/





































































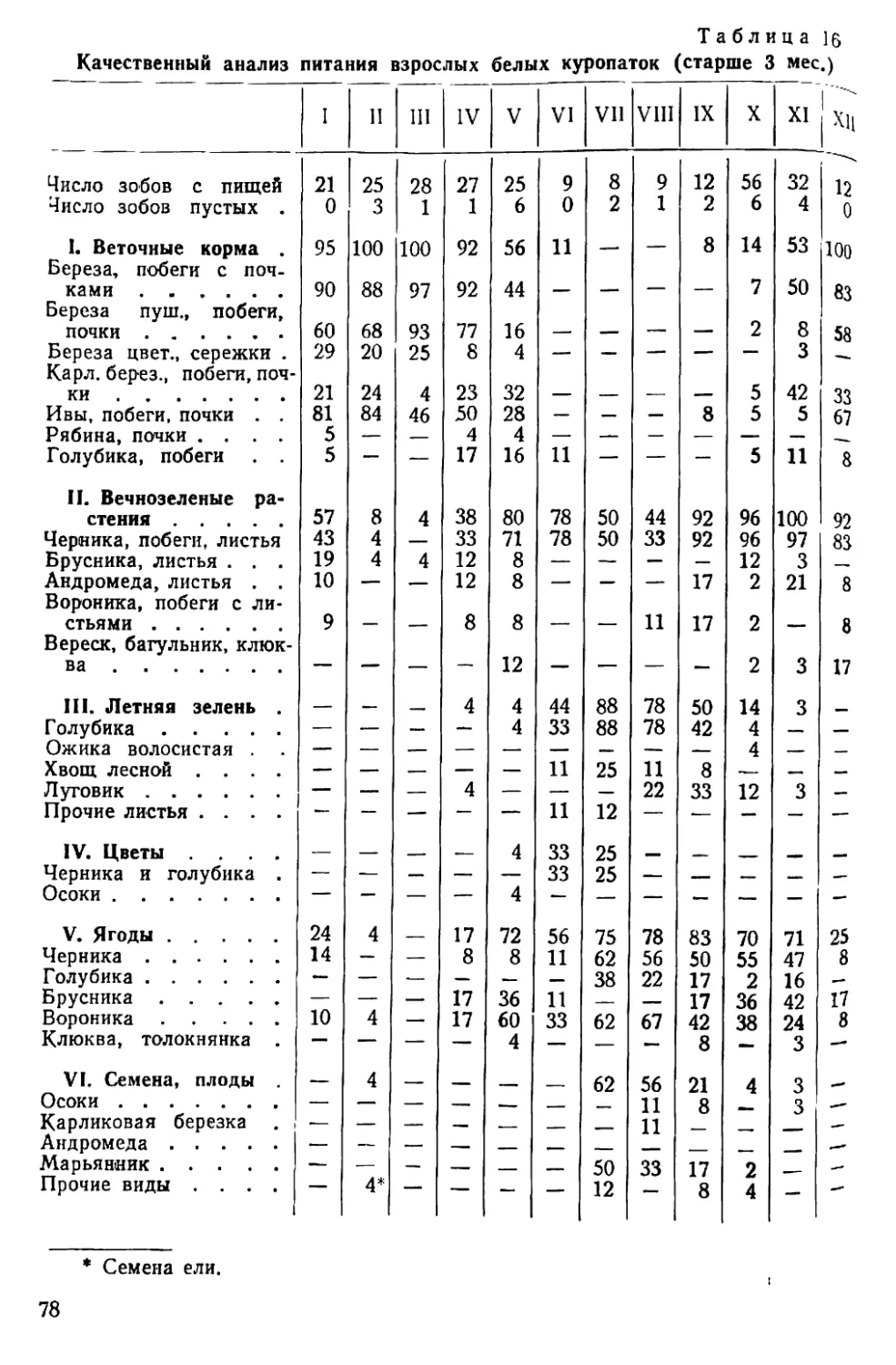
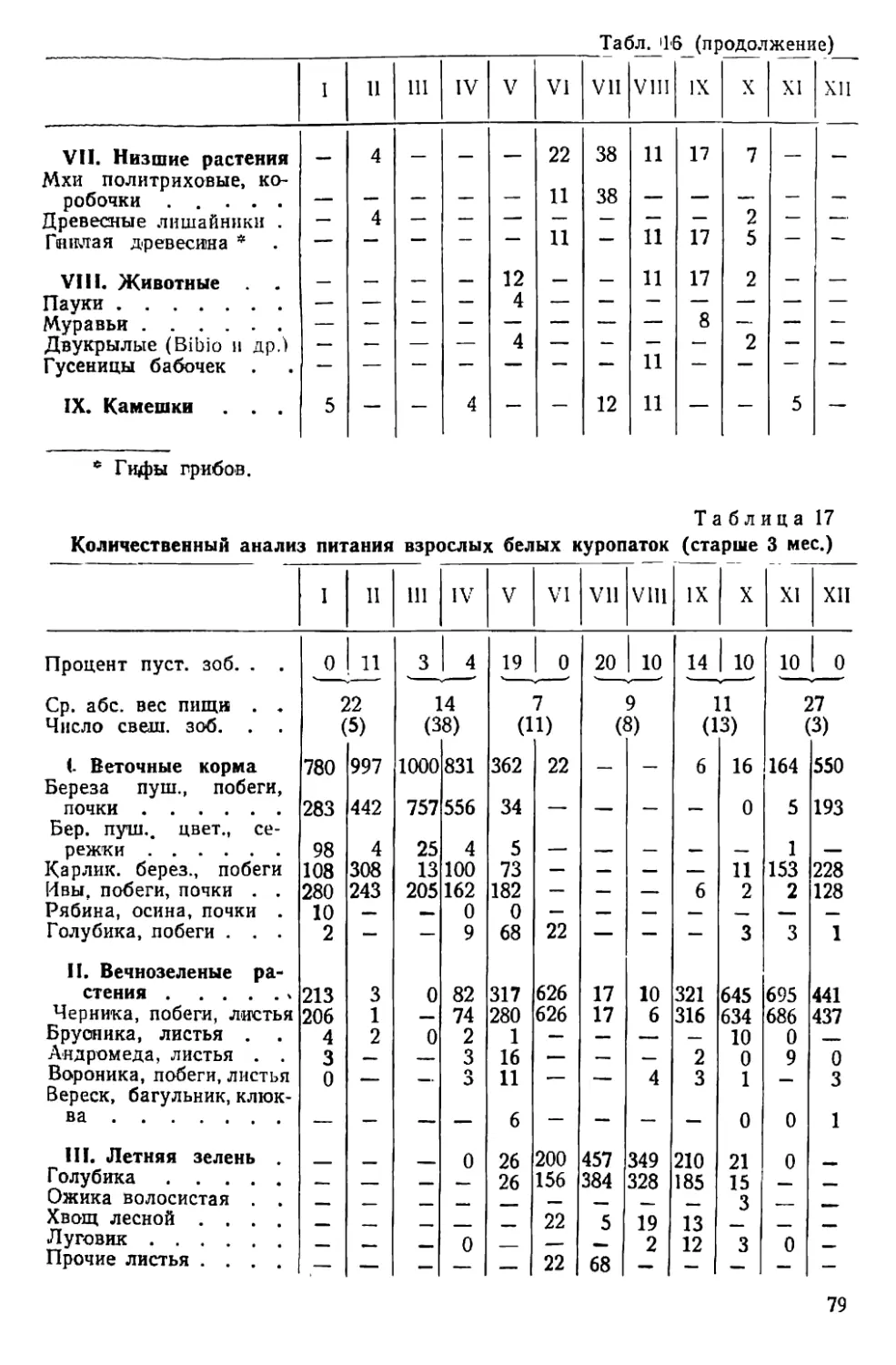
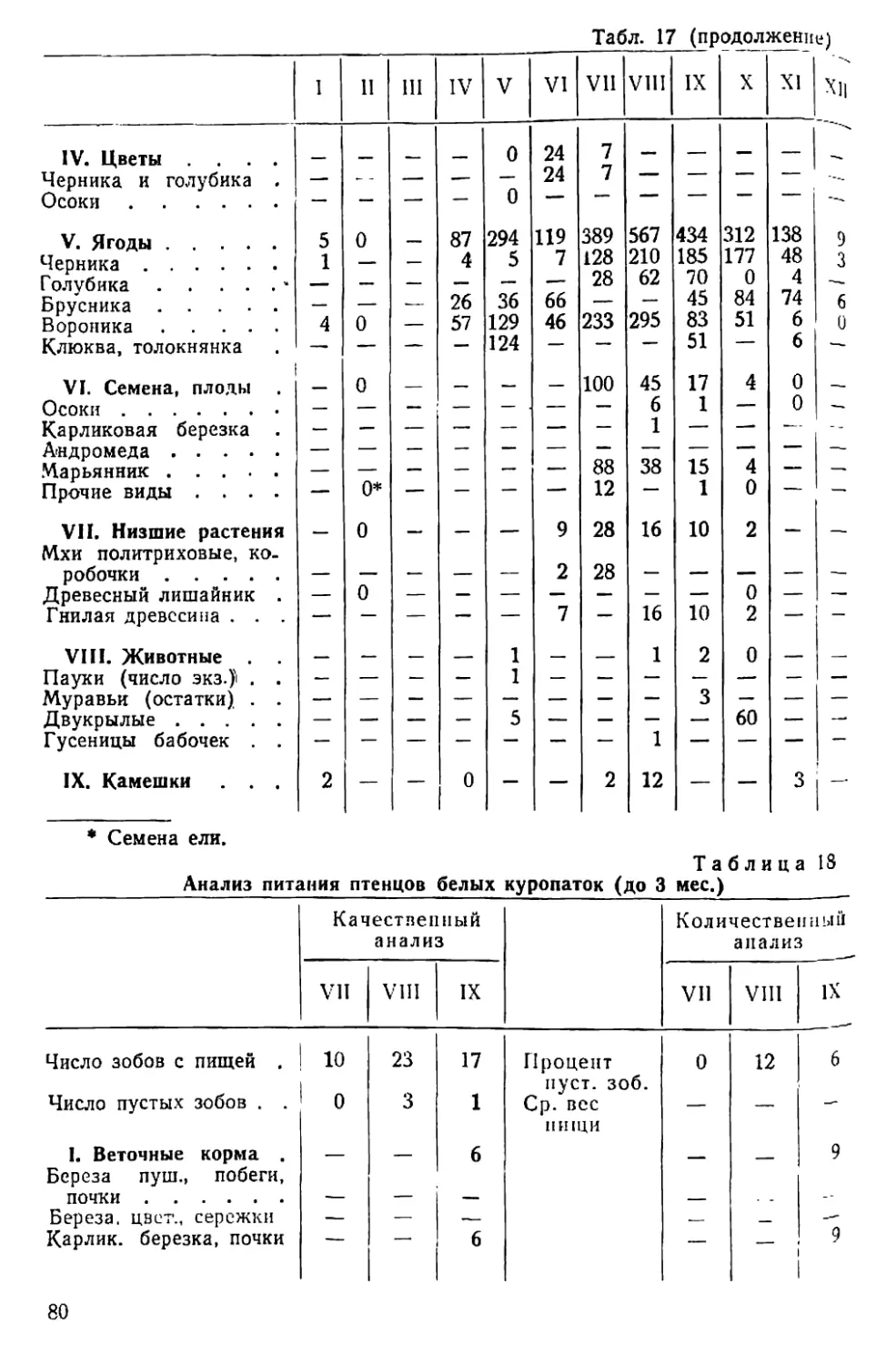
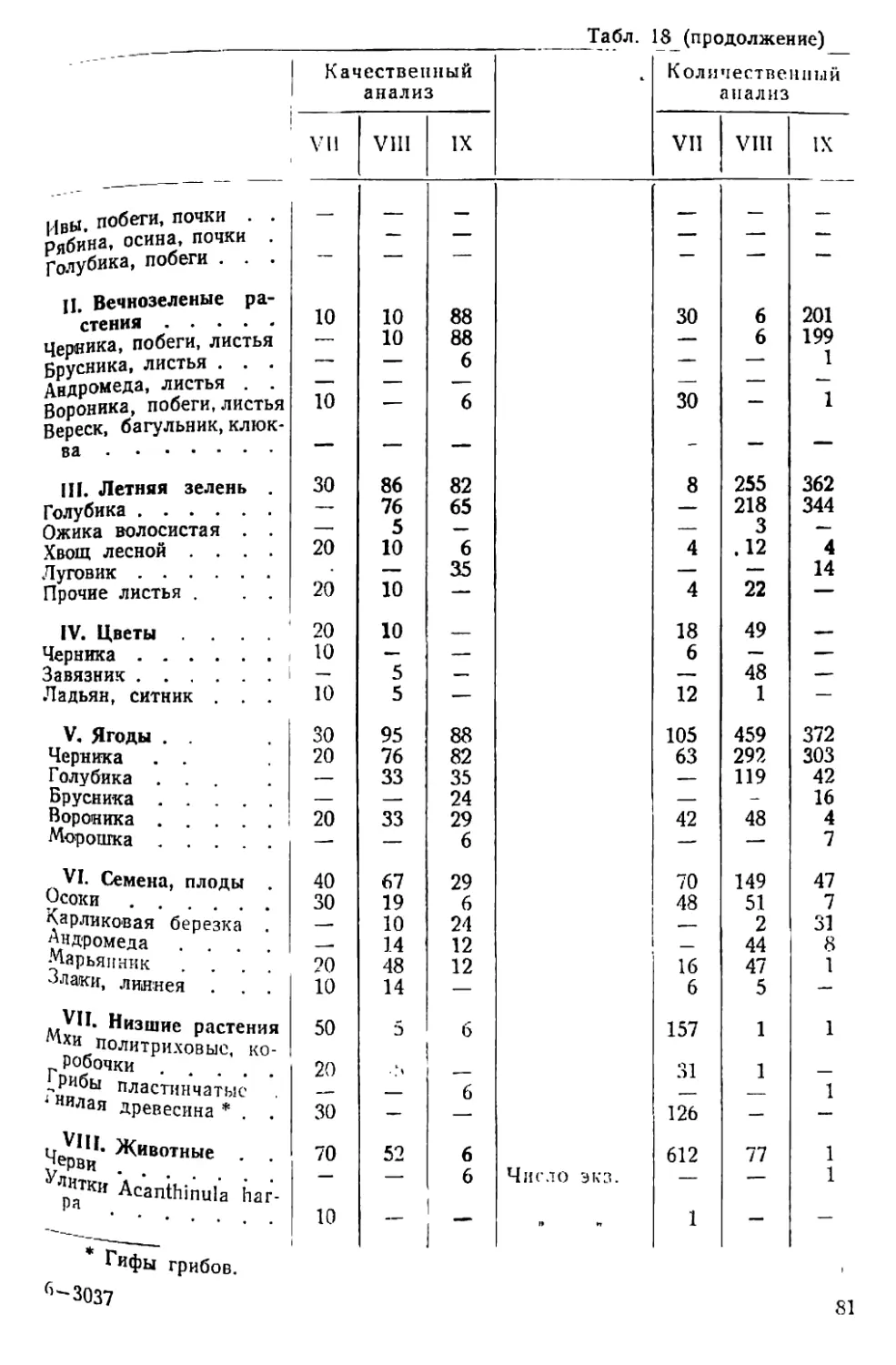






















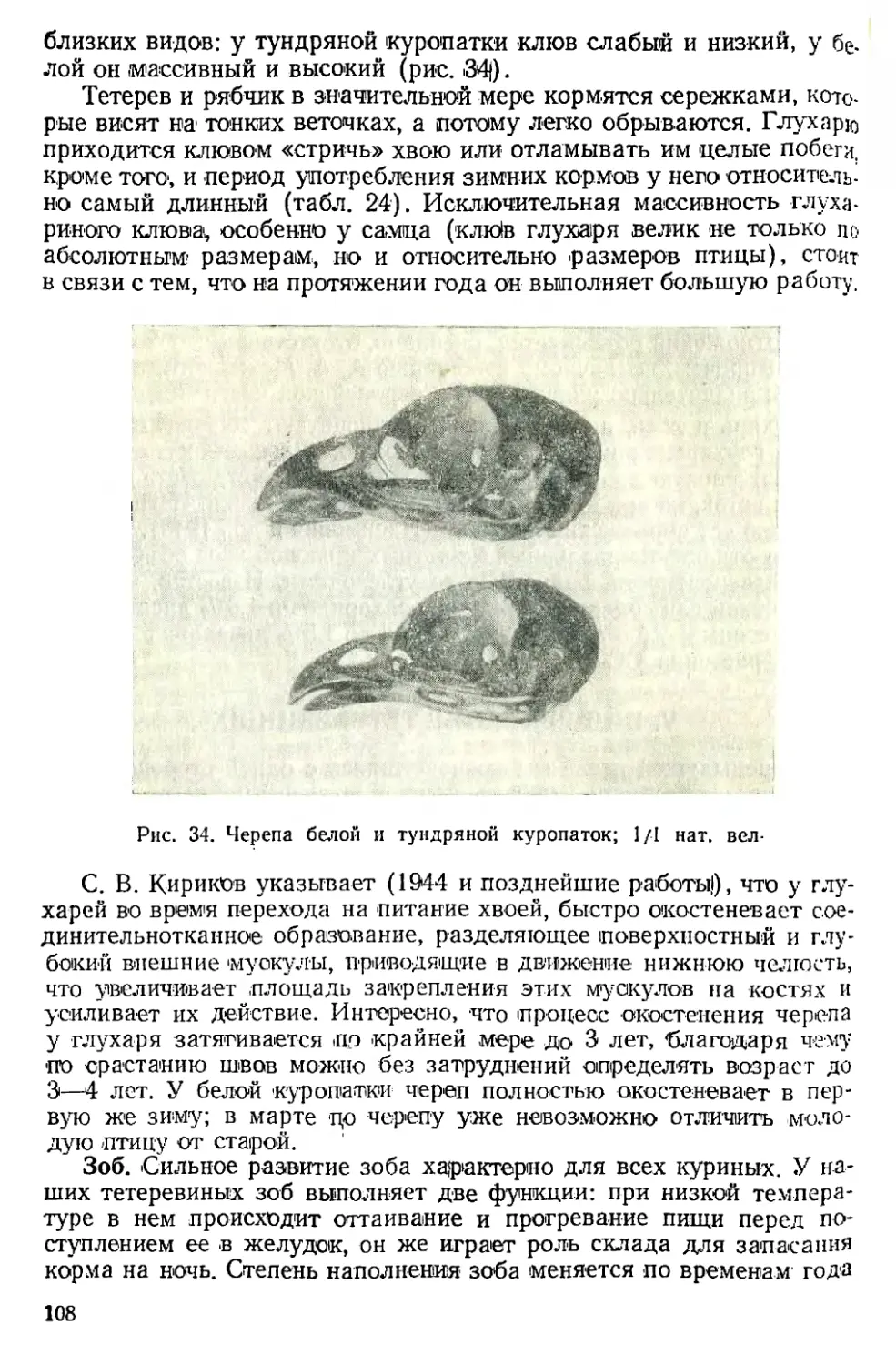







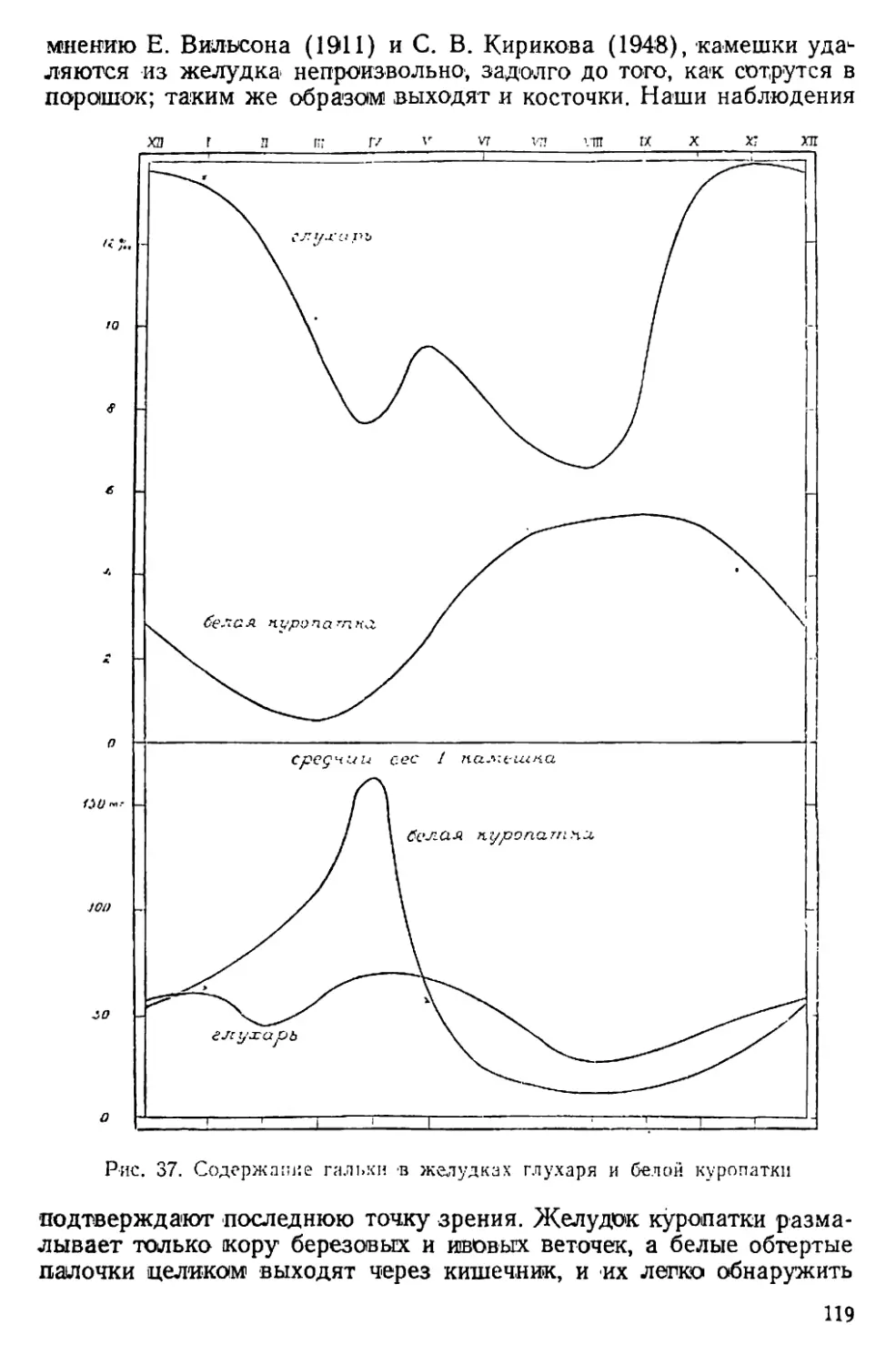
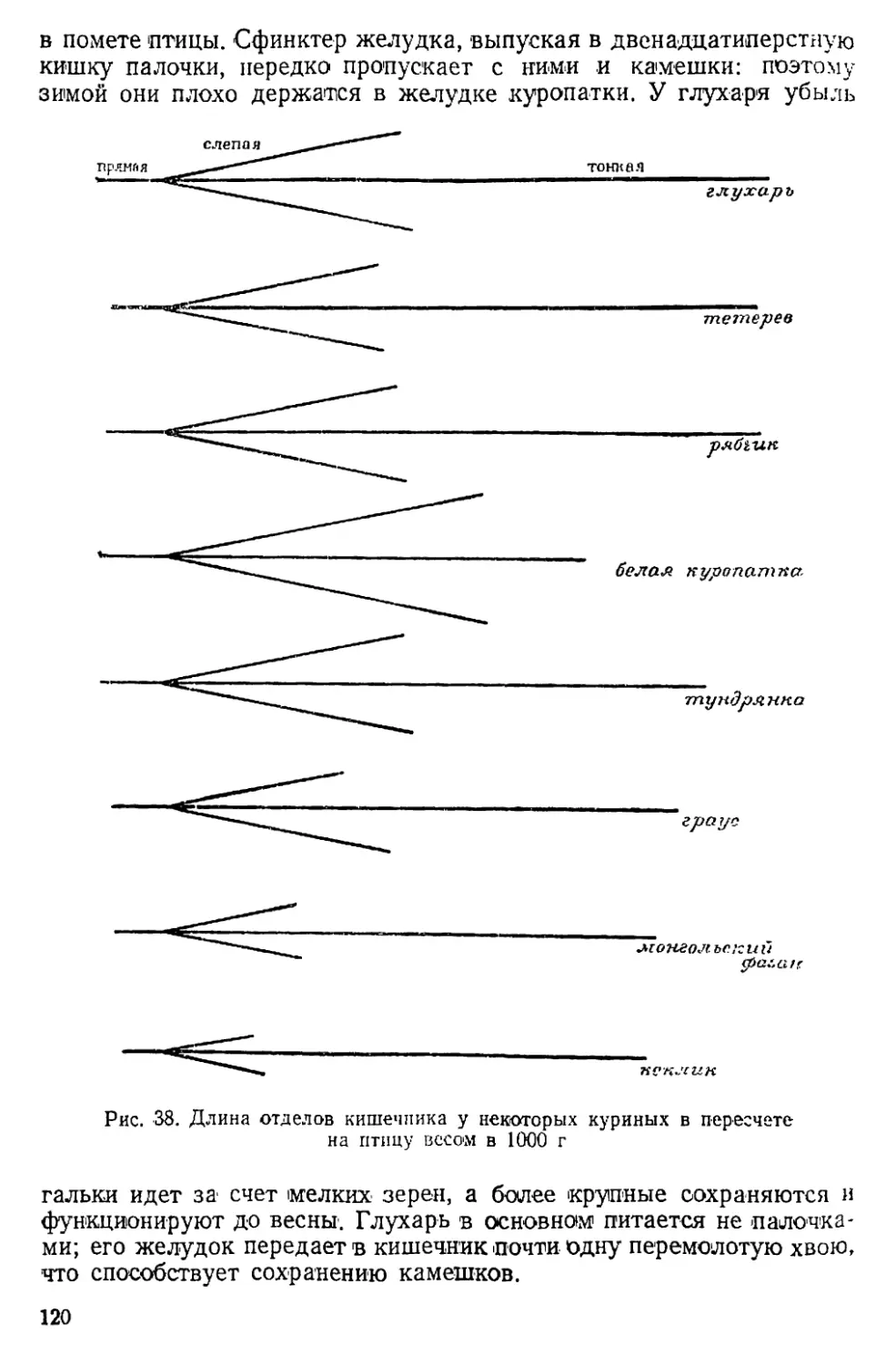

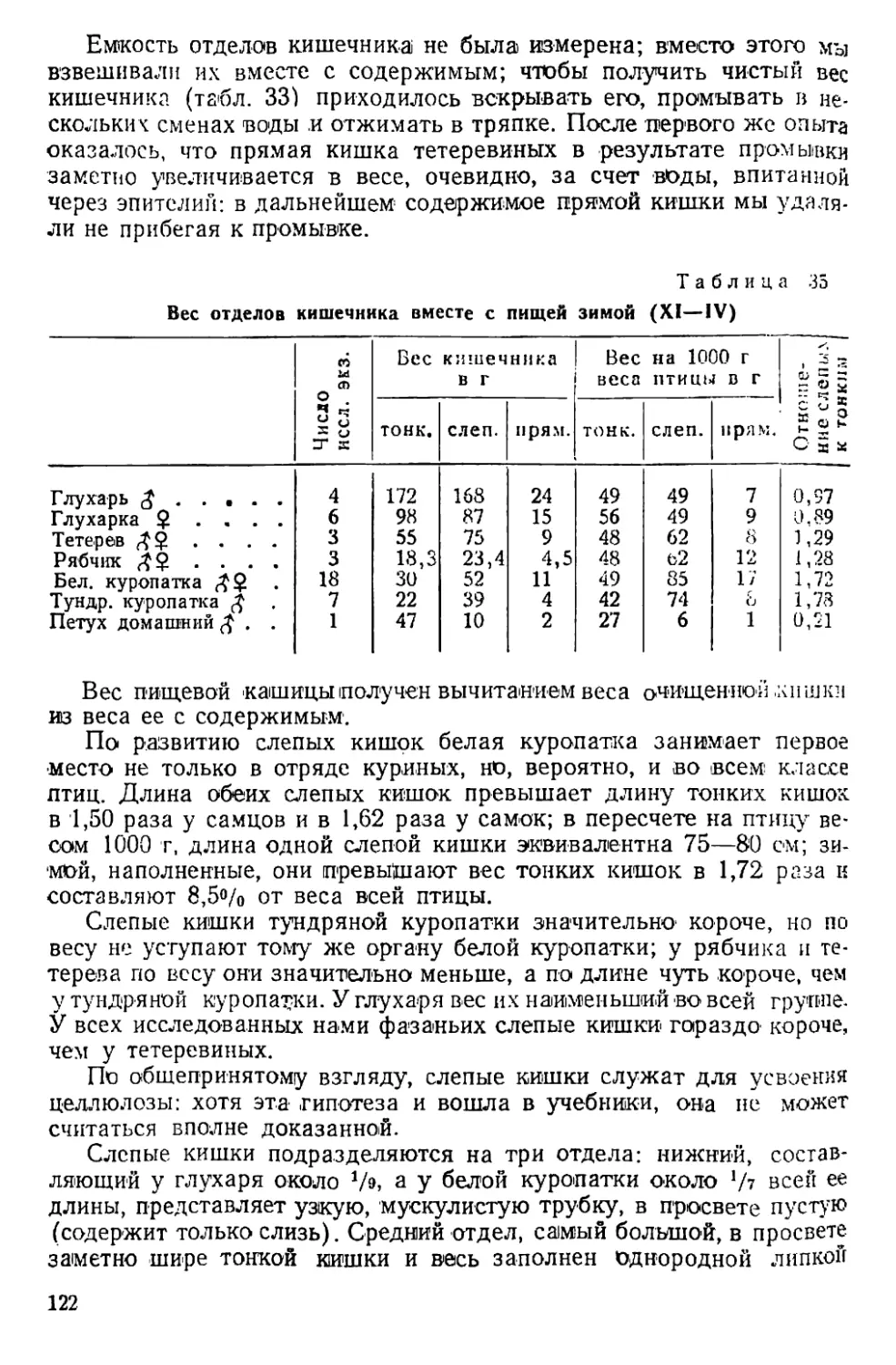

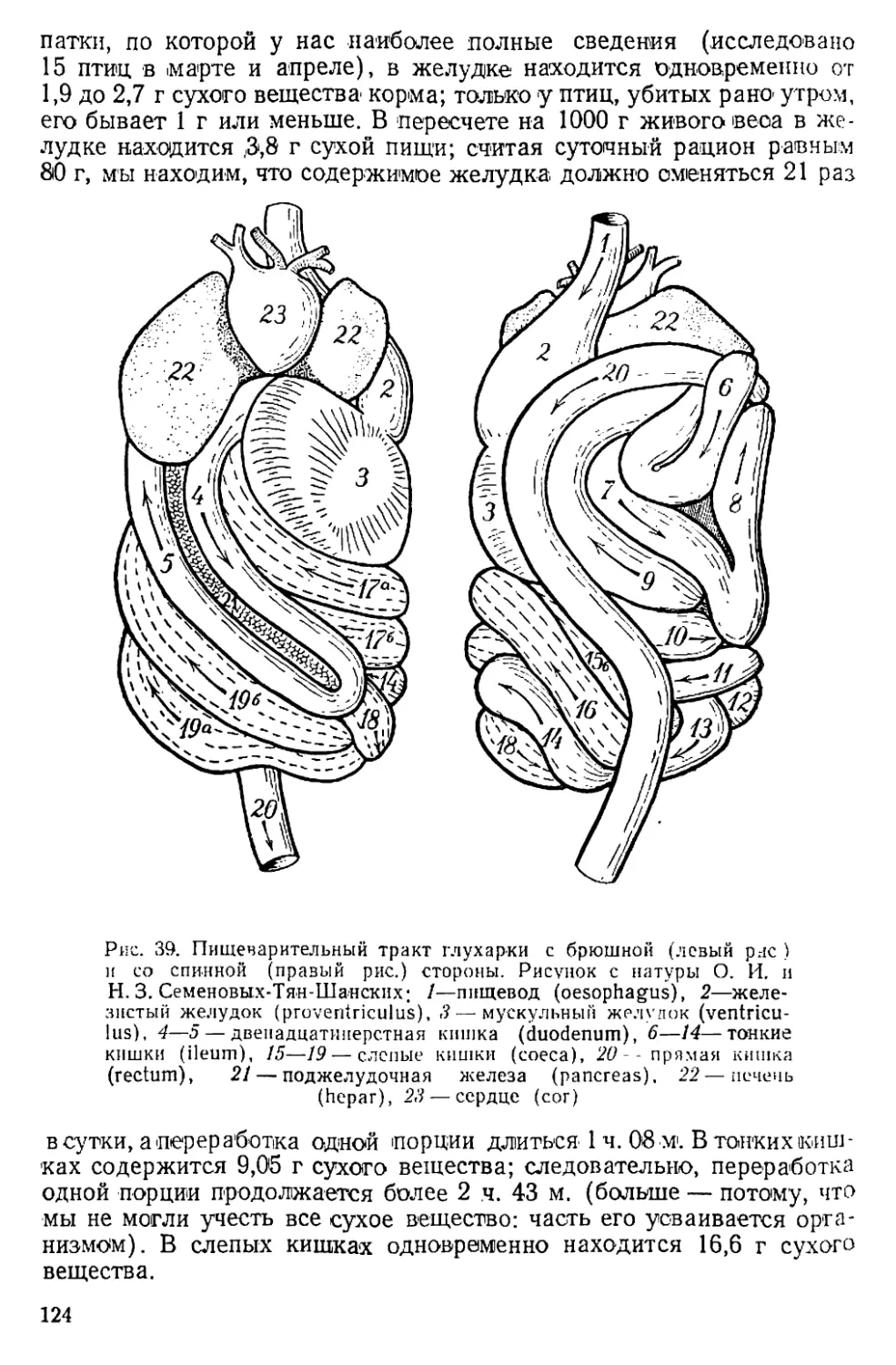
































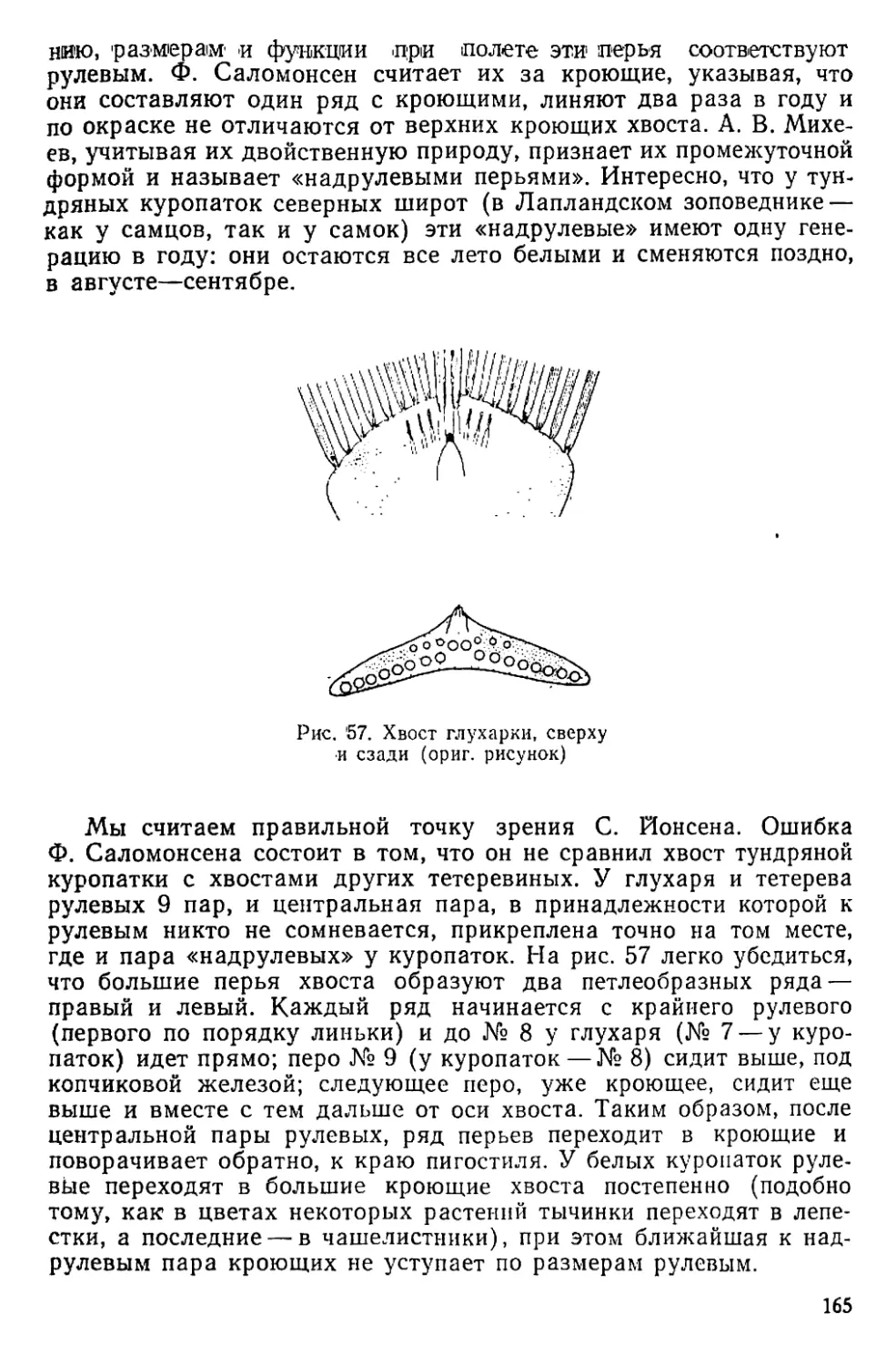


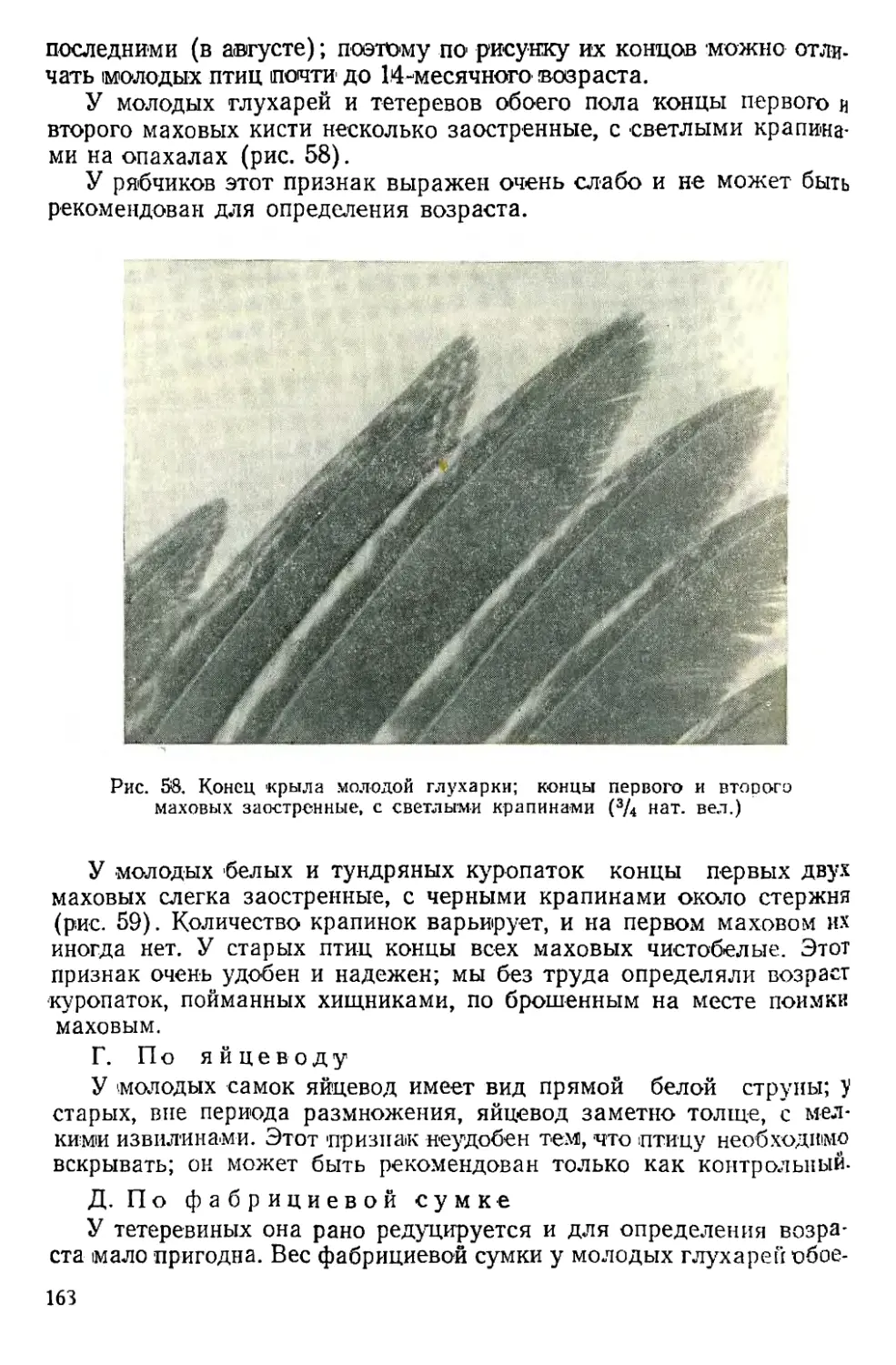

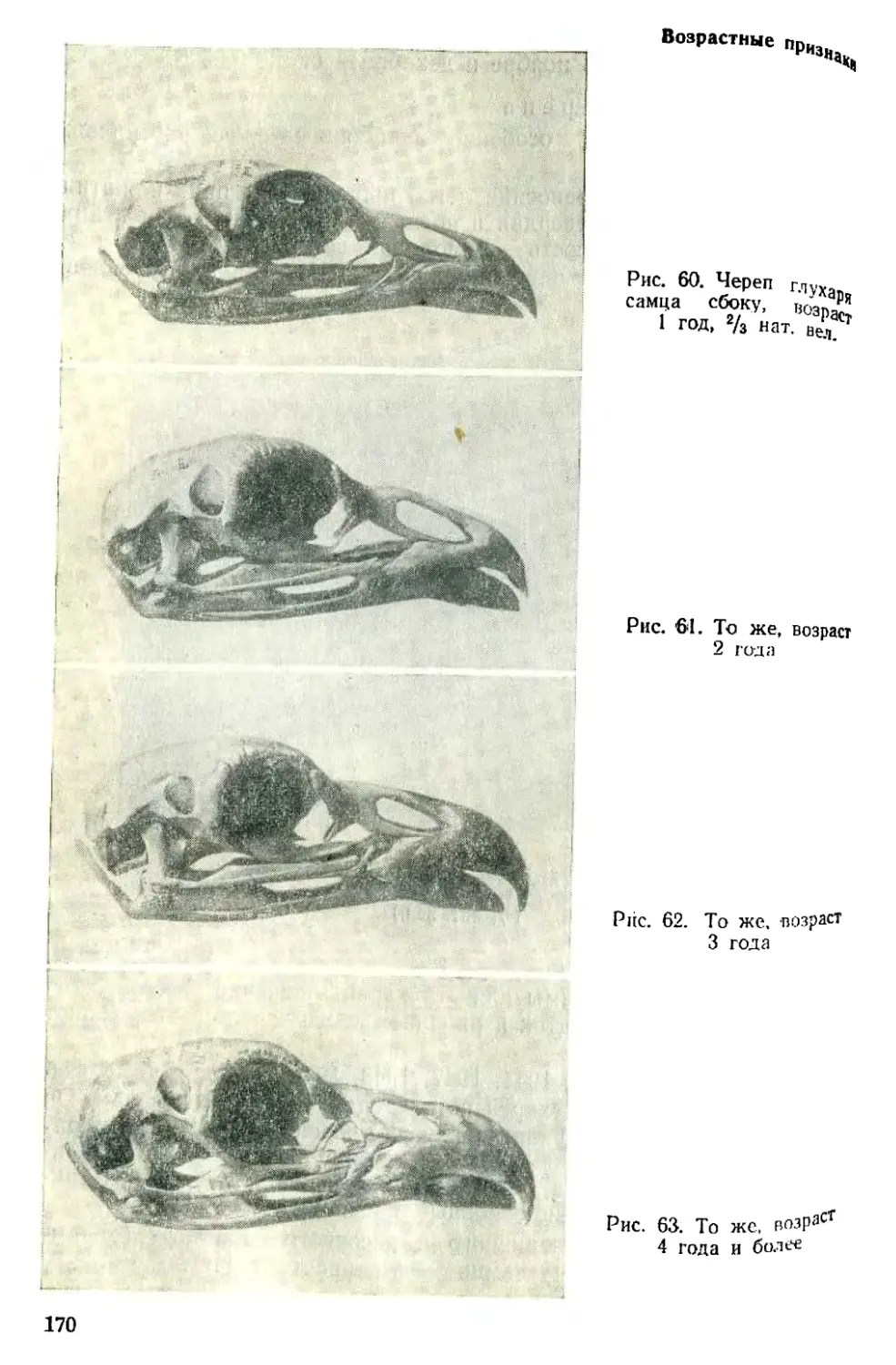




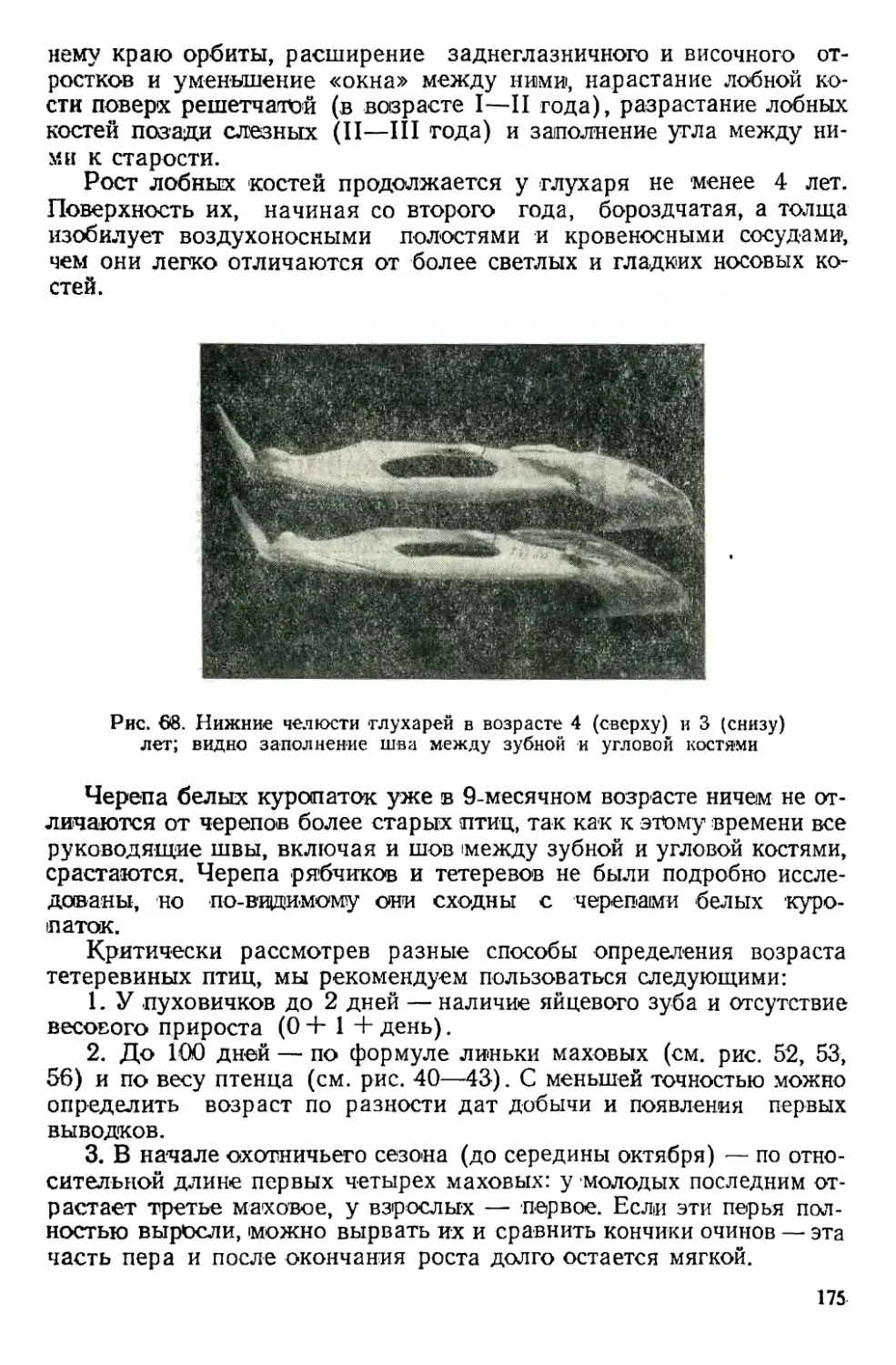




























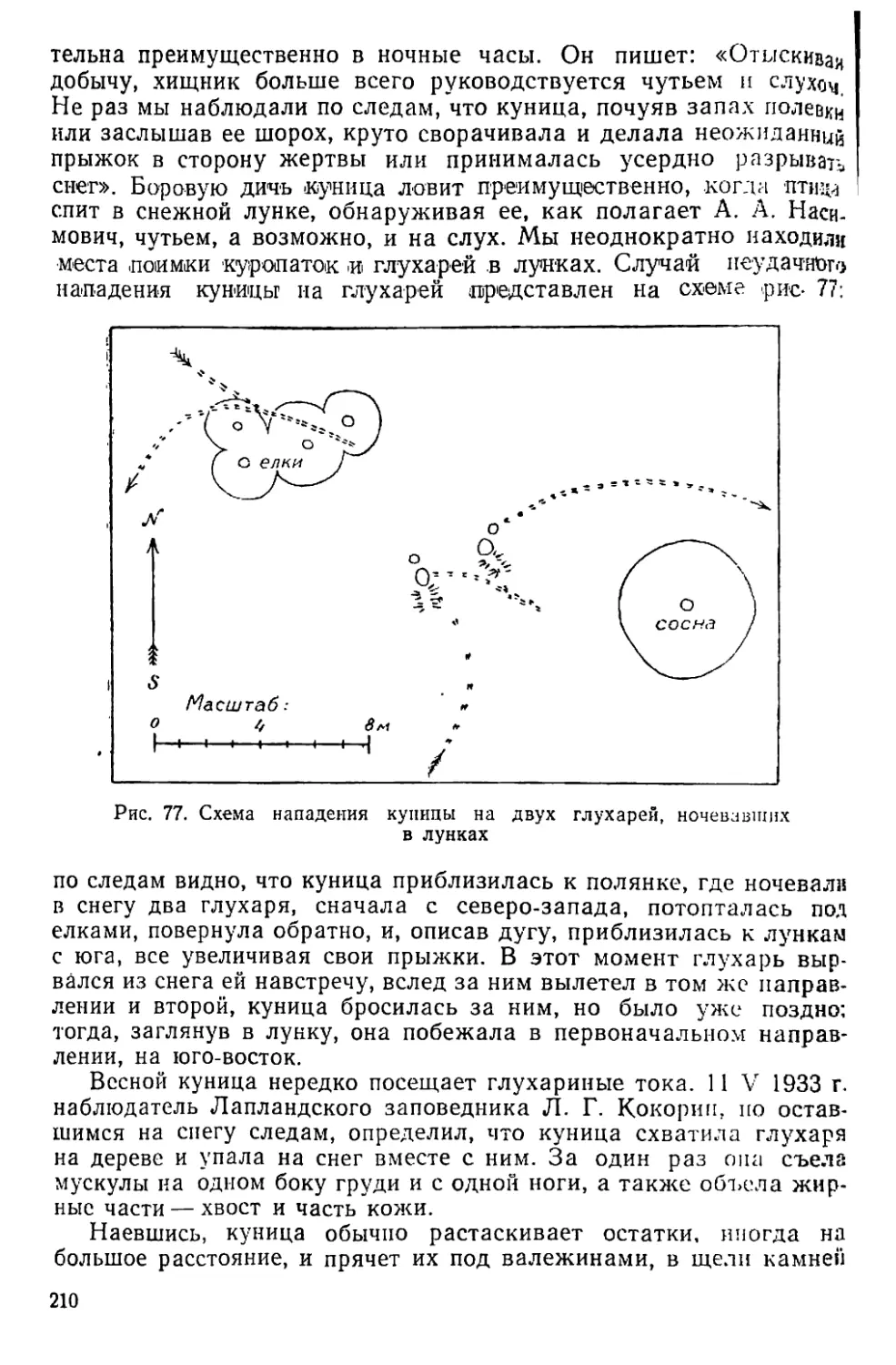







































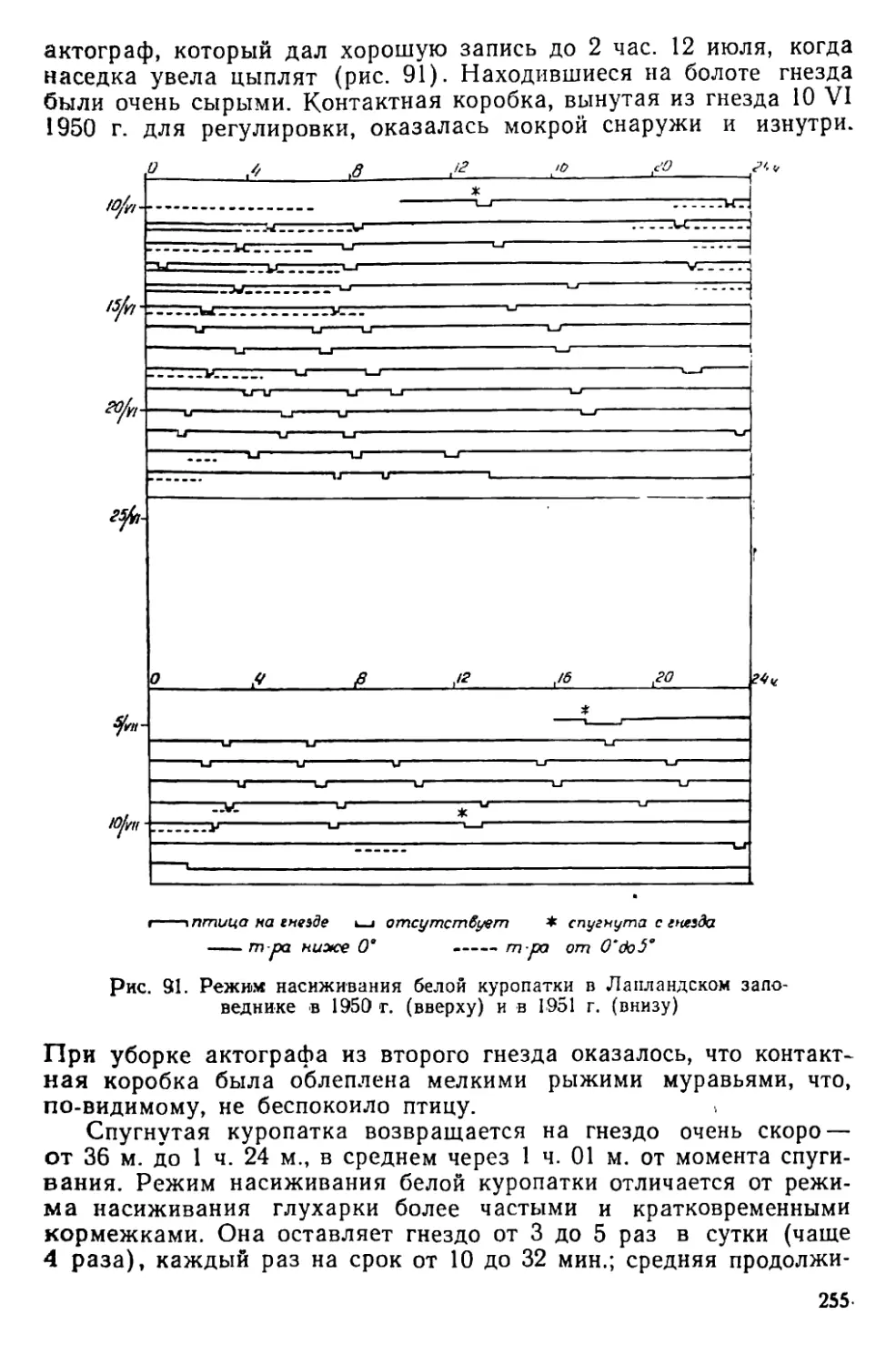








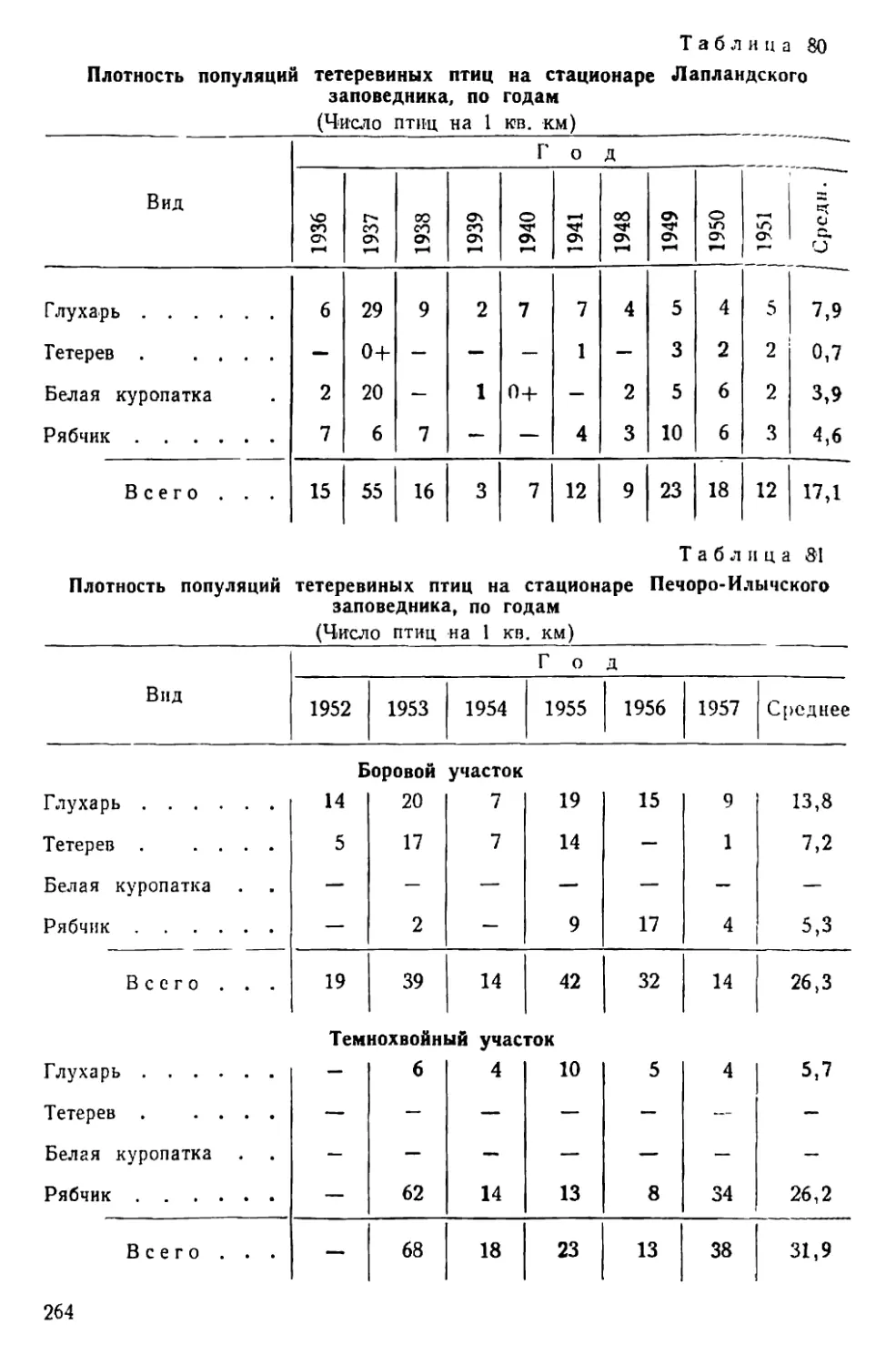



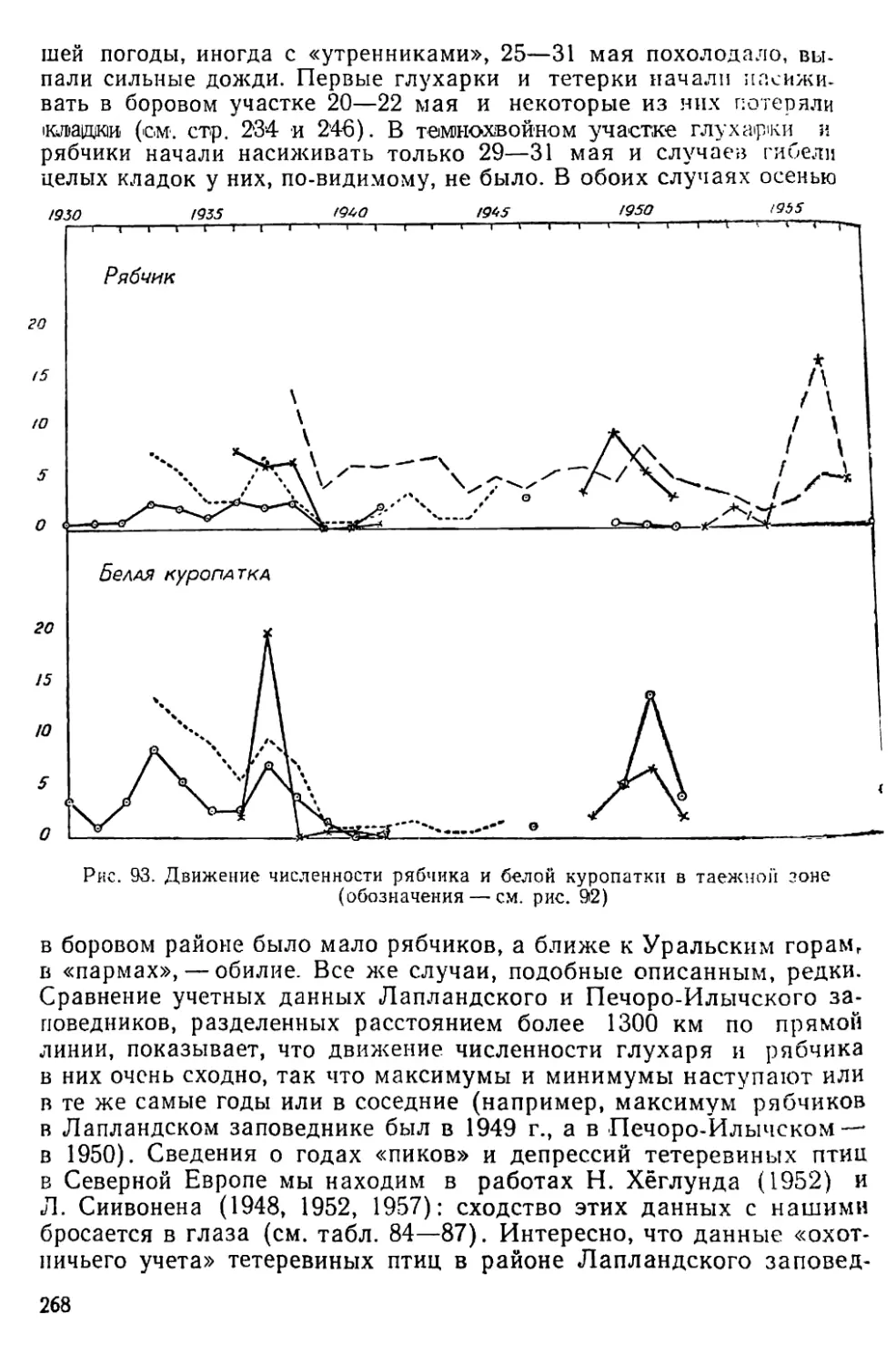

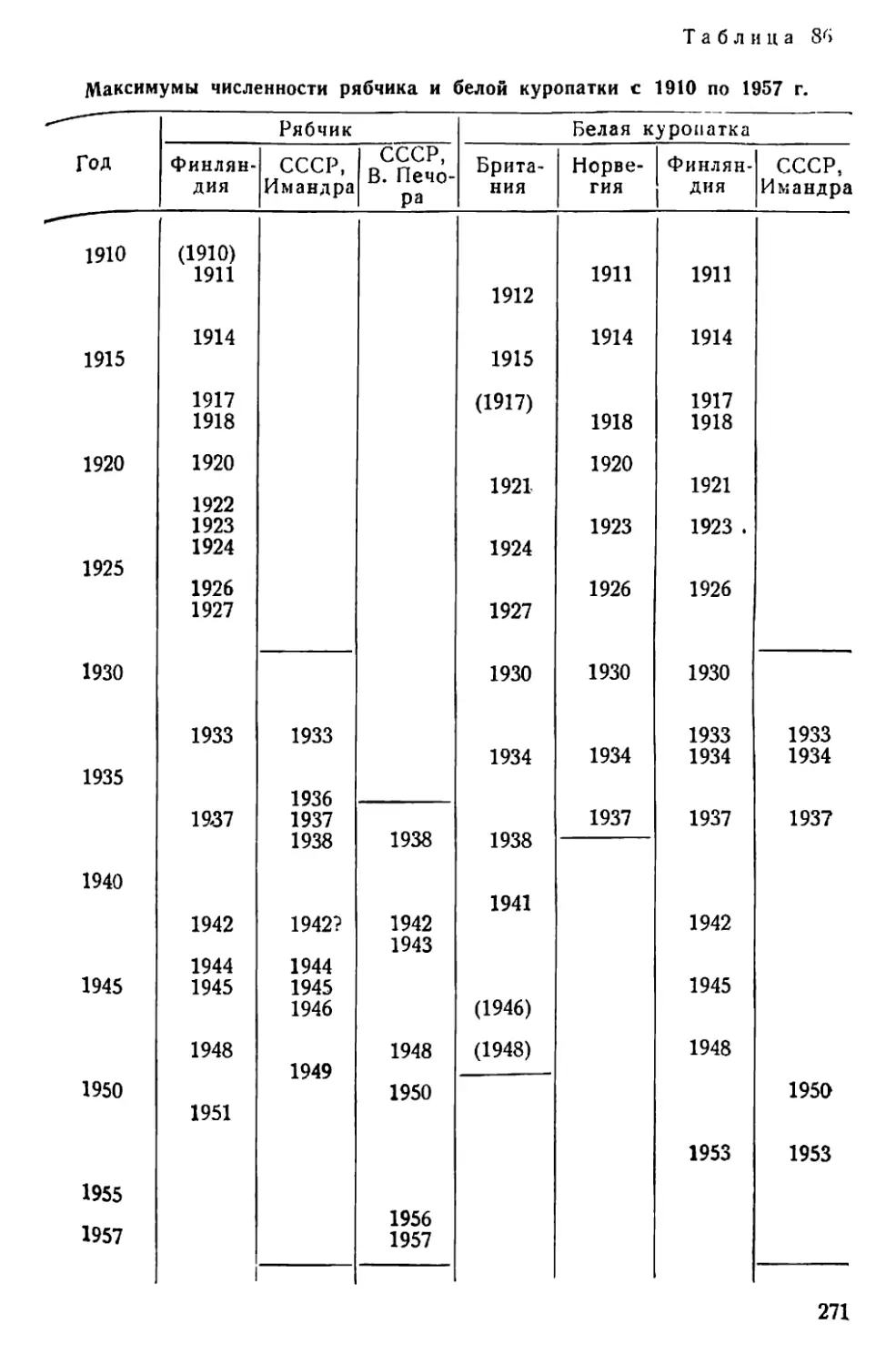




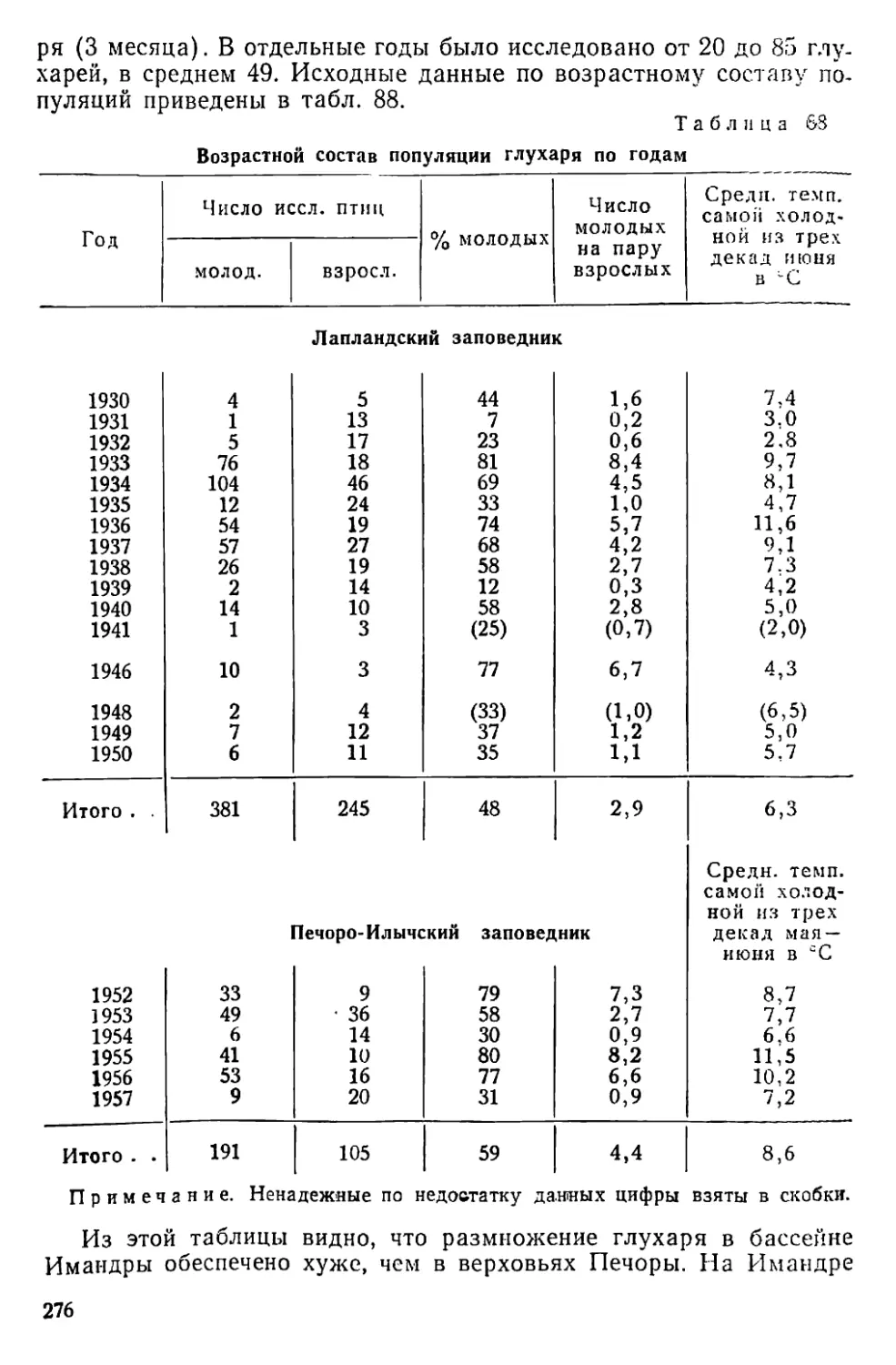




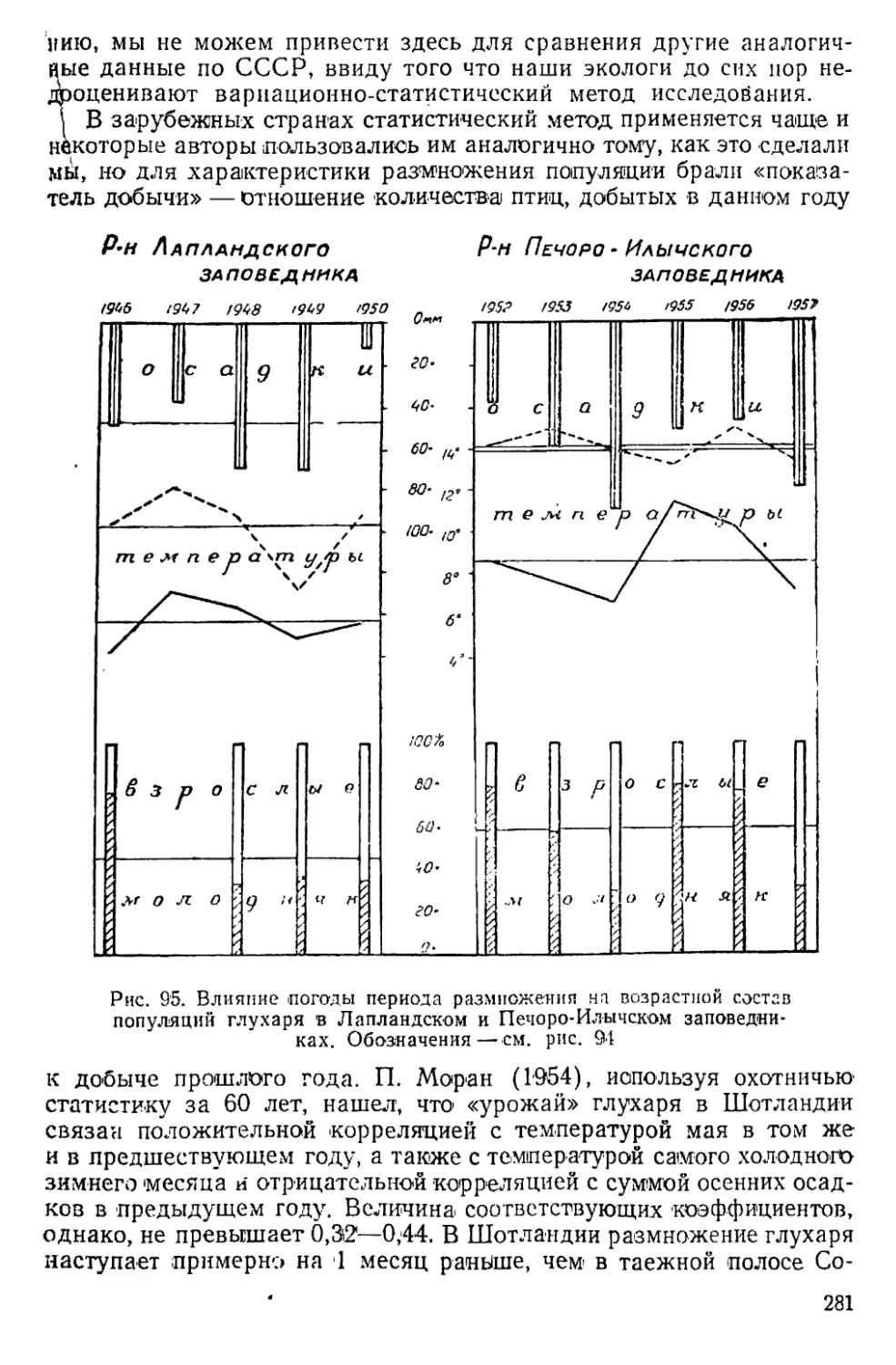





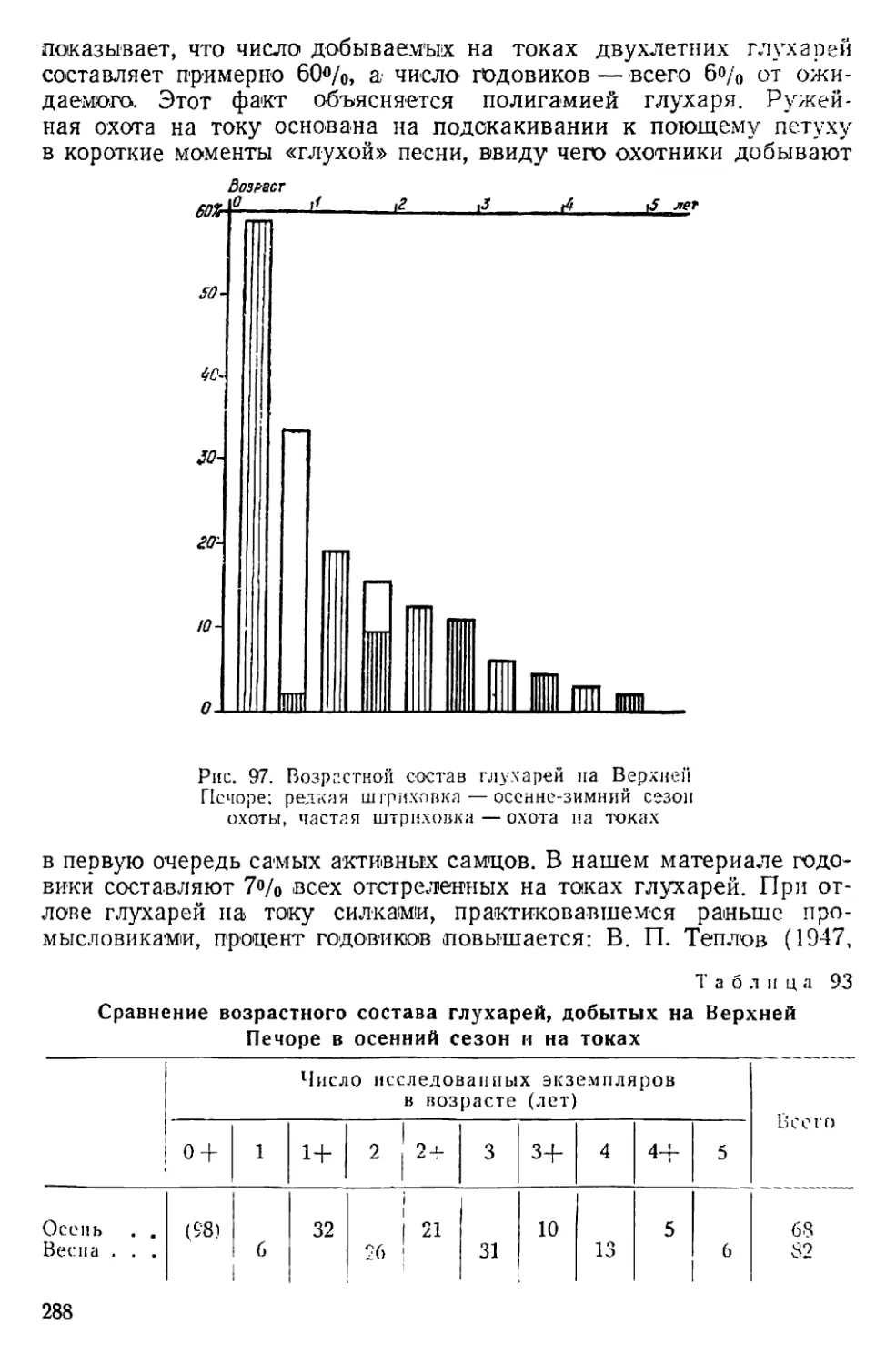
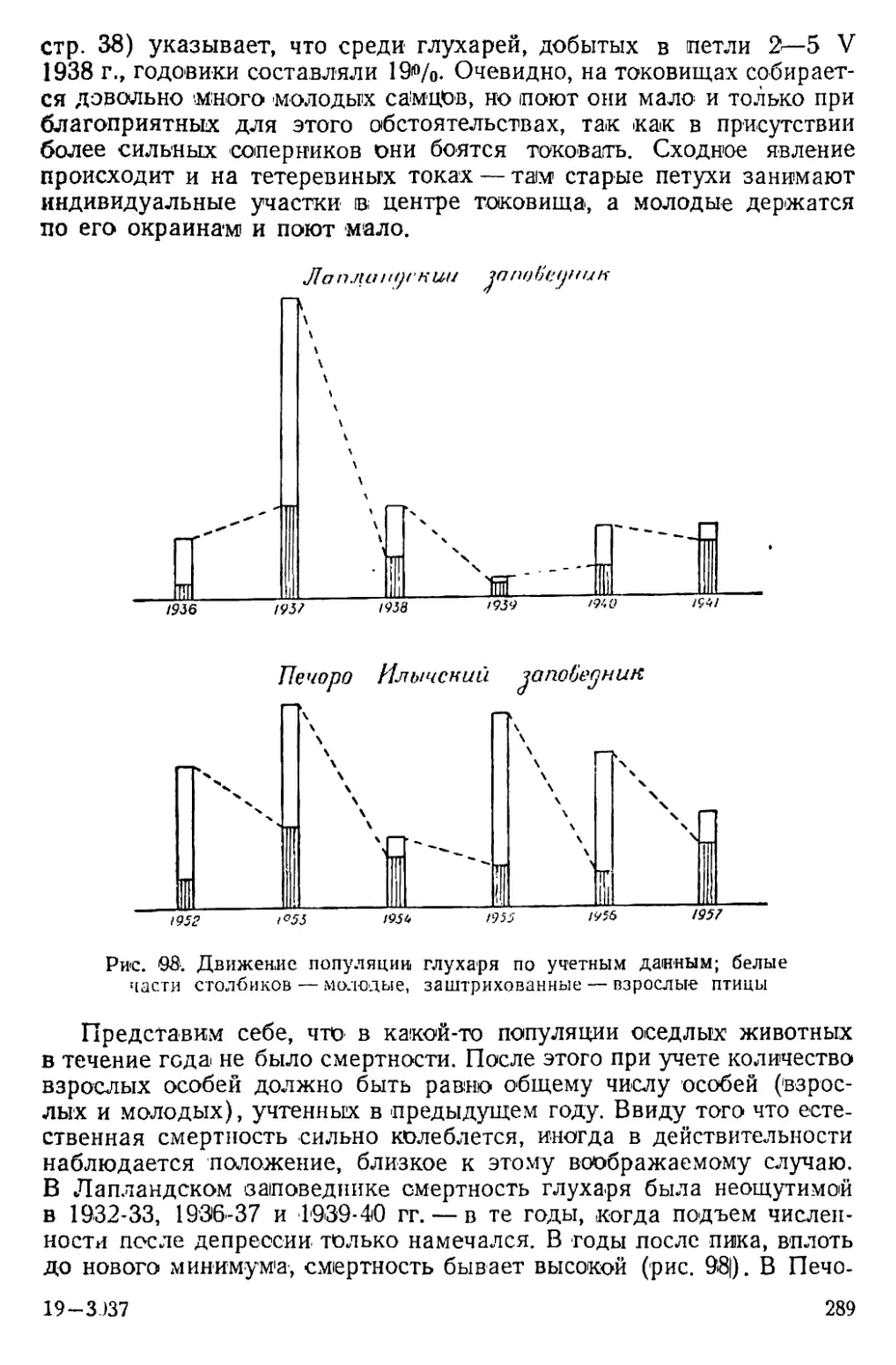
































