/























































































































Text
»
А. А. ЛОГИНОВ
ОЧЕРКИ
ПО ОБЩЕЙ
ФИЗИОЛОГИИ
Основные свойства
и закономерности
живых систем
Издательство «Вышэйшая школа»
Минск 1976
57.04
Л69
УДК 612.0
Логинов А. А.
Л69 Очерки по общей физиологии. Основные свойства
и закономерности живых систем. Минск, «Вышэйш.
школа», 1976.
240 с, с ил.
Очерки познакомят читателя с такими интересными проблемами,
как едмистио. взаимосвязанность и взаимообусловлен кость, всеобщ-
ность и универсальность основных жизненных свойств и процессов на
всех уровнях проявления жизни.
Большое внимание уделяется единству и взаимосвязи живых орга-
низмов с окружающей средой, а также приспособительному поведе-
нию живых объектов в системе организм — среда.
Очерки рассчитаны на широкий круг читателей, имеющих доста-
точную общебнологическую подготовку (студенты биологических, ме-
дицинских и философских факультетов), а также на лиц, интересую-
щихся филоссфскнми проблемами современного естествознания, учи-
телей средних школ, слушателей народных университетов естествен-
нонаучных знании высшей ступени сложности.
21000—112
Л ----------- 44—76
М 304(05)—76
57.04
(Б) Издательство «Вышэйшая школа», 1976
ПРЕДИСЛОВИЕ
Предлагаемая читателю книга весьма своеобразна как по
форме, так и по содержанию. Эта книга — размышление. Раз-
мышление автора, в которое исподволь, но активно вовлека-
ется читатель. Размышление об общих, наиболее фунда-
ментальных и злободневных проблемах современной биологии.
Размышление подчас глубоко философское и в то же время не
отрывающееся от конкретного материала.
Автор назвал свою книгу «Очерки». Это слово ассоциируется
с разрозненными фрагментами, этюдами, вырванными из общей
картины, системы наших знаний о жизни и ее закономерностях.
На самом деле это не так. Книга имеет внутренний логический
стержень, авторское кредо и авторская позиция отчетливо
прослеживаются через весь текст. Внутренняя «монистичность»
книги — одна из ее сильных сторон. В центре внимания — го-
меостические, регуляторные, взаимодействия и взаимоотноше-
ния в системе организм — окружающая среда и принцип «ус-
тойчивой неравновесности» во множественных их проявлениях
и уровнях значимости.
Книга выходит за рамки и второй части своего официально-
го названия «...по общей физиологии». Ее автор с единых по-
зиций неназойливо и по мере надобности знакомит читателя с
многими вопросами из области термодинамики равновесных и
неравновесных систем, биофизики, кибернетики, биохимии,
цитологии, эмбриологии, генетики, экологии, геронтологии и
лаже геологии—амплитуда, посильная далеко не каждому ав-
тору. В этом — третья сильная сторона книги.
И, наконец, автор постоянно чувствует и не дает забывать
читателю об идеях развития, об органической эволюции, ее
факторах и определяющей роли для ныне существующих форм
8
жизни, структурно-функционального устройства и поведения
организмов. Эволюционные аспекты последовательно развива-
ются на страницах книги. Диапазон представленного материа-
ла чрезвычайно широк — от молекулярного до биосферного
уровня. По вполне понятным причинам, однако, автор сосре-
доточивает основное внимание на проблемах физиологии жи-
вотных и человека.
Приходится удивляться, каким образом сравнительно
небольшая по размерам книга вмещает в себя такое обилие
информации и идей, каким образом в ней постоянно поддержи-
вается волнующая мыслительно-эмоциональная напряженность.
В значительной мере это определяется тем, что автор умело
разграничивает самое важное и второстепенное и находит
удачные ракурсы в подаче и компоновке материала.
Хочу подчеркнуть, что в эпоху информационного взрыва и
Сурной дифференциации наук остро ощущается необходи-
мость в научно-популярной литературе для самих научных
работников, поскольку в пределах биологии, не говоря уже о
других науках, представители различных ее разделов разучи-
лись понимать друг друга и мало осведомлены о состоянии
вопроса у «соседей».
В «Очерках» А. Л. Логинова содержатся некоторые дискус-
сионные положения, авторская постановка новых вопросов,
авторские гипотезы, классификация процессов, не прошедшие
апробации научной общественности. По моему убеждению, та-
кого права нельзя лишать автора научно-популярной книги,
если его гипотезы н предложения не противоречат фундамен-
тальным законам природы.
Несмотря на отдельные неточности книги, подчас связан-
ные с научно-популярным жанром, она, без сомнения, ока-
жется полезной широкому кругу читателей, будет встречена
с большим интересом и может послужить «острой» затравкой
для дискуссий средн специалистов, особенно в отношении про-
блем «термодинамика и биология», «организм и внешняя
среда».
проф. С. Конев
I. ОРГАНИЗМ И СРЕДА
Очерк 1. ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ.
ОБЩИЕ ПОЛОЖЕНИЯ
Обязательным процессом, без которого немыслима жизнь,
является обмен веществ. Ои возможен только при условии
трат свободной энергии, т. е. при совершении работы. Под ра-
ботой в широком смысле следует понимать «изменение формы
движения материи, рассматриваемое с количественной стороны»
(Ф. Энгельс). Обмен веществ — это одновременно протекаю-
щие, но не всегда равные по интенсивности процессы ассимиля-
ции и диссимиляции. Ассимиляция приводит к накоплению в
организме пластических веществ, идущих на формирование раз-
личных тканей (массы тела), и энергетических, необходимых
для осуществления всех жизненных процессов. За счет дисси-
миляции происходит распад химических соединений, разруше-
ние отживших и поврежденных элементов тканей и высвобож-
дение энергии. Оба процесса осуществляются в организме при
условии поступления, переработки и усвоения белков, жиров,
углеводов, витаминов, минеральных веществ, микроэлементов
из внешней среды в виде продуктов питания и удаления их в
виде продуктов распада. Принципиальная схема обмена ве-
ществ в организме представлена па рис. I.
Процесс обмена веществ подчиняется всеобщему закону
сохранения материи, который гласит; при всех явлениях при-
роды видоизменяется только форма вещества, количество же
его остается постоянным, или — данная масса материи незави-
сима от изменений, происходящих в ней.
В справедливости этого закона в отношении живых объектов
можно убедиться на следующем.
1. На примере суточного потребления и выделения веществ
(в граммах) организмом человека, находящегося в состоянии
максимального покоя — основного обмена;
б
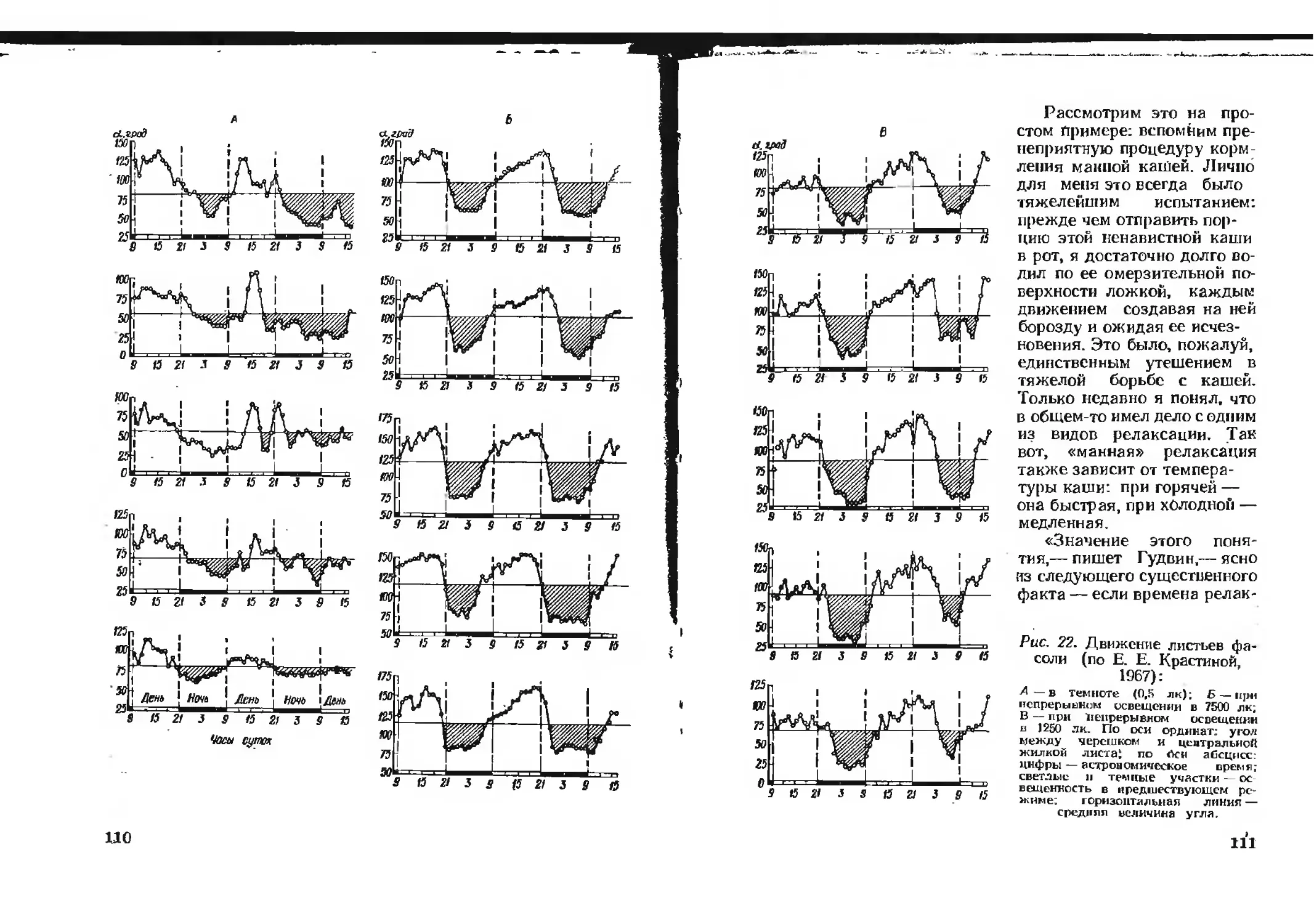
Рис. 1. Три стадии катаболизма и анаболизма
(по А. Ленинджеру, 1974):
жирные стрелки — катаболические пути, светлые—анаболические. Ста-
дия III амфиболическая. На этой стадии завершается разрушение
пищевых молекул до СОг и восстанавливаются низкомолекулярные
предшественники для анаболических процессов.
Потребление
Кислорода через легкие 800
Воды через пищеварительный аппарат 2800
Солей минеральных 30
Органических веществ (100 г белков, 60 г жиров, 500 г
углеводов) 660
Итого , . .4290
Выделение
Углекислоты через легкие 900
Воды через легкие 900
Воды через кожу (пот) 800
Воды с мочой 1500
Воды с калом 100
Солей с мочой 25
Органических веществ с мочой 35
Органических и неорганических веществ с калом . 30
, Итого . . .4290
2. На примере включения живых организмов (их обмена
веществ) в общий круговорот веществ в природе и поддержа-
ния его постоянства (рис. 2).
Согласно закону сохранения энергии, общая сумма энергии
материальной системы остается постоянной независимо от
изменений в самой системе, или — энергия не исчезает и не
творится вновь, а только переходит из одной формы в другую.
Так, при полном превращении 1 ккал (4,18 Дж) в механическую
работу ее всегда совершается 427 кГм (4162 Дж). Это и есть
механический эквивалент тепла. При превращении 1 ккал в
электрическую энергию получается такое ее количество, кото-
рое будучи превращено в механическую работу даст те же
427 кГм.
Приложимость этого закона к живым организмам была до-
казана Робертом Майером. Простота и изящество, с каким это
было сделано, являются примером поразительной наблюдатель-
ности и научной прозорливости исследователя. Находясь в
7
Рис. 2. Схема круговорота веществ в природе
в общих чертах существование биосферы основано на использовании солнеч-
углерода; причем образуются, во-первых, органические соединения и, во-
некоторых других
должности судового врача, по прибытии на остров Ява Майеру
пришлось с лечебной целью провести кровопускание нескольким
заболевшим матросам. При этом он заметил, что венозная
кровь, которая обычно значительно темнее богатой кислородом
артериальной, почти не отличалась по цвету. Это натолкнуло
Майера на мысль, что в условиях жаркого климата тропиков
для поддержания температуры тела 37° и совершения работы
организм нуждается в менее интенсивном обмене веществ, чем
в условиях холодного климата, а следовательно, и в меньшем
потреблении пищи. Так была установлена связь между пита-
8
(из Дж. Хатчинсона, 1972):
ной энергии для фотосинтетического восстановления атмосферной двуокиси
вторых, молекулярный кислород. На схеме изображен также круговорот
важных элементов.
пием и окружающей температурой. Естественны и связи ин-
тенсивности окислительных процессов с теплообразованием;
образования тепла в теле человека с теплопотерями; теплоты
с произведенной работой. Эти умозаключения привели Майера
к выводу о том, что механическая работа, кинетическая энергия
и теплота могут превращаться друг в друга, т. с. все это раз-
личные формы энергии. Так Майер в 1842 году пришел к фор-
мулировке закона сохранения и превращения энергии. По су-
ществу он его переоткрыл после Ломоносова (1748—1756) и
Карно (1824).
9
Поскольку материя и энергия неотделимы друг от друга,
следует говорить о единстве рассматриваемых законов, т. е. о
законе сохранения н превращения материи и энергии. Поэтому
для понимания протекания основных жизненных процессов
весьма важно познать сущность проявления этого закона в
жизнедеятельности живых объектов. Рассмотрим в общем виде
схему круговорота материи и энергии на примере основной
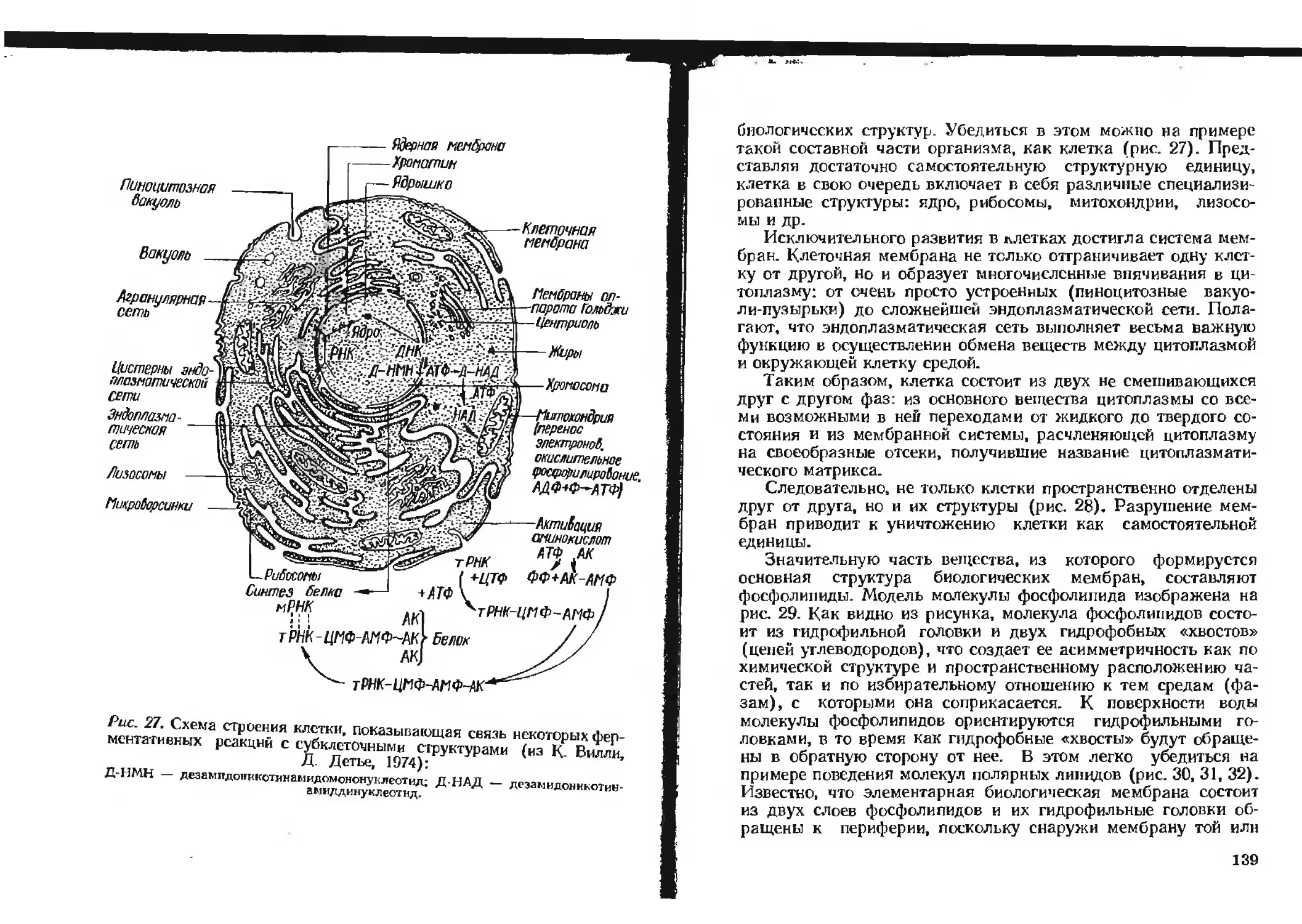
функциональной единицы организма — клетки (рис. 3).
Рис. 3. Поток энергии в клетке
(из А. Лёви и Г. Сиксвица, 1971).
По особенности потребления энергии растения и животные
сходны друг с другом, за исключением конечного источника
ее. Растения — это аутотрофы; они способны путем использо-
вания световой энергии (например, солнца) потреблять неор-
ганические соединения и синтезировать из них углеводы, жиры
и белки, т. е. осуществлять фотосинтез. Животные — гетеро-
трофы, так как потребляют органические вещества в готовом
виде и превращают их в углеводы, жиры и белки, специфиче-
ские для собственного организма.
10
Углеводы, жиры или белки, а также продукты их расщеп-
ления не могут непосредственно служить источником энергии
для клеточных процессов. Эту роль выполняет АТФ (аденозин-
трифосфат), или, как принято говорить, энергия макроэргиче-
ских фосфатных связей ~Ф. Расщепление пищевых веществ с
высвобождением энергии, используемой для синтеза АТФ, про-
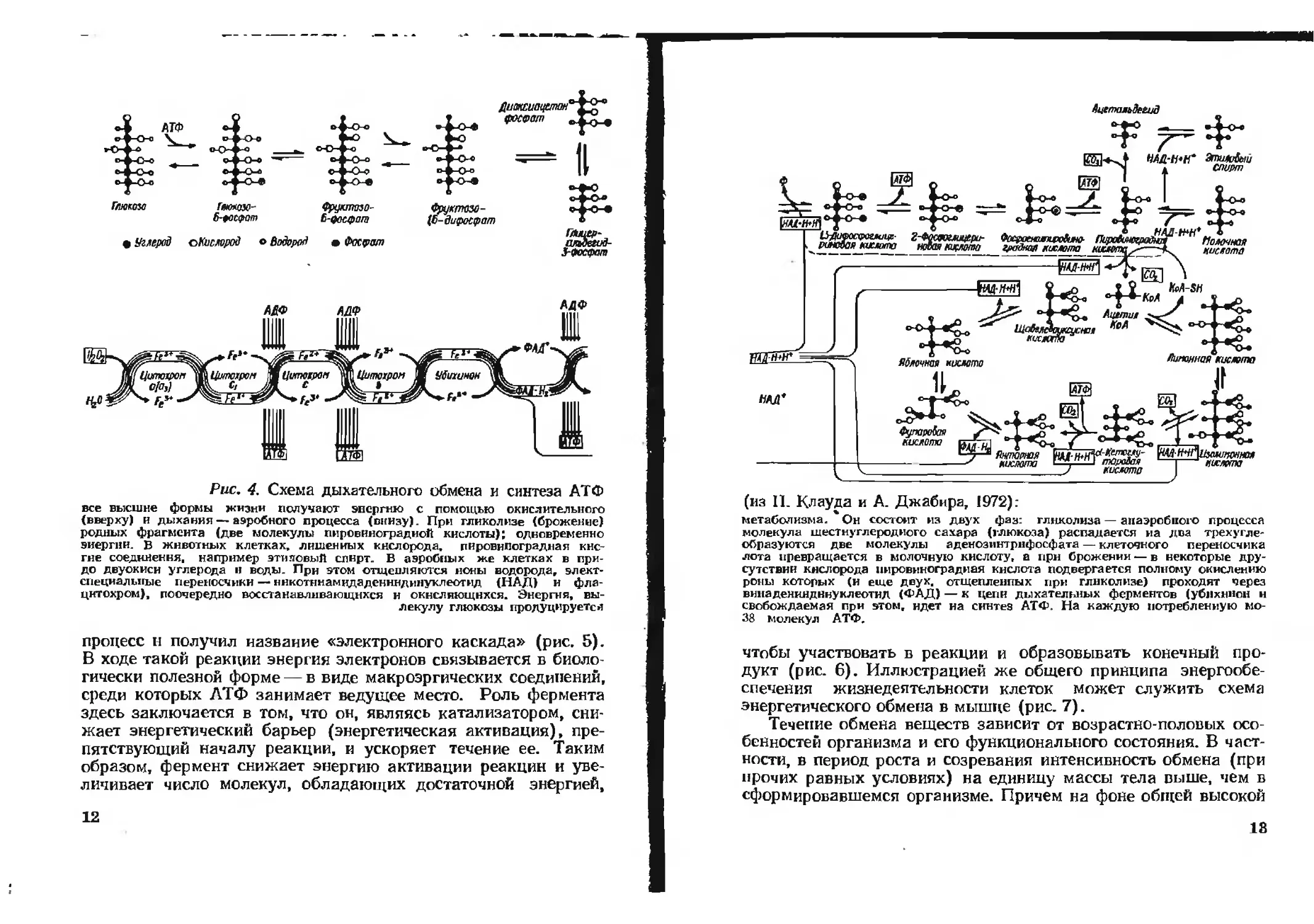
исходит в результате дыхательного обмена (рис. 4). Этот обмен
поставляет не только энергию, но и химические вещества, не-
обходимые для синтеза новых углеводов, жиров и белков. Во
всех клеточных процессах обычно происходит преобразование
одного вида энергии в другой. Подобные процессы можно рас-
сматривать как различные формы работы, выполняемые клет-
кой (см. рис. 3): а) химическая работа, использующая энергию
АТФ на химические превращения, связанные с обменом угле-
водов, .жиров, белков и нуклеиновых кислот; на рост и размно-
жение клетки; на синтез и передачу наследственного материала;
б) осмотическая работа, приводящая к накоплению в клетке
веществ, присутствующих в окружающей среде в малых коли-
чествах, н выделению веществ, которых в окружающей среде
больше, чем в клетке; в) электрическая работа, обусловлива-
ющая возникновение и поддержание разности потенциалов
наружной и внутренней поверхностей клеточной мембраны;
г) механическая работа, лежащая в основе всех форм движе-
ния (сокращение мышечных клеток, биение ресничек, течение
цитоплазмы и др.); д) регуляторная работа, упорядочивающая
клеточные процессы, связанные с использованием энергии АТФ,
соответственно конкретным условиям существования клетки в
данный момент. О том, каким образом осуществляется синтез
АТФ в клетках, можно составить представление на основании
вышеуказанной схемы (см. рис. 4).
В последнее время было установлено, что в процессах био-
синтеза принимают участие, помимо АТФ, и другие нуклеотиды
с макроэргическими фосфатными связями, например цито-
зинтрифосфат (ЦТФ), уридинтрифосфат (УТФ) и гуанидин-
трнфосфат (ГТФ). Процессы их образования катализируются
группой ферментов — нуклеозиддифосфокиназами. Однако пер-
вичным источником энергии, заключенной в этих нуклеозид-
трифосфатах, служит АТФ. Главным источником макроэргиче-
скнх связей в клетке является перенос электронов от одного
акцептора к другому подобно каскаду водопадов, за что данный
11
Глюкоза Гйюнюо- Фруктао-
6-foc<?am £-^осфат
• Углерод оКистрод о Водород • Фосфат
Рис. 4. Схема дыхательного обмена и синтеза АТФ
все внешне формы жизни получают энергию с помощью окислительного
(вверху) и дыхания—аэробного процесса (внизу). При гликолизе (брожение)
родных фрагмента (две молекулы пировиноградной кислоты); одновременно
энергии, в животных клетках, лишенных кислорода, пировиноградная кис-
гне соединен ня, например этиновый спирт. В аэробных же клетках в при-
до двуокиси углерода и воды. При этом отщепляются ноны водорода, элект-
специалыгые переносчики — ннкотннамцдаденнндннуклеотид (НАД) и фла-
цнтохром), поочередно восстанавливающихся и окисляющихся. Энергия, вы-
лекулу глюкозы продуцируется
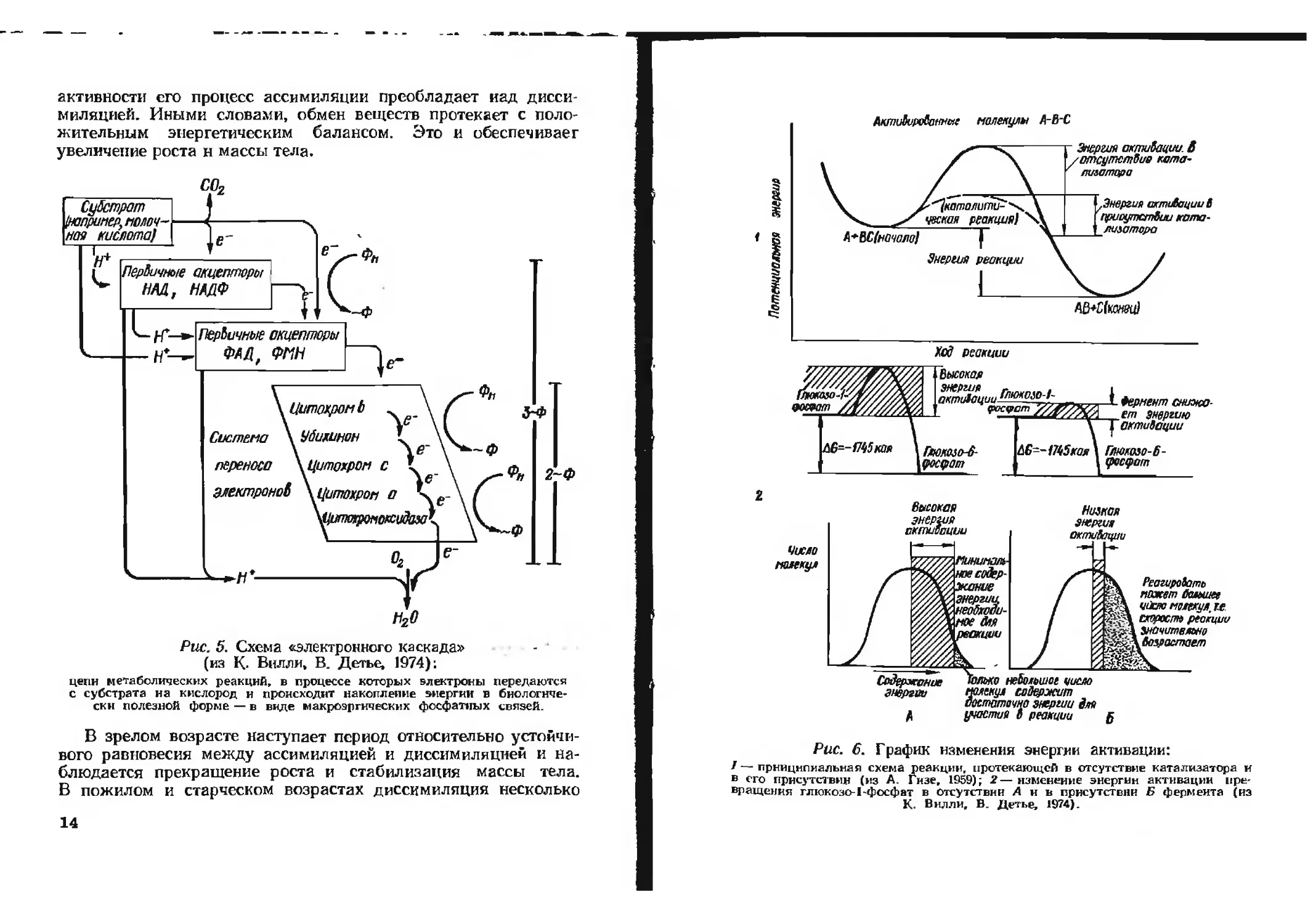
процесс н получил название «электронного каскада» (рис. 5).
В ходе такой реакции энергия электронов связывается в биоло-
гически полезной форме — в виде макроэргических соединений,
среди которых АТФ занимает ведущее место. Роль фермента
здесь заключается в том, что он, являясь катализатором, сни-
жает энергетический барьер (энергетическая активация), пре-
пятствующий началу реакции, и ускоряет течение ее. Таким
образом, фермент снижает энергию активации реакции и уве-
личивает число молекул, обладающих достаточной энергией,
12
Ацетальдегид
(из II. Клауда и А. Джабира, 1972):
метаболизма. Он состоит из двух фаз: гликолиза — анаэробного процесса
молекула шестнуглеродного сахара (глюкоза) распадается на два трехугде-
образуются две молекулы аденозинтрнфосфата — клеточного переносчика
лота превращается в молочную кислоту, а при брожении — в некоторые дру-
сутствни кислорода пировиноградная кислота подвергается полному окислению
роиы которых (и еще двух, отщепленных при гликолизе) проходит через
випадениядннуклеотнд (ФАД) — к цепи дыхательных ферментов (ублхпнон и
свобождаемая при этом, идет на синтез АТФ. На каждую потребленную мо-
38 молекул АТФ.
чтобы участвовать в реакции и образовывать конечный про-
дукт (рис. 6). Иллюстрацией же общего принципа энергообе-
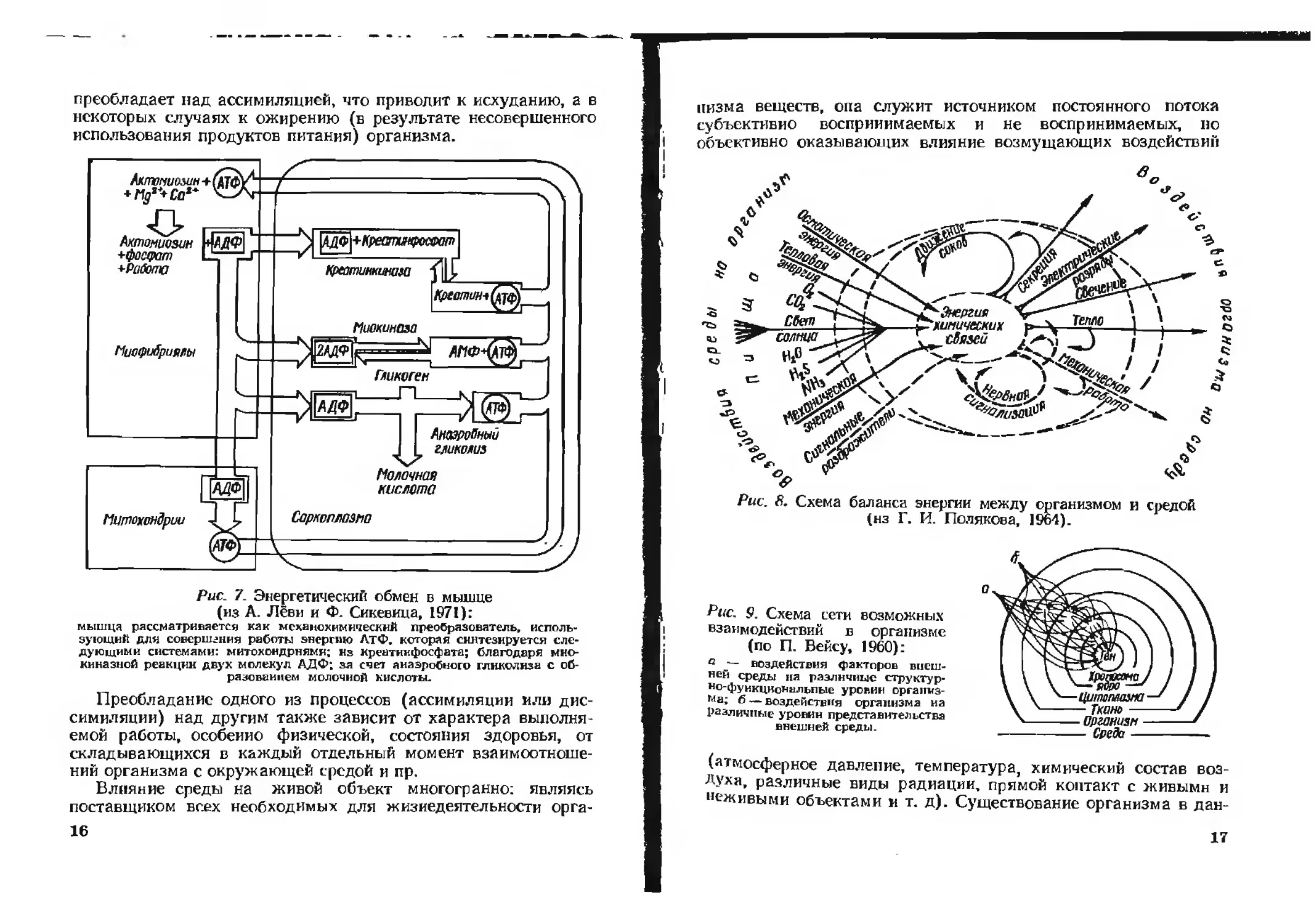
спечения жизнедеятельности клеток может служить схема
энергетического обмена в мышце (рис. 7).
Течение обмена веществ зависит от возрастно-половых осо-
бенностей организма и его функционального состояния. В част-
ности, в период роста и созревания интенсивность обмена (при
прочих равных условиях) на единицу массы тела выше, чем в
сформировавшемся организме. Причем на фоне общей высокой
18
активности его процесс ассимиляции преобладает над дисси-
миляцией. Иными словами, обмен веществ протекает с поло-
жительным энергетическим балансом. Это и обеспечивает
увеличение роста и массы тела.
Рис. 5. Схема «электронного каскада»
(из К- Вилли, В. Детье, 1974):
цепи метаболических реакций, в процессе которых электроны передаются
с субстрата на кислород и происходит накопление энергии в биологиче-
ски полезной форме — в виде макроэргических фосфатных связей.
В зрелом возрасте наступает период относительно устойчи-
вого равновесия между ассимиляцией и диссимиляцией и на-
блюдается прекращение роста и стабилизация массы тела.
В пожилом и старческом возрастах диссимиляция несколько
14
АктиЪироВаннае молекулы А-В-С
^^католить \
чвская реакция) X
Л * ВС(ночало) I”
Энергия реакции
^Энергия астиВацииВ
присутствии ката-
лизатора
- Энергия активации. В
/отсутствие ката-
' лилатора
AQ*CfKOMffcd
Хед реакции
90Ж№
Высокая
энергия г„1Пип,л,
пктЛтуш *люкою-1-
фосуст
АВ--Р^5кая
Глюкоза-6-
фосфат
—L Фермент снижа-
ет энергию
октиВоции
Глмколо-6-
, фосфат
Число
молекул
Высокая
энергия
октиопции
Codfpxn^ue
эмергпи
для
Низкая
энергия
октиВации
ное содер-
жание
о небольшое цисло
молекул содержит
достаточно энергии для
участия В реакции
Реагировать
пакет большее
ч&т молекул,и.
схорост реакции
Значительно
Возрастает
В
Рис. 6. График изменения энергии активации:
f — принципиальная схема реакции, протекающей в отсутствие катализатора и
в его присутствии (из А. Гизе, 1959); 2— изменение энергии активации пре-
вращения глюкозе-1-фосфат в отсутствии Ань присутствии Б фермента (из
К. Вилли, В. Детье, 1974).
преобладает над ассимиляцией, что приводит к исхуданию, а в
некоторых случаях к ожирению (в результате несовершенного
использования продуктов питания) организма.
Рис 7. Энергетический обмен в мышце
(из А. Лёви и Ф. Сикевица, 1971):
мышца рассматривается как мсханохнмнческий преобразователь, исполь-
зующий для совершения работы энергию ЛтФ. которая синтезируется сле-
дующими системами: митохондриями; из креатиифосфата; благодаря мио-
киназной реакции двух молекул АДФ; за счет анаэробного гликолиза с об-
разованием молочной кислоты.
Преобладание одного из процессов (ассимиляции или дис-
симиляции) над другим также зависит от характера выполня-
емой работы, особенно физической, состояния здоровья, от
складывающихся в каждый отдельный момент взаимоотноше-
ний организма с окружающей средой и пр.
Влияние среды на живой объект многогранно: являясь
поставщиком всех необходимых для жизнедеятельности орга-
16
пизма веществ, она служит источником постоянного потока
субъективно воспринимаемых и не воспринимаемых, но
объективно оказывающих влияние возмущающих воздействий
!чд a d S
^пногЛО 8
9. Схема сети возможных
взаимодействий в организме
(по П. Вейсу, 1960):
а ~~ воздействия факторов внеш-
ней среды ня различные структур-
но-фуикциональпые уровни организ-
ма; б — воздействия организма на
различные уровни представительства
внешней среды.
(атмосферное давление, температура, химический состав воз-
Духа, различные виды радиации, прямой контакт с живыми и
неживыми объектами и т. д). Существование организма в дан-
17
ных условиях возможно, если он на все эти воздействия точно
и своевременно реагирует приспособительными реакциями:
изменением обмена веществ, работы органов кровообращения,
Na'
к----------------
Сог*-------------
Л/у2+------------
СГ------------—
нсо;-------------
фоароты----------
SO}'-------------
Глюкоза----------
Аниншислоты
Холестерин
Фоаролипиди
Нейтральные Жиры
Р«г--------------
------0,5г %
scot
pH
Внеклеточная
жидкости
-----137нзкВ/л
-------5нэкВ!л -
-------Знаке/п -
-------ЗнзкВ/л —
-----Ю5нжВ/л
------2ВнэкВ/л
-------9нзк6/л
--------1нзкВ/е
------90нг%
------ ЗОнг%
Внутриклеточная
жидкость
— ЮнзкВ/л
- МнзкВ/л
-ОнзкВ/л
-62нэкВ/л
----АнзкВ/л
----ЮнзкВ/л
----75нэкВ/л
------2нзкВ/л
0-20нг 7,
— 200нг % ?
— 2-95г %
— 35нн рт ст —|—--------20ннртст?
— 50 нн рт ст. ?
— 7,1?
----Wntipm ст
-----jit-------
Рис. 10. Наиболее типичный состав внутриклеточных и внеклеточных
(тканевых) жидкостей у человека (из К- Вилли, В. Дет1>е, 1974).
дыхания, пищеварения, аппарата движения, деятельности ре-
гулирующих систем (нервной и желез внутренней секреции).
При этом изменения организма не должны выходить за рамки
допустимых пределов физиологических колебаний, иначе насту-
18
пит нарушение его нормальной жизнедеятельности — заболе-
вание и даже смерть.
Живые объекты представляют собой своеобразные «физио-
логические маятники», в пределах амплитуды колебания кото-
рых возможны отклонения тех или иных функций от какой-то
средней величины.
В равной мере воздействия организма на среду, приводящие
к нарушению сложившихся в ней взаимоотношений, могут
создать условия, неблагоприятные для жизни этого организма.
Таким образом, между живыми объектами и окружающим их
внешним миром, органической частью которого они сами явля-
ются, должен сложиться оптимальный баланс обмена вещест-
вом и энергией, обеспечивающий их взаимное благополучие
(рис. 8). Подобные двусторонние взаимоотношения прослежи-
ваются на всех структурно-функциональных уровнях, из кото-
рых складываются живые организмы и окружающая их среда,
включая и живые объекты (рис. 9). Вот почему животные и
растительные организмы в процессе эволюции не только при-
обретают новые наследственно фиксируемые качества, но и
удерживают относительное постоянство химического состава и
функций, т. е. сохраняют неповторимую биохимическую и
функциональную индивидуальность максимально независимо от
воздействий внешней среды. Это свойство получило название
гомеостаз. В самом деле животные и растения и по химическо-
му составу и концентрации циркулирующих в них жидкостей
(кровь, лимфа, тканевая жидкость) и по температурному ре-
жиму окружающей среды (рис. 10) не только отличаются, но
и, невзирая на эту вещественно-температурную разницу, тре-
бующую от организмов, согласно законам физики и, в част-
ности второму закону термодинамики, уравновешивания,
сопротивляются ему, сохраняя свое неравновесное состояние.
Элементарный анализ «термодинамического» поведения биоло-
гических систем подтверждает это положение.
Очерк 2. «ТЕРМОДИНАМИЧЕСКИЙ» АНАЛИЗ
ВЗАИМООТНОШЕНИИ В СИСТЕМЕ ЖИВОЙ
ОРГАНИЗМ — ОКРУЖАЮЩАЯ СРВДА
Термодинамика — наука, изучающая взаимные превраще-
ния различных видов энергии, связанные с переходом ее при
19
различных физических и химических процессах от одних тел
к другим в форме теплоты и работы. В основе термодинамики
лежат два закона.
Первый по существу есть закон сохранения энергии. Он
устанавливает, что количество теплоты ДО, полученное какой-
либо системой* тел, затрачивается на повышение ее внутренней
энергии Дб/ и на работу ДЛ, совершаемую ею против действия
внешних снл:
ДО=ДО+ДЛ.
Значит работа, совершаемая системой, равна разности между
количеством сообщаемой ей теплоты и изменением ее внутрен-
ней энергии:
ДЛ=ДО- ДГЛ
Внутренняя энергия системы может уменьшаться, переходя в
окружающее пространство в виде тепла, или увеличиваться за
счет его поступления.
В первом случае изменение внутренней энергии будет отри-
цательным:
-Дб?=ДО+ДД.
Таким образом, убыль внутренней энергии равна сумме выде-
ленного тепла и произведенной работы.
Во втором случае изменение внутренней энергии будет по-
ложительным:
+Дб/=Д(2-ДД.
Следовательно, увеличение внутренней энергии равно количе-
ству поглощенной извне теплоты за вычетом произведенной си-
стемой работы.
Из первого закона термодинамики следует, что работа со-
вершается за счет изменения внутренней энергии системы или
за счет сообщения ей некоторого количества тепла. В живой
системе работа осуществляется не за счет притока тепла извне,
т. е. живой организм не может работать, как тепловая маши-
* В термодинамике системой называется всякий материальный
объект или совокупность взаимодействующих объектов, мысленно
обособляемых от окружающей среды.
20
на. Установлено, что в тепловой машине количество тепла,
превращенного в работу, зависит от разности абсолютных тем-
ператур нагревателя Ti и холодильника Та по Кельвину (Т=
=1+273°), причем максимальная работа тем больше, чем
больше температурный перепад между Ti и Та. Коэффициент
полезного действия тепловой машины вычисляется по формуле:
Предположим, что мышца работает как тепловая машина при
температуре +25° С и к. п. д. 30% (1/3). Подставляя эти дан-
ные в формулу, получим
71 — 298___1_
Tt ~ 3 ’
71 = 447° К, или 174° С (447-273=174).
Значит, если бы мышца работала как тепловая машина, то
при заданных условиях она нагрелась бы до 174° С, т. е. до
температуры, ие совместимой с жизнью, так как белки денату-
рируют при 40—60°.
Следовательно, в живом организме работа совершается за
счет изменения внутренней энергии системы в результате про-
текающих в ней биохимических процессов с участием фермен-
тов или за счет изменения энтропии.
Приложимость первого закона термодинамики к биологиче-
ским объектам была доказана Лавуазье и Лапласом в 1780 го-
ду. Они измерили количество выделенного тепла и углекислоты
морской свинкой, помещенной в ледяной калориметр, и при
прямом сжигании продуктов питания, получаемых этой свин-
кой. Результаты совпали. Аналогичная зависимость была уста-
новлена между количеством кислорода, пошедшего на окисле-
ние пищевых веществ, и тепла, выделенного организмом.
Таким образом, потребление кислорода и выделение угле-
кислоты животным тесно коррелирует с теплопродукцией. Это
привело к заключению о существовании эквивалентности хими-
ческой энергии, высвобождающейся в процессе обмена веществ,
и теплотой, выделяемой во внешнюю среду. Следовательно, по
количеству поглощенного кислорода и выделенной углекислоты
можно судить о состоянии энергетического обмена организма.
21
В частности, установлено, что на оскисление в организме 1 г
белков, жиров, углеводов требуется различное количество
кислорода. Соответственно этому выделяется разное количество
тепла (табл. I).
Стандарты пересчета
Таблица 1
При окислении I г вещества*
Питательное вещество поглощается кислорода, дм3 выделяется углекислоты, дм5 освобождается тепловой энергии, кДж Калорический эк- вивалент кисло- рода Дыхательный коэффициент
Белки Жиры Углеводы 0,965 1,990 0,830 0,775 1,420 0,830 17,18 (4,1 ккал) 38,97 (9,3 ккал) 17,18 (4,1 ккал) 17,60(4,2 ккал) 19,69(4,7 ккал) 21,16(5,05 ккал) 0,8 0,7 1,0
Количество тепла, высвобождающееся при поглощении
i дм3 кислорода или выделении углекислоты, называется кало-
рическим эквивалентом кислорода или углекислого газа. Зная
общее количество кислорода, потребленное организмом в еди-
ницу времени, можно вычислить его энергетические траты, если,
конечно, известны вещества, подвергшиеся окислению. Они опре-
деляются по величине дыхательного коэффициента, т. е. по
отношению объемов выделенной углекислоты и поглощенного
за то же время кислорода. При смешанной пище этот коэффи-
циент равен 0,85—0,9.
Описанный метод оценки энерготрат организма получил
название непрямой калориметрии. Для расчета энерготрат дан-
ным методом существуют специальные таблицы. Однако более
точным является метод прямой калориметрии. Суть его состоит
в непосредственном определении количества выделенного орга-
низмом тепла, углекислоты, азота, мочевины и поглощенного
кислорода. Это возможно при помещении объекта исследования
* Для жиров и углеводов приводятся данные, получаемые при
окислении до СОг и НгО, для белков — до мочевины, как это имеет
место в организме.
22
(человек, животное) в специальную калориметрическую ка-
меру.
Если человек в сутки получил 56,8 г белка, 140 г жира и
79,9 г углеводов, то по табл. 1 можно рассчитать количество
тепловой энергии, высвобождающейся при окислении этих
продуктов: белка — 974 кДж (233 ккал); жира-—5442 кДж
(1302 ккал); углеводов — 1371 кДж (328 ккал); всего 7787 кДж
(1863 ккал). В соответствии с первым законом термодинамики
следует ожидать, что в течение суток выделится столько тепло-
вой энергии, сколько поступило. И в самом деле, теплоты было
выделено 5743 кДж (1374 ккал); калорический эквивалент
выдыхаемых газов оказался равным 180 кДж (43 ккал); кало-
рийность выделенного кала и мочи — 95 кДж (23 ккал); испа-
рения через дыхание — 757 кДж (181 ккал); через кожу —
949 кДж (227 ккал); различные поправки составили 46 кДж
(11 ккал); всего 7770 кДж (1859 ккал).
Такой энергетический баланс говорит о том. что живой орга-
низм не является источником новой энергии, т. е. происходя-
щие в нем процессы полностью подчиняются первому закону
термодинамики. Однако подобный баланс возможен, если не
выполняется значительная работа, не происходит накопления
массы и не совершаются и другие процессы, связанные с повы-
шенным распадом нли накоплением энергии, т. е. в условиях
так называемого основного обмена. Прн совершении работы к
энергетическим тратам основного обмена прибавляются допол-
нительные траты, что составляет общий обмен веществ, кото-
рый значительно выше основного. Его интенсивность зависит
преимущественно от мышечной деятельности н сопровождаю-
щей ее работы внутренних органов. Энергия, расходуемая при
мышечной работе, называется моторной энергией (моторные
калории). Общий обмен веществ идет со сдвигом энергетиче-
ского баланса в сторону превосходства расхода, который ком-
пенсируется приемом пищи. Именно поэтому составление пи-
щевых рационов по энергетической полноценности продуктов
должно базироваться на знании энергетической стоимости той
или иной формы работы (физической, умственной, комбиниро-
ванной), которая определяется методом прямой и непрямой
калориметрии.
Таким образом, теплоту, выделенную организмом в данный
момент, можно разделить на первичную (основную) и вторич-
23
ную (активную). Первичная теплота — это теплота, постоянно
высвобождающаяся в ходе основного обмена веществ. Вторич-
ная теплота выделяется при совершении организмом любой ра-
боты за счет запаса энергетических соединений, накопившихся
в процессе метаболизма. В обычных условиях существования
оба вида теплоты находятся в состоянии относительно устой-
чивого равновесия: уменьшение производства первичной ком-
пенсируется увеличением вторичной и наоборот. Отдача первич-
ной теплоты в окружающую среду — неизбежное следствие
термодинамических взаимоотношений организма со средой.
Рассеивание первичной теплоты происходит, даже если темпе-
ратура окружающей среды более высокая, чем тела. Это обе-
спечивается механизмом теплорегуляции, в частности благода-
ря усиливающемуся потоотделению и испарению. В противном
случае избыточное поглощение тепла извне может приве-
сти к катастрофическим последствиям — перегреву. При
переохлаждении же, т. е. в условиях избыточных теплопотерь,
поддержание необходимого уровня выделения первичной теп-
лоты обеспечивается за счет увеличения доли вторичной тепло-
ты (усиление двигательной активности вплоть до появления
непроизвольной дрожи), а также усиления теплопродукции
путем перехода па менее экономный способ ее производ-
ства.
Итак, само существование и все процессы жизнедеятельно-
сти организмов связаны с изменением веществеиио-энергетиче-
ского баланса в системе живой организм — окружающая среда,
т. е. со всеобщим законом сохранения и превращения материи
и энергии.
Следовательно, первый закон термодинамики позволяет оп-
ределить количественные соотношения различных форм энергии,
принимающих участие в данном процессе. Однако ои ничего
не говорит о возможности протекания этого процесса и о пре-
имущественном направлении его. Так, например, исходя из пер-
вого закона термодинамики, можно допустить переход тепла
в любом направлении, от менее нагретой системы к более на-
гретой, лишь бы общее количество теплоты оставалось постоян-
ным. Между тем знание преимущественной направленности
превращения энергии имеет огромное практическое значение,
так как позволяет в рамках закона сохранения управлять про-
цессом. Решить все эти вопросы можно на основе второго закона
24
термодинамики, который налагает определенные ограничения
на взаимные переходы энергии.
Первое ограничение: тепло невозможно перевести от более
холодной системы к более горячей без затраты энергии извне.
Это означает, что самопроизвольно протекают только процессы,
связанные с переносом тепловой энергии от более высокого
уровня к более низкому, т. е. по градиенту. Чем выше градиент
взаимодействующих систем, тем интенсивнее осуществляется
перенос энергии, т. е. работа.
Второе ограничение: любая форма энергии может практи-
чески. полностью перейти в тепловую, но последняя не полно-
стью переходит в иную форму и частично остается в виде теп-
лоты. Это означает, что внутренняя энергия системы неодно-
родна. Одна часть ее способна к превращениям и к производ-
ству полезной работы, другая же при данных условиях этого
не может. Первая часть получила название свободной энергии
/•', вторая — связанной энергии G (по Гельмгольцу). Следова-
тельно, внутренняя энергия системы есть сумма свободной и
связанной энергии:
U=F+G.
Связанная энергия тем больше, чем меньше разность тем-
ператур в системе, и в отношении выполнения полезной рабо-
ты как бы обесценена. Эту' непроизводительную энергию на-
зывают энтропией S. Для того чтобы в определенных условиях
уменьшить энтропию, т. е. поддержать высокий жизненный
потенциал, живые организмы должны постоянно заряжаться
энергией извне и притом с некоторым запасом, подобно тому
как заряжается аккумулятор. Находясь «в долгу» у окружа-
ющей природы и тем самым снижая свою энтропию, живые
организмы увеличивают общую энтропию системы организм —
среда. Есть все основания полагать, что если бы на Земле не
было жизни, ее энтропия была бы значительно меньше. О при-
роде малой энтропии живых существ очень образно сказал
Э. Шредингер: «Отрицательная энтропия — вот то, чем орга-
низм питается. Или, чтобы выразить это менее парадоксально,
существенно в метаболизме то, что организму удается осво-
бождать себя от всей той энтропии, которую он вынужден про-
изводить, пока жив».
25
Временное прекращение поступления энергии извне (прекра-
щение питания) приводит к снижению уровня жизненной актив-
ности организма. Эта своеобразная приспособительная реакция
преследует основную цель сократить прирост энтропии при
недостаточной компенсации ее свободной энергией извне. Сле-
довательно, живые организмы, безусловно, находятся в сфере
действия второго закона термодинамики. Наука располагает
возможностью измерить как величину энтропии системы, так и
интенсивность антиэнтропийных процессов, необходимых для
сохранения данной системы, например живых организмов. Так,
связанная энергия и энтропия отражают одно и то же явление.
Всякий раз, когда в системе совершается какой-либо само-
произвольный процесс, осуществляющийся по градиенту', про-
исходит не только перераспределение различных форм энергии,
но и увеличение энтропии, которое является своеобразной пла-
той за каждое использование энергии. Следовательно, величина
энтропии системы в данный момент &S равна разности энтро-
пии системы в начале Si и в конце St процесса:
AS=5j—Sj.
Если процесс обратим, т. е. допускается возвращение систе-
мы в первоначальное состояние без каких-либо изменений
в окружающей среде, то энтропия этой системы, естественно,
равна нулю. При необратимых процессах энтропия системы
больше нуля, т. е. она увеличивается. Таким образом, в общем
виде величина энтропии может быть выражена формулой
„ Л<2
AS=-^->0,
где &Q — количество тепла, поглощенного из среды; Т — абсо-
лютная температура.
Все реальные процессы, осуществляющиеся в окружающем
иас мире, необратимы. Это прежде всего касается живой при-
роды. Примером может служить однонаправленность и необ-
ратимость возрастных изменений человека в течение жизни
(рождение — детство — отрочество — юность — зрелость — ста-
рость— смерть). Такое явление объясняется тем, что источни-
ком жизнедеятельности живых организмов является химическая
энергия, переходящая в работу и тепло, большая часть которого
рассеивается в результате теплоотдачи. Кроме того, в соответ-
26
ствии со вторым законом термодинамики полное превращение
тепла в работу невозможно.
Следовательно, энтропия — это характеристика необратимо-
сти процесса такой формой энергии, из которой она самопро-
извольно не может переходить в другие формы. Можно пред-
полагать, что количество тепла, высвободившееся в процессе
превращения всех мыслимых видов энергии и переданное от
более горячей системы к более холодной, будет систематически
превращаться в связанную энергию последней, т. е. будет уве-
личивать ее энтропию. Такой процесс может идти до тех пор,
пока между системами не установится равенство температур,
т. е. пока энтропия их не станет максимальной. Это значит, что
произойдет качественное изменение энергии взаимодействую-
щих систем при неизменном общем количестве ее. Наступит
момент, когда в системах сохранится только один вид энергии —
энергия теплового движения молекул, атомов и элементарных
частиц (связанная энергия); исчезнет температурный гради-
ент — температурная асимметрия между ними; установится
термодинамическое равновесие, т. е. наступит тепловая смерть
систем, а шире — тепловая смерть вселенной, так как исчезнет
возможность самопроизвольного возникновения процессов в
связи с потерей способности энергии к превращениям.
Признание второго закона термодинамики как абсолютно-
го— вселенского — равно признанию некоего начала и возмож-
ного конца вселенной, что явно абсурдно. Следовательно, вто-
рой закон термодинамики не универсальный, а частный, т. е.
вероятностный. Это было доказано Т. Сведбергом, М. Смолу-
ховским, Л. Больцманом. В частности, Сведберг, исследуя броу-
новское движение частиц коллоидного золота, заметил, что ка-
кое-то количество их под влиянием толчков молекул окружаю-
щей среды перемещается против градиента силы тяжести (снизу
вверх). Значит покоящееся тепло среды, т. е. связанная энергия
системы, превращается в работу вопреки второму закону тер-
модинамики. Статистическая вероятность данного перемещения
частиц коллоидного золота оказалась весьма малой величиной.
Однако это не значит, что она отсутствует вообще. Отсюда
можно сделать заключение, что второй закон термодинамики в
земных условиях отражает лишь вероятностную преимуществен-
ность протекающих в соответствии с ним процессов, а не абсо-
лютную закономерность.
27
В условиях безграничного космоса, т. е. в условиях абсо-
лютного господства закона сохранения материи и энергии, вто-
рой закон термодинамики еще менее приложим. Само понятие
бесконечности делает все возможные процессы равновероятны-
ми. Именно это и позволило Больцману высказать положение
о том, что во вселенной есть области, где второй закон термо-
динамики не имеет силы, и области, где энтропия не увеличи-
вается, а, напротив, уменьшается. В последних, в противопо-
ложность земным условиям, вероятность антиэнтропийных про-
цессов более высокая, чем энтропийных.
