
































































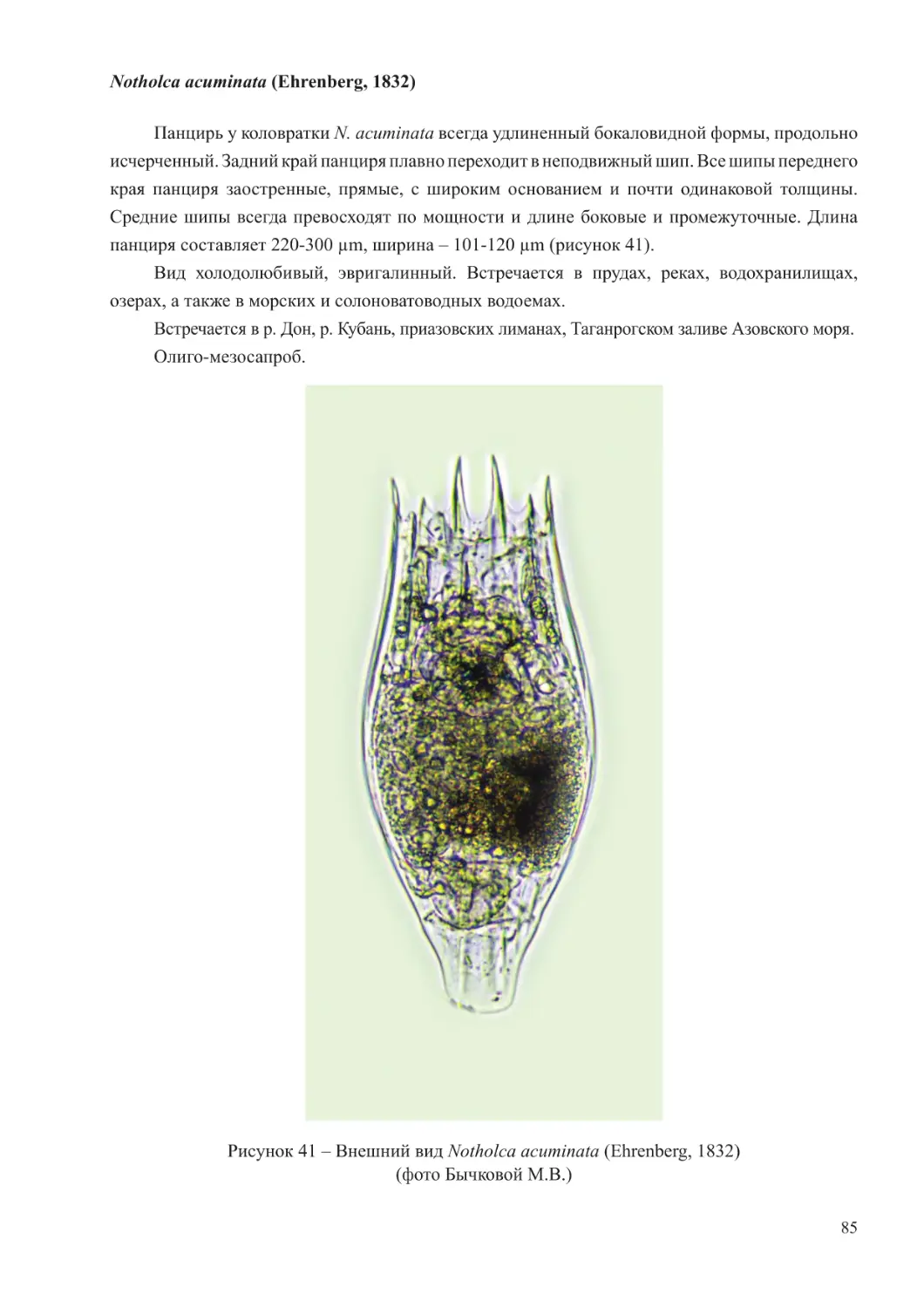

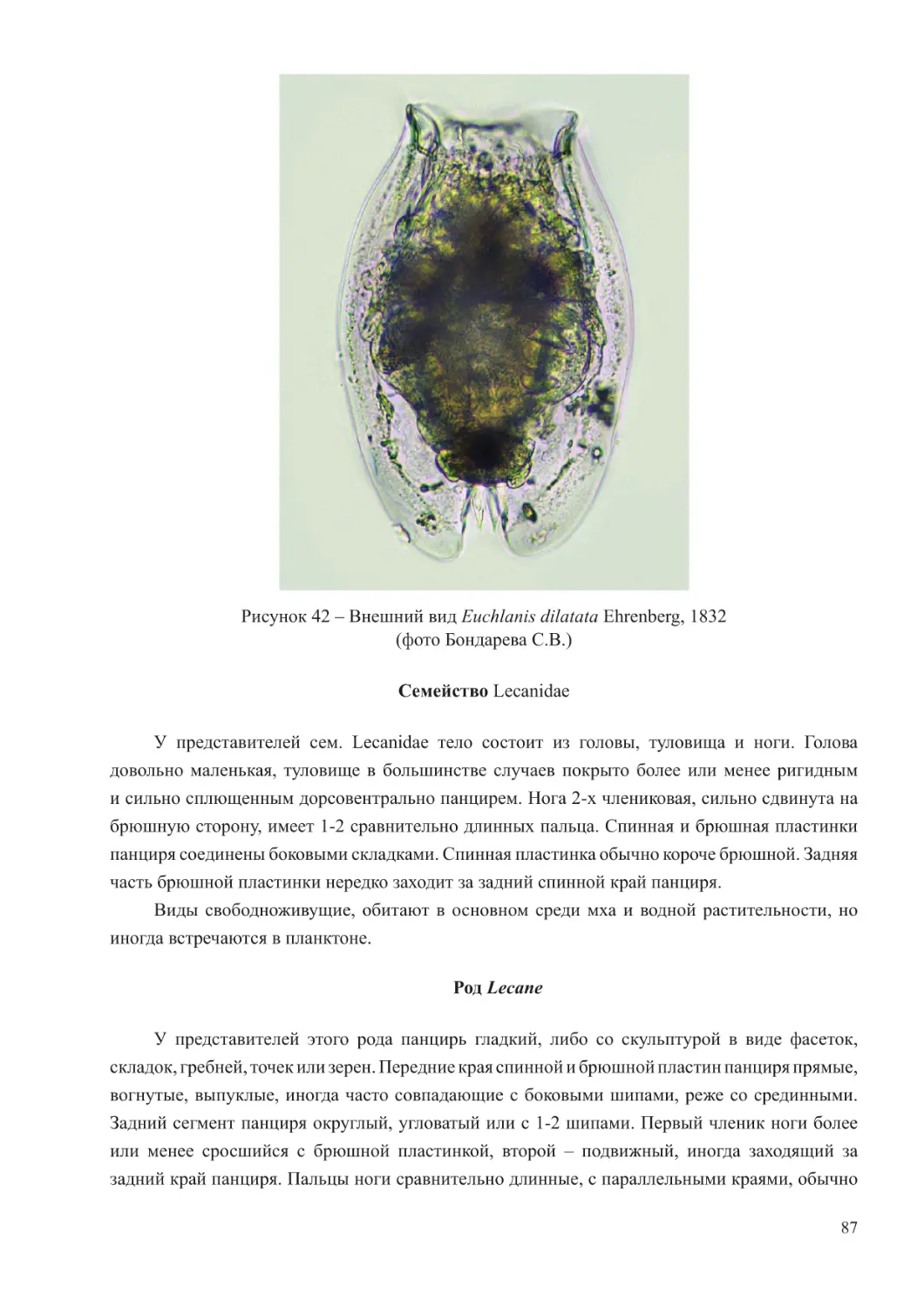

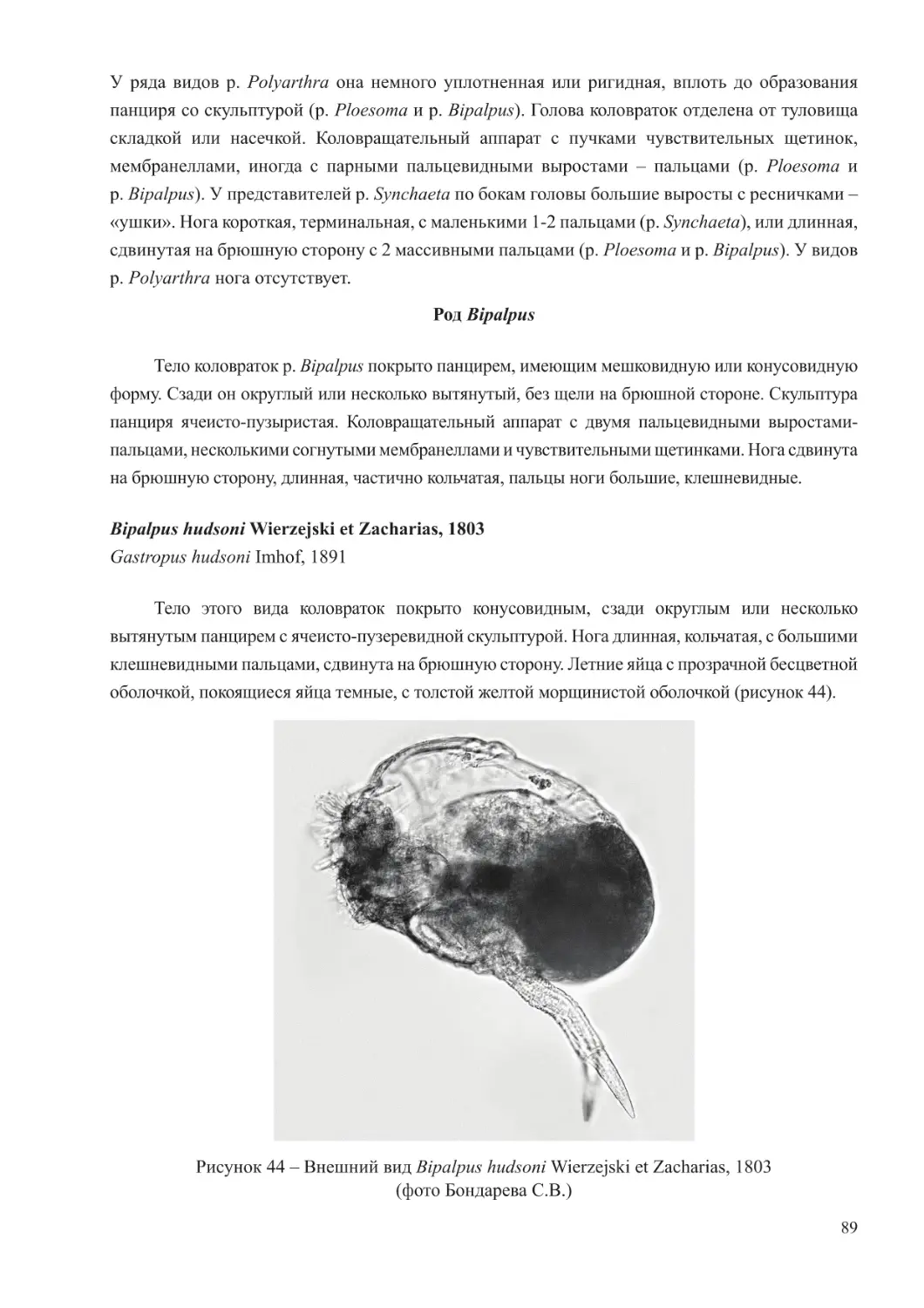

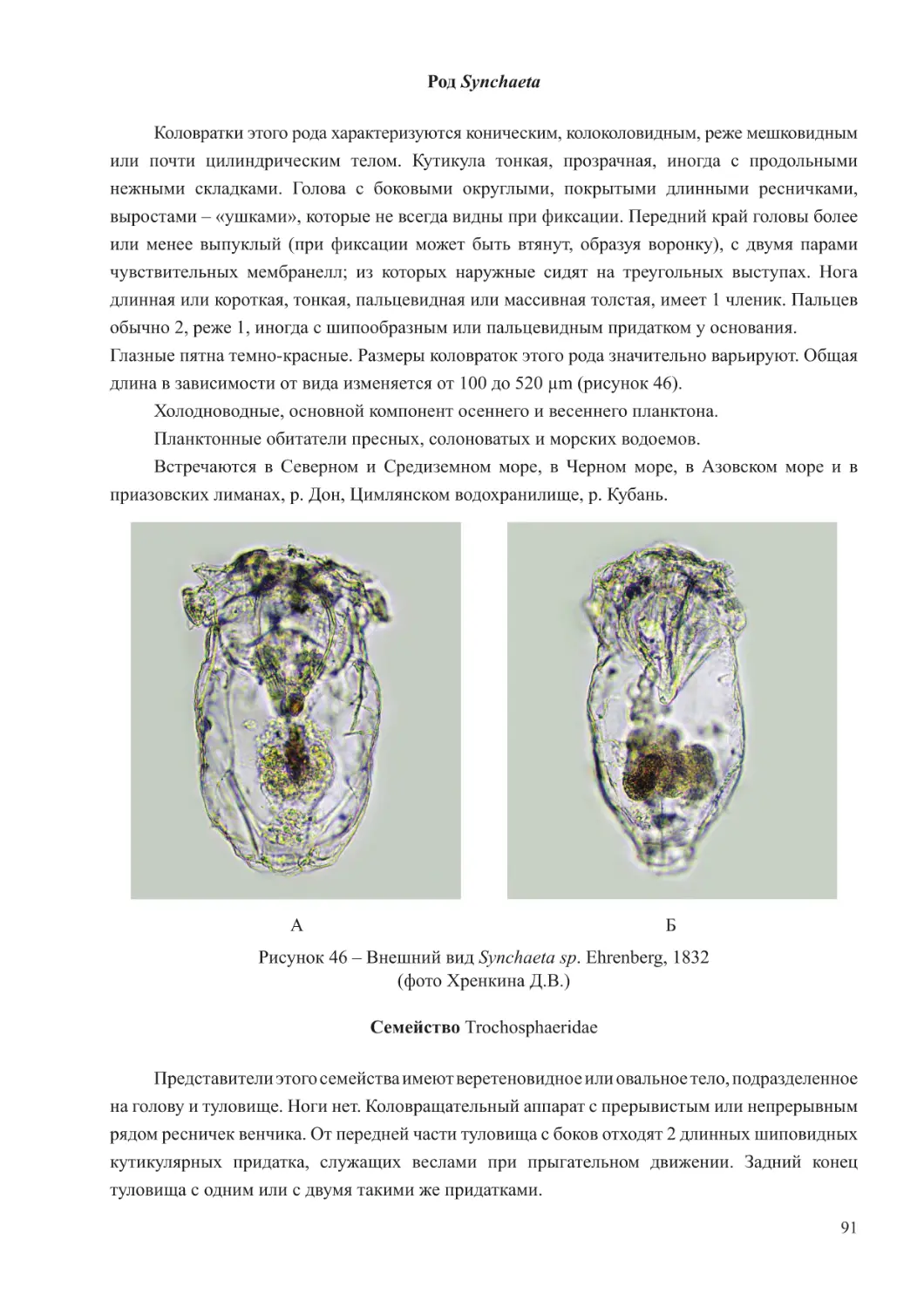













Author: Афанасьев Д.Ф. Хренкин Д.В. Мартынюк М.Л. Мирзоян З.А. Бычкова М.В. Шляхова Н.А. Кожурин Е.А.
Tags: общая экология биоценология гидробиология биогеография иудаизм ашкенази география зоология подводный мир
ISBN: 978-5-904063-32-0
Year: 2020




ФГБНУ «Всероссийский научно-исследовательский институт
рыбного хозяйства и океанографии»
Азово-Черноморский филиал
Афанасьев Д.Ф., Хренкин Д.В., Мартынюк М.Л., Мирзоян З.А.,
Бычкова М.В., Шляхова Н.А., Кожурин Е.А.
Иллюстрированный атлас массовых
видов зоопланктона Азовского
и Черного морей
Ростов-на-Дону
2020
УДК 574.583(035)(262.5+262.54)
Издается по решению Ученого совета Азово-Черноморского филиала ФГБНУ «ВНИРО»
(«АзНИИРХ»)
Рецензенты:
кандидат биологических наук А.Д. Губанова
кандидат биологических наук, доцент С.И. Решетников
Научный редактор:
кандидат биологических наук, доцент Д.Ф. Афанасьев
Афанасьев, Д.Ф.
А 94 Иллюстрированный атлас массовых видов зоопланктона Азовского и Черного морей /
Д.Ф. Афанасьев, Д.В. Хренкин, М.Л . Мартынюк, З.А . Мирзоян, М.В. Бычкова, Н.А . Шляхова,
Е.А. Кожурин.
–
Ростов-на-Дону: Азово-Черноморский филиал ФГБНУ «ВНИРО»
(«АзНИИРХ»), 2020. – 112 с.: цв. ил.
ISBN 978-5-904063-32-0
В иллюстрированном атласе массовых видов зоопланктона, встречающихся в Азовском
и Черном морях, приводятся описания и фотографии 55 видов. Кратко характеризуется их
распространение, отношение к солености, температуре и другим абиотическим факторам.
Атлас предназначен для научных сотрудников, работающих в области рыбного хозяйства,
аспирантов и студентов биологических специальностей.
Ключевые слова: зоопланктон, Азовское море, Черное море, копеподы, кладоцеры,
коловратки, меропланктон.
ISBN 978-5-904063-32-0
© Коллектив авторов, 2020
© АзНИИРХ, 2020
3
DDC 574.583(035) (262.5+262.54)
Reviewers:
A.D. Gubanova, PhD of Biology
S.I. Reshetnikov, PhD of Biology, Associate Professor
Scientific editor:
D.F. Afanasyev, PhD of Biology, Associate Professor
ISBN 978-5 -904063-32-0
Afanasyev D.F., Khrenkin D.V., Martynyuk M.L., Mirzoyan Z.A., Bychkova M.V., Shlyakhova N.A.,
Kozhurin E.A.
Illustrated Atlas of mass species of zooplankton of the Azov and Black Seas.
In the illustrated atlas of mass species of zooplankton, found in the Sea of Azov and the Black
Sea, descriptions and photographs of 55 species are given. Their distribution, relation to salinity,
temperature and other abiotic factors are briefly characterized. The atlas is intended for researchers
working in the field of fisheries, graduate students and students of biological specialities.
Key words: zooplankton, Sea of Azov, Black Sea, copepods, cladocerans, rotifers, meroplankton.
ISBN 978-5-904063-32-0
DDC 574.583(035)(262.5+262.54)
© Groupe of authors, 2020
© AzNIIRKH, 2020
4
ПРЕДИСЛОВИЕ
Одним из основных факторов, определяющих полноценный рост и развитие промысловых
рыб, особенно на ранних стадиях онтогенеза, является трофность водоема, степень развития
кормовой базы. Таким образом, оценка количественных показателей развития и идентификация
кормовых организмов является важнейшей рыбохозяйственной задачей. Следует отметить
малочисленность современных пособий, в том числе хорошо иллюстрированных атласов,
для обучения молодых специалистов-гидробиологов. Настоящим атласом массовых видов
зоопланктона Азовского и Черного морей мы надеемся частично заполнить этот пробел.
Цифровая фотосъемка массовых видов зоопланктонных организмов была проведена
коллективом лаборатории гидробиологии Азово-Черноморского филиала ФГБНУ «ВНИРО»
(«АзНИИРХ») в течение весны, лета и осени 2019 г. в рамках проведения прикладных научных
исследований по теме «Разработка и совершенствование инструментальных методов и
технических средств учета численности и управления поведением гидробионтов» для создания
базы цифровых изображений гидробионтов Черного и Азовского морей в целях разработки
средств для оценки характеристик мезозопланктона in situ».
Распространение видов и их описание приводится как по собственным данным, так и
по литературным источникам (Рылов, 1930, 1948; Бенинг, 1941; Боруцкий, 1960; Мордухай-
Болтовской, 1968, 1969, 1972; Кутикова, 1970, 1977; Набережный, 1984; Боруцкий, Степанова,
Кос, 1991 и др.). Названия видов приведены согласно World Register of Marine Species (WoRMS)
(http://www.marinespecies.org/).
5
ВВЕДЕНИЕ
Черное море – внутриконтинентальный, глубоководный, соленый водоем, относящийся
к бассейну Атлантического океана. Среднее значение солености воды составляет 21,9 ‰,
в поверхностных водах она находится на уровне 17-18 ‰, что почти вдвое ниже таковой в
поверхностных водах Мирового океана (Черное..., 2019). Изолированные объемы почти пресной
воды формируются вблизи устьев рек в период паводков, высокосоленые средиземноморские
воды проникают с нижнебосфорским течением. Большие глубины моря, ограниченный
водообмен с Мировым океаном, интенсивный приток пресных вод, наличие сероводородной
зоны определяют главную особенность моря – устойчивую вертикальную стратификацию
вод и отсутствие жизни на глубинах более 200 м, за исключением ряда анаэробных бактерий
(Проект «Моря СССР», 1992; Черное..., 2019).
Азовское море расположено на юге Европейской части Российской Федерации,
является полузамкнутым внутренним водоемом, входящим в Средиземноморскую систему
морей. Мелководность этого водоема (средняя глубина составляет 8,5 м, а максимальная –
14 м) обуславливает динамичность его термического режима. Температура воды изменяется
от отрицательных значений в зимний период до 25-26 oС в июле-августе. Для этого моря
характерны резкие колебания температуры воды во времени: в течение нескольких суток она
может снизиться или возрасти сразу на несколько градусов (Бронфман и др., 1979; Воловик,
1985; Гребневик..., 2000).
Азовское море – солоноватоводный водоем. Колебания его солевого режима определяются
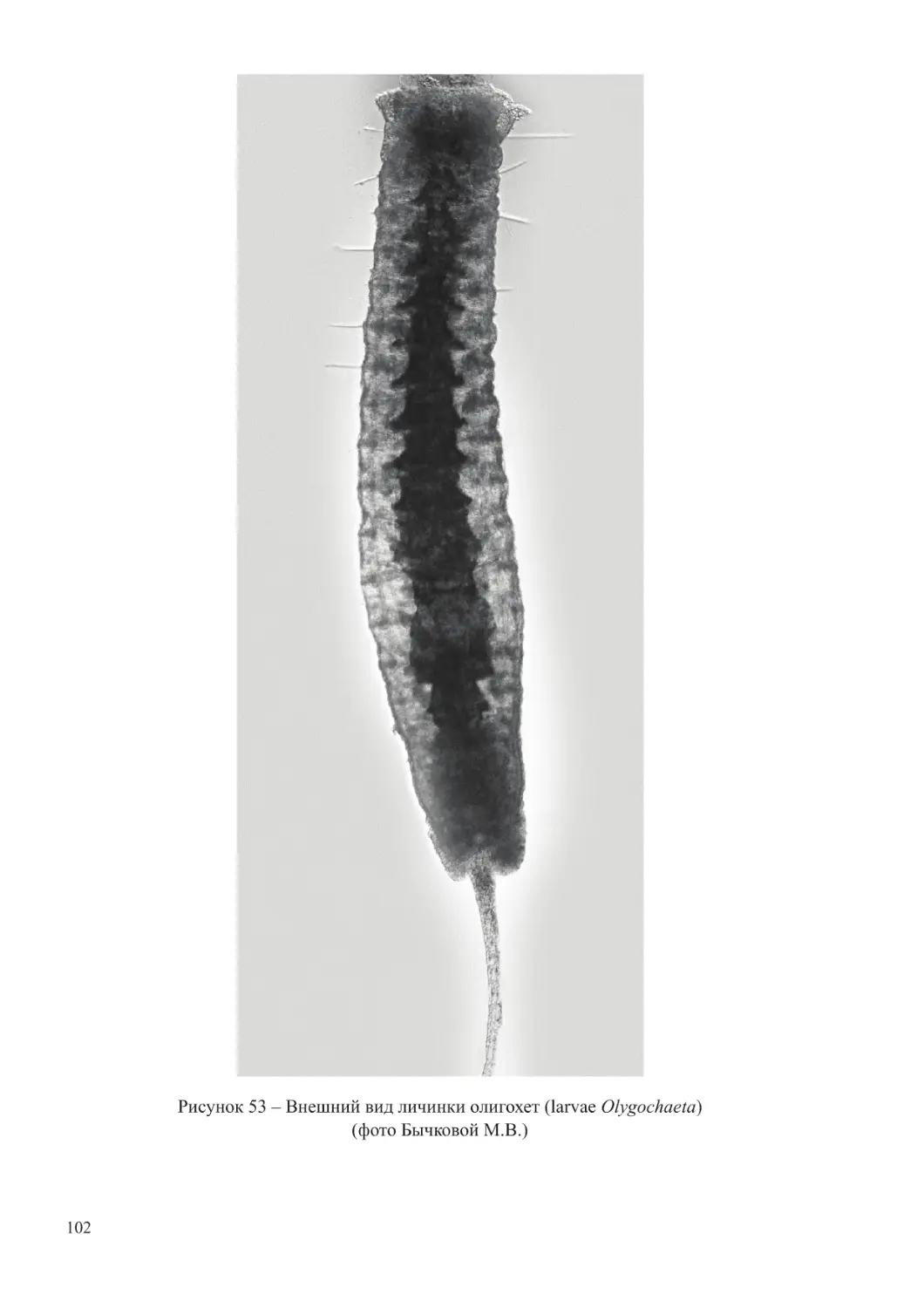
увлажненностью Азовского бассейна и флюктуациями речного стока. В годы продолжительной
многоводной фазы средняя соленость воды в Азовском море невысокая. В собственно море она
составляет около 11 ‰, в Таганрогском заливе – 6,5 ‰. При недостаточной обеспеченности
материковым стоком соленость Азовского моря заметно повышается: в открытой части моря
она возрастает в среднем до 13 ‰, в Таганрогском заливе – до 9,5 ‰. В отдельные годы
маловодных фаз соленость может быть несколько выше этих значений (Бронфман и др., 1979;
Жукова, Дубинина, 2018).
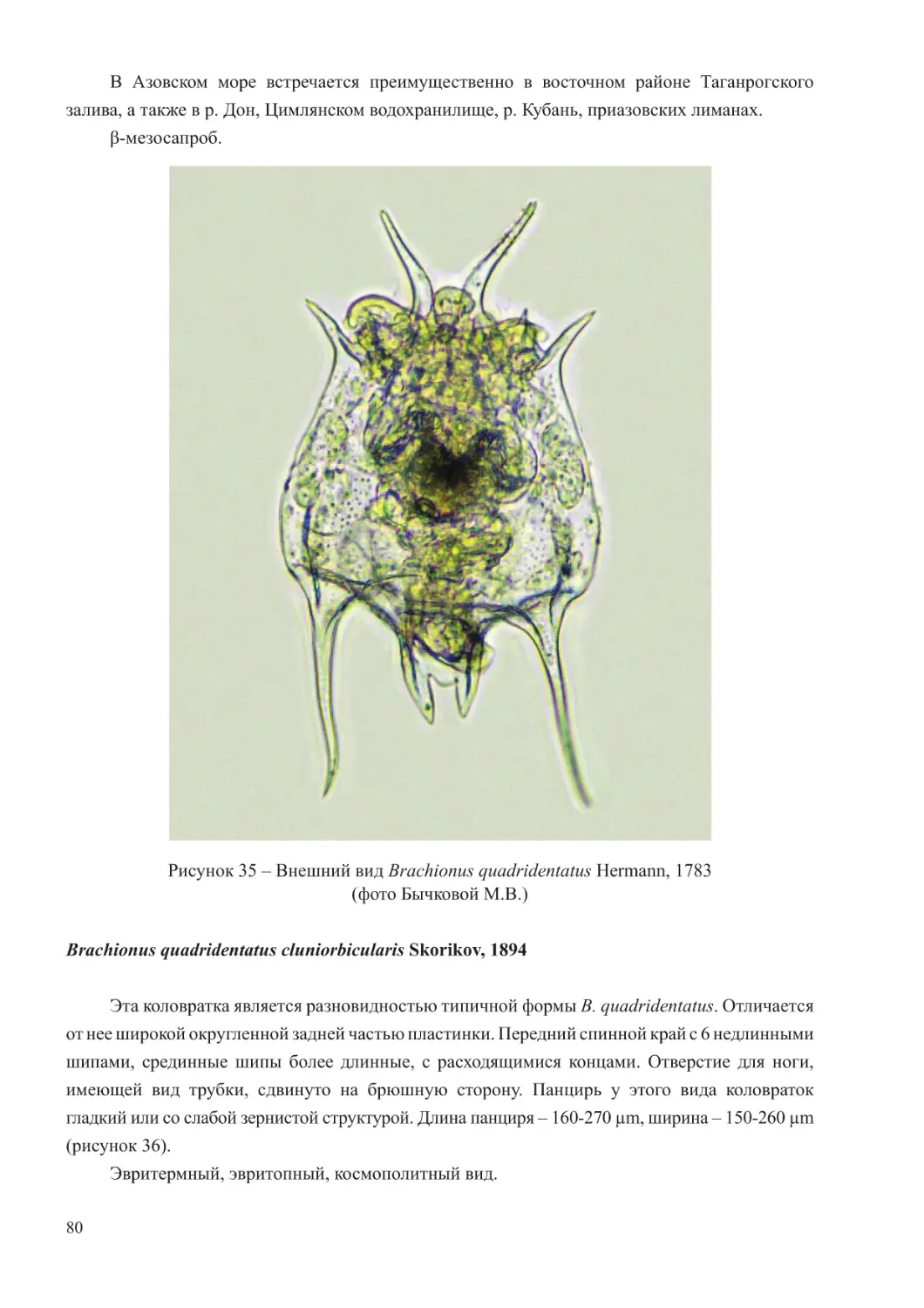
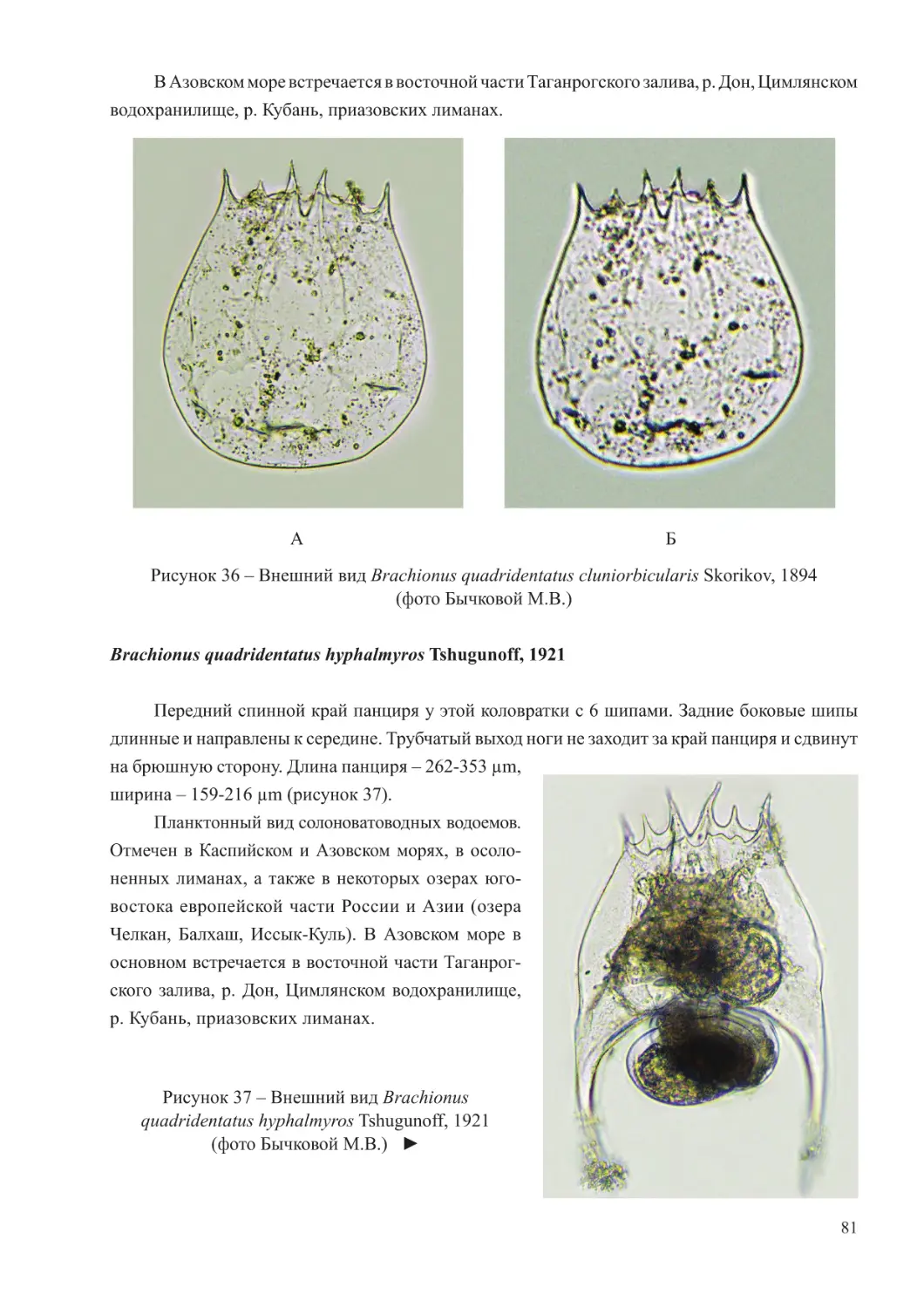
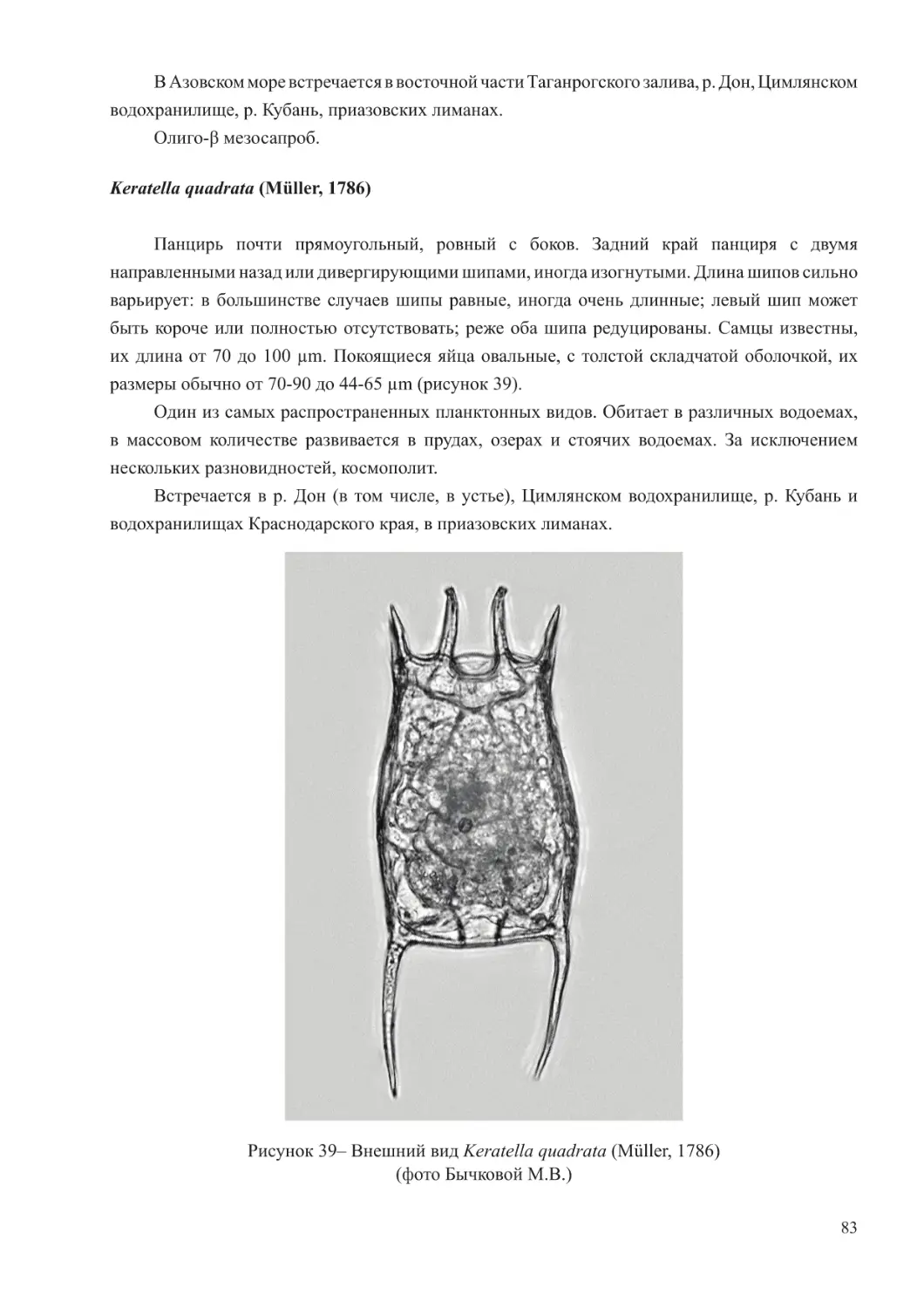
Краткая характеристика зоопланктона Черного и Азовского морей
Для зоопланктона Черного моря характерно относительно невысокое биологическое
разнообразие. Его состав насчитывает около 100 видов эупланктонных организмов,
значительная часть которых имеет ограниченное распространение, приуроченное к
районам с меньшей соленостью, нежели основные воды этого водоема (Грезе, 1979;
Ковалев, 1991).