Text
ГЛАВНОЕ УПРАВЛЕНИЕ ОХОТНИЧЬЕГО ХОЗЯЙСТВА И ЗАПОВЕДНИКОВ
ПРИ СОВЕТЕ МИНИСТРОВ РСФСР
ЛАПЛАНДСКИЙ ГОСУДАРСТВЕННЫЙ ЗАПОВЕДНИК
О. И. СЕМЕНОВ-ТЯН-ШАНСКИЙ
экология
ТЕТЕРЕВИНЫХ
ПТИЦ
МОСКВА — 1!№и
ГЛАВНОЕ УПРАВЛЕНИЕ ОХОТНИЧЬЕГО ХОЗЯЙСТВА И ЗАПОВЕДНИКОВ
ПРИ СОВЕТЕ МИНИСТРОВ РСФСР
ЛАПЛАНДСКИЙ ГОСУДАРСТВЕННЫЙ ЗАПОВЕДНИК
О. И. СЕМЕНОВ-ТЯН-ШАНСКИЙ
экология
ТЕТЕРЕВИНЫХ
ПТИЦ
МОСКВА—1959
ГЛАВНОЕ УПРАВЛЕНИЕ ОХОТНИЧЬЕГО ХОЗЯЙСТВА И ЗАПОВЕДНИКОВ
ПРИ СОВЕТЕ МИНИСТРОВ РСФСР
ТРУД ы
ЛАПЛАНДСКОГО
ГОСУДАРСТВЕННОГО ЗАПОВЕДНИКА
Выпуск V
О. И. СЕМЕНОВ-ТЯН-ШАНСКИИ.
ЭКОЛОГИЯ
ТЕТЕРЕВИНЫХ ПТИЦ
МОСКВА — 1959
РЕДАКЦИОННАЯ КОЛЛЕГИЯ:
доктор биолог, наук Н. Л. АВРОРИН,
доктор географ, наук А. А- НАСИМОВИЧ (редактор),
канд. биолог, наук О. И- СЕМЕНОВ-ТЯН-ШАНСКИЙ,
канд. биолог, наук Р- Н. ШЛЯКОВ
ВВЕДЕНИЕ
В нашей стране тетеревиные птицы имеют громадное экономи-
ческое значение. Величину охотничьей добычи, к сожалению, труд-
но определить, потому что учета ее мы до сих пор не имеем, а циф-
ры заготовок не дают полного представления о количестве добы-
ваемой дичи по той причине, что большая часть ее потребляется на
месте и в заготовки не поступает. В связи с этим у нас шйроко рас-
пространено мнение, что- пушной промысел имеет гораздо большее
значение в экономике, чем дичной; между тем в действительности
дело обстоит, вероятно, как раз наоборот. Необходимо помнить, что
дичь, съедаемая семьями охотников, практически также входит
в народнохозяйственный бюджет, так как использование ее эконо-
мит потребление других продуктов населением.
Среди всех наших промысловых птиц А. Н. Формозов (1940,
стр. 789—794) ставит диких птиц отряда куриных на первое место
по хозяйственному значению. Размер добычи можно приблизи-
тельно определить косвенным путем. По данным приполярной пе-
реписи 1926—1927 г. товарный выход боровой дичи составлял
у рябчика —12,5% от всего количества добытых птиц, у глуха-
ря— 1,Оо/о, остальные виды занимали промежуточное положение.
Допустим, что в среднем в заготовки поступает не более 10% всей
добычи охотников. В период с 1928 по 1956 г., исключая годы вой-
ны, заготовки боровой дичи в Советском Союзе составляли от
4*58 000 до 4 755 000 штук в год (Колосов и Шибанов, 1957); от-
сюда всю добычу куриных (в основном — из семейства тетереви-
ных) можно оценить от 5 до 50 миллионов штук в год, что соответ-
ствует 3000—30 000 тонн. В Финляндии добыча этих птиц в уро-
жайные годы достигает 1,3 млн. голов («Финляндия — страна озер
и лесов», 1954); в Канаде средняя ежегодная добыча боровой ди-
чи оценивается в 1400 тонн (Коуан, 1955). <
Относительное значение тетеревиных птиц в охотничьем хозяй-
стве ближайшего будущего, по всей вероятности, возрастет. У них
есть важные преимущества перед другими видами дичи: во-пер-
вых оседлость, упрощающая задачу охраны и рациональной экс-
плуатации стада; во-вторых — особый характер питания, благо-
5
даря чему тетеревиные не конкурируют ни с одним из домашних
животных.
В зонах тундры и лесотундры, в частности на Кольском полу-
острове, наибольшее экономическое значение имеет белая куро-
патка. В старину она входила в число основных средств существо-
вания саамов. В. В. Чарнолуский (1930, стр. 26) передает со слов
иокангских саамов: «Досельные» * лопари были знаменитые охот-
ники и промышляли больше всего дикарей **, мясом которых пи-
тались, еще брали куропаток и рыбу». Есть указание, что некто
Богдан Никулин 23 IX 167 года (1659 г. нашей эры) уплатил 20 ал-
тын и 2 деньги пошлины за вывезенные им с Поноя 54 пуда куро-
пачьего пера (Андреев, 1930, стр. 16). Учитывая, что вес оперения
куропатки зимой составляет около 42 г, а за вычетом маховых,
рулевых и оперения лапок, которые в дело не идут, — 33 г, легко
рассчитать, что в тот сезон в районе села Поной было добыто не
менее 26 800 куропаток.
В 1952( г. в Кожвинском и Усть-Усинском районах Коми АССР
было заготовлено 267 тыс. куропаток, причем добыча отдельных
охотников достигала! 5000 штук (Маслов, 1953).
Практическое значение тетеревиных птиц вполне оправдывает
то внимание, которое им уделялось за последние 30 лет. Интерес-
ные исследования, были проведены в ряде заповедников — Алтай-
ском, Башкирском, Беловежской пуще, Кавказском, Краснояр-
ском, Лапландском, Наурзумском, Печоро-Илычском и других.
Предлагаемая работа отличается от всех ранее опубликованных
прежде всего длительностью исследования. Автор этих строк рабо-
тал в Лапландском заповеднике с первого года его существования
(1930) и систематически собирал материал по тетеревиным птицам
все время, включая и годы, когда был занят, в основном, другой
тематикой.
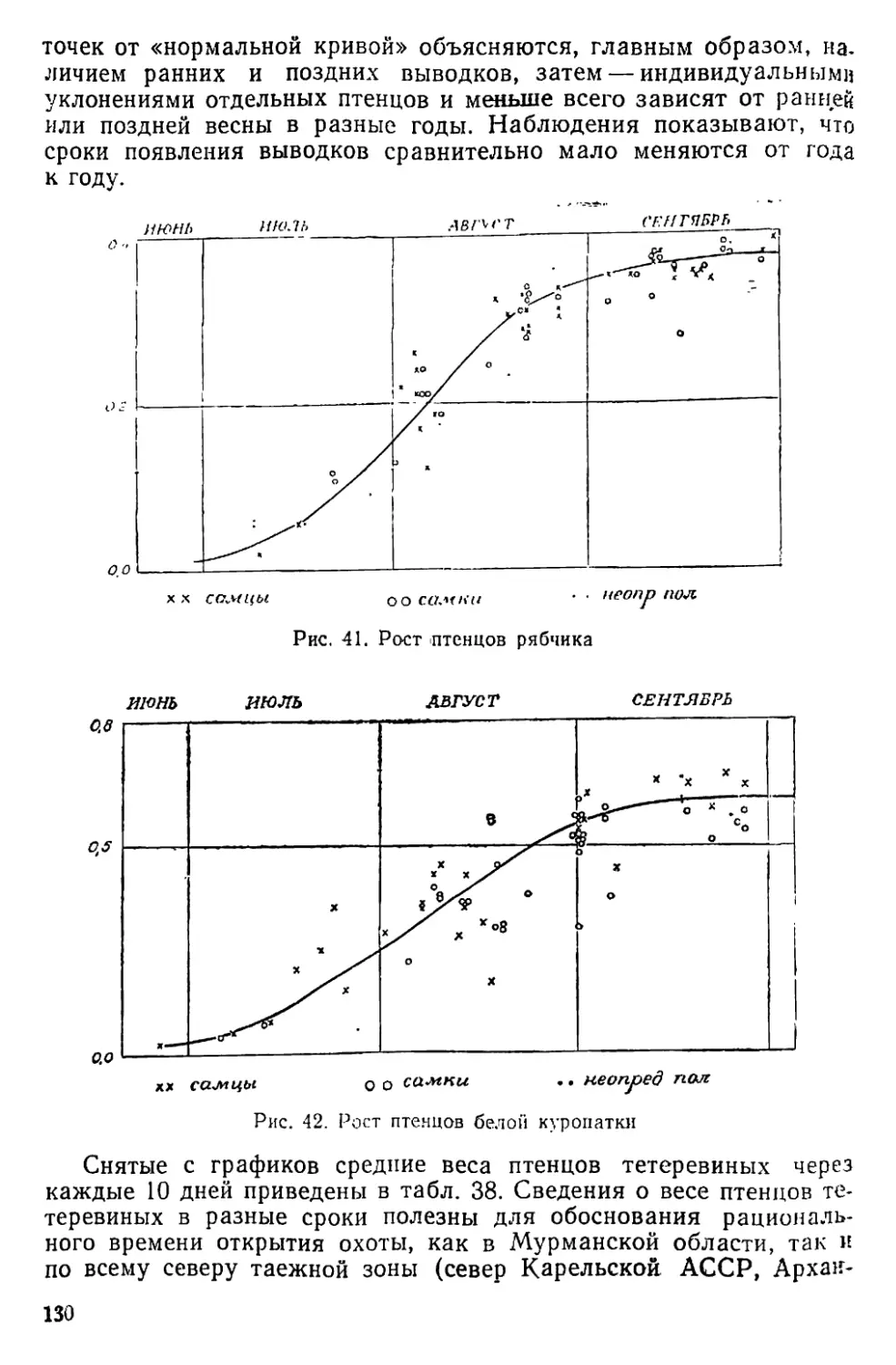
Осенью 1931 г. меня заинтересовало странное явление почти
полного отсутствия молодняка у глухарей. Обсуждая этот вопрос
с проф. Г. Г. Доппельмаиром, я узнал от него, что то же самое на-
блюдалось и в Карелии; из этого факта был даже сделан поспеш-
ный вывод, что преобладание в популяции старых птиц является
показателем недопромысла глухаря в крае. Согласиться с таким
объяснением было невозможно, но оно способствовало тому, что
я обратил особое внимание па выяснение причин «неурожаев» те-
теревиных птиц, и в первую очередь глухаря. Прежде всего нужно
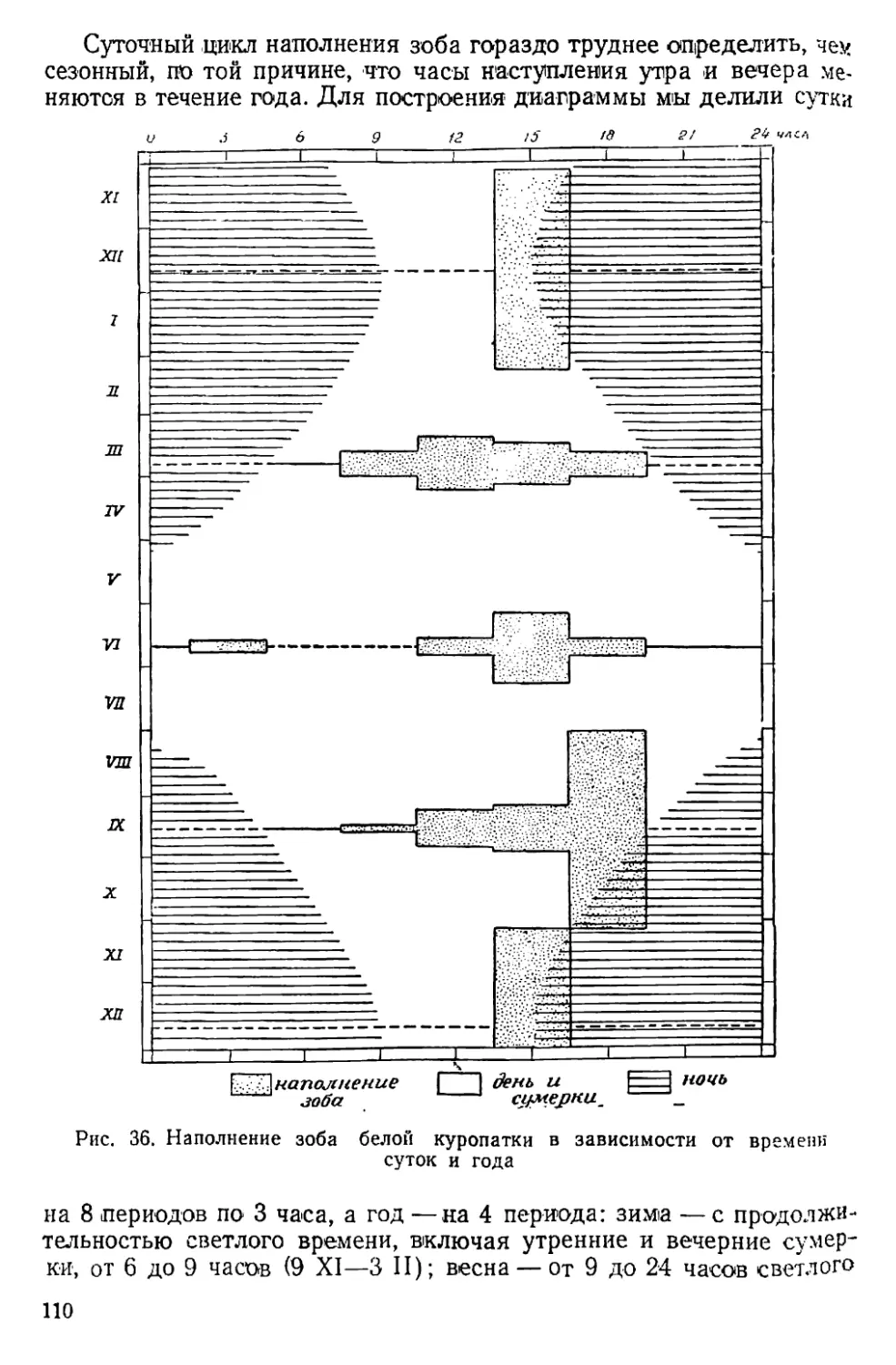
было найти способ выразить величину «урожая» дичи в числах:
эта задача была решена путем определения возрастного состава
популяции в каждый охотничий сезон отдельно. Собрав такой мате-
риал за 7 лет, я сопоставил его с метеорологическими условиями
сезона размножения и получил поразительный результат — оказа-
лось, что величина «урожая» глухаря почти в точности опреде-
* Прежние, старинные.
♦♦ «Дикарь» — дикий северный олень.
6
ляется температурой воздуха в июне, вернее в первой декаде этого
месяца (Семенов-Тян-Шанский, 1938, стр. 296—297; 1937, стр.
23—24). Примененный при этом метод корреляции устанавливает
только формальную связь двух рядов чисел; причинная сторона на-
блюдаемого явления по-прежнему оставалась неясной. Чтобы разо-
браться в этом вопросе, надо было хорошо знать режим насижива-
ния яиц, а также характер действия низких температур на яйца
и на пуховых птенцов. Визуальные наблюдения тут мало чем
могли помочь. Знакомство с метеорологическими самописцами на-
толкнуло меня на мысль снабдить обычный термограф вторым
пером, связанным с якорем электромагнитной катушки, а провод
от батареи пропустить через гнездо и там поместить приспособле-
ние, включающее ток в то время, когда птица сидит на гнезде.
Таким образом перо будет отмечать на ленте моменты ухода птицы
с гнезда и возвращения на него; тут же рядом будет записываться
и температура воздуха. В то время (1944 г.) я находился в Совет-
ской действующей армии; построить прибор и применить его на
практике мне удалось только в 1950 году.
В 1951 г. Лапландский заповедник был закрыт, и я перешел на
работу в Печоро-Илычский заповедник, где начал обработку при-
везенных с собой материалов, одновременно продолжай полевые
исследования на новом ‘месте. Законченные в начале 1953 г. первые
пять глав настоящей работы, так же как и написанные несколько
позднее главы VI—VIII, составлены только по материалам Лап-
ландского заповедника. Разделы, посвященные вопросам размно-
жения, смертности, возрастному составу и движению численности
популяций, построены на базе материалов, собранных мною в обоих
заповедниках, с использованием картотек и «Летописей природы»
Печоро-Илычского заповедника за прошлые годы. Эти ценнейшие
материалы тщательно собирал с 1938 по 1951 г. В. П. Теплов. Ра-
бота писалась в течение семи лет (1952—1959), чем и объясняется
неоднородность ее состава. Материал оказался так обширен, что
я не мог включить его полностью. По той же причине и литература
использована в меньшей мере, чем хотелось бы. Мы надеемся, что
отмеченные недочеты компенсируются обилием новых оригиналь-
ных материалов.
Материал для работы, как полевой, так и литературный, соби-
рался постепенно в течение 29 лет. За это время я пользовался
консультацией и советами многих зоологов, в числе которых сле-
дует назвать П. А. Мантейфеля, А. А. Насимовича, Г. А. Новикова,
А. Н. Формозова, Б. К. Штегмапа и ныне покойных Г. Г. Доппель-
маира, И. В. Кожанчикова, Г. М. Крепса, А. Н. Промптова и
В. Ю. Фридолина. В сборе материалов, проведении учетов и т. д.
большую поддержку оказал весь персонал Лапландского и Печоро-
Илычского заповедников, особенно А. Б. Васильев, А. Н. Жуков,
Е. П. Кнорре, Т. В. Кошкина, П. С. Мальчевский, А. А. Меженный,
В. С. Поярков, Г. А. Предтеченский, А. С. Сергин, В. П. Теплов,
JO. п. Язан и другие. В устройстве самописцев, работе с иимп и
7
проведении опыта искусственной инкубации яиц большое участие
приняли М. И. Владимирская, 3. И. Кондратьева, Р. Е. Оплеснина
и В. П. Чижевский. Л. А. Портенко, К. А. Юдин и Ю. Г. Гаев лю-
безно просмотрели рукопись и внесли ряд ценных поправок. Всем
этим лицам я искренне благодарен за дружеское отношение и по-
мощь в работе.
В заключение необходимо отметить, что осуществление этой ра-
боты было возможно только в заповеднике. Наши заповедники
являются идеальной базой для проведения всех экологических ра-
бот, требующих длительного времени и соблюдения относительного
постоянства условий. Эту возможность оценили и ею охотно поль-
зуются многие сотрудники высших учебных заведений и научно-
исследовательских институтов. Наиболее плодотворной формой ра-
боты являются все же исследования, сосредоточенные в самих запо-
ведниках и выполняемые силами их постоянных сотрудников. Их
задача состоит в том, чтобы полностью оправдать возможности,
предоставляемые им наличием заповедных территорий. Я не могу
судить о том, в какой мере мне удалось с этой задачей справиться,
но я глубоко убежден в том, что экологические исследования запо-
ведников создадут прочную основу для будущего развития у нас
охотничьего дела, а также других отраслей хозяйства, использую-
щих ресурсы дикой природы.
I. РАСПРОСТРАНЕНИЕ ТЕТЕРЕВИНЫХ ПО ТЕРРИТОРИИ
; МУРМАНСКОЙ ОБЛАСТИ
Как мы уже упоминали, в Мурманской области шире всего рас-
пространена белая куропатка. Опа населяет все пространство от
Печенги на северо-западе до горла Белого моря на юго-востоке
и от морского побережья до верхней границы леса в горах и даже
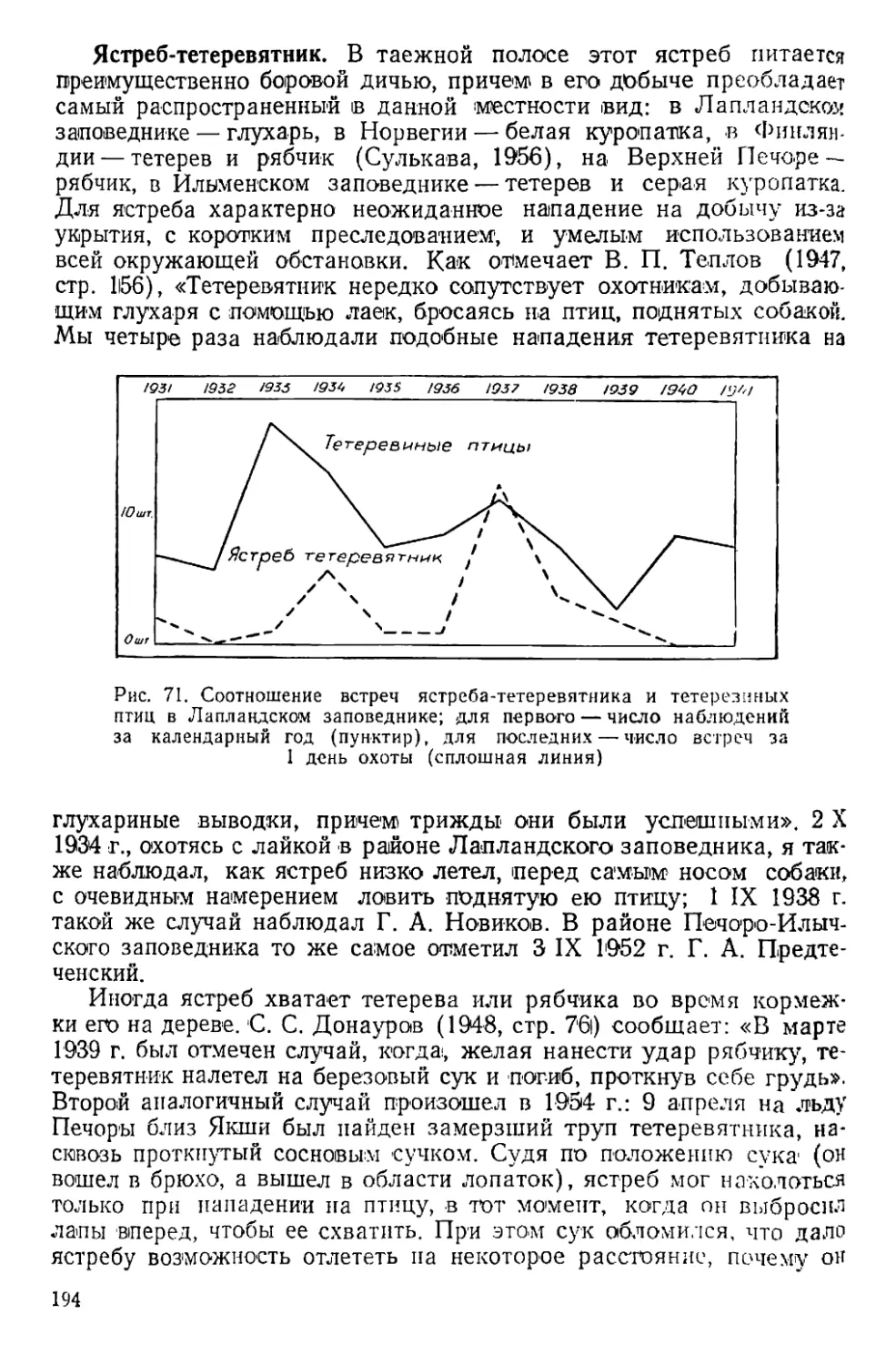
выше, примерно до 600 м абс.
Внешне очень похожая на нее тундряная куропатка ограничена
в своем распространении горными тундрами (выше верхней гра-
ницы леса, проходящей, обычно, на высоте около 380 м абс.) и ка-
менистыми участками равнинной тундры вдоль берега Варенцова
моря. В Лапландском заповеднике этот вид по общей численности
стоит на последнем месте. Тундряные куропатки встречаются не
только на больших скалистых горных массивах, как Хибинские
горы, Чуна-, Монче- и Волчья тундры, но и на невысоких плоских
тундрицах, иногда совсем незначительных по площади. В коллек-
циях Зоологического института АН СССР есть экземпляр тундря-
ной куропатки, добытый на тундрице Ерм-острова 7 VII 1927 г.
Л. О. Белопольским. Эта тундрица, безлесная площадь которой не
превышает нескольких гектаров, отделена от ближайшего горного
массива (Кыма-тундра) расстоянием в 14 км, включая озерный
пролив.
Распространение глухаря в Мурманской области совпадает
с распространением хвойных лесов. Глухарь обычен в бассейне
Нотозера и р. Туломы, по системе р. Колы, по всему бассейну
-оз. Имандры'И р. Умбы- Встречается от на Ловозере и по р. По-
ною, от истоков вниз до пос. Каневки (67°09' с. ш., 39°40' в. д.).
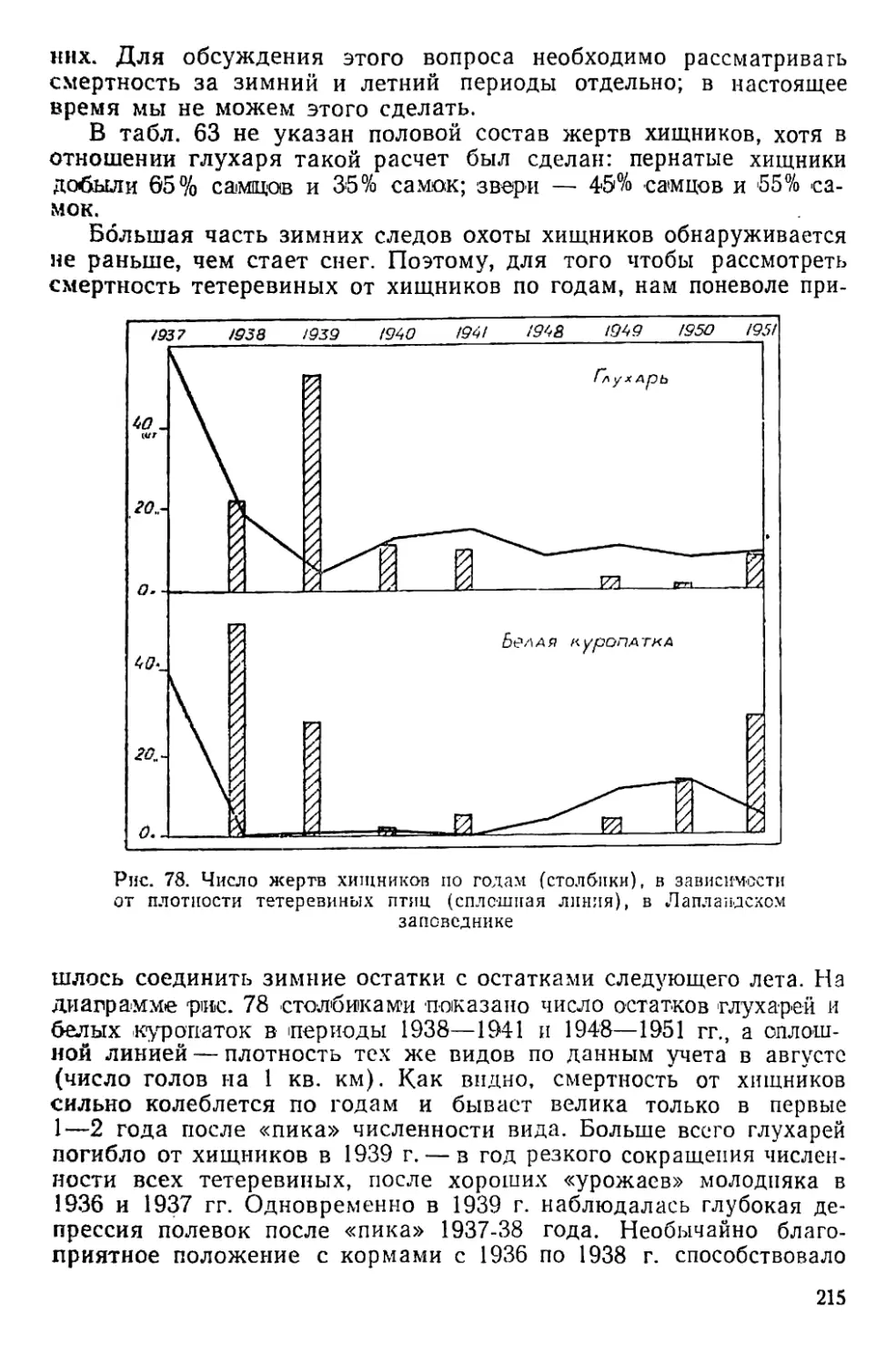
На юго-востоке полуострова он указан для окрестностей с. Пя-
лицы: здесь глухарь, по-видимому, не достигает даже полярного
круга, тогда как на западе, по Кольскому заливу, он поднимался
До 69°04' с. ш. (Семенов-Тян-Шанский, 1938, стр. 223). '
Рябчик в нашей области более редок, чем глухарь, н распро-
странен не так далеко к северу. На Поное он доходит до пос. Крас-
нощелье (67с25' с. ш., 37°00' в. д.; устное сообщение Е. Г. Чер-
нова). По Туломе он доходил почти до устья (68°5СГ с. ш.), но после
9
вырубки лесов вдоль р. Колы и нижнего течения Туломы граница
распространения рябчика и глухаря здесь отступила к югу.
Ареал тетерева, напротив, расширяется. А. Ф. Миддендорф
(1840), а за ним и Ф. Д. Плеске (1880) проводили северную гра-
ницу тетерева по южному берегу оз. Имандры. В первые годы на-
шей работы (1930—1935) тетерева были обыкновенны вдоль ли-
нии Кировской ж. д. к северу до станции Имандра; к западу от
оз. Большой Имандры они попадались значительно реже и токо-
вали только1 поодиночке. В Лапландском заповеднике тетерев
встречался лишь на юго-восточной окраине. Нотозерским саамам
тетерев был неизвестен до 30-х годов: когда 7 V 1935 г. под восточ-
ным склоном Сальной тундры (68°20'с. ш.) был убит один косач —
определил его русский охотник. В восточной части полуострова
Е. Г. Чернов проводит северную границу тетерева от южной оконеч-
ности Умбозера на восток, по водоразделу Поноя с Паной и до
слияния последней с Варзугой, дальше она идет вверх по Варзуге
и Стрельне, выходя к Белому морю у с. Тетрина. В этих преде-
лах тетерев приурочен к долинам рек и местами, например, вокруг
Сергозера, совсем не встречается.
Интересно отметить, что за последние 30 лет численность тете-
рева в Лапландском заповеднике возрастала из года в год. «Удель-
ный вес» тетерева по данным отстрела боровой дичи* близ южной
и восточной границ Лапландского заповедника с 1930 по 1951 г.
был равен (в %):
1930-1933 гг. 1934-1937 гг. 1938-1941 гг. 1948-1951 гг.
1,8 2,2 8,1 11,9
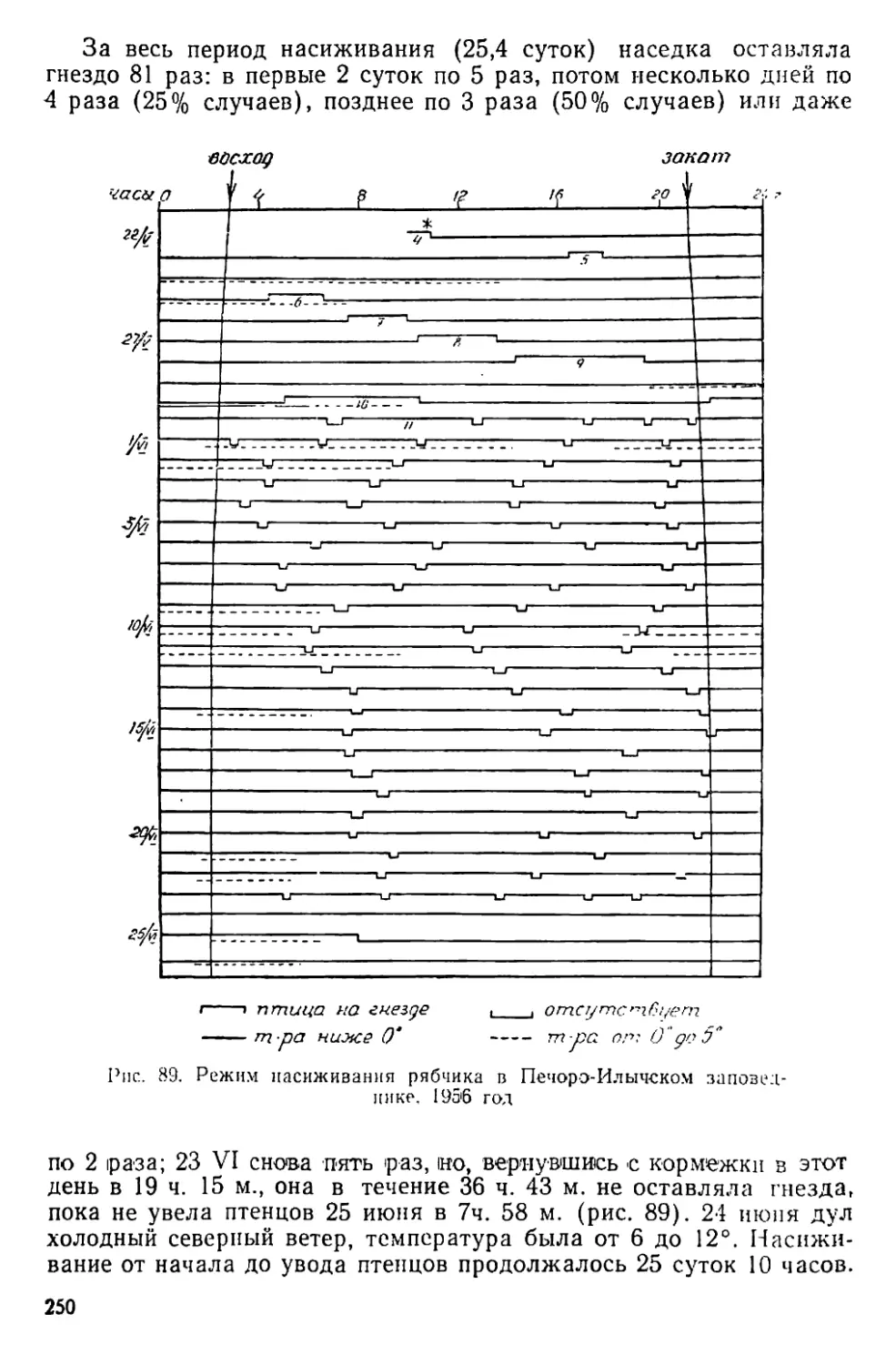
Данные относительного и абсолютного учетов тетеревиных птиц
и просмотр карточек биологических наблюдений по тетереву (осо-
бенно в период токования) подтверждают, что численность этого
вида сильно увеличилась за годы нашей работы в заповеднике;
одновременно тетерев все дальше продвигался на север.
Е. Ф. Бартольд (1930), обследовавший в 1929 г. территорию
намечавшегося тогда заповедника, отметил, что в долине р. Вите
тетерев не встречается. В 1937 г. на оз. Островском, в долине этой
реки был построен кордон. 26 мая 1941 г. наблюдатель охраны
М. С. Тютин писал: «...на льду Островского озера токует косач.
В прошлые весны я ни разу не встречал косачей в этом районе».
16 мая 1942 г. он же писал, что «тетеревов появилась в 1942 г.
масса. Токуют на Островском озере, а также на оз. Вите посере-
дине».
Еще севернее, в окрестностях Мончегорска, выводок тетеревов
впервые был встречен в 1937 г. (сообщение С. Стрелкова и Л. Пир-
мапа). В последние годы тетерев дошел почти до Мурманска:
А. Б. Васильев сообщил, что в 1957 г. он видел тетерева на запад-
ной стороне Кольского залива (68°55' с. ш.).
* Всего 1745 птиц.
10
К сожалению, мы не смогли выяснить положение северной гра-
ницы тетерева в прошлые эпохи: исторические материалы по пти-
цам и костные остатки их гораздо скуднее, чем аналогичные дан-
ные по крупным зверям. Мы полагаем, что быстрое расширение
ареала тетерева в Мурманской области не имело прецедентов в
прошлом; причиной его является вырубка хвойных лесов, замеще-
ние их березовыми и успешное продвижение земледелия на север.
В подтверждение можно указать, что на саамском языке нет сво-
его, оригинального названия тетерева. В районе нашего исследо-
вания распостранены следующие названия птиц:
Международное Русское Саамское
Tetrao urogallus £ глухарь чуухч
Tetrao urogallus $ кбппала куаппель
Lyrurus tetrix ft косач, тетерев кбссач
Lyrurus tetrix $ тетёрка тетёрка
Tetrastes bonasia рябчик, рябок пынг
Lagopus lagopus куропатка, куропать иногда — лесная куропатка (прилаг. «куропачий») рехпь •
Lagopus lagopus (весной) — лякстак
Lagopus lagopus (весной) — коамгынч
Lagopus mutus тупдрянка, горная куропатка кйёрымь
Очевидно, что название тетерева саамы заимствовали с рус-
ского языка, перенеся только ударение на первый слог.
Не все русские охотники зпают и отличают тундряную куро-
патку от обыкновенной белой (транскрипцию «тундреная» мы счи-
таем неправильной, так как при обычном в разговоре переносе уда-
рения на предпоследний слог в нем ясно слышится звук «я»: туид-
рянка, тундряная куропатка). Напротив, тундровые саамы пре-
красно знают эту птицу. От саамов Нотозерского погоста я слы-
шал поговорку-определение: «кйёрымь — чаппись нюн сиёрымь»
(буквально: «тундряпка — черные пос и ноздри»).
О соотношении видов тетеревиных в районе Лапландского запо-
ведника в известной мере можно судить по числу карточек на добы-
тых птиц, использованных для этой работы:
В шт. В %
Глухарь . '807 46
Тетерев 60 1 3
Рябчик . 241 14
Белая куропатка . 550 32
Тундряная куропатка . 87 5
11
II. СРЕДА ОБИТАНИЯ ТЕТЕРЕВИНЫХ
Северная граница леса проходит примерно в 120—130 км к се-
веро-востоку от Чунозера, в районе которого собрана большая
часть нашего материала; в связи с этим и верхняя граница хвой-
ного леса проходит тут невысоко, в среднем не выше 250 м над
самыми низкими отметками окружающей равнины (рис. 1).
Рис. 1. Верхняя граница леса в горах Чуиа-тундры зимой
Лапландские леса своеобразны даже по внешнему виду. Пре-
обладающая порода — ель, именно Picea fennica, по всем призна-
кам более близкая к сибирской Р. obovata, чем к европейской
Р. excelsa. Северные ельники отличаются от южных прежде всего
редким древостоем: в связи с этим самоочищения от сучьев обыч-
но не происходит, и охвоенные ветви спускаются по стволу до са-
мой поверхности земли. Узкие остроконечные кроны елей напоми-
нают шпили готических построек (рис. 2). Рост елей достигает 24,
но обычно не превышает 15—18 м.
В ельниках обязательна примесь березы Betula pubescens (по
новейшей номенклатуре — В. subarctica). В отличие от лесов более
южных широт, на Кольском полуострове береза далеко не дости-
гает роста елей и всегда остается во втором ярусе. У основания
стволов нередка корневая поросль. В небольшом количестве встре-
чаются рябина, козья и двуцветная ивы и можжевельник. Как пра-
вило, древостой ельников очень разновозрастный, и обновляется он
12
постепенно. Старые деревья засыхают на корню или валятся вет-
ром. Валежник гниет и постепенно затягивается моховым покро-
вом. Иногда мох затягивает живые ветви елей, которые потом уко-
реняются и образуют любопытные «коврики» из еловых лапок. Об-
наженную землю можно найти только под корнями валежин или
под густыми ветками вокруг стволов елей. Лес мало затеняет поч-
ву, которая поэтому покрыта сплошным зеленым ковром, состоя-
Рис. 2. Ельник с покровом из зеленых мхов, черники
и вороники
щим, в массе, из вечнозеленых форм. Особенно характерны два
вида зеленых мхов (Pleurozium Schreberi, Hylocomium splendens),
черника * (Vaccinium myrtillus), брусника (Vaccinium vitis idaea),
багульник (Ledum palustre), грушанка (Pirola secunda), два плауна
(Lycopodium annotinum, L. anceps), линиея (Linnaea borealis), во-
роника (Empetrum hermaphrodilum). Летнезеленйе растения по
числу видов многочисленнее вечнозеленых, но далеко уступают им
по массе. Из них в ельниках наиболее постоянны: марьянник (Мс-
lampyrum pratense), луговик (Deschampsia flexuosa), золотая роз-
* Черника осенью теряет лист; однако ее толстые граненые побеги оста-
ются всю зиму зелеными. Как мы увидим, это один из главнейших зеленых
кормов боровой дичи.
13
га (Solidago virga aurea), голубика (Vaccinium uliginosum), швед-
ский дерен (Cornus suecica), папоротничек трехраздельный (Gym-
nocarpium dryopteris), лесная герань (Geranium silvaticum), лесной
хвощ (Equisetum silvaticum), морошка (Rubus chamaemorus). По-
следние виды характеризуют более влажные местообитания.
В сосновых лесах обычна примесь березы и ели. Как и в ель-
никах, деревья в борах ниже ростом, чем в умеренных широтах»
Рис. 3. Бор с чернично-ягельным покровом
стоят они реже и возобновление их происходит исподволь
(рис. 3). Наличие смолистых сушин, нередко обугленных (следы
пожара, прошедшего иногда более сотни лет назад), выворочен-
ных ветром с корнями деревьев («кокоры»), валежника и под-
роста, а также примесь елей и берез — все это делает боры Лап-
ландского заповедника очень живописными и обогащает их есте-
ственными укрытиями. Настоящего '.подлеска в борах не бывает,
попадаются только отдельные деревца козьей ивы (Salix саргеа),
можжевельник (Juniperus sibirica) и маленькие шарообразные ку-
сты ивы Salix xerophila. Основу напочвенного покрова состав-
ляет либо ягель (Cladonia alpestris), либо зеленый мох (Pleuro-
14
zium Schreberi); чаще они комбинируются, образуя пятнистый мо-
заичный покров. Кустарничково-травяной ярус состоит из черники,
брусники, вороники, иногда вереска (Calluna vulgaris) и толок-
нянки (Arctostaphylos uva ursi), марьянника, луговика — в общем
почти из тех же видов, что и в ельниках. <
В августе и сентябре в лесах бывает много грибов, особенно
красных, березовиков, моховиков и различных сырбежек. Хороший
урожай грибов бывает обычно через год. Грибы служат кормом
многим животным: их едят северные олени, белки, лемминги и по-
левки, глухари и кукши, слизни и, наконец, личинки многочисленных
двукрылых *, от которых грибы становятся червивыми.
Из насекомых в лесах севера множество муравьев: муравейники
с годами достигают высоты 1,5 м при диаметре основания до 5 м.
Бабочек и их гусениц в тайге мало; большее значение имеют ли-
чинки пилильщиков (Tenthnedinidae), науки и мелкие саранчовые.
В середине лета леса и болота изобилуют кровососущими насеко-
мыми— комарами (преимущественно относящимися к роду Aedes),
мошками-кровососками (Simulium sp.) и мокрецом (Culicoides sp.).
Указания старых авторов, что птенцы белой куропатки будто бы
питаются комарами или личинками, как мы убедились, не со-
ответствуют действительности. В жизни птиц эти кровососущие на-
секомые играют только отрицательную роль.
Благодаря прохладному лету и недостаточному испарению ме-
стность богата водой. Грунтовые воды находятся очень неглубоко,
ручейки попадаются часто. Рельеф местности сильно расчленен,
и все понижения более или менее заболочены. Очень распростра-
нены осоковые болота с карликовой березкой (Betula папа). Не-
редко в заболоченной лощине образуется сложная сеть более или
менее поперечных перемычек, играющих роль плотин и задержи-
вающих сток воды, отчего поверхность болота становится ступенча-
той. Эти «плотины» покрыты зарослью карликовой березки, осокой
(Carex lasiocarpa), молинией (Molinia coerulea), завязником (Ро-
tentilla erecta). Между «плотинами» в мокрое лето — более или ме-
нее открытая водная поверхность, в сухое — растрескавшийся ило-
ватый грунт;
Сфагновые болота обычно представлены комплексным типом.
Торфяные бугры покрыты сфагновыми мхами, нередко ягелями,
карликовой березкой, багульником, голубикой, вороникой, андро-
медой (Andromeda polifolia), вереском, морошкой, мелкоплодной
клюквой (Oxycoccos microcarpa) и пушицей влагалищной (Eriopho-
rum vaginatum). На буграх растут редко разбросанные сосны, вы-
сотой до 5—8 м, с жидкой, не дающей тени кроной. В «ямах» между
буграми, на топком иловато-торфяном грунту растут осоки (Carex
irrigua, С. rariflora, С. pauciflora и другие), пушина узколистная
* По любезному указанию А. А. Штакельберга они принадлежат к семей-
ствам Fungivoridae, Clythiidae и Helomyzidae.
15
(Eriophorum angustifolium), шейхцерия (Scheuchzeria palustris)
и некоторые мхи.
Летом белые куропатки держатся преимущественно на болотах,
а зимой глухари охотно кормятся на низких болотных соснах.
Окраины болот и влажных лощин — излюбленное место обитания
всех видов боровой дичи. Глухарь, тетерев, рябчик и белая куро-
патка хорошо используют кормовые ресурсы и все природные осо-
бенности тайги. Даже такие детали ландшафта, как наличие в лесу
выворотов, молодых или угнетенных елочек, укромных «беседок»
под большими елями, играют определенную роль в жизни этих
птиц.
Горные тундры занимают около 28% всей площади Лапланд-
ского заповедника, образуя несколько массивов, самый значитель-
ный из которых — Чуна-тундра — расположен в восточной части
заповедника и на севере смыкается с Монче-тундрой. Оба хребта
каменисты, расчленены ущельями, а вершины их достигают 900—
1 100 м абс. высоты. В западной части заповедника находится не-
сколько массивов меньшего размера; очертания их сглаженные,
округлые, а высшие точки большею частью не превышают 600 м.
В нижних частях альпийского пояса гор на сухих участках об-
щий фон дают ягельники, с низкой, почти не возвышающейся над
ягелем карликовой березкой и кустарничками — вороникой, брус-
никой, голубикой, миниатюрной азалией (Loiseleuria procumbens),
«гром'-ягодой» * (Arctons alpina). Характерны также'.плауны (Lyco-
podium alpinum, L. selago), осоки (Carex Bigelowii, C. vaginata),
ситник (Juncus trifidus), куропачья трава (Dryas octopetala, D. pun-
ctata), альпийская ястребинка (Hieracium alpinum) и т. д. Увлаж-
ненные долины покрыты зарослями различных видов ив и карли-
ковой березки с купальницей, лесной геранью, речным гравила-
том, грушанками, медвежьей дудкой, бодяком и другим разно-
травьем.
Выше по склонам растительный покров редеет, заменяясь рос-
сыпями камней и выходами скал, поверхность которых покрыта
только накипными лишайниками. Снежные сумёты сохраняются
здесь до середины лета, а иногда и до выпадения нового снега.
Для влажных площадок у снежников характерны карликовые виды
ив (Salix herbacea, S. polaris), кассиопа (Cassiope hypnoides), сиб-
бальдия (Sibbaldia procumbens), гречишник (Polygonum vivipa-
rum), полярный щавель с почковидными листьями (Oxyria digyna),
сердечник (Cardamine bellidifolia) и другие виды.
Тундряная куропатка — единственная оседлая птица наших гор-
ных тундр. Помет ее мы встречали даже на самых вершинах голь-
* Это растение не имеет народного русского названия, а книжное «аль-
пийская толокнянка» — неудачно/мякоть ягоды толокнянки в самом деле по-
хожа на толокно, а ягоды «альпийской толокнянки» такие же сочные и чер-
ные, как ягоды вороники, только значительно крупнее. Предложенное нами
в 1937 г. название «гром-ягода» — буквальный перевод саамского «тиярмес-
мубрьх»; оно начинает прививаться в научной литературе (Фолитарек, 1939)
16
цов. Другие виды тетеревиных только залетают в горы. В конце
зимы на первых проталинах и выдувах выше границы леса (рис. 4)
часто встречаются белые куропатки, а иногда тетерева и даже глу-
хари.
Климат нашего района характеризуется коротким и довольно
прохладным летом; заморозки и даже снегопады возможны почти
во все месяцы (правда, в июле заморозки очень редки, а выпаде-
ние снега наблюдалось только на горных тундрах). Солнце со-
Рис. 4. Местообитание тундряных куропаток на западном склоне
Чуна-тундры (апрель)
всем не заходит в течение приблизительно 48 суток, а вместе с по-
рой «белых ночей» весь период непрерывного света достигает
102 дней. В то время, когда стоят светлые ночи, росы почти не бы-
вает, потому что суточная амплитуда температуры уменьшается.
Отсутствие росы, широкое распространение ягельников, захламлен-
ность лесов и нередкие летом грозы способствуют возникновению
и распространению лесных пожаров. Значительная часть лесной
площади Мурманской области представлена в настоящее время га-
рями разных возрастов.
Иногда уже в конце августа при прохождении холодного фрон-
та циклона горы покрываются снегом. В начале октября нормально
в горах устанавливается снежный покров, а в лесном поясе в это
время идет первый снег. В последних числах октября (крайние
даты — 8 X и 16 XI) снежный покров устанавливается здесь на
зиму. Приблизительно в это же время замерзают мелкие озера и
тихие участки рек. Большие и глубокие озера становятся поздно,
нормально в середине ноября, изредка даже в декабре (1938 г.).
2—3037
17
Зимой преобладает пасмурная погода, часто с ветром. Для Мур-
манской области характерно, что в это время северные ветры при-
носят сравнительно теплый воздух с поверхности океана. В это
время нередки оттепели- Январь in февраль сравнительно редко про-
ходят совсем без оттепелей: максим1альн!а1Я температура месяца
чаще бывает выше 0°, чем ниже 0°. Сильные ,морозы наступают
только при ясной безветренной погоде. В таких случаях, как пра вило,
наблюдается температурная инверсия и на горных тундрах бывает
теплее, чем в лесной зоне.
Около времени зимнего солнцестояния солнце на протяжении
20—30 суток совсем не восходит. Время первого появления солнца
в разные годы варьирует между 1—5 января, в зависимости от ве-
личины рефракции в атмосфере. Период «зимней ночи» не так мра-
чен, как может показаться, потому что в это время утренние сумер-
ки сливаются с вечерними и продолжаются в общей сложности 6 ча-
сов. К тому же снежный покров хорошо отражает свет; когда небо
пасмурно — отсутствия солнца не чувствуется.
В середине зимы преобладает сплошная низкая облачность, ви-
димость плохая, горы обычно окутаны облаками. Самый холодный
месяц — февраль или январь; впрочем, минимальная температура
редко и далеко не каждый год достигает —40°. От изморози и сне-
га на кронах деревьев лес становится как будто седым. Накопле-
ние «кухты» на ветвях продолжается обыкновенно до февраля или
марта, но нередко оттепель или сильный ветер очищают лес от
кухты еще в середине зимы.
Март по температуре вполне зимний месяц; правда, день в это
время длинный и настолько светлый, что с 1 марта снег начинает
на солнце таять, образуя под крышами сосульки. Снежный покров
продолжает нарастать и обычно достигает годового максимума
в последней декаде марта. До этого времени он остается с поверх-
ности мягким, а на глубине уже в январе становится зернистым и
сыпучим.
В апреле наступает перелом на весну. Снег дает осадку, поверх-
ность его уплотняется (образуется наст); сначала на склонах гор,
на берегах рек и озер, позднее—на буграх болот, а потом и вокруг
стволов деревьев появляются небольшие проталины. В реках вода
начинает прибывать, промывая местами лед. Снегопады и метели
становятся реже. Яркие солнечные дни, короткие ночи, обычно мяг-
кая температура, хороший лыжный путь, чистые очертания гор
в сухом прозрачном воздухе—все это делает апрель лучшей по-
рой года. В утренние и полуденные часы нижние слои воздуха
сильно нагреваются солнцем и отраженной от поверхности снега
радиацией, отчего в ясные дни влажность сильно падает, иногда
до 20%; при этом снег и изморозь понемногу «растворяются» в воз-
духе, без таяния (сублимация). По этой же причине следы живот-
ных на снегу быстро стареют и деформируются, а шишки хвойных,
семенные сережки ольх и берез рассыхаются и освобождают се-
мена, цветочные почки козьей ивы сбрасывают чешуйки и обра-
18
зуют пушистые «барашки». В апреле появляются первые проталины
на обдуваемых ветром местах. На снегу появляются насекомые,
чаще выбегают на поверхность полевки.
С 1 мая наступают белые ночи и температура становится поло-
жительной. Начинается общее таяние снега — как на горных скло-
нах, так и на равнине. К 20 мая нормально уже больше половины
поверхности в лесу освобождается от снега, а 5 июня этот момент
наступает и на горных тундрах. Повсюду шумят ручьи; поверх-
ность льда, покрывшаяся в конце апреля слоем воды, обсыхает на
короткое время перед вскрытием озер; у берегов появляются за-
береги.
Около 20 мая распускаются первые цветы — сережки серой
ольхи и козьей ивы, колоски пушицы влагалищной и мелкие цве-
точки вороники. Пробуждаются разные насекомые — муравьи,
шмели, крупные комары — теобальдия. В мае прилетает большин-
ство перелетных птиц и некоторые начинают гнездиться.
Июнь — месяц незаходящего солнца. В первых числах июня
вскрываются озера, исчезают остатки снега (в лесной зоне), зеле-
неют березы и другие лиственные деревья. В середине месяца за-
цветают черника, морошка, андромеда и другие растения; в это
время обычно прекращаются заморозки. В двадцатых числах начи-
нается нудная «комарная пора».
В этих высоких широтах случается, что в июне в зеленом уже
лесу начинается снежная буря, выпадает слой снега (иногда боль-
ше 10 см) и лежит несколько суток, пока северный ветер не стих-
нет и погода не переменится к лучшему. За 2© лет, с 1930 по 194И и
с 1945 по 19511, только шесть весен прошли без июньских снегопа-
дов (1936, 1937, 1938, 1939, 1948 и 1950). Сильная снежная буря
с северным ветром была 2 июня 1935 года. В 1940 г., когда березы
полностью зазеленели и зацвели, 9 июня внезапно пошел снег, вы-
пал слоем до 10 см и лежал до 12 VI. Самое позднее выпадение
снега было в 1941 г. — с 10 по 16 июня. В 1949 г. снег выпал
4 июня и лежал до 6-го, а в 1951 г. 1 июня выпало не менее 10 см
снега, правда, на следующий день этот снег стаял. На вершинах
гор снегопады бывают еще позже. В 1949 г. снег выпал даже
14 июля, а в 1958 г.— 13 июля.
Заморозки у поверхности почвы в июне бывают почти каждый
год (только в 1936 г. последний утренник был 30 мая), но они
кратковременны и повреждают, главным образом, цветы морошки:
поэтому она редко дает хороший урожай. В 1948 г. заморозок был
4 VII, а в 1941 г. —даже 17 VII.
Северный ветер, по местному «моряпка», летом, как правило,
приносит с собой ненастную погоду на несколько дней. Типичный
случай «морянки» был 5—8 июля 1950 г.; дождь при сильном ле-
денящем северо-восточном ветре шел без перерыва с 19 час.
6 VII до 9 час. 8 VII; температура воздуха в это время была около
4—5°. В лесу буря повалила множество деревьев. В колониях кра-
чек на островках погибли от холода почти все птенцы.
2
19
Летом даже в хорошую погоду редко бывает жарко: абсолют-
ный максимум температуры за год обычно бывает между 25 и 30°.
Вот в общих чертах та обстановка, в которой протекает жизнь
тетеревиных птиц в исследованной нами местности.
III. ОБРАЗ ЖИЗНИ ТЕТЕРЕВИНЫХ ПТИЦ ПО СЕЗОНАМ
По плану настоящей работы каждая глава излагает одну из
сторон экологии нескольких видов тетеревиных птиц параллельно.
При такой системе нельзя было обойтись без предварительного
Рис. 5. Ветки сосны, общипанные кормившимся на ней
глухарем
очерка образа жизни каждого вида в отдельности. Во избежание
частых повторений, эти очерки, по возможности, сокращены.
Глухарь. В самый холодный период зимы — в январе и фев-
рале— большую часть времени птица проводит, зарывшись в снег.
Глухарь выходит на поверхность только для того, чтобы наполнить
зоб сосновой или можжевеловой хвоей, составляющей в это время
20
года его единственную пищу. Зимой самцы и самки живут раз-
дельно, поодиночке или, чаще, небольшими стайками от двух до
шести голов, иногда больше. Самцы всю зиму предпочитают опи-
санные выше верховые сфагновые болота с низкими, редко разбро-
санными соснами, или же сосновые редколесья по склонам возвы-
шенностей. Глухарки встречаются в борах с более частым древо-
стоем. Изредка глухари (самцы или самки) встречаются среди
обширных еловых массивов, где сосны совсем нет. Как показали
анализы зобов убитых птиц, в этих случаях они питаются хвоей
можжевельника и ели.
Во время непрерывной полярной ночи глухарь кормится два
раза в сутки — утром и вечером; если погода мягкая и тихая, он
остается на деревьях всю светлую часть суток, но чаще среди дня
прячется в снег или садится под елку, под прикрытие ее ветвей. Под
соснами, на которых птица кормилась, видна осыпавшаяся хвоя,
а если снег плотный, то и помет. Если глухаря никто не тревожит,
он долго- живет на одном) месте, жируя на одних и тех же
соснах, после чего убыль хвои становится легко заметной на глаз
(рис. 5)-
В 1931 г. я поднял в одном и том же месте 3 глухарок 22 марта
и 4 апреля; в 1941 г. один глухарь самец наблюдался йа болотце
недалеко от Управления заповедника на кормежке последователь-
но 6, 8 и 10 января, 12 февраля и 8 марта — на протяжении более
двух месяцев. На месте такого «жительства» глухаря можно на-
считать с десяток и более лунок различной свежести; так, напри-
мер, 17 III 1941 г. А. Б. Васильев, подняв из снега 2 глухарок, на-
шел на этом месте 32 лунки.
Зимой глухарь в полном смысле слова оседлая птица. Если бы
удалось провести хроном»етраж его времяпрепровождения в течение
нескольких суток, оказалось бы, что около 80% времени, если не
больше, птица проводит, зарывшись в толщу снега, занятая только
пищеварением. У глухаря и прочих тетеревиных больше, чем у дру-
гих птиц, проявляется зимнее понижение активности, сходное с та-
ким же понижением жизнедеятельности лося (Семенов-Тян-Шан-
ский, 1948). Такой путь приспособления к зимним условиям суще-
ствования— отчасти аналогичен явлению зимней спячки.
Отдельные наблюдения показывают, что в сильные морозы глу-
хари избегают взлетать на сосны, а предпочитают бродить по сне-
гу, обклевывая побеги можжевельника (27 XII 1930 г. при темпе-
ратуре воздуха —33,5°), или взлетают на низкие сосенки, переходя
от одной к другой пешком (21—23 I 1933 г. при температуре —36,5°
и ниже).
В оттепель, когда снег сырой, глухарь ночует на дереве, а в слу-
чае наста он не пытается пробивать его и спит на поверхности сне-
га, иногда под елкой, иногда открыто.
В марте глухари заметно оживляются. Как только снег начнет
таять и испаряться на солнце, обнажая корни вывороченных ветром
деревьев, глухари наведываются в такие места, разыскивая и за-
21
глатывая камешки, застрявшие в переплете корней (см. табл. 1).
12 III 1939 г. А. Пономарев нашел, что по краю обтаявшего берега
ручья снег был истоптан глухарями, клевавшими тут гальку.
Вскоре глухари начинают «чертить» крыльями: утром в сол-
нечную погоду они бродят и кружатся по снегу, опустив крылья
так, что концы маховых бороздят снег. В это время глухари еще
Рис. 6. Взрыв снежной пыли при вылете глухаря из
лунки
не поют и даже не поднимают хвост веером; характерные «набро-
ды» с бороздами не приурочены обязательно к токовищам.
Весной у петухов проявляется склонность путешествовать. В ап-
реле следы их попадались нам даже на горных тундрах. 23 IV
1931 г. я нашел след глухаря на западном склоне Чуна-тундры,
метров на 200 (по вертикали) выше границы леса; 17 V 1935 г. на
южном склоне тундры был встречен совершенно свежий след глу-
харя, поднимавшегося вверх по склону; 20 IV 1940 г. на западном
склоне Чуна-тундры (абс. выс. 675 м) найдены остатки глухаря,
добытого здесь каким-то хищником.
В конце апреля (27 IV в среднем за 15 лет) глухари начинают
токовать. В это время снег тает днем и подмерзает ночью; на про-
22
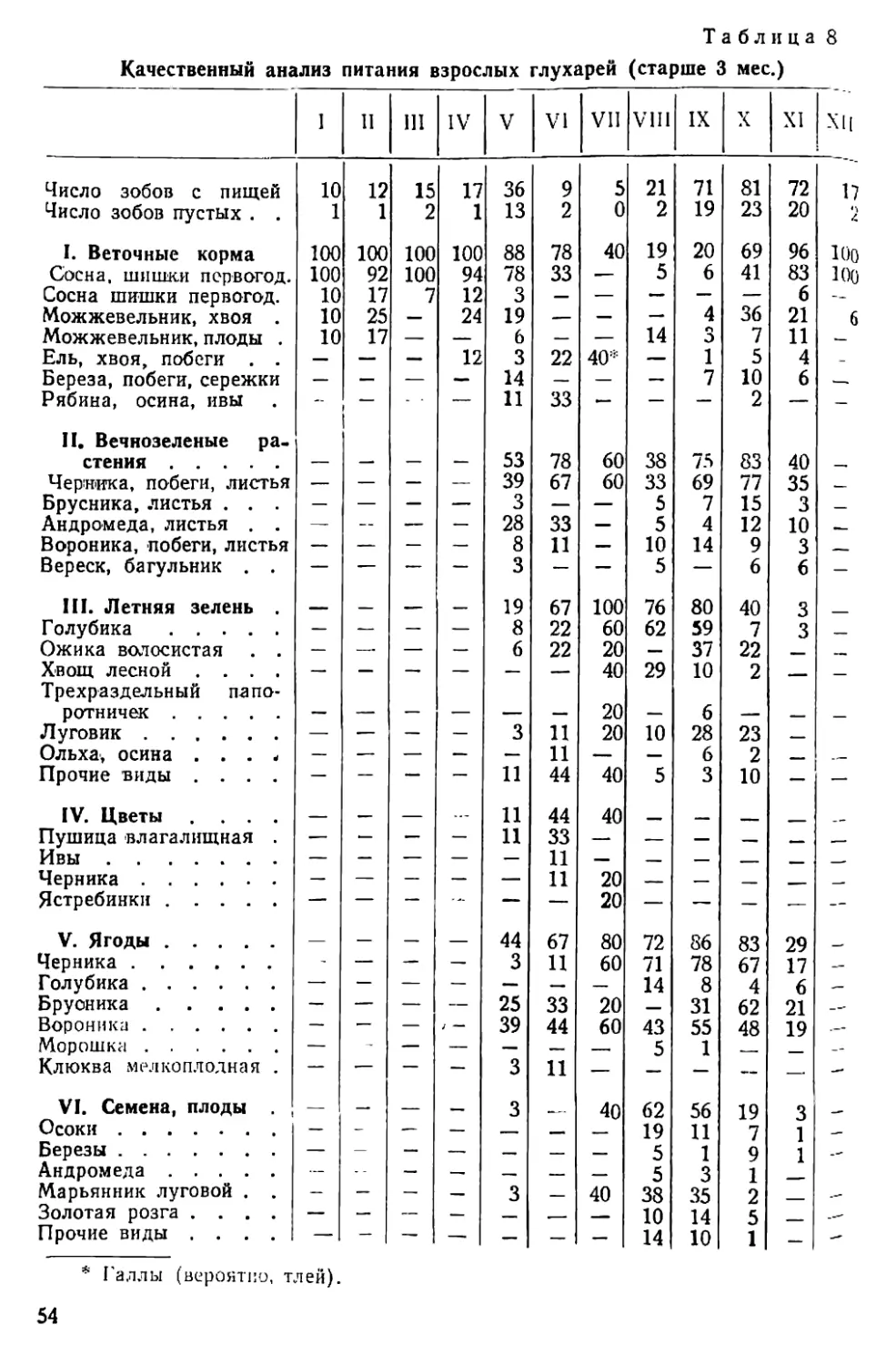
Таблица 1
Периодические явления у глухаря
Год Первые поиски гальки Начало чир- канья крылья- ми Начало токован. (песня) Послед- нее токо- вание Первый выводок Вылеты на гальку
первый последи.
1931 15 IV 28 IV 31 V 18 VI 18 IX 4 XI
1932 20 III 15 III 16 IV 9 VI 1 VII 3 X 1 XI
1933 — 22 ill 8 V 4 VI 6 VII 27 IX 29 X
1934 — 23 III 16 IV 6 VI 25 VI 8 IX 7 XI
1935 — 18 III 14 V 7 VI — 25 IX 1 XI
1936 4 IV 41V 201V 20 V 20 VI 23 IX 22 X
1937 — до 3 1V 24 1V — 19 VI — 8 XI
1938 — 10 Ill 27 IV — 7 VII 26 IX 2 XI
1939 12 III 11IV — — — —
1940 — 8 111 4 V — 17 VI — —
1941 — 25 HI 2 V 20 VI — — —
1942 14 IV — — — — —
1943 — — — — — —
1944 — — — — — — —
1945 — — 16 IV — 28 VI — —
1946 — — -— —- — — —
1947 — — 15 IV — — —
1948 10 IV 28 IV — — — —
1949 — 10 III 28 IV — 24 VI — —
1950 23 III 24 III — — 21 VI — —
1951 — 19 HI 9 V 25 V 29 VI — —
Среднее: 22 III 26 III 27 IV 4 VI 25 VI 23 IX 2 XI
талинах становится доступным подножный корм, но, в основном,
глухарь кормится все еще хвоей. В связи с этим преобладающая
древесная порода на токовищах — сосна; характерен редкий разно-
возрастный древостой с наличием сушин и примесью ели и березы.
Иногда ток находится на склоне тундры или большой Бараки (ме-
стное название возвышенности, облесенной до самой вершины; тер-
мин саамского происхождения), иногда на холмах или пригорках
у края сфагнового болота или на самом болоте. Возможно, что
глухарей привлекают на такие места проталины, рано появляю-
щиеся на местах с расчлененным рельефом. В частом однородном
бору на равнине глухари не токуют.
Если глухарей не преследуют, они токуют у самых домов, почти
на усадьбе. В первоначальном поселке Лапландского заповедника
(так называемая «старая база», 1930—1939 гг.) мы нередко слы-
шали песню глухаря с крыльца дома, несмотря на то, что в поселке
днем было достаточно шумно (колка дров, громкий говор, лай со-
бак). На месте «новой базы», построенной в 1936—1939 гг., раньше
23
глухари не токовали, но в 1951 г. в 7г км от усадьбы наметился
небольшой ток. В предшествующие годы отмечено несколько слу-
чаев, когда глухарь пел на самой усадьбе заповедника. Подобное
маленькое токовище было и у кордона Нявка: 29 V 1941 г. наблю-
датель охраны А. С. Сергин записал, что глухарь токовал на земле,
в 15 шагах от собак, сидевших на цепи и лаявших.
В районе Лапландского заповедника тока невелики и на них
собирается немного птиц: мы наблюдали, самое большее, 8—12 пе-
тухов, а обычно меньше, от 2 до 5. При плохой погоде (особенно
неблагоприятны ветер и метель) глухарки не вылетают па ток, и
в этих случаях петухи почти совсем не поют, а сидят на деревьях,
изредка пробуя «тёкать».
Токование обнимает весь период таяния и заканчивается, когда
снег в лесу исчезнет, а лиственные породы деревьев зазеленеют.
В это время стоят белые ночи. Глухари слетаются на ток, как толь-
ко начнет смеркаться. Они начинают петь между 21 и 23 час. и кон-
чают между 2 и 3 ч. 30 м. утра *. После этого они еще некоторое
время сидят на току, а с наступлением полного дня перелетают на
кормежку. Иногда глухари поют всю ночь напролет, иногда около
полуночи наблюдается небольшой перерыв. В начале токов глу-
харь иногда поет и днем, на солнце (28 IV 1931 г. до 11 час. утра).
Петухи начинают петь, сидя на деревьях. Во время перерывов
они понемногу кормятся хвоей. Входя в азарт, они спускаются на
землю и поют, бродя по проталинам или по снегу. В момент «глу-
хой песни» глухарь иногда подпрыгивает, с шумом взмахивая
крыльями. В Лапландском) заповеднике драк между петухами нам
ни разу не пришлось наблюдать и ни на одном из убитых глухарей
не было замечено ран или шрамов от боев с соперниками; в Печо-
ро-Илычском, напротив, такие случаи были нередки. Активность
петухов на току всецело зависит от поведения глухарок: чем их
больше и чем они крикливее, тем азартнее поют глухари. В начале
токов токуют только вполне взрослые птицы; позднее, примерно
с половины мая, поют и молодые самцы по первому году, которых
не трудно отличить на слух, потому что песня их еще не выработа-
лась и не приобрела необходимой отчетливости. При ружейной
охоте на токах их добывают мало и преимущественно в такие ночи,
когда стариков па току нет или они поют вяло. В районе Лапланд-
ского заповедника из 52 петухов, убитых на току, молодых было
только 9 (17<>/о), причем два из них, как отмечено на карточках,
не пели. На Верхней Печоре из 82 добытых на токах глухарей было
6 молодых (7%); из них трое заведомо не пели.
В конце мая глухарки уже оплодотворены, несутся и перестают
посещать тока. Мы не будем останавливаться на гнездовании, так
как этот вопрос рассмотрен ниже. Заметим только, что гнезда глу-
харок всегда расположены недалеко от токовища. Поэтому распо-
ложение токовищ в конечном итоге сказывается и на размещении
* Здесь и дальше указывается местное среднее солнечное время.
24
выводков. Первые выводки глухарей встречаются, в среднем,
27 июня (см. табл. 1).
В середине лета глухари редко встречаются в бору. Как вывод-
ки, так и взрослые петухи предпочитают ельники, но не сплошные
массивы елового леса, а скорее лощины среди боров, занятые ель-
ником, долины рек и ручьев, переходный лес у окраин болот. Изме-
нение мест обитания объясняется тем, что во время токов глухари
Рис. 7. Глухарка, слетающая с дерева
переключаются с древесного питания на подножный корм. Весь
бесснежный период года они ведут наземный образ жизни и только
спугнутые садятся на дерево. По-видимому, иногда они и спят на
дереве. Однажды летом я наблюдал одинокую глухарку, сидевшую
открыто на сушине среди старой гари: создалось впечатление, что
там она искала защиты от кровососущих насекомых. В случае силь-
ного дождя, а осенью — при выпадении мокрого снега глухарь пря-
чется под елку.
Размещение глухарей летом зависит не столько от распределе-
ния кормов, сколько от наличия необходимых им в этот период за-
щитных условий: чистые боры не обеспечивают укрытием ни вывод-
25
ков, ни взрослых птиц. В случае опасности маленькие глухарята за-
таиваются в траве, а матка, сгорбившись и опустив крылья, бежит
и перепархивает, стараясь отвлечь за собою врага. В возрасте 2—
3 недель глухарята летят сколько сил хватит, а потом садятся
в траву и затаиваются, не давая следа. Практика показывает, что
после такого маневра собака редко находит цыпленка. 11 VII
1938 г. я поднял выводок на берегу р. Чуны: один птенец полетел
Рис. 8. Глухарь подходит к «порхалищу»
через реку, но израсходовал силы раньше, чем добрался до берега,
упал в воду и остаток пути проплыл.
Летом глухарята страдают от кровососущих насекомых и пухо-
едов. В зобах птенцов нередко можно встретить одиночных кома-
ров, очевидно, пойманных на себе. Птичья кровососка (см. ниже,
стр. 1'83) — довольно крупная, хорошо бегающая, плоская, зеле-
новато-серая муха — часто попадается на птицах, добытых после
25 июля; пухоеды встречаются круглый год. Птицы борются с ними,
купаясь в сухой пыли. Свежие «купалки», или «порхалища», глуха-
рей с оброненными перышками и другими признаками птицы, мож-
но встретить, как только сойдет снег и обсохнет поверхность земли:
6
первое наблюдение — 31 V 1949 г., последнее — 24 IX 1949 г. В опе-
рении убитых летом птиц легко обнаружить песок.
Глухари купаются в песке под свежими выворотами (пока об-
наженный грунт не зарастет), на торных оленьих тропах, иногда на
кострищах, крышах промысловых лесных избушек (такие крыши
строятся из плах и засыпаются землей или песком), даже в насы-
пях песка перед челом медвежьих берлог (наблюдения 29 VI
1935 г. и 22 VII 1951 г.). На вершине Бабьей горы есть заброшен-
Рис. 9. Глухарь купается в песке
Снимки рис. 8 и 9 сделаны в Печор о-Илы чеком заповеднике аппаратом
«Любитель* с приспособлением для самосъемки
ная избушка. 24 VII 1951 г. лаборант Т. К- Рыбина, ничего не подо-
зревая, подошла к избушке, и внезапно из дверного проема на-
встречу ей вылетел взрослый глухарь. На месте очага в избе ока-
залось много глухариного помета: птица не раз купалась там в зо-
ле. Глухари купаются также в сухой древесной трухе от сгнивших
валежин.
В июле молодых петушков можно отличить от самочек по опе-
рению шеи, которое у петушков нейтрально-серое с мелкими попе-
речными пестринами, а у курочек — рябое, со сравнительно круп-
ными охристыми пестринами. Около 10 августа на шее и зобу пе-
тушков пробиваются первые черные перья. Линька продолжается
1 месяц и заканчивается в середине сентября. Одновременно
с линькой в петушье перо молодые самцы отделяются от вывод-
ков. Самочки остаются с маткой: возможно, что те стайки глуха-
рок, которые мы встречаем зимой, представляют остатки выводков.
27
В осенней жизни глухарей характерны вылеты на берега за
галькой. В сущности, для пополнения запаса камешков в желудке
глухарям нет необходимости выходить на пляжи по берегам озер,
потому что на Кольском полуострове, где почва состоит из плохо
отсортированного моренного материала, гальку можно найти под
любым выворотом или на другом обнажении грунта. Заметим, что
Рис. 10. Глухарка в месячном возрасте
(Печоро-Илычский заповедник)
белая куропатка и рябчик, в желудках которых также есть гастро-
литы, никогда не вылетают «на гальку».
Глухари вылетают из леса на плоские намывные берега озер
еще в утренние сумерки и несколько часов бродят по берегу. Про-
езжая на лодке в тихую погоду, можно наблюдать на открытом
берегу птиц, иногда до 9 штук сразу (18 X 1937 г., И. Яницкий).
В эту пору днем можно заметить, что глухари концентрируются пре-
имущественно вдоль берегов,—в- «сузёмке» (лесном массиве), по-
дальше от берега их и с собакой не найдешь. Охотники полагают,
что глухари перекочевывают на время к источникам гальки, ио
может быть и обратное: глухари появляются на берегах в связи
с происходящими местными миграциями.
28
Осеннее токование глухаря сам я наблюдал только один раз.
20 IX 1935 г. в 8 час. утра.
В конце октября или начале ноября петухи собираются стая-
ми, садятся открыто на вершины деревьев и становятся очень
«строги1М1и». Глухарь, поднятый летом или зимой, как правило, не
летит далеко и редко поднимается выше деревьев. В октябре —
Рис. П. Глухаренок самец в месячном возрасте.
Снимки рис. 10 и 11: сделаны в один день; обратите
внимание на разницу в позах
ноябре, после окончания линьки, глухари находятся в расцвете
сил и охотно летают высоко над землей, по-видимому, без всякой
необходимости. Иногда удается наблюдать форменный перелет:
так, 4 XI 1931 г. я видел стаю штук в 20 самцов и самок, летевшую
высоко над лесом на северо-запад; 13 XII 1934 г. стая глухарей про-
летела высоко над лесом на северо-восток. Одиночные глухари
осенью часто перелетают через озера шириной 1 км и больше,
а иногда летят над горной тундрой на такой высоте, что трудно
даже сообразить, что это за птица, особенно, если глухарь несется
не шевеля крыльями, так, что ветер свистит в перьях. Однажды,
29
12 XI 1937 г., мы застрелили молодого глухаря, ходившего по от-
крытой тундре и кормившегося побегами можжевельника.
В октябре глухари переходят с летних кормов на зимние, древес-
ные; в это же время у них заканчивается линька, продолжавшаяся
5 месяцев. Эти факты говорят о том, что в октябре в жизни глухаря
происходит такой же перелом, как и в марте — апреле. Важно от-
метить, что в оба переходных периода глухари переселяются из
одних стаций в другие; популяция при этом частично перемеши-
вается.
Вопрос о том, на какое расстояние перелетают глухари, можно
решить только кольцеванием: имеющиеся данные говорят о том,
что расстояние это невелико. Колхозник оленевод А. Герасимов
рассказал нам, что летом 1934 г. он поймал глухаренка близ де-
ревни Падун (по р. Туломе) и выпустил его, привязав к ноге крас-
ную тряпочку; в конце зимы глухарка с тряпочкой была убита
почти на том же месте.
После выпадения снега посещение галечников прекращается.
Глухари не раскапывают снег в поисках пищи и берут только то,
что находится на виду. По этой причине они переходят полностью
на древесный корм раньше, чем другие представители семейства
тетеревиных (см. следующую главу).
Пока тепло, глухари нормально спят на дереве. 15 VIII 1950 г.
в 22 ч. 45 м. я застрелил глухарку, ночевавшую на сосне; 13 X
1949 г. отметил, что глухари спят на деревьях; 19 XI 1934 г., в мо-
мент, когда сумерки так сгустились, что луна бросала уже тень от
деревьев, собака облаяла на сосне двух глухарок, одну из которых
я убил, а другая слетела; в ту же ночь я поднял! двух глухарей,
ночевавших на снегу; 13 XI 1937 г., когда мне пришлось ночью, при
густо шедшем мокром снеге, рубить сушник для костра, я поднял
с сосен более десятка глухарей и глухарок. Если мокрый снег идет
днем, глухари прячутся под кроны елей (24 IX 1939 г.). В декабре,
пока снега мало, глухари ночуют в полулунке — открытой снежной
ямке. При высоте снега 30 см глухарка не может в него зарыться,
хотя и делает попытки; для ночлега под снегом самкам нужен слой
не менее 40 см, самцам — около 50 см.
В начале зимы глухари самцы иногда собираются на открытых
болотах большими стаями: так, 8 XII 1934 г. на болоте к югу от
Чунозера с одного места я насчитал 26 петухов; на некоторых сос-
нах сидело по 2—3 птицы. 5 II 1939 г. наблюдатель охраны
И. Ф. Зайков наблюдал на соснах по северному берегу Пасма-лам-
бины 25 глухарей на кормежке и отметил, что они жили в этом
районе с осени. Самки ведут себя иначе — они живут в борах и на
кормежке прячутся в ветвях или у ствола. Оперение глухарки ря-
бое, большая часть перьев темно-серая с охристыми пятнами и бе-
ловатым краем, ио область зоба представляет большое одноцвет-
ное яркое охристо-рыжее пятно. Когда глухарка сидит в кроне
сосны лицом к наблюдателю, яркий цвет ее зоба совершенно сли-
вается с тоном коры верхней половины соснового ствола и боль-
30
ших ветвей. Существование такой узкоспециальной защитной
окраски, полезной лишь в одном определенном случае, подтвер-
ждает тесную связь всей истории развития глухаря с одним хвой-
ным деревом, хвоя которого составляет его корм в течение большей
части года.
Тетерев. По образу жизни зимой тетерева сходны с глухарями,
но, в отличие от них, держатся смешанными стаями, самки вместе
Рис. 12. Лунка тетерки
с самцами. Они подвижнее глухарей, что, должно быть, зависит
и от рода пищи: глухарь довольствуется одной хвоей, а в рацион
тетерева входят молодые шишки сосен и цветочные сережки берез.
Это сравнительно дефицитные корма: если тетерева покормились
на определенной березе один раз — то вторично в ту же зиму они
не найдут на ней сережек. Поэтому стаи тетеревов нередко летают
на кормежку далеко.
Тетерева — более общественные птицы, чем глухари, и на ноч-
легах лунки тетеревов располагаются гораздо теснее, чем лунки
глухарей. Наблюдатель охраны заповедника А. С. Сергин 10 XII
1940 г. видел на березах стаю тетеревов в 39 голов, а 7 II 1949 г. —
стаю в 34 шт. на лету.
31
Тетерева, как и глухари, зимой кормятся два раза в сутки —.
утром и вечером, а около полудня чаще их можно поднять из лу.
нок: им достаточно и 30 см снега, чтобы в нем скрыться.
На склонах горных тундр бугры и гребни вытаивают весной
раньше, чем появятся проталины в лесу. На горных проталинах
бывает много перезимовавших ягод вороники, брусники и «гром,
ягоды». По этой причине ранней весной тетерева иногда кормятся
выше верхней границы леса. 11 IV 1950 г. на Чуна-тундре мы под,
няли с такой проталины на высоте 390 м абс. 5 косачей, прилетев,
ших сюда на жировку, а в следующем году 17 IV нашли на том же
месте свежие следы тетеревов на снегу.
Тетерев начинает токовать раньше, чем глухарь, токует гораздо
энергичнее и кончает петь позднее глухаря; правда, токование те-
терева легче обнаружить, его «бормотание» разносится примерно
до 4 км, и, чтобы установить самый факт, нет надобности ходить на
ток. По этой причине наблюдения получаются полнее и точнее.
Таблица 2
Сроки токования тетерева
Весеннее токование Осеннее токование
Год ф о <D О к О ° s= w О ф о ф 3 * О к оя£§
СП t; S со S3 £ о s и s се я *=! ’S q Е
o Ф О E H и Q. Ф Ф Q.E к Ф Ф £ -
ф o ф Ф о s о ca Ф О Ф Ф о ЕЕ пз
с e ~ C t- « sr я о м С Е Е Е н Я е ех ф а
1931 25 IV
1932 3 V — —
1933 25 IV — —
1934 1 V — —
1935 10 V — — —
1936 1 V — —
1937 13 IV 7 VI 55 — 16 IX (23IX) 2
1938 27 IV 14 VI 48 14 — —
1939 29 IV 23 VI 55 18 — __ __
1940 27 IV 17 VI 51 6 — 2 XI 2
1941 28 IV 13 VI 46 11 23 IX 10 XI 48 2
1942 — — —
1943 — — ___
1944 24 IV — — .
1945 27 IV — —
1946 4 IV — —
1947 24 IV 2 VI 39 И X 1
1948 31 III 2 VI 63 16 19 IX 15 XI 57 6
1949 10 IV 16 VI 67 29 19 VIII 3 XI 76 7
1950 26 III 16 VI 82 16 15 VIII 2 X 48 5
1951 12 IV 16 VI 65 24 5 IX (29 IX) 3
Среднее 21 IV 13 VI 57 17 6 IX 28 X 57 4
Как мы уже упоминали, численность тетеревов за годы нашей
работы значительно увеличилась. Ввиду этого сильно изменилась
вся картина токов. В 1936 г. мы описывали ее такими словам^
32
«В районе Чунозерской базы заповедника песню тетерева можно
слышать, в среднем за 6 лет, с 6 мая по 23 мая, причем токует он
в одиночку; но там, где он многочисленнее (Хибины, Апатиты, За-
шеек), токование начинается примерно на неделю раньше и растя-
гивается на более продолжительный срок, причем на токах наблю-
дается по несколько петухов». (Семенов-Тян-Шанский, 1938,
стр. 247).
За прошедшие с тех пор 15 лет тетеревиные тока в районе запо-
ведника сильно выросли и в пространстве, и во времени, и по числу
участвующих в них птиц. В табл. 3, для того чтобы даты были срав-
нимы, нам пришлось день первого токования в 1931—1936 гг. взять
не по Чунозеру, а по Хибинам или Апатитам.
Т а б л и ц а
Число дней наблюдений токования тетерева
(За все годы вместе, по декадам)
Март Апрель Май Июнь Июль Всего весной
И III 1 11 Ill 1 11 111 1 II III I II III.
— 3 4 7 13 20 29 25 17 15 1 2 — — 136
Август Сентябрь Октябрь Ноябрь Всего осенью
Декада I 11 Ill 1 11 III I II 111 1 II III
— 2 — 1 6 7 4 2 1 2 1 — 26
Когда тетерев только начинает токовать (конец марта), он «бор-
мочет» днем: так, в 1950 г. первую в году песню я слышал 26 III
в 12час. дня, а 28 III — в 8 час. утра. В апреле токование передви-
гается на более ранние часы (4—7 час. утра), но тетерева по-преж-
нему поют преимущественно на деревьях. В конце апреля тетерева
с вечера, обыкновенно в 18—20 час., вылетают один за другим из
леса на лед озера, на 100—500 м от берега, и тут токуют всю ночь
до 4—7 Час. утра, Если ночь морозная, тетерева после полуночи
замолкают и прячутся где-нибудь на берегу; но едва солнце пока-
жется из-за гор, как тетерева вылетают на лед и ток возобнов-
ляется с новой энергией.
Каждый петух токует сам по себе, в промежутках между пес-
нями перебегая с места на' место. Когда петухи распоются, тетер-
ки, до того сидевшие на деревьях около берега, также вылетают
на лед, ближайшие косачи устремляются к самке и вступают в дра-
ку. Бои бывают довольно ожесточенными: днем 20 V 1951 г. я за-
стрелил на побережье Чунозера взрослого (старше года) косача,
у которого на затылке и зобу были опухшие, но уже заживающие
3—3037
33
поверхностные раны и не хватало перьев. Место тока тетеревов
можно узнать по следам на снегу с характерными бороздами от
опущенных крыльев по бокам, попадаются и черные перышки, вьк
рванные в драке.
С одного места можно насчитать до десятка и более птиц, из-
дали похожих на черные шарики, катающиеся в разных направ-
лениях. Подойти к ним на выстрел совершенно невозможно: токую-
щий тетерев подпускает открыто идущего человека метров на 200,
не ближе, а потом сразу улетает.
Рис. 13. Токующие тетерева (Печоро-Илычский заповедник,
16 V 1954 г.)
В мае, когда ток в полном разгаре, в благоприятную погоду
(тихую и не очень морозную), проходя по озеру или наблюдая
с берега в бинокль, можно бывает насчитать на льду десятки тете-
ревов. Наблюдатель охраны А. С. Сергин, проезжая 11 V 1951 г. по
оз. Пиренга, насчитал в разных местах 98 косачей, а до 20—30 птиц
разные лица отмечали неоднократно. Характерно полное отсут-
ствие обособленных токовищ.
Судя по наблюдениям токования, в глубине Лапландского запо-
ведника сравнительно мало тетеревов; напротив, к югу от него,
в обширном «сузёмке» Чайцкл-туйпл, окруженном цепью озер
(Имандра, Воче-ламбина, Чунозеро, Охтозеро, Пиренгское озеро
и р. Пиренга), их много. Из этого лесного массива тетерева выле-
тают токовать на все окружающие озера, причем довольно много их
токует внутри «сузёмка», на озерах (Осиновое, Куропачье, Мерьт-
явр) и обширных болотах. Внутри заповедника по нескольку коса-
чей токовало на озерах Островском, Кензис-явре, Чингльс-явре н
34
Румель-явре. Косачи токуют на льду даже тогда, когда он придет
в движение. После очищения озер от льда они поют на деревьях
и перестают токовать только в середине июня, когда у них начнет-
ся линька. Как видно из табл. 3, разгар тока приходится на май.
В это время у птиц уже летний режим питания, о чем можно су-
дить по «травяному» составу находимого на льду помета.
Линька начинается рано: у самца, добытого 20 V 195П г., все
перья, кроме вырванных в драке, были целы; у самца 1 VI 1949 г.
выпали 3 внутренние пары первостепенных маховых и уже появи-
лись 2 пары пеньков; 27 VI 1950 г. поднятый в кустах тетерев поте-
рял при взлете одно маховое.
Рис. 14. Лунка рябчика
Тетерки выводят птенцов в одно время с глухарками (24 VI
1937 г.; 27 VI 1949 г.; 27 VI 1951 г.). Линька молодых петушков
во взрослое перо происходит у обоих видов почти одновременно.
В отличие от глухарей тетерева предпочитают летом более от-
крытые места — болота, гари и лесосеки.
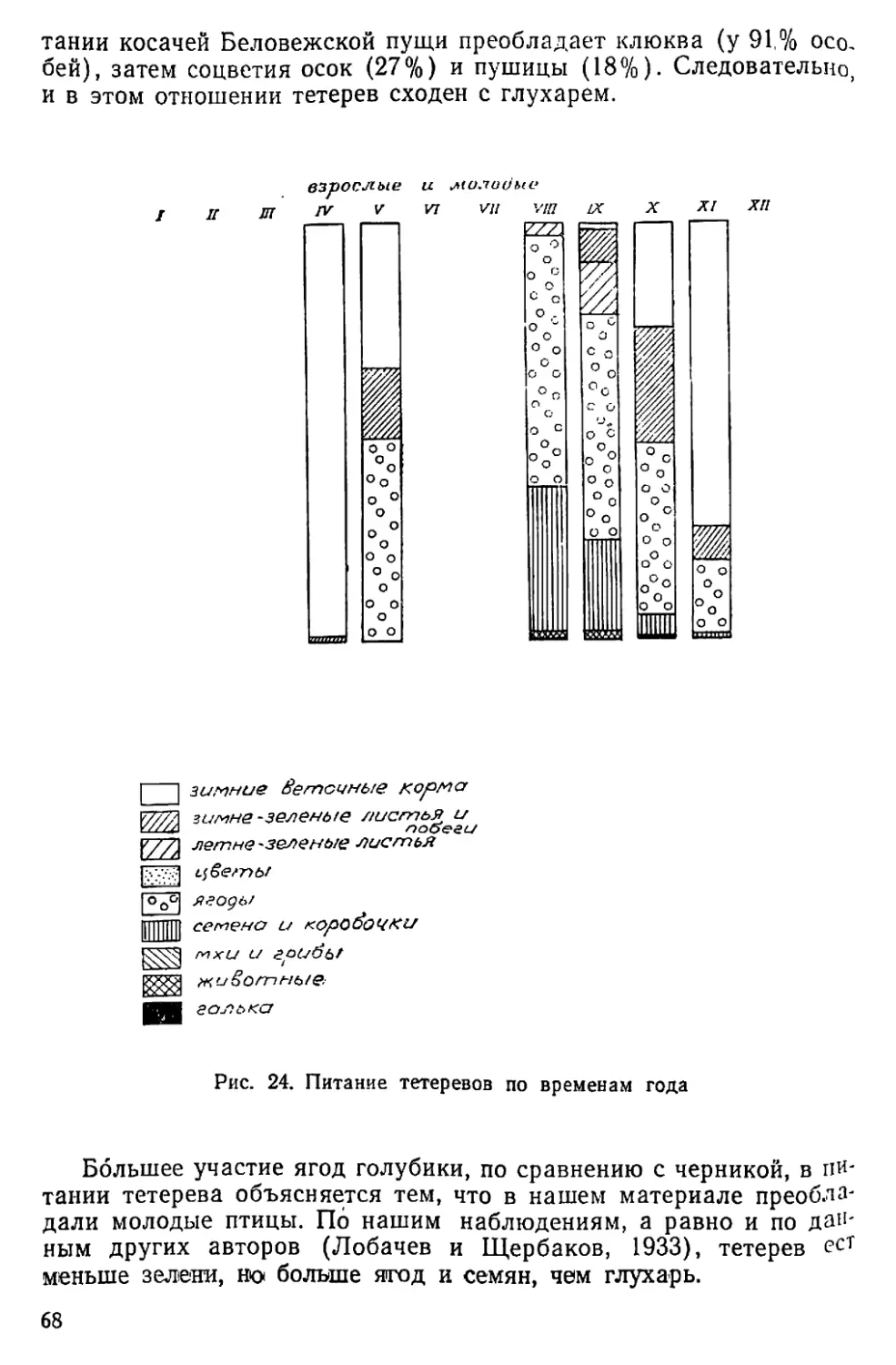
Осеннее токование у тетерева наблюдается гораздо чаще, чем
у глухаря: оно начинается иногда уже в августе (19 VIII 1949 г.;
15 VIII 1950 г.), достигает наибольшего развития в конце сентября
и, постепенно затихая, заканчивается в середине ноября (см.
табл. 2). Косачи поют обычно на деревьях, чаще всего рано утром,
в августе с 4 час. утра, в сентябре — с 6—8 час. Бормотание моло-
дых петушков можно слышать и среди дня (18 IX 1950 г. в 16 ч.
45 м.). Иногда на току собирается до 8 птиц. После замерзания
озер тетерева, как и весною, вылетают на лед: 10 XI 1941 г.
А. С. Сергин наблюдал на льду Охтозера большую стаю, косачи
бегали и гоняли тетерок; 15 XI 1948 г. там же два наблюдателя ви-
дели на току 24 тетерева.
3* 35
Перелеты тетеревов стаями, иногда на большой высоте, мы от-
мечали в разное время — с конца сентября до конца марта. В об-
щем, образ жизни тетерева меньше меняется по сезонам, чем об-
раз Жизни глухаря.
Рябчик. Зимой рябчиков легче всего встретить в долинах ручьев
или в лощинах с зарослями серой ольхи, сережки и почки которой
составляют их любимую зимнюю пищу. Кроме того, они нередки
в ельниках, особенно в густых и захламленных. Рябчику достаточно
очень маленького участка, если там есть все необходимые ему усло-
вия жизни. Поэтому они попадаются и среди боров в подходящих
уголках — еловых лощинах или долинах ручьев, где есть ольха.
Чаще они держатся парами, реже — поодиночке.
При удобном случае рябчик и зимой добывает зелень и ягоды,
очевидно, выкапывая их из-под снега в местах, где слой его тонок,
например, под ветками елей, как это наблюдал по следам А. А. На-
симович 2 и 8 января 1939 г. В марте и апреле, как только пока-
жутся проталины на косогорах и под камнями, на них можно встре-
тить рябчиков.
Ввиду очень скрытного образа жизни рябчика, сведения о гнез-
довании его в Лапландском заповеднике скудны: они приведены
ниже, в главе о размножении.
Первые встречи выводка рябчика отмечены в следующие числа:
1937 г. 22 VI 1945 г. 2 VII
1938 г. 4 VII 1946 г. —
1089 г. — 1947 г. —
1940 г. 29 VI 1948 г. —
19411 г. — 1949 г. 27 VI
Г942 г. — 1950 г. 29 VI
194'3 г. — 1961 г. —
1944 г. —• Среднее: 29 VI
Выводки держатся обычно в долинах ручьев или сыроватых ло-
щинах в лесу; попадались они и в травянистых березняках по бере-
гам рек- Нередко при выводке бывает не только матка, но и самец.
Наравне с глухарями и тетеревами рябчики любят летом «порхать-
ся» в песке или древесной трухе. Рябчик избегает вылетать на откры-
тые места (тундра, берег озера, открытое болото) и необходимый
ему гравий находит в лесу, на обнажениях грунта под выворотами.
Рябчик в особенности любит ягоды брусники. По-видимому»
именно брусника привлекает его в ягельные -боры, где его иногда
можно встретить осенью, перед выпадением снега, и рано весной.
Для рябчика характерны резкие изменения численности: в неко-
торые годы он был очень многочислен (1936—1938 и, особенно»
1944—1946 гг.), в другие почти полностью исчезал (1939—1941 гг.)-
Белая куропатка. Если глухарь, тетерев и рябчик по спо-
собу добывания корми зимой в основном древесные птицы, то бе-
36
лая и тундряная куропатки — наземные круглый год. Русское наз-
вание белых •куропаток, сближающее их с серой куропаткой, под-
черкивает общие свойства этих птиц—наземный образ жизни и
бег как главное средство передвижения.
Зимой белая куропатка питается веточками разных видов ив и
берез; род пищи определяет и характер занимаемых ею стаций. В
снежное время она держится преимущественно по берегам рек и
озер, особенно там, где есть заросли ивняков, по окраинам болот и
в заболоченных долинах — в таких местах бывает много кустов.
Кроме того, ее можно встретить в заболоченных еловых и сосновых
лесах, реже — в глубине елово-березовых лесов, вдали от опушек.
Следы по -белой тропе показывают, что куропатка живет там же,
где и заяц-беляк.
Куропатки обычно встречаются маленькими стайками (от
двух птиц до десятка), которые, вероятно, представляют остатки
выводков; состав их смешанный, самцы и самки держатся вместе,
Дневная деятельность у куропаток начинается раньше, чем у
других птиц. Иногда, выйдя из снежных,лунок, куропатки тут же на-
чинают кормиться, иногда они некоторое время перелетают с места
на место, причем самцы, садясь, выкрикивают свою брачную песню.
Эти перелеты происходят в сумерках, иногда так рано, что шум
крыльев слышен, а птиц еще не видно по причине слабого освеще-
ния.
На жировке куропатки перебегают от куста к кусту, обламывая
кончики побегов. По-видимому, если их никто не тревожит, они мо-
гут кормиться весь день, не поднимаясь на крыло: длина их набро-
дов иной раз превышает 1 км. Перед тем как зарыться в снег на
отдых, куропатки обязательно перелетают на новое место: поэтому
при троплении куропаток нет шансов поднять птиц из лунок — мож-
но или застать их на жировке, или же следы кончаются взлетом. В
середине зимы куропатки редко зарываются в снег днем и отды-
хают под елкой, валежиной или другим укрытием. У куропатки
почти идеальная защитная окраска, и заметить ее на фоне снега,
пока она неподвижна, невозможно. Только приглядевшись, можно
заметить, что оттенок белого оперения куропатки, чуть чуть кремо-
вый, теплее голубоватого тона снега.
Неоднократно указывалось, что белый цвет оперения куропатки
способствует лучшему сохранению тепла, потому что белое тело
теряет меньше тепла через излучение. Йонсен (1929) указал также,
что белые перья менее теплопроводны, чем окрашенные, потому что
белый цвет их зависит от мельчайших пузырьков воздуха в роговой
ткани. Может быть, эти соображения и справедливы, но с физиче-
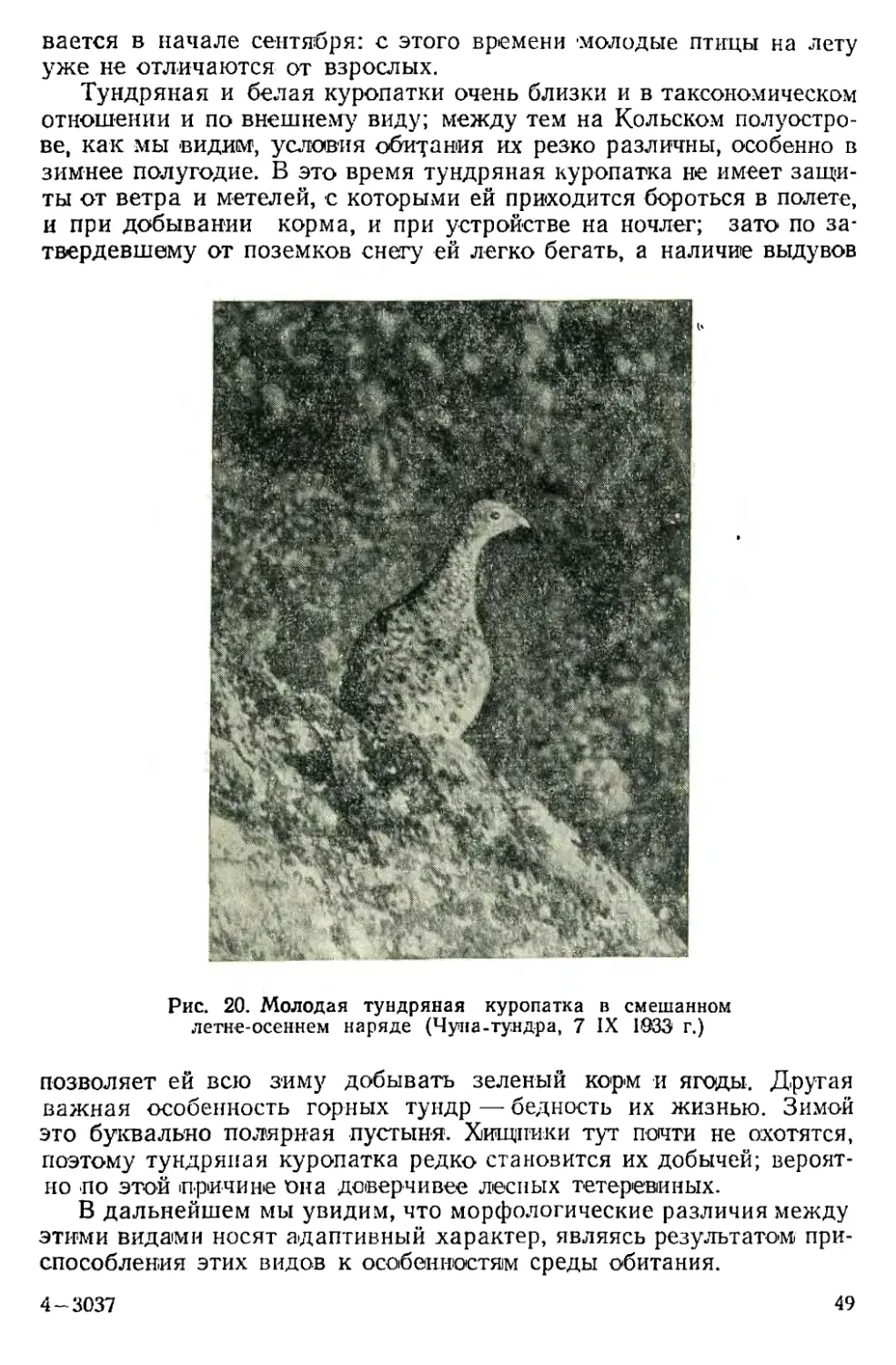
ской стороны они еще не доказаны. У куропатки, как и у всякой
другой птицы, потеря тепла идет не с поверхности перьев, а с по-
верхности кожи; перья играют роль экрана на пути тепловых лу-
чей, а какой экран лучше сберегает тепло, темный или светлый —
вопрос довольно сложный, учитывая, что речь идет не о световых
лучах, а о тепловых, отличающихся большой длиной волны.
37
Хотя мы и считаем куропатку наземной, бегающей птицей, иног-
да она кормится на березах, точно тетерев. Куропатка кормится не
столько почками, как палочками ивы и березы, отламывая их ку-
сочек за кусочком от тонких веток. Тем же кормом питаются и зай-
цы. В конце концов запас тонких, наиболее «съедобных» веточек
истощается. Видимо, именно по этой причине во вторую половину
Рис. 15. Следы белых куропаток на кормежке
зимы куропатки охотнее всего кормятся у берез, кроны которых
пригнулись к земле под тяжестью снега, объедают веточки с ив
и берез, поваленных бобрами, а также взлетают на ветки берез. В
первой половине зимы каждый большой снегопад закрывает исполь-
зованный куропатками ярус кустарниковой поросли и переносит их
деятельность на этаж выше. Позднее, в период максимальной высоты
снега поверхность его долго остается на одном уровне. В это время
все низкие кусты (карликовая березка, большинство ив) полностью
скрыты в снегу, а .потому «продовольственный вопрос» у куропаток
обостряется.
Веточками ив и берез питаются также лоси, лемминги и полев'
ки: однако их конкуренция не играет роли потому, что первые еры*
38
вают ветки на высоте 1—2 м от поверхности снега, а вторые
используют только ветки, находящиеся под снегом.
Как и рябчик, куропатка не упускает случая достать зеленый
корм и ягоды из-под снега. В малоснежные зимы, к которым отно-
сились зимы 1930-31 и 1938-391 гг., в январе и феврале мы не раз
наблюдали, как куропатки разрывали тонкий снег. Ранней весной
куропаток легче всего встретить на проталинах — по бере-
гам озер и рек, на кочках бугристых болот, на склонах горных
тундр.
В предвесеннее время куропатки становятся деятельнее, хотя
у них и нет такого явного перелома в поведении, как у глухаря.
В марте они забегают под вывороты в поисках гравия; по утрам все
чаще они летают стаями, все громче и задорнее кричат самцы.
День начала токования куропаток невозможно определить. В
сущности, у них и нет токования в .смысле соревнования или борь-
бы самцов за самку. Весной куропатки разбиваются на пары, и их
ток — не более как брачная игра, аналогичная той, какую мы наб-
людаем весной у гагар, сов и многих других птиц.
Перед началом брачного периода куропатки меняют.места оби-
тания. Они уходят из районов с глубоким и ровным снежным! по-
кровом, где долго не образуется проталин, частью на болота, где
рано вытаивают кочки, частью к верхней границе леса. Пояс бере-
зового криволесья обеспечивает их обильньим веточным кормом, а
все остальное они находят на проталинах в альпийском поясе, где
встречаются с коренным жителем этих мест — тундряной куропат-
кой.
Во время ежегодных оленных поездок для учета диких оленей
на горных тундрах в некоторые годы (1937, 1949) мы поднимали с
проталин десятки белых куропаток. Они встречались на высотах до
500—600 м абс.: разумеется, не высота места ограничивает их за-
леты, а распределение проталин, которые рано появляются в ниж-
ней части альпийского пояса, в то время, когда в гольцовой области
гор ледянистый, состоящий наполовину из изморози, снежный пок-
ров еще не начал подтаивать.
В это время (в среднем 17 IV) у самцов начинается линька. За-
мечательно, что в тех местах, где снег сходит рано, и у куропаток
линька протекает раньше; напротив, в многоснежных районах ку-
ропатки дольше сохраняют зимний наряд. Первые весенние перья
появляются на горле. Скоро «красное» горловое пятно расширяется,
охватывая всю шею и голову; после этого темп линьки замедляется,
и петушок долго ходит в ярком брачном наряде, с коричнево-крас-
ной головой и шеей и белоснежными туловищем и крыльями. В то
же время у него сильно развиваются брови —участки голой кожи
светлого суриково-красного цвета, отороченные сверху тонким гре-
бешком. Птица может произвольно поднимать и опускать этот гре-
бешок; когда он опущен, кроющие перья темени свисают на бровь
и почти совсем ее закрывают.
39
Днем куропаток можно встретить парочками на кормежке пре-
имущественно на проталинах. Как только начнет смеркаться, самцы
начинают токовать и токуют всю ночь напролет: ночи в конце апре-
ля — мае уже светлые. .
Петушок, ухаживая за самкой, расширяет брови, вытягивает
вверх шею и поднимает хвост, но не распускает его веером так, как
делает глухарь или тетерев. Он перебегает и перелетает с места на
место, время от времени громко выкрикивая свою странную брач-
ную песню. Буквами ее можно передать приблизительно так:
« Кэррр -эр-эр -эр-эр -эр -эр -эр -э •..».
«Кабао, кабао, кабао, кабао, кабао... кабао... кабао... кабао...
(Сначала часто, потом вое реже; затем1 после паузы): кав... кав...
кав...».
В этом варианте он начинает песню на лету и заканчивает ее уже
сидя на земле. Если он поет не взлетая, то песня звучит иначе, сна-
чала редко, потом чаще:
«Ко.... ко... ко-ка-ка-ка-ка-ка-ка-какакёррр...»
Потом следует «кабао», а иногда песня обходится и без этого
колена.
Раскатистые брачные крики белых куропаток — характерная чер-
та природы Лапландии. В фольклоре саамов крик белой куропатки
заменяет крик петуха в русских сказках: и тот и другой являются
предвестниками утра, и обоим сказки приписывают магическую
силу (Харузин, 1890).
Призывный крик курочек звучит: «м-е... ме...» или «мя... мя...»;
подражая ему, нетрудно подманить петуха. Весной куропачи ста-
новятся, как говорят, «дурными» и теряют обычную осторожность.
Мне случалось белой ночью ехать по озеру на оленях. Из лесу вы-
летает куропач и с криком садится впереди. Олени бегут на него
рысью, но он не пугается. Кончается тем, что олени сворачивают в
сторону, косясь на птицу. Человека куропачи подпускают не так
близко, но все же на десять метров подойти к ним удается.
Во время брачных игр самцы любят садиться на возвышенные
предметы—кочки, пни, валежины, крыши избушек, камни а неред-
ко и на березы. Несколько раз в мае и июне я наблюдал, как самец
садился с брачным криком на большие ели. 21 VI 1951 г. М. И. Вла-
димирская видела, как самец куропатки прилетел на маленький
островок среди озера и сел на елку. На островке гнездились две
пары крачек: одна из крачек с криком пикировала на куропача, ста-
раясь ударить его в голову.
Самец не принимает участия в насиживании, но все время дер-
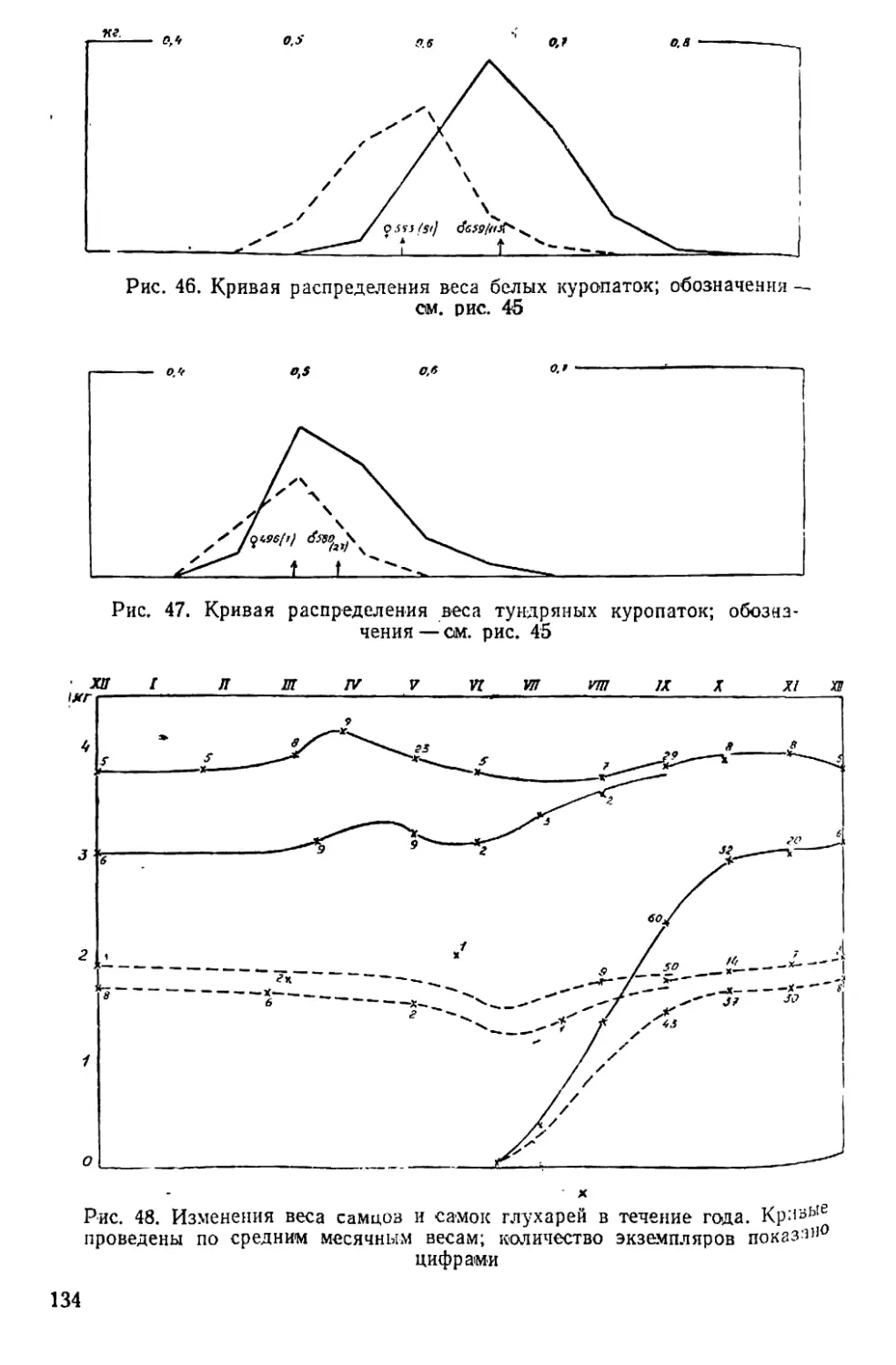
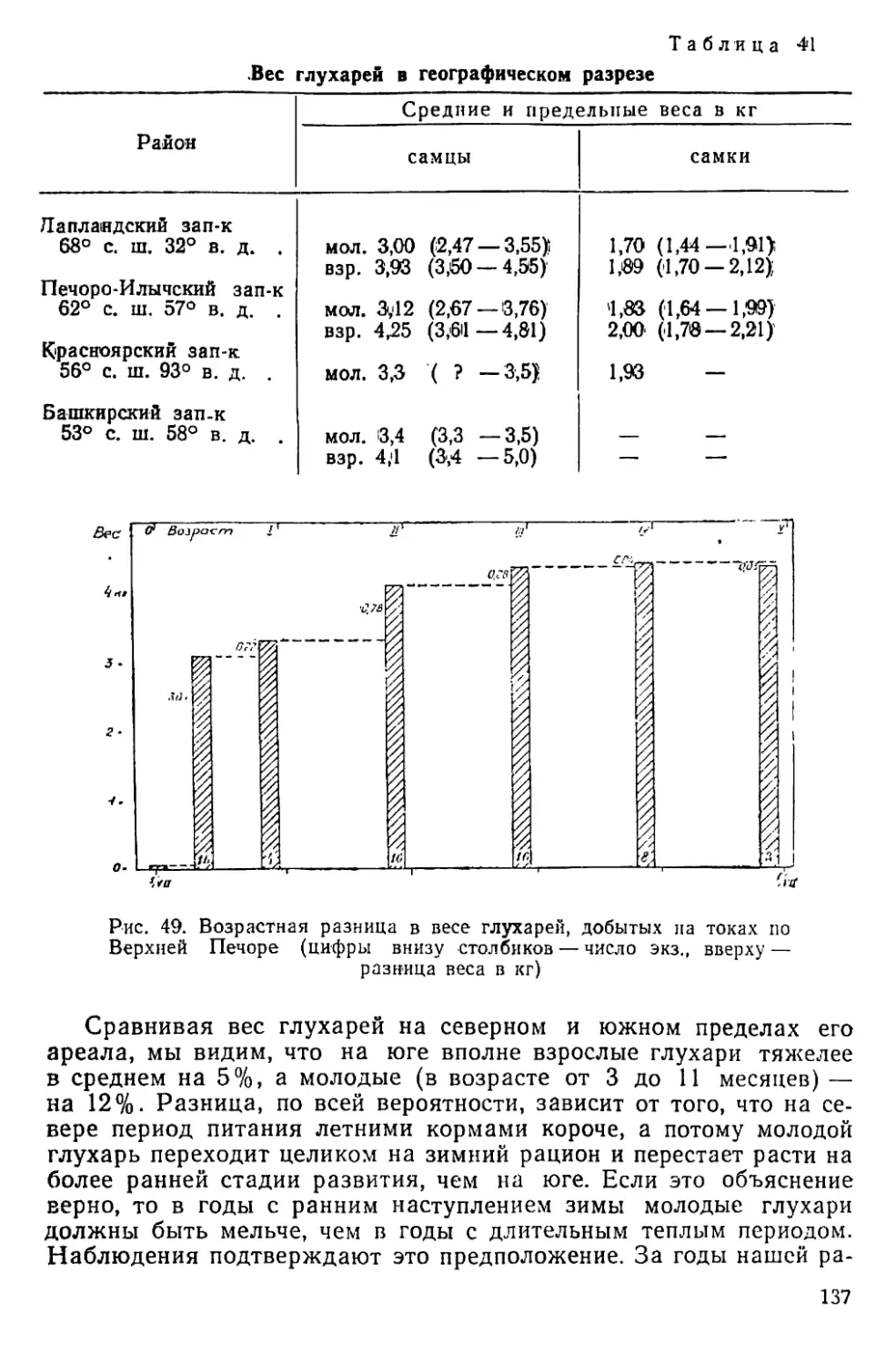
жится около гнезда и, вероятно, способствует его охране, отвлекая
на себя внимание хищников. Пока самка насиживает, у самца за-
канчивается весенняя линька. Если петух не погибает к моменту вы-
лупления птенцов, он так и остается при выводке. Куропатка выво-
дит на неделю позже, чем глухарка, в среднем за 13 лет —3 июля
(табл. 4).
40
Таблица 4
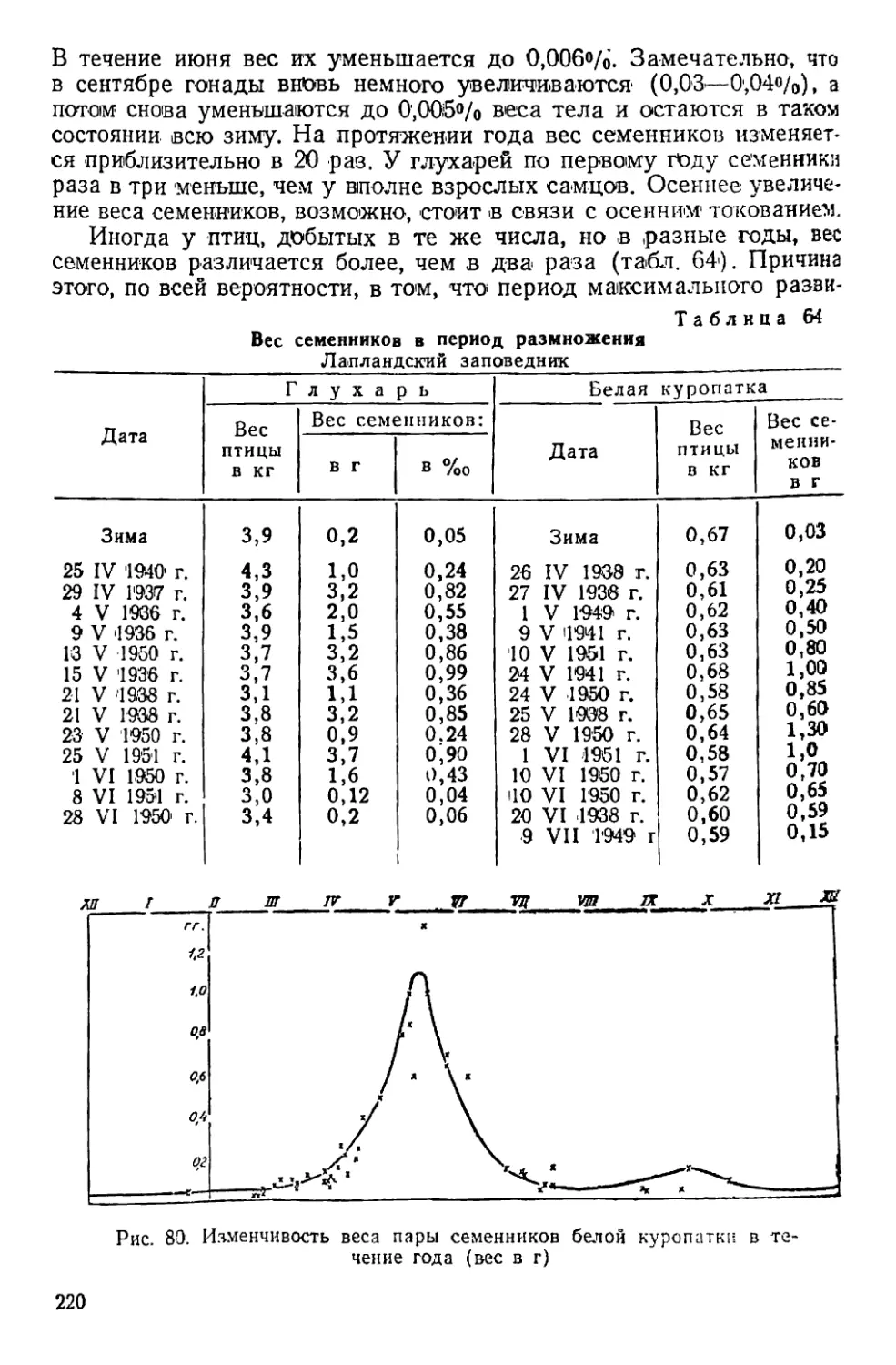
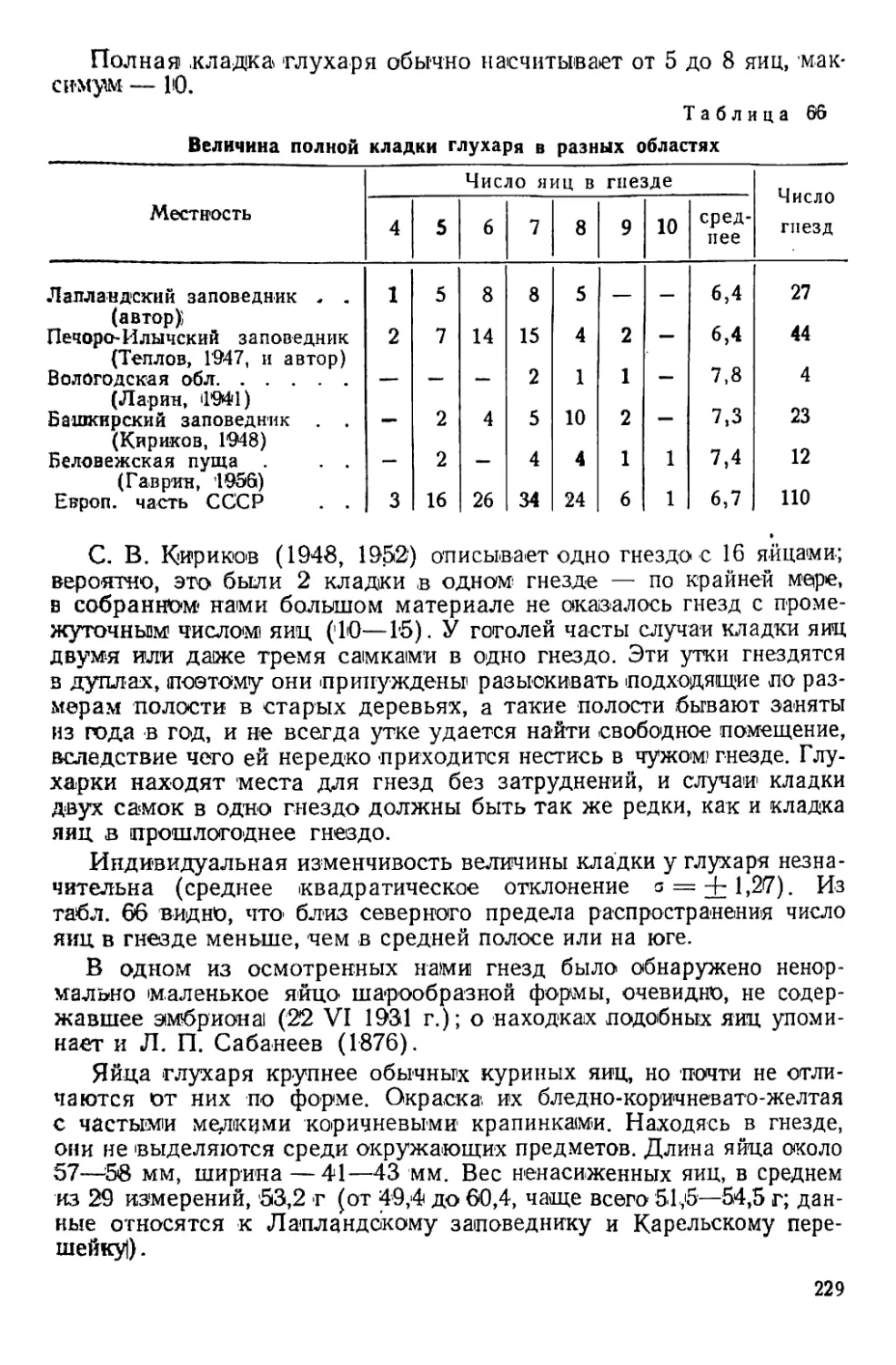
Периодические явления у белой куропатки
Год Красное пятно на горле самца Начало весенней линьки самки Начало летней линьки самца Первый выводок Начало зимней линьки Первая в зимнем наряде
1930 28 X
1931 25 IV 21 V 25 V — — 24 X
1932 15 IV 6 V 1 VI — 4 X 22 X
1933 24 IV 12 V 17 V 19 VII 28 IX 19 X
1934 29 IV 23 V 23 V 26 VI 27 IX 1 XI
1935 29 IV 10 V 27 V 7 VII 30 IX 1 XI
1936 20 IV — 20 V 4 VII 22 IX 19 X
1937 17 IV 9 V 11 V 20 VI 23 IX 19 X
1938 17 IV 8 V 25 V 8 VII 16 IX 23 X
1939 — 1 VI 9 VI — 18 IX 12 X
1940 17 IV 28 V 28 V 9 VII 15 IX 15 X
1941 9 IV — 23 V 5 VII — 17 X
1942 — — — — — —
1943 — — — — — —
1944 7 IV •— — — — —
1945 16 IV — — 4 VII — —
1946 — — — — — 20 X
1947 13 IV — — — 23 IX 19 X
1948 10 IV 26 IV 17 V 22 VI 26 1£ 15 X
1949 14 IV 1 V 28 V 3 VII 25 IX 27 X
1950 12 IV 19 V 20 V 23 VI 29 IX 26 X
1951 15 IV 20 V 20 V 12 VII 20 IX 20 X
1952 14 IV — — — — —
Среднее: 17 IV 14 V 24 V 3 VII 24 IX 22 X
Выводки кормятся чаще всего на болотах — описанных выше
«ступенчатых», осоково-пушицевых или сфагновых, преимуще-
ственно у окраин, где защитные условия лучше; затем в черничных
и травянистых ельниках, а также на гарях с почвенным покровом
из политриховых мхов. Выводки куропаток можно встретить по
склонам гор вплоть до верхней границы лесной растительности.
Иногда два выводка объединяются и пасутся вместе, что обна-
руживается по наличию при птенцах 3 или 4 взрослых птиц; число
птенцов при этом достигает 15—20 штук. Такие выводки мы наблю-
дали, например, 13 VII 1932, 19 VIII 1933, 27 VII 1934, 14 VII 1938,
19 VII 1949, 25 VII 1950 г. Эта своеобразная черта, отличающая ку-
ропатку от других тетеревиных, впервые была отмечена, насколько
мне известно, охотничьим писателем XVIII века Василием Левши-
ным (1779, стр. 132), который объяснил ее таким образом:
«Примечено, что самец сего рода держит у себя по две самки,
которые несутся обе в одном гнезде, и одна из них общие яйца вы-
41
сиживает; а цыплят водят обе вместе, и вероятно потому, что когда
случится найтить их с молодыми, то бывает от двадцати до тридцати
вместе и две самки старых в одном стаде».
Куропатки купаются в пыли, как и остальные тетеревиные. Са~
мые ранние находки свежих ямок с взбитой пылью и потерянными
белыми перышками были сделаны 25 V 1937 и 22 V 1949 г.; самые
поздние — 24 IX 1949 и даже 4 X 1934 г.
Если к выводку приближается враг, старики с криком вылетают
ему навстречу и стараются отвести от птенцов; при этом самец дер^
Рис. ’1'6. Молодая белая куропатка в ягельном бору.
Фото М. Владимирской, Печоро-Илычский заповедник.
жится осторожнее, чем самка. Через некоторое время один или оба,
смотря по обстоятельствам, возвращаются и начинают скликать по-
прятавшихся птенцов. Птенцы постарше разлетаются в разные сто-
роны с писком и, отлетев как можно дальше, молча затаиваются в
траве; когда все вокруг затихнет, они начинают перебегать и пи-
щать, разыскивая друг друга. 2l2 VII 1’950 г. мы подняли выводок
из 7 портков и 2 взрослых, самки и самца. Заметив место, где сел
один из птенцов, мы подняли его вторично, он сделал короткий пере-
лет и сел под елку; ©третий раз он не полетел, а бегал вокруг елки,
пока его не поймали руками. Птенец весил 244 г, ему было около
20 дней. Этот пример показывает, как быстро птенцы тетеревиных
истощают силы в полете.
Летом самцы куропаток почти не кричат. Ближе к осени поведе-
ние их 'меняется. 28—2’9 VII 1950 -г. мы записали: «Ночью куропачи
кричали, точно в апреле», а 1—2 VIII 1938 г.: «В полночь было из-
42
рядно темно, и впервые за лето слышал токовый крик куропатки».
Осенью спугнутый самец обычно взлетает с криком.
В начале сентября все серые маховые молодой куропатки (кро-
ме внутренних на предплечье) заменяются белыми. Теперь молодых
трудно отличить на лету от взрослых, тем более, что и летают они
хорошо. 9 IX 1950 г. на рассвете мы наблюдали выводок белых -ку-
ропаток, перелетавших через озеро Воче-ламбину шириной около
1 км. Во второй половине сентября выводки переселяются с летних
местообитаний — осоковых и сфагновых болот — на сухие и возвы-
шенные участки елового леса. Это перемещение, по всей вероятно-
сти, связано с одновременно происходящей переменой питания:
осенью куропатки -кормятся в основном побегами черники и только
после выпадения снега постепенно переключаются на зимний корм—
веточки ив и берез.
С этого времени на рассвете можно слышать такие же пере-
клички самцов куропатки, как весной.
В начале зимы куропатки иногда залетают даже в населенные
пункты. 19 X 1930 г. А- Б. Васильев застрелил куропатку с крыльца,
лесничества в поселке Хибины; 21 XI 193*1 г. в tomi же поселке
Г. Д. Рихтер застрелил 3 куропатки. В метель молодые куропатки
иногда бывают так смирны, что случается застрелить • несколько
птиц прежде, чем оставшиеся полетят; 29 X 1935 г. на берегу р. Ниж-
ней Чуны одну куропатку убили даже палкой. Нормально зимой
куропатки взлетают еще издали, раньше, чем успеешь их заметить
на снегу.
Стайки куропаток иногда ночуют в снежных лунках на льду рек
и озер. Пока снега мало, лунки представляют простую ямку в снегу;
позднее птицы целиком зарываются в снег.
Численность куропаток сильно меняется по годам. В период
с 1930 по 1951 г. ('22 года) их было много <в 1933 и 1934, в 1937,
в il9(4© и 1950 гг., мало в 1939, 1940 и 1941 и, по рассказам, дальше
вплоть до 1945 года. В тундровой зоне колебания численности куро-
паток обычно связываются с миграциями. Наиболее полная сводка
по этому вопросу дана Михеевым (1948).
На Кольском полуострове, по словам Е. Г. Чернова, массовая
миграция наблюдалась зимой 1938-39 года. Куропатки летели с Тер-
ского берега через горло Белого моря и сотнями замерзали на льду.
Особенно много их было между Пялицей и Сосновкой. При таких
миграциях происходит концентрация птиц в прибрежной полосе,
чем промысловые охотники и пользуются для ловли сетями и сил-
ками.
Замечательно, что в районе Лапландского заповедника мы ни
разу не наблюдали перелетов куропаток на большие расстояния,
чем перелеты глухарей и тетеревов. Нам кажется, что в гористой
местности так и должно быть: если бескормица или другие неблаго-
приятные причины вынуждают куропаток равнинных тундр к пере-
селению, то в горах местные условия гораздо разнообразнее и птице
нет надобности лететь далеко.
43
Тундряная куропатка. В исследованном нами районе это?
вид населяет только горные тундры, никогда не попадается
лесу и в этом отношении может быть противопоставлен остальным
тетеревиным. В первую половину зимы, самую темную, тундряная
куропатка — единственная птица горных тундр. Если исключить
норвежского лемминга и некоторых полевок, ведущих подснежный
образ жизни, и принять во внимание, что дикий северный олень в
это время пасется в лесной зоне, то окажется, что тундрянка вооб-
ще единственное живое существо, которое можно в это время встре-
тить в горах. Недостаток света и низкая зимняя облачность, вслед-
ствие чего тундра почти ежедневно окутана туманом, крайне за-
трудняют зимние экскурсии в горы. Поэтому зимняя жизнь тундря-
ной куропатки нам мало известна.
В это- время они живут в нижней части альпийского пояса гор,
на высоте от 300 до 600 м абс. Тундряные куропатки раскапывают
тонкий снег на обдутых местах по буграм и гребням горных скло-
нов или в воронках выдува вокруг отдельных камней. Корм их со-
стоит из зеленых побегов, почек и ягод альпийских растений. Куро-
патка может разгребать только свежевыпавший снег; перевеваемый
постоянными ветрами, он скоро так затвердевает, что иногда даже
след не печатается на его поверхности. Ввиду этого распределение
куропаток по массиву зависит, в основном, от наличия выдувов, ко-
торых больше на наветренном склоне гор. Зимой на Кольском полу-
острове преобладают северные ветры муссонного характера. Поэто-
му в снежные зимы тундряные куропатки перебираются на север-
ный конец Монче-тундры, где их истребляют охотники из г. Монче-
горска. Охотник 14. Малодушев сообщил нам, что в апреле 1938 г.
он один убил здесь около 50 тундряных куропаток.
Нормально тундряные куропатки держатся стайками до 6 голов,
а то и поодиночке и тогда легко подпускают на выстрел, но иногда
собираются стадами до 30—35 штук и тогда бывают очень осторож-
ны, взлетают дружно и с шумо!М, точно стая белых голубей (27 X
1930, .22 X 1934, 21 XII 194В г.).
Во второй половине зимы дикие олени переходят из ягельных
боров, где снег становится глубок, на горные тундры. Как и куро-
патки олени пасутся не по всей площади, а только на малоснежных
участках, но их сила и особая форма копыт позволяет им разбивать
довольно крепкую снежную корку. В этот период тундряные куро-
патки охотно кормятся на оленьих «тегорах» (местах жировки),
где снег разбит копытами и потому птицам легко найти корм. Нам
приходилось наблюдать в бинокль, как они бегали в стаде диких
оленей буквально между ногами животных (23 IV 1931, 22 IV
1949 г.). Эта форма комменсализма — использование птицами в
снежное время года мест пастьбы крупных копытных животных —
описана в различных вариантах и для разных местностей рядом ав-
торов (Бируля, 1907, стр. 20'; Чарнолуский, 1930-а, стр. 29; Рома-
нов, 1934; Тугаринов и ТоЛхмачев, 1934, стр. 20).
44
У тундряной куропатки клюв меньше и слабее, чем у белой; хотя
она и ломает им веточки тундровых ив и берез, но одними ими не
может прокормиться. Только после снегопадов, пока ветер не сметет
снег с бугров, тундрянки спускаются в пояс березового криволесья и
тут либо обклевывают почки с кустов, бегая по снегу, либо взлетают
кормиться на кроны берез (наблюдения 15 IV—16 IV 1932 г. и
7 IV —8 IV 193® г.).
Рис. 17, Тундряные куропатки в ветреную погоду (Чуна-тундра)
В ветреную погоду тундряная куропатка сидит всегда головой
навстречу ветру — в положении наилучшего обтекания воздушной
струей. В метель она садится всегда у камней; если она сядет с под-
ветренной стороны, то ее быстро заносит снегом, так что она оказы-
вается в снежной лунке. Возможно, это происходит просто случай-
но, но не исключено, что птица выработала такой прием вместо то-
го, чтобы активно зарываться в снег. В сильный ветер они бывают
очень смирны и подпускают на 4 метра (рис. 17).
Брачные игры тундряных куропаток можно наблюдать с середи-
ны или конца апреля до начала июня. Они, как и белые куропатки,
токуют напролет всю светлую северную ночь. Самцы перебегают по
снегу, подняв яркие брови, останавливаясь для исполнения брачной
песни. В первый момент самец опускает хвост и шею так, что вместе
с линией спины они образуют пологую дугу, а кончая песню,
птица поднимает шею и принимает обычную позу. Крик тун-
дрянки не похож на крик белой куропатки, он скорее напоминает
скрип коростеля. Звукоподражательно его можно изобразить так:
«Кррраааэ-крэ-кэ-крэээ-э-э-э-э...»
45
Петух тундряной куропатки кричит так не только во время брач-
ных игр, но и в другое время года. По мнению саамов, зимой крик
тундряной куропатки (предвещает-метель. Лучше 'всего они токуют в
тихие и сравнительно теплые ночи. Призывный крик самки тундрян-
ки не отличается от соответствующего крика белой куропатки. Во
время токования курочки держатся очень тихо и их трудно даже за-
метить. Как и у белой куропатки, токование имеет характер брач-
ной игры. Тундряные куропатки разбиваются на пары около 1 мая.
Если к парочке приблизится посторонний петух, куропач немедлен-
но гонит его и преследует с криком, напоминающим голос кряквы.
Рис. 18. Тундряная куропатка на взлете (Чуна-тундра)
Днем тундряные куропатки не токуют; самец обычно сидит на вы-
ступающем камне, прижавшись к нему грудью, головой на ветер;
курочка прячется у подножья камня. При подходе самец взлетает и
садится тут же рядом, не решаясь оставить самку; последняя же не
имеет причины взлетать, пока не увидит человека. Как только она
поднимется, петух присоединяется к ней, и оба улетают вдаль. Вес-
ной самца легко отличить от самки по манере приподнимать на бегу
шею и полу распущенный хвост (курочка на бегу горбится). Весной
тундряные куропатки мало осторожны, особенно самцы, и нередко
подпускают метров на 10 (рис. 18).
Весенняя линька у самцов задерживается на время токования,
как и у белых куропаток. Существенное различие только в том, что
линька у них начинается не с горла, а с затылка и притом сравни^
телъно поздно, так что первые весенние перья появляются на виду
уже после начала брачных игр. Соответственно и пауза в ходе линь'
46
ки наступает на более ранней стадии, чем у белой куропатки. Весь
период токов, до начала или середины июня, петух остается в белом
зимнем наряде, с небольшим числом черноватых весенних перьев на
затылке и темени. У самки весенняя линька проходит быстро и за-
Рис. 19. Самец тундряной куропатки во время весенней
линьки
канчивается за неделю до начала линьки туловища у петухов. Бла-
годаря такой разнице в сроках линьки разных полов бывает «вре-
мя, когда самец весь белый (чернота затылка издали не видна), а
самка с ним в паре — в летнем наряде, без белого на спинной сто-
роне (наблюдения 24 V 1936 и 30 V 1938 г.).
В начале насиживания еще можно встретить самцов в районе
гнезда, но ко времени вывода птенцов петухи поднимаются в гольцы,
к вершинам гор, где и заканчивают линьку (рис. 19) .Там их можно
встретить поодиночке или стайками до 5 птиц; вместе с ними могут
47
быть и самки, потерявшие кладку. Места, где тундряные куропатку
купались ® леске («порхалища»), мы встречали с конца мая
(в I960 г. 28 V).
Несмотря на то, что на горных тундрах снег дольше задержи-
вается, чем в лесу, тундряная куропатка гнездится и выводит птец.
цов приблизительно одновременно с белой куропаткой, живущей
лесу. В табл. 5 указаны даты встреч только что вылупившихся цып-
лят тундряной куропатки.
Таблица 5
Периодические явления у тундряной куропатки
Год Начало брачн. линьки Начало весен- ней линьки 65 Начало весен- ней линьки dd Вывод птенцов Начало зимней линьки Первая в зим- нем наряде
1930 1 1 26 VI 5 VII 6 IX
1931 10 V 19 V — — —
1932 — 20 V — — — 22 X
1933 6 V — — 3 VII — 14 X
1934 1 V 18 V 17 VI 24 VI — 22 X
1935 — — 22 VI — 15 X
1936 24 V — 17 VI ( 30 VI 1 4 VII — —
1937 11 V 12 V — 22 VI — —
1938 — 27 V 8 VI — 16 IX —
1939 20 V — 4 VI — — —
1940 — — — 30 VI 19 IX 21 X
1948 12 V — 18 VI — — 6 X
Среднее 12 V 19 V 16 VI 30 VI 14 IX 17 X
Матка самоотверженно защищает цыплят: стараясь отвести от
выводка, она волочит крылом и бьется иногда в 2—3 м от человека.
Самец тундряной куропатки, как не раз отмечалось, не водит цып-
лят с самкой. Только один раз, 16 VIII 1933 г., я встретил выво-
док, при котором были вместе самка и самец; через год, 25 VI
1934 г., на том же месте петух тундряной куропатки на глазах у ме-
ня отводил собаку, по-видимому, от гнезда.
Выводки встречаются на различных высотах — от верхней гра-
ницы леса примерно до 800 м абс. Места обитания их разнообраз-
ны — то сыроватые луговины с низкой травой, то кустарниковые за-
росли в долинах горных ручьев, то скалистые ущелья, где долго за-
леживается снег, то тундровые гари, заросшие кукушкиным льном,
иногда ягельники, если поблизости есть вода или залежи снега.
Летний наряд тундряной куропатки поразительно подходит
рисунку и тональности к поверхности выветрившихся, покрытых на-
кипными лишайниками, камней. Пользуясь этим обстоятельством,
она часто прячется на открытом месте среди камней, избегая взлс1'
тать (рис. 20). Смена серых «детских» маховых на белые заканчИ'
48
вается в начале сентября: с этого времени молодые птицы на лету
уже не отличаются от взрослых.
Тундряная и белая куропатки очень близки и в таксономическом
отношении и по внешнему виду; между тем на Кольском полуостро-
ве, как мы видим, условия обидания их резко различны, особенно в
зимнее полугодие. В это время тундряная куропатка не имеет защи-
ты от ветра и метелей, с которыми ей приходится бороться в полете,
и при добывании корма, и при устройстве на ночлег; зато по за-
твердевшему от поземков снегу ей легко бегать, а наличие выдувов
Рис. 20. Молодая тундряная куропатка в смешанном
летнс-осеннем наряде (Чуна-тундра, 7 IX 1933 г.)
позволяет ей всю зиму добывать зеленый корм и ягоды. Другая
важная особенность горных тундр — бедность их жизнью. Зимой
это буквально полярная пустыня. Хищники тут почти не охотятся,
поэтому тундряная куропатка редко становится их добычей; вероят-
но по этой причине она доверчивее лесных тетеревиных.
В дальнейшем мы увидим, что морфологические различия между
этими видами носят адаптивный характер, являясь результатом при-
способления этих видов к особенностям среды обитания.
4 — 3037
49
IV. ПИТАНИЕ
Материалы по питанию тетеревиных в районе Лапландского запо.
ведника собраны преимущественно мною (1930—1951 гг.), отчасти
Г. А. Новиковым (19'37—1939 гг.)„ П. С. Мальчевским (1937 г.),
А. А. Насимовичем (1946—1948 гг.) и другими сотрудниками. Часть
этого материала была обработана нами в 1935 (Семенов-Тян-Шап-
ский, 1938-а) и 1936 гг. (Семенов-Тян-Шанский, 1938). Питание ряб-
чика позднее обработано Новиковым1 (1952); в его работе приведены
также, к сожалению, в сыром1 виде, некоторые данные по питанию
белой куропатки, глухаря и тетерева. Предлагаемый обзор питания
тетеревиных — вполне самостоятельная работа, выполненная по пер-
воисточникам. Все сотрудники заповедника пользовались при сборе
и обработке материала почти одинаковой методикой. Содержимое
зоба добытых птиц исследовалось, как правило, в свежем виде, что
облегчало точное определение растений. В большинстве случаев раз-
бор зоба и определение кормов проводилось лицом, добывшим пти-
цу. Содержимое желудков подробно не исследовалось, ввиду недо-
статочной репрезентативности этих данных и трудности определения
измельченных частиц.
В последнее время в практику экологических работ прочно (вошел
метод подсчета «процента встреч» отдельных кормов в исследуемых
объедках, зобах, желудках, погадках и экскрементах: этот прием
удобен тем, что применим к любым материалам от различных живот-
ных. К сожалению, процент встречаемости не всегда пропорционален
