на терри-
тории города также связано с отдельными промзонами и наиболее загруженными
магистралями. В северо-западной части города их расположение коррелирует с
промзонами “Трикотажная”, “Братцево”; на западе - с промзонами “Строгино”,
“Октябрьское поле”, “Кунцево”; на юго-западе - с промзоной “Очаково”; на севе-
ро-востоке - с промзонами “Северянин”, “Осташковская ул.”, “Свиблово”, “Ка-
лошино”, с территориями ТЭЦ, пересечениями ряда крупных автомагистралей с
МКАД.
Данных инструментальных измерений концентрации загрязнителей в возду-
хе автор не имеет, однако по аналогии с подобными наблюдениями в сходном по
размерам и численности населения городе Париже (Deruelle, Guilloux, 1993), с
поправкой на географическое положение г. Москвы, можно полагать, что соче-
танное загрязнение в пределах “лишайниковой пустыни” соответствует воздей-
ствию более 180 мкг/м3 SO2
Выдел картосхемы с высоким уровнем загрязнения (ИРЭЛ =11-30) занимает
около 21% территории города. Вероятно, концентрация загрязнителей в воздухе
здесь соответствует воздействию 130-180 мкг/м3 SO2 Эпифитные лишайники
встречаются во всех квадратах, но их трудно обнаружить, и число видов невели-
ко (наряду с двумя названными выше здесь также можно встретить слоевища
Parmelia sulcata, Scoliciosporum chlorococcum и другие). Основная часть этого
выдела примыкает к “лишайниковой пустыне” в основном с запада, частично - с
востока, а также встречается отдельными участками разного размера на всей об-
следованной территории. На формирование выдела влияют как крупные промзо-
ны с обширной сферой воздействия, так и мелкие производства, автохозяйства и
котельные. Наличие здесь таких крупных автомагистралей как Кутузовский и
Ленинский проспекты, Профсоюзная улица, Можайское, Волоколамское, Ленин-
градское, Ярославское, Щелковское шоссе, Волгоградский и Рязанский проспек-
ты также оказывает значительное воздействие на экологическую ситуацию этих
районов города.
10. Лихеноиндикация качества воздуха в г. Москве
233
На участки с умеренно высоким уровнем загрязнения (ИРЭЛ= 31-70) прихо-
дится почти 27% территории города. Вероятно, концентрация загрязнителей в
воздухе здесь соответствует воздействию 80-130 мкг/м3 SO2. Лишайники обнару-
жены во всех растрах с числом видов в квадрате до 8. К упомянутым выше добав-
ляются виды Lecanora, Opegrapha, Hypogymnia physodes и другие. Представите-
ли массовых в городе видов обнаруживаются сравнительно легко, поскольку встре-
чаются относительно часто и достаточно заметны на стволах деревьев.
Сосредоточены такие участки ближе к МКАД практически во всех частях города.
Как правило, это либо территории, занимаемые лесопарками и парками (Кузь-
минский, Кусковский, Измайловский, Сокольники, Останкино, Царицыно и дру-
гие), либо относительно недавно построенные жилые массивы (Ясенево, Теплый
Стан, Строгино и другие).
Участки со средним уровнем загрязнения (ИРЭЛ = 71-150) занимают в Мос-
кве около 13% территории (к этому выделу отнесены и два квадрата, включаю-
щие Востряковское кладбище, с максимальными на обследованной местности
величинами ИРЭЛ, превышающими 150-176 и 182). Вероятно, концентрация заг-
рязнителей в воздухе здесь соответствует воздействию 60-80 мкг/м3 БО2.Число
видов эпифитных лишайников в квадрате здесь достигает 18, на участках выдела
встречаются представители большинства обнаруженных в Москве видов, а са-
мые массовые из них достаточно заметны на стволах деревьев. Самые крупные
массивы выдела приурочены к Лосиноостровскому, Серебряноборскому, Битцев-
скому лесопаркам. Филевский парк и Лесная опытная дача ТСХА также выделя-
ются развитием эпифитных лишайников.
Сравнение административных округов по среднему значению ИРЭЛ в одном
квадрате (см. рис. 10.17) показывает, что наихудшие экологические условия - в
Центральном округе (среднее значение ИРЭЛ здесь 7 ), тогда как в Западном ок-
руге эта величина равна 49 и превышает среднюю для Москвы (31). В последнем
округе находятся и квадраты с максимальными для Москвы значениями индекса.
По средней величине ИРЭЛ в одном квадрате для округа можно разделить 9 ад-
министративных округов Москвы натри группы: 1) округа с опасно неблагопри-
ятной экологической ситуацией (средняя величина ИРЭЛ в одном квадрате = 7-
10) - Центральный и Юго-Восточный; 2) округа с неблагоприятной экологичес-
кой ситуацией (средняя величина ИРЭЛ = 23-27) - Южный, Северный,
Северо-Восточный; 3) округа с умеренно неблагоприятной экологической ситуа-
цией (средняя величина ИРЭЛ = 40-49) - Западный, Юго-Западный, Восточный,
Северо-Западный.
Опасно неблагоприятная экологическая ситуация в Центральном округе
определяется прежде всего сильнейшим воздействием автотранспорта как по лу-
чевым (улицы Тверская, Новый Арбат, Бол.Якиманка, Бол.Полянка, Люсиновс-
кая, Марксистская, Нижегородская, Мясницкая, Краснопрудная, Новослободс-
кая, Сретенка, проспект Мира), так и кольцевым магистралям (Садовое кольцо,
234 10. Лихеноиндикация качества воздуха в г. Москве
Бульварное полукольцо), а также по набережным рек Москва и Яуза. В пределах
Садового кольца расположено множество автохозяйств, на территории округа
находится большинство железнодорожных вокзалов города, что обуславливает
наличие здесь подъездных путей к ним, зон безопасности и т.д. Имеются здесь и
ТЭЦ, и опасные промзоны типа Павелецкой и другое. Это самый старый и наибо-
лее застроенный городской район с незначительной долей озелененных площа-
дей (Центральный парк культуры и отдыха им. А.М.Горького с Нескучным садом
на правом берегу реки Москвы, Лужнецкая пойма, Краснопресненский парк, парк
Дома армии, небольшой Ботанический сад МГУ в начале проспекта Мира, не-
сколько кладбищ - на левом берегу), но основное число деревьев представлено в
посадках вдоль транспортных магистралей, улиц, внутри дворов, в скверах. На
территории округа обнаружены представители 10 видов лишайников в 45% квад-
ратов.
Юго-Восточный округ также пересекают крупные автомагистрали с интен-
сивным движением транспорта, в котором весьма заметна роль грузовых крупно-
тоннажных автомобилей (Волгоградский и Рязанский проспекты), однако эколо-
гическая ситуация определяется наличием здесь таких крупных загрязнителей
как промзоны “завод Серп и молот”, “Карачарово”, “Чагино-Капотня”, “Курья-
ново”, “Грайвороново”, “ Люблино-Перерва” на левом берегу реки Москвы, не-
скольких ТЭЦ. Застроен он несравнимо меньше, чем Центральный округ, но
названные промзоны занимают крупные плошали с большим радиусом воздей-
ствия на среду. Озелененные площади представлены Парком окружного дома
офицеров на левом берегу реки Яузы, Кузьминским лесопарком и одноименным
парком культуры и отдыха с прудами. На них и представлены участки с несколь-
ко более благоприятной экологической ситуацией в сравнении с основной пло-
щадью округа, хотя его территория и примыкает к реке Москве, что с позиции
здравого смысла должно было бы улучшать, а не ухудшать комфортность жиз-
ни людей. На территории округа обнаружены представители 16 видов в 31%
квадратов.
Территория Южного округа входит в группу округов с несколько более бла-
гоприятной ситуацией, чем в двух названных выше. Южный и Юго-Восточнй
округа на большей части разделены рекой, и проблемы территорий, к ней примы-
кающих, здесь общие. Именно у реки сосредоточены здесь наиболее обширные и
мощные промзоны, такие как “ЗИЛ”, “Южный порт”, “Нагатино”, “Коломен-
ское”, которые вместе с такими промзонами как “Бирюлево”, “Котляково”, “Вер-
хние Котлы”, “Царицыно”, расположенного неподалеку (с другой стороны МКАД)
Московского коксогазового завода, а также крупными автомагистралями - Вар-
шавским и Каширским шоссе - и несколькими ТЭЦ, негативно воздействуют на
состояние воздушного бассейна. Наряду с районами весьма плотной застройки в
северной части округа по обоим берегам реки Москвы, на юге здесь нередко мож-
но встретить пустыри, овраги, долины небольших рек - Чертановки, Горобни и
10. Лихеноиндикация качества воздуха в г. Москве
235
других, пруды. Озелененные площади представлены Царицынским и Коломенс-
ким парками, небольшим лесным массивом западнее Варшавского шоссе у МКАД,
несколькими кладбищами, остатками промышленного сада совхоза им. Ленина.
На территории округа обнаружены представители 29 видов лишайников в 65%
квадратов.
Северный округ, отнесенный ко второй группе округов, с запада выходит к
Химкинскому водохранилищу и каналу “Москва-Волга”. Основные автомагист-
рали - Ленинградский проспект с одноименным шоссе и Дмитровское шоссе.
Это густо населенный округ с такими крупными промзонами как “Магистраль-
ная ул.”, “Боткинский проезд”, “ул. Правды”, “Коптево” с чугуно-литейным заво-
дом, “Коровино” с мощнейшей ТЭЦ, “Вагоноремонтная ул.”, “Бескудниково”,
“Химки-Ховрино”, “Савеловский вокзал”, также оказывающими сильнейшее воз-
действие на прилегающие территории Центрального и Северо-Западного окру-
гов. Озелененные площади представлены Лесной опытной дачей Тимирязевской
сельхозакадемии, заложенной более 100 лет назад, парком Дружбы, лесным мас-
сивом между Октябрьской железной дорогой, каналом и МКАД, территорией ста-
диона “Динамо”, небольшими лесными островками на ул. Куусинена и западнее
платформы Марк, кладбищами. На территории округа обнаружены представите-
ли 18 видов лишайников в 56% квадратов.
Через Северо-Восточный округ, также входящий во вторую группу округов,
проходят автомагистрали проспект Мира с Ярославским шоссе и Алтуфьевское
шоссе. Южная часть округа - старая плотно застроенная городская территория
со множеством промзон, таких как “Сущево”, заводы “Калибр” и “Станколит”,
“Огородный проезд”, “Алексеевская ул.”, “Митьковская ветка”. Северная часть-
это и территория некогда самостоятельного города Бабушкин, и районы недавне-
го интенсивного жилищного строительства - Бибирево, Владыкино, Лианозово,
Медведково, Отрадное - с промзонами “Алтуфьевское шоссе”, “Дегунино”, “Ос-
ташковское шоссе”, “Окружная”, “Медведково”, “Северянин”. Долины реки Яузы
и ее притоков Лихоборки, Чермянки, Ички, пустыри, овраги, многочисленные
пруды также свойственны центральной и северной частям округа. Озелененные
площади представлены территориями Главного ботанического сада РАН и Все-
российского выставочного центра, старинным Останкинским парком, парком в
Лианозово, лесными массивами восточнее платформы Лианозово, остатками са-
довых участков, несколькими кладбищами. На территории округа обнаружены
представители 29 видов лишайников почти в 60% квадратов.
Восточный округ отнесен к третьей группе округов с умеренно неблагопри-
ятной экологической ситуацией. Это наименее застроенная часть города, здесь
сосредоточены крупнейшие лесные массивы - Лосиноостровский и Измайловс-
кий лесопарки, парки Сокольники, Кусково. Много прудов, территорию на отрез-
ке около 6 км пересекает река Яуза и ее притоки. Негативное воздействие на эко-
логическую ситуацию в округе оказывают находящиеся в застроенных районах
236
10. Лихеноиндикация качества воздуха в г. Москве
такие промзоны как “Кирпичная ул.”, “Соколиная гора”, “завод Прожектор”, “Пе-
рово”, “Хроматрон”, ТЭЦ на Амурской улице и другие. Округ пересекают шоссе
Энтузиастов и Щелковское с крупнейшим в городе автовокзалом. Ощущается здесь
и влияние крупных производств соседних Мытищ. На территории округа обна-
ружены представители 27 видов лишайников в 79% квадратов.
Северо-Западный округ, пожалуй, наиболее “водный” в городе, поскольку
здесь протекают реки Сходня, Химка и Москва со старицами и излучинами, про-
рыто несколько каналов, а также находится западная половина Химкинского во-
дохранилища. Значительна в округе доля озелененных площадей - часть Сереб-
ряноборского лесничества, лесной массив в Братцево, парки Покровское-Глебо-
во и Покровское-Стрешнево и другие. Загрязнителями среды здесь являются
промзоны “Силикатный проезд”, “Октябрьское поле”, “Тушино-Трикотажная”,
несколько ТЭЦ, автомагистраль Волоколамское шоссе. На территории округа об-
наружены представители 31 вида лишайников в 89% квадратов.
Естественной границей Западного округа с северо-востока является река
Москва. Здесь же протекает Сетунь с притоками Раменка-Очаковка-Самородин-
ка. Много прудов. Основные автомагистрали - Кутузовский проспект с Можайс-
ким шоссе, проспекты Вернадского и Мичуринский, Рублевское шоссе. В округе
также имеется несколько промзон, крупнейшие из которых-“Бережковская набе-
режная” с Киевским вокзалом, “Очаково”, “Кунцево”, “Фили”, “ул.Барклая”, ТЭЦ.
Озелененные площади - значительная часть Серебряноборского лесничества с
обеих сторон Рублевского шоссе, парки Победы, Фили, Тропарево,50-летия Ок-
тября и другие, спорткомплекс в Крылатском, высокий правый берег реки Моск-
вы от впадения Сетуни до проспекта Вернадского (Воробьевы, или Ленинские
горы), территория МГУ с Ботаническим садом, ряд крупных кладбищ. В округе
довольно много пустырей, оврагов. Здесь обнаружены представители 32 видов
лишайников почти в 85% квадратов, в одном из которых отмечено максимальное
для Москвы число видов (18).
Юго-Западный округ с севера также выходит к реке Москве, но всего лишь
на отрезке около 1 км. На остальной части округа, в основном, на юге протекают
речки Коршуниха, Котловка, Чертановка, Очаковка. Северная половина террито-
рии - район весьма плотной застройки с ТЭЦ на улице Вавилова. В округе нахо-
дятся также промзоны “Черемушки”, “Воронцово”, “Теплый стан”, однако их
нагрузка на среду относительно невелика. Основные магистрали с интенсивным
движением автотранспорта - Ленинский проспект и Профсоюзная улица. Озеле-
ненные площади представлены крупным Битцевским лесопарком, массивом между
Ленинским проспектом, Профсоюзной улицей и МКАД, парком в Воронцове,
частью Воробьевых гор, территорией Дворца пионеров и школьников. Число об-
наруженных здесь видов лишайников сравнительно невелико (22), но они отме-
чены почти в 92% квадратов и представители отдельных видов развиты здесь
лучше.
10. Лихеноиндикация качества воздуха в г. Москве
237
10.5. РЕЗУЛЬТАТЫ НАБЛЮДЕНИЙ
ЗА ТРАНСПЛАНТИРОВАННЫМИ В ГОРОД
СЛОЕВИЩАМИ ЛИШАЙНИКА
Ранее (гл. 7) было указано, что в центре города между бывшим домом Пашко-
ва и Боровицкой башней Кремля, а также в массиве Лесной опытной дачи ТСХА
проводились наблюдения над трансплантированными слоевищами Hypogymnia
physodes. Они подтверждают высказанное выше заключение, что в городе нет
участков с нормальным чистым воздухом, поскольку в местах экспонирования
талломов достаточно быстро возрастала доля отмерших водорослевых клеток и
происходило накопление ряда металлов (см. табл. 7.4 и 7.5).
Так, в центре города, в “лишайниковой пустыне” (выдел 0-10) содержание
меди с декабря по апрель увеличилось на 220-520% (по август - 350-590%), свинца
- на 130-140% (по август - на 180-190%). На участке Лесной опытной дачи,
относящемуся к лучшему в Москве выделу (71-150), также шло достаточно ин-
тенсивное накопление металлов - содержание меди увеличилось на 110-280%, а
свинца - на 45-80%. Все это - свидетельства низкого качества воздуха в Москве.
Метод трансплантации - достаточно нагляден, но не надежен, поскольку в
условиях густо населенных территорий из-за вандализма отдельных представи-
телей населения опрометчиво надеяться на возможность проведения длительных
сравнимых наблюдений и получение репрезентативных результатов. Хотя, воз-
можно, к кому-то судьба будет благосклонна.
11. ЗАКЛЮЧЕНИЕ:
ЛИШАЙНИКИ И ДИАГНОЗ ЗДОРОВЬЯ ЭКОСИСТЕМ
Мы являемся свидетелями проникновения слова “экология” в лексику пред-
ставителей разных сфер человеческой деятельности - наряду с профессионала-
ми, применяющими этот термин в строго научном его смысле, а именно, для обо-
значения отношений живых организмов со средой их существования, в том числе
и с другими организмами, это слово в разных значениях используют журналис-
ты, политики, писатели, филологи, оппозиционеры, лица, желающие улучшить
свои жилищные условия и другие. В непрофессиональной среде под экологией в
лучшем случае, понимают изучение только гигиенических аспектов состояния
окружающей среды, а нередко и просто уровень ее техногенного загрязнения (Боль-
шаков и др., 1996). Можно услышать и о “плохой (?) экологии”, и об “экологии
языка”, и об “экологии культуры”, и об “экологии литературы” и множество дру-
гих сочетаний, режущих слух специалиста-биолога. Однако, есть мнение, что ши-
рокое внедрение этого слова отражает назревшую необходимость экологизации
разных отраслей знаний (Розенберг, 1999). В то же время происходит и обратное
- социологизация экологии, проникновение в понятийный аппарат экологии тер-
минов, обычно используемых для обозначения явлений человеческого общества.
Давно известен пример “фитоценология” - “фитосоциология”. Ныне говорят и
пишут о “здоровье” экосистем. На страницах научных изданий одни полагают,
что это, возможно, новая научная парадигма (Rapport, 1989,1995; Shrader-Frechette,
1994; Rapport et al., 1995, 1998), другие утверждают, что понятие “здоровье эко-
систем” - всего лишь метафора, и оно не имеет научного смысла (Sutter, 1993;
Wicklum, Davies, 1995). Тем не менее, издается журнал “Ecosystem Health”, функ-
ционирует научное общество с одноименным названием, проводятся междуна-
родные научные форумы по этой проблеме. Все это свидетельствует о том, что в
эту идею вложены немалые деньги.
Используют этот термин и применительно к лишайникам как индикаторам
состояния среды (Hawksworth, 1991, Biazrov, 1994b, 1995а, b; DePriest, 1994; Бяз-
ров, 1995, 1996в), особенно в исследованиях по программе “Мониторинг здоро-
вья лесов” (McCune, Dey, 1994; Бязров, 19986; McCune, 2000).
Я полагаю, что если рассматривать словосочетание “здоровье экосистем”, пред-
ставляющее объединение терминов двух разных наук, даже как метафору, то она
11. Заключение: лишайники и диагноз здоровья экосистем
239
довольно удачна для обозначения загрязненных территорий, хотя сами понятия
“экосистема” и “здоровье” неконкретные. Экосистема-это внеранговая террито-
риальная единица, которая включает все живые организмы данного объема про-
странства и взаимодействующую с ними физическую среду. Экосистема может
быть небольшой как крохотное помещение, и громадной как биосфера. Понятия
“здоровье”, “здоровый” также не имеют четких границ даже по отношению к
человеку. Здоровый, значит, не больной в данное время. Общепризнанных коли-
чественных критериев “здоровой” экосистемы пока нет. В общем виде индекс
здоровья экосистемы принимается как функция (i) энергии, выражением которой
являются продуктивность и метаболизм, (ii) организации (структура, биоразно-
образие) и (iii) лабильности или эластичности (Ramade, 1995). Я понимаю под
“здоровьем экосистемы” степень соответствия выявленных интегральных при-
знаков изучаемой экосистемы ожидаемым в данном месте ее идеальным свой-
ствам, представление о которых складывается на основе либо личного опыта эк-
сперта, либо результатов других исследований аналогичных экосистем. Диагноз
может ставиться путем сравнения широкого комплекса показателей обследован-
ной экосистемы с признаками другой, идеализированной, экосистемы. Несом-
ненно, использование экологических индикаторов, в частности лишайников, мо-
жет существенно помочь установлению верного диагноза, оценке возможного риска
для экосистемы и планированию действий по уменьшению риска (Eduljee, 2000).
Вероятно, поэтому программой “Мониторинг здоровья лесов” (FHM), наряду с
другими индикаторами, предусмотрено обследование и эпифитных макролишай-
ников (McCune, Dey, 1994; Ferretti, 1997). Анализ публикаций позволяет заклю-
чить, что лишайники чаще всего применяют для выявления степени нарушенно-
сти лесов, качественной оценки загрязнения воздушного бассейна и оценки ра-
диоактивного загрязнения территорий.
11 Л. ЛИШАЙНИКИ - ИНДИКАТОРЫ НЕНАРУШЕННЫХ ЛЕСОВ
В обжитых районах нашей планеты теперь довольно трудно встретить нетро-
нутые девственные леса. Однако их ищут, ибо эти сообщества представляют ин-
терес как эталоны природы, как здоровые экосистемы. Для суждения о степени
нарушенности лесов, наряду с другими признаками, используют и лишайники.
Уместно отметить, что в разных природных зонах Земли экспертные оценки в
основном базируются на особенностях видового состава эпифитных лишайни-
ков в экосистемах (Горбач, 1965; Rose, 1976; Hyvarinen et al., 1992; Turk, 1993;
Goward, 1994; Kuusinen, 1994; Sillett, 1994; Kuusinen, Siitonen, 1998; Trass et al.,
1999; Norden, Appelqvist, 2001; Rees, Juday, 2002; и другие). Привлекаются и дру-
гие показатели, например, соотношение размеров популяций разных видов, ин-
дексы, характеризующие экологию и видовой состав лишайников и другое (Rose,
240
//. Заключение: лишайники и диагноз здоровья экосистем
Trass et al., 1999). Однако необходимо отметить, что состав лишайников в
сообществе зависит не только от происхождения и степени нарушенное™ после-
днего, но и от особенностей структуры, типа леса, состава древесных пород,
географического положения территории (табл. 11.1).
Таблица 11.1
Количественные показатели эпифитных лишайников
в некоторых типах леса Северной Евразии
Показатель Тип леса, местоположение, возраст
Querceto-Piceetum oxalidoso-asperu- losum, Россия, Смоленская обл., около 200 лет Abieto-Piceetum oxalidosum, Россия, Республика Мари Эл, около 200 лет Pineto-Laricetum mixherbosum, Монголия, Хангайский хребет, около 200 лет
Число видов 49 55 27
Спектр жизненных форм*, % Eph 2
Сг 61 71 27
L1 4 4 4
SI 6 9 23
С1 2 2 8
Sc 8 4 19
Lfp 2 - -
Fp 15 10 19
Масса, кг/га, сух. вес 700 450 720
* - обозначения жизненных с юрм соответствуют табл. 1.1.
Опубликованные материалы свидетельствуют, что для ненарушенных лесах,
как правило, характерно значительное видовое и морфологическое разнообразие
встреченных на стволах и ветвях деревьев лишайников (табл. 11.2). Индикатора-
ми ненарушенных лесов называют представителей порошкоплодных калициевых
лишайников, виды семейства уснеевых, рода Lobaria, лишайники с цианобакте-
риальным фотобионтом и другие (Lesica et al., 1991; Karstrom, 1992; Tibell, 1992;
Anonby, 1994; Gauslaa, 1994,1997; Selva, 1994; Holien, 1996; Kuusinen, 1996; Rolstad,
Rolstad, 1999; Gauslaa, Solhaug, 2000). Для таких лесов характерны относительно
большой запас формируемой эпифитными лишайниками массы, частая встреча-
емость на поверхности почвы опавших с деревьев слоевищ (Бязров, 1970; Esseen,
Renhom, 1996; Arseneau et al., 1998). При этом необходимо отметить, что универ-
сальных видов-индикаторов, видимо, нет, ибо они достаточно специфичны для
каждого континента, для каждой природной зоны или горного пояса, для каждой
лесной формации.
11. Заключение: лишайники и диагноз здоровья экосистем
241
Таблица 11.2
Виды лишайников, которые используются как индикаторы естественных старых лесов
Европы (номенклатура таксонов - согласно цитируемым источникам)
Вид Великобритания (Rose, 1976) Швеция (Karstrom, 1992; Tibell, 1992)
Alectoria sarmentosa ssp. sarmentosa 4-
Arthonia didyma 4-
A. leucope Haena +
A. vinosa - +
Arthopyrenia cinereopruinosa +
Bacidia igniarii +
Biatora epixanthoidiza +
B. sphaeroides +
Bryoria capillaris +
B. lanestris 4-
B. nadvornikiana +
B. schaereri 4-
Calicium adaequatum +
Catinaria atropurpurea +
Cavernularia hultenii +
Chaenotheca chi ore ll a + 4-
C chrysocephala 4- 4-
C. gracillima 4- 4-
C. laevigata 4- 4-
C. phaeocephala 4- 4-
C. subroscida 4- 4-
Chaenothecopsis папа 4-
C. viridialba 4-
Cladonia parasitica 4-
Collema curtisporum 4-
C. flaccidum 4-
C. fragrans 4-
C. furfur aceum 4-
C. nigrescens 4-
C. occultatum 4-
Conotrema populorum 4-
Cybebe gracilenta 4-
Cyphelium inquinans 4-
C. karelicum 4-
C. pinicola 4-
C. tigillare 4-
Degelia plumbea 4-
Dimerella lutea 4-
D. pineti 4-
Enterographa crassa 4-
Evernia divaricata 4-
242
11. Заключение: лишайники и диагноз здоровья экосистем
таблица 11.2 (продолжение)
Е. mesomorpha +
Е. prunastri +
Heterodermis speciosa +
Нуросепотусе friesii +
Hypogymnia austerodes +
H. bitteri +
H. vittata +
Lecanactis premnea +
Lecanora ochrococca +
Leptogium teretiusculum +
Lithographa flexella +
Lobaria amplissima +
L. pulmonaria + +
L. virens +
Loxospora elatinum +
Micarea globulosella +
Microcalicium ahlneri +
M. arenarium
Mycoblastus affinis +
Nephroma laevigatum +
Opegrapha lyncea +
Pachyphiale carneola +
Pannaria conoplea
P. mediterranea +
Parmeliella triptophylla +
Parmotrema crinita +
Peltigera collina +
P. horizontalis +
Pertusaria pupillaris +
Phaeocalicium populneum
P. praecedans +
Platismatia norvegica +
Porina leptalea +
Punctelia ruddenda +
Pyrenula nitida +
Pyrrhospora cinnabarina +
P. elabens +
Ramalina dilacerata +
R. farinacea +
R. thrausta +
Rinodina isidioides +
Schismatomma pericleum +
Sclerophora coniophaea +
Stenocybe septata +
II. Заключение: лишайники и диагноз здоровья экосистем
243
таблица 11.2 (окончание)
Sticta limbata +
S. sylvatica +
Thelopsis rubella +
Thelotrema lepadinum +
Usnea extensa +
Vulpicida juniperinus +
11.2. ОЦЕНКА ЗАГРЯЗНЕНИЯ
ВОЗДУШНОГО БАССЕЙНА
Эта проблема обсуждалась в предшествующих разделах книги. В связи с рас-
сматриваемым понятием “здоровье экосистем” считаю необходимым подчеркнуть,
что экосистемы крупных городов, в частности, Москвы, а также территории, на-
ходящиеся под воздействием различных источников загрязнения, здоровыми на-
звать нельзя. Они, несомненно, больны, и лишайники здесь полезны для оценки
тяжести заболевания, а не степени здоровья.
11.3. ОЦЕНКА РАДИОАКТИВНОГО
ЗАГРЯЗНЕНИЯ ТЕРРИТОРИИ
Наличие в экосистеме искусственных радионуклидов, поступление которых в
среду связано с термоядерными взрывами различного назначения, с работой раз-
личных энергетических установок и деятельностью ряда предприятий, несомнен-
но, следует считать одним из серьезнейших симптомов нездоровья экосистемы.
Наблюдения в одном из районов Брянской области, загрязненных в результате
аварии на Чернобыльской АЭС, показали, что при относительно невысоких уров-
нях радиоактивного загрязнения серьезные нарушения здоровья выявлены у пред-
ставителей всех исследованных таксономических групп, от растений до млеко-
питающих (Последствия Чернобыльской катастрофы, 1996). “Выздоровление”
загрязненных радионуклидами территорий, которое прежде всего зависит от пе-
риода полураспада изотопов, может продолжаться сотни и многие тысячи лет.
Некоторые аспекты использования лишайников в индикации радиоактивного заг-
рязнения уже обсуждались ранее (гл. 9).
Несомненно, лишайники могут быть полезны для постановки диагноза при
обследовании состояния экосистемы. Однако не следует абсолютизировать зна-
чение лишайников как индикаторов среды. Их видовой состав практически не
меняется при поступлении в экосистему значительного количества радионукли-
244
11. Заключение: лишайники и диагноз здоровья эко систем
дов, и только инструментальное измерение активности и состава радионуклидов
позволяет получить представление о масштабах опасности загрязнения и его осо-
бенностях. С другой стороны, такие загрязнители как двуокись серы и ряд ток-
сичных металлов поражают многие виды лишайников, но исчезновение чувстви-
тельных видов не позволяет судить о количественных показателях загрязнителей.
Использование показателей лишайников для диагноза состояния экосистемы тре-
бует всестороннего учета специфики реагирования их на действие факторов сре-
ды, что, в свою очередь, предполагает привлечение к такой работе компетентных
специалистов.
***
Итак, лихеноиндикация представляет собой часть биоиндикации - обширно-
го прикладного раздела экологии. Проблема, решаемая специалистами в области
биоиндикации - по свойствам представителей живых организмов установить те
или иные особенности среды, в которой эти организмы обитают и с которой вза-
имодействуют. Соответственно, теоретической и методической основой биоинди-
кации является экология, наука о взаимоотношениях организмов и среды. Иссле-
дователь, решившийся по показателям той или иной группы организмов устано-
вить какие-то свойства среды, должен хорошо ориентироваться в экологии этих
организмов. Лишайник - комплекс двух или трех организмов, представляющих
два-три царства органического мира, которые находятся в сложных отношениях
друг с другом, скорее всего, гриб паразитирует на зеленой водоросли или циано-
бактерии, или сразу на представителях обеих таксономических групп. И распро-
странение этой сложно устроенной ассоциации организмов зависит как от взаи-
моотношений между бионтами лишайниковой ассоциации, так и от комплекса
экологических факторов. Загрязнение воздуха - всего лишь одно из множества
свойств среды, сказывающихся на встречаемости лишайников. Основным мето-
дом лихеноиндикационных исследований является градиентный анализ, и глав-
ная проблема-обеспечение при сборе материала по какому-либо градиенту прин-
ципа “при прочих равных условиях”.
Технология любого исследования, в особенности связанного с измерением
тех или иных величин, характеризующих изучаемый объект, требует гарантии
качества работы и его контроля . Гарантия качества мониторингового исследова-
ния -это система процедур, обеспечивающих наличие документации, показыва-
ющей, что работа проведена согласно оговоренным с заказчиком методам, обес-
печивающим соответствующий времени стандарт для данного типа исследова-
ний, а измерения показателей получены с известной степенью точности (Shampine,
1993). Это предполагает последовательное документирование процесса планиро-
вания исследования, полевых и лабораторных процедур, методов анализа дан-
ных. Если исследование проводится в связи с экспертизой какого-либо проекта,
11. Заключение: лишайники и диагноз здоровья экосистем
245
реализация которого может отрицательно сказаться на состоянии здоровья насе-
ления или стать причиной необратимых изменений природной среды, необходи-
мо предусмотреть проведение анализов в разных лабораториях или внешнюю
проверку данных анализов и эталонных образцов лишайников.
Контроль качества определяется как процедура по установлению, что поле-
вой или лабораторный методы являются статистически контролируемыми в пре-
делах известных уровней вероятности и обеспечивают декларируемую точность,
приемлемую для данного периода времени. Контроль качества предполагает воз-
можность проверки полевых коллекций, фактических данных лабораторных ана-
лизов, результатов анализов возможных дублирующих учетов в поле другим кол-
лектором, анализ повторных и стандартных эталонных материалов, экспертную
оценку сделанных по полученным результатам выводов. Гарантия качества обес-
печивается также соответствующей квалификацией исполнителей работы, про-
ведением химических анализов в лабораториях, имеющих сертификат на опреде-
ление интересующих веществ в биологических объектах. Выполнение условий,
обеспечивающих необходимый уровень сбора материала, его анализа, интерпре-
тации - важнейшее требование при выполнении проекта, связанного с лихеноин-
дикацией. Так, в Италии установлено, что точность выявления числа видов эпи-
фитных лишайников у разных исполнителей варьирует от 52 до 88% (Brunialti
et al., 2002). Еще ниже была точность идентификации видов в поле - от 29
до 61%. Значительная вариабельность показателей наблюдалась и при проверке
данных качественной оценки признаков лишайников, сделанной разными испол-
нителями.
При всей привлекательности лишайников как биоиндикаторов считаю необ-
ходимым предостеречь от переоценки их индикаторного значения. Здесь уместен
умеренный скептицизм. Прежде всего, на территориях с высоким уровнем заг-
рязнения образуются “лишайниковые пустыни”, где лишайники отсутствуют.
Нередко это значительные по размерам участки (в Москве, например, это более
чем треть обследованной площади города). Здесь как бы порог условий для жиз-
ни лишайников, и мы лишены возможности сравнивать по показателям лишай-
ников такие участки между собой, хотя они явно неравноценны. Кроме того, нельзя
ожидать, что картографирование распространения лишайников по той или иной
методике, даже с применением интегральных оценочных индексов, даст сведения
о конкретных загрязнителях и величинах их концентрации в среде. Эмпирически
установленные индексы и корреляции с концентрациями имеют локальное значе-
ние, и закономерности, установленные в Англии или Альпах, не всегда можно
экстраполировать на Европейскую Россию. Мало еще известно о действии одних
фитотоксикантов при наличии в среде других или о трансформации токсикантов
под воздействием факторов среды. Например, выбрасываемые органические со-
единения могут образовать 16-20 продуктов трансформации, среди которых мо-
гут быть более токсичные или опасные, чем исходное вещество (А. Малышева,
246
11. Заключение: лишайники и диагноз здоровья экосистем
1997). Однако с биологической точки зрения и с позиций охраны природы факти-
ческие концентрации загрязнителей в среде малоинформативны. Важнее биоло-
гический эффект многовариантного действия различных фитотоксикантов, ин-
тегрируемый в реакции лишайников. Такие влияния, особенно кислотное загряз-
нение, можно выявить с помощью лишайников довольно рано и надежно. Если
не настаивать на широко распространенном требовании очень точной информа-
ции о конкретном загрязнителе и фактической концентрации его в среде, лишай-
ники могут быть полезными для оценки действия на обследуемой территории
кислых загрязнителей и относительных изменений в условиях загрязнения воз-
духа. Результаты картографирования распространения лишайников могут быть
полезны при планировании мероприятий по оздоровлению среды, адресного об-
следования населения для предупреждения и выявления некоторых заболеваний,
например, рака легких. Так что для жителей городов или промышленных регио-
нов исчезновение лишайников из привычных для них местообитаний должно быть
тревожным симптомом, сигналом опасности. С другой стороны, если на поверх-
ности какого-либо субстрата (кора деревьев, древесина, камни и другое) появи-
лись представители лихенобиоты, которые постепенно расширяют освоенные ими
площади, это означало бы, что в данной местности произошло снижение уровня
загрязнения воздуха и есть надежда, что качество жизни как лишайников, так и
людей, улучшится. Но не надо забывать, что значительное видовое разнообразие
и обилие эпифитных лишайников в интересующем вас районе свидетельствуют о
чистоте воздуха, однако бедный состав видов и их слабое развитие не всегда яв-
ляются признаками нездоровья экосистемы.
ЛИТЕРАТУРА И АВТОРСКИЙ ИНДЕКС
(цифры курсивом в круглых скобках означают номера страниц текста,
где содержится ссылка на литературный источник)
Аблаева З.Х. Лихеноиндикационное картирование Лапландского заповедника //
Природа Севера и ее охрана. Мурманск, 1981а. С.38-44. (102)
Аблаева З.Х. Эпифитные лихеносинузии естественных и антропогенных ланд-
шафтов на территории Лапландского заповедника И Брио-лихенологичес-
кие исследования высокогорных районов и Севера СССР. Апатиты, 19816.
С.73-74. (118)
Адамова Л.И., Бязров Л.Г. Тяжелые естественные радионуклиды в лишайниках
ряда экосистем Западного Кавказа // Биоиндикация и биомониторинг.