Черноморскую планктонную фауну формируют простейшие, коловратки, ракообразные
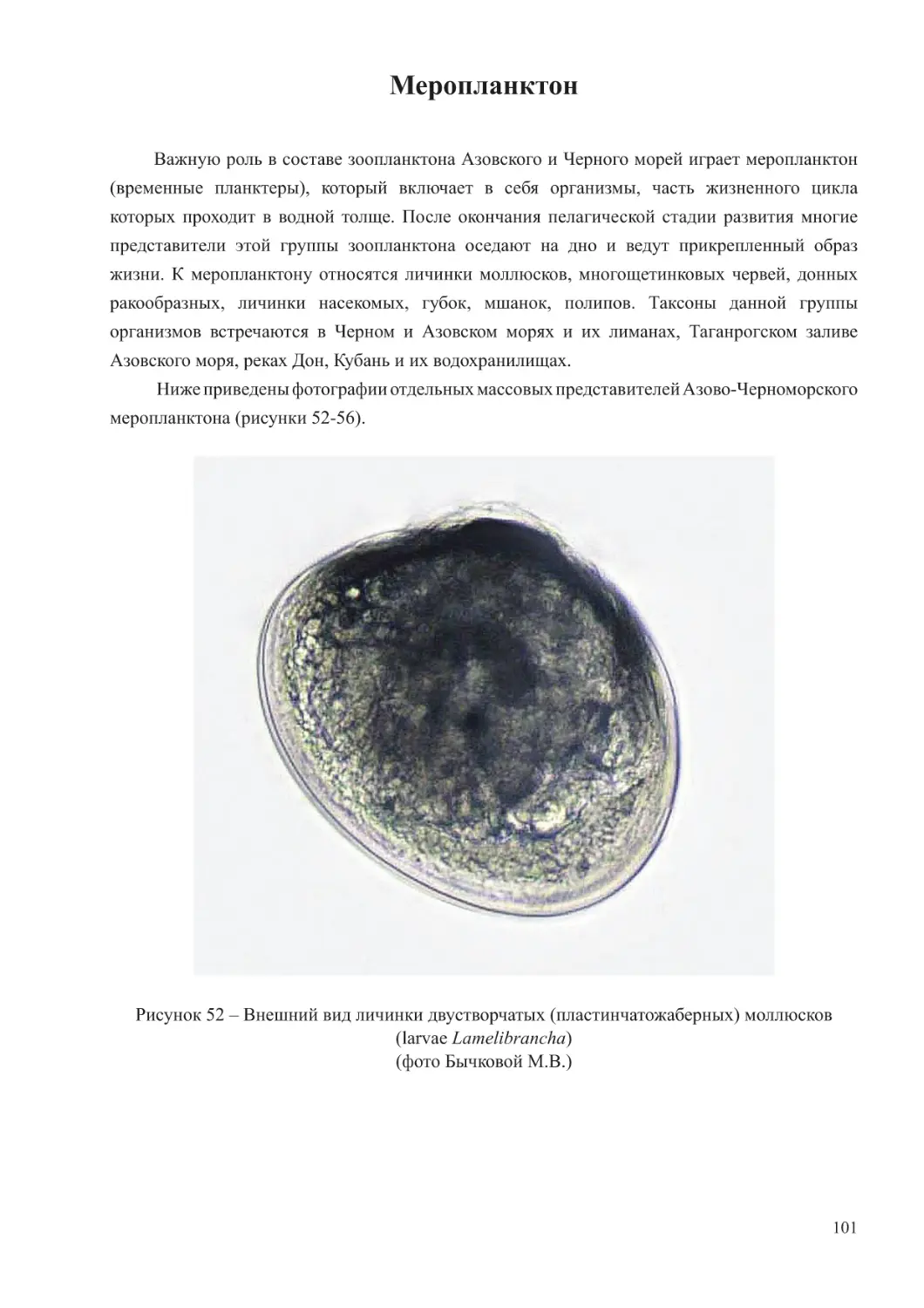
(копеподы и кладоцеры), щетинкочелюстные, аппендикулярии, меропланктон, а также
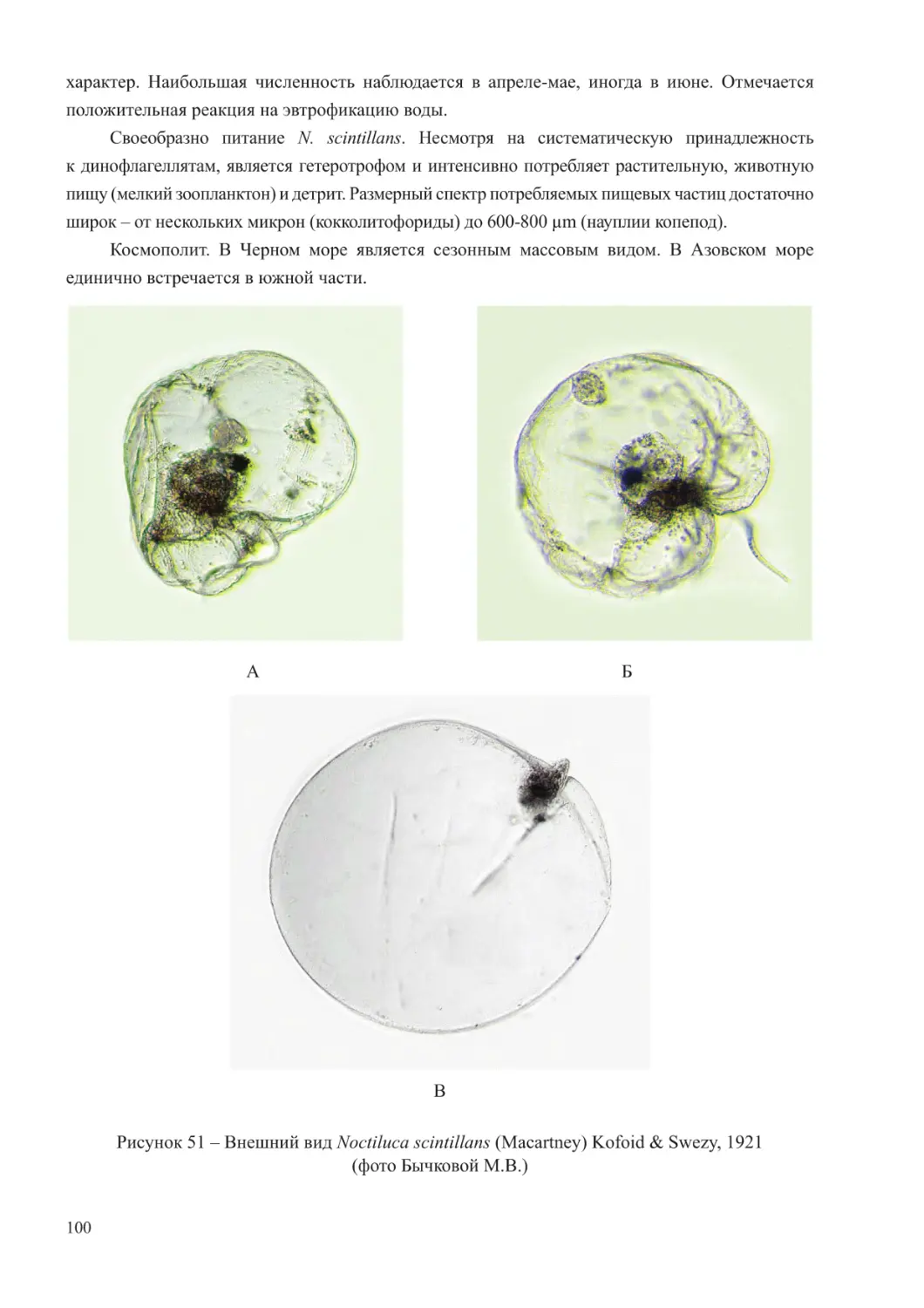
гребневики и медузы. Гетеротрофный вид динофлагеллят Noctiluca scintillans из-за крупного
размера и всеядного типа питания также рассматривается как компонент мезозоопланктона
(Грезе, 1979; Арашкевич и др., 2015).
6
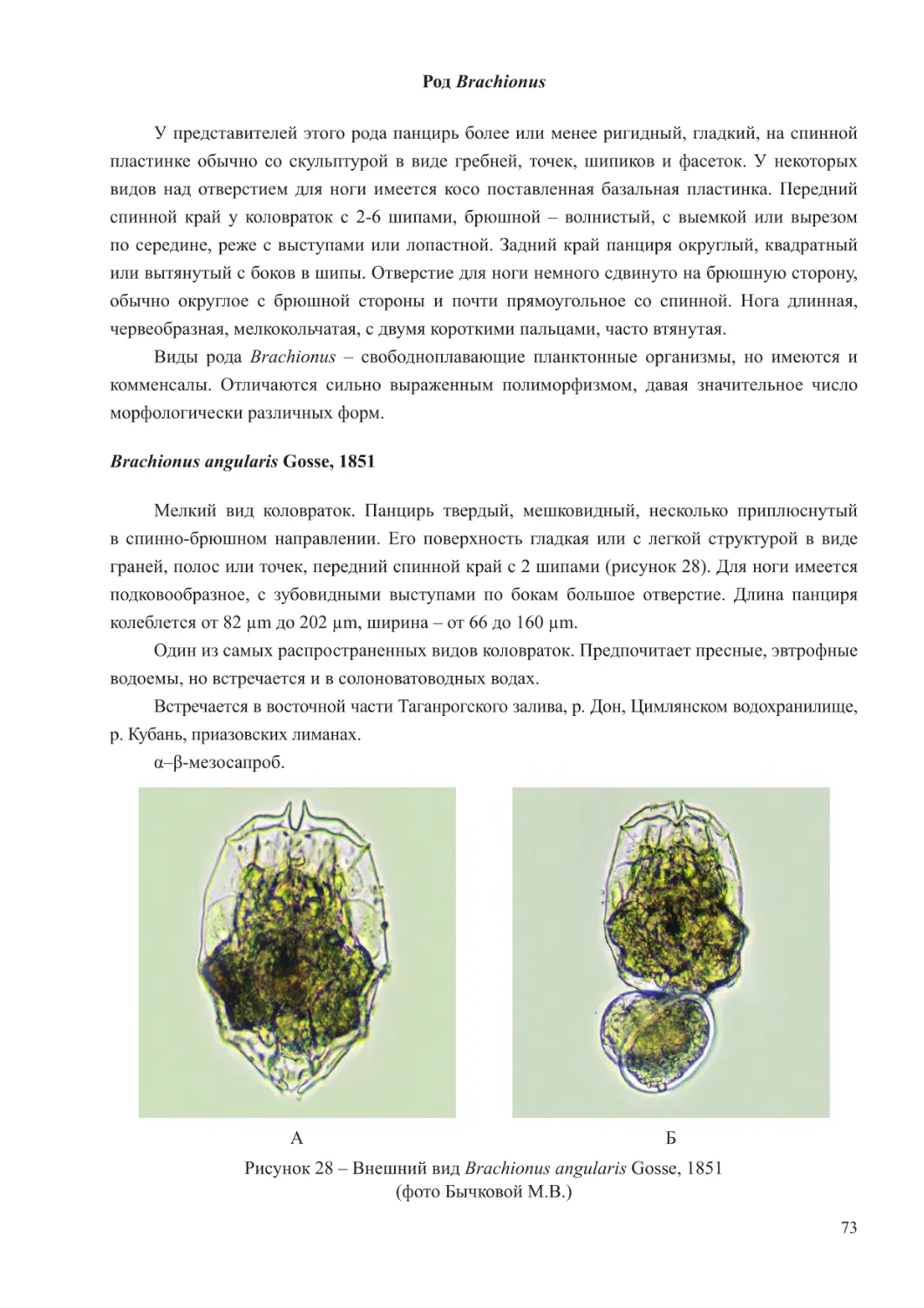
В составе планктона Черного моря выделяются три экологических комплекса видов –
холодоводный, тепловодный и эвритермный. К первой группе относятся копеподы: Calanus
euxinus, Pseudocalanus elongatus, Oithona similis, населяющие не только более холодные
глубинные слои моря, но обитающие также и в поверхностных горизонтах в холодное
время года. Теплолюбивый комплекс планктона включает такие виды кладоцер, как Penilia
avirostris, Pseudevadne tergestina, Evadne spinifera, Pleopis polyphemoides и копепод –
Pontella mediterranea, Labidocera brunescens, Centropages ponticus, Oithona davisae, Acartia
(Acanthacartia) tonsa, которые в массе развиваются в теплое время. Ряд видов проявляют
черты эвритермности и встречается в море во все сезоны года. К таковым относятся
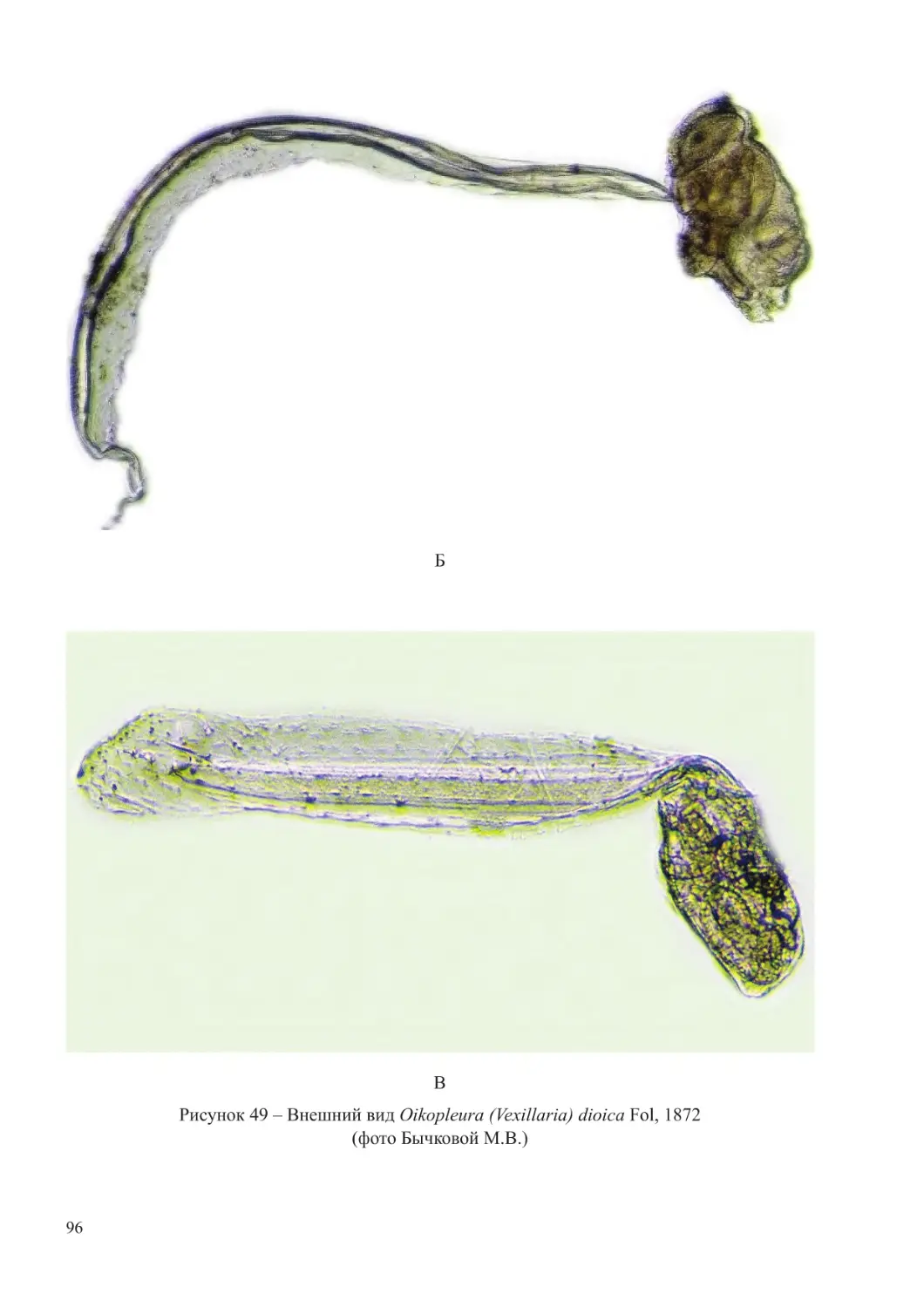
Paracalanus parvus, Acartia (Acartiura) clausi, Oithona nana, Oikopleura (Vexillaria) dioica,
Parasagitta setosa (Грезе, 1979; Губанова, 2000; Прусова и др., 2002; Губарева и др., 2016;
Серегин и др., 2016).
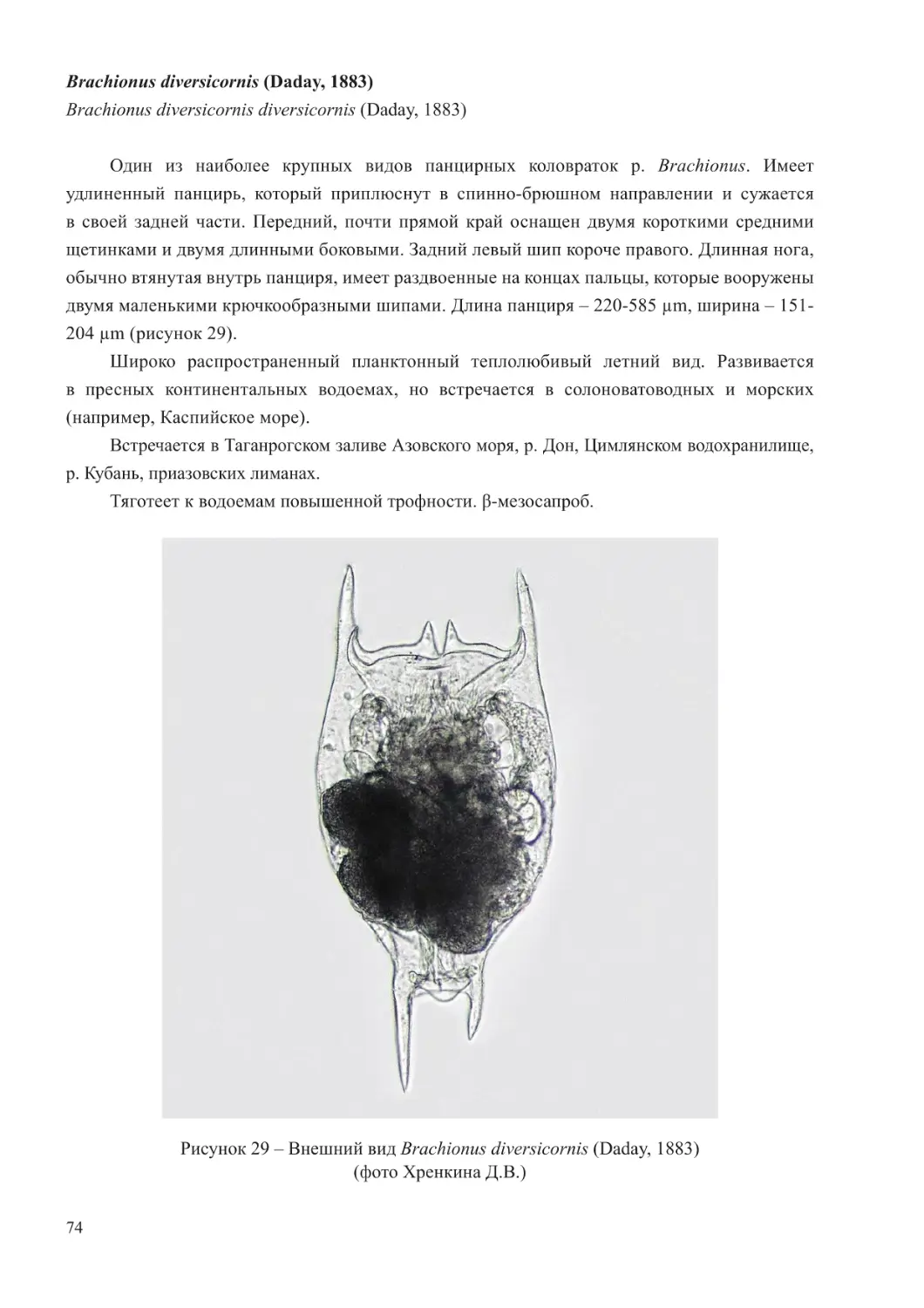
Состав и численность зоопланктона в Черном море претерпевают существенные
изменения в течение года, связанные с сезонной динамикой в развитии видов. В зимний
период в составе сообщества отмечается небольшое число таксонов холодоводного комплекса,
формирующих достаточно высокую численность. Весной интенсивность развития этой группы
значительно снижается, на смену ей появляются эвритермные виды. В летнее время видовое
разнообразие зоопланктона значительно возрастает. В планктоне появляются теплолюбивые
виды, продолжают достаточно активно развиваться эвритермные, отмечается массовое
размножение большинства бентосных животных и появление в планктоне их личиночных
стадий. В результате в прибрежных районах наибольшие показатели развития зоопланктона
отмечаются в августе-сентябре. Значительную роль в формировании этой вспышки играет
развитие популяции кладоцеры Penilia avirostris, а в последние годы – и нового для Черного
моря вида копепод O. davisae. С понижением температуры воды исчезают теплолюбивые
формы планктона и остаются круглогодичные, к которым присоединяются холодолюбивые
животные (Коваль, 1968; Грезе, 1979).
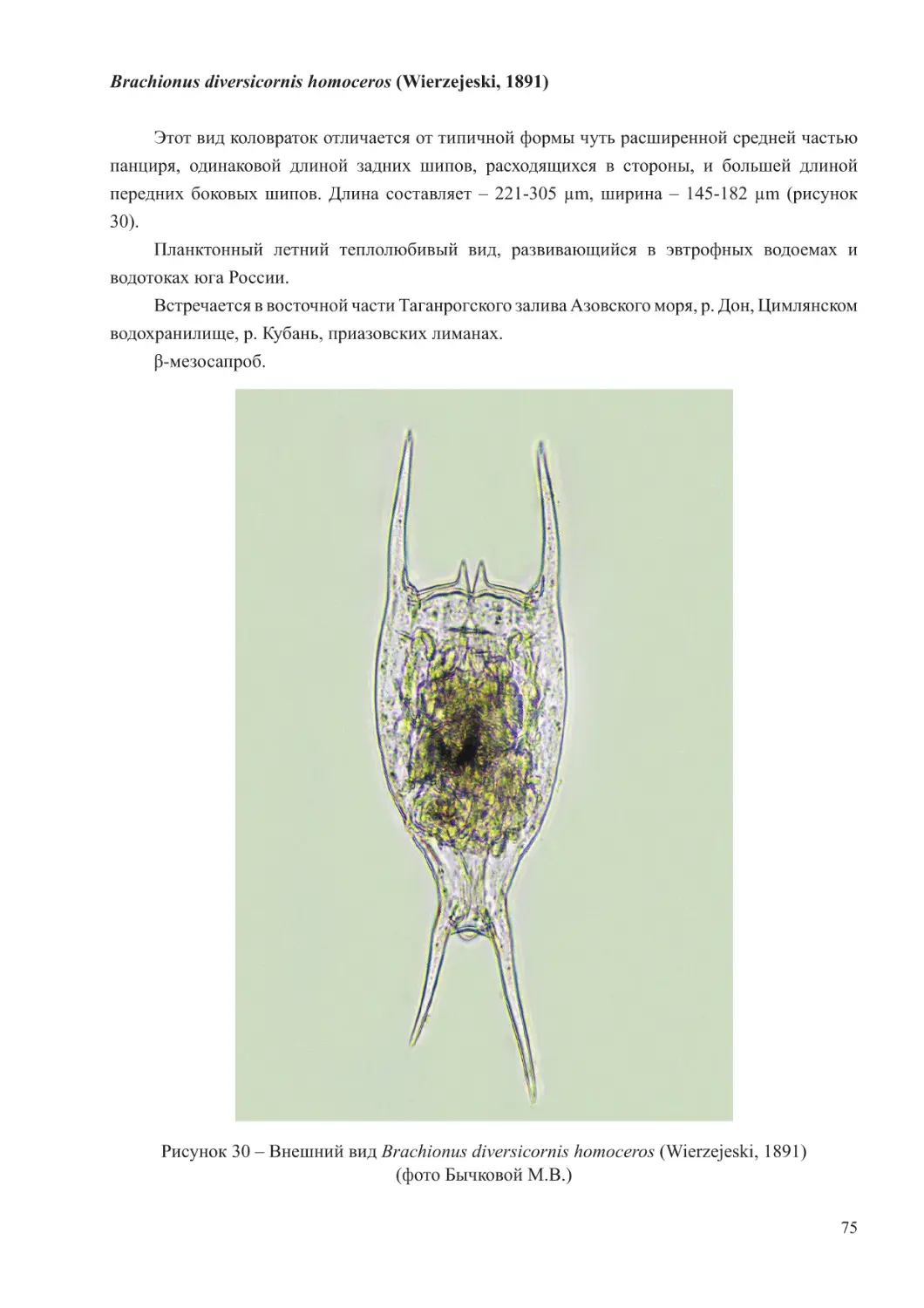
Зоопланктон Азовского моря имеет пресноводное, солоноватоводное и морское
происхождение. При оптимальном водном режиме моря состав сообществ включал около 150
видов. В настоящее время, в условиях зарегулирования стока, здесь встречается немногим
более 70 таксонов (Воловик, 1985). В это число входят свободноживущие представители
членистоногих (Arthropoda), относящиеся к двум классам ракообразных (Branchiopoda и
Hexanauplia), коловратки (Eurotatoria), инфузории (Protozoa), кишечнополостные (Coelenterata),
гребневики (Ctenophora) и пелагические личинки моллюсков и червей. Щетинкочелюстные
(Chaetognatha) и хордовые (Hordata) в Азовском море отмечаются в единичных экземплярах
при высокой солености в южной части (Мирзоян, 2000).