весу или объему данного корма. Поэтому при обработке питания жи-
вотного по методу встречаемости иногда трудно выделить важные
корма от случайных примесей.
Чтобы получить количественные показатели, Г. А. Новиков и мы,
начиная с 1936 г., по возможности взвешивали содержимое зобов по
отдельным фракциям, либо подсчитывали количество частиц (ягод,
кусочков побегов, экземпляров насекомых и т. д.). Еще раньше была
•введена балловая оценка кормов на карточках, причем Новиков при-
менял пятибалльную шкалу, а мы — четырехбалльную, в подража-
ние геоботанической шкале обилия Друде. Значение этих баллов сле-
дующее:
IV — зоб наполнен исключительно или преимущественно одним
данным кормом;
III. — корма ’много, но наряду с другими видами кормов;
II — в небольшом количестве;
I — единично.
Для возможно более полного использования собранного материа-
ла я счел возможным на всех карточках проставить приблизительньго
веса кормов, на основе имевшихся на карточках указаний. Известно,
что одна ягода черники в зобу куропатки весит, в среднем1, 0,2—0,3 г,
кусочек березовой ветки — 0,02 г и так далее. Для перевода в вес
балловых оценок мы использовали 184 зоба, содержимое который
50
было взвешено по фракциям, и рассчитали весовое значение каж-
дого балла по каждому виду птиц в отдельности (см. табл. 6|).
Таблица 6
Весовое значение балловой оценки кормов тетеревиных птиц
Вид птицы Число контроль- ных птиц Весовое значение бал- ловой оценки в г Переводи, коэффиц. (приблиз.)
IV III II I
„ 1 Глухарь, самец . . . 30 126,0 15,0 1,75 0,18 9
Глухарка 24 34,5 9,4 1,10 0,20 6
Тетерев 14 — 9,6 0,90 0,08 10
Рябчик 15 — 7,3 0,50 0,10 8
Белая куропатка . . . 43 18,1 4,5 0,62 0,07 6
Тундряная куропатка . 8 — 4,8 0,60 0,09 7
Из просмотра цифр- этой таблицы видно, что низший балл у всех
наших птиц имеет почти одинаковое значение, а последующие можно
получить последовательным умножением, предыдущего балла на не-
который коэффициент, приблизительная величина которого» показана
в последнем столбце. Мы видим, что баллы глазомерной оценки стоят
друг к другу в отношении геометрической прогрессии. Поэтому нет
смысла вычислять из баллов встреч кормов арифметическое среднее
и давать его в таблицах питания (как это сделано нами в работе
1938 года). Важно то обстоятельство, что при высшем балле количе-
ство корма в 200—750 раз больше, чем при низшем балле, хотя «про-
цент встреч» может быть одинаковым в обоих случаях. Отсюда сле-
дует, что при серьезной работе совершенно необходим количествен-
ный анализ питания. Именно поэтому, рискуя получить упрек в субъ-
ективном подходе к материалу, я решил последовательно провести
грубый количественный анализ питания птиц путем перевода баллов
в весовые единицы, но параллельно с этим даю и обычные «качест-
венные» таблицы питания. Все таблицы построены по месяцам;
число данных указано в подзаголовке «качественной» таблицы (в
«количественной» ,оно не повторяется); встречаемость кормов дана в
«качественных» таблицах в целых процентах, причем пустые зобы
не приняты в расчет. В подзаголовке «количественных» таблиц дан
процент пустых зобов и средний вес содержимого одного зоба, вы-
численный только по фактически взвешенным, без поправки на пу-
стые зобы. Питание птенцов (с момента рождения до 1 октября, т. е.
до трех'месячного возраста) дается особо. Начиная с L октября все
птицы, молодые и старые, рассматриваются вместе. Число птенцов,
добытых в июне, так мало, что их не стоило» выделять, и они вклю-
чены в июльский столбец. В тех случаях, когда за месяц было добы-
то Ь—2 птицы, я /присоединял их к тому из соседних месяцев, куда
4*
51
они больше подходили по датам, что в каждом отдельном случае
указано стрелкой.
Таким образом, по каждому виду птиц мы даем три таблицы:
1) качественную таблицу питания взрослых—здесь дается ветре,
чаемость кормов в целых процентах; за ЮО принято число исследо
ванных зобов, не считая пустых;
2) количественную таблицу 'питания взрослых птиц — дается
«удельный вес» корма по месяцам в целых промиллях; здесь сумма
по столбцу равна 10*00;
3) качественную и количественную таблицы питания птенцов за
3 месяца.
Большинство авторов делят корма куриных на несколько групп,
каждая из которых необходимо- входит в состав пищи в определенное
время года: грубые древесные корма, зеленые травяные, ягодные,
животные и минеральные. Мы детализировали эту схему, разбив
вторую из перечисленных групп на 4 самостоятельных. В наших таб-
лицах приняты 9 групп кормов, по каждой из которых мы даем
отдельной строкой итого1вый процент встречи — «удельный вес».
Чтобы не перегружать таблицы, мы не даем в них полного списка
всех встреченных растений и животных: дополнительный перечень
более редких и случайных кормов дан в тексте. Количественные таб-
лицы составлены следующим' образом: в карточках на добытых птиц
проставлены действительные или приближенные -веса кормов в све-
жем виде; веса каждого корма в отдельности складываются по меся-
цам; сумма их делится на общий вес всех кормов во всех зобах за
Таблица 7
Число птиц, использованных в таблицах питания
Вид Возраст Число зобов
всего с кормом пустых
абс. 1 В %
Глухарь Взрослые 452 366 86 19
Птенцы 131 105 26 20
Тетерев > Взрослые 26 24 2 8
Птенцы 27 22 5 18
Рябчик Взрослые 136 128 8 6
Птенцы 74 61 13 18
Белая куропатка . . Взрослые 296 270 26 9
Птенцы 54 50 4 7
Тундряная куропатка . Взрослые 64 58 6 9
Птенцы 21 19 2 10
Итого: 1281 1103 178 —
52
данный месяц; частное от деления ,выраженное в промиллях, вносит-
ся в соответствующую графу таблицы. Таким образом, сумма в каж-
дой колонке должна быть равна 1000, если складывать отдельные
корма; итог сумм по группам кормов также равен 1000. Исключение
сделано только для группы животных кормов — здесь для отдельных
отрядов дан не вес, а число найденных особей: «удельный вес» жи-
вотных кормов показан только в заголовке этой группы кор'мо>в.
Глухарь (табл. 8—10, рис. 21).
В питании глухаря группа зимних кормов играет исключительно
важную роль. В Лапландском заповеднике в течение 5 месяцев (с
декабря по апрель) глухарь питается исключительно хвоей с при-
месью веток, шишечек сосны и плодов можжевельника; в небольшом
количестве он ест их и летом. Хвоя преобладает в пище глухаря с 28 X
до20 V (1204 дня в году). В годовом цикле питания первое место- за-
нимает сосно!вая хвоя. Глухарь или отстригает ее острыми краями
клюва, как ножницами, или отльпмывает клювом концевой побег
вместе с хвоей и верхушечной почкой. Хвою молодых сосенок глу-
хари иногда щиплют стоя в снегу, но обычно они кормятся на кро-
нах взрослых сосен. Неоднократно указывалось, что у глухарей бы-
вают любимые сосны, на которых они питаются преимущественно
перед другими (Лобачев и Щербаков, 1'936; Семенов-Тяй-Шанский,
1938; Кирилов, 1952; Линдрют и Линдгрен, 1950). Ветви таких сосен
бывают сильно общипаны (см. рис. 5|). Зимой 1940-41’ г. мы заме-
тили, что глухари охотно кормились на погибающих соснах в зоне
затопления Охтозера, уровень которого был поднят в связи со строи-
тельством Нивской гидроэлектростанции. Можно предполагать, что
глухарь выбирает те сосны, у которых почему-либо нарушен пра-
вильный обмен веществ и продукт фотосинтеза (углеводы) задер-
живается в хвое.
Известно, что семена сосны созревают не через 3—4 месяца после
цветения, как у ели и пихты, а только через 16 месяцев. Зимой на
сосне бывают шишки двух родов — взрослые, со спелыми семенами,
и (молодые, опыленные, но с еще неоплодотворенными яйцеклетками.
Эти молодые шишечки глухарь также ест, хотя и реже, чем хвою. В
зобах двух глухарок были встречены и целые всходы сосны, 25 IX
1939 г.—2 экз., 27 X 1938 г. 8 экз.
Охвоенные веточки и плоды можжевельника — любимая пища
глухаря, когда он кормится на поверхности снежного покрова.
Еловую хвою глухари обычно не трогают и едят ее только в слу-
чае зимовки в обширных еловых массивах, где совсем1 нет сосны.
Таких птиц нам случалось добывать только весною, отсюда и уве-
личение встреч еловой хвои в апреле—июне. Летом 1в двух зобах
была встречена собственно говоря не хвоя, а галлы с еловых веток
(6 VII 1937 г. — 39 галлов и 24 VII 1940 г. — 4 галла).
В целом в снежное полугодие (XI—IV) сосна составляет 9Оо/о
рациона глухаря, можжевельник—8°/0, а все «подснежные» корма
в совокупности—2о/о. В летнее полугодие (V—X), при обильном под-
ножном корме, сосна составляет Ii6o/G, можжевельник — 2»/0, ель —
53
Таблица 8
Качественный анализ питания взрослых глухарей (старше 3 мес.)
I 11 Ill IV V VI VII VIII IX X XI XII
Число зобов с пищей 10 12 15 17 36 9 5 21 71 81 72 17
Число зобов пустых . . 1 1 2 1 13 2 0 2 19 23 20 4
I. Веточные корма 100 100 100 100 88 78 40 19 20 69 96 100
Сосна, шишки первогод. 100 92 100 94 78 33 — 5 6 41 83 100
Сосна шишки первогод. 10 17 7 12 3 — — — — — 6 --
Можжевельник, хвоя . 10 25 — 24 19 — — —- 4 36 21 6
Можжевельник, плоды . 10 17 — — 6 — — 14 3 7 11 —
Ель, хвоя, побеги . . —. — — 12 3 22 40* — 1 5 4 -
Береза, побеги, сережки — — — — 14 — — — 7 10 6 —
Рябина, осина, ивы - — — И 33 — — — 2 — —
П. Вечнозеленые ра.
стения — —> — — 53 78 60 38 75 83 40
Черника, побеги, листья — — — — 39 67 60 33 69 77 35
Брусника, листья . . . — — — — 3 — — 5 7 15 3 —.
Андромеда, листья . . —- — — — 28 33 — 5 4 12 10
Вороника, побеги, листья — — — — 8 11 — 10 14 9 3 —
Вереск, багульник . . — — — — 3 — — 5 — 6 6 —
HI. Летняя зелень — — — 19 67 100 76 80 40 3
Голубика — — — — 8 22 60 62 59 7 3 —
Ожика волосистая . . — — — — 6 22 20 —- 37 22 —
Хвощ лесной .... Трехраздельный папо- — — — — — — 40 29 10 2 — —
ротничек — — — — — — 20 — 6 — — —
Луговик — — — — 3 11 20 10 28 23 —
Ольха, осина . . . - — — — — — 11 — — 6 2
Прочие виды .... — — — — 11 44 40 5 3 10 — —
IV. Цветы .... — — — 11 44 40 — —
Пушица влагалищная . -— •— — — 11 33 — — — — — .—
Ивы — — — — — 11 — — — — —-
Черника — — — — — 11 20 — — — — —
Ястребинки — — — ... — — 20 — — — — —
V. Ягоды — — — — 44 67 80 72 86 83 29 —
Черника - — — — 3 11 60 71 78 67 17 —
Голубика — — — — — — — 14 8 4 6 —
Брусника — — — — 25 33 20 — 31 62 21 —-
Вороника — — — i — 39 44 60 43 55 48 19
Морошка — - — — — — — 5 1 — —
Клюква мелкоплодная . — — — — 3 11 — — — — —• —
VI. Семена, плоды — — — — 3 — 40 62 56 19 3
Осоки — — -— — — — 19 11 7 1 —
Березы — — — — — — — 5 1 9 1
Андромеда — — — — — — 5 3 1
Марьянник луговой . . — — — — 3 — 40 38 35 2
Золотая розга .... — — — — — — — 10 14 5 ---г
Прочие виды .... — — — — — — — 14 10 1 —
* Галлы (вероятно, тлей).
54
Табл. 8 (продолжение)
; । । II 111 1 1 IV V VI VII VIII IX X XI XII
VIL Низшие растения — — — 3 20 10 18 6 1 —
Грибы пластинчатые . — 16 1 — —
Гнилая древесина . . — — 5
Мхи политриховые . . — — — — — — 20 5 3 4 1 ’—
Ягель, древ. лиш. . . — — — — 3 — — — 3 2 — —
VIII. Животные . . — — — — 3 11 20 24 11 1 — —
Слизни и улитки ... — — 5 3 — — —
Пауки —* — — . — — — 20 — 1 — — —
Прямокрылые (кобылки) — —
Жуки и их личинки . . — -— — — 3 11 20 5 1 — -— —
Личинки пилильщиков . — — — — — — 20 — — — — —
Муравьи и их коконы . — — — — — — 20 14 6 — — —
Двукрылые, их личинки — — — — — — — 5 — 1 — —
Гусеницы бабочек . . — — — — — 20 •— 1 — — —
Ящерица живородящая —
IX. Камешки ... — — — — т- — 20 — 10 4 1 —
Та б л и ц а 9
Количественный анализ питания глухарей (старше 3 мес.)
I 11 Ill IV V VI VII VIII IX X XI XII
Пустые зобы, % . . . 8 12 6 27 18 0 _9 21 22 22 11
Ср. вес зоба на 1 кг ве- са птицы 27 20 1 1 1 2 10 29
Число взвеш. зобов . . (7) ( 9) (И) (5) (36) (17)
I. Веточные корма . 1000 1000 1000 1000 684 260 102 9 40 421 877 1000
Сосна, хвоя, побеги . . 880 933 999 887 431 176 — 1 28 352 691 962
Сосна, шишки первогод- ние 27 5 1 4 0 — 5
Можжевельник, хвоя . 93 ^8 — 107 18 — — — 1 56 137 38
Можжевельник, плоды . 0 4 — — 1 — — 8 6 10 41 —
Ель, хвоя, галлы . . . — 2 115 53* 102* — 0 1 1
Береза, побеги, сережки — — — 116 — — — 5 2 2 —
Рябина, осина, ива . . — — — — 3 31 — — — 0 — —
II. Вечнозеленые ра- стения 119 347 32 5 148 169 84
Черника, побеги, листья — — — — 86 335 32 4 146 111 76
Брусника, листья . . . — — — — 0 —• — 0 0 9 1
Андромеда, листья . . — — — 33 12 — 0 1 44 6
Вороника, побеги, листья — — — — 0 0 — 1 1 1 0
Вереск, багульник . . — — — — 0 — — 0 — 4 1 —
* Галлы
55
Табл. 9 (продолжение)
1 II III IV V VI VII VIII IX X XI XII
III. Летняя зелень . 3 90 553 359 331 59 0
Голубика .— — — — 0 60 337 314 210 6 0 —
Ожика волосистая . . — — — 2 3 14 — 95 40 — —
Хвощ лесной .... — — — — — — 13 39 13 0 — —
Трехраздельный папо- ротничек — — — 160 — 4 — — —
Луговик — — — — 0 0 1 1 4 10 — —
Ольха, осина .... — — — — — 3 — — 4 0 — —
Прочие виды .... — — — — 1 24 28 3 1 3 — —
IV. Цветы .... — — — — 90 142 38 — — — — —
Пушица влагалищная . — — — — 90 138 — — — — — —
Ивы 3
Черника — — — — — 1 11 — — — — —
Ястребинки 27
V. Ягоды — — — 104 160 91 472 373 332 37 —
Черника — — — — 2 0 76 234 260 119 6 —
Голубика — 52 4 3 1 —
Брусника — — — — 42 18 2 — 20 98 18 —
Вороника — — — — 35 138 13 163 86 112 12 —
Морошка — — — — — — — 23 3 — — —
Клюква мелкоплодная — — — — 25 4 — — — — — —
VI. Семена и плоды — 0 — 3 144 70 7 2
Осоки — 26 12 2 1 —
Марьянник (с листьями^ — — — — 0 — 3 64 33 0 —. —
Андромеда — 5 4 1 — —
Золотая розга .... — 1 16 0 — —
Березы — 0 1 4 1 —
Прочие виды .... — 48 4 0 — —
VII. Низшие растения — — — — 0 — 0 1 35 3 0 —
Грибы пластинчатые . — — — — — — —- — 35 3 —. —
Гнилое дерево ... - 0
Мхи политриховые . . — — — — — — 0 1 0 0 0 —
Ягели, древ, лишайники — — — — 0 — — — 0 0 — —
VIII. Животные . . — — — 0 0 170 10 1 9
Слизни и улитки (число экз.) 4 2
Пауки — — — — — — 1 — 1 — —
Прямокрылые . . . «
Жуки и их личинки . — — — — 1 1 2 67 1 —
Пилильщики, личинки . 303
Муравьи и их коконы . — — — — — — 1 3 6 — —
Двукрылые, их личинки — — — —. — — — 1 — 187 —
Гусеницы бабочек . . — — — — — — 1 — 1 — —
Живородящая ящерица — — — — —
IX. Камешки . . . — 1 — — — — 11 — 2 1 0 —
56
Таблица 10
Анализ питания птенцов глухарей (до 3 мес.)
Качествен- Количест-
пый венный
VII VIII IX VII VIII IX
Число зобов с пищей . 12 22 71 % пустых зобов . . . 8 21 21
Число пустых зобов . . 1 6 19 Ср. абс. вес пищи . . 2 6 33
Число взвешен, зобов . 3 1 8
I. Веточные корма . — 5 18 — 8 28
Сосна, хвоя, побеги . . — 5 6 — 0 2
Сосна, шишки первогод. Можжевельник, побеги — — — — — —
с хвоей — —— 3 — — 0
Можжевельник, плоды . — 5 8 — 8 17
Ель, побеги с хвоей . — — 4 — — 0
Береза, побеги, сережки — — 6 — — 9
Рябина, осина, ива, поч- ки .
П. Вечнозеленые раст. 42 14 53 14 8 48
Черника, побеги, листья 25 14 38 10 7 36
Брусника, листья . . — — 8 — — 1
Андромеда, листья . . 8 — 14 ’ 3 — 9
Вороника, побеги, листья 8 5 17 1 1 2
Вереск, толокнянка . . — 5 1 — 0 0
III. Летняя зелень . 33 68 92 51 490 500
Голубика 33 59 65 50 386 347
Ожика волосистая . . — — 49 — — 91
Хвощ лесной .... — 41 20 — 104 38
Трехраздельный папо- л
ротничек Луговик 8 — 1 41 *7 1 — О 17
Береза, осина, листья . — — / 12 — 1
Прочие виды .... — — 6
IV. Цветы .... 8 — — 10 — —
Пушица влагалищная . — — — —
Ивы — —
Черника 8 10 —
Ястребинки 85 78 18 28
V. Ягоды . . • • • Черника Голубика 67 25 42 8 68 50 32 634 48 67 8 209 53 101 304 166 30
Брусника 58 32 45 511 36 oZ 53 1 В
Вороника . • • • • 9 3 18
Морошка . • ’ ’ ' Толокнянка, клюк , 5 3 1 л
смородина • • • VI. Семена и плоды . 67 17 77 64 63 23 74 28 183 112 4 90
Осоки 50 55 27 46 57 •JU 19 *7
Марьянник • • * * ‘ — 6
Андромеда • * \ ’ — — 17 — __ / Q
Золотая розга • • ’ . Березы . • ‘ . — 5 18 10 13 3 11 У 15 А
Прочие виды •
Табл. 10 (продолжение)
Качествеп- н ы й Количест- венный
VII VIII IX VII VIII IX
VII. Низшие растения 17 18 14 2 2 7
Грибы пластинчатые . — — 6 4
Гнилое дерево .... - — — о о —
Мхи политриховые . . 17 14 6 1 2 г\ 3
Ягели, древ, лишайники — 5 3 и 0
VIII. Животные . . 83 73 42 199 96 21
Слизни и улитки . . . 8 9 16 4 1 с 6 246
Пауки 33 23 4 16 19 4
Прямокрылые (кобыл-
ки) — 5 — — 5 —
Жуки и личинки . . . 8 18 1 1 5 1
Пилильщики, личинки . 42 — — 73 — —-
Муравьи и их коконы . 33 32 25 14 180 83
Двукрылые и их личинки 25 18 1 8 11 1
Гусеницы бабочек . . 58 32 7 60 10 11
Ящерица живородящая — 9 — — 2 —
IX. Камешки . . . 8 14 13 16 4 2
4<Vo, лиственные породы — Зю/о рациона. Важно отметить, что по ;всей
области распространения глухаря зимой он питается преимуществен,
но сосновой хвоей (см. ниже о питании глухаря в географическом
разрезе); исключения наблюдаются только на южной окраине ареа-
ла (Кириков, 1952, — на Южном Урале; Вурм, 1885,—в Южном
Европе|).
Вечнозеленые кустарнички, преимущественно зеленые веточки
черники, играют большую роль в питании глухаря дважды в году:
весной — от момента появления первых проталин до начала роста
трав, и осенью — вплоть до установления снежного покрова. Летом
глухарь иногда ест листья черники (хотя они и опадают на зиму, но
для упрощения обработки мы объединили их с веточками черники).
Листья и другие части летнезеленых растений составляют основу
питания глухаря летом. Особенно важны листья голубики, затем
волосистая ожика (листья и стебли с плодами), лесной хвощ, и трех-
раздельный папоротничек. Листья луговика встречаются часто, но
всегда в малом количестве. Находили мы также листья осины, не-
которых ив, ольхи, березы и многих травянистых растений: осоки
(Carex sp. sp.), трихофорум1а (Trichophorum bracteatum), купальни-
цы (Trollius europaeus), сабельника (Comarum palustre), завязника,
лабазника1 (Filipendula ulmaria), морошки, герани, шведского дер^'
на, грушанок (Pirola sp. sp.|), марьянника, линнеи, ястребинкп
(Hieracium sp. sp.), бодяка (Cirsium heterophyllum) и золотой розги.
Судя по- составу поедаемых растений, летом, глухари охотно жор'
мятся по» берегам! ручьев, окраинам болот и другим влажным местам*
Цветы. В этой группе кормив наибольшее значение имеет пушин^
колоски которой появляются рано* весной на вытая!вших от снега
58
кочках сфагновых болот (рис. 22). В зобу у глухарей, добытых на
току, сйбычно обнаруживается сосновая хвоя, которую они клюют
тут же, с дерева; но у застреленных днем зоб иногда буквально на-
бит колосками пушицы (самец 15 V 1938 г. — 165 колосков общим
зимние веточные корма
зим не зеленые листья и
nodeeu
летне - зеленые листья
ягооь/
семена и коробочки
мхи и грибы
еалъко
молодые
Рис. 21. Питание глухарей по временам года
весом 17,9 г; самка 6 VI 1939 г.— 112 колосков; кроме того, еще
у 4 самцов и 1 самки). Все эти птицы добыты в период максималь-
ной половой активности, обе самки — в период откладки яиц.
В. П. Теплой (1'947) обнаружил цветы пушицы в зобах :2!8°/о глу-
харей, добытых на Верхней Печоре в Miae. Замечательно, что и на
59
Южном Урале С. В. Кириков (1952) нашел, что «в весеннем питание
глухарей очень большое значение имеют цветы», особенно оон-травц
(до 21<>/о), хохлатки, дуба, осины и ивы; он же ссылается на А. Чер-
касова (1867|), указавшего, что цветы сон-травы — один из главных
весенних кормов глухаря в Восточной Сибири. В Беловежской путце,
как указывает В. Ф. Гаврин (1956), в апреле и мае глухари едят
бутоны сон-тра!вы, соцветия пушицы и другие цветы.
Рис. 22. Пушица влагалищная в цвету; 7/в нат. вел.
Сведения об употреблении цветов в разных местах ареала глу-
харя показывают, что весной, как и летом, глухарь не зависит от
каких-либо определенных видов растений, а легко заменяет цветы,
листья или ягоды одних видов растений другими: выбор кормовых
растений определяется составом флоры местности. Вместе с тем глу-
харь не может обойтись без одной из (групп кормов — например, без
ягод. Зимой дело обстоит совсем иначе: © это время существование
его тесно связано с хвойными породами, в частности с сосною.
Питание глухарей цветами в период размножения может -объяс-
няться тем, что в это время очень мало свежей зелени, но вполне
возможно, что в цветах 'содержатся вещества, необходимые для нор-
мального хода размножения.
60
Ягоды. Весной, на проталинах, глухарь наьходит перезимовав-
шие ягоды брусники и (вороники. На Кольском полуострове последние
сохраняются до поспевания новых в таком количестве, что живот-
ные не поспевают их съесть. В связи с этим мы иногда находили в
одном зобу ягоды вороники двух генераций (свежие и прошлогод-
ние). У черники глухарь ест сначала цветы, потом завязи; с момента
поспевания ягод (в среднем за 20 лет 28 VII) до выпадения снега
черника — любимая ягода глухаря. За черникой идут по порядку: во-
Рис. 23. Марьянник с семенами; немного увеличено
роника, брусника и на последнем месте голубика. Морошка рано схо-
дити редко дает хороший урожай, вероятно, потому глухари мало
ее едят. Остальные ягоды (клюква, толокнянка, красная смороди-
на) — случайный корм.
Замечательно, что глухарь никогда не наполняет зоб одними яго-
дами: вместе с ними, мы (всегда находим зелень и другие корма. В
августе, сентябре и октябре глухарь питается преимущественно яго-
дами; в этот период все содержимое кишечного тракта и помет птиц
окрашены черникой в темно-синий цвет. В целом за летнее полугодие
ягоды составляют около 25о/о рациона.
61
Семена и коробочки разных растений поедаются в небольшом ко.
личестве (4<>/о за летнее полугодие), но очень регулярно, особенно
белые семечки марьянника (рис. 23), которые можно отнести к числу
самых любимых кормов. Марьянник растет всюду, и в борах и в ель.
никах, но всегда рассеянно: на той же площади он производит в де.
сятки раз меньше плодов и зеленой массы, чем черника. Глухарь
ест не только семена марьянника, но вместе с ними чашечки, листья
и стебли. Семена по вкусу напоминают лесной орех и, вероятно,
очень питательны.
Из осок в зобах отмечены следующие виды: Carex Buxbaumii,
С. canescens, С. flava, С. irrigua, С. lasiocarpa, С. limosa, С. раись
flora, С. rariflora, С. rotundarta, С. vaginata (некоторые определения
неточны|). Нередко встречались коробочки подбела, побуревшие кор-
зинки золотой розги с плодами, семенные сережки берез — пушистой
и карликовой и единично плоды грушанок, лесной герани, линнси,
завязника, купальницы, корзинки ястребинок, колоски душистого
колоска, ситников и плоды шейхцерии.
Грибы — сезонный осенний корм. Глухарь заглатывает грибы
М'елкими кусочками, почему определить их до вида нам не удава-
лось: обычно они относятся к пластинчатым. Политриховые мхи,
клочки ягелей и древесных лишайников почти не имеют значения в
питании и, возможно, являются случайной примесью.
Животные в пище глухаря принадлежат к трем типам; они
играют заметную роль с июля по сентябрь.
Моллюски представлены слизнями рода Arion, многочисленными
в лесах Кольского полуострова, и мелкими наземными улитками,
наиболее похожими на Succinea и Goniodiscus; членистоногие —
пауками, кобылками, муравьями (коконы и взрослые), личинками
пилильщиков, гусеницами бабочек, жуками и их личинками (Curcu-
lionidae, Chrysomelidae др.), мухами и их личинками; позвоноч-
ные— только живородящей ящерицей: хвосты этого вида встречены
в зобах двух молодых птиц в августе.
Животные корма могут преобладать над растительными только
у маленьких птенцов. Приведем данные о нескольких зобах, содер-
жавших наибольшее число животных:
6 VII 1937 г. взр., вес 3325 г — 303 личинки пилильщиков, 1 паук.
3 гусеницы, 5 муравьев, 1 долгоножка Tipula, 3 жука (Sericeus
brunneus, Otiorrhynchus sp.). Вес животной пищи —15,5 г, расти-
тельной — 37,6.
15 VII 1938 г., 2мол., вес 122 г — 24 гусеницы бабочек, 8 личинок пи-
лильщиков, 7 пауков, 1 жук, 3 муравья, 1 кокон паука, 4 улитки;
растительной пищи пет.
17 VII 1950 г., 2 мол., вес 283 г —26 личинок пилильщиков, 5 муравьев.
Вес животной пищи 1,3 г, растительной — 1,35 г.
28 VII 1950 г., $ мол., вес 193 г, 22 личинки пилильщиков, 2 остатка
не опр. насекомых. Вес животной пищи —1,05 г, раститель-
ной — 1,7 г.
5 VIII 1935 г., 2 мол., вес 528 г — преобладают муравьи и их коконы.
3 паука, 5 их коконов, 1 куколка, часть хвоста ящерицы; живот-
ной и растительной пищи приблизительно поровну.
62
5VIII 1940 г., $ мол., — коконы муравьев, 1 хвост живородящей ящерицы;
растительный корм преобладает.
15VIII 1950 г., $ взр., вес 1895 г — 67 жуков сем. слоников; вес их 2,9 г,
растительной пищи — 78.1 г.
14 IX 1933 г., J мол., вес 2052 г —около 200 слизней Arion subfuscus
(опр. Е. С. Раммельмсйср); растительная пнш,а преобладает.
4 X 1950 г., о мол., вес 1732 г—187 личинок Bibio sp. (опр. А. А.
Штаксльберг); вес их 16,2 г, растительной пищи — 45,6 г.
Камешки и песок составляют большую часть содержимого желуд-
ка, но в зобах встречены только у 12 взрослых птиц и 13 птенцов,
в период с 5 VII по 26 IX. По приблизительному подсчету, они со-
ставляют 0,lo/о всего рациона взрослых птиц за год и О’,7о/б рациона
птенцов (за 3 месяца). К вопросу о камешках мы вернемся в сле-
дующей главе.
Питание птенцов глухаря. По пуховым птенцам у нас нет мате-
риала. Представленные в таблице данные за июль относятся к птен-
цам весом от 100 до 500 г. Глухарята едят те же корма, что и взрос-
лые, только в несколько ином соотношении.
Животные корма у птенцов встречаются приблизительно в 4 раза
чаще, чем у взрослых, а по количеству — раз в 10 больше *. Пре-
обладание насекомых в корме птенцов известно и для многих певчих
птиц со смешанным питанием.
Птенцы поедают также больше семян, чем взрослые: так, напри-
мер, семена осок за 3 месяца составили: у взрослых— 1°/о, у моло-
дых птиц — 6^/о всего рациона. Судя по диаграмме (рис. 21) в июле
птенцы едят больше ягод, чем взрослые, но этот вывод не подтверж-
дается на других птицах и должен быть приписан случайности: дело
в том, что все птенцы были добыты- во второй половине июля, когда
ягоды уже поспевают, а из 5 взрослых 3 добыты в первых числах
месяца — в это время ягодники только цветут. В августе и сентябре
взрослые птицы в большей мере питаются ягодами, а молодые — зе-
леными частями растений, особенно листьями голубики (26о/0 рацио-
на взрослых птиц и 3i7<>/o рациона молодых за эти месяцы).
К ягодам голубики птенцы относятся иначе, чем взрослые: в.
июле—сентябре у первых черника составила 19о/о рациона, а голу-
бика — 2%, т. е. в 10 раз меньше; у вторых черника — 9о/о, голуби-
ка— 7о/о, разница только в 1,3 раза. В свое время мы высказали
предположение, что «молодые едят ее по неопытности, не отличая от
черники, а взрослые избегают ее есть» (Семенов-Тян-Шанский,
1938). Интересно, что значение голубики закономерно уменьшается
по мере роста птенцов. Как мы видели, в пище молодых глухарей
первое место по количеству занимают листья голубики; кормясь у
кустов голубики, птенцы вместе с листьями срывают и ягоды, почему
•в первый месяц их жизни голубика преобладает над черникой, хотя
ягоды последней и сахаристее и сытнее. Постепенно птенец научает-
ся различать эти ягоды и к концу ягодного сезона ест голубику
* Помещенные в табл. 9 данные о поедании животных кормов в июле не
показательны потому, что было исследовано только 5 взрослых птиц, одна из ко-
торых съела много жуков.
63
только тогда, когда кустики черники скроются под снегом: поэтому в
ноябре значение голубики вновь немного возрастает.
Из зимних кормов глухарята прежде начинают есть плоды мо>к.
жевельника (5 VIII 1935 г.), а потом уже сосновую хвою (12 IX
1936 г.; 27 IX 1937 г.). Питание хвоей требует новых навыков -,
умения кормиться на дереве: молодые птицы приобретают этот на,
вык в возрасте 3—4 месяцев.
В заключение перечислим работы по питанию глухаря в СССР,
учитывая лишь те, где материал охватывает большую часть года.
Район Широта Долгота Автор и год опубликования
Лапландский заповедник . . Печоро-Илычский заповедник РСФСР, ряд областей . . . Башкирский заповедник . . 68° с. ш. 62° , „ 53о? , 32° в. д. 57° , „ 58о г в Семенов-Тян-Шанский (1938) Теплов (4'944) Лобачев и Щербаков (1'936) Кириков (4952 и др.)
Тетерев (табл. 11—12, рис. 24).
Питание тетерева изменяется в течение года таким же образом,
как и питание глухаря. Не повторяя того, что уже сказано по пово-
ду глухаря, остановимся на отличительных особенностях тетерева.
Зимний рацион его, очевидно, более разнообразен. К сожалению,
судить об этом приходится по 9 птицам, добытым частью в начале
зимы (XI—ХП|), частью в конце ее (III—IV).
Береза, по нашим данным, главный зимний корм: тетерев ест
как мужские сережки, так и кусочки веток с почками (недаром
его и зовут местами березовиком). Сосна занимает второе место.
Тетерев ест не столько хвою, сколько уже упомянутые нами шишки
по первому году, число которых в одном зобу иногда достигало
166—176 шт. (6 IV 1948 г., ?, вес шишек 24,0 г; 12 XI 1938 г.д?).
Как это отметил и Теплов (1947), тетерев ест шишечки не каждую
зиму, а только в те годы, когда их много, что бывает за год до уро-
жая семян. Мы находили их в пище тетерева зимой 1938—39,
1947—48 и 19481—49 г., но не встречали зимой 1'9134—35 г. В 1939 г.
был хороший урожай сосновой шишки, а в 1935 г. — низкий (Вла-
димирская, 1948-а, стр. 129).
Можжевельник тетерев ест приблизительно в тех же размерах,
как и глухарь. С. В. Лобачев (1933) считал ягоды можжевельника
важнейшим кормом тетерева, от которого зависит благополучие
его популяции. Из текста автора трудно понять, хорошо ли этот
вывод обоснован на его материале; впрочем, и А. Н. Дубровский
(1930) считал, что в Новгородской области можжевельник имеет
для тетерева не меньшее значение, чем береза. А. С. Фетисов (1934)
для Прибайкалья, С. В. Кириков (1952) для Южного и В. П. Теи-
лов (1947) для Северного Урала, В. Ф. Гаврин (1956) для Бел°'
вежской пущи указывают тот же состав зимних кормов, как и мы*
даже кавказский тетерев, по наблюдениям Ю. В. Аверина (1938)»
64
Таблица 11
Качественный анализ питания тетеревов (взрослых и молодых)
1 II ш IV V VI VII VIII IX X XI XII
1
Число зобов с пищей . Число зобов пустых . . — — - 2 1 U1 1 1Н - 4 3 22 2 6 1 5-
I Веточные корма — — — 100 50 — — 0 18 83 100 —
Сосна, хвоя .... Сосна, шишки первогод. Можжевельник, побеги — 100 67 — — — 33 33 67 17 —
с хвоей 5 17 —
Можжевельник, плоды 100 50 — 5 17 17 —
Березы, побеги, почки . —• — — — —- — 5 67 50 —
березы, цвет, сережки Ивы, побеги, почки . . — — — 33 25 — — — 5 33 17 —
17
П. Вечнозеленые ра-
стения — — — 33 100, — — 20 59 83 50 —
Черника, побеги, листья — — — — 100 — — 20 46 67 50 —
Брусника, листья . . . — — — 33 — — — — 5 .— — —
Андромеда, листья . . — — — — 25 — — — 14 33 33 —
Вороника, побеги, листая Вереск, побеги с листья- — — — 33 — — — — 5 — — —
ми — — — — — — — — 14 — — —
III. Летняя зелень 40 86 33
Голубика — —. — — — — — 20 50 —
Хвощ лесной — — — — — — — 9
Луговик 55 33 —
Березы — — — — — — — 20 9 —
Герань лесная . . . — — 20
Ястребинки .... — 20 5 — — —
IV. Цветы 25
Андромеда — - — — 25 — — — — — — —
V. Ягоды ... — — — 75 — — 40 91 100 33
Черника — — 20 82 83 17
Голубика Брусника .... * Вороника ... * * ДеРен, клюква мелко- — — — — — —. — 20 23 17 17 —
— — — — 75 — — — 50 83 33 —
— — — — 50 — — 40 36 17 — —
плодная — — — — — — — — 5 — 17 —
ОсокиСеМе"а “ плоды • 1,Рочце виды . ’. — 80 68 50 17 —
— — — — — — — 60 27 33 17 —
— — — — 20 40 20 14 36 14 33 17 17 17
Низшие растения политриховые 5 17
— 5 17 — —
5
3037
65
Табл. 11 (продолжение)
1 11 1П IV V VI VII VIII IX X XI XII
VIII. Животные . < — 25 — — 60 64 17 — —
Пауки — — - — — — — — 5 17 — —
Прямокрылые (кобылки) Жуки — — — — — — *— 20 5 9 17 —
Пилильщики-личинки . — — — — — — — — 5 — — —
Муравьи и их коконы . — — — — 25 — — 40 59 — — '—
Двукрылые * . . . . — — — — — — — 20 — — — —
Гусеницы бабочек . . 5
IX. Камешки . . . । - • — — — — — — 20 9 33 — -
* Bibio sp.
Таблица 12
Количественный анализ питания тетеревов (взрослых и молодых)
I 11 Ш IV V VI VII V111 IX X XI XII
Процент пустых зоб. . — — — 0 20 — — 37 8 I 14 0 - V
Ср. вес пищи в зобу . — — 75 21 13 15 67
Число свеш. зобов . . — — ( :2) 1 :з) i :2) (10) ( :п
I. Веточные корма . — — — 990 344 — — — 14 251 729 —
Сосна, хвоя .... — — — 255 — — — — — 8 38 —
Сосна, шишки первогод. Можжевельник, побеги — — — 130 — — — — — — 245 —
с хвоей 0 1 31 —
Можжевельник, плоды — 10 1 56 —
Березы, побеги, почки . — — — 600 8 — — — 0 28 45 —
Березы, цвет, сережки — — — 5 336 — — — 4 203 314 .—
Ивы, побеги .... — — — — — — — — — 10 — —
II. Вечнозеленые ра-
стения — — — 10 170 — — 2 83 279 83 —
Черника, побеги, листья — — — —• 168 — 2 51 76 45 —
Брусника, листья . . — — — 5 — — — 10 —
Андромеда, листья . . — — — — 2 — — — 4 203 38 —
Вороника, побеги, листья Вереск, побеги с листь- — — — 5 — — — — 0 — — —-
ями — — — — — — — — 18 — — —
III. Летняя зелень . 27 124 1
Голубика — — — — 7 92
Хвощ лесной .... — — — — 4
Луговик — — — — 18 1 .
Березы — — —, — — 1 7
Герань лесная .... — — .— 3 -
Ястребинки — — — — — — — 16 3 — — 1 —
66
Табл. |J2 (продолжение)
1 II 111 IV V VI VII VIII IX X XI XII
IV. Цветы .... 2 — — — —
Андромеда — — — — 2 — — — — — — —
V. Ягоды — — — — 484 — — 605 542 413 176 —
Черника — 80 348 136 75 —
Голубика — 517 105 183 3 —
Брусника — — — — 420 — — — 63 86 97 —
Вороника Дерен, клюква мелко- — — — 64 — — 8 26 8 — —
плодная 0 —• 1 —
VI. Семена и плоды . 348 215 47 12 —
Осоки — — — — — — — 216 76 7 6 —
Береза пушистая — 63 9 40 6 —
Марьянник .... — 56 127 0 — —
Прочие виды .... — 13 3 0 — —
VII. Низшие растения — 0 2 — —
Мхи политриховые . . 0 2 — —
VIII. Животные . . — — — — 0 — — 15 22 0 — —
Пауки — — — — — — — — 3 2 — —
Прямокрылые (кобылки) Жуки — — — — — — — 1 1 2 1 — —
Пилильщики, личинки . — 1
Муравьи — — — — 1 — — 15 220 — — —
Двукрылые * . . . . — 6
Гусеницы бабочек . . — 8
IX. Камешки . . . 3 0 7
* iBibio sp.
питается зимой в первую очередь березой, затем можжевельником
и другими растениями.
Как видно из рис. 24, период преимущественного употребления
зимних кормов у тетерева меньше, чем у глухаря. По карточкам
биосъемки, составленным на всех добытых птиц, мы определили
его продолжительность в 194 дня (с 31 X по 13 V). По недостатку
материала потребление цветов в период токования не нашло отра-
жения в наших таблицах и диаграмме. А. С. Фетисов (1934) нашел
цветы сон-травы* у 70% тетеревов, добытых в мае в юго-восточном
Прибайкалье. В. Ф. Гаврин (1956) сообщает, что в весеннем пи-
* Pulsatilla patens.
5*
67
тании косачей Беловежской пущи преобладает клюква (у 91,% осек
бей), затем соцветия осок (27%) и пушицы (18%). Следовательно,
и в этом отношении тетерев сходен с глухарем.
зимние Неточные корма
гимне-зеленые f/истья и
поблей
летне -зеленые листья
| о ос>] ягоды
ШШ №ММО
и коробочки
мхи и гаибы
Рис. 24. Питание тетеревов
по временам года
Большее участие ягод голубики, по сравнению с черникой, в пи-
тании тетерева объясняется тем, что в нашем материале преобла-
дали молодые птицы. По нашим наблюдениям, а равно и по дан-
ным других авторов (Лобачев и Щербаков, 1933), тетерев ест
меньше зелени, но больше ягод и семян, чем глухарь.
68
Приводим приблизительные координаты пунктов, где было изу-
чено питание тетерева:
Район Широта Долгота Автор и год опубликования
Лапландский заповедник . . 68° с. ш. 32° в. д. Семенов-Тян-Шанский (1'938}
Печоро-Илычский заповедник 62о . . 57° > „ Теплов (1947-а)
РСФСР, ряд областей . . . — — Лобачев и Щербаков (11933)
Польский р-н Новгор. обл. . 58° „ , 32о. , Дубровский 01930}
Беловежская пуща .... 53». . 240 . , Гаврин (1966).
Бапткирский заповедник . . 530 , „ 580 . , Кириков (il'952J)
Юго-восточ. Прибайкалье . 52о , . 108о . Фетисов (11934)
Кавказский заповедник . . . 44о , . 40» , * Аверин (1936)
Рябчик (табл. 13—15, рис. 25).
Зимнее питание рябчика обеспечивают, в основном, два расте-
ния: береза и серая ольха; у них он ест веточки, листовые и цветоч-
ные почки и, в особенности, ‘мужские цветочные сережки. К этому
присоединяются в небольшом количестве веточки и почйи осины, ив
(козья ива и другие виды) и рябины; осенью — также карликовой
березки.
Из хвойных шород рябчик поедает только плоды можже-
вельника и галлы с веточек 'ели; ест он также и галлы с ивовых ве-
точек.
Все древесно-кустарниковые корма в сумме составляют около
90о/о рациона в снежное полугодие и 14о/о!— в бесснежное. Зимние
корма преобладают в питании приблизительно с 19 октября до
1 мая, т. е. 194 дня, одинаково с тетеревом.
Наряду с веточным кормом рябчик всю зиму добывает из-под
снега в небольшом количестве и зеленый корм (веточки черники
и др.), а в начале и конце зимы — также ягоды брусники и дру-
гих растений. Наблюдения показывают, что он может разрывать
только мелкий снег, например под ветками елей, где они спуска-
ются до земли. В марте рябчик разыскивает первые проталинки
в скалистых местах на южных склонах тундр и варак. Наземные
корма начинают преобладать в питании рябчика раньше, чем
у других тетеревиных. Рябчики кормятся на деревьях и после того,
как стает снег, но в это время они едят не веточный корм, а цветы
ив (наблюдения 14 V 1938 г. и 26 V 1938 г.). Позднее они очень
охотно едят цветы черники, а отчасти и других растений — голу-
бики, незабудки и линнеи.
В летнем питании рябчика главную роль играют ягоды, лю-
бит он также семена, особенно осок, а зелень ест менее охотно,
чем глухарь.
Животные в питании рябчика те же, что у глухаря и другой бо-
ровой дичи: слизни Agriolimax sp. и Arion sp., улитки Goniodiscus
ruderatus и Acanthinula harpa; один раз — многоножка; пауки и
69
Таблица 13
Качественный анализ питания взрослых рябчиков (старше 3 мес.)
1 II III IV V VI VII VIII IX X XI XII
Число зобов с пищей . 2- <-2 5 4 5 2Н 1-12 10 10 54 13 9
Число зобов пустых . 0 0 0 0 1 1 0 0 0 4 2 0
I. Веточные корма . 100 — 100 75 40 21 0 30 85 100 100
Можжевельник, плоды 25 — — — 7 — — 4 — -
Береза, побеги, почки . 50 80 50 20 — — 20 32 54 67
Береза, цвет, сережки Ольха серая, побеги, поч- 75 80 50 20 7 — 10 50 31 33
ки 100 20 50 — — — 10 20 69 100
Ольха цвет, сережки . Проч, виды, побеги, поч- — — 25 —• — — — 6 54 33
ки 50 20 25 20 — — — 30 31 78
Ивы и ель, галлы . . — — 25 — 7 — — — — —
11. Вечнозеленые ра-
стения 25 — 20 25 80 — 21 20 20 59 69 89
Черника, побеги, листья 25 20 25 80 21 20 10 57 69 89
Брусника, листья . . — —— —— — — — — — 8 11
Вороника, побеги, листья — — — 20 — — 10 2 23 22
Линнея, листья . . . — 20 — — — — — 2 — 11
III. Летняя зелень — — — 50 20 9
Голубика, листья . < — — — — 36 — —
Иван-чай, листья . . Ивы, осина, ольха, ря- — — — — — — 10 4 — —
бина — — — — 21 — 20 2
Прочие листья . . . — — — — 21 — — 7 — —
IV. Цветы .... — 40 64
Ивы — — — 40 —
Черника, голубика . . — — — — 50 — — —
Ястребинка, незабудка — — — — 21 — — — — —•
V. Ягоды — — — 25 80 29 100 90 89 62 11
Черника — — — — 21 70 70 76 31
Голубика — — — — 14 10 4 8
Брусника — — 25 80 — 30 60 56 31 11
Вороника Смородина, рябина, — — 25 20 7 30 22 15
клюква — — — — 7 — — 6 — —
VI. Семена и плоды . — — — 21 60 70 20 8
Осоки — — —. 7 10 30 2 —
Береза — — — 7 30 20 17 .
Герань лесная . . . — — — — 20
Марьянник — — — 20 10
Прочие семена . . . — — — — 7 20 20 4 8 . —
VII. Низшие растения — — 7 7
Мхи политриховые . . —. — 7 5
Древесные лишайники . — — — — — — 2 — —
70
Табл. 113 (продолжениej
1 II III IV V VI VII VIII IX V XI XII
VIII. Животные . . ,— 43 40 30 । 2
Слизни — — — — 7 20 10 — — —
Пауки — — — — 7 — 10 2 — —
Многоножки .... — — — — — 10 — — — —
Жуки — — — — 14 — — — — —
Пилильщики, личинки — — — — 14 10 — — — —•
Муравьи — — — — — 10 — 2 — —
Наездники и проч. . . — — — — — 10 — 2 — —
Двукрылые .... — — — — 14 — 10 — — —
Гусеницы бабочек . . — — — — 21 20 — — — —
IX. Камешки . . . — 2 8 11
Таблица 14
Количественный анализ питания взрослых рябчиков (старше 3 мес.)
1 II Ill IV V VI VII VIII IX X XI XII
Процент пустых зобов . । 0 0 0 17 7 _0 0 7 13 0
. '
Ср. вес пищи в зобу . — — 9,7 — — 4,1 9,3 40,0
Число свеш. зобов . . — — 3 — — 4 22 7
I. Веточные корма 845 — 996 877 79 24 0 117 590 797 937
Можжевельник, плоды 2 — 2 — — 3 — —
Береза, побеги, почки . ’ 111 567 148 36 — — 11 61 69 143
Береза, цвет, сережки . 421 393 316 36 11 — 4 292 111 189
Ольха, побеги, почки . 200 11 123 , — 102 112 177 438
Ольха, цвет, сережки . — 52 — — 72 408 38
Прочие, побеги, почки . 111 25 94 7 — — 50 32 129
Ивы и ель, галлы . . — — 144 11 — — — — —
II. Вечнозеленые ра-
стения 155 — 4 10 300 13 7 108 96 168 63
Чериика, побеги, листья 155 3 10 264 13 7 102 94 165 62
Брусника, листья . . — 0 0
Вороника, побеги, листья Линнея, листья . . — 1 36 6 1 1 3 1 0
III. Летняя зелень Голубика, листья . — — — — — — 302 — 29 15 — —
Иван-чай, листья — — — 144 — 1 о
Ивы, осина, ольха, ря- —- — — — — 4 и
бина ..... 25
Прочие листья . — —- — 119 — 0 — —
— 39 2
71
Табл. (14 (продолжение)
I 11 ш IV V VI V11 VIII IX X XI All
IV. Цветы .... - — 364 — 310 — — — — —
Ива козья (и др.) . . — — — 364 — — — — — —
Черника, голубика . . Ястребинка, незабудка, — — — —• 294 —
линнея — — — — 14 —
V. Ягоды из 257 278 675 568 267 34 0
Черника — — — •— 52 588 413 120 8 —
Голубика — — — — 45 16 — 5 1 —•
Брусника — — 103 236 — 41 155 110 24 0
Вороника Смородина, рябина, — — 10 21 30 151 30 — 26 1
клюква — — — — — — 6 —
VI. Семена и плоды . — — — — — — 6 285 130 32 0 —
Осоки — — — — 5 230 13 0 — —
Береза — — — — 0 36 0 27 — —
Герань лесная .... — — — — — — 106 — — '—
Марьянник — — — — — 13 1 — — —
Прочие семена .... — — — — 1 6 10 5 0 —
VII. Низшие растения — — — — — 2 — 0 — —
Мхи политриховые . . — — — — 2 — — 0 — —
Древесные лишайники — — — — — — — 0 — —
VIII. Животные . . — — — — 67 33 48 0 — —
Слизни (число экз.) . — — — — 2 4 11 — —
Пауки .— — — 2 1 1 — —
Многоножки .... — — — — — 1 — —
Жуки — — — 2 — — — —
Пилильщики, личинки . — — — — 14 1 — —
Муравьи — — — — — 1 — 1 — —
Наездники и проч. . . — — — — — 2 — 1 — —-
Двукрылые — — — — 12 — 1 — — —
Гусеницы бабочек . . । 1 — — — — 11 2 — — — —
IX. Камешки . . . — 0 1 0
Таблица 15
Анализ питания птенцов рябчиков (до 3 мес.)
Качественный анализ Количественный анализ
VII VIII IX VII VIII IX
Число зобов с пищей . 6 28 27 % пустых зоб. 25 18 16
Число пустых зобов . . 2 6 5 Ср. абс. вес пищи Число взве- шен. зобов. • 10 1
72
Табл. 15 (продолжен не)
' Качественный анализ Количественный анализ
VII VIII IX VII VIII IX
—
I Веточные корма . можжевельник, плоды Береза, побеги, почки . Береза, Цвет, сережки . Ольха серая, побеги, ПОЧКИ Ольха, цвет, сережки . — 7 26 — 7 46
— 4 4 7 7 4 — 2 5 7 7 14
— — — — — —
Пр. ДеР-’ побеги* почки — — 11 — — 18
Р1вы и ель, галлы . . —
11. Вечнозеленые ра-
стевия — — 26 — — 4
Черника, побеги, листья — — 18 — — 3
Брусника, листья . . — 4 «7 — — 0
Багульник и зимн. хвощ 7 — — 1
Вороника и линнея . . — •— — ——
III. Летняя зелень . — 4 22 — 2 23
Голубика ..... — 4 11 — 2 • 2
Иван-чай Ивы, осина, ольха, ря- — — — —
бина — — — — — —
Прочие листья .... — — 11 — — 21
IV. Цветы . . . — — — — — —
V. Ягоды 50 86 100 667 716 647
Черника 50 57 82 667 408 386
Голубика — 39 4 252 35
Брусника — 14 48 . 12 150
Вороника — 14 4 27 9
Шведский дерен Клюква, морошка, то- — — 4 — — 43
локнянка, рябина . . — — И — — 24
VI. Семена и плоды . — 68 56 152 188
Осоки — 14 22 39 86
Береза — 36 22 53 84
Герань лесная .... Марьянник Прочие виды 21 7 11 — 47 5 4
— 11 15 — 13 9
^уп. Низшие растения п *и п°литриховые . . цРевесные лишайники . — 4 4 5 0
— 4 4 — 5 0
Животные . . Па^и’ У™ . . . 50 36 52 333 113 87
17 14 18 Число экз. 3 20 11
Мн°гоновди ’. *. *. ; ; — 11 4 п » W 9 — 14 5
73
Табл. 15 (продолжение)
Качественный анализ Количественной анализ
VII VIII IX VII VIII IX
Цикадки Auchenorhyncha 17 — — Число экз. 2 — —
Тли Aphididae .... 17 — 4 я W 7
Клопы, личинки Stenodema . 17 — — 1» л 1 — —
Жуки Galerucella . . . — 11 7 в я — 7 о 215
Пилильщики, лич. . . 33 11 4 л я 7 3 1
Муравьи — — 22 п я — — 6 3
Орехотворки, наездники — — 4 я я — —
Двукрылые — — 7 и л — — 4
Гусеницы бабочек; 1 моль ..... 33 22 22 я - 4 12 8
Прочие насекомые . . — 7 26 и л — 2 21
IX. Камешки . . . — 4 11 п л — 5 с
мелкие красные клещики; зеленые тли с берез; цикадки; жуки, пре-
имущественно листоеды Chrysomelidae (в зобу молодого рябчика,
добытого 4 IX 1908 г., Г. А. Новиков нашет 205 листоедов Galeru-
cella sp.); гусеницы бабочек, в частности пядениц; одиночные му-
равьи, наездники и орехотворки; личинки (лжегусеницы) пилиль-
щиков; мухи.
В таблицах питания (табл. 13—15) перечислено лишь 14 видов
сосудистых растений. В рубрику «прочие» попали отчасти другие
органы упомянутых ранее 14 видов (так, например, у ив рябчик
поедает не только цветы и галлы, но также побеги, почки и листья),
а кроме того: трехраздельный папоротничек, колоски зимнего хво-
ща (Equisetum hiemale), семена перловника (Melica nutans), листья
сизой ивы (Salix glauca) и осины (Populus tremula), почки и по-
беги карликовой березки, луковички гречишника живородящего,
плодики лютика (Ranunculus propinquus), ягоды красной сморо-
дины, ягоды, почки и листья рябины, листья и ягоды морошки,
семена фиалок, ягоды шведского дерена, плоды и листья кругло-
листной грушанки (Pirola rotundifolia), листья багульника, ягоды
толокнянки и мелкоплодной клюквы, плоды седмичника (Trienta-
lis сигораеа), цветы незабудки (Myosotis sp.), листья болотной
мытника (Pedicularis palustris), листья золотой розги, цветы 11
листья ястребинок.
Из низших растений в зобах рябчиков найдены мхи PolytrichuH1
sp. sp. и лишайник Bryopogon chalybeiforme. Из упомянутых здесь
и в таблицах 38 видов растений 5 видов в сумме дают около 83%
всей пищи рябчика за год, 9 видов дают 15%, а на долю остал^'
ных 24 видов приходится менее 2о/о рациона: очевидно, некоторы6
74
из них поедаются только случайно. По этой причине скрупулезное
пополнение списка кормовых растений является, в сущности,
данью рутине.
А-ООЛ7С7
| | зитние CemoQHbfQ
У//Л зычне- зеленые листья и
;побеги
[//J летне-зеленые ласты/
цёеты
[о~^о| ягоды
ГПППТП сетено и коробочки
тки и ер и бы
молодые
VII Vlll IX
Рис. 25. Питание рябчиков по временам года
Питание птенцов характеризуется увеличением, по сравнению
со взрослыми птицами, роли насекомых, моллюсков и ягод (моло-
дые рябчики, как и глухарята, едят больше голубики, чем взрос-
лые) и уменьшением роли зелени.
75
Питание рябчика в годовом разрезе изучено в следующИх
пунктах СССР начиная! с севера:
Район Широта Долгота Автор и год опубликования
Лапландский заповедник и
бассейн Имандры .... 68° с. ш. 32° в. д. Семенов-Тян-Шанский (1938)
ы Новиков (1952):
Печоро-Илычский зап-к . . • 62° . , 57° . „ Донауров (1947)
Шарьинский р-н Костр. обл. 58° „ , 46° , . Формозов (1934)
Башкирский заповедник в . 53° . . 58° . . Кириков (1962)
Алтайский „ 51», , 88° , , Цвеленьев (19318)
Сопоставив эти же материалы, собранные на громадном про-
странстве (17° по широте и 56° по долготе), С. В. Кириков (1952)
отмечает:
«Зимой рябчики кормятся исключительно на деревьях. Виды
зимних древесных кормов во всех переименованных выше местно-
стях немногочисленны; сережки и почки березы и ольхи — главней-
ший зимний корм. Летом рябчики кормятся на земле. Группы лет-
них кормов в различных физико-географических областях одина-
ковы. Одно из самых важных мест занимает группа ягод...»
В действительности и зимой рябчик отчасти кормится на земле,
а летом — на деревьях (обратите внимание на роль древесных
листьев в питании рябчика — табл. 13 и 14). В остальном мы со-
гласны с Кириковым. Замечательно, что видовой состав зимних
кормов рябчика во всех исследованных пунктах одинаков* и вме-
сте с тем отличается от кормов других тетеревиных. Иначе обстоит
дело летом: возьмем, для примера, ягодную группу кормов. На
Имандре рябчик ест** чернику, бруснику, воронику, голубику,
реже другие виды; на Печоре — бруснику, чернику, шиповник, ко-
стянику и др.; в Костромской области — чернику, малину, бруснику
и др.; на Южном Урале — костянику, шиповник, черемуху, ма-
лину, клубнику и др.; на Алтае — рябину, красную смородину, лес-
ную землянику, малину, костянику и др. виды.
В различных областях рябчик питается ягодами самых различ-
ных растений, и вместе с тем всюду ягоды — его важнейший летне-
осенний корм. В этот период на ягодниках кормятся также тетерев,
глухарь и белая куропатка, причем в одной и той же местности
разные виды тетеревиных едят ягоды тех же самых растений. От-
сюда -можно заключить, что летом питание отдельных видов тетере-
виных не специфично и состав его определяется, в основном, харак-
тером растительного покрова местности, потому что питание тете-
ревиных строится, как правило, на массовых видах кормов. Зимой?
напротив, каждый вид тетеревиных специализируется на использсн
* С. С. Донауров относит сережки и почки ольхи к случайным кормам»
считая, что печорский рябчик зимой кормится почти одной березой.
** Перечисляем в нисходящем порядке.
76
вании своего, определенного вида корма. Летом по содержимому
зоба или желудка почти невозможно сказать, какого вида птице
он принадлежит; зимой в большинстве случаев это легко сделать
не только по желудку, но даже по экскрементам.
Белая куропатка, (табл. 16—18, рис. 27). Белая куропатка по
способу питания весь год остается наземной птицей, чем и отли-
чается от глухаря, тетерева или рябчика, которые зимой кормятся
на деревьях.
Зимой белая куропатка ест, главным образом, палочки, которые
отламывает клювом от веток ив и берез, бегая по снегу от куста
к кусту. Длина каждой палочки около 8 мм, вес (в свежем ви-
де) — 20—30 мг. В сосновых, еловых лесах и на гарях Мурманской
прочие Сеточные норма бысопт снежного
I. . ' покрова
береза триигтол '
nap.t и нова я оерёзка
Рис. 26. Веточные корма в питании белой куропатки в зависимости от
высоты снежного покрова
области куропатки общипывают, главным образом, прикорневую
поросль берез, а по берегам рек и ручьев «подстригают» ивовые
кусты; с осени, пока мало снега, они охотно объедают карликовую
березку (рис. 26). По мере увеличения высоты снежного покрова
«ярус» кормежки куропаток повышается, потому что ранее объ-
еденные побеги покрываются снегом, зато становятся доступными
выше расположенные веточки. В период максимальной высоты сне-
га (март — апрель) уровень его поверхности на некоторое время
стабилизируется, вследствие чего кормовой фонд куропаток заметно
сокращается. Вот почему в этот период куропатки нередко кор-
мятся сидя на березах, точно тетерева, а также обклевывают ветви
берез, сломанных или согнутых тяжестью снега; на берегах рек
они не упускают случая подстригать концы веток с деревьев, сре-
занных бобрами.
77
Таблица ]&
Качественный анализ питания взрослых белых куропаток (старше 3 мес.)
--
I И III IV V VI VII VIII IX X XI Хц
Число зобов с пищей 21 25 28 27 25 9 8 9 12 56 32 12
Число зобов пустых . 0 3 1 1 6 0 2 1 2 6 4 0
I. Веточные корма . Береза, побеги с поч- 95 100 100 92 56 И — — 8 14 53 50 100
ками 90 88 97 92 44 — — — — 7 83
Береза пуш., побеги,
почки 60 68 93 77 16 — — — — 2 8 58
Береза цвет., сережки . Карл, берез., побеги, поч- 29 20 25 8 4 — — — — — 3 42 —-
ки 21 24 4 23 32 — — •— — 5 33
Ивы, побеги, почки . . 81 84 46 50 28 — — — 8 5 5 67
Рябина, почки .... 5 — — 4 4 — — — — — — —-
Голубика, побеги . . 5 — — 17 16 11 — — — 5 11 8
II. Вечнозеленые ра- 96
стения 57 8 4 38 80 78 50 44 92 100 92
Черника, побеги, листья 43 4 — 33 71 78 50 33 92 96 97 83
Брусника, листья . . . 19 4 4 12 8 — — — — 12 3 —
Андромеда, листья . . Вороника, побеги с ли- 10 — — 12 8 8 — — — 17 2 21 8
стьями 9 — — 8 — — 11 17 2 — 8
Вереск, багульник, клюк- 12
ва — — — — — —- — —- 2 3 17
III. Летняя зелень . — — 4 4 44 88 78 50 14 3
Голубика — — — — 4 33 88 78 42 4 — —
Ожика волосистая . . 4
Хвощ лесной .... — — — — — 11 25 11 8 — —
Луговик — — — 4 — — — 22 33 12 3 —
Прочие листья .... — — — — — 11 12 — — — — —
IV. Цветы .... — — — — 4 33 25 — — — — —
Черника и голубика . — — — — — 33 25 — — — — —
Осоки 4
V. Ягоды 24 4 — 17 72 56 75 78 83 70 71 25
Черника 14 — — 8 8 11 62 56 50 55 47 8
Голубика — — — — — — 38 22 17 2 16 —
Брусника — — — 17 36 11 — — 17 36 42 17
Вороника 10 4 — 17 60 33 62 67 42 38 24 8
Клюква, толокнянка . — — — — 4 — — — 8 — 3 —
VI. Семена, плоды — 4 — — — — 62 56 21 4 3 —
Осоки — — — — — — — 11 8 3
Карликовая березка — — — — — — — 11 — — —
Андромеда
Марьянник — — — — — — 50 33 17 2 .—
Прочие виды .... — 4* — — — — 12 — 8 4 —
* Семена ели.
78
VII. Низшие растения
Мхи политриховые, ко
робочки .....
Древесные лишайники .
Гнилая древесина *
VIII. Животные . .
Пауки ..............
Муравьи.............
Двукрылые (Bibio и др.)
Гусеницы бабочек .
IX. Камешки . . .
* Гифы грибов.
Та бл. I '6_( п р од о л ж ен и е)
I 11 111 IV V VI VII VIII IX X X! XII
— 4 — — — 22 38 11 17 7 — —
— — — — 11 38 — — — — —
4 — — — — — — 2 — —
— — - — 11 — 11 17 5 — —
12 и 17 2 — —
4
8
— — — 4 —— — — — 2 —
— 11
5 — — 4 — — 12 11 — — 5 —
Таблица 17
Количественный анализ питания взрослых белых куропаток (старше 3 мес.)
I 11 111 IV V VI VII VIII IX X XI XII
Процент пуст. зоб. . . Ср. абс. вес пищи . . jr Г |_11 >2 3 1 4 14 19 1_о 7 20 Но 9 14 1 10 L1 10 | 0 27
Число свет. зоб. . . ( 5) (3 8) (1 1) С ?) (1 3) ( 3)
1 Веточные корма Береза пуш., побеги, 780 997 1000 831 362 22 — — 6 16 164 550
почки Бер. пуш.. цвет., се- 283 442 757 556 34 — — — — 0 5 193
режки 98 4 25 4 5 — — — — — 1 —
Карлик, берез., побеги 108 308 13 100 73 — — — — 11 153 228
Ивы, побеги, почки . . 280 243 205 162 182 — — — 6 2 2 128
Рябина, осина, почки . Голубика, побеги . . . II. Вечнозеленые ра- 10 — 0 0 — — —. — — — —
2 — — 9 68 22 — — — 3 3 1
стения ч 213 3 0 82 317 626 17 10 321 645 695 441
Черника, побеги, листья 206 1 74 280 626 17 6 316 634 686 437
Брусника, листья . 4 2 0 2 1 — — — — 10 0
Андромеда, листья . . 3 — 3 16 — — — 2 0 9 0
Вороника, побеги, листья Вереск, багульник, клюк- 0 — — п О И — — 4 3 1 — 3
ва — — — — 6 — — — — 0 0 1
Ш. Летняя зелень 0 26 200 457 349 210 21 0
Голубика — 26 156 384 328 185 15
Ожика волосистая . . 3
Хвощ лесной .... 22 5 19 13
Луговик 0 — 2 12 3 0 —
Прочие листья .... — — — — — 22 68 — — — — —
79
Табл. 17 (продолжение)
I 11 III IV V VI VII VIII IX X XI Х11
IV. Цветы .... . 0 24 7 — — — z
Черника и голубика . — — — — 24 7 — — —
Осоки — — — — 0 — — — — — —
V. Ягоды 5 0 87 294 119 389 567 434 312 138 9
Черника 1 — 4 5 7 128 210 185 177 48 3
Голубика ' — — — — — — 28 62 70 0 4 —
Брусника — — — 26 36 66 — — 45 84 74 6
Вороника 4 0 — 57 129 46 233 295 83 51 6 0
Клюква, толокнянка — — — — 124 — — — 51 — 6 —
VI. Семена, плоды — 0 — — — — 100 45 17 4 0 —.
Осоки — — — — — . — — 6 1 — 0 —
Карликовая березка . — — — — — — — 1 — —• — --
Андромеда 88 38
Марьянник — — — — — — 15 4 — —-
Прочие виды .... — 0* — — — — 12 — 1 0 — —
VII. Низшие растения — 0 — — — 9 28 16 10 2 — —
Мхи политриховые, ко- робочки — — — 2 28 — — —
Древесный лишайник . — 0 — — — — — — — 0 — —
Гнилая древесина . . . — — — — — 7 — 16 10 2 — —
VIII. Животные . . — — — — 1 — — 1 2 0
Пауки (число экз.)| . . 1
Муравьи (остатки) . . 3
Двукрылые — — — — 5 — — — —> 60 — —
Гусеницы бабочек . . — 1
IX. Камешки . . . 2 — — 0 — — 2 12 — — 3 Г'
* Семена ели.
Таблица 18
Анализ питания птенцов белых куропаток (до 3 мес.)
Качественный анализ Количественный анализ
VII VIII IX VII VIII IX
Число зобов с пищей . I 10 23 17 Процент 0 12 6
Число пустых зобов . . 0 3 1 пуст. зоб. Ср. вес — —
I. Веточные корма . 6 пищи 9
Береза пуш., побеги, почки 1
Береза, цвет., сережки — — — —
Карлик, березка, почки — — 6 — 9
80
Табл. 18 (продолжение)
Качественный Количестве ними
анализ анализ
VII VIII IX VII VIII IX
Ппы побеги, почки . . — — — — — —
рябина, осина, почки , Голубика, побеги . . . — — — — — —
ц Вечнозеленые ра- 10 10 10 88 88 г 30 201 199 1
стения Черника, побеги, листья О' О'
Брусника, листья . . . о 1
Андромеда, листья . . Вороника, побеги, листья Вереск, багульник, клюк- 10 — 6 30 — 1
— — — - — —
III. Летняя зелень . 30 86 82 8 255 362
Голубика — 76 65 — 218 344
Ожика волосистая . . —• 5 — — 3
Хвощ лесной .... 20 10 6 4 .12 4
Луговик Прочие листья . . . 20 10 35 4 22 14
IV. Цветы .... 20 10 — 18 49 —
Черника । 10 — — 6 — —
Завязник 1 — 5 —-” — 48 —
Ладьян, ситник . . . 10 5 — 12 1 —
V. Ягоды . . 30 95 88 105 459 372
Черника . . 20 76 82 63 292 303
Голубика ... — 33 35 — 119 42
Брусника — 24 — — 16
Вороника 20 33 29 42 48 4
Морошка — — 6 — — 7
VI. Семена, плоды . 40 67 29 70 149 47
Осоки 30 19 6 48 51 7
Карликовая березка 10 24 2 31
Андромеда .... 14 12 44 8
Марьянник Злаки, линнея . . . 20 48 12 16 47 1
10 14 — 6 5 —
VII. Низшие растения 1ХИ политриховые, ко- 50 5 6 157 1 1
робочки Гмибы пласгинчатыс 20 • ,и* 31 1
6 1
нилая древесина * . 30 — 126 —
и Животные V рви - ЛЛТКИ Acanthinula har- р <1 70 52 6 612 77 1
— — 6 Число экз. — — 1
10 — 1* н 1 — —
~~—'— -
Гифы грибов.
*’"3037
________________________________Табл. 18 (продолжение!
Качественный анализ Количественны анализ
VII VIII IX VII VIII 1\
Пауки, сенокосцы, кле- 19 ЧИ-.ТО -;КЗ.
щи 40 — 18 14 —
Кобылки — 5 — — 2 —
Гл и 10 — — .. 4 •—
Цик а ди. Th a mno 1 el t ix sp. /Куки-слоники и личин- 20 10 — п и 2 —
ки листогрызов 20 — It 4 13 -
Муравьи 10 10 — " j> 1 4 •-
Личинки пилильщиков • 40 10 — V n 93 2 --
Двукрылые, комары и др. 30 10 — я n 4 2
Гусеницы бабочек . . 30 24 — fl Л 18 19 —
IX. Камешки . . . — 5 6 — 1 1
Способ питания куропаток требует большей затраты времени
по сравнению с древесным питанием других тетеревиных птиц.
В связи с этим, куропатка проводит на ногах значительную часть
дня; много бегая, куропатка имеет возможность найти в каком-
нибудь укромном месте подснежную зелень или ягоды. За 6 зимних
месяцев (XI—IV) «надснежные» веточные корма составляют око-
ло 72% рациона куропаток, вечнозеленые кустарнички — 24 % и
ягоды — 4о/0. В летнее полугодие веточный корм играет ничтожную
роль, всего 7%. Период веточного питания у куропатки короче,
чем у рассмотренных выше птиц: зимние корма преобладают
с 15 XII до 2 V, всего 138 дней.
Весной, до начала вегетации трав, и осенью, после ее оконча-
ния, главнейший и необходимый корм куропатки — веточки черни-
ки. Она стрижет их на такие же мелкие кусочки, как и березовые
веточки. По подсчетам Г. А. Новикова, количество частиц в одном
зобу достигает 6600 (Новиков, 1952): если допустить, что куро-
патка на жировке затрачивает на один кусочек только секунду,
и то для наполнения зоба ей потребуется два часа непрерывной
работы.
Побеги черники птица ест круглый год и они занимают первое
место в годовом рационе; другие вечнозеленые растения попа да'
ются только как примесь. В середине лета, пока куропатки живут
на болотах, их главный зеленый корм — листья голубики; лисгь*1
других травянистых растений, а равно и цветы, не играют большой
роли в питании этого вида. Вместе с тем куропатка очень охотно
ест ягоды вороники, черники, брусники, реже — голубику и клюкву
Из семян растений главную роль играет марьянник. На гарях кУ
ропатки охотно едят коробочки политриховых мхов; плодовые тела
iрибов они почти не едят (один случай), но сравнительно нередко
заглатывают кусочки гнилой древесины или коры. Животный ьоГм
82
в питании взрослых белых куропаток принимает самое небольшое
участие.
взрослые
I П Ш IV V VI VII Vlll IX X XI XI/
зимние Неточные корма
зимне 'Зеленые листья и
побеги
летне 'Зеленые листья
ягоды
семена и коробочки
тли и грабь/
ниботнся?
галька
куропаток по временам года
Рис. 27. Питание белых
Список кормов белой куропатки меньше, чем у других тетереви-
ных. Шесть растений — черника, пушистая береза, карликовая бе-
резка, ива (вернее, несколько видов ив), голубика и вороника
в сумме дают 91 % годичного рациона; всего же нами обнаружено
35 видов сосудистых растений. Кроме перечисленных в таблицах,
в зобах куропаток найдены веточки селагинеллы (Selaginella sela-
6*
83
ginoides), семена ели, соцветия ситников, плоды тофильдии И ceci-
dia pusilia), цветы и стебли ладьяна (Coralliorhiza trifida), луко.
вички живородящего гречишника, листья василистника (Thalictruni
alpinum), росянки (Drozera anglica), лесной герани, шведского де,
рена, грушанок и багульника, ягоды толокнянки, веточки вереска,
плоды линнеи, листья барчии (Bartsia alpina), золотой розги ц
ястребиною
Рис. 28. Побеги черники, обклеванные тетеревиными
птицами
В питании птенцов, особенно в первый месяц их жизни, боль-
шую роль играют беспозвоночные. В пище куропаток встречены:
мелкие черви (Oligochaeta) и улитки (Acanthinula harpa) по одно-
му разу; пауки и клещики; кобылки, тли, цикадка (Thamnoicttis
sp.), жуки сем. Curculionidae и Chrysomelidae. муравьи, личинки
пилильщиков, комары Aedes и Tipula, мухи и их личинки, в тон
числе Bibio sp. (imago), моль, гусеницы пядениц (з том числе
Eupithecia sp.), медведиц, совок и других бабочек.
84
Как и у других тетеревиных, молодые куропатки едят больше
семян, но меньше ягод, чем взрослые.
Несмотря на широкое распространение белой куропатки в СССР,
захватывающее три географические зоны, питание ее еще мало
изучено, особенно скудны сведения по зимнему периоду.
Рис. 29. Ягоды вороники (Empetrum nigrum); 2/з нат. вел.
Опубликованные материалы относятся к следующим районам:
Район Широта Долгота Автор и год опубликования
Новосибирские острова Южный Ямал Тимаиская тундра . . , . Зона т 75° с. hi. 6£° я . 68° . „ у н д р ы 140° в. д. 68». . 50». . Бируля (4*907), Городков и Короткевич (1957) Дунаева и Кучерук (1941) Михеев (1948)
Зопа тайги
Лапландский заповедник и Хибинские горы . . . , . 68° . . 32». . Семенов-Тян-Шанский (1938) и Новиков (1952)
Псчоро-Илычский заповедник 62» , . 57». Теплов ('1947-а)
Зона степей
Бараба и Кулунда .... W . . 78» . . 1 1 Юрлов ('1958)
Наурзумский заповедник . . 51» . . 65» . . Ульянин (191319)
Для тундровой зоны есть кое-какие сведения и по другим райо-
нам, но -мы не будем 'их рассматривать, поскольку такой обзор
85
сделан недавно А. В. Михеевым (1948). Надо заметить, что глаз,
ная масса белых куропаток тундры на зиму откочевывает в лес^
тундру, а остающиеся питаются, судя по отрывочным наблюди
ниям, сходно с тундряной куропаткой, то есть не столько неточны^,,
кормом (запасы его в тундре очень скудны), сколько ягодах^
семенами и зелеными частями растений, добываемыми из-под сне.
га. В связи с этим в разных местах отмечен факт жировки бель^
куропаток на оленьих копанках.
Летнее питание куропаток тундры изучено достаточно подробно
Элементы его не отличаются от описанных нами (зеленые чаш и
растений, цветы, ягоды, семена|) и находятся почти в том же
соотношении. Список поедаемых растений определяется составом
флоры. На Новосибирских островах, например, главные кормовые
растения — Salix polaris, Oxyria digyna, Papaver polare, Saxifra^a
sp. sp. и другие тундровые виды. Все исследователи отмечают пре.
обладание зелени и цветов над ягодами и очень редкое нахождение
насекомых в зобах взрослых птиц. Городков и Короткевич нашли
в зобах 25 куропаток только одну гусеницу и несколько жуков
(Chrysomela).
Сведения по питанию куропаток лесной полосы Европы, опубли-
кованные Нордхагеном (1928) и Тепловым (1947-а), еще ближе
к нашему описанию. И в Норвегии и на Верхней Печоре куропатки
зимой питаются, в основном, побегами и почками березы и различ-
ных видо® ив; кроме того, в их рацион входят побеги черники, а на
западе — также и вереска. Нордхаген подчеркивает большое зна-
чение стеблей черники, а для лета упоминает, кроме листьев трав,
луковички живородящего гречишника и коробочки политриховых
мхов, указываемые и русскими авторами для зоны тундр.
Режим питания белой куропатки в степи, па южном предел'’
ее ареала, описан Ульяниным. Его материал охватывает весь год.
но по объему он невелик, дан без разделения по сезонам и, по-внди-
мому, с пропусками. В Барабннской и Кулундинской степях Юр.юз
исследовал только птиц, добытых в теплое полугодие (с апреля
октябрь), а зимнее питание описывает по визуальным наблюде-
ниям. В степи, как и в тайге, веточные корма представлены ы^1-
ми, березой, а в Наурзумском заповеднике также и осиной. Меле-
ные корма целиком представлены степными растениями из родов
Vicia, Lathyrus, Potentilla, Spiraea, Filipendula, Artemisia и т. Д-;
ягоды — шиповником (Rosa cinnamomea) и костяникой (Rubus гД
xatilis). Поедаются также зерна пшеницы, овса, овсюга, семена
различных гречишников. Насекомых Юрлов нашел у 18п/0 иссл^
доваиных им куропаток. Ульянин отмечает, как и я, что «бсльД
куропатки имеют по сравнению с тетеревом более узкий и 11<?'
сколько своеобразный круг кормов».
Для белой куропатки очень характерно то, что зимой она и11
тается веточным кормом, приспособление к чему бросается в глаз*1
при сравнении ее с близким видом — тундряной куропаткой. Э?'1
особенность могла выработаться только в зонах тайги и леС°
86
тундры, которые и надо считать родиной белой куропатки. От-
сюда она распространилась впоследствии на север и на юг; при
этом в тундрах она стала, в общем, перелетной птицей. Широ-
кому распространению куропатки способствовали ее наземный об-
раз жизни и наземное питание. Напротив, глухарь, тетерев и ряб-
чик, которые зимой только спят в снегу, а кормятся на деревьях,
к северу не могли выйти за пределы лесной зоны, а к югу от нее,
в лесостепь, выходит только тетерев.
Тундряная куропатка (табл. 19—22, рис. 30).
Питание этой птицы в зимний период изучено нами недоста-
точно: за 4 месяца (XI—II) не было добыто ни одного экземпляра,
в марте — один и только в апреле—17. Поэтому нам приходится
описывать зимнее питание по данным за март и апрель. На гор-
ных тундрах в апреле еще полная зима, а так как в это время chci
бывает наиболее плотным, то ошибка при экстраполировании ап-
рельских данных на всю зиму может быть только в сторону пре-
увеличения роли веточных кормов: это тем более вероятно, что
в октябре птица их совсем не потребляет.
Таблица 19
Качественный анализ питания взрослых тундряных куропаток (старше 3 нес.)
1 1 1 11 Ill г v 1 VI vn Vlli 'X x XI Xll
Число зобов с пищей . 1 1 -16 8 13 7 3 5 5
Число зобов пустых . — — 1 — — 1 2 2 — — —
I. Веточный корм . . — — 94 75 — —
Можжевельник, плоды . i 12 —
Береза, побеги, почки i 6 — — — —
Береза, цвет, сережки ' 18
Карлик, березка, побеги 76 62 i ~ '
Ивы, побеги, почки . . I 12
Голубика, побеги . . । 21 38 — ' — — — —
II. Вечнозеленые ра-
стения Черника, побеги . . Брусника, листья . . Андромеда, листья . Гром-ягода, листья, поч- — 88 12 24 29 50 12 25 25 62 31 8 86 43 14 33 33 60 40 100 60 — —
ки Вороника, побеги, листья Куропачья трава, листья Прочие виды .... 24 59 18 12 38 12 15 15 8 15 43 14 33 60 20 20 40 40
Ш. Летняя зелень 92 43 67
Г олубика . — — — 60 20 — —
Карлик, березка . — — 23 14 — 60 —
Ива травянистая и др’ Василистник альпий- — — 38 — 33 — —•
— — 62 28 33 — 20
ский * . . .
Ястребинка альпийская — — — 14 — — —
Прочие виды . — — — — 33 — —
’ Thalictrum alpinum. — — 23 29 — -— —
87
Т а б л и ц aJlG i (продолжении ।
111 IV V VI Vll Vill IX X XI ; X|J
( 1 IV. Цветы .... — 1 — - - - 38 57 - — — —
Черника, гол\и:.’:.: . . | - - 15 43 - — —
Андромеда . . . । - - — 14 - — — ।
Гром-ягода । — - 8 — — — —
Кошачья лапка* . . . — — 8 — — — —
Ястрсбинка альпийская . — — - 29 - — —
Вероника альп., осоки . - - 15 14 -
V. Яг ды . . . । — — - 82 88 85 — 100 100 80 - 1
Черника ... . . 6 — — — 33 - )0
голубика 6 — — — — 20 40
Брусника ...... 35 62 62 - 33 20 20
Гром-ягода 18 62 31 - - ~ 1
Толокнянка — — 15 — —- — — 1
Вороника < 65 50 24 — 33 80 20 ।
VI. Семена, плоды . - — — - — - 29 67 40 80 - 1 -
Эсоки (назв. в тексте, ; 1
стр. 93) .... _ _ _ 14 67 20 20
Ситник ** — — — — — 40 40
Ожика *** - - - — 33 — —
Карлик, березка . . . - - - - 33 — 60
Живород. гречишник,
луковички . ... — — — 14 — — —
Ива травянистая . . — —14 — — —
VII. Низшие растения — — - - - 31 14 33 — — — —
Мхи политриховые, ко- i
робочки - - 31 14 33 - -
Ягели — - 8 - —
VIII. Животные . л — — - — — 8 — - — — — —
Муравьи . . ... - - 8 - - — —
IX. Камешки . . — — _ 6 12 I - - - 20 - [ - -
* Antennaria sp.
** Juncus trifidus.
*** Luzula spicata.
Таблица 2U
Количественный анализ питания взрослых тундряных куропаток (старше 3 «ес
] 11 111 | IV V VI Vll Vlll IX X XI | x"
Пустые зобы, % . . . — — - 6 0 0 12 40 29 0 - -
Ср. абс. вес пищи . . — — - 5 - — — 15
Число взвешен, зобов . — — - 7 - - - 1 — —. —
I. Веточные корма . — — - 385 127---
Можжевельник, плоды . 4 — - — — —. —
Береза, побеги, почки . 6 - - - - —- —
Береза, цвет., сережки . 148 - - - — — —
8S
Табл. 20 (продолжение)
1 11 111 IV v v‘l 1 Vli | V111 IX X XI XII
_ 1
Карл- берез., побеги, 1 210 121
поЧКН • Ивы побеги, почки . . Голубика, побеги . . • 3 14 6 — — — — — । .
Л. Вечнозеленые ра- 328 633 86 177 71 202 117 I
стения Черника, побеги, листья Брусника, листья Андромеда, листья • 15 13 55 2 19 230 30 1 67 4 7 36 81
Гром-ягода, листья, поч- 15 24 .—. — —
ки • • • • • • • Вороника, побеги, листья Куропачья трава, листья Прочие виды . - . . 178 50 2 380 2 2 23 6 64 42 64 154 6 6 18 18
Ill, Летняя зелень — — — — .— 338 217 28 157 254 —
Голубика Карликов, березка . . Ивы — — 52 40 199 105 106 4 157 254
Василистник альпийский — — -— 1 — — —
Ястребинка альпийская — — — 21 — —
Прочие виды .... — —• 47 5 — — —
IV. Цветы ... « — — — — — 52 409 — — — — ! —
Черника, голубики . . — — 5 240 — — — 1
Андромеда — — — 105 — — —
Гром-ягода — — 23 — — — —
Кошачья лапка . . . — — 23 — — — — 1
Ястребинка альпийская — — — 63 — — —
Вероника алыт., осоки — — 1 1 — — —
V. Ягоды — — 280 229 449 — 644 528 588 —
Черника 1 — — -— 177 — 496 । —
Голубика 6 — — — — 18 20
Брусника 6 । 70 286 — 4 148 8
Гром-ягода ... 48 98 144 — — — —
Толокнянка ... — — 11 — — — —
Вороника 165 61 8 — 463 362 64
VI. Семена плоды — 88 169 ПО 41
Осоки м Ситник Ожика Карлик, березка . . . речишник, луковички . ^ва травянистая . . — — — 4 46 Г> Г 18 92 3 5 ।
— 35 88 — 33
— — — 42 — — — ।
— — 42 — — —
VII. Низшие растения политри ховые, ко- — — — — — 75 109 88 — — — —
Робочки . Ягеля 1 — 75 0 109 88 — ।
^у7тиЖ"В“ТНЫе. •_: — — — 0 1 — — — —
Камешки . — — — 7 И — — — 3 — — —
89
Таблица 2 ]
Анализ питания птенцов тундряной куропатки (до 3 нес.)
Качественный анализ Количественны!, анализ
VI VII VIII IX VII VIII IX
Число зобов с пищей 1 _ 1 -^7 5 6 Процент 20 0 0
пуст. зоб.
Число зобов пустых . 1* 1 0 0 Ср. вес — —
пищи в зоб.
I. Веточные корма —. — 17 —- — 19
Можжевельник, плоды — — — — — —
Береза, побеги, почки — — — — —
Карл, березка, побеги.
почки — — 17 — — 3
Карл, березка, цвет, се-
режки — — 17 — — 16
Ивы, побеги, почки . . — — — — — —-
Голубика, побеги . . . — — — — — —
II. Вечнозеленые ра-
стения 11 80 17 15 166 13
Черника, побеги, листья 20 17 — 8 5
Брусника, листья . . ; - — — — — —
Андромеда, листья . . i — — — — —
Гром-ягода, листья . . — — — — — —
Вороника, побеги, листья — 17 11 — <8
Куропачья трава, листья 1 - — — — — —
Камнеломка розовая . 1 - 60 — — 158 —
III. Летняя зелень . ' 44 100 67 51 214 164
Голубика 80 — — 166 —
Ива травянистая, сизая
и ДР । 11 40 — 15 32 —
Карл, березка . . . . 1 — — — — — —
Василистник альпийский 22 — — 11 — —
Вероника альпийская . | 11 — 17 7 3
Ястребинка альпийская — 20 50 5 155
Проч, и неопр. виды . ( 22 40 33 18 11 6
IV. Цветы .... 55 — 170 —
Черника, голубика . . । 55 — — 148
Андромеда 22 — — 22 —
Гром-ягода | — — — — __ —
Кошачья лапка . . . । > — —. —-
Ястребинка, воро-л ... 1
осоки . .... ' - — — — — -
1 I V. Ягоды । ‘ 22 100 100 126 348 367
Черника — 40 *3 185 217
Голубика — 60 — 158 —-
Брусника 1 22 — 50 111 137
* Желудок включен в обработку.
<>0
Табл. 21 (продолжение)
Качественный анализ Коли чествеин ый анализ
VI VII VIII IX VII VIII IX
Гром ягода — — — — — —
Толокнянка — — — — — —
Вороника 11 20 33 15 5 13
VI. Семена и плоды . 55 40 83 237 130 365
Осоки — 40 33 — 56 322
Ситник — - 33 — — 19
Ожика — — — — — —
Карлик, березка . . . — 20 33 — 66 24
Гречишник, луковички . 55 — — 237 — —
Линнея, седмичник и пр. — 40 — — 8 —
VII. Низшие растения Мхи политриховые, ко- j 33 20 33 156 156 105 8
робочки 33 20 17 105 5
Ягели [ — — 17 — — 3
VIII. Животные . . i 66 20 17 2 8 29 8
Слизни — 20 — — 10 —
Пауки — — 17 — — 3
Клещики 22 — — 33 —— —
Жуки Лжегусеницы пилильщи- 44 20 — 27 1 —
ков 11 — — 2 — —
Муравьи 11 — — 2 — —
Мухи, комары .... 11 20 — 5 3 —
Гусеницы бабочек . . 22 — — 11 — —
Неопред, насекомые 1 33 — — 13 — —
IX. Камешки и земля 11 20 50 37 8 56
Сравнивая питание белой и тундряной куропаток в апреле, мы
видим, что у первой подснежные корма (ягоды вороники, брус-
ники и стебли черники) составляют 17% рациона, а у второй —
62%, причем состав их заметно отличается. В это время тундрянка
кормится, в основном, облиственными веточками и ягодами воро-
ники, а также зимующими ягодами брусники и «гром-ягоды»,
листьями куропачьей травы, андромеды, а на Хибинских тундрах —
обычной там розовой камнеломкой (Saxifraga oppositifolia). Все
5то она находит на малоснежных участках, разгребая мелкий снег.
Веточный корм тундряная куропатка ест, очевидно, только в край-
ности, когда на тундрах нет ни выдувов, ни проталин. Тогда она
ломает самые тонкие веточки карликовой березки и голубики,
а иногда спускается по склонам до верхнего предела лесной рас-
тительности, садится на кроны берез и поедает мужские сережки,
91
а отчасти листовые почки и веточки. Слабый клюв не позволял-
ей кормиться одними ветками, как это делает белая куропатка.
Ргодь/
мхи и гриЗы
ъаЗогппь/е
еалько
молодые
Неточные
ХОрПО
оберег
IX
Рис. 30. Питание тундряных куропаток по временам года
Тундряная куропатка приспособилась к жизни в другой геогра*
фической среде — в открытой местности с сильными ветрами, сме'
тающими снег: в этом и состоит главное экологическое различи1’
обоих видов.
92
Летом пища обоих видов куропаток состоит из тех же элемен-
тов— зелени, цветов и ягод; по сравнению с белой, тундряная ку-
ропатка ест относительно меньше зелени, но больше цветов, ягод
и семян. Веточные корма в летнее полугодие (V—X) составляют
только 2% (рациона. Взрослая тундряная куропатка -почти совсем
не ест животных: из 64 исследованных зобов только в одном был
найден муравей.
Рис. '31. «Гром-ягода» весиою; 2/з нат. вел.
Приводим названия кормов тундряной куропатки, не вошедшие
в таблицы 19—21: печеночный мох Lophozia lycopodioides, коро-
бочки верхоплодных мхов Polytrichum juniperinum, Р. pilifcrum,
Р. strictum, обрывки ягелей, плоды и колоски осок (Carex Ьгип-
nesoens, С. Bigelowii, С. vaginala), почки -и листья сизой ивы,
листья сиббальдии, распускающиеся почки морошки, бутоны иван-
чая (Chamacnerium angustifolium), незрелые ягоды шведского де-
рена, почки багульника, побеги с листьями азалии (Loiseleuria рго-
curnbens) и филлодоце (Phyllodocc coerulea), плоды седмичника,
цветы и листья альпийской вероники, листья и плоды линией. Всего
в пище тундряной куропатки найдено 32 вида сосудистых растений.
В питании птенцов важный элемент—беспозвоночные живот-
ные. Найдены те же формы, что и у других тетеревиных: слизни,
пауки, клещики, жуки (в частности, листогрызы и щелкуны), му-
равьи, лжегусеницы пилильщиков, некоторые двукрылые, гусеницы
бабочек. По сравнению со взрослыми птицами, птенцы едят боль-
ше семян (осок и других растений), а также разных цветов и лу-
ковичек из соцветий живородящего гречишника.
93
Тундряная куропатка питается разнообразнее, чем белая, и
ее никогда не бывает наполнен чем-нибудь одним. Ни один :Нг.(
корма не имеет для нее такого исключительного значения, как с0:.
новая хвоя для глухаря или березовые палочки и стебли чернц^.
для белой куропатки. При подсчете «удельного веса» отдельных Pjjl
дов за весь год на первом месте оказалась вороника (около
Рис. 32. Куропачья трава (дриада) в цвету;
2/з нат. вел.
Тундряная куропатка распространена циркумполярно, по горам
и отчасти равнинным тундрам Европы, Азии и Америки, от Грен-
ландии, Шпицбергена и Земли Франца Иосифа на севере (81° с. ш )
до Пиренеев и гор острова Хонсю на юге (изолированная колония
па 36° с. ш.). Условия жизни ее на этом обширном пространстве ие
могут быть одинаковыми, но всюду она живет в условиях откры-
того ландшафта, каменистого, с неровным рельефом и неравномер-
ным снежным покровом. Нигде она не бывает обильной, и мож'1°
думать, что общая численность этого вида в СССР в десятки р"‘ч
меньше численности белой куропатки. Опа не делает таких нере'
летов, как белая куропатка, что отметил для Ямала еще Житко0
(1913) и недавно подтвердил Саломонссн (1940, стр. 153).
94
Тундряная куропатка мало изучена. Из старых авторов
М. А. Мензбир (1895) описывает ее верно, но поверхностно, а \
Брема (1912) мы находим баснословный рассказ о том, что «они
без труда выкапывают себе в снегу длинные ходы, ведущие их
к корму», и что «они устраивают себе зимние жилища в виде глу-
боких ям в снегу,поблизости от собранных в кучи запасов пищи» л
В недавнее время в Данин п Норвегии вышли 3 интересные ра-
боты, посвященные тундряной куропатке — Ф. Саломонсена (1940)
п С. Йонсена (1941) о линьке и П. Гельтинга (1937) о питании
Последняя работа дает нам возможность сравнить пищу этой к\
ропатки в разное время года в трех горных местностях арктики
и Субарктики. Питание тундряной куропатки на Северном Таймыре
описано В. М. Сдобниковым.
Район Широта | Долгота Автор и год опубликования
Восточная Гренландия . . . 74° с. ш. 22° з. д. Гельтинг (1937)
Северный Таймыр .... 75й . . 102ов. д. Сдобников (1957)
Лапландский заповедник и Хибинские горы 680 _ „ 32’ . . Семенов-Тян-Шанскин
Иотунгейм, Норвегия . . . 62’ , , 8» » . (Г938-а) Лид - (4927)
Кроме того, у перечисленных авторов мы находим обзоры
if других опубликованных материалов по питанию тундряной ку-
ропатки (большей частью отрывочных — серий от 2 до 18 птиц,
собранных за короткий срок).
В Восточной Гренландии список кормовых растений, по ана-
лизу зобов 78 птиц, включает 52 вида сосудистых растений; там,
как и у нас, зимой тундряная куропатка кормится на выдувах,
разгребая тонкий снег. На северном побережье Таймырского озе-
ра у 199 исследованных в теплую половину года (V—IX) тундря-
нок найден 51 вид растений. В горах Южной Норвегии список
кормов 21 исследованной птицы (I—III и VI—VIII) включает
30 видов сосудистых растений. Для наглядности сопоставим глав-
ные корма в этих местах и их «удельный вес» по полугодиям
(табл. 22). В Гренландии наибольшее значение имеет куропачья
|рава и арктическая ива, а летом живородящий гречишник;
в Лапландском заповеднике зимой — вороника и два вида берез,
а летом — разные ягодные кустарнички; в Южной Норвегии зи-
мой— вороника и береза, а летом — ивы; на Таймыре весной и
летом — куропачья трава и полярная ива, составляющие по общ-
ему 94% пищи в V, 88% в VI—VII и 47% в VIII—IX. В Русской
* Эти наблюдения принадлежат Арнольду Пайку, зимовавшему .на Шпиц-
бергене в 1888—1889 г.; Ф. Д. Плеске в свое время усомнился в прав-ильностп
этого сообщения (Йонсен, 1941).
о-;
Лапландии тундряная куропатка ест больше ягод, чем в других
местах, но в общем характер ее питания везде одинаковы й, и,
в отличие от белой куропатки, в ее пище веточный корм не преоб-
ладает нигде и никогда.
На Кавказе тундряной куропатки нет; между тем там, ца
альпийских лугах, она была бы вполне обеспечена кормами круг,
лый год. Мы не рекомендуем акклиматизацию тундряной куро-
патки на Кавказе только потому, что не предвидим от этого меро-
приятия какого-либо практического эффекта: трудно ожидать,
чтобы эта птица, всюду распространенная с малой плотностью,
сильно размножилась на новом месте.
Таблица 22
«Удельный вес» главных кормов в питании тундряной куропатки в разных
областях (в
Зимнее полугодие
(XI—IV)
Летнее полугодие
(V—X)
Виды
Salix arctica
Salix herbacea
Salix lapponum, S. hastata
glauca
Betula nana
Betula pubescens (s. 1.)
Polygonum viviparum .
Dryas octopetala, punctata
Saxifraga oppositifolia .
Empetrum nigrum (s. 1.)
Arctous alpina ....
Vaccinium vitis idaea
Vaccinium uliginosurn
Vaccinium myrtillus
Сумма no 13 видам
26
2
65
6
0
0
3
21
15
5
34
6
7
2
2
95
39
— 0 5 45
4 — 5 20
1 — 5 8
30 - • — —
— 31 1 1
— 9 2 2
— 2 — 4
60 — 26 1
— 1 5 1
3 12 —
2 7
— — 16 —
98 84 84 S2
S
Питание пуховых птенцов. Мы рассматривали, в основном, пищу
взрослых и подрастающих птиц, по возможности проводя сравне-
ние режима питания в географическом разрезе. По питанию пухо-
вых птенцов у нас мало материала. Учитывая сходство кормов всех
тетеревиных, особенно молодняка, летом, мы рассмотрим питание
маленьких птенцов, расположив их не по видам, а по возрасту.
Данные относятся только к тундряной, белой куропатке и рябчику
(•пуховичков глухаря и тетерева в нашем распоряжении не
было).
Возраст птенцов определен по весу.
С6
Таблица 23
Питание пуховичков рябчика, белой и тундряной куропаток
Возрасти в^днях \ Вид ПТИЦЫ Пол Дата Вес в г В зобу В желудке
0 Тундр. с? 22 VI 1'937 г. 13 Пустой Коробочки политри- хума; кусочки хити- на; песчинка
1 Тундр. — 30 VI 1936 г. 14 1 цветок черники Цветы черники; не- изв. почка; камешков нет
2 Бел. к. д’ 26 VI 1934 г. 20 2 цветка черники; 7 гусениц 1 семя вороники; колоски злака; зе- лень; неокатанные ка- мешки
6 Рябч. д' 10 VII 1906 г. 21 3 улитки; 2 цикад- ки; 3 лич. пилильщи- ков; 1 клоп Stenode- ma Остатки пищи; пе- сок
10 Бел. к. $ 6 VII 1936 г. 40 1 паук крестовик Цветы черники; 1 яг. и семя вороники; семя марьянника; ве- точка Bryopogon; му- равей; камешки
11 Бел. к. 8 VII 1934 г. 50 Коробочки политри- хума —
12 Бел. к. — 6 VII 1937 г. — 6 завязей черники; побеги вороники; ку- сочек коры (сосн.?) Завязи и листья чер- ники; семена и листья вороники; камешки
12 Бел. к. — 28 VII 1933 г. 54 Плодики и чешуи осок; 1 лич. пилиль- щика Семена осок; семе- на вороники; камеш- ки
13 1А Бел. к. 2 13 VII 1938 г. 66 Паук сем. Tomisi- dae; клещик; 2 тли; 45 лич. пилильщиков; лич. мухи; гусеница совки Eupithecia
14 1/1 Бел. к. д’ 14 VII 1938 г. 76 Улитка Acanthinula harpa; паук; 2 цикад- ки; 2 Tipula; 46 лич. пилильщ. 7 гусениц (6 —совки Anarta); куколка бабочки; моль Семена вороники; камешков нет.
14 Рябч. Рябч. — 17 VII 1938 г. 53 Много зеленых тлей; 4 лич. пилиль- щиков; 1 моль —
а 17 VII 1937 г. 54 Пустой Семена вороники; зе- лень; песчинки
7
3037
97
Табл. 23 (продолжение)
Возрасс в днях Вид птицы Пол Дата Все В зобу В желудке
15 Тундр. d 1’4 VII 1938 г. 59 Цветы голубики, андромеды; соцв. гре- чишника; листья ва- силистника; клещики; 1 лич. пилильщика; 3 жука; 7 гусениц (1 — Zygaena); ок. . 10 неоп. насеком. и 1 лич.
16 Тундр. 2 19 VII 1934 г. 56 Короб, политриху- ма; бутоны иваи-чая; куски неопр. листьев; 2 жука; 1 перепонча- токрылое
16 Тундр, (из того же вы- водка) с? 19 VII 1934 г. 64 Короб, политриху- ма; 9 прошл. ягод брусники; 1 прошл. яг. вороники; не опр. листья; мошки и др. насекомые
17 Тундр. 3 17 VII 119318 г. 67 Цветок голубики; 3 лук. гречишника; лист, василистника; кусок неизв. листа
17 Тундр, (из того же вы- водка). с? 17 VII 11938: г. 70 Цветок голубики; 2 цветка андромеды; соцв. с луковичк. гре- чишника
17 Тундр. а 1'6 VII 1938 г. 70 33 цв. голубики; 3 яг. брусники; 2 короб, политрихума; 3 кле- щика; 2 муравья; 17 жуков; (1 щелкун, 2 листогрыза); 4 гусе- ницы; 8 камешков
Рассматривая питание птенцов, мы приходим к следующим вы-
водам:
1. С первого дня жизни птенцы едят, наряду с животной пищей,
которая преобладает в рационе, и растительный корм.
2. Состав беспозвоночных у пуховиков тот же, что и у старших
птенцов, исключая лишь слизней; учитывая размеры добычи, глав-
ную роль играют гусеницы бабочек и личинки пилильщиков, затем
тли и некоторые жуки.
3. Муравьиных «яиц», комаров и их личинок, которыми, по ука-
занию некоторых авторов, в основном кормятся птенцы куропаток
и других тетеревиных (Брем, 1912; Тугаринов и Портенко, 1950)
они у нас совсем не едят.
4. Растительный корм пуховичков состоит из цветов и завязей
черники, голубики и андромеды; цветов и луковичек живородящего
98
гречишника; перезимовавших ягод вороники и брусники, коробо-
чек политрихума; зеленых листьев разных растений.
5. Камешки и песок появляются в желудках с первого дня
жизни; отсутствие их в отдельных случаях проще всего объяснить
тем, что у пуховичков они легко проскакивают из желудка в ки-
шечник, почему птенцу приходится постоянно их возобновлять.
6. С ростом птенцов количество съедаемых растительных кор-
мов все увеличивается, а животных — остается примерно на одном
уровне (возможность найти насекомое не увеличивается с возра-
стом птенца), почему «удельный вес» животных падает. В октябре
из животных попадаются только пауки и насекомые, но и те редко.
7» Взрослые птицы едят меньше животных, чем птенцы, в сред-
нем в 3 раза по частоте встреч или в 5 раз по массе.
8. Взрослые птицы едят немного больше ягод и меньше семян,
чем птенцы; рябчик представляет исключение из этого правила.
Основные особенности питания взрослых птиц. Зная состав пищи
тетеревиных птиц Лапландского заповедника по месяцам, можно
определить относительное значение отдельных кормов или групп
кормов в течение всего года. Для этой цели предпочтительно взять
таблицы количественного анализа питания. Помещенные в них ма-
териалы по глухарю и белой куропатке охватывают все 12 месяцев,
по рябчику— 10, по тундряной куропатке—7, по тетереву — толь-
ко 6 месяцев. Состав пищи за недостающие месяцы можно прибли-
зительно определить путем интерполяции. Для рябчика в II и VI,
а также для тетерева в VI и VII мы берем среднее за 2 соседних
месяца. Для тундряной куропатки, учитывая, что на горных тунд-
рах зима продолжается долго и наземные корма доступны все это
время благодаря наличию выдувов, условно принимаем, что ап-
рельские материалы характеризуют питание за весь период с нояб-
ря по март (см. стр. 87). Для тетерева пришлось сделать допуще-
ние, что в декабре пища его состоит на 90% из веточных кормов, на
9% из вечнозеленых растений и на 1% из ягод, а в I—III он пи-
тается, как глухарь, исключительно веточным кормом. Получен-
ные этим путем значения отдельных групп кормов в питании раз-
ных видов приведены в табл. 24.
Важнейшее значение в питании тетеревиных имеют веточные
корма. Роль их особенно велика у глухаря, у остальных видов она
убывает в той же последовательности, как стоят они в таблице.
Второе место занимают или зеленые части растений (у глухаря
и белой куропатки) или ягоды (у тетерева и рябчика). У тундряной
куропатки первое и второе место делят зелень и ягоды, а древес-
ные веточки занимают только третье.
Всем тетеревиным свойственно весь год питаться самыми обык-
новенными растениями тех мест, где они живут: в этом легко убе-
диться, если сопоставить характеристики растительного покрова
различных стаций и таблицы питания отдельных видов (табл.
8—21). Замечательно, что вегетативные части растений, особенно
хвою, ветки, сережки и почки деревьев, составляющие основную
7*
99
i Таблица 24
Годовой спектр питания тетеревиных птиц (в %)____
Типы кормов ! Глухарь Тетерев Рябчик Белая куро- патка Тундря- ная ку- ропатка
Веточные 62 53 52 39 20
Вечнозеленые .... 7 9 10 28 27
Летнезеленые .... 12 5 4 11 8
Цветы л , ... 2 2 8 0 4
Ягоды ....... 13 24 21 20 35
Семена 2 6 4 1 4
Мхи, грибы ... . 0 0 0 1 2
Животные 2 1 1 0 0
Период преимуществ, пи- тания веточными кор- мами (в долях года} 0,56 0,53 0,53 0,38 0,00
массу годового рациона тетеревиных птиц (81 % —у глухаря), дру-
гие птицы вообще не едят, так что конкуренция по этой линии воз-
можна только с некоторыми млекопитающими. Общее представле-
ние об использовании лесных кормовых фондов Лапландского за-
поведника различными дикими животными дает табл. 25.
Таблица 25
Использование кормовых фондов лесов Лапландского заповедника
Еловая хвоя . . .
Сосновая хвоя . . .
Можжевеловая хвоя
Береза, ветки . . .
Береза, муж. сережки
Карл, берез., ветки .
Ольха, ветки . .
Ольха, муж. сережки
Ивы, ветки ....
Черника, стебли . .
Вороника, побеги
Пушица, цветы . .
Летняя зелень . . .
Ягоды .............
Марьянник, семена .
Мхи................
Ягели..............
Грибы . . . . - .
Условные обозначения:
++ важный или излюбленный корм.
+ мало поедается.
— не поедается.
100
Самый факт наличия общих кормов у глухаря и лося (хвоя
сосны), у белой куропатки, лося, зайца-беляка и бобра (веточки
ив и берез), у глухаря, северного оленя, белки, лемминга, полевок
и кукши (грибы), у всех тетеревиных, полевок, леммингов и лося
(побеги черники) еще не дает права считать этих животных кон-
курентами. Если производство данного корма во много раз боль-
ше потребления, не может быть и речи о конкуренции. В настоящее
время мы не в состоянии дать полный расчет такого рода, но неко-
торые материалы можно использовать в качестве наглядного при-
мера.
В 1939 и 1940 гг. ботаник Т. П. Некрасова взяла в девяти типах
ельников Лапландского заповедника серию укосов зеленой массы.
В среднем, запас ее оказался равным 2,57 т на 1 га (см. табл. 26).
Таблица 26
Запас зеленой массы растений в ельниках Лапландского заповедника
Виды растений Сырой вес в ц/га „Удельный вес“ в % в годовом . рационе
глухаря бел. куроп.
Черника Vaccinium myrtillus . . Вороника Empetrum nigrum . . . Брусника Vaccinium vitis idaea . . Голубика Vaccinium uliginosum . Луговик Deschampsia fiexuosa Шведский дерен . . . Cornus suecica Трехразд, папоротничек Gymnocarplum dryopteris Золотая розга .... Solidago vlrga aurea . . Марьянник Melampyrum pratense . Герань Geranium silvaticum . . Хвощ лесной .... Equisetum silvaticum . . Ожика Luzula pilosa ..... Прочие виды .... 15,8 4,7 1,6 1,3 0,4 0,4 0,2 0,2 0,1 0,1 } 0,9 6,7 0,0 0,1 7,7 0,1 0,0 1,4 0,1 0,4 0,3 0,5 1,3 3,5* 27,5 0,2 0,2 10,0 0,1 0,0 0,6 0,5 0,0 1,1*
Всего 3,6 22,1% 40,2%
* Главным образом болотные виды (пушица, морошка, андромеда).
По данным П. А. Богданова (1952), относящимся к Лисинскому
лесхозу Ленинградской области, средний возраст стеблей черни-
ки—15 лет, брусники — 8 лет; листья черники возобновляются
ежегодно, а листья брусники — в течение 2—3 лет. На этом осно-
вании запас зеленой массы этих двух видов, не считая опадающих
листьев, можно принять равным приблизительно десятилетнему их
приросту. Отсюда продукцию зеленой массы черничников и брус-
101
ничников можно определить в 0,22 т с 1 га в год; урожай летне,
зеленых растений равен их запасу. Общий прирост зеленой массы
травяно-кустарничкового яруса в еловых лесах Лапландского за-
поведника должен быть порядка 60 т на 1 кв. км (не считая мхов
и лишайников). На 1 кв. км в среднем приходится около 17 птиц
семейства тетеревиных общим весом около 26 кг. Суточное потреб-
ление пищи на 1 кг живого веса птицы мы определили от 110 до
200 г, что дает приблизительно 1,4 т с 1 кв. км в год, из которых
21%, или 0,3 т приходится на зеленые корма. Таким образом, те-
теревиные птицы тайги, при данной плотности популяций, распо-
лагают практически неисчерпаемым запасом зеленых кормов.
В правой части табл. 26 показано соотношение отдельных видов
зеленых кормов в пище глухаря и белой куропатки — самых «тра-
воядных» из. тетеревиных птиц. Сравнение этих цифр с величинами
запасов показывает, что глухарь предпочитает есть голубику, па-
поротничек, марьянник, ожику и лесной хвощ, а куропатка — чер-
нику, голубику, марьянник и хвощ.
Таблица 27
Урожай ягод черники, вороники и семян марьянника в ельнике (кг/га)
Вид 1938 1939 1940 1949 1950 1951 Среднее
Черника 128 121 132 138 66 134 120
Вороника 62 76 178 76 21 34 75
Марьянник . — — — 13 — 25 19
В табл. 27 показан урожай главнейших ягод в чернично-ягель-