Надо полагать, что соблюдение закона сохранения материи
и энергии во вселенной возможно при количественном и каче-
ственном балансе (равнозначности, симметричности) энтро-
пийных и антиэнтропийных областей. Однако этот баланс при-
менителен к вселенской бесконечности. Каждая взятая отдельно
область ее не сбалансирована, термодинамически неравновес-
на, она или преимущественно эптропийна или антиэнтропийна,
т. е. негэнтропийна (процесс обратной энтропии). В целом же
все области обмениваются как энтропией, так и негэнтропией,
обеспечивая всеобщий термодинамический баланс бесконечной
вселенной.
Полемизируя со сторонниками приложимости второго зако-
на термодинамики к окружающей природе, а следовательно, и
тепловой смерти вселенной, Энгельс нарисовал замечательную
картину вечно движущейся и превращающейся материи: «Вот
вечный круговорот, в котором движется материя,— круговорот,
который завершает свой путь лишь в такие промежутки вре-
мени, для которых наш земной год уже не может служить до-
статочной единицей измерения; круговорот, в котором время
наивысшего развития, время органической жизни и, тем более,
время жизни существ, сознающих себя и природу, отмерено
столь же скудно, как и то пространство, в пределах которого
существует жизнь и самосознание; круговорот, в котором каж-
дая конечная форма существования материй — безразлично,
солнце или туманность, отдельное животное или животный вид,
химическое соединение или разложение — одинаково прехо-
дяща и в котором ничто не вечно, кроме вечно изменяющейся,
вечно движущейся материи и законов ее движеиня и изменения.
Но как бы часто и как бы безжалостно ни совершался во вре-
мени и пространстве этот круговорот; сколько бы миллионов
28
солнц и земель ни возникало и ни погибало; как бы долго
ии длилось время, пока в какой-нибудь солнечной системе и
только на одной планете не создались условия для органической
жизни; сколько бы бесчисленных органических существ ни
должно было раньше возникнуть и погибнуть, прежде чем из
их среды разовьются животные со способным к мышлению моз-
гом, находя на короткий срок пригодные для своей жизни
условия, чтобы затем быть тоже истребленными без милосер-
дия,— у нас есть уверенность в том, что материя во всех своих
превращениях остается вечно, одной и той же, что ни одни
из ее атрибутов никогда не может быть утрачен и что поэтому
с той же самой жизненной необходимостью, с какой она когда-
нибудь истребит на Земле свой высший цвет — мыслящий дух,
она должна будет его снова породить где-нибудь в другом месте
и в другое время». (Маркс К-, Энгельс Ф. Соч. Изд. 2-е. Т. 20,
с. 362—363).
Итак, волею сложившихся обстоятельств Земля н все су'щее
на ней находится в той части бесконечной вселенной, где пре-
обладает энтропийный процесс, т. е. царит второй закон термо-
динамики.
Для более ясного представления о термодинамических взаи-
моотношениях материальных тел в рамках второго закона тер-
модинамики, исходя из особенностей взаимодействия с окру-
жающей средой, принято различать три типа систем:
I — изолированные системы, не обменивающиеся со сре-
дой пи веществом, ни энергией;
II — замкнутые системы, обменивающиеся со средой
энергией, ио не обменивающиеся веществом;
III — открытые системы, обменивающиеся со средой и
энергией, и веществом.
Два первых типа относятся к закрытым системам. Однако,
так как общей закономерностью материального мира, основной
формой бытия движущейся материи является всеобщая связь
и взаимообусловленность предметов, процессов и явлений, к по-
нятиям изолированных и замкнутых систем следует относиться
лишь как к удобным для познания абстракциям, не существую-
щим в реальном мире. Единственной замкнутой системой может
быть только сама вселенная, замкнутая, как это ни парадок-
сально, в своей бесконечности. Бесконечность в силу своего
свойства бесконечности не может быть полностью охвачена ни
20
мысленно, ни опытом. Следовательно, в реальном мире мысля-
щий человек в своей познавательной деятельности всегда имеет
дело с открытыми системами, мерой «открытости» которых яв-
ляется интенсивность взаимного обмена их энергий и веществ.
Системы могут находиться в термодинамически равновес-
ном, стационарном и неравновесном состояниях.
Термодинамическое равновесие — это такое состояние, при
котором способность системы производить работу равна нулю.
Из этого состояния система не может выйти без траты энергии,
поступившей извне. Такое состояние может вечно поддержи-
ваться за счет внутренней энергии системы при обязательном
условии — неизменности окружающей среды. У систем, нахо-
дящихся в подобном состоянии, свободная энергия равна нулю,
а энтропия максимуму. Из условий, обеспечивающих термо-
динамическое равновесие, следует, что система, находящаяся
в этом состоянии, имеет постоянные неизменяющиеся основные
параметры характеристик. Естественно, что в состоянии термо-
динамического равновесия могут находиться лишь идеальные
закрытые системы.
Стационарное состояние — это состояние, при котором сис-
тема, подобно термодинамически равновесной системе, сохра-
няет во времени свои основные параметры неизменными при
необязательном постоянстве внешней среды. Свободная энер-
гия такой системы не равна нулю. Это означает, что данная
система, будучи выведенной из стационарности под влиянием
изменений в окружающей среде, способна производить работу,
направленную против нарушения этой стационарности, т. е. на
восстановление ее. Таким образом, если при термодинамиче-
ском равновесии энтропия системы максимальна и приращение
ее практически невозможно, т. е. оно равно нулю, то при ста-
ционарном состоянии скорость создания и величина энтропии
равны интенсивности обмена ее с окружающей средой. Значит
скорость приращения энтропии в стационарной системе по-
стоянна и выше нуля. Наличие свободной энергии, характерное
для стационарного состояния, обеспечивает системе возмож-
ность совершения работы и как следствие роста энтропии. Од-
нако избыточный рост энтропии может привести в конце концов
открытую стационарную систему к «насыщению» энтропией до
уровня, имеющегося в окружающей среде, и в результате к пре-
кращению процессов обмена энергии и вещества между этой
30
системой и средой, т. е. к поглощению системы средой, а для
живого организма — к смерти.
Удержание стационарного состояния, следовательно, воз-
можно при обязательном наличии относительно уравновешиваю-
щих друг друга потоков вещества и энергии из окружающей
среды в систему и обратно. Отличие приращения энтропии в
открытой стационарной системе от нуля говорит об отсутствии
полного равенства этих потоков. Таким образом, стационарное
состояние — это динамическое состояние, под которым следует
понимать способность открытой системы в течение определен-
ного времени относительно стабильно удерживать свои основ-
ные параметры, т. е. стремиться к минимуму прироста энтро-
пии в единицу времени В каждый отдельный момент система
может иметь соответствующее взаимодействиям со средой ста-
ционарное состояние. На основании изложенного приходим к
выводу о существовании двух видов стационарного состояния
живых организмов. Одно из иих характерно для каждого дан-
ного момента времени, другое - конечное, к которому система
приближается в процессе развития, роста и старения (А. И. Зо-
тин, 1974).
Принцип механизма, поддерживающего стационарное со-
стояние в открытой системе, и отличие его от такового в закры-
тых системах рассмотрим на примере двух гидродинамических
моделей (рис. II, 12), которые дают возможность составить
представление о двух важнейших свойствах открытых стацио-
нарных систем.
Первое свойство заключается в том, что открытые стацио-
нарные системы чрезвычайно чувствительны к притоку и оттоку
вещества и энергии. В самом деле, даже если из крапа / в си-
стему начнет поступать на одну молекулу воды больше или
меньше при неизменном положении кранов 2 и 3, стационарное
состояние в сосудах начнет сейчас же изменяться в сторону
последовательного увеличения или уменьшения в единицу вре-
мени уровня жидкости в них на одну молекулу. Этот процесс
будет идти до тех пор. пока вновь не восстановится прежний
приток или отток молекул воды. Иначе говоря, открытые ста-
ционарные системы практически не имеют порога, т. е. той
минимальной дискретной величины воздействия, ниже которой
они бы не реагировали на изменения среды. Для открытых
стационарных систем, к которым относятся и все живые орга-
81
1
Рас. It. Равновесное
состояние жидкости
в закрытой системе сосудов
(по 3. П. Беликовой
и Р. С. Павлову, 1969):
жидкость переливается из верх-
него сосуда в нижний со ско-
ростью. зависящей от того, на-
сколько открыт край, в резуль-
тате устанавливается равновесие,
так как в этих условиях систе-
ма изолирована от окружающей
среды (испарение исключается).
Естественно, что подобное со-
стояние для биологических си-
стем несовместимо с понятием
жизнь, основным свойством кото-
рой является постоянный об-
мен веществ-
Рис. 12. Стационарное
состояние жидкости в
открытой системе сосудов
(по 3. П. Беликовой
и Р. С. Павлову, 1969):
краны /, 3 моделируют константы
переноса веществ в системе, огра
инчивающие интенсивность поступ-
ления и удаления последних; кран
2 — константу скорости химических
реакций в открытой системе. При
изменении «концентрации катализа-
тора» (поворот крана 2) устанавли-
вается ИОнЫЙ уровень жидкости и
создается новое стационарное со-
стояние. В данных условиях проис-
ходит постоянное обновление жидко
сти в сосудах и среде. Для орга-
низма — это обмен веществ и энер-
гий со средой, обновление его со-
става при сохранении стационарного
состояния — динамического равнове-
сия.
низмы, нет безразличных воздействий внешней среды, какими
бы, незначительными они формально ни казались.
Второе свойство органически вытекает из первого и сводится
к тому, что открытые стационарные системы практически инди-
скретно изменяют свое стационарное состояние, так как для
них не существует пороговой (минимальной) разницы между
двумя или несколькими следующими друг за другом или одно-
временными воздействиями внешней среды, па которые бы они
не отреагировали. Если в условиях нарушенного стационарного
состояния в сосудах многократно изменять положение крана
1 так, чтобы каждый раз приток воды увеличивался на одну
молекулу, то можно наблюдать еще более интенсивное изме-
нение стационарного состояния системы. Следовательно, для
живых организмов вообще не существует безразличных собы-
тий во внешней и внутренней среде, как бы малы они ни были
и в какой бы пространственной и временной последователь-
ности ни воздействовали на эти организмы.
Высокая чувствительность открытых систем, и в первую
очередь живых организмов, к изменениям в окружающей среде
предполагает возможность существования для них бесчислен-
ного множества стационарных состояний в пределах допусти-
мых для сохранения жизни колебаний функций органов, систем
органов и всего организма (в зависимости, например, от актив-
ности организма, от времени суток и года, возраста, пола, роста
и развития, питания, места жительства и т. д.).
Стационарное состояние может относительно стабильно со-
храняться в течение какого-то времени. Стабильность не явля-
ется жестко фиксируемым неподвижным состоянием, подобным
термодинамическому равновесию. Такого в природе быть не
может, поскольку в соответствии с законом сохранения и пре-
вращения материи и энергии идет постоянный обмен веществом
и энергией между различными объектами, входящими в систе-
му, называемую Природой, а шире—Вселенной.
Все события совершаются во времени. Вероятность абсо-
лютного совпадения во времени двух или нескольких событий,
полностью уравновешивающих друг друга, в любом ограничен-
ном пространстве ничтожна. Поэтому под стационарным со-
стоянием следует понимать колебания параметров того или
иного явления или процесса около какой-то «идеальной» кон-
станты, наиболее соответствующей данным условиям взаимоот-
2 А. А. Логинов 33
ношений открытой системы (организма) со средой. Например,
частота дыхания в атмосфере, обедненной кислородом и насы-
щенной углекислотой, и наоборот, будет изменяться в сторону
учащения в первом случае и урежения во втором. Однако как
учащение, так и урежение дыхания имеет пределы, выше кото-
рых частота дыхания без нарушения функций организма изме-
няться ие может. Так, частота дыхания у человека 1—2 раза
в минуту несовместима с жизнью. Это справедливо для всех
констант показателей функционального состояния организма:
для пределов колебаний скорости двигательной реакции на
раздражение, для уровня сахара или pH крови и т. д. Медико-
биологические исследования в области оценки состояния здо-
ровья преследуют цель — выяснить, насколько измеряемый па-
раметр укладывается в пределы возможных для нормального
существования организма колебаний и каков резерв для опти-
мального развития данной функции. Именно в рамках крайних
пределов этих колебаний и может быть сколь угодно много
различных уровней стационарных состояний. Определенный
уровень стационарного состояния (динамического равновесии)
в открытой системе поддерживается не потому, что свободная
энергия сведена до минимума, как это имеет место при термо- ;
динамическом равновесии, а потому, что система получает из
среды свободную энергию в количестве, достаточном для ком-
пенсации потерь в соответствии с особенностями ее взаимоот- j
ношений со средой. ,
Не менее важным для возникновения и поддержания уровня
стационарного состояния является и скорость протекания ре- ,
акций в системе. При формировании того или иного стацио- ‘
нарного состояния скорость реакции в одном из направлений j
больше, чем в другом. Сохранение данного уровня стационар- j
нести во времени зависит от длительности удержания нового 1
баланса прихода и расхода. J
Так, сохранение постоянства веса тела возможно при сба 1
лансированном приходе и расходе вещества и энергии в орга- 1
низме. Однако при желании снизить вес до определенного |
уровня, т. е. создать новый уровень стационарного состояния, 1
следует изменить соотношение между поступлением и расходом ]
вещества и энергии. При этом необходимо различать устойчи- I
вые и неустойчивые стационарные состояния. В качестве при- 3
мера биологических моделей относительно устойчивого стацио- я
84 I
।
нарного состояния может служить мембранный потенциал по-
коя, а неустойчивого — потенциал действия.
Сущность же проявления основных жизненных свойств та-
кой открытой системы, как организм, заключается в том, что
под влиянием внешней среды наблюдается постоянный переход
одного стационарного состояния в другое, обеспечивающее уста-
новление оптимальных для данных условий отношений со сре-
дой. Он осуществляется по различным переходным кривым,
представленным в упрощенном виде на рис. 13.
V
Рис. 13. Основные типы перехода
одного стационарного
состояния А в другое В:
I — переход с избыточным отклоне-
нием (типичная кривая при возбужде-
нии); II — переход по типу «экспонен-
циального приближения» (наиболее эко-
номный переход); III — переход с
«ложным стартом».
Возбуждение, возникновение потенциала действия, сокра-
щение мышцы и другие подобные процессы являются приме-
рами перехода одних стационарных уровней в другие. Способ-
ность открытых систем сохранять допустимо возможное ста-
ционарное состояние или переходить с одного уровня на другой
предполагает одновременно и вещественно-термодинамическую
иеравновесность открытых систем относительно среды. Однако
описанные механизмы поддержания и изменения уровня того
или иного стационарного состояния только объясняют само су-
ществование стационарного состояния, но не позволяют выя-
вить факторы, его определяющие, т. е. факторы, обусловливаю-
щие иеравновесность стационарных состояний открытых систем
с окружающей средой. Удовлетворительный ответ на этот во-
прос дает теория Онзаге.ра (1931; 1945).
Неравновесное состояние, по Онзагеру, определяется нали-
чием сил и вызываемых этими силами потоков. Силы — это все
2* ЗБ
виды существующих в природе градиентов и потенциалов (диф-
фузионные и температурные градиенты, химические и электри-
ческие потенциалы и Др.). Для биологических систем основной
силой является химическая активность, т. е. активность метабо-
лических процессов. Силы вызывают появление потоков. Наи-
более важный для биологических систем — диффузионный по-
ток вещества, вызываемый протеканием химических реакций.
В результате того или иного химического процесса концентра-
ция веществ в месте произошедшей реакции уменьшается или
увеличивается. В обоих случаях возникает диффузионный (кон-
центрационный) градиент, благодаря которому данное вещество
поступает из внешней среды в биосистему или наоборот. При-
мером может быть возникновение и поддержание потенциала
покоя, местного потенциала и потенциала действия. Химические
реакции (экзо- и эндотермические, экзер- и эндергеннческие)
создают в организме не только диффузионные потоки, но и по-
токи тепла и энергии.
Итак, благодаря силам и потокам создаются условия не
только для возникновения и поддержания того или иного уровня
стационарного состояния, но и для неравновесного распреде-
ления вещества и температуры между биологической системой
и средой.
Подобно тому как понятие «стационарное состояние» пред-
полагало существование неравновесного состояния, так нерав-
новесное состояние предполагает существование у открытых
систем свойства сохранять относительно постоянный состав и
энергетический потенциал. Специфика этого свойства в свое
время была определена принципом Ле Шателье относительно
термодинамического равновесия: «Если система находится в
состоянии равновесия, то при действии на нее сил, вызывающих
нарушение равновесия, система приходит в такое состояние, в
котором эффект внешнего воздействия ослабляется». Так, если
химическая реакция протекает с уменьшением числа молей (а
соответственно и объема), то давление благоприятствует пря-
мой реакции и препятствует обратной (например, Кг+ЗНгч*
s±2NHs, где уменьшение объема обусловлено тем, что вместо
4 молей исходных веществ образуется только 2 моля аммиака).
Впоследствии принцип Ле Шателье был применен для от-
крытых систем в условиях стационарного состояния. В этом
случае он обрел следующее содержание: всякое превращение
36
(процесс), приводящее к нарушению стационарного состояния
(динамического равновесия), тормозит причину возмущения.
Необходимо помнить, что все процессы носят необратимый
характер. Существующая же обратимость явлений относительна
уже хотя бы потому, что процессы протекают во времени, а вре-
мя необратимо, однонаправленно, асимметрично. Это имеет
самое непосредственное отношение не только к индивидуальным
биологическим системам (отдельные особи), но и к более слож-
ным (популяция, вид и т. д.). Однонаправленность программы
их существования жестко предопределена: возникновение (за-
рождение) — развитие — гибель.
Для биологических систем особое значение имеет взаи-
модействие протекающих в них необратимых процессов. Однако
поскольку в биологической системе, как и в любой другой от-
крытой системе, процесс есть производное потоков I, то речь
должна идти о взаимодействии потоков. Взаимодействие двух
потоков в открытой системе описывается двумя феноменологи-
ческими уравнениями:
где Л-!, Z2.2 — коэффициенты с однозначными индексами,
характеризующие отдельный процесс (диффузия, теплопровод-
ность, электропроводность и др.); Л.2- Д-i ~~ коэффициенты
с неоднозначными индексами, отражающие взаимодействие не-
обратимых процессов (термодиффузня, электроосмос, термо-
электричество и др.). Эти коэффициенты называются феноме-
нологическими. Онзагер (1931) установил, что в области дей-
ствия феноменологических уравнений коэффициенты, отражаю-
щие взаимодействие процессов, равны между собой 1ш=1ъ
(i, A=l, ..., л). Это равенство (равенство взаимоотношений
Онзагера) утверждает, что если поток необратимого
процесса X испытывает влияние силы необратимого процесса
К(хь), то и поток процесса К также испытывает влияние силы X
посредством того же самого коэффициента.
На основании этого принципа Н. Пригожин сформулировал
и доказал главное свойство стационарного состояния — в ста-
ционарной неравновесной системе скорость возрастания энтро-
37
JO
nuu (или скорость рассеяния свободной энергии) ^21, обу-
dt
словленная потоками необратимых процессов, имеет по-
ложительное и минимальное (но не нулевое) из возможных
значение.
При достаточной стройности и логической строгости изло-
женного следует, однако, помнить, что принцип Ле Шателье
буквально гласит: «Всякое изменение одного из факторов рав-
новесия видоизменяет систему в том направлении, в котором
рассматриваемый фактор испытывает изменение, противопо-
ложное первоначальному». Это значит, что «принцип Ле Ша-
тслье относится к системам, находящимся в равновесии, и из-
менение состояния, т. е. реакции системы, которую требует
принцип при изменении окружающей среды, ведет именно к
ожидаемому при данной окружающей среде равновесию, иначе
говоря, принцип указывает, при каком именно направлении ре-
акции при данной новой окружающей среде наступит равнове-
сие» (Бауэр, 1935). Короче, изменение состояния системы при
измеиепии окружающей среды приводит к вещественно-термо-
динамическому равновесию между ними. Между тем, как уже
было сказано, все открытые системы, и тем более биологиче-
ские, стационарны и неравновесны с окружающей средой. Фи-
гурально выражаясь, состояния возможного в земных услови-
ях вещественно-термодинамического равновесия, т. е. максиму-
ма энтропии, биологические системы могут достичь после
гибели, да и то не сразу, а только в процессе окончательной
минерализации и растворения в среде.
Таким образом, распространение принципа Ле Шателье на
открытые стационарные системы и особенно на биологические,
в которых все процессы необратимы, недостаточно корректно.
Формально «спасательным кругом», позволяющим применять
принцип Ле Шателье ко всем открытым стационарным систе-
мам, является уже рассмотренная нами теорема Пригожина.
Однако несмотря на меньшую по сравнению с теоремой Приго-
жина математическую оснащенность, в сущностном отношении
правильно было бы обратиться к открытому замечательным и
незаслуженно забытым мыслителем и исследователем Эрвином
Симоновичем Бауэром всеобщему закону биологии, который
утверждает, что «все и только живые системы никогда не бы-
вают в равновесии и исполняют за счет своей свободной энергии
Зв
Ш 7 ,
постоянную работу против равновесия, требуемого законами
физики и химии при существующих внешних условиях». Этот
закон Бауэр обозначил как «принцип устойчивого неравновесия
живых систем». «Этот принцип имеет ясный термодинамический
смысл: так же как устойчивое равновесие, характеризуется тем,
что, будучи нарушено, всегда наступает вновь (или устанавли-
вается на новом уровне в соответствии с принципом Ле Ша-
телье.— А. Л.), так и в организмах, в живой материи, нерав-
новесное состояние сохраняется постоянно и обладает всеми
признаками устойчивости... Мы утверждаем, что живая система
всегда превращает свою свободную энергию в работу против
ожидаемого равновесия... Прежде всего из этого выражения
следует, что живая система вообще не исполняет никакой ра-
боты, кроме работы против равновесия» (Бауэр, 1935).
Фундаментальность положения Бауэра столь значима в об-
щетеоретическом, философско-методологическом и прикладном
отношениях, что правильнее было бы его назвать именем авто-
ра — принципом Бауэра, принципом устойчивого неравновесия
живых систем.
Между принципами Ле Шателье н Бауэра имеется внешнее
сходство, «заключающееся в том, что оба они содержат обшее
указание, в каком направлении будет происходить реакция, т. е.
изменение состояния системы при каком-либо изменении со-
стояния окружающей среды. Кроме того, оба принципа говорят,
что изменение состояния системы направлено в некотором
смысле против изменения состояния окружающей среды». Вмес-
те с тем «физический смысл в обоих случаях, однако, совер-
шенно различный и не имеет друг к другу никакого отношения»
(Бауэр, 1935). Смешение принципа Ле Шателье с принципом
Бауэра означает непонимание принципа устойчивого неравно-
весия живых систем.
Кардинальное различие между ними состоит в том, что
принцип Ле Шателье относится к системам, находящимся в рав-
новесии, принцип Бауэра — к живым системам, не находящимся
в равновесии. В первом случае изменение состояния системы
при изменении окружающей среды приводит к их равновесию.
Во втором — живая система отвечает работой, направленной
против тенденции к равновесию, т. с. против тех изменений,
которые могли бы произойти согласно принципу, сформулиро-
ванному Ле Шателье.
89
Однако равнозначно ли изложенное отказу от приложения
принципа Ле Шателье к живым системам вообще? Разумеется,
нет! Вот что по этому поводу говорит Бауэр (1935): «Принцип
Ле Шателье является собственно следствием второго начала
термодинамики и может быть из него выведен. Он может быть
применен и для живых систем, как и второе начало, и не про-
тиворечит нашему принципу живых систем. Наш принцип
исходит из действительности второго начала и принципа Ле
Шателье (выделено нами.— А. Л.), и именно при их помощи
возможно заранее сказать, какие изменения состояния долж-
ны были произойти и при каком состоянии должно было на-
ступить равновесие, следовательно, какая работа против него
должна быть исполнена живой системой и за счет каких при-
способлений и механизмов, существующих в ней. Ко второму
началу и к принципу Ле Шателье паш принцип непосредствен-
ного отношения не имеет (выделено нами.— А. Л.); он уже
предполагает их действительность и в сущности дает некото-
рые указания относительно условий структуры и приспособлений
живых систем».
Исходя из принципа Бауэра, становится понятным одно из
наиболее основных свойств живого — способность самопроиз-
вольно, активно осуществлять самые разнообразные приспосо-
бительные реакции на любые возмущающие воздействия с ми-
нимально возможными в этих условиях изменениями своего
устойчивого неравновесия, т. е. гомеостаза.
Убедиться в справедливости этого положения можно на
примере динамики экономизации энергетических трат при вы-
полнении физических упражнений по мере совершенствования
мастерства спортсмена. Эта экономизация реализуется не толь-
ко за счет физических качеств приспособления и совершенст-
вования биохимических процессов и различных функций орга-
низма, но и технико-тактического мастерства. Это обстоятель-
ство имеет чрезвычайное значение для того свойства
стационарного состояния, которое можно назвать
аутостабилизацией. Аутостабилизация стационарного состояния
лежит в основе обратной связи, обязательной для всех
существующих в природе форм регуляции и саморегуляции.
Она обеспечивает как стационарное, так и вещественное и
термодинамическое неравновесное состояние открытых систем,
т. е. гомеостаз. Например, концентрация тех или иных веществ
40
в открытых системах (в данном случае в живых организмах,
см. рис. 10) существенно отличается от таковой в окружающей
среде (в том числе и в поглощаемой пище). Причем концентра-
ция веществ в организме достаточно постоянна, несмотря на
весьма существенные изменения ее в окружающей среде. Та-
кое свойство называется эквифинальностыо. Удивительным
примером эквифинальности открытых неравновесных живых
систем является факт довольно длительного сохранения в них
нормального содержания и соотношения различных химических
веществ не только в условиях ограничения или избытка пита-
ния, но и при полном голодании. Нарушение состава химичес-
ких веществ развивается сравнительно медленно, а для белко-
вого обмена существенное нарушение наступает практически
накануне гибели биологической системы. Следовательно, для
того чтобы открытая система нарушила свою эквифинальность,
вещественно-термодинамическую неравновеспость, асиммет-
ричность со средой, недостаточно просто воздействия внешних
сил. Необходимо, чтобы эти силы достигли такого критического
уровня, при котором система не могла бы сопротивляться воз-
действиям, т. е. утратила бы свойство неравповесности, экви-
финалыюсти.
На основании изложенного можно определить важнейшее
свойство неравновесных систем — пороговость. Оно заключа-
ется в том, что для изменения состояния неравновесное™ си-
стемы в отличие от стационарного состояния необходима крити-
ческая сила — порог воздействия. Под влиянием порогового и
тем более сверхпорогового воздействия неравновесное состоя-
ние биосистемы может быть сведено к нулю, что означает ги-
бель ее; может также возникнуть и качественно новое, иногда
прямо противоположное, неравновесное состояние. Например,
потенциал действия, при котором соотношение ионов К+ и
Na+ на поверхностях мембраны становится обратным соотно-
шению, характерному для условий потенциала покоя. По
существу потенциал действия — это трансформированный под
влиянием порогового раздражения потенциал покоя. Смена
состояний неравновесное™ ионов имеет пространственно-вре-
менные промежуточные стадии. Для приведенного примера
такой стадией будет местный потенциал.
Противоречит ли свойство пороговое™ неравновесных со-
стояний свойствам беспороговости и индискретиости стацио-
41
парных состояний открытых систем? Разумеется, пет. Эти свой-
ства тесно взаимосвязаны и взаимопереходящи (потенциал по-
коя — местный потенциал — потенциал действия — местный
потенциал — потенциал покоя). Между ними имеются качест-
венно-количественные различия в динамике развития какого-
либо процесса, возникающего или существующего во взаимо-
отношениях открытой системы со средой.
Резюмируя, приходим к следующим выводам.
1. Все закрытые системы, кроме вселенной, идеальны.
2. Все реальные системы, кроме вселенной, открытые.
3. Все открытые системы стационарны.
4. Все открытые системы неравновесны.
5. Для каждой открытой системы присуща специфическая
стационарность и вещественно термодинамическая неравно
вескость, отличающая ее от всех окружающих, в том числе н
от себе подобных, систем.
6. Стационарные состояния, которые способна приобретать
открытая система, характеризуют возможности ее адаптировать
свое поведение во взаимоотношениях со средой.
7. Во всех случаях открытая стационарная неравновесная
система стремится к минимально возможному приращению
энтропии.
Противоречат ли свойства неравновесное™ и стационарно-
сти открытых систем второму закону термодинамики, требую-
щему всеобщего уравновешивания и максимума энтропии?
Формально противоречат, так как открытые неравновесные
стационарные системы и особенно биологические ведут себя
внешне явно аптиэнтронийпо. Следует ли из этого, что живые
организмы не накапливают энтропию, а напротив, негэнтро-
пичны, т. е. являются накопителями, а возможно и продуцента-
ми свободной энергии? Нет, не следует: как было отмечено,
живые организмы постоянно находятся в «долгу» у природы,
поддерживая свой высокий энергетический потенциал, и это
заставляет их сопротивляться фатальной силе второго закона
термодинамики. Однако сопротивляться — не значит не подчи-
няться. Живые организмы не могут уйти из сферы влияния
второго закона термодинамики. Они лишь в состоянии про-
длить или сократить свое существование. В этом отношении
огромную роль на уровне человека играют социально-экономи-
ческие факторы, и среди них физическая культура и спорт
42
занимают нс последнее место. «Индивидуальная жизнь, ее
зарождение, детство, юность, зрелость — это как бы перерывы
в непрерывности действия принципа энтропии, но старость и
смерть все же отражают именно этот принцип, его индивиду-
альный вариант. Старость — это рассеивание или нейтрализа-
ция энергии, невозможность метаболических или трансформи-
рованных процессов на высоком биологическом уровне. Ста-
рость — это начало процесса индивидуальной энтропии,
завершение этого процесса — биохимические превращения и
молекулярный распад — является необходимой прелюдией
развития новой жизни» (И. В. Давыдовский, 1966). Как, в
каком темпе и качестве совершится эта непременная цепь
событий для человека — дело разума самого человека!
Итак, с позиции физики, в частности с позиции второго за-
кона термодинамики, все живое действительно ие только
подчиняется, но и сопротивляется этому закону. Чем дольше
сохраняется такая способность, тем более продолжительна п
относительно «независима» жизнь животных и растений в не-
благоприятных условиях. Таким образом, живые существа
действительно подчиняются всеобщему закону биологии —
принципу устойчивого неравновесия живых систем, основным
выражением которого является способность живых объектов
сохранять свой гомеостаз.
Очерк 3. ГОМЕОСТАЗ. ОБЩИЕ ПОЛОЖЕНИЯ
В 1857 году Клод Бернар впервые высказал и обосновал
важную идею о внутренней среде организма и постоянстве ее
как необходимом условии жизни. (Позже, в 1939 году В. Кен-
нон это постоянство внутренней среды обозначил термином
«гомеостаз».)
Почти одновременно с Бернаром в лекции «О значении так
называемых растительных актов в животной жизни» И. М. Се-
ченов (1861) указывал: «Говоря же вообще, совокупное дейст-
вие всех растительных актов в организме дает в результате
Для зрелого человека количественный и качественный status guo
всех частей тела; для ребенка — прибывание массы тела с тон
же неизменностью его физических свойств». И. М. Сеченов ви-
дел в status gup (т. е. в гомеостазе.— Л. Л.) «...сущность и
48
л.
значение в жизни животного тела всех растительных прогрес-
сов, взятых вместе».
В свою очередь Бернар утверждал : «Все физиологические
механизмы, сколь бы различны они ни были, имеют только
одну цель — сохранение постоянства условий жизни во внут-
ренней фазе». Это выражение понималось Бернаром не как аб-
солютное, а как относительное, поскольку в противном случае
ие нужны были бы «физиологические механизмы», обеспечи-
вающие достижение одной цели — сохранение постоянства
внутренней среды. Сеченов же утверждает: «Регуляция возмож-
на только между известными пределами».
Нам представляется, что трактовка явления поддержания
постоянства внутренней среды Бернаром и Сеченовым выража-
ет идею саморегуляции и гомеостаза. У многих исследователей
в результате буквального перевода термина сформировалось
мнение, что гомеостаз — это состояние абсолютной стабильно-
сти показателей внутренней среды организма. Крайнее выраже-
ние такая точка зрения получила у Р. Гольдекра (1960), счи-
тающего возможным существование в организме одного из
двух альтернативных состояний — «состояния поддержания
ритма» или «состояния гомеостазиса». Но если организм может
быть в двух состояниях, то вполне естественно полагать, что
это осуществляется путем их взаимопревращения. Однако чет-
кой теоретически обоснованной границы перехода обнаружить
не удается, вследствие чего Гольдекру приходится прибегать к
обозначению удобной в гносеологическом отношении точки от-
счета: с одной стороны, состояние расценивается как гомеоста-
тическое, с другой — ритмическое, осцилляторное. Он считает,
что, когда «сдвиг фаз» увеличивается от 0 до 2л, ритмические
колебания должны исчезнуть.
Несоответствие буквального содержания термина «гомео-
стаз» реально существующему явлению приводит некоторых
исследователей к мысли о необходимости вообще отказаться
от этого термина, заменив его другими, например «гомеокинез»
(постоянство движения), «гомеорезис» (постоянство течения).
Гомеокинез неадекватно характеризует рассматриваемое явле-
ние сохранения относительного постоянства внутренней среды.
Сущность его раскрывается только с одной стороны за счет
различных форм движения. Параметрическая же характери-
стика этим термином ие отражается.
44
Термин «гомеорез», предложенный У. Уоддингтоном, более
близок к истине. В отличие от термина «гомеостаз», под кото-
рым Уоддингтон понимает поддержание «постоянства некото-
рых параметров физиологических процессов», гомеорез рас-
сматривается нм шире и предусматривает поддержание на
постоянном уровне не какого-то одного параметра, а «протяжен-
ного во времени процесса изменения». В это понятие
вкладывается представление о неизбежности развития
(процесса, потока) по определенному и достаточно жестко
запрограммированному пути (каналу), несмотря на помехи,
создаваемые внешними условиями и генетической средой.
В связи с этим Уоддингтоном введен термин «канализация»,
основное содержание которого акцентируется на конечном
продукте процесса во времени.
Многие авторы предпочитают пользоваться термином «го-
меостаз развития». Данный термин также предусматривает
параметр времени. Этот параметр предполагает и формула
Бернара: «Постоянство внутренней среды есть основа свободной
жизни», т. е. основа процесса, имеющего временную протяжен-
ность — жизнь. Кеннон считал, что явление поддержания по-
стоянства внутренней среды обусловливается двумя факторами.
Первый сводится к резервированию и мобилизации веществ;
второй — к регуляции скорости (ускорение или замедление) и
интенсивности (уменьшение или увеличение) процессов. Таким
образом, понятие «гомеостаз», по Кеннону, со всей очевидно-
стью включает параметр времени. Поэтому понятие «гомеорез»,
введенное для характеристики устойчивости процесса развития,
соответствует понятию «гомеостаза», устойчиво изменяющегося
во времени.
Следует помнить, что для биологических систем существуют
два вида стационарных состояний: в каждый данный момент
времени и конечное, к которому эта система, согласно второму
закону термодинамики, неизбежно приближается в процессе
развития, роста и старения. В обоих случаях стремление си-
стемы может быть направлено как к стационарному состоянию,
так и на уклонение от него. Однако смысл и механизм подоб-
ного поведения системы для каждого из двух стационарных
состояний будет разным.
Подробно эту систему рассмотрел Зотип (1974). Уклоне-
ние— приближенно организмов к конечному стационарному
45
состоянию получило название конститутивного, а временное
уклонение к текущему стационарному состоянию — индуцибиль-
ного уклонения. Индуцибильное уклонение живого организма
к новому стационарному состоянию может происходить под
влиянием возмущающих воздействий без возвращения доста-
точно длительное время к исходному состоянию. Такой процесс
назван ипдуцибильно-адаптивным. Если же живая система
быстро возвращается к прежнему состоянию, процесс будет
индуцибильно-импульсным*. По мнению Зотина, примером ии-
дуцибильно-адаптивного уклонения является переход организ-
мов на новый уровень теплопродукции и дыхания при изменении
внешней среды (температуры, газового или солевого состава,
давления и т. д.) или прн длительных изменениях внутренних
факторов (патологических, гормональных и др.). Примером
иидуцибильно-импульсного уклонения может быть любое уси-
ление теплопродукции на кратковременные воздействия среды.
По существу это интерпретация известной теории Приго-
жина— Виам, которая утверждает, что в процессе развития
и жизни животные в каждый данный момент находятся в ста-
ционарном состоянии, характеризующемся минимальной поте-
рей энергии (диссипация) при непрерывном эволюционировании
в направлении снижения ее. Значит развитие, рост и старение
организма представляют собой абсолютизированный процесс
приближения к конечному стационарному состоянию.
Таким образом, ортодоксальное понимание стационарности
(гомеостатичности) как стабильного состояния не выдерживает
сколько-нибудь серьезной критики. Понятия «гомеорез» и
«гомеостаз» не только имеют точки соприкосновения, но и
взаимозаменяемы, так как гомеорез характеризует устойчивость
процесса развития, что соответствует гомеостазу, явлению, ус-
тойчиво изменяющемуся во времени.
* Понятия «индуцибильно-адаптивиый» и «индуцибильно-им-
пульсный» следует рассматривать в относительном, а не абсолютном
смысле, так как идеального возвращения живой системы к исходно-
му состоянию не может быть в силу: а) необратимости любых про-
цессов, тем более в живых системах; б) практической невероятности
идеального повторения обстоятельств, вызвавших даииое изменение
стационарного состояния; в) скорее эмоционального, чем рациональ-
ного восприятия понятия длительности времени в отношении наблю-
даемых процессов.
46
Поддержание гомеостаза как частного (для отдельных пе-
ременных), так и суммарного (для целостной жилой системы)
обеспечивается множественностью причинно-следственных отно-
шений между различными процессами, совершающимися в
системе. Конечным результатом этих процессов является
стремление возвратить систему в лоно предопределенной для
нее траектории (канал) развития, носящего название креод.
Такое толкование гомеостаза позволяет с полным правом рас-
пространить и па неге соображения Уоддингтона относительно
гомеорезиса, траектория притяжения которого параллельна
оси времени. «Во многих креодах, наблюдаемых нами у реаль-
ных животных,— пишет Уоддингтон (1970),— естественный
отбор контролирует как величину, так и степень стабилизации.
Например, Робертсон в опытах по селекции на размеры тела
у дрозофилы обнаружил существование буферности в отноше-
нии окончательных размеров тела» (выделено нами.— А. Л.),
т. е. контролирует относительную гомеостатичность. Еще при-
мер: «Введение в относительно нормальный генотип какого-
нибудь мутанта с сильным эффектом... часто приводит к обра-
зованию фенотипа не только в высшей степени аномального, но
и весьма изменчивого, т. е. дестабилизирует креод. Однако
это, как мне кажется,— продолжает Уоддингтон (1970),— не
исключает возможности, что существование креода того или
иного рода необходимо... Если какие-либо взаимодействия меж-
ду компонентами (какой-либо системы.— А. Л.) нелинейны...,
то появление каких-то креодов (забуференности — гомеостатич-
нссти.— А. Л.) неизбежно». Другой механизм забуференности
реализуется обратной связью, обеспечивающей возвращение
того или иного процесса, отклоненного возмущающим воздейст-
вием, к возможно близкому исходному состоянию. Такое по-
нимание гомеостаза и будет отправным для последующих рас-
суждений.
Закон сохранения и превращения материи и энергии явля-
ется основополагающим и абсолютным, хотя «уже имеющиеся
скудные факты говорят о том, что эти данные могут привести к
противоречию с законом сохранения энергии (и вещества) в его
современной форме, ограниченной известными нам формами
энергии» (В. А. Амбарцумян, 1962). Имеется в виду противоре-
чие не вообще со вселенским законом сохранения, а только в
рамках известных его форм. Таким образом, классическое по-
47
а —*
нимание закона сохранения и превращения материн и энергии
в сущности не меняется и должно рассматриваться как тенден-
ция сохранения, неразрывно связанная с тенденцией измене
ния, т. е. характеризовать не только сохранение, но и превра-
щение, изменение.
Даже если принять океаническое или любое иное земное
происхождение жизни как абсолютное, то уже в изначальный
момент она обладала какими-то механизмами, поддерживаю-
щими ее неравновесное (гомеостатическое) состояние относи-
тельно среды. Если же предположить, что жизнь есть явление
космическое и представляет собой один из способов существо-
вания материи с безграничными возможностями миграции во
вселенной, а следовательно, и па Землю, то проблема выбора
способа существования ее здесь (как и вообще во вселенной)
не может быть альтернативной (следовать за изменениями со-
левого состава океана, «перестраивая закрепленную в ряде
поколений адаптацию, или создать органы, дающие возмож-
ность пройти через геологические эпохи, не изменив привычных
условий существования клеток организма». А. Г. Гинецинский,
1963). Вряд ли во вселенной существует проблема «или, или»,
более конструктивно и реально «и то, и другое». Каким бы
способом пи возникла (появилась) жизнь на Земле, она неми-
нуемо должна была воспользоваться и действительно восполь-
зовалась принципом «и то, и другое», т. е. и консервативным
принципом сохранения и принципом адаптивного структурно-
функционального эволюционирования. Оба принципа нашли
отражение в эволюции животного мира не врозь, а в диалекти-
ческом единстве, двойственности и противоречивости всех явле-
ний и процессов. Именно поэтому они не только выявляются
друг через друга, но перекрывают и совершенствуют друг дру-
га во всех формах проявления жизни. С этой позиции гомеостаз
следует рассматривать более широко — как состояние-процесс,
позволяющее любой материальной единице возможно дольше
сохранять свое относительное (возможно, минимально изме-
няемое) структурно-функциональное постоянство.
Гомеостаз — по существу частный случай закона сохране-
ния материи и энергии. Следовательно, под гомеостазом надо
понимать единственно возможный способ существования лю-
бых материальных систем, характеризующихся нсравновесно-
стью (иеществешюй и термодинамической эквифиналыюстыо)
48
с окружающей средой, неограниченным числом стационарных
состояний различных параметров и переходом их из одного
состояния в другое во времени и в пределах возможных (со-
храняющих достаточно высокую специфичность иеравновесно-
сти), как правило, нелинейных изменений существенных пере-
менных.
Таким образом, даже элементарный анализ термодинамиче-
ских взаимоотношений в системе организм — среда убеждает
в универсальной применимости этой области знаний к явлениям
в живой природе.
Принципиальные механизмы поддержания гомеостаза (точ-
нее, адаптивного гомеостаза) в живых и неживых объектах,
вероятно, сходны, но не всегда тождественны. Мера же исполь-
зования этих механизмов разная. В данном случае речь идет
в первую очередь о мере активного использования структурно-
функцно'нальных возможностей живых и неживых объектов.
Подобная активность, очевидно, может проявляться во многих
видах, однако из них следует выделить два как наиболее
существенные: 1) структурно-функциональная (пассивная) со-
противляемость возмущающим воздействиям, заложенная в са-
мой материальной природе объекта, и 2) структурно-функцио-
нальная (активная) сопротивляемость возмущающим воздей-
ствиям, определяющаяся степенью активности таких его свойств,
как самоорганизация, управление, регулирование, репродукция,
сохранение и передача наследственной информации, воспроиз-
ведение и обновление составляющих его элементов в результате
переработки и усвоения веществ, поступающих извне.
Разумеется, обе активности не взаимоисключают, но и не
взаимозаменяют друг друга. Они, сосуществуя в единстве, про-
являются степенью выраженности. В объектах неживой приро-
ды преобладает первый, а живой — второй вид активности
механизмов гомеостаза.
Таким образом, под системой следует понимать такую це-
лостность, которая способна удерживать гомеостатическое
единство составляющих ее частей (элементов) и обладает ин-
тегративными качествами, не свойственными этим элементам.
Элементом же является любая часть, не обладающая интегра-
тивными свойствами той системы, в которую опа входит, но в
свою очередь способная представлять собой какую то новую
систему.
49
В неорганической природе подобное представление о систе-
ме, видимо, может распространяться на любую материальную
субъединицу вплоть до атомов и элементарных частиц.
Данное понятие системы приложимо и к органической при-
роде, в том числе и к биологическим объектам. Однако для ор-
ганической природы минимальной субъединицей, которую при
сегодняшнем нашем знании органического мира можно рас-
сматривать как систему, следует считать молекулу, в биологи-
ческом же мире—минимальную совокупность молекул, отве-
чающую требованиям активного поддержания, сохранения и
воспроизведения ее. Подобной совокупностью, т. е. наиболее
элементарным гомеостатическим уровнем биологических си-
стем, следует считать организменный, независимо от его струк-
турно-функциональной сложности и эволюционного положения,
ибо только организм как реальная биологическая целостность
способен к активным взаимодействиям со средой для поддер-
жания и сохранения индивидуальности — гомеостатичности (не-
равновесное™). Основой этого уровня гомеостаза является
генетически зафиксированная специфика обмена веществ или
уже — контроль за генетическим постоянством — геностазис
клеток организма или клетки (если организм одноклеточный
или доклеточный, например вирус).
Таким образом, элементом биологической системы является
любая ее часть, лишенная не только интегративных свойств
системы, по и возможности поддерживать вне ес свое гомео-
статическое состояние и индивидуальное существование.
Эта формулировка закономерна в отношении и естествепно-
временно переживающего органа или ткани (в условиях кли-
нической смерти) н в условиях искусственно поддерживаемой
культуры тканей. Однако она не распространяется на часть,
отделенную от биологической системы, но способную к само-
стоятельной регенерации целостности даже формально дефект-
ной, но адаптивно-гомеостатически активной, что наблюдается,
например, у кишечнополостных, червей и др.
Элемент биологической системы является ее составной ча-
стью только до тех пор, пока играет в ней определенную роль.
Генетически запрограммированное (например, смена молочных
зубов и др.) или возникшее под влиянием различных факторов
выпадение элемента, а также любое изменение его, в том числе
фенотипическое и генотипическое, влечет за собой как «поло-
50
жительные», так и «отрицательные» отклонения в деятельности
всей системы. Во всех случаях, сохраняя свою гомеостатиче-
скую сущность и принципиальную специфику, биосистема бу-
дет находиться на одном из возможных уровней стационарно-
сти.
Итак, адаптивный гомеостаз — это способность организма
удерживать постоянство внутренней среды, или шире — поддер-
живать оптимальные взаимоотношения с окружающим внеш-
ним миром. Это единственный способ существования любой
биологической системы независимо от ее эволюционного совер-
шенства. Справедливость данного положения подтверждается
тем, что только путем ограничения зависимости от окружающей
среды (именно ограничения, а не полной независимости) жи-
вые существа относительно стабильно фиксируют и передают
потомству наследственную информацию, сохраняя тем самым
данный вид.
Одни организмы, следуя за изменениями среды, перестраи-
вают закрепленную в ряде поколений адаптацию и приобрета-
ют приспособления, позволяющие перейти на максимально
низкий уровень функциональной активности вплоть до анабио-
за, вернее, гипобиоза. Несмотря на весьма широкое распростра-
нение такого способа приспособления к изменившимся усло-
виям среди живых организмов, в эволюционном отношении ои
недостаточно надежен, так как не гарантирует 100%-ную вы-
живаемость всех впавших в гипобиоз особей. Обстоятельства
могут сложиться так, что восстановление благоприятных жиз-
ненных условий произойдет после утраты организмом
способности вернуться к активной жизнедеятельности.
Перестройка адаптаций идет нс далее стабилизации генетически
зафиксированной специфики обмена веществ, характеризую-
щегося предельно минимальной интенсивностью. Другого не
дано! До тех пор пока в данных условиях эта специфика обме-
на сохраняется, зависимость живого существа от окружающей
среды снижена, причем целостность и жизнеспособность его не
утрачивается.
Нарушение специфики обмена неминуемо приводит к распа-
ду системы. Образование же новой биологической системы в
этом случае ни теоретически, ни практически представить не-
возможно. Следовательно, распад системы может завершиться
только ее термодинамическим уравновешиванием со средой.
51
Между тем не вызывает сомнения справедливость вышеупомя-
нутого закона Бауэра (1935): «Все и только живые системы
никогда не бывают в равновесии и исполняют за счет своей
свободной энергии постоянно работу против равновесия, тре-
буемого законами физики и химии при существующих внеш-
них условиях».
Несмотря на то что перестройка адаптации в результате
«стабильно» изменившихся условий внешней среды, казалось
бы, не отражается на биохимико-генетическом гомеостазе, тем
не менее она не проходит бесследно для него. Это лежит в
основе эволюции вначале иа уровне фено-, а затем и геноти-
па*, который представляет собой так называемый пассивный
способ приспособления к окружающей среде. У других пред-
ставителей живой природы под действием неблагоприятной
среды формируются приспособления, позволяющие искать бо-
лее подходящие для сохранения и поддержания гомеостаза
условия существования. Поиск осуществим только в том слу-
чае, если живая система приобретает способность перемещать-
ся в пространстве. Различают трн вида подобного приспособле-
ния: 1) пассивное перемещение—перенос воздушными и
водными потоками, активно подвижными существами малопо-
движных или неподвижных форм (семена, паразиты и др.);
2) активное перемещение (индивидуальные и массовые мигра-
ции); 3) смешанное (пассивное и активное) перемещение в
пространстве.
Естественно, что пассивное перемещение менее надежно га-
рантирует выживание, т. е. сохранение гомеостаза особи. Ви-
димо, потому большинство пассивно перемещающихся форм
живых существ обладает высокой репродуктивной способностью,
имеет промежуточные стадии развития и может длительно на-
ходиться в состоянии различной степени гипобиоза.
Активно подвижные формы благодаря способности произ-
вольно перемещаться в пространстве обладают более высокой
выживаемостью — гомеостатичностью. В данном случае даже
можно говорить о еще большем снижении зависимости живых
организмов от условий окружающей среды. Одиако это лишь
* Генотип — фактор, лимитирующий фенотипическую изменчи-
вость, однако воздействия внешней среды, вызывающие генетически
фиксируемые мутации, способствуют эволюции генотипа.
52
формально. В действительности же она (зависимость) возрас-
тает, в чем можно убедиться на примере перелетных птиц. По
существу смысл преодоления огромных расстояний (до десяти
и более тысяч километров в один конец, например с севера иа
юг) перелетной птицей заключается в том, чтобы в течение
года находиться в условиях относительно стабильного темпера-
турного режима, т. е. в условиях зафиксированного эволюцией
термодинамического баланса со средой. Разумеется, здесь не
исключаются и другие жизнеобеспечивающие факторы — пи-
тание и др. Естественно, что поведение живых существ, направ-
ленное на пребывание в течение всей жизни в максимально
стабильных условиях, снижает возможность формирования
фенотипических изменений. Это в свою очередь в определенной
мере замедляет и генотипическую эволюцию данного вида.
Коротко говоря, этот способ приспособления делает более
жесткой зависимость гомеостаза внутренней среды живых орга-
низмов от гомеостаза окружающих условий существования.
Формируется замкнутый контур взаимоотношений: гомеостаз
среды гомеостаз организма — гомеостаз среды. Однако и в
этом случае неизбежны, хотя и менее выражены, эффекты дав-
ления отбора, фено- и генотипические изменения В1гутренней
природы гомеостаза.
Наконец, у многих живых существ развились механизмы
активного вмешательства в окружающий мир для радикального
преобразования его в соответствии со своими гомеостатическими
потребностями. Высшего совершенства подобная форма при-
способления достигла у человека в виде его сложнокоордини-
рованной двигательной активности, связанной с трудовой де-
ятельностью, которая является прямым следствием и причиной
дальнейшего совершенствования двигательной активности.
Здесь движение выступает уже не просто как средство переме-
щений в пространстве, а как тончайший механизм реализации
всех форм трудовой, творческой, преобразующей деятельности.
Таким образом, движение во всем многообразии своего выра-
жения становится наиболее совершенным способом приспособ-
ления и активного воздействия на окружающую среду. Этот
способ приспособления по праву может быть назван способом
активного преобразования.
По существу все специализированные органеллы и органы,
в том числе органы кровообращения, дыхания, пищеварения и
68
другие, а также основной механизм управления и регулирова-
ния деятельностью организма — нервная система — в конечном
счете необходимы для осуществления движений, т. е. для адап-
тивно-гомеостатического поведения животного. Естественно,
что при этих обстоятельствах предельно ограничены возможно-
сти изменения адаптаций и максимально жестко фиксирован
гомеостаз. Столь же естественно и то, что в этих условиях не
только фено-, но и тем более генотипическая изменчивость ми-
нимальна.
Таким образом, вполне очевидна двойственность как пере-
стройки закрепленных в ряде поколений адаптаций, так и го-
меостаза. Оба эти явления, существуя в единстве и перекрывая
друг друга в проявлении, выполняют функцию и сдерживающих,
и движущих биологическую эволюцию начал.
Изложенное позволяет представить себе три основные воз-
можности осуществления оптимальных отношений организма
со средой: 1) специализация зависимости, реализующаяся
путем снижении уровня функциональной активности вплоть
до гипобиоза; 2) специализация максимального приспособления
путем поиска адекватных условий существования; 3) специа-
лизация активного преобразования. Во всех случаях речь идет
не об абсолютности, а о преимуществениости данной специали-
зации у живых организмов, находящихся на самых разных
уровнях эволюционного развития.
Если же ставить вопрос об эволюции совершенствования
механизмов поддержания гомеостаза внутренней среды орга-
низма, то оно шло путем узкой специализации оргапелл, орга-
нов и систем. Специализация в свою очередь требовала макси-
мально жестких параметров (кислотность, температура и др.)
циркулирующих в организме жидкостей (окружающая среда).
Следовательно, функциональная активность механизмов гомео-
стаза, как и вообще всего организма, находится в прямой за-
висимости от характера и интенсивности обмена веществ объе-
диненного в целом организме спецификой изменения состава
омывающих его жидкостей, опосредующих обменные взаимо-
отношения между элементами живой системы.
Подводя итог, можно сказать, что па заре формирования
жизни на Земле живое выделилось из окружающей среды я
стало от нее отличаться благодаря приобретению способности
относиться избирательно как к поступлению веществ из этой
64
среды, так и к выделению продуктов жизнедеятельности на
ружу.
Живое стало живым благодаря активному проявлению
свойств, обеспечивающих вещественную и термодинамическую
неравновеспость и стационарность (гомеостаз). Это снизило за-
висимость живых существ от внешней среды. Однако, предельно
освободившись от такого «плена», живые существа оказались
практически в абсолютной зависимости от своей внутренней
среды. Бывшая, эволюционно наиболее древняя, форма взаи-
моотношений биологической системы с внешними условиями
стала ее внутренним состоянием и определяющим способом
существования. Наиболее экономное сохранение внутреннего
гомеостаза возможно только при максимальной стабилизации
и ограничении колебаний параметров среды обитания, т. е. при
целенаправленной, преобразующей природу деятельности би-
ологической системы. Для человека — трудовой. В противном
случае это неминуемо должно привести к еще большей зависи-
мости системы от внешних условий.
Таким образом, принципиально верный тезис Бернара
(1878) «постоянство внутренней среды есть условие свободной
независимой жизни» в крайнем выражении явно противоречит
самому себе. Животные со сверхгомсостазироваиной внутрен-
ней средой обречены на вымирание из-за неспособности творче-
ски и активно преобразовывать внешнюю среду соответственно
споим гомеостатическим потребностям. Примером может быть
сумчатый зверек коала, приспособившийся к питанию листья-
ми только двух видов эвкалипта.
Неизмеримо более «свободен» в этом отношении человек.
Однако и он может попасть в своеобразный «цейтнот» при не-
померном стремлении приспосабливать среду к себе, ограни-
чивая тем самым свои возможности приспособления к ней.
Из сказанного, разумеется, не следует делать вывод о роли
цивилизации (фактора, явно способствующего жесткому го-
меостазированию внутренней среды организма) как «могиль-
щика» биологического вида человека. Напротив, «социально-
организованный опыт живых существ есть производное от фи-
зической природы, результат долгого развития ее, развития из
такого состояния физической природы, когда пи социальности,
ни организованности, ни опыта, ни живых существ не было и
не могло быть» (Ленив В. И. Поли, собр, соч. Изд. 5-е. Т. 18,
65
с. 241), и уж если человек в процессе становления достиг по-
нимания сущности и понимания, если пока не космического,
во всяком случае земного и околоземного значения собствен-
ного «социально-организованного опыта», то именно эта соци-
альная сущность человека мыслящего должна использовать
возможности разума на создание оптимальных взаимоотноше-
ний в системе человек — среда для параллельной и прогрессив-
кой биологической и социальной эволюции вида Homo sapiens.
Очерк 4. УРОВНИ И ВИДЫ ГОМЕОСТАЗА
БИОЛОГИЧЕСКИХ СИСТЕМ
Как уже было сказано, наиболее элементарным гомеоста-
тическим уровнем биологических систем следует считать орга-
низменный. Основой его является эволюционно-зафиксирован-
ный в высшей степени специфичный обмен веществ, свойствен-
ный как для отдельной особи, так и для вида в целом.
Утверждение это справедливо, несмотря на то что при всем
индивидуальном и видовом многообразии проявления жизни на
Земле природа не была щедра на разнообразие принципов и
механизмов реализации обмена веществ. Здесь уместен афоризм
Сент-Дьерди: «Природа создала все разнообразие жизни из
очень небольшого количества кирпичиков».
Таким образом, в качестве фундамента всей иерархии уров-
ней гомеостаза заложен эволюционно зафиксированный обмен-
ный гомеостаз. Значит, обменный гомеостаз — это единственная
реально существующая основа и суть всех уровней и видов био-
логического гомеостаза.
Организменный уровень складывается из: а) цитогенетиче-
ского гомеостаза; б) соматического геностаза; в) соматическо-
го гомеостаза; г) функционального (физиологического) гомео-
стаза и д) онтогенетического гомеостаза.