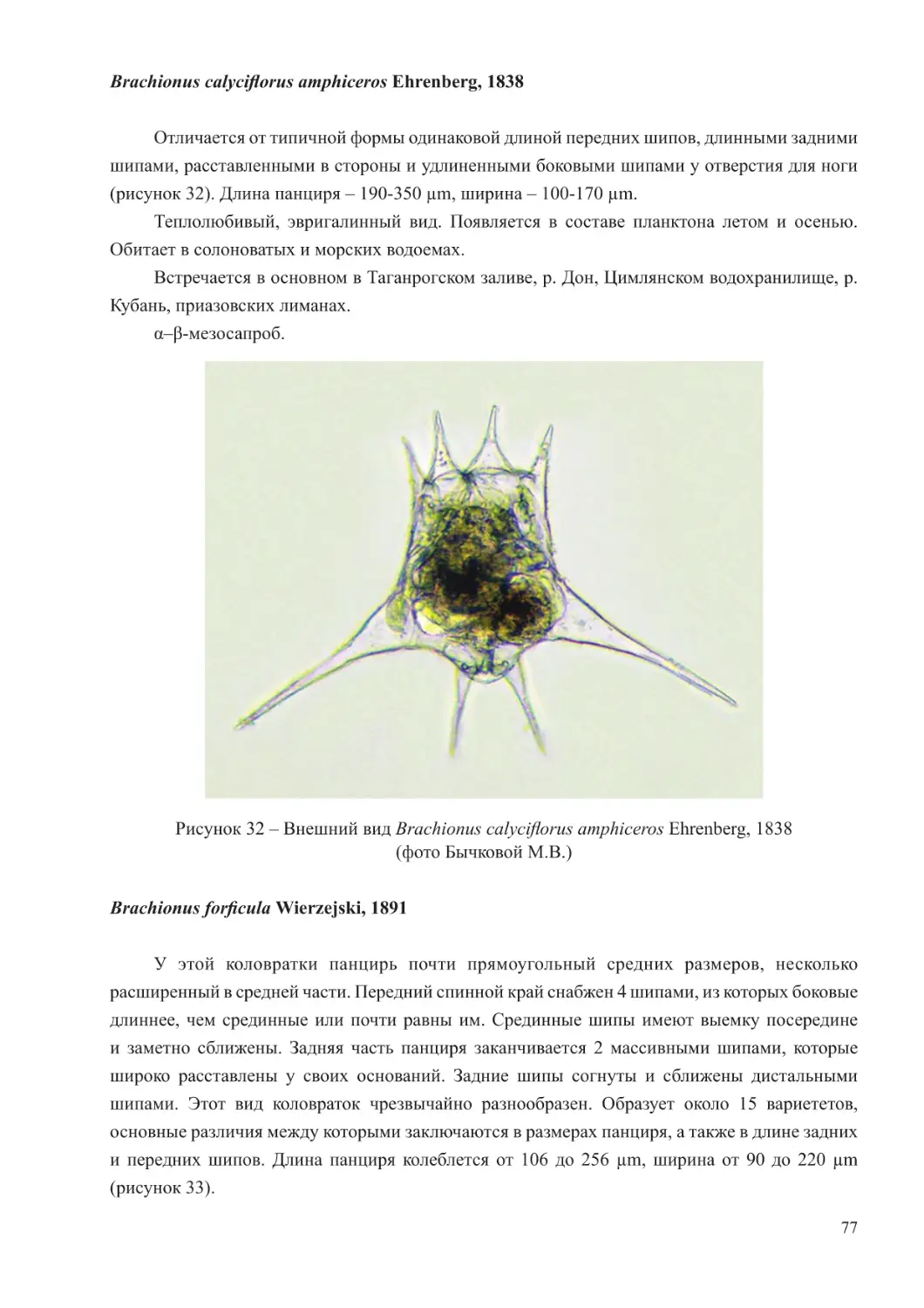
Планктонная фауна Азовского моря весьма разнообразна в районах, отличающихся
соленостью. Так, Таганрогский залив под влиянием стока р. Дон имеет пониженную соленость,
которая возрастает в направлении от устья реки Дон до выхода в открытую часть моря
(Бронфман и др., 1979). Такая особенность солевого режима определяет в заливе сложный
состав зоопланктона. По мере продвижения в собственно море наблюдается снижение числа
7
видов пресноводного комплекса, таких, как Cyclops strenuus, C. vicinus, Mesocyclops leucarti,
Cornigerius maeoticus, Bosmina longirostris, Chydorus sphaericus, коловраток р. Brachionus, и
появление эвригалинных и морских форм, основными представителями которых являются
виды копепод родов Eurytemora, Calanipeda, Acartia.
Собственно море осваивают в основном эвригалинные виды копепод морского комплекса,
например, относящиеся к р. Acartia, коловратки р. Synchaeta, временные обитатели пелагиали
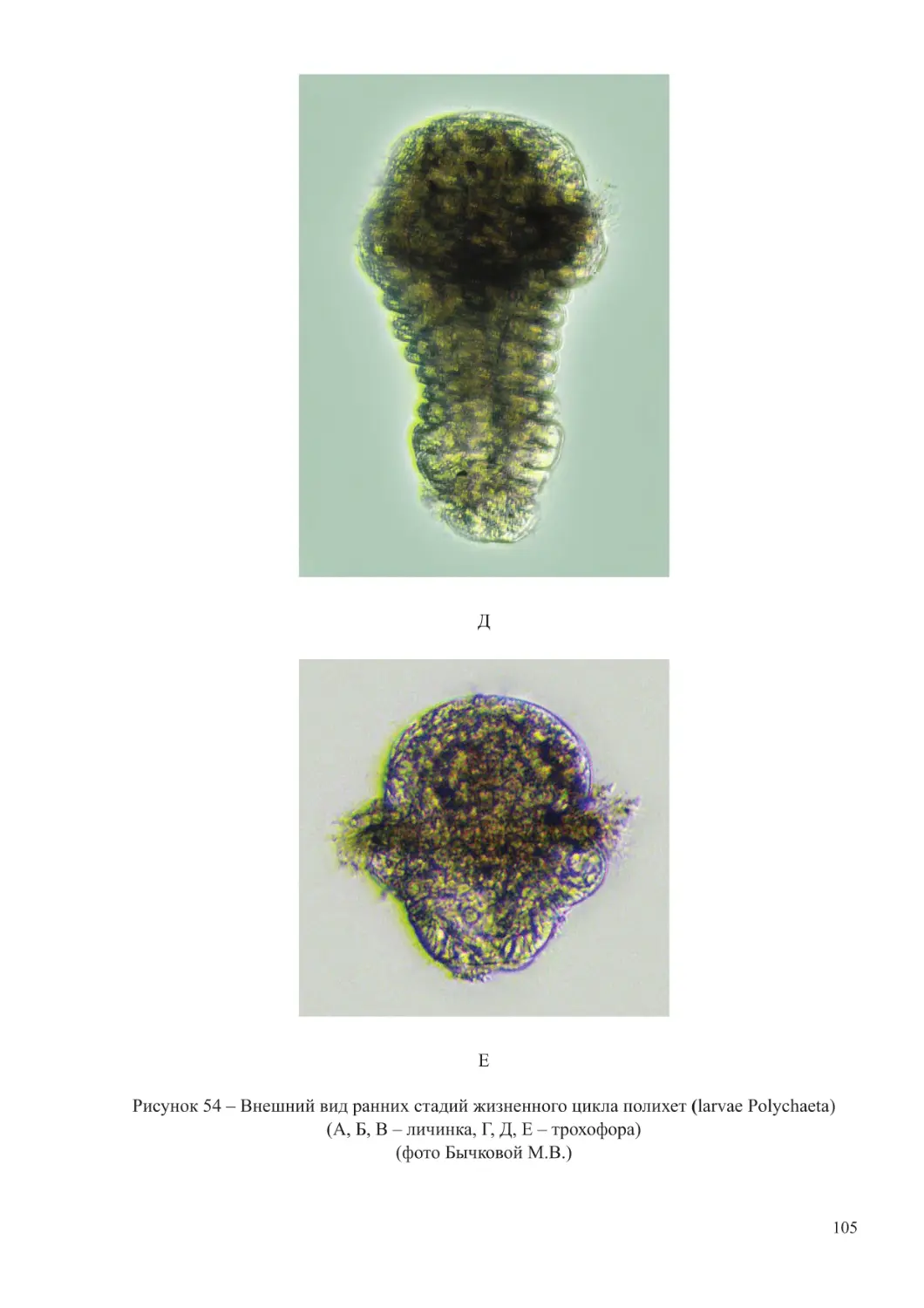
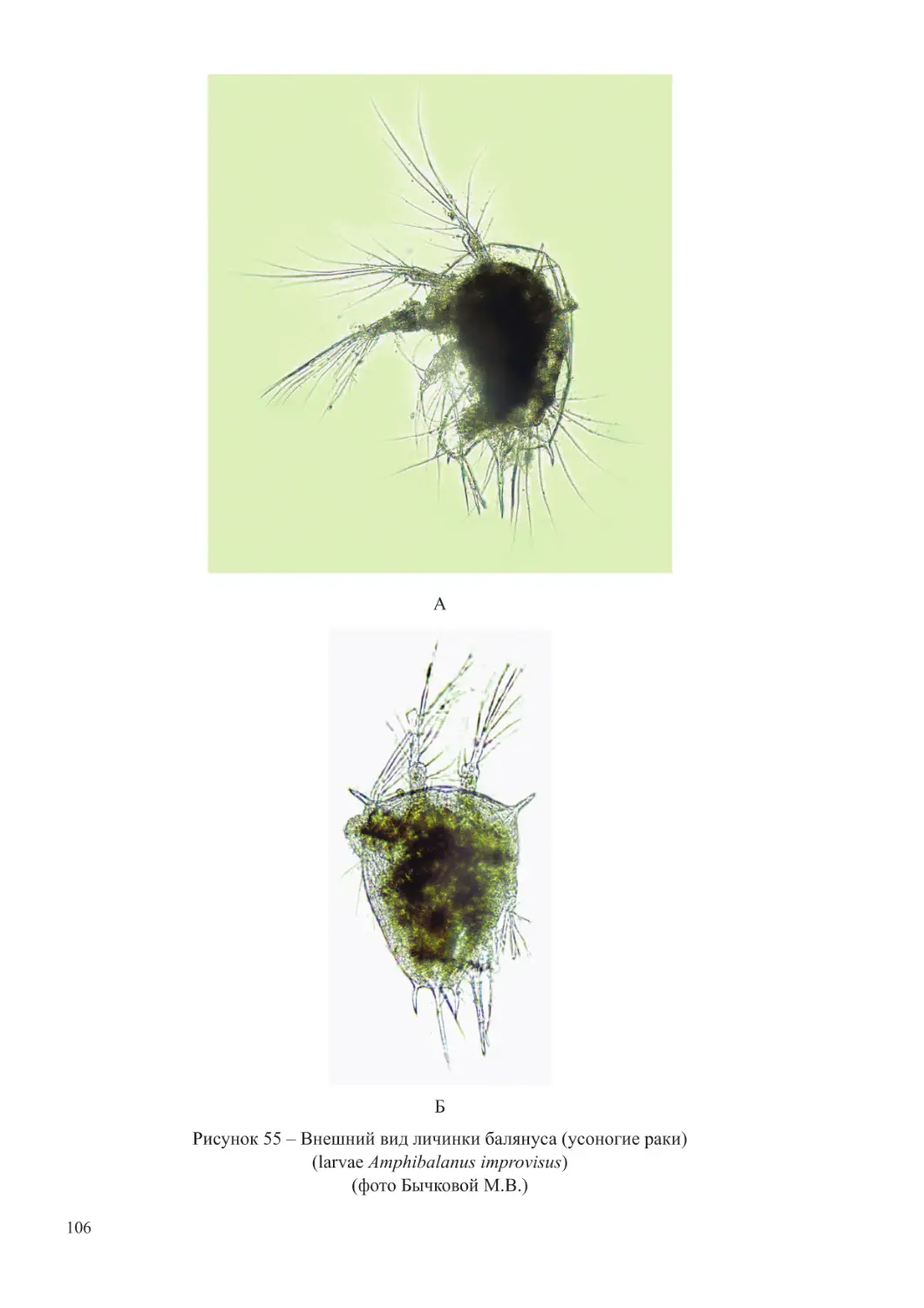
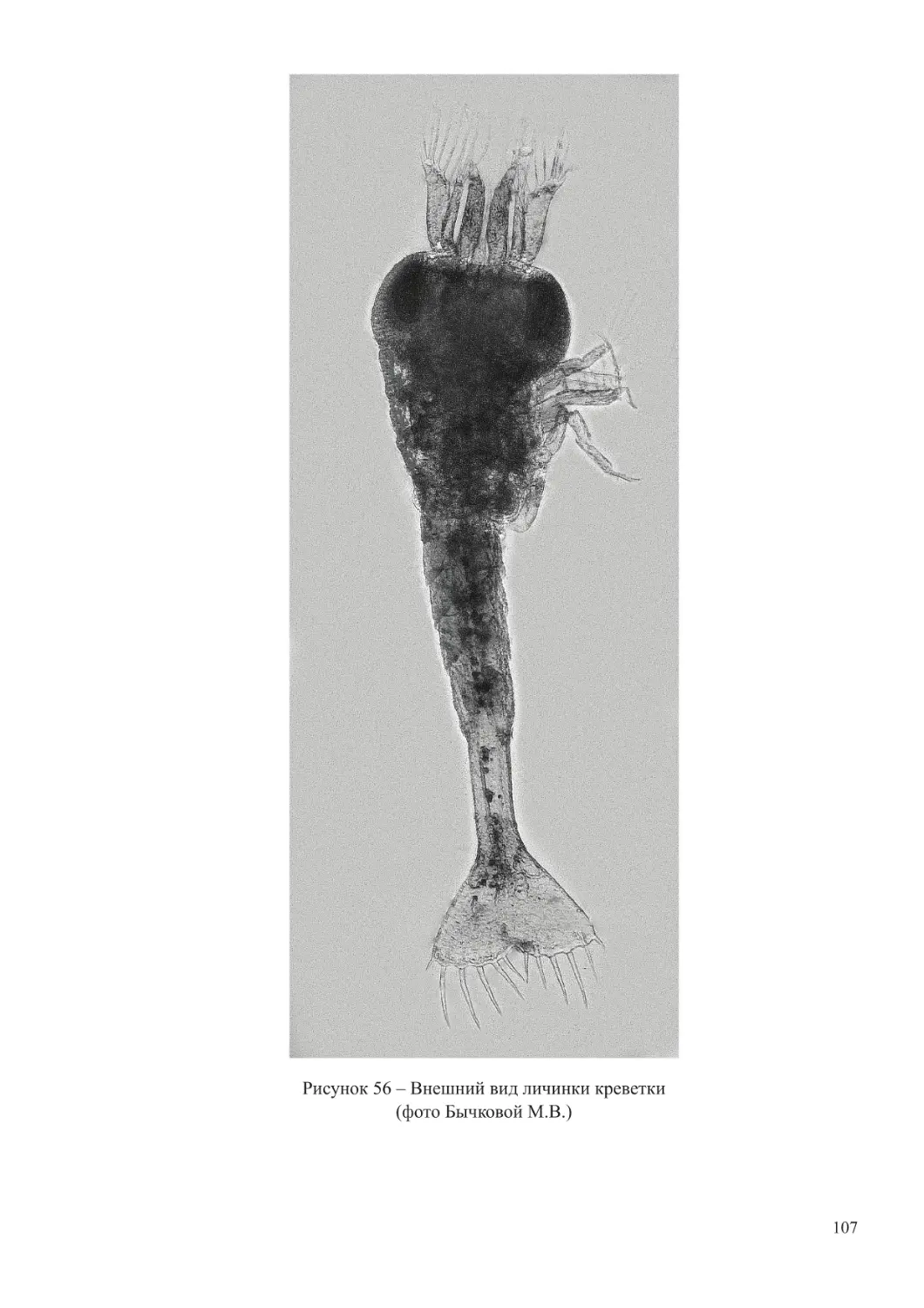
(личинки морских видов усоногих ракообразных, полихет, моллюсков). При повышении
солености в южной части моря появляются чисто морские иммигранты из Черного моря
(Алдакимова и др., 1977; Воловик, 1985).
Зоопланктону Азовского моря свойственны достаточно большие качественные и
количественные изменения, связанные с сезонностью развития организмов. В собственно
море зимой планктон очень беден, состоит из зимующих форм копепод и коловраток. Весной
сохраняется такой же видовой состав, но более многочисленными становятся коловратки.
В конце весны – начале лета активизируется размножение копепод, интенсивно развивается
меропланктон. С появлением в конце 80-х гг. ХХ века в Азовском море хищного гребневика
Mnemiopsis leidyi второй летний копеподный пик развития зоопланктона более не наблюдается,
а сезонная динамика его биомассы в последние десятилетия имеет вид одновершинной
кривой. Осенью в составе азовского планктона вновь возрастает доля холодолюбивых
коловраток.
Общая характеристика морфологии ракообразных и коловраток
В разделе приводится описание характерных черт строения основных групп планктонных
животных, почерпнутое из различных источников (Рылов, 1948; Мануйлова, 1964; Мордухай-
Болтовской; 1969; Кутикова, 1970; Алексеева, Цалолихина, 2010).
Ракообразные – наиболее богатая видами группа планктонных животных в Черном и
Азовском морях, внешний вид которых очень разнообразен.
Cladocera (ветвистоусые ракообразные) имеют короткое тело с плохо заметной или
совсем незаметной сегментацией. Большинство из них имеют двустворчатую хитиновую
раковинку, закрывающую только туловище и конечности. У некоторых видов раковинка по-
крывает только спинную сторону тела, а иногда совсем отсутствует. Голова имеет различные
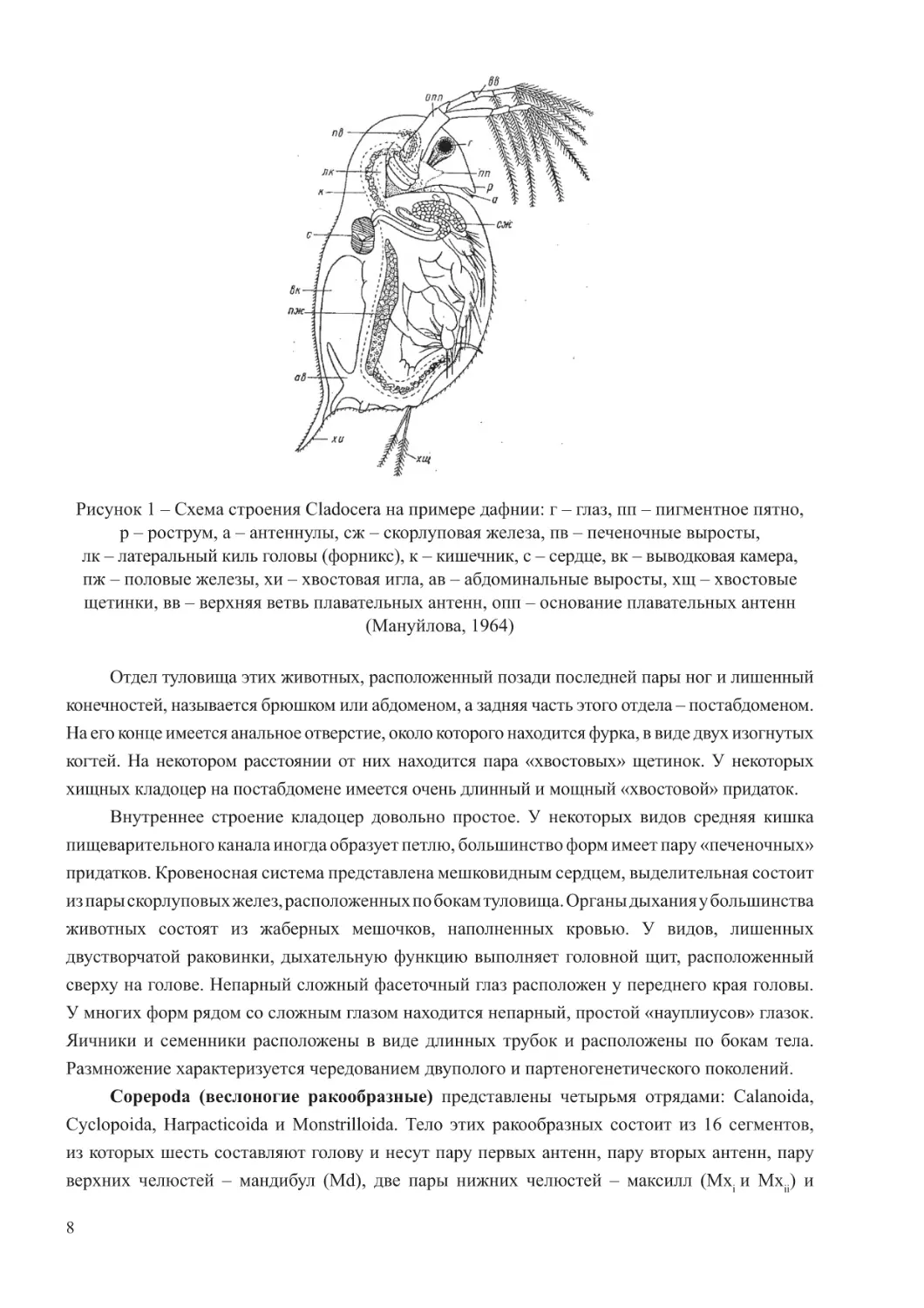
выросты и придатки (рострум, шлем, рога), глаза, две пары антенн и ротовые органы (рисунок 1).
Антенны I обычно короткие, с чувствительными образованиями. Антенны II сильно развиты и
служат органом передвижения (плавания). Туловище несет 4-6 пар конечностей листовидной
или палочковидной формы. У разных семейств кладоцер строение листовидных конечностей
в деталях различно, но во всех случаях они обеспечивают животным дыхание и питание.
Своим движением они вызывают сильный ток воды с питательными частицами, которые
отфильтровываются щетинками и продвигаются к ротовому отверстию. Палочковидные
конечности имеются лишь у хищных кладоцер и служат для схватывания и передачи к ротовому
отверстию пойманной добычи.
8
Рисунок 1 – Схема строения Cladocera на примере дафнии: г – глаз, пп – пигментное пятно,
р – рострум, а – антеннулы, сж – скорлуповая железа, пв – печеночные выросты,
лк – латеральный киль головы (форникс), к – кишечник, с – сердце, вк – выводковая камера,
пж – половые железы, хи – хвостовая игла, ав – абдоминальные выросты, хщ – хвостовые
щетинки, вв – верхняя ветвь плавательных антенн, опп – основание плавательных антенн
(Мануйлова, 1964)
Отдел туловища этих животных, расположенный позади последней пары ног и лишенный
конечностей, называется брюшком или абдоменом, а задняя часть этого отдела – постабдоменом.
На его конце имеется анальное отверстие, около которого находится фурка, в виде двух изогнутых
когтей. На некотором расстоянии от них находится пара «хвостовых» щетинок. У некоторых
хищных кладоцер на постабдомене имеется очень длинный и мощный «хвостовой» придаток.
Внутреннее строение кладоцер довольно простое. У некоторых видов средняя кишка
пищеварительного канала иногда образует петлю, большинство форм имеет пару «печеночных»
придатков. Кровеносная система представлена мешковидным сердцем, выделительная состоит
из пары скорлуповых желез, расположенных по бокам туловища. Органы дыхания у большинства
животных состоят из жаберных мешочков, наполненных кровью. У видов, лишенных
двустворчатой раковинки, дыхательную функцию выполняет головной щит, расположенный
сверху на голове. Непарный сложный фасеточный глаз расположен у переднего края головы.
У многих форм рядом со сложным глазом находится непарный, простой «науплиусов» глазок.
Яичники и семенники расположены в виде длинных трубок и расположены по бокам тела.
Размножение характеризуется чередованием двуполого и партеногенетического поколений.
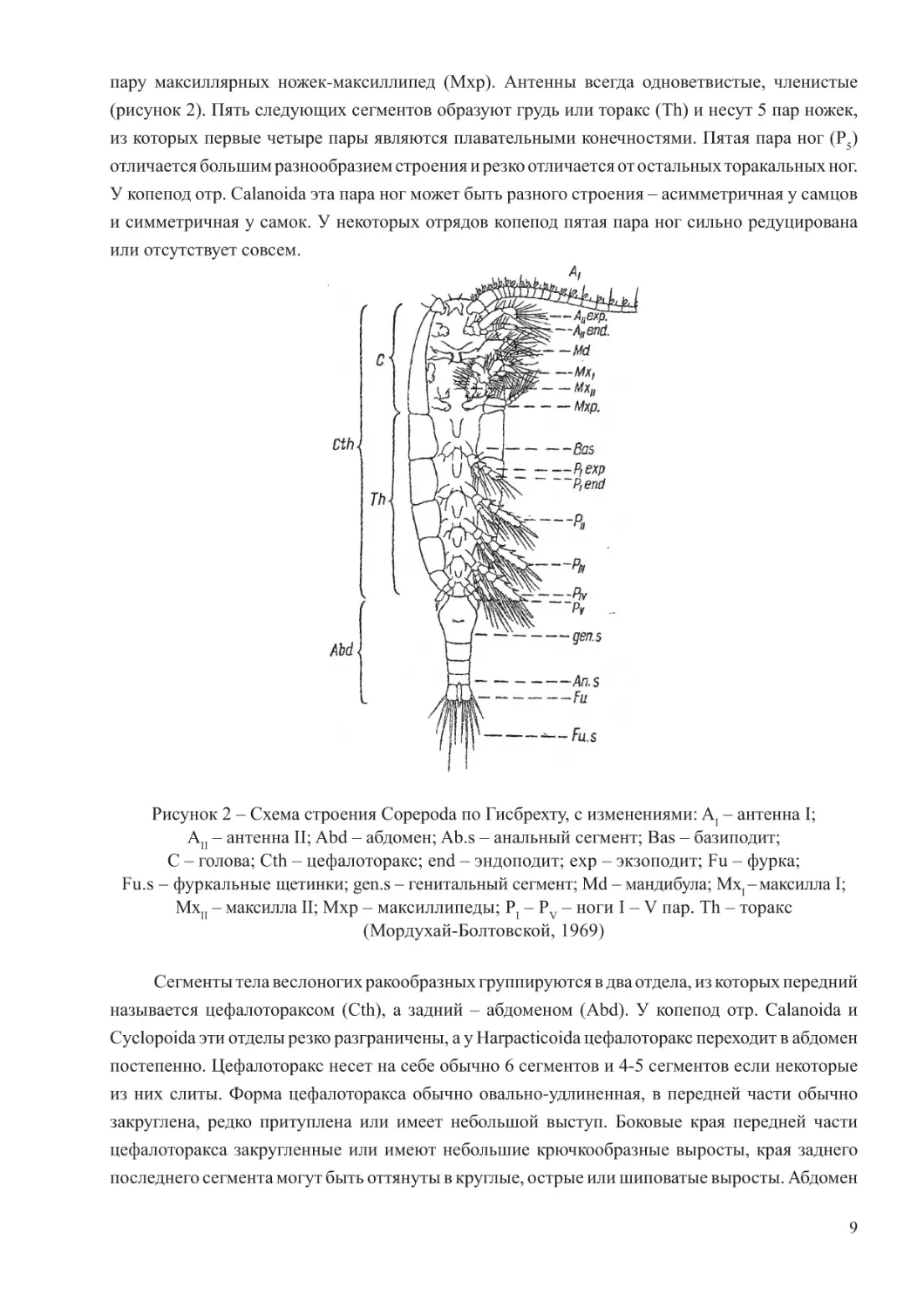
Copepoda (веслоногие ракообразные) представлены четырьмя отрядами: Calanoida,
Cyclopoida, Harpacticoida и Monstrilloida. Тело этих ракообразных состоит из 16 сегментов,
из которых шесть составляют голову и несут пару первых антенн, пару вторых антенн, пару
верхних челюстей – мандибул (Md), две пары нижних челюстей – максилл (Mxi и Mxii) и
9
пару максиллярных ножек-максиллипед (Mxp). Антенны всегда одноветвистые, членистые
(рисунок 2). Пять следующих сегментов образуют грудь или торакс (Th) и несут 5 пар ножек,
из которых первые четыре пары являются плавательными конечностями. Пятая пара ног (P5)
отличается большим разнообразием строения и резко отличается от остальных торакальных ног.
У копепод отр. Calanoida эта пара ног может быть разного строения – асимметричная у самцов
и симметричная у самок. У некоторых отрядов копепод пятая пара ног сильно редуцирована
или отсутствует совсем.
Рисунок 2 – Схема строения Copepoda по Гисбрехту, с изменениями: AI
–
антенна I;
AII
–
антенна II; Abd – абдомен; Ab.s – анальный сегмент; Bas – базиподит;
C – голова; Cth – цефалоторакс; end – эндоподит; exp – экзоподит; Fu – фурка;
Fu.s – фуркальные щетинки; gen.s – генитальный сегмент; Md – мандибула; MxI
– максилла I;
MxII
–
максилла II; Mxp – максиллипеды; PI
–
PV
–
ногиI–Vпар.Th–торакс
(Мордухай-Болтовской, 1969)
Сегменты тела веслоногих ракообразных группируются в два отдела, из которых передний
называется цефалотораксом (Cth), а задний – абдоменом (Abd). У копепод отр. Calanoida и
Cyclopoida эти отделы резко разграничены, а у Harpacticoida цефалоторакс переходит в абдомен
постепенно. Цефалоторакс несет на себе обычно 6 сегментов и 4-5 сегментов если некоторые
из них слиты. Форма цефалоторакса обычно овально-удлиненная, в передней части обычно
закруглена, редко притуплена или имеет небольшой выступ. Боковые края передней части
цефалоторакса закругленные или имеют небольшие крючкообразные выросты, края заднего
последнего сегмента могут быть оттянуты в круглые, острые или шиповатые выросты. Абдомен
10
у самок 2-4 сегментный, у самцов – 5 сегментный. К первому сегменту абдомена (генитальному)
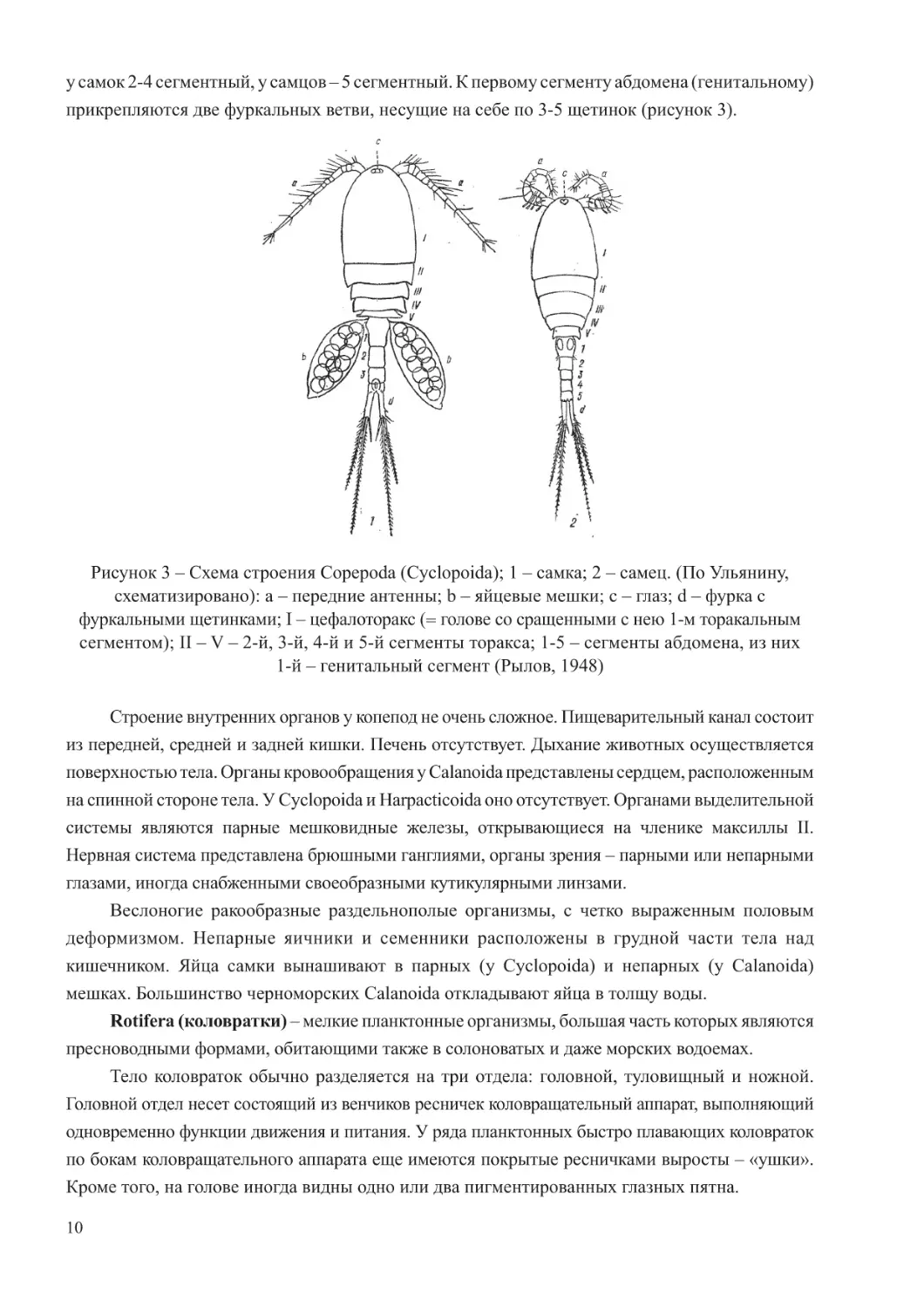
прикрепляются две фуркальных ветви, несущие на себе по 3-5 щетинок (рисунок 3).
Рисунок 3 – Схема строения Copepoda (Cyclopoida); 1 – самка; 2 – самец. (По Ульянину,
схематизировано): a – передние антенны; b – яйцевые мешки; c – глаз; d – фурка с
фуркальными щетинками; I – цефалоторакс (= голове со сращенными с нею 1-м торакальным
сегментом); II – V
–
2-й, 3-й, 4-й и 5-й сегменты торакса; 1-5
–
сегменты абдомена, из них
1-й
–
генитальный сегмент (Рылов, 1948)
Строение внутренних органов у копепод не очень сложное. Пищеварительный канал состоит
из передней, средней и задней кишки. Печень отсутствует. Дыхание животных осуществляется
поверхностью тела. Органы кровообращения у Calanoida представлены сердцем, расположенным
на спинной стороне тела. У Cyclopoida и Harpacticoida оно отсутствует. Органами выделительной
системы являются парные мешковидные железы, открывающиеся на членике максиллы II.
Нервная система представлена брюшными ганглиями, органы зрения – парными или непарными
глазами, иногда снабженными своеобразными кутикулярными линзами.
Веслоногие ракообразные раздельнополые организмы, с четко выраженным половым
деформизмом. Непарные яичники и семенники расположены в грудной части тела над
кишечником. Яйца самки вынашивают в парных (у Cyclopoida) и непарных (у Calanoida)
мешках. Большинство черноморских Calanoida откладывают яйца в толщу воды.