ных ельниках Лапландского заповедника за 6 лет. В 1938—1940 гг.
учеты проводила Т. П. Некрасова на 20 площадках по 10 кв. м ка-
ждая; в 1949—1951 гг.— О. И. Семен ов-Тян-Шанокий на одной
площадке в 10 кв. м. При учетах спелые ягоды и завязи подсчи-
тывались отдельно, после чего число завязей множилось на сред-
ний вес спелой ягоды и прибавлялось к общему весу ягод. Эта
предосторожность необходима ввиду того, что ягоды поспевают
неодновременно. Учеты 1949—1951 гг. показали, что однократный
учет в начале сентября дает только 0,69 полного урожая черники,
0,94 урожая вороники и около половины урожая марьянника. Сред-
ний урожай ягод составляет, круглым числом, 20 т на 1 кв. км.
Общую годовую потребность тетеревиных птиц в кормах мы опре-
делили в 1,4 т с 1 кв. км. Поскольку наиболее многочисленный и
крупный представитель, глухарь, ест сравнительно мало ягод, об-
щую долю последних в годовом рационе можно принять в 18%; это
составляет 0,25 т с 1 кв. км — величина ничтожная по сравнению
с запасами. Ввиду этого, даже при низком урожае ягод, их хватает
102
для всех потребителей, что дает возможность тетеревиным кор-
миться ими и весной, вплоть до поспевания нового урожая. Лучше
и дольше всех ягод сохраняется на стеблях вороника. Очевидно,
именно по этой причине она играет большую роль в питании птиц,
чем можно было ожидать по величине ее урожаев:
Вид Урожай ц/га „Удельный вес* (в %) в рационе
глухаря белой куропатки
Черника 1,2 5,8 6,4
Вороника ...... 0,7 4,7 7,5
Марьянник 0,2 0,4 0,6
Ввиду того что тетеревиные птицы питаются только массовыми
кормами, нельзя ожидать больших отклонений в составе их пищи
по годам. Вместе с тем, ввиду большой сезонной изменчивости ра-
циона, доказать это предположение трудно: нужен большой мате-
риал. Наибольшее количество глухарей было исследовано нами
в сентябре, с 1932 по 1938 г. Встречаемость главных кормов в сен-
тябре в разные годы показана в табл. 28. Просматривая ее можно
обнаружить некоторые отклонения от нормального рациона (как.
например, отсутствие ягод брусники в 1932 г., семян осок в 1936 г.,
семян марьянника в 1938 г.), однако они наблюдались как раз в те
годы, когда материала было меньше; поэтому мы вправе считать
эти отклонения случайными и недостоверными.
Таблица 28
Встречаемость главных кормов в питании глухаря в сентябре за разные
годы (%)
Корм . 1932 1933 1934 1935 1936 1937 1938 Средн, за все годы
Побеги и листья черники 45 56 43 55 42 71 62 54
Листья голубики . . 45 67 66 50 83 35 88 62
Листья ожики . . . 27 50 60 27 67 47 50 43
Хвощ лесной . м , 9 33 17 5 8 0 50 15
Листья луговика « . . 18 61 34 23 42 47 38 34
Ягоды черники < . . 82 83 69 86 100 71 88 78
Ягоды брусникц . . . 0 33 29 14 58 53 38 30
Ягоды вороники . . . 27 83 34 59 67 29 62 50
Семена осок .... Семена и листья марь- 27 28 17 9 0 6 38 17
янника 64 33 20 50 33 29 0 31
Грибы 27 17 3 14 25 6 12 11
Беспозвоночные . 18 39 29 18 33 18 50 27
Число данных . . 11 18 35 22 12 17 8 142
103
Летнее питание всех лесных тетеревиных почти одинаково по
видовому составу, разница наблюдается только в количественно^
соотношении отдельных компонентов. Наиболее отличается питание
тундряной куропатки, но только по причине своеобразия флоры
Рис, 33. Значение кормовых растений в питании тетеревиных в годовом
аспекте. Зимние веточные корма охвачены наружной дугой
местообитаний. Особенности питания отдельных видов мы отме-
чали при описании питания данного вида и возвращаться к этому
вопросу не будем. Из всего списка кормовых растений (у глухаря —
51 вид сосудистых растений, у тетерева — 22, у рябчика — 38, у бе-
104
лой куропатки — 35 и у тундряной — 32) большое значение имеют
только немногие виды, причем они частично совпадают у разных
птиц. Пять или шесть видов растений в сумме дают 83—90% всех
кормов, съедаемых за год этой группой птиц. Рис. 33 показывает
роль главных кормовых растений в питании отдельных видов тете-
ревиных: в этой диаграмме помещены все растения, дающие не ме-
нее 5% годового рациона хотя бы у одного из исследованных видов.
Летом все наши тетеревиные кормятся почти исключительно на
поверхности земли. Очень характерна их манера срывать ягоды,
цветы, семена, листья и насекомых с растений на корню. Замечено,
что при содержании в клетке или вольере тетеревиных трудно кор-
мить потому, что они не в состоянии оторвать кусочки корма ог
брошенных им веток, если последние не .пр-И1К|реплены (Вильсон и
Лесли, 1911). По той же причине возникают трудности при воспи-
тании тетеревят под наседкой-курицей, которая не может научить
их брать корм (Хватов, 1860; Ларин, 1941).
Если бы зимой совсем не было снега, тетеревиные, очевидно,,
круглый год кормились бы на поверхности земли, как это и на-
блюдается в отдельных случаях (граус в Великобритании, белая
куропатка на побережьях Норвегии). У нас рябчик и оба вида
куропаток в течение всей зимы разгребают снег, где он тонок, в по-
исках корма. Однако норы в снегу они роют только для ночлега,
а не для добывания пищи, как писали старые авторы. Глухарь бе-
рет корм, только находящийся на виду, и не пытается искать его
под снегом.
Питание тетеревиных птиц зимою на деревьях веточными кор-
мами дает им возможность жить оседло при наличии толстого
снежного покрова и могло возникнуть только как адаптация к су-
ровым зимам таежной зоны. Как мы увидим дальше, это приспо-
собление имело следствием глубокие изменения в строении и функ-
циях пищеварительных органов. При этом каждый вид приспосо-
бился к питанию своим, специфичным для него кормом. Разница
в питании отдельных видов так велика, что по зимнему помету
легко определить, какой птице он принадлежит. При однообразии
диеты в течение продолжительного времени (6—7 месяцев) тетере-
виные птицы потребляют в течение года значительно больше зим-
них кормов, чем летних; вместе с тем запасы зимних кормов срав-
нительно невелики, а главное—их возобновление происходит толь-
ко летом. Массовое объедание цветочных и листовых почек
деревьев в течение зимы угнетает дерево и может привести к его
гибели. По всем этим причинам в совокупности продовольственный
вопрос зимой становится серьезнее, чем летом.
Глухарь лучше обеспечен зимними кормами, чем остальные ви-
ды тетеревиных. По данным А. А. Молчанова (1953), запас хвои
в полных сосновых насаждениях 150-летиего возраста составляет
8,9 т/га. В условиях Лапландского заповедника средняя полнота
сосновых лесов около 0,3, следовательно, запас хвои не превышает
3 т/га; поскольку на побегах сосны одновременно находится хвоя
105
5—6 генераций, ежегодный прирост ее составляет 0,5—0,6 т/га, или
50—60 т на 1 кв. км. На этой площади нормально проживает 4 глу,
харя, средним весом по 3,5 кг, и 4 глухарки, по 1,7 кг (всего20,8 кг).
Годовая потребность их в пище, считая по НО г в сутки на 1 кг
живого веса, составляет 835 кг, из которых на зимние корма (сое.
новую хвою) приходится 420 кг — около 1»/о всего прироста. Ввиду
того что глухарь кормится не на всех соснах подряд, а только па
определенных «любимых», глухари могут объесть у них от 5 д0
50% всей хвои с кроны (Линдрот и Линдгрен, 1950).
В зимнем питании тетерева важны мужские цветочные сережки
и веточки березы, первогодние шишечки и хвоя сосны, плоды и
хвоя можжевельника. Шишечки сосны бывают в большом количе.
стве только в зиму перед хорошим урожаем семян и могут счи-
таться дефицитным кормом; плоды можжевельника также доступ,
ны только в некоторые годы. Плодоношение березы более стабиль-
но, и цветочные сережки каждое лето образуются в достаточном
количестве, однако общий запас их на 1 кв. км невелик, так что
если тетеревов много — они могут уничтожить за зиму значитель-
ную часть запаса.К сожалению, интересный вопрос о влиянии птиц
на плодоношение древесных пород еще совершенно не изучен.
При обледенении ветвей тетеревам приходится заглатывать се-
режки и другой корм со льдом; оттаивание его требует большой
затраты тепла и в результате птица голодает. По сообщению охот-
ника К. Е. Собяника из деревни Курьи на Верхней Печоре, такой
случай был у них в начале декабря 1953 г. Две тетерки, добытые
им после 20 XII, были истощены. Впоследствии лед исчез с ветвей
и зимовка прошла благополучно: по крайней мере, мертвых птиц
в ту зиму не встречали. Аналогичное явление, происшедшее в на-
чале 30-х годов, было причиной массовой гибели рябчиков. К- Е. Со-
бянин находил тогда в день до 2—3 мертвых рябчиков со смерз-
шимся в зобу кормом.
Зимняя пища рябчика состоит из сережек, цветочных и листовых
почек и веточек березы и ольхи. В отношении обеспеченности зимниь
ми кормами рябчик сходен с тетеревом.
Белая куропатка отличается тем, что зимой кормится не столько
почками, как именно веточками ив и берез и рвет их с молодои по-
росли, бегая по поверхности снега. Ее питание приурочено к узкому
ярусу, положение которого зависит от высоты снежного покрова в
данный момент. Скрывая низкие кусты, снежный покров сильно со-
кращает кормовые фонды куропатки к концу зимы. А. В. Михеев
(1948, стр. 150) указывает, что в Калининской области для прокорм-
ления одной куропатки в течение зимы требуется около 4—9 га ку-
старниковых зарослей. Поэтому зимняя кормовая база является для
нее «узким местом». На Крайнем Севере напряженное положение
с кормами вынуждает куропаток к массовым перекочевкам. По той
же причине конкуренция со стороны зайца-беляка имеет для куро-
патки реальное значение (оба вида питаются теми же кормами и в
том же ярусе); напротив, лось использует ветки на недоступной ку-
106
роиатке высоте (зимой главным образом в 1—3 м над землей) и не
может считаться конкурентом.
Тундряная куропатка мало приспособлена к ританию веточным
кормом и всю зиму достает из-под снега ягоды и зеленые части ран
стений.
По степени использования зимних веточно-хвойных кормов наши
тетеревиные образуют экологический ряд, представленный в табл. 24
и на рис. 33.
Благодаря использованию древесных кормов тетеревиные птицы
зимой лучше обеспечены пищей, чем другие лесные птицы, и голо-
дают только в исключительных случаях. Слабое развитие у них жи-
ровых отложений объясняется, очевидно, отсутствием необходимости
в них. Впрочем, по личному сообщению А. А. Меженного, в Якутии
глухари значительно жирнее, чем. в европейской части Союза.
Глухарь и лось, питаясь хвоей, предпочитают сосновую (в При-
уралье глухарь, кроме того, ест кедровую, а лось — пихтовую), но
избегают еловую хвою. Белка в голодные зимы «стрижет» кончики
еловых веток, но при этом ест не хвою, а только почки. Изредка ело-
вую хвою ест норвежский лемминг (Насимович и др., 1948). Отрица-
тельное отношение различных животных к еловой хвое объясняется,
по всей вероятности, бедностью ее углеводами. Известно, что хвоя
европейской ели содержит 0,8% моносахаридов и 1,8% дисакаридов,
а хвоя сосны — до 5% моносахаридов и до 1,5% дисахаридов. («По-
лезные растения СССР», 1951).
V. ПИЩЕВАРЕНИЕ ТЕТЕРЕВИНЫХ
В предыдущей главе мы познакомились с одной стороной пита-
ния — составом пищи отдельных видов в зависимости от времени
года и возраста птицы. Мы убедились, что они питаются, в основном,
массовыми кормами, несвойственными птицам и мало используемы-
ми другими животными. Своеобразие кормов вызывает соответствую-
щие изменения в физиологии пищеварения и строении пищеваритель-
ного аппарата. Поскольку это необходимо для правильного понима-
ния связей организма со средой обитания, нам необходимо коснуть-
ся и этой стороны питания тетеревиных.
Клюв. Тетеревиные не подбирают корм с земли, как настоящие
куры, а рвут его. Чтобы сорвать ягоду или лист, нет необходимости
в особом устройстве клюва; напротив, срезание сосновой хвои и от-
ламывание веточек сосны, ив и берез требует значительного усилия
и наличия на клюве режущей кромки.
Наименьшую работу выполняет клюв тундряной куропатки, так
как веточный корм составляет не более 20<% ее годового рациона,
а веточки черники — не более 3%. Лесная белая куропатка на 40о/0
питается ветками деревьев и кустарников и на 27% — веточками
черники. Эта разница в питании объясняет происхождение главного
диагностического признака, используемого для различения этих двух
107
близких видов: у тундряной 'куропатки клюв слабый и низкий, у бе.
лой он массивный и высокий (рис. 34|).
Тетерев и рябчик в значительной мере кормятся сережками, кото*
рые висят на тонких веточках, а потому легко обрываются. Глухарю
приходится клювом «стричь» хвою или отламывать им целые побеги,
кроме того-, и период употребления зимних кормов у него относитель-
но самый длинный (табл. 24). Исключительная массивность глуха-
риного клюв'а, особенно у самца (клю!в глухаря велик не только по
абсолютным- размерам, но и относительно -размеров птицы), стоит
в связи с тем, что на протяжении года он выполняет большую работу.
Рис. 34. Черепа белой и тундряной куропаток; 1/1 нат. Вел-
C. В. Кириков указывает (19*44 и позднейшие работы!), что у глу-
харей во время перехода на питание хвоей, быстро окостеневает сое-
динительнотканное образование, разделяющее поверхностный и глу-
бокий внешние ‘мускулы, приводящие в движение нижнюю челюсть,
что увеличивает площадь закрепления этих мускулов на костях и
усиливает их действие. Интересно, что процесс окостенения черепа
у глухаря затягивается по -крайней мере до 3 лет, благодаря чему
-по срастанию швов можно без затруднений определять возраст до
3—4 лет. У белой куропатки череп полностью окостеневает в пер-
вую же зиму; в марте цо черепу уже невозможно отличить моло-
дую птицу от старой.
Зоб. Сильное развитие зоба характерно для всех куриных. У на-
ших тетеревиных зоб выполняет две функции: при низкой темпера-
туре в нем происходит оттаивание и прогревание пищи перед по-
ступлением ее в желудок, он же играет роль склада для запасания
корма на ночь. Степень наполнения зоба меняется по временам года
108
и по часам суток. Средний вес содержимого зоба и процент пустых
зобов по месяцам.приведены в табл. 9—22. Зимой зоб содержит в не-
сколько раз боль|ше корма, чем в бесснежное время года: у глухаря
Рис- 35. Наполнение зоба глухаря в зависимости от времен суток и года
в 6 pas, у рябчика почти в 10 рав, у белой куропатки в 4 раза. Пу-
стые зобы реже всего встречаются в темные месяцы (XII—I); в
остальные месяцы1 процентное отнюрхение пустых зобов не обнаружи-
вает определенной закономерности.
109
Суточный цикл наполнения зоба гораздо труднее определить, чем
сезонный, по той причине, что часы наступления утра и вечера ме-
Рис. 36. Наполнение зоба белой куропатки в зависимости от времени
суток и года
на 8 периодов по 3 часа, а год —на 4 периода: зима —с продолжи-
тельностью светлого времени, включая утренние и вечерние сумер-
ки, от 6 до 9 часов (9 XI—3 II); весна — от 9 до 24 часов светлого
110
времени (4 II—5V); лето — 24 часа светлого времени (6 V—7 VIII);
осень — от 24 до 9 часов светлого времени (8 VIII—8 XI).
Вес зоба мы берем в процентах от веса птицы (с опорожненным
зобом) и вводим поправку на процент пустых зобов за тот же период.
К сожалению, число птиц с указанием часа добычи и» веса зоба было
невелико (158 куропаток и 71 глухарь); поэтому полученные нами
данные имеют значение ориентировочных (рис. 35 и 36|). При всех
расчетах за начало' и конец сумерек мы принимаем момент, когда
центр солнца находится на 7° ниже горизонта.
Из диаграмм видно, что летом у глухаря и белой куропатки нет
определенных часов кормежки и зоб почти круглые сутки содержит
небольшое количество корми, в среднем около 1°/0 веса птицы. Вес-
ной и осенью, в связи с сокращением долготы дня, на рассвете зобы
оказываются совершенно пустыми, а после заката они всего полнее.
У белой куропатки осенью хорошо видно, что наполнение зоба, про-
должается весь день; глухарь наполняет зоб гораздо быстрее, почему
зимой у него заметно наличие двух кормежек — предварительной,,
утренней, и главной, вечерней.
Таблица* 29
Максимальный вес пищи в зобу тетеревиных
Назв. птицы и состав пищи Дата и время добычи (в часах и мину- тах) Продол, ночи (в часах) Вес птицы с зобом (в г) Абс. вес; пищи (в г) То же в % от веса птицы Сут. рацион (в % от веса птицы)-
свеж. сух.
Глухарь* £ мол., сосн. 25 XI 1937 г. 15-20 16,8 242 8,3 118 48
хвоя Глухарь £ мол., сосн. (сумер.) 24 XI 1938 г. — 16,8 2810 194 7,4 106 44
хвоя и др.
Глухарь мол., сосн. 3 XII 1934г. су- 17,3 3039 200 7,1 98 40
хвоя Глухарка $ мол., сосн. мер. 30 XI 1952 г. 15-38 16,0 1922 148 8,3 124 51
хвоя
лУхарка $ взр , ягоды, .5 VIII I960 г. 22-45 3,2 1895 81 4,2 315 70
листья лухарка $ мол., сосн. хвоя етерев £ взр., сосн. хв.» вЛрез- ветки Бел Kvn’ ?’ берез- веТф ЛйЛУ₽> мол- ЯГ<ЭДЫ. (ночь) 19 XI 11934 г. су- мер. 10 IV 1949 г. 17-10 16,3 10,2 1818 1437 80 95 4,6 7,0 68 164 28 97
(день): 167 87
3 XII 19134 г. — 17,3 692 74 * 12,0
I X 1949 г. 18-15 10,8 645 58 10,0 222 49
Ро«СТЬя Чйк $, ольха, береза (сумер.); 31 X 1933 г. су- 14,0 461 58 14,4 246 145
Ф ольха> береза ?ябчак ф ольха и др. мер. 12 XI 1934 г. 16-00 16,0 470 55 13,3 200 118
17 XII 1938 г. — 18,0 420 48 12,9 177 105 по
₽ябчик О’ Ольха> береза 22 XII '1934 г. — 18,0 450 47 11,6 155 92 11 с
* • береза, ольха 22 XII 1934 г. — 18,0 424 54 > 14,6 195 11Э
111
В диаграммах наполнение зоба выражено в процентах от веса
птицы. Заметно, что маленькая птица ест относительно больше кор-
ма, чем крупная. Картина еще яснее, если сопоставить максималь-
ные веса зобов (табл. 29). Вместимость полного зоба глухаря со-
ставляет около 7—8°/о веса птицы (без зоба), белой куропатки 10 —
12%, рябчика 13—15%. Наиболее полные .зобы встречены в ноябре
и декабре, в вечерние сумерки; обычно они принадлежат молодым
птицам. Мы не имели полных зобов тетеревов и тундряных куропа-
ток, удовлетворяющих, этим1, условиям, но по аналогии надо думать,
что максимальный вес пищи в зобу у них 'близок к белой куропатке
(у тетерева — меньше 12%, у тундрянки, возможно, больше).
Суточная активность. Для определения времени суточной актив-
ности глухаря, я на экскурсиях отмечал по часам вое встречи дея-
тельных птиц в сумерках. В отношении белой куропатки задача ре-
шается проще, так как время пробуждения самцов легко узнать по
крику. Время окончания дневной деятельности и утреннее пробужде-
ние должны быть связаны с определенной величиной освещения. При
наблюдениях я замечал время по точным! часам, а потом, выбрав
эфемериду солнца на этот день, определял по известным формулам
высоту центра солнца Ьр;в момент наблюдения (в сумерки она имеет
отрицательный знак). Величина освещения в сумерках зависит, глав-
ным образом, от глубины погружения солнца под горизонт: наличие
или отсутствие снега и характер облачности сравнительно мало
влияют на освещенность. («Таблицы для расчета природной осве-
щенности», 1945).
Установленное таким образом время начала и окончания днев-
ной активности глухаря указано в табл. 30.
Таблица 30
(Время начала и окончания дневной активности глухаря *
Часы и Высота
Дата солнца Что наблюдалось
минуты в град.
5 VIII 1937 г. 22—40 -3,8° £ мол. кормился на земле
15 VIII 1950 г. 22-45 -7,0 Добыта $ на сосне во время ночлега
18 IX 1’950 г. 19-15 -5,8 $ поднята с земли на дерево, кокала
18 IX 1950 г. 19-18 -6,1 £ слетел с дерева, «скйркая»
26 IX 193i8 г. 5-30 -2,9 1 и 2 $ $ искали гальку по берегу
1 X 11949 г. 18—29 -6,6 2 усевшиеся на ночлег, слетели с сосен
29 X 193'6 г. 8—05 -0,0 $ бродила по земле и кормилась
25 XI 1936 г. 15-00 -5,1 Добыта $, только что севшая на ночлег
30 XI 1952 г. 15-38 -5,0 1 $ еще кормилась на сосне
6 II 1951 г. 16-32 -5,2 2 $ $ подняты с кормежки на соснах
28 IV 1937 г. 22-56 -7,4 «Тёка.нье» $ на току.
29 IV 1937 г. 0-46 -7,6 То же
29 IV 1937 г. 1-36 -6,2 2 глухаря распелись
Здесь, как и во всей работе, указано местное среднее солнечное время.
112
Таблица 3'1
Время начала и окончания дневной активности белой куропатки
Дата Часы и минуты Высота солнца в град. Что наблюдалось
9 IX 1950 г. 3-57 -5,9 Стая перелетела через озеро
27 IX 1937 г. 19—04 —6,0 Первый крик самца.
28 IX 1'937 г. 4—25 -10,0 То же
29 IX 1930 г. 5-00 -7,0 То же
30 IX 1938 г. 4-50 -8,0 То же,
30 IX 1937 г. 5-05 -7,0 То же
1 X 1938 г. 4-53 -8,1 Первый крик самца
1 X 1949 г. 18—15 —5,2 Самец перелетел
7 X 1'949 г. 18-13 -7,2 Самец взлетел с криком
10 X 11939 г. 5-23 -8,0 Крик самца
18 X 1950 г. 17-17 -6,2 2 куропатки подняты с кормежки
20 X 1946 г. 6—20 -7,0 Крик самца
9 XI 1937 г. 7-00 —8,0 То же.
12 XI 1937 г. 7-20 -7,0 То же.
12 XI 1937 г. 15-55 -7,0 То же
13 XI ’1'9137 г. 7-15 -8,0 То же
7 XII 1'930 г. 8-15 -6,5 Перелетала стайка куропаток
8 XII 1938 г. 18—30 -20,0 3 куропатки спугнуты с ночлега на льду
14 XII 11938 г. 8—30 -10,0 Поднята стайка
14 XII '1'938' г. 9-30 -7,0 Поднято 6 куропаток
10 I >1951 г. 15-40 -7.7 3 куропатки еще на кормежке
23 I 1949 г. 8-12 -7,3 Крик самца
20 III (1949 г. 5-00 -6,7 То же
3 IV 1938 г. 9 IV 1'951 г. 12 IV 1951 г. 20-55 3-26 3-15 1 1 1 . - о о Первый крик самца в горной тундре То же в лесу То же
13 IV 1948 г. 2-47 -7,9 То же
14 IV 1949 г. 14 IV 1'949 г. 14 IV 4950 г. 2—52 -7,2 То же в горной тундре
20—54 2-50 —6,0 -7,2 Последний крик самца в горной тундре Первый крик самца в горной тундре
15 IV 1951 г. 3—05 -6,0 1 Начало и конец утреннего тока, горн, тунд-
1'6 IV 1950 г. 5—03 +3,8 J ра.
21-00 21-44 -5,8 1 -8,4 / Вечернее токование в горной тундре
}? JV 1950 г. 2-25 -7,8 Первый крик утром, горная тундра
' IV 1949 г. 2—28 -7,7 Первый крик в горной тундре
7 IV 1'951 г. 2-30 7,5 То же
18 IV 1949 г. 21—00 —5,1 ) Начало и конец вечернего тока в горах;
22-10 -9,0 /
21 m 1949 г- 1-05 -10,4 Первый крик в горной тундре
2 w 1949 г- 1—45 -8,3 Первый крик утром в горной тундре
‘У 1950 г. 22 IV 1950 г. 1—45 2—00 -8,3 —7,7 ) Первый крик утром в горной тундре
23 IV 1938 г. 2-30 1—05 -6,1 | —7.0 1 Начало и конец утреннего тока в горах
1 V 2-05 -5,0 J Начало и конец утреннего тока в лесу
* V 0-00 -7,5 С 1 мая куропатки токуют всю ночь
8"3037
Из табл. 30 можно заключить, что глухарь кончает кормиться и
устраивается на ночлег, когда солнце опустится на 5—6° ниже гори*
зонта; утром он выходит приблизительно при таком же освещении
и к восходу солнца успевает уже поесть. Большинство лесных птиц,
в частности дятлы, кукша и все певчие, нуждаются в свете больше,
чем 1лухарь; их деятельность обычно начинается, когда солнце на-
ходится на 2—3° ниже горизонта.
В Лапландском заповеднике глухари токуют в период белых но-
чей: поэтому наблюдения на токах не дают возможности определить
их потребность в свете. Только в раннюю весну 1937 г. я мог заме-
тить, что в середине ночи, когда солнце было ниже горизонта на (5°,
глухари только «тёкали», но не пели по-настоящему.
Белая куропатка еще непритязательнее к свету. В лесном поясе
она активна при положении солнца выше —7°, —8°, а на горных
тундрах, где освещение усиливается открытой местностью, с —8°,
—10°.
Указанным положениям солнца приблизительно соответствуют
следующие величины освещенности горизонтальной плоскости на
открытом месте:
Момент восхода или заката 750—250 люксов
hQ -5°, -6°. . 10- 1
h0. -7°, —8° . . 1-0,1
h^-8°, —!0° . 0,1-0,01 ,
0
Сила света зависит также от облачности и наличия или отсут-
ствия снежного покрова.
Для упрощения дальнейших расчетов мы принимаем, что наши
тетеревиные активны в продолжении периода, когда солнце нахо-
дится выше —7°: при этом мы немного преувеличиваем активность
глухаря и преуменьшаем' активность куропатки. В течение года на
широте Лапландского заповедника длина этого периода изменяется
от 6 часов в момент зимнего солнцестояния до 24 часов в течение
3 летних месяцев (см. рис. 35 и 36). В темный период зимы ночь
в нашем районе продолжается в среднем 17 часов, максимум
18 часов.
Величина суточного рациона. Калорийность кормов тетеревиных
птиц нам неизвестна. Чтобы привести равные корма по возможности
к одному знаменателю, мы ведем расчет на воздушно-сухое вещество.
Взвешивая корма непосредственно из зобов птиц и потом вторично»
после 1—2-недельного просушивания на печке в сухой комнате, мы
получили следующие показатели содержания сухого вещества в
Основных осенних и зимних корм'ах (табл. 32).
С. В. Лобачев и Ф. А. Щербаков (1936|) определили величину су~
точного рациона глухаря по максимальному количеству корма в зобу
и получили 40 г сухого вещества хвои на 1 кг живого веса
птицы.
114
Таблица 32
Содержание сухого вещества в различных кормах
Корм Птица Сухое вещ. в % Число проб
Сосновая хвоя с веточками Глухарь 41 7
Березовые веточки Белая куропатка . . . 52 12
Березовые сережки, почки, ве- точки Рябчик, тетерев . . , 59 3
Ягоды и зелень в смеси Глухарь 22 9
То же Белая куропатка . . . 22 7
Стебли черники Ягоды вороники и брусники Глухарь и куропатка . 30 5
весной Глухарь и куропатка . 17 5
Ягоды черники осенью Зелень и веточки в смеси, зи- Глухарь и рябчик . . 14 4
мой Тундряная куропатка . 46 3
Попробуем рассчитать суточный рацион более 'сложным1 путем,
исходя из следующих (Предпосылок:
1) желудок тетеревиных работает равномерно и непрерывно круг-
лые сутки;
2) запасенный в зобу с вечера корм полностью расходуется толь-
ко к (моменту выхода птицы на утреннюю кормежку;
3) птица активна от h о — —7° утром до h о — —7° вечером.
Если обозначить суточную потребность в пище на 1 кг живого
веса через С, отношение веса полного зоба в конце вечерней кормеж-
ки к весу тела через А и число часов от окончания вечерней кор-
межки до начала утренней через В получим:
С = 24 — .
В
Подставив соответствующие величины для глухаря и темного
периода зимы получаем:
С -24 A2L =0,113 кг.
17
В переводе на сухое вещество 113 г X 0,41 = 46 г/кг живого веса,
что хорошо согласуется с определением Лобачева и Щербакова. Вы-
численные этим путем величины рациона для нескольких конкрет-
ных случен приведены в двух последних столбцах табл. 29; здесь
даем обобщенные показатели на 1 кг живого веса для разных птиц:
Глухарь............110 г свежей хвои, или 46 г на сухой вес
Рябчик............ 200 » сережек, почек и
веток, или 115 „ ,
Белая куропатка . 116 „ ивовых и березо-
вых веточек, или 80 „ »
8*
115
Можно допустить, что суточная норма свежего корма остается
весь год одинаковой; bi этом нет большой натяжки, так как хотя лет*
ние корма содержат больше воды, чем зимние, но и потребность в
пище в это время меньше. С этой оговоркой мы можем! определить
годовую потребность птицы в пище, а, зная состав рациона по сезо-
нам (табл. 9, 14, 17) или в целом за год (табл. 24), прибли-
зительно рассчитать расход каждого корма в отдельности. Пользуясь
данными учетов тетеревиных птиц нетрудно определить эти величи-
ны не па птицу, а на I1 кв. км, как это и сделано нами.
Желудок. У тетеревиных птиц желудочный мускул очень сильно
развит. Его внутренняя кератиновая оболочка крепкая, довольно
толстая, легко отделяемая от мускула, что дает возможность взве-
шивать мускульный и железистый желудки отдельно от кератиновой
капсулы с наполняющими ее пищей и галькой. В табл. 33 приведен
средний вес различных органов тела тетеревиных птиц без разделе-
ния по сезонам. Все цифры даны 1в пересчете на 1 кг живого веса
птицы с опорожненным зобом,, другими словами, вес органов выра-
жен в промиллях от веса птицы. У рябчика отделы кишечника взве-
шивались только с пищей: разделение на вес тканей кишечника и
пищевую кашицу сделано по аналогии с другими птицами, вследст-
вие чего цифры даны с вопросительным знаком.
Вес мускульного и железистого желудка (без кератиновой кап-
сулы с пищей|) наибольший у глухарки (4«/0 веса птицы), наимень-
ший у тетерева (2,5«/о); самый большой желудок из всех известных
нам птиц оказался у лысухи — птицы по преимуществу травоядной
(6°/о веса птицы)). Вес желудка (в основном желудочного мускула)
меняется на протяжении года.
Количество и качество гальки в желудках также меняется по се-
зонам. У глухаря больше всего камешков в начале зимы, причем у
самки их меньше не только по абсолютному, но и по относительно му
весу. К весне запас камешков понемногу уменьшается, а средний
вес одного гастролита увеличивается: это показывает, что уменьше-
ние веса гальки происходит не за счет истирания камешков, а за счет
потери их вследст1вие проскальзывания с пищей в кишечник. В мар-
те — мае гастролиты представлены только крупными, ровными, хоро-
шо окатанными кварцевыми зернами, средний вес которых около
80 мг у самца и 40 мг у самки; число их в одном желудке от 230 до
800, а общий вес — у самца 22—60 г, у самки 10—30 г. После окон-
чания токов глухарь снова заглатывает гальку, о чем, можно судить
по появлению мелких, угловатых камешков: в результате средний вес
зерна уменьшается, но общий вес запаса все лето остается почти
на одном' уровне (7—8%о от веса птицы). В октябре—ноябре содер-
жание гальки достигает максимума, от 630 до 1230 зерен, средним
весом по 50 мг у самца и по 30 мг у самки. Суммарный вес — у самца
12—57 г, у самки 17—39 г, относительный вес достигает 20%и от
веса птицы, а в среднем равен 14°/оо-
Запас гальки у молодых птиц летом приблизительно та'К*о!1 же,
как и у взрослых.
116
Таблица 33
Средний вес частей тела тетеревиных в %о от веса птицы
Глухарь Тетерев Ряб- чик, л 2 Б. куро- патка Тунд- рян- ка,
а>1 г. ? * $ л 2
Число экз 12 17 5 3 7 18 12 6
Вес без зоба .... 3722 1760 1273 928 344 616 587 531
Оперение 67,7 70,5 61,5 74,9 74,7 65,7 69,8 70,4 29,7
Голова 34,5 27,8 29,5 28,6 36,5 32,0 28,6
Ножки ощип Тушка с легкими и поч- 16,3 14,2 15,6 16,6 16,4 13,7 12,0 10,0
ками 696,0 664,3 688,8 684,2 13,5 669,0 663,0 14,4 629,1 647,4
Сердце 10,8 12,4 12,3 5,1 14,5 20,5
Селезенка 0.7 0,8 0,6 1,7 0,4 0,7 18,6 0,35 0,2
Печень 16,3 18,7 18,8 25,7 18,0 17,6 23,4
Поджелуд. железа . . 1.5 1,9 1,8 2,5 2,7 0,2 2,3 2,3 2,4
Гонады 0.2 0,15 0,5 0,2 0,7 0,25 0,3
Зоб + пищевод . . . 7i6 8,9 7,1 7,0 7,4 6,4 7,4 7,9
Желудок без капсулы . 29,0 39,0 25,6 26,3 26,9 28,2 32,4 28,3
Тонкие кишки . . . 7,4 9,3 7.5 7,3 7,5? 7,5 8,8 1 9,1
Слепые „ ... 7,7 7,8 7,4 6,6 7,5? 10,1 14,1 14,8
Прямая „ ... 2,9 3,4 2,3 3,0 3,0? 4,4 5,3 3,7
Сумма 898,6 879,2 879,3 898,1 875,3 867,7 842,5 868,1
Содерж. зоба .... 7,3 14,6 26,6 0,0 16', 6 16.1 19,3 10,6
„ жел. + капе. . . 18,1 26,0 17,0 14,3 18,1 15,1 16,2 15,1
„ тонк. киш. . . 34,2 36,5 37,4 38,3 39,1? 39,1 37,7 32,4
„ слеп. „ ... 32,2 34,5 43,5 28,3 42,1? 56,3 67,4 59,5
»> прям. ,,.... 2,8 3,2 3,8 1,5 6,0? 6,6 9,8 4,5
Содерж. пищеварительн. тракта без зоба . . 87,3 100,2 3,9 2,8 101,7 82,4 105,3 117,1 131,1 111,5
Сух. содерж. зоба . . ,, капе, желуд. . . 2,2 1)9 7,7 1.7 0,0 6,2 6,3 1,8 8,9 2,4 4,2 1,9
Галька в желуд. 8,5 8,9 1,8 — — 2,8 0,9 0,5
Сух. пища желуд. . . 1,7 4,1 4,0 — — 2,9 4,1 4,7 6,8
„ тонк. кищ. 7,2 7,7 7,8 8,0 — 8,2 7,9 16,2
„ „ слеп. киш. 7,7 8,3 10,4 6,8 — 13,5 14,3
„ „ прям. киш. 0.7 0,8 0,9 0,4 — 1,7 2,5 1.1
Сух. вещ. пищи . . . 17,3 20,9 23,1 — — 26,3 30,7 26,9
Вода в пище .... 70,0 79,3 78,6 — — 90,8 100,4 84,6
Желудок у тетерева в марте и апреле почта лишен гальки (10—
13 зерен общим весом около 1 г, или О,8%о от веса те ла). В мае часть
птиц пополняет запас, так что число гастролитов достигает 250 при
суммарном весе 5 г; в октябре число камешков доходит до 7001 шт.,
а вес достигает 10 г, или 8%о от веса самой птицы. Как и у глухаря,
средний вес отдельных гастролитов выше всего в конце зимы (у сам-
ца в апреле 85 мг), летом и осенью он уменьшается (12—24 мг) за
счет заглатывания мелких камешков.
' По рябчику данные скудны и относятся только к августу и октя-
брю. В августе запас гальки от 0,5 до 1,7 г (80—1801 шт.), в октя-
бре — От 0,8 до 2,7 г (от 2 до 8%о от веса птицы).
117
В желудках белой куропатки во второй половине зимы почти нет
камешков: в период с 1 февраля до середины апреля мы находили
в среднем по 3 гастролита, причем в 14% желудков их совсем не
было. Интересно, что в это время абсолютный размер гастролитов
у белой куропатки больше, чем у глухаря, а сами они не окатаны
(средний вес ПО—160 мг). Наблюдения показывают, что начиная
с 20 «марта куропатки забегают под торчащие из снега корпи вале-
жин, очевидно, в поисках гальки. Число камешков и их общий вес
возрастают до июля — августа, но средний размер одного камешка
при этом уменьшается, и в июле он в 10 раз меньше, чем в апреле.
В октябре запас камешков по весу почти такой же, как и в июле
(до 7у от веса птицы, в среднем >, или 3 т). В течение года числе
камешков в желудке (исследовано 35 экз.) изменяется от 0 до
300 штук.
У тундряной куропатки, как и у белой, в апреле в желудке нет
или почти нет камешков, хотя, кормясь на выдувах, она находит их
без всяких затруднений.
Резюмируя данные по динамике запаса гальки в желудках тете-
ревиных птиц, мы можем 'сказать, что число камешков — минималь-
ное в конце зимы (III—IV), максимальное в конце лета (VII). Общий
вес гастролитов — наибольший перед наступлением зимы (X), наи-
меньший в конце ее (III); размер зерен — наибольший в апреле, наи-
меньший — с июля по сентябрь. В это время камешки почти не ока-
таны и наиболее разнообразны по минеральному составу (большая
примесь полевого шпата).
У глухаря камешки в желудке не переводятся весь год: (вес их не
бывает меньше 5%о от веса птицы и не превышает 2О%о. Гастроли-
ты, как известно, способствуют измельчению пищи в желудке и функ-
ционально заменяют птицам зубы. Замечательно, что у тетерева,
белой и тундряной куропаток желудок почти совсем лишен камешков
во второй половине зимы, когда они, по-видимому, всего нужнее.
Е. Вильсон (1911, стр. 117) указывал, что обладая запасом камеш-
ков в желудке, белая куропатка легко перерабатывает самый дере-
вянистый корм. А. В. Михеев (1948|) в Тиманской тундре находил
камешки в 100% желудков белых куропаток в марте и апреле,
а летом 16—20% желудков совсем1 не содержали камешков. Эти дан-
ные полностью противоречат нашим. Чтобы объяснить такое расхож-
дение, остается предположить, что наличие или отсутствие гальки
зависит не столько ОТ потребности в ней в тот или другой сезон,
сколько от возможности ее нахождения в природе и условий со*
хранности в желудке птицы.
Различные исследователи не раз отмечали, что, если в желудках
рябчиков или белых куропаток много ягодных косточек ('майника,
шиповника, костяники и др.), в них мало> камешков, и наоборот. При
этом одни (Формозов, 1'904; Михеев, 1948) допускали, что косточки
выполняют функцию камешков, другие (Фолитарек, 1938) считали
косточки вредной примесью, неспособной до перемалывания выйти
в кишечник и бесполезно сокращающей рабочий объем желудка. По
118
мнению Е. Вильсона (1911) и С. В. Кирикова (1948), камешки уда^
ляются из желудка непроизвольно, задолго до того, как сотрутся в
порошок; таким же образом! выходят и косточки. Наши наблюдения
Рис. 37. Содержа пне гальки -в желудках глухаря и белой куропатки
подтверждают последнюю точку зрения. Желудок куропатки разма-
лывает только кору березовых и ивовых веточек, а белые обтертые
палочки целикам выходят через кишечник, и их легко обнаружить
119
в помете птицы. Сфинктер желудка, выпуская в двенадцатиперстную
белая куропатка
Рис. 38. Длина отделов кишечника у некоторых куриных в пересчете
на птицу весом в 1000 г
гальки идет за счет мелких зерен, а более крупные сохраняются и
функционируют до весны. Глухарь в основном питается не палочка-
ми; его желудок передает в кишечник почти одну перемолотую хвою,
что способствует сохранению камешков.
120
Таким образом, наблюдаемое явление просто и без натяжек
объясняется механическими причинами. Здесь мы видим интересный
пример несовершенной адаптации. Заглатывание камешков—при-
способление, способствующее лучшей переработке грубого корма;
между тем это полезное приспособление отказывает как раз в тот
период, когда оно всего нужнее.
Динамика запаса гальки в желудках глухаря и белой куропатки
показана на рис. 37. Ввиду большой разницы в размерах самцов и
самок глухарей, по ним мы даем не абсолютный вес камешков, а от-
носительный (на 1000 г живого веса птицы).
Кишечник. Кишечник тетеревиных замечателен исключительным
раэвитиехМ слепых кишок, которые у них значительно больше, чем
у друпи-х 'Куриных. Ввиду разницы в размерах отдельных видов,
длину кишечника полезно привести к условному весу птицы = 1000 г;
для этого непосредственно измеренная длина кишки делится на ко-
рень кубичный из веса птицы (см. рис. 38 и табл. 34). А. А. Межен-
ный сообщил нам длину отделов кишечника и вес двух самцов фаза-
нов с р. Или; аналогичные данные по граусу заимствованы из рабо-
ты Е. Вильсона (1911|); две самки кеклика измерены мною в долине
Терека; сведения об американском дымчатом тетереве (Dendrogapus
obscurus) заимствованы из статьи Дж. Бира (1955), остальные дан-
ные получены в Лапландском заповеднике и представляют средние
по измерениям 17 глухарей, 4 тетеревов, 7 рябчиков, 19 белых и
6 тундряных куропаток. Обычно одна из слепых кишок немного длин-
нее другой: для рисунка мы вычислили средние для более короткой
и длинной отдельно.
Таблица 34
Развитие кишечника у различных куриных
Вид птицы Число иссл. экз. Длина кишечника абс. (в см) Длина на 1000 г. веса птиц От нош. слепых К ТОЦКИМ
тонк. обе слеп. прям. тонк. обе слеп. прям.
Сем. тетеревиных Глухарь, самец . . . 8 215 184 22 144 123 14 0,86
„ самка . . . 9 173 136 18 144 113 15 0.78
Тетерев, оба пола . . 4 146 ПО 12 141 106 12 0,75
Рябчик, оба пола . . 7 101 86 10 138 114 14 0,85
Бел. куроп. самец . . 11 86 129 13 101 152 16 1,50
„ „ самка 8 82 132 13 98 158 16 1,62
Тундр, куроп. самец . 6 104 96 12 129 120 15 0,92
Граус, оба пола . . . — 104 84 12 121 98 14 0,81
Дымчат, тетерев, самец 10 110 74 14 103 69 13 0,67
„ самка . 31 93 63 11 97 66 12 0,68
Сем. фазаньих Серая куроп., самец . 2 66 30 7 89 40 10 0,46
Кек лик, самка . . . 2 82 35 7 115 48 10 0 43
Фазан монгол., самец 2 132 78 14 117 68 12 0,59
Петух домаш., самец . 1 136 33 8 112 27 6 0,25
121
Емкость отделов кишечника не была измерена; вместо этого мы
взвешивали их вместе с содержимым; чтобы получить чистый вес
кишечника (табл. 33) приходилось вскрывать его, промывать в не-
скольких сменах воды и отжимать в тряпке. После первого же опыта
оказалось, что прямая кишка тетеревиных в результате промывки
заметно увеличивается в весе, очевидно, за счет воды, впитанной
через эпителий: в дальнейшем1 содержимое прямой кишки мы удаля-
ли не прибегая к промывке.
Таблица 35
Вес отделов кишечника вместе с пищей зимой (XI—IV)
Число нссл. экз. Вес кишечника в г Вес на 1000 г веса птицы в г Отноше- ние слепы> к тонким
тонк. слеп. прям. тонк. слеп. прям.
Глухарь $ 4 172 168 24 49 49 7 0,97
Глухарка $ . , . . 6 98 87 15 56 49 9 0.89
Тетерев Д9 • • . . 3 55 75 9 48 62 8 1,29
Рябчик .... 3 18,3 23,4 4,5 48 ь2 12 1,28
Бел. куропатка 18 30 52 11 49 85 17 1,72
Тундр, куропатка £ 7 22 39 4 42 74 i'j 1,78
Петух домашний £ . . 1 47 10 2 27 6 1 0,21
Вес пищевой кашицы получен вычитанием веса очищенной кишки
из веса ее с содержимым.
По развитию слепых кишок белая куропатка занимает первое
•место не только в отряде куриных, но, вероятно, и во всем; классе
птиц. Длина обеих слепых кишок превышает длину тонких кишок
в 1,50 раза у самцов и в 1,62 раза у самок; в пересчете на птицу ве-
сом 1000 г, длина одной слепой кишки эквивалентна 75—80 ом; зи-
мой, наполненные, они превышают вес тонких кишок в 1,72 раза к
составляют 8,5% от веса всей птицы.
Слепые кишки тундряной куропатки значительно короче, но но
весу не уступают тому же органу белой куропатки; у рябчика и те-
терева по весу они значительно меньше, а по длине чуть короче, чем
у тундряной куропатки. У глухаря вес их наименьший во всей группе-
У всех исследованных нами фазаньих слепые кишки гораздо короче,
чем у тетеревиных.
По общепринятому взгляду, слепые кишки служат для усвоения
целлюлозы: хотя эта гипотеза и вошла в учебники, она не может
считаться вполне доказанной.
Слепые кишки подразделяются на три отдела: нижний, состав-
ляющий у глухаря около %, а у белой куропатки около всей ее
длины, представляет узкую, мускулистую трубку, в просвете пустую
(содержит только слизь). Средний отдел, самый большой, в просвете
заметно шире тонкой кишки и весь заполнен однородной липкой
122
массой, напоминающей по консистенции и цвету столовую горчицу.
Зимою она интенсивно-охристая (особенно у глухаря); в мае, с пере-
ходом на травяное питание, становится темно-бурой, а в августе и
сентябре, в период ягодного питания, окра'шена в фиолетово-черный
цвет. На внутренней поверхности кишки есть невысокие продольные
складки, числом 8—9 у глухаря, 7—9 у белой куропатки; они не
вполне правильны, местами какая-нибудь из них выклинивается, а
другая раздваивается вильчато. С внешней стороны кишки они вы-
глядят как белая связка или шов. Верхний конец—червеобразный
отросток; длина его у глухаря около 6 мм, у белой куропатки —
-1 мм *.
Большие железы пищеварительного тракта. Поджелудочная же-
леза и печень хорошо развиты. Последняя замечательна полным от-
сутствием желчного пузыря, хорошо развитого у фазаньих. По ука-
занию Ф. Греббельса (1932), желчного пузыря нет у страусов, нанду,
цесарок, голубей, попугаев, кукушек, туканов и колибри. Странно,
что ни Греббельс, ни позднейшие авторы (Дементьев, 1940, стр. 234',
639; «Птицы Советского Союза», 1952, стр. 3) не включают тетереви-
ных в число этих птиц. Причины редукции желчного пузыря у неко-
торых птиц до сих пор не выяснены. Вполне возможно, что> отсутст-
вие этого органа в разных случаях зависит от разных причин. Инте-
ресно, что среди млекопитающих желчного пузыря нет у оленей,
группы до некоторой степени аналогичной тетеревиным1 (приспособ-
ление к питанию веточным кормом зимой)).
Печень и желудок тетеревиных расположены у вентральной
стенки брюшной полости, причем одним из сухожильных «зеркал»
желудок прилегает к ней. Выходное отверстие его обращено, как
обычно, вправо, и петля двенадцатиперстной кишки также прилегает
к брюшной стенке. Петли тонких кишок расположены в глубине
брюшной полости, а слепые кишки ниже их, местами касаясь наруж-
ной стенки-брюха (рис. 39).
Пищеварение. Пищеварение у тетеревиных происходит, по наше-
му представлению, таким образом.
Зоб является только резервуаром для хранения запаса корма, а
зимой также и для подогрева' его. Корм в нем остается почти сухим
и не подвергается ни механической, ни химической обработке. Желе-
зистый желудок нормально бывает пустым; по-видимому, выделяе-
мый его железами сок стекает в кератиновую капсулу, выстилающую
изнутри мускульный желудок. Рабочий объем последнего отдела не-
велик и к тому же частично заполнен камешками и . балластом,
обтертыми палочками — зимой, косточками ягод — летом. Корм по-
ступает сюда (маленькими порциями и довольно быстро перетирается
в результате работы мощного желудочного мускула. У белой куро-
* Ф. Нансен (1937, русское издание) сообщает, что гренландские эскимосы
больше всего ценят в тундряной куропатке кишки с их содержимым, которые они
едят сырыми, прямо из птицы. «Это единственное гренландское блюдо, которого
я не попробовал», — добавляет Нансен. По-видимому, он имел в виду именно
слепые кишки.
123
патки, по которой у нас наиболее полные сведения (исследовано
15 птиц в марте и апреле), в желудке находится одновременно от
1,9 до 2,7 г сухого вещества корма; только у птиц, убитых рано- утром,
его бывает 1 г или меньше. В пересчете на 1000 г живого веса в же-
лудке находится Д8 г сухой пищи; считая суточный рацион равным
80 г, мы находим, что содержимое желудка должно сменяться 21 раз
Рис. 39. Пищеварительный тракт глухарки с брюшной (левый ряс )
и со спинной (правый рис.) стороны. Рисунок с натуры О. И. и
Н. 3. Семеновых-Тян-Шанских; 1—пищевод (oesophagus), 2—желе-
зистый желудок (proventriculus), 3— мускульный желудок (ventricu-
lus), 4—5 — двенадцатиперстная кишка (duodenum), 6—14—тонкие
кишки (ileum), 15—19 — слепые кишки (соеса), 20 - - прямая кишка
(rectum), 21— поджелудочная железа (pancreas), 22—печень
(hepar), 23— сердце (сог)
в сутки, а переработка одной порции длиться 1 ч. 08 ml В тонких киш-
ках содержится 9,05 г сухого вещества; следовательно, переработка
одной порции продолжается более 2 ч. 43 м. (больше — потому, что
мы не могли учесть все сухое вещество: часть его усваивается орга-
низмом). В слепых кишках одновременно находится 16,6 г сухого
вещества.
124
У глухаря (10 птиц с ноября ,по М1арт) на 1000 г живого веса в
желудке находится 2,5 г сухого вещества, в тонких кишках — 9,3
в слепых — 9,9; считая суточный рацион равным 46 г, находим, что
переработка в желудке занимает 1 ч. 18 м., в тонких кишках — более
2 ч. 01 м.
Содержимое тонких кишок неоднородно; среди жидкой пищевой
кашицы находится множество березовых палочек (у белой куропат-
ки) или измятых волокон хвои (у глухаря) и других неусваиваемых
частиц. На границе тонкой и прямой кишок происходит сортировка:
слепые кишки, действуя как пипетки, отсасывают жидкую пульпу,
а все твердые частицы проходят мимо их входных отверстий в пря-
мую кишку, где они формируются в экскременты, выделяемые по
мере накопления как днем, так и ночью. Как мы уже упоминали,
слизистая оболочка прямой кишки жадно впитывает влагу: видимо,
здесь отбросы пищеварения отдают остатки свободной воды.
Усвоение пищи в слепых кишках занимает много времени. Кал
из них, насколько известно (Брем, 1911; Вильсон, 1911; Крутовская,
1953; Вурм, 1885 и др.), выделяется один раз в сутки, на рассвете,
при пробуждении птицы. При этом она выделяет не более половины
всего содержимого обеих слепых кишок. У 14 белых куропаток, до-
бытых в марте и апреле в разные часы дня, от 7 ч. 20 м. до 16 ч. 30 м.,
вес содержимого слепых кишок колебался от 32,1 (8 ч. 24 м.) до
61,9 г (14 ч. 20 iM.), или, в переводе на сухое вещество, от 7,1 до
12,6 г. У ГГ глухарей, добытых в октябре—декабре между 9 ч. 43 м.
и 15 ,ч. 38 м., — от 40,1 г (12 ч. 00 м.) до 80,2 г (15 ч. 38 м.), а в
переводе на сухое вещество—от 9,6 до 19,2 г. У 4 глухарей, добы-
тых в январе — марте, от 107,5 г до 166,4 г, или от 25,8 до 40,0 г
сухого вещества.
Когда птица ночует в снегу, кал слепых кишок обычно обливает
экскременты прямой кишки, и разделить их тогда невозможно. По-
этому у нас мало данных по раздельному весу помета, и они отно-
сятся почти только к глухарю. Свежим весом пользоваться невоз-
можно, потому что к калу примерзает много снега. Воздушно-сухой
вес ночного помета из слепых кишок:
Глухарь, 29 I 1949 г................. . 32,5 г
Глухарь, 20 III 1951 г....................40,0 „
Глухарь, 23 III 1951 г.................. 17,1 „
Глухарка, 19 III 1951 г...................3,7 „
Тетерев, 23 XI 495Й г....................1'4,8 „
Из приведенных данных видно, что после утренней дефекации
в слепых кишках глухаря остается порция приблизительно равная
выделенному помету, и к следующему утру количество содержимого
увеличивается вдвое. Отсюда мы заключаем], что переваривание од-
ной порции пищевой массы в слепых кишках продолжается около
2 суток и содержимое их в середине дня приблизительно соответст-
вует тому количеству, какое отсасывается из тонкой кишки за 24 часа
(или даже за больший срок).
125
Зимний суточный -рацион глухаря мы определили в 46 г сухого
вещества на 1000 г веса птицы, вес содержимого слепых кишок со.
стайзляет 39 г (табл. 36), или, в переводе на сухое вещество, 9,4 г
(табл. 37), т. е. около 20% величины суточного рациона. Эти 2Оо/0
представляют «концентрат» пищевой кашицы, прошедшей через тон-
кие кишки за одни—двое суток. Все остальное удаляется из кишечни.
ка без задержки. Таким образом, значительное развитие слепых
Таблица 36
Изменения веса кишечника тетеревиных птиц по сезонам
(вес в граммах на 1000 г живого веса птицы}
Сезон Число птиц Тонкие Слепые Прямая
Общ. вес Ткань Содер- жимое Общ. вес Ткань Содер- жимое Общ. Ткань Содер- 1 жимое 1
Глухарь
1—IV 4 49,2 (39,0) 48,8 (39,5) 6,9 (3,4)
V —VIII 6 40,8 7,3 33,5 39,4 7,2 32,2 5,5 5,4 3,0 2,5
IX-X 6 40,4 8,9 31,5 39,6 8,7 30,9 2,7 2,7
Глухарка
XI —111 8 54,4 10,6 43,8 47,8 9,3 38,5 8,9 4,0 4,9
VIII 3 40,7 7,8 32,9 31,5 6,4 25,1 33,5 6,4 3,2 3,2
1Х-Х 7 43,3 9,0 34,3 42,2 8,7 4,9 3,1 1,8
Бел. кур.
Ш-IV 19 49,3 8,3 41,0 85,6 12,4 73,2 16,8 5,3 11,5
V-V1 6 39,5 7,0 32,5 51,1 9,6 41,5 5,2 7,2 3,4 1,8
VII —VIII 2 48,2 (41,2) 46,8 (37,2) (3,8)
1X-XI 3 42,5 9,1 33,4 57,4 12,6 44,8 6,7 4,1 2,6
Таблица 37
Содержание воздушно-сухого вещества в пробах из разных отделов кишечника
тетеревиных птиц (в %)
Вид птицы Дата | Тонкие | Слепые | Прямая
Глухарка мол 30 XI 1952 г. 20 35 25
Тетерка мол 21 IX 1949 г. 20 24 16
Бел. куропатка $ взр. . . . 17 III 1950 г. 21 20 28
„ „ Д мол.. . . 28 III 1951 г. 20 22 27
„ $ МОЛ. . . . 1 IV 1949 г. 20 26 30
„ м Л взр. . . . 2 IV 1949 г. 25 20 29
„ „ л взр. . . . 13 IV 1947 г. 22 22 17
„ „ $ взр. • • . 1 V 1949 г. 21 23 26
„ „ А взр. . . . 1 V 1949 г. 22 23 27
„ „ л МОЛ.. . . 24 V 1950 г. 21 26 27
Среднее . . . 21 24 25
126
кишок у тетеревиных позволяет им: продлить процесс кишечного пи-
щеварения почти в 20 раз (60 часов вместо 2—3 часов), при быстрой
разгрузке кишечника от неусвояемых кусочков древесины. Мы не
беремся судить, происходит ли в слепых кишках усвоение целлюло-
зы или в них только полнее используются крахмал и сахар, нахо-
дящиеся в тканях веточек и хвое, — решить этот вопрос невозможно
без специального исследования. Заметим, что слепые кишки рабо-
тают с полной нагрузкой лишь зимой. Как видно из табл. 37, у куро-
паток летом в них приблизительно вдвое, а у глухаря — в полтора
раза меньше содержимого, чем: зимой.
Наш расчет суточного рациона глухаря был приблизительно про-
верен учетом кала на местах ночлега птиц: эти данные не представ-
ляют особого интереса и здесь не приводятся. Укажем только, что
сухой вес выделений слепых кишок составляет приблизительно 20о/о
сухого веса всего кала за сутки.
Заметки о строении и весе некоторых органов. Мы познакомились
с особенностями строения зоба, желудка и слепых кишок тетереви-
ных и пытались объяснить их, исходя из функций этих органов.
Мы отметили также факт отсутствия у тетеревиных желчного
пузыря. По-видимому, единственная попытка объяснить эту анатоми-
ческую особенность принадлежит Гадову (по Греббельсу, 1923); он
полагал, что развитие желчного пузыря стоит в связи с питанием
жирной пищей. К тетеревиным это объяснение неприменимо, так как
их пища не содержит жиров. Можно предложить другое, более про-
стое объяснение. Пищеварение у тетеревиных идет беспрерывно,
днем и ночью, от первого и до последнего дня жизни птицы. Поэтому
желчь поступает в кишечник все время, без задержек, а при таких
условиях желчный пузырь был бы излишним.
Это соображение до некоторой степени аналогично приведенному
выше объяснению слабого развития жировых отложений у тетереви-
ных умеренных широт: они круглый год обеспечены пищей и не
имеют приспособлений к перенесению голодовок. Из этого правила
есть исключения. С. Ионсен (1941, стр. 20), указывая, что на Шпиц-
бергене у тундряных куропаток под кожей имеется жировой слой,
добавляет: «Я был поражен, заметив это на экземплярах, прислан-
ных замороженными в Бергенский музей, потому что не -видел ничего
подобного ни у норвежских белой и тундряной куропаток, ни у
исландской тундряной». Это явление отмечено и другими авторами
(Кейльхау, 1831; Пайк, 1889; Бируля, 1907; Мюнстерхьельм; 1911,
цит. по Йонсену).
Эти факты не противоречат нашему объяснению, а скорее допол-
няют его. Жирные куропатки наблюдались на Шпицбергене, распсх
ложенном к северу от 77° с. ш., и в других местах Крайнего Севера.
В период зимнего солнцестояния там не хватает света даже для кор-
межки белых куропаток, потому что 22 XII в полдень солнце там
находится на 10° ниже горизонта. На материке куропатки отлетают
из таких местностей на юг, а на островах им некуда уйти, и в это
время они, очевидно, недоедают. Запас жира дает им возможность
127
благополучно перенести полярную ночь. Необходимость этой адап-
тации не подлежит сомнению. Удивительно только, как способность
к ожирению развилась у вида, в норме не имеющего этого свойства,
притом без существенной перестройки всего организма.
Рассматривая табл. 33, мы можем отметить, во-первых, что из
всех внутренних органов сильно различается по развитию
у разных видов только один—сердце и, во-вторых, что у всех заметна
небольшая, но определенная разница в развитии отдельных опганоз
у птиц разного пола. По относительному весу сердца тетеревиные
располагаются в таком порядке (в %о):
Рябчик, З4'?
Глухарь: —
Тетерев: $ —
Бел. куропатка: —
Тундр, куроп.: Д —
1,5
10,8 $ — 12,4
12,3 $ —11'3,5
14,4 $ — 14,5
20,5
По нашим описаниям образа жизни этих видов можно заметить,
что подвижность их и способность к полету стоят в той же последо-
вательности. Замечательно, что индекс сердца рябчика приближает-
ся к самому (низкому в (классе птиц (альбатрос — 5,4%о, фазан —
4,2°/оо, по Греббельсу, 1932, стр. 169 и след.), а у тундряной куропат-
ки — к высшему, так как более крупное сердце имеют только неко-
торые колибри и кулики (перевозчик — 24,4%п, Греббельс). Оче-
видно, что тундряная куропатка, будучи горной птицей, летает го-
раздо чаще, чем остальные виды, и при этом' выполняет большую
работу (большие подъемы, борьба с сильным1 ветром').
Половая разница в весе органов выражается в tomi, что у самцов
относительно массивнее голова, лапы, костяк со скелетной мускула-
турой, а у самок — оперение, сердце, печень, поджелудочная железа,
зоб, желудок и кишечник (отдельные исключения объясняются, по-
видимбму, сезонной изменчивостью веса органов и неравномерным
распределением исследованных нами самцов и самок по временам
года). В сравнении с самками у самцов избыточно развиты скелет-
ная мускулатура и костяк.
Наибольшие индивидуальные различия дает вес печени и селе-
зенки, что и попятно, так как вес этих паренхиматозных органов ме-
няется в зависимости от наполнения их кровью. О сезонных измене-
ниях веса желудка упоминалось выше. Добавим, что параллельно
с :желудком меняется вес зоба и кишечника — максимум зимой, мп-
нимум—в мае—июне (см. табл. 36). Иначе меняется вес печени —
максимум ссенью, минимум —в конце зимы. Интересно, что вес опе-
рения летом почти не уменьшается, а у глухаря даже увеличивается,
несмотря на то что' во время линьки число перьев бывает меньше,
чем зимой: объясняется .это тем, что мы взвешивали перья свежими,
а летом они тяжелее, так как в период роста содержат в очинах
много влаги.
128
VI. РОСТ ПТЕНЦОВ И ДИНАМИКА ВЕСА ВЗРОСЛЫХ ПТИЦ
ПО СЕЗОНАМ
Для изучения роста птенцов и сезонных изменений веса взрос-
лых нами исследовано следующее количество птиц:
Глухарь: самцов 290, самок 250
Тетерев: „ 26, „ 20
Рябчик: „ 87, „ 88
Белая куропатка; „ Г85, „ 120
Тундряная куропатка: „ 48, „ 19
Всего ЫЗЗ птицы
В нашей работе «Экология боровой дичи Лапландского заповед-
ника» (Семенов-Тян-Шанский, 1938) приведены графики роста
молодых глухарей за 1933, 1934 и 1936 гг., белых куропаток за
1934 г. и рябчиков за 1936 г. За последние годы мы не имели доста-
точного материала для составления таких же графиков. Здесь да-
ются только обобщенные графики роста птенцов глухаря, рябчика,
белой и тундряной^куропаток за все годы вместе (рис. 40—43). Все
4 диаграммы построены по одному типу: по оси абсцисс отложено
время, по оси ординат — вес; каждая добытая птица обозначена на
этой сетке крестиком (самец), кружком (самка) или точкой (если
пол не был определен). Кривые проводились от руки с таким рас-
четом, чтобы точки располагались по обе стороны ее возможно сим-
метричнее. Эти линии дают представление о средней скорости роста
птенцов соответствующих видов. Большие отклонения отдельных
9—3037 129
точек от «нормальной кривой» объясняются, главным образом, на*
личием ранних и поздних выводков, затем — индивидуальными
уклонениями отдельных птенцов и меньше всего зависят от ранней
или поздней весны в разные годы. Наблюдения показывают, что
сроки появления выводков сравнительно мало меняются от года
к году.
Рис. 41. Рост птенцов рябчика
Снятые с графиков средние веса птенцов тетеревиных через
каждые 10 дней приведены в табл. 38. Сведения о весе птенцов те-
теревиных в разные сроки полезны для обоснования рациональ-
ного времени открытия охоты, как в Мурманской области, так и
по всему северу таежной зоны (север Карельской АССР, Архан-
130
Рис. 43. Рост птенцов тундряной куропатки
не могут обходиться без матки, вследствие чего охота в это время
носит истребительный характер, не представляя никакого спортив-
ного интереса. Качество добытой дичи низкое; к тому же при теп-
лой погоде ее невозможно сохранить даже несколько дней. Наи-
Таблица 38
Средний вес птенцов тетеревиных птиц в южной части Мурманской области
по декадам (в г}
Дата Глухарь Тетерев Рябчик Бед. ку- ропатка Тундр, ку- ропатка
с? 2 2 А ? 2 A S
"РИ рождении . 36 26 — 14 13
2* ЧЮля . . . 1 • • • jJ Wycra . . . 2||.” ; ; ; ||‘ййтября , . ” ... 1|Ь*Ября . . . (X-V) 200 400 690 1070 154fr 2030 2370 2620 2800 170 340 580 870 ИЗО 1350 1470 1560 1630 1000* 1200* 750* 900* 35 80 150 240 305 340 360 370 370 60 140 240 350 440 520 570 590 600 40 по 210 310 375 430 460 485 500
молод. . взр. . - 3001 3930 1698 1888 1242 1426 957 978 | 398 398 659 583 580 496
* Интерполировано по нескольким экземплярам.
131
более удобным сроком открытия охоты на боровую дичь во всей
северной полосе надо считать 1 сентября. В это время молодняк
тетеревиных птиц достигает двухмесячного возраста; петушки глу.
хари и тетерева линяют во взрослое перо и легко отличаются о?
курочек по окраске и величине; выводки начинают разбиваться;
вес птенцов рябчиков и белых куропаток уже приближается к весу
взрослых птиц.
В годы «неурожая» тетеревиных птиц следует сокращать продол,
жительность охотничьего сезона, начиная его позже или заканчи.
вая раньше. В целях охраны дичи в тяжелый для нее год целесо-
образно сокращать срок охоты именно в начале сезона, потому что
в это время добывается больше всего глухарей, тетеревов и рябчи-
ков. Впоследствии дичь становится осторожнее, и зимняя охота
мало отражается на ее численности; вместе с тем зимой боровая
дичь — продовольственный ресурс для охотника на промысле пупь
пого зверя. Только промысловая охота на белую куропатку интен-
сивно ведется до конца зимы, особенно в полосе лесотундры.
Поскольку в октябре вес молодняка тетеревиных уже стабили-
зируется, для определения «нормального» веса птиц мы останови-
лись на периоде с 1 октября по 31 мая (срок окончания весенней
охоты на токах). Средний вес вычислен для каждого пола отдель-
но. В октябре весовой прирост молодых самцов глухарей не пре-
кращается, а только приостанавливается до следующего лета. По-
этому вес глухарей первогодков и вполне взрослых дается раз-
дельно. Особи, явно задержавшиеся в росте («заморыши»), при
обработке исключались. Результаты подсчетов представлены
в табл. 39 и на рис. 44—47.
Таблица 39
Вес тетеревиных в период с октября по май (в г}
Вид и пол Средний вес и ср. ошибка Среднее квадр. отклон. Пределы веса Число особей
Лапландский заповедник
Глухарь £ взрослый .... 3930+28 +229 3500-4548 66
Глухарь молодой .... 3001+23 +205 2471-3546 76
Глухарка $ взрослая # . . 1888±21 +108 1700-2121 26
Глухарка £ молодая ’ . , 1698±10 + 94 1468-1910 80
Глухарка £ все возрасты . 1748+11 + 127 1440—2121 122
Тетерев взрослый .... 1426±21 + 43 1364-1462 4
Тетерев £ молодой .... 1242+29 + 81 1120-1400 9
Тетерка £ все возрасты . . 968+12 ± 40 900-1056 12
Рябчик $ все возрасты . . 398+ 6 + 34 310- 485 36
Рябчик £ все возрасты . . 398+ 4 + 29 352— 465 42
Белая куропатка £ ... . Белая куропатка £ . . . . Тундряная куроп. (с 1' IX); Тундряная куроп. £ (с 1 IX} 659+ 5 583+ 5 530± 9 496+16 + 49 + 45 ± 46 ± 43 536 - 795 475- 692 459— 640 442- 580 114 80 27 7
132
Вид и пол Средний вес и ср. ошибка Среднее квадр. отклон. Пределы веса Число особей
Печоро-Илычский заповедник
Глухарь $ взрослый .... 4254±35 ±267 3610- 4810 57
Глухарь $ молодой .... 3123±64 +270 2665—3760 20
Глухарка $ взрослая . . . 2004±29 ±123 1780- 2210 18
Глухарка $ молодая . . . 1834+28 ±112 1637—1993 16
Вес взрослых птиц обнаруживает определенную сезонную из-
менчивость. Для тетеревиных это явление впервые описано Виль-
соном (1911). Он нашел, что вес самки грауса в Англии выше все-
Рис. 44. Кривая распределения веса глухарей: прерывистая линия —
самки, тире с тачками — молодые самцы, сплошная линия —
взрослые самцы
го в апреле — мае, в период кладки яиц; во время насиживания вес
быстро падает и в июне — июле достигает минимума (85% макси-
Рис. 45. Кривая распределения веса рябчиков: прерывистая линия —
самки, сп тошная линия •— самцы
мального веса). У самцов максимальный вес наблюдается в де-
кабре, минимальный — в марте (80% максимального). В мае — ию-
ле вес самок уменьшается, а вес самцов увеличивается. Некоторые
133
Рис. 46. Кривая распределения веса белых куропаток; обозначения —
см. рис. 45
Рис, 47. Кривая распределения веса тундряных куропаток; обозна-
чения — см. рис. 45
Рис. 48. Изменения веса самцов и самок глухарей в течение года. Кривее
проведены по средним месячным весам; количество экземпляров показан
цифрами
134
данные о сезонной изменчивости веса приводят и советские иссле-
дователи (Фолитарек и Дементьев, 1938, и Донауров, 1947, для ряб-
чика; Семенов-Тян-Шанский, 1938, и Ульянин, 1939, для белой ку-
ропатки), причем картина получается сходная с описанной Вильсо-
ном. На наших диаграммах показана динамика веса взрослых глу-
харей (рис. 48), рябчиков (рис. 50) и белых куропаток (рис. 51).
Основой для проведения кривых служили средние веса птиц по ме-
сяцам, показанные крестиками (самцы) и точками (самки): цифры
при них показывают число использованных данных. Местами, где
данных было мало, средний вес отнесен не к середине месяца,
а к средней дате добычи соответствующих экземпляров. Ввиду не-
достаточного количества данных, особенно по самкам, диаграммы
рис. 48—51 следует считать ориентировочными. Рассмотрим сведе-
ния о росте птенцов и динамике веса взрослых птиц по видам.
Глухарь. Появление первых выводков наблюдалось между 17 VI
и 6 VII, в среднем за 12 лет — 25 июня. Эта дата, по-видимому,
близка к моменту массового вывода, так как встретить самый ран-
ний выводок и притом в первый же день по оставлении им гнезда
мало шансов.
Точные данные по росту птенцов в условиях вольерного содер-
жания приводят С. А. Ларин (1941) и Е. А. и Е. В. Крутовские
(1953). Наши данные взяты с графика роста птенцов (рис. 40).
У Ларина и Крутовской возраст птиц определен точно, но данные
относятся к небольшому числу особей, находившихся притом в ис-
кусственных условиях содержания; мы определяли возраст ориен-
тировочно и даем обобщенные цифры. Учитывая различия в мето-
дике, лучшего согласования результатов мы не вправе были и ожи-
дать. Данные по росту глухаря в Печоро-Илычском заповеднике
приводит В. П. Теплов (1947), однако его цифры плохо согласу-
ются с другими источниками, возможно, по причине неточности
в оценке возраста птиц.
Вес 3 новорожденных глухарят, по нашим измерениям, был 36,
36 и 37 г. В первые двое суток, пока глухаренок живет, главным
образом, за счет желточного мешка, вес его не увеличивается (по
Ларину — на второй день даже уменьшается примерно на 10%).
На третий день глухаренок начинает расти, и темп роста в даль-
нейшем все увеличивается, достигая у самок в возрасте 40—50 дней
28—29 г., а у самцов в возрасте 50—70 дней 43—47 г. в сутки.
После того прирост постепенно уменьшается, у самок быстро, у сам-
цов медленно, вследствие чего кривые роста самцов и самок, при-
мерно с 25 августа, сильно расходятся.
В возрасте около 120 дней рост прекращается у обоих полов.
Небольшое увеличение веса в ноябре — декабре (рис. 48) объяс-
няется тем, что в это время зобы убитых птиц содержат больше
корма, чем осенью. К весне вес самцов немного увеличивается.
В июне у молодых глухарей возобновляется прерванный на зиму
рост, и к сентябрю, в возрасте 14 месяцев, они по весу почти срав-
ниваются со стариками (см. табл. 40 и рис. 48).
135
Молодые самки уже в трехмесячном возрасте мало отличаются
от взрослых, и зимой их средний вес составляет 90% веса взрос-
лых, тогда как молодые петухи достигают в это время только 76%
веса взрослых (см. табл. 40).
Таблица 40
Вес лапландских глухарей по месяцам
(в г; в скобках число экз.)
Месяц Самцы мод. Самцы взр. Самки мол. Самки взр.
Январь .... 13343 (3) 3798 (5)
Февраль . . . . Март 3947 (8) 1 1666 (6) 1784 (2)
Апрель .... 3025 (7) 4180 (9) — —
Май 3170 (9) 3901 (23) 1 1720 (3)
Июнь 3092 (2) 3776 (5) f JL /Х/V/ у*"*/
Июль . . . . • 3361 (3) — 1400 (1)
Август .... 3552 (2) 3718 (7) 1754 (9)
Сентябрь . . . 2317 (60) 3829 (29) 1488 (43) 1784 (50)
Октябрь .... 2916 (32) 3873 (8) 1678 (37) 1878 (14)
Ноябрь .... 2978 (20) 3968 (8) 1710 (30) 1925 (7)
Декабрь . . . « 3062 (6) 3788 (5) 1718 (8) 1942 (3)
Питание глухарят во время самого быстрого роста (в августе)
состоит в основном из зелени, ягод, семян растений и животных
кормов. В этот период они поедают больше насекомых и моллюсков
(даже в абсолютных величинах), а также семян растений (в про-
центах ко всему рациону), чем в другие месяцы. В зимний период
приостановки роста глухари кормятся почти исключительно хвоей
и веточками деревьев.
В отличие от рябчиков и белых куропаток, вес глухарок в пе-
риод кладки яиц, по-видимому, не увеличивается: три глухарки
с функционирующим яичником (возраст не определен) весили
1440, 1690 и 2030 г. Во время насиживания и выкармливания цып-
лят они тощают. Самая легковесная взрослая глухарка, добытая
24 VII 1940 г., была без выводка, но имела наседное пятно: она ве-
сила 1400 г, т. е. 75% веса взрослой самки зимою. Вес самок уве-
личивается уже в августе и в декабре достигает максимума.
Вес самцов выше всего в апреле (на 6% больше среднего зим-
него). У глухаря, в отличие от мелких видов тетеревиных, пониже-
ние веса наступает у обоих полов одновременно. Представление
о возрастной разнице в весе глухарей, добытых на токах, дает диа-
грамма, составленная по материалам, собранным нами в Псчоро-
Илычском заповеднике (рис. 49).
Вес глухарей из различных местностей трудно сравнивать по
той причине, что в большинстве случаев не проводится разделение
на взрослых и молодых. В табл. 4L мы приводим только те данные,
где это требование было учтено (Кириков, 1948; Крутовская, 1953).
136
Таблица 41
Вес глухарей в географическом разрезе
Район Средние и предельные веса в кг
самцы самки
Лапландский зап-к 68° с. ш. 32° в. д. . Печоро-Илычский зап-к 62° с. ш. 57° в. д. . Красноярский зап-к 56° с. ш. 93° в. д. . Башкирский зап-к 53° с. ш. 58° в. д. . мол. 3,00 (2,47 — 3,55) взр. 3,93 (3,50 — 4,55) мол. 3,42 (2,67 — 3,76) взр. 4,25 (3,611—4,81) мол. 3,3 ( ? — 3,5) мол. 3,4 (3,3 —3,5) взр. 4,4 (3,4 —5,0) 1,70 (1,44 — 1,01) 1,89 (1,70 — 2,12); 4,83 (1,64 — 1,99) 2,00' (1,718 — 2,21) 1,93 —
Рис. 49. Возрастная разница в весе глухарей, добытых па токах по
Верхней Печоре (цифры внизу столбиков — число экз., вверху —
разница веса в кг)
Сравнивая вес глухарей на северном и южном пределах его
ареала, мы видим, что на юге вполне взрослые глухари тяжелее
в среднем на 5%, а молодые (в возрасте от 3 до И месяцев) —
на 12%. Разница, по всей вероятности, зависит от того, что на се-
вере период питания летними кормами короче, а потому молодой
глухарь переходит целиком на зимний рацион и перестает расти на
более ранней стадии развития, чем на юге. Если это объяснение
верно, то в годы с ранним наступлением зимы молодые глухари
должны быть мельче, чем в годы с длительным теплым периодом.
Наблюдения подтверждают это предположение. За годы нашей ра-
137
боты в Печоро-Илычском заповеднике самая поздняя весна и самая,
ранняя зима были в одном и том же 1952 году. Замечательно, что
в охотничий сезон 1952 г. молодые глухари были мельче, чем во все
остальные годы. Средний вес четырех исследованных нами в ту
осень самцов оказался равным 2655 г, или 86 % от нормального
веса их в это время, а средний вес пяти молодых глухарок был
равен 96% нормального.
В Мурманской области молодые глухари в первую зиму дости-
гают 76%, а в Башкирии — 83% веса старых петухов. Вес молодой
глухарки составляет 56% веса молодого самца, а вес старой глу-
харки— 48% веса старого глухаря. Глухарь представляет один из
самых выдающихся примеров полового диморфизма в классе птиц.
Тетерев. По недостатку данных сведения о росте тетеревят, при-
веденные в табл. 38, очень приблизительны, и мы их не комменти-
руем. Заметим только, что молодые косачи в первую зиму дости-
гают 87% веса стариков, а молодые тетерки 98% веса взрослых;
отношение веса самки к весу самца у молодых птиц равно 77%,
у старых — 69%.
Сведения о весе тетеревов в разных географических условиях
скудны. В Печоро-Илычском заповеднике вес тетерева меньше, чем
в Лапландском, и в промысловый период составляет в среднем:
1225 г —самцы и 910 г — самки.
Рябчик. Вследствие небольшого размера рябчика, птенцы его
растут и развиваются быстрее, чем глухарята. Самый быстрый рост
Рис. 50. Изменения веса самцов и самок рябчика в течение года
(9 г в сутки) происходит в первой декаде августа. В сентябре сред-
ний вес взрослых рябчиков равен 374 г, молодых — 354 г, или 95%
веса взрослых. В октябре отличить молодых от старых уже труд-
но, и сравнить вес тех и других мы не смогли.
138
Динамика веса взрослых рябчиков показана на рис. 50 и в
табл. 42. Аналогичные сведения приводят С. С. Фолитарек и
Г. П. Дементьев (1938) и С. С. Донауров* (1947). Имеющиеся дан-
ные относятся к Лапландскому, Печоро-Илычскому и Алтайскому
заповедникам. Несмотря на огромные расстояния между ними,
всюду вес рябчиков изменяется сходным образом. В ноябре и де-
Таблица 42
Вес рябчиков, белых и тундряных куропаток по месяцам
(в г; в скобках число экз.)
Месяц Рябчик Белая куропатка Тундряная куропатка
£2 £2 $2 | 22
Январь . . . 384 (2) 727 (6) 628 (5)
Февраль . . — — 648 (6). 554 (8) — —
Март . . . . 396 (1) 377 (3) 662 (13) 606 (9) — —
Апрель . . . 339 (1) — 656 (14) 572 (8) 519 (12) 498 (2)
Май .... 310 (1) 406 (4) 615 (21) 618 (4) 508 (8) —
Июнь . . . 348 (2) — 597 (7) — 487 (5) —
Июль . . . . 346 (5) 316 (3) 597 (11) 464 (1) — 455 (3)
Август . . . 352 (1) 336 (8) 631 (7) 541 (42) 519 (3) 446 (2)
Сентябрь . . 377 (5) 370 (3) 670 (13) 603 (5) 557 (5) 509 (3)
Октябрь . . 386 (22) 396 (22) 653 (37) 567 (23) 625 (2) 475 (2)
Ноябрь . . . 427 (5) 405 (7) 701 (14) 581 (17) — —
Декабрь . . 437 (6) 429 (4) 718 (3) 601 (6) — —
кабре он достигает максимума; в это время вес одинаков у обоих
полов. Во второй половине зимы вес немного уменьшается, при-
чем у самцов он продолжает падать и дальше, достигая в мае
(в Печоро-Илычском з-ке — в июне) минимального уровня за год,
равного 71—80% от максимального. Вес самок в мае возрастает,
образуя второй максимум. В это время самец легче самки — в Лап-
ландском заповеднике вес его равен 76%, на Алтае — 80%, на Пе-
чоре— 84% среднего веса самки в той же местности и в то же
время. Июньских самок мы, к сожалению, не имели, но в июле,
когда они водят птенцов, вес их самый низкий за весь год. Самцы
в это время уже поправляются и весят больше самок — в Лаплан-
дии на 10%, на Печоре — на 6%, на Алтае — на 7%.
Ввиду того, что в некоторые месяцы самцы легче самок, а в дру-
гие— тяжелее, в среднем за весь год оба пола по весу почти в точ-
ности равны (табл. 39 и 42). Приводим вес рябчиков (в г) в раз-
ных географических областях (в скобках указано число особей):
* Донауров дает не вес рябчиков, а частное от деления веса (в г) на длину
птицы (в см); причина выбора такого странного индекса не ясна.
139
Район Самец Самка Период
Лапландский заповедник 68° с. ш. 32° в. д. 398 (36) 398 (42) С X по V
Печоро-Илычский заповедник 62° с. ш. 57° в. д. 435 (32) 422 (24) Поздняя осень и зима
Алтайский заповедник 51° с. ш. 88° в. д. 381 (72) 393 (45) IV, V, IX, X
Белая куропатка. Первые выводки были встречены между 20 Vi
и 19 VII, в среднем за 13 лет — 3 июля, на 8 дней позже появления
глухариных выводков. Вес только что вылупившегося птенца, по
литературным указаниям (Ульянин, 1939; Дунаева и Кучерук
1941; Михеев, 1948), от 13,5 до 16 г.
Суточный прирост птенцов увеличивается, пока в первой декаде
августа не достигнет 11 г в день (при возрасте птенцов 30—35
дней), потом рост замедляется и после 1 октября почти совсем пре-
кращается. Молодые куропатки в первую зиму по весу не отлича-
ются от взрослых (табл. 43).
Таблица 43
Соотношение веса белых куропаток (с X по V) по полу и возрасту
__________________(в в скобках число особей)__________________
— Возраст Самцы Самки Отношение веса самок к весу самцов в %
Взрослые 665 (23) 597 (6) 90
Молодые . 649 (12) 566 (19) 90
Отношение веса молодых к ве-
су взрослых в % .... 98 98
Все куропатки 659 (114) 583 (80) 89
Изменения веса белых куропаток по сезонам (рис. 51) сходны
с тем, что мы видели у рябчиков (рис. 50). По нашим данным,
наибольший вес у обоих полов наблюдается ib январе (самец — 727,
самка — 628 г); после этого вес самок сначала уменьшается, а в мае
вновь поднимается, причем на короткое время самки сравниваются
с самцами (618 г). Во время насиживания кладок и вождения птен-
цов вес самок падает до минимума1, составляя около 525 г, или 82%
от максимального за год. Вес самцов достигает минимума в июне-
июле в среднем 597 г, или 82«/0 от максимального за год.
На южной границе ареала белой куропатки, в Наурзумском за-
поведнике (Ульянин, 1939), минимальный вес самцов наблюдался
в мае (в среднем 514 г), максимальный, вероятно, в феврале, когда
был добыт самый крупный самец (872 г). У шотландской куропатки,
как мы уже видели, амплитуда изменений веса примерно такая же.
но сроки наступления минимумов более ранние, особенно у самца
(март).
140
При сравнении веса куропаток из различных географических
районов самыми крупными оказываются южные, степные, самыми
•мелкими — лесные, например с верховьев Печоры, а тундровые за-
няли промежуточное положение *. Приводим средний вес куропаток
из разных местностей, группируя их по> зонам (Фолитарек, 1939;
Михеев, 1948; Теплов, 1947-а; Ульянин, 1939):
Зона и район Самцы Самки Период
Тундра Мурманск, побережье 67—68° с. ш. 40—41°в. д. 652 605 1Х-Х
Тиманская тундра 68° с. ш. 50° в. д. Тайга 662 (15) 636 (19) IV-V
Лапландский заповедник 68° с. ш. Зй° в д. 659 (114) 583 (80) X-V
Печоро-Илычский заповедник 62° с. ш. 57° в. д. Степь 650 (15) 550 (15) IX—IV
Наурзумский заповедник 52° с. ш. 64° в. д. 711 (12) 653 (2) IV и IX
Тундряная куропатка. Первые выводки отмечены в среднем за
6 лет 2 июля (24 VI—12 VII|), следовательно, не позднее, чем у белой
куропатки. Вес пуховичков в первый день жизни, по нашим1 измере-
ниям, равен 10—14 г. Из сравнения рис. 42 и 43 можно убедиться,
что птенцы обоих видов развиваются одинаково и что наибольшая
* Научные названия этих форм Lagopus lagopus major Lorenz, L. 1. rossicus
Serebrowsky, L. I. koreni Thayer et Bangs.
141
прибыль в весе (10 г в сутки) наблюдается в первой декаде августа.
К сожалению, наш материал для тундряной куропатки недостато-
чен— всего 75 птиц, и не охватывает всего года; к тому же весной
добывались почти одни самцы.
Вес самцов выше всего в октябре или, может быть, несколько
позже, а к апрелю они заметно тощают (83о/о осеннего веса); в июне
вес достигает минимума (78о/о осеннего). Таким образом, амплитуда
колебаний веса у тундряной куропатки больше, чем у белой и поху-
дание за зиму—значительнее. У самок, как и у белой куропатки,
минимальный вес в июле и августе, а максимальный, вероятно, позд-
ней осенью (см. табл. 42).
Определить разницу в весе молодых и старых птиц осенью не
удалось по той причине, что большая часть экземпляров добыта в
1930—1934 гг., когда мы еще не различали возраста куропаток. Вес
самок (с IX по V) составляет 94о/о веса самцов; разница меньше,
чем1 у белой куропатки.
Данные по весу тундряных куропаток из разных частей ее ареала
скудны и относятся к разным сезонам, а потому трудно сравнимы
(Йонсен, 1941; Гельтинг, 1937):
Раной Самцы Самки Период
Шпицберген 738 (9) 701 (11) 1Х-Х
77—80° с. ш. 11—25° в. д.
Вост. Гренландия 502 (5) Vll—VIII
74° с. ш. 22° з. д.
Лапландский заповедник 576 (7) | I 495 (5) 1Х-Х
68° с. ш. 32° в. д. 476 (8) Vll—VIII
йотунгейм, Норвегия 439 (18) I —III и
52° с. ш. 8° в. д. VI—VIII
Средние веса для Лапландского заповедника подсчитаны таким
образом, чтобы их можно было сравнить с данными для Шпицбер-
гена и восточной Гренландии. Вес северных тундряных куропаток
значительно больше веса южных. Шпицбергенская куропатка * ве-
сит приблизительно на 35% больше типичной, а гренландская — на
6°/0 больше. Норвежская тундряная куропатка с гор йотунгейма
по крайней мере на 8% легче лапландской. Географическая измен-
чивость белой куропатки оказывается более сложной, но и этот вид,
вообще говоря, не становится крупнее к северу («Птицы Советского
Союза», т. IV, стр. 11—42). Попытки найти универсальный закон,
ставящий величину птиц разных популяций в зависимость от широты
* Lagopus mutus hyperboreus Sundev. (название типичной формы L.. m. mulus
(Mont.).
142
местности («правило Бергмана»}), очевидно, обречены на неудачу.
Разницу в росте отдельных популяций надо искать в каждом отдель-
ном случае в конкретных особенностях взаимоотношений организма
со средой обитания. Один пример таких отношений мы уже рассмот-
рели.
Мы уже упоминали о жировых отложениях шпицбергенской тунд-
ряной куропатки. А. Пайк (цит. по Ионсену, 1941) сообщает, что
добытые 5 X 1868 г. на Шпицбергене три куропатки весили ровно
1 кг каждая и были очень жирными. По-видимому, разница в весе
разных популяций зависит не только от размеров птиц, но и от их
упитанности, которая, в свою очередь, связана с условиями питания
и добывания корма.
В связи с этим1, возникает вопрос: если у тетеревиных вес регу-
лярно меняется на протяжении года и амплитуда этих колебаний
составляет от 10 до 30% максимального веса, то за счет каких тканей
он меняется? Для освещения этого вопроса, в 1949—1951 гг. мы
исследовали около 90 птиц, определяя вес каждого органа в отдель-
ности. Общее число взвешенных по сезонам птиц значительно пре-
вышает число исследованных на вес органов. Зная, что индивидуаль-
ные колебания веса птиц гораздо больше его сезонных изменений,
мы не могли непосредственно сравнивать веса органов небольшого
числа птиц, добытых в разные времена года. С целью свести к мини-
муму влияние индивидуальных отклонений веса птиц на вес их орга-
нов, мы вычислили в отдельности, по> двухмесячным периодам, сред-
ние веса птиц (без содержимого зоба) и средние индексы веса раз-
личных их органов. Помножив средний вес птицы на индекс соответ-
ствующего органа в тот же период, мы получили средние веса орга-
нов по сезонам. Они дают представление о динамике веса отдель-
ных частей тела «нормальной» особи на протяжении года. При по-
добных расчетах большая точность не нужна, вследствие чего все
цифры округлены до целых граммов. Здесь приведен расчет только
для самцов белой куропатки, по которым 1мы имели больше материа-
ла (табл. 44). В правом столбце таблицы дана разница в весе органа
в период его наибольшего и наименьшего развития — в граммах и
в процентах от максимального веса1, показанного в предыдущих ко-
лонках таблицы.
Анализируя табл. 44, мы приходим ,к следующим выводам:
1. Резче всего меняется по сезонам среднее содержание корма
в зобу — оно колеблется от 4% веса птицы в V—VI до 4^/0 в темный
период зимы (XI—XII; см. также стр. 109).
2. Вес содержимого желудка (с гастролитами и кератиновой кап-
сулой) и кишечника изменяется аналогичным образом; но с той раз-
ницей, что максимума он достигает в марте, в период, когда куро-
патка питается единственно веточками ив и берез, кормом, по-види-
мому, плохо усваиваемым.
3. Вес желудочного мускула в мае—июне приблизительно на 40о/0
меньше, чем в ноябре—декабре. Это объясняется, видимо, тем, что
летом желудок работает с небольшой нагрузкой, вследствие чего
143
Таблица 44
Динамика веса частей тела самца белой куропатки по временам года (в г)
Части тела III—IV V-V1 VII—VIII 1Х-Х XI—XII Разность макс. — мин,
абс. в %
Средний вес самца . . (688) 659 606 614 662 710 104 1 15
Вес корма в зобу . , (21) 13 6 8 10 25 19 76
Вес птицы без зоба . Содержимое пищевари- (667) 646 600 606 652 685 85 12
тельного тракта . . — 91 54 61 72 73 37 41
Желудок муск. и жел. — 21 14 21 20 24 10 42
Пищев., зоб и кишки . — 21 15 17 18 26 11 42
Печень — 10 11 13 17 16 7 41
Сердце — 9 9 8 10 11 3 27
Тушка с гол. и ногами —' 432 445 428 456 469 41 1 9
Оперение Число экз., у которых — 45 38 41 39 51 13 25
взвеш. органы 0 19 6 3 2 1 — —
мускул временно редуцируется. Вес остальных частей пищеваритель-
ного тракта (пищевод, зоб и кишечник|) изменяется параллельно
весу желудка.
4. Амплитуда изменений веса печени такая же, как и частей пи-
щеварительного тракта (41—42о/о максимального веса|), однако
сроки наступления максимума (IX—XII) и минимума (III—IV, то
есть в конце зимы) иные. Поскольку в печени сосредоточены энерге-
тические ресурсы, понижение ее веса может быть показателем пло-
хого питания организма. Известно, что- в марте белая куропатка пи-
тается скудно и однообразно. С другой стороны, упитанность куро-
паток в начале зимы наибольшая.
5. Вес сердечного мускула достигает минимума в мае—июне,
максимума — в ноябре—декабре; амплитуда — около 25<*/0 наи-
большего веса сердца.
6. Вес ощипанной и выпотрошенной птицы (скелетная мускула-
тура, костяк, нервная система, кожа, органы дыхания и почки) изме-
няется параллельно с весом сердца. Замечательно, что амплитуда
колебаний веса тушки составляет только 9°/0, тогда как амплитуда
живот веса птицы равна 1бо/о. Отсюда следует, что скелетная муску-
латура— наиболее стабильная часть тела куропатки. Если бы куро-
патки осенью жирели, амплитуда веса тушки была бы гораздо
большей.
7. Вес оперения сравнительно мало меняется. Летнее оперение
заметно уступает зимнему и по числу перьев и по их размеру, но
разница отчасти компенсируется тем, что в период роста перья тя-
желее.
Динамика веса частей тела других видов тетеревиных, в общем,
совпадает с этой схемой, но сроки наступления максимумов и мини-
144
мумов иногда отличаются. Так, у глухарей наибольший вес печени
бывает в августе и сентябре, минимальный — с января по март; наи-
больший вес желудка — также с января по март, а минимальный
(на 19% меньше максимального) — в мае—июне. Как правило, са-
мой стабильной частью птицы является скелетная мускулатура и
костяк, затем оперение и сердце.
Явления сезонной изменчивости организма наших птиц и зверей,
к сожалению, почти не изучены, а потому и значение их обычно не-
дооценивается. В литературе есть некоторые данные только по до-
машнему северному оленю (Иванов* и Николаевский, 1936). Авторы
исследовали гистологические срезы костей, щитовидной железы,
семенников, печени и селезенки и пришли к выводу, что, например,
кости оленя находятся в полноценном состоянии только в октябре —
феврале; с апреля до половины июня происходит перестройка кост-
ной ткани, преимущественно ее разрушение: в это время кости оленя
наиболее хрупки (нередки случаи переломов ног и позвоночника);
с половины июня до сентября перестройка кости идет преимущест-
венно в сторону ее формирования. Полноценное состояние печени,
селезенки и лимфатических узлов наблюдалось в июле—августе и
в октябре, атрофические процессы более выражены в апреле — на-
чале мая.
В общем, фазы сезонной изменчивости северного оленя, установ-
ленные путем изучения тканей под микроскопом, довольно хорошо
согласуются с описанными здесь фазами изменчивости белой куро-
патки и глухаря. Этого и следовало ожидать, зная, что наши тетере-
виные обитают в тех же географических зонах, что и северный олень,
вследствие чего сходен и годичный цикл их питания.
Обобщая приведенные здесь факты, мы можем сказать, что се-
зонная изменчивость организма не сводится только к разнице в его
упитанности. Поскольку величина отдельных внутренних органов ме-
няется в гораздо большей степени, чем* вес животного в целом, глав-
ную роль в этом вопросе играют физиологические явления, стоящие
в тесной связи с сезонным циклом питания.
VII. ЛИНЬКА И МЕТОДЫ ОПРЕДЕЛЕНИЯ ВОЗРАСТА
ТЕТЕРЕВИНЫХ
Белая и тундряная куропатки сменяют на протяжении года не-
сколько хорошо различимых нарядов (одни авторы насчитывают
три, другие даже четыре наряда). Аналогию белому зимнему наряду
куропаток мы находим только у некоторых млекопитающих (зайцы,
горностай, ласка, песец, копытный лемминг и лемминговидная полев-
ка!), но у других птиц нет ничего подобного. Резкая разница в пиг-
ментации и рисунке перьев разных нарядов белых куропаток облег-
чает исследование их линек на обычном материале — музейных
шкурках. По этим причинам линька этих двух видов, по сравнению
с другими тетеревиными, изучена очень подробно. В частности линь-
ка белой куропатки исследована Йонсеном (1929—на норвежском'
10-3037
145
языке с английским резюме) и Михеевым (1939, 1948), линька тунд-
ряной куропатки — Саломонсеном (1940 — монография на англий-
ском языке), линька всех наших ©идов тетеревиных—Снигиревским
(I960).
Мы не занимались специально исследованием линьки. Здесь
дается краткое описание линьки тетеревиных птиц только по собст-
венным наблюдениям. Ценность их в том, что они относятся к одно-
му определенному пункту — Лапландскому заповеднику. Наше вни-
мание было обращено преимущественно на установление сроков ли-
нек и связи их с условиями среды обитания, а с другой стороны —
на способы определения возраста по перьям.
Глухарь. Рост миховых первой генерации1 * начинается с рожде-
ния птенца. 21 VI 1950 г. мы поймали трех; глухарят в возрасте менее
2 суток (у них еще сохранился «яйцевой зуб», который, по указанию
Е. А. Крутовской (1953), отпадает к концу второго дня жизни), и
у них были хорошо заметны пенечки маховых перьев. У глухаренка
приблизительно 4-дневного возраста, найденного 29 VI 1951 г. в гнез-
де зимняка, на крыльях были растущие маховые с эмбриональным
пухом на вершине стержня. ^Благодаря быстрому росту ювенильных
маховых птенцы способны перепархивать в возрасте уже нескольких
дней, а достигнув веса 200 г (приблизительно в 16 дней) —даже
взлетать на деревья.
Контурные перья появляются почти в одно время на зобу и на
спине. У птенцов весом 170—200 г (возраст 15 дней) туловище, в
Общем, прикрыто перьями, растущими на спине, крыльях и зобу; го-
лова, шея, середина груди по сторонам киля, брюхо и голени еще
одеты эмбриональным пухом. Хвост состоит из 9 пар рулевых пер-
вой генерации, причем внутренние из них еще короткие, с синим ство-
лом; ювенильные маховые на разных ступенях развития, но три ма-
ховых кисти и семь маховых предплечья уже закончили рост; в то же
время растут первые маховые второй генерации, а именно самые
внутренние из маховых предплечья (рис. 52 и 53).
При весе 300 г (возраст 22 дня) вся поверхность тела покрыта
перьями, но пух заметен между перьями головы, груди и голеней.
Замечательно, что на этих перьях, вырастающих на третьей педеле
жизни глухаренка, проявляется половой диморфизм — у самцов они
(особенно кроющие уха и темени) сероватые, с мелкими поперечны-
ми полосками, у самок охристые, с черными поперечными перевязя-
ми, очень похожие на рябые перья взрослых самок. Е. А. Крутов-
ская (1953) указывает, что по этому признаку можно определить пол
глухаренка с 15—20-дневного возраста **. Теперь уже 4 маховых
• Они принадлежат к так называемому «гнездовому» наряду: этот термин
в применении к выводковым птицам звучит странно, и мы предпочитаем назы-
вать первый наряд тетеревиных ювенильным.
♦♦ По Н. Хеглунду (195<8) пол глухаренка после отпадения яйцевого зуба
можно определить по цвету клюва: у самцов он темный, у самок светлый, с тем-
ными продольными полосками. В возрасте 6—12 дней есть разница в окраске
маховых и их кроющих: у самца по переднему краю опахала есть белая кромка,
у самки — только белые пятна.
146
кисти и от 8 до 14 маховых предплечья ювенильного наряда закон-
чили рост и имеют твердый белый очин.
Около 1 августа при весе самки 580 г и самца 690 г (возраст
35 дней) все ювенильные рулевые, кроме двух центральных пар, уже
выпали и на месте их, начиная с крайних, растут рулевые второй ге-
нерации, которые будут служить до следующего лета. Маховые сме-
няются медленнее, особенно в отделе предплечья, где больше поло-
вины присутствующих маховых принадлежит еще к первой генера-
ции. Параллельно с ростом маховых кисти первой генерации
(№ 6—10, считая от внешнего края крыла) происходит рост двух
дистальных маховых, которые хотя и принадлежат фактически к пер-
вой генерации, но уже не сменяются до конца следующего лета: это
явление впервые отмечено О. Хейнротом в 1898 г. и подробно описа-
но Ф. Саломонсеном (1940/).
В случае ранней весны уже в первых числах августа, а чаще--
с 10 VIII, когда глухарятам исполнится 45 дней и самцы достигнут
веса 1 кг, на зобу и шее у них появляются первые черные, с зеленым
металлическом блеском., контурные перья второй генерации. В это
время маховые первой генерации представлены 5 перьями на кисти
(№ 1—5, из них первые три еще растут) и 11 перьями на*предплечье
(начиная с № 8).
Развитие петушиных перьев начинается с краев птерилий. Снача-
ла появляются две продольные черные полосы по бокам зоба и ниж-
ней стороны шеи до подбородка; когда выпадут ювенильные перья
внизу шеи, обнаруживаются две продольные полосы черных перьев
по бокам груди а между ними полоса ювенильных перьев; сзади она
расширяется и охватывает почти все брюхо. Одновременно начинает-
ся линька верхних кроющих крыла, плечевых и спины, включая над-
хвостье; у сидящего петушка становится заметным белое «зеркаль-
це» под плечевым суставом. Омена ювенильного серого оперения го-
ловы на более темный наряд петуха начинается с «бородки» и теме-
ни, распространяясь далее на затылок, заднюю сторону шеи, крою-
щие ушей и заканчивается надбровной областью, щеками и ветвями
нижней челюсти.
Около 1 сентября, в возрасте 65 дней и при весе 2 кг, петушок
бывает снизу черноватым. К этому времени из рябых ювенильных
перьев сохраняются 5 последних маховых предплечья, характерных
своей ланцетовидной формой, часть средних кроющих предплечья и
плеча и перья щек и горла вокруг «бородки», заходящие немного
и на шею.
В половине сентября курочки достигают веса 1,5 кг, петушки —
2,5 кг; в это время они получают полный наряд молодого самца, ру-
левые заканчивают рост и имеют в длину около 25 см. На крыльях
молодых птиц обоих полов продолжается рост маховых кисти № 3
и № 4, а также маховых предплечья № 1 и № 14—19.
В середине октября у молодых птиц затвердевают стволы махо-
вого кисти № 3 и маховых предплечья № 17—18; однако и после