Д.А.Криволуцкий - ред. М.: Наука, 1991. С.125-129. (179, 189, 195)
Аксенова О.И., Волкова И.Ф., Ефимов М.В., Корниенко А.П. Здоровье населения
города Москвы И Гигиена и санитария. 1997. № 3. С.44-46. (212)
Алексахин Р.М., Равикович М.М. О естественной радиоактивности различных ком-
понентов лесных биогеоценозов, связанной с 40К И Радиоактивные изотопы
в почвах и растениях. Л.: Колос, 1969. Т. 16. С.123-133. (192, 195)
Алехин В.В. Растительность и геоботанические районы Московской и сопредель-
ных областей. М.: МОИП, 1947. 79 с. (201)
Андреев В.Н. Некоторые данные о делихенизации тундры И Растительность и
почвы субарктической тундры. Новосибирск, 1980. С. 201-205. (10, 28)
Аржанова В.С., Скирина И.Ф. Значение и роль лихеноиндикационных исследо-
ваний при эколого-геохимической оценке состояния окружающей среды //
География и природные ресурсы. 2000. № 4. С.33-40. (179, 184)
Баденкова С.В., Княжева Л.А., Качур А.Н., Кононова И.Ф., Седова В.М. Лишай-
ники как индикаторы загрязнения атмосферы вблизи металлургического
завода (Южное Приморье) // Лихеноиндикация состояния окружающей сре-
ды. Таллин, 1978. С. 125-128. (184)
Баденкова С.В., КняжеваЛ.А., Кононова И.Ф. Опыт лихеноиндикации загрязне-
ния ландшафтов восточного Сихотэ-Алиня И Сихотэ-Алинский биосфер-
ный район: принципы и методы экологического мониторинга. Владивос-
ток: ДВНЦАНСССР, 1981. С.128-135. (66, 184)
248
Литература и авторский индекс
Бадтиев Ю.С., Кулемин А.А. Методика биоиндикации окружающей природой
среды И Экологический вестник России. 2001. № 4. С.27-29. (10)
Байбаков Э.И. Лихеноиндикационное картографирование атмосферных загряз-
нений городов Волжско-Камского края // Современные проблемы миколо-
гии, альгологии и фитопатологии. Сборник трудов Международной конфе-
ренции, Москва, апрель 1998 г. М.: МГУ. 1998. С.366-367. (101)
Барашкова Э.А. Питание кладонии оленьей через мертвую часть подециев И Во-
тан. журн. 1963. Т. 48. №4. С.588-591. (30)
Баумгертнер М.В. Лишайники - биоиндикаторы загрязнения окружающей сре-
ды юга Кемеровской области // Сибирский экологический журнал. 1998.
Т.5. №2. С. 191-196. (65)
Безуглая Э.Ю., Расторгуева Г.П., Смирнова И.В. Чем дышит промышленный го-
род. Л.: Гидрометеоиздат, 1991.255 с. (201, 202)
Беккер А.А. Воздух Москвы // Природа. 1989. №8. С.30-40. (213, 232)
Беспамятное Г.П., Кротов Ю.А. Предельно допустимые концентрации химичес-
ких веществ в окружающей среде. Л.: Химия. 1985. 528 с. (57)
Блюм О.Б. Водный режим лишайников // Украинский ботан. журн. 1965. Т.22.
№2. С. 3-14. (25)
Блюм О.Б. Влияние газообразных атмосферных загрязнителей на лишайники //
Международная школа по лихеноиндикации, Таллин, 22-25 июня, 1982.
Таллин, 1984. С.35-51. (50)
Блюм О.Б., Тютюнник Ю.Г. Исторический биомониторинг содержания свинца в
атмосфере с помощью лишайников //ДАН УССР. Сер. Б: геол., хим. и биол.
науки. 1985. №10. С.53-55. (182)
Блюм О.Б., Тютюнник Ю.Г. Исторический аспект регионального мониторинга
тяжелых металлов в атмосфере, осуществляемый методом биогеохимиче-
ской лихеноиндикации (на примере Украинской ССР) // Проблемы эколо-
гического мониторинга и моделирования экосистем. Л.: Гидрометеоиздат,
1989. Т. 12. С.73-87. (182)
Бобрицкая М.А. Поглощение литофильной растительностью минеральных эле-
ментов из массивно-кристаллических пород // Тр. Почвенного института.
М, 1950. Т.34. С.5-27. (31, 34, 35, 179)
Богатов С.А., Боровой А.А., Дубасов Ю.В., Ломоносов В.В. Форма и характерис-
тика частиц топливного выброса при аварии на Чернобыльской АЭС //
Атомная энергия. 1990. Т.69. Вып. 1. С.36-40. (192)
Бойко А.В., Киселев В.Н., Чубанов К.Д, Бортник А.В., Пикулик Н.И., Базыленко
ГГ Лихеноиндикация рассеивания серосодержащих техногенных эмиссий
в зеленой зоне Минска// Вестник АН БССР. Сер. биол. 1981. №1. С.23-26.
(43)
Бойко А.В., Киселев В.Н., Чубанов КД., Пикулик Н.И. Фитоиндикация рассеива-
ния некоторых тяжелых металлов в зеленой зоне Минска И Тяжелые метал-
Литература и авторский индекс
249
лы в окружающей среде и охрана природы. Материалы 2 Всесоюзной кон-
ференции, 28-30 декабря 1987. М., 1988. 4.2. С.342-345. (184)
Бойко А.В., Лукашев В.К., Киселев В.Н., Чубанов КД., Мазо Е.И., Пикулик Н.И.
Лихеноиндикация рассеивания микроэлементов в зеленой зоне Минска //
ДАН БССР. 1984. Т.28. С.451-454. (184)
Большаков В.Н., Криницин С.В., Кряжимский Ф.В., Мартинес Р.Х.П. Проблемы
восприятия современным обществом основных понятий экологической на-
уки // Экология. 1996. №3. С. 165-170. (238)
Борзилов В.А., Клепикова Н.В., Костриков А.А., Хваленский Ю.А., Троянова Н.И.
Метеорологические условия дальнего переноса радиоактивных продуктов
аварии на Чернобыльской атомной станции // Метеорология и гидрология.
1989. №11. С.5-11. (193)
Боровой А.А., Гагаринский А.Ю. Выбросы радионуклидов из разрушенного бло-
ка Чернобыльской АЭС И Атомная энергия. 2001. Т.90. Вып. 2. С.137-145.
(192)
Буткевич Н. А. Эпифитная флора в районе опытной станции лесоводства Мос-
ковской сельскохозяйственной академии И Проблемы экологического мо-
ниторинга и моделирования экосистем. Л.: Гидрометеоиздат, 1985. Т.8. С.72-
74. (205)
Бучельников М.А., Григорьев Ю.С. Применение замедленной флуоресценции хло-
рофилла при проведении активной лихеноиндикации воздушной среды го-
рода (на примере г. Красноярска) И Ботанические исследования в Сибири.
1996. №5. С. 11-14, 99. (144, 150)
Бязров Л.Г. Влагоемкость коры некоторых древесных пород // Лесное хозяйство.
1969а. №11. С.23-24. (22)
Бязров Л.Г Синузии эпифитных лишайников в широколиственно-еловых
лесах Подмосковья И Ботан. журн. 19696. Т.52. №2. С.239-249. (22, ПО,
212)
Бязров Л.Г. Синузии эпифитных лишайников некоторых типов лесных биогеоце-
нозов Смоленской области // Бюлл. МОИП. Отд. биол. 1969в. Т.74. №6.
С. 115-124. (119, 212)
Бязров Л.Г Эпифитные лишайниковые синузии в лесных биогеоценозах подзо-
ны широколиственно-еловых лесов. Автореф. дис. канд. биол. наук. Л.: БИН
АН СССР, 1970. 26 с. (22, 25, 31, 49, 96, 118, 240)
Бязров Л.Г. Сукцессии эпифитных лишайников в сосняках Подмосковья // Лесо-
ведение. 1972. №5. С.62-68. (27)
Бязров Л.Г. Лишайниковые синузии в лиственничнике разнотравном (Хангай-
ский хребет, МНР) // Ботан. журн. 1974. Т.59. №10. С. 1425-1438. (23)
Бязров Л.Г. Об использовании лишайников как индикаторов степени пастбищ-
ной нагрузки // Индикация природных процессов и среды. Вильнюс, 1976.
С.38-39. (10)
250
Литература и авторский индекс
Бязров Л.Г. О водном режиме лишайников горной лесостепи Восточного Хангая
// География и динамика растительного и животного мира МНР. М.: Наука,
1978а. С. 119-123. (25)
Бязров Л.Г. Эпифитизм как форма консортивных отношений // Материалы 6 Кон-
ференции по споровым растениям Средней Азии и Казахстана. Душанбе,
19786. С.281. (21)
Бязров Л.Г. Беспозвоночные животные в эпифитных лишайниках разных жиз-
ненных форм в лесах Подмосковья И Биология почв Северной Европы. М.:
Наука, 1988а. С. 149-154. (28)
Бязров Л.Г. Линейный прирост эпилитных лишайников И Тезисы докладов 11
симпозиума микологов и лихенологов Прибалтийских республик и Бело-
руссии. Таллин: АН ЭССР, 19886. С. 121-124. (27, 188)
Бязров Л.Г. Лишайники в сухих степях Восточного Хангая // Бюлл. МОИП. Отд.
биол. 1988в. Т.93. №3. С.66-80. (188)
Бязров Л.Г. Лишайниковые синузии и структура биогеоценоза // Журн. общей
биологии. 1990. Т.51. №5. С.632-641. (18, 122, 125)
Бязров Л.Г. Некоторые результаты использования лихеноценологических иссле-
дований при мониторинге состояния среды в Подмосковье И Биоиндика-
ция в городах и пригородных зонах. М.: Наука, 1993а. С.55-72. (123, 125,
128, 129, 134)
Бязров Л.Г. Эпифитные лишайниковые синузии в березовых лесах Восточноу-
ральского радиоактивного следа// Экологические последствия радиоактив-
ного загрязнения на Южном Урале. М.: Наука, 19936. С.134-155. (23, 123,
125, 187)
Бязров Л.Г. Видовой состав и распределение эпифитных лишайников в лесных
насаждениях Москвы // Лесоведение. 1994. №1. С. 45-54. (92, 96, 109, 200,
205, 209)
Бязров Л.Г. (Biazrov L.G.). Ecosystem health as a new paradigm for ecological
assessment and some aspects of lichens use for ecosystem state control // Пробле-
мы изучения биологического разнообразия водорослей, грибов, мохообраз-
ных Арктики. Международная конференция, Санкт-Петербург, 12-16 де-
кабря, 1995 г. Программа и Тезисы докладов. СПб, 1995. С.35-36. (238)
Бязров Л.Г. Видовое разнообразие лишайников Москвы // Бюлл. МОИП. Отд.
биол. 1996а. Т.101. №3. С.68-77. (95, 200, 210)
Бязров Л.Г. Исчезновение лишайников - сигнал опасности // Наука в России.
19966. №4. С.64-68. (98, 114, 200)
Бязров Л.Г. Концепция “здоровья экосистем” в отношении радиоактивно загряз-
ненных территорий и использование лишайников для диагноза состояния
экосистем // Пятая международная научно-техническая конференция “Чер-
нобыль-96”: итоги 10 лет работы по ликвидации последствий аварии на
ЧАЭС. Сборник тезисов. Н.П.Архипов-ред. Зеленый Мыс, 1996в. С.360. (238)
Литература и авторский индекс
251
Бязров Л.Г. Биоиндикация качества воздуха в Москве по картированию распрос-
транения эпифитных лишайников // Экология и промышленность России.
1998а. №7. С.27-31. (110, 114, 200, 212)
Бязров Л.Г. Изучение реакции эпифитных макролишайников на многолетнее эк-
спериментальное внесение сульфата аммония на лесную экосистему // Ле-
соведение. 19986. №2. С.22-29. (124, 238)
Бязров Л.Г. Использование индекса развития эпифитных лишайников для оцен-
ки качества воздуха в Москве И Тезисы докладов Международной конфе-
ренции “Проблемы ботаники на рубеже 20-21 веков”, С.-Петербург, 26-
30 мая 1998 г. СПб, 1998в. Т.2. С.62. (ПО, 200)
Бязров Л.Г. Оценка качества воздуха г. Москвы по распространению лишайников
// Современные проблемы микологии, альгологии и фитопатологии. Сбор-
ник трудов Международной конференции, Москва, апрель 1998 г. М.: МГУ,
1998. С.368-369. (НО, 200)
Бязров Л.Г. Индекс развития эпифитных лишайников и оценка состояния воз-
душного бассейна территории г. Москвы И Бюлл. МОИП. Отд. биол. 1999а.
Т.104. №6. С.30-40. (65, ПО, 114, 200, 212)
Бязров Л.Г. Лишайники эпицентра ядерного взрыва на Семипалатинском ядер-
ном полигоне // Биоиндикация радиоактивных загрязнений. Д.А.Криволуц-
кий - ред. М.: Наука, 19996. С.336-339. (187)
Бязров Л.Г. Синузии эпифитных лишайников Рудных гор (Германия) И Бюлл.
МОИП. Отд. биол. 2001. Т. 106. №5. С.93-97. (123, 125)
Бязров Л.Г, АдамоваЛ.И. Тяжелые металлы в лишайниках Кавказского и Рицин-
ского заповедников // Заповедники СССР - их настоящее и будущее. Тезисы
докладов Всесоюзной конференции. Новгород, 1990. 4.1. С.338-339. (179)
Бязров Л.Г, Ганболд Э., Губанов И. А., Улзийхутаг Н. Флора Хангая. Л.: Наука,
1989. 191 с. (26)
Бязров Л.Г, Максимова В.Ф., Рухадзе Е.В. Использование эпифитных лишайни-
ков для индикации загрязнения воздушной среды в Москве И Вестник МГУ.
Сер. 5. География. 1997. №4. С.32-36. (98, 114, 200, 210)
Бязров Л.Г, Штанчаева У.Я. Некоторые особенности радиоэкологии лишайни-
ков как среды обитания беспозвоночных животных на территории радио-
активного загрязнения И Биоиндикация радиоактивных загрязнений.
Д.А.Криволуцкий - ред. М.: Наука, 1999. С.169-176. (171)
Вайнштейн Е.А. Некоторые вопросы физиологии лишайников. I. Дыхание // Бо-
тан. журн. 1972. Т.57. №7. С.832-840. (11, 86)
Вайнштейн Е.А. Некоторые вопросы физиологии лишайников. П. Фотосинтез //
Боган, журн. 1973. Т.58. №3. С.454-464. (11, 86)
Вайнштейн Е.А. Некоторые вопросы физиологии лишайников. III. Минеральное
питание// Боган, журн. 1982. Т.67. №5. С.561-571. (11,32, 33, 34, 35, 38, 39,
46, 48)
252
Литература и авторский индекс
Вайнштейн Е.А. Применение флуоресцентного метода для оценки состояния ли-
шайников И Новости систематики низших растений. СПб.: Наука, 1996.
Т.31. С.94-97. (81, 84, 88)
Василевич В.И. Статистические методы в геоботанике. Л.: Наука, 1969. 232 с.
(167)
Ветров В.А., Алексеенко В.А., Пословин А.Л., Толоконников А.В. Чернобыльские
радионуклиды в компонентах экосистемы Байкала как метка дальнего пе-
реноса атмосферных загрязнений // Мониторинг и оценка состояния Бай-
кала и Прибайкалья. Л.: Гидрометеоиздат, 1991. С.108-113. (193)
Викторов С.В. Лишайники как индикаторы литологических и геохимических
условий в пустыне И Вестник МГУ. Сер. естеств. науки. 1956. №5. С.115-
119. (10, 30)
Викторов С.В. Лишайники пустыни Устюрт и их связь с некоторыми свойствами
почвы и горных пород // Вопросы индикационной геоботаники. М., 1960.
С.53-56. (10)
Викторов С.В., Джураева З.Д. Индикационное значение лишайниковых сооб-
ществ в пустынях // Бюлл. МОИП. Отд. биол. 1979. Т.84. №5. С.130-134.
(Ю)
Второва В.Н., Маркерт Б. Мультиэлементный анализ растений лесных экосис-
тем Восточной Европы // Изв. РАН. Сер. биол. 1995. №4. С.447-454. (179)
Галич Б. Полеофобия лишайников И Научные чтения памяти М.Г.Попова. Ир-
кутск, 1972. №12/13. С.3-11. (89)
Глазовский Н.Ф., УчватовВ.П., Злобина А.И. Некоторые геохимические особен-
ности экологии эпифитов в южной части Московской области //Лихеноин-
дикация состояния окружающей среды. Таллин, 1978. С.57-60. (179)
Глуздаков С.И. Внутривидовые отношения у лишайников // Проблемы внутриви-
довых отношений организмов. М., 1962. С. 144-145. (27)
Голубкова Н.С. Очерк флоры лишайников Московской области и смежных райо-
нов//Ботан. журн. 1959. Т.44. №2. С. 153-161. (22)
Голубкова Н.С. Определитель лишайников средней полосы Европейской части
СССР. М.- Л.: Наука, 1966. 256 с. (200, 203)
Голубкова Н.С. Анализ флоры лишайников Монголии. Л.: Наука, 1983. 248 с.
(18)
Голубкова Н.С. К вопросу о происхождении и путях эволюции лишайникового
симбиоза // Новости систематики низших растений. СПб.: Наука, 1993. Т.29.
С.84-104. (11, 13, 14)
Голубкова Н.С., Бязров Л.Г. Жизненные формы лишайников и лихеносинузии И
Ботан. журн. 1989. Т.74. №6. С.794-805. (18, 19, 125)
Голубкова НС., Малышева НВ. Влияние роста города на лишайники и лихено-
индикация атмосферных загрязнений г. Казани // Ботан. журн. 1978. Т.63.
№8. С. 1145-1154. (95, 101)
Литература и авторский индекс
253
Горбанов Ст., Хаджиатанасов Д. Върху акумуляцията на микоелементи и есте-
ствени радиоактивни елементи от някои видове мъхове и лишен, распрост-
ранени в България // Научн. тр. Внеш, сельскост. ин-т “В. Коларов”. Плов-
див, 1973. Т.22. №4. С.28-37. (34, 179)
Горбач НВ. К вопросу об индикаторной роли лишайников в лесных сообществах
// Проблемы изучения грибов и лишайников. Тарту, 1965. С.177-181. (239)
Горбач Н.В., КобзарьН.Н. Возможности использования Hypogymniaphysodes (L.)
Nyl. в качестве индикатора загрязнения атмосферного воздуха SO2 // Эко-
логия и биология низших растений. Тезисы докладов Всесоюзного симпо-
зиума микологов и лихенологов. Минск, 1982. С.224-225. (43)
Горбач Н.В., Ходасевич Э.В., Мельникова Л.М., Кобзарь Н.Н. Изменение количе-
ственных характеристик фонда хлорофиллов в лишайниках под воздействи-
ем загрязнения воздуха SO, И ДАН БССР. 1982. Т. 11. №9. С.850-852. (73, 88)
Гордеева М.М., Толпышева Т.Ю. О реакции эпифитных лишайников на внесение
удобрений //Лихеноиндикация состояния окружающей среды. Таллин, 1978.
С.41-43. (10)
Горшков В.В. Эпифитные лихеносинузии сосновых лесов Кольского полуострова
(формирование, экология, влияние антропогенных факторов). Автореф. дис.
канд. биол. наук. Л.: БИН АН СССР, 1986. 21 с. (58, 59, 123)
Горшков В.В. Влияние атмосферного загрязнения окислами серм на эпифитный
лишайниковый покров северотаежных сосновых лесов // Лесные экосисте-
мы и.атмосферное загрязнение. Л.: Наука, 1990. С.144-159. (58, 74, 91)
Горшков В.В. Разрушение эпифитного лишайникового покрова стволов сосен в
сосновых лесах Кольского полуострова под действием атмосферного заг-
рязнения // Экология. 1991. №4. С.20-27. (58, 59)
Грейг-Смит П. Количественная экология растений. М.: Мир, 1967. 359 с. (167)
Григорьев Ю.С., Бучельников М.А. Трансплантационная лихеноиндикация заг-
рязнения воздушной среды на основе замедленной флуоресценции хлоро-
филла // Экология. 1997. №6. С.465-467. (144, 150, 155)
Данилов-Данильян В. И. (ред.). О состоянии окружающей природной среды Рос-
сийской Федерации за 1988-1998 гг. (аналитическая записка). Приложение
2 к Государственному докладу “О состоянии окружающей природной сре-
ды Российской Федерации в 1998 г.”. Госкомитет РФ по охране окружающей
среды: Гос. центр экологических программ. М., 1999. 135 с. (53, 54, 64, 65)
Джураева 3. Индикационное значение лишайников при определении некоторых
литологических и инженерно-геологических условий в экосистемах арид-
ных территорий И Лихеноиндикация состояния окружающей среды. Тал-
лин, 1978. С. 114-117. (10)
Джураева 3. Лишайниковые группировки в основных растительных ассоциаци-
ях такыров и такыровидных почв // Изв. АН ТуркмССР. Сер. биол. 1979.
№3. С.37-43. (47)
254
Литература и авторский индекс
Добровольский И.А., Гаевая Н.В. Лишайниковая растительность искусственных
лесных насаждений степи и ее индикаторная роль // Вопросы степного ле-
соведения, научные основы лесной рекультивации земель. Днепропетровск,
1985. С.92-97. (29)
Дылис Н.В. Межбиогеоценозные связи, их механизмы и изучение // Проблемы
биогеоценологии. Е.М.Лавренко, Т.А.Работнов-ред. М.: Наука, 1973. С.71-
79. (48)
Еленкин А.А. Лихенологические заметки. 1. // Изв. Импер. СПб Ботан. Сада. 1901.
Т.1.№З.С 117-123.(25/
Ермолаева-Маковская А.П., Литвер Б.Я. Свинец-210 и полоний-210 в биосфере.
М.: Атомиздат, 1978. 130 с. (195)
Жизнь растений. М.: Просвещение, 1977. Т.З. 488 с. (15)
Зайцев ГН. Методика биометрических расчетов. М.: Наука, 1973. 256 с. (168)
Закутнова В.И. Эпифитные лишайники как индикаторы загрязнения атмосфер-
ного воздуха И Изв. АН Азерб. ССР. Сер. биол. 1988. №4. С. 122-128. (102)
Залиханов А.М. Природная индикация экологических модификаций в районе
Тырнаузского горно-металлургического комбината И Экологическая моди-
фикация и критерии экологического нормирования: Труды Международно-
го симпозиума, Нальчик, 1-12 июня 1990 г. Л., 1991. С.368-372. (60)
Золотарева Б.Н., Скрипниченко И.И., Мартин Ю.Л. Лишайники - индикаторы
загрязнения среды тяжелыми металлами И Природа. 1981. №1. С.86-88.
(179)
Иванов Е.А., Рамзина Т.В., ХамьяновЛ.П., Васильченко В.Н., Коротков В.Т., Но-
совский А.В., Осколков Б.Я. Радиоактивное загрязнение окружающей сре-
ды Ат-241 вследствие аварии на Чернобыльской АЭС // Атомная энергия.
1994. Т.77. №2. С.140-145. (199)
Иванов М.А. Эпигейные лишайники в проблеме индикации окружающей среды
Воркутинского промышленного комплекса//Тр. Коми научного центра УрО
РАН. 1993. Т.135. С.52-58. (65)
Иванова Н.В. Структура и эволюция генов рРНК у лишайников семейства
Umbilicariaceae. Автореф. дис. канд. биол. наук. М.: МГУ, 1998. 22 с. (17)
Израэль Ю.А., Филиппова Л.М., Ровинский Ф.Я. Программа экологического мо-
ниторинга в биосферных заповедниках И Биосферные заповедники. Труды
2 советско-американского симпозиума. Л.: Гидрометеоиздат, 1982. С.128-
141. (9)
Инсаров Г.Э., Инсарова ИД. Лишайники в условиях фонового загрязнения ат-
мосферы двуокисью серы // Проблемы экологического мониторинга и мо-
делирования экосистем. Л.: Гидрометеоиздат, 1986. Т.9. С.242-258. (177)
Инсаров Г.Э., Пчелкин А.В. Сравнение различных методов учета лишайников-
эпифитов // Проблемы экологического мониторинга и моделирования эко-
систем. Л.: Гидрометеоиздат, 1983. Т.6. С.90-101. (94)
Литература и авторский индекс
255
Инсаров Г.Э., Пчелкин А.В. Количественные характеристики состояния эпифит-
ной лихенофлоры Курильского заповедника. М., 1988. 174 с. (94)
Инсарова ИД. Влияние сернистого газа на лишайники // Проблемы экологичес-
кого мониторинга и моделирования экосистем. Л.: Гидрометеоиздат, 1982.
Т.5. С.33-48. (177)
Инсарова И.Д. Влияние тяжелых металлов на лишайники // Проблемы экологи-
ческого мониторинга и моделирования экосистем. Л.: Гидрометеоиздат,
1983. Т.6. С. 101-113. (164, 177)
Инсарова И.Д., Инсаров Г.Э. Анализ роста лишайников // Проблемы экологичес-
кого мониторинга и моделирования экосистем. Л.: Гидрометеоиздат, 1987.
Т.10. С.87-114. (80)
Инсарова ИД., Инсаров Г.Э. Сравнительные оценки чувствительности эпифит-
ных лишайников различных видов к загрязнению воздуха // Проблемы эко-
логического мониторинга и моделирования экосистем. Л.: Гидрометеоиз-
дат, 1989. Т.12. С. 113-175. (74, 89, 90, 91, 210)
Исаев А.А. Экологическая климатология. М.: Научный мир, 2001.456 с. (53, 55,
56, 57, 178, 193, 195)
Использование эпифитных лишайников для индикации атмосферного загрязне-
ния (методические рекомендации). Составитель В.В. Горшков. Апатиты:
Институт промышленной экологии Севера КНЦ АН СССР, 1990.48 с. (123)
История Москвы. С.С.Хромов и др. -ред. М.: Наука, 1978. 543 с. (201, 202)
Казачевский ИВ., Ветров В.А., Трасс Х.Х. Нейтронно-активационный анализ
лишайников, хвои кедра и пихты для контроля загрязнения атмосферного
воздуха в Южном Прибайкалье // Ядерно-физические методы анализа в
контроле окружающей среды. Труды 3 Всесоюзного совещания, Томск 21-
23 мая 1985 г. Л.: Гидрометеоиздат, 1987. №4. С.91-102. (177, 179)
Квасникова Е.В., Стукин Е.Д., Голосов В.Н. Неравномерность загрязнения цези-
ем- 137 территорий, расположенных на большом расстоянии от Чернобыль-
ской АЭС // Метеорология и гидрология. 1999. №2. С.5-11. (193)
Киселев В.Н., ЧубановК.Д., Бойко А.В., Лукашев В.К, Суровая Т.П., Бортник А.В.,
Пикулик Н.И. Лихеноиндикация загрязнения воздушной среды зеленых зон
промышленных центров Белоруссии // Экология. 1986. №2. С.30-35. (43, 184)
Киселев В.Н., Чубанов К.Д., Бойко А.В., Пикулик Н.И. Анализ техногенного заг-
рязнения соединениями серы зеленой зоны Минска по содержанию суль-
фат-ионов в лишайнике Hypogymniaphysodes И ДАН БССР. 1983. Т.27. № 12.
С.1109-1111. (184)
Клауснитцер Б. Экология городской фауны. М.: Мир, 1990. 246 с. (202)
Климат Москвы за последние 30 лет. М.: МГУ, 1989. 94 с. (201)
Кондратюк С.Я., Кучерявый В.А., Крамарец В.А. Сравнительное лихеноиндика-
ционное картирование городов Украины И Укр. ботан. журн. 1993а. Т.50.
№4. С.74-83. (102, 114)
256
Литература и авторский индекс
Кондратюк С.Я., Кучерявый В.А., Крамарец В.А., Зинько Ю.В., Сиренко И.Н.
Лихеноиндикация загрязнения воздуха в г. Львове // Укр. ботан. журн. 1991.
Т.48. №2. С.72-76. (102, 105, 114)
Кондратюк С.Я., Навроцкая И.Л., БруньГ.А., Люгин В.А. Содержание радионук-
лидов в лишайниках Украинского Полесья (1990-1991) // Укр. ботан. журн.
19936. Т.50. №3. С.13-22. (192)
Котлов Ю.В. Лишайники, собранные на птичьей колонии в северо-западной ча-
сти острова Большевик (Северная Земля) // Ботан. журн. 1993. Т.78. №8.
С.34-36. (28)
Котлов Ю.В. О моделировании эволюции основных жизненных форм лишайни-
ков И Ботан. журн. 1995. Т.80. №3. С.26-30. (18)
Кравчук Л. А. Лихеноиндикация загрязнения атмосферного воздуха городов Бела-
руси. Автореф. дис. канд. геогр. наук. Минск: БГУ, 2001.21 с. (123)
Красная книга города Москвы. М., 2001.622 с. (208)
Криволуцкий Д.А. Почвенная фауна в экологическом контроле. М.: Наука, 1994.
272 с. (10)
Крючков В.В., Сыроид НА. Лишайники как биоиндикаторы качества окружаю-
щей среды в северной тайге И Экология. 1990. №6. С.63-66. (160)
Курсанов А.Л., Дьячков НИ. Лишайники и их практическое использование. М.-
Л.: АН СССР, 1945. 55 с. (179)
Кучерявый В.А., Кондратюк С.Я., Крамарец В.А., Зинько Ю.В, Сиренко И.Н.
Лихеноиндикационная оценка степени загрязненности атмосферного воз-
духа в г. Львов И ДАН УССР. Сер. Б: геол., хим. и биол. науки. 1989. №11.
С.67-72. (102, 105, 114)
Лапицкая С.К., Свириденко В.Г., Поломарчук А.С., Закриничная С.П., Шахрай
О.П. Содержание микроэлементов в лишайниках Припятского заповедни-
ка (БССР) // Растительные ресурсы. 1979. Т.15. №4. С.584-587. (179)
Лебедева А.Ф., СаванинаЯ.В., БарскийЕ.Л., ГусевМ.В. Устойчивость цианобак-
терий и микроводорослей к действию тяжелых металлов: роль металлосвя-
зывающих белков // Вестник МГУ. Сер. 16. Биология. 1998. Вып. 2. С.42-
49. (46)
Левин Ф.И Роль лишайников в выветривании известняков и диоритов И Вестник
МГУ. 1949. №9. С.149-159. (31, 51)
Левина В. И. Определение массы ежегодного опада в двух типах соснового леса
на Кольском полуострове//Ботан. журн. 1960. Т.45. №3. С.418-423. (31, 79)
Левицкая К. И. Зольность доминантных видов основных фитоценозов района гео-
графической станции “Железо” (Ленинградская область) // 27 Герценовс-
кие чтения. Биология (научные доклады). Л.: ЛГПИ им. А.И.Герцена, 1975.
С.49-55. (31,179)
Лекарственно-техническое сырье. Сборник стандартов. М.: Стандарты, 1958.
431 с. (173)
Литература и авторский индекс
257
Лийв С.Э. Лихеноиндикация состояния загрязнения воздуха в г. Вал га // Флора и
группировки низших растений в природных и антропогенных экстремаль-
ных условиях среды. Таллин, 1984. С. 187-202. (102)
Лийв С.Э. Лихеноиндикация загрязнения воздуха в городах южной Эстонии. Авто-
реф. дис. канд. биол. наук. Свердловск: ИЭРЖ УрО АН СССР, 1988.16 с. (102)
ЛийвС.Э., МартинЛ.Н. Сравнениеполеотолерантностиэпифитныхлишайников
в городах Эстонии // Экологические особенности низших растений Совет-
ской Прибалтики. Вильнюс, 1977. С.130-132. (102)
Локинская М.А. Некоторые данные о составе зольных элементов в лишайниках
рода Cladonia северо-востока Сибири И Водоросли и грибы Сибири и Даль-
него Востока. Новосибирск: Наука, 1970. 4.1(3). С.230-232. (35, 179)
Магомедова М.А. Лишайники как индикаторы ландшафтных различий в горных
тундрах Северного Урала // Охрана и рациональное использование биоло-
гических ресурсов Урала (информационные материалы). 4. Флора и фауна.
Свердловск, 1980. №4. С.55-57. (10)
Магомедова М.А. Возможности лихеноиндикации в высокогорьях // Седьмая кон-
ференция по споровым растениям Средней Азии и Казахстана: тезисы док-
ладов. Алма-Ата, 1984. С.343-344. (10)
Магомедова М.А. Лишайники как компонент северных экосистем и объект мони-
торинга // Проблемы экологического мониторинга и моделирования экоси-
стем. СПб.: Гидрометеоиздат, 1996. Т.16. С.105-121. (10)
Магомедова М.А., Корытин Н.С., Нифонтова М.Г., Ендукин А.Ю. Влияние вы-
паса оленей на лишайниковый покров сосновых лесов // Рациональное ис-
пользование и охрана растительного мира Урала. Свердловск, 1991. С.72-
78. (10, 78)
Максимова В.Ф., Бязров Л.Г. Оценка загрязнения воздуха по данным о распрост-
ранении лишайников [карта] // Экологический атлас Москвы. М.: АБФ, 2000.
С.66-67. (98, 114, 200, 210)
Малышева А.Г. Закономерности трансформации органических соединений в ок-
ружающей среде И Гигиена и санитария. 1997. №3. С.5-10. (17, 245)
Малышева Н.В. Лишайники Старо-Петергофского парка Биологического инсти-
тута Санкт-Петербургского государственного университета и изменение
лихенофлоры парка за 65 лет И Вестник СПбУ Серия 3. 1992. №17. С.33-
38. (95)
Малышева Н.В. Лишайники окрестностей Ленинграда. 1. Изменение видового
состава лишайников в окрестностях станции Ольгино (Ленинградская об-
ласть) за 72 года И Новости систематики низших растений. СПб.: Наука,
1993. Т.29 С.119-124. (95)
Малышева Н.В. Об экологической патоморфологии лишайников в окрестностях
Санкт-Петербурга // Новости систематики низших растений. СПб: Наука,
1995. Т.30. С.78-85. (79)
258
Литература и авторский индекс
Малышева Н.В. Биоразнообразие лишайников и оценка экологического состоя-
ния парков окрестностей Санкт-Петербурга // Новости систематики низ-
ших растений. СПб.: Наука, 1996а. Т.31. С.135-137. (95)
Малышева Н.В. Лишайники Санкт-Петербурга. 1. Современная лихенофлора и
ее анализ И Ботан. журн. 19966. Т.81. №6. С.23-30. (95)
Малышева Н.В. Лишайники Санкт-Петербурга. 2. Изменения лихенофлоры за270
лет // Ботан. журн. 1996в. Т.81. №7. С.55-72. (95)
Малышева Н.В. Морфолого-анатомическое строение накипных лишайников в
условиях загрязнения окружающей среды И Новости систематики низших
растений. СПб.: Наука. 1996г. Т.31. С. 130-134. (79)
Малышева Н.В. Лишайники Санкт-Петербурга. 3. Влияние городских условий и
лихеноиндикация атмосферного загрязнения И Ботан. журн. 1998. Т.83. №9.
С.39-45. (65, 95, 102, 105)
Малышева Н.В. Лишайники городов Ивановской области // Ботан. журн. 1999.
Т.84. №2. С.59-67. (65)
Малышева Н.В., Булыгина НЕ. Эпифитные лишайники арборетума Санкт-Пе-
тербургской лесотехнической академии и выяснение их индикаторной роли
И Изв. ВУЗов. Лесной журнал. 1996. №6. С. 13-18. (95)
Малышева Т.В. Использование напочвенного лишайникового и мохового покрова
для индикации стадий рекреационной дигрессии сосняков Подмосковья //
Лихеноиндикация состояния окружающей среды. Таллин, 1978. С.38-40. (10)
Малышева Т. В., Толпышева Т.Ю. Влияние вытаптывания на восстановление на-
почвенного покрова // Биогеохимические аспекты криптоиндикации. Тал-
лин, 1982. С.52-53. (10, 28, 205)
Манаков К.Н Продуктивность и биологический круговорот в тундровых биогео-
ценозах Кольского полуострова. Л.: Наука, 1972. 147 с. (179)
Маркерт Б., Второва В.Н. Кадастры концентраций химических элементов рас-
тений лесных экосистем Восточной Европы И Изве. РАН. Сер. биол. 1995.
№5. С.545-553. (179)
Маркина Н.В., Рязанов Д.К., Павлов В.В., Личадеев В.В., Тэн Э.В., Музалева О.В.
Использование инструментального нейтронно-активационного анализа для
исследования элементного состава проб растительного и животного мира
//Атомная энергия. 1995. Т.79. №3. С.201-203. (177)
Мартин Л.Н. Лихеноиндикационное картирование г. Таллина // Лихеноиндика-
ция состояния окружающей среды. Таллин, 1978. С. 134-139. (102)
Мартин Л.Н. Лихеноиндикация в условиях различного загрязнения воздуха. Ав-
тореф. дис. канд. биол. наук. Свердловск: ИЭРЖ УНЦ АН СССР, 1984.
17 с. (36)
Мартин Л.Н, Каннукене Л., Костенчук НА. Лишайники и мхи как показатели
состояния окружающей среды в Приокско-Террасном заповеднике // Лихе-
ноиндикация состояния окружающей среды. Таллин, 1978. С.49-56. (66)
Литература и авторский индекс
259
Мартин Л.Н., Нильсон Э.М. Распространение эпифитных лишайников в городе
Кохтла-Ярве И Человек, растительность, почва. Тарту, 1983. С.62-80. (65)
Мартин Ю.Л. Формирование лишайниковых синузий на моренах ледников По-
лярного Урала. Автореф. дис. канд. биол. наук. Свердловск: ИЭРЖ УФ АН
СССР, 1967. 22 с. (10, 27)
Мартин Ю.Л. Лихенометрическая индикация времени обнажения каменистого
субстрата // Экология. 1970. №5. С. 16-24. (10)
Мартин Ю.Л. Биогеохимическая индикация загрязнения среды при помощи
споровых растений // Изв. АН ЭССР. Сер. биол. 1985. Т.34. №1. С.1-15.
(164)
Мартин Ю.Л. Биогеохимические аспекты криптоиндикации И Биогеохимичес-
кий круговорот веществ в биосфере. М.: Наука, 1987. С.95-101. (164)
Медведев Ж.А. Чернобыльские радионуклиды за пределами СССР. Европейский
континент// Радиобиология. 1991. Т.31. №6. С.771-793. (193)
Михайлова И.Н. Возможности использования характеристик эпифитных лишай-
никовых группировок для индикации аэротехногенного загрязнения // Спо-
ровые растения Крайнего Севера России. Сыктывкар, 1993. С.72-83. (60,
118)
Михайлова И.Н. Сообщества эпифитных лишайников // Экологическое нормиро-
вание техногенных загрязнений наземных экосистем. Е.Л.Воробейчик,
О.Ф.Садыков, М.Г.Фарафонтов - ред. Екатеринбург: Наука, 1994. С.210-
216. (60, 118)
Михайлова И.Н. Эпифитные лихеносинузии лесов Среднего Урала в условиях
аэротехногенного загрязнения. Автореф. дис. канд. биол. наук. Екатерин-
бург: ИЭРЖ УрО РАН, 1996. 24 с. (60, 118)
Михайлова И.Н., Воробейчик Е.Л. Эпифитные лихеносинузии в условиях хими-
ческого загрязнения: зависимости доза - эффект // Экология. 1995. №6.