Rotifera (коловратки) – мелкие планктонные организмы, большая часть которых являются
пресноводными формами, обитающими также в солоноватых и даже морских водоемах.
Тело коловраток обычно разделяется на три отдела: головной, туловищный и ножной.
Головной отдел несет состоящий из венчиков ресничек коловращательный аппарат, выполняющий
одновременно функции движения и питания. У ряда планктонных быстро плавающих коловраток
по бокам коловращательного аппарата еще имеются покрытые ресничками выросты – «ушки».
Кроме того, на голове иногда видны одно или два пигментированных глазных пятна.
11
Форма туловищного отдела различается у разных экологических групп (рисунок 4).
У планктонных видов коловраток туловище округлое, мешковидное или колоколовидное,
у ползающих – чаще червеобразное. Туловище коловраток многих групп покрыто тонкой ложной
кутикулой, представляющей собой внутриклеточное уплотнение покровов. Однако чаще
развивается эндокутикула в виде волокнистой двухслойной пластинки, которая образует панцирь.
Тело панцирных коловраток часто имеет сложную форму, несет разнообразные выросты и шипы.
У планктонных видов от туловища иногда отходят подвижные придатки, служащие для
плавания.
Рисунок 4 – Схема строения Rotifera (по Wulfert, 1969): I – дорсально; II – латерально.
1 – передние (лобные) глаза; 2 – мозг (ганглий); 3 – субцеребральные железы;
4 – ретроцеребральный мешок; 5 – протонефридии; 6 – яичник; 7 – желточник; 8 – боковое
чувствительное щупальце; 9 – мочевой пузырь; 10 – нога; 11 – пальцы ноги; 12 – ножное
чувствительное щупальце; 13 – железы ноги; 14 – анальное отверстие; 15 – кишечник;
16 – мерцательная клетка протонефридиев; 17 – желудок; 18 – желудочные железы;
19 – слюнные железы мастакса; 20 – мастакс; 21 – глаза на ганглии; 22 – спинное
чувствительное щупальце; 23 – ушки с ресничками; 24 – рот; 25 – труба глотки;
26 – резервуар ножных желез; 27 – пищевод (Алексеева, 2010)
Ножной отдел (нога) наиболее развит у ползающих, прикрепленных и колониальных
коловраток, а у планктонных часто отсутствует. Нога обычно оканчивается выростами –
пальцами, реже венчиком ресничек или стебельком с прикрепленной пластинкой. Пальцы
у большинства коловраток веретеновидные или цилиндрические, реже игловидные.
Для коловраток известно половое размножение, но более характерно партено-
генетическое – бесполое, при котором самки откладывают неоплодотворенные яйца, из
которых развиваются только самки. Самцы в популяции встречаются редко, а для многих
видов неизвестны; они мельче и устроены проще, чем самки.
12
Crustacea (ракообразные)
Класс Branchiopoda (листоногие ракообразные)
Виды, относящиеся к этому классу, имеют листовидные туловищные конечности,
несущие жабры (эпиподы) и недоразвитые максиллы, из которых II пара отсутствует,
а I – рудиментарная. Лишь у небольшой группы хищных форм ноги имеют другое
строение. Обитают эти ракообразные преимущественно в пресных водах и делятся на
несколько отрядов.
Надотряд Cladocera
Отряд Anomopoda
У представителей этого отряда тело разнообразно по форме. Голова может иметь головной
щит или быть без него. Створки хорошо развитой раковинки полностью закрывают туловище
и грудные конечности. Верхняя ветвь плавательной антенны трех или четырехчлениковая,
нижняя ветвь всегда трехчлениковая. Каждая из ветвей оснащена не более чем пятью
оперенными щетинками, сидящими только на концах члеников. Каждая из пяти или шести пар
листовидных, нечленистых грудных конечностей имеет особое строение, шестая пара всегда
сильно упрощенная.
В Черном и Азовском морях из этого отряда планктонной фауны обитают представители
двух семейств – Bosminidae и Chydoridae.
Семейство Bosminidae
Виды этого семейства имеют очень укороченное тело. Задне-нижний край створок
обычно с шипом или с серией выростов, около него имеется одинокая щетинка. Глазок
и абдоминальные выросты отсутствуют. Хоботообразные антеннулы самки, сросшиеся
с рострумом, неподвижны. Антеннулы самца подвижные, служат для удержания самки
при копуляции. Все представители этого семейства имеют пять-шесть пар конечностей,
из которых последняя рудиментарна. Эфиппиум слабо хитинизирован, с продольными
килями. Постабдомиальные щетинки короткие. Являются планктонными фильтраторами.
Латентные яйца откладывают в примитивном эфиппии.
Из двух родов этого семейства в Черном и Азовском морях встречается только один –
Bosmina.
13
Род Bosmina
Тело самки этого рода в основном округлое. На задне-нижнем углу имеется один
шип, иногда редуцированный. Постабдомен сильно сплющен с боков, дистально прямо
обрубленный. Постабдомиальные коготки с гребнем тонких шипиков, расположенных при
основании. Базальные части антенн самки несросшиеся. Одна из ветвей плавательных антенн
трехчлениковая, другая – четырехчлениковая. Грудных конечностей шесть пар.
В Черном и Азовском морях встречается один вид – Bosmina (Bosmina) longirostris,
обитающий в опресненных районах указанных водоемов.
Bosmina (Bosmina) longirostris (O.F. Müller, 1785)
Bosmina (Bosmina) longirostris cornuta (Jurine, 1820)
Bosmina (Bosmina) longirostris curvirostris Fischer, 1854
Bosmina (Bosmina) longirostris pellucida Stingelin, 1895
Bosmina (Bosmina) longirostris similis (G.O. Sars, 1890)
Bosmina africanaaustralis Methuen, 1911
Bosmina avrostris Schiklejew, 1930
Bosmina brevicornis Hellich, 1877
Bosmina curvirostris Fischer, 1854
Bosmina japonica Poppe & Richard, 1890
Bosmina pelagica Stingelin, 1895
Bosmina pellucida Stingelin, 1895
Bosmina stuhlmanni Weltner, 1898
Garbinia adriani Grochowski, 1910
Lynceus longirostris O. F. Müller, 1785
Monoculus cornuta Jurine, 1820
Вид имеет очень укороченное тело. У самки край головы и створок почти полукруглой
формы, слегка выпуклый в передней части. Спинной край створок равномерно выпуклый,
брюшной почти прямой. Задний верхний угол створок тупой, нижний вытянут в небольшой
тонкий шипообразный вырост, впереди которого имеется хорошо развитая щетинка Курца.
Передний край головы равномерно выпуклый, глаз небольшой. Передние антенны тонкие,
короткие, изогнутые. Постабдомен имеет вид прямоугольной удлиненной пластинки. Его
верхний край слегка вогнутый, оканчивается небольшим лопастьевидным выступом с тремя
рядами мелких щетинок. Нижний край постабдомена вытянут в конический вырост, несущий
на конце тонкие, слегка изогнутые коготки. Длина самки – 0,25-0,62 мм.
Самец несколько мельче самки. В отличие от нее спинной край створок самца спереди
горбообразно – выпуклый, сзади прямой. Верхний задний угол почти прямой. Передние антенны
длинные, утолщенные, слегка изогнутые. Постабдомен самца короткий, сходный по форме
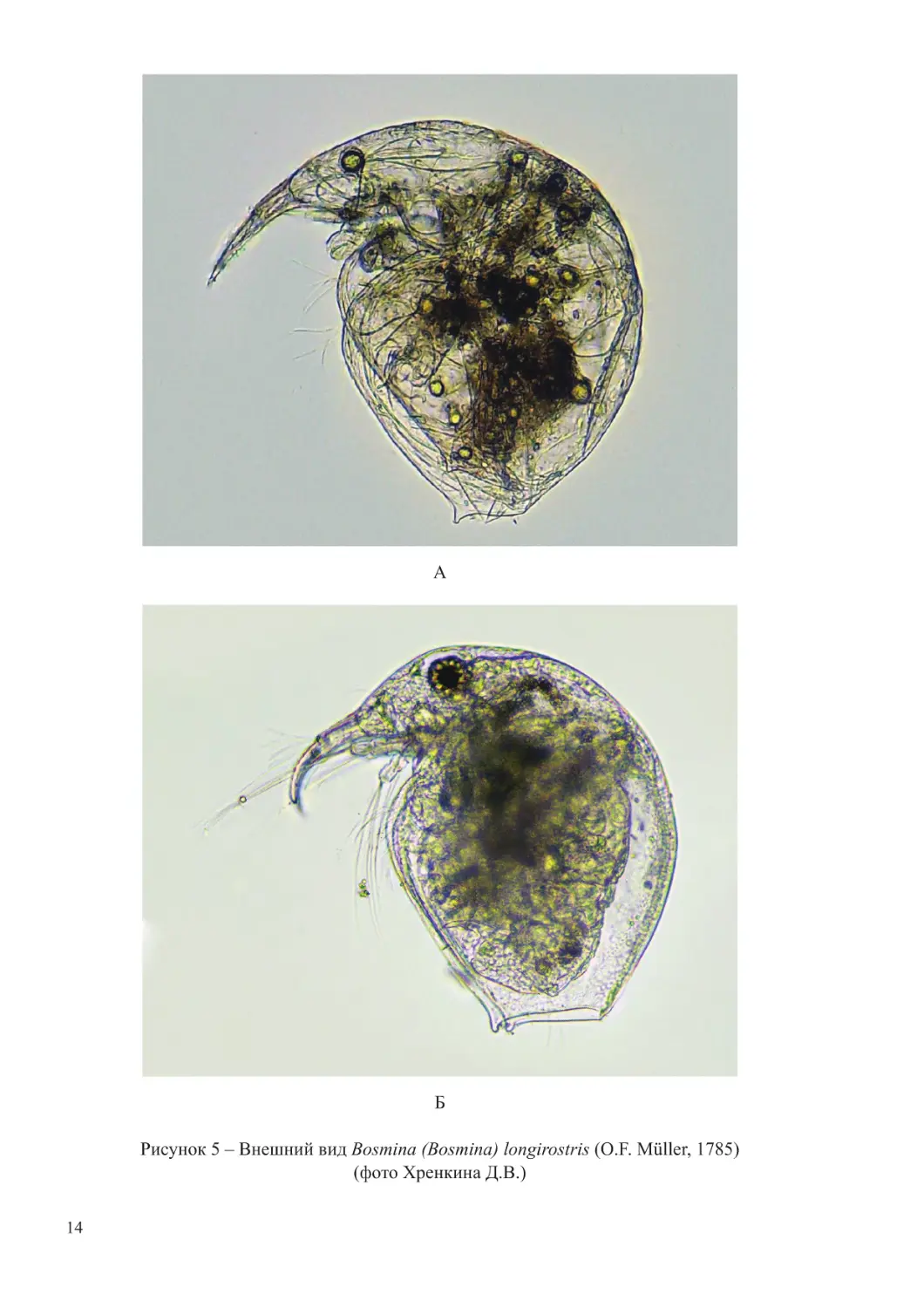
с таковым у самки. Коготки тупые, короткие, гладкие. Длина самца 0,25-0,44 мм (рисунок 5).
14
А
Б
Рисунок 5 – Внешний вид Bosmina (Bosmina) longirostris (O.F. Müller, 1785)
(фото Хренкина Д.В.)
15
У самок можно встретить до 7 яиц, самцы и эфиппиумы появляются обычно со второй
половины лета и осенью.
B. (B.) longirostris – вероятно, комплекс криптических видов. Является космополитом.
Населяет главным образом прибрежные районы крупных водоемов, бухт и заливов, в мелких
водоемах осваивает всю пелагиаль. Типичная форма прудового зоопланктона, где нередко
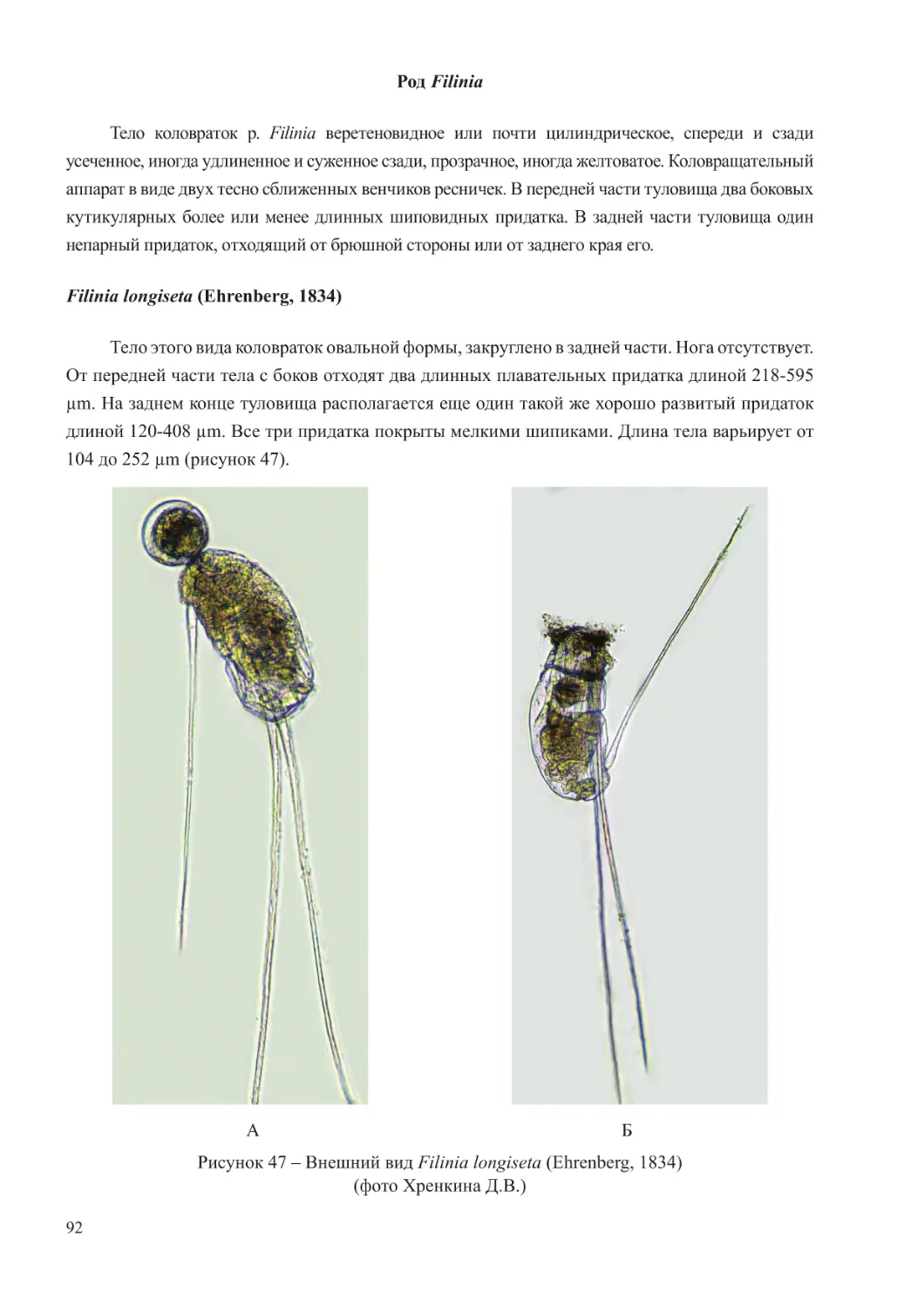
развивается в огромных количествах. В водоемах с соленостью до 5-8 ‰.
Встречается в р. Дон, р. Кубань, Таганрогском заливе Азовского моря, в лиманах Азовского
моря. Наибольшая интенсивность развития отмечается в апреле.
ß-мезосапроб.
Семейство Chydoridae
Представители этого семейства имеют округлое, почти сферическое или овальное, сжатое
с боков тело. Головной щит с хорошо выраженным рострумом. Сложный глаз маленький, у
большинства видов имеется глазок. Выводковая сумка не замкнутая, подавляющее большинство
представителей откладывает не более двух яиц. Абдомиальные выросты отсутствуют,
постабдомен разнообразной формы. Антеннулы у самок короткие, подвижные, отходят от
основания рострума и обычно не достигают его конца. Плавательные антенны короткие, обе
их ветви трехчлениковые. Пять или шесть пар грудных конечностей имеют веер маленьких
фильтрующих щетинок. Антеннулы самцов короче и толще чем у самок и не участвуют в
удержании самки при копуляции.
Виды р. Chydorus встречаются только в Азовском море.
Род Chydorus
Для видов этого рода характерно шарообразное тело. Нижний край створок очень выпуклый.
Скульптура створок может быть представлена в виде многоугольников, бугорков или структур,
напоминающих соты (у тропических и американских форм). Рострум короткий, либо средней
длины, прямой или слабо изогнутый. Киль имеет обычную форму, без зубцов на переднем крае.
Постабдомен короткий или средней длины, с закругленным дистальным и сильно выступающим
преданальным углом. Зубцы постабдомена мелкие, боковые щетинки короткие, коготок короткий,
имеются два базальных шипа умеренной длины. Наиболее распространенным видом является
Chydorus sphaericus. Встречается в Азовском море, в Черном море отсутствует.
Chydorus sphaericus (O.F. Müller, 1776)
Chydorus arcticus Røen, 1987
Chydorus coelatus Werestchagin, 1913
Chydorus lynceus Langhans, 1911
Chydorus mutilus Kreis, 1921
Lynceus sphaericus O.F. Müller, 1776
Monoculus infusorius Schrank, 1781
16
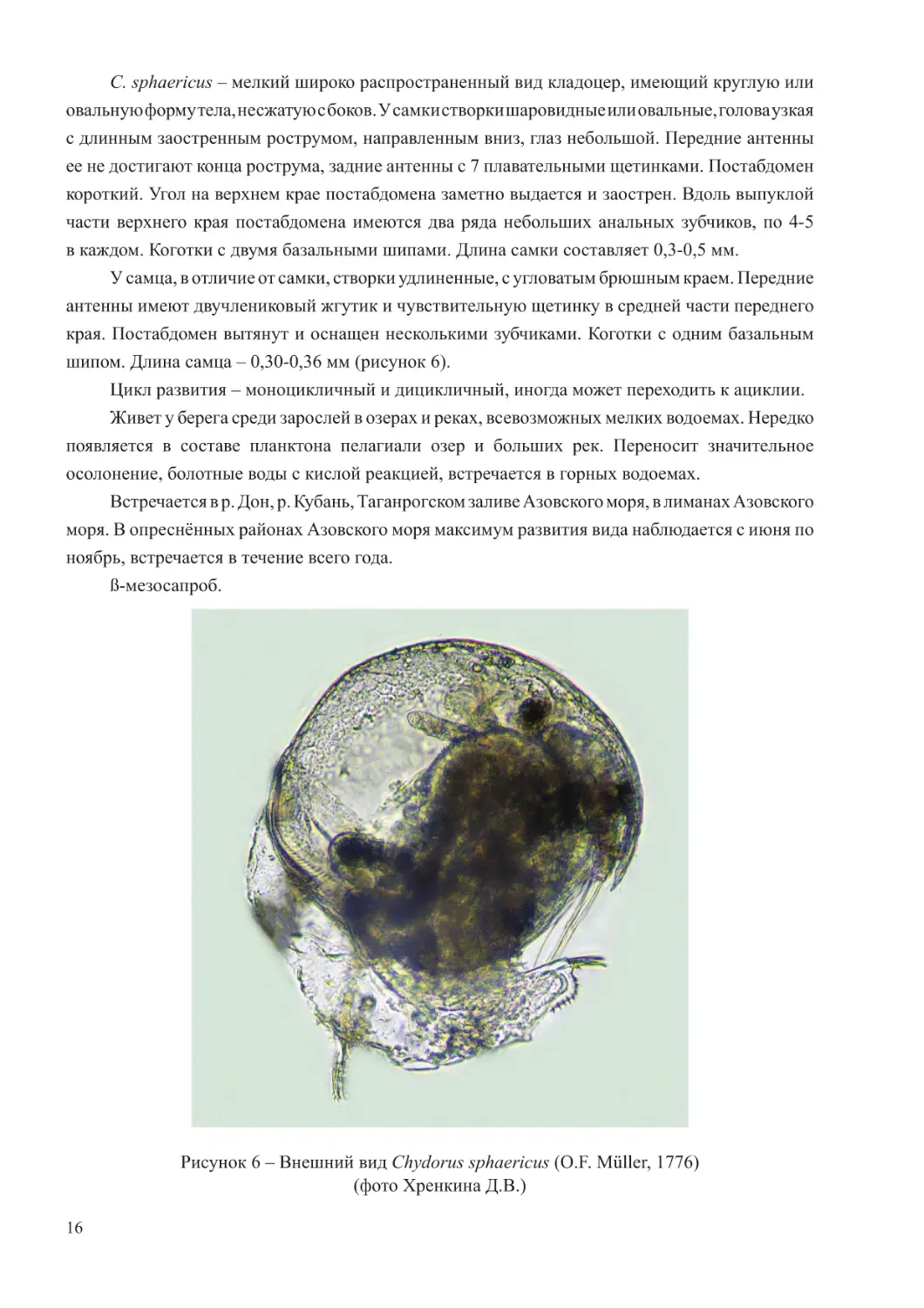
C. sphaericus – мелкий широко распространенный вид кладоцер, имеющий круглую или
овальную форму тела, несжатую с боков. У самки створки шаровидные или овальные, голова узкая
с длинным заостренным рострумом, направленным вниз, глаз небольшой. Передние антенны
ее не достигают конца рострума, задние антенны с 7 плавательными щетинками. Постабдомен
короткий. Угол на верхнем крае постабдомена заметно выдается и заострен. Вдоль выпуклой
части верхнего края постабдомена имеются два ряда небольших анальных зубчиков, по 4-5
в каждом. Коготки с двумя базальными шипами. Длина самки составляет 0,3-0,5 мм.
У самца, в отличие от самки, створки удлиненные, с угловатым брюшным краем. Передние
антенны имеют двучлениковый жгутик и чувствительную щетинку в средней части переднего
края. Постабдомен вытянут и оснащен несколькими зубчиками. Коготки с одним базальным
шипом. Длина самца – 0,30-0,36 мм (рисунок 6).
Цикл развития – моноцикличный и дицикличный, иногда может переходить к ациклии.
Живет у берега среди зарослей в озерах и реках, всевозможных мелких водоемах. Нередко
появляется в составе планктона пелагиали озер и больших рек. Переносит значительное
осолонение, болотные воды с кислой реакцией, встречается в горных водоемах.
Встречается в р. Дон, р. Кубань, Таганрогском заливе Азовского моря, в лиманах Азовского
моря. В опреснённых районах Азовского моря максимум развития вида наблюдается с июня по
ноябрь, встречается в течение всего года.
ß-мезосапроб.
Рисунок 6 – Внешний вид Chydorus sphaericus (O.F. Müller, 1776)
(фото Хренкина Д.В.)
17
Отряд Onychopoda
Представители этого отряда имеют округлое или удлиненное тело, большую голову,
отчетливо отделенную от туловища. В передней части головы находится большой сложный
глаз, глазок отсутствует. Раковинка имеет разные размеры, но во всех случаях она не прикрывает
грудные ноги. Сегменты грудного отдела между собой слиты. Абдомен у некоторых видов
бывает сильно редуцирован или хорошо развит. Постабдомен имеет вид закругленных
бугорков, несущих заостренные коготки различной формы и длины. Может быть вытянут в
виде хвостового придатка или иметь хвостовую иглу (у полифемид и церкопагид). Антеннулы
короткие, либо подвижно сочленены с небольшим выступом головы, либо неподвижны и слиты
между собой. Плавательные антенны мощные. Эфиппиум отсутствует.
Все представители этого отряда являются в основном обитателями пелагиали пресных,
солоноватых водоемов, открытых зон Мирового океана и внутренних морей.
Семейство Podonidae
У представителей ракообразных этого семейства тело овальное, более или менее
вытянутое. Раковинка большая, полусферическая или коническая, занимает спинную сторону
тела, оставляя конечности открытыми. Абдоминальный отдел сильно укорочен, сразу за IV
парой торакальных конечностей располагается постабдомен с хорошо развитыми коготками.
Хвостового придатка нет, но есть пара хвостовых щетинок на бугорках (подставках). Антеннулы
маленькие, неподвижные, приросшие к голове.
Род Cornigerius
Представители этого рода ветвистоусых ракообразных имеют продолговатую раковину,
вершина которой закруглена или заострена. На голове располагаются крупные выросты,
идущие от одного ствола и расходящиеся в разные стороны. Постабдоминальные коготки
чрезвычайно развиты, сильно расходятся в стороны, загнуты и более тонкие на концах. Обе
ветви плавательной антенны несут по 6 щетинок.
Cornigerius maeoticus (Pengo, 1879)
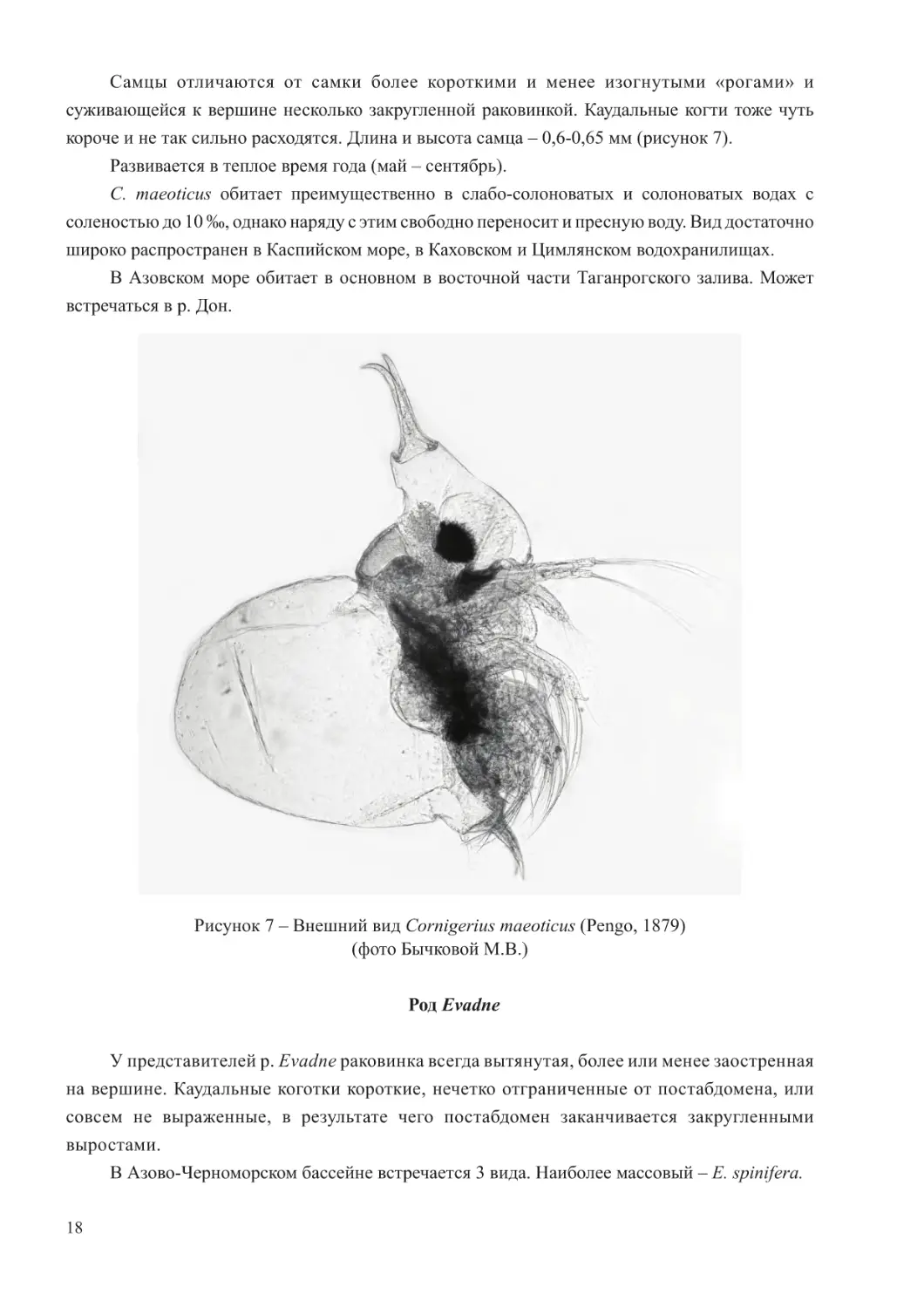
Форма тела у этого вида подобна типичной форме тела представителей семейста, т.е.
голова отделена от туловища четко выраженной выемкой. На передней поверхности головы
расположены два изогнутых отростка, так называемые «рога», расходящиеся от вершины
общего ствола. Они почти всегда сильно пигментированы. У самки раковина эллиптическая,
на вершине закруглена. Каудальные когти чрезвычайно длинные, тонкие, сильно изогнутые и
резко расходятся в самом начале. Длина самки (без изогнутых «рогов») – 0,5-0,8 мм, высота –
0,6-0,9 мм.
18
Самцы отличаются от самки более короткими и менее изогнутыми «рогами» и
суживающейся к вершине несколько закругленной раковинкой. Каудальные когти тоже чуть
короче и не так сильно расходятся. Длина и высота самца – 0,6-0,65 мм (рисунок 7).
Развивается в теплое время года (май – сентябрь).
C. maeoticus обитает преимущественно в слабо-солоноватых и солоноватых водах с
соленостью до 10 ‰, однако наряду с этим свободно переносит и пресную воду. Вид достаточно
широко распространен в Каспийском море, в Каховском и Цимлянском водохранилищах.
В Азовском море обитает в основном в восточной части Таганрогского залива. Может
встречаться в р. Дон.
Рисунок 7 – Внешний вид Cornigerius maeoticus (Pengo, 1879)
(фото Бычковой М.В.)
Род Evadne
У представителей р. Evadne раковинка всегда вытянутая, более или менее заостренная
на вершине. Каудальные коготки короткие, нечетко отграниченные от постабдомена, или
совсем не выраженные, в результате чего постабдомен заканчивается закругленными
выростами.
В Азово-Черноморском бассейне встречается 3 вида. Наиболее массовый – E. spinifera.
19
Evadne spinifera P.E. Müller, 1867
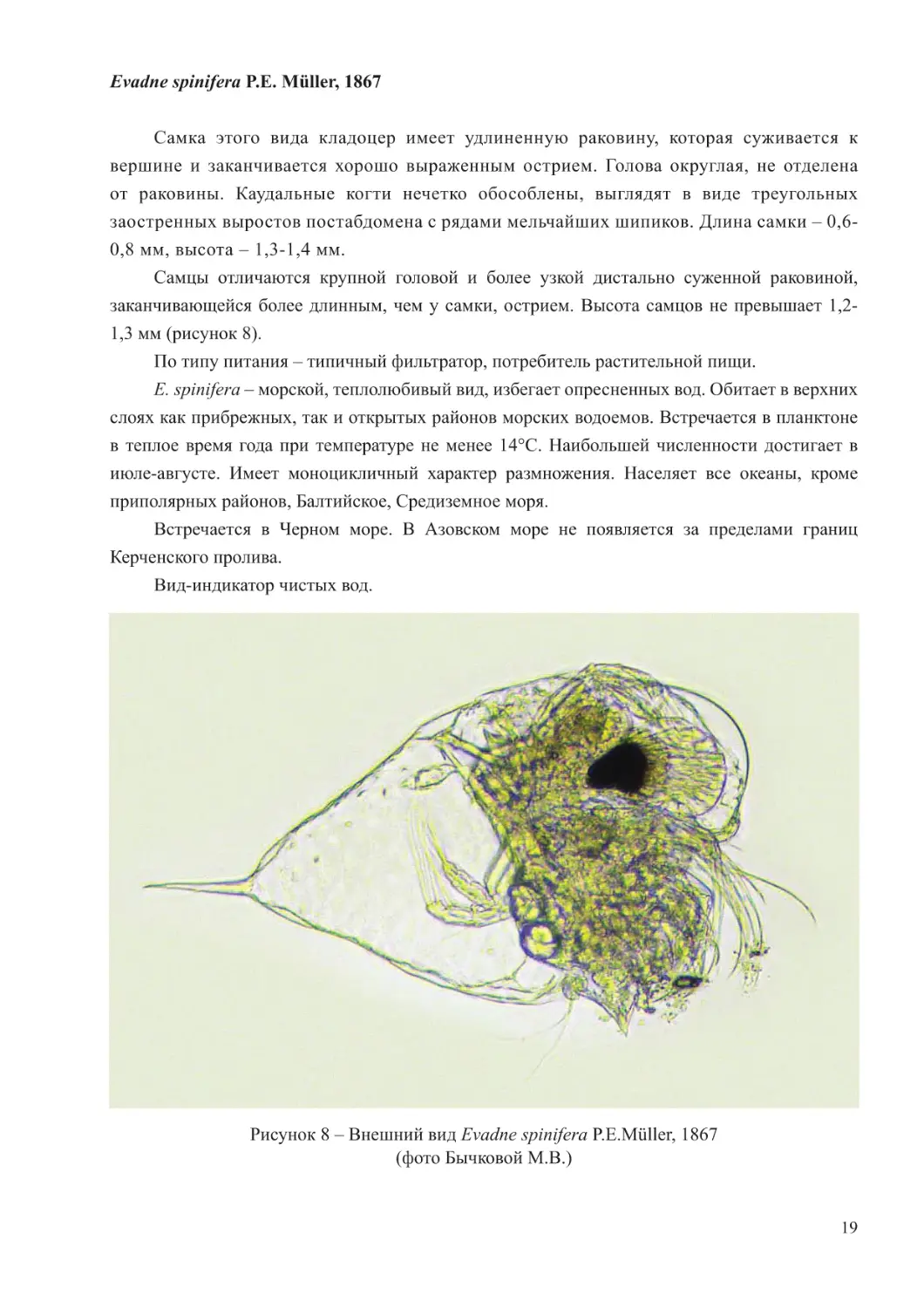
Самка этого вида кладоцер имеет удлиненную раковину, которая суживается к
вершине и заканчивается хорошо выраженным острием. Голова округлая, не отделена
от раковины. Каудальные когти нечетко обособлены, выглядят в виде треугольных
заостренных выростов постабдомена с рядами мельчайших шипиков. Длина самки – 0,6-
0,8 мм, высота – 1,3-1,4 мм.