окончания линьки маховых, на теле, особенно на спине, можно найти
10*
147
Таблица 45
Линька маховых у молодых глухарок
Второстепенные Первостепенные
18 17 16 15 14 13 12 11 10 9 8 7 6 5 4 3 2 1 10 9 8 7 6 5 4 3 2 1
28 VII 1950 г. 0 0 0 0 0 0 0 0 0 0 — 0 0 0 — — — 0 0 0 0 0
17 VII 1950 г. 0 0 — 0 - — 0 0 0 — — — —- 0 0 0
1 VIII 1950 г. 0 0 0 0 0 0 0 0 - — 0 0 0 0 0 — — 0 0 0
5 VIII 1935 г. — — 4 4 4 - — 4 + 0 0 • — — 0 0 0
4 VIII 1984 г. — 0 0 0 0 4 + - — 4 4 + 0 0 0 — — 0 0
9 VIII 1937 г. — • 0 0 0 0 0 tr 4 - — 4 + 4 4 0 0 0 — 0 0
5 VIII 1937 г — 0 0 0 0 0 0 - — 4 4 4 0 0 0 — — 0 0
15 VIII 1936 г. — — — — — — • 0 0 0 0 0 4 4 4 4 0 — 4 + + 0 0 0 0 — 0 0
15 VIII 1936 г. — 0 0 0 0 4 4 4 4 4 0 — 4” 4 4 _!_ 0 0 0 — 0 0
27 VIII 1936 г — — — • 0 0 0 0 0 0 4 4 4 4 4 4- 0 — 4 4 4 4 0 0 • 4 +
6 IX 1936 г. — — — 0 0 0 0 0 0 4 4 4 4 4 + 4 0 — 4 -L 4 + 4 0 0 0 4 +
8 IX 1938 г. — — — 0 0 0 0 0 + 4 + + 4 + 4 4 0 — ч- 4 4 4 + 0 0 0 4 4
8 IX 1938 г. — — 0 0 0 0 0 4 4 + + + 4 4 + 4 4 0 + 4 4 Ь + 4 0 0 4 4
6 IX 1936 г. — — • 0 0 0 + 4 4 4 + 4 4 4 4 4 + 0 4 -г 4 4 4 + 0 0 4 +
16. IX 1949 г. \ — 0 0 0 0 О 0 -1- -Г 4- 4 4- 4- 4- 4 4- 4 0 4 / 4 4 4 -i- 0 0 + +
Второстепенные
Табл. 45. (5\род.олж.енме>
Первостепенные
18 17 16 15 14 13 12 11 10 9 8 7 6 5 4 3 2 1 10 9 8 7 6 5 4 3 2 1
27 IX 1936 г.
10 X 1938 г.
12 IX 1936 г.
1 X 1936 г.
10 X 1938 г.
4 X 1950 г.
1 X 1936 г.
10 X 1938 г.
8 X 1937 г.
26 X 1937 г.
14 XI 1949 г.
— — 0 0 4 4 + 4 + 4 * 4 + 4 4 + 4 4 4 4 + 4 + 4 + 0 4 4
— — 0 4 4 4 4 4 + 4 + 4 + 4 4 + 0 + 4 4 + 4 + 4 + 0 4 4
— 0 0 4 4 4 4 4 + 4 + 4 + 4 4 + 0 + 4 4 + 4 + 4 + 0 4 4
— 0 0 4 4 4 + 4 + 4 + 4 + 4 4 + 4 + 4 4 + 4 + 4 4 0 4 4
0 0 4 + 4 4 + 4 + 4 + 4 + 4 4 + 0 + 4 4 + 4 + 4 4 0 4 4
0 0 4 4 4 4 + 4 + 4 + 4 + 4 4 + 4 + 4 4 4 4 + 4 4 0 4 4
0 0 4 4 4 4 + 4 4 4 + 4 + 4 4 + 4 + 4 4 4 4 + 4 4 4 4
4 4 4 4 4 4 + 4 4 4 "Г 4- + 4 4 + 4 + 4 4 4 4 + 4 + 0 4 4
4 4 4 4 4 4 + 1 4 4 4 4 + 4 4 + 4 + 4 4 4 4 + 4 + 4 + +
4 4 4 4 4 4 + + 4 4 4 4 + 4 4 + 4 + 4 4 4 4 + 4 + 4 4~ 4
4 4- 4 4- 4 4 + 4 4- 4 4 + 4 4 + 4 + 4 4 4 4 + 4 4 + +
Пояснения к таблице: • перо выпало
0 перо растет
— вполне развитое перо 1 генерации
I вполне развитое перо II генерации
пеньки или контурные перья с мягким стволом. Только в середине
ноября линька полностью заканчивается.
Порядок и сроки линьки маховых представлены в табл. 45, со-
ставленной непосредственно по карточкам обработки добытых птиц
и потому вполне документальной. Чтобы расположить птиц в поряд-
150
ке развития линьки, пришлось местами нарушить правильную по-
следовательность дат. Аналогичные таблицы были составлены для
молодых самцов, взрослых самцов и самок глухарей, а также для
молодых и взрослых белых куропаток. В целях экономии места они
не приводятся. На основе этих таблиц составлены схематические
диаграммы линьки маховых (рис. 52 и 53), образец которых заим-
ствован из книги Саломонсена. Слева показаны календарные даты,
справа — возраст птенцов, считая, что в среднем они выводятся
25 VI. Фактически добытые птицы принадлежали и к ранним и к
поздним выводкам. Для того чтобы выровнять данные наблюде-
ний, пришлось карточки на добытых птиц расположить в последова-
тельности фаз линьки и перенумеровать. После этого даты с тех же
карточек были выписаны в хронологическом порядке и также пере-
нумерованы, а затем дата № 1 приписана! к фазе № 1 и так далее,
до конца ряда. Этот прием позволил выровнять картину линьки без
риска систематической ошибки в определении возраста птенцов. Эти
диаграммы можно использовать для приблизительного определения
возраста глухаря, причем результаты должны быть точнее, чем при
определении по весу (рис. 40).
Изучение линьки молодых глухарей приводит нас к следующим
выводам’:
1. Рост маховых 1 генерации (ювенильные) начинается с рожде-
ния цыпленка, а в возрасте 20 дней некоторые из них успевают не
только вырасти, но и выпасть, заменяясь маховыми II генерации.
2. Самый короткий цикл развития у ювенильных маховых близ-
ких к кистевому сгибу крыла (№ 6—10 на кисти и № 3—8 на пред-
плечье) — от 10 до 20 дней. Самые внутренние маховые заканчивают
рост к 40 дням, после третьего Махового кисти.
3. Маховые кисти первой генерации № 1 и 2 представляют исклю-
чение; рост их начинается в возрасте 20—30 дней и заканчивается
к 50—63 дню; они не сменяются до следующей осени.
4. Ювенильные маховые предплечья № 1 и 2 представляют исклю-
чение: они начинают расти, вероятно, с 17—18 дней и заканчивают
к 30—33 дню.
5. Развитие маховых второй генерации начинается с появления
десятого махового кисти (в возрасте приблизительно 19 дней|), за-
тем следуют девятое и восьмое, а вместе с ними, или чуть позже,
третье и четвертое маховые предплечья; далее линька идет в обе
стороны, пока не сменятся третье маховое кисти и все маховые пред-
плечья до конца, что наступает в возрасте около ПО дней.
6. Первые два маховых предплечья сменяются по' очереди, снача-
ла второе (в возрасте 50—55 до 70—75 дней), потом первое (с 75—
80 до 100 дней).
7. Рост отдельного махового (от появления пенька до полного за-
твердения ствола) продолжается от 13 до 41 дня; быстрее всех ра-
сту7? рано сменяющиеся перья у кистевого сгиба, дольше всех — пер-
вые четыре маховых кисти.
151
8. Рост новых маховых происходит в трех или четырех разобщен-
ных участках крыла одновременно. При постъювенильной линьке,
обычно одновременно растут 3 смежных маховых кисти и 4—5 смеж-
ных маховых предплечья. Одновременно на крыле может развивать-
ся до 10—11 маховых, причем все время птица сохраняет способность
полета.
9. В зимнем оперении молодой птицы первое и второе маховые
кисти принадлежат к I генерации; они начинают расти очень рано,
когда цыпленку исполнится 20—.30 дней. В связи с этим они хорошо
отличаются по форме и рисунку, как от соседних с ними маховых,
так и от первого и второго маховых взрослых птиц: опахала их на
конце заострены и имеют светлые пестринки, отсутствующие у взрос-
лых (ом. рис. 58).
Линька взрослых глухарей. Линька петухов растягивается на
очень длительный срок. У глухаря, добытого на току 13 V 1950 г.,
оперение подбородка было изрежено (возможно, вследствие дра-
ки?), однако пеньков новых перьев еще нигде не было. У другого,
добытого на току 20 V 1934 г., в верхней части шеи показались пень-
ки. У глухарей, добытых 8 VI 1951 г. и 18 VI 1934 г., много пеньков,
частично» развернувшихся кисточкой на конце, на голове, шее и верх-
ней стороне крыльев; у первого из них отмечено, что 5 внутренних
маховых кисти выпали, причем № 9 и 10 уже заменены вполне вырос-
шими новыми. В это время глухари уже не токуют. Нередко в лесу
можно найти свежевыпавшие внутренние маховые кистевого отдела
глухарей, по которым можно судить о начавшейся линьке (19 VI
1'950 г. и 18 VI 1951 г.).
У глухарей, добытых 26 VI 1934 и 28 VI 1950 г., линька в полном
ходу, все тело в пеньках; верхние кроющие крыльев и хвоста уже вы-
росли: роговые бахромки на пальцах полностью исчезли: две или че-
тыре внешние пары рулевых выпали и на месте их подрастают но-
вые; выпало по 6 маховых кисти и по- 3—4 — предплечья на каждом
крыле, на месте которых подрастают новые.
В дальнейшем темп линьки замедляется. 5 VIII 1935 г. отмечено,
что у глухаря на голове и шее нет свежих пеньков, следовательно,
линька этих частей закончена. Продолжается смена маховых кисти,
в направлении от внутренних к дистальным, и маховых предплечья,
начиная с № 3 к туловищу. Около 1 августа выпадают последние,
центральные, рулевые, а новые периферические заканчивают в это
время свой рост. В середине месяца у годовалых птиц выпадает
«детское» второе маховое кисти, а к 1 IX — и первое. К этому вре-
мени линька контурного пера затихает, и при ощипывании птицы на
ней оказывается мало пеньков. В октябре отрастают на зиму опере-
ние лапок и роговые бахромки по краям пальцев. Линька полностью
заканчивается только в начале ноября.
Таким образом, взрослый петух линяет почти полгода. От выпа-
дения десятого1 махового кисти (третья декада мая|) до затвердения
ствола первого махового (третья декада октября) проходит 5 меся-
152
цев; линька контурных перьев начинается на декаду раньше, а за-
канчивается примерно на декаду позже линьки маховых.
Смена рулевых начинается в середине июня и заканчивается в се-
редине сентября: самое раннее окончание линьки хвоста—у самца
3 IX 1990 г. (все рулевые окрепли), самое позднее — у самца 21 IX
1951 г., имевшего еще мягкие очины центральных рулевых. В общем,
линька хвоста занимает 3 месяца. i
Рис. 54. Схема линьки маховых взрослых глухарей; сплошные стрел-
ки — маховые самца в стадии роста, прерывчатые стрелки — маховые
самки
У старых птиц каждое отдельное маховое развивается медленнее,
чем у молодых, у них одновременно! растут обычно, по 3 смежных
маховых предплечья, а не по 4—5, как у птенцов. Порядок смены
первого и второго маховых предплечья такой же, как у молодых
птиц, но внутренние маховые начинают линять с № il6 в обе стороны,
Одновременно с выпадением' № 3. Впрочем, линька внутренних ма-
ховых идет не в строгой последовательности; создается впечатление,
что выпадение того или другого пера в этой группе зависит от слу-
чайных причин. Остальные маховые предплечья линяют также не
153
вполне последовательно, почему невозможно точно подобрать
экземпляры по порядку линьки: часто на правом крыле сменяются
одни перья, а на левом — другие. Для диаграммы линьки (рис. 54)
данные пришлось, по необходимости, округлять.
Неравномерность хода линьки еще резче выражена у самок.
Глухарки начинают линять по окончании периода насиживания. В
дальнейшем течении их линьки, вероятно, большую роль играет на-
личие или отсутствие выводка (вывела она птенцов или бросила
кладку). Выпадение того или другого пера нередко зависит от слу-
чая. Так, 21 VI 1950 г. матка, отводившая от однодневных цыплят,
была схвачена за крыло: маховые предплечья держались так слабо,
что несколько перьев осталось в руке наблюдателя, отделившись без
всякого усилия. В общем, порядок линьки крыла у взрослых самок
тот же, что и у самцов; важнейшие различия следующие:
1. Линька маховых начинается, по-видимому, в первой декаде
июля, на полтора месяца позже, чем у самцов.
2. Смена маховых и отчасти рост новых протекает быстрее, а по-
тому линька крыльев, а всиестес тем1 и линька контурного пера и под-
отеки у обоих полов заканчиваются приблизительно в одно время
(начало ноября).
3. Линька маховых предплечья у самок протекает еще неправиль-
нее, чем у самцов.
4. Линька маховых предплечья идет в двух направлениях, от № 3
и от № 17 навстречу друг другу; до № 12 и № 13 очередь доходит
с обеих сторон одновременно. У самцов между линькой этих участ-
ков заметен разрыв: № 14—47 сменяются с середины июня до сере-
дины августа, а № 10—13 только в сентябре.
Хвост у глухарки, добытой 24 VII 1940 г., еще не начинал линять;
у самки 15 VIII 'I960 г. все старые рулевые выпали и на месте их
росли новые, причем внешние 3 пары достигли полного ростам по еще
не окрепли. У глухарки Ы IX 1936 г. центральные 4 пары рулевых
еще не окрепли, у одной, добытой 10 IX 1936 г., они не вполне вы-
росли, а у другой — закончили рост и окрепли. Таким образом, у са-
мок хвост начинает линять позже, но конец линьки у обоих полов
наступает одновременно в середине сентября.
Тетерев. Наш материал по линьке тетерева настолько невелик,
что рассматривать его здесь не имеет смысла. Заметим только, что
молодые самцы перелинивают в наряд петуха в той же последова-
тельности и в те же сроки, что и молодые глухари. Взрослые тетерева
по срокам линьки едва ли отличаются от глухарей; так. у самца
20 V 1951 г. линька маховых не начиналась, а у самца 1 VI 1949 г.
маховые предплечья не начали линять, но три внутренних маховых
кисти выпали и на месте № 9 и 10 появились пеньки; бородки под-
отеки сохранялись на концах пальцев, но отпали у основания их.
Материал по линьке рябчика также недостаточен. Укажем только,
что линька оканчивается раньше, чем у глухарей: у молодых — пер-
вое и второе маховые кисти прекращают рост в середине августа,
а третье кистевое и последние маховые предплечья — к концу сен-
154
тября. У взрослых птиц линька заканчивается в середине октября,
когда, окрепнет первое маховое кисти.
Белая куропатка. Весенняя линька у белой куропатки начинается
очень рано. В начале апреля, а иногда даже в последних числах
марта (28 III 1950 г., 2 IV 1949 г.) у петуха на горле, под белыми
перьями, можно найти пеньки или кисточки подрастающих красно-
коричневых перышек брачного наряда: при этом можно заметить,
что белое оперение тут изрежено. Вскоре новые перья пробиваются
на поверхность, образуя на горле маленькое темное пятно, хорошо
видное при наблюдении птицы в природе: этот момент наступает,
в среднем', 17 апреля, с небольшими отклонениями по годам (см.
табл. 4). «Красное» горловое пятно быстро расширяется и через не-
делю охватывает всю шею, распространяясь и на голову. На этой
фазе дальнейшее течение линьки замедляется почти до приостановки.
Задержка в ходе линьки продолжается около месяца, совпадая
с периодом «токования» белых куропаток и разбивкой их на пары.
Самка в это время сохраняет белое зимнее оперение и поэтому резко
отличается от самца по внешности. Описанную здесь первую фазу
линьки самца белой куропатки (с частично или полностью «крас-
ными» шеей и головой, а остальным оперением — белым, как зимой)
можно называть брачным нарядом!.
Первые летние кроющие на плечах, а чуть позднее — на крестце,
появляются, в среднем, 24 мая; последние петухи в «брачном наря-
де», без летних перьев на туловище, встречены 30 мая. Линька спи-
ны, кроющих крыльев и хвоста идет уже без задержек, заканчиваясь
через месяц (в среднем к 23 июня).
В период интенсивной линьки в летнее перо в лесу часто попа-
даются выпавшие белые перья куропаток. Это явление настолько
характерно, что существует даже постоянный потребитель этого
«утил-я», ia именно пеночка-весничка (Phylloscopus trochilus) *.
Одновременно с окончанием линьки кроющих спины начинает-
ся смена маховых и рулевых. Прежде всего выпадают внутренние
маховые предплечья, начиная с № 13, 14, 15 или 16 — до конца
(№ 17 или 18). Сменяющие их летние перья по рисунку и расцветке
похожи на кроющие. Затем (в середине июня) выпадают самые
внутренние маховые кисти, начиная с десятого; когда настанет
очередь шестого, вместе с ним выпадает и третье маховое пред-
плечья, после чего линька идет в двух противоположных направ-
лениях: маховые кисти сменяются от внутренних к наружным,
предплечья — от внешних внутрь. Первые два маховые предплечья
сменяются поздно, сначала второе, потом первое. Во второй поло-
вине сентября заканчивается рост последних новых маховых, пер-
вого и второго кисти и № 1, 12 и 13 предплечья. В октябре выпа-
* Сотрудники Лапландского заповедника нашли и описали не один десяток
гнезд веснички и все они, без исключения, были устланы белыми перьями куро-
яатки. Весничка строит гнездо на поверхности почвы, во мху, и закрывает
гнездо сводчатой крышей с боковым входным отверстием: благодаря этому
белая подстилка не может демаскировать гнездо.
155
дают летние (рябые) внутренние маховые предплечья, заменяясь
зимними (белыми): окончание роста белых маховых № 15 и jf,
совпадает с окончанием линьки контурных перьев. В общем линька
маховых занимает четыре с половиной месяца: за это время около
пяти (у разных особей число это варьирует) внутренних маховых
сменяется дважды (рис. 55).
Рис. 55. Схема линьки маховых взрослой белой куропатки; топкие
стрелки — белые маховые в стадии роста, толстые — пигментирован-
ные маховые
Линька хвоста протекает быстрее. Центральная белая пара ру-
левых («надрулевые», по терминологии А. В. Михеева и некоторых
других авторов) выпадает около 1 июня, вслед за белыми верхними
кроющими хвоста. Вместо них вырастают летние рябые перья.
Черные рулевые сохраняются до середины июля; они сменяются от
крайних к средним такими же черными рулевыми, рост которых
заканчивается к середине сентября. Приблизительно в это время
выпадают и летние «надрулевые», вместо которых появляются зим-
ние, белые. В сроках линьки средних рулевых и самых больших
кроющих наблюдаются большие индивидуальные различия, ио в
середине октября линька хвоста заканчивается у всех особей.
В августе — начале сентября перелинивают белые перья нижней
стороны тела и вообще все белые кроющие, сменяющиеся один раз
в году. К моменту выпадения эти белые перья сильно обнашивают-
ся и пачкаются. Поэтому с появлением на месте их новых, пышных
156
и чистых, вся куропатка заметно белеет; области, покрытые летними
перьями, к этому времени сокращаются.
Около 21 сентября на спине, среди летних перьев, показываются
первые белые перья (крайние сроки— 15 IX 1940 г. и 29 IX 1950 г.).
Белое перо пробивается одновременно всюду — на голове, на шее,
зобу, спине, плечах и кроющих предплечья. Полный зимний наряд
куропатки получают, в среднем за 18 лет, 22 октября (крайние
сроки—12 X 1939 г. и 1 XI в 1934 и 1935 гг.). Единичные рыжие
перышки, чаще на голове, попадаются до 1 ноября.
Рис. 56. Схема линьки маховых молодой белой куропатки (условные
обозначения как на рис. 55}
Густое оперение лапок куропатки выпадает в начале июля, и
летом пальцы почти голые. Когти в июне — начале июля (а иногда
и до начала августа) еще длинные и широкие, как зимой; после
спадения они вновь начинают отрастать в конце августа, а в тече-
ние сентября лапки опушаются по-зимнему.
Осенне-зимняя линька происходит почти одновременно у моло-
дых птиц и старых, у самцов и у самок. Весенняя линька, напротив,
у самок начинается позднее, чем у самцов, но протекает без всякой
задержки, а потому заканчивается раньше: начинаясь, в среднем,
14 мая (26 IV — 1 VI), она оканчивается 29 мая (28 V — 3 VI).
Единичные летние перышки у самок появляются, иногда, очень
рано, например, таких самок я наблюдал 1 IV 1949 г., 15 IV 1932 г.,
15 IV 1940 г. и 28 IV 1933 г. Это явление объясняется преждевре-
менной потерей отдельных перьев зимнего наряда. Б. Г. Новиков
(1939) нашел, что контурные перья, выщипанные у белой куропатки
в январе—феврале, заменяются уже не белыми, а пигментирован-
157
ными. Эффект наблюдается как у самцов, так и у самок, в те же
сроки. Потенциальная способность производить пигментированные
перья наступает у обоих полов одновременно, но при нормальном
ходе линьки у самцов белые перья начинают выпадать на опреде.
ленных местах (шея) гораздо раньше, чем у самок.
Птенцы белой куропатки получают сначала наряд, сходный с
нарядом взрослой самки, с той разницей, что маховые первой гене,
рации* у них серые, как у рябчика. Появление белых маховые
второй генерации начинается с десятого махового кисти, в возрасте
около 10 дней. Смена серых «детских» маховых идет в той же
последовательности, как и смена прошлогодних маховых у взрос-
лых птиц. Последнее серое маховое кисти (№ 3) выпадает в воз-
расте 55 дней, но на внутреннем краю крыла серые маховые со-
храняются до начала октября, то есть до возраста 90 дней. Ди-
аграммы линьки маховые (рис. 56) белой куропатки построены
так же, как и диаграммы линьки глухарей, с той разницей, что
средним сроком вывода птенцов принято 3 июля. Чтобы нагляднее
отличить на рисунке пигментированные маховые от белых, первые
начерчены толстыми стрелками.
Тундряная куропатка. В отличие от белой куропатки у самца
тундряной весенняя линька начинается с затылка и прилегающей
к нему части шеи. Первые пигментированные перья появляются
здесь около 9 V (IV — 20 V); они темно-серые, почти без пестрин.
Линька затылка начинается после наступления белых ночей, при
отрицательных температурах воздуха, и при почти сплошном снеж-
ном покрове на горных тундрах. Как только почернеет затылок,
дальнейшее движение линьки задерживается на месяц. Самки на-
чинают линять чуть позднее самцов, но у них весенние перья
пробиваются одновременно на голове, шее и плечах и линька идет
безостановочно, так что к 5 июня на спине не остается белых перьев
и курочка получает защитный летний наряд, под цвет и рисунок
поверхности камней (рис. 20). Кроющие перья весенне-летнего
наряда темно-серые, с довольно крупными бледно-охристыми пят-
нышками или поперечными перевязками.
У самцов линька кроющих спинной стороны туловища начи-
нается около 11 июня и заканчивается к 8 июля. Поэтому в период
брачных игр (конец мая — начало июня) полы резко различаются
по окраске — в это время самец почти совсем белый, а самка тем-
ная, рябая, и только на лету заметно, что крылья у нее белые.
Во время летней линьки самцы, как уже упоминалось, живут
обособленно от самок, выше в горах. Даже по окончании линьки
тундряная куропатка выглядит довольно пестрой, что происходит
оттого, что опа в это время одета перьями трех разных нарядов.
Часть перьев линяет один раз в году и остается весь год белой:
сюда принадлежат:
1) маховые кисти и предплечья, за исключением самых внутрен-
них;
151 За исключением первого и второго.
158
2) все нижние и большая часть верхних кроющих крыла (осо-
бенно малые и большие); (
3) рулевые, из них 7 пар боковых черные с белыми кончиками
и основаниями, восьмая, центральная, пара белая;
4) перья ног, включая и оперение лапок, брюха и нижней части
груди;
5) часть нижних кроющих хвоста и кроющих боков.
Остальное оперение линяет дважды или трижды, а по мнению
некоторых старых авторов — даже четыре раза в год. Представле-
ние о многократной линьке тундряной и белой куропаток про-
изошло, без сомнения, от наличия перьев трех или четырех типов
окраски, принадлежащих к разным нарядам.
Действительно, в летнем и осеннем оперении петуха мы нахо-
дим, кроме перечисленных выше белых перьев и черных рулевых,
пигментированные перья двух резко различных типов. К первому
типу (летний наряд) принадлежат кроющие головы, почти всей шеи
и отдельные перья в верхней части спины, на плечах и локтях.
Фон этих перьев почти черный, с редкими, сравнительно крупными,,
охристыми или беловато-охристыми пятнышками.
Ко второму типу (осенний наряд) относится большая час;ть кро-
ющих спины, надхвостья и хвоста (сверху), зоба и передней части
груди, часть кроющих боков под крыльями, подхвостье и сменяю-
щиеся на лето перья крыла. К последним относятся внутренние
маховые предплечья, большие кроющие плеча, средние кроющие
плечевого отдела и предплечья. В целом эта группа перьев обра-
зует на белом крыле темную косую полосу, которая идет от внут-
реннего угла крыла, постепенно суживаясь, к кистевому сгибу.
Перья осеннего наряда заметно светлее летних, так как темно-
серый фон их подавлен частыми светло-охристыми крапинками,
сливающимися в неправильные струйчатые поперечные полоски.
Перья летнего и осеннего нарядов удобно различать, лишь имея
птицу в руках. Первые экземпляры с наличием осенних перьев
добыты: взрослый самец — 23 VI 1939 г., взрослая самка— 12 VII
1935 г., птенец —23 VII 1938 г.
Части тела взрослых птиц, одетые преимущественно перьями
летнего наряда, сохраняют его до зимней линьки. Птицы, добытые
в сентябре и первых числах октября, сильно запестревшие белым
пером, имели перья обоих нарядов примерно в том же соотноше-
нии, как у июльских и августовских экземпляров. У взрослого сам-
ца 14 X 1932 г. оперение было в основном белое, с остатками
летнего пера в области затылка и двумя осенними перьями на пра-
вом плече. Таким образом, замены перьев летнего наряда пигмен-
тированными же перьями осеннего наряда не происходит (по край-
ней мере в районе нашего исследования).
Зимняя линька тундряных куропаток происходит приблизительно
одновременно у молодых и старых птиц обоих полов и на неделю
раньше, чем у белых куропаток (табл. 46). Экскурсии в горы —
единственное местообитание тундряной куропатки — невозможно
159
было проводить регулярно, а потому даты фенологичных явлений
у нее часто отмечались с запозданием. Чтобы избежать вредной
систематической ошибки в определении «нормальных» сроков сме
ны нарядов, здесь применен особый прием: за начало линьки при.
пято арифметическое среднее между датами встреч — первой пти-
цы, начавшей линять, и последней птицы, не начинавшей линять.
Когда наблюдений много, первая дата бывает более ранней, чем
вторая, когда наблюдений недостаточно — наоборот; однако, и в
том и другом случае наш прием достигает цели.
Таблица 46
Средние сроки линьки куропаток за 17 лет наблюдений
Наряды и линька Тундряная Белая
С аж цы Самки Самцы Самки
16 X 16 X 22 X 22 X
Зимнее оперение, дней 205 208 178 201
9 V — 18 IV —
Брачный наряд, дней 33 — 39 —
11 VI 12 V 27 V 11 V
Летняя линька, дней 27 24 27 18
8 VII 5 VI 23 VI 29 V
Летне-осенний наряд, 65 98 90 115
дней
11 IX 11 IX 21 IX 21 IX
Зимняя линька, дней 35 35 31 31
16 X 16 X 22 X 22 X
Смена маховых у тундряной куропатки идет в общем по той же
схеме, что и у белой. Мы не будем рассматривать линьку взрослых
детально и совсем не касаемся линьки птенцов по той причине,
что старые материалы уже давно опубликованы (Семенов-Тян-
Шанский, 1938-а), а новых собрано- немного.
Линька белых куропаток описывалась подробно и неоднократно,
но тем не менее отдельные вопросы до сих пор не вполне ясны. На
некоторых из них мы остановимся.
Число линек. Современные авторы, описывая наряды куропаток,
обычно обходят вопрос о числе линек (йонсен, 1929; Михеев, 1939).
В сущности, главной причиной путаницы с числом линек у рода
белых куропаток является нечеткость самого определения линьки.
Некоторые авторы, как, например, С. И. Снигиревский (1950.
стр. 215), принимают фенологическую терминологию линек: «Одно-
временно растущие перья относятся к одному наряду. Таким обра-
зом устанавливаются наряды — весенний, летний, осенний и зимний
и соответствующие линьки — весенняя, которая приводит к весен-
нему наряду...» (и т. д.). При таком понимании число линек будет
равно числу нарядов, которые можно различить и описать (исклю-
чая,. разумеется, наряды, приобретаемые без линьки, путем обна-
шивания перьев). Мы не согласны с таким определением. Наличие
160
двух линек в году предполагает, что все перьевые сосочки или
часть их производят за этот период две генерации перьев. В свое
время я утверждал, что у тундряной куропатки только две линьки
в году (Семенов-Тян-Шанский, 1938-а, стр. 193). В настоящее время
можно привести дополнительные соображения в пользу того, что
двумя линьками можно легко объяснить все наряды тундряной ку-
ропатки в течение года.
Среди млекопитающих известны факты, когда определенная
линька, при изменении тех или других условий среды, дает меховой
покров (несвойственного этой линьке типа. Бельчата летних выводков
родятся в летней шерсти, между тем как бельчата ранних выводков,
рожденные белкой в зимнем меху, выходят из гнезд также в зимнем
сером меху и только после линьки получают летний рыжий мехо-
вой покров (Гольцмайер, 1935; Салмин, 1938; Огнев, 1951).
Всем нашим тетеревиным свойственен резко выраженный сезон-
ный цикл питания и состояния органов усвоения пищи. Поэтому
и условия роста пера в разные месяцы должны быть резко раз-
личными. Процесс линьки тетеревиных птиц, примем ли мы две,
три или четыре линьки в году — безразлично, продолжается все
теплое полугодие без перерывов. Явление линьки усложняется еще
и тем, что часть перьев начинает расти не сразу по выпадении
старого пера, а через какой-то промежуток времени.
Ф. Саломонсен (1940, стр. 46) указывает, что белые перья на
середине груди тундряной куропатки «выпадают в начале лета, но
не возмещаются никакими новыми перьями в летнем наряде; места
их остаются незанятыми до осенней линьки. Также дорзальные,
части боков, прикрытые крыльями, голы в летнем наряде».
С. И. Снигиревский (1950) также нашел, что при частичной лет-
ней линьке тетеревиных птиц на некоторых участках перья выпа-
дают без следующей замены, и указал, что существуют, по крайней
мере, два таких места — голова и бока тела под крыльями. Он
пишет: «Зимою перья расположены на теле куропаток гуще, чем
летом, так как на летний период многие перья выпадают и времен-
но не заменяются вплоть до осенней и зимней линек» (там же,
стр. 228). И далее: «Перьевые фолликулы на какой-то срок впа-
дают в состояние покоя. Значение этой линьки заключается в
изреживании оперения на теплый период года» (там же, стр. 232).
Если на месте одних выпавших перьев новые развиваются сразу,
а на месте других — только после длительной паузы, естественно
предположить, что и скорость роста и внешний вид этих перьев
будут резко различными. Поэтому мы допускаем, что одна линька
(назовем ее летней) дает в начале и конце перья различного рисун-
ка и окраски, хотя они и принадлежат к одной генерации.
Выпадение зимнего пера у тундряной и белой куропаток растя-
гивается на долгий срок. Из табл. 46 видно, что от внешнего появ-
ления первых пигментированных перьев до выпадения последних
белых на спине у петуха тундряной куропатки проходит 60 дней,
у петуха белой — 66, у самок этих видов соответственно — 24 и
11 3037
161
18 дней. Можно приблизительно рассчитать, сколько времени про.
ходит от выпадения первых белых перьев до появления их преем-
ников на поверхности оперения. Ф. Саломоноен указывает (по лите-
ратурным источникам), что после удаления у домашней птицы
пера, новый пенек выходит на поверхность кожи через 9—11 дней.
У самцов белой куропатки мы нашли на горле пеньки пигментиро-
ванных перьев в первый раз 2 IV 1949 г. и 28 III 1950 г., а первое
появление «красных» перьев на поверхности оперения наступило
14 IV в 1949 и 12 IV в 1950 году. Отсюда следует, что мелкое перо
подрастает не менее 12—15 дней, а от выпадения старого пера до
появления кончика нового на поверхности оперения проходит не
менее 21—26 суток. Весь процесс выпадения зимнего пера на спин-
ной стороне куропаток обоих видов занимает 80—95 дней у самцов
и около 40—50 дней у самок.
Внешние условия за это время сильнейшим образом изменя-
ются. У самца белой куропатки выпадение белых перьев начинается
около 24 III, при сплошном снежном покрове, температуре воздуха
около —8° и продолжительности светлого времени суток 15 часов;
кончается около 23 VI, при полном отсутствии снега в лесах, тем-
пературе около + 11° и круглосуточном дне. У самца тундряной
куропатки линька начинается около 14 IV, в период появления
первых проталин, при температуре около —4° и 18—19 часах свет-
лого времени в сутки; при окончании ее (около 8 VII) только на
вершинах гор сохраняются клочки слежавшегося снега, темпера-
тура воздуха в местах обитания птицы около + 11° и стоит непре-
рывный полярный день. Выпадение пера начинается в период пи-
тания зимними кормами, конец — при очень разнообразном летнем
рационе.
У самок период выпадения зимнего пера вдвое короче, вслед-
ствие чего разница условий в начале и конце его не так велика:
по температуре она составляет 8—9°, а не 15—19°, как у самцов.
Соответственно и другие условия изменяются меньше. Мы пола-
гаем, что именно по этой причине разница во внешнем виде перьев
летнего и осеннего нарядов у самок слабее выражена, чем у сам-
цов. На летних перьях курочек светлые охристые пятна образуют
довольно правильные поперечные полоски, концы перьев часто с
беловатой каемкой; на осенних перьях — струйчатые пестрины ши-
ре, чем у самцов. Это подтверждает наш взгляд, что разница в
аспекте перьев летнего и осеннего нарядов определяется разницей
во времени и условиях их формирования, а не принадлежностью их
к разным генерациям.
Можно представить дело так, что до какого-то определенного
срока на месте выпавших зимних перьев вырастают перья летнего
наряда, после этого срока — перья осеннего наряда, независимо от
пола птицы, а только под влиянием внешних условий в период
роста пера. Для взрослых птиц на горах Кольского полуострова
поворотный пункт наступает около 1 июня. В таком случае у птен-
цов, которые появляются на свет около 1 июля, ювенильный наряд
16'
должен бы состоять из одних перьев осеннего наряда. В действи-
тельности их первое оперение комбинируется из перьев летнего и
осеннего нарядов, причем на голове и шее, как и у взрослых птиц,
преобладают летние, а на спине, плечах и надхвостье они состав-
ляют */< — 1/з общего числа, остальные являются осенними. При
этом оба наряда выражены менее типично, чем даже у самок, и
большая часть перьев имеет переходный характер.
Из приведенных фактов можно заключить, что развитие того
или другого наряда определяется сезонной цикличностью орга-
низма. Смена отдельных стадий этого цикла зависит как от перио-
дических изменений среды обитания, так и от возраста птицы.
Большое значение должно иметь описанное Ф. Саломонсеном и
С. И. Снигиревским явление задержки развития некоторых перьев.
После периода покоя соответствующий сосочек даст перо уже не
летнего, а следующего за ним наряда, что создает иллюзию выпа-
дения летних перьев и замены их осенними. Затронутые здесь во-
просы линьки рода белых куропаток могут быть окончательно раз-
решены на живых птицах, содержимых в вольере, путем мечения
перьев и учета выпадающих при линьке.
Связь зимнего наряда со снежным покровом. Белый зимний
наряд куропаток — классический пример «покровительственной
окраски». Интересно, что средние сроки появления и выпадения
белого пера довольно близки к средним датам выпадения снега
осенью и таяния его весною. От выпадения перьев ста-
рого наряда до формирования нового проходит много дней: поэ-
тому мы не можем ожидать такого же хорошего совпадения дат
линьки и изменений снежного покрова по отдельным годам. Попыт-
ка найти коррелятивную зависимость сроков линьки по годам от
средних месячных температур и некоторых фенологических явле-
ний предшествующих данной фазе линьки, дала следующие резуль-
таты.
Коэффициент корреляции между появлением «красного» пятна
па горле самца куропатки и
появлением первых проталин .... г = 4-0,31 ±0,24 (14 лет)
средней температурой марта . . . г =4-0,23 + 0,24 (16 лет)
средней температурой апреля . . . г=—0,27 ±0,23 (16 лет)
между первой встречей куропатки в полном зимнем наряде и
выпадением первого снега...........г =4~0,08±0,23 (18 лет)
средней температурой сентября . . . г =4~0,24±0,23 (16 лет)
средней температурой октября . . . г =-4^0,46 ± 0,20 (16 лет)
Все коэффициенты малы и к тому же статистически недосто-
верны, из этого можно заключить, что прямой связи между сроками
линьки и температурой предшествующего периода нет. Любопытно,
что амплитуда колебаний сроков линьки по годам меньше, чем
сроков выпадения и таяния снега: это обстоятельство показывает,
11*
163
что ход линьки вообще меньше зависит от погоды, чем наступление
гидро-фенологических явлений.
Таблица 47
Ход линьки белой куропатки и гидро-фенологические явления в природе
Явления (в последовательном по- рядке) Средняя дата Амплиту- да коле- баний в днях Среднее квадра- ги чн. отклоне- ние (с)
Появление первых проталин . . . 9 IV (16 л.) 59 ± П,0
Появление красного пятна на горле 22 ± 6,4
с? белой куропатки 17 IV (18 л.)
Половина поверхности очистилась 19 V (20 л.) ± 8,7
от снега . * • 31
Появление летних перьев на спине
£ белой куропатки 24 V (15 л.) 29 ± 5,5
Появление белых перьев на спине
белой куропатки 24 IX (14 л.) 19 ± 5,5
Выпадение первого снега в лесу . 6 X (21 л.) 37 ± Ю,5 ± 5,8
Первые вполне белые куропатки . 24 X' (18 л.) 20
Установление снежного покрова 30 X (21 л.) 39 ± 12,1
Из всех приведенных нами фактов можно заключить, что сроки
смены пигментированного оперения на белое и белого на пигменти-
рованное являются приспособительными к среде обитания в такой
же мере, как и возникновение самого белого зимнего наряда. Вме-
сте с тем сроки линьки не стоят под непосредственным контролем
условий погоды. Вероятнее всего, что изменения температуры воз-
духа, появление и таяпие снежного покрова, изменения условий
освещения и другие перемены ассимилируются организмом в це-
лом и путем нервной или гуморальной регуляции вызывают выпа-
дение старых перьев и рост новых. Развитие перьев того или иного
внешнего вида регулируется более или менее сходным путем, ко-
торый, однако, 1не связан полностью с первым. Благодаря этому
в зависимости от пола и возраста линяющих особей в одно и то
же время могут развиваться перья, принадлежащие к разным
нарядам. Интересные факты такого рода приводит Ф. Саломонсен,
который сам склоняется к мысли, что основным контролирующим
линьку органом является щитовидная железа. Попытки связать
линьку белых куропаток с внутренней секрецией гонад не увен-
чались успехом. Б. Г. Новиков (1939) показал на опыте, что каст-
рация не изменяет сроков линьки и характера нарядов ни у самцов,
ни у самок белой куропатки.
Число рулевых у куропаток. У белой и тундряной куропаток
7 пар черных рулевых с белой каемкой па конце и белым основа-
нием, которые линяют один раз в году (с середины июля до сере-
дины августа). Восьмая, центральная, пара — спорная: С. Ионсеп
причисляет ее к рулевым на том основании, что по местоположе-
164
нию, 'размерам* и функции при полете эти перья соотв1етствуют
рулевым. Ф. Саломонсен считает их за кроющие, указывая, что
они составляют один ряд с кроющими, линяют два раза в году и
по окраске не отличаются от верхних кроющих хвоста. А. В. Михе-
ев, учитывая их двойственную природу, признает их промежуточной
формой и называет «надрулевыми перьями». Интересно, что у тун-
дряных куропаток северных широт (в Лапландском зоповеднике —
как у самцов, так и у самок) эти «надрулевые» имеют одну гене-
рацию в году: они остаются все лето белыми и сменяются поздно,
в августе—сентябре.
Рис. '57. Хвост глухарки, сверху
и сзади (ориг. рисунок)
Мы считаем правильной точку зрения С. йонсена. Ошибка
Ф. Саломонсена состоит в том, что он не сравнил хвост тундряной
куропатки с хвостами других тетеревиных. У глухаря и тетерева
рулевых 9 пар, и центральная пара, в принадлежности которой к
рулевым никто не сомневается, прикреплена точно на том месте,
где и пара «надрулевых» у куропаток. На рис. 57 легко убедиться,
что большие перья хвоста образуют два петлеобразных ряда —
правый и левый. Каждый ряд начинается с крайнего рулевого
(первого по порядку линьки) и до № 8 у глухаря (№ 7 — у куро-
паток) идет прямо; перо № 9 (у куропаток —№ 8) сидит выше, под
копчиковой железой; следующее перо, уже кроющее, сидит еще
выше и вместе с тем дальше от оси хвоста. Таким образом, после
центральной пары рулевых, ряд перьев переходит в кроющие и
поворачивает обратно, к краю пигостиля. У белых куропаток руле-
вые переходят в большие кроющие хвоста постепенно (подобно
тому, как в цветах некоторых растений тычинки переходят в лепе-
стки, а последние — в чашелистники), при этом ближайшая к над-
рулевым пара кроющих не уступает по размерам рулевым.
165
Хвост рябчика сходен с хвостом куропаток в том, что восьмая
пара рулевых у него резко отличается от остальных по окраске.
Определение возраста тетеревиных. Возрастный анализ состава
популяций за1вое®ыв1ает все большее значение в экологии животных.
Он уже давно и прочно вошел в практику ихтиологических работ.
Определение возраста млекопитающих представляет гораздо боль-
шие трудности. В основе его лежит порядок смены молочных зубов
постоянными и стирание коронок последних. Для каждого вида
приходится разрабатывать особую схему определения возраста.
Хуже всего обстоит дело у птиц. Не имея в виду разбирать этот во-
прос детально, я опишу только проверенные нами и использованные
в настоящем исследовании методы.
А. П о весу.
Из рис. 44 и табл. 39 видно, что в сезон охоты только у сам-
цов глухарей и отчасти тетеревов возраст можно определить по ве-
су. Глухарь в возрасте до 11 месяцев весит менее 3,5 кг (обычно
2,8—3,2 кг), вполне взрослый — более 3,5 кг (обычно 3,7—4,3 кг).
Молодой тетерев весит до 1,40 кг, взрослый — более 1,3*5 кг. Эти
цифры относятся к Мурманской области; в других популяциях гра-
ница между молодыми и старыми петухами может быть несколько
иной.
Б. П о размерам.
Возраст самцов глухарей и косачей легко определить по хвосту.
У молодых глухарей длина его 22—27 см, рулевые узкие, округлен-
ные на конце. У старого глухаря длина хвоста 30—36 см, рулевые
широкие, на конце срезанные прямо, с небольшим выступом у стерж-
ня.
У молодых косачей длина выпрямленных крайних рулевых IS-
IS см, изгиб пера незначителен. У старых косачей длина крайних
рулевых 20—23 ом и концы их круто загнуты в стороны.
У остальных тетеревиных возрастная разница в длине хвоста
незначительна (у глухарки она равна, в среднем, 9 мм) и гораздо
меньше индивидуальных колебаний. У молодой глухарки концы ру-
левых округлые; у старой—они срезаны в виде фигурной скобки
благодаря выступающему стержню; признак этот, однако, не очень
отчетлив.
У самцов белой и тундряной куропаток хвост заметно длиннее,
чем у самок: у первого вида — на 12 мм, у второго вида — на 10 мм.
Если у белой куропатки хвост длинее 121 мм, это почти всегда са-
мец, если меньше — самка: в среднем ошибка получается в одном
случае из девяти. У тундряной куропатки длина хвоста у самца
обычно больше, а у самки меньше ПО мм: ошибка — в одном слу-
чае из восьми.
В. По маховым перьям.
Как уже упоминалось, зимой у молодых тетеревиных первое и
второе маховые кисти принадлежат к первой генерации, а все
166
остальные — ко второй; первые два маховых начинают расти рано,
уже на; первом месяце жизни птенца1. В связи с этим, их концы (са-
мая «детская» часть пера в онтогенезе) хорошо отличаются как от
концов махового № 3 и следующих за ним, так и от № 1 и № 2
взрослых птиц. В ходе линьки взрослых птиц эти маховые выпадают
Таблица 48
Размеры тетеревиных птиц Лапландского заповедника
(в мм)
Название птицы Длина тела Крыло
Средняя и ср. ошибка Пределы^ Сред, квадр. откл. Число экз. Среднее и ср. ошибка Пределы Средн, квадр. откл. Число экз.
Глухарь мол, 804+3 745-870 ±25 63 374+1 330-395 ±П 66
Л взр. 903±2 860-950 ±18 74 388±1 361—410 ± 9 78
600+1 560-650 ±16 124 295+1 275—315 ± 8 124
Тетерев £ мол. 569+5 550-595 ±14 7 268+2 262-274 + 4 6
rf взр. 616+9 600-640 ±17 4 272±2 270-277 ± з 4
472±6 440—495 ±16 8 240±1 236-249 ± 4 9
Рябчик $ . . 378+1 355 -390 + 8 32 167±1 152-174 ± 5 27
370+8 350—395 ±12 35 165±1 152—173 ± 4 32
Бел. куроп. $ 417+1 380- 445 ±13 118 210+0 196-220 ± 5 126
.. $> • • 392+2 350 - 420 ±14 81 197±1 180-207 ± 6 86
Тундр, куроп. X 383+2 360—400 ±И 26 204±1 197-210 ± з 26
368+4 360—384 ± 9 6 198±2 192-202 ± 4 6
Название птицы Хвост
Средняя и ср. ошибка Пределы Средн, квадр. откл. Число экз.
Глухарь мол. 247+1 225-270 ±ю 67
взр. 321 + 1 300-360 ±12 81
„ ? • • 186+1 160-210 X 9 128
Тетерев £ мол. 174+3 165-180 ± 7 7
взр. 216+6 203—232 ±13 4
126+2 113-130 ± 6 9
Рябчик Л . . 123±1 110—131 ± 5 36
$ • • 116+1 110-125 ± 4 36
Бел. куроп. $ 127+0 115-138 5 123
115±0 105-125 ± 4 84
Тундр, куроп.Д 115+1 105-124 ± 4 26
$ • 105±1 100-108 X 3 6 1
Примечание: Использованы птицы добытые в период X—V, тундряные
куропатки — в IX—V; крыло выпрямлялось по масштаб-
ной линейке; хвост измерен от основания копчиковой же-
лезы.
167
последними (в августе); поэтому по1 рисунку их концов можно отли-
чать молодых птиц почти до 14-месячного возраста.
У молодых глухарей и тетеревов обоего пола концы первого и
второго маховых кисти несколько заостренные, с светлыми крапина-
ми на опахалах (рис. 58).
У рябчиков этот признак выражен очень слабо и не может быть
рекомендован для определения возраста.
Рис. 58. Конец крыла молодой глухарки; концы первого и второго
маховых заостренные, с светлыми крапинами (3/4 нат. вел.)
У молодых белых и тундряных куропаток концы первых двух
маховых слегка заостренные, с черными крапинами около стержня
(рис. 59). Количество крапинок варьирует, и на первом маховом их
иногда нет. У старых птиц концы всех маховых чистобелые. Этот
признак очень удобен и надежен; мы без труда определяли возраст
куропаток, пойманных хищниками, по брошенным на месте поимки
маховым.
Г. По яйцеводу
У молодых самок яйцевод имеет вид прямой белой струны; у
старых, вне периода размножения, яйцевод заметно толще, с мел-
кими извилинами. Этот ’признак неудобен тем, что птицу необходимо
вскрывать; он может быть рекомендован только как контрольный.
Д. По фабрициевой сумке
У тетеревиных она рано редуцируется и для определения возра-
ста мало пригодна. Вес фабрициевой сумки у молодых глухарей обое-
163
го пола составлял: в июле — около О,25о/о веса птицы, в сентябре и
октябре — около О,1о/о, в ноябре и декабре — около 0,05о/о.
Е. По костям черепа
Простейшие способы, особенно популярные среди охотников,
следующие:
У молодых птиц переносица легко прогибается при нажатии
пальцем, у старых она твердая и не поддается пальцу; если птицу
поднять за нижнюю челюсть — у молодой ветви ее согнутся под
весом птицы, у старой — нет (Семенов-ТянШанский, 1938; Теплов,
1947).
Рис. 59. Концы маховых белых куропаток в возрасте до 11 года и
старше года; у молодой маховое № 2 с черными крапинами
(3/4 нат. вел.)
Оба способа применимы для глухарей в течение всего охот-
ничьего сезона. У куропаток и рябчиков кости черепа затвердевают
уже в начале зимы.
С. В. Кириков (11939, 1944, 1'947, 1948, 1952) разработал способ
определения возраста глухарей по отпрепарированным черепам. Он
основан на том, что у глухаря некоторые черепные швы поздно сра-
стаются, а кости продолжают расти в течение нескольких лет, увели-
чивая массивность черепа, особенно в надглазничной области. Кири-
ков предложил следующие основные признаки:
1. Срастание заднеглазничного и височного отростков (у сам-
ки— в возрасте 1 года, у самцов — в возрасте 2 лет).
169
Возрастные
приз«ач
Рис. 60. Череп глуХа.
самца сбоку, Ii03 Р
1 год, % нат. в4
Я
т
Рис. 61. То же, возраст
2 года
Рис. 62. То же, возраст
3 года
Рис. 63. То
4 года
же, В03РаСГ
и более
170
в черепах глухарей, добытых на току.
I Заднеглазничный (processus postorbitalis posterior) и височный (proces-
sus zygomaticus) отростки разделены щелью, шириной около 1 мм; задние
концы носовых костей (оо. nasalia) лежат в выемках лобных костей (оо fron-
talia), не срастаясь с ними. Вес очищенного черепа с нижней челюстью и рам-
фотекой около 20 г: ширина межглазничного промежутка позади слезных ко-
стей около 25 мм. ।
II Продолжение височного отростка срастается с заднеглазничным, но
спереди между ними остается выемка; задние концы носовых костей врастают
в лобные, но, ввиду различия в поверхности и цвете этих костей, граница
между ними хорошо заметна; вес черепа около 26 г; межглазничный проме-
жуток около 31 мм.
III Заднеглазничный и височный отростки срослись на всем протяжении
участка смыкания; лобные кости нарастают поверх задних концов носовых на
5—7 мм, но по сторонам остаются швы, а разница в структуре этих костей
отчетливо заметна; спиральный шов, отделяющий зубную (os dentale) кость
от угловой (os angulare) отчетлив на всем протяжении; вес черепа около
28 г; межглазничный промежуток около- 34 мм.
IV Место сращения заднеглазничного и височного отростков спереди за-
круглено; структура носовых и лобных костей становится сходной, и место
сращения их сглаживается, но щелки по сторонам еще заметны; спиральный
шов на ветвях нижней челюсти в виде неглубокой бороздки, зарастающей
спереди и сзади; вес черепа около 30 г; межгла эпичный промежуток 36 мм.
V Носовые кости незаметно переходят в лобные, наружные стороны места
смыкания закругленные, без признаков шва; спиральный шов на нижней че-
люсти едва прослеживается или совсем зарос; вес черепа около 33 г; меж-
глазничный промежуток около 38 мм. ,
171
Возрастные
Рис. 64. Черепа
реи самцов сверл« Пг-
раст 1 год, 2/3 На- *оз-
• аЧ,
Рис. 65. То же, возраст
2 года
Рис. 67. То же, п°зраСТ
4 года и бодее
Рис. 66. Тоже, возраст
3 года
172
глухарей в осенне-зимний сезон
, iePena*
Самки
Самцы
Заднеглазничные и височ-
(Н отростки разделены щелью;
пние концы носовых костей
33 бодно налегают на лобные.
04- Между заднеглазничными и ви-
сочными отростками большой про-
межуток; носовые кости свободно
налегают на лобные; вес черепа око-
ло 11 г; мел^глазничный промежуток
ок. 23 мм.
Заднеглазничные отростки
срастаются с височными, но
спереди может оставаться
щель; носовые кости начинают
срастаться с лобными, но гра-
ница между ними заметна на
всем протяжении; вес черепа
ок. 5,9 г; межглазничный про-
межуток ок. 17 мм.
14- Заднеглазничные и височные от-
ростки срастаются сзади, разделены
щелью спереди; носовые кости нале-
гают на лобные, без сращения; по-
верхность носовых гладкая, светлая,
лобных — бороздчатая, темная; вес
черепа около 17 г; межглазничный
промежуток ок. 28 мм.
П4- Заднеглазничные отростки II
сращены с височными; концы
носовых костей слились с лоб-
ными, но по бокам остаются
щелки; спиральный шов меж-
ду зубной и угловой костями
отчетлив на всем протяжении;
вес черепа ок. 6,4 г; межглаз-
ничный промежуток ок. 17,5 мм.
4" Заднеглазничные и височные от-
ростки срастаются на всем участке
прилегания; концы носовых костей
врастают в лобные, но структура их
поверхностей различна и швы хоро-
шо заметны по бокам; вес черепа
около 24 г; межглазничный промежу-
ток ок. 32 мм.
U+
Швы по сторонам носовых
костей исчезают; спиральный
Шов на нижней челюсти исчез;
вес черепа ок. 6,8 г; межглаз-
аичный промежуток 18—20 мм.
Ill + Носовые кости по структуре
сходны с лобными, внутренние швы
между ними хорошо заметны, внеш-
ние исчезают; спиральный шов на
нижней челюсти явственный до пе-
реднего конца фонтанели; вес чере-
па ок. 27 г, межглазн. промежуток
ок. 35 мм.
1V4- Граница между носовыми и лоб-
ными костями неуловима; спираль-
ный шов на нижней челюсти в виде
слабой ложбинки или исчез; вес че-
репа ок. 29 г; межглазпичный проме-
жуток ок. 37—38 мм.
^Рена^оле 4 а Н И е’ ^се измерения даны по черепам с Печоры; в Лапландии
пР0МежАЫХ самцов примерно на 6 г, старых — на 3 г легче, а межглазнич-
жУток примерно на 4 мм меньше. Черепа самок по размерам почтя
173
2. Срастание носовых костей с лобными (у самцов на третье?*
году).
3. Зарастание шва между зубной и угловой костями нижней че-
люсти (у самцов старше 3 лет).
4. Расширение с возрастом межглазничного промежутка.
5. Увеличение веса черепа.
Проверка этих признаков на коллекционном материале, собран-
ном нами в Лапландском и Печоро-Илычском заповедниках, пока-
зала, что признаки эти вполне реальны, но на севере последователь-
ность срастания костей иная. Если серия собрана за сравнительно
короткий промежуток времени, черепа распределяются по возрастам
довольно легко и случаи затруднений в отнесении экземпляра к той
или другой возрастной группе редки. Между типичными черепами
каждой группы возрастная разница выражена отчетливо.
Исследовав почти двести черепов глухарей и глухарок старше
одного года, я составил определительную таблицу, не вполне совпа-
дающую с таблицей С. В. Кирилова (Кириков, 1948 и 1952|). Из на-
шей таблицы видно, что срастание определенных черепных костей
у самок происходит приблизительно на год раньше, чем у самцов.
При сравнении с данными Кирикова, у лапландских и североураль-
ских глухарей срастание носовых костей с лобными наступает на
год раньше, чем у южноуральских.
При разработке определительной таблицы возраст птиц до 14 ме-
сяцев определялся по крылу или хвосту, с учетом даты добычи:
таким образом первая возрастная группа была выделена вполне
объективно. Контрольных экземпляров старших возрастов мы на
имели. Черепа были разделены по возрастам только на основании
признаков, указанных С. В. Кириковым. Обработка черепов, добы-
тых на токах и в осенне-зимний сезон охоты, проводилась раздельно,
что обеспечивало разрыв в возрасте двух соседних групп не менее
9 месяцев. Несмотря на индивидуальные отклонения в отдельных
признаках, при учете их в совокупности почти не возникало сомне-
ний в правильности отнесения отдельного черепа к данной группе.
Своеобразной проверкой объективности определения возраста глу-
харей оказалась равномерная убыль числа особей с увеличением
возраста птиц (см. ниже, рис. 97), а также закономерное изменение
веса птиц с возрастам; (см. выше, рис. 49).
Приводим здесь две таблицы для определения возраста. Первая
составлена только по материалам, собранным в Печоро-Илычском
заповеднике, на токах; вторая относится к осенне-зимнему сезону
охоты и включает самцов и самок, добытых как на Верхней Печоре,
так и в Лапландии. Возрастные признаки хорошо видны на наших
фотографиях (рис. 60—68). Для модели подобраны летние экземп-
ляры из коллекции Лапландского заповедника. На снимках замет-
ны не только упоминаемые в таблицах признаки, но и некоторые
другие, как, например, утолщение лобных и слезных костей по верх-
174
нему краю орбиты, расширение заднеглазничного и височного от-
ростков и уменьшение «окна» между ними, нарастание лобной ко-
сти поверх решетчатой (в возрасте I—II года), разрастание лобных
костей позади слезных (II—III года) и заполнение угла между ни-
ми к старости.
Рост лобных костей продолжается у глухаря не менее 4 лет.
Поверхность их, начиная со второго года, бороздчатая, а толща
изобилует воздухоносными полостями и кровеносными сосудами,
чем они легко отличаются от более светлых и гладких носовых ко-
стей.
Рис. 68. Нижние челюсти глухарей в возрасте 4 (сверху) и 3 (снизу)
лет; видно заполнение шва между зубной и угловой костями
Черепа белых куропаток уже в 9-месячном возрасте ничем не от-
личаются от черепов более старых птиц, так как к этому времени все
руководящие швы, включая и шов между зубной и угловой костями,
срастаются. Черепа рябчиков и тетеревов не были подробно иссле-
дованы, но по-видимому они сходны с чер.епа1ми белых куро-
паток.
Критически рассмотрев разные способы определения возраста
тетеревиных птиц, мы рекомендуем пользоваться следующими:
1. У пуховичков до 2 дней — наличие яйцевого зуба и отсутствие
весового прироста (0 + 1 + день).
2. До 100 дней — по формуле линьки маховых (см. рис. 52, 53,
56) и по весу птенца (см. рис. 40—43). С меньшей точностью можно
определить возраст по разности дат добычи и появления первых
выводков.
3. В начале охотничьего сезона (до середины октября) — по отно-
сительной длине первых четырех маховых: у молодых последним от-
растает третье маховое, у взрослых — первое. Если эти перья пол-
ностью выросли, можно вырвать их и сравнить кончики очинов — эта
часть пера и после окончания роста долго остается мягкой.
175
4. Зимой, весной и летом (до июля) у самцов глухарей и коса-
чей самый простой возрастной признак—длина хвоста (табл. 48 и
стр. 166): он П'р1ИМ0Н1Игм и при наблюдении живых птиц в при-
роде.
5. Зимой, весной и летом (до августа) молодых птиц можно от-
личить от старых по светлым (глухарь, тетерев) или черным (белая
и тундряная куропатки) крапинам на конце второго махового (рис.
58—59).
6. В период зимней охоты (до апреля) — у самок рябчиков воз-
раст определяется по внешнему виду яйцевода (стр. 168). Для сам-
цов падежного способа не найдено.
7. В возрасте старше 14 месяцев возраст распознается только у
глухарей, по срастанию швбв и другим черепным признакам, у са-
мок— до 3+ лет; у самцов — до 4-f-, а весной—до 5 лет (см.
стр. 170—173).
VIII. ПАРАЗИТЫ ТЕТЕРЕВИНЫХ
В Лапландском заповеднике не проводилось паразитологических
исследований, но у части добытых птиц кишечник обследовался на
крупных паразитических червей; всего было вскрыто 287 птиц, что
составляет 7*—Vr> всех исследованных экземпляров. В двенадцати-
перстной кишке, слепых и прямой мы не встречали паразитических
червей, но в тонкой кишке часто наблюдались аскариды и крупные
ленточники из рода Raillietina, ширина стробилы которых достигает
3 мм.
Ближайшая к заповеднику местность, где изучены кишечные па*
разиты белой куропатки и тетерева, — это Норвегия (см. Хуус, 1928;
Хольт, 1952; Лунд, 1952). При исследовании 517 экз. белых куропа-
ток Хуус нашел два вида нематод и два вида цестод. Ascaridia compar
(Schrank) найдена у 37% куропаток, в тонкой кишке; Capillaria
longicollis (Rudolphi) —редка, встречена в двенадцатиперстной
кишке у трех птиц; Raillietina urogalli (Modeer) —найдена у Г1%
птиц, в тонкой кишке; Weinlandia microps (Diesing) —у 12% ис-
следованных птиц. У куропаток в южной Норвегии Хольт нашел в
15% всех исследованных кишечников цестод и в 45% —аскарид, в
основном Ascaridia compar. Лунд описывает паразитов тетерева сле-
дующим образом': Capillaria caudinflata — у 25% птиц в двенадцати-
перстной кишке; Ascaridia compar — у 27%, во всех отделах кишеч-
ника, исключая только слепые кишки; Raillietinai urogalli — у 12%
птиц, в нижней части тонких кишок; R. globocaudata—у 5%* птиц,
в двенадцатиперстной кишке (ширина до 1,1 мм); R. retiisa —у од-
ной птицы; Hymenolepis (Weinlandia) microps — у одной птицы в
двенадцатиперстной кишке; Heterakis gallinarum — у Двух экз., в
слепых кишках; Subulura sp. —у одного экз.
Учитывая распространение этих паразитов и их размеры (диа-
метр тела Capillaria caudinflata и Hymenolepis microps едва дости-
гает 0,1—0,2 мм) мы решили, что имели дело только1 с Ascaridia
176
compar— из нематод и с представителями рода Raillietina— из це-
стод.
Нормальное местонахождение аскарид — в верхней части соб-
ственно тонкой кишки, цестод — в нижних двух третях протяжения
того же отдела. Через несколько часов после смерти птицы аскарид
можно встретить не только в двенадцатиперстной кишке, но даже
в желудке и в полости тела, куда они выходят, прободая стенку ки-
шечника. Обычно в кишечнике бывает от одной до пяти аскарид, но
иногда несколько десятков. У молодой самки куропатки, добытой
20 X 1934 г., было около 40 аскарид, у другой (7 XI 1949 г.) 22 экз.
общим весом 2,2 г; у молодого тетерева 20 IX 1951 г. найдено 14 экз.,
но у глухарей я не находил более 4 экз. В Печоро-Илычском запо-
веднике В. П. Теплов (1947) находил у глухарей до 6 экз. аскарид,
преимущественно Ascaridia cylindrica (Biome), изредка — A. galli
(Schrank). Из цестод он встречал Ralllietina globooaudata (Cohn.),
изредка R. urogalli и Davainea tetraoensis (Fuhrmann). Заражен-
ность тетеревиных птиц аскаридами «и цестодами всюду имеет се-
зонный характер (табл. 49|).
Таблица 49
Годовой ход встречаемости кишечных паразитов глухаря и белой куропатки в Лапландском заповеднике (в %)
I 11 III IV V VI VII VIII IX X XI XII
Глухарь
Аскариды 0 0 0 0 0 0 0 31 6 4 0 0
Цестоды 0 50 63 89 67 60 20 15 6 7 0 0
Без глистов .... 100 50 37 11 33 40 80 61 88 89 100 100
Число птиц .... 2 4 8 9 6 5 5 13 36 27 3 4
Белая куропатка ,
Аскариды ! 1 0 0 0 15 33 0 20 69 52 40 —
Цестоды — 100 68 88 85 100 100 80 85 57 0 —
Без глистов ..... — 0 32 12 8 0 0 20 8 19 60 —
Число птиц 0 2 22 17 13 3 4 5 13 21 15 0
Примечание: Цестоды учитывались только по наличию развитой стробилы.
Аскариды встречены мною исключительно в летнее полугодие,
самые ранние даты 20 V 1951 г. и 28 V 1950 г., самые поздние 19 и
16 X 1934 г. (у белых куропаток). На диаграмме (рис. 69) хорошо
видно, что в сезонном ходе зараженности белых куропаток аскари-
дами как в Норвегии, так и в Мурманской области наблюдаются
два «ма'Коимума: в июне и в сентябре. В Норвегии зимой аскариды
редки, в Л апл а ндс колг заповеднике совсем пе встречаются. Жизнен-
ный цикл Ascaridia compar, по-видимому, мало известен; поэтому по-
12—3037
177
лезно предварительно ознакомиться с жизненным циклом хорошо
изученной A. galli, паразита тонких кишок домашних кур и других
фазановых.
У кур заражается, в основном, молодняк до одного года. Источ-
ником являются инвазированные птицы и перезимовавшие во внеш-
ней среде яйца аскарид. Личинка выходит из заглотанного птицей
спелого яйца уже в желудке, спускается в кишечник, внедряется
в просвет либеркюновой железы, где развивается в течение 19 дней,
потом выходит в просвет кишечника и на 35—58-й день с момента
заражения достигает половой зрелости. Продолжительность жиз-
ни куриной аскариды 9-14 месяцев (Антипин, Ершов и др., 1956,
стр. 128—131).
Рис. 69. Сезонные изменения зараженности аскаридами. Сплошная
линия — Лапландский заповедник, прерывистая — Печоро-Илычский
заповедник, тире с точкой — Норвегия
Аскариды куропатки и глухаря, видимо, менее долговечны; как
только выпадет снег и новое заражение станет невозможным —
они постепенно исчезают. В Мурманской области они могут перези-
мовать в стадии яиц, сохраняющихся под снежным покровом. Новая
инвазия начинается очень рано, вскоре после появления больших
проталин. При обсуждении условий заражения надо иметь в виду,
что тетеревиные не ищут корм в земле, как куры, а обрывают ягоды,
побеги и другой корм прямо с живого растения; поэтому у них нет
никаких шансов проглотить яйцо аскариды с кормом. К тому же
при невысокой плотности боровой дичи, порядка 15—30 птиц разных
видов на 1 кв. км, зараженность /местности яйцами не может быть
значительной. Наиболее вероятный путь передав паразитов (не
имеющих промежуточного хозяина) от одной особи к другой — это
«порхалища» боровой дичи *. В лесах со сплошным растительным
покровом мало мест с обнаженным грунтом, почему каждое такое
место привлекает птиц; на Верхней Печоре охотники ловили глухарей
* Такое же предложение высказано П. Г. Ефремовым (1940).
178
слопцами и силками исключительно на обнажениях грунта, так как
ни на какую другую приманку они не идут. В разрытом» птицей песке
или древесной трухе постоянно попадаются не только выпавшие при
линьке перья (по ним можно судить, какие птицы посещали это ме-
сто), но и помет. Сопоставление дат появления аскарид в кишечни-
ках птиц с некоторыми другими периодическими явлениями, наблю-
давшимися в Лапландском заповеднике, говорят в пользу того, что
они заражаются именно через «порхалища»:
1949 1950 1951
Массовое появление проталин в лесу 26 IV 18 IV 23 IV
Последний день с снежным покровом в ель-
никах 17 V 10 V 20 V
Первые «пыльные ванны> боровой дичи 22 V 13 V —
Первые аскариды в кишечниках — 28 V 20 V
Первый (июньский) «пик» встречаемости аскарид является ре-
зультатом заражения взрослых птиц перезимовавшими яйцами.
Второй «пик» (в сентябре) можно объяснить массовым заражением
Рис. 70. Сезонные изменения в зараженности цестодами. Сплошная
линия — Лапландский, прерывистая — Печоро-Илычский заповедник
птенцов боровой дичи, которое происходит, вероятно, также через
«порхалища». В Лапландском заповеднике аскариды встречались у
глухарей реже, чем у 'куропаток; возможно, по этой причине мы не
наблюдали у них двух максимумов аскарид. Однако в Печоро-
Илычском заповеднике, где глухари заражены значительно сильнее
(Теплой, 1947, стр. 58—60), у них также наблюдается один макси-
мум (в июне, см. рис. 70).
12*
179
Насколько можно судить по нашему небольшому материалу, те-
терев и рябчик менее подвержены аскаридозу, чем« белая куропат-
ка, но больше, чем глухарь (табл. 50).
Таблица .50
Сравнение глистных инвазий тетеревиных птиц в периоды наибольшего
заражения
(в %)
(В скобках — число исследованных кишечников)
Вид хозяина । Аскариды 1 VIII - IX 1 Цестоды II — VI Нес годы, весь год
Глухарь 12 (49) 69 (32) 23 (122)
Тетерев 38 (13) 50 ( 4) 20 ( 20)
Рябчик 20 ( 5) 0 ( 3) 0 ( 12)
Белая куропатка 56 (18) 61 (57) 67 (115)
Тундряная куропатка . - ( 1) 8 (13) И ( 18)
Продолжительность жизни отдельной аскариды, по-видимомуч
не превышает 3 месяцев; возможно, что и период от момента зара-
жения до развития взрослой аскариды у нее соответственно короче,
чем у аскариды домашних кур.
Аскариды встречаются приблизительно одинаково часто у обоих
полов, но молодняк в пять—шесть раз больше подвержен аскари-
дозу, чем взрослые птицы. Ввиду того что весной аскариды почти
не попадаются, а в числе отстреленных птиц преобладали самцы,
сравнение целесообразно провести только за вторую половину года.
Количество птиц с аскаридами оказалось следующим1 (в %; в скоб-
ках— общее число исследованных экземпляров):
Глухари самцы........12 (32) Белые куропатки самцы . 44 (33)
„ самки ............5 (56) „ » самки . 55 (22)
Глухари, моложе Va года 13 (46) „ моложе >/2 года . . 72 (18)
„ старше 1 года . 2 (42) „ старше 1 года ... 14 (14)
Странно, что в Печоро-Илычском заповеднике, по данным
В. П. Теплова (1947, стр. 60), молодые глухари заражены круглы-
ми червями значительно меньше, чем взрослые: этот вывод противо-
речит не только данным Лапландского заповедника, но и тому, что
известно об аскаридозе домашних кур.
Определить статистическим путем потерю веса птиц в результате
глистных инвазий довольно трудно: с одной стороны, зараженность
меняется в зависимости от сезона и возраста птицы, с другой сто-
роны, и вес меняется по сезонам, полу и возрасту птицы. Наличный
материал приходится дробить на много групп, а при малом числе
экземпляров в> каждой результаты получаются недостоверными. В
частности, выводы по этому вопросу в нашей первой работе по тете-
ревиным (Семенов-Тян-Шанский, 1938, стр. 292) неубедительны.
Есть местности, где дикие куриные свободны от аскаридоза; к
ним относится все морское побережье Норвегии (Хуус, 1928). Холь-
180
rep (1952) обратил внимание на то обстоятельство, что там, где
нет аскарид, куриные страдают от паразита! слепых кишок Heterakis
gallinarum, и наоборот; по-видимому, 'между этими видами сущест-
вует антагонизм. Причины этих особенностей распространения ас-
каридоза неясны.
Цестоды встречаются в кишечниках тетеревиных птиц большую
часть года; только в период с 21 октября по 22 февраля мы не встре-
тили их ни у одной из 41 исследованных за это время птиц. Наибо-
лее заражены белые куропатки; за 8 месяцев (II—IX) цестоды были
найдены у 82% исследованных экземпляров, без существенных от-
клонений в отдельные месяцы; даты последних встреч — 20 X
1934 г. и 19 X 1906 г. В конце февраля — марте цестоды снова на-
блюдаются у 71 % исследованных птиц. Глухарь и тетерев меньше по-
ражены цестодами; еще реже они попадаются у рябчика и тундря-
ной куропатки (табл. 50). В Лапландском заповеднике цестод в ряб-
чиках мы не находили, но они были обнаружены в кишечнике ряб-
чика, добытого 20 V 1943 г. близ Повенда в Карелии. В отличие от
белой куропатки, у глухарей и тетеревов зараженность сокращается
уже летом’ (VII—VIII); последняя встреча была 10 X 1938 г: Новое
появление наблюдается также в конце февраля. Иногда, особенно у
белых куропаток, цестоды заполняют почти все сечение кишечника,»
просвечивая через его стенку. Наибольшее количество, 16 экз., об-
наружено у самца белой куропатки 11 III 1941 г.
Оба пола приблизительно одинаково заражены цестодами. В
отношении возраста — молодые куропатки заражены меньше, чем
старые, а молодые глухари, напротив, больше. Поскольку заражен-
ность гельминтами, с одной стороны, и половые и возрастные соот-
ношения добытых птиц, с другой, меняются по сезонам, весь мате-
риал, для удобства сравнения, полезно разбить по крайней мере на
два периода — первое и второе полугодие.
Зараженность цестодами (в %)'
Первое полугодие Второе полугодие
Глухарь Белая куропатка Глухарь Белая куропатка
Самцы . . . .65(26) 86 (37) 9 (32) 38 (34)
Самки . . . 62 ( 8) 70 (20) 7 (56) 41 (22)
Молодые . . 70 (10) 80 (20) 15 (46) 56 (18)
Взрослые , . . 62 (24) 90 (19) о (42) 93 (14)
Сходным образом меняется по сезонам зараженность цестодами
и в других местах. Так, например, в Норвегии, кишечники белых ку-
ропаток свободны от цестод с ноября по март (Хуус, 1928), а на
Верхней Печоре, у глухарей, с ноября по февраль (Теплов, 1947).
Судя по крупным размерам' стробилы и локализации паразита в
нижней части тонких кишок, мы — имели дело с Raillietina urogali
(возможно, и с другими видами того же рода). Промежуточный хо-
зяин этого вида нам неизвестен, но для близких видов того же рода
указываются муравьи и некоторые напочвенные жуки — мелкие жу-
181
желицы и навозники (Снигиревский, 1947; Антипин, Ершов и др.,
1956).
Давно уже высказывалось мнение, что освобождение кишечни-
ков тетеревиных птиц от цестод объясняется глистогонным действием
веточных кормов, что веточные корма служат не только пищей, но
и лекарством, дающим птице возможность освободиться от парази-
тов на зиму — период, по-видимому, наиболее для них тяжелый
(Вольтер, 1930). Однако массовое появление цестод в конце зимы,
когда вся боровая дичь питается еще исключительно зимними кор-
мами, противоречит этому объяснению. В 1936 г. мы писали по этому
поводу: «Если исключить совершенно невероятную возможность но-
вой инвазии в течении зимы,'нам остается предположить, что осенью
птицы действительно освобождаются от цестод, причем из кишечни-
ка удаляется большая часть тела червя, сохраняется только голов-
ка, а при вскрытиях она легко может остаться незамеченной. Вре-
мя обрыва цестод и удаления их из кишечника приурочено к
моменту перехода на зимние корма... Впоследствии остаток цестоды
приспособляется к изменившейся среде (к пищевой кашице, напол-
няющей кишечник птицы зимой), начинает расти и в феврале дости-
гает полного развития» (Семенов-Тян-Шанский, 1938, стр. 291—292).
В 1940—1942 гг. А. В. Федюшин, изучая цестод тетерева и бе-
лой куропатки в Омской области, столкнулся с тем же явлением.
Исследование свыЩе 300 зимних экземпляров диких куриных из
Омской области «показало зараженность только ленточными чер-
вями тетеревов на 20-о/о, белых куропаток на 5О«/о, рябчиков на
13,5о/о и серых куропаток на 68,9«/о- Все паразитирующие в это вре-
мя у тетеревиных птиц ленточные черви дестробилируют, т. е. отбра-
сывают всю свою стробилу, и сохраняются в слизистой кишечника
в течение всей зимы в виде сколексов с 5—8 передними члениками...
с сентября, наряду с полными экземплярами ленточных червей,
встречались и сколексы (21»/0)... Но уже с X—XI, после выпадения
снега и наступления морозов, у тетеревов, белых куропаток и рябчи-
ков мы находили исключительно сколексы (100о/0)... Таким обра-
зом, осенью у тетеревиных, несмотря на переход их на грубые дре-
весные корма (хвою, почки), полной дегельминтизации кишечника
все же не происходит... В конце зимы начинается отрастание новых
стробил от зимовавших в кишечнике головок и уже добытые в это
время и ранней весной птицы сильно заражены ленточными червя-
ми» (Федюшин, 1946, 101—105).
Явление дестробилизации Федюшин рассматривает как истори-
чески сложившуюся реакцию со стороны паразита на неблагоприят-
ные условия среды — изменение химизма пищи и отсутствие условий
для развития личиночных стадий. Дальнейшая дискуссия по вопросу
о дестробилизации не внесла в это дело ясности. Представленные
новые фактические материалы в общем совпадали с прежде опуб-
ликованными, а новые соображения нельзя считать более убеди-
тельными, чем прежние. М. Н. Дубинина (1950) считает непосред-
ственной причиной отпада стробилы голодание хозяина в связи с
182
малой питательностью его зимних кормов. Между тем в феврале
состав пищи, как мы показали выше, еще не меняется, и для нового
роста стробилы в это время, по-видимому, нет причин. И. М. Олигер
(1050) полагает, что дестробиляцию и .вторичный рост цестод мож-
но объяснить происходящими зимой изменениями в содержании
крахмала в древесных породах. Насколько это предположение
обосновано фактами — судить трудно. Интереснее другое указание
Олигера, что период дестробиляции совпадает со значительным
снижением активности у тетеревиных птиц. П. И. Феоктистов (1953)
наблюдал дестробиляцию Drepanidotaenia lanceolata у домашних
гусей при неизменном' кормовом рационе и потому полагает, что это
явление вообще не зависит от питания хозяина. Он не учитывает,
что питание организма зависит не только от наличия кормов, но и от
активности животного. В начале работы (стр. 21 и 40) я упоминал,
что уже в марте глухари и куропатки заметно оживляются, а это не
может не отразиться на обмене веществ. Период полного отсут-
ствия цестод в кишечниках начинается за 62 дня до зим-
него солнцестояния, а кончается также через 62 дня, следовательно,
при тех же точно условиях освещения. Долгота дня в Лапландском
заповеднике 2)1 X и 22 II составляет одинаково 8 ч. 45 м.; вероятно,
это не случайное совпадение.
Вероятность инвазии зависит от зараженности окружающей сре-
ды яйцами или личинками соответствующего паразита, от численно-
сти и зараженности промежуточных хозяев. Естественно предполо-
жить, что в годы обилия дичи глистные инвазии возрастают, а в го-
ды депрессии они соответственно уменьшаются. Для проверки этого
предположения надо иметь достаточно большой и сравнимый мате-
риал за несколько лет подряд: при малых пробах добавление одной
птицы уже заметно меняет результат.
В 1939 г. в Мурманской области численность всех видов тетере-
виных была минимальной. Хотя после того она и возрастала, но до
195.1 г. не было ни одного настоящего «пика». Поэтому, по общему
уровню плотности популяций период 1933—1938 гг. удобно противо-
Таблица 51
Зараженность гельминтами в периоды с разным уровнем численности
тетеревиных
1933-1938 гг. 1939-1951 г.
Среднее число тетеревиных яа 100 га 34 12
% птиц с аскаридами:
глухарь .... . . 10 (63 экз.) 2 (59 экз.)
белая куропатка 41 (59 . ) П (55 . )
% птиц с цестодами:
глухарь ... - . 32 (63 , ) 1! <59 • >
белая куропатка I 58 (59 „ ) 78 (55 . )
183
поставить периоду 1939—1901 гг. Встречаемость паразитических
червей в эти периоды, действительно, была различной (табл. 51):
после 1939 г. аскариды у обоих видов встречались в 4—5 раз реже,
чем раньше; цестоды у глухарей в 2 раза реже, но у куропаток, на-
против, в полтора раза чаще. По-видимому, только аскарид нал инва-
зия определенно зависит от плотности популяции хозяина.
Эктопаразиты, На тетеревиных часто встречаются пухоеды, а ле.
том, кроме того, птичья кровососка Ornithomyia chloropus. Самые
ранние встречи кровососок на добытых птицах отмечены: 22 VII
1934 г. — на взрослой тундряной куропатке, 25 VII I960 г. на (мо-
лодом глухаре, 216 VII 1938 г. на молодой белой куропатке. Больше
всего их бывает в августе. Самые поздние встречи: 1 X 1935 г. на мо-
лодой белой куропатке, 29 IX 1949 г. на молодом тетереве и 20 IX
1951 г. на нескольких тетеревах. На одной птице мы не встречали
больше <5—6 кровососок. Они легко бегают по телу птицы под опе-
рением, а при обработке ее переходят на человека, забираясь под
белье, но не делая попыток проколоть кожу.
Пухоеды встречаются в течение круглого года и на большей ча-
сти особей: из 42 птиц, добытых ® 1949—1951 гг. они не были найде-
ны только на 19. Пухоедов легче всего обнаружить среди мелких
перьев вокруг основания клюва. Количество их на птице обычно не-
велико, около десятка или даже меньше; обилие пухоедов служит
верным признаком больной птицы: у нее оказывается либо травма,
либо большая зараженность гельминтами, либо вес ее значительно
ниже нормы. Один взрослый и внешне вполне здоровый глухарь,
пойманный при взлете с земли собакой, оказался сильно заражен-
ным пухоедами.
Все виды куриных очень любят купаться в пыли. Эта привычка,
по-видимому, является средством борьбы с пухоедами. Иначе труд-
но объяснить, почему пухоеды несколько чаще наблюдаются зимой,
когда птицы не могут «порхаться». В мае-сентябре они найдены на
11 птицах из 23 обследованных (48о/о), а в октябре-апреле—на 15
птицах из 22 (68»/о).
IX. ВРАГИ ТЕТЕРЕВИНЫХ
Для того чтобы правильно оценить роль тетеревиных птиц в био-
ценозе тайги, необходимо иметь ясное представление о численном
соотношении отдельных экологических групп видов и об их био-
массе. Для Лапландского заповедника такой расчет можно прове-
сти по материалам исследований Г. А. Новикова (1956): в его книге
приведены показатели численности птиц еловых лесов заповедника
в пересчете на 1 час экскурсии, для теплого и холодного полугодий
отдельно (там же, стр. 25—26]). Веса птиц мы выбрали из картотек
Лапландского и Печоро-Илычского заповедников, а по отдельным
видам — заимствовали из сводки «Птицы СССР» (1951 —1954).
При наличии большой разницы в весе полов мы принимаем вое пти-
цы равным среднему между самцом и самкой. Результаты подсчета
даны в табл. 52.
184
Таблица 52
биомассы птиц в еловых лесах Лапландского заповедника
(1937—1939 гг., по Г. А. Новикову)
расчеТ
Лето Зима
ВИДЫ птиц Вес число индекс число индекс
(в г) встреч био- встреч био-
на 1 час массы на 1 час массы
Глухарь • 2620 0,82 2150 0,11 288
1100 — — 0,02 22
Тетерев • • • 400 0,48 192 0,25 100
Рябчик • Белая куропатка . . . Тетеревиные в сумме . . . 620 0,57 1,87 353 2695 0,25 0,63 155 565
Кречет Дербник Ястреб тетеревятник . . 1420 0,01 14 —
190 1140 0,01 0,01 3 17 0,03 34
Ястреб-перепелятн. . . . Орлан-белохвост . . . Зимняк Дневные хищные в сумме . 200 4450 990 0,01 0,01 0,02 0,07 2 44 20 100 0,03 1 1 1 £
Воробьиный сыч .... 70 0,01 1 — —
Ястребиная сова .... 320 0,01 3 0,05 16
Совы в сумме — 0,02 4 0,05 16
Большой пестр, дятел . . 95 0,03 3 0,04 4
Малый пестр, дятел . . 23 — — 0,01 О
Черный дятел 330 0,04 13 0,02 7
Трехпалый дятел .... 68 0,12 8 0,18 12
Дятлы в сумме — 0,19 24 0,25 23
Ворон 1230 0,04 49 0,06 74
Кунша 85 0,64 54 0,21 18
Чечетка . 13 3,32 43 3,66 48
снегирь 30 0,23 7 0,06 2
Й*УР Клесты (3 вида} .... 53 0,07 4 0,75 40
41 0,54 22 6,57 269
восковка 9 0,02 0
г±<а черноголовая свХ9 _•; : 10 0,34 3 0,16 2
12 56 0,43 0,16 5 9 0,21 0,08 3 4
^иж . зяблик ’ Свая1 (реМез и камыше- й^зкАбелая • • • • ^Ролек СН' и луг') • 12 23 22 0,05 0,01 3,55 1 0 78 — —
19 22 0 03 0,02 1 0 —
20 6 0,54 0,01 11 0 — —
185
Табл. 52 _(п р о до л жени е)
Виды птиц Вес (в г) Лето Зима
число встреч на 1 час индекс био- массы число встреч на 1 чае । иидекг био 1 массы 1
Мухоловка серая . . . 17 0,19 3
Мухоловка пеструшка . . 13 0,03 0 — —
Пеночка теньковка . . . 10 0,15 2 —
Пеночка весничка . . . 9 5,80 52 — —
Пеночка таловка . . . 10 0,20 2 — —
Дрозд деряба 103 0,01 1 — —
Дрозд певчий 65 0,51 33 — —
Дрозд белобровик . . . 61 2,16 132 — —
Дрозд рябинник .... 93 0,13 12 — —
Синехвостка 13 0,01 0 — —
Горихвостка 16 0,26 4 — —
Варакушка 20 0,01 0 —- —
Зарянка 16 0,12 2 — —
Завирушка 20 0,01 0 — —
Крапивник 10 0,01 0 — —
Воробьиные в сумме . 19,58 530 11,78 469
Кукушка ....... 110 0,07 8 — —
Сумма индексов птиц . . . — 21,80 3361 12,74 1098
При летних маршрутных учетах Г. А. Новикова 4 вида тетереви-
ных дали около 80% биомассы всех встреченных птиц. Ввиду тогог
что часть певчих птиц и хищников, в сумме дающих около 12<>/0 био-
массы, отлетает на зиму, казалось бы, «удельный вес» тетеревиных
зимой должен еще более возрасти. Данные Новикова показывают
обратное — зимой тетеревиные дали только 52% массы птиц.
Это несоответствие объясняется тем, что Новиков работал в лесу
только одну зиму 1938-39 г., когда боровой дичи было мало и на-
блюдалось массовое появление клестов и чечеток (по случаю обиль-
ного урожая семян ели и березы). С другой стороны, зимой тетере-
виные прячутся в снегу даже днем, отчего они становятся менее за-
метными на экскурсиях. Данные Новикова относятся к еловым ле'
сам, в борах «удельный вес» тетеревиных птиц должен быть
выше.
В Печоро-Илычском заповеднике подсчет биомассы птиц по эко-
логическим группам проведен Е. Н. Тепловой (1957|); для Кондо*
Сосвинского заповедника я сделал аналогичный расчет, пользуясь
данными В. В. Раевского (1952). Птиц тайги мы делим на следую-
щие экологические группы:
186
1. Тетеревиные.
2. Птицы, питающиеся зимой преимущественно семенами и яго-
дами деревьев и кустарников; численность этих птиц на зимовке за-
висит от «урожая» их основных кормов. Сюда относятся клесты,
большой пестрый дятел, чечетка, свиристель, щур и снегирь.
3. Насекомоядные оседлые птицы с относительно устойчивой
кормовой базой — все синицы, дятлы (кроме большого пестрого),
пищуха' и поползень.
4. Всеядные птицы из семейства вороновых — ворон, кукша,
кедровка и сойка.
5. Дневные хищные птицы и совы.
6. Перелетные певчие птицы и кукушка.
Для каждой группы даются два показателя — численности (чис-
литель!) и массы (знаменатель). В табл. 53 они даны в процентах
от общего числа учтенных птиц и от их общей массы.
Таблица 53
Численность и 'биомасса птиц в заповедниках
по экологическим группам
Экологические группы птиц Ла пл а идс к ип, 68° с. ш., Печоро- Илычский, 62° с. ш. зима Копдо-Со- свинский, 62° с. ш., зима
лето зима 1
Тетеревиные — 4 вида . . . 9/80 5/52 23/89 5/70
Семеноядные зимующие — 8 видов 20/3 87/33 41/4 43/15
Насекомоядные зимующие — 10 видов 4/1 5/2 28'1 47;6
Вороновые — '3 вида . . . 3/3 2/8 6/4 5/8
Хищные и совы — ГЗ видов 1/3 1/5 1/2 0+/1
Перелетные певчие — 24 вида 63 10 — —
Приведенные здесь цифры надо считать грубо приблизительными
(у разных наблюдателей методика могла быть не вполне одинако-
вой, не учтены особенности состава фауны в отдельные зимы и так
далее), но все же из них видно, что тетеревиные, составляющие по
числу особей 5—25% всего птичьего населения тайги, по весу со-
ставляют около 75% его.
Среди млекопитающих тайги первое место (по биомассе) могут
оспаривать две группы: с одной стороны, олени, с другой — мыше-
видные грызуны. Учеты разных млекопитающих проводятся различ-
ными способами, и «привести их к одному знаменателю» очень труд-
но. Попытаемся сделать расчет биомассы хотя бы для двух главных
групп.
В Лапландском заповеднике абсолютный учет дикого северного
оленя и лося проводился на всей его территории (1042 кв. км,). За
187
13 лет число учтенных оленей колебалось в пределах от 99 до 970
голов. Принимая средний вес оленя в 80 кг, получаем общий вес
стада от 8 до 78 т, что дает От 5 до 47 кг на 1 кв. км1. Число учтен-
ных лосей за 5 лет менялось от 32 до 141; при среднем весе лося
около 2176 кг это дает от 5 до 24 кг живого веса на 1 кв. км. В Печо-
ро-Илычском заповеднике, по условиям местности, абсолютный
учет копытных не проводился.
Учет мышевидных млекопитающих в обоих заповедниках прово-
дился одним1 способом — путем облова в августе-сентябре линии,
длиной около 1 км 100 ловушка-ми в течение 5 суток. Чтобы отнести
данные этих учетов к определенной площади необходимо знать ши-
рину облавливаемой полосы и отношение числа пойманных зверьков
к числу оставшихся: эти величины можно приблизительно опреде-
лить.
Леса таежной полосы заселены, в основном, тремя видами ры-
жих полевок (Clethrionomys glareolus, CI. rutilus, CI. rufocanus).
Случаи повторной поимки полевок, у которых раныше ловушкой был
отбит хвост или повреждена лапка, а иногда—распределение по
ловчей линии членов одной семьи полевок (например, матки и мо-
лодых европейской рыжей полевки в год, когда этот вид редок) поз-
воляют считать, что осенью диаметр индивидуального1 или семейно-
го участка рыжих полевок не превышает 25—40 м. Учитывая, что
к ловчей линии полевки подходят с обеих сторон, ширину облавли-
ваемой полосы можно принять, круглым числом, в 50 м? При от-
лове без перемещения ловушек в течение 5 суток попадаемость
день за днем снижается (это в среднем, так как в отдельных слу-
чаях она зависит от погоды). Обработка данных учета в Печоро-
Илычском заповеднике за 5 лет показала, что в среднем улов за
вторые сутки составляет 0,8 улова первых суток, за третьи сутки —
0,8 улова вторых суток и так далее. Предел суммы членов такого
ряда в 1,5 раза больше суммы первых 5 его членов. Зная, что длина
учетной линии в Лапландском заповеднике составляет 1 км, мы при-
нимаем облавливаемую площадь равной 0,05 кв. км, а общее коли-
чество пойманных на ней мелких млекопитающих увеличиваем в 1,5
раза.
Необходимо иметь в виду, что стационарные учетные линии не
захватывают заведомо бедных зверьками участков (например, мок-
рых болот); поэтому, распространяя результаты наших подсчетов
(табл. 54) на большую площадь, мы неизбежно получаем несколько
преувеличенные числа.
Землеройки мельче полевок и попадаются реже их, поэтому по
весу они составляют около 5% всех учтенных мелких млекопитаю-
щих.
* А. А. Насимович (1948, стр. 8) приводит сходный по результатам расчет,
хотя принятую им ширину учетной полосы и процент облова он не обосновывает
расчетом.
188
Серые полевки * попадаются еще реже; наибольшее значение
имеют рыжие полевки.
Учет боровой дичи проводится в августе на ленте определенной
ширины и длиною несколько десятков километров: он непосредст-
венно дает запас птиц на данной площади. Однако и тут надо учи-
тывать, что лента нс захватывает горных тундр и других участков,
бедных дичью, поэтому для заповедника в целом эти данные будут
преувеличенными.
Таблица 54
Приблизительный расчет массы важнейших позвоночных в заповедниках
(в кг на 1 кв. км )
Группы животных Миним. величина Средн, величина Максим, величина Число лет учета
Полевки и землеройки (ель-
ник) ......................
Копытные...................
Тетеревиные птицы . . . .
Лапландский заповедник
5 88 290 ’ 12
10 29 71 13
6 26 93 10
Печоро-Илычский заповедник
Полевки и землеройки (ель-
ник) ............................. 7
То же, в бору..................... 5
Тетеревиные птицы .... 23
I
88
26
42
150
38
57
5
5
4
Из таблицы видно, что запасы тетеревиных птиц, полевок и оле-
ней представляют, в общем, близкие величины. Для трофических
связей внутри биоценоза это обстоятельство должно иметь большое
значение. Отражается оно и на специализации хищников. Копытны-
ми питаются медведь, волк, росомаха; полевками — горностай, зим-
ник и большинство видов сов; боровой дичью — ястреб-тетеревят-
ник и кречет. Наша задача — показать значение группы тетереви-
ных ® питании различных хищников по материалам' заповед-
ников.
Пернатые хищники. Материалы по питанию хищных птиц Лап-
ландского заповедника опубликованы в работе М. И. Владимирской
(1948|). Данные, помещенные в табл. 55 переработаны по картоте-
кам заново, с включением дополнительных материалов. Обработан-
ный материал состоит из погадок, собранных под гнездами, остатков
* Microtus agrestis, М. oeconomus.
189
добычи (как под гнездами, так и на местах Охоты) и содержимого
желудков добытых хищников; не использованы только визуальные
наблюдения погони хищника за животным (если оно не было тут
же добыто). Для удобства сравнения проценты вычислены от обще,
го числа определенных объектов: при такой системе общая сумма
должна быть равна 100о/0-
Таблица 55
Характеристика питания хищных птиц Лапландского заповедника
(68° с. пг.)
Дербник* Кречет Ястреб тетеревят- ник Ястреб перепелят- ник Орлан £ ct о- о О о ч 1 Зимняк | 1 Ястреби- ная сова
Общее число данных 16 125 8 7 127 94 270 28
Млекопитающие . . . 19 28 — — 42 52 82 100
Копытные (падаль) — — — — 2 13 — —
Хищные . . . . . — — — — — 15 — —
Грызуны крупные . Грызуны мелкие и — — — — 25 16 — —
землеройки . . . 19 28 — — 15 8 82 100
Птицы 25 66 100 100 37 47 9
Тетеревиные . . . — 51 88 — 13 37 4 —
Водоплавающие . . — 3 — — 22 6 — —
Хищники, совы и др. — 2 12 14 — 4 1 —
Певчие, дятлы . . 25 10 — 86 2 — 4 —
Рептилии и амфибии . — — — — 0,5 — 1 —
Рыбы — — — — 18 1 — —
Насекомые 56 4 — — 2 — 1 —
Ягоды — 2 — — 0,5 — 7 —
* Включены и материалы Печоро-Илычского заповедника.
Из табл. 55—56 видно, что самое большое значение тетеревиные
имеют в питании ястреба-тетеревятника, затем — кречета, беркута,
а зимой также филина. Гораздо меньшее значение они имеют для
орлана белохвоста, подорлика и зимняка. В материалах по питанию
чеглока, ястреба-перепелятника, канюка и осоеда из тетеревиных
птиц был встречен только рябчик. В пище дербника, пустельги, ско-
пы, коршуна и ястребиной совы птицы этой группы не были встре-
чены.
190
По-видимо му, значение тетеревиных в питании того же самого
вида хищника на юге меньше, чем на севере, что может зависеть от
большего разнообразия стаций и, соответственно, более разнообраз-
ной авифауны (больше уток, болотных птиц и т. д.). Для правиль-
ной оценки значения отдельных хищников необходимо учесть не
только состав их пищи, но и относительную численность, а также
Таблица 56
Характеристика питания хищных птиц Печоро-Илычского заповедника
(62° с. ш.)
По материалам С. С. Донаурова (1948): и Е. Н. Тепловой
(11957; последние отмечены звездочкой}
Виды птиц Чеглок* Пустельга Ястреб тетере - вятник ос 0.0 о сГ Орлан белохвост Орел беркут Канюк Обь; к нов. Осоед Скопа* Филин*
Лето 1 •>? cd S СП
Общее число данных 66 129 57 52 82 92 33 439 34 16 404
Млекопитающие . . . 3 82 26 4 13 39 53 74 — 6 74
Копытные (падаль) — — — — — 1 — — — — —
Хищные 1 — 2 — — — — 0-г — — 04-
Грызуны крупные . — 7 2 4 — 34 53 5 — — 61
Грызуны мелкие и землеройки . . . 2 75 22 — 13 4 — 69 — 6 13
Птицы 45 6 74 96 76 40 47 23 6 6 23
Тетеревиные . . « ' 7 — 27 87 14 11 43 9 3 — 16
Водоплавающие . . — — 2 4 — 24 — 0 (- — 6 5
Хищники, совы и др. 3 — 3 — 3 — — 1 — — 2
Певчие, дятлы . . 35 6 42 5 59 5 4 13 3 — —
Рептилии и амфибии . — 2 — — — — — 1 30 — 1,5
Рыбы — — — — — 20 — — — 88 0,5
Насекомые .... 52 57 — — 11 1 — 2 64 — 1
Ягоды — — — — -— — — — — — —
—
Примечание. Значение кормов в %; подробности см. в тексте.
191
Таблица 57
Характеристика питания хищных птиц Ильменского заповедника
(55° с. ш.)
По Ушкову, 1949 и 4949-а
480 400 300 180
Общее число данных
Млекопитающие . . .
Копытные (падаль)
Хищные..............
Грызуны крупные .
Грызуны мелкие и
землеройки . . .
Птицы...............
Куриные ..........
Водоплавающие * **
Хищники, совы, кули-
ки................
Певчие, дятлы . .
Рептилии и амфибии .
Рыбы................
Насекомые...........
Ягоды...............
400
20
5
15
80
14
15
23
28
301
61
6
55
39
3
5
5
26
56 81
140
44
3
41
48
4
16
2
26
8
0+
41
7
1
0 +
* Сюда же отнесены лысуха и гагары.
** Встречены, но автором в таблицу не введены.
размеры. Эти сведения собраны в табл. 58. В качестве показателя
размера хищника взят его вес (в кг, среднее арифметическое веса
самца и самки). Показатель численности в Лапландском и Ильмен-
ском заповедниках означает среднее число встреч данного вида
одним наблюдателем) в год (для Печоро-Илычского заповедника
даны условные показатели). Перемножив три показателя — числен-
ность хищника, его размер и «удельный вес» тетеревиных в его< пита-
нии, мы получаем более близкую к действительности оценку роли
данного хищника, как -врага тетеревиных птиц. Результаты по-
казаны в табл. 59: для удобства сравнения они приведены к такому
виду, что сумма их в каждохМ столбце равна 100о/0. В следующем
обзоре хищные птицы размещены в порядке их значения для боро-
вой дичи.
192
Таблица 58
Относительная численность хищных птиц и их размеры___________
Вид хищной птицы Средний вес в кг < 2 ) Заповедники
Лапланд- ский Печоро- Илычский Ильмен- ский
Сапсан 0,9 — 1,0 2,6
Кречет 1,4 1,1 — —
Чеглок 0,2 — 4,7 —-
Дербник 0,2 5,3 0,2 —
Пустельга 0,2 1,7 1,0 —
Кобчик 0,2 — 0,4 —
Ястреб-тетеревятник .... 1,1 2,5 7,4 8,8
Ястреб-перепелятник . 0,2 1,8 6,2 6,2
Лунь луговой 0,3 — 0,2 —
Лунь полевой 0,4 — 0,4 —
Лунь болотный 0,5 — — 5,6
Коршун черный 1,0 0,2 7,1 22,0
Орлан белохвост 4,4 4,6 4,3 16,2
Орел беркут 3,4 1,6 1,6 9,6
Орел могильник 3,0 ? — — 1,6
Подорлик большой .... 2,2 — — ‘ 12,2
Канюк малый - 0,7 — 31,4* 15,0
Зимник 1,0 9,8 0,2 —
Осоед 0,8 0,1 2,3* —
Скопа 1,5 5,5 4,1 2,4
Филин 2,1 0,3 . 2,5 9,4
Ястребиная сова 0,3 8,3 — —
Прочие совы — 2,9 — —
