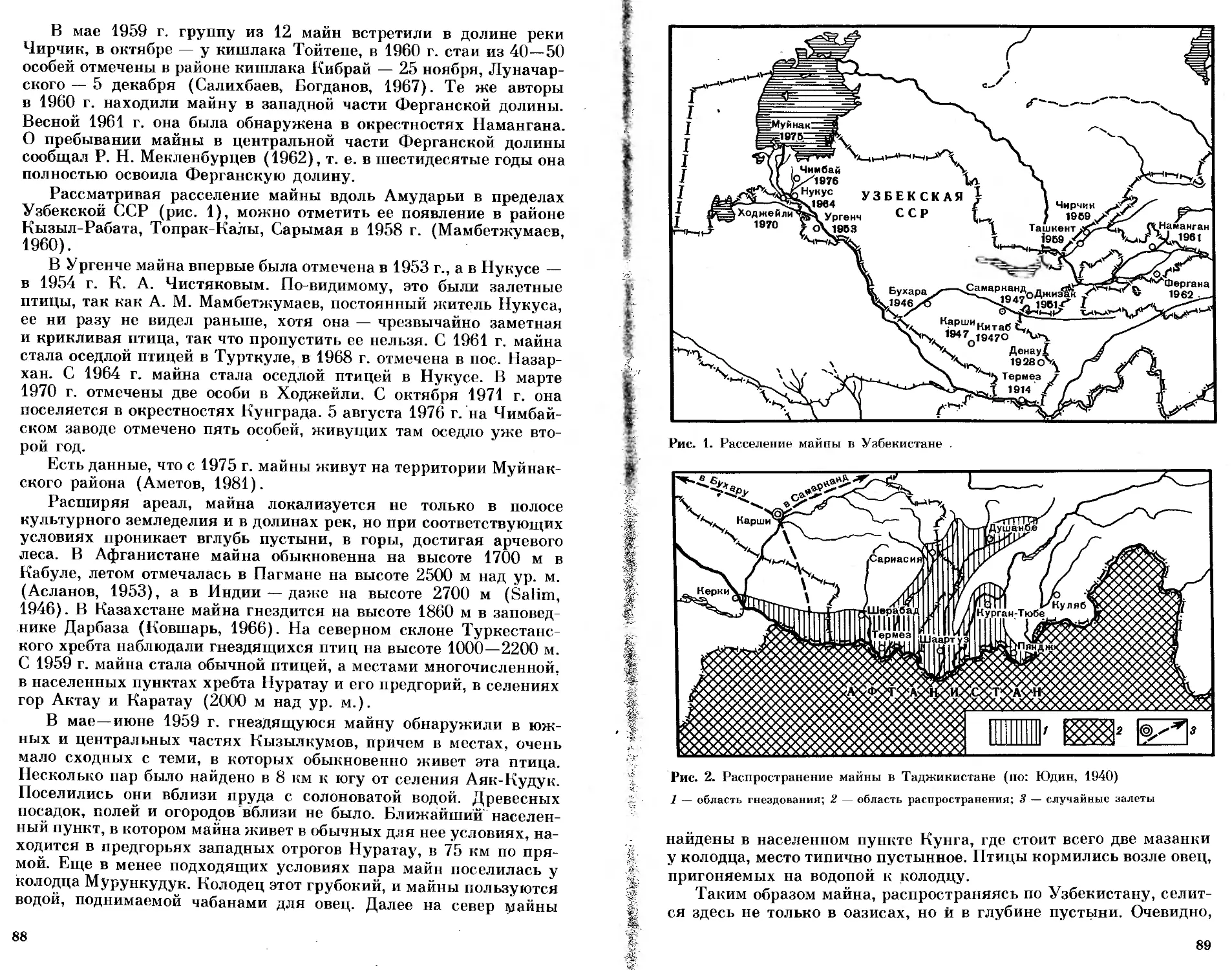
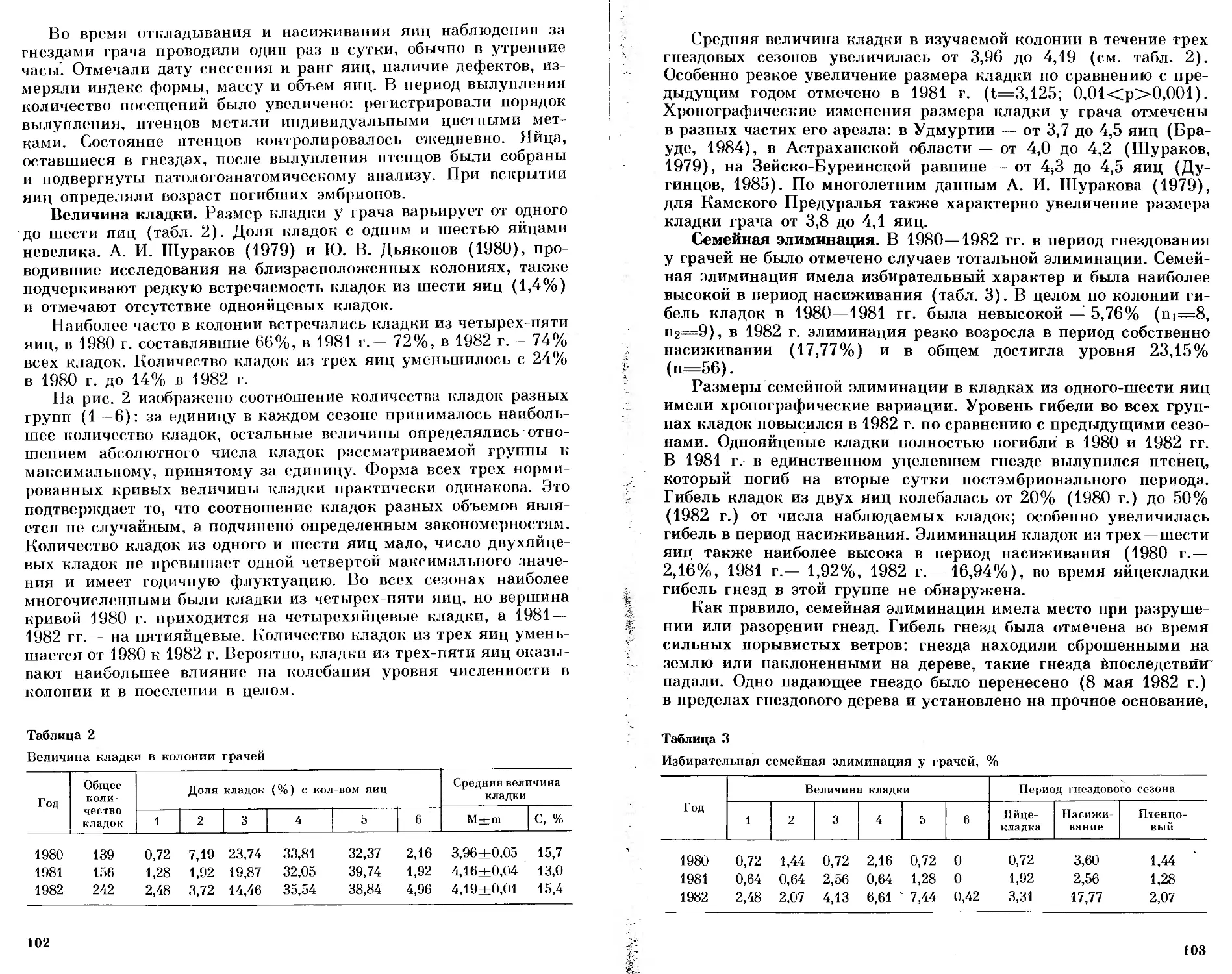
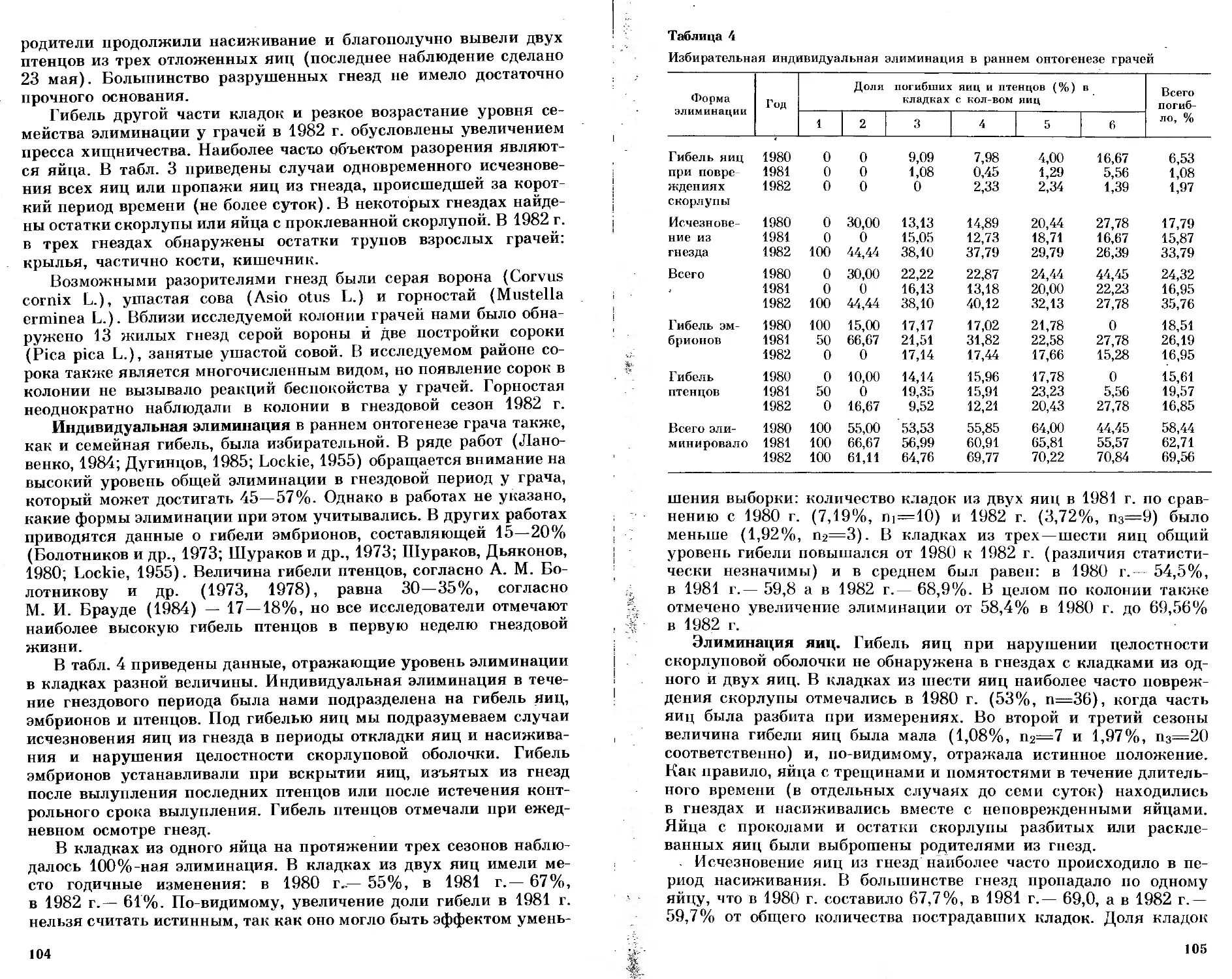
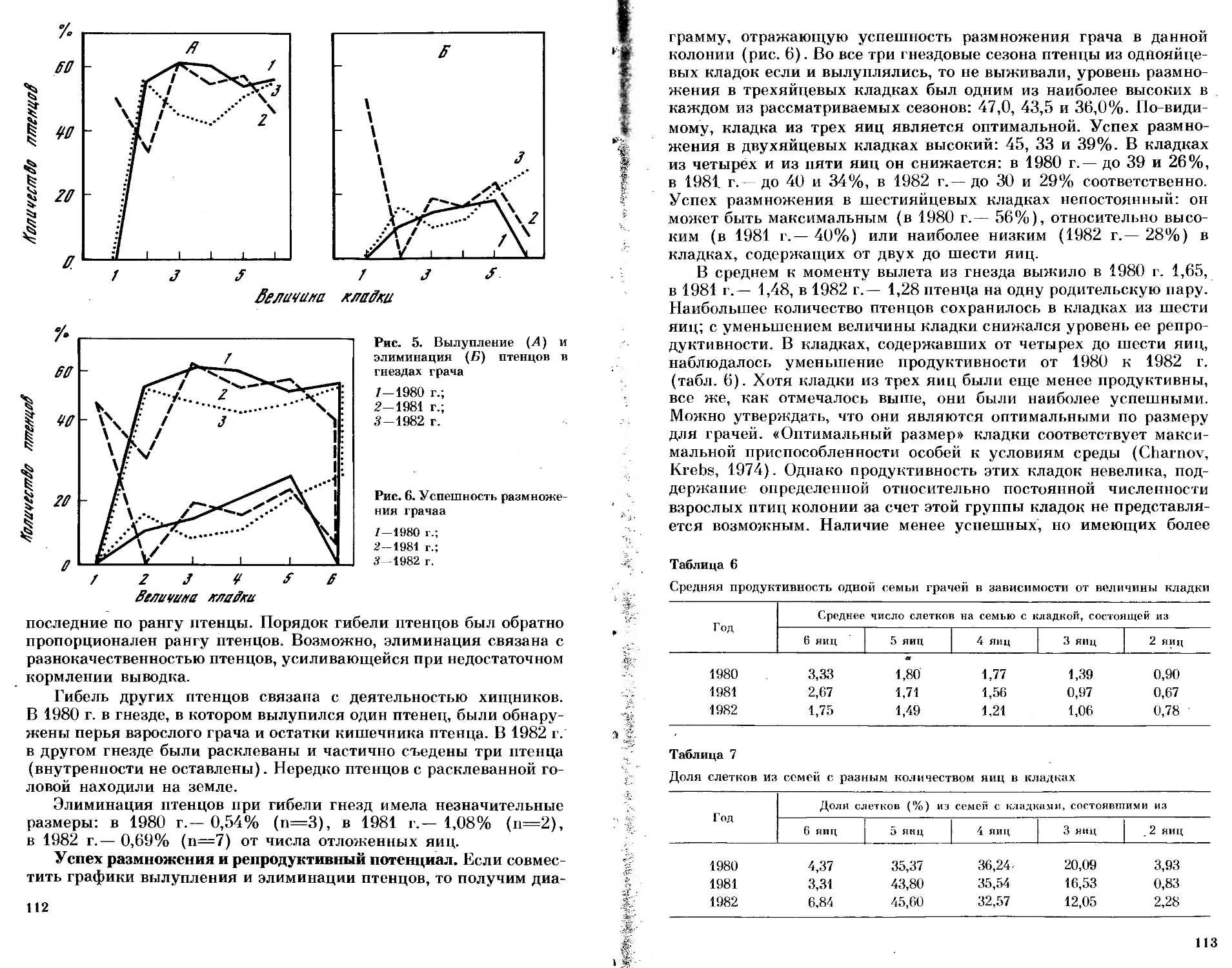
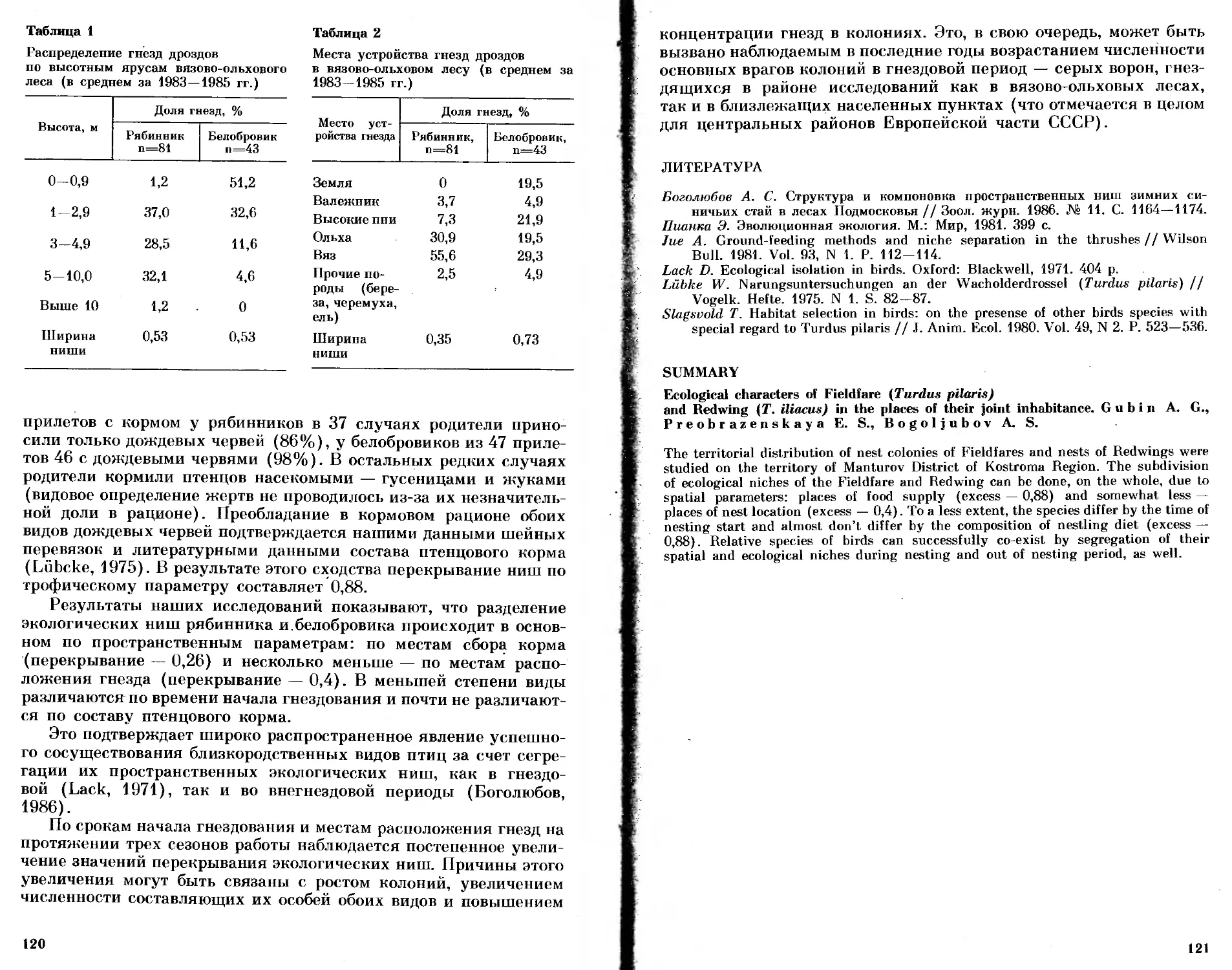
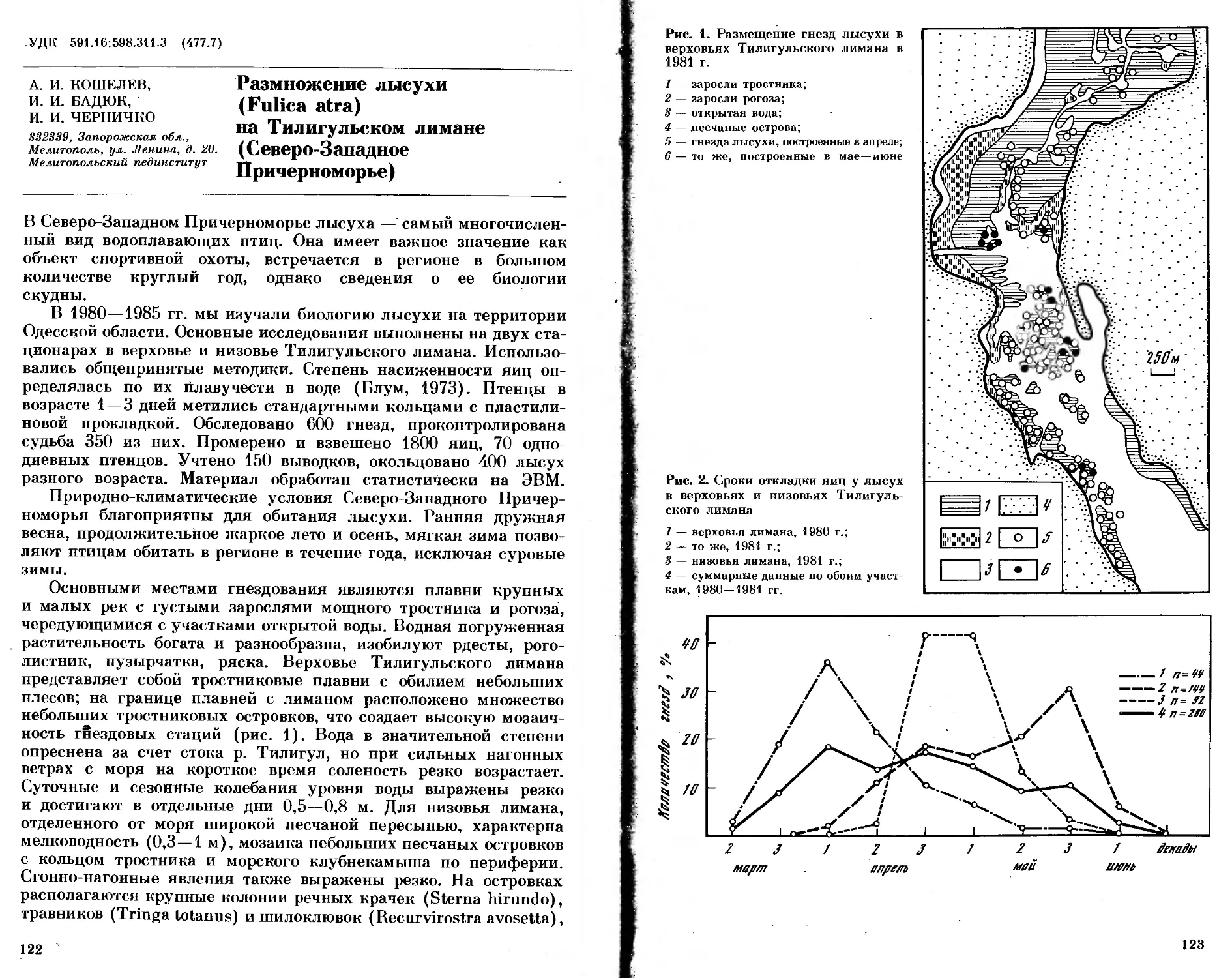
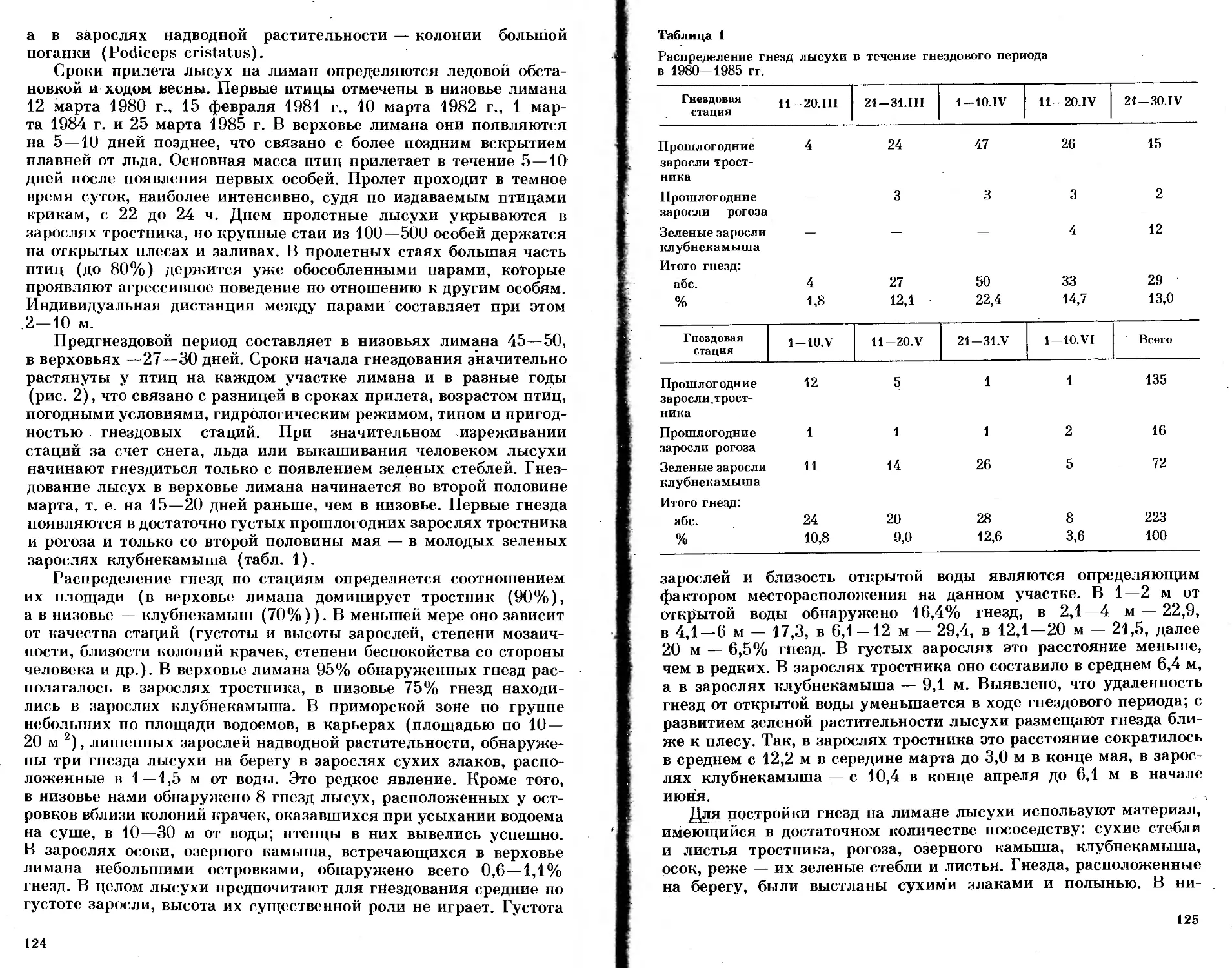
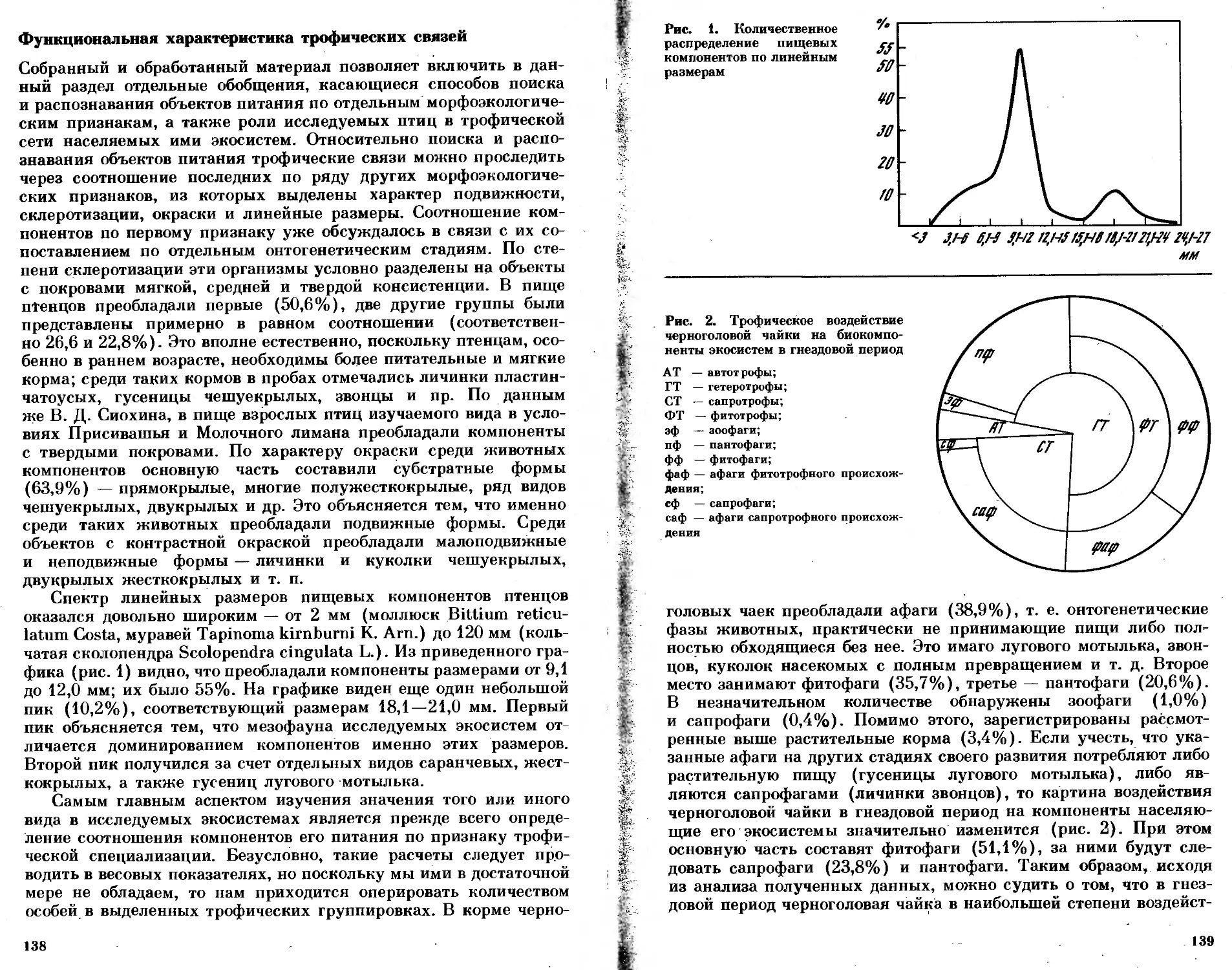
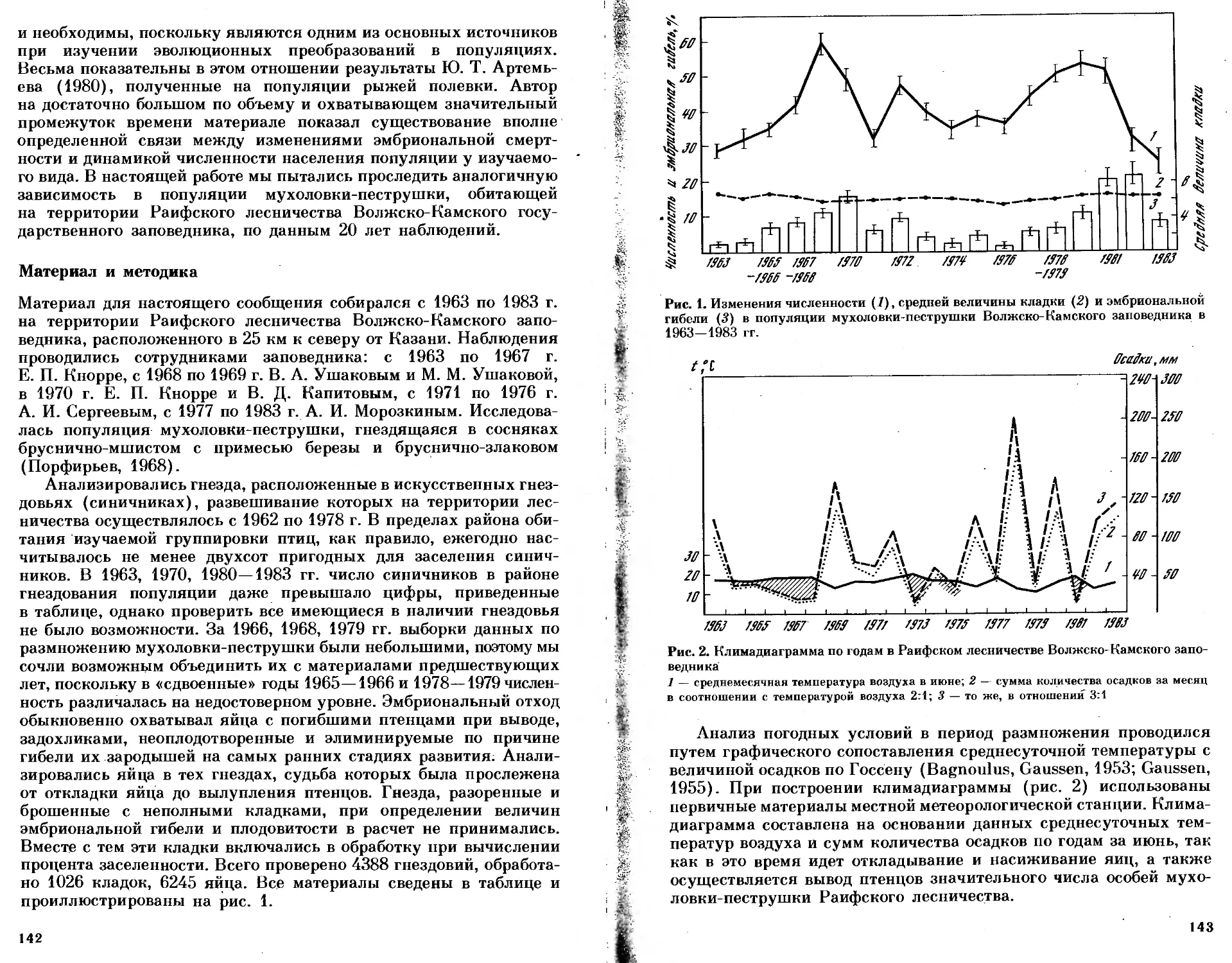
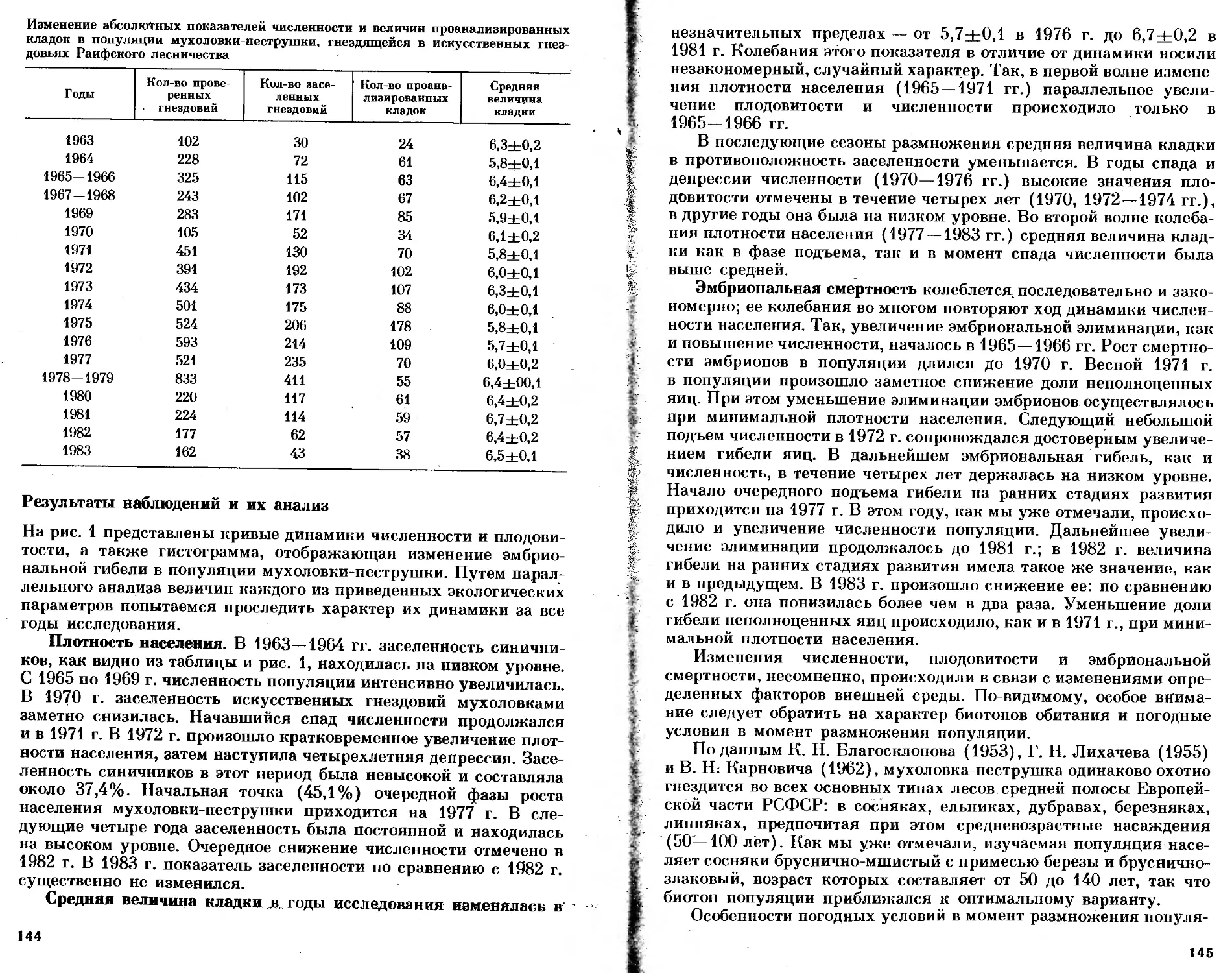
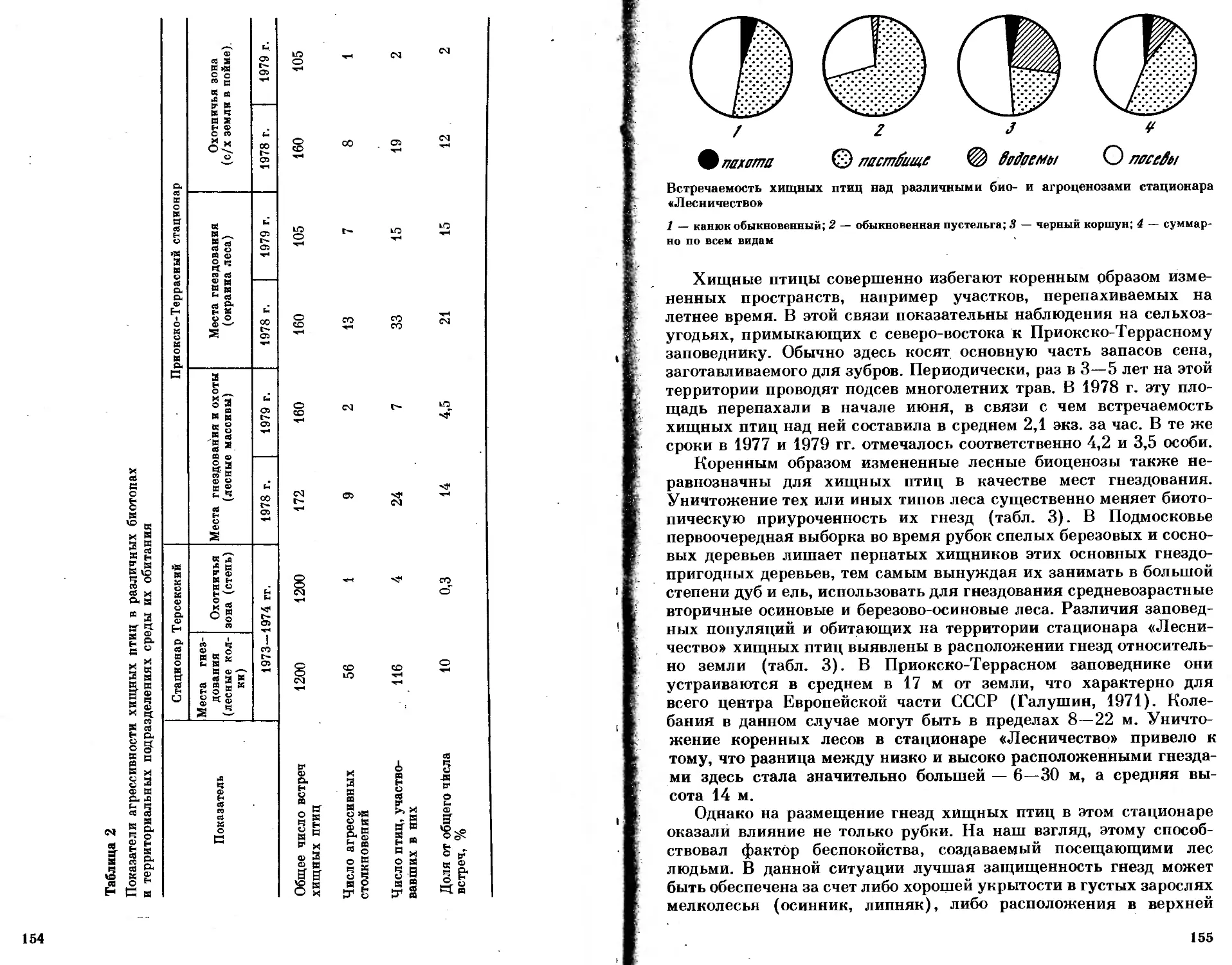
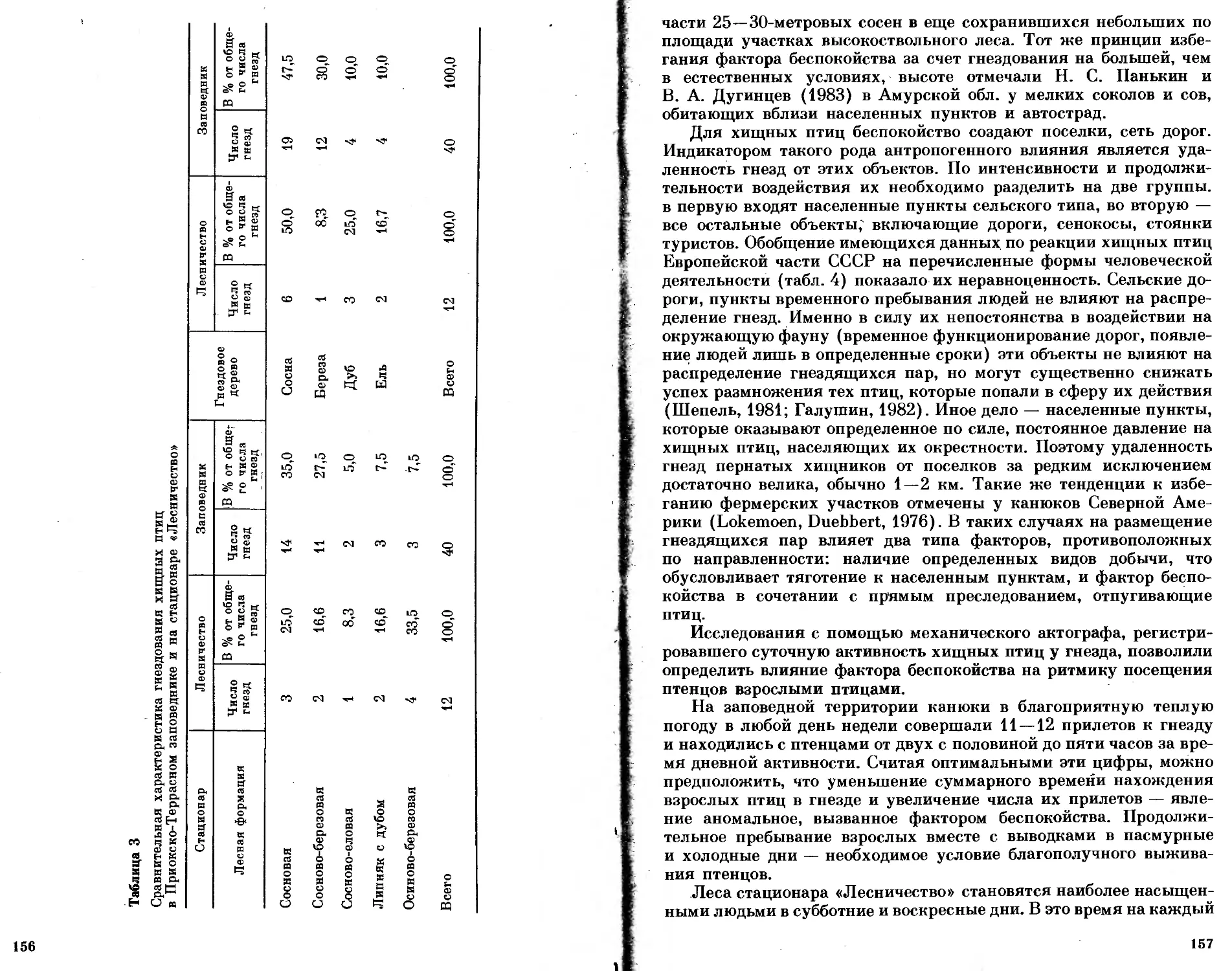
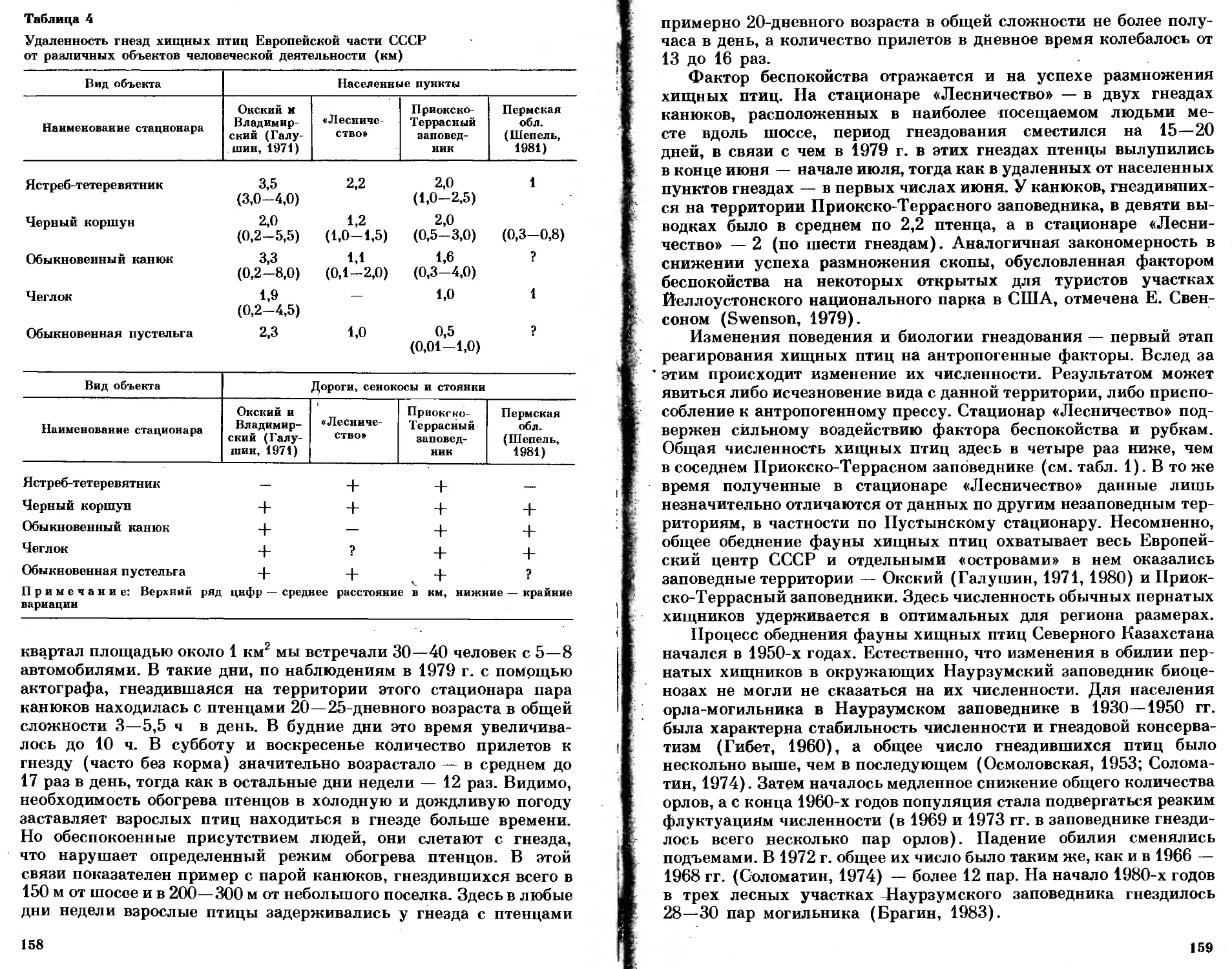
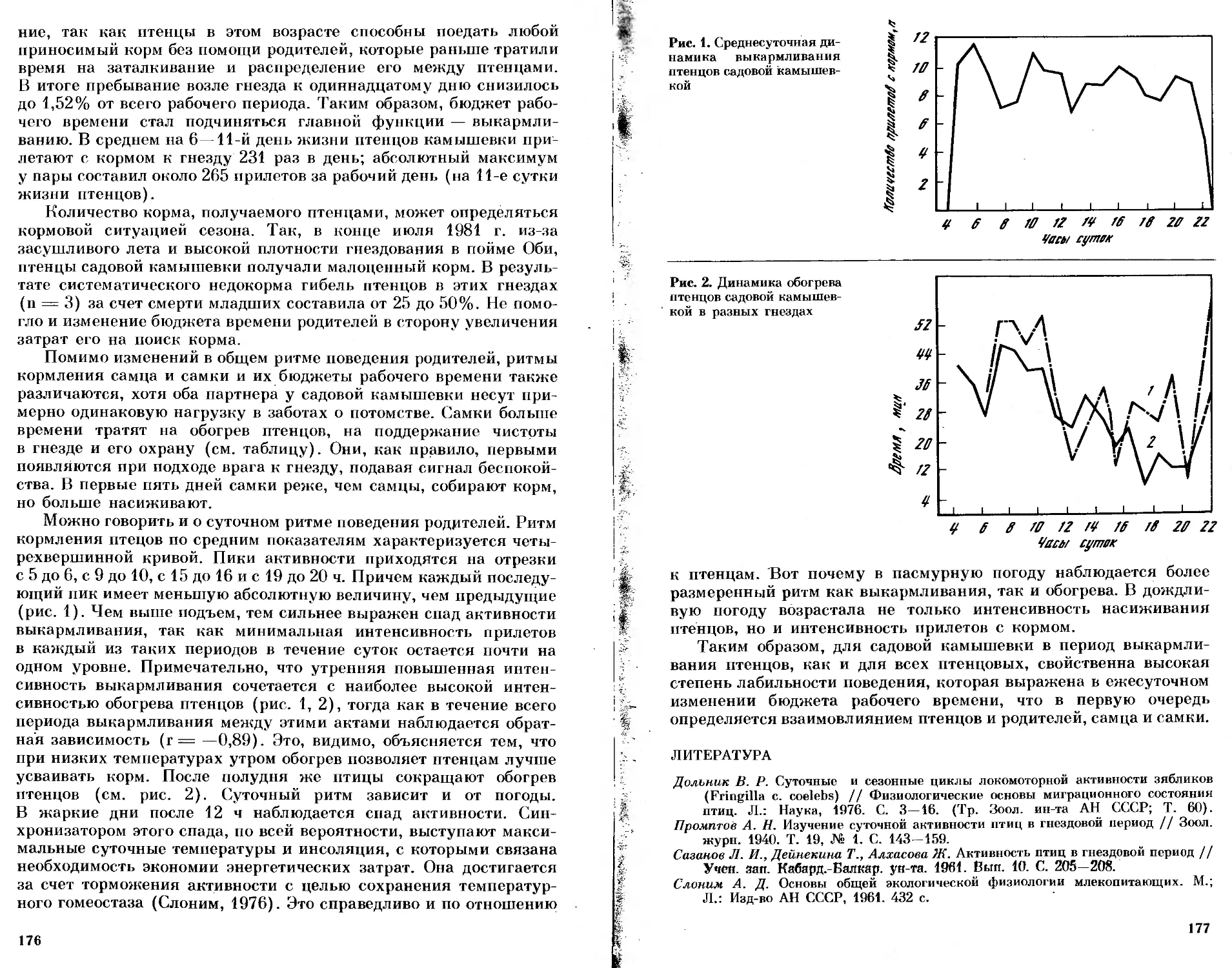
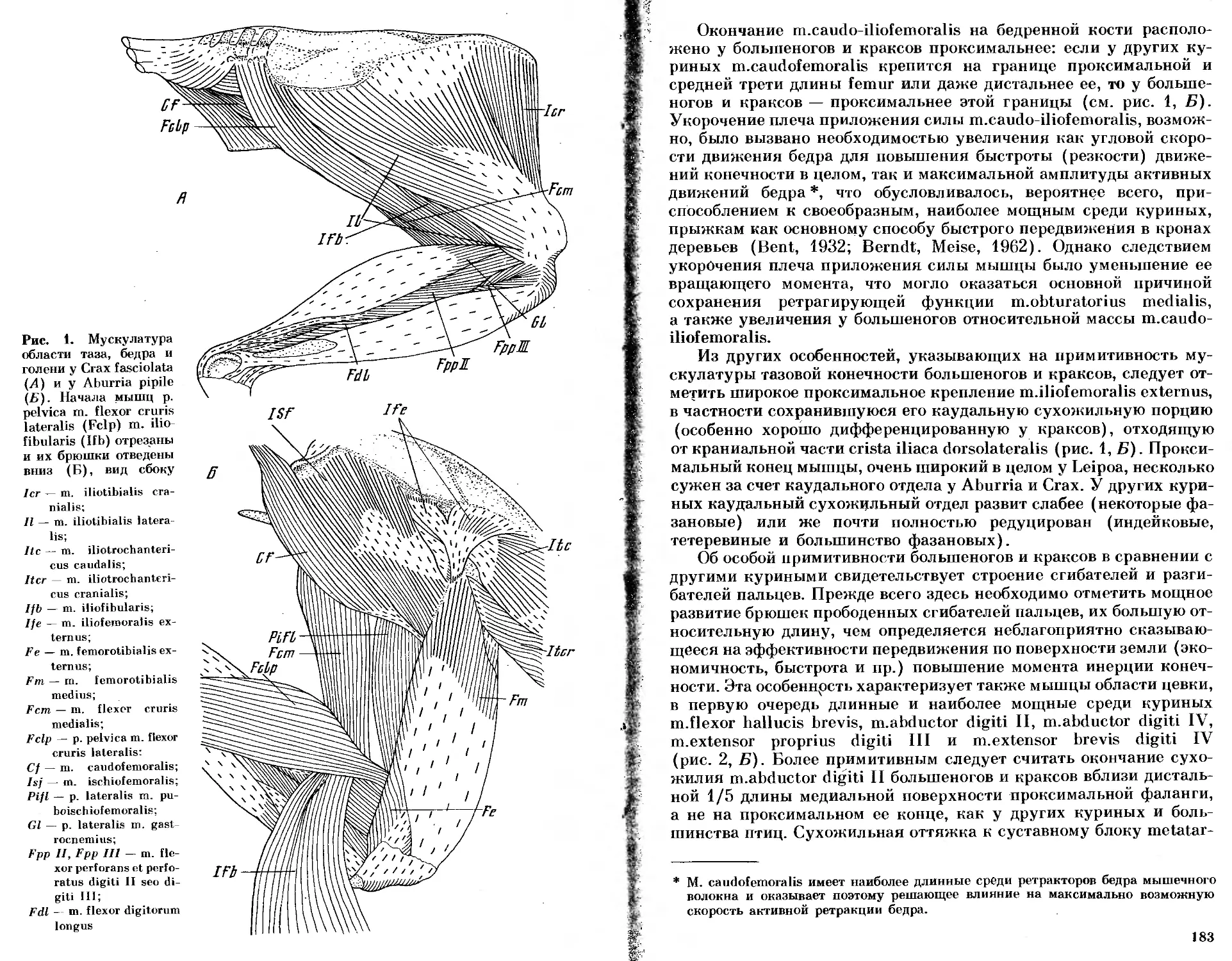
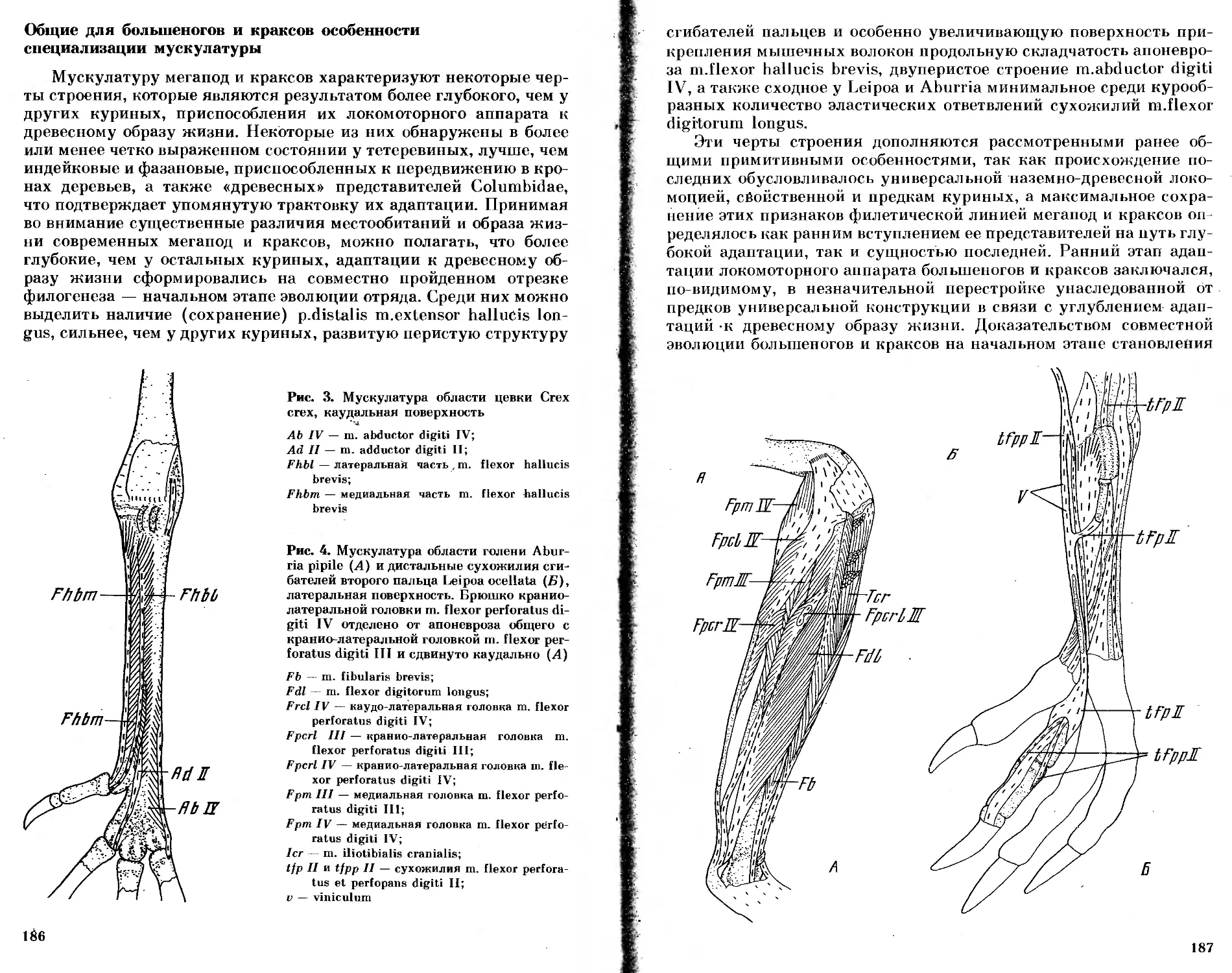
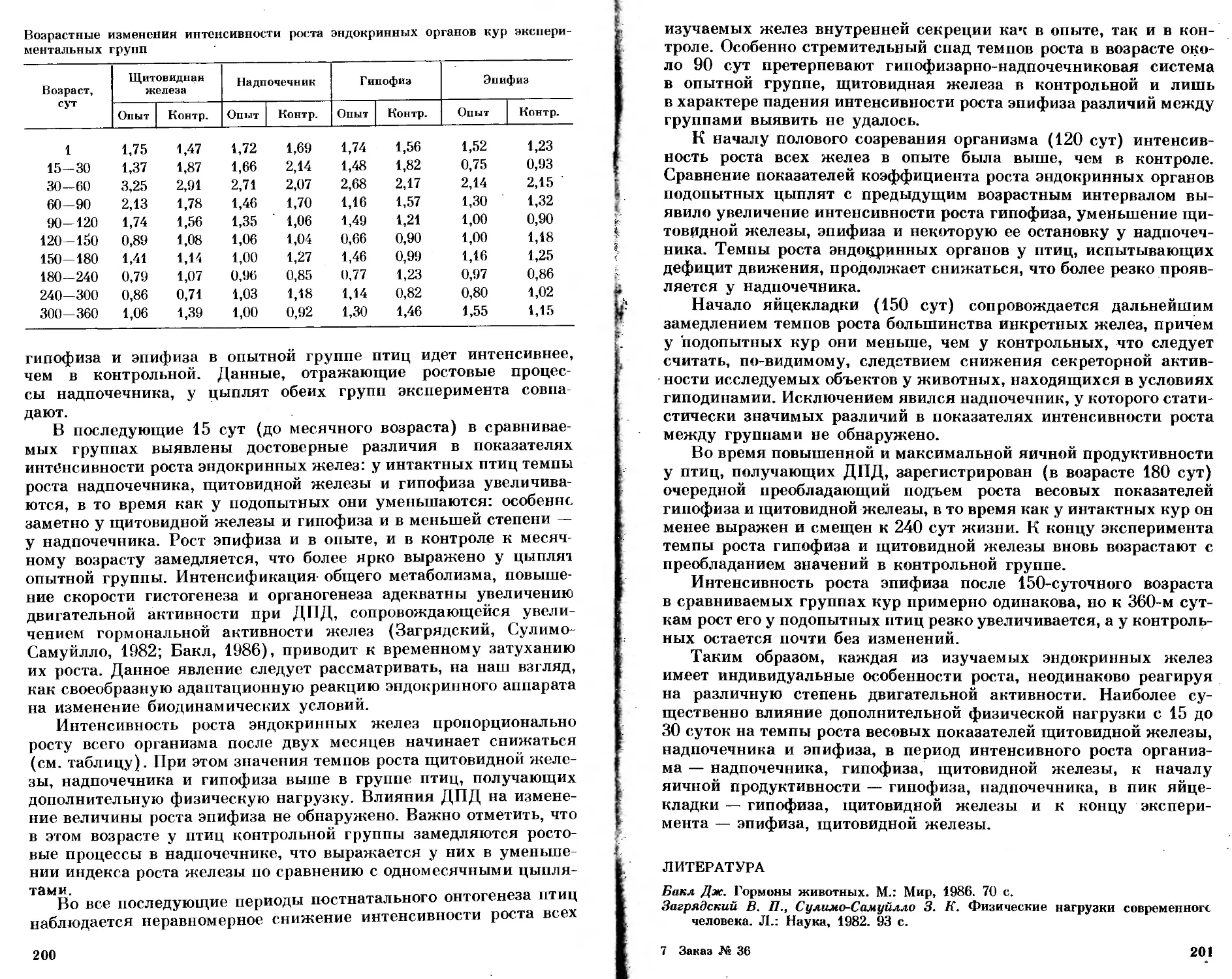
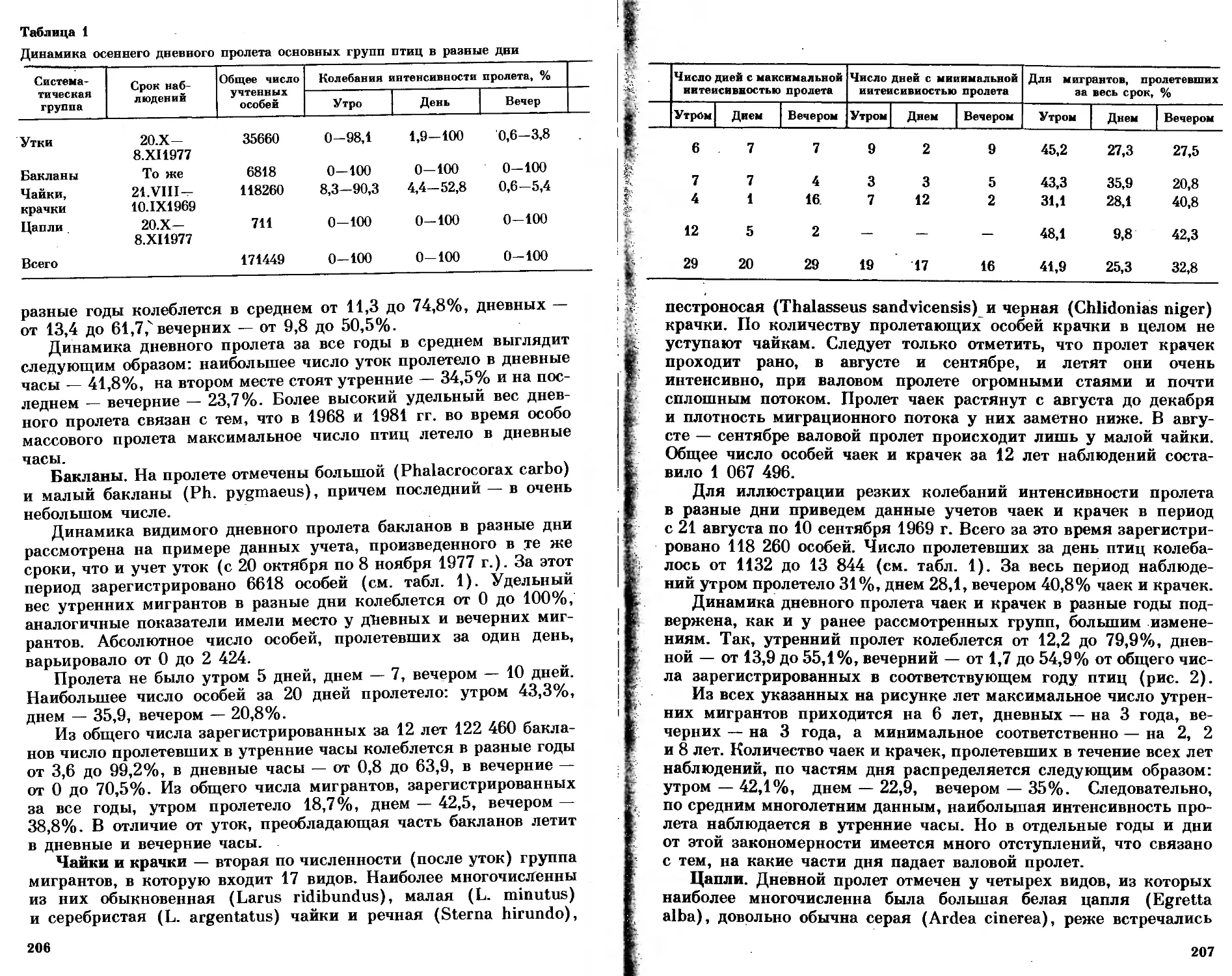
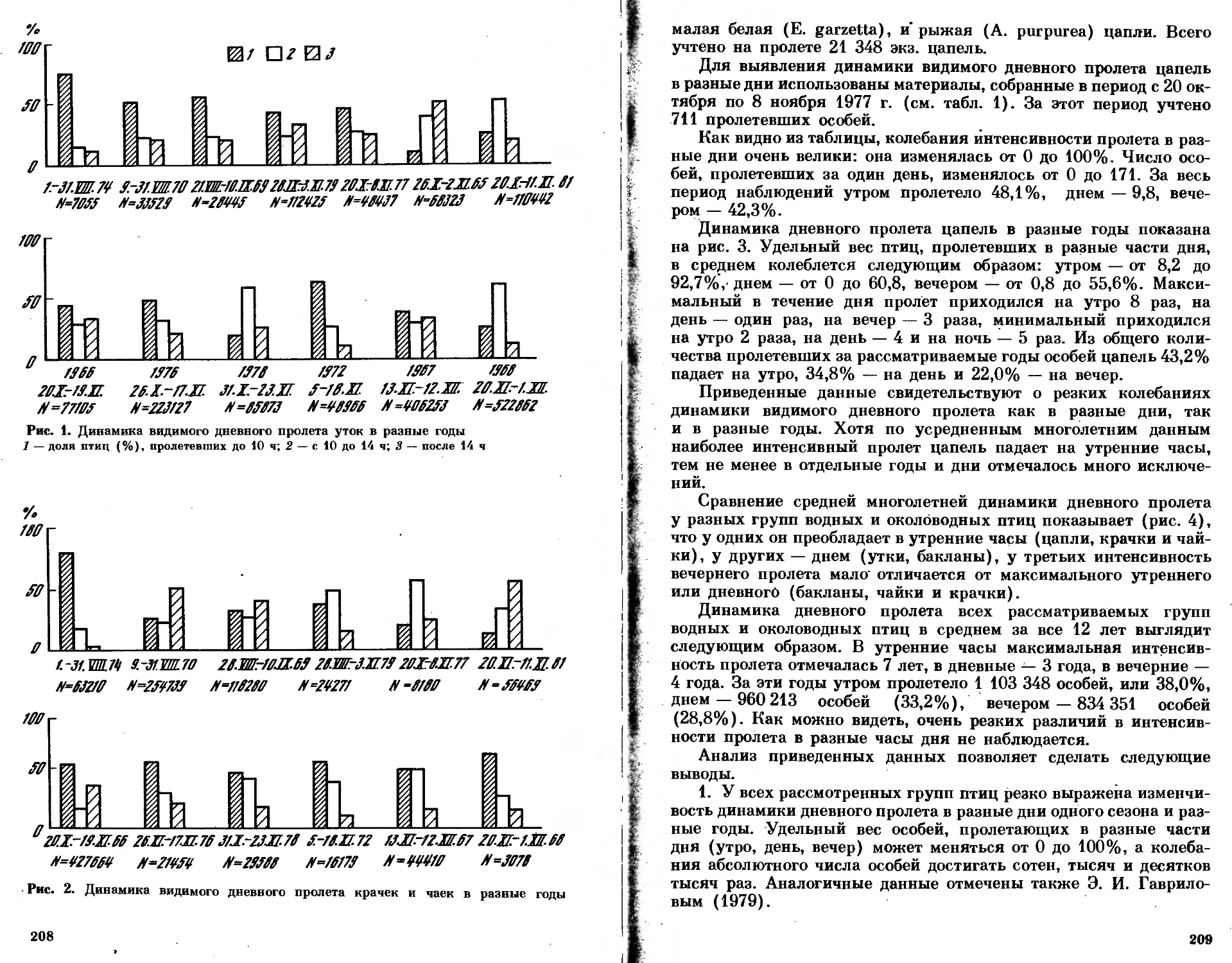
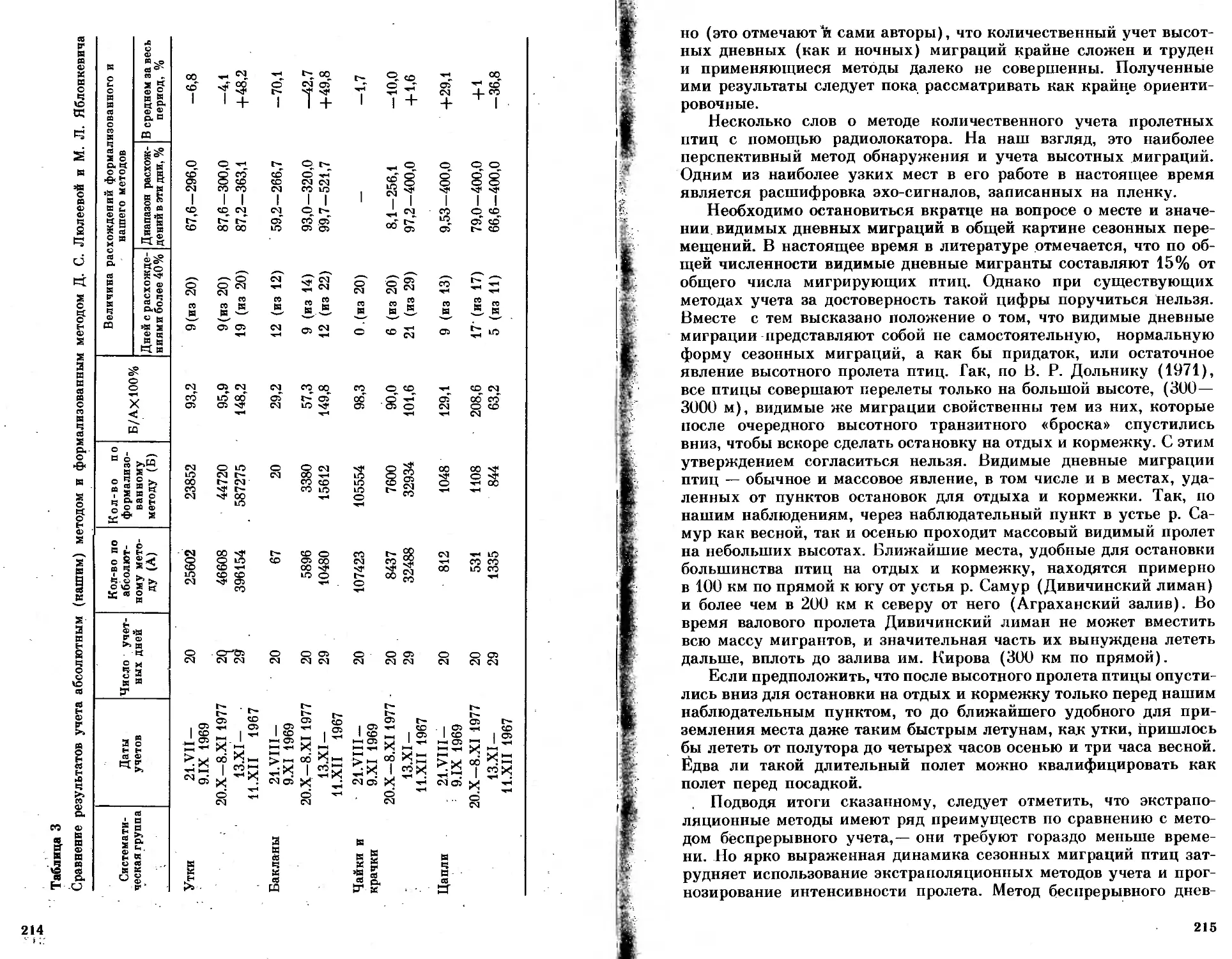
/





































































































Author: Курочкин Е.Н. Правдивая Е.Ф.
Tags: aves птицы в целом орнитология зоология птицы
ISBN: 5-02-005877-7
Year: 1990




Text
СОВРЕМЕННАЯ
ОРНИТОЛОГИЯ
1990
USSR ACADEMY
OF SCIENCE
USSR
ORNITHOLOGICAL
SOCIETY
MODERN
ORNITHOLOGY
wo
Collected papers
of the USSR
Ornithological Society
Editor-in-chief
E. N. KUROCHKIN
MOSCOW
NAUKA PUBLISHERS
1990
АКАДЕМИЯ НАУК СОВРЕМЕННАЯ
ВСЕСОЮЗНОЕ
орнитологическое \^гПР114/1 Plrl
ОБЩЕСТВО
1990
Сборник
научных трудов
Ответственный редактор
кандидат биологических наук
Е. Н. КУРОЧКИН
МОСКВА
«НАУКА»
1990
УДК 598.219: 591.5
Современная орнитология 1990.— М.: Наука, 1990.—
264 с.- ISBN 5-02-005877-7.
Книга составлена из статей орнитологов СССР по различным во-
просам орнитологии: экологии и адаптации птиц к среде обитания,
методам охраны птиц, динамике полового и возрастного состава неко-
торых видов, в том числе редких и включенных в Красные книги.
Исследуются биология птиц в антропогенных комплексах и влияние
человека на состав авифауны, функциональная морфология птиц
и методика их изучения. Сборник содержит статьи, посвященные па-
мятным датам, юбилеям, хронике, а также рецензии.
Редакционная коллегия:
Я. А. ВИКСНЕ, О. С. ГАБУЗОВ, В. М. ГАЛУШИН,
В. Р. ДОЛЬНИК, В. Д. ИЛЬИЧЕВ, В. В. ИВАНИЦКИЙ,
А. А. КАРХУ, Е. Н. КУРОЧКИН (ответственный редактор),
К. Е. МИХАЙЛОВ, В. А. ОСТАПЕНКО,
Е. Ф. ПРАВДИВАЯ (ответственный секретарь)
Рецензенты:
канд. биол. наук Е. П. ПИВОВАРОВА,
канд. биол. наук К. В. АВИЛОВА
Modem Ornithology 1990.— Moscow: Nauka, 1990.
The papers of the Soviet ornithologists on the ecology and adapta-
tions of birds, the methods of their conservation, dynamics of sexual and
age composition, including rare and Red data books species compile this
collected volume. Some papers investigate the biology of birds in anthro-
pogenic landscapes, a human influence on avifaunas, a functional mor-
phology and the methods of birds study. The chapters on the memorial
dates, chronicles and reviews contain separate personal notes, important
ornithological meetings etc.
Editorial board:
J. A. VIKSNE, O. S. GABUZOV, V. M. GALUSHIN,
V. R. DOLNIK, V. D. ILIJCHEV, V. V. IVANITSKY,
A. A. KARKHU, E. N. KUROCHKIN (Editor-in-Chief),
С. E. MIKHAILOV, V. A. OSTAPENKO,
E. F. PRAVDIVAJA (Responsible Secretary).
C
1907000000-303
042(02)-90
575-90, II пол.
ISBN 5-02-005877-7
© Издательство «Наука», 1990
ВОПРОСЫ
экологии
УДК 591.524(98)
А. В. АНДРЕЕВ
685010, Магадан, ул. К. Маркса,
д. 24. Институт биологических
проблем Севера ДВО АН СССР
Экологическая
энергетика
арктических птенцов
Известно, что период появления на свет и последующий рост
птенцов у тундровых птиц в целом синхронизирован с годовым
максимумом температуры. Этот момент характеризуется и наибо-
лее интенсивной вегетацией растений, и наивысшей активностью
беспозвоночных.
Хотя температурные и пищевые условия в середине лета
благоприятны, воспитание нового поколения — всегда напряжен-
ный период в жизни птиц. Потери популяции в это время
могут быть значительны. Многие признаки репродуктивной био-
логии тундровых видов, такие, например, как выраженность в
популяции территориального поведения, свойственный виду
характер брачных отношений, выбор особями гнездовых участков,
обнаруживают связь с последующим периодом роста птенцов
и направлены на повышение их выживаемости (Андреев, 1984).
С другой стороны, эти признаки как будто соответствуют плотно-
сти, запасам и размещению пищевых ресурсов популяции в период
размножения (Хлебосолов, 1984, 1986).
Поскольку выживаемость птенцов существенно влияет на
последующую динамику популяций, а через нее и на другие
аспекты естественной истории видов, целесообразно рассмотреть
различные аспекты экологии роста более внимательно.
Росту и развитию птиц посвящена обширная литература, одна-
ко морфологические и физиологические аспекты роста изучены
гораздо лучше, чем вопросы экологии птенцов в этот период.
Для нас же представляют интерес именно последние. В данной ра-
боте рассмотрены условия, которые обеспечивают нормальный
рост и выживание птенцов на примере нескольких видов птиц,
обитающих в тундрах северо-восточной Якутии. В связи с очерчен-
ной проблемой наиболее важными представляются следующие
вопросы.
1) Каковы темпы роста массы различных видов птиц и насколь-
ко велики различия между птенцами, выращенными в лаборатории
и выросшими в природе?
5
2) Каков уровень метаболизма растущих птенцов и как он
зависит от видовой принадлежности, возраста, размеров, скорости
роста и температурных условий?
3) Каковы соотношения между интенсивностью метаболизма,
скоростью роста и пищевыми потребностями у растущих птиц?
4) Каким образом, в какой мере и за счет каких ресурсов
тундровые птицы обеспечивают свои ростовые потребности?
5) Каких поведенческих или эколого-физиологических страте-
гий должны придерживаться особи и семьи, чтобы достигнуть
наибольшего соответствия между своими потребностями и ресурс-
ными возможностями среды при наименьшей смертности молодых
индивидуумов?
Сведения, позволяющие более или менее подробно ответить
на поставленные вопросы, собраны автором в период с 1978 по
1986 г. на полевых базах Института биологических проблем
Севера, расположенных в Нижнеколымских тундрах, однако
последние два вопроса выходят за рамки настоящей работы. Суще-
ственную помощь в сборе полевых и экспериментальных данных
оказали мои коллеги — сотрудники лаборатории эксперименталь-
ной экологии ИБПС ДВНЦ АН СССР — Е. И. Хлебосолов, Е. Р. По-
тапов, Л. В. Задорина, В. Н. Хлесткин, а также студенты В. В. Гав
рилов (МГУ) и А. И. Пинчук (ЛГУ). Считаю приятным долгом
выразить здесь всем им свою признательность.
Материал и методика
В качестве объектов исследования были выбраны несколько видов
птиц, воспитывающих своих птенцов по выводковому, полувывод-
ковому и птенцовому способам и доминирующих в ландшафтах
нижнеколымских тундр. В их числе гуменник (Anser fabalis),
зимняк (Buteo lagopus), белая куропатка (Lagopus lagopus), се-
ребристая чайка (Larus argentatus), розовая чайка (Rhodostethia
rosea), полярная Крачка (Sterna paradisaea), турухтан (Philoma-
chus pugnax) и дутыш (Calidris melanotos). В целях сравнения
были использованы данные по другим полярным и восточно-
азиатским видам, опубликованные в печати или любезно представ-
ленные сотрудниками нашей лаборатории. Куликов и куропаток
выращивали из яиц, взятых в гнездах за три-четыре дня до вылуп-
ления и помещенных в инкубатор. В других случаях птенцов
брали из гнезд в возрасте одного-двух дней. Температурные усло-
вия в период роста поддерживали в оптимальных пределах (15—
22°). В первые дни жизни птенцов содержали в садках, где они
могли пользоваться искусственным обогревом. Позднее, в дневное
время при сухой и теплой погоде птенцов содержали в уличных
загонах, защищенных от хищников мелкоячеистой сеткой. В холод-
ную погоду их снова переносили в обогреваемое помещение.
Таким образом, в экспериментах птенцов выращивали при тем-
пературах, не достигавших термонейтральности (30—33°) и при
избытке пищи. Ростовые процессы в этом случае должны достигать
6
найлучшего выражения (Kleiber, Dougherty, 1933). В качестве
корма птенцам куликов и чаек давали хироном ид, водных рако-
образных и моллюсков, вареное яйцо и рыбу. Куропаткам с первых
дней наряду с животными, давали зеленые корма (мятлик, щавель,
звездчатку, иву), гречневую крупу и комбикорм. Гусей содержали
на полувольном выпасе, где они кормились хвощами, арктофилой
и мятликом, получая в качестве дополнения некоторое количество
увлажненного комбикорма. Птенцов зимняка выкармливали мыше-
видными грызунами (Потапов, 1986).
Птенцов ежедневно взвешивали и обмеряли. Параллельно про-
водили сбор данных о скорости роста маркированных птенцов
в природе. Для массовых видов (белая куропатка, дутыш, турух-
тан) скорость роста в естественных условиях устанавливали также
по статистическим данным. Для каждого вида строили кривые
роста (рис. 1). Обычно изменение веса тела во время роста харак-
теризуют величиной привеса (dW) за известный промежуток вре-
мени, либо логарифмическими константами роста, позволяющими
сравнить относительный рост у различных по размеру животных
(Мина, Клевезаль, 1976). Для наших целей важны также и вре-
менные характеристики роста, поскольку в реальной среде про-
исходят непрерывные климатические и фенологические изменения,
влияющие на пищевые потребности птенцов и доступность корма.
В целях анализа весь период роста (Т) оказалось удобным раз-
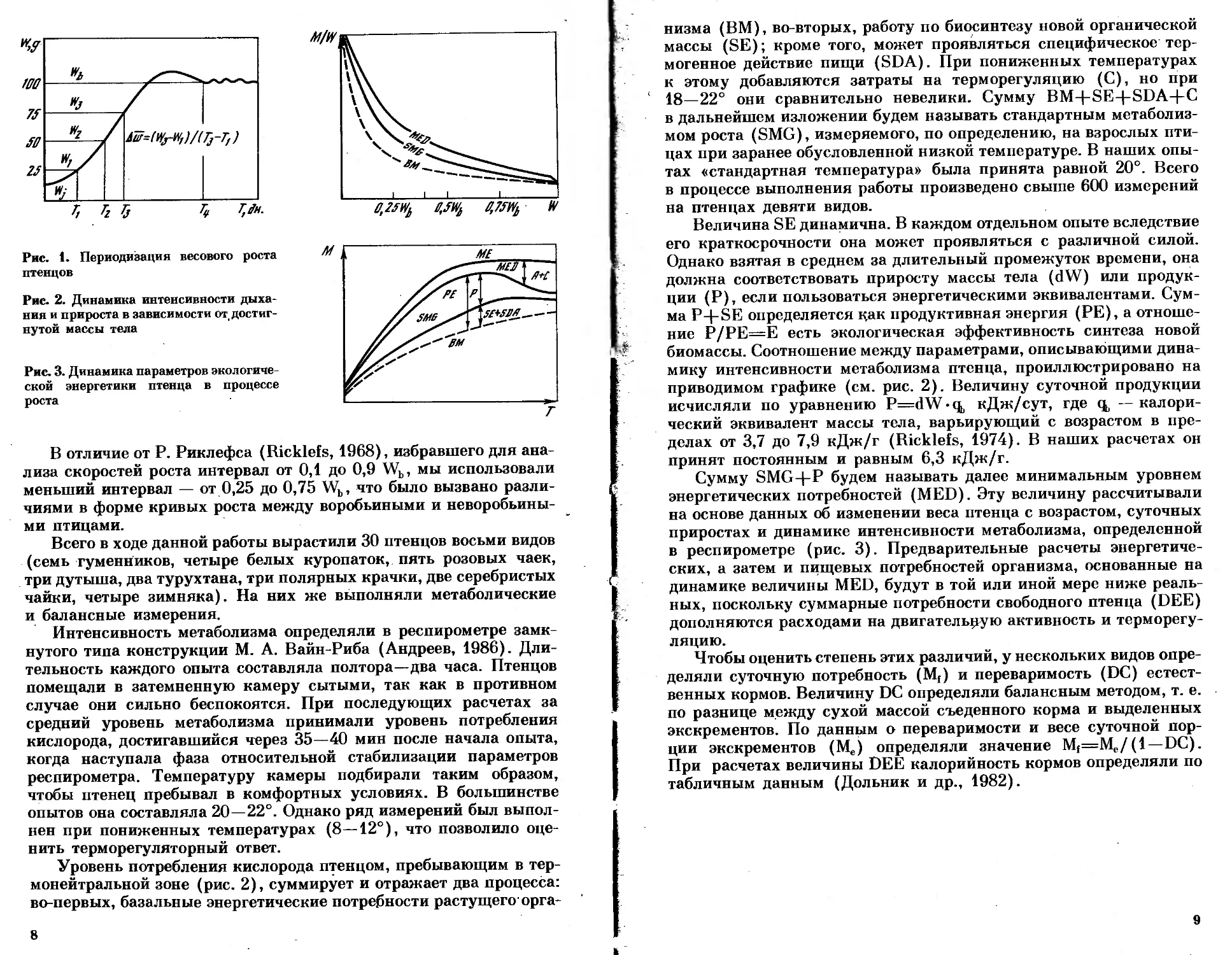
бить на четыре периода (см. рис. 1):
1) начальная фаза, когда привесы еще невелики, терморегуля-
ция неустойчива, рост оперения выражен слабо или не выражен
вовсе. В этой фазе достигается 15—25% конечной массы тела;
2) фаза быстрого роста. В течение ее достигается 75—90% ко-
нечной массы, отрастает контурное перо, растет и развивается
осевой скелет и конечности. Уже в начале этой фазы терморегу-
ляция становится устойчивой;
3) фаза замедленного роста: весовой и линейный рост тормозит-
ся, достигается 90—100% конечной массы, заканчивается рост кон-
турного оперения;
4) фаза стабилизации; относительно устойчивое состояние,
в котором отклонения обычно не превышают ±3% от некоторого
среднего значения конечной массы (Wb).
Соответственно такому подразделению из большого возможного
числа ростовых характеристик были выбраны шесть, которые пред-
ставляются наиболее удобными и информативными для дальней-
шего анализа и сравнений:
Wj — стартовый вес, средний вес птенца в течение первых
6—8 часов жизни;
Wh — конечный вес (определен как средний вес в четвертом
периоде);
dW — скорость линейного роста массы, или средний привес
массы в интервале от 1/4 до 3/4 конечной массы;
Т|, Тг, Тз — периоды, в течение которых достигается соответ-
ственно четверть, половина и три четверти конечной
массы.
7
Рис. 1. Периодизация весового роста
птенцов
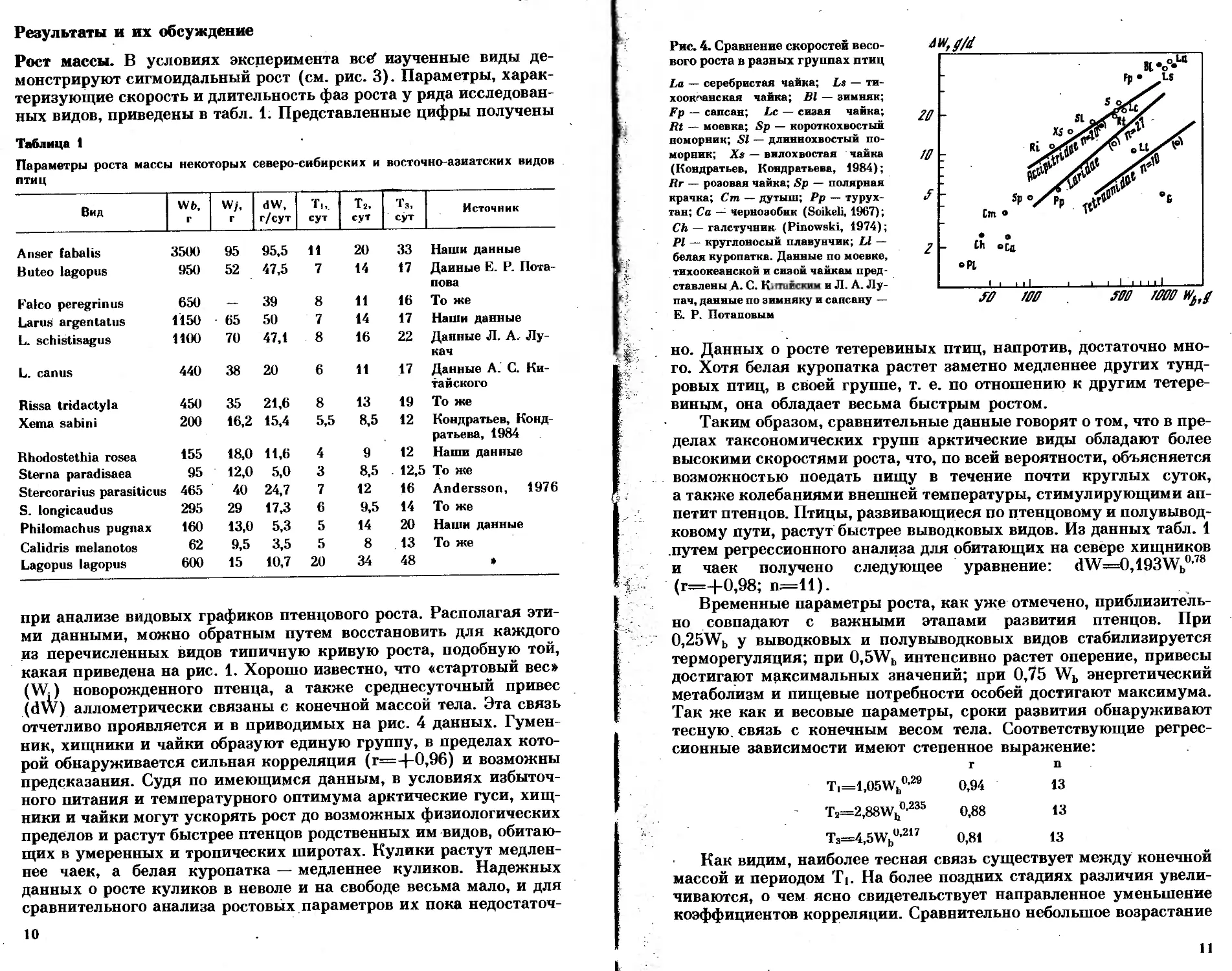
Рис. 2. Динамика интенсивности дыха-
ния и прироста в зависимости от, достиг-
нутой массы тела
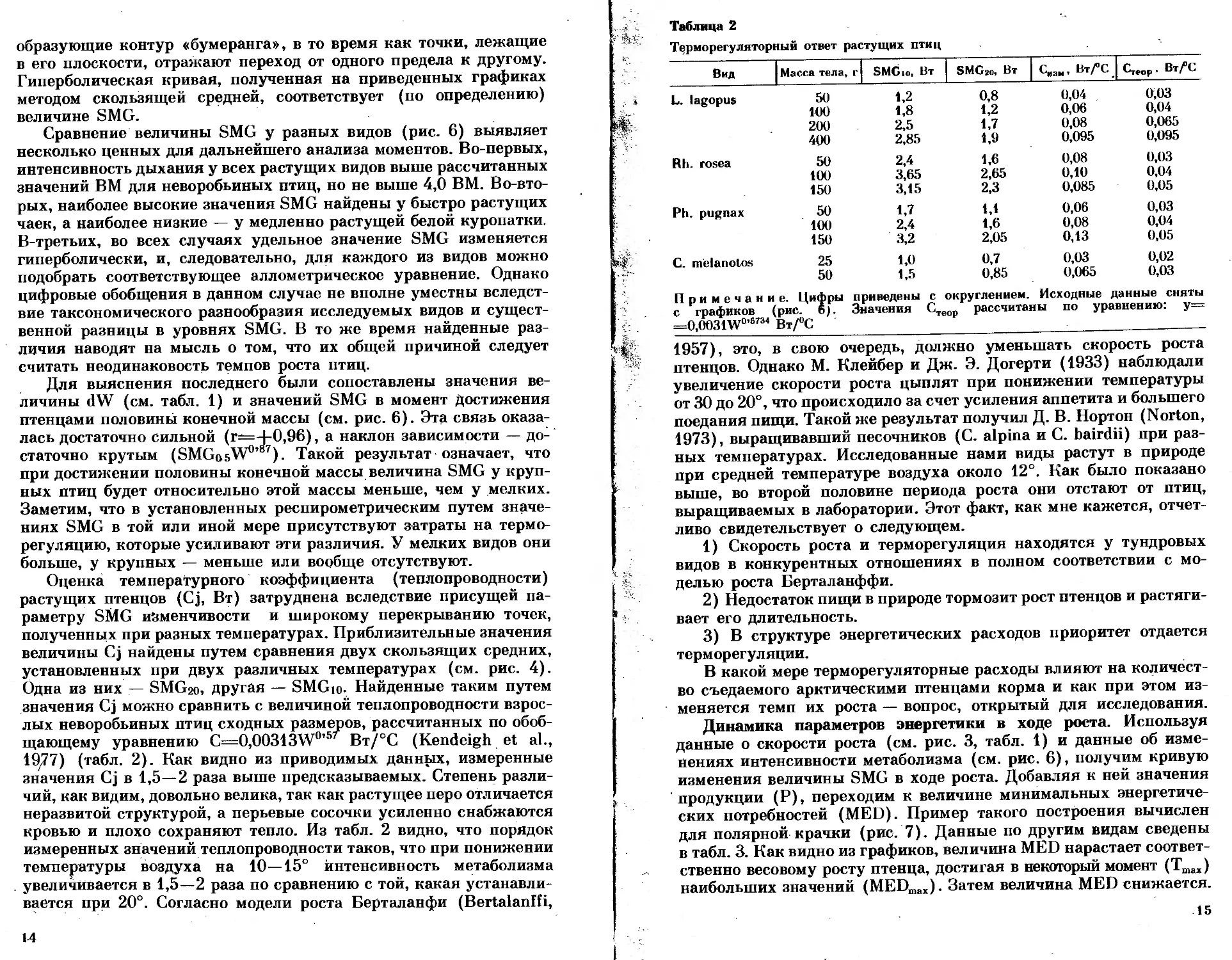
Рис. 3. Динамика параметров экологиче-
ской энергетики птенца в процессе
роста
В отличие от Р. Риклефса (Ricklefs, 1968), избравшего для ана-
лиза скоростей роста интервал от 0,1 до 0,9 Wb, мы использовали
меньший интервал — от 0,25 до 0,75 Wb, что было вызвано разли-
чиями в форме кривых роста между воробьиными и неворобьины-
ми птицами.