Самцы отличаются крупной головой и более узкой дистально суженной раковиной,
заканчивающейся более длинным, чем у самки, острием. Высота самцов не превышает 1,2-
1,3 мм (рисунок 8).
По типу питания – типичный фильтратор, потребитель растительной пищи.
E. spinifera – морской, теплолюбивый вид, избегает опресненных вод. Обитает в верхних
слоях как прибрежных, так и открытых районов морских водоемов. Встречается в планктоне
в теплое время года при температуре не менее 14°С. Наибольшей численности достигает в
июле-августе. Имеет моноцикличный характер размножения. Населяет все океаны, кроме
приполярных районов, Балтийское, Средиземное моря.
Встречается в Черном море. В Азовском море не появляется за пределами границ
Керченского пролива.
Вид-индикатор чистых вод.
Рисунок 8 – Внешний вид Evadne spinifera P.E.Müller, 1867
(фото Бычковой М.В.)
20
Род Pleopis
Представители этого рода имеют эллиптическую или полусферическую раковинку, всегда
закругленную на вершине. Голова в большей или меньшей степени отграничена от раковинки
выемкой, иногда слабо выраженной. Каудальные когти острые, тонкие, часто сильно изогнутые
и значительно длиннее хвостовых щетинок, всегда хорошо обособлены.
Pleopis polyphemoides (Leuckart, 1859)
Evadne polyphemoides Leuckart, 1859
Pleopis minutus G.O. Sars, 1862
Podon mecznikovi Chernyavskii, 1868
Podon polyphemoides (Leuckart, 1859)
P. polyphemoides – самый мелкий вид подонид. У самки раковинка небольшая,
полусферическая или почти шаровидная при наличии зрелых зародышей. На голове имеется
присоска, вторые антенны короткие. Хвостовые щетинки маленькие, хвостовой стебелек почти
рудиментирован. Длина зрелой самки не превышает 0,4-0,45 мм, высота примерно такая же.
Раковинка самца маленькая, менее округлая и несколько сужена сзади. Голова и глаз
крупнее, чем у самки. Длина самца не более 0,4 мм (рисунок 9).
Рисунок 9 – Внешний вид Pleopis polyphemoides (Leuckart, 1859)
(фото Бычковой М.В.)
21
Тип питания – фильтрация, в качестве пищи использует детрит.
Это морской, эвригалинный, умеренно тепловодный вид, выносит понижение солености
до 6-8 ‰. Встречается в поверхностных слоях морских водоемов, наибольшее развитие получает
в акваториях со слабой интенсивностью водообмена и высокой степенью эвтрофированности.
Из всех видов кладоцер P. polyphemoides первым появляется в планктоне. Температурный
оптимум его развития находится в пределах 16-20°С.
Населяет все океаны, за исключением тропической зоны, Балтийское, Средиземное,
Каспийское моря.
Встречается в Черном море. В Азовском море при невысокой солености обитает в
основном в открытой части водоема, при осолонении заходит в Таганрогском заливе.
Род Podonevadne
У представителей р. Podonevadne раковинка конической формы с заостренной или
закругленной вершиной. Голова крупная, между нею и раковинкой всегда есть более или менее
заметная выемка. Постабдоминальные коготки острые, прямые, или загнутые. Обе ветви
плавательной антенны несут по 6 щетинок.
В Азовском море встречается 1 вид – Podonevadne trigona.
Podonevadne trigona (G.O. Sars, 1897)
Evadne intermedia G.O. Sars, 1902
Evadne pusilla G.O. Sars, 1902
Evadne trigona G.O. Sars, 1897
Evadne trigonoides G.O. Sars, 1902
Некрупный вид кладоцер. У самки передняя часть головы округленная, слегка отделенная
от задней. Раковинка различной формы – от треугольной до эллиптической или яйцевидной.
Между раковинкой и головой имеется иногда незначительная выемка. Выводковая камера
у самки сильно расширена в базальной части, имеет почти треугольную форму, на конце
сильно заострена. Постабдомен заканчивается двумя острыми каудальными коготками.
Они длинные, узкие, прямые или лишь незначительно изогнутые, параллельные друг другу,
хорошо обособлены от постабдомена. Длина самки – 0,35-0,50 мм. Самец отличается от самки
суживающейся к вершине почти округленно-треугольной раковиной и более крупным глазом.
Длина самца около 0,4 мм (рисунок 10).
P. trigona обитает в солоноватых и слабосолоноватых водоемах, интенсивно развивается
преимущественно летом. Переносит соленость до 10 ‰, но вместе с тем может встречаться и
в пресной воде.
В Азовском море при естественном солевом режиме иногда встречается в открытой
части водоема и достаточно часто в Таганрогском заливе. При повышении солености этот
вид отмечается только в Таганрогском заливе, главным образом, в его опресненной восточной
части. Широко распространен в Каспийском море.
22
Рисунок 10 – Внешний вид Podonevadne trigona (G.O. Sars, 1897)
(фото Бондарева С.В.)
Род Pseudevadne
Представители этого рода кладоцер имеют раковинку эллиптической или яйцевидной
формы, слегка суживающуюся к вершине. Выемка за головой часто слабо выражена. Выводковая
камера округленно-яйцевидная. Хвостовые щетинки очень короткие, отходящие от маленького
бугорка. Абдомиальные коготки хорошо развиты.
Pseudevadne tergestina (Claus, 1877)
Evadne tergestina Claus, 1864
Pleopis schoedleri Chernyavskii, 1868
Pleopis tergestina (Claus, 1877)
Podon schoedleri Caporiacio, 1938
Этот вид кладоцер довольно крупный. У самки голова в большей или меньшей степени
отделена от раковины слабо выраженной, почти незаметной выемкой. Раковинка эллиптическая
или яйцевидная, слегка суживается к вершине. Для обоих полов вида характерна легкая окраска
раковинки. Длина самки доходит до 0,7 мм, высота – до 1,1 мм.
23
Самец несколько мельче, чем самка. Отличается более крупной головой и заметно
суживающейся к вершине, раковинкой. Высота самца составляет не более 0,8 мм.
По типу питания является фильтратором, потребляет растительную пищу.
P. tergestina – морская, тепловодная, сравнительно стеногалинная кладоцера, которая не
встречается при солености менее 16 ‰. Живет в поверхностных слоях воды (до 20 м), обычно
недалеко от берега. Встречается в летне-осенний период (июнь-ноябрь), наиболее активно
размножается в конце лета.
Районами обитания этого вида являются теплые воды всех океанов между 40° с.ш. и
40° ю.ш., Средиземное и Черное моря. Может появляться в Азовском море при его осолонении.
Отряд Ctenopoda
У представителей отр. Ctenopoda раковинка либо полностью закрывает ноги, или
несколько укорочена. Все тело животных покрыто массивной студенистой оболочкой,
покрывающей все тело. Членики верхней ветви плавательных антенн оснащены щетинками,
сидящими как на концах этих члеников, так и на их боковой стороне. Имеется шесть пар
листовидных, нечленистых грудных конечностей, из которых пять передних сходного строения.
Покоящиеся яйца откладываются без эфиппиума.
Семейство Sididae
Представители этого семейства имеют крупную голову без головного щита, отчетливо
отделенную от туловища. Раковина продолговатая, полностью закрывает 6 пар грудных
конечностей, из которых пять передних сходны по своему строению и вооружены мощными
веерами из длинных фильтрующих щетинок. Плавательные антенны длинные, массивные с
двумя ветвями. Верхняя ветвь оснащена многочисленными щетинками, сидящими как на конце,
так и на боковой стороне члеников. Антеннулы самцов часто имеют копулятивные придатки.
Постабдомен имеет хорошо развитые коготки с добавочными зубчиками, хвостовые щетинки
сидят на сильно развитых подставках. Латентные яйца откладываются без эфиппия.
Все виды этого семейства являются фильтраторы, среди которых есть как планктонные,
так и зарослевые и донные формы, потребляющие микроскопические водоросли, тонкий
детрит и бактерии.
Род Diaphanosoma
У представителей этого рода голова у обоих полов внизу без выростов, достаточно
отчетливо отделена от туловища, различается по размерам и форме. Антеннулы самок
маленькие, самцы имеют длинные антеннулы с копулятивными придатками разной формы.
Плавательные антенны мощные и длинные. Брюшной край створок раковинки загибается
внутрь, образуя полость различной глубины и формы.
24
В составе этого рода известно более 30 видов, но для европейской части России достоверно
указаны только 4 таксона.
Diaphanosoma brachyurum (Liévin, 1848)
Diaphanosoma brandtianum Fischer, 1850
Diaphanosoma perarmatum Brehm, 1933
Sida brachyura Liévin, 1848
Имеет удлиненное тело. У самки узкая, удлиненная голова отделена от створок туловища
выемкой. Верхний край головы слегка выпуклый, нижний почти прямой. Рострум отсутствует.
Большой глаз находится в переднем нижнем углу головы. Передние антенны маленькие, с
палочковидным основанием. Створки более высокие в задней части. Брюшной и спинной край
створок слегка выпуклые, задний край прямой. Середина брюшного края створок усажена
5-6 довольно длинными щетинками, задняя часть – 6-7 короткими щетинками. Между ними
помещаются многочисленные шипики. Такие же шипики имеются и на нижней половине его
заднего края. Постабдомен небольшой, почти конической формы. Крупные, сильно изогнутые
коготки являются непосредственным его продолжением. Вогнутый край коготков вооружен
тремя зубцами. Длина самки – 0,8-1,3 мм.
У самцов форма туловища и головы напоминает таковую у самки, но у самца она
несколько уже. Первые антенны очень длинные, значительно длиннее, чем у самки, заходят
за середину створок. Первая пара ног вооружена крючком. Длина самца – 0,75-0,80 мм.
Особи обоих полов D. brachyurum очень прозрачны (рисунок 11).
Это широко распространенный пресноводный вид, выносящий осолонение. Обитает
во всевозможных внутренних водоемах, водохранилищах, в крупных и мелких озерах,
прудах, реках. Населяет поверхностные слои водной толщи, встречается как в открытой
части водоемов, так и в зарослях. Вид с четко выраженными суточными миграциями.
Обычная массовая форма кислых и солоноватых водоемов.
D. brachyurum – теплолюбивый вид. В планктоне появляется при температуре воды
около 12°С, массовое развитие отмечается в летний период. Имеет моноцикличный
характер размножения. Самка вынашивает до 8 яиц. Самцы и эфиппиальные самки
встречаются при охлаждении воды до 17-16°С. Покоящиеся яйца попадаются со второй
половины лета.
В России обитает повсеместно, кроме крайнего севера.
Встречается в р. Дон, р. Кубань, р. Маныч, Цимлянском водохранилище, в Таганрогском
заливе Азовского моря до солености около 6-8 ‰. В слабосолоноватых лиманах нередко
развивается в массовом количестве.
Олиго-мезосапроб.
25
Рисунок 11 – Внешний вид Diaphanosoma brachyurum (Liévin, 1848)
(фото Бондарева С.В.)
Род Penilia
У представителей р. Penilia голова низкая, у самки она вытянута в два выроста с
шипами, направленных книзу, у самца она округлая. На краях створок по всей длине находятся
шиповидные зубчики, между которыми расположены короткие щетинки. Каудальные когти
очень тонкие и длинные, с двумя шипиками у основания и мелкими щетинками или зубчиками
с вогнутой стороны. Хвостовые щетинки длинные (приблизительно равны длине тела), сидят
на двух отдельных подставках. Постабдомен конический, невооруженный.
Род включает один вид, обитающий в Черном море – Penilia avirostris.
Penilia avirostris Dana, 1849
Penilia orientalis Dana, 1852
Penilia pacifica Kraemer, 1895
Penilia schmackeri Richard, 1895
26
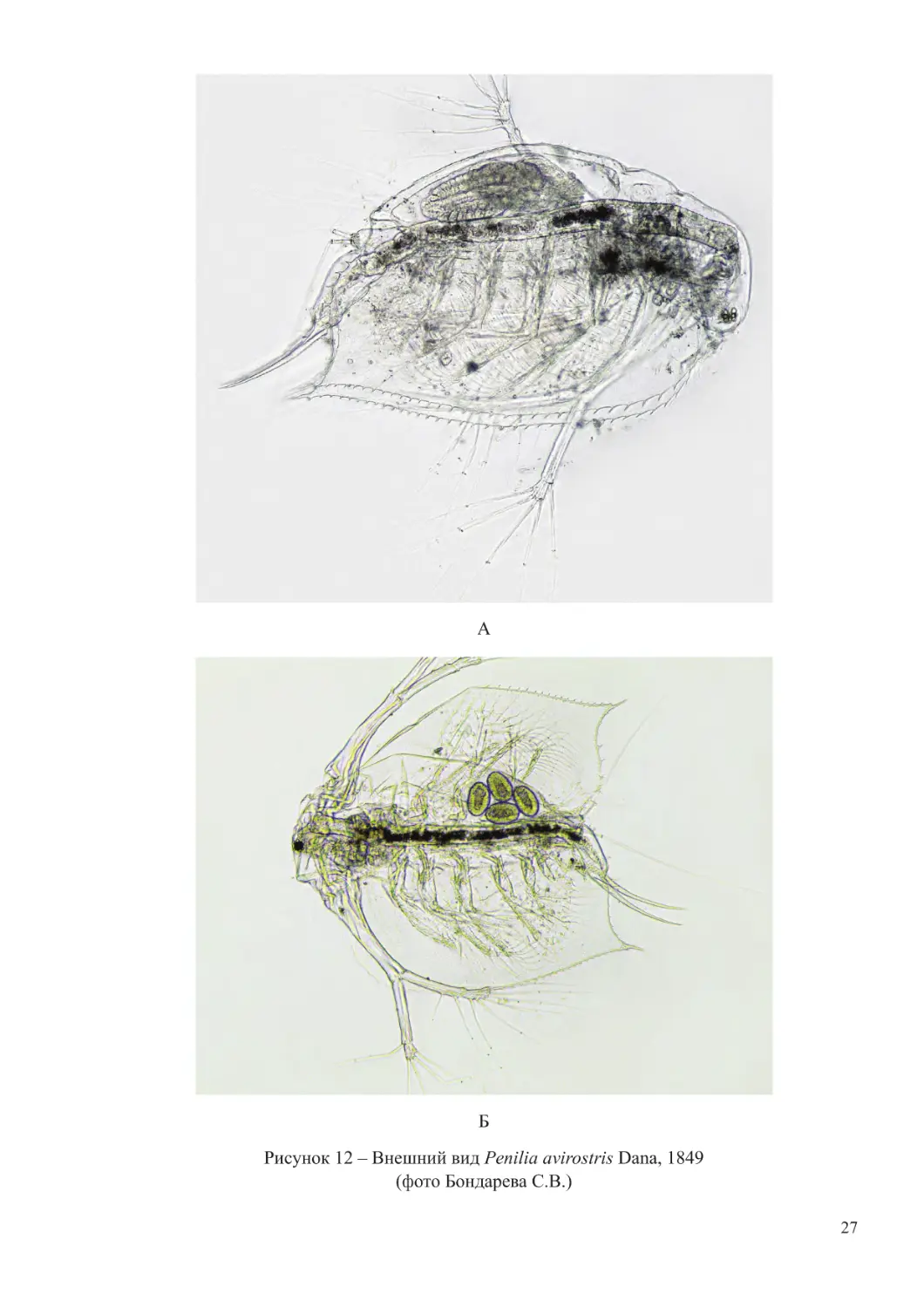
Кладоцера имеет удлиненное тело. У самки голова вытянута в два направленные
книзу выроста с шипами. Антенны I короткие с длинной чувствительной щетинкой,
антенны II имеют две двучленистых ветви. По всей длине краев створок находятся
шиповидные зубчики. Длина самки – 0,8-1,0 мм.
У самца голова округлая, слегка отделена от туловища. Антенны очень длинные,
доходят до конца раковины. Длина самца – 0,65-0,80 мм.
Оба пола этого вида имеют общие морфологические признаки: небольшой глаз,
довольно длинное острие в месте перехода нижнего края створок в задний, длинные и
тонкие абдоминальные коготки, а также очень длинные, приблизительно равные длине
тела, хвостовые щетинки (рисунок 12).
Партеногенетические самки достигают размеров 0,91-0,98 мм и дают в среднем
по 8 яиц. К осени количество яиц в выводковых камерах снижается до 3-4. Развиваются
в планктоне в течение 3-4 месяцев в году. Осенью откладывают покоящиеся (зимующие)
яйца, которые обеспечивают воспроизводство популяции в следующем году. Максимальная
продолжительность жизни около 20-25 суток. Наибольшая численность вида отмечается
в августе-сентябре.
По типу питания является тонким фильтратором, потребляет детрит, бактерии и
мелкие жгутиковые водоросли размером менее 8 μm.
Это чисто морской вид, обитающий в теплых и умеренных прибрежных водах.
Держится в верхних слоях воды, выносит сильные колебания солености.
Обитает в Атлантическом, Тихом и Индийском океанах, в Средиземном и Черном
морях. В Азовском море встречается редко в южной части, попадая сюда с адвекцией
черноморских вод.
Очень чувствителен к загрязнению, в связи с чем может служить индикатором
чистых вод.
27
А
Б
Рисунок 12 – Внешний вид Penilia avirostris Dana, 1849
(фото Бондарева С.В.)
28
Класс Hexanauplia
Подкласс Copepoda
Тело представителей подкласса Copepoda состоит из 16 сегментов, в том числе 6 головных,
5 сегментов формирующих грудь (торакс) и 5 конечных сегментов, образующих брюшко
(абдомен).
Форма цефалоторакса (головогруди) у веслоногих ракообразных овально-удлиненная, в
передней части обычно закруглена, иногда притуплена либо имеет небольшой выступ. Боковые
края передней части цефалоторакса закругленные или с небольшими крючкообразными
выростами, края заднего последнего сегмента могут быть оттянуты в круглые, острые или
шиповатые выросты. Абдомен у самок 2-4 сегментный, у самцов – 5 сегментный. К первому
сегменту абдомена (генитальному) прикрепляются две фуркальных ветви, несущие на себе по
3-5 щетинок.
Веслоногие ракообразные раздельнополые организмы, с четко выраженным половым
деформизмом. Непарные яичники и семенники расположены в грудной части тела над
кишечником. Самки представителей отряда Cyclopoida вынашивают яйца в парных мешках, а
самки отряда Calanoida – в непарных.
Подкласс Copepoda характеризуется двухэтапным постэмбриональным развитием.
Науплиальная личинка имеет яйцевидно-округлую форму. В начале своего развития она
несегментирована, в конце приобретают слабую сегментацию, один глаз и 2– 4-х членистые
антенны I с развитыми головными конечностями. Копеподиты имеют «имагинальный» вид,
тело состоит из цефалоторакса и абдомена. Торакальные плавательные ножки двуветвистые,
их число после каждой линьки увеличивается. У разных отрядов копепод бывает пять-шесть
науплиальных стадий и шесть копеподитных, включая взрослую.
Отряд Calanoida
У представителей этого отряда цефалоторакс заметно шире и длиннее абдомена, форма
тела яйцевидно-овальная со срезанным задним краем различного очертания. Передний край
цефалоторакса закруглен, однако у некоторых видов посредине он слегка оттянут вперед.
Рострум имеет форму нитей или глубоко вырезанных пластинок.
Тело самок разных видов отр. Calanoida состоит из 8-11 сегментов, самцов – из 7-10.
Торакальные ноги хорошо развиты. Первые четыре пары плавательного типа. Пятая пара ног с
признаками полового диморфизма, симметричная у самок, асимметричная у самцов, может быть
одноветвистой или двуветвистой. Строение пятой пары ног у обоих полов служит хорошим
диагностическим признаком.
Фуркальные ветви разной длины, чаще короткие, оснащены веерообразно рас-
положенными фуркальными щетинками, число которых составляет от трех до пяти
29
штук. У разных таксонов длина щетинок неодинакова, но в пределах одного вида
больших различий в их длине нет.
К отряду Calanoida относятся морские, солоноватоводные и пресноводные свободно-
живущие веслоногие ракообразные.
Семейство Acartiidae
Ракообразные этого семейства имеют длинное, узкое тело, у которого цефалоторакс
значительно превышает длину абдомена, соотношение составляет примерно 1:4.
Головной сегмент не сливается с первым торакальным, последние грудные сегменты
между собой слиты. Антенны самки относительно длинные, немного заходят за задний
край генитального сегмента. Абдомен у самки состоит обычно из 3, а у самца – из 4-5
сегментов. Пятая пара ног у обоих полов одноветвистая, у самцов с загнутыми внутрь
концами.
Род Acartia
Голова у представителей этого рода без рострума и без боковых крючков, последний
торакальный сегмент закруглен. Абдомен состоит из 3 сегменов, все фуркальные щетинки
имеют одинаковую толщину. В Черном и Азовском морях встречаются два вида – Acartia
(Acartiura) clausi и Acartia (Acanthacartia) tonsa.
Acartia (Acartiura) clausi Giesbrecht, 1889
Acartia clausi Giesbrecht, 1889
Представители вида имеют очень стройное, удлиненное, прозрачное, непигментированное
тело, один непарный глаз и заметно притупленный передний край головы. Рострум
отсутствует, антенны достигают конца генитального сегмента. Абдомен и фуркальные
ветви несколько длиннее, чем у Acartia (Acanthacartia) tonsa. Фуркальных щетинок
пять, они довольно длинные и широко расставлены. Пятая пара ног симметричная и
очень маленькая. Длина самки – 0,7-1,75 мм. У самца пятая пара ног асимметричная, на
внутренней поверхности левой ноги имеется два довольно больших округлых выступа.
Размеры самца составляют 0,7-1,5 мм (рисунок 13).
Встречается A. ( A.) clausi в течение всего года, совершает незначительные вертикальные
миграции. В Черном море наиболее интенсивно размножается весной и в начале лета (май-
июнь), в Азовском море – в середине лета. Нередко определяет общую динамику развития
планктона указанных морей. В течение года наблюдается 8-9 генераций, количество яиц в
кладке составляет 12-22 штуки. Общая продолжительность развития от оплодотворенного
яйца до созревания взрослой особи составляет 30-33 суток, продолжительность жизни
особи – около 60 дней.
30
А
Б
31
В
Г
32
Д
Е
33
Ж
34
З
Рисунок 13 – Внешний вид Acartia (Acartiura) clausi Giesbrecht, 1889
(А, Б, В – науплиус, Г – копеподит, Д, Е, Ж – самка, З – самец)
(фото Хренкина Д.В.)
A. (A.) clausi – эврифаг, имеет смешанный тип питания, Наряду со способностью питаться
путем фильтрации, существенную роль играет хищничество. В качестве корма использует
фитопланктон, животную пищу и детрит.
A. (A.) clausi – эвритермный, полигалобный, морской вид, широко распространенный
вид в умеренной зоне Атлантического океана, в Тихом и Индийском океанах, а также в
Северном и Средиземном морях. Наиболее распространенный вид в планктоне Черного
и Азовского морей. Для него свойственны широкая экологическая валентность и
положительная реакция на эвтрофирование водоема. Нижняя граница солености, при
которой встречается эта копепода, составляет около 5 ‰. Интенсивное развитие она
получает при солености выше 12,5 ‰, в связи с чем при осолонении Азовского моря
становится массовым видом. В Черном море повсеместно. Встречается в устьевых зонах
приазовских лиманов.
35
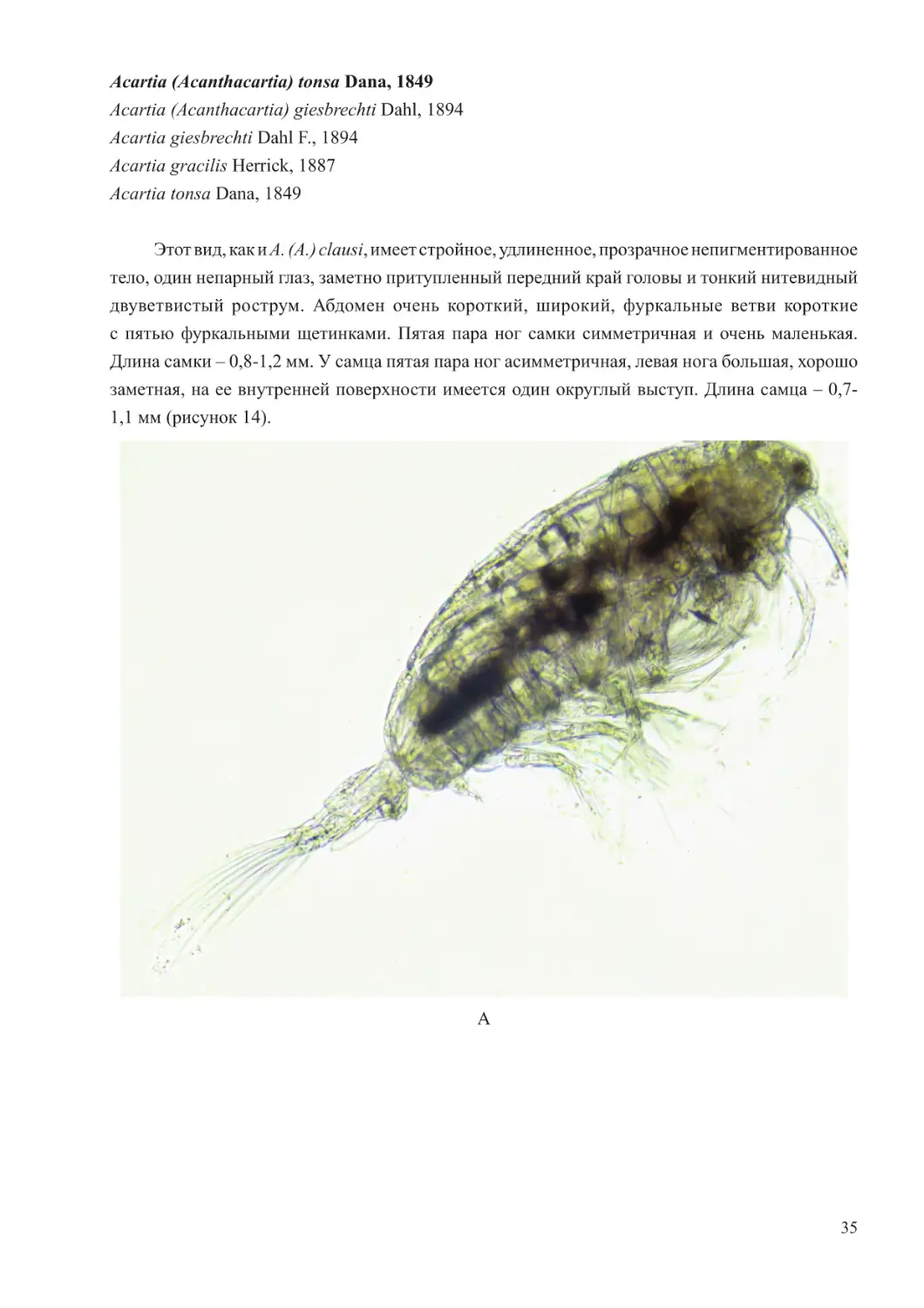
Acartia (Acanthacartia) tonsa Dana, 1849
Acartia (Acanthacartia) giesbrechti Dahl, 1894
Acartia giesbrechti Dahl F., 1894
Acartia gracilis Herrick, 1887
Acartia tonsa Dana, 1849
Этот вид, как и A. (A.) clausi, имеет стройное, удлиненное, прозрачное непигментированное
тело, один непарный глаз, заметно притупленный передний край головы и тонкий нитевидный
двуветвистый рострум. Абдомен очень короткий, широкий, фуркальные ветви короткие
с пятью фуркальными щетинками. Пятая пара ног самки симметричная и очень маленькая.
Длина самки – 0,8-1,2 мм. У самца пятая пара ног асимметричная, левая нога большая, хорошо
заметная, на ее внутренней поверхности имеется один округлый выступ. Длина самца – 0,7-
1,1 мм (рисунок 14).
А
36
Б
В
37
Г
Рисунок 14 – Внешний вид Acartia (Acanthacartia) tonsa Dana, 1849
(А,Б,В –самка,Г –самец)
(фото Хренкина Д.В.)
Вид отличается высокой плодовитостью. Наиболее интенсивно развивается при
температуре выше 16°С. Уровень ее развития даже при оптимальных условиях подвержен
значительным колебаниям, связанным с трофическими условиями и выеданием. При
температуре 10°С вид практически перестает размножаться. Откладывает покоящиеся яйца,
которые могут сохраняться в илах в течение нескольких лет. При наступлении благоприятных
условий яйца развиваются до нормальных взрослых особей.
Как и A. (A.) clausi, имеет смешанный тип питания, является эврифагом, потребляет
фитопланктон, животную пищу и детрит.
Неритический, типичный стенотермный, теплолюбивый вид. Может обитать как в
слабосоленой, так и в океанической воде.
38
Встречается в водах тихоокеанского и атлантического берегов Америки, в Индийском
океане, в районе Малайского архипелага, Средиземном, Балтийском и Каспийском морях.
В южных морях России A. (A.) tonsa – вид-вселенец. В Черном и Азовском морях впервые
был обнаружен в 1970-х годах, в настоящее время является массовым видом планктонной
фауны этих водоемов в теплое время года. В Черном море была обнаружена раньше, чем в
Средиземном, и, следовательно, в Азово-Черноморский бассейн, возможно, была занесена с
балластными водами из другого района Мирового океана. Относится к тепловодному комплексу
черноморских видов, отсутствует в планктоне в холодный период года, устойчив к загрязнению
и эвтрофированию водоема. Приурочен к бухтам, эстуариям и заливам. В Азовском море
A. (A.) tonsa в настоящее время является доминирующим таксоном в сообществе животного
планктона в летний период.
Семейство Calanidae
У представителей семейства Calanidae тело крупных размеров, длиной от 2,7 мм до
4,5 мм. Форма тела удлиненная, овальная. Головной сегмент четко отделен от I грудного,
последние торакальные сегменты не слиты. Абдомен у самки состоит из четырех, у самца – из
пяти сегментов. Пятая пара ног у обоих полов плавательного типа и по-своему по строению
мало отличается от остальных торакальных ног. Исключительно морское семейство, состоит
из пяти родов.
В Черном море обитает только один род.
Род Calanus
Половой деформизм у представителей этого рода выражен слабо. Отмечаются лишь
незначительные различия в строении антенн I и пятой пары торакальных ног. Абдомен и фурка
симметричны, пять пар хорошо развитых фуркальных щетинок располагаются веерообразно.
В Черном море обитает один вид – Calanus euxinus.
Calanus euxinus Hulsemann, 1991
Calanus ponticus Karavaev, 1894
Сalanus helgolandicus Claus, 1863
C. euxinus – крупный морской вид, имеющий узкое, стройное тело. На переднем конце
явно отчлененного головного сегмента имеется небольшой тупой выступ, который несколько
заметнее выражен у самки. Антенны тонкие, длинные, несколько превышают длину тела и
широко расставлены в стороны. Абдомен и фурка симметричны, пять пар хорошо развитых
фуркальных щетинок располагаются веерообразно. Пятая пара ног у обоих полов двуветвистая,
от первых четырех пар отличается характером оперения. На генитальном сегменте самки
имеется вентральный выступ, который хорошо виден сбоку. Длина самки – 2,7-4,5 мм, длина
39
самца – 2,35-3,2 мм. За счет жировых включений C. euxinus в зимнее время приобретает
красноватую окраску.