• В оригинале канюк и осоед показаны вместе; они разделены нами
ориентировочно.
Таблица 59
_____________Значение тетеревиных птиц в добыче хищников_______________
Вид хищной птицы Заповедники
Лапланд- ский Печоро- Илычский Ильмен- ский
Кречет Falco gyrfalco 10 __
Чеглок Falco subbuteo — 1 —
Ястреб-тетеревятник Accipiter gen- tiiis . 29 37 24
Ястреб-перепелятник Accipiter nisus 1 1 0 +
Коршун Milvus korschun ... — 6
Орлан белохвост Haliaeetus albicilla 31 16 —.
Орел беркут Aquila chrysaetus . . 23 17 17
Орел могильник Aquila heliaca . . — — 2
Подорлик большой Aquila clanga . — — 21
Канюк Buteo buteo vulpinus . . . Зимняк Buteo lagopus 4 16 2
Осоед Pernis apivorus — 0-р —
Филип Bubo bubo 2 6 34
Сумма показателей .... 100 100 100
13-3037
193
Ястреб-тетеревятник. В таежной полосе этот ястреб питается
преимущественно боровой дичью, причем в его добыче преобладает
самый распространенный в данной местности вид: в Лапландское
заповеднике — глухарь, в Норвегии — белая куропатка, в Финлян-
дии— тетерев и рябчик (Сулькава, 1956), на Верхней Печоре —
рябчик, в Ильменском заповеднике — тетерев и серая куропатка.
Для ястреба характерно неожиданное нападение на добычу из-за
укрытия, с коротким преследованием, и умелым использованием
всей окружающей обстановки. Как отмечает В. П. Теплев (1947,
стр. 1156), «Тетеревятник нередко сопутствует охотникам, добываю-
щим глухаря с помощью лаек, бросаясь на птиц, поднятых собакой.
Мы четыре раза наблюдали подобные нападения тетеревятника на
Рис. 71. Соотношение встреч ястреба-тетеревятника и тетеревиных
птиц в Лапландском заповеднике; для первого — число наблюдений
за календарный год (пунктир), для последних — число встреч за
1 день охоты (сплошная линия)
глухариные выводки, причем трижды они были успешными». 2 X
1934 г., охотясь с лайкой в районе Лапландского заповедника, я так-
же наблюдал, как ястреб низко летел, перед самым носом собаки,
с очевидным намерением ловить поднятую ею птицу; 1 IX 1938 г.
такой же случай наблюдал Г. А. Новиков. В районе Печоро-Илыч-
ского заповедника то же самое отметил 3 IX 1952 г. Г. А. Предте-
ченский.
Иногда ястреб хватает тетерева или рябчика во время кормеж-
ки его на дереве. С. С. Донауров (1948, стр. 76|) сообщает: «В марте
1939 г. был отмечен случай, когда:, желая нанести удар рябчику, те-
теревятник налетел на березовый сук и погиб, проткнув себе грудь».
Второй аналогичный случай произошел в 1954 г.: 9 апреля на льду
Печоры близ Якши был найден замерзший труп тетеревятника, на-
сквозь проткнутый сосновым сучком. Судя по положению сука1 (он
вошел в брюхо, а вышел в области лопаток), ястреб мог наколоться
только при нападении на птицу, в тот момент, когда он выбросил
лапы вперед, чтобы ее схватить. При этом сук обломился, что дало
ястребу возможность отлететь па некоторое расстояние, почему он
194
I! оказался на середине реки. Эти факты показывают, как стреми-
тельно ястреб наносит свой удар жертве.
В Лапландском заповеднике тетеревятник попадался во все ме-
сяцы года, кроме декабря; встречаемость была, в общем, пропор-
циональна численности тетеревиных птиц в том же или предыду-
щем году (рис. 71); гнездо с яйцами найдено один раз 20 V 1*951 г.
Сулькава (1956) отмечает, что в Финляндии, где тетеревятник так-
же в основном питается боровой дичью (77о/о добычи), доля прочих
животных в его добыче обратно пропорциональна численности тете-
ревиных в том? же году.
В Печоро'-Илычском заповеднике этот ястреб, по мнению
Е. Н. Тепловой (1957), на самое темное время (XII—I) отлетает на
юг. В этой работе питание ястреба исследовано методически непра-
вильно (по наблюдениям охоты ястреба, сделанным преимущест-
венно над рекой, а не в лесу); поэтому в табл. 58 приведены более
старые сведения С. С. Донаурюва (Г948|), основанные на анализе
погадок и остатков добычи под гнездами.
Орел беркут. По своей величине и мощному вооружению лап
(рис. 72) беркут — самый опасный для крупных птиц хищник. В
Рис. 72. Лапа орла беркута; 0,6 нат. вел.
Лапландском заповеднике питание беркута изучено по остаткам пи-
щи и погадкам. Гнездо беркута (рис. 73) было найдено в лесу в до-
лине р. Вите 14 VI 1939 г; в нем было 2 пуховых птенца, позднее
погибших. Среди остатков добычи первое место принадлежало глу-
харю (остатки 12 самцов и 2 самок), второе — белой куропатке
(5 костяков и 5 встреч в погадках). Замечательно, что в числе
13*
195
жертв оказалось много хищников-—3 лисы, 2 куницы, 1 горностай
и 2 ястребиные совы. Объясняется это тем», что 1939 г. был годом
полного «неурожая» выводков боровой дичи, совпавшим с резкой
депрессией численности полевок. После обильных кормам и 1937 и
1938 гг. хищникам пришлось голодать, и сильнейшие из них истреб-
ляли более слабых. Возьможно, что лисиц орел поймал тут же у
Рис. 73. Гнездо беркута близ Мясного озера в Лапландском
заповеднике
гнезда, куда они подходили в поисках костей и других остатков. В
снежное время года беркут в значительной мере питается па-
далью —остатками северных оленей и лосей, погибших от волков
и медведей. При наличии такого корма беркут на зиму не отлетает.
В Лапландском заповеднике три беркута попались в капканы, по-
ставленные с целью уничтожения волков и росомах на остатках их
добычи (1-6 II и 22 IV 1941 г. и 3 IV 1951 г.).
196
3 IV 1986 г. мы наблюдали по следам, как взрослый глухарь
спрятался от орла в малоснежном пространстве под елкой с низко-
опущенными и засыпанными снегом ветками. Орел сел на снег, обе-
жал елку кругом, бросился на глухаря сверху через ветки, вырвав
при этом хвост и много перьев, вытащил его на поверхность паста
и расклевал. За один раз он объел мясо с шеи, спины, одной ноги,
вплоть до таза, с хвоста и отчасти с плечевых костей; съедено было
и сердце. Остатки были еще теплые, из чего мы заключили, что на-
падение произошло около 16 часов. Хищник был определен по сле-
ду, имевшему Г5—18 см в длину.
В Печоро-Илычском заповеднике одно гнездо было найдено в
горном районе у р. Егро-ляги. Среди остатков добычи преобладали
заяц и белка, затем глухарь (6 самцов и 2 самки), рябчик, тетерев
и куропатка (Донауров, 1948). Орлы здесь зимой охотно кормятся
трупами копытных (Теплова, 11967).
В Ильменском заповеднике (Ушков, 1949, стр. 14’3—1631) ® пище
беркута первое место занимала косуля (падаль, отчасти телята),
затем мелкие млекопитающие. Тетерев, глухарь и другие куриные
попадались в пище беркута, но редко; еще меньшее значение они
имеют для большого подорлика.
В Англии, где беркут охраняется, он питается преимущественно
кроликами, зайцами и шотландскими куропатками («Редкие хищные
птицы», 1967).
В СССР беркут всюду, за исключением некоторых заповедников,
относится к числу самых редких хищников, а потому причиняемый
им ущерб не так велик, как можно бы ожидать на основании табл.
59/
Орлан белохвост встречается у нас значительно' чаще, чем бер-
кут. Орлан массивнее беркута, но вооружен слабее его. В таежной
полосе он является регулярной перелетной птицей: в Лапландский
заповедник он прилетает, в среднем, 16 IV и улетает 29 IX. По ро-
ду-питания, орлан связан с водоемами. Материал, собранный в Лап-
ландском заповеднике у гнезд орланов, показывает, что в период
размножения он добывает ондатру, водяную крысу, уток, щуку, ха-
риуса и т. д. Тетеревиные птицы, хотя и не относятся к основным
кормам, попадаются нередко (встречено 9 глухарей, 5 белых куро-
паток и 1 тетерев). Ранней весной большое значение имеет для ор-
лана падаль.
На верхней Печоре (Донауров, 1948) в гнездовой период орлан
питается в основном белками, утками и рыбой, в первую очередь
хариусом; реже берет глухаря и других тетеревиных (преимущест-
венно» взрослых особей). Осенью, перед отлетом, главная его добы-
ча— отнерестившаяся семга.
В Ильменском) заповеднике (Ушков, 1949) в питании орлана
первое место заним'ает рыба, особенно щука, затем водяная крыса
и падаль, гораздо' реже — водоплавающие птицы; остатков куриных
в его добыче не было обнаружено.
197
Ястреб-перепелятник—настоящий орнитофаг, но питается прей,
мущественно певчими птицами. Однажды <в сентябре 1932 г. в Лап-
ландском заповеднике перепелятник был спугнут у выводка рябчи-
ков, которых он, очевидно, караулил. На Печоре перепелятник не-
редко ловит птенцов рябчиков, а один раз в добыче его отмечен глу.
харенок (Донауров, 194®). В большом1 материале по питанию пере-
пелятника в Ильменском заповеднике оказался только один взрос-
лый рябчик (Уников, 1949). В таежной полосе перепелятник — пере-
летная птица.
В пище чеглока два главных элемента—певчие птицы и круп-
ные насекомые: стрекозы, жуки и другие. В Печоро-Илычском запо-
веднике из куриных он добывает только* 'молодых рябчиков. Чеглок
поздно прилетает и рано улетает. Заметного* вреда боровой дичи он
не может принести.
Дербник в Лапландском заповеднике — обыкновенная гнездя-
щаяся птица, в Печоро-Илычском заповеднике преимущественно
пролетная (гнездится только па Уральских горах|). Питание его не-
достаточно изучено, но случаев поедания им тетеревиных птиц не
установлено. В добыче пустельги, коршуна, скопы и мелких сов те-
теревиные также не найдены.
По наблюдениям Лапландского, Кандалакшского, Омского,
Астраханского и некоторых других заповедников известно, что во-
рон и ворона в гнездовой период уничтожают яйца птиц. По нашим
наблюдениям в Лапландском заповеднике (Семенов-Тяп-Шанский,
1060), ворон чаще всего разоряет гнезда крохалей, вероятно высле-
живая утку при ее уходе на кормежку и возвращении на гнездо.
Крохали устраивают гнезда по берегам водоемов, непосредственно
у воды, а потому находить их довольно легко. Гнезда тетеревиных
не привязаны к определенным ориентирам, и разыскивать их гораз-
до труднее. В Лапландском и Печоро-Илычском заповедниках не
было случаев разорения гнезд тетеревиных птиц вороном или воро-
ной. По наблюдениям В. П. Теплова и И. С. Турова (1956) в Окском
заповеднике, 70% яиц, уничтоженных серой вороной, принадлежали
уткам и только* 2% — куриным птицам (тетерев).
Все хищные птицы обрабатывают пойманных ими птиц сходны-
ми приемами — выщипывают часть перьев, а потом обклевывают
мясо вместе с кожей по кусочкам. На месте остаются перья и ске-
лет или части его с костями конечностей, соединенными связками.
Тонкие кости, как грудина и таз, нередко бывают обклеваны по
краям. Тут же можно найти кишечник, содержимое желудка и зоба
жертвы, выброшенный струей белый помет хищника, а иногда и его
погадку. Совы в первую очередь съедают голову жертвы, дневные
хищники, по-видимому, ее оставляют. Остатки часто, особенно зи-
мой, доедают лиса или куница. При таком комбинированном исполь-
зовании жертвы трудно бывает определить, кем она была добыта,
а в тех случаях, когда на месте остались только перья, задача ока-
зывается и неразрешимой.
200
Хищные звери. В Лапландском заповеднике питание хищных
млекопитающих было подробно изучено А. А. Касимовичем (горно-
стай, лисица, куница и росомаха — Касимович, 1948, 1948-а, б, в;
медведь — Касимович и Семенов-Тян-Шанский, 1951; выдра —
Владимирская, Лебедев и Касимович, 1953). Печоро-Илычским за-
поведником опубликованы данные о питании лисы (Теплова, 1947),
росомахи (Теплов, 1955), медведя (Теплов, 1953|) и выдры (Теплов,
1953-а). Питание куницы и горностая изучалось в сезон промысло-
вой охоты по желудкам, полученным от охотников, а летом — по
экскремента1м; для Летописи природы заповедника сводки составля-
лись каждый год отдельно. По горностаю мною испльзованы
здесь данные Летописи за все годы, по кунице — только с зимы
1950-51 г. Табл. 60 и 61 составлены по образцу помещенных выше
таблиц, характеризующих питание хищных птиц; с этой целью ори-
гинальные материалы авторов пришлось в некоторых случаях пере-
работать.
Таблица 60
Характеристика питания хищных зверей Лапландского заповедника
________________(68° с. ш.; по Насимовичу, в %)_______________
Горностай Куница Ро- сома- ха Лиса Мед- ведь
лето зима лето зима лето зима лето
зима
Общее число данных 137 133 485 214 113 358 433 150
Млекопитающие . . . Копытные (падаль)) 1 2 52 2 9 10
Хищные ..... — — — — 3 0 + 1 —
Грызуны крупные . 1 1 1 8 4 7 4 —
Полевки и землеройки 37 90 54 36 15 48 56 7
Птицы Тетеревиные . . 5 3 18 19 8 21 0,5
Прочие 4 1 6 6 — 6 2 2
Яйца птиц .... 1 — 2 7 1 2 1 0,5
Рептилии и амфибии . 3 — 5 1 — 2 — —
Рыбы 2 1 0 + 1 1 1 1 о+
Насекомые . . . - 15 1 12 6 — 9 0 + 25
Ягоды 37 1 16 15 1 15 3 35
Травянистые части раст. — — — — — — — 20
Отбросы . ... — — — — 4 — 2 —
201
Таблица 61
Характеристика питания хищных зверей Печоро-Илычского заповедника
(62° с. ш.: по Теплову, Тепловой и рукописным фондам, в %j
Горно- стай Куница Росо- маха Лиса Мед- ведь
зима лето зима зима лето зима лето
Общее число данных 546 58 577 810 392 1031 400
Млекопитающие . . .
Копытные (падаль) 1 — 1 75 1 3 12
Хищные — — — 2 — —
Грызуны крупные . 5 5 15 10 21 45 i 4
Полевки и землеройки 72 47 37 2 33 15
Птицы
Тетеревиные . . . 8 5 15 6 14 24 1 1
Прочие птицы . . . 10 6 6 1 И 6
Яйца птиц .... — 2 4 — — * — * —
Рептилии и амфибии . 2 — 2 — * * —
Рыбы 1 —. — о+ * —
Насекомые .... 1/2 11 1 — 14 0 + 43
Растительные объекты .
Кедровый орех . . Ягоды 4 19 9 6 ) 2 6 6 !
Травян. части . . . 0,5 — 1 — — 23
Отбросы — — 3 3 — — —
* В цитируемой работе Тепловой и Теплова (1947) эти корма не выделены;
в более старой работе Тепловой (1947), где питание рассматривается без разде-
ления по сезонам, встречаемость яиц определена в 2,5%, рептилий и амфибий —
в 1%, рыб — в 0'+ % (от числа данных)’.
Лиса. В зимнем питании лисы главную роль играют грызуны
(в Лапландском заповеднике — полевки, в Печоро-Илычском—зай-
цы и затем полевки), а группа тетеревиных занимает второе или
третье место. Летом боровая дичь меньше страдает от лисы. В
зависимости от численности тетеревиных, меняется по годам и зна-
чение их в добыче лисы. Так, например, в Лапландском заповед-
нике летом 1938 г. они были встречены в 33% всех данных по пи-
танию лисицы, а в неурожайном на боровую дичь 1939 г. — только
в 6%.
Роль отдельных видов птиц в питании лисы определяется И5
численным соотношением в природе. В Лапландском заповеднике
202
на первом месте стоит глухарь, на втором — белая куропатка, ре-
же попадаются тетерев и рябчик; в Печоро-Илычском заповеднике
большое значение имеют рябчик и глухарь, реже — тетерев и белая
куропатка.
По мнению А. А. Касимовича (1948-а, стр. 55), «большинство
тетеревиных лисица добывает не самостоятельно, а в виде остат-
ков на следах лесной куницы». На Верхней Печоре замечено, что
лисица иногда поедает остатки боровой дичи, убитой ястребом-те-
теревятником (Теплова, 1947, стр. 248). По мнению того же автора,
лиса питается боровой дичью больше всего в глубокоснежный
период, с декабря по февраль, когда остатки куриных птиц най-
дены в 24% всех данных по ее питанию. В это время тетеревиные
проводят значительную часть времени в снегу, а рыхлый снежный
покров облегчает охоту тем, что заглушает шаги зверя и дает ему
возможность лучше чуять сквозь снег. 27 XI 1934 г. на моих глазах
охотничья лайка поймала таким способом тетерку, зарывшуюся в
рыхлый снег глубиной около 50 см. Нередко собаки ловят тетере-
виных (особенно белых куропаток) и в бесснежный период.
Поршни боровой дичи летают бойко только по внешнему впечат-
лению: в действительности они быстро утомляются и после одного-
двух перелетов уже не могут подняться, и тогда их можно поймать
руками — важно только «наглядеть» затаившегося птенца. В рай-
оне Лапландского заповедника, в период с 6 VII по 20 VIII отме-
чено 5 случаев поимки молодых куропаток собаками. Три раза я
был свидетелем, как лайка ловила вполне взрослых здоровых птип.
1 X 1949 г. в вечерних сумерках перелетавший куропач сел в за-
росль карликовой березки на берегу озера в нескольких шагах
от собаки и был схвачен ею прежде, чем успел взлететь. 7 X 1949 г.
в 16 час. лайка наскочила на взрослого глухаря (4+года), кормив-
шегося на черничнике в бору и успела поймать его на взлете. В
районе Печоро-Илычского заповедника 11 IX 1955 г. в 11 час. та
же лайка опять поймала взрослого глухаря (2+ года), чему спо-
собствовал мелкий еловый подрост в бору, затруднявший взлет
птицы. У обоих глухарей при поимке был вырван весь хвост и зна-
чительная часть оперения спины. Я остановился на этих случаях
потому, что они помогают уяснить приемы лисы на охоте. В янва-
ре 1939 г. А. А. Насимович наблюдал по следам на снегу, как глу-
харь опустился на косогор, неподалеку от лисицы, которая в два-
три прыжка настигла его, схватила и скатилась вместе с ним по
склону под небольшую елочку: тут глухарю удалось вырваться и
па месте осталось только немного крови и вырванных перьев (На-
симович, 1948-а, стр. 63). 2 V 1941 г. в районе Лапландского заповед-
ника был добыт сидевший под елкой глухарь с вырванным хвостом:
очевидно, незадолго до того он подвергся нападению лисицы.
Для лисы характерно, что она уносит свою добычу от места
поимки целиком и съедает ее в другом месте, иногда на льду реки
или озера (куропатка 24 IV 1935 г.; глухарка 23 II 1935 г.; глухарь
14 XI 1937 г; глухарка 8 V 1938 г.; глухарь 25 И 1940 г.). После
203
лисы от куропатки и глухаря обычно остаются только содержимое
зоба и мускульного желудка и перья, причем у маховых очини
бывают обгрызены. Иногда по следам можно убедиться, что лис^
выкопала из-под снега остатки чужой добычи и доела ее (28 VI[
1934 г.; 8 II 1935 г.). Она охотно ходит по следам других хищиц,
ков—куниц, росомах и лис—отчасти в поисках объедков, а отчасти
и потому, что ей трудно передвигаться по рыхлому снегу и всякие
чужой след облегчает ей путь. В тайге с ее высоким и рыхлым
снежным покровом лисам трудно сводить концы с концами. В годы
«неурожая» полевок и боровой дичи они голодают, подъедаю?
отбросы на свалках у окраин населенных пунктов и становятся
легкой добычей охотников-капканщиков (в Мурманской области —
зимы 1935-36, 1939-40, 1947-48, 1951-52 гг.).
Куница. В зимнем рационе куницы, как в Лапландском, так и
в Печоро-Илычском заповеднике, боровая дичь занимает второе
место (после группы мелких млекопитающих). Летом тетеревиные
птицы встречаются в пище куницы реже, чем зимой: в Лапланд-
ском заповеднике — в 6 раз, в Печоро-Илычском — в 3 раза.
Об охоте куницы зимой А. А. Насимович (1948-6) пишет: «В
противоположность другим хищникам — росомахе, лисице и горно-
стаю— куница редко ходит по чужим следам, охотясь самостоя-
тельно. Куница не «нахлебник», а настоящий «добытчик».
Различие в манере охоты куницы и лисы объясняется их приспо-
собленностью к передвижению по снегу. След куницы по площади
приблизительно равен следу лисы: по Насимовичу (1948-а), площадь
следа всех четырех лап у лисы равна 148 кв. см; а у самца куни-
цы —140 кв. см. Между тем даже крупная лапландская куница
(вес самца около 1,68 кг, самки— 1,03 кг) весит втрое меньше лисы
(в Лапландском заповеднике вес ее около 4,3 кг). Поэтому куница
имеет возможность бегать по рыхлому снегу, а лисица в нем вяз-
нет и старается облегчить путь, проходя по льду озер и рек, по
зимним дорогам, лыжням, а также по следам других животных.
Хотя процент встреч тетеревиных птиц в зимнем питании обоих
хищников приблизительно одинаков, куница, как активный хищ-
ник, для них опаснее. Случаев голодовки всей популяции у куниц,
по-видимому, не бывает. В Печоро-Илычском заповеднике попада-
лись отдельные истощенные или даже погибшие от голода особи
(например, в 1951 г. две—10 II и 10 V), но большинство добытых
куниц всю зиму сохраняет достаточную упитанность. Из приведен-
ных фактов можно заключить, что лиса добывает боровую дичь
практически только в некоторые зимы с благоприятными кормовы-
ми условиями, а куница — во все зимы, в том числе и тогда, когда
в лесу очень мало птицы.
В Лапландском заповеднике и, особенно, в Печоро-Илычском»
питание куницы изучалось систематически из года в год. В Лап-
ландском заповеднике желудки и экскременты куниц с 1938 — по
1941 г. исследованы А. А. Насимовичем (1948-6), в 1948—1951 гг.
Т. В. Кошкиной, в 1958 г. — мною (картотека заповедника); в Пе-
204
чоро-ИпычскоМ' заповеднике с 1937 по 1951 г. П. Б. Юрвенсоном,
В. П. Тепловым и Е. Н. Тепловой, после 1952 г. А. А. Меженным,
Г. А. Предтеченским, Ю. П. Язаном, В. П. Поярковым и мною
(Летопись природы и картотека заповедника). Имея в руках такой
обширный материал по питанию куницы, целесообразно рассмот-
реть его подробнее, используя как основу для понимания отношений
хищника и жертвы в природе.
Куница достаточно сильна, чтобы справиться с любой птицей
из боровой дичи, включая и взрослых глухарей самцов; поэтому
состав ее жертв приблизительно соответствует отношению отдель-
ных видов тетеревиных в стациях куницы. В изученных нами мате-
риалах эти отношения оказались следующими (табл. 62, в %).
Таблица 62
Заповедник Глухарь Тетерев Б. куро- патка ^Рябчик Число случаев
Лапландский 81 1 16 2 135
Печоро-Илычский .... 17 13 2 68 270
Диаграммы рис. 74 и 75 наглядно показывают изменения соста-
ва пищи 'Куницы по годам, в зависимости от 'конъюнктуры. В обоих
заповедниках и летом и зимой основным кормом куницы являются
полевки (и отчасти землеройки), причем «удельный вес» полевок
непосредственно зависит от численности их в соответствующем го-
ду. Поэтому коэффициент корреляции между показателем учета
полевок (при отлове на стационаре в августе) и встречаемостью
их в материалах по питанию куницы очень высок: летом, в Лап-
ландском заповеднике, г = +0,91 +0,06 (за 8 лет), зимой, в Печо-
ро-Илычском, г =+0,86+ 0,10 (7 лет). Коэффициент корреляции
между данными учета тетеревиных птиц и встречаемостью их в
зимнем питании куницы на Печоре оказался гораздо ниже,
г = +0,43 + 0,31*. Если взять только рябчика (учет проводился
в начале октября с помощью манка) г = +0,38 + 0,32. Не лучшее
соответствие получается между учетом белки в ноябре и ее значе-
нием в питании куницы: г = + 0,19 + 0,37.
Отсюда можно заключить, что основной объект охоты куницы в
тайге — полевки, а остальных животных она добывает только по-
путно. При недостатке полевок кунице приходится пробегать боль-
шее пространство, что увеличивает шансы не только поимки поле-
вок, но и встречи с боровой дичыо. Подтверждение этому выводу
можно найти и в наших диаграммах: на них хорошо видно, что
амплитуда показателя учета полевок гораздо больше, чем ампли-
туда их значения в питании куницы. Потверждается он и статисти-
ческим анализом питания: корреляция между процентом полевок
* Определение коэффициента корреляции для летнего питания боровой дичью
не имеет смысла, так как в это время года она является случайной и редкой
добычей (г = — 0,46 ± 0,28 за 8 лет).
205
в рационе кунины, с одной стороны, и процентом тетеревиных
том же рационе — с другой, дают коэффициент г = — 0,81 + 0,Ifj.
Корреляция между процентом полевок и процентом белок в пишл
куницы дает коэффициент г = —0,74 + 0,17. Оба коэффициента
велики и имеют отрицательный знак (чем меньше полевок, тем
больше тетеревиных птиц и белок). Только коэффициент корреля-
ции между процентом полевок и процентом землероек в питании
куниц оказался положительным: г = +0,43+ 0,31 —доказательст-
во того, что землеройки служат не заменой полевок при недостатке
последних, а скорее «приловом» при мышковании. Судя по рис. 74,
Рис. 74. Состав пищи куницы в Лапландском заповеднике, истом;
черные столбики — показатели учета полевок и землероек (основная
пища куницы)
летом в Лапландском заповеднике, при недостатке полевок, куница
возмещает их, главным образом, ягодами и насекомыми (осы,
шмели и их гнезда, отчасти жуки и др.). Вообще ягоды — обяза-
тельная составная часть рациона куницы: полное отсутствие их в
пище куницы летом 1941 г. объясняется тем, что весь исследован*
ный материал был собран в самом начале лета, когда ягод еще не
было. Связь между процентом полевок и процентом насекомых в
питаний куницы Лапландского заповедника выражается коэффи-
циентом г = — 0,92 + 0,05, а между встречаемостью полевок, с од-
ной стороны, и ягод — с другой, г = —0,84 + 0,10.
В Лапландском заповеднике и летом и зимой растительные
корма куницы ограничиваются почти одними ягодами — вороникой,
черникой, голубикой, рябиной и другими. Зимой потребление их
сокращается по мере увеличения высоты снежного покрова. В Пе-
чоро-Илычском заповеднике в желудках куницы находят, главным
образом, кедровые орешки и плоды рябины: и те и другие отно-
сятся к излюбленным кормам, доступным только в годы урожая.
Поэтому между встречаемостью полевок и растительных кормов
здесь нет связи (г = +0,15 + 0,37).
206
Все приведенные материалы показывают, что основным факто-
ром, определяющим состав пищи куницы, является урожай главных
кормов. При обилии основного животного корма (полевок) суточ-
ный пробег куницы сокращается, вследствие чего вероятность
встречи ею на охоте других животных снижается. Наоборот, при
снеурожае» полевок куница вынуждена увеличить длину охотничь-
его поиска и компенсировать недостаток полевок другими кормами.
В таких случаях, в таежной полосе на первое место могут выйти
тетеревиные птицы, как это и наблюдалось в Печоро-Илычском
заповеднике в зимы 1941-42, 1950-51 и 1953-54 гг. В другие годы
главную роль играет белка (зимы 1938-39, 1940-41, шесть зим с 1942
по 1948 г. и 1954-55 г.).
На значении птичьих яиц в питании куницы необходимо остано-
виться. Кроме куницы, яйца птиц поедают и другие хищники (лиса,
росомаха, горностай и медведь; см. табл. 60 и 61), но в питании
куницы они играют большую роль. Интересно, что в зимних мате-
риалах яйца встречаются чаще, чем в летних; А. А. Насимович
(1948-6) объясняет это тем, что летом хищник, выпив яйцо, бросает
скорлупу на месте, а зимой ему достаются испорченные, засохшие
яйца, которые приходится есть вместе с оболочками и скорлупой.
По мере увеличения высоты снежного покрова куница перестает
добывать яйца. Распространено мнение, что куница достает из-под
снега яйца, припрятанные ею же или другими хищниками с лета.
Нам неизвестно, насколько это объяснение обосновано фактами:
по крайней мере, ни Насимович, ни я не находили спрятанных яиц.
Обычно хищный зверек, найдя гнездо, выпивает яйца тут же, на
месте. 6 VI 1936 г. в Лапландском заповеднике было найдено гнез-
до глухарки со скорлупами 4 яиц, выпитых, вероятно, куницей.
8 VII 1951 г. на одном из островов Чунозера было разорено гнездо
крохаля с 8 яйцами; через несколько дней скорлупы 4 выпитых яиц
207
были найдены запрятанными под елками, валунами и другие
укрытиями на расстоянии от 1,5 до 5 м от гнезда. Судя по следам
зубов на скорлупе, они были выпиты горностаем. 1 VI 1956 г. в
Печоро-Илычском заповеднике из гнезда глухарки, в котором на-
кануне я установил самописец для записи режима насиживания,
бесследно исчезла вся кладка из 7 яиц: очевидно, они были ута-
щены целиком. На следующий день на песчаном берегу реки в
160 м от гнезда я нашел свежий след куницы, что дает основание
предполагать, что яйца были унесены ею и где-то съедены или
спрятаны. Доказательств устройства куницей складов мы не имеем.
Гораздо больше оснований считать, что зимой куница достает
яйца, оставшиеся в гнездах. В Лапландском заповеднике зимой
1938-39 г. А. А. Насимович нашел яйца птиц в 3% данных по
питанию куницы, а в следующие две зимы — в 14 и 13% данных
Эту разницу он объясняет массовой гибелью кладок тетеревиных
во время июньских холодов 1939 и 1940 гг. (Насимович, 1948-6.
стр. 95). В Печоро-Илычском заповеднике за 20 лет В. П. Тепло-
вым и нами собран уникальный материал по питанию куницы, что
дало возможность составить интересную диаграмму (рис. 76),
наглядно показывающую, что значение яиц в зимнем пи-
тании куницы обратно пропорционально кол^
честву молодняка в популяции глухаря. Эта зако-
номерность наблюдается как в долине Печоры, так и в окрестностях
оз. Имандры. Ниже мы увидим, что соотношение молодых и старых
глухарей в популяции в сезон охоты зависит от условий погоды
в период инкубации. Таким образом, статистический метод под-
тверждает, что основным источником яиц для куницы (а равно и
для лисицы и других хищников) служат не заранее запасенные
ею яйца, а найденные в течение зимы брошенные кладки или
отдельные испорченные яйца — «болтуны».
В Печоро-Илычском заповеднике однажды мы нашли такую
брошенную кладку. 31 V 1952 г. близ Якши подняли глухарку,
по-видимому с гнезда, которое, однако, не было обнаружено; 24 VI
гнездо было разыскано, и в нем оказались 4 совершенно свежих
яйца, сверху выцветших до полного побеления. Яйца пролежали
в лесу, никем не тронутые, почти месяц: очевидно, они могли сохра-
ниться так и до зимы. Весной 1953 г. одна глухарка в течение
31 дня насиживала яйца с погибшими эмбрионами: если бы -мы их
не забрали — рано или поздно их нашел бы какой-нибудь хищник
Летом хищники лучше обеспечены кормами и, вероятно, не так
подвижны, как зимой, когда они вынуждены тщательнее обследо-
вать свой охотничий участок. Куница находит яйца, очевидно, по
запаху, проникающему сквозь снег. На поверхности снега в пас-
мурную погоду температура близка к температуре воздуха, а в
ясные ночи она бывает на несколько градусов ниже, между тем
как под снегом температура всю зиму остается немного ниже нуля.
Таким образом на расстоянии 0,5—1 м существует большой темпе-
ратурный градиент, способствующий циркуляции воздуха в направ-
208
лении перпендикулярном слою снега. Человек хорошо слышит про-
никающий сквозь снег запах болота, хотя обоняние у него развито
слабо; северный олень чутьем разыскивает под снегом ягель; таким
же образом белка находит под снегом и лесной подстилкой олений
трюфель.
Из диаграммы рис. 76 видно, что встречаемость яиц в зимнем
питании куницы увеличивается при недостатке полевок; следова-
тельно, нахождение яиц подчинено тому же правилу обратной зави-
ZPJ7
19*2
1947 195?
80%
бо-
со-
го-
о-
/О %
.г -
о-
ьо-
60-
80'
Рис. 76. Значение яиц в зимнем рационе куницы, в зависимости от
эмбриональной смертности у глухаря и от доступности полевок.
Сплошная линия — Печоро-Илычский, прерывистая — Лапландский
заповедник
симости от урожая полевок, как добывание тетеревиных птиц и
других дополнительных кормов, что мы объясняем увеличением
длины охотничьего поиска куницы в год неурожая полевок. Коэф-
фициент корреляции между встречаемостью полевок, с одной сто-
роны, и яиц — с другой, в зимнем питании куницы за 13 лет (1944—
1956 гг.) оказался равен —0,68 + 0,15. Эта закономерность не
прослеживается в период 1937—1943 гг., что объясняется, по-види-
мому, тем, что в те годы главным кормом куницы была белка.
В пище лисы яйца имеют меньшее значение, но изменения их
«удельного веса» по годам носят тот же характер, как и в пище
куницы. Насимович (1948-а) отмечает, что яйца поедались в боль-
шем количестве при нехватке основных кормов.
Охотничьи приемы куницы хорошо описаны А. А. Насимовичем.
Он отмечает, что даже к северу от полярного круга куница дея-
14-3037
209
тельна преимущественно в ночные часы. Он пишет: «Отыскивая
добычу, хищник больше всего руководствуется чутьем и слухом.
Не раз мы наблюдали по следам, что куница, почуяв запах полевки
или заслышав ее шорох, круто сворачивала и делала неожиданный
прыжок в сторону жертвы или принималась усердно разрывать
снег». Боровую дичь -куница ловит преимущественно, когда птида
спит в снежной лунке, обнаруживая ее, как полагает А. А. Наси-
мович, чутьем, а возможно, и на слух. Мы неоднократно находили
места .поимки куропаток <и> глухарей в лунках. Случай пеудачн&го
нападения куницы на глухарей представлен на схеме рис 77:
Рис. 77. Схема нападения куницы на двух глухарей, ночевавших
в лунках
по следам видно, что куница приблизилась к полянке, где ночевали
в снегу два глухаря, сначала с северо-запада, потопталась под
елками, повернула обратно, и, описав дугу, приблизилась к лункам
с юга, все увеличивая свои прыжки. В этот момент глухарь выр-
вался из снега ей навстречу, вслед за ним вылетел в том же направ-
лении и второй, куница бросилась за ним, но было уже поздно;
тогда, заглянув в лунку, она побежала в первоначальном направ-
лении, на юго-восток.
Весной куница нередко посещает глухариные тока. 11 V 1933 г.
наблюдатель Лапландского заповедника Л. Г. Кокорин, по остав-
шимся на снегу следам, определил, что куница схватила глухаря
на дереве и упала на снег вместе с ним. За один раз опа съела
мускулы на одном боку груди и с одной ноги, а также объела жир-
ные части — хвост и часть кожи.
Наевшись, куница обычно растаскивает остатки, иногда на
большое расстояние, и прячет их под валежинами, в щели камней
210
и по другим укромным местам. Впоследствии куница разыскивает
и доедает припрятанное; однако нередко ее опережают другие хищ-
ники— лиса, росомаха и горностай.
Характерные признаки птицы, съеденной куницей — целость всех
крупных костей (коракоиды, плечевые и бедренные); грудина и таз
обгрызаются только по краям; очины маховых перьев иногда по-
грызены, а иногда и целы; остатки растаскиваются от места поимки
и на них можно найти помет куницы.
Соболь заселяет восточную часть прежней (до 1951 г.) террито-
рии Печоро-Илычского заповедника, но и там, по-видимому, пре-
обладают его гибриды с куницей. В Коми АССР соболь давно уже
находится под полной охраной, но кидусы (гибриды) принимаются
в заготовки, а следовательно, и тушки их поступают в заповедник
от охотников. Материалы по питанию кидусов и куниц обрабаты-
вались совместно.
В северо-западной части Свердловской области питание соболя
исследовано В. А. Залекером и Н. Б. Полузадовым (1955): оно ока-
залось очень сходным с питанием куницы по нашим данным за те
же годы, с той лишь разницей, что соболь в большей мере питался
полевками (это различие особенно заметно зимой 1952-53 г.). Зимой
1951-52 г. тетеревиные птицы найдены в 19% исследованных собо-
линых желудков, а яйца птиц — в 3%; в 1952-53 г. они встречены
соответственно в 3 и 1 % желудков.
В. В. Раевский (1947), описывая питание соболя в Кондо-Сое-
винскюм заповеднике, относит группу тетеревиных птиц к основ-
ным зимним кормам соболя и указывает, что «в зимнее время,
когда пернатых хищников в заповеднике почти нет, именно соболь
представляет основного врага куриных» (там же, стр. 126). В снеж-
ный период года тетеревиные найдены им в 11% данных, летом —
в 8 ,%; яйца птиц — зимой в 3.%, летом — в 4 %.
Горностай питается главным образом полевками и может с пол-
ным правом считаться стенофагом. В зимнем питании горностая
Лапландского заповедника А. А. Насимович (1948) встретил тете-
ревиных в 5% данных: при разборе 133 зимних экскрементов, же-
лудков и остатков еды глухарь встречен 3 раза, белая куропатка
4 раза и прочие птицы 1 раз. Автор сопровождает эти данные сле-
дующим комментарием: «Как показали зимние тропления горно-
стая, почти все встречи в его экскрементах остатков крупных птиц
следует отнести за счет поедания хищником объедков чужой добы-
чи— куницы или лисы. Лично нам ни разу не пришлось наблюдать
следы успешной охоты горностая на боровую дичь. Изредка можно
было отметить лишь попытки скрадывать птиц, тщательный осмотр
лунок белых куропаток и выискивание птиц по свежим жировочным
следам» (там же, стр. 17).
В летнем питании горностая тетеревиные не обнаружены; яйца
их, напротив, встречены только летом (2 раза в 137 данных); одно
наблюдение, сделанное уже после опубликования работы Насимо-
вича, приведено выше.
14*
211
В Печоро-Илычском заповеднике за 13 зим исследовано 473
желудка и 73 экскремента горностая (картотека заповедника),
причем рябчик встречен 32 раза, тетерев 3 раза и куропатка 1 раз,
что в сумме составляет 8% от всех найденных объектов. В зимней
пище горностая яйца не были встречены. Хотя в Печоро-Илычском
заповеднике горностая специально не тропили, в картотеке есть
отдельные наблюдения за приемами горностая на охоте. 3 XII
1938 г. наблюдатель охраны Е. Т. Пашин видел горностая, раста-
скивавшего задавленную им глухарку. Подобные единичные наблю-
дения не дают оснований причислять горностая к активным врагам
тетеревиных птиц.
Росомаха. Как в Лапландском заповеднике, так и в Печоро.
Илычском изучено только зимнее питание росомахи (Насимович,
1948-в; Теплов, 1955). Этот хищник в основном питается дикими
копытными (лось и северный олень); в Лапландском заповедники
тетеревиные птицы стоят на втором месте (19% от всех встречен-
ных объектов); на Печоре второе место занимает заяц-беляк (9%),
а тетеревиные только третье (6%). Среди птиц росомаха чаше
добывает глухаря (15 встреч на 113 данных в Лапландском запо-
веднике и 28 из 749 в Печоро-Илычском), затем белую куропатку
(в Лапландском заповеднике 11 встреч) и рябчика (в Печоро-
Илычском заповеднике 18 встреч рябчиков и куропаток вместе
взятых). Один раз в пище росомахи встречены яйца: по мнению
А. А. Насимовича, росомаха выкопала из-под снега погибшую
прошлым летом кладку аналогично тому, как это делают лисица
и куница.
Относительно способа добывания птиц В. П. Теплов сообщает
лаконично: «Тетеревиных птиц она ловит во время пребывания их
в снегу; нередко посещает глухариные и тетеревиные тока, скра-
дывая токующих на земле самцов». А. А. Насимович, пройдя около
100 км по следам росомах, пришел к другому заключению: «Сама
росомаха зимой обычно не ловит птиц, а довольствуется остатками
добычи других хищников, которые она подбирает либо прямо на
следах хищников, либо утаскивает из мест, где они бывают припря-
таны».
20 I 1957 г., в Печоро-Илычском заповеднике, пройдя 2 км по
следу двух росомах, мы нашли место, где они выкопали из-под
60-сантиметрового слоя снега остатки глухарки, но не стали их есть:
по-видимому, они недавно кормились на падали и были сыты. В
пушных заготовках Мурманской области одна росомаха приходит-
ся, в среднем, на 11 куниц, а в Коми АССР — на 16 куниц (Произ-
водительные силы Коми АССР, 1953). Часть росомах добывается
в тундровой зоне, где куница не встречается; поэтому в тайге соот-
ношение между этими видами изменяется еще больше в пользу
куницы. Так, например, в Троицко-Печорском районе, на террито-
рии которого находится Печор о-Илычский заповедник, одна зап>
товленная росомаха приходится на 30 куниц. Росомаха приблизи-
тельно в 8 раз крупнее куницы. Перемножив соответствующие
212
показатели, нетрудно подсчитать, что на Верхней Печоре росомахи
должны съесть за зиму почти в 10 раз меньше тетеревиных птиц,
чем куницы. В Лапландском заповеднике росомаха относительно
многочисленнее и в большей мере питается птицами: но тут сле-
дует иметь в виду, что значительную часть боровой дичи она не
ловит сама, а подъедает за другими хищниками.
Медведь. В Лапландском заповеднике, по А. А. Насимовичу
и О. И. Семенову-Тян-Шанскому (1951), встречи птиц в пище мед-
ведя составили около 3% всех данных: в их числе отмечены птенцы
крупных хищных птиц (орлан, беркут) на взлете, тетеревиные, а
также яйца птиц. Встреча в экскрементах остатков яиц показывает,
что медведи разоряют птичьи гнезда. Замечено, что весной медведи
нередко посещают глухариные тока: предполагается, что им удает-
ся иногда поймать токующего на земле петуха. В Печоро-Илычском
заповеднике, по Е. Н. Тепловой (1957), птицы составляют 1,2% всех
данных по питанию медведя; обнаружены глухарь, певчие птицы,
а также яйца тетеревиных.
В списках кормов медведя тетеревиные птицы и их яйца зани-
мают самое последнее место; как враг тетеревиных он не имеет
значения.
Выдра. Питание выдры детально изучено в Лапландском и Пе-
чоро-Илычском заповедниках. Птицы занимают довольно большое
место в ее рационе, -но ей не приходится ловить боровую дичь. В
Лапландском заповеднике в 414 исследованных экскрементах и
остатках ее еды пти-цы встречены 28 раз, преимущественно летом
Н5 уток, 8 певчих птиц, остальные не определены; Владимирская,
Лебедев и Насимович, 1953). В исследованном1 'Материале Печоро-
Илычского заповедника (1530 данных) обнаружено 159 уток, 11 ку-
ликов, 7 оляпок и 1 белая куропатка (0,06%; Теплов, 1953-а).
Волк в обоих заповедниках встречался редко и нерегулярно,
почему питание его не могло быть изучено.
Рысь в Лапландском заповеднике не найдена, а в Печоро-Илыч-
ском она редка и даже следы ее встречаются не каждую зиму. По
литературным данным (Дулькейт, 1953; Кончиц, 1937; Новиков,
1956), рысь, в отличие от росомахи, питается почти исключительно
свежей добычей, чаще всего зайцами и другими грызунами, места-
ми— мелкими копытными, нередко ловит и птиц. В Центральном
лесном заповеднике (Калининская, Псковская и Смоленская обла-
сти) остатки птиц были встречены в 13% исследованных экскре-
ментов рыси. В Алтайском заповеднике в числе 56 остатков добычи
рыси был 1 глухарь, а среди 30 остатков пищи росомахи — 2 глу-
харя. По аналогии с другими хищниками, в питании которых на
севере боровая дичь имеет большее значение, чем на юге, можно
ожидать, что и рысь на северном пределе своего ареала чаще ловит
птиц; впрочем здесь она так редка, что с приносимым ею вредом
можно не считаться.
Статистика находок жертв хищников дает возможность оценить
величину отхода по годам, а также выяснить, какие хищники важ-
213
нее — пернатые или четвероногие. В 1941 г. А. А. Насимович обра-
ботал в этом плане материал картотек Лапландского заповедника
и любезно передал его в наше распоряжение; нами сделан допол-
нительный подсчет за 1948—1951 гг. В каждой колонке табл. 63 пер-
вое число означает число находок за 1931—1941 гг., а второе за
1948—1951 гг.; проценты даны за все годы вместе.
Таблица 63
Остатки тетеревиных птиц, погибших от хищников в Лапландском заповеднике
за 1931—1941 и 1948—1951 гг.
Виды жертв Хищп. звери Хищн. птицы Всего* Соотно- шение в отстре- лах в %
абс. В % абс. В % абс. В %
Глухарь ...... 68+ 3 52 43+ 8 37 Ш + 15 40 47
Тетерев 3+ 6 6 7+ 7 10 10+17 8 3
Рябчик 4+ 1 4 24- 0 1 6+1 2 14
Белая куропатка . . 40+11 37 57+11 49 97+53 47 31
Тундряная куропатка . 0+ 1 1 4+ 0 3 7+ 2 3 5
Всего . . . 115+22 100 113+26 100 231 + 88 100 100
* Вместе с остатками, для которых хищник не был определен.
Признаки, которыми мы пользовались при отнесении жертвы
на счет пернатых или четвероногих хищников, упомянуты выше.
Если на месте оставались одни перья, определение хищника обыч-
но было невозможно. А. А. Насимович отнес 51 % всех исследован-
ных им остатков жертв на счет хищных зверей и 49% на счет птиц.
В обработанных мною данных от зверей погибло 26%, от птиц 29%,
хищник не определен в 45% случаев. Отсюда следует, что потери
тетеревиных от птиц и от зверей почти одинаковы; при этом перна-
тые хищники добывают относительно больше куропаток (оба вида)
и тетеревов, то есть птиц сравнительно открытого ландшафта, а
хищные звери — больше глухарей и рябчиков. Известно, что птицы
в поисках пищи руководствуются зрением, а хищные млекопитаю-
щие— обонянием. Лиса и куница зимой ловят куропаток, когда они
отдыхают, зарывшись в снег, и в этих условиях цвет их оперения
не дает им никаких преимуществ перед другими птицами. Теоре-
тически в снежное полугодие куропатки должны нести меньше
потерь от хищных птиц, чем от млекопитающих. Между тем из
табл. 63 нельзя сделать такого заключения. В целом за год белые
куропатки имеют относительно большее значение в добыче хищных
птиц, чем темноокрашенные глухари, тетерева и рябчики. Зимний
наряд белых куропаток не предохраняет их от нападения хищных
птиц, но, вероятно, способствует некоторому снижению потерь от
214
них. Для обсуждения этого вопроса необходимо рассматривать
смертность за зимний и летний периоды отдельно; в настоящее
время мы не можем этого сделать.
В табл. 63 не указан половой состав жертв хищников, хотя в
отношении глухаря такой расчет был сделан: пернатые хищники
добыли 65% самцов и 35% самок; звери — 45% самцов и 55% са-
мок.
Большая часть зимних следов охоты хищников обнаруживается
не раньше, чем стает снег. Поэтому, для того чтобы рассмотреть
смертность тетеревиных от хищников по годам, нам поневоле при-
от плотности тетеревиных птиц (сплошная линия), в Лапландском
заповеднике
шлось соединить зимние остатки с остатками следующего лета. На
диаграмме рис. 78 столбиками показано число остатков глухарей и
белых куропаток в периоды 1938—1941 и 1948—1951 гг., а оплош-
ной линией — плотность тех же видов по данным учета в августе
(число голов на 1 кв. км). Как видно, смертность от хищников
сильно колеблется по годам и бывает велика только в первые
1—2 года после «пика» численности вида. Больше всего глухарей
погибло от хищников в 1939 г. — в год резкого сокращения числен-
ности всех тетеревиных, после хороших «урожаев» молодняка в
1936 и 1937 гг. Одновременно в 1939 г. наблюдалась глубокая де-
прессия полевок после «пика» 1937-38 года. Необычайно благо-
приятное положение с кормами с 1936 по 1938 г. способствовало
215
увеличению численности хищников, после чего в 1939 г. наступила
катастрофа, сопровождавшаяся усилением конкуренции между
ними. Характерно, что добыча, которую орел принес в 1939 г. своим
птенцам, на 18% состояла из хищников. Глухарь дольше сохранял
высокий уровень численности, чем мелкие представители тетере,
виных, ввиду чего число погибших в сезон 1938-39 г. оказалось
выше, чем год назад. У белых куропаток наибольшая смертность
была в сезоны 1937-38 г. и 1950-51 г., непосредственно после макси,
мумов численности 1937 и 1950 гг. Много куропаток погибло также
зимой 1934-35 г. В другие годы смертность от хищников была в
5—25 раз меньше, чем в зимы после «пика». /
Таким образом, хищники сильно снижают численность тетере-
виных птиц при большой плотности популяции, но не препятствуют
ее восстановлению после депрессии. Однако, ввиду того что мак-
симумы и минимумы смертности от хищников отстают от макси-
мумов и минимумов численности жертвы приблизительно на один
год, деятельность хищников не смягчает амплитуды колебаний чис-
ленности, а скорее даже усиливает ее.
Простые случаи отношений хищника и жертвы описаны и объяс-
нены уже давно (рысь и заяц — Сетоном, песец и лемминг, белая сова
и лемминг — Элтоном и др. авторами). В этих случаях изме-
нение «нагрузки» со стороны хищника объясняется изменением
численных соотношений популяций хищника и жертвы. При этом
рост популяции хищника может происходить не только вследствие
размножения ее при благоприятных условиях, но и за счет имми-
грации. Примером таких взаимоотношений, характерных для хищ-
ников стенофагов, может служить ястреб-тетеревятник. Хищники
эврифаги более гибки в выборе кормов и состав их пиши отражаег
кормовые условия данного сезона. Мы уже видели, что куница при
«неурожае» главного корма сравнительно легко компенсирует не-
хватку за счет других кормов. Хорошо приспособленная к зимним
условиям тайги, она обычно успешно переживает голодные зимы.
Куница опасна тем, что не снижает «нагрузку» на боровую дичь
даже в те зимы, когда запас ее очень мал (если в это время мало
полевок и белок). Другие эврифаги, как, например, лиса, в неблаго-
приятные зимы (бескормные или очень снежные) голодают, отко-
чевывают или гибнут, а следовательно, снижают «нагрузку» на
тетеревиных птиц такта же путем, как это происходит со стено-
фагами.
В заключение необходимо рассмотреть встречи травмированных
и мертвых птиц. Имея целью только пополнить наши представле-
ния о причинах смертности боровой дичи, мы не останавливаемся
на встречах подранков и жертв браконьерства.
Некоторые случаи объясняются скорее всего нападением хищ-
ника, почему-либо не смогшего затем воспользоваться своей
добычей.
Например, 27 V 1934 г. к берегу Чунозера волной прибпло ку-
ропатку с рваной раной на боку и небольшим кровоподтеком над
216
глазом: ее могла выронить хищная птица, если на нее напал другой
хищник. Утром 5 XI 1936 г. на Чунозере, в воде, среди образую-
щейся ледяной пленки нашли еще живого взрослого самца глухаря.
Ночь перед тем была ясная и тихая, температура на восходе солн-
ца— 8°, а следующие 8 дней были пасмурные и теплые. В один из
этих дней на том же месте нашли еще глухаря, уже мертвого,
прибитого волной к берегу. Ширина озера в этом месте достигает
3 нем1; поэтому возможно, что глухарь, перелетая через озеро пр«и
полном штиле, не видел поверхности воды и снизился настолько,
что попал в воду. Более вероятно, что глухаря сбила в воду хищ-
ная птица.
8 I 1950 г. был добыт глухарь с большой заживающей поверх-
ностной раной на плече; вокруг нее и на хвосте было несколько
пеньков перьев, которые могли вырасти только на месте вырванных
перьев. 9 X 1933 г. и 6 XI 1933 г. добыты две глухарки, у которых
ранее была вырвана часть рулевых и на месте их росли новые.
2 V 1941 г. добыт взрослый глухарь, у которого были вырваны все
рулевые и значительная часть оперения спины. Все эти птицы, оче-
видно, пострадали от нападения лисы или куницы.
Иногда на птицах обнаруживаются следы старых ранений ког-
тями хищников или дробью. 23 X 1936 г. добыта крайне истощен-
ная самка белой куропатки (вес 386 г) с задержавшейся зимней
линькой. Стенка «мускульного желудка была прободена, и- из нее
выдавался обросший живой тканью комок пищи. На грудных мыш-
цах были затвердевшие рубцы, а вся птица издавала тяжелый за-
пах. В кишечнике ее никаких паразитарных заболеваний не оказа-
лось (исследован на кафедре проф. В. А. Догеля в Лея. гос. уни-
верситете). У самки рябчика, добытой 16 VIII 1937 г., в области
зоба, под кожей, найдено образование размером с семя фасоли,
внутри которого оказались две незрелые ягоды черники и часть
колоска осоки. Очевидно, у рябчика был прорван зоб, причем часть
пищи вышла из ранки под кожу и инкапсулировалась. 26 IV 1938 г.
добыт самец белой куропатки с большим шрамом на грудных
мышцах и костной мозолью на киле. Вес птицы нормальный (626 г),
но в оперении было много пухоедов, а в кишечнике — цестод. 18 IV
1939 г. на Чуна-тундре добыт самец тундряной куропатки с комоч-
ком обросших тканью древесных побегов в стенке зоба.
21 VIII 1939 г. был добыт самец белой куропатки, подпустив-
ший на 5 шагов и не взлетевший. Птица была истощена (вес 489 г),
заражена цестодами и нематодами, но признаков ранения не имела;
ее состояние объясняется, вероятно, хронической болезнью.
Отдельные травмы могут быть следствием несчастного случая.
3 IX 1934 г. на р. Чуне в воде была найдена мертвая глухарка со
сломанной ногой. 2 XI 1933 г. я добыл глухарку с вытекшим левым
глазом. 7 IX 1932 г. добыта взрослая глухарка, у которой на одной
лапе не было 2-го и 3-го пальцев. У взрослой самки рябчика, до-
бытой 19 X 1950' г., отсутствовал второй палец левой лапы. Можно
предполагать, что в обоих случаях пальцы отпали в результате
217
сухой гангрены, развившейся от ранения или перетяжки пальца
птенца волокном. У самца куропатки 2 IV 1949 г. на левой лапе
были отломаны или повреждены три когтя, что могло произойти,
например, от попадания птицы в капкан.
Другого рода несчастный случай — удар летящей птицы о про-
вод или ветку.
Вовремя августовского учета дичи-в 1937 г. взлетевшая мо-
лодая тетерка на наших глазах ударилась о дерево и упала, после
чего была поймана руками. Возможно, таким же образом было сло-
мано крыло у молодой куропатки, пойманной 19 VIII 1935 г. около
поселка Хибины. 23 X I960 г. под телефонной линией был найден
труп разбившейся о провод белой куропатки. Утром 23 I 1937 г. во
дворе одного из домов по Чкаловскому проспекту в Ленинграде
(б. Геслеровский переулок) был найден разбившийся о провод ко-
сач. В этом случае трудно объяснить, каким образом косач залетел
так далеко вглубь большого города. Судя по содержимому зоба
(плоды рябины, сережки и почки Ольхи, листья андромеды), птица
была дикая и незадолго перед тем кормилась вне города.
В Лапландском заповеднике сравнительно редкая причина
травм—драки самцов на току: только один раз, 2(0 V 1951 г., мы
добыли взрослого косача с многочисленными заживающими ранками
и нехваткой перьев на затылке и в области зоба.
Все описанные выше случаи заимствованы из картотеки Лапланд-
ского заповедника (117'55 карточек на добытых и исследованных те-
теревиных птиц). Несколько подобных случаев обнаружено и в Пе-
чоро-Илычском заповеднике, при просмотре более 500 карточек за
1952—1957 гг.
9 XI 1952 г. добыта молодая глухарка, у которой были вырваны
все рулевые. У рябчика, добытого 11 V 1953 г., не хватало четырех
,рулевых, па месте которых росли новые перья. И IX 1956 г. добыта
молодая глухарка нормальной упитанности с одной ногой; на месте
другой ноги был затянувшийся шрам. 7 X 1956 г. добыт молодой
глухарь с сильно поврежденной ногой — вырвана часть мускулов и
нога высохла. 7 X 1953 г. у молодого рябчика отмечены шрамы на
3-м и 4-м пальцах ноги.
Эти травмы можно приписать с большей или меньшей вероятно-
стью нападению какого-нибудь хищника. В нескольких случаях
можно предполагать огнестрельные ранения.
13 X 1952 г. добыт молодой глухарь с фистулой зоба, проникаю-
щей и сквозь кожу. 27 IV 1953 г. добыт глухарь с хорошо сросшимся
переломом правой бедренной кости. 7 V 1954 г. добыт взрослый глу-
харь со сросшимся повреждением левой теменной кости. 11 X 1956 г
застрелен молодой глухарь с вытекшим глазом.
Следы драк на току замечены на двух взрослых глухарях. 17 V
1954 г. добыт довольно тощий глухарь трех лет с заживающей по-
верхностной рапой на подбородке, свежей на крыле и в двух местах
на спине. 6 V 1955 г. у трехлетнего глухаря, добытого па току, в обла-
сти плеч обнаружено несколько заживающих ссадин.
218
Учет характера трасвм .позволяет судить и о причинах элиминации
среди тетеревиных. Описанные здесь случаи мы отнесли за счет:
'1 . Неудачное нападение хищника . • • П случаев
2. Старые огнестрельные ранения ... в
3. Собственно несчастные случаи .... 9 г
4. Половой отбор....................'З ,,
5. Хроническая болезнь..............1
Рассмотренные в этой и предыдущей главах материалы приводят
нас к выводу, что дикие тетеревиные погибают главным! образом на-
сильственной смертью — в когтях хищников или от выстрела охот-
ника. Часть птиц, погибших от несчастных случайностей или от вы-
стрелов, также достается хищникам. Смертность от болезней, судя
по всем данным, очень «мала, а драки на токах имеют результатом
только отстранение слабейших самцов от участия в размножении,
без серьезных физических повреждений.
По своим крупным размерам и сравнительно! высокой численно-
сти тетеревиные птицы являются одним из главных источников жи-
вотной пиши для обитателей леса. Для некоторых хищников тетере-
виные птицы служат главным источником существования, а для
большинства — важным дополнительным кормом. В такой обстанов-
ке у глухаря, тетерева, куропатки или рябчика нет почти никаких
шансов умереть от старости или болезни. Правильность этого1 вывода
подтверждается и исследованием возрастного состава популяции
глухаря.
X. РАЗМНОЖЕНИЕ
Признаком наступления брачного периода служит увеличение се-
менников у самцов и развитие яйцевых фолликул у самок.
Рис. 79. Изменчивость веса пары семенников глухаря в течение гота
(вес в г)
У глухаря семенники заметно увеличиваются уже в апреле, а
около 20 мая они достигают максимального развития: в это время
вес пары семенников составляет от 0,085 до 0,1°/0 веса самой птицы *.
♦ Если семенники не были взвешены на весах, можно определить их объем
по длине и наибольшему диаметру, по формуле, предложенной для измерения
объема яйцевидных тел (Дементьев, 1940, стр. 339): если G объем в мм3 (рав-
ный весу в мг), А длинная и В короткая оси яйца, то G = 0;5 (В2А).
219
В течение июня вес их уменьшается до 0,006%. Замечательно, что
в сентябре гонады вновь немного увеличиваются (0,03'—0,04%), а
потом снова уменьшаются до 0,005% веса тела и остаются в таком
состоянии всю зиму. На протяжении года вес семенников изменяет-
ся приблизительно в 20 раз. У глухарей по первому году семенники
раза в три меньше, чем у вполне взрослых самцов. Осеннее увеличе-
ние веса семенников, возможно, стоит в связи с осенним токованием.
Иногда у птиц, добытых в те же числа, но в разные годы, вес
семенников различается более, чем в два раза (табл. 64'). Причина
этого, по всей вероятности, в том, что период максимального разви-
Таблица 64
Вес семенников в период размножения
________ Лапландский заповедник __________ ________
Дата Глухарь Белая куропатка
Вес птицы в кг Вес семенников: Дата Вес птицы в кг Вес се- менни- ков в г
в г в %0
Зима 3,9
25 IV '1940' г. 4,3
29 IV 1937 г. 3,9
4 V 1986 г. 3,6
9 V 4936 г. 3,9
13 V 1950 г. 3,7
15 V 1936 г. 3,7
21 V 4938 г. 3,1
21 V 1938 г. 3,8
23 V 1950 г. 3,8
25 V 19'54 г. 4,1
1 VI 1950 г. 3,8
8 VI 1951 г. 3,0
28 VI 1950' г. 3,4
0,2 0,05 Зима 0,67 0,03
1,0 0,24 26 IV 1938 г. 0,63 0,20
3,2 0,82 27 IV 1938 г. 0,61 0,25
2,0 0,55 1 V 1949 г. 0,62 0,40
1,5 0,38 9 V 4941 г. 0,63 0,50
3,2 0,86 10 V 1951 г. 0,63 0,80
3,6 0,99 24 V 1941 г. 0,68 1,00
1,1 0,36 24 V 1950 г. 0,58 0,85
3,2 0,85 25 V 19138 г. 0,65 0,60
0,9 0.24 28 V 1950 г. 0,64 1,30
3,7 0,90 1 VI 1951 г. 0,58 1,0
1,6 0,43 10 VI 1950 г. 0,57 0,70
0,12 0,04 НО VI 1950 г. 0,62 0,65
0,2 0,06 20 VI 4938 г. 0,60 0,59
i 9 VII 1'949 г 0,59 0,15
Рис. 80. Изменчивость веса пары семенников белой куропатки в те-
чение года (вес в г)
220
тия гонад очень короток и, в зависимости от хода весны, в разные
годы приходится на разные числа. Подтверждение этому можно
найти в таблице. Максимальный вес семенников глухарей (3,2 —-
3,7 г) наблюдался раньше всего в 1937 г., когда была самая ранняя
весна за все годы исследования, и позже всего в 1938 и 1950 гг.,
при относительно поздней весне.
У самца белой куропатки момент максимального развития гонад
наступает около 30 мая, дней на 10 позже, чем у глухаря. Вес семен-
ников изменяется в течение года от 0,05 до 1,3. г, т. е. приблизительно
в 25 раз. Замечательно, что у белой куропатки около 1 октября так-
же наблюдается временное увеличение их размеров до 0,1—0,15 г.
У тетерева' зарегистрированы наименьший вес семенников 0,04 г,
наибольший 1,9 г (19 V 1951 г.). Максимальный вес семенников ряб-
чика в Лапландском заповеднике не определен.
Сведения о развитии яичников приведены в табл. 65. У самок
темп роста гонад более быстрый, чем’ у самцов. В Лапландском за-
Таблица 65
Увеличение яичников в период размножения
Вид Дата Вос птицы в кг Вес яичн. в г Наиболь- шее яйцо 1 Примечания
вес в г диам. в мм
Глухарка Зимой 1,75 0,23 — — В среднем
14 V 4508 г. 1,44 0,9 0,065 5
44 V 4938 г. 1,69 4,5 1,8 15
19 V 1938 г. — — 16,0 32
21 V 1938 г. — — 2,3 17
6 VI 1938 г. 2,03 21,3 16,0 32 Яйцо в скорлупе
и 2 снесены
Рябчик Зимой 0,40 0,07 — — В среднем
10 V 1936 г. 0,39 0,39 0,033 4
44 V 1938 г. 0,36 0,55 0,033 4
22 V 1934 г. 0,42 — — — Яйцо в скорлупе
26 V 1'938 г. 0,45 — 5,9 23
23 V 1942 ♦ г. — — — — То же
Б/ куропат- Зимой 0,58 0,15 — — В среднем
ка 11 V 1939 г. 0,60 — 0,014 3
15 V 1938 г. 0,61 0,7 0,034 4
23 V 1934 г. 0,61 —. 8,0 25
26 V 1938 г. 0,68 21,7 8,0 25
27 V 1934 г. — — — — Яйцо в скорлупе
3 VI 1948 г. — — 8,0 25 Яйцо в скорлупе
и 3 снесены
* Добыта на северном побережье Онежского озера.
221
поведнике кладка яиц у глухарок, рябчиков и белых куропаток п-ро-
исходит в последней декаде мая — первой декаде июня.
Брачные игры и токование тетеревиных птиц были вкратце описа.
ны выше (глава Ш|), и возвращаться к этому вопросу мы не будем,
В дальнейшем описании гнездования тетеревиных птиц, наряду с
собственными материалами, использованы и литературные, что дает
возможность осветить некоторые вопросы в географическом раз-
резе. 1
Русские авторы давно замечали, что после холодной весны -вы-
водки у боровой дичи, в частности у глухаря, бывают малочислен-
ными. Так, К. Л. Сабанеев (1904) писал про лето 1903 г., в районе
Свердловска: «Гнездистых выводков ныне совсем! не встречалось ни
у какой птицы, так как кладка яиц прерывалась холодом и яйца пер-
вой кладки позябли, вторая же кладка была невелика».
В свое время мы исследовали этот вопрос статистическим мето-
дом (Семенов-Тян-Шанский, 1938 и 1947) и нашли, что возрастной
состав стада -в период охоты изменяется по годам, -причем' «удель-
ный вес» -молодняка связан тесной корреляцией с температурой июня.
Удалось установить, что наибольшее значение имеет температура
первой декады июня — начального периода насиживания. Поэтому
мы предположили, что при низкой температуре воздуха яйца засты-
вают в гнезде в то время, когда глухарка уходит на кормежку или
«отводит» приблизившегося к гнезду хищника. При температуре воз-
духа ниже критической эмбрионы погибают от холода. Это объясне-
ние вызвало дискуссию, к которой мы вернемся -в следующей главе,
Вполне отдавая себе отчет в том, что предложенная нами гипотеза
требует проверки и доказательства, мы решили исследовать режим
насижвания глухаря и других тетеревиных точными методами и со-
брать возможно более полные сведения по эмбриональной смертно-
сти тетеревиных птиц.
В 1944 г. была намечена в общих чертах принципиальная схема
прибора для автоматической регистрации поведения наседки, В
1946 г. я консультировался по этому вопросу у А. Н. Промлтова; он
познакомил меня с устройством как своих приборов сходного назна-
чения, так и актографов с пневматической передачей системы Ганике,
которые применялись -в Физиологическом институте им. И. П. Пав-
лова для опытов с грызунами. Эти приборы, созданные для работы
в лаборатории, казались мало пригодными для наших целей. Для
установки в лесу на’м нужен был портативный недорогой прибор-, на-
дежный в действии при всякой погоде и не требующий частой регули-
ровки. Мы взяли в основу испытанный и широко распространенный
метеорологический инструмент — термограф. В начале 1950 г. два
комплекта актографа были пострены по нашей схеме конструктором
Центрального Бюро поверки метеорологических приборов В. П. Чи-
жевским (Семенов-Тян-Шанский, 1952). В процессе работы та часть
прибора, которая ставится в гнездо, была значительна усовершен-
ствована. Здесь мы приводим описание прибора в том виде, какой
он получил к весне 1954 года. Гнездовой актограф пашей системы
222
состоит из контактной коробки, комбинированного самописца, прово-
да и батареи питания.
Контактная коробка (рис. 81) делается из двух дисков или таре-
лочек из пластмассы или другого диэлектрика, причем нижний яв-
ляется основой, на которой крепятся контакты, клеммы и три спи-
ральные пружины, поддерживающие верхний диск (крышку короб-
ки). Близ края основного диска прикреплен двумя болтиками пру-
жинящий контакт — изогнутая горбом вверху полоска тонкой ла-
туни или алюминия. На противоположном краю диска выше и ниже
пружинящего контакта укреплены два неподвижных контакта, изо-
лированных друг от друга и разделенных промежутком в 3 мм. Вы-
ступающие концы крепежных болтиков с гайками служат клеммами
для подключения проводов.
-резина,
www- па астмасса ,
Рис. 81. Контактная коробка в разрезе (0,65 нат. вел.)
—металл
Подвижный контакт 77, в силу своей упругости прижатый к верх-
нему неподвижному контакту В, при нажиме сверху отходит от него
и прикасается к нижнему контакту Н. К крышке контактной коробки
приклеена гайка (она может быть сделана из текстолита), через ко-
торую сверху пропущен короткий винтик, головка которого укрыта
в отверстии, просверленном <в центре крышки. Выпуская конец винта
длиннее или короче, можно регулировать величину критической на-
грузки, приводящей в движение пружинный контакт П.
Спиральные пружины вставлены в гнезда, высверленные в кусоч-
ках пластмассы, приклеенных по окружности одного из дисков на рас-
стоянии Г20° один от другого; в другой диск, точно против центров
гнезд, вклеены деревянные или пластмассовые шпеньки, которые
входят внутрь соответствующих спиралей. В собранном приборе
крышка держится только на пружинах, и при нажатии опускается
нз 5—10 мм, пока не ляжет на опарные гнезда пружин. Если прибор
поставить в гнездо в таком1 виде, то в промежутки между дисками
попадет мусор и крышка потеряет подвижность. Для предотвраще-
ния этого, на собранную контактную коробку надевается чехол, в
форме берета, который можно вырезать из резинового пузыря для
льда или сшить из толстой материи (см. рис. 81|).
Внешние размеры контактной коробки, рассчитанной для уста-
новки под глухарку, следующие: диаметр 150—180 мм, высота 20—
25 мм, вес с чехлом около 250 г (без чехла — 1’50 г). Для куропатки
диаметр контактной коробки может быть уменьшен до 120 мм.
223
В основании прибора просверливается отверстие для выпуска
двойного, а еще лучше тройного изолированного провода. Если про-
вод двойной, один подключается к клемме контакта 77, второй к кон-
такту В: тогда цепь будет замкнута, пока контактная коробка сво-
бодна или стоит под легкой нагрузкой (яйца), и разомкнута все вре
мя, пока птица сидит на гнезде. Если прибор ставится для записи
откладки яиц, выгоднее второй провод подключить к контакт]/
чтобы цепь замыкалась только при посещении гнезда птицей. Если
провод тройной, провода подключаются ко всем- трем контактам,
что дает возможность изменять характер записи, не подходя
к гнезду.
Постоянная нагрузка на крьйпку контактной коробки в рабочем
ее положении равна весу яиц и гнездовой подстилки. Глухариная
кладка из 6 яиц весит 6 X ’53 = 318 г, а с подстилкой — около 350,
но не более 400 г. Вес глухарки — около' 1700 г. Для более надежного
действия переключение должно происходить при нагрузке в половит;
веса птицы плюс постоянная нагрузка (гнездо); для глухарки это
составит: 1700/2 + 350 = 1200 г. Наши контактные коробки устрое-
ны таким образом, что передвижением верхнего винтика можно ре-
гулировать критическую нагрузку в пределах 500—1500 г, благодаря
чему их можно использовать для птиц разного веса.
Изоляция провода должна быть рассчитана на прокладку его в
сырой среде, а иногда и в воде. Удобнее всего мягкий многожиль-
ный медный шнур с поливиниловой или резиновой изоляцией, при-
меняемый для комнатной электропроводки, но можно воспользовать-
ся и одножильным звонковым! проводом, хотя он жестковат. Телефон-
ные провода со стальными жилами не годятся, виду своей упругости
и большого сопротивления. Длина провода берется с таким расчетом,
чтобы птица не слышала тиканья часового механизма и спокойно
оставалась на гнезде, когда наблюдатель подходит к самописцу; наы
опыт показывает, что достаточно иметь 20 м провода.
Актограф (рис. 82) представляет переделанный термограф, по
возможности — с суточным'оборотом барабана. Запись температуры
воздуха сохраняется. В период насиживания температура опускает-
ся только немного ниже 0: поэтому нижняя половина ленты остается
чистой и может быть использована для записи насиживания. Для
этой цели нужно установить пониже температурного пера второе
перо, рычаг которого присоединен к якорю маленького электромаг-
нита, (можно сделать его самому или использовать катушку от элек-
трического звонка или телеграфного аппарата). Рычаг пера должен
быть снабжен слабой пружинкой, вытягивающей якорь из катушки
на 1—3 мм каждый раз, когда ток разомкнут; полезно предусмот-
реть регулировку натяжения этой пружинки. Необходимо пристроить
винтик для регулировки прижима конца пера к ленте, так, как это
сделано на всех метеорологических самописцах. Дополнительные
приспособления крепятся на корпусе термографа с помощью винтов
с гайками. Перо, чернила и ленты берутся стандартные, принятые
для всех метеорологических самописцев.
224
Если рычаг и электромагнитная катушка сделаны аккуратно1, без
излишнего трения, и хорошо отрегулированы, для приведения их в
действие достаточно тока от одного элемента в 1,6 в. Ток будет за-
мыкаться на сравнительно короткое время и для питания цепи при-
бора в течение сезона достаточно одного-двух телефонных элемен-
тов марки ЗС-Л-ЗО; можно использовать также любой аккумулятор
накала от радиоприемника.
Рис. 82. Гнездовой актограф, переделанный из термографа; справа
контактная коробка с проводом и сухим элементом
Чтобы изготовить контакную коробку и пристроить к термографу
электромагнитную катушку и второй рыча-г с пером, нужно иметь
элементарные навыки в слесарном! деле и набор необходимых инстру-
ментов (дрель со сверлами' по металлу, плоскогубцы, напильник,
отвертка, пинцет и другие мелочи). Необходимые материалы — су-
точная модель термюграфа, электромагнитная катушка, винты с гай-
ками, обрезки листовой латуни или алюминия (толщиной 0,2 —
0,5 мм)), стальная струнная проволока диаметром около 0,5 мм для
навивки пружин, кусочки текстолита и пластмассы, две тарелочки
диаметром 15—18 см из пластмассы, резиновый пузырь для льда,
перья и чернила для метеорологических самописцев, запас лент для
суточного термографа, 20 м двойного провода, 1—2 телефонных эле-
мента. Вес и приблизительная стоимость этих предметов следующие:
Термограф суточный.........3,00 кг 220 р.
Дополнительные части к нему . . 0,10 „ 25 „
Контактная коробка...........0^5 „ 15 „
20 м двойного изолир. провода 0,50 „ 20 „
Элемент ЗС-Л-ЗО...................0,65 „3 „ 30 к.
Итого . . . 4,50 кг 283 р.
15-3037 225
Установка прибора требует особого внимания, так как потрево-
женная наседка легко бросает гнездо. Если гнездо найдено по взле-
тевшей птице, лучше отложить установку прибора до следующего
дня и взять с собой помощника. Предварительно надо проверить и
отрегулировать работу контакта и катушки и даже провести репети-
цию установки — это важно, чтобы не задерживаться лишнее время
у гнезда. Самописец надо ставить с той стороны, откуда будет под-
ходить наблюдатель и, по возможности, в условиях, сходных с рас-
положением гнезда. После этого к клеммам подключается батарея
питания и провода; около приемника температуры кладется конт-
рольный термометр; прокладывается1 провод. Провод следует пря-
тать, пропуская под валежинами и маскируя мхом. Перед установ-
кой контактной коробки следует переложить яйца так, чтобы они не-
стыли, аккуратно вынуть всю подстилку на лист бумаги и выкопать
под гнездом ямку по размерам контактной коробки. Важно, чтобы
она устойчиво держалась в горизонтальном положении, иначе яйца
будут скатываться в одну сторону. Подстилка! и яйца укладываются
поверх коробки и все приводится в прежний вид. Для проверки дей-
ствия контакта в гнездо следует положить какой-либо предмет, при-
близительно равный половине веса птицы, и посмотреть .положение
пера. Остается завести, механизм!, наложить на барабан ленту, отме-
тить на ней дату, время по часам и температуру по контрольному
термометру, записать в книжке те же данные, ввести в углубления
перьев по' капле чернил, сделать контрольные {штрихи перьями и не-
медленно уйти. Для облегчения обработки данных рекомендуется
сменять ленту и ставить контрольные штрихи на ней в «круглые»
часы, для которых есть отметка на ленте (у недельного прибора
линии проведены через два часа, у 'Суточного — через четверть
часа).
При экскурсиях специально' с целью поиска гнезд надо брать с
собой контактную коробку и провод, чтобы установку их можно- было
выполнить сразу же и больше не тревожить птицу; тогда останется
только принести на место и подключить к свободным концам прово-
дов элемент и самописец: положение пера покажет, сидит ли птица
на гнезде или нет.
Ленту на суточных барабанах надо менять ровно через 2Ф часа,
в крайнем’ случае с опозданием на один час: после этого перья перей-
дут через планку, закрепляющую концы ленты, и старая запись пере-
путается с новой. Обработка термограммы ведется по указаниям
инструкции для метеорологических станций, с введением1 поправок на
ход часового механизма и на температуру воздуха. «Гнездовое» перо
ведет горизонтальную линию пока цепь разомкнута, а когда, ток замк-
нется, оно делает скачок вниз и чертит ниже, пока цепь снова не
разомкнется (см. рис. 82). Если контактная коробка слишком! мала,
вес птицы частично ложится на края гнезда, вследствие чего перо
чиркает каждый раз, когда птица шевелится на гнезде; того же мож-
но добиться, если отрегулировать контакт на больший критический
вес, чем следует по расчету.
226
Отпечатанная на ленте разметка часов рассчитана только на одно
перо, ось которого совпадает с центром дуговых линий. Обработка
записи режима насиживания ведется отдельно с помощью милли-
метровой линейки, которая накладывается на ленту параллельно
черте второго пера так, чтобы ноль совпал с начальным контрольным
Рис. 83. Актограф в рабочем положении у гнезда глухарки
штрихом. Расстояние от контрольного штриха до всех последующих
отсчитывается с точностью до О J1 мм (с помощью лупы). Зная рас-
стояние между контрольными штрихами -и время, прошедшее между
запуском> и снятием ленты, легко определить цену одного часа в мил-
лиметрах и перевести все измеренные расстояния в часы, после чего
доли часа перевести © минуты. Вычисления удобнее всего вести с
помощью счетной линейки. Приведем) для примера обработку одной
из лент (28—29 V 1953 г., длина 1 часа=11,24 im>mi):___________
Отсчет в мм Часы Часы и минуты Момент собы- тия Отсутство- вала (часы и мин.)
Ушла Вернулась 25.0 28;9 2,22 2,57 2-13 2-34 17-00 17-21 } 0-21
Ушла Вернулась 133,4 136,0 11,87 12,10 11-52 12-06 2-39 2-63 | 0-14
Ушла Вернулась 188,0 191,8 16,73 17,06 16-44 17-04 7-31 7-51 | 0-20
Конец записи 269,7 24,00 24-00 14-47
15*
227
На ленте недельного самописца длина одного часа равна прибли-
зительно 1,6 мм. Первое время я пользовался недельными приборами,
но запись получалась слишком (мелкой и неудобной для обработки.
Единственное преимущество недельного прибора в том, что его мож-
но проверять не ежедневно.
Переходим к описанию результатов наших исследований.
Размножение глухаря. Иногда глухарка гнездится на территории
токовища, а чаще — по соседству с ним, не далее 1 км. Примерно
в половине случаев гнездо устраивается под елкой, реже — у осно-
вания сосны или под березовым кустом:, иногда — под валежиной или
корневым выворотом.; одно гнездо находилось под прикрытием кам-
ня. Мы 1никопда не находили гнезд, устроенных на открытом -месте,
без какого-либо прикрытия. В Лапландском заповеднике ветки елок
обычно свешиваются до- земли, поэтому ель хорошо закрывает гнездо
и от дождя и от посторонних глаз. Стволы сосен и кусты берез, без-
листные в период кладки яиц, не дают никакой защиты от непогоды;
это почти «символическое» укрытие, но оно может иметь значение
ориентира для птицы.
В трех случаях (10% гнезд) гнездо находилось на берегу реки
или озера, не далее 10 м от воды; в пяти случаях — на краю болота,
на прикорневой кочке сосны или же под прикрытием крошечной
елочки, кустов багульника и других растений. В восьми случаях
(28о/о) гнездо было найдено рядом с тропой или дорогой, иногда по
соседству с жилыми или нежилыми постройками. Так, например,
I1! VI 1932' г. гнездо глухарки было найдено в L50 м от кордона, при
котором содержались две лайки. Другое гнездо было» найдено 8 VI
1951 г. в 400 !м от жилых домов управления Лапландского заповед-
ника; третье — 31 V 1953 г., в 20 м от нежилой лесной избушки в
Печоро-Илычском заповеднике. В двух из этих гнезд цыплята выве-
лись благополучно, а одно гнездо было брошено наседкой. Hai срав-
нительно частые случаи находки глухариных гнезд у людных дорог
и лесных тропинок обращали внимание и некоторые старые авторы
(Гейер, по Брему, 1911, т. VII; Романов, по Сабанееву, 1876, стр.34).
Гнездо имеет вид небольшого углубления в напочвенном покрове
со скудной подстилкой из сухих листьев березы, обрывков мхов,
хвоинок и перьев наседки; если вокруг растут -злаки или осоки, в
подстилке бывают их листья и стебли. Почва под гнездом всегда
сухая. Л. П. Сабанеев (1870) и Е. А. и Е. В. Крутовские (1953) ука-
зывают, что глухарка, сходя с гнезда, прикрывает яйца травой, мхом
или сухим листом. Нам не раз приходилось осматривать гнездо глу-
харки во время ее кормежки, но яйца- ничем не были прикрыты, так
же как и в гнездах, с которых птица была спугнута.
В любому уголке тайги можно найти сколько угодно подходящих
для глухариного гнезда мест. Поэтому глухарка, как 'правило, не
возвращается на следующий год на старое место. Единственный
известный нам случай гнездования глухарки в старом гнезде наблю-
дался-в Красноярском заповеднике «Столбы» в 1940 г. Е. А. Крутов-
ской (письм. сообщ.) .
228
Полная .кладка глухаря обычно насчитывает от 5 до 8 яиц, ‘мак-
симум — 10.
Таблица 66
Величина полной кладки глухаря в разных областях
Местность Число яиц в гнезде Число гнезд
4 5 6 7 8 9 10 сред- нее
Лапландский заповедник , . 1 5 8 8 5 6,4 27
(автор) Печоро-Илычский заповедник 2 7 14 15 4 2 — 6,4 44
(Теплов, 1947, и автор) Вологодская обл 2 1 1 7,8 4
(Ларин, 11941) Башкирский заповедник . . 2 4 5 10 2 — 7,3 23
(Кириков, 1948) Беловежская пуща . . 2 4 4 1 1 7,4 12
(Гаврин, 1956) Европ. часть СССР . . 3 16 26 34 24 6 1 6,7 ПО
С. В. Кириков (1948, 1952) описывает одно гнездо с 16 яйцами;
вероятно, это были 2 кладки .в одном гнезде — по крайней мере,
в собранном нами большом материале не оказалось гнезд с проме-
жуточным числом яиц ('10—15). У гоголей часты случаи кладки яиц
двумя или даже тремя самками в одно гнездо. Эти утки гнездятся
в дуплах, поэтому они 1припуждены разыскивать подходящие по раз-
мерам полости в старых деревьях, а такие полости бывают заняты
нз года в год, и не всегда утке удается найти свободное помещение,
вследствие чего ей нередко приходится нестись в чужом? гнезде. Глу-
харки находят места для гнезд без затруднений, и случаи кладки
двух самок в одно гнездо должны быть так же редки, как и кладка
яяц в прошлогоднее гнездо.
Индивидуальная изменчивость величины кладки у глухаря незна-
чительна (среднее квадратическое отклонение □ = + 1,27). Из
табл. 66 видно, что близ северного предела распространения число
яиц в гнезде меньше, чем в средней полосе или на юге.
В одном из осмотренных на>ми гнезд было обнаружено ненор-
мально маленькое яйцо- шарообразной формы, очевидно, не содер-
жавшее эмбрион al (22 VI 1931 г.); о находках подобных яиц упоми-
нает и Л. П. Сабанеев (1876).
Яйца глухаря крупнее обычных куриных яиц, но почти не отли-
чаются от них по форме. Окраска, их бледно-коричневато-желтая
с частыми медкцми коричневыми крапинками. Находясь в гнезде,
они не выделяются среди окружающих предметов. Длина яйца около
57—58 мм, ширина — 41—43 мм. Вес ненасиженных яиц, в среднем
из 29 измерений, 53,2 г (от 49,4 до 60,4, чаще всего 51,5—54,5 г; дан-
ные относятся к Лапландскому заповеднику и Карельскому пере-
шейку!) -
229
А. И. Хватов (1861; 2 кладки) и Е. А. Крутовская (1953; 6 кла-
док), наблюдавшие кладку яиц глухарками в неволе, указывают, что
они несутся не ежедневно, а через день; точнее, в описании Крутов-
ской, (минимальный (Промежуток между откладкой яиц был 2 суток
(13 случаев), реже 3 суток (7 случаев) и в остальных 7 случаях —
от 4 до 7 суток. С, В. Кириков (1948) приводит одно наблюдение
откладки яиц в природе: за 11 суток в гнезде прибавилось 5 яиц.
М. А. Мензбир (1900—-*1902, стр. 155) приводит противоречивые дан-
ные: по Л. П. Сабанееву, глухарка несется ежедневно, ранним утрам,
а по Л. Ушкову — через день, между 5—6 часами вечера.
Мы наблюдали кладку яиц глухаркой один раз, в Печоро-Илыч-
ском заповеднике. Гнездо было найдено утром1 21 V 1966 г., по взле-
тевшей глухарке: в нем было одно яйцо, очевидно только что сне-
сенное. 22 мая между 121—18 час. глухарка снова была поднята и
в гнезде оказалось два яйца. 24' мая в 16 час. при установке самопис-
ца для записи насиживания, глухарки не оказалось, а в гнезде было
3 холодных яйца. Четвертое яйцо, согласно записи прибора, было
снесено 25 мая между 6—46 час, а в 22 час. в тот же день глухарка
начала насиживать (рис. 85|). Последнее, пятое, яйцо> снесено 26 мая.
Таким образом пять яиц были снесены в течение 6 суток. Промежу-
ток между откладкой двух яиц больше 24 часов, вследствие чего
время откладки яиц сдвигается к вечеру; после этого птица пропу-
скает один день (в данном случае «днем отдыха» было, по- всей ве-
роятности, 24 мая) и следующий цикл начинается опять с утра. Ве-
роятно, этот порядок кладки и является нормальным (см. ниже, о
рябчике), но при менее благоприятных условиях кладка растягивает-
ся на более долгий срок.
В Лапландском! заповеднике гнезда глухарок были найдены: са-
мое раннее 25 V 1937 г., самое позднее 24 VI 1933 г.; средняя дата,
по 28 гнездам, 7 июня. Ввиду того, что кладки могли быть найдены
на разных стадиях инкубации, начало насиживания отсюда нельзя
определить. Исходя из того1, что средняя многолетняя дата встречи
первого выводка — 25 июня, продолжительность инкубации состав-
ляет около 25 суток, средняя величина кладки 6—7 яиц и откладка
их занимает 7—9 дней, мы находим, что нормально глухарки начи-
нают нестись около 22—24 мая. К этому времени поверхность земли
в лесу в основном очистилась от снега, некоторые растения начинают
цвести, летают многие насекомые, но березы начинают зеленеть уже
после начала насиживания.
Продолжительность насиживания разные авторы оценивают от
20 до 30 суток. Обратившись к первоисточникам, мы нашли такие
определения:
Л. Ушков (1887), окрестности Свердловска . . • . 20—24 сут.
С. В. Кириков (1948), Башкирский заповедник, 11 случай . 22—23 „
С. В. Кириков (1952), тот же случай, в другой интерпретации 24—25 „
к. Крюденер (Kriidener, 1928), Германия............... 27—28
К. Кронау (1904)', Германия (инкубатор) « 28 „
Л. П. Сабанеев (1876), Свердловская область .... 28—30 „
Е. А. Крутовская (19513], Красноярский заповедник . . . 29—30 ,,
230
Авторы сводок на русском языке следуют указаниям1 либо Ушко-
ва (Мензбир, 1900—11912; Иванов и др., 1951; Доппельмаир, Маль-
чевский и др., 1951), либо Сабанеева (Мензбир, 1895; Холодковский,
1901). К сожалению, теперь невозможно установить, каким спосо-
бом определяли продолжительность насиживания Сабанеев и Ушков.
Кириков, располагая наблюдениями только над одним) гнездом1, в
двух работах определил срок насиживания с разницей в 2 дня. Если
04 8 is is го гъь
26/v *
1 о f
Ufa
-------
5/А ' ** ~~~~ Ж
— —
' 1—1—
...
Ю/vi ... . W. 1—J-
—
—
•—i —
15/<!
. . - 1—1
» _ _ ~ ~ J
2Ofij 1——> >—• 1 1
1—1 L.
... - . в
птица на гнезде
Ж
отсутствует
спугнута с гнезда
___ т ра ниже О*
___т ра от 0* до 5*
Рис. 84. Режим насиживания глухарки в Лапландском заповеднике,
1950 г.
принять, что глухарка неслась точно через день, то из приводимых
им дат откладки можно заключить, что последнее яйцо было снесено
15 V 41, цыплята вывелись 9 VI, и насиживание продолжалось 25 су-
ток. Е. А. Крутовская, по нашему запросу, разъяснила, что она на-
блюдала два случая: в первом инкубация длилась 29—30 дней, во
вторам — 25—26 дней. По А. М. Грюнеру (1951), в Висимском за-
поведнике глухарята вывелись через 24 дня после находки гнезда.
Рассмотрим наши наблюдения. В Лапландском заповеднике гнез-
до с 5 яйцами было найдено 26 V 1950 г. в 11 час.; 27 V в 13 час.
231
в нем было уже 6 яиц (оба раза птица была поднята с гнезда);
21 VI в 2 ч. 30 м. цыплята начали выводиться, а 22 VI в б ч. 30 м,
оставили гнездо; продолжительность инкубации, считая с дня от-
кладки последнего' яйца до ухода цыплят с гнезда, — 26 суток.
Глухариные яйца из гнезда, найденного на Карельском перешей-
ке утром 10 V 1952 г. (см. ниже|), были взяты нами для искусствен-
ной инкубации в экологической лаборатории Зоологического инсти-
тута АН СССР 11 мая в 14 час. Насиживание началось, очевидно,
ДО' 10 мая, так как птица оба раза была поднята с гнезда, а на сле-
дующий день число яиц в кладке не увеличилось. Яйца были поме-
щены в термостат и обогрев включен 12 мая в И ч. 15 м. Развитие
яиц продолжалось, и :5 июня в 17 час. был отмечен первый проклев,
однако птенцы погибли, не выбравшись из скорлупы. От момента
находки гнезда до проклева прошло 26 суток, но насиживание было
прервано в самом начале на 1 сутки. Продолжительность инкубации
до вылупления птенцов была бы 25—26 суток.
В Печоро-Илычском заповеднике днем 31 V 1952 г. спугнули глу-
харку с гнезда, в котором было 7 яиц. При установке самописца
21 июня в гнезде было 8 яиц, следовательно, одно было снесено уже
после 31 мая. 24 июня в 9 ч. 14 м. цыплята начали выводиться, а
25 июня в 8 ч. 24 *м. матка их увела. Продолжительность инкубации,
считая от находки гнезда до увода птенцов, оказалась 25 суток.
Насиживание кладки, найденной 21 V 1956 г. (ом. выше), нача-
лось 25 мая в 22 ч. 06 м., а 20 июня в 12 ч. 40 м. глухарка увела
цыплят с гнезда. Насиживание продолжалось 25 суток и 1472 часов.
Во всех четырех случаях инкубация продолжалась от 25 до 26 су-
ток: это и есть нормальный срок инкубации глухаря, установленный
с такою точностью, какую допускает определение начала и конца
насиживания.
Из диаграмм рис. 84 и 85 видно, что тетеревиные птицы начинают
насиживание постепенно: по-видимому, откладка яйца занимает не-
много времени (вероятно, около получаса), но птица и после того
некоторое время остается на гнезде, сначала на несколько минут,
но с каждым разом все дольше, пока не дойдет до нескольких часов;
так, например, 25 мая глухарка просидела 9 ч. 43 ;м. (рис. 85). Пере-
ломным моментом следует считать установление характерного режи-
ма насиживания, с короткими отлучками для кормежки. Этот момент
наступает, примерно, за полсуток до откладки последнего яйца. За
начало инкубации можно принять один из этих двух моментов, но
следует предпочесть первый.
Время вывода птенцов труднее определить, тем более, что они
выходят из скорлупы неодновременно. Практически лучше всего
измерять период насиживания до- увода птенцов с гнезда: этот мо-
мент, по крайней мере, поддается точному определению. Отдельные
случаи, когда глухарка насиживала 219—30 дней, можно объяснить
только резкими уклонениями в режиме инкубации, например, дли-
тельными перерывами или недостаточно высокой температурой яиц.
Почти всегда гнезда глухарок находят по взлетевшей птице, по-
232
этому находки неполных кладок очень редки. Если из полной, но
ненасиженной кладки взять часть яиц, глухарка не дополняет кладку,
а или насиживает оставшиеся яйца (наблюдение Г .Г. Шубина
в окрестностях Лапландского заповедника в 1936 г.), или бросает
гнездо (наблюдение Е. А. Крутовской в Красноярском- заповеднике
I---\ птице/ на гнездо ----т-ра
лнтгутгтпг/от ___т - pci от 0" до J
спугнута с гнезда
Рис. 85. Режим насиживания глухарки в Печоро-Илычском заповед-
нике, 1956 год
В Лапландском заповеднике за два сезона (I960 и 1'951 гг.|) мы
ставили самописцы в три глухариных гнезда. Первое мы нашли 26 V
1950 г. на берегу озера, в 1,3 км от усадьбы заповедника; 221 июня
в 6 ч. 30 м. глухарка увела цыплят. На этом гнезде получены четыре
недельные ленты записи, охватывающие весь период насиживания,
но, вследствие неполадок в работе прибора, первая лента оказалась
непригодной для обработки (рис. 84). Два других гнезда были най-
233-
дены в один день, 8 VI I1951i г., одно — в 1,5 им от усадьбы, у края
малохожей тропинки на заболоченном южном! склоне Чуна-тундры;
другое—в 0,4 км от жилых домов, под густою елкой на краю поляны.
Первую глухарку пришлось несколько раз тревожить для регулиров-
ки контактной коробки, вследствие чего уже на третий день она бро-
сила гнездо. Вторая также бросила гнездо через три дня в связи
с начатым нами опытом по измерению температуры внутри яйца;.
В Печоро-Илычском заповеднике за шесть сезонов (1952—
1957 гг.) исследовано шесть гнезд. Первое обнаружено 31 V 1952 г.
в бору-брусничнике на краю болота, в 3,5 км от поселка Якша. При-
бор был поставлен 21 июня, а 25 VI глухарка увела птенцов. Полу-
чены 4 суточные ленты с хорошей записью.
Второе найдено в бору-брусничнике на другой стороне р. Печоры,
в 4 км от Якши. 25 мая мы. установили самописец и за все время
получили 17 хорРших суточных лент. Глухарка сидела до 23 июня,
и, когда стало ясно, что из яиц ничего не выйдет, аппаратура была
убрана. Во всех 6 яйцах эмбрионы не развивались или погибли на
ранней стадии развития. 21—22 мая, накануне дня находки гнезда
(и, по-видимому, до начала инкубации) в продолжение 9 часов был
мороз до —5,3°. Вероятнее всего, что яйца были подморожены;
не исключено, также, что они вообще не имели эмбрионов (напри-
мер, не были оплодотворены|).
Третье гнездо было найдено в 1,5 км от Якши, в заболоченном
сосновом лесу 24 V 1955 г. Запись велась с 27 мая по 10 июня, когда
в 16 час. глухарка увела цыплят. В этом'гнезде впервые была постав-
лена описанная выше усовершенствованная контактная коробка. Все
14 суток прибор работал безотказно.
Четвертое гнездо, найденное 21 V 1956 г. в 2,5 км> от Якши, в за-
болоченном бору за рекой, оказалось самым интересным — в нем
нам впервые удалось наблюдать откладку яиц и начало насижива-
ния (см. рис. 85). 30 V 1956 г., по пути к этому гнезду, в сухом бору-
брусничнике было обнаружено еще одно гнездо. Самописец был
установлен 31 мая в 14 час. за несколько минут до наступления
сильной грозы с ливнем1. Глухарка вернулась и села на яйца еще до
того, как пошел дождь, но 1 июня в 8 ч. 07 м. ее согнал и уничтожил
яйца какой-то хищник, по-видимому, куница.
19 V 1957 г. в 13 ч. 26 м. не далее 0,2 км от гнезда 1952 г. мы
увидели ^спокойно сидевшую на гнезде глухарку. Контактная короб-
ка была установлена сразу-же, al самописец принесен и подключен
в 17 ч. 30 м. Наседка вернулась и села на гнездо только в 19 ч. 44 м.;
в дальнейшем насиживание и запись его шли отлично до 10 июня,
когда в 16 ч. 28 м. матка увела птенцов.
Из 9 гнезд, в которые ставился актограф, 2 кладки были брошены
в результате причиненного нами беспокойства, одно гнездо разорено
хищником независимо от нашей работы, а в другом эмбрионы по-
гибли еще до установки прибора. Остальные пять глухарок благо-
получно вывели цыплят. По' всем девяти гнездам мы получили в сум-
ме 98 суток записи насиживания. Это вполне достаточный материал,
234
чтобы судить о режиме инкубации глухаря (не говоря уже о том,
что пока он является единственным в мире).
Мы уже рассматривали порядок откладки яиц и вопрос о продол-
жительности инкубации. Переходим собственно к режиму насижи-
вания яиц.
Глухарка оставляет гнездо для прогулки до 4 рш в сутки, чаще
всего 2 раза (61°/о всех записанных прибором дней|), реже три (22o/J
случаев) или один (14о/о случаев). В период вылупления птенцов
глухарка иногда не сходит с гнезда более суток. По диаграммам на-
сиживания (рис. 84 и 85) видно, что глухарка уходит с гнезда при-
близительно в то же время, что и накануне; Однако постепенно сроки
кормежки смещаются в ту или другую сторону, и в целом за< весь
период насиживания их трудно приурочить к определенным часам.
Периодические прогулки непродолжительны, чаще всего от 10 до
35 минут. Использовав все записи гнездовых самописцев в Лапланд-
ском п Печоро-Илычском заповедниках, мы составили сводную таб-
личку продолжительности отлучек насиживающих глухарок
(табл. 67).
Таблица, 67
Число отлучек различной продолжительности глухарки с гнезда
Название заповедника Продолжительность в мин. Число случаев