Всего в ходе данной работы вырастили 30 птенцов восьми видов
(семь гуменников, четыре белых куропаток, пять розовых чаек,
три дутыша, два турухтана, три полярных крачки, две серебристых
чайки, четыре зимняка). На них же выполняли метаболические
и балансные измерения.
Интенсивность метаболизма определяли в респирометре замк
нутого типа конструкции М. А. Вайн-Риба (Андреев, 1986). Дли-
тельность каждого опыта составляла полтора—два часа. Птенцов
помещали в затемненную камеру сытыми, так как в противном
случае они сильно беспокоятся. При последующих расчетах за
средний уровень метаболизма принимали уровень потребления
кислорода, достигавшийся через 35—40 мин после начала опыта,
когда наступала фаза относительной стабилизации параметров
респирометра. Температуру камеры подбирали таким образом,
чтобы птенец пребывал в комфортных условиях. В большинстве
опытов она составляла 20—22°. Однако ряд измерений был выпол-
нен при пониженных температурах (8—12°), что позволило оце-
нить терморегуляторный ответ.
Уровень потребления кислорода птенцом, пребывающим в тер-
монейтральной зоне (рис. 2), суммирует и отражает два процесса:
во-первых, базальные энергетические потребности растущего1 орга-
8
низма (ВМ), во-вторых, работу по биосинтезу новой органической
массы (SE); кроме того, может проявляться специфическое тер-
могенное действие пищи (SDA). При пониженных температурах
к этому добавляются затраты на терморегуляцию (С), но при
18—22° они сравнительно невелики. Сумму BM-|-SE-|-SDA-|-C
в дальнейшем изложении будем называть стандартным метаболиз-
мом роста (SMG), измеряемого, по определению, на взрослых пти-
цах при заранее обусловленной низкой температуре. В наших опы-
тах «стандартная температура» была принята равной 20°. Всего
в процессе выполнения работы произведено свыше 600 измерений
на птенцах девяти видов.
Величина SE динамична. В каждом отдельном опыте вследствие
его краткосрочности она может проявляться с различной силой.
Однако взятая в среднем за длительный промежуток времени, она
должна соответствовать приросту массы тела (dW) или продук-
ции (Р), если пользоваться энергетическими эквивалентами. Сум-
ма P-(-SE определяется как продуктивная энергия (РЕ), а отноше-
ние Р/РЕ=Е есть экологическая эффективность синтеза новой
биомассы. Соотношение между параметрами, описывающими дина-
мику интенсивности метаболизма птенца, проиллюстрировано на
приводимом графике (см. рис. 2). Величину суточной продукции
исчисляли по уравнению P=dW-qb кДж/сут, где q —калори-
ческий эквивалент массы тела, варьирующий с возрастом в пре-
делах от 3,7 до 7,9 кДж/г (Bicklefs, 1974). В наших расчетах он
принят постоянным и равным 6,3 кДж/г.
Сумму SMG-pP будем называть далее минимальным уровнем
энергетических потребностей (MED). Эту величину рассчитывали
на основе данных об изменении веса птенца с возрастом, суточных
приростах и динамике интенсивности метаболизма, определенной
в респирометре (рис. 3). Предварительные расчеты энергетиче-
ских, а затем и пищевых потребностей организма, основанные на
динамике величины MED, будут в той или иной мере ниже реаль-
ных, поскольку суммарные потребности свободного птенца (DEE)
дополняются расходами на двигательную активность и терморегу-
ляцию.
Чтобы оценить степень этих различий, у нескольких видов опре-
деляли суточную потребность (Mf) и переваримость (DC) естест-
венных кормов. Величину DC определяли балансным методом, т. е.
по разнице между сухой массой съеденного корма и выделенных
экскрементов. По данным о переваримости и весе суточной пор-
ции экскрементов (Ме) определяли значение Mf=Me/(l—DC).
При расчетах величины DEE калорийность кормов определяли по
табличным данным (Дольник и др., 1982).
9
Результаты и их обсуждение
Рост массы. В условиях эксперимента все” изученные виды де-
монстрируют сигмоидальный рост (см. рис. 3). Параметры, харак-
теризующие скорость и длительность фаз роста у ряда исследован-
ных видов, приведены в табл. 1. Представленные цифры получены
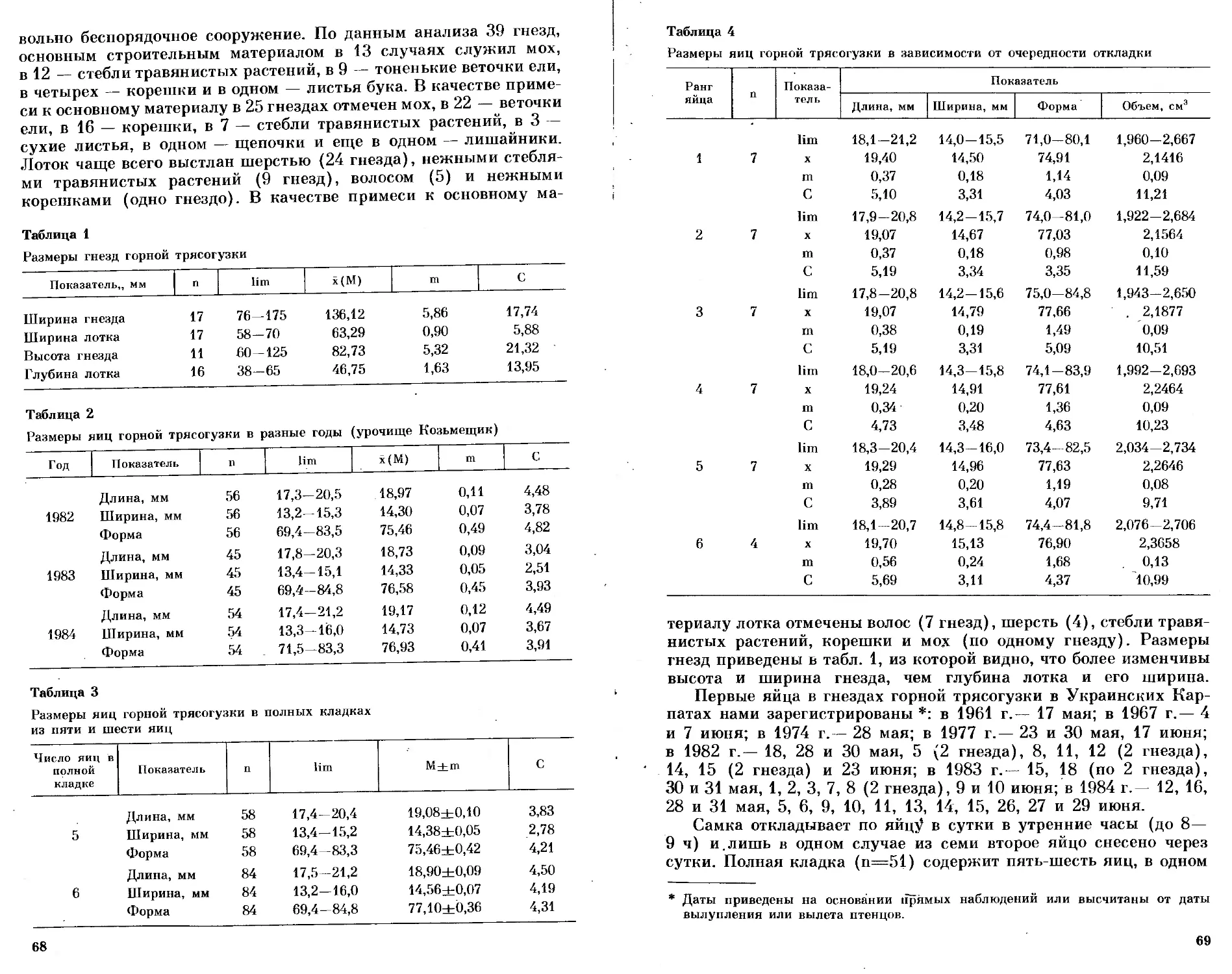
Таблица 1
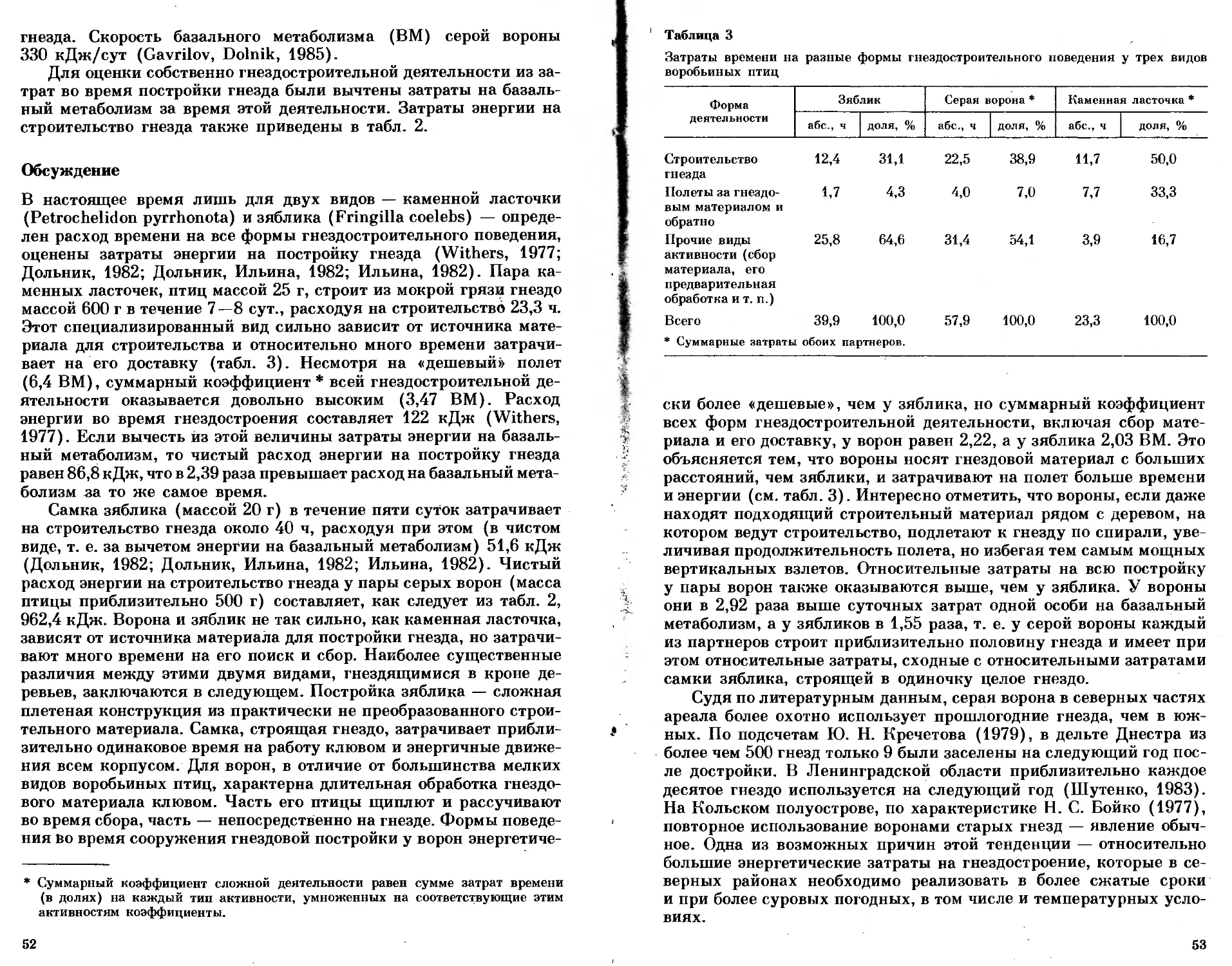
Параметры роста массы некоторых северо-сибирских и восточно-азиатских видов
птиц
Вид Wfe, г w>, Г dW, г/сут T„. сут т2, сут Тз, сут Источник
Anser fabalis 3500 95 95,5 11 20 33 Наши данные
Buteo lagopus 950 52 47,5 7 14 17 Данные Е. Р. Пота- пова
Falco peregrin us 650 — 39 8 11 16 То же
Larus argenlatus 1150 65 50 7 14 17 Наши данные
L. schistisagus 1100 70 47,1 8 16 22 Данные Л. А. Лу- кач
L. can us 440 38 20 6 11 17 Данные А. С. Ки- тайского
Rissa tridactyla 450 35 21,6 8 13 19 То же
Хеша sabini 200 16,2 15,4 5,5 8,5 12 Кондратьев, Конд- ратьева, 1984
Rhodostethia rosea 155 18,0 11,6 4 9 12 Наши данные
Sterna paradisaea 95 12,0 5,0 3 8,5 12,5 То же
Stercorarius parasiticus 465 40 24,7 7 12 16 Andersson, 1976
S. longicaudus 295 29 17,3 6 9,5 14 То же
Philomachus pugnax 160 13,0 5,3 5 14 20 Наши данные
Calidris melanotos 62 9,5 3,5 5 8 13 То же
Lagopus lagopus 600 15 10,7 20 34 48 »
при анализе видовых графиков птенцового роста. Располагая эти-
ми данными, можно обратным путем восстановить для каждого
из перечисленных видов типичную кривую роста, подобную той,
какая приведена на рис. 1. Хорошо известно, что «стартовый вес»
(W=) новорожденного птенца, а также среднесуточный привес
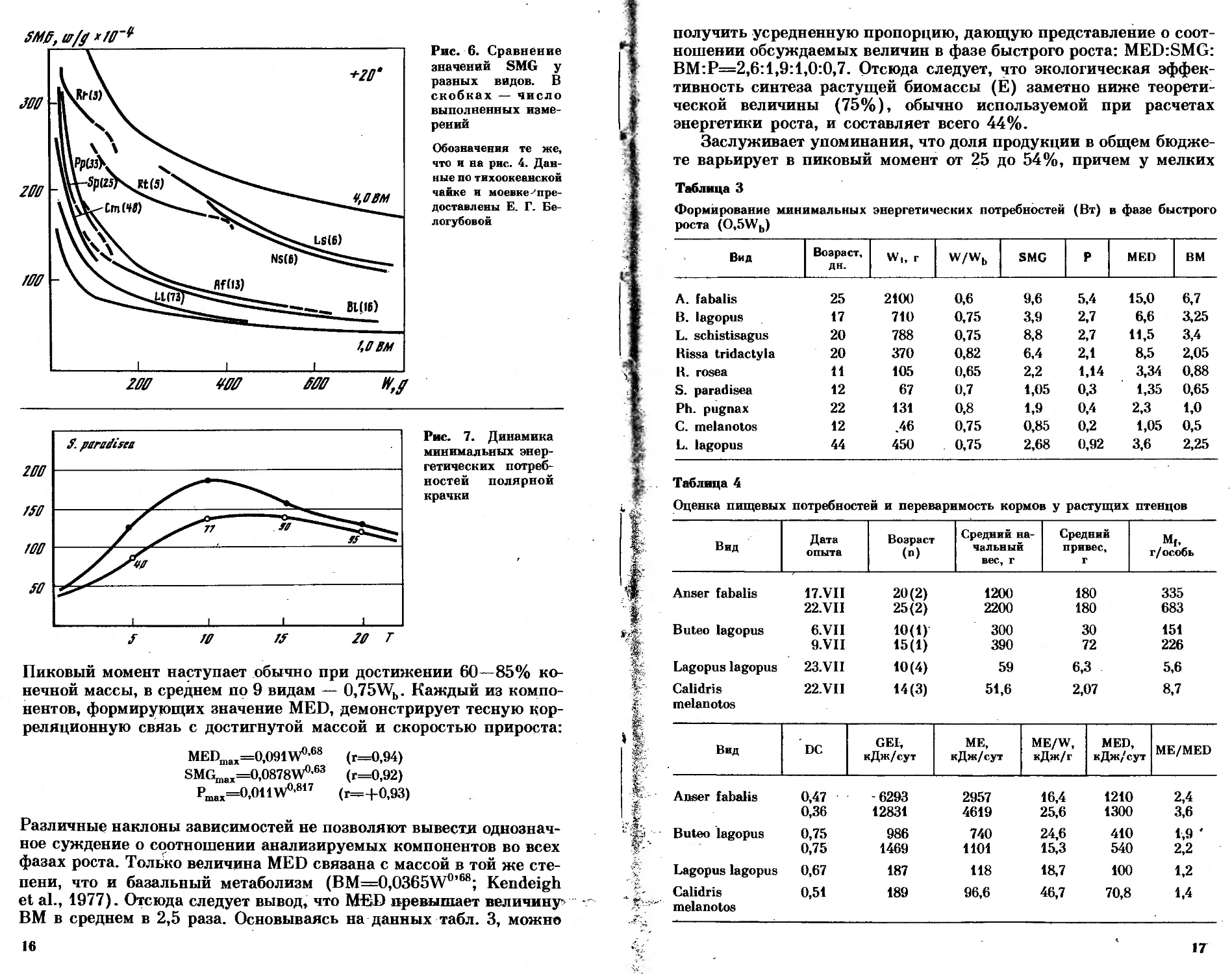
(d\V) аллометрически связаны с конечной массой тела. Эта связь
отчетливо проявляется и в приводимых на рис. 4 данных. Гумен-
ник, хищники и чайки образуют единую группу, в пределах кото-
рой обнаруживается сильная корреляция (г=-|-0,96) и возможны
предсказания. Судя по имеющимся данным, в условиях избыточ-
ного питания и температурного оптимума арктические гуси, хищ-
ники и чайки могут ускорять рост до возможных физиологических
пределов и растут быстрее птенцов родственных им видов, обитаю-
щих в умеренных и тропических широтах. Кулики растут медлен-
нее чаек, а белая куропатка — медленнее куликов. Надежных
данных о росте куликов в неволе и на свободе весьма мало, и для
сравнительного анализа ростовых параметров их пока недостаточ-
10
Рис. 4. Сравнение скоростей весо-
вого роста в разных группах птиц
La — серебристая чайка; Ls — ти-
хоокеанская чайка; В1 — зим ня к;
Fp — сапсан; Lc — сизая чайка;
Rt — моевка; Sp — короткохвостый
поморник; SI — длиннохвостый по-
морник; Xs — вилохвостая чайка
(Кондратьев, Кондратьева, 1984);
Rr — розовая чайка; Sp — полярная
крачка; Ст — дутыш; Рр — турух-
тан; Са — чернозобик (Soikeli, 1967);
Ch — галстучник (Pinowski, 1974);
Pl — круглоносый плавунчик; LI —
белая куропатка. Данные по моевке,
тихоокеанской и сизой чайкам пред-
ставлены А. С. К^гтиесклм и Л. А. Лу-
пач, данные по зимняку и сапсану —
Е. Р. Потаповым
но. Данных о росте тетеревиных птиц, напротив, достаточно мно-
го. Хотя белая куропатка растет заметно медленнее других тунд-
ровых птиц, в своей группе, т. е. по отношению к другим тетере-
виным, она обладает весьма быстрым ростом.
Таким образом, сравнительные данные говорят о том, что в пре-
делах таксономических групп арктические виды обладают более
высокими скоростями роста, что, по всей вероятности, объясняется
возможностью поедать пищу в течение почти круглых суток,
а также колебаниями внешней температуры, стимулирующими ап-
петит птенцов. Птицы, развивающиеся по птенцовому и полувывод-
ковому пути, растут быстрее выводковых видов. Из данных табл. 1
.путем регрессионного анализа для обитающих на севере хищников
и чаек получено следующее уравнение: dW=O,193Wb0,78
(г=+0,98; п=11).
Временные параметры роста, как уже отмечено, приблизитель-
но совпадают с важными этапами развития птенцов. При
0,25Wb у выводковых и полувыводковых видов стабилизируется
терморегуляция; при 0,5Wb интенсивно растет оперение, привесы
достигают максимальных значений; при 0,75 Wb энергетический
метаболизм и пищевые потребности особей достигают максимума.
Так же как и весовые параметры, сроки развития обнаруживают
тесную, связь с конечным весом тела. Соответствующие регрес-
сионные зависимости имеют степенное выражение:
г п
T^l.OSWj,0’29 0,94 13
Т2=2,88Wh°’235 0,88 13
T3=4,5Wb°’2n 0,81 13
Как видим, наиболее тесная связь существует между конечной
массой и периодом Ть На более поздних стадиях различия увели-
чиваются, о чем ясно свидетельствует направленное уменьшение
коэффициентов корреляции. Сравнительно небольшое возрастание
11
периодов роста (пропорциональное в среднем корню четвертой
степени из конечной массы) говорит о том, что крупные птицы как
бы обладают способностью сокращать период своего роста по срав-
нению с более мелкими видами.
Небольшой пример, рассчитанный из вышеприведенных урав-
нений, может дать представление о типичных скоростях роста двух
видов, взрослые особи которых имеют массу 100 и 1000 г.:
Wb т, т2 Тз
100 4 8 12
1000 8 15 20
Как видим, с увеличением конечной массы в 10 раз период ро-
ста удлиняется только вдвое. Поскольку относительный рост более
крупных птиц не такой быстрый, как у мелких, этот результат
можно истолковать в том смысле, что эффективность процессов
роста у крупных птиц более высокая, а кормовая ситуация ни-
чуть не менее напряженная, чем у мелких.
Сравнение данных о скорости роста птенцовых видов в природе
и лаборатории показывает, что выкармливаемые родителями птен-
цы чаек растут в общем так же быстро, как и птенцы в неволе или
даже быстрее. В отношении птенцов выводковых видов ситуация
иная. Здесь обнаружено два варианта. У белой куропатки часть
птенцов растет с максимальной скоростью, а часть — с отстава-
нием, которое становится заметным уже с первых дней жизни
и усиливается по мере роста. У дутыша в первом и начале второго
периода скорость роста птенцов в природе и в лаборатории одина-
ковы. Однако в дальнейшем намечаются и затем последовательно
усиливаются различия, так что дикие птицы отстают в росте и до-
стигают конечного веса заметно позднее лабораторных. Замедле-
ние темпов роста у части птенцов дутыша из выводков, выращен-
ных на огороженных участках тундры, отмечал Е. И. Хлебосолов
(1983). По его мнению, в этом обнаруживаются конкурентные
отношения между особями внутри выводка из-за недостатка пищи..
При этом некоторые птенцы сохраняли высокий (максимальный)
темп роста, в то время как у других он снижался; теряя иерар-
хический ранг, эти последние птенцы погибали. Эти результаты,
как и данные ростовой статистики, можно объяснить напряженной
кормовой ситуацией у части свободно живущих особей, которая
ускоренно выявляется в загородках. В целом же замедленный
рост выводковых птиц, по-видимому, связан с повышенным расхо-
дованием энергии на двигательную активность и терморегуляцию
в естественной среде.
Интенсивность метаболизма. Интенсивность дыхания, рас-
считанная на единицу массы (VO2 мл/мин-г), гиперболически
снижается по мере возрастания массы тела (рис. 5). Характерно
размещение экспериментальных точек на графиках: практически
во всех случаях они очерчиваются контуром, напоминающим
по форме бумеранг. Разброс значений параметра VO2 в первой
12
*М~*1ил/гмии
Рис. 5. Динамика интен-
сивности дыхания в зави-
симости от возраста у раз-
личных видов
и четвертой фазах роста минимальный; в промежуточных перио-
дах разброс точек велик и достигает 25—30% от среднего значе-
ния скользящей средней.
С моей точки зрения, такую картину можно объяснить суще-
ствованием у растущего организма двух чередующихся состояний.
Одно из них можно назвать «инертным», второе — «продуктив-
ным». В первом случае процессы роста в целом заторможены и не
выражаются в дыхании. «Продуктивное» состояние характери-
зуется наибольшей скоростью биосинтетических процессов, соот-
ветственно чему возрастает й темп потребления кислорода птен-
цом. В каждом отдельном опыте в силу его сравнительной крат-
ковременности фиксируется либо одно из крайних состояний, либо
переход между ними. Если изложенная точка зрения верна, то двум
предельным состояниям соответствуют гиперболические кривые,
13
образующие контур «бумеранга», в то время как точки, лежащие
в его плоскости, отражают переход от одного предела к другому.
Гиперболическая кривая, полученная на приведенных графиках
методом скользящей средней, соответствует (по определению)
величине SMG.
Сравнение величины SMG у разных видов (рис. 6) выявляет
несколько ценных для дальнейшего анализа моментов. Во-первых,
интенсивность дыхания у всех растущих видов выше рассчитанных
значений ВМ для неворобьиных птиц, но не выше 4,0 ВМ. Во-вто-
рых, наиболее высокие значения SMG найдены у быстро растущих
чаек, а наиболее низкие — у медленно растущей белой куропатки.
В-третьих, во всех случаях удельное значение SMG изменяется
гиперболически, и, следовательно, для каждого из видов можно
подобрать соответствующее аллометрическое уравнение. Однако
цифровые обобщения в данном случае не вполне уместны вследст-
вие таксономического разнообразия исследуемых видов и сущест-
венной разницы в уровнях SMG. В то же время найденные раз-
личия наводят на мысль о том, что их общей причиной следует
считать неодинаковость темпов роста птиц.
Для выяснения последнего были сопоставлены значения ве-
личины dW (см. табл. 1) и значений SMG в момент Достижения
птенцами половины конечной массы (см. рис. 6). Эта связь оказа-
лась достаточно сильной (г=-|-0,96), а наклон зависимости — до-
статочно крутым (SMGftsW0’87). Такой результат означает, что
при достижении половины конечной массы величина SMG у круп-
ных птиц будет относительно этой массы меньше, чем у мелких.
Заметим, что в установленных респирометрическим путем значе-
ниях SMG в той или иной мере присутствуют затраты на термо-
регуляцию, которые усиливают эти различия. У мелких видов они
больше, у крупных — меньше или вообще отсутствуют.
Оценка температурного коэффициента (теплопроводности)
растущих птенцов (Cj, Вт) затруднена вследствие присущей па-
раметру SMG изменчивости и широкому перекрыванию точек,
полученных при разных температурах. Приблизительные значения
величины Cj найдены путем сравнения двух скользящих средних,
установленных при двух различных температурах (см. рис. 4).
Одна из них — SMG20, другая — SMGio. Найденные таким путем
значения Cj можно сравнить с величиной теплопроводности взрос-
лых неворобьиных птиц сходных размеров, рассчитанных по обоб-
щающему уравнению C=0,00313W°’57 Вт/°С (Kendeigh et al.,
1977) (табл. 2). Как видно из приводимых данных, измеренные
значения Cj в 1,5—2 раза выше предсказываемых. Степень разли-
чий, как видим, довольно велика, так как растущее перо отличается
неразвитой структурой, а перьевые сосочки усиленно снабжаются
кровью и плохо сохраняют тепло. Из табл. 2 видно, что порядок
измеренных значений теплопроводности таков, что при понижении
температуры воздуха на 10—15° интенсивность метаболизма
увеличивается в 1,5—2 раза по сравнению с той, какая устанавли-
вается при 20°. Согласно модели роста Берталанфи (Bertalanffi,
14
Таблица 2
Терморегуляторный ответ растущих птиц
Вид Масса тела, г SMGio, Вт SMG2o, Вт СЙЗМ, Вт/°С Стеор • ВтА’.
L. lagopus 50 1,2 0,8 0,04 0,03 100 1,8 1,2 0,06 0,04 200 2,5 1,7 0,08 0,065 400 2,85 1,9 0,095 0,095 Rh. rosea 50 2,4 1,6 0,08 0,03 100 3,65 2,65 0,10 0,04 150 3,15 2,3 0,085 0,05 Ph. pugnax 50 1,7 1,1 0,06 0,03 100 2,4 1,6 0,08 0,04 150 3,2 2,05 0,13 0,05 C. melanotos 25 1,0 0,7 0,03 0,02 50 1,5 0,85 0,065 0,03 Примечание. Цифры приведены с округлением. Исходные данные сняты с графиков (рис. 6). Значения Стеор рассчитаны по уравнению: у= =0,0031W°’5734 Вт/°С
1957), это, в свою очередь, должно уменьшать скорость роста
птенцов. Однако М. Клейбер и Дж. Э. Догерти (1933) наблюдали
увеличение скорости роста цыплят при понижении температуры
от 30 до 20°, что происходило за счет усиления аппетита и большего
поедания пищи. Такой же результат получил Д. В. Нортон (Norton,
1973), выращивавший песочников (С. alpina и С. bairdii) при раз-
ных температурах. Исследованные нами виды растут в природе
при средней температуре воздуха около 12°. Как было показано
выше, во второй половине периода роста они отстают от птиц,
выращиваемых в лаборатории. Этот факт, как мне кажется, отчет-
ливо свидетельствует о следующем.
1) Скорость роста и терморегуляция находятся у тундровых
видов в конкурентных отношениях в полном соответствии с мо-
делью роста Берталанффи.
2) Недостаток пищи в природе тормозит рост птенцов и растяги-
вает его длительность.
3) В структуре энергетических расходов приоритет отдается
терморегуляции.
В какой мере терморегуляторные расходы влияют на количест-
во съедаемого арктическими птенцами корма и как при этом из-
меняется темп их роста — вопрос, открытый для исследования.
Динамика параметров энергетики в ходе роста. Используя
данные о скорости роста (см. рис. 3, табл. 1) и данные об изме-
нениях интенсивности метаболизма (см. рис. 6), получим кривую
изменения величины SMG в ходе роста. Добавляя к ней значения
продукции (Р), переходим к величине минимальных энергетиче-
ских потребностей (MED). Пример такого построения вычислен
для полярной крачки (рис. 7). Данные по другим видам сведены
в табл. 3. Как видно из графиков, величина MED нарастает соответ-
ственно весовому росту птенца, достигая в некоторый момент (Ттах)
наибольших значений (MEDmax). Затем величина MED снижается.
15
Рис. 6. Сравнение
значений SMG у
разных видов. В
скобках — число
выполненных изме-
рении
Обозначения те же,
что и на рис. 4. Дан-
ные по тихоокеанской
чайке и моевке ^пре-
доставлены Е. Г. Бе-
логубовой
zoo
zoo
ZOO
Рис. 7. Динамика
минимальных энер-
гетических потреб-
ностей полярной
крачки
Пиковый момент наступает обычно при достижении 60—85% ко-
нечной массы, в среднем по 9 видам — 0,75Wb. Каждый из компо-
нентов, формирующих значение MED, демонстрирует тесную кор-
реляционную связь с достигнутой массой и скоростью прироста:
MEDmax=0,091W°’68 (г=0,94)
SMGmax=0,0878W°’63 (г=0,92)
Pmax=0.011W°-817 (г=+0,93)
Различные наклоны зависимостей не позволяют вывести однознач-
ное суждение о соотношении анализируемых компонентов во всех
фазах роста. Только величина MED связана с массой в той же сте-
пени, что и базальный метаболизм (BM=0,0365W°’68; Kendeigh
et al., 1977). Отсюда следует вывод, что MED превышает величину-
ВМ в среднем в 2,5 раза. Основываясь на данных табл. 3, можно
16
получить усредненную пропорцию, дающую представление о соот-
ношении обсуждаемых величин в фазе быстрого роста: MED:SMG:
ВМ:Р=2,6:1,9:1,0:0,7. Отсюда следует, что экологическая эффек-
тивность синтеза растущей биомассы (Ё) заметно ниже теорети-
ческой величины (75%), обычно используемой при расчетах
энергетики роста, и составляет всего 44%.
Заслуживает упоминания, что доля продукции в общем бюдже-
те варьирует в пиковый момент от 25 до 54%, причем у мелких
Таблица 3
Формирование минимальных энергетических потребностей (Вт) в фазе быстрого
роста (O,5Wb)
Вид Возраст, дн. W,, г w/wb SMG P MED BM
A. fabalis 25 2100 0,6 9,6 5,4 15,0 6,7
В. lagopus 17 710 0,75 3,9 2,7 6,6 3,25
L. schistisagus 20 788 0,75 8,8 2,7 11,5 3,4
Bissa tridactyla 20 370 0,82 6,4 2,1 8,5 2,05
R. rosea 11 105 0,65 2,2 1,14 3,34 0,88
S. paradisea 12 67 0,7 1,05 0,3 1,35 0,65
Ph. pugnax 22 131 0,8 1,9 0,4 2,3 1,0
C. melanotos 12 .46 0,75 0,85 0,2 1,05 0,5
L. lagopus 44 450 0,75 2,68 0,92 3,6 2,25
Таблица 4
Оценка пищевых потребностей и переваримость кормов у растущих птенцов
Вид Дата опыта Возраст (n) Средний на- чальный вес, г Средний привес, г Mf, г/особь
Anser fabalis 17.VII 20(2) 1200 180 335 22. VII 25(2) 2200 180 683 Buteo lagopus 6.VII 10(1) 300 30 151 9. VII 15(1) 390 72 226 Lagopus lagopus 23.VII 10(4) 59 6,3 5,6 Calidris 22. VII 14(3) 51,6 2,07 8,7 melanotos
Вид DC GEL, кДж/сут ME, кДж/сут ME/W, кДж/г MED, кДж/сут ME/MED
Anser fabalis 0,47 - 6293 2957 16,4 1210 2,4 0,36 12831 4619 25,6 1300 3,6 Buteo lagopus 0,75 986 740 24,6 410 1,9 ' 0,75 1469 1101 15,3 540 2,2 Lagopus lagopus 0,67 187 118 18,7 100 1,2 Calidris 0,51 189 96,6 46,7 70,8 1,4 melanotos
17
чаек и куликов, растущих с наибольшей скоростью, она как раз
самая низкая. У более крупных видов соотношение обратное. Это,
по-видимому, опять-таки говорит о более высокой эффективности
биосинтеза у крупных видов и существовании у них резервов мощ-
ности, которые могут быть использованы на комфортные актив-
ности.
Определение суточных расходов энергии. Изменение величины
MED с возрастом определяет и динамику минимальных пищевых
потребностей (Mfmin). Однако реальное потребление корма (М()
должно обеспечивать весь спектр энергетических расходов расту-
щего птенца (DEE), куда, помимо стандартного метаболизма ро-
ста и продукции, входит еще терморегуляция (С) и двигательная
активность (А). Определение величины DEE возможно двумя спо-
собами: либо путем добавления к величине MED рассчитанных
надбавок С и А, либо путем измерения валовой (GEI) и метабо-
лизированной энергии (ME) в естественных или близких к ним
условиях.
В табл. 4 представлены результаты измерений, выполненных по
второму способу. Данные о переваримости нищи имеют вспомога-
тельное значение и дополняют уже опубликованные сведения на
этот счет (Дольник и др., 1982). Несмотря на фрагментарность дан-
ных табл. 4, можно видеть, что величина ME в 1,2—3,3 раза выше
MED. У выводковых видов различия увеличиваются с возрастом
(гуменник) и наиболее велики в пиковый момент роста (0,75Wb).
У птенцовых (сапсан, канюк) и полувыводковых (чайки) соотно-
10
20 30 ta
Рис. 8. Модель динамики параметров энергетики в ходе роста
18
шение ME/MED не столь велико (1,2—1,4), что хорошо соот-
ветствует их крупным размерам и малоподвижному образу
жизни.
Затраты метаболизированной энергии на продукцию 1 г новой
массы варьируют в ходе быстрого роста в пределах от 15
до 20 кдж/г, в среднем 16,6 кдж/г. Более высокие цифры относят-
ся либо к началу, либо к концу периода быстрого роста. Поскольку
плотность энергии во вновь создаваемой массе приблизительно
равна 6,3 кдж/г, экологическая эффективность трансформации
энергии пищи в рост (Р/МЕ) оказывается в среднем равной
38%.
Таким образом, экологически значимую величину — общие
и суточные пищевые потребности растущего птенца — можно удов-
летворительно предсказать, основываясь на данных о скорости
роста и соотношениях, выведенных в ходе вышеизложенного ана-
лиза (рис. 8). При этом значения величины SMG в точках дости-
жения четверти, половины и трех четвертей конечной массы уста-
навливаются по уравнениям SMG=f (w—), продукция — по усред-
ненным за 3—5 дней данным о величине dw, а метаболизирован-
ная энергия может принимать следующие ориентировочные значе-
ния (в долях MED, см. табл. 4):
0,25 0,5 0,75
Водоплавающи е 1,5 2,5 3,5
Хищники 1,1 1,2 1,2
Чайки 1,1 1,4 1,4
Кулики 1,5 2 2,5
Тетеревиные 1,5 2 3
Заключение
Выполненный анализ показывает, что параметры экологической
энергетики растущих птиц определяются в поле по меньшей мере
четырех аргументов: возраста, скорости роста, температуры среды,
концентрации, распределения и наличия пищи в окружающей
среде.
Соотношение и динамика параметров, вытекающие из нашего
анализа, схематически изображены на рис. 6, который развивает
и уточняет двухмерную схему энергетики роста Клейбера—
Догерти, а также терморегуляционную модель Шоландера (Scho-
lander et al., 1950).
Как видно из объемной модели, при температурах ниже кри-
тических рост и терморегуляция находятся в конкурентных отно-
шениях, причем наиболее острый период приходится на момент
достижения 0,5—0,75Wb. Поддержанию скорости роста мы должны
отводить приоритетную роль, рассматривая ее в качестве важней-
шей эволюционной доминанты. Однако в условиях низких темпе-
ратур воздуха потребности, связанные с поддержанием гомеостаза,
также оказываются необходимым условием выживания. Величина
19
MED формируется в плоскости свода, образующего третий «этаж»
модели. В пространстве, разделяющем поверхности SMG и MED,
привес и терморегуляция находятся в конкурентных отношениях.
Поскольку с течением времени среднее значение ta возрастает,
направление изменения внешних условий благоприятствует росту.
В пространстве между третьим (MED) и четвертым (ME) «эта-
жами» в действие включаются другие экологические факторы,
такие, например, как необходимое время и интенсивность кормовой
и прочей двигательной активности.
«Крыша» DEA демонстрирует уровень доступной энергии, т. е.
то ее количество, которое особь может получить (разыскать, добыть
и поглотить) за период кормовой активности. Изменчивые очерта-
ния поверхности DEA отражают неравномерный характер разме-
щения и доступности ресурсов. Она формируется в сфере взаимо-
действия видовых адаптивных свойств и особенностей среды, т. е.
как бы символизирует экологическую нишу в количественном
аспекте.
Высоту пространства, разделяющего поверхности DEA и ME,
можно рассматривать и как запас устойчивости, и как меру адапти-
рованности. Чем шире это пространство, тем выше вероятность
индивидуального выживания.
Первые четыре «этажа» модели связаны предсказуемыми коли-
чественными соотношениями. Закономерности формирования
последнего этажа пока не выяснены. Эта задача хотя и более
сложная, но достойная углубленных исследований.
ЛИТЕРАТУРА
Андреев А. В. Энергетика особи и видовые адаптивные признаки // Адаптация
организмов к условиям Крайнего Севера, Таллин: Изд-во АН ЭССР, 1984.
С. 15—20.
Андреев А. В. Инструментальные методы в исследованиях по экологической
энергетике птиц // Экспериментальные методы в изучении северных птиц
и результаты их применения. Владивосток: ДВНЦ АН СССР, 1986. С. 59—
83.
Дольник В. Р., Дольник Т. В., Постников С. В. Калорийность и усвояемость
объектов питания птиц // Бюджеты времени и энергии у птиц в природе.
Л.: Наука, 1982. С. 143—153. (Тр. Зоол. ин-та АН СССР; Т. ИЗ).
Кондратьев А. Я., Кондратьева Л. Ф. Рост и развитие птенцов вилохвостой чай-
ки // Орнитология. М.: Изд-во МГУ, 1984. Вып. 19. С. 81—88.
Мина М. В., Клевезаль Г. А. Рост животных. М.: Наука, 1976. 291 с.
Потапов Е. Р. Экспериментальное изучение и экологическая интерпретация пере-
варимости корма у птенцов зимняка // Экспериментальные методы в изучении
северных птиц и результаты их применения. Владивосток: ДВНЦ АН СССР,
1986. С. 114-119.
Хлебосолов Е. И. О трофической избирательности некоторых тундровых кули-
ков //Зоол. журн. 1983. Т. 62, № 6. С. 963—965.
Хлебосолов Е. И. Видовое разнообразие и адаптивная стратегия в жизни птиц
тундры (на примере куликов) // Адаптация организмов к условиям Крайнего
Севера. Таллинн: Изд-во АН ЭССР, 1984. С. 181 — 183.
Хлебосолов Е. И. Огораживание территории как метод исследования биологии
тундровых куликов в постэмбриональный период // Экспериментальные мето-
ды в изучении северных птиц и результаты их применения. Владивосток,
1986. С. 108-113.
20
Andersson M. Population ecology of long tailed skua (Stercorarius longicaudus
Vteill.) 11 Journ. Aniin. Ecol. 1976. Vol. 45. N 2. P. 537-559.
Bertalanjfi L. Quantitative laws in metabolism and growth // The Quaterly Review
of Biology. 1957. Vol. 32, N 3. P. 217-231.
Kendeigh S. Ch., Dolntk V. R., Gavrilov V. M. Avian energetics // Granivorous
birds in ecosystems. Cambridge: Univ, press, 1977. P. 127—204.
Kleiber M., Dougherty J. E. The influence of environmental temperature on the
utilisation of food energy in baby chicks //J. Gen. Physiology. 1933. Vol. 17.
P. 701-726.
Norton D. W. Ecological energetics of Calidrine Sandpipers breeding in Northern
Alaska: Ph. D. Thesis. Fairbanks, 1973. 163 p.
Ricklefs R. E. Patterns of growth in birds // Ibis. 1968. Vol. 110. P. 419—451.
Ricklefs R. E. Energetics of reproduction in birds // Anian Energetics / R. A. Paynter
(ed.). 1974. P. 152-297.
Scholander P. F., Hock R., Walters V. et others. Heat regulation in some arctic
and tropical mammals and birds//Biol. Bull. 1950. Vol. 99. P. 237—258.
SUMMARY
Ecological Energetics of Arctic nestlings. AndreievA. V.
Various aspects of the growth of 8 Arctic birds species were studied in Lower Kolyma
tundras in 1978—1986: Bean Goose, Rough-legged Hawk, Willow Grouse, Herring
Gull, Ross’s Gull, Arctic Tern, Ruff, Pectoral Sandpiper. One—three day nestlings
were bred in the artificial conditions at t°=15—22° C. The data analysis showed that
the ecological energetic parameters of growing nestlings can be determined in the
field, at least, by 4 arguments: age, growth rate, environmental temperature, concentra-
tion, distribution and food supply in the surroundings. The volumetric model which,
develops and verifies 2—D Kleiberg and Dogerti (1933) scheme of growth energetics
and also Sholander’s (1955) thermoregulation model, was constructed.
УДК 591.543.43(470.36)
Л. В. СОКОЛОВ,
В. П. ДЬЯЧЕНКО,
Т. А. ИЛЬИНА,
Д. С. ЛЮЛЕЕВА,
М. Л. ЯБЛОНКЕВИЧ
199164, Ленинград, Универси-
тетская набережная, д. 1. Зоо-
логический институт АН СССР
Проверка возможности
образования зимовки
у молодых зябликов,
задержанных
на Куршской косе
Широко распространено представление о том, что привязанность
перелетных птиц к постоянным зимовкам выражена значительно
слабее, чем к местам гнездования. Считается, что строгая привя-
занность птиц к одним и тем же местам зимовки становится не
адаптивной в условиях резких колебаний кормового и погодных
факторов (Михеев, 1962; Паевский, 1976, 1985; Jenni, 1982). Это
подтверждается данными о значительных колебаниях зимнего
ареала у ряда видов в зависимости от суровости зим и урожая
растений, обеспечивающих их кормом, а также находками околь-
цованных особей в разных местах ареала в течение одного или
21
нескольких сезонов (Finlayson, 1980; Jenni, 1982; Troy, 1983;
Yunick, 1983). Отсутствие постоянства зимовки обычно характерно
для так называемых кочующих видов, питающихся семенами и
плодами различных растений (Дольник, 1975; Finlayson, 1980;
Носков, 1981; Паевский, 1985). Однако у этих видов непостоян-
ство мест зимовки может быть относительным. Например, у юрка
(Fringilla montifringilla), вида, известного своими широкими
кочевками в пределах зимнего ареала (в связи со спецификой
питания буковыми орешками), и у которого одни и те же особи
могут зимовать в течение одной и нескольких зим в разных частях
ареала, была также обнаружена четкая связь птиц с определен-
ными местами зимовки (Haar, 1976; Schenker, 1976; Jenni, 1982).
Кольцевание и отлов юрков на одном и том же месте (в ФРГ) в те-
чение нескольких зимних сезонов показали, что птицы на сле-
дующий год после кольцевания могут зазимовать в другой части
ареала, а через год или более вновь вернуться на место кольцева-
ния (Schenker, 1976).
В противоположность этому у дальних мигрантов обнаружи-
вается строгая привязанность к постоянным местам зимовки. С по-
мощью кольцевания доказано, что значительная часть птиц у мно-
гих перелетных видов возвращается в прежний район зимовки,
при этом они могут занимать прежние индивидуальные участки
и проявлять типичное поведение, связанное с охраной террито-
рии (Moreau, 1969; Brosset, 1971; Zahavi, 1974; Pearson 1972, 1981;
Mewaldt, 1976; Price, 1981; O. Johnson, P. Johnson, 1983). Такие ви
ды показывают хорошую способность к зимнему хомингу при заво-
зах как на близкие, так и на дальние расстояния (Schwartz, 1963;
Mewaldt, 1964; Ralph, Mewaldt, 1976; Benvenuti, loale, 1980). Вер-
ность прежнему месту зимовки дает особи ряд преимуществ, свя-
занных со знанием территории, повышает ее конкурентоспособ-
ность, а соответственно и выживаемость (Brosset, 1971; Ralph,
Mewaldt, 1975; Fuller, Youngman, 1979).
Относительно времени формирования связи птиц с местом зи-
мовки высказывались разные предположения. Э. Зауэр (Sauer,
1963), а затем С. Фретвел (Fretwell, 1969) считали, что связь
с зимовочной территорией может устанавливаться на протяже-
нии почти всей зимы. П. Шварц (Schwartz, 1963) и Л. Мэдуей
(Medway, 1973) предполагали, что это происходит непосредственно
перед началом весенней миграции. Однако в работе американских
исследователей Дж. Ральфа и Р. Мевальдта (Ralph, Mewaldt, 1975)
было убедительно показано с помощью перевозок молодых и взрос-
лых белогорлых (Zonotrichia leucophrys) и пестроголовых (Z. atri-
capilla) зонотрихий в районе их основной зимовки (штат Калифор-
ния) , что образование прочной связи с конкретным местом зимовки
происходит в ограниченный период непосредственно после оконча-
ния первой осенней мйграции (до 1 января). Позже было установ-
лено, что у пальмовой древесницы Dendroica palmarum связь
с территорией зимовки у молодых птиц образуется уже к началу
декабря (Stewart, Connor, 1980).
22
Тот факт, что конкретные места зимовки у перелетных птиц не
являются генетически фиксированными, а запечатлеваются ими
в определенный период жизни после попадания в них в процессе
первой осенней миграции, дает возможность проводить экспери-
менты с завозами птиц в новые районы зимовки. Такие опыты,
проведенные на зонотрихиях (Mewaldt, 1964; Ralph, Mewaldt,
1975), показали, что птицы возвращаются в новое место зимовки
(даже если оно находится в нескольких сотнях километров от тра-
диционного района зимовки) только в том случае, если они были
перевезены молодыми, до начала периода становления терри-
ториальных связей. Взрослых птиц в этих экспериментах не
удалось осадить в новых районах зимовки более чем на один
сезон.
Однако все эти эксперименты проводились с птицами, которые
уже прибыли в район зимовки, а не были задержаны на трассе
осенней миграции. Опыты по созданию мест зимовки у каких-либо
видов непосредственно на трассе их миграции, насколько нам
известно, не проводились. В связи с этим мы решили провести
такой эксперимент на зяблике (Fringilla coelebs), который в боль-
шом числе мигрирует осенью через Куршскую косу Балтийского
моря.
Зяблик является внутриконтинентальнЫм мигрантом, зи-
мующим преимущественно в юго-западной части Франции и на
Пиренейском полуострове (Шумаков, Соколов, 1982). Климат
в этих районах переходный от умеренного к средиземноморскому,
зима чаще мягкая (средние температуры января — 5—12 °) с боль-
шим количеством осадков. Снег не образует устойчивого покрова,
реки обычно не замерзают. В горных областях зима более суровая,
с морозами и сильными снегопадами (Люлеева, 1982). Основным
кормом для зяблика являются семена дикорастущих и культур-
ных растений (Дольник, 1982), поэтому для него важно наличие
свободных от снега участков для успешного добывания корма. По
данным П. Марлера (Marler, 1956) и И. Ньютона (Newton, 1972),
континентальные зяблики, зимующие в Британии, кормятся круп
ними стаями на вспаханных полях, открытых участках, на скло-
нах холмов, опушках леса и т. п. Достоверных сведений о постоян-
стве мест зимовки у зяблика мало. Только Дж. Финлайсон (Fin-
layson, 1980), кольцующий зимующих птиц в районе Гибралтара
(Испания), обнаружил на следующий год одного окольцованного
им зяблика (из шести помеченных особей) в том же месте. Правда,
имеются косвенные данные, полученные А. Пердеком (Perdeck,
1958) в экспериментах по смещению пролетных молодых и взрос-
лых зябликов с миграционной трассы осенью. Они свидетель-
ствуют о том, что взрослые птицы в отличие от молодых обладают
знанием района зимовки, поскольку вносят поправку на смещение,
проявляя способность к навигации. В пользу этого говорят и экспе-
рименты с задержанными осенью на Куршской косе зябликами,
которые показали, что у взрослых птиц в отличие от молодых
наблюдается пролонгация миграционного состояния, вероятно,
23
из-за отсутствия необходимой для завершения миграционного
полета информации о достижении ими знакомого района зимовки
(Дольник, Паевский, 1965; Шумаков, Виноградов, 1971; Дольник,
1975; Соколов, 1976, 1978). О прекрасных навигационных способ-
ностях зябликов свидетельствуют и данные по возвращению их
в узколокальный район рождения и гнездования (Соколов, 1981;
1982; Соколов и др., 1984).
Основная цель данного исследования состояла в выяснении
возможности образования зимовки у молодых зябликов, задер-
жанных в разгар осенней миграции на Куршской косе, находя-
щейся примерно на расстоянии 1500 км от традиционных районов
их зимовки. Предполагалось выяснить следующие вопросы:
1) останутся ли молодые зяблики зимовать в районе выпуска;
2) вернутся ли они в этот район на следующий год; 3) в какой
именно период у зябликов устанавливается связь с зимовочной
территорией.
Материал и методика
Птицы для эксперимента были пойманы большими ловушками на
территории полевого стационара биостанции «Фрингилла» в пе-
риод с 20 сентября по 3 октября 1979 г., когда через Куршскую
косу пролетают преимущественно зяблики из Финляндии (Паев-
ский, 1971; Шумаков, Соколов, 1982). Было взято 470 молодых
самцов. Их перевезли в пос. Рыбачий, расположенный в 12 км се-
веро-восточнее стационара, и посадили в две большие вольеры —
комнатную, находящуюся в помещении станции, и уличную, распо-
ложенную снаружи. В комнатную вольеру поместили 270 особей,
в уличную — 200. В обоих случаях фотопериод был естественным
для района исследования. Температура в уличной вольере не отли
чалась от температуры внешнего воздуха, в комнатной вольере
она была в среднем на 11° выше. Птицы содержались на обычной
зерновой смеси (подсолнечник, конопля, просо) и мягком корме
(куриное яйцо с морковью). Систематически давался пророщен-
ный овес.
Выпуск птиц на свободу производился из уличной вольеры
партиями по 40—60 особей при хороших погодных условиях
(отсутствии осадков, сильного ветра) примерно один раз в две
недели начиная с 1 ноября (см. таблицу). После выпуска в осво-
божденную уличную вольеру переводились птицы из комнатной
вольеры, чтобы они могли адаптироваться к наружной темпера
туре.
Постепенный выпуск птиц на протяжении всей зимы был
запланирован нами с целью выяснения сроков формирования у них
связи с местом зимовки. В случае, если бы птицы вернулись в район
выпуска на следующий год, можно было бы определить, в какой
именно период им важнее всего быть на свободе, чтобы снять
координаты зимовки.
24
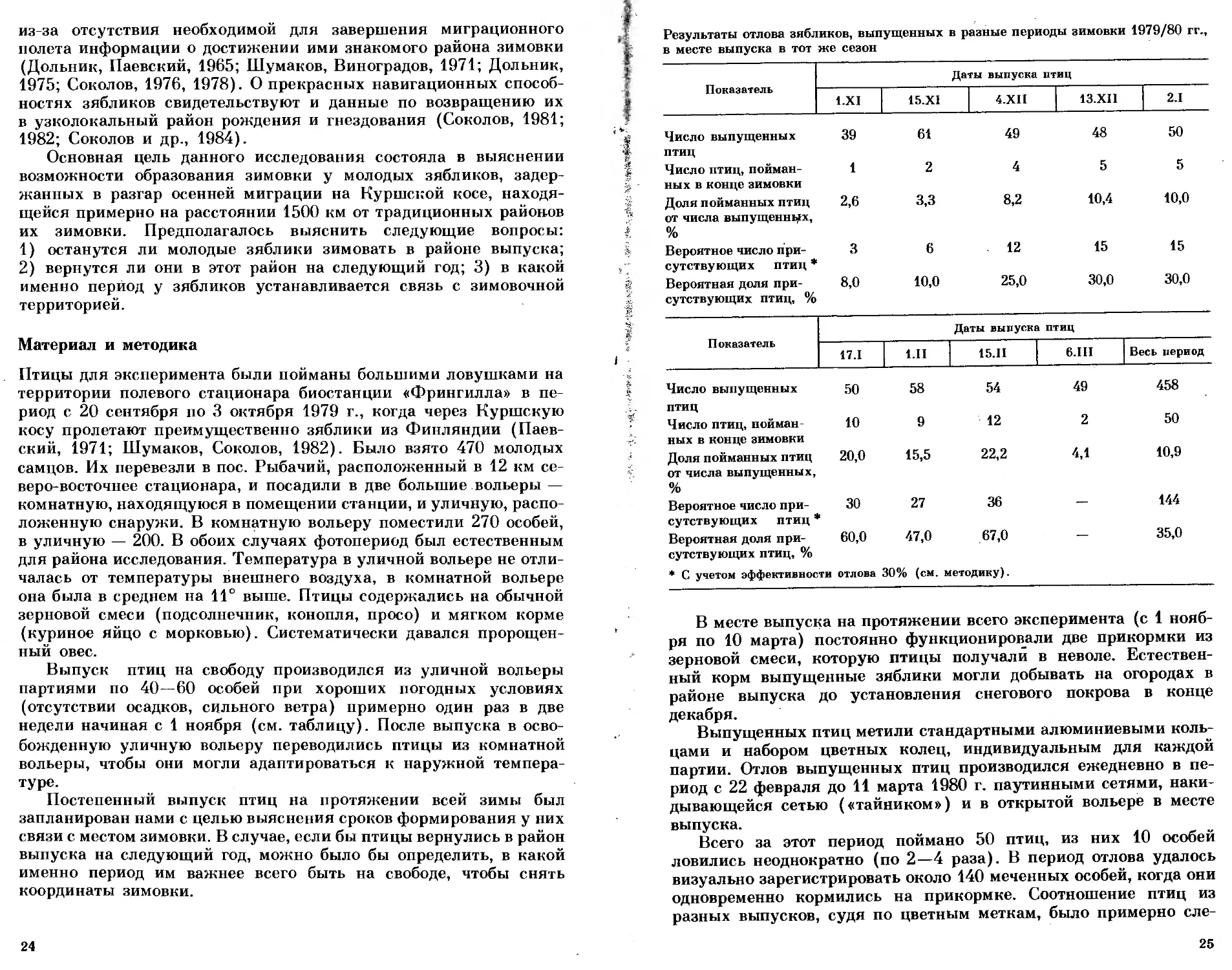
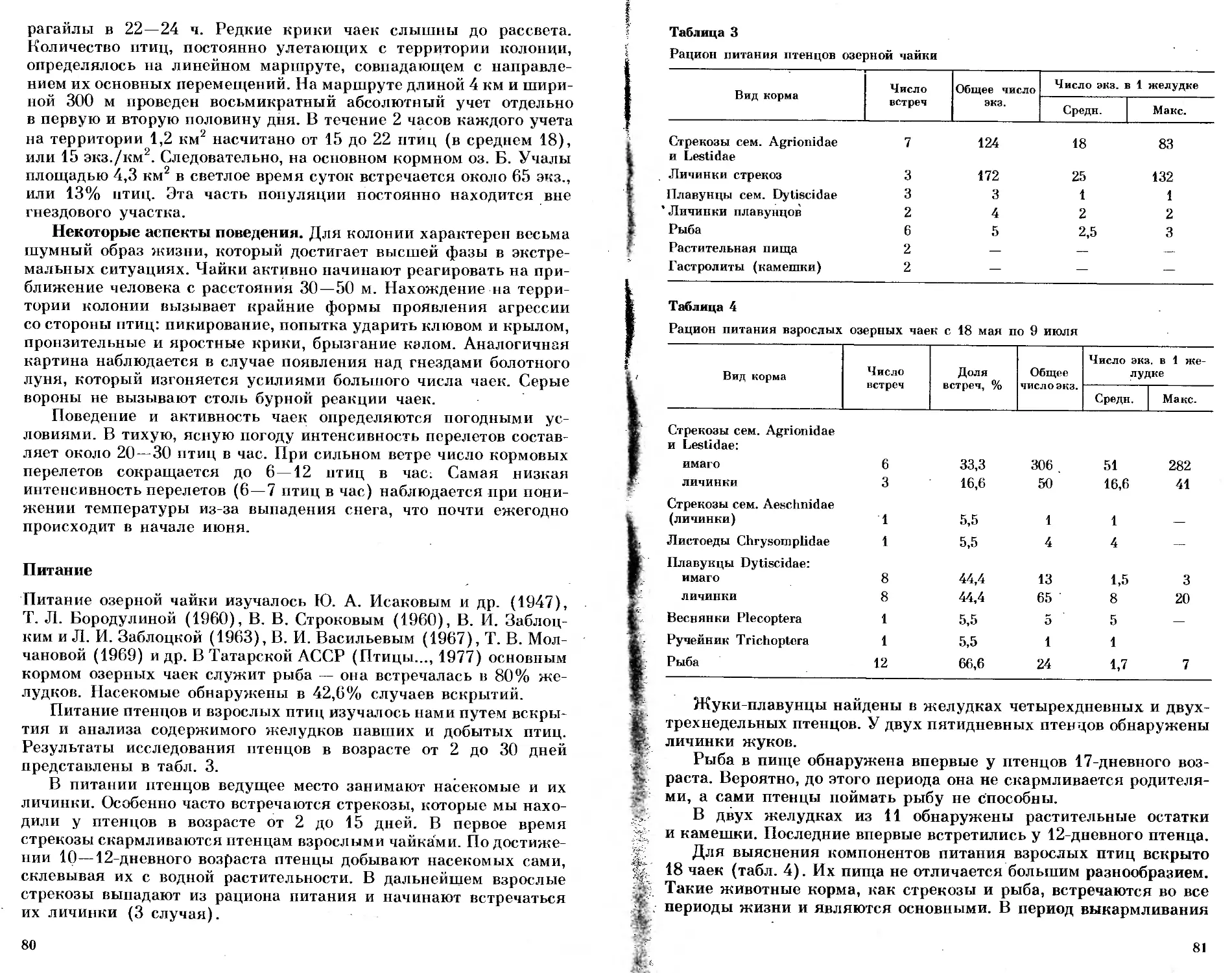
Результаты отлова зябликов, выпущенных в разные периоды зимовки 1979/80 гг.,
в месте выпуска в тот же сезон
Показатель Даты выпуска птиц
1.XI 15.XI 4.XII 13.XII 2.1
Число выпущенных 39 61 49 48 50
птиц
Число птиц, пойман- ных в конце зимовки 1 2 4 5 5
Доля пойманных птиц от числа выпущенных, % 2,6 3,3 8,2 10,4 10,0
Вероятное число при- сутствующих птиц * 3 6 12 15 15
Вероятная доля при- сутствующих птиц, % 8,0 10,0 25,0 30,0 30,0
Показатель Даты выпуска птиц
17.1 1.П 15.11 6.П1 Весь период
Число выпущенных 50 58 54 49 458
ПТИЦ Число птиц, пойман- 10 9 12 2 50
ных в конце зимовки Доля пойманных птиц 20,0 15,5 22,2 4,1 10,9
от числа выпущенных, % Вероятное число при- 30 27 36 144
сутствующих птиц * Вероятная доля при- 60,0 47,0 67,0 — 35,0
сутствующих птиц, %
* С учетом эффективности отлова 30% (см. методику).
В месте выпуска на протяжении всего эксперимента (с 1 нояб-
ря по 10 марта) постоянно функционировали две прикормки из
зерновой смеси, которую птицы получали в неволе. Естествен-
ный корм выпущенные зяблики могли добывать на огородах в
районе выпуска до установления снегового покрова в конце
декабря.
Выпущенных птиц метили стандартными алюминиевыми коль-
цами и набором цветных колец, индивидуальным для каждой
партии. Отлов выпущенных птиц производился ежедневно в пе-
риод с 22 февраля до 11 марта 1980 г. паутинными сетями, наки-
дывающейся сетью («тайником») и в открытой вольере в месте
выпуска.
Всего за этот период поймано 50 птиц, из них 10 особей
ловились неоднократно (по 2—4 раза). В период отлова удалось
визуально зарегистрировать около 140 меченных особей, когда они
одновременно кормились на прикормке. Соотношение птиц из
разных выпусков, судя по цветным меткам, было примерно сле-
25
дующим: 10 особей из ноябрьского выпуска, 30 — из декабрьского,
40 — из январского, 60 — из февральского. Поймано птиц с этими
метками было соответственно 3:9:15:21 особь (см. таблицу).
Следовательно, эффективность отлова составляла примерно 30%
(48 из 140 особей, присутствующих во время отлова).
С целью обнаружения вернувшихся в район выпуска зябликов
с 1 по 15 ноября 1980 г. и с 3 по 15 января 1981 г. проводились
визуальные наблюдения и контрольный отлов птиц на прикормке,
организованной на прежнем месте. Прикормка функционировала
постоянно с 1 ноября по 15 января. Визуальные наблюдения
проводились ежедневно (по 2—3 ч) непосредственно в месте вы-
пуска и в окрестностях пос. Рыбачий. Отлов птиц на прикормке
производился ежедневно с10до17чс помощью паутинных сетей
и «тайником».
На протяжении всего эксперимента у птиц, содержавшихся
в комнатной и уличной вольерах, производилось измерение массы
тела (в утренние часы до начала кормовой активности) и опреде-
лялся уровень жировых резервов по визуальной методике
Т. И. Блюменталь и В. Р. Дольника (1962). Эти показатели регист-
рировались также при отловах у выпущенных птиц. Параллельно
регистрировались наружная температура воздуха и температура
в комнатной вольере, а также погодные условия (осадки, направ-
ление и сила ветра).
Оценка разности между долями повторных поимок птиц из
разных выпусков производилась по критерию у 2. Проведен кор-
реляционный и регрессионный анализ по Г. Ф. Лакину (1973)
зависимости величины массы тела птиц от температуры внеш-
него воздуха.
Результаты
Анализ отловов в конце февраля и начале марта показал, что
наименьшая доля поимок (3,0%) приходится на птиц, выпущен-
ных на свободу в ноябре (см. таблицу). Она достоверно ниже,
чем доля пойманных птиц из выпущенных в январе — 15% (% 2=
=7,4 для Р=0,01). Доля птиц декабрьского выпуска (9,3%) ниже,
чем доля птиц январского выпуска (15,0%), однако разница между
этими долями недостоверна (/ 2=1,0 для Р=0,05). Доля птиц,
выпущенных-в феврале, составила 18,8%. Незначительную долю
(4,1%) среди отловов составили птицы из последнего выпуска
6 марта.
Если рассматривать вероятное число присутствующих птиц
во время отлова, которое примерно в три раза больше числа пой-
манных (см. методику контроля), то получается, что в месте
отлова в конце февраля находилось около 10% зябликов, выпу-
щенных в ноябре, около 30% — выпущенных в декабре, около
40% — выпущенных в январе и около 60% — выпущенных в
феврале.
26
Средняя продолжительность пребывания выпущенных в раз-
ные сроки зябликов в районе выпуска составила:
срок выпуска продолжительность кол-во особей
1 ноября 114 сут 1
15 ноября 104 сут 2
4 декабря ' 86 сут 4
13 декабря 77 сут 5
2 января 54 сут 5
17 января 41 сут 10
1 февраля 28 сут 9
15! февраля 12 сут 12
6 марта 4 сут 2
В период отлова наибольшее число особей (46 из 50) было пой-
мано с 23 февраля по 2 марта. Позже удалось поймать только че-
тыре особи, поскольку резко снизилось число птиц, державшихся
в месте прикормки.
Обычно при плохих погодных условиях (низкой температуре,
снегопаде, сильном ветре) зяблики практически в течение всей
светлой части суток держались непосредственно возле прикормки.
Ночевали они обычно за пределами кормовой территории. Во
время оттепели птицы, как правило, рассредотачивались на боль-
шей территории. В целом зима 1979/80 гг. на Куршской косе была
довольно теплой: средняя температура декабря составляла
+2,2±1,4О, средняя температура января —4,6±0,6° и средняя
температура февраля------4,4±0,7 °C. Однако снеговой покров
был выше обычного — 50—-60 см. Это делало для зябликов прак-
тически невозможным добывание естественного корма с начала
января до конца февраля. Лишь во второй декаде марта снег начал
интенсивно таять. К этому времени, как показали наблюдения,
выпущенные зяблики всё реже посещали прикормку и чаще
стали встречаться за пределами района выпуска.
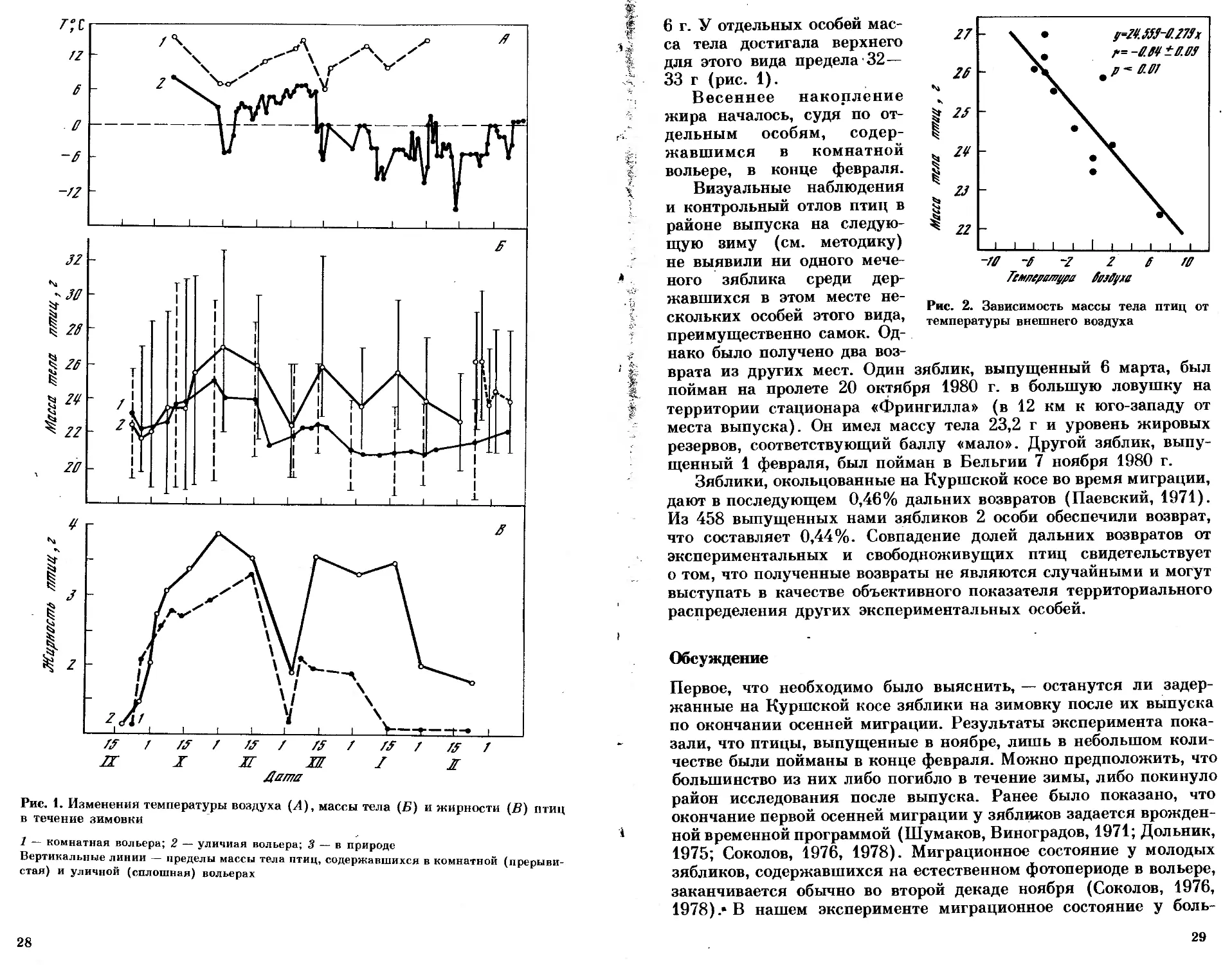
Анализируя изменения массы тела и уровня жировых резер-
вов у зябликов, мы обнаружили, что осеннее миграционное состоя-
ние у птиц, содержавшихся как в уличной, так и в комнатной
вольере, закончилось, судя по резкому уменьшению жироотло-
жений и снижению массы тела, к 20 ноября (рис. 1), т. е. в сроки,
обычные для молодых птиц этого вида (Соколов 1976; Шумаков,
Соколов, 1982). После этого у зябликов началось зимнее ожирение,
которое достигло максимальной величины в середине декабря.
В целом у птиц, живущих на улице, средняя масса тела (24,9±
±0,4 г) и средняя величина жировых резервов (2,66±0,20 г)
были достоверно выше, чем у содержавшихся в помещении (22,1 ±
±0,30 и 1,90±0,18 г соответственно). Эти различия связаны
с тем, что температура в уличной вольере была в среднем на 110
ниже, чем в комнатной (см. рис. 1). Обнаружена высокая отри-
цательная корреляция (г=—0,84) между массой тела у зябликов,
отловленных на свободе и живших в уличной вольере, и темпера-
турой внешнего воздуха (рис. 2).
В период с наиболее низкой температурой средняя масса тела
у зябликов на улице достигала 26—27 г, а жировые резервы —
27
Дата
Рис. 1. Изменения температуры воздуха (Л), массы тела (Б) и жирности (В) птиц
в течение зимовки
i
1 — комнатная вольера; 2 — уличная вольера; 3 — в природе
Вертикальные линии — пределы массы тела птиц, содержавшихся в комнатной (прерыви-
стая) и уличной (сплошная) вольерах
28
6 г. У отдельных особей мас-
са тела достигала верхнего
для этого вида предела 32—
33 г (рис. 1).
Весеннее накопление
жира началось, судя по от-
дельным особям, содер-
жавшимся в комнатной
вольере, в конце февраля.
Визуальные наблюдения
и контрольный отлов птиц в
районе выпуска на следую-
щую зиму (см. методику)
не выявили ни одного мече-
ного зяблика среди дер-
Рис. 2. Зависимость массы тела птиц от
температуры внешнего воздуха
жавшихся в этом месте не-
скольких особей этого вида,
преимущественно самок. Од-
нако было получено два воз-
врата из других мест. Один зяблик, выпущенный 6 марта, был
пойман на пролете 20 октября 1980 г. в большую ловушку на
территории стационара «Фрингилла» (в 12 км к юго-западу от
места выпуска). Он имел массу тела 23,2 г и уровень жировых
резервов, соответствующий баллу «мало». Другой зяблик, выпу-
щенный 1 февраля, был пойман в Бельгии 7 ноября 1980 г.
Зяблики, окольцованные на Куршской косе во время миграции,
дают в последующем 0,46% дальних возвратов (Паевский, 1971).
Из 458 выпущенных нами зябликов 2 особи обеспечили возврат,
что составляет 0,44%. Совпадение долей дальних возвратов от
экспериментальных и свободноживущих птиц свидетельствует
о том, что полученные возвраты не являются случайными и могут
выступать в качестве объективного показателя территориального
распределения других экспериментальных особей.
Обсуждение
Первое, что необходимо было выяснить, — останутся ли задер-
жанные на Куршской косе зяблики на зимовку после их выпуска
по окончании осенней миграции. Результаты эксперимента пока-
зали, что птицы, выпущенные в ноябре, лишь в небольшом коли-
честве были пойманы в конце февраля. Можно предположить, что
большинство из них либо погибло в течение зимы, либо покинуло
район исследования после выпуска. Ранее было показано, что
окончание первой осенней миграции у зябликов задается врожден-
ной временной программой (Шумаков, Виноградов, 1971; Дольник,
1975; Соколов, 1976, 1978). Миграционное состояние у молодых
зябликов, содержавшихся на естественном фотопериоде в вольере,
заканчивается обычно во второй декаде ноября (Соколов, 1976,
1978).* В нашем эксперименте миграционное состояние у боль-
29
шинства зябликов также закончилось во второй декаде ноября.
Следовательно, птицы, выпущенные в ноябре (особенно 1 ноября),
находились еще в миграционном состоянии, что могло стимулиро-
вать их к отлету из района выпуска. Если бы основной причиной
низкой численности ноябрьских птиц в конце зимовки была смерт-
нось, то тогда не наблюдалось бы существенных различий в отловах
ноябрьских и декабрьских птиц (см. табл.), поскольку и те и
другие находились после выпуска практически в одинаковых
условиях. Визуальные наблюдения подтверждают, что до де-
кабря в районе выпуска задержалось только 15 зябликов (из 100
выпущенных в ноябре), причем большинство из этих птиц
(12 особей) было выпущено 15 ноября.
В отличие от птиц ноябрьского выпуска большинство зябликов,
выпущенных в декабре, вероятно, осталось на зимовку, судя по
доле повторных поимок их в конце февраля. Эта доля достоверно
не отличается от доли поимок птиц, выпущенных в январе. Ранее
Г. А. Носковым (1970) было показано, что зяблики, выпущенные
из вольеры в декабре в отличие от чижей и чечеток остаются
в месте выпуска, даже если их перестают кормить.
Зяблики, выпущенные в январе и феврале, также остались
зимовать в месте выпуска. Они держались там вплоть до первой
декады марта. Визуальные наблюдения показали, что в конце зи-
мовки в месте выпуска держалось около 130 зябликов, выпущен-
ных после 1 декабря. Это составляет примерно 40% от общего
числа (309) выпущенных в этот период птиц. Судьба остальных
60% зябликов неизвестна. Значительная часть их могла погибнуть,
другая часть птиц могла быть нами не обнаружена.
На следующую зиму в районе исследования эксперименталь-
ных зябликов обнаружено не было, несмотря на активный их
поиск и контрольный отлов. Этому можно дать несколько объяс-
нений: 1) выпущенные птицы не дожили до следующей зимы;
2) зяблики вообще не склонны проявлять верность прежнему
месту зимовки; 3) птицы не смогли установить связь с необычным
местом зимовки, поэтому и не смогли вернуться; 4) птицы верну-
лись в район выпуска, но не были обнаружены; 5) птицы устано-
вили связь с местом зимовки, но не остались в нем зимовать на
следующий год по причине значительного несоответствия геогра-
фического положения зимовки и эндогенной миграционной про-
граммы длительности полета.
Рассмотрим, насколько вероятно каждое из этих объяснений.
Первое предположение следует отвергнуть, поскольку в конце
осени 1980 г. (20 октября и 7 ноября) были пойманы две особи.
Кроме того, точно было известно, что первую зимовку благополучно
пережили по крайней мере 140 особей. Вероятность, что все они
погибли за весенне-летний период, крайне мала.
Второе предположение противоречит в первую очередь данным
Дж. Финлайсона (Finlayson, 1980) и экспериментам А. Пердека
(Perdeck, 1958), приведенным нами во введении. Поэтому оно так-
же должно быть отвергнуто.
30
Относительно третьего объяснения следует заметить, что вы-
пущенные нами зяблики должны были вести себя точно так же,
как если бы они оказались в нормальном для них районе зимовки,
поскольку у птиц нет врожденного знания местоположения зимов-
ки. Птицы, выпущенные в ноябре и декабре, т. е. в период наиболее
вероятного запечатлевания зимовочной территории, должны были
установить связь с местом выпуска, поскольку они находились
в нем (имеются в виду особи, оставшиеся на зимовку) достаточное
для этого время. Птицы, выпущенные в январе и позже, т. е. после
окончания предполагаемого периода запечатлевания зимовочной
территории, были поставлены в условия, с которыми они обычно
не сталкиваются в природе. Неизвестно даже в отношении запе-
чатлевания гнездовой территории, могут ли птицы установить
связь с территорией после окончания «чувствительного» периода
в том случае, если они были лишены возможности сделать это
в срок. Если могут, то тогда зяблики, выпущенные в январе и фев-
рале, также должны были установить связь с местом первой зимов-
ки. Поэтому нет оснований считать, по крайней мере для птиц, вы-
пущенных до января, что они не смогли установить связь с местом
выпуска.
Предположение о том, что птицы вернулись в район выпуска,
но не были обнаружены, нам кажется маловероятным. Зима
1980/81 гг. была достаточно снежной и холодной, чтобы вынудить
вернувшихся и оставшихся на зимовку зябликов сконцентриро-
ваться в районе прикормки — единственном месте с постоянным
источником корма для птиц этого вида на большой территории.
Даже если бы вернулись только птицы, выпущенные в ноябре и де-
кабре и благополучно пережившие зимовку — не менее 40 особей
(или около 20 особей, если допустить, что половина из них погибла
к моменту контрольного отлова), — то все равно мы их обязательно
обнаружили бы на прикормке в январе, когда вся коса была по-
крыта снегом.
Наиболее вероятным кажется нам последнее, пятое объяснение.
Мы предполагаем, что зяблики, по крайней мере те особи, которые
были выпущены в наиболее вероятный период запечатлевания зи-
мовочной территории (до 1 января), установили связь с местом вы-
пуска. На следующую осень они, вероятно, мигрировали из своих
гнездовых районов, куда они должны были вернуться весной после
вынужденной зимовки на Куршской косе, в направлении первой
зимовки и достигли ее, видимо, в конце сентября—октябре, когда
обычно идет пролет зябликов финляндских популяций. Доказа-
тельством тому служит поимка зяблика 20 октября 1980 г., о кото-
рой уже было сказано ранее. В первую миграцию (в 1979 г.) он был
пойман на пролете 26 сентября, т. е. примерно на три недели рань-
ше. Возможно, что это объясняется обычным индивидуальным от-
клонением в сроках миграции, известным для зябликов, пролета-
ющих через косу (Паевский, 1962), но не исключена возможность,
что эта птица на некоторое время задержалась в районе своей быв-
шей зимовки, расположенной северо-восточнее, прежде чем мигри-
31
ровать дальше. Поимка другой птицы 7 ноября 1980 г. в Бельгии,
на традиционной трассе миграции, подтверждает это предположе-
ние. Если бы мы проводили свой эксперимент не в середине,
а в конце миграционного пути, когда птицы уже завершают мигра-
цию, то тогда, возможно, удалось бы осадить их в новом районе
зимовки более чем на один сезон.
Итак, результат эксперимента по созданию зимовки у молодых
зябликов, задержанных на Куршской косе во время осенней миг-
рации, показали, что: 1) пролетных молодых зябликов можно вы-
нудить зимовать на трассе миграции, даже если этот район распо-
ложен на значительном расстоянии (около 1500 км) от традици-
онных мест зимовки, если их задержать до 1 декабря. Большинство
птиц, выпущенных в ноябре, покидает район выпуска, поскольку
еще находится в миграционном состоянии; 2) при наличии посто-
янной прикормки значительная часть птиц (не менее 40%), выпу-
щенных после окончания осенней миграции, успешно переживает
зимовку и держится до марта, после чего покидает район выпуска;
3) следующую зиму задержанные зяблики не проводят в районе
вынужденной зимовки, находящейся примерно на середине их про-
летного пути, а предпочитают мигрировать в направлении естест-
венных зимовок.
Авторы статьи приносят благодарность К. В. Большакову
и В. А. Паевскому за помощь, оказанную ими при постановке экспе-
римента, и критические замечания.
ЛИТЕРАТУРА
Блюменталь Т. И., Дольник В. Р. Оценка энергетических показателей птиц в поле-
вых условиях // Орнитология. М.: Изд-во МГУ, 1962. Вып. 4. С. 394—407.
Дольник В. Р. Миграционное состояние птиц. М.: Наука, 1975. 398 с.
Дольник В. Р., Паевский В. А. О пределах способности мелких птиц к ориентации //
Бионика. М.: Наука, 1965. С. 364—371.
Дольник Т. В. Пищевое поведение, питание и усвоение пищи зябликом // Попу-
ляционная экология зяблика. Л.: Наука, 1982. С. 18—41. (Тр. Зоол. ин-та
АН СССР; Т. 90).
Лакин Г. Ф. Биометрия. М.: Высш, шк., 1973. 344 с.
Люлеева Д. С. Зимовки зябликов // Популяционная экология зяблика. Л.: Наука,
1982. С. 162-164. (Тр. Зоол. ин-та АН СССР; Т. 90).
Михеев А. В. О закономерностях размещения зимовок и пролетных путей попу-
ляций у птиц // Зоол. журн. 1962. Т. 41, № 8. С. 1210—1219.
Носков Г. А. Об особенностях миграционного поведения кочующих видов вьюрко-
вых в зимний период // Материалы 7-й Прибалт, орнитол. конф. Рига: Зинатне,
1970. Т. 1. С. 61-64.
Носков Г. А. Тип миграционной активности и структура местного населения во-
робьиных птиц // Тез. докл. 10-й Прибалт, орнитол. конф. Рига, 1981. Т. 1.
С. 156-160.
Паевский В. А. Постоянство индивидуальных сроков и путей сезонных передвиже-
ний птиц // Материалы 3-й Всесоюз. орнитол. конф. Львов, 1962. Т. 2.
С. 131-132.
Паевский В. А. Атлас миграций птиц по данным кольцевания на Куршской косе //
Экологические и физиологические аспекты перелетов птиц. Л.: Наука,
1971. С. 3-110. (Тр. Зоол. ин-та АН СССР; Т. 50).
Паевский В. А. Популяционно-демографические аспекты миграций птиц / / Итоги
науки и техники. Зоология позвоночных. М.: ВИНИТИ. 1976. Т. 9. Миграции
птиц. С. 8—60.
32
Паевский В. А. Демография птиц. Л.: Наука, 1985. С. 285. (Тр. Зоол. ин-та
АН СССР; Т. 125).
Соколов Л. В. Временная и энергетическая регуляция продолжительности осеннего
миграционного состояния у молодых и взрослых зябликов // Исследования по
биологии птиц. Л.: Наука, 1976. С. 80—86. (Тр. Зоол. ин-та АН СССР; Т. 65).
Соколов Л. В. Влияние фотопериода на сроки окончания осеннего миграционного
состояния у молодых и взрослых зябликов. // Тез. докл. 2 Всесоюз. конф, по
миграциям птиц. Алма-Ата, 1978. Ч. 2. С. 299—300.
Соколов Л. В. «Чувствительный» период в процессе формирования связи с терри-
торией будущего гнездования у зяблика (Fringilla coelebs) на Куршской ко-
се // Зоол. журн. 1981. Т. 60, № 6. С. 887—893.
Соколов Л. В. Послегнездовые перемещения и постоянство мест гнездования у зяб-
лика на Куршской косе // Популяционная экология зяблика. Л.: Наука, 1982.
С. 215-228. (Тр. Зоол. ин-та АН СССР; Т. 90).
Соколов Л. В., Большаков К. В., Виноградов Н. В. и др. Проверка способности моло-
дых зябликов запечатлевать и находить территорию будущего гнездования //
Зоол. журн. 1984. Т. 63, №11. С. 1671-1681.
Шумаков М. Е., Виноградова Н. В. Формирование и длительность осеннего мигра-
ционного состояния у молодых и взрослых зябликов Куршской популяции //
Анализаторные системы и ориентационное поведение птиц: Материалы к сим-
поз. М.: Изд-во МГУ, 1971. С. 115-116.
Шумаков М. Е., Соколов Л. В. Миграция зябликов на Куршской косе // Популя-
ционная экология зяблика. Л.: Наука, 1982. С. 144—161. (Тр. Зоол. ин-та
АН СССР; Т. 90).
Benvenuti S., loale Р. Homing experiments with birds displaced from their wintering
ground // J. Ornithol. 1980. Vol. 121, № 3. P. 281-286.
Brosset P. A. Territorialisme et de’fence du territoire chez les migrateurs palearctiques
hivernan au Gabon // Alauda. 1971. Vol. 39, № 2. P. 127—131.
Finlayson J. C. The recurrence in winter quarters at Gibraltar of some scrub passeri-
nes // Ring and Migration. 1980. Vol. 3, N 1. P. 32—34.
Fretwell S. Ecotypic variation in the non-breeding season in migratory populations:
A study of tarsal lenght in some Fringillidae // Evolution. 1969. Vol. 23.
P. 406-420.
Fuller R. J., Youngman R. E. The utilisation of farmland by golden plovers wintering
in southern England // Bird Study. 1979. Vol. 26, N 1. P. 37—46.
Haar H. Der Bergfink (Fringilla montifringilla) als Durchziigler und Wintergast in der
Oststeiermark (Aves) // Jahrb. naturwiss. Abt. Joanneum Gras. 1976. S. 393—402.
Jenni L. Schweizerische Ringfunde von Bergfinken Fringilla montifringilla: Ein Beit-
rag zum Problem der Masseneinfliige // Ornithol. Beob. 1982. Bd. 79, N 4.
S. 265-272.
Johnson O. W., Johnson P. M. Plumage-moultage relationships in «oversummering»
and migratory lesser golden Plovers // Condor. 1983. Vol. 85, N 4. P. 406—419.
Marler P. Behaviour of the chaffinch, Fringilla coelebs // Behaviour. 1956. Vol. 5.
184 p.
Medway L. A ringing study of the migratory Broun Shrike in West Malaysia // Ibis.
1973. Vol. 115, N 1. P. 60-86.
Mewaldt L. R. California Sparrows return from displacement to Maryland // Science.
1964. Vol. 146. P. 941-942.
Mewaldt L. R. Winter philopatry in White-crowned Sparrows (Zonotrichia leucoph-
rys) // North Amer. Bird Band. 1976. Vol. 1, N 1. P. 14—20.
Moreau R. E. The recurrence in winter quarters (Ortstreue) of trans-Saharan mi-
grants // Bird Study. 1969. Vol. 16, N 2. P. 108-110.
Newton I. Finches. London: Collins, 1972. 288 p.
Pearson D. I. The wintering and migration of paleartic passerines at Kampala, Sou-
thern Uganda // Ibis. 1972. Vol. 114, N 1. P. 43—60.
Pearson D. J. The wintering and moult of ruffs Philomachus pugnax in the Kenyan
rift valley // Ibis. 1981. Vol. 123, N 2. P. 158-182.
Perdeck A. C. Two types of orientation in migrating starlings Sturnus vulgaris L. and
chaffinches Fringilla coelebs L., as revealed by displacement experiments //
Ardea. 1958. Vol. 46, N 1/2. P. 1-37.
2 Заказ № 36
33
Price T. The ecology of the Greenish Warbler Phylloscopus trochiloides in its winter
quarters // Ibis. 1981. Vol. 123, N 2. P. 131 —144.
Ralph C. J., Mewaldt L. R. Timing of site fixation upon the wintering grounds in spar-
rows // Auk. 1975. Vol. 92, N 4. P. 698- 705.
Ralph C. J., Mewaldt L. R. Homing success in wintering sparrows // Auk. 1976. Vol.
93, N 1. P. 1 — 14.
Sauer E. G. F. Migration habits of Golden Plovers // Proc. 13-th Intern. Ornithol.
Congr. Helsinki, 1963. P. 454-467.
Schenker R. Winterplatztreue beim Bergfinken (Fringilla monti fringilla) // Vogel-
warte. 1976. Bd. 28, N 4. S. 313-314.
Schwartz P. Orientation experiments with Northern Watertrushes wintering in Vene-
suela) // Proc. 13-th Intern. Ornithol. Congr. Helsinki, 1963. P. 481—484.
Stewart P. A., Connor H. A. Fixation of wintering Palm Warblers to a specific site //
J. Field Ornithol. 1980. Vol. 51, N 4. P. 365-367.
Troy D. M. Recaptures of redpolls: movements of an irruptive species // Ibid. 1983.
Vol. 54, N 2. P. 145-151.
Yinick R. P. Winter site fidelity of some northern finches (Fringillidae) // Ibid. 1983.
Vol. 54, N 3. P. 254-258.
Zahavi A. The social behavior of the White Wagtail (Motacilla alba) wintering in
Israel // Ibis. 1974. Vol. 113, N 2. P. 203-211.
SUMMARY
Test of possible wintering of young chaffinches, kept off on Kurshskaya Kosa.
Sokolov L. V., Diachenko V. P., Ilyina T. A., Liuleieva D. S.,
YablonkevichM. L.
The possibility of wintering of young chaffinches, kept off during their autumn mi-
gration on Kurshskaya Kosa, situated 1500 km far from their traditional wintering
territories, was tested. The young males, caught for the experiment, were kept and
released in groups of 40—60 specimens every 2 weeks during November 1 — March 6
on the territory of Ribachi Biostation (in all, 450 specimens were released). The results
of the experiment showed: 1) most of the birds, released in December—February, stayed
for wintering in the region of their release; 2) a considerable part (40%) of wintering
birds, if constantly fed up, can survive their wintering well and stay in the region of
their release till March 10; 3) the next winter chaffinches don’t stay in the region of the
forced wintering, but they preffer to migrate to their natural wintering places.
34
УДК 574.24: 591: 598.841
А. В. БАРДИН
199164, Ленинград, Универси-
тетская набережная, д. 1. Зоо-
логический институт АН СССР
Динамика полового
и возрастного состава
и жировые резервы
больших синиц
(Parus m. major)
в зимний период
В северных частях своего ареала большие синицы (Parus m. major
L.) могут зимовать в значительной мере лишь благодаря использо-
ванию кормов антропогенного происхождения (Snow, 1952; Ulf-
strand, 1962; Haartman, 1969; Носков, 1976; Бианки, Шутова, 1978;
Носков и др., 1981; Бардин, 1983). В зоне тайги большинство особей
этого вида уходят на зиму к поселениям человека. Согласно
О. П. Смирнову и Г. А. Носкову (1975), в Ленинградской области
около 87% больших синиц проводят зиму в населенных пунктах.
Наиболее оптимальные условия они находят в поселках сель-
ского и дачного типов, а также в небольших озелененных го-
родах.
Таким образом, на Северо-Западе СССР населенные пункты
являются важнейшей стацией большой синицы в зимний период.
Цель данного сообщения — описать динамику полового и возраст-
ного состава больших синиц в подобных стациях на протяжении
осенне-зимнего сезона и в разные годы, а также характер пребыва-
ния птиц в городе и масштабы их перемещений в зимнее время.
С целью сравнения состояния особей разных половозрастных
групп, зимующих в одних условиях, исследовали суточную дина-
мику их жировых резервов.
Район исследования, материал и методика
Работу проводили в г. Печоры Псковской области в 1968—1983 гг.
Печоры — небольшой город с населением порядка 3 тыс. чел., боль-
шим количеством садов, парков, скверов, окруженный населен-
ными пунктами сельского типа. Многие его жители подкармливают
синиц в зимнее время. Кормушки есть почти у каждого дома.
Поэтому постоянные прикормки, на которых ловили синиц
с целью кольцевания, существенно не изменяли кормовых условий.
Всего в пределах города было 16 пунктов прикормки. Ежегодно
ловили синиц в 1 —12 точках. Пойманным птицам, кроме стандарт-
ного алюминиевого кольца с номером, надевали цветные кольца
или окрашивали оперение, чтобы их можно было узнать на расстоя-
нии. Получены данные наблюдений за 1920 окольцованными осо-
бями.
Прижизненную оценку жировых запасов проводили по методу
Т. И. Блюменталь и В. Р. Дольника (1962). Выделяли 4 основных
(«нет», «мало», «средне», «много») и 6 промежуточных, т. е. всего
2*
35
/Пасса meat
Рис. 1. Масса тела самцов
и самок большой синицы
разной жирности
Точки — среднее арифмети-
ческое значение массы, пря-
моугольники — 95%-ный до-
верительный интервал сред-
него, вертикальные линии —
вариационный размах.
Обозначение баллов жирно-
сти:
1 — нет;
2 — больше,
3 — меньше,
4 — мало;
5 — больше,
6 — меньше,
7 — средне;
8 — больше,
9 — меньше,
10 ~ много;
11 — больше,
чем нет;
чем мало;
чем мало;
чем средне;
чем средне;
чем много;
чем много
11 баллов жирности. При этой полуколичественной оценке количе-
ство жира у особей с определенным баллом жирности в среднем вы-
ше, чем с предыдущим баллом, и ниже, чем с последующим, но на-
сколько — для большинства видов неизвестно. У мелких воробьи-
ных количества липидов у особей 4 основных классов жирности
соотносятся в среднем как 1:3:5:14 (Блюменталь, 1967, 1971).
Масса тела птиц частично отражает количество депонированного
жира. На рис. 1 представлена средняя масса больших синиц раз-
ных классов жирности в осенне-зимний период. Масса самцов
с баллами жирности «мало», «средне» и «много» отличается от то-
щей массы в среднем на 1,6; 2,2 и 3,2 г соответственно. У самок с те-
ми же баллами жирности превышение массы составляет в среднем
1,1; 1,9 и 2,7 г. При одинаковой массе тощих больших синиц масса
жирных особей зимой выше, чем в период осенней миграции. По
целому ряду причин, прежде всего из-за большой изменчивости
массы тела, использование полуколичественного метода определе-
ния жирности предпочтительнее косвенного весового (Блюмен-
36
таль, 1967; Дольник, 1975). Поэтому в данной работе используется
именно этот метод. Баллы жирности образуют порядковую шкалу.
К ней применим ряд методов непараметрической статистики,
в частности критерий U Манна—Уитни, критерий Краснела—Уол-
лиса, метод множественных сравнений, основанный на суммах ран-
гов Краскела—Уоллиса.