Цитогенетический гомеостаз — морфологическая и функци-
ональная приспособляемость — есть выражение непрерывной
перестройки организмов соответственно условиям существова-
ния. Устойчивость организмов к действию различных факторов
определяется тем, что перестройка осуществляется под контро-
лем регулирующего клеточного механизма. Прямо или косвен-
но функции такого механизма выполняет наследственный
аппарат клетки (гены). В основе подвижной устойчивости
66
живых существ лежит регуляция физиологических и морфоге-
нетических процессов адаптации на уровне клеточной организа-
ции. т. е. цитогенетический гомеостаз. Именно на этом уровне до-
стигает своего полного завершения интеграция наследственных
факторов любой, даже наиболее сложной, особи (И. И. Шмаль-
гаузен, I960).
Соматический геностаз — это контроль за генетическим по-
стоянством соматических клеток, составляющих конкретный
индивидуальный организм. В чем смысл подобного контроля?
Продолжительность жизни клетки одноклеточного или много-
клеточного организма сравнительно невелика. Так, клетка тон-
кого кишечника живет 24 часа, печени — 180 суток при средней
продолжительности жизни человека 70 лет. Следовательно,
сохранение столь длительной жизни целого организма воз-
можно только при условии конвейерного воспроизведения •—
рождения новых клеток взамен погибших, причем не только
воспроизведения, но и изменения их суммарного количества в
процессе роста и развития индивидуума и поддержания относи-
тельно постоянного количества во взрослом организме. Мало
того, изменение и восстановление клеточного состава должно
быть специфическим для каждого органа или ткани (например,
печеночная клетка должна воспроизвести только себе подобную,
а не какую-либо другую). Такая непрерывная поточная смена
клеточных популяций тант в себе опасность: во время деления
клеток под влиянием тех или иных факторов неизбежны ошиб-
ки в точности воспроизведения того или иного гена, т. е. про-
исходит мутация. Накопление мутантных клеток, естественно,
ведет к перерождению данной ткани, органа и в результате к
утрате обычной для них функции; и чем больше клеток в орга-
низме, чем больше специализированы отдельные популяции
их (почки, печень, эпителиальная, соединительная ткани и др.),
тем больше вероятность возникновения мутации. К примеру,
для организма человека, состоящего приблизительно из 10н
клеток, из которых 1С"—1012 ежесуточно возобновляются, риск
возникновения мутаций достаточно велик. Если предположить,
что мутация возникает у одной из миллиона воспроизводимых
клеток, то следует ожидать возникновения около 106 мутантных
клеток в сутки. Даже если какая-то часть мутаций (как правило,
очень небольшая) имеет для организма положительное значе-
ние, шанс на заболевание слишком велик. Однако реальная
67
частота заболеваний вследствие мутаций значительно ниже ста-
тистически ожидаемой. В чем же дело? Надо полагать, что
большинство мутантных клеток по какой-то причине не может
реализовать свои пагубные для организма возможности. Сле-
довательно, должен сущестновать какой-то механизм, органи-
Р А К
Рис. 14. Две иммунные линии обороны (по В. Г. Галактионову, 1972):
«активный иммунитет» (слева) н «алогенпая ингнбнцня» (справа), защищаю-
щие организм от мутантных клеток тела
чиваюшин эти возможности, механизм, контролирующий со-
хранение постоянства нормального генотипа. Суть его заклю-
чается в образовании антител.
Разумеется, сказанным не исчерпываются средства контро-
ля за генетическим постоянством клеток. При всей простоте
и логичности механизма поддержания соматического геностаза
есть много спорных вопросов и неопределенностей, хотя бы в
отношении фагоцитирования мутантных клеток лимфацитами.
Однако факт существования соматического геностаза не вызы-
58
вает сомнений (рис. 14). Отрицать его из-за неполноты пони-
мании механизма неразумно. Как сказал Хевисайд, «вряд ли
следует отказываться от обеда только потому, что и до конца
не знаю, как происходит переваривание пищи».
Соматический гомеостаз является результирующим выра-
жением цитогенетического гомеостаза и соматического гено-
стаза. Соматический гомеостаз направляет суммарные сдвиги
функциональной активности организма на установление наибо-
лее оптимальных отношений данной особи со средой. Под этим
следует понимать, что любая реакция организма в ответ на вся-
кое воздействие адаптивна по смыслу и гомеостатична по со-
держанию. Это значит, что она проявляется наилучшим при-
способительным эффектом при наименьших затратах для его
достижения. Классическим примером, иллюстрирующим это
положение, может быть экономизация (до определенного пре-
дела) энергетических трат на выполнение того или иного физи-
ческого упражнения по мере совершенствования мастерства
спортсмена.
Функциональный гомеостаз — это оптимальная функцио-
нальная активность различных органов (органелл), их систем
и взаимоотношений и, наконец, деятельности всего организма
н конкретных условиях среды. Реализация этих функций должна
осуществляться с минимальными тратами и отклонениями той
или иной активности от состояния покоя и с наибольшим при-
способительным эффектом. Функциональный гомеостаз вклю
чает: обменный гомеостаз, обеспечивающий относительное по-
стоянство специфики и интенсивности обмена веществ, концен-
трации продуктов метаболизма в циркулирующих жидкостях
и т. д.; системный (органный) гомеостаз, поддерживающий отно-
сительную стабильность деятельности органов и систем (уровень
кровяного давления, скорость кровотока, интенсивность дыха-
ния и др.). Поэтому можно выделять дыхательный гомеостаз,
пищеварительный, выделительный, циркуляторный, двигатель-
ный, сенсорный, сенсомоторный, психомоторный, психологиче-
ский, регуляторный (обеспечивает оптимальный уровень нервно-
гуморальной регуляции в данных условиях) гомеостаз. В сферу
функционального гомеостаза может также входить: информа-
ционный гомеостаз, обеспечивающий оптимальную реакцию
организма на поступающую информацию, ее отбор и переработ-
ку; пространственный гомеостаз, определяющийся поведением
Б©
организма при выборе оптимального пространства для реали-
зации данного действия; временной гомеостаз, реализующийся
способностью организма осуществлять данное действие в опти-
мальное время; временно-пространственный, объединяющий в
себе два последних гомеостаза, и др.
Онтогенетический гомеостаз — это индивидуальное развитие
особи от образования зародышевой клетки (гаметы, зиготы,
споры) до смерти или до прекращения существования в преж-
нем качестве (в случае деления клетки).
Условно онтогенетический гомеостаз любого индивидуума
можно расчленить на две составляющие.
1. Развертывание и реализация наследственной информации,
заложенной в зародышевых клетках (или в наследственном ве-
ществе клеток, размножающихся делением), т. е. развертыва-
нием генотипа. Оно осуществляется по строго предопределен-
ному пути и проходит через все этапы становления индивида
до смерти- Причем генетический механизм обеспечивает фор-
мирование не точных родительских копий, а систем, которые в
известных пределах изменчивости способны развиваться в ор-
ганизмы родительского типа.
Эта составляющая онтогенетического гомеостаза может быть
названа генотипическим индивидуальным гомеостазом разви-
тия.
2. Процесс фенотипических изменений, представляющий
собой результат действия двух сил:
а) генотипического компонента, который определяется взаи-
модействием наследственного материала (генотипа) половых
клеток обоих родителей;
б) компонента, образующегося под влиянием материнской
цитоплазмы зиготы на развертывание находящегося в ней ге-
нетического материала (материнской среды, где развивается
яйцо или зародыш), а также всех внутренних и внешних фак-
торов' на фенотипическое проявление наследственных признаков
в течение жизни особи.
При действии этих факторов фенотипической изменчивости
возможны отклонения от генетически предопределенного эво-
люцией пути развития индивидуума. Однако гомеостатическая
сущность онтогенеза заключается в том, что система постоянно
стремится аннулировать, ослабить отклонения и возвратиться
в ложе предопределенного канала развития. Это проявляется
60
в таких крайних выражениях, как стремление стабилизировать
ход онтогенеза и возвратиться к развитию по намеченной кри-
вой. Примером может быть замедление с последующим уско-
рением развития животных (особенно молодых) во время за-
болеваний, голода и после них. Надо полагать, что в реализации
этих процессов ведущая роль принадлежит морфогенетическому
механизму. Здесь мы с благодарностью должны вспомнить
Уоддингтона, обратившего особое внимание на подобный факт,
названный им канализированием, или гомеорезом.
Вторая составляющая онтогенетического гомеостаза может
быть также названа фенотипическим индивидуальным гомео-
стазом развития или морфогенетическим гомеостазом. Благо-
даря функциональным перестройкам он обеспечивает форми-
рование нормального фенотипа, несмотря на помехи со стороны
внешней среды или нарушения развития. Онтогенетический
гомеостаз может быть назван также индивидуальным гомеоре-
зом или индивидуальным гомеостазом развития.
Уже упоминалось о том, что биологическими системами яв-
ляются не только отдельные особи, по и надындивидуальные
совокупности. К ним могут быть отнесены: семья, колония, ста-
до, популяция, вид и т. д., в которых каждая особь занимает
самостоятельное положение не только в «социальном», но и в
функциональном отношении (например, семья пчел, колонии
муравьев и т. д.). Это обстоятельство определяет необходимость
общефизиологического осмысления явления надорганизменного
гомеостаза.
В пределах задач нашего обсуждения следует остановиться
на таких надындивидуальных биосистемах, от которых зависит
перспектива морфофункционального эволюционирования вида.
Наиболее полно этому требованию соответствует популяция.
В популяции ясно проявляется гетерогенность состава, устойчи-
вая подвижность структуры, достаточная непрерывность суще-
ствования во времени, обеспечивающая преемственность жизни
и эволюции.
Можно было бы предположить, что гомеостатичность попу-
ляции определяется простым суммированием онтогенетических
гомеостазов входящих в нее особей. Такое соображение могло
родиться из известного факта, что процессы развития специфи-
ческой формы (организмов) канализованы. Это, естественно,
приводит к фенотипической однородности (внешнему сходству)
«1
особей в данной популяции при достаточных генетических раз-
личиях между ними- В связи с тем что фенотипическая измен-
чивость является высокоприспособительной, генетическая может
вызвать лишь минимальное снижение приспособленности в дан-
ный момент. При этих обстоятельствах очевидна важность ге-
нетической изменчивости в отдаленной перспективе. Таким
образом, и онтогенетический гомеостаз и тем более гомеостаз
популяции отвечают принципу сохранения и развития.
В основе достаточной устойчивости популяции лежит гене-
тический гомеостаз. Понятие это было введено Лернером (1954)
в связи с осмыслением любопытного факта — способности по-
пуляции изменять под давлением отбора исторически сложив-
шийся фенотип (по одному или нескольким признакам), а затем
после прекращения действия факторов достаточно быстро вос-
станавливать его. Так, снятие воздействия факторов направлен-
ного отбора в пользу увеличения количества щетинок или раз-
меров тела дрозофилы или количества и размеров яиц у до-
машних птиц приводило в скором времени к восстановлению
исходных показателей.
Лернер определил это явление как «свойство популяции
сохранять равновесие своего генетического состава и противо-
стоять внезапным изменениям». Генетический гомеостаз харак-
теризуется следующим.
1. Существующий в природе фенотип является продуктом
генотипа, подвергавшегося в течение длительного времени
отбору на максимальную приспособленность к данной среде.
2. Любой отбор на новый фенотип (по одному или несколь-
ким признакам), возникающий при изменении условий суще-
ствования, ведет к перестройке ранее сформировавшегося гено-
типа, т. е. к снижению его приспособленности. Это происходит
либо путем накопления рецессивных гомозигот, либо возникно-
вения дисгармонии между новыми «выгодными» генами и ос-
тавшейся неизменной основной частью генотипа.
3. Смысл генетического гомеостаза заключается в том, что
путем ослабления давления отбора на новый фенотип в резуль-
тате преимущественно гетерозиготных комбинаций создается
возможность (если не полностью, то частично) восстановить
исторически сложившийся генотип, обладающий максимальной
приспособленностью. При этом в популяции обязательно сохра-
няются некоторые фенотипические черты предшествовавшего
62
отбора, что объясняется либо закреплением определенных го-
мозигот, либо, что более вероятно, переходом ее на новый мор-
фофункциональиый адаптивно-гомеостатический уровень. Таким
образом, этому уровню свойственны два качества — он соответ-
ствует первоначальному генотипу в отношении общей приспо-
собленности и одновременно превосходит его по конкретному
фенотипическому признаку, на который было направлено дав-
ление отбора.
Более того, генетический гомеостаз определяет, в какой
степени данный генофонд может отвечать на отбор. Здесь
весьма важен тот факт, что чем меньше какая-либо черта фе-
нотипа приспособлена к среде, тем легче она будет реагировать
на специальный отбор. Примером этому может быть форми-
рование устойчивости к токсическому веществу.
Следовательно, внутреннее морфофункциональное содер-
жание генетического гомеостаза позволяет объяснить такие
явления, как эволюционная косность, генетическая инертность,
явление стабилизирующего отбора и др. Собственно, термин
«стабилизирующий отбор», предложенный Шмальгаузеном
(1949), близкие ему по смыслу «центростремительный» (Симп-
сон, 1953) и «нормализующий» (Уоддингтон, 1949) сводятся
к представлению, что изменения в популяции с высокой геноти-
пической изменчивостью неизбежно приводят к возникновению
большого количества особей, отклоняющихся от среднего фе-
нотипа и значит обладающих пониженной жизнеспособностью.
Исключение этих особей из оборота в популяции и есть функция
генетического гомеостаза. Отсюда следует, что все перечислен-
ные типы отбора могут быть определены одним понятием —
генетический гомеостатический отбор. Такая формулировка ха
рактеризует не только процессуальную сторону явления, по и
его природу. (Выживаемость и эволюционная перспективность
популяции определяется не только генетическим гомеостазом,
обеспечивающим оптимальную фенотипическую структуру, но
и многими другими обстоятельствами.)
Итак, функциональное значение гомеостаза такой элемен-
тарной надындивидуальной биологической системы, как попу-
ляция, весьма велико. Реализация гомеостаза популяции нахо-
дится в прямой зависимости от среды обитания, т. е. от качества
и разнообразия возмущающих воздействий. Следовательно,
судьба особей, как и в целом популяции, зависит от адекват-
63
ности их морфофункциональных отношений со средой. Поэтому
важно понимание таких явлений, как биоценотический, био-
сферный и филогенетический гомеостазы, отражающие гло-
бальные общефизиологические аспекты биологических систем
в целом.
Биоценотический гомеостаз обеспечивает относительно устой-
чивое постоянство различных видов и популяций в биоценозе
и значительно реже—в биотопе. Главным фактором, поддер-
живающим гомеостатическую целостность биоценоза, являются
сложные пищевые взаимозависимости между населяющими его
организмами. Основное значение имеет прямой тип взаимо-
отношений хищник — жертва и косвенный — пищевая кон-
куренция.
В качестве примера рассмотрим взаимоотношения хищник —
жертва. В простейшем виде это выглядит так: увеличение чис-
ленности популяции ведет к усиленному потреблению и в
результате к уменьшению количества объектов питания (живот-
ные, растения), т. е. к ухудшению питания хищников, что сни-
жает их плодовитость, численность и, следовательно, создает
благоприятные условия для восстановления численности попу-
ляции жертвы. Так возникает замкнутый цикл регуляций. При
любом усложнении (путем включения других циклов) принцип
и суть гомеостатической саморегуляции пе изменяется. Однако
следует обратить внимание на одну особенность этой саморегу-
ляции. Так как процесс саморегуляции реализуется в первую
очередь посредством размножения, то эффективность его про-
является в следующем поколении, т. е. с опозданием. Это ска-
зывается на популяции, находящейся в фазе увеличенной или
уменьшенной численности: происходит либо сложение, либо
вычитание эффектов воздействия, что приводит к существенным
периодическим колебаниям — от вспышек массового размноже-
ния до почти полного уничтожения популяции. Таким образом,
биоценотический гомеостаз, как и все прочие, имеет осцилля-
торную колебательную природу.
Математическое описание подобных ритмов представлено
правилами Вольтерры, носящими имя автора, впервые сформу-
лировавшего их в 1926 году.
1. Период осцилляции зависит от коэффициентов увеличения
(уменьшения) элементов каждого вида и от первоначального
количества каждого из них.
64
2. Среднее количество элементов обоих видов стремится к
постоянной величине, каким бы ни было первоначальное коли-
чество.
3. Уничтожение поедающих элементов (условно — акулы)
ускоряет колебания; уничтожение поедаемых элементов (ус-
ловно — рыбы) замедляет колебания.
4. Увеличение степени защиты поедаемых элементов ведет
к увеличению количества элементов обоих видов.
5. При одновременном и однообразном уничтожении двух
видов соотношение амплитуд колебаний поедающего и поедае-
мого видов имеет тенденцию к увеличению.
6. Если попытаться уничтожить оба вида однообразно и
пропорционально имеющемуся их количеству, то среднее коли-
чество элементов поедаемого вила увеличивается, а поедающе-
го — уменьшается.
Итак, мы пришли к заключению, что механизмы биоцено-
тического гомеостаза такие же, как и гомеостаза популяции
и индивида. В этом убеждаемся даже на примере любой клет-
ки. Можно провести аналогию между взаимоотношениями
«акул» и «рыб» и ядра и цитоплазмы. Ядро «питается» за счет
цитоплазмы, а цитоплазма за счет окружающей среды (мы
исключаем возможность получения цитоплазмой вещества ядра,
так как в конечном счете это вещество получено из него). Если
экспериментально удалить из клетки какое-то количество цито-
плазмы (например, отрезав часть цитоплазмы у амебы), то
можно наблюдать сморщивание ядра, достигающее максимума
через несколько часов (запаздывание!). Это классический при-
мер событий, реализующихся механизмами биоценотического
гомеостаза.
Биосферный гомеостаз — поддержание постоянства живого
вещества («количества жизни») в биосфере в исторически обо-
зримые периоды времени. В 1875 году профессор Венского
университета Э. Зюсс ввел в науку представление о биосфере
как особой оболочке земной коры, охваченной жизнью. Строй-
ное учение о биосфере создал наш замечательный ученый ака-
демик В. И. Вернадский. Ему же принадлежат и чрезвычайно
интересные идеи, развивающие представление высоко почитае-
мого им. Ж. Бюффона о том, что «фонд» Живых субстанций
на Земле неизменен: «Если взять все организмы вообще, то в
целом количество жизни всегда то же».
8 А. А, Логшюв
65
Удивительно сходство биосферного гомеостаза с биоцено-
тическим А стоит ли этому удивляться? Разумеется, нет! Ведь
ь космическом представлении жизни как способа существования
материи (а в том, что жизнь явление не уникальное и присущее
не только Земле, сейчас мало кто сомневается) Земля не что
иное, как один из космических биоценозов. Законы же биоце-
нозов, вероятно, всюду' одинаковы!
Представления Бюффона об органических «молекулах», из
которых образуются живые вещества, веют некоторым мисти-
цизмом, так как, по его мнению, они бессмертны и неизменны.
Идея же Вернадского о постоянстве массы живого вещества
на Земле обретает достаточно строгое научное содержание:
«Изучая историю Земли, мы наталкиваемся на факт огромной
важности, последствия которого обыкновенно не замечаются:
факт постоянства химического облика земной коры в течение
всего геологического времени. Совершенно несомненно, что ми-
нералы, образующиеся в течение геологического времени, всегда
одинаковы. Всюду и всегда, не только со времени кембрийской
эры, но и со времени архейских эр, образуются те же минералы;
пет никакого изменения. Не только нет изменений в самих
минералах, но и их парагенезис остается тот же и их взаимные
количества во все времена кажутся тождественными.
Отсюда необходимо заключить, что геохимические явления
не изменились заметным образом со времени архейских эр. Из
этого также следует, что средние количества и состав живого
вещества оставались приблизительно одинаковыми в течение
всего этого непостижимого по длительности времени. Иначе,
ввиду значения организмов в геохимической истории всех хи-
мических элементов, ни минералы, ни их соединения не могли
бы оставаться все время тождественными.
Таким образом, количество живого вещества, по-видимо.му>
является планетной константой со времени архейской эпохи,
т. е. все деление геологического времени.
В тех сложных динамических равновесиях,— пишет далее
Вернадский,— какие мы видим в биосфере, говоря о постоян-
стве явления, очевидно, отнюдь нельзя считать, что данное яв-
ление не меняется е своем числовом значении (выделено на-
ми.— А- Л.). Можно только утверждать, что пределы колебаний
не меняются. Мы привыкли к такой форме постоянства в со-
ставе воздуха или в солевом составе океана. Их постоянство,
66
т. е. неизменность пределов колебаний (относительная,- -
А. Л.), указывает, что биосфера находится в очень устойчивом
состоянии.
Сейчас можно ставить только как подлежащую проверке
очень вероятную гипотезу, что все эти постоянные пределы
колебаний геологически вечны, т. е. неизменны в геологическом
времени, составляют основные черты структуры биосферы, т. е.
планеты (выделено нами,—Л. Л.)».
Чем это не описание классического механизма биоценоти-
ческого гомеостаза в объеме всей планеты? Отсутствует только
сам термин. Подтверждением мысли о существовании биосфер-
ного гомеостаза служит общеизвестный факт, что биологическая
эволюция по сравнению с геологической более консервативна.
В целях доказательства этого положения мы позволим себе в
какой-то мере по-своему интерпретировать замечательную
мысль Гинецинского (1963) о происхождении и развитии жизни.
Жизнь, несомненно, возникла в воде. Столь же несомненно,
что первые живые существа появились не в пресной воде, а в
растворе солей натрия, калия, кальция и магния. Иначе нельзя
объяснить тот факт, что клетки всех живых существ от самых
простых до самых сложных, какова бы ни была среда их оби-
тания, имеют в своем составе эти вещества и погибают, когда
они отсутствуют.
В настоящее время никто не сомневается в том, что жизнь
возникла в океане архейской эпохи, имеющем определенные
соотношения одновалентных и двухвалентных катионов, кото-
рые с полным основанием могут быть названы биологическими.
У примитивных прототипов животного мира, так же как у
современных кишечнополостных, морская вода является одно-
временно и внешней и внутренней средой организма. При этих
условиях и произошло приспособление клеток к пропорции
ионов, свойственной воде древних морей. Укрепляясь необозри-
мые периоды времени, эта пропорция была возведена в степень
абсолютной необходимости, вне которой благополучная жизнь
невозможна.
Биологическая эволюция шла под воздействием непрерыв-
ных изменений неживой природы. Возрастала суммарная соле-
ность оксана. Изменялся его ионный состав. Морская вода
утрачивала соли калия и кальция ( они постепенно оседали па
дно океана) и обогащалась солями натрия и магния, приноси-
3*
67
мыми речными потоками. Живым существам, обитавшим в из-
меняющемся по солевому составу океане, предоставлялся вы-
бор: или следовать за этими изменениями, перестраивая за-
крепленные в ряде поколений приспособления, или создать
органы, дающие возможность пройти через геологические эпо-
хи, не изменив привычных условий существования. Природа
не решила эту проблему альтернативно. Она оказалась мудрее,
использовав и тот и другой путь с различной степенью участия
альтернатив. Наивысшего развития достигла ветвь, идущая
преимущественно по консервативному гомеостатическому пути
сохранения-изменения. В результате, противопоставив эволю-
ции океана (геологической эволюции) свою собственную эво-
люцию, живые существа замкнули морскую воду в системе
кровеносных сосудов. Они превратили внешнюю среду во внут-
реннюю и выработали физиологические механизмы, охраняю-
щие ее постоянство (гомеостаз).
Захваченная «в плен» морская вода превратилась в кровь,
циркулирующую по всему телу. Выйдя миллионы лет тому на-
зад из пучин океана на сушу, наши древние предки — амфибии
унесли «в своих сердцах», а затем передали нам по наследству
частицу прародины— морскую воду, преобразованную в кровь.
Животные, создавшие замкнутую систему кровообращения,
сохранили в ней соотношение ионов, которое было присуще
среде обитания их предков. А это значит, что по соотношению
калия, натрия, кальция, магния и других веществ высших жи-
вотных и человека можно судить о составе воды океана в эпоху,
когда возникли предки позвоночных. Вот почему тщетны по-
пытки найти полное тождество солевого состава крови совре-
менных животных и вод ныне существующих океанов.
Подвергшийся геологическим воздействиям первобытный
океан, став современным, резко отличается по концентрации
солей от своего прародителя. Оказывается, внутренняя среда
организма — кровь более консервативна, чем тысячевековые
геологические события, преобразовавшие облик Земли и ее
недра.
Необходимо отдать должное огромной мощи мышления
Вернадского, идея которого о том, что и в образовании всех
земных сфер и пород роль живого вещества весьма высока,
доказана. Даже столь фантастическая мысль, что гранитная
оболочка Земли образовалась за счет переплавления осадочных
68
пород и что граниты — это «былые биосферы», получила се-
годня достаточное подтверждение.
В громадности геологической силы человечества, способного
преобразовывать не только геологический, но и геохимический
лик Земли, мы каждый день убеждаемся на примере произ-
водственной деятельности человека. Что человек?! Огромна
геологопреобразующая функция животных благодаря способ-
ности концентрировать те или иные химические элементы (на-
пример, залежи селитры биогенной природы в Чили).
Суммарный вещественный состав планеты достаточно ста-
билен (гомеостатичен), значительно более неустойчива его
комбинаторика и дислокация. Это, пожалуй, относится и к го-
меостатическим колебаниям вещества нашей планеты. Правда,
нужно добавить, что и вещественный состав планеты в опреде-
ленной степени изменчив за счет получения из космоса и отдачи
ему тех или иных веществ. Но это уже выходит за пределы
обсуждения проблемы биосферного гомеостаза. Хотя автор
настоящих очерков не прочь пофантазировать и предложить
идею о существовании зависимости количества живого веще-
ства на Земле от ее массы, т. е. о своеобразном лимите накоп-
ления массы живого вещества (критическая масса) и массы
планеты. В этом случае биосферный гомеостаз, любой его уро-
вень, приобретет еще одно очень важное качество: ему будет
свойствен не только колебательный характер внутри постоянной
массы живого вещества, но и количественно-качественная эво-
люция в планетарном масштабе.
Так идея биосферного гомеостаза родила идею планетарного
гомеостаза, в котором живая материя также занимает почетное
место как более активная часть механизма, поддержи-
вающего его.
Собственно, не произнеся формулы «биосферный гомеостаз»,
Вернадский по существу создал ее. Ведь именно ему принад-
лежит мысль о том, что «необходимо сейчас пересмотреть пред-
ставление о характере обмена — движения атомов—внутри
организма, об устойчивости «протоплазмы».
Может быть, существует связь между идеями К. Бернара и
К. М. Бэра относительно «бережливости» жизни и установлен-
ного геохимией факта — постоянства количества живого веще-
ства в биосфере. Возможно, что все эти идеи относятся к одному
и тому же явлению, а именно к «неизменности (относитель-
69
ной.— А. Л.) протоплазматических образований в биосфере
в течение геологических периодов» (не вообще, а геологиче-
ских.— А. Л.).
Филогенетический гомеостаз — историческое развитие орга-
низмов, сопровождающееся тенденцией сохранения-изменения.
В силу того что существуют так называемые филогенетические
запреты, ограничивающие возможность организмов (даже в
высшей степени легко приспосабливаемой эволюционной линии)
к беспредельному освоению любых адаптивных зон — биотопов,
возможности эволюционного процесса небесконечны. Рассмот-
рим один из многих примеров. Под влиянием окружающей
среды грызуны приспособились к подземному, древесному, реч-
ному и другому образу жизни. Однако несмотря на практически
неистощимую кормовую растительную базу морей, нет ни одно-
го вида морских грызунов. Случайность ли это? Нет, в силу
генетически закрепленных признаков строения и функции гры-
зунов зона эта для них запретна.
Таким образом, можно говорить, что эволюционный процесс
связан не только с приобретением новых особенностей, расши-
рением возможности освоения новых биотопов, но и с появле-
нием системы запретов. Запреты не позволяют на данном
морфофункциональном генотипическом и фенотипическом уров-
не организации осваивать определенные среды. Важен и другой
факт. Для большого таксона в целом (например, отряд грызу-
нов) число запретов сравнительно небольшое. Как уже было
сказано, таким универсальным запретом для всех представите-
лей отряда грызунов является обитание в морской среде. Однако
чем ниже ранг таксона — надсемейство, семейство, вид, тем
выше функциональная специализация его форм и, следователь-
но, тем больше для него запретов. Как, например, не существует
кротов, ведущих речной или древесный образ жизни, так нет
и белок, живущих под землей. Система запретов лежит и в
основе эволюции того или иного таксона, обеспечивая ее опре-
деленную направленность и степень перспективности. Так, пи-
щевой запрет, «наложенный» па коалу, практически поставил
животное не только в абсолютную зависимость от среды оби-
тания, но и вообще перед проблемой возможности его дальней-
шего эволюционирования. Таким образом, филогенетическое
развитие таксона предоставляет ие только широкие возможно-
сти для формирования разнообразия фенотипов и генотипов и
70
их приспособляемости, но и накладывает на этот процесс опре-
деленные ограничения, т. е. канализирует его. Следует вспом-
нить Уоддингтона, предложившего для характеристики такого
процесса термин «гомеорез». Правда, применение его здесь
менее удачно, чем термина «гомеостаз развития», так как он
основное внимание акцентирует на конечном продукте, дости-
жение которого осуществляется в рамках довольно жесткого
канала, траектории. Это делает эволюцию в какой-то степени
обозримой во времени, конечной. Между тем даже предполо-
жить подобное вряд ли возможно. Следовательно, если гово-
рить об эволюции живого вообще, то канализированность по-
зволяет лишь предвидеть возможный путь ее, по не конечный
результат. В отношении же отдельных эволюционных линий
результат можно предсказать, тем более, что истории известны
примеры эволюционного конца многих из них (всевозможные
завры и др.). Здесь термин «гомеостаз развития», обозначающий
по существу, но не обязательно предусматривающий достиже-
ние однозначного конечного результата, следует заменить на
филогенетический гомеостаз. Этот термин включает представ-
ление о гомеостазе как канализированном, исторически (эволю-
ционно) развивающемся процессе в силу семантического со-
держания слова филогенез.
Предлагаемая система гомеостазов условна и не вполне
совершенна. Здесь есть еще над чем и работать, и спорить. Но
не вызывает никакого сомнения факт, что любая морфофунк-
циональная гомеостатическая устойчивость относительна во
времени и пространстве в пределах размаха допустимых откло-
нений от идеальной средней. За пределами допустимого про-
странственно-временнбго и морфофункциональиого размаха жи-
вая система разрушается, вымирает. Собственно, она развивает-
ся,но отстает от эволюции среды. Это приводит к дезадаптации
биологической системы, на смену ей приходит новая зародив-
шаяся (ответвившаяся, трансформировавшаяся и т. д.) в
недрах старой. Следовательно, совсем не парадоксальным будет
следующий окончательный вывод.
Эволюция живых систем — это саморегулирующийся про-
цесс, в основе которого лежит устойчивая, специфическая, ди-
намическая неравновесность этих систем с окружающей средой-
Специфичсская неравновесность обеспечивает отличительные
характеристики данной эволюционной линии живых систем от
71
других линий. Устойчивая неравновесность гарантирует воз-
можно длительное существование данной неравновесной спе-
цифичности (форма, функция, сохранение и передача наслед-
ственной информации, воспроизведение себе подобных). Дина-
мическая неравновесность обусловливает ту или иную форму
эволюции (прогрессивная или регрессивная, медленная или
быстрая).
Итак, эволюция — процесс, канализированный по форме,
гомеостатичный по содержанию и адаптивный по сущности;
процесс, суть которого в полной мерс раскрывается философ-
ской трактовкой развития, данной В. И. Лениным: «Развитие
есть «борьба» противоположностей. Две основные (или две
возможные? или две в истории наблюдающиеся?) концепции
развития (эволюции) суть; развитие как уменьшение и увели-
чение, как повторение, и развитие, как единство противополож-
ностей (раздвоение единого на взаимоисключающие противо-
положности и взаимоотношение между ними).
При первой концепции движения остается в тени само дви-
жение, его двигательная сила, его источник, его мотив
(или сей источник переносится ео вне — бог, субъект etc.). При
второй концепции главное внимание устремляется именно на
познание источника и «с а м о»движения.
Первая концепция мертва, бледна, суха. Вторая — жизнен-
на. Только вторая дает ключ к «самодвижению» всего су-
щего» (Ленин В. И. Поли. собр. соч. Изд. 5-е. Т. 29, с. 317).
Удивительно убедительным по глубине философского и на-
учного содержания и поэтическим по эмоциональной красоте
примером является кровь как элемент внутренней среды орга-
низма, обеспечивающей ее постоянство. Ведь в самом деле жи-
вотные, обладающие обособленной внутренней средой, избрали
такой путь, который привел их к высшему совершенству. Став
свободными от влияния геологических превращений, существа
с независимой внутренней средой пережили периоды опреснения
и увеличения солености океана и смогли наконец выйти на
сушу. Они обрели способность поддерживать привычное и ве-
ками закрепленное постоянство внутренней среды, обеспечи-
вающее наиболее благоприятные условия для существования
всех без исключения клеток своего тела, постоянство, сохраняю-
щееся даже при чрезвычайных изменениях окружающего внеш-
него мира. Степень этой свободы и независимости дала орга-
72
низмам ту внутреннюю силу, которая обеспечила их индивиду-
альное совершенствование в борьбе за существование, за
сохранение своего вида и его эволюцию.
Основной силой, управляющей процессом сохранения и эво-
люцией данного вида, является естественный отбор.
Некоторые раздумья возникают в отношении человека. Ведь
естественный отбор в самом общем понимании представляет
собой такой процесс, при котором происходит или закрепление
в поколениях приспособлений, что ведет не только к адаптации,
но и образованию нового вида, или исключение этой эволюци-
онной линии из биологического оборота. И тот и другой ва-
риант— следствие возникновения определенной специализа-
ции. В отношении же человека этого в прямом смысле сказать
нельзя. Так, по крайней мерс, па сегодня нет никаких ни тео-
ретических, ни фактических данных о процессе видообразования
на уровне человека. По образному выражению Э. Майера
(1950), это, по-видимому, произошло вследствие того, что «че-
ловек, так сказать, специализировался к деспециализации...
занимает более разнообразные экологические ниши, чем любое
из всех известных животных. Если один вид—человек — ус-
пешно занимает все пиши, открытые для человекоподобных су-
ществ (и легко перемещается из одной ниши в другую.—А. Л.),
то ясно, что процесс видообразования происходить не может».
Согласно Д. Роза (1899), существует гипотеза об утрате
изменчивости или «пластичности» филогенетически вымираю-
щими линиями вследствие чрезмерной специализации — сверх-
специализации, т. е. гипотеза об «уменьшении приспособляе-
мости с увеличением специализации». Предполагается, что
сверхспециализация резко снижает темпы и ограничивает мас-
штабы эволюционных изменений и тем самым ставит подверг-
шиеся ей филогенетические линии перед опасностью вымира-
ния. Таким образом, специализация сама по себе не есть при-
чина вымирания, но она создает для этого благоприятные
предпосылки.
Однако с 1950 года в социально-преобразующей деятель-
ности человека произошли весьма существенные изменения, ко-
торые еще более увеличили способность его к экологической
деспециализации. Но это лишь формально. В действительности
же человек сверхспециализировался в искусственно созданной
нм самим и жестко застабилизированной экологической среде.
73
Точнее, современный человек не просто существует в искус-
ственно созданной им стабильной среде, а живет по принципу,
omnia mea mecum porto («все мое ношу с собой»). Это жилье,
одежда, питание и другие средства поддержания постоянства
и индивидуального микроклимата окружающей среды каждой
человеческой особи. Таким образом, человек действительно не
столько деспециализирован, сколько сверхспециализнрован к
существованию в весьма узких и систематически суживающихся
параметрах искусственно создаваемой им среды обитания.
С биологической точки зрения это обстоятельство может иметь
несколько аспектов и один из них проявляется в неминуемом
снижении фенотипической, да и генотипической, пластичности,
изменчивости, сверхспециализированных (сверхгомеостазиро-
ванных) биологических систем. Это явно не способствует не
только видообразованию на уровне человека, по и принципи-
альному использованию возможностей прогрессивной феноти-
пической изменчивости, заложенной в генотипе индивида. Рас-
пространение подобной сверхспецнализацни к искусственной
и в общем упрощенной среде па все географические зоны оби-
тания человека — перспектива не особенно утешительная в свя-
зи с весьма возможным проявлением теломорфоза и ката-
морфоза.
Понятия тсломорфоз и катаморфоз обычно пе используются
в биологии человека, по имеют к ней непосредственное отно-
шение. Теломорфоз—специализация организма, связанная с
переходом от более общей среды к частной, более ограниченной.
При этом происходит одностороннее развитие некоторых орга-
нов и частичное обратное развитие (редукция) других. Прогрес-
сивная дифференциация распространяется на тс части организ-
ма, которые связывают его со специфичным для данной частной
среды условиями. Организация в целом остается на исходном
уровне или испытывает даже некоторое упрощение. Катамор-
фоз — регрессивные изменения организма, связанные с перехо-
дом к более простым взаимоотношениям со средой и с общим
понижением жизнедеятельности. Примерами катаморфоза мо-
гут служить все явления перехода от более активной жизни к
менее активной, от активного питания к пассивному, от жизни
в открытых пространствах к скрытой, защищенной жизни в
1рунте, от самостоятельного движения к сидячему образу жизни
н т. п. (Шмальгаузсн, 1969).
74
Именно это обстоятельство городило идею о генетическом
вырождении человека. Согласно этой идее, человечество в ре-
зультате создания собственной среды и благодаря успехам
медицины уклонилось от действия естественного отбора и тем
самым возложило на себя повышенный груз давления со сто-
роны накапливающихся мутаций. Не исключая динамичности
генетики популяции человека, в том числе и негативной, сле-
дует сказать, что обычные процессы сдвига концентрации генов
идут медленно. Все они в настоящее время отступают перед
двумя мощными процессами, оказывающими оздоравливающее
влияние на генетику человека. Первый из них — это громадный
рост численности людей на Земле, захватывающий весь гене-
тический потенциал человечества. Второй — смешение рас в на-
правлении создания единого, внутренне разнообразного чело-
вечества. Этот процесс интеграции генетической системы чело-
века облегчается почти полным исчезновением географической
изоляции между различными человеческими популяциями бла-
годаря совершенствованию средств передвижения и др.
Н. П. Дубинин (1971) считает, что благодаря влиянию этих
мощных оздоравливающих факторов генетическим основам
человечества как биологического вида ничто не угрожает:
«Идущие в них изменения так малы, что в течение тысячелетий
эти основы полностью сохраняют свои мощные биологические
потенциалы. Это генетическое строение может быть нарушено,
если в среде возникает резко увеличенный фон мутагенных
факторов. Однако дело регуляции среды обитания человека
целиком зависит от него самого».
По данным В. Гласс и К- Ли, негры, проживающие в Север-
ной Америке, в результате смешивания с белым населением
сейчас в своем генотипе несут 30% генов белых людей. Если
учесть, что негры были привезены в Новый свет три с половиной
века тому назад, то скорость этого процесса указывает, что
через 75 поколений (2000 лет) установится полное генетиче-
ское равновесие, т. е. объединение генного пула негров и белых
в единой популяции. Однако процесс смешения популяций че-
ловека ставит ряд серьезных проблем Соотношение численно-
сти представителей белых, черных и желтых людей очень нерав-
номерно. По подсчетам Хацера, общая популяция людей на
Земле к 2000 году достигнет 6267 миллионов человек. Из них
по разным областям планеты будет жить: в Африке — 517 мил-
75
лионов человек, в Северной Америке—312, в Центральной и
Южной Америке — 529, в Азии — 3870, в Европе (включая всю
территорию Советского Союза) — 947, в Океании — 29 миллио-
нов человек.
Очевидно, простое объединение этих столь разных по чис-
ленности популяций может изменить то биологически хорошее
разнообразие (дифференциация на расы), которое характери-
зует современное человечество. Надо представить себе, какой
человек может возникнуть, если стихийно перемешать генные
пулы всех существующих рас. Следовательно, процесс объеди-
нения всех популяций несет людям будущего серьезные пробле-
мы, обдумывание которых надо начинать уже сейчас.
Таким образом, возникает не только биологический, но и
социальный аспект специализации человека к созданной им
искусственной среде обитания. Частным выражением его явля-
ется разумное «расшатывание» сложившейся сверхспециализа-
ции путем привлечения в искусственную среду элементов, адек-
ватных эволюционно определившимся биологическим (морфо-
функциональным) запросам человеческой особи. К ним отно-
сятся эмоциональное и информационное насыщение обедненных
в этом отношении современных монотонных производственных
процессов (конвейеры, высокая механизация и др ), увеличение
двигательной активности регламентированным введением физи-
ческой культуры и спорта во все возрастно-половые и производ-
ственные группы людей, организация труда и отдыха и т. д.
П.ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ
ВЗАИМООТНОШЕНИИ В СИСТЕМЕ ОРГАНИЗМ —
СРЕДА
Очерк 1. ГОМЕОСТАТИЧЕСКИЕ ПАРАМЕТРЫ
БИОЛОГИЧЕСКИХ СИСТЕМ
Рассматривая уровни и виды гомеостаза биологических си-
стем, мы постоянно обнаруживали в его сущностном содержа-
нии две противоположные пространственно-временные качест-
венные тенденции: 1) стабильность и 2) динамичность.
Пространственно-временная качественная стабильность раз-
личных компонентов гомеостаза проявляется в активном про-
тиводействии системы влияниям возмущающих факторов окру-
жающей среды. Это противодействие осуществляется множест-
вом различных по природе и функциональному назначению
механизмов, удерживающих биологическую систему в рамках
минимальных (или жизненно допустимо максимальных) откло-
нений от уровня функциональной активности, свойственной
идеальному .состоянию бносистемы с минимальной энтропией
в широком смысле.
Пространственно-временная качественная динамичность, из-
менчивость, напротив, неминуемо ведет биологическую систему
по достаточно жестко запрограммированному каналу к термо-
динамическому равновесию ее со средой (максимуму энтропии),
т. е. к фатальному финалу.
Таким образом, гомеостаз (процесс-состояние) условно из-
меняется в лонгитудинальной (продольной) и трансеерзальной
(поперечной) пространственно-временной протяженности. Пер-
вая является параметром, ограничивающим общую или этапную
протяженность существования биосистемы и реализацию гене-
ральной программы ее гомеостатического развития «от — до»
(онтогенетический гомеостаз). Это — протяженность траекто-
рии развития бносистемы во времени.
Трансверзальная протяженность является параметром про-
странственно-качественного состояния гомеостаза биосистемы
77
в данный момент и определяет форму( отлогая, крутая и т. д.),
общую лонгитудинальную объемность траектории, объемность
морфофункциональных пуффов (здесь — временное ускорение
и активизация процессов) или, напротив, временное уменьшение
относительно нормы «толщины» данного участка траек-
тории.
Так, например, для общей лонгитудинальной жизнен-
ной траектории человека характерна строго определенная по-
следовательность достаточно четко обозначенных периодов его
онтогенеза. В начале этой траектории (нуль—точка отсчета)
находится одиночная оплодотворенная женская яйцеклетка.
В стадии зрелого возраста уже насчитывается до 1012—10” кле-
ток— результат по меньшей мере 38-1012 ~ 238 клеточных деле-
ний и увеличения общего размера по отношению к одиночной
клетке не менее чем в 1О10 раз. Несмотря на почти фантастиче-
скую неправдоподобность указанных величин, они — объектив-
ная реальность. Причем реален не просто процесс количествен-
ного изменения клеточного состава организма, но и еще более
сложный процесс дифференцировки клеток на специфические
тканевые и органные структуры, завершающийся в конце кон-
цов формированием до предельной точки развития (восходя-
щая часть траектории) целостного организма. Затем наступает
деградация биологической системы и смерть (нисходящая
часть).
Геронтология в настоящее время располагает обоснованны-
ми теоретическими данными о возможности нормальной жизне-
деятельности человека как естественного процесса до 150 лет.
Более короткая продолжительность жизни и принимаемая сей-
час за нормальную среднюю продолжительность 70—72 года —
результат помех (болезнь, социальные и другие неблагополу-
чия), ограничивающих проявление биологических жизненных
возможностей человека. Однако примем эту длительность жиз-
ни за оптимальную. Но принимаемая условно оптимальная дли-
тельность жизни содержит в себе возможность вариации дли-
тельности как всей жизни, так и ее отдельных этапов. Вариации
длительности этапов определяют форму траектории жизни как
в целом, так и по этапам. Однако изменение длительности эта-
пов, т. е. их «кривизна» во времени, зависит от событий, раз-
вивающихся в траисверзальных параметрах, т. е. от отклонений
гомеостатических характеристик данного этапа от «идеального»
78
уровня. Эти изменения не только влияют на кривизну всей
траектории жизни и ее этапов, но и могут быть факторами, ли-
митирующими ее развитие вообще. Именно поэтому траектория
жизни Может быть оборвана в любой точке развития. Напри-
мер, данные статистики говорят, что на 100 зачинающихся в
результате оплодотворения девочек, зачинается 140 мальчиков.
Реально же на 1000 новорожденных приходится 513 мальчиков
(элиминация мужских особен происходит на очень ранних ста-
диях развития, еше до появления самых первых признаков
беременности). Это не говоря уже о самопроизвольных абор-
тах, возникающих по тем или иным обстоятельствам, мертво-
рождаемости, смертности по разным причинам, в том числе
в связи с несчастными случаями и т. д.
Однако вернемся к реальной средней жизненной траектории
человека протяженностью во времени 70 лет. Попробуем рас-
смотреть лонгитудинальные характеристики каждого этапа ее.
Здесь сразу обращает на себя внимание замечательный факт.
Оказывается, что наиболее точно выдерживаются временные
параметры этапов внутриутробного развития. Причем чем бли-
же данный этап к нуль-точке (оплодотвореная яйцеклетка),
тем точнее его временная характеристика. Так, например,
оплодотворение яйцеклетки происходит в течение 24 часов
после овуляции- В течение 72 часов (!) оплодотворенная
яйцеклетка, пройдя все стадии дробления (до образования бла-
стоцисты) попадает в матку, на 6-й день после овуляции бла-
стоциста прикрепляется к эндометрию, на 7-й — имплантиру-
ется в него. К этому времени на вентральной стороне зароды-
шевого диска различается энтодерма (в виде тонкого слоя
клеток) и появляется полость амниона. Все! Наступил второй
период развития — зародышевый. Начало его определяется не
только структурными усложнениями в результате дробления,
но и в первую очередь переходом на новый способ питания.
В это время питание зародыша осуществляется за счет веществ,
проникающих из эндометрия путем диффузии. С 7-го по 14-й
день происходят процессы образования плаценты.
Следует обратить особое внимание па поразительную вре-
менную точность всех событий и особенно тех, которые связа-
ны с дроблением яйцеклетки до момента образования бласто-
цисты. События разворачиваются буквально с точностью до
минут и часов (особенно отчетливо это можно видеть на при-
79
мере дробления яйца одного из видов лягушки — Rana sylva
tica, рис. 15).
В чем же дело? Да, видимо, в том, что работает предельно
жесткая генетическая программа, отклонение от которой как
Рис. 15. Развитие лягушки. Рапа sylvaiica (по У. Бодсмеру, 1971).
Л — неоплодотворенное яйцо; Б — образование серого серпа; Ь — Е — первое
второе, третье и четвертое деления; Ж — позднее дробление; 3 — ранняя;
It — поздняя бластулы; К — ранняя. Л — средняя, М — поздняя гаструлы;
1 — аномальное полушарие; 2 — вегетативное полушарие; 3 — микромеры,
— микромеры; б — мелкие клетки; б — крупные клетки; 7 — спинная губа
бластопора; 8 — желточные клетки.
во времени, так и по существу смерти подобно. Материальное
обеспечение этой программы происходит за счет веществ опло-
дотворенной яйцеклетки. В этом случае любое изменение ско-
рости формирования бластоцисты может сказаться на матери-
альных (обменных) возможностях дробящейся яйцеклетки.
Другое дело, когда имплантирования бластоциста переходит на
80
«довольствие» матери. Здесь уже могут быть несколько большие
отклонения в скорости дифференциации яйцеклетки, ставшей
зародышем. Однако и в данном случае они не должны быть
значительными во времени. На этой стадии зародыш еще не-
достаточно «активен» в своих запросах к матери, во многом
«полагается» на самого себя и не особенно нарушает времен-
ную программу развития. И все же, если до сих пор речь шла
о часах и даже минутах временных отклонений, то после 14-го
дня развития уже можно говорить о сутках Так, примерно (!)
на 16-й день у зародыша появляется первичная полоска. Да-
лее временная амплитуда еще более увеличивается. С 16-го по
21-й день образуется нервная пластинка, которая затем пре-
вращается в нервную трубку. Период между 21-м и 23-м днями
после овуляции характеризуется прогрессирующей сегментаци-
ей и началом органогенеза.
По мере дифференциации зародыша увеличивается степень
его зависимости от матери: свои запасы уже давным-давно ис-
черпаны, а расти и развиваться надо. Между тем поступление
питательных веществ во многом зависит от особенностей отно-
шений матери со средой, которые не так уж просты и не всегда
удовлетворяют ее потребности. На генетическую программу,
заложенную в схеме развития зародыша, с каждым днем все
большее влияние оказывает окружающая среда, т. е. мать. Вот
почему с началом третьего —• плодного периода временная
«раскачка» этапов развития будущего человека увеличивается.
Несмотря на то что в общем этапы пренатального онтоге-
неза человека соблюдаются довольно строго и по времени, и
по содержанию, все же можно прийти к чрезвычайно существен-
ному выводу. Смысл его в том, что по мере увеличения зави-
симости развития плода от условий среды временные и качест-
венные параметры генетической программы организма претер-
певают все большие изменения. Этот закон еще более четко
выражен в процессе постнатального развития человека. Имен-
но поэтому временные и качественные характеристики периодов
(«идеальные» параметры) детства, отрочества, юности, зрело-
сти и т. д. вплоть до умирания более подвижны, чем периоды
пренатального развития. Соответственно увеличивается актив-
ность сил противодействия «идеальной» жизненной траектории
(от простейших автоматических до осознанных, творческих, со-
циальных форы).
81
К началу просесса умирания, т. е. в условиях практически
полного прекращения активных форм противодействия фаталь-
но развивающимся событиям, последние вновь начинают
«вкладываться» в жесткие временные параметры. Здесь, по-ви-
димому, «работают» механизмы, сходные с механизмами
начальных этапов развития, но, подобно пущенной в противо-
положном направлении киноленте, в обратном порядке. Если
на начальных этапах генетическая программа разворачивалась
прогрессивно в направлении формирования данной системы, то
в процессе умирания происходит деградация ее. В соответствии
с законом двойственности и внутренней противоречивости яв-
лений эта деградация заложена в самой программе. Она
начинает реализовываться с начала формирования биосистемы
и завершает ее существование.
Итак, первый и основной параметр гомеостатического пове-
дения биосистемы — качало и конец ее существования.
Диапазон индивидуальной длительности существования
биосистем может быть достаточно широк даже в пределах
одной популяции или вида, но никогда не длительнее, чем это
установлено генотипом и предельными фенотипическими вари-
ациями его проявления. То, что родилось,— умрет!
Временной параметр существования распространяется не
только на биологическую, но и на любую материальную систему.
Это всеобщий закон материального мира, в котором вечность
материи складывается из бесконечного многообразия конечных
составляющих.
Тезис Энгельса о том, что «жизнь всегда мыслиться в со-
отношении со своим необходимым результатом, заключающим-
ся в ней постоянно в зародыше,— смертью» замечателен тем,
что он подразумевает ограниченность существования биосисте-
мы в одном состоянии, т. е. динамичность и конечность ее го-
меостатических свойств как в целом, так и на отдельных
временных этапах существования.
Начало, т. е. рождение данной индивидуальной биологиче-
ской системы, в определенном смысле, явление случайное. Как
говорится, «дети не выбирают себе родителей». Вместе с тем
случайность появления данного индивида не исключает неслу-
чайности проявляемых им особенностей. В частности, сходство
с родительской особью неслучайно. Оно определено генетически.
Это второй параметр, параметр гомеостатического сходства не
82
только поколений родственных особей, по и внутри популяции
и вида. Шире — биологическая форма движения материи, от-
личающая живое от неживого, есть также общий для всего
живого гомеостатический параметр.
В качестве частного примера этого генетического параметра
гомеостаза биосистемы может быть упомянут уже известный
нам соматический геностаз или еще более конкретное его
выражение — генетическая специфичность (геностатичность)
соматических клеток (цитогенетический гомеостаз). Известно,
что все соматические клетки независимо от их морфофупкцио-
нальной специфичности содержат в себе набор генетического
материала, достаточный для воспроизведения целого организ-
ма, т. е. обладают сходным геномом (возможно, и генотипом
в целом). Доказательством этого являются блестящие эксиери
менты с воспроизведением растения, например моркови из од-
ной клетки, нли лягушки из яйцеклетки с удаленным ядром,
на место которого пересажено ядро соматической клетки (ядро
взято из клетки слизистой кишечника взрослой лягушки). Сле-
довательно, соматические клетки являются единственным
классом «живых существ» с избыточной наследственной ин-
формацией, не имеющей прямого отношения к их приспособлен-
ности к самостоятельному существованию (Ю. Б. Вахтин,
1974). Это значит, что в обычных условиях существования со-
матической клетки происходит не полное использование воз-
можностей заключенного в ней генетического материала.
Исследования показали, например, что в клетках печени мы-
ши в неактивном состоянии находится около 80% ядерной
ДНК, т. е. генома клетки.* Поэтому морфофункциональная
специфичность и репродукция их только из клеток печени опре-
деляется активностью 20% ее нерепрессированного генетическо-
го материала.
Есть основание полагать (Бахтин, 1974), что генетический
материал в соматических клетках может находиться по крайней
мере в трех состояниях: облигатно активном, факультативно
* Прямых доказательств того, что репрессия приводит к полно-
му прекращению геном синтеза информационной РНК (и-РНК) в со-
матических клетках- в настоящее время нет; возможно, что синтез
РНК при репрессии лишь снижается настолько, что его ие удается
выявить
83
активном и облигатно неактивном. Таким образом, параметром,
определяющим морфофункциональную специфичность (гомео-
статичность) соматической клетки, является специфичный для
пее цитогенетический гомеостаз (клеточный геностаэ), имею-
щий определенное количественное выражение облигатно и фа-
культативно активного генетического материала.
Соматические клетки представляют собой «организмы», ли-
шенные как полового процесса, так и каких-либо иных меха-
низмов (в обычных условиях) обмена генетической информа-
цией. Поэтому популяции соматических клеток представляют
собой совокупность клеток, специфика каждой из которых
(наследственная структура) может быть воспроизведена в
потомстве. Генетическая целостность популяций соматических
клеток обусловлена генотипической и эпигенетической * из
менчивостью. Размерность наследования генотипических и
эпигенетических изменений на клеточном уровне определяется
устойчивостью генотипа клетки к воздействиям и является
параметром тканевого и органного, а также системно-органно-
го геностаза.
Следовательно, специфические морфофункциональные (го-
меостатические) свойства ткани проявляются лишь в том слу-
чае, если она состоит из клеток с эпигенетической наследствен-
ностью, реализуемой одинаковым образом. Это значит, что эпи-
генетические различия клеток ткани не выходят за пределы ее
«объема потенции». По мнению Вахтина (1974), эпигенотип
ткани в целом «богаче», чем эпигенотип любой из составляю-
щих ее клеточных субпонуляций. Возможно, он даже «богаче»,
чем совокупность эпигеиотипов всех ее субпопуляций, реализу-
ющихся в онтогенезе. Этот «эпигенотипический резерв» может
быть использован в патологических ситуациях как один из
механизмов поддержания тканевого и органного морфофункци-
онального гомеостаза и выступить в качестве одного из его
параметров.
Надо полагать, что так называемый эволюционно фиксиро-
ванный консерватизм генетических возможностей биосистемы,
* Эпигеном — совокупность наследственных факторов, обуслов-
ливающих фенотипическую изменчивость клеток при действии внеш-
ней среды. Эпигенетическая изменчивость клеток в отличие от гене-
тической не связана с глубокой перестройкой генома.
84
а также наследование его генотипической и эпигенетической
(фенотипической) изменчивости лежит в основе параметриче-
ских характеристик функциональной активности всех уровней
соматического гомеостаза. Рассмотрим это положение на одном
из проявлений функционального гомеостаза — обменном гомео-
стазе. В качестве модели возьмем состояние кислотно-щелочно-
го баланса крови (активная реакция крови). В настоящее время
этот баланс оценивается по трем показателям кровн: pH, рСОа
и сдвигу буферных соединений (бикарбонаты, фосфаты, белки).
У здоровых людей величина pH крови колеблется в пределах
7,36—7,44. Отклонения, совместимые с жизнью, составляют
примерно ±6,4 (от 7,0 до 7,8). Много это или мало?
Когда речь идет о параметрах гомеостаза внутренней среды
или любого другого гомеостаза, такие понятия, как «много»
или «мало», отступают на второй план и не потому, что это
относительные понятия. Суть в том, чтобы определить, насколь-
ко эволюционно фиксированные пределы изменения тех или
иных параметров удерживаются достаточно строго и постоян-
но. Поэтому сложившееся представление об исключительной
чувствительности тканей к изменению концентрации водородных
ионов в омывающих жидкостях достаточно формально. В са-
мом деле в диапазон величин pH от 7,0 до 7,8 собственно вхо-
дят концентрации водородных ионов от 100 н-экв/л (в 2,5 раза
превышает нормальную) до 16 н-экв/л (0,4 нормальной).