С.455-460. (60)
Михайлова И.Н., Воробейчик Е.Л. Размерная и возрастная структура популяций
эпифитного лишайника Hypogymniaphysodes (L.) Nyl. в условиях антропо-
генного загрязнения // Экология. 1999. №2. С. 130-137. (60)
Моисеева Е.Н. Биохимические свойства лишайников и их практическое значе-
ние. М.-Л.: АН СССР, 1961. 82 с. (179)
Москва. План города. Масштаб 1:38000. М.: ГУГК СССР, 1989. 1 лист. (109)
Нильсон Э.М., Мартин Л.Н. Эпифитные лишайники в условиях кислотного и
щелочного загрязнения // Взаимодействие лесных экосистем и атмосфер-
ных загрязнителей. Таллин, 1982. 4.2. С.88-100. (36)
Нифонтова М.Г. Экспериментальные исследования накопления цезия-137 лишай-
никами // Материалы Всесоюзного симпозиума “Теоретические и практи-
ческие аспекты действия малых доз ионизирующей радиации, 1973”. Сык-
тывкар, 1973. С.167-168. (198)
260
Литература и авторский индекс
Нифонтова М.Г. Накопление Sr-90 и Cs-137 лишайниками в условиях экспери-
мента// Экология. 1976. №1. С.89-92. (198)
Нифонтова М.Г., Куликов Н.В. О накоплении Sr-90 и Cs-137 лишайниками в при-
родных условиях // Экология. 1977. №3. С.93-96. (198)
Нифонтова М.Г., Куликов Н.В., Таршис Г.И., Дьяченко А.П. Радиоэкологическое
изучение природных экосистем в зоне атомных электростанций // Эколо-
гия. 1988. №3. С.40-45. (193)
Нифонтова М.Г., Лебедева А.В., Куликов Н.В. Накопление Sr-90 и Cs-137 живы-
ми и мертвыми лишайниками И Экология. 1979. №1. С.94-97. (30)
Норин Б.Н., Ярмишко В.Т.- ред. Влияние промышленного атмосферного загряз-
нения на сосновые леса Кольского полуострова. Л.: БИН АН СССР, 1990.
195 с. (58, 59, 60, 184)
Обухов А.М. Контроль чистоты воздушного океана // Город, природа, человек. М.:
Мысль, 1982. С.91-108. (210)
Окснер А.Н. Взаимоотношения между лишайниками в литотических группиров-
ках степных заповедников Украины // Укр. ботан. журн. 1961. Т.18. №6.
С.64-73. (26, 27)
Окснер А.Н. Морфология, систематика и географическое распространение И Опре-
делитель лишайников СССР. Л.: Наука, 1974. Вып.2. С.1-283. (11, 16,17,18)
Отнюкова Т.Н. Диагностика загрязнения атмосферы по состоянию ягельного
лишайника // Доклады Российской академии сельскохозяйственных наук.
1997а. №3. С.21-22. (66, 75, 79)
Отнюкова Т.Н. Морфологическое состояние Cladina stellaris (Cladoniaceae,
Lichenes) как показатель атмосферного загрязнения // Ботан. журн. 19976.
Т.82. №3. С.57-66. (66, 75, 79)
Оценка и регулирование качества окружающей природной среды. М.: НУМЦ
Минприроды России, изд. дом “Прибой”, 1996. 350 с. (53, 54, 55)
Пауков А.Г. Лихенофлора урбоэкосистем. Автореф. дис. канд. биол. наук. Екате-
ринбург: ИЭРЖ УрО РАН. 2001. 18 с. (123)
Перельман А. И., Касимов НС. Геохимия ландшафтов. М.: Астрея-2000, 1999.
768 с. (42, 48)
Петрушина М.Н Геохимические особенности лишайника Caloplaca elegans в
сфере воздействия горно-промышленного производства// Влияние промыш-
ленных предприятий на окружающую среду. М.: Наука, 1987. С. 182-189.
(60, 184)
Плакунова О.В., Плакунова В.Г., Гусев М.В. Физиология эпигейных лишайников
в связи с ранней индикацией загрязнения окружающей среды // Изв. АН
СССР. Сер. биол. 1984. №6. С.888-896. (82)
Покаржевский А.Д. Геохимическая экология наземных и почвенных животных
(биоиндикационные и радиоэкологические аспекты). Автореф. дис. канд.
биол. наук. М.: ИЭМЭЖ РАН, 1993. 40 с. (10)
Литература и авторский индекс
261
Полынов Б.Б. Первые стадии почвообразования на массивно-кристаллических
породах И Почвоведение. 1945. №7. С.325-339. (31, 42, 51, 179)
Последствия Чернобыльской катастрофы: здоровье среды / В.М.Захаров,
Е.Ю.Крысанов-ред. М.: Центр экологической политики, 1996. 169 с. (243)
Поташева М.А. Влияние аэротехногенных выбросов Костомукшского горно-обо-
гатительного комбината на эпифитный лишайниковый покров сосны И
Новости систематики низших растений. СПб.: Наука, 1995. Т.30. С.85-89.
(67, 62)
Предварительная программа по геосистемному мониторингу в биосферных за-
поведниках. М.: ИГ АН СССР, 1985. 96 с. (9)
Пристяжнюк С.А. Жизненные формы лишайников субарктических тундр полу-
острова Ямал. 1. Система жизненных форм // Ботан. журн. 1996. Т.81. №3.
С.34-42. (18)
Пчелкин А.В. Лихеноиндикационное картирование территории парка “Сокольни-
ки” И Современные проблемы микологии, альгологии и фитопатологии.
Сборник трудов Международной конференции (Москва, апрель 1998 г.).
М.: МГУ, 1998. С.379-380. (105)
ПярнА.И., Трасс Х.Х. Эпифитные лишайники горных лесов хребта Хамар-Дабан
(Прибайкалье) // Ботан. журн. 1990. Т.75. №3. С.358-368. (66)
Пярн А., Трасс X., Цобель К. Лихеноиндикация степени загрязненности атмос-
ферного воздуха в Прибайкалье // Биогеохимические аспекты криптоинди-
кации. Таллин, 1982. С.37-38. (66)
Работное Т.А. Экологические наблюдения над лишайниками в Южной Якутии
// Советская ботаника. 1936. №6. С. 149-153. (21)
Работное Т.А. Фитоценология. М.: МГУ, 1983. 296 с. (128)
Работное Т.А., Говорухин В.С. Lichenes- лишайники // Кормовые растения сено-
косов и пастбищ СССР. И.В.Ларин - ред. М.-Л.: Гос. изд. сельскохоз. лите-
ратуры, 1950. Т.1. С.54-98. (31, 179)
Родин Л. Е., Базилевич Н. И. Динамика органического вещества и биологический
круговорот в основных типах растительности. М.-Л.: Наука, 1965. 253 с.
(179)
Родин Л.Е., Ремезов Н.П., Базилевич Н.И. Методические указания к изучению
динамики и биологического круговорота в фитоценозах. Л.: Наука, 1968.
143 с. (31)
Родникова И.М., Скирина И.Ф., Христофорова Н.К. Оценка воздушной среды в
Лазовском заповеднике (Приморский край) методами лихеноиндикации //
Ботан. журн. 1998. Т.83. №5. С.48-56. (66, 102)
Розенберг ГС. Анализ определений и понятия “экология” И Экология. 1999. №2.
С.89-98. (238.)
Савоскул О.С. Лихенометрический метод датирования голоценовых отложений
//Изв. РАН. Сер. географ. 1995. №5. С.94-101. (10)
262
Литература и авторский индекс
Сает Ю.Е., Ревич Б.А., Янин Е.П., Смирнова Р.С., Башаркевич И.Л., Онищенко
Т.Л., Павлова Л.И, Трефилова Н.Я., Ачкасов А.И., Саркисян С.Ш. Геохи-
мия окружающей среды. М.: Недра, 1990. 335 с. (56)
Сафонова М.Ю., Саканян Е.И., Лесиовская Е.Е. Cetraria islandica (L.) Ach.: хи-
мический состав и перспективы применения в медицине // Растительные
ресурсы. 1999. Т.35. №2. С. 106-115. (179)
Седельникова Н.В. Лишайники - индикаторы горных пород // Нетрадиционные
методы в исследованиях растительности Сибири. Новосибирск: Наука, 1982.
С.35-40. (10.)
Седельникова Н.В. Лишайники Горного Алтая и Кузнецкого нагорья. Автореф.
дис. докт. биол. наук. Новосибирск: СБС СО АН СССР, 1994. 33 с. (26, 122,
124)
Седельникова Н.В., Седельников В.П. Роль лишайниковых синузий в высокогор-
ных фитоценозах северной части Алтае-Саянской горной области И Ботан.
журн. 1979. Т.64. №5. С.671-679. (125)
Седельникова Н.В., Черемисин Д.В. Использование лишайников для датировки
петроглифов //Сибирский экологический журнал. 2001. №4. С.479-481. (10)
Скирина И.Ф., Качур А.Н. Накопление тяжелых металлов в лишайниках как ин-
дикатор состояния окружающей среды (на примере горно-металлургиче-
ского и химического производств ПО “Дальполимерметалл”) // Сихотэ-
Алинский биосферный район: производственно-природные отношения.
Владивосток: ДВНЦ АН СССР, 1991. С. 109-117. (60, 184)
Скрипниченко И.И., Золотарева Б.Н., Мартин Ю.Л. Мхи и лишайники как ин-
дикаторы содержания ртути в окружающей среде // Лихеноиндикация со-
стояния окружающей среды. Таллин, 1978. С.72-74. (179)
Слепян Э.И., Шейнерман Н.А., Кольчевский А.Г Бенз(а)пирен в Hypogymnia
physodes (L.) Nyl. и Parmelia sulcata TayL, произрастающих на коре Betula
pendula Roth., Populus tremula L. и Pinus sylvestris L. в лесопарковой зоне
Ленинграда, лихеноиндикация и экологический мониторинг // Проблемы
фитогигиены и охраны окружающей среды. Л.: ЗИН АН СССР, 1981. С. 145-
152. (178)
Слука 3. А., Абрамова Л.И. Мхи и лишайники лесопарка “Лосиный остров” И
Вестник МГУ. Сер. 16, Биология. 1984. №2. С. 11-19. (205)
Соколов В.С., Бочаров Б.В., Криволуцкий Д.А. Экотоксикология и проблемы за-
щиты окружающей среды от загрязнений // Экотоксикология и охрана при-
роды. Д.А.Криволуцкий, Б.В.Бочаров-ред. М.: Наука, 1988. С.4-19. (74)
Соколов Л.С., Астрахан Е.Д. Загрязнение территории Москвы металлами И При-
рода. 1993. №7. С.68-73. (232)
Солдатенкова Ю.П. Некоторые данные о составе зольных элементов в Hypogymnia
physodes (L.) Nyl. // Лихеноиндикация состояния окружающей среды. Тал-
лин, 1978. С.98-101. (179)
Литература и авторский индекс
263
Стебаева СК, Седелъникова Н.В. Население коллембол (Hexapoda, Collembola)
лишайниковых консорций нагорья Сангилен // Сибирский экологический
журнал. 1999. №5. С.509-513. (28)
Стебаева С.К., Седельникова Н.В., Андриевский В.С., Волонихина И.И. Сообще-
ства микроартропод под лишайниками на хребте Восточный Танну-Ола
(Тува) И Зоол. журн. 2001. Т.80. №2. С. 170-182. (28)
Сукачев В.Н. Основные понятия лесной биогеоценологии // Основы лесной био-
геоценологии. М.: Наука, 1964. С.5-49. (48, 118)
Таисаев Т.Г., Константинова ИМ. Концентрация золота в ягеле на золоторудных
полях гольцовых ландшафтов И ДАН СССР. 1988. Т.302. №3. С.706-709.
(10, 30)
Тищенко Н. Ф. Охрана атмосферного воздуха. Расчет содержания вредных веществ
и их распределение в воздухе. Справочник. М.: Химия, 1991.368 с. (53, 54)
ТрассХ.Х. Некоторые вопросы фитоценологического изучения лишайников //Уч.
зап. Латвийского университета. 1966. Т.74. №2. С.122-133. (117, 125)
Трасс Х.Х. Анализ лихенофлоры Эстонии. Автореф. дис. докт. биол. наук. Л.:
БИН АН СССР, 1968. 80 с. (89, 101, 121)
Трасс Х.Х. Полеотолерантность лишайников // Материалы 6 симпозиума ми-
кологов и лихенологов Прибалтийских республик. Рига, 1971. Т. 1. С.66-70.
Трасс Х.Х. Биоиндикация состояния атмосферной среды городов // Экологиче-
ские аспекты городских систем. Минск: Наука и техника, 1984. С.96-109.
(101)
Трасс Х.Х. Классы полеотолерантности лишайников и экологический мониторинг
И Проблемы экологического мониторинга и моделирования экосистем. Л.:
Гидрометеоиздат, 1985а. Т.7. С.122-137. (90, 101, 135)
ТрассХ.Х. Трансплантационные методы лихеноиндикации // Проблемы экологи-
ческого мониторинга и моделирования экосистем. Л.: Гидрометеоиздат,
19856. Т.8. С. 140-144. (137, 139, 143, 149, 153)
Трасс XX. Лихеноиндикационные индексы и SO2 // Биогеохимический кругово-
рот веществ в биосфере. М.: Наука, 1987. С.111-115. (106)
ТрассХ.Х., ПярнА.И., Цобелъ К.Р. Лихеноиндикационная оценка степени загряз-
ненности атмосферной среды Южного Прибайкалья // Уч. зап. Тартуского
университета. 1988. №812. С.32-46. (66, 102, 105)
Троицкая М.Н., Рамзаев П.В., Моисеев А.А., Нижников А.И., Бельцев Д.И., Иба-
туллин М.С., Литвер Б.Я., Дмитриев ИМ. Радиоэкология ландшафтов
Крайнего Севера// Современные проблемы радиоэкологии. М., 1971. Т.2.
С.325-353. (198)
Турманина В. И. Исследование эволюции ледника Джанкуат фитоиндикационны-
ми методами // Материалы гляциологических исследований. 1971а. №18.
С. 106-109.(70)
264
Литература и авторский индекс
Турманина В. И. Перспективы применения фитоиндикационных методов в гляци-
ологии // Фитоиндикационные методы в гляциологии. М., 19716. С.5-19.
(10)
Турманина В.И., Баденкова С.В. Тяжелые металлы в эпифитных лишайниках
Приэльбрусья // Биогеохимические аспекты криптоиндикации. Таллин, 1982.
С.43-44. (179)
Тютюнник Ю.Г Лишайники как индикаторы загрязнения окружающей среды
свинцом // Укр. ботан. журн. 1987. Т.44. №5. С.73-75,112. (182)
Урбанавичюс Т.П. Лихеноиндикация современных и палеоклиматических усло-
вий Южного Прибайкалья // Изв. РАН. Сер. географ. 2002. №1. С.81-90.
Финни Д. Введение в теорию планирования экспериментов. М.: Наука, 1970.
287 с. (168)
ЧерриР.Д. Альфа-активность планктона// Вопросы радиоэкологии. М.: Атомиздат,
1968. С.252-262. (195)
Шапиро И.А. Азотный обмен у лишайников и его регуляция // Ботан. журн. 1986.
Т.71. №7. С.841-850. (11, 84)
Шапиро И.А. Влияние сернистого ангидрида на содержание азота и пероксидаз-
ную активность у лишайников // Ботан. журн. 1993. Т.78. №6. С.66-72. (11,
84)
Шапиро И.А. Физиолого-биохимические изменения у лишайников под влиянием
атмосферного загрязнения И Успехи современной биологии. 1996. Т.П6.
№2. С. 158-171. (11, 72, 82, 88)
Шапиро И.А., Нифонтова М.Г. Действие сернистого газа и гамма-излучения на
нитроредуктазную активность у лишайника Lobariapulmonaria (L.) Hoffm.
// Экология. 1991. №3. С.47-51. (84)
Шенников А.П. Экология растений. М.: Советская наука, 1950. 376 с. (24)
Шуберт Р. - ред. Биоиндикация загрязнений наземных экосистем. М.: Мир, 1988.
348 с. (100)
Щелкунова Р.П. Прирост кормовых лишайников и делихенизация растительного
покрова на Енисейском Севере И Сибирский вестник сельскохозяйствен-
ной науки. 1978. №2. С.36-43. (28)
Эйзенбад М. Радиоактивность внешней среды. М.: Атомиздат, 1967. 332 с. (195)
Юницкий А. О значении в лесоводстве лихенологии и о коллектировании лишай-
ников в связи с производством лихенологических наблюдений. СПб, 1909.
8 с. (23)
Якушев Б.И., Будкевич Т.А., Заболотный А.И., Миронов В.П., Кудряшов В.К.,
Ярошевич О.И., ЖукИ.В. Америций-241 в растительности природных ком-
плексов и агроценозов на территории Беларуси, загрязненной аварийными
выбросами ЧАЭС И Доклады Национальной АН Беларуси. 2000. Т.44. №2.
С.73-76. (199)
Литература и авторский индекс
265
Ярилова Е.А. Роль литофильных лишайников в выветривании массивно-крис-
таллических пород И Почвоведение. 1947. Т.42. №9. С.533. (31, 51, 179)
Ackermann I. Flechtenkartietung in Raum Leipzig // Flechtenmonitoring - ein
kommunales Kontrollinstrument. P.Scholz-ed. Halle: Unabhangingen Institutes
fur Umweltfragen, 1993. S.21-34. (100)
Adamson E., SeppeltR.D. A comparison of airborne alkaline pollution damage in selected
lichens and mosses at Casey Station, Wilkes Land, Antarctica // Antarctic
ecosystems. Ecological change and conservation / K.P.Kerry, G.Hempel - eds.
Berlin: Springer-Verlag, 1990. P.347-353. (79)
Addison P.A. Quantification of branch dwelling lichens for the detection of air pollution
impact// Lichenologist. 1984. V.16. P.297-304. (140, 149)
Ahmadjian V. The lichen symbiosis. New York: John Wiley & Sons, Inc., 1993. 250 p.
(12, 15, 33)
Ahti T, Vitikainen O. Bacidia chlorococca, a common toxitolerant lichen in Finland //
Memoranda Soc. Fauna Flora Fennica. 1974. V.49. P.95-100. (93)
Alebic-Juretic A., Arko-Pijevac M. Air pollution damage to cell membranes in lichens -
results of simple biological test applied in Rijeka, Yugoslavia // Water, Air, and
Soil Pollution. 1989. V.47. Nl-2. P.25-33. (85)
Alstrup V. Cryptogams on imported timber in West Greenland // Lichenologist. 1977.
V.9.N2. P. 113-117. (29)
Alstrup V., Hansen E.S. Three species of lichens tolerant of high concentrations of cooper
// Oikos. 1977. V.29. N2. P.290-293. (35, 40)
AnonbyJ.E. Epifyttisk lavflora i vestnorsk furuskog - sammenligning av en naturskog
og en kulturskog // Blyttia. 1994. V.52. N2. P.81-87. (240)
vonArb C. Photosynthesis and chlorophyll content of the lichen Parmelia sulcata Taylor
from locations with different levels of air pollution // Progress and problems in
lichenology in the eighties. E.Peveling - ed. J. Cramer, Berlin-Stuttgart. 1987.
P. 343-345. (143)
von Arb C., BrunoldC. Lichen physiology and air pollution. 1. Physiological responses
of in situ Parmelia sulcata among air pollution zones within Biel, Switzerland 11
Canad. J. Bot. 1990. V.68. NI. P.35-42. (139, 143, 158)
von Arb C., Mueller C., Ammann K., Brunold C. Lichen physiology and air pollu-
tion.II. Statistical analysis of the correlation between SO,, N2, N and O3, and
chlorophyll content, net photosynthesis, sulphate uptake and protein synthesis
of Parmelia sulcata Taylor H New Phytologist. 199Q. V.115. P.431-437. (83,
86,89)
Armaleo D., Clerc P Lichen chimeras - DNA analysis suggests that one fungus
forms two morphotypes // Experimental Mycology. 1991. V.15. NI. P.1-10.
Armstrong R.A. The descriptive ecology of saxicolous lichens in an area of South
Merionethshire, Wales//J. Ecology. 1974. V.62. NI. P.33-45. (26)
266
Литература и авторский индекс
Armstrong R.A. The influence of calcium and magnesium on the growth of the lichens
Parmelia saxatilis and Xanthoria parietina on slate substrates // Environ.
Experiment. Bot. 1990. V.30.NI. P.51-57. (143, 154)
Armstrong R.A. Competitive interactions between four foliose lichens on north and
south facing rock surfaces // Environ. Experiment. Bot. 1991. V.31. N1. P.51 -58.
(26, 79)
Armstrong R.A. The growth of six saxicolous lichens transplanted to lime-rich and
lime-poor substrates in South Gwynedd, Wales 11 Symbiosis. 1993. V.15. N3.
P.257-267. (143, 154)
Armstrong R.A. The influence of bird droppings on the growth of lichen fragments
transplanted to slate and cement substrates // Symbiosis. 1994. V.17. NI. P.75-
86. (143)
Arseneau M.-J., OuelletJ-P., Sirois L. Fruticose arboreal lichen biomass accumulation
in an old-growth balsam fir forest // Canad. J. Bot.. 1998. V.76. N10. P.1669-
1676. (240)
Ausmus R.S. An argument for ecosystem level monitoring // Environ. Monit. Assess.
1984. V.4. N3. P.275-293. (114)
Bacci E., Calamari D., Fanelli R., Focardi S, Gaggi C., Moros ini M. Chlorinated
hydrocarbons in lichen and moss samples from the Antarctic Peninsula //
Chemosphere. 1986. V.15.N6. P.747-754. (43, 178, 179)
Baddeley M.S., Ferry B.W.. Finegan E.J. A new method of measuring lichen respi-
ration: response of selected species to temperature, pH and sulphur dioxide //
Lichenologist. 1971. V.5. NI. P.18-25. (87)
de Bakker A. J. Effects of ammonia emission on epiphytic lichen vegetation // Acta Bot.
Neerlandica. 1989. V.38. N3. P.337-342. (115)
Bargagli R. Determination of metal deposition patterns by epiphytic lichens // Toxicol.
Environ. Chem. 1989. V.18. N4. P.249-256 . (174)
Bargagli R. Assessment of metal air pollution by epiphytic lichens: the incidence of
crustal materials and of the possible uptake from substrate barks // Studia
Geobotanica. 1990a. V.10. P.97-103. (171)
Bargagli R. Mercury emission in an abandoned mining area: assessment by epiphytic
lichens 11 Encyclopedia of Environmental Control Technology. Vol. 4. Hazardous
Waste Containment and Treatment. P.N.Cheremisinofif - ed. Houston: Gulf
Publishing Co., 1990b. P.613-640. (184.)
Bargagli R., Barghigiani C. Lichen biomonitoring of mercury emission and deposition
in mining, geothermal and volcanic areas of Italy // Environ. Monit. Assess. 1991.
V.16.N32. P.265-275. (10, 53, 182, 183)
Bargagli R., Barghigiani C., Siegel B.Z., Siegel S.M. Accumulation of mercury
and other metals by the lichen, Parmelia sulcata, at an Italian mine site
and a volcanic area // Water, Air, and Soil Pollution. 1989. V.45. N3-4. P.315-
327. (182)
Литература и авторский индекс
267
Bargagli R., Barghigiani С., Siegel B.Z., Siegel S.M. Trace metal anomalies in sur-
face soils and vegetation on two active island volcanoes: Stromboli and Vulcano
(Italy)// Sci. Total Environ. 1991. V.102. P.209-222. (182)
Bargagli R., Battisti E., Focardi S, Formichi R Preliminary data on environmental
distribution of mercury in northern Victoria Land, Antarctica // Antarctic Science.
1993. V.5.N1. P.3-8. (179, 180)
Bargagli R., Cateni D., Nelli L., Olmastroni S, Zagarese B. Environmental impact of
trace element emissions from geothermal power plants // Arch. Environ.
Contamin. Toxicol. 1997. V.33. N2. P.172-182. (183, 184)
Bargagli R„ Iosco F.P., Barghigiani C. Assessment of mercury dispersal in an abandoned
mining area by soil and lichen analysis // Water, Air, and Soil Pollution. 1987a.
V.36. N1-2. P.219-225. (184)
Bargagli R., Iosco F.P., D'Amato M.L. Zonation of trace metal accumulation in three
species of epiphytic lichens belonging to the genus Parmelia // Cryptogamie,
Bryologie et Lichenologie. 1987b. V.8. N4. P.331-337. (167, 170)
Bargagli R, Monad F, Borghini F, Bravi F, Agnorelli C. Mosses and lichens as
biomonitors of trace metals. A comparison study on Hypnum cupressiforme and
Parmelia caperata in a former mining district in Italy // Environ. Pollut. 2002.
V.116.N2. P.279-287. (184)
Bargagli R., Sanchez-Hernandez J.C., Monad F Baseline concentrations of elements in
the Antarctic macrolichen Umbilicaria decussata // Chemosphere. 1999. V.38.
N3. P.475-487. (179, 180)
Barghigiani C., Bargagli R., Gioffre D. Mercury in the environment of the Mt. Etna
volcanic area// Environmental Technology Letters. 1988a. V.9. N2. P.239-244.
Barghigiani C., Bargagli R., Siegel B.Z., Siegel S.M. Source and selectivity in the
accumulation of mercury and other metals by the plants of Mt. Etna // Water, Air,
and Soil Pollution. 1988b. V.39. P.395-408. (182)
Barghigiani C., Bargagli R., Siegel B.Z., Siegel S.M. A comparative study of mercury
distribution on the Aeolian Volcanos, Vulcano and Stromboli // Water, Air, and
Soil Pollution. 1990. V.53.N1-2. P.179-188. (53, 182)
Bari A., Rosso A., Minciardi M.R., Troiani F., Piervittori R. Analysis of heavy metals in
atmospheric particulates in relation to their bioaccumulation in explanted
Pseudeverniafurfuraceath&Wx H Environ. Monit. Assess. 2001. V.69.N3. P.205-
220. (143)
Barkman J. J. Phytosociology and ecology of cryptogamie epiphytes. Assen: van Gorcum,
1958. 628 p. (20, 21, 22, 23, 24, 27, 35, 89, 117)
Barkman J., Moravec J., Rauschert S. Code of phytosociological nomenclature. 2nd ed.
// Vegetatio. 1986. V.67. N3. P.145—195. (117)
Bartok K. Heavy metal distribution in several lichen species in a polluted area // Rev.
Rom. Biol. Biol. Veget. 1988. V.33. N2. P. 127-134. (143, 146)
268
Литература и авторский индекс
Bartok К., Bercea V. Effectul poluantilordin zona industrial a de juliu asupra continutului
de pigmenti asimilatori al speciilorde licheni Physcia aipolia si Xanthoriaparietina
11 Study si cercetari de biologie, Seria biologie vegetalia. 1991. V.43. Nl-2. P.63-
69. (144)
Bartok K., NicoaraA., Bercea V., Tibor O. Biological responses in the lichen Xanthoria
parietina transplanted in biomonitoring stations // Rev. Rom. Biol. Biol. Veget.
1992. V.37. N2. P.135-142. (145)
Bartok K., Osvath T. Influence of pollutants emanated from the Dej cellulose
manufactures, over lichens // Contributii Botanice. Cluj-Napoca. 1990, P.67-72.
(146, 184)
Bates J. W., Bell J.N.B., Farmer A.M. Epiphyte recolonization of oaks along a gradient
of air pollution in South-East England, 1979-1990//Environ. Pollut. 1990. V.68.
Nl-2. P.81-99. (94.)
Bates J.W., Bell J.N.B., Massara A.C. Loss of Lecanora conizaeoides and other
fluctuations of epiphytes on oak in SE England over 21 years with declining SO2
concentrations // Atmosph. Environ. 2001. V.35. N14. P.2557-2568. (93)
Beckett P.J., Boileau L.J.R., Padovan D., Richardson D.H.S., Nieboer E. Lichens
and mosses as monitors of industrial activity associated with uranium mining in
northern Ontario, Canada. Part 2: Distance dependent uranium and lead
accumulation patterns // Environ. Pollut. (Series B). 1982. V.4. N2. P.91-107.
(40, 197.)
Beckett R.P., Brown D.H. Natural and experimental-induced zinc and copper resistance
in the lichen genus Peltigera // Ann. Bot. 1983. V.52. P.43-50. (44, 87)
Beckett R.P., Brown D.H. The control of cadmium uptake in the lichen genus Peltigera
ПЗ. Experim. Bot. 1984a. V.35. N156. P.1071-1082. (38, 46)
Beckett R.P., Brown D.H. The relationship between cadmium uptake and heavy metal
tolerance in the lichen genus Peltigera // New Phytologist. 1984b. V.97. P.301-
311. (38, 46)
Beekley P.K., Hoffman G.R. Effects of sulfur dioxide fumigation on photosynthesis,
respiration, and chlorophyll content of selected lichens // Bryologist. 1981. V.84.
N3. P.379-390. (83, 87)
BelnapJ. Effects of air pollutants on cold-desert cyanobacterial-lichen crusts and rock
lichens: chlorophyll degradation, electrolyte leakage and nitrogenase //
Measurement of toxic and related air pollutants. Proceedings of the 1990 EPA/A
& WMA International Symposium. 1990. P.661-667. (84)
BelnapJ., Harper К. T. Effects of a coal fired power plant on the rock lichen Rhizoplaca
melanophthalma - chlorophyll degradation and electrolyte leakage 11 Bryologist.
1990. V.93. N3. P.309-312. (26, 83, 85)
BelnapJ., Sigal L., Moir W., Eversman S. Identification of sensitive species 11 Lichens
as bioindicators of air quality. General technical report RM-224. K.Stolte et al. -
eds. Fort Collins, CO: US department of agriculture, forest service, Rocky
Литература и авторский индекс
269
Mountain Forestand Range Experiment Station. 1993. P.67-88. (74, 75, 76, 77,
78, 85)
Benedict J.В. Lichen mortality due to late-lying snow: results of a transplant study //
Arctic and Alpine Research. 1990a. V.22. NI. P.81-89. (144)
Benedict J.B. Winter frost injury to lichens - Colorado Front Range // Bryologist. 1990b.
V.93.N4. P.423-426. (99, 145)
Benedict J.B. Experiments on lichen growth. II. Effects of a seasonal snow cover //
Arctic and Alpine Research. 1991. V.23. N2. P. 189-199. (144, 146, 154)
Benedict J.B. Effects of changing climate on game animal and human use of the Colorado
High Country (USA) since 1000-BC // Arctic and Alpine Research. 1999. V.31.
NLP. 1-15. (145)
Benedict J.B., Nash Т.Н. Radial growth and habitat selection by morphologically similar
chemotypes of Xanthoparmelia!'! Bryologist. 1990. V.93. N3. P.319-327. (144,
146)
Bennett J.P. Abnormal chemical element concentrations in lichens of Isle Royale National
Park// Environ. Experim. Bot.. 1995. V.35. N3. P.259-277. (179, 180)
Bennett J.P, Dibben M.J., Lyman K.J. Element concentrations in the lichen Hypogymnia
physodes (L.) Nyl. after 3 years of transplanting along Lake Michigan // Environ.
Experim. Bot.. 1996. V.36. N3. P.255-270. (142)
Bennett J.P, Wetmore C.M. Chemical element concentrations in four lichens on a transect
entering Voyageurs National Park // Environ. Experim. Bot. 1997. V.37. N2-3.
P. 173-185. (179)
Bennett J.P, Wetmore C.M. Changes in element contents of selected lichens over 11
years in Northern Minnesota, USA // Environ. Experim. Bot. 1999a. V.41. NI.
P.75-82. (181)
Bennett J.P, Wetmore C.M. Geothermal elements in lichens of Yellowstone National
Park, USA // Environ. Experim. Bot. 1999b. V.42. N3. P.191-200. (183)
Bennett J.P., Wetmore C.M. 16-Year trends in elements of lichens at Theodore Roosevelt
National Park, North Dakota // Sci. Total Environ. 2000. V.263. Nl-3. P.231-
241. (179, 181)
Beschel R. Eine Flechte als Niederschlagsmesser 11 Wetter und Leben. 1954. Bd.6.
N3-4. S.56-60. (25, 27)
Biazrov L.G. Air pollution damage to algal cells of the lichens and accumulation of the
metals by transplanten lichen thalli - the results of lichens use for active monitoring
of air pollution in Moscow // Fourth SETAC-EUROPE Congress towards
sustainable environmental management. Abstract book. Brussels, Belgium, 11-
14 April 1994. 1994a. P.31. (142)
Biazrov L.H. Lichens as indicators of ecosystem health I I Abstracts of 1st Internatio-
nal Symposium on Ecosystem Health and Medicine: Integrating Science,
Policy and Management. June 19-23 1994, Ottawa, Ontario, Canada. 1994b.
P.10. (238)
270
Литература и авторский индекс
Biazrov L.G. Radionuclide content in lichen thallus in the forests adjacent to the Chernobyl
atomic power plant// Sci. Total Environ. 1994c. V.157. P.25-28. (187, 189)
Biazrov L.G. The radionuclides in lichen thalli in Chernobyl and East Urals areas after
nuclear accidents // Phyton (Hom, Austria). 1994d. V.34. Fasc.l. P.85-94. (187,
192, 193)
Biazrov L. G. The use of lichens for diagnosis of ecosystem health П Radiation Research
1895-1995. Tenth Inter. Congress of Radiation Research, Wurzburg, Germany,
August 27 - September 1., 1995. V.l: Congress Abstracts. U.Hagen, H.Jung,
C.Streffer-eds. 1995b. P.258. (238)
Biazrov L. G. Conception of “ecosystem health’’ with regard to radioactive contaminated
areas and use of lichens for diagnosing of ecosystem state // Abstracts of First
practical conference sustainable development: Environ. Pollut. and ecological
safety. Dniepropetrovsk, Ukraine, December 4-8, 1995. Dniepropetrovsk.
V.F.Prisnyakov-ed. 1995a. V.l. P.144. (238)
Biazrov L.G., Adamova L.I. The toxic metals in lichens of trans-polar Europe //
Programme and Abstracts of the International Nordic Symposium on Chemicals
in the Arctic-boreal Environment, Marina Congress Center, Helsinki, Finland,
May 12-14,1993. M.Lutuola, E. Nurmi-eds. Helsinki: Marina Congress Center,
1993. P.57. (179, 184)
Biazrov L.G., Mineyeva N.Y. The environmental indication of area of the factory for
processing and storage of low-radioactive waste products with help of the lichens
// Abstracts of 3d European Conference on Ecotoxicology, Zurich, Switzerland,
August 28-31, 1994. 1994. P. 5.13. (184)
Bitter G. Ueber die Variability einiger Laubflechten und bber den EinfluB ausserer
Bedingungen auf ihr Wachstum //Jahrb. f. wissensch. Bot. 1901. Bd. 36. S. 421-
492. (22, 23)
Boileau L.J.R., Beckett P.J., Lavoie P, Richardson D.H.S., Nieboer E. Lichens
and mosses as monitors of industrial activity associated with uranium mining in
northern Ontario, Canada. Part 1: Field procedures, chemical analysis and
interspecies comparisons // Environ. Pollut. Series B. 1982. V.4. P.69-84. (40, 197)
Boileau L.J.R, Nieboer E., Richardson D.H.S. Uranium accumulation in the lichen
Cladonia rangiferina. Part 1. Uptake of cationic, neutral, and anionic forms of
the uranyl ion // Canad. J. Bot. 1985a. V.63. N3. P.384-389. (197.)
Boileau L.J.R., Nieboer E., Richardson D.H.S. Uranium accumulation in the lichen
Cladonia rangiferina. Part 2. Toxic effects of cationic, neutral, and anionic forms
of the uranyl ion // Canad. J. Bot. 1985b. V.63. N3. P.390-397. (197)
Boonpragob K., Fox C.A., Nash Т.Н. Seasonal deposition patterns of acidic ions and
ammonium to the lichen Ramalina menziesii Tayl. in Southern California// Environ.
Experim. Bot. 1989. V.29. N2. P. 187-197. (37, 43)
Boonpragob K., Nash Т.Н. Seasonal variation of elemental status in the lichen Ramalina
menziesii Tayl. from two sites in Southern California: evidence for dry deposition
Литература и авторский индекс
271
accumulation//Environ. Experim. Bot. 1990. V.30. N4. P.415-428. (141, 160, 172)
Boonpragob K., Nash Т.Н. Physiological responses of the lichen Ramalina menziesii
Tayl. to the Los Angeles urban environment // Environ. Experim. Bot. 1991. V.31.
N2. P.229-238. (141, 157)
Bosserman R. W., Hagner J.E. Elemental composition of epiphytic lichens from
Okefenokee Swamp// Bryologist. 1981. V.84. NI. P.48-58. (167)
Boucher V.L., Nash Т.Н. Growth patterns in Ramalina menziesii in California: coastal
vs. island populations // Bryologist. 1990. V.93. N3. P.295-302. (141)
ter Braak C.J.F., Prentice I.C. A theory of gradient analysis // Advances in Ecological
Research. 1988 V.18. P.101-146. (75)
Branquinho C., Catarino E, Brown D.H., Pereira M.J., Soares A. Improving the use of
lichens as biomonitors of atmospheric metal pollution // Sci. Total Environ. 1999.
V.232. N1-2. P.67-77. (184)
Brady P. V., Dorn R.I., Brazel A.J., Clark J., Moore R.B., Glidewell T. Direct measure-
ment of the combined effects of lichen, rainfall, and temperature on silicate
weathering // Geochimica et Cosmochimica Acta. 1999. V.63. N19-20. P.3293-
3300. (50)
Brodo I.M. Transplant experiments with corticolous lichens using a new technique //
Ecology. 1961. V.42.N4. P.838-841. (137, 139, 144, 146, 152)
Brodo I.M. Field studies of the effects of ionizing radiation on lichens // Bryologist.
1964. V.67.N1. P.76-87. (187)
Brodo I.M. Lichen growth and cities: a study on Long Island, New York // Bryologist.
1966. V.69. P.427-449. (99, 144, 150, 152)
Brodo I.M. Lichens and cities // International symposium on identification and
measurement of Environ. Pollut. B. Westley - ed. Ottawa: Nat. Res. Con., 1972.