При анализе данных по половому и возрастному составу и его
зависимостей от сезона и года использовали дисперсионный анализ
для качественных данных, критерий %2 и, в случае небольших выбо-
рок, критерий точной вероятности Фишера. Границы доверитель-
ного интервала доли вычисляли с помощью «р-преобразования Фи-
шера с поправкой Йейтса на непрерывность (Sokal, Rohlf, 1969;
Лакин, 1973; Холлендер, Вулф, 1983).
Результаты
Характер пребывания в городе зимующих больших синиц. По дан-
ным кольцевания, большие синицы, наблюдающиеся зимой в Печо-
рах, представлены следующими фракциями: 1) взрослые особи,
размножавшиеся в городе и проводящие здесь зиму; 2) молодые,
родившиеся в городе или осевшие здесь после окончания расселе-
ния и оставшиеся здесь на зиму; 3) взрослые, гнездившиеся в ок-
рестных лесах и переселившиеся на зиму в город; 4) взрослые,
гнездящиеся и зимующие в лесах, но регулярно прилетающие в го-
род на кормежку; 5) молодые, родившиеся или осевшие в лесах
и переселившиеся на зиму в город; 6) молодые, живущие в лесу
и регулярно прилетающие в город; 7) взрослые и молодые, приле-
тевшие на зиму из других географических районов. Повторные от-
ловы дают примеры всех указанных типов, однако не позволяют
оценить доли отдельных фракций среди зимующих в городе птиц.
Предположительно среди молодых преобладают фракции вторая
и седьмая, а среди взрослых — первая, третья и четвертая. Кроме
того, в сентябре — начале октября и во второй половине февраля —
марте через город пролетает большое количество мигрирующих
больших синиц.
В течение одного зимнего сезона состав больших синиц в городе
несколько меняется, но все же сохраняет известную степень посто-
янства. В последующий сезон контингент зимующих особей значи-
тельно обновляется, в среднем на 89%.
В зимнее время у некоторых больших синиц наблюдаются регу-
лярные перемещения из леса в город и обратно. Более детально та-
кое поведение прослежено у 13 окольцованных особей. В лесу пти-
цы держались на определенных участках обитания диаметром не
более 500 м. В городе они также ограничивали свои перемещения
участком 200—300 м в диаметре. Одна особь сохраняла привязан-
ность к постоянным участкам в лесу и городе на протяжении 5 лет,
другая — 4 лет. Расстояния между лесными и городскими участ-
ками были 1,2 км (одна), 1,4 (три), 1,6 (две), 2,1 (одна), 2,2 (две),
2,5 (две), 3,0 (одна), 4,0 км (одна особь). Среди этих птиц было
37
9 взрослых (5 самцов и 4 самки) и 4 молодых (3 самца и 1 самка).
Птицы могли перемещаться из леса в город и обратно ежедневно
или какое-то время постоянно жили в городе. Между своими лес-
ными и городскими участками обитания синицы не наблюдались
ни разу. Вероятно, они быстро и целенаправлено пролетали это
расстояние. Для многих, особенно молодых синиц, характерна се-
зонная смена лесных летних и городских зимних участков обита-
ния.
Масштабы перемещений больших синиц внутри города в целом
невелики ’(табл. 1). По-видимому, встречи особей далее 0,5 км сви-
детельствуют не о большом участке обитания, а о смене участка.
В марте значительная часть зимовавших в городе больших си-
ниц исчезала и в дальнейшем не наблюдалась.
Таблица 1
Масштабы перемещений больших синиц в г. Печоры в зимний период по данным
кольцевания
Расстояние между наиболее удаленными точками встреч, км Количество особей
абс. % 95%-ный доверительный интервал (%)
0,0-0,3 357 80,4 76,6-4-84,0
0,3-0,5 56 12,6 9,74-15,9
0,5-0,7 15 3,4 1,94-5,3
0,7-0,9 12 2,7 1,44-4,4
0,9-1,1 2 0,5 0,44-1,3
1,1-1,3 1 0,2 0,04-0,9
>1,3 1 0,2 0,04-0,9
Всего 444 100,0
Половой и возрастной состав зимующих больших синиц в раз-
ные годы. В декабре и январе индивидуальный состав встречаю-
щихся в городе синиц наиболее постоянен. Соотношение взрослых
и молодых в этот период в разные годы показано на рис. 2. Доля
взрослых значимо менялась год от года (дисперсионный анализ для
качественных данных, Р<0,01). Она колебалась от 13 до 57%
и в среднем составляла 32±1% (п=1727).
В целом за все годы количество самцов среди всех особей превы-
шало количество самок (%2=5,33, Р<0,05) и составило 53±1%.
При этом соотношение полов среди взрослых птиц не отличалось
значимо от 1:1 (%2=0,06), в то время как среди молодых самцов бы-
ло значимо больше: 55±1% (%2=6,76, Р<0,01).
При данном объеме материала дисперсионный анализ не выя-
вил значимых изменений по годам в соотношении полов среди об-
щей выборки и среди молодых. Однако для взрослых птиц разли-
чия в соотношении полов оказались значимыми (Р<0,05), доля
самцов менялась от 42 до 71%.
Как видно из рис. 3, изменения в соотношении полов по годам
38
Рис. 2. Возрастной состав больших синиц в декабре—январе в г. Печоры в разные
годы
Вертикальные линнн — 95%-ный доверительный интервал. Значения, значимо отличаю-
щиеся от средней за все годы (Р<0,05) обведены. Числа на поле графика — количество
обследованных особей
Рис. 3. Соотношение полов в декабре—январе среди молодых (светлые кружки)
и взрослых (темные кружки) больших синиц в г. Печоры в разные годы
При отсутствии значимых различий в доле самцов между молодыми и взрослыми птицами
соответствующие значения соединены вертикальными линиями. При наличии значимых
различий в соотношении полов от 1:1 соответствующие значения обведены. Горизонтальны-
ми линиями показано соотношение полов в общих выборках
у молодых и взрослых не однонаправлены. Значимая корреляция
между долей взрослых и соотношением полов у последних отсутст-
вует. Полученные материалы также не выявили значимой корреля-
ции между половым и возрастным составом зимующих больших си-
ниц и условиями зимы (средней температурой воздуха и состоя-
нием снежного покрова).
39
Сравнение полового и возрастного состава больших синиц в ле-
су и в городе. В зимний период в лесах встречается гораздо меньше
больших синиц, чем в населенных пунктах. Достаточно указать, что
зимой в городе отловлено 1727 особей, а в больших лесных масси-
вах — только 61, хотя основная работа велась именно в лесу и было
окольцовано более 1000 синиц других видов. Следует заметить, что
большие синицы, наблюдающиеся в лесу и в населенных пунктах,
не представляют изолированные группировки. Напротив, и здесь
и там мы можем видеть одних и тех же особей. Возможно, число
птиц с двумя «местами жительства» достаточно велико в местно-
сти, где в пределах 10 км всегда можно найти поселение человека.
Вместе с тем половой и возрастной состав зимующих или регуляр-
но появляющихся в лесных массивах больших синиц существенно
отличается от того, что наблюдается в городе, где зимует много осо-
бей, не посещающих в этот сезон лес. Как видно из табл. 2, в лесу
значимо преобладают взрослые птицы и самцы.
Таблица 2 ,
Половой и возрастной состав больших синиц в г. Печоры и окрестных больших
лесных массивах в декабре—январе
Возрастные н половые группы Город (п=1727) Смешанный лес с преобла- данием сосны и ели (п—61)
% 95%-ный дове- рительный ин- тервал, % % 95%-ный дове- рительный ин- тервал, %
Взрослые самцы 16 14-4-18 41 294-54
Взрослые самки 16 14-4-18 26 164-38
Молодые самцы 37 34-4-40 30 194-42
Молодые самки 31 284-33 3 0-4-9
Все взрослые 32 304-35 67 554-79
Все самцы 53 51-4-56 70 584-81
Доля самцов среди взрослых 51 46-4-56 61 454-76
Доля самцов среди молодых 55 51-4-58 90 724-99
Динамика полового и возрастного состава больших синиц в го-
роде в течение осенне-зимнего периода. Изменение соотношения
взрослых и молодых птиц с октября по март представлено на рис. 4.
Материал, собранный в марте, невелик; собирался он лишь в первой
декаде месяца. В общем наблюдается тенденция увеличения доли
взрослых в городе ко второй половине зимы и резкое уменьшение
ее в марте. Эта тенденция в разные зимы выражена в различной
степени. В годы с большим количеством молодых она практически
не прослеживается. По средним данным за все годы, она выявляет-
ся, однако статистически не значима. В зимы со средней и выше
средней долей взрослых описываемая тенденция наиболее выраже-
на и значима (дисперсионный анализ для качественных данных,
Р<0,01). По всей видимости, изменение соотношения возрастных
групп в течение сезона связано с более частым появлением в горо-
40
Рис. 4. Изменение возрастного состава больших синиц в г. Печоры по месяцам
1 — в среднем за все годы; 2 — в годы со средней и ниже средней долей взрослых в де-
кабре— январе; 3 — в годы со средней и выше средней долей взрослых в декабре—январе.
Числа над столбцами гистограммы — количество обследованных особей
Рис. 5. Изменение соотношения по-
лов среди молодых и взрослых боль-
ших синиц в г. Печоры по месяцам
Обозначения те же, что и на рис. 3
де и переселением сюда взрослых птиц из леса в наиболее суровый
период. В годы с большим количеством молодых, обитающих в пре-
делах города, появление здесь лесных птиц, количество которых
гораздо меньше, не вызывает существенного изменения в соотно-
шении взрослых и молодых птиц.
Изменение соотношения полов по месяцам показано на рис. 5.
Полученный материал не позволяет выявить какой-либо опреде-
ленной тенденции. Доля самцов максимальна в феврале как у моло-
дых, так и у взрослых, в обоих случаях значимо превышая 50%: для
молодых 59±4% (%2=4,89, Р<0,05), для взрослых 63±5% (%2=
=5,13, Р<0,05).
Жировые резервы зимующих больших синиц исследованы на
материале 689 визуальных определений жирности больших синиц
в городе во второй половине декабря — январе. Сравнивали жир-
ность птиц в разное время дня, при разной температуре воздуха
и между разными половыми и возрастными группами.
Наиболее значительными были изменения жирности в зависи-
мости от времени суток. Изменения были значимыми и сходными
при температуре воздуха от 0 до -f-3° и при —1 10°. Анализ
с помощью критерия Краснела — Уоллиса показал большую зна-
чимость различий (Р<0,001). Следует заметить, что после 15 ч
(время местное солнечное) часть птиц отлетает к местам ночлега,
41
а ловиться продолжают лишь менее жирные особи, что завышает
долю последних. Наибольшее количество птиц с жирностью
«меньшей чем мало» отмечено не на рассвете, а на полтора—два
часа позже, между 11 и 12 ч. Особи без видимых подкожных отло-
жений жира встречались крайне редко. Медианное значение балла
жирности неуклонно повышалось на протяжении дня и составляло
после 9 ч «мало», в 10 ч — «меньше среднего», в 11 и 12 ч —
«средне», в 13 и 14 ч — «меньше чем много», в 15 ч — «много».
Значимых различий в жирности синиц при разной температуре
воздуха в одинаковое время суток обнаружить не удалось, возмож-
но, из-за незначительного диапазона температур, при которых об-
следовано большинство особей.
В каждый часовой интервал дневного времени проведен анализ
различий в жирности птиц четырех групп: взрослых самцов, взро-
слых самок, молодых самцов и молодых самок. На имеющемся ма-
териале значимые различия выявлены только в интервалах 10—
11 ч (критерий Краскела—Уоллиса Нк =29,77, Р<0,05) и 15—
16 ч (Нк=85,47, Р<0,001). Последующее множественное сравне-
ние на основе сумм рангов Краскела—Уоллиса показало, что жир-
ность самцов была меньше, чем самок, как среди взрослых, так
и среди молодых птиц.
Обсуждение
Зимнее население больших синиц в Печорах формируется в конце
октября — начале ноября и распадается в начале марта. Большин-
ство зимовавших особей исчезает и в дальнейшем не отмечается
ни в городе, ни в окружающих его лесных массивах. Сходная кар-
тина наблюдается в Вильнюсе (Шяштокас, Идзелис, 1972). Появ-
ление в городе большого количества новых птиц в конце осени и их
исчезновение в конце зимы дает основание предполагать, что в кон-
тингенте зимующих больших синиц заметную долю составляют
особи, пришедшие из других географических районов. Это подтвер-
ждают три случая, когда молодые синицы, окольцованные зимой
в Печорах, были найдены взрослыми в более северных и восточ-
ных районах: самец в Новгородской области (на расстоянии
210 км), самки в Финляндии (400 км) и Карелии (440 км). В то же
время какая-то часть особей местного происхождения улетает
осенью из района исследования. Об этом свидетельствует находка
23 октября в Польше молодой птицы, окольцованной слетком
в Печорах 9 августа (780 км).
Соотношение взрослых и молодых в разные годы варьирует до-
вольно значительно. Доля взрослых изменяется от 13 до 57%
и в среднем составляет 32%. В соседних регионах картина весьма
сходная: в Южной Финляндии — от 20 до 52%, в среднем 45%
(Hilden, 1978), в поселках Ленинградской области — в среднем
26% (Смирнов, Носков, 1975). Возрастной состав зимующих боль-
ших синиц в существенной мере определяют пришельцы. При
этом чем больше последних, тем выше доля молодых. Интересно,
42
что вариации возрастного состава не проявляют заметной связи
с условиями зимы. Аналогичный вывод делает О. Хильден (Hilden,
1978). Он обнаружил, что доля молодых положительно коррелиру-
ет с успешностью размножения в предшествующее лето. Для
Печор отсутствуют полноценные данные о гнездовании этого вида.
Можно только предполагать, что указанная зависимость и здесь
в известной мере определяет соотношение возрастных групп зимой.
Соотношение полов среди молодых не имеет значимых разли-
чий по годам, в то время как среди взрослых такие различия про-
слеживаются. Так как они не связаны с условиями зимы, они, види-
мо, представляют собой результат различий в смертности половых
групп, варьирующей год от года.
По сравнению с городом в лесу ситуация проще. Зимой здесь
остается лишь небольшая часть гнездившихся взрослых и осевших
после расселения молодых. Пришельцы из других географических
районов в лесу, по всей видимости, отсутствуют — они проводят
зиму в поселениях человека. В зимний сезон в лесах существенно
преобладают взрослые и самцы.
Динамика жировых резервов у особей разных половозрастных
групп, зимующих в городе, оказалась практически одинаковой. Как
известно, у этого вида самцы доминируют над самками, получая
преимущественный доступ к пище при лимитированных кормовых
ресурсах (Hinde, 1952; Kluyver, 1957; De Laet, 1983). Однако
в нашем случае жирность самцов утром и вечером оказалась даже
ниже, чем у самок. Возможно, это объясняется большей активно-
стью самцов и их большим риском в поддержании энергетического
баланса, оправдываемым их более доминантным положением.
Полученные данные свидетельствуют о том, что в городе с его бога-
той кормовой базой условия зимовки особей разных половозраст-
ных групп заметно не различаются. Исключение могут составлять
особенно суровые периоды, по которым данные отсутствуют из-за
их редкости.
Следует отметить, что у больших синиц наблюдается очень
сильные суточные колебания уровня жировых резервов. Используя
результаты визуальных определений жирности и эмпирическую
зависимость прироста массы от жирности в баллах, получаем, что
как самцы, так и самки накапливают зимой за светлую часть суток
в среднем 1,6 г жира, большая часть которого используется следую-
щей ночью и в первые утренние часы. Эта величина хорошо согла-
суется с данными исследований суточной динамики массы тела
больших синиц зимой (Лехикойнен, 1980). По суточной динамике
жировых резервов у свободноживущих мелких воробьиных зимой
в литературе имеется гораздо меньше данных, чем по сезонной.
У всех изученных видов, как и у большой синицы, наблюдается
быстрое накопление жира днем с последующей его тратой во время
ночевки, причем в высоких широтах суточные колебания жирности
более резкие (Newton, 1969;.Blem, 1976). Для ряда видов показана
негативная корреляция количества депонированного жира с темпе-
ратурой воздуха ниже определенной «критической величины»,
43
выше которой корреляция отсутствует (Odum, 1949; King, Farner,
1966; Helms, 1968). Для большой синицы этого не удалось показать,
что, возможно, связано с небольшим диапазоном температур, когда
обследовалось большинство особей (-|-3--10°), хотя отдельных
птиц осматривали и при —20—28°. Может быть, однако, что у боль-
шой синицы вообще слабо выражена или отсутствует реакция
жироотложения на температуру среды как сигнальный фактор.
Во всяком случае, у ряда видов ее также не удалось выявить (King,
1972). П. Р. Эванс (Evans, 1969) обнаружил, что вечерний уровень
жировых резервов у обыкновенной овсянки Emberiza citrinella
сильнее коррелирует со средними многолетними температурами,
Чем с конкретной температурой во время исследования.
До сих пор однозначно не решен вопрос, что представляют собой
ежегодно наблюдающиеся осенние и весенние миграции большой
синицы. Г. А. Носков и О. П. Смирнов (1981) считают, что эти ми-
грации представляют собой в основном процесс биотопического
перераспределения особей. Перемещения географического масшта-
ба, по их мнению, свойственны лишь незначительной части боль-
ших синиц, прежде всего молодых.
Согласно другой точке зрения, большим синицам на Северо-
Западе СССР свойственна частичная перелетность (Вилкс, 1966;
Паевский, 1971; Ефремов, 1975; Липсберг, Руте, 1975; Вильбасте,
1975; Руте, 1981). Поданным, X. Вильбасте (1975), в Эстонии пере-
летность наиболее характерна для больших синиц, живущих в ле-
сах, в то время как в урбанизированном ландшафте они преимуще-
ственно оседлы.
Если бы в окрестностях Печор большие синицы совершали толь-
ко местные биотопические перемещения, то зимой в городе при
столь значительном преобладании самцов в лесу должны были пре-
обладать самки. Этого, однако, не наблюдается.
Известно, что соотношение полов среди слетков больших синиц
может значимо отличаться от 1:1 (Dhondt, 1970; Drent, 1984), одна-
ко смещение всегда относительно невелико. В целом долю молодых
самцов в популяции можно принять равной 0,5. На осеннем пролете
среди молодых больших синиц доля самцов составляет примерно
0,4 (38—39%, см. Ефремов, 1975; Липсберг, Руте, 1975), в зимнем
лесу — 0,9. Исходя из этих показателей, можно грубо оценить, ка-
кая часть лесных молодых синиц улетает на зиму. Если к — доля
улетевших, (1—к) — доля оставшихся, р — доля самцов на проле-
те, q — доля самцов среди оставшихся в лесу, то
ь 0,5—q 0,5—0,9 R
К_ р—q ~ 0,4—0,9 —и,в-
Таким образом, около 80% молодых больших синиц на зиму
улетает из леса, причем не столько к ближайшему жилью человека,
сколько в более удаленные регионы.
У взрослых птиц соотношение полов может сильнее отличаться
от 1:1 из-за различной смертности полов (Kluyver, 1951; Bulmer,
Perrins, 1973; Orell, Ojanen, 1979; Hilden, 1982), поэтому для них
подобная оценка может дать большое смещение.
44
Сроки осенней миграции и сроки формирования зимнего насе-
ления в городе не совпадают. Прилет новых синиц в город начина-
ется в конце или после окончания периода миграции. Таким обра-
зом, в биотопических перемещениях могут принимать участие
только те местные особи, которые не приняли участие в миграции,
а также особи из других регионов, закончившие миграцию в данном
месте.
Следует еще раз подчеркнуть, что в наблюдавшихся условиях
большие синицы ведут зимой преимущественно оседлый образ жиз-
ни как в лесу, так и в городе. Некоторые (возможно, многие) особи
имеют не один, а два или более участков обитания и совершают
между ними сезонные, не периодические и даже суточные пере-
мещения. В первом случае их поведение сходно с перелетностью
и фактически отличается от нее лишь расстоянием между двумя
сезонными участками обитания. Подобная картина полностью соот-
ветствует выводам, сделанным в результате многолетнего кольцева-
ния синиц в окрестностях г. Стренчи в Латвии (Вилкс, Вилкс,
1961; Вилкс, 1966). Как в Стренчи, так и в Печорах не получено
данных, свидетельствующих о кочевках больших синиц в зимнее
время.
Заключение
Межгодовую и сезонную динамику полового и возрастного состава
больших синиц, зимующих в городе, в принципе, можно объяснить
суммарным действием четырех причин: общей успешностью раз-
множения на достаточно большой территории; дифференциальной
смертностью членов разных половозрастных групп; широкими
осенними и весенними перемещениями особей; попеременным ис-
пользованием участков обитания некоторыми особями в зимнее
время.. Различия в половом и возрастном составе больших синиц
в городе и лесных массивах невозможно объяснить только пере-
мещениями части лесных птиц в город. Необходимо допустить, что
значительная часть больших синиц улетает из леса в другие гео-
графические регионы. Это заставляет присоединиться к мнению
о том, что части больших синиц, обитающих на Северо-Западе
СССР, свойственна типичная перелетность. Более того, существуют
свидетельства того, что в последние десятилетия перелетность
больших синиц принимает более массовый характер (Руте, 1981).
Очевидно, это связано с расширением северной границы ареала этого
вида и проникновением его в таежные леса.
ЛИТЕРАТУРА
Бардин А. В. Семейство Синицы — Paridae //А. С. Мальчевский, Ю. Б. Пукин-
ский. Птицы Ленинградской области и сопредельных территорий. История,
биология, охрана. Л.: Изд-во ЛГУ, 1983. Т. 2. С. 269—299.
Бианки В. В., Шутова Е. В. К экологии большой синицы в Мурманской области //
Бюл. МОИП. Отд. биол. 1978. Т. 83, № 2. С. 63—70.
Блюменталь Т. И. Изменение энергетических запасов (жирности) у некоторых
45
воробьиных птиц Куршской косы в связи с участием их в миграции // Мигра-
ции птиц Прибалтики. Л.: Наука, 1967. С. 164—202. (Тр. Зоол. ин-та АН СССР;
Т. 40).
Блюменталь Т. И. Формирование осеннего миграционного состояния в природе
у некоторых воробьиных птиц (биоэнергетический аспект) // Экологические
и физиологические аспекты перелетов птиц. Л.: Наука, 1971. С. 111 — 182.
(Тр. Зоол. ин-та АН СССР; Т. 50).
Блюменталь Т. И., Дольник В. Р. Оценка энергетических показателей птиц в поле-
вых условиях // Орнитология. М.: Изд-во МГУ, 1962. Вып. 4. С. 394—407.
Вилкс Е. К. Миграции и территориальное поведение латвийских синиц и пополз-
ней но данным кольцевания // Миграция птиц Латвийской ССР. Рига: Зипат-
не, 1966. С. 69-88.
Вилкс К. А., Вилкс Е. К. Сезонное размещение синиц и поползня в Латвийской
ССР и их зимняя подкормка // Экология и миграции птиц Прибалтики. Рига:
Зинатне, 1961. С. 151-160.
Вильбасте X. Формирование осенней миграции большой синицы в Эстонской
ССР // Материалы Всесоюз. конф, по миграциям птиц. М., 1975. Кн. 1. С. 162 —
163.
Дольник В. Р. Миграционное состояние птиц. М.: Наука, 1975. 398 с.
Ефремов В. Д. Возрастной и половой состав больших синиц (Parus major L.), про-
ходящих Куршскую косу в период осенней миграции // Сообщ. Прибалт,
комиссии по изучению миграций птиц. Тарту: АН ЭССР, 1975. № 9. С. 83—90.
Лакин Г. Ф. Биометрия. М.: Высш, шк., 1973. 343 с.
Лехикойнен Е. Характер изменения веса у некоторых воробьиных птиц, зимующих
в северной умеренной зоне // Адаптация животных к зимним условиям. М.:
Наука, 1980. С. 44 - 57.
Липсберг Ю. К., Руте Ю. Я. Осенняя миграция синиц семейства Paridae и ополов-
ника на юго-западном побережье Латвии в 1967—1971 гг. // Сообщ. Прибалт,
комиссии по изучению миграций птиц. Тарту; АН СССР, 1975. № 9. С. 105—
122.
Носков Г. А. К вопросу об эволюции территориального поведения так называемых
«оседлых» видов воробьиных птиц // Биоценотические отношения организмов.
Л.: Изд-во ЛГУ, 1976. С. 163 176.
Носков Г. А., Зимин В. Б., Резвый С. П. и др. Птицы Ладожского орнитологического
стационара и его окрестностей // Экология птиц Приладожьн. Л.: Изд-во ЛГУ,
1981. С. 3-86.
Носков Г. А., Смирнов О. П. Территориальное поведение и миграции большой сини-
цы (Parus m. major L.) // Экология птиц Приладожья. Л.: Изд-во ЛГУ, 1981.
С. 100-130.
Паевский В. А. Атлас миграций птиц по данным кольцевания на Куршской косе //
Экологические и физиологические аспекты перелетов птиц. Л.: Наука, 1971.
С. 3-110 (Тр. Зоол. ин-та АН СССР; Т. 50).
Руте Ю. Я. Изменение в характере осеннего пролета большой синицы (Parus major)
//Экологические и поведенческие исследования позвоночных животных
в Прибалтике. Рига: Латв. гос. ун-т, 1981. С. 52—69.
Смирнов О. П., Носков Г. А. Структура популяции большой синицы в Ленинград-
ской области // Экология. 1975. № 6. С. 79—83.
Холлендер М., Вулф Д. А. Нелараметрические методы статистики. М.: Финансы
и статистика, 1983. 518 с.
Шяштокас И., Идзелис Р. К вопросу оседлости и миграций большой синицы //
Тез. докл. VIII Прибалт, орнитол. конф. Таллин, 1972. С. 111 — 112.
Blem С. В. Patterns of lipid storage and utilization in birds // Amer. Zool. 1976. Vol.
16. P. 671-684.
Bulmer M. G., Perrins С. M. Mortality in the Great Tit Parus major // Ibis. 1973. Vol.
115. P. 277-281.
De Laet J. Ecological implications of social dominance of the Great Tit Parus major L.
during winter // Proc. 18-th Intern. Ethol. Conf. Brisbane, 29 Aug.— 6 Sept.
1983. S. 1., [1983]. P. 78.
Dhondt A. A. The sex ratio of nestling Great Tits // Bird Study. 1970. Vol. 17. P. 282—
286.
Drent P. J. Mortality and dispersal in summer and its consequences for the density of
46
Great Tils Parus major at the onset of autumn // Ardea. 1984. Vol. 72. P. 127 —
162.
Evans P. R. Winter fat deposition and overnight survival of yellow buntings (Emberiza
citrinella L.) // 3. Anim. Ecol. 1969. Vol. 38. P. 415—423.
Haarlman L. The nesting habits of Finnish birds. 1. Passeriformes // Comment. Biol.
Soc. Sci. fenn. 1969. Vol. 32. P. 1 — 187.
Helms C. W. Food, fat, and feathers//Amer. Zool. 1968. Vol. 8. P. 151 —167.
Hilden O. Ika- ja sukupuolijakauma eraassa talitiaisen Parus major talvipopulaatiossa
Etela-Suomessa // Ornis fenn. 1978. Vol. 55. P. 120—125.
Hilden O. Proportion of juveniles as a measure of adult mortality in a breeding popula-
tion of Great Tits Parus major and Blue Tits P. caeruleus // Ornis fenn. 1982.
Vol. 59. P. 149-151.
Hinde R. A. The behaviour of the Great Tit (Parus major) and some other related spe-
cies // Behaviour. Leiden, 1952. Suppl. 2. P. 1—202.
King J. R. Adaptive periodic fat storage by birds // Proc. XVth Intern. Ornithol.
Congr. 1972. P. 200-217.
King J. R., Earner D. S. The adaptive role of winter fattening in the White-crowned
Sparrow with comments of its regulation // Amer. Nalur. 1966. Vol. 100. P. 403—
418.
Kluyver H. N. The population ecology of Great Tit, Parus m. major L. // Ardea. 1951.
Vol. 39. P. 1-135.
Kluyver H. N. Roosting habits, sexual dominance and survival in the great tit // Cold
Spring Harb., Symp. quant, biol. 1957. Vol. 22. P. 281— 285.
Newton I. Winter fattening in the Bullfinch // Physiol. Zool. 1969. Vol. 42. P. 96—107.
Odum E. P. Weight variations in wintering White-throated Sparrows in relation to
temperature and migration//Wilson Bull. 1949. Vol. 61. P. 3—14.
Orell M., Ojanen M. Mortality rates of the Great Tit Parus major in a northern popula-
tion //Ardea. 1979. Vol. 67. P. 130-133.
Snow D. W. The winter avifauna of Arctic Lapland // Ibis. 1952. Vol. 94. P. 133—143.
Sokal R., Rohlj F. J. Biometry. The principles and practice of statistics in biological
research. San Francisco: Freeman and Co, 1969. 776 p.
Uljstrand S. On the nonbreeding ecology and migratory movements of the Great Tit
(Parus major} and the Blue Tit (Parus caeruleus) in Southern Sweden [Kpbn-
havn], 1962. 146 p. (Var. Fagelvarld; Suppl. 3).
SUMMARY
The dynamics of sexual and age composition and fatty reserves of Great Titmice (Parus
m. major) in winter time. В a r d i n A. V.
In 1968—1983 in the Town Pechor in Pskov Region Great Titmice were caught and
painted in different colours and were birdbanding with standard aluminium rings.
The fatty reserves were estimated by Blumenthal and Dolnik Method (1962). The
data on 1920 specimens were obtained. Among wintering birds the specimens which
migrate from the other regions, form a considerable part. Some part of local birds
migrate from the studied region in autumn. The correlation of adult and young birds
varies in different years. The part of the adults changes from 13 to 57%, on the average
it is 32%. About 80% of young birds migrate from the forest for winter time to the other
geographical regions. The annual and seasonal dynamics of sexual and age composition
of Great Titmice, wintering in the town, depends on the general successfulness of
their reproduction on a rather large territory, differential death rate of members of
various sexual and age groups, wide autumn and spring migrations of specimens, the
alternate use of habitat zones by some specimens in winter time.
47
УДК 591.563:598.816
Т. А. ИЛЬИНА
119899, Москва,
Ленинские горы.
МГУ им. М. В. Ломоносова.
Биологический факультет.
Кафедра зоологии позвоночных
Расход времени и энергии
на гнездование
у серой вороны
(Corvus cornix)
Изучение затрат времени и энергии на различные формы репро-
дуктивного поведения увеличивает возможность сравнения страте-
гий размножения разных видов, разных географических популя
ций одного вида и т. д. Этот подход, в частности, очень перспекти-
вен при изучении гнездостроительного поведения. Продолжитель-
ность постройки гнезда в отличие от большинства других фаз пери-
ода размножения (откладки яиц, насиживания и т. п.) может очень
сильно варьировать в зависимости от даты ее начала, погодных ус-
ловий и т. п. О необходимости количественно охарактеризовать
процесс постройки гнезда часто свидетельствуют встречающиеся
в орнитологических работах описания типа: «строят долго и мед-
ленно», «сначала строят медленно, а к концу строительства темпы
увеличиваются», «строят очень интенсивно» и т. п. В рамках част-
ного исследования такие субъективные характеристики иногда
оказываются вполне достаточными, но для объективных коли-
чественных сравнений необходимы такие оценки, как расход
времени и энергии за какой-либо определенный период деятель-
ности.
Материал и методика
Согласно многолетним данным о фенологии гнездового периода
ворон в Ленинграде и Ленинградской области, самые ранние даты
начала строительства гнезд— 15—17 марта, а наиболее поздние
даты окончания — двадцатые числа мая (Мальчевский, 1959; Шу-
тенко, 1983).
Наблюдения за серыми воронами, строящими гнезда, проведены
в Ленинграде в период с 29 марта по 28 апреля 1986 г. Температура
воздуха во время наблюдений была в среднем -|-2о и колебалась
от —5 до -|-8о. Во время сеансов наблюдений продолжительностью
от 0,5 до 2 ч, а также во время одного наблюдения, проведенного
5 апреля в течение всей светлой части суток (с 7 до 21 ч), в общей
сложности в течение 40 ч, осуществлено хронометрирование пове-
дения четырех пар ворон. Для этой цели использовались секундо-
меры и метроном. Последний особенно удобен при измерении дли-
тельности кратковременных, часто чередующихся друг с другом
поведенческих актов, например перемещения в кроне дерева, вы-
сматривания веток, пригодных для гнезда, и т. п. На основе данных
хронометрирования определены средние затраты времени на раз-
личные формы гнездостроительного поведения и по ним рассчита-
48
ны затраты энергии на гнездостроение. Четыре гнезда после выле-
та птенцов были сняты, разобраны на компоненты и взвешены по
частям на весах с точностью до 0,5 г.
Результаты наблюдений за строительством гнезд
Гнездовая постройка ворон состоит из наружного слоя толстых
веток, слоя земли и верхних слоев, образованных тонкими веточ-
ками, кусочками древесной коры, луба и разнообразных мягких
материалов, выстилающих лоток (шерсти, перьев и т. п.). Масса
гнездовой постройки в среднем равна 1843 г (1200—2280 г).
Основная ее доля приходится на ветки (1175 г), число которых
в разобранных нами гнездах составило 242 (186—310), слой земли
(300 г), кусочки коры и луба (225 г).
Продолжительность постройки может варьировать от трех дней
до трех недель (Мальчевский, 1959; Грабовский, 1983). Наиболее
растянута постройка ранних гнезд, наиболее сжата постройка позд-
них.
Средняя продолжительность строительства, по данным раз-
ных авторов, составляет: 6—7 дней (Чмутова, 1953), 10—15 (Шка-
тулова, 1959), 9 (Дорофеев, Звездина, 1972), 7—8 (Сметана, 1978).
По нашим наблюдениям за четырьмя гнездами, строящимися в се-
редине периода гнездостроения, продолжительность их строитель-
ства равнялась 8—9, 7—8, 7 и 6—7, в среднем 7,4 дням. По нашим
наблюдениям за гнездами, строившимися при благоприятных по-
годных условиях в «средние» сроки, оба партнера поочередно при-
летали к гнезду в течение всей светлой части суток (четырнадцати
часов) в среднем 8,63 раза в час (максимум до 18 прилетов в час).
Таким образом, за время постройки одного гнезда партнеры совер-
шают в общей сложности 895 прилетов.
При хронометрировании гнездостроительной деятельности мы
выделяли следующие формы поведения.
Поиск гнездового материала — полет от места расположения
гнездовой постройки к месту сбора гнездового материала, переме-
щения по земле или по ветвям деревьев (ходьба и перепархивание),
высматривание подходящих объектов. Основную часть гнездового
материала вороны собирают на гнездовом участке. За время наблю-
дений лишь в пяти случаях гнездовой материал был принесен
с расстояния 100 м и более, а средняя дистанция сбора составила
42±1,9 м. Расстояние между гнездом и местами сбора замеряли
при помощи рулетки.
Добывание объекта — отламывание веток, отрывание кус-
ков луба, коры, рассучивание и щипание материала для внутрен-
них слоев гнезда. Большую часть гнездового материала (3/4) воро-
ны собирают на деревьях, а остальную — на земле.
Т ране портировка добытого материала к гнезду включа-
ет как все типы перемещения с гнездовым материалом в клюве
(полет, перепархивание, ходьба), так и остановки, связанные
с перехватыванием объекта. Во время интенсивной постройки гнез-
49
Таблица 1
Средние затраты времени (с), необходимые для доставки в гнездо одной порции
строительного материала
Форма поведения Доставка объекта с дерева Доставка объекта с земли
М±т lim М±т lim
Поиск гнездового материала полет 6,60+0,63 2-21 5,88+0,73 2-15
перепархивание 1,08±0,64 0-30 0,94±0,54 0-7
ходьба 5,59+1,57 0-69 145,30+31,02 21-660
высматривание 21,00±5,66 0-180 27,29+15,70 0-315
Отламывание ветки, сдирание 34,22±4,30 5-145 — —
коры, луба Транспортирование материа- ла к гнезду полет 7,21+0,62 3-24 9,12±1,04 4-20
перепархивание 1,37+0,54 0-20 3,81+2,88 0-60
ходьба 1,73+0,60 0-18 — -—
перехватывание, щипание, 25,08+4,60 0-108 19,67+11,46 3-240
сидение в позе готовности
да птица, уложив принесенную порцию гнездового материала,
целенаправленно летит за новой, но очень часто, слетев с гнезда,
переключается на какую-либо другую деятельность, не связанную
непосредственно с гнездостроением, и лишь через некоторое время
начинает добывать гнездовой материал. В таких ситуациях затраты
на поисковый полет выделить невозможно, поэтому мы условно
приравнивали их к затратам, необходимым на безостановочный по-
лет от места постройки гнезда к месту сбора материала.
Затраты времени, необходимые для поиска, добывания и транс-
портировки одного объекта, приведены в табл. 1. В некоторых слу-
чаях (14,6%) вороны прилетали к гнезду без материала и занима-
лись укладыванием принесенных ранее порций. Такие прилеты мы
также включали в гнездостроительную деятельность.
Продолжительность обработки и укладывания принесенного
материала варьирует от 12 до 280 с, составляя в среднем 90,7+5,2 с.
Из них 82 с птица работает преимущественно клювом и толь-
ко в течение 8,7 с совершает энергичные движения всем кор-
пусом.
Реальные затраты времени на строительство гнезда выше
затрат, которые можно было бы рассчитать, исходя из приведен7
ных в табл. 1 данных и числа прилетов к гнезду с материалом
и без него, так как не весь добытый материал вороны доставляют
к месту строительства. Иногда это происходит без видимых причин,
а иногда из-за нападения соседей. Так, например, в отдельные дни
мы регистрировали по 3—4 конфликта в час, общая продолжитель-
ность которых доходила до 4 мин, из которых 58 с приходилось на
драки и преследования в воздухе. На наш взгляд, затраты на поиск
50
и добывание материала, который оказывается утерянным по раз-
ным причинам, необходимо включать в затраты на гнездостроение.
Доля таких порций значительна: по наблюдениям за сбором гнездо-
вого материала, из 106 порций, собранных на деревьях, на месте
сбора было брошено 28, а из 32 порций, собранных на земле, было
брошено 7. В табл. 2 приведены затраты времени на постройку все-
го гнезда с учетом затрат на поиск и добычу объектов, не доставлен-
ных в гнездо, а также на полеты к гнезду без строительного мате-
риала.
По данным о расходе времени на перечисленные выше формы
поведения при помощи универсальных коэффициентов пересчета
(Дольник, 1980) рассчитаны затраты энергии на строительство
Таблица 2
Полные затраты времени и энергии на постройку гнезда
Форма поведения Затраты времени, ч Коэффициент пересчета Затраты энергии, кДж за вычетом затрат на ВМ
Поиск объектов на деревьях
полет 1,40 12,0 211,75
перепархивание 0,23 6,0 15,81
ходьба, прыжки 1,19 1,6 9,76
высматривание 4,46 1,3 18,43
Добывание объектов на деревьях 7,26 1,6 59,95
Транспортировка объектов, добытых на
деревьях
полет 1,49 12,0 225,36
перепархивание 0,28 6,0 19,25
ходьба, прыжки 0,36 1,6 3,03
балансирование, перехватывание 5,17 1,3 21,31
и т. п.
Поиск объектов на земле
полет 0,38 12,0 57,48
перепархивание 0,06 6,0 4,13
ходьба 9,28 1,6 76,59
высматривание 1,74 1,3 7,15
Транспортировка объектов, добытых на
земле
полет 0,46 12,0 69,58
перепархивание 0,19 6,0 13,06
перехватывание, щипание, рассучи- 0,98 1,3 3,99
вание
Строительство гнезда
преимущественно клювом 20,39 1,3 84,15
всем корпусом 2,15 1,6 17,74
Прилеты без гнездового материала 0,29 12,0 43,86
Всего 57,76 962,38
51
гнезда. Скорость базального метаболизма (ВМ) серой вороны
330 кДж/сут (Gavrilov, Dolnik, 1985).
Для оценки собственно гнездостроительной деятельности из за-
трат во время постройки гнезда были вычтены затраты на базаль-
ный метаболизм за время этой деятельности. Затраты энергии на
строительство гнезда также приведены в табл. 2.
Обсуждение
В настоящее время лишь для двух видов — каменной ласточки
(Petrochelidon pyrrhonota) и зяблика (Fringilla coelebs) — опреде-
лен расход времени на все формы гнездостроительного поведения,
оценены затраты энергии на постройку гнезда (Withers, 1977;
Дольник, 1982; Дольник, Ильина, 1982; Ильина, 1982). Пара ка-
менных ласточек, птиц массой 25 г, строит из мокрой грязи гнездо
массой 600 г в течение 7—8 сут., расходуя на строительство 23,3 ч.
Этот специализированный вид сильно зависит от источника мате-
риала для строительства и относительно много времени затрачи-
вает на его доставку (табл. 3). Несмотря на «дешевый» полет
(6,4 ВМ), суммарный коэффициент * всей гнездостроительной де-
ятельности оказывается довольно высоким (3,47 ВМ). Расход
энергии во время гнездостроения составляет 122 кДж (Withers,
1977). Если вычесть йз этой величины затраты энергии на базаль-
ный метаболизм, то чистый расход энергии на постройку гнезда
равен 86,8 кДж, что в 2,39 раза превышает расход на базальный мета-
болизм за то же самое время.
Самка зяблика (массой 20 г) в течение пяти суток затрачивает
на строительство гнезда около 40 ч, расходуя при этом (в чистом
виде, т. е. за вычетом энергии на базальный метаболизм) 51,6 кДж
(Дольник, 1982; Дольник, Ильина, 1982; Ильина, 1982). Чистый
расход энергии на строительство гнезда у пары серых ворон (масса
птицы приблизительно 500 г) составляет, как следует из табл. 2,
962,4 кДж. Ворона и зяблик не так сильно, как каменная ласточка,
зависят от источника материала для постройки гнезда, но затрачи-
вают много времени на его поиск и сбор. Наиболее существенные
различия между этими двумя видами, гнездящимися в кроне де-
ревьев, заключаются в следующем. Постройка зяблика — сложная
плетеная конструкция из практически не преобразованного строи-
тельного материала. Самка, строящая гнездо, затрачивает прибли-
зительно одинаковое время на работу клювом и энергичные движе-
ния всем корпусом. Для ворон, в отличие от большинства мелких
видов воробьиных птиц, характерна длительная обработка гнездо-
вого материала клювом. Часть его птицы щиплют и рассучивают
во время сбора, часть — непосредственно на гнезде. Формы поведе-
ния во время сооружения гнездовой постройки у ворон энергетиче-
Суммарный коэффициент сложной деятельности равен сумме затрат времени
(в долях) на каждый тип активности, умноженных на соответствующие этим
активностям коэффициенты.
52
Таблица 3
Затраты времени на разные формы гнездостроительного поведения у трех видов
воробьиных птиц
Форма деятельности Зяблик Серая ворона * Каменная ласточка *
абс., ч доля, % абс., ч доля, % абс., ч доля, %
Строительство гнезда 12,4 31,1 22,5 38,9 11,7 50,0
Полеты за гнездо- вым материалом и обратно 1,7 4,3 4,0 7,0 7,7 33,3
Прочие виды активности (сбор материала, его предварительная обработка и т. п.) 25,8 64,6 31,4 54,1 3,9 16,7
Всего 39,9 100,0 * Суммарные затраты обоих партнеров. 57,9 100,0 23,3 100,0
ски более «дешевые», чем у зяблика, но суммарный коэффициент
всех форм гнездостроительной деятельности, включая сбор мате-
риала и его доставку, у ворон равен 2,22, а у зяблика 2,03 ВМ. Это
объясняется тем, что вороны носят гнездовой материал с больших
расстояний, чем зяблики, и затрачивают на полет больше времени
и энергии (см. табл. 3). Интересно отметить, что вороны, если даже
находят подходящий строительный материал рядом с деревом, на
котором ведут строительство, подлетают к гнезду по спирали, уве-
личивая продолжительность полета, но избегая тем самым мощных
вертикальных взлетов. Относительные затраты на всю постройку
у пары ворон также оказываются выше, чем у зяблика. У вороны
они в 2,92 раза выше суточных затрат одной особи на базальный
метаболизм, а у зябликов в 1,55 раза, т. е. у серой вороны каждый
из партнеров строит приблизительно половину гнезда и имеет при
этом относительные затраты, сходные с относительными затратами
самки зяблика, строящей в одиночку целое гнездо.
Судя по литературным данным, серая ворона в северных частях
ареала более охотно использует прошлогодние гнезда, чем в юж-
ных. По подсчетам Ю. Н. Кречетова (1979), в дельте Днестра из
более чем 500 гнезд только 9 были заселены на следующий год пос-
ле достройки. В Ленинградской области приблизительно каждое
десятое гнездо используется на следующий год (Шутенко, 1983).
На Кольском полуострове, по характеристике Н. С. Бойко (1977),
повторное использование воронами старых гнезд — явление обыч-
ное. Одна из возможных причин этой тенденции — относительно
большие энергетические затраты на гнездостроение, которые в се-
верных районах необходимо реализовать в более сжатые сроки
и при более суровых погодных, в том числе и температурных усло-
виях.
53
ЛИТЕРАТУРА t
Бойко И. С. Экология и практическое значение серой вороны в Мурманской области:
Автореф. дис. ... канд. биол. наук. М., 1977. 16 с.
Грабовский В. И. Социальная структура популяции серых ворон (Corvus cornix) на
Ленинских горах в Москве // Зоол. журн. 1983. Т. 62, № 3. С. 389 - 398.
Дольник В. Р. Коэффициенты для расчета расхода энергии свободноживущими
птицами по данным хронометрирования их активности // Орнитология. М.:
Изд-во МГУ, 1980. Вып. 15. С. 63-72.
Дольник В. Р. Продуктивная энергия и продукция взрослых зябликов // Популя-
ционная экология зяблика. Л.: Наука, 1982. С. 64—86.
Дольник В. Р., Ильина Т. А. Бюджет энергии свободноживугцих зябликов в гнездо-
вой период // Там же. 1982. С. 229—239.
Дорофеев А. М., Звездина Л. Ф. Экология серой вороны в Белорусском Поозерье //
Животный мир Белорусского Поозерья. Минск: Изд-во Белорус, ун-та, 1972. J
Вып. 2. С. 25-38.
Ильина Т. А. Бюджет времени и поведение зяблика в гнездовой период // Популя-
ционная экология зяблика. Л.: Наука, 1982. С. 191—214.
Кречетов Ю. И. Гнездование серой вороны в дельте Днестра // Экология гнездова-
ния птиц и методы ее изучения. Самарканд: Самарканд, ун-т., 1979. С. 116—
117.
Мальчевский А. С. Гнездовая жизнь певчих птиц. Л.: Изд-во ЛГУ, 1959. 281 с.
Сметана Н. М. Фенология серой вороны в Наурзумском заповеднике // Вторая , >
Всесоюз. конф, по миграциям птиц: Тез. докл. Алма-Ата: Наука, 1978. Ч. 2. S
С. 140-142. 1
Чмутова А. И. Особенности развития и размножения птиц (серая ворона) в различ- ;
ных географических зонах СССР // Бюл. МОИН. Отд. биол. 1953. Т. 58, вып. 6. J
С. 27-34. !
Шкатулова А. П. Экология серой вороны и ее практическое значение: Автореф. дис.
... канд. биол. наук. М., 1959. 24 с.
Шутенко Е. В. Серая ворона // Птицы Ленинградской области и сопредельных тер-
риторий: история, биология, охрана. Л.: Изд-во ЛГУ, 1983. Т. 2. С. 439—449.
Gavrilov V. М., Dolnik V. В. Basal metabolic Rate, Thermoregulation and Existence
Energy in Birds: World data // Acta XVIII Congressus Internationalis Ornitholo-
gicus, Moscow I 16—25 August, 1982. Moscow: Nauka, 1985. Vol. I. P. 421—466.
Withers P. C. Energetic aspects of reproduction by the Cliff Swallow // Auk. 1977.
Vol. 94, N 4. P. 718-725.
SUMMARY
Time and energy expenditure for nest building in the Hooded Crow (Corvus cornix).
11 у i n a T. A.
From the data of visual observations of 4 pairs of the Hooded Crows in Leningrad in
April 1986 the time budget was determined. Based on it, the energy expenditure for
nest building was calculated. The duration of the nest building period is 7—8 days in
average. During this period a pair of crows spends the next amount of time and energy
(except the value of basal metabolism) on different forms of nest building behaviour: ,
nest construction — 22,5 hours and 101 kJ; flights for nest material and its transporta-
tion — 4,0 hours and 608,0 kJ; the collection of nest material and its advance prepara-
tion — 31,4 hours and 252,5 kJ. The energetic cost of building of one nest is 2,92 times
higher than the value of individual’s daily basal metabolism (dBM). It is higher than
a relative cost, known for nests of smaller passerines: it is 2,39 dBM in a pair of Cliff
Swallow (Petrochelidon pyrrhonota) (Withers, 1977) and 1,55 dBM in Chaffinch fema-
le (Fringilla coelebs) (Dolnik, 1982; Dolnik, Ilyina, 1982).
54
УДК 591.525:598.412.2
В. Г. ПАПЧЕНКОВ
420041,
Казань, ул. Тактам, д. 114.
Волжско-Камское отделение
ВНИИОЗ
Характер
разногодичных изменений
условий обитания утиных
на мелководьях водохранилища
с переменным уровнем
наполнения
Большие и малые водохранилища стали неотъемлемой частью
ландшафтов нашей страны. Особенно много их создано в европей-
ской части СССР. Помимо глубоководной рабочей зоны, подавляю-
щее большинство водохранилищ имеет обширные зоны мелководий
с глубиной до 2 м, занимающих обычно от 12 до 46% площади
водохранилища (Успенский, 1974). В целом по стране их площадь
равна примерно 1 млн га (Синявская, Лисицкий, 1974). Обычно
мелководья зарастают высшей водной растительностью, что создает
благоприятные условия для обитания здесь многих животных
водно-болотного комплекса, в том числе различным представите-
лям водоплавающих птиц.
Продуктивность мелководий определяется прежде всего ста-
бильностью их обводнения и защищенностью от волнобоя. В связи
с этим все водохранилища могут быть подразделены на водоемы с
относительно стабильным уровнем воды по сезонам года и между
годами, как это имеет место, например, на Иваньковском и Киев-
ском водохранилищах, мелководья которых зарастают соответ-
ственно на 36 и 95% (Успенский, 1974), и водоемы с сезонным
и межгодовым изменением водного уровня на 1,5 м и более,
мелководья которых зарастают слабо (Голубева, Шпак, 1984;
Экзерцев, Лисицина, 1984). По степени защищенности от волно-
боя с учетом работы В. М. Синявской и Н. Т. Лисицкого (1974)
мелководья можно разделить на 4 типа: 1) автономные мелководья,
соединяющиеся с чашей водохранилища нешироким проливом,
который легко может быть перекрыт дамбой, в результате чего
образуются замкнутые водоемы с относительно независимым
гидрорежимом, подобные озерам пойм крупных рек; 2) широкие
заливы, достаточно защищенные от ветра (высота возникающих
волн не более 0,5 м); 3) узкие меандровые заливы и протоки;
4) широкие открытые заливы, мелководья у островов и берегов
основного плеса водохранилища, подверженные сильному
волновою.
Первый тип мелководий наиболее продуктивен на водохранили-
щах как с постоянным, так и с переменным уровнем наполнения;
четвертый тип наименее продуктивен. Продуктивность второго
и третьего типов зависит от постоянства уровня воды и имеет
средние и низкие значения.
55
Для конкретизации этих наиболее общих закономерностей
и непосредственно мониторинга природных процессов, происходя-
щих в биогеоценозах разных типов мелководий с переменным уро-
венным режимом, нами в 1977 г. начаты стационарные наблюдения
на водоемах острова-заказника Мизиновского охотничьего хо-
зяйства (Татарская АССР), расположенного в зоне мелководий
Свияжского отрога Куйбышевского водохранилища.
Программа работ на стационаре наряду с другими вопросами
включала в себя выявление запасов растительных кормов речных
уток и относительный учет их численности перед открытием охоты.
Согласно литературным источникам (Краевский, 1954; Приклон-
ский, Самарина, 1974; Рыбакова, 1983; Виноградов, Чернявская,
1984; и др.), в качестве растительных кормов речных уток рас-
сматривались семена и плоды таких водных и воздушно-водных
растений, растущих в изучаемых угодьях, как различные виды
рдестов (Potaniogeton lucens L., Р. pectinatus L., P. perfoliatus L.,
P. gramineus L., P. compressus L., P. berchtoldii Fieb.), горец
земноводный Polygonum amphibium L., манник большой Glyceria
maxima (C. Hartm.) Holmb., камыш озерный Scirpus lacustris L.,
двукисточник тростниковый Phalaroides arundinacea (L.) Rau-
schret, ежеголовники (Sparganim erectum L., S. emersum Rehm.),
стрелолист обыкновенный Sagittaria sagittifolia L., различные вре-
менники (Rorippa amphibia (L.) Bess., Oenanthe aquatica (L.)
Poir., Rumex maritimus L. и др.) и осоки (Carex acuta L. и др.).
Кроме того, в качестве кормов учитывались все запасы рясковых
(Lemna minor L., L. trisulca L., Spirodela polyrhiza (L.) Schleid).,
луковички стрелолиста, вегетативные почки и бутоны телореза
алоэвидного Stratiotes aloides L., молодые побеги рдестов, рого-
листника темно-зеленого Ceratophyllum demersum L. и элодеи
канадской Elodea canadensis Michx., запасы которых принимались
равными трети надземной биомассы этих растений. Материалы
по семенной продуктивности макрофитов Куйбышевского водо-
хранилища нами опубликованы (Папченков, 1984).
Описание угодий
Площадь острова-заказника с заливами и отделяющей его от
коренного берега протокой равна 229 га. Из них при нормальном
подпорном уровне (НПУ-53,0 абс. м) 174 га приходится на вод-
ную поверхность и 55 га — на сушу. Водные угодья представлены
автономными, искусственно отделенными от основной части
мелководий земляными плотинами озеровидными водоемами:
оз: Большим — 36 га, оз. Малым — 4 га (до 1983 г.— 8 га, затем
было перекрыто дополнительной плотиной), оз. Лосиным — 4 га,
а также защищенным от сильного волнобоя заливом с тремя лопа-
стями — 77 га, протокой — 42 га и более мелкими, заливаемыми
водой низинами, суммарная площадь которых около 11 га. Водоемы
в разной степени зарастают водной растительностью, в которой
56
I --------- ---------- ---------- ---------- ----------- ---------
I
преобладают сообщества с доминированием рогоза узколистного
Typha angustifolia L., тростника обыкновенного Phragmites austra-
Ilis (Cav.) Trin. ex Steud., манника большого, рдеста блестящего
Potamogeton lucens L., роголистника темно-зеленого, горца земно-
водного, а в отдельные годы — пузырчатки обыкновенной Utri-
cullaria vulgaris L. и урути мутовчатой Myriophyllum verticil-
latum L. Основную часть суши (43 га) занимает дубняк с осиной
Populus tremula L. и липой Tilia cordata Mill., остальные 12 га —
луга и ивняки. При горизонте воды ниже НПУ к ним добавляются
площади затопленных лугов и обсыхающие обнаженные отмели,
площади же водоемов, естественно, сокращаются. Особенно сильно
; (более чем наполовину) усыхают залив и протока, а также озера
Малое и Лосиное, тогда как площадь оз. Большого почти не сокра-
щается даже при очень слабой обводненности мелководий.
I Общая схема изучаемых угодий и поперечные профили через
них, выполненные при разных уровнях воды, показаны на рис. 1.
Промеры глубин на профилях показали, что на протоке их вели-
ь чины достигают 3,9 _м, в заливе — 3,8 м, на оз. Лосином — 2,4 м,
на оз. Большом — 1,9 м, на оз. Малом — 1,7 м. Но глубины эти
являются величинами очень непостоянными, они зависят от уров-
ня воды в водохранилище. В маловодные годы глубина озер не
J превышает 70—120 см, а в незарегулированных заливе и протоке
J вода остается только в наиболее глубоких ямах и ложбинах,
i Остров с юго-запада отделен от коренного берега 80—100-метровой
протокой, на северо-востоке к нему прилегают основные мелково-
дья с рядом похожих островов и грив. На коренном берегу
небольшие лесные участки чередуются с полями и небольшими за-
болоченными лугами. Остров является частью заказника охотни-
чьего хозяйства. Он сравнительно мало посещается людьми
(исключением являются одна-две недели июля, когда обкаши-
вается часть лугов), на нем не ведется охота. Водоемы острова-
заказника, и прежде всего зарастающие водной растительностью
озера (особенно оз. Большое), используются водоплавающими
птицами как кормовые угодья. По их берегам, а также в биотопах
коренного берега гнездятся обычные здесь кряква Anas platyr-
hynchos и чирки (A. crecca, A. querquedula), реже — широконоска
A. clypeata и шилохвость A. acuta. Помимо речных уток в изучае-
мых угодьях постоянно гнездятся одна-две пары лысухи Fulica
atra, две-три пары чомги Podiceps cristatus, обитает серая цапля
Ardea cinerea и другие птицы водно-болотного комплекса.
Динамика угодий
Исследования, проведенные нами в 1977 —1985 гг., показали, что
мелководья Куйбышевского водохранилища как места обитания
утиных птиц претерпевают значительные изменения по годам,
и вызваны они прежде всего характером гидрорежима водохрани-
лища в весенне-летний период.
57
Рис. 1. Профили (Л и Б) и схема их расположения в изучаемых угодьях (В)
1 — лес; 2 — ивняки; 3 — рогоз узколистный; 4 — тростник обыкно- рдесты и другие погруженные растения, 10 — луг; сплошная горизоя-
венный; 5 — двукисточник тростниковый; 6 — манник большой; 7 — тальная линия — высокий уровень воды, август 1978 г., пунктирная
стрелолист обыкновенный; 8 — роголистник темно-зеленый; 9 — линия — пониженный уровень, август 1977 г.
58
i
Начало наблюдений пришлось на последний маловодный год
в их длительном ряду (1972 — 1977 гг.). В середине вегетационных
периодов засушливых 1972, 1973, 1975 гг. уровень воды в водохра
нилище находился на 2,5—3,5 м ниже НПУ и многие мелководья
находились в обсохшем состоянии (Голубева, Шпак, 1984). В 1976
и 1977 гг. среднелетний горизонт воды был меньше нормы на 120
и 70 см соответственно, обводненность зоны мелководий составля-
ла примерно 50%. Многие отмели по-прежнему оставались над
водой. В протоке они представляли собой свободные от раститель-
ности грязевые пространства. Обсохшие отмели залива и замкну-
тых водоемов были заняты плотными зарослями двукисточника
тростникового и (ближе к воде) полевицы побегообразующей
Agrostis stolonifera L., увлажненные и топкие места — зарослями
манника большого. Обводненные части озер имели хорошо разви-
тую водную и особенно воздушно-водную растительность, в которой
доминировали сообщества рогоза узколистного, двукисточника и
манника. Заметную биомассу здесь давали также тростник, рдест
блестящий, ежеголовник прямой и уруть мутовчатая, но аспекти-
рующим видом был, несомненно, рогоз узколистный. Особенно
мощно он был развит на оз. Большом, где его сообщества росли и об-
ширными купаками, и кольцеобразными и дугообразными линиями
и стенками, чередуясь с небольшими плесами, занятыми ценозами
рдеста блестящего и урути мутовчатой. Это создавало прекрасные
условия для обитания утиных и других водоплавающих птиц. Сте-
пень зарастания озер, т. е. отношение площади зарослей к площади
водоема, составляла 75—100% (табл. 1), интенсивность зараста
ния, под которой мы понимаем запас сырой надземной массы
макрофитов, приходящийся на 1 м2 водоема (Папченков, 1979,
1982, 1985), была в пределах 4,08—7,05 кг/м2 (табл. 2). Согласно
нашей шкале (Папченков, 1979, 1982), это соответствует сильному
и Очень сильному зарастанию водоемов.
Достаточно обильными были растительные корма речных уток.
Из-за мелководности водоемов в них были хорошо развиты и обиль-
но плодоносили ежеголовник прямой и стрелолист обыкновенный.
Их плоды и семена составляли более 90% запасов этой группы
кормов. Среди зеленого корма уток наиболее велики были запасы
рясок. Их дополняли запасы побегов рдеста блестящего, роголи-
стника темпо-зеленого и луковиц стрелолиста. В августе угодья
были заполнены утками. Большинство из них (около 600 птиц)
держалось на оз. Большом. Численность речных уток в угодьях в
этот год была максимальной за все время наблюдений (рис. 2).
С 1978 г. начинается новый, многоводный, период в жизни
обитателей мелководий Куйбышевского водохранилища. Характер
гидрорежима этого года внес резкие перемены в сложившиеся и
адаптировавшиеся к «малой» воде биоценозы этой зоны. Весенний
паводок 1978 г. был по среднемноголетним данным нормальным
по величине, хотя и большим, чем в предыдущие маловодные годы
(на 70 см выше НПУ). Начался он на 12 дней раньше срока, за-
кончился на 6 дней раньше. Своего пика половодье достигло
59
Таблица 1
Динамика степени зарастания водоемов, %
Водоем Годы
1977 1978 1979 1980 1981 1982 1983 1984 1985
Оз. Большое 82 100 30 13 26 34 46 67 81
Оз. Малое 75 75 26 9 15 46 80 16 94
Оз. Лосиное 100 100 15 7 16 34 36 100 94
Протока 16 15 5 3 9 18 21 0 25
Залив 9 12 8 6 11 20 17 5 23
Прочие водоемы 95 98 9 3 20 25 24 — 54
В целом 49 56 12 7 14 24 26 26 40
Таблица 2
Динамика интенсивности зарастания водоемов, кг/м2
Водоем Годы
1977 1978 1979 1980 1981 1982 1983 1984 1985
Оз. Большое 4,8 6,8 1,3 0,5 1,3 2,0 3,2 4,5 6,0 Оз. Малое 4,1 4,2 0,9 0,3 0,7 1,4 1,8 0,5 5,2 Оз. Лосиное 7,1 4,2 0,4 0,1 0,4 1,4 1,0 4,3 5,5 Протока 0,8 0,7 0,3 0,1 0,5 1,0 1,0 0,0 1,4 Залив 0,3 0,6 0,3 0,2 0,6 1,1 1,2 0,1 1,6 Прочие водоемы 3,8 5,3 0,2 0,2 1,0 0,7 0,4 — 1,9 В целом 2,0 2,4 0,5 0,2 0,7 1,2 1,5 1,6 2,7
в третьей декаде апреля; с началом мая началось интенсивное
падение водного уровня, и в первой декаде июня он уже был на
1,4 м ниже НПУ. На мелководьях сложилась обстановка, типичная
для маловодных лет. Несмотря на неустойчивую погоду, вода
прогрелась до 12—15°. В ней и на обнажившихся отмелях нача-
лась интенсивная вегетация водных и околоводных растений, в тех
же биотопах, что и в предшествующие годы, загнездились водопла-
вающие птицы. Но 30 мая была закрыта водосливная плотина
Волжской ГЭС. В условиях частых и обильных дождей приток
воды в водохранилище стал заметно превышать ее сброс. Начался
новый быстрый подьем водного уровня, который в первой декаде
июля был уже на 0,5 м выше НПУ. Вода залила все понижения,
водоемы залили все близлежащие низины, захватили большие
участки лугов, кустарников и леса, некоторые из них на больших
участках береговой линии слились друг с другом. Повышение
уровня воды привело к тому, что большая часть утиных кладок
была затоплена и погибла. В результате к осени в водоемах острова-
заказника насчитывалось лишь 20—30% от прошлогоднего числа
речных уток.
60
Рис. 2. Динамика численности речных уток и запасов растительных кормов в угодьях острова-заказника
1 — ежеголовник прямой; 2 — частуха подорожниковая; 3 — стрело- надская; 7 — ряски; 8 — рдесты; 9 — телорез алоэвидный; 19
лист обыкновенный; 4 — манник большой; 5 — осоки; б — злодея ка- голистник; 11 — малообильные виды в сумме
61
На водной растительности такое колебание уровня воды ска
залось весьма положительно. Воздушно-водные растения начали
вегетацию при слабой обводненности водоемов, последовавшее за
тем повышение водного уровня как бы подгоняло их рост. К кон-
цу лета рогоз, тростник,' манник, стрелолист были сильно развиты.
Целиком из водных фитоценозов выпал ежеголовник прямой,
не осталось полевицы. Но эти потери в общих запасах биомассы
растений были многократно восполнены необычно сильным разви
тием в озерах погруженных макрофитов, которые образовали слож-
ные многокомпонентные фитоценозы с преобладанием пузырчатки
обыкновенной, урути мутовчатой, различных рдестов и рясковых.
На оз. Большом они заполнили весь объем плесов, повысив степень
зарастания водоема до 100% (см. табл. 1), а интенсивность за-
растания— до 6,8 кг/м2 (см. табл. 2). Обилие рясок повысило
запасы кормов речных уток, но при этом из-за выпадения из
угодий ежеголовника снизились запасы семян и плодов водных
растений (см. рис. 2).
1979 г.— второй высоководный год — выделился очень высоким
паводком, достигшим пика на уровне отметки 55,4 м в начале
мая. При этом максимальном за все время существования водо-
хранилища уровне практически весь остров был залит водой.
Летняя сработка воды шла очень медленно. В начале июня водный
горизонт находился на отметке 54,0 м, НПУ был достигнут лишь
в конце августа. Такая гидрологическая обстановка способство-
вала задержке в развитии водных растений, лишила водопла
вающих птиц привычных мест гнездования. В этих условиях
началось изреживание зарослей рогоза и отмирание его на наиболее
глубоких местах. Обильные ранее манник и двукисточник сохрани-
лись только в виде небольших сплавин. Исчезли огромные прошло-
годние заросли пузырчатки, урути и узколистных рдестов. Их
место занял роголистник темно-зеленый. Мощное развитие полу-
чила зона осок, поднявшаяся на луга среднего уровня. Повсеме-
стно вдоль берегов распространился горец земноводный. В целом
запасы растительных кормов речных уток резко снизились (с 46,7
до 6,6 т абсолютно сухого вещества). Их основу составили побеги
роголистника, дополняемые небольшими запасами рдеста блестя-
щего, рясок и стрелолиста. Очень низка в этот год была семенная
продуктивность водных растений. Общий запас семян и плодов
не превышал 300 кг, среди них преобладали семена осок (см.
рис. 2). Численность уток в угодьях еще более снизилась.
Характерной особенностью 1980 г. была ранняя, но затяжная,
с частыми возвратами холодов, весна и прохладное лето. Уровень
воды в водохранилище, упавший за зиму до отметки 47,5 м, подни-
мался медленно и достиг своего максимума (53,6 м) лишь в третьей
декаде мая. В июне уровень воды был повышенным, в июле и авгу-
сте — нормальным. В этих условиях продолжилась начавшаяся в
прошлом году тенденция снижения степени и интенсивности за-
растания водоемов. Их величины упали до минимальных из отме-
ченных за время наблюдений. Больше стало только горца земно-
62
водного, который в этот год был аспектирующим видом на водое-
мах. Запасы растительных кормов речных уток были незначи-
тельны, численность птиц осталась на низком уровне.
В 1981 г. уровень воды в водохранилище достиг максимальной
высоты (54,0 м) в начале мая. Затем началось его равномерное,
со скоростью примерно 0,5 м в месяц, падение. В середине июля
водный уровень был на отметке НПУ, в конце августа — на 1 м
ниже. Лето было жарким и сухим, с большим количеством солнеч-
ных дней. В условиях сильной инсоляции основные доминанты
водных фитоценозов начали интенсивное восстановление своих
запасов. Почти в 2,5 раза по сравнению с предыдущим годом увели-
чились площади, занятые рогозом узколистным. Более чем в три
раза возросли запасы манника большого, осок, горца земноводного
и рдеста блестящего. Столь же заметно увеличились запасы семян
и вегетативных частей растений, поедаемых речными утками.
Одновременно с улучшением кормовых и защитных условий в уго-
дьях повысилась и численность утиных птиц (см. рис. 2). Таким
образом, летом 1981 г. наметилась адаптация биоценозов мелково-
дий к условиям подобного многоводного гидрорежима.
1982 год, подобно 1978 г., имел два пика наполнения водо-
хранилища. Первый, весенний пик, пришелся на первую декаду
мая. Он был низким и на 40 см не достиг НПУ. К концу мая уровень
воды понизился до отметки 52,1 м. Обнажились многие отмели;
в условиях теплой и солнечной погоды на них стали быстро раз
виваться и зацвели временники. Затем началось новое поднятие
водного уровня, который в конце июня приблизился к НПУ, в июле
держался около него, а в конце августа был уже на 1 м ниже НПУ.
Затопленные временники продолжили свое развитие в воде, развив
при этом большую биомассу и вытеснив местами манник большой.
На общем фоне увеличения зарастания водоемов особенно замет-
ным было возрастание площади сообществ рдеста блестящего,
молодые зеленые побеги и плоды которого составили около 80%
всех запасов растительных кормов речных уток (см. рис. 2).
Утки, в условиях многоводья последних лет ставшие гнездиться в
биотопах коренного берега, от двойного паводка не пострадали,
и их численность вновь стала достаточно большой и составила
примерно 60—70% от максимально отмеченной в 1977 г.
1983 г. по гидрорежиму в общих чертах сходен в 1981 г.,
но имел несколько более низкий паводок, а летний уровень более
длительное время держался у НПУ. Хорошо развилась водная
растительность. Полностью восстановилась площадь поредевших
после 1979 г. сообществ рогоза узколистного. Продолжилось расши-
рение площади сообществ рдеста блестящего, увеличилась его про-
дуктивность. Запасы растительных кормов речных уток поднялись
почти до уровня 1977 г. Численность утиных продолжала увели-
чиваться (см. рис. 2).
1984 год прервал серию лет с высоким и нормальным обводне-
нием мелководий Куйбышевского водохранилища. В ряду наших
наблюдений это был самый маловодный год. Площадь водного зер-
63
кала в угодьях сократилась со 174 до 91 га. Примерно на половину
уменьшились залив, протока, оз. Малое и оз. Лосиное. Освободив-
шиеся из-под воды обширные отмели быстро затянулись зарослями
разнообразных временников, среди которых наиболее обильны бы-
ли горец щавелелистпый Polygonum lapathifolium L. и щавель
приморский Rumex maritimus L. Только оз. Большое, сильно
обмелевшее, но потерявшее лишь 12% площади, полностью сохра-
нило свое значение как кормовой и выводковый водоем для водо-
плавающих птиц. Степень его зарастания увеличились по сравне-
нию с прошедшим годом с 46 до 67%, а интенсивность — с 3,2 до
4,5 кг/м 2. Произошло это преимущественно за счет массового
развития роголистника темно-зеленого, стрелолиста обыкновенного
и частухи подорожниковой. Обильное плодоношение последних
заметно обогатило кормовую базу речных уток, численность кото-
рых в угодьях снова стала очень большой. Около десятка пар
утиных, преимущественно кряквы, вновь загнездились в пределах
острова-заказника.
В 1985 г. летний уровень воды вновь был высоким. В отли-
чие от 1979 г., в этот раз не было катастрофического весеннего
паводка, хотя он был все же очень высок и в середине мая достигал
отметки 54,3 м. В летние месяцы водный горизонт находился в
пределах 53,5—53,3 м. Так же, как и в 1978 г., высокая вода после
года маловодья привела к значительному усилению интенсивно-
сти зарастания озер (см. табл. 2). Запасы растительных кормов
речных уток возросли более чем в 2 раза. Еще сильнее был развит
и обильно плодоносил рдест блестящий, очень большой урожай
семян и плодов дали вновь появившийся в больших количествах
ежеголовник прямой и стрелолист обыкновенный. Среди запасов
зеленой массы, помимо рдеста блестящего, доля которого была бо-
лее 60%, большую роль играли запасы роголистника темно-зелено-
го, рясок, элодеи канадской, телореза алоэвидного. Численность же
утиных вновь стала меньше и составляла 60—70% от максимума.
Причиной этого может быть как высокое и позднее весеннее напол-
нение водохранилища, заставившее птиц искать места для гнездо-
вий снова вне острова, так и меньший, чем в прошлые годы, возврат
птиц с мест зимовок, что было отмечено нами в других местах Куй-
бышевского водохранилища и на Нижнекамском водохранилище,
отличающемся более стабильным режимом уровня.
Такова картина изменения угодий и колебания в них числен-
ности речных уток за десятилетний ряд наблюдений. Думается,
что она присуща подобным типам мелководий водохранилищ с
переменным уровнем наполнения. Исследования позволяют заклю-
чить, что такие угодья интенсивнее используются речными утками
в маловодные годы и слабее в годы с уровнями воды, превышаю-
щими НПУ. Сокращение численности уток связано с двойным
весенне-летним подъемом воды, а также с запоздалыми высокими
паводками. Медленное нарастание численности речных уток в
1981 —1983 гг. шло синхронно росту запасов их растительных
кормов, что позволяет предполагать, что численность птиц на этом
64
этапе была обусловлена кормностыо угодий. Сопоставление дина-
мики запаса кормов с динамикой численности уток показывает, что
для обеспечения кормами отмеченного в угодьях максимума птиц
(около 800 шт.) достаточно 30 т абс. сух. вещества семян, плодов
и молодых вегетативных частей водных растений.
ЛИТЕРАТУРА
Виноградов В. В., Чернявская С. И. О некоторых связях птиц с растительностью
дельты Волги // Экология. 1984. № 1. С. 70—73.
Голубева И. Д., Шпак Т. Л. Продуктивность высшей водной растительности на
мелководьях Куйбышевского водохранилища // Продуктивность островных
и прибрежно-мелководных экосистем Куйбышевского водохранилища.
Казань, 1984. С. 5-21.
Краевский И. М. Гигрофиты как источник питания промысловых водоплавающих
птиц // Тр. Моск, пушно-мехов. ин-та. 1954. Т. 5. С. 77—91.
Папченков В. Г. К методике изучения продуктивности водной растительности
в средних и малых реках // Растительные ресурсы. 1979. Т. 15, вып. 3.
С. 454-459.
Папченков В. Г. Характеристика высшей водной растительности рек Среднего
Поволжья: Автореф. дис. ... канд. биол. наук. Свердловск, 1982. 20 с.
Папченков В. Г. Растительные корма утиных, их продуктивность и динамика
запасов на мелководья Куйбышевского водохранилища // Современное
состояние ресурсов водоплавающих птиц: Тез. Всесоюз. семинара, 20—23
октября 1984 г. М.: ВНИИприрода, 1984. С. 319—321.
Папченков В. Г. Зарастание рек Среднего Поволжья и связь его с условиями
среды // Экология. 1985. № 3. С. 20—27.
Приклонский С. Г., Самарина Б. Ф. Опыт оценки утиных угодий в охранной зоне
Окского заповедника//Тр. Окск. гос. зап-ка. 1974. Вып. 10. С. 174 — 194.
Рыбакова Т. И. Сезонная динамика питания некоторых видов пластинчатоклювых
срединного региона СССР // Экология животных и фаунистика. Тюмень:
Тюмен. ун-т, 1983. С. 72—76.
Синявская В. М., Лисицкий Н. Т. Некоторые аспекты сельскохозяйственного
использования мелководий водохранилищ // Мелиорация земель в зоне влия
ния равнинных водохранилищ. М.: Колос, 1974. С. 5—21.
Успенский С. М. Мелководья и подтопленные земли — мелиоративный фонд для
сельского хозяйства //Там же. 1974. С. 198—211.
Экзерцев В. А., Лисицына Л. И. Изучение растительных ресурсов водохранилищ
волжского каскада // Биологические ресурсы водохранилищ. М.: Наука,
1984. С. 89-99.
SUMMARY
The character of environmental changes of waterfowls in different years on the
shallow water basins with changeable water filling level. Papchenkov V. G.
The stationary investigations in 1977 — 1985 showed that among the various types
of shallow water basins with changeable water filling level the most suitable for
dwelling of waterflows are limnetic water basins with a developed aquatic vegetation.
The maximum number of river ducks in such areas can be observed in arid years.
The reduction of birds population is connected with the double spring—summer water
rise (1978), that brought to the flooding and destruction of their egglayings, and
with the late high floods which deprived waterfowls of their habitual nest places
(1975 and 1985). Followed one after another, some full-flowing years cause, at first,
a big reduction of fodder and protective characteristics of the territories (1979 —
1980) by oppression of the aquatic vegetation; then on the 4th full-flowing year
adaptation and rise of productivity of shallow waters biocenoses occurred in 1981 —
1983. The complete restoration of land characteristics was marked on the 6th full-
flowing year in 1983. The river ducks population raised to the former high positions
only in the arid year 1984.
65
3 Заказ № 36
УДК 591.524.23:598.529
В. С. ТАЛПОШ 282011, Тернополь, ул. Ленина, д. 144, кв. 86 О биологии европейской горной трясогузки (Motacilla cinerea cinerea) в Украинских Карпатах
Материалом для данной работы послужили наблюдения автора
в 1962 — 1987 гг. главным образом на Черногоре (урочище Козьме-
щик) и на Свидовецком горном массиве (Раховский р-н Закар-
патской обл.).
На Украине горная трясогузка гнездится в Карпатах и Крым
ских горах. Вне Украинских Карпат мы нашли ее на гнездовье
только в районе с. Скоморохи Бучацкого р-на Тернопольской обл.,
вблизи турбазы «Лесная» (в каньоне р. Стрыпа). В других районах
западных областей УССР она отмечалась изредка лишь во время
миграций или в качестве залетной птицы.
По литературным данным (Портенко, 1950; Кштяшвський, 1950;
Страутман, 1954, 1963 и др.) и личным наблюдениям, горная
трясогузка в Украинских Карпатах населяет и северные, и южные
склоны гор по речным долинам в поясе тайги и широколиственных
лесов.
По горным ручьям она поднимается высоко в горы, в пояс
криволесья, и выходит на субальпийские луга (полонины), где
иногда ее можно встретить у источников на высоте 1500—1800 м
над ур. м.
Гнездится она по берегам горных речек, ручьев и их
притоков, поросших лесом и кустарником, по берегам водо-
хранилищ, горных озер и т. п.; вдоль речек и ручьев, протекающих
через населенные пункты (поселки Ясиня, Усть-Черная), попа-
даются весьма доверчивые особи, подпускающие людей, подобно
воробьям, на расстояние до 0,5 м. Иногда горная трясогузка гнез-
дится в стороне от воды (пос. Ясиня).
В поясе карпатской еловой тайги это обыкновенная, а местами
довольно многочисленная птица. Например, в урочище Козьмещик
(высота 800—1000 м над ур. м.) на 1 км маршрута в 1983
и 1984 гг. гнездилось в среднем около 5 пар. На нижних участках
гор, в поясе криволесья и на субальпийских лугах она встречается
значительно реже.
На западе УССР это перелетная, изредка в небольшом количест-
ве зимующая на южных предгорьях Карпат, обращенных к Закар-
патской низменности, птица. В Украинские Карпаты горная трясо-
гузка прилетает в конце, марта — начале апреля. Вначале она
держится в нижней части- гор и постепенно, по мере таяния
снега, поднимается вверх, вглубь лесов и к их верхней границе
(Страутман, 1963). В конце февраля 1966 г. отмечена в окрест-
ностях с. Среднее Ужгородского р-на (сообщение Ю. И. Крочко).
66
24 марта 1965 г. самца и самку мы наблюдали на р. Тиса в
окрестностях г. Хуст в Закарпатье, а 12 апреля 1987 г. двух сам-
цов — на р. Серет вблизи очистных сооружений Тернополя. 20—
25 марта 1972 г. пару этих птйц отметил В. Н. Кравчук на р. Иква
вблизи с. Сапанов на Терйополыцине. Средние участки гор трясо-
гузки обычно покидают в середине- октября, редко — в начале
ноября и даже позже. К. А. Татаринов (1973) наблюдал несколько
особей теплой осенью (27 октября) 1968 г. вблизи полонины
Брецкул.
Горная трясогузка гнездится обособленными, не очень близко
располагающимися парами. В урочище Козьмещик в мае — июле
1982—1984 гг. четырнадцать ее гнезд располагались на расстоянии
от 55 до 420 м, в среднем — 222 м. Поисками места для гнезда
занимается самка при участии самца. В это время самец часто
издает призывный крик и время от времени совершает токовый
полет. Призывный крик чаще всего звучит как звонкое и быстрое
«цицицицици», реже как «цицицицицици-си», «циви-циви-циви-
цицицицицици» или «ци-ци-ци-ци-цицицицицицицици». Звонкое
и быстрое «цицицицицици» чаще всего бывает 6—8-сложным,
реже — 5 или 9—10-сложным. В последнем случае два концевые
слога «цици» произносились очень быстро и смазывались. Описан-
ную в субвысокогорье Тянь-Шаня демонстративную песню горной
трясогузки, напоминающую нечто вроде «цзиль-цзиль-цзиль»
(Ковшарь, 1979), как и подпесню (тихое бормотание самца),
в Карпатах нам слышать не приходилось. Совершая токовый полет,
самец сначала набирает высоту, а потом по косой (параболе),
мелко тряся крыльями, опускается вниз, реже — планирует. При
спуске хвост приподнят примерно на 45° по отношению к телу,
а не вертикально, как это характерно для трясогузок на Тянь-
Шане. Сидя на возвышенном месте (дереве), токующая птица
высоту может не набирать, а сразу опускается вниз по параболе.
Призывные звук «цицицицицици» токующие самцы издают, хотя
и не всегда, и при токовом полете, что не отмечено на Тянь-Шане
(Ковшарь, 1979).
Свои гнезда горная трясогузка устраивает на земле или невы-
соко над ней, в пещерках и щелях береговых обрывов речек
и ручьев (нами отмечено 27 гнезд), на полочках прибрежных
скал (12 гнезд), под корягами елей у ручьев (7 гнезд), в корягах
вывернутых деревьев (9), под упавшими стволами (2 гнезда),
реже — в углублениях пней, среди травы, между бревнами дере-
вянного моста и на бетонной полке заброшенного полутемного под-
вала (по одному гнезду). В большинстве случаев гнезда защищены
сверху от дождя выступающим камнем, свисающим пучком тра-
вянистых растений, мхом, дерном и хорошо замаскированы. Только
в шести случаях из 61 они были открыты сверху, а в одном —
издали хорошо заметно. Три близкие к завершению гнезда строили
самки в сопровождении самцов (в одно гнездо с 18 до 20 ч самка
три раза приносила строительный материал, а в другое с 10
до 14 ч — 14 раз). Гнездо представляет собой чашеобразное, до-
з»
67
вольно беспорядочное сооружение. По данным анализа 39 гнезд,
основным строительным материалом в 13 случаях служил мох,
в 12 — стебли травянистых растений, в 9 — тоненькие веточки ели,
в четырех — корешки и в одном — листья бука. В качестве приме-
си к основному материалу в 25 гнездах отмечен мох, в 22 — веточки
ели, в 16 — корешки, в 7 — стебли травянистых растений, в 3 —
сухие листья, в одном — щепочки и еще в одном — лишайники.
Лоток чаще всего выстлан шерстью (24 гнезда), нежными стебля-
ми травянистых растений (9 гнезд), волосом (5) и нежными
корешками (одно гнездо). В качестве примеси к основному ма-
Таблица 1
Размеры гнезд горной трясогузки
Показатель,, мм п lim х(М) m С
Ширина гнезда 17 76-175 136,12 5,86 17,74
Ширина лотка 17 58-70 63,29 0,90 5,88
Высота гнезда 11 60-125 82,73 5,32 21,32
Рлубина лотка 16 38-65 46,75 1,63 13,95
Таблица 2 Размеры яиц горной трясогузки в разные годы ^урочище Козьмещик)
Год Показатель п Him х(М) m С
Длина, мм 56 17,3-20,5 18,97 0,11 4,48
1982 Ширина, мм 56 13,2-15,3 14,30 0,07 3,78
Форма 56 69,4-83,5 75,46 0,49 4,82
Длина, мм 45 17,8-20,3 18,73 0,09 3,04
1983 Ширина, мм 45 13,4-15,1 14,33 0,05 2,51
Форма 45 69,4—84,8 76,58 0,45 3,93
Длина, мм 54 17,4-21,2 19,17 0,12 4,49
1984 Ширина, мм 54 13,3-16,0 14,73 0,07 3,67
Форма 54 71,5-83,3 76,93 0,41 3,91
Таблица 3
Размеры яиц горной трясогузки в полных кладках
из пяти и шести яиц
Число яиц в полной кладке Показатель п lim С
Длина, мм 58 17,4-20,4 19,08+0,10 3,83
5 Ширина, мм 58 13,4-15,2 14,38+0,05 2,78
Форма 58 69,4-83,3 75,46+0,42 4,21
Длина, мм 84 17,5-21,2 18,90+0,09 4,50
6 Ширина, мм 84 13,2-16,0 14,56+0,07 4,19
Форма 84 69,4-84,8 77,10+0,36 4,31
68
Таблица 4
Размеры яиц горной трясогузки в зависимости от очередности откладки
Ранг яйца п Показа- тель Показатель
Длина, мм Ширина, мм Форма Объем, см3
lim 18,1-21,2 14,0-15,5 71,0-80,1 1,960-2,667
1 7 X 19,40 14,50 74,91 2,1416
in 0,37 0,18 1,14 0,09
с 5,10 3,31 4,03 11,21
lim 17,9-20,8 14,2-15,7 74,0-81,0 1,922-2,684
2 7 X 19,07 14,67 77,03 2,1564
m 0,37 0,18 0,98 0,10
с 5,19 3,34 3,35 11,59
lim 17,8-20,8 14,2-15,6 75,0-84,8 1,943-2,650
3 7 X 19,07 14,79 77,66 . 2,1877
m 0,38 0,19 1,49 0,09
с 5,19 3,31 5,09 10,51
lim 18,0-20,6 14,3-15,8 74,1-83,9 1,992-2,693
4 7 X 19,24 14,91 77,61 2,2464
m 0,34 0,20 1,36 0,09
c 4,73 3,48 4,63 10,23
lim 18,3-20,4 14,3-16,0 73,4- 82,5 2,034-2,734
5 7 X 19,29 14,96 77,63 2,2646
m 0,28 0,20 1,19 0,08
c 3,89 3,61 4,07 9,71
lim 18,1-20,7 14,8-15,8 74,4-81,8 2,076-2,706
6 4 X 19,70 15,13 76,90 2,3658
m 0,56 0,24 1,68 0,13
c 5,69 3,11 4,37 10,99
териалу лотка отмечены волос (7 гнезд), шерсть (4), стебли травя-
нистых растений, корешки и мох (по одному гнезду). Размеры
гнезд приведены в табл. 1, из которой видно, что более изменчивы
высота и ширина гнезда, чем глубина лотка и его ширина.
Первые яйца в гнездах горной трясогузки в Украинских Кар-
патах нами зарегистрированы *: в 1961 г.— 17 мая; в 1967 г. — 4
и 7 июня; в 1974 г,— 28 мая; в 1977 г.— 23 и 30 мая, 17 июня;
в 1982 г.— 18, 28 и 30 мая, 5 (2 гнезда), 8, 11, 12 (2 гнезда),
14, 15 (2 гнезда) и 23 июня; в 1983 г.— 15, 18 (по 2 гнезда),
30 и 31 мая, 1, 2, 3, 7, 8 (2 гнезда), 9 и 10 июня; в 1984 г. 12, 16,
28 и 31 мая, 5, 6, 9, 10, 11, 13, 14, 15, 26, 27 и 29 июня.
Самка откладывает по яйцУ в сутки в утренние часы (до 8—
9 ч) и.лишь в одном случае из семи второе яйцо снесено через
сутки. Полная кладка (п=51) содержит пять-шесть яиц, в одном
Даты приведены на основании прямых наблюдений или высчитаны от даты
вылупления или вылета птенцов.
69
Таблица 5
Бюджет поведения горной трясогузки при воспитании потомства
№ гнез- да Дата наб- людения Число яиц(я) или птен- цов (п) в гнезде Возраст птенцов в днях Время, ч мин
Длитель- ность «рабоче- го дня» Обогрева птенцов или яиц в гнезде за «рабочий день,»
б 9 б + 9
2 19.06.1982 6я - 14,51 - 11,15 11,15 7 2.07.1984 5я - 14,24 3,51 8,17 12,08 6 23.06.1984 5п-|-1я -1 15,29 4,01 10,05 14,06 2 1.07.1982 6п 2-3 15,39 1,07 5,43 6,50 3 22.06.1982 5п 3-4 15,45 1,01 7,22 8,23 6 26.06.1984 6п 3+ 16,05 1,39 5,51 7,30 2 3.07.1982 6п 4-5 16,02 4 29.06.1982 6п 7-8 16,02 0„09 0,14 0,23 6 29.06.1984 6п 64- 15,45 — - - 7 23.06.1984 6п 10-11 16,25 - - - 1 21.06.1977 6п * 16,23 - - - * Наблюдения проводились за два дня до их вылета из гнезда.
случае было отмечено четыре яйца; в среднем — 5,51. Яйца желто-
вато-белого, бледно-серого или палевого цвета с охристыми и — ре-
же — рыжими, серовато-бурыми или рыжевато-бурыми пестрина-
ми или пятнышками, часто более сконцентрированными на тупом
конце, где образуют почти сплошное пятно, реже — равномерно
разбросаны по всему яйцу. Интенсивность опятнения бывает раз-
личной и в пределах одной кладки, вследствие чего одно-два яйца
выглядит светлее. В одной кладке четыре из шести яиц на тупом
конце имели по одной темно-бурой каракуле. Изменчивость длины,
ширины и формы яиц довольно низкая, причем длина яйца более
изменчива, чем его ширина (табл. 2). Имеются, видимо, различия
по ширине (t=2,34) и форме (t=2,96) яиц в зависимости от их
числа в полной кладке (табл. 3). Ширина, форма и объем яиц
из кладок, содержащих шесть яиц, больше, чем в кладках, содер-
жащих пять яиц (объем равен 2,098 и 2,066 см3 соответственно).
Размеры яиц изменчивы и в пределах одной кладки (С. У.=0,25—
3,44%), причем различия по длине в 18 кладках были больше,
чем по ширине, в 9 —. наоборот, в одной — одинаковы. В 28 гнездах
разница по длине яиц в пределах кладки колебалась в пределах
0,1 —1,9 мм (в среднем 0,80), а по ширине — 0,1 —1,1 мм (в сред-
нем 0,45). Размеры яиц в пределах кладки, видимо, зависят от
очередности их снесения. Они постепенно увеличиваются от перво-
го к шестому яйцу, что хорошо заметно по средним показателям
(X) ширины и объема яиц * (табл. 4). Выявлены достоверные раз-
* Объем яйца У высчитан по формуле: У=О,5236 Rr2, где R — длина яйца,
г — ширина его.
70
/ Без обогрева птенцов или яиц за «рабочий день» Число
кормлений выносов фекальных капсул обогревов птенцов или яиц за «рабочий день»
б 9 б+р б 9 б + р б 9 б+р
3,36 — — — — — — — 27 27
2,16 — — — — — — 22 20 42
1,23 27 31 58 1 6 7 • 33 39 72
8,49 33 но 143 18 48 66 16 73 89
7,22 90 46 136 39 16 55 9 61 70
8,35 95 113 208 44 42 86 20 61 81
2 201 203 — 69 69
15,39 157 123 280 32 28 60 9 8 17
15,45 179 186 365 41 46 87 — — —
16,25 225 185 410 61 39 100 — — —
16,23 49 75 124 12 37 49 — — —
личия в размерах (ширине, длине) и форме (ширина X 100) /
длина яиц, отложенных в разные годы. В частности, яйца, отло-
женные в 1984 г., оказались больше, чем снесенные в 1982
и 1983 гг. (см. табл. 2). Создается впечатление, что при повышен-
ной влажности и пониженной температуре горная трясогузка в
Карпатах откладывает более крупные яйца. Объем яиц в 1984 г.,
отличавшемся наиболее влажной, прохладной и поздней весной,
составил 2,178 см3; в 1982 г. он составлял 2,031 см3, а в 1983 г.—
2,014 см3.
Насиживание чаще всего начинается после откладки предпо-
следнего яйца, реже — четвертого или последнего (пятого или
шестого) и длится 13—14 сут. Доля участия партнеров в насижи-
вании у разных пар неодинакова. В большинстве случаев насижи-
вают, видимо, и самец и самка (самка больше), но бывают
случаи, когда самец участия в насиживании не принимает
(табл. 5). Насиживающая самка время от времени привстает и ме-
няет ориентацию яиц в гнезде клювом. Слетает самка с кладки, как
правило, тогда, когда вблизи гнезда появляется самец, реже — в его
отсутствие. Во всех случаях кладка некоторое время не обогре-
вается, хотя плотность насиживания со временем возрастает, дости-
гая максимума, вероятно, в день вылупления птенцов.
Как правило, один птенец вылупляется на сутки позже ос-
тальных. В двух полных кладках, содержащих по шесть яиц,
вылупление происходило в течение трех суток (4 —1 —1), а в одной,
содержащей пять яиц,— в течение одного дня. Птенцы находятся
в гнезде 13—16 и даже 18 сут (п=11), но при опасности они
могут покинуть его еще совсем неспособными к полету, в 11 —
71
12-суточном возрасте, что иногда приводит к их гибели (падают
в горные ручьи и тонут). Кормят птенцов и чистят гнездо и самец
и самка, хотя доля участия партнеров в этом носит индивидуаль-
ный характер (в шести гнездах более активной была самка,
а в трех — самец). Пока птенцы маленькие, самка при сравнитель-
но небольшом участии самца довольно часто и долго обогревает их.
По мере роста птенцов интенсивность обогрева падает (см. табл. 5),
хотя она зависит и от погодных условий. Интенсивность корм-
ления птенцов и величина пищевых порций с их возрастом
увеличивается, но перед вылетом заметно снижается. Судя по ос-
реднениым данным девяти суточных наблюдений у шести гнезд,
родители в течение рабочего дня кормят птенцов более-менее
равномерно — 11—15 раз в час (в среднем 13,4 кормлений на
гнездо или 2,5 кормлений в один час на одного птенца). За время
пребывания птенцов в гнезде самец и самка прилетают к ним
с кормом около 3000 раз. Более-менее равномерно проходит и
вынос фекальных капсул. Корм для птенцов трясогузки собирают
на земле, часто на дорогах вдоль горных ручьев и речек или на
выступающих камнях среди них. Нередко птицы «бродят» по
мелководью и корм собирают прямо из воды, добывая его из коряг
и иного мусора. Время от времени птицы подпрыгивают за ле-
тающими насекомыми, ловят их на лету на высоте 0,5 —1,0 м от
земли, реже — выше (до 5 м) или нагоняют летящее насекомое
пешком и резким броском хватают его; иногда ловят насекомых
на лету, слетая с веток деревьев, выделывая при этом довольно
сложные пируэты. При поисках корма для птенцов горная трясо-
гузка издает отрывистые и четкие одно-двусложные звуки «ци»
или «ци-ци». Такие же звуки она произносит и при опасности,
но при этом добавляет высокий протяжный печальный писк
«ииить». Звук «ци» при опасности чаще всего бывает дву-, трех-,
реже одно- или четырехсложным.
После вылета птенцов из гнезда их примерно неделю сопро-
вождают взрослые и подкармливают вне гнезда. Если самка вскоре
приступает ко второй кладке, то эта функция в основном ложится
на самца.
В урочище Козьмещик Раховского р-на в 1982 и 1984 гг.