Таким образом, концентрация водородных ионов колеблется
в пределах от 40 до 250% величины, которая поддерживается
в нормальных условиях. Это значительно больше совместимого
с жизнью диапазона изменений калия, натрия и других жизнен-
но важных электролитов (однако это не значит, что клетки
обладают принципиально малой чувствительностью к измене-
нию pH омывающих их жидкостей).
Концентрации водородных ионов цельной крови, плазмы и
межклеточной жидкости равны. Это значит, что кислотно-ще-
лочной баланс всех жидкостей, образующих внешнюю среду
клеток, постоянен, одинаково гомеостатичен. Концентрация
водородных ионов во внутриклеточной жидкости — цитоплазме
ниже, чем во внеклеточной (pH около 6,9). Такое соотношение
pH внешней и внутриклеточной среды неслучайно и несет в
себе гомеостатическое содержание. В частности, это имеет
отношение к поддержанию необходимого постоянства концен-
85
трации внутри и вне клетки таких электролитов, как К+, Na+
и др. Особенно гомеостатическая функция pH проявляется в
том, что кислотно-щелочной баланс артериальной, капиллярной
и венозной крови практически и по общепринятым средним ве-
личинам, а также по средним величинам и размаху в каждом
определенном исследовании одинаков. Однако наиболее важным
является факт высокой стабильности величины pH всех ви-
дов крови на протяжении всего онтогенеза.
Следовательно, говоря о параметрической гомеостатичности
того или иного функционального показателя биоснстемы, необ-
ходимо сосредоточить внимание не на субъективно (пожалуй,
даже эмоционально) воспринимаемом представлении о «боль-
шом» или «малом» размахе его, а на предельно возможном
сохранении постоянства этого размаха как в данный момент
и при данной ситуации, так и на протяжении длительных от-
резков времени.
После рассмотрения конкретного примера, имеющего четко
выраженные параметрические гомеостатические ограничения - -
pH крови (кстати, можно было использовать любой другой
химический показатель внутренней среды организма — концен-
трацию калия, натрия, сахара; функцию какого-либо органа —
частоту сокращений сердца, быстроту двигательной реакции,
скорость проведения возбуждения и т. д.), есть смысл обсу-
дить вообще функционально-параметрические возможности го-
меостатического поведения организма в целом или какой-либо
его системы. Эта проблема не так проста, как может показаться
па первый взгляд. Для решения ее вновь необходимо догово-
риться о терминах. Основная сложность здесь заключается в
том, что каждая биосистема при различных обстоятельствах
ведет себя адаптивно по сути и гомсостатично по содержанию.
Это значит, что биосистема, адаптируясь к любым условиям
существования, стремится не изменить своего гомеостатического
состояния, а если даже и изменить, то возможно минимально.
В этом рассуждении нет особой новизны. При анализе термо-
динамических аспектов жизни оно было рассмотрено на при-
мере фундаментального закона природы о том, что под влияни-
ем внешнего воздействия в системе происходят процессы,
ослабляющие данное воздействие. Напомним это на упрощен-
ном примере: каждый из нас немного присядет, если тяжелая
рука ляжет ему на плечо, и напротив, если кто-либо попытает-
86
ся насильно приподнять нас за руки, мы станем на носки. Ко-
роче говоря, при любых ситуациях биосистема выбирает наибо-
лее оптимальный режим своего поведения в отношении
возмущающих воздействий. Понятие «оптимально» восприни-
мается легко и просто. Еще легче оно воспринимается при
дословном переводе этого слова: оптимальный — значит наи-
более благоприятный, наиболее соответствующий. Как будто
все ясно. Это соблюдается далее в том случае, когда побеждает
болезнь. Ведь мысль, что при заболевании организм стремится
не улучшить, а ухудшить свое состояние, усугубить течение бо-
лезни, абсурдна. Она абсурдна не только с позиции здравого
смысла, но и биологии- Если бы дело обстояло действительно
так, то жизнь погибла бы, не успев развиться. Точнее, жизни
вообще бы не возникла! Следовательно, при любых обстоятель-
ствах и в соответствии с ними бносистемы в меру имеющихся
возможностей реализуют свое поведение оптимально. Таким
образом, понятие оптимальности, казалось бы, утрачивает свое
реальное значение как параметр адаптивно-гомеостатического
поведения живых систем в окружающей среде и теряет свой
смысл.
В действительности это не так. Анализ накопленных клас-
сической физиологией данных позволяет найти принципиаль-
ную общность функциональной направленности сдвигов, про-
исходящих в организме спортсменов под влиянием тренировки,
со сдвигами в деятельности организма нетренированного чело-
века или животного в ответ па возмущающее воздействие раз-
личной длительности и интенсивности. Однако количественно-
качественная характеристика этих сдвигов у спортсмена и нс
занимающегося спортом различна. Так, сравнение величины н
характера функциональных сдвигов, возникающих в организме
спортсменов, показало, что по мере повышения спортивного
мастерства временная, а по многим показателям и амплитуд-
ная характеристика сдвигов, необходимых для осуществления
одного и того же (или более сложного )действия, последова-
тельно снижается. Это обеспечивает более экономную по срав-
нению с имевшейся ранее оптимизацию функций как в услови-
ях покоя, так и активной деятельности, и, следовательно, удер
жание существенных переменных организма у допустимо
минимальных (функционально оптимальных в каждом случае)
границ индивидуальных возможностей.
87
Переносимо ли подобное рассуждение на использованный
выше пример с заболеванием? Разумеется. Это и явление при-
обретенного иммунитета, ослабляющее течение повторного за-
болевания или вообще предотвращающего его, и большая
сопротивляемость закаленного организма заболеваниям и т. д.
Итак, понятие оптимальности в биологическом смысле на
чинает обретать достаточно четкие очертания, но еще не имеет
размерности.* Приобретет ли оно вообще размерность? Веро-
ятно, да, если будут определены параметры, в рамках которых
сможет осуществляться та или иная функция без нарушений.
Дело в том, что возможности выражения ее без нарушений ог-
раничены предельными гомеостатическими возможностями.
Для решения этого вопроса необходимо ввести по меньшей
мере три новых понятия: размах (предел) функции, размах
оптимальности функции и резерв оптимальности функции.
Рассмотрим это на конкретном примере. Верхняя частотная
граница сокращений сердца в ответ на максимальную нагруз-
ку практически одинакова по абсолютному показателю как
для тренированного, так и нетренированного организма, но от-
личается предельной длительностью ее удержания и, что еще
важнее, уровнем пропорциональности роста частоты сокраще-
ний сердца (ЧСС) росту систолического выброса крови, т. е.
критической частоте сокращений сердца (в том числе и нижней
границы) в пользу тренированного. Это явные преимущества
тренированного сердца в смысле совершения предельной рабо-
ты в оптимальном режиме. Его временная и качественно-функ-
цинальная характеристика выше. Частотный же оптимальный
уровень покоя при этом ниже, чем у нетренированного (боль-
шая величина систолического выброса и др.).
Таким образом, размах оптимальности (гомеостатичности)
у тренированного значительно шире. Значит, разница в разма
хе оптимальности между тренированным и нетренированным
сердцем будет выражать резерв оптимальности по величине,
мощности, интенсивности и т. д.
В формализованном виде сказанное в отношении ЧСС
может быть представлено следующим образом:
* Автор не склонен безоговорочно переносить трактовки понятия
оптимальности, существующие в технике, кибернетике и других об-
ластях знаний, на биологические объекты.
88
1. Разм. ЧСС=Макс. ЧСС-Мин. ЧСС,
где Разм. ЧСС— размах (предел) функции; Макс, и Мин.— до-
пустимые частотные пределы, при которых сердце выполняет
функции на уровне, достаточном для поддержания жизни;
2. Разм. опт. ЧСС=КЧС.С-ПЧСС,
где Разм. опт. ЧСС — размах (предел) оптимальности функ
ции; КЧСС — критическая ЧСС; ГТЧСС — ЧСС в покое;
3. Рез. опт. ЧСС=ЧСС2-ЧСС1,
где Рез. опт. ЧСС — резерв оптимальности функции; индекс
1 — величина, принятая за исходную; индекс 2—сравниваемая
величина.*
Здесь в формализованном виде выражены количественно-
качественные показатели состояния данной функции. Однако в
эти формулы можно ввести и параметр времени. Тогда речь
будет идти о пределе и резерве времени для достижения тех
или иных качественно-количественных показателей функции.
1. Временной резерв достижения критического состояния
(предел оптимума) функции:
Рез. /<?КЧСС=«КЧСС2 - «КЧСС|,
где t — время, д — знак достижения;
2. Временной резерв удержания функции в пределах крити-
ческой оптимальности.
Рез. tyK4CC=h/K4CC2-Z{/K4CCi,
где у — знак удержания;
3. Временной резерв восстановления состояния покоя функ-
ции:
Рез. /вЧСС=/вЧСС2-<вЧСС1,
где в — знак восстановления.
Следует полагать, что такие показатели, как длительность
удержания критической (желательнее докритической) частоты
и тем более максимальной частоты сердечных сокращений с
полноценным и оперативным восстановлением уровня покоя,
после прекращения выполнения работы будут характеризовать
* Размах оптимальности функции с индексом 2 может рассмат-
риваться как характеристика изменившейся величины (относительно
размаха с индексом 1) или сравниваемых объектов (1 и 2).
89
не только состояние оптимальности функции, но и уровень
большей или меньшей прочности, надежности, биологической
системы (в данном случае организма спортсмена), включая и
расширение возможности чисто биологической выживаемости
в экстремальных, предельных условиях.
Представляется также справедливым перенести приведенные
здесь рассуждения вообще на все показатели функциональных
возможностей биосистем, особенно если известны их должные
и предельно возможные (верхние и нижние) оценочные харак-
теристики, например основного обмена, показателей внешнего
дыхания и т. д.
Таким образом, мы пришли к реальной количественной и
качественной оценке оптимальных адаптивных возможностей
биологических систем в допустимых границах гомеостатическо-
го «разброса» функций. Однако встречаются такие жизненные
ситуации, когда к функции предъявляются требования за пре-
делами реальных возможностей оптимальной реализации ее.
Нечто в этом роде мы имеем в случае увеличения частоты сер-
дечных сокращений выше критической. Подобные ситуации
могут вызвать перенапряжение адаптивно-гомеостатических
возможностей биосистсмы и вывести ее за грань допустимых
гомеостатических пределов со всеми вытекающими отсюда по-
следствиями. В частности, это поддержание функций за счет
компенсаторных, суперкомпенсаторных и субкомпенсаторных
механизмов в условиях декомпенсации — компенсируемой и
пекомпенсирусмой (выпадение функции данного органа(ов) и
его (их) компенсирующих и, наконец, полное прекращение
функций — смерть. Все стадии, за исключением некомпенсируе-
мой декомпенсации и, естественно, смерти, адаптивно-гомеоста-
тичны для биосистемы. Однако резерв их адаптивно-гомеоста-
тических возможностей последовательно убывает.
Следует сказать, что в зоне гомеостатической оптимальности
функций могут быть выделены подуровни. Например, оптималь-
ные и субоптималыгые уровни функционирования. В качестве
отправной точки отсчета следует брать параметрическую ха-
рактеристику состояния активности «здоровой» биосистемы в
условиях максимального покоя (основного обмена).
По существу изложенным можно ограничить принципиаль-
ное обсуждение проблемы гомеостатических параметров биоло-
гических систем организменного (индивидуального) уровня,
60
памятуя при этом, что в нх основе лежат генотипические воз-
можности индивида.
Естественно, что гомеостатические параметры надоргапиз-
мсниых (надындивидуальных) уровней гомеостаза живых си-
стем —• популяций, вида — также в первую очередь определя-
ются генетическим фоктором — генофондом. Однако обсуждение
этого вопроса выходит за рамки задач настоящих очерков.
Читателям, интересующимся данной проблемой, автор реко-
мендует обратиться к специальной литературе (Э. Майр, 1974;
А. А. Гимельфарб, Л. Р. Гинзбург и др., 1974; П. Ф. Рокицкий,
1974; М. Эйген, 1973).
Очерк 2. НЕПАРАДОКСАЛЬНЫЕ ПАРАДОКСЫ
ГОМЕОСТАЗА
Изменяемость лонгитудинальных н трансверзальных парамет-
ров онтогенетического гомеостаза позволяет говорить о колеба-
тельной природе как жизненной траектории в целом, так и ее
отдельных этапов или любых временных отрезков. В более
широком смысле это следует понимать как свойство гомеостаза
вообще. Более того, неосциллирующий гомеостаз — явление не-
реальное. Собственно, эта идея явно или не явно сопутствует
нашим рассуждениям с самого начала — с того момента, как
было установлено, что все биологические системы являются
открытыми, находятся в неравновесном стационарном сотоянии
с окружающей их средой и постоянно затрачивают свободную
энергию на поддержание этой неравповесности.
Одиако что такое колебание? Воспользуемся образным
примером А. М. Жаботинского. Колебание — «это когда нечто
исчезает в одном месте и появляется в другом, потом исчезает
и в этом другом месте и возвращается в прежнее-—и так да-
лее». Но так как все в пашем мире существует в пространстве,
времени и взаимодействии с окружающим внешним миром, то
нс может «нечто исчезающее в другом месте возвращаться в
прежнее», поскольку «прежнего места» уже не будет, оно из-
менится качественно, изменится и само «нечто», ибо «все течет,
все изменяется». Следовательно, колебания, совершающиеся
во времени и пространстве, да и еще имеющие какие-либо связи
с соседними частицами, вовлекаемыми в колебательное движе-
ние,— это уже не просто колебания, а волны со своей собствен-
91
ной временной и амплитудной характеристикой, относительной
повторяемостью даже в случае незатухающих автоколебаний,*
Подобное рассуждение в отношении биологических систем не
должно нас слишком удивлять. Ведь возникновение живой ма-
терии— это лишь следствие качественной эволюции неоргани-
ческой материи. Особенностью же материального мира, его ат-
рибутивным свойством является движение. Движение в своем
изначальном материальном обличии — микрочастице имеет
колебательную природу.** Отсюда вывод: «каков фундамент,
таков и дом», т. е. все процессы, протекающие в живых систе-
мах, должны иметь колебательную природу. Это в первую оче-
редь имеет отношение к колебательной природе физических,
химических, физико-химических процессов, совершающихся в
биосистемах и лежащих в основе их биохимических событий.
Факт колебательной природы биохимических процессов (си-
стем) установлен на примере периодических изменений актив-
ности ферментов поджелудочной железы, слюны, уреазы, при
гидролизе мочевины и т. д. Было также установлено, что неко-
торые биохимические колебания могут протекать с периодиче-
ски меняющейся скоростью и интенсивностью. В этом можно
убедиться на так называемых «спонтанных» колебаниях како-
го-либо одного компонента крови. Например, уровень сахара
в крови одного и того же животного от минуты к минуте, не
говоря уже о часах и днях, претерпевает такие резкие колеба-
ния, что концентрация его порой достигает то гипергликемиче-
ской величины, то снижается до резкой степени гипогликемии.
* Автоколебательная система содержит источник энергии, за
счет которой поддерживаются колебания. Происходящие в таких
системах колебания называются автоколебаниями. Если источник
энергии постоянно пополняется, то автоколебания становятся неза-
тухающими.
** Французский физик Лун де Бройль в 1942 году выдвинул идею
волновых свойств материи. Согласно его гипотезе, каждой частице,
обладающей определенной массой и движущейся с определенной
скоростью, соответствует волна, длина которой зависит от массы и
скорости частиц. Опыты подтвердили это положение и показали, что
все микрочастицы, независимо от их природы и строения, обладают,
кроме корпускулярных, еще и волновыми свойствами. Важность этой
теории столь велика, что физики, зная о происхождении Луи де
Бройля из старинного королевского рода Бурбокон, придумали шу-
точный афоризм: «Напоследок Бурбоны создали короля — физика».
92
Самым важным здесь является временной, периодический, ха-
рактер колебаний концентрации того или иного вещества, уста-
навливающийся на переносимую организмом длительность.
Таким образом, гомеостатичпость колебаний — это не только
частотно-амплитудная характеристика, по и допустимая дли-
тельность удержания ее при данных обстоятельствах.
Рис. 17. Колебания концентра
ции четырехвалентного церия
(по А. И. Жаботинсксму, 1974):
Со — равновесная реакция: С, - -
верхний порог концентрации в от-
сутствие восстановителя; Cz—
нижний порог концентрации в
присутствии восстановителя.
Рис. 16. Схема периодиче-
ской реакции (по А. И. Жй-
ботинскому, 1974):
ВгВ — Хромированный восста-
новитель; НВгО — бромид,
образующийся в результате
реакций с, б, е. тормозит ре-
акцию.
Рассмотрим одну из возможных моделей самопроизвольного
колебательного процесса в гомогенной химической среде —
реакцию Б. П. Белоусова — А. И. Жаботинского. Суть ее
заключается в том, что при окислении лимонной кислоты
бромноватокислым калием в присутствии ионов церия (ката-
лизатор) отмечаются периодические изменения цвета раствора.
В ходе этой работы было установлено, что в гомогенном раство-
ре действительно наблюдаются незатухающие колебания (авто-
колебания) концентрации (рис. 16, 17). Причем подобная
химическая система практически может генерировать все воз-
можные в электрических и механических устройствах типы
колебаний. Если в 6 мл воды растворить 2 г лимонной кислоты,
0,2 г KBr2Os, добавить 0,7 мл концентрированной HsSO4, затем
довести общий объем раствора до 10 мл, добавляя воду и тща-
93
тельно перемешивая его, то немного терпения — и любители
эксперимента могут убедиться в существовании периодических
(автоколебательных) реакций в гомогенной среде.
Итак, периодические биохимические колебания в настоящее
время являются твердо установленным фактом. Даже можно
говорить о механизмах этих колебаний. По предложению
С. Э. Шноля (1966), следует различать два механизма перио-
дических колебаний в макромолекулах; «кинетический» (регу-
ляционный) и «конформационный». К первому относятся про-
цессы, периодический ход которых обусловлен кинетическими
механизмами, т. е. временем химических превращений моле-
кулы в отдельных промежуточных стадиях, активирующих или
тормозящих предшествующие.
Если учесть, что константы скорости тех или иных биохи-
мических реакций в какой-то мере отражают время ковалентной
модификации (химической релаксации молекулы) или их- от-
дельных этапов и измеряются (особенно при образовании фер-
мент-субстратных комплексов) величинами 10—®—10~8 секун-
ды, то можно представить эту фантастическую частотную ха-
рактеристику статистических колебательных процессов на мо-
лекулярном уровне. Еще большей скоростью характеризуются
колебания электронпо-возбужденпых молекул— 10s—10 11 с.
Конформационный механизм, согласно Шнолю, характери-
зуется следующим; «Представим себе большую молекулу бел-
ка — фермента, окруженную молекулами воды, субстратов,
коферментов, катионами и анионами. В такой системе осуще-
ствляется множество колебательных процессов. Электронные
колебания — переход электронов с одного энергетического уров-
ня на другой — обусловливают поглощение света в ультрафио-
летовой области или при наличии системы сопряженных связей и
хромофорных групп поглощение и в видимой области оптическо-
го спектра. Вращательные, деформационные колебательные дви-
жения атомов и атомных групп в молекулах обусловливают
поглощение в инфракрасной области. Вращательные и прочие
движения молекул обусловливают поглощение электромагнит-
ных волн в диапазоне от сотен микрон до миллиметров и санти
метров. Сложные многоатомные макромолекулы могут сущест-
вовать в различных конформациях, почти не отличающихся
друг от друга по уровню энергии... Разность энергетических
уровней при конформационных переходах может быть значи-
94
тельно меньше величин активационных барьеров, препятствую-
щих осуществлению таких периодов. В этом случае частости ре-
ально осуществляемых переходов будут определяться внешни-
ми причинами и прежде всего температурой среды... Для
белков-ферментов можно оценить вероятную среднюю частоту
(частость) релаксационных конформационных колебаний в
ходе катализируемых ими реакций. Известно, что при некото-
рой концентрации субстрата скорость катализируемой фермен-
том реакции достигает максимума. В биохимии принято
характеризовать эту максимальную скорость «числом оборо-
тов» — числом молекул субстрата, превращенных в продукт
молекулой фермента в 1 минуту. Каждый оборот включает
соединение фермента с субстратом, осуществление соответству-
ющего превращения... каждый оборот (выделено нами.—
А. Л.)—это конформационный цикл». Таким образом, кон-
формационный механизм является одним из механизмов, опре-
деляющих скорость каталитических процессов.
Ферментативные же процессы — основа жизни. Вне их
понятие «жизнь» превращается в беспочвенную абстракцию.
Важность ферментных систем в жизнедеятельности биосистем
столь значительна, что многие исследователи справедливо по-
лагают, что ферментно-каталитические процессы лежат в осно-
ве возникновения жизни. Как показали исследования, количест-
во оборотов большинства ферментов соответствует частотным
характеристикам слышимого и инфразвукового диапазона и
только у нескольких ферментов — ультразвукового (табл. 2).
На основании данных табл. 2 можно сделать вывод о том,
что особенно интенсивно функционируют преимущественно ка-
тализаторы универсальных биохимических процессов, таких,
как гликолиз, цикл Кребса, транспорт электронов и ионов,
фосфорилирование. Наибольшая частость конформационных
изменений у ферментов, катализирующих «узкие места» мно-
гократных превращений (например, фосфофруктокиназа при
гликолизе). Есть основание утверждать, что именно эти фермен-
ты «задают тон», т. е. лимитируют скорость процесса в целом.
Идеи о роли конформационных изменений в определении
колебательной природы биохимических процессов позволили
высказать Шнолю интересное соображение: «Конформация сво-
бодной молекулы фермента отличается от конформации белка
в фсрмснт-субстратном комплексе. Макромолекула белка
96
Таблица 2
Частоты оборотов некоторых ферментов
(по С. Э. Шнолю, 1966)
фермент Кол-во об/cat фермент Кол-во об/сек
Пепсин 0,001 Конденсирующий фермент
Пируват-карбоксилаза 13 (цикл Кребса) 450
Альдолаза 33 Дегидрогеназа молочной
АТ Ф-азамнозиназа 104 КИСЛОТЫ 1215
Миокиназа 166 Фосфорилаза 1600
Эиолаза 150 Фумараза 77000
Дегидрогеназа триозофос- Каталаза 510s
фата 166 Карбоангидраза 1,6-10®
Гексокиназа 215 Ацетилхолинэстераза 310е
Фосфоглюкомутаза 280
устроена так, что после того как первый акт превращения
субстрата осуществился (после случайной тепловой флуктуа-
ции), выделяющаяся энергия переводит молекулы фермента в
особое неравновесное состояние (выделено нами.— А. Л.) — в
состояние термодинамически невыгодной конформации, в кото-
ром по кинетическим причинам (большой активационный
барьер) макромолекула может находиться достаточно большое
время... Неравновесная в результате этого макромолекула мо-
жет перейти в более вероятное состояние (равновесное.— А. Л.)
лишь при контакте с субстратом (катализируемый объект.—
А. Л.). Получается забавный, но вполне реальный парадокс
(вернее один из непарадоксальпых парадоксов механизма го-
меостаза, в данном случае метаболического.— А. Л.): субстрат
катализирует переход фермента в равновесное состояние. Зато
сам субстрат претерпевает необходимые превращения, акти-
вационный барьер которых понижается за счет конформацион-
ной энергии макромолекулы фермента. Ясно, что при этом
необходимо специальное устройство полипептидных цепей —
ограниченное число степеней свободы * их взаимного движения,
* Степень свободы — число независимых видов движений, кото-
рые может совершать тело.
96
большая амплитуда их подвижности в небольшом числе направ-
лений. Особый смысл поэтому может иметь и определенная
последовательность аминокислот в «активных центрах» и при-
легающих к ним областях макромолекул ферментов— В свете
сказанного о механизме ферментативного акта не кажется столь
удивительным сходство последовательности аминокислот в
активных центрах совершенно различных ферментов — фосфо-
глюкомутазы, трипсина, тромбина и др.». По утверждению
Шноля, один н тот же каталитический двигатель может рабо-
тать с различными «навесными» приспособлениями — перено-
сить группы, разрушать пептидные связи и пр. Автор даже
считает возможным существование «универсальной каталити-
ческой конформационно-подвижной (ритмически подвижной.—
А. Л.) конструкции» макромолекул ферментов.
А теперь — кажущееся отступление от основной темы.
В Лондонском институте инженеров-электриков 11 февраля
1969 года профессором Уилки была прочитана лекция. Лекцию
предварял анонс, написанный в лучших традициях классичес-
кого англо-саксонского юмора: «В серийном производстве. Ли-
нейный двигатель характеризуется прочностью и надежностью.
Конструкция проверена и оптимизирована в ходе испытаний,
проводившихся в течение очень длительного времени и в гло-
бальном масштабе. Все модели высокоэкономичны (использу-
ется тот же механизм преобразования энергии, что н в топлив-
ном элементе) и работают на общедоступных видах топлива.
В состоянии покоя мощность почти не потребляется, но
двигатель может мгновенно включаться без добавления топлива
и за несколько миллисекунд развивать удельную мощность до
1 кВт на 1 кг веса. Конструкция модульная, имеется большой
набор субблоков, что дает возможность в каждом конкретном
случае получить наивысшую экономическую эффективность.
Два режима управления.
1. Управление с внешним запуском. Универсальный ре-
жим, пригодный на все случаи жизни. Управление цифровое от
импульсов с энергией в несколько пикоджоулей. Хотя входная
мощность ничтожна, достигается весьма высокое отношение
сигнал/шум. Усиление мощности — около 10е. Механические
характеристики (при толщине модуля 1 см): скорость плавно
регулируется от 0,1 до 100 мм/с; развиваемое давление
(2—5) 10— И/м2.
4 А. А. Логинов
97
2. Автономный режим с внутренней, автогенерацией. При-
меняется главным образом для повторяющихся однотипных
движений (типа насоса). Есть модули, удобные по частотным
и механическим характеристикам для перекачки:
а) твердых и полужидких веществ (0,01—1 Гц);
б) жидкостей (0,5—5 Гц); срок службы 2,6- 109 (тип, зна-
чение), 3,6-10° (макс, значение) — независимо от частоты;
в) газов (50—1000 Гц).
Имеется большое число дополнительных функциональных
блоков: встроенные следящие системы (с позиционным и ско-
ростным регулированием) на случай, когда требуется точная
подстройка. Прямая подача кислорода. Генерация термо-э. д. с.
Может служить пищей».
Какова же тема лекции? Чему она посвящена?
Тема лекции — живая мышца!
В чем же дело? Для чего столь много интригующих заявок?
Да просто потому, что мышца того заслуживает, и в первую
очередь благодаря актомиозиновому комплексу, составляюще-
му ее основную массу.
В 1939 году В. А. Энгельгардт и М. IT. Любимова обнару-
жили, что выделенный из мышцы белок — миозин обладает
ферментативной активностью. В присутствии ионов кальция
миозин катализирует гидролиз концевой фосфатной группы
АТФ — основного источника энергии для сокращения мышцы —
с образованием АДФ и неорганического фосфата. Каталитиче-
ская активность миозина проявляется также и в отношении
ГТФ, ИТФ, ЦТФ, т. е. практически всех основных макроэрги-
ческнх соединений. Важным свойством миозина служит его
способность связывать актин — второй главный белковый ком-
понент мышцы—и образовывать так называемый актомиози-
новый комплекс — материальную основу обеспечения сократи-
тельной функции мышцы.
Отмечен также еше один интереснейший факт: в растворах
белков актомиозинового комплекса обнаружены синхронные
колебания таких показателен, как АТФ-азная активность, ад-
сорбционная способность по отношению к красителям и др. Как
оказалось, эти колебания синхронны конформационным коле-
баниям макромолекул актомиозинового комплекса. Естествен
вывод, что конформационные колебания белков актомиозиново-
го комплекса могут иметь прямое отношение вообще ко всем
98
видам движения в живых системах. Кстати, об этом свидетель-
ствуют опыты на моделях — нитях актомиозина, проявляю-
щих ритмическую сократительную автоматичность под воздей-
ствием АТФ. Выше было упомянуто. Что число оборотов боль-
шинства известных ферментов соответствует акустическйм по-
лям слышимого диапазона. В связи с этим предлагается инфор-
мация для размышления. Первое — еще в 1883 году Гельмгольц
установил существование вибрационных колебаний мышечных
волокон в звуковой частоте. (Микровибрация мышечных воло-
кон наблюдается от рождения до смерти любого теплокровно-
го организма при всех физиологических состояниях как при
полном мышечном расслаблении, так и во время сна в состоя-
нии наркоза; она сохраняется в течение 50—70 минут даже
после смерти.) Второе — физико-химическая среда в мышце
и протоплазме соответствует условиям, максимально благопри-
ятным длн перехода из одной конформации в другую. Тре-
тье — почему же эти конформационные изменения не заметны
при наблюдении за целой мышцей и какая связь между первым
и вторым?
Конформационные изменения в целой мышце незаметны
не потому, что их нет, а потому, что они не регистрируются
слишком инерционными приборами, используемыми для этих
целей. Более совершенные методы исследования позволили
зарегистрировать в самой мышце ритмическое скольжение ни-
тей актина и актомиозина относительно друг друга. Есть до-
статочно оснований полагать, что эти шаги отражают конфор-
мационные изменения актомиозиновых комплексов всех мио;
фибрилл целой мышцы или группы мышц и' происходят
неодновременно, ступенчато, дискретными шагами. Значит, в
качестве микровибраций регистрируются, по-видимому, синхро-
низированные конформационные изменения какой-то ограни-
ченной, ио достаточно многочисленной группы или rpytin
актомиозиновых комплексов. Именно поэтому частотная харак-
теристика спонтанных конформационных изменений одиночного
актомиозинового комплекса может не совпадать с таковой
группового, которая в свою очередь ие совпадает с микровибра-
цией группы мышечных волокон или же целой мышцы. Видимо,
это может быть объяснено следующим образом.
1. Принципом большинства: наблюдателем регистрируются'
конформационные изменения в виде сокращений й расслабления
4
99
актомиозиновых комплексов какого-то количества синхронно
конформирующих элементов.
2. Пейсмекерным эффектом, при котором некоторому коли-
честву элементов навязывается ритм «водителя».
3. Способностью некоторого количества элементов подклю-
чаться к ритму «водителя», т. е. способностью усваивать его
ритм.
Таким образом, связь между первым и вторым, по-видимо-
му, есть; по крайней мере с позиции здравого смысла правиль-
нее предполагать ее, чем отрицать.
В качестве иллюстрации роли конформационных изменений,
обусловливающих ритмическую двигательную активность, мо-
жет служить один из самых распространенных видов движе-
ния — биение жгутиков и ресничек. Как известно, это наиболее
древний аппарат автоматизированных движений. Он сохранил-
ся не только морфологически в сократительном аппарате
произвольной мускулатуры фибрилл (91 2), но и, по нашему
мнению, функционально в виде неуправляемой спонтанной
микровибрации, лежащей в основе ультра микроструктуры
(тремор, микротремор) как произвольной, так и непроизвольной
двигательной активности.
Мы рассмотрели колебательную характеристику биохими-
ческих, кинетических и конформационных процессов, обеспе-
чивающих относительное постоянство метаболизма на молеку-
лярном уровне. Эти периодические процессы сказываются и на
«внешнем» выражении той или иной функции (например, со-
кратительной на определенном уровне и в течение определенно-
го отрезка времени), т. е. проявляют активность па одном из
возможных уровней стационарности в пределах допустимого
гомеостатического размаха.
Подобная схема гомеоритмических колебаний какого-либо
процесса гомеостаза прослеживается от молекулярного уровня
до органа и системы в целом.
Это убедительно демонстрируется на примере структуры и
функции молекулы гемоглобина.
Однако рассмотрим вначале вообще молекулу белка, и
прежде всего ее структурную организацию. Белки являются
высокомолекулярными соединениями. Пр уровню организации
молекула белка состоит из «-последовательностей «-аминокислот
и по структуре может быть первичной, вторичной, третичной и
100
четвертичной. Первичная структура — это последовательное
расположение аминокислотных остатков в полипептидной цепи
(рис. 18); вторичная — вид ее организации (право- или лево-
закругленная спираль, рис. 19); третичная —способ укладки
спиральных структур в частицах (глобулах) белков (рис. 20),
который определяет внешнюю форму молекулы (нарушение
этой структуры, как правило, лишает белки их биологической
активности); четвертичная структура характеризуется еще бо-
лее сложной пространственной конфигурацией, где молекула
(или агрегат молекул) состоит из субъединиц, каждая из ко-
торых имеет третичную структуру Примером может быть
гемоглобин. Что же в нем особенно замечательного? Молекула
гемоглобина (рис. 21) — эффективнейший переносчик кисло-
рода. Она способна постепенно связывать четыре молекулы
его, причем каждая предыдущая связь повышает сродство ге-
моглобина к следующим молекулам кислорода. Поскольку
атомы железа в четырех гемгруппах находятся друг от друга на
расстоянии по крайней мере 25 А, можно сделать вывод, что
взаимное влияние четырех кислородосвязывающих центров
осуществляется через конформационные изменения в пептид-
ных цепих. Это предположение нашло подтверждение в иссле-
дованиях М. Перутца (1964). В частности, им показано, что
после связывания кислорода субъединицы гемоглобина повора-
чиваются относительно друг друга так, что расстояние между
атомами железа в ₽-цепях уменьшается на 6,5 А, а в о-цепях
увеличивается па 1 А-
Оказывается, молекула гемоглобина «дышит») Только тут
соотношения обратные: «вдох» молекулы сопровождается сжа-
тием, «выдох» — расширением. В. Л. Энгельгардт так оц^ил
это открытие: «Думается, что этим достигнут предел глубины
нашего проникновения в молекулярные механизмы такого фун-
даментального явления, каким является кислородоперенося-
шая функция гемоглобина».
Таким обазом, самая замечательная особенность молекулы
гемоглобина — ее конформационные периодические изменения,
определяющие кислородную насыщенность и определяемые
ею. Периодичность этих изменений находится в прямой зави-
симости от кислородных запросов тканей, их поглотительной
способности, а также и от скорости использования ими кис-
лорода.
101
4Й «7 46 45 44 43 42 41 40 59 38 37 36 35 34
Рис. 18. Первичная структура молекулы белка инсулина:
I, S, 3. .... >М — сокращенные наименования аминокислот.
Рис. 19. Вторичная структура (а=спираль) молекулы белка
(по А. Ленииджеру, 1974):
Л -- правая спираль (плоскости жестких пептидных связей расположены
параллельно длинной оси); Б — модель а=спирали, на которой покаяаны
внутри цепочные связи (шаг а= спирали и смещение в расчете на один ами-
нокислотный остаток соответствует большому (5,4А) и малому (I.5A) перио-
дам); Й— средине размеры с—спирали; Г — пространственная модель а*=спн-
ралн.
Рис. 20. Третичная структура молекулы гемоглобина (по М. Пе-
рутцу, 1966):
п-цепь (слева — белая) н fJ-цепь (справа — черная). Кривой линией показано
направление главной цепи. Частично видна группа гема (дисковидная струк-
тура).
Рис. 21. Четвертичная структура молекулы
гемоглобина (по М. Перутцу, 1964).
Если колебательная природа кинетических (химических, ме-
таболических), в том числе и конформационных, изменений дик-
тует формирование программы оптимального поведения в био-
системе, если это обусловлено факторами среды и «личностны-
ми» свойствами всех структурно-функциональных микроуровней
(атомно-молекулярные), то любопытно было бы проследить за
подобным проявлением адаптивного поведения на уровне клетки
и целого организма.
Клетка в этом отношении представляет особый интерес. Ведь
в живой природе клетка представительствует, так сказать, в
двух ипостасях: как самостоятельный одноклеточный организм
и как мельчайшая специализированная морфофункциональпая
единица многоклеточных.
Грешным делом, автор настоящих очерков склонен крамольно
думать, что если не у всех, то, по крайней мере, у большинства одно-
клеточных или в отдельных клетках многоклеточных организмов
некоторые органеллы (митохондрии, хлоропласты, рибосомы и др.)
не являются продуктом дифференциации живого вещества первичной
клетки, бывшего некогда однородным. Набравшись дерзновенной
смелости и фантазии, можно думать, что как ныне существующие
одноклеточные, так и клетки специализированных органов и тканей
многоклеточных в прошлом представляли собой многоклеточные (ве-
роятно, колониальные) образования неспециализированных клеток
(или обособленных доклеточных структур). В процессе эволюции ко-
лониальных образований отдельные структуры, обретая ту или иную
специализацию, утрачивали самостоятельность н объединялись в об-
разования, ныне называемые клеткой, в качестве узкоспециали-
зированных органелл. Одним из доказательств справедливости тако-
го рассуждения является достаточно обоснованное представление о
бактериальной природе митохондрий, в какой-то степени еще сохра-
нивших относительную внутриклеточную самостоятельность (актив-
но функционирующая ДНК, отличная по составу и продолжительно-
сти жизни от ДНК ядра; большее сходство ее с нуклеоплазмой
бактерий, чем с ядерной ДНК; отсутствие сформированных мито-
хондрий; наличие митохондриальных мутантов, нс дающих расщеп-
ления в потомстве; автономный синтез белка, чувствительного к
ингибиторам бактериального синтеза, и т. д.). Возможно, сказанное
относится и к другим менее самостоятельным органеллам клетки, не
говоря уже об облигатном внутриклеточном паразитизме вирусов.
Таким образом, ныне существующие клетки можно рассматри-
вать не как изначальные структуры, в отношении которых постулат
«Omnis cellule е cellula» («каждая клетка от клетки») абсолютен, а
105
как продукт длительной эволюции и специализации примитивных
клеточных или доклеточных образований колониальных структур,
утративших широкую адаптивную универсальность, базирующуюся,
папримср, на таком примитивном механизме регуляции как зависи-
мость скорости реакции от концентрации реагентов и др.
Усложнение взаимоотношений со средой, естественно, должно
было вынудить живые организмы прибегнуть к Осуществлению
основного принципа — обеспечению надежности через множество
дублирующих и менее надежных элементов, т. е. к образованию
клетки.
Первым обстоятельством, не столь удивляющим, сколь под-
тверждающим сложившееся представление, является факт су-
ществования колебательно-волновой периодичности в движении
цитоплазмы, причем не просто периодичности, а синхронизации
колебаний всех органелл и клетки в целом. Как считает Шноль,
«такая синхронизация может осуществляться как при взаимо-
действии посредством акустического или электромагнитного
полей, так и посредством «химического поля», т. е. при взаимо-
действии через «общие метаболические фонды», что в данном
случае кажется более вероятным (выделено нами,— А. Л.). Так,
кинетический (метаболический.— А. Л.), колебательный про-
цесс в клетке может синхронизировать колебания форм и раз-
меров (объема) всех органелл в данной клетке».
По мнению Д. Ц. Спаннера (1959), органеллы клетки, на-
пример митохондрии, благодаря протекающим в них периоди-
ческим осцилляторным химическим реакциям могут быть цент-
рами двух видов излучений — электромагнитного и «эластич-
ного». К какому же выводу иа основании этого приходит Спан-
нер? «Фактически, митохондрии могут говорить по собственному
каналу радиовещания» (III). Пока ограничимся лишь тремя
(хотя идея заслуживает значительно больше) восклицатель-
ными знаками.
Исследователи, изучающие колебательные процессы в био-
логических системах считают, что конформационные колебания
молекул, клеточных органелл и клеток в целом вместе с кине-
тическими колебаниями биохимических процессов могут созда-
вать такое колебательное поле, которое в состоянии обеспечить
не только индивидуальность каждого элемента, но и целост-
ность — гомеостатичность, в том числе и морфогенетическую,
всей клетки как единой морфофуикциональной структуры.
106
Вот несколько соображений по этому поводу. Посредством
акустических и электромагнитных полей, существующих в клет-
ке благодаря конформационным колебаниям макромолекул,
могут взаимодействовать не только молекулы, но и все состав-
ляющие клетку, а также клеток друг с другом. Так, собственно
акустические колебания, как и прочие, по сути и есть передви-
жение частиц в пространстве с определенной частотно-ампли-
тудной размерностью — скоростью. Согласно мнению Шноля,
«градиенты акустического поля, создаваемые метаболическими
и механохимическимп процессами, могут быть причиной «пра-
вильного» расположения структурных элементов клеток (орга-
нелл и др.) и самих клеток в заключающем их пространстве.
Да и не только клеток, но и пространственных отношений вну-
триклеточных образований, в том числе и во время митоза.
Важным является то, что синхронизированные конформацион-
ные колебания макромолекул и градиентов создают условия
для оптимальных внутриклеточных пространственно-гомеоста-
тических взаимоотношений, способных обеспечить ритмическую
гомеостатичность биохимических превращений (например, роль
конформационных изменений молекулы гемоглобина в выпол-
нении его кислородотранспортных функций). Такие колебатель-
ные метаболические процессы в клетке в равной степени необ-
ходимы для поддержания всех возможных стационарных
состояний — от различных уровней покоя, активности до мито-
тического деления.
Итак, колебательная природа гомеостатического поведения
биосистсм рассмотрена на молекулярном, клеточном и частично
органном уровнях. Для того чтобы еще более убедиться в ко-
лебательной природе гомеостатического поведения функцио-
нальной активности отдельных органов, достаточно вспомнить
хотя бы авторитмичность деятельности сердца, систем орга-
нов — ритмическую синхронность деятельности сердца и пери-
ферического сосудистого русла, межсистемных отношений —
ритмическую активность сердечно-сосудистой системы и син-
хронизированную в определенном отношении с ней систему ор-
ганов дыхания (например, частота сокращения сердца и дыха-
ния в покое в отношении 4—5:1) н т. д. Причем во всех случаях
только при строго определенных частотно-амплитудных соот-
ношениях возможно поддержание в течение необходимого вре-
мени стационарно-гомеостатического функционального состоя-
107
пня окислительно-восстановительных процессов в органах и
тканях.
Вернемся вновь к Шнолю. «Подволя некоторые итоги рас-
смотрения кинетических (химических, метаболических.— А. Л.)
колебаний и синхронизированных конформационных колеба-
ний в клетках, мы можем задать вопрос: что полезного для
живых организмов в колебательном режиме указанных двух
типов? Кинетическим и конформационным колебательным ре-
жимам живые организмы обязаны: 1) согласованным (адаптив-
но-гомеостатичным.— А. Л.) во времени течением различных
биохимических процессов; 2) наличием механизма биологиче-
ских часов: 3) всеми видами движения — от биения ресничек,
жгутиков и движения протоплазмы до ритмического сокращения
сердца и работы поперечнополосатых мышц; взаимодействием
клеток в процессе морфогенеза; 5) основным механизмом ра-
боты ферментов».
Безграничность, сложность и неисчерпаемость внутри и вне
организменных связей столь велики, что приводить далее ана-
логии этому — труд более чем неблагодарный.
Все циклично в этом мире!
Вращение Земли неравномерно, циклично: в августе оно бы-
стрее, а в марте медленнее. Разница, по нашим понятиям, не-
большая, всего 0,0025 секунды, ио она есть! Эта цикличность
вызвана Солнцем, временами года. Вместе с тем Солнце имеет
свою цикличность, которая зависит от цикличности других кос-
мических тел и т. д. до бесконечности. Естественно, что все эти
формы н виды циклов и тем более циклов земных и солнечных
не могли и не могут не сказаться па событиях, свойствах, каче-
ствах, динамике активности биологических систем. Надо пола-
гать, что ритмический колебательный характер биохимических
процессов и, следовательно, вообще всех форм проявления жи-
знедеятельности биологических систем имеет свою причинную
обусловленность, которая заложена в сущностном содержании
закона о всеобщей связи и взаимообусловленности всех явлений.
Физические ритмы — электромагнитные, световые, темпера-
турные, ритмы волн морских приливов и других изменений
среды — должны быть усвоены живой материей, чтобы не ока-
заться с ними в слишком большом противоречии. Теоретический
анализ значения пространственно-временных законов физиче-
ского мира для возникновения условнорефлекторных механиз-
108
мои и регулирования функций привел академика П. К. Анохина
(1968) к важнейшему общебиологическому обобщению. Смысл
его — в утверждении пространства и времени как абсолютных
факторов приспособления живой природы к внешнему миру
и в том, что «движение материи по последовательным ритми-
чески повторяющимся фазам является универсальным законом,
определившим основную организацию живых существ на пашей
планете». Усвоение этих ритмов приводит к формированию в
биологических системах собственных биоритмов (смена сна
и бодрствования, степени активности, форм поведения и т. д.) и
способности «следить» за временем, т. е. создает так называе-
мые биологические часы (рис. 22). Фазы периодических процес-
сов, совершающихся на всех структурно-функциональных уров-
нях биологических систем, могут сдвигаться и изменяться.
В связи с тем что колебательный периодический процесс
является свойством живой материи, биосистема должна обла-
дать авторитмичностыо. Причем не вообще авторитмичностыо,
а такой, которая по своим частотно-амплитудным характерис-
тикам не выходит за рамки эволюционно-фиксированных экст-
ремумов, т. е. она должна обладать достаточно высокой гомео-
статичиостью.
Видимо, для правильного понимания соотношения понятий
устойчивости, гомеостатичиости и ритмичности, осцилляторно-
сти биологических процессов следует воспользоваться пред-
ставлением Уоддингтона (1956) о трех главных уровнях рас-
пределения активности во времени: биохимическом, морфогене-
тическом (эпигенетическом) и эволюционном. Именно это
деление Гудвин (1966) положил в основу разграничения
различных систем, предложив рассматривать три вида их:
метаболический, эпигенетический и генетический.
Следуя Гудвину, в целях формализации наших представле-
ний введем понятие релаксации, точнее, времени релаксации.
Время релаксации в широком смысле—это время, необходи-
мое для того, чтобы те или иные переменные (например, откло-
нение в течении биохимических реакций, частоты сердечных
сокращений и т. д.) после некоторого возмущающего воздей-
ствия на биосистему возвратились к своим стационарным со-
стояниям, к исходным колебаниям. Оно зависит от темпера-
туры в момент и после воздействия возмущающего фак-
тора.
109
110
Рассмотрим это на про-
стом Примере: вспомним пре-
неприятную процедуру корм-
ления манной кашей. Лично
для меня это всегда было
тяжелейшим испытанием:
прежде чем отправить пор-
цию этой ненавистной каши
в рот, я достаточно долго во-
дил по ее омерзительной по-
верхности ложкой, каждым
движением создавая на ней
борозду и ожидая ее исчез-
новения. Это было, пожалуй,
единственным утешением в
тяжелой борьбе с кашей.
Только недавно я понял, что
в общем-то имел дело с одним
из видов релаксации. Так
вот, «манная» релаксация
также зависит от темпера-
туры каши: при горячей —
она быстрая, при холодной —
медленная,
«Значение этого поня-
тия,— пишет Гудвин,— ясно
из следующего существенного
факта — если времена релак-
Рис. 22. Движение листьев фа-
соли (по Е. Е. Крастиной,
1967):
А~ в темноте (0,5 лк); Б — при
непрерывном освещения в 7500 лк;
В — при непрерывном освещении
в 1250 лк. По оси ординат: угол
между черешком и центральной
жилкой листа! по Леи абсцисс:
цифры — астрономическое время;
светлые и темные участки — ос
вещенность в предшествующем ре-
жиме; горизонтальная линия —
средняя величина угля.
111
сации в двух системах различаются очень сильно (например, в
100 раз), то во время изменения переменных «медленной» сис-
темы (с большим временем релаксации) «быструю» систему (с
малым временем релаксации) можно рассматривать как систе-
му, находящуюся в стационарном состоянии.
Тогда, с одной стороны, в динамические уравнения медлен-
ной системы войдут лишь стационарные Значения переменных
быстрой системы и число уравнений резко уменьшится. С дру-
гой стороны, переменные «медленной» системы войдут в урав-
нения «быстрой» системы как параметры, а не как переменные.
Вообще говоря, эти параметры будут медленно изменяться,
вызывая медленные изменения быстрой системы во времени;
однако при изучении динамики быстрой системы на малых от-
резках времени переменные медленной системы можно рассмат-
ривать как параметры среды. Таким образом, временной кри-
терий, позволяющий отличить одну систему от другой, будет
достаточно строгим только в том случае, если времена релак-
сации этих двух систем существенно различны» (выделено
нами.— А. Л.).
Вернемся к системам. Метаболическая система (в данном
случае клетки) включает основные процессы, определяющие
скорость изменения се строения. Это пассивный и активный
перенос веществ, интенсивность и скорость биохимических пре-
вращений, в том числе и ферментативные взаимодействия и
превращения «малых» молекул (не макромолекул), а также
взаимодействия между малыми молекулами и макромолеку-
лами (например, активация и ингибирование ферментов и др.).
Синтез макромолекул ие входит в метаболическую систему, так
как считается, что концентрация макромолекул играет в ней
роль постоянных или очень медленно меняющихся параметров
среды. Разумеется, это своеобразная абстракция, облегчающая
(и, кстати, не слишком нарушающая) принципиальную строй
ность и правомочность рассуждений.
Установление стационарного состояния в метаболической
системе определяется главным образом скоростью превращений
субстрата под влиянием ферментов, т. е. числом оборотов в
единицу времени. Однако, учитывая, что внутри клетки (или
любой биосистемы) происходит множество фсрмент-субстрат-
ных реакций, протекающих в определенной последовательности
и в сложнейших циклических взаимоотношениях и имеющих
112
собственный временной параметр протекания, конечный резуль-
тат оборачиваемости, необходимый для сохранения суммарной
метаболической стационарности, будет измеряться средне-обоб-
щенным временем, а не быстротой отдельного цикла. Причем,
чем короче это время, тем более стационарной будет данная
биосистема. Это рассуждение особенно существенно, потому что
оно имеет прямое отношение к широкому классу динамических
процессов обмена веществ, при которых концентрации опреде-
ленных метаболитов подвержены незатухающим периодическим
изменениям. Именно эти процессы как раз и играют централь-
ную роль в динамической организации не только метаболиче-
ской, по и вообще всех видов биосистем.
П. Л. Спавдглер, Ф. М. Шелл показали, что колебания
могут возникнуть в системе, где два фермента сопряжены
взаимным ингибированием. Вот что по этому поводу пишет
Гудвин: «Существование метаболических осцилляторов услож-
няет анализ временной организации клетки, так как в этих
условиях необходимо учитывать влияние осцилляторов на ди-
намику эпигенетической системы. И в этом случае ситуация
зависит от соотношения времени релаксации метаболической
и эпигенетической систем... Таким образом, даже если период
метаболического осциллятора равен I или 2 минутам и... нахо-
дится за пределами эпигенетической системы, явление деления
частоты может дать колебания с любым периодом — от 5 до
30 минут и более.
Эти верхние значения уже близки к собственным временам
эпигенетической системы, и, таким образом, метаболит, уча-
ствующий в относительно медленных колебаниях, может стать
элементом эпигенетических процессов... Итак, рассматривая
колебательные явления в метаболической системе, мы будем
считать, что их частоты достаточно велики и что они лежат
вне временного диапазона эпигенетической системы. В этом
случае можно будет использовать допущение об относительной
стационарности (гомеостатичности.— А. Л.) метаболической
системы». Гудвин прав, говоря, что «мы должны быть готовы
к тому, что такой подход окажется совершенно неадекватным
поставленной задаче, т. е. что динамическое разделение мета-
болической и эпигенетической системы внутри одной клетки (а
возможно, и вообще биосистсмы.— А. Л.) не имеет смыс-
ла».
113
В эпигенетической системе клетки следует рассматривать
осцилляции, связанные с процессами достаточно длительными.
Это и биосинтез, и диффузия, и взаимодействие макромолекул.
Так, время, затрачиваемое бактерией на синтез одной молекулы
белка, равно 5 секундам, у высших организмов оно возрастает
до нескольких минут. По достаточно грубым расчетам, синтез
одной молекулы РНК у бактерий продолжается 1 секунду, у
высших организмов — около 1 минуты. Для обсуждаемой нами
проблемы весьма полезны данные, полученные П. Фейгельсоном
и О. Грипгардом (1962). Они установили следующий факт.
После внутривенного введения крысам триптофана синтез трип-
тофанпиролазы в печени начинается только через 2 часа. При-
чем первый этап, связанный с превращениями, не включающими
синтез макромолекул, продолжается несколько минут, т. е.
столько, сколько свойственно релаксациям, совершающимся
в пределах «компетенции» метаболической системы, так как это
характерная «метаболическая» реакция, не включающая синтез
макромолекул (в данном случае ацетил-КоА). Для деградации
избыточно введенного триптофана в ацетил-КоА необходим до-
полнительный синтез фермента триптофанпиролазы. Он проис-
ходит приблизительно через 2 часа благодаря индуцирующему
действию триптофана. Таким образом, синтез фермента de novo
осуществляется во временных параметрах эпигенетической си-
стемы. При этом, по мнению исследователей, происходит и син-
тез гомологической информационной РНК- Итак, эксперимен-
таторам удалось достаточно четко разделить по времени мета-
болические и эпигенетические реакции. Причем самым важным
здесь является то, что эпигенетическая реакция не может про-
явиться до тех пор, пока в метаболической системе не уста-
новится допустимо возможное стационарное состояние. На-
пример, время реакции эпигенетической системы клеток печени
должно составлять 2 часа... Можно предположить, что время
релаксации эпигенетической системы клетки лежит в диапазоне
102—10* секунд» (от 1,5 минуты—продолжительность метабо-
лической реакции—до 3 часов — продолжительность эпигене-
тической реакции.— А. Л.). Однако с позиции, рассматриваю-
щей гомеостаз как осцилляторный процесс, еще большее зна-
чение приобретает заключение Гудвина, что «...эпигенетическую
систему одиночной клетки можно рассматривать как часть
среды, в которой находится се метаболическая система, прн
114
условии, что разница между временами релаксации этих двух
систем достаточно велика.
Согласно нашим сценкам, обычно это условие выполняется,
и, следовательно, концентрации макромолекул можно рассмат-
ривать как постоянные или медленно меняющиеся параметры
на отрезке времени, необходимом для метаболической реакции
на малые стимулы. Однако, если стимулы велики и длительны,
то сначала может последовать быстрая реакция метаболической
системы, а затем более медленное и, вероятно, более сильное
изменение метаболических переменных, вызванное изменением
концентрации макромолекул. Во время этой второй фазы реак-
ции «параметры» метаболической системы изменяются и время
наблюдения должно быть увеличено по сравнению с тем, ко-
торое необходимо для регистрации метаболической реакции.
Таким образом, в наших терминах этот процесс следует опи-
сывать как реакцию эпигенетической системы клетки».
Вот мы и добрались до самой сути осцилляторной природы
гомеостатического поведения биологических систем — соотно-
шения осцилляторных характеристик метаболической и эпиге-
нетической систем; эпигенетическая система содержит в себе
метаболическую систему в том смысле, что все переменные по-
следней входят в первую.
Если идти дальше, то с помощью понятия времени релак-
сации в одиночной клетке (бактериальной или у простейших)
можно выделить и генетическую систему.
Гудвин не выделяет время релаксации генетической систе-
мы. И это не случайно, так как часто приходится сталкиваться
с чрезвычайными сложностями, суть которых в первую очередь
в недостаточности наших знаний. Однако в том, что существуют
генетические релаксации, сомнений нет. Поэтому в математи
ческом выражении эти системы представляют ряд, связанный
соотношениями включений;
sic:S2c:Ssc...Sn
где Si— метаболическая система; Ss— эпигенетическая система,
S3 — генетическая система и т. д. вплоть до таких образований
высокого порядка, как экологическая система, и др.
Изложенная концепция временной организации биологиче-
ских систем, разумеется, лишь попытка построения теории.
Однако «лучше синица в руках, чем журавль в небе». Наиболее
115
уязвимым местом теории является исходная позиция, опираю-
щаяся на понятие «релаксация», т. е. па понятие времени, необ-
ходимого для того, чтобы те или иные смещения (колебания)
возвратились к своим исходным стационарным состояниям
после некоторого возмущения.
Этого-то в реальной природе, мертвой и тем более живой
ие бывает. Здесь релаксация осуществляется всегда с опреде-
ленным сдвигом относительно исходной величины и, что самое
скверное, не в «положительную» cTopoity, особенно для био-
снетем, в силу фатальной обреченности ее онтогенетического
гомеостаза. Поэтому временная релаксационная концеппция
Гудвина почти полностью (если ие считать необратимость со-
бытий, происходящих в открытых системах) удовлетворяет
понятию метаболической системы на данном и трудно учиты-
ваемом отрезке времени. Менее удовлетворительна она для
эпигенетической системы, для которой характерна тенденция к
затухающим автоколебаниям (в широком смысле слова).
В генетической (в филогенетическом плане) системе в ко-
нечном результате (учитывая зарождение генотипа, его расцвет
и т. д.) релаксации как таковой фактически нет. В принципе
нет даже затухающих колебаний. Есть только конец. Не слу-
чайно существует понятие «вымирание живого», причем как
характеристика естественного явления для данного вида или
типа. Вымирание, разумеется, не следует понимать как некий
недетерминированный рок. Напротив, это естественное след-
ствие накопления генотипической необратимости и нарастания
рассогласования между темпами генетической и средово-геоло-
гической эволюции, завершающегося биологической катастро-
фой для таксона. Так по крайней мере было во все времена
существования жизни на Земле. Будет ли так далее? Это, ви-
димо, в какой-то степени зависит от человека, обретшего спо-
собность не только приспосабливаться к среде, но и приспосаб-
ливать ее к себе. От того, насколько социально разумно будет
его поведение в этом отношении, настолько адекватно будет
релаксационное, да и вообще колебательное проявление био-
логического гомеостаза на нашей планете.