P.325-328. (89)
Brodo I.M. Substrate ecology // The Lichens. V.Ahmadjian, M.E.Hale -eds. New York,
London: Academic Press, 1973. P.401-441. (32, 35)
Bronstein J.L. Our current understanding of mutualism //Quarterly Review of Biology.
1994. V.69. NI. P.31-51. (21)
Brown D.H., Beckett R.P. Uptake and effect of cations on lichen metabolism //
Lichenologist. 1984. V.16.N2. P.173-188. (38, 44)
Brown D.H., Beckett R.P. The role of the cell wall in the intracellular uptake of cations
by lichens // Lichen physiology and cell biology. D.H.Brown - ed. New York,
London: Plenum Press, 1985. P.247-258. (38)
Brown D.H, Slingsby D.R. The cellular location of lead and potassium in the lichen
Cladonia rangiformis (L.) Hoffm. // New Phytologist. 1972. V.71. P.297-305.
(35, 38, 41)
Brunialti G., Giordani P, Ispcrono D., Loppi S. Evaluation of data quality in lichen
biomonitoring studies: the Italian experience // Environ. Monit. Assess. 2002.
V.75. P.271-280. (98, 245)
272
Литература и авторский индекс
BruteigLE. The epiphytic lichen Hypogymnia physodes as a biomonitor of atmospheric
nitrogen and sulphur deposition in Norway // Environ. Monit. Assess. 1993b.
V.26. NI. P.27-47. (43, 185)
Bruteig I.E. Large scale survey of the distribution and ecology of common epiphytic
lichens on Pinus silvestris in Norway H Ann. Bot. Fennici. 1993a. V.30. N3.
P. 161-179. (114)
Biidel B„ Wessels D.C.J. Parmelia hueana Gyeln., a vagrant lichen from the Namib
Desert, SWA/Namibia. 1 Anatomical and reproductive adaptations // Dinteria.
1986. V. 18. P.3-12. (18)
Butin H. Physiologisch-okologische Untersuchungen liber den Wasserhaushalt und die
Photosynthese bei Flechten // Biol. Zentralb. 1954. Bd.73, N9-10. S.459-502. (23)
Cairns J. Are single species toxicity tests alone adequate for estimating environmental
hazard? // Environ. Monit. Assess. 1984. V.4. N3. P.259-273. (114)
CalliariL, Caniglia G., NardiS., Tollardo A.M., CallegaroR. EDXRS study of lichens
as biomonitors and effect of washing procedure on element concentrations //
X-Ray Spectrometry. 1995. V.24. N3. P.143-146. (143)
Calvelo S., Baccala N., Arribere M.A., Ribeiro Guevara S., Bubach D. Concentration
of biological relevant elements of foliose and fruticose lichens from Nahuel Haupi
national park (Patagonia), analyzed by IN AA H Sauteria. 1998. Bd.9. S.263-272.
(179)
Campo G., Orsi M., Badino G., Giacomelli R., Spezzano P. Evaluation of motorway
pollution in a mountain ecosystem. Pilot project: Susa Valley (Northwest Italy)
years 1990-1994 // Sci. Total Environ. 1996. V.190. P. 161-166. (43)
Canas M.S., Orellana L., Pignata M.L. Chemical response of the lichens Parmotrema
austrosinense and Parmotrema conferendum transplanted to urban and non-
polluted environments I I Ann. Bot. Fennici. 1997. V.34. NI. P.27-34. (144, 158)
Canas M.S., Pignata M.L. Temporal variation of pigments and peroxidation products in
the lichen Parmotrema uruguense (Kremplh.) Hale transplanted to urban and
non-polluted environments // Symbiosis. 1998. V.24. NI. P. 147-161. (144)
Caniglia G., Calliari I., Celin L, Tollardo A.M. Metal determination by EDXRF in
lichens. A contribution to pollutants monitoring // Biological Trace Element
Research. 1994. V.43-45. P.213-221. (140)
Caniglia G., Tassoni T, Carboni C. Thallus variations of Parmelia caperata exposed
in suburbs and industrial area (Venice, North-East Italy) // Sauteria. 1998. Bd.9.
S.95-102. (143, 146, 154)
Carignan J., Gariepy C. Isotopic composition of epiphytic lichens as a tracer of the
sources of atmospheric lead emissions in southern Quebec, Canada // Geochimica
et Cosmochimica Acta. 1995. V.59. N21. P.4427-4433. (42)
Carlberg G.E., Ofstad E.B., Drangsholt H., Steinnes E. Atmospheric deposition of
organic micropollutants in Norway studied by means of moss and lichen analysis
//Chemosphere. 1983. V.12. N3. P.341-356. (43, 178)
Литература и авторский индекс
273
Carreras Н.А., Gudino G.L., Pignata M.L. Comparative biomonitoring of atmospheric
quality in five zones of Cordoba city (Argentina) employing the transplanted lichen
Usnea sp. // Environ. Pollut. 1998. V.103. N2-3. P.317-325. (140)
Carreras H.A., Pignata M.L. Comparison among air pollutants, meteorological
conditions and some chemical parameters in the transplanted lichen Usnea
amblyoclada // Environ. Pollut.. 2001. V. 111.N1. P.45-52. (140)
Carreras H.A., Pignata M.L. Biomonitoring of heavy metals and air quality in Cordoba
city, using transplanted lichens//Environ. Pollut. 2002. V.l 17. NI. P.77-87. (140)
Case J. W. The influence of three sour gas processing plants on the ecological distribution
of epiphytic lichens in the vicinity of Fox Creek and Whitecourt, Alberta, Canada
// Water, Air, and Soil Pollution. 1980. V.l4. P.45-68. (102, 103, 114, 115)
Case J. W. Lichen biomonitoring networks in Alberta // Environ. Monit. Assess. 1984.
V.4.N3. P.303-313. (112)
Cercasov V., PantelicaA., Salagean M.,.Caniglia G., ScarlatA. Comparative study of
the suitability of three lichen species to trace-element air monitoring // Environ.
Pollut. 2002. V.l 19. P. 129-139. (140, 167)
ChettriM.K., Sawidis T, Karataglis S. Lichens as a tool for biogeochemical prospecting
// Ecotoxicology and Environmental Safety. 1997a. V.38. N3. P.322-335. (30)
Chettri M.K., Sawidis T, Zachariadis G.A., Stratis J.A. Uptake of heavy metals by
living and dead Cladoniathalli // Environ. Experim. Bot. 1997b. V.37. NI. P.39-
52. (30)
Chiarenzelli J.R., Aspler L.B., Ozarko D.L., Hall G.E.M., Powis K.B., Donaldson J.A.
Heavy metals in lichens, Southern District of Keewatin, Northwest Territories,
Canada//Chemosphere. 1997. V.35.N6. P.1329-1341. (180)
Christ R., Turk R. Die Indikation von Luftverunreinigungen durch CO2-Gaswech-
selmessungen an Flechtentransplantaten // Mitteilungen der Forstlichen
Bundesversuchsanstalt Wien. 1981. Bd.137. S.145-150. (143, 157)
Christ R., TurkR. CO2-Gaswechselmessungen an Flechtentransplanten zur Indikation
der SO2-Belastung im Stadtgebiet von Salzburg // Schriftenr. Luftguteunter-
suchung. Amt der Salzburger Landesregierung. 1982. Bd.7. S.36-77. (86, 143,
157)
CislaghiC, NimisP.L. Lichens, air-pollution and lung cancer//Nature (London). 1997.
V.387. N6623. P.463-464. (9)
Coccaro D.M.B., Saiki M., Vasconcellos M.B.A., Marcelli M.P. Evaluation of trace
elements in different species of lichens by neutron activation analysis // J.
Radioanal. Nuclear Chem. 2000. V.244. NI. P. 141-145. (177)
Collins C.R., Farrar J.F. Structural resistances to mass transfer in the lichen Xanthoria
parietina// New Phytologist. 1978. V.81. P.71-83. (39)
Connor M., Dempsey E., Smyth M.R., Richardson D.H.S. Determination of some metal
ions using lichen modified carbon paste electrodes // Electroanalysis. 1991. V.3.
N4-5. P.331-336. (177)
274
Литература и авторский индекс
Conti М.Е., Cecchetti G. Biological monitoring: lichens as bioindicators air pollution
assessment-a review//Environ. Pollut. 2001. V.114. P.471-492. (73, 164)
Crete M., Lefebvre M.A., Zikovsky L., Walsh P. Cadmium, lead, mercury and cesium-
137 in fruticose lichens of Northern Quebec // Sci. Total Environ. 1992. V.121.
P.217-230. (180)
Culberson C.F., Culberson W.L., Johnson A. Thermally induced chemical artifacts in
lichens//Phytochemistry. 1977. V.16. P.127-130. (183)
Culberson W.L. The corticolous communities of lichens and bryophytes in the upland
forests of northern Wisconsin // Ecol. Monogr. 1955. V.25. N2. P.215-231.
(22)
Culberson W.L., Egan R.S., Esslinger T.L. Recent literature on lichens. 2002. http://
www.nhm.uio/botanisk/bot-mus/lav/sok_rll.htm. [Presented on the Web by
E.Timdal. First posted 1997.04.14, latest update 2002.06.27]. (11)
Сипу D., van Haluwyn C, Pesch R. Biomonitoring of trace elements in air and soil
compartments along the major motorway in France // Water, Air, and Soil Pollution.
2001. V. 125. N1-4. P.273-289. (143, 184)
Czehura S.J. A lichen indicator ofcoppermineralization, Lights Creek district, Plumas
County, California // Econ. Geol. 1977. V.72. P.796-803. (45)
Davies F.B.M., Notcutt G. Accumulation of fluoride by lichens in the vicinity of Etna
Volcano // Water, Air, and Soil Pollution. 1988. V.42. N3-4. P.365-371. (182)
Davies F.B.M., Notcutt G. Accumulation of volcanogenic fluorides by lichens // Fluoride.
1989. V.22.N2. P.59-65. (182)
Dayan F.E., RomagniJ.G. Lichens as potential source of pesticides // Pesticides outlook.
2002. V.12. N6. P.229-232. (28)
Dempsey E., Smyth M.R, Richardson D.H.S. Application of lichen-modified carbon
paste electrodes to the voltammetric determination of metal ions in multi-element
and speciation studies // Analyst. 1992. V.l 17. N9. P.1467-1470. (177)
Denison R, CaldwellB., Bormann B., EldredL., Swanberg C, Anderson S. The effects
of acid rain on nitrogen fixation in western Washington coniferous forests //
Water, Air, and Soil Pollution. 1977. V.8. P.21-34. (84)
Denison W.C. Life in tall trees // Scientific American. 1973. V.228. N6. P.75-80. (84)
Denison W.C. Culturing the lichens Lobaria oregana and Lobariapulmonaria on nylon
monofilament//Mycologia. 1988. V.80.N6. P.811-814 . (145)
DePriest P. T. Lichenology challenges and opportunities // Amer. J. Botany. 1994. V.81.
N6. Suppl. P.4-5. (238)
Deruelle S. Influence de la pollution atmospherique sur la vegetation lichenique des
arbres isoles dans la region de Mantes (Yvelines) // Rev. Bryol. Lichenol. 1977.
V.43.N2. P. 137-158. (99)
Deruelle S. Etude comparee de la sensibilite de trois methodes d’estimation de la pollution
atmospherique, en utilisant les lichens comme indicateurs biologiques, dans la
region de Mantes (Yvelines)// Rev. Bryol. Lichenol. 1978a. V.44. P.429-441. (93)
Литература и авторский индекс
275
Deruelle S. Les lichens et la pollution atmospherique // Bulletin d’Ecologie. 1978b. V.9.
P.87-128. (106)
Deruelle S„ Guilloux F. Evolution de la vegetation lichenique en region Parisienne
entre 1981 et 1991, en relation avec la qualite de 1’air// Bulletin d’informations de
1’Association Francaise de lichenologie. Memoire. 1993. N2. P.23-44. (66,115, 232)
DeSloover J., LeBlanc F. Mapping of atmospheric pollution on the basis of lichen
sensitivity И Proceedings of the symposium in recent advances in tropical ecology.
International Society for tropical ecology, Banaras Hindu University, Varansi.
R.Misra, B.Gopal -eds. 1968. P.42-56. (102)
Dibben M.J. Whole lichen culture in a phytotron // Lichenologist. 1971. V.5. P.1-10. (48)
Dillman K.L. Use of the lichen Rhizoplaca melanophthalma as a biomonitor in relation
to phosphate refineries near Pocatello, Idaho H Environ. Pollut. 1996. V.92. NI.
P.91-96. (184)
Djalali B. Flechtenkartierung und Transplantatuntersuchungen in Stadtgebiet von
Stuttgart// Hohenheim. Arb. Schriftenr. Univ. Hohenheim. 1973. Bd.74. S.15-
30. (142)
van Dobben H.F. Vegetation as a monitor for deposition of nitrogen and acidity [Vegetatie
als monitor voor depositie van stikstof en zuur], Rijksuniversiteit te Utrecht, Utrecht.
1993. 214 p. (33, 43, 64)
van Dobben H.F, ter Braak C.J.F. Effects of atmospheric NH3 on epiphytic lichens in
the Netherlands - the pitfalls of biological monitoring // Atmosph. Environ. 1998.
V.32. N3. P.551-557. (43)
van Dobben H.F., ter Braak C.J.F. Ranking of epiphytic lichen sensitivity to air pollution
using survey data: a comparison of indicator scales H Lichenologist. 1999. V.31.
NI. P.27-39. (89)
van Dobben H.F., Wolterbeek H.T., Wamelink G.W.W., ter Braak C.J.F. Relationship
between epiphytic lichens, trace elements and gaseous atmospheric pollutants //
Environ. Pollut. 2001. V.112. P. 163-169. (45)
Dodds W.K., Gudder D.A., Mollenhauer D. The ecology of Nostoc // J. Phycology.
1995. V.31.NI. P.2-18. (82)
Dongarra G., Ottonello D., Sabatino G., Triscari M. Preliminary data on heavy metal
content in lichens from North Eastern Sicily // Mineralogia Petrographica Acta.
1994. V.37. P.141-153. (185)
Dongarra G., Varrica D. The presence of heavy metals in air particulate at Vulcano
Island (Italy) // Sci. Total Environ. 1998. V.212. NI. P.1-9. (182)
Doyle P, Fletcher W.K., Brink V.C. Trace element content of soils and plants from the
Selwyn Mountains, Yukon and Northwest Territories // Canad. J. Bot. 1973. V.54.
N3. P.421-427. (32, 179)
Du Rietz G.E. Zur Vegetationsokologie der Ostschwedischen Kiistenfelsen //
Beihefte zum Botanischen Centralblatt. 1932. Bd.49, Erganzungsband. S.61-
112. (21)
276
Литература и авторский индекс
Du Rietz G.E. От fattigbark - och rikbarsamhallen // Svensk Bot. Tidskr. 1945. V.39.
NI. P. 147-150. (20)
Eckl P, Hofmann W., TiirkR Uptake of natural and man-made radionuclides by lichens
and mushrooms // Radiat. Environ. Biophys. 1986. V.25. NI. P.43-54. (189, 193,
195)
Eduljee G.E. Trends in risk assessment and risk management // Sci. Total Environ.
2000. V.249. P. 13-23. (239)
van der Eerden L., de Vries W., van Dobben H. Effects of ammonia deposition on forests
in the Netherlands // Atmosph. Environ. 1998. V.32. N3. P.525-532. (33)
Egger R., Schlee D., TiirkR. Changes of physiological and biochemical parameters in
the lichen Hypogymnia physodes (L.) Nyl. due to the action of air pollutants - a
field study // Phyton [Austria]. 1994. Bd.34. N2. P.229-242. (142, 157)
Eldridge D.J. Ecology and management of biological soil crusts: recent developments
and future challenges // Bryologist. 2000. V.103. N4. P.742-747. (47)
Eldridge D.J., Zaady E., Shachak M. Infiltration through three contrasting biological
soil crusts in patterned landscapes in the Negev, Israel 11 Catena. 2000. V.40. N3.
P.323-336. (47)
Elkin B.T., Bethke R.W. Environmental contaminants in caribou in the Northwest
Territories, Canada// Sci. Total Environ. 1995. V.l60/161. P.307-321. (178)
Ellis K.E., Smith J.N. Dynamic model for radionuclide uptake in lichen // J. Environ.
Radioactivity. 1987. V.5.N3. P.185-208. (198)
Epstein E., Sagee O., Cohen J.D., Garty J. Endogenous auxin and ethylene in the lichen
Ramalina duriaei I I Plant Physiology. 1986. V.82. N4. P.l 122-1125. (140, 159)
Erbisch F.H. Lichens of the Enterprise radiation forest // The Enterprise, Wisconsin,
radiation forest: pre-irradiation ecological studies. T.D.Rudolph - ed. USAEC
Report TID-26113. 1974. P.47-53. (187)
Erbisch F.H. Effects of chronic gamma irradiation on the lichen Parmelia sulcata Tayl
// The Enterprise, Wisconsin, radiation forest: radioecological studies. LZavitkov-
ski-ed. USAEC Report T1D-26113-2. 1977. P.25-34. (187)
Erbisch F.H. Effect of chronic gamma radiation on Parmelia subaurifera in the Enterprise
radiation forest// Bryologist. 1978. V.81. NI. P.137-143. (187)
Erbisch F.H, Ligon J.B., Mathena D.E. Measurement of tensile strength of Cladonia
podetia // Bryologist. 1977. V.80. N3. P.524 -527. (187)
Erdman J.A., Gough L.P. Variation in the element content of Parmelia chlorochroa
from the Powder River Basin of Wyoming and Montana I I Bryologist. 1977.
V.80. N2. P.292-303. (34, 179)
Erdman J.A., Gough L.P, White R. W. Calcium oxalate as source of high ash yields in
terricolous lichen Parmelia chlorochroa 11 Bryologist. 1977. V.80. N2. P.334-
339. (31, 179)
Ernst W.H.O. Sampling of plant material for chemical analysis // Sci. Total Environ.
1995. V.176.N1-3. P.15-24. (31, 169)
Литература и авторский индекс
277
Ertl L. Uber die Lichtverhaltnisse in Laubflechten // Planta. 1951. Bd.39. N3. S.245-
270. (23)
EsseenP.-A., Renhom K.-E. Epiphytic lichen biomass in managed and old-growth boreal
forests: effect of branch quality // Ecological Applications. 1996. V.6. N1. P.228-
238. (240)
Esslinger T., EganR.S. A sixth checklist of the lichen-forming, lichenicolous, and allied
fungi of the continental United States and Canada // Bryologist. 1995. V.98. N4.
P.467-549. (200)
Evans C.A., Hutchinson T.C. Mercury accumulation in transplanted moss and lichens at
high elevation sites in Quebec // Water, Air, and Soil Pollution. 1996. V.90. P.475-
488. (142)
Eversman S. Effects of low-level SO2 on Usnea hirta and Parmelia chlorochroa //
Bryologist. 1978. V.81. P.368-377. (81, 87)
Eversman S. Preliminary distributional study of the epiphytic macrolichens of Montana
// Proceedings of the Montana Academy of Science. 1979. V.38. P.60-62. (87)
Eversman S., Sigal L.L. Ultrastructural effects of peroxyacetyl nitrate (PAN) on two
lichen species // Bryologist. 1984. V.87. N2. P. 112-118. (81)
Eversman S., Sigal L.L. Ultrastructural effects of gaseous pollutants and acid precipitation
on lichens // Canad. J. Bot. 1987. V.65. P.1806-1818. (81, 82)
Fahselt D. Lichen substances of transplanted thallus segments of Parmelia cumberlandia
// Canad. J. Bot. 1979. V.57. NI. P.23-25. (144)
Fahselt D. Secondary compounds following north-south transplantation of three lichen
species // Lichenologist. 1981. V.13. N3. P.253-258. (144)
Fahselt D. Growth form and reproductive character of lichens near active fumaroles in
Japan//Symbiosis. 1995. V.18.N3. P.211-231. (182)
Fahselt D., WuT.W., MottB. Trace element patterns in lichens following uranium mine
closures // Bryologist. 1995. V.98. N2. P.228-234. (184)
Falla J., Laval-Gilly P., Henryon M, Morlot D., Ferard J.F. Biological air quality moni-
toring: a review // Environ. Monit. Assess. 2000. V.64. N3. P.627-644. (73, 164)
Farkas E„ Lokos L., Verseghy K. Lichens as indicators of air pollution in the Budapest
agglomeration. I. Air pollution map based on floristic data and heavy metal
concentration measurements //Acta Botanica Hungarica. 1985. V.31. N1 -4. P.45-
68. (142, 146, 153)
Farmer A.M., Bates J. W., Bell J.N.B. The transplantation of four species of Lobaria
lichens to demonstrate a field acid rain effect // Acidification research, evaluation
and policy application. T.Schneider-ed. Elsevier Science Publishers, 1992. P.295-
300. (145)
Farrar J.F. The lichen as an ecosystem: observation and experiment // Licheno-
logy: progress and problems. D.H.Brown, D.L.Hawksworth, R.H.Bailey -
eds. Academic Press: London, New York, San Francisco, 1976a. P.385-406.
(12, 48)
278
Литература и авторский индекс
Farrar J.F. Ecological physiology of the lichen Hypogymnia physodes. I. Some effects
of constant water saturation // New Phytologist. 1976b. V.77. P.93-103. (25, 48)
Farrar J.F. Ecological physiology of the lichen Hypogymnia physodes. II. Effects of
wetting and drying cycles and the concept of “physiological buffering” // New
Phytologist. 1976c. V.77. P.105-113. (25)
Farrar J.F. The uptake and metabolism of phosphate by the lichen Hypogymnia physodes
//New Phytologist. 1976d. V.77. P. 127-134. (37, 88)
Farrar J.F., Smith D.C. Ecological physiology of the lichen Hypogymnia physodes. III.
The importance of the rewetting phase 11 New Phytologist. 1976. V.77. P.l 15-
125. (48)
Feige G.B. Niedere Pflanzen - speziell Flechten - als Bioindikatoren // Decheniana,
Beihefte (Bonn). 1982. Bd.26. S.23-30. (80)
Feige G.B., Niemann L., Jahnke S. Lichens and mosses - silent chronists of the Chernobyl
accident // Contributions to Lichenology in Honour of A. Henssen. H.M.Jahns -
ed. J. Cramer, Berlin-Stuttgart, 1990. P.63-77. (192)
Ferrara R, Maserti B.E., Bargagli R. Mercury in the atmosphere and in lichens in a
region affected by a geochemical anomaly // Environmental Technology Letters.
1988. V.9. N7. P.689-694. (183)
Ferretti M. Forest health assessment and monitoring: Issues for consideration // Environ.
Monit. Assess. 1997. V.48. P.45-72. (239)
Ferris J.M., Jeffree R.A., Prince K.E., Hoffman E., HeijnisH. Plutonium immobilization
by lichens at Meralinga, a former nuclear test site in semi-arid South Australia И
Environmental Impact Radioact. Release: Proc. Int. Symp., Vienna, 8-12 May,
1995. Vienna. 1995. P.710-712. (189)
Ferry B.W., Baddeley M.S., Hawksworth D.L (eds.). Air pollution and lichens. Toronto:
University of Toronto Press, 1973. 389 p. (73)
Ferry B.W., Coppins B.J. Lichen transplant experiments and air pollution studies //
Lichenologist. 1979. V.11.N1. P.63-73. (88, 139, 141)
Fields R. Physiological responses of lichens to air pollution fumigations // Lichens,
bryophytes and air quality. T.H.Nash, V. Wirth - eds. J. Cramer, Berlin-Stuttgart,
1988. P.l75-200. (77, 83, 84, 86)
Fields RD., St.Clair L.L. A comparison of methods for evaluating SO2 impacton selected
lichen species: Parmelia chlorochroa, Collemapolycarpon and Lecanora muralis
11 Bryologist. 1984a. V.87. N4. P.297-301. (87)
Fields RD., St. Clair L.L. The effects of SO2 on photosynthesis and carbohydrate transfer
in the two lichens: Collema polycarpon and Parmelia chlorochroa // Amer. J.
Bot. 1984b. V.71. P.986-998. (85, 87)
FigueiraR, Catarino F, Pacheco A.M.G., Sousa A.J. In situ studies on sea-salt uptake
by epiphytic lichens // Sauteria. 1998a. V.9. P.l43-150. (140, 183)
Figueira R., Pacheco A.M.G., Sousa A.J., Branquinho C, Catarino F. Assessment of
two epiphytic lichens as saltfall biomonitors: calibration of transplants // Ecotoxico-
Литература и авторский индекс
279
logy and Environmental Restoration. 1998a. V.l. N2. P.61-69. (140, 183)
FigueiraR, Pacheco A.M.G., Sousa A.J., CatarinoF. Biological monitoring of airborne
salinity through epiphytic lichens // Revista de Biologia (Lisbon). 1999a. V.l7.
NI. P.3-12. (183)
Figueira R, Sousa A.J., Brown D.H., Catarino F., Pacheco A.M.G. Natural levels of
saline elements in lichens - determination of cellular fractions and their importance
as saline tracers//Lichenologist. 1999b. V.31. N2. P. 183-196. (183)
FigueiraR., Sousa A. J., Pacheco A.M.G., CatarinoF. Saline variability at ground level
after kriging data from Ramalina spp. biomonitors // Science of the Total
Environment. 1999c. V.232. P.3-11. (183)
Figueira R., Sousa A.J., Pacheco A.M.G., Catarino F. Space-time geostatistical
modelling: a case study of sea-salt measured on lichens // geoEnv 11 - Geostatistics
for Environmental Applications. J.Gymez-HemGndez, A.Soares, R.Froidevaux-
eds. Kluwer Academic Publishers, Dordrecht, The Netherlands, 1999d. P.53-64. (183)
FigueiraR, Sousa A.J., Pacheco A.M.G., Catarino F. Variability of sea-salt deposition
assessed by lichen monitoring in the south-west Portuguese coast // Air Pollution
VII. C.A.Brebbia, M.Jacobson, H.Power-eds. WIT Press, Boston, 1999e. P.599-
607. (183)
Flora C.J., Nieboer E. Determination of nickel by differential pulse polarography at a
dropping mercury electrode // Analytical Chemistry. 1980. V.52. N7. P.1013-
1020. (177)
Focardi S, Gaggi C, Chemollo G., Bacci E. Organochlorine residues in moss and lichen
samples from two Antarctic area // Polar Research. 1991. V.27. P.241 -244. (178)
Folkeson L. Impact of airborne copper, iron and zinc pollution on lichen and bryophyte
vegetation near a brass foundry // Silva Fennica. 1981. V. 15. N4. P.446-449. (174)
France R, Coquery M. Lead concentrations in lichens from the Canadian High Arctic
in relation to the latitudinal pollution gradient // Water, Air, and Soil Pollution.
1996. V.90. N3-4. P.469-474. (180)
Franzle S., Markert B. The biological system of the elements (BSE). Part II: a theoretical
model for establishing the essentiality of chemical elements. The application of
stochiometric network analysis to the biological system of the elements // Sci.
Total Environ. 2000. V.249. Nl-3. P.223-241. (32)
Freitas M.C. Heavy metals in Parmelia sulcata collected in the neighborhood of a coal-
fired power station // Biological Trace Element Research. 1994. V.43-45. P.207-
212. (184)
Freitas M.C. Application of radioanalytical techniques to environmental studies -
elemental bioaccumulators in air pollution studies // J. Radioanal. & Nuclear
Chem. - Articles. 1995. V. 192. N2. P. 171 -181. (184)
Freitas M.C., Reis M.A., Alves L.C., Wolterbeek H.T. Distribution in Portugal of some
pollutants in the lichen Parmelia sulcata // Environ. Pollut. 1999. V.l06. N2.
P.229-235. (185)
280
Литература и авторский индекс
Freitas М.С., Reis М.А., Alves L.C., Wolterbeek H.T., Gouveia M.A. Elemental
accumulation in lichen transplants in the neighborhood of thermal power stations
//Nuclear instrumental methods: Proc. 7th Intem.Conf.on P1XE and its analytical
applications, Padova, May 26-30, 1995. 1995. P. 117-118. (143, 168)
Freitas M.C., Reis M.A., Alves L.C., Wolterbeek H.T., Verburg T, Gouveia M.A.
Biomonitoring of trace element air pollution in Portugal: qualitative survey // J.
Radioanal. & Nuclear Chem. 1997. V.217.N1. P.21-30. (184, 185)
Freitas M.C., ReisM.A., Marques A.P., Wolterbeek H.T. Dispersion of chemical elements
in an industrial environment studied by biomonitoring using Parmelia sulcata /
I J. Radioanal. Nuclear Chem. 2000. V.244. NI. P. 109-113. (143, 185)
Frenzel R.W, Witmer G.W., Starkey E.E. Heavy metal concentrations in a lichen of
Mt.Rainier and Olympic National Park, Washington, USA // Bull. Environ.
Contamin. Toxic. 1990. V.44. NI. P. 158-164. (179)
Friedl T, Biidel B. Photobionts 11 Biology of lichens. T.Nash - ed. Oxford: Acad.Press,
1996. P.8-23. (15, 16)
Friedmann Е.1. Endolithic microorganisms in the Antarctic cold desert// Science. 1982.
V.215. P. 1045-1053. (50)
Fuchs C., Garty J. Elemental content on the lichen Ramalina duriaei (De Not.) Jatta at
air quality biomonitoring stations// Environ. Experim. Bot. 1983. V.23. P.29-43.
(62, 141, 161)
Gaare E. The Chernobyl accident: can lichens be used to characterize a radiocesium
contaminated range? // Rangifer. 1987. V.7. N2. P.46-50. (189)
Gailey F.A. Y., Lloyd O.L. Spatial and temporal patterns of airborne metal pollution - the
value of low technology sampling to an environmental epidemiology study //
Sci. Total Environ. 1993. V.133. N3. P.201-219. (142)
Gailey F.A.Y., Smith G.H., RintoulL.J., LloydO.L. Metal deposition patterns in central
Scotland, as determined by lichen transplants// Environ. Monit. Assess. 1985.
V.5. P.291-309. (142)
Galun M., Garty J., Ronen R. Lichens as bioindicators of air pollution // Webbia. 1984.
V.38. P.371-383. (11)
Galun M., Ronen R. Interactions of lichens and pollutants // Handbook of Lichenology
I Ed. M. Galun. Boca Raton: CRC Press, 1988. V.3. P.55-72. (11, 88, 158)
Galvan J., Rodriguez C., Ascaso C. The pedogenic action of lichens in metamorphic
rocks//Pedobiologia. 1981. V.21. P.60-73. (51)
Garty J. Metal amounts in the lichen Ramalina duriaei (De Not) Bagl. transplanted at
biomonitoring sites around a new coal-fired power station after 1 year of operation
//Environ. Res. 1987. V.43.N1. P. 104-116. (62, 141, 161)
Garty J. Comparisons between the metal content of a transplanted lichen before and
after the start-up of a coal-fired power station in Israel // Canad. J. Bot. 1988.
V.66.N4. P.668-671. (141, 161, 162)
Garty J. Re-establishment of lichens on chalk rocks after a forest fire // Fire in Ecosystem
Литература и авторский индекс
281
Dynamics. J.G.Goldammer, MJ Jenkins - eds. The Hague: SPB Acad. Publ.,
1990. P.77-84. (188)
Garty J. Lichens as biomonitors for heavy metal pollution H Plants as biomonitors //
Indicators for heavy metals in the terrestrial environment. B.Markert - ed.
Weinheim etc.: VCH, 1993. P.193-263. (32, 35, 164, 177)
Garty J. Environment and elemental content of lichens И Trace elements - their
distribution and effects in the environment. B.Markert, K.Friese - eds. Elsevier
Science B.V. 2000a. P.245-276. (35, 164, 167, 177)
Garty J. Trace metals, other chemical elements and lichen physiology: research in the
nineties // Trace elements - their distribution and effects in the environment.
B.Markert, K.Friese - eds. Elsevier Science B.V. 2000b. P. 277-322. (83, 84,
164, 177)
Garty J., Ammann K. The amounts ofNi, CR, Zn, Pb, Cu, Fe and Mn in some lichens
growing in Switzerland // Environ. Experim. Bot.. 1987. V.27. N2. P.l27-138.
(168)
Garty J., Cohen Y, Kloog N. Airborne elements, cell membranes, and chlorophyll in
transplanted lichens // J. Environ. Quality. 1998a. V.27. N4. P.973-979. (141, 159)
Garty J., Cohen Y, Kloog N., Karnieli A. Effects of air pollution on cell membrane
integrity, spectral reflectance and metal and sulfur concentrations in lichens //
Environ. Toxicol. Chem. 1997a. V.16. N7. P.l396-1402. (141, 158, 159)
Garty J., Fuchs C. Heavy metals in lichen Ramalina duriaei transplanted in biomoni-
toring stations // Water, Air, and Soil Pollution. 1982. V.17. P.l75-183. (62,141)
Garty J., Fuchs C, Zisapel N., Galun M. Heavy metals in lichen Caloplaca aurantia
from urban, suburban and rural regions in Israel. A comparative study H Water,
Air, and Soil Pollution. 1977. V.8. P.l71-188. (174)
Garty J., Galun M., Kessel M. Localization of heavy metals and other elements
accumulated in the lichen thallus // New Phytologist. 1979. V.82. N2. P. 159-168.
(33, 39, 41)
Garty J., Hagemeyer J. Heavy metals in the lichen Ramalina duriaei transplanted at
biomonitoring stations in the region of a coal fired power plant in Israel after three
years of operation // Water, Air, and Soil Pollution. 1988. V.38. N3-4. P.311-323.
(141, 161)
Garty J., Harel Y, Steinberger Y The role of lichens in the cycling of metals in the
Negev desert//Arch. Environ. Contamin. Toxicol. 1995a. V.29. N2. P.247-253. (179)
Garty J., Karary Y, Harel J. Effect of low pH, heavy metals and anions on chlorophyll
degradation in the lichen Ramalina duriaei (De Not.) Bagl. // Environ. Experim.
Bot. 1992. V.32. N3. P.229-241. (141)
Garty J., Karary Y, Harel J. The impact of air pollution on the integrity of cell
membranes and chlorophyll in the lichen Ramalina duriaei (De Not.) Bagl.
transplanted in industrial site in Israel // Arch. Environ. Contamin. Toxicol. 1993a.
V.24. N4. P.455-460. (84, 85, 141, 158, 159)
282
Литература и авторский индекс
Garty J., Karary К, Harel J., Lurie S. Temporal and spatial fluctuations of ethylene
production and concentrations of sulfur, sodium, chlorine and iron on in the thallus
cortex in the lichen Ramalina duriaei (De Not) Bagl. // Environ. Experim. Bot.
1993b. V.33. N4. P.553-563. (88, 141, 159)
Garty J., KardishN., Hagemeyer J., Ronen R. Correlations between the concentration
of adenosine tri phosphate, chlorophyll degradation and the amounts of airborne
heavy metals and sulfur in a transplanted lichen // Arch. Environ. Contamin.
Toxicol. 1988. V.17.N5. P.601-611. (141, 159)
Garty J., Karnieli A., Wolfson R., Kunin P, Garty-Spitz R. Spectral reflectance and
integrity of cell membranes and chlorophyll relative to the concentration of airborne
elements in a lichen//Physiologia Plantarum. 1997a. V.101.N2. P.257-264. (141,
158, 159)
Garty J., Kauppi M., Kauppi A. Differential responses of certain lichen species to sulfur
containing solutions under acidic conditions as expressed by the production of
stress ethylene // Environ. Res. 1995b. V.69. N2. P.132-143. (141)
Garty J., Kauppi M., Kauppi A. Accumulation of airborne elements from vehicles in
transplanted lichens in urban sites // J. Environ. Quality. 1996. V.25. N2. P.265-
272. (141, 162)
Garty J., Kauppi M., Kauppi A. The influence of air pollution on the concentration of
airborne elements and on the production of stress ethylene in the lichen Usnea
hirta (L) Weber em Mot. transplanted in urban sites in Oulu, N. Finland // Arch.
Environ. Contamin. Toxicol. 1997c. V.32. N3. P.285-290. (141, 162)
Garty J., Kauppi M., Kauppi A. The production of stress ethylene relative to the
concentration of heavy metals and other elements in the lichen Hypogym-
nia physodes //Environ. Toxicol. Chem. 1997d. V.l6. Nil. P.2404-2408.
Garty J., KloogN., Cohen X Integrity of lichen cell membranes in relation to concentration
of airborne elements//Arch. Environ. Contamin. Toxicol. 1998b. V.34. N2. P.136-
144. (141, 149, 158, 159)
Garty J., Kloog N„ Cohen Y, Wolfson R, Karnieli A. The effect of air pollution on the
integrity of chlorophyll, spectral reflectance response, and on concentrations of
nickel, vanadium, and sulfur in the lichen Ramalina duriaei (De Not.) Bagl. //
Environ. Res. 1997e. V.74.N2. P. 174-187. (141, 149, 158, 159)
Garty J., Kloog N., Wolfson R„ Cohen Y, Karnieli A., Avni A. The influence of air
pollution on the concentration of mineral elements, on the spectral reflectance
response and on the production of stress ethylene in the lichen Ramalina duriaei
//New Phytologist. 1997f. V.137.N4. P.587-597. (141, 149, 158, 159)
Garty J., Perry A.S., Mozel J. Accumulation of polychlorinated biphenyls (PCBs)
in the transplanted lichen Ramalina duriaei (De Not.) Jatta at air qua-
lity biomonitoring experiments // Nordic J. Bot. 1982. V.2. N6. P.583-586.
Литература и авторский индекс
283
Garty J., Ronen R., Galun M. Correlation between chlorophyll degradation and the
amount of some elements in the lichen Ramalina duriaei (De Not.) Jatta // Environ.
Experim. Bot. 1985a. V.25. NI. P.67-74. (83, 141)
Garty J., Weissman L., Cohen Y., KarnieliA., Orlovsky L. Transplanted lichens in and
around the Mount Carmel National Park and the Haifa Bay industrial region in
Israel: physiological and chemical responses И Environ. Res. 2001. V.85. N2.