летные птенцы в сопровождении взрослых стали попадаться со вто-
рой декады июня, хотя в это время находили строящиеся гнезда,
гнезда с кладками, реже — с птенцами. В 1983 г. летные птенцы
наблюдались уже с 30 мая. В последнем случае у сопровождающей
выводок пары наблюдали токовый полет и копуляцию. Выводки,
токовые полеты и копуляция отмечены мною также 31 мая, 5 и
6 июня 1983 г. Аналогичную картину я наблюдал здесь и с 30 июня
по 8 июля 1983 г., когда ежедневно в сопровождении взрослых
встречались от 2 до 10 выводков (короткохвостых или с более-
менее отросшими хвостами).
Сравнение наблюдений 1982—1984 гг. свидетельствует о том,
что период размножения горной трясогузки в Украинских Карпа-
тах зависит от высоты местности над уровнем моря и характера
72
погодных условий. В частности, в условиях поздней весны 1982
и 1984 гг. в урочище Козьмещик горные трясогузки приступили
к размножению на 10—12 дней позже, чем в 1983 г., и лишь дважды
выводили птенцов, в то время как в 1983 г. часть птиц имела, ве-
роятно, три кладки.
Судя по имеющимся в нашем распоряжении фактическим дан-
ным, на средних участках гор (урочище Козьмещик) в зависимости
от характера хода весны к первой кладке трясогузки приступают
в третьей декаде апреля — начале второй декады мая, а ко вто-
рой — с конца мая до второй декады июня. В благоприятные годы
часть птиц, возможно, приступает к третьей кладке начиная
с первой декады июля, хотя, несомненно, более вероятны три клад-
ки на нижних участках гор. У верхней границы леса (1200 —
1400 м над ур. м., окрестности приюта «Драгобрат» Раховско-
го р-на) часть трясогузок тоже дважды за лето выводит птенцов,
о чем свидетельствуют находки гнезд с кладками и птенцами
в июне, а также наблюдение выводков, токовых полетов и гнездо-
строения 26 и 28 июня 1974 и 1975 гг.
Успех размножения горной трясогузки в Карпатах довольно
высок. Вылупляемость птенцов достигает 96,00% (в 28 гнездах
из 150 яиц вылупилось 144 птенца, 5,04 птенца на гнездо).
В 22 гнездах птенцы вылупились из всех яиц, в четырех было
по одному болтуну, еще в двух — по одному яйцу с задохликом.
Из 20 гнезд, в которых вылупилось 106 птенцов, благополучно
вылетело 94 птенца (88,68% от числа вылупившихся, или 4,70
птенца на гнездо). Таким образом, из одного гнезда вылетает 3,42
птенца на гнездо (около 62% от числа отложенных яиц).
Естественными врагами горной трясогузки в Карпатах являют-
ся сойка, сони (полчек и лесная), горностай, муравьи и, воз-
можно, гадюка обыкновенная, уничтожающие кладки и птенцов.
Однажды зарегистрировано паразитирование кукушки. Неблаго-
приятное влияние оказывают фактор беспокойства (гибнут кладки
и падают в воду потревоженные еще неспособные к полету
птенцы) и резкая смена уровня воды в горных ручьях. Яйца
в одной кладке оказались прогрызенными голым слизнем.
ЛИТЕРАТУРА
К1стяк1всъкий О. Б. Птахи Закарпатсько! облает! // Тр. 1н-ту зоол. Ки!в: Вид-во
АН УРСР, 1950. Т. 4. С. 3-77.
Ковшарь А. Ф. Певчие птицы в субвысокогорье Тянь-Шаня. Алма-Ата: Наука,
1979. 312 с.
Портенко Л. А. Очерк фауны птиц Западного Закарпатья // Сб. памяти акад.
П. П. Сушкина. М.; Л.: Изд-во АН СССР, 1950. С. 301-359.
Страутман Ф. И. Птицы Советских Карпат. Киев: Изд-во АН УССР, 1954. 332 с.
Страутман Ф. И. Птицы западных областей УССР. Львов: Изд-во Львов, ун-та,
1963. Т. 2. 183 с.
Татаринов К. А. Фауна хребетних заходу Украши. Льв1в: Вид-во Льв!в. ун-ту,
1973. 257 с.
73
SUMMARY
On biology of European Grey Wagtail (Motacilla cinerea cinerea)
in the Ukrainian Carpathians. TalposhV. S.
The information on the population distribution, biotopic distribution, phenology,
biology and reproduction of Grey Wagtail in the Ukrainian Carpathians is given.
Eggs’ sizes and their coloration, sizes and construction material, etc. are described.
Digital indices are processed by using the method of variation statistics. Big changeabi-
lity is established. The nest height and width, the eggs’ length are varied more
than the depth and width of the nest cup and the egg’s width. The sizes of the eggs,
laid in different years, depend on the number of the eggs, as a whole, and on the
rank of an egg.
УДК 598.422.1(470.57)
А. Ф. МАМАТОВ Озерная чайка
450074, Уфа ул. Фрунзе, д. 32. БашГУ. Кафедра зоологии и дарвинизма (Larus ridibundus) в Башкирии
Озерная чайка в различных частях ареала служила объектом
исследования многих орнитологов (Модестов, 1943; Исаков и др.,
1947; Бородулина, 1960; Строков, 1960; Creutz, 1963; Заблоцкий,
Заблоцкая, 1963; Михлин, 1963, 1966; Wundsch, 1963; Klima, 1964;
Молчанова, 1969; и др.). В последнее время изучение колониаль-
ных птиц успешно ведется на уровне мелких структур внутри
популяции (Зубакин, 1976; Виксне, 1968; Харитонов, 1981а, б;
1983а, б; Харитонов, Зубйкин, 1984). В Башкирии экология озерной
чайки специально не изучалась. В работах П. П. Сушкина (1897),
С. В. Кирикова (1952), М. Г. Баянова (1977), Птицы... (1977),
В. Д. Ильичева, В. Е. Фомина (1979) можно найти краткие
сведения об этой птице. Некоторые вопросы экологии озер-
ной чайки рассматривались ранее автором (Маматов, 1975 а, б;
1979).
Материалы собраны на оз. Карагайлы Учалинского р-иа Баш-
кирской АССР в 1963, 1973, 1978, 1979, 1980, 1982, 1986 гг. Здесь
расположено около десяти озер, где чайки постоянно держатся,
а два-три озера являются местами массового гнездования и кормеж-
ки озерных чаек, малых чаек (Larus minutus), речной (Sterna
hirundo) и черной (Chlidonias nigra) крачек, красноголового ныр-
ка (Aythya ferina), хохлатой чернети (A. fuligula), лысухи (Fulica
atra), чомги (Podiceps cristatus), красношейной (Р. auritus),
черношейной (Р. nigricollis) и серощекой (Р. griseigena) по-
ганок.
74
Предгнездовой период
Озерные чайки на водоемы учалинской группы прилетают с зимо-
вок примерно в середине апреля. Сроки прилета колеблются и
зависят от метеорологической ситуации конкретного года. Основ-
ная масса птиц отмечается в местах будущего гнездования не
позднее 20—22 апреля, а пролет полностью завершается в первой
неделе мая. Сначала (13—15 апреля, самая ранняя дата прилета
10 апреля) появляются единичные особи, которые держатся по
высоким местам и на полях, где раньше сходит снег. В дневное
время неоттаявшие озера птицами почти не посещаются, но вече-
ром они собираются на озерах, устраиваясь ночевать на сплавинах
или на льду. Пары у них еще не формируются, чайки держатся
на земле поодиночке или группами.
Образование пар завершается преимущественно к началу мая.
Процесс формирования пар зависит от условий гнездования. В годы
с высоким уровнем воды площади, пригодные для гнездования,
значительно сокращаются, и нехватка гнездовых территорий ощу-
щается остро. Уровень паводка может играть решающую роль
в жизни популяции. В 1979 г. уровень воды на оз. Карагайлы
был высоким и глубина была 1 м. В результате подтопления
низинных частей колонии они не заселялись птицами, а более
приподнятые стали местом поселения поганок, занимавших ранее
нижний ярус. Для чаек подходящих гнездовых участков не оста-
лось. Птицы продолжали держаться в районе бывшего гнездования
до начала июня, после чего исчезли. В первой декаде июня
на оз: Чупликуль, в- 12 км от оз. Карагайлы на тростниковых
заломах они образовали колонию в 450—500 особей. После раз-
множения общее количество озерных чаек там достигло 1000 экз.
По материалам учета 1986 г., численность озерной чайки была
максимальна среди чайковых. Общая экстраполяционная оценка
достигает 20 000 экз., т. е. превосходит численность серебристой,
малой чаек, речной, черной, белокрылой крачек в 7—20 раз.
Морфометрия. В числе 18 птиц, добытых в период с 15 мая
по 10 июля с целью гельминтологического вскрытия, оказалось
12 самцов и 6 самок. Из табл. 1 видно, что самцы превосходят
Таблица 1
Размеры озерных чаек из Башкирии
Показатель Самцы, п=12 Самки, п—6
М±т lim М+т lim
Вес, г 285+4,7 233-312 257+16,6 216-311
Общая длина, мм 402±6,8 362-453 368+8,2 355-410
Размах крыльев, мм 961+19,6 855-1030 899+7,4 875-920
Длина крыла, мм 312+2,2 292-320 297+3,5 281-300
Длина клюва, мм 37+0,6 34-40 34+1,3 28-36
Длина цевки, мм 54 + 1,0 49-60 49+1,1 45-50
75
самок по всем морфометрическим характеристам. При р=0,05 раз-
личия в показателях, кроме веса, достоверны. Учитывая морфо-
метрические отличия, а также общий габитус, ритуальное поведе-
ние и др., в природе возможно определение пола птицы.
Наши данные по среднему весу заметно отличаются от данных
Г. П. Дементьева и др. (1951): самцы из нашей выборки мельче,
чем самцы из других популяций, на 33 г, а самки — на 19 г.
Гнездование
Гнездовые биотопы. Для гнездования чайки выбирают малодоступ-
ные места среди бордюра прибрежной надводной травянистой
растительности или селятся на островах с твердым субстратом и
на плавающих временных скоплениях отмершей растительности.
В наших условиях, избегая беспокойства, они отдают предпочтение
плавучим островкам, где сосредоточивается до 95% всех кладок.
На оз. Карагайлы на протяжении многих лет существует смешан-
ная колония, состоящая из восьми видов водоплавающих птиц —
озерной чайки, речной крачки, чомги, красношейной и черношей-
ной поганок, хохлатой чернети, красноголового нырка, лысухи.
Гнездовой островок длиной 50—60 м и шириной 30 м, сужи-
вающийся к концам, представляет собой массу из отмерших стеб-
лей тростника, рогозов (в основном узколистного), находящуюся
на плаву. Этот своеобразный понтон фиксируется на одном месте
проросшими насквозь стеблями тех же растений, растущих на
глубине 1 —1,5 м. Летом остров густо зарастает тростником,
создавая ремизы для всех его обитателей. В июле уровень воды
снижается, обнажаются дно, камни, травянистая растительность.
Гнезда распределяются по острову неравномерно. По периферии,
где полегший старый тростник образует плотную массу, гнезда
располагаются почти вплотную или на 30—40 см отстоят друг
от друга (индекс плотности гнездования К (Зубакин, 1976)
составлял 0,43). Ближе к центру плотность гнездования ниже,
расстояние между гнездами достигает 2—2,5 м. Однако в некоторые
годы для чаек характерно равномерно плотное гнездование. При
этом нет одиночных кладок, а среднее расстояние между гнездами
составляет 30—60 см (в 1981 г. К был равен 0,31).
Форма и размеры гнезд. Постройка гнезд идет вслед за таянием
льда на сплавинах. Вначале заселяются более сухие участки,
а по мере возрастания количества гнезд чайки занимают влажные
места.
По характеру постройки и размерам можно выделить три основ-
ных типа гнезд (табл. 2). По конструкции наиболее совершенны
гнезда, расположенные в местах с повышенной влажностью или
в зоне волнобоя. Их форма конусовидная, лоток бысоко приподнят
над водой и не подмокает. Они отличаются аккуратностью и плот-
ностью исполнения, лоток имеет высокие и хорошо оформленные
борта. Гнезда, помещенные в сухих местах плавней, представляют
собой небольшую, беспорядочно набросанную массу из раститель-
76
Таблица 2
Форма и размеры (см) гнезд чаек
Тип гнезда Диаметр основания Высота бортов Лоток
Глубина Диаметр
Высокий, конусообразный 36—50 21-30 4-5,5 14-16
Плоский, плохо оформленный 21-24 4-12 2,5-5 16-18
Переходный 32-34 13-15 4-5 18-19
ной ветоши. Их лоток почти не имеет бортов и опирается дном
на землю или едва приподнят над грунтом. Между этими типами
крайне редко встречаются промежуточные формы, которые отли-
чаются в деталях.
При постройке любых типов гнезд используется однотипный
строительный материал, собранный вблизи постройки: стебли и
листья тростника (Phragmites communis), рогозов (Typha angusti-
folia, T. latifolia), камыша (Scirpus lacustris), хвоща болотного
(Equisetum palustre), прутья и т. п. Свежие листья и стебли при-
меняются чайками при дополнительной надстройке или строитель-
стве поздних гнезд. Основная масса птиц строит гнезда первого
типа, так как гнездится в основном на острове в зоне значитель-
ного волнобоя.
Сроки откладки яиц непостоянны из года в год даже внутри
одной колонии. В условиях Башкирского Зауралья озерные чайки
приступают к откладке яиц 6—10 мая. Период яйцекладки обычно
растягивается примерно на месяц и дольше. Самые ранние кладки
приходятся на первые числа мая, поздние — на середину июня.
К 20 мая в 70% гнезд идет насиживание, как правило, полных
кладок из трех яиц. Наряду с этим значительное количество гнезд
еще содержит одно-два яйца или же только построены. Но еще
в десятых числах июня, когда на плавнях большое количество
птенцов, имеются кладки чаек на разной стадии насиживания.
В 1979 г., когда, как мы уже отмечали, уровень воды на оз. Кара-
гайлы был очень высок, чайки гнездились на другом водоеме с
опозданием от обычных сроков гнездования почти на месяц. Все
это говорит о том, что четкой синхронности сроков откладки яиц
в колонии не обнаруживается. Можно утверждать, что порядок
откладки яиц повторяет очередность заселения гнездовых участков
в границах колонии.
В абсолютном большинстве гнезд количество яиц не превышает
трех. Кладки с четырьмя яйцами составляют менее 1 % от общего
их количества. В двух гнездах из 250 обнаружено по пять яиц,
форма, размеры и окраска которых не отличались от типичных.
Видимо, в таких гнездах кладка принадлежит двум самкам.
Размеры и форма яиц колеблются в следующих пределах:
длина — от 46,3 до 55,0 мм, ширина — от 36,0 до 40,5 мм. Средние
длина и ширина, по данным измерений 47 яиц, составляет 49,7±
77
0,37x37,4+0,37 мм. Вес яиц изменяется от 33,0 до 38,0 г, сред-
нее — 35,9 г.
Наиболее типична более или менее эллиптическая форма,
составляющая 85% просмотренных яиц. Яйца округлой формы
с яйцами равномерно-эллиптической формы в кладках всегда соче-
таются, а удлиненно-эллиптические яйца бывают только в виде
отдельных кладок.
Отмечены и цветовые аномалии яиц. Обычно они имеют грязно-
зеленоватый или зеленовато-коричневатый основной фон, по кото-
рому рассыпано большое количество пятен разнообразной конфи-
гурации и размеров бурого или оливкового цвета. Наряду с такими
обнаружены, начиная с 1971 г., яйца светлой окраски, слабо пиг-
ментированные основным фоном. Такие яйца встречались лишь в
одном или двух гнездах в виде полной кладки или ее части.
Дальнейшая судьба нетипичных яиц неизвестна.
Сформированность гонад. В литературе вопрос о состоянии
половой системы чаек по этапам репродуктивного цикла освещен
слабо. По нашим данным, сроки ярко выраженной половой актив-
ности приходятся у самцов на май, когда семенники имеют наи-
большие размеры и массу. В середине июня семенники сохра-
няют крупные размеры, однако их масса в 2—3 раза меньше, чем
в мае. У самок картина несколько иная. Максимальные размеры
яйцеклеток и массы яичника отмечаются во второй декаде мая
и резко снижаются в первой декаде июня.
Сравнение состояния половых желез в разные сроки дает
интересные результаты. Во время массового размножения (в мае)
масса семенников возрастает по сравнению с периодом покоя
(июль) в 22—25 раз. Аналогичное явление наблюдается и у самок.
В мае масса яичника достигает 10,6 г, но к середине июня она
снижается до 0,40 г, т. е. уменьшается в 26,5 раз.
Сроки насиживания определялись по пяти контрольным гнез-
дам с полными кладками. Насиживание в них началось 18 —19 мая
и продолжалось 23—24 дня. Выклев птенцов в первом гнезде
отмечен 9 июня. За последующие два дня все птенцы покинули
скорлупу яиц. Однако 9—11 июня нельзя считать самым ранним
сроком вылупления, так как в других гнездах (помимо контроль-
ных) птенцы появились 5—6 июня. Если принять, что насижива-
ние в них длилось столько же .времени, то его начало приходится
на 14 15 мая. В дальнейших наблюдениях мы получили подтверж-
дение того, что начало насиживания в благоприятные годы прихо-
дится на 10—12 мая.
Хотя в насиживании принимают участие оба партнера, самка
сидит на гнезде примерно 75% времени. Особенно плотно она
обогревает кладку в последнюю треть времени инкубации. Самец
в это время занят добыванием корма. Он возвращается к гнезду
каждые 40—50 мин, принося мелкую рыбу, стрекоз и их личинок,
листоедов, плавунцов, веснянок, ручейников и т. д. В период
насиживания самки тоже могут посещать негнездовые водоемы в
поисках корма. По состоянию половой системы вскрытых самок
78
установлено, что на озерах, где чайки не гнездятся, встречаются
~ самки или не начавшие яйцекладку (поздно гнездящиеся), или
потерявшие ее в результате разорения хищниками или людьми.
Остальные птицы, добытые в мае—июне, были самцами.
Вылупление птенцов. Раньше всего птенцы появляются в сухих
местах плавней, т. е. порядок вылупления птенцов повторяет
*. очередность гнездования внутри колонии. Обсыхание птенца
происходит через 2—3 ч. Потребность в корме возникает спустя
12 —15 ч, когда птенцы активно выпрашивают пищу. Двухдневные
птенцы способны реагировать на опасность: они разбегаются и
прячутся в зарослях. До этого времени они остаются в гнезде
• при приближении человека, но уже затаиваются. В раннем возрасте
наблюдается самая высокая смертность птенцов. Так, с 10 по
12 июня 1980 г. обнаружено 30 мертвых чайчат, что составляет
2,5% от общего зарегистрированного в 1980 г. количества птенцов.
Плавать чайчата начинают в возрасте 4 — 5 дней. В 8—10 дней
птенцы переселяются в заросли густой растительности, где каждый
выводок держится отдельно. При появлении родителей с кормом
У они выбегают на открытое место и поедают принесенную пищу.
! К двадцатому дню жизни птенцы образуют группы в 20—30 особей,
которые самостоятельно бродят по острову в поисках пищи. Чайча-
та собирают сидящих на стеблях растений или на земле насекомых.
Самки подкармливают их лишь изредка, прилетая один-два раза
в день с кормом, который отрыгивают на землю или на мелко-
водье. Ночь птенцы проводят в зарослях вместе с обоими родите-
,( ’ лями или только с матерью. С наступлением активного функциони-
рования копчиковой железы (примерно в 25 дней) выводок может
ночевать на воде, что представляет для чаек обычное явление.
До недельного возраста птенцы прибавляют в весе сравнительно
медленно; позже он увеличивается быстрыми темпами:
возраст, дней вес, г возраст, дней вес, г
2 33,2 12 114,5
3 34,3 15 157,5
4 43,2 17 174,1
5 46,1 20 229,7
6 49,8 30 263,6
Молодые чайки становятся лётными в возрасте пяти недель.
Полет их в это время тяжелый, маломаневреиный и птицы,
пролетев 50—60 м, садятся на воду. Шестинедельные чайки уже
летают легко и стремительно. Они набирают массу взрослых,
но отличаются от родителей цветом оперения.
Суточная активность. Чайки активны 19—20 ч в сутки. Первые
полеты начинаются в 4 ч утра в виде поискового полета над зерка-
лом озера. Около 5 ч начинаются перелеты по направлению к
другим водоемам. Чайки часто встречаются на озерах, находящих-
ся в 10—15 км от гнездовий. Полеты по кормовым маршрутам
продолжаются до 19—20 ч. Отдельные особи курсируют над оз. Ка-
79
рагайлы в 22—24 ч. Редкие крики чаек слышны до рассвета.
Количество птиц, постоянно улетающих с территории колонии,
определялось на линейном маршруте, совпадающем с направле-
нием их основных перемещений. На маршруте длиной 4 км и шири-
ной 300 м проведен восьмикратный абсолютный учет отдельно
в первую и вторую половину дня. В течение 2 часов каждого учета
на территории 1,2 км2 насчитано от 15 до 22 птиц (в среднем 18),
или 15 экз./км2. Следовательно, на основном кормном оз. Б. Учалы
площадью 4,3 км2 в светлое время суток встречается около 65 экз.,
или 13% птиц. Эта часть популяции постоянно находится вне
гнездового участка.
Некоторые аспекты поведения. Для колонии характерен весьма
шумный образ жизни, который достигает высшей фазы в экстре-
мальных ситуациях. Чайки активно начинают реагировать на при-
ближение человека с расстояния 30—50 м. Нахождение на терри-
тории колонии вызывает крайние формы проявления агрессии
со стороны птиц: пикирование, попытка ударить клювом и крылом,
пронзительные и яростные крики, брызгание калом. Аналогичная
картина наблюдается в случае появления над гнездами болотного
луня, который изгоняется усилиями большого числа чаек. Серые
вороны не вызывают столь бурной реакции чаек.
Поведение и активность чаек определяются погодными ус-
ловиями. В тихую, ясную погоду интенсивность перелетов состав-
ляет около 20—30 птиц в час. При сильном ветре число кормовых
перелетов сокращается до 6 —12 птиц в час. Самая низкая
интенсивность перелетов (6—7 птиц в час) наблюдается при пони-
жении температуры из-за выпадения снега, что почти ежегодно
происходит в начале июня.
Питание
Питание озерной чайки изучалось Ю. А. Исаковым и др. (1947),
Т. Л. Бородулиной (1960), В. В. Строковым (1960), В. И. Заблоц-
ким и Л. И. Заблоцкой (1963), В. И. Васильевым (1967), Т. В. Мол-
чановой (1969) и др. В Татарской АССР (Птицы..., 1977) основным
кормом озерных чаек служит рыба — она встречалась в 80% же-
лудков. Насекомые обнаружены в 42,6% случаев вскрытий.
Питание птенцов и взрослых птиц изучалось нами путем вскры-
тия и анализа содержимого желудков павших и добытых птиц.
Результаты исследования птенцов в возрасте от 2 до 30 дней
представлены в табл. 3.
В питании птенцов ведущее место занимают насекомые и их
личинки. Особенно часто встречаются стрекозы, которые мы нахо-
дили у птенцов в возрасте от 2 до 15 дней. В первое время
стрекозы скармливаются птенцам взрослыми чайками. По достиже-
нии 10—12-дневного возраста птенцы добывают насекомых сами,
склевывая их с водной растительности. В дальнейшем взрослые
стрекозы выпадают из рациона питания и начинают встречаться
их личинки (3 случая).
80
Таблица 3
Рацион питания птенцов озерной чайки
Вид корма Число встреч Общее число экз. Число экз. в 1 желудке
Средн. Макс.
Стрекозы сем. Agrionidae и Lestidae 7 124 18 83
Личинки стрекоз 3 172 25 132
Плавунцы сем. Dytiscidae 3 3 1 1
Личинки плавунцов 2 4 2 2
Рыба 6 5 2,5 3
Растительная пища 2 — — —
Гастролиты (камешки) 2 — — —
Таблица 4
Рацион питания взрослых озерных чаек с 18 мая по 9 июля
Вид корма Число встреч Доля встреч, % Общее число экз. Число экз. в 1 же- лудке
Средн. Макс.
Стрекозы сем. Agrionidae и Lestidae: имаго 6 33,3 306 . 51 282
ЛИЧИНКИ 3 16,6 50 16,6 41
Стрекозы сем. Aeschnidae (личинки) 1 5,5 1 1 —
Листоеды Chrysomplidae 1 5,5 4 4 —
Плавунцы Dytiscidae: имаго 8 44,4 13 1,5 3
личинки 8 44,4 65 8 20
Веснянки Plecoptera 1 5,5 5 5 —
Ручейник Trichoptera 1 5,5 1 1
Рыба 12 66,6 24 1,7 7
Жуки-плавунцы найдены в желудках четырехдневных и двух-
трехнедельных птенцов. У двух пятидневных птенцов обнаружены
личинки жуков.
Рыба в пище обнаружена впервые у птенцов 17-дневного воз-
раста. Вероятно, до этого периода она не скармливается родителя-
ми, а сами птеицы поймать рыбу не способны.
В двух желудках из 11 обнаружены растительные остатки
и камешки. Последние впервые встретились у 12-дневного птенца.
Для выяснения компонентов питания взрослых птиц вскрыто
18 чаек (табл. 4). Их пища не отличается большим разнообразием.
Такие животные корма, как стрекозы и рыба, встречаются во все
периоды жизни и являются основными. В период выкармливания
81
Таблица 5
Вид и возраст рыб, добываемых озерными чайками
Вид и возраст рыб Число встреч Доля встреч, % Общее число экз. Число экз. в 1 желудке
Средн. Макс.
Карась 1 1 1 5,5 2 2 —
Карась 2 5 27,7 9 1.В 3
Карась 3+ 6 33,3 7 U 2
Карась 4+ 1 5,5 2 2 —
Карась 5 1 5,5 1 1 —
Плотва 3 1 5,5 3 3 —
птенцов обычными кормами служат взрослые и личиночные формы
плавунцов, равнокрылые стрекозы. Лишь у одной чайки найдена
личинка разнокрылой стрекозы большое коромысло (Aeschna
grandi). Обычной пищей в это время служит рыба.
В летний период основной пищей чаек являются личинки и
имаго плавунцов и стрекоз. Их число в одном желудке может
отличаться на 1 — 2 порядка. Так, в момент вылета равнокрылых
стрекоз в одном желудке было обнаружено 283 стрекозы. Макси-
мальное число личинок жука-плавунца достигало 20 экз.
В одном желудке мы находили от 1 до 7 экз. рыбы. Чайки
ловили плотву и карасей возраста I*-—3+. Наиболее частой их до-
бычей оказывались двух-трехлетние караси. Выяснилось, что такие
мелкие караси наиболее часто заражены ремнецами.
Вид и возраст рыб, встречающихся в пище чаек, представлены
в табл. 5.
Озерные чайки проявляют клептопаразитизм, характерный
для некоторых чайковых (Бузун, 1984), по отношению к крачкам.
Попытки отнять рыбу чайкам удаются редко: из 18 случаев, заме-
ченных нами, только в одном преследование было успешным.
Роль озерной чайки в распространении лигулидозов
В Башкирии чайки играют заметную роль в распространении
заболеваний, вызываемых ремнецами. М. Г. Баяновым (1976)
проведено вскрытие 121 экз. озерных чаек. Их гельминтофауна
оказалась наиболее богатой — 51 вид, из которых около трети
они приобретают через рыб. Эти заболевания широко известны
у рыб в различных регионах республики, в том числе и в Зауралье.
Ремнецы паразитируют на плотве, золотом и серебряном карасе
и др. Известно, что окончательными хозяевами паразитов служат
рыбоядные птицы более чем 40 видов. В числе этих птиц значится
и озерная чайка.
При вскрытии 30 экз. взрослых чаек ремнецы найдены у
15 птиц. Результаты вскрытия показаны в табл. 6. По данным
табл. 10, интенсивность инвазии колебалась преимущественно от 1
до 3 экз. Лишь в трех случаях отмечено более трех ремнецов.
82
Таблица 6
Зараженность озерных чаек ремнецами
Место добычи (озеро) Количе- ство ремнецов Digramma interrupta Ligula intestinalis
Общий вес, г Полово- зрелые Неполо- возрелые Общий вес, г Полово- зрелые Неполо- возрелые
Аслыкуль 2 — — — 1 1
Аслыкуль 2 — — — 3,9 — —
Карагайлы 1 1,35 1 — — — —
Карагайлы 3 7,78 1 2 — — —
Ургун 13 — — — 9,7 — —
Б. Учалы 1 0,34 — 1 — — —
Карагайлы 1 2,2 1 — — — —
Б. Учалы 7 2,87 — 7 — — —.
Карагайлы 1 — — — — — —
Калкан 2 1,8 1 1 — — —
Б. Учалы 7 2,8 — 7 — — —
Карагайлы 2 — — — — — —
Карагайлы 1 0,18 1 — — — —
Карагайлы 1 — 1 — —
Б. Учалы 2 — 2 — . — —
Среди вскрытых птиц было 17 птенцов и молодых чаек в воз-
расте от 2 до 60 дней; гельминты обнаружены у шести из них,
или у 28,3%. Наиболее ранние случаи заражения зарегистрирова-
ны у 15-дневного птенца (19 июня).
Найденные ремнецы относятся к двум видам. Ligula intestila-
lis L., паразитирует главным образом на плотве, Digramma inter-
rupta Rud.— на обоих видах карасей. Общий вес паразитов дости-
гал у одной чайки 9,7 г, а максимальная длина гельминта 286 мм.
Оба вида ремнецов одновременно ни у одной чайки не встречены,
хотя водоемы, в которых имеет место лигулез плотвы (оз. Ургун)
и диграммоз карасей (озера Б. Учалы и Карагайлы), находятся
друг от друга на расстоянии 1 — 3 км и птицы часто перелетают
с одного озера на другое.
Другие гельминты, преимущественно цестоды, обнаружены у
88,8% взрослых и у 55% птенцов. Наиболее ранние случаи
заражения отмечались у птенцов 12-дневного возраста.
Значение чаек в биоценозах
В условиях современных антропогенных изменений среды обита-
ния чайки испытывают многосторонние негативные воздействия
хозяйственной деятельности человека. Озерная чайка — экологи-
чески пластичный вид, но все же надо отметить, что численность
и состояние ее колоний зависят от уровня антропогенного пресса,
поэтому характерны периодические колебания численности птиц
по годам.
83
В Башкирии для создания оптимальных условий для коло-
ниальных птиц необходимо следующее.
Усилить разъяснительную работу о необходимости охраны ко-
лоний.
Установить режим заказности на водоемах и болотистых
угодьях, где концентрируется на гнездовье большое количество
колониальных птиц (смешанные и чистые колонии чаек, крачек,
уток, поганок, пастушков и других птиц).
Запретить посещение островов, плавней, где гнездится много
птиц, в гнездовой период.
С целью привлечения птиц организовать микрозаказники, зоны
покоя на водоемах, где имеются условия для гнездования, но
сильно сказывается фактор беспокойства.
ЛИТЕРАТУРА
Баянов М. Г. Гельминты чаек на Южном Урале.//Видовой состав и экология
животных Башкирии. 1976. Ден. ВИНИТИ, № 1974—78 Деп. С. 44—56.
Баянов М. Г. Класс Птицы // Животный мир Башкирии. Уфа, 1977. С. 259—312.
Бородулина Т. Л. Биология и хозяйственное значение чайковых птиц южных во-
доемов СССР//Тр. ин-та морфологии животных им. Северцова. 1960.
Вып. 32.
Бузун В. А. Клептопаразитизм у чаек и степень агрессивности в Ответах крачек //
Зоол. журн. 1984. Т. 62, № 6. С. 874-881.
Васильев В. И. К питанию Larus ridibundus L., Sterna hirundo L., Colymbus
cristatus L. и их роль в рыбном хозяйстве Азербайджана // Изв. .АН Аз.ССР.
Сер. биол. наук. 1967. № 6. С. 60—65.
Виксне В. А. О структуре гнездовой популяции озерной чайки (Larus ridibundus L.)
на приморских озерах Латвии // Орнитологические исследования. Рига: Зи-
натне, 1968. № 5. С. 207 -228.
Дементьев Г. П., Гладков Н. А., Спангенберг Е. И. Отряд Чайки //Птицы Со-
ветского Союза: М.: Сов. наука, 1954. Т. 6. С. 372—603.
Заблоцкий В. И., Заблоцкая Л. И. Эколого-фаунистический обзор чайковых птиц
Юго-Западного Каспия и их рыбохозяйственное значение//Тр. Астрахан.
зап-ка. 1963. Вып. 8. С. 309—348.
Зубакин В. А. Сравнительная экология колониального гнездования чайковых птиц:
Автореф. дис. ... канд. биол. наук. М., 1976. 22 с.
Ильичев В. Д., Фомин В. Е. Орнитофауна Башкирии и ее изменения в XX веке //
Орнитология. М.: Изд-во МГУ, 1979. Вып. 14. С. 83—96.
Исаков Ю. А., Крумина М. К., Распопов М. И. Материалы по экологии обыкновен-
ной чайки // Очерки природы Подмосковья и Московской области. М.: Изд-во
МОИП, 1947. С. 104-187.
Кириков С. В. Птицы и млекопитающие в условиях ландшафтов Южной оконеч-
ности Урала. М.; Л.: Изд-во АН СССР, 1952. 412 с.
Маматов А. Ф. Воздействие человека на структуру гнездовой колонии оз. Кара
гайлы // Колониальные гнездовья околоводных птиц и их охрана. М.: Наука,
1975а. С. 20-21.
Маматов А. Ф. Гнездовая колония околоводных птиц оз. Карагайлы // Там же.
19756. С. 146.
Маматов А. Ф. Условия гнездования водоплавающих птиц в Башкирском Зау-
ралье // Тез. Всесоюз. конф. «Экология гнездования птиц и методы ее изуче-
ния». Самарканд: Самарканд, ун-т, 1979. С. 135—136.
Михлин В. Е. Изучение охотничьих районов озерной чайки //Учен. зап. Горьк.
ун-та. 1963. Вып. 63. С. 11 — 14.
Михлин В. Е. О гнездовой площади обитания озерных чаек Заболотновской коло-
нии // Там же. 1966. Вып. 75. С. 63—66.
84
Модестов Б. М. К экологии гнездового периода чаек в дельте Волги // Зоол. журн.
1943. Т. 22, № 2. С. 92—102.
Молчанова Т. В. Очерки по биологии озерной чайки на озере Тайпшно Курган-
ской области // Вопр. зоологии. Челябинск, 1969. Вып. 1. С. 87—91.
Птицы Волжско-Камского края. Неворобьиные. М.: Наука, 1977. 296 с.
Строков В. В. Обыкновенная чайка в ближнем Подмосковье // Охрана природы
и озеленение. М., 1960. Вып. 4. С. 77—94.
Сушкин П. П. Птицы Уфимской губернии. М.: Кушнерев, 1897. XI, 331 с. (Ма-
териалы к познанию фауны и флоры Российской империи; Вып. 4).
Харитонов С. П. О формировании микроколоний у озерной чайки (Larus ridibun-
dus L.) //Зоол. журн. 1981а. Т. 60, № 4. С. 540—547.
Харитонов С. П. 'Взаимоотношения озерных чаек (Larus ridibundus) в локальной
группировке в пределах колонии //Там же. 19816. Т. 60, № 6. С. 871—877.
Харитонов С. П. Индивидуальное использование мест кормежки озерными чайками
(Larus ridibundus L.) // Там же. 1983а. Т. 62, № 5. С. 748— 754.
Харитонов С. П. Структура колоний и динамика переселений озерных чаек
(Larus ridibundus L.) в сезон размножения // Там же. 19836. Т. 62. № 7.
С. 1068-1076.
Харитонов С. П., Зубакин В. А. Процесс формирования пар у озерных чаек //
Там же. 1984. Т. 63. № 1. С. 95-104.
Creutz G. Ernahrungsweise und Aktionsradius der Lachmowe (Larus ridibundus L.) //
Beitr. Vogelkunde. 1963, Bd. 9, N 1/2, S. 3—58.
Klima M. Prispevek k hnizdni bionomii racka chechtaveho (Larus ridibundus L.) //
Zool. listy, 1964. Sv. 13, N 2. S. 111-124.
Wundsch H. H. Die Steilung der Lachmowe (Larus ridibundus) in des teichwirts-
chaftlichen Production (Karpfenzucht) // Beitr. Vogelkunde. 1963. Bd. 9, N 3.
S. 191-199.
SUMMARY
Black-headed Gull in Bashkiria. MamatovA. F.
Black-headed Gull forms a colony, numbering to 250 pairs in Karagaili Lake in
Bashkirian Transuralia. The information on phenology of the arrival and morphometric
data are given. The terms and characteristics of nesting were observed, the form
and sizes of nests and eggs were described. In the food of nestlings 63,6% are
adult dragonflies, 27,3% larvae of dragonflies and diving beetles, 18,2% — beetles
larvae, 54,6% — fish (golden silver crucians, roach). In the food of the adult birds
in 66,6% of cases is fish, 44,4% are diving beetles, 33,3% are damselflies, etc.
Gulls promote considerably spreading fish diseases (digrammosis, ligulydosis) which
had infected 30% of adult birds and 28,3% of gull chicks. Black-headed Gull is of
great use feeding with insects, including pests of fishery and some little value
fishes. The joint nesting of gulls and ducks is useful for the letters.
85
УДК 591.91: 598.822 (5744-575)
А. К. САГИТОВ,
Л. Э. БЕЛЯЛОВА,
С. Э. ФУНДУКЧИЕВ
703004, Самарканд,
бульвар М. Горького, д. 3.
Самаркандский
государственный университет.
Биологический факультет
Расселение майны
(Acridotheres
tristis tristis)
в Средней Азии
и Казахстане
Майна, или индийский скворец, является единственным предста-
вителем рода Acridotheres, встречающимся в пределах Советского
Союза. Прочие шесть видов этого рода населяют Южную и Юго-
Восточную Азию.
На территорию СССР майна проникла из Афганистана. Но
400 лет назад в Кабуле и Кабульской провинции майны не было.
Захириддин Бабур отмечает ее лишь для области Ламнагат (юго-
восточнее Кабула). Бабур пишет: «... одна из птиц — шарак (аф-
ганцы и сейчас называют майну «шаро»). Много его в Ламнагате.
Южнее же часто встречается во всем Индостане» (цит. по
М. П. Асланову (1953)).
Путешествуя по южному, западному и северному Афганистану
в октябре—ноябре 1936 г., М. П. Асланов (1953) видел майну в
Кандагаре, Ферахе, Герате, Меймене, Акче, Мазари-Шерифе,
Аибике, т. е. вокруг Гиндукуша.
В Герате майна появилась лет 100 назад, так как Эчисон,
посетивший северо-западный Афганистан в 1884—1885 гг., ее не
указывает (Асланов, 1953). Герат находится'от советско-афганской
границы всего в 100 км, а Меймене и Мазари-Шериф расположены
еще ближе. По-видимому, для майны нетрудно было перелететь
через Амударью — из Меймене в Керки, из Мазари-Шерифа в Тер-
мез и т. д. Расселение майны по поймам среднеазиатских рек,
в частности вдоль культурной полосы по Амударье, бесспорно. Этот
факт проливает свет на происхождение индийских (южноазиат-
ских) элементов в фауне Средней Азии.
М. А. Мензбир (1914) полагал, что такие элементы в фауне
Туркестана являются местными и древними, т. е. реликтами
прошлой фауны Средней Азии. С другой стороны, Г. П. Дементье-
вым (1938), выдвинута гипотеза, согласно которой такие виды,
как нитехвостая ласточка (Ilirundo smithii L.), полосатая тимелия
(Garrulus lineatus Vigors), синий дрозд (Myophonus caeruleus
Scop.), райская мухоловка (Terpsiphone paradisi L.), представляют
новейшее наслоение в среднеазиатской фауне. К. А. Воробьев
(1944) объединяет противоположные позиции М. А. Мензбира и
Г. П. Дементьева. Согласно ему, появление южноазиатских элемен-
тов в Средней Азии шло различными путями в различное время.
По своим биогеографическим связям майна относится к группе
видов южноазиатского происхождения, которых М. А. Мензбир
86
считал за реликтов, и если бы расселение майны произошло 50 лет
тому назад, то можно было высказать предположение о принадлеж-
ности ее к третичным реликтам туркестанской фауны. Между тем
4 заселение майной территории Средней Азии происходит на наших
? глазах и может рассматриваться как аргумент в пользу взгляда о
недавнем появлении других южноазиатских элементов в Средней
< Азии (Рустамов, 1946).
Впервые появившись на территории СССР в начале столетия,
| майна за последние 70 лет продвинулась от южных районов
| Узбекистана за его северные границы, освоила многие районы
Казахстана и продолжает распространяться на север, представляя
тем самым исключительный пример стремительного расширения
£ ареала.
Расселение майны в Узбекистане
На территории СССР майна впервые отмечена С. И. Билькевичем
V в 1907 г. у Камар-Сарая.
: В 1912 г. залетные птицы были встречены Н. А. Зарудным в
районе Термеза. Там же 11 июня 1914 г. Н. А. Бобринским добыты
С гнездовые птицы. Н. Ю. Юдин в 1927 г. добыл взрослых птиц в
S районе Термеза и Джар-Кургана, в 1928 г.— в районе Денау,
. в 1929 г. было найдено гнездо в кишлаке Юрчи, пара майн была
Э обнаружена в районе Сары-Асия (Юдин, 1940). В дальнейшем
майна в долине р. Суржан становится обычной и к 1953 г. ареал,
расширение которого шло исключительно по речным долинам,
V. продвинулся по долине р. Сурхан по Денау и дальше по Гис-
сарской долине, а по р. Шарабаддарье поднялся почти до Дер-
бента.
В Чарджоу майна появилась в 1939 г. (Рустамов, 1946) и
отсюда проникла в Зарафшанскую долину (Богданов, 1952).
В 1940 г. добыча в районе Каракуля (Богданов, 1952), а в 1946—
£•' 1947 гг. встречалась в Бухаре, Гиждуване, еще через год — в Ну-
рате; в это же время обнаружена в Карши, Чиракчи, Китабе
(Юдин, 1940).
Несколько позднее, в 1947 —1949 гг., майна появилась в Самар-
кандской области. И. М. Ананьев добыл двух самцов в 15 км
от Каттакургана, а стайки в 20—30 особей часто встречал в самом
городе (Сагитов и др., 1957). 15 марта 1947 г. майна была добыта
на территории Пайарыкского района в 40 км к северо-западу от
Самарканда, осенью 1949 г.— в 12 км к северу от Самарканда.
На железнодорожной станции Джума в 25 км к западу от города
пару майн видели 5 июня 1951 г. (Мекленбурцев, 1962). В 1951 г.
залетная птица была добыта в окрестностях Джизака.
В начале 1953 г. майна отмечена в Зааминском районе (50 км
северо-восточнее Джизака), а в районе станции Милютинской
(90 км к северо-востоку от Самарканда) видели майну на гнездо-
вании (Богданов, 1952).
87
В мае 1959 г. группу из 12 майн встретили в долине реки
Чирчик, в октябре — у кишлака Тойтепе, в 1960 г. стаи из 40—50
особей отмечены в районе кишлака Кибрай — 25 ноября, Луначар-
ского — 5 декабря (Салихбаев, Богданов, 1967). Те же авторы
в 1960 г. находили майну в западной части Ферганской долины.
Весной 1961 г. она была обнаружена в окрестностях Намангана.
О пребывании майны в центральной части Ферганской долины
сообщал Р. Н. Мекленбурцев (1962), т. е. в шестидесятые годы она
полностью освоила Ферганскую долину.
Рассматривая расселение майны вдоль Амударьи в пределах
Узбекской ССР (рис. 1), можно отметить ее появление в районе
Кызыл-Рабата, Топрак-Калы, Сарымая в 1958 г. (Мамбетжумаев,
1960).
В Ургенче майна впервые была отмечена в 1953 г., а в Нукусе —
в 1954 г. К. А. Чистяковым. По-видимому, это были залетные
птицы, так как А. М. Мамбетжумаев, постоянный житель Нукуса,
ее ни разу не видел раньше, хотя она — чрезвычайно заметная
и крикливая птица, так что пропустить ее нельзя. С 1961 г. майна
стала оседлой птицей в Турткуле, в 1968 г. отмечена в пос. Назар-
хан. С 1964 г. майна стала оседлой птицей в Нукусе. В марте
1970 г. отмечены две особи в Ходжейли. С октября 1971 г. она
поселяется в окрестностях Кунграда. 5 августа 1976 г. на Чимбай-
ском заводе отмечено пять особей, живущих там оседло уже вто-
рой год.
Есть данные, что с 1975 г. майны живут на территории Муйнак-
ского района (Аметов, 1981).
Расширяя ареал, майна локализуется не только в полосе
культурного земледелия и в долинах рек, но при соответствующих
условиях проникает вглубь пустыни, в горы, достигая арчевого
леса. В Афганистане майна обыкновенна на высоте 1700 м в
Кабуле, летом отмечалась в Пагмане на высоте 2500 м над ур. м.
(Асланов, 1953), а в Индии — даже на высоте 2700 м (Salim,
1946). В Казахстане майна гнездится на высоте 1860 м в заповед
нике Дарбаза (Ковшарь, 1966). На северном склоне Туркестанс-
кого хребта наблюдали гнездящихся птиц на высоте 1000—2200 м.
С 1959 г. майна стала обычной птицей, а местами многочисленной,
в населенных пунктах хребта Нуратау и его предгорий, в селениях
гор Актау и Каратау (2000 м над ур. м.).
В мае—июне 1959 г. гнездящуюся майну обнаружили в юж-
ных и центральных частях Кызылкумов, причем в местах, очень
мало сходных с теми, в которых обыкновенно живет эта птица.
Несколько пар было найдено в 8 км к югу от селения Аяк-Кудук.
Поселились они вблизи пруда с солоноватой водой. Древесных
посадок, полей и огородов вблизи не было. Ближайший населен-
ный пункт, в котором майна живет в обычных для нее условиях, на-
ходится в предгорьях западных отрогов Нуратау, в 75 км по пря-
мой. Еще в менее подходящих условиях пара майн поселилась у
колодца Мурункудук. Колодец этот грубокий, и майны пользуются
водой, поднимаемой чабанами для овец. Далее на север майны
88
Рис. 1. Расселение майны в Узбекистане .
Рис. 2. Распространение майны в Таджикистане (по: Юдин, 1940)
1 — область гнездования; 2 — область распространения; 3 — случайные залеты
найдены в населенном пункте Кунга, где стоит всего две мазанки
у колодца, место типично пустынное. Птицы кормились возле овец,
пригоняемых на водопой к колодцу.
Таким образом майна, распространяясь по Узбекистану, селит-
ся здесь не только в оазисах, но й в глубине пустыни. Очевидно,
89
с дальнейшим освоением пустыни майна станет таким же обычным
видом Кызылкумов, каким она за последние годы стала в долине
Зарафшана (Ишунин, 1960).
Расселение майны в Таджикистане
Расселение майны в Таджикистане шло в основном по главным
водным артериям — вдоль Пянджа и его притоков (рис. 2).
В значительном количестве майны обнаружены в поливной зоне
р. Кафирниган, в окрестностях Шаартуза и Кабодиена еще в 1929 г.
(Ахмедов, 1953).
В 1935—1936 гг. майны найдены на гнездовье восточнее Шах-
ринава (Иванов, 1940). В 1936 г. три птенца добыты в Курган-
Тюбе, гнездовья были обнаружены также в районе Джиликуля,
а в окрестностях Душанбе зарегистрировано семь гнездящихся пар.
До 1934 г. их здесь не встречали. В 1934 г. появилась первая пара,
и с тех пор число птиц с каждым годом увеличивалось (Юдин,
1940).
Расселение майны происходит и по Гиссарскому хребту,
в Варзобской долине по хр. Ромит и Харангон. По Вахшскому
хребту майна проникла до Кангурта, Бальджувона, Ховалинга.
Если в 1934—1935 гг. она была гнездящейся птицей лишь в ни-
зовьях Пянджа, то в 1961 г. майны впервые отмечены у подно-
жия Дарвазского хр. около селения Висхара (Абдусалямов, 1973).
На гнездовье майна теперь встречается в предгорьях север-
ного склона Туркестанского хребта от Воруха, Исфары, Пшемыша,
включая Ура-Тюбе, Шахристан, Окташ. Кроме основной долины,
майны на гнездовье заходят в населенные пункты вдоль большин-
ства притоков Зерафшана.
Майна — обычная гнездящаяся птица по склонам Курамин-
ского хребта, гор Могол-Тау, во всей долине Сырдарьи от урочища
Сыры-Камыш до Фархадского водохранилища, включая К'айрак
Кум, Ленинабад, Пролетарск, Новую Матчу (Иванов, 1969).
В 1961 г. пара майн отмечена у Канибадама (Мекленбурцев,
1962).
Расселение майны в Туркмении
В Туркмении расселение майны первоначально строго ограничи-
валось долиной Амударьи (рис. 3).
К. А. Воробьев (1941) отмечал этих птиц только для юго-вос-
точной части Туркмении. Весной 1939 г. он нашел майн в значи-
тельном количестве по долине Амударьи в районе станции Чар-
шанга, в горах встретил этих птиц в районе аула Кугитанг (70 км к
северу от Амударьи). По словам местных жителей, первые птицы
появились в Кугитанге только в 1933 г. А. И. Иванов (1940) от-
мечал майну для станции Мукры (Керкинский район) в 1934 г.
В 1936 г. она была встречена в Карабекауле и добыта севернее
Чарджоу, в ауле Денау (Гизенко, 1946).
90
Рис. 3. Расселение майны в Туркмении
Зоологическая экспедиция Туркменского филиала АН СССР
добыла майну 24 апреля 1942 г. в ауле Гайнук по Амударье
(65 км севернее Чарджоу; см.: (Гизенко, 1946)). В Кабаклы пер-
вое появление майн отмечено в 1945 — 1946 гг., а к 1953 г. они стали
обычными на гнездовье. Северная граница ареала к этому времени
продвинулась от Кабаклы на 40—55 км к тугаям Гогорчинлы
(Дементьев, 1953). В некоторые зимы стайки майн появлялись
тогда и севернее Гогорчинлы — в районном центре Дарган-Ата
(Клюшкиц, 1959).
Посетив в октябре-ноябре 1957 г. г. Дарган-Ату и прилегающие
к нему тугаи Карайгыр и Джербент, расположенные на 25—35 км
севернее, А. М. Мамбетжумаев (1960) повсеместно видел стайки
майн.
Расселение шло не только с юга на север, по долине Амударьи,
но и с востока на запад, вдоль Каракумского канала. В мае 1955 г.
майна была обнаружена в 80 км восточнее Керки (Нургельдиев,
Секунова, 1955). В 1961 г. они были обычными птицами в поселке
Карамет-Нияз (Ташлиев, 1961).
Первое появление майны в Репетеке отмечено В. А. Стальмако-
вой в 1945 г. (Рустамов, 1946). Затем они там не появлялись до
мая 1953 г. (Клюшкин, 1959).
Достоверные сведения о появлении майны в долине Мургаба
относятся к 1954 г., когда в Тахта-Базаре были добыты две
птицы из стайки численностью около 40 особей. В марте 1954 г.
пара майн в Тахта-Базаре искала место для устройства гнезда
(Сухинин, 1956). В мае—июне 1955 г. майны постоянно держались
91
в Ташкепри и в 1956 г. найдены здесь на гнездовье А. К. Рустамо-
вым (1958). С 1954 по I960 г. майны расселились к северу от Тахта-
Базара до колхоза «Ленинград», расположенного севернее г. Мары
(Ташлиев и др., 1964).
На сегодняшний день пункты нахождения этого скворца нахо-
дятся в 200 км к востоку от Ашхабада и ограничиваются долиной
р. Теджен, где приходилось наблюдать майн в сентябре—декабре
1983 и 1984 гг. (Рустамов, Эминов, 1985).
Расселение майн в Казахстане
К концу 1950-х — началу 1960-х годов майна, освоив территории
Туркмении и Таджикистана, достигла северной границы Узбеки
стана и проникла в Казахстан.
За 10 лет майна заселила значительную часть Южного Ка-
захстана. Расселение ее шло от Ташкента предгорными районами
Западного Тянь-Шаня на северо-восток и восток, по долине Сыр-
дарьи — на северо-запад.
В Казахстане, в прилегающих к Ташкенту районах, первых
майн отметили в 1959 г. 30 сентября 1961 г. Д. Ю. Кашкаров добыл
майну в долине Келеса (Мекленбурцев, 1962).
Одна пара майн обнаружена А. Ф. Ковшарем (1966) в ноябре
1961 г. южнее Кузгурта. Через несколько дней он же встретил ее
в Туркестане. В 1962 г. майны гнездились в Чимкенте, в 1963 г.
продвинулись дальше на восток по долине р. Арыс. 19 января в
селе Высоком встречена стайка из 11 майн. Кроме того, майны поя-
вились и на кордонах заповедников Джагаблы (1400 м) и Дарбаза
(1860 м) (Ковшарь, 1966).
По сообщению Р. И. Малышевского (1965), в мае-июне 1962 г.
пара майн обнаружена в Яны-Курганском районе в 12 км запад-
нее станции Беш-Арык, позднее там найдено гнездо с птенцами.
К 1963 г. майна заселила значительную часть Чимкентской об-
ласти, распространяясь в двух основных направлениях: по долине
Сырдарьи и вдоль северных предгорий Таласского Ала-Тау, где
достигла границы Чимкентской и Джамбульской областей.
В 1964 г. гнездо майны обнаружено на станции Тимур (Варшав-
ский, 1968).
В 1962 г., не преследуя каких-либо определенных целей, по
инициативе Главного управления охотничьего хозяйства и запо-
ведников Казахской ССР через Зооцентр было получено 200 птиц,
и 24 марта их выпустили на территории Алма-Атинского заповед-
ника. Разлет майн был невелик. Поступившие возвраты окольцо-
ванных птиц свидетельствуют о том, что к 1965 г. они расселились
не далее 30—40 км от Алма-Аты.
В 1965 г. одиночная птица была встречена в ауле Сары-Булак
в 80 км восточнее Казалинска (Ротшильд, 1965).
В августе 1968 г. майна была встречена в селе Михайловка
в 12 км северо-восточнее Джамбула (Сема, 1970). В то же время
10 птиц были отмечены в 45 км восточнее Джамбула. Отмеченное
92
Рис. 4. Расселение майны в Казахстане
ранее А. Ф. Ковшарем (1966) гнездование майны в селе Высоком
(верховье р. Арысь), в 1964 г. было самым северо-восточным.
Таким образом, за четыре года майна расселилась на 100 км, прео-
долев при этом Чокпакский перевал, имеющий высоту 1200 км над
ур. м.
18 октября 1968 г. А. П. Моисеев добыл заканчивавшую линьку
самку в долине р. Чу в поселке М. Камкалы.
К 1981 г. в Сырдарьинском Каратау майна стала обычной,
а местами многочисленной по всем населенным пунктам, как в
юго-восточной, так и в северо-западной его части (Ковшарь, 1984).
Особенно высока ее численность по долинам рек Арысь, Чаян,
Бугунь, Боролдай. Она обычна в поселках Сузак и Чулак-Курган,
в низовьях и в среднем течении р. Чу, а на юго-западной окраине
Бетпак-Далы — в поселке Жуван-Толбе (в 75 км к северу от Су-
зака). Гнездится она во всех населенных пунктах от села Фурма-
новка до совхоза Байтал. В Фурмановке она появилась в 1978 г.,
в Байтале — в 1980 г. В низовьях Сарысу в 1982 г. майны приле-
тели 31 марта, и небольшие стайки регистрировались весь апрель
(Сема, Гисцов, 1984).
В настоящее время майну можно встретить в пустынях Южного
Прибалхашья, куда она приникла по долинам Или и Каратала.
93
В низовьях Или, в поселке Баканас, майна гнездится с 1981 г.
Две пары майн появились в поселке Караой в 1982 г. В том же году
несколько раз одиночные птицы были встречены в окрестностях
Караоя — в урочище Ушкудук в 20 км к северу от Караоя, в уро-
чище Жагуйген в 40 км к востоку от Караоя, в урочище Карадон в
30 км к юго-востоку от Караоя. По-видимому, встречи майн
вне населенных пунктов свидетельствуют об активном поиске
ими новых мест для гнездования. По долинам Каратала майна
проникла на 95 км к северу от станции Уш-Тобе (рис. 4). Видимо,
майна, осваивая новые территории, активно проникает в пустын-
ные районы по долинам крупных рек.
Заключение
Анализируя расселение майны в пределах Средней Азии и Казах-
стана, можно сказать, что этот вид продвигался на север по куль-
турной полосе — по пойме Амударьи, постепенно заселяя ее право-
бережные притоки. В Шахрисябзе майна стала гнездиться с
1942 г., в Карши, Чиракчи первых майн встретили только в 1946—
1947 гг. Возможно, в Карши майна проникла из Керков по Каршин-
скому каналу, а в Шахрисябз — по притокам Сурхандарьи и Ка-
шкадарьи. Определенную роль в расселении майны сыграло освое
ние пустыни, строительство каналов.
Говоря о скорости расселения майны, можно отметить, что
распространение ее шло неравномерно как в отдельные годы,
так и в различных местностях (рис. 5). А. К. Рустамов (1946)
приводит следующие данные: по Амударье за 35 лет (1912—
1946 гг.) она распространилась на 500 км — от Термеза до
Кабаклы. С 1907 по 1947 г. прошла по пойме Амударьи от Сарай-
Камара до Кабаклы на 760 км (Юдин, 1940). Как видно из этих
примеров, средняя скорость распространения составляет в первом
случае 16,6 во втором — 19 км в год.
По данным О. П. Богданова (1952), расселение происходило
значительно быстрее, так как майна впервые отмечается в Чард-
жоу в 1939 г., а в 1949 г. она достигла Самарканда, покрыв рассто-
яние, превышающее 300 км, за 10 лет, т. е. со средней скоростью
30 км в год. По А. М. Мамбетжумаеву (1960) с 1952 по март 1959 г.
майна продвинулась на север по Амударье на расстояние свыше
150 км — от Кабаклы, где в это время проходила северная граница
гнездования майн, до Питняка. Если же включить сюда и пос. Ха-
зарасп (на территории которого майна считалась в то время залет-
ной птицей), то расстояние увеличивается до 190 км и средняя
скорость возрастает от 25 до 31 км в год.
От Гойнука до Кабаклы майна продвинулась за три года на
60—65 км, а от Кабаклы до Гогорчинлы более чем за девять лет —
на 50 км (Рустамов, 1958). На основании этих данных можно
говорить о некотором замедлении темпов расселения майны на
север по сравнению с предыдущими годами.
Причиной неравномерного расселения майны может быть пре-
94
Рис. 5. Расселение майны в Средней Азии и Казахстане
1 — в 1920-е годы; 2 — 1930-е годы; 3 — 1940-е годы; 4 — 1950-е годы; 5 — 1960-е годы;
6 — 1970-е годы
рывистость подходящих для нее биотопических условий (Дементь-
ев, 1953).
Майна тяготеет к культурному ландшафту. Обычно расселе-
ние ее идет вдоль рек, вдоль железных дорог, рядом с населенными
пунктами. Но в последнее время майна отмечена на гнездовьях в
пустынных районах, далеко от населенных пунктов, куда она по-
падает, преодолев огромные пустынные территории.
Причиной расширения ареала майны, как считают одни авторы,
может быть увеличение площадей культурного ландшафта.
Г. П. Дементьев (1953) связывает это явление с каким-то повы
шением численности майны, отчасти обусловленной их высокой
плодовитостью. М. П. Асланов (1953) считает, что расселению май-
ны способствуют рост населения, увеличение числа населенных
пунктов, развитие ирригации, постройка высоких зданий, кото-
рые они используют для ночевок.
По мнению А. К. Рустамова (1946), развитие культурного
ландшафта не является непосредственной причиной расселения
майн. А. К. Рустамов считает, что в расселении майны можно ви
деть результат пульсации границ ареалов, обусловливаемой бла-
гоприятными для вида экологическими условиями, в первую оче-
95
рёдь трофическими, определяющими, по-видимому, высокую
плодовитость. В расселении майн в Средней Азии он склонен
видеть результат перенаселения основной гнездовой области.
ЛИТЕРАТУРА
Абдусалямов И. А. Птицы. Душанбе: Дониш, 1973. 396 с. (Фауна Таджик-
ской ССР; Т. 19, ч. 2).
Аметов М. Птицы Каракалпакии и их охрана. Нукус; Каракалпакстан,
1981. 137 с.
Асланов М. П. Распространение индийских птиц в Афганистане // Тр. Ин-та
зоологии и паразитологии АН УзССР. 1953. Т. 2. С. 84—89.
Ахмедов К. Р. Биология и хозяйственное значение майны в Таждикиста-
не // Изв. АН ТаджССР. Отд-ние естеств. наук. 1953. № 4. С. 80—84.
Богданов О. П. Новые данные о распространении майны Acridotheres tristis L. //
Изв. АН УзССР. 1952. № 6. С. 104-105.
Варшавский С. И. К расселению майны в среднем течении Сыр-Дарьи // Ор-
нитология. М.: Изд-во МГУ, 1968. Вып. 9. С. 340—341.
Воробьев К. А. Новые данные по распространению и биологии птиц Южной
Туркмении//Докл. АН СССР. 1941. Т. 33, № 5. С. 381-384.
Воробьев К. А. Индо-гималайские элементы в орнитологической фауне
Русского Туркестана // Там же. 1944. Т. 45, № 9. С. 381—384.
Гизенко А. И. О распространении майны Acridotheres tristis tristis // Изв.
Туркм. филиала АН СССР. 1946. № 3 / 4. С. 483—485.
Дементьев Г. П. К вопросу о взаимоотношениях палеарктической и восточной
зоогеографических областей на примере Туркестанской и Индийской ави-
фаун // Бюл. МОИП. Отд. биол. 1938. Вып. 5 / 6. С. 343—355.
Дементьев Г. П. Новые материалы по распространению и биологии майны
в Туркмении // Изв. АН ТССР. Сер. биол. наук. 1953. № 3. С. 20—24.
Зарудный И. А. Заметки по орнитологии Туркестана // Орнитол. вестник.
1912. № 1. С. 16-30.
Иванов А. И. Птицы Таджикистана//Тр. Тадж. базы АН СССР. 1940 Т. 10.
С. 1-229.
Иванов А. И. Птицы Памиро-Алая. Л.: Наука, 1969. 447 с.
Ишунин Г. И. Майна в Кызылкумах // Природа. 1960. № 11. С. 132.
Илюшкин Е. А. О майне в Кара-Кумах (Репетек) // Тр. Ин-та зоологии
и паразитологии АН ТССР. 1959. Т. 4. С. 150—155.
Ковшарь А. Ф. Птицы Таласского Алатау. Алма-Ата: Кайнар. 1966. 436 с.
Ковшарь А. Ф. Майна у северных пределов Средней Азии//Тез. докл.
VIII Всесоюз. зоогеогр. конф. М., 1984. С. 72—74.
Малышевский Р. И. К расселению майны в Казахстане // Орнитология.
М.: Изд-во МГУ, 1965. Вып. 7. С. 479.
Мамбетжумаев А. М. О расселении обыкновенной майны на юге Каракал-
пакской АССР, Хорезмской области и северо-западе Бухарской области
Узбекской ССР//Зоол. журн. 1960. Т. 34, вып. 5. С. 781—783.
Мекленбурцев Р. И. Новые данные о распространении некоторых птиц
в горах Средней Азии // Орнитология. М.: Изд-во МГУ, 1962. Вып. 5.
С. 211-214.
Мензбир М. А. Зоологические участки Туркестанского края и вероятное
происхождение последнего. М., 1914. 144. с.
Нургельдыее О. И., Секунова С. А. Новые сведения о распространении
майны на территории Туркменистана // Изв. АН ТССР. Сер. биол. наук. 1955,
вып. 5. С. 87.
Ротшильд Е. В. Залет майны в Северные Кызылкумы // Новости орнитоло-
гии. Алма-Ата: Наука, 1965. С. 326.
Рустамов А. К. Расселение майны в бассейне Аму-Дарьи // Природа. 1946.
№ 2. С. 89—92.
Рустамов А. К. О гнездовой фауне птиц и практическом значении наземных
позвоночных животных Ташаузской области (Северо-Восточный Туркме-
нистан) //Учен. зап. Туркм. ун-та. 1958. Вып. 4. С. 95—129.
96
Рустамов Э. А., Эминов А. Майна в Ашхабаде // Изв. АН ТССР. Сер. биол.
наук. 1985. № 2. С. *59-61.
Сагитов А. К., Калужина М. В., Киселева Е. В. Индийская майна в Зараф
шанской долине // Тр. УзГУ. Н. С. 1957. № 67. С. 73—83.
Салихбаев X. С., Богданов А. И. Фауна Узбекской ССР. Т. 2, ч. 4. Птицы.
Ташкент, 1967. С. 56—59.
Сема А. М. К расселению майны в Казахстане // Материалы 2-й науч. конф,
молодых ученых АН КазССР. Алма-Ата, 1970. С. 376.
Сема А. М., Гисцов А. П. Расселение майны в Казахстане//Орнитология.
М.: Изд-во МГУ, 1984. Вып. 19. С. 211-212.
Сухинин А. Н. Материалы о распространении некоторых птиц в Туркме-
нии // Зоол. журн. 1956. Т. 35. № 5. С. 612—613.
Ташлиев А. О. Состав и распределение авифауны трассы Каракумского канала
первой очереди//Изв. АН ТССР. Сер. биол. наук. 1961. № 6. С. 68 — 75.
Ташлиев А. О., Эминов А., Сухинин А. Н. Новые данные о распространении
некоторых птиц в Туркмении // Там же. 1964. № 1. С. 83—86.
Юдин Н. М. Майна или афганский скворец — Acridotheres tristis L. Его
биология и распространение в Средней Азии // Тр. Узбекистан, зоол. сада.
1940. Т. 2. С. 35-43.
Юдин Н. М. Расселение майны//Природа. 1955. № 10. С. 151.
Salim A. The book of Indian Birds. Bombay: Bombay Natural History Society,
1946. 437 p.
SUMMARY
Distribution of Common Mynah (Acridotheres tristis tristis) in Middle Asia and
Kazakhstan. Sagitov. A. K., Fundukchiev S. E., Beljalova L. E.
Common Mynah in the USSB was firstly marked by S. I. Bilkevich in 1907 near
Kamar-Sarai. By the end of the 1950’s — beginning of the 1960’s the Common Mynah
was distributed on the territories of Turkmain, Tadjikistan, reached the northern
borders of Uzbekistan and spread within the Kazakhstan. Common Mynah settled a
considerable part of the Southern Kazakhstan over 10 years. Its distribution began
from Tashkent along the foothills regions of West Tjan-Shan, to the north-east and the
east, and along the Valley of Syrdaria Biver to the north-west. The distribution of the
Common Mynah proceeded unevenly, both during different years and in different
localities. The average annual rate of the distribution, according to the data of some
authors, ranges from 16,6 to 30 km.
4 Заказ № 36
97
УДК 574.2 3: 591: 598.816
Л. п. БЫКОВА
614000, Пермь,
ул. Коммунистическая, д. 23.
Пермский сельско-
хозяйственный институт.
Кафедра анатомии
Элиминация
в раннем онтогенезе
и величина кладки у грача
(Corvus frugilegus)
Элиминация с точки зрения эволюции представляет собой выпа-
дение из генофонда популяции части генов и их сочетаний (Се-
верцов, 1981), которые не принимают участия в дальнейших
преобразованиях популяций. Репродуктивный потенциал по-
пуляций очень велик. У птиц величина кладки, характеризующая
репродуктивные возможности популяции, реализуемые в опреде-
ленных условиях внешней среды, варьирует довольно в широких
пределах. Например, у хохлатой чернети в кладке 6—12 яиц
(Брауде, 1983), у обыкновенной сороки — 4—10 (Блинов, 1981;
Дугинцов, 1985), у скворца (Шкарин, Родимцев, 1982; Ну-
меров, 1984) и серой вороны (Блинов, 1981; наши данные) —
3-7.
В то же время численность большинства видов птиц довольно
постоянна, соотношение максимального и минимального ее зна-
чений редко превышает 2—6 раз (Лэк, 1957). Кроме других фак-
торов, незначительность колебаний численности обусловлена вы-
соким уровнем элиминации.
На существование определенного равновесного соотношения
между рождаемостью животных и смертностью указывают многие
авторы (Одум, 1975; Риклефс, 1979; Солбриг О., Солбриг Д.,
1982; и др.). Рождаемость и смертность отражают взаимодействие
особи с окружающей ее средой: способность организма направить
имеющиеся ресурсы на размножение и на избегание хищников,
голода или действия неблагоприятных факторов. Однако меха-
низмы, обусловливающие равновесие между рождаемостью и эли-
минацией, не выявлены.
Как подчеркивал И. И. Шмальгаузен (1982), нельзя пред-
полагать, что естественный отбор действует изолированно на
отдельные стадии развития организма. В борьбе за существование
побеждают те организмы, которые в течение всего онтогенеза
приспособлены к среде таким образом, что, достигая взрослого
состояния, дают наибольшее потомство (Рубайлова, 1981). Анализ
интенсивности элиминации на разных стадиях онтогенеза и вы-
явление обусловивших ее факторов представляют возможность
оценить как качественную, так и количественную стороны слож-
ного и многогранного процесса борьбы за существование. Мы оста-
новимся на раннем онтогенезе как. периоде с наиболее высокой
смертностью птиц.
98
Массовая, или общая, элиминация (здесь и далее классифи-
кация приведена по И. И. Шмальгаузену (1939, 1946,- 1969))
происходит редко: гибель птенцов-слетков воробья во время
летнего града описана А. Н. Хохловым и Л. В. Афонасовой (1985),
ежегодную гибель гнезд чаек при весеннем подъеме уровня воды
наблюдали Ю. И. Мельников (1982), А. М. Болотников и Л. П. Бы-
кова (1984). Тотальная элиминация представляет собой случай
крайне интенсивного отбора, при котором гибель не обусловлена
индивидуальными свойствами особи.
Семейная элиминация проявляется у птиц при активном
хищничестве или разрушении гнезд. Вероятность гибели гнезд
различна у разных видов и зависит от степени защищенности
(надежности) гнезда и активности поведения родителей (Гаври-
лов, Родионов, 1965; Ковшарь, 1981; Ash, 1970). Известны случаи
семейной гибели птенцов от паразитических насекомых (Бак-
кал, 1980). В некоторых биотопах резко увеличивается количество
брошенных и разоренных кладок вследствие сильного антропо-
генного воздействия (Myrberget, 1983; Orell, Ojanen, 1983).
Индивидуальная элиминация у птиц встречается наиболее
часто, однако полных сведений об элиминации на всех стадиях
раннего онтогенеза мало (Семенов-Тян-Шанский, 1983; Нумеров,
1985; Orell, Ojanen 1983). В просмотренной нами литературе мы
не нашли достаточно полного анализа причин гибели птиц в гнез-
довой период. За исключением статьи Р. А. Зацепиной (1981),
нам не известны исследования по диким птицам, в которых бы
авторы пользовались возрастным и патолого-анатомическим
анализами погибших особей для установления причин элими-
нации.
В настоящей статье мы попытались провести анализ элими-
нации грача (Corvus frugilegus L.) в раннем онтогенезе, опреде-
лить причины, вызывающие гибель особей на разных стадиях
развития, а также выяснить взаимоотношения элиминации и ве-
личины кладки.
Материал и методика
Исследования проводились в 1980 —1982 гг. в одном из наиболее
крупных поселений (около 1000 гнезд) грача в Камском Пред-
уралье, расположенном в защитной лесополосе, вытянутой вдоль
железнодорожного полотна. В состав грачевника входят три
колонии, гнездовые участки которых разделены молодыми лесо-
посадками (Шураков, 1982).
Изучаемая колония расположена в западной части поселения
и занимает площадь 0,56 га (длина — 331 м, ширина — 17 м).
Среди древесных пород на участке преобладают ели и березы. Гра-
чи предпочитают гнездиться на елях, число построек на березах
незначительно, и появляются они позднее. Количество занятых
грачами деревьев увеличилось с 84 в 1980 г. до 103 в 1982 г., хотя
численность колонии изменялась незначительно: в 1980 г. было
4*
99
учтено 280 гнездящихся пар грачей, в 1981 г.— 252, в 1982 г, - 251.
Наиболее высокая плотность размещения гнезд — 3,3 гнезда на
дерево — отмечена в 1980 г. В 1981 и в 1982 гг. среднее количество
гнезд, приходящихся на одно дерево, было почти одинаково
(2,5 и 2,4 соответственно) и с высокой степенью достоверности
(t=6,25, Р<0,001) отличалось от данных 1980 г. (табл. 1).
Таблица 1
Распределение гнезд в колонии грача
Год Доля деревьев (%) с числом гнезд Всего занято деревьев Среднее кол-во гнезд на Г дерево
1 2 3 4 >4 М+т С, %
1980 9,5 29,76 15,48 23,81 21,43 84 3,3+0,1 28
1981 30,39 30,39 16,67 11,76 10,78 102 2,5+0,08 32
1982 27,18 33,01 22,33 8,74 8,74 103 2,4+0,07 29
Большинство заселенных деревьев имело по два гнезда. Ко-
личество деревьев с одним гнездом резко увеличилось в 1981 г.
(30,39%, п=31) по сравнению с предыдущим годом (9,5%,
п=8). Доля деревьев с тремя гнездами возросла от 15,48%
(п=13) в 1980 г. до 22,33% (п=23) в 1982 г. За изучаемый период
число деревьев с большим количеством гнезд (4 и более) сокра-
тилось в 2 раза (см. табл. 1).
В 1981 использование для гнезд старых построек составило
65,8%, а в 1982 г. грачами было занято 87,2% гнезд предыдущего
года. Причиной переселения грачей в новое гнездо может быть
гибель гнезда, гибель гнездового дерева, неуспешность размно-
жения в предыдущем сезоне. Разрушение гнездовых постро-
ек — естественный процесс, который усиливается с увеличением
массы гнезда. В. В. Корбут (1985) указывает на значение влаж-
ности для сохранности гнезд грачей. В исследуемой колонии при
сильном порывистом ветре в мае 1981 г. мы наблюдали гибель
большого количества птенцов и разрушение гнезд, построенных на
березах, растущих на южной (обращенной к полю) границе ле-
сополосы. На следующий год гнезда на этих деревьях не были
обнаружены.
Таким образом, ежегодно в пределах колонии при сохранении
относительно постоянной численности птиц происходит частичное
перераспределение гнездовых участков, которое обеспечивается
механизмом саморегуляции численности и структуры популяции
и обусловливается влиянием изменяющихся условий внешней
среды.
Ежегодно наблюдения за гнездованием грача начинались в
период яйцекладки. Как указывают многие исследователи (Дья-
конов, 1980; Брауде, 1984; и др.), откладка яиц у грача начина-
ется при повышении среднесуточных температур до -|-6° и при
интенсивном таянии снежного покрова. В 1981 г. откладка яиц
100
Рис. 1. Интенсивность откладки
яиц (Л) и вылупления птенцов
(Б) в колонии грача
2—1980 г.;
2-1981 г.;
3—1982 г.
Рис. 2. Нормированная по мак-
симуму встречаемость кладок
разной величины у грача
2—1980 г.;
2-1981 г.;
3—1982 г.
началась раньше/чем в 1980 и 1982 гг. (рис. 1), и имела растяну-
тый характер. Пик массовой яйцекладки в 1981 г. был отмечен
12 апреля. Откладка яиц в 1980 и 1982 гг. происходила в более
сжатые сроки с пиками 19—20 и 17 —19 апреля соответственно.
Появление первых птенцов в гнездах ежегодно наблюдалось
26—28 апреля. Появление птенцов во все три гнездовые сезона
было менее растянуто, чем откладка яиц (см. рис. 1). Массовое
вылупление в 1980 г. отмечено 7 —11 мая (пик — 8 мая),
в 1981 г.— 1—4 мая (2 мая) в 1982 г,— 5—8 мая (6 мая). Небла-
гоприятные условия гнездового сезона 1981 г. (резкие понижения
температуры воздуха, длительные снегопады) обусловили увели-
чение числа возобновленных кладок и растянутость сроков
вылупления.
101
Во время откладывания и насиживания яиц наблюдения за
гнездами грача проводили один раз в сутки, обычно в утренние
часы. Отмечали дату снесения и ранг яиц, наличие дефектов, из-
меряли индекс формы, массу и объем яиц. В период вылупления
количество посещений было увеличено: регистрировали порядок
вылупления, птенцов метили индивидуальными цветными мет-
ками. Состояние птенцов контролировалось ежедневно. Яйца,
оставшиеся в гнездах, после вылупления птенцов были собраны
и подвергнуты патологоанатомическому анализу. При вскрытии
яиц определяли возраст погибших эмбрионов.
Величина кладки. Размер кладки у грача варьирует от одного
до шести яиц (табл. 2). Доля кладок с одним и шестью яйцами
невелика. А. И. Шураков (1979) и Ю. В. Дьяконов (1980), про-
водившие исследования на близрасположенных колониях, также
подчеркивают редкую встречаемость кладок из шести яиц (1,4%)
и отмечают отсутствие однояйцевых кладок.
Наиболее часто в колонии встречались кладки из четырех-пяти
яиц, в 1980 г. составлявшие 66%, в 1981 г.— 72%, в 1982 г.— 74%
всех кладок. Количество кладок из трех яиц уменьшилось с 24%
в 1980 г. до 14% в 1982 г.
На рис. 2 изображено соотношение количества кладок разных
групп (1—6): за единицу в каждом сезоне принималось наиболь-
шее количество кладок, остальные величины определялись отно-
шением абсолютного числа кладок рассматриваемой группы к
максимальному, принятому за единицу. Форма всех трех норми-
рованных кривых величины кладки практически одинакова. Это
подтверждает то, что соотношение кладок разных объемов явля-
ется не случайным, а подчинено определенным закономерностям.
Количество кладок из одного и шести яиц мало, число двухяйце-
вых кладок не превышает одной четвертой максимального значе-
ния и имеет годичную флуктуацию. Во всех сезонах наиболее
многочисленными были кладки из четырех-пяти яиц, но вершина
кривой 1980 г. приходится на четырехяйцевые кладки, а 1981 —
1982 гг.— на пятияйцевые. Количество кладок из трех яиц умень-
шается от 1980 к 1982 г. Вероятно, кладки из трех-пяти яиц оказы-
вают наибольшее влияние на колебания уровня численности в
колонии и в поселении в целом.
Таблица 2
Величина кладки в колонии грачей
Год Общее коли- честно кладок Доля кладок (%) с кол-вом яиц Средняя величина кладки
1 2 3 4 5 6 М±т С, %
1980 139 0,72 7,19 23,74 33,81 32,37 2,16 3,96+0,05 15,7
1981 156 1,28 1,92 19,87 32,05 39,74 1,92 4,16+0,04 13,0
1982 242 2,48 3,72 14,46 35,54 38,84 4,96 4,19+0,01 15,4
102
Средняя величина кладки в изучаемой колонии в течение трех
гнездовых сезонов увеличилась от 3,96 до 4,19 (см. табл. 2).
Особенно резкое увеличение размера кладки по сравнению с пре-
дыдущим годом отмечено в 1981 г. (t=3,125; 0,01<р>0,001).
Хронографические изменения размера кладки у грача отмечены
в разных частях его ареала: в Удмуртии — от 3,7 до 4,5 яиц (Бра-
уде, 1984), в Астраханской области — от 4,0 до 4,2 (Шураков,
1979), на Зейско-Буреинской равнине — от 4»3 до 4,5 яиц (Ду-
гинцов, 1985). По многолетним данным А. И. Шуракова (1979),
для Камского Предуралья также характерно увеличение размера
кладки грача от 3,8 до 4,1 яиц.
Семейная элиминация. В 1980—1982 гг. в период гнездования
у грачей не было отмечено случаев тотальной элиминации. Семей-
ная элиминация имела избирательный характер и была наиболее
высокой в период насиживания (табл. 3). В целом по колонии ги-
бель кладок в 1980—1981 гг. была невысокой — 5,76% (щ=8,
п2=9), в 1982 г. элиминация резко возросла в период собственно
насиживания (17,77%) и в общем достигла уровня 23,15%
(п—56).
Размеры семейной элиминации в кладках из одного-шести яиц
имели хронографические вариации. Уровень гибели во всех груп-
пах кладок повысился в 1982 г. по сравнению с предыдущими сезо-
нами. Однояйцевые кладки полностью погибли в 1980 и 1982 гг.
В 1981 г. в единственном уцелевшем гнезде вылупился птенец,
который погиб на вторые сутки постэмбрионального периода.
Гибель кладок из двух яиц колебалась от 20% (1980 г.) до 50%
(1982 г.) от числа наблюдаемых кладок; особенно увеличилась
гибель в период насиживания. Элиминация кладок из трех—шести
яиц также наиболее высока в период насиживания (1980 г.—
2,16%, 1981 г,— 1,92%, 1982 г,— 16,94%), во время яйцекладки
гибель гнезд в этой группе не обнаружена.
Как правило, семейная элиминация имела место при разруше-
нии или разорении гнезд. Гибель гнезд была отмечена во время
сильных порывистых ветров: гнезда находили сброшенными на
землю или наклоненными на дереве, такие гнезда йпоследствшг
падали. Одно падающее гнездо было перенесено (8 мая 1982 г.)
в пределах гнездового дерева и установлено на прочное основание,
Таблица 3
Избирательная семейная элиминация у грачей, %
Год Величина кладки Период гнездового сезона
1 2 3 4 5 6 Яйце- кладка Насижи- вание Птенцо- вый
1980 0,72 1,44 0,72 2,16 0,72 0 0,72 3,60 1,44
1981 0,64 0,64 2,56 0,64 1,28 0 1,92 2,56 1,28
1982 2,48 2,07 4,13 6,61 7,44 0,42 3,31 17,77 2,07
103
родители продолжили насиживание и благополучно вывели двух
птенцов из трех отложенных яиц (последнее наблюдение сделано
23 мая). Большинство разрушенных гнезд не имело достаточно
прочного основания.
Гибель другой части кладок и резкое возрастание уровня се-
мейства элиминации у грачей в 1982 г. обусловлены увеличением
пресса хищничества. Наиболее часто объектом разорения являют-
ся яйца. В табл. 3 приведены случаи одновременного исчезнове-
ния всех яиц или пропажи яиц из гнезда, происшедшей за корот-
кий период времени (не более суток). В некоторых гнездах найде-
ны остатки скорлупы или яйца с проклеванной скорлупой. В 1982 г.
в трех гнездах обнаружены остатки трупов взрослых грачей:
крылья, частично кости, кишечник.
Возможными разорителями гнезд были серая ворона (Corvus
cornix L.), ушастая сова (Asio otus L.) и горностай (Mustella
erminea L.). Вблизи исследуемой колонии грачей нами было обна-
ружено 13 жилых гнезд серой вороны й две постройки сороки
(Pica pica L.), занятые ушастой совой. В исследуемом районе со-
рока также является многочисленным видом, но появление сорок в
колонии не вызывало реакций беспокойства у грачей. Горностая
неоднократно наблюдали в колонии в гнездовой сезон 1982 г.
Индивидуальная элиминация в раннем онтогенезе грача также,
как и семейная гибель, была избирательной. В ряде работ (Лано-
венко, 1984; Дугинцов, 1985; Lockie, 1955) обращается внимание на
высокий уровень общей элиминации в гнездовой период у грача,
который может достигать 45—57%. Однако в работах не указано,
какие формы элиминации при этом учитывались. В других работах
приводятся данные о гибели эмбрионов, составляющей 15—20%
(Болотников и др., 1973; Шураков и др., 1973; Шураков, Дьяконов,
1980; Lockie, 1955). Величина гибели птенцов, согласно А. М. Бо-
лотникову и др. (1973, 1978), равна 30—35%, согласно
М. И. Брауде (1984) — 17 —18%, но все исследователи отмечают
наиболее высокую гибель птенцов в первую неделю гнездовой
жизни.
В табл. 4 приведены данные, отражающие уровень элиминации
в кладках разной величины. Индивидуальная элиминация в тече-
ние гнездового периода была нами подразделена на гибель яиц,
эмбрионов и птенцов. Под гибелью яиц мы подразумеваем случаи
исчезновения яиц из гнезда в периоды откладки яиц и насижива-
ния и нарушения целостности скорлуповой оболочки. Гибель
эмбрионов устанавливали при вскрытии яиц, изъятых из гнезд
после вылупления последних птенцов или после истечения конт-
рольного срока вылупления. Гибель птенцов отмечали при ежед-
невном осмотре гнезд.
В кладках из одного яйца на протяжении трех сезонов наблю-
далось 100%-ная элиминация. В кладках из двух яиц имели ме-
сто годичные изменения: в 1980 г,— 55%, в 1981 г,— 67%,
в 1982 г.— 61%. По-видимому, увеличение доли гибели в 1981 г.
нельзя считать истинным, так как оно могло быть эффектом умень-
104
Таблица 4
Избирательная индивидуальная элиминация в раннем онтогенезе грачей
Форма элиминации Год Доля погибших яиц и птенцов (%) в кладках с кол-вом яиц Всего погиб- ло, %
1 2 3 4 5 6
Гибель яиц 1980 0 0 9,09 7,98 4,00 16,67 6,53
при повре- 1981 0 0 1,08 0,45 1,29 5,56 1,08
ждениях 1982 0 0 0 2,33 2,34 1,39 1,97
скорлупы
Исчезнове- 1980 0 30,00 13,13 14,89 20,44 27,78 17,79
ние из 1981 0 0 15,05 12,73 18,71 16,67 15,87
гнезда 1982 100 44,44 38,10 37,79 29,79 26,39 33,79
Всего 1980 0 30,00 22,22 22,87 24,44 44,45 24,32
1981 0 0 16,13 13,18 20,00 22,23 16,95
1982 100 44,44 38,10 40,12 32,13 27,78 35,76
Гибель эм- 1980 100 15,00 17,17 17,02 21,78 0 18,51
брионов 1981 50 66,67 21,51 31,82 22,58 27,78 26,19
1982 0 0 17,14 17,44 17,66 15,28 16,95
Гибель 1980 0 10,00 14,14 15,96 17,78 0 15,61
птенцов 1981 50 0 19,35 15,91 23,23 5,56 19,57
1982 0 16,67 9,52 12,21 20,43 27,78 16,85
Всего эли- 1980 100 55,00 53,53 55,85 64,00 44,45 58,44
минировало 1981 100 66,67 56,99 60,91 65,81 55,57 62,71
1982 100 61,11 64,76 69,77 70,22 70,84 69,56
шения выборки: количество кладок из двух яиц в 1981 г. по срав-
нению с 1980 г. (7,19%, ш=10) и 1982 г. (3,72%, пз=9) было
меньше (1,92%, п2=3). В кладках из трех—шести яиц общий
уровень гибели повышался от 1980 к 1982 г. (различия статисти-
чески незначимы) и в среднем был равен: в 1980 г.— 54,5%,
в 1981 г.— 59,8 а в 1982 г.— 68,9%. В целом по колонии также
отмечено увеличение элиминации от 58,4% в 1980 г. до 69,56%
в 1982 г. ‘
Элиминация яиц. Гибель яиц при нарушении целостности
скорлуповой оболочки не обнаружена в гнездах с кладками из од-
ного и двух яиц. В кладках из шести яиц наиболее часто повреж-
дения скорлупы отмечались в 1980 г. (53%, п=36), когда часть
яиц была разбита при измерениях. Во второй и третий сезоны
величина гибели яиц была мала (1,08%, п2=7 и 1,97%, п3=20
соответственно) и, по-видимому, отражала истинное положение.
Как правило, яйца с трещинами и помятостями в течение длитель-
ного времени (в отдельных случаях до семи суток) находились
в гнездах и насиживались вместе с неповрежденными яйцами.
Яйца с проколами и остатки скорлупы разбитых или раскле-
ванных яиц были выброшены родителями из гнезд.
. Исчезновение яиц из гнезд наиболее часто происходило в пе-
риод насиживания. В большинстве гнезд пропадало по одному
яйцу, что в 1980 г. составило 67,7%, в 1981 г.— 69,0, а в 1982 г.—
59,7% от общего количества пострадавших кладок. Доля кладок
105
Рис. 3. Нормированная по
максимуму элиминация
эмбрионов в гнездах грача
7-1980 г.;
2-1981 г.;
3—1982 г.
Рис. 4. Элиминация эм-
брионов грача (нормиро-
ванная по максимуму) в
зависимости от возраста
эмбрионов
1 - 1980 г.,
2 — 1981 г.,
3 - 1982 г.
с двумя исчезнувшими яйцами меньше: в 1980 г.— 27,7%, в
1981 г. — 19,0, в 1982 г.— 33,3%. Исчезновение трех яиц из одной
кладки встречалось редко: в 1980 г.— 4,6% (п=7), в 1981 г.— 12,1
(п=5), в 1982 г.— 6,9% (п=3). Характер нормированной по мак-
симуму кривой гибели яиц в 1982 г. указывает на отсутствие кор-
реляции между размертми кладки и величиной гибели яиц. За
исключением резкого скачка кривой в области однояйцевых кла-
док, который, возможно, вызван малочисленностью таких кладок
(п=6), соотношение количества погибших яиц почти постоянно во
всех группах кладок, т. е. величина кладки не оказывала заметного
влияния на выбор объекта хищником.
В 1980—1981 гг. пропажа яиц происходила приблизительно
в 2 раза реже (17,79 и 15,87% от числа отложенных яиц), чем
106
в 1982 г. (33,79%). Количество исчезнувших яиц колебалось
в кладках разной величины от 13,13 до 30,0% в 1980 г., от 12,73
до 18,71% в 1981 г., однако эти колебания также не зависели от
величины кладки. По-видимому, сохранность яиц во время инку-
бации у грачей обеспечивается комплексом поведенческих реак-
ций родительской пары.
Основной причиной появления повреждений скорлупы явля-
ется хищничество пернатых, которое, кроме непосредственного
уничтожения яиц, вносило сильное беспокойство в ритм жизни
колонии, особенно в период насиживания.
Исчезновение яиц из кладок также обусловлено действием
пресса хищничества, резкое усиление которого было отмечено
в 1982 г. В этот сезон мы наблюдали неоднократное появление
в колонии горностая и белок (Sciurus vulgaris L.), отмечали ак-
тивные реакции грачей на серую ворону, ушастую сову, ястреба-
перепелятника (Accipiter nisus L.).
Элиминация эмбрионов. Уровень гибели эмбрионов был наибо-
лее низким в 1982 г. — 16,95% (п=172), в 1980 г. он составил —
18,51 (п=102), а в 1981 г.— 29,19% (п=170). Повышение уровня
элиминации в 1981 г. было характерно для всех групп кладок, но
особенно резко он возрастал в кладках из одного и двух яиц. Высо-
кая гибель эмбрионов и яиц, а также немногочисленность во всех
сезонах кладок из одного (щ=1, п2=2, пз=6) и двух (п1=10,
п2=3, Пз=9) яиц позволяют утверждать, что роль этих кладок в
регуляции численности колонии минимальна.
Кривые нормированной по максимуму элиминации эмбрионов
(рис. 3) в 1980 и 1982 гг. имеют одинаковую форму в той части
графика, которая отражает уровень гибели в кладках из трех—
шести яиц, что свидетельствует о наличии определенных внутрен-
них причин гибели эмбрионов. Кривая гибели 1981 г.— лома-
ная линия, не выявляющая четкой зависимости элиминации от
величины кладки.
Возрастная дифференцированность погибших эмбрионов от-
ражена в табл. 5, в которой использована классификация ста-
дий развития воробьиных птиц, разработанная А. И. Шураковым
(1981). Нами дополнительно введена графа, указывающая элими-
нацию эмбрионов в ранний зародышевый период (первые—пятые
Таблица 5
Возраст погибших эмбрионов грача
Период развития Возраст эмбриона Элиминация эмбрионов, %
Сутки Стадии 1980 г. 1981 г. 1982 г.
Зародышевый 1-8 1-31 65,06 64,00 50,00
Всего В том числе 1-5 1-24 60,24 32,00 40,48
предплодный 9-12 32-37 21,69 12,77 23,81
плодный 13-17 38-43 13,25 24,00 26,19
107
сутки развития), когда аллантоис не превышает размеров за-
родыша.
Более половины эмбрионов погибло в зародышевый период
инкубации (50—65%). Особенно велика была элиминация в пер-
вые пять суток развития: в 1980 г.— 60,24%, в 1982 г.— 40,48% от
общего числа погибших.
Исследуя распределение погибших эмбрионов в зависимости
от возраста (рис. 4), можно выделить несколько периодов повы-
шенной гибели. Для 1980 и 1982 гг. это 1—5-е, 10—13-е и 16—17-е
сутки развития. Наличие первых двух пиков эмбриональной ги-
бели подтвердилось при вскрытии 76 яиц, собранных в 1980 г. при
уничтожении гнезд грачей на опорах ЛЭП в пригороде Перми.
Выявление третьего периода было затруднено малым количеством
эмбрионов, имевших возраст 15—17 сут.
Возраст эмбрионов, погибших в 1981 г., варьировал сильнее,
и на графике можно выявить пять пиков смертности: 1-е, 4—7-е,
9-е, 13-е и 16—17-е сутки. Два последних пика совпадают по
времени с пиками, отмечавшимися в 1980 и 1982 гг. Доля 6—9 су-
точных эмбрионов увеличилась до 50%, тогда как в 1980 г. они со-
ставляли 15,7%, а в 1982 г.— 10,64%.
Сравнивая даты гибели эмбрионов в 1981 г. и кривую средне-
дневных температур воздуха, мы обнаружили, что появление вто-
рого пика повышенной гибели обусловлено понижением темпера-
туры воздуха до —|—1°, которое отмечалось в течение пяти дней
и сопровождалось обильными снегопадами. Элиминация эмбри-
онов зависела от степени насиженности яиц к моменту выпадения,
снега (Быкова, 1981). Гибель эмбрионов на девятые сутки разви-
тия, возможно, является последствием воздействия длительного
низкотемпературного охлаждения. А. И. Шураков и Ю. В. Дья-
конов (1980) также отмечают три периода повышенной гибели
у грача: в первые двое суток, на 5—13-е и 16—17-е сутки раз-
вития.
Повышенную элиминацию на ранних стадиях развития обна-
ружили Р. А. Зацепина (1981) у полевого воробья, А. И. Мороз-
кин (1984) у мухоловки-пеструшки, М. Джилбертсон и Р. Хейл
(Gilbertson, Hale, 1974) у серебристой чайки. А. Ф. Ковшарь
(1981), на основе анализа результатов размножения воробьиных
птиц указывает, что 84,6% отходов инкубации составляют «бол-
туны», т. е. яйца с эмбрионами, погибшими в ранний зародыше-
вый период еще до развития кровеносной системы.
Таким образом, в эмбриональном развитии у диких птиц, как
и у домашних (Отрыганьев и др., 1964; Третьяков, Крок, 1978;
Орлов и др., 1982), имеются три критических периода. Наиболее
значимым является первый, приходящийся на первые пять суток
развития; в это время развивающиеся зародыши испытывают
минимальное воздействие внешней среды и уровень гибели, по-
видимому, определяется действием внутренних механизмов регу-
ляции численности популяции и отражает степень загруженности
особей летальными генами.
108
При патологоанатомическом вскрытии яиц нами установлено,
что наиболее часто у погибших грачей в предплодный и плодный
периоды наблюдаются изменения в системе кровоснабжения.