C. euxinus – круглогодичный, морской, холодолюбивый, батипелагический вид. Совершает
четко выраженные вертикальные миграции в летнее время. Младшевозрастные группы этих
животных (копеподиты I-III стадий) держатся в основном в верхнем перемешанном слое,
старшевозрастные особи (копеподиты IV-VI стадии), как правило, не поднимаются выше
термоклина. C. euxinus наиболее активно развивается в открытых водах на больших глубинах,
в прибрежных районах встречается единично (рисунок 15).
А
40
Б
Рисунок 15 – Внешний вид Calanus euxinus Hulsemann, 1991 (науплиус)
(фото Хренкина Д.В.)
Имеет 4-6 генераций, наибольшее число яиц (до 20 шт.) откладывает при температуре
10°С. При повышении температуры (до 18-22°С) количество яиц в кладке снижается до 13-
14 шт. Общая продолжительность развития от оплодотворенного яйца до созревания взрослой
особи составляет 33-38 суток.
По типу питания этот вид является грубым пассивным фильтратором, фитофагом. Основу
пищи составляют водоросли размером от 16-20 μm до 500 μm.
Обитает в Северной части Атлантики, Северном, Норвежском, Средиземном и Черном
морях. В Азовское море при повышении солености иногда встречаются копеподиты C. euxinus,
попадающие сюда с адвекцией черноморских вод.
Семейство Centropagidae
Представители этого семейства имеют стройное тело, с заметно расширенным в средней
части цефалотораксом. У самки общее число сегментов равно 9, у самца – 10-11. Головной
сегмент отделен от I торакального. Антенны I самки длинные, почти достигают конца
фуркальных ветвей. Антенна самца петлеобразно изогнута в конечном отделе. Пятая пара ног
самок двуветвистая, плавательного типа. У самца левая нога также сохраняет тип плавательной
конечности, у правой ноги плавательным является эндоподит, последний членик экзоподита
превращен в клешню.
41
Представители этого семейства являются морскими и солоноватоводными видами. В
Черном и Азовском море живет только один род – Centropages.
Род Centropages
У представителей этого рода цефалоторакс с оттянутыми боковыми углами, 6-сегментный,
все сегменты почти одинаковой длины. Генитальный сегмент самки по бокам с небольшими
кнопкообразными выростами, покрытыми группой мелких шипиков. Фуркальные ветви
симметричные, относительно длинные, с оперенными внутренними краями. Фуркальные
щетинки густо оперены.
В Черном и Азовском морях массовым видом является Centropages ponticus.
Centropages ponticus Karavaev, 1895
Centropages kroyeri pontica Karavaev, 1895
Тело крепкое, коренастое, четко пигментированное. Абдомен значительно короче
цефалоторакса. Последний сегмент цефалоторакса имеет четко выраженные оттянутые
боковые углы. Пятая пара ног самки плавательного типа. Длина самки составляет 1,25-1,65 мм.
У самца пятая пара ног асимметричная, двуветвистая. Левая ветвь плавательного типа, а правая
выглядит в виде большой клешни. Длина самца – 1,4 мм (рисунок 16).
C. ponticus – морской, теплолюбивый, летне-осенний вид копепод, однако единичные
экземпляры встречаются в зимнее время и весной. Обитает в основном в прибрежной зоне.
Совершает сезонные миграции: летом держится в верхних горизонтах, зимой опускается
глубже. Нижняя граница солености, при которой встречается эта копепода в Азовском море,
составляет 8,5 ‰. Интенсивное развитие C. ponticus получает при солености 12,4-13,0 ‰,
в результате чего при осолонении водоема становится достаточно распространенным
видом. В Черном море соленость воды не лимитирует его развитие, и он является массовой
формой.
Этот вид отличается высокой плодовитостью, одна самка дает до 14 генераций. В течение
4-5 месяцев наблюдается 3-4 генерации. Наибольшее число яиц в кладке (20-25 шт.) отмечается
при температуре воды 23-25°С. При снижении температуры до 18-22 °С количество яиц в кладке
уменьшается до 10-12 шт. Продолжительность науплиального периода развития составляет 10
суток, копеподитного – 27 суток, взрослой стадии – 58 суток. После периода размножения
летними яйцами (июнь-август) самки откладывают зимующие формы яиц (август-сентябрь),
которые обеспечивают воспроизводство популяции в следующем году.
Копепода C. ponticus обладает истинным смешанным типом питания, так как у него
наряду с фильтрацией водорослей иногда наблюдается активное захватывание животной пищи.
Размер животных и растительных пищевых объектов колеблется от 1,5-3,0 μm до 0,4-1,0 мм.
Вид встречается в умеренной зоне Атлантического океана, в Индийском океане, в
Средиземном, Черном и Азовском морях.
42
А
Б
43
В
Рисунок 16 – Внешний вид Centropages ponticus Karavaev, 1895
(А, Б – науплиус, В – самец)
(фото Назирова М.А.)
Семейство Clausocalanidae
У представителей этого семейства цефалоторакс яйцевидной формы, несколько расширен
к средней части. Головной сегмент слит с первым торакальным, последние торакальные
сегменты слиты. Абдомен самки состоит из 4-х, а самца – из 5 сегментов. Общее число
сегментов у самки составляет 8, у самца – 9. Пятая пара ног у самки одноветвистая, 3-х
членистая, иногда полностью или частично редуцированная до небольшой щетинки. Пятая
пара ног самца одноветвистая, ноги тонкие, длинные.
В Черном море обитает один род – Pseudocalanus.
44
Род Pseudocalanus
Имеет признаки семейства. В Черном море встречается один вид – Pseudocalanus
elongatus.
Pseudocalanus elongatus (Boeck, 1865)
Calanus clausi Brady, 1865
Clausia elongata Boeck, 1865
Pseudocalanus minutus elongatus (Boeck, 1865)
У P. elongatus тело удлиненное, стройное, прозрачное, иногда розоватое или красноватое
в области рта и на конце абдомена, Верхняя часть головы овально закруглена. Антенны у обоих
полов длинные, достигают середины узкого симметричного абдомена. Генитальный конец
самки с сильно выступающим вентральным утолщением, длиннее, чем у самца. Ноги пятой
пары самца почти одинаковой длины. Длина самки – 1,3-1,8 мм, самца – 1,0 мм.
Это холодолюбивый, батипелагический, морской вид. Зимой встречается от поверхности
до сероводородной зоны, летом обычно отсутствует в слое 0-25 м. Совершает суточные
вертикальные миграции, связанные со степенью прогрева верхних слоев воды. P. elongatus
более многочисленен в открытых водах, нежели в прибрежных районах. Вспышки численности
младших копеподитных стадий в прибрежье связаны с привносом их из открытой части моря
во время сгонно-нагонных ветров.
В Черном море P. elongatus размножается в течение всего года, давая 5-6 генераций.
Наиболее интенсивное размножение этого вида отмечается в зимний и весенний периоды.
Максимальное количество яиц (27-30 шт.) он откладывает при температуре 7-15°С. Общая
продолжительность развития от оплодотворенного яйца до созревания взрослой особи
составляет 34-38 суток.
Этот вид копепод является грубым фильтратором, фитофагом. Основу пищи составляют
развивающиеся в массе водоросли – жгутиковые, динофлагелляты, диатомовые размером от
16 до 160 μm.
Является широко распространённым, массовым видом северной части Атлантического,
Тихого океанов и Черного моря. В Азовском море встречаются только копеподитные стадии,
которые проникают в юго-восточный район при повышении солености.
Семейство Diaptomidae
У представителей семейства Diaptomidae тело широкое, укороченное. Головной сегмент
цефалоторакса отчленен от первого торакального. Последние торакальные сегменты или
полностью расчленены, или след их расчленения в виде насечки заметен по бокам тела.
Абдомен самки состоит из 3, самца – из 5 сегментов. Общее число их у самки составляет 9,
самца – 11.
45
Для этого семейства характерна асимметрия в строении последнего торакального
и анального сегментов, а также наличие у самки крыловидных лопастей на последнем
торакальном сегменте. Пятая пара ног у обоих полов двуветвистая, у самки – симметричная, у
самца – асимметричная.
Виды данного семейства являются пресноводными и солоноватоводными.
Род Eudiaptomus
У представителей этого рода тело чаще стройное, удлиненное, 7-8-сегментное у самки
и 10-сегментное у самца. Головной сегмент сливается с первым торакальным, а четвертый
и пятый торакальный четко не разделены, по краю на границе этих сегментов имеются
лишь более или менее заметные насечки. Нижние углы последнего торакального сегмента
у самок вытянуты в лопасти. У разных видов они различно устроены. В середине заднего
края последнего торакального сегмента имеется глубокий вырез. Генитальный сегмент
самки симметричный, в средней части заметно расширен, по бокам вооружен небольшими
коническими зубчиками, длина его превышает ширину. У самца генитальный сегмент тоже
симметричный, но значительно короче, чем у самки, более прямоугольный, без зубчиков по
бокам. Фуркальные ветви короткие, широкие, каждая вооружена пятью короткими оперенными
щетинками, расположенными веерообразно. Пятая пара ног у обоих полов не плавательная,
двуветвистая, у самки симметричная, у самца асимметричная. У самки один яйцевой мешок.
Встречаются в лиманах и эстуариях рек, впадающих в Азовское и Черное моря. В этих
водоемах обитают два вида – Eudiaptomus gracilis и E. salinus.
Eudiaptomus gracilis (Sars G.O., 1863)
Diaptomus gracilis Sars G.O., 1863
Diaptomus westwoodii Lubbock, 1864
Вид имеет стройное, удлиненное тело. У обоих полов нижние углы последнего
сегмента цефалоторакса с широкими лопастями, по краям вооружены шипами. У самки
цефалоторакс сужен в нижней части и наиболее широкий – в средней. Передние антенны очень
длинные, обычно длиннее всего туловища. Абдомен 2-3 сегментный, генитальный сегмент
симметричный, длинный, в средней части заметно расширен. Фуркальные ветви короткие,
широкие, каждая вооружена пятью короткими оперенными щетинками, расположенными
веерообразно. Ноги пятой пары самки с одночленистыми эндоподитами палочковидной
формы и с тремя придатками на конце. Экзоподит трехчленистый, первый членик которого
длинный, без придатков, второй – небольшой, а третий очень маленький, квадратный, вооружен
длинной щетинкой и шипиком. Правая пятая нога самца с двухчленистым экзоподитом первый
членик которого имеет квадратную форму с маленьким шипиком. Второй членик удлиненно-
овальный, с мощным шипом на внешнем крае и слабоизогнутым концевым когтем. Эндоподит
колбасовидной формы. Левая нога пятой пары самца с двухчленистым экзоподитом, первый
46
членик которого удлиненный с маленькой гиалиновой пластинкой, второй – грушевидной
формы с пучком тонких волосков. Длина самца составляет 1,0-1,2 мм. Длина самки составляет
1,0-1,5 мм (рисунок 17).
Рисунок 17 – Внешний вид Eudiaptomus gracilis (Sars G.O., 1863) (самка)
(фото Бычковой М.В.)
Обычно этот вид каляноид дицикличен. Имеет два сезонных максимума – в теплое
летнее время года и зимой, однако в некоторых водоемах он моноцикличен. В одних случаях
встречается только летом (Боденское озеро), в других (оз. Кабана в Татарстане) доминирует в
зимнее время.
E. gracilis – эвритермный вид, характерный для пелагического планктона озер, лиманов и
слабосоленых морских водоемов. В более мелких водоемах (пруды) встречается гораздо реже,
а в лужах только в виде исключения, попадая в них случайно при разливе рек.
47
Встречается в самой распресненной оконечности восточной части Таганрогского залива,
р. Дон, Цимлянском водохранилище, р. Кубань и водохранилищах Краснодарского края, в
приазовских лиманах.
Обитает в олиготрофных и эвтрофных водоемах и отсутствует в дистрофных озерах.
Олиго-мезасапроб.
Семейство Paracalanidae
У представителей этого семейства головной сегмент не отчленен от первого торакального,
последние грудные сегменты не расчленены. Общее число сегментов тела самки 6-8, самца –
9. Пятая пара ног у обоих полов одноветвистая, не плавательного типа.
В Черном и Азовском морях выявлен только один род – Paracalanus.
Род Paracalanus
Имеет признаки семейства. В Черном и Азовском морях встречается один вид.
Paracalanus parvus (Claus, 1863)
согласно WoRMS – Paracalanus parvus parvus (Claus, 1863)
Calanus parvus Claus, 1863
Paracalanus lagunaris (Grandori, 1912)
Paracalanus parvus (Claus, 1863)
Piezocalanus lagunaris Grandori, 1912
Scolecithrix ancorarum Oliveira, 1946
Вид имеет удлиненно-овальное, крепкое, коренастое тело. Головной конец
цефалоторакса плавно закруглен у самки и незначительно выступает у самца. В верхней
части цефалоторакса самца имеется горб, хорошо различимый при рассмотрении его
сбоку. Антенны достигают середины абдомена. Фуркальные ветви симметричные,
короткие, на них расположены четыре пары фуркальных щетинок почти одинаковой
длины. На генитальном сегменте самки имеется вентральный выступ, хорошо видимый
сбоку. Ноги пятой пары у обоих полов одноветвистые. У самки пятая пара ног маленькая.
Пятая пара ног самца асимметричная, правая нога очень маленькая, левая значительно
длиннее (приблизительно в 3 раза). Тело самки иногда с синеватым оттенком, самцы
светло-желтые, иногда с красными пятнышками на голове. Длина самки – 0,8-1,0 мм.
Длина самца достигает 1,0 мм (рисунок 18).
P. p. parvus наиболее интенсивно размножается ранней весной и в начале лета. Количество
генераций достигает 7-8, а число яиц в кладке при температуре 18-22°С насчитывает 13 шт.
Общая продолжительность развития от оплодотворенного яйца до созревания взрослой особи
составляет 29-38 суток.
48
Является пассивным, тонким фильтратором, фитофаг. Спектр питания у этого вида
состоит из мелких водорослей (размер от 5-8 до 16 μm), простейших и бактериопланктона.
Морской, эвритермный, эвригалинный вид. Температурный оптимум обитания находится
в пределах 10-20°С. Относится к олиготрофным формам планктофауны.
Широко распространен в водах Атлантического, Тихого и Индийского океанов, Северного,
Балтийского, Норвежского, Средиземного, Адриатического, Красного и Черного морей. В
Черном море встречается до глубины 150-175 м, но предпочитает обитать в поверхностном
горизонте. В Азовском море встречается только в периоды его осолонения, проникая сюда с
компенсационными потоками черноморской воды.
А
49
Б
Рисунок 18 – Внешний вид Paracalanus parvus (Claus, 1863) (самка)
(фото Бычковой М.В.)
Семейство Pontellidae
Тело узкое стройное. Характерно наличие крупных глаз. Головной сегмент отделен от
первого торакального, последние торакальные сегменты слиты или разделены. Абдомен самки
состоит из двух-трех, самца – из пяти сегментов. Общее число сегментов самки восемь-девять,
самца – девять-десять. Длина рачков – 2-3 мм.
Антенны I самки по длине почти равны цефалотораксу. Эндоодиты торакальных ног
двухчленистые. Пятая пара ног у самок одно-двуветвистая, у самцов – одноветвистая, правая
нога обычно с клешней. Обитатели морей. В Черном и Азовском морях встречается три рода.
50
Род Anomalocera
Рачки крупные, легко распознаются по наличию двух глазных линз. Цефалоторакс
шестисегментный, трехсегментный абдомен самки слабо асимметричен, в строении абдомена
самца резко выражена асимметрия. В Черном море живет только один вид – Anomalocera
patersonii Templeton, 1837.
Anomalocera patersonii Templeton, 1837
Anomalocera patersoni Templeton, 1837
Anomalocera splendidus (Goodsir, 1843)
Irenaeus patersonii Claus, 1866
Irenaeus splendidus Goodsir, 1843
Pontella eugeniae Leuckart, 1859
Pontia pattersonii (Templeton, 1837)
На нижнем углу правой стороны генитального сегмента самки небольшая щетинка. Фуркальные
ветви слабо асимметричны. У самца правый нижний угол последнего торакального сегмента и правый
угол генитального сегмента крючкообразно оттянуты назад. Пятая пара ног сапки двуветвистая.
Пятая пара ног у самца одноветвистая. Правая нога четырехчленистая. Третий членик вздутый, с
вытянутым нижним внутренним углом, вместе с четвертым крючкообразно направленным наружу
члеником образует клешню. Левая нога трехчленистая, с сильно удлиненным вторым члеником.
Длина самки 3,5-4,2 мм, самца – 3 -4 мм (рисунок 19). A. patersonii имеет фиолетовую окраску.
Эвритермная круглогодичная форма Черного моря, в массовых количествах обычно не
встречается.
Рисунок 19 – Внешний вид науплиуса Anomalocera patersonii Templeton, 1837 (?)
(фото Бычковой М.В.)
51
Семейство Pseudodiaptomidae
У представителей этого семейства тело стройное, удлиненное. Головной сегмент
отделен от грудного, последние грудные сегменты не слиты. Абдомен у самки имеет 4, а у
самца – 5 сегментов. Общее число сегментов тела самки 10, самца – 11. Пятая пара ног самки
одноветвистая, самца – двуветвистая. Семейство состоит из одного рода – Calanipeda.
Род Calanipeda
Имеет признаки семейства. В Черном и Азовском морях обитает один вид – Calanipeda
aquaedulcis.
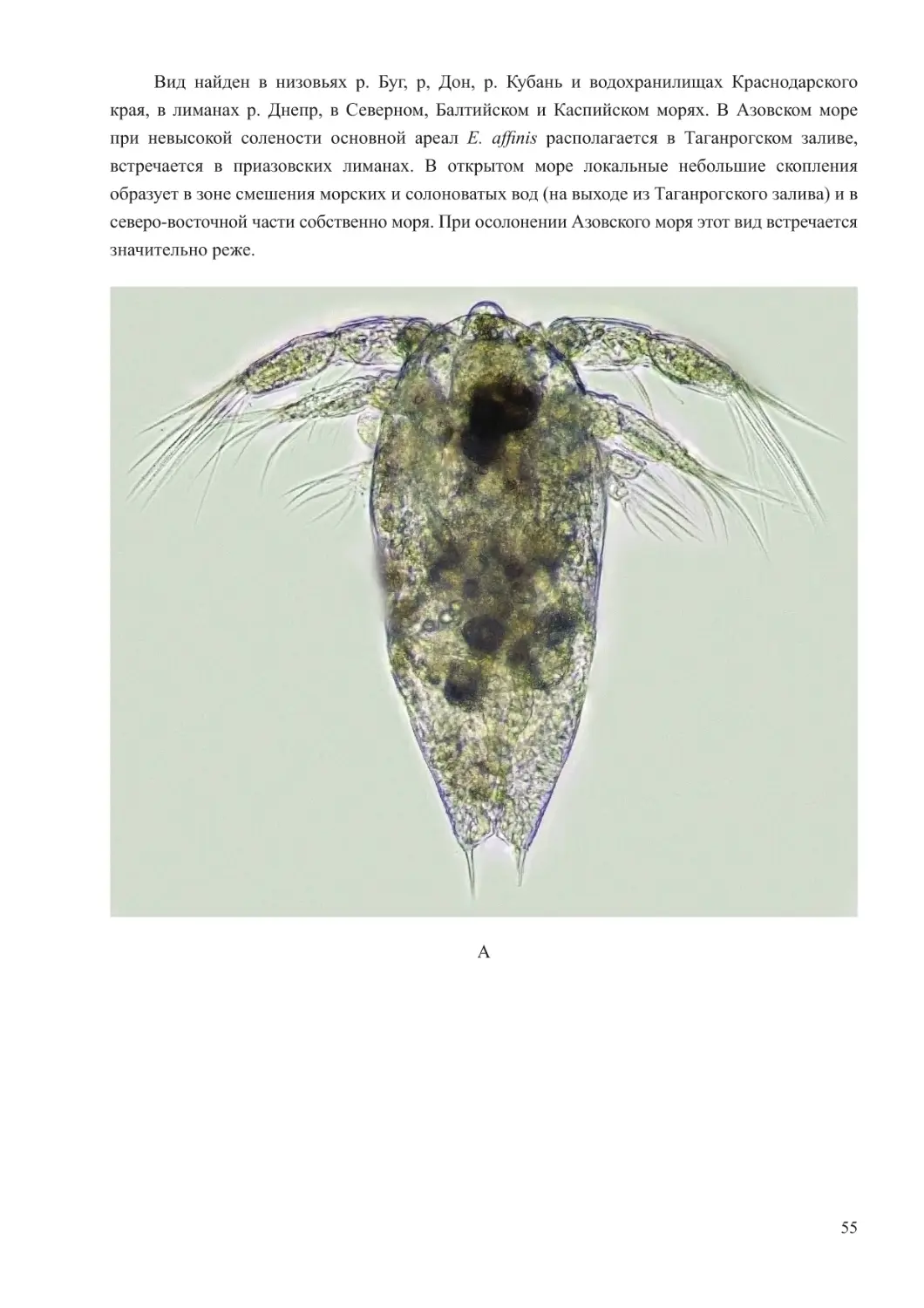
Calanipeda aquaedulcis Krichagin, 1873
Poppella guernei Richard, 1888
Siatella durbini Labbé, 1927
Этот вид имеет очень стройное, удлиненное тело, равномерно сужающееся к переднему
и заднему концам цефалоторакса. Антенны у обоих полов широко расставлены в стороны.
Асимметричный, генитальный сегмент самки значительно длиннее остальных, на левой стороне
снабжен крючковидным придатком. Генитальный сегмент самца симметричный. Тонкие, длинные,
фуркальные ветви имеют 5 пар коротких оперенных щетинок. Пятая нога самки одноветвистая.
У самца пятая пара ног резко асимметричная, обе ноги двуветвистые, левая нога гораздо короче
правой. Характерной особенностью строения 5 пары ног самца является наличие на правой ноге
серповидного когтя. Длина самок – 1,20-1,45, самцов около – 1,0 мм (рисунок 20).
Этот вид копепод по типу питания является фильтратором, питается в основном
растительной пищей, потребляя фитопланктон (в основном мелкие и средние водоросли
размером 3-30 μm и 30-50 μm и детритные частицы размером 20-150 μm.
C. aquaedulcis является солоноватводным, пелагическим видом, но активно развивается
также в прибрежных районах и лиманах. Границы солености, при которых вид встречается,
находятся в пределах 1-13 ‰. Соленость порядка 5-7 ‰ является оптимальной для массового
развития популяции. В пресной воде C. aquaedulcis живет продолжительное время, но
становится менее устойчивой к неблагоприятным факторам среды. Переносит соленость более
13 ‰ при невысокой температуре воды, но погибает при температуре 24-25 °С.
Обитает в Каспийском и Азовском морях, в устьях их рек, лиманах и заливах. При
невысокой солености Азовского моря ареал C. aquaedulcis распространяется почти на всю
его акваторию. В Таганрогском заливе с начала лета по середину осени образует большие
скопления. При осолонении Азовского моря ареал заметно сужается и ограничивается восточной
частью Таганрогского залива. Встречается в р. Дон, Цимлянском водохранилище, р. Кубань и
водохранилищах Краснодарского края. В Черном море встречаемость этого вида приурочена к
сильно опресненным участкам (лиманам, устьям рек), в самом море он отсутствует.
52
А
Б
В
Г
53
Д
Рисунок 20 – Внешний вид Calanipeda aquaedulcis Krichagin, 1873
(А, Б – науплиус, В, Г – самка, Д – самец)
(фото Бычковой М.В.)
Семейство Temoridae
Тело у представителей семейства Temoridae различной формы: у одних видов оно
стройное, удлиненное, у других – расширенное, укороченное. Головной сегмент отделен от
первого торакального, последние грудные сегменты могут быть разделены, или иметь только
следы слияния. Абдомен самки состоит из 3-х, самца – из 5 сегментов. Обще число сегментов
тела у самки 9, у самца – 11. Пятая пара ног у обоих полов одноветвистая, иногда на левой ноге
самца сохраняется рудимент эндоподита.
Представители этого семейства – пресноводные и солоноватоводные формы. В Азовском
море представлено двумя родами – Eurytemora и Heterocope.
54
Род Eurytemora
У представителей этого рода цефалоторакс незначительно шире спереди, имеет 6
сегментов, последние торакальные сегменты отелены друг от друга. Головной конец у обоих
полов закруглен, последний торакальный сегмент самки с оттянутыми углами, у самца углы
закруглены. Абдомен симметричный у обоих полов, у самки он в 1,5 раза короче цефалоторакса.
Генитальный сегмент самки мало отличается от последующих. Фуркальные ветви у разных
видов различной длины, у самца они обычно несколько длиннее. Фуркальных щетинок пять,
они короче фуркальных ветвей и расположены веерообразно. Ноги пятой пары одноветвистые
у обоих полов: у самки симметричные, у самца – асимметричные.
В лиманах Черного моря и в Азовском море встречаются два вида – Eurytemora affinis и
Eurytemora velox.
Eurytemora affinis (Poppe, 1880)
Eurytemora affinis hirundoides (Nordquist, 1888)
Eurytemora hirundo Giesbrecht, 1881
Eurytemora hirundoides (Nordquist, 1888)
Eurytemora inermis (Boeck, 1865)
Temora affinis Poppe, 1880
Temora inermis Boeck, 1865
Temorella affinis Poppe, 1885
Temorella affinis hirundoides Nordquist, 1888
Temorella inermis (Boeck, 1865)
Этот вид ракообразных имеет широкое, укороченное тело. У самки цефалоторакс заметно
сужается к нижней его части. Последний сегмент цефалоторакса имеет большие треугольные
лопасти, внешние края которых вооружены мелкими шипиками. Вершины этих лопастей
направлены в стороны. Генитальный сегмент самки крупный, более широкий в верхней своей
половине, посередине имеются направленные вниз выступы. Фуркальные ветви длинные,
тонкие с пятью щетинками, которые густо оперены. Наружная поверхность фуркальных ветвей
и почти всего анального сегмента покрыта мелкими шипиками. Пятая пара ног самки и обе
пары ног самца трехчленистые. У самки второй членик пятой пары ног с двумя большими
шипами на внешнем крае. У самца последний членик правой ноги пятой пары пальцеобразный
и на конце закруглен. Последние членики обеих ног самца обычно образуют перекрест. Длина
самки – 1,5-1,8 мм, самца – 1,2-1,5 мм (рисунок 21).
По типу питания E. affinis является тонким фильтратором. В качестве пищи использует в
основном мелкие и средние водоросли размером от 3-30 μm до 30-50 μm.
Характерной чертой вида является его эвритермность. Он широко распространен в
солоноватых водах, в опресненных прибрежных участках морских водоемов (лиманы и устья
рек), встречается в пресной воде. Обитает как в условиях пелагической зоны глубоководья, так
и в прибрежных районах.
55
Вид найден в низовьях р. Буг, р, Дон, р. Кубань и водохранилищах Краснодарского
края, в лиманах р. Днепр, в Северном, Балтийском и Каспийском морях. В Азовском море
при невысокой солености основной ареал E. affinis располагается в Таганрогском заливе,
встречается в приазовских лиманах. В открытом море локальные небольшие скопления
образует в зоне смешения морских и солоноватых вод (на выходе из Таганрогского залива) и в
северо-восточной части собственно моря. При осолонении Азовского моря этот вид встречается
значительно реже.
А
56
Б
В
57
Г
Д
Рисунок 21 – Внешний вид Eurytemora affinis (Poppe, 1880)
(А – науплиус, Б, В – самка, Г, Д – самец)
(фото Назирова М.А.)
Eurytemora velox (Lilljeborg, 1853)
Eurytemora adleri Schiklejew, 1931
Eurytemora clausi Brady, 1891
Eurytemora clausi (Hoek, 1876)
Temora clausii Hoek, 1878
Temora velox Lilljeborg, 1853
Temorella clausi Claus, 1881
Temorella lacinulata Sars G.O., 1897
58
Вид имеет укороченное, коренастое тело у самки и более стройное у самца. Задний
сегмент цефалоторакса самки несет угловые, направленные назад выросты с тонкими шипами
с боков. Фуркальные ветви короче двух последних сегментов абдомена вместе взятых.
Внутренние края ветвей опушены волосками. Фуркальных щетинок пять, четыре из которых
прикреплены на концах ветвей, а одна в нижней части наружного края. Шетинки значительно
короче фуркальных ветвей, густо оперены волосками. Пятая пара ног самки трехчленистая.
Второй членик пятой пары ног имеет один небольшой шип на внешнем крае.
У самца на заднем закругленном сегменте цефалоторакса выростов нет. Правая пятая
нога самца четырехчленистая, изогнутая, левая – трехчленистая, в месте сочленения образует
изгиб. Длина фуркальных ветвей равна длине двух последних сегментов абдомена. Длина
самца – 1,2-1,8 мм. Длина самки – 1,3-2,2 мм.
E. velox – эвригалинный вид, широко распространен в пресных и слабосоленых водах.
Обитает в лиманах Днепра и Буга, в низовьях Дона и Кубани, и водохранилищах Краснодарского
края, Цимлянском водохранилище. Встречается в Северном, Балтийском и Каспийском морях
(вблизи устьев рек). В Азовском море ареал E. velox при невысокой солености занимает почти
весь Таганрогский залив, а при осолонении только его восточную часть, присутствует в
кубанских лиманах.
Род Heterocope
У представителей этого рода цефалоторакс 6 - сегмнтный, два последних торакальных
сегмента нечетко разделены. Углы последнего торакальнго сегмента у обоих полов закруглены.
Абдомен у самки состоит из 3-х, у самца – из 5 сегментов. Конечных фуркальных щетинок три.
Пятая пара ног у самки симметричная, у самца – асимметричная.
В Азовском море встречается один вид – Heterocope caspia.
Heterocope caspia Sars G.O., 1897
Вид имеет удлиненное, стройное тело с наибольшей шириной в области переднего
сегмента. Цефалоторакс 6-сегментный, с нечетким разделением двух последних сегментов.
Передние антенны почти достигают заднего конца брюшка. Углы последнего сегмента у
обоих полов закруглены. Абдомен стройный, удлиненный, генитальный сегмент расширен в
его переднем отделе. Каудальные ветви стройные, почти параллельные, с тремя относительно
длинными щетинками на каждой ветви. Ноги пятой пары самки одноветвистые, трехчленистые.
Правая пятая нога самца одноветвистая, левая – двуветвистая, с одночленистым эндоподитом
в виде серповидного отростка. Длина самцов – около 1,5 мм. Длина самок (с каудальными
щетинками) 1,7-1,9 мм (рисунок 22).
H. caspia является эвригалинным, планктическим видом, предпочитает районы с низкой
соленостью, но может жить и в пресной воде, хотя здесь не достигает значительного
количественного развития. Является характерным видом для Каспийского моря и достаточно
59
многочисленным в восточной части Таганрогского залива при солености 0,3-3,0 ‰. В открытом
море не встречается. Обитает в р. Дон, Цимлянском водохранилище, р. Кубань и водохранилищах
Краснодарского края, лиманах Азовского моря.
А
60
Б
Рисунок 22 – Внешний вид Heterocope caspia Sars G.O., 1897
(А – науплиус, Б – копеподит)
(фото Бычковой М.В.)
61
Отряд Cyclopoida
У представителей этого отряда цефалоторакс яйцевидной формы, абдомен
значительно уже цефалоторакса. Голова слита с первым торакальным сегментом (сем.
Cyclopinidae и Cyclopidae) или отделена от него (сем. Oithonidae). Задние углы последнего
сегмента цефалоторакса закруглены или оттянуты. Цефалоторакс у самок включает 4
сегмента, самцов, за редким исключением, 5 сегментов. Общее число сегментов тела
у самок составляет 9-10, у самцов – 10-11. Фуркальные ветви разной длины и формы,
оснащены одной боковой и одной спинной щетинкой и четырьмя конечными. Средние
конечные щетинки значительно длиннее крайних. Ноги пятой пары у обоих полов
имеют одинаковое строение. Они резко отличаются от плавательных торакальных ног,
маленькие, чаще одноветвистые, расчлененность ветвей различна. Самка вынашивает
яйца в двух яйцевых мешках, расположенных по бокам генитального сегмента. Самки
крупнее самцов, иногда значительно.
Представители отряда Cyclopoida – свободноживущие и паразитические формы,
большей частью пресноводные виды, но есть солоноватоводные и морские.
Семейство Cyclopidae
У представителей этого семейства головной сегмент слит с I торакальным. Вместе с ним
может превышать длину всех остальных сегментов цефалоторакса (род Euryte). Общее число
сегментов у тела самки 9, у самца – 10. У самок некоторых видов последний торакальный
сегмент образует боковые выросты (род Cyclops). Фуркальные ветви имеют разную длину, у
некоторых видов они могут быть очень короткими. Ноги пятой пары, обычно состоящие из
1-2, редко 3 члеников, достаточно развиты.