P. 159-176. (141)
GartyJ., Weissman L, TamirO., BeerS., Cohen Y, KarnieliA., OrlovskyL. Comparison
of five physiological parameters to assess the vitality of the lichen Ramalina lacera
exposed to air pollution // Physiologia Plantarum. 2000. V. 109. N4. P.410-418. (141)
Garty J., Ziv 0., Eshel A. The effect of coating polymers on accumulation of airborne
heavy metals by lichens// Environ. Pollut., Series A. 1985b. V.38. P.213-220. (141)
Gaskell S.J., Eglinton G., Bruun T Hydrocarbon constituents of three species of
Norwegian lichen: Cetraria nivalis, C. crispa, Siphula ceratites I'/ Phytochemistry.
1973. V.12. P.1174-1176. (775)
GasparoD., Castello M., BargagliR Biomonitoraggiodell’inquinamento atmosferico
tramite licheni. Studio presso un inceneritore (Macerata) H Studia Geobotanica.
1989. V.9. P. 152-250. (175, 184)
Gauslaa Y Lungenever, Lobariapulmonaria, som indikator pe arstrike kontinuitetsskoger
[Lobaria pulmonaria, an indicator of species-rich forests of long ecological
continuity] // Blyttia. 1994. V.52. N3. P.l 19-128. (240)
Gauslaa Y Population structure of the epiphytic lichen Usnea longissima in a boreal
Picea abies canopy // Lichenologist. 1997. V.29. N5. P.455-469. (240)
Gauslaa Y, Solhaug KA. High light intensity damage to the foliose lichen Lobaria
pulmonaria within a natural forest - the applicability of chlorophyll fluorescence
methods И Lichenologist. 2000. V.32. N3. P.271-289. (145, 240)
Geebelen W., Hoffmann M. Evaluation of bioindication methods using epiphytes by
correlating with SO2-pollution parameters H Lichenologist. 2001. V.33. N3. P.249-
260. (72, 89, 114, 115)
Geiser L.H., Derr C.C., Dillman K. Air quality monitoring on the Tongass national
forest. Methods and baselines using lichens. USDA Forest Service, Tongass
National Foresty Stikine Area. R10-TB-46. 1994. 85 p. (67, 179)
Getty S.R., Gutzler D.S., Asmerom Y, Shearer C.K., Free S.J. Chemical signals of
epiphytic lichens in southwestern North America; natural versus man-made
sources for airborne particulates // Atmosph. Environ. 1999. V.33. N30. P.5095-
5104. (72)
Gilbert O.L. A biological scale for the estimation of sulphur dioxide pollution // New
Phytologist. 1970. V.69. P.629-634. (115)
Gilbert O.L. The effect of airborne fluorides // Air Pollution and Lichens. B.W.Ferry,
M.S.Baddeley, D.L.Hawksworth - eds. Toronto: University of Toronto Press,
1973. P.176-191. (73)
284
Литература и авторский индекс
Gilbert O.L. Colonization by Parmeliasaxatilis transplanted onto a suburban wall during
declining SO, pollution // Lichenologist. 1988. V.20. N2. P. 197-198. (144)
Gilbert O.L. A successful transplant operation involving Lobaria amplissima //
Lichenologist. 1991. V.23. NI. P.73-76. (145, 146)
Giordani P, Brunialti G., Modenesi P. Applicability of the lichen biodiversity (L.B.) to
a Mediterranean area (Liguria, NW Italy) // Cryptogamie, Mycologie. 2001. V.22.
N3. P.l93-208. (98, 118)
Giordani P, Brunialti G., Alleteo D. Effects of atmospheric pollution on lichen
biodiversity (LB) in a Mediterranean region (Liguria, Northwest Italy) // Environ.
Pollut. 2002. V.118. NI. P.53-64. (98)
Glenn M.G., Gomez-Bolea A., Lobello R. Metal content and community structure
of cryptogam bioindicators in relation to vehicular traffic in Montseny Bio-
sphere Reserve (Catalonia, Spain) // Lichenologist. 1995. V.27. N4. P.291-304.
(184)
Glenn M.G., Orsi E. V., Hemsley M.E. Lichen metal contents as correlates of air filter
measurements // Grana. 1991. V.30. P.44-47. (184)
Glime J.M., Iwatsuki Z. Niche characteristics of Cladonia lichens associated with
geothermal vents in Japan // Ecological Research. 1990. V.5. P.131-141. (188)
Glotfelty D.E., Seiber J.N., Liljedahl L.A. Pesticides in fog // Nature. 1987. V.325.
P.602-605. (74)
Gombert S., Asta J. The effect of refuse incinerator fumes on the lead and cadmium
content of experimentally exposed corticolous lichens // Water, Air, and Soil
Pollution. 1998. V.104. Nl-2. P.29-40. (145)
Gonzalez CM., Casanovas S.S., Pignata M.L. Biomonitoring of air pollutants from
traffic and industries employing Ramalina ecklonii (Spreng.) Mey. and Flot. in
Cyrdoba, Argentina//Environ. Pollut. 1996. V.91.N3. P.269-277. (140, 158,159)
Gonzalez CM., Orellana L.C., Casanovas S.S., PignataM.L. Environmental conditions
and chemical response of a transplanted lichen to an urban area // J. Environ.
Manag. 1998. V.53. P.73-81. (140, 158, 159)
Gonzalez C.M., Pignata M.L. The influence of air pollution on soluble proteins,
chlorophyll degradation, MDA, sulphur and heavy metals in a transplanted lichen
//Chemistry and Ecology. 1994. V.9. P.105-113. (144)
Gonzalez C.M., Pignata M.L. Chemical response of the lichen Punctelia subrudecta
(Nyl.) Krog transplanted close to a power station in an urban-industrial
environment // Environ. Pollut. 1997. V.97. N3. P.195-203. (144, 158)
Gonzalez C.M., Pignata M.L. Effect of pollutants emitted by different urban-industrial
sources on the chemical response of the transplanted Ramalina ecklonii (Spreng.)
Mey. & Flot.// Toxicol. Environ. Chem. 1999. V.69. P.61-73. (140)
Gonzalez C.M., Pignata M.L. Chemical response of transplanted lichen Canomaculina
pilosa to different emission sources of air pollutants // Environ. Pollut. 2000.
V.110. N2. P.235-242. (144)
Литература и авторский индекс
285
Gordon С.А., Herrera R„ Hutchinson TC. The use of a common epiphytic lichen as a
bioindicator of atmospheric inputs to two Venezuelan cloud forests // J. Tropical
Ecology. 1995. V.l 1. NI. P.1-26. (114, 144)
Gorham E. A comparison of lower and higher plants as accumulators of radioactive fall-
out // Canad. J. Bot. 1959. V.37. N2. P.327-329. (189)
Gouaux P, Vincent J.P. Evidence and control of pollution action on lichens (Peltigera
canina) using infrared color film // Sci. Total Environ. 1990. V.95. P. 181-190. (151)
Gough L.P, Jackson L.L., Sacklin J.A. Determining baseline element composition of
lichens .2. Hypogymnia enteromorpha and Usnea spp at Redwood National Park,
California // Water, Air, and Soil Pollution. 1988a. V.38. Nl-2. P.169-180. (40,
175, 179)
Gough L.P, Severson R.C., Jackson L.L. Determining baseline element composition of
lichens . 1. Parmelia sulcata at Theodore Roosevelt National Park, North Dakota
// Water, Air, and Soil Pollution. 1988b. V.38. Nl-2. P.157-167. (175, 179)
Gough L.P, Severson R.C., Jackson L.L. Baseline element concentrations in soils and
plants, Bull Island, Cape Romain National Wildlife Refuge, South Carolina, USA
11 Water, Air, and Soil Pollution. 1994. V.74. Nl-2. P.1-17. (175)
Goward T. Notes on oldgrowth-dependent epiphytic macrolichens in inland British
Columbia, Canada // Acta Bot. Fennica. 1994. V.l50. P.31-38. (239)
Goyal R., Seaward M.R.D. Lichen ecology of the Scunthorpe heathlands.II.Industrial
metal fallout pattern from lichen and soil assays // Lichenologist. 1981a. V.13.
N3. P.289-300. (30)
Goyal R., Seaward M.R.D. Metal uptake in terricolous lichens. I. Metal localization
within the thallus // New Phytologist. 1981b. V.89. N4. P.631-645. (174)
Goyal R., Seaward NL.R.D. Metal uptake in terricolous lichens.II.Effects on the
morphology of Peltigera canina and Peltigera rufescens // New Phytologist.
1982a. V.90. NI. P.73-84. (30, 79)
Goyal R., SeawardM.R.D. Metal uptake in terricolous lichens.IILTranslocation in the
thallus of Peltigera caninaI'/New Phytologist. 1982b. V.90. NI. P.85-98. (30)
Grasso M.F., Clocchiatti R., Carrot F., Deschamps C., VurroF. Lichens as bioindicators
in volcanic areas: Mt. Etna and Vulcano Island (Italy) // Environmental Geology.
1999. V.37. N3. P.207-217. (182)
Gries C. Lichens as indicators of air pollution // Lichen Biology. T.Nash - ed. Oxford:
Acad. Press, 1996. P.240-254. (80, 82, 86, 87, 158)
Gries C, Nash Т.Н., Kesselmeier J. Exchange of reduced sulfur gases between lichens
and the atmosphere // Bidgeochemistry. 1994. V.26. NI. P.25-39. (50)
Gries C, RomagniJ.G., Nash Т.Н., Kuhn U., Kesselmeier J. The relation of H2S release
to SO2 fumigation of lichens // New Phytologist. 1997. V. 136. N4. P.703-711. (50)
Gries C., Sanz M.-J., Nash Т.Н. The effect of SO2 fumigation on CO, gas exchange,
chlorophyll degradation in different lichen species from western North America
// Cryptogamie Botany. 1995. V.5. N3. P.239-246. (73)
286
Литература и авторский индекс
Griininger W., Monge-Najera J. Use of the temperate lichen Hypogymnia physodes
(Parmeliaceae) to evaluate air pollution in the tropics // Revista de Biologia
Tropical. 1988. V.36. N2B. P.545-547. (139, 142, 146)
Guillitte O., Kirchmann R., van GelderE., Hurtgen Ch. Radionuclides fallout on lichens
and mosses and their leaching by rain in a forest ecosystem // Transfer of
radionuclides in natural and semi-natural environments. G.Desmet, P.Nassimbeni,
M.Belli - eds. London and New York: Elsevier Applied Science, 1990. P.110-
117. (199)
Haapala H, Goltsova N., Seppala R., Huttunen S., Kouki J., Lamppu J., Popovichev
B. Ecological condition of forests around the eastern part of the Gulf of Finland
// Environ. Pollut. 1996. V.91. N2. P.253-265. (184)
Haas J.R., Bailey E.H., Purvis O. W. Bioaccumulation of metals by lichens - uptake of
aqueous uranium by Peltigera membranacea as a function of time and pH //
Amer. Mineralogist. 1998. V.83. NI 1-12. P.1494-1502. (197, 199)
Haffner E., Lomsky B., Hynek V., Hallgren J.E., Batic E, Pfanz H Air pollution and
lichen physiology. Physiological responses of different lichens in a transplant
experiment following an SO2-gradient // Water, Air, and Soil Pollution. 2001.
V. 131. N1 -4. P. 185-201. (82, 140, 157)
Hale M.E. The lichens of Aton Forest, Connecticut // Bryologist. 1950. V.53. N3. P. 181 -
213. (139)
Hale M.E. Phytosociology of cryptogams in the hardwoods of southern Wisconsin //
Bull. Ecol. Soc. America. 1952. V.33. N3. P.59. (22)
Hale M.E. First report on lichen growth rate and succession at Aton Forest, Connecticut
// Bryologist. 1954. V.57. N3. P.244-247. (139)
Hale M.E. Phytosociology of corticolous cryptogams in the upland forests of southern
Wisconsin // Ecology. 1955. V.36. NI. P.45-63. (22)
Hale M.E. Studies on lichen growth rate and succession // Bull. Torrey Bot. Club.
1959. V.86. N2. P. 126-129. (139)
Hale M.E- The biology of the lichens. 2d editon. Edward Arnold, Baltimore, 1974.
181 p. (17)
Hallbauer D.K., Jahns H.M. Attack of lichens on quartzitic rock surfaces //
Lichenologist. 1977. V.9. P. 119-122. (50)
Hallbom L., Bergman B. Effects of inorganic nitrogen on C,H2 reduction and CO2
exchange in the Peltigera praetextata-Nostoc and Peltigera aphthosa-
Coccomyxa-Nostoc symbioses // Planta. 1983. V.157. P.441-445. (84)
Hallgren J.E., Nyman B. Observations on trees of Scots pine (Pinus silvestris L.) and
lichens around a HF and CO2 emission source // Stud. For. Suecica. 1977. V. 137.
P.1-40. (84, 87)
Halonen P, Hyv'arinen M., Kauppi M. Emission related and repeated monitoring of
element concentrations in the epiphytic lichen Hypogymniaphysodes in a coastal
area, W. Finland // Ann. Bot. Fennici. 1993. V.30. N4. P.251-261. (181)
Литература и авторский индекс
287
Handley R., Overstreet R. Uptake of carrier-free Cs-137 by Ramalina reticulata //Plant
Physiol. 1968. V.43. P.1401-1405. (38)
Hanson W.C. Radioecological concentration processes characterizing Arctic ecosystems
// Radioecological concentration processes. B.Aberg, F.P.Hungate - eds. Oxford
and other: Pergamon press, 1967. P.l83-191. (189)
Harris E., MackR., Ku M.S.B. Death of steppe cryptogams under the ash from Mount
St. Helens // Amer. J. Bot. 1987. V.74. N8. P.l249-1253. (182)
Harris G.P. The ecology of corticolous lichens. 3. A simulation model of productivity as
function of light intensity and water availability // J. Ecology. 1972. V.60. NI.
P.l9-40. (48)
Hart R., Webb P.G., Biggs R.H., Portier K.M. The use of lichen fumigation studies to
evaluate the effects of new emission sources on class-I areas // JAPCA. 1988.
V.38. N2. P.l44-147. (85)
Hauck M., Hesse V., JungR., Zoller T, Runge M. Long-distance transported sulphur as
limiting factor for the abundance of Lecanora conizaeoides in montane spruce
forests // Lichenologist. 2001a. V.33. N3. P.267-269. (21)
Hauck M., JungR., Runge M. Does water holding capacity of bark have an influence on
lichen performance in dieback affected spruce forests // Lichenologist. 2000. V.32.
N4. P.407-409. (22, 90)
Hauck M., Jung R., Runge M. Relevance of element content of bark for the distribution
of epiphytic lichens in a montane spruce forest affected by forest dieback И Environ.
Pollut.. 2001b. V. 112. N2. P.221 -227. (21)
Hawksworth D.L. The variety of fungal - algal symbioses, their evolutionary significance,
and the nature of lichens // Botan. J. Linnean Soc. 1988. V.96. NI. P.3-20. (15)
Hawksworth D.L. The fungal dimension of biodiversity - magnitude, significance, and
conservation // Mycological Research. 1991. V. 95. P. 641-655. (238)
Hawksworth D.L., Coppins B.J., Rose F. Changes in the British lichen flora // The
Changing Flora and Fauna of Britain. D.L.Hawksworth - ed. London, New York:
Academic Press, 1974. P.47-78. (93)
Hawksworth D.L., Hill D.J. The lichen-forming fungi. Blackie, Glasgow and London,
1984. 158 p. (17)
Hawksworth D.L, McManus P.M. Lichen recolonization in London under conditions of
rapidly falling sulphur dioxide levels, and the concept of zone skipping // Bot. J.
Linnean Soc. 1989. V.100. NI. P.99-109. (65, 66, 209)
Hawksworth D.L., Rose F. Qualitative scale for estimating sulphur dioxide air pollution
in England and Wales using epiphytic lichens // Nature (London). 1970. V.227.
P.l45-148. (66. 89, 98, 115)
Hawksworth D.L., Rose F. Lichens as pollution monitors И Studies in Biology. 1976.
N66. P.l-60. (10, 11, 137)
Hawksworth D.L., Rose E, Coppins B.J. Changes in the lichen flora of England and
Wales attributable to pollution of the air by sulphur dioxide // Air Pollution and
288
Литература и авторский индекс
Lichens. B.W.Ferry, M.S.Baddeley, D.L.Hawksworth-eds. Toronto: University
of Toronto Press, 1973. P.330-367. (93)
Heber I., Heber W, TiirkR. Aktives monitoring mit Flechten in der Landeshauptstadt
Linz (Oberosterreich, Osterreich)// VDI Berichte. 1991.N901. S.143-148. (142)
Heber L, Heber W., Turk R. Die Luftqualitat in der Stadt Linz (Oberosterreich) von
Oktober 1990 bis Oktober 1991 Festgestellt anhand von Flechtenexponaten //
Naturkundliches Jahrbuch der Stadt Linz. 1994. Bd.37-39. S.491-552. (142)
Henderson-Sellers A., Seaward M.R.D. Monitoring lichen reinvasion of ameliorating
environments//Environ. Pollut. 1979. V.19. P.207-213. (94, 114)
Henriksson E., Pearson L.C. Nitrogen fixation rate and chlorophyll content of the lichen
Peltigera canina exposed to sulfur dioxide // Amer. J. Bot. 1981. V.68. P.680-
684. (83, 84)
Henriksson L.E., da Silva E.J. Effects of some inorganic elements on nitrogen fixation
in blue-green algae and some ecological aspects of pollution // Zeitschr. f.
Allgemeine Mikrobiologie. 1978. Bd.18. S.487-494. (84)
Herzig R.,Urech M. Flechten als Bioindikatoren // Bibliotheca Lichenologica. 1991.
Bd.43. S.1-283. (102, 104, 105, 114)
Hickmott M. Lichens on lead II Lichenologist. 1980. V.12. N3. P.405-406. (19)
Hill D.J. The nature of the symbiotic relationship in lichens // Endeavour. 1994. V.18.
N3. P.96-103. (12)
Hoffman G.R. The influence of a paper pulp mill on the ecological distribution of epiphytic
cryptogams in the vicinity of Lewiston, Idaho and Clarkson, Washington //
Environ. Pollut. 1974. V.7. P.283-301. (102, 103)
Hofmann W., Attarpour N., Tiirk R. Verteilung von Caesium-137 in Wald-Okosystemen
im Bundesland Salzburg (Osterreich) // FlW-Symposium 1988: Waldsterben in
Osterreich-Theorien, Tendenzen, Therapien. 27, und 28. Oktober 1988. E.Fiihrer,
F.Neuhuber-eds. Universitat fur Bodenkultur, Wien, 1988. S.xl-x3. (193)
Holien H Influence of site and stand factors on the distribution of crustose lichens of
the Caliciales in a suboceanic spruce forest area in central Norway //
Lichenologist. 1996. V.28. N4. P.315-330. (240)
Holopainen J.K., Mustaniemi A., Kainulainen P, SatkaH, OksanenJ. Conifer aphids
in an air polluted environment. 1. Aphid density, growth and accumulation of
sulfur and nitrogen by Scots pine and Norway spruce seedlings // Environ. Pollut.
1993. V.80.N2. P. 185-191. (142)
Holopainen T Alterations in the ultrastructure of epiphytic lichens Hypogymnia physodes
and Alectoria capillaris caused by air pollution // Silva Fennica. 1981. V.15.
P.469-474. (81)
Holopainen T Comparison of the ultrastructural changes in epiphytic lichens at different
distances from two industrial pollution sources И Savonia. 1982. V.5. P.l 5-30. (81)
Holopainen TH. Cellular injuries in epiphytic lichens transplanted to air polluted areas
//Nordic J. Bot. 1984a. V.4. P.393-408. (141, 155, 156)
Литература и авторский индекс
289
Holopainen Т. Types and distribution of ultrastructural symptoms in epiphytic lichens
in several urban and industrial environments in Finland // Ann. Bot. Fennici.
1984b. V.21. P.213-219. (79, 81, 146, 156)
Holopainen T, Karenlampi L Injuries to lichen ultrastructure caused by sulphur dioxide
fumigations//New Phytologist. 1984. V.98. P.285-294. (82)
Holopainen T, Kauppi M. A comparison of light, fluorescence and electron microscopic
observations in assessing the SO2 injury of lichens under different moisture
conditions // Lichenologist. 1989. V.21. N2. P.l 19-134. (81)
Honegger R. The lichen symbiosis - what is so spectacular about it? // Lichenologist.
1998. V.30. N3. P.l93-212. (15)
Honegger R. The symbiotic phenotype of lichen-forming Ascomycetes // The Mycota
IX. B.Hock-ed. Fungal Associations. Springer-Verlag, Berlin, Heidelberg, 2001.
P.l65-188. (13, 14, 15)
Horvat M., Jeran Z., Spiric Z., Jacimovic R., Miklavcic V. Mercury and other elements
in lichens near the INA Naftaplin gas treatment plant, Molve, Croatia // J. Environ.
Monitoring. 2000. V.2. N2. P.l39-144. (142)
Hrynkiewicz A.Z., Szymczyk S„ Kajfosz J., Olech M. PIXE and NRA environmental
studies by means of lichen indicators П Nuclear Instruments and Methods. 1980.
V. 168. N1-3. P.517-521. (177, 184)
Huebert D.B., L’Hirondelle S.J., Addison P.A. The effects of sulphur dioxide on net
CO2 assimilation in the lichen Evernia mesomorpha Nyl. // New Phytologist.
1985. V.l00. P.643-651. (86)
Hujnagel G., TiirkR Monitoring of air pollutants by exposed lichens in Salzburg (Austria)
// Sauteria. 1998. Bd.9. S.281-288. (142, 153, 157)
Hultengren S., Martinsson P.O., Steinstrom J. Lavar och luftfororenigar kanslighets-
klassing och indexberaknine av epifytisca lavar // Swedish Environmental
Protection Agency, Soina. Report 3967. 1991. 58 p. (24)
HuneckS. New results on the chemistry of lichen substances // Fortschritte der Chemie
organischerNaturstoffe. W.Herz, H.Falk, G.W.Kirby, R.E.Moore-eds. Springer:
Wien, New York, 2001. V.81. P. 1-276. (12)
HurkaH, Winkler S. Statistische Analyse der rindenbewohnenden Flechtenvegetation
einer Allee Tubingens // Flora. 1973. Bd.162. S.61-80. (118)
Hutchinson T.C., Scott M., Soto C., Dixon M. The effect of simulated acid
rain on boreal forest floor feather moss and lichen species // Effects of
Atmospheric Pollutants on Forests, Wetlands and Agricultural Ecosystems.
T.C.Hutchinson, K.M.Meema - eds. Berlin: Springer-Verlag, 1987. P.411-426.
(82)
Hyvarinen M., Crittenden P.D. Growth of the cushion forming lichen, Cladonia
portentosa, at nitrogen polluted and unpolluted heathland sites // Environ. Experim.
Bot. 1998. V.40. NI. P.67-76. (145)
290
Литература и авторский индекс
Hyvarinen М, Halonen Р, Kauppi М. Influence of stand age and structure on the epiphytic
lichen vegetation in the middle-boreal forests of Finland // Lichenologist. 1992.
V.24. N2. P.l65-180. (239)
Ikonen S., Karenlampi L. Physiological and structural changes in reindeer lichens
transplanted around a sulphite pulp mill И Proceedings of the Kuopio meeting on
plant damages caused by air pollution. L.Karenlampi - ed. University of Kuopio,
Kuopio, 1976. P.37-45. (145, 154, 157)
Jaakola T. Fe-55 and stable iron in some environmental samples in Finland //
Radioecological Concentration Processes. B.Aberg, F.P.Hungate - eds. Peigamon
Press: London, 1967. P.247-251. (34)
Jaakkola T, Heinonen O.J., Keinonen M., Salmi A., Miettinen JK. Use ofPb-206/Pb-
204 isotope ratio in lichen, air filter, incinerator ash and gasoline samples as
pollution source indicator// International J. of Mass Spectrometry and Ion Physics.
1983. V.48. P.347-350. (184)
Jackson L.L., Engleman E.E., PeardJ.L Determination of total sulfur in lichens and
plants by combustion-infrared analysis // Environ. Sci. Technol. 1985. V. 19. P.437-
441. (174, 177)
Jackson L.L., Ford J., Schwartzman D. Collection and chemical analysis of lichens for
biomonitoring H Lichens as bioindicators of air quality. General technical report.
K.Stolte et al. - eds. U.S. Department of Agriculture, Forest Service, Rocky
Mountain Forest and Range Experiment Station, Fort Collins, Colorado. 1993.
P. 96-115. (169,173, 175, 176, 177, 179)
Jacquiot L, Daillant O. Bioaccumulation des metaux lourds et d’autres elements traces
par les lichens. Revue bibliographique // Bulletin de 1’Observatoire Mycologique,
2000. V.12. P.2-31. (177)
Jahns H.J. Fame-Moose-Flechten Mittel-, Nord- und Westeuropas. BLV Verlagsgesell-
schaft, Munchen, Wein, Zurich. 1980. 256 S. (18)
Jalkanen L, Makinen A., Hasanen E., Juhanoja J. The effect of large anthropogenic
particulate emissions on atmospheric aerosols, deposition and bioindicators in
the eastern Gulf of Finland region // Science of the Total Environment. 2000.
V.262.N1-2. P.123-136. (185)
Jaros W. The influence of cavernous microclimate on the variability of the lichen Lepra-
ria crassissima // Acta Biol. Cracow. Ser. Bot. 1964. V.7. N1. P.89-105. (23)
Jenkins D.A., Davies R.I. Trace element content of organic accumulations // Nature.
1966. V.210. P.l296-1297. (33)
Jeran Z., Byrne A.R., Batic F. Transplanted epiphytic lichens as biomonitors of air
contamination by natural radionuclides around the Zirovski vrh uranium mine,
Slovenia// Lichenologist. 1995. V.27. N5. P.373-385. (142)
Jeran Z., Smodis B., Jacimovic R. Multielemental analysis of transplanted lichens
{Hypogymnia physodes, L. Nyl.) by instrumental neutron activation analysis //
Acta Chimica Slovenica. 1993. V.40. N4. P.289-299. (142, 161)
Литература и авторский индекс
291
Jezierski A., Bylinska Е., Seaward M.R.D. Electron paramagnetic resonance (EPR)
investigations of lichens. 1. Effects of air pollution H Atmosph. Environ. 1999.
V.33. N28. P.4629-4635. (177)
Jezierski A., Czechowski F, Jerzykiewicz M., Golonka I., Drozd J., Bylinska, E Chen
Y., Seaward M.R.D. Quantitative EPR study on free radicals in the natural
polyphenols interacting with metal ions and other environmental pollutants //
Spectrochimica Acta. Part A. 2002. V.58. N6. P.l293-1300. (177)
John E.A. An assessment of the role of biotic interactions and dynamic processes in the
organization of species in a saxicolous lichen community // Canad. J. Bot. 1989a.
V.67. N7. P.2025-2037. (122)
John E.A. The saxicolous lichen flora of Jonas Rockslide, Jasper National Park, Alberta
11 Bryologist. 1989b. V.92. N1. P. 105-111. (122)
John E., Dale M.R.T. Environmental correlates of species distribution in saxicolous
lichen community // J. Veget. Sci..1990. V.l. N3. P.385-392. (26, 122)
Johnsen I. Problems in relation to the use of plants as monitors of air pollution with
metals // Proceedings of Kuopio meeting on plant damages caused by air pollution.
L. Karenlampi -ed. University of Kuopio, Kuopio, Finland, 1976. P.l 10-114. (75)
Johnsen I., Sochting U. Influence of air pollution on the epiphytic lichen vegetation and
bark properties of deciduous trees in the Copenhagen area // Oikos. 1973. V.24.
P.344-351. (153)
Jones D. Lichens and pedogenesis // CRC Handbook of Lichenology. M.Galun - ed.
CRC Press, Inc., Boca Raton, 1988.V.3. P.l09-124. (31, 50)
Jones D., Wilson M.J., Laundon J.R. Observations on the location and form of lead in
Stereocaulon vesuvianumll Lichenologist. 1982. V.14. N3. P.281-286. (39)
Jorgensen RM. Difficulties in lichen nomenclature // Mycotaxon. 1991. V.40. P.497-
501. (14)
Jovanovic S, Carrot F, Deschamps C., Deschamps N., Vukotic P. A study of the air
pollution in the surroundings of an aluminium smelter, using epiphytic and
litophytic lichens H J. of Trace and Microprobe Techniques. 1995. V.13. N4.
P.463-471. (184)
Jozwik Z. Heavy metals in tundra plants of Bellsund area, Spitsbergen // Polish Polar
Research. 1990. V.l 1. P.401-409. (180)
Juinchang R., Freedman B., Coles C., Zwicker B., Holzbecker J., Chatt A. Vanadium
contamination of lichens and tree foliage in the vicinity of 3 oil-fired power
plants in Eastern Canada // J. of the Air and Waste Management Association.
1995. V.45. N6. P.461-464. (184)
Kandler O. Lichen and conifer recolonization in Munich’s cleaner air// Air pollut. and
ecosyst.: Proc. Int. Symposium,Grenoble, 18-22 May, 1987. Dordrecht etc.:
D. Reidel Publ. Co. 1988. P.784-790. (66, 137)
Kandler O., PoeltJ. Wiederbesiedlung der Innenstadt von Miinchen durch Flechten //
Naturwissensch. Rundschau. 1984. Bd.37. S.90-95. (66)
292
Литература и авторский индекс
Kansanen Р.Н., Venetvaara J. Comparison of biological collectors of airborne heavy
metals near ferrochrome and steel works // Water, Air, and Soil Pollution. 1991.
V.60. N3-4. P.337-359. (184)
Kappen L., Lange O.L. Die Kaltresistenz einiger Makrolichenen // Flora. 1972. Bd. 161.
S.l-29. (99)
Kardish N., Ronen R., Bubrick P, Garty J. The influence of air pollution on the
concentration of ATP and on chlorophyll degradation in the lichen, Ramalina
duriaei (De Not.) Bagl. // New Phytologist. 1987. V.106. N4. P.697-706. (83,
141, 159.)
Karenlampi L. Distribution of chlorophyll in the lichen Cladonia alpestris II Rept. Kevo
Subarctic Res. Station. 1970. V.7. P.1-8. (77, 79, 83)
Karenlampi L. On methods for measuring and calculating the energy flow through lichens
// Rept. Kevo Subarctic Res. Station. 1971. V.7. P.40-46. (79)
Karstrom M. The project One step ahead - a presentation // Svensk Bot. Tidskr. 1992.
V.86. N3. P.103-114. (240, 241)
Kauppi M. Fruticose lichen transplant technique for air pollution experiments // Flora.
1976. V. 165. P.407-414. (159)
Kauppi M. Fluorescence microscopy and microfluorometry for the examination of
pollution damage in lichens//Ann. Bot. Fennici. 1980. V.17.N2. P. 163-173. (33,
81, 84, 142, 155)
Kauppi M. The gathering of lichens as a trade // Aquilo, Ser. Bot. 1993. V.31. P.89-91. (28)
Kauppi M., Halonen P. Lichens as indicators of air pollution in Oulu, Northern Finland
//Ann. Bot. Fennici. 1992. V.29. NI. P.1-9. (184)
Kauppi M., Kauppi A. Infrared color photography for the examination of lichens used in
pollution damage experiments // J. Biolog. Photograph. Ass. 1978. V.46. P.105-
107. (151)
Kauranen P, Miettinen J.K. Po-210 and Pb-210 in environmental samples in Finland
// Radioecological Concentration Processes. B.Aberg, F.P.Hungate-eds. Pergamon
Press: London, 1967. P.275-280. (195)
Kelly B.C., Gobas A.P.C. Bioaccumulation of persistent organic pollutants in lichen -
caribou - wolf food chains of Canada’s central and western Arctic // Environ.
Sci. Technol. 2001. V.35. P.325-334. (43, 178)
Kershaw KA. Physiological ecology of lichens. London: Camridge University Press,
1985. 293 p. (198)
Kershaw K.A., Rose W.R. Studies on lichen-dominated systems. l.The water relations
of Cladonia alpestris in spruce-lichen woodland in northern Ontario // Canad. J.
Bot. 1971. V.49. P.1389-1399 (47)
Kirschbaum U., Windisch U. Beurteilung der lufthygienischen Situation Hessens mittels
epiphytischer Flechten // Umweltplanung, Arbeits- und Umweltschutz.
Schriftenreihe der Hessischen Landesanstalt fur Umwelt, Wiesbaden. 1995. Heft
171. 150 S. (65, 107, 109)
Литература и авторский индекс
293
Kirschbaum U., Wirth V. Flechten erkennen Luftgiite bestimmen. Eugen Ulmer GmbH
& Co.: Stuttgart, 1995. 128 S. (72)
KiszkaJ Wplyw emisji miejskich i przemyslowych na flore porostow (Lichenes) Krakowa
i puszczy Niepolomickiej // Prace monogarficzne wyzszej szkoly pedagogicznej
w Krakowie. 1977. T.19. P.5-133. (72)
Klement O. Der okologische Zeigerwert der Flechten in the Forstwirtschaft // Forstarchiv.
1951.N9-10. S.138-140. (23)
Klement O. Prodromus der mitteleuropaischen Flechtengesellschaften // Feddes Repert.
Beih. 135 (Beitrage zur Vegetationskunde). 1955. S.5-194. (22, 117)
Knops J.M.H., Nash Т.Н., Boucher V.L., Schlesinger W.H. Mineral cycling and epiphytic
lichens - implications at the ecosystem level // Lichenologist. 1991. V.23. N3.
P.309-321. (33, 49, 50)
Knops Nash Т.Н., Schlesinger W.H. The influence of epiphytic lichens on the
nutrient cycling of an oak woodland I I Ecol. Monogr. 1996. V.66. N2. P. 159-179.
(31, 49)
KockM., Schlee D. Effect of sulphite on adenine nucleotides of the green alga Trebouxia
// Phytochemistry. 1981. V.20. P.2089-2092. (88)
Kong EX., Hu W, Chao S.Y., Sang W.L., Wang L.S. Physiological responses of the
lichenXanthoparmelia mexicana to oxidative stress of SO2 // Environ. Experim.
Bot. 1999. V.42. N3. P.201-209. (82)
Korte E, Kvesitadze G., Ugrekhelidze D., Gordeziani M., Khatisashvili G., Buadze O.,
Zaalishvili G., Coulston F. Organic toxicants and plants // Ecotoxicology and
Environmental Safety. Sect. B. 2000. V.47. NI. P.1-26. (43)
KortesharjuJ., Savonen K, Saynatkari T. Element contents of raw humus, forest moss
and reindeer lichens around a cement works in Northern Finland // Ann. Bot.
Fennici. 1990. V.27. N3. P.221-230 . (184)
Koskinen A. Uber die Kryptogamen der Baume, besonders die Flechten, im
Gewassergebiet des Paijanne sowie an den Flussen Kalajoki, Lestijoki und
Pyhajoki. Floristische, soziologische und okologische Studie I. - Privately pub-
lished, printed by Mercatoprin Kirjapaino, Helsinki, 1955. 176 p. (20, 22, 27)
Kral R., Kryzova L., Liska J. Background concentrations of lead and cadmium in the
lichen Hypogymnia physodes at different altitudes // Sci. Total Environ. 1989.
V.84. P.201-209. (184)
Kreuzer W., Schauer T. The vertical distribution of Cs-137 in Cladonia rangiferina and
C. silvaticall Svensk Bot. Tidskr. 1972. V.66. N3. P.226-238. (198)
Kubin E. A survey of element concentrations in the epiphytic lichen Hypogymnia
physodes in Finland in 1985-86 // Acidification in Finland. P.Kauppi, P.Anttila,
K.Kenttflmies-eds. Springer-Verlag, Berlin, 1990. P.421-446. (185)
Kubin E., Lippo H, Karhu J., Poikolainen J. Environmental specimen banking of
nationwide biomonitoring samples in Finland //Chemosphere. 1997. V.34. N9-
10. P.l939-1944. (185)
294
Литература и авторский индекс
Kuusinen М. Epiphytic lichen diversity on Salix caprea in old-growth southern and
middle boreal forests of Finland // Ann. Bot. Fennici. 1994. V.31. N2. P.77-92. (239)
Kuusinen M. Cyanobacteria! macrolichens on Populus tremula as indicators of forest
continuity in Finland // Biological Conservation. 1996. V.75. NI. P.43-49. (240)
Kuusinen M., Siitonen J. Epiphytic lichen diversity in old-growth and managed Picea
abies stands in southern Finland //J. Veget. Sci. 1998. V.9. N2. P.283-292. (239)
Kuziel S. The ratio of К to Ca in thalli of several species of lichens occurring on various
trees//Acad. Soc. Bot. Poloniae. 1973. V.42. P.63-71. (35)
Kwapulinski J., Seaward M.R.D., Bylinska E.A. l37Caesium content of Umbilicaria
species, with particular reference to altitude // Sci. Total Environ. 1985.V.41. N2.
P.125-133. (189)
Kytomaa A., Nieminen S., Thuneberg P, Haapala H., Nuorteva P. Accumulation of
aluminum in Hypogymnia physodes in the surroundings of a Finnish sulfite cellulose
factory // Water, Air, and Soil Pollution. 1995. V.81. N3-4. P.401-409. (184)
Laaksovirta K, Olkkonen H. Effect of air pollution on epiphytic lichen vegetation and
element contents of a lichen and pine needles at Valkeakoski, S. Finland // Ann.
Bot. Fennici. 1979. V.l6. N4. P.285-296. (40, 184)
Laaksovirta K, Olkkonen H, Alakuijala P. Observation on the lead content of lichen
and bark adjacent to a highway in southern Finland // Environ. Pollut. 1976. V. 11.
P.247-255. (40, 184)
Lackovicova A., KubinskaA. The fluorine accumulation in transplanted soil mosses and
lichen in flood plain forests close to Bratislava (Southwestern Slovakia) // Biologia
[Bratislava]. 1991. V.46. N9. P.781-789. (145)
Lang G.E., Reiners W.A., Heier R.K. Potential alteration of precipitation chemistry by
epiphytic lichens П Oecologia. 1976. V.25. P.229-241. (35)
Lange O.L. Hitze- und Trockenresistenz der Flechten in Beziehung zu ihrer Verbreitung
11 Flora. 1953. Bd.140. S.39-97. (24)
Lange O.L. Einige Messungen zum Warmehaushalt poikilohydrer Flechten und Moose
//Arch. Meterol., Geophysik. und Bioklimatol. Ser. B: Allg. und biol. Klimatol.