Гиперемия кожных покровов, вызываемая усиленным притоком
крови, была отмечена в 1980 г. у 6% вскрытых эмбрионов,
в 1981 г,— у 55, в 1982 г,— у 56%. Множественные кровоизлия-
ния в кожных покровах, особенно на крыльях, в головном мозге
и на стенках брюшной полости также наиболее часто встречались
в 1982 г.— у 45%; в 1981 г.— у 35, а в 1980 г. — у 6% вскрытых
эмбрионов. У многих эмбрионов одновременно с гиперемией кожи
отмечено обесцвечивание внутренних органов, вызванное оттоком
крови: в 1980 и 1981 гг.— у 30%, в 1982 г.— у 48%. Гиперемия
органов и тканей, кровоизлияния могут быть вызваны различными
причинами, от нарушения режима инкубации до инфекционных
заболеваний, поэтому необходимо рассматривать их в комплексе с
другими патологическими признаками.
Отек шеи, вызванный увеличением жидкости в тканях, во всех
сезонах отмечали в сочетании с гиперемией кожных покровов и
анемией внутренних органов: в 1980 г. его имели 22%, в 1981 г.—
10 и в 1982 г.— 30% эмбрионов. Отеки органов и кожных покровов
обычно возникают при нарушении кровообращения, нарушении
обмена веществ и общем голодании.
Неправильное положение тела эмбриона (расположение головы
под левым крылом, опускание клюва или конечностей в желточный
мешок) является характерным признаком длительной гипоксии
или асфиксии, но может проявляться и при нарушении режима
поворачивания яиц. В 1980 г. неправильное положение тела имели
2% погибших, в 1982 г. —19%, в 1981 г. такие отклонения от нор-
мы не обнаружены.
Изменение формы клюва у эмбрионов (перекрещивание челю-
стей, недоразвитие верхней челюсти) для нашей колонии — яв-
ление редкое: в 1980 г.'оно не наблюдалось, в 1981 г. было обнару-
жено у 10% и в 1982 г.— у 7% просмотренных эмбрионов. Урод-
ства клюва в естественных условиях обычно вызываются леталь-
ными генетическими факторами, но при искусственной инкубации
получены при перегреве в начальный период инкубации (Отры-
ганьев и др., 1981).
В 1982 г. у части обследованных яиц на подскорлуповых обо-
лочках в районе воздушной камеры обнаружены сильно разросши-
еся колонии плесени.
Кроме перечисленных признаков, были отмечены уплотнение
подскорлуповых оболочек, переполнение кишечника (иногда
и брюшной полости) жидкостью, эктопия, изменение цвета желтка
на желто-зеленый, прилипание зародыша к подскорлуповой обо-
лочке.
В 1980 г. гиперемия кожных покровов и множественные крово-
излияния сочетались с увеличением кровенаполнения сосудов
аллантоиса и анемией внутренних органов, что свидетельствует
о нарушениях в системе кровоснабжения у части эмбрионов.
109
Однако мы не можем говорить о направленном воздействии внеш-
ней среды, так как каждый отдельный эмбрион не имел преобла-
дающего большинства или всех указанных признаков. Элимина-
ция эмбрионов, вероятно, была вызвана разными причинами.
Наблюдавшиеся в 1981 г. у эмбрионов переполнение сосудов,
особенно аллантоиса, кровью, гиперемия кожных покровов с одно-
временным оттоком крови от внутренних органов, множественные
кровоизлияния могли быть обусловлены, по Ю. Исаеву и др.
(1979), гипоксией, острой гипер- или гипотермией. Гипоксия
была возможна при закрывании кладок, снегом или при частичной
закупорке пор скорлупы грязью или-содержимым разбитых яиц.
Однако наиболее характерный признак длительной гипоксии —
неправильное положение тела — отсутствовал у эмбрионов этого
года. Кроме того, отмечены два случая погружения эмбрионов
в желток, что происходит обычно при сильно пониженных темпе-
ратурах. На влияние низких температур указывает уменьшение
количества эмбрионов с отеками шеи до 10%: отеки встречаются
редко при снижении температуры насиживания до 22° и ниже
(Отрыганьев и др., 1981). По-видимому, большое число исследо-
ванных нами особей испытало воздействие низких температур
и погибло во время или до снегопада.
В 1982 г. мы отметили увеличение количества эмбрионов,
имевших неправильное положение тела (19%). У 30% эмбрионов
наблюдался отек шеи и у 26% — переполнение кишечника жид-
костью. Массовые кровоизлияния (45%), гиперемия кожных пок-
ровов (56%), анемия внутренних органов (48%) сочетались с уп-
лотнением подскорлуповых оболочек (22%) и увеличением напол-
нения кровеносных сосудов аллантоиса (15%). У 4% эмбрионов
желток был желто-зеленого цвета, у 11% зарегистрирована экто-
пия.
Перечисленные изменения могут возникать при постоянных
(длительных), но неглубоких недогревах, но не исключено, что ги-
бель их была обусловлена целым рядом факторов, действие которых
усиливается при активной деятельности хищников.
Таким образом, из возрастного и патологоанатомического ана-
лиза следует, что наиболее высока гибель эмбрионов в ранний заро-
дышевый период; элиминация была обусловлена наследственными
факторами. Величина гибели в предплодный и плодный периоды
более подвержена колебаниям. На уровень элиминации в этн пе-
риоды оказывает влияние ряд факторов внутренней и внешней
среды.
Элиминация птенцов. Гибель птенцов грача в период от вы-
лупления до вылета из гнезда в течение трех лет наблюдений была,
как и эмбриональная гибель, высокой. В 1980 г. она составляла
15,5%, в 1981 г, —19,6, а в 1982 г, —16,9% от общего количества
птенцов (см. табл. 4). В 1980 г. элиминация птенцов зарегистри-
рована в 29,6% находившихся под наблюдением гнезд, в 1981 г.
она охватывала 51,3, а в 1982 г,— в 44,2% гнезд.
Гибель всех вылупившихся птенцов произошла в 10—11,5%
ПО
семей. Чаще 100%-ная элиминация наблюдалась в небольших
выводках:
с одним птенцом с двумя птенцами
1980 г. 64,3% 35,7%
1981 г. 33,3% 33,3%
1982 г. 40,0% 36,0%
Гибель половины птенцов и более отмечена в 1980 г. в 16,6%,
в 1981 г.— в 22,4, в 1982 г.— в 22,3% случаев.
Наиболее высокая гибель птенцов наблюдалась в кладках с
большим количеством яиц. Выживаемость выводков имела хроно-
графические изменения, например, гибель птенцов в кладках из
трех яиц варьировала от 9,52% в 1982 г. до 19,35% в 1981 г., из
пяти яиц — от 17,78% в 1980 г. до 23,23% в 1981 г.
При сравнении кривых элиминации и вылупления птенцов в
кладках разной величины (рис. 5) нетрудно заметить идентичность
их формы: величина элиминации пропорциональна количеству
птенцов в выводке. Наиболее высокая выводимость в 1980—1981 гг.
обнаружена в кладках из трех яиц (60,61—62,37%), в 1982 г.— из
двух и из шести яиц (55,55 и 56,94%). В 1980 г. количество вылу-
пившихся птенцов было приблизительно одинаково в кладках,
содержавших от двух до шести яиц (nmax —nmin =6,83%), в 1981 г.
выводимость в четырех- и пятияйцевых кладках была относительно
высокой, а в кладках из шести и двух яиц она снизилась. В 1982 г.
количество птенцов уменьшалось последовательно в кладках из пя-
ти, трех и четырех яиц.
Наиболее часто среди погибших птенцов мы отмечали выпав-
щих из гнезда. Падение птенцов может происходить при сильных
порывистых ветрах. Так, 11 мая 1981 г. после урагана была обнару-
жена гибель 19 птенцов (14,96% от общего числа погибших),
которая превысила суточный уровень элиминации в предыдущие
дни в 3,8 раза. Кроме того, падение птенцов может наблюдаться
без вмешательства извне: отмечены случаи нахождения птенцов на
краю гнезда (в одном гнезде — один птенец, в другом — три) и
падения птенцов.
Значительную часть от числа погибших составили птенцы, об-
наруженные в гнезде мертвыми: в 1980 г,—31,40% (п=27), в
1981 г.—31,50% (п=40), в 1982 г.—17,54% (п=30). Трупы не
имели внешних повреждений и находились либо на краю лотка,
либо под живыми птенцами, лишь в четырех случаях из 101 — на
краю гнезда. Увеличение количества мертвых птенцов в два раза
зафиксировано 9 мая 1981 г. после ночной грозы, сопровождавшей-
ся сильным ветром и резким понижением температуры воздуха от
20° в 20 ч 8 мая до 6° в 8 ч 9 мая (в ночное время температура возду-
ха не измерялась). В последующие сутки температура колебалась
в пределах 5—9°.
В обычных условиях мы наблюдали гибель птенцов в гнезде
поодиночке, одновременное нахождение двух трупов в одном
гнезде встречалось редко. Как правило, первыми погибали в гнезде
11)
Рис. 5. Вылупление (Л) и
элиминация (Б) птенцов в
гнездах грача
7—1980 г.;
2—1981 г.;
3-1982 г.
Рис. 6. Успешность размноже-
ния грачаа
I—1880 г.;
2-1981 г.;
3-1982 г.
последние по рангу птенцы. Порядок гибели птенцов был обратно
пропорционален рангу птенцов. Возможно, элиминация связана с
разнокачественностью птенцов, усиливающейся при недостаточном
кормлении выводка.
Гибель других птенцов связана с деятельностью хищников.
В 1980 г. в гнезде, в котором вылупился один птенец, были обнару-
жены перья взрослого грача и остатки кишечника птенца. В 1982 г.
в другом гнезде были расклеваны и частично съедены три птенца
(внутренности не оставлены). Нередко птенцов с расклеванной го-
ловой находили на земле.
Элиминация птенцов при гибели гнезд имела незначительные
размеры: в 1980 г.— 0,54% (п=3), в 1981 г,— 1,08% (п=2),
в 1982 г.— 0,69% (п=7) от числа отложенных яиц.
Успех размножения и репродуктивный потенциал. Если совмес-
тить графики вылупления и элиминации птенцов, то получим диа-
112
грамму, отражающую успешность размножения грача в данной
[колонии (рис. 6). Во все три гнездовые сезона птенцы из однояйце-
вых кладок если и вылуплялись, то не выживали, уровень размно-
жения в трехяйцевых кладках был одним из наиболее высоких в
каждом из рассматриваемых сезонов: 47,0, 43,5 и 36,0%. По-види-
мому, кладка из трех яиц является оптимальной. Успех размно-
J жения в двухяйцевых кладках высокий: 45, 33 и 39%. В кладках
t из четырёх и из пяти яиц он снижается: в 1980 г.— до 39 и 26%,
| в 1981 I. до 40 и 34%, в 1982 г,— до 30 и 29% соответственно.
i Успех размножения в шестияйцевых кладках непостоянный: он
5 может быть максимальным (в 1980 г,— 56%), относительно высо-
ким (в 1981 г.— 40%) или наиболее низким (1982 г,— 28%) в
кладках, содержащих от двух до шести яиц.
В среднем к моменту вылета из гнезда выжило в 1980 г. 1,65,
в 1981 г.— 1,48, в 1982 г,— 1,28 птенца на одну родительскую пару.
Наибольшее количество птенцов сохранилось в кладках из шести
яиц; с уменьшением величины кладки снижался уровень ее репро-
дуктивности. В кладках, содержавших от четырех до шести яиц,
наблюдалось уменьшение продуктивности от 1980 к 1982 г.
(табл. 6). Хотя кладки из трех яиц были еще менее продуктивны,
все же, как отмечалось выше, они были наиболее успешными.
Можно утверждать, что они являются оптимальными по размеру
для грачей. «Оптимальный размер» кладки соответствует макси-
мальной приспособленности особей к условиям среды (Charnov,
; Krebs, 1974). Однако продуктивность этих кладок невелика, под-
держание определенной относительно постоянной численности
взрослых птиц колонии за счет этой группы кладок не представля-
ется возможным. Наличие менее успешных, но имеющих более
Таблица 6
Средняя продуктивность одной семьи грачей в зависимости от величины кладки
Год Среднее число слетков на семью с кладкой, состоящей из
6 яиц 5 яиц 4 яиц 3 яиц 2 яиц
1980 3,33 1,80 1,77 1,39 0,90
1981 2,67 1,71 1,56 0,97 0,67
1982 1,75 1,49 1,21 1,06 0,78
Таблица 7
Доля слетков из семей с разным количеством яиц в кладках
Год Доля слетков (%) из семей с кладками, состоявшими из
6 яиц 5 яиц 4 яиц 3 яиц | 2 яиц
1980 4,37 35,37 36,24- 20,09 3,93
1981 3,31 43,80 35,54 16,53 0,83
1982 6,84 45,60 32,57 12,05 2,28
113
высокий репродукционный потенциал кладок из четырех — ше-
сти яиц позволяет восполнять потери колонии и популяции в
целом.
Из табл. 7 видно, что доля слетков из семей, в которых было
по четыре—пять яиц в кладке, наиболее велика, а участие в попол-
нении популяции семей, имевших по три яйца, значительно ниже.
Вклад в популяцию семей, величина кладок которых сильнее всего
отклонялась от среднего, минимален.
Выводы
Средняя величина кладки в колонии грача в 1980—1982 гг. соста-
вляла 4,16 яиц. В исследуемый период размер кладки увеличился с
3,96 яиц в 1980 г. до 4,19 в 1982 г. Во все сезоны большая часть кла-
док (89—95%) состояла из 3—5 яиц.
Элиминация в раннем онтогенезе грача возросла с 63% в 1980 г.
до 73% в 1982 г., в среднем —67%. Семейная элиминация охваты-
вала 6—23% кладок, имела значительные годовые вариации и была
наиболее высокой в одно- и двухяйцевых кладках. Индивидуальная
элиминация увеличилась от 58% в 1980 г до 70% в 1982 г. Наибо-
лее низкий уровень гибели имели кладки из трех яиц. В кладках из
одного яйца отмечена 100%-ная элиминация.
Элиминация яиц (исчезновение и нарушение целостности скор-
лупы) не зависит от величины кладки и увеличивается при усиле-
нии пресса хищничества. В среднем она составляет 25,7% (ко-
лебания от 10 до 36%).
Эмбриональная гибель поддерживается на уровне 16—17%,
увеличиваясь при резком изменении условий внешней среды. Ги-
бель большого числа зародышей (32—60%) в первые пять суток
развития свидетельствует, вероятно, о значительной степени загру-
женности популяции летальными генами. Высокая смертность
эмбрионов отмечена в кладках из одного-двух яиц.
Элиминация птенцов составляла 16 — 20%, в среднем 18%.
Порядок гибели птенцов обратно пропорционален их рангу.
Наиболее высокий непродуктивный потенциал имели кладки из
четырех-шести яиц, а успешность размножения была самой высо-
кой в кладках из трех яиц. Средняя продуктивность колонии —1,47
птенца на одну родительскую пару.
ЛИТЕРАТУРА
Баккал С. Н. О гибели птенцов воробьиных птиц от паразитических мух // Вести.
ЛГУ. Биология, 1980. № 9. С. 106—108.
Блинов В. Н. Материалы по экологии гнездового периода сороки, серой вороны
и галки в пойме Верхней Оби // Экология и биоценотические связи перелетных
птиц Западной Сибири. Новосибирск: Наука, 1981. С. 109—127.
Болотников А. М., Быкова Л. П. Динамика численности и межвидовые отношения
сизой и серебристой чаек в условиях ограниченной площади гнездова-
ния //Вид и его продуктивность в ареале. Свердловск, 1984. Ч. 2. Млекопи-
тающие и птицы. С. 58—59.
114
Болотников А. М., Скрылаев Л. Ф., Тарасов В. А., Ангальт В. 3. О разнокачествен-
ности яиц одной кладки и выживаемости птенцов грача // Экология. 1978. № 2.
С. 86—88.
Болотников А. М., Соколова Т. И., Чащин С. П. Циклы яйцекладки, плодови-
тость и выживаемость птенцов у грача // Учен. зап. Перм. педин-та. Пермь,
1973. С. 39- 42.
Брауде М. И. Материалы к гнездовой биологии хохлатой чернети в низовьях
Оби // Структура населения птиц Европейского Северо-Востока СССР.
Сыктывкар, 1983. С. 10—16. (Тр. Коми фил. АН СССР; № 62).
Брауде М. М. К вопросу о гнездовой биологии грача в условиях Приуралья // Жи-
вотные — компоненты экосистем Европейского Севера и Урала. Сыктывкар,
1984. С. 89-94.
Быкова Л. П. Элиминация у грача в условиях естественного охлаждения яиц //
Гнездовая жизнь птиц. Пермь: ПГПИ, 1981. С. 104—107.
Гаврилов Э. И., Родионов Э. Ф. О гибели птичьих гнезд//Новости орнитологии.
Алма-Ата: Наука, 1965. С. 81—82.
Дугинцов В. А. Биология врановых птиц Зейско-Буреинской равнины-и их хо-
зяйственное значение: Автореф. дис. ... канд. биол. наук. М., 1985. 18 с.
Дьяконов Ю. В. Гетерогенность яиц и размер кладок грача /'/ Гнездовая жизнь
птиц. Пермь: ПГПИ, 1980. С. 6-15.
Зацепина Р. А. О методике оценки эмбриональной смертности у птиц // Микроэво-
люция. Казань: Казан, фил. АН СССР, 1981. Вып. 1. С. 75—79.
Исаев Ю., Владимирова Ю., Никитченко Л. Дифференциальная диагностика
эмбриональной смертности при аварийных нарушениях режима инкубации //
Достижения науки в области повышения продуктивности сельскохозяйствен-
ной птицы и улучшения качества птицепродуктов: Тез. докл. конф.,
19—22 июня 1979, г. Одесса. М., 1979. С. 70—79.
Ковшарь А. Ф. Особенности размножения птиц в субвысокогорье. Алма-Ата:
Наука, 1981. 260 с.
Корбут В. В. Распределение и структура колоний грача в окрестностях Звениго-
родской биостанции МГУ. Орнитология. М.: Изд-во МГУ, 1985. Вып. 20.
С. 186-189.
Лановенко Е. Н. Экология и хозяйственное значение вороновых птиц в различных
районах Узбекистана: Автореф. дис. ... канд. биол. наук. М., 1984. 18 с.
Лэк Д. Численность животных и ее регуляция в природе. М.: Изд-во иностр, лит.,
1957. 404 с.
Мельников Ю. И. Адаптивные черты колониального гнездования в нестабильных
условиях среды // XVIII Междунар. орнитол. конгр.: Тез. докл. и стендовых
сообщ. М.: Наука, 1982. С. 195-196.
Морозкин А. И. О связи эмбриональной гибели с динамикой популяции мухоловки-
пеструшки // Отражение достижений орнитологической науки в учебном
процессе средних школ и ВУЗов и народном хозяйстве. Пермь, 1984. С. 63—64.
Нумеров А. Д. Взаимосвязь числа яиц в кладке и других показателей успешности
размножения обыкновенного скворца // Труды Окского гос. заповедника.
1984. Вып. 15. С. 229-231.
Нумеров А. Д. Возрастные особенности размножения скворца в районе Окского за-
поведника // Вести, зоологии. 1985. № 2. С. 46—51.
Одум Ю. Основы экологии: М.: Мир, 1975. 740 с.
Орлов М. В., Быховец А. У., Злочевская К. В. Инкубация. М.: Колос, 1982. 223 с.
Отрыганьев Г. К., Бессарабов Б. Ф., Исаев Ю. В. Болезни эмбрионов птиц. М.:
Россельхозиздат, 1981. 136 с.
Отрыганьев Г. К., Хмыров В. А., Колобов Г. М. Инкубация. М.: Колос, 1964. 288 с.
Риклефс Р. Основы общей экологии. М.: Мир, 1979. 424 с.
Рубайлова Н. Г. Формирование и развитие теории естественного отбора. М.: Наука,
1981. 196 с.
Северцов С. А. Введение в теорию эволюции. М.: Изд-во МГУ, 1981. 318 с.
Семенов-Тян-Шанский О. И. Биология размножения тетеревиных птиц на Севере.
М.: Наука, 1983. 61 с.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. М.: Мир, 1982.
488 с.
115
Третъяков Н. П., Крок Г. С. Инкубация с основами эмбриологии. М.: Колос,
1978. 304 с.
Хохлов А. Н., Афанасова Л. В. Массовая гибель домового и полевого воробьев в
Ставропольском крае // Орнитология. М.: Изд-во МГУ, 1985. Вып. 20. С. 200.
Шкарин В. С., Родимцев А. С. Материалы по размножению скворца на юге Кеме-
ровской области // Гнездовая жизнь птиц. Пермь, 1982. С. 20—25.
Шмалъгаузен И. И. Пути и закономерности эволюционного процесса. М.; Л.:
Изд-во АН СССР. 1939.
Шмалъгаузен И. И. Факторы эволюции. М.; Л.: Изд-во АН СССР. 1946. 396 с.
Шмалъгаузен И. И. Проблемы дарвинизма. Л.: Наука, 1969. 494 с.
Шмалъгаузен И. И. Организм как единое целое в индивидуальном и историче-
ском развитии. М.: Наука, 1982. 383 с.
Шу раков А. И. Экологические аспекты изменчивости величины кладки некоторых
колониальных птиц//Гнездовая жизнь птиц. Пермь, 1979. С. 12—19.
Шураков А. И. Стадии развития воробьиных птиц // Там же. Пермь, 1981. С. 3—22.
Шу раков А. И., Дьяконов Ю. В. Особенности развития зародышей грача в основ-
ных и возобновленных кладках на севере ареала // Гнездовая жизнь птиц.
Пермь, 1980. С. 122-127.
Шураков А. И., Соколова Т. И., Швецов А. А. К экологии размножения грача в
Пермской области//Учен. зап. Перм. педин-та. Пермь, 1973. С. 54—68.
Шураков С. А. Динамика величины Ергачинской колонии грачей // Гнездовая
жизнь птиц. Пермь, 1982. С. 92—95.
Ash J. S. Observation on a decreasing population of Redbacked Shrikes // Brit, birds.
1970. Vol. 6. P. 225-239.
Charnov E. L., Krebs J. B. On clutch-size and fitness // Ibis. 1974. Vol. 116, N 2.
P. 217-219.
Gilbertson M., Hale R. Early embrionic mortality in a Herring Gull colony in Lake On-
tario // Canad. Field-Natur. 1974. Vol. 88, N 3. P. 354-356.
Lockie J. D. The breeding and feeding of Jakdaws and Rooks, with notes on Carrion
crows and other Corvidae// Ibis, 1955. Vol. 97. P. 341—369.
Myrberget S. Desertion of nests by Willow Grouse Lagopus lagopus // Fauna norv.
1983. Жол. 2. П. 109-113.
Orell M., Ojanen M. Breeding success and population dinamics in a northern great tit
Parus major population//Ann. Zool. fenn. 1983. Vol. 20, N 2. P. 77—98.
SUMMARY
Elimination in the early ontogenesis and the clutch size
in Rook (Corvus frugilegus). Bykova L. P.
The increase of the average size of Rook’s clutch from 3,96 (1980) to 4,19 (1982) was
marked in Kama Pre-Urals. 89—95% of clutches consisted of 3—5 eggs. The eli-
mination level in the early ontogenesis (before leaving the nests by nestlings) ranged
ftom 63% in 1980 to 73% in 1982. 100% elimination is characteristic for one egg-
clutch nests. The lowest destruction of eggs was in the nests with 3 eggs. The average
beproduction of the colony was 1,47 nestlings per each parents couple. Clutches of
4—5 eggs are leading ones in the regulation of colony population. The nestlings
from these clutches form 72—78% of the total number of fledgelings for a colony.
Family elimination spread over 6—23 clutches, and as well as the eggs’ destruction
(17—36% on the average — 25,7%) it was increasing with the intensification of pre-
dation pressure. The average embryonal death rate in the colony was 16—17% (the
highest death rate-during 1—5 days). The elimination of nestlings was 16—20%
(on the average 18%). The reasons causing the elimination on different stages of the
early ontogenesis are discussed.
116
УДК 591.557:598.842.7
А. Г. ГУБИН,
Е. С. ПРЕОБРАЖЕНСКАЯ,
А. С. БОГОЛЮБОВ
117071, Москва, Ленинский
проспект, д. 33. Институт
эволюционной морфологии
и экологии животных
им. А. Н. Северцова АН СССР
Экологические особенности
дроздов рябинника
(Turdus pilaris)
и белобровика (Т. iliacus)
в местах совместного обитания
Объектами подробных экологических исследований часто и вполне
оправдано служат группы совместно обитающих близкород-
ственных видов. Выявление условий их сосуществования тради-
ционно включает анализ трех основных компонентов их экологиче-
ских ниш: пространственного, временного и трофического (Пиан-
ка, 1981).
Одним из таких объектов служат дрозды: рябинник Tur-
dus pilaris и белобровик Т. iliacus. Достаточно полно изучены
в отдельности их трофические (Liibcke, 1975) и пространственные
ниши (Jue, 1981), а также территориальные взаимоотношения этих
видов (Slagsvold, 1980). Задача нашей работы заключалась в уста-
новлении условий сосуществования дроздов рябинника и белобро-
вика путем одновременного анализа пространственного, временно-
го и трофического компонентов их экологических ниш в гнездовой
период.
Работа проводилась в 1983—1985 гг. на территории биостанции
Института эволюционной морфологии и экологии животных
им. А. Н. Северцова АН СССР (Мантуровский район Костромской
области). Изучали территориальное распределение гнездовых ко-
лоний рябинников и гнезд белобровиков.
Сравнивались также четыре параметра экологических ниш этих
видов:
1) время прилета и начала гнездования;
2) место сбора корма;
3) характер расположения гнезда;
4) состав корма птенцов в местах совместного обитания.
Размещение гнездовых участков и колоний дроздов изучали
вскоре после прилета, в мае—июне, методами маршрутных учетов
и картирования гнезд на участке перелесков, граничащих с полями,
площадью около 2 км2 (в том числе площадь леса около 1,4 км2).
Был составлен план размещения колоний рябинников и участ-
ков самцов белобровиков. Часть этой работы в 1985 г. была вы-
полнена И. Ю. Губенко, которому авторы выражают благодар
ность.
Характер расположения гнезд, места сбора и состав корма,
сроки начала гнездования изучались на площадке в вязово-ольхо-
вом лесу (площадью 33 га). Здесь также проводили многократные
(5—6 раз в период начала гнездования) учеты и картирование мест
117
встреч поющих самцов белобровика и их гнезд и колоний рябинни-
ков.
При описании гнезда указывались высота его расположения
и порода дерева, на котором оно находится, содержимое гнезда и
возраст птенцов. После вылета птенцов или опустошения гнезда
по другой причине гнездо уничтожалось. Всего обследовано 81
гнездо рябинников и 43 гнезда белобровиков.
Состав птенцового корма дроздов определяли способом шейных
.перевязок и путем наблюдения за гнездом в моменты прилета ро-
дителей с кормом из скрадка, установленного на расстоянии 2—
5 м от гнезда. Для установления мест сбора корма птицами было
заложено несколько маршрутов, которые проходили по лесу и
опушкам полей. Во время учета на маршруте регистрировали
птиц, встреченных в лесу, летящих в лес с кормом и из леса за кор-
мом, а также птиц, встреченных на полях. По полям с этой целью
пройдено 8,4 км, по лесу — 13,6 км.
Ширина экологических ниш рассчитана с использованием ин
декса полидоминантности Симпсона и нормирована по рангам из-
менчивости каждого параметра
где В — ширина ниши, В' — индекс полидоминантности Симпсона,
N — число градаций данного параметра.
Перекрывание ниш рассчитали по индексу Пианки.
Распределение по территории
На изученной территории лиственных перелесков (площадь —
5 км2) оба вида дроздов предпочитают гнездиться в массивах вязо-
во-ольхового леса, причем рябинники образуют здесь колонии.
В случае конкурентного вытеснения гнёзда белобровиков чаще на-
ходились бы вне колоний рябинников. Результаты майских
учетов показали, однако, следующее. На площади вязово-оль-
ховых лесов (1,4 км) зарегистрировано 34 гнездовых участка бе-
лобровиков. ПлоЩадь колоний рябинников занимает 18 га. При
равномерном распределении белобровиков на 18 га их пришлось
бы 4 пары, в действительности же на этой площади обнаружено
12 участков белобровиков, что в 3 раза больше ожидаемого при
отсутствии предпочтения территорий колоний.
Аналогичная система анализа была применена к данным
картирования гнезд на площадке вязово-ольхового леса площадью
33 га. Здесь гнезда белобровиков также оказались приурочены к
колониям рябинников. Их концентрация в местах колоний в 4,5 ра-
за выше ожидаемой при равномерном распределении. Представ-
ленные данные свидетельствуют об отсутствии избегания одним
видом территорий, занятых другим, а при условии структурной од-
нородности среды и избытка мест для гнездования — о тяготении
белобровиков к колониям рябинников. Тяготение ряда видов (в
118
том числе дроздов-белобровиков) к гнездовым колониям рябин-
ников отмечено также для лесов Норвегии (Slagsvold, 1980).
Т. Слагсвольд высказал предположение о том, что гнездование
ряда неколониальных видов в колониях рябинников снижает воз-
можность разорения их гнезд хищниками, в частности серыми во-
ронами. Наши исследования также свидетельствуют о значитель-
ной гибели кладок обоих видов (от 20 до 50% за период гнездо-
вания) на изучаемой территории в результате разорения их серыми
воронами и мелкими куньими.
Разделение экологических ниш
Время прилета и начала гнездования. Рябинники и белобровики
прилетают одновременно, обычно в середине апреля, и держатся
до начала гнездования в смешанных стаях. К гнездованию обычно
приступают в начале мая, причем у белобровиков этот период
растянут на месяц, что обусловливает различие между видами в
сроках и продолжительности гнездования. Расчет перекрывания
ниш по этому параметру производили по пятидневкам, при этом
сильных различий в сроках не отмечено. Среднее перекрывание по
всем годам — 0,69; различия по годам существенны: в 1983 г,—
0,47, в 1984 г.-0,67, в 1985 г.—0,92.
Места сбора корма. В естественных условиях для большинства
видов характерно совпадение гнездовых и кормовых территорий.
Исключение составляют колониальные виды, которые могут
гнездиться в одном биотопе, собирая корм в других. Так, два изу-
чаемых нами вида, гнездящихся в одном и том же биотопе, имеют
различные места сбора корма. Рябинники для сбора корма пред-
почитают поля (89% встреч кормящихся птиц), белобровики же,
наоборот, кормятся в основном в лесу (79% встреч). Все рябин-
ники, кормящиеся в лесу, по нашим данным, оказались одино-
кими родителями. Перекрывание ниш по этому параметру наи-
меньшее — 0,26.
Места расположения гнезд. Анализ высоты расположения гнезд
рябинников и белобровиков показывает, что рябинники главным
образом гнездятся в среднем высотном ярусе леса, а белобровики —
в нижнем (табл. 1). При этом рябинники в основном используют
лиственные деревья, а белобровики — в большей степени землю и
высокие пни. Рябинники при гнездовании используют больший
диапазон высот, но при этом меньший, чем у белобровиков, набор
мест устройства гнезда (табл. 2). Перекрывание по месту устрой-
ства гнезда велико — 0,81, меньше по высоте его расположения —
0,50. Среднее суммарное перекрывание по местам расположения
гнезд — 0,4, при этом в 1983 г.—0,34, в 1984 г. — 0,38, в 1985 г.—
0,49.
Состав корма птенцов. Основным методом выяснения состава
корма было наблюдение за птицами в момент кормления птенцов.
За рябинниками наблюдали 11 ч 45 мин, за белобровиками — 9 ч.
По составу корма изученные виды почти не различаются. Из 43
119
Таблица 1
Распределение гнезд дроздов
по высотным ярусам вязово-ольхового
леса (в среднем за 1983—1985 гг.)
Высота, м Доля гнезд, %
Рябинник п=81 Белобровик п=43
0-0,9 1,2 51,2
1—2,9 37,0 32,6
3-4,9 28,5 11,6
5-10,0 32,1 4,6
Выше 10 1,2 0
Ширина ниши 0,53 0,53
Таблица 2
Места устройства гнезд дроздов
в вязово-ольховом лесу (в среднем за
1983-1985 гг.)
Место уст- ройства гнезда Доля гнезд, %
Рябинник, п—81 Белобровик, п=43
Земля 0 19,5
Валежник 3,7 4,9
Высокие пни 7,3 21,9
Ольха 30,9 19,5
Вяз 55,6 29,3
Прочие по- роды (бере- за, черемуха, ель) 2,5 4,9
Ширина ниши 0,35 0,73
прилетов с кормом у рябинников в 37 случаях родители прино-
сили только дождевых червей (86%), у белобровиков из 47 приле-
тов 46 с дождевыми червями (98%). В остальных редких случаях
родители кормили птенцов насекомыми — гусеницами и жуками
(видовое определение жертв не проводилось из-за их незначитель-
ной доли в рационе). Преобладание в кормовом рационе обоих
видов дождевых червей подтверждается нашими данными шейных
перевязок и литературными данными состава птенцового корма
(Liibcke, 1975). В результате этого сходства перекрывание ниш по
трофическому параметру составляет 0,88.
Результаты наших исследований показывают, что разделение
экологических ниш рябинника и.белобровика происходит в основ-
ном по пространственным параметрам: по местам сбора корма
(перекрывание — 0,26) и несколько меньше — по местам распо-
ложения гнезда (перекрывание — 0,4). В меньшей степени виды
различаются по времени начала гнездования и почти не различают-
ся по составу птенцового корма.
Это подтверждает широко распространенное явление успешно-
го сосуществования близкородственных видов птиц за счет сегре-
гации их пространственных экологических ниш, как в гнездо-
вой (Lack, 1971), так и во внегнездовой периоды (Боголюбов,
1986).
По срокам начала гнездования и местам расположения гнезд на
протяжении трех сезонов работы наблюдается постепенное увели-
чение значений перекрывания экологических ниш. Причины этого
увеличения могут быть связаны с ростом колоний, увеличением
численности составляющих их особей обоих видов и повышением
120
концентрации гнезд в колониях. Это, в свою очередь, может быть
вызвано наблюдаемым в последние годы возрастанием численности
основных врагов колоний в гнездовой период — серых ворон, гнез-
дящихся в районе исследований как в вязово-ольховых лесах,
так и в близлежащих населенных пунктах (что отмечается в целом
для центральных районов Европейской части СССР).
ЛИТЕРАТУРА
Боголюбов А. С. Структура и компоновка пространственных ниш зимних си-
ничьих стай в лесах Подмосковья // Зоол. журн. 1986. № 11. С. 1164—1174.
Пианка Э. Эволюционная экология. М.: Мир, 1981. 399 с.
Jue A. Ground-feeding methods and niche separation in the thrushes // Wilson
Bull. 1981. Vol. 93, N 1. P. 112-114.
Lack D. Ecological isolation in birds. Oxford: Blackwell, 1971. 404 p.
Liibke W. Narungsuntersuchungen an der Wacholderdrossel (Turdus pilaris) //
Vogelk. Hefte. 1975. N 1. S. 82-87.
Slagsvold T. Habitat selection in birds: on the presense of other birds species with
special regard to Turdus pilaris //J. Anim. Ecol. 1980. Vol. 49, N 2. P. 523—536.
SUMMARY
Ecological characters of Fieldfare (Turdus pilaris)
and Redwing (T. iliacus) in the places of their joint inhabitance. Gubin A. G.,
Preobrazenskaya E. S., Bogoljubov A. S.
The territorial distribution of nest colonies of Fieldfares and nests of Redwings were
studied on the territory of Manturov District of Kostroma Region. The subdivision
of ecological niches of the Fieldfare and Redwing can be done, on the whole, due to
spatial parameters: places of food supply (excess — 0,88) and somewhat less —
places of nest location (excess — 0,4). To a less extent, the species differ by the time of
nesting start and almost don’t differ by the composition of nestling diet (excess —
0,88). Relative species of birds can successfully co-exist by segregation of their
spatial and ecological niches during nesting and out of nesting period, as well.
121
УДК 591.16:598.311.3 (477.7)
А. И. КОШЕЛЕВ,
И. И. БАД ЮК,
И. И. ЧЕРНИЧКО
332339, Запорожская обл.,
Мелитополь, ул. Ленина, д. 20.
Мелитопольский пединститут
Размножение лысухи
(Fulica atra)
на Тилигульском лимане
(Северо-Западное
Причерноморье)
В Северо-Западном Причерноморье лысуха — самый многочислен-
ный вид водоплавающих птиц. Она имеет важное значение как
объект спортивной охоты, встречается в регионе в большом
количестве круглый год, однако сведения о ее биологии
скудны.
В 1980—1985 гг. мы изучали биологию лысухи на территории
Одесской области. Основные исследования выполнены на двух ста-
ционарах в верховье и низовье Тилигульского лимана. Использо-
вались общепринятые методики. Степень насиженности яиц оп-
ределялась по их плавучести в воде (Блум, 1973). Птенцы в
возрасте 1—3 дней метились стандартными кольцами с пластили-
новой прокладкой. Обследовано 600 гнезд, проконтролирована
судьба 350 из них. Промерено и взвешено 1800 яиц, 70 одно-
дневных птенцов. Учтено 150 выводков, окольцовано 400 лысух
разного возраста. Материал обработан статистически на ЭВМ.
Природно-климатические условия Северо-Западного Причер-
номорья благоприятны для обитания лысухи. Ранняя дружная
весна, продолжительное жаркое лето и осень, мягкая зима позво-
ляют птицам обитать в регионе в течение года, исключая суровые
зимы.
Основными местами гнездования являются плавни крупных
и малых рек с густыми зарослями мощного тростника и рогоза,
чередующимися с участками открытой воды. Водная погруженная
растительность богата и разнообразна, изобилуют рдесты, рого-
листник, пузырчатка, ряска. Верховье Тилигульского лимана
представляет собой тростниковые плавни с обилием небольших
плесов; на границе плавней с лиманом расположено множество
небольших тростниковых островков, что создает высокую мозаич-
ность гнездовых стаций (рис. 1). Вода в значительной степени
опреснена за счет стока р. Тилигул, но при сильных нагонных
ветрах с моря на короткое время соленость резко возрастает.
Суточные и сезонные колебания уровня воды выражены резко
и достигают в отдельные дни 0,5—0,8 м. Для низовья лимана,
отделенного от моря широкой песчаной пересыпью, характерна
мелководность (0,3—1 м), мозаика небольших песчаных островков
с кольцом тростника и морского клубнекамыша по периферии.
Сгонно-нагонные явления также выражены резко. На островках
располагаются крупные колонии речных крачек (Sterna hirundo),
травников (Tringa totanus) и шилоклювок (Recurvirostra avosetta),
122
Рис. 1. Размещение гнезд лысухи в
верховьях Тилигульского лимана в
1981 г.
1 — заросли тростника;
2 — заросли рогоза;
3 — открытая вода;
4 — .песчаные острова;
5 — гнезда лысухи, построенные в апреле;
6 — то же, построенные в мае—июне
Рис. 2. Сроки откладки яиц у лысух
в верховьях и низовьях Тилигуль-
ского лимана
1 — верховья лимана, 1980 г.;
2 — то же, 1981 г.;
3 — низовья лимана, 1981 г.;
4 — суммарные данные по обоим участ-
кам, 1980—1981 гг.
123
а в зарослях надводной растительности — колонии большой
поганки (Podiceps cristatus).
Сроки прилета лысух на лиман определяются ледовой обста-
новкой и ходом весны. Первые птицы отмечены в низовье лимана
12 марта 1980 г., 15 февраля 1981 г., 10 марта 1982 г., 1 мар-
та 1984 г. и 25 марта 1985 г. В верховье лимана они появляются
на 5—10 дней позднее, что связано с более поздним вскрытием
плавней от льда. Основная масса птиц прилетает в течение 5 —10
дней после появления первых особей. Пролет проходит в темное
время суток, наиболее интенсивно, судя по издаваемым птицами
крикам, с 22 до 24 ч. Днем пролетные лысухи укрываются в
зарослях тростника, но крупные стаи из 100—500 особей держатся
на открытых плесах и заливах. В пролетных стаях большая часть
птиц (до 80%) держится уже обособленными парами, которые
проявляют агрессивное поведение по отношению к другим особям.
Индивидуальная дистанция между парами составляет при этом
.2 — 10 м.
Предгнездовой период составляет в низовьях лимана 45—50,
в верховьях —27—30 дней. Сроки начала гнездования значительно
растянуты у птиц на каждом участке лимана и в разные годы
(рис. 2), что связано с разницей в сроках прилета, возрастом птиц,
погодными условиями, гидрологическим режимом, типом и пригод-
ностью гнездовых стаций. При значительном изреживании
стаций за счет снега, льда или выкашивания человеком лысухи
начинают гнездиться только с появлением зеленых стеблей. Гнез-
дование лысух в верховье лимана начинается во второй половине
марта, т. е. на 15—20 дней раньше, чем в низовье. Первые гнезда
появляются в достаточно густых прошлогодних зарослях тростника
и рогоза и только со второй половины мая — в молодых зеленых
зарослях клубнекамыша (табл. 1).
Распределение гнезд по стациям определяется соотношением
их площади (в верховье лимана доминирует тростник (90%),
а в низовье — клубнекамыш (70%)). В меньшей мере оно зависит
от качества стаций (густоты и высоты зарослей, степени мозаич-
ности, близости колоний крачек, степени беспокойства со стороны
человека и др.). В верховье лимана 95% обнаруженных гнезд рас-
полагалось в зарослях тростника, в низовье 75% гнезд находи-
лись в зарослях клубнекамыша. В приморской зоне по группе
небольших по площади водоемов, в карьерах (площадью по 10 —
20 м 2), лишенных зарослей надводной растительности, обнаруже-
ны три гнезда лысухи на берегу в зарослях сухих злаков, распо-
ложенные в 1 — 1,5 м от воды. Это редкое явление. Кроме того,
в низовье нами обнаружено 8 гнезд лысух, расположенных у ост-
ровков вблизи колоний крачек, оказавшихся при усыхании водоема
на суше, в 10—30 м от воды; птенцы в них вывелись успешно.
В зарослях осоки, озерного камыша, встречающихся в верховье
лимана небольшими островками, обнаружено всего 0,6—1,1%
гнезд. В целом лысухи предпочитают для гйездования средние по
густоте заросли, высота их существенной роли не играет. Густота
124
Таблица 1
Распределение гнезд лысухи в течение гнездового периода
в 1980-1985 гг.
Гнездовая стация 11-20.1П 21-31.111 1-10.IV И-20.1 V 21-30.IV
Прошлогодние заросли трост- ника 4 24 47 26 15
Прошлогодние заросли рогоза 3 3 3 2
Зеленые заросли клубнекамыша Итого гнезд: — — — 4 12
абс. 4 27 50 33 29
% 1,8 12,1 22,4 14,7 13,0
Гнездовая стация 1-10.V 11-20.V 21-31.V 1-10. VI Всего
Прошлогодние за рос ли. трост- ника 12 5 1 1 135
Прошлогодние заросли рогоза 1 1 1 2 16
Зеленые заросли клубнекамыша Итого гнезд: И 14 26 5 72
абс. 24 20 28 8 223
% 10,8 9,0 12,6 3,6 100
зарослей и близость открытой воды являются определяющим
фактором месторасположения на данном участке. В 1—2 м от
открытой воды обнаружено 16,4% гнезд, в 2,1—4 м—22,9,
в 4,1—6 м — 17,3, в 6,1—12 м — 29,4, в 12,1—20 м — 21,5, далее
20 м — 6,5% гнезд. В густых зарослях это расстояние меньше,
чем в редких. В зарослях тростника оно составило в среднем 6,4 м,
а в зарослях клубнекамыша — 9,1 м. Выявлено, что удаленность
гнезд от открытой воды уменьшается в ходе гнездового периода; с
развитием зеленой растительности лысухи размещают гнезда бли-
же к плесу. Так, в зарослях тростника это расстояние сократилось
в среднем с 12,2 м в середине марта до 3,0 м в конце мая, в зарос-
лях клубнекамыша — с 10,4 в конце апреля до 6,1 м в начале
июня.
Для постройки гнезд на лимане лысухи используют материал,
имеющийся в достаточном количестве пососедству: сухие стебли
и листья тростника, рогоза, озерного камыша, клубнекамыша,
осок, реже — их зеленые стебли и листья. Гнезда, расположенные
на берегу, были выстланы сухими злаками и полынью. В ни-
125
зовьях лимана близость курортной зоны обусловливает значитель-
ное загрязнение водоема бумагой, ветками, досками, кусками поли-
этиленовой пленки. Эти материалы также изредка встречаются
в стенках гнезд лысухи (8 случаев) В верховьях лимана, однако,
95% гнезд изготовлено из тростника. В низовьях 40% гнезд было
выстроено из клубнекамыша, что связано с его обилием здесь,
остальные 60% гнезд были выстроены из тростника. Размеры гнезд
лысухи на Тилигульском лимане не отличаются от приводимых
для других районов СССР (Блум, 1973; Кошелев, 1984; и др.).
Диаметр гнезд (п=120) составляет 28—44 см (среднее — 35 см),
диаметр лотка 11 —19 (17,5) см, высота надводной части гнезда
5— 35 (18,5) см, глубина лотка 3 — 13 (7) см. Длина сходен 30—7Q
(45) см, их ширина 20—50 (25) см. Сходни обнаружены у 70%
осмотренных гнезд, причем 3% гнезд было с двумя сходнями,
0,5% — с тремя. Установлено, что гнезда со сходнями были на
21,1% выше, чем гнезда без сходен. Некоторые гнезда, жестко
закрепленные между стеблями тростника, оказались при падении
уровня воды в висячем положении, на высоте 20—50 см от поверх-
ности воды. В зарослях клубнекамыша гнезда имели сходни в 80%,
а в тростниковых зарослях — в 65%. Характерной особенностью
гнезд, построенных в конце мая — июне, является устройство над
ними своеобразной крыши из надломленных и переплетенных зе-
леных Листьев и стеблей тростника, рогоза или клубнекамыша.
Крыши обнаружены у 25—45% поздних гнезд, причем чаще в за-
рослях клубнекамыша (30%) и рогоза (18%), чем в тростнике
(8%). Вблизи основного гнезда, служащего для откладки яиц,
лысухи часто строят одну-две платформы, используемые для отды-
ха свободным партнером. Платформы встречены у 20% гнезд,
причем в зарослях тростника чаще (32% гнезд), чем в зарослях
клубнекамыша (16%). Это связано с наличием островков, отмелей
и близостью берега к гнездам в низовье лимана, охотно использу-
емых птицами для отдыха вместо платформ. В зарослях тростника
платформы размещаются в 1—15 м в среднем в 5,5 м от гнезд,
а в зарослях клубнекамыша — в 1—8 (3,7) м.
Плотность гнездования лысух на обоих участках лимана до-
вольно высокая. Соседние гнезда располагались в 1—45 м друг от
друга (в среднем — в 14,5 м), причем в зарослях тростника эта
дистанция в среднем была меньше (14,2 м), чем в зарослях
клубнекамыша (16,2 м). По расстоянию друг от друга гнезда
распределялись следующим образом: 1—7 м — 10,2%, 7,1 —
15 м — 32,2, 15,1—21 м — 42,4, 21,1—30 м — 11,8, свыше 30 м —
3,4% случаев. Видимо, столь высокая плотность гнездования
связана с нехваткой пригодных для гнездования мест. Косвенно
об этом свидетельствует заселение лысухами вновь образовавших-
ся зеленых зарослей рогоза и клубнекамыша в конце мая — июне,
повторное занятие участков новыми парами при уходе выводков.
Такой популяционный «резерв» из поздно гнездящихся или не-
гнездящихся пар был особенно значителен в низовье — до 35—50%
от общего числа учтенных птиц. Не участвующие в размножении
126
пары держались на плесах вблизи зарослей, их активно преследова-
ли и отгоняли от зарослей территориальные птицы. При отсутствии
возможности загнездиться такие пары присоединялись в конце
июня к скоплениям линных неполовозрелых лысух и приступали
к линьке.
По соседству с гнездами лысух нами обнаружены гнезда семи
видов птиц. В 1 — 15 м (в среднем — в 6,5 м) от гнезд лысухи
встречено 91 гнездо большой поганки, причем лысухи иногда от-
кладывает яйца в ее гнезда (6 случаев). Серощекая поганка
(Podiceps grisegena) гнездится только в верховье лимана, 95% ее
гнезд располагалось в 0,5—5 м от построек лысух (65 случаев).
Гнезда кряквы (Anas platyrhynchos), расположенные в искусствен-
ных гнездовьях, были обнаружены в 1 — 15 м от гнезд лысух
(25 случаев). Гнезда камышницы (Gallinula chloropus) найдены
в 5—20 м от гнезд лысух (15 случаев). Вблизи от гнезд лысух
найдено также 7 гнезд серой цапли (Ardea cinerea) 60 гнезд речной
крачки, 3 гнезда гнездящейся в густых зарослях тростника сороки
(Pica pica). Несмотря на агрессивный характер лысухи в период
размножения (Блум, 1973; Кошелев, 1984), случаев столкновений
между ней и другими птицами-соседями на данном водоеме мы
не отмечали. Это связано, по-видимому, с разобщенностью гнездо-
вых и кормовых микростаций разных видов, несмотря на близкое
расположение их гнезд. Отсутствие конфликтов с камышницей
объясняется, видимо, тем, что камышницы не показывались на
открытой воде, где в основном осуществляется патрулирование
границ лысухами. Взаимному избеганию птиц-соседей способству-
ет и развитая система их звуковой сигнализации.
У лысух, гнездящихся в Тилигульском лимане, кладка состоит
из 4 —15, чаще из 7—8 яиц, в среднем из 7,38 яиц (табл. 2).
Выявлено, что величина кладки у птиц, гнездящихся на разных
участках лимана, статистически не различается. Это позволило
объединить данные о размерах кладок: имеющиеся различия
перекрываются изменчивостью данного показателя в пределах
каждого участка. Величина кладки у лысух, гнездящихся в ап-
реле — начале мая в зарослях тростника в верховье лимана, со-
Таблица 2
Величина кладки лысух в 1980—1985 гг.
Год п Количество кладок с числом яиц Среднее
4 5 6 7 8 9 10 11 11 М + ш С, %
1980 127 4 10 14 30 37 16 11 6 0 7,63+0,14 21,3 1981 75 4 8 14 13 16 И 5 2 1 7,31+0,23 27,0 1982 46 0 678 12 543 1 7,70+0,18 25,5 1983 23 0 3 7 8 3 2 0 0 0 6,7+0,15 23,0 1984 23 1 4 9 4 5 0 0 0 0 6,3+0,20 18,7 1985 25 0 05937100 7,6+0,14 21,5 Всего 319 9 31 56 72 76 41 21 И 2 7,38+0,25 23,0
127
ставила 7,62±0,18 яиц, в низовье — 7,54±0,22 яиц. В конце
мая — июне величина кладки у птиц, гнездящихся в зарослях
клубнекамыша, составила 6,0±0,30 яиц. Величина кладки последо-
вательно уменьшается в ходе гнездового периода. В конце марта
она составила 8,2 яйца, в конце апреля — 7,8, в конце мая — 6,1,
в первой половине июня — 5,0 яиц. Мы связываем это с более позд-
ним гнездованием птиц младших возрастных групп (одно-двугодо-
валых), откладывающих меньшее число яиц, а также с включением
в анализ повторных кладок, отложенных самками взамен утрачен-
ных. Различия в размерах кладки по годам незначительны. Не
выявлено также достоверных различий по данному показателю
у птиц, населяющих разные микростации, гнездящихся в колони-
ях крачек и куликов и вне колоний.
Сроки размножения лысух на обоих участках лимана значи-
тельно растянуты и составляют в верховье 95—100, а в низовье —
75—80 дней (от появления первых яиц до вылупления последних
птенцов). Как отмечалось выше, это связано с разновременностью
гнездования птиц разных возрастных групп, поселением в разных
стациях, откладкой повторных кладок (иногда у отдельных пар
до 2—4 раз), участием в размножении резервных пар. Пик массо-
вой откладки яиц приходится в разные годы на начало апреля —
начало мая, что зависит от погодных условий весны.
Первые яйца лысухи откладывают как в полностью отстроенное
гнездо (90% кладок), так и недостроенное, достраиваемое в про-
цессе откладки яиц и насиживания. По окраске преобладают яйца
со светло-песочным фоном, с редким крапом из темно-коричневых,
мелких пятен (65%), реже встречаются яйца со светло-глинистым
фоном и более крупным крапом (35%). У птиц, гнездящихся
в зарослях клубнекамыша, окраска яиц заметно темнее, чем у тех,
что населяют заросли тростника. Аномальная окраска яиц (почти
белые, с едва заметным крапом) встречается исключительно редко
(2 яйца). Самка откладывает ежедневно по одному яйцу в утренние
часы. Размеры яиц варьируют в широких пределах.. Длина яиц
составляет 42,0—63,0 мм, ширина — 32,0—41,2 мм, масса — 30,0—
53,8 г. Выявлены достоверные различия размеров и массы яиц
у птиц, гнездящихся на разных участках лимана (табл. 3).
Эти различия свидетельствуют о значительной разнокачествен-
ности лысух, с одной стороны, и о более оптимальных условиях
размножения в верховьях, с другой. Меньшие размеры яиц у птиц
в низовьях лимана связаны, вероятно, с участием в размножении
птиц младших возрастных групп и, возможно, с высоким популя-
ционным давлением в виде резерва гнездящихся пар. В среднем
размеры яиц на Тилигульском лимане составили 53,31±0,08Х
Х36,38±0,04 мм, масса — 38,60±0,18 г, т. е. они не отличаются
от показателей из других частей ареала (Блум, 1973; Кошелев,
1984).
Размеры яиц последовательно уменьшаются в течение гнездово-
го периода на обоих участках: в марте они составляли 53,8Х
Х37,0 мм, в апреле — 53,4X36,5 мм, в мае — 53,1X35,9 мм и в
128
I
I
i
Таблица 3 Размеры и масса яиц лысух в 1980—1985 гг. а С if «Г С с с с о- С С 4 150 Таблица 4 Успешность гнездования лысух в 1981 — 1985 гг. Доля погибших, % 11—20 марта 15 0 100,0 1 — 10 мая 65 60 7,7 21-31 марта 77 33 57,1 11-20 мая 61 50 18,0 1-10 апреля 215 181 15,8 21-31 мая 149 43 71,1 11—20 апреля 154 142 7,8 1 — 10 июня 19 1 94,7 21 — 30 апреля 92 83 9,8 Всего 847 593 30,0
Масса, г M±m lim > °9. Г се ю 1 се се О 00 СМ СО " о -н
Вылупилось птенцов
550 38,: 480 35,' Всего яиц
е
Ширина, мм М±т Пт С с 1 с о СГ с\ •V xj с СГ с 60 Срок вылупления
S о o' o' -н -н о о со 1Q со се Доля погибших, %
Длина, мм — М±т Пт 53.54-0.09 42.0—01.0 с СТ О 1Г О“ Vt 76 Вылупилось птенцов
52,7±0,13 5, Всего яиц
Участок лимана Верховья Низовья Критерий Стьюдента Срок вылупления
5 Заказ № 36
129
июне — 51,5x35,8 мм (объем выборки — по 100—250 экз. для
каждого периода). В каждой кладке яйца, отложенные первыми,
были несколько крупнее и тяжелее, чем отложенные последними.
С увеличением размеров кладки размеры и масса каждого яйца
возрастали и составили для кладки из трех-четырех яиц 52,6Х
Х36,2 мм, для кладок из пяти-шести яиц 53,2X36,3 мм, для кладок
из семи-восьми яиц 53,6X36,4 мм, для кладок из девяти-десяти
яиц 54,2x36,7 мм, для кладок из 11—12 яиц 55,3x37,2 мм.
Возможно, эти различия определяются возрастом самок: известно,
что чем старше самки, тем больше откладывают яиц и тем крупнее
яйца. В ходе насиживания масса яиц ежедневно уменьшается на
0,28 г и составляет к моменту вылупления птенцов 82,6% от перво-
начальной. Поэтому для анализа были взяты только свежие яйца.
Насиживание кладки длится 21 —22 дня (п=15), первые птен-
цы отмечены в начале апреля — начале мая. Общий период вылуп-
ления птенцов составил в верховье 70—75, в низовье 45—50 дней.
В зависимости от величины кладки и времени начала насиживания
яиц, которое чаще начинается после откладки третьего-четвертого
яйца, птенцы выводятся в одном гнезде в течение 2—8 сут. Вес
обсохших однодневных птенцов равен 21,8—30,0 г, в среднем
25,77+0,28 г, что составляет 66,8% от массы свежих яиц. Из более
крупных яиц вылупляются более крупные и тяжелые птенцы.
Масса птенцов в верховье лимана была несколько выше, чем в
низовье (25,97+0,32 и 25,61+0,45 г соответственно). Птенцы об-
сыхают под наседкой в течение 10—24 ч, после чего уводятся от
гнезда вторым партнером. При опасности птенцы обычно прячутся
в основании гнезда, реже — быстро отплывают, затаиваются или
заныривают, выставив на поверхность только голову.
Птенцы лысухи вылуплялись в среднем из 70% яиц, но в начале
и конце периода размножения доля погибших значительно выше
30% (табл. 4).
Основные причины гибели яиц: яйца выпали из гнезда во вре-
мя волнобоя и разрушения гнезда (53,5%), уничтожены болотным
лунем Gircus aeruginosus (18,0%) и сорокой (7,6%), выкатились
из гнезда при поспешном бегстве самки (8,7%), взяты человеком
(4,3%), уничтожены ондатрой (2%), замурованы птицей в стен-
ках и дне лотка (1,2%); неоплодотворенными оказались 2,7% яиц,
эмбрионы погибли в 2,0% яиц. В 97 полных успешных кладках
отход составил 0,69 яйца на кладку, а клев птенцов — 89,9%.
Установлено, что выклев птенцов в гнездах, расположенных в за-
рослях тростника, составлял 80,3%, в зарослях клубнекамыша —
46,6% из-за более высокой гибели целых кладок. Доля гибели
их изменялась в течение сезона: в середине марта погибло 100%,
во второй декаде марта — 75%, в первой декаде апреля — 11,5%,
во второй декаде апреля — 4,6%, в третьей декаде апреля — 6,3%,
в третьей декаде мая — 56%, в первой декаде июня — 80%
учтенных в июне кладок. Всего погибло 27,6% гнезд (п=37) из
134 исследованных.
Причины гибели гнезд и кладок лысухи следующие: разру-
130
Таблица 5
Величина выводка лысух в 1980—1985 гг.
Участок лимана Количество выводков с числом птенцов Сред- нее Выживае- мость птенцов, %
3 4 5 6 7 8 9 11
Верховья 6 24 27 27 21 7 2 114 5,7 75,0
Низовья 6 4 12 5 5 2 0 36 5,0 68,5
Всего 12 28 39 32 26 9 2 150 6,0 81,3
шено волнобоем 32,4%, разорено болотным лунем и сорокой 40,6%,
оставлено птицами вследствие частого беспокойства человеком
10,8%, разорено ондатрой 8,1%, перевернулось при падении
уровня воды 8,1% гнезд. Гибель гнезд в зарослях тростника
достигала 10%, в зарослях клубнекамыша — 38,1 %, что связано
с худшими защитными условиями зарослей клубнекамыша и
сильным антропогенным прессом (выпас скота, частые выходы
на остров рыбаков, подростков и отдыхающих) в низовьях. В пери-
од яйцекладки погибло 10 гнезд, в период насиживания — 26,
во время вылупления птенцов — одно гнездо.
Выводки лысух с подросшими птенцами в возрасте 2,5 недель
и старше состоят, по нашим наблюдениям, из трех—одиннадцати,
в среднем из шести птенцов (табл. 5).
Птенцы, особенно в возрасте до десяти дней, гибнут от пере-
охлаждения, иногда затаптываются родителями на гнезде (5% об-
щей гибели). В верховьях лимана основным врагом птенцов
является болотный лунь, а в низовьях главной причиной их гибели
является холодная ветренная погода, сопровождаемая волнобоем
и нагоном воды. Каждый выводок лысух обитает на постоянном
выводковом участке, протяженность границ которого вдоль кромки
зарослей и открытой воды составляет 20—50 м. Границы участков
охраняют оба партнера. В случае резкого ухудшения условий
(усыхание зарослей, вытравливание их скотом) выводки лысух
уходят в другое место; это явление известно и на юге западной Си-
бири (Кошелев, 1984).
Во второй половине июля — начале августа выводки с вырос-
шими лётными молодыми выходят из плавней на акваторию
лимана, образуют вначале рыхлые, а позднее плотные скопления
сеголеток. Взрослые лысухи уходят на линьку в густые заросли
тростника, где держатся чрезвычайно скрытно группами по 30—
100 особей. В низовьях лимана нет обширных густых зарослей,
поэтому лысухи линяют там небольшими группами по 30 —10 осо-
бей. Возможно, часть птиц линяет на выводковых участках при
выводке, совмещая линьку с размножением. Кроме того, на обоих
участках лимана ежегодно формируются скопления из линных
холостых и неполовозрелых лысух, до 400—500 особей на каждом
участке. В верховье лимана они держатся в июне—июле на грани-
це плеса и зарослей рогоза и тростника, а в низовье — в густых
5»
131
зеленых зарослях клубнекамыша, вырастающего на мелководьях.
В 1979—1980 гг. в низовье 26—29 июня было отловлено 12 лысух,
не имевших маховых перьев, только у трех птиц на крыльях
были второстепенные и третьестепенные маховые (личное сообще-
ние М. Жмуда). Отлов линных птиц не представляет на мелко-
водье трудностей (поймано и окольцовано 90 экз.). Отпущенные
лысухи вплавь уходили в другое место за 1—3 км, где отлавли-
вались повторно. С отрастанием первостепенных маховых перьев
на треть их нормальной длины линные лысухи выходят из зарослей
на открытую воду, мелкие группы объединяются в скопления
по 100—500 особей. При преследовании они убегают по воде
вплавь, реже скрываются в зарослях.
Кочевки и отлет лысух из плавней лимана начинается с с.ере-
дины августа и ускоряется интенсивной охотой. Лысухи уходят
на акваторию лимана, где держатся в 1—3 км от берега. К берегу
они приближаются только в безопасных местах в дни, когда
не ведется охота. Сеголетки держатся на лимане скоплениями
из 100—10 000 особей до конца ноября — конца декабря (до ледо-
става), а в теплые зимы часть птиц остается зимовать на лимане.
Однако основная масса сеголетков, по данным кольцевания (Блум,
Лицбарский, 1982), отлетает на зимовки в Румынию и страны
Средиземноморья. Осенний пролет проходит также только в темное
время суток. Среди зимующих птиц обнаружены только сеголетки
обоих полов (просмотрено свыше 200 экз. погибших птиц, более
20 тыс. прижизненно с помощью бинокля). Взрослые лысухи
по окончании линьки отлетают в конце августа—сентября отдель-
но от молодых и не остаются на зимовку в гнездовой области.
Итак, Тилигульский лиман населяет перелетная местная по-
пуляция лысух, представленная двумя пространственно изолиро-
ванными в гнездовое время группировками; в послегнездовой
период молодые птицы частично смешиваются на акватории ли-
мана. Для лысух данной популяции характерны полуоседлость
вплоть до оседлого образа жизни части птиц в особенно теплые
зимы, длительный срок пребывания в гнездовой области (до 9 —
И мес) короткий миграционный путь, значительный интервал
между прилетом и началом размножения (от двух до 10 недель),
растянутые сроки размножения, наличие значительного числа
половозрелых резервных особей. Размещение гнезд на открытых
(мелководья, берег) местах связано с отсутствием или малочис-
ленностью пернатых и наземных хищников. Каждая пара совер-
шает за сезон только один цикл размножения, предположение для
соседних районов Молдавии о двух успешных циклах кладки
(Мунтяну, 1970) нашими исследованиями не подтверждается.
Лысухи занимают гнездовые стации в зависимости от их при-
годности для гнездования, что позволяет гнездиться им с большой
плотностью. При этом существенное значение имеет нехватка
пригодных для гнездования мест, особенно в низовье, а также
большая протяженность благоприятного для размножения пе-
риода.
132
Гнездование и размножение у лысух данной популяции доволь-
но успешны, несмотря на значительную гибель гнезд, кладок
?; и яиц. Гибель первых кладок компенсируется откладкой повтор-
’ ных. Высокой выживаемости птенцов способствует участие в их
й воспитании и кормлении обоих родителей, активная защита от
пернатых хищников, пребывание выводков в густых зарослях
I первые полторы-две недели жизни, разделение выводка на се-
мейные группы между родителями (Кошелев, 1984), гибкая, под-
вижная система использования территории (смена гнездовых
участков на выводковые, образование скоплений, кочевки вы-
водков) .
Значение лысухи в охотничьих хозяйствах возрастает. В вер-
„ховьях Тилигульского лимана, на территории Тилигульского
£ охотхозяйства ВВОО гнездится ежегодно 200—250 пар, осенние
запасы составляют 2,5—4 тыс. лысух. Из них добывается осенью
400—500 экз., что дает 35—40% от общего числа добываемых
водоплавающих птиц (Кошелев, Шулим, 1984). Площадь вод-
£ ных угодий в хозяйстве превышает 2800 га, т. е. потенциальная
f емкость угодий велика. Однако нехватка подходящих для гнез-
f дования мест сдерживает рост численности лысух. Они избегают
| заселять сплошные массивы тростниковых зарослей в плавнях
и густые потравленные скотом заросли рогоза на прибрежных
мелководьях. Большинство гнезд располагается в островках трост-
ника и вдоль искусственных плесов и проходов к ним. Расширяя
£ их сеть и протяженность, можно значительно увеличить гнездо-
| вую емкость угодий. Однако в охотхозяйстве на охоту па лысуху
f обращают мало внимания, биотехнические работы направлены
только на увеличение численности кряквы путем установки искус-
ственных гнездовий.
В низовье лимана на территории Коблевского охотхозяйства
Николаевского облсовета УООР ежегодно гнездится от 50
Д до 120 пар лысух, осенью добывается 450 — 500 экз., что Состав-
S' ляет 30—35% общего числа добытых водоплавающих птиц. Даль-
й нейший рост численности лысух сдерживается нехваткой гнез-
довых стаций. Осуществление биотехнических мероприятий зат-
руднительно. Для сохранения зарослей тростника и клубнекамыша
следует ограничить и регулировать выпас скота, желательна вы-
садка надводных солеустойчивых растений и расширение их пло-
К щади вокруг всех островов. Следует учитывать, что на многочис-
ленных островках в низовье лимана располагаются уникальные
для региона гнездовые колонии крачек и куликов, в том числе ред-
ких и исчезающих видов, гнездятся многие утиные птицы (Кор-
зюков, Черничке, 1977). Проводятся многолетние исследования
по изучению миграций и популяционной экологии птиц препо-
давателями и студентами Одесского университета. Благодаря
их инициативе в низовье лимана (на территории Одесской обла-
й сти) в 1983 г. организован областной сезонный орнитологический
заказник «Тилигульская пересыпь» площадью 1200 га. Охрана
; гнездовий птиц ведется силами студентов.
133
Интенсивное сельское хозяйство, расширение рекреационных
зон в приморской зоне обусловливают необходимость особенно бе-
режного, заботливого отношения к немногим охраняемым участкам
естественных ландшафтов степной зоны и побережья моря,
где еще уцелели редкие виды растений и животных. Существую-
щая в регионе сеть заказников и заповедников явно недостаточна,
нуждается в пересмотре и расширении. Поэтому большая приро-
доохранная роль отводится приписным охотничьим хозяйствам
УООР и ВВОО, которые обязаны уделять внимание не только охот-
ничьим животным, но и редким и исчезающим видам. Следует
также срочно пересмотреть отношение к так называемым второ-
степенным видам дичи, включая лысуху, шире применять, к ним
специальные биотехнические мероприятия и искусственное раз-
ведение с последующим выпуском молодняка в природу.
ЛИТЕРАТУРА
Блум П. Н. Лысуха в Латвии. Рига: Зинатне, 1973. 156 с.
Блум П. И., Лицбарский X. Лысуха // Миграции птиц Восточной Европы и Север-
ной Азии: Хищные—журавлеобразные. М.: Наука, 1982. С. 209—273.
Корзюков А. И., Черничке И. И. Тилигульский лиман — место концентрации во-
доплавающих и околоводных птиц // Фауна и биология гусеобразных птиц.
М.: Наука, 1977. С. 57-58.
Кошелев А. И. Лысуха в Западной Сибири. Новосибирск: Наука, 1984. 176 с.
Кошелев А. И., Шулим А. С. Тилигульское охотхозяйство // Охота и охотничье
хоз-во. 1984. № И. С. 6-8.
Мунтяну А. И. Экология лысухи в Молдавии // Изв. АН МССР. Сер. биол. и хим.
наук. 1970. № 1. С. 29-35.
SUMMARY
Reproduction of the Coot on the Tiiigul Liman in the north-west Black Sea Region.
Koshelev A. I., В a d i u k I. I., Chernichko I. I.
Over 500 clutches, 1800 eggs of the Coot were studied and over 150 broods were counted
up in 1980—1984 in two points of the Tiiigul Liman. The dates of egg laying lasted
since the second decade of March till the second decade of June. One clutch consists
of 5—12 eggs, on the average 7,5+0,12 eggs. The eggs’ sizes: 53,31+0,08X36,38+
+0,04 mm, mass 38,60+0,18 g. In broods of 3—11 nestlings, on the average 6,0 nest-
lings, their survival is 81,3%. The coefficient of their successful nesting reaches 72,4%.
There were no true distinctions in the reproduction between two groupings of Coots,
which nested on the Liman, 60 km from each other. They form one local population. The
expanded reproduction terms and other periodical phenomena, the high reproduction
potential, a short migration way are characteristic for the Coots of the given population.
134
УДК 591.3: 598.422
Т. Б. АРДАМАЦКАЯ, Трофические связи
В. И. ВАКАРЕНКО, А. А. ПЕТРУСЕНКО черноголовой чайки
326240, Херсонская об л.. в гнездовой период
Голая Пристань, в условиях
Днепровская ул. д. 1. Черноморский государственный Черноморского заповедника
заповедник АН СССР Сообщение II. Пространственно- временные характеристики трофических связей *
В условиях заповедника черноголовая чайка гнездится на остро-
вах. В поисках корма взрослые птицы посещают разнообразней-
шие биотопы, о чем свидетельствует наличие в пищевых пробах
птенцов представителей не менее чем 10 группировок организмов.
Большинство составляли степные элементы (46,1%). Это прямо-
крылые Calliptamus barbarus Costa, Myrmeleotettix sp., Euchort-
hippus pulvinatus F.-W., Oedipoda caerulescens L., маврская че-
репашка; жужелицы Amara similata Gyll., Zabrus tenebrioides Gz.,
пластинчатоусые Amphicoma vulpes F., Anisoplia austriaca Hbst.,
A. deserticola F.-W., чернотелки Anatolica eremita Stev., Pedinus
borysthenicus Rchdt., долгоносики Mesagroicus obscurus Bohm.,
муравьи Messor clivorum Ruzs., Cataglyphis aenescens Nyl., Formica
cunicularia Forst., Polyergus rufescens Latr. и др. Довольно значи-
тельным числом представлены политопные (ряд видов прямо-
крылых, полужесткокрылых, пластинчатоусых, щелкунов,коровок),
различных двукрылых, особенно среди звонцов (27,4%), и луговые
(некоторые жужелицы, долгоносики, чешуекрылые, в частности
луговой мотылек, двукрылые) формы. На долю представителей
других группировок (соленоводных, пресноводных, лесных, га-
лофильных, болотных, пойменнолесных и супралиторальных)
в целом приходится всего лишь 1,2%. Отсюда можно сделать
вывод, что птицы собирают корм главным образом в степных био-
топах и агроценозах. Довольно охотно они посещают и пойменно-
луговые биотопы. С другой стороны, не исключена возможность
попадания луговых элементов на плакорные участки поливных
земель.
Согласно визуальным наблюдениям, в предгнездовой период
чайки пролетают за кормом до 115 км. В гнездовое время, особенно
после появления птенцов, радиус полетов сокращается до 50—
60 км, часто и меньше — до 5—45 км. При этом, согласно литера-
турным данным (Клименко, 1950), зона полетов охватывает пло-
щадь в 4000 км2, а в исследуемый период — видимо, значительно
больше (до 6000 км2). Вполне естественно, что такая территория
* Сообщение I «Пространственно-временные характеристики трофических связей»
опубликовано в кн.: Экология и поведение птиц (М.: Наука, 1988).
135
включает значительное количество мозаично перемежающихся
биотопов. Этим и объясняется присутствие различных экотопи-
ческих элементов в пищевых пробах. Тем не менее в каждой отдель-
но взятой пробе чаще всего присутствуют представители одной—
двух, реже трех и лишь изредка четырех и более экотопических
группировок. По-видимому, взрослые птицы собирают один пи-
щевой комок, как правило, из одного биотопа.
Другим не менее важным фактором, определяющим способ
добывания корма, является ярусная приуроченность объектов пи-
тания, отражающая особенности вертикального размещения орга-
низмов. По данному признаку четко выделяется девять их. груп-
пировок, причем доминировали обитатели растительных ярусов,
или фитобионты,— 65,2%. В довольно значительном количестве
обнаружены беспозвоночные, обитающие как в растительных
ярусах, так и в опаде, или стратофитобионты,— 22,9%. В меньшем
числе отмечены организмы, связанные с поверхностью почвы,
или эпигеобионты,— 5,6%; с растительным опадом, или страто-
бионты,— 4,0%; активно прокладывающие ходы в почве,
или роющие геобионты,— 1,6% и в незначительном — беспозво-
ночные, попеременно обитающие в почве и опаде, или геострато-
бионты,— 0,1%. Из сказанного следует, что основную часть ком-
понентов корма птицы собирают с поверхности растений, а также
с поверхности почвы, куда они, будучи потревоженными, часто
падают. Что касается роющих геобионтов, геостратобионтов и стра-
тобионтов, ведущих скрытый образ жизни, следует иметь в виду,
что эти организмы часто оказываются на поверхности при прове-
дении культивации и других сельхозработ. Кроме того, в пасмур-
ную погоду они часто активны и днем. Из водных обитателей по-
падались нектонты, планктонты и бентонты, вместе составив-
шие 0,6% что еще раз подтверждает известный вывод о том,
что черноголовая чайка питается практически почти полностью
на суше.
В пространственном отношении трофические связи животных
тесно связаны и с характером распределения популяций предста-
вителей отдельных их видов. Поэтому компоненты питания черно-
головой чайки были условно разделены на две группы — обра-
зующие различные скопления в виде колоний, роев, при массовом
выплоде и т. д. и равномерно-диффузно распределяющиеся по всей
поверхности субстрата, с которой собирается корм. Доминирующее
положение в пище птенцов исследуемого вида заняли предста-
вители первой группы — саранчевые (Calliptamus italicus L.,
С. barbatus Costa), пластинчатоусые родов Gymnopleurus Ill.,
Onthophagus Latr., Aphodius Ill., Oniticellus Serv., имаго и гусе-
ницы лугового мотылька, муравьи и двукрылые из сем. Chirono-
midae (всего 17 видов), вместе составивших 67,0% от общего ко-
личества обнаруженных пищевых объектов.
К временным аспектам питания относится потребление живот-
ного корма разных онтогенетических стадий составляющих его
компонентов, возраст потребителей, а также характер суточной
136
активности тех и других. Нами обнаружено подавляющее домини-
рование взрослых (имагинальных) форм (75,5%). Личинки и ку-
колки обнаружены в значительно меньшем количестве (9,6%),
остальное приходится на беспозвоночных, у которых возрастные
стадии определить трудно (малощетинковые черви, моллюски,
ракообразные, паукообразные и др.— 11,6%) и растительные
корма. Вероятно, это связано с преобладанием в корме подвижных
компонентов (63,2%), большую часть которых как раз и составили
имагинальные формы насекомых. Это большинство жуков-жуже-
лиц, щелкуны, чернотелки, муравьи и др., среди которых сравни-
тельно небольшим количеством представлены летающие фор-
мы (23,0%) — большинство прямокрылых, чешуекрылые, ряд
видов перепончатокрылых, двукрылые. Довольно значительной
была доля (36,9%) сравнительно малоподвижных и неподвижных
объектов — малощетинковые черви, моллюски, пластинчатоусые
жуки, листоеды, долгоносики и др. Очевидно, бегающие и ле-
тающие формы беспозвоночных чайкам трудно доступны, о чем
свидетельствует их незначительная (4,6%) доля в рационе.
Для характеристики возрастных стадий потребителей мы рас-
полагаем пищевыми пробами, взятыми у птенцов трех возрастных
групп: младшего (6 — 10 дней), среднего (11—20 дней) и стар-
шего (21—35 дней). Количество наименований компонентов пи-
тания в этих группах колебалось от 51 до 128; причины таких ко-
лебаний не выявлены. Обнаружено только, что в 1974 г. в пищевых
пробах птенцов старшего возраста было значительное количество
растительных кормов, тогда как в 1975 и 1977 гг. они не были от-
мечены. Среди кормов животного происхождения в 1974 г. у птен-
цов старшего возраста преобладали прямокрылые, жуки-чернотел-
ки, муравьи, в меньшем количестве попадались чешуекрылые
и двукрылые. В 1975 г. количество муравьев, полужесткокрылых
и жесткокрылых резко снизилось. Указанные обстоятельства тре-
буют дополнительных исследований. Противоречивы данные и от-
носительно наполненности пищевых проб. Так, среднее количество
компонентов на одну пробу у птенцов младшего возраста в 1974 г.
составляло 130 экз., среднего — 208, старшего — 70, тогда как
в 1975 г,— 73, 60 и 287 соответственно.
Весьма неожиданным было подавляющее доминирование
(50,3%) в собранных днем пробах компонентов с сумеречно-ноч-
ным типом активности, прежде всего массовых видов — лугового
мотылька, звонцов и др., собираемых птицами с поверхности ра-
стений, где эти насекомые проводят дневное время суток. Другие
беспозвоночные с подобным типом активности, как уже отмечалось,
часто бывают доступны птицам при проведении сельхозработ.
Беспозвоночные с дневной активностью в рационе птенцов соста-
вили 29,4%, а с круглосуточной — 20,3%.
137
Функциональная характеристика трофических связей
Собранный и обработанный материал позволяет включить в дан-
ный раздел отдельные обобщения, касающиеся способов поиска
и распознавания объектов питания по отдельным морфоэкологиче-
ским признакам, а также роли исследуемых птиц в трофической
сети населяемых ими экосистем. Относительно поиска и распо-
знавания объектов питания трофические связи можно проследить
через соотношение последних по ряду других морфоэкологиче-
ских признаков, из которых выделены характер подвижности,
склеротизации, окраски и линейные размеры. Соотношение ком-
понентов по первому признаку уже обсуждалось в связи с их со-
поставлением по отдельным онтогенетическим стадиям. По сте-
пени склеротизации эти организмы условно разделены на объекты
с покровами мягкой, средней и твердой консистенции. В пище
птенцов преобладали первые (50,6%), две другие группы были
представлены примерно в равном соотношении (соответствен-
но 26,6 и 22,8%). Это вполне естественно, поскольку птенцам, осо-
бенно в раннем возрасте, необходимы более питательные и мягкие
корма; среди таких кормов в пробах отмечались личинки пластин-
чатоусых, гусеницы чешуекрылых, звонцы и пр. По данным
же В. Д. Сиохина, в пище взрослых птиц изучаемого вида в усло-
виях Присивашья и Молочного лимана преобладали компоненты
с твердыми покровами. По характеру окраски среди животных
компонентов основную часть составили субстратные формы
(63,9%) — прямокрылые, многие полужесткокрылые, ряд видов
чешуекрылых, двукрылых и др. Это объясняется тем, что именно
среди таких животных преобладали подвижные формы. Среди
объектов с контрастной окраской преобладали малоподвижные
и неподвижные формы — личинки и куколки чешуекрылых,
двукрылых жесткокрылых и т. п.
Спектр линейных размеров пищевых компонентов птенцов
оказался довольно широким — от 2 мм (моллюск Bittium reticu-
latum Costa, муравей Tapinoma kirnburni К. Arn.) до 120 мм (коль-
чатая сколопендра Scolopendra cingulata L.). Из приведенного гра-
фика (рис. 1) видно, что преобладали компоненты размерами от 9,1
до 12,0 мм; их было 55%. На графике виден еще один небольшой
пик (10,2%), соответствующий размерам 18,1—21,0 мм. Первый
пик объясняется тем, что мезофауна исследуемых экосистем от-
личается доминированием компонентов именно этих размеров.
Второй пик получился за счет отдельных видов саранчевых, жест-
кокрылых, а также гусениц лугового мотылька.
Самым главным аспектом изучения значения того или иного
вида в исследуемых экосистемах является прежде всего опреде-
ление соотношения компонентов его питания по признаку трофи-
ческой специализации. Безусловно, такие расчеты следует про-
водить в весовых показателях, но поскольку мы ими в достаточной
мере не обладаем, то нам приходится оперировать количеством
особей в выделенных трофических группировках. В корме черно-
138
Рис. 1. Количественное
распределение пищевых
компонентов по линейным
размерам
7*
Я
Л7
40
J0
20
/О
З.Н 0,М 0,М2 1ZtHf 70,НО 70,1-21Z1,HV Z4J-Z7
мм
Рис. 2. Трофическое воздействие
черноголовой чайки на биокомпо-
ненты экосистем в гнездовой период
АТ — автотрофы;
ГТ — гетеротрофы;
СТ — сапротрофы;
ФТ — фитотрофы;
зф — зоофаги;
пф — пантофаги;
фф — фитофаги;
фаф — афаги фитотрофного происхож-
дения;
сф — сапрофаги;
саф — афаги сапротрофного происхож-
дения
головых чаек преобладали афаги (38,9%), т. е. онтогенетические
фазы животных, практически не принимающие пищи либо пол-
ностью обходящиеся без нее. Это имаго лугового мотылька, звон-
цов, куколок насекомых с полным превращением и т. д. Второе
место занимают фитофаги (35,7%), третье — пантофаги (20,6%).
В незначительном количестве обнаружены зоофаги (1,0%)
и сапрофаги (0,4%). Помимо этого, зарегистрированы рассмот-
ренные выше растительные корма (3,4%). Если учесть, что ука-
занные афаги на других стадиях своего развития потребляют либо
растительную пищу (гусеницы лугового мотылька), либо яв-
ляются сапрофагами (личинки звонцов), то картина воздействия
черноголовой чайки в гнездовой период на компоненты населяю-
щие его экосистемы значительно изменится (рис. 2). При этом
основную часть составят фитофаги (51,1%), за ними будут сле-
довать сапрофаги (23,8%) и пантофаги. Таким образом, исходя
из анализа полученных данных, можно судить о том, что в гнез-
довой период черноголовая чайка в наибольшей степени воздейст-
139
вует на гетеротрофов (фитофаги, зоофаги, пантофаги — 72,7%),
в меньшей степени — на сапротрофов и в незначительной сте-
пени — на автотрофов.
Практическое значение
Из всего списка объектов питания черноголовой чайки животного
происхождения (295 наименований) около половины (150) из-
вестны как вредители и переносчики заболеваний культурных
растений, и в количественном отношении они составили лишь
немногим менее половины (45,0%) общего объема проанализиро-
ванных проб. Из них основную часть (26,0%) составили чешуе-
крылые, главным образом за счет лугового мотылька (24,0%).
Из других вредителей более или менее многочисленными оказа-
лись прямокрылые (4,2%), пластинчатоусые (3,5%) и долгоно-
сики (1,0%). Из отдельных видов в большом количестве отмечены
маврская черепашка (7,0%) жужелицы родов Amara Bon. et Zab-
rus Clrv. (5,5%), кузьки Anisoplia Latr. (2,7%).
Если учесть, что колония черноголовой чайки в Черноморском
заповеднике насчитывает более 330 тыс. пар и добавить к ним при-
мерно 9—25% холостых птиц, то число добытых ими вредителей
превысит 100 млн особей за один день и 5 млрд за гнездовой период.
Из этого количества более 1,5 млрд особей приходится на долю
вредителей зерновых культур, среди которых основное место за-
нимают вредители пшеницы. Из обнаруженных в пищевых пробах
вредителей к ним относятся хлебная и некоторые другие жу-
желицы, клопы-черепашки и кузьки; кроме того, черепашки и ли-
чинки хлебных жужелиц часто препятствуют колошению хлебов.
Заключение
Данное сообщение не претендует на полное раскрытие рассмат-
риваемого вопроса. Главная его задача заключается в поисках эф-
фективных методик, направленных на возможно более детальное
изучение экологии черноголовой чайки через анализ сопоставле-
ния потребляемых им объектов по выделенным качественно-
количественным и морфоэкологическим признакам. В частности,
обнаружено, что при весьма обширном спектре корма (около 300
названий), свидетельствующем о широкой экологической пластич-
ности черноголовой чайки, более или менее обычными оказались
компоненты всего 16 названий, а массовыми — только 8.
Обобщение материала по признакам ярусно-биотопической
приуроченности объектов питания, с одной стороны, отразило
мозаику биотопов региона, а с другой — преимущественный сбор
корма из степных экосистем и агроценозов. Кроме того, оно фак-
тически подтвердило результаты визуальных наблюдений, касаю-
щихся сбора пищи большей частью с поверхности растений, в мень-
шей степени — из растительного опада, поверхности почвы и в нез-
начительной — из верхних слоев почвы и с водоемов. Ряд не менее
140
I; интересных и уже приведенных выводов сделан при сопоставлении
* компонентов по другим признакам — характеру распределения
популяций, численности, подвижности, онтогенетическим стадиям
суточной активности, окраске и линейным размерам.
.<.) Соотношение объектов по пищевой специализации наглядно
показало роль черноголовой чайки в трофической сети исследу-
5 емых экосистем. Прежде всего отмечено, что в гнездовой период
данный вид наибольшее воздействие оказывает на представителей
блока гетеротрофов, прежде всего на консументов первого порядка
(фитофагов). В практическом отношении около половины его ра-
циона составили вредители и переносчики заболеваний культур-
ных растений.
ЛИТЕРАТУРА
Клименко М. И. Материалы по фауне птиц Черноморского заповедника//Тр.
Черномор, гос. заповедника. 1950. № 1. С. 8—15.
SUMMARY
Trophic relations of the Mediterranean Gull during the nesting period at the Black
Sea State Reserve. Report II. The spatial-temporal charasteristics of trophic relations.
Ardamatskaya T. В., V a k а г e n k о V. I., P e t г u s e n k о A. A.
Based on the long-term studies at the Black Sea State Reserve, the materials on the
characters of the layer-biotopical confinement of food places, which had expressed
the mosaic of the regional biotopes and the principal food from the steppe ecosystems
and agrocenoses. During the nesting period near half of the diet of the Mediterranean
Gull contained pests and carriers of cultural plant deseases.
УДК 591. 525: 598. 813
А. И. МОРОЗКИН,
В. Д. КАПИТОВ
422523, Татарская АССР,
Зеленодольский район.
Волжско-Камский
государственный заповедник
Изменение
эмбриональной смертности
в связи с многолетней
динамикой численности
у мухоловки-пеструшки
(Ficedula hypoleuca)
Экология мухоловки-пеструшки представлена достаточно большим
количеством исследований, в которых анализируются средняя ве-
личина кладки, фенология гнездования, интенсивность размно-
жения, возрастная, пространственная структура популяций.
Однако в этих работах крайне редко приводятся данные, связанные
с многолетней динамикой популяций (Карпович, 1962; Зимин,
1972; Лихачев, 1978; Jarvinen, 1980; Jarvinen, Ulimaunu, 1984;
Virolainen, 1984). Исследования, выполненные на других видах
животных организмов, говорят о том, что такие сведения интересны
4
141
и необходимы, поскольку являются одним из основных источников
при изучении эволюционных преобразований в популяциях.
Весьма показательны в этом отношении результаты Ю. Т. Артемь-
ева (1980), полученные на популяции рыжей полевки. Автор
на достаточно большом по объему и охватывающем значительный
промежуток времени материале показал существование вполне
определенной связи между изменениями эмбриональной смерт-
ности и динамикой численности населения популяции у изучаемо-
го вида. В настоящей работе мы пытались проследить аналогичную
зависимость в популяции мухоловки-пеструшки, обитающей
на территории Раифского лесничества Волжско-Камского госу-
дарственного заповедника, по данным 20 лет наблюдений.
Материал и методика
Материал для настоящего сообщения собирался с 1963 по 1983 г.
на территории Раифского лесничества Волжско-Камского запо-
ведника, расположенного в 25 км к северу от Казани. Наблюдения
проводились сотрудниками заповедника: с 1963 по 1967 г.
Е. П. Кнорре, с 1968 по 1969 г. В. А. Ушаковым и М. М. Ушаковой,
в 1970 г. Е. П. Кнорре и В. Д. Капитовым, с 1971 по 1976 г.
А. И. Сергеевым, с 1977 по 1983 г. А. И. Морозкиным. Исследова-
лась популяция мухоловки-пеструшки, гнездящаяся в сосняках
бруснично-мшистом с примесью березы и бруснично-злаковом
(Порфирьев, 1968).
Анализировались гнезда, расположенные в искусственных гнез-
довьях (синичниках), развешивание которых на территории лес-
ничества осуществлялось с 1962 по 1978 г. В пределах района оби-
тания изучаемой группировки птиц, как правило, ежегодно нас-
читывалось не менее двухсот пригодных для заселения синич-
ников. В 1963, 1970, 1980—1983 гг. число синичников в районе
гнездования популяции даже превышало цифры, приведенные
в таблице, однако проверить все имеющиеся в наличии гнездовья
не было возможности. За 1966, 1968, 1979 гг. выборки данных по
размножению мухоловки-пеструшки были небольшими, поэтому мы
сочли возможным объединить их с материалами предшествующих
лет, поскольку в «сдвоенные» годы 1965—1966 и 1978—1979 числен-
ность различалась на недостоверном уровне. Эмбриональный отход
обыкновенно охватывал яйца с погибшими птенцами при выводе,
задохликами, неоплодотворенные и элиминируемые по причине
гибели их зародышей на самых ранних стадиях развития. Анали-
зировались яйца в тех гнездах, судьба которых была прослежена
от откладки яйца до вылупления птенцов. Гнезда, разоренные и
брошенные с неполными кладками, при определении величин
эмбриональной гибели и плодовитости в расчет не принимались.
Вместе с тем эти кладки включались в обработку при вычислении
процента заселенности. Всего проверено 4388 гнездовий, обработа-
но 1026 кладок, 6245 яйца. Все материалы сведены в таблице и
проиллюстрированы на рис. 1.
142
-ff66 ~/Ш -fw
Рис. 1. Изменения численности (У), средней величины кладки (2) и эмбриональной
гибели (3) в популяции мухоловки-пеструшки Волжско-Камского заповедника в
1963-1983 гг.
Рис. 2. Климадиаграмма по годам в Раифском лесничестве Волжске- Камского запо-
ведника
1 — среднемесячная температура воздуха в июне; 2 — сумма количества осадков за месяц
в соотношении с температурой воздуха 2:1; 3 — то же, в отношений 3:1
Анализ погодных условий в период размножения проводился
путем графического сопоставления среднесуточной температуры с
величиной осадков по Госсену (Bagnoulus, Gaussen, 1953; Gaussen,
1955). При построении климадиаграммы (рис. 2) использованы
первичные материалы местной метеорологической станции. Клима
диаграмма составлена на основании данных среднесуточных тем-
ператур воздуха и сумм количества осадков по годам за июнь, так
как в это время идет откладывание и насиживание яиц, а также
осуществляется вывод птенцов значительного числа особей мухо-
ловки-пеструшки Раифского лесничества.
143
Изменение абсолютных показателей численности и величин проанализированных
кладок в популяции мухоловки пеструшки, гнездящейся в искусственных гнез-
довьях Раифского лесничества
Годы Кол-во прове- ренных гнездовий Кол-во засе- ленных гнездовий Кол-во проана- лизированных кладок Средняя величина кладки
1963 102 30 24 6,3±0,2
1964 228 72 61 5,8±0,1
1965-1966 325 115 63 6,4±0,1
1967-1968 243 102 67 6,2±0,1
1969 283 171 85 5,9±0,1
1970 105 52 34 6,1 ±0,2
1971 451 130 70 5,8±0,1
1972 391 192 102 6,0±0,1
1973 434 173 107 6,3±0,1
1974 501 175 88 6,0±0,1
1975 524 206 178 5,8±0,1
1976 593 214 109 5,7±0,1
1977 521 235 70 6,0±0,2
1978-1979 833 411 55 6,4±00,1
1980 220 117 61 6,4±0,2
1981 224 114 59 6,7±0,2
1982 177 62 57 6,4±0,2
1983 162 43 38 6,5±0,1
Результаты наблюдений и их анализ
На рис. 1 представлены кривые динамики численности и плодови-
тости, а также гистограмма, отображающая изменение эмбрио-
нальной гибели в популяции мухоловки-пеструшки. Путем парал-
лельного анализа величин каждого из приведенных экологических
параметров попытаемся проследить характер их динамики за все
годы исследования.
Плотность населения. В 1963—1964 гг. заселенность синични-
ков, как видно из таблицы и рис. 1, находилась на низком уровне.
С 1965 по 1969 г. численность популяции интенсивно увеличилась.
В 1970 г. заселенность искусственных гнездовий мухоловками
заметно снизилась. Начавшийся спад численности продолжался
и в 1971 г. В 1972 г. произошло кратковременное увеличение плот-
ности населения, затем наступила четырехлетняя депрессия. Засе-
ленность синичников в этот период была невысокой и составляла
около 37,4%. Начальная точка (45,1%) очередной фазы роста
населения мухоловки-пеструшки приходится на 1977 г. В сле-
дующие четыре года заселенность была постоянной и находилась
па высоком уровне. Очередное снижение численности отмечено в
1982 г. В 1983 г. показатель заселенности по сравнению с 1982 г.
существенно не изменился.
Средняя величина кладки jb. годы исследования изменялась в
144
незначительных пределах — от 5,7±0,1 в 1976 г. до 6,7±0,2 в
1981 г. Колебания этого показателя в отличие от динамики носили
незакономерный, случайный характер. Так, в первой волне измене-
ния плотности населения (1965—1971 гг.) параллельное увели-
чение плодовитости и численности происходило только в
1965-1966 гг.
В последующие сезоны размножения средняя величина кладки
в противоположность заселенности уменьшается. В годы спада и
депрессии численности (1970—1976 гг.) высокие значения пло-
довитости отмечены в течение четырех лет (1970, 1972 — 1974 гг.),
в другие годы она была на низком уровне. Во второй волне колеба-
ния плотности населения (1977 —1983 гг.) средняя величина клад-
ки как в фазе подъема, так и в момент спада численности была
выше средней.
Эмбриональная смертность колеблется, последовательно и зако-
номерно; ее колебания во многом повторяют ход динамики числен-
ности населения. Так, увеличение эмбриональной элиминации, как
и повышение численности, началось в 1965—1966 гг. Рост смертно-
сти эмбрионов в популяции длился до 1970 г. Весной 1971 г.
в популяции произошло заметное снижение доли неполноценных
яиц. При этом уменьшение элиминации эмбрионов осуществлялось
при минимальной плотности населения. Следующий небольшой
подъем численности в 1972 г. сопровождался достоверным увеличе-
нием гибели яиц. В дальнейшем эмбриональная гибель, как и
численность, в течение четырех лет держалась на низком уровне.
Начало очередного подъема гибели на ранних стадиях развития
приходится на 1977 г. В этом году, как мы уже отмечали, происхо-
дило и увеличение численности популяции. Дальнейшее увели-
чение элиминации продолжалось до 1981 г.; в 1982 г. величина
гибели на ранних стадиях развития имела такое же значение, как
и в предыдущем. В 1983 г. произошло снижение ее: по сравнению
с 1982 г. она понизилась более чем в два раза. Уменьшение доли
гибели неполноценных яиц происходило, как и в 1971 г., при мини-
мальной плотности населения.