Род Cyclops
К этому роду относятся крупные циклопы длиной 1,15-2,32 мм. Цефалоторакс животных
ступенчатого вида. Первый сегмент головогруди слегка суживается к переднему краю,
остальные сегменты со свободными боковыми углами. Внутренние края фуркальных ветвей
оперены волосками. Ноги пятой пары двучленистые, членики расположены один на другом.
Яйцевые мешки округлые или овальные, яйца круглые.
Род очень богат видами, содержит около 20 таксонов. В Азовском море в опресненных
районах встречаются два вида – Cyclops vicinus vicinus и Cyclops strenuus.
Cyclops vicinus vicinus Uljanin, 1875
Cyclops uljanini Sovinsky, 1887
Cyclops vicinus Uljanin, 1875
62
Этот вид циклопод имеет стройное тело. У самки первые три сегмента цефалоторакса без
оттянутых углов. Предпоследний грудной сегмент очень крупный, последний – значительно
уже, оба имеют сильно развитые, оттянутые, заостренные лопасти. Передние антенны у этих
циклопов доходят примерно до середины второго сегмента цефалоторакса. Генитальный сегмент в
верхней части расширен, имеет небольшие выступы по бокам. Фуркальные ветви длинные, узкие,
обычно несильно расходящиеся, со сплошным рядом волосков на внутренних краях. Из крайних
фуркальных щетинок внутренние щетинки приблизительно вдвое длиннее внешних. Средние
щетинки хорошо развиты, по длине незначительно отличаются друг от друга. Самка имеет два
яйцевых мешка овальной формы, тесно прилегающих к абдомену. Длина самки составляет 1,25-
2,18 мм (рисунок 23).
Рисунок 23 – Внешний вид Cyclops vicinus vicinus Uljanin, 1875 (самка)
(фото Бычковой М.В.)
63
У самцов тело стройнее, чем у самки. Два задних сегмента цефалоторакса без выростов
сзади и с боков. Ноги шестой пары вооружены очень длинной внешней щетинкой, длина самца
1,15-1,50 мм.
В эвтрофных водоемах средней Европы половозрелые особи встречаются весь год,
однако максимум их количества отмечается зимой (декабрь) и весной (апрель). Зимние самки
несколько мельче летних и содержат меньшее количество яиц в мешках.
C. v. vicinus – пресноводный, эвритермный вид, выносит некоторое осолонение, интенсивно
развивается в летний период. Обитает в пелагиали эвтрофных озер, а также в мелких стоячих
водоемах и медленно текучих реках с рН 5,9-9,1. Избегает значительно дистрофированные
водоемы.
Исключая водоемы субтропиков, обитает в пределах всех ландшафтных зон, в том числе
и в лиманах Черного и Азовского морей, р. Дон, р. Кубань, Цимлянском водохранилище.
C. v. vicinus относится к ß-мезосапробам.
Род Mesocyclops
У представителей этого рода тело большей частью стройное, удлиненное, с резко
дифференцированными передним и задним отделами. Генитальный сегмент удлиненный,
равномерно суживающийся кзади. Фуркальные ветви относительно короткие, их длина обычно
в 2,5-3,5 раза (редко в 4-5 раз) более их ширины. Лишь у немногих видов на внутренних краях
фуркальных ветвей имеются ряды волосков. Род представлен 6 видами, из которых в Азовском
море в сильно опресненных районах встречается Mesocyclops leuckarti leuckarti.
Mesocyclops leuckarti leuckarti (Claus, 1857)
Cyclops leewenhoekii Hoek, 1878
Cyclops leuckarti Claus, 1857
Cyclops leuckarti setosa Wierzejski, 1893
Cyclops longicaudatus Poggenpol, 1874
Cyclops scourfieldi Brady, 1891
Mesocyclops leuckarti (Claus, 1857)
Тело самки M. l. leuckarti относительно широкое. Передние антенны одноветвистые,
обычно доходят до заднего конца второго сегмента цефалоторакса или немного длиннее
его. Генитальный сегмент стройный, удлиненный, спереди расширенный и несколько
суживающийся кзади. Фуркальные ветви либо слабо расходятся, либо почти параллельны
друг другу. Их длина в 3,0-3.5 раза превышает ширину, внутренние края ветвей без волосков.
Длина самки – 0,9-1,3 мм. Яйцевые мешки обычно крупные, расходящиеся по бокам, парные, с
различным количеством яиц – от 4 до 30 шт. в каждом. У самцов, как и у самок, тело стройное,
удлиненное. Передние антенны, в связи с их функцией при копуляции, сильно изменены в
специальный хватательный орган. Длина самца – 0,8-1,0 мм (рисунок 24).
64
А
Б
Рисунок 24 – Внешний вид Mesocyclops leuckarti leuckarti (Claus, 1857)
(А – самка, Б – самец)
(фото Хренкина Д.В.)
M. l. leuckarti – истинный планктонный вид. Обитает в самых различных водоемах –
от мелких луж и болот до крупных озер. В озерах является характерным компонентом не
только пелагического планктона, но и прибрежной зоны. В пелагиали озер обитает в зоне
65
эпилимниона, совершая активные суточные вертикальные миграции, доходящие в некоторых
глубоких озерах до 40 м. В мелких водоемах и ключах встречается гораздо реже, но бывает
многочисленным в составе планктона рек и прудов.
Это теплолюбивый вид, развивается в теплое время года и исчезает зимой. При
температуре от 15-20°C до 25°C размножается особенно интенсивно. Более низкая температура
воды порядка 4°C вызывает гибель науплиусов. Кислородный оптимум, вероятно, лежит в
пределах, близких к нормальному насыщению. Оптимальный интервал рН воды равен 6,0-8,5,
более низкое значение (4,5) является критическим, а около 4,0 – летальным.
M. l. leuckarti – космополит. Встречается в водоемах крайнего Севера, южного Кавказа,
Сахалина и бассейна р. Амур. В зоне тундры встречается гораздо реже и отсутствует в районе
арктических островов. Неоднократно указывался для планктона горных озер, расположенных
не выше 1240 м над уровнем моря.
В Азовском море встречается в опресненных районах восточной части Таганрогского
залива, р. Дон, р. Кубань, приазовских лиманах.
Олиго-мезосапроб.
Семейство Oithonidae
Ракообразные семейства Oithonidae имеют стройное тело. Антенны I у самки длинные,
тонкие. Мандибулярные щупики хорошо развиты, мандибулы с сильными когтеобразно
загнутыми шипами. Максилярные ноги удлиненные. Ветви торакальных ног чаще 3-членистые.
Ноги пятой пары слабо развиты, редуцированы.
В Черном и Азовском морях встречается только один род – Oithona.
Род Oithona
У представителей рода Oithona тело узкое, стройное, передний конец цефалоторакса у
самок заострен или закруглен, у самцов всегда закруглен. Рострум может быть у обоих полов,
либо только у самки, либо отсутствовать у обоих полов.
В Черном и Азовском морях обитают два вида – Oithona davisae и Oithona similis.
Oithona davisae Ferrari F.D. & Orsi, 1984
По форме тела и размерам вид близок к O. nana. У самок передний отдел короткий, чуть
длиннее заднего. Передняя часть головы срезана прямолинейно, имеется довольно длинный
остроконечный рострум с резким изгибом. Антенны достигают начала III торакального
сегмента. На передней части головы обычно хорошо заметен темный глаз. Длина самки
составляет 0,48-0,56 мм (рисунок 25).
У самца передняя часть головы широкая, рострум отсутствует. Торакс немного сужается
по направлению к нижней своей части. Хватательные антенны обычного строения, но несколько
сжаты. Длина самца – 0,46-0,50 мм.
66
По типу питания этот вид является хищником, его рацион составляют мелкие
гетеротрофные жгутиковые.
O. davisae – эпипланктонный теплолюбивый, эвригалинный эндемик умеренных
прибрежных вод Восточной Азии. В южных морях России – вид-вселенец. В 2001 г.
в Севастопольской бухте был обнаружен новый для Черного моря вид копепод, который
вначале определили как O. brevicornis. На протяжении последующих 4 лет его не встречали,
а в 2005 г. было зарегистрировано несколько экземпляров чужеродного вида циклопоидной
копеподы, который также идентифицировали как O. brevicornis. Позже было установлено,
что этот вид морфологически близок к виду O. davisae, который с 2006 г. стал доминировать
по численности в летне-осенний период в Севастопольской бухте (Губанова и др., 2018). С
2009 г. O. davisae широко распространилась в прибрежных районах Черного моря. В северо-
восточной части моря этот вид в массовых количествах был обнаружен в 2010 г.
В больших количествах развивается в прибрежных эвтрофированных районах при
температуре воды не ниже 18-20°С и солености от 5 до 55 ‰. Цикл развития этой циклопоиды
от яйца до взрослой особи в Черном море длится 25±5 дней, число яиц в кладке доходит
до 18 штук. Совершает незначительные вертикальные миграции. В светлое время суток
наибольшую численность формирует в слое 10-25 м, в темное – в слое 0-10 м.
В настоящее время является массовый формой зоопланктона Черного и Азовского морей,
заняв экологическую нишу ранее многочисленного вида O. nana.
А
67
Б
В
Рисунок 25 – Внешний вид Oithona davisae Ferrari F.D. & Orsi, 1984
(А – науплиус, Б, В – самка)
(фото Хренкина Д.В.)
68
Oithona similis Claus, 1866
Oithona helgolandica (Claus, 1863)
Oithona pygmaea Boeck, 1865
Oithona spinifrons Boeck, 1865
Oithonella helgolandica Claus, 1863
Вид имеет узкое, стройное тело. Цефалоторакс самки одинаково широкий на всем его
протяжении. Верхняя часть цефалоторакса прямо срезана или слегка округлена, последний
сегмент маленький, округлой формы, Рострум острый, направлен вниз. Антенны не очень
густо оперены щетинками, по длине достигают половины генитального сегмента. Генитальный
сегмент очень длинный и почти равен суммарной длине трех последующих. Фуркальные ветви
такой же длины, как и анальный сегмент. Две средние щетинки фурок длинные, а две крайние
очень маленькие. Ноги пятой пары слабо развиты. Средняя длина самки 0,78 мм с вариациями
от 0,64 до 1,04 мм (рисунок 26).
У самца цефалоторакс несколько расширен в нижнем отделе. Его верхняя часть широко
закруглена, с вентральной стороны имеет тупой вырост, рострум отсутствует. Фуркальные
ветви маленькие. Пятая пара ног двухчленистая, первый членик имеет форму бугорка, второй
членик маленький, удлиненный. Длина самца – 0,6-0,7 мм. Для обоих полов характерна
сезонная изменчивость размера, который увеличивается от середины лета к зиме.
Для вида характерно хищничество. Она активно захватывает одиночные клетки
гетеротрофных флагеллят и инфузорий средних размеров.
O. similis – морской, эвригалинный вид, относится к холодоводному комплексу
морского зоопланктона. Температурный оптимум развития вида находится в пределах
7-15 °С. Размножается в течение всего года, но наиболее интенсивно в холодный период.
С мая по ноябрь предпочитает более глубокие горизонты, в остальное время держится в
поверхностных слоях. Число генераций составляет 6-7, количество яиц в кладке – 14-18 штук.
Общая продолжительность развития от оплодотворенного яйца до созревания взрослой особи
составляет 33-43 суток.
Вид чрезвычайно широко распространен в полярных морях, в Антарктике, в Атлантическом
(умеренный пояс), Тихом и Индийском океанах, Средиземном, Адриатическом, Красном и
Черном морях. В Азовском море эта циклопоида начинает развиваться при солености более 9
‰, проникая через Керченский пролив.
69
А
Б
В
Рисунок 26 – Внешний вид копеподитов Oithona similis Claus, 1866
(фото Бычковой М.В.)
Класс Eurotatoria
Подкласс Monogononta
Отряд Ploima
Представители этого отряда коловраток свободно передвигающиеся, прикрепленных
форм нет. Нога с 1-2 пальцами; у пелагических форм может полностью отсутствовать.
Семейство Asplanchnidae
У представителей этого семейства тело крупное, обычно мешковидное, прозрачное,
покрытое тонкой кутикулой. Оно разделено на голову, туловище (р. Asplanchna) и нередко ногу
(р. Asplanchnopus, р. Harringia) с короткими пальцами. На голове находится коловращательный
аппарат типа Asplanchna с редуцированными буккальным и большим аппикальным полем,
окруженным циркумапикальным венчиком ресничек, колебательное движение которых создает
70
около коловратки вихревые потоки. Туловище часто вздутое, иногда с боковыми выростами.
Нога, если имеется, может быть длинной (р. Harringia) или короткой, сильно сдвинутой на
брюшную сторону (р. Asplanchnopus). Мастакс (глотка) инкудатного типа, иногда с признаками
малеатного. Глазное пятно, если имеется, обычно 1 церебральное. Самки живородящие или
яйцекладущие. Все виды семейства планктонные обитатели.
Род Asplanchna
Тело у представителей этого рода мешковидное, колоколовидное, иногда с боковыми
выступами, прозрачное, покрыто нежной кутикулой. Коловращательный аппарат типа
Asplanchna с большим апикальным полем и простым венчиком ресничек. Мастакс большой,
инкудатного типа, характерный для вида. Нога с пальцами отсутствует. Амиктические самки
этого рода живородящие, а миктические откладывают покоящиеся яйца. В Азовском море
обитает несколько видов, из которых массовый – A. priodonta.
Asplanchna priodonta Gosse, 1850
Asplanchna priodonta priodonta Gosse, 1850
A. priodonta – беспанцирный вид коловраток, имеющий удлиненно-мешковидное,
прозрачное тело, несколько суженное в передней части. Нога отсутствует. Коловращательный
аппарат с большим аппикальным полем и простым венчиком ресничек. Самки живородящие
или яйцекладущие. Яйценосные самки появляются при температуре воды 12-15°С, а наиболее
интенсивное размножение отмечается при температуре 20-22°С. Покоящиеся яйца имеют
толстую оболочку, внутри которой просматриваются жировые капли Длина самок –
250-1500 μm, ширина – 300-500 μm. Размер самцов изменяется в пределах 200-500 μm
(рисунок 27).
A. priodonta – планктонный, эвритермный вид. Типичный представитель планктофауны озер
и прудов, при этом хорошо приспосабливается к солоноватым и даже морским водоемам. Обычно
живет в верхних слоях воды, однако в глубоких озерах встречается и на глубине 25-50 м.
По типу питания – это хищная коловратка. Зубы на внутреннем крае рамусов позволяют
перетирать ей пищевые объекты. Излюбленным кормом являются другие коловратки и мелкие
планктонные ракообразные, в случае их недостатка она переходит на каннибализм и питание
водорослями.
Вид-космополит. В России встречается повсеместно.
В Азовском море основным районом обитания A. priodonta является Таганрогский
залив. При невысокой солености она расселяется почти на всей его акватории, формируя
максимальную численность в восточной части. При повышении солености ареал заметно
сужается, ограничиваясь опресненными районами. Встречается в р. Дон, Цимлянском водо-
хранилище, р. Кубань, приазовских лиманах.
β-мезосапроб.
71
А
Б
72
В
Рисунок 27 – Внешний вид Asplanchna sp. Gosse, 1850
(фото Хренкина Д.В.)
Семейство Brachionidae
Тело коловраток семейства Brachionidae состоит из головы, туловища, а у двух
родов (Brachionus и Platyias) еще и ноги. На голове находится коловращательный аппарат
типа Euchlanis с некоторыми особенностями в строении парацингулума и буккального
поля. Туловище всегда покрыто панцирем, который чаще всего имеет более или менее
выраженную скульптуру в виде фасеток, ограниченных гребнями, сеточки, бугорков,
точек и зернистости. Панцирь у большинства видов дорсовентрально сильно сплющен
и образован брюшной и спинной пластинками, плотно прижатыми краями друг к другу.
Передний спинной, реже брюшной, а также задний край панциря обычно с шипами. Нога
длинная, втягивается, мелкокольчатая или членистая, с 2 короткими пальцами. Мастакс
маллеатного типа.
Представители этого семейства коловраток ведут планктонный образ жизни и лишь
изредка встречаются в планктонобентосе.
73
Род Brachionus
У представителей этого рода панцирь более или менее ригидный, гладкий, на спинной
пластинке обычно со скульптурой в виде гребней, точек, шипиков и фасеток. У некоторых
видов над отверстием для ноги имеется косо поставленная базальная пластинка. Передний
спинной край у коловраток с 2-6 шипами, брюшной – волнистый, с выемкой или вырезом
по середине, реже с выступами или лопастной. Задний край панциря округлый, квадратный
или вытянутый с боков в шипы. Отверстие для ноги немного сдвинуто на брюшную сторону,
обычно округлое с брюшной стороны и почти прямоугольное со спинной. Нога длинная,
червеобразная, мелкокольчатая, с двумя короткими пальцами, часто втянутая.
Виды рода Brachionus – свободноплавающие планктонные организмы, но имеются и
комменсалы. Отличаются сильно выраженным полиморфизмом, давая значительное число
морфологически различных форм.
Brachionus angularis Gosse, 1851
Мелкий вид коловраток. Панцирь твердый, мешковидный, несколько приплюснутый
в спинно-брюшном направлении. Его поверхность гладкая или с легкой структурой в виде
граней, полос или точек, передний спинной край с 2 шипами (рисунок 28). Для ноги имеется
подковообразное, с зубовидными выступами по бокам большое отверстие. Длина панциря
колеблется от 82 μm до 202 μm, ширина – от 66 до 160 μm.
Один из самых распространенных видов коловраток. Предпочитает пресные, эвтрофные
водоемы, но встречается и в солоноватоводных водах.
Встречается в восточной части Таганрогского залива, р. Дон, Цимлянском водохранилище,
р. Кубань, приазовских лиманах.
α–β -мезосапроб.
А
Б
Рисунок 28 – Внешний вид Brachionus angularis Gosse, 1851
(фото Бычковой М.В.)
74
Brachionus diversicornis (Daday, 1883)
Brachionus diversicornis diversicornis (Daday, 1883)
Один из наиболее крупных видов панцирных коловраток р. Brachionus. Имеет
удлиненный панцирь, который приплюснут в спинно-брюшном направлении и сужается
в своей задней части. Передний, почти прямой край оснащен двумя короткими средними
щетинками и двумя длинными боковыми. Задний левый шип короче правого. Длинная нога,
обычно втянутая внутрь панциря, имеет раздвоенные на концах пальцы, которые вооружены
двумя маленькими крючкообразными шипами. Длина панциря – 220-585 μm, ширина – 151-
204 μm (рисунок 29).
Широко распространенный планктонный теплолюбивый летний вид. Развивается
в пресных континентальных водоемах, но встречается в солоноватоводных и морских
(например, Каспийское море).
Встречается в Таганрогском заливе Азовского моря, р. Дон, Цимлянском водохранилище,
р. Кубань, приазовских лиманах.
Тяготеет к водоемам повышенной трофности. β-мезосапроб.
Рисунок 29 – Внешний вид Brachionus diversicornis (Daday, 1883)
(фото Хренкина Д.В.)
75
Brachionus diversicornis homoceros (Wierzejeski, 1891)
Этот вид коловраток отличается от типичной формы чуть расширенной средней частью
панциря, одинаковой длиной задних шипов, расходящихся в стороны, и большей длиной
передних боковых шипов. Длина составляет – 221-305 μm, ширина – 145-182 μm (рисунок
30).
Планктонный летний теплолюбивый вид, развивающийся в эвтрофных водоемах и
водотоках юга России.
Встречается в восточной части Таганрогского залива Азовского моря, р. Дон, Цимлянском
водохранилище, р. Кубань, приазовских лиманах.
β-мезосапроб.
Рисунок 30 – Внешний вид Brachionus diversicornis homoceros (Wierzejeski, 1891)
(фото Бычковой М.В.)
76
Brachionus calyciflorus calyciflorus Pallas, 1766
Brachionus calyciflorus f. amphiceros Ehrenberg, 1838
У этого вида коловраток панцирь очень тонкий, овальный, неразделенный на спинную
и брюшную пластинки. На переднем крае спинной пластинки хорошо видны 4 сравнительно
длинных, почти равных по длине заостренных шипа. Боковые края отверстия для ноги немного
выступают. Длина панциря – 150-570 μm, ширина – 124-300 μm (рисунок 31).
Активная пищевая избирательность отсутствует.
Эвритермный, эвригалинный вид. Встречается в солоноватых водоемах и морях. В
динамике его развития наблюдаются два пика – первый в мае-июне, второй в сентябре-октябре.
Предпочитает водоемы со щелочной средой. Обитает в водоемах различного типа южной и
средней полосы, реже встречается в северных широтах.
Встречается в р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах,
Таганрогском заливе Азовского моря.
α–β -мезосапроб.
Рисунок 31 – Внешний вид Brachionus calyciflorus calyciflorus Pallas, 1766
(фото Бычковой М.В.)
77
Brachionus calyciflorus amphiceros Ehrenberg, 1838
Отличается от типичной формы одинаковой длиной передних шипов, длинными задними
шипами, расставленными в стороны и удлиненными боковыми шипами у отверстия для ноги
(рисунок 32). Длина панциря – 190-350 μm, ширина – 100-170 μm.
Теплолюбивый, эвригалинный вид. Появляется в составе планктона летом и осенью.
Обитает в солоноватых и морских водоемах.
Встречается в основном в Таганрогском заливе, р. Дон, Цимлянском водохранилище, р.
Кубань, приазовских лиманах.
α–β -мезосапроб.
Рисунок 32 – Внешний вид Brachionus calyciflorus amphiceros Ehrenberg, 1838
(фото Бычковой М.В.)
Brachionus forficula Wierzejski, 1891
У этой коловратки панцирь почти прямоугольный средних размеров, несколько
расширенный в средней части. Передний спинной край снабжен 4 шипами, из которых боковые
длиннее, чем срединные или почти равны им. Срединные шипы имеют выемку посередине
и заметно сближены. Задняя часть панциря заканчивается 2 массивными шипами, которые
широко расставлены у своих оснований. Задние шипы согнуты и сближены дистальными
шипами. Этот вид коловраток чрезвычайно разнообразен. Образует около 15 вариететов,
основные различия между которыми заключаются в размерах панциря, а также в длине задних
и передних шипов. Длина панциря колеблется от 106 до 256 μm, ширина от 90 до 220 μm
(рисунок 33).
78
Широко распространенный теплолюбивый, южный вид озер, прудов, водохранилищ и рек.
Встречается в р. Дон (в том числе в устье реки), Цимлянском водохранилище, р. Кубань,
приазовских лиманах.
β-мезосапроб.
Рисунок 33 – Внешний вид Brachionus forficula Wierzejski, 1891
(фото Бычковой М.В.)
Brachionus plicatilis longicornis Fadeev, 1786
Панцирь у этого вида удлиненно-овальный, чуть приплюснутый в спинно-брюшном
направлении, без четкого разделения на спинную и брюшную пластинки. Передний спинной
край панциря имеет четыре хорошо выраженных выступа, отделенных друг от друга
неглубокими вырезами, и снабжен 6 заостренными шипами, крайние из которых самые
маленькие. Задний край округлый, лишен шипов, вырез отверстия для ноги четко обозначен.
Длина панциря изменяется в пределах 125-315 μm, ширина – 105-260 μm (рисунок 34). B. p.
longicornis является одним из вариететов типичной формы Brachionus plicatilis и встречается
с ней совместно.
Эвригалинный, эвритермный вид, широко распространен в солоноватых, морских
водоемах, но встречается и в пресных.
Обитает в прибрежных районах Черного, Азовского (Таганрогский залив) и Каспийского
морей, р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах.
β-мезосапроб.
79
Рисунок 34 – Внешний вид Brachionus plicatilis longicornis Fadeev, 1786
(фото Бычковой М.В.)
Brachionus quadridentatus Hermann, 1783
Этот вид имеет панцирь почти четырехугольной формы. Передний край брюшной
пластинки имеет слабую волнистость, спинной снабжен 6 шипами. Срединные шипы, с
расходящимися концами, самые длинные, а самые короткие – промежуточные. Задний край
панциря с шипами, направленными вниз. Характерным признаком вида является трубчатая
форма отверстия для ноги и скульптура панциря в виде гребней, точек, ровных линий или
узоров. Длина панциря – 156-353 μm, ширина – 103-216 μm (рисунок 35). Это один из самых
распространенных и изменчивых видов коловраток р. Brachionus.
B. quadridentatus – планктонный эвригалинный вид, космополит. Встречается в водоемах
с кислой средой, требователен к повышенному содержанию органических веществ. Обитает в
пресных, соловатоводных и прибрежных морских водоемах.
80
В Азовском море встречается преимущественно в восточном районе Таганрогского
залива, а также в р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах.
β-мезосапроб.
Рисунок 35 – Внешний вид Brachionus quadridentatus Hermann, 1783
(фото Бычковой М.В.)
Brachionus quadridentatus cluniorbicularis Skorikov, 1894
Эта коловратка является разновидностью типичной формы B. quadridentatus. Отличается
от нее широкой округленной задней частью пластинки. Передний спинной край с 6 недлинными
шипами, срединные шипы более длинные, с расходящимися концами. Отверстие для ноги,
имеющей вид трубки, сдвинуто на брюшную сторону. Панцирь у этого вида коловраток
гладкий или со слабой зернистой структурой. Длина панциря – 160-270 μm, ширина – 150-260 μm
(рисунок 36).
Эвритермный, эвритопный, космополитный вид.
81
В Азовском море встречается в восточной части Таганрогского залива, р. Дон, Цимлянском
водохранилище, р. Кубань, приазовских лиманах.
А
Б
Рисунок 36 – Внешний вид Brachionus quadridentatus cluniorbicularis Skorikov, 1894
(фото Бычковой М.В.)
Brachionus quadridentatus hyphalmyros Tshugunoff, 1921
Передний спинной край панциря у этой коловратки с 6 шипами. Задние боковые шипы
длинные и направлены к середине. Трубчатый выход ноги не заходит за край панциря и сдвинут
на брюшную сторону. Длина панциря – 262-353 μm,
ширина – 159-216 μm (рисунок 37).
Планктонный вид солоноватоводных водоемов.
Отмечен в Каспийском и Азовском морях, в осоло-
ненных лиманах, а также в некоторых озерах юго-
востока европейской части России и Азии (озера
Челкан, Балхаш, Иссык-Куль). В Азовском море в
основном встречается в восточной части Таганрог-
ского залива, р. Дон, Цимлянском водохранилище,
р. Кубань, приазовских лиманах.
Рисунок 37 – Внешний вид Brachionus
quadridentatus hyphalmyros Tshugunoff, 1921
(фото Бычковой М.В.) ►
82
Род Keratella
Представители этого рода коловраток имеют ложкоподобный или прямоугольный
панцирь. Поверхность спинной пластинки панциря обычно с более или менее выраженной
скульптурой в виде фасеток или полей, покрытых мелкой сеточкой, шипиками или точками.
Брюшная пластинка полностью, либо на 1/3 от переднего края покрыта точками или шипиками.
По переднему спинному краю расположены 6 шипов, из которых срединные шипы более
массивные и длинные, промежуточные меньше боковых. Передний брюшной край с небольшой
выемкой посередине. На заднем крае 1-2 шипа, или же они вовсе отсутствуют.
Keratella cochlearis (Gosse, 1851)
Этот вид коловраток имеет ложкоподобную форму панциря, у которого спинная пластинка
выпуклая, а брюшная плоская. Спинная пластинка панциря покрыта сеточкой, шипиками или
точками. На брюшной пластинке точки располагаются только на первой трети ее поверхности.
Задний край коловратки посередине переходит в длинный шип. Мелкий вид, длина панциря –
130-196 μm, ширина – 52-82 μm (рисунок 38).
А
Б
Рисунок 38 – Внешний вид Keratella cochlearis (Gosse, 1851)
(фото Бычковой М.В.)
K. cochlearis – один из наиболее распространенных видов коловраток, способен жить
в воде с различной кислотностью. Эвритермный, круглогодичный, пресноводный организм,
переносящий соленость до 5 ‰.
83
В Азовском море встречается в восточной части Таганрогского залива, р. Дон, Цимлянском
водохранилище, р. Кубань, приазовских лиманах.
Олиго-β мезосапроб.
Keratella quadrata (Müller, 1786)
Панцирь почти прямоугольный, ровный с боков. Задний край панциря с двумя
направленными назад или дивергирующими шипами, иногда изогнутыми. Длина шипов сильно
варьирует: в большинстве случаев шипы равные, иногда очень длинные; левый шип может
быть короче или полностью отсутствовать; реже оба шипа редуцированы. Самцы известны,
их длина от 70 до 100 μm. Покоящиеся яйца овальные, с толстой складчатой оболочкой, их
размеры обычно от 70-90 до 44-65 μm (рисунок 39).
Один из самых распространенных планктонных видов. Обитает в различных водоемах,
в массовом количестве развивается в прудах, озерах и стоячих водоемах. За исключением
нескольких разновидностей, космополит.
Встречается в р. Дон (в том числе, в устье), Цимлянском водохранилище, р. Кубань и
водохранилищах Краснодарского края, в приазовских лиманах.
Рисунок 39– Внешний вид Keratella quadrata (Müller, 1786)
(фото Бычковой М.В.)
84
Keratella tecta (Gosse, 1851)
Keratella cochlearis tecta (Gosse, 1851)
Форма панциря у этого вида коловраток аналогична типичной форме. Отличие заключается
в отсутствии заднего шипа. Вид имеет укороченные передние шипы, самые длинные средние
и совсем короткие промежуточные. Размер меньше, чем у типичной формы. Длина панциря –
82-123 μm, ширина – 46-66 μm (рисунок 40).
Теплолюбивый вид, развивается обычно в теплое время года. Обитает в планктоне мелких
и крупных полупроточных пресных и солоноватоводных водоемов.
Встречается в восточной части Таганрогского залива, р. Дон, Цимлянском водохранилище,
р. Кубань, приазовских лиманах.
А
Б
Рисунок 40 – Внешний вид Keratella tecta (Gosse, 1851)
(фото Бычковой М.В.)
Род Notholca
У коловраток р. Notholca панцирь имеет широкую - удлиненно-овальную или бокаловидную
форму, выпуклую спинную пластинку, сплющенную дорсовентрально. Скульптура панциря обычно
хорошо выражена лишь на спинной пластинке в виде продольной штриховатости, точек, зерен
или альвеол. На переднем спинном крае панциря хорошо видны 4-6 парных шипа. Брюшной край
панциря – волнистый, с выступами, чаще со срединной выемкой. Задний край панциря округлый
или с 1-2 шипами, которые иногда прикреплены с боков панциря или на спинной пластинке.
Большинство видов – холодолюбивые типичные планктонты. Обитают в пресных,
солоноватых водах и в прибрежных районах морей и океанов.
85
Notholca acuminata (Ehrenberg, 1832)
Панцирь у коловратки N. acuminata всегда удлиненный бокаловидной формы, продольно
исчерченный. Задний край панциря плавно переходит в неподвижный шип. Все шипы переднего
края панциря заостренные, прямые, с широким основанием и почти одинаковой толщины.
Средние шипы всегда превосходят по мощности и длине боковые и промежуточные. Длина
панциря составляет 220-300 μm, ширина – 101-120 μm (рисунок 41).
Вид холодолюбивый, эвригалинный. Встречается в прудах, реках, водохранилищах,
озерах, а также в морских и солоноватоводных водоемах.
Встречается в р. Дон, р. Кубань, приазовских лиманах, Таганрогском заливе Азовского моря.
Олиго-мезосапроб.
Рисунок 41 – Внешний вид Notholca acuminata (Ehrenberg, 1832)
(фото Бычковой М.В.)
86
Семейство Euchlanidae
Тело коловраток этого семейства подразделено на голову, туловище и ногу. Голова хорошо
отделена от туловища небольшим шейным сужением. Туловище стекловидно- прозрачное,
покрыто обычно гладким, чаще дорсовентрально сплющенным панцирем. Панцирь более
или менее овальный, состоит из двух или трех пластинок. Пластинки соединены тонкой
перепонкой, обычно лежащей в более или менее глубоких бороздах, которые в некоторых
случаях отсутствуют. Спинная пластинка цельная, выпуклая, иногда с высоким срединным
килем, брюшная пластинка плоская или немного вогнутая. Передние края панциря чаще с
вырезами, задний край закругленный, иногда угловатый, часто со срединным вырезом. Нога
хорошо обособленная, с 2-3 длинными на конце заостренными пальцами.
Род Euchlanis
У коловраток этого рода панцирь туловища имеет продолговатую, яйцевидную или
округлую форму. Панцирь очень прозрачный, составлен из спинной и заметно меньшей
брюшной пластинок. Пластинки обычно соединены гибкой перепонкой, образующей
продольные боковые и задние борозды. Реже перепонка редуцируется, и брюшная пластинка
боковыми краями сливается со спинной. Спинная пластинка округлая, сводчатая, либо с
острым продольным килем. Задний край ее часто имеет более или менее гибкий V- или
U-образный вырез или только чуть заметное углубление. Брюшная пластинка плоская и
сзади округлая. Передние края обеих пластинок утончаются до эластичных пленок, имеющих
широкие срединные выемки. Нога с неясно выраженными 2-3-члениками, с двумя длинными
или короткими, ровными или веретеновидными, тонкими или толстыми пальцами.