Но отбросим печальные размышления и будем более опти-
мистичными. Попробуем привести взаимоотношения между
метаболической и эпигенетической системами к общему, форма-
лизованному выражению, учитывая при этом, что в отличие от
116
всех других специалистов биолог имеет дело со сложно орга-
низованной системой и его подход должен быть совершенно
отличным от подхода инженера. Он не может изолировать
даже условно часть, содержащую лишь несколько переменных,
не нарушив общего поведения всей системы. Он не может
произвести «линеаризацию», не утратив важнейших динамиче-
ских свойств системы, ибо все явления в биологических систе-
мах, в том числе и результирующие (индивидуальный онтоге-
нез), нелинейны. И наконец, он ие может исследовать проблему
регулирования так, как это делает инженер, т. е. пользуясь
некоторыми известными критериями, искать сигнал обратной
связи, обеспечивающей оптимальное поведение системы. Биолог
не всегда может установить критерий оптимального протекания
регуляторных или адаптивных процессов в клетке. Поэтому
будем пока довольствоваться лишь некоторыми достаточно кор-
ректными математическими расчетами Гудвина.
Предположим, что концентрация метаболита определя-
ется концентрацией белка Ъ- Значит, Г; представляет собой
переменную, ограничивающую скорость синтеза Л1(. Рассмотрим
случай, когда М/, являясь конечным продуктом реакционной
цепи, поступает в общий метаболический фонд. В этом случае
получается простейшая кинетическая схема, которую можно
представить в виде уравнения:
dM,
dt
ryi — SiMi,
где Sf— константа скорости распада которая имеет первый
порядок; rt — сложная константа, зависящая от констант
скорости элементарных стадий, концентрации субстрата и кон-
станты Михоэлиса (Кт) для реакции, продуктом которой явля-
ется Mi.
Вспомним, что У/ и Mi принадлежат к разным системам:
первая — к эпигенетической, вторая — к метаболической. Поэ-
тому различия между изменениями их скоростей очень велики
(вспомним также идею о временных релаксациях). Следова-
тельно, можно считать, что Mt находится в стационарном со-
стоянии во время т. е. можно написать;
117
dM,
dt
Динамическое поведение метаболических фондов в клетке,
где работают колебательные регуляторные механизмы, осложня-
ется тем, что размеры этих фондов сами колеблются. Это сле-
дует из уравнения, показывающего, что при рассмотрении от-
резков времени, характерных для эпигенетической системы,
Ml ведет себя по существу так же, как и Yt На более коротких
отрезках времени поведение Mt будет определяться также
взаимодействиями внутри метаболической системы, но они мо-
гут рассматриваться как «шумы» по отношению к эпигенети-
ческим процессам, если принять предположение о наличии су-
щественной разницы в величинах релаксаций. Однако так назы-
ваемый метаболический шум отнюдь не помеха, а основа про-
явления релаксационных свойств эпигенетической системы. В
связи с тем что и метаболические, и эпигенетические процессы
имеют колебательную природу, естественно, что используемая
при этом свободная энергия G также носит колебательный ха-
рактер и соответственно названа Гудвином таландической.
Так, можно представить, что некоторое число соединений V
не изолировано от остальных компонентов клетки и обменива-
ется с ними таландической энергией n— V. Если п очень вели-
ко, то для подавляющего большинства компонентов канониче-
ского ансамбля Gt будет довольно близко к каноническому
среднему G.
Это значит: 1) величина G распределяется по всей системе
благодаря взаимодействию эпигенетических компонентов через
общие генетические фонды; 2) так как интенсивность метабо-
лических осцилляций находится в зависимости от температуры
© (таландическая), последняя в свою очередь служит мерой
возбуждения системы, находящейся «в полном покое» относи-
тельно устойчивого уровня (стационарного состояния). Следо-
вательно, первый случай является материально-энергетической
основой возникновения и поддержания гомеостатического со
стояния биосистемы, второй — определяет меру возможных ста-
ционарных состояний ее в пределах допустимых гомеостатиче-
ских колебаний, а вместе они оба выражают адаптивно-гомео-
статические возможности этой системы в конкретных ситуациях.
Отсюда «...клетка (биосистема.— А. Л.), в которой эпигенети-
ческая система находится на высоком G-уровие, имеет зиачитель-
118
ное преимущество в смысле адаптации перед клеткой (биоси-
стемой.— А. Л.) с очень низким G-уровнем (если обе они поме-
щены в периодически изменяющиеся внешние условия, что
обычно и происходит на нашей планете). Кроме того, для до-
стижения устойчивых упорядоченных во времени взаимоотно-
шений между происходящими в клетке биохимическими про-
цессами и приспособления ее к периодическим изменениям
внешней среды требуется значительная нелинейность... (!! —
А. Л.). Эпигенетическая система, находящаяся на высоком
G-уровие, должна совершать меньше таландической работы,
чтобы адаптироваться, чем система, находящаяся в состоянии
с очень малым в» (Гудвии, 1966). Следовательно, надо пола-
гать, что для такого утверждения есть достаточные основания.
Итак, в эпигенетической системе (процессах) клетки роль
переменных играют концентрации т-РНК н белка: т-РНК
является переносчиком аминокислот, необходимых для синтеза
белка; количество же синтезируемого белка по принципу об-
ратной связи регулирует концентрацию т-РНК. С позиции
представлений непосвященного человека многие процессы со-
вершаются весьма быстро. Например, некоторые бактериальные
клетки синтезируют до 1000 молекул протеинов в 1 секунду.
У млекопитающих белок клеток обновляется в среднем за 1 час
на 1%, а белки некоторых ферментов на 10% (у человека ве-
сом 70 кг в 1 час обновляется около 100 г белка, т. е. «полное»
белковое обновление происходит менее чем за трое суток). Не-
смотря на это, данные процессы следует отнести к эпигенети-
ческим, так как оии «вызывают появление макроскопических
(морфогенетических.— А. Л.), т. е. иерархически более высоко
расположенных свойств клетки» (Гудвин). Вывод сводится к
тому, что для клетки, а в широком смысле и для любой био-
логической системы характерна, временная (осцилляторная)
организация трех видов (возможно, и больше): метаболическая,
эпигенетическая и эволюционная. Понятие «эволюционная си-
стема» рассматривается широко, в том числе и па уровне клет-
ки, в виде деградации клеточных структур при раковом и других
заболеваниях или врожденных дефектах с проявлением эво-
люционно более (менее) примитивных морфофункциональных
изменений.
И еще один вывод—ритмический колебательный характер
метаболических процессов имеет свою причинную обусловлен
119
ность. Она в первую очередь заложена в сущностном содержа-
нии закона о связи и взаимообусловленности всех явлений.
Итак, опять приходим к тому, что все циклично в этом мире.
Степень гомеостатичности колебательных и тем более авто-
колебательных процессов биосистем находится в сложной за-
висимости от амплитудно-частотных характеристик, в которые
мы вкладываем следующее содержание.
1. Амплитуда гомеостатического параметра — абсолютная
величина его максимального смещения в данных условиях.
2. Частота гомеостатического параметра — количество функ-
циональных циклов (замкнутых процессов), совершающихся
в единицу времени.
3. Амплитудно-частотная интенсивность гомеостатического
параметра — это энергетическая «стоимость» функционального
цикла в единицу времени в принятых для данного процесса
единицах измерения.
Читатель должен учитывать некоторую условность опреде-
ления этих понятий, так как в биосистеме нет изолированных
функциональных циклов. Мало того, одно и то же функциональ-
ное событие реализуется множеством совершающихся в стро-
гой иерархической последовательности процессов. Каждое пре-
дыдущее звено (микроцикл) вовлекает следующее (мезоцикл),
которое в свою очередь усложняется подключением новых
звеньев (макроцикл), и циклы завершаются целостной реак-
цией биосистемы на возмущающее воздействие. Все микроциклы
и особенно обменио-энергетические более «быстрые», чем мезо-
и тем более макроциклы. Чем быстрее возникает целостная от-
ветная реакция на возмущение, тем более адаптивно-гомеоста-
тичной она будет по своему результату. Отсюда явно вытекает,
что чем более высокочастотным и низкоамплитудным будет
процесс, тем меньше он отклоняется от некой гипотетической
идеальной траектории как по своим лонгитудинальным, так и
трансверзальным параметрам, тем более стационарным, устой-
чивым, адаптивно-гомеостатичным будет данный процесс.
Это один из важнейших непарадоксальных парадкосов го-
меостаза: чем подвижнее, тем стабильнее. Его в свое время
подметил А. А. Ухтомский: «Высоколабильный в физиологи-
ческом смысле — высокоработоспособный и надежный, ибо он
наименьше изменчив (в формально регистрируемом внешнем
проявлении.— А. Л.) в своей деятельности».
120
Попробуем убедиться в этом, минуя промежуточные «ин-
станции», на примере колебаний общего центра тяжести (ОЦТ)
при сохранении вертикального положения тела у человека
в различных условиях. Установлено (В. А. Цаун, 1971), что
при поддержании вертикальной позы, т. е. с нашей точки зре-
ния одного из стационарных состояний двигательного гомеоста-
за, стоя иа земле, количество колебаний ОЦТ с частотой от
1 до 10 Гц в пересчете на 1 минуту равно 53,6±9,8 (X±SjT)
При помещении человека на высоту 3 метров с ограниченной
площадью опоры количество этих колебаний достоверно возрас-
тает до 77,2± 14,84 (па 44%). Количество же колебаний с час-
тотой ниже 1 Гц, вызванных переносом центра тяжести с левой
стороны стопы на правую, напротив, уменьшается с 14,0±3,58
на земле до 10,6±2,34 колебаний в минуту иа высоте, что со-
ставляет 17,1% (различие достоверно).
Обратим внимание — в условиях, опасных для жизни и спо-
собствующих смещению центра тяжести тела, частотная харак-
теристика колебаний увеличивается) При анализе же расстоя-
ний между верхними и нижними экстремумами стабилограммы
обнаружено, что на земле амплитуда смещения равна 19,0±
±2,53 мм, а на высоте 3 м — 14,8±2,65 (разница достоверна).
Следовательно, амплитуда колебаний уменьшается в более
сложных условиях. Итак, увеличение частоты колебаний ОЦТ
и уменьшение их амплитуды свидетельствуют о стремлении
(непроизвольном!) уменьшить колебания тела и не допустить
смещения в область, граничащую с потерей равновесия. Пове-
дение явно адаптивно-гомеостатическое. Еще более любопытен
в этом отношении тот факт, что максимально воспроизводимая
частота произвольных движений у подростков практически ос-
тается прежней и на высоте 10 м, а у взрослых даже снижается.
Это можно рассматривать как эффект переноса адаптивно-го-
меостатической регуляции двигательной активности (сохране-
ние наиболее безопасной позы в экстремальных условиях) из
произвольной сферы в непроизвольную — автоматическую. Л
возрастные отличия объясняются большим совершенством этих
механизмов у взрослых. Тут речь идет об изменении частотно-
амплитудной характеристики поддержания двигательного го-
меостаза.
Особенно демонстративен подобный механизм поддержания
гомеостатического уровня тех или иных параметров внутренней
121
среды В первую очередь это уже упомянутый выше pH крови.
Необычное постоянство pH есть следствие высочайшей чув-
ствительности организма к смещению концентрации ионов
водорода.
Весьма любопытно проследить изменения колебаний содер-
жания сахара в крови. В условиях покоя они имеют для опре-
деленной биосистемы индивидуальный профиль, рисунок, как
говорят иногда, «сахарное зеркало». Нанесение раздражения
вызывает значительное и длительное изменение концентрации
сахара в крови. Эти изменения имеют волнообразный характер.
Они вначале высокоамплитудные и низкочастотные, причем
столь значительные, что почти полностью перекрывают (сти-
рают) колебания, регистрировавшиеся за более короткие про-
межутки времени в условиях покоя. Вызванные возмущением
колебания содержания сахара в крови постепенно затухают
и приобретают особенности исходного ритма физиологического
покоя. В случае достаточно полной компенсации сдвига наблю-
дается восстановление исходного уровня обмена веществ. При
неполной компенсации концентрация сахара экспоненциально
или логистически переходит на новый уровень, на котором вос-
станавливается исходный «спонтанный» ритм, или последний
может приобрести новую, более адекватную (оптимальный,
стационарный уровень) возникшим условиям характеристику.
Переход биосистемы с одного стационарного уровня на другой,
т. е. к новому гомеостатическому «потолку», лается недешево.
На это расходуется энергия и не малая. Так, при изменении
колебаний ОЦТ на различных высотах одновременно регистри-
ровались существенные сдвиги в частоте сердечных сокращений
и дыхания, т. е. в работе системы энергообеспечения. Читатель,
разумеется, представляет себе, что в этом случае даже формаль-
но незначительные сдвиги обмена веществ по своему функцио-
нальному содержанию значимы. Ведь энергия расходуется и на
реализацию приспособительного действия (например, движе-
ния), и па минимально допустимое (экономичное) в этих усло-
виях изменение состава внутренней среды. «Этот процесс про-
ходит не слишком гладко. Прежде чем рост достигнет средней
точки (оптимума, нового стационарного состояния для данных
условий), он начинает скакать и вертеться,— пишет Прайс,—
менять, подобно проказливым духам, формы и определения с
тем, чтобы обойти этот ужасный потолок. Или говоря в менее
122
антропоморфных аналогиях, здесь устанавливается кибернети-
ческий эффект поиска, и кривая испытывает резкие колебания».
Колебательной природе гомеостаза присущ еще один удиви-
тельный механизм, работающий в соответствии с известным нам
принципом устранения или максимального ограничения дей-
ствия внешних и внутренних факторов, нарушающих относи-
Рис. 23. Концентрация глюкозы в крови у кошки:
I — динамика концентрации в условиях внутривенного введе-
ния глюкозы; //-—динамика концентрации в условиях вве-
дения инсулина (штриховая линия — те же услсння, но при
болевом раздражении редел торов прямой кишкн повышением
давления в момент максимальной концентрации). Глюкоза и
инсулин вводились из расчета влияния на концентрацию глю-
козы в крови в течение равных отрезков времени.
123
тельное динамическое постоянство биосистемы. Одно из частных
выражений его, как отметил Гудвин, проявляется следующим
образом: «Направление смещения колебательных траекторий
(от идеального стационарного состояния или к нему.— А. Л.)
под влиянием импульса будет зависеть от того, в какой части
траектории находилась система в начале воздействия. Если она
находилась на части траектории, расположенной иод стацио-
нарным состоянием, тогда импульс сместит систему «вверх»,
к нему, уменьшив тем самым амплитуду колебаний (вот он,
«укротитель проказливых духов» Прайса.— А Л). Если же
в начале возмущения концентрация белка превосходила ста-
ционарную (автор иллюстрирует это положение на примере
эпигенетической системы клетка — синтез белка.— А. Л.), то
в результате сдвига система будет удаляться от стационарного
состояния и амплитуда колебаний возрастет». Это еще один
непарадоксальный парадокс гомеостаза. Он выявляет одно из
принципиальных свойств механизма управления и регулирова-
ния в биосистеме. Это результатирующее свойство осциллято-
ров, сводящееся к тому, что колебания, происходящие в дан-
ных регуляционных системах, явно асимметричны.
И, наконец, еще один непарадоксальный парадокс, на ко-
тором мы остановимся. Он выражает следующий тезис: смысл
процесса авторегуляции сводится к тому, что само отклонение
от оптимально гомеостатического состояния покоя (нормы)
служит стимулом восстановления нарушенного состояния. Это
нашло отражение в ряде теоретических представлений и экспе-
риментальных факторов, что позволило сформулировать, если
так можно выразиться, гомеостатический закон гомеостаза —
закон исходной величины. По этому закону, «чем сильнее воз-
буждение вегетативных нервов, степень напряжения деятель-
ности вегетативного органа, тем слабее их возбудимость в от-
ношении возбуждения и тем сильнее их реактивность в отно-
шении угнетающих факторов» (Вильдер, 1931). Закон этот
может быть распространен вообще на все функции. Мы проил-
люстрируем его примером изменения направления обмена ве-
ществ (уровень сахара в крови) при возмущающих воздействи-
ях, поступающих в восходящую или нисходящую фазу откло-
нения его от уровня колебаний физиологического покоя
(рис. 23).
124
Очерк 3. ГОМЕОСТАЗ И КОНЕЧНОСТЬ
РАЗВИТИЯ БИОСИСТЕМ. ЕЩЕ ОДНА ПОПЫТКА
ИЗМЕРЕНИЯ ГОМЕОСТАЗА
В предыдущем очерке мы пришли к выводу, что наиболее
стабильное сохранение одного из предельного множества со-
стояний биосистемы осуществимо лишь при соблюдении обяза-
тельного условия — стационарность должна иметь колебатель-
ную, ритмическую природу. Эту мысль с полным правом можно
возвести в ранг общебиологического закона, закона биологи-
ческой адаптации, ибо постоянство живой системы свидетель-
ствует о том, что в пей действуют или готовы действовать аген-
ты, поддерживающие его. Значит, гомеостатическое состояние
не лишено способности к развитию, т. е. оно адаптивно. Именно
в силу этих обстоятельств мы и предпочитаем пользоваться
понятием «адаптивный гомеостаз», подчеркивая тем самым,
что адаптация биосистемы представляет собой процесс, в ходе
которого уровень гомеостаза непрерывно подвергается измене-
нию. Кроме того, чем более функционально специализирована
биосистема, тем более высокие у нее требования к поддержанию
строгости гомеостатического состояния, т. е. тем меньше по
амплитуде и больше по частоте должны быть колебания пара-
метров ее внутренней среды (а во многих случаях и функций
в целом).
Чтобы представить себе это положение достаточно четко,
рассмотрим, например, такую сложную биосистему, как орга
низм человека. Из левого желудочка сердца в аорт)', а затем
но всей артериальной системе сосудов к органам и тканям при-
текает кровь, практически однородная по составу. Между тем
организм человека состоит из множества специализированных
тканей и органов, каждый из которых специфичен не только
по строению и функции, но и в первую очередь по химическому
составу, т. е. по обмену веществ. Отсюда следует единственно
возможный вывод: проницаемость микрососудов избирательна
как для компонентов, притекающих с кровью, так и для посту-
пающих из клеток продуктов метаболизма. В результате сохра-
няется адаптивно-гомеостатическая специфичность внешней
среды (межклеточная жидкость). Таким образом, каждая клет-
ка благодаря практически неповторимой биохимической инди-
видуальности имеет свой внутри- и внеклеточный гомеостати-
125
ческий профиль, отличающийся, хотя и незначительно, от по-
добных ей клеток. Группа однородных клеток с весьма сходным
гомеостатическим профилем образует орган, характеризую-
щийся морфофункциопальной специфичностью. По этому прин-
ципу строится система органов и организма в целом. В резуль-
тате каждому организму присуща его собственная, сугубо ин-
дивидуальная иерархия адаптивно-гомеостатических отношений.
На основании изложенного читатель, естественно, пришел
к выводу: чем специфичнее морфофункциональные особенности
данной биологической системы (клетка — ткань — орган — си-
стема— системы — организм), тем жестче должны быть адап-
тивно-гомеостатические механизмы поддержания этой специ-
фичности. Приобретение и сохранение такого качества возможно
при условии, если биосистема приобретает способность реаги-
ровать с максимальной быстротой иа любое минимальное от-
клонение от свойственного ей адаптивно-гомеостатического
оптимума функции. Уровень чувствительности биоскстемы к
подобным отклонениям лежит в основе амплитудно-частотной
характеристики отклонений ее гомеостатических параметров.
Проиллюстрируем это соображение. В ходе эволюционного
развития животного мира нервные клетки (особеиио мозга)
приобретают наиболее совершенную специализацию в проявле-
нии свойств живой материи. Вот почему именно мозг функцио-
нирует в живом организме в условиях максимального гомео-
стаза. Разве только сердце может конкурировать с мозгом, для
которого характерны минимальные изменения внутримозгового
кровообращения даже при обильной потере крови, травмах,
гравитационных воздействиях; мозгу присуще сохранение нор-
мального метаболизма вплоть до наступления смерти в усло-
виях полного голодания и т. д. Нервные клетки более, чем
какие-либо другие, специализированы к восприятию возмущаю-
щих воздействий внешней или внутренней среды как организ-
ма, так и самой клетки. Именно это свойство поставило мозг
во главу механизмов регуляции общеорганизмениого гомеостаза
и обеспечило ему наиболее совершенные возможности регули-
рования собственного гомеостаза. В результате мозг приобрел
способность реагировать на самые минимальные изменения
внутренней среды. Возможно, процесс шел и в обратном по-
рядке или имели место одновременно оба процесса, выражая
тем самым диалектическое единство сущности адаптивного
1?6
гомеостаза. Каковы возможности дальнейшего совершенство-
вания мозга в этом отношении? Беспредельны! Совершенство
регуляции, осуществляемой мозгом (нервной системой), будет
расти пропорционально его способности оценивать изменения
внутренней или внешней среды. Количество же этих факторов
по мере повышения чувствительности нервной системы будет
беспредельно возрастать. Именно беспредельно! Однако при-
оритетное положение нервной системы относительно. Мы склон
иы думать, что в биосистеме в таком положении находятся
вообще все составляющие ее элементы. Подтверждением тому
служат безнервные бносистемы.
Вероятно, настоящее рассуждение читатель не воспримет
как отречение от ранее доказанного тезиса, что живое конечно.
Ведь там речь шла о конечности индивидуального проявления
жизни. Даже конечность жизни в планетарном—земном объе-
ме— это еще ие конечность жизни в принципе. Поэтому эво-
люция жизни в принципе не ограничена. Ограниченность же
индивидуального проявления жизни в первую очередь зависит
от временного использования выделенных ему природой ресур-
сов. О временной лонгитудинальной судьбе особи речь шла при
обсуждении онтогенетического гомеостаза; трансверзальиой —
при обсуждении колебательной природы гомеостаза.* Однако
до сих пор не ясны пределы этих параметров индивида в бес-
предельности общего проявления жизни и в бесконечности ее
эволюционирования. Правда, первая попытка была предприня-
та в предыдущем очерке; надо думать, и вторая будет лишь
очередным приближением.
На наш взгляд, правильно считает В. К- Прохоренко (1969).
что основные моменты процесса развития системы вообще, в том
числе и биосистемы, можно определить следующим: нетожде-
ственным обменом веществ между биосистемой и окружающей
средой (практически это равноценно идее динамического не-
равновесия, стационарности и эквифинальностн биосистемы с
окружающей средой) и его кумулятивным эффектом. Кумуля-
тивный эффект нетождественного обмена имеет как позитивное
значение в том отношении, что именно благодаря ему происхо-
дит накопление веществ в системе, ее рост и развитие, так и
* Напомним, что разделение на лонгитудинальные и трансвер-
зальные гомеостатические характеристики в данном случае условно.
127
21 {Предел насши.
t, tCp t2 Врет
Рис. 24. Логистическая кривая по-
ведения систем (по Д. Прайсу).
негативное — существование биосистемы в режиме нетожде-
ственного обмнлв приводит к противоречиям между внешним
и внутренним Обменом и в результате к уничтожению биосисте-
мы. Последнее может произойти при достижении биосистемой
предела насыщения отрицательными эффектами в процессе
сопротивления накоплению энтропии, т. е. удержания нетожде-
ственного обмена. Предел насыщения может возникнуть вне-
запно (трансверзально) или
развиваться медленно (лон-
гитудиально).
Как было отмечено, мно-
гие события, совершающиеся
в биосистемах, реализуются
по закону развития логисти-
ческой кривой (рис. 24). Про-
анализируем ее. Например,
параметр z начиная от ис-
ходного значения («пол» ло-
гистической кривой), растет
по экспоненте. Экспоненци-
альный рост принято харак-
теризовать периодом удвоения, т. е. временем, достаточным
для удвоения исходного значения измеряемого параметра.
Обычно через несколько периодов удвоения логистическая кри-
вая отрывается от экспоненты, и далее лараментр 2 растет
медленнее последней. Замедление роста прогрессирует, и при-
мерно через три периода удвоения после отрыва от экспоненты
логиста_ческая кривая выходит в точку перегиба (в момент вре-
мени i). Затем рост с насыщением представляется правой
ветвью кривой, симметричной левой. Пределом насыщения
роста является «потолок» логистической кривой, завершающий
конечное развитие.
За логистическим потолком кривой, описывающей всю жиз-
ненную «траекторию» биосистемы, нет жизненных перспектив.
Это конец, структура системы расформировывается. Если же
логистическая кривая описывает частный процесс, например
динамику изменения силы мышцы в единицу времени, то здесь
возможен «взрыв» логистического потолка и новое экспонен-
циальное восхождение, когда, как говорит Прайс, «подобно
фениксу из пепла старой логистической кривой возникает новая
128
кривая, имеет место так называемая эскалация». Для биологи-
ческих систем это явление нормальное. Убедител дм примером
может служить описание достижения предела насыщения сило-
вых качеств спортсмена на данном уровне развития их (спор-
тивная квалификация, тренированность и т. л.). Оно представ-
лено логистической кривой с пределом насыщения, потолком,
на базе которого впоследствии с ростом спортивного мастерства
может произойти не взрыв потолка и расформирование систе-
мы, а переход на новый не чисто экспоненциальный, а логисти-
ческий рост. Разумеется, и этому рано или поздно наступает
конец. Однако деградация функций чаще совершается с «эска
лаиией наоборот» — движение экспонент и логистических кри-
вых вниз (например, возрастная утрата ритма в энергетическом
гомеостазе). В частности, возрастное увеличение вероятности
возникновения таких заболеваний, как ожирение, предиабет,
сахарный диабет тучных, атеросклероз, рак, понижение сопро-
тивляемости к инфекционным заболеваниям, В. М. Днльмаи
(1972) с достаточно высокой научной обоснованностью рас-
сматривает как нарушение ритма, т. е. колебательных, экспо-
ненциальных и логистических характеристик энергетического
i гомеостаза. «Возможность свести все разнообразие возрастной
патологии... к одной лишь причине— нарушению ритма в энер
i гетическом гомеостазе — является самым универсальным и по-
тому поразительным по своей простоте ответом на вопрос,
почему так единообразны и закономерны проявления возраст-
ной патологии современного человека».
Мы в принципе разделяем эту' концепцию, по поводу кото-
| рой Дильман далее пишет: «Энергетический гомеостат пред-
। ставляет собою открытую систему. Это выражается в том, что
нормальный ритм его работы связан с поступлением энергети-
I ческих субстратов с пищей извне. В более широком смысле
I энергетический гомеостат открыт для других влияний внешне-
, то мира, что проявляется, например, изменением интенсивности
. его работы под влиянием стресса (любого возмущающего
воздействия.— А. .П.) или в связи с необходимостью его им-
мунологической защиты.. Одним из основных элементов энер-
гетического гомеостата является взаимодействие между
гормоном роста, инсулином и глюкозой. Когда человек голода-
ет, увеличивается выделение гормона роста, который обладает
• мощным жиромобилизующим эффектом. Это выражается в
5 А. А. Логинов
129
расщеплении жира в жировой ткани, вследствие чего в ток
крови поступают жирные кислоты, обладающие высокой кало-
рийностью. Этим объясняется похудание человека при голода-
нии, когда энергетические процессы обеспечиваются в основном
за счет сгорания жирных кислот после их мобилизации из
жировых депо. С другой стороны, когда сахар или глюкоза
поступают с пищей, происходит, как показали Глик и его со-
трудники, торможение секреции гормона роста. Такой эффект
вполне целесообразен, так как если энергетический материал
берется из внешнего мира, то организм не должен использовать
собственных запасов, и поэтому происходит уменьшение уров-
ня жиромобилизующего гормона роста под влиянием глюкозы.
Само тормозящее влияние глюкозы осуществляется через со-
ответствующий гипоталамический центр, который контролиру-
ет выделение гормона роста из гипофиза. Таким образом,
глюкоза участвует в механизме отрицательной обратной связи
энергетического гомеостата, контролируя, подобно рабочему
гормону, уровень секреции гормона роста — гормоиа-регуля-
тора. Так совершаются ритмические колебания в энергетиче-
ском гомеостате в зависимости от периодов поступления пищи.
Одновременно глюкоза стимулирует выход из поджелудочной
железы инсулина — гормона, который обеспечивает сгорание
глюкозы в тканях. В результате в условиях голодания основ-
ным источником энергии служат жиры собственных запасов
организма, а при поступлении пищи — углеводы. И в этом
проявляется внутренний ритм смены источников энергии в те-
чение суток.
Однако столь необходимый для жизнедеятельности ритм
при старении и возрастной патологии выходит из строя. Дей-
ствительно, как можно видеть на рис. 25, построенном на
основании полученных в нашей лаборатории данных, стандарт-
ная доза глюкозы вызывает (в соответствии со своей ролью в
энергетическом гомеостазе) падение уровня гормона роста в
крови у молодых лиц, но не дает этого важнейшего эффекта ни
у мужчин среднего возраста, ни при такой характерной возраст-
ной патологии, как рак молочной железы и рак тела матки у
женщин. Это означает, что гипоталамус проявляет резистент-
ность к тормозящему гомеостатическому действию глюкозы.
Именно наличие резистентности создает нарушение ритма в
смене источников энергии при поступлении в организм углево-
130
дов. Можно видеть, что в ответ на введение глюкозы не только
не происходит снижения продукции гормона роста у лиц с
возрастными изменениями, по не снижается и концентрация в
крови жирных кислот. Иными словами, несмотря на поступле-
ние пищи извне и увеличение в крови концентрации глюкозы.
Рис. 25. Эндокринный гомеостаз (по В. М. Дильману, 1972).
концентрация жирных кислот тоже остается высокой, т. е.
происходит нарушение ритма в работе энергетического гомео-
стата».
Попытки привести все сказанное к каким-то количественным
оценкам предпринимались неоднократно. Но цель настоящих
очерков не сказать все, а попробовать сказать обо всем в са-
мом обобщенном, принципиальном виде.* Поэтому следует
познакомиться с одной из наиболее общих моделей процесса
конечного развития по Прохоренко (рис. 26).
* Автор приносит свои извинения за чрезмерную самонадеян-
ность.
5*
131
1. Кумулятивный обмен обеспечивает накопление субстра-
та, измеренного по какому-то параметру z, в системе по логи-
стическому закону. На участке молодого экспоненциального
роста, простирающегося от <=0 до i~li, будем иметь:
г(/) = гу?*,
где zo#=O и означает величину параметра г при 1=0, когда на-
чинается экспоненциальный рост; а — постоянная экспоненци-
ального роста.
2. Мощность внешнего обмена «(1) в соответствии с основ-
ным свойством кумулятивности определяется количеством на-
Рис. 26. Кривые мощности обмена
систем (по В. К- Прохоренко,
1969).
личного субстрата в системе
н в каждый момент времени
пропорциональна ему.
н(0 = рг(0.
где Р — удельная мощность
внешнего обмена.
3- Поставляемый внеш-
ним обменом субстрат реали-
зуется внутренним обменом
по двум существенно различ-
ным каналам: истинный об-
мен (обновление наличного
субстрата) с мощностью
и‘(0> также определяемой
количеством наличного суб-
страта и пропорциональной
последнему:
щ(О=уг(0,
где у—постоянная удельная мощность истинного обмена и,
во-вторых, обмен роста (кумулятивное накопление субстрата)
системы с мощностью «2(9, равной скорости роста параметра
z и выражающейся через производную функцию z по времени:
В стадии экспоненциального роста последнее уравнение
дает;
132
и постоянная а приобретает смысл удельной мощности роста.
Записываем баланс мощностей обмена:
ч (0 (0+М0.
который должен выполняться по крайней мере в период ранне-
го развития. Подставляя значения мощностей щ, иг, иа, получим:
Р=а+у.
Кривые мощности обмена строим следующим образом. До-
пустим Р=1, а <х=у, тогда кривая внешнего обмена и воспро-
изводит график z. Дифференцируя графически логистическую
кривую и, находим мощность роста Щ. Кривую мощности ис-
тинного обмена Hi строим, удовлетворяя баланс мощностей об-
мена на участке экспоненциального роста. Суммируя
кривые ш и иг, получим мощность полного внутреннего обмена
и,+иг как функцию времени. Графическим дифференцировани-
ем кривой иг находим ускорение роста системы и2.
Обсудим полученные кривые. Мощности внешнего обмена
и и внутреннего истинного обмена «1 воспроизводят логистиче-
скую кривую в различных масштабах. Кривая мощности роста
"2, представляющая также скорость роста, приближенно имеет
гауссову форму (нормальное распределение): мощность се
сначала растет по экспоненте, затем отрывается от последней
при достигает максимума при t—icp и убывает вправо
симметрично левой ветви. Кривая и (или z) по отношению к иг
будет интегральной, фиксирующей кумулятивное действие об-
мена роста. Полная мощность внутреннего обмена ш+щ в на-
чальной стадии совпадает, как и было предусмотрено, с мощ-
ностью внешнего обмена. Этим демонстрируется единство
внешнего и внутреннего обмена. Мощность ш+иг в точке k от-
рывается от кривой и, сравнительно быстро достигает насыще-
ния и в пределе стремится к кривой истинного обмена. Чрез-
вычайно важным является то обстоятельство, что с прекраще-
нием роста внутренний обмен сохраняет лишь один канал
потребления нового субстрата — истинный обмен. Кривая
ускорения роста вначале идет экспоненциально вверх, дости-
гает максимума при Z=A, после чего убывает к нулевому зна-
чению при t—fcp и симметрично левой ветви уходит в отрица-
тельную область (область замедленного роста).
133
Обратимся теперь к точке k, в которой, по эмпирическим
данным, рассмотренным выше, наступает кризис развития.
В этой точке, кроме того, что рост параметра отклоняется or
экспоненциального, ускорение его достигает максимума, а
мощность внутреннего обмена отрывается от мощности внеш-
него. Таким образом, в точке k впервые появляется противоре-
чие: внешний обмен развивает такой поток субстрата, который
внутренний обмен не в состоянии реализовать. Разрыв в мощ-
ностях прогрессирует. Система вынуждена либо «заморажи-
вать производительные силы» и искусственно снижать реальную
мощность внешнего обмена, либо накапливать в виде балласта
вновь произведенный субстрат. Но ни одни из этих путей не
спасает ее, поскольку противоречие между производством и
потреблением субстрата такими способами не разрешается.
Внутренняя структура системы вообще не имеет никаких шан-
сов на спасение (стационарная система с непрерывным нетож-
дественным обменом невозможна). Она погибает вместе с
системой, если последняя ие находит возможностей к внутрен-
ней радикальной трансформации (так умирает развитой орга-
низм), или расформировывается для воплощения в новую
структуру, обеспечивающую свободную кумулятивную дея-
тельность нетождественного обмена, (ароморфозы А. Н. Север-
цова в эволюции жизни и т. д.).
Очерк 4. БИОЛОГИЧЕСКИЕ СТРУКТУРЫ
И ПРОЦЕССЫ ОБМЕНА ВЕЩЕСТВ
Наступил момент поставить перед собой новый вопрос: что
же является той материальной основой, па которой разыгрыва-
ются все сложные перипетии адаптивно-гомеостатического по-
ведения биологических систем? Ответ может быть лишь один —
все формы приспособительного поведения возможны только на
базе обмена веществ, свойственного биосистемам как индиви-
дуального, так и надындивидуального уровней.
Оптимальное течение его определяется строгой упорядочен
ностью химических процессов. Эта упорядоченность зависит
как от соотношения скоростей регулируемых ферментами хи-
мических реакций, так и от степени структурной организованно-
сти всех элементов живого организма.
В связи с этим возникает новый вопрос о соотношении ме-
134
жду структурой и функцией биосистемы. При огромном разно-
образии точек зрения относительно системы, структуры, функ-
ции вряд ли возможно в рамках настоящих очерков широкое
обсуждение их. Если обратиться к философскому словарю, то
«структура — строение и внутренняя (выделено нами.— А. Л.)
форма организации системы, выступающая как единство устой-
чивых взаимосвязей между ее элементами... Функция — внешнее
проявление (необязательно формально внешнее, регистриру-
емое, а принципиально проявляемое для структуры любой
сложности.— А. Л.) свойств какого-либо объекта в дайной си-
стеме отношений...»
Читатель, по-виднмому, уже почувствовал единство связи
н взаимозависимости структуры и функции в любых, и тем бо-
лее биологических, системах. Это позволяет с достаточной уве-
ренностью утверждать, что функция есть направленное, изби-
рательное воздействие структуры на определенные стороны
внешней среды.
Вероятно, следует согласиться с тем, что «...необходимо
обобщить понятие функции, и проще всего это сделать, но-вн-
димому, на базе элементно-структурных представлений. Для
этой цели н необходимо ввести понятие функциональной струк-
туры (выделено нами.-—А. Л.), которую можно определить
следующим образом. Функциональная структура представляет
собой закон связи между функционально определенными эле-
ментами (частями) исследуемого объекта» (Ф. 3. Меерсон,
1964). В качестве примера можно рассмотреть структуру моле-
кулы ДНК, ее двуспиральность и последовательность располо-
жения нуклеотидов, определящие функцию только данной мо-
лекулы. Любое структурное изменение в ней, например замена
нуклеотида, неминуемо приводит к изменению функции (в
смысле возможности воспроизведения себе подобной, а также
информационной и-РНК для синтеза молекулы белка) или к
полному исчезновению ее.*
Одиако о единстве структуры и функции следует говорить
как о противоречивом единстве, в котором функция выполняет
определенные структурообразующие обязанности, а структура
не является пассивным следствием свойств функции н играет
активную роль в становлении и развитии ее.
* Исключение составляет мутагенная ДНК, участки которой, по
данным сегодняшнего дня, ничего ие кодируют.
13Б
На примере специализированных педелящихся клеток
взрослого организма можно выявить несколько основных про-
цессов, неразрывно связанных между собой и имеющих свою
преимущественную локализацию в определенных клеточных
структурах: 1) процесс возбуждения (восприятие регуляторных
стимулов и регулирование функции клетки в условиях целост-
ного организма), протекающий в основном на клеточных мем-
бранах и образованиях, связывающих мембраны со специали-
зированными внутриклеточными структурами; 2) специфиче-
ская физиологическая функция клетки (сокращение, секреция
и т. д.), осуществляемая расположенными в цитоплазме спе-
циализированными органеллами — миофибриллами, секретор-
ными гранулами и т. д.; 3) образование энергии в форме АТФ,
локализованной главным образом в митохондриях и необходи-
мой для осуществления физиологических функций синтеза бел-
ка в клетке; 4) синтез белков, образующих митохондрии, мем-
браны и другие структурные элементы, локализованный в ге-
нетическом аппарате и рибосомах клетки. Противоречивость
структурно-функциональных взаимоотношений ие ограничи-
вается изложенным: если геологические (условно структура)
преобразования более динамичны, чем внутренние преобразова
ния биосистем или чем их эволюционно-приспособительные воз-
можности (условно функция), то «функции организма, регу-
лируемые нервно-зндокринным аппаратом, несоизмеримо бо-
лее динамичны, чем осуществляющие их структуры» (Меерсон,
1964). Здесь консервативность структур значительно выше кон-
сервативности функций. Однако не следует делать «тупикового»
вывода о безнадежности понимания соотношения структуры и
функции.
Мы более ортодоксальны в своих п[»едставлениях по этому
поводу и считаем, что любое событие (функция), происходя-
щее в биосистеме, реализуется в условиях структурных изме-
нений системы (хотя бы конформационных или иных, нам еще
неизвестных), равно как любые структурные изменения, сопро
вождаются изменением функции (возможно, и в неизвестном
нам проявлении), т. е. нет функциональных сдвигов без орга-
нических (структурных) изменений, как нет «органики» без
соответствующего изменения функции. Структура и функция —
неразрывные характеристики любых материальных тел.
Опыт показывает, что интенсификация функции под влиянн-
136
ем тех или иных воздействий осуществляется за счет возмож-
ностей, заложенных в обеспечивающих эти функции структу-
рах. В свою очередь интенсификация влияет на активность
аппарата синтеза белка и через его посредство на структуру.
В случае возникновения со стороны возмущающих воздейст-
вий (внешней среды) «запросов» к интенсификации функции
выше экстремумов, заложенных в реализующих ее структурах,
процесс может выйти за рамки гомеостатических пределов
структурных изменений вплоть до гипертрофии и дегенерации
органов и тканей, т. е. патологического изменения структур.
Предложенная философско-теоретическая предпосылка по-
зволяет перейти к более частному рассмотрению проблемы —
биологическим структурам и процессам обмена вещетв. В этом
отношении значение биологических структур для живого орга-
низма и его адаптационно-гомеостатического поведения, соглас-
но А. Г. Пасынскому (1968), заключается в том, что они
обеспечивают:
1) пространственное отделение живого организма от внеш-
ней среды (форму организма), без чего было бы невозможно
существование его как самостоятельного целого (биологиче-
ской системы);
2) внутреннюю организацию живого организма, обусловли-
вающую выполнение специализированными органами и тканями
различных функций, необходимых для жизнедеятельности (воз-
буждения, движения, пищеварения и др.);
3) полезное использование энергии в форме механической,
осмотической, электрической, химической и других работ, что
возможно только при наличии специализированных структур.
Без них энергия биохимических процессов приводила бы лишь
к бесполезному выделению и рассеянию теплоты;
4) регулирование ферментативных процессов путем изме-
нения условий взаимодействия ферментов с субстратами, из-
менения действующих концентраций ферментов, обеспечения
определенных пространственных условий последовательного
протекания и чередования реакций в организме;
5) взаимосвязь биологических структур и биохимических
процессов.
В живом организме структуры, регулирующие ход биохи-
мических процессов, порождаются этими процессами. При на-
рушении или прекращении их происходит быстрый распад
137
Пиноцитозная
Вакуоль
------(верная мембрана
-----Хроматин
— Ядрышко
Вакуоль
Агранулярная
сеть
Цистерны эндо
плазматической
сети
Эндоплазма-
тическая
сет
Лизосомы
Жикроборсинки
—Клеточная
мембрана
-Жиры
Хромосома
Ш
Мембраны on-
-порото Гольджи
-Центриоль
—Рибосомы
Синтез белка
мРНК
оюндрия
{перенос
зпектроноб.
окислительное
фоарорилиробоние
АДФлф~АТФ)
-Активация
аминокислот
гРНК
( +ЦТФ ФФ+АКАНФ
дк-| ^тРНК-ЦПФ-АПф
т'Р'НК-ЦМФ-АЖФ-Ак}- Белок
AKJ
тРНК-ЦНФ-АНФ-АК
Рис. 27. Схема строения клетки, показывающая связь некоторых фер-
ментативных реакций с субклеточными структурами (из К Вилли
Д. Детье, 1974):
Д-НМН — дезампдопикотинамидомононунлеотнд; ДНАД — демнидоннкотнн-
амидд ннукл еот ид.
биологических структур. Убедиться в этом можно иа примере
такой составной части организма, как клетка (рис. 27). Пред-
ставляя достаточно самостоятельную структурную единицу,
клетка в свою очередь включает в себя различные специализи-
рованные структуры: ядро, рибосомы, митохондрии, лизосо-
мы и др.
Исключительного развития в клетках достигла система мем-
бран. Клеточная мембрана не только отграничивает одну клет-
ку от другой, но и образует многочисленные впячивания в ци-
топлазму: от очень просто устроенных (пиноцитозные вакуо-
ли-пузырьки) до сложнейшей эндоплазматической сети. Пола-
гают, что эндоплазматическая сеть выполняет весьма важную
функцию в осуществлении обмена веществ между цитоплазмой
и окружающей клетку средой.
Таким образом, клетка состоит из двух не смешивающихся
друг с другом фаз: из основного вещества цитоплазмы со все-
ми возможными в ней переходами от жидкого до твердого со-
стояния и из мембранной системы, расчленяющей цитоплазму
на своеобразные отсеки, получившие название цитоплазмати-
ческого матрикса.
Следовательно, не только клетки пространственно отделены
друг от друга, но и их структуры (рис. 28). Разрушение мем-
бран приводит к уничтожению клетки как самостоятельной
единицы.
Значительную часть вещества, из которого формируется
основная структура биологических мембран, составляют
фосфолипиды. Модель молекулы фосфолипида изображена на
рис. 29. Как видно из рисунка, молекула фосфолипидов состо-
ит из гидрофильной головки и двух гидрофобных «хвостов»
(цепей углеводородов), что создает ее асимметричность как по
химической структуре и пространственному расположению ча-
стей, так и по избирательному отношению к тем средам (фа-
зам), с которыми она соприкасается. К поверхности воды
молекулы фосфолипидов ориентируются гидрофильными го-
ловками, в то время как гидрофобные «хвосты» будут обраще-
ны в обратную сторону от нее. В этом легко убедиться на
примере поведения молекул полярных липидов (рис. 30, 31, 32).
Известно, что элементарная биологическая мембрана состоит
из двух слоев фосфолипидов и их гидрофильные головки об-
ращены к периферии, поскольку снаружи мембрану той или
1зе
a
8
140
Рис. 28. Различные моде-
Ципоплазт
тт-<яш
ЖЖ
ШИШ
ЖГЖЖ
ШАШШ
ли молекулярного строе-
ния биологических мем-
бран:
I. В основу моделей поло-
жены; двухслойная липид-
ная структура (а — е):
глобулярная организация
{ж — к); а — белок в 0-фор-
ме; б — а-спираль; в — гло-
булярный белок; г — асим-
метрия в расположении
белка; д— канальцы и по-
ры, частично пронизываю-
щие белковый слой; е —
белок внутри двойного слоя
липидов; ж—липидные ми-
целлы с 0-белком; з — ли-
пидные мицеллы с глобу-
лярным белком; и, к — пре-
вращение глобулярной орга-
низации в двухслойную. ВО
всех случа ях фосфолипид-
ные белковые слои поверх-
ностей мембран не симмет-
ричны друг другу, еще бо-
лее асимметрично по соста-
ву н свойствам расположе-
ние ферментных и других
функциональных групп (по
Э. де Робертасу, В. Новин-
скому. Ф. Саэсу, 1973).
II. Схема строения мембра-
ны по Валлаху (из С. В. Конева и др., 1973): а — схематический вид поверх-
ности мембраны (’/ — ориентированные липиды; 2—белки); б — продольный
разрез мембраны по ее средней гидрофобной части (/—липидный цилиндр.
2 — водный канал, 3 — белковая палочка). ///. А — непрерывная мембрана;
Б— начало образования прерывистой мембраны; В—-конец образования пре-
рывистой мембраны (из А. Поликара. 1972). IV. Модель большой мембраны,
включающей наружную поверхность оболочки клетки (из Э. де Робертиса,
В. Новинского, Ф. Саэса. 1973).
141
иной клетки окружает межклеточная жидкость — внешняя вод-
ная фаза, а внутренняя — цитоплазма также представляет со-
бой водную фазу. В аналогичных условиях находятся и все
1тк
Рис. 29. Полная молекулярная модель фосфолипида (молекулы
фосфатидилэтаноламина):
Д|. Д®—углеводородные цело олеиновой и ояльметиновой кислот; ХГ —
характеристическая группа, определяющая принадлежность к данному
виду фосфолипидов; ФГ — группа, соединяющая характеристическую
группу с двумя углеводородными цепями;---------одинарная связь, = —
двойная связь между атомами углерода; О — группы, участвующие в
водородной связи.
органеллы клеток. Следовательно, биологические мембраны
отделяют друг от друга две различные по составу и свойствам
водные фазы.
Помимо фосфолипидов, структуру клеточных мембран обра-
зуют и белки. Масса их несколько больше. Однако, если учесть
молекулярные массы, то на одну молекулу белка приходится
142
flocm
boia
Рис. 30. Схема расположения полярных липидов (из Ж- Хаггиса,
Д. Михи н др., 1967);
с — расположение липидов на поверхности раздела масло — вода; б — сфе-
рические мицеллы в масляной н водной фазах.
Рис. 31. Схема образования молекулярных пленок (из Э. Де Робсрти-
са, В. Новинского, Ф. Саэса, 1973):
А ~ молекула стеариновой кислоты с полярной группой, погруженной в во-
ДУ: расположение молекул: Б — при слабом сдавливании; В — при сильном
сдавливании. Степень упорядоченности молекул зависит ог их количестве,
приходящегося на площадь поверхности соприкосновения с водой и в большей
мере — от выраженности их асимметрии (чем менее они симметричны, тем
более упорядочено их расположение в иоиослое), так как по мере увеличе-
ния последней усиливаются гидрофильные свойства отдельных частей мо
лекул; Г- при погружении стеклянной пластинки с моиомолекулярной плен-
кой стеарата бария второй молекулярный слой соединяется с первым при
помощи ыеполяриык целей и образуется бимолекулярная пленка. Справа
показано образование нескольких бимолекулярных слоев стеарата бария на
стеклянной пластинке после ее многократных погружений в воду.
144
около 70 молекул липидов. Предполагают, что белки включа-
ются в липидный слой или покрывают его.
Вероятно, слои фосфолипидов и белков не статичны, а пе-
ремещаются относительно друг друга, сохраняя при этом функ-
циональную взаимосвязь .
Рис. 32. Образование бимолекулярного слоя путем выталкивания
воды из толстой пленки (по Томпсону и А. Поликару, 1972).
До сих пор не выяснено, существуют поры и каналы в
мембране постоянно или же они образуются во время тех или
иных воздействий каждый раз заново.
Проблема строения клеточной мембраны не исчерпывается
изложенным. Предложена так называемая «модель большой
мембраны», которая включает не только липопротеидную струк-
туру, но и оболочку на ее наружной поверхности.
Таким образом, проблема построения клеточных мембран
не решена, но совершенно очевидно, что они являются сложно
устроенными и активно функционирующими образованиями
клеток животных и растительных организмов. От них зависит
характер и особенности протекания всех клеточных процессов.
Благодаря избирательной способности мембраны являются не
только механическими барьерами, но и регуляторами обмена
145
веществ между клеткой и окружающей ее средой. Различают
пассивный и активный транспорт веществ через мембраны.
Пассивный транспорт осуществляется практически без су-
щественных затрат энергии обмена веществ клетки и идет по
градиенту (от большей величины к меньшей) до установления
равенства концентраций по обе стороны мембраны. Это возмож-
но только в том случае, если мембраны полностью проницаемы
лишь для немногих веществ (полупроницаемы), что зависит в
первую очередь от размеров молекул веществ. Если они пре-
восходят поры мембраны и не растворяются в ее липидной
фазе, то транспорт частиц через них путем пассивного переноса
вообще невозможен, несмотря на градиент концентрации.
Пассивный перенос через клеточную мембрану зависит так-
же от электрического градиента. Движение ионов по электри-
ческому градиенту происходит в том случае, если они переме-
щаются против градиента концентрации или градиента потен-
циала.
Перенос веществ через клеточную мембрану может осущест-
вляться и за счет осмоса, т. е. осмотического градиента.
Пассивный перенос осуществляется легко в отношении ве-
ществ, хорошо растворимых в жирах. Они передвигаются че-
рез мембрану по законам диффузии. Сложнее обстоит дело с
веществами, которые растворимы не в жирах, а в воде. Слож-
ность заключается в том, что гидрофильные вещества, чтобы
пройти через мембрану, должны преодолеть водоотталкиваю-
щее — гидрофобное действие липидов мембраны. А это невоз-
можно. Однако через мембрану дрожжевой клетки проходит, и
весьма энергично (300 млн. молекул за один час), такое водо
растворимое вещество, как глюкоза. И это далеко не единствен-
ный пример. Существует множество теорий, пытающихся объ-
яснить этот «казус».
Так, Эйринг пассивный транспорт гидрофильных веществ
через мембрану рассматривает как ряд переходов одного рав
новссного состояния в другое. Однако это утверждение мало
что объясняет и не слишком убедительно.
Согласно другой точке зрения, основанной на теории «вре-
менных дырок», потоки гидрофильных веществ устремляются
через временные «пустоты» клеточных мембран.
По наиболее распространенной теории, проникновение во-
дорастворимых веществ происходит через постоянно существу-
146
ющие в биологической мембране поры, как через сито, и зави-
сит от их концентрации по обе стороны мембраны. Однако
через поры молекулярного сита в обе стороны движутся встреч-
ные потоки множества самых разнообразных водорастворимых
веществ. Естественно, в этом случае должна возникнуть свое-
образная «борьба» молекул. Значит, транспорт каждого веще-
ства здесь зависит не только от собственного градиента концен-
трации, но и от концентрации других веществ, стремящихся
проникнуть через пору. Перенос может осуществляться также
и благодаря тому, что свободно проникающая через пору вода
проталкивает молекулы растворенного в ней вещества, и диф-
фузия его через мембрану будет более интенсивной. Простота
и убедительность такого объяснения переноса веществ через
клеточную мембрану весьма привлекательна. Однако пока не
удалось с высокой достоверностью доказать существование
мембранных пор. Вероятно, они есть, но в незначительном ко-
личестве.
До сих пор речь шла о пассивном переносе нейтральных
молекул. Многие же водорастворимые вещества имеют электри-
ческий заряд. Рассмотрим подробно транспорт ионов через
клеточную мембрану.
При изучении проницаемости клеточных мембран было за-
мечено, что многие вещества проходят через них значительно
быстрее, чем это может обеспечить концентрационный гради-
ент, а иногда они проникают через мембрану клетки вообще
против градиента. Естественно, при этом должна затрачивать-
ся энергия. Транспорт вещества через клеточную мембрану с
затратой энергий обмена веществ клетки получил название
активного транспорта. С его помощью обеспечивается доставка
в клетку необходимых для ее жизнедеятельности веществ, а
также удаление ненужных продуктов обмена веществ.
Считают, что одним из механизмов активного переноса ве-
ществ через мембрану являются «ионные насосы». Как извест-
но, в условиях физиологического покоя и нормальной темпера-
туры внеклеточная среда богата ионами натрия и бедна ионами
калия. Так, в цитоплазме эритроцитов, и мышечных и нервных
волокон ионов калия в 30—50 раз больше, чем в лимфе и меж-
клеточной жидкости, а натрия в 8—10 раз меньше. Объяснить
подобное резко асимметричное распределение ионов натрия и
калия по обе стороны мембраны действием только механизма
147
пассивного переноса (диффузия) нельзя. Предполагают, что в
мембране существует специальный «натрий-калнсвый насос»,
также принимающий участие в поддержании ионной асиммет-
рии. В условиях физиологического покоя он обеспечивает
«откачивание» натрия из клетки и «подсасывание» калия
внутрь ее. Происходит это при помощи особых химических
«переносчиков». Ионы натрия захватываются ими у внутренней
Рис. 33. Предполагаемый механизм «калий-натриевого насоса»
с одним переносчиком (из Лёви и Ф. Сикевпц, 1973):
— X ~ Ф. NaH~ — X ~ Ф — комплексы кали» и натрия с фосфо-
рилмрованным белком — ферментом.
поверхности мембраны и переносятся иа наружную, где от-
щепляются и таким образом выводятся из клетки. На наруж-
ной же поверхности х переносчику присоединяются ионы калия,
которые вместе с ним транспортируются на внутреннюю поверх-
ность мембраны и внутрь клетки (рис. 33). Работа эта осу-
ществляется с затратой энергии, высвобождающейся при рас-
щеплении АТФ прн участии фермента аденозинтрифосфатазы.
Ионные насосы известны ие только для натрия и калия, но
И для других веществ. Способность этих насосов «перекачи-
вать» ионы против градиента концентрации просто фантастич-
на. Так, сказалось, что изолированная кожа лягушки перека-
чивает поваренную соль из раствора, омывающего ее наруж-
ную поверхность, в раствор, соприкасающийся с внутренней
поверхностью, даже тогда, когда концентрация ее во внутреи-
148
нем растворе в 10 000 раз больше, чем в наружном. Еще при-
мер — через 1 смг мембраны нерва кальмара перекачивается
10 млрд, ионов натрия в секунду. На эту работу затрачивается
около 20% всей энергии, производимой клеткой. Коэффици-
ент же полезного действия таких насосов достигает 50-—95Уо.
Такого к. п. д. не имеет ни одна самая совершенная машина,
созданная человеком.
Рис. 34. Предполагаемый механизм «калий-иатриевого
насоса» с двумя переносчиками X и У (из К. Вилли,
Е. Детье, 1974).
Есть предположение, что существует не общий переносчик
ионов натрия и калия, а для каждого свой собственный или
один, ио способный превращаться то в переносчик ионов ка-
лия, то натрия (рис. 34).
Г1редполагается также, что активный перенос ионов через
биологические мембраны осуществляется за счет биохимичес-
ких процессов и благодаря особенностям расположения в мем-
149
браке биохимических ансамблей белков, ферментов и др. При-
мером может быть дыхательный ансамбль мембраны мито-
хондрий. Эта мощная «машина» способна не только к образо-
ванию АТФ, но и к перемещению ионов к конформационным
изменениям мембран. Последние, вероятно, связаны и с КОН-
fl
Рис. 35. Предполагаемая схема воздействия электрического поля на
контакт «жестких» белковых глобул в мембране:
А — наиболее устойчивое постулируемое состояние мембранных белков в от-
сутствие трансмембранного поля; Б — состояние белков при появлении элект-
рического поля; с, б — глобулы, расходящиеся (обозначено стрелками) под
действием электростатическою притяжения разноименных зарядов; в. г —
глобулы, приходящие в контакт в результате отталкивания одноименных за-
рядов белков и поверхностей мембраны; д. е—глобулы. разъединяющиеся
(яс, з —соединяющиеся) из-за мшрацип одной из них (е и ос).
формационными изменениями углеводородных цепей фосфоли-
пидов, и с движением белков в мембранах (рис. 35). Сущест-
вуют данные, что все перечисленные активности являются
«векторными». Для этого должна быть соответствующая струк-
турная основа, т. е. асимметричная организация индивидуаль-
ных ферментов или целых ансамблей в мембране. Если мембра-
на сама по себе асимметрична (к примеру, одна из ее поверх-
150
ностей более гидрофильна), то перенос
молекул будет носить векторный харак-
тер. На рис. 36 изображена модель мем-
браны, где фиксируются молекулы фер-
мента с асимметричным активным цент-
ром. Дыхательные ансамбли также ха-
рактеризуются асимметричной организа-
цией (рис. 37).