Изменения численности, плодовитости и эмбриональной
смертности, несомненно, происходили в связи с изменениями опре-
деленных факторов внешней среды. По-видимому, особое внима-
ние следует обратить на характер биотопов обитания и погодные
условия в момент размножения популяции.
Поданным К. Н. Благосклонова (1953), Г. Н. Лихачева (1955)
и В. Н; Карновича (1962), мухоловка-пеструшка одинаково охотно
гнездится во всех основных типах лесов средней полосы Европей-
ской части РСФСР: в сосняках, ельниках, дубравах, березняках,
липняках, предпочитая при этом средневозрастные насаждения
(50—100 лёт). Как мы уже отмечали, изучаемая популяция насе-
ляет сосняки бруснично-мшистый с примесью березы и бруснично-
злаковый, возраст которых составляет от 50 до 140 лет, так что
биотоп популяции приближался к оптимальному варианту.
Особенности погодных условий в момент размножения популя-
145
ции показаны на рис. 2. На основании июньских данных средне-
суточной температуры и суммы количества осадков все годы
наблюдения можно подразделить на пять групп: 1) 1966 — 1968,
1973 и 1981 — засушливые годы (экстремальные условия);
2) 1964, 1965 и 1975 — годы с незначительным понижением влаж,-
ности (условия, близкие к экстремальным); 3) 1969, 1978, 1980 —
1983 — годы с повышенной влажностью (экстремальные условия);
4) 1963, 1972, 1976—1982 — годы с незначительным понижением
влажности (условия, близкие к экстремальным); 5) 1970, 1971,
1974 и 1977 — годы, когда сумма количества осадков и средне-
суточная температура июня были близки к средним многолетним
(нормальные условия).
Большинство исследователей (Кнорре, 1947; Тауринып, 1951;
Тауринып, Тима, 1953; Лихачев, 1953; Данилов, 1959; Езерскас,
1959; Jarvinen, Vaisanen, 1984; Virolainen, 1984) указывали на то,
что чем ближе к экстремуму значения температуры и влажности,
тем хуже обеспечены условия размножения мухоловки-пеструшки.
Наряду с этим в работах В. Н. Карповича (1962), Т. Ю. Хохловой
и В. И. Головань (1981) приведены сведения, которые, на наш
взгляд, могут служить показателем того, что не всегда успешность
размножения популяции согласуется с ходом среднесуточных
температур воздуха и количества осадков в период репродукции.
Если принять за основу первую точку зрения, наиболее распро-
страненную среди экологов, тогда в нашем случае при неблаго-
приятных условиях может быть выше величина гибели эмбрионов
и ниже число самок, приступивших к репродукции.
Для выяснения возможности такой зависимости сравним между
собой кривую динамики численности и гистограмму эмбриональ-
ной гибели с климадиаграммой (см. рис. 2). В качестве примера
возьмем 1969 и 1970 гг. Июнь 1969 г. выдался холодным и дождли-
вым, осадков выпало в 2,5 раза выше нормы, температура была на
3° ниже средней многолетней. В ночное время температура часто
падала до -4-3, +4°. В июне 1970 г. количество осадков (58 мм)
и среднесуточная температура воздуха (16,5°) были очень близки к
средней многолетней норме (55 мм и 16,8°). Следовательно, по-
годные условия периода размножения в 1970 г. были более благо
приятны, чем в 1969 г.
Однако, вопреки ожиданию, в 1970 г. эмбриональная смертность
оказалась выше, а численность особей ниже. Погодные условия
июня 1971 г, также были оптимальными. Тем не менее 1971 г. от-
личался наименьшими численностью и уровнем гибели яиц. Если
мы попытаемся описать дальнейший ход кривых на рис. 1 и 2,
то получим результат, подобный приведенным примерам. Все это,
очевидно, говорит об отсутствии прямых связей между абиотиче-
скими факторами среды (среднесуточная температура воздуха,
сумма количества осадков) и динамикой численности и колеба-
ниями эмбриональной гибели мухоловки-пеструшки.
146
Обсуждение
Как показал анализ результатов, колебания численности и эмбрио-
нальной смертности не имеют выраженной зависимости от погод-
ных условий в период размножения. Вместе с тем известно, что
в среднем гибель взрослых мухоловок в течение одного между-
гнездового периода на большей части ареала вида составляет 55%
(Карпович, 1962; Nilsson, 1984). Все это свидетельствует, видимо,
о том, что не погодные условия в момент размножения, а особен-
ности осенних и весенних миграций, а также характер протекания
зимовок являются основными факторами, на фоне которых форми-
руются приспособительные возможности популяции. Когда пере-
леты и зимовки проходят в благоприятных условиях, ослабляется
действие естественного отбора. В итоге выживают особи с понижен-
ной жизнеспособностью, отягощенные различными отрицательны-
ми наследственными факторами. Напротив, при неблагоприятных
условиях в течение года особи с пониженной жизнеспособностью
должны вымирать. Популяция после этого может состоять из
особей с высокой жизнеспособностью, отличаться минимальным
содержанием летальных наследственных факторов. Эти генети-
ческие особенности должны теоретически отражаться в измене-
ниях величины эмбриональной гибели (Артемьев, 1981). Исходя из
этого, колебания численности и гибели яиц можно объяснить сле-
дующим образом. В 1963—1964 гг. численность была низкой. Это,
видимо, было связано со значительной гибелью птиц во время
перелетов. Популяция, вероятно, состояла из особей с высокой
жизнестойкостью. Ей соответствовала низкая гибель яиц. С 1965
по 1969 г. численность нарастала. Это говорит о возможном ослаб-
лении естественного отбора и накоплении в популяции за счет
действия поступательности популяционного размножения (Арте-
мьев, 1980, 1981) особей с наследственными нарушениями. Дей-
ствительно, в эти годы наблюдается последовательное нарастание
эмбриональной смертности. Судя по величине эмбриональных
потерь, в 1970 г. популяция обладала самой низкой жизнеспособ-
ностью в первой волне численности. Возможно, этим объясняется
тот факт, что после зимовок 1969/70 и 1970/71 гг. к размножению
в 1971 г. приступило в 2,1 раза меньше особей, чем в 1969 г. Мало-
жизнеспособная часть птиц, вероятно, погибла при перелетах. При
анализе особенностей популяции выяснилось, что она в период раз-
множения весной 1971 г. состояла из птиц, имевших низкие величи-
ны эмбриональных потерь. В 1972 г. произошло новое повышение
численности, что говорит о возможном улучшении условий пере-
летов и зимовок 1971 — 1972 гг. Наблюдаемое ослабление действия
отбора дало возможность поступательности размножения за ко-
роткий срок несколько повысить концентрацию наследственных
аномалий в популяции. Подтверждением этому служит отмеченная
весной 1972 г. повышенная гибель яиц. В следующие четыре года
численность и эмбриональная гибель были крайне низкими. Вполне
допустимо, что значительная элиминация взрослой части населе-
147
ния птиц была связана с возможными экстремальными условиями
зимовок и перелетов. Очередное повышение численности насе-
ления произошло в 1977 г. Подобные перемены были, по-видимому,
связаны с улучшением условий зимовок и перелетов, что привело
к ослаблению отбора. Его ослабление создало предпосылки для
последовательного накопления в популяции особей с пониженной
устойчивостью к отрицательному влиянию условий существования
под действием поступательности размножения. Это наглядно про-
явилось через интенсивное повышение в эти годы эмбриональной
смертности. В 1981 и 1982 гг., судя по максимальному количеству
погибших яиц, популяция состояла из птиц, жизнеспособность
которых была самой низкой за все годы наблюдения. Очевидно
по причине невысокой приспособленности, после зимовок 1981/82
и 1982/83 гг. к размножению в 1983 г. приступило в 2 раза меньше
особей, чем в 1981 г. Недостающая часть птиц с низким адаптивным
потенциалом, вероятно, погибла при перелетах. При оценке эколо-
гических особенностей популяции выяснилось, что она в момент
размножения в 1983 г. состояла из птиц, которые имели невы-
сокий показатель гибели на ранних стадиях развития, т. е., видимо,
обладали повышенной жизнеспособностью.
Таким образом, популяция мухоловки-пеструшки в течение
20 лет была охвачена двумя волнами динамики численности. В ходе
исследования выяснилось, что динамика ее численности не связана
с ходом изменения среднесуточных температур воздуха и количе-
ства осадков в момент размножения. Вместе с тем параллельное
изучение величины гибели на ранних стадиях развития, отражаю-
щей приспособительные возможности популяции, показывает, что
этот экологический показатель очень часто совпадает с изменения-
ми численности. Как видно из рис. 1, повышение или понижение
численности, как правило, сопровождается противоположными
изменениями в показателях жизнеспособности. Отсюда следует,
что ранняя элиминация, как и показатель плотности, очевидно, мо-
жет быть использована в качестве дополнительной характеристики
при изучении популяционной динамики мухоловки-пеструшки.
ЛИТЕРАТУРА
Артемьев Ю. Т. Биология размножения. Казань: Изд-во Казан, ун-та, 1980. 87 с.
Артемьев Ю. Т. Метод зародышевых деталей при микроэволюционном изучении
природных популяций млекопитающих//Микроэволюция. Казань, 1981.
Вып. 1. С. 5-30.
Благосклонов К. Н. Особенности гнездования птиц-дуплогнездников в искусствен-
ных гнездовьях // Перелеты птиц в Европейской части СССР. Рига: Изд-во
АН ЛатвССР, 1953. С. 10-11.
Данилов Н. Н. Размножение мухоловки-пеструшки на Среднем Урале//Вторая
Всесоюз. орнитол. конф.: Тез. докл. М., 1959. Т. 2. С. 106.
Езерскас Л. И. Птицы-дуплогнездники юго-восточной части Литвы / Там же.
1959. Т. 2. С. 110.
Зимин В. Б. Экология размножения мухоловки-пеструшки в Южной Карелии //
Экология. 1972. № 5. С. 23-29.
Карпович В. В. Экология массовых обитателей искусственных гнездовий (скворца,
мухоловки-пеструшки) в районе Окского заповедника//Тр. Окского гос.
заповедника. 1962. Вып. 4. С. 65—176.
148
Кнорре Е. II. Опыт привлечения в очаги сосновой пяденицы насекомоядных птиц
как мера борьбы с этим вредителем // Научно-методические записки. М.:
Гл. упр. по заповедникам, 1947. Вып. 9. С. 25—26.
Лихачев Г. Н. Наблюдения над размножением мухоловки-пеструшки в искусствен-
ных гнездовьях // Бюл. МОИП. Отд. биол. 1953. Т. 8, вып. 2. С. 15—17.
Лихачев Г. Н. Мухоловка-пеструшка (Ficedula hypoleuca L.) и ее связь с гнездовой
территорией // Тр. Бюро кольцевания. М„ 1955. Вып. 8. С. 40—43.
Лихачев Г. Н. Размножение и численность мухоловки-пеструшки на юге Москов;
ской обл. // Птицы и пресмыкающиеся: (Исследования по фауне Советского
Союза). М., 1978. С. 18-19.
Порфирьев В. С. Растительность Райфы // Тр. Волж.-Кам. заповедника. 1968.
Вып. 1. С. 106—136.
Тауринъш Э. Материалы по биологии мухоловки-пеструшки Ficedula hypoleuca
на искусственных гнездовьях в сосновом бору в условиях Латвийской ССР //
Вторая экол. конф, по пробл. «Массовые размножения животных и их прогно-
зы»: Тез. докл. Киев, 1951. Т. 3. С. 101 —104.
. Тауриныи Э. Я., Тима Ч. Б. Наблюдения по биологии некоторых видов птиц,
гнездящихся в искусственных гнездовьях в условиях Латвийской ССР // Пере-
леты птиц в Европейской части СССР. Рига: Изд-во АН ЛатвССР, 1953. С. 70—72.
Хохлова Т. Ю., Головань В. И. К биологии размножения мухоловки-пеструшки в
Южной Карелии//Экология наземных позвоночных северо-запада СССР.
Петрозаводск, 1981. С. 50—62.
Bagnouls F., Gaussen Н. Saison seche et indice xerothermique //Docum. carles pro-
duct. veget. Ser. gen. 1953. T. 3, Vol. 1, art. 8. P. 1—48.
Gaussen H. Experession des milieux par des formules ecologiques: leur representation
cartographique//Colloq. intern. CNRS. 1955. Vol. 59. P. 257—269.
Jiirvinen A. Population dynamics in the Pied Flycatcher, Ficedula hypoleuca at
subarctic Kilpisjarvi Finnish Lapland // Ornis fenn. 1980. Vol. 57, N 1. P. 17—25.
larvinen A., Ulimaunu G. Significance of egg size on the growth of nestling Pied
Flycatcher Ficedula hypoleuca//Ann. Zool. Fenn. 1984. Vol. 21. P. 213—216.
larvinen A., Vaisanen A. Reproduction of Pied Flycatcher Ficedula hypoleuca in good
and bad breeding seasons in a Northern marginal area//Auk. 1984. Vol. 101.
P. 439-450.
Nilsson S. G. The evolution of nest-site selection among holenestling birds: the
importance of nest predation and competition // Ornis Scand. 1984. Vol. 15.
P. 167-175.
Virolainen M. Breeding biology of the Pied Flycatcher Ficedula hypoleuca in relation
to population density//Ann. Zool. Fenn. 1984. Vol. 21. P. 187 — 197.
SUMMARY
The change of the embryonal mortality in connection with the long-term dynamics
of the Pied Flycatcher (Ficedula hypoleuca). Morozkin A. I., Kapi-
t о v V. D.
It is an attempt, undertaken for tracing the long-term changes of egg’s destruction
on the background of fluctuations of the Pied Flycatcher population in Raif Forestry
of Volga—Kama State Reserve during 20 years (1963—1983). The index of eggs
destruction included: elimination on the early stages of development,
unfertilized eggs, eggs with chokes, death of broods. The size of the population was
determined by counting the number of Titmice nesting pairs living man-made hollows
from the number of suitable for nesting man-made hollows as a whole. At this time
there were two cycles of size fluctuation and embryonal mortality in the population.
It was succeeded to find a well-expressed connection between changes in the popula-
tion and embryonal mortality. The density of the population grew, the embryonal
mortality increased, but during the depression years it stayed on the minimum level.
It follows, evidently, that the peculiarities of the early ontogenesis are closely connected
with the population dynamics, and the researches on the population level should be
carried out with regard to the population phase.
149
УДК 574.91:598.422
п. С. томкович
103009, Москва,
ул. Герцена, д. 6.
Зоологический музей
МГУ им. М. В. Ломоносова
К вопросу
о внегнездовых перемещениях
белых чаек
(Pagophila eburnea)
Сведения о миграциях белых чаек исключительно скудны, посколь-
ку эти птицы почти круглый год придерживаются паковых льдов
незамерзающих северных морей. Кольцевание белых чаек в Грен-
ландии, близ Лабрадора и в 1930-х годах на Земле Франца-Иосифа
дало несколько возвратов, которые позволили заключить о суще-
ствовании контактов между белыми чайками, посещающими Лаб-
радор, Восточную Гренландию, Шпицберген и Землю Франца-
Иосифа (Дементьев, 1951; The birds..., 1983). Перемещения белых
чаек в пределах приатлантического сектора Арктики подтверждаю-
тся еще двумя возвратами колец, имеющимися в Центре кольце-
вания АН СССР. Чайки, помеченные птенцами 25 июля и 3 августа
1960 г. на о-ве Виктория (между Шпицбергеном и Землей Франца-
Иосифа) обнаружены: первая — 3 марта 1965 г. на р. Кия на п-ове
Канин, вторая — 8 августа 1966 г. на Земле Александры в архипе-
лаге Земля Франца-Иосифа. Вместе с тем до настоящего времени
остается невыясненной популяционная принадлежность белых
чаек, зимующих в заметном числе в Беринговом море (Косыгин,
1975).
В 1981 г. нам удалось отловить и окольцевать 46 взрослых бе-
лых чаек в период предгнездовых кочевок на о-ве Греэм-Белл, Зем-
ля Франца-Иосифа (Томкович, 1986), от одной из этих птиц по-
лучен неожиданный возврат. Самец, окольцованный 17 июня 1981 г.
(он имел большие формирующиеся наседные пятна) отловлен в
январе 1984 г. возле протоки из лагуны Кэйнгыпильгин в Берин-
говском районе Чукотского автономного округа. Охотник-про-
мысловик В. В. Радивилов любезно сообщил, что в январе того года
во время тайфуна у побережья Анадырского залива появилось
большое число чаек (особенно серебристых Larus argentatus);
в капканы на песцов на участке побережья от мыса Гинтера до мы-
са Гека оказались пойманными около 400 чаек, в том числе упомя-
нутая белая чайка с кольцом. Этот факт, во-первых, доказывает
возможность широких кочевок белых чаек не только в пределах
приатлантического сектора Арктики, но по всей Арктике и, во-
вторых, отчасти проясняет вопрос о происхождении птиц, зи-
мующих в Беринговом море. Насколько регулярно белые чайки
из Европы оказываются на зимовках в Беринговом море, пока
неясно, однако можно предположить, что эти чайки «оттесняются»
наступающими зимними условиями к югу в северные части либо
Атлантического, либо Тихого океанов, в зависимости от того, где
птицы были застигнуты во время своих позднеосепних кочевок
по Арктике.
150
ЛИТЕРАТУРА
Дементьев Г. П. Отряд Чайки // Птицы Советского Союза. М.: Сов. наука, 1951.
Т. 3. С. 372-602.
Косыгин Г. М. Белая чайка на Чукотском полуострове // Колониальные гнез-
довья околоводных птиц и их охрана. М.: Наука, 1975. С. 176.
Томкович П. С. Материалы по биологии белой чайки на о-ве Греэм-Белл (Земля
Франца-Иосифа) //Актуальные проблемы орнитологии. М.: Наука, 1986.
С. 34-49.
The birds of the Western Palearctic. Oxford; London; New York, 1983. Vol. 3. 913 p.
SUMMARY
On the question of out-of-nest movements of Ivory Gulls (Pagophila eburnea).
Tomkovich P. S.
Banding of 46 adult specimens of Ivory Gulls was carried out in 1981 during the
period of pre-nesting flights to Gream Bell Island, Franz Iosif Land. The second
recover of a male in January 1984 is from the Bering District of the Chukotka Auto-
nomous Okrug. This fact shows the possibility of wide movements of Ivory Gulls
all over the Arctics.
УДК 591.543.43:598.9
В. И. ПЕРЕРВА 142790, Московская обл., Ленинский р-н, n/о ВИЛ АР. С адки-Знаменс кое. ВНИИ охраны природы и заповедного дела Госкомприроды СССР Изменение экологии хищных птиц под влиянием антропогенных факторов
Одной из форм экологического мониторинга является сравнение
экологии пернатых хищников, обитающих в биоценозах, макси-
мально приближенных к естественным (например, в заповедни-
ках), и на территориях, подверженных разнообразным антропо-
генным преобразованиям.
Рассмотрены реакции хищных птиц на хозяйственное преобра-
зование их кормовых и гнездовых биотопов, а также на фактор
беспокойства. Наблюдения проводились в двух регионах: в Север-
ном Казахстане и в Европейском центре СССР в 1972 — 1979 гг.,
где были избраны четыре стационара — Терсекский заповедный
(площадь 60 км2) в первом; Пустынский (50 км2, Горьковская
обл.), Приокско-Террасный заповедный (50 км2) и «Лесничество»
(40 км2, оба на юге Московской обл.) во втором. Начальным эта-
пом экологических исследований являются учеты численности,
которые проводились методом картирования встреч, поиска гнезд
и описания гнездовых территорий (Галушин, 1971). В Северном
Казахстане в колковых сосновых борах было проведено сплошное
обследование лесных участков с поиском гнезд, а в ряде случаев —
151
Таблица 1
Численность хищных птиц исследованных стационаров
(число гнездящихся пар на 100 км2 всех угодий)
152
вылетевших выводков. За время наблюдений описано более 300
гнезд и зарегистрировано 2,6 тыс. встреч хищных птиц 12 видов.
Материалы учетов позволили сделать пересчет на площадь в
100 км2 всех угодий (табл. 1). Суточная активность и частота
приноса добычи хищниками определялись во время наблюдений
у гнезд (9 сут) и механическим автографом (Перерва, 1977), кото-
рый работал в общей сложности 70 сут на нескольких гнездах
пяти видов птиц.
Одной из первых реакций хищных птиц на воздействие антро-
погенных факторов является изменение поведения, обусловленное,
в частности, различным их отношением к гнездовой территории при
обитании в сплошных лесах или в колках, окруженных открытыми
пространствами. Охрана границ всей гнездовой территории, выра
жающаяся в облете ее периметра, конфликтах в зоне соприкосно-
вения территорий соседствующих пар, имеет место в обширных
лесных массивах. Однако в мозаичных биотопах, возникающих
после рубок, хищные птицы ведут себя по-другому. Создается си-
туация, аналогичная взаимоотношениям птиц со средой обитания
в островных лесах в Северном Казахстане (Перерва, 1984) или
прериях Северной Америки (Schmutz et aL, 1980). При дефи-
ците пригодных для гнездования биотопов, где к тому же повы-
шается плотность гнездящихся пар, окружающие их открытые
пространства становятся единой охотничьей зоной. Здесь одна или
даже обе птицы пары уже не в состоянии обеспечить сохранность
достаточно обширного кормового участка от вторжения соседей.
В связи с этим снижается агрессивность птиц в охотничьей зоне,
но сохраняется в местах гнездования, т. е. вблизи гнезд (табл. 2).
Агрессивность хищных птиц была определена как процентное
соотношение числа птиц, участвовавших в конфликте, к общему
количеству всех встреч за весь период наблюдений. Судя по полу-
ченным результатам, в охотничьих зонах; как вокруг колков в
Северном Казахстане, так и вдоль края леса в Московской области
над сельскохозяйственными угодьями агрессивность хищных птиц
была значительно ниже, чем над гнездовыми биотопами. Такой же
результат был получен Ф. Трильмихом (Trillmich, 1969) в усло-
виях антропогенного ландшафта близ г. Хильдесгейма в ФРГ.
Расположение заповедных территорий вблизи различных
сельскохозяйственных угодий дало возможность сравнить прёдпо-
читаемость этих биоценозов хищными птицами. На Терсекском
стационаре в утренний период подъема суточной активности пер-
натых хищников за час над целиной встречалось в среднем 14,2
птиц, а над соседними полями пшеницы — 8,8. В остальное время
суток над угодьями первого типа встречалось 7,6, а второго — 7,1
экз. Однако различные типы угодий неодинаково привлекательны
для хищных птиц (см. рисунок): на стационаре «Лесничество»
над пастбищами пернатые хищники, особенно пустельга, встре-
чались чаще. Посевы многолетних трав по предпочитаемости лишь
немногим уступают пастбищам, они даже более привлекательны
для канюка и черного коршуна.
153
Таблица 2
Показатели агрессивности хищных птиц в различных биотопах
и территориальных подразделениях среды их обитания
S
к
и
3
в
S'
S
и
154
/ z 3 V
Ъ па хата © пастбище % бсбаемь/ О посети
Встречаемость хищных птиц над различными био- и агроценозами стационара
«Лесничество»
1 — канюк обыкновенный; 2 — обыкновенная пустельга; 3 — черный коршун; 4 — суммар-
но по всем видам
Хищные птицы совершенно избегают коренным образом изме-
ненных пространств, например участков, перепахиваемых на
летнее время. В этой связи показательны наблюдения на сельхоз-
угодьях, примыкающих с северо-востока к Приокско-Террасному
заповеднику. Обычно здесь косят основную часть запасов сена,
заготавливаемого для зубров. Периодически, раз в 3—5 лет на этой
территории проводят подсев многолетних трав. В 1978 г. эту пло-
щадь перепахали в начале июня, в связи с чем встречаемость
хищных птиц над ней составила в среднем 2,1 экз. за час. В те же
сроки в 1977 и 1979 гг. отмечалось соответственно 4,2 и 3,5 особи.
Коренным образом измененные лесные биоценозы также не-
равнозначны для хищных птиц в качестве мест гнездования.
Уничтожение тех или иных типов леса существенно меняет биото-
пическую приуроченность их гнезд (табл. 3). В Подмосковье
первоочередная выборка во время рубок спелых березовых и сосно-
вых деревьев лишает пернатых хищников этих основных гнездо-
пригодных деревьев, тем самым вынуждая их занимать в большой
степени дуб и ель, использовать для гнездования средневозрастные
вторичные осиновые и березово-осиновые леса. Различия заповед-
ных популяций и обитающих на территории стационара «Лесни-
чество» хищных птиц выявлены в расположении гнезд относитель-
но земли (табл. 3). В Приокско-Террасном заповеднике они
устраиваются в среднем в 17 м от земли, что характерно для
всего центра Европейской части СССР (Галушин, 1971). Коле-
бания в данном случае могут быть в пределах 8—22 м. Уничто-
жение коренных лесов в стационаре «Лесничество» привело к
тому, что разница между низко и высоко расположенными гнезда-
ми здесь стала значительно большей — 6—30 м, а средняя вы-
сота 14 м.
Однако на размещение гнезд хищных птиц в этом стационаре
оказали влияние не только рубки. На наш взгляд, этому способ-
ствовал фактор беспокойства, создаваемый посещающими лес
людьми. В данной ситуации лучшая защищенность гнезд может
быть обеспечена за счет либо хорошей укрытости в густых зарослях
мелколесья (осинник, липняк), либо расположения в верхней
155
5
6
ф
А © .? - о 5 5'
и К я ь Я ф О Я Я ;
Q д
t£
Я я Ф № СО
я S С
еч Я ео § 4
с « S з
X ш з 5" a 2 З4
К о
5& Н «
<л Я ь я о о № от об ЧИС л гнезд
2 к О Ф £
° S J-J Я Р* S CQ
го т К ф
£ « я я £ С 1 Q ч Числе 1 гнезд
я J
а
“ я
я о я
Я я я
а © я
Я я а «
X « S
X и а
Я С о о
я ь я я я е
Л о W «
со Ч к Сб
GJ О о Я
Я ь я Ф
Я Я о Ф
я я я ES
ч я а
ко я Е а^1
н О п
Сосна I-1 ~ S I £ й i g « и с э 5 а Q
156
1
части 25—30-метровых сосен в еще сохранившихся небольших по
площади участках высокоствольного леса. Тот же принцип избе-
гания фактора беспокойства за счет гнездования на большей, чем
в естественных условиях, высоте отмечали Н. С. Панькин и
В. А. Дугинцев (1983) в Амурской обл. у мелких соколов и сов,
обитающих вблизи населенных пунктов и автострад.
Для хищных птиц беспокойство создают поселки, сеть дорог.
Индикатором такого рода антропогенного влияния является уда-
ленность гнезд от этих объектов. По интенсивности и продолжи-
тельности воздействия их необходимо разделить на две группы,
в первую входят населенные пункты сельского типа, во вторую —
все остальные объекты, включающие дороги, сенокосы, стоянки
туристов. Обобщение имеющихся данных по реакции хищных птиц
Европейской части СССР на перечисленные формы человеческой
деятельности (табл. 4) показало их неравноценность. Сельские до-
роги, пункты временного пребывания людей не влияют на распре-
деление гнезд. Именно в силу их непостоянства в воздействии на
окружающую фауну (временное функционирование дорог, появле-
ние людей лишь в определенные сроки) эти объекты не влияют на
распределение гнездящихся пар, но могут существенно снижать
успех размножения тех птиц, которые попали в сферу их действия
(Шепель, 1981; Галушин, 1982). Иное дело — населенные пункты,
которые оказывают определенное по силе, постоянное давление на
хищных птиц, населяющих их окрестности. Поэтому удаленность
гнезд пернатых хищников от поселков за редким исключением
достаточно велика, обычно 1—2 км. Такие же тенденции к избе-
ганию фермерских участков отмечены у канюков Северной Аме-
рики (Lokemoen, Duebbert, 1976). В таких случаях на размещение
гнездящихся пар влияет два типа факторов, противоположных
по направленности: наличие определенных видов добычи, что
обусловливает тяготение к населенным пунктам, и фактор беспо-
койства в сочетании с прямым преследованием, отпугивающие
птиц.
Исследования с помощью механического актографа, регистри-
ровавшего суточную активность хищных птиц у гнезда, позволили
определить влияние фактора беспокойства на ритмику посещения
птенцов взрослыми птицами.
На заповедной территории канюки в благоприятную теплую
погоду в любой день недели совершали 11 —12 прилетов к гнезду
и находились с птенцами от двух с половиной до пяти часов за вре-
мя дневной активности. Считая оптимальными эти цифры, можно
предположить, что уменьшение суммарного времени нахождения
взрослых птиц в гнезде и увеличение числа их прилетов — явле-
ние аномальное, вызванное фактором беспокойства. Продолжи-
тельное пребывание взрослых вместе с выводками в пасмурные
и холодные дни — необходимое условие благополучного выжива-
ния птенцов.
Леса стационара «Лесничество» становятся наиболее насыщен-
ными людьми в субботние и воскресные дни. В это время на каждый
157
Таблица 4
Удаленность гнезд хищных птиц Европейской части СССР
от различных объектов человеческой деятельности (км)
Вид объекта Населенные пункты
Наименование стационара Окский и Владимир- ский (Галу- шин, 1971) «Лесниче- ство» Приокско- Террасный заповед- ник Пермская обл. (Шепель, 1981)
Ястреб-тетеревятник 3,5 2,2 2,0 1 (3,0-4,0) (1,0-2,5) Черный коршун 2,0 1,2 2,0 (0,2-5,5) (1,0-1,5) (0,5-3,0) (0,3-0,8) Обыкновенный канюк 3,3 1,1 1,6 ? (0,2-8,0) (0,1-2,0) (0,3-4,0) Чеглок 1,9 — 1,0 1 (0,2-4,5) Обыкновенная пустельга 2,3 1,0 0,5 ? (0,01-1,0)
Вид объекта Дороги, сенокосы и стоянки
Наименование стационара Окский и Владимир- ский (Галу- шин, 1971) «Лесниче- ство» Приокско- Террасный заповед- ник Пермская обл. (Шепель, 1981)
Ястреб-тетеревятник — Черный коршун -|- -(- _|_ _|_ Обыкновенный канюк -|- — _|_ _|_ Чеглок -|- ? _|_ । Обыкновенная пустельга -|- _|_ _|_ ? Примечание: Верхний ряд цифр — среднее расстояние в км, нижние — крайние
вариации
квартал площадью около 1 км2 мы встречали 30—40 человек с 5—8
автомобилями. В такие дни, по наблюдениям в 1979 г. с помощью
актографа, гнездившаяся на территории этого стационара пара
канюков находилась с птенцами 20—25-дневного возраста в общей
сложности 3—5,5 ч в день. В будние дни это время увеличива-
лось до 10 ч. В субботу и воскресенье количество прилетов к
гнезду (часто без корма) значительно возрастало — в среднем до
17 раз в день, тогда как в остальные дни недели — 12 раз. Видимо,
необходимость обогрева птенцов в холодную и дождливую погоду
заставляет взрослых птиц находиться в гнезде больше времени.
Но обеспокоенные присутствием людей, они слетают с гнезда,
что нарушает определенный режим обогрева птенцов. В этой
связи показателен пример с парой канюков, гнездившихся всего в
150 м от шоссе и в 200—300 м от небольшого поселка. Здесь в любые
дни недели взрослые птицы задерживались у гнезда с птенцами
158
примерно 20-дневного возраста в общей сложности не более полу-
часа в день, а количество прилетов в дневное время колебалось от
13 до 16 раз.
Фактор беспокойства отражается и на успехе размножения
хищных птиц. На стационаре «Лесничество» — в двух гнездах
канюков, расположенных в наиболее посещаемом людьми ме-
сте вдоль шоссе, период гнездования сместился на 15—20
дней, в связи с чем в 1979 г. в этих гнездах птенцы вылупились
в конце июня — начале июля, тогда как в удаленных от населенных
пунктов гнездах — в первых числах июня. У канюков, гнездивших-
ся на территории Приокско-Террасного заповедника, в девяти вы-
водках было в среднем по 2,2 птенца, а в стационаре «Лесни-
чество» — 2 (по шести гнездам). Аналогичная закономерность в
снижении успеха размножения скопы, обусловленная фактором
беспокойства на некоторых открытых для туристов участках
Йеллоустонского национального парка в США, отмечена Е. Свен-
соном (Swenson, 1979).
Изменения поведения и биологии гнездования — первый этап
реагирования хищных птиц на антропогенные факторы. Вслед за
этим происходит изменение их численности. Результатом может
явиться либо исчезновение вида с данной территории, либо приспо-
собление к антропогенному прессу. Стационар «Лесничество» под-
вержен сильному воздействию фактора беспокойства и рубкам.
Общая численность хищных птиц здесь в четыре раз ниже, чем
в соседнем Приокско-Террасном заповеднике (см. табл. 1). В то же
время полученные в стационаре «Лесничество» данные лишь
незначительно отличаются от данных по другим незаповедным тер-
риториям, в частности по Пустынскому стационару. Несомненно,
общее обеднение фауны хищных птиц охватывает весь Европей-
ский центр СССР и отдельными «островами» в нем оказались
заповедные территории — Окский (Галушин, 1971, 1980) и Приок-
ско-Террасный заповедники. Здесь численность обычных пернатых
хищников удерживается в оптимальных для региона размерах.
Процесс обеднения фауны хищных птиц Северного Казахстана
начался в 1950-х годах. Естественно, что изменения в обилии пер-
натых хищников в окружающих Наурзумский заповедник биоце-
нозах не могли не сказаться на их численности. Для населения
орла-могильника в Наурзумском заповеднике в 1930—1950 гг.
была характерна стабильность численности и гнездовой консерва-
тизм (Тибет, 1960), а общее число гнездившихся птиц было
нескольно выше, чем в последующем (Осмоловская, 1953; Солома-
тин, 1974). Затем началось медленное снижение общего количества
орлов, а с конца 1960-х годов популяция стала подвергаться резким
флуктуациям численности (в 1969 и 1973 гг. в заповеднике гнезди-
лось всего несколько пар орлов). Падение обилия сменялись
подъемами. В 1972 г. общее их число было таким же, как и в 1966 —
1968 гг. (Соломатин, 1974) — более 12 пар. На начало 1980-х годов
в трех лесных участках Лаурзумского заповедника гнездилось
28—30 пар могильника (Брагин, 1983).
159
В 1938 г. на территории Наурзумского заповедника обитало
более 20 пар балобанов (Дементьев, 1951), а к середине 1960-х —
началу 1970-х годов — 15 пар и менее (Соломатин, 1974). В 1972 г.
на одном небольшом участке Наурзумского бора балобаны совсем
не гнездились, тогда как в 1946 и 1947 гг. здесь обитало 5—8 пар
(Тибет, 1960). В Терсекском бору их количество исчислялось 8 па-
рами в 1968 г., одной — в 1969 г., а в 1973 и 1974 гг.— 11 — 12 пара-
ми. Общая численность вида в заповеднике в 1980-е годы вновь
оценивается в 20—22 пары (Брагин и др., 1983). Динамика числен-
ности балобанов в Наурзумском заповеднике отражает общую тен-
денцию к сокращению популяции в 1960—1970-х годах в целом
по Северному Казахстану, но с той лишь разницей, что она менее
значительна в сравнении с незаповедными территориями.
Необходимо отметить одинаковую реакцию орлов-могильников
и балобанов на антропогенное воздействие, а именно смену периода
стабильности популяций снижением их численности, а затем появ-
ление резких флуктуаций. Можно предположить, что такой харак-
тер динамики численности обусловлен временным негнездованием
части населения видов в первые годы начавшегося постепенного
снижения их количества. При сохранении дестабилизирующих
условий может произойти исчезновение птиц из данного района.
Планомерная реализация природоохранных мероприятий, в пер-
вую очередь сохранение заповедного режима, стабилизирует со-
стояние популяций хищных птиц в заповедниках. Выявленные ко-
лебания численности других обычных видов пернатых хищников
обусловлены естественными факторами и не меняют общую карти-
ну стабильности всего фаунистического комплекса хищных птиц
Наурзумского заповедника. В этой связи можно говорить о его
значении для Северного Казахстана как резервата, способствующе-
го сохранению и расселению хищных птиц.
Таким образом, в результате сравнительного анализа поведения
и экологии хищных птиц в заповедниках и на территориях общего
пользования выявились определенные тенденции в реагировании
на антропогенные факторы. Относительно слабые преобразования
ландшафтов вызывают достаточно быструю адаптацию птиц.
Вырубка лесов, перепашка угодий влекут качественное изменение
способов использования хищными птицами таких ценозов. Это
сказывается на их поведении, характере гнездования, ритмике
насиживания, выкармливания, обогрева птенцов и т. д. Вслед за
изменением поведения и наиболее лабильных экологических пара-
метров дестабилизируется вся система «хищник—жертва», в ре-
зультате чего происходит падение численности пернатых хищни-
ков. Значительно снижается численность наиболее уязвимых
видов, к которым относятся в первую очередь стенобионтные, узко
специализированные хищники. При продолжающемся прессе
антропогенных факторов они могут исчезнуть с данной территории.
Результирующим всех описанных процессов является формиро-
вание обедненных фаунистических комплексов хищных птиц,
представляющих собой набор видов, легко приспосабливающихся
160
к современной антропогенной среде. Однако интенсификация хо-
зяйственной деятельности, отсутствие соответствующих мер охра-
ны может привести к их исчезновению. В данном случае особенно
велика роль охраняемых территорий и в первую очередь запо-
ведников.
ЛИТЕРАТУРА
Брагин Е. А. Экология могильника в островных лесах Кустанайских степей // Ох-
рана хищных птиц. М„ 1983. С. 92—97.
Брагин Е. А., Сметана Н. М., Сметана Н. Г., Гордиенко Н. С. Наурзум. Алма-Ата:
Кайнар, 1983. 218 с.
Галушин В. М. Численность и территориальное распределение хищных птиц Евро-
пейского центра СССР // Тр. Окского гос. заповедника. 1971. Вып. 8. С. 5—132.
Галушин В. М. Современное состояние численности дневных хищных птиц в евро-
пейской части СССР // Экология, география и охрана птиц. М.: Наука, 1980.
С. 156-167.
Галушин В. М. Адаптация хищных птиц к современным антропогенным воздей-
ствиям // Зоол. журн. 1982. Т. 61, № 7. С. 1088—1096.
Гибет Л. А. Численность хищных птиц и ее связь с грызунами в степной зоне
Северного Казахстана // Орнитология. М.: Изд-во МГУ, 1960. Вып. 3.
С. 278-291.
Дементьев Г. II. Отряд Хищные птицы // Птицы Советского Союза. М.: Сов. наука,
1951. Т. 1. С. 70-341.
Осмоловская В. И. Географическое распределение хищных птиц равнинного
Казахстана и их значение в истреблении вредителей // Материалы по
биогеографии СССР. М.; Л.: Изд-во АН СССР, 1953. Вып. 1. С. 219—307.
(Тр. Ин-та географии; Т. 54).
Панькин Н. С., Дугинцев В. А. О заселяемости гнезд врановых птиц пернатыми
хищниками в антропогенном ландшафте Зейско-Буреинской равнины // Ох-
рана хищных птиц. М.: Наука, 1983. С. 63—65.
Перерва В. И. Опыт использования механического актографа для изучения трофи-
ческой активности чеглока и обыкновенной пустельги в Наурзумском запо-
веднике // Орнитология. М.: Изд-во МГУ, 1977. Вып. 13. С. 173—176.
Перерва В. И. Особенности стабилизации численности хищных птиц // Журн. общ.
биологии. 1984. Т. 45, № 5. С. 644—652.
Соломатин А. О. Орел-могильник и балобан на Тургайском плато и природная
среда // Бюл. МОИП. Отд. биол. 1974. Т. 79, вып. 3. С. 40—51.
Шепель А. И. Характер пребывания, территориальное распределение и особенности
гнездования хищных птиц и сов юга Пермской области//Гнездовая жизнь
птиц. Пермь: ПГПИ, 1981. С. 66-70.
Lokemoen J. Т., Duebbert Н. F. Ferruginous hawk nesting ecology and raptor popu-
lations in Northern South Dakota // Condor. 1976. Vol. 78, N 4. P. 464—470.
Schmutz J. K., Schmutz S. M., Boag D. A. Coexistence of three species of hawks
(Buteo spp.) it the prairie-parkland ecotone// Canad. J. Zool. 1980. Vol. 58,
N 6. P. 1075-1089.
Swenson J. E. Factors affecting status and reproduction of Osprey in Yellowstone
National Park //J. Wildlife Manag. 1979. Vol. 43, N 3. P. 595-601.
Trillmich F. Zur Siedlungsdichte von Rotmilan (Milvus milvus) und Mansebussard
(Buteo buteo) bei Hiblesheim // Vogelwelt. 1969. Bd. 90, N 3.
'/26 Заказ № 36
161
SUMMARY
Ecological changes of birds of prey, influenced by anthropogenic factors.
Pererva V. I.
The information on the specific behaviour and ecological reactions of birds of prey
to the economical transformations of biocenoses and to anxiety factor is presented.
The creation of the mosaic landscapes, combining woodlands and farmlands, makes
birds of prey change the character of protection their food biotopes, in connection
with which the aggressive relations between the neighbouring pairs
remain only near the nests. The woodcuttings change the composition of trees and
cause redistribution of nests both according to biotopes suitable for nests and to the
location of buildings relative to the earth. The anxiety factor decreases, to a certain
extent, the successful reproduction of birds of prey on the unprotected territories.
The destabilization of the population of the whole complex of birds of prey in that
or another district up to the disappearance of some species is the result of the
influence of the named anthropogenic factors.
ПОВЕДЕНИЕ
УДК 591.556: 598.412
М. А. ТАРХАНОВА,
Л. Я. КУРИЛОВИЧ
123820, Москва,
ул. Б. Грузинская, д. 1.
Московский зоопарк
О взаимоотношениях
некоторых видов уток
в выводковый период
Работа проводилась на о-ве Лодейном, расположенном в Канда-
лакшском заливе на территории Кандалакшского государственного
заповедника. В отличие от большинства островов Северного архи-
пелага, о-в Лодейный имеет сильно изрезанную береговую линию
с глубоко вдающимися заливами и несколько внутренних озер,
удобных для выводков. На самом большом озере Бабье море (пло-
щадью около 10 га), где проводилась большая часть наблюдений,
ежегодно встречаются выводки нескольких видов речных и нырко-
вых уток. Обычно там держится по два-три выводка гоголя, турпа-
на, кряквы, шилохвости, свиязи, по одному-два — среднего кроха-
ля, чирка-свистунка и хохлатой чернети, некоторое время (до двух
недель) на озере проводят три-четыре выводка морской чернети.
Часть самок (в частности, практически все гоголи) гнездится на
о-ве Лодейном, часть приводит птенцов с близлежащих луд или
даже других островов. Вероятно, привлекательность этого озера
для выводков связана с тем, что, как и на любом внутреннем во-
доеме, там не так чувствуются изменения погоды, нет волнобоя
(что особенно важно для маленьких птенцов) и реже охотятся се-
ребристые чайки. Несмотря на то, что на о-ве Лодейном есть еще
несколько озер, наибольшая численность и видовое разнообразие
уток наблюдается именно на Бабьем море. Очевидно, это объясняет-
ся тем, что это — единственное на острове слабосоленое озеро (ос-
тальные— пресные), что может сказываться на составе беспозво-
ночных и на растительности, создавая лучшие кормовые условия не
только для нырковых, но и для речных уток. Последние питаются
рдестом, обильно встречающемся на мелководье, из прибрежных
растений используют некоторые виды осок и ситняг (на пресных
озерах состав растительности другой). Небольшая глубина озера
позволяет кормиться птенцам всех видов уток. Таким образом,
в летний период оз. Бабье море становится местом обитания девяти
видов уток — образуется смешанная популяция или «межвидовая
ассоциация» (Иваницкий, 1986).
‘/26*
163
Основной задачей данной работы является соотнесение особен-
ности поведения каждого вида уток с наличием межвидовой кон-
куренции и механизмов, снижающих ее.
Визуальные наблюдения за выводками уток на о-ве Лодейном
велись в летние периоды 1978—1980 и 1983—1986 гг. Особое вни-
мание обращали на поведение самок гоголей и турпанов с вывод
ками. Регистрировались их взаимодействия со всеми видами уток.
У гоголя и турпана были выделены следующие формы агрессив-
ного поведения.
Гоголь
1. Угрожающая поза (шея вытянута и
почти касается воды).
2. Движение по направлению к объекту
со значительного расстояния (более
10—15 м).
3. Угрожающий рывок (то же, что и
.№ 2, но с большей скоростью и на
короткое расстояние).
4. Ныряние в сторону объекта на зна-
чительное расстояние.
5. Ныряние с последующим ударом из-
под воды.
6. Драка.
7. Полет к объекту (за 50—100 м), при
этом утка старается сесть на воду как
можно ближе к нему.
8. Полет-преследование (утка летит за
объектом, иногда нанося удары клю-
вом) .
9. Демонстрация вдоль границ (птицы
плавают на расстоянии 5—20 м друг
от друга в угрожающих позах, пери-
одически ныряя).
Турпан
1. Угрожающая поза (шея втянута, клюв
прижат к груди и касается воды).
2. Угрожающий рывок (быстрое пере-
мещение в сторону объекта на не-
большом расстоянии в угрожающей
позе).
3. Нападение (птица бросается на объект
с вытянутой шеей и раскрытым клю-
вом).
4. Драка.
Все взаимодействия, в которых птицы демонстрировали хотя
бы один из указанных видов активности, считались агрессивными.
Сравнительный материал по агрессивности гоголя и турпана соб-
ран в основном в 1986 г. (за каждым видом проведено около 100 ч
наблюдений). Практически все самки гоголя с выводками опоз-
навались индивидуально, так как были помечены цветными коль-
цами и пикриновой кислотой. Самки турпана опознавались по ин-
дивидуальным особенностям окраски оперения (форме и размерам
белых пятен на голове), числу и размеру птенцов в выводках.
164
Взаимоотношения разных видов уток
Большая часть акватории оз. Бабье море распределена между
несколькими выводками гоголя — единственного территориального
вида (Savard, 1982; Тарханова и др., 1983) уток из наблюдавшихся.
Границы участков гоголей достаточно четки, выводки большую
часть времени проводят внутри своих территорий и редко наруша-
ют границы. Самки остальных видов охраняют только небольшие
индивидуальные участки (3—5 м) вокруг своих птенцов и переме-
щаются по всему озеру, т.е. их территории как бы накладываются
на территории гоголей. Это и позволяет разным видам использовать
одну и ту же площадь. Вероятно, присутствие еще нескольких тер-
риториальных видов сделало бы невозможным совместное обитание
около 20 выводков довятИ" видов уток на одном небольшом озере.
Однако распределение выводков гоголя оказывает определенное
влияние на распределение выводков других видов, в частности тур-
пана. Взаимоотношения самок гоголя и турпана могут складывать-
ся по-разному, и в большой степени это, вероятно, зависит от инди-
видуальности самок турпана. Более робкие (возможно, молодые)
самки после первых же нападений гоголей покидают их тер-
ритории.
Выводки остальных видов уток использут всю акваторию озера,
но по-разному. Так, хохлатые и морские чернети, кормящиеся на
каком-либо участке территории гоголя, при первом же нападении
самки гоголя обычно покидают его. Однако позже, когда самка за-
нята чем-то другим, отдыхает на суше или находится в другой час-
ти своей территории, чернети опять появляются на том же участке
и кормятся там до нового нападения.
Длиннохвостые крохали и речные утки менее других подверже-
ны воздействию гоголя. Выводки речных уток кормятся в заливах
часами, не вызывая агрессии гоголей. Выводки крохалей, двигаясь
с большой скоростью вдоль берегов всего озера, не избегая и терри-
тории гоголей, тоже относительно редко подвергаются их нападе-
ниям. Выводки всех этих видов конфликтуют между собой гораздо
реже и могут подолгу кормиться на небольшом расстоянии друг от
друга. Таким образом, разный характер использования территории
делает возможным взаимное существование всех этих видов.
Популяция гоголя организована на основе жесткой террито-
риальности, популяции остальных видов имеют аморфную струк-
туру. Похожая ситуация описана В. В. Иваницким (1980) для сме-
шанной популяции птиц на юге Тувы. Автор отмечает, что виды
с жесткой территориальностью обычно обладают и более высокой
агрессивностью. Это справедливо и для нашего случая. Во взаи-
моотношениях гоголя со всеми остальными видами уток, оби-
тающими на острове, обращает на себя внимание асимметрия:
подавляющее большинство межвидовых конфликтов инициируют
самки гоголя, препятствующие проникновению на свою террито-
рию других уток. Этому способствует и то, что из всех перечислен-
ных видов нырковых уток гоголь первый приводит выводки на озе-
6* Заказ № 36
165
ро, остальные же как бы пытаются внедриться на уже занятую
акваторию.
Высокая агрессивность обыкновенного гоголя не раз отмечалась
в литературе (Savard, 1982, 1984). Данных об агрессивности тур-
пана нам обнаружить не удалось, и мы можем ориентироваться
только на свои наблюдения. Количество внутривидовых конфлик-
тов у обоих видов сравнимо с количеством межвидовых, но пре-
восходит их, составляя 60—70% от всех столкновений. Средняя
частота внутривидовых конфликтов у гоголя несколько выше, чем
у турпана: у гоголя — 2,4 конфликта в час, у турпана — 2. Мак-
симальная частота конфликтов у обоих видов—около 10 в час.
Гораздо ярче проявляются различия в продолжительности кон-
фликтов: внутривидовые конфликты у гоголя продолжаются в
среднем 3 мин, у турпана — одну (п=174). При этом конфликты,
длящиеся не более 10 с, у турпана составляют около половины
отмеченных случаев (54%), у гоголя —15%; 5—7 мин —5% у тур-
пана; столкновения, продолжающиеся более 7 мин, составляют
у гоголя 10%, а у турпана их не наблюдалось.
При установлении территорий между самками гоголя иногда
происходят длительные антагонистические взаимодействия, для-
щиеся до 20 мин. Для турпана характерна относительная бедность
форм агрессивного поведения, особенно в сравнении с гоголем.
Практически все формы агрессивного поведения, характерные для
гоголя и отсутствующие у турпана, связаны с территориальным
поведением. Многие формы агрессивного поведения у гоголя рас-
считаны на воздействие на объект, находящийся на значительном
расстоянии, у турпана же оно не превышает нескольких метров. У
обоих видов для внутривидовых взаимоотношений используются те
же формы агрессии, что и для межвидовых. Однако у гоголя есть и
специфически внутривидовая «демонстрация вдоль границ». Эта
форма агрессивного поведения становится характерной после того,
как соседние выводки познакомятся друг с другом, с ее появлением
драки становятся редкими. Разнообразие поведения гоголя отража-
ется и в том, что в каждом агрессивном акте чаще всего использую-
тся последовательно сразу несколько форм. По предварительным
данным, не более 20% от всех взаимодействий гоголя содержат в
себе одну форму агрессии; у турпана же таких взаимодействий
85%. Таким образом, по целому ряду показателей: большой про-
должительности конфликтов, преобладанию активных форм агрес-
сии, разнообразию и экспрессивности агрессивного поведения,
большему расстоянию, на котором объект может вызвать агрессию,
агрессивность гоголя выше, чем у турпана.
Известно, что у многих видов животных отмечена корреляция
между внутри- и межвидовой агрессивностью (Иваницкий, 1986).
Как же ведут себя гоголь и турпан в межвидовых взаимодействиях?
Как уже было упомянуто, и здесь гоголь гораздо агрессивнее,
он — инициатор почти всех межвидовых конфликтов. На эти
взаимодействия у обоих видов приходится 30—40% всех стол-
кновений. Средняя частота межвидовых конфликтов у гоголя нес-
166
колько выше (1,3 конфликта в час), чем у турпана (0,8). Средняя
продолжительность межвидовых конфликтов у гоголя равна 1 мин,
у турпана —40 с. Интересно, что конфликты между гоголем и тур-
паном часто бывают достаточно длительными (максимально до
20 мин), в среднем их продолжительность 2 мин, в то время как
конфликты с остальными видами уток длятся не более 10 с. По со-
отношению разных форм агрессии у гоголя межвидовые конф-
ликты мало отличаются от внутривидовых, поскольку практи-
чески все они инициируются самками гоголя, применяющими при
этом те же формы агрессии. У турпанов же дело обстоит совсем ина-
че: в межвидовых конфликтах сильно возрастает доля драк, их в
12 раз больше, чем во внутривидовых! Связано это с тем, что у них
две трети всех межвидовых конфликтов происходит с самками гого-
ля (чаще всего это нападения самок гоголя, которые нередко за-
канчиваются драками). В антагонистических взаимоотношениях с
другими видами уток инициатором чаще всего бывает самка турпа-
на. У самок гоголя примерно половина межвидовых конфликтов
происходит с турпанами. С другими видами уток конфликты у
гоголя и турпана происходят реже: с крохалями происходит 14%
всех межвидовых конфликтов у турпана, 17% — у гоголя, с черне-
тями —10% у турпана, 13%— у гоголя. С речными утками у гоголя
конфликтов почти в два раза больше, чем у турпана (17 и 9,4%;
данные 1986 г.). Вероятно, частота конфликтов с этими видами
может колебаться в зависимости от их численности (так, в 1983 г.
36% конфликтов у самок гоголя приходилось на длинноносого кро-
халя, чья численность была велика). Расстояние, на котором самка
турпана реагирует на другие виды, практически никогда не превы-
шает нескольких метров. Для гоголей это расстояние значительно
больше и сильно зависит от видовой принадлежности объекта
агрессии. Наиболее велико оно для чернетей, меньше всего — для
речных уток. Чтобы реагировать на разные виды дифференци-
рованно, гоголи, безусловно, должны опознавать их визуально.
Судя по всему, и другие виды уток опознают гоголей. По-видимому,
в этом смешанном поселении утки обучаются распознавать более
агрессивные виды (Иваницкий, 1986). Особенно ярко это прояв-
ляется у чернетей обоих видов, которые менее всех способны на
ответную агрессию. Чернети хорошо отличают гоголей от всех
других уток и часто уже при первых признаках агрессии со стороны
самок гоголя, даже если те находятся на большом расстоянии,
стараются как можно быстрее ретироваться.
Территориальность и агрессивность гоголя
и их «относительность»
Территория выводка гоголя представляет собой залив моря или
озера, участок акватории, вытянутый вдоль береговой линии или
озеро целиком, если оно небольшое. Но о-ве Лодейном гоголь пред-
почитает два залива озера Бабье море и оз. Островное. На Бабьем
море эти заливы занимают в первую очередь выводки, первыми
6**
167
спустившиеся на воду, более поздние занимают участки вдоль севе-
ро-восточного берега. Чем объясняется это предпочтение — не
совсем ясно, но в годы с высокой плотностью выводков выживае-
мость птенцов из заливов выше (Тарханова, Курилович, 1986).
Возможно, это связано с тем, что самке легче защищать вход в
замкнутый залив, чем длинную, вытянутую вдоль берега террито-
рию, и у них меньше конфликтов (количество конфликтов отрица-
тельно сказывается на выживаемости птенцов). К тому же в зали-
вах серебристым чайкам охотиться менее удобно, чем посередине
озера. Самка гоголя с выводком занимает территорию примерно в
3 га. Внутри территории у выводка есть постоянные места дневного
отдыха (чаще всего это торчащие из воды камни), кормится же
он практически по всей площади. Степень использования террито-
рии выводком зависит от плотности населения. Так, в 1985 г. при
появлении на Бабьем море четвертого выводка количество конф-
ликтов между самками резко возросло, но через несколько дней
уменьшилось, поскольку новый выводок занял небольшой участок
на территории одной из самок, которая стала посещать этот участок
очень редко.
Территории выводков неоднородны по степени запрета дру-
гих самок. Так, в 1985 г. одна из самок занимала два залива Бабьего
моря, но в первом из них она проводила гораздо больше времени.
Туда редко заходили другие выводки, а если это случалось, она
была к ним максимально нетерпима. Совсем по-другому исполь-
зовался второй залив: самка делила его с другой самкой, причем
они редко появлялись там одновременно. За все время наблюде-
ний там было зафиксировано только два конфликта. Поочередное
использование части территории, несомненно, сильно сокращает
количество конфликтов. Иногда за счет такого общего участка
становится возможным внедрение нового выводка. Наличие таких
«буферных» участков уменьшает число конфликтов. При росте
плотности они исчезают в первую очередь.
Заняв определенные территории, самки хорошо представляют
себе их границы, птенцы же усваивают их постепенно. Часто,
кормясь вдоль берега, они заходят на чужую территорию, а самка
следует за ними на некотором расстоянии. Если самка-резидент
находится достаточно далеко, они могут кормиться там некоторое
время, но как только она замечает их и идет в атаку, вторгшийся
выводок без всякого сопротивления немедленно покидает чужую
территорию. Время от времени самки появляются на чужих тер-
риториях одни, без птенцов. Создается такое впечатление, что
они проверяют их занятость. Если по какой-либо причине один
из участков освобождается, через некоторое время он начинает
использоваться другим выводком. Однако при низкой плотности
выводков он может пустовать значительное время.
Предварительно мы выделили четыре этапа освоения выводком
своей территории. В течение первого этапа выводок сразу после
спуска на воду довольно много перемещается, нигде подолгу не
задерживаясь, «ищет место». Однако так ведут себя не все вы-
168
водки. Возможно, «поиски места» вызваны тем, что самка не знако-
ма с акваторией озера. Если же она хорошо знакома с местностью,
этот этап может отсутствовать. Для второго этапа характерна «ма-
лая подвижность выводка». В это время птенцы кормятся на не-
большой площади. Такое поведение объясняется в первую очередь
возрастом птенцов (на этом этапе птенцам обычно 3—5 сут.) . Одна-
ко такая же «малая подвижность» может наблюдаться и у выводка
после его переселения на новую территорию, что наблюдалось нами
у выводка с птенцами в возрасте по крайней мере десяти дней.
По-видимому, эта стадия связана с освоением новой территории.
Для следующего, третьего этапа характерно расширение первона-
чальной территории и установление ее окончательных границ.
Он может продолжаться около месяца; затем самка уводит птенцов
в море (четвертый этап). Впрочем, переход подросших птенцов на
море наблюдается не у всех выводков.
На о-ве Лодейном из-за наличия большого количества искус-
ственных гнездовий не все загнездившиеся самки гоголей имеют
возможность обладать территорией — площадь водоемов, пригод-
ных для обитания выводков, ограничена. В годы с высокой плот-
ностью выводков между ними существует жесткая конкуренция
за жизненное пространство. Практически все птенцы из нетерри-
ториальных выводков погибают (Тарханова, Курилович, 1986).
Однако агрессивность и территориальное поведение гоголя ха-
рактерны для него не всегда. Е. Н. Панов (1983) отмечает, что у
особей одного и того же вида территориальное поведение может
проявляться или не проявляться в зависимости от целого ряда
обстоятельств — пола, возраста, социального опыта, сезона и
даже плотности популяции. Это справедливо и для такого вы-
сокоагрессивного и жестко территориального вида, как гоголь.
В брачный период агрессивны как самцы, так и самки, но ос-
новную нагрузку по охране территории несет самец (по наблюдени-
ям в Московском зоопарке). При появлении другой пары самец
проявляет гораздо большую агрессивность и прогоняет ее, в то вре-
мя как самка в его присутствии может продолжать кормиться, не
реагируя на другую пару. Очевидно, это имеет место и в природе.
В середине инкубационного периода ситуация меняется на
противоположную: самцы теряют агрессивность и улетают с мест
размножения, а между самками, если они одновременно вылетают
из гоголятников, наблюдаются жестокие драки. В дальнейшем,
в отличие от большинства других видов уток, агрессивность самок
гоголя очень долго остается на высоком уровне (Тарханова, Кури-
лович, 1984), Вероятно, даже после того, как птенцы становятся са-
мостоятельными, самки продолжают держаться на Бабьем море,
охраняя небольшие «кормовые участки» и отгоняя и чужих, и соб-
ственных птенцов. Птенцы гоголя рано начинают проявлять агрес-
сивность. На о-ве Лодейном агрессивное поведение гоголенка по от-
ношению к птенцу турпана было впервые отмечено в возрасте около
месяца. В Московском зоопарке уже двухнедельные птенцы под-
плывали к местам, где посетители кормили уток хлебом, и, совер-
169
шая резкие броски, разгоняли крякв, значительно превосходящих
их по размеру. Однако молодые гоголи (первогодки) держатся
стайно и не проявляют никаких признаков агрессии и территори-
ального поведения.
Самки, потерявшие птенцов, остаются агрессивными один-два
дня, продолжая держаться на той же территории, где были с птен-
цами; позже их вытесняют другие выводки.
Агрессивность самки зависит и от степени ее знакомства и
продолжительности владения территорией. Самка, плохо знакомая
с территорией, обычно держится настороженно и робко, а по мере
ознакомления становится более агрессивной. Это подтверждается
нашими наблюдениями за самкой, обитавшей в 1986 г. на оз. Пить-
евом. Несколько раз она пыталась переселиться на оз. Бабье море.
Первые две попытки закончились неудачно: державшаяся там сам-
ка с выводком была значительно агрессивнее и инициировала
практически все столкновения. При третьей попытке ситуация из-
менилась: агрессивнее стала вселявшаяся самка. Несколько раз
обитавшая на Бабьем море самка пыталась уклониться от кон-
фликта и отступала, но новая самка преследовала ее и иницииро-
вала драки. Она была гораздо агрессивнее старой и по отношению
к турпанам: за час семь раз дралась с самками двух выводков тур-
пана и добилась того, что они ушли. Вероятно, причиной измене-
ния поведения было постепенное ознакомление самки с акваторией
залива Бабьего моря, где происходили описываемые события.
После третьей попытки она осталась в заливе.
Территориальное поведение гоголя зависит от плотности по-
пуляции. В условиях высокой плотности практически вся акватория
Бабьего моря распределена между выводками гоголя, перемещение
каждого выводка ограничено пределами его участка, при низкой
плотности выводки перемещаются более свободно. В 1986 г. в
результате хищничества чаек выводков осталось мало. С 21 июня
по 5 июля на Бабьем море чайки уничтожили пять выводков гоголя,
причем это происходило настолько быстро, что выводки соседство-
вали не более одного-двух дней. За это время каждый из них не
успевал закрепить за собой четкой территории, и практически не
было жесткой конкуренции. Ни один из уцелевших выводков не
ушел с озера, и все они смогли стать территориальными. Каждый
выводок занял определенную территорию, однако в отличие от
прошлых лет она не оставалась неизменной в течение длительного
срока.
Заключение
Летом на оз. Бабье море гнездится девять видов уток. Конф-
ликты внутри группы нырковых уток (гоголь, турпан, мор-
ская и хохлатая чернети) происходят значительно чаще, чем между
нырковыми утками и длинноносыми крохалями, а также между
речными и нырковыми утками. В то же время, судя по литератур-
ным данным, внутри группы нырковых уток существуют значи-
170
тельные перекрытия в питании и способе добычи корма. Это позво-
ляет предположить зависимость степени агрессивности (числа
конфликтов) между представителями разных видов от наличия или
отсутствия конкуренции между ними. Частота агрессивных взаи-
модействий между разными видами нырковых уток свидетельству-
ет о том, что конкуренция между ними существует. Однако они мо-
гут обитать совместно, поскольку эта конкуренция снижается за
счет различий в использовании территории, сопряженных с разным
поведением при межвидовых конфликтах (гоголь — доминант,
турпан — субдоминант, еще ниже статус двух видов чернетей) и
«выучивания» своего места в структуре смешанной популяции.
Авторы выражают глубокую благодарность сотрудникам Канда-
лакшского заповедника и особенно В. В. Бианки за предостав-
ленную возможность сбора материала и всестороннюю помощь.
ЛИТЕРАТУРА
Иваницкий В. В. Межвидовые отношения симпатрических видов каменок // Зоол.
журн. 1980. Т. 59, № 5. С. 739-749.
Иваницкий В. В. Социальная организация смешанных популяций близких видов
животных // Итоги науки и техники. Зоология позвоночных. М.: ВИНИТИ,
1986. Т. 14. С. 127—180.
Панов Е. Н. Поведение животных и экологическая структура популяций. М.: Наука,
1983. 423 с.
Тарханова М. А., Еурилович Л. Я. Некоторые аспекты поведения выводков турпана
и гоголя в Кандалакшском заливе // Докл. МОИП: Зоология и ботаника. М.,
1984. С. 35.
Тарханова М. А., Еурилович Л. Я. Выживаемость птенцов гоголя на о. Лодейный //
Тез. докл. IX Всесоюз. орнитол. конф. Л., 1986. Ч. 2. С. 272.
Тарханова М. А., Еурилович Л. Я., Харитонова И. А. Поведение выводков гоголя,
турпана и длинноносого крохаля в Кандалакшском заливе // Материалы
III Всесоюз. конф, по поведению животных. М., 1983. Т. 2. С. 280.
Handbook of behavioral neurobiology: Social behavior and communication / Ed.
P. Marler, J. G. Vandenbergh. N. Y., 1979.
Savard J. P. Intra- and interspecific competition between Bucephala islandica and
B. albeola 11 Canad. J. Zool. 1982. Vol. 60. P. 3439 3446.
Savard J. P. Territorial behaviour of Common Goldeneye and buffleaded in areas
of sympatry // Ornis. Scand. 1984. Vol. 15, N 4. P. 211—216.
SUMMARY
On relations of some duck species during hatching period.
Tarkhanova M. А., Kurilovich L. Ya.
The behaviour of females with the broods of the Goldeneyes and Valvet Scoters
was compared during the summers of 1978—1980 and 1983—1986 on the Lodeiny
Island in the Kandalaksha Bay of the White Sea. The Goldeneye happened
to be more aggressive — it was the initiator of almost all conflicts between the species
(in both species — 30—40% of conflicts). The conflicts within the group of ducks
(Goldeneye, Valvet Scoter, Tufted Duck and Create Scaup) occur considerably oftener
than those between ducks and Mergansers, and between river and sea ducks. The
available completition between the sea ducks can be reduced at the expense of the
differences in the use of the territory, conjugated with the different behaviour during
the interspecific conflicts and the place «learning» in the mixed populations.
171
УДК 591.51:591.568: 598.842:3
С. с. москвитян, Т. Л. АНАНЬИНА 634010, Томск, ул. Ленина, д. 21. Томский государственный университет, Зоологический музей Ритмика поведения и бюджет рабочего времени садовой камышевки (Acrocephalus dumetorum) в период выкармливания птенцов
Одной из видовых и популяционных характеристик у животных
является ритмика поведения. Количественное выражение ее — хо-
роший показатель взаимоотношения организма со средой. К сожа-
лению, по этому вопросу региональные работы на птицах отсут-
ствуют. Поэтому мы сделали попытку показать на примере широко
распространенного насекомоядного вида — садовой камышевки —
норму поведенческих реакций в условиях южной тайги Западной
Сибири.
Для этого мы использовали наблюдения за поведением роди-
телей при выкармливании птенцов, так как именно в этот период
мы имеем дело с наиболее разнообразным и «эволюционно выверен-
ным» действием птиц, ибо размножение — самый ответственный
и напряженный период в их жизни, от которого зависит существо-
вание и вида, и популяции. При наблюдении из укрытий за 12 гнез-
дами фиксировалась частота кормлений, время, затрачиваемое
на скармливание пищи, обогрев птенцов, отдых, сторожевые функ-
ции, поддержание санитарного состояния в гнезде и т. д. О ритмике
других стандартных поведенческих актов, таких, как кормодобыва-
ние, питание, пространственные перемещения, отдых вне гнезда,
можно судить лишь косвенно по ритму и частоте выкармливания.
Гнезда были с птенцами почти всех возрастов, но для получения
более точных сравниваемых данных использовались лишь те,
которые имели по четыре птенца. Общий объем наблюдений за
ними составил 230 ч.
Для рассматриваемого вида в период выкармливания птенцов,
в отличие от периода миграций и активного пения, характерен
строго дневной ритм активности. Этот период приходится на июль,
когда наблюдается убывание светового дня. Время активной дея-
тельности продолжалось от 17 ч 30 м в начале месяца до 16 ч 35 м
в конце его, или от 72,9 до 69,2% длительности суток. Кормление
начиналось через 28—30 мин после восхода солнца и кончалось че-
рез 30—35 мин после захода. Таким образом, освещенность, с ко-
торой связана и возможность обнаруживать корм, выступает в ка-
честве синхронизатора ритма. Это согласуется и с литературными
материалами, согласно которым активность и обмен веществ у птиц,
ведущих дневной образ жизни, связаны с освещенностью (Пром-
птов, 1940; Сазанов и др., 1961; Слоним, 1961).
172
В процессе роста птенцов ритмика и суммарное время, затра-
чиваемое на основные регистрируемые акты деятельности, ме-
няются (см. таблицу). Эти изменения по дням имели как общие
закономерности, так и особенности, связанные с поведением роди-
телей, иногда погодой, кормностью территории и т. д. Общий
ритм выкармливания имел тенденцию усиливаться до шестого
дня включительно, а затем до конца пребывания птенцов в гнезде
поддерживалась относительная стаэильность. Число кормлений
несколько увеличилось лишь в последний день. Минимальная
за весь гнездовой период (между первыми и одиннадцатыми
сутками) средняя суточная интенсивность выкармливания отлича-
лась от максимальной в 5,4 раза. Приход же энергии в единицу
времени в результате кормления возрастал не только за счет увели-
чения числа прилетов с кормом, но и за счет некоторого укрупнения
массы приносимого корма.
Одновременно с увеличением активности выкармливания бюд-
жет непищевой активности имел тенденцию к уменьшению. Напри-
мер, в первые сутки обогрев птенцов составил 55,7% времени
активного периода, на шестые сутки он сократился в 3,6 раза,
на восьмые — в 11,2 раза, а на девятые полностью прекратился,
исключая ночное время. Переломными явились седьмые сутки,
когда резко уменьшилось время, затрачиваемое на обогрев, что
связано с переходом птенцов от пойкилотермии к гомотермии
(Слоним, 1976). Поэтому речь, видимо, должна идти об эволю-
ционно закрепленном ритме обогрева. Однако синхронизатором
выкармливания и, вероятно, обогрева выступает еще и двигатель-
ная активность птенцов, взаимодействие ритмов самца и самки.
В результате этого мы - наблюдаем изменение бюджета времени
с ростом птенцов. Слабая двигательная активность птенцов до чет-
вертого дня жизни и небольшие потребности их в корме позволяют
родителям много времени проводить в гнезде. Дальнейший рост
двигательной активности, сопровождающийся усилением конку-
рентности отношений между птенцами, поддерживается звуковыми
сигналами и влияет на активацию родителей. Проголодавшиеся
птенцы буквально выталкивают из гнезда обогревающих родите-
телей, побуждая их к полету за кормом. Поддержанию ритма ак-
тивности выкармливания способствует существующая очередность
кормления птенцов, поэтому в выводке наблюдается снижение
активности накормленных и повышение активности голодных
птенцов. Это ведет к их постоянному, обычно круговому перемеще-
нию в гнезде. В течение суток все они получают почти одинаковое
количество корма. Конечно, активность движения связана не
только с ощущением голода, она дополняется и тем, что сам рост
стимулирует двигательный аппарат и мышечную систему (Слоним,
1976). С возрастом птенцы все больше и действеннее мешают
обогреву, а их поведение по выпрашиванию корма становится все
более сильным раздражителем и способствует усилению кормо-
добывающей деятельности родителей.
Следует добавить, что ритм задается не только этим. Практи-
173
Возрастная динамика интенсивности выкармливания птенцов садовой камышевкой
Воз- раст, сут Дли- тель- ность наб люде- НИЙ, ч Среднее кол-во при летов с кормом в час Затраты времени, %
на кормление на обогрев
б 9 д+2 <5 9 д+9 д 9 д+9
1 12 1,83 1,67 3,50 0,75 0,66 1,41 23,5 32,25 55,75 4 19,5 - - 7,56 - - 2,88 - - 38,87 5 37 6,20 4,13 10,33 2,40 1,60 4,00 2,20 12,90 15,10 6 35 7,61 7,11 14,72 2,94 2,70 5,64 1,94 13,4 15,34 7 35 - - 13,45 - - 5,18 - - 4,09 8 28 4,53 7,94 11,47 1,77 2,71 4,48 1,76 3,76 5,52 9 15 13,07 - - 5,06 0 0 0 10 30 - - 13,11 - - 3,00 0 0 0 11 18,5 - - . 15,57 - - 1,52 0 0 0 Всего в 230 - - 11,42 - - 3,68 — — 22,45
среднем
чески каждый сход птицы с гнезда заканчивается приносом корма.
Поэтому прилет с кормом к гнезду одного родителя ведет к тому,
что второй, который в это время обогревает птенцов, уступает
ему место и, в свою очередь, летит за кормом. Таким образом, рит-
мы выкармливания самцом и самкой в известной степени синхро-
низируются. Такое взаимодействие было проверено нами и с по-
мощью специального вспугивания птицы. Вспугнутая с гнезда, она
возвращалась, как правило, с кормом. Особое значение активи-
зация одного партнера другим имеет для начального периода вы-
кармливания, когда птицы много обогревают’птенцов. В этом мы
убедились на примере самки рябинника, которая из-за отсутствия
самца не могла «расстатюя» с доминантой насиживания. В резуль-
тате этого маленькие птенцы от постоянного недокорма не только
слабо росли, но и начали погибать друг за другом. Самка кормила
их с нормальной частотой лишь тогда, когда мы ее регулярно
вспугивали. Если партнер исчезает не на самых начальных стадиях
кормления, а при подросших птенцах, то наблюдается повышение
интенсивности выкармливания оставшейся птицей. Причем как в
случаях с камышевкой, так и на примере с другим гнездом рябин
ника видно, что активность одиночной птицы превышает суммар-
ную частоту приноса корма двумя родителями. Абсолютный макси-
мум частоты приносов корма садовой камышевкой за все время
наших наблюдений пришелся на одиночного родителя и состав-
лял 32 раза в час. Он же установил своеобразный рекорд, накормив
птенцов за весь рабочий день 370 раз, в то время как при наличии
обоих родителей в этом возрасте птенцы получали корм около
265 раз. Видимо, при исчезновении одного партнера в период ин-
тенсивного выкармливания птенцы до такой степени стимулируют
родителя-одиночку, что в его поведенческом ритме начинает доми-
нировать кормление. Такой родитель обычно не проявляет беспо-
койства у гнезда, обращая мало внимания на появление внешних
раздражителей. Такое подчинение ритма главной задаче — выкар-
174
и бюджет рабочего времени взрослых птиц
Затраты времени, %
на поддержание санитар- ного состояния на отдых у гнезда общий объем времени нахожде- ния птиц у гнезда
г 9 б +9 б . 9 б +9 б 9 6+9
0 1,15 1,15 1,67 1,75 3,42 25,92 35,81 61,73 - - 0,95 2,00 2,13 4,13 - - 46,83 0,35 1,45 1,80 0,90 3,30 4,20 5,85 19,25 25,10 0,57 3,13 3,70 2,75 4,85 7,60 Й,20 24,08 32,28 - - 2,35 - - 4,90 — - 16,52 1,34 2,96 4,30 5,8 3,60 9,40 10,67 13,03 23,71 - - 2,11 - - 0 - - 7,17 - - 0,96 - - 0,78 — - 4,74 — - 0- — 0 — - 1,52 - - 2,16 - - 4,92 - - 24,4
мливанию — имеет особый биологический смысл, связанный с вос-
производством.
Обычно частота прилетов родителей с кормом к гнезду значи-
тельно ниже (см. таблицу). Этот средний уровень определяется
не только потребностями птенцов и ритмом поведения роди-
телей, но и энергетическими возможностями родителей. Начи-
ная с шестых суток затраты у родителей на локомоторную актив-
ность резко возрастают, так как увеличивается дальность их поле-
тов (Дольник, 1976), дополнительно растет частота перепархива-
ний, зависаний в воздухе и т. д. Увеличивается расход энергии
(Яблонкевич, 1976) за счет повышения нервной нагрузки, связан-
ной как с охраной птенцов, так и с развитием инстинкта материн-
ства. Об этом, по нашему мнению, убедительно говорит тот факт,
что у самок камышевки, на которых ложится основная нагрузка
по охране гнезда и которые летают за кормом на меньшее (в сред-
нем), чем самцы, расстояние, вес в период выкармливания умень-
шился в среднем, по нашим наблюдениям, на 8,3% (п = 25),
тогда как у самцов — на 4,4% (н=28) по сравнению с периодом
насиживания яиц. Да и сам факт относительной стабилизации
интенсивности выкармливания птенцов в возрасте 6—10 сут,
несмотря на их дальнейший рост и развитие, свидетельствует о до-
стижении высокой напряженности энергетических усилий.
Эта напряженность объясняется не только возможностями
родителей, но и изменениями кормовой ситуации, особенно вблизи
гнезда, где возникает дефицит предпочитаемых объектов корма.
Поэтому поддержание стабильности и даже некоторое увеличение
интенсивности выкармливания, наблюдающееся на одиннадцатый
день, достигалось в известной мере за счет перераспределения
в бюджете рабочего времени. Во второй период выкармливания
резко сокращается время на обогрев птенцов, в меньшей сте-
пени — на отдых у гнезда и поддержание чистоты в гнезде. С девя-
тых суток сокращаются затраты времени и на собственное кормле-
175
ние, так как птенцы в этом возрасте способны поедать любой
приносимый корм без помощи родителей, которые раньше тратили
время на заталкивание и распределение его между птенцами.
В итоге пребывание возле гнезда к одиннадцатому дню снизилось
до 1,52% от всего рабочего периода. Таким образом, бюджет рабо-
чего времени стал подчиняться главной функции — выкармли-
ванию. В среднем на 6—11-й день жизни птенцов камышевки при-
летают с кормом к гнезду 231 раз в день; абсолютный максимум
у пары составил около 265 прилетов за рабочий день (на 11-е сутки
жизни птенцов).
Количество корма, получаемого птенцами, может определяться
кормовой ситуацией сезона. Так, в конце июля 1981 г. из-за
засушливого лета и высокой плотности гнездования в пойме Оби,
птенцы садовой камышевки получали малоценный корм. В резуль-
тате систематического недокорма гибель птенцов в этих гнездах
(п = 3) за счет смерти младших составила от 25 до 50%. Не помо-
гло и изменение бюджета времени родителей в сторону увеличения
затрат его на поиск корма.
Помимо изменений в общем ритме поведения родителей, ритмы
кормления самца и самки и их бюджеты рабочего времени также
различаются, хотя оба партнера у садовой камышевки несут при-
мерно одинаковую нагрузку в заботах о потомстве. Самки больше
времени тратят на обогрев птенцов, на поддержание чистоты
в гнезде и его охрану (см. таблицу). Они, как правило, первыми
появляются при подходе врага к гнезду, подавая сигнал беспокой-
ства. В первые пять дней самки реже, чем самцы, собирают корм,
но больше насиживают.
Можно говорить и о суточном ритме поведения родителей. Ритм
кормления птецов по средним показателям характеризуется четы-
рехвершинной кривой. Пики активности приходятся на отрезки
с 5 до 6, с 9 до 10, с 15 до 16 и с 19 до 20 ч. Причем каждый последу-
ющий пик имеет меньшую абсолютную величину, чем предыдущие
(рис. 1). Чем выше подъем, тем сильнее выражен спад активности
выкармливания, так как минимальная интенсивность прилетов
в каждый из таких периодов в течение суток остается почти на
одном уровне. Примечательно, что утренняя повышенная интен-
сивность выкармливания сочетается с наиболее высокой интен-
сивностью обогрева птенцов (рис. 1, 2), тогда как в течение всего
периода выкармливания между этими актами наблюдается обрат-
ная зависимость (г = —0,89). Это, видимо, объясняется тем, что
при низких температурах утром обогрев позволяет птенцам лучше
усваивать корм. После полудня же птицы сокращают обогрев
птенцов (см. рис. 2). Суточный ритм зависит и от погоды.
В жаркие дни после 12 ч наблюдается спад активности. Син-
хронизатором этого спада, по всей вероятности, выступают макси-
мальные суточные температуры и инсоляция, с которыми связана
необходимость экономии энергетических затрат. Она достигается
за счет торможения активности с целью сохранения температур-
ного гомеостаза (Слоним, 1976). Это справедливо и по отношению
176
Рис. 1. Среднесуточная ди-
намика выкармливания
птенцов садовой камышев-
кой
Рис. 2. Динамика обогрева
птенцов садовой камышев-
кой в разных гнездах
I I I I | I I ।_______I--
V й й Ю 22 М /6 /й 20 22
Vaaf суток
к птенцам. Вот почему в пасмурную погоду наблюдается более
размеренный ритм как выкармливания, так и обогрева. В дождли-
вую погоду возрастала не только интенсивность насиживания
птенцов, но и интенсивность прилетов с кормом.
Таким образом, для садовой камышевки в период выкармли-
вания птенцов, как и для всех птенцовых, свойственна высокая
степень лабильности поведения, которая выражена в ежесуточном
изменении бюджета рабочего времени, что в первую очередь
определяется взаимовлиянием птенцов и родителей, самца и самки.
ЛИТЕРАТУРА
Дольник В. Р. Суточные и сезонные циклы локомоторной активности зябликов
(Fringilla с. coelebs) // Физиологические основы миграционного состояния
птиц. Л.: Наука, 1976. С. 3—16. (Тр. Зоол. ин-та АН СССР; Т. 60).
Промптов А. Н. Изучение суточной активности птиц в гнездовой период // Зоол.
журн. 1940. Т. 19, № 1. С. 143-159.
Сазанов Л. И., Дейнекина Т., Алхасова Ж. Активность птиц в гнездовой период //
Учен. яап. Кабард.-Валкар. ун-та. 1961. Вып. 10. С. 205—208.
Слоним А. Д. Основы общей экологической физиологии млекопитающих. М.;
Л.: Изд-во АН СССР, 1961. 432 с.
177
Слоним А. Д. Среда и поведение. Л.: Наука, 1976. 240 с.
Яблонкевич М. Л. Годовые и суточные ритмы веса печени и составляющих его
компонентов у зябликов (Fringilla с. coelebs) // Физиологические основы
миграционного состояния птиц. Л.: Наука, 1976. С. 63 — 76. (Тр. Зоол. ин-та
АН СССР; Т. 60).
SUMMARY
Behavioral rythms and time budget during the hatching period of Blyth’s Reed Warbler
(Aerocepkalus dumetorum). M о s k v i t i n S. S., A n a n j i n a T. L.
The observations after 12 nests of 4 Blyth’s Reed Warbler nestlings were carried out.
The total volume of observations came to 230 hours. The feeding rythm of the nestlings
by lease indices was characterized by a 4-peaked curve. The activity peaks fall
on the sections 5—6 a. m., 9—10 a. m., 3—4 p. in., 7—8 p. m. Every next peak
has its smaller absolute value than the previous ones. On hot days the decrease
of the activity was marked. The high degree of behaviour lability is typical for the
Blyth’s Reed Warbler, expressed in a daily change of time budget, that can be
determined, in the first place, by the mutual influence of the nestlings and parents,
the male and female.
МОРФОЛОГИЯ
УДК 591.473:598.6
Б. Ф. СЫЧ
432600, Ульяновск,
бульвар Новый Венец, д. 1.
Ульяновский сельско-
хозяйственный институт
Морфологические
особенности мускулатуры
тазовой конечности
большеногое и краксов
(Galliformes;
Megapodiidae, Gracidae)
Среди представителей отряда куриных (Galliformes) особый инте-
рес для теоретической орнитологии, в первую очередь филогенети-
ки и систематики, представляет наиболее четко обособленная
группа, сорных кур или болыпеногов (Megapodiidae) и краксов
(Cracidae), таксономический статус которых в сравнении с дру-
гими представителями отряда определяется большинством систе-
матиков однозначно (Мауг, Amadon, 1951; Wetmore, 1960; Brod-
korb, 1964; Vaurie, 1968; Howard, Moore, 1980; и др.). Непосред-
ственным объектом интереса являются как специфические и даже
уникальные для отряда и класса особенности экологии предста-
вителей этих семейств, так и признаваемая рядом авторов (Brod-
korb, 1964; Holman, 1964; Hudson et al., 1966; Delacour, Amadon,
1973; Роме, 1978; и др.) примитивность их организации и большее,
чем у остальных куриных, сходство с предполагаемой предковой
для отряда формой.
Хотя эта точка зрения и выглядит на первый взгляд убедитель-
ной, тем не менее продолжает оставаться недостаточно обосно-
ванной и аргументированной данными глубоких специальных
исследований.
Успешно использовав морфобиологический метод при изучении
челюстного аппарата куриных и, в частности, одного из предста-
вителей краксов пенелопы (Penelopa jacucaca) и проанализировав
с учетом данных по экологии соответствующих видов его морфо-
функциональные особенности, Ф. Я. Дзержинский (1977, 1980)
пришел к оригинальному заключению об архаичности конструкции
челюстного аппарата краксов и показал возможный путь его даль-
нейшей перестройки у представителей других семейств куриных.
В связи с этими выводами возрос интерес к посткраниальному от-
делу скелетно-мышечной системы куриных и особенно к локомо-
торному аппарату, так как передвижение представляет одну из
179
наиболее существенных сторон жизни птиц, а различные его спо-
собы имели большое значение в эволюции класса в целом (Позна-
нии, 1978).
Посвященные мускулатуре грудных и тазовых конечностей ку-
риных работы Г. Хадсона с соавторами (Hudson et al., 1959; Hud-
son, Lanzillotti, 1964), располагавших обширным материалом для
анатомического исследованияя, к сожалению, не восполнили про-
бел в морфологическом изучении отряда. Главная ценность этих
работ заключается, пожалуй, в том, что они являются поучитель-
ным образцом низкорезультативной утилизации ценного в морфо-
логическом отношении материала. Задавшись целью получить как
можно большее количество цифровых данных для математической
обработки и последующего нумерического систематического ис-
следования, авторы ограничились субъективной оценкой в баллах
таких характеристик, как форма, длина, ширина и толщина брю-
шек мышц, тканевой состав начального отдела мышцы, длина сухо
жилий и наличие внутрисухожильных сесамоидов и пр. Вне поля
зрения авторов остались практически все необходимые для морфо-
функционального анализа аппарата морфологические особенности.
Поэтому основная морфологический ценность их работ свелась
лишь к обстоятельным данным по топографии мускулатуры до-
машней курицы и дымчатого тетерева, избранных авторами
в качестве модельных объектов исследования.
Учитывая упомянутое состояние изученности морфологии ку-
риных, в том числе большеногой и краксов, мы воспользовались лю-
безностью проф. X. Сика (Бразильский музей естественной исто-
рии) и Ш. Паркера (Южно-Австралийский музей естественной
истории), предоставивших в наше распоряжение фиксированные
трупы пипиле (Aburria ptpile), глазчатого кракса (Стах fasciolata)
и глазчатой курицы (Leipoa ocellata) и провели исследование
мускулатуры их тазовых конечностей. В ходе исследования строе-
ние мускулатуры болыпеногов и краксов сравнивалось со строени-
ем мускулатуры ранее изученных нами видов семейства фазано-
вых, а также птиц других отрядов (Anseriforines, Gruiformes,
Charadriiformes и Columbiformes). Исходя из наиболее аргументи-
рованного фактическим материалом представления о филогенезе
куриных и близкородственных им отрядов птиц (Fiirbringer, 1888),
мы на основе сравнительно-анатомического анализа установили
предковые морфологические особенности аппарата двуногой локо-
моции болыпеногов и краксов, утраченные остальными куриными,
по сохранившиеся в отдельных случаях у журавлеобразных и в
меньшей степени у гусеобразных и ржанкообразных. При сравни-
тельной оценке в рассматриваемом ряду преобразований мышц,
состоящих из нескольких головок или частей, мы руководствова-
лись созданным на основе результатов нашего исследования пред-
ставлением о примитивности универсальной наземно-древесной
локомоции предковой для отряда куриных формы. На последую-
щем этане адаптации тазовых конечностей к преимуществен-
нодревесной (Cracidae, Megapodiidae) или наземной (Phasianidae)
180
локомоции функция мускулатуры тазовых конечностей в целом
упростилась в связи с более выраженной стереотипностью движе-
ний. Результатом взаимосвязанного преобразования морфологии
тазовых конечностей явилось слияние порций и головок мышц или
их полная редукция, отмеченные в более яркой форме у предста-
вителей других, в том числе близкородственных куриным отрядов
птиц, представители которых обладают узко специализированными
тазовыми конечностями.
Примитивные (архаичные) особенности мускулатуры,
сохранившиеся у современных представителей большеногое
и краксов
Мускулатура тазовой конечности болыпеногов и краксов характе-
ризуется наиболее примитивным по сравнению с индейковыми,
тетеревиными и фазановыми строением. Так, у представителей
обоих семейств сохранились наиболее дифференцированные m.fe-
morotibialis medius и m.femorotibialis externus, вторичное сращение
которых менее выражено, чем у других куриных. Обширную об-
ласть отхождения, включая латеральную поверхность processus
dorsolateralis ilium, имеет m. flexor cruris lateralis (рис. 1, Л), кото-
рый у других куриных сужается и «оттесняется» в каудальном
направлении мощно развитым каудальным отделом m.iliotibialis
lateralis и начинается поэтому только от двух краниальных свобод-
ных хвостовых позвонков, каудальной поверхности ischium и
мембраны между хвостовыми позвонками, ischium и ilium.
M.iliotibialis lateralis, усиление которого отличает адаптации
тазовой конечности фазановых к быстрому передвижению по по-
верхности земли, развит слабо у болыпеногов и краксов, как у ин-
дей-ковых и тетеревиных. Однако, если ослабление мышцы у тетере-
виных, вероятно, результат более поздней перестройки их ло-
комоторного аппарата (Сыч, 1986), то отмеченная особенность
большеногой, краксов и индейковых, на наш взгляд, примитивная,
унаследованная от предковой формы, обладавшей слабо адапти-
рованным к быстрому передвижению по поверхности земли аппа-
ратом двуногой локомоции.
Большеногой и краксов отличает короткая область отхождения
m.iliofibularis, каудальный отдел которого начинается лишь от
краниальной половины (Стах), или большей части (Leipoa, Abur-
ria) длины crista iliaca dorsolateralis. Наиболее дистально, причем в
сравнении не только с другими курообразнымн, но и исследован-
ными представителями гусеобразных, журавлеобразных, ржанко-
образных и голубеобразных, расположено у болыпеногов и краксов
окончание m.obturatorius medialis на бедренной кости. Локализа-
ция окончания дистальнее оси вращения головки бедра создает воз-
можность участия мускула в ретракции и аддукции бедра. По-
скольку у остальных куриных и многих других птиц он узко спе-
циализирован как супинатор, его полифункциональность у мегапод
и краксов мы склонны считать унаследованной от предков особен-
ностью примитивной универсальной коиечйости.
181
Рис. 1. Мускулатура
области таза, бедра и
голени у Стах fasciolata
(Л) и у Aburria pipile
(Е). Начала мышц р.
Fdb
Gf
Fcbp
pelvica m. flexor cruris
lateralis (Fclp) m. ilio-
fibularis (Ifb) отрезаны
и их брюшки отведены
вниз (Б), вид сбоку
Icr — m. iliotibialis cra-
nialis;
Il — m. iliotibialis latera-
lis;
Itc — m. iliotrochanteri-
cus caudalis;
Iter — m. iliotrochanteri-
cus cranialis;
Ifb — m. iliofibularis;
Ife — m. iliofemoralis ex-
tern us;
Fe — m. femorotibialis ex-
tern us;
Fm — m. femorotibialis
medius;
Fem — m. flexor cruris
medialis;
Fclp — p. pelvica m. flexor
cruris lateralis:
Cf — m. caudofemoralis;
Isf — m. ischiofemoralis:
Pifl — p. lateralis m. pu-
bo isch iof e moral is;
Gl — p. lateralis m. gast-
rocnemius;
Fpp II, Fpp III — m. fle-
xor perforans et perfo-
ratus digiti II seo di-
giti III;
Fdl — m. flexor digitorum
longus
Fem
FppUL
FppI
Окончание m.caudo-iliofemoralis на бедренной кости располо-
жено у большеногов и краксов проксимальнее: если у других ку-
риных m.caudofemoralis крепится на границе проксимальной и
средней трети длины femur или даже дистальнее ее, то у больше-
ногов и краксов — проксимальнее этой границы (см. рис. 1, Б).
Укорочение плеча приложения силы m.caudo-iliofemoralis, возмож-
но, было вызвано необходимостью увеличения как угловой скоро-
сти движения бедра для повышения быстроты (резкости) движе-
ний конечности в целом, так и максимальной амплитуды активных
движений бедра *, что обусловливалось, вероятнее всего, при-
способлением к своеобразным, наиболее мощным среди куриных,
прыжкам как основному способу быстрого передвижения в кронах
деревьев (Bent, 1932; Berndt, Meise, 1962). Однако следствием
укорбчения плеча приложения силы мышцы было уменьшение ее
вращающего момента, что могло оказаться основной причиной
сохранения ретрагирующей функции m.obturatorius medialis,
а также увеличения у большеногов относительной массы m.caudo-
iliofemoralis.
Из других особенностей, указывающих на примитивность му-
скулатуры тазовой конечности большеногов и краксов, следует от-
метить широкое проксимальное крепление rn.il iofemoralis externus,
в частности сохранившуюся его каудальную сухожильную порцию
(особенно хорошо дифференцированную у краксов), отходящую
от краниальной части crista iliaca dorsolateralis (рис. 1, Б). Прокси-
мальный конец мышцы, очень широкий в целом у Leipoa, несколько
сужен за счет каудального отдела у Aburria и Стах. У других кури-
ных каудальный сухожильный отдел развит слабее (некоторые фа-
зановые) или же почти полностью редуцирован (индейковые,
тетеревиные и большинство фазановых).
Об особой примитивности большеногов и краксов в сравнении с
другими куриными свидетельствует строение сгибателей и разги-
бателей пальцев. Прежде всего здесь необходимо отметить мощное
развитие брюшек прободенных сгибателей пальцев, их большую от-
носительную длину, чем определяется неблагоприятно сказываю-
щееся на эффективности передвижения по поверхности земли (эко-
номичность, быстрота и пр.) повышение момента инерции конеч-
ности. Эта особеннрсть характеризует также мышцы области цевки,
в первую очередь длинные и наиболее мощные среди куриных
m.flexor hallucis brevis, m.abductor digiti II, m.abductor digiti IV,
m.extensor proprius digiti III и m.extensor brevis digiti IV
(рис. 2, Б). Более примитивным следует считать окончание сухо-
жилия m.abductor digiti II большеногов и краксов вблизи дисталь-
ной 1/5 длины медиальной поверхности проксимальной фаланги,
а не на проксимальном ее конце, как у других куриных и боль-
шинства птиц. Сухожильная оттяжка к суставному блоку metatar-
* М. caudofemoralis имеет наиболее длинные среди ретракторов бедра мышечного
волокна и оказывает поэтому решающее влияние на максимально возможную
скорость активной ретракции бедра.
183
Рис. 2. Мускулатура области голени и цев-
ки Leipoa ocellata;
А — каудо-латеральная поверхность голе-
ни; Б — Краниальная; В — каудальная по-
верхности цевки
Ab II — m. abductor digiti II; Ab IV — т.
abductor digiti IV; Ad II — т. abductor digiti II;
aif — ansa m. iliofibularis; Eb IV — m. extensor
brevis digiti IV; Ehld — p. distalis m. extensor
hallucis longus; Ehlp — p. proximalis m. exten-
sor hallucis longus; Ep III — m. extensor prop-
rius digiti III; Fb — m. fibularis brevis; Fl — m.
fibularis longus; Fdl — m. flexor digitorum lon-
gus; Fpcrl III — кранио-латеральная головка
m. flexor perforatus digiti III; Fpcl IV — каудо-
латеральная головка m. flexor perforatus digiti
IV; Fpcrl IV — латеральная головка m. flexor
perforatus digiti IV, Fpm III — медиальная
головка m. flexor perforatus digiti III, Fpm IV —
медиальная головка m. flexor perforatus digiti
IV; Icr —,m. iliotibialis cranialis; tarn — ди-
стальное сухожилие m. ambiens
FpcrLlU
EhLp
FpcLIZ
FL
Lam
Fpm IK
ЕртШ
*~FpcrLJZ
FPL
Fhb
РЫТ
Ehbd
EbLd
184
sale III (см. рис. 2, Б), ограничивающая абдуцирующий эффект
мускула, очевидно, благоприятствовала укорочению дистального
сухожилия.