1954. Bd.5. N2. S. 182-190. (24)
Lange O.L. Pflanzenleben unter Stress: Flechten als Pioniere der Vegetation an
Extremstandorten der Erde. Rostra Universitatis Wirceburgensis. 1992. 59 S. (25,
26, 47)
Lange O.L., Kilian E., Ziegler H. Photosynthese von Blattflechten mit hygroskopischen
Thallusbewegungen bei Befeuchtung durch Wasserdampf oder mit fliissigem
Wasser // Contributions to Lichenology. In honour of A.Henssen. H.M.Jahns -
ed. J. Cramer, Berlin-Stuttgart, 1990. S.311-323. (19)
Lange O.L., Ziegler H. Der Schwermetallgehalt von Flechten aus dem Acarosporetum
sinopicae auf Erzschlackenhalden des Harzes. I. Eisen und Kupfer// Mitteil. der
Florist-soziologische Arbeitsgemeinschaft.N.F. 1963. Bd.10. S.156-183. (35,40, 45)
Larson D. W., Kershaw KA. Studies on lichen-dominated systems. XVIII. Morphological
Литература и авторский индекс
295
control of evaporation in lichens//Canad. J. Bot. 1976. V.54. N17. P.2061-2073.
(48)
Laundon J.R. A study of the lichen flora of London // Lichenologist. 1967. V.3. P.277-
327. (66, 93)
Laundon J.R. London’s lichens // London Naturalist. 1970. V.49. P.20-69. (66)
LawreyJ.D. Trace element accumulation by plant species from a coal strip-mining area
in Ohio // Bull. Torrey Bot. Club. 1977a. V.104. P.368-375. (40)
Lawrey J.D. X-ray emission microanalysis of Cladonia cristatella from a coal strip-
mining area in Ohio // Mycologia. 1977b. V.69. N4. P.855-860. (177)
LawreyJ.D. Lichens as monitors of pollutant elements at permanent sites in Maryland
and Virginia// Bryologist. 1993. V96. N3. P.339-341. (181)
Lawrey J.D., Hale M.E. Retrospective study of lichen lead accumulation in the
northeastern United States // Bryologist. 1981. V.84. N4. P.449-456. (174, 181)
Lawrey J.D., Hale M.E. Lichen evidence for changes in atmospheric pollution in
Shenandoah National Park, Virginia// Bryologist. 1988. V.91. NI. P.21-23. (64,
181, 182)
Lawrey J.D., Rudolph E.D. Lichen accumulation of some heavy metals from acidic
surface substrates of coal mine ecosystems in southeastern Ohio // Ohio J. Sci.
1975. V.75. P.113-117. (34)
LeBlanc F. Quotient d’epiphytisme des arbes du Sud du Quebec И Rev. Canad. Biol.
1963. V.22. NI. P. 19-25. (21)
LeBlanc F. Possibilities and methods for mapping air pollution on the basis of lichen
sensitivity П Mitt. Forstlich. Bundes-Veruchsantalt Wein. 1971. Bd.92. S. ЮЗ-
126. (112, 114)
LeBlanc E, DeSIoover J. Relation between industrialization and the distribution and
growth of epiphytic lichens and mosses in Montreal // Canad. J. Bot. 1970. V.48.
P.1485-1496. (102, 103, 112)
LeBlanc C, Rao D.N. Reaction de quelques lichens et mousses epiphytiques a 1’anhydride
sulfureux dans le region de Sudbury, Ontario // Bryologist. 1966. V.69. P.338-
346. (143)
LeBlanc E, Rao D.N. Effects of sulphur dioxide on lichens and moss transplants //
Ecology. 1973a. V.54. P.612-617. (83, 143)
LeBlanc F., Rao D.N. Evaluation of the pollution and drought hypotheses in relation to
lichens and biyophytes in urban environments // Bryologist. 1973b. V.76. NI.
P.1-19. (11)
LeBlanc E, Rao D.N. Effects of air pollutants on lichens and bryophytes // Responses
of plants to air pollution. J.B.Mudd, T.T.Kozlowski - eds. New York: Academic
Press, 1975. P.237-272. (102, 143)
LeBlanc E, Rao D.N., Comeau G. The epiphytic vegetation of Populus balsamifera and
its significance as an air pollution indicator in Sudbury, Ontario // Canad. J. Bot.
1972a. V.50. P.519-528. (103, 106)
296
Литература и авторский индекс
LeBlanc Е, Rao D.N., Comeau G. Indices of atmospheric purity and fluoride pollution
pattern in Arvida, Quebec // Canad. J. Bot. 1972b. V.50. P.991-998. (102, 115)
LeBlanc E, Robitaille G., Rao D.N. Ecophysiological response of lichen transplants to
air pollution in the Murdochville Gaspe Copper Mines area, Quebec // J. Hattori
Bot. Lab. 1976. V.40. P.27-40. (143)
Lechowicz M.J. Ecological trends in lichen photosynthesis И Oecologia. 1982. V.53.
P.330-336. (87)
Lesica P., McCune B., Cooper S. V., Hong W.S. Differences in lichen and bryophyte
communities between old-growth and managed second-growth forests in the Swan
Valley, Montana // Canad. J. Bot. 1991. V.69. N8. P. 1745-1755. (122, 140)
Letrouit-Galinou M.A., Seaward M.R.D., Deruelle S. A propos du return des lichens
epiphytes dans le Jardin du Luxembourg (Paris) // Bull. Soc. Bot. France. 1992.
V.139. Letters bot.(2). P. 115-126. (66, 94, 209)
Levi H. W. Radioactive deposition in Europe after the Chernobyl accident and its long-
term consequences 11 Ecol. Res. 1991. V.6. P.201-216. (192, 193)
Levin A.G., Pignata M.L. Ramalina ecklonii as a bioindicator of atmospheric pollution
in Argentina // Canad. J. Bot. 1995. V.73. N8. P.l 196-1202. (140, 159)
Liden K, Gustafsson M. Relationships and seasonal variation of Cs-137 in lichen, reindeer
and man in northern Sweden 1961-1965 // Radioecological Concentration Pro-
cesses. B.Aberg, F.P.Hungate- eds. Pergamon Press, Oxford, 1967. P.l93-208.
W
Lindahl P. O. The taxonomy and ecology of some Peltigera species, P. canina (L.) Willd.,
P. refescens (Weis) Humb., P. praetextata (Flk.) Vain. // Svensk Bot. Tidskr. 1953.
V.47. NI. P.94-106. (139)
Lindsay D.C. Probable introductions of lichens to South Georgia // British Antarctic
Survey Bulletin. 1973. V.33-34. P.l69-172. (29)
LinkS.O., Nash Т.Н. An analysis of an Arctic lichen community with respect to slope
onsiliciousrocksat AnaktuvukPass, Alaska//Bryologist. 1984. V.87.N1. P.162-
166. (26, 78)
Lippo H, Poikolainen J., Kubin E. The use of moss, lichen and pine bark in the
nationwide monitoring of atmospheric heavy metal deposition in Finland // Water,
Air, and Soil Pollution. 1995. V.85. N4. P.2241-2246. (185)
Lodenius M. Regional distribution of mercury in Hypogymnia physodes in Finland //
AMB1O. 1981. V. 10. N4. P.l83-184. (185)
Lodenius M., Kumpulainen J. Cd, Fe and Zn content of the epiphytic lichen Hypogymnia
physodes in a Finnish suburb // Sci. Total Environ. 1983. V.32. P.81-85. (184)
Lodenius M., Laaksovirta K. Mercury content of Hypogymniaphysodes and pine needles
affected by a chlor-alkali works at Kuusankoski, SE Finland // Ann. Bot. Fennici.
1979. V.16. NI. P.7-10. (40, 184)
Longton RE. Biology of polar bryophytes and lichens. Cambridge: Cambridge University
Press, 1988. 391 p. (28)
Литература и авторский индекс
297
Looney Kershaw К.А., Nieboer Е., Webber С., Stetsko P.L The distribution of
uranium and companion elements in lichen heath associated with undisturbed
uranium deposits in the Canadian Arctic // Lichen Physiology and Cell Biology.
D.H.Brown-ed. Plenum Press, New York and London, 1985. P.l93-209. (197)
Looney Webber C.E., Nieboer E., Stetsko P.L, Kershaw K.A. Interrelationships
between concentrations of Cs-137 and various stable elements in three lichen species
И Health Physics. 1986. V.50. NI. P. 148-152. (184)
LoppiS. Biomonitoraggio della qualita dell’aria tramite licheni nell’ area geotermica di
Travale Radicondoly // Geotermal in Toscana: Ambiente e Sviluppo. Ricerca
Scientificae Technologica. 1993. V.l. P.12-19. (53, 183)
Loppi S. Lichens as bioindicators of geothermal air pollution in Central Italy // Bryologist.
1996. V.99. NI. P.41-48. (53, 183)
Loppi S. Monitoring of arsenic, boron and mercury by lichen and soil analysis in the Mt.
Amiata geothermal area (central Italy) 11 Geothermal Resources Council
Transactions. 1997. V.21. P.137-140. (53, 183)
Loppi S. Environmental distribution of mercury and other trace elements in the geo-
thermal area of Bagnore (Mt. Amita, Italy) // Chemosphere. 2001. V.45. P.991-
995. (53)
Loppi S., Bargagli R. Lichen biomonitoring of trace elements in a geothermal area (Central
Italy) // Water, Air, and Soil Pollution. 1996. V.88. Nl-2. P.l77-187. (183)
Loppi S., Bonini I. Lichens and mosses as biomonitors of trace elements in areas with
thermal springs and fumarole activity (Mt. Amiata, Central Italy) // Chemosphere.
2000. V.41. N9. P.l333-1336. (183)
Loppi S., Cenni E., Bussotti F, Ferretti M. Epiphytic lichens and tree leaves as
biomonitors of trace elements released by geothermal power plants // Chemistry
and Ecology. 1997a. V. 14. NI. P.31-38. (183)
Loppi S, Cenni E., Bussotti E, Ferretti M. Biomonitoring of geothermal air pollution
by epiphytic lichensand forest trees//Chemosphere. 1998a. V.36. N4-5. P.l 079-
1082. (183)
Loppi S., Chiarucci A., Bonini I. Epiphytic lichen communities as indicators for
atmospheric pollution by geothermal emission // Giomale Botanico Italiano.
1991a. V.l25. N3. P.354. (183)
LoppiS., Chiarucci A., Boninil. Preliminary data on the useoflichens as bioindicators
of air pollution by geothermal emission // Intern. Symposium Environmental impact
assessment situations and perspectives in Europe. Genova. Part III. 1991b. P.455-
456. (183)
Loppi S, De Dominicis V. Lichens as long-term biomonitors of air quality in Central
Italy // Acta Bot. Neerlandica. 1996. V.45. N4. P.563-570. (181)
LoppiS., Giomarelli B., BargagliR. Lichensand mosses as biomonitors of trace elements
in a geothermal area (Mt. Amiata, Central Italy) // Cryptogamie, Mycologie. 1999.
V.20. N2. P.l 19-126. (183)
298
Литература и авторский индекс
Loppi S., Ivanov D., Boccardi R Biodiversity of epiphytic lichens and air pollution in the
town of Siena (Central Italy) // Environ. Pollut. 2002. V. 116. N1. P. 123-128. (98)
Loppi S., Malfatti A., Sani M., Whitehead N.E. Lichens as biomonitors of geothermal
radionuclide pollution // Geothermics. 1997b. V.26. N4. P.535-540. (183)
Loppi S., Nascimbene J. Lichen bioindication of air quality in the Mt. Amiata geothermal
area (Tuscany, Italy) // Geothermics. 1998. V.27. N3. P.295-304. (183)
Loppi S., Nelli L., AncoraS., Bargagli R Accumulation of trace elements in the peripheral
and central parts of a foliose lichen thallus // Bryologist. 1997c. V. 100. N2. P.251 -
253. (170)
Loppi S., Pacioni G., Olivieri N., Digiacomo E Accumulation of trace-metals in the
lichen Evernia prunastri transplanted at biomonitoring sites in Central Italy //
Bryologist. 1998b. V.101.N3. P.451-454. (140)
Loppi S., Putorti E., Pirintsos S.A., De Dominicis V. Accumulation of heavy metals in
epiphytic lichens near a municipal solid waste incinerator (Central Italy) // Environ.
Monit. Assess. 2000. V.61. N3. P.361-371. (184)
Loppi S„ Putorti E., Signorini C., Fommei S., Pirintsos S. A., De Dominicis V. A
retrospective study using epiphytic lichens as biomonitors of air quality - 1980
and 1996 (Tuscany, Central Italy) П Acta Oecologica. 1998c. V.19. N4. P.405-
408. (95, 181)
Lounamaa J. Trace elements in plants growing wild on different rocks in Finland.
A semi-quantitative spectrographic survey H Ann. Bot. Soc. Zool. Bot. Fenn.
“Vanamo”. 1956. V.39. N4. P.l-196. (32)
Lounamaa K.J. Studies on the content of iron, manganese and zinc in macrolichens //
Ann. Bot. Fennici. 1965. V.2. P.l27-137. (34, 35)
Lumbsch H. T, Kothe H. W. Resurrection of the lichen genus Pleurosticta Petrak
(Parmeliaceae'. Ascomycotyna) // Mycotaxon. 1988. V.33. P.447-455. (18)
Lupsina V., Horvat M., Jeran Z., Stegnar P Investigation of mercury speciation in
lichens // Aanalyst. 1992. V.117. N3. P.673-675. (184)
LutzoniF., Pagel M„ Reeb V. Major fungal lineages are derived from lichen symbiotic
ancestor // Nature. 2001. V.411. N6840. P.937-940. (13)
MacNair M.R., Bert V., Huitson S.B., Saumiton-Laprade P, Petit D. Zinc tolerance and
hyperaccumulation are genetically independent characters // Proc. Royal Soc.
London. Ser. B. 1999. V.266. P.2175-2179. (47, 166)
Mahaney W.C., Wilson E., Boyer M.G.. Hancock R.G. V. Marginal bleaching of thalli of
Rhizocarpon as evidence for acid rain in the Norra Storfjallet, Sweden // Environ.
Pollut. 1995. V.87. NI. P.71-75. (79, 83)
Mahon D. C. Uptake and translocation of naturally-occurring radionuclides of the uranium
series // Bull. Environ. Contamin. Toxicol. 1982. V.29. P.697-703. (197)
Makholm M.M., Bennett J.P Mercury accumulation in transplanted Hypogymnia
physodes lichens downwind of Wisconsin chloralkali plant // Water, Air, and Soil
Pollution. 1998. V.102. N3-4. P.427-436. (142)
Литература и авторский индекс
299
Malhotra S.S., Kahn A. A. Sensitivity to SO, of various metabolic processes in an epiphytic
lichen, Evernia mesomorpha 11 Biochemie und Physiologie der Pflanzen. 1983.
Bd.178. S.121-130. (88)
ManninenS., HuttunenS., TorvelaH. Needle and lichen sulfur analyses on two industrial
gradients // Water, Air, and Soil Pollution. 1991. V.59. Nl-2. P.153-163. (184)
Markert B. Presence and significance of naturally occurring chemical elements of the
periodic system in the plant organisms and consequences for future investigations
of inorganic environmental chemistry in ecosystems H Vegetatio. 1992. V.103.
NI. P.l-30. (32)
Markert B. (ed.). Plants as biomonitors. Indicators for heavy metals in the terrestrial
environment. VCH. Weinheim. 1993.644 p. (169)
Markert B. The biological system of the elements (BSF) for terrestrial plants (glycophytes)
// Sci. Total Environ. 1994a. V.155. N3. P.221-228. (32)
Markert В. (ed.). Environmental sampling fortrace analysis. VCH Verlagsgesellschaft
mbH, Weinheim. 1994b. 524 p. (31, 169)
Markert B. Sample preparation (cleaning, drying, homogenization) for trace elements
analysis in plant matrices // Sci. Total Environ. 1995. V. 176. N1 -3. P.45-61. (31,169)
Martin L., Martin J. Comparison of two epiphytic lichen community indices reflecting
air pollution // Folia Cryptogamica Estonica. 1974. Fasc 6. P.47-48. (102, 106)
Martin M.H., Coughtrey P. J. Biological monitoring of heavy metal pollution. Applied
Science Publishers. London, 1982. 476 p. (164)
Mathis P.M., Tomlinsoon G. Lichens: bioassay for air pollution in a metropolitan area
(Nasheille, Tennessee)//}. Tennessee Acad. Sci. 1972. V.47. NI. P.67-73. (102,
114, 115)
Matschullat J., Ottenstein R., Reimann C. Geochemical background - can we calculate
it? // Environmen. Geol. 2000. V.39, N9. P.990-1000. (178,179)
Matthes-Sears U., Nash Т.Н., Larson D. W. The ecology of Ramalina menziesii. IV. In
situ photosynthetic patterns and water relations of reciprocal transplants between
two sites on a coastal-inland gradient // Canad. J. Bot. 1986. V.64. P. 1183-1187. (141)
Matthews KM. The use of lichens in a study of geothermal radon emissions in New
Zealand // Environ. Pollut. Ser. A. 1981. V.24. P.l05-116. (183)
Mattick F. Bodenreaktion und Flechtenverbreitung // Beihefte zum Botanisch.
Centralblatt. 1932. Bd. 49. Abt. 2. Erg. Bd. S.241-271. (21)
Mattsson L.J.S. Deposition, retention and internal distribution of the l55Eu, l44Ce, l25Sb,
l06Ru, 95Zr, 54Mn and 7Be in the reindeer lichen Cladonia alpestris, 1961-1970//
Health Physics. 1975. V.29. NI. P.27-41. (189, 198)
McCarroll D., Viles H. Rock weathering by the lichen Lecidea auriculata in an Arctic
alpine environment // Earth Surface Processes and Landsforms. 1995. V.20. N3.
P.l99-206. (50)
McCune B. Lichen communities along O3 and SO2 gradients in Indianapolis // Bryologist.
1988. V.91. N3. P.223-228. (121, 123)
300
Литература и авторский индекс
McCune В. Lichen communities as indicators of forest health // Bryologist. 2000. V. 103.
N2. P.353-356. (238)
McCune B., AntosJ.A. Epiphyte communities of the Swan Valley, Montana// Bryologist.
1982. V.85. NI. P.1-12. (122)
McCune B., Derr C.C., Muir PS., Shirazi A., Sillett S.C., Daly W.J. Lichen pendants
for transplant and growth experiments // Lichenologist. 1996. V.28. N2. P.l 61-
169. (140, 149, 154)
McCune B., Dey J. P Chapter 7. Lichen Communities // Forest Health Monitoring.
Field Methods Guide. N.G.Tallent-Halsell - ed. U.S. Environmental Protection
Agency, Washington D.C. EPA/620/R94/027. 1994. P.7.1-7.14. (124, 238, 239)
McCune B., Lesica P The trade-off between species capture and quantitative accuracy
in ecological inventory of lichens and bryophytes in forests in Montana// Bryologist.
1992. V.95. N3. P.296-304. (119, 120)
McLean J., Purvis O. W, Williamson B.J., Bailey E.H. Role for lichen melanins in uranium
remediation //Nature (London). 1998. V.391. N6668. P.649-650. (197, 198)
Mendez O.L, Fournier L.A. Los liquenes сото indicatores de la contaminacion
atmosferica en el area metropolitana de San Jose, Costa Rica И Revista de Biologia
Tropical. 1980. V.28. NI. P.31-39. (143, 150)
Mezger U. Flechtenkartierungen und Exponierung von Hypogymniaphysodes in Berlin
und dem Umland // P. Scholz - ed. Flechtenmonitoring - ein kommunales
Kontrollinstrument. Halle: Unabhangingen Institutes fur Umweltfragen, 1993.
S.77-94. (100)
Mietelski J. W., Gaea P, Olech M.A. Radioactive contamination of lichens and mosses
collected in South Shetlands and Antarctic Peninsula // J. Radioanal. & Nuclear
Chem. 2000. V.245. N3. P.527-537. (197, 199)
Migaszewski Z.M. Determining organic compound ratios in soils and vegetation of the
Holy Cross Mts, Poland // Water, Air, and Soil Pollution. 1999. V.l 11. Nl-4.
P.l23-138. (43, 178)
Migaszewski Z.M., Galuszka A., Swiercz A., Kucharzyk J. Element concentrations in
soils and plant bioindicators in selected habitats of the Holy Cross Mountains,
Poland // Water, Air, and Soil Pollution. 2001. 2001. V.l29. Nl-4. P.369-386. (179)
Mikhailova I.N., Scheidegger C. Early development of Hypogymniaphysodes (L.) Nyl.
in response to emissions from a copper smelter//Lichenologist. 2001. V.33. N6.
P.527-538. (80)
Monad F, Moni E, Lanciotti E., Grechi D., Bargagli R. Biomonitoring of airborne
metals in urban environments - new tracers of vehicle emission, in place of lead
// Environ. Pollut. 2000. V.l07. N3. P.321-327. (184)
Monitoring with lichens - monitoring lichens: Proceeding of the NATO advanced research
workshop on lichen monitoring, Wales, United Kindom, 16-23 August, 2000.
P.L.Nimis, Ch.Scheidegger, P.A.Wolseley - eds. Kluwer Academic Publ.:
Dordrecht ets, 2002. 408 p. (9)
Литература и авторский индекс
301
Моппа Е, AiuppaA., Varrica D., Dongarra G. Pb isotope composition in lichens and
aerosols from eastern Sicily: insights into the regional impact of volcanoes on the
environment// Environ. Sci. Technol. 1999. V.33. N15. P.2517-2523. (182)
Morgan-Huws D.L, Haynes EN. Distribution of some epiphytic lichens around an oil
refinery at Fawley, Hampshire// Air Pollution and Lichens. B.W.Ferry
M.S.Baddeley, D.L.Hawksworth - eds. University Press, London, 1973. P.89-
108. (63)
Morosini M., SchreitmvJler J., Reuter U., Ballschmiter K. Correlation between C-6/C-
14 chlorinated hydrocarbons levels in the vegetation and in the boundary layer of
the troposphere // Environ. Sci. Technol. 1993. V.27. N8. P.1517-1523. (178)
Moser T.J., Nash Т.Н. Photosynthetic patterns of Ce'traria cucullata (Bell.) Ach. at
Anaktuvuk Pass, Alaska//Oecologia. 1978. V.34. NI. P.37-43. (77, 83)
Moser T.J., Nash Т.Н., Clark W.D. Effects of a long-term sulfur dioxide fumigation on
arctic caribou forage lichens // Canad. J. Bot. 1980. V.58. P.2235-2240. (83)
Moser T.J., Swafford J.R., Nash TH. Impact of Mount St. Helens’ emissions on two
lichen species of south-central Washington // Environ. Experim. Bot. 1983. V.23.
P.321-329. (77, 83, 85, 182)
MuirD.C.G., Segstro M.D., WelbournP.M., ToomD., EisenreichS.J., MacDonaldC.R.,
Whelpdale D.M. Patterns of accumulation of airborne organochlorine
contaminants in lichens from the Upper Great Lakes region of Ontario // Environ.
Sci. Technol. 1993. V.27. N6. P.1201-1210. (43, 178)
Muir P.S., McCune B. Lichens, tree growth, and foliar symptoms of air-pollution - are
the stories consistent // J. Environ. Quality. 1988. V.17. N3. P.361-370. (121,
123)
Muir P.S., ShiraziA.M. Effects of formaldehyde-enriched mists on Pseudotsuga menziesii
(Mirbel) Franco and Lobariapulmonaria (L.) Hoffm. // Environ. Pollut. 1996.
V.94. N2. P.227-234. (43, 178)
Muir P.S., Shirazi A.M., Patrie J. Seasonal growth dynamics in the lichen Lobaria
pulmonaria//Bryologist. 1998. V.100. N4. P.458-464. (145, 146, 154)
Mukherjee A.B., Nuorteva P. Toxic metals in forest biota around the steel works of
Rautaruukki Oy, Raahe, Finland // Sci. Total Environ. 1994. V.151. P. 191-204.
(184)
Murphy K.J., Alpert P, Cosentino D. Local impacts of a rural coal-burning generating
station on lichen abundance in a New England forest // Environ. Pollut. 1999.
VI05. N3. P.349-354. (60)
Narewski U., Werner G., Schulz H, Vogt C. Application of laser ablation inductively
coupled mass spectrometry (LA-ICP-MS) for the determination of major, minor,
and trace elements in bark samples // Fres. J. Anal. Chem. 2000. V366. N2.
P.l67-170. (177)
Nash Т.Н. Simplification of the Blue Mountain lichen communities near a zinc factory
// Bryologist. 1972. V.75. N3. P.315-324. (44, 73)
302
Литература и авторский индекс
Nash Т.Н. Sensitivity of lichens to sulphur dioxide//Bryologist. 1973. V.76. N3. P.333-
339. (83)
Nash Т.Н. Influence of effluents from a zinc factory on lichens // Ecol. Monogr. 1975.
V.45.N2. P.l83-198. (40, 44)
Nash TH. Sensitivity of lichens to nitrogen dioxide fumigations H Bryologist. 1976.
V.79. NI. P.l03-106. (83)
Nash Т.Н. Correlating fumigation studies with field effects // Lichens, Bryophytes and
Air Quality. T.H.Nash, V.Wirth - eds. J. Cramer, Berlin-Stuttgart, 1988. P.201-
216. (73, 77)
Nash TH Metal tolerance in lichens // Heavy metal tolerance in plants: evolutionary
aspects. A.J.Shaw - ed. CRC Press, Inc., Boca Raton, 1989. P.l 19-131. (36, 42)
Nash Т.Н. (ed.). Lichen Biology. Cambridge: University Press, 1996.303 p. (11, 31, 33,
36, 40, 42, 45, 47, 49, 84)
Nash Т.Н., Boucher V.L., Gebauer R., Larson D.W. Morphological and physiological
plasticity in Ramalina menziesii'. studies with reciprocal transplants between a
coastal and inland site // Contributions to Lichenology in Honour of A. Henssen.
H.M.Jahns - ed. J. Cramer, Berlin-Stuttgart. 1990. P.357-365. (146, 183)
Nash Т.Н., Dibben M.J. On the ethics of group lichen collecting // International
Lichenological Newsletter. 1979. V.12. NI. P.1-3. (28)
Nash TH, Gries C. Lichens as indicators of air pollution// The Handbook of
Environmental Chemistry. V.4, part C . O.Hutzinger - ed. New York: Springer-
Verlag, 1991. P.l-29. (74, 86)
Nash Т.Н, Gries C. The response of lichens to atmospheric deposition with an emphasis
on the Arctic // Sci. Total Environ. 1995a. V.161. P.737-747. (180)
Nash Т.Н, Gries C. The use of lichens in atmospheric deposition studies with an emphasis
on the Arctic // Sci. Total Environ. 1995b. V.161. P.729-736. (180)
Nash Т.Н, Lange O.L. Responses of lichens to salinity - concentration and time-course
relationships and variability among Californian species //New Phytologist. 1988.
V.109. N3. P.361-367. (183)
Nash Т.Н, Moser T.J. Vegetational and physiological patterns of lichens in North
American deserts // J. Hattori Bot. Labor. 1982. V.53. P.331-336. (25)
Nash Т.Н., Moser T.J., LinkS.O. Nonrandom variation of gas exchange within arctic
lichens // Canad. J. Bot. 1980. V.58. N10. P.l 181-1186. (77, 83)
Nash TH, Sommerfeld M.R. Elemental concentrations in lichens in the area of the Four
Comers power plants, New Mexico//Environ. Experim. Bot. 1981. V.21. P. 153-
162. (184)
Nazarov V. M., Frontasyeva M. V., Peresedov V.F., Chinaeva V.P., Ostrovnaya T.M.,
Gundorina S.F., Nikonov V.V., LoukinaN. V. Epithermal neutron activation analysis
of moss, lichen and pine needles in atmospheric deposition monitoring // Краткие
сообщения: Объединенный институт ядерных исследований. Дубна, 1995.
N3. Р.25-34. (177)
Литература и авторский индекс
303
Nejedly Z, Kral J., Voltr J., Krejci R., Swietlicki E., Cerny J., Kubik P, Svejda J.
Quality assurance of environmental P1XE analysis in Prague //Nuclear Instruments
and Methods in Physics Research. Sect. B. 1998. V.138. P.981-985. (177)
Nevstrueva M.A., Ramzaev P. V., Moiseev A.A., Ibatullin M.S., Teplykh LA. The nature
of Cs-137 and Sr-90 transport over the lichen-reindeer-man food chain //
Radioecological Concentration Processes. B.Aberg, F.P.Hungate-eds. Pergamon
Press: London, 1967. P.209-215. (198)
Newberry G. The influence of a sulfate-process paper mill on corticolous lichens //
Bryologist. 1974. V.77. N4. P.561-576. (60)
Nieboer E., Ahmed H.M., Puckett KJ., Richardson D.H.S. Heavy metal content of
lichens in relation to distance from a nickel smelter in Sudbury, Ontario //
Lichenologist. 1972. V.5.N3-4. P.292-304. (174, 184)
Nieboer E., Lavoie P, Sasseville R.L.P., Puckett K.J., Richardson D.H.S. Cation-
exchange equilibrium and mass balance in the lichen Umbilicaria muhlenbergii
// Canad. J. Bot. 1976a. V.54. N8. P.720-723. (38)
Nieboer E., Padovan D., Lavoie P, Richardson D.H.S. Anion accumulation by lichens.
2. Competition and toxicity studies involving arsenate, phosphate, sulphate and
sulphite //New Phytologist. 1984. V.96. NI. P.83-93. (37)
Nieboer E., Puckett K.J., Grace B. The uptake of nickel by Umbilicaria muhlenbergii:
a physicochemical process // Canad. J. Bot. 1976b. V.54. N8. P.724-733. (38)
Nieboer E., Puckett K.J., Richardson D.H.S., Tomassini F.D., Grace B. Ecological and
physiological aspects of the accumulation of heavy metals and sulphur in lichens
// Proceedings of International Conference on Heavy Metals in the Environment,
Toronto, Ontario, Canada, Oct. 27-31, 1975. 1977. V.2. Part 1. P.331-351. (38)
Nieboer E., Richardson D.H.S. The replacement of the nondescript term “heavy metals”
by a biologically and chemically significant classification of metal ions // Environ.
Pollut. Ser. B. 1981. V.l. NI. P.3-26. (32, 36, 37, 42)
Nieboer E., Richardson D.H.S., Boileau L.J.R., Beckett P.J., Lavoie P, Padovan D.
Lichens and mosses as monitors of industrial activity associated with uranium
mining in northern Ontario, Canada - Part 3: Accumulation of iron and titanium
and their mutual dependence 11 Environ. Pollut. Ser. B. 1982. V.4. P.l 81-192.
(184, 197)
Nieboer E., Richardson D.H.S., Tomassini ED. Mineral uptake and release by lichens:
an overview // Biyologist. 1978. V.81. N2. P.226-246. (30, 35, 36, 39, 40)
Nieminen M., Rantataro M. Mineral and trace element contents of lichens in Finnish
reindeer herding area// Rangifer. 1990. Special issue 4. P.56. (184)
Nimis P.L., Andreussi S., Pittao E. The performance of two lichen species as
bioaccumulators of trace metals // Sci. Total Environ. 2001. V.275. Nl-3. P.43-
51. (167, 170, 174)
Nimis P.L., Castello M., PerottiM. Lichens as bioindicators of heavy metal pollution: a
case studia at La Spezia (N. Italy) // Plants as Biomonitors: Indicators for heavy
304
Литература и авторский индекс
metals in the terrestrial environment. B.Markert-ed. VCH, Weinheim etc., 1993.
P.265-284. (167, 174, 184, 185)
Nimis P.L., Lazzarini A., Lazzarini G., Gasparo D. Lichens as bioindicators of air
pollution by SO2 in the Veneto region (NE Italy) // Studia Geobotanica. 1991.
V.ll. NI. P.3-76. (64, 65, 102, 105)
Nimis P.L., Lazzarini G., Lazzarini A., Skert N. Biomonitoring of trace elements with
lichens in Veneto (NE Italy) // Sci. Total Environ. 2000. V.255. N1 -3. P.97-111.
(185)
Norden B., Appelqvist T. Conceptual problems of ecological continuity and its
bioindicators // Biodiversity and Conservation. 2001. V.10. N5. P.779-791. (239)
Noske O., Lauchili A., Lange O.L., Vieweg G.H., Ziegler H. Konzentration und
Lokalisieruing von Schwermetallen in Flechten der Erzschlackenhalden des Harzes
// Vbrtrage aus dem Gesamtegebiet der Botanik, N.F. [Deutsch. Bot. Ges.]. 1970.
Bd. 4. S.67-79. (39, 41, 45)
NotcuttG., Davies E Dispersion of gaseous volcanogenic fluoride, Island ofHawaii//J.
of Volcanology and Geothermal Research. 1993. V.56. P.125-131. (182)
Notcutt G., Davies E Biomonitoring of volcanogenic fluoride, Furnas Caldera, Sao
Miguel, Azores // J. of Volcanology and Geothermal Research. 1999. V.92. Nl-2.
P.209-214. (182)
Nyangababo J. T. Lichens as monitors of aerial heavy metal pollutants in and around
Kampala// Bull. Environ. Contamin. Toxicol. 1987. V.38. NI. P.91-95. (184)
Nygard S, Harju L. A study of the short range pollution around a power plant using
heavy fuel oil by analysing vanadium in lichens // Lichenologist. 1983. VI5.
P.89-93. (184)
Nylander W. Les lichens du jardin du Luxembourg // Bull, de la Soc. Bot. France. 1866.
V.13.N1. P.1-19. (10)
Oksanen L. On the use of the Scandinavian type class system in coverage estimation /
/Ann. Bot. Fennici. 1976. V.13. P.149-153. (121)
Olech M. Preliminary observations on the content of heavy metals in thalli of Usnea
antarctica Du Rietz (Lichenes) in the vicinity of the “H. Arctowski” Polish
Antarctic station // Polish Polar Research. 1991. VI2. NI. P.129-131. (180)
Olmez L, Gulovali M.C., Gordon G.E. Trace element concentrations in lichens near a
coal-fired power plant//Atmosph. Environ. 1985. V. 19. P.l663-1669. (40,171,184)
Orsi E. V., Glenn M.G. Lichens as microenvironmental markers of air quality: relative to
topography, wind direction and vehicular traffic patterns // Grana. 1991. V.30.
P.51-58. (184)
Qystedal D.O., Lewis Smith R.I. Lichens of Antarctica and South Georgia: a guide to
their identification and ecology. Cambridge: Cambridge University Press, 2001.
411 p. (26)
Owczarek M., Guidotti M., Blasi G., De Simone C., De Marco A., Spadoni M. Traffic
pollution monitoring using lichens as bioaccumulators of heavy metals and
Литература и авторский индекс
305
policyclic aromatic hydrocarbons//Fres. Environ. Bull. 2001. V.10. NI. P.42-45.
(184)
Pakarinen P Nutrient and trace metal content and retention in reindeer lichen carpets
of Finnish ombrotrophic bogs // Ann. Bot. Fennici. 1981a. V.18. P.265-274.
(W
Pakarinen P Regional variation of sulphur concentrations in Sphagnum and Cladonia
lichens in Finnish bogs//Ann. Bot. Fennici. 1981b. V.18. P.275-279. (185)
Pakarinen P, Makinen A., Rinne R.J.K. Heavy metals in Cladonia arbuscula and
Cladonia mitis in eastern Fennoscandia// Ann. Bot. Fennici. 1978. V.15. P.281-
286. (174, 184)
Palmer H.E., Hanson W.C, Griffin B.J., Roesch W.C. Cesium-137 in Alaskan Eskimos
// Science. 1963. V.142. N3588. P.64-66. (186)
Palomaki V., Tynnyrinen S., Holopainen T. Lichen transplantation in monitoring fluoride
and sulfur deposition in the surroundings of a fertilizer plant and a strip mine at
Siilinjarvi // Ann. Bot. Fennici. 1992. V.29. NI. P.25-34. (142)
Pearson L.C. Varying environmental factors in order to grow intact lichens
under laboratory conditions И Amer. J. Bot. 1970. V.57. N6. Part 1. P.659-664.
(48, 78)
Pearson L.C. Air pollution damage to cell membranes in lichens. 1. Development of a
simple monitoring test //Atmosph. Environ. 1985. V.19. P.209-21’2. (85)
Pearson L.C. Active monitoring // Lichens as bioindicators of air quality. General
Technical Report RM-224. K.Stolte - ed. Fort Collins, CO: US Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station, 1993. P.89-95. (137, 138, 146, 149, 151)
Pearson L.C., BensonS. Laboratory growth experiments with lichens based on distribution
in nature// Bryologist. 1977. V.80. N2. P.317-327. (78)
Pearson L.C., Rogers G.A. Air pollution damage to cell membranes in Lichens.3. Field
experiments // Phyton (Austria). 1982. Bd.22. S.329-337. (85, 137, 138, 159)
Peciar V. Epiphytische Moosgesellschaften der Slowakei // Acta Fac. Rer. Nat. Univ.
Comeniae. Botanika. 1965. Bd.9. N12. S.371-470. (21, 22)
Peck J. E., Ford J., McCune B., Daly W. Tethered transplants for estimating biomass
growth rates of the Arctic lichen Masonhalea richardsonii II Bryologist. 2000.
V.103. N3. P.449-454. (144)
Pentecost A. Aspect and slope preference in a saxicolous lichen community //
Lichenologist. 1979. V. 11. N1. P.81 -83. (26)
Pentecost A., Rose F. Changes in the ciyptogam flora of the Wealden sandrocks, 1688-
1984 // Bot. J. Lin. Soc. 1985. V.90. N3. P.217-230. (28)
Perkins D.F. Relationship between fluoride contents and loss of lichens near an aluminium
works // Water, Air, and Soil Pollution. 1992. V.64. P.503-510. (184)
Persson R.B.R. 210-Pb atmospheric deposition in lichen carpets in northern Sweden
during 1961-1969//Tellus. 1970. V.22. P.564-571. (195)
306
Литература и авторский индекс
Pfeiffer H.N., Barclay-Estrup Р. The use of a single lichen species, Hypogymnia physodes,
as an indicator of air quality in Northwestern Ontario // Bryologist. 1992. V.95.