Таким образом, действительно суще-
ствуют механизмы («насосы») активно-
го переноса через мембрану нс только
ионов калия и натрия, но и ионов водо-
рода и гидроксильной группы, т. е. ки-
слоты.
Более сложны и менее изучены меха-
низмы активного переноса других ве-
ществ, например сахаров, аминокислот.
Что касается сахаров, то они почти не
растворяются п липидах, а размеры их
молекул достаточно велики и испытыва-
ют затруднения для свободного прохож-
дения через мембрану. Несмотря на это,
сахара довольно легко транспортируют-
ся через нее. Предполагают, что в дан-
ном случае происходит пассивный пе-
ренос молекул сахаров благодаря их об-
легченой диффузии (рис. 38). (При об-
легченной диффузии вещество передви-
гается по градиенту концентрации с
более высокой скоростью, чем по клас-
сическим законам диффузии). Большин
Рис. 36. Модель векторной реакции (из
А. Ленииджсра, 1974):
верхняя схема — асимметричное образование по-
пов Н^" и ОН— в процессе переноса электронов;
средняя схема — мембранная АТФ-аза. способ-
ная к асимметричному образованию (связыва-
нию) и ОН— (или фосфолипидных групп);
нижняя схема — модификация. объясняющая
асимметричность образования ионов и ОН
и АТФ в каждом пункте сопряжения.
ЦыпЗныи центр
со сбязыСсющипи
Учтали конпленемтюрныш ас В
д----В
'Субстрат
Ъенброна
внутри Сцаружь
Ченброна
151
центр
Рис. 37. Асимметричное расположение активных центров
АТФ-аз и переносчиков электронов Н+ и ОН— в мембранной
структуре относительно непроницаемой для ионов Н+ (схема
механизма переноса ионов) (по А. Ленинджеру, 1966):
А — асимметричная АтФ-аза. создающая градиент pH; Б — разделе-
ние ионов Н"•* и ОН ' в процессе окисления Нг-субстрата кислородом,
обусловленное асимметричным н противоположным расположением
флавопротеида н цитохромоксидазы в мембране; В— ДТФ-аза, ответ-
етвеииая за окислительное фосфорилирование АДФ с одновременным
разделением попов Н"^ и ОН"-"I — белковый слой, П — липидный
слой.
152
стпо веществ, обладающих этим свойством, растворимы в воде.
Однако интенсивность «облегчения» зависит и от многих дру-
гих свойств, в частности от структуры веществ. Так, например,
для зеркальных изомеров известны случаи, когда один из них
переносится при помощи облегченной диффузии, а другой вооб-
ще не диффундирует. Это еще одно доказательство избира-
тельного отношения клеточных мембран к веществам.
Наружная
среда
Мендрша
/ Внутренняя
/ среда
Рис. 38. Схема облегченной диффузии глюкозы через
мембрану (из К. Вилли ,В. Детье, 1974):
G—молекула глюкозы; X—пептид пли белок (переносчик).
В настоящее время явление облегченной диффузии объясня-
ют существованием специальных веществ — хорошо раствори-
мых в мембранах переносчиков. Эту роль, видимо, могут вы-
полнять липиды и белки, входящие в состав самой мембраны,
или же посторонние вещества.*
Для переноса через мембрану одновременно нескольких ви-
дов сахаров предложена схема, представленная на рис. 39.
Установлено, что перенос сахаров (возможно, и аминокислот)
зависит от присутствия ионов натрия. Это так называемый
натрий-зависимый перенос (рис. 40).
С активностью клеточных мембран тесно связаны также про-
цессы непосредственного поглощения клеткой твердых и жид-
* В последнее время установлено, что переносчиками щелочных
металлов могут быть вводимые извне антибиотики — так называе-
мые циклодепсипептиды.
153
Рис. 39. Перенос сахаров через мембрану (из А. Лёви и Ф Сиксвица
1971):
S, и Sa —сахара; X — подвижный переносчик (если S2 добавить к среде по-
еле того, как между переносчиком Л и Si устанавливается равновесие с об-
разованием S|A, то S2 вытесняется против градиента концентрации; это мож-
но объяснить, предположив, что S2 конкурирует с Si, вытесняет последний и
соединяется с X),
Рис. 40. Натрий-зависимый перенос сахаров (из А. Лёви и Ф. Си-
кевица, 1971):
S — сахар.
ких веществ. Это фагоцитоз и пиноцитоз. Так как процессы
фагоцитоза и пиноцитоза принципиально сходны друг с дру-
гом, их объединяют под общим понятием эндоцитоз и связы-
Эндоцитог
Рибосомы
\ * V/
Фагосомы
яите-
кжуоль
Гранула
накопления
таптное
тельце
тая
мембрана
Аутофагирую
щая бакуоль
Экскреция Плеточная
мембрана
~ сеть
Рис. 41. Предполагаемый механизм переработки веществ, поступив-
ших в клетку в результате эндоцитоза (из Э. де Робсртиса, В. Но-
винского. Ф. Саэса, 1973).
Кислые
гидролазы
Лизосонь
вают с понятием экзоиитоз. Эндоцитоз наблюдается лишь у
некоторых клеток (лейкоциты и др.). Он осуществляется
благодаря подвижности клеточных мембран и их способности
перемещаться и образовывать выступы. Гипотетический меха-
низм эндоцитоза схематически изображен на рис. 41.
155
Итак, активный перенос в отличие от пассивного является
чисто структурно-биохимическим процессом. Следовательно,
мембрана не столько механический барьер, отделяющий одну
клетку от другой, сколько сложная функциональная структу-
ра, где совершаются важнейшие ферментативные, окислитель-
но-восстановительные метаболические реакции, определяющие
судьбу обмена веществ, а вместе с тем н организма в целом.
in. ВИДЫ И ФОРМЫ ПРОЯВЛЕНИЯ
ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ живых
СИСТЕМ
Очерк 1. ФУНКЦИОНАЛЬНАЯ АКТИВНОСТЬ —
ЕДИНСТВЕННО ВОЗМОЖНОЕ СОСТОЯНИЕ
ЖИВЫХ СИСТЕМ
Как мы уже знаем, абсолютным свойством живого является
обмен веществ, поддерживающий динамически стационарное
и термодинамически и вещественно неравновесное состояние
его относительно окружающей среды, что возможно только при
условии трат свободной энергии системы, т. е. при соверше-
нии работы. Такое состояние живых систем следует именовать
функциональной активностью. Поэтому, перефразируя всеоб-
щий закон биологии Бауэра, можно сказать, что все и только
живые системы всю свою свободную энергию расходуют на
постоянное поддерживание состояния функциональной актив-
ности, уровень которой определяется конкретными формами
взаимодействий их (систем) со средой. В каждом отдельном
случае он всегда максимально адекватен характеру этих вза-
имодействий и энергетически допустимо минимален, т. е- гоме-
остатичеи. Даже предельные критические уровни активности
(нижний — анабиоз, вернее, гипобиоз и верхний — максималь-
ная интенсификация) адаптивны по характеру поведения;
оптимальны для данных конкретных условий и гомеостатичны
по содержанию.
Из-за отсутствия в настоящее время достаточно объектив-
ных методов регистрации исследователь нс оценит момент на-
чала или прекращения активности биосистемы, не впав при
этом в присущий ему антропоморфизм. Невозможность реги-
страции электрических характеристик бносистемы, а также
иных выражений ее активности, например при глубоком гипо
бибзе или других состояниях, именуемых торможением, факт
экспериментальный, а не биологический. Единственным и пока
157
методически недостижимым критерием оценки может быть ре-
гистрация момента обратимости (или необратимости) состоя-
ния активности. Нам представляется, что понимание процес-
сов возбуждения и торможения их стадий развития и т. д. долж-
но рассматриваться как выражение различных уровней актив-
ности адаптивно-гомеостатических механизмов биосистемы.
Возникновение адаптивно-гомеостатической реакции возможно
в случае, если на это есть побудительные причины. К таковым
относятся факторы внешней среды — раздражитель, стимул,
возмущающее воздействие.
Являясь по своей энергетической природе химическими,
электрическими, механическими, температурными, радиацион-
ными и другими, по биологическому значению раздражители
могут быть «адекватными» и «неадекватными». Адекватные —
это раздражители, на воздействие которых возбудимое обра-
зование (рецептор, клетка) в естественных условиях специаль-
но приспособлено реагировать изменением функциональной
активности при весьма малой энергии раздражения. Например,
для чувствительных клеток сетчатки глаза адекватным раздра-
жителем будет видимая часть спектра электромагнитных волн
(световые излучения) с энергией всего в несколько квантов,
для мышцы — нервный импульс и т. д. Все остальные раздра-
жители для данных систем будут неадекватными. Адекватность
раздражения определяется силой и длительностью действия
раздражителя, а также количеством элементов, вовлекаемых
в ответную реакцию. Исключение хотя бы одной из составляю-
щих частично или полностью устраняет адекватность и лишает
биологическую систему способности оптимально реагировать
на раздражитель. Таким образом, биологически наивыгодней-
шая «отзывчивость» биосистемы возникает только на раздра
жители, адекватные по всем показателям. Способность же жи-
вого отвечать на действие раздражителя изменениями своих
физиологических свойств и функциональных отправлений, как
известно, есть возбудимость, а характер и величина этих изме-
нений — эффективность, или дееспособность.
В живых объектах следует выделить два вида функциональ-
ной активности — физиологический покой и возбуждение. Со-
стояние физиологического покоя характеризуется отсутствием
внешне выраженных проявлений специфической деятельности,
168
например сокращение, секреция и др. Вместе с тем это ие
бездеятельное состояние. Оно является той исходной основой,
без которой специфическая активность не может проявиться.
В условиях физиологического покоя процессы обмена веществ
протекают на уровне, необходимом для поддержания и сохра-
нения жизнедеятельности организма и обеспечения готовности
его реагировать на воздействия, требующие проявления специ-
физичсской активности. Поэтому правильнее говорить не просто
о состоянии физиологического покоя, а о состоянии относитель-
ного физиологического покоя, в котором, и только в котором,
может находиться живое существо, не проявляющее «активной»
деятельности. В сущности, состояние физиологического покоя —
это способ реагирования (уровень активности) на те воздейст-
вия внешней среды, которые нс требуют проявлений приспосо-
бительных реакций.
Возбуждение же, как известно, это процесс, возникающий
под влиянием того или иного воздействия и сопровождающийся
переходом биосистемы на более высокий уровень активности.
Первое (при существующих методах регистрации) охватывает
сравнительно ограниченный участок вблизи от места нанесения
раздражения, второе распространяется на большее расстояние,
вовлекая в ответную реакцию биосистему в целом. С достаточ-
ной степенью условности проявления возбуждения можно ква-
лифицировать как не специфические, принципиально общие для
всех живых объектов (например, изменения процесса обмена
веществ, связанные с высвобождением энергии под влиянием
раздражения), и специфические, в виде физиологической функ-
ции, свойственной только данному органу или ткани (сокра-
щение мышцы, секреция железы и др.). Специфическая реак-
ция — это следствие дальнейшего развития местного возбужде-
ния.
Для более удобной оценки уровней функциональной актив-
ности биосистем необходимо применить понятие «торможение».
Его следует рассматривать как характеристику такой формы
активности биосистемы, которая проявляется в ослаблении или
прекращении текущей функциональной активности (переход
биосистемы на более низкий уровень относительно предшест-
вующего) .
Очерк 2. МЕМБРАННЫЙ ПОТЕНЦИАЛ —
ОДНА ИЗ ОСНОВНЫХ ХАРАКТЕРИСТИК
ЖИВОЕ СИСТЕМЫ
Мембранный потенциал характеризует разность потенциа-
лов поверхностей мембраны в результате избирательного пере-
носа катионов и анионов. Различают потенциал покоя, мест-
ный потенциал и потенциал действия.
Еще в 1838 году Маттеучи обнаружил, что мышца на по-
верхности заряжена положительно, а внутри отрицательно.
Позже это явление было открыто у большинства клеток живот-
ных и растений. Такая поляризация электрических зарядов
характерна и для поверхностей клеточной мембраны, когда
клетка находится в состоянии физиологического покоя. Эта раз-
ность потенциалов получила название мембранный потенциал
покоя (потенциал покоя).
Какова природа его возникновения и поддержания? Теорий,
пытающихся ответить на этот вопрос, несколько. Рассмотрим
одну из них, получившую наибольшее признание. Она исхо-
дит из того, что избирательно проницаемая (полупроницаемая)
мембрана разделяет два раствора, содержащие ионы,— цито-
плазму, находящуюся внутри клетки, и межклеточную жид-
кость, ее омывающую. Известно, если в растворе находится
какое-то количество катионов, то и анионов в нем должно быть
столько же.
Электронейтральность цитоплазмы клетки могла бы под-
держиваться неорганическими анионами (НРО,~. НРО^-,
ИаНСз ) и крупными органическими анионами (А-— белки,
аминокислоты и др ), накапливающимися в ней в процессе
внутриклеточного обмена. Однако подобная электронейтраль-
иость по обе стороны клеточной мембраны не удерживается по
одной из основных причин — в условиях покоя проницаемость
клеточной мембраны для ионов калия достаточно высокая,
а для ионов натрия — низкая, так как гидратированные ионы
последнего крупнее, что и затрудняет прохождение их через
поры мембраны.
Всякая несущая электрический заряд частица (ион или поляр-
ная молекула), находящаяся в растворе, окружается сольватной
оболочкой. Если растворитель вода, оболочка будет называться гид-
ратной, а процесс — гидратацией. Степень гидратации различных
160
ионов и молекул неодинакова и зависит от размеров частиц и их за-
рядов. Чем выше удельная плотность заряда, тем сильнее гидрата-
ция (рис. 42). Поэтому ион калия, несмотря на большой радиус, н
гидратированной форме меньше гидратированного иона натрия. Со-
гласно теории Муллииза, однократно гидратированные ионы могут
проникать через клеточную мембрану только тогда, когда их радиу-
сы близки радиусу поры. Если же радиус такого иона значительно
Рис. 42. Гидратация и стабилизация (из А. Гизе, 1959):
Л—модель и дипольная схема молекулы воды; К— схема гидратации
электрически нейтральной частицы: В — гидратация положительно заря-
женной коллоидной частицы.
меньше радиуса поры, то свободная энергия гидратации препятству-
ет этому. Данная гипотеза при многих недостатках удобна для
объяснения того факта, что проницаемость покоящейся мембраны
(например, нервных и мышечных волокон) для ионов калия значи-
тельно выше, чем для иоиов натрия.
При таких условиях благодаря существующему концентра-
ционному градиенту какое-то количество ионов калия выходит
из клетки через поры мембраны, создавая тем самым на се
наружной поверхности избыток положительно заряженных
частиц. На внутренней же поверхности возникает избыток
крупных молекул органических анионов, оставшихся без ней-
трализующих их калий-положительных ионов. Благодаря элек-
тростатическим силам вышедшие катионы калия нс могут
далеко удалиться от наружной поверхности мембраны (поло-
6 А. А. Логинов
161
жительно заряженные частицы, находящиеся вне клетки, при-
жимают их к мембране, а скопившиеся на внутренней поверх-
ности отрицательно заряженные частицы стремятся «втащить»
их внутрь клетки). Однако высокий концентрационный гради-
ент для калия препятствует этому. В результате на поверхно-
стях клеточной мембраны и возникает разность потенциалов —
Рис. 43. Образование мембранного потенциала за счет градиента кон-
центрации (по Вудсбери, 1962):
размерами символов Na"K Cl , в других обозначена степень отно-
сительной концентрации соответствующих ионов.
потенциал покоя (рис. 43). Разумеется, калий играет здесь,
так сказать, основную, ведущую роль. Вместе с тем по гради-
енту концентрации в клетку поступает и какое-то количество
катионов натрия. Но положительный заряд потока натрия зна-
чительно меньше противоположного потока ионов калия. Более
того, некоторое количество анионов хлора, захватываемых
натрием и поступающих вместе с органическими анионами,
162
также участвует в образовании слоя отрицательных зарядов
на внутренней поверхности мембраны, а оставшиеся в избыт-
ке катионы натрия и калия — слоя положительных зарядов па
наружной поверхности мембраны. (Надо сказать, что в воз-
никновении и поддержании потенциала покоя существенную
роль играет и активный перенос, т. е. работа «калий-натриево-
го насоса».)
На образование двойного электрического слоя идет очень
немного ионов. Поэтому концентрационный градиент сохраня-
ется высоким. Следовательно, диффузия ионов должна продол-
жаться. Однако в условиях уже существующего электрическо-
го потенциала диффундирующие ионы будут испытывать пре-
пятствие со стороны электрических зарядов на поверхности
мембраны: положительные ограничивают дальнейшее поступ-
ление нз клетки катионов калия, а отрицательные — приток в
клетку анионов. В результате возникает состояние, при котором
силы диффузии (концентрационного градиента) и электроста-
тического отталкивания уравновешиваются: количество ионов,
проходящих через мембрану по градиенту концентрации, ста-
новится равным количеству ионов, проходящих в обратном
направлении под влиянием сил электростатического отталкива-
ния. Уровень этого электрохимического равновесия и будет
определять как ионную асимметрию по обе стороны мембраны
(клетка — среда), так и величину мембранного потенциала.
Следует обратить особое внимание иа земечательный факт
возникновения и поддержания потенциала покоя как активно-
го саморегулирующегося процесса. Вот почему потенциал по-
коя специфичен для различных тканей и колеблется, например,
в клетках скелетных мышц в пределах 80—90 мВ, а эпители-
альной ткани — 20—30 мВ. Потенциал покоя одной и той же
ткани претерпевает изменения в зависимости от возраста, со-
стояния обмена веществ и т. д., а также определяется особен-
ностями проницаемости клеточной мембраны, изменения кон-
центрации ионов внутри и вне клетки и др.
Проницаемость мембраны в свою очередь зависит от харак-
тера отношений в ней водных и липидных фаз. В зове между
этими фазами ионы преодолевают силы гидратации. Запас сво-
бодной энергии ионов зависит от разности энергии их при пе-
реходе из одной фазы в другую. Таким образом, при миграции
частиц через биологическую мембрану возникает определенный
6*
163
энергетический барьер. За счет этого барьера и создается из-
бирательная проницаемость и высокое электрическое сопро-
тивление клеточных мембран.
Регулятором, определяющим пропускную способность мем-
бран и разделение встречных потоков анионов и катионов, яв-
ляется электрический заряд поры мембраны. Привлекающая
своей простотой и ясностью мембранная теория возникновения
элсктропотеициалов клетки встречает значительные трудности
при объяснении многих вопросов, связанных с выяснением их
природы.
Сорбционная, или так называемая фазовая, теория *, в от-
личие от мебранной теории, рассматривающей клетку и окружа-
ющую ее среду как два различных водных раствора, разделен-
ные друг с другом мембраной, заключается в сущности в том.
что цитоплазма и омывающая клетку жидкость представляют
собой две фазы одного и того же раствора, отличающиеся друг
от друга способностью сорбировать электролиты. Но этой тео-
рии различная проницаемость клеток обусловлена разной раст
воримостью веществ в цитоплазме и в окружающей их среде.
Эта теория полагает, что основная часть электролитов, и в пер-
вую очередь фосфатов калия, находится в цитоплазме в свя
занном состоянии, образуя электролитно-белковые комплексы.
Значит, в клетке, находящейся в покое, не может быть электри-
ческого потенциала, так как потенциал возникает при условии
существования свободных положительно и отрицательно заря-
женных ионов и может возникнуть лишь при разрушении элек-
тролитно-белковых комплексов. Это происходит при поврежде-
нии клетки, частным случаем которого, по мнению авторов
сорбционной теории, является нанесение раздражения, доста-
точного для возникновения возбуждения. Однако сорбционная
теория полностью нс игнорирует биологических свойств мем-
бран. I
Теория редокспотенциалов объясняет возникновение элект-
родвижущих сил (электрических потенциалов) в клетке, пред-
ставляющей собой редокс-систему (окислительно-восстанови-
тельная система), разным уровнем окислительно-восстанови-
тельных процессов. Согласно этой теории, источником возникно-
вения потенциалов на поверхностях биологических мембран
* Эта теория в настоящее время большинством исследователей
ие признается.
164
(потенциала покоя) являются высвобождающиеся при окисли-
тельно-восстановительных процессах электроны. Вследствие
повышения интенсивности обмена веществ, под влиянием раз-
дражений окислительные процессы усиливаются и потенциал
покоя может перейти в местный потенциал и потенциал дейст-
вия. Следует обратить внимание на то, что эта теория ставит
возникновение электрических потенциалов в зависимость от
состояния метаболических процессов в клетке, а не только от
обмена веществ мембраны, обеспечивающей активный тран-
спорт.
Близка по идее к теории редокспотенциалов теория протон-
но-химических процессов, которая считает возникновение мем-
бранного потенциала результатом переноса положительно
заряженных частиц — протонов.
Этими теориями не исчерпываются попытки научного объ-
яснения природы электрических потенциалов клетки. В частно-
сти, С. В. Коневым с сотрудниками (1970) развивается ориги-
нальная концепция о роли свойств клетки как кооперативной
системы в возникновении всех видов потенциалов. В коопера-
тивной системе состояние одного элемента зависит от состояния
другого. Высокие степени кооперативности присущи лишь
упорядоченным кристаллическим или квазикристаллическим
телам, к которым относится клетка в целом и тем более ее
мембраны. По мнению авторов, кооперативные свойства гене-
раций потенциала действия сводятся по существу к межбелко-
вым взаимодействиям, когда изменения одного промотора *
облегчают взаимный переход в соседних. Так, рецепторы аце-
тилхолина, содержащиеся в мембране, белковой природы. Этот
рецепторный белок может находиться в двух конформацион-
ных состояниях, переходящих друг в друга: Р-^Д. Форма Р
соответствует поляризованному, т. е. покоящемуся, исходному,
состоянию мембраны, Д — деполяризованному, возбужденному.
Скачкообразным изменениям проницаемости мембраны для
ионов натрия и калия, обусловливающим возникновение элек-
трических потенциалов клетки, согласно С. В. Коневу (1970),
должны предшествовать определенные перестройки в мембра-
нах, которые протекают по кооперативному закону. Одним из
механизмов его реализации являются конформационные изме-
* Промотор — рецепторный белок (терминология Шаиже).
16S
нения не только углеводородных цепей фосфолипидов мембра-
ны, но и конформационные (в широком смысле) изменения
мембраны в целом. В частности, показано, что простой механи-
ческой деформацией мембраны можно изменить ее электрофи-
зиологические свойства.
Нам представляется правомочным высказать еще одно пред-
положение о возникновении потенциала действия на базе
пьезоэлектрического эффекта в условиях конформационных
изменений клеточной мембраны, если рассматривать ее как
кристаллическое и квазикристаллическое образование. Так,
влияние растяжения аксона на потенциал покоя и скачкообраз-
ное изменение потенциала действия в опытах Спириопоулоса
(1968), по-видимому, можно объяснить не только кооперативно-
конформационными эффектами в их обычном понимании.
Предлагаемые теории при всех недостатках не лишены и
достоинств. Поэтому правильнее решать вопрос комплексной
природы проницаемости клетки и ее электрических потенциа-
лов. Какие бы дискуссионные бури ие возникали вокруг того
или иного объяснения, отправной точкой для каждого из них
должен быть основной жизнеобеспечивающий процесс — про-
цесс обмена веществ. Именно такой «метаболический» подход
позволил установить точки соприкосновения противоположных
друг другу теорий. В связи с этим электрические потенциалы
(потенциал покоя, местный потенциал и потенциал действия)
следовало бы называть метаболическими независимо от частно-
го механизма возникновения каждого из них.
Классическим доказательством проявления метаболических
процессов при развитии потенциала действия может быть опыт
Хилла (1963) по изучению теплопродукции нерва при ответной
реакции на раздражение. Оказалось, что одиночное раздраже-
ние нерва конечности краба при 0° вызывает общее повышение
температуры примерно на 2-10~6 градуса. Это повышение тем-
пературы — суммарный результат двухфазного процесса: пер-
вая фаза -— повышение температуры па 10 • 10~6 градуса во
время прохождения импульса и вторая—понижение темпера-
туры на 8 • 10—° градуса в течение нескольких десятых долей
секунды после проведения импульса. Хилл отмечает, что выде-
ление тепла в фазе положительной теплопродукции, «вероят-
но, носило бы характер одиночной резкой «вспышки», совпа-
дающей по времени с прохождением импульса. Фаза отрица-
166
тельной теплопродукции, однако, занимает около 300 мс—-
значительно больше времени, чем любой известный процесс,
связанный с самим импульсом».
Однако такой подход к роли метаболизма в возникновении
И поддержании всех видов потенциалов еще не получил доста-
точного распространения. Поэтому в литературе можно встре-
тить и другую терминологию, как например, потенциал градиен-
та основного обмена, называемый также «истинным потенциа-
лом» или метаболическим. Выделение их в самостоятельную
группу весьма условно. Эти потенциалы генерируются в процес-
се основного обмена, т. е. в состоянии максимально допустимого
покоя биологических систем. По характеру протекания потен-
циалы основного обмена очень вариабельны, но в большинстве
случаев постоянны по полярности и медленно колеблемы по
величине. При этом участки с более интенсивным обменом
электроотрицательны по отношению к окружающим элемен-
там биосистемы.
Применяется формулировка метаболические потенциалы в
иной трактовке. По А. В . Латманизовой (1972), например, это
«стойкие, неизменные во времени разности потенциалов, су-
ществующие между противоположными поверхностями от-
дельных тканевых структур (например, кожи лягушки и др.)
или между различными частями организма, расположенными
вдоль его продольной оси. В биологическом отношении здесь
речь идет, по-видимому, о примерах суммарных различий в
величинах потенциалов покоя отдельных структурных звеньев
исследуемой сложной морфологической системы».
Таким образом, состояние так называемого физиологиче-
ского покоя, хотя внешне и не проявляет специфической дея-
тельности (например, движение, секреция и др.), в действи-
тельности пе бездеятельно. Состояние относительно устойчи-
вого физиологического покоя живой клетки находится в пря-
мой зависимости от относительно устойчивого значения метабо-
лизма и потенциала покоя. Сохранение последнего и есть один
из основных показателей состояния ее физиологического покоя.
Снижение потенциала покоя до нуля свидетельствует о ката-
строфическом нарушении проницаемости клеточной мембраны,
в результате чего концентрация одноименных ионов на наруж-
ной и внутренней поверхности становится одинаковой. Возни-
кает ионная симметрия мембраны. Это говорит о потере мем-
167
браной способности к избирательной проницаемости, о полном
уравновешивании внутреннего содержимого клетки с окружа-
ющей средой, поглощении ее средой, т. е. о смерти. Вот почему
потенциал покоя является одной из основных характеристик
живой клетки, а следовательно, и организм в целом.
Если воспользоваться современными методами, то можно
заметить, что на одном и том же участке клеточной мембраны
потенциал покоя то уменьшается (гипополяризация), то увели-
чивается (гиперполяризация). Как известно, величина потен-
циала покоя зависит от разности концентрации иоиов на по-
верхностях мембраны. Чем больше эта разность, тем больше
поляризована мембрана и выше потенциал покоя. Эта разность
поддерживается' избирательной проницаемостью мембраны и
процессами активного транспорта. В тех участках мембраны,
где изменяется обмен веществ (в результате изменения состава
межклеточной жидкости, вдыхаемого воздуха, температурных
и других воздействий), наблюдается изменение проницаемости
ее. Это сказывается на состоянии всей клетки и может быть
зарегистрировано в виде колебания потенциала покоя. Так, по
мере роста силы раздражения до определенного предела в месте
нанесения его происходит градуально (соответственно увели-
чению силы раздражения) нарастающее уменьшение потенциа-
ла покоя (рис. 44). Это возможно только при уменьшении уров-
ня поляризации поверхностей клеточной мембраны, что осуще-
ствимо лишь при условии увеличения ее проницаемости для
всех потенциалобразующих ионов и в первую очередь натрия.
Проникающие внутрь клетки катионы натрия уменьшают вели-
чину положительного заряда на наружной поверхности мембра-
ны и в целом потенциала покоя. Прекращение раздражения
ведет к восстановлению его исходной величины. В связи с тем
что эти изменения распространяются от места своего возник-
новения, постепенно ослабляясь вплоть до исчезновения, а в
мышечной и нервной ткани вообще ограничены небольшим
участком нанесения раздражения, они и получили название
местного потенциала. Важнейшим свойством местного потен-
циала является градуальная зависимость от силы раздражения,
какой бы малой она ни была. Благодаря этому биоснстема
находится в состоянии постоянной готовности к реагированию
на всякое воздействие. Следовательно, даже самые незначи-
тельные события небезразличны для живых организмов. Это
168
свойство находится в
полном соответствии
с термодинамическими
свойств а м и откр ы тых
стационарных и неравно-
весных систем.
Другой не менее су-
щественной особенностью
местного потенциала яв-
ляется отсутствие латент-
ного периода при ответе
на любое роздействие,
что обеспечивает макси-
мальную оперативность
в установлении наиболее
оптимальных отношений
организма с внешней сре-
дой и, следовательно, со-
хранение гомеостаза. В
этом отношении замеча-
телен тот факт, что для
клеток соединительной и
эпителиальной тканей,
т. е. тканей, из которых в
основном образованы
почти все наши внутрен-
ние органы, единственной
формой электрического
ответа на воздействия
извне являются градуаль-
ные изменения потеициа-
Рис. 44. Изменение мембранного по-
тенциала в зависимости от силы на-
носимого раздражения длительностью
2 мс (по Б. Катцу, 1968 и Е. К. Жу-
кову, 1969):
1—5 изменение мембранного потенциа-
ла. Внизу—относительная сила наносимо-
го раздражения (электрический ток). Го-
ризонтальные линии: сплошная — уровень
потенциала покоя; штрих пунктирна я — кри-
тический уровень местного потенциала
(подпороговая область), т. е. момент воз-
никновения потенциала действия; штрихо-
вая— уровень перехода деполяризации в
инверсию знака потенциала действия (над-
пороговая область); а, б — местный потен-
циал, включающий локальный потенциал
возбуждения б; с — потенциал действия.
ла покоя, достижение критического уровня (вероятно, близкого
по своей природе к явлению местного подпорогового ответа нерв-
ной и мышечной тканей) которого сопровождается возникнове-
нием выраженной функциональной активности биосистемы
(для железы, например, секреция). Да как может быть по-
другому, если благодаря работе именно внутренних органов
обеспечивается строгое поддержание постоянства внутренней
среды (состав плазмы, лимфы, межклеточной жидкости). Ес-
тественно, что ткани, их образующие, практически не могут
169
находиться в состоянии физиологического покоя. Они должны Щ
быть постоянно в деятельности, в «готовности № 1!». Неу диви- W
тельно, что местная реакция на раздражение вызвана местным В
возбуждением, где местный потенциал будет одним из выра-
женин его, а величина потенциала в определенной степени—
мерой возбуждения, т. е. мерой функциональной активности В
тканей. Причем под ростом величины возбуждения следует W
понимать не только и не столько внешнее его проявление (на- Ч
пример, секреция), сколько усиление интенсивности обмена |
веществ. 11
В ходе развития местного возбуждения в отличие от рас-
пространяющегося возбуждения отсутствует состояние так на-
зываемой рефрактериости — снижения возбудимости. В этом
заложен большой биологический смысл. Ведь местное возбуж-
дение в большинстве случаев скорее характеризует готовность
живой системы к ответу на раздражение, чем его непосредст-
венное осуществление. Это относится и к тканям, для которых
местный потенциал является единственной электрической ха-
рактеристикой состояния возбуждения. В данном случае мест-
ный потенциал, достигнув критического уровня (возможно,
сходного с локальным потенциалом возбуждения, свойственным
нервной и мышечной тканям), вызывает специфическую ответ-
ную реакцию органов, состоящих из эпителиальной, железистой
и соединительной тканей. Рефрактерность. свойственная оди-
ночному циклу (волна) распространяющегося возбуждения,
дает возможность полноценной реализации ответной реакции J
на пороговое и надпороговое раздражение. Присутствие ре- 3
фрактерности в местном возбуждении так называемых возбу-
димых тканей (нервные клетки, мышцы) лишило бы биологи-
ческую систему способности вообще реагировать специфиче-
ской ответной реакцией на раздражения.
Наконец, еще одним важным свойством местного возбуж-
дения является ограниченность его распространения, т. е. де-
крементность. Говоря о местном характере возбуждения, мы
имеем в виду лишь количественную сторону его, да и то в
пределах возможностей измерительной аппаратуры. Теорети-
чески нераспространяющихся и ограниченно распространяю-
щихся событий в биологической системе пет в силу ее морфо-
функционалыюй целостности. При изучении электрических
свойств мышечной и нервной тканей было замечено, что зави-
170
симость местного потенциала от силы раздражителя при до-
стижении определенной величины нарушается (см. рис. 44).
Как только местный потенциал достигает некоторой критиче-
ской (пороговой) величины, он превращается в потенциал
Рис. 45. Механизм возникновения потенциала действия (по Е. К. Жу-
кову, 1969; Ходжии и Хаксли, 1952):
J. А—раздражающий толчок, Б — возникновение потенциала действия;
90 — уровень потенциала покоя; 55 мВ — уровень порогового (критиче-
ского) потенциала; 90—55 мВ-местный потенциал; стрелка и ПД —момент
возникновения потенциала действия; В — иопные потоки; нар.— направлен-
ные из клетки, пн.— внутрь; Г — суммарный ионный поток через мембрану.
Вертикальные линии связывают одновременные точки на кривых ионных
потоков. О — «нуль-точка» суммарною потока. //. ‘ ~ кР11Вые измене-
ния проницаемости мембраны для ионов калия и натрин во время развитая
потенциала действия; ПД — потенциал действия, мВ; Е^, — уровни по-
тенциалов равновесия для ионов натрия и калия. ///. Вертикальная стрел-
ка — момент порогового сдвига проницаемости.
действия, что говорит о возникновении распространяющегося
возбуждения.
Потенциал действия есть следствие дальнейшего развития
ионных пертурбаций на поверхностях мембраны под влиянием
возросшей силы раздражения (рис. 45). Так, в подпороговой
области приблизительно в течение 2/3 времени (Z, А, а) от
момента нанесения порогового раздражения развитие местного
потенциала (I, Б) идет в условиях усиления выходящего по-
тока ионов калия из клетки (/, В, Л<) из-за снижения мем-
171
бранного потенциала Ен относительно потенциала равновесия*
для калия (//, £к). «Входящий» же поток ионов натрия в
клетку (/, В. Juz) еще незначителен, так как проницаемость
мембраны для них (//, ?№) пока невелика; в этих условиях
суммарный поток ионов калия и натрия (/, Г) направлен из
клетки наружу. По мере усиления деполяризации проницае-
мость мембраны для ионов натрия растет. Рост этот начина-
ется в последней трети времени действия порогового раздраже-
ния (/, А, б); когда местный потенциал достигает критического
(порогового) уровня (/, Б, ПД), встречные потоки ионов натрия
и калия уравновешиваются (/, В) и суммарный поток ионов
становится равным пулю (/, Г, О)**.
Нуль-точка суммарного потока (I, Г, О) ионов—* это момент
возникновения неустойчивого равновесия, т. е. неустойчивого
стационарного состояния. В этих условиях даже незначитель-
ная дальнейшая деполяризация мембраны (обязательная при
действии порогового раздражения) нарушает равновесие. Про-
ницаемость мембраны для ионов натрия повышается в сотни
раз (II, <zNa). В результате поток их в клетку усиливается
настолько, что начинает превышать поток нопов калия из клет-
ки (7, В). Вследствие этого суммарный поток меняет свое на-
правление — теперь он входит в клетку (/, Г) и практически
не изменяется под влиянием раздражений любой свсрхпорого-
вой силы. Местный же потенциал круто преобразуется в вос-
ходящее колено потенциала действия (/, Г, Д, П, ПД). Объяс-
нение этому можно найти, если принять предположение, что
здесь регенеративный процесс идет за счет собственных заря-
дов мембраны клетки. Стремительно проникающий поток ионов
натрия в клетку способствует и ускорению деполяризации мем-
браны. В результате в определенный момент мембранный по-
тенциал становится равным нулю. Однако такое состояние
(если оно практически вообще возможно) сохраняется очень
* Величина мембранного потенциала, уравновешивающая диф-
фузию иона, называется потенциалом равновесия.
** Возможность абсолютного совпадения в дайной временной
точке полного равенства противоположных потоков ионов маловеро-
ятна; реально под «нуль-точкон» следует повимать момент возникно-
вения наименьшей разницы между разнонаправленными потоками
ионов.
172
короткое, не поддающееся учету время. Дело в том, что про-
должающееся поступление ионов натрия в клетку ие только
«гасит» избыток имеющихся там отрицательно заряженных
анионов, но и приводит к накоплению положительно заряжен-
ных катионов на внутренней поверхности мембраны и к недо-
статку их на наружной. В итоге в месте нанесения раздражения
на наружной поверхности мембраны заряд меняется с положи-
тельного на отрицательный, а на внутренней — наоборот, с от-
рицательного па положительный, т. с. происходит изменение
(инверсия) знака потенциала мембраны. При достижении по-
тенциалом действия максимума (77, 77//), находящегося в пря-
мой зависимости от количества ионов, перешедших с наружной
поверхности мембраны на внутреннюю, поток ионов в клетку
ослабляется, так как точка максимума потенциала действия
близка по величине к потенциалу равновесия для ионов натрия
(7/, 7?ыа). Одновременно для этих ионов начинает снижаться
проницаемость мембраны (7, в, 77, <7Na), что способствует
ослаблению суммарного потока ионов (7, Г).
В связи с ослаблением потока ионов натрия в клетку и
продолжением роста потока ионов калия из клетки роль первых
в возникновении потенциала действия снижается, а вторых
в возникновении (восстановлении) потенциала покоя — увели-
чивается. Этим и объясняется крутое начало нисходящего ко-
лена (фазы) потенциала действия (7/, ПД), т. е. первой стадии
реполяризации мембраны — реверсии знака потенциала. По
мере приближения потенциала действия (в нисходящей фазе)
к величине потенциала равновесия для ионов калия (7/, Ец)
ноток их ослабевает (7, В, П, ?Na). Это сопровождается за-
медлением процесса реполяризации — восстановления потен-
циала покоя (зона следового отрицательного потенциала).
Дальнейшее превышение потока ионов калия из клетки над
потоками ионов натрия в клетку приводит не только к восста-
новлению исходного потенциала покоя мембраны, ио и к неко-
торой ее гиперполяризации (зона следового положительного
потенциала). Окончательное восстановление исходного потен-
циала покоя соответствует моменту достижения потоком ионов
калия величины потенциала равновесия.
Все эти события совершаются преимущественно за счет
пассивного переноса ионов по градиенту концентрации. Однако,
если бы в процессе возникновения и тем более затухания по-
173
тенциала действия происходил только пассивный перенос ио-
нов, то очень скоро установилось бы стойкое равенство содер-
жания их на внутренней и наружной поверхностях мембраны.
Между тем этого в действительности не происходит, так как
здесь включается (особенно в фазе реполяризации) механизм
активного переноса — калий-натриевый пасос. В результате
восстанавливается потенциал покоя с асимметричной концен-
трацией ионов натрия и калия на поверхностях мембраны.
Особо следует отметить еще один чрезвычайно существенный
факт: потенциал действия, как и местный потенциал, может
возникнуть лишь при обязательном условии — при наличии
ионов кальция во внешней среде, т. е. на наружной поверхности
мембраны, поскольку переход гидратированных ионов натрия
в цитоплазму зависит нс только от их размеров, но и от содер-
жания в окружающей среде ионов кальция. Полагают, что
ионы кальция адсорбированы на белковых молекулах, высти-
лающих канал натриевой поры мембраны. В силу одноимен-
ности зарядов ионов кальция и натрия при их встрече возни-
кают электростатические силы отталкивания, препятствующие
прохождению натрия через пору. При уменьшении концентра-
ции ионов кальция в окружающей среде проницаемость мем-
бран для натрия увеличивается. Возможно также, что здесь
играют роль и конкурентные отношения ионов кальция и на-
трия в силу их высокого химического сродства.
Процесс деполяризации и инверсии знака потенциала дей-
ствия при его возникновении в нервных и мышечных клетках
развивается очень быстро (0,1—0,5 мс); реполяризация про-
текает значительно медленнее (0,5—4,5 мс). Общая продол-
жительность потенциала действия равна 1—5 мс.
Таким образом, приходим к выводу, что в основе механизма
возникновения и поддержания потенциала покоя ведущая роль
действительно принадлежит ионам калия, а потенциала дей-
ствия— ионам натрия. Поэтому для уровня потенциала покоя
существенное значение имеет концентрация в окружающей
клетку среде ионов калия, а для величины потенциала дейст-
вия — ионов натрия.
Однако структура и природа потенциалов возбуждения
несколько сложнее. Если вернуться к рис. 44, то можно увидеть,
что как только сила раздражения (4) оказывается достаточной
для деполяризации мембраны (в данном случае нервного во-
174
локна), при которой величина местного потенциала равна —65—
60 мВ, на кривой появляется дополнительный «горб» (4, б).
Эта маленькая волна деполяризации между а и б, так же как
и предыдущая а, локальна и носит название местного подпо-
рогового ответа. Он является следующим этапом развития
местного потенциала и выражением первой активной реакции
возбудимой системы на раздражение. Именно поэтому он на-
зван локальным потенциалом возбуждения. Уровень раздра-
жения, при котором местный потенциал переходит в потенциал
действия (4, в),— критический или пороговый уровень. При
значительных силах раздражений (5) развитие местного по-
тенциала, включая локальный потенциал возбуждения, идет
столь быстро, что они практически могут быть не зарегистри-
рованы. В этом случае создается впечатление, что потенциал
действия (распространяющееся возбуждение) возникает сразу
в ответ на раздражение, минуя стадию развития местного по-
тенциала (местного возбуждения).
Потенциал действия отличается от местного потенциала тем,
что он возникает на строго определенную критическую, порого-
вую, силу раздражения и только через определенный латентный
период, равный длительности достижения местным потенциалом
критической величины. Возбуждение, сопровождающееся воз-
никновением потенциала действия (величина его практически
не зависит от любого изменения силы раздражения, т. е. он гра-
дуально независим от надпороговой силы), в отличие от воз-
буждения, сопровождающегося местным потенциалом, способно
иррадиировать без ослабления (декремента) на большие рас-
стояния от места возникновения. (Распространяющееся воз-
буждение в отличие от местного также обладает свойством
рефрактерности.) Эти свойства потенциала действия опреде-
ляют и биологический смысл характеризуемых им явлений —
включение систем, обеспечивающих проявление и регулирование
непроизвольных моторных (работа сердца, сократительная
функция пищеварительного аппарата и др.) и особенно произ-
вольных сложнокоординированных (движение, ходьба, бег,
трудовая, спортивная деятельность и др.) двигательных реак-
ций, лишь при необходимости, т. е. тогда, когда факторы внеш-
ней среды достигают значений, вызывающих существенные
отклонения в жизнедеятельности организма. В противном слу-
чае эти системы находятся в состоянии физиологического покоя
176
или близком к нему. В первую очередь это имеет отношение
к аппарату произвольных движений — скелетной мускулатуре.
Таким образом, местный потенциал и потенциал действия —
наиболее общие выражения состояния возбуждения, т. е. функ-
циональной активности тканей, органов и организма в целом.
Величина и характер местного потенциала и потенциала дей-
ствия, так же как и потенциала покоя, находится в прямой
зависимости от состояния обмена веществ в данный момент.
Именно поэтому все регистрируемые электрические явления
живых образований следует рассматривать как одно из прояв
лений жизнедеятельности биосистемы, источник которой зало
жен в обмене веществ.
Очерк 3. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ
И СВОЙСТВА ВОЗБУДИМОСТИ ЖИВЫХ СИСТЕМ
Для того чтобы биологическая система могла перейти из
состояния физиологического покоя в состояние возбуждения,
недостаточно одного контакта с раздражающим фактором.
Сила действующего раздражения при этом должна достичь
критической величины. Та минимальная сила раздражения, при
воздействии которой возникает минимальная по величине и ин-
тенсивности специфическая ответная реакция, называется
порогом возбуждения. Как известно, момент появления мини-
мальной ответной реакции во многих случаях практически
невозможно точно зарегистрировать. Например, очень трудно
зафиксировать момент возникновения минимального сокраще-
ния целой скелетной мышцы. Да и вообще понятие минималь-
ная ответная реакция весьма неопределенно. Одпако известно,
что возникновение критического местного потенциала для ор-
ганов, образованных из эпителиальных тканей (железы и др.),
и потенциала действия для нервной и мышечной тканей явля-
ется верным признаком возникновения возбуждения. Поэтому
регистрация этих потенциалов — наиболее удобный прием для
выявления момента возникновения минимальной специфической
ответной реакции.
Сила раздражения ниже пороговой называется подпорого-
вой (в ответ на нее возникает только местное возбуждение,
местный потенциал, не сопровождающийся специфической ре-
акцией), выше—над- или сверхиороговой. Чем больше сила
176
раздражения по сравнению с пороговой, тем сильнее (до опре-
деленного предела) у возбудимой системы ответная реакция.
При достижении этого предела дальнейшее увеличение силы
раздражения вызывает только увеличение (а иногда и сниже-
ние) интенсивности обмена веществ и не обусловливает сокра-
Рис. 46 Схема зависимости ответ-
ной реакции биосистемы от силы
раздражения (по Л. В. Латмани-
зовой, 1972):
А — подпороговая сила; Б — порого-
вая; Б — субмаксимальная: Г — мак-
симальна и сила; штриховая линия —
величина суммарных энергетических
сдвигов; сплошные вертикальные ли-
нии — величина функционального от-
вета; по оси абсцисс — сипа раздра-
жения, по осн ординат — величина от-
вета в условных единицах.
Рис. 47. Схема зависимости
ответной, реакции от дли-
тельности раздражения (по
Л. В. Латманизовой, 1972):
А —- пороговая длительность раз-
дражения; Б — максимальная
длительность раздражения; пун-
ктирная линия — величина сум-
марных энергетических сдвигов;
сплошные вертикальные линии —
величина функционального от-
вета; по оси абсцисс — дли-
тельность раздражения, по осн
ординат — величина ответа в ус-
ловных единицах.
щепия мышцы. Сила раздражения, вызывающая наибольшую
ответную реакцию, называется максимальной, а выше макси-
мальной— супермаксимальиой. Все силы, располагающиеся
между пороговой и максимальной, будут субмаксимальными
(рис. 46). Чем ниже порог силы, тем выше возбудимость, т. е.
способность реагировать па минимальные по силе раздражения.
Однако ясно, что для возникновения возбуждения педоста
точно только действия на живой объект силы раздражения.
Необходимо, чтобы это раздражение длилось какое-то время
7 А. А. ЛогинО!
177
(рис. 47). Та минимальная сила раздражения, на которую при
практически неограниченно большой длительности ее действия
возникает минимальная ответная реакция, называется реобазой.
Наименьшая длительность раздражения силой в одну реобазу,
необходимая для возникновения возбуждения, называется по-
лезным временем, так как дальнейшее увеличение длительности
раздражения при тех же условиях
ие оказывает регистрируе-
мого влияния иа величи-
ну ответной реакции
(рис. 48). Для возникно-
вения возбуждения на
раздражение по силе вы-
ше реобазы также тре-
буется какое-то мини-
мальное (пороговое)
время его действия. Одна-
ко величина ответной ре-
акции на неизмеияющую-
ся силу выше одной
реобазы, до определен-
ного предела, находится
в прямой зависимости от
увеличения длительности
порогового времени разд-
ражения. Это значит, чем
длительнее раздражение, тем сильнее (до предела) ответная
реакция живого объекта (см. рис. 47). Таким образом, фактор
времени играет существенную роль для величины ответной
^JOO
i
§
I
60
60
40
£ 0 0! 0.6 0.5 0.7 Q5 Ю
Ввепп, нс
Рис. 48. Кривая зависимости си-
лы — длительности:
О-Р— реабаза; О-В — минимальное
время, необходимое для возникновения
возбуждения при действии силы раз-
дражителя в одну реабазу.
реакции иа раздражение.
В громадном разнообразии взаимоотношений живого объек-
та со средой только закон силы или только закон длительности
раздражения практически ие проявляются. Это объясняется
тем, что любое возмущающее воздействие одновременно обла-
дает двумя неотделимыми друг от друга свойствами — силой
и длительностью действия. Следовательно, помимо порога воз-
буждения необходимо выделять еще и порог времени возбуж-
дения. Чем больше сила раздражения, тем меньше требуется
времени для возникновения возбуждения. Таким образом, для
каждой силы раздражения существует свой порог времени
возбуждения. Итак, возникновение возбуждения находится в
178
прямой зависимости от неразрывного единства силы-длитель-
ности действия раздражения. Поэтому правильнее говорить
о существовании единого закона силы-длительиости раздраже-
ния. При изучении соотношения силы и длительности раздра-
жения, исходя из закона их обратной зависимости, Гоорверг,
Вейс и Лапик построили кривую на основании положения, что
усиление раздражения приводит к укорочению времени, необ-
ходимого для возникновения возбуждения, ио ие беспредельно
(см. рис. 48). Отправными точками были взяты величины рео-
базы и полезного времени. Из рисунка видно, что при очень
короткой длительности раздражения, как бы велико оно ни
было по силе, возбуждение ие возникает (восходящая ветвь
кривой параллельна ординате). Следовательно, возникновение
возбуждения в зависимости от силы-длительности раздраже-
ния выявляется лишь в определенных пределах.
В сложных взаимоотношениях организма со средой ему
приходится сталкиваться не только с изменением силы н дли-
тельности действия факторов, но н с быстротой их изменений,
т. е. чем выше градиент раздражения (быстрота нарастания
силы раздражения в единицу времени), тем быстрее (до из-
вестных пределов) возникает реакция возбуждения. Более того,
при медленном нарастании силы раздражения, даже при до-
стижении ею чрезвычайных значений, специфическая ответная
реакция не возникает. Так, например, иа быстро возросшую
от нуля до порогового значения силу раздражения мышечное
волокно отвечает сокращением. На силу же, во много раз пре-
восходящую пороговую, но нарастающую во времени очень
медленно, сокращение не наступает. Сдвиги в мышце ограни-
чиваются развитием местного возбуждения — местного потен-
циала (рис. 49).
Биологическое значение закона градиента раздражения
чрезвычайно велико. Быстро нарастающие во времени раздра-
жения требуют от живой системы развития срочной приспосо-
бительной реакции, проявляющейся в виде возбуждения (со-
кращение, секреция и др.). Напротив, медленно нарастающее
раздражение не вызывает внешне выраженного ответа. Это
говорит о том, что отдельная клетка, орган или в цепом орга-
низм сумели «справиться» с возмущающим воздействием «внут-
ренними», «местными» перестройками, т. е. произошло приспо-
собление к раздражителю—аккомодация (адаптация). Оиа
Г 179
очень важна, так как открывает биосистеме возможность в этих
условиях реагировать на действие других раздражителей, тре-
бующих в отношении себя оперативных, срочных специфических
ответов.
Установление оптимальных отношений биосистемы со сре-
дой определяется не только ее способностью быстро реагировать
па возможно минимальные
по силе раздражения или
адаптироваться к медлен-
но нарастающим большим.
Часто иа живой объект дей-
ствует серия одновременно
или последовательно посту-
пающих раздражений, каж-
дое из которых требует
соответствующей ответной
реакции. Живой объект дол-
жен их различать не только
Рис 49. Влияние крутизны на-
растания силы раздражения на
возникновение потенциала дей-
ствия и его величины (по
Е. Б. Бабскому и др, 1972):
I — раздражение прямоугольным
(«мгновенным») толчком тока; 2—5 —
линейно нарастающие силы раздра-
жений. Горизонтальным и стрелка-
ми отмечен критический (порого-
вый) уровень деполяризации мем-
браны при дайкой крутнзпе. Верти-
кальными стрелками отмечена дли-
тельность нарастания силы раздра-
жения от нуля до возникновения по-
тенциала действия. При снижении
крутизны нарастания силы раздра-
жения ниже градиента (кривая 5)
возникает только местный потен-
циал.
по качественным показате-
лям, по и по силе и длитель-
пости действия. Поэтому
существует еще одна мера
оценки функциональных
возможностей живой систе-
мы — порог разности, т. е. та
минимальная разность силы
длительности действия меж-
ду двумя раздражениями,
которая достаточна для воз-
никновения нового уровня
активности биосистемы. Чем
меньше эта разность, тем
легче и оперативнее осуще-
ствляется переход иа новый
уровень активности или поддерживается прежний. Так, для
выполнения какого-либо упражнения спортсмен должен посто-
янно оценивать местоположение центра тяжести тела и чем
точнее, тем меньше смещение его, тем легче удерживается нуж-
ное положение. Можно сказать, что именно разностная чувст-
вительность биосистемы к силам возмущающих воздействий
180
лежит в основе той или иной
колебательной характеристи-
ки адаптивно-гомеостатическо-
го поведения биосистем.
Как известно, во взаимоот-
ношениях со средой биосистема
никогда не сталкивается с дей-
ствием лишь одного фактора.
Одновременно или в какой-то
последовательности иа иее вли-
яют не только одинаковые, но
и разные раздражители. Всегда
ли живая система внешне реа-
гирует иа все раздражения.
Для этого надо знать, какова
возбудимость живого объекта,
т. е. какова способность его
отвечать на раздражение, по-
ступающее в различные фазы
одного и того же цикла воз-
буждения. Если принять состо-
яние возбудимости в условиях
физиологического покоя за
100%, то в ходе развития одно-
го цикла возбуждения (рис. 50)
оио изменяется как в сторону
повышения (экзальтация), так
и понижения (рефрактерпость).
В результате анализа дина-
мики изменения возбудимости
от момента нанесения раздра-
жения до завершения одиночно-
го цикла возбуждения удалось
(фаз). В период развития местного возбуждения (местного по-
тенциала) возбудимость несколько повышается. Каждое допол-
Рис. ЬО. Схема изменения воз-
будимости при протекании оди-
ночного цикла возбуждения:
Л— электрическая реакция /потен-
циал действия); С — изменение воз-
будимости (уровень состояния по-
коя HXJ%); 1—начальное повыше-
ние возбудимости (развитие мест-
ного потенциала); 2— фаза абсо-
лютной рефрактерное™ (пик потен-
циала действия — деполяризация и
инверсия — восходящее колено); 3 —
фаза относительной рефрактерное™
(восстановление потенциала покоя —
реверсия и реполярнзацня — быст-
рая и часть медленной, начальная
часть отрицательного следового по-
тенциала); 4 фаза экзальтации
(продолжение восстановления по-
тенциала покоя — отрицательный
следовый потенциал); 5—фаза
субнормальной возбудимости (поло-
жительный следовый потенциал). В
скобках — фаза развития потенциа-
ла действия, соответствующая дан-
ному состоянию возбудимости.
выявить несколько периодов
нительное раздражение, нанесенное в этот период, даже ниже
порогового по силе, ускоряет развитие местного возбуждения.
Как только оно достигает критической величины и переходит в
распространяющееся возбуждение (высоковольтный потенциал
действия — пик, его восходящее колено — деполяризация и
181
инверсия), возбудимость быстро снижается и становится прак-
тически равной нулю (особенно в верхней точке пика). Время, в
течение которого происходит это снижение, н будет абсолютной
рефрактерной фазой, а само снижение возбудимости — абсо-
лютной рефрактерностыо. Раздражение любой сверхпороговой
силой, нанесенное в этот период, практически не влияет на
развитие текущего возбуждения.
В процессе угашения возбуждения и восстановления состоя
ния физиологического покоя (восстановление потенциала покоя
нисходящее колено потенциала действия — реверсия и репо-
ляризация) возбудимость последовательно восстанавливается.
Время восстановления возбудимости до исходного значения —
относительная рефрактерная фаза. Состояние же, в котором
находится живой объект при этих же условиях, представляет
собой относительную рефрактериость. Раздражение, нанесенное
в этот период, может вызвать усиление возбуждения только
в том случае, если оно будет надпороговым. Длительность отно-
сительной рефрактерной фазы значительно больше, чем абсо-
лютной. Она захватывает начальную часть отрицательного сле-
дового потенциала действия.
Вслед за периодом относительной невозбудимости наступает
фаза экзальтации — фаза повышенной возбудимости. В этой
фазе биосистема способна отвечать повторной волной возбуж-
дения даже на раздражения, которые в состоянии физиологи-
ческого покоя ее были подпороговыми. Фаза экзальтации сов-
падает по времени с фазой окончания отрицательного и начала
развития положительного следового потенциала действия.
В чем же биологический смысл фазового изменения возбу-
димости в ходе развития одиночной волны возбуждения? На-
чальная фаза повышения возбудимости обеспечивает условие,
при котором каждый дополнительный раздражитель ускоряет
процесс подготовки (местное возбуждение) для перехода си-
стемы к специфической приспособительной реакции. Состояние
абсолютной рефрактер кости позволяет «без помех» осущест-
вить текущую приспособительную реакцию. Если бы в этих
условиях возбудимость была нормальной, то дополнительное
раздражение, вызвав дополнительное возбуждение, могло бы
исказить эту реакцию, превратив ее в чрезмерную или недо-
статочную для данных условий. Абсолютная рефрактериость
защищает биосистему от чрезмерных энергетических трат.