Наряду с рассмотренными выше общими для большеногов и
краксов примитивными чертами строения мускулатуры аппарата
двуногой локомоции выявлены специфические, характерные толь-
ко для Megapodiidae или. же только для представителей Cracidae
примитивные особенности. В частности, у Leipoa отмечены более
дистальное окончание ротаторов бедра — mrn.iliotrochanterici и
m.ischiofemoralis (у большинства птиц, в том числе у других ку-
риных, преобразование шло по пути смещения места фиксации в
проксимальном направлении), а также наиболее длинные среди
куриных брюшки хорошо развитых m.ambiens и m.fibularis longus.
Более дистальное прикрепление mm.ischiofemoralis и iliotrochante-
rici создает возможность для возникновения абдуцирующего эф-
фекта, компенсирующего слабое развитие m.iliotibialis lateralis.
Только у Aburria сохранилась дистальная ветвь сухожилия m.fibu-
laris longus, слитая в области цевки с сухожилием in.flexor perfora-
tus digiti III. Прободенные сгибатели третьего и четвертого пальцев
у Leipoa имеют хорошо обособленные кранио-латеральные, а также
каудо-латеральную (m.flexor perforatus digiti IV) головки, которые
у других птиц (Cracidae, Anseriformes, Charadriiformes, Columbifor-
mes) подверглись частичной редукции или полному осухожилива-
нию и по-разному выраженному слиянию в единое брюшко с меди-
альной головкой (см. рис. 2, А). М. plantaris хорошо развит как у
Aburria, так и у Стах; брюшко его простирается за пределы прок-
симальной половины голени (у других птиц он ослаблен, и его дли-
на лишь у курообразных достигает трети длины tibiotarsus).
Заслуживает внимания строение m.flexor hallucis brevis у Lei-
poa: в брюшке, медиальнее его средины, расположен ориентиро-
ванный в парасагиттальной плоскости внутренний апоневроз, от
противоположных поверхностей которого отходят мышечные во-
локна, формирующие двуперистую структуру. Дистальные концы
волокон прикрепляются к внутренней поверхности наружного апо-
невроза каудальной поверхности брюшка, переходящего в дисталь-
ное сухожилие. Последнее разделено за пределами брюшка
на две ветви, оканчивающиеся обособленными расширениями на
плантарной поверхности основания hallux (см. рис. 2, В). Так как
у исследованных пастушковых, ржанковых и бекасовых рассмат-
риваемый мускул представлен двумя самостоятельными однопе-
ристыми брюшками, оканчивающимися обособленными сухожили-
ями (рис. 3), его структуру у Leipoa мы склонны рассматривать как
сохранившую следы предшествовавшего ей двумышечвого состо-
яния. В сравнении с прикрепляющейся единым сухожилием
мышцей других куриных m.flexor hallucis brevis Leipoa имеет бо-
лее примитивную структуру.
185
Общие для болыпеногов и краксов особенности
специализации мускулатуры
Мускулатуру мегапод и краксов характеризуют некоторые чер-
ты строения, которые являются результатом более глубокого, чем у
других куриных, приспособления их локомоторного аппарата к
древесному образу жизни. Некоторые из них обнаружены в более
или менее четко выраженном состоянии у тетеревиных, лучше, чем
индейковые и фазановые, приспособленных к передвижению в кро-
нах деревьев, а также «древесных» представителей Columbidae,
что подтверждает упомянутую трактовку их адаптации. Принимая
во внимание существенные различия местообитаний и образа жиз-
ни современных мегапод и краксов, можно полагать, что более
глубокие, чем у остальных куриных, адаптации к древесному об-
разу жизни сформировались на совместно пройденном отрезке
филогенеза — начальном этапе эволюции отряда. Среди них можно
выделить наличие (сохранение) p.distalis m.extensor hallucis lon-
gus, сильнее, чем у других куриных, развитую перистую структуру
Рис. 3. Мускулатура области цевки Crex
сгех, каудальная поверхность
Ab IV — m. abductor digiti IV;
Ad II — т. adductor digiti II;
Fhbl — латеральная часть zm. flexor hallucis
brevis;
Fhbm — медиальная часть m. flexor hallucis
brevis
Рис. 4. Мускулатура области голени Abur-
ria pipile (А) и дистальные сухожилия сги-
бателей второго пальца Leipoa ocellata (Б),
латеральная поверхность. Брюшко кранио-
латеральной головки ш. flexor perforatus di-
giti IV отделено от апоневроза общего с
кранио-латеральной головкой m. flexor per-
foratus digiti III и сдвинуто каудально (А)
Fb — m. fibularis brevis;
Fdl — m. flexor digitorum longus;
Frcl IV — каудо-латеральная головка m. flexor
perforatus digiti IV;
Fpcrl III — кранио-латеральная головка m.
flexor perforatus digiti III;
Fpcrl IV — кранио-латеральная головка m. fle-
xor perforatus digiti IV;
Fpm III — медиальная головка m. flexor perfo-
ratus digiti III;
Fpm IV — медиальная головка m. flexor perfo-
ratus digiti IV;
Icr — m. iliotibialis cranialis;
tjp II и tfpp II — сухожилия m. flexor perfora-
tus et perfopans digiti II;
v — viniculum
186
сгибателей пальцев и особенно увеличивающую поверхность при-
крепления мышечных волокон продольную складчатость апоневро-
за ш.flexor hallucis brevis, двуперистое строение m.abductor digiti
IV, а также сходное у Leipoa и Aburria минимальное среди курооб-
разных количество эластических ответвлений сухожилий m.flexor
digrtorum longus.
Эти черты строения дополняются рассмотренными ранее об-
щими примитивными особенностями, так как происхождение по-
следних обусловливалось универсальной наземно-древесной локо-
моцией, свойственной и предкам куриных, а максимальное сохра-
нение этих признаков филетической линией мегапод и краксов оп-
ределялось как ранним вступлением ее представителей на путь глу-
бокой адаптации, так и сущностью последней. Ранний этап адап-
тации локомоторного аппарата болыпеногов и краксов заключался,
по-видимому, в незначительной перестройке унаследованной от
предков универсальной конструкции в связи с углублением адап-
таций -к древесному образу жизни. Доказательством совместной
эволюции болыпеногов и краксов на начальном этапе становления
187
отряда, а также существования длительного этапа их обособленного
развития от другой фазианоидной филетической линии куриных
являются общие для первых черты строения мускулатуры конеч-
ности, отсутствующие у всех исследованных представителей по-
следних (Meleagridinae, Tetraoninae, Phasianinae). Оценка их
адаптивного значения у современных мегаиод и краксов затрудне-
на. В числе таковых можно отметить проксимальное сращение
между собой трех порций m.gastrocnemius и локализацию хрящево-
го утолщения не в латеральной, как у других курообразных, а в
медиальной ветви сухожилия m.flexor perforatus digiti И, а также
наличие перемычки от суставного блока metatarsale III, ограничи-
вающей абдуцирующий эффект m.abductor digiti II.
Адаптивные особенности, приобретенные на этапе
обособленной эволюции большеногов и краксов
Черты узкой адаптации аппарата двуногой локомоции, обличаю-
щие мегапод и краксов друг от друга, возникли на этапе их обособ-
ленного филогенеза. У краксов приспособление к древесному об-
разу жизни, отразившееся в комплексе морфологических и
экологических особенностей, включая кормление, гнездование и
передвижение, в значительной степени предопределило эволюцию
аппарата локомоции. С одной стороны, это развитие способности к
планированию, с другой — к быстрому передвижению прыжками
с ветки на ветку за счет поочередных толчков конечностями с
целью достижения верхушки деревьев как отправной точки плани-
рующего полета (Bent, 1932; Berndt, Медее, 1962). Эти экологи-
ческие особенности формировались в неразрывной взаимосвязи и
обусловили ряд специфических для краксов перестроек как скелета
(сужение таза, удлинение конечности и особенно цевки), так и
мускулатуры тазовой конечности.
Среди особенностей мускулатуры краксов, отличающих их от
большеногов, в первую очередь необходимо отметить продолже-
ние медиальной порции сухожилия m.gastrocnemius до основания
проксимальной фаланги первого пальца, осухожиливание одной из
головок m.flexor perforans et perforatus digiti II, ослабление (у
Aburria) либо полную редукцию (у Стах) кранио-латеральной го-
ловки m.flexor perforatus digiti IV (рис. 4, А), редукцию короткого
латерального ответвления сухожилия m.flexor perforatus digiti IV
к плантарной поверхности основания четвертой фаланги четвертого
пальца, максимальную среди курообразных выраженность струк-
тур фиксирующего механизма на сухожилиях m.flexor digitorum
longus и т.flexor hallucis longus, двуперистую структуру m.exten-
sor hallucis longus, m.extensor brevis digiti IV, мощное развитие
сгибателей пальцев и m.piantaris, ослабление, в свою очередь,
m.iliotibialis lateralis и пронаторов бедра.
Нами обнаружены также специфические для Aburria черты
строения мускулатуры тазовой конечности, ставящие под сомнение
выводы Ч. Нори (Vaurie, 1968) о большей примитивности Penelopi-
188
ni в сравнении с более специализированными Cracini: .максималь-
ное развитие промежуточного апоневроза m.iliofemoralis и полное
разделение брюшка на каудальный и краниальный мышечные от-
делы, развитие мощного, начинающегося от crista cnemialis latera-
lis, цроксимального апоневроза m.flexor perforans et perforatus di-
giti II, сращение брюшка кранио-латеральной головки m.flexor per-
foratus digiti II с брюшком медиальной головки и сохранение в
обособленном виде только сухожильного начала первой, а также
обнаруженное только у Aburria полное сращение дистального сухо-
жилия m.ambiens с каудальной поверхностью сухожилия patella.
В связи с этим сухожилие m.ambiens не продолжается, как у других
курообразных, в проксимальное сухожилие кранио-латеральных
головок прободенных сгибателей. Начало их у Aburria расположено
не на латеральной поверхности дистального эпифиза femur, как у
других куриных, а смещено дистально и находится на латеральной
поверхности fibula в конце проксимальной четверти голени (см.
рис. 4, А). Поэтому влияние сокращения кранио-латеральных го-
ловок на амплитуду перемещения сухожилия прободенных сгиба-
телей не зависит от положения в коленном суставе *.
Исходя из нашего представления об аппарате двуногой локомо-
ции предковой для отряда куриных формы, можно считать, что у
видов рода Стах, способного дольше и более эффективно передви-
гаться по земле, чем у Aburria (Bent, 1932; Delacour, Amadon,
1973), двуногая локомоция более универсальная**. Поэтому
Cracini по строению аппарата двуногой локомоции, на наш взгляд,
менее удалены от предковой формы, чем представители трибы
Penelopini, вопреки мнению Ч. Вори (Vaurie, 1968), полагавше-
го, что у краксовых адаптация к древесному образу жизни первич-
на, а к наземному — относительно вторична.
Специализация многих представителей Megapodiidae на этапе
обособленного от Cracidae филогенеза включала адаптацию к пре-
имущественно наземному образу жизни и становление комплекса
морфологических и экологических особенностей, обеспечивающих
специфику размножения большеногов, в частности «инкубацию»
яиц (Clark, 1964а,b). С эволюцией размножения неразрывно вза-
имосвязана интенсификация функции разгребания тазовыми ко-
нечностями у мегапод. С представлениями Г. Кларка о примитив-
ных и специализированных современных болыпеногах, в частно-
сти с отнесением к первым кустарниковых индеек (Alectura, Tale-
galla, Aepypodius и представителя рода Leipoa), а ко вторым —
более адаптированных к наземному образу жизни представителей
родов Megapodius и Macrocephalon, в целом согласуется предлагае-
мая схема развития адаптаций аппарата двуногой локомоции пред-
* При максимально согнутом положении в коленном суставе сохраняется возмож-
ность максимального сгибания пальцев, необходимого для охватывания тон-
ких ветвей.
** По данным А. Бента (Bent, 1932), другой представитель Penelopini (род
Penelopa) весьма неуклюже передвигается по поверхности земли.
189
ков современных болыпеногов. В ней мы выделяем следующие
этапы: 1) примитивная универсальная наземно-древесная локо-
моция; 2) углубление адаптаций преимущественно к древесной
локомоции; 3) развитие адаптаций к преимущественно наземной
локомоции и наиболее интенсивному среди куриных разгре-
банию.
Среди характерных только для мегапод особенностей мускула-
туры локомоторного аппарата следует выделить удлиненное брюш-
ко и более каудальное начало m.ischiofemoralis, укороченную ме-
диальную порцию сухожилия ш.gastrocnemius и формирование
бугорка в области дистальной 1 /5 медиальной поверхности tarsome-
tatarsus, с которым срастается дистальный конец упомянутого су-
хожилия, двуперистую (в отличие от одноперистой у других ку-
риных) структуру M.flexor hallucis brevis, относительно более
мощное развитие на фоне других сгибателей пальцев m.flexor digi-
torum longus, усиление m.tibialis cranialis, более выраженную, чем
у краксов, дифференцировку (обособленность) трех головок
m.flexor perforatus digiti IV.
Уникальна перестройка сгибателей второго пальца. Брюшко
медиальной головки m.flexor perforatus digiti II у Leipoa развито
слабо и начинается от единого апоневроза медиальных головок на-
иболее дистально. Дистальное сухожилие мышцы, миновав канал
hypotarsus, становится окостеневшим, причем основание сесамоида
расширено до такой степени, что лишает последний возможности
входить в канал (см. рис. 4, Б). Продолжением короткого сесамои-
да является эластическая связка, с. которой сращен vinculum су-
хожилия m.flexor perforans et perforatus digiti II. Дистальнее этого
места сращения оно содержит сесамоид, который затем переходит в
типичное сухожильное окончание на проксимальном конце основ-
ной фаланги. Это окончание функционально подчинено уже m.fle-
xor perforans et perforatus digiti II, тогда как m.flexor perforatus
digiti II превратился, по сути, в разгибатель цевки — синергист
m.gastrocnemius. С этим, очевидно, взаимосвязано и его ослабление,
в наибольшей степени отразившееся на медиальной головке. Таким
образом, типичная схема окончания m.flexor perforans et perforatus
digiti II усложнена наличием упомянутого vinculum, посредством
которого этот мускул соединил в себе функции обоих сгибателей
второго пальца.
Заключение
Изложенные особенности морфологии мускулатуры тазовой конеч-
ности позволяют заключить, что исходная для отряда куриных
предковая форма, очевидно, обладала универсальным (лазающе-
ходящим) аппаратом двуногой локомоции, обеспечивавшим
сочетание слабо специализированного древесного и наземного пе-
редвижения при отличающемся незначительной двигательной ак-
тивностью в целом оседлом образе жизни. Примитивные черты уни-
версальной конструкции аппарата двуногой локомоции предковой
190
для отряда формы сохранились в более или менее выраженном виде
у всех современных куринообразных. Для всех исследованных
нами семейств отряда характерно наибольшее для птиц число
мышц тазовой конечности, их дифференцированных головок и пор-
ций, мощное развитие мускульной массы в целом, особенно в об-
ласти голени и цевки, примитивные особенности топографии
m.iliotibialis lateralis, m.flexor cruris medialis, m.flexor cruris latera-
lis, m.fibularis brevis, m.adductor digiti II и m.abductor digiti IV,
а также структуры и дистального прикрепления m.flexor hallucis
longus, m.flexor hallucis brevis, m.flexor perforans et perforatus digi-
ti III, m.flexor perforatus digiti IV и m.extensor digitorum longus.
Большеноги и краксы образуют единую, резко обособленную
от других куриных филетическую линию, начальный этап отделе-
ния которой происходил в связи с углублением адаптаций к древес-
ному образу жизни. Это подтверждается их сходными, отсутствую-
щими у других куриных морфологическими особенностями, явля-
ющимися в своем большинстве результатом более глубокого при-
способления к древесной локомоции. Сохранение менее измененной
в сравнении с другими куриными предковой для отряда конструк-
ции локомоторного аппарата при достаточно глубокой адаптации
тазовых конечностей у современных представителей как большено-
гое, так и краксов позволяет предположить, что каждая из этих
групп раньше других куриных вступила на путь специализации.
Перестройка в направлении приспособления к обитанию в кронах
деревьев и выработки специфической формы передвижения мощ-
ными прыжками в тропическом лесу у краксов или
частичной утраты ряда особенностей, связанных с древесным
образом жизни, и приспособление к более продолжительному
пребыванию и передвижению по поверхности земли в связи с
формированием экологических особенностей у современных боль-
шеногов отражена в отмеченных различиях строения мускулатуры,
которые подтверждают обоснованность рассмотрения каждой
группы в ранге семейства. Наконец, в свете изложенных данных не
вызывает сомнений филогенетическая древность Megapodiidae и
Cracidae в сравнении с более молодыми Phasianidae (Meleagridinae,
Tetraoninae и Phasianinae).
ЛИТЕРАТУРА
Дзержинский Ф. Я. О значении биомеханического анализа в реконструкции фило-
гении птиц//Адаптивные особенности и эволюция птиц. М.: Наука, 1977.
С. 53-60.
Дзержинский Ф. Я. Адаптивные преобразования челюстного аппарата в эволюции
куриных // Морфологические аспекты эволюции. М.: Наука, 1980. С. 148—158.
Познании Л. П. Экологические аспекты эволюции птиц. М.: Наука, 1978. 148 с.
Сыч В. Ф. Морфоэкологические особенности аппарата двуногой локомоции кури-
ных в связи с возможными путями эволюции отряда // Изучение птиц СССР,
их охрана и рациональное использование. Л., 1986. Ч. 2. С. 268—269.
Bent А. С. Life histories of North American Gallinaceous birds // Smithsonian Inst.
US Nat. Mus. Bull. 1932. Vol. 162. P. 1-490.
Berger-Dell’mour H. Her Ubergang von Echse zu Schleiche in der Gattung Tetradacty-
lus, Merrem // Zool. Jb. Anat. 1983. Bd. 110, N 1. S. 1 —152.
191
Berndt В., Meise W. Naturgeschichte der Vogel. Stuttgart: Kosmos, 1962. Bd. 2. 673 S.
Brodkorb P. Catalogue of fossil birds. Pt 2. Anseriformes through Galliformes // Bull.
Fla. Slate Mus. 1964. Vol. 8, N 3. P. 195-335.
Clark G. A. Life histories and the evolution of Megapodes//Living Bird. 1964a.
P. 149-167.
Clark G. A. Ontogeny and evolution in the Megapodes (Aves: Galliformes) // Postilla.
1964b. Vol. 78. P. 1-37.
Delacour J., Amadon D. Corassows and related birds. New York: Amer. Mus. Natur.
Hist., 1973. 247 p.
Fiirbringer M. Untersuchungen zur Morphologic und Systematik der Vogel. Amster-
dam; Jena, 1888. Bd. 1/2. 1751 S.
Holman /. A. Osteology of Gallinaceous birds // Quart. J. of Fla. Sci. 1964. Vol. 27,
N 3. P. 230-252.
Howard R., Moore A. A complete checklist of the birds of the world. Oxford, etc.,
1980. 701 p.
Hudson G. E., Lanzilotti P. I. Muscles of the pectoral limb in Galliform birds // Amer.
Midland Natur. // 1964. Vol. 71, N 1. P. 1-113.
Hudson G. E., Lanzilotti P. I., Edwards P. Muscles of the pelvic limb in Galliform
birds // Ibid. 1959. Vol. 61, N 1. P. 1—67.
Hudson G. E., Parker H. A., Berge I. V., Lanzilotti P. I. A numerical analysis of the
modifications of the appendicular muscles in various genera of Gallinaceous
birds // Ibid. 1966. Vol. 76, N 1. P. 1-73.
Mayr E., Amadon D. A classification of recent birds//Amer. Mus. Novit. 1951.
N 1496. P. 1-42.
Rome A. Considerazioni sulla sistematica del galliformi // Nature. 1978. Vol. 69, N 3/4.
P. 146-152.
Stolpe M. Physiologisch-anatornische Untersuchungen uber die hintere Extremitat //
.1. Ortithol. 1932. Bd. 2. S. 162-247.
Vaurie Ch. Taxonomy of the Cracidae (Aves) // Bull. Amer. Mus. Natur. Hist. 1968.
Vol. 138, N 4. P. 135 -256.
Wetmore A. A classification for the birds of the world // Smithsonian MiscelL Coll.
1960. Vol. 139, N 11. P. 1-37.
SUMMARY
Morphological peculiarities of the pelvic musculature of megapods and cracids
(Galliformes; Megapodiidae, Cracidae).
Sytch V. F.
The comparative-morphological study of the pelvic musculature showed the simila
rity of the bipedal locomotion of representatives of Megapodiidae and Cracidae, and
its essential distinctions from those of Pasianidae (Meleagridinae, Tetraoninae,
Phasianinae). In spite of the adaptation specifics of the recent representatives, the lo-
comotion apparatus of Megapods and Cracids had preserved a bigger archaism than
that of the other Galliforms, and the similarity with the universal climbing-walking
apparatus of the ancestral for the order form. It is confirmed that the Megapods and
Cracids form the oldest phylogenetic branch of Galliforms. It is assumed that the iso-
lation of this branch at the early stage of the order evolution was connected with the
deeper development of the «wood» adaptations complex than that of other Galliformes.
192
УДК 591.473:598.826.1
Е. А. ДЯДИЧЕВА,
А. Н. ЦВЕЛЫХ
332339, Запорожская обл.,
Мелитополь, ул. Ленина, д. 20.
Педагогический институт,
Орнитологическая станция
Морфо-экологический
анализ
органов полета
трех видов
вьюрковых
Сравнительные исследования органов полета близких видов птиц с
учетом различий в образе жизни позволяют выявить тонкие разли-
чия в морфологии, обусловленные их экологической специализа-
цией. Имеется очень немного работ, в которых сравнительные мор-
фологические исследования органов полета проводились бы на ви-
дах с хорошо изученной экологией, близких систематически и не
очень различающихся размерами. Последнее условие крайне важ-
но, так как многие морфологические показатели зависят от абсо-
лютных размеров птиц. Игнорирование этого обстоятельства часто
приводило к ошибочным выводам.
Объектом нашего исследования были выбраны три достаточно
близких и достаточно изученных вида семейства вьюрковых: зяб-
лик (Fringilla coelebs), юрок (F. montifringilla) и чиж (Spinus
spinus).
В этой работе мы попытались проверить, насколько морфологи-
ческие различия в органах полета у близких видов птиц соответ-
ствуют различиям их экологии.
Материал и методика
Материал для данного исследования собирался в середине сентяб-
ря — октябре 1981—1984 гг. в период массовой миграции воробь-
иных птиц вдоль левого берега Киевского водохранилища. В иссле-
довании использованы промеры птиц, отлавливавшихся для пара-
зитологических исследований или погибших при случайном стол-
кновении с ловушкой (в этом месте действует большая ловушка
рыбачинского типа, принадлежащая Институту зоологии АН
СССР). Для сравнения органов полета исследуемых птиц использо-
вались индексы, наиболее часто применяемые в подобных исследо-
ваниях (Гладков, 1949). Для характеристики степени заостренно-
сти вершины крыла использовался индекс заостренности крыла
(Цвелых, 1983). С целью исключения возможного влияния пола
и возраста исследованных птиц на результаты работы нами мето-
дом случайного подбора было отобрано по 18 птиц каждого вида
так, чтобы в каждой из групп половозрастные соотношения были
идентичны. Пол и возраст устанавливались по определителю (Ви-
ноградова и др., 1976).
193
Результаты и обсуждение
Сравнение индексов органов полета вьюрковых (см. таблицу) пока-
зывает, что у зяблика наиболее широкие крылья, наиболее удли-
ненный хвост, максимально развитое крылышко, наименьшее уд-
линение и наименьшая острота крыла (различия достоверны).
Кроме того, зяблик имеет максимально развитую мускулатуру
крыла и минимальный индекс сердца. При сравнениимежду собой
чижа и юрка оказалось, что индексы удлинения и крылышка у них
одинаковы (различия недостоверны), но чиж имеет существенно
более короткий хвост, узкие и острые крылья, индекс сердца у него
максимален, а индекс развития летательной мускулатуры минима-
лен среди трех исследованных видов.
Увеличение нагрузки на крылья обычно требует и увеличения
скорости полета птицы. В нашем случае сравниваемые виды имеют
сходные, достоверно не различающиеся величины нагрузки. Не
отличаются они и по относительному размаху крыльев. Таким об-
разом, эти показатели не должны оказывать влияния на летные
различия исследуемых видов. Укорочение хвоста характерно для
птиц, имеющих быстрый полет, в то время как относительно более
длинохвостые птицы более маневренны (Бородулина, 1953). Удли-
нение характеризует относительную' ширину и относительный
размах крыльев. Но более важно то, что увеличение этого показате-
ля ведет к снижению индуктивного сопротивления крыльев, а это
способствует повышению аэродинамического качества летательно-
го аппарата. Удлинение крыльев зяблика существенно меньше,
чем у чижа и юрка, у которых индексы удлинения отличаются мало
(различия недостоверны). Однако если сравнить эти виды по ин-
дексу ширины крыла, то чижи оказываются достоверно более узко-
Средние значения (М+m) относительных показателей (в %) органов полета
трех видов вьюрковых
Отношение Чиж Юрок Зяблик
ширины крыла * к его длине 63,75+0,25 67,74+0,20 71,82+0,31
длины хвоста к длине крыла 65,64+0,46 74,55+0,59 80,33+0,51
длины крылышка к длине крыла 28,57+0,30 28,71+0,24 29,79+0,34
размаха крыльев к длине тела без длины хвоста 295,38+2,79 306,54+5,31 301,35+3,64
размаха крыльев к длине корпуса 617,52+14,12 631,92+16,40 616,12+15,89
Удлинение 5,63+0,10 5,54+0,10 5,12+0,08
веса сердца к весу тела 1,50+0,04 1,36+0,02 1,25+0,03
веса больших грудных мышц к весу тела 19,13+0,37 21,56+0,24 22,47+0,34
веса подключичных мышц к весу больших грудных мышц 9,34+0,26 9,19+0,16 9,60+0,21
Острота крыла + 1,26+0,180 —0,507+0,283 —2,58+0,300
* Ширина крыла измерялась от кистевого сгиба до махового. вершины первого второстепенного
194
крылыми. Степень снижения индуктивного сопротивления крыла
сильно зависит и от формы концов крыльев птицы. В нашем слу-
чае чиж существенно более острокрыл, чем юрок, а зяблик имеет
достоверно менее заостренные крылья, чем юрок. Известно также,
что незаостренные крылья имеют птицы, которым часто приходит-
ся круто взлетать и садиться, т. е. летать при больших углах атаки
(Гладков, 1949). Поскольку и крылышко обычно функционирует
при таких режимах полета, то оно более развито у птиц с тупым
крылом. В нашем случае более острокрылые чиж и юрок имеют
достоверно меньший индекс крылышка, чем зяблик. Однако тупое
и широкое крыло создает большее сопротивление при горизонталь-
ных полетах.
Таким образом, птицам, которым часто приходится взлетать и
садиться, выгодно иметь менее заостренные и широкие крылья.
Однако при полетах на дальность энергетически выгоднее иметь
крылья более заостренными и узкими. В связи с этим можно вспом-
нить известное эко-географическое правило — правило Сибома,
связывающее форму крыла птиц с дальностью перелетов. Согласно
этому правилу, у близких видов и подвидов, а в некоторых случаях
и у различных популяций одного вида (Потапов, 1967) более за-
остренные крылья характерны для тех групп птиц, протяжен-
ность миграционных путей которых больше. Кроме того, более
острокрылые птицы имеют и большую скорость полета (Якоби,.
1966).
Таким образом, сравнительный анализ органов полета исследо-
ванных нами видов (на индексах летательной мускулатуры и серд-
ца мы остановимся позже) позволяет нам предполагать, что у чи-
жей скорость полета выше, а протяженность миграционных путей
больше, чем у юрков, которые, в свою очередь, должны превосхо-
дить по этим показателям зябликов.
Остановимся теперь на некоторых чертах биологии вьюрковых,
которые могут быть причиной обнаруженных различий в строении
органов полета. Наши наблюдения за осенней миграцией вьюрко-
вых вдоль берега Киевского водохранилища показывают, что у чи-
жей скорость полета выше, чем у юрков, а у юрков она больше, чем
у зябликов. Наблюдения проводились за относительной скоростью
стай, летящих параллельным курсом в одном направлении.
Аналогичные наблюдения сделаны и в других местах (Дольник,
1975). Многочисленные измерения скорости полета зябликов и юр-
ков, произведенные в идентичных ветровых условиях (Gatter,
1979), показали, что скорость полета у юрков (55,8 км/ч) сущест-
венно больше, чем у зябликов (51,5 км/ч). Интересны также заме-
чания В. Гаттера о том, что только отдельные зяблики в состоя-
нии продолжительное время лететь в стае юрков, а наблюдаемые
иногда летящие смешанные стаи являются временными. Дж. Ти-
неманн (Tienemann, 1910), измерявший скорости полета птиц во
время миграций, определил, что в сходных ветровых условиях сме-
шанные стаи чижей и чечеток летели быстрее, чем смешанные стаи
зябликов и юрков (скорость 55,8 и 52,6 км/ч соответственно).
195
Юрки совершают значительно более дальние перелеты, чем зяб-
лики: протяженность миграционных путей юрков — 3—5 тыс. км,
а зябликов — 1,7—2,5 тыс. км (Дольник, 1967). Длина сезонных
миграционных путей чижей не больше, чем у юрков. Однако для
чижей характерны массовые летние миграции, отсутствующие у
других вьюрковых. Чижи совершают продолжительные по времени
и дальности летние перемещения в северном направлении и, таким
образом, в отличие от зябликов и юрков, трижды пересекают зна-
чительные пространства Европы (Паевский, 1971). В итоге сум-
марная протяженность сезонных перелетов чижей оказывается все
же наибольшей в рассматриваемой группе видов.
Интересно также рассмотреть летную активность вьюрковых на
зимовках. Юрок, в отличие от зяблика, более специализированный
в питании вид. Прямые наблюдения и повторные находки околь-
цованных птиц свидетельствуют о значительной пульсации границ
их зимовок и значительных перемещениях птиц на зимовках в за-
висимости от урожая семян бука (Лэк, 1957; Eriksson, 1970). Ана-
логичные массовые перемещения характерны и для чижей (Ме-
женный, 1963). Кроме того, зимой чижи редко подолгу задержива-
ются на одном месте, совершая длительные суточные перемещения.
В отличие от зябликов и юрков, кормящихся на земле или на
крупных ветвях в нижнем ярусе леса, чижи обычно держатся в
верхнем ярусе. Тем самым они, хотя и обитают в густом лесу, реже,
чем юрки и зяблики, используют полет при больших углах атаки.
Таким образом, экологические различия рассматриваемых ви-
дов не противоречат результатам, полученным на основании ана-
лиза органов полета этих видов.
Особый интерес представляет анализ степени развития лета-
тельной мускулатуры и сердца. Б. К. Штегман (1962) считал, что
увеличение сердечного индекса характерно для видов, приспособ-
ленных к продолжительному активному полету, а увеличение от-
носительного веса летательной мускулатуры — для видов, приспо-
собленных к значительным кратковременным летным нагрузкам.
Иными словами, виды, более приспособленные к длительному
полету, имеют относительно большую сердечную мышцу на фоне
относительно менее развитой летательной мускулатуры. Эти вы-
воды сделаны в основном на основании изучения органов полета
довольно сильно отличающихся по образу жизни групп куликов
(ржанок и бекасов), поэтому представляет значительный интерес
сравнение этих индексов у исследованных нами видов вьюрковых.
Наибольший индекс сердца и наименьший индекс мускулатуры
имеет чиж, наименьший сердечный индекс и наиболее развитую
мускулатуру крыла имеет зяблик. Юрок занимает промежуточное
положение. Эти различия (они достоверны) вполне соответству-
ют описанным выше различиям в степени подвижности и другим
особенностям экологии исследованных видов. Однако увеличение
относительного веса мускулатуры, возможно, связано и с формой
крыла. Ведь крыло худшего аэродинамического качества требует
затраты больших мышечных усилий на полет, создает большее ло-
196
бовое сопротивление. В отряде куриных, например, виды, имеющие
широкие, наименее заостренные разрезные крылья, имеют и более
развитую летательную мускулатуру (Штегман, 1950); в семействе
ястребиных наиболее ширококрылые и тупокрылые представите-
ли — ястребы — имеют значительно более развитую летательную
мускулатуру, чем виды сходных размеров, но имеющих более узкие
и заостренные крылья, а грудная мускулатура более острокрыло-
го и узкокрылого ястреба — тетеревятника значительно менее
мощная, чем у близкого вида — перепелятника (Якоби, 1960).
Можно считать, что в данном случае степень развития летательной
мускулатуры носит компенсаторный характер.
Соотношение веса мышц-антагонистов — большой грудной и
подключичной — часто используется для решения вопроса о раз-
личиях в степени активности подъема крыла. Однако в нашем слу-
чае достоверные отличия по относительному весу подключичной
мышцы у исследованных видов отсутствуют.
Выводы
Из трех сравниваемых видов вьюрковых чиж имеет наиболее острое
и узкое крыло, укороченный хвост и наименьшее крылышко.
Юрок по этим показателям занимает промежуточное положение.
Эти различия обусловлены более быстрым полетом чижа, менее
быстрым полетом юрка и наименее стремительным полетом зябли-
ка, а также соответствующими различиями в степени летной ак-
тивности исследованных видов. Различия в индексах сердца и ле-
тательной мускулатуры подчиняются закономерности увеличения
сердечного индекса при специализации к длительному полету и
увеличению индекса летательной мускулатуры при специализации
к интенсивным кратковременным нагрузкам. Увеличение относи-
тельного веса летательной мускулатуры связано также с формой
крыла. Управление крылом худшего аэродинамического качества
требует большего развития летательной мускулатуры, поэтому бо
лее острокрылые и узкокрылые птицы имеют относительно мень-
шую массу полетных мышц.
ЛИТЕРАТУРА
Бородулина Т. Л. Морфо-функциональное исследование хвоста птиц // Работы по
морфологии и экологии птиц и млекопитающих. М.; Л.: Изд-во АН СССР,
1953. С. 76—91. (Тр. Ин-та морфологии животных АН СССР; Вып. 9).
Виноградова Н. В., Дольник В. Р., Ефремов В. Д., Паевский В. А. Определение
пола и возраста воробьиных птиц фауны СССР. М.: Наука, 1976. 192 с.
Гладков Н. А. Биологические основы полета птиц. М.: МОИН, 1949. 248 с.
Дольник В. Р. Годовые циклы биоэнергетических приспособлений к условиям
существования у 16 видов Passeriformes // Миграции птиц Прибалтики. 1967.
С. 115-163. (Тр. Зоол. ин-та АН СССР; Т. 40).
Дольник В. Р. Миграционное состояние птиц. М.: Наука, 1975. 398 с.
Лэк Д. Численность животных и ее регуляция в природе. М.: Изд-во иностр, лит.,
1957. 404 с.
Меженный А. А. Результаты кольцевания некоторых видов птиц за 1957—1960 гг.
на Биологической станции ЗИН АН СССР в Рыбачьем // Сообщ. Прибалт,
комис. по изуч. миграций птиц. 1963. № 7. 2. С. 3—25.
197
Паевский В. А. Особенности перелета чижей в Европе // Материалы VII Прибалт,
орнитол. конф. Рига, 1970. Кн. 2. С. 68—76.
Потапов Р. Л. Зависимость формы крыла птицы от дальности перелета // Миграции
птиц Прибалтики. 1967. С. 213—230. (Тр. Зоол. ин-та АН СССР; Т. 40).
Цвелых А. Н. Форма вершины крыла птиц и ее оценка // Вести, зоологии. 1983.
№ 6. С. 52-58.
Штегман В. К. Исследования о полете птиц // Памяти академика П. П. Сушкина.
М.; Л.: Изд-во АН СССР, 1950. С. 135-208.
Штегман Б. К. Основные направления в развитии летательного аппарата кули-
ков // Зоол. журн. 1962. Т. 41, № 2. С. 125—131.
Якоби В. Э. Морфо-функциональные исследования полета птиц семейства ястре-
биных //Работы по экологической морфологии птиц и млекопитающих. М.;
Л.: Изд-во АН СССР, 1960. С. 14—214. (Тр. Ин-та морфологии животных АН
СССР; Вып. 32).
Якоби В. Э. Морфо-экологические приспособления к скоростному полету птиц //
Механизмы полета и ориентации птиц. М.: Наука, 1966. С. 64—81.
Eriksson К. Wintering and autumn migration ecology of the bramling, Fringilla monti-
fringilla //Sterna. 1970. N 9. P. 77—90.
Gatter W. Unterschiedliche Zuggaschwingkeit nahe verwandter Vogelarten //
J. Ornithol. 1979. Bd. 120. S. 221-225.
Tienemann J. Untersuchungen iiber die Schnelligkeit des Vogelflugels // Ibid. 1910.
Bd. 58. S. 46-62.
SUMMARY
Morpho-ecological analysis of flying system of three species of Fringillidae.
Diadicheva E. A., Tsvelych A. N.
The degree of correspondence of the morphological distinctions of birds flying system
of Cardullinae birds to their ecological specialization is studied in three species of the
Fringillidae. The true distinctions in the structure of the flying system of the Chaffinch,
Brambling and Siskin are caused by differences in flight speed, summer activity and
other ecological peculiarities. The cardiac index of the Fringillidae increases under the
specialization for a long flight. The increase of the relative weight of the flying
musculature is connected with the adaptation to the intensive short-term loadings
and also corresponds to the deterioration of the airdynamic quality of the wing.
198
УДК 591.351: 636.551
Е. В. ГРИШИНА
109472, Москва,
ул. акад.-Скрябина, д. 23.
Московская ветеринарная
академия им. К. И. Скрябина.
ДОС-2. Кафедра зоологии
Динамика роста
эндокринных органов
домашней курицы
в зависимости
от различной степени
двигательной активности
Одним из критериев оценки физиологического состояния орга-
низма и его продуктивности является экдокринный аппарат,
определяющий уровень приспособительных возможностей орга-
низма к необычным условиям обитания, что оказывает сущест-
венное влияние на продуктивные качества животных. Между
ростом, размерами важнейших эндокринных органов и интенсив-
ностью их гормональной деятельности выявлена положительная
корреляция (Селье, 1960; Шварц и др., 1968; и др.), поэтому в
качестве морфофизиологического индикатора активности инкрет-
ных желез были взяты характеристики изменения роста их весовых
показателей.
Задача настоящего исследования — выяснить закономерности
ростовых процессов желез внутренней секреции кур постнаталь-
ного онтогенеза при различной степени двигательной активности.
Для реализации поставленной задачи в условиях Томилинского
производственного птицеводческого объединения (Константи-
новская птицефабрика) был поставлен эксперимент на двух
группах племенных кур породы белый леггорн, сформированных
по принципу аналогов. Опытная группа получала ежедневно
возрастающую дозу принудительного движения (ДПД) по раз-
работанной схеме (Хрусталева, Криштофорова, 1986); контроль-
ная группа находилась в стандартных условиях того же птичника.
У экспериментальных птиц (всего 120) однодневного, 15-, 30-, 60-,
90-, 120-, 150-, 180-, 240-, 300- и 360-дневного возраста для морфо-
логического исследования извлекали щитовидную железу, над-
почечник, гипофиз и эпифиз.
Изучаемые железы анатомировали, измеряли и затем опреде-
ляли рост весовых показателей по Н. П. Чирвинскому (1949).
Обработку данных проводили на электронно-цифровой вычисли-
тельной машине с использованием стандартных программ матема-
тической статистики.
На основании сопоставления полученных данных (см. таблицу)
прослежена возрастная динамика интенсивности роста органов
внутренней секреции кур с учетом биомеханики их двигательного
поведения (гиподинамия и применение ДПД). Установлено, что
в первые 15 сут жизни, в период напряженной адаптации орга-
низма к новым условиям существования, рост щитовидной железы,
199
Возрастные изменения интенсивности роста эндокринных органов кур экспери-
ментальных групп
Возраст, СУТ Щитовидная железа Надпочечник Гипофиз Эпифиз
Опыт Контр. Опыт Контр. Опыт Контр. Опыт Контр.
1 1,75 1,47 1,72 1,69 1,74 1,56 1,52 1,23
15-30 1,37 1,87 1,66 2,14 1,48 1,82 0,75 0,93
30-60 3,25 2,91 2,71 2,07 2,68 2,17 2,14 2,15
60-90 2,13 1,78 1,46 1,70 1,16 1,57 1,30 1,32
90-120 1,74 1,56 1,35 1,06 1,49 1,21 1,00 0,90
120-150 0,89 1,08 1,06 1,04 0,66 0,90 1,00 1,18
150-180 1,41 1,14 1,00 1,27 1,46 0,99 1,16 1,25
180—240 0,79 1,07 0,96 0,85 0,77 1,23 0,97 0,86
240-300 0,86 0,71 1,03 1,18 1,14 0,82 0,80 1,02
300-360 1,06 1,39 1,00 0,92 1,30 1,46 1,55 1,15
гипофиза и эпифиза в опытной группе птиц идет интенсивнее,
чем в контрольной. Данные, отражающие ростовые процес-
сы надпочечника, у цыплят обеих групп эксперимента совпа
дают.
В последующие 15 сут (до месячного возраста) в сравнивае-
мых группах выявлены достоверные различия в показателях
интенсивности роста эндокринных желез: у интактных птиц темпы
роста надпочечника, щитовидной железы и гипофиза увеличива
ются, в то время как у подопытных они уменьшаются: особенно
заметно у щитовидной железы и гипофиза и в меньшей степени —
у надпочечника. Рост эпифиза и в опыте, и в контроле к месяч-
ному возрасту замедляется, что более ярко выражено у цыплят
опытной группы. Интенсификация- общего метаболизма, повыше-
ние скорости гистогенеза и органогенеза адекватны увеличению
двигательной активности при ДПД, сопровождающейся увели-
чением гормональной активности желез (Загрядский, Сулимо-
Самуйлло, 1982; Бакл, 1986), приводит к временному затуханию
их роста. Данное явление следует рассматривать, на наш взгляд,
как своеобразную адаптационную реакцию эндокринного аппарата
на изменение биодинамических условий.
Интенсивность роста эндокринных желез пропорционально
росту всего организма после двух месяцев начинает снижаться
(см. таблицу). При этом значения темпов роста щитовидной желе-
зы, надпочечника и гипофиза выше в группе птиц, получающих
дополнительную физическую нагрузку. Влияния ДПД на измене-
ние величины роста эпифиза не обнаружено. Важно отметить, что
в этом возрасте у птиц контрольной группы замедляются росто-
вые процессы в надпочечнике, что выражается у них в уменьше-
нии индекса роста железы по сравнению с одномесячными цыпля-
тами.
Во все последующие периоды постнатального онтогенеза птиц
наблюдается неравномерное снижение интенсивности роста всех
200
изучаемых желез внутренней секреции как в опыте, так и в кон-
троле. Особенно стремительный спад темпов роста в возрасте око-
ло 90 сут претерпевают гипофизарно-надпочечниковая система
в опытной группе, щитовидная железа в контрольной и лишь
в характере падения интенсивности роста эпифиза различий между
группами выявить не удалось.
К началу полового созревания организма (120 сут) интенсив-
ность роста всех желез в опыте была выше, чем в контроле.
Сравнение показателей коэффициента роста эндокринных органов
подопытных цыплят с предыдущим возрастным интервалом вы-
явило увеличение интенсивности роста гипофиза, уменьшение щи-
товидной железы, эпифиза и некоторую ее остановку у надпочеч-
ника. Темпы роста эндокринных органов у птиц, испытывающих
дефицит движения, продолжает снижаться, что более резко прояв-
ляется у надпочечника.
Начало яйцекладки (150 сут) сопровождается дальнейшим
замедлением темпов роста большинства инкретных желез, причем
у подопытных кур они меньше, чем у контрольных, что следует
считать, по-видимому, следствием снижения секреторной актив-
ности исследуемых объектов у животных, находящихся в условиях
гиподинамии. Исключением явился надпочечник, у которого стати-
стически значимых различий в показателях интенсивности роста
между группами не обнаружено.
Во время повышенной и максимальной яичной продуктивности
у птиц, получающих ДПД, зарегистрирован (в возрасте 180 сут)
очередной преобладающий подъем роста весовых показателей
гипофиза и щитовидной железы, в то время как у интактных кур он
менее выражен и смещен к 240 сут жизни. К концу эксперимента
темпы роста гипофиза и щитовидной железы вновь возрастают с
преобладанием значений в контрольной группе.
Интенсивность роста эпифиза после 150-суточного возраста
в сравниваемых группах кур примерно одинакова, но к 360-м сут-
кам рост его у подопытных птиц резко увеличивается, а у контроль-
ных остается почти без изменений.
Таким образом, каждая из изучаемых эндокринных желез
имеет индивидуальные особенности роста, неодинаково реагируя
на различную степень двигательной активности. Наиболее су-
щественно влияние дополнительной физической нагрузки с 15 до
30 суток на темпы роста весовых показателей щитовидной железы,
надпочечника и эпифиза, в период интенсивного роста организ-
ма — надпочечника, гипофиза, щитовидной железы, к началу
яичной продуктивности — гипофиза, надпочечника, в пик яйце-
кладки — гипофиза, щитовидной железы и к концу экспери-
мента — эпифиза, щитовидной железы.
ЛИТЕРАТУРА
Бакл Дж. Гормоны животных. М.: Мир, 1986. 70 с.
Загрядский В. П., Сулимо-Самуйлло 3. К. Физические нагрузки современного
человека. Л.: Наука, 1982. 93 с.
7 Заказ № 36
201
Селье Г. Очерки об адаптационном синдроме. М.: Сов. наука, 1960.
Хрусталева И. В., Криштофорова Б. В. Развитие скелета у кур-несушек//Птице-
водство. 1986. № 12. С. 29—32.
Чирвинский Н. П. Изменение сельскохозяйственных животных под влиянием
обильного и скудного питания в молодом возрасте//Избр. соч. М.: Сельхоз-
гиз, 1949. Т. 1. С. 125-143.
Шварц С. С., Добринский Л. Н., Смирнов В. С. Метод морфофизиологических
индикаторов в экологии наземных позвоночных//Тр. Ин-та экологии рас.ений
и животных. Свердловск: УФ АН СССР, 1968. Вып. 58. С. 386.
SUMMARY
The dynamics of the growth of domestic hen endocrine organs in dependence upon
the various degree of the locomotion activity. Grishina E. V.
For the elucidation of the laws of hens endocrine glands growth processes under a
different degree of the locomotion activity, there was an experiment, carried out
on two groups of the white leghorn hens, being formed on the principle of analo-
gues. The experimental group got daily an increasing dose of forced locomotion
(DFL) according to the worked out scheme (Khrustaleva, Krishtoforova, 1986).
The thyroid gland, adrenal gland, hypophysis and epyphysis were periodicallj
derived from 120 studied birds at the age of 1—360 days. The additional physica.
loading from 15 to 30 days exercises the most essential influence on the growth
rates of the weight indices of the thyroid gland, adrenal gland and the epyphysis,
and during the period of the intensive growth — the adrenal gland, hypophysis ant
the thyroid gland, by the beginning of eggs productivity — the hypophysis, the adre-
nal gland,, during the peak period of egg laying — the hypophysis, the thyroid gland,
and by the end of the experiment — the epyphysis and thyroid gland.
МЕТОДИКА
ОРНИТОЛОГИЧЕСКИХ
ИССЛЕДОВАНИЙ
УДК 598.2.08
А. В. МИХЕЕВ
129243, Москва,
ул. Кибальчича, д. 6, корп. 5.
МГПИ им. В. И. Ленина.
Биологический факультет
Дневные видимые
перелеты птиц
и методы
их количественного учета
Сезонные миграции птиц привлекают к себе большое внимание ор-
нитологов, что связано с разработкой важных проблем как теорети-
ческого, так и прикладного характера. При решении ряда из них
необходимы данные о численности и динамике пролета птиц пс
тем или иным миграционным путям, в тех или иных географических
регионах. Достоверность этих данных зависит от совершенства
методов количественного учета птиц на пролете. В последнее время
разработке этих методов орнитологи уделяют большое внимание.
Помимо видимых дневных, усиленно исследуются ночные и
дневные высотные перелеты.
До недавнего времени усилия орнитологов были направлены
на изучение перелетов отдельно взятыми методами, каждый из
которых не мог дать представления об общей картине перелетов.
Впервые в СССР и за рубежом эксперимент по одновременному
комплексному применению разных методов учета миграций в
одном и том же регионе (Куршская коса, Прибалтика) с целью
проверки их пригодности для обнаружения и количественного
учета всего пролетающего в течение суток потока мигрантов на
разных высотах провела в 1977 г. Биологическая станция Зоологи-
ческого института АН СССР под руководством В. Р. Дольника
(19816). Эксперимент дал весьма интересные результаты и пред-
ставляется единственным способом изучения вопроса о величине
и распределении миграционного потока во времени и пространстве.
Для получения истинной картины перелетов желательна
максимальная точность каждого метода учета, и не случайно, что в
упомянутом эксперименте большая часть проверяемых методов
была забракована. Однако и рекомендованные методы, на наш
взгляд, еще недостаточно совершенны и нуждаются в дальнейшей
разработке.
Собранный нами фактический материал позволяет показать
7*
203
основные закономерности динамики видимого дневного пролета
и на этой основе высказать ряд соображений по поводу приме-
няемых методов их количественного учета. Этим двум вопросам
и посвящена настоящая статья.
Динамика видимых дневных миграций водных и околоводных
птиц
Наблюдения велись с 1964 по 1981 г. с постоянного наблюдательно-
го пункта, расположенного на западном побережье Каспийского
моря в устье р. Самур (юг Дагестана). Примыкающая в этом
районе к побережью моря полоса полупустыни непригодна для
остановок водных и околоводных птиц. Тяготея к берегу моря,
они летят здесь транзитом, представляя удобный объект для
визуального количественного учета. Генеральное направление
пролета осенью — юго-восточное, весной — северо-западное,
т. е. параллельно береговой линии. Иные направления свойственны
лишь небольшому числу некоторых сухопутных видов и в приво-
димых в статье данных не фигурируют.
В указанный период в устье р. Самур было совершено 12 осен-
них экспедиционных выездов продолжительностью от 10 до
37 дней. Ими охвачен весь сезон осеннего пролета, с начала августа
до середины декабря. В некоторые периоды повторность наблюде-
ний в разные годы — четырех- и пятикратная.
Мы применили тот же метод учета, который в свое время был
применен В. Ф. Гавриным (1964): учет численности пролетных
птиц проводили в течение дня с рассвета до темноты. При обработке
материала ни к какой экстраполяции не прибегали. Для учета
ежедневно выделяли четырех дежурных наблюдателей, сменяв-
ших друг друга каждые 3 ч. При валовом пролете каждую смену
дежурили два человека, один из которых определял виды и под-
считывал летящих птиц, другой записывал данные. Ширины учет-
ной полосы для птиц крупных и средних размеров (гуси, цапли,
утки, чайки и др.) составляла 2 км (1 км над сушей, 1 км над
морем), для мелких (крачки, мелкие чайки, утки и кулики
и др.) — 800 м — 1 км. В основу статьи вошел материал осеннего
пролета, собранный за 12 лет в течение 349 полных учетных дней;
за эти дни зарегистрировано около 3 млн мигрантов.
Основные особенности динамики дневного пролета мы покажем
на следующих четырех модельных группах птиц: утки (Anatini,
Aythyni и др.), бакланы (Phalacrocoracidae), в основном большой
(Phalacrocorax carbo), чайки (Larinae) и крачки (Sterninae),
цапли (Ardeidae). Сравнение динамики дневного пролета столь
разных экологических групп имеет несомненный интерес.
Учитывая, что существующие методы количественного учета
дневных миграций базируются, как правило, на показателях ин-
тенсивности пролета в утренние и вечерние часы, мы для полу-
чения сравнимых данных при анализе дневной ритмики пролета
всю светлую часть суток разбили на три части: утро — до 10 ч,
204
день — с 10 до 14 ч, вечер — после 14 ч. В октябре здесь рассве-
тает примерно в 6 ч, темнеет в 18 ч и на каждую выделенную
нами часть светового дня падает по 4 ч. В августе и в сентябре
утро и вечер длиннее, в ноябре и декабре — короче, чем день.
В среднем за весь осенний сезон пролета эти три части примерно
равны.
Анализ наших данных направлен в первую очередь на выявле-
ние особенностей динамики видимого дневного пролета как в раз-
ные части суток и дни одного сезона, так и в разные годы. Первые
два вопроса мы рассматриваем на примере первичных данных еже-
дневных учетов за конкретный отрезок времени года (он опреде-
лялся сроками полевых работ в данном году). Выводы по второму
вопросу базируются на средних показателях дневной динамики
пролета за каждый из 12 лет наблюдения и за все годы в целом.
Утки — самая многочисленная группа птиц на пролете пс
западному побережью Каспийского моря. Пролет уток растянут с
августа до декабря. В августе—сентябре летят чирок-свистунок
и широконоска, массовый пролет остальных падает преимущест-
венно на вторую половину ноября — первую половину декабря.
Ритмика видимого дневного пролета уток в разные дни показана
на примере данных, полученных путем учета мигрантов в течение
20 полных дней в период с 20 сентября по 8 ноября 1977 г. (табл. 1).
Всего за этот период зарегистрировано 45 660 птиц. В разные
дни этого периода число пролетных птиц колебалось: утром (де
10 ч утра) — от 0 до 98,1% от всех пролетевших за данный день
птиц, днем (с 10 до 14 ч) — от 1,9 до 100%, вечером (после 14 ч) —
от 0 до 63,8%. Общее число пролетевших за день особей за 2С
дней колебалось от 2 до 11 683.
Приведенные показатели свидетельствуют о значительных
колебаниях в ритмике дневного пролета в разные дни. Они нь
дают каких-либо оснований говорить о преобладании интенсив-
ности миграций в утренние часы. В то же время сопоставление
абсолютного числа мигрантов, пролетевших за весь период наблю-
дений в разные части дня, свидетельствуют об обратном. За 20 дней
наблюдений в утренние часы зарегистрировано 45,2%, в дневные —
27,3 и в вечерние — 27,5%, пролетавших уток. Большой удельный
вес утренних мигрантов следует отнести на счет валового пролета,
проходившего преимущественно в утренние часы 22 и 23 октября и
4 ноября. В эти дни до 10 ч пролетело три четверти всех уток,
зарегистрированных в утренние часы в течение всего 20-дневногс
периода наблюдений. Этот пример лишний раз показывает, сколь
велики могут быть различия в динамике миграций в разные
дни.
Динамика видимого пролета в разные годы показана на
основе данных наблюдений за 12 лет, в течении которых за 349
учетных дней зарегистрировано 1 770 842 особи уток. Анализ этих
данных показывает, что ритмика дневного пролета птиц подвер-
жена таким же резким изменениям по годам, как и по дням одно-
го и того же года (рис. 1). Удельный вес утренних мигрантов ь
205
Таблица 1
Динамика осеннего дневного пролета основных групп птиц в разные дни
Система- тическая группа Срок наб- людений Общее число учтенных особей Колебания интенсивности пролета, %
Утро День Вечер
Утки 20.Х- 8.XI1977 35660 0-98,1 1,9-100 0,6-3,8
Бакланы То же 6818 0-100 0-100 0-100
Чайки, 21.VIII — 118260 8,3-90,3 4,4-52,8 0,6-5,4
крачки 10.1X1969
Цапли 20.Х- 8.XI1977 711 0-100 0-100 0-100
Всего 171449 0-100 0-100 0-100
разные годы колеблется в среднем от 11,3 до 74,8%, дневных —
от 13,4 до 61,7,'вечерних — от 9,8 до 50,5%.
Динамика дневного пролета за все годы в среднем выглядит
следующим образом: наибольшее число уток пролетело в дневные
часы — 41,8%, на втором месте стоят утренние — 34,5% и на пос-
леднем — вечерние — 23,7%. Более высокий удельный вес днев-
ного пролета связан с тем, что в 1968 и 1981 гг. во время особо
массового пролета максимальное число птиц летело в дневные
часы.
Бакланы. На пролете отмечены большой (Phalacrocorax carbo)
и малый бакланы (Ph. pygmaeus), причем последний — в очень
небольшом числе.
Динамика видимого дневного пролета бакланов в разные дни
рассмотрена на примере данных учета, произведенного в те же
сроки, что и учет уток (с 20 октября по 8 ноября 1977 г.). За этот
период зарегистрировано 6618 особей (см. табл. 1). Удельный
вес утренних мигрантов в разные дни колеблется от 0 до 100%,
аналогичные показатели имели место у дневных и вечерних миг-
рантов. Абсолютное число особей, пролетевших за один день,
варьировало от 0 до 2 424.
Пролета не было утром 5 дней, днем — 7, вечером — 10 дней.
Наибольшее число особей за 20 дней пролетело: утром 43,3%,
днем — 35,9, вечером — 20,8%.
Из общего числа зарегистрированных за 12 лет 122 460 бакла-
нов число пролетевших в утренние часы колеблется в разные годы
от 3,6 до 99,2%, в дневные часы — от 0,8 до 63,9, в вечерние —
от 0 до 70,5%. Из общего числа мигрантов, зарегистрированных
за все годы, утром пролетело 18,7%, днем — 42,5, вечером —
38,8%. В отличие от уток, преобладающая часть бакланов летит
в дневные и вечерние часы.
Чайки и крачки — вторая по численности (после уток) группа
мигрантов, в которую входит 17 видов. Наиболее многочисленны
из них обыкновенная (Larus ridibundus), малая (L. minutus)
и серебристая (L. argentatus) чайки и речная (Sterna hirundo),
206
Число диен с максимальной интенсивностью пролета Число дней с минимальной интенсивностью пролета Для мигрантов, пролетевших за весь срок, %
Утром Дием Вечером Утром Днем Вечером Утром Днем Вечером
6 7 7 9 2 9 45,2 27,3 27,5 7 7 4 3 3 5 43,3 35,9 20,8 4 1 16 7 12 2 31,1 28,1 40,8 12 5 2 — — — 48,1 9,8 42,3 29 20 29 19 17 16 41,9 25,3 32,8
пестроносая (Thalasseus sandvicensis) и черная (Chlidonias niger)
крачки. По количеству пролетающих особей крачки в целом не
уступают чайкам. Следует только отметить, что пролет крачек
проходит рано, в августе и сентябре, и летят они очень
интенсивно, при валовом пролете огромными стаями и почти
сплошным потоком. Пролет чаек растянут с августа до декабря
и плотность миграционного потока у них заметно ниже. В авгу-
сте — сентябре валовой пролет происходит лишь у малой чайки.
Общее число особей чаек и крачек за 12 лет наблюдений соста-
вило 1 067 496.
Для иллюстрации резких колебаний интенсивности пролета
в разные дни приведем данные учетов чаек и крачек в период
с 21 августа по 10 сентября 1969 г. Всего за это время зарегистри-
ровано 118 260 особей. Число пролетевших за день птиц колеба-
лось от 1132 до 13 844 (см. табл. 1). За весь период наблюде-
ний утром пролетело 31%, днем 28,1, вечером 40,8% чаек и крачек.
Динамика дневного пролета чаек и крачек в разные годы под-
вержена, как и у ранее рассмотренных групп, большим измене-
ниям. Так, утренний пролет колеблется от 12,2 до 79,9%, днев-
ной — от 13,9 до 55,1%, вечерний — от 1,7 до 54,9% от общего чис-
ла зарегистрированных в соответствующем году птиц (рис. 2).
Из всех указанных на рисунке лет максимальное число утрен-
них мигрантов приходится на 6 лет, дневных — на 3 года, ве-
черних — на 3 года, а минимальное соответственно — на 2, 2
и 8 лет. Количество чаек и крачек, пролетевших в течение всех лет
наблюдений, по частям дня распределяется следующим образом:
утром — 42,1%, днем — 22,9, вечером — 35%. Следовательно,
по средним многолетним данным, наибольшая интенсивность про-
лета наблюдается в утренние часы. Но в отдельные годы и дни
от этой закономерности имеется много отступлений, что связано
с тем, на какие части дня падает валовой пролет.
Цапли. Дневной пролет отмечен у четырех видов, из которых
наиболее многочисленна была большая белая цапля (Egretta
alba), довольно обычна серая (Ardea cinerea), реже встречались
207
//=77/03 M=223/27 3=03673 N=7SS£6 3=306233 /7=322062
Рис. 1. Динамика видимого дневного пролета уток в разные годы
1 — доля птиц (%), пролетевших до 10 ч; 2 — с 10 до 14 ч; 3 — после 14 ч
Рис. 2. Динамика видимого дневного пролета крачек и чаек в разные годы
208
малая белая (Е. garzetta), и* рыжая (A. purpurea) цапли. Всего
учтено на пролете 21 348 экз. цапель.
Для выявления динамики видимого дневного пролета цапель
в разные дни использованы материалы, собранные в период с 20 ок-
тября по 8 ноября 1977 г. (см. табл. 1). За этот период учтено
711 пролетевших особей.
Как видно из таблицы, колебания интенсивности пролета в раз-
ные дни очень велики: она изменялась от 0 до 100%. Число осо-
бей, пролетевших за один день, изменялось от 0 до 171. За весь
период наблюдений утром пролетело 48,1%, днем — 9,8, вече-
ром — 42,3%.
Динамика дневного пролета цапель в разные годы показана
на рис. 3. Удельный вес птиц, пролетевших в разные части дня,
в среднем колеблется следующим образом: утром — от 8,2 до
92,7%, днем — от 0 до 60,8, вечером — от 0,8 до 55,6%. Макси-
мальный в течение дня пролет приходился на утро 8 раз, на
день — один раз, на вечер — 3 раза, минимальный приходился
на утро 2 раза, на день — 4 и на ночь — 5 раз. Из общего коли-
чества пролетевших за рассматриваемые годы особей цапель 43,2%
падает на утро, 34,8% — на день и 22,0% — на вечер.
Приведенные данные свидетельствуют о резких колебаниях
динамики видимого дневного пролета как в разные дни, так
и в разные годы. Хотя по усредненным многолетним данным
наиболее интенсивный пролет цапель падает на утренние часы,
тем не менее в отдельные годы и дни отмечалось много исключе-
ний.
Сравнение средней многолетней динамики дневного пролета
у разных групп водных и околоводных птиц показывает (рис. 4),
что у одних он преобладает в утренние часы (цапли, крачки и чай-
ки), У других — днем (утки, бакланы), у третьих интенсивность
вечернего пролета мало отличается от максимального утреннего
или дневного (бакланы, чайки и крачки).
Динамика дневного пролета всех рассматриваемых групп
водных и околоводных птиц в среднем за все 12 лет выглядит
следующим образом. В утренние часы максимальная интенсив-
ность пролета отмечалась 7 лет, в дневные — 3 года, в вечерние —
4 года. За эти годы утром пролетело 1 103 348 особей, или 38,0%,
днем — 960 213 особей (33,2%), вечером — 834 351 особей
(28,8%). Как можно видеть, очень резких различий в интенсив-
ности пролета в разные часы дня не наблюдается.
Анализ приведенных данных позволяет сделать следующие
выводы.
1. У всех рассмотренных групп птиц резко выражена изменчи-
вость динамики дневного пролета в разные дни одного сезона и раз-
ные годы. Удельный вес особей, пролетающих в разные части
дня (утро, день, вечер) может меняться от 0 до 100%, а колеба-
ния абсолютного числа особей достигать сотен, тысяч и десятков
тысяч раз. Аналогичные данные отмечены также Э. И. Гаврило-
вым (1979).
209
Рис. 3. Динамика видимо-
го дневного пролета цапель
в разные годы
Рис. 4. Средние многолет-
ние показатели динамики
пролета разных групп птиц
2. По средним многолетним данным, у всех групп водных
и околоводных птиц интенсивность пролета максимальна утром,
меньше — днем, минимальна — вечером, хотя в среднем резких
различии интенсивности в разные части дня не надблюдается.
У цапель, крачек и чаек проявляется тенденция к наиболее ин-
тенсивному пролету в утренние часы, а у уток и особенно у бак- 1
ланов — в дневные. Однако в разные годы наблюдается много
самых разнообразных исключений. Решающее влияние на характер I
показателей динамики дневного пролета имеет время валового про-
лета; от него зависит, в какую часть дня пролетает абсолютное
большинство мигрантов.
210
।
О применяемых методах количественного учета
видимых дневных перелетов птиц
Все современные методы количественного учета видимых дневных
перелетов птиц относятся к экстраполяционным: непосредствен-
ные подсчеты пролетающих через наблюдательный пункт птиц
производятся в течение только части светлого дня в полосе опре-
деленной для разных по размеру птиц ширины, а затем получен-
ные показатели экстраполируются на весь день, сезон и единый
для всех мигрантов фронт пролета.
Показанная выше изменчивость характера динамики видимого
дневного пролета в разные дни и разные годы осложняет приме-
нение экстраполяционных методов и ограничивает точность их
результатов. Для выяснения степени точности этих методов мы по-
пытались сравнить результаты метода беспрерывного дневного
учета с данными, полученными при помощи двух наиболее заслу-
живающих внимание экстраполяционных методов.
Первый метод разработан Э. И. Гавриловым (1974, 1979)
и широко распространен среди орнитологов Казахстана и Средней
Азии. Другой метод (Люлеева, Яблонкевич, 1981) применяется
в Прибалтике. Поскольку эти методы относятся к абсолютным
методам учета и рекомендуются для применения к разным группам
птиц и в разных условиях, мы сочли правомерным сравнение
наших данных с данными упомянутых методов.
Суть сравнения сводится к следующему. Из данных нашего
учета мы сделали выборки показателей числа пролетевших птиц
в те часы дня, которые приняты каждым экстраполяционным ме-
тодом, а затем по способу каждого из них вычисляли общее число
пролетевших за день и весь период наблюдений. Экстраполяцию
на ширину фронта пролета мы не проводили. Такого рода срав-
нение вполне правомерно, поскольку оно касается не разных
экологических групп, с которыми имели дело авторы сравнивае-
мых методов, а только тех, с которыми имели дело мы. По сущест-
ву, описанная методика сравнения дает нам ответ на вопрос:
какие результаты количественного учета мигрантов мы получили
бы на своем стационаре, применяя упомянутые экстраполяцион-
ные методы. Полученные данные правомерно сравнивать с нашими
данными.
Метод Э. И. Гаврилова сводится к тому, что ежедневные коли-
чественные учеты пролетающих птиц проводятся с постоянного
наблюдательного пункта в течение 2 ч утром и 2 ч вечером,
а один раз в пять дней проводятся полные учеты птиц в течение
всего дня. Мелких воробьиных птиц учитывают в полосе шириною
100 м, птиц средних размеров — 500 м, крупных — около 2000 м.
В процессе обработки материала применяется экстраполяция по-
лученных данных на единую для всех птиц ширину пролетной
полосы в 500 м и на весь световой день. В первом варианте число
зарегистрированных мелких воробьиных птиц умножается на 5,
число крупных птиц делится на 4, число средних остается неизмен-
211
ным; во втором производятся более сложные расчеты. Сначала под-
считывается число птиц, пролетевших за все дни полного учета
и определяется, какая доля (в %) из него падает на утренние
и вечерние часы (6—8 и 16—18 ч). Затем подытоживается общее
число зарегистрированных в эти часы птиц за весь период ежеднев-
ных наблюдений и с учетом ранее установленных коэффициен-
тов, высчитывается суммарное число птиц, которое должно проле-
теть за весь период наблюдений в светлое время суток.
Сделав выборки из наших данных и проведя их экстраполя-
цию по способу, указанному в описанном методе, мы сравнили
полученные результаты с результатами учета нашим методом
(табл. 2). Из нее видно, что данные, полученные рассматривае-
мым экстраполяционным методом, отклоняются от фактических
в 1,5—4 раза. Экстраполяция числа мигрантов на единую ширину
фронта пролета усиливает величину расхождений ввиду простран-
ственной неравномерности плотности пролета.
Таблица 2
Сравнение результатов учета абсолютным методом и методом экстраполяции
Э. И. Гаврилова
Виды и группы Даты учета Число учет- ных дней Кол-во по абс. методу (А) Кол-во по ме- тоду экстра- поляции (Б) Б/АХЮО%
Кряква 20.Х—19.XI1966 30 13103 13800 100,5
Чайки 20.Х-8.XI1977 20 8180 16100 196,8
Кряква 20.IX-23.XI1976 58 8662 11433 132,0
Бакланы 9.XI-8.ХП1976 31 20854 93290 447,3
Утки 20.Х—8.XI1977 20 45660 30024 65,8
Кряква То же 20 12316 8770 71,2
Чайки 20.Х—19.XI1966 30 427664 254818 59,4
Бакланы 20.Х—8.XI1977 20 6118 1642 24,1
Всего 229 541254 888340 164,1
Метод Д. С. Люлеевой и М. Л. Яблонкевича (1981), названный
авторами «формализованным», состоит в том, что наблюдения
с постоянного наблюдательного пункта начинаются за 30 мин до
восхода солнца и заканчиваются через 30 мин после его захода.
Каждый час учет производится только в течение первых 15 мин.
Полученные данные экстраполируются на один час (умножаются
на 4). Учет мелких птиц проводится в полосе 150 м, крупных —
500 м. Для определения плотности пролета по фронту шириной
1 км за один час число мелких птиц умножалось на 7, а крупных —
на 2. В итоге коэффициент пересчета для крупных птиц составлял
8 (4X2), для.мелких — 28 (4X7). Полученные таким образом дан-
ные затем суммировались за весь день.
Для сравнения результатов этого метода с нашими мы произве-
ли выборку из наших данных и обработали ее в соответствии
с описанной методикой. Полученные таким образом экстраполя-
ционные данные «фнвамааднсь © даивями каянеги методе ыошюго
212
дневного учета. В качестве модельных мы взяли те же четыре груп-
пы птиц, а сезон — начало осенного пролета, его середину и ко-
нечную стадию. Результаты этих сравнений показаны в табл. 3.
Оценивая рассматриваемый метод, следует сказать, что вели-
чина его ошибки в наших примерах меньше, чем предыдущего.
Максимальная ошибка формализованного метода достигает по
отдельным группам 120—200, за весь период наблюдений по всем
группам — около 1,3 крат. За некоторые отрезки времени по не-
которым группам (чайкам, крачкам и уткам) средние показатели
достаточно близки к нашим. Высокая достоверность данного ме-
тода связана с небольшой протяженностью отрезков времени,
отводимых на экстраполяцию. Вместе с тем формализованный
метод страдает известными изъянами.
Так, количество учетных дней с расхождениями в 40% и более
превышает 50% всего периода наблюдений.. При этом величина
расхождений колеблется от 66,6 до 400%. Даже у крачек и чаек,
суммарные данные учета которых близки к нашим, доля дней
с расхождениями в 40% и более равна 40%, с расхождениями
от 200 до 400% — 20%. Наибольший удельный вес расхождений
падает на период валового пролета, а по группам — на бакланов
и цапель.
Кроме того, формализованный метод может искажать картину
волнообразности пролета. Регистрируя большинство волн, в ряде
случаев он не улавливает их или, наоборот, показывает их в дни,
когда их фактически нет. Так, например, в период с 13 ноября
по 11 декабря 1967 г. этот метод не позволил обнаружить две
волны из шести у бакланов и дал две волны у бакланов, три —
у чаек и крачек и две — у уток, которых фактически не было.
Наконец, ошибки формализованного метода, как й метода Э. И. Гав-
рилова, могут возрастать за счет экстраполяции числа мигрантов
на единый фронт пролета в 1 км, поскольку далеко не всегда и не
везде птицы летят по территории равномерно.
Средние расхождения показателей методов беспрерывного
и экстраполяционных количественных учетов видимых дневных
миграций как будто невелики и для выявления общих крупномас-
штабных закономерностей и теоретических построений вполне
пригодны. Однако при решении более детальных вопросов допус-
каемый масштаб расхождений (в 2—4 раза) значителен: при вало-
вом пролете на путях массовых миграций это может составлять
десятки и сотни тысяч птиц в день. Поэтому необходима дальней-
шая работа по усовершенствованию экстраполяционных методов
и выявлению близких к истине показателей дневного видимого
пролета за более короткие отрезки времени.
Все вышесказанное позволяет сделать некоторые замечания по
поводу методов учета ночных и высотных миграций, применяю-
щих приемы экстраполяции в неизмеримо больших масштабах.
Так, например, в методе учета М. Л. Яблонкевича и Д. С. Люлее-
вой (1981) дневных высотных миграций в 8-кратный телескоп
коэффициент экстраполяции достигает 280 и даже 560. Несомнен-
213
20.X-8.XI 1977 20 531 1108 208,6 17' (из 17) 79,0-400,0
13.XI- 29 1335 844 63,2 5 (из И) 66,6-400,0
11.XII 1967
но (это отмечают и сами авторы), что количественный учет высот-
ных дневных (как и ночных) миграций крайне сложен и труден
и применяющиеся методы далеко не совершенны. Полученные
ими результаты следует пока рассматривать как крайне ориенти
ровочные.
Несколько слов о методе количественного учета пролетных
птиц с помощью радиолокатора. На наш взгляд, это наиболее
перспективный метод обнаружения и учета высотных миграций.
Одним из наиболее узких мест в его работе в настоящее время
является расшифровка эхо-сигналов, записанных на пленку.
Необходимо остановиться вкратце на вопросе о месте и значе-
нии видимых дневных миграций в общей картине сезонных пере-
мещений. В настоящее время в литературе отмечается, что по об-
щей численности видимые дневные мигранты составляют 15% от
общего числа мигрирующих птиц. Однако при существующих
методах учета за достоверность такой цифры поручиться нельзя.
Вместе с тем высказано положение о том, что видимые дневные
миграции представляют собой не самостоятельную, нормальную
форму сезонных миграций, а как бы придаток, или остаточное
явление высотного пролета птиц. Гак, по В. Р. Дольнику (1971),
все птицы совершают перелеты только на большой высоте, (300—
3000 м), видимые же миграции свойственны тем из них, которые
после очередного высотного транзитного «броска» спустились
вниз, чтобы вскоре сделать остановку на отдых и кормежку. С этим
утверждением согласиться нельзя. Видимые дневные миграции
птиц — обычное и массовое явление, в том числе и в местах, уда-
ленных от пунктов остановок для отдыха и кормежки. Так, по
нашим наблюдениям, через наблюдательный пункт в устье р. Са-
мур как весной, так и осенью проходит массовый видимый пролет
на небольших высотах. Ближайшие места, удобные для остановки
большинства птиц на отдых и кормежку, находятся примерно
в 100 км по прямой к югу от устья р. Самур (Дивичинский лиман)
и более чем в 200 км к северу от него (Аграханский залив). Во
время валового пролета Дивичинский лиман не может вместить
всю массу мигрантов, и значительная часть их вынуждена лететь
дальше, вплоть до залива им. Кирова (300 км по прямой).
Если предположить, что после высотного пролета птицы опусти-
лись вниз для остановки на отдых и кормежку только перед нашим
наблюдательным пунктом, то до ближайшего удобного для при-
земления места даже таким быстрым летунам, как утки, пришлось
бы лететь от полутора до четырех часов осенью и три часа весной.
Едва ли такой длительный полет можно квалифицировать как
полет перед посадкой.
Подводя итоги сказанному, следует отметить, что экстрапо-
ляционные методы имеют ряд преимуществ по сравнению с мето-
дом беспрерывного учета,— они требуют гораздо меньше време-
ни. Но ярко выраженная динамика сезонных миграций птиц зат-
рудняет использование экстраполяционных методов учета и прог-
нозирование интенсивности пролета. Метод беспрерывного днев-
215
ного учета более точен, но трудоемок и для широкого применения
не может быть рекомендован. Его можно рекомендовать для спе-
циальных исследований, а также для проверки старых и разработ-
ки новых экстраполяционных методов. Внедрение последних
в практику желательно после их апробации.
Особое внимание должно быть уделено методам учета ночного
и высотного пролетов и в первую очередь с помощью радиолока-
торов. Надежный количественный учет и распределение всего
потока мигрантов во времени и пространстве могут быть осущест-
влены только комплексом хорошо отработанных методов.
ЛИТЕРАТУРА
Большаков К. В. Реконструкция полной картины ночного пролета и эффективность
обнаружения ее разными методами // Методы обнаружения и учета миграций
птиц. Л„ 1981. С. 95-123. (Тр. Зоол. ин-та АН СССР; Т. 104).
Большаков К. В., Булюк В. И., Жалкявичус М. М. и др. Сравнение результатов
учета ночного пролета осенью 1977 г. на Куршской косе семью методами наб-
людений //Там же. 1981. С. 79—95.
Гаврилов Э. И. О двух формах миграций у птиц // Изв. АН КазССР. Сер. биол.
1974. № 5. С. 55-58.
Гаврилов Э. И. Сезонные миграции птиц на территории Казахстана. Алма-Ата:
Наука, 1979. 254 с.
Гаврин В. Ф. Экология шилохвости в Казахстане // Охотничьи птицы Казахстана.
Алма-Ата: Изд-во АН КазССР, 1964. С. 5—58. (Тр. Ин-та зоологии АН КазССР;
Т. 24).
Губин Б. М. Пролет птиц в Таласском Алатау (Западный Тянь-Шань) // Миграции
птиц в Азии. Алма-Ата: Наука, 1976. С. 49—69.
Дольник В. Р. Энергетика перелетов птиц // Вопросы информации. М.: ВИНИТИ,
1971. Вып. 1. С. 52-81.
Дольник В. Р. Миграционное состояние птиц. М.: Наука, 1975. 398 с.
Дольник В. Р. Динамическая модель прогноза миграции птиц // Методы обнару-
жения и учета миграций птиц. Л., 1981а. С. 123—133. (Тр. Зоол. ин-та
АН СССР; Т. 104).
Дольник В. Р. Программа «Эксперимент»: сравнение методов наблюдения пролета
птиц // Там же. 19816. С. 3—7.
Дольник В. Р., Большаков Б. В., Жалкявичус М. М. Реконструкция полной
картины дневного пролета и эффективность обнаружения ее разными мето-
дами // Там же. 1981. С. 70—79.
Кумари Э. В. Инструкция Для изучения миграций птиц. Тарту, 1955. 28 с.
Люлеева Д. С. Весенние миграции птиц на Куршской косе по данным наблюде-
ний 1959—1960 гг. // Миграции птиц Прибалтики. Л.: Наука, 1967. С. 25—50.
Люлеева Д. С., Жалкявичус М. М., Шумаков М. Е. и др. Сравнение результатов
учета дневного пролета птиц осенью 1977 г. на Куршской косе пятью мето-
дами наблюдений // Методы обнаружения и учета миграций птиц. Л., 1981.
С. 57-70. (Тр. Зоол. ин-та АН СССР; Т. 104).
Люлеева Д. С., Яблонкевич М. Л. Визуальные наблюдения формализованным ме-
тодом дневного пролета птиц осенью 1977 г. на Куршской косе // Там же.
1981. С. 24-36.
Меженный А. А. Миграции птиц на Куршской косе и некоторые задачи их изу-
чения // Экология и миграции птиц Прибалтики. Рига: Изд-во АН ЛатвССР,
1961. С. 235-240.
Яблонкевич М. Л., Люлеева Д. С. Оптические наблюдения дневного высотного
пролета птиц осенью 1977 г. на Куршской косе // Методы обнаружения
и учета миграции птиц. Л., 1981. С. 37—44. (Тр. Зоол. ин-та АН СССР; Т. 104).
216
SUMMARY
Day-time visible migrations of birds and the methods of their quantitative registration.
Mikheev A. V.
The main objective regularities of the day-time dynamics of the visible migrations of
birds are considered. In the light of these data the applied methods of the quantitative
registration of immigrants were estimated. The materials of the daily registrations
of migrating birds during the whole light day for 249 days (about 3 mln of specimens
registered) were considered. On the example of the model groups of birds a big chan-
geability of arrival intensity was shown in the morning, day-time and evening hours
on different days of one season in different years. It’s shown that the methods of the
quantitative registration which resort to the extrapolation in time and space, allows
to obtain the data, suitable for the determination of a common big-scale phenomena and
the theoretical conclusions. The used methods need a further elaboration.
УДК 598.2.08:598.412
Н. П. КОЛОМИЙЦЕВ 692532, Приморский край, Уссурийский район, поселок Каменушка. Уссурийский государственный заповедник К методике учета чешуйчатого крохаля (Mergus squamatus) и мандаринки (Aix galericulata)
Для контроля за состоянием популяций редких и легко уязвимых
видов первостепенное значение имеет регулярный сбор сведений
об их численности и пространственном распределении. Однако
из-за большой трудоемкости и ограниченного числа специалистов
такого рода исследования часто оказываются неосуществимы, и
необходимы дальнейшая разработка, упрощение и стандартизация
методических приемов проведения учетов. Некоторый положитель-
ный опыт в этом направлении был накоплен нами в 1980—1986 гг.
при изучении экологии чешуйчатого крохаля и мандаринки на
реках восточных склонов Южного Сихотэ-Алиня.
Что касается чешуйчатого крохаля, то к настоящему времени
уже имеются попытки объективной оценки его численности. В ряде
работ авторы указывали либо абсолютную численность птиц (вы-
водков, брачных пар) для некоторых сравнительно небольших
участков рек (Яхонтов, 1976; Смиренский, 1977; Елсуков, 1979),
либо усредненные данные, пересчитанные на 1 или 10 км протя-
женности русла (Шибнев, 1976). Отнесение результатов учетов
не к площади, а к линейной протяженности русла для этого вида
вполне оправдано, так как только река да узкая полоса прибрежной
растительности, а не площадь поймы или долины в целом, опреде-
ляют собой стацию его обитания.
Как показали серии учетов и стационарные наблюдения, у
чешуйчатого крохаля весенний пролет протекает в сжатые сроки
и к концу второй декады апреля в основном заканчивается.
217
В третьей декаде апреля уже начинается насиживание наиболее
ранних кладок. Учитывая, однако, то, что общая доля таких кладок
незначительна, учеты по брачным парам и группам (последние,
кроме взрослых самца и самки, обычно включают еще молодых
птиц) с небольшими погрешностями можно проводить всю вторую
и третью декады апреля.
Занятие гнездовых дупел у многих пар происходит почти сра-
зу же после прилета (в одном случае даже 31 марта), к в дальней-
шем птицы держатся преимущественно в районе своих гнезд.
В период откладки яиц самки тоже очень привязаны к местам
размножения и продолжают встречаться в парах. Поэтому там,
где птиц не беспокоят, учеты в весеннее время могут дать
довольно точные сведения не только о численности, но и о прост-
ранственном распределении вида. При обработке данных этих уче-
тов брачную группу любого состава, а также одиночных самок
следует принимать за одну размножающуюся пару, так как из
нескольких самок, входящих в состав некоторых групп, в дальней-
шем к размножению приступает, как правило, только одна. Другие
птицы к началу насиживания собираются в обособленные стайки,
но часть самок первого года жизни продолжает держаться около
гнезд до появления птенцов и позже. Укажем здесь, что гнезда
чешуйчатого крохаля отличаются от гнезд близких видов цветом
пуховой обкладки, которая у него светло-серая стального оттенка,
внешне очень похожая на гнездовой пух мандаринки.
Некоторые сложности при проведении учетов в ранневесенние
сроки возникают в связи с приходящимся на это же время проле-
том большого крохаля (Mergus merganser L.). Во избежание оши-
бок в определении самцов основное внимание следует обращать
на наличие у них на голове хохла удлиненных перьев, который
у чешуйчатого крохаля длинный, почти касается спины, и заметен
в бинокль на большом расстоянии. Самки хорошо различаются
окраской нижней части шеи и грудки. У самок большого крохаля
этот участок спереди почти белый и хорошо виден издали в виде
резко отграниченного пятна. У самок же чешуйчатого крохаля он
не отличим по цвету от боков грудки. С более близкого расстояния
у самок большого крохаля заметно также белое горло. Кроме того,
весной и для самцов, и для самок чешуйчатого крохаля характер-
на ярко-красная (алая) окраска клюва. У большого крохаля окрас-
ка клюва темнее, и на расстоянии он кажется черным. В полете
чешуйчатые крохали узнаются по трехраздельному у самцов и
двухраздельному у самок зеркалу крыла (Бутурлин, Дементьев,
1935).
Что касается определения в природе возраста птиц, то самцы
чешуйчатого крохаля на первом году жизни отличаются от старых
особей коричневым оттенком оперения головы и распространением
мелкочешуйчатого рисунка на бока передней части тела и низ шеи.
Молодых самок можно узнать по поведению: они держатся обычно
небольшими группами, более доверчивы и не так привязаны к опре-
деленным местам.
218
Учеты выводков лучше всего проводить в третьей декаде
июня — первых числах июля, когда птенцы в них уже немного
подрастают, но еще сильно привязаны к участкам своего обитания.
Активны выводки бывают в течение всего светлого времени,
но там, где беспокойство птиц со стороны людей носит системати-
ческий характер, суточная активность может изменяться — они
держатся более скрытно и от них уже на ранних стадиях начинают
отбиваться отдельные молодые. Вследствие малочисленности вида
недоучет в таких условиях бывает весьма значительным, до 15% и
более. Во избежание этого учеты необходимо повторять.
Из других причин, влияющих на точность учетов выводков,
укажем присущую самкам этого вида склонность к гнездованию
поблизости друг от друга, что при незначительных различиях
в количестве и возрасте молодых существенно затрудняет иденти-
фикацию отдельных семей. Следует учитывать и то, что чешуйчато-
му крохалю свойственно в ряде случаев образование объединен-
ных семейных групп во главе с одной старой птицей. Такие
группы далеко не всегда можно с полной уверенностью отличить
от больших выводков, которые, если судить по максимальным
размерам кладок, могут включать до 14 пуховичков.
При тщательном выполнении работ учеты брачных пар, самок
в период откладки яиц и выводков чешуйчатого крохаля дают
сопоставимые результаты. В сочетании с дневной активностью и
заметностью птиц это открывает возможность ежегодного обследо-
вания значительных территорий. Экстраполяцию полученных дан-
ных следует производить только на те участки таежных горных рек,
где они протекают в зоне долинных кедрово-широколиственных
лесов. При этом надо иметь в виду, что на совсем маленьких
реках протяженностью около 30 км и менее чешуйчатый крохаль,
как правило, вообще не гнездится.
. На мелководных загроможденных завалами участках рек лучше
передвигаться пешком, спускаясь вниз по течению и внимательно
осматривая в бинокль открывающиеся плесы й отмели. Регистри-
ровать следует только тех птиц, которые остались сзади. Там, где
условия позволяют, можно использовать надувную резиновую лод-
ку-
Мандаринка, в отличие от чешуйчатого крохаля, в период
гнездования и воспитания потомства держится скрытно и активна
преимущественно в сумерки. Она более пластична в выборе мест
обитания. Находить ее гнезда и искать выводки приходится на
горных реках и пойменных озерах, по небольшим ручьям и даже
просто у лесных луж. Не избегает мандаринка и освоенных
человеком мест, поселяясь вблизи дорог, у окраин полей и посел-
ков. В то же время общее распределение этой утки на гнездовании
в бассейнах рек, как это было замечено еще Е. П. Спангенбер-
гом (1952), весьма неравномерно, и на многих притоках она может
вообще не Отмечаться.
В силу сказанного учеты мандаринки на большой площади
весной после распределения пар по местам гнездования или
219
по выводкам в конце июня очень затруднительны. Из-за общей
малой численности обитания на пересеченной лесной местности
и трудности поиска гнезд для оценки численности мандаринки
оказываются малопригодными традиционные методы, применяе-
мые для водоплавающих птиц. Нельзя учитывать мандаринку
и подобно каролинской утке (Aix sponsa L.) на зорях (Hain, 1966),
так как локальных мест кормежки и ночевок, а следовательно,
и более-менее массовых суточных перелетов у нее нет.
В процессе изучения фенологии и пространственного распреде-
ления мандаринки нам удалось установить, что с момента прилета
и до начала откладки яиц она в. основной своей массе концентри-
руется на реках и активна большую часть суток. В какой-то мере
птиц принуждают к этому еще не полностью оттаявшие ручьи,
тихие протоки и лесные озерки, но и в годы с ранней и дружной
весной мандаринка в период прилета и брачных игр явно тяготеет
к более крупным водоемам. Это время, особенно третью неделю
апреля, когда транзитных птиц уже мало, можно рекомендовать
для проведения учетов.
Фенология размножения вида на разных участках ареала,
по всей видимости, различается незначительно. Гем не менее на
севере Приморского края и в Приамурье учет, скорее всего, нужно
проводить в несколько более поздние сроки. Так, для р. Бикин
Ю. Б. Шибнев (1985) в качестве наиболее подходящего времени
проведения учетов указывает конец апреля.
Пересчет и экстраполяцию полученных данных следует произ-
водить только на облесенные участки долин рек и низовьев ручьев,
исключая площади населенных пунктов и больших полей. Для
небольшой территории нельзя пересчитывать плотность гнездова-
ния на всю площадь горных лесов, так как материал, представлен-
ный в таком виде, не будет отражать ни действительного состоя-
ния местной популяции, ни усредненной плотности гнездования
вида на данном участке ареала.
Для обеспечения необходимой объективности учеты чешуйчато-
го крохаля и мандаринки нужно проводить либо на всем протяже-
нии обследуемой реки, либо на разных участках ее течения.
В последнем случае длина каждого маршрута должна быть не менее
15—20 км, а общая их протяженность 50 км и более. В апреле,
когда горные реки еще полноводны, один-два человека, используя
надувную резиновую лодку, могут осуществить такой учет
за 2—3 дня.
ЛИТЕРАТУРА
Бутурлин С. А., Дементьев Г. П. Полный определитель птиц СССР. М.; Л.: КОИЗ,
1935. Т. 2. 280 с.
Елсуков С. В. К биологии чешуйчатого крохаля Mergus squamatus // Биология
птиц юга Дальнего Востока СССР. Владивосток: ДВНЦ АН СССР, 1979.
С. 82-86.
Смиренский С. М. Новый район гнездования чешуйчатого крохаля // VII Всесоюз.
орнитол. конф.: Тез. докл. Киев, 1977. Ч. 2. С. 245—246.
220
Спангенберг Е. П. Материалы по экологии мандаринки//Охрана природы. М-,
1952. Вып. 15. С. 105-118.
Шибнев Б. К. Краткое сообщение о чешуйчатом крохале на р. Бикине // Редкие,
исчезающие и малоизученные птицы СССР. Рязань: Моск, рабочий, 1976.
С. 73—74. (Тр. Окск. гос. заповедника; Вып. 13).
Шибнев Ю. Б. О современном состоянии мандаринки и чешуйчатого крохаля
на р. Бикине // Редкие и исчезающие птицы Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 1985. С. 95-99.
Яхонтов В. Д. Краткое сообщение о чешуйчатом крохале в районе залива
Счастья / / Редкие, исчезающие и малоизученные птицы СССР. Рязань: Моск,
рабочий, 1976. С. 72—73. (Тр. Окск. гос. заповедника; Вып. 13).
Hain D. Float counts and flight counts as indices to abundance of nesting wood
ducks // J. Wildlife Manag. 1966. Vol. 30, N 1. P. 13-16.
SUMMARY
About methods of the counts of the Chinese Merganser (Mergus squamatus) and the
Mandarin Duck (Aix galericulata).
Kolomiitsev N. P.
The count methods of the Chinese Merganser and the Mandarin, based on the study
of the ecology of these species in 1980—1986 on the rivers of the eastern slopes
of the Southern Sikhote-Alin are recommended. The counts of the Chinese Merganser
during the period of arrival and egg laying (from the beginning of the second decade
and to the end of the third decade of April) and of broods (in the third decade
of June — the beginning of July) give some comparative results. It enables to
investigate annually the large territories. The field characters of the adult males,
females and birds of the 1st year are given. The down in the nest of the Chinese
Merganser is of light-grey colour, tinged with steel shadow. The third decade
of April is the most suitable time for the count of the Mandarin Duck when there are
few transit birds and most of ones are concentrated on the rivers, and they
are very active almost the whole day.
ПАМЯТНЫЕ ДАТЫ
УДК 598.20(092)
В. Е. СОКОЛОВ,
В. Д. ИЛЬИЧЕВ
117071, Москва, Ленинский
проспект, д. 33. Институт
эволюционной морфологии
и экологии животных
им. А. И. Северцова АН СССР
Выдающийся
советский орнитолог
и деятель охраны природы
К 90-летию
со дня рождения Г. П. Дементьева
(1898—1969)
Отечественная наука по праву гордится именами Г. А. Кожевнико-
ва, Н. М. Кулагина, Б. М. Житкова и других выдающихся зоологов,
чья научная и организаторская деятельность уже в первой четвер-
ти нашего столетия заложила основы нового направления, значение
которого в наши дни все более возрастает,— охраны природы.
В этой плеяде замечательных ученых профессор Московского
университета Георгий Петрович Дементьев (1898—1969) занимает
особое место как представитель следующего поколения советских
ученых, принявших эстафету от своих учителей, успешно развив-
ший идеи охраны природы применительно к новым экологическим
и хозяйственным условиям — периоду бурного развития экономи-
ки и освоения природных ресурсов, возрастающего загрязнения
окружающей среды, интенсивного промысла, сокращения числен-
ности и вымирания многих видов животных под влиянием челове-
ка.
В истории советской науки Г. П. Дементьев оставил значитель-
ный след как один из выдающихся деятелей международного
движения охраны природы, инициатор международных природо-
охранных программ с участием советских ученых, активный про-
пагандист достижений нашей страны в области охраны природы,
способствовавший повышению ее научного престижа в тяжелые
послевоенные годы.
Именно в 1950—1960-е годы Г. П. Дементьев был признанным
лидером отечественной школы охраны природы, неизменно пред-
ставительствовал на всех международных форумах, занимал руко-
водящие посты самого высокого ранга в международных организа-
циях, таких, как Международный союз по охране природы, Между-
народный совет по охране птиц и др. Он был создателем первого
научно-координирующего органа по охране природы в систе-
ме АН СССР — Комиссии по охране природы и долгие годы воз-
главлял ее в качестве председателя.
222
Возникнув в системе Академии наук СССР, эта Комиссия уже
в ранге головного научно-исследовательского института перешла
в ведение сначала Министерства сельского хозяйства СССР, а те-
перь Госкомприроды СССР.
Г. П. Дементьев родился в Петергофе 5 июля 1898 г. Его дед
по отцовской линии — Николай Федорович — был ветеринаром.
Интерес ко всему живому в семье всячески поддерживался от-
цом — Петром Николаевичем —врачом по профессии, высокообра-
зованным человеком, имевшим глубокие познания в области
биологии. Рано пробудившийся интерес к собиранию насекомых
у среднего сына Георгия (два его брата, кстати, также выбрали
себе профессию зоолога) он всячески поддерживал. Увлечение
энтомологией в детские годы у Георгия Петровича было серьез-
ным — собранные им по всем правилам коллекции позднее посту-
пили в Зоологический музей Московского университета. Приобре-
тенные в детстве навыки Георгий Петрович широко использовал
в монгольских и туркменских экспедициях, собирая насекомых
во время наблюдений за птицами.