NI. P.38-41. (114, 184)
Piervittori R., Usai L., Alessio E, Maffei M. The effect of simulated acid rain on surface
morphology and n-alkane composition of Pseudeverniafurfuracea // Lichenologist.
1997. V.29. N2. P.191-198 . (78, 79)
PilegaardK. Heavy metals in bulk precipitation and transplanted Hypogymniaphysodes
and Dicranoweisia cirrata in the vicinity of a Danish steelworks // Water, Air,
and Soil Pollution. 1979. V.ll. NI. P.77-91. (142)
Pilegaard K, Rasmussen L, GydeseuH Atmospheric background deposition of heavy
metals in Denmark monitored by epiphytic cryptogams // J. Applied Ecology.
1979. V.16. N3. P.843-853. (174)
Pirintsos S.A., Vokou D., Diamantopoulos J., Galloway D.J. An assessment of the
sampling procedure for estimating air pollution using epiphytic lichens as
indicators// Lichenologist. 1993. V.25. N2. P.165-173. (169)
Pisut L, Jelinkova E. Anmerkungen bber die Immissionseinflusse auf die Variabilitnt
der Flechte Lecanora varia (Ehrh.) Ach. im Gebiet von Bratislava // Biologia
[Bratislava]. 1973. V.28. P.279-287. (145)
Plummer G.L. Color infrared photography, land-use patterns and plant sciences // Bull.
Georgia Acad. Sci. 1968. V.26. P.23-32. (151)
Poblet A., Andrade S., Scagliola M., Vodopivez C., Curtosi A., Pucci A., Marcovec-
chio J. The use of epilithic antarctic lichens {Usnea aurantiacoatra and
Usnea antartica) to determine deposition patterns of heavy metals in the Shet-
land Islands, Antarctica // Sci. Total Environ. 1997. V.207. N2-3. P.l87-194.
(180)
Prussia C.M., Killingbeck К. T. Concentrations of ten elements in two common foliose
lichens - leachability, seasonality, and the influence of rock and tree bark substrates
// Bryologist. 1991. V.94. N2. P.135-142. (172)
Puckett KJ. The effect of heavy metals on some aspects of lichen physiology // Canad.
J. Bot. 1976. V.54. P.2695-2703. (85)
Puckett K.J. Bryophytes and lichens as monitors of metal deposition // Lichens,
Bryophytes and Air Quality. T.H.Nash, V. Wirth - eds. J. Cramer, Berlin-Stuttgart,
1988. S.231-267. (30, 41, 88, 177)
Puckett K.J., Burton M.A.S. The effect of trace elements on lower plants // Effect of
heavy metal pollution on plants. N.W.Lepp-ed. London: Applied Science Publ.,
1981. V.2. P.213-238. (40)
Puckett KJ., Finegan E.J. An analysis of the element content of lichens from the Northwest
Territories, Canada// Canad. J. Bot. 1980. V.58. P.2073-2089. (42, 184)
Puckett K.J., Nieboer E., Gorzynski M.J., Richardson D.H.S. The uptake of metal ions
by lichens: a modified ion-exchange process//New Phytologist. 1973. V.72.N2.
P.329-342. (36, 38)
Литература и авторский индекс
307
Pullum Р.А., Erbisch ЕН. Effects of gamma radiation on the lichen Cladonia verticillata
(Hoffm.) Schaer.// Bryologist. 1972. V.75. NI. P.48-53. (187)
Purvis O. W The occurrence of copper oxalate in lichens growing on copper sulphide-
bearing rocks in Scandinavia // Lichenologist. 1984. V.16. N2. P.l97-204. (45,
46)
Purvis O. W., Broomhead J. A., ElixJ.A., Jones G.C. The occurrence of copper norstictic
acid in lichens from cupriferous substrata // Lichenologist. 1987. V. 19. N2. P. 193-
203. (46)
Purvis O. W., Halls C. A review of lichens in metal-enriched environments // Lichenologist.
1996. V.28.N6. P.571-601. (35, 45, 177)
Pustelniak L. Application of the transplantation method in studies on the influence of the
urban environment upon vitality of Hypogymniaphysodes (L.) Nyl. thalli // Zeszyty
Naukowe Uniwersytetu Jagiellonskiego. Prace Botaniczne. 1991. V.997. P. 193-
20\.(142)
Pyatt F.B., Grattan J.P., Lacy D., Seaward M.R.D. Comparative effectiveness of
Tillandsia usneoides L. and Parmotrema praesorediosum (Nyl.) Hale as bio-
indicators of atmospheric pollution in Louisiana (USA) // Water, Air, and Soil
Pollution. V.111.N1-4. P.317-326. (177)
Quevauviller P. Conclusions of workshop - improvements of trace element
determinations in plant matrices// Sci. Total Environ. 1995.V.176. Nl-3. P.141-
148. (177)
Quevauviller P, Herzig R., Muntau H Certified reference material of lichen (CRM-
482) for the quality control of trace element biomonitoring // Sci. Total Environ.
1996. V.187. N2. P.l43-152. (168, 173, 174, 175, 177)
Quevauviller P, van Renterghem D., Muntau H, GriepinkB. Intercomparison to improve
the quality of trace element determination in lichens // Intern. J. Environ. Anal.
Chem. 1993. V.53. N3. P.233-242. (168)
Rabe R., Wiegel H. Wiederbesiedlung des Ruhrgebietes durch Flechten zeigt
Verbesserung der Luftqualitat an // Staub Reinhalt. d. Luft. 1985. Bd. 45. N3.
S. 124-126. (66)
Ramade E Qualitative and quantitative criteria defining a ’’healthy ecosystem” //
Evaluating and monitoring the health of large-scale ecosystems. NATO AS1 Series.
Ser.l. Global environment change. Springer-Verlag: Berlin, Heidelberg, 1995. V.28.
P.43-61. (239.)
Rao D.N., Le Blanc F. Influence of an iron sintering plant on corticolous epiphytes in
Wawa, Ontario // Bryologist. 1967. V.70. N2. P.141-157. (98, 99)
Rapport D.J. What constitutes ecosystem health? // Perspectives in Biology and Medicine.
1989. V.33.N1. P.l20-132. (238)
Rapport D.J. Ecosystem health: an emerging integrative science // Evaluating and
monitoring the health of large-scale ecosystems. NATO ASI Series. Ser. 1. Global
environment change. Springer-Verlag: Berlin, Heidelberg, 1995. V.28. P.3-31. (238)
308
Литература и авторский индекс
Rapport D.J., Costanza R., McMichael A. J. Assessing ecosystem health // Trends in
Ecology and Evolution. 1998. V.13. N10. P.397-402. (238)
Rapport D.J., Gaudet C.L., Calow P. (eds.). Evaluating and monitoring the health of
large-scale ecosystems. NATO AS1 Series. Ser. 1. Global environment change.
Springer-Verlag: Berlin, Heidelberg, 1995. V.28. 454 p . (238)
Rees D.C., Juday G.R Plant species diversity on logged versus burned sites in central
Alaska // Forest Ecology and Management. 2002. V.155. Nl-3. P.291-302.
(239)
Reis M.A., Alves L.C., Freitas M.C., van Os B., Wolterbeek H.T Lichens Parmelia
sulcata time response model to environmental elemental availability // Sci. Total
Environ. 1999a. V.232. Nl-2. P.l05-115. (143)
ReisM.A., Freitas M.C., AlvesL.C., Marques A.P., Costa C. Environmental assessment
in an industrial area of Portugal // Biological Trace Element Research. 1999b.
V.71-72. P.273-280. (143)
Renhom K-E, Esseen P.-А. Biomass growth in five alectorioid lichen epiphytes //
Conservation Biology of Lichenised Fungi. Mitteilungen der Eidgenossischen
Forschungsanstalt fur Wald, Schnee und Landschaft, Birmensdorf, Switzerland.
C.Scheidegger, P.A.Wolseley, G.Thor-eds. 1995. P.l33-140. (140, 146, 154)
Renhom, K-E., Esseen, P-А., Palmqvist K, SundbergB. Growth and vitality of epiphytic
lichens. I. Responses to microclimate along a forest edge-interior gradient //
Oecologia. 1997. V.109. NI. P.l-9. (144, 146, 154)
Richardson D.H.S. The transplantation of lichen thalli to solve some taxonomic problems
in Xanthoriaparietina (L.) Th. Fr. // Lichenologist. 1967. V.3. P.386-391. (145)
Richardson D.H.S. Understanding the pollution sensitivity of lichens // Bot. J. Linnean
Soc. 1988. V.96. NI. P.31-43. (41)
Richardson D.H.S. Metal uptake in lichens // Symbiosis. 1995. V.18.N2. P.l 19-127.
(36, 38, 177)
Richardson D.H.S., KiangS., Ahmadjian V., Nieboer E. Lead and uranium uptake by
lichens // Lichen physiology and cell biology. D.H.Brown - ed. Plenum Press,
New York, London. 1985. P.227-246. (197)
Richardson D.H.S., Nieboer E., Lavoie P, Padovan D. The role of metal-ion binding in
modifying the toxic effects of sulfur dioxide on the lichen Umbilicaria
muhelenbergii.W. l4C-fixation studies//New Phytologist. 1979. V.82. N3. P.633-
643. (46)
Richardson D.H.S., Nieboer E., Lavoie P, Padovan D. Anion accumulation by lichens
1. The characteristics and kinetics of arsenate uptake by Umbilicaria muhlenbergii
//New Phytologist. 1984. V.96. NI. P.71-82. (37.)
Riga-Karandinos A.N., Karandinos M.G. Assessment of air pollution from a lignite
power plant in the plain of Megalopolis (Greece) using as biomonitors three species
of lichens - impacts on some biochemical parameters of lichens // Sci. Total
Environ. 1998. V.215. Nl-2. P.167-183. (60)
Литература и авторский индекс
309
Riget Е, Asmund G., Aastrup Р. The use of lichen (Cetraria nivalis) and moss
(Rhacomitrium lanuginosum) as monitors for atmospheric deposition in Greenland
// Sci. Total Environ. 2000. V.245. Nl-3. P.137-148. (180)
Rissanen K., Rahola T. Radiocesium in lichens and reindeer after the Chernobyl accident
// Rangifer. 1990. Special Issue. N3. P.55-61. (189, 198)
Roberts B.A., Thompson L.K. Lichens as indicators of fluoride emission from a
phosphorous plant, Long Harbour, Newfoundland, Canada // Canad. J. Bot. 1980.
V.58. P.2218-2228. (60)
Rolstad J., Rolstad E. Does tree age predict the occurrence and abundance of Usnea
longissima in multi-aged submontane Picea abies stands // Lichenologist. 1999.
V.31.N6. P.613-625. (240)
Ronen R., Galun M. Pigment extraction from lichens with dimethyl sulfoxide (DMSO)
and estimation of chlorophyll degradation // Environ. Experim. Bot. 1984. V.24.
N3. P.239-245. (83)
Rope S.K., Pearson L.C. Lichens as air pollution biomonitors in a semiarid environment
in Idaho// Bryologist. 1990. V.93. NI. P.50-61. (184)
Rose C.I., Hawksworth D.L. Lichen recolonization in London’s cleaner air // Nature
(London). 1981. V.289.N5795. P.289-292. (65, 66)
Rose E Lichenological indicators of age and environmental continuity in woodland //
Lichenology: Progress and Problems. Systematics Association Spec. D.H.Brown,
D.L.Hawksworth, R.H.Bailey - eds. Academic Press, London and New York,
1976. V.8. P.279-307. (239, 240, 241)
Rosentreter R. Indicator value of lichen cover on desert shrubs // Proceedings-Symposium
on Cheatgrass Invasion, Shrub Die-off, and Other Aspects of Shrub Biology and
Management. General Technical Report INT-276. E.D.McArthur, E.M.Romney,
S.D.Smith & P.T.Tueller - eds. Intermountain Research Station, U.S. Forest
Service, U.S.D.A., Ogden, Utah. 1990. P.282-289. (10)
RoserD.J., MelickD.R., Seppelt R.D. Reductions in the polyhydric alcohool content of
lichens as an indicator of environmental pollution // Antarctic Science. 1992. V.4.
N2. P.l85-189. (88)
Ross L.J., Nash Т.Н. Effect of ozone on gross photosynthesis of lichens // Environ.
Experim. Bot. 1983. V.23. NI. P.71-77. (87)
Rossbach M., Jayasekera R., Kniewald G., Nguyen Huu Thang. Large scale air
monitoring - lichen vs. air particulate matter analysis // Sci. Total Environ. 1999.
V.232. Nl-2. P.59-66. (169, 180)
Roux C. Etude ecologique et phytosociologique des peuplements licheniques saxicolec-
calcicoles du sus-est de la France // Bibliotheca Lichenologica. Vaduz, 1981. Bd.
15. 558 s. (34)
RydzakJ. Rozmieszczenie i ekologia porostow miasta Lubina// Ann. Univer. M. Curie-
Sklodowska (Lublin). Sect. C. 1953. V.8(9). P.233-356. (11)
310
Литература и авторский индекс
RydzakJ. Lichens as indicators of the ecological condition of the habitat //Ann.Univer.
M.Curie-Sklodowska (Lublin). Sect. C. 1968. V.32. P.l31-164. (11)
Saeki M., Kunii K., Seki T, Sigiyama K., Suzuki T, Shishido S. Metal burden of urban
lichens // Environ. Res. 1977. V.13. N2. P.256-266. (184)
Saeki M., Kunii K, Seki T, Suzuki T A lichen {Parmelia conspersa) surviving with
elevated concentrations of lead and cooper in the center of Sendai city // Bull.
Environ. Contamin. Toxicol. 1975. V.14. N6. P.726-730. (184)
SalemaaM., Vanha-Majamaa I., DeromeJ Understorey vegetation along a heavy-metal
pollution gradient in SW Finland//Environ. Pollut. 2001. V.l 12.N3. P.339-350.
(185)
Salisbury F.B., Sporner G.G., Sobral M., Ward R.T. Analysis of an alpine environment
// Bot. Gaz. 1968. V.l29. P.l6-32. (35)
Salomon H. Uber das Vorkommen und die Aufnahme einiger wichtiger Naahrsalze bei
den Flechten //Jahrb. f. wissensch. Bot. 1914. Bd. 54. S.309-354. (33, 39)
Sansone U., Danesi P.R., Barbizzi S, Belli M., Campbell M. Gaudino S, Jia G.G.,
Ocone R., Pali A., Rosamilia S., Stellato L. Radioecological survey at selected
sites hit by depleted uranium ammunitions during the 1999 Kosovo conflict // Sci.
Total Environ. 2001. V.281. Nl-3. P.23-35. (10, 197)
Santesson R. The lichens and lichenicolous fungi of Sweden and Norway. Lund. 1993.
240 p. (200)
Sanz M.J., Gries C., Nash Т.Н. Dose-response relationships for SO2 fumigations in the
lichens Evernia prunastri (L.) Ach. and Ramalina fraxinea (L.) Ach. // New
Phytologist. 1992. V.l22. P.313-319. (86)
SarkelaM., NuortevaP. Levels of aluminium, iron, zinc, cadmium and mercury in some
indicator plants growing in unpolluted Finnish Lapland I I Ann. Bot. Fennici. 1987.
V.24. P.301-305. (179)
SarretG., Manceau A., CunyD., vanHaluwyn C., Deruelle S., Hazemann J.-L., Soldo
Y, Eybert-BurardL., MenthonnexJ.-J. Mechanisms oflichen resistance to metallic
pollution // Environ. Sci. Technol. 1998a. V.32. N21. P.3325-3330. (47, 177)
Sarret G., Manceau A., Spadini L., Roux, J.-C., Hazemann J.-L., Soldo Y, Eybert-
BiirardL., Menthonnex J.-J. Structural determination of Zn and Pb binding sites
in Penicillium chrysogenum cell walls by EXAFS spectroscopy // Environ. Sci.
Technol. 1998b. V.32. P.1648-1655. (177)
Scerbo R., Possenti L., Lampugnani L., Ristori T, Barale R„ Barghigiani C. Lichen
(Xanthoria parietina) biomonitoring of trace element contamination and air quality
assessment in Livorno province (Tuscany, Italy) // Sci. Total Environ. 1999. V.241.
Nl-3. P.91-106. (185)
Scerbo R., Ristori T, Possenti L., Lampugnani L., Barale R., Barghigiani C. Lichen
(Xanthoria parietina) biomonitoring of trace element contamination and air quality
assessment in Pisa province (Tuscany, Italy) // Sci. Total Environ. 2002. V.286.
Nl-3. P.27-40. (185)
Литература и авторский индекс
311
Scheidegger С. Early development of transplanted isidioid soredia of Lobaria pulmonaria
in an endangered population // Lichenologist. 1995. V.27. N5. P.361-374. (145,
146)
SchimperA.F.W PflanzengeographieaufphysiologischerGrundlage. Jena, 1898. (21)
Schipperges B., Kappen L., Sonesson M. Intraspecific variations of morphology and
physiology of temperate to Arctic populations of Cetraria nivalis // Lichenologist.
1995. V. 27, N 6. P. 517-529. (144, 146)
SchmelingR., AltF., Klockenkamper R Multielement analysis by total reflection X-ray-
fluorescence spectrometry for the certification of lichen research material // Fres.
J. Anal. Chem. 1997. V.357. N8. P.l024-1044. (169)
Schaller H. Veranderungen der Flechtenflora und Flechtenvegetation im Frankfurter Raum
seit 1800//Cour. Forsch. Inst. Senckenberg. Frankfurt a.M. 1995. Bd. 186. S. 149-
168. (95)
Scholz P. Kartierung epiphytischer Flechten und Luftgute in Sachsen-Anhalt //
Flechtenmonitoring - ein kommunales Kontrollinstrument. P.Scholz-ed. Halle:
Unabhangingen Institutes fur Umweltfragen, 1993. S.l 19-128. (100)
Schanbeck H. Einfluss von Luftverunreinigungen (SO2) auf transplantierte Flechten //
Naturwussenschaften. 1968. Bd. 55. N9. S.451-452. (139, 142)
Schanbeck H. Eine Methode zur Erfassung der biologischen Wirkung von
Luftverunreinigungen durch transplantiert Flechten // Staub- Reinhaltung der
Luft. 1969. Bd. 29. NI. S. 14-18. (139, 142)
Schanbeck H., Holte W. Zum Nachweis von Luftverunreinigungen durch die
transplantierte Biattflechte Parmelia physodes //Zentralbl. Bakteriol., Parasitenk.,
Infektionskrankh. Hygiene, Erst Abteil., Originale. 1970. Bd. 212. S.356-357.
Schuster G. Die Jugendentwicklung von Flechten // Bibliotheca Lichenologica. 1985.
Bd. 20. S.3-206. (80)
Schutte J.A. Chromium in two corticolous lichens from Ohio and West Virginia //
Bryologist. 1977. V.80. N2. P.279-283. (170, 181, 182)
Schwartzman D., KasimM., StieffL., Johnson J.H. Quantitative monitoring of airborne
lead pollution by a foliose lichen // Water, Air, and Soil Pollution. 1987. V.32.
N3-4. P.363-378. (171)
Schwartzman D. W., Stieff L., Kasim M., Kombe E., Aung S, Atekwana E., Johnson J.,
Schwartzman K. An ion-exchange model of Pb-210 and lead uptake in a foliose
lichen - application to quantitative monitoring of airborne lead fallout // Sci. Total
Environ. 1991. V. 100. P.319-336. (171)
Scott M.G., Hutchinson T.C. The use of lichen growth abnormalities as an early
warning indicator of forest dieback//Environ. Monit. Assess. 1990. V.15. P.213-
218. (79)
SeawardM.R.D. Lichen ecology in the Scunthorpe heathlands. 1. Mineral accumulation
// Lichenologist. 1973. V.5. P 423-433. (39)
312
Литература и авторский индекс
Seaward M.R.D. The use of lichens as bioindicators of ameliorating environments //
Bioindikation auf der Ebene der Individuen / Eds.R. Schubert, J. Schuh. Martin-
Luther-Universitat, Halle-Wittenberg, 1980. S.17-23. (114)
Seaward, M.R.D. Lichen flora of the West Yorkshire Conturbation-supplement 11(178-
1980)//Naturalist, Hull. 1981.V.106. P.89-95. (94)
Seaward, M.R.D. Lichen ecology and changing urban environments // Urban Ecology.
The Second European Ecological Symposium, Berlin, 8-12 September 1980.
R.Bomkamm, J. A.Lee, M.R.D.Seaward-eds. Blackwell Sceintific Publications,
Oxford, London, Edinburgh, Boston, Melbourne, 1982. P.l81-189. (94)
Seaward M.RD. Contribution of lichens to ecosystems // CRC Handbook of Lichenology.
M.Galun - ed. CRC Press Inc., Boca Raton, 1988. V.2. P.l07-129. (39)
Seaward M.R.D. Lichens, silent witnesses of the Chernobyl disaster. Bradford: University
of Bradford, 1992. 14 p. (189, 192)
Seaward M.R.D., Letrouit-Galinou M.A. Lichen recolonization of trees in the Jardin du
Luxembourg, Paris // Lichenologist. 1991. V.23. N2. P.181-186. (66)
Selva S.B. Lichen diversity and stand continuity in the northern hardwoods and spruce-
fir forests of northern New England and western New Brunswick // Bryologist.
1994. V.97. N4. P.424-429. (240)
Senhou A., El-Khoukhi T, ChouakA., ElMoursili CherkaouiR., El-Yahiaoui A., Lferde
M. Study of the accumulation of air pollution by the biological indicators, using
14 MeV neutron activation // Radiation Physics and Chemistry. 2001. V.61. N3-
6. P.701-703. (177)
Sernander R. Studier ofver lafvames biologi. 1. Nitrofila lafvar // Svensk Bot. Tidskr.
1912. V.6. P.803-883. (20)
Shampine W.J. Quality assurance and quality control in monitoring programs // Environ.
Monit. Assess. 1993. V.26. P.143-151. (168, 244)
SheardJ. W. Distribution of uranium series radionuclides in upland vegetation of northern
Saskatchewan. I. Plant and soil concentration // Canad. J. Bot. 1986a. V.64. Nil.
P2446-2452 . (197)
SheardJ. W. Distribution of uranium series radionuclides in upland vegetation of northern
Saskatchewan. II. Patterns of accumulation among species and location // Canad.
J. Bot. 1986b. V.64. Nil. P.2453-2463. (197)
Sheppard M.L, Thibault D.H. Natural uranium concentrations of native plants over a
low-grade ore body // Canad. J. Bot. 1984. V.62. P.l069-1075. (197)
Sheridan R.P. Impact of emissions from coal fired electricity generating facilities on
N2-fixing lichens П Bryologist. 1979. V.82. N1. P.54-58. (84)
Shirazi A.M., Muir PS., McCune B. Environmental factors influencing the distribution
of the lichens Lobaria oregana and L. pulmonaria II Bryologist. 1996. V.99. N1.
P.l2-18. (145, 146)
Showman R.E. Lichens as indicators of air quality around a coal-fired power generating
plant И Bryologist. 1975. V.78.N1. P.1-6. (61, 114)
Литература и авторский индекс
313
Showman R.E. Cold injury to lichens in southern Ohio // Bryologist. 1979. V.82. N4.
P.620-621. (99)
Showman R.E. Lichen recolonization following air quality improvement // Bryologist.
1981. V.84. N4. P.492-497. (61, 94, 98, 114)
Showman R.E. Mapping air quality with lichens, the North American experience //
Lichens, bryophytes and air quality. T.H.Nash, V. Wirth - eds. J. Cramer, Berlin-
Stuttgart, 1988. P.67-90. (96, 99, 100, 114)
Showman R.E. Lichen recolonization in the Upper Ohio River Valley // Bryologist. 1990.
V.93. N4. P.427-428. (94, 114)
Showman R.E. Continuing Lichen Recolonization in the Upper Ohio River Valley //
Bryologist. 1997. V.l00. N4. P.478-481. (114)
Shrader-Frechette K.S. Ecosystem health: a new paradigm for ecological assessment?
// Trends in Ecology and Evolution. 1994. V.9. N12. P.456-457. (238)
Sigal L.L. The relationship of lichen and bryophyte research to regulatory decisions in
the United States // Lichens, Bryophytes and Air Quality. T.H.Nash, V. Wirth -
eds. J. Cramer, Berlin-Stuttgart, 1988. P.269-287. (116)
Sigal L.L., Johnston W.J. Effects of acidic rain and ozone on nitrogen fixation and
photosynthesis in the lichen Lobaria pulmonaria (L.)WoWm. // Environ. Experim.
Bot. 1986. V.26. P.59-64. (84, 87)
SigalL.L., Nash Т.Н. Lichen communities on conifers in southern California mountains:
an ecological survey relative to oxidant air pollution // Ecology. 1983. V.64.
P.l345-1354. (72, 73, 79, 80, 143, 153)
Silberstein L., Siegel B.Z., Siegel S.M., MukhtarA., Galun M. Comparative studies on
Xanthoriaparietina, a pollution resistant lichen, and Ramalina duriaei, a sensitive
species. 1. Effects of air pollution on physiological processes // Lichenologist.
1996a. V.28. N4. P.355-365. (141)
Silberstein L., Siegel B.Z., Siegel S.M., MukhtarA., Galun M. Comparative studies on
Xanthoriaparietina, a pollution resistant lichen, and Ramalina duriaei, a sensitive
species. 2. Evaluation of possible air pollution-protection mechanisms //
Lichenologist. 1996b. V.28. N4. P.367-383. (141)
Sillett S.C. Growth rates of two epiphytic cyanolichen species at the edge and in the
interior of a 700-year-old Douglas fir forest in the western Cascades of Oregon
// Bryologist. 1994. V.97.N3. P.321-324. (145, 146, 154, 239)
Sillett S.C., McCune B. Survival and growth of cyanolichen transplants in Douglas-fir
forest canopies//Bryologist. 1998. V.101.N1. P.20-31. (145, 146, 154)
Sillett S.C., McCune B., Peck J.E., Rambo T.R., Ruchty A. Dispersal limitations of
epiphytic lichens result in species-dependent on old-growth forests // Ecological
Applications. 2000. V.10. N3. P.789-799. (145)
Silva-Pando F.J., Ascaso C. Modificaciones ultraestructurales de liquenes epifitos
transplantados a zonas urbanas de Madrid // Collectanea Botanica. 1982. V.13.
P.351-374. (146, 156)
314
Литература и авторский индекс
Sipman H.J.M., Aptroot A. Where are the missing lichens? // Mycological Research.
2001. V.105. N12. P.1433-1439. (17)
Skacel E, PekarekJ. Monitoring of lead, cadmium and mercury in environmental samples
at the regional station of the integrated background monitoring network of Gems
in Czechoslovakia// Sci. Total Environ. 1992. V.l 15. N3. P.261-276. (179)
Skye E. Luftfororeningars inverkan pa busk- och badlavfloran kring skifferoljeverket i
Narkes Kvamtorp // Svensk Bot. Tidskr. 1958. V.52. NI. P.l33-190. (137)
Skye E. Lichens and air pollution. A study of cryptogam ic epiphytes and environment in
the Stockholm region // Acta Phytogeogr. Suecica. 1968. V.52. P.1-123. (36)
Skye E. Lichens as biological indicators of air pollution //Annual Review of
Phytopathology. 1979. V.l7. P.325-341. (137)
SloofJ.E. Lichens as quantitative biomonitors for atmospheric trace-element deposition,
using transplants // Atmospheric Environment. 1995a V.29. NI. P.11-20. (143,
146, 161)
SloofJ.E. Pattern recognition in lichens for source apportionment // Atmosph. Environ.
1995b. V.29. N3. P.333-343. (185)
SloofJ.E., WolterbeekH. T. National trace element air pollution monitoring survey using
epiphytic lichens // Lichenologist. 1991a. V.23. N2. P.139-165. (185)
SloofJ.E., Wolterbeek H.T. Patterns in trace elements in lichens // Water, Air, and Soil
Pollution. 1991b. V.57-58. P.785-795. (185)
SloofJ.E., Wolterbeek В. T. Interspecies comparison of lichens as biomonitors of trace-
element air pollution//Environ. Monit. Assess. 1993a. V.25.N2. P.149-157. (185)
SloofJ.E., Wolterbeek B.T. Substrate influence on epiphytic lichens // Environ. Monit.
Assess. 1993b. V.25. N3. P.225-234. (185)
Smith A.L. A Handbook of the British Lichens. London: British Museum, 1921. 160 p.
(51)
Smith D.C. Studies in the physiology of lichens. 1. The effects of starvation and of
ammonia absorption upon the nitrogen content of Peltigerapolydactyla // Ann.
Bot. 1960a. V.24. N93. P.52-62. (139)
Smith D.C. Studies in the physiology of lichens. 2. Absorption and utilization of some
simple organic nitrogen compounds by Peltigerapolydactyla!I Ann. Bot. 1960b.
V.24. N94. P.l72-185. (139)
Smith D.C. Studies in the physiology of lichens. 3. Experiments with dissected disc of
Peltigerapolydactyla// Ann. Bot. 1960c. V.24. N94. P.l86-199. (139)
Smith D.C. The biology of lichen thalli//Biol. Revs Cambridge Philos. Soc. 1962. V.37.
N4. P.537-550. (37)
Smith D.C. Symbiosis and the biology of lichenised fungi // Symposia Soc. Exper. Biol.
[University Press, Cambridge]. 1975. V.29. P.373-405. (35, 48)
Smith D. C. Comparison between the lichen sumbiosis and other symbioses // D.H.Brown,
D.L.Hawksworth, R.H.Bailey-eds. Lichenology: Progress and Problems. London:
Academic Press, 1976. P.497-513. (12, 48)
Литература и авторский индекс
315
Smith D.C. Symbiosis research at the end of the millennium // Hydrobiologia. 2001.
V.461. P.49-54. (15)
Smith D.C., Douglas A.E. The biology of symbiosis. London: Edward Arnold.
Contemporary Biology series, 1987. 302 p. (12)
Smith D.C., Molesworth S. Lichen physiology. XIII. Effects of rewetting dry lichens //
New Phytologist. 1973. V.72. P.525-533. (48)
Sochting U. Injured reindeer lichens in Danish lichen heaths // Graphis Scripta. 1987.
V.1.N4. P.l03-106. (145)
Sochting U., Johnsen I. Lichen transplants as biological indicators of SO2 air pollution
in Copenhagen // Bull. Environ. Contam. Toxicol. 1978. V.19. NI. P.l-7. (142,
152, 154)
Solberg Y.J. Studies on the chemistry of lichens. XX. The element concentration of the
lichen species Alectoria fremontii and its associated bark substrate of Pinus
silvestris // Zeitschrift fur Naturforschung, Part C. 1979. Bd. 34. S.1275-1277.
(77/;
Sommerfeldt M., John V. Evaluation of a method for the reassessment of air quality by
lichen mapping in the city of Izmir, Turkey // Turkish J. Bot. 2001. V.25. N2.
P.45-55. (109)
Spencer S. Effects of coal dust on species composition of mosses and lichens in an arid
environment//J. Arid Environ. 2001. V.49. N4. P.843-853. (99)
Spencer S, Tinnin R. Effects of coal dust on plant growth and species composition in
an arid environment // J. Arid Environ. 1997. V.37. N3. P.475-485. (99)
SteinkampA., KreebK-H, Weinmann-Kreeb R Flechtentransplantante zur bberwachung
der Luftqualitnt in Bremen //Abh. Naturwiss. Ver. Bremen. 1999. Bd. 44, H. 2-3.
S.737-746. (143, 153)
Steinnes E., Krog H. Mercury, arsenic and selenium fall-out from industrial complex
studied by means of lichen transplants // Oikos. 1977. V.28. N2-3. P.160-164.
SteubingL., Klee R., Kirschbaum U. Beurteilung der lufthygienischen Bedingungen in
der Region Untermain mittels niederer und hoherer Pflanzen // Staub - Reinhaltung
der Luft. 1974. Bd. 34. N6. S.206-209. (142)
SteubingL., Vogel A. Biomonitoring von Immissionswirkungen durch verkehrsbedingte
Schwermetalle in einem Heideokosystem //Umweltwissenschaften und Schadstoflf
Forschung. 1998. Bd. 10, H.6. S.333-338. (184)
Stolte K., MangisD., DotyR., Tonnessen K, Huckaby L.S. (eds.). Lichens as bioindicators
of air Quality. General technical report, U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station, Fort Collins,
Colorado. 1993. 131 p. (68, 70, 74, 78. 99, 119, 121, 122, 123, 124, 168, 172, 178)
Stone S.F., Freitas M.C., ParrR.M., ZeislerR Elemental characterization of a candidate
lichen research material - IAEA-336 // Fres. J. Anal. Chem. 1995. V.352. Nl-2.
P.227-231. (168)
316
Литература и авторский индекс
Stringer Р. W., Stringer M.H.L. Air pollution and the distribution of epiphytic lichens
and bryophytes in Winnipeg, Manitoba// Bryologist. 1974. V.77. N3. P.405-426.
(102, 103)
Sundberg B., Palmqvist K, Esseen PA., Renhorn K.E. Growth and vitality of epiphytic
lichens .2. Modeling of carbon gain using field and laboratory data // Oecologia.
1997. V.l09. NI. P.l0-18. (144)
Sutter G.W. A critique of ecosystem health concepts and indexes // Environ. Toxicol.
Chem. 1993. V.12. N9. P.1533-1539. (238)
Swieboda M., Kalemba A. The lichen Parmelia physodes (L.) Ach. as indicator for
determination of the degree of atmospheric air pollution in the area contaminated
by fluorine and sulphur dioxide emission // Acta Societatis Botanicorum Poloniae.
1978. V.47. Nl-2. P.25-40. (142)
Swieboda M., Kalemba A. The thallus of the Hypogymnia physodes (L.) Nyl. as biotest
// Международная школа по лихеноиндикации, Таллин, 22-25 июня, 1982.
Таллин, 1982. С.65-74. (142)
Syers J.K., Iskandar I.K. Pedogenic significance of lichens // The Lichens. V.Ahmad-
jian, M.E.Hale-eds. Academic Press: New York, London, 1973. P.225-248. (34,
50)
Takala К, Kauranen P, Olkkonen H. Fluorine content of two lichen species in the
vicinity of a fertilizer factory // Ann. Bot. Fennici. 1978. V.15. P.158-166. (40,
60)
Takala K, Olkkonen H. Lead content of the lichen Pseudevernia furfuracea in the
urban area of Kuopio, central Finland // Proceedings of the Kuopio Meeting on
Plant Damages Caused by Air Pollution. L.Karenlampi - ed. University of Kuopio,
Kuopio, Finland. 1976. P.64-67. (184)
Takala K, Olkkonen H. Lead content of an epiphytic lichen in the urban area of Kuopio,
east central Finland // Ann. Bot. Fennici. 1981. V.l 8. N2. P.85-89. (184)
Takala K, Olkkonen H. Titanium content of lichens in Finland // Ann. Bot. Fennici.
1985. V.22. P.299-305. (185)
Takala K, Olkkonen H, Ikonen J., Jaaskelainen J., Puumalainen P. Total sulphur
contents of epiphytic and terricolous lichens in Finland //Ann. Bot. Feninici. 1985.
V.22. N2. P.91-100. (40, 43, 185)
Takala K, Olkkonen H, Jaaskelainen J., Selkainaho К Total chlorine content of epiphytic
and terricolous lichens and birch bark in Finland //Ann. Bot. Fennici. 1990. V.27.
N2. P.131-137. (185)
Takala K, Olkkonen H, Krouse HR. Sulphur isotope composition of epiphytic and
terricolous lichens and pine bark in Finland // Environ. Pollut. 1991. V.69. P.337-
348. (185)
Takala K, Olkkonen H, Salminen R. Iron content and its relations to the sulphur and
titanium contents of epiphytic and terricolous lichens and pine bark in Finland //
Environ. Pollut. 1994. V.84. N2. P.131-138. (185)
Литература и авторский индекс
317
Takala К., Salminen R., Olkkonen Н. Geogenic and anthropogenic zinc in epiphytic and
terricolous lichens in Finland// J. Geochem. Explor. 1998. V.63. P.57-66. (185)
Tarhanen S. Ultrastructural response of the lichen Bryoriafuscescens to simulated acid
rain and heavy metal deposition // Ann. Bot. 1998. V.82. N6. P.735-746. (81)
Tarhanen S., Holopainen T, OksanenJ. Ultrastructural changes and electrolyte leakage
from ozone fumigated epiphytic lichens // Ann. Bot. 1997. V.80. N5. P.611-621.
/87;
Tarhanen S., Holopainen T, Poikolainen J., OksanenJ. Effect of industrial emissions
on membrane permeability of epiphytic lichens in Northern Finland and the Kola
Peninsula industrial areas // Water, Air, and Soil Pollution. 1996. V.88. Nl-2.
P.l89-201. (85)
Tarhanen S, Metsarinne S., Holopainen T, OksanenJ. Membrane permeability response
of lichen Bryoria fuscescens to wet deposited heavy metals and acid rain //
Environ. Pollut. 1999. V.l04. NI. P.121-129. (85)
Tarhanen S., Poikolainen J., Holopainen T, Oksanen J. Severe photobiont injuries of
lichens strongly associated with air pollution // New Phytologist. 2000. V.l47.
N3. P.579-590. (81)
Tehler A. Systematics, phylogeny and classification // Lichen Biology. T.H.Nash - ed.
Cambridge: University Press, 1996. P.217-239. (17)
Thomas W., Ruhling A., Simon H. Accumulation of airborne pollutants (PAN, chlorinated
hydrocarbons, heavy metals) in various plant species and humus // Environ. Pollut.
Ser. A. 1984. V.36. P.295-310. (178)
Thomson J. W. The lichen genus Physcia in North America // Beih. Nova Hedwigia.
1963. Bd. 7. S.1-172. (29)
Thrower S.L. Air pollution and lichens in Hong Kong // Lichenologist. 1980. V. 12,
N 3. P. 305-311. (99)
Tibell L. Crustose lichens as indicators of forest continuity in boreal coniferous forests
//Nordic J. Bot. 1992. V.l2. N4. P.427-450. (240, 241)
Tomassini F.D., Puckett K. J, Nieboer E., Richardson D.H.S., Grace B. Determination
of copper, iron, nickel, and sulphur by X-ray fluorescence in lichens from the
Mackenzie valley, Northwest Territories, and the Sudbury district, Ontario // Canad.