5* 10 15 20 25 30 35 40 и более
Лапландский .... 1 4 9 8 12 11 12 57
Печоро-Илычский 2 15 72 44 23 6 7 5 174
Отлучки продолжительностью от 3 до 7 мин. считаются за 5 мин. и т. д.
Продолжительность кормежек глухарки мало изменяется с те-
чением времени. Если откинуть первые сутки инкубации (когда ре-
жим еще не установился) и последние сутки (во время вывода птен-
цов наседка не покидает гнезда), а остальной период разделить на
три части, продолжительность времени, когда наседки нет на гнез-
де, составит в долях всего времени инкубации:
Начало Сере- дина Конец
Глухарка 4956 г Глухарка 1957 г 0,031 0,021 0,028 0,022 0,030 0,032
Температура воздуха влияет на продолжительность отлучек на-
седки, но незначительно. В теплую погоду глухарка кормится
дольше, чем в холодную: соответствующий коэффициент корреля-
235
ции оказался равным: в 1956 г. г = 4-0,29 + 0,19; в 1957 г. г —
= +0,52 + 0,16.
Начало вывода птенцов характеризуется усиленным чирканьем
пера актографа и прекращением очередных кормежек глухарки: если
она и сходит в этот период, то на самое короткое время (не более
11 мин.), вероятно, только для дефекации. От последней кормежки
глухарки до увода птенцов с гнезда в 1950 г. прошло 34 часа,
в 1952 г. —27, в 1955 г. —31, в 1956 г. —34 и в 1957 г. — 23 часа.
От начала вывода птенцов до ухода их с гнезда в 1950 г. прошло
Рис. 86. Гнездо глухарки в момент вылупления птенцов; Лапланд-
ский заповедник, (16 VI 1940 г.
28 часов, в 1952 г. — 23, в 1957 г.— 11 час. Выводок покидает
гнездо в различные часы дня, самое раннее в 6 ч. 30 м. (1950 г.),
самое позднее в 16 ч. 30 м (1957 г.).
16 VI 1940 г. в 10 ч. 30 м. мы спугнули глухарку с гнезда, в кото-
ром было 7 наклюнутых яиц. Птенцы, прорезав «яйцевым зубом»
поясок по краю воздушной камеры, отделяли крышку и выбира-
лись на волю (рис. 86). Через 50 мин. вышли 3 глухаренка, но
гнездо тем временем остыло, движения птенцов стали вялыми и
в следующие 30 мин. не вывелся ни один. В это время на птенцов
напали муравьи. Глухарка, очевидно, вернулась не сразу, в ре-
зультате чего 3 птенцов закусали'муравьи, 3 погибли, не выйдя
из яиц, и матка увела с собой, пО-видимоМу, только одного.
Первое время выводок движется очень медленно. В 1957 г. че-
рез 42 мин. после оставления гнезда глухарята были найдены в 5 м.
от него (скорость движения около 7 м в час). Осматривая поки-
нутое гнездо, мы иногда обнаруживали примерно в 1 м от него боль-
236
шой ком помета матки. Нормально пищеварение у тетеревиных идет
равномерно круглые сутки, но в период насиживания кал надолго
задерживается в прямой кишке, вследствие чего объем ее сильно
увеличивается (Вильсон, 1911, стр. ПО). Возможно, что накопле-
ние кала является непосредственной причиной, побуждающей на-
седку сойти с гнезда. Исходя из этого, можно понять, почему глу-
харка в первый день насиживания часто оставляет гнездо (рис. 85)
и приспосабливается к двухразовым кормежкам только постепенно.
Иногда, как наблюдалось в гнезде 1952 г., она кормится только
один раз в сутки.
В Лапландском заповеднике глухарки насиживают в то время,
когда солнце совсем не заходит; в связи с этим у них нет ни отчет-
ливого суточного цикла деятельности, ни определенного периода
ночного покоя. Печоро-Илычский заповедник расположен на 5° юж-
нее; там1 в период с 22 V по 23 VI солнце заходит каждую ночь и
в течение 2 часов находится на 4—8° ниже горизонта, что создает
эффект «белой ночи». В самые темные часы (23 ч. — 1 ч. по ме-
стному среднему солнечному времени) глухарки не оставляют гнез-
да: вернее, наблюдалась одна отлучка в первые часы после начала
насиживания. Кроме того, сравнительно редко глухарка оставляет
гнездо между 22—2 и 11 —17 часами.
Таблица 68
Суточный цикл насиживающей глухарки
(Суммарная продолжительность отлучек в промиллях от всего времени
насиживания)
Часы суток ф 3 -
Название „ х
заповедника 23 1 3 5 7 9 И 13 15 17 19 21 23 °
с: о
Лапландский . . 16 94 30 129 44 30 116 120 52 53 16 29 61
(486 час. набл.) Печоро-Илычский (1857 час. набл.)' 7 47 60 12 23 13 7 5 4 44 71 28 27
Таблица 69
Температура воздуха во время насиживания и отлучек глухарки (В % от числа всех случаев)
Название заповедника Пределы температур в °C Средн, т-ра в °C
-10 —5 0 5 10 15 20 25 30 35
Лапландский . . 485 час. насиж. 53 кормежки — 2 2 23 28 40 40 20 17 11 4 4 7 0 + 2 — 9,0 8,7
Печоро-Илычский Г888 час. насиж. 0 + 4 17 31 26 14 7 1 о+ 10,2
161 кормежка 1 6 16 39 24 9 4 1 8,9
237
Глухарка нередко оставляет гнездо во время суточного мини,
мума температуры воздуха, который наступает около 2 час. утра
в Лапландском заповеднике и около 3—4 час. утра в Печоро-
Илычском. В связи с этим, вероятность кормежек при низких тем-
пературах воздуха несколько выше, чем при высоких. Глухарки, как
и другие тетеревиные, уходя на кормежку, ничем не прикрывают
яйца (в отличие от уток). В Печоро-Илычском заповеднике заре-
гистрировано 8 случаев, когда яйца оставались без обогрева в те-
чение 13—19 мин. при температуре воздуха от —1,1 до —5,5=;
один раз гнездо оставалось при температуре —1,0е в течение
69 мин., а другой раз при —0,8° в течение 42 мин., что, однако, не
отразилось на жизнеспособности эмбрионов. 27 V 1953 г. около
3 час. утра глухарка уходила кормиться при 0° и наличии снеж-
ного покрова, образовавшегося ночью и лежавшего до 10 час. утра.
При сильном и длительном дожде глухарка остается на гнезде,
пропуская очередную кормежку, как это наблюдалось, например,
8 VI 1953 г, 29 V 1955 г., 23 V 1957 г. и 31 V 1957 г. утром и 5 VI
1956 г. вечером. При этом птица сидела на гнезде непрерывно
около 24 час., а иногда и больше.
Возвращаясь после кормежки на гнездо, глухарка некоторое
время усаживается и переворачивает яйца: актограф отмечает это
утолщенным штрихом на ленте. Сидя на гнезде, она также время
от времени переворачивает яйца. В 1950 г. глухарка делала это,
в среднем, 26 раз в сутки, в 1953 г. — 32 раза, в 1957 г. — 22 раза.
Примерно так же часто переворачивает яйца и домашняя курина
(Третьяков, 1951). В ненастную погоду количество движений на-
седки сокращается: так, например, в 1957 г. в хорошую погоду
(или даже с ливневыми осадками) в среднем глухарка перевора-
чивала яйца 26 раз в сутки (максимальное число 38 было в са-
мый жаркий день за весь период), в дождливую — в среднем
13 раз, а в самые ненастные дни только 6—9 раз. В один из таких
дней (31 V 1957 г.) мы видели в бинокль, что перья на спине
наседки намокли и слиплись. Глухарка не может голодать больше
полутора суток: в случае длительного дождя ей неизбежно придется
уйти на кормежку под дождем. Очевидно, что, не просушив опере-
ние, она не сможет возобновить насиживание и кладка будет
брошена. Мы не можем подтвердить эти соображения фактами,
так как за 8 сезонов работы с самописцами не наблюдалось об-
ложных дождей, продолжавшихся более 12 час.
В отдельных случаях глухарка оставляет насиженную кладку
на несколько часов без видимых причин. 12 VI 1950 г. в 5 ч. 24 Nt-
глухарка ушла с гнезда и вернулась к нему только в 12 ч. 55 м.,
то есть через 7*/г час. В этот период гнездо было осмотрено п
яйца оказались холодными. Температура воздуха была 5—9°, и эм-
брионы от такого охлаждения не пострадали.
Насиживающая глухарка обычно подпускает человека метров
на пять и почти с места взлетает, не пытаясь «отводить»; некоторые
подпускают и ближе, иногда до 2 м. Однажды глухарка, сойдя
238
с гнезда для кормежки, услыхала наше приближение и сейчас же
вернулась (отсутствовала только 3 минуты); только через 2 ч. 40 м.
она снова вышла на кормежку.
Спугнутая глухарка возвращается на гнездо в большинстве слу-
чаев через 1—3 часа, а в среднем из 19 случаев — через 2 ч. 23 м.
(максимум 6 ч. 18 <м., и только один раз через Юмин.). Приповтор-
ных спугиваниях глухарка нередко бросает гнездо. В 1951 г. спуг-
нутая в третий раз за трое суток глухарка не вернулась к кладке,
которую насиживала около 10 дней. 12 VI 1951 г. другая глухарка,
разбив яйцо, оборудованное для измерения температуры в нем,
приклеилась перьями к другим яйцам, вследствие чего разбросала,
разбила их и бросила кладку на седьмой день инкубации. В других
случаях, наоборот, глухарка продолжает насиживание, несмотря на
явную опасность. 20 VI и 22 VI 1932 г. одну глухарку дважды пы-
тались поймать на гнезде сачком (чтобы закольцевать): оба раза
ей удалось избежать поимки, но гнезда она не бросила и вывела
птенцов. 10 V 1952 г. близ станции Горьковской на Карельском пе-
решейке глухариное гнездо с полной кладкой было найдено в
20 шагах от проезжей дороги, на поляне, под валежиной. Глухарка
взлетала каждый раз, когда кто-нибудь проходил. Кто-то привалил
с другой стороны гнезда сосновую вершинку и в образовавшемся
узком проходе поставил несколько силков. Несмотря на грубое на-
рушение окружающей обстановки, глухарка вернулась к гнезду
и попала в силки. Проходя 11 мая в 14 ч. мы увидели, как глухарка
вскочила и забилась в силках. Нам ничего не оставалось, как осво-
бодить птицу и использовать яйца для опыта с искусственной инку-
бацией (см. выше).
Нередко глухариные кладки гибнут при лесных пожарах. 21 VI
1936 г. на пожаре мы нашли обгоревшее гнездо и дважды подни-
мали около него глухарку. Другое сгоревшее гнездо было найдено
на лесном пожаре 21 VI 1950 г. Накануне около того же места, на
кочке мокрого болота, к краю которого подходил огонь, нашли еще
одно гнездо глухарки. Несмотря на то что огонь не мог дойти до
гнезда, а люди сразу ушли и больше не беспокоили птицу, гнездо
было брошено и прошедшие половину инкубации яйца погибли.
Сообщения некоторых авторов, что глухарка будто бы выкиды-
вает из гнезда яйцо с погибшим зародышем и что она не сидит на
гнезде лишних дней, если птенцы не выводятся в срок, — неверны.
В гнезде 1953 г. глухарка в течение 31 дня насиживала мертвые
яйца и, вероятно, просидела бы еще дольше, если бы наша сотруд-
ница не убрала яйца и аппаратуру. Если гнездо расположено на
косогоре, то при поспешном бегстве глухарки яйцо иногда откаты-
вается: наседка не возвращает яйцо на место, и оно погибает. Та-
кие случаи мы наблюдали 20 VI 1932 г., 22 VI 1932 г. и 15 VI 1953 г.
Возможно, что подобные наблюдения дали повод предположить, что
глухарка выбрасывает испортившиеся яйца.
А. М. Грюнер (1951) в Висимском заповеднике измерял темпе-
ратуру в гнезде глухарки с помощью двух термометров — макси-
239
мального и минимального. Отсчеты их производились через не*
сколько дней. В период с 13 V по 6 VI 1950 г. максимальные темпе*
ратуры оказались равны 37,7; 38,7; 37,0; 40,9; 43,6; 34,8; 36,3°.
Минимальные температуры были: 14,6; 20,2; 18,8; 23,7; 22,7° и три
раза выше предела шкалы (выше 24°). Температуры выше 40° мы
считаем сомнительными, так как они превышают температуру по*
верхности тела птицы (по крайней мере, у домашних птиц темпе-
ратура в клоаке равна40—41°, а между перьев — только 38,5°). По
всей вероятности, резервуар термометра находился выше капил-
лярной трубки, и при толчках наседки ртуть проскакивала через
сужение капилляра в трубку. Хотя в период наблюдений были ча-
стые заморозки, температура в гнезде почти не опускалась ниже
20° и только один раз была ниже 15°. Между тем в ту же весну
в Висимском заповеднике было найдено несколько частично замерз,
ших кладок тетеревиных птиц. Грюнер объясняет эмбриональную
смертность тем, что похолодания захватывали период кладки и пер-
вые дни жизни птенцов.
С целью получить представление о ходе остывания яиц на воз-
духе мы провели весной 1951 г. опыт с глухариными яйцами, взя-
тыми из двух брошенных кладок. Вечером 15 июня, когда ожидался
ночной заморозок, 3 глухариных и 2 гоголиных яйца (примерно оди-
накового веса) были нагреты в воде при температуре 40°, осушены
полотенцем, перенесены в лес в сосуде Дюара и уложены в пустое
глухариное гнездо. В двух глухариных яйцах были просверлены
дырки, в которые были введены на глубину 10—20 мм резервуары
маленьких термометров: таким образом температура измерялась
там, где в живом яйце находится эмбрион. Опыт был начат в 23 ч.
00 м. при температуре воздуха; 2,6° и закончен в 1 я. ОО м. при тем-
пературе воздуха —1,1°. Термометры отсчитывались через 5 минут.
В начале опыта температура внутри яиц была 37,2 и 37,0°; через
1 час 19,9° в яйце, находившемся в середине гнезда, и 17,6° в яйце
на периферии; по истечении 2 часов температура яиц была 11,4
и 9,6°. Опыт был проведен в условиях очень близких к естествен-
ным, с той лишь разницей, что эмбрионы были мертвы и сами не
производили тепла. Обработка полученных цифр показала, что сни-
жение температуры яиц в единицу времени пропорционально раз-
нице температур яйцо — воздух. Если обозначить температуру воз-
духа через а внутреннюю температуру глухариного яйца через
Го, Г5, 7\ (соответственно в начале опыта, через 5 мин., через
10 мин. и т. д.), мы получим ряд уравнений вида:
Т3-Т* К (Го - Гв );
Г10-Гв = № (Го -Гв );
7'15 — Гв =/<3 (Го — Гв ); и т. д.
«Коэффициент остывания» К для центрального яйца оказался
равным 0,9585, для периферийного 0,9526 (при отсчетах через ка-
ждые 5 мин.). Формула дает возможность вычислить температуру
240
внутри яйца глухарки в любой момент, если известны исходные ве-
личины— температуры воздуха и яйца: последняя, судя по литера-
турным данным, должна быть близка к 37°. Далее, как показал по-
ставленный нами опыт по изучению скорости остывания глухари-
ных яиц в гнезде, разность температур «яйцо — воздух» умень-
шается:
с 35 до 30'° через 17 мин., или, от начала опыта, через 17 мин.
„ зо „ 25° 20 37 ..
„ 25 „ 20° '25 „ 1 и. 02
„ 20 „ 15° 31 1 .. 33
„ 15 „ ю° 45 2 „ 18 ..
„ 10 „ 5° 76 ,, э „ 34
м Э „ 1° ... 176 6 „ 30
Поверхность яиц уже через 20 мин. после начала опыта была
на ощупь холодной, хотя внутренняя температура их в это время
была около 28—29°. Из приведенных цифр следует, что при темпе-
ратуре воздуха 17° за получасовую кормежку глухарки яйца осты-
вают не больше, чем на 5°, а при температуре 0° — только на 10°.
Небольшие периодические охлаждения яиц необходимы для нор-
мального развития зародыша, так как они способствуют вентиля-
ции воздуха внутри яйца через поры скорлупы. В настоящее .время
охлаждение применяется при промышленной инкубации яиц до-
машних птиц. Н. П. Третьяков указывает, что «охлаждение яиц во-
доплавающей птицы (уток, гусей) является необходимым меро-
приятием, и без него трудно получить хорошие показатели выводи-
мости... в инкубаторах» (Третьяков, 1951, стр. 197).
Яйца, оставленные на холоду на срок более 6 час., практически
принимают температуру окружающего воздуха. При температуре
•вьтое 0° яйца хорошо переносят даже длительные перерывы в на-
сиживании. При искусственной инкубации семи глухариных яиц
в термостате на двенадцатые сутки два яйца были вынесены нами
в погреб и оставлены там на 16 час. при температуре 8,0°. Впо-
следствии оказалось, что в одном из яиц эмбрион погиб еще до
опыта, а в другом он продолжал нормально развиваться до конца
инкубации.
Наблюдения не подтвердили нашего предположения о том, что
в холодные весны яйца тетеревиных погибают от холода во время
обычных кормежек наседки; в то же время выяснилось, что яйца
могут замерзнуть до начала насиживания или во время насижи-
вания, если оно прерывается на длительное время. Мы наблюдали
случай, когда тетерка оставила гнездо па 40 часов, в течение кото-
рых был сильный заморозок (см. ниже). Нередко глухарки бро-
сают кладки во время насиживания; эта аномалия также связана
с погодой, так как только в холодные весны эмбриональная смерт-
ность у тетеревиных птиц принимает массовый характер. В такие
годы большое число погибших кладок дает возможность кунице
и другим хищникам кормиться яйцами птиц даже зимой.
Замечательно, что реакция глухарки (а также самок других
тетеревиных птиц) на неблагоприятные условия погоды полезна
16-3037
241
только для данной особи, но не способствует обеспечению размно-
жения популяции при всякой обстановке. В результате этого раз-
множение вида в отдельные годы может быть подавлено, и тогда
популяция состоит, главным образом, из старых особей (см. ниже).
Это явление — основная причина колебаний численности тетереви-
ных в северных широтах.
В холодную и позднюю весну 1951 г. не только глухарки и те-
TepiKiw, но >и ужи бросали гнезда. В мае мы наблюдали, как гоголи,
снесшие полные кладки в двух дуплянках, начали-было «рассижи-
ваться», а потом бросили гнезда. По-видимому, причиной этого был
возврат холодов, сопровождавшийся выпадением снега 1 и 2 июня,
задержавший вскрытие озер. Позднее еще в нескольких дуплянках
были найдены брошенные ненасиженные кладки гоголей.
Мы полагаем, что инстинкт размножения в большей мере кон-
тролируется высшей нервной деятельностью, чем принято думать.
Птица живет в определенной географической среде и реагирует на
все изменения в ней, в частности на изменения погоды и аспекта
местности, что, вероятно, и определяет все сезонные периодические
явления в жизни птицы. Необходимым условием для насижива-
ния, помимо наличия гнезда, является наступление определенного
состояния психики (и организма в целом), без чего птица, отло-
жившая яйца, или совсем не приступает к насиживанию, или же
сидит не крепко и легко бросает гнездо по такой причине, которая
в другой год не имела бы никакого значения.
Эмбриональная смертность. В Лапландском заповеднике заре-
гистрировано 34 глухариных гнезда с 197 яйцами; дальнейшая
судьба 17 гнезд с 97 яйцами, не прослежена. В одиннадцати гнез-
дах, содержавших 63 яйца, выведено 53 птенца, семь погибли при
выводе (из них шесть в одном гнезде остыли и были закусаны му-
равьями), одно случайно разбито, два погибли на ранней стадии
развития (так называемые «болтуны»). Шесть кладок с 32 яйцами
погибли целиком: две с 11 яйцами сгорели в пожарах, одна
с 4 яйцами была разорена, по-видимому, куницей, три с 17 яйцами
брошены маткой. 1
Число брошенных кладок в действительности, возможно, и вы-
ше, потому что почти все гнезда были найдены по взлетевшей на-
седке, следовательно, уже после начала насиживания. Неполные
кладки гораздо труднее обнаружить, а среди них процент брошен-
ных должен быть гораздо больше, чем среди насиженных. Те яйца,
которые добывают из-под снега куницы и лисы, относятся либо
к брошенным кладкам, либо являются «болтунами», оставшимися
в гнездах после вывода птенцов. Большое значение яиц в зимнем
питании куницы косвенно подтверждает, что эмбриональная смерт-
ность является главной причиной «неурожая» тетеревиных птиц.
В Печоро-Илычском заповеднике в 1952—1958 гг. прослежена
судьба 10 гнезд, содержавших 62 яйца; еще 5 гнезд с 31 яйцом были
найдены уже после вывода птенцов. В одиннадцати гнездах из
69 яиц вывелось 59 птенцов, 1 яйцо случайно выкатилось из гнез-
242
да, в девяти эмбрионы погибли на ранней стадии. Остальные четыре
кладки погибли целиком, две из них с 14 яйцами были разорены
хищниками, одно с 4 ненасиженными яйцами брошено маткой (или
она погибла?), в одной кладке все 6 яиц погибли до насиживания
или в начале его.
Суммируя наблюдения по эмбриональной смертности у глу-
харя, мы получаем следующие цифры:
Лапландский заповедник Печоро-Илыч - ский запо- ведник Всего
Птенцы уведены с гнезда 56 % 64 % 59 %
Птенцы погибли при выводе 7 „ — 4 „
Эмбрионы погибли на ранней
стадии 2 „ 16 „ 9 „
Яйцо разбито или выкатилось 1 . 1 . 1 ,
Кладка уничтожена хищником 4 „ 15 „ 10 „
„ сгорела при лесном по-
жаре 12 „ — 6 „
„ брошена наседкой 18 . 4 „ и „
Общее число яиц, шт. . . . 95 93 188
Число уведенных цыплят определялось нами по числу скорлуп
в гнезде, так как разыскать выводок обычно не удается.
Рис. ‘87. Гнездо тетерки в Лапландском заповеднике, 8 VI 1951 г.
Размножение тетерева. В отличие от глухарки тетерка иногда
устраивает гнездо среди поляны или на открытой кочке в заболо-
ченном лесу, в стороне от деревьев. Тетеревиные яйца отличаются
16*
243
от глухариных меньшей величиной и более крупными и яркими
крапинами по основному фону (рис. 87): обычно фон имеет более
теплый оттенок, а крапинки — кирпично-бурые, а не коричневые,
как у глухаря. Вес яиц из двух кладок, найденных в Печоро-Илыч-
ском заповеднике (одна из них неиасиженная, другая —на второй
день инкубации), — от 30 до 38 г, в среднем 36 г в первом гнезде
и 32,2 г во втором. В Северном Казахстане, по Н. С. Ульянину
(1949), вес яиц от 28 до 35 г. Одно яйцо из окрестностей Ленин-
града весило 35 г. Чаще всего кладка насчитывает 8 яиц; геогра-
фической изменчивости в ее величине не обнаружено (табл. 70).
Таблица 70
Величина полной кладки тетерева в разных областях
Местность 4 5 Число яиц в гнезде Число гнезд
6 7 8 9 10 11 сред- нее
Лапландский зап-к, 68° с. ш. '32° в. д. (автор). — — — 1 2 1 — — 8,0 4
Печоро-Илыч. зап-к, 62° с. ш. 57° в. д, (автор) Печоро-Илыч. зап-к (Тепло©, Г947, всего 117 яиц) (1) (2) 7,0 7,3 3 16
Вологодская обл., 60° с. ш. 40° в. д. (Ларин, 1*94-1) 1 1 — 2 3 4 1 — 7,8 12
Башкирский зап-к, 53° с. ш. 58° в. д. (Кириков, 1*952) — 1 3 2 3 — 1 1 7,5 11
Беловежская пуща, 33° с. ш. 24° в. д. (Гаврин, 1956) — 1 2 — 3 3 1 2 8,3 12
Наурзумский зап-к, 52° с. ш. 64° в. д. (Ульянин, 4949) 2 — — 2 3 1 1 — 7,3 9
Прибайкалье, 52° с. ш. 108° в. д. (Фетисов, 4934) — 1 — — 2 1 — — 7,5 4
По всем пунктам 3 5 5 5 17 9 7 4 7,7 71
Одно гнездо найдено 5 VI 1951 г. на побережье Охтозера,
в 15 км от управления Лапландского заповедника, в редком забо-
лоченном сосняке с напочвенным покровом из багульника, воро-
ники и брусники, на кочке (без всякого прикрытия); оно содер-
жало 7 яиц. Обнаружено по тетерке, которая прилетела и села
около гнезда. 18 VI, когда мы установили самописец, в гнезде ока-
залось только 6 яиц. Прибор проверялся только один раз в не-
делю, в результате чего запись оказалась почти непригодной для
обработки. Три тетеревенка вывелись, по-видимому, 26 июня,
а 28-го 'Б 1 ч. 3'3 м. матка увела их от гнезда. 24 июня одно из яии
было найдено около гнезда разбитым, а в остальных двух эмбрио-
ны погибли на разных стадиях развития.
244
В Печоро-Илычском заповеднике мы исследовали два гнезда.
Одно было найдено 19 V 1953 г. на захламленной вырубке близ
границы заповедника, под густой молодой ёлочкой, по слетевшей
тетерке; в нем оказалось 7 яиц. При установке самописца в 13 ч.,
20 мая тетерка снова была поднята с гнезда, и в нем было уже
8 ненасиженных яиц (дальнейшая история этого гнезда описана
ниже). Другое гнездо с 8 насиженными яйцами было найдено
и отсутствует —__т-ра от 0*до5°
Ж спугнута с гнезда
Рис. 88. Режим насиживания тетерки в Печоро-Илычском заповед-
нике, 1957 год
24 V 1957 г. в 1 км от управления заповедника, в редком ягельном
бору, близ края посадочной площадки, под молодой сосенкой. Са-
мописец был установлен на следующий день; насиживание продол-
жалось нормально, и 16 июня в 16 ч. 30 м. матка увела птенцов.
По этим двум гнездам мы получили более 31 суток записи насижи-
вания: режим его сходен с режимом насиживания глухарки.
Тетерка оставляет гнездо от 1 до 4 раз в сутки, чаще два раза
(50% случаев) или три раза (37%). Продолжительность отлучек
чаще всего от 10 до 30 мин.: самая длинная наблюдалась в первые
245
часы насиживания. Число и продолжительность отлучек наседки
характеризуются следующими цифрами:
Продолжитель- ность отлучки в мин. Число случаев
5 3
10 11
15 29
20 16
25 12
30 7
35 —
40 —
45 1
Средн. 18 мин.
Общая продолжительность отлучек составляла: у первой тетер-
ки 5,3%, у второй 2,4% всего времени насиживания, причем с те-
чением времени отлучки становились все более и более короткими.
У второй тетерки в первую неделю они составляли 2,6%, во вто-
рую— 2,4%, в третью — 2,1% всего времени. Продолжительность
«прогулок» наседки не зависит от температуры воздуха (г = —
— 0,18 + 0,20). Несмотря на то что в период насиживания в Пе-
чоро-Илычском заповеднике стоят белые ночи — у тетерок хорошо
выражен период ночного покоя , от 2Й до 1 час. по местному
среднему солнечному времени; она редко оставляет гнездо также
между 13 и 21 час. Вечерняя кормежка приурочена к 21—23 час.,
первая утренняя кормежка происходит обычно в 1—3 час., вто-
рая— в разное время между 5 и 13 час. (табл. 71).
T а б л и ц а 71
Суточный цикл насиживания тетерки в Печоро-Илычском заповеднике (в %0 от всего времени наблюдений — 751 час.)
Часы суток 23 1 3 5 7 9 11 13 15 17 19 21 23 Полные сутки
— 71 23 39 31 28 45 8 5 8 5 120 32
Хотя тетерка оставляет гнездо при различных температурах
воздуха, но при 0—5° несколько чаще, а при температурах выше
15° сравнительно редко (табл. 72). Первая утренняя кормежка
происходит приблизительно во время суточного минимума темпе-
ратуры воздуха, иногда при заморозке. Так, например, 24 V 1953 г.
тетерка оставила гнездо на 19 мин. при —2,0°, а 27 V 1957 г. на
11 мин. при —1,5°. Как мы уже упоминали, 20 V 1953 г. в 13 час.
тетерка была спугнута с гнезда для установки в нем актографа.
В тот же вечер наступило ненастье, около суток дул северный ве-
тер, сопровождавшийся дождем или снегом, но 21 мая к ночи про-
яснило и наступил мороз, достигавший у гнезда —5,3°. Тетерка не
246
появлялась на гнезде в течение 40 час. и вернулась только 22 мая
в 5 ч. 10 м., после почти 8-часового заморозка. В эту ночь яйца
.должны были пострадать от мороза, но проверить это предполо-
жение, к сожалению, не удалось, так как 1 июня в 9 час. кладка
была целиком уничтожена каким-то хищником. Однако в том же
районе 23 мая было найдено гнездо глухарки, в котором все яйца
оказались «болтунами», то есть с мертвыми эмбрионами.
Таблица 72
Температура воздуха (в °C) во время насиживания и отлучек тетерки
(в % от общего числа случаев)
В ночь на 27 V 1953 г. выпал снег, однако тетерка вышла на
кормежку в обычное время, несмотря на снежный покров. В 1957 г.
тетерка при дождливой погоде обычно пропускала очередную кор-
межку (28 V — утреннюю, 2, 4 и 5 VI — вечернюю). В четырех
других случаях пропуск кормежки нельзя было объяснить небла-
гоприятной погодой.
В 1953 г. тетерка переворачивала яйца от 18 до 34 раз, в сред-
нем 25 раз в сутки, как и глухарка. Движения наседки распределя-
лись по часам довольно равномерно, но от 0 до 6 ч. несколько реже,
от 6 до 12 ч. чаще. В 1957 г. тетерка в конце насиживания шевели-
лась на гнезде в один день 38, в другой — 39 раз.
Спугнутая с гнезда тетерка возвращается через такое же время,
как и глухарка (1 ч. 19 м. и 2 ч. 53 м. в Лапландском заповеднике,
2 ч. 34 м. и 0 ч. 58 м. в Печоро-Илычском). Во время вывода птен-
цов матка в течение I8V2 часов не оставляла гнезда. Птенцы начали
выводиться в 22 ч. 39 м., в 5 ч. 42 м. наседка успокоилась и пере-
стала шевелиться, а в 16 ч. 30 м. 16 VI 1957 г. увела цыплят. Через
2 ч. 35 м. после этого выводок был обнаружен в 26 м от гнезда
(средняя скорость 'передвижения 10 м/час).
Продолжительность инкубации яиц в 1951 г. была не менее
22 суток, в 1957 г. — не менее 23 суток. Литературные указания на
этот счет расходятся: А. А. Хватов (1860) определил ее в 28 дней,
С. А. Ларин (1941) —в 24 дня, Н. С. Ульянин (1949) — в 23 дня,
С. И. Шпаковский (1937) —от 19 до 22 дней. По мнению М. А.
Мензбира (1895, 1900—1912), ,в средней полосе тетерка насиживает
19—22 дня, а на севере 23—25 дней. Это предположение мало ве-
роятно, так как у других видов тетеревиных не обнаружено геог-
рафической изменчивости периода инкубации.
247
В. П. Теплов (1947) нашел, что в Печоро-Илычском заповед-
нике «первые выводки глухаря появляются в среднем на 12 дней
раньше выводков тетерева» (первые — 21 VI, вторые — 3 VI1). По
данным, собранным тем же автором, средний размер выводка поч-
ти нс меняется по годам, а также и в географическом разрезе:
у глухаря в среднем 3,4 птенца, у тетерева — 4,5, у рябчика5,3.
Размножение рябчика. Л. П. Сабанеев (1876) справедливо за-
метил, что «найти гнездо рябчика очень трудно, так как оно всегда
бывает в чаще и вообще в малодоступных местах и потому, хотя
рябчики ©езде многочисленнее глухарей, но яйца последних попа-
даются гораздо чаще. На Урале/например, я нашел всего только
2 гнезда рябчика, между тем как яйца кополухи видел раз 12—15,
если не больше». Сабанеев указывает далее, что самка несется ка-
ждый день и, положив 8—10 яиц, садится высиживать их; иногда
кладки достигают 14—15 яиц; насиживание продолжается 3 не-
дели и начинается несколько позже, чем у тетеревов; самец, по-
видимому, не принимает участия в насиживании, хотя и держится
около гнезда и позднее водит птенцов вместе с маткой и даже за-
меняет ее в случае гибели. До последнего времени к этим сведениям
мало что можно было добавить. Определение продолжительности
инкубации в 21 день приводится почти во всех книгах без указания
источника; только А. Брем (1911, т. VII) упоминает, что это на-
блюдение принадлежит Сабанееву.
Интересные новые данные о размножении рябчика сообщает
А. Пюннонен (1954). В районе Йоэнсу (восточная Финляндия) он
наблюдал кладку яиц и насиживание их в двух гнездах. Одна
самка рябчика за 15 дней снесла 9 яиц (май 1950 г.), у другой от-
кладка 10 яиц продолжалась 13 или 14 дней (весна 1952 г.). Пол-
ная кладка содержит от 7 до 12 яиц, чаще всего 8—10. Пюннонен
упоминает также О' гнездах с 15 и даже с 19 яйцами. Откладка
последнего яйца происходит уже после того, как самка начнет
насиживать. В одном гнезде инкубация продолжалась 27 суток,
в другом — 25. Показания Пюннонена противоречат данным Саба-
неева только в отношении периодичности откладки яиц и продол-
жительности периода насиживания.
Точных данных о размножении рябчика в Советском Союзе
мало. В Лапландском заповеднике было найдено только одно
гнездо, 31 V 1947 г.: в нем было 7 яиц и при нем находились как
самка, так и самец. Птенцы у рябчика появляются немного позже,
чем у глухарки. Сравнительно много гнезд рябчика найдено в Пе-
чоро-Илычском заповеднике (Донауров, 1947): в табл. 73 эти дан-
ные пополнены позднейшими наблюдениями по картотеке заповед-
ника.
Подробно исследовано одно гнездо рябчика, найденное нами
в елово-пихтовом лесу на берегу Печоры 22 V 1956 г., в 11 час., под
старой еловой валежиной, в углублении почвы. Я собирался поста-
вить туда мышиный капканчик и нагнулся к выходу, в этот момент
самка рябчика вылетела из-под валежины и под ней оказалось
248
Таблица 73
Величина полной кладки рябчика на Урале и в Белоруссии
Название заповедника
Лапландский, 68° с. ш.
32° в. д....................
(автор)
Печоро-Илычский, 62° с. ш.
57° в. д....................
(Донауров, '1'947 и автор)
Башкирский, 53° с. ш. 58° в. д.
Ильменский, 55° с. ш. '60° в. д.
(Кириков, 1948, Ушков,
И949)
Беловежская пуща, 53° с. ш.
24° в. д....................
(Гаврин, 1'956)
По всем пунктам:
Число яиц в гнезде
4 5 6
1 2 1
1 2 4
1
22
5
17
45
4 холодноватых яйца. При установке актографа па следующий
день в 12 ч. 15 м. матки на гнезде не было, но она была на нем
раньше, так как яйца были слегка прикрыты опавшими березо-
выми листьями; время кладки яиц показано на диаграмме рис. 89.
30 мая утром матка снесла десятое яйцо и в тот же день в 22 час.
начала насиживание. После этого она снесла последнее (одиннад-
цатое) яйцо, что было установлено только 17 VI, во время кор-
межки наседки (за весь период наблюдений ее не спугивали ни
разу). Вес ненасиженных яиц был от 16,5 до 17,9 г, в среднем
17,2 г (свешано 9 яиц утром 29 мая). Промежуток между отклад-
кой двух яиц у рябчика оказался от 26 до 30 часов, причем каждый
следующий раз птица проводила на гнезде все большее время —
видимо, уже после того, как снесла яйцо. Время кладки яиц посте-
пенно сдвигается с утра на вечер; после этого птица пропускает
день и новый цикл кладки начинается опять с утра. У нашего ряб-
чика днями пропуска были 24 и 29 мая, а также, по-видимому,
19 мая.
Кладка яиц у рябчика
(в скобках данные экстраполяции)
Таблица 74
№ яйца
1|2|3|4|5|6| 7 | 8 9 | 10 | 11
День откладки (май)'
Часы суток (прибл.)
Интервал (в часах)
Проведено на гнезде,
минут...............
(18) (20)
(16) (5)
(21)
(8)
22 23
11 17
30
25 26
5 8
27 27
27 28 30 (31)
11 15 6 (8)
28 39 26
79 130 138 186 308 328 318
249
За весь период насиживания (25,4 суток) наседка оставляла
гнездо 81 раз: в первые 2 суток по 5 раз, потом несколько дней по
4 раза (25% случаев), позднее по 3 раза (50% случаев) или даже
I--> птица на гнезде t____j отсутствует
----т-ра ниже 0* ---т-ра or? О"до 5*
Рис. 89. Режим насиживания рябчика в Печоро-Илычском заповед-
нике, 1956 год
по 2 раза; 23 VI снова пять раз, но, вернувшись с кормежки в этот
день в 19 ч. 15 м., она в течение 36 ч. 43 м. не оставляла гнезда,
пока не увела птенцов 25 июня в 7ч. 58 м. (рис. 89). 24 июня дул
холодный северный ветер, температура была от 6 до 12°. Насижи-
вание от начала до увода птенцов продолжалось 25 суток 10 часов.
250
Число и продолжительность отлучек наседки характеризуются
следующими цифрами:
Продолжитель-
ность отлучки
в мин.
Число
случаев
5 —
10 —
15 17
20 34
25 20
30 6
35 3
40 —
45 1
Среди. 22 мин.
В большинстве случаев самка рябчика отсутствовала от 15 до
25 мин. Утром 17 июня мы подкараулили, когда она слетела на
кормежку, и осмотрели гнездо; она возвратилась только , через
47 мин. Общая продолжительность кормежек составила 5,2% соб-
ственно периода насиживания (без времени вывода и обогревания
цыплят). Как и у других тетеревиных, время кормежек с течением
времени немного сокращается: в первые 26 часов — 7,1 %, в следую-
щие 8 суток — 5,9%, в средние 7 суток — 4,8%, в последние 8 су-
ток— 4,5%, в течение 32 часов вывода и обогрева птенцов — 0%.
Период ночного покоя хорошо выражен: с 22 ч. 15 м. до 2 ч. 56 м.,
то есть в течение 4,7 час., наседка ни разу не оставляла гнезда.
Утром она уходила на кормежку в разное время, от 3 час. в на-
чале насиживания до 8 час. в конце его. Вечерняя кормежка обыч-
но наблюдалась между 19 и 21 ч. (табл. 75).
Таблица 75
Суточный цикл насаживающего рябчика на Верхней Печоре
(в %0 от всего времени наблюдений — 610 час.)
Часы суток Полные
23 1 3 5 7_9 11 13 15 17 19 21 23 СУТКИ
1 37 56 106 45 19 57 65 59 90 56 49
В отличие от глухарки и тетерки, у рябчика средняя темпера-
тура моментов кормежки немного выше температуры всего периода
инкубации, включая и вывод птенцов: объясняется это тем, что
рябчик чаще кормится среди дня, а ночной отдых у него продол-
жительнее. Температура воздуха не влияет на продолжительность
кормежки: коэффициент корреляции г = — 0,14 + 0,20 (при вы-
числении коэффициента первый и последний дни насиживания от-
кинуты). Во время насиживания рябчика в 1956 г. температура
251
воздуха ни разу не опускалась ниже 0, но перед его началом, ночью
29—30 мая, в течение 7 часов был мороз до —1,8°; яйцам он не
повредил.
Табл и ц а 76
Температура воздуха (в °C) во время насиживания и отлучек рябчика
___________ (в % от общего числа случаев)__________________________
Пределы температур Средн, темпера- тура
-5 0 5 10 15 20 25 30
610 час. насижи- вания ... — 14 32 31 19 4 — 10,9
81 кормежка . . — 11 27 40 18 4 — 11,4
Переворачивание яиц актограф отмечал только в последние
10 дней работы: оно производилось 27—30 раз в сутки, а в послед-
ние четыре дня от 33 до 47 раз; после того, как птенцы вывелись
(24 VI в 21 ч.), движения на гнезде прекратились. Во время клад-
ки яиц рябчик шевелится на гнезде чаще, чем во время насижива-
ния, в среднем 4—5 раз в час.
Самец рябчик не принимает участия в насиживании, по дер-
жится около гнезда. При смене лент мы несколько раз слышали
его голос, а 2 VI видели его на земле в 27 м от гнезда.
Размножение белой куропатки. В Лапландском заповеднике бе-
лые куропатки гнездятся либо в ельниках, либо на сфагновых или
осоково-злаковых болотах. Сравнительно редко гнездо устраи-
вается под елью или другим деревом (3 случая), чаще под крохот-
ными, угнетенными елочками с грибообразной кроной и не выше
0.5 м ростом (4 случая) или на кочке среди болота, открыто или
под защитой кустиков карликовой березки (6 случаев); иногда
гнездо располагается на прогалине, совсем открыто или около мо-
лодой сосенки или побега березы, служащего ориентиром (4 слу-
чая). В сходных условиях белая куропатка гнездится и в других
географических зонах — тундровой и лесостепной.
В гнезде куропатки более глубокий лоток, чем в гнезде глу-
харки и в нем больше подстилки: она состоит из стеблей и листьев
осок и злаков (напр., молинии) с примесью перьев матки, а иногда
и сухих березовых листьев. Даты находок гнезд следующие: 25 V
1937 г., 5 VI 1950 г., 8 VI 1938 г. (все три в начальной стадии инку-
бации); 14 VI 1933 г., 15 VI 1939 г., 18 VI 1934 г., 20 VI 1934 г.;
30 VI 1950 г. (птенцы постукивают); 4 VII 1936 г., 5 VII 1951 г.
(в последнем птенцы вывелись 18 VII). Средняя дата всех 10 на-
ходок 17 июня, на 10 дней позже средних дат находок гнезд глу-
харок и тетерок.
Яйца куропатки пигментированы сильнее, чем тетеревиные; по
общему бледно-желтоватому фону они густо испещрены довольно
крупными темно-красно-бурыми пестринами. Вес девяти свежих
252
яиц от 19,7 до 21,0 г, в среднем 20,3 г; сходные величины приводят:
для тундровой зоны А. В. Михеев (19—22г) и для степной —
Рис. 90. Гнездо белой куропатки в Лапландском заповеднике,
10 VI 1950 г.
Н. С. Ульянин (16—21 г.). Сведения о количестве яиц в гнезде при-
ведены в табл. 77.
Таблица 77
Величина полной кладки белой куропатки в разных областях
Местность Чис л о яиц Число гнезд
6 7 8 9 10 11 12 13 14 15 Сред- не
Южный Ямал, 68° с. ш.
&8° в. д — . — 8,2 13
(Дунаева, Кучерук. 1941)
Тим а иска я тунд. ’68° с. ш.
50° в. д (Михеев, 1948) — — 1 1 3 2 5 — 1 1 11,3 14
Лапландск. зап-к. 68° с. ш.
32° в. д (автор) 2 3 6 2 1 1 — — — — 8,0 15
Наурзумск. зап-к 52° с. ш.
64° в. д (Ульянин, 1'939, 1949) — — — 2 3 7 6 1 — — ПЛ 19
По всем пунктам . . . Юкон, Аляска, 63° с. ш. 2 3 7 5 7 10 11 1 1 1 9,7 61
1*63° 3. Д. . . . . . (Бент, 1932) — 3 2 1 3 1 — — — — 8,7 10
253
Обычная величина кладки — от 8 до 12 яиц; среднее число —
в одних местах 8, в других—11, без определенной географической
закономерности. Необходимо учитывать, что величина кладки мо-
жет быть разной не только в отдельных популяциях, но и в зави-
симости от условий данного года, а возможно, и от возраста пти-
цы: впрочем, указания старых авторов, что молодые птицы откла-
дывают меньше яиц, чем старые, нельзя принимато на веру, если
нет упоминания о способе определения возраста этих птиц. Клад-
ка шотландской куропатки, по А. Лесли (1911), чаще всего содер-
жит 7 или 8 яиц.
По наблюдениям Н. С. Ульянина, самка белой куропатки несется
утром и притом ежедневно. Продолжительность инкубации лам не
удалось определить, но она во всяком случае больше 18 суток.
А. В. Михеев (1948) наблюдал в Тиманской тундре вывод птенцов
через 19 дней после находки гнезда (3 случая); Н. С. Ульянин
(1939) нашел, что насиживание продолжается 18 дней, но в двух
гнездах оно длилось 20 дней. Другие авторы указывают большие
сроки: К. Вестеркув (1956) в Норвегии 20*/2—22 дня, А. Бент
(1932) в Аляске около 22 дней, А. Лесли (1911) в Англии 23—
24 дня, В. Ушаков (1904) в «Сибири» 27—30 дней. Последнее опре-
деление мы считаем ошибочным и склонны думать, что инкуба-
ция у белой куропатки продолжается 21—22 дня.
Белая куропатка сидит на гнезде очень крепко. Нам удавалось
осторожно приблизиться к гнезду для фотосъемки до 2 м, что легко
было установить по метражной шкале объектива зеркальной ка-
меры; при этом фотограф стоял во весь рост. Бент описывает инте-
ресный случай, когда наблюдатель накрыл гнездо куропатки сво-
ей курткой: наседка нашла кладку, сделала новое гнездо в 30 см
от старого, перенесла в него яйца и продолжала насиживание,
Ульянин сгонял одну куропатку 25 раз на протяжении всего пе-
риода насиживания, и несмотря на это она все же вывела цыплят.
У другой самки вырвали все перья хвоста, но через 25 минут она
снова сидела на яйцах. Спугнутая с гнезда собакой, куропатка
обычно убегает, подпархивая и крича, и уводит собаку за собой;
спугнутая человеком, иногда бежит, а иногда улетает. В большин-
стве случаев около гнезда находится и самец, который также уча-
ствует в демонстрации тревоги, но при этом держится осторожнее,
чем самка.
Запись поведения насиживающей куропатки проведена нами
в двух гнездах. Первое было найдено 5 VI 1950 г. в 21 час. на сфаг-
новой кочке среди мокрого осоково-сфагнового болота. В нем было-
8 яиц, самка насиживала, самец находился поблизости. Актограф
был установлен около 14 час. 6 июня, но только с 13 час. 10-го по-
лучена хорошая запись, продолжавшаяся до увода птенцов с гнез-
да 23 VI в 13 ч. 30 м. Второе найдено 5 VII 1951 г. не далее 1 км от
первого, у края мокрого осокового болота, на сфагновой кочке, под
прикрытием двух крохотных (0,5 м) елочек: в нем было 7 яиц,
самца при нем не было. В тот же день в 17 час. был установлен
254
актограф, который дал хорошую запись до 2 час. 12 июля, когда
наседка увела цыплят (рис. 91). Находившиеся на болоте гнезда
были очень сырыми. Контактная коробка, вынутая из гнезда 10 VI
1950 г. для регулировки, оказалась мокрой снаружи и изнутри.
г- птица на гнезде u_i отсутствует * спугнута с гнезда
--------тра ниже 0° --— тра от О'доУ
Рис. 91. Режим насиживания белой куропатки в Лапландском запо-
веднике в 1950 г. (вверху) и в 1951 г. (внизу)
При уборке актографа из второго гнезда оказалось, что контакт-
ная коробка была облеплена мелкими рыжими муравьями, что,
по-видимому, не беспокоило птицу.
Спугнутая куропатка возвращается на гнездо очень скоро —
от 36 м. до 1 ч. 24 м., в среднем через 1 ч. 01 м. от момента спуги-
вания. Режим насиживания белой куропатки отличается от режи-
ма насиживания глухарки более частыми и кратковременными
кормежками. Она оставляет гнездо от 3 до 5 раз в сутки (чаще
4 раза), каждый раз на срок от 10 до 32 мин.; средняя продолжи-
255
тельность кормежки—19 мин. Суммарная продолжительность
всех кормежек составляет 4,5% всего времени насиживания. В обо-
их гнездах оно происходило в период незаходящего солнца. Как
и у других тетеревиных, при сходстве часов кормежки в соседние
дни они со временем смещаются таким образом, что распределя-
ются по всем часам суток (рис. 91 и табл. 78). Наседка наиболее
активна утром, от 7 до 9 ч., наименее — в полуденные часы, с 10 до
16 ч., период ночного покоя на 68° с. ш. полностью отсутствует.
Число и продолжительность отлучек наседки с гнезда характеризу-
ются следующими цифрами:
Продолжительность Число
отлучки в мин. случаев
5
10
15
20
25
30
35
40
45
14
12
28
2
8
Среди. 19 мин.
Таблица 78
Суточный цикл насиживающей белой куропатки в Лапландском
заповеднике
(в %0 от всего времени наблюдений — 464 час.)
Часы суток Полные сутки
23 1 3 5 7 9 11 13 15 17 19 21 23
23 54 68 . 58 124 29 14 23 55 27 16 37 45
Таблица 79
Температура воздуха (в ОС) во время насиживания и отлучек
белой курапатки
______________(в % от общего числа случаев)___________
Пределы температур Средн- темпера- тура
-5 0 5 10 15 20 25 30
464 час. насижи- вания .... 65 кормежек . . 2 2 12 18 43 40 24 28 13 9 6 3 — 10,1 9,2
256
Кормежка при отрицательной температуре наблюдалась толь-
ко один раз, 13 VI 1950 г., в течение 18 мин.
В период вывода птенцов поведение белой куропатки меняется
так же, как и у глухарки. 22 VI 1950 г. в 12 ч. 17 м. самка вернулась
с кормежки и не оставляла гнезда в течение 19 часов; 23 июня она
оставляла гнездо на короткое время в 7 ч. 18 м. и в 9 ч. 13 м.;
в U31 ч. 29 м. матка увела птенцов с гнезда, а через 46 мин. мы разы-
скали их в И м> от гнезда (скорость движения 14 mi в час). Удалось
обнаружить только 5 птенцов из 8. Самец в это время беспокойно
бегал по другой стороне болотца. Позднее там же было найдено не-
сколько перьев, потерянных шм или в драке с другим петухом, или
при нападении хищника. Непосредственно у гнезда найдена боль-
шая куча помета наседки.
10 VII 1951 г. в 12 ч. 23 м. мы спугнули куропатку с гнезда для
его осмотра: в это время птенцы в яйцах еще не стучали. Через
43 мин. наседка вернулась и не сходила с гнезда на протяжении
34 час., только один раз «чиркнув» пером на ленте. 11 июля между
23 и 24 ч. она отсутствовала в течение 28 мин. (возможно, пыталась
увести выводок?), а 12-го в 1 ч. 24 м. увела цыплят окончательно.
Через 10 час. после этого выводок уже не удалось разыскать. В 1 м
от гнезда была большая куча свежего помета. В гнезде не оказалось
скорлуп: очевидно, они были кем-то съедены. Таким образом, во
время вывода птенцов белая куропатка не кормится и не оставляет
гнезда около суток; после того она может один или два раза сойти
на несколько минут и вскоре уводит птенцов, что в обоих наблюдав-
шихся нами случаях происходило утром.
А. Лесли (1911) указывает, что наседка грауса (в неволе) иног-
да, без видимых причин, покидает гнездо па несколько дней, потом
возвращается и высиживает цыплят. Весной 1908 г. наблюдались
случаи, когда гнезда (в природе, на верещатниках) были несколько
дней покрыты снегом, причем в некоторых из них даже яйца обес-
цветились, и все же из ’них вывелись здоровые цыплята1. * Основной
причиной гибели кладок Лесли считает то, что самка грауса остав-
ляет гнездо, когда суровая погода делает насиживание невыноси-
мым. Приостановка инкубации в начальном ее периоде не приносит
вреда, ио, когда в яйцах началось кровообращение, их уже нельзя
остуживать. «Если самка принуждена оставить гнездо вследствие
сильного дождя, есть опасность, что она совсем бросит кладку —
для этого достаточно трех дней мокрой погоды... Другая опасность,
которой подвержены яйца грауса, это гибель их от мороза во время
отсутствия птицы с гнезда. Эта опасность наиболее велика в период
* Приводимые Снигиревским (4947) со ссылкой на Лесли показания очевид-
цев о том. что птенцы выводились из яиц, примерзших к гнездовой подстилке
или переживших мороз в 25°, мы считаем сомнительными; невероятно, чтобы
на Британских островах в апреле мороз мог достигать —25° по Фаренгейту
(—82° С) или даже—25° по Цельсию. Подлинное выражение автора «25 degrees
of frost» — по всей вероятности надо понимать как 4-25° по Фаренгейту, т. е.
—4° С.
17—3037
257
до окончания полной кладки, ибо, после того, как насиживание на-
чалось, курочка неохотно оставляет свое гнездо во время мороза на
сколько-нибудь продолжительное время» (Лесли, 1911, стр. 10).
Наши выводы близки к заключению Лесли.
В Лапландском заповеднике найдено было 15 кладок белой ку-
ропатки во время насиживания и 5 старых гнезд со скорлупами.
Прослежена судьба девяти гнезд с 73 яйцами. Одна неполная
кладка из 5 яиц была уничтожена хищником (7% яиц); япце погиб-
шими эмбрионами1 не было отмечено; таким образом, птенцы вы-
велись из 93% яиц. Одно гнездо сгорело в пожаре, но произошло
это до вывода птенцов или после — установить не удалось.
Тундряная куропатка. Размножение тундряной куропатки мало
изучено. В литературе упоминаются только случайные находки
гнезд и не всегда можно разобрать, были ли они с насиженной или
неполной кладкой.
В Лапландском заповеднике одно гнездо, с 4 яйцами, было
встречено 4 VII 1931 г.; другое гнездо, с 2 яйцами и одним только
что выведшимся цыпленком, было найдено 3 VII 1933 г. на высоте
около 500 м абс.; третье, с 7 яйцами, насиженными примерно не-
делю, найдено 23 VI 1939 г., на такой же высоте, как и предыду-
щее. Все три гнезда были обнаружены по вспорхнувшей наседке;
они были расположены в одном районе, на юго-восточной оконеч-
ности Чуна-тундры. Кроме того, в июне 19*34 г. были обнаружены
две прошлогодние кладки: в одной было 3 яйца с оперенными
птенцами и еще скорлупы от 1—2 яиц, другая была брошена
с 8 насиженными яйцами.
С. йонсен (1941) описывает 4 гнезда, найденных в Финмаркене
и на Шпицбергене; они содержали 2, 5, 11 и 13 яиц. Г. П. Де-
ментьев и др. (1952) упоминают пять гнезд, с 4, 5, 9, 11 и 13 яйца-
ми. В. М. Сдобников (1957) нашел на Северном Таймыре в 1947
и 1948 гг., между 22 VI и 7 VII, шесть гнезд, содержавших 3, 3,
4, 6, 7 и 11 яиц.
Судя по этим данным, средняя величина кладки тундряной
куропатки — 6,7 яиц, причем большие (11 —13 яиц) и малые
(2—3 яйца) кладки встречаются одинаково часто. По Лапланд-
скому заповеднику среднее число яиц 5,3, а средняя дата находки
гнезд — 30 июня, позже, чем у других видов тетеревиных.
Гнездо располагается в открытой тундре, на ягельнике с кар-
ликовой березкой и вороникой или на чистом вороничнике, чаще —
между скалистых россыпей; подстилка лотка скудная. Яйца сходны
с яйцами лесной белой куропатки: размеры одного — длина 42 мм,
ширина 30 мм, вес 19,5 г (приблизительно на шестой день инку-
бации).
Срок насиживания дается только в научно-популярных изда-
ниях (Брем, 1911, — «около 3 недель»; Иванов, 1951, — 24—26 дней;
Тугаринов и Портенко, 1950, — 30 дней), без указания источников
и способа определения: эти данные не могут претендовать на точ-
ность.
258
Замечательно, что из 26 или 27 яиц, найденных на Чуна-тунд-
ре, одиннадцать (41—42%) были прошлогодними, с эмбрионами,
погибшими в процессе инкубации или в конце ее. Судя по этим
небольшим данным, эмбриональная смертность у тундряной куро-
патки выше, чем у других тетеревиных, но причины ее тс же, что
и у них,— прекращение насиживания или гибель наседки в этот
период, а также гибель птенцов во время вылупления.
Все 5 взрослых самок тундряных куропаток, добытые в июле
и первой половине августа, имели наседные пятна. Одна из них
(12 VIII 1930 г.) была с выводком в 6 птенцов, другая (9 VIII
1935 г.) поймана браконьером в силок и наличие птенцов при ней
не установлено; остальные три (22 VII 1933 г., 12 VII 1935 г.,
14 VII 1938 г.) были заведомо без выводков. Случаи полной гибели
кладок или выводков нередки и у других тетеревиных; так, на-
пример, 24 VII 1940 г. нами была добыта одинокая глухарка с от-
четливым наседпым пятном.
Приведенные факты подтверждают наше представление о боль-
шой роли эмбриональной смертности в колебаниях численности те-
теревиных птиц.
XI. УЧЕТ И ДВИЖЕНИЕ ЧИСЛЕННОСТИ
Способы учета. Обзоры способов учета куриных птиц даны
в работах С. В. Кирикова, А. В. Михеева и Е. П. Спангепберга
(1952, стр. 260—275) и Г. А. Новикова (1953, стр. 189—192), однако
без критической их оценки. Едва ли в лучшем положении находится
и методика учета какой-либо другой группы животных. П. В. Те-
рентьев указывает: «К сожалению, математическая теория количе-
ственного учета пока еще не только не разработана, но большин-
ство исследователей даже не отдает себе ясного отчета в том, что
именно представляют собой полученные ими цифры» (Новиков,
1953, стр. 171). Между тем для изучения движения численности по-
пуляций животных теоретическая разработка основ учета совер-
шенно необходима; без этого мы не знаем, в какой мере данные
наших учетов пригодны для решения поставленных перед исследо-
вателем задач. *
Не имея возможности углубиться в теорию вопросов учета, мы
попытаемся разобраться в рекомендуемых в настоящее время спо-
собах его только с практической стороны.
В качестве основного способа учета различных куриных птиц
Кириков, Михеев и Спангенберг выдвигают учет самцов весной,
либо на токах (глухарь, тетерев), либо по гнездовым территориям,
обнаруженным по брачным крикам или визуально (рябчик, белая
куропатка, различные представители семейства фазановых).
С. В. Кириков пишет: «Учет глухарей на токах есть достаточно точ-
ная перепись глухарей определенного участка или района, а по ко-
личеству глухарок, вылетающих на токовище в разгар токового пе-
риода можно с большой доле?! вероятности предсказать (раз-
259
рядка паша. — О. С.-Т.-Ш.) количество возможных глухариных вы-
водков» (там же, стр. 261). Как для хозяйственных целей, так и
для изучения движения численности животных важнее всего знать
величину популяции не перед началом сезона размножения, а после
его окончания, когда период детской смертности уже миновал и мо-
лодняк по всем признакам приблизился к взрослым птицам или
сравнялся с ними, то есть непосредственно перед началом охотничь-
его сезона. Весенние учеты не дают возможности определить «про-
дукцию» популяции за прошедший сезон потому, что к сроку про-
ведения учета она сильно сократилась в результате продолжавших-
ся несколько месяцев потерь от охотников, хищников, стихийных
и других причин, а величину этих потерь трудно определить. С дру-
гой стороны, по учетам на току едва ли возможно дать удовлетво-
рительный прогноз количества глухариных и тетеревиных вывод-
ков, ввиду того что эмбриональная смертность сильно меняется от
года к году. Провести учет выводков осенью и проще и надежнее,
чем пытаться предсказать их количество весной *.
У весенних учетов есть и другая отрицательная сторона — боль-
шая трудоемкость. Кириков указывает, что при учете глухарей
«очень большие тока следует разбивать на 2—3 участка и учет ве-
сти должны 2—3 лица. На каждом току глухарей следует учиты-
вать в течение 2—5 зорь подряд (нельзя учитывать глухарей в вет-
реное холодное утро, когда поет только небольшая часть самцов,
да и то плохо)» (там же, стр. 261). Из приводимых тем же авто-
ром цифр учета видно, что среднее число самцов на току в разных
местностях колеблется от 2 до 12. Легко подсчитать, что в среднем
1 работник в 1 ночь сможет учесть всего 1—2 глухарей. Если при-
нять во внимание предварительную работу по разысканию токо-
вищ, малую продолжительность периода энергичного токования и
необходимость считаться с погодой, нам станет ясно, почему спо-
соб учета на токах, несмотря на его несложность, не привился
в практике заповедников и охотничьих хозяйств. Многолетних учет-
ных данных, собранных по этому способу, у нас, по-видимому, во-
обще нет **.
Весенний учет воротничкового рябчика по брачным крикам ши-
роко применяется в Сев. Америке, однако и там стараются заме-
нить его более экономными способами. В недавно вышедшей статье
Дорни и др. (1958) подсчитано, что затрата труда для учета токую-
щих самцов на площади 800 га составляет 500 человеко-часов. Ме-
жду тем для проведения на той же площади зимнего учета вспар-
хивающих рябчиков достаточно затратить только 3 человеко-часа.
Таким образом весенние учеты принадлежат к самым дорогим спо-
собам учета. В тех местах, где водится несколько видов тетереви-
* «Цыплят по осени считают» — вошло в пословицу.
** Данные Дица для Гатчинской охоты, использованные Северцовым (1944)
и Кириловым и др. (il.9i52), мы считаем неубедительными. За .15 лет число
учтенных глухарей изменялось менее, чем в два раза: такая малая амплитуда
изменений численности глухаря неправдоподобна. По всей вероятности, учет
проводился недобросовестно.
260
ных птиц, учет каждой из них весной приходится проводить от-
дельно, что делает результаты этих учетов трудно сравнимыми.
По нашему мнению, наиболее универсальным из предложенных
до настоящего времени способов учета тетеревиных надо считать
учет на ленточных пробах, при условии проведения его ранней
осенью, когда выводки еще не разбились и ведут наземный образ
жизни. Учет состоит в регистрации всех птиц, поднятых цепью за-
гонщиков на определенном прямолинейном маршруте, пересекаю-
щем разнообразные стации: благодаря этому он пригоден для пере-
расчетов на большую площадь. В Лапландском заповеднике учет
по этому способу применяется с 1936 года. Средний учетчик (он же
ведет и запись) идет по лесоустроительной визирке, а два помощ-
ника идут параллельно ему в 20 м справа и слева. Дичь учиты-
вается как в полосе между учетчиками, так и по сторонам от край-
них до 7,5 м: таким образом, полная ширина учетной ленты равна
55 м. За все годы учета не было случая, чтобы птица поднялась
позади загонщиков; ввиду этого пропуск какой-то части птиц вслед-
ствие затаивания их в полосе учета мало вероятен.
Маршрут состоит из 4 участков, общим протяжением 41,3 км,
что дает учетную площадь в 227 га. Важно следить, чтобы загон-
щики соблюдали установленную дистанцию и шли фронтом, не от-
ставая от товарищей и не забегая вперед. Ввиду этого скорость
учетного хода невелика, обычно около 45—50 м- в минуту, или
2,7—3,0 км в час; проведение учета занимает 4 дня. В Печоро-
Илычском заповеднике этот способ учета введен с 1952 года: до
того (с 1938 г.) В. П. Теплов проводил только маршрутные учеты,
притом в более поздние сроки. В средней полосе тайги, где распо-
ложен Печоро-Илычский заповедник, лес гуще и труднее прохо-
дим, чем на крайнем севере таежной зоны. Поэтому здесь мы не
добивались сохранения определенного расстояния между загонщи-
ками, но зато измеряли расстояние от места взлета каждой птицы
до визирки по перпендикуляру: таким образом, вопрос о том, под-
нята ли птица в ленте или вне ее, решался уже после взлета
птицы. С целью сокращения холостых ходов маршруты были сде-
ланы в форме буквы П, что дало нам возможность за 8 рабочих
часов отрабатывать более 18 километров, включая в это время
необходимые остановки для записей и отдыха, но не считая перехо-
дов к начальному пункту учета и возвращения от конечного пункта
домой. При ширине учетной ленты 50 м три учетчика отрабатывают
за день 91 га, а в среднем за ряд лет на этом пространстве реги-
стрировались 24 особи тетеревиных птиц разных видов, то есть для
учета 8 птиц требуется 1 человеко-день.
Хотя вопрос о репрезентативности данных учетов до сих пор не
исследован, полезно привести некоторые соображения об особен-
ностях ленточного учета. Форма пробных площадей длинная и
узкая, они пересекают разнообразные стации, в различной степени
заселенные птицами, и потому должны дать более объективное
представление о средней плотности популяций, чем учет птиц на
261
местах концентрации их (токовища, долины ручьев и т. д.). Однако
в тех случаях, когда пробные ленты намечены с расчетом избежать
заведомо бедных участков (обширные болота, горные тундры
ц т: д.), пересчет данных на всю площадь хозяйства даст преувели-
ченные показатели. При обработке материалов следует избегать по-
казной точности, не оправдываемой условиями учета. Очевидно, что
если мы проведем контрольный учет на том же маршруте в другой
день, результаты первого и второго учетов не совпадут. Поэтому
нет смысла вычислять плотность населения до десятых и сотых
долей, за верность которых мы не можем ручаться. Если общая
площадь пробы была от 100 до 500 га, данные учета следует при-
водить к 1 кв. км (100 га) и давать их в целых числах; только
в том случае, если учетом охвачено около 1000 га или больше, це-
лесообразно вести расчет на 10 кв. км (1000 га).
Приблизительное представление о численности дичи, пригодное
для характеристики движения численности по годам, можно полу-
чить и без специальных учетов. Достаточно систематически реги-
стрировать всех птиц, встреченных на охоте или на экскурсиях,
учитывая длину или продолжительность пройденных маршрутов.
Расчет можно вести на 100 километров или на 100 часов хода,
в крайнем случае — на 1 день. Таким путем, пользуясь старыми
дневниками, мы получили количественные показатели встречае-
мости тетеревиных птиц за первые 6 лет существования Лапланд-
ского заповедника, до организации учета на ленточных пробах.
Для характеристики колебаний численности тундряной куропатки
мы пользовались только этим способом. В. П. Теплов успешно при-
менял его в течение многих лет в Печоро-Ильичжом1 заповед-
нике.
Главное преимущество этого примитивного способа учета--на-
зовем его «охотничьим учетом» — в том, что он *пе требует пи спе-
циальной затраты времени, ни какой-либо подготовки учетной пло-
щади и потому особенно пригоден для массового применения. При
сравнении полученных этим путем данных с результатами ленточ-
ных учетов выясняется, что «охотничий учет» имеет тенденцию
сглаживать колебания численности, преуменьшая их амплитуду
(см. табл. 80 и 81). В период 1936—1941 гг. оба способа учета оди-
наково определяют самый богатый по суммарной численности
дичи год (1937) и самый бедный (1939); однако плотность дичи по
данным ленточного учета в 1939 г. была в 18 раз меньше, чем
в 1937 г., а по данным «охотничьего учета» — только в 4,5 раза.
Причина такого расхождения, по-видимому, в том, что при охоте
с лайкой каждая птица отнимает у охотника сколько-то времени на
подход и отстрел или наблюдение; когда дичи много, охотник и
его собака не могут обнаружить всех глухарей и тетеревов по
маршруту потому, что времени на это не хватает. В год бедный
дичью, наоборот, охотник тратит на подходы меньшее время и мо-
жет обследовать на том же маршруте большее пространство, чем
в урожайный на птицу год.
262
Один из главных недостатков «охотничьего учета» состоит в том,
что величину обследованной площади трудно определить. Учетные
данные, полученные в начале и в конце охотничьего сезона, нерав-
ноценны: в начале сезона птица, как говорится, «смирнее», обнару-
живается на меньшем расстоянии и легче определяется, чем позднее.
По всем этим причинам запись птиц на охотничьих экскурсиях не
может заменить настоящий учет. Гораздо лучший показатель чис-
ленности промысловых животных мог бы дать полный учет всей
дичи, добываемой охотниками по отдельным районам или областям.
В некоторых странах проводится сбор и обработка таких статисти-
ческих данных. Приведенные ниже сведения о движении числен-
ности тетеревиных птиц в Финляндии основаны частью на стати-
стике добычи охотничьих клубов (с 1906 г.) и сведениях о добыче
дичи по разрешениям на право охоты (с 1933 г.), а частью на ан-
кетном материале*.
Подобных данных по Советскому Союзу мы до сих пор не
имеем. Приводимые изредка в литературе цифры заготовок дичи
мало показательны, так как величина заготовок, пожалуй, в боль-
шей степени зависит от приемных цен на дичь и других конъюнк-
турных условий, чем от состояния ее запасов в природе. Очевидно,
что количество заготовленной дичи составляет лишь незначитель-
ную часть всей добычи охотников, однако о соотношении этих вели-
чин мы имеем самое смутное представление. Казалось бы, никакое
хозяйство не может действовать без учета своих фондов и продук-
ции— одно только охотничье хозяйство до сих пор как-то обходится
без учета; в результате этого мы почти ничего не знаем о количе-
стве добываемой дичи, распределении ее по областям, видовом со-
ставе и изменениях величины добычи по годам.
Результаты учета тетеревиных в двух таежных заповедниках.
В Лапландском заповеднике учет на ленточных пробах ведется
с 1936 года. Учетные линии находятся в южной части заповедника,
в районе преобладания сосновых лесов. Они не захватывают гор-
ных тундр, хотя и поднимаются по западному склону Чуна-туидры
до границы леса. Сосновые и смешанные леса с напочвенным по-
кровом из ягелей, зеленых мхов и ягодников занимают около 64%
площади стационара (в том числе гари — 6%),еловые леса — 22%,
березовые — 3%, сфагновые и осоковые болота— 11%. При ширине
учетных лент 55 м и суммарной длине их 41,3 км общая площадь
стационара равна 227 га. Учет проводится в период с 1 по 22 авгу-
ста (средняя дата 12 VIII) и занимает обычно 4 дня. Наибольшее
число птиц было учтено в 1937 г. — 67 глухарей, 46 белых куропа-
ток, 14 рябчиков и 1 тетерев; наименьшее в 1939 г. — 5 глухарей и
1 куропатка. Результаты учета по годам, в пересчете на 1 кв. км,
указаны в табл. 80.
*» В финляндских анкетах величина популяции оценивается по 3-балльной
шкале: 3 — выше средней, 2 — средняя, 1 — :ниже средней, О'—вид отсутствует.
263
Таблица 80
Плотность популяций тетеревиных птиц на стационаре Лапландского
заповедника, по годам (Число птиц на 1 кв. км)
Год
Вид
Глухарь 6 29 9 2 7 7 4 5 4 5 7,9
Тетерев . .... - 0+ - - - 1 - 3 2 2 0,7
Белая куропатка 2 20 - 1 0+ - 2 5 6 2 3,9
Рябчик 7 6 7 - — 4 3 10 6 3 4,6
Всего . . . 15 55 16 3 7 12 9 23 18 12 17,1
Таблица 81
Плотность популяций тетеревиных птиц на стационаре Печоро-Илычского
заповедника, по годам
(Число птиц на 1 кв. км)
Год
Вид 1952 1953 1954 1955 1956 1957 Среднее
Боровой участок
Глухарь 14 20 7 19 15 9 13,8
Тетерев . .... 5 17 7 14 - 1 7,2
Белая куропатка — — — — — — —
Рябчик - 2 9 17 4 5,3
Всего . . . 19 39 14 42 32 14 26,3
Темнохвойный участок
Глухарь - 6 4 10 5 4 5,7
Тетерев . .... — — — — — — —
Белая куропатка . . — — — — — — —
Рябчик - 62 14 13 8 34 26,2
Всего . . . — 68 18 23 13 38 31,9
264
В Печоро-Илычском заповеднике ленточный учет введен с 1952 г.,
маршруты установились только с 1955 г. Первый участок ста-
ционара находится близ поселка Якша, в районе преобладания бо-
пов-брусничников; есть тут также старые гари и заболоченные сос-
ново-еловые леса на мокрой почве, с вахтой, сабельником и бело-
крыльником. Маршрут длиной 18,2 км имеет форму буквы П, начи-
наясь и заканчиваясь на берегу р. Печоры; площадь его, при ши-
рине ленты 50 м, равна 91 га. Другие два участка расположены
приблизительно в 70 км от первого и в другой ландшафтной зоне,
почему результаты учетов даются по ним отдельно. Здесь преобла-
дают елово-пихтовые леса с примесью березы и кедра, с наземным
покровом из черники, кислицы, борца, папоротников и других рас-
тений (местное название этого типа тайги «парма»); есть и уча-
стки сосново-лиственничных лесов с ягельно-черничным покровом.
Длина второго участка — 8,5 км, третьего — 18,1 км, их общая пло-
щадь равна 133 га. Учет проводился между 4 и 26 августа (в сред-
нем 17 августа) за три рабочих дня, не считая времени на переезды.
В те годы, когда учет на ленточных пробах не проводился,
о движении численности тетеревиных птиц можно судить по учету
встречаемости птиц на охоте и экскурсиях в сентябре —декабре
(табл. 82—83). На рис. 92—93 движение численности тетеревиных
Таблица 82
«Охотничий учет» дичи в районе Лапландского заповедника
(Число встреч на 100 часов хода в IX—XII) *
Год Глухарь Тетерев Б. куроп. Рябчик Всего Пройдено часов
1930 74 3 34 3 114 121
1931 98 — 11 5 114 104
1932 69 1 36 7 113 115
1933 219 3 84 24 330 145
1934 165 5 54 19 243 155
1935 108 3 26 7 144 98
1936 95 2 26 27 150 58
1937 103 2 70 20 195 177
1938 57 2 39 24 122 156
1939 25 1 16 1 43 146
1940 50 94 «_ — 144 18
1941 43 80 3 18 144 33
1946 39 22 9 30 100 46
1949 71 44 52 6 173 111
1950 69 45 135 2 251 49
1951 29 32 39 — 100 59
Среднее 93 11 44 13 161 100
* Годы максимумов напечатаны курсивом (большая численность тетерева
в 1940 г. сомнительна, так как основана на малом материале).
265
показано для обоих заповедников параллельно, причем даны как
цифры учета на пробных лентах, так и учет встреч на охоте. В боль-
шинстве случаев обе характеристики численности изменяются в
том же направлении. Резкие различия наблюдались только у тете-
рева: в Лапландском заповедник1е — в 1940 г., в Печоро-Илыч-
ском— в 1952 , 1953 и 1955 гг. Объяснение этому надо искать в ме-
стных перекочевках тетерева в предзимний период: в одних случаях
Рис. 92. Движение численности глухаря и тетерева в таежной зоне. Обозначения:
линия е кружками — Лапландский заповедник, число встреч па 10 часов; линия
с крестиками — там же, учет на ленте (число птиц па 100 га); пунктир — Финская
Лапландия, учет добытых в тысячах; прерывистая линия—Печоро-Илычскии
заповедник, число встреч па 25 км; то же с крестиками — там же, учет на ленте
(число птиц на 100 га)
они могли налететь из других мест (1940, 1952 гг.), в других, наобо-
рот, откочевать (1953, 1955 гг.). Между временем проведения лен-
точного учета и регистрацией встреч на охоте проходит 1—4 ме-
сяца, причем работа проводится, хотя и на близких, но различных
участках. Замечательно, что в Печоро-Илычском заповеднике нет
прямой связи между изменением численности рябчиков в боровом
и темнохвойном участках заповедника (см. табл. 82). Разница
в движении численности рябчика на этих участках объясняется, по
266
Таблица 83
«Охотничий учет» дичи в районе Печоро-Илычского заповедника
(до 1951 г. В. П. Теплов *, с 4952 г. О. И. С.-Т.-Ш.),
Год Глухарь Тетерев Б. куроп. Рябчик Всего Пройдено километров
1937 112 58 не учит. 94
1938 88 40 137 265 102
1939 22 12 40 74 ПО
1940 48 2 58 108 166
1941 52 35 58 145 230
1942 92 42 67 201 110
1943 88 42 69 199 959
1944 110 15 35 160 175
1945 58 20 46 124 213
1946 42 10 43 95 165
1947 62 32 54 148 413
1948 125 118 60 303 225
1949 158 55 43 256 310
1950 120 38 81 239 •155
1951 102 82 52 236 205
1952 65 69 3 39 176 187
1953 179 21 0 60 260 64
1954 31 33 0 17 81 104
1955 56 13 1 50 120 198
1956 115 3 0 21 139 92
1957 41 12 0 47 100 190
Среди, за 6 л. 81 25 1 39 146 139
* В. П. Теплов относил встречаемость птиц к 100 км; для сравнимости ре-
зультатов мы пересчитали эти данные (1937—ЮЗ! гг.) на 100 час. экскурсий,
принимая скорость хода охотника в 2,5 км в час. Учет рябчиков В. П. Теплов
проводил отдельно от других видов, в IX—X, с помощью манка.
всей вероятности, различием метеорологических условий в критиче-
ский период гнездования. Весна 1957 г. была неблагоприятной для
размножения тетеревиных в боровом участке. С 19 IV по 15 V была
устойчивая теплая солнечная погода, снег в это время сошел и пти-
цы начали гнездиться; судя по срокам появления птенцов, первые
глухарки начали насиживать 15 мая, а рябчики — не позднее
25 мая. 16 мая настала холодная и дождливая погода, продолжав-
шаяся до 5 июня; окончание кладки яиц и насиживание происхо-
дили при очень неблагоприятных условиях, вследствие чего в этом
районе было мало выводков. В темнохвойном районе заповедника
снег всегда сходит позже, чем в боровом, вследствие чего запазды-
вает и размножение птиц. Очевидно, что в 1957 г. рябчики (а воз-
можно, и глухарки) приступили там к гнездованию уже после окон-
чания неблагоприятного периода. Аналогичное явление и с теми же
последствиями наблюдалось и весной 1953 г.: после периода хоро-
267
шей погоды, иногда с «утренниками», 25—31 мая похолодало, вы-
пали сильные дожди. Первые глухарки и тетерки начали насижи-
вать в боровом участке 20—22 мая и некоторые из них потеряли
1кл>адии (см. стр. 234 и 246). В темнохвойном участке глухарки и
рябчики начали насиживать только 29—31 мая и случаев гибели
целых кладок у них, по-видимому, не было. В обоих случаях осенью
Рис. 93. Движение численности рябчика и белой куропатки в таежной эоне
(обозначения — см. рис. 92)
в боровом районе было мало рябчиков, а ближе к Уральским горам,
в «пармах», — обилие. Все же случаи, подобные описанным, редки.
Сравнение учетных данных Лапландского и Печоро-Илычского за-
поведников, разделенных расстоянием более 1300 км по прямой
линии, показывает, что движение численности глухаря и рябчика
в них очень сходно, так что максимумы и минимумы наступают или
в те же самые годы или в соседние (например, максимум рябчиков
в Лапландском заповеднике был в 1949 г., а в Печоро-Илычском —
в 1950). Сведения о годах «пиков» и депрессий тетеревиных птиц
в Северной Европе мы находим в работах Н. Хёглунда (1952) и
Л. Сиивонена (1948, 1952, 1957): сходство этих данных с нашими
бросается в глаза (см. табл. 84—87). Интересно, что данные «охот-
ничьего учета» тетеревиных птиц в районе Лапландского заповед-
268
Таблица 8-1
Максимумы численности глухаря с 1910 по 1957 г.
--— Год Шотлан- дия Норве- гия Швеция, Хельсин- гланд Финлян- дия СССР, Имандра СССР, Верхняя Печора
1910 (1910)
1911 1911
1914 1914
1915 1917 1917
1918 1919 1918
1920 1922 1920
1923
1925 1926 1926 (1926)
1930 1930 1930 1930
1931 (1931) (1931)
1933 1933
1934 1934 1934
1935 1936
1937 1937 1937 1937 1937 1937
1940 1941 1940 1942 1942
1943
1944 1944 1944
1945 1946 1945
1948 1949 1949
1950 1950 1950
1953 1953 1953
1955 1955
1956
1957
Примечание: I "оды пло> со выраж! енных ма: ксимумов взяты в скобки.
269
Таблица
Максимумы численности тетерева с 4910 по 1957 г.
Год Шотлан- дия Норве- гия Швеция Финлян- дия СССР, Имандра 1 СССР |В- HeU 1 Ра
1910 1910 (1910) 1911
1915 1915 1914 (1913) 1914
1918 (1917) 1918
1920 1920 1921 1920 (1920)
1925 1923 1926 (1923) 1926 1924 1927
1930 1930 1930 1930
1935 1933 1934 1933 1933 (1934) (1934)
1937 1937 (1937) 1937
1940 1945 1941 1942 1944 1945 1941 1942 1943
1950 1948 1951 1949 1948 1951
1955
1957
270
Таблица 8r>
Максимумы численности рябчика и белой куропатки с 1910 по 1957 г.
Год Рябчик Белая куропатка
Финлян- дия СССР, Имандра СССР, В. Печо- ра Брита- ния Норве- гия Финлян- дия СССР, Имандра
1910 (1910)
1911 1912 1911 1911
1914 1914 1914
1915 1917 1915 (19П) 1917
1918 1918 1918
1920 1920 1921 1920 1921
1922
1923 1923 1923 .
1924 1924
1925 1926 1926 1926
1927 1927
1930 1930 1930 1930
1933 1933 1933 1933
1934 1934 1934 1934
1935 1936
1937 1937 1938 1938 1938 1937 1937 1937
1940 1942 1942? 1942 1941 1942
1943
1944 1944
1945 1945 1945 1945
1946 (1946)
1948 1949 1948 (1948) 1948
1950 1951 1950 1950
1953 1953
1955
1957 1956
1957
1
271
Таблица
Минимумы численности тетеревиных птиц (в целом) с 1910 по 1957 г
Год Шотлан- дия Норве- гия Швеция, Хель- сингланд Финлян- дия СССР, Имандра СССР, в- Heoi. Ра
—
1910 1912 1912
1915 1916 1916 1915 1916
1920 (1919) 1919
(1922) 1922 1922
1925 1925 1925 1925
1928 1929 1932 1928 1928 1
1930 1932 1933 1932 1931 1932
1935 1935 1935 1935 1935
1940 1939 1940 1939 1938 1939 1939 1940 1939 (1940) 1939 1940
(1943) 1943 (1943)
1945 1945 1947 1946 (1947) 1948 1951 1946
1950 1949
1955 19:4
1957 1957 1957
272
ника почти совпадают со статистикой добычи тех же видов птиц
в финской Лапландии, что дает нам возможность использовать
финские данные для интерполяции движения численности тетере-
виных Лапландского заповедника в те годы, когда он нс функцио-
нировал (1910—1929, 1942—1945 и 1952—1957).
Учет плотности популяций тетеревиных птиц важен нс только
в охотничьем хозяйстве, но и при проведении экологических иссле-
дований, например, при определении степени использования кормо-
вых фондов, анализе отношений хищника и жертвы или паразита
и хозяина. Учеты животных являются исходным материалом для
исследований колебаний численности животных, их периодичности
н причин.
Зарубежная литература о колебаниях численности диких живот-
ных, преимущественно охотничье-промысловых, очень велика. К со-
жалению, скандинавские, английские и американские зоологи в
большинстве случаев суживают эту проблему, ограничиваясь опре-
делением цикличности популяций животных и рассмотрением свя-
зей между циклами различных животных, но почти не затрагивают
непосредственных причин роста или убыли популяций. Обзор ис-
следований «циклического направления» можно найти в•недавно
вышедшей книге Д. Лэка (1957) и статье Г. Вильямса (1954).
При первой попытке выяснить зависимость размножения глу-
харя от переменных условий среды обитания мы установили только
статистическую связь двух коротких рядов чисел (Семенов-Тян-
Шанский, 1938). Наша работа отличалась от других исследований
такого типа в трех отношениях: вместо цифр количественного учета
был использован показатель размножения, так сказать, в чистом
виде; температуры воздуха были взяты за короткие периоды, что
дало возможность показать плавное увеличение, а затем уменьше-
ние величин коэффициентов корреляций; корреляция между темпе-
ратурой критического периода и показателем размножения глухаря
оказалась очень высокой (г = +0,93+ 0,05, в 20 раз больше ве-
роятной ошибки). Предварительные выводы в дальнейшем были
подтверждены нами на новом материале (Семенов-Тян-Шанский,
1947). Они вызвали дискуссию, которая, к сожалению, имела в зна-
чительной мере спекулятивный характер, так как никто из совет-
ских экологов не использовал при этом данных по возрастному со-
ставу популяций, аналогичных нашим.
В. П. Теплов (1947, стр. 57 и 140) полностью принял наше пред-
ставление о зависимости размножения глухаря от температур пе-
риода насиживания, но лучшим индексом неблагоприятных метео-
рологических условий считает сумму отрицательных температур
в июне (по всей вероятности подразумевается сумма минимальных
температур ниже 0 в термометрической будке, хотя в тексте такой
расшифровки нет). Этот вывод не подкреплен математической об-
работкой связей «урожаев» глухаря с температурными условиями.
Полемизируя с нами и с Тепловым одновременно, С. В. Кириков
пишет в одной из своих работ по глухарю (Кириков, 1952, стр. 93):
18-3037
273
«По-моему, в гибели глухариных яиц во время насиживания боль,
шее значение, чем утренние и ночные заморозки, должна иметь дли-
тельная холодная и ненастная погода, хотя бы температура и не
падала ниже нуля» (Кириков, 1952, стр. 93). Изложив два одиноч-
ных наблюдения, сделанных в Башкирском заповеднике в 1941
и 1946 гг., Кириков развивает свою мысль следующим образом:
«Можно предположить, что если в первые дни после выхода глуха-
рят из яиц в течение нескольких дней будет стоять холодная дожд-
ливая погода, то такая погода может отразиться на маленьких глу-
харятах более губительно, чем утренники. Ио непосредственных,
прямых наблюдений в природе над губительным действием на глу-
харей заморозков и продолжительной холодной ненастной погоды
нет, и поэтому выводы о том, что указанные метеорологические яв-
ления могут быть причиной высокой ранней «детской смертности»,
нельзя оценивать иначе, как возможные предположения» (там же,
стр. 94).
За границей над проблемой причинной зависимости выживае-
мости молодняка глухаря от условий погоды в последнее время ра-
ботал Н. Хёглунд (1952). Из непосредственных наблюдений, прове-
денных в Швеции, он выяснил, что активность птенцов глухаря
и других тетеревиных определяется температурой внешней среды,
вследствие чего в холодную погоду они голодают. Нормальное раз-
витие птенцов в первые дни их жизни зависит от максимальных
температур воздуха в этот период, причем критическая максималь-
ная температура близка к 19°. Автор приводит график максималь-
ных температур и количества осадков за соответствующий период
на протяжении 20 лет в связи с численностью глухаря в следующую
осень, однако без вариационно-статистической обработки этих
данных.
Новую точку зрения выдвинул недавно Л. Сиивонен (1957). Со-
поставляя движение численности глухаря и тетерева в Финляндии
по годам с температурой отдельных периодов, определяемых вре-
менем размножения этих птиц (до кладки, во время кладки, при
насиживании яиц и после вылупления 'Птенцов), он нашел, что дви-
жение численности птиц определяется главным образом темпера-
турой апреля, то есть периода непосредственно перед кладкой. Ав-
тор объясняет найденную им зависимость, на основе известных
в настоящее время фактов, следующим образом. Зимнее питание
тетеревиных птиц неполноценно, вследствие чего вес птиц к весне
падает. У самцов он продолжает падать и в период токования, но
у самок он в это время резко увеличивается, что предполагает уси-
ленное питание их перед началом кладки. В случае позднею появ-
ления проталин подкормка глухарок и тетерок зеленью, цветами
и ягодами становится невозможной, что неблагоприятно сказы-
вается на результатах сезона размножения. Слабое место в теории
Сиивонена в том, что величину урожая и неурожая он определяет
по анкетным данным, где они характеризуются только качественно
(прирост — плюс, убыль популяции за год — минус). В приводимых
274
автором 11-летних рядах наблюдений, вслед за высокими темпера-
турами периода перед кладкой (3—4°) происходил прирост попу-
ляций, а за низкими (0—2°) —их убыль. Пока эта закономерность
не подтвердится на новом материале из других мест, ее нельзя счи-
тать доказанной.
Наш обзор новейших взглядов на давно известную зависимость
успеха размножения тетеревиных птиц от погоды весны приводит
к странному результату. Все цитированные здесь авторы (Теплов,
Кириков, Хёглунд, Сиивонен и автор этих строк) согласны в том,
ито зависимость эта вполне реальна, но в отношении того, какие
элементы погоды играют роль, какой период является критическим
и каким путем погода влияет на размножение птиц, мнения расхо-
дятся. Сиивонен полагает, что успех размножения зависит от тем-
пературы воздуха перед началом кладки и причиной «неурожая»
является истощенное состояние самок во время кладки; Теплов свя-
зывает неурожай с заморозками во время насиживания и причиной
гибели яиц считает замерзание их в это время; Кириков полагает,
что играют роль не заморозки, а затяжные дожди и гибнут не яйца,
а новорожденные птенцы; Хёглунд находит, что птенцы гибнут от
недоедания, если температура воздуха среди дня недостаточно вы-
сока. Не пытаясь оспаривать мнение других исследователей, заме-
тим только, что наши выводы основаны на более обширных и раз-
носторонних материалах, чем тс, которыми располагали остальные
авторы. Для дальнейшего обсуждения вопроса необходимо иметь
новые, систематически собранные материалы, пригодные для мате-
матической обработки. По нашему мнению, развитому в предыду-
щей главе, главной причиной большой эмбриональной смертности
в некоторые годы является прекращение обогрева яиц наседкой при
неблагоприятных условиях погоды, а также замерзание яиц при
длительных перерывах в насиживании или перед началом его.
Переходим к рассмотрению корреляционных связей между от-
дельными метеорологическими факторами и показателями размно-
жения глухаря. В качестве такого показателя мы используем отно-
шение количества молодых птиц по первому году к общему коли-
честву глухарей, добытых в осенне-зимний охотничий сезон. Мож-
но было бы взять другой показатель — число молодых птиц на одну
пару взрослых; однако при хороших урожаях этот показатель воз-
растает очень быстро, вследствие чего он оказался менее удобным
при вычислении корреляционных связей.
В районе Лапландского заповедника отстрел (с исследователь-
скими целями) проводился нами в течение всего года, но общее
количество добытых птиц было обычно невелико; поэтому для опре-
деления возрастного состава мы использовали всех глухарей, добы-
тых с 1 сентября по 30 апреля, т. е. за 8 месяцев. Материал коле-
бался от 9 до 150 особей в год, в среднем — 44. На Верхней Пе-
чоре научный отстрел нами не проводился, а осенняя охота там
заканчивается уже *к 1 декабря; ввиду этого возрастной состав
определялся нами по результатам отстрела с 1 сентября по 30 нояб-
18*
275
ря (3 месяца). В отдельные годы было исследовано от 20 до 85 глу-
харей, в среднем 49. Исходные данные по возрастному составу по-
пуляций приведены в табл. 88.
Таблица 63
Возрастной состав популяции глухаря по годам
Год Число И( молод. :сл. птиц взросл. % молодых Число молодых на пару взрослых Среди, темп, самой холод- ной из трех декад июня в LC
Лапландский заповедник
1930 4 5 44 1,6 7.4
1931 1 13 7 0,2 3.0
1932 5 17 23 0,6 2.8
1933 76 18 81 8,4 9.7
1934 104 46 69 4,5 8,1
1935 12 24 33 1,0 4,7
1936 54 19 74 5,7 11,6
1937 57 27 68 4,2 9,1
1938 26 19 58 2,7 7.3
1939 2 14 12 0,3 4,2
1940 14 10 58 2,8 5,0
1941 1 3 (25) (0,7) (2,0)
1946 10 3 77 6,7 4,3
1948 2 4 (33) (1,0) (6,5)
1949 7 12 37 1,2 5,0
1950 6 И 35 1,1 5,7
Итого . . 381 245 48 2,9 6,3 Средн, темп, самой холод- ной из трех
1 Печоро-Илычс :кий запо вед ,ник декад ман- ию ня в сС
1952 33 9 79 7,3 8,7
3953 49 • 36 58 2,7 7,7
1954 6 14 30 0,9 6.6
1955 41 10 80 8,2 п;5
1956 53 16 77 6,6 10,2
1957 9 20 31 0,9 7,2
Итого . . 191 105 59 4,4 8,6
Примечание. Ненадежные по недостатку данных цифры взяты в скобки.
Из этой таблицы видно, что размножение глухаря в бассейне
Имандры обеспечено хуже, чем в верховьях Печоры. Иа Имандре
276
урожаи были в 1933, 1934, 1936, 1937 и 1946 гг. — примерно один
раз в три года; на Верхней Печоре — в 1952, 1955 и 1956 гг., следо-
вательно, один раз в два года. Урожайными мы считаем годы, когда
количество приплода приближается к максимально возможному и
составляет от 2/3 до 4/5 всей популяции. Откинув годы, когда воз-
растный состав популяции был определен по недостаточному мате-
риалу (1941 и 1948), мы получаем по Лапландскому заповеднику
14 пригодных для исследования лет (в том числе непрерывный ряд
в 11 лет), а по Печоро-Илычскому заповеднику — 6 лет. Сведения
о температурах воздуха и осадках взяты с ближайшей к заповед-
нику метеорологической станции, в первом случае это Хибины
(28 км к востоку от заповедника), во втором случае — Якша (при
управлении заповедника). Для сравнения были использованы сред-
ние температуры, абсолютный минимум температуры и суммы
осадков по месяцам; средние декадные температуры за 3 месяца
н самая холодная из трех декад июня (Хибины) или из 3 декад,
включая последнюю декаду мая и две первые декады июня (Якша),
а также число холодных дней за этот период. Соответствующие
коэффициенты корреляции с их вероятными ошибками приведены
в табл. 89.
В Лапландском заповеднике размножение глухаря зависит,
главным образом, от температур июня (средней месячной или ми-
нимальной). Рассмотрение коэффициентов по отдельным декадам
показывает, что наибольшее значение имеет первая декада июня,
период, во время которого при нормальном ходе весны глухарки
начинают насиживание яиц. Предыдущая и последующие декады
имеют меньшее значение: ввиду большой величины вероятных оши-
бок наличие корреляции даже сомнительно. В Печоро-Илычском
заповеднике из средних месячных температур наибольший коэффи-
циент дала температура июля; однако отрицательный знак коэффи-
циента показывает, что урожай связан с низкими температурами
июля, что мало вероятно. Ввиду большой вероятной ошибки мы
предполагаем тут случайное совпадение чисел, вполне возможное
при небольшой длине ряда наблюдений. Корреляция между возра-
стным составом популяции и температурой воздуха по декадам дает
в обоих заповедниках одинаковый результат с разницей во времени
па одну декаду. В годы нашего исследования первый выводок глу-
харя наблюдался в Лапландском заповеднике 25 июня (в среднем
за 12 лет), в Печоро-Илычском — 16 июня (среднее за 6 лет), то есть
на 9 дней раньше. Продолжительность инкубации составляет 25—
26 суток, отсюда следует, что насиживание начинается в Лапланд-
ском заповеднике около 30 мая, а в Печоро-Илычском — 21 мая.
В обоих случаях «критической» оказывается температура первой
декады насиживания. Если весна поздняя, в «критическую» декаду
глухарки еще не насиживают, а несут яйца. Низкие температуры
опасны не только в начале инкубации: плохие выводки наблюда-
лись и в тех случаях, когда холода наступили в середине или конце
насиживания, а начало его проходило при благоприятной темпера-
277
Таблица
Корреляционные связи возрастного состава популяции глухаря
с метеорологическими условиями сезона размножения
Заповедни к
Лапландский, 14 лет Лапландский, 11 лет Печоро-Или'ц скин, 6 лет
Ср. т-ра апреля . . . — 0,08+0,27 +0,06+0,30 —0,32+0,37
мая .... +0,22+0,25 +0,31+0,27 -0,32+0,37
„ июня .... ±0,67±0,15 +0,7<9±О,12 +0,20+0.39
„ июля .... +0,32+0,24 +0,24+0,28 —0,60+0,26
Абс. мин. апреля —0,18+0,26* -0,17+0,29 —0,17±0,40
„ мая . . . +0,26+0,25 +0,34+0,26 —0,24+0,38
„ июня .... +М2±0,09 +0,7<9±О,12 —0,14+0,40
„ июля .... +0,61+0,17 +0,52±0,21 —0,65+0,26
Сумма осадк. апреля . —0,20+0,26 —0,08+0,30 —0,14±0,40
„ мая .... —0,61+0,17 —0,52+0,22 +0,15+0,40
„ июня . . —0,36+0,23 -0,43+0,24 -0,96+0,03
„ июля .... + 0,13+0,27 + 0,20+0,29 —0,23+0,30
Ср. т-ра III дек. апреля — — —0,25+0,38
м I дек. мая +0,32+0,24 +0,26±0,28 -0,58+0,27
„ II дек. мая . -0,05+0,27 +0,07±0,30 -0,31 ±0,37
III дек. мая . +0,17±0,2б + 0,17+0,29 + 0,00+0.15
„ I дек. июня . +0,77+0,13 +0,7<$±ОД2 +0,13±0,40
„ II дек. июня . ±0,30±0,24 +0,30+0,27 -0,35+0,36
„ III дек. июня + 0,29+0,24 +0,33+0,27 ±0,14+0,40
„ I дек. июля ±0,06+0,27 —0,07±0,30 —
Сам. холоди, из трех декад 4-0,74+0,12 + 0,92+0,05 + 0,&5±0Д1
Чщсло дней ниже -|- 5° за три декады . . — 0,67+0,15 —0,77+0,13 -0,76+0,17
День зацв. пушицы . —0,37+0,25 — 0,43+0,25 4-0.44±0,33
День зелен, берез . . — 0,45+0,22 —0,46+0,24 4-0,42+0,
Дата встречи перв. вывод, глухаря +0,22+0,30 — ±0,10+°>40
278
туре. В Лапландском заповеднике примером может служить 1938 г.,
когда урожай глухаря был снижен сравнительно холодной второй
декадой июня. В Печоро-Илычском заповеднике то же, но в более
отчетливой форме, произошло в 1954 г.: к сожалению в тот сезон
нам не представилось случая использовать гнездовые самописцы.
По этой причине лучший результат дает сопоставление возра-
стного состава популяции не с одной определенной декадой, а с са-
мой холодной из трех декад, в течение которых происходит наси-
живание. При этом условии коэффициент корреляции в Лапланд-
ском заповеднике достигает +’0,92 + 0,05 (за 1'1 лет непрерывных на-
блюдений), а в Печоро-Илычском +0,85 + 0,11 (за 6 лет).
Таблица 90
Средние декадные температуры (в °C) в период размножения глухаря
Год VI V2 V’5 VI' Vl= , VI-5 VII1 Колич. молодых в популяц. В %
Лапландский заповедник
1930 5,0 7,0 7,4 12,1 9,3 17,4 44
1931 7,4 5,0 3,0 7,5 8,9 10,9 18,0 7
1932 3,3 1.8 2,8 12,2 13,1 23
1933 0,9 4^5 9,7 13.2 10,8 15,4 81
1934 5,9 5.3 8,4 8,1 13,5 11,3 69
1935 1,8 3,2 4,7 12,0 11,6 7,9 33
1936 6,7 4,9 13,3 18,3 11,6 12,7 74
1937 6,7 5.0 9,1 13.4 18.0 16,8 68
1938 1,8 8,0 10,7 7,3 12.6 13,1 58
1939 1,9 5,8 4,2 15,4 10,8 13,9 12
1940 6,4 3.7 5,0 18,3 21,6 14,2 58
'Средн. 4.3 4.9 7,1 12,8 13.0 14,0 48
Печоро-Илычский заповедник
1952 -4,4 2,6 13,2 8,7 14,2 19,4 — 79
1953 1,5 11,2 7,7 18,0 14,3 12.5 — 58
1954 4,4 8,2 8,6 6,6 18,3 17,2 — 30
1955 4,5 10,4 11,5 11,6 16,3 11,8 — 80
1956 4,9 9.3 10,2 11,5 16,0 17,2 — 77
1957 10,9 10,6 7,2 12,3 14,9 13,1 — 31
Средн. 3,6 8,7 9,7 11,4 15,7 15,2 — 59
Средняя температура периода 1 инкубации в Лапландском заповеднике
(I VI—25 VI) 10;б°.
Средняя температура периода инкубации в Печоро-Илычском заповеднике
(21 у—15 VI) -1'1,6°.
Температура самой холодной из трех декад насиживания напечатана
курсивом.
Табл. 90 и рис. 94 и 95 дают возможность определить критиче-
ские для глухаря пределы температур. Нормальное количество при-
плода (свыше 2/з популяции) бывает в годы, когда средняя темпе-
ратура всех трех декад насиживания была выше 8°: неурожай на-
279
ступает, когда средняя температура одной из этих декад окажется
ниже 5° (Лапландский заповедник) #или ниже 7,5° (Печоро-Илыч-
скип заповедник). Ввиду быстрого {ютепления воздуха в мае —
июне температуры ниже критических чаще всего случаются именно
в начале насиживания (примерно в 70% случаев): именно этим
Рис. 94. Влияние погоды периода размножения на возрастной состав популяции
глухаря в Лапландском заповеднике. Обозначения: столбики вверху—сумма
осадков за июнь, пунктирная линия — средняя т-ра воздуха в июне, сплошная
линия — т-ра самой холодной из трех декад июня; столбики внизу — возраст-
ной состав популяции (заштрихованная часть — молодые)
и объясняется, почему первая декада дает лучшую корреляцию
с количеством молодняка у глухаря. Температура воздуха не един-
ственный метеорологический элемент, влияющий на размножение.
В Печоро-Илычском заповеднике осадки имеют, по-видимому, еще
большее значение (коэффициент корреляции +0,96 + 0,03); В Лап-
ландском заповеднике осадки такой роли не играют.
Приведенные здесь статистические данные, собранные в двух
заповедниках, разделенных расстоянием свыше 1300 км, убеди-
тельно показывают, что возрастный состав популяции глухаря тесно