Очень рано у Г. П. Дементьева возникла особая привязанность
к птицам. Ребенком он начал рисовать птиц и вести дневник
наблюдений, в тринадцатилетнем возрасте написал свою первую
научную работу, посвященную соколам. В 1911 г. поступил сразу
в четвертый класс гимназии — так хорошо он был подготовлен
дома. В гимназические годы интересовался историей, особенно
Древним Римом, Грецией, временем Петра I, интенсивно учил
языки — в совершенстве знал латинский и греческий, немецкий
и французский (позднее изучил английский, научился переводить
с итальянского и шведского, говорить на польском и монгольском).
Но, конечно, больше всего внимания уделял птицам — изучал их
повадки, вел дневники, много экскурсировал по территории тог-
дашней Петербургской губернии и Финляндии. Другим увлече-
нием была юриспруденция — здесь, видимо, сказалось влияние
деда по материнской линии — известного адвоката. Влияние это
оказалось столь значительным, что, окончив гимназию с золотой
медалью в 1915 г., Георгий Петрович поступил на юридический
факультет Петроградского университета. Первоначально занятия
биологией и птицами представлялись ему как бы второй профес-
сией, однако позднее это стало главным делом его жизни. Уже
в 1920 г. он покинул Петроград и вместе с матерью переехал
в Москву. Работая в Наркомате социального обеспечения, в тече-
ние 10 лет он одновременно изучал и обрабатывал коллекции
птиц Зоологического музея Московского университета, сначала
под наблюдением С. А. Бутурлина, а позднее самостоятельно.
В 1931 г. произошло решающее событие его жизни. Он перешел
на работу в Московский университет и с тех пор не покидал
его стен до самых последних дней жизни. Здесь он прошел путь
от хранителя орнитологических коллекций до профессора кафедры,
заведующего межкафедральной лабораторией, стал всемирно из-
вестным ученым, гордостью Московского университета. И все это
223
произошло в удивительно короткие сроки — к 1940 г. он уже был
в ряду ведущих орнитологов страны, профессором Московского
университета. Вместе с С. А. Бутурлиным он написал сводку
«Птицы СССР» (1934—1941 гг.) в пяти томах; в 1936 г. в Париже
вышла его книга «Systema avium Rossicarum» (т. 1), а в 1940 г.—
шестой том «Руководства по зоологии (Птицы)».
В 1935 г. Георгий Петрович после Б. М. Житкова возглавил
орнитологическую секцию Всероссийского общества охраны приро-
ды, объединявшую свыше 200 орнитологов. Руководить этой
секцией было тем более почетно, что она носила имя М. А. Мензби-
ра, влияние которого на жизнь и деятельность Георгия Петровича
было особенно велико.. В середине 1940-х годов начались регуляр-
ные «дементьевские субботы» в Зоологическом музее — семинары,
сыгравшие огромную роль в развитии отечественной орнитологии,
по своему значению для нее сравнимые со знаменитыми «налич-
никами» физиков. «Дементьевские субботы» работали еженедель-
но при жизни Георгия Петровича тридцать лет (за исключением
военного периода) и существуют по настоящее время. Участниками
«суббот» были практически все московские орнитологи; многие
приезжали из других городов — специально для этой цели.
Тяжелые годы войны Георгий Петрович провел вместе с уни-
верситетом в Ашхабаде и Свердловске, не прерывал своей научной
и педагогической деятельности, много экскурсировал, собирал ма-
териал для монографии «Птицы Туркменистана», увидевшей свет
в 1952 г., завершая другую свою книгу — «Сокола-кречеты»
(1951). Обе монографии отразили комплексные подходы, разрабо-
танные Георгием Петровичем еще в предвоенные годы специально
для изучения фаун и отдельных систематических групп птиц.
Развивая эти подходы, большая группа учеников Георгия Пет-
ровича — академик АН Туркменской ССР А. К. Рустамов, профес-
сора А. В. Михеев, М. А. Воинственский, Ф. И. Страутман
и Ю. М. Кафтановский с В. М. Модестовым заложили основы,
позволившие позднее создать шеститомную сводку «Птицы Совет-
ского Союза» (1951 —1954). Инициатором и душой всей работы,
ведущим автором и руководителем авторского коллектива был
Георгий Петрович. «Птицы Советского Союза» заняли заметное
место в мировой орнитологии. Вскоре эта работа была переведена
на английский язык и издана за рубежом. «Птицы Советского
Союза» были удостоены Государственной премии СССР.
Высоко оценивая научное значение этой сводки, необходимо
подчеркнуть, что она появилась в результате реализации обще-
союзной программы изучения птиц, объединившей усилия десят-
ков орнитологов, работающих в различных регионах нашей страны.
И хотя эта программа носила, так сказать, «неофициальный»
характер и во многом поддерживалась личным обаянием и высо-
ким научным авторитетом Георгия Петровича, она явилась прооб-
разом общегосударственных программ по орнитологии 1970—
1980 гг.— Миграционной программы, учрежденной специальным
постановлением ГКНТ СССР в 1973 г., Сводного плана научно-
224
исследовательских работ по теоретической и прикладной орнитоло-
гии 1982—1985 гг. и, наконец, программы развития региональных
исследований в области изучения и охраны птиц, принятой Отделе-
нием общей биологии Президиума АН СССР в 1987 г. Следует
отметить, что научная и научно-организационная деятельность
Георгия Петровича создала предпосылки для «официального»
оформления и последующего практического осуществления этих
программ, при этом инициаторами и координаторами их стали
его ученики.
Следует особо подчеркнуть, что Георгий Петрович придавал
большое значение развитию региональной орнитологии, справед-
ливо полагая, что именно регион как исторически и социально-
экономически сложившаяся ландшафтно-географическая единица
должен всегда быть объектом самостоятельного, пристального и
постоянного внимания орнитологов, притом не только представи-
телей центров, но и, так сказать, периферийных научных уч-
реждений.
Считая, что комплексное изучение фауны «своего» региона
является первоочередной задачей последних, он всячески способст-
вовал становлению и развитию орнитологических коллективов в
Туркмении, Казахстане, Украине, Молдавии, Белоруссии, Тад-
жикистане. Возглавившие эти коллективы ученики Георгия Пет-
ровича к настоящему времени подготовили уже своих учеников
и создали свои научные школы, являющиеся как бы «внучатым»
поколением по отношению к Георгию Петровичу,— это ли не сви-
детельство плодотворности его научных идей, его выдающихся
успехов в воспитании научной смены.
Следуя этим подходам, советские орнитологи комплексно изу-
чают отдельные наиболее интересные в лаучном отношении ре-
гионы, в первую очередь малоизученные, но подлежащие интен-
сивному хозяйственному освоению в ближайшие годы. Особое
внимание уделяется и освоенным регионам, длительное время
изучаемым орнитологами. Тем самым реализуется на практике
мысль академика П. С. Палласа о том, что все регионы нашей
огромной территории должны быть хорошо изученными в фауни-
стическом отношении и каждый из них должен иметь свою регио-
нальную монографию. В реализации этой мысли огромный практи-
ческий вклад вложила дементьевская школа во главе со своим
основателем.
Георгий Петрович считал необходимым всесторонне и в первую
очередь в экологическом и систематическом аспекте изучать всех
представителей орнитофауны СССР. И хотя главными любимцами
его были хищные птицы, все остальные группы никогда не выпада-
ли из его поля зрения. Многочисленные статьи и главная сводка
«Птицы Советского Союза» отразили и обобщили весь этот огром-
ный материал в виде видовых очерков, каждый из которых ценен
и как часть фундаментального труда и в тоже время представляет
вполне самостоятельный интерес. Естественно, осознавая всю
сложность поставленной задачи, Георгий Петрович рассматривал
225
эти видовые очерки как прообраз будущих монографий, обобщаю-
щих все сведения о конкретных, сначала наиболее массовых,
хозяйственно важных, интересных в научном и практическом отно-
шении видах, а затем и вообще всех, составляющих нашу фауну.
Сегодняшние зоологи, интенсивно и плодотворно работающие
над академической серией видовых монографий, не должны забы-
вать, что одним из авторов этой идеи был Георгий Петрович.
Используя птиц как природоохранительную модель, Геор-
гий Петрович рассматривал их взаимоотношения с человеком не
только с экологических позиций. Его интересовали и социально-
экономические, и исторические аспекты, и даже лингвистические.
Он является одним из основоположников нового направления на
стыке орнитологии с лингвистикой — экологической биолингви-
стики птиц, занимающейся ролью птичьего голоса в формирова-
нии человеческой речи.
Именно на орнитологической модели он выдвинул и обосновал
оригинальные представления о редких видах животных и их эколо-
гическом статусе в фауне, использовании охраняемых территорий,
в частности заповедников, в охране и восстановлении исчезающих
видов. Его взгляды на охрану природы носили комплексный
характер, и их правильность подтвердило все последующее разви-
тие экологической проблематики. И хотя Георгий Петрович не
оставил обобщающей монографии по охране природы и его идеи
рассеяны по журналам и сборникам, время сохраняет нам облик
гражданина, беззаветно любящего природу и жизнь во всех ее
проявлениях и формах.
И хотя еще при жизни Георгий Петрович был отмечен высокими
наградами, главной своей наградой он считал признательность
и любовь коллег, своих товарищей по работе, всех кто его знал.
Трудно представить себе другого человека, имевшего так много
друзей и так уважаемого и любимого ими. Георгий Петрович
всем этим обладал в избытке.
Заключая статью об этом замечательном человеке, авторы
выражают чувство глубокого уважения к его памяти и ко всему
тому доброму и прекрасному, что он оставил после себя людям
и птицам.
SUMMARY
A Famous Soviet ornithologist and conservationist of nature.
To the 90th Birthday of G. P. Dementiev (1898—1969).
Sokolov V. E., IlijchevV. D.
The main periods of life and scientific activity of Professor G. P. Dementiev are
described. The contribution of G. P. Dementiev to the creation of the first scientific-
coordinative organ on nature protection within the system of the USSR Academy
of Sciences, called the Committee for Nature Conservation, to the development of the
regional ornithology, into the leadership of the All-Union programme of study
of birds as a result of which was the publication of 6 Volumes of «Birds of the'
Soviet Union», awarded with the State Prize of the USSR and was published abroad,
is shown. The contribution of G. P. Dementiev to the bringing up the galaxy
of the Soviet ornithologists is great.
226
УДК 598.20(092)
А. В. МИХЕЕВ
129243, Москва,
ул. Кибальчича, д. 6, корп. 5.
МГПИ им. В. И. Ленина.
Биологический факультет
Г. П. Дементьев
и изучение
миграций птиц
в СССР
С Георгием Петровичем Дементьевым мне посчастливилось не-
посредственно работать на протяжении шести лет, с 1934 по 1940 г.
В дальнейшем дружеские контакты с ним сохранялись у меня
до последних дней его жизни.
По окончании биологического факультета МГУ я был направ-
лен в Зоологический музей, перешедший впоследствии в веде-
ние МГУ, где работал вначале экскурсоводом, а затем младшим
научным сотрудником. Георгий Петрович, ведавший орнитологи-
ческим отделом, да и весь коллектив отдела — Николай Алек-
сеевич Гладков, Ангелина Михайловна Судиловская, Евге-
ний Павлович Спангенберг, директор музея Сергей Сергеевич Ту-
ров — оставили неизгладимый след на всю мою жизнь.
Георгий Петрович был очень скромный, обаятельный и весьма
благожелательный к людям человек. В обращении он был прост,
улыбчив; среди сотрудников музея и биологического факультета
пользовался огромным уважением. В отделе царила дружная,
спокойная, рабочая обстановка.
Бросалась в глаза одна особенность: к сотрудникам отдела,
и особенно к Георгию Петровичу, систематически приходило боль-
шое количество разных по возрасту и своему положению лиц за
консультациями. Среди них немало было орнитологов с периферии,
приезжавших в Москву и считавших необходимым воспользоваться
возможностью выяснить у Г. П. Дементьева возникшие в работе
вопросы. Георгий Петрович всегда приветливо встречал посетите-
лей и исчерпывающе отвечал на вопросы. Это у него занимало
много времени, но он никогда не давал даже малейшего намека
на то, что это отрывает его от текущих научных дел.
Г. П. Дементьев был глубоко эрудированным биологом, на
орнитологическом материале разрабатывавшим общие проблемы
эволюции, экологии, биогеографии и др. Его интересы были
многоплановыми, что определялось широтой и глубиной его науч-
ного кругозора. И каким бы вопросом он ни начинал заниматься,
разрабатывал он его глубоко, фундаментально.
Большой вклад Г. П. Дементьев внес в изучение миграции
птиц.
Эта проблема была в его поле зрения на протяжении большей
части творческой деятельности. Из работ по миграциям прежде
всего следует отметить большую главу «Перелеты и их место
в биологии птиц», в его замечательном капитальном труде «Пти-
цы», 6-го тома «Руководства по зоологии» (1940). В нашей литера-
227
туре это была наиболее полная и фундаментальная сводка новей-
ших для того времени отечественных и зарубежных данных
о перелетах. В ней рассматриваются, по существу, все основные
аспекты перелетов. О широте вопросов, охватываемых сводкой,
можно судить по перечню подзаголовков: перелетные, оседлые
и кочующие формы; непереодические выселения из гнездовой
области; перелеты птиц в тропической и жаркой-зонах; миграции
океанических птиц; ход перелетов, значение гнездовой территории
(родина); места зимовок; направление и пути перелетов; ланд-
шафтные факторы; способы и виды передвижения перелетных
птиц; скорость, высота и продолжительность перелетов; фенологи-
ческие и метеорологические факторы перелетов; непосредственные
причины перелетов; вероятное происхождение перелетов; ориента-
ция; история изучения перелетов.
В сводке использовано большое количество отечественных и
особенно иностранных литературных источников. При этом надо
отметить то, что Г. П. Дементьев не просто изложил взгляды
тех или иных авторов, а давал оценку этим взглядам, высказывал
свою точку зрения по обсуждаемому вопросу. Замечательно, что
абсолютное большинство высказанных им в то время положений
остается верным и в настоящее время.
Многое сделано Г. П. Дементьевым по организации изучения
сезонных миграций птиц в СССР, и в первую очередь через
Центральное Бюро кольцевания. С начала 1940-х годов, на протя-
жении почти 30 лет, т. ,е. почти до конца жизни, он был факти-
ческим руководителем и идейным вдохновителем деятельности
Центрального Бюро кольцевания, переименованного впоследствии
в Центр кольцевания. Непосредственное руководство Центральным
Бюро кольцевания он осуществлял в 1943—1946 гг. Несмотря на
все трудности военной обстановки, работа Бюро не только не
прерывалась, но и несколько расширилась по своему содержанию.
«Проблема миграций птиц... стала изучаться на более широкой
основе, как один из элементов общей биологической проблемы
периодических циклов жизнедеятельности животных в разных
географических условиях» *. Такая постановка вопроса нашла свое
отражение в содержании двух выпусков Трудов Бюро (V и VI),
подготовленных в эти годы Г. П. Дементьевым и опубликованных
в 1947 г. Большинство статей в этих сборниках было написано им.
Указанное направление исследований прослеживается в VII
(1948 г.), VIII (1959 г.) и в ряде последующих выпусков Тру-
дов Бюро кольцевания. VII выпуск Трудов был посвящен Геор-
гию Петровичу в связи с его 50-летием.
В 1956 г. Бюро кольцевания, в связи с упразднением Управле-
ния по заповедникам и охотничьему хозяйству Министерства лес-
ного хозяйства СССР, в состав которого оно входило, по инициативе
и при личном содействии Г. П. Дементьева было передано в' Ко-
* Дементьев Г. П. От редакции // Тр. Центр, бюро кольцеваний. М. 1947.
Вып. VI. С. 5.
228
миссию по охране природы АН СССР, председателем которой
был Г. П. Дементьев.
В феврале 1957 г. постановлением бюро Отделения биологи-
ческих наук АН СССР, опять-таки по. инициативе Георгия Петро-
вича, был создан Совет по изучению миграций животных при
Комиссии по охране природы. В состав Совета вошли крупнейшие
орнитологи и маммологи страны. Общее руководство работой Со-
вета и контроль за его деятельностью были возложены на Комис-
сию по охране природы АН СССР.
Период с 1956 по 1961 г., когда общее руководство работой
Бюро кольцевания, по существу, осуществлял Г. П. Дементьев,
был наиболее плодотворным в работе этой организации. Перед ней
были поставлены новые задачи, что привело к необходимости
применения метода кольцевания для изучения более широкого
круга вопросов биологии птиц. Если раньше кольцевание применя-
лось в основном для изучения сезонного размещения и миграций
птиц, то теперь оно используется для решения вопросов популя-
ционной экологии: выяснения средней продолжительности жизни
особей вида, привязанности их к гнездовой территории, расселения
молодых, причин гибели птиц в природе и т. д. Перед кольцеванием
были поставлены и вопросы прикладного значения, в частности
выявление степени воздействия охоты на численность как промы-
словых, так и непромысловых видов птиц.
За этот период резко возросло и количество помеченных птиц.
Если в 1956 г. в СССР было окольцовано 116 тыс. птиц, то
в 1958 г. помечено 172 тыс. птиц, в 1959 г.— около 210 тыс.,
а в 1960 г.— более 260 тыс.
Весьма плодотворным было участие Георгия Петровича в изда-
нии Трудов Бюро кольцевания. В период с 1937 по 1956 г., когда
Бюро кольцевания находилось в ведении Главного Управления
по заповедникам при Совете Министров РСФСР, а затем при
Министерстве лесного хозяйства СССР, было издано девять номе-
ров сборника «Труды Центрального Бюро кольцевания». С переда-
чей дела кольцевания в систему Академии наук СССР «Труды»
' были заменены сборниками «Миграции животных». С 1959
по 1968 г. вышло пять выпусков этого сборника. Г. П. Дементьев
был идейным вдохновителем и тех и других сборников, оказал
влияние на их теоретическую направленность. Абсолютное боль-
шинство сборников вышло при непосредственном активном его
участии как редактора или как автора помещенных в них статей.
Кроме сборников, под редакцией Георгия Петровича были изданы
два выпуска информационных бюллетеней «Кольцевание и мече-
ние животных» (в 1961 и 1964 гг.), в которых подводились итоги
кольцевания птиц в нашей стране.
Проблема сезонных перелетов птиц интересовала Г. П. Демен-
тьева на протяжении почти всей его творческой деятельности.
Первая публикация по этой проблеме появилась в 1935 г., послед-
няя — в 1966 г. Всего специальных публикаций по миграциям
птиц было пятнадцать, не считая многочисленных предисловий
к сборникам и книгам, посвященным миграциям.
229
Из важнейших работ Г. П. Дементьева этого периода следует
упомянуть, помимо отмеченной выше главы о миграциях птиц,
также брошюру «Сезонное размещение и миграция чаек по данным
кольцевания в СССР» (М., 1947), написанную в соавторстве с
умершим к тому времени В. Н. Вучетичем, статьи «Сезонное
размещение и миграции кречета», «Результаты кольцевания чи-
стиковых» в VI выпуске «Трудов Центрального бюро кольцевания»
(1947 г.) и «О перелетах чернозобой гагары» (VII вып., 1948 г.).
Это были первые обобщения накопленного материала по кольцева-
нию птиц в СССР, представлявшие, несомненно, большой науч
ный интерес.
Придавая важное значение знакомству орнитологов с зарубеж
ной литературой о сезонных миграциях птиц, Г. П. Дементьев
проявил инициативу в осуществлении издания на русском языке
двух книг: «Перелеты птиц и их изучение» И. Штейнбахера
(М.: Изд-во иностр, лит., 1956) и «Перелеты птиц» Д. Гриффина
(М.: Мир, 1966). Обе книги вышли под его редакцией, причем
к первой он написал предисловие, а ко второй — введение. Кроме
того, им была опубликована рецензия на книгу А. Д. Дирксена
«Перелеты птиц» (Новые книги за рубежом. Сер. биол. 1962. № 5).
Г. П. Дементьев придавал большое значение пропаганде работ
по кольцеванию и миграциям птиц вообще. Под его руководством
сотрудники Бюро кольцевания проводили большую популяриза-
торскую работу, выступая со статьями на страницах журналов
«Охота и охотничье хозяйство», «Природа» и др; Будучи крупным
ученым, не пренебрегал этой стороной дела и сам Георгий Петро-
вич. В качестве примера можно привести следующие его публика-
ции.
В 1935 г. в журнале Боец-охотник им были напечатаны две
статьи: «Перелет птиц» (№ 3) и «Еще о птичьих перелетах»
(№ 11); в 1937 г. в сборнике «Биология в школе» (№ 6) — статья
«Ориентация у птиц». В 1956 г., в восьмом номере журнала «Охота
и охотничье хозяйство» появилась статья «Перелеты птиц». Раз-
дел с таким же названием, написанный совместно с С. С. Гуровым,
включен в четвертый том Детской энциклопедии (М.: Учпедгиз,
1960).
Почти за полувековой период кольцевания в нашей стране
накопился огромный новый материал, что сыграло большую роль
в изучении сезонных миграций птиц. Большая заслуга в этом
Г. II. Дементьева.
Огромное влияние на подготовку орнитологических кадров,
повышение их квалификации, на развитие орнитологии в нашей
стране оказала деятельность Орнитологической секции, которой
начиная с 1930-х годов на протяжении нескольких десятилетий
руководил Георгий Петрович. На заседания этой секции собира-
лись как старшие, так и начинающие орнитологи-москвичи, а не-
редко и представители периферии. Среди вопросов, обсуждающих-
ся на заседаниях, заметное место занимали сезонные миграции
птиц. В целом эта секция, душой которой был Г. П. Дементьев
230
и в которой систематически работали многие орнитологи, сыграла
важную роль в активизации и повышении научного уровня иссле-
дований в области орнитологии вообще и перелетов птиц в част-
ности.
Наконец, нельзя не подчеркнуть большого влияния на разви-
тие орнитологических исследований, в том числе и по перелетам
птиц, упоминавшихся уже личных контактов Георгия Петровича
с орнитологами, его консультаций, которые он давал очень многим
московским и периферийным коллегам, обращавшимся к нему
устно и письменно по вопросам личной работы.
Георгий Петрович Дементьев был постоянно в курсе современ-
ных достижений мировой и отечественной орнитологии, хорошо
представлял наиболее желательные пути ее развития. Будучи
человеком весьма скромным и отзывчивым, он снискал себе
большой авторитет, любовь и уважение среди широких кругов
отечественных орнитологов. Вся долголетняя деятельность
Г. П. Дементьева явилась выдающимся вкладом в развитие нашей
отечественной орнитологии, в том числе и одного из его разде-
лов — сезонных миграций птиц.
SUMMARY
G. Р. Dementiev and the study of birds migrations in the USSR.
A. V. Mikheev
G. P. Dementiev practically was a leader in the USSR for the works on ringing
and migrations of birds. He published some important books and papers on migrations
of birds, organized the researches on these problems in the country. Many years
he was as a Chief of the Central Ringing Bureau, in 1957 he created the Council
on study of animals migrations in Academy of Sciences. G. P. Dementiev was as
editor-in-chief and author for the numerous volumes of «Transactions of Central
Ringing Bureau», «Migrations of Animals» and «Ringing and Marking of Animals».
Many popular papers by G. P. Dementiev was published in non-fiction magazines.
231
УДК 598.20(092)
В. Е. СОКОЛОВ, Все силы —
В. Д. ИЛЬИЧЕВ изучению и охране птиц
117071, Москва, Ленинский проспект, д. 33. Институт Анверу Кеюшевичу
эволюционной морфологии и экологии животных Рустамову —
им. А. Н. Северцова АН СССР 70 лет
В 1950 г. в одной из аудиторий Московского университета за-
щищалась докторская диссертация, посвященная птицам Турк-
менистана. По единодушному мнению членов Ученого Совета, эта
работа внесла выдающийся вклад в изучение орнитофауны мало-
изученного региона. Сама работа не вызывала никаких сомнений,
однако необычен был возраст диссертанта — 33 года — редчайший
случай в зоологии тех лет. Этим диссертантом был Анвер Кеюше-
вич Рустамов.
Прошло около 40 лет и 17 декабря 1987 г. он — крупный ученый
и деятель охраны природы, педагог и организатор науки — встре-
тил свое семидесятилетие уже как действительный член Академии
наук Туркменской ССР, профессор и ректор Туркменского сельско-
хозяйственного института, заслуженный деятель науки республи-
ки. Среди многочисленной армии орнитологов и любителей птиц
нашей страны А. К. Рустамов приобрел широкую известность
как вице-президент Всесоюзного орнитологического общества, ав-
тор многих научных трудов и популярных книг, посвященных
птицам Средней Азии. И хотя он занимался также млекопитаю-
щими и рептилиями, основным объектом его внимания всегда
были и остаются птицы.
Во многом это связано с тем, что в становлении А. К. Руста-
мова как ученого ведущую роль сыграл выдающийся советский
орнитолог Г. П. Дементьев. Отношения между ними были не толь-
ко отношениями ученика и учителя. Долгие годы, вплоть до кончи-
ны Г. П. Дементьева, их связывали узы тесной дружбы и това-
рищества, сердечной теплоты. Точно так же в формировании
личности А. К. Рустамова — уроженца Ашхабада, выпускника
Туркменского педагогического института — большое значение
имело его длительное, многолетнее пребывание в стенах Москов-
ского университета в качестве стажера и докторанта. Связь
А. К. Рустамова с этим старейшим университетом страны не
прерывается до сих пор.
Именно в стенах Московского университета в постоянном обще-
нии с А. Н. Формозовым, С. С. Туровым, В. Г. Гептнером,
Е. П. Спангенбергом, Е. С. Птушенко и другими выдающимися
зоологами старшего поколения вырабатывались оригинальные под-
ходы к комплексному изучению орнитофауны Туркменистана,
экологии и поведения пустынных птиц. Для А. К. Рустамова
особое значение приобрели слова, часто повторяемые его учителем:
«изучать птиц как модель общебиологических проблем». Следуя
232
Анвер Кеюшевич Рустамов
за обработкой коллекции птиц
этому девизу, он сосредоточил основное внимание на выяснении
адаптаций пустынных птиц и, в частности, на изучении экологи-
ческих, физиологических и зоогеографических механизмов этих
адаптаций. Общебиологическое значение имеют работы А. К. Ру-
стамова, раскрывающие адаптивные явления в жизни отдельных
наиболее интересных по своей биологии пустынных обитателей.
К числу наиболее важных обобщений и гипотез, высказанных
А. К. Рустамовым, следует отнести оригинальные представления
об адаптивной природе эколого-географических параллелизмов
и феномене эко-географического изоморфизма в отношении пу-
стынных птиц, адаптивной трактовке понятия «жизненная форма»,
антропогенным факторам в изменении региональной фауны и роли
человека в формировании культурного ландшафта в условиях
Средней Азии.
Одновременно с этим А. К. Рустамов уделяет пристальное вни-
мание изучению региональной фауны, используя комплексные под-
ходы и методы зоогеографии, систематики и экологии. Именно ему
принадлежит оригинальная трактовка понятия «фаунистический
комплекс», именно он одним из первых поставил вопрос о воз-
можности управления региональными фаунами. Эти направления
нашли свое отражение в фундаментальной монографии А. К. Ру
8 Заказ № 36
233
стамова «Птицы Туркменистана» (1958), значение которой как
исторической точки отсчета в динамике среднеазиатской фауны,
своеобразного эталона ее состояния на период середины века
все более возрастает в наши дни.
Огромный фактический материал, собранный Л. К. Рустамовым
и его многочисленными учениками, вошел составной частью в фун-
даментальную монографию «Птицы Советского Союза» (1951 —
1956) и был обобщен ими уже на общесоюзном уровне. Как извест-
но, эта монография стала первой орнитологической работой, удо-
стоенной Государственной премии СССР. Участие в этой моно-
графии, как и его эколого-фаунистические исследования по пти-
цам Туркмении, позволили Л. К. Рустамову войти в круг ведущих
орнитологов страны, обеспечили высокий научный авторитет и
признание коллег.
Экологическими аспектами охраны природы А. К. Рустамов
заинтересовался с первых шагов своей научной деятельности под
влиянием Г. П. Дементьева. Всесторонне изучая проблемы охраны
природы Туркменистана, падение численности отдельных видов,
последствия антропогенных изменений, он формулирует важные
представления об экологических механизмах влияния человека
на отдельные виды и фауну в целом, месте и значении редких
видов в зоогеографических обобщениях, урбанизации как экологи-
ческом факторе в жизни пустынных птиц. Эти исследования слу-
жат ему теоретической основой для осуществления (под его
руководством и при непосредственном участии) важных практи-
ческих мероприятий по охране природы Туркмении — создании
Красной книги Туркменской ССР (1985) и коллективного труда
«Охрана природы Туркменистана» (1978), организации сети запо-
ведников и заказников в республике, а также специализированных
питомников для разведения редких видов.
Особое место в деятельности А. К. Рустамова всегда занимала
и занимает подготовка научных кадров, в том числе и высшей
квалификации. Преподавая в Туркменском педагогическом инсти-
туте, Туркменском университете и в Туркменском сельскохозяйст-
венном институте, он создал школу учеников, работающих ныне
во многих вузах и практических зоологических учреждениях.
Учениками А. К. Рустамова с гордостью называют себя 15 докто-
ров и кандидатов наук. А. К. Рустамов воспитывает у своих учени-
ков широкие экологические взгляды на сущность природоохранных
задач, высокую ответственность за их решение. Как ректор Турк-
менского сельскохозяйственного института, он делает все для повы-
шения уровня экологизации мышления будущих специалистов
сельского хозяйства республики. Влияние А. К. Рустамова как
воспитателя и педагога распространяется во многие республики
Средней Азии и Кавказа, с орнитологами которых он давно и тесно
сотрудничает.
А. К. Рустамов пользуется широкой международной извест-
ностью, активно участвует в деятельности Международного союза
охраны природы как член его руководящего органа. В проведении
234
впервые в нашей стране XIV Генеральный ассамблеи Международ-
ного союза охраны природы в 1978 г. он выполнял наиболее
трудньГе и ответственные функции Генерального секретаря.
В большом успехе этого важного мероприятия есть значительная
доля его труда.
В краткой статье невозможно коснуться всей многогранной
деятельности А. К. Рустамова, в том числе и его вклада в развитие
герпетологии и териологии. Конечно, как ученый он един в своих
достижениях и поисках, и Рустамов-герпетолог и териолог в чем-то
дополняет Рустамова-орнитолога. Но еще большее значение имеет
этот союз в общественной деятельности А. К. Рустамова, выпол-
няющего ответственные функции председателя республиканского
общества охраны природы и Комиссии по охране природы Турк-
мении.
Участие А. К. Рустамова в Великой Отечественной войне,
его плодотворная научная и общественная деятельность отмечены
многими правительственными наградами, Почетными грамотами
Верховного Совета республики. Но что особенно приятно отметить
в юбилейную дату — это его высокие человеческие качества, тепло-
ту и внимательность к людям. Вместе со всеми авторы статьи
желают А. К. Рустамову новых творческих успехов, здоровья
и бодрости вперед на долгие годы.
SUMMARY
All activity for study and protection of birds
(The 70th Anniversary of Anver Keushevich Rustamov).
S о k о 1 о v V. E., 1 1 i j c h e v V. D.
A. K. Rustamov, the follower of G. P. Dementiev, is a great scientist and nature
conservationist, teacher and organizer in the science. He is the author of the
fundamental study «Birds of Turkmenistan» (1958), one of the authors of 6-Volumed
«Birds of the Soviet Union» (1951 —1956); he supervised the publication of «The
Red Data Book of the Turkmain SSR» (1985) and the collective work «The Nature
Conservation in Turkmenistan» (1978). A. K. Rustamov took a direct participation
in the organization of the system of sanctuaries, reserves and specialized nurseries
for breeding of rare species of animals. In 1978 he performed the function of the
General Secretary of the XIV General Assembly of the International Union for
Nature Conservation. 15 Doctor’s and Master’s Thesises were defended under his
supervision.
8*
235
УДК 598.20(092)
в. и. гаранин В. А. Попов
245018, Казань, ул. Ленина, д. 1. -j.
КГУ им. В. И. Ленина. Н ©U-ЛеТИЮ
Биологический факультет. СО ДНЯ рО5КДСНИЯ
Кафедра охраны природы г
Виктор Алексеевич Попов (1910—1980) был одним из учеников
А. А. Першакова (1875—1942), основавшего в 1920—1930-х годах
казанскую школу экологов. В научной деятельности В. А. Попова
можно выделить три основных направления: изучение промысло-
вых животных; фауна и экология животных; охрана природы. Хотя
он и считался в большей степени териологом, изучение птиц
являлось составной частью каждого из упомянутых направлений.
Первыми орнитологическими работами В. А. Попова были за-
метка о перспективах развития дичного охотхозяйства, опублико-
ванная в журнале «Боец-охотник» (1935), и статья «К экологии
лысухи» *, основанная на материалах, собранных в 1933 г. в Зау-
ралье и отчасти в Татарии. Работа по лысухе включает общую
характеристику вида, географическое распространение, биотопи-
ческое размещение, сезонную динамику, данные по ее размноже-
нию и питанию. Статью отличает обстоятельность изложения ма-
териала, типичная для автора (хотя это только третья его печат-
ная работа).
После длительного перерыва, связанного с работой над канди-
датской диссертацией, посвященной американской норке, и года-
ми Великой Отечественной войны, выходит написанная вместе
с А. В. Лукиным книга «Животный мир Татарии» (1949 г.) —
первая популярная книга о фауне позвоночных республики, где
соответствующее место отведено птицам. В послевоенные годы,
когда в Татарии возобновилась работа Всероссийского общества
охраны природы, в республиканских газетах часто стали появлять-
ся заметки В. А. Попова о природе, в частности, посвященные
Дню птиц. Орнитологическая тематика разрабатывалась и в ходе
исследований, проводившихся под руководством В. А. Попова в
Казанском филиале АН СССР по изучению зоны затопления Куй-
бышевской ГЭС. Эти работы завершились коллективной моногра-
фией, содержащей уникальные данные по фауне поймы Средней
Волги и Нижней Камы, залитой водами Куйбышевского водохра-
нилища **. В 1950—60-е годы основное внимание уделялось изуче-
нию богатой авифауны Волжско-Камского края — географии, эколо-
гии, фенологии, морфологии, использованию и охране птиц, чему
посвящен ряд статей и докладов ученого на конференциях. Орни-
тологические исследования, сочетавшие стационарные наблюде-
ния, в основном в Волжско-Камском заповеднике, организованном
* Тр. О-ва естествоиспытателей при Казан, ун-те. 1938. Т.55, вып. 3/4.
** Попов В. А., Попов Ю. К., Приезжаев Г. П. и др. Результаты изучения животного
мира зоны затопления Куйбышевской ГЭС // Тр. Казан, фил. АН СССР. Сер.
биол. 1954. Вып. 3.
236
в 1960 г., и экспедиционные работы на территории 12 областей
и республик, завершились изданием двухтомной монографии
«Птицы Волжско-Камского края» (1977—1978). В. А. Поповым
были написаны «Введения» обоих томов и 11 очерков. В сводке
подведены итоги изучения птиц Волжско-Камского края за 200 лет.
В монографии приняли участие 28 авторов из Казани, Москвы,
Ленинграда, Горького, Ижевска, Йошкар-Олы, Кирова, Чебоксар,
Окского заповедника. Продолжалось изучение промысловых птиц,
преимущественно водоплавающих, что получило отражение в ряде
работ и выступлений, в том числе на Международных конгрессах
биологов-охотоведов в Москве, Хельсинки и Стокгольме. Значи-
тельное место уделялось вопросам охраны природы вообще, охране
и использованию животных, в том числе птиц. С этим связаны
статьи в сборниках «Природа Татарии и ее охрана» (1963, 1971),
доклад на соискание ученой степени доктора биологических наук,
защищенный в Московском университете (1964), статьи об истории
исследований на территории Волжско-Камского заповедника
(1968), о роли человека в формировании фауны птиц (1971 г.),
второе издание «Животного мира Татарии» (1971). В более
общей форме эти вопросы затрагиваются в популярной книге
«Биосфера и вопросы ее охраны» (1975), в описании памятников
природы Татарии (1977), в работе о некоторых общих закономер-
ностях формирования прибрежных акваторий и береговых био-
геоценозов водохранилищ (1978), в книге «Редкие и исчезающие
виды животных Татарии» (1978) и в уже несостоявшемся докладе
на научной конференции, посвященной 20-летию Волжско-Камско-
го заповедника (1980). Последняя из вышедших посмертно работ
В. А. Попова касается также орнитологии: это очерк об истории
изучения птиц Волжско-Уральского края в первом томе сводки
«Птицы СССР» (1982).
В настоящее время исследования по экологии птиц и их охране
продолжаются в отделе экологии Казанского филиала АН СССР,
на кафедре зоологии позвоночных Казанского университета и ка-
федре зоологии Казанского пединститута, а также учениками
Виктора Алексеевича Попова разных поколений, работающими
во многих точках Советского Союза — от Белоруссии до Приморья.
SUMMARY
V. A. Popov. For the 80th birthday anniversary. V. I. Garanin.
Victor Alexeevitch Popov (1910—1980) was famous ornithologist, theriologist and
conservationist of nature. He was born, lived and worked in Kazan. His main
scientific researches were dedicated to animal world of Tataria and to birds of
Volga—Kama region. The problems of nature conservation took a marked place
in his investigations.
237
ХРОНИКА
УДК 598.2
Международная сотая сессия
Немецкого Орнитологического общества
С 25 сентября по 1 октября 1988 г. в г. Бонне (Федеративная
Республика Германии) проходила Юбилейная годичная сессия
Немецкого Орнитологического Общества, посвященная 100-летию
со дня его основания. Она была задумана и проведена как между-
народное собрание, что было отражено и в официальном назва-
нии ее: «Международная сотая годичная сессия Немецкого Орни-
тологического Общества». Помимо ученых из ФРГ, для участия
в этом мероприятии было приглашено около 50 наиболее известных
орнитологов из США, Великобритании, Канады, Польши, Швей-
царии, Австрии, Германской Демократической Республики, Да-
нии, Голландии, Бразилии. Получили приглашение и приняли
участие в работе сессии орнитологи СССР. В делегацию Советско-
го Союза вошли проф. В. Д. Ильичев, Л. С. Степанян, А. А. Наза-
ренко.
Немецкое Орнитологическое общество — одна из старейших в
мире и одна из наиболее авторитетных орнитологических органи-
заций.
В разное время в его состав входили все крупнейшие орни-
тологи мира. Отечественная наука в прошлом была представ-
лена академиком П. П. Сушкиным, Г. П. Дементьевым, Л. А. Пор-
тенко, Б. К. Штегманом. Общество насчитывает около 3000 ин-
дивидуальных и большое число коллективных членов — различ-
ных научных учреждений и организаций. Помимо профессиональ-
ных ученых, в Общество входит большое число орнитологов-
любителей.
Общество проводит обширную научно-исследовательскую рабо-
ту, а также координирует многие орнитологические изыскания как
в Федеративной Республике Германии, так и за ее пределами.
Основной печатный орган Общества — «Journal fiir Ornithologies;
в 1988 г. вышел его 129-й том.
Международная юбилейная сессия Общества включала офи-
циальные приветственные мероприятия, научную часть (26 сен-
тября — 1 октября), экскурсии (2—3октября), прощальные меро-
приятия. Научная часть была необыкновенно насыщенной. Состоя-
238
лось пять пленарных заседаний, прошло восемь симпозиумов:
Энергетика и терморегуляция, Роль эндогенных и экзогенных фак-
торов в биологии размножения и плотности населения хищных
птиц, Годовой цикл и гормональная регуляция, Телеметрия, Эколо-
гия поведения и выбор местообитаний, Охрана природы — особен-
но воздействие врановых на редкие виды птиц, Экология морских
птиц, Ориентация и навигация. Вторая половина дня дважды
выделялась для постерных демонстраций. Советская делегация,
кроме того, провела круглый стол на тему о современном состоянии
орнитологической науки в СССР и по вопросам охраны птиц
в нашей стране. В конце каждого рабочего дня выделялось время
для показа орнитологических фильмов и докладов, посвященных
истории орнитологии и отражению роли птиц в искусстве и духов-
ной жизни человека. Все без исключения демонстрировавшиеся
фильмы были технически безупречны и содержали большой объем
познавательного материала (например, великолепная картина по
экологии колибри, снятая К.-Л. Шухманом).
Составной частью программы сессии были анималистические
выставки (рисунки птиц, выполненные для научных монографий,
сводок и определителей; высокохудожественные живописные
произведения с изображением птиц), выставки научной и научно-
популярной орнитологической литературы, показ орнитологи-
ческих и оологических коллекций. Работала богатая выставка-про-
дажа орнитологической литературы и богатейшая выставка-прода-
жа оптических приборов, фото- и кинотехники, адаптированных
для наблюдений и съемок птиц.
Все мероприятия проводились в помещениях Музея Александ-
ра Кёнига и в залах дома Конрада Аденауэра — официального
здания, предназначенного для наиболее важных государственных
и общественных мероприятий. К открытию сессии были опубли-
кованы и розданы участникам программа, список участвующих
лиц, тезисы докладов, а также многочисленные вспомогательные
информационные материалы, касающиеся деятельности Общества,
истории и современности города Бонна и др.
Сессия открылась 26 сентября приветственным словом Прези-
дента Немецкого Орнитологического общества проф. К. Шмидт-
Кёнига. Затем собравшиеся заслушали приветствия мэра г. Бонна,
представителя партии «зеленых», министра Земли Рейн-Вестфа-
лия.
Юбилейный характер сессии наложил отпечаток на ее научную
программу. По замыслу организаторов, она должна была отразить
достижения орнитологической науки в историческом аспекте и,
с другой стороны, дать абрис современного состояния и перспектив
развития. Эта цель была полностью достигнута.
В настоящем сообщении нет возможности охарактеризовать весь
спектр интересных и весьма содержательных докладов и сообще-
ний по всем разделам изучения птиц, зачитанных на сессии.
Существенно отметить, что классические направления орнитоло-
гии, такие, как систематика, орнитогеография, эволюция, морфо-
239
логия, нашли достойное отражение в программе. Значительное
место было уделено вопросам экологии, ориентации и навигации,
поведения, физиологии, биоакустики, охраны птиц. Среди методи-
ческих приемов наибольшее внимание было уделено телеметрии
й биоакустическим методам. Подчеркнуты контакты орнитологии
со смежными науками.
Здесь нет возможности дать обзор всей научной программы
сессии, можно лишь упомянуть авторов наиболее интересных док-
ладов. С весьма большим вниманием были выслушаны доклады
Э. Майра (Гарвардский университет, США), В. Бока с соавторами
(Колумбийский университет, США) | Ю. Хаффера (Эссенекий уни-
верситет, ФРГ), П. Гранта (Принстонский университет, США),
Л. Шорта и Д. Горн (Американский музей естественной истории,
США, и Естественно-исторический музей Найроби, Кения),
Г. Гедульдига и И. Мартенса (университет Майнца, ФРГ), Ф. Ву-
лимье (Американский музей естественной истории, США), Г. Узле
(Оттавский национальный музей, Канада). Кроме того, очень
интересные доклады и сообщения сделали П. Бертольд, Е. Новак,
Г.-И. Кюнне, Я. Пиновский, В. Эльстродт, Г.-Г. Бергман, Г. Не-
мечкал, А. Эльцен, Р. Эльцен, П. Бюлер, Д. Хомбергер, Р. Прин-
цингер, Р. Дрент, К. Шмидт-Кёниг, П. Даум-Бенц, Р. Вильчко,
Р. П. Балда, В. Вильчко, Д. У. Ганцорн. Советские участники
сессии В. Д. Ильичев и А. А. Назаренко сделали два доклада.
Юбилейная сотая годичная сессия Немецкого Орнитологическо-
го Общества стала крупной вехой в развитии мировой орнитологии.
Широта охвата научных проблем, самый высокий уровень не толь-
ко пленарных, но и симпозиальных докладов, наконец, широкое
международное представительство участников делают прошедшую
сессию большим международным научным событием.
Мы желаем дальнейших успехов в плодотворной научной
и организационной деятельности Немецкому Орнитологическому
обществу и благодарим организаторов Юбилейной сессии за при-
глашение участвовать в ее работе.
В. Д. ИЛЬИЧЕВ, Л. С. СТЕПАНЯН
УДК 598.2
Четвертые чтения
памяти академика М. А. Мензбира
12—13 мая 1986 г. в Туле на кафедре зоологии Тульского го-
сударственного педагогического института им. Л. Н. Толстого про-
ходили IV чтения памяти академика М. А. Мензбира (1855—1935),
приуроченные к 130-летию со дня его рождения. В них приняли
участие президент Всесоюзного орнитологического общества проф.
В. Д. Ильичев (Москва), проф. Е. Н. Дерим-Оглу (Орехово-Зуево),
проф. И. Д. Миллер (Тула) и многие орнитологи и зоологи Москвы,
Перми; Свердловска, Фрунзе, Харькова и других городов страны.
Чтения были организованы Всесоюзным орнитологическим об-
ществом АН СССР, Орнитологическим комитетом СССР и Туль-
ским государственным педагогическим институтом. Первые чтения
памяти академика М. А. Мензбира проходили в Москве в 1978 г.,
вторые — в Ленинграде, третьи — в Киеве, четвертые чтения со-
стоялись на родине выдающегося орнитолога.
Проф. В. Д. Ильичев в обстоятельном докладе «М. А. Мензбир
и общебиологическое направление в изучении птиц» показал
основополагающую роль М. А. Мензбира в развитии многих
направлений орнитологии. Именно Мензбир впервые наладил коор-
динацию фаунистических исследований в различных регионах Рос-
сии и опубликовал результаты этой работы в 15 томах «Мате-
риалов к познанию фауны и флоры России». Обобщение огромного
материала позволило ему подготовить фаунистические сводки «Ор-
нитологическая география Европейской России» (1882) и «Птицы
России» (1895). Михаил Александрович составил карту перелетов
птиц и объяснил причины миграций. Анатомические исследования
привели его к серьезным филогенетическим выводам, касающимся
происхождения пингвинов и их места в системе птиц. М. А. Менз-
бир занимался охотничьими птицами и вопросами их практическо-
го использования, посвятив этому специальную монографию.
В одной из своих крупных работ — «Орнитологические участки
Туркестанского края и вероятное происхождение фауны послед-
него» (1914) М. А. Мензбир произвел анализ истории фауны
Средней Азии в связи с современными геологическими данными.
Значение М. А. Мензбира как учителя, создавшего школу зоо-
географов, систематиков, эволюционистов, а также фаунистов,
обследовавших огромные просторы России, исключительно ве-
лико.
Проф. И. Д. Миллер выступил с докладом «Тульский край —
родина М. А. Мензбира». Михаил Александрович родился 4 нояб-
ря 1855 г. в Туле в небогатой дворянской семье. С детства
он увлекался изучением животных, собирая коллекции беспозво-
ночных в различных уездах Тульской губернии. В 1869 г. М. А. Мен-
збир поступает в Тульскую гимназию, которую заканчивает с сере-
241
бряной медалью. С 1874 г. он — студент Естественного отделения
физико-математического факультета Московского университета,
который закончил с золотой медалью в 1878 г. Вся его дальнейшая
жизнь была связана с Московским университетом. В 1917 г.
Мензбир стал первым выборным ректором МГУ, а в 1929 г.—
академиком АН СССР. Велики заслуги Михаила Александровича
перед мировой наукой. А. В. Луначарский отмечал: «Высоко
ценя Ваши научные заслуги, создавшие Вам мировую известность,
и Вашу многолетнюю профессорскую деятельность, коллегия Нар-
компроса на заседании своем от 7-го апреля сего 1924 года, постано-
вила отметить 40-летний юбилей Вашей профессорской деятель-
ности. Ваше имя уже давно заняло одно из самых выдающихся
мест среди европейских орнитологов и зоогеографов».
Проф. К. А. Татаринов подчеркнул, что М. А. Мензбир был
орнитологом — энциклопедистом, и посвятил свое выступление
экологии и охране редких видов птиц Карпатского региона. Док-
ладчик отметил дальнейшее снижение численности хищных птиц,
занесенных в Красную книгу Украинской ССР. Постепенно воз-
растает лишь численность черного аиста, у которого намечается
тенденция перехода к синантропному образу жизни, что подтвер-
ждает пребывание этих птиц на пашнях совместно с белыми
аистами.
Проф. Е. Н. Дерим-Оглу рассказала о способности к обучению
некоторых мелких воробьиных птиц, в частности методе проб
и ошибок обыкновенной горихвостки и мухоловки-пеструшки. Эти
птицы адекватно реагируют на цвет и игнорируют форму.
Широкое обсуждение вызвал доклад сотрудников Палеонтоло-
гического института АН СССР Е. Н. Курочкина и А. С. Рутиана
«Структура филогенеза i происхождение птиц», а также сообще-
ние доц. Б. Г. Петрова йз Пермского университета «Экология
раннего онтогенеза птиц». Отмеченные выступления радовали
своей оригинальностью, новизной и дискуссионностью, привлече-
нием математических методов и определенных философских умо-
заключений.
Проведение IV чтений памяти академика М. А. Мензбира на
его родине — знаменательное событие. Продолжая традиции вели-
кого орнитолога, преподаватели и студенты кафедры зоологии
Тульского пединститута проводят мониторинговые исследования
фауны Ясной Поляны, которые обеспечат охрану природной среды
Ясной Поляны и сопредельных территорий Тульской области,
дадут материалы для составления Всесоюзного кадастра животно-
го мира СССР.
В память о замечательном земляке на здании бывшей Тульской
гимназии по ул. Менделеевской, д. 7, установлена мемориальная
доска с надписью: «Здесь учился выдающийся русский зоолог
М. А. Мензбир с 1868 по 1874 год». Участники чтений возложили
живые цветы у этой доски.
К. А. ТАТАРИНОВ
242
УДК 598.20: 59.006
Орнитологические исследования в зоопарках
(К 125-летию Московского зоологического парка)
В настоящее время ни у кого не вызывает сомнений необходи-
мость проведения научных исследований на базе зоопарков и пи-
томников. Возрастает роль зоопарков и зоосадов, ведущих свои
исследования в области пропаганды идей охраны природы и естест-
венно-научных знаний среди населения, разрабатывающих науч-
ные основы воспроизводства редких и ценных для народного
хозяйства видов животных, в том числе и птиц. Из 8600 видов
птиц мировой фауны около половины содержится в зоопарках,
питомниках и частных коллекциях. О характере и сроках размно-
жения, линьки, о поведении и т. д. многих из них, особенно
редких и малоизученных, данные получены только в неволе. Целый
ряд видов спасен от исчезновения исключительно благодаря
зоопаркам. Возросло число видов, находящихся в угрожаемом
состоянии и требующих срочного вмешательства природоохранных
органов. Возникает необходимость разработки технологии искус-
ственного разведения птиц различных таксонов с целью создания
генетически полноценных воспроизводящихся групп птиц в нево-
ле — своего рода искусственных «популяций». Разведение редких
видов предполагает возможность их реинтродукции и интродук-
ции в природные местообитания. Таким образом, разведение
птиц и других групп животных — основная задача зоопарков, она
подчиняет себе другие направления научных исследований
здесь.
Зоопарки сейчас обладают огромным опытом по содержанию
и разведению многих видов птиц. Особенно ценен опыт узко-
специализированных зоопарков. Например, Слимбриджский
зоопарк в Англии содержит крупнейшую коллекцию гусеобразных
мира. Трест по разведению фазанов в этой же стране имеет
большую коллекцию фазанов и большой опыт по их разве-
дению.
В Берлинском зоопарке (ГДР) содержится крупная коллекция
хищных птиц и сов, он является методическим центром по .их
разведению. В орнитопарке Вальсроде (ФРГ) содержится большая'
коллекция попугаев. Многие отечественные и зарубежные зоопар-
ки занимаются разведением птиц из отрядов курообразных и гусе-
образных. Многие представители этих отрядов имеют огромную
народнохозяйственную ценность. Так, Московский зоопарк со дня
своего основания в 1864 г. специализируется на разведении птиц
из семейства утиных, а в последние Годы поле его деятельности
расширилось и зоопарк приобрел новые направления научных
исследований в области разведения журавлеобразных, хищных
птиц и курообразных. Алма-Атинский зоопарк имеет свой питом-
ник, где разводятся некоторые редкие веды хищных птиц.
243
Достаточно актуальны сейчас работы по изучению методов
разведения охотничье-промысловых видов птиц. Кроме видов, ши-
роко использующихся в дичеразведении, необходимо вовлекать
перспективные виды, обладающие достаточно ценными свойствами,
экологически пластичные. Это поможет обогатить нашу фауну
и уравновесить численное соотношение птиц различных видов,
нарушенное в периоды массового выпуска в природу птиц из
дичеферм. Например, кроме кряквы, можно будет с успехом вы-
пускать различных нырковых уток и гусей, а также виды, ставшие
редкими в конкретных местах выпуска.
С целью обмена информацией сведения о разведении различных
видов птиц публикуются в Международном ежегоднике зоопарков
(Лондон), в выпусках трудов отдельных зоопарков, тематических
сборниках и специальных журналах. Анализ этих данных показал,
что зоопарки добились больших успехов в разведении птиц многих
систематических групп. Остановимся на группах, являющихся
объектом специализации Московского зоопарка.
Гусеобразные. Во многих зоопарках мира успешно разводят
различные виды гусей, казарок, огариных, речных, древесных,
нырковых уток, однако имеется лишь несколько случаев раз-
множения таких «проблемных» видов, как гага-гребенушка, каме-
пушка, морянка, синьга, горбоносый турпан, луток, средний и боль-
шой крохали, савка. Особенно хорошие результаты по разведе-
нию водоплавающих получены в Слимбридже, зоопарках Берлина,
Хельсинки и Филадельфии. В Московском зоопарке к настоящему
времени содержится коллекция, включающая до 70 видов и подви-
дов водоплавающих птиц. На четырех прудах общей площадью
2,5 га созданы условия, приближенные к естественным. Строятся
стационарные и плавучие острова, на берегах имеются каменные
горки с пещерками, устанавливаются домики различной конфигу-
рации и размеров, на деревьях подвешиваются дуплянки для уток-
дуплогнездников. Создана инкубаторная станция, обрабатывающая
полторы—две тысячи яиц в год. За время существования зоопарка
получен и выращен приплод от черных и тундровых лебедей,
кликуна, шипуна и трубача, сухоноса и горного гуся, большого
и малого белых гусей, серого гуся и гуменников, белощекой,
канадской и краснозобой казарок. Магелланова и нильского гусей,
различных видов огарей, многих видов уток. В зоопарке много лет
ведется работа по созданию свободноживущих групп гусеобразных.
Положительные результаты достигнуты по крякве, гоголю, хохла-
той чернети, красноголовому и красноносому ныркам, огарю и пе-
ганке. В разное время на прудах жили отдельные особи и пары
летных черных лебедей, шипунов, горного и белого гусей, серого
гуся, белоглазого нырка и свиязи. Миграционное состояние сохра-
няется только у нырковых уток, а все остальные ведут оседлый
образ жизни. Такая работа способствует обогащению не только
фауны города, но и пригородной зоны.
Особое природоохранное значение получили работы по разведе-
нию редких и исчезающих видов птиц, внесенных в Международ-
244
ную и региональные Красные книги. Из них в Московском зоопарке
размножались лайсанский чирок, кубинская свистящая утка, бело-
щекая и краснозобая казарки, сухонос и горный гусь, а также
тундровый лебедь и трубач.
Курообразные представляют интерес не только для использова-
ния в народном хозяйстве, но и с природоохранных позиций.
Так, во многих зоопарках мира хорошо размножаются кеклики,
серые куропатки, перепела, обыкновенные фазаны, а также ряд
экзотических форм фазановых, цесарок, индеек. Сложнее обстоит
дело с разведением тетеревиных птиц ввиду специфичности их
биологии и питания. Они в условиях неволи в большей мере
подвержены различного рода заболеваниям, особенно паразитар-
ным. Наибольшего успеха зоопарки достигли при разведении глу-
харя, в то же время белая куропатка размножалась лишь в семи-
восьми зоопарках мира, в том числе в Московском (в 1978—
1979 гг.) и Ленинградском. Лишь в четырех зоопарках размно-
жалась тундряная куропатка, около десяти зоопарков разводили
тетерева, в том числе Ленинградский и Новосибирский, и лишь
в трех получили приплод от рябчика.
В последние годы в Московском зоопарке создана коллекция
редких видов фазанов, среди которых успешно разводятся фазаны
Свайно, Микадо, Эллиота, Хьюма, получено потомство от хохлато-
го, белого и синего ушастых и непальского фазанов. Содержится
несколько подвидов обыкновенного фазана: зеравшанский, тад-
жикский, семиреченский, амударьинский, маньчжурский, тайвань-
ский и зеленый. Интерес представляет разведение гололицего гок-
ко, гималайского монала и кокласа. Есть опыт по содержанию
каспийского улара, турача, пустынной куропатки, ряда тетереви-
ных птиц.
Журавлеобразные. Журавлей часто содержат в зоопарках, од-
нако случаи успешного разведения их довольно редки. Координа-
той такой работы является Международный фонд охраны журав-
лей (шт. Висконсин, США), а в СССР — Питомник редких видов
журавлей Окского государственного заповедника. Наибольших ус-
пехов зоопарки мира достигли в разведении японского, индийского
и венценосных журавлей, значительно реже размножаются серый,
даурский, черный и другие виды журавлей. Хорошие результаты
по разведению журавлей и аистов получены в зоопарках Тама
(Япония), Франкфурта-на Майне (ФРГ) и орнитопарке Вальсро-
де (ФРГ). В Московском зоопарке в настоящее время содержатся
такие редкие журавли, как стерх, даурский, японский и черный —
всего 10 видов. Успешно размножаются даурские, японские, ин-
дийские журавли. Для стимуляции размножения используется
метод искусственного осеменения, освоены и методы искусствен-
ного инкубирования и выращивания молодых птиц. Немаловажную
роль в успешном разведении журавлей играет содержание отдель
ных пар взрослых птиц на больших площадях с домиками для
зимовки. В теплые зимние дни птицы имеют возможность находить-
ся на открытом воздухе, что способствует повышению резистент-
245
ности организма птиц. Важен и правильный подбор фотопериодов
в зимнее и весеннее время.
Соколообразные и сокообразные. Если по некоторым группам
птиц собрана большая информация и разведение в неволе осущест-
вляется довольно давно, то разведением хищных птиц зоопарки
занимаются лишь последние десятилетия. В то время как питом-
ники хищных птиц специализируются на разведении различных
соколов, используемых для ловчей охоты, в зоопарках начали
целенаправленно разводить крупных хищных птиц — орлов, орла-
нов, грифов Старого и Нового Света. Особенно успешно разводятся
беркуты, белоголовые орланы и некоторые грифы. Зоопарки
США — Сан-Диего и Лос-Анжелесский — пытаются сохранить ис-
чезнувшего в природе калифорнийского кондора. В зоопарках Ин-
сбрука (Швейцария) и Вены осуществляется программа по раз-
ведению и выпуску в Альпах бородача, исчезнувшего здесь до-
вольно давно.
В последние годы хищные птицы стали чаще размножаться
и в советских зоопарках. В Московском, Ленинградском и Таш-
кентском зоопарках размножались андские кондоры, в Алма-Атин-
ском и Московском успешно разводятся бородачи, в Николаевском
зоопарке — белоголовые орланы, в Свердловском — орланы-бело-
хвосты, а в Московском — кумаи, беркуты, степные орлы, белопле-
чие и белоголовые орланы, яйца без насиживания откладывали
орланы-белохвосты и орлы-могильники.
Многие виды сов размножаются в Берлинском зоопарке (ГДР),
в Лейпцигском успешно разводят белых сов. Серая неясыть и
ушастая сова размножались в Ленинградском зоопарке, а филин
и сипуха — в Московском. В зоопарках отрабатываются методы
искусственного и естественного выращивания птенцов хищных
птиц и сов. Применяется метод изъятия первой кладки для
искусственного инкубирования, а повторную оставляют под птица-
ми. К сожалению, нередко птицы бросают или съедают собствен-
ные кладки и птенцов, что вызывается прежде всего стрессовой
ситуацией — реакция на посетителей, а также неправильным
кормлением.
Таким образом, выявление основных закономерностей, влияю-
щих на процессы размножения птиц в искусственных условиях,
приведет к успехам в важном деле — разработке технологии раз-
ведения отдельных видов или целых эколого-систематических
групп. Зоопарки здесь призваны сыграть одну из главных ролей.
Богатый опыт практической деятельности зоопарков и наличие
базы в совокупности с развивающимися научными исследованиями
помогут повысить их природоохранное и народнохозяйственное
значение.
В. А. ОСТАПЕНКО
246
УДК 636.58
Проблемы и перспективы развития
любительского птицеводства
(певчие и декоративные птицы)
Наш народ исстари содержал в своем жилище певчих птиц. Однако
первое упоминание об организации любителей птиц относится
только к 1901 г., когда впервые на территории нашей страны был
создан Московский кружок любителей певчей и другой вольной
птицы. Этот кружок вел довольно активную деятельность — вы-
пускал свой печатный сборник, организовывал конкурсы и вы-
ставки (был проведен даже конкурс пения соловьев с использо-
ванием телефона), осуществлял обследование разводческих хо-
зяйств любителей, организовывал помощь Московскому зоо-
парку.
На начало нашего столетия приходится пик расцвета в нашей
стране канароводства. Именно в этот период сложилась особая
порода — русская канарейка овсяночного напева. Наши деды
и прадеды постарались вложить в песню этой канарейки наиболее
звучные и мелодичные строфы отечественных птиц: овсянки, юлы,
весенний бой большой синицы, посвист московки, а также милые
сердцу русского человека звуки поддужного колокольчика, ра-
достное серебро кузнечного отбойного молоточка. На разведении
канареек овсяночного напева специализировались жители целых
сел — Полотняного Завода, Павлова-на-Оке. Отдельным люби-
телям удавалось создать песни, отличающиеся особым напевом —
«ходом». До сих пор известны канарейки рябовского и музланов-
ского «ходов», названных по фамилиям канароводов. Не меньше
было развито и любительское содержание птиц отечественной
фауны.
Революционные события, а затем и Великая Отечественная
война нанесли большой урон любительским птицеводческим хо-
зяйствам. Однако постепенно, с возрастанием благосостояния на-
рода, начала возрастать и потребность в певчей птице. Удовле-
творению спроса весьма способствовало и образование Зооцентра,
а затем и Зоообъединения, которые в послевоенные годы стали
завозить в нашу страну большие партии экзотических птиц, цвет-
ных канареек и Канареек дудочного напева. Яркие краски экзоти-
ческих птиц и их неприхотливость в содержании привлекли вни-
мание многих любителей природы. Вместе с тем завоз цветных
канареек и канареек дудочного напева при отсутствии культуры
разведения этих птиц, бессистемном скрещивании с русской кана-
рейкой привели к практической потере уникальной породы и сни-
зили интерес к нашей отечественной птице.
Обеспокоенные сложившейся ситуацией, любители-канаро-
воды Москвы с целью восстановления породы создали в 1947 г. при
247
НИИ птицеводства секцию любителей канареек овсяночного на-
пева. Впоследствии преобразованная в клуб секция приняла к себе
любителей птиц отечественной фауны, а затем и экзотических
птиц. Со временем в различных городах Советского Союза начали
возникать аналогичные любительские самодеятельные организа-
ции. Надо сказать, что при содержании птиц отечественной фауны
наши любители ценили лишь способность птицы к пению. Со-
ответственно с этой целью отлавливались только самцы, отлов
самок считался неэтичным и наносящим ущерб и до сегодняшнего
дня. Это стало серьезным препятствием на пути развития люби-
тельского птицеводства. Во многих странах из-за серьезной опас-
ности антропогенного пресса на птиц законодательно ограничены
отлов и содержание птиц, обитающих на территории их стран.
Вслед за этим среди любителей распространилось движение за
создание клеточных «популяций» птиц отечественной фауны. Эту
же задачу поставила перед любителями Международная конфе-
дерация орнитологов-любителей (СОМ), объединяющая в насто-
ящее время любительские организации большинства развитых
стран Европы и Америки (из социалистических стран в нее входят
Болгария, Чехословакия, Югославия, Румыния и Куба). С целью
охраны птиц путем их репродукции в клеточных условиях СОМ
принял решение о допустимости демонстрации только тех птиц,
которые были выведены любителями. По положению СОМ к вы-
ставкам и конкурсам не допускаются птицы, не имеющие не-
разъемного номерного кольца.
Вопрос об организации единого союзного координационного
органа орнитофильских самодеятельных клубов и секций впервые
был поднят в 1981 г. на Ленинградской клубной выставке птиц
с участием иногородних любителей. Однако объективные условия
в те времена позволили создать только Всероссийский республи-
канский координационный совет клубов любителей певчих и де-
коративных птиц, учрежденный Центральным советом Всерос-
сийского общества охраны природы в 1986 г. Тем не менее за пе-
риод с 1981 г. инициативной группе и Всероссийскому совету
клубов удалось провести несколько совещаний и семинаров по
проблемам любительского содержания птиц, принять ряд осново-
полагающих документов — типовое положение о клубе, положе-
ние о выставке птиц, организовать проведение четырех всесоюз-
ных выставок и чемпионатов птиц (в Москве, Харькове, Минске,
Ереване), установить связь с зарубежными любительскими орга-
низациями аналогичного профиля и принять участие в зарубеж-
ной выставке. Совет установил и поддерживает связь почти с 60
клубами и секциями нашей страны. Однако различная ведомствен-
ная подчиненность клубов и секций, республиканский статус Ко-
ординационного совета, его подведомственность республиканско-
му обществу охраны природы создавали значительные трудности
при координации деятельности клубов, не позволяли войти ни
в один из международных союзов любителей птиц. Координацион-
ный совет не оставлял попыток создания единого всесоюзного ор-
248
гана. В 1988 г. любителям птиц была предоставлена возможность
создания такого органа отделом любительских объединений Все-
союзного научно-методического центра народного творчества
и культурно-просветительной работы (ВНМЦ НТ и КНР) Мини-
стерства культуры СССР. Модель структуры всесоюзного органа,
разработанная ВНМЦ, позволяла объединить клубы и секции раз-
личных республик независимо от их ведомственной принадлеж-
ности и преодолеть таким образом все трудности, стоящие перед
ВКСК. В соответствии с данной моделью Всесоюзное объедине-
ние клубов любителей певчих и декоративных птиц является
юридическим лицом и получает право на организацию своих хоз-
расчетных подразделений для обеспечения своей деятельности.
Объединение функционирует на принципах хозрасчета, само-
окупаемости и самофинансирования. Учредителем Объединения
выступает ВНМЦ НТ и КПР, соучредителем — Всесоюзное орни-
тологическое общество, взявшее на себя вопросы методического
руководства деятельностью любительских клубов. Гибкая струк-
тура модели позволит использовать методики и разработки Мини-
стерства культуры СССР в части организации любительских объе-
динений и профессиональное руководство Орнитологического об-
щества в области разработки стратегических направлений раз-
вития клубов любителей певчих и декоративных птиц.
С созданием Всесоюзного объединения появились перспективы
широкого привлечения любителей птиц к участию в орнитоло-
гических исследованиях, к тесному сотрудничеству профессио-
налов и любителей. Успешная деятельность всего Объединения
должна базироваться в первую очередь па экономически сильных
и активно действующих клубах. Поэтому совет Объединения одну
из первых задач видит в необходимости развития материальной
базы клубов, достижения их устойчивого финансового положения,
внедрения в деятельность клубов принципов хозрасчета и само-
финансирования. С этой целью Совет планирует проведение регу-
лярных, школ лидерского актива, семинаров но обмену опытом.
Другой насущной задачей является повышение профессиональ-
ного уровня наших любителей. В этом направлении Объединение
намерено осуществить целую программу и, в частности, издание
отечественных и перевод некоторых зарубежных книг, отражаю-
щих современный уровень содержания, разведения и селекции
певчих и декоративных птиц. Издание «Стандартов экзотических
птиц» является необходимым условием выхода советских люби-
телей на международные соревнования разводчиков певчих и деко-
ративных птиц. Другим условием является внедрение в практику
советских любителей кольцевания приплода и ведение родослов-
ных документов на выводимую птицу. Дальнейшее развитие оте
явственного любительского птицеводства напрямую зависит от
решения проблемы изготовления неразъемных колец. Кольцо,
надетое на ножку семи-восьмидневного птенца, уже через два-три
дня становится практически песнимаемым и служит на протя-
жении всей жизни своеобразным паспортом птицы. Информация,
249
нанесенная на наружную поверхность кольца, позволяет в любой
момент определить клуб и даже разводчика, получившего дан-
ную птицу. Такие кольца должны распределяться по клубам цен-
трализованно, на основании их заявок.
В отношении любителей птиц отечественной фауны ставится
задача переориентации их увлечения на Содержание с целью раз-
ведения и создания одомашненных самовозобновляющихся «по-
пуляций» и дальнейшей селекции с целью получения разноокрашен
ных птиц, птиц с ярковыраженными певчими способностями.
К сожалению, пальма первенства разведения многих отечествен-
ных птиц уже находится у зарубежных разводчиков. Безусловно,
отсутствие опыта, многолетние заблуждения • относительно
трудностей разведения отечественных видов, поддерживаемые не-
которыми нашими уважаемыми орнитологами, долго еще будут
сказываться на успехах любителей. Всесоюзное объединение клу-
бов любителей певчих и декоративных птиц ждет от Орнитологи-
ческого общества активной помощи любителям и всемерного по-
ощрения их работы.
Таким образом, речь идет об изменении психологии наших
любителей птиц отечественной фауны от содержания к разведе-
нию. Хотелось бы, чтобы достижение этой цели проходило красной
нитью через все популярные и специальные издания, предназна-
ченные для любителей природы. Большим подспорьем в нашей
работе должны стать специальные разработки для разводчиков
птиц по основам генетики, практической селекции, ветеринарии.
Тесное взаимодействие любителей с профессиональными орни-
тологами, всемерная помощь клубов и Всесоюзного орнитологи-
ческого общества, связь с государственными и общественными ор
типизациями и, конечно, инициатива самих членов Совета и дру-
гих-выборных органов Объединения смогут принести положитель-
ные результаты.
Е. В. МАЛАХАНОВ
РЕЦЕНЗИИ
В. Д. Ильичев и В. Е. Фомин
Орнитофауна и изменение среды. М.: Наука, 1988
Фаунистические исследования сегодняшнего дня предполагают
выявление тенденций изменения фауны под влиянием человека.
И хотя этот процесс значительно ускорился, в последнее время
орнитолог-фаунист вынужден тратить долгие годы на то, чтобы
установить общую динамику региональной фауны, определяющие
ее основные факторы, состояние популяций, отдельных видов, по
крайней мере, наиболее интересных с эколого-хозяйственной или
природоохранной точки зрения. Здесь орнитологу, помимо соб-
ственных услилий, могут помочь далекие предшественники, если,
конечно, региональная фауна долгие годы была предметом их
внимания.
Именно такая ситуация сложилась в Южно-Уральском реги-
оне, авифауна которой всесторонне и полно изучалась еще со
времен П. С. Палласа, а позднее ее исследователями были такие
выдающиеся специалисты, как Э. А. Эверсманн, П. П. Сушкин,
С. В. Кириков, С. А. Снигиревский и другие. Последние 30 лет
авифауну региона изучал В. Д. Ильичев, а в 1974 —1976 гг. здесь
работал экспедиционный отряд ИЭМЭЖ АН СССР (начальник
отряда — В. Е. Фомин, научный руководитель — В. Д. Ильичев).
Результаты этих исследований отражены в недавно опубликован-
ной монографии В. Д. Ильичева и В. Е. Фомина «Орнитофауна
и изменение среды», посвященной современному состоянию орни-
тофауны региона, тенденциям ее изменений под влиянием челове-
ческой деятельности.
Эта монография продолжает своеобразный фаунистический
мониторинг, и это мы рассматриваем как ее главное положитель-
ное качество, осуществляющееся традиционно в течение около
200 лет. Сопоставление фаунистических обзоров, выполненных за
этот период, показывает, какие глубокие изменения претерпела
фауна региона и как значительно они усилились и ускорились в
последние годы. Авторы выявляют и рассматривают роль различных
факторов (как отрицательных, так и положительных в этом процес-
се) — вырубки лесов, распашка целины, рекреационной нагрузки,
с одной стороны, урбанизации и расширении сети охраняемых тер-
риторий — с другой. Этот анализ дается как в общем виде, так
и по отношению к отдельным представителям фауны, в первую
очередь редким и исчезающим с территории региона. Важно под-
251
черкнуть, что в качестве приложения авторы подготовили пере-
чень редких видов, оформленный по стандартам Красных книг,
что крайне полезно для практики охраны природы.
Таким образом, рецензируемая монография не только обоб-
щает большой фактический материал, характеризующий совре-
менное состояние фауны региона, но является и своеобразным
образцом исследований, отражающих динамику фауны и ее
мониторинг. Авторы считают разработку таких подходов одной из
главных своих задач, справедливо полагая, что это открывает пер-
спективы использования региональных фаун в качестве крупно-
масштабного индикатора состояния среды.
Конечно, это пока еще первые шаги в разработке индикатор-
ных подходов к изучению фауны, как всякие первые шаги — не
очень уверенные, встречающие много трудностей. Однако важ-
ность и нужность таких подходов ни у кого сомнений не вызывает.
Авторы, кажется, впервые рассматривают урбанизированную
фауну как автономный компонент региональной фауны, вступа-
ющий с последней в сложные и противоречивые отношения. Заслу-
живает внимания формула этих отношений, предложенная в моно-
графии (урбанизированная фауна как активный фильтр регио-
нальной), а также большой фактический материал, собранный
В. Е. Каревым — автором главы, посвященной фауне г. Уфы. По-
лезным является и дусторонний взгляд на роль человека в изме-
нении фауны: характеризуются не только отрицательные воз-
действия, но и положительное значение деятельности человека,
в частности, за счет создания охраняемых территорий — заповед-
ника и национального парка.
Оценивая монографию в целом, следует еще раз подчеркнуть
не только региональное, но и общее ее значение, выразившееся
в использовании региона как «модельной» территории для реше-
ния более широких проблем, связанных с экологией и охраной
природы. Этот аспект приобретает в настоящее время все большее
значение и в педагогической науке, и как одна из эффективных
форм экологического воспитания населения. Поэтому нам кажется,
что книга найдет себе широкого читателя и среди студентов и
преподавателей педагогических институтов, школьников старших
классов и учителей. .
В. М. КОНСТАНТИНОВ
В. Gensbol. Greifvogel. Munchen, 1986.
(Б. Генсбол. Хищные птицы. Мюнхен, 1986)
В последние годы хищным птицам уделяется особое внимание
специалистов-орнитологов и любителей птиц. Издается большое
количество литературы, посвященной различным проблемам био-
логии, хозяйственного и культурного использования хищных
птиц, их практического значения. В подавляющем большинстве
252
это книги зарубежных авторов. Отечественной литературы по наз-
ванным вопросам в настоящее время почти нет. В связи с этим
особый интерес представляет рецензируемая нами книга немец-
кого орнитолога Б. Генсбола «Хищные птицы».
Книга состоит из трех частей: общей, повидовой и определи-
тельной. В общую часть включены главы, в которых анализируется
биотопическое распределение хищных птиц, характер питания
и способы добывания корма, пути пролета и места концентрации
птиц в периоды миграционных перемещений и зимовки. Хорошо
представлена биология гнездования и другие вопросы экологии.
Особый интерес вызывают главы, в которых описывается антропо-
генное влияние на хищных птиц, приводящее к резкому сокраще-
нию их численности в природе. Это прежде всего загрязнение
среды различными химическими веществами — пестицидами,
азотосодержащими удобрениями, отходами различных производств.
Вредные вещества не только губительны для самих птиц, но и вли-
яют на их репродуктивные способности, на толщину скорлупы
яиц и пр. Большую опасность для птиц представляют изменения
природных биотопов человеком. В результате обеднения естест-
венных сообществ исчезают исконные места обитания хищных
птиц. Так, сведение высокоствольных лесов лишило птиц удобных
для строительства гнезд деревьев. В книге приводятся формы пря-
мого преследования птиц человеком — ружейная охота, сбор яиц
коллекционерами, изъятие из гнезд птенцов любителями соко-
линой охоты, от которых страдают в основном крупные и средние
соколы, а также ястребы-тетеревятники, а местами и перепелят-
ники. Перечисляются также и другие опасности для хищных
птиц — большая рекреационная нагрузка естественных местооби-
таний, повышенный интерес орнитологов, приводящий к тому, что
хищные птицы становятся особенно пугливыми и нередко поки-
дают гнезда в период гнездования, оставляя яйца и птенцов.
В книге даны рекомендации, как следует правильно подходить
к гнездам, как устраивать засидки и т. д. Приводится статистика
гибели хищных птиц на линиях электропередач, на автомобиль-
ных дорогах, от столкновений со зданиями, налетевших на окна
и витрины, прожекторы, запутавшихся в колючей проволоке.
Особый отрицательный эффект достигнут в результате много-
векового преследования хищных птиц охотниками. Планомерное
истребление пернатых хищников началось 200—300 лет тому на-
зад. Охотникам выплачивались премии за добытых ими соколов,
ястребов, орлов и орланов. Все эти сведения, приведенные в книге
в интересной и доступной широкому читателю форме, являются
великолепным природоохранным материалом, призывающим лю-
бить природу во всем ее многообразии.
Во второй части книги помещены повидовые очерки о хищных
птицах, обитающих в западной части Палеарктики: в Европе,
Северной Африке и на Аравийском полуострове. Для большинства
видов даны сведения о современной численности, иногда в виде
таблиц. В разделе, посвященном распространению, имеются кар-
253
ты гнездового и зимовочного ареалов, что хорошо иллюстрирует
материал. Приводятся сведения по динамике численности по-
пуляций, биотопическому распределению птиц, местам и характеру
пролета в периоды миграционных перемещений, дано описание
голосов, гнездовой биологии, питания и способов охоты хищ-
ных птиц. Особенно следует отметить качество и обилие иллюстра-
тивного материала. Имеются хорошие цветные фотоснимки и штри-
ховые рисунки. Иллюстрации хорошо раскрывают содержание
книги и делают ее особенно привлекательной для читателя.
Третья часть посвящена определению хищных птиц. В ней даны
сведения, позволяющие довольно точно определить птицу в при-
роде. Приведены рисунки силуэтов птиц в различных положениях,
что очень облегчает определение и экономит время, что немало-
важно в полевых условиях. Даны повидовые описания птиц, вклю-
чающие описания внешнего вида, пропорций тела, молодых, из-
вестные вариации в окраске, вид в полете, манеры и способы поле-
та, типичное поведение. Дается описание близких видов, с которы-
ми можно спутать данный вид в природе, их отличительные
особенности. В конце книги приводится список литературы.
Высоко оценивая научную, научно-практическую и природо-
охранную значимость рецензируемой книги, мы рекомендуем ее
для перевода и издания на русском языке. В связи с тем, что
хищные птицы в настоящее время в Европе испытывают сильное
воздействие со стороны человека и их численность резко сокраща-
ется (большинство их уже внесено в Красные книги различных
рангов), публикация справочника у нас в стране будет иметь важ-
ное значение. Важно учесть и то, что многие виды хищных птиц
широко используются в качестве индикаторов загрязнения среды,
в частности пестицидами. Учитывая катастрофическое положение
ряда видов хищных птиц во всем мире, в том числе и в СССР, пред-
принимаются крупномасштабные эксперименты по их разведе-
нию в специальных питомниках и зоопарках с последующим вы-
пуском в природу. Внимание международной общественности
приковано к охоте с ловчими птицами, возрождение которой свя-
зано также и с проблемами отпугивания птиц от аэродромов, где
с помощью хищных птиц удается рассеивать массовые самолето-
опасные скопления птиц (в мире ежегодно около 4 тыс. самолетов
сталкивается с птицами, из них около 300 на территории нашей
страны). Наконец, существен еще один аспект, касающийся ме-
тодов определения хищных птиц в природе. Естественно, что без
соответствующих навыков и руководств по методам определения
видов никакая охрана хищных птиц невозможна. Рецензируемая
книга удовлетворяет всем этим требованиям и поставленным за-
дачам. Она рассчитана как на биологов различного профиля, за-
нимающихся птицами, так и на широкий круг любителей природы.
В. А. ОСТАПЕНКО
Информация
Редколлегия «Современной орнитологии» — ежегодника Всевоюзного ор-
нитологического общества АН СССР — приглашает членов Общества ак-
тивнее печататься в нашем издании. Мы ждем проблемные, обзорные и
критические статьи по орнитологии, а также материалы, сообщающие или
обобщающие конкретные факты или результаты исследований, получен-
ные в поле или в лабораториях. Приветствуются также материалы, подго-
товленные непрофессиональными орнитологами — членами Общества.
Обращаем внимание всех авторов, что представляемые рукописи дол-
жны полностью удовлетворять «Правилам для авторов», - опубликованным
в первом (1985), втором (1986) и пятом (1989) выпусках «Информацион-
ных материалов ВОО». Рукописи необходимо готовить предельно аккурат-
но, использовать только белую бумагу (не желтую или желтоватую!), пе-
репечатывать на машинке с чистыми литерами, латинские названия впеча-
тывать на машинке, а не вписывать от руки. Требуются оригиналы рисун-
ков (в первом экземпляре), рукописи, иллюстрированные ксеро- или фото-
копиями, рассматриваться не будут. Цифровые таблицы (если они не со-
ставляют Приложения) необходимо помещать по тексту внутри статьи, же-
лательно на отдельных страницах, и считать их в числе страниц текста.
Как и вся рукопись, таблицы должны перепечатываться через 2 интервала.
Просим учесть, что «портфели» ежегодника «Современная орнитоло-
гия» и сборника «Орнитология» теперь разделены, и дело самих авторов
решать, куда направлять рукопись.
Редколлегия оставляет за собой право редактирования и сокращения
рукописей, направления их на рецензирование. Непринятые рукописи
возвращаются авторам.
Для своевременного формирования содержания ежегодника нам необ-
ходимо получать Ваши рукописи не позднее 20 ноября текущего года.
В связи с включением в ежегодник резюме статей на английском язы-
ке просим авторов тщательно готовить тексты резюме для своих статей, ко-
торые должны отражать существо содержания и основные результаты,
сообщаемые в основном тексте. Текст резюме не может превышать полови-
ны машинописной страницы. Он должен быть пригоден для перевода на ан-
глийский язык или сразу представлен на нем с приложением русского па-
раллельного текста.
Рукописи (в 2 экземплярах) можно направлять по любому из двух ад-
ресов: 117674, Москва, Профсоюзная ул., д. 123, Палеонтологиче-
ский институт АН СССР, Ответственному редактору'Е. Н. Курочкину;
109240, Москва, 1-й Котельнический пер. д. 10, Всесоюзное орнитологиче-
ское общество, Ответственному секретарю редколлегии Е. Ф. Правдивой.
РЕДКОЛЛЕГИЯ
СОДЕРЖАНИЕ
ВОПРОСЫ
экологии
А. В. АНДРЕЕВ
Экологическая энергетика арктических птенцов 5
Л. В. СОКОЛОВ, В. П. ДЬЯЧЕНКО,
Т. А. ИЛЬИНА, Д. С. ЛЮЛЕЕВА, М. Л. ЯБЛОНКЕВИЧ
Проверка возможности образования зимовки у моло-
дых зябликов, задержанных на Куршской косе 21
А. В. БАРДИН
Динамика полового и возрастного состава и жиро-
вые резервы больших синиц (Parus m. major) в зим-
ний период 35
Т. А. ИЛЬИНА
-— Расход времени и энергии на гнездостроение у серой
вороны (Corvus cornix) 48
В. Г. ПАПЧЕНКОВ
Характер разногодичных изменений условий обита-
ния утиных на мелководьях водохранилища с пере-
менным уровнем наполнения 55
В. С. ТАЛПОШ
— О биологин европейской горной трясогузки (Mota-
cilla cinerea cinerea) в Украинских Карпатах 66
А. Ф. МАМАТОВ
— Озерная чайка (Larus ridibundus) в Башкирии 74
А. К. САГИТОВ, Л. Э. БЕЛЯЛОВА,
С. Э. ФУНДУКЧИЕВ
Расселение майны (Acridotheres tristis tristis) в Сред-
ней Азии и Казахстане 86
Л. П. БЫКОВА
— Элиминация в раннем онтогенезе и величина кладки
у грача (Corvus frugilegus) 98
А. Г. ГУБИН, Е. С. ПРЕОБРАЖЕНСКАЯ,
А. С. БОГОЛЮБОВ
Экологические особенности дроздов рябинника (Tur-
dus pilaris) и белобровика (Т. iliacus) в местах
совместного обитания 117
А. И. КОШЕЛЕВ, И. И. БАДЮК,
И. И. ЧЕРНИЧКО
Размножение лысухи (Fulica atra) на Тилигульском
лимане (Северо-Западное Причерноморье) 122
Т. Б. АРДАМАЦКАЯ, В. И. ВАКАРЕНКО,
А. А. ПЕТРУСЕНКО '
— Трофические связи черноголовой чайки в гнездовой
период в условиях Черноморского заповедника. Сооб-
цение II. Пространственно-временные характерис-
тики трофических связей 135
256
А. И. МОРОЗКИН, В. Д. КАПИТОВ Изменение эмбриональной смертности в связи с мно-
голетней динамикой численности у мухоловки-пес- трушки (Ficedula hypoleuca) 141
П. С. ТОМ КО ВИЧ
К вопросу о внегнездовых перемещениях белых чаек (Pagophila eburnea) 150
В. И. ПЕРЕРВА
Изменение экологии хищных птиц под влиянием антропогенных факторов 151
ПОВЕДЕНИЕ М. А. ТАРХАНОВА, Л. Я. КУРИЛОВИЧ О взаимоотношениях некоторых видов уток в вывод- ковый период С. С. МОСКВИТИН, Т. Л. АНАНЬИНА 163
Ритмика поведения и бюджет рабочего времени садо- вой камышевки (Acrocephalus dumetorum) в период 172
вскармливания птенцов
МОРФОЛОГИЯ В. Ф. СЫЧ
Морфологические особенности мускулатуры тазовой конечности большеногое и краксов (Galliformes; Me- gapodiidae, Cracidae) 179
Е. А. ДЯДИЧЕВА, А. Н. ЦВЕЛЫХ Морфо-зкологический анализ органов полета трех видов вьюрковых 193
Е. В. ГРИШИНА
Динамика роста эндокринных органов домашней ку- рицы в зависимости от различной степени двигатель- ной активности 199
МЕТОДИКА А. В. МИХЕЕВ
ОРНИТО- Дневные видимые перелеты птиц и методы их ко-
ЛОГИЧЕСКИХ личественного учета 203
ИССЛЕДОВАНИЙ Н. П. КОЛОМИЙЦЕВ К методике учета чешуйчатого крохаля (Mergus squamatus) и мандаринки (Aix galericulata) 217
ПАМЯТНЫЕ В. Е. СОКОЛОВ, В. Д. ИЛЬИЧЕВ
ДАТЫ Выдающийся советский орнитолог и деятель охраны природы. К 90-летию со дня рождения Г. П. Дементь- ева (1898-1969) 222
А. В. МИХЕЕВ Г. П. Дементьев и изучение миграций птиц в СССР 227
В. Е. СОКОЛОВ, В. Д. ИЛЬИЧЕВ Все силы — изучению и охране птиц (Анверу Кею- шевичу Рустамову — 70 лет) 232
В. И. ГАРАНИН В. А. Попов. К 80-летию со дня рождения 236
ХРОНИКА В. Д. ИЛЬИЧЕВ, Л. С. СТЕПАНЯН Международная сотая сессия Немецкого Орнитологи- ческого общества 238
К. А. ТАТАРИНОВ
Четвертые чтения памяти академика М. А. Мензбира 241
257
В. А. ОСТАПЕНКО
РЕЦЕНЗИИ
Орнитологические исследования в зоопарках (К 125-
летию Московского зоологического парка) 243
Е. В. МАЛ АХАНОВ
Проблемы и перспективы развития любительского
птицеводства (певчие и декоративные птицы) 247
В. Д. Ильичев и В. 1'1 Фомин. Орнитофауна и изме-
нение среды. М.: Наука, 1988 (В. М. Константинов) 251
В. Gensbol. Greifvogel. Munchen, 1986 (Б. Генсбол.
Хищные птицы. Мюнхен, 1986) (В. А. Остапенко) 252
CONTENTS
PROBLEMS
OF ECOLOGY
A. V. ANDREEV
Ecological Energetics of Arctic nestlings
L. V. SOKOLOV, V. P. DIACHENKO,
T. A. ILYINA, D. S. LIULEIEVA,
M. L. YABL0NKEV1CH
Test of possible wintering of young chaffinches, kept
off on Kurshskaya Kosa
A. V. BARDIN
The dynamics of sexual and age composition and fatly
reserves of Great Titmice (Parus m. major) in winter
time
T. A. ILYINA
Time and energy budget for nest building by the
Hooded Crow (Corvus comix)
V. G. PAPCHENKOV
Tlie character of environmental changes of waterfowls
in different years on the shallow water basins with
changeable water filling level
V. s. talposh
On biology of European Grey Wagtail (Motacilla
cinerea cinerea) in the Ukrainian Carpathians
A. F. MAMATOV
Black-headed Gull (Larus ridibundus) in Bashkiria
A. K. SAGITOV, S. E FUNDUKCHIEV,
L. E. BELJALOVA
Distribution of Common Mynah (Acridotheres tristis
tristis) in Soviet Central Asia and Kazakhstan
L. Р. BYKOVA
Elimination in the early ontogenesis and clutch size in
Rook (Corvus frugilegus)
A. G. GUBIN, E. S. PREOBRAZENSKAYA,
A. S. BOGOLJUBOV
Ecological characters of Fieldfare (Turdus pilaris)
and Redwing (T. iliacus) in the places of their joint
inhabitance
A. I. KOSHELEV, I. I. BADIUK,
1. I. TCHERNICHKO
Reproduction of the Coot (Fulica atra) on the Tiiigul
Liman in the north-west Black Sea region
T. B. ARDAMATSKAYA, V. I. VAKARENKO,
A A. PETR USENKO
Trophic relations of the Mediterranean Gull during
the nesting period at the Black Sea State Reserve
.Report П. The spatial-temporal characteristics of
trophic relations
21
34
47
54
65
74
85
97
116
121
134
141
259
Л. I. MOROZKIN, V. D. KAPITOV Hie change of the embryonal mortality in connection with the long-term dynamics of the Pied Flycatcher (Ficedula hypoleuca) 149
P. S. TOMKOVICH On the question of out-of-nest movements of Ivory Gulls (Pagophila eburnea) 151
V. I. PERERVA Ecological changes of birds of prey, influenced by anthropogenic factors 162
BEHAVIOUR M. A. TARKHANOVA, L. YA. KURILOVICH
On relations of some duck species during hatching period 171
S. S. MOSKVITIN, T. L. ANANJINA Behavioral rythms and time budget during the hatching 1
period of Blyth’s Reed Warbler (Acrocephalus dumeto- rum) 178
MORPHOLOGY V. F. SYTCH
Morphological peculiarites of the pelvic musculature of megapods and cracids (Galliformes; Megapodiidae, Cracidae) 192 ’
E. A. DIADICHEVA, A. N. TSVELYCH Morpho ecological analysis of flying system of three species of Fringillidae 198 ;
E. V. GRISHINA The dynamics of the growth of domestic hen endocrine organs in dependence upon the various degree of the locomotion activity 202
METHODS A. V. MIKHEEV
OF INVESTI- Day-time visible migrations of birds and the methods of
GATIONS their quantitative registration 217
N. P. KOLOMIITSEV About methods of the counts of the Chinese Merganser (Mergus squamatus) and the Mandarin Duck (Aix galericulata) J 221 i
MEMORIAL V. E. SOKOLOV, V. D. ILIJCHEV I
DATES A Famous Soviet ornithologist and conservationist
of nature. To the 90th Birthday of G. P. Dementiev (1898—1969) 226
A. V. MIKHEEV G. P. Dementiev and the study of birds migrations in the USSR 231
V. E. SOKOLOV, V. D. ILIJCHEV All activity for study and protection of birds (The 70th years Birthday of Anver Rustamov) 235
V. I. GARANIN V. A. POPOV. For the 80th birthday anniversary 237 '
CHRONICLE V. D. ILIJCHEV, L. S. STEPANJAN
Centenial International Meeting of the German Ornitho- logical Society 238
К. A. TATARINOV
Fourth Meeting in the memory of Academician
M. A. Menzbier 241
260
REVIEWS
V. A. OSTAPENKO
Ornithological studies in the zoological gardens (To the
125th Anniversary of the Moscow Zoological Park)
E. V. MALAKHANOV
Problems and perspectives of the development of the
amateur poultry farming (singinig and decorative
birds)
V. I). Ilijchev and V. E. Fomin. Ornithofauna and
environmental changes. M.: Nauka, 1988 (Konstan
tinov V. M.)
B. Gensbol. Birds of Prey. Munchen, 1986 (Osta-
penko V. A.)
243
247
251
252
Научное издание
СОВРЕМЕННАЯ
ОРНИТОЛОГИЯ
1990
Утверждено к печати
Всесоюзным
Орнитол огич ески м
обществом
Редактор издательства А. В. Бирюков
Художник Э. А. Смирнов
Художественный редактор Н. Н. Михайлов
Технический редактор Н. Н. Кокина
Корректоры А. Б. Васильев, К. П. Лосева
ИБ № 47029
Сдано в набор 19.04.90
Подписано к печати 10.08.90
Формат 60Х90’/1б
Бумага офсетная № 1
Гарнитура обыкновенная новая
Печать офсетная
Усл. печ. л. 16,5. Усл. кр. отт. 17.
Уч.-изд. л. 18.
Тираж 1900 экз. Тип. зак. 36.
Цена 3 р. 80 к.
Ордена Трудового Красного Знамени
издательство «Наука»
117864, ГСП-7, Москва, В-485,
Профсоюзная ул., 90.
2-я типография издательства «Наука»
121099, Москва, Г-99, Шуби некий пер., 6.
Приглашаем
РЕКЛАМУ
Редколлегия ежегодника «Современная орнитология» приг-
лашает советских и иностранных производителей товаров и из-
дателей книг, которые могут заинтересовать орнитологов, по-
мещать свою рекламу в нашем издании.
С предложениями обращаться по адресу:
СССР, 117647, Москва, Профсоюзная ул., д. 123,
Палеонтологический институт АН СССР
Александру Альфредовичу Карху
К иностранным читателям
«Современной орнитологии»
За Пределами СССР ежегодник «Современная орнитология»
распространяется по заказам через:
В/О «Международная книга»
СССР, 113095, Москва,
ул. Димитрова, д. 39
или
Секретариат Всесоюзного орнитологического общества АН СССР
готов обменивать наше издание на соответствующую орнитоло-
гическую литературу из других стран:
СССР, 109240, Москва,
1-й Котельнический пер., д. 10,
Всесоюзное орнитологическое общество.
Вице-президенту Общества
Евгению Николаевичу Курочкину
дДЮЛОГду,
3 р. 80 коп.
Сборник содержит статьи,
посвященные различным
вопросам орнитологии: экологии
и адаптации птиц к среде
обитания, методам их охраны,
динамике полового и возрастного
состава некоторых видов, в том
числе редких и включенных в
Красные книги. Исследуются
биология птиц в антропогенных
комплексах, функциональная
морфология, методика изучения.
Освещается ряд памятных дат
и др.
Белая чайка (Pagophila
eburnea). Земля Франца-
Иосифа, о-в Греэм-Белл,
конец июля 1981 г.
Фото П. С. Томковича
На передней сторонке
переплета: Белые гуси
(Chen caerulescens). О-в
Врангеля, начало июня
1986 г. Фото С. П. Харитонова