182
Сходную роль играет и относительная рефрактериость, с той
лишь разницей, что в данном случае система в состоянии реа-
гировать иа раздражения, требующие срочного специфического
ответа. Именно поэтому большинство тканей, работающих не-
прерывно и не обладающих длительными периодами физиоло-
гического покоя (например, сердце), имеют по сравнению со
скелетной мускулатурой более длительную рефрактериость.
Наконец, состояние экзальтации обеспечивает готовность
живого объекта к ответу ие только на повторное раздражение
прежней силы, но и даже на более слабые, бывшие ранее под-
пороговыми. Раздражение, поступившее в эту фазу, вызывает
ответную реакцию с большей легкостью и меньшими энергети-
ческими тратами.
Описанная динамика состояния возбудимости в одиночном
цикле возбуждения позволяет сделать чрезвычайно важный
вывод — биоснстема и в первую очередь нервная и мышечная
ткань проявляют на возмущающие воздействия две взаимосвя-
занные формы возбуждения: постоянную, непрерывную, в виде
местного возбуждения различной интенсивности (в том числе
и физиологический покой) и ритмическую (распространяющееся
возбуждение способно воспроизводиться в данных условиях
с интервалом, равным длительности рефрактерного состояния).
Из изложенного очевидно, что чем короче длительность
рефрактерных фаз, тем с большей частотой биосистема в со-
стоянии отвечать на повторные раздражения и наоборот. Эта
быстрота протекания процессов возбуждения, названная
Н. Е. Введенским функциональной подвижностью (лабильно-
стью), измеряется максимальным количеством циклов возбуж-
дений, воспроизводимых в единицу времени (в секунду). Функ-
циональная подвижность не является абсолютно постоянной.
В зависимости от обстоятельств она может то увеличиваться,
то уменьшаться. Особенно четко выражена эта изменчивость
у малолабильных образований (например, мышца сердца). В
процессе жизнедеятельности наблюдается соиастроение, «усвое-
ние ритма», т. е. система, изменяя свою лабильность, в состоя-
нии реагировать (усваивать) как на более высокие, так и на
более низкие ритмы действующих раздражений (А. А. Ухтом-
ский).
Снижение лабильности системы уменьшает способность ее
к ритмической работе. А это не менее важно, чем повышение
183
лабильности. Обычно снижение наступает в результате дли-
тельной или слишком интенсивной работы, приводящей к из-
быточным энергетическим тратам и утомлению, и ослабляет
и лаже предотвращает последние, выполняя при этом защит-
ную — охранительную, гомеостатическую, роль.
В обычных условиях практически ни одна биосистема не
проявляет максимальной функциональной подвижности. В
частности, в естественном состоянии скелетная мышца «рабо-
тает» в частоте, не превышающей 150—200 импульсов в секунду.
Эта же мышца в эксперименте воспроизводит до 1000 им-
пульсов за то же время. Такое поведение мышцы говорит
о том, что в опыте действует фактор, не имеющий места в ес-
тественных условиях, и поэтому выявляется наличный «запас
прочности», который обычно ие используется, та? как во взаи-
моотношениях со средой почти никогда не возникает необхо-
димость в эксплуатации этого резерва биосистемы. Мало того,
в пределах шкалы функциональной подвижности обнаружи-
вается относительно постоянный диапазон ритма, где работа
оказывается максимально эффективной по производительности
и энергетической экономичности и может продолжаться значи-
тельно более длительное время, чем при поступлении раздра-
жений с частотой за пределами этого диапазона. Это так назы-
ваемый собственный, или оптимальный, ритм возбуждения.
Например, для одиночных волокон скелетной мышцы лягушки
максимальный ритм равен 50— 120 импульсам в секунду, опти-
мальный— 10—20 (резерв 40—100 импульсов). Следовательно,
ритм функциональной лабильности имеет две меры — макси-
мальную, характеризующую предельные возможности ритмиче-
ской активности, и оптимальную — ритм работы в естественных
условиях.
Н. Е. Введенский в опытах на нервно-мышечном препарате
лягушки обнаружил, что если наносить раздражения одной и
той же силы, но последовательно увеличивать частоту подавае-
мых сигналов в единицу времени, то ответная реакция также
последовательно нарастает. Достигнув максимальной величины,
опа, несмотря на продолжающийся рост частоты наносимых
раздражений, не только не увеличивается, но постепенно умень-
шается и даже прекращается вовсе. Иначе говоря, частота
раздражений, вызывающая максимальный функциональный
ответ, будет оптимальной, а приводящая к уменьшению отве-
184
та — пессимальной; само же явление соответственно носит на-
звание оптимум и нессимум частоты. Здесь максимальная от-
ветная реакция определяется тем, что каждое следующее
раздражение попадает в фазу экзальтации, наилучшей готов-
ности к ответу. При последовательном повышении частоты
часть следующих раздражений попадает сперва в относи-
тельную, а затем и абсолютную рефрактерную фазу. Естествен-
но, ответная реакция в этих условиях прогрессивно уменьшается
вплоть до нуля, так как в фазах рефрактерности возбудимость
снижается в направлении от относительной рефрактерности к
абсолютной.
Аналогичная последовательность изменения ответной реак-
ции была выявлена на целой мышце и при применении возрас-
тающей силы раздражений, наносимых с постоянной частотой.
Сила раздражений, вызывающая максимальный функциональ-
ный ответ, была названа оптимальной, а само явление — опти-
мумом силы; сила же, ведущая к уменьшению ответа,— песси-
мальной, а явление — пессимумом силы. Одним из механизмов
оптимума силы может быть постепенное вовлечение все боль-
шего количества функциональных единиц (например, мышеч-
ных волокон), имеющих разные пороги возбудимости, в ответ-
ную реакцию биосистемы.
Природа возникновения псссимума силы вытекает из закона
силы раздражения. В частности, после достижения оптимума
снижение ответной реакции на более значительное раздраже-
ние, т. е. развитие пессимума силы, может иметь в своей основе
не усиление, а угнетение интенсивности обмена веществ
(Л. В. Латманизова, 1972).
В естественных условиях на живые объекты обычно дей-
ствуют одновременно и силовые и частотные раздражители.
Поэтому правильнее говорить о существовании оптимума
и пессимума частоты-силы.
Что же все-таки представляет собой пессимум, каков его
биологический смысл, является ли он состоянием прекращения
активности? Оказывается пет! Установлено, что если в условиях
развивающегося пессимума уменьшить силу или частоту раз-
дражения до оптимальных значений, ответная реакция сейчас
же восстанавливается (рис. 51). Следовательно, пессимум —
особая форма активной реакции, направленной против чрезмер
ных по силе и частоте раздражений, т. е. пессимум — одна из
186
форм проявления торможения. Это ио существу обратная сто-
рона возбуждения, достигшего критического уровня и превра-
тившегося в свою функциональную противоположность. Таким
образом, мы снова приходим к заключению, что возбуждение
и торможение — это различные уровни единственно возможно-
го состояния биологических систем — активности.
Сопоставляя два рода тор-
можения — синаптическое и
пессималыюе, можно видеть,
что оба они являются выраже-
нием активной деятельности
биосистемы. Торможение, воз-
никающее вследствие возбуж-
дения тормозящих синапсов,—
первичное торможение, а пере-
ход возбуждения в торможение
без участия особых тормозя-
щих структур — вторичное. Са-
мое важное здесь то, что воз-
буждение и торможение не
Рис. 51. Оптимум и пессимум
частоты раздражения скелет-
ной мышцы лягушки (по
Н. Е. Введенскому).
просто состояния, сменяющие друг друга, а саморегулирующи-
еся процессы, необходимые для установления оптимального
уровня отношений организма со средой.
Из изложенного выше видно, что большая частота при отно-
сительно малой силе раздражения может вызвать такой же
тормозящий — пессимальный эффект, как и малая частота при
достаточно большой силе раздражения. Хотя закон оптимума
и пессимума частоты силы был обнаружен в опытах на изоли-
рованной мышце, он имеет равное отношение к деятельности
всего организма. Только в этом случае проявление его будет
выражением суммарных (интегративных) частотно-силовых
возможностей организма в целом.
Своеобразие взаимосвязи и взаимоперехода состояния воз-
буждения и торможения на примере оптимума и пессимума
побудило Введенского заняться разработкой этого вопроса.
В частности, им было обнаружено, что на различные воздейст-
вия нерв отвечает однотипной местной реакцией, где можно
различить несколько последовательно протекающих стадий.
Обнаружение этих стадий проводилось иа нервно-мышечном
препарате лягушки (рис. 52). Вблизи и вдали от мышцы иа
нерв накладывались электроды, с помощью которых наноси
лнсь раздражения с нарастающей частотой и силой —- от поро-
говых и выше (условно — слабые, средние, сильные), вызыва-
ющие сокращения мышцы в соответствии с законом силы
раздражения. Если же на небольшой участок нерва между
электродами а и б воздействовать каким-либо химическим
Г
Рис. 52. Схема постановки опытов Введенского по изучению па-
рабиоза (по Л. В. Латманизовой, 1972):
1 — раздражение током малой частоты; 2— раздражение током сред-
ней частоты; 3 —* раздражение током высокой частоты; вертикальные
линии — величина ответной реакции в различные стации парабиоза:
Л — норма; £ —уравнительная стадия; В — парадоксальная стадия;
Г—схема нервио-мышсчного препарата; ц, б—места наложения
электродов на нерв. Участок нерва, подвергающийся химическому,
температурному н другим воздействиям, заштрихован; в — икронож-
ная мышца.
(эфир, хлороформ и др.) или физическим (температурным
и др.) фактором, изменяющим функциональное состояние
нерва, то при повторном раздражении с электрода б ответная
реакция не изменяется, с электрода же а — меняется. Это и
понятно, так как возбуждение, возникающее в точке а, должно
пройти через функционально измененный участок нерва. При
этом вначале проявляется уравнительная (трансформационная)
стадия — на все три силы интенсивности раздражения возни-
кает равная по величине ответная реакция мышцы. Это говорит
187
о том, что в участке нерва, на который оказано воздействие,
прежде всего повышается возбудимость и проводимость. Не-
сколько позже эффект раздражения в точке а становится еще
более необычным: возникает парадоксальная стадия — слабые
раздражения вызывают более сильный эффект, чем средние, а
средние — более значительный, чем сильные. Это в первую
очередь результат снижения лабильности в участке нерва,
подвергшегося воздействию тех или иных факторов, а следова-
тельно, и уменьшения его пропускной способности для сильных
высокочастотных раздражений. Через некоторое время, когда
под влиянием воздействия возбудимость и лабильность участ-
ка нерва еще более снизится, наступает тормозящая стадия:
при раздражении нерва в точке а мышца «ие реагирует» иа
раздражения любой силы. Возбуждение, возникающее в этой
точке, доходит до зоны измененной функциональной активности
и далее не распространяется. Таким образом, мы вновь встре-
тились с фактом последовательного развития состояния тормо-
жения из возбуждения.
Тормозящая стадия обратима, но в случае углубления оиа
переходит в необратимую—наступает смерть. Именно поэто-
му обнаруженное явление было названо парабиозом, а после-
довательные стадии его получили название парабиотического
процесса. Наиболее важно то, что три стадии функциональных
парабиотических сдвигов обязательно проявляются в реакции
нерва на самые разнообразные воздействия извне. Это нор-
мальная ответная реакция на раздражение любой биологиче-
ской системы.
Парабиоз — действительно динамическая характеристика
всеобщего свойства живых систем, яркий пример взаимообу-
словленности и взаимосвязи саморегулирующихся процессов
возбуждения и торможения.
IV. УПРАВЛЕНИЕ И РЕГУЛИРОВАНИЕ ФУНКЦИЙ
Очерк 1. ОБЩИЕ ПОЛОЖЕНИЯ ОБ УПРАВЛЕНИИ
И РЕГУЛИРОВАНИИ ФУНКЦИЙ
Самоорганизации и саморегулирование биосистемы любой
сложности направлено на оптимальное взаимодействие состав-
ляющих ее элементов, процессов, обеспечивающих поддержа-
ние целостности и уровней активности, необходимых для
установления адекватных отношений со средой. Процесс
саморегулирования осуществляется на всех уровнях живого;
молекулярном, клеточном, органном, системном (система
органов кровообращения, дыхания и др.), организменном, а так-
же межмолекулярном, межклеточном, межоргапном, межсис-
темном, межорганизменном и средовом. Саморегулирование реа-
лизуется механизмами, заложенными в самой сущности биологи-
ческой формы движения, т. е. жизни. Под самоорганизацией,
управлением и регулированием следует понимать также соойст
ва системы, которые обеспечивают ее сохранение — гомеоста-
тичность. Можно выделить три механизма регулирования —
генный, гуморальный и нервный. Изучение генного механизма
регулирования находится в компетенции молекулярной генети-
ки. Гуморальный и нервный на уровне высокоорганизованных
живых существ объединены в единый нейрогуморальпый ме-
ханизм. Кроме того, различают четыре основных принципа
механизма управления и регулирования; рассогласования —
неравновестности градиента; замкнутого контура; прогнозиро-
вания; целостности.
Наиболее древний в эволюционном отношении гуморальный
механизм регулирования функций. Он осуществляется за счет
химических веществ циркулирующих в организме жидкостей
(кровь, лимфа, тканевая жидкость, цитоплазма). Химическими
регуляторами могут быть и вещества, поступающие извне
вместе с продуктами питания, при дыхании, через покровы
189
Тела; неспецифическиё продукты обмена ВСщеСтВ органов и
тканей (например, накопление углекислоты в крови возбуждает
центр дыхания); некоторые специфические продукты обмена
веществ клеток, например медиаторы синапсов (ацетилхолин,
норадреналин и др.); гормоны. Гормоны можно вообще
назвать важнейшими специализированными химическими ре-
гуляторами жизнедеятельности организма благодаря их влия-
нию на течение обмена веществ, формирование органов и тка-
ней, способности «запускать» деятельность различных органов
и, наконец, коррсгировать интенсивность функций организма.
Характерно, что химический регулятор, попадая в кровоток
или другие циркулирующие жидкости, поступает к каждому
органу независимо от того, участвует ли он в регуляции его
функций или нет. Только избирательное отношение тканей к
химическим веществам определяет успешность включения гор-
монов в процесс регуляции. Следует отметить: скорость рас-
пространения химического регулятора от места образования
до места приложения невысокая и соответствует скорости дви-
жения переносящих его жидкостей.
Между различными химическими регуляторами, особенно
между гормонами, часто проявляется принцип саморегуляции.
Так, если гормон поджелудочной железы — инсулин способст-
вует снижению уровня сахара в крови, то гормон мозгового
слоя надпочечников — адреналин — увеличению. В случае, ког-
да продукция инсулина становится избыточной, сразу же уси-
ливается продукция адреналина и наоборот. Это касается и
других желез.
Нервный механизм регуляции эволюционно более молодой.
Он отличается от гуморального тем, что его сигналы распро-
страняются по нервным путям с достаточно большой скоростью
(в зависимости от характера нерва) —от 0,5 до 80—120 м/с —
к строго определенным органам или частям организма. Нервные
механизмы регуляции, так же как и гуморальные, работают
ио принципу саморегуляции, в основе которого лежат сменяю-
щие друг друга возбуждение и торможение.
Основным нервным механизмом регуляции функций являет-
ся рефлекс, реализующийся по рефлекторной дуге. Простейшая
рефлекторная дуга включает минимум два нейрона — аффе-
рентный и эфферентный (рис. 53). В большинстве случаев
рефлекторные дуги многонейронны.
190
Помимо рефлекторных дуг, замыкающихся в центральной
нервной системе, есть так называемые местные, или внутрнор-
ганные, рефлекторные дуги. Они также состоят из двух нейро-
нов, но замыкаются внутри тканн органа (сердце, стенка киш-
ки, сосуда и т. д.). Эти рефлекторные дуги осуществляют
«местную» рефлекторную регуляцию функций (например, ме-
Рис. 53. Схема двухнейронной (А) и трехиейроннои (Б) дуг егшино-
мозгового рефлекса:
Р — рецепторный (афферентный) нейрон; М — мотонейрон (эфферентный).
стную регуляцию просвета мелких сосудов и др.). В некоторых
случаях функцию рефлекторной дуги может выполнять одна
нервная клетка. Это может быть в том случае, когда ее аксон
на конце одной ветви имеет чувствительное окончание, а дру-
гая ветвь иннервирует, например, мышечные волокна в стенке
сосуда. Рецепторное окончание, находящееся на внутренней
поверхности стенки сосуда, воспринимая изменение химичес-
кого состава протекающей крови или ее температуры, возбуж-
дается, возбуждение передается на эффекторную ветвь, в
результате сосуд либо суживается, либо расширяется. Такая
«местная» регуляция называется ак«он-рефлексом.
191
В механизме нервной регуляции функций различают два
вида рефлексов: безусловные—врожденные, наследственно пе-
редающиеся и свойственные всем представителям вида; и услов-
ные — приобретенные, характерные для индивида (у одних
представителей вида могут быть, у других — отсутствуют).
Весьма важным признаком условных рефлексов является
их сигнальный характер. И. П. Павлов назвал эти рефлексы
«предупредительными». Вспомним его знаменитые опыты на
собаке, когда неподкрепленпый безусловным раздражителем
звонок вызывает у нее рефлекс пищевого поведения, т. е. «пре-
дупреждает» животное о возможности получения пищи, к ко-
торому ей следует подготовиться.
Нервная регуляция функций складывается из сложнейших
взаимоотношений безусловных и условных рефлексов.
Рассмотрев врозь гуморальную и нервную регуляции, мы
в какой-то степени нарушили принцип целостного понимания
организма. Однако такое разделение условно. Это только при-
ем, позволяющий правильно представить каждый из механиз-
мов регуляции, действующих в комплексе. В самом деле, пе-
редатчиком нервного возбуждения служит медиатор, хими-
ческое физиологически активное вещество. Значит, неотделимой
частью нервной регуляции является гуморальный компонент.
В это же время деятельность многих желез внутренней секреции
«запускается» нервным импульсом. Следовательно, органиче-
ской частью гуморальной (гормональной) регуляции будет
нервный компонент. Мало того, есть особые нейросекреторные
клетки, занимающие промежуточное положение между «чисто»
нервными и железистыми клетками.
И наконец, любая рефлекторная реакция обязательно со-
провождается гуморальными сдвигами, равно как и гумораль-
ный сдвиг вызывает изменение рефлекторной регуляции. Так,
при нанесении раздражения на рецептор в качестве эффектора
может выступить множество органов. В этом случае мышца
сократится, сердце изменит свою работу', а железы выделят
гормоны, которые по принципу обратной связи (см. рис. 54)
могут оказать на мышцу, сердце и другие органы либо усили
вающее, либо ослабляющее воздействие. Таким образом, у ор-
ганизмов, имеющих нервную систему, действительно существу-
ет единая нейро-гуморальиая регуляция, способствующая уста
повлепию оптимальных отношений их с окружающей средой.
192
Очерк 2. ПРИНЦИП РАССОГЛАСОВАНИЯ —
НЕРАВНОВЕСНОСТИ ГРАДИЕНТА
Уже высказывалась мысль о том. что гомеостаз как способ
существования любой материальной индивидуальности (откры-
той системы), и в первую очередь биологической, предполагает
многоуровневую и бесконечно разнообразную градиентпость
энтропийных и негэнтропийных процессов. Следовательно, со-
вершенно неизбежно рассогласование — «конфликтность» меж-
ду «индивидуально-эгоистическими» формами поведения био-
систем и средой. Это рассогласование, в принципе абсолютное
и постоянное, меняется по степени в зависимости от антиэнтро-
пийной активности биосистемы. Значит, управление и регули-
рование - атрибутивные свойства биосистем. Их выраженность
пропорциональна степени рассогласования взаимоотношений
биосистема — среда.
В основе поддержания активности механизмов управления и
регулирования лежит также практическая инлискретность
поступления возмущений на вход и развитие событий во вре-
мени. Это обусловливает поступление выходной информации о
степени рассогласования на вход системы (см. рис. 54) в мо-
мент действия нового возмущающего фактора, требующего
адекват'/эго ответа, независимо от предшествующих событий.
Это делает невозможным идеальное согласование не только во
взаимоотношениях биосистема — среда, но и внутри самой
биосистемы, между выходными параметрами и входными «за-
просами». Вот почему качество управления и регулирования,
качество стабильности поддержания стационарного состояния,
зависит не от вещественно-термодинамической перавновесности
системы вообще, а в первую очередь от способности ее контро-
лировать степень рассогласования между «агрессивностью»
возмущающего воздействия и возможностью сопротивляться
ему. Вместе с тем любая система во взаимоотношениях со
средой неуклонно движется к своему наиболее вероятному со-
стоянию — к равновесию со средой, т. с. к переходу от более
упорядоченного состояния к менее упорядоченному.
Таким образом, смысл управления и регулирования двой-
ствен и внутренне противоречив. В самом деле, рано или поздно
биосистема обязательно приходит к равновесному со средой
состоянию. Следовательно, управление и регулирование на-
193
правлено на ликвидацию рассогласования в системе биосисте-
ма— среда, вход — выход. Достижение же столь идеального
эффекта — приведение рассогласования к нулю — аннулирует
сам процесс управления и регулирования, так как создает усло-
вия для термодинамического и вещественного равновесия био-
системы со средой — смерти.
Однако реальное положение вещей позволяет утверждать,
что все системы, и тем более биосистемы, не просто сохраняют
свою неравновесность, а активно борются за нее. Значит, управ-
ление и регулирование противодействует установлению согла-
сования — равновесия. Конечное же равновесие — это сниже-
ние возможностей управления и регулирования противостоять
возмущающим воздействиям, каждый раз оставляющим после
себя необратимый «энтропийный» след.
Итак, рассогласование в системе организм — среда действи-
тельно представляет собой движущую силу управления и регу-
лирования (самоуправления и саморегулирования), смысл
которых заключается в приведении этого рассогласования к
оптимальному уровню, обеспечивающему биосистеме макси-
мальную выживаемость. Подобно максимальному согласова-
нию, приводящему к эффекту «зарегулирования», максимальное
рассогласование «ломает» механизмы регулирования и ведет
к разрегулированию с одним и тем же результатом в обоих
случаях.
Является ли этот принцип управления и регулирования
теоретической догадкой или он имеет под собой реальное ма-
териальное основание? Все вышеизложенное о взаимонеравно-
весиости и временной стационарности материальных объектов
исключает мысль об умозрительной природе принципа рассо-
гласования. Тогда закономерен вопрос, что же лежит в мате-
риальной основе нс только межобъектных неравновесных, но
и внутренних отношений систем. Прежде всего можно предпо-
лагать как внутрисистемную межэлементную (в том числе и
«межподсистемную») взаимосвязь, так и взаимонеравновесность
(вещественную и термодинамическую градиентность) био-
систем.
Видимо, первым, кто обратил на это внимание, был Чайльд
(1924), развивший представления о краниокаудальном (про-
дольно-осевом) морфофуикциопальном градиенте. Как извест-
но, понятие градиент тесно связано с представлением о физио-
194
логическом Доминировании и подчиненности. Чайльд рассмат-
ривал проблему градиентов преимущественно с позиции после-
довательного функционального ослабления их от головного кон-
ца тела. Однако правильнее подходить к этой проблеме шире.
Высшая точка градиента — это область самого интенсивного
обмена, наибольшей активности. Она влияет на физиологиче-
ское состояние других уровней сильнее, чем те могут влиять
на нее, т. е. можно говорить о градиенте взаимоуправляюших и
взаиморегулирующих влияний различных уровней. Видимо,
следует выделять не только продольно-осевую, но и фронталь-
ную градиентность, градиентность горизонтальной морфофунк-
циональной асимметрии как материальной основы феномена
управления и регулирования.
Вообще проблема асимметрии — симметрии прямо или кос-
венно давно находится в сфере интересов науки. Правда, в
большинстве случаев она рассматривается в обратном поряд-
ке: симметрия — асимметрия даже тогда, когда утверждается
примат последней. Асимметрия — единственное, что действи-
тельно в доступном нашим чувственным и умозрительным воз-
можностям мире, за исключением, видимо, только вселенной,
симметричной в своей бесконечности и «растворяющей» сим-
метрию до бесконечности. Говоря о симметрии, люди принима-
ют желаемое за действительное. Хорошо по этому поводу ска-
зал А. И. Вейник (1973): «...Кинеата наносит сокрушительный
удар абсолютности симметрии мира. Мие лично такой мир ка-
жется менее красивым, стройным и симметричным». Введение
понятия симметрии как основополагающего отправного момен-
та познания позволило упростить объект исследования, осво-
бодиться от не поддающихся никакому учету переменных,
существующих в реальном асимметричном мире. Поэтому нет
ничего удивительного в том, что все законы, открытые в «сим-
метричном мире», являются постоянным источником новых
законов, их нарушающих. Вряд ли стоит удивляться тому, что
вполне естественно и лишь нарушает наши традиционные пред-
ставления, поражая воображение сокрушительной мощью по-
трясения основ. При обратном порядке познания (симметрия —
асимметрия) люди ничего не могли бы познать или в лучшем
случае заблудились бы в косности сущностного содержания,
вкладываемого в понятие «симметрия». Познание мира шло бы
по пути роста гносеологической энтропии, а не наоборот.
195
При всей магнетической притягательности мифической сим-
метрии человек в конечном счете объективно видит преиму-
щество реально существующей асимметрии не только в силу ее
естественного проявления, но и в эстетическом отношении. Этот
как будто парадоксальный феномен оригинально сформулиро-
вал Д. Н. Узнадзе (1966): «.. Человек скорее настроен воспри-
нимать окружающее асимметрично, чем наоборот, и ...вообще
он психически склонен больше к явлениям асимметрии, чем
симметрии».
Таким образом, остается справедливым принцип Р. Фейн-
мана о том, что «из елки (асимметрии.— А. Л.) можно сделать
палку (симметрию.— А. Л.), а из палки не сделаешь елки».
И это оправдано тем простым обстоятельством, что асиммет-
рия — это градиептность, а наличие градиента предполагает и
наличие процесса, действия, динамики, управления и регулиро-
вания на всех уровнях сложности материальных систем, и тем
более биологических.
Поэтому абсолютно целесообразно в основе представлений
об управлении и регулировании принимать положение II. Кюри
(1894), что «некоторые элементы симметрии могут сосущество-
вать с некоторыми явлениями, но это не обязательно. Необхо-
димо. чтобы некоторые элементы симметрии отсутствовали. Это
и есть та дисимметрия, которая творит явление» (цит. по
А. В. Шубникову и В. А. Копцик, 1972). Совершенно справед-
ливо в связи с. этим замечание Шубникова и Копцик. «что
условия симметрии в силу своей абстрактной природы только
необходимы, но недостаточны для реализации явления».
Следовательно, в объектах живой природы поддержание
динамичности возможно благодаря совершенству механизмов
управления и регулирования, т. е. механизмов обеспечение
оптимальной дисимметричности* внутри биологической системы
* Термины «дисимметрия» и «дисимметризация», «симметрия»
и «симметризация» нами рассматриваются только как удобные в об-
ращении, а не отражающие действительность хотя бы потому, что
они предполагают реальность симметрии, дли оценки меры которой
используются наши антропоморфные возможности. Единственной
симметрией может быть симметрия типа доннановского равновесия,
допускаемая по аналогии и в космосе, где функцию «полупроницае-
мой мембраны» могут выполнять поля и взаимоотношения между
ними.
196
и в сс отношении с окружающей средой. Активность поведений
любой системы определяется количеством степеней свободы,
которыми она располагает для оптимального управления и ре-
гулирования веек форм взаимоотношений внутри и вне себя.
И в самом деле, «молекулы высокой симметрии достаточно
часто образуют кристаллы, в которых эти молекулы занимают
частные положения с пониженным числом степеней свободы
и даже без степеней свободы» (А. И. Китайгородский, 1971).
В этом случае имеет место максимум энтропии системы и ми-
нимум энергии, в том числе и колебательной ее части. Все же
живое складывается из асимметричных молекул даже в том
случае, если внешне биосистема высоко морфофункционально
симметрична.
Именно в этом внутрисистемном рассогласовании и заложе-
на природа активности биосистемы. Причем чем выше это
рассогласование, тем, по-видимому, активнее асимметрично-мо-
лекулярные механизмы регулирования адаптивной гомеостатич-
ности внутренней среды и тем выше результирующее (ие обя-
зательно позитивное для биосистемы) выражение его во внеш-
ней активности биосистемы.
Попробуем проследить правильность этого соображения на
примере эволюции живого. Вероятно, есть смысл миновать
предбиологическис стадии. Важно, что первичные клеточные
организмы, построенные из асимметричных молекул и, следо-
вательно, асимметричной протоплазмы и внутриклеточных об-
разований, внешне были или максимально симметричны (сфе-
ричны или близки к этому), или полностью асимметричны
(амебы и амебоподобные одноклеточные). Первые в силу «пол-
ной симметрии внешних форм» практически были лишены всех
степеней свободы, а значит, и избирательного (преимуществен-
ного) реагирования той или иной части тела на возмущающие
воздействия, т. е. лишены возможности выбора направления
перемещения в пространстве, активного движения в окружаю-
щей среде. Поэтому они могли поддерживать индивидуальное
адаптивно-гомеостатическое состояние внутренней среды либо
за счет колоссальной интенсивности внутриклеточного обмена
(энергия обмена ныне существующих неподвижных бактериаль-
ных и микробных форм), либо за счет перехода в гипобиотиче-
ское состояние, либо за счет того и другого одновременно. Под-
держание же видового гомеостаза у многих малоподвижных
197
форм, как известие, происходит благодаря фантастической
интенсивности размножении и малой продолжительности жизни
индивида.
Напротив, у простейших с полной асимметрией внешних
форм (анаксоиты) любая часть тела, подвергшаяся воздей-
ствию среды, могла стать ведущей или ведомой при переме-
щении, направленном в сторону возмущающего фактора или
от него. В этом случае живые существа получали явное
преимущество в установлении оптимальных адаптивно-гомео-
статических отношений со средой (по сравнению с «полпосим-
метричными») и имели возможность перейти на более эко-
номную энергетику обмена, приобрести большую продолжи-
тельность жизни индивида и меньшую интенсивность воспро-
изведения потомства. Вместе с тем можно предположить, что
отсутствие достаточно жесткой фиксации ведущей и ведомой
частей тела послужило источником осложнения в выборе наи-
более адекватно-адаптивного поведения при одновременном
воздействии на апаксонт достаточно сильных и биологически
равных или разных по актуальности возмущений. Надо пола-
гать, что такие ситуации нередки. Именно поэтому должны
были определиться ведущие и ведомые продольно-осевые гра-
диентные отношения между частями тела (передне-задними),
т. е. должна была произойти наиболее вероятная симметриза-
ция. Такой могла быть только билатеральная симметризация.
Итак, приходим к выводу, что анаксонты совершенствовали
активно-адаптивные отношения со средой, направленные на
снижение зависимости от нее, путем ослабления неограничен-
ной асимметрии при соответствующей фиксации некоторых
степеней свободы. Здесь примером может быть приобретение
Amoeba proteus под влиянием гальванотаксиса почти двусто-
ронне-симметричной формы. Сферические формы для подобной
активизации должны были избрать прямо противоположный
путь — путь ослабления симметрии и высвобождения чрезмер-
но-фиксированных степеней свободы.
Разная степень «освобождения» от чрезмерной симметрии
выразилась в появлении всех полиаксоиных форм вплоть до
моноаксониой гетерополярной, т. е. билатерально-симметрич-
ной. Надо полагать, что все изложенное выше в равной мере
приложимо не только к простейшим. Так, у иглокожих, пере-
двигающихся путем ползания, несмотря на радиально-симмет-
198
ричную форму в моторике выступают принципы билатераль-
ной симметрии. Морские звезды передвигаются по прямой,
которая условно делит их тело на две симметричные половины.
При этом движении вперед перемещаются один или два луча,
называемые ведущими. (Ведущим может быть любой из
лучей в зависимости от места нанесения раздражения.) У
офиур передвижение идет также по принципу билатеральной
симметрии: локомоторными являются только две пары лучей;
пятый, непарный, располагающийся спереди или сзади, фор-
мально в движении участия не принимает. По-видимому, он
выполняет функцию «правила», определяющего направление
движения.
Подытоживая изложенное, можно утверждать, что системы
как с «полной», так и с отсутствующей симметрией в равной
степени малопригодны в отношении активно-приспособитель-
ного поведения. В первом случае из-за бесконечного количества
элементов симметрии «нельзя найти никаких градиентов
свойств пи в одном направлении, кроме направления от центра
к периферии», во втором случае «не существует постоянных
градиентов свойств ни в одном направлении, кроме направле-
ния от глубины к поверхности» (В. Н. Беклемишев, 1944).
Весьма интересна еще одна мысль В. Н. Беклемишева*:
«...между асимметрией амеб и инфузорий — колоссальное рас-
стояние: у первых асимметрия неопределенности: симметрия
нарушается здесь тем, что в любой точке и в любое время
возникают новые и непостоянные образования, а сложные
формы, которые мы находим у отдельных особей в тот или
иной момент времени, нс представляют ни правильности, ии
повторяемости; асимметрия амеб есть отсутствие определен-
ного плана строения. Асимметрия Hypotridia или Dinoflagellate,
наоборот,— проявление сложного плана, полной дифференци-
ровки, где симметрическое повторение частей исключается
чрезвычайной специализацией и точным распределением их по
определенным местам, т. е. животные, по общему уровню ор-
ганизации стоящие выше, обладают и менее богатой симмет-
рией... вплоть до установления билатеральной симметрии».
Однако установление билатеральной симметрии внешних
форм биосистемы не сопровождается столь же выраженной
* Б. II. Беклемишев придерживается примата симметрии.
199
симметризацией формы и расположения внутренних органов
(аппарат пищеварения, сердце), в том числе и парных (легкие,
почки и др.) Причину большей морфофункциональной асим-
метрии последних можно видеть в чрезвычайной загомеоста-
зированпости внутренней среды (циркулирующие жидкости —
кровь, лимфа и тем более локальная межклеточная жидкость)
вследствие исключительной специализации органов и тканей.
Лишь высокая гомеостатичность ее могла обеспечить доста-
точно жесткое автоматическое регулирование деятельности
этих органов и «позволила» организму многие из них иметь
в единственном числе. Дублирующие же билатерально-сим-
метричные внутренние органы в большинстве своем эволюци-
онно более молоды и менее совершенны, несмотря на то что
в высшей степени специализированы.
Правильность высказанного подтверждается огромным ко-
личеством фактов, говорящих о том, что по мере повышения
специализации и ее совершенства парные — билатеральные
органы — становятся в морфофункциональном отношении все
более и более асимметричными. В первую очередь это имеет
отношение к морфофункциональной дисимметризации цент-
ральной нервной системы. Асимметрия больших полушарий
столь явно выражена, что в литературе по этому поводу даже
поднят вопрос, один мозг у «полушарных» животных или два.
Так, П. Линдсей и Д. Норман (1974) склонны два симмет-
рично расположенных полушария рассматривать как самостоя-
тельные образования, способные не только работать врозь, но
и выполнять разные (асимметричные) функции. Однако это
крайняя точка зрения. В самом деле, как показывают данные
науки, «правополушарный» человек более склонен к восприятию
мира в его конкретном вещественном выражении. Он, так ска-
зать, лишен абстрактного мышления, в силу чего не в состоянии
установить логической связи между конкретно воспринимае-
мыми явлениями, соотнести их с определенными категориями.
Это богатый фактами бедняк, неспособный воспользоваться
своими богатствами. Человек в «левополушарном» варианте —
полный антипод «правополушарному». Его «интеллектуально-
абстрактная» обогащенность — та же бедность, поскольку он
не располагает конкретным материалом, который можно было
бы обобщать и анализировать. Только при совместной работе
обоих полушарий возможно осуществление всех функций. Это
200
верно даже при условии, что каждое полушарие «приторма-
живает», ослабляет деятельность противоположного полуша-
рия. Вот почему правилен тезис — полушарий два, а мозг один!
Билатеральная симметрия — единственный вид симмет-
рии, способный обеспечить при перемещении в пространстве
точный выбор направления движения вправо — влево, так же
как наличие продольной оси тела обусловливает точность вы-
бора движения в направлении вперед—назад, а спинно-брюш-
ная асимметрия — вверх и вниз. Здесь обращает на себя вни-
мание тот факт, что Движения вперед — назад и вверх — вниз
осуществляются скорее благодаря передне-задней и спинно-
брюшной асимметрии, а влево —- вправо — билатеральной сим-
метрии. Однако симметрия, как уже было сказано выше, огра-
ничивает активность, закрепощая степени свободы. Между
тем актуальность выбора направления влево — вправо ничуть
не меньшая, чем вперед — назад или вверх — вниз. Более того,
движения почти всегда комбинированы и значительно чаще
направлены влево—вправо, чем вперед — назад — вверх или
вперед — назад—вниз, хотя бы потому, что переход от дви-
жения вперед — назад и наоборот, за крайне редким исклю-
чением, осуществляется благодаря полному лево-правому
повороту. Следовательно, идеальная билатеральная симметрия,
гарантирующая жесткую равнозначность, вероятность движе-
ния вправо и влево, может поставить биосистему в затрудни-
тельное и даже катастрофическое положение при выборе на-
правления перемещения в условиях «равнозначной простран-
ственной монотонности» возмущающих воздействий внешней
среды. Подобную ситуацию можно сравнить с образным «фе-
номеном Буриданова* осла». Последний, находясь между
двумя совершенно одинаковыми по виду и качеству охапками
сена, т. е. будучи подвержен действию двух абсолютно равных
факторов, обречен якобы на абсолютное бездействие, на го-
лодную смерть.
Однако в действительности при нормальных взаимоотно-
шениях биосистемы со средой этот феномен не имеет места.
Отсутствие его объясняется биологической неравнозначностью
даже идеально симметрично расположенных (пространственно
и временно) и физически равных возмущений, обусловленной
Буридан—французский философ (умер около 1360 г.).
201
морфофункциональной (например, пороговая) асимметрич-
ностью (дисимметричностью) биосистемы. Внешне билате-
рально-симметричные биосистемы ведут себя во взаимоотно-
шениях со средой как асимметричные. Эта асимметричность
поведения (функции) имеет материальную базу — асиммет-
ричность структуры, формы.
Весьма важно вникнуть в сущность понятия билатераль-
ная симметрия, симметрия типа гп, образующая эиантиаморф-
ную пару, т. е. «существование» (сосуществование.— A.JJ.)
зеркально равных форм одной и той же симметрии: в природе
симметрия т осуществляется, конечно, только приблизительно»
(А. В. Шубников, В. А. Копцик, 1972). «Всякое же нарушение
эпантиаморфности, или асимметрия, может, по геометрическим
представлениям, осуществляться в двух вариантах, которые в
свою очередь составляют энаптиаморфпую пару, а любые фи-
гуры, которые могут быть представлены в энантиаморфных
модификациях,— асимметричные» (В. Б. Касииов, 1971).
В асимметричности правой или левой половины тела био-
системы с билатеральной симметрией можно убедиться на
бесчисленном количестве фактов. У человека, например, нет
ни одной мышцы, симметричной или состоящей из энантиа-
морфной пары. Все мышцы асимметричны по форме, размерам,
местам прикрепления, так как каждая из них является членом
^энантиаморфной пары. Формы же с симметрией m не имеют
энантиаморфных модификаций, так как «правизна» и «левизна»
содержится в самих формах. Следовательно, пи одна мышца
не в состоянии создавать симметричной тяги. В этом великий
биологический смысл — асимметричность развиваемой тяги
позволяет осуществлять нестандартные движения, что, веро-
ятно, объясняет явление, привлекшее внимание Н. А. Бернштей-
на (1945, 1961, 1963): «Строгая стандартность формы и мет-
рики циклических навыков движений никогда не реализуется
сама собой и никогда не бывает самоцелью. Ее приходится
вырабатывать, и мозг идет на это не только в тех случаях или
в тех деталях нли звеньях двигательного акта, где такая стан-
дартность существенно необходима».
Итак, билатеральная симметрия — это пара ие только
дисимметричных относительно друг друга, но и максимально
асимметричных энантиаморфов (энантиомеров, антимеров, ан-
типодов), это тот оптимальный минимум симметрии, точнее
202
максимум асимметрии, который обеспечивает возможность
асимметричного (в широком смысле) активно-нриспособитель
ного поведения биосистем в окружающем мире. Ибо, как из-
вестно из принципа П. Кюри, «когда несколько различных
явлений природы накладываются друг на друга, образуй одну
систему, дисимметрии их складываются. В результате оста-
ются лишь те элементы симметрии, которые являются общими
для каждого явления, взятого отдельно» (цит. по Шубникову,
Копцик. 1972), т. е. увеличивается асимметрия.
Учитывая абсолютную невероятность идеальной симметрии
не только двух явлений, но и взятого отдельно явления внутри
себя, можно утверждать, что во взаимоотношениях организма
со средой из всех симметрий билатеральная оптимально огра-
ничивает интенсивность роста энтропии.
Рассматриваемая нами билатеральная симметрия, безу-
словно, является материальной основой феномена управления
и регулирования, так как несет в себе лево-правый (плюс-ми-
нус и др.) градиент свойств. Отсюда ясно, что существование
генетической предопределенности лево-правой асимметрии на
всех уровнях живых существ, имеющих билатерально-энап-
тйаморфное тело, не вызывает сомнений. Суть этого утвержде-
ния не умаляется фактом, отмеченным Г. Вейлем (1968), «что
все регенеративные ткани (или иные формы компенсации.—
А-Л.), асимметричные в потенции, могут производить обе
формы... однако при нормальном (выделено нами.— А. Л.)
развитии всегда развивается только одна из них — левая или
правая. Какая именно—это определяется генетическими осо-
бенностями...». Это натолкнуло нас на мысль о том, что асим
метрия функции и функциональная асимметрия — самостоя-
тельные понятия.
Асимметрия функции — генетически предопределенное пре-
имущественное участие в процессе одного из симметричных
органов (частей тела) при равной возможности выбора их
(равная нагрузка, симметричные требования).
Функциональная же асимметрия — деятельность симметрич-
ных органов или частей тела под влиянием асимметричной
нагрузки — формируется вследствие как усиления асимметрии
функции путем направленной тренировки генетически ведуще-
го в этом процессе органа или части тела, так и ослабления
и даже «инверсии» ее.
203
Разумеется, второй вариант и особенно инверсия — явле-
ние нежелательное, во всяком случае биологически не оправ-
данное, поскольку приводит иногда к совершенно неожидан-
ным последствиям. Так, у занимающихся футболом мальчиков
в возрасте 9- 10 лет, у которых тренировочный процесс в те-
чение 6—9 месяцев был построен на «работу» генетически не
ведущей ноги, обнаружено статистически достоверное замед-
ление роста по сравнению с тренировавшимися не по этому
принципу. Прекращение эксперимента способствовало энергич-
ному росту, и в результате эти дети за три месяца догнали
своих сверстников.
Итак, работа билатерально-симметричных органов и час-
тей тела осуществляется в общем асимметрично, даже тогда,
когда внешне это не выявляется. Например, при одновремен-
ных движениях обеими ногами или руками никогда не бывает
одновременности, и даже при выполнении их с максимальной
скоростью уже в течение первых пяти секунд регистрируется
существенное рассогласование между количеством ударов
правой и левой руки в пользу той, которая является генетиче-
ски ведущей.
Если асимметричны движения, естественно, должно быть
асимметричным и их вегетативное обеспечение. Да и подчи-
няясь закону билатеральной градиентности, вегетативная, в
том числе и метаболическая, асимметрия должна рассматри-
ваться как естественная, что подтверждается обширными дан-
ными. Например, температурная и сосудистая асимметрия
зависит от возраста и функционального состояния организма
(Л. А. Логинов, 1955—1961, 1963). Вегетативная асимметрия —
столь необычно сильное проявление билатеральной функции,
что не только наблюдается, но даже в конечном эффекте уси-
ливается при таком казалось бы не имеющем строгой литера-
лизации явлении, как одностороннее удаление или раздраже-
ние околощитовидных желез. Между тем И. Л. Скобским
(1969) показано изменение асимметрии электролитного состава
крови, оттекающей от правой и левой пог собаки за счет сдвигов
па той стороне, где было удаление (раздражение) железы.
Им же показано усиление асимметрии коллоидного состава,
свертывания, РОЭ, состава лейкоцитарной формулы и других
показателей крови у людей при заболеваниях нервной системы,
особенно при центральных параличах и парезах. Как полагает
204
Скобский, подобные сдвиги начинают формироваться в докли-
нической стадии развития болезни. Более того, в последнее
время установлено, что если крыс-левшей научить брать пишу
правой лапкой, то в цитоплазме, кариоплазме и ядрышках их
нейронов изменяется содержание РНК. Это весьма существенно.
Ведь насильственное и тем более форсированное переучивание
генетически предопределенной двигательной асимметрии, т. е.
создание функциональной асимметрии путем подавления асим-
метрии функции, по своей биологической сути может быть рав-
ноценно сильному стрессорному воздействию. Необычайно
мощный экстеро- и тем более проприоцептивный поток импуль-
сации с генетически не ведущей части тела и резкое ослабле-
ние его с генетически ведущей при насильственном переучи-
вании справа налево и наоборот требует инверсии функцио-
нального состояния симметричных нервных центров, т. е. воз-
никновения генетически неадекватной асимметрии.
Подобное переучивание — переучивание ие только перифе-
рии, а и периферией нервных центров. Вероятность влияния
периферии на состояние нервных центров предполагал
И. П. Павлов, отмечавший, что развитие функций обеих рук
обеспечивает развитие «центров» речи в обоих полушариях.
М. М. Кольцова (1973), ссылаясь на электроэнцефалографи-
ческие данные Л. А. Папащспко, пришла к выводу, что тре-
нировка пальцев (пассивное сгибание и разгибание, массаж
кисти) в течение 1—2 месяцев у детей в возрасте 6 недель на
два с половиной месяца ускоряет процесс функционального
созревания полушария, контрлатеральвого тренируемой руке.
В этом исследовании не учитывалось, является ли тренируемая
рука генетически ведущей. Автор исходила из представления
об эквипотенциальности полушарий у детей раннего возраста,
формально подтвержденной ею электроэнцефалографически.
Между тем известно, что формальная эквипотенциальность
еще не доказательство ее истинности, так как даже без тре-
нирующих воздействий генетическая латерализация обязатель-
но себя проявит. Кроме того, генетически предопределенная
правизна и левизна существует у животных, и здесь вряд ли
можно говорить о социально управляемом воспитании право-
или левосторонней направленности действий.
Вот почему необходимо более сдержанно относиться к ре-
комендациям Кольцовой в отношении того, что «по-видимому.
205
в играх следует равным образом развивать тонкие движении
пальцев обеих рук» и стимулировать, т. е. ускорять их
регламентированной проприоцептивной импульсацией функ-
циональное созревание мозга. Напротив, положительно сле-
дует относиться к предложению «как-то распределять выпол-
нение действий с предметами в обиходе». Точнее «не как-то»,
а в соответствии с генетически предопределенной асимметрией
функций. Кроме того, было бы правильным постулировать
необходимость не «стимуляции», а нормохропичной естествен-
ному ходу онтогенетического развития ребенка функциональ-
ной тренировки. Гетерохропичная естественному онтогенезу и
генетически неадекватно латерализованная не только стиму-
ляция, но и тренировка — процессы по всем показателям, в том
числе и энергетическим, достаточно дорогие и нуждающиеся
в существенных метаболических тратах. Не в этом ли скрыта
разгадка замедления роста юпых футболистов?
Из сказанного отнюдь не следует вывод о том, что нужно
полностью передать «судьбе» формирование того или иного
двигательного навыка. Напротив, как было показано нами
(А. А. Логинов, В. М. Лебедев, 1972, 1973), при свободном
выборе поведения и реализации заданного действия превали-
рует генетический фактор, получающий дополнительную «за-
грузку» за счет непроизвольно формирующейся направленной
тренировки. На начальных этапах обучения морфофункцио-
пальная асимметрия нарастает, предъявляя тем самым повы-
шенные требования к регуляторным механизмам «выравнива-
ния» возникшего рассогласования. Своевременное включение
этих механизмов приводит к стабильному и «хорошо» осуще-
ствляемому действию с минимальной асимметрией. Дальней-
шее совершенствование реализации действия вновь сопровож-
дается некоторым ростом асимметрии (вторичная асимметрия).
Если она оказывается ниже исходной и выше минимальной, то
это будет характеризовать степень оптимальности асимметрии,
необходимой для высоконадежного и совершенного осущест-
вления действия, в том числе и сугубо асимметричного.
Именно поэтому в процессе приобретения двигательного
навыка целесообразно пользоваться правом свободного выбора
ведущей стороны для выполнения заданного действия. После
же достижения в этих условиях лучшего его осуществления
полезно ввести в обучение элемент нагрузки на неведущую
206
сторону (выравнивание асимметрии) с последующим повтор-
ным (при достижении достаточного успеха) свободным выбо-
ром ведущей стороны. Такое искусственное «подтягивание»
неведущей стороны особенно важно, когда замедляется естест-
венное, что происходит, если обучение, особенно «асимметрич-
ным видам движений» (метание, гребля па каноэ), строится
на преобладании в нем узкоспециализированных упражнений.
В этом случае дисимметрия, согласно правилу П. Кюри, может
привести к нарушению всех видов коррелятивно-координацион-
ных отношений между ведущей и неведущей частями тела с
конечным эффектом перерегулирования и разрушения сфор-
мированного (иногда с большим трудом) двигательного навы-
ка, да и не только навыка, а и функционального благополучия
организма в целом.
В данном случае приложимо соображение Г. Вейля (1968)
о том, что «действие» факторов филогенетической эволюции,
стремящихся вызвать в организме наследственное различие
(выделено нами.— А. Л.} между левым и правым, тормозится,
вероятно, за счет тех преимуществ, которые животное извле-
кает из зеркальносимметричпого (относительно.— А. Л.) рас-
положения своих органов движения — ресничек или мышц и
конечностей; в случае асимметричного (чрезмерно.— А. Л.) их
развития, естественно, получилось бы винтовое, а не прямо-
линейное движение. Это может нам объяснить, почему паши
конечности подчиняются закону симметрии более строго, чем
внутренние органы».
Действительно, в отношении внутренних органов феномен
«закручивания» выражен наиболее явно (например, кишечник
и тем более сердце млекопитающих являются асимметричным
винтом). В. Людвиг (1932) все эти явления рассматривал как
вторичные филогенетические эффекты нарушения симметрии.
Однако на основании уже сказанного выше представляется,
что это скорее вторичные филогенетические эффекты роста
дисимметрии, обусловленные высокой специализацией пнут
ренпих органов в условиях сверхгомеостазированной внутрен-
ней среды (жидкости) организма. В перспективе эволюционно-
филогенетического развития феномен «закручивания» может,
видимо, стать генотипически фатальным и для органов движе-
ния и частей тела в результате их чрезмерной специализации
(например, краб-скрипач и др.).
207
Таким образом, морфофункциональная симметрия дейст-
вительно является материальной основой управления и регу-
лирования вообще и двигательной активности в частности.
Причем в каждом отдельном случае мерой оптимального уп-
равления и регулирования будет оптимальная для данного
случая морфофункциональная асимметрия.
Согласно Б. Г. Ананьеву (1968), «в билатеральном регули-
ровании субординационные отношения выступают в виде ла-
терального доминирования, которое, однако, иосит относи-
тельный, многозначный и временный характер». Более того,
«доминантность одновременно выступает как структурный и
как функционально-динамический принцип их (полушарий,—
А. Л.) деятельности, как безусловно рефлекторное, наследст-
венно-врожденное и индивидуально приобретенное свойство
индивида».
Как уже было сказано выше, в целях большей точности
целесообразно различать наследственную и врожденную лате-
рализацию, т. е. асимметрию функции, и индивидуально при-
обретенную латерализацию, в том числе викарную, компенса-
торную и другие, т. е. функциональную асимметрию.
Функциональная асимметрия может формироваться по трем
типам: I — на базе усиления наследственной и врожденной
латерализации направленной тренировкой; II — на базе на-
сильственной латерализации стороны, противоположной на-
следственной и врожденной латерализации; III — как следст-
вие функционально-динамической латерализации, возникающей
по принципу доминанты Ухтомского. В повседневной жизни
мы имеем дело со всеми тремя видами функциональной асим-
метрии (табл. 3).
Функциональная асимметрия I типа — это функционально
усиленная истинно латерализованная асимметрия, являющаяся
первичной основой «горизонтальной» (термин Ананьева) си-
стемы регулирования, работающей по жесткой, преимуществен-
но генетически детерминированной программе. Говоря о пре-
валировании одного из симметричных механизмов регулиро-
вания, здесь, вероятно, правильнее пользоваться понятием «нс
доминантное», а «ведущее полушарие» (ведущий центр).
Функциональная симметрия II типа — принудительно лате-
рализованпая асимметрия, возникающая благодаря морфофунк-
циопальпой пластичности нервных структур. Это вторичная
208
Виды горизонтальной системы регулирования
Таблица 3
Асимметрия функций (на- следственная и в рожденна я ла- те ралиаацня) Функциональней асимметрия (индивидуально-приоб- ретенная латерализации, в том числе иикяриая, компенсаторная к Другие)
истинная фун- кционально латерализо- еаннаи принудительно латерализованная функционально латералгизопаниая
Морфо- функцио- нальная структура видов асимме- трии Первичная основа гори- зонтальной системы регу- лирования Природа: генетически пре- допределенная морфо- функциональная латера- лизация нервных цент- ров (регуляторов) Механизм: «жесткая», пре- имущественно генети- чески детерминирован- ная программа Регулятор: ведущее полу- шарие, центр и др. Вторичная осно- ва горизонталь- ной системы ре- гулирования Природа: морфо- фуикциональ- ная пластич- ность нервных центров (регу- ляторов) Механизм: «по- лу жесткая», преимущест- венно функци- онально детер- минированная программа Регулятор: фун- кционально-ве- дущее полу- шарие, центр и др. Третичная осно- ва горизонталь- ной системы ре- гулирования Природе: пре- имущественно функциональ- ная пластич- ность нервных центров (регу- ляторов) Механизм: € гиб- ка я» функци- онально детер- минированная программа Регулятор: фун- кционально- доминантное полушарие, центр и др.
Способы формиро- вания За счет на- следственной и врожден- ной морфо- функпиональ- ной литерали- зации нерв- ных центров (регулято- ров) За счет уси- ления наслед- ственной и врожденной литерализа- ции, направ- ленной трени- ровкой-воспи- танием За счет насиль- ственной трени- ровки стороны, противополож- ной наследствен- ной и врожден- ной литерализа- ции За счет функпи- оналызо-Дина ми- ческой латерали- зации по прин- ципу доминанты А. А Ухтомско- го
8 А. А. Логинов
209
основа «горизонтальной» системы регулирования, работающей
по «полужесткой», преимущественно функционально-детерми-
нированной программе. В этом случае, говоря о превалирова-
нии того или иного механизма регулирования, лучше пользо-
ваться понятием «функционально-ведущее полушарие». Такое
представление функциональной асимметрии этого типа опреде-
ляется известными фактами о ее «сбое» в экстремальных
условиях и включении механизмов управления функциональ-
ной асимметрии 1 типа даже при ее меньшей функциональной
готовности. Одним из примеров может быть известный случай
с летчиком, рожденным левшой. «Переучившись» на правшу,
он погиб в условиях аварийной ситуации, безуспешно пытаясь
достать кольцо и раскрыть парашют левой генетически веду-
щей рукой.
Функциональная асимметрия HI типа — наиболее функ-
ционально-лабильная латерализованная асимметрия, форми-
рующаяся по принципу доминанты Ухтомского. Это третичная,
преимущественно фуикционалыю-коррекциониая основа гори-
зонтальной системы регулирования, работающей по гибкой
функционально-детерминированной программе. При этом, го-
воря о временном превалировании право- и левостороннего
механизма регулирования, видимо, тоже точнее пользоваться
термином «функционально-доминантное полушарие».
Опыт показывает, что при определенных обстоятельствах
функционально-доминантное состояние, возникшее в неведущем
или функционально-неведущем полушарии, может не только
ослабить, но и полностью подчинить себе ведущие или функ-
ционально-ведущие механизмы управления и регулирования.
Именно в подобной интерпретации следует понимать мысль
Н. Винера (1958): «...функции мозга и доминирующая роль
одной из рук могут быть не связаны между собой». Кстати,
проблема латерализации всех видов значительно более слож-
на, чем это может показаться на первый взгляд. Основная
сложность заключается в том, что ни один центр, как и целиком
полушарие, никогда не является одновременно ведущим,
функционально-ведущим и тем более функционально-доми-
нантным по всем возможным параметрам. Например, глаз,
ведущий по остроте зрения, может быть неведущим по при-
цельной способности или величине поля зрения; при малом
угле зрения у большинства людей в роли ведущего по при-
210
цельной способности выступает правый глаз, при увеличении
же угла зрения ведущим становится левый. Многообразность
и динамичность функциональной асимметрии установлена и в
отношении других анализаторов. Об этом говорит и фундамен-
тальный факт динамической реципрокности нервных механиз-
мов управления и регулирования локомоторикой да и вообще
координацией движения. Из-за асимметрии реципрокных отно-
шений, по всей вероятности, в нервных центрах практически
невозможно осуществить абсолютно одновременное действие
пары эиантиаморфов (прыжок с места и тем более с разбега,
с отталкиванием двумя ногами и т. д.). Эта асимметричность
латералцзации «веса» управляющей и регулирующей функции
симметричных центров является источником их парной работы.
Истинным проявлением парности, пожалуй, служит ие столько
полная синхронизация (симметричность) и, следовательно, в
какой-то мере дублирование, сколько оптимальная десинхро-
низация — дисимметричность работы механизмов управления
и регулирования как по временным, так и по количественно-
качественным показателям. Такое распределение обязанностей
между симметричными центрами не только не разобщает их, а
напротив, объединяет, обеспечивает установление оптимальных
координационных отношений.