связан с метеорологическими условиями весны и может быть опре-
делен по ним за два месяца до начала охоты. К большому сожале-
280
нию, мы не можем привести здесь для сравнения другие аналогич-
ные данные по СССР, ввиду того что наши экологи до сих нор не-
^оценивают вариационно-статистический метод исследования.
1 В зарубежных странах статистический метод применяется чаще и
некоторые авторы пользовались им аналогично тому, как это сделали
мы, но для характеристики размножения популяции брали «показа-
тель добычи» — отношение количества птиц, добытых в данном году
Рис. 95. Влияние погоды периода размножения на возрастной состав
популяций глухаря в Лапландском и Печоро-Илычском заповедни-
ках. Обозначения— см. рис. 94
к добыче прошлого года. П. Моран (1'964), используя охотничью
статистику за 60 лет, нашел, что «урожай» глухаря в Шотландии
связан положительной корреляцией с температурой мая в том же
и в предшествующем году, а также с температурой самого холодного
зимнего месяца и отрицательной корреляцией с суммой осенних осад-
ков в предыдущем году. Величина соответствующих коэффициентов,
однако, не превышает 0,Э2—0,44. В Шотландии размножение глухаря
наступает примерно на 1 месяц раньше, чем- в таежной полосе Со-
281
ветского Союза: поэтому вывод Морана в отношении температуры
мая совпадает с нашим. Коэффициенты корреляции «урожаев» грау-
са и тундряной куропатки с условиями погоды оказались, по Мора-
ну, более низкими и статистически недостоверными. Автор не касает-
ся вопроса о том, каким образом температура и осадки влияют
на размножение тетеревиных птиц. HaiM' кажется, что осенние осадки
и зимние температуры могут влиять только на выживание произво-
дителей; возможно также, что осенние осадки и зимние температуры
каким-то образом связаны с температурой и осадками весны, а по-
следние непосредственно влияют на размножение птиц.
В. Шелфорд и Р. Ииттер (1955) нашли связь между данными
учета токующих самцов лугового тетерева в США и осадками апреля
в сочетании с показателем излучения солнца в ультрафиолетовой
области спектра, а также с температурой и осадками июня (в по-
следний перед токованием' сезон). Использование актинометриче-
ских данных объясняется общим представлением о влиянии света на
деятельность гонад. Однако авторы не разъясняют, как они пред-
ставляют себе это влияние.
Еще более отвлеченный подход к вопросу о влиянии света на
размножение тетеревиных мы находим в статье Л. Сиивонспа и
Ю- KooKniMineca о колебаниях численности и лунном цикле (1955).
Известно, чтс фазы луны сдвигаются вперед на 10,9 суток в год. По-
этому в определенный небольшой календарный промежуток (поло-
жим, в последнюю декаду апреля) полнолуние наступает периоди-
чески через несколько лет и длина периода будет тем больше, чем
короче выбранный нами промежуток. Подобрав соответствующую
длину и положение этого промежутка, можно добиться совпаде-
ния годов наступления полнолуния в этот фиксированный календар-
ный промежуток с любым событием, правильно повторяющимся че-
рез несколько лет, например.с «пиками» лемминга и других живот-
ных (если ряды наблюдений сравнительно коротки). Хотя авторы
и ссылаются на стимулирующую роль света, мы не можем себе пред-
ставить, какое значение может иметь лунный свет для тетеревиных
пгиц, одинаково недеятельных и в безлунные и в лунные ночи. К то-
му же весной дни настолько длинные и светлые, что в сравнении
с ними лунное освещение играет ничтожную роль.
Ларсеи и Лейхи (1958) сопоставили результаты весенних учетов
ворстничковогс рябчика на севере США за 29 лет с различными эле-
ментами погоды предшествующего года, взятыми за короткие про-
межутки, и нашли, что высокой численности рябчика предшествуют
повышенные дневные температуры летних месяцев (IV—IX) и пони-
женные температуры зимних (XI—XII?). Однако коэффициенты кор-
реляции здесь очень малы, а причинные связи явлений авторами не
анализируются. Ларсен и Лейхи приходят к выводу, что определен-
ных результатов по этому вопросу не достигнуто ввиду неизученно-
сти конкретных причин смертности тетеревиных птиц и подмены не-
достающих фактов спекулятивными построениями.
282
Очевидно, что нахождение связи двух явлений вариационно-ста-
тистическим методом — только первый этап исследования; в даль-
нейшем необходимо найти и подтвердить все последовательные
звенья этой связи. Снивонен (1957) видит недостаток нашей первой
работы (Семенов-Тян-Шанский, 1938) в том, что, объясняя неурожай
глухаря гибелью кладок от холода, мы не привели фактов, доказы-
вающих эту гибель. Собрать такие сведения очень трудно, так как
случаи находки гнезд без наседок слишком редки. Хищные звери
справляются с этой задачей легче, чем человек: поэтому мы сделали
попытку попользовать многолетние материалы по питанию куницы
для подтверждения нашей гипотезы о «гибели- кладок (см1. стр. 207—
209)“. Этот пример (показывает, какими окольными путями прихо-
дится иногда пользоваться экологу для разрешения своих задач.
Колебания численности тетерева, рябчика и белой куропатки мы
не будем здесь рассматривать, так как по возрастному составу их
популяций собрано слишком мало материала. Остается сказать не-
сколько слово о тундряной куропатке. Мы учитывали ее только пу-
тем регистрации встреченных на экскурсиях птиц с пересчетом либо
на 1<0 км пути, либо на •! день работы в тундре. Материалы, собран-
ные до 1936 г., были опубликованы более 20 лет тому назад (Семе-
пов-Тян-Шаяский, 1938-а)’. В качестве предварительного вывода мы
указывали, что главным фактором, вызывающим убыль популяции,
я?вляется снеговой режим зимы. Численность тундряной куропатки
была высокой в малоснежные зимы 1930-31 и 1933-34 гг. и в следую-
щее за ними лето« (1931 и 1934 гг.). Популяция резко сократилась
в 1931-32 и 1934-35 гг. — в зимы с высоким и .плотным снежным по-
кровом. Возрастной состав популяции определялся в течение б лет
по карточкам регистрации встреч птиц в июле и августе. При этом
выяснилось, что количество молодняка меняется по годам так же
резко, как и у глухаря, а годы «урожаев» (1930, 19341) и «неурожаев»
(*1901, 1935) этих птиц в общем совпадают. Предположение, что
причина плохого раэмно-жения тундряной куропатки в отдельные
годы та же, что и у глухаря, подтверждается добычей трех самок
с наседн'ыми пятнами, но без птенцов, а также находкой двух прош-
логодних кладок с погибшими во время инкубации яйцами (18 VI
1934 и 27 VI 1934 г.).
После 1936 г. мы не имели возможности регулярно посещать гор-
ные тундры в различные сезоны и новых наблюдений над тундряной
куропаткой собрано мало. Мы встречали ее главным образом при
объездах горных тундр заповедника на оленьих упряжках в конце
зимы, с целью учета диких оленей. Только за этот период (апрель)
можно было получить сравнимые учетные данные для всех лет на-
блюдений: они представлены на рис. 96. После 1936 г. депрессии
тундряной куропатки были в 1939-40, 1947-48 и «1949-50 гг.; хорошо
выраженных «пиков» в эти годы не было.
В «1908 г. мы писали: «В экологии тундряной куропатки есть два
«узких места»: летние волны холода, вызывающие гибель яиц или
цыплят в раннем возрасте, и многоснежные зимы, особенно если им
283
сопутствуют Оттепели и гололедица; тундрянка голодает, часть птиц
гибнет (или мигрирует?), а уцелевшие плохо размножаются». Важно
было выяснить, что происходит с тундряными куропатками в снеж-
ные зимы и влияет ли плохая зимовка на условия размножения, тем
более, что почти то же предполагает и Л. Сиивонен в отношении
глухаря и тетерева.
Поскольку материалов по- этому вопросу еще не собрано, нам не-
чего добавить к высказанным' ранее положениям.
Количество молодняка, учтенное перед началом охотничьего се-
зона, представляет то, что осталось после потерь от эмбриональной
и ранней детской смертности. Если бы в этот момент потери полно-
Рис. 96. Маршрутный учет тундряной куропатки на Чуна-тундре
в апреле; показано число птиц на 1 ходовой день
стью прекратились, популяция должна была бы из года в год увели-
чиваться: в Лапландском заповеднике на 48^/0 в год (в среднем),
в Печоро-Илычском — на 59%. При таком темпе прироста через
10 лет численность глухарей должна бы увеличиться при-
мерно в 50 раз (в первом случае) или в 100 рав (во втором слу-
чае}) . В действительности, рост численности компенсируется убылью,
если не в Тот же год, то в следующие, и на протяжении достаточно
долгого времени средняя величина популяции остается на одном
уровне. Из этого следует, что средняя величина смертности за год
должна быть равна среднему количеству приплода. Учитывая, что
практически все тетеревиные гибнут либо от хищников, либо в
результате охоты, мы можем определить продуктивность охотничьих
угодий по этой группе птиц: для этого нужно знать плотность засе-
ления угодий и процент молодняка в популяциях. В Лапландском
заповеднике в среднем учитывается 79 глухарей на 1000 га, а молод-
няк составляет 48% всей популяции: отсюда средняя продуктивность
будет около1 38 глухарей с 1000 га в год. Печоро-Илычский заповед-
ник расположен южнее, природные условия там более благоприятны
для размножения боровой дичи, вследствие чего продуктивность (в
284
боровом районе) оказалась вдвое выше — около 81 глухаря с 1000 га.
Охотники могли бы добывать такое количество глухарей без наруше-
ния основного запаса, но только при условии полного отсутствия по-
терь от хищников (условие это невыполнимо); наоборот, при отсутст-
вии охоты весь приплод поглощается хищниками. Судя по низкому
проценту возврата колец с окольцованных тетеревиных птиц, потери
их от хищников всюду значительно больше потерь от охотников (см*,
ниже).
Допустив, в порядке первого приближения, что смертность оди-
накова во1 всех возрастах, мы «можем рассчитать вероятный возраст-
ной состав популяции. В табл. 91 приведены подобные расчеты для
глухаря в сравнении с двумя видами млекопитающих — красной
полевкой * (пример крайне быстро размножающегося и недолговеч-
ного животного) и усатой ночницей ** (медленно размножающийся
и долговечный вид). Возрастной состав популяции красной полевки
определен нами в Печоро-Илычском заповеднике по ежегодным осен-
ним учетам, проводившимся в течение 5 лет (Семенов-Тян-Шанский,
1969). Сведения о возрастном составе популяции усатой ночницы
заимствованы из работы Слейтера, Хердта и Беземи (1956): они
относятся к южной Голландии и являются результатом1 систематиче-
ской 20-летпей работы по кольцеванию летучих мышей в пещерах.
Таблица 91
Возрастной состав популяций различных по плодовитости животных
Число лет При условии равномерной смертности из 1000 особей выживает:
красная полевка, В. Печора глухарь, В. Печора глухарь, Лапл. зап-к усатая ночница, Ю. Голландия
0 1000 1000 1000 1000
1 77 410 520 795
2 6 168 270 632
3 0,5 0,04 69 141 502
4 28 73 399
5 — 12 38 317
6 5 20 252
7 2 10 200
8 — 0,8 5 159
9 — 0,3 3 126
10 — 0,1 1 100
20 — — — 10
Выживаемость за 1 год . . 0,077 0,41 0,52 0,795
♦ Clethrionomys rutilus.
♦♦ Myotis mystacinus.
Из таблицы видно, что из 100 особей остается в живых одна —
у красной полевки через 210 месяцев, у глухаря — через 5 или 7 лет.
у ночницы —только через 20 лет. Структуры популяций этих живот-
ных резко различны. Ясно, что популяции такого типа, как у летучих
мышей, мало производительны и не могут стать основой каких-либо
цепей питания в биоценозе; приплод подобных видов может компен-
сировать только смертность от несчастных случаев или от старости.
Этот простой пример показывает, что выяснение возрастной струк-
туры популяций промысловых животных важно как для развития
экологии, так и для охотничьего хозяйства.
По черепу можно определять возраст глухарок до 3 лет, самцов
глухарей — до 4—5 лет. В принципе это открывает возможность
определить смертность в каждой возрастной группе отдельно. В
194'9—1951 гг. в Лапландском заповеднике нами исследовано 40 глу-
хариных черепов (21 самка и 19 самцов), в 1962—196*8 гг. в Печоро-
Илычском— 452 экз. (42 самки и 140 самцов, из числа! которых 82
добыты на токах). Черепа птиц по первому году не препарировались,
так как возраст этих особей проще было узнать по перьям1. Черепа
были исследованы не у всех старых птиц, а с другой стороны часть
черепов получена от охотников без учета количества добытых ими
молодых птиц. По этим причинам возрастной состав популяции нель-
зя получить простым подсчетом карточек на добытых птиц. Прихо-
дится прибегать к более сложному приему: отношение числа моло-
дых и взрослых птиц определяется по одной пробе, а возрастной
состав взрослых птиц — по другой, после чего общее число взрослых
птиц, найденное при анализе первой пробы, делится по возрастам
пропорционально результатам исследования второй пробы. Получен-
ные нами результаты представлены в табл. 92.
В районе Лапландского' Заповедника часть птиц была добыта в
закрытое для охоты время: чтобы использовать весь материал, надо
было установить, с какого дня считать птицу на один год старше;
таким моментом принято 1 июля. Исследование возрастного состава
самок и самцов в отдельности мы откладываем на будущее. Возраст-
ной состав глухарей, добытых на токах, рассматривается
отдельно.
Смертность и выживаемость определены путем логарифмирова-
ния численности отдельных возрастных групп (табл. 92).
Средняя выживаемость глухаря от первого до второго охот-
ничьего сезона составляет: в Лапландском* заповеднике 0,455, в Пе-
чоро-Илычском 0,326; коэффициент выживания в последующие годы
составляет, соответственно, 0,585 и 0,540. С помощью этих коэффи-
циентов вычислены числа в соответствующих колонках табл. 92' (ци-
тированные выше голландские авторы составили таблицу выживае-
мости усатой ночницы аналогичным способом)).
Результаты, полученные в обоих заповедниках, близко сходны.
Смертность глухаря в каждый из последующих лет жизни состав-
ляет, в среднем, 0,70'—0,75 смертности его за первую зиму и второе
лето жизни.
286
Таблица 92
Возрастной состав глухарей в окрестностях
Лапландского и Печоро-Илычского заповедников
Возраст птиц, лет Лапландский заповедник Печоро-Илычский заповедник
Число экз. Фактич. состав популяции (В %) То же по вычи- слению (В %) Число осенне- зимних экз. Фактич. состав популяции (в %) То же по вычи- слению (в Ул)
0+ (22)* 48 47,9 (98)* 59 58,8
1 + 10 22 21,8 32 19 19,1
2+ 8 17 12,7 21 13 10,3
н- 4 9 7,4 10 6 5,6
4+ 2 4 4,3 5 3 3,0
5+ — — 2,5 — — 1,6
6+ — — 1,5 — — 0,8
7+ — — 0,8 — — 0,4
8+ — —. 0,5 — — 0,2
9+ — — 0,3 — — 0,1
10+
н старше — — 0,3 — — од
Всего 24 100 100,0 68 100 100,0
Результат логарифмирования
Разности Разности
0 1g 22=1,343 lg 98=1,992
0,343 0,487
1 1g 10=1,000 lg 32 = 1,505
0,097 0,183
2 1g 8=0,903 lg 21=1,322
0,301 • 0,233 0,322 0,269
3 lg 4=0.602 lg 10-1,000
0,301 0,301
4 lg 2=0,301 lg 5=0,699
* Вычислено по среднему за ряд лет проценту молодых птиц.
Возрастной состав глухарей, добытых на токах в районе Печоро-
Илычского заповедника, на первый взгляд противоречит результа-
там анализа популяции в осенне-зимний период: основную ‘.массу
токовиков составляют самцы 3 лет и старше — они дают 61% всех
добытых птиц; между тем в сентябре—ноябре, то есть 6—8 месяцев
назад, глухари в возрасте 2 -f- лет и старше составляли только 22%
популяции. Замечательно, что численное соотношение старших поко-
лений весной и осенью одинаково; число птиц годом старше умень-
шается приблизительно вдвое. На диаграмме рис. 97 масштаб подо-
бран таким образом, чтобы уменьшение численности возрастных
групп шло по одной плавной кривой и осенью и весной. Диаграмма
287
показывает, что число добываемых на токах двухлетних глухарей
составляет примерно 60%, а число годовиков —всего 6% от ожи-
даемого. Этот факт объясняется полигамией глухаря. Ружей-
ная охота на току основана на подскакивании к поющему петуху
в короткие моменты «глухой» песни, ввиду чего охотники добывают
Рис. 97. Возрастной состав глухарей па Верхней
Печоре; редкая штриховка—осенне-зимний сезон
охоты, частая штриховка—охота па токах
в первую очередь самых активных самцов. В нашем материале годо-
вики составляют 7% всех отстреленных на токах глухарей. При от-
лове глухарей на току силками, практиковавшемся раньше про-
мысловиками, процент годовиков повышается: В. П. Теплов (1947,
Т а б л н ц а 93
Сравнение возрастного состава глухарей, добытых на Верхней
Печоре в осенний сезон и на токах
Число исследованных экземпляров в возрасте (лет) Всего
0 + 1 1+ 2 [2- 3 3+ 4 4+ 5
Осень . . Веспа . . . (98) 6 1 32 '21 26 I 31 10 13 5 6 68 82
288
стр. 38) указывает, что среди глухарей, добытых в петли 2—5 V
1938 г., годовики составляли 19i°/o- Очевидно, на токовищах собирает-
ся довольно много молодых са’м-цов, но поют они мало; и только при
благоприятных для этого обстоятельствах, так как в присутствии
более сильных соперников они боятся токовать. Сходное явление
происходит и на тетеревиных токах — там старые петухи занимают
индивидуальные участки в центре токовища, а молодые держатся
по его окраинам и поют мало.
Рис. 93. Движение популяции глухаря по учетным данным; белые
части столбиков — молодые, заштрихованные — взрослые птицы
Представим себе, что в какой-то популяции оседлых животных
в течение года не было смертности. После этого при учете количество
взрослых особей должно быть равно общему числу особей (взрос-
лых и молодых), учтенных в предыдущем году. Ввиду того что есте-
ственная смертность сильно колеблется, иногда в действительности
наблюдается положение, близкое к этому воображаемому случаю.
В Лапландском заповеднике смертность глухаря была неощутимой
в 1932-33, 1936-37 и 1'939-40 гг. — в те годы, когда подъем числен-
ности после депрессии только намечался. В годы после пика, вплоть
до нового минимума, смертность бывает высокой (рис. 98|). В Печо-
19—3)37
289
ро-Илычском заповеднике незначительная смертность была в
1964-55 г. Для дифференцированного определения смертности по
отдельным годам необходимо иметь увереннность в том, что ошибка
учетных цифр невелика (не выше, положим, 10о/0). Применявшаяся
нами методика учетов и малый размер пробных площадей не обеспе-
чивают необходимой для таких расчетов точности. По- этой причине
количество старых птиц иногда оказывалось даже больше количе-
ства всех птиц в предыдущем году.
Сокращение смертности в периоды депрессий объясняется изме-
нением численных соотношений популяций хищников и жертв. Воз-
можно, что имеет значение и изменение возрастного состава тетере-
виных птиц, так как в моменты депрессий популяция состоит пре-
имущественно из старых птиц, обладающих большим жизненным
опытом: нам удалось показать, что смертность у них в самом деле
меньше, чем у молодых птиц по первому году.
XII. МИГРАЦИИ
Обсуждая вопрос о движении численности тетеревиных птиц, мы
не принимали во внимание возможности изменений ее в результате
перекочевок, хотя такое представление распространено не только
среди охотников, но« и в литературе. Для рассмотрения этого вопроса
наших собственных наблюдений недостаточно; новейшие и наиболее
объективные данные по нему дает метод кольцевания — их рассмот-
рением мы и ограничимся.
В Советском Союзе кольцуют очень мало куриных, в связи с тем,
что взрослых сравнительно трудно ловить, а птенцов нельзя метить
ножными кольцами, так как со временем эти кольца станут малы и
искалечат лапу. На русском языке нет ни одной работы о результа-
тах кольцевания тетеревиных птиц. Для развития дела кольцевания
куриных необходимо перейти на другие способы мечения; попытки
в этом направлении делаются как за границей, так и у нас. В Швеции
и в Финляндии с 1945 г. широко применяют крыловую метку, сде-
ланную по принципу английской булавки: ее закрепляют на переднем
краю перепонки, соединяющей предплечье и плечо*. Введение этого
способа мечения позволило резко увеличить количество кольцуемых
тетеревиных за счет маленьких птенцов, однако количество возвра-
тов при этом снизилось, что объясняется высокой птенцовой смертно-
стью у куриных (Койвисто, 19*56).
В Коми АССР А. Н. Романовым в 1955—1957 гг. успешно испы-
тан способ автоматического кольцевания тетеревиных специальными
силками из толстой капроновой лесы, с застежкой в виде карабин-
чика или чеки. Установленная в воротцах ловчей изгороди петля со-
стоит из двух частей, одна из которых застегивается на шее птицы
и образует прочный ошейник с номером, а другая обрывается, как
только ошейник застегнется и птица рванется. Преимущество этого
* Propatagium.
290
способа в том, что отлов и кольцевание происходят автоматически и
одновременно, причем метятся только взрослые или полувзрослые
птицы; недостаток — в том, что не всегда можно определить по сле-
дам, какая птица окольцована, и невозможно установить число
окольцованных птиц: одна птица может иметь на себе несколько
меток, а учет ведется только по количеству исчезнувших петель
с метками. К сожалению, результаты работы автором еще не опубли-
кованы; описана только методика (Романов, 1956).
В Швеции кольцевание проводит Шведский охотничий союз; от-
четы опубликованы Н. Хёглундом: (1956, 1957|). В Финляндии мече-
нием руководит Управление охраны дичи; отчеты и некоторые резуль-
таты опубликованы Ю. Коскимиесом (1953), Нордстрёмом (1954)
и И. Койвисто (1956). В этих странах и у нас, в Печоро-Илычском
заповеднике, окольцовано следующее количество тетеревиных (в
числителе — число возвратов, в знаменателе — число окольцованных
птиц):
Вид Швеция Финляндия СССР Возврат в %
Глухарь 20/1165 20/718 0/12 2,1
Тетерев 22/473 14/624 6/30 3,7
Рябчик 6/378 6/401 1/3 1,7
Белая куропатка ...» 12/625 3/175 0/0 1,9
Тундряная куропатка . . . 0/39 0/2 0/0 0,0
По опубликованным шведским и финляндским материалам с при-
соединением и небольших материалов Печоро-Илычского заповед-
ника мы получили следующие расстояния между пунктами мечения
и возврата:
Вид Расстояние в км Всего случаев
0-2,5 2,5-5 5-10 10—20 20-40
Глухарь , » « 19 2 6 9 4 40
Тетерев . . ...... 28 2 2 5 2 39
Рябчик 7 3 1 — — 11
Белая куропатка .... 7 3 1 2 2 15
19*
291
Приблизительно, половина возвратов поступила с расстояния не
более 2,5 км от места мечения. Максимальные расстояния зарегист-
рированы: у глухаря (Швеция) — 38 км, у глухарки (Финляндия) —
25 км, у тетерева (Финляндия;)—26' км, у тетерки (Швеция)—27 км,
у рябчика (СССР) —б км, у белой куропатки (Финляндия) — 30 км.
В использованные нами сводки не попал описанный Л. Сиивоненом
('1943) случай с куропаткой, окольцованной летом 1913 г. ® Финской
Лапландии на широте 68°, и добытой в Куусамо, в 240 км к югу.
Койвисто (1956), обработав финляндские и шведские данные
по кольцеванию глухаря, пришел к выводу, что молодые самки отко-
чевывают сравнительно далеко; от места мечения (6—22 км), тогда
как самцы добывались обычно не далее 21 км. Самки старше года, по-
видимому, не кочуют. По мнению Койвисто, расселение глухаря про-
исходит только за счет самок, а петухи остаются оседлыми, и этим
объясняется известное постоянство токовищ. У тетерева это явление
выражено слабо.
Данные кольцевания дают основания считать всех тетеревиных
таежной полосы оседлыми. Наблюдавшиеся нами в Лапландском
заповеднике перелеты глухарей поздней осенью (см-, стр. 29),пред-
ставляют, вероятно, местное явление, связанное с переменой мест
обитания при наступлении зимы, а может быть — и с расселением.
Тетерев не отличается от глухаря по1 масштабу своих перелетов *.
Белые куропатки, гнездящиеся в тундре, регулярно мигрируют,
отлетая зимовать в лесотундру, а иногда (вероятно, при недостатке
кормов или вследствие больших снегопадов) залетают далеко
вглубь лесной зоны. В Коми АССР зимний промысел белой куро-
патки сосредоточен в северных районах — Усть-Цилемском, Ижем-
ском, Усть-Усинском и Кожвинском. В Печоро-Илычском заповед-
нике белая куропатка довольно редка и. встречается поодиночке
или стайками по 2—4 птицы. Всего за пять зимних месяцев сотруд-
ники заповедника отмечают обычно не более 100 птиц. Зимой
1956—57 г. в картотеке наблюдений заповедника было зарегистри-
ровано около 700 встреч белых куропаток. В предгорьях Урала стаи
куропаток по 10—35 птиц появились уже в декабре и держались
там до конца марта. Очевидно, это был случай необычайно глубо-
кого залета куропаток из тундр в область тайги.
Тундряная куропатка в горах Лапландского заповедника жи-
вет, по-видимому, почти оседло, по при выпадении глубоких снегов
перелетает на те склоны, где есть выдувы. В апреле 193'8 г. на севе-
ро-восточных склонах Монче-тундры, по сообщению наблюдателя
охраны заповедника М. Малодушева, держалась стая куропаток
около 80 штук; вскоре ее перестреляли охотники. Множество тунд-
ряных куропаток в ту зиму наблюдалось около метеорологической
станции Юкспор на Хибинских горах (сообщение Париенко). На
горах Чуна-тундры в ту зиму не было выдувов, и при учете диких
оленей в апреле мы видели сравнительно мало тундряных куропа-
* Случаи залета глухарей и тетеревов в города, о которых иногда сообщают
в журналах, могут объясняться местными перелетами на расстояние свыше 10 км.
292
ток. Этот пример показывает, что местные миграции могут создать
превратное представление о численности куропаток.
По всей вероятности, зимовка тундряных куропаток на Шпиц-
бергене, Земле Франца Иосифа и других островах протекает при-
мерно так же, как и в Лапландском заповеднике; однако тундряные
куропатки с берегов Таймырского озера уже в сентябре — октябре
отлетают на зимовки, возвращаясь с юга в апреле—мае (Сдобни-
ков, 1957).
Резюмируя сказанное, мы приходим к заключению, что при ис-
следовании колебаний численности белой и тундряной куропаток
необходимо учитывать возможность массовых миграций, регуляр-
ных в тундровой зоне, эпизодических в тайге или на горах. Измене-
ния численности глухаря, тетерева и рябчика, если они охватывают
значительное пространство, нельзя объяснить перелетами.
Возвратимся еще раз к материалам кольцевания, поскольку по
ним можно судить о долголетии птиц. Обычно при кольцевании
большая часть возвратов поступает в первом же году. В рассмот-
ренном нами материале (90 возвратов) 71 % меток вернулся менее
чем через 1 год, 19%—на втором году после мечения,t 8 %—на
третьем и 2% через большие сроки. Из этих двух меток одна снята
через 6 лет с остатков погибшей глухарки (Швеция), другая —
с рябчика, застреленного в северной Швеции в возрасте 7 лет и
4 месяцев. Этот случай можно считать рекордным, поскольку воз-
вратов по четвертому, пятому и шестому году не было.
ВЫВОДЫ
Материал для работы собирался в 1930—1941 и 1949—1951 гг.
в Лапландском заповеднике и в 1952—1957 гг. в Печоро-Илычском
заповеднике: материалы последнего использованы только в главах
IX—XII. Длительность исследования дала возможность сосредото-
чить внимание на движении численности тетеревиных птиц, в пер-
вую очередь глухаря, и на причинах, от которых оно зависит.
Распространение. Белая куропатка заселяет всю Мурманскую
область. Тундряная куропатка приурочена к горным тундрам и ка-
менистым участкам равнинных тундр. Ареал глухаря и рябчика
ограничен областью распространения хвойных пород. Распростра-
нение тетерева в годы исследования сильно расширилось в связи
с вырубкой первичных хвойных лесов, заменой их лиственными
молодняками (частично по гари) и значительным расширением
площади возделанных земель. В 1957 г. тетерев достиг Кольского
залива. На южной границе заповедника относительная численность
тетерева увеличилась за 18 лет почти в 7 раз.
Питание. Тетеревиные, в отличие от других птиц, питаются в ос-
новном вегетативными частями растений: в этом отношении они
сходны с некоторыми млекопитающими (копытные, грызуны). Ле-
том все виды тетеревиных питаются почти одинаковой пищей, со-
стоящей из листьев, цветов, ягод и сухих плодов самых распростра-
293
ненных растений местной флоры. Различия в питании отдельных
видов незначительны. В Мурманской области для глухаря, напри-
мер, характерно употребление колосков пушицы весной, грибов и
листьев волосистой ожики— осенью; тетерев и рябчик в большей
мере питаются ягодами и семенами, чем остальные виды (рябчик
особенно предпочитает ягоды брусники); основной корм белой ку-
ропатки весной и осенью — стебли черники, а осенью и ягоды ее.
В питании птенцов особое значение имеет животный корм, осо-
бенно личинки пилильщиков и гусеницы бабочек; вместе с тем
с первого дня они поедают цветы, ягоды и другие части растений,
а также заглатывают камешки. В августе и сентябре в питании
взрослых птиц животный корм и семена играют меньшую роль,
а ягоды — большую, чем у молодых. Зимой лесные виды тетереви-
ных питаются веточным кормом, причем состав зимних кормов
у разных видов настолько специфичен, что даже по помету легко
определить, кому он принадлежит. Глухарь питается хвоей и веточ-
ками сосны, отчасти можжевельника, изредка — ели: хвойное пита-
ние у него преобладает в течение 0,56 года. Тетерев кормится муж-
скими сережками и ветками березы, первотоднимм шишечками и
хвоей сосны, плодами и хвоей можжевельника. Рябчик ест преиму-
щественно мужские сережки, женские и листовые почки серой ольхи
и березы. У обоих видов зимние корма преобладают в течение
0,53 года. Белая куропатка в продолжение 0,38 года питается, в ос-
новном, ветками различных видов ив и берез: вместе с тем она (как
и рябчик) всю зиму добывает из-под снега зелень и ягоды. Тундря-
ная куропатка всю зиму кормится плодами и зелеными частями
ягодных кустарничков и других альпийских растений, выкапывая
их из-под снега: веточные корма не играют большой роли в ее пи-
тании. 1
Приблизительный расчет показывает, что при средней плотности
популяций тетеревиных они съедают в год около 0,5 % годового при-
роста зеленой массы напочвенного покрова и 1 — 1,5% урожая
ягод. Летом тетеревиные в изобилии обеспечены кормом, но зимой
кормовой вопрос обостряется. Глухари, например, съедают в год
около 1% прироста сосновой хвои. Запасы зимних кормов других
видов тетеревиных значительно меньше, вследствие чего нехватка
пищи в некоторые зимы является реальной угрозой. Вследствие
специфичности зимнего питания отдельных видов внутривидовая
конкуренция должна проявляться при этом раньше, чем межви-
довая.
Пищеварение. При максимальном наполнении зоб глухаря содер-
жит корма до 7—8% веса тела птицы, зоб белой куропатки 10—
12%, зоб рябчика 13—15%. Это количество пищи переваривается
в течение ночи. Глухарь активен в сумерках при положении центра
солнца до 5—6° ниже горизонта, белая куропатка до 7—8° (в лесу)
или до 8—10° (в тундре). Суточное потребление зимних кормов
определено в ПО г корма (сырой вес) на 1 кг живого веса птицы —
для глухаря; 160 г — для белой куропатки и 200 г — для рябчика.
294
Анатомо-физиологические особенности тетеревиных птиц явля-
ются результатом приспособления их именно к зимним условиям
питания. Характерно сильное развитие клюва и наличие на нем ре-
жущей кромки — приспособления для отстригания или отламывания
веток. Необычайно развит зоб, служащий резервуаром пищи на
ночь и для прогревания ее. Мускульный желудок очень развит, до-
стигая 4% веса птицы. Летом, когда птица питается менее грубой
пищей, вес желудка уменьшается: у куропаток в V—VI он на 33%
меньше, чем в II—IV и на 42% меньше, чем у одного экземпляра
в декабре. Полное отсутствие желчного пузыря связано, по всей ве-
роятности, с тем, что пища поступает из желудка в кишечник бес-
прерывно. Необычайное развитие слепых кишек дает возможность
продлить процесс усвоения пищи при быстрой разгрузке кишечника
от непереваримых грубых частиц, составляющих, по сухому весу,
около 80% всей поступающей в кишечник массы корма. Пища в же-
лудке измельчается с помощью гастролитов, количество которых
меняется по сезонам, достигая максимума в октябре и минимума
в марте. Сокращение их количества зимой объясняется невозмож-
ностью возобновления запаса в это время.
Рост птенцов и вес взрослых птиц. Молодые глухари прибавляют
в весе до момента перехода на зимний режим питания, что проис-
ходит в октябре. Вес молодых самок зимой равен 90% веса взрос-
лых, но молодые самцы достигают только 76% веса взрослых сам-
цов. Рост их возобновляется на следующее лето, прибавка в весе
наблюдается и на третье лето. Вес взрослых глухарей изменяется
по сезонам, у самцов он достигает максимума в апреле, минимума
в июне — августе. Вес молодых тетеревов в первую зиму равен 87%
веса взрослых, вес молодых самок — 98%. Рябчики и белые куро-
патки в возрасте 4 месяцев по весу не отличаются от взрослых.
У последних видов хорошо выражены сезонные изменения веса.
Максимум у обоих полов бывает в ноябре — январе, минимум —
у самцов рябчика в мае, у самцов куропатки в июне — июле; у са-
мок в период кладки (май) наблюдается второй максимум веса,
а годовой минимум приходится на июль. Амплитуда сезонных
колебаний веса достигает 10—30% от максимального за год. При
сезонных колебаниях веса в наибольшей мере меняется вес пище-
варительных органов и их содержимого.
Линька. У взрослых тетеревиных птиц она начинается в ап-
реле— мае и заканчивается в октябре. Смена маховых, например
у глухаря, продолжается 5 месяцев. У молодых птиц при постъю-
венильной линьке первые два маховых кисти не сменяются. Они
заметно отличаются от остальных маховых наличием светлых
(глухарь, тетерев) или темных (белая и тундряная куропатки)
крапинок на концах опахал и заостренной их формой. Эти призна-
ки удобны для определения возраста птиц. Молодых самцов глу-
харя и тетерева легко отличить по длине рулевых, которые на
20—25% короче, чем у взрослых. У глухарей старше года возраст
можно определить по срастанию определенных швов на черепе
295
(способ С. В. Кирикова). Возраст самок различается до 3 лет, сам.
нов — до 4—5. Для остальных тетеревиных птиц этот способ не
применим.
Паразиты. Встречаемость аскарид и крупных ленточников из
рода Raillietina подчинена определенному сезонному циклу. Аска-
риды встречаются только летом, причем у глухаря наблюдается
один максимум (август), у белой куропатки их два в (июне и
сентябре). Цестоды встречаются с февраля по октябрь. Время исчез-
новения их из кишечников совпадает с моментом перехода птицы
на зимний кормовой режим, однако регенерация стробил наступает
задолго до перехода на летние корма. Период полного отсутствия
стробил цестод совпадает с периодом, когда долгота дня менее
8 ч. 45 м. Предполагается, что дестробиляция связана с зимним
снижением активности птиц, а регенерация с повышением ее в кон-
це зимы.
Враги. Тетеревиные составляют приблизительно 80% всей био-
массы птиц в тайге; они уступают по значению только двум груп-
пам диких млекопитающих — копытным и полевкам. Поэтому тете-
ревиные играют важную роль в питании многих хищников, осо-
бенно ястреба-тетеревятника, кречета, беркута, орлана, куницы,
лисы и росомахи. Изменения состава пищи куницы по годам опре-
деляются, в основном, урожаем главного корма (полевки). Недо-
статок полевок заставляет куницу увеличить длину суточного про-
бега, и это дает ей возможность компенсировать недостаток основ-
ного корма второстепенными. Хищные звери уничтожают прибли-
зительно столько же тетеревиных птиц, как и хищные птицы; при
этом глухарь и рябчик больше страдают от зверей, а тетерев и оба
вида куропаток — от птиц. Смертность от хищников сильно ме-
няется по годам и бывает наибольшей в годы падения численности
жертвы, то есть в годы, когда соотношение численности жертв и
хищников изменяется в пользу последних.
Размножение. Амплитуда сезонных изменений веса семенников
достигает 20—25 крат, причем максимум наступает на очень корот-
кое время. В сентябре — октябре наблюдается вторичное увеличе-
ние веса семенников в 2—3 раза.
Режим насиживания изучался в Лапландском и Печоро-Илыч-
ском заповедниках с 1950 г. с помощью актографов особой кон-
струкции, регистрировавших наличие или отсутствие наседки на
гнезде, а также и ее движения. Рябчик несет яйца с интервалом
в 26—30 часов, так что время кладки постепенно сдвигается с утра
на вечер; после этого птица пропускает один день, а затем с утра
начинается новый цикл. Одинаковым образом несется и глухарка.
Насиживание начинается вечером, накануне откладки послед-
него яйца. В первые сутки наседка отлучается чаще и на большие
сроки; нормальный режим устанавливается на второй день. Глу-
харки и тетерки оставляют гнездо 2—3 раза в день, рябчики
3—5 раз, белая куропатка 3—5 раз. Продолжительность каждой
отлучки в Лапландском заповеднике 10—40 мин., в Печоро-Илыч-
296
ском 10—25 мин. Суммарная продолжительность отлучек состав-
ляет в Лапландском заповеднике у глухарки 0,061, у куропатки
0,045 всего времени насиживания; в Печоро-Илычском заповед-
нике .у глухарки 0,027, у тетерки 0,032, у рябчика 0,05-2 времени на-
сиживания. Наседки переворачивают яйца в среднем 22—33 раза
в день, в жаркую погоду чаще, в дождливую — реже. Продолжи-
тельность насиживания, включая и обогревание птенцов в гнезде
до ухода выводка, у глухарки колебалась в пределах 25—26 суток,
у рябчика была равна 25,4 суток. В период вывода птенцов наседка
совсем не оставляет гнезда в продолжении 23—37 часов и только
в одном случае—18 часов. Спугнутая с гнезда наседка обычно
долго не возвращается. В отдельных случаях гнездо оставалось
без наседки до 6—7,5 час., однако без вредных последствий * для
эмбрионов. В опыте яйца оставались 17 час. при температуре 8°,
после чего развитие эмбриона продолжалось. Замерзание яиц в
период инкубации крайне мало вероятно. Главными причинами эм-
бриональной смертности мы считаем то, что наседки нередко бро-
сают кладки во время насиживания или до его начала, а также за-
мерзание яиц в период кладки. Замечательно, что в зимы после
«неурожая» выводков глухаря в питании куницы увеличивается
значение птичьих яиц: хищник добывает их из-под снега, обнару-
живая чутьем.
Учет и движение численности. Обсуждается методика учетов и
приводятся данные учета на ленточных пробах в августе, а также
учетов встреч на экскурсиях в период осенне-зимней охоты. Боль-
шие пики и депрессии численности тетеревиных птиц охватывают
огромные пространства тайги от Норвегии до Урала, как это на-
блюдалось при депрессиях 1932, 1935 и 1939 гг. или при пиках
1937 и 1953 гг. Причиной колебаний численности является взаи-
модействие двух переменных факторов — успешности размножения
и величины смертности. Хорошим показателем успешности размно-
жения может служить возрастной состав популяции осенью, полу-
ченный путем исследования птиц, добытых на осенней охоте. У глу-
харя процент молодых птиц колебался в районе Лапландского за-
поведника от 7 до 81 %; на Верхней Печоре — от 30 до 80%. Между
этими показателями, с одной стороны, и температурой и осадками
периода насиживания—с другой, существуют тесные коррелятив-
ные связи. В Лапландском заповеднике насиживание происходит,
в среднем, с 30 V по 25 VI, в Печоро-Илычском — с 21 V по 16 VI.
Величины соответствующих коэффициентов корреляции были от
0,67 + 0,15 до 0,96 + 0,03. При таких высоких показателях связи
можно уже 1 июля давать вполне обоснованный прогноз возра-
стного состава, а следовательно, и численности глухаря в сезон
охоты. /Лолодняка много в те годы, когда средняя температура
каждой из трех декад насиживания была выше 8°. В Лапланд-
ском заповеднике «неурожай» наступает, если средняя температура
одной из этих трех декад окажется ниже 5°; в Печоро-Илычском
заповеднике — если сумма июньских осадков окажется больше
70 мм.
297
Средняя величина смертности в популяции равна среднему
многолетнему проценту молодых птиц в ней. Основных причин
смертности две — хищники и охота. В Лапландском заповеднике
средняя плотность популяции глухаря в годы исследования была
79 птиц на 1000 га, из них 48% молодых, откуда средняя продук-
тивность лесов учетного стационара равна 38 шт. с 1000 га. В Пе-
чоро-Илычском заповеднике плотность равна 138 шт., молодняка —
59%, а продуктивность популяции около 81 шт. с той же площади.
Определение возрастного состава популяции глухаря по возра-
стным признакам дало возможность дифференцировать величину
смертности по возрастам. За первую зиму и второе лето выжи-
вает около 46% в Лапландии и 33% на Верхней Печоре, а в по-
следующие годы — соответственно 59 и 54%. Таким образом, смерт-
ность старших возрастных групп составляет только 70—75% смерт-
ности годовиков. Среди глухарей, убитых на Верхней Печоре на
токах, преобладают особи от трех лет и старше (61 % всех добытых
самцов), двухлетков всего 32%, годовиков — 7%. Объясняется это
тем, что старые петухи не дают токовать молодым.
Миграции. По данным кольцевания (главным образом финским
и шведским), более половины возвратов меток с тетеревиных посту-
пает от птиц, добытых не далее 2,5 км от места мечения. Макси-
мальные зарегистрированные расстояния 25—38 км и только у бе-
лой куропатки отмечен один возврат за 240 км. Миграции лесных
видов тетеревиных имеют местный характер; они не могут быть
причиной значительных колебаний численности.
Практические выводы. Излагаем их в виде тезисов:
1. Осенняя охота на боровую дичь должна открываться не ра-
нее начала рассеивания выводков: таким путем можно избежать
непроизводительного истребления маток и птенцов. Для всей таеж-
ной полосы Союза следует установить стандартный срок открытия
охоты на боровую дичь 1 сентября, а в годы неурожая на выводки
переносить начало охоты на 1—3 недели позже.
2. Весенняя охота на токах вполне допустима в хозяйствах, где
можно обеспечить проведение учета добычи охотников и соблюде-
ние определенной нормы отстрела. На токах отстреливаются в пер-
вую голову более старые самцы, а половозрелые молодые вполне
могут обеспечить оплодотворение самок. Отстрел самцов полигам-
ных птиц улучшает соотношение полов в популяции. В местах, где
глухарь и тетерев редки, целесообразно запретить осеннюю охоту,
но сохранить весеннюю, как это и практикуется в Средней Европе.
Весенняя охота может быть разрешена на срок не более двух пе-
дель, причем необходимо установить предельные нормы добычи.
3. Мероприятия по истреблению хищников целесообразны толь-
ко в периоды снижения численности дичи, когда соотношение чис-
ленности хищника и жертвы наиболее неблагоприятно. Отстрел та-
ких редких птиц, как кречет, беркут или орлан, не должен допу-
скаться; целесообразно контролировать только ястреба-тетеревят-
ника и филина. В хозяйствах, имеющих задачей обеспечение спор-
298
тивной охоты, целесообразно контролировать и численность пушных
хищников — лисы и куницы.
4. Во всех таежных заповедниках необходимо организовать
•службу учета плотности популяций тетеревиных птиц, включая про-
ведение ежегодного учета на лентах общей площадью не менее
200 га, и определение возрастного состава наиболее распростра-
ненных видов. Эти данные должны использоваться для прогноза и
назначения рациональных сроков охоты, а также для учета движе-
ния численности промысловых птиц на обширном пространстве.
5. Совершенно необходимо организовать статистический учет
добываемой охотниками дичи. Это прямая обязанность Управлений
охотничьего хозяйства союзных республик. Работу целесообразно
проводить через Добровольные общества охотников. Никакое хо-
зяйство, в том числе и охотничье, не может существовать без учета
своих фондов и продукции.
Semenov-Tian-Shansky, Oleg
ON ECOLOGY OF TETRAONIDS
Summary
The present study was carried out in Lapland Reserve (district of
Murmansk) in 1930—1951, and partly in Petshora-Ylytsh Reserve
(Republic of Komi), in 1952—1958; both Reserves are situated’in taiga
region.
Chapter 1 deals with the distribution of Tetraonids in Murmansk
district. Capercaillie and Hasel grouse areas are limited to the northern
range of forests. The distribution of Blackgame, owing to progress of
agriculture and wood-cutting, greatly increased in last decades; being
limited up to 1900 to the south coast of Imandra Lake, its area now
reaches Kola fiord. Willow grouse is widely distributed on the whole
district, but Rock ptarmigan is restricted mainly to the tundra region
of mountain ranges.
In Ch. II plant cover and some physical-geography features of
Lapland Reserve are described.
In Ch. Ill habits of our Tetraonids and their seasonal changes are
described. In tables 1—5 some phenologic data are recorded.
Food habits are described in Ch. IV. In tables 8—21 food items
found in crops are recorded for months separately. They are classified
as follows: I — browse food, forming the bulk of winter diet; II — ever-
green plants; III — leaves of deciduous plants; IV — blossoms; V — ber-
ries; VI — capsules and seeds; VII — mushrooms, mosses and lichens;
VIII — animal food; IX — grit. For each species three tables are given,
in the first — frequency of occurrence of different food items in adult
birds (per cent) is recorded, in the second — the approximate quantity
of them (per mille); in the third — frequency and quantity of corres-
ponding food items of juvenile birds. In summer all grouse are mostly
herbivorous and their food habits have much in common. Animal food
(mainly insects) is consumed chiefly by juveniles. In winter all grouse
forage on trees and each species has its own specific diet. The brow-
sing season is the longest in Capercaillie (0,56 of year), then follow
301
Blackgame and Hazel grouse (0,53); snow cover interferes less with
willow grouse feeding; it forages on birch and willow twigs 0.38 of
year only. Rock ptarmigan gets berries and evergreen leaves all win-
ter long on hillocks, wind swept of snow.
Ch. V deals with digestion-tract structure und function, including
also dependence of grouse activity upon day light. Seasonal changes
of grit contents in gizzards are discussed. The absence of vesica fellea
and extraordinary size of coeca are the principal anatomic characters
of Tetraonids (tabic 34). Size of heart in various Tetraonids greatly
differs, being the least in Hazel grouse (5»/Oo of body-weight) and the
greatest in Rock ptarmigan (2O»/oo). In Capercaillie and Blackgame
the heart is comparatively larger in females and smaller in males.
In Ch. VI growth of juveniles and seasonal changes in weight of
adult birds are described. Weight growth of young birds ceases with
the change of their diet to winter ligneous food. Weight of immature
Capercaillie females the first winter is about 90—91 % of that of adult
females; weight of immature males does not exceed 73—76% of that
of adult males (table 39); thus the growth of Capercaillie continues
next summers (table 40 and fig. 49). Weight, of Willow grouse and
Rock ptarmigan hens (and probably other hens also) rises in time of
mating and egg-laying, but decreases when they raise chickens. Weight
of males reaches its maximum in XII-I and minimum in VI—VII
(table 42). Weight of gizzard muscles changes in the course of year
most of all: the difference makes about 20% in Capercaillie and 40%
in Willow grouse; size of liver changes likewise (table 44).
In Ch. VII moulting and methods of age determination of Tetraonids
are considered. Aging of juveniles can be carried out by their body-
weight or by the state of primaries moulting. Immature Capercaillie
and Blackgame males can be distinguished from adults by their tails
being shorter. The best method for aging both males and females is
by the two outermost primary feathers: tips of these primaries are
pointed and speckled in immature birds, but rounded and coloured
uniformly with others in adults (fig. 58 and 59). S. Kirikov proposed
a method for the estimation of the age of Capercaillies by a definite
order in effacing certain fissures in cranium; this method is effectual
up to 4 or 5 years in males and 3 years in females (pp. 171 —173.
fig. 60—68).
Ch. VIII — Parasites of Tetraonids. Frequency of occurrence of
cestodes (Raillietina sp.) and ascarides (Ascaridia sp.) is subject to
distinct seasonal changes (table 49, fig. 69—70). In general, Willow
grouse are intensely invaded with these gelminths; Hazel grouse and
Rock ptarmigan — least of all (table 50).
Ch. IX considers predators and their activity. In both Reserves the
stock of Tetraonids amounts to 26—42 kg for 1 sq. km approximately:
being the largest forest birds, they make the bulk of total birds-mass.
In northern forests only voles (mainly Clethrionomys sp.) and deer
(elk and reindeer) are comparable with Tetraonids in their capacity as
302
«кеу-industry» (table 54). Lists of birds of prey with data of their food
composition are recorded in tab!. 55—57; analogous in tabl. 60—61
for beasts of prey. Fig 74—75 demonstrate, how the role of Tetraonids
in marten’s diet differs from year to year, according to accesibility of
voles, constituting their principal food item. In winter martens (as well
as loxes) often get eggs of Tetraonids from under the snow. These
eggs are the result of embryonic mortality in course of brooding; for
this reason their occurrence in winter diet depends upon the age ratio
of Capercaillie population, and, on the other hand, upon the accessibility
of voles (fig. 76).
Ch. X — Breeding habits. Brooding regime studies with actographs
of original construction (fig. 81—83) are considered here. Registered
by actographs brooding regimes of Capercaillie (fig. 84, 85), Blackgame
(fig. 88), Hazel grouse (fig. 89) and Willow grouse (fig. 91) are partly
represented graphically. Egg-laying in Capercaillie and Hazel grouse-
daily takes place later and then, after passing one day, the new cycle-
begins in the morning (fig. 85, 89). Brooding begins in the evening,
before the last egg is laid. On the first day of brooding the hen leaves
nest more frequenty and for a longer time than later. While chick-
hatching, during about 23—37 hours, the brooding hen .does not
abandon her nest at all. Going from the nest away, the covey makes
only 7—14 m in an hour.
Brooding regime can be characterized as follows:
Capercaillie Black- grouse Petsh. Hazel grouse Petsh. Willow grouse Lapl.
Lapl. Petsh.
Duration of incubation ♦, days . . Ordinary numb, of walks, in a day . 26 25-26 >23 25,4 >18
2-3 2-3 2-3 3-4 3—5
Duration of each walk, min 20-40 10-25 10-25 15-25 10-30
Complete dur. of walks, part of a day 0,061. 0,027 0,032 0,049 0,045
Egg turnings, in a day . . ; . . 26 22-32 25 33 ?
Air temp, of incub. period, av. . . . 9,0° 10,2° 9,5° 7,8° 10,9° 10,1°
„ „ at time or hens absence . 8,7° 8,9е 11,4° 9,2°
Minim, temp. „ „ „ . . Time of abs., when hen is driv., hours 0,9° -5,5° -2,0° 0,3° -0,9°
1-3 1-3 1—3 7 1
Max. time of absence, hours .... Time on nest, when chicks hatch., 7,5 6,3 (40)** 0,8 1,4
hours 34 23-34 18 37 34
Number of nests studied 1 5 2 1 2
* From beginning of brooding up to departure of the covey.
** Perhaps the eggs were frozen (nest later destroyed).
The eggs of Tetraonids can not be frozen during the absence of
brooding female, because their cooling is a comparatively slow process.
Losses of eggs in cold springs take place partly if they freeze before
303
brooding, partly when brooding hens abandon their nests in cold or
rainy weather.
Ch. XI — Counts and population fluctuations. In tabl. 80—81 are
cited census figures of Tetraonids for 1 sq. km, and in tabl. 82—83-^
counts for 100-hours trip. In tabl. 84—87 «peak»-years and depression,
years of Tetraonids in Scandinavian countries and USSR are compa-
red. Sometimes, as in 1932, 1935, 1937, 1939 and 1953, peak years or
low years spread on immense space of forest region, from Norway to
Ural. Population fluctuations are the result of interaction of two
changeable factors — success of breeding and rate of mortality. When
the meaning of separate meteorological factors in population fluctua-
tions must be revealed, census figures are but of little use; age ratios
in population (table 88) make better index for such studies. Correlation
months or 10-days periods and ratios of immature birds in Capercaillie
coefficients between air temperatures (or precipitation sums) for
populations are listed in table 89. In Lapalnd Reserve best results are
received with the mean temperature of June (r = + 0,67 + 0Л5),
with first 10 days of June (r = + 0»71 + 0,13), with the coldest 10-days
period of June (r == + 0,74 + 0,12) and minimum temperature in June
(r = + 0,82 + 0,09). In Petshora-Ylytsh Reserve — with precipitation
sum in June (r = — 0,96 + 0,03), with mean temperature in 10 last
days of May (r = + 0,80 + 0,15) and with the coldest of 3 ten-days
periods of incubation (r = + 0,85 + 0,11). Thus the close connection
of breeding success in Capercaillie with weather conditions in brooding
time is well demonstrated.
In table 92 the age composition of Capercaillie populations in both
Reserves is calculated. For age determination two methods were used,
immature birds were distinguished by pinion tips or tail length, the age
of adults was estimated by cranium fissures. The survival rate in the
first winter and second summer of life is about 46% near Lapland,
and 33% near Petshora-Ylytsh Reserves; in each of further years —
about 59% and 54% correspondingly. Thus the mortality rate for adult
Capercaillies is about 70—75% of that of yearlings. The age composi-
tion in Capercaillie cocks shot on mating grounds is quite remarkable.
In table 93 the number of Capercaillies shot in open season and on
mating grounds, c'onsidcrnig their age, is recorded and the figures are
compared (see also fig. 97). On mating grounds 1- and even 2- years-old
cocks are suppressed in display by males of three years and older; for
this reason elder birds prevail in the bag.
In Ch. XII migrations of Tetraonids are discussed. Marking records
suggest that forest species of Tetraonids are almost stationary birds:
about 58% of marked birds were recorded not farther than 2.5 km
(or 1,5 miles) from the point of marking and none farther than 40 km.
Willow grouse and Rock ptarmigans of tundra regions pass winter by
the northern timber-line and their migrations reach some hundreds
kilometers; in fluctuation studies of these birds, migrations should be
taken into consideration.
304
EXPLANATIONS FOR ILLUSTRATIONS
1. Timberline on slopes of Tshuna-tundra in winter.
2. Spruce forest with Pleurozium Schreberi, Vaccinium myrtillus
and Empetrum nigrum in the floor cover.
3. Pine forest with Cladonia alpestris and Vaccinium myrtillus in
the floor cover.
4. The habitat of the Rock ptarmigan on the west slope of Tshuna-
tundra.
5. Pine branches, browsed by the Capercaillie in foraging.
6. An explosion of snow-dust, caused bu the Capercaillie starting
from under the snow.
7. Capercaillie female in fly.
8. Capercaillie male approaching his dust-bath.
9. Capercaillie male dusting. Phot. 8 & 9 are made with a self-
photo-snapshot device, in Petshora-Ylytch Reserve.
10. Capercaillie female of a month age.
11. Capercaillie male of a month age. Phot. 10 and 11 are made on
the same day; note the difference in postures, owing to sex.
12. Snow burrow of a Blackgame female.
13. Blackgame males displaying (Petshora-Ylytsh Reserve).
14. Snow burrow of a Hazel Grouse.
15. The tracks of Willow Grouse foraging.
16. Willow Grouse juvenile (Petshora-Ylytsh Reserve).
17. Rock ptarmigans in windy weather (Tshuna-tundra).
18. Rock ptarmigan flying up (Tshuna-tundra).
19. Rock ptarmigan male at spring moulting.
20. Rock ptarmigan juvenile in mixed summer-autumn plumage.
21. Seasonal changes of food composition in Tetrao urogallus
(adults above, juveniles below). Explications, from above: winter brow-
sing forage (white); evergreen forage (frequent oblique hatched);
summer-green forage (sparse oblique hatched) blossoms (dotted);
berries (circles); seeds and capsules (vertical hatched); mosses and
mushrooms (oblique hatched); animals (cross hatched); grit (black).
22. Cottonweed (Eriophorum vaginatum) in bloom.
23. Melampyrum pratense with seeds.
24. Seasonal changes in food composition of Lyrurus tetrix (for
explications — see above, fig. 21).
25. Seasonal changes in food composition of Tetrastes bonasia
(for explications— see above, fig. 21).
26. Alternations in browsing food of Lagopus lagopus, in connetion
With snow cover height.
27. Seasonal changes in food composition of Lagopus lagopus (for
explications — see above, fig. 21)
28. Bilberry (Vaccinium myrtillus) stalks, browsed by Tetraonids.
29. Crowberry (Empetrum nigrum) with fruits.
30. Seasonal changes in food composition of Lagopus mutus (for
explications — see above, fig. 21).
20 -3037
305
31. Arctous alpina — the «Thunder-berry» in Lapp.
32. Dryas punctata — the «Ptarmigan grass» in Russian.
33. Evaluation of Tetraonids principal food plants in annual aspect.
Names of food plants listed (from above) are: Pinus silvestris, Junipe-
rus sibirica (dotted), Salix sp. sp., Betula pubescens and B. verrucosa
(horizontal hatched), Alnus borealis (cross hatched), Betula nana
(vertical hatched), Empetrum nigrum (blackened), Arctous alpina,
Vaccinium vitis idaea, Vaccinium uliginosum (circles), Vaccinium
myrtillus, other plants (white).
34. Skulls of Lagopus lagopus (above) and Lagopus mutus (below),
in comparison.
35. Daily and seasonal changes in crop contents of Tetrao uroga-
llus. Explications: day and twilight period (white), dark period
(hatched), weight of crop contents (dotted).
36. Daily and seasonal changes in crop contents of Lagopus lagopus.
For explications — see above, fig. 35.
37. Grit contents in stomachs of Tetrao urogallus and Lagopus
lagopus; above — weight ratio per mille, below — average weight
of one stone.
38. Comparison of intestine parts in some Gallinaceous birds;
all measures calculated for 1000 g body weight. From above: Tetrao
urogallus, Lyrurus tetrix, Tetrastes bonasia, Lagopus lagopus,
L. mutus, L. scoticus, Phasianus mongolicus, Alectoris graeca.
39. Digestive tract of Tetrao urogallus female, from abdominal
side (left) and from back side (right).
40. Growth of Tetrao urogallus juv.; xx — males, oo — females.
41. Growth of Tetrastes bonasia juveniles; for explications — see
fig. 40.
42. Growth of Lagopus lagopus juveniles; for explications — see
fig. 40.
43. Growth of Lagopus mutus juveniles; for explications — see
fig. 40.
44 .Distribution of Tetrao urogallus body weights; dash line —
females, dash-and dot-line — immature males, solid line — adult
males.
45. Distribution of Tetrastes bonasia body weights; dash line —
females, solid line — males.
46. Distribution of Lagopus lagopus body weights; for explica-
tions — see fig. 45.
47. Distribution of Lagopus mutus body weights; for explica-
tions — see fig. 45.
48. Seasonal alternations of Tetrao urogallus body weights. Figu-
res indicate number of specimens used for building the line.
49. Age difference of Capercaillie males weight in spring. Expli-
cations: figures below the columns indicate the quantity of specimens,
figures above — addition in weight in kg.
306
50. Seasonal alternations of Tetrastes bonasia body weights.
51. Seasonal alternations of Lagopus lagopus body weights.
52. Moulting diagram of primaries and secondaries feathers in
Capercaillie juveniles (males).
53. Moulting diagram of primaries and secondaries feathers in
Capercaillie juveniles (females).
54. Moulting diagram of primaries and secondaries feathers in
Capercaillie adults; solid arrows — male feathers in state of growth,
break-off arrows — those of females.
55. Moulting diagram of primaries and secondaries in Willow
Grouse adults; thin arrows indicate white feathers, thick arrows —
coloured secondary feathers.
56. Moulting diagram of primaries and secondaries in Willow
Grouse juveniles. For explications see fig. 55.
57. The structure of Capercaillie female tail from above and from
behind.
58. Wing’s end of an immature Capercaillie female. Primaries
NN 1 and 2 are pointed, tops light-spotted.
59. Primaries feathers of Willow Grouse, juvenile (left) and
adult (right). The primary N 2 of juvenile bird is at the top pointed,
black-spotted.
60— 63. Skulls of Capercaillie males from the side. Note the
course of processus postorbitalis posterior and proc, zygomaticus
joining together. Fig. 60—1 year old, 61—2 years, 62 — 3 years,
63 — 4 years or more.
64— 67. Skulls of Capercaillie males from above. Note the course
of oo. nasalici and oo. frontalia joining together. Fig. 64— 1 year old,
65 — 2 years, 66 — 3 years, 67 — 4 years or more.
68. Mandibulae of Capercaillie males, 4 years old (above) and
3 years old (below). Note the seam between os dentale and os angu-
lare filling up.
69. Seasonal changes in Ascaridia invasion of Capercaillie and
Willow Grouse. Solid line — in Lapland Reserve, dash-and dot-
line — in Norway, dash line — in Petshora-Ylytsh Reserve.
70. Seasonal changes in Gestoda invasion of Capercaillie and
Willow Grouse. For explications see fig. 69.
71. Number of Goshawks met in a year (dash line) and of
Tetraonids met during one hunting day (solid line); both in Lapland
Reserve.
72. Golden eagles claw (Aquila chrysaetus).
73. Golden eagles aerie in Lapland Reserve.
74. Food composition of marten in Lapland Reserve in summer.
Explications: in broad columns, from above — berries (circles),
insects (cross hatched), frogs (dotted), eggs (white), various birds
(vertical hatched), Tetraonids (sparse hatched), various mammals
(oblique hatched), voles and shrews (sparse hatched). Black arrow
columns — census indices of voles and shrews.
20*
307
75. Food composition of marten in Petshora-Ylytsh Reserve. For
explications see fig. 74 (cedar nuts are indicated together with
berries).
76. Value of eggs in martens winter diet, in comparison with
embryonic mortality in Capercaillie (above) and accessibility of voles
(below). Solid line — Petshora-Ylytsh Reserve, dash line — Lapland
Reserve.
77. Plan of martens attack on two Capercaillies, hidden in snow
burrows.
78. Figures of Capercaillies (columns above) and Willow Grouse
(columns below) destroyed by predators, in connection with
values of their populations (solid line).
79. Alternations in Capercaillie testiculae weight in course of
year (grams).
80. Alternations in Willow Grouse testiculae weight in course
of year (grams).
81. Switch box of a nesting recorder (in section).
82. Thermograph with nest-recording device; on the right — the
switch box with double wire and a dry pile.
83. Actograph in work by the Capercaillie nest.
84. Brooding regime of a Capercaillie female in Lapland Reserve.
Explications: articulate line on high level indicates the fowl being on
nest, on low level — fowl being absent; solid'line (under the articulate
line) — air temperature below zero; dash line — air temperature
from 0° up to 5°C; asterisk — fowl driven away.
85. Brooding regime of Capercaillie female in Petshora-Ylytsh
Reserve. For explications — see fig. 84.
86. Capercaillie nest in time of hatching the chickens.
87. Blackgame nest in Lapland Reserve.
88. Brooding regime of Blackgame female in Petshora-Ylytsh
Reserve. For explications — see fig. 84.
89. Brooding regime of Hazel Grouse female in Petshora-Ylytsh
Reserve. For explications — see fig. 84.
90. Willow Grouse nest in Lapland Reserve.
91. Brooding regime of Willow Grouse female in Lapland Reserve,
in 1950 (above) and in 1951 (below). For explications — see fig. 84.
92. Capercaillie and Blackgame fluctuations in numbers in taiga-
zone. Explications: line with rings — Lapland Reserve, number of
specimens for 10-hour trip; line with crosses — Lapland Reserve,
census figures for 1 sq. km; dot line — Finnish Lapland, bag in
thousands; dash line — Petshora-Ylytsh Reserve, number of specimens
for 25 km trip; dash line with crosses — census figures for 1 sq. km.
93. Hazel Grouse and Willow Grouse fluctuations in numbers in
taiga zone. For explications — see fig. 92.
94. Weather of breeding period and age ratios in Capercaillie
population in Lapland Reserve. Explications: columns above —
precipitation sums in June, dash line — average air temperatures of
308
June, solid line — average temperature of the coldest 10-days period
of June; columns below — age ratios of population (hatched part —
juveniles, white part — adults).
95. Weather of breeding period and age ratios in Capercaillie
populations in Lapland (left) and Petshora-Ylytsh Reserve (right).
For further explications see fig. 94 and the summary.
96. Counts of Rock Ptarmigan on Tshuma-tundra in April; number
of specimens for one day trip.
97. Age composition of Capercaillie population in Upper Petshora
district; sparse hatched — in open season, dense hatched — in spring
hunting season.
98. Dynamics of Capercaillie population based on census figures;
white columns — immature birds, hatched columns — the adults.
All photographs, excluding N 16, are made by 0. Semenov-Tian-
Shansky.
ЛИТЕРАТУРА
Аверин Ю. В., 1938. Кавказский тетерев. Тр. Кавк. гос. заповеди., в. 1, М.,
стр. 57—86.
Андреев А. М., 1930. Исторические материалы о Кольском полуострове, хра-
нящиеся в Ленинграде. Сб. мат. по ист. Кольск. п-ва в XVI—XVII вв Изд.
АН СССР.
Антипин Д. Н., Ершов В. С., Золотарев И. А., Саляев В. А., 1956.
Паразитология и инвазионные болезни сельскохозяйственных животных. М.
Бартольд Е. Ф., 1930. Краткий отчет о< рекогносцировочной поездке в запо-
ведник Чуна-тундра. Тр. по лесн. оп. делу ЦЛОС, в. 7, М., стр. 151—156.
Б и рул я А. А., 1907. Очерки из жизни птиц полярного побережья Сибири. Изд.
Ак. наук. СПб.
Богданов П. А., 1952. Биология и динамика травяного и мохового покрова
ельника-черничника. Бот. журн., т. 37, в. 6, стр. 864—868.
Болезни птиц, т. Л, 1951. М.
Брем А., 1911. Жизнь животных, т. VII, изд. 4, СПб.
Владимирская М. И., 1948. Птицы Лапландского заповедника. Тр. Л апл.
гос. заповеди., в. 3, М., стр. 11711—245.
Владимирская М. И., 1948-а. Экология белки на северном пределе ее рас-
пространения. Там же, стр. 1125—470.
Владимирская М. И., Лебедев В. Д., Насимович А. А., 4953. Новые
данные по экологии выдры. Бюлл. Моск, о-ва исп. прир., отд. биол., т. 58,
в. 3, стр. 14—24.
Вольтер Ю., 11933'. К вопросу о сезонных кормах диких птиц из отр. куриных.
Бюлл. зоопарков и зоосадов, № 2, М.
Воропай И., 1891. Записки об охоте в Шенкурском уезде. Пермь.
Гаврин В. Ф., '1956. Экология тетеревиных птиц Беловежской пущи. Авторе-
ферат канд. дисс., Алма-Ата, стр. *1—46.
Гольцмайер Г. К.„ 4935. Белка телеутка. М.
Городков Б. Н., Короткевич Е. С., 1937. Питание белой куропатки в зо-
не арктических пустынь. Зоол. журн., т. 36, № 9, стр. 1382—1284*.
Грюнер А. М., 4951. Предельные температуры в гнезде глухаря в период
насиживания. Природа, №5, стр. 68—69.
Дементьев Г. П„ 4940. Птицы. Руководство по зоологии^ т. VI, М.
Добровольский Н., 4956. Лесные гости. Огонек, № 47.
Дон аур ов С. С., 41947. Рябчик в Печорско-Ылычском заповеднике. Тр. Печ.-
Ыл. заповеди., в. 4, ч. 4, М., стр. 77—422.
Д о н а у р о в С. С., 1948. Распространение и питание дневных хищных птиц в
Печорско-Ылычском заповеднике. Тр. Печ.-Ыл. гос. заповеди., .в. 4, ч. 2,
М„ стр. 67—87.
Доп пельм аир Г. Г., Мальчевский А. С., Новиков Г. А., Фаль-
кенштейнБ. Ю., 1951. Биология лесных зверей и птиц.
Дубинина М. Н., '1950. Дестробиляция у ленточных червей и причины ее
вызывающие. Зоол. журн., т. 29, в. 2, стр. 147—>151.
311
Дубровский А. Н., 11*9130. Материалы к изучению питания тетерева в Поль
ском р-не Новгородской юбл. Труды по лесному оп. делу ЦЛОС, в. 5.
Дулькейт Г. Д.» 49513; Значение рыси и росомахи как хищников в природном
комплексе Алтайской тайги. Сб. Преобразование фауны позвоночных нашей
страны, М., стр. 147—>152.
Дунаева Т. Н., Куче рук В. В., 194'1. Материалы по экологии наземных
позвоночных тундры южного Ямала. Мат. к позн. фауны и фл. СССР, в. 4
изд. Моск, о-ва исп. прир., М.
Ефремов П. Г., 1910. Летние наблюдения над, глухарем и его промыслом в
Кайском р-не Кировской обл. Уч. зап. Марийск. гос. пед. ин-та, т. 1, i
Йошкар-Ола.
Житков Б. П., 1913. Полуостров Ямал. Зап. И. Рус. геогр. о-ва, т. 49, СПБ.
3 а л е к е р В. А., По л у з а д о в Н. Б., 1955. Плодовитость и зимнее питание
соболя Ивдельского р-на Свердловской обл. Тр. Н.-и. ин-та ох, пром., в, 14,
стр. 145—152.
Иванов А. И., Козлова Е. В., Пор те нк о Л. А., Тугаринов А. Я.,
1’951. Птицы СССР. ч. И, Изд. АН СССР.
Иванов Б. Г., Николаевский Л. Д., ,1936. Явления изменчивости в орга-
низме северного оленя по сезонам года. Сов. оленев., в. 7, изд. ГСМП, Л.,
стр. 57—84.
К а л а б у х о в Н. И., 1947. Динамика численности наземных позвоночных.
Зоол. журн., т. 26, в. 6, стр. 503—520.
Кириков С. В., 1937. Сравнительно-экологический очерк южноуральского дуб-
равного и борового глухаря. Изв. АН СССР, сер. биол. № Й.
Кириков С. В., 4939. О возрастных изменениях глухарей и возрастном соста-
ве их на токах. Научно-мет. записки Ком. по заповеди., в. 2, М., стр. 98—
1109,
Кириков С. В., 1944. Возрастные изменения жевательной мускулатуры и че-
репа у глухарей. Зоол. журн., т. 23, в. 4, стр. 161—25.
Кириков С. В., 1947. Токование и биология размножения южноуральского
глухаря. Зоол. журн., т. 26, в. 1, стр. 71—84.
Кириков С. В., 11948. Охотничьи птицы Южного Урала, М.
Кириков С. В., 1949. Очередные вопросы изучения экологии тетеревиных птиц
в заповедниках. Научно-мет. записки Гл. упр. по заповеди., в. 13, М.,
стр. 47—56.
Кириков С. В., 1952. Птицы и млекопитающие в условиях ландшафтов южной
оконечности Урала. Изд. АН СССР, М. .
Кириков С. В., Мих е ев А. В., С п а н г е н б е р г Е. П., '1952. Учет куриных
птиц. Сб. Методы учета числ. и геогр. распр. наземн. позв., Изд. АН СССР,
М.
Колосов А. М., Шибанов С. В., *1957. Боровая дичь, ее промысел и заго-
товка. Изд. Центросоюза, М.
Косенков С. А., 4936. Веточный корм в лесной промышленности. Гослес-
техиздат, М.
Крепе Г. М., Семено в-Т я н-Ш а н с к и й О. И., 1934. Очерк биологии дико-
го северного оленя в Лапландии. Сб. Промысловая фауна и ох. хоз.,
Изд. КОИЗ, Л., стр. 27—49.
Кронау К., 1904. Разведение глухарей в неволе. Псовая и руж. охота, № 18,
стр. 287—4289.
Крутовская Е. А. 11941. Материалы по птицам Красноярского гос. заповед-
ника «Столбы» (рукопись).
Крутовская Е. А., Крутовская Е. В., 4953. Опыт одомашнивания и по-
лувального разведения глухаря. Сб. Преобраз, фауны позв. 'нашей страны,
Изд. Моск, о-ва исп. прир., стр. 201—234.
Л а р и н И. В. (и др.), 1950. Кормовые растения сенокосов и пастбищ СССР»
т. 1, Сельхоз. ГИЗ, М—Л.
Ларин С. А., 1941. Выращивание глухарей и тетеревов в искусственных усло-
виях. Тр. Мос. зоот. ин-та, т. 1, М., стр. 466-^181. <
312
Дев шин В., 1779L Совершенный Егер или знание о всех принадлежностях к
ружейной и прочей полевой охоте, СПБ.
Д о б а ч е в С. В., Щ е р б а к о в Ф. А., 1933. Естественные корма тетерева-коса-
ча. Бюлл. Моск, о-ва исп. прир., отд. биол. т. 42, в. 1, стр. 42—61.
Лобачев С. В., Щербаков Ф. А., 1936. Естественные корма тетерева-глу-
харя в годовом цикле жизни (и т. д.). Зоол. журн., т. 40, в. 2, М.
Дэк Д., Г957. Численность животных и ее регуляция в природе. Изд. иностр,
лит., М.
Маслов В., 1953. Белая куропатка. Газета За Новый Север, № 248, 20 XII 1953.
Мензбир М. А., 1'805, Птицы России, т. 1.
Мензбир М. А., Г900—1'912. Охотничьи и промысловые птицы Евр. России и
Кавказа, т. II, М.
Михеев А. В., !l*909l Линька и изменчивость белой куропатки восточной Пале-
арктики. Сб. тр. гос. Зоол. муз. МГУ, т. 5, стр. 65^108.
Михеев А. В., Г9418. Белая куропатка. Изд. Гл. упр. зап-ков, М.
Младенов 3. М., 1955. Некоторые физиологические особенности птиц в связи
с полом и временем года. Журн. общ. биол., т. 16, № 4, стр. 285—290.
Молчанов А. А., 1953. Сосновый лес и влага. Изд. АН СССР, М.
Нансен Ф., 1937. На лыжах через Гренландию. Собр. соч., т. 1, изд. ГСМП, Л.
Нансен Ф., 4937. Жизнь эскимосов. Собр. соч., т. 1, изд. ГСМП, Л.
Насимович А. А., 1939. Зима в жизни копытных Зап. Кавказа. Сб. Вопр.
экол. и биоцен., в. 7, Л., стр. 3—*91.
Насимович А. А., 1948. Очерк экологии горностая в Лапландском заповед-
нике. Тр. Лапл. гос. заповеди., в. 3, М., стр. ’3—138.
Насимович А. А., 4948-а. Экология лисицы в Лапландском заповеднике.
Тр. Лапл. гос. заповеди., в. 3, М., стр. 39—79.
Насимович А. А., 1948-6. Экология лесной куницы. Тр. Лапл. гос. заповеди.,
в. 3, М., стр. 81—106.
Насимович А. А., 1948-в. Новые данные по экологии росомахи в Лапланд-
ском заповеднике. Тр. Лапл. гос. заповеди., в. 3, М., стр. 107—424.
Насимович А. А., Новиков Г. А., С е м ен ов-Тя н-Ш а нск и й О. И.,
4'948. Норвежский лемминг. Сб. Фауна и? экология грызунов, в. 0, изд. Моск,
о-ва исп. прир., стр. 203;—262.
Насимович А. А., Семе но в-Т я н-Ш а н с к и й О. И., 1951. Питание бурого
медведя и оценка роли его как хищника в Лапландском заповеднике. Бюлл.
Моск, о-ва исп. прир., отд. биол., т. 56, в. 4, стр. 3-42.
Новиков Б. Г., 1939. К анализу механизма сезонной изменчивости у Lagopus
lagopus (L.J. Докл. АН СССР, т. 25, № 6, стр. 554—556.
Новиков Г. А., 1939. Из опыта биоценотического изучения млекопитающих и
птиц в еловых лесах Лапландского заповедника. Научно-мет. записки Гл.
Упр. заповеди., в. 5, М, стр. 51—58.
Новиков Г. А., 1952. Материалы по питанию лесных птиц Кольского полу-
острова. Тр. Зоол. ин-та АН СССР, в. 9, стр. 1455—1198.
Новиков Г. А., '1953. Полевые исследования по экологии наземных позвоноч-
ных. 2-ое изд., Сов. Наука, стр. 189—492.
Новиков Г. А., 1956. Еловые леса как среда обитания и роль в их жизни
млекопитающих и птиц. Сб. Роль жив. в жизни леса, изд. МГУ, стр. 6—‘165.
Новиков Г. А., 1956-а. Хищные млекопитающие фауны СССР. Изд. АН СССР,
М—Л., стр. 252—257.
Нордстрём, 1955. Обзор кольцевания птиц в Финляндии в 1913—1952 гг.
Реф. журн. Биология, № 21, 59369.
Огнев С. И., 1951. Экология млекопитающих. Изд. Моск. О-ва исп. прир. •
Олигер И. М„ 1940. Паразитические простейшие и их роль в колебаниях чис-
ленности рябчика (Tetrastes bonasia L.) на севере Горьковской обл. Докл.
АН СССР, т. 28, № 5.
Олигер И. М., 1950. Паразитофауна тетеревиных птиц лесной зоны Европ. ча-
сти РСФСР. Автореферат канд. дисс. Л.
Олигер И. М., 1950»а. Причины дестробиляции цестод тетеревиных птиц. Докл.
АН СССР, т. 24, № 4, стр. 869—872.
313
Олигер И. М., Немцев В. В., 1950. Изменения в биологии некоторых жи-
вотных Дарвинского зап-ка под влиянием изменений условий существе^
ния. Природа, № 2, стр. 42—43.
Осмоловская В. И., 1948. Экология хищных птиц полуострова Ямала.
Ин-та Геогр., в 41:, изд. АН СССР.
Пидопличко И, Г., 1951. О ледниковом периоде. В. 2, изд. АН Укр. ССР,
Киев.
Плеске Ф. Д., 1387. Критический обзор птиц и млекопитающих Кольского
полуострова. СПб.
Полезные растения СССР, т. 1, 1951.
Подсосов Л. А., 1947. Изменения в семенниках глухаря и вальдшнепа
в период весенних токов и тяги. Тр. Уральск, фил. АН СССР, в. 2, Сверд-
ловск, стр. 26—29.
Раевский В. В., 1947. Жизнь Кондо-Сосвинского соболя. Изд. Гл. Упр. по
заповеди. М.,
Раевский В. В., 1952. Опыт количественной характеристики зимней фауны
птиц тайги Западной Сибири. Сб. Методы учета числ. и геогр. распр. наземн.
позв., изд. АН СССР, М., стр. 294—303.
Птицы Советского Союза, т. IV, 1952. Изд. Сов. Наука, М.
Рольни к В., 1939. Температурный режим естественной инкубации у нанду.
Сб. Вопр. экол. и биоцен., № 5—6, Л., стр. 236—262.
Романов А. П., 1934. О белой куропатке Ленско-Хатангского района.
Тр. Аркт. ин-та, т. I, в. 11.
Романов А. Н., 1956. Автоматическое кольцевание диких животных и перс-
пективы его применения. Зоол. журн., т. 35 № 12, стр. 1902—1905.
Рудакове к ий В. Я., Насимович А. А., 1933. Боровая дичь. Заготовка,
обработка, экспорт.
Сабанеев Л. П., 1876. Рябчик. Сб. Природа, кн. 4, М., стр. 91—128.
Сабанеев Л. П., 1876. Глухой тетерев. Охотничья монография. М.
Сабанеев К. Л., 1904. Лето и осень 1903 г. Псовая и руж. ох. № 5, стр. 87.
С а л м и и Ю. А., 1938. К биологии маньчжурской или уссурийской белки. Тр.
Сихотэ-Алинск. гос. заповеди., в. 2, М.
Сдобников В. М., 1957. Сибирская тундровая куропатка (L. mutus kellogae
Grin.} на северном пределе своего ареала. Тр. Аркт. н.-и. ин-та, в. 205,
стр. 63-71.
С е в е р ц о в С. А., 1932. Материалы по биологии размножения Tetraonidae.
Тр. лабор. прикл. зоол. АН СССР, Л.
С ев ер цо в С. А., 1935. К познанию экологии размножения. Зоол. журн., т. 14,
в. 2.
С ев ер цо в С. А., 1941. Динамика населения и приспособительная эволюция.
Изд. АН СССР.
С е м е н о в-Т я н-Ш а н с к и й О. И., 1937. Лапландский государственный запо-
ведник. Изд. Ком. по заповеди, М., стр. 13—122.
Семене в-Т я н-Ш а н с к и й О. И., 1938. Экология боровой дичи Лапландского
зап-ка. Тр. Лапл. гос. заповеди., в. 1, М. стр. 217—>306.
С е м е н о в-Т я н-Ш а н с к и й О. И. 1938-а. Экология тундряной куропатки в
Лапландском заповеднике. Сб. Вопр. экол. и биоц., в. 4, Л., стр. 173—207.
С е м е н о в-Т я н-Ш а н ск и й О. И., 1947. Опыт учета боровой дичи в Лапланд-
ском заповеднике. Научно-мет. записки Гл. Упр. по заповед., в. 9, М.«
। стр. 55—59.
С е м е н о в-Т я н-Ш а н ск и й О. И., 1948. Дикий северный олень на Кольском
полуострове. Тр. Лапл. гос. заповеди, в. 2, М.» стр. 3—90.
Семене в-Т я н-Ш а н с к и й О. И., 1948-а. Лось на Кольском полуострове-
Там же, стр. 91—162.
С е м е н о в-Т я н-Ш а н с к и й О. И., 1952. Изучение инкубации тетеревиных
птиц в природных условиях. Бюлл. Моск, о-ва исп. прир., отд. биол. т. 57,
в. 6, стр. 35—41. ।
314
с е м е н о в-Т я н-Ш а н ски й О. И., 1960. Изучение поведения диких птиц
на гнезде с помощью самописцев. Тр. пробл.-тем. совещ. Зоол. ин-та. Изд.
АН СССР (в печати}.
Сем еко в-Т я н-Ш а н с к и й О. И., Значение анализа возрастного со-
става популяций для охотничьего хозяйства. Тр. Уральск, фил. АН СССР,
Свердловск (в печати).
Снигиревский С. И., 1947. Обзор семейства тетеревиных. Канд. дисс. Л.
С н и ги р ев ск и й С. И., 1950. Линька тетеревиных. Сб. Памяти ак. П. П. Суш-
кина. Изд. АН СССР, стр. 215—236. >
Таблицы для расчета природной освещенности и видимости, 1945. Изд.
АН СССР.
Те пл о в В. П., 1947. Глухарь в Печорско-Ылычском заповеднике. Тр. Печ.-Ыл.
гос. заповеди., в. 4, ч. 1, М., ст]р. 3—76.
Теп лов В. П., 1947-а. К экологии боровой дичи Печорско-Ылычского заповед-
ника. Там же, стр. 123—467.
Тепло в В. П., 1948-6. Материалы по экологии филина, в Печорско-Ылычском
заповеднике. Тр. Печ.-Ыл. гос. заповеди., в. 4, ч. 2, М.
Т е п л о в В. П., 1950. К изучению питания и динамики численности популяций
обыкновенной лисицы. Иаучн.-мет. записки Гл. Упр. по заповеди., в. 13, М.
Теп лов В. П., 1953. Опыт оценки хищнической деятельности бурого медведя.
Сб. Преобр. фауны позв. нашей страны, изд. Моск, о-ва исп. прир.,
стр. 153—166. ।
Те пл о.в В. П., 1953-а. Речная выдра в районе Печоро-Илычского заповедника.
Бюлл. Моск, о-ва исп. прир., отд. биол. т. 58, в. 6, стр. 7—16.*
Теп лов В. П., 1955. К зимней экологии росомахи в районе Печоро-Илычского
заповедника. Там же, т. 60. в. 1, стр. 3—И.
Теп лов В. П., Туров И. С., 1956. О значении серой вороны в пойменных
охотничьих угодьях среднего течения р. Оки. Зоол. журн., т. 35, в. 5,
стр. 735—757.
Тепло ва Е. Н., Теп лов В. П., 1947. Значение снежного покрова в экологии
млекопитающих и птиц Печорско-Ылычского заповедника. Тр. Печ.-Ыл. гос.
заповеди., в. 4, М., стр. 181—254.
Теплое а Е. Н., 1947. Питание лисицы в Печорско-Ылычском заповеднике.
Тр. Печ.-Ыл. гос. заповеди., в. 5, М., стр. 245—260.
Теп лов а Е. Н., 1957. Птицы района Печоро-Илычского заповедника Тр.
Печ.-Ил. гос. заповеди., в. 6, стр. 5—115. Сыктывкар.
Третьяков Н. П., 1951. Инкубация яиц сельскохозяйственных птиц. Сельхоз-
гиз, М.
Третьяков Н. П., 1954. Длительное глубокое охлаждение эмбрионов кур на
разных стадиях развития. Реф. журн. Биология, № 7, 12729.
Тугаринов А. Я, Толмачев А. И., 1934. Материалы для авифауны восточ-
ного Таймыра. Изд. АН СССР.
Тугаринов А. А., Портенко Л. А., 1950. Атлас охотничьих и промысло-
вых птиц и зверей СССР, т. 1, Птицы. Изд. АН СССР.
Ульянин Н. С„ 1949. К экологии тетерева, белой и серой куропаток Северного
Казахстана. Тр. Наурзумск. гос. заповеди., в. 2, М., стр. 5—57.
Ульянин Н. С., 1939. К биологии белой куропатки Северного Казахстана.
Сб. тр. гос. Зоол. муз. МГУ, в. 5, стр. 109—126.
Ушаков В., 1904. Белые куропатки Сибири. Ж. Псов, и руж. ох., № 2,
стр. 35—37.
Ушков Л., 1887. Краткий обзор распространения и жизни глухаря и тетерева
и охота на них в окрестностях Екатеринбурга. Екатеринбург.
Ушков С., 1929. Перелеты глухарей над Пермью. Ж. Уральск, ох., № 10,
стр. 4611.
Ушков С. Л., 1949. Материалы по изучению роли пернатых хищников в усло-
виях заповедности. Тр. Ильменск. гос. заповеди, им. В. И. Ленина, в. 4,
Челябинск, стр. 1111—181.
Ушков С. Л.. 1949-а, Орел могильник в Ильменском заповеднике. Там же»
стр. 182—201. г
315
Ушков С. Л., 1949-6. Некоторые примеры приспособления птиц к окружаю^»
их обстановке. Там же, стр. 202—208.
Федюшин А. В., 1946. Новая форма сезонной адаптации у цестод оссЦЛы
птиц. Зоол. журн., т. 25, № 2, стр. 101—105.
Феоктистов П. И., 1953. Дестробиляция и другие формы сезонной адаптацИи
у ленточных червей. Зоол. журн., т. 32, в. 1, стр. 49—52.
Фетисов А. С., 1934. Материалы по питанию тетерева в юго-восточной части
Прибайкалья. Зоол. журн., т. 13, в. 2, стр. 369—396.
Фол и тарек С. С., 1938’. Некоторые данные по питанию рябчика. Сб. ВоПр
экол. и биоц., в. 4, Л., стр. 89—112.
Ф о л и т а р е к С. С., Д е ме и т ь е в Г. П., 1938. Птицы Алтайского гос. заповед.
ника. Труды Алтайск. гос. заповеди., в. 1,'М., 7—91.
Ф о л и т а р ек С. С., 1939'. Материалы по питанию белой и тундряной куропаток
Научно-мет. записки Гл. Упр. по заповеди., в. 5, М„ стр. 130—1'38.
Формозов А. Н., 1934. Материалы к биологии рябчика по наблюдениям на
севере Горьковского края. Бюлл. Моск, о-ва исп. прир., отд. бнол., т. 43. в. ]
Формозов А. Н., 1935. Колебания численности промысловых животных. П3д*
КОИЗ, М.-Л.
Формозов А. Н., 1940. Практическое значение птиц. В книге Рук. по зоол.,
т. 6, М., стр. 762—801.
Формозов А. Н., 1946. Снежный покров как фактор среды, его значение в
жизни млекопитающих и птиц СССР. Изд. Моск, о-ва исп. прир.
ФормозовА. Н., 1952. Спутник следопыта. Изд. Моск, о-ва исп. прир.
Ха рузин Н., 1890. Русские лопари.
Хватов А. И., 1'860. 18-летние опыты приручения и одомашнения тетеревов-
березовиков. Акклиматизация, т. 1, в. 4, стр. 146—153.
Хватов А. И., 1861. Приручение и одомашнение тетеревов глухих. Акклимати-
зация, т. 2, в. 1, стр. 105—ПО.
Хватов А. И., 1861-а. Лечение тетеревов березовым углем. Акклиматизация,
т. 2, стр. 162—163.
X о л о д к о в с к и й Н. А., 1901. Атлас птиц Европы.
Ц веленье в Л. A., 11'9138. Материалы по питанию рябчика на Алтае. Тр. Ал-
тайск. гос. заповеди., в. 1, М., стр. 263—294.
Ч а р н о л у с к и й В. В., 1930. Материалы по быту лопарей. Л.
Чарнолуский В. В., 1930-а. Заметки о пастьбе и организации стада у лопа-
рей. Кольский сборник.
Черкасов А., '1'8*84. Записки охотника Восточной Сибири, изд. 2, СПб.
Шпаковский С. И. 1937. К вопросу об одомашнивании тетерева. Тр. Ново-
сиб. зоос., т. 1. । I
Ште гм ан Б. К-, 1938. Основы орнитогеографического деления Палеарктики.
Фауна СССР, Птицы, т. 1, в. 2, изд. АН СССР.
Штег м ан Б. К., 1950. Исследования о полете птиц. Сб. Памяти ак. П. П. Суш-
кина, изд. АН СССР, стр. 237—265.
Ште г м а н Б. К-, 1950-а Функциональное значение особеннностей строения
грудины у куриных птиц. Там же, стр. 129—134.
Юр ген сон П. Б., 1951. Эколого-географические аспекты в питании лесной ку-
ницы и географическая изменчивость эколого-морфологическх адаптаций ее
жевательного ^аппарата. Зоол. журн., т.е30, в. 2, стр. 172—185.
Юрлов К. Т., 1956. Материалы по питанию белой куропатки в Барабе и Кулун*
де. Тр. Биол. ин-та Зап.-Сиб. фил. АН СССР, в. 1, стр. 87—97.
Beer J. R., 1955. Some gross morphology of the blue grouse digestive tract.
Murrelet, v. 36, № 2, pp. 22—25.
Bent A. C., 1932. Life histories of North American Gallinaceous birds. Smith-
Inst., Un. St. Nat. Mus., Bull. 162, Washington.
Boker H., 1935—1937. Einfiirung in die vergleichende Biologische Anatomic def
Wirbeltiere. В. I, II, Jena.
Breckenridge W. J., 1956. Nesting study of wood ducks. J. Wildlife Manag-»
v. 20, № 1, pp. 16—21.
Brinkmann A., 1926. Coccidiosen hos lirypen. Berg. Mus. Aarbok, H. 2, Bergen-
316
rowan J. Me T., 1955. Wildlife conservation in Canada. J. Wildlife Manag.,
G v. 19» № 2, pp. 161—176.
norney R. S., Thompson D. D., Hale J. B., Wendt R. F., 1958. An eva-
V luation of Ruffed Grouse drumming counts. J. Wildlife Manag., v. 22, № 1,
pp. 35—40.
Finland —a country of lakes and wilderness, 1954. Helsinki.
Gel ting P., 1937. Studies on the food of the East-Greenland Ptarmigan especially
in its relation to vegetation and snow-cover. Kobenhavn.
Groebbels Fr., 1932. Der Vogel, b. I Atmungswelt und Nahrungswelt. Berlin.
Gurr L., 1955. A pneumatic nest-recording device. Ibis, v. 97, № 3, pp. 584—586.
Holt Adv., 1952. Lirypas parasitter. Riistatieteellisia Julkaisuja, № 8, Helsinki,
pp. 142—149.
Hoglund Nils, 1952. Capercaillie reproduction and climate. Riistatieteellisia
Julkaisuja, № 8, pp. 78—80.
Hoglund Nils, 1956. Pieten metsopoikasten sukupuolen maarittatriinen.
Suomen Riista, № 10, Helsinki, pp. 150—155.
Hoglund Nils, 1956. Swedish Sportsmen’s Association’s marking of game
1945—1954. Viltrevy, v. I. № 2, Stockholm, pp. 162—224.
Hu us J., 1928. Darmparasiten des norwegischen Moorschneehuhns. Bergens Mus.
Aarbok, H. 2, Bergen.
Johnsen S., 1929. Draktskiftet hos lirypen i Norge. Berg. Mus. Aarbok, H. 2,
Bergen.
Johnsen S., 1941. Remarks on the Swalbard Ptarmigan (Lagopus mutus
hyperboreus Sundev.). Berg. Mus. Aarbok, H. 3, № 8, pp. 1—30.
Klogan E. D., Coleman I. А., К о z i с к у E. L., 1956., A pheasant nest
activity recording instrument J. Wildlife Manag., v. 20, № 2, pp. 173—177.
Krudener A., 1928. Auer wild. Naturgeschichte Jagd.
Koivisto Ilkka, 1956. Metsokannoissa tapahtuvista siitymisista. Suomen
Riista, № 10, pp. 179—184.
Koivisto Ilkka, 1956. Riistantutkimuslaitoksen siipimerkinta vv. 1953—55.
Suomen Riista. № 10, pp. 166—175.
Kos ki mi es Jukka. 1953. Suomen Riistanhoito-saation siipimerkinta 1950—52.
Suomen Riista, № 8, pp. 165—174.
Larsen J. A., Lahey J. F., 1958. Influence of weather upon a ruffed grouse
population. J. Wildlife Manag., v. 22, № 1, pp. 63—70.
Leslie A. S., 1911. The Life History of the Grouse. The Grouse in health and
in disease, v. 1, London, pp. 5—28.
Lindroth H., Lindgren L., 1950. Metson hakomisen metsanhoidollisesta
merkityksesta. Suomen Riista, № 5, pp. 60—81.
Linkola Pentti, 1956. Kanahaukkakannan romahdus vuonna 1956. Luonnon
tutkija, v. 61, № 2, pp. 49—58.
Lund H j., 1952. Entoparasitter hos 136 orrfugl Lyrurus t. tetrix (L.), fra Norge.
Riistatieteellisia Julkaisuja, № 8, Helsinki, pp. 150—154.
Madsen H., 1952. Uber Parasitose jagdbarer Hiihnervogel. Riistatieteellisia
Julkaisuja, № 8, Helsinki, pp. 154—158.
Marples B. J., Gurr L., 1943. A mechanism for recording automatically the
nesting habits of birds. The Emu, v. 43, pp. 67—71.
Moran P. A. P., 1954. The statistical analysis of Game-bird records. J. Animal
Ecol., v. 25, № 1, pp. 35—37.
Nordhagen Rolf, 1928. Rypeaar og Baeraar. Berg. Mus. Aarbok. Naturv.
rekke, № 2, Bergen.
Rynnonen Alpi, 1954. Beitrage zur kenntnis der Lebensweise des Haselhuhns,
Tetrastes bonasia (L.). Riistatieteellisia Julkaisuja, v. 12, Helsinki, pp. 1—90.
Salomonsen F., 1940. Moults and sequence of plumages in the Rock Ptarmi-
gan (L. mutus). Copenhague.
Shelf ord V. E., Y eat ter R. E., 1955. Some suggested relations of prairie
chicken abundance to physical factors, especially rainfall and solar radiation.
J. Wildlife Manag., v. 19, № 2, pp. 233—242.
317
Siivonen L., 1943. Vaeltavatko kanalintumme? Metsastys ja Kalastus, r
Siivonen L., 1948. Decline in numerous mammal and bird populations in north
western Europe during the 1940. s. Riistatieteellisia Julkaisuja, № 2,
pp. 1—26.
Siivonen L., 1952. On the reflection of short-term fluctuations in numbers jn
the reproduction of tetraonids. Riistatieteellisia Julkaisuja, № 9, Helsinki
pp. 1—43.
Siivonen L., 1954. Some essential features of short-term population fluctuation
J. Wildlife Manag., v. 18, № 1, pp. 38—45.
Siivonen L., 1954. On the short-term fluctuations in numbers of Tetraonids.
Riistatieteellisia Julkaisuja, № 13, Helsinki, pp. 1—10.
Siivonen L., 1957. The problem of the short-term fluctuations in numbers in
tetraonids in Europe. Riistatieteellisia Julkaisuja, № 19, Helsinki, pp. 1—44.
Siivonen L., Koskimies J., 1955. Population fluctuations and the lunar
cycle. Riistatieteellisia Julkaisuja, № 14, Helsinki, pp. 1—22.
Sluiter J. W., Heerdt P. F., Bezem J. J., 1956. Popualtion statistics of the
bat Myotis mystacinus, based on the markingrecapture method. Arch. Neerl.
zool., v. 12, № 1, pp. 63—88.
Sulkava S., 1956. Kanahaukan pesimisaikaisesta ravinnosta. Suomen Riista,
№ 10, Helsinki, pp. 44—62.
The rarer birds of prey. Their status in the British Isles, 1957. British Birds, v. 50,
№ 4, pp. 129—155.
Wes ter s ко v К a j, 1956. Age determination and dating nesting events in
the willow ptarmigan. J. Wildlife Manag., v. 20, № 3, pp. 274—279.
Williams G. R., 1954. Population fluctuations in some Northern Hemisphere
Game birds (Tetraonidae). J. Animal Ecol., v. 23, № 1, pp. 1—34.
Wilson E. A., 1911. Physiology and Anatomie of the Red Grouse. The Grouse
in health and in disease. London, pp. 100—129.
Wilson E. A., Leslie A. S., 1911. Food of the Red Grouse. Ibid.
Wurm W., 1885. Das Auerwild. Wien.
ОГЛАВЛЕНИЕ
Введение..............................................................5
I. Распространение тетеревиных по территории Мурманской области 9
II. Среда обитания тетеревиных....................................12
III. Образ жизни тетеревиных по сезонам . 20
IV. Питание........................................................50
V. Пищеварение тетеревиных....................................107
VI. Рост птенцов и динамика веса взрослых птиц по сезонам .... 129
VII. Линька и методы определения возраста тетеревиных.145
VIII. Паразиты тетеревиных.........................................176-
IX. Враги тетеревиных.............................................184
X. Размножение...................................................219
XI. Учет и движение численности . 259
XII. Миграции...................................... .............290
Выводы.................... . . . . . 293-
Резюме (на англ, яз.) . . . . . . 301
Литература .... . . . . . 311
Редактор А. А. Насимович Техн, редактор С. П. Житов
ПН 00002 Подпис. к печ. 5/1 1960 г. 20 печ. л. (24 уч.-изд. л.)
Бумага 60 X 92 Зак. 3037 Тир. 3.000
Типография, проезд Сапунова, 2.