J. Bot. 1976. V.54. N14. P.l591-1603. (35, 38, 39, 40, 177)
Трасс X.X. (Trass H.). Cryptoindication of the level of atmospheric air pollution and
ecological monitoring //Уч. зап. Тартуского университета. 1988. №812. С.25-
31. (106)
Trass Н, VellakK., IngerpuuN. Floristical and ecological properties for identifying of
primeval forests in Estonia//Ann. Bot. Fennici. 1999. V.36.NI. P.67-80. (239, 240)
Tretiach M., Ganis P. Hydrogen sulphide and epiphytic lichen vegetation: a case study
on Mt. Amiata (central Italy) // Lichenologist. 1999. V.31. N2. P. 163-181. (182)
Trotet G. Recherches sur la nutrition des lichens. Premiers resultants // Rev. Bryol. et
Lichenol. 1968-1969 (1970). V.36. N3-4. P.733-736. (35)
318
Литература и авторский индекс
Triimpener Е. Uber die Bedeutung der Wasserstoffionenkonnzentration fur die Verbreitung
von Flechten // Beihefte zur Botanisch. Centralblatt. 1926. Bd. 42. N3. S.321-
354. (21)
Tschermak-Woess E. The algal partner // CRC Handbook of Lichenology. M.Galun -
ed. Boca Raton: CRC Press, Inc., 1988. V.l. P.39-92. (14)
Tuba Z., Csintalan Z. Bioindication of road motor traffic caused heavy metal pollution
by lichen transplants // Plants as biomonitors: indicators for heavy metals in the
terrestrial environment. B.Markert-ed. VCH, Weinheim, 1993. P.329-341. (145,
150, 162)
Tuba Z., Csintalan Z., Badacsonyi A., Proctor M.C.F. Chlorophyll fluorescence as an
exploratory tool for ecophysiological studies on mosses and other small poikilo-
hydric plants // J. Bryol. 1997. V.19. N3. P.401-407. (145)
Tuba Z., Csintalan Z., Nagy Z., Szente K, Takacs Z. Sampling of terricolous lichen
and moss species for trace element analysis, with special reference to bioindi-
cation of air pollution // Environmental Sampling for Trace Analysis. B.Mar-
kert-ed. VCH Verlagsgesellschaft mbH. Weinheim, 1994. P.415-434. (170)
Tuominen К Studies on the strontium uptake of the Cladonia alpestris thallus // Ann.
Bot. Fennici. 1967. V.4. NI. P.l-28. (38)
Tiirk R. Nationalparke und Flechten - Aufgaben, Chancen, Folgerungen // Salzburger
Geographische Materialien. 1993. Heft 19. S.53-55. (239)
Tiirk R., Christ R. Untersuchungen des CO2-Gaswechsels von Flechtenexplanta-
ten zur Indikation von SO2-Belastung im Stadtgebiet von Salzburg // Bioindika-
tion auf subzellularen und zellular Ebene (Bioindikation 2). R.Schubert,
J.Schuh-eds. Martin-Luther-Universitflt, Halle-Wittenberg, 1980. S.39-45. (143,
157)
Tiirk R., Wirth V. The pH dependence of SO2 damage to lichens // Oecologia. 1975.
V.19. P.285-291. (86)
Tyler G. Uptake, retention and toxicity of heavy metals in lichens - a brief review //
Water, Air, and Soil Pollution. 1989. V.47. N3-4. P.321-333. (177)
Tyler G., PahlssonA.M.B., Bengtsson G., Baath E., TranvikL. Heavy metal ecology of
terrestrial plants, microorganisms and invertebrates. A review // Water, Air, and
Soil Pollution. 1989. V.47. N3-4. P.l89-215. (177)
Tynnyrinen S., Palomaki V., Holopainen T, Karenlampi L. Comparison of several
bioindicator methods in monitoring the effects on forest of a fertilizer plant and a
strip mine // Ann. Bot. Fennici. 1992. V.29. N1. P.l 1-24. (184)
Upreti D.K., Pandev V. Determination of heavy metals in lichens growing on different
ecological habitats in Schirmacher Oasis, East Antarctica// Spectroscopy Letters.
2000. V.33. N3. P.435-444. (180)
Vaeck L. van, Poels K, de Nollin S., Hachimi A., Gijbels R. Laser microprobe mass
spectrometry - principle and applications in biology and medicine // Cell Biology
International. 1997. V.21.N10. P.635-648. (177)
Литература и авторский индекс
319
Vagts I., Kinder М., Muller J. The effect of agrochemicals on the growth of Cladonia
furcata // Lichenologist. 1994. V.26.N1. P.73-82. (145, 146, 154)
Varrica D., AiuppaA., Dongarra G. Volcanic and anthropogenic contribution to heavy
metal content in lichens from Mt. Etna and Vulcano island (Sicily) // Environ.
Pollut. 2000. V.l08. N2. P.l53-162. (182)
VDI (Verein deutscher lngenieure)-Richtlinien Messen von Immissionswirkungen,
Ermittlung und Beurteilung phytotoxischer Wirkungen von Immissionen mit
Flechten. Verfahren der standardisierten Flechtenexposition 11 VDI 3799. 1991.
Blatt 2/Part 2. S.l-14. (146, 147, 148, 149, 150)
VDI (Verein deutscher lngenieure)-Richtlinien Messen von Immissionswirkungen,
Ermittlung und Beurteilung phytotoxischer Wirkungen von Immissionen mit
Flechten. Flechtenkartierung zur Ermittlung der Luftgtitewertes (LGW) // VDI
3799, 1995. Blatt 1/Part 1. S.l-25. (96, 106, 107, 108, 109)
Vestergaard N.K., Stephansen U., Rasmussen L, Pileegard K. Airborne heavy metal
pollution in the environment of Danish steel plant // Water, Air, and Soil Pollution.
1986. V.27. P.363-377. (184)
VezdaA. Lesni, mechorosty a lisejniky jako indikatory kyselosti svrchnich vrstev lesnich
pud // Sbomik Vysoke Skoly Zemedelske a Lesnicke fakulty v Bme. 1955. V.4C.
P.l87-190. (21)
Villeneuve J.P., Cattini C., Holm E. Transfer of chlorinated hydrocarbons in the
food-chain lichen-reindeer-man //Chemosphere. 1985. V. 14. N11 -1. P. 1651 -1658.
(77S)
Villeneuve J.P, Fogelqvist E., Cattini C. Lichens as bioindicators for atmospheric
pollution by chlorinated hydrocarbons // Chemosphere. 1988. V.l7. N2. P.399-
403. (178)
Villeneuve J.P, Holm E. Atmospheric background of chlorinated hydrocarbons studied
in Swedish lichens//Chemosphere. 1984. V. 13. P.l 133-1138. (178)
Vogel S. Niedere “Fensterpflanzen” in der siidafrikanischen Wiiste. Eine okologische
Schilderung//Beitr. Biol. Pflanzen. 1955. Bd. 31. NI. S.45-135. (23)
Vokou D., Pirintsos S.A., Loppi S. Lichens as bioindicators of temporal variations in air
quality around Thessaloniki, Northern Greece//Ecol. Res. 1999. V.14.N2. P.89-
96. (105)
Wadleigh M.A., Blake D.M. Tracing sources of atmospheric sulfur using epiphytic lichens
// Environ. Pollut. 1999. V.l06. N3. P.265-271. (66, 140, 183)
Wagner G. Basic approaches and methods for quality assurance and quality control in
sample collection and storage for environmental monitoring// Sci. Total Environ.
1995. V.l76. Nl-3. P.63-71. (168)
Walker C.H. Organic pollutants: an ecotoxicological perspective. London: Taylor &
Francis, 2001.282 p. (43, 74.)
Walker C.H, Hopkin S.P, SiblyRM., PeakailD.B. Principles of ecotoxicology. London:
Taylor & Francis, 2001. 309 p. (32, 43, 74)
320
Литература и авторский индекс
Walther D.A., Ramelow G.J., Beck J.N., Young J.С., Callahan J.D., Marcon M.F.
Distribution of airborne heavy metals as measured in the lichens Ramalina
stenospora and Parmotremapraesorediosum in Baton-Rouge, Louisiana // Water,
Air, and Soil Pollution. 1990a. V.50. N3-4. P.l89-200. (185)
Walther D.A., Ramelow G.J., Beck J.N., Young J.C., Callahan J.D., Marcon M.F.
Temporal changes in metal levels of the lichens Parmotremapraesorediosum and
Ramalina stenospora, Southwest Louisiana // Water, Air, and Soil Pollution.
1990b. V.53. Nl-2. P.l89-200. (180)
WaringR.H., Schlesinger W.H. Forest ecosystems. Concept and management. Orlando:
Academic Press, 1985. (48)
Webster M., Brown D.H. Preliminary observations on the growth of transplanted Peltigera
canina under seminatural conditions // Lichenologist. 1997. V.29. NI. P.91-96.
(145)
Weinmann-Kreeb R., KreebK.H. In-vivo-FluoreszenzbeiChloroplasten: Modellierungs-
ansatz und Anwendung in der Bioindikation //Archiv fur Hydrobiologie Beihefte
Ergebnisse der Limnologie. 1987. Bd. 29. S.131-141. (142)
Werner A. Aktives Biomonitoring mit der Flechte Hypogymniaphysodes zur Ermittlung
der Luftqualitat in Hannover // Bibliotheca Lichenologica. J. Cramer, Berlin,
Stuttgart, 1993. Bd. 49. 113 S. (86, 142, 153)
Werner A., Niemeyer R. Aktives Biomonitoring mit der Flechte Hypogymnia physodes.
Ein integriertes Programm zur Erfassung der biologischen Wirkung von
Luftbelastungen // Staub-Reinhaltung der Luft. 1994. Bd. 54. S.313-320. (142,153)
Wetmore CM. Lichens and air quality in Big Bend National Park,Texas // Bryologist.
1981a. V.84. N3. P.426-433 (94)
Wetmore C.M. Lichens of Voyageus National Park, Minnesota // Bryologist. 1981b.
V.84. N4. P.482-491. (94)
Wetmore C.M. Lichen decomposition in a black spruce bog // Lichenologist. 1982. V. 14.
N3. P.267-271. (93)
Wetmore C.M. Lichens of Theodore Roosevelt National Park // Mycotaxon. 1985. V.23.
Nl-4. P.241-249. (94)
Wetmore C.M. Lichens and air quality in Indiana Dunes National Lakeshore // Mycotaxon.
1988a. V.33. P.25-39. (94)
Wetmore C.M. Lichen floristics and air quality // Lichens, bryophytes and air quality.
T.H.Nash, V.Wirth - eds. J. Cramer, Berlin-Stuttgart, 1988b. S.55-65. (84, 93, 210)
Wetmore C.M. Lichens and air quality in Cuyahoga Valley National Recreation Area,
Ohio // Bryologist. 1989. V.92. N3. P.273-281. (92, 94)
Wicklum D., Davies R. W. Ecosystem health and integrity? // Canad. J. Bot. 1995. V.73.
N7. P.997-100. (238)
WiersmaG.B., BrunsD.A., Finley K., McAnulty L., Whitworth C, Boelcke C. Elemental
composition of lichens from a remote Nothofagus forest site in Southern Chile //
Chemosphere. 1992. V.24. N2. P.155-167. (179)
Литература и авторский индекс
321
Wietschorke G.M., Liihmann H.J., Kreeb К.Н. Erfassung kleinraumiger Immissions-
belastung durch transplantierte Flechten // Verh. Ges. Okologie. 1985. Bd. 13.
S.653-656. (142)
Will-Wolf S. Effects of a “clean” coal-fired power generating station on four common
Wisconsin lichen species // Bryologist. 1980a. V.83. N3. P.296-300. (81, 137)
Will-Wolf S. Structure of corticoious lichen communities before and after exposure to
emissions from a “clean” coal-fired generating station I I Bryologist. 1980b. V.83.
N3. P.281-295. (124, 137)
Will-WolfS. Quantitative approaches to air quality studies 11 Lichens, Bryophytes and
Air Quality. T.H.Nash, V.Wirth - eds. J. Cramer,Berlin-Stuttgart. 1988. P.l 09-
140. (137)
Williams R.N., Casellas R.C., Mangelson N.F., Rees L.B., St.Clair L.L., Schaalje G.B.,
Swalberg K.D. Elemental analysis of lichens from the intermountain western
USA using PIXE//Nuclear Instruments and Methods in Physics Research. Sect.
B. 1996. V.109. P.336-340. (177)
Winner W.E., Atkinson C.J., Nash Т.Н. Comparison of SO2 absorption capacities of
mosses, lichens, and vascular plants in diverse habitats // Lichens, bryophytes and
air quality. T.H.Nash, V.Wirth - eds. J. Cramer, Berlin-Stuttgart, 1988. S.217-
230. (33, 50)
Wirth V. Die Silikatflechten-Gemeinschaften in ausseralpinen Zentraleuropa //
Dissertationes Botanicae. Cramer, Lehre, 1972. Bd. 17. S. 1-306. (34)
Wirth V The influence of water relations on lichen SO2-resistance // Progress and
Problems in Lichenology in the Eighties. E.Peveling - ed. J. Cramer, Berlin-
Stuttgart, 1987. S.347-350. (86)
Wirth V. Phytosociologicai approaches to air pollution monitoring with lichens // Lichens,
bryophytes and air quality. T.H.Nash, V.Wirth - eds. J. Cramer, Berlin-Stuttgart,
1988. P.97-107. (84, 93, 121, 124, 125, 127)
Wirth V. Zeigewerte von Flechten // Scripta Geobotanica. 1991. Bd. 18. S.215-237. (74,
89, 90, 210)
Wirth V., Turk R. Zur SO2-Resistenz von Flechten verschiedener Wuchsform // Flora.
1975. Bd. 164. N2/3. S.133-143. (89)
Wiseman R.D., Wadleigh M.A. Lichen response to changes in atmospheric sulphur:
isotopic evidence // Environ. Pollut. 2002. VI16. N2. P.235-241. (72, 140)
Wit T. de. Flechtenverbreitung in den Niederlanden und ihre AbhSngigkeit von der
Luftverunreinigung // Schriftenr. Vegetationsk. 1976. N10. S. 169-175. (115,
116)
Wittig R. General aspects of biomonitoring heavy metals by plants I I Plants as biomonitors.
B.Markert - ed. CH Verlagsges., Weinheim, etc., 1993. P.3-27. (31, 32)
WolterbeekH.T., Bode P. Strategies in sampling and sample handling in the context of
large-scale plant biomonitoring surveys of trace element air-pollution 11 Sci. Total
Environ. 1995. V.176. Nl-3. P.33-43. (169)
322
Литература и авторский индекс
Wonitzki С., Coburger К. Flechtenkartierung in der Stadt Greiz // Flechtenmonitoring -
ein kommunales Kontrollinstrument. P.Scholz-ed. Halle: Unabhangingen Institutes
fur Umweltfragen. 1993. S.129-136. (65)
Woodwell G.M., Gannutz T.P. Effects of chronic gamma irradiation on lichen communities
of a forest//Amer. J. Bot. 1967. V.54. N10. P.1210-1215. (187)
Woodwell G.M., Whittaker R.H. Effects of chronic gamma irradiation on plant
communities //Quarterly Review of Biology. 1968. V.43. NI. P.43-55. (187)
YevseevA. V., Krasovskaya T.M. Spatial and temporal patterns in pollutant accumulation
in the ecosystems of the Kola Peninsula // Aquilo. Ser. Bot. 1994. V.34. P.65-68.
W
Zakshek E.M., Percy K.E., Puckett K.J. Lichen sulfur and lead levels in relation to
deposition patterns in Eastern Canada // Water, Air, and Soil Pollution. 1986.
V.30.N1-2. P.l61-169. (184)
Zambrano A., Nash Т.Н. Lichen responses to short term transplantation in desierto de
Los Leones, Mexico city // Environ. Pollut. 2000. V.107. N3. P.407-412. (140)
Zambrano A., Nash Т.Н., Gries C. Physiological effects of the Mexico city atmosphere
on lichen transplants on oaks // J. Environ. Quality. 1999. V.28. N5. P.l 548-
1555. (140)
Zanini E., Bonifacio E., Cuttica G.C. Heavy metals in soils near a steel-making industry
- a case study in a complex valley situation in Italy // J. Environ. Sci. & Health.
Part A. 1992. V.A27. N8. P.2019-2036. (184)
Zobel К. (Цобель К.) Use of epiphytic lichens as bioindicators of atmospheric pollution
in the Khamar-Daban Mountains (Lake Baikal region) // Уч. зап. Тартуского
гос. университета. 1988. №812. С.47-66. (106)
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ТАКСОНОВ,
УПОМЯНУТЫХ В ТЕКСТЕ
Abies nordmaniana 197
Acarospora 45
A. clauzadeana 34
A. fuscata 44
A. gwynnii 26
A. impressula 188
A. rugulosa 46
A. sinopica 40, 41
A. smaragdula 39, 41, 46
Acer platanoides 107
A. pseudoplat anus 107, 196
Acrocordia gemmata 90
Alectoria implexa 203
A. sarmentosa 67, 90, 140, 141, 179, 183
A. sarmentosa ssp. sarmentosa 241
Alectoria sp. 140
Alnus glutinosalb, 107
A. incana 20
Anaptychia ciliaris 90, 104, 127, 145, 203
Arthonia byssacea 90
A. cinnabarina 90
A. didyma24\
A. leucopellaena 90, 241
A. patellulata 131, 132, 133, 134
A. radiata 91
A. spadicea 91
A. vinosa 90, 241
Arthopyrenia cinereopruinosa 241
A. punctiformis 90
Arthothelium ruanum 91
Arthrosporum populorum 130, 132, 133, 134
Ascomycota 13, 17
Aspicilia 45
A. cinerea 44
A. desert or um 188
A. esculenta 18
324
Указатель латинских названий таксонов, упомянутых в тексте
Bacidia arceutina 90
В. beckhausii 129, 130, 134
В. globulosa 90
В. igniarii 241
В. rubella 90
В. subincompta 90
Bactrospora dryina 90
Baeomyces roseus 203, 205
Basidiomycota 13, 17
Betula 36
B. pendula 107
B. platyphylla 194
B. verrucosa 20
Biatora epixanthoidiza 241
B. helvola 129, 131, 132, 134
B. sphaeroides 241
B. symmicta 204
Biatorella moriformis 204
Bryoria 205
Bryoria bicolor 90
B. capillaris 90, 92, 141, 146, 155, 156, 241
B. fremontii 91, 141, 183
B. furcellata 92
B. fuscescens 90, 91, 140, 141, 157, 208
B. implexa 203
B. lanestris 92, 241
B. nadvornikiana 241
B. osteola 92
B. pseudofuscescens 92
B. schaereri 241
B. simplicior 91
Bryoria sp. 128, 140
B. trichodes 92
Buellia alboatra 134
B. disciformis 90, 129, 130, 132, 133, 134
B. frigida 26
B. geophila 134
B. grisea 26
B. griseovirens 91
B. punctata 91, 132, 133, 188, 203
Caliciales 13
Calicium adaequatum 241
C. adpressum 90
C. lichenoides 132, 133, 135
C. salicinum 90
C. viride 90
Указатель латинских названий таксонов, упомянутых в тексте
325
Calicium sp. 92
Caloplaca 188
С. aurantia 41
С cerina 90, 188, 203, 206, 211, 218
С. citrina 203, 205, 206
С. geophila 188
С. herbidella 90
С holocarpa 203, 206, 211, 219
С. pyracea 203
С. vitellina 188
Candelaria concolor 90, 104, 203
Candelariella 115
C. efflorescens 91
C. reflexa 90
C vitellina 188, 203, 206, 211, 220
C. vitellina var. xanthostigma 203
C. xanthostigma 91, 203
Canomaculina pilosa 144
Carbonea vorticosa 26
Castanea sativa 196
Catinaria atropurpurea 130, 134, 241
Cavernularia hultenii 241
Cetraria 140
C. chlorophylla 91
C. cucullata9\, 143
C. halei 140
C islandica9\, 140, 157, 167, 203, 205, 208
C. nivalis 144
C. pinastri 90, 91, 140, 204
C. sepincola 90, 91
Cetrelia cetrarioides 90, 196
C. olivetorum 203
Chaenotheca brunneola 90
C. chlorella 241
C. chrysocephala 90, 126, 135, 241
C. ferruginea 91
C. furfur ace a 90, 132, 135
C. gracillima 241
C. laevigata 241
C. phaeocephala 90, 241
C. subroscida 241
C. trichialis 90
Chaenothecopsis папа 241
C. viridialba 241
Chondropsis semiviridis 18
Chrysothrix candelaris 90
326
Указатель латинских названий таксонов, упомянутых в тексте
Cladina 30, 34, 59, 145, 187, 205
С. arbuscula 91
С. mitis 91, 154, 157, 190, 191
С. rangiferina 33, 38, 60, 67, 91, 142, 154, 157, 179, 181, 193, 198
С. stellaris 32, 34, 41, 59, 60, 92, 141, 154, 157
Cladonia 34, 45, 59, 79, 187
С. alcicornis 26
С alpestris 145, 154, 157
С. amaurocraea 139, 143
С. arbuscula 203, 208
С. bacillaris 203
С. bacilliformis 91
С. botrytes 91, 208
С. caespitica 206, 211,215
С. cariosa 203, 205, 206, 208
С. cenotea 91
С. cervicornis ssp. verticillata 187
C. chlorophaea 92, 139, 144, 203, 206, 211,218
C. coccifera 91
C. coniocraea 92, 126, 129, 130, 131, 132, 133, 134, 135, 194, 203, 206, 211, 221
C. convoluta 30, 145, 146, 162
C. cornuta 91
C. cornutoradiata f. cornutoradiata 203
C. crispata9\, 190, 191
C. cristatella 44
C. cryptochlorophaea 92
C. cyanipes 92
C. deformis 91
C. digitataVX, 126, 206,211,215
C. ecmocyna 139, 143
C fimbriata 92, 129, 130, 131, 132, 133, 203, 206, 211, 222
C foliacea 126
C furcat a 130, 145, 146, 154, 170, 193,208
C. gracilis 91, 190
C. impexa ssp. spumosa 203
C. macilenta 205, 208
C. macilenta ssp. macilenta 203
C. merochlorophae 91
C. mitis 145
C. ochrochlora 132, 206, 211, 223
C. parasitica 203, 205, 241
C. pityrea 203
C. pleurota 91, 126
C. pocillum 24
C. portentosa 145, 203
C. pyxidata 126, 203, 205
Указатель латинских названий таксонов, упомянутых в тексте
327
С. ramulosa 203, 205
С. rangiferina 145, 203
С. rangiformis 30, 145
С. rei 92
С. silvatica 203
Cladonia sp. 44
С. squamosa 203
С. stellaris 208
С. subulata 203, 205
С. sulphurina 91
С. turgida 144, 203, 208
С. uncialis 44, 191
Cliostomum griffithii 129, 130, 132, 134
Coenogonium 15
Collema 16, 44
C. curtisporum 241
C. flaccidum 90, 241
C. fragrans 241
C. furfuraceum 241
C. nigrescens 90, 241
C. occultatum 241
Conotrema populorum 241
Cybebe gracilenta 90, 241
Cyphelium inquinans 241
C. karelicum 241
C. pinicola 241
C. tigillare 241
Cystocoleus 15
Degelia plumbea 241
Dermatocarpon 189
Deuteromycota 13, 17
Dibaes baeomyces 203, 205
Dimelaena oreina 50
Dimerella lutea 241
D. pineti 91, 241
Diploica canescens 91
Diploschistes muscorum 47
Dirinaria picta 99'
Endocarpon subfoliaceum 188
Enterographa crassa 241
E. hutchinsiae 90
Ephebe 15
Evernia 140
Evernia divaricata 90, 241
E. mesomorpha 92, 140, 180, 181, 208, 242
328
Указатель латинских названий таксонов, упомянутых в тексте
Е. prunastri 41, 63, 91, 115, 140, 146, 156, 167, 169, 191, 194, 203, 206, 208, 209, 211,
215, 242
Everniastrum neocirrhatum 140
Flavocetraria (Cetraria) cucullata 139
E (Cetraria) nivalis 146
Flavoparmelia baltimorensis 64, 172, 182
E (Pseudoparemlia) baltimorensis 171
E caperata 44, 61, 114, 127, 194, 196, 203
E (Parmelia) caperata 139, 146, 150, 152, 154, 167, 170, 172, 181, 183
Fraxinus 36, 124
E excelsior 107
Fuscidea cyathoides 90
E viridis 91
Gleocapsa 16
Graphis scripta 9\, 129, 130, 131, 132, 133, 134, 144, 206, 208, 209, 211, 215
Gyalecta ulmi 90
Heder a colchica 196
Heterodermis speciosa 242
Hyperphyscia adglutinata 90
Hypocenomyce caradocensis 91
Hfriesii 92, 242
H. scalaris 44, 91, 126, 135, 206, 211, 223
Hypogymnia 140
H austerodes 92, 242
H bitteri 92, 242
H. enteromorpha 67, 79, 143, 153, 179
H. farinacea 91
H physodes 22, 23, 24, 37, 43, 48, 59, 63,71,79, 80, 89, 91, 115, 126, 129, 130, 131, 132,
133, 134, 139, 140, 141, 142, 143, 145, 146, 152, 153, 154, 155, 156, 157, 160,
167, 180, 181, 190, 191, 192, 193, 194, 196, 197, 200, 203, 206,211,217, 233
H tubulosa 91, 92, 208
H. vittata 242
Hypotrachyna partita 183
Imshaugia aleurites 90
Juglans regia 107
Lasallia papulosa 44
L. pustulata 51
Lecanactis amylacea 90
L. premnea 242
Lecania cyrtella 90
L. dimera 204
L. dubitans 204
Lecanora 45, 233
L. albella 129, 134, 203
L. allophana 90, 134, 143, 153, 203, 206, 211, 215
L. argentata 90
Указатель латинских названий таксонов, упомянутых в тексте
329
L. carpinea 91, 203
L. chlarotera9\
L. chloropolia 92
L. coilocarpa 92
L. conizaeoides 63, 91, 93, 98, 100, 104, 105, 108, 115, 126, 128
L. expaliens 91
L. fuscescens 91
L. hagenii 89, 91, 188, 206, 211, 224
L. impudens 90, 91
L. intumescens 90, 203
L. leptyrodes 139, 144
L. muralis 39, 205, 206, 208
L. ochrococca 242
L. pallida 90
L. pallida.NZX. cinerella 203
L. piniperda 204, 211,215
L. pulicaris9\, 130, 132
L. rugosella 204
L. saligna 91
L. sambuci 90
L. subfusca 143, 146
L. symmicta 90, 129, 132, 133, 204, 206, 211,215
L. umbrina 204, 205, 206
L. varia909 145, 206,211,215
L. vinetorum 45
Lecidea 45
L. canciformis 26
L. erythrophaea 206, 211,216
L. flexuosa 91
L. glomerulosa 204
L. lactea 46
L. plebeja 92
L. sphaerella 126
L. symmicta 204
L. the io des 46
Lecidella elaeochroma 91, 129, 130, 132, 133, 134, 143, 153
L. euphorea 204, 206, 211,216
Lepraria 23, 44, 99
L. aeruginosa 204
L. incana 91, 126, 129, 130, 131, 132, 133, 134, 135, 204, 206,211,225
L. lobificans 90
Leptogium saturninum 90, 194
L. teretiusculum 242
Letharia divaricata 145
L. vulpina 183
Lichenes 10
330
Указатель латинских названий таксонов, упомянутых в тексте
Lichina 44
Lithographa flexella 242
Lobaria 44, 146, 240
L. amplissima 90, 145, 242
L. oregana 67, 140, 145, 154, 179
L. pulmonaria 2"!, 43, 90, 140, 144, 145, 154, 196, 242
L. scrobiculata 145
L. virens 145, 242
Loxospora cismonica 90
L. elarina 90
L. elatinum 242
Malus sp. 107
Masonhalea richardsonii 144
Melanelia exasperata 207, 209, 211,216
M. exasperatula 197, 207, 211, 215
M. olivacea 153, 194, 204
M. (Parmelia) glabratula ssp. fuliginosa 154
M. (Parmelia) olivacea 153
M. subaurifera 187
Menegazzia terebrata 90
Micarea adnata 90
M. denigrata 91
M. globulosella 242
M. melaena 91
M. peliocarpa 90
M. prasina 90
M. trisepta 44
Microcalicium ahlneri 242
M. arenarium 242
Mycobilimbia tetramera 133, 134
Mycoblastus affinis 242
M. sanguinarius 92
M. sterilis 91
M. tornoensis 91
Neckera sp. 194, 195, 196
Nephroma 44
N. arcticum 139, 143
N. bellum 90
N. laevigatum 90, 242
N. parile 90
N. resupinatum 90
Neuropogon acromelanus 26
Normandina pulchella 90
Nostoc 16, 82, 84
Ochrolechia androgyna 90, 91
O. arborea 90, 92
Указатель латинских названий таксонов, упомянутых в тексте
331
О. pallescens 90
О. turneri 90
Opegrapha 233
О. atra9\, 129, 130, 132, 133, 134, 207,211,224
О. diaphora 204
О. lyncea 242
О. puli car is 134
О. rufescens 90, 207, 211,215
О. varia 130, 132, 133, 204
О. vermicellifera 91
О. viridis 90
Pachyphiale carneola 242
Л fagicola 90
Pannaria conoplea 90, 242
P. mediterranea 242
Parmelia 3\, 143, 156, 167
P. acetabulum 91, 141
P caperata 63, 90, 141, 143, 144, 145, 203
P. centrifuga 91
P. cetrarioides 203
P chlorochroa 31
P coniocraea 90
P. conspersa 143
P contorta 90
P. cumberlandia 144
P elegantula 90
P exasperata 90
P exasperatula 91
P flaventior 91
P glabratula 63, 91
P glabratula ssp. fuliginosa 143
P. melanothrix 35
P mougeotii 91
P. olivacea 143, 204
P pastilifera 90
P. perlata 63, 141
P physodes 142
P revoluta 91
P. saxatilis 91, 143, 144, 154, 167, 170, 204
P. scortea 204
P septentrionalis 92
P sinuosa 90
P squarrosa 143
P stenophylla 144, 150, 155
P. subargentifera 90
P subrudecta 63, 91
332
Указатель латинских названий таксонов, упомянутых в тексте
Р sulcata29, 63, 89,91, 128, 129, 130, 131, 133, 134, 135, 140, 141, 143, 145, 146, 153,
157, 158, 161, 167, 170, 179, 181, 185, 187, 190, 191, 194, 204, 207, 209,211,217, 232
Р tiliacea 91, 204
Р vagans 18
Parmeliella plumbea 90
Р triptophylla 90, 242
Parmelina (Parmelia) quercina 146, 156
P. (Parmelia) tiliacea 51
P tiliaceae 204
Parmeliopsis aleurites 91
P ambigua 91, 93, 126, 204, 207, 208, 211, 220
P hyperopta 91, 204, 207, 211, 216
Parmotrema austrosinense 144, 158
P chinense 194
P conferendum 144, 158
P crinita 242
P madagascariaceum 144
P. praesorediosum 180
P tinctorum 99
P. uruguense 144
Peltigera 30, 44, 46, 79, 205
P. aphthosa f. variolosa 204
P. canina 82, 143, 145, 146, 204, 208
P. canina f. subcanina 204
P collina 90, 242
P. didactyla 204
P horizontalis 44, 242
P leucophlebia 204
P. membranacea 194, 197, 198
P polydactyla 204
P praetextata 91, 204
P. rufescens 39, 204, 208
P spuria f. spuria 204
P venosa 204
Peltula 16
P. impressula 188
Pertusaria 205
P albescens 91, 132, 135, 204
P amara 9\, 130, 131, 132, 133
P coccodes 9\, 132, 134
P coronata 91
P. flavida 91
P globulifera 204
P. hemisphaerica 91
P. hymenea 90
P leioplaca9\, 132, 135
Указатель латинских названий таксонов, упомянутых в тексте
333
Р pertusa 91
Р pupillaris 242
Р. trachythallina 90
Phaeocalicium populneum 242
P praecedans 242
Phaeophyscia ciliata 204, 207, 211,219
P. endophoenicea 90
P. nigricans 207, 211, 223
P. orbicularis 91, 204, 207, 209, 211, 213, 214
P (Physcia) orbicularis 154
Phlyctis agelaea 90
P. argena 91, 143, 153
Physcia 29, 115, 167
P adscendens 91, 145
P aipolia 91, 127, 144, 146, 204, 207, 211, 215
P caesia 207, 211, 215
P. ciliata 204
P dubia 204
P grisea 204
P. hispida 204
P millegrana 143
P. orbicularis 143
P pulverulenta 204
P stellaris 91, 127, 204, 207, 208, 211, 213, 214
P. tenella9\, 143, 154, 204
P teretiuscula 204
P. tribacia 204, 207,211,221
P. virella 204
Physconia distorta 90, 127, 134, 194, 204
R enteroxantha 90
R grisea 91, 133, 134, 204, 207,211,222
P perisidiosa 91
Picea 36
P. abies 20,21, 125
Pinus 36
P. kochiana 196
P rigida 186
P sylvestris 20, 123
Platismatia glauca 91, 92, 144, 154
P. norvegica 242
Populus balsamifera 123
Populus sp. 107
P tremula 20
Porina aenea 91
P leptalea 242
Porpidia 45
334
Указатель латинских названий таксонов, упомянутых в тексте
Protococcus 156
Prunus avium 107
Р domestica 107
Pseudeverniafurfuraceal9,9\, 143, 169, 174
Pseudocyphellaria rainierensis 140, 145, 154
Punctelia ruddenda 242
P (Parmelia) rudecta 182
P. subrudecta 144, 158
Pyrenula nitida 91, 242
P nitidella 91
Pyrrhospora cinnabarina 242
P. elabens 133, 134, 242
Pyrus communis 107
Pyxine cocoes 99
Quercus 124, 172
Q. alba 172, 186
Q. borealis 172
Q. coccinea 186
Q. petraea 107
Q. robur20, 63, 107
Q. suber 35
Racodium 15
Ramalina 183
R. calicaris 35, 140
R. dilacerata 92, 242
R. duriaei 62, 140, 141
R. (duriaei) lacera 158, 161
R. ecklonii 140, 158, 159
R. farinacea 9\. 140, 141, 153, 167, 196, 208, 242
R. fastigiata 90, 141
R. fraxinea 90
R. lacera 62, 141
R. menziesii 49, 141, 157, 160
R. pollinaria 91, 204
R. protecta 51
R. stenospora 180
Ramalina sp. 63, 140
R. thrausta 242
Rhizocarpon 45
R. oederi 40
Rhizocarpon sp. 144, 145
Rhizoplaca melanophthalma 26
Rinodina colobina 90
R. exigua 91
R. isidioides 242
R. pyrina 90
Указатель латинских названий таксонов, упомянутых в тексте
335
R. sophodes 90
Robinia pseudoacacia 107
Saccomorpha icmalea 91
Salix sp. 20
Sarcogyne privigna 26
Schismatomma pericleum 90, 242
Sclerophora coniophaea 242
S. nivea 90
Scoliciosporum chlorococcum 44, 91, 93, 129, 130, 131, 135, 207, 211, 216, 232
Scytonema 16
Sorbus aucuparia 20
Sphaerophorus globosus 90
Stenocybe major 90
S. pullatula 90
S. septata 242
Stereocaulon 45
S. glareosum 194
S. saxatile 144
S. tomentosum 204, 205
Sticta 44
S. limbata 243
S. sylvatica 90, 243
Strangospora moriformis 204
S. pinicola 91
Taxus baccata 197
Thamnolia subuliformis 42
T vermicularis 139, 143
Thelopsis rubella 243
Thelotrema lepadinum 90, 243
Tilia 36
T cordata 20, 107
T platyphyllos 107
Trapelia granulosa 91
T. involuta 198
T. pseudogranulosa 91
Trebouxia 14, 15, 33, 88, 156
T arboricola 14
T decolorans 14
T irregularis 14
Tremolecia 45
Trentepohlia 16
Tucker manops is chlorophylla 207, 211,215
T sepincola 207,211,219
Ulmus carpinifolia 107
Umbilicaria muehlenbergii 38
Usnea 128, 167, 205
U. amblyoclada 140
U. bailey 99
U. ceratina9&, 140
U. extensa 243
U. filipendula 90
U. florida 90, 99
U. fulvoreagens 90
U. hirta 91, 92, 141, 162, 204, 208
U. longissima 140
U. sphacelata 79
Usnea sp. 63, 140, 145, 179, 180, 196, 197
U. subfloridana 90
Vaccinium myrtyllus 125
Verrucaria nigrescens 44
Vulpicida juniperinus 243
V. pinastri 129, 130, 134, 204, 207, 211, 220
Xanthomaculina convoluta 18
Xanthoparmelia camtchadalis 18
X. chlorochroa 181
X. (Parmelia) conspersa 51, 146, 154
X. decipiens 154
X. lineola 144, 154
X. som^nsis 150, 155, 194
X. subdecipiens 144
Xanthoria 115, 167
X. candelaria 91, 204
X. elegans 188, 194
X.fallax 91, 207, 211,216
X. lychnea 204
X parietina 14, 23, 39, 47, 91, 104, 141, 143, 144, 145, 146, 154, 170, 204, 207, 211, 225
X. polycarpa 91, 204, 207, 211, 223
Научное издание
Бязров Лев Георгиевич
ЛИШАЙНИКИ В ЭКОЛОГИЧЕСКОМ МОНИТОРИНГЕ
«Научный мир», Телефакс (007) (095) 291-2847
E-mail: naumir@ben.irex.ru. Internet: http://195.178.196.201/N_M/n_m.htm
Лицензия ИД № 0322 i аг 10.11.2000.
Подписано к печати 13.08.2002. Формат 70*100/16
Гарнитура Таймс. Печать офсетная. Печ. л. 21. Усл. печ. л 27.3
Тираж 700 экз. Заказ 199
Издание отпечатано в типотрафии ООО -Галлея-Принт”
Москва, 5-я Кабельная, 26