Итак, феномен управления и регулирования действительно
имеет в своей материальной основе как вертикальную, так и
горизонтальную градиентность (асимметричность). И если до
сих пор обсуждались преимущественно функциональные ас-
пекты ее, а также глобально выраженная билатеральная энан-
тиаморфность, то теперь назрела необходимость обратить осо-
бое внимание на чрезвычайно важное широко распространен-
ное свойство билатеральных биологических систем, роль кото-
рого до сих пор еще не оценена. Речь идет об общеизвестном
феномене перекрещивания, и особенно неполного перекрещи-
вания, восходящих и нисходящих проводящих путей нервной
системы, в том числе и на уровне периферических нервных
сплетений. Достаточно вдуматься в сущность этого явления,
чтобы прийти к заключению, что перекрест проводящих путей
усиливает не только структурно-морфологическую асимметрию
нервных образований на всех уровнях нервной системы, но,
что еще более важно, создает дополнительные возможности
для усиления функциональной асимметрии. Наиболее ярким
8»
211
примером, иллюстрирующим это положение, является несим-
метричное представительство половин сетчатки (латеральной —
правого глаза, медиальной — левого в правом полушарии; ла-
теральной— левого глаза, медиальной — правого—в левом)
в полушариях головного мозга. При бинокулярности зрения эф-
фект центральной диспаратности, стереоскопичности и прост-
ранственности восприятия должен быть сохранен даже при
идеальном корреспондировании изображения на сетчатке. Зна-
чит, должен быть сохранен и оптимальный зрительный контроль
за пространственным расположением частей тела при беге,
ходьбе и т. д. как относительно друг друга, так и по отношению
к объектам окружающей среды.
Это распространяется и на другие анализаторные системы.
Принцип оценки локализации источника звука осуществляется,
например, благодаря бинауральности слуха. Причем у живот-
ных, которым требуется точное определение местоположения
источника звука, адаптивная локомоторика зависит от работы
слухового анализатора; бинауральная асимметрия слухового
восприятия дополнительно обеспечивается трехмерной асим-
метрией структуры ушных раковин и асимметричностью
движения при локации звука.
Наконец, следует сказать, что ритмические молекулярные
явления в клетке, связанные, например, с синтезом белка, реа-
лизуются в соответствии с уравнением, выведенным Гудвином
и показывающим, что в этом процессе переменные Xi (коли-
чество РНК) и (фермент) имеют ббльшие отклонения от
стационарных значений вверх, чем вниз. При возрастании та-
ландической температуры эта асимметрия также возрастает,
т. е. положительные отклонения еще более превышают отри-
цательные.
Итак, принцип внутрисистемного и внесистемного рассог
ласования, градиентности, конфликтности действительно явля-
ется принципом управления и регулирования поведения био-
системы. Основа феномена управления и регулирования —
морфофункциональная асимметрия биосистемы и внутри, и по
отношению к окружающей среде — также материальна. Смысл
же управления и регулирования заключается в достижении
оптимального уровня асимметрии, необходимого для сохране-
ния способности активного поддержания гомеостатичности
бносистемы. Это возможно в том случае, если биосистема в
212
состоянии оценивать степень рассогласования внутри и вне
ее, что осуществимо только при наличии важнейшего элемента
управления и регулирования—обратной связи.
Таким образом, морфофункциональная асимметрия заведо-
мо предполагает наличие в живой природе обратной связи,
т. е. принципа замкнутого контура управления и регулиро-
вания.
Очерк 3. ПРИНЦИП ЗАМКНУТОГО КОНТУРА
РЕГУЛИРОВАНИЯ И ПРОГНОЗИРОВАНИЯ
Принцип замкнутого контура регулирования вытекает из
способности живого сопротивляться разрушающим воздейст-
виям среды: Поэтому биосистеме недостаточно ответить па
раздражение той или иной реакцией Необходимо еще оценить,
насколько эта реакция на выходе соответствует требованиям
входного раздражения и приспособительна для поддержания
оптимальных жизненных свойств системы в данных условиях.
Одну из попыток проанализировать общие принципы орга-
низации управляющих систем осуществил Ф. Гродинз (1966).
В частности, он выделил в управляющем устройстве две под-
системы— детектор ошибки и собственно регулятор (рис. 54).
Первая оценивает рассогласование, т. е. величину ошибки
kye, равную разности между выходным у0 и задающим
сигналами, а также генерирует сигнал ошибки. Этот сигнал
поступает в регулятор, который выдает на объект управления
(на выход) скорректированную управляющую команду. В ре-
зультате система уменьшает величину рассогласования между
входом и выходом, т. е. приспосабливается к существованию
в конкретных условиях.
Подобное поведение бносистемы возможно при одном обя-
зательном условии — прн наличии канала связи, по которому
на вход управляющего устройства поступает информация о со-
бытиях, происходящих на выходе системы. Такие каналы
получили название обратной связи. Функцию их в биосистемах
выполняют любые пути (нервные проводники, циркулирующие
в организме жидкости и др.). Часто обратная связь устанав-
ливается вне биосистемы. Например, при осуществлении зри-
тельного контроля (внешняя обратная связь) за перемещением
звеньев тела в пространстве по заданной программе.
213
Все это позволяет сделать чрезвычайно важное заключе-
ние: бносистемы с обратной связью (а других не бывает) в
реализации отношений со средой преследуют цель поддержи-
вать выходной сигнал, равным (симметричным) задающему.
Однако подобное было бы возможно, считает Ф. Гродинз,
«если бы существовала уверенность в том, что па вход объ-
возпдщенив ит
таш
Рис. 54. Блок-схема системы с обратной связью (по Ф. Гродинзу,
1966).
екта управления не поступает никаких других сигналов, кроме
сигнала управления Fe, и что свойства этого объекта остаются
абсолютно неизменными во времени». Такой уверенности не
только нет, но и шансы на ее появление совершенно невероят-
ны. Ведь в этом случае «регулятор мог бы неявным образом
«знать» о результатах своих управляющих воздействий на
{/о, и обратная связь, снабжающая регулятор информацией,
об этих результатах, в явном виде не требовалось бы». Пред-
положить возможность отсутствия обратной связи в механиз-
мах управления и регулирования, самоуправления и саморе-
гулирования в живых системах — идея более чем несостоя-
тельная. Это понятно хотя бы потому, что все биосистемы
открытые и находятся в неравновесном стационарном состоя-
нии со средой, а его необходимо постоянно поддерживать.
Следовательно, объективно эти системы беспороговые в от-
ношении реагирования на любые события, происходящие в
беспрестанно изменяющихся условиях. Мало того, изменения,
происходящие в биосистеме, реализующей ответную реакцию
на события внешней среды, не полностью обратимы. Значит,
в каждое мгновение состояние биосистемы отличается от
214
предыдущего не только в силу естественно развивающихся
онтогенетических изменений, по и благодаря следам преды-
щих взаимодействий со средой. Ясно, что даже в маловероятных
условиях неизменности сигналов, поступающих на вход, биоло-
гическая значимость их каждый раз будет иная, иным будет
и выходной сигнал, информация о котором при любой
скорости достижения входа будет постоянно поступать на
функционально иной вход. Кроме того, -биологические систе-
мы существуют в условиях не только избирательно задающих
сигналов управления Л-, по и возмущающих воздействий
F<i, не предусмотренных программой поведения биосистемы.
Отсюда вывод: выходной сигнал, т. е. управляемая величина,
всегда будет существенным образом отличаться от запросов
входного, т. е. от основного задающего сигнала и шума, созда-
ваемого дополнительными возмущающими воздействиями, ко-
торые в свою очередь также являются задающими сигналами
и иногда даже могут меняться ролями с основным.
Поэтому главная задача саморегулирующейся системы за-
ключается в том, чтобы в реализации реакции поддерживать
управляющую величину (выходной сигнал) не столько равной
основному задающему сигналу, сколько оптимально отлича-
ющейся от него в условиях действия возмущения. Единствен-
ный способ установить наличие таких возмущений состоит в
наблюдении их действия на выходной сигнал с помощью обрат-
ной связи, т. е. с помощью замкнутого контура регулирования.
Необходимо указать еще на одно важное свойство обрат-
ной связи — на эффект запаздывания. Суть его в том, что все
события развиваются во времени. Следовательно, от момента
поступления задающего сигнала на управляющее устройство
до момента поступления управляющего сигнала по прямым
связям на управляемую величину (выход) должно пройти ка-
кое-то время. Время требуется и для поступления сигнала по
обратной связи. Значит, какова бы ни была скорость «обора-
чиваемости» потока информации в замкнутом контуре регули-
рования, сигнал обратной связи всегда будет запаздывать.
Поэтому всегда будет запаздывать и его коррегирующее воз-
действие на управляющее и управляемое устройство. Так как
компенсирующий эффект сигналов обратной связи пропорци-
онален возмущению, их вызвавшему, то за время, в течение
которого происходит включение коррегирующих механизмов.
215
продолжающееся действие возмущающего фактора приводит
к накоплению ошибок. Вот почему новая реакция системы на
следующее возмущение реализуется в «неблагоприятных»
условиях систематического запаздывания. Это, разумеется,
умножает величину и интенсивность накопления ошибок.
В результате возникают колебания в реакциях системы. Она
самовозбуждается. Соответственно степени самовозбуждения
изменяется эксплуатация механизмов управления и регулиро-
вания, в том числе и каналов обратной связи. Здесь возможно
оптимальное регулирование и перерегулирование. Последнее
бывает в двух формах: перерегулирование в сторону чрезмер-
ного сглаживания (загомеостазирование амплитудно-частот-
ных характеристик реакций биосистемы на возмущение при
повышенной относительно оптимальной оборачиваемости про-
цессов в замкнутом контуре регулирования) и перерегулиро-
вание в сторону выхода амплитудно-частотных характеристик
за пределы зоны насыщения (разгомеостазирование). В осно-
ве механизма разгомеостазирования, по-видимому, лежит
преимущественно замедление оборачиваемости процессов в
контуре регулирования.
Однако подобные грубые отклонения гомеостатического
оптимума в биосистемах — явление чрезвычайное. Оно почти
всегда сопровождается возникновением патологических со-
стояний. Устойчивость биосистем при этом объясняется прин-
ципиальной, особенно в отношении обратных связей, нелиней-
ностью их ответов на возмущения, а также «эффектом упраж-
нения». Именно нелинейность и упражняемость—меры,
эволюционно выработанные для поддержания колебаний на
уровне оптимальности при контакте биосистемы со средой.
Необходимо обратить внимание, что в многоконтурных си-
стемах регулирования содружественность взаимодействий це-
цей обратных связей в конечном итоге определяется не только
их формальным содружеством, функциональной однонаправ-
ленностью, но и функциональной разнонаправленностью отно-
сительно друг друга.
Теоретически в живых объектах можно выделять одно-,
дву-... и-контурные системы управления и регулирования как
функций отдельных элементов, так и организма в целом.
Практически живые объекты имеют функционально взаимо-
связанные многоконтурные системы автоматического регули-
216
рования. Всегда на любые возмущающие воздействия реаги-
руют все контуры с формально разной степенью активности,
фактически необходимой в каждом отдельном случае.
Принцип действия замкнутого контура состоит из следу-
ющих этапов.
1. Па вход (например, воспринимающие приборы-рецеп
торы) объекта управления — регулирования (например, ске-
летная мышца) поступает возмущающее воздействие (хими-
ческое раздражение — молочная кислота, в избытке образовав-
шаяся в результате длительной работы этой мышцы и
вызывающая болевое, ощущение).
2. На выходе системы (мышцы) — управляемая величина
(сокращение).
3. Сильное сокращение сопровождается усилением болевых
ощущений, возникает защитная реакция, сигнал с сокращаю-
щейся мышцы по обратной связи поступает в управляющее
устройство (функции его выполняют соответствующие отделы
центральной нервной системы), затем происходит оценка
качества поступившего сигнала, его биологического значения,
и в результате управляющее устройство оказывает на объект
управления тормозящее воздействие, используя известные
способы торможения.
Вследствие этого управляемая величина (сокращение)
на выходе уменьшается, благодаря чему снижается болевое
ощущение, о чем по обратной связи информируется управля-
ющее устройство. Таким образом, саморегуляция действитель-
но осуществляется по принципу работы замкнутого контура.
Данному принципу подчиняется и работа рефлекторной дуги.
Здесь описана отрицательная обратная связь. Она называется
так потому, что вызывает ослабление управляемой величины
(работы органа). Если же обратная связь усиливает работу
того или иного органа, то она будет положительной. Отрица-
тельные обратные связи, как правило, сохраняют устойчивое,
стабильное, гомеостатическое состояние биосистемы. Это, если
можно так выразиться, преимущественно функционально ща-
дящие связи, так как благодаря им первоначальное отклонение
в управляемой величине, вызванное возмущающим воздейст-
вием, уменьшается. Положительные же обратные связи в
большинстве своем приводят к неустойчивой работе системы
в целом.
217
Помимо обратных, следует выделять еще и прямые связи,
по которым управляющее устройство включает объекты управ-
ления в ту или иную форму активности.
Замкнутый контур регулирования обусловливает биосисте-
ме работу в компенсационном режиме и режиме слежения.
Суть первого режима заключается в том, что система постоян-
но стремится (максимально допустимо в конкретных услови-
ях) уменьшить рассогласование между заданным состоянием
и состоянием, сложившимся в результате воздействия. Откло-
нение от заданного состояния есть ошибка системы регулиро-
вания, величина которой определяет качество компенсацион-
ного регулирования, т. е. способность биосистемы удерживать
заданное (необходимое в данных условиях) стационарное
(гомеостатическое) состояние. Примером компенсационного
режима может быть сужение и расширение зрачка при разной
освещенности для поддержания оптимальной «засветки» сет-
чатки глаза или изменение просвета сосудов прн изменении
кровяного давления с целью восстановления его исходного
уровня и т. д.
В случае необходимости регулирования той или иной функ-
ции по изменяющейся программе (в отличие от стабильной
программы в условиях компенсационного режима) использу-
ется режим слежения. Программа может задаваться как внут-
ренними, так и внешними условиями существования биосисте-
мы. Примером здесь может служить суточная периодика
функций.
Как заметил читатель, существование двух режимов работы
системы довольно условно, поскольку н тот и другой — в
сущности разновидности режима слежения за отклонением от
программы, качественно и количественно постоянной во вре-
мени (компенсационный режим) и изменяющейся (режим
слежения).
Регулирование в компенсационном (по заданному уровню)
режиме и режиме слежения (по программе) потребовало вве-
дения новых терминов — саморегулирование (самонастраива-
ние), самоприспособление, самообучение. Именно самообуче-
ние наиболее тесно связано с таким свойством биосистемы,
как способность к самосовершенствованию, самоулучшению.
Причем самым важным является то, что биосистема способна
к самообучению даже при условии получения неполной инфор-
218
мации о том или ином событии. Она использует накопленный
ею опыт и восполняет недостаток информации некоторыми
общими сведениями, сохраняющимися в ее «запоминающих»
и «логических» блоках. Таким образом, биосистема способна
к прогнозированию своего поведения при проявлении неявных,
предстоящих событий. В этом процессе роль обратных связей
не только не снижается, а, напротив, увеличивается, поскольку
благодаря именно им возможно получение максимума инфор-
мации, в том числе и о приближающемся событии, и умень-
шение информационной энтропии о нем. Здесь нельзя не
вспомнить замечательную мысль Н. Винера (1958) о том, что
обратная связь — «свойство, позволяющее регулировать буду-
щее поведение прошлым выполнением приказов». Одновремен-
но следует отметить, что прогноз только оценивает возможное
развитие системы, но не влияет на него.
Материальной основой элементарного прогноза является
повторяемость событий. Классическим примером может слу-
жить условный рефлекс. Как правило, большинство этих реф-
лексов образуется в условиях максимума информации о на-
мечающемся событии и минимума информационной энтропии.
Значительно сложнее реализуется принцип прогнозирования в
регуляции деятельности биосисгемы, связанной с рассудочной.
Одним из наиболее постоянных периодических процессов,
определяющих формирование прогностического поведения всех
уровней сложности, является смена дня и ночи, позволяющая
на основе опыта поведения в течение суток в прошлом прогно-
зировать его на будущее различной временной отдаленности.
Однако смена дня и ночи — это только принципиальная, а не
идеальная, абсолютная повторяемость событий, так как каж-
дый новый день и новая ночь отличается от предыдущих по
множеству параметров (в том числе и насыщенностью собы-
тий). В силу этого, используя прошлый опыт как модель бу-
дущего поведения, биосистема всегда сталкивается с чем-то
новым. Следовательно, прогнозируется только принципиальное
поведение, в которое реальное явление вносит свои поправки.
Принципиальный прогноз обеспечивает основу предупредитель-
ной регуляции — настройки на ожидаемое явление, а встреча
с ним включает механизмы коррегирующей регуляции. Так,
например, при приобретении двигательных навыков формиро-
вание и сохранение в памяти идеальной (основной, принципи-
219
алыюй) модели, а также некоторый набор моделей движений
для обеспечения возможностей оптимального выбора их в
конкретных условиях — одно из важных обстоятельств осу-
ществления наиболее выгодной двигательной активности.
Однако мы не склонны рассматривать ритмические автома-
тизмы (биологические ритмы) в деятельности животных и
растений как проявление прогностического поведения, хотя за
последние годы накапливается все больше данных о порази-
тельной чувствительности их к событиям во внешней среде,
определяющим возникновение или изменение биоритмов, но
не воспринимаемым человеком и всем арсеналом его исследо-
вательской техники. Поэтому проявление или изменение био-
ритма в большинстве случаев — это не столько прогноз пред-
стоящего события, сколько прямая реакция на реально дейст-
вующие возмущения. Тем более затруднительно отнести к
прогностическому поведению «включение» биологических ча-
сов однократной засветкой растения, находящегося в темноте.
Как показывают опыты, такое растение в течение длительного
времени ведет себя так, как если бы оно находилось в естест-
венных условиях (опускает и подымает листья соответственно
нормальным суточным ритмам смеиы света и тьмы), чего не
делало до засветки. (Скорее до засветки растение вело себя
в соответствии с принципом прогнозирования.)
Таким образом, в иерархии принципов управления и регу-
лирования принцип прогнозирования является ведущим в осу-
ществлении наиболее сложных форм поведения биосистем.
Очерк 4. ПРИНЦИП ЦЕЛОСТНОСТИ
В настоящем очерке прежде всего необходимо рассмотреть
в общем виде некоторые особенности категорий «часть» и
«целое» применительно к биологическим объектам.
Следует признать, что биологической системой любой
сложности, как уже говорилось, может быть только такая це-
лостность, которая удерживает гомеостатическое единство со-
ставляющих ее частей и обладает интегративными качествами,
не свойственными образующим систему элементам. Элемент
биологической системы является ее составной частью только
до тех пор, пока он играет какую-то совершенно определенную
и обязательную роль в общем ансамбле элементов. Именно
220
.поэтому биологическая система па любое воздействие отвеча-
ет целостной реакцией с разной (необходимой в данной ситуа-
ции) степенью участия в ней составляющих элементов. Совер-
шенно справедливое утверждение У. Эшби (1962) в отношении
мозга — «даже изменение одной молекулы нельзя заранее
счесть неэффективными» — вполне может быть распространено
на все биологические системы как системы открытые, находя-
щиеся в динамически стационарном и неравновесном состоянии
относительно среды. Методическая невозможность регистрации
малых изменений в деятельности системы или ее отдельных
элементов не столько наша вина, сколько беда. Формально же
малое не означает наименее значимое для реализации той или
иной функции.
Вряд ли можно согласиться с Анохиным (1970), предпола-
гающим, что система «только такой комплекс избирательно
(выделено нами.— А. Л.) вовлеченных компонентов, у кото-
рых взаимодействие и взаимоотношение приобретают характер
воздействия компонентов на получение фокусированного по-
лезного действия». Стоит ли удивляться такой обедненной
формулировке понятия «система», если автор считает, что
«конкретным механизмом взаимодействия компонентов явля-
ется освобождение их от избыточных степеней свободы, ненуж-
ных (выделено нами.— А. Л.) для получения конкретного ре-
зультата, и, наоборот, сохранение всех тех степеней свободы,
которые способствуют получению результата». С философской
и тем более биологической точки зрения подобное утверждение
не выдерживает критики, так как ненужных степеней свободы
в любой и особенно в биологической системе нет и быть не
может. Все имеющиеся в системе степени свободы всегда нуж-
ны и используются в зависимости от обстоятельств: либо как
свободные, либо как фиксированные, поскольку для реализа-
ции функции фиксация одних степеней свободы ничуть не ме-
нее существенна, чем высвобождение других.
Согласно Анохину, логическим следствием упрощенного
представления о функциональной системе является открытие
нереальной закономерности «минимального обеспечения функ-
циональной системы» числом элементов, реализующих данную
функцию. Следует ли доказывать, что в целостности не может
быть ни минимума, ии максимума участников действия, так
как мы имеем дело с целостностью, ответная реакция которой
221
Ий воздействие всегда тотальна и качественно определяется не
количеством, а степенью участия в ней каждого элемента био-
системы, включая и случаи реципрокных отношений между
элементами, и формально регистрируемое отсутствием реакции.
Отсутствие — это форма присутствия, без которой реакция
системы, адекватная создавшейся ситуации, была бы невоз-
можной.
Итак, любой организм представляет собой целое, т. е.
живую биологическую систему, способную удерживать гомео-
статическое единство составляющих ее частей. Естественно, что
для поддержания этого единства все элементы организма
(клетки, органы, системы органов) должны иметь возмож-
ность общаться друг с другом, обмениваться информацией,
продуктами жизнедеятельности, т. е. пространственно взаимо-
действовать. Только на основе подобного взаимодействия
между органами и тканями устанавливается оптимальный
уровень активности организма в целом. Например, чтобы под-
нять правую руку необходимо такое перераспределение взаимо-
отношений органов, которое позволило бы осуществить только
данное движение. Короче, поднятие руки — это суммарная,
очень сложнокоординированная работа всего организма.
Наиболее древним способом пространственного взаимодей-
ствия элементов для взаимообмена, координации и объедине-
ния системы в единое целое является рассмотренный выше гу-
моральный механизм. При помощи гуморальной системы ин-
теграции внешние и внутренние воздействия передаются, по
образному выражению Ухтомского, «всем, всем, всем», мно-
жеству клеток организма, вызывая более обобщенные диффуз-
ные реакции, ограничителем которых является избирательное
отношение тканей к гормонам.
Относительно эволюционно молодой системой интеграции
является нервная система, обеспечивающая, по сравнению с
гуморальной, более точно и избирательно адресованные по-
сылки от места возникновения возбуждения до места назна-
чения, где они приводят в действие соответствующую часть
организма и притормаживают активность других. Как проис-
ходит подобное распространение возбуждения, легче всего
понять на примере продвижения потенциала действия вдоль
нервного волокна, а также иа примере передачи возбуждения
через синапсы с одной нервной клетки на другую или с нерв-
222
ной клетки на мышечное волокно. Предполагается, что возник-
ший в месте нанесения раздражения потенциал действия явля-
ется не только показателем возбуждения, но и источником
раздражения соседнего невозбужденного участка. По своей
силе потенциал действия в 5—10 раз превышает силу раздра-
жения, необходимую для возбуждения рядом расположенного
участка. Благодаря этому последовательно возникающие по-
тенциалы действия создают как бы ток действия. Ток дейст-
вия в отличие от местного потенциала распространяется по
миелинизированному нервному волокну без декремента, а по
безмякотному — с некоторым затуханием.
Так же как и возбуждение, проведение не сводится к чисто
физическим схемам (процессам), в частности к току действия.
Известно, что в клеточной мембране совершаются метаболиче-
ские процессы с накоплением и высвобождением энергии.
В связи с этим проведение также имеет химическую природу,
и, следовательно, распространение возбуждения — это бегу-
щая по биологическому объекту метаболическая волна с
энергией, достаточной для создания состояния возбуждения,
т. е. возникновения специфической реакции (сокращение, сек-
реция и др.). Вот почему биоэлектрические явления (потенциал
покоя, местный потенциал, потенциал действия и ток покоя)
скорее не причина, а следствие функционального состояния
(обмена веществ) живого объекта.
Пространственные отношения при помощи синапсов осу-
ществляются благодаря передаче возбуждения с одной нервной
клетки на другую или с нервного волокна на иннервируемую
им клетку — мышечную, железистую и др. В нещелевых синап-
сах возбуждение передается с пресинаптической мембраны на
постсинаптическую и обратно. Это, как полагают, осуществля-
ется электрическим путем. Однако в организме наземных жи-
вотных большинство синапсов щелевые. В них возбуждение
распространяется в одном направлении (от пресинаптической
к постсинаптической мембране) с помощью особых химических
веществ — медиаторов (ацетилхолин и норадреналин). Разли-
чают холинэргические и адренергические синапсы. В последнее
время накоплены убедительные данные, свидетельствующие
о том, что медиаторную функцию выполняют многие вещества,
в том числе и АТФ. Есть сведения, что в холинэнергических
синапсах наряду с большим количеством пузырьков, содержа-
228
1цих ацетилхолин, есть некоторое количество заполненных
норадреналином, а в адренергических наоборот. Возможно,
именно поэтому при исчерпании основного медиатора холин-
эргические синапсы могут выполнять функцию адренергиче-
ских, а адренергические — холинэргических. Если в постсинап-
тическую щель поступило медиатора больше, чем это было
необходимо для возникновения возбуждения, избыток его раз-
рушается (инактивируется) специальными ферментами (напри-
мер, ацетилхолинэстеразой). Недостаточность медиатора не
обеспечивает передачи возбуждения, но создает более благо-
приятные условия для его возникновения при поступлении сле-
дующего импульса. Именно поэтому в синаптической щели
всегда находится некоторое количество медиатора, поддержи-
вающее состояние готовности постсинаптической мембраны к
передаче возбуждения.
Есть синапсы, в которых под влиянием возбуждения в си-
наптическую щель поступают медиаторы, вызывающие не
деполяризацию, а гиперполяризацию постсинаптической мем-
браны (например, гамма-аминомасляная кислота). Естественно,
при этом возбудимость постсинаптической мембраны снижает-
ся, что служит препятствием для продвижения возбуждения.
Это так называемый тормозный постсинаптический потенциал.
Синапсы, работающие по этому принципу, называются тормо-
зящими (тормозными).
Если учесть, что при проведении возбуждения срабатывают
и возбуждающие, и тормозящие синапсы, значит, существует
возможность усиления деятельности одних органов и систем
и торможения других, т. е. в зависимости от обстоятельств ор-
ганизм избирательно использует различные части тела в при-
способительной реакции иа то или иное воздействие. Например,
сгибание в любом суставе обязательно обусловлено возбужде-
нием, возникающим в центрах сгибателей, и торможением —
в центрах разгибателей. Итак, действие может быть выполнено
не только благодаря «непосредственным» исполнителям, но и
потому, что «не участвующие» части тела «не мешают» этому.
Следовательно, в любой ответной реакции организма не участ-
вующих элементов нет.
Значит, распространение возбуждения (в широком смысле)
один из основных механизмов целостной приспособительной
реакции организма на воздействие внешней среды. Л само
224
возбуждение — это процесс, связанный не только с освобожде-
нием энергии, необходимой для совершения действия, но и с
доставкой информации, па основании которой целенаправлен-
но осуществляется именно это, а не какое-либо иное действие.
В последнее время начинает все более утверждаться пред-
ставление о множественности механизмов интеграции органи-
зма. К ним можно отнести механизмы межмолекулярного и
межатомного переноса энергии (диффузионный перенос макро-
эргических молекул, перенос электронов через зоны проводи-
мости, резонансный перенос электронного возбуждения,
комплексный перенос заряда, кооперативные и конформацион-
ные процессы, пьезоэлектрические эффекты и, возможно,
такие диффузионно-кристаллические эффекты, как замещение
вакансий, дислокационные процессы и др.).
Весьма привлекательна идея о значении электромагнитных
полей (ЭМП) в организме для регуляции (интеграции) про-
цессов жизнедеятельности. Речь идет не об известных способах
передачи информации по нервам с помощью биоэлектрических
импульсов, а о своеобразной радиосвязи между различными
элементами и системами. Конечно, термин «радиосвязь» усло-
вен, так как связи предполагаются не только в радиочастотном
диапазоне, но и с помощью ЭМП низких и даже высоких
частот.
В 1959 году была высказана гипотеза о таком управлении,
в котором молекулы ДНК рассматриваются как генераторы
радиочастотных сигналов, РНК — как усилители, а ферменты
и аминокислоты—как исполнители сигналов, кодированных
в различных участках спектра; оболочке клетки приписыва-
ется роль шумового фильтра. В связи с этим примечательна
мысль Сент-Дьерди (1960): «...две молекулы, электроны ко-
торых могут одинаковым образом возбуждаться, способны
вести себя как связанные осцилляторы. В этом случае нет
необходимости в наличии материальной связи между ними,
так как их связывает электромагнитное поле, если только
расстояние между ними слишком велико (мало по сравнению
с длиной волны)». Отсюда, по мнению А. С. Пресмана (1968).
«при оценке возможного частотного диапазона электромаг-
нитных взаимосвязей между клетками следует учитывать два
обстоятельства: с одной стороны, экспериментальные данные
указывают на чувствительность клеток к ЭМП самых различ-
225
пых частот, с другой — проникновение ЭМП внутрь клетки
должно зависеть от частоты, так как от нее зависит импеданс
мембраны и поглощение, обусловленное полярными молекула-
ми воды. Очевидно, что компромиссным может быть диапазон
от десятков до 1000 мГц, в котором влияние клеточных мем-
бран становится уже незначительным, а полярные свойства
воды еще не сказываются. Иначе говоря, в этом диапазоне
взаимосвязь между внутренними элементами различных кле-
ток может быть частотно-независимой. При широкой полосе
частот и соответствующем способе кодирования может быть
обеспечена высокая надежность межклеточной взаимосвязи,
которая осуществляется, по-видимому, и через субклеточные
структуры и на молекулярном уровне» (выделено.— А. Л.).
Как оказалось, возникающее ЭМП проявляется не только
внутри биообъекта, но и вокруг него. Так, ныне существующие
методы измерения позволили зарегистрировать ЭМП изолиро-
ванного нерва лягушки на расстоянии 25 см (напряжение
равнялось 1 мВ), изолированной мышцы и сердца-—на рас-
стоянии 14 см, сердца и мышцы человека — на расстоянии
10 см. Зарегистрированы ЭМП и целых организмов, например,
при полете шмеля, комара и др.
Весьма интересным является факт регистрации напряже-
ния магнитной составляющей электромагнитного поля вблизи
возбужденного нерва даже раньше, чем регистрировался
электрический сигнал, проходящий по нему. Это значит, что
реагирующий на возбуждение объект подготовлен к его при-
ему заранее (до того, как возбуждение достигло объекта).
Учитывая практическую пространственную неограниченность
и, как было показано выше, беспрепятственность распростра-
нения ЭМП по биообъекту, следует полагать, что в состоянии
готовности к ответу даже на строго адресованную посылку
оказывается вся биосистема, а не только та ее часть, куда
поступает «классическое» возбуждение. Такой тип связи подо-
бен тому, который рассматривал II. Винер (1958) как сигнал
пожарной сирены, называя его сигналом «тем, кого это каса-
ется». По существу, это иная редакция принципа «всем, всем,
всем», распространенного Ухтомским только на гуморальный
механизм регуляции.
В связи с этим Пресман (1968) выдвинул предположение
о существовании системы «аварийной» сигнализации одновре-
226
к
менно всем органам, не связанной с нервной сетью. По его
мнению, такая сигнализация могла бы обеспечить практически
мгновенную общую мобилизацию организма (особенно при
эмоциональных состояниях). В этих случаях, вероятно, цен-
тральная нервная система передает электромагнитные сигаа-
лЫ" «тем, кого это касается», по всему организму па «аварий-
ной» частоте. Исходя из способности биосистем, в том числе
и безнервных (это касается и растений), продуцировать воз-
буждение под влиянием возмущающих воздействий, свойство
сигнализации и интеграции при помощи ЭМП может быть
распространено и на эти системы. В частности, исследованиями
В. П. Казначеева, Л. П. Михайлова и С. П. Шурина показано,
что если поместить в два сосуда из кварцевого стекла культу-
ры нормальных живых клеток, а затем одну из них заразить
вирусом, то почти одновременно клетки в другом сосуде, на-
ходящиеся лишь в оптическом контакте с зараженными, вос-
производят похожий патологический процесс. Так, авторы
травили культуру клеток в одном сосуде сулемой, облучали
смертельными дозами ультрафиолетового возбуждения, одна-
ко погибала и культура клеток, не подвергавшаяся воздействи-
ям, несмотря на то что сосуды соприкасались только доныш-
ками.
По формуле регистрации этого открытия клеточную струк-
туру можно определить «как детектор модуляционных особен-
ностей электромагнитных излучений». Следовательно, электро-
магнитное поле биологических объектов может быть источником
не только межобъектного обмена информацией, но и внутри-
объектной интеграции элементов биосистемы в единое целое.
Отдавая дань справедливости, следует отметить, что прин-
ципиально, в общем виде, на биологическое поле как средство
внутри- и межсистемных взаимодействий впервые указал
А. Г. Гурвич (1944) на основании открытого им явления мито-
генетического излучения *. Под биологическим полем клетки
* Митогенетическое излучение—ультрафиолетовое излучение
широкого диапазона малой интенсивности, возникающее при экзо-
термических реакциях, протекающих в живых системах. Митогенети-
ческое излучение впервые было обнаружено в опытах по изучению
влияния активно митотически делящихся клеток корешка лука на
усиление размножения расположенных в непосредственной близости
культуры дрожжей.
227
Гурвич понимал силы, которые поддерживают «в живых си-
стемах специфическую для них Молекулярную упорядочен-
ность». «Понятие молекулярной упорядоченности в нашем
смысле,— пишет Гурвич,— всякое пространственное распре-
деление молекул, не вытекающее из их химической структуры
или состояния равновесия, т. е. химических связей, ван-дер-
ваальсовых сил и т. п. В силу этого молекулярная упорядочен-
ность, вообще говоря, не равновесна». Таким образом, биоло-
гическое поле является фактором, определяющим основное
свойство биосистем — направленность, векторпость, всех про-
явлений жизнедеятельности этих систем. В частности, анали-
зируя зависимость элементов от целого во время формообра-
зования, Гурвич пришел к выводу о необходимости создания
пространственно-временной конструкции, облегчающей понима-
ние последовательного развертывания формообразовательных
процессов. Для этого были сделаны следующие допущения: к
каждой фазе развития (например, зародыша) следует отнести
понятие целого, имеющего свою пространственно-временную
динамику; наряду с переменными параметрами допускается
существование постоянного члена — инвариантного принципа
действия целого на элементы; ход формообразования является
функцией от инвариантной слагаемой.
На основании таких представлений и подтверждающих их
фактов (анализ распределения митозов и др.) Гурвич выде-
лил две категории взаимоотношений между клеточным уровнем
и уровнем целого: а) если все факторы, определяющие данную
фазу процесса, «жестко» связаны между собой, то целое «де-
терминирует» поведение элементов; б) если хотя бы один фак-
тор из группы относительно «независим» от других, то возник-
новение процесса приобретает характер случайного события,
т. е. целое «нормирует» поведение элементов. Понятие «нор-
мировка» шире понятия «детерминация», так как охватывает
все процессы взаимодействия (в том числе и случайные, ве-
роятностные), где нужно говорить о различных степенях
свободы.
Таким образом, понятие нормировки, возможно, даже более
определенно, чем понятие регуляции. С этой точки зрения
ядро, а в некоторых случаях при наличии цитохроматина и
тело клетки, следует рассматривать как систему, включающую
в себя множество элементарных источников полей. Действие
228
совокупности элементарных полей подчиняется вследствие
векторного характера явлений гомеостатическому сложению.
Результирующий вектор в свою очередь характеризует более
сложное поле—ядерное или клеточное, которое не ограничи-
вается пределами клетки и проявляется уже за ее пределами,
в частности, в векторной характеристике соседних клеток.
Геометрическое сложение векторов клеточных плоей вместе с
тем создает так называемое «актуальное иоле», характеризу-
ющее совокупность клеток и т. д.
Нам кажется, что на основании общетеоретических поло-
жений Гурвича о биологическом поле, даже без достаточного
знания его природы, можно представить в общем виде все
варианты межклеточных, тканевых (клеточная и тканевая ин-
теграция и дифференциация, например органная, органно-тка-
невая и др.) и, вероятно, организменных отношений.
Не в прямой связи с теорией биологического поля, но с
учетом ее следует рассматривать процессы интеграции и диф-
ференциации иммуннобиологической природы (система анти-
ген— антитело, изоиммунные реакции — аутоантитела и др.).
Появление в наших рассуждениях наряду с понятием ин-
теграции понятия дифференциации при рассмотрении пробле-
мы целостности не случайно. Клеточно-тканевая и органная
дифференциация, обеспечивая функциональную специализацию
органов и тканей, не только не приводит (в естественных усло-
виях) к расформированию биосистемы на элементы, но и еще
более увеличивает межэлементную взаимозависимость и пот-
ребность в интеграции.
Таким образом, принцип целостности как принцип регуля-
ции адаптивно-гомеостатического поведения биосистем приоб-
ретает еще большую актуальность именно в силу внутренней
противоречивости и двойственности материальной основы, за
счет которой он осуществляется.
Итак, мы рассмотрели общие свойства и закономерности
живых систем. Эти свойства и закономерности являются
фундаментальными, присущими всей живой природе, даже
тогда, когда у биосистемы то или иное свойство проявляется
при помощи иных механизмов.
ПРИЛОЖЕНИЕ
К РАЗДЕЛУ «ОРГАНИЗМ И СРЕДА»
МЕХАНИЗМЫ ИНДИВИДУАЛЬНЫХ УРОВНЕЙ
И ВИДОВ ГОМЕОСТАЗА
1. Цитогенетический гомеостаз: клеточный регулирующий
механизм (ядро, митохондрии, вакуоли, лизосомы, мембраны
и в первую очередь генетический аппарат),
2. Соматический геностаз: противомутационный, иммуно-
логический, программно-генетический (регрессионный) кон-
троль.
2.1. Тканевый и органный геностаз: противомутационный,
иммунологический, программно-генетический (релрессионный)
контроль, механизмы поддержания тканевого гомеостаза
(микроциркуляция и др.).
2.2. Системно-органный геностаз: см. 2.1.
3. Соматический гомеостаз: механизмы координационных
отношений цитогенетического гомеостаза, соматического гено-
стаза; все механизмы управления и регулирования функций,
интегрирующих биологическую систему в единую целостность.
3.1. Функциональный (физиологический) гомеостаз: вну-
триклеточные, межклеточные, органные; межорганные, сис-
темные, межсистемныс, организменные генетические, гумо-
ральные (биохимические внутриклеточные), интегрирующие
(внутри- и межорганные) гуморальные и нейрогуморальные
механизмы управления и регулирования.
3.2. Обменный (организменный) гомеостаз: генетический
контроль специфики обмена органелл, клеток, органов, тканей
и организма в целом; все виды циркуляции; гуморальный и
нейрогуморальиый механизмы управления и регулирования.
3.2.1. Клеточно-тканевый (органный) гомеостаз: см. 3.2.
3.2.2. Гуморальный гомеостаз: см. 3.2.
3.3. Системный (органный) гомеостаз: генетический функ-
циональный контроль специфики обменного гомеостаза; вну-
280
триклеточные и внутрисистемные интегрирующие гуморальные
и нейрогуморальные механизмы управления и регулирования.
3.3.1. Дыхательный гомеостаз: механизм внутриклеточно-
го тканевого дыхания (аэробное и анаэробное биологическое
окисление); все механизмы управления и регулирования обме-
на газов между организмом и средой.
3.3.2. Пищеварительный гомеостаз: механизмы поглоще-
ния (фагоцитоз, пиноцитоз, пищеварительные вакуоли, полос-
ти, пищеварительный тракт; внешнее переваривание, фермен-
тативное расщепление, внутреннее переваривание — пищевари-
тельные вакуоли, лизосомы, полостное и пристеночное); все
виды всасывания, в том числе и перенос через клеточные мем-
браны; гуморальная и нейрогуморальная регуляция.
3.3.3. Выделительный гомеостаз: физико-химические меха-
низмы (эффект поверхностного натяжения, явление капилляр-
ности, диффузии); пассивный и активный перенос веществ че-
рез клеточные мембраны; внутриклеточные специализирован-
ные механизмы, выделительные органы и системы (железы
и др.), мочевыделительная система и ее аналоги, вспомога-
тельные системы выделения — дыхательная и пищеваритель-
ная; гуморальные и нейрогуморальные механизмы регуляции
выделительных функций.
3.3.4. Циркуляторный гомеостаз: физико-химические меха-
низмы (эффект поверхностного натяжения, явление капилляр-
ности, диффузии и др.); перенос жидкостей через клеточную
мембрану, внутриклеточные специализированные механизмы
управления движением цитоплазмы — циклос (сократительные
вакуоли и пр.); специализированные системы циркуляции
(системы органов крово- и лимфообращения и др.); гумораль-
ные и нейрогуморальные механизмы регуляции всех видов
циркуляции.
3.3-5. Двигательный (моторный) гомеостаз: все способы
перемещения в пространстве, в том числе за счет водных
и воздушных потоков и некоторых существ (семена, парази-
тирующие организмы, молодые особи популяции и др.); рас-
пространение активно не перемещающихся клеток, включая
и опухолевые и «захват» территории за счет размножения и
увеличения размеров органа или ткани, в состав которых они
входят; движение цитоплазмы, обусловленное активным
передвижением некоторых одноклеточных организмов (аме-
231
боидное движение), а также движение, не сопровождающееся
перемещением клетки в пространстве, но обеспечивающее вну-
триклеточное перемещение органелл ее. Передвижение одно-
клеточных организмов при помощи жгутиков, ресничек, унду-
лирующих мембран; частичное смещение в пространстве по-
верхностей фиксированных клеток, снабженных ресничками;
движение при помощи специализированных сократительных и
других структур, образующих сложные опорно-двигательные
аппараты (скелет, мышцы, связки); физико-химические, биохи-
мические, гидравлические и другие; гуморальные, нейрогумо-
ральиые механизмы управления и регулирования двигательной
активностью.
3.3.6. Сенсорный гомеостаз: все уровни рецепирования —
принятие и переработка информации (в том числе и биосис-
тем, ие располагающих дифференцированной сенсорикой); все
уровни специализации (химические, физико-химические меха-
низмы получения и переработки информации); гуморальные
и нейрогуморальные механизмы регуляции деятельности сен-
сорных систем.
3.3.7. Сенсомоторный гомеостаз: координационно-функцио-
нальные механизмы двигательного и сенсорного гомеостаза
(для всех уровней биосистем, в том числе и реализующих
двигательную активность путем таксисов).
3.3.8. Психомоторный гомеостаз: все механизмы обеспече-
ния нервной и психонервной деятельности; для низших форм —
механизм условнорсфлекторной и инстинктивной деятельности.
3.3.9. Психологический гомеостаз: механизмы обеспечения
высших форм психического поведения (механизмы психонерв-
ной деятельности).
3.3.10. Регуляторный гомеостаз: механизмы всех перечис-
ленных гомеостазов — от генного до гуморального, нейрогумо-
рального и психонервного.
4. Онтогенетический гомеостаз (подразделяется на пре- и
постнатальный и др.): механизмы витогенетического гомео-
стаза, соматического геносгаза, соматического гомеостаза, фа
нотипической п риспособлениости.
МЕХАНИЗМЫ ПОПУЛЯЦИОННОГО
(НАДЫНДИВИДУАЛЬНОГО) ГОМЕОСТАЗА
1. Генетический гомеостаз: отбор (в основном стабилизи-
рующий), благоприятствующий поддержанию сбалансирован-
ного «максимально приспособленного» генотипа.
1.1. Гомеостаз оптимальности (см. от 1.1 до 1.2): под-
держивает оптимальную общую и структурно-функциональ-
ную численность (организацию) популяции в определенных
условиях существования.
1.1.1. Гомеостаз плодовитости (фертильный гомеостаз):
скорость размножения организмов в зависимости от скорости
полового созревания, величины приносимого приплода и часто-
ты размножения; обеспечивает поддержание оптимального по-
полнения численности популяции.
11.2. Сексуальный гомеостаз: плодовитость популяции и все
факторы, обеспечивающие рождение мужских и женских осо-
бей; обеспечивает соотношение мужских и женских особей,
необходимое для оптимальной рождаемости в популяции.
1.1.3. Сексуальнорепродуктивный гомеостаз: плодовитость
популяции и все факторы, исключающие особи, находящиеся
в активном репродуктивном возрасте; обеспечивает соотноше-
ние мужских и женских особей, находящихся в активном ре-
продуктивном возрасте, необходимое для поддержания опти-
мальной рождаемости.
1.1.4. Ювенальный гомеостаз: плодовитость популяции и
все факторы, исключающие особи, не достигшие репродуктив-
ного возраста; обеспечивает оптимальное соотношение мужских
и женских особей, не достигших репродуктивного возраста и
составляющих резерв сексуальнорепродуктивпого гомеостаза.
1.1.5. Климактерический гомеостаз: плодовитость популя-
ции и все факторы, исключающие особи климактерического
возраста; обеспечивает оптимальное соотношение мужских
и женских особей в возрасте старше активно-репродуктивного
(лимитирующий фактор рождаемости).
1.2. Витальный гомеостаз: плодовитость популяции и все
факторы, исключающие особи из биологического «оборота»
популяции или ограничивающие эту возможность; обеспечивает
оптимальную биологическую жизненность популяции.
•233
1.2.1. Ёиталонгитудиналъный гомеостаз: все факторы, опре-
деляющие длительность жизни особей в популяции; обеспечи-
вает оптимальную среднюю продолжительность жизни особи
(суммарно и по полу).
1.2.2. Морталитный гомеостаз: все факторы, определяющие
смертность особей в популяции; обеспечивает оптимальную
(суммарно и по полу) смертность особей.
1.2.2.1. Морталитно-ювенальный гомеостаз: все факторы,
определяющие смертность (суммарно и по полу) особей, на-
ходящихся в ювенальном возрасте; поддерживает оптимальный
сексуальнорепродуктивный резерв популяции.
1.2.2.2. Морталитно-репродуктивный гомеостаз: все факто-
ры, определяющие смертность особей в репродуктивном воз-
расте; обеспечивает оптимальную смертность (суммарно и по
полу) особей, находящихся в репродуктивном возрасте.
1.2.2.3. Функционально-репродуктивный гомеостаз: все фак-
торы, определяющие смертность особей в биологически и фун-
кционально непродуктивном возрасте; обеспечивает оптималь-
ную смертность (суммарно, по полу и возрасту) особей в
биологически и функционально («социально») непродуктив-
ном возрасте.
1.2.2.4. Аффективный гомеостаз: заболеваемость и болез-
ненность и все другие факторы, исключающие дефектные осо-
би из популяции; обеспечивает оптимальное количество (сум-
марно, по полу и возрасту) больных и дефектных особей.
1.3. Структурно-функциональный гомеостаз: плодовитость
популяции, гено- и фенотипические особенности популяции,
все факторы, регулирующие избыток или недостаток внутрипо-
пуляционной специализации (в том числе фактор конкурентной
борьбы за самку и др.); обеспечивает оптимальное соотноше-
ние количества (суммарно и по полу) особей, выполняющих
специализированные функции (временно или постоянно) в
«социальной» структуре популяции.
1.4. Внутрипопуляционный комфортный гомеостаз: гено- и
фенотипические особенности данной особи, определяющие ее
положение в популяции (лидер и др.); обеспечивает особи, вхо-
дящей в популяцию, оптимальные условия существования (ин-
дивидуальная среда обитания, питание, право на воспроизведе-
ние потомства и т. д.).
1.5. Крмфортно-биоценотический гомеостаз: механизмы,
234
обеспечивающие популяции высокую конкурентоспособность в
данном биоценозе (гено- и фенотип популяции и др ); обеспе-
чивает популяции комфортное существование в биоценозе.
1.5.1- Лдаптивно-комфортный гомеостаз: гено- и фенотипи-
ческие механизмы адаптации; обеспечивает популяции опти-
мальную приспособляемость к тем или иным природно-клима-
тическим условиям существования.
1.5.2. Конкурентно-комфортный межпопуляционный гомео-
стаз: гено- и фенотипические механизмы адаптации; обеспе-
чивает оптимальную конкурентоспособность популяции за
рраво существования в биотопе
ЛИТЕРАТУРА
Учебники и учебные пособия
Ашмарин И. П. Молекулярная биология. Л., «Медицина», 1974.
Байер В. Биофизика. М., «Иностр, литература», 1962.
Бергельсон Л. Д. Биологические мембраны. М., «Наука», 1976.
Биологическая кибернетика. Под рсд. А. В. Когана. М., «Высш,
школа», 1972.
Биофизика. Под ред. Б. Н. Тарусова, О. Р. Кольсе. М., «Высш,
школа», 1968.
Боген Г. Современная биология. М., «Мир», 1970.
Вилли К., Детье В. Биология. М., «Мир», 1974.
Готт В. С. Философские вопросы современной физики. М., «Высш,
школа», 1967
Еремин Е Н. Основы химической термодинамики. М., «Высш,
школа», 1974.
Пост. X. Физиология клетки. М., «Мир», 1975.
Коган А. Б. Электрофизиология. М., «Высш, школа», 1969.
Кузнецов В. В. Физическая и коллоидная химия. М., «Высш,
школа», 1968.
Латманизова Л. В. Очерк физиологии возбуждения. М., «Высш,
школа», 1972.
Ленинджер А. Биохимия. М., «Мир», 1974.
Леви А., Сикевиц Ф. Структура и функции клетки. М„ «Мир»,
1971.
Линдсей П.. Норман Д. Переработка информации у человека.
М.. «Мир», 1974.
Пасынский А. Г. Биофизическая химия. М., «Высш, школа», 1968.
Проссер Л., Браун Ф. Сравнительная физиология животных. М..
«Мир», 1967.
Робертис де С., Новинский В., Саэс Ф. Биология клетки. М.,
«Мир», 1973.
Рокецкий П. Ф. Введение в статистическую генетику. Минск,
«Вышэйш. школа», 1974.
Фридрихсберг Д. А. Курс коллоидной химии. Л.. «Химия», 1974.
236
Специальная литература
Адольф Э Развитие физиологических регуляций. М., «Мир», 1971.
Ананьев Б. Г. Человек как предмет познания Л.. Изд-во Ле-
нингр. ун-та, 1969.
Анохин П. К. Биология и нейрофизиология условного рефлекса.
М., «Наука», 1968.
Бауэр Э. С. Теоретическая биология. М.— Л.. Изд-во ВИЭМ,
1935.
Бреслер С. Е. Молекулярная биология. Л., «Наука», 1973.
Бюннмнг Э. Ритмы физиологических процессов. М., «Иностр- ли-
тература», 1961
Бахтин Ю. Б. Генетика соматических клеток. Л., «Наука», 1974.
Бейль Г. Симметрия. М., «Мир», 1968.
Вернадский В. М. Биосфера. М., «Мысль», 1967.
Винер Н. Кибернетика. М., «Советское радио», 1968.
Гарднер А. Ф. Этот правый и левый мир. М, «Мир», 1967.
Гаузе А. Ф. Асимметрия протоплазмы. М.— Л., 1940.
Гельфер Я. М. Законы сохранения. М., «Наука», 1967.
Гинсдорф П., Пригожин И. Термодинамическая теория структу-
ры, устойчивости и флуктуации. М., «Мир», 1973.
Гродинз Ф. Теория регулирования и биологические системы. М.,
«Мир», 1966.
Гудвин Б. Временная организация клетки. М., «Мир», 1966.
Гурвич А. Г. Теория биологического поля. М., «Советская нау-
ка». 1944.
Гурвич А. А., Еремеев В. Ф., Карабчевскцй Ю. А. Энергетические
основы митогенетического излучения и его регистрация на фотоэлект-
ронных умножителях. М., «Медицина», 1974.
Зетин А. И. Термодинамический подход к проблемам развития,
роста и старения. М., «Наука», 1974.
Карпинская Г. С. Философские проблемы молекулярной биоло-
гии. М., «Мысль», 1971.
Касиное В. Б. Биологическая изомерия. Л., «Наука», 1973.
Катц Б. Нерв, мышца и синапс. М., «Мир», 1968.
Конев С. В., Аксенцев С. Л., Черницкий Е. А. Кооперативные
переходы белков в клетке. Минск, «Наука и техника», 1970.
Майр Э. Популяции, виды и эволюция. М, «Мир», 1974.
На пути к теоретической биологии. 1. Пролегомены. Под ред.
Б. А. Астаурова. М., «Мир», 1970.
Петрушенко Л. А. Самодвижение материи в свете кибернетики.
М„ «Наука», 1971.
Поликар Л. Поверхность клетки и ее микросреда. М., «Мир»,
1975.
237
Пресман Л. С. Электромагнитные поля и живая природа. М.,
«Наука», 1968.
Прохоренко В. К- Методологические принципы общей динамики
систем. Минск, Изд-во Бслор. ун-та, 1969.
Садовский В. Н. Основы общей теории систем. М., «Наука», 1974.
Скулачев В. П. Трансформация энергии в мембранах. М., «Нау-
ка», 1972.
Тесаки И. Нервное возбуждение. М., «Мир», 1971.
Тимофеев-Ресовский Н. В., Воронцов Н. В.. Яблоков Н. В- Крат-
кий очерк теории эволюции. М., «Наука», 1968.
Уорд Р. Живые часы. М., «Мир», 1974.
Урманцев Ю. А. Симметрия природы и природа симметрии. М.,
«Мысль», 1974.
Харрисон Дм., Уайнер Дж., Таппер Дж., Барникот Н. Биология
человека. М., «Мир», 1968.
Ходоров Б. М. Проблема возбудимости. Л., «Медицина», 1969.
Чижевский Э. Л. Земное эхо солнечных бурь. М., «Мысль», 1973.
Шафрановский И. И. Симметрия в природе. Л., «Недра», 1968.
Шмалвгаузен И. И. Факторы эволюции. М., «Наука», 1968.
Шмальгаузен И. И. Проблемы дарвинизма. Л., «Наука», 1969.
Шредингер Э. Что такое жизнь? М., «Атомиздат», 1972.
Шубников А. В., Цопцик В. А. Симметрия в науке и искусстве.
М., «Наука», 1972.
СОДЕРЖАНИЕ
Предисловие........................................ °
I. ОРГАНИЗМ И СРЕДА
Очерк. 1. Обмен веществ и энергии. Общие положения . . 5
Очерк 2. «Термодинамический» анализ взаимоотношений
в системе живой организм — окружающая среда . 19
Очерк 3. Гомеостаз. Общие положения...............43
Очерк 4. Уровни н виды гомеостаза биологических систем 56
II. ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ ВЗАИМООТНОШЕНИЙ
В СИСТЕМЕ ОРГАНИЗМ - СРЕДА
Очерк 1. Гомеостатические параметры биологических сис-
тем ............................................ 77
Очерк 2. Непарадоксальиые парадоксы гомеостаза .... 91
Очерк 3. Гомеостаз и конечность развития биосистем. Еще
одна попытка измерения гомеостаза................125
Очерк 4. Биологические структуры и процессы обмена ве-
ществ ...........................................134
III. ВИДЫ И ФОРМЫ ПРОЯВЛЕНИЯ ФУНКЦИОНАЛЬНОЙ
АКТИВНОСТИ живых СИСТЕМ
Очерк 1. Функциональная активность — единственно воз-
можное состояние живых систем ...................157
Очерк 2. Мембранный потенциал — одна из основных ха-
рактеристик живой системы........................160
Очерк 3. Основные закономерности и свойства возбудимо-
сти живых систем.................................176
239
IV- УПРАВЛЕНИЕ И РЕГУЛИРОВАНИЕ ФУНКЦИЙ
Очерк /. Общие положения об управлении и регулировании
функций...................................189
Очерк 2. Принцип рассогласования — иеравновесности гра-
диента ............................................193
Очерк 3 Принцип замкнутого контура регулирования и
протезирования ............................... . . .213
Очерк 4. Принцип целостности..................... .220
ПРИЛОЖЕНИЕ к РАЗДЕЛУ «ОРГАНИЗМ И СРЕДА» 230
Механизмы индивидуальных уровней и видов гомеостаза 230
Механизмы популяционного (надындивидуального) гоме-
остаза .............,............................ .233
Литература ..... . . ..... .236
Лнаголий Алексеевич Логинов
ОЧЕРКИ ПО ОБЩЕЙ ФИЗИОЛОГИИ.
ОСНОВНЫЕ СВОЙСТВА И ЗАКОНОМЕРНОСТИ ЖИВЫХ СИСТЕМ
Редактор Л. В. Харитонович. Худож. оформление В. В. Бесо-
нова. Худож. редактрр В. И. Валентович. Техн, редактор М. Н.
Кислякова. Корректор А. А. Савицкая
АТ 11168. Сдано в набор 12/1 1976 г. Подписано к печати 26/V 1976 г.
Бумага 70X108'/3S ТНПОгр. № 1. Печ. л 7,5+0,125ф. (10,5+0,175ф). Уч,-
нзд. л. 13,17. Изд. № 75-158. Тип. зак. 2942. Тираж 10000 экз. Цена
94 коп.
Издательство «Вышэйшая школа» Государственною комитета Совета
Министров БССР по делам издательств, полиграфии и книжной торгов-
ли. Редакции литературы по естествознанию. 220600. Минск, ул. Киро-
ва, 24.
Полиграфкомбинат нм. J1. Коласа Государственною комитета Совета
Министров БССР по делам издательств, полиграфии и книжной тор-
говли. Минск, ул Красная, 28.