/























































































































































































































































































































































































































































































































































































Author: Беритов И.С.
Tags: медицина физиология практическая медицина физиология человека мышцы издательство академия наук ссср
Year: 1947




Text
Б-Ц8
і ^*.і
АКАДЕМИЯ НАУК СОЮЗА СОР
Академик И. С. БЕРИТОВ
ОБЩАЯ ФИЗИОЛОГИЯ
МЫШЕЧНОЙ И НЕРВНОЙ
СИСТЕМ
ТЕОРЕТИЧЕСКОЕ И ПРАКТИЧЕСКОЕ
РУКОВОДСТВО
ТОМ I
(МЫШЕЧНАЯ СИСТЕМА;
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА:
СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ)
Второе, переработанное издание
ыо№
БИБЛИОТЕКА
; -,«»а
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва
1947
Ленинград
9
ПРЕДИСЛОВИЕ КО ВТ ОРОМУ И ЗДАНИЮ
Новое издание «Общей физиологии мышечной и нервной систем»
выходит в совершенно переработанном виде. Многие главы написаны
почти заново, и добавлено несколько новых. Почти все главы
переработаны по новейшим литературным источникам. Из этого можно,
пожалуй, сделать вывод, что первое издание страдало определенными
недостатками, которые с дальнейшим развитием физиологии стали
особенно заметны.
За последние десять лет многие проблемы общей физиологии
подверглись чрезвычайно углубленному и всестороннему исследованию во
многих лабораториях нашего Союза и за границей. Так случилось
с 'вопросами о происхождении мышечного сокращения, об эластических
и пластических свойствах мышцы, о передаче возбуждения в синапсах,
о распространении возбуждения по нервному волокну, о
физиологических особенностях вегетативной нервной системы, о координации
движений у беспозвоночных животных, о центральном торіможении, об
автоматической деятельности центральной нервной системы, о
биотоках стволовой части головного мозга и коры большого мозга, об
отрицательном действии посторонних раздражений на индивидуально
приобретенные рефлексы и т. д. Эти проблемы в последние годы
изучались большей частью совершенно новыми методами, как
микрофизиология, изучающая отдельные мышечные и нервные волокна, как
осциллография, дающая возможность изучить биотоки от одного
мышечного или нервного волокна. Отсюда совершенно понятна
необходимость основательной переработки многих глав на основании
нового фактического материала и новых идей.
Уже в первом издании «Общей физиологии» я старался излагать
функции той или другой ткани или органа вместе с их структурой,
с целью выяснить их связь друг с другом и взаимную обусловленность.
В новом издании структурный материал излагается значительно
обширнее, и лучше выявлены его взаимоотношения с функциональной
деятельностью. Это опять-таки непосредственно связано с
многочисленными за последние годы открытиями в области тончайшей структуры
мышечных и нервных клеток и тканей. Особенно много сделано по
вопросам общей структуры живой системы, гладкой /мускулатуры,
а также тончайшего строения центральной нервной системы. Новый
структурный материал еще не нашел отражения в современных
учебниках и руководствах гистологии, а потому мне пришлось привести
в новом издании целый ряд новых гистологических рисунков.
Изучение биохимических и биофизических процессов также далеко
шагнуло вперед за последние десять лет, причем оно проводилось
8 связи с структурными и функциональными изменениями. Это в осо-
VI
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
бенности касается тех процессов, которые протекают внутри живой
протоплазмы. На этом основании возникло новое понимание
биологических процессов в живой клетке, новое понимание сократительного
процесса и биоэлектрических явлений. Естественно, что и в отношении
этих процессов мне пришлось многое изменить.
Сильно подвинулось вперед за последние годы изучение общих
свойств и функций мышечной и нервной систем на эмбриональном и
постэмбриональном материале позвоночных, а также на сравнительном
материале беспозвоночных животных. Это изучение дало много нового
и интересного. В первом издании уже приводились результаты,
добытые в этом направлении. В настоящем издании пришлось значительно
расширить этого рода сведения и использовать их для целого ряда
обобщений с точки зрения филогенеза и онтогенеза функций.
Трудно найти такую проблему, которая разрабатывалась бы под
одним и тем же углом зрения во всех лабораториях, где ее изучают.
Очень часто сильно разнятся не только теоретические подходы, но и
добытые при этом фактические результаты ив особенности
формулированные на основании их теоретические толкования. Я не імог не
учесть.этого обстоятельства и довольствоваться изложением данных
только одной какой-либо лаборатории. Как и в первом издании, я
старался беспристрастно изложить то существенное, что дала . каждая
лаборатория по данной проблеме, а затем пытался дать всему этому
то научное понимание, какое напрашивается на основании всего
собранного по данной проблеме фактического материала.
В результате существенных изменений и дополнений, в новом