Прибрежные обитатели, живущие среди водной растительности, иногда в пелагиали.
Euchlanis dilatata Ehrenberg, 1832
Этот вид имеет продолговатый панцирь яйцевидной или округлой формы. Спинная
пластинка панциря низкая, выпуклая, без киля. По бокам видны широкие, четко выраженные
борозды. На переднем крае спинной пластинки хорошо заметен глубокий и закругленный
вырез, на заднем крае – U-образный. Нога короткая с двумя длинными, тонкими, почти
параллельными или слегка веретеновидными пальцами. Длина спинной пластинки составляет
155-320 μm, ширина – 131-225 μm, брюшной – 130-255 μm и 110-172 μm соответственно.
Летние яйца прикрепляются к стеблям растений (рисунок 42).
Один из самых распространенных видов. Обитает в пресных, солоноватых и морских
водоемах среди прибрежной растительности. Гораздо реже находится в планктоне.
Встречается в р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах, в
восточной части Таганрогского залива Азовского моря.
Олиго-β мезосапроб.
87
Рисунок 42 – Внешний вид Euchlanis dilatata Ehrenberg, 1832
(фото Бондарева С.В.)
Семейство Lecanidae
У представителей сем. Lecanidae тело состоит из головы, туловища и ноги. Голова
довольно маленькая, туловище в большинстве случаев покрыто более или менее ригидным
и сильно сплющенным дорсовентрально панцирем. Нога 2-х члениковая, сильно сдвинута на
брюшную сторону, имеет 1-2 сравнительно длинных пальца. Спинная и брюшная пластинки
панциря соединены боковыми складками. Спинная пластинка обычно короче брюшной. Задняя
часть брюшной пластинки нередко заходит за задний спинной край панциря.
Виды свободноживущие, обитают в основном среди мха и водной растительности, но
иногда встречаются в планктоне.
Род Lecane
У представителей этого рода панцирь гладкий, либо со скульптурой в виде фасеток,
складок, гребней, точек или зерен. Передние края спинной и брюшной пластин панциря прямые,
вогнутые, выпуклые, иногда часто совпадающие с боковыми шипами, реже со срединными.
Задний сегмент панциря округлый, угловатый или с 1-2 шипами. Первый членик ноги более
или менее сросшийся с брюшной пластинкой, второй – подвижный, иногда заходящий за
задний край панциря. Пальцы ноги сравнительно длинные, с параллельными краями, обычно
88
к концу суженные, реже веретеновидные, вздутые. Они могут быть разъединены до основания,
либо сросшиеся на 1/3 – 1/2 у основания, или полностью слиты в единое целое.
Lecane luna (Müller, 1776)
Вид имеет широко-округлый, гладкий панцирь. Задний край панциря гладкий, без
придатков, передний спинной край с двумя срединными углами и насечками. Брюшной
передний край панциря с боковыми заостренными углами. Пальцы с параллельными краями,
коготок с острым плечиком. Длина коловратки – 140-220 μm (рисунок 43).
Эвригалинный, эвритермный, хорошо приспособленный к различным гидрохимическим
условиям среды обитания вид коловраток. Широко распространен в прибрежной зоне пресных
и солоноватых водоемов. В пелагиали встречается реже.
Обитает р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах, выноситься
токами пресной воды в море.
Олиго-β-мезосапроб.
Рисунок 43 – Внешний вид Lecane luna (Müller, 1776)
(фото Бондарева С.В.)
Семейство Synchatidae
Тело коловраток этого семейства у представителей р. Synchaeta имеет конусовидно-
цилиндрическую форму. У видов р. Ploesoma и р. Bipalpus отмечается мешковидная, либо
прямоугольная форма, сплющенная с боков, у коловраток р. Polyarthra она сплющена
дорсовентрально. Для представителей р. Synchaeta характерна нежная, складчатая кутикула,
89
У ряда видов р. Polyarthra она немного уплотненная или ригидная, вплоть до образования
панциря со скульптурой (р. Ploesoma и р. Bipalpus). Голова коловраток отделена от туловища
складкой или насечкой. Коловращательный аппарат с пучками чувствительных щетинок,
мембранеллами, иногда с парными пальцевидными выростами – пальцами (р. Ploesoma и
р. Bipalpus). У представителей р. Synchaeta по бокам головы большие выросты с ресничками –
«ушки». Нога короткая, терминальная, с маленькими 1-2 пальцами (р. Synchaeta), или длинная,
сдвинутая на брюшную сторону с 2 массивными пальцами (р. Ploesoma и р. Bipalpus). У видов
р. Polyarthra нога отсутствует.
Род Bipalpus
Тело коловраток р. Bipalpus покрыто панцирем, имеющим мешковидную или конусовидную
форму. Сзади он округлый или несколько вытянутый, без щели на брюшной стороне. Скульптура
панциря ячеисто-пузыристая. Коловращательный аппарат с двумя пальцевидными выростами-
пальцами, несколькими согнутыми мембранеллами и чувствительными щетинками. Нога сдвинута
на брюшную сторону, длинная, частично кольчатая, пальцы ноги большие, клешневидные.
Bipalpus hudsoni Wierzejski et Zacharias, 1803
Gastropus hudsoni Imhof, 1891
Тело этого вида коловраток покрыто конусовидным, сзади округлым или несколько
вытянутым панцирем с ячеисто-пузеревидной скульптурой. Нога длинная, кольчатая, с большими
клешневидными пальцами, сдвинута на брюшную сторону. Летние яйца с прозрачной бесцветной
оболочкой, покоящиеся яйца темные, с толстой желтой морщинистой оболочкой (рисунок 44).
Рисунок 44 – Внешний вид Bipalpus hudsoni Wierzejski et Zacharias, 1803
(фото Бондарева С.В.)
90
Вид широко распространен в планктоне пресных и солоноватых водоемов.
Встречается в восточной части Таганрогского залива.
Олиго-мезосапроб.
Род Polyarthra
Тело коловраток р. Polyarthra прямоугольное либо цилиндрическое. Часто оно
дорсовентрально сплющено, прозрачное, бесцветное или может иметь желтоватый,
фиолетовый, иногда синеватый оттенок. Голова от туловища отделена насечкой.
Коловращательный аппарат в виде венчика ресничек. На теменной стороне 3 пары
конусовидных или цилиндрических выростов с чувствительными ресничками. Ноги нет.
Туловище типичных форм со спинными и брюшными парными боковыми узлами. В каждом
узле по три плавательных придатка, плавника, соединенных с мощными мышцами, идущими
по бокам туловища. Некоторые виды с маленькими брюшными плавничками, отходящими
несколько ниже ротового отверстия.
Polyarthra vulgaris Carlin, 1943
Тело коловратки P. vulgaris имеет прямоугольную форму. Голова от туловища отделена
насечкой, ноги нет. Плавники широкоперистые или ланцетовидные, зазубренные по краям. На
них отчетливо видна срединная жилка и поперечно-косая штриховатость. Брюшные плавнички
узкие, умеренно длинные с зазубренными краями. Боковые щупальца расположены недалеко
от заднего края. Длина тела – 100-205 μm, длина плавников – 120-150 μm (рисунок 45).
Представитель планктона.
Вид эвригалинный, пере-
носит соленость до 6-8 ‰.
Обитатель медленно текучих
малых, больших рек, водо-
хранилищ, пресных и солоно-
ватых водоемов.
В Азовском море встре-
чается в восточной части
Таганрогского залива, в р. Дон,
Цимлянском водохранилище,
р. Кубань, приазовских лиманах.
β-мезосапроб.
Рисунок 45 – Внешний вид
Polyarthra vulgaris Carlin, 1943
(фото Хренкина Д.В) ►
91
Род Synchaeta
Коловратки этого рода характеризуются коническим, колоколовидным, реже мешковидным
или почти цилиндрическим телом. Кутикула тонкая, прозрачная, иногда с продольными
нежными складками. Голова с боковыми округлыми, покрытыми длинными ресничками,
выростами – «ушками», которые не всегда видны при фиксации. Передний край головы более
или менее выпуклый (при фиксации может быть втянут, образуя воронку), с двумя парами
чувствительных мембранелл; из которых наружные сидят на треугольных выступах. Нога
длинная или короткая, тонкая, пальцевидная или массивная толстая, имеет 1 членик. Пальцев
обычно 2, реже 1, иногда с шипообразным или пальцевидным придатком у основания.
Глазные пятна темно-красные. Размеры коловраток этого рода значительно варьируют. Общая
длина в зависимости от вида изменяется от 100 до 520 μm (рисунок 46).
Холодноводные, основной компонент осеннего и весеннего планктона.
Планктонные обитатели пресных, солоноватых и морских водоемов.
Встречаются в Северном и Средиземном море, в Черном море, в Азовском море и в
приазовских лиманах, р. Дон, Цимлянском водохранилище, р. Кубань.
А
Б
Рисунок 46 – Внешний вид Synchaeta sp. Ehrenberg, 1832
(фото Хренкина Д.В.)
Семейство Trochosphaeridae
Представители этого семейства имеют веретеновидное или овальное тело, подразделенное
на голову и туловище. Ноги нет. Коловращательный аппарат с прерывистым или непрерывным
рядом ресничек венчика. От передней части туловища с боков отходят 2 длинных шиповидных
кутикулярных придатка, служащих веслами при прыгательном движении. Задний конец
туловища с одним или с двумя такими же придатками.
92
Род Filinia
Тело коловраток р. Filinia веретеновидное или почти цилиндрическое, спереди и сзади
усеченное, иногда удлиненное и суженное сзади, прозрачное, иногда желтоватое. Коловращательный
аппарат в виде двух тесно сближенных венчиков ресничек. В передней части туловища два боковых
кутикулярных более или менее длинных шиповидных придатка. В задней части туловища один
непарный придаток, отходящий от брюшной стороны или от заднего края его.
Filinia longiseta (Ehrenberg, 1834)
Тело этого вида коловраток овальной формы, закруглено в задней части. Нога отсутствует.
От передней части тела с боков отходят два длинных плавательных придатка длиной 218-595
μm. На заднем конце туловища располагается еще один такой же хорошо развитый придаток
длиной 120-408 μm. Все три придатка покрыты мелкими шипиками. Длина тела варьирует от
104 до 252 μm (рисунок 47).
А
Б
Рисунок 47 – Внешний вид Filinia longiseta (Ehrenberg, 1834)
(фото Хренкина Д.В.)
93
Вид приспособлен к различным гидрохимическим условиям среды обитания. Переносит
колебания pH воды от 6,6 до 9,0, соленость от 6,3 до 12,7 ‰, повышенное загрязнение до
α-мезосапробного состояния и значительные перепады температуры воды, в связи с чем
встречается на протяжении всего года.
Обитает в лужах, прудах, лиманах, водохранилищах.
В Азовском море встречается в Таганрогском заливе, в основном в опресненной восточной
части, р. Дон, Цимлянском водохранилище, р. Кубань, приазовских лиманах.
β-мезосапроб.
Класс Sagittoidea
К этому классу относятся морские планктонные животные, имеющие удлиненное,
двухсторонне симметричное, прозрачное тело. На голове расположен ловчий аппарат,
состоящий из системы челюстных крючковидных щетинок. Тело покрыто однослойным
эпителием, который за головой образует многослойное утолщение. По бокам тела расположены
две пары боковых плавников, заканчивается тело непарным хвостовым плавником. Имеются
два глаза, каждый из которых состоит из пяти пигментных клеток. Животные – гермофродиты,
Развитие прямое, проходит без личиночной стадии.
Отряд Aphragmophora
Семейство Sagittidae
Род Parasagitta
Представители этого рода имеют вытянутое тело с двумя парами боковых плавников.
Хвостовой плавник округло-треугольный. Хватательные крючья серповидные, подвижный
кончик их прямой. По сторонам рта в два ряда расположены многочисленне зубчики. Кишечник
часто с боковыми выпячиваниями.
Parasagitta setosa (J. Müller, 1847)
Sagitta batava Biersteker & Van der Spoel, 1966
Sagitta euxina Moltschanoff, 1909
Sagitta setosa J. Müller, 1847
У этого вида щетинкочелюстных вытянутое тело с передними, задними и хвостовыми
плавниками, последние имеют округло-треугольную форму. Длина тела достигает 26 мм.
P. setosa – морской вид, проявляет черты эвритермности, совершает незначительные
вертикальные миграции. За год может давать до 11 генераций. Число яиц в кладке составляет
94
12-17 штук, особи первых весенних генераций более плодовиты. По типу питания является
хищником - зоофагом. В пищевом спектре вида преобладают копеподы. Отмечено явление
каннибализма (рисунок 48).
Встречается в Атлантическом океане, Средиземном море.
Обитает в Черном море. В Азовском море при осолонении появляется в южной части.
А
Б
В
Г
Рисунок 48 – Внешний вид Parasagitta setosa (J. Müller, 1847)
(фото Бычковой М.В.)
Класс Appendicularia
К этому классу относятся морские, одиночные, пелагические оболочники мелких
размеров. Взрослые особи имеют хорду. Тело делится на туловище и хвост. Передняя часть
туловища покрыта железистым эпителием – ойкопластом, секретирующим тунику, называемую
«домиком», который в той или иной степени покрывает животное. Иногда он значительно
превышает размеры тела, представляя собой сложное фильтровальное устройство. Хвост
плоский и длинный, отходит от вентральной стороны туловища и является основным органом
движения. Терминальный рот ведет в глотку, переходящую в пищевод, за которым следует
желудок и кишечник.
95
Отряд Copelata
Семейство Oikopleuridae
Род Oikopleura
Oikopleura (Vexillaria) dioica Fol, 1872
Appendicularia coerulescens Gegenbaur, 1855
Oikopleura flabellum Traustedt, 1880
Oikopleura malmii Hartmann, 1878
Vexillaria flabellum Müller, 1846
Vexillaria speciosa Eisen, 1874
Это морские, одиночные, мелкие планктонные животные. Тело делится на туловище
длиной до 1 мм и хвост. Хвост плоский, длиннее туловища примерно в четыре раза и является
основным органом движения. В хвосте помещается хорда (рисунок 49).
Особи этого вида раздельнополые. Размножаются только половым путем. Крупные
самки содержат в гонаде до 100 яиц. Летом самки приступают к размножению при длине тела
0,65 мм, весной – не менее 0,8 мм. Сразу после размножения они гибнут. Основные периоды
размножения – май-июнь и август-сентябрь. Развитие особей до стадии половой зрелости
весной составляет 15-17 суток.
O. (V.) dioica является фильтратором, потребляет растительную пищу. В качестве кормовых
объектов использует мелких жгутиковых, перидиней, диатомовых, кокколитофорид.
O. (V.) dioica – эвритермный и эвригалинный вид. Обитает в широком диапазоне солености
и температуры. Встречается во все сезоны, обитает обычно в прибрежных акваториях.
Обитает в теплых и умеренных водах многих морей Атлантического, Тихого и Индийского
океанов,
В Черном море, иногда появляется в южной части Азовского моря при адвекции
черноморских вод.
А
96
Б
В
Рисунок 49 – Внешний вид Oikopleura (Vexillaria) dioica Fol, 1872
(фото Бычковой М.В.)
97
Класс Oligotrichea
Отряд Choreotrichida
Семейство Codonellidae
Домики трубчатые, шарообразные или чашеобразные, с шейкой или без шейки, если
шейка имеется, она не спиральная и не гиалиновая. Задний конец закрыт, иногда заканчивается
стебельком. На поверхности домика многочисленны посторонние тельца. Стенки домика
имеют первичную, вторичную или даже третичную структуру.
Род Tintinnopsis
Наиболее распространенный род тинтинноидей. Представители рода широко распространены,
как в Черном и Азовском морях, так и в пресных водах. Домик очень разнообразен по форме,
инкрустирован у некоторых форм блестящими частицами и посторонними телами (рисунок 50).
Задний конец домиков закрыт и иногда переходит в узкий вырост – стебелек. Стенки
однослойные, тонкие, покрытые многочисленными посторонними тельцами. Первичная
структура тонкая, вторичная – грубая. Один-два макронуклеуса, один-два микронуклеуса и
20-24 мембранеллы.
А
98
А
Б
Рисунок 50 – Внешний вид представителей рода Tintinnopsis Stein, 1867
(фото Бычковой М.В.)
Класс Dinophyceae
К классу Dinophyceae относятся микроскопические одноклеточные, подвижные
одиночные, реже колониальные или нитчатые организмы. Клетки имеют разнообразную
форму, но в большинстве случаев округленную, дорсовентрального строения. Два жгута
разной конструкции и функций прикрепляются в передней, редко в задней или в центральной
части клетки с брюшной стороны. В последнем случае они лежат в углублениях (бороздах),
одна из которых опоясывает клетку, как бы деля ее на две части (верхнюю и нижнюю), а другая
располагается продольно на брюшной стороне.
Отряд Noctilucales
Организмы весьма разнообразные по форме и величине, обычно крупные или
средних размеров. Поясок, борозда и жгутики сильно варьируют в зависимости от родов,
иногда видоизменены, редуцированы или отсутствуют. Оболочка деформируемая, нередко
«голая», без явного тубалирования. Сложная ротоглоточная система, расположенная
на пересечении пояска и борозды, включает жгутики, служащие для захвата добычи,
щупальце, цитостом и глотку. У некоторых видов имеется велум или прозрачная
раковинка. Цикл развития весьма сложный, полностью известный у единичных родов.
Отряд исключительно морской.
99
Семейство Noctilucaceae
Клетки крупные, шарообразные. Поясок рудиментарный или отсутствует, борозда
присутствует, но аберрантная. Имеется щупальце. Оболочка мягкая, деформируемая, без
велума, панциря или раковики, но с хорошо развитым пелликулярным слоем.
Род Noctiluca
Клетки крупные, в виде пузыря, сферические или почковидные в начале вегетативного
деления, 200-2000 μm в диаметре. Эписома и гипосома неразличимы, т.к. поясок практически
отсутствует, его остаток находится в ротовой складке. Тут же располагается редуцированный
до торчащего подвижного зубца поперечный жгутик. Борозда короткая, в форме ротовой
складки, из заднего конца которой выходит длинное поперечно исчерченное щупальце,
лопатообразное на конце и служащее для отлова добычи. Продольный жгутик короткий, он не
приводит клетку в движение, а выполняет функцию чувствительного волоска (щупика). Тело
заполнено гидростатическими вакуолями. Жизненный цикл сложный, с чередованием половой
и бесполой стадий. В отличие от других динофлагеллят, вегетативная планктонная стадия р.
Noctiluca является диплоидной, хлоропласты отсутствуют, типичная гетеротрофия.
Noctiluca scintillans (Macartney) Kofoid & Swezy, 1921
Mammaria scintillans Ehrenberg, 1834
Medusa marina Slabber, 1771
Medusa scintillans Macartney, 1810
Noctiluca marina Ehrenberg, 1834
Noctiluca miliaris Suriray, 1816
Noctiluca omogenea Giglioli, 1870
Noctiluca pacifica Giglioli, 1870
Noctiluca punctata Busch, 1851
N. scintillans рассматривается как компонент мезозоопланктона из-за своего крупного
размера и всеядности. Является некормовым компонентом планктона.
N. scintillans имеет вид крупных клеток в виде пузыря размером 200-2000 μm в диаметре.
В центре клеток находится ядро, оболочка эластичная и прозрачная, хлоропласты отсутствуют.
В начале вегетационного деления клетки сферические или почковидные. Для этого организма
свойственно интенсивное свечение отдельных частей тела. Размножение осуществляется
бесполым и половым путем. В первом случае вегетативная клетка делится пополам, во втором
в процессе мейоза возникают одножгутиковые гаметы, при слиянии которых образуется зигота,
дающая начало взрослой особи
N. scintillans – морской, неритический, холодолюбивый вид (рисунок 51). Образует
обширные «цветения воды» у берегов. Развитие этого вида имеет хорошо выраженный сезонный
100
характер. Наибольшая численность наблюдается в апреле-мае, иногда в июне. Отмечается
положительная реакция на эвтрофикацию воды.
Своеобразно питание N. scintillans. Несмотря на систематическую принадлежность
к динофлагеллятам, является гетеротрофом и интенсивно потребляет растительную, животную
пищу (мелкий зоопланктон) и детрит. Размерный спектр потребляемых пищевых частиц достаточно
широк – от нескольких микрон (кокколитофориды) до 600-800 μm (науплии копепод).
Космополит. В Черном море является сезонным массовым видом. В Азовском море
единично встречается в южной части.
А
Б
В
Рисунок 51 – Внешний вид Noctiluca scintillans (Macartney) Kofoid & Swezy, 1921
(фото Бычковой М.В.)
101
Меропланктон
Важную роль в составе зоопланктона Азовского и Черного морей играет меропланктон
(временные планктеры), который включает в себя организмы, часть жизненного цикла
которых проходит в водной толще. После окончания пелагической стадии развития многие
представители этой группы зоопланктона оседают на дно и ведут прикрепленный образ
жизни. К меропланктону относятся личинки моллюсков, многощетинковых червей, донных
ракообразных, личинки насекомых, губок, мшанок, полипов. Таксоны данной группы
организмов встречаются в Черном и Азовском морях и их лиманах, Таганрогском заливе
Азовского моря, реках Дон, Кубань и их водохранилищах.
Ниже приведены фотографии отдельных массовых представителей Азово-Черноморского
меропланктона (рисунки 52-56).
Рисунок 52 – Внешний вид личинки двустворчатых (пластинчатожаберных) моллюсков
(larvae Lamelibrancha)
(фото Бычковой М.В.)
102
Рисунок 53 – Внешний вид личинки олигохет (larvae Olygochaeta)
(фото Бычковой М.В.)
103
А
Б
104
В
Г
105
Д
Е
Рисунок 54 – Внешний вид ранних стадий жизненного цикла полихет (larvae Polychaeta)
(А, Б, В – личинка, Г, Д, Е – трохофора)
(фото Бычковой М.В.)
106
А
Б
Рисунок 55 – Внешний вид личинки балянуса (усоногие раки)
(larvae Amphibalanus improvisus)
(фото Бычковой М.В.)
107
Рисунок 56 – Внешний вид личинки креветки
(фото Бычковой М.В.)
108
Благодарности
Авторы считают своим долгом выразить благодарность всем сотрудникам Азово-
Черноморского филиала ФГБНУ «ВНИРО» («АзНИИРХ»), которые помогали отбирать пробы
зоопланктона в Азовском и Черном морях, определять таксономическую принадлежность
и проводить цифровую фотосъемку организмов. Особую благодарность авторы выражают
Н.С . Елфимовой и М.А. Назирову за цифровую обработку фотографий, Г.В . Ермолаевой
за корректуру рукописи и рецензентам за замечания, позволившие значительно улучшить
манускрипт.
109
Список литературы
Арашкевич Е.Г., Луппова Н.Е., Никишина А.Б., Паутова Л.А. и др. Судовой экологический
1.
мониторинг в шельфовой зоне Черного моря: оценка современного состояния пелагической
экосистемы // Океанология. –
2015. – Т.55. – No 6. – С.964-970.
Алдакимова А.Я., Некрасова М.Я., Студеникина Е.И. Гидробиологический режим
2.
Азовского моря и его изменения в связи с преобразованием речного стока.
–
Л., 1977.
–
С. 90-103.
Бенинг А.Л. Кладоцера Кавказа. – Тбилиси: Грузмедгиз, 1941. – 384 с.
3.
Бродский К.А Веслоногие рачки Calanoida дальневосточных морей СССР и полярного
4.
бассейна. – М.-Л.: Изд-во АН СССР, 1950. – 442 с.
Боруцкий Е.В. Определитель свободноживущих пресноводных веслоногих раков СССР
5.
и сопредельных стран по фрагментам в кишечниках рыб. – М.: Изд-во АН СССР, 1960. –
218 с.
Боруцкий Е.В., Степанова Л.А., Кос М.С. Определитель Calanoida пресных вод СССР //
6.
Определители по фауне СССР, издаваемые зоологическим институтом АН СССР. Т. 157. –
СПб.: Наука, 1991. – 503 с.
Бродский К.А., Вышкварцева Н.В., Кос М.С., Мархасева Е.Л. Веслоногие ракообразные
7.
(Copepoda: Calanoida) морей СССР и сопредельных вод. Т. 1. – Л.: Изд-во «Наука», 1983. –
358 с.
Бронфман А.М., Дубинина В.Г., Макарова Г.Д. Гидрологические и гидрохимические основы
8.
продуктивности Азовского моря. – Москва: Пищевая промышленность, 1979. – 288 с.
Воловик С.П. Продуктивность и проблемы управления экосистемой Азовского моря:
9.
дисс. на соиск. уч. степ. д-ра биол. наук. – Ростов-на-Дону, 1985. – 563 с.
Гребневик
10.
Mnemiopsis leidyi (A. Agassiz) в Азовском и Черном морях: биология и последствия
вселения // Под ред. С.П. Воловика. – Ростов-на-Дону: БКИ, 2000 – 500 с.
Грезе В.Н. Основы биологической продуктивности Черного моря.
–
Киев: «Наукова
11.
думка», 1979. – С. 143-164.
Губанова А.Д.
12.
Acartia tonsa Dana в Черном море: появление, сезонная динамика и
размерная структура // Экология моря. – 2000. – Вып. 51. – С. 55-58.
Губанова А.Д., Загородняя Ю.А., Фенева И.Ю.
13.
Oithona davisae Ferrari F.D . et Orsi,
1984 // Самые опасные инвазионные виды России (ТОП-100) / ред.: Ю.Ю . Дгебуадзе,
В.Г. Петросян, Л.А. Хляп. – Москва: Т-во науч. изд. КМК, 2018. – С. 414-419.
Губарева Е.С., Светличный Л.С. Копеподы Oithona similis и Oithona davisae – две стратегии
14.
эколого-физиологической адаптации в Черном море // Океанология. – 2016. – Т. 56. – No 2. –
С. 258-265.
Жукова С.В., Дубинина В.Г. Государственный мониторинг среды обитания водных
15.
биологических ресурсов Азовского моря // Водные ресурсы России: современное состояние
и управление: сб. матер. Всерос. науч.-практ. конф. (г. Сочи, 8-14 октября 2018 г.).
–
Новочеркасск: Лик, 2018. – Т. 1. – С. 275-284.
110
Киселев И.А. Панцирные жгутиконосцы (DINOFLAGELLATA) морей и пресных вод
16.
СССР. – М.-Л.: Изд-во АН СССР, 1950. – 279 с.
Ковалев А.В. Структура зоопланктонных сообществ Атлантики и Средиземного морского
17.
бассейна. – Киев: Наук. думка, 1991. – 144 с.
Коваль Л.Г. Значение биологических факторов в сезонной динамике зоопланктона
18.
северо-западной части Черного моря // Биологические исследования Черного моря и его
промысловых ресурсов. – М.: Наука, 1968. – С. 79-86.
Коновалова Г.В . Динофлагелляты (Dinophyta) дальневосточных морей России и
19.
сопредельных акваторий Тихого океана. – Владивосток: Дальнаука, 1998. – 297 с.
Краткий определитель беспозвоночных пресных вод центра Европейской России /
20.
М.В. Чертопруд, Е.С. Чертопруд. – М.: Центр Москва, 2010. – 185 с.
Крахмальный А.Ф. Динофитовые водоросли Украины (иллюстрированный определитель).
–
21.
Киев: Альтерпресс, 2011. – 444 с.
Кутикова Л.А. Бделоидные коловратки фауны России. – М.: Т-во научных изданий КМК,
22.
2005. – 315 с.
Кутикова Л.А. Коловратки фауны СССР (Rotatoria). – Л.: Изд-во «Наука», 1970. – 744 с.
23.
Мануйлова Е.Ф . Ветвистоусые рачки (Cladocera) фауны СССР. – М . - Л .: Наука, 1964. –
24.
327 с.
Мирзоян З.А. Изменения структуры и продуктивности сообщества зоопланктона при
25.
вселении гребневика / Гребневик Mnemiopsis leidyi (A. Agassiz) в Азовском и Черном
морях: биология и последствия вселения // Под ред. С.П. Воловика.
–
Ростов-на-Дону:
БКИ, 2000 – С.189-208.
Набережный А.И. Коловратки водоемов Молдавии. –
Кишинев, изд-во «Штиинца»,
26.
1984. – 326 с.
Никишина А.Б., Дриц А.В., Васильева Ю.В., Тимонин А.Г., Соловьев, Ратькова Т.Н.,
27.
Сергеева В.М. Роль популяции Noctiluca scintillans в трофодинамике планктона Черного
моря в весенний период // Океанология. – 2011. – Т. 51. – No 6. – С. 1090-1101.
Определитель безпозвоночных России и сопредельных стран. Т. 2. Ракообразные / Под
28.
ред. С.Я. Цалолихина. – СПб.: зоол. ин-т РАН, 1995. – 627 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1.
29.
Зоопланктон / Под ред. В.Р. Алексеева, С.Я. Цалолихина.
–
М.: Товарищество научных
изданий КМК, 2010. – 495 с.
Определитель фауны Черного и Азовского морей. Свободноживущие беспозвоночные / Под
30.
ред. Ф.Д. Мордухай-Болтовского. Том 1. – Киев, 1968. – 440 с.
Определитель фауны Черного и Азовского морей. Свободноживущие беспозвоночные / Под
31.
ред. Ф.Д. Мордухай-Болтовского. Том 2. – Киев, 1969. – 536 с.
Определитель фауны Черного и Азовского морей. Свободноживущие беспозвоночные / Под
32.
ред. Ф.Д. Мордухай-Болтовского. Том 3. – Киев, 1972. – 340 с.
Определитель пресноводных беспозвоночных Европейской части СССР / Под ред.
33.
Кутиковой Л.А., Старобогатова Я.И. – Л.: Гидрометеоиздат, 1977. – 511 с.
111
Петипа Т.С. Питание планктонных ракообразных в Черном море. // Биологические
34.
исследования Черного моря и его промысловых ресурсов. – М.: Наука, 1968. – С. 67-71.
Петипа Т.С. Трофодинамика копепод в морских планктонных сообществах. – Киев: Наук.
35.
думка, 1981. – 245 с.
Прусова И.Ю., Губанова А.Д., Шадрин Н.В., Курашова Е.К., Тиненкова Д.Х.
36.
Acartia tonsa
(COPEPODA, CALANOIDA) новый вид в зоопланктоне Каспийского и Азовского морей //
Vestnik zoologii. – No 36 (5). – 2002. – С. 65-68.
Проект «Моря СССР». Гидрометеорология и гидрохимия морей СССР. Том IV. Черное море.
37.
Вып. 2. Гидрохимические условия и океанологические основы формирования биологической
продуктивности. – Санкт-Петербург: Гидрометеоиздат, 1992. – С. 175-178.
Рылов В.М. Пресноводные Calanoida СССР // Определители организмов пресных вод
38.
СССР. А. Пресноводная фауна. Вып. 1. – Л.: Изд-во АН СССР, 1930. – 288 с.
Рылов В.М. Cyclopoida пресных вод. – М. -Л.: Изд-во АН СССР, 1948. – 319 с.
39.
Серегин С.А., Попова Е.В. Многолетняя динамика численности копеподы-вселенца,
40.
Oithona davisae, в прибрежных водах Черного моря // Российский Журнал Биологических
Инвазий. – 2016. – No 3. – С. 90-100.
Темных А.В., Токарев Ю.Н., Мельников В.В., Загородняя Ю.А. Суточная динамика и
41.
вертикальное распределение пелагических COPEPODA в открытых водах у юго-западного
Крыма (Черное море) осенью 2010 г. // Морський екологiчный журнал. – Т. XI. – No 2. –
2012. – С. 75-84.
Черное и Азовское моря. Экологический атлас. Москва: Фонд «НИР». 2019. – 464 с.
42.
Научное издание
Д.Ф. Афанасьев, Д.В. Хренкин, М.Л . Мартынюк,
З.А. Мирзоян, М.В. Бычкова, Н.А. Шляхова, Е.А. Кожурин
Иллюстрированный атлас массовых видов
зоопланктона Азовского и Черного морей
Подписано в печать 23.11.2020 г. Формат 60x84/8.
Печать цифровая. Бумага офсетная. Объем 14,0 печ. л.
Тираж 500. Заказ No 4323.
ФГБНУ «ВНИРО». Азово-Черноморский филиал ФГБНУ «ВНИРО» («АЗНИИРХ»).
344002, Россия, г. Ростов-на-Дону, ул. Береговая, 21в.
Отпечатано в типографии ООО «Диапазон-Плюс».
344011, г. Ростов-на-Дону, пер. Островского, 124.
Лиц. ПЛД No 65-116 от 29.09.1997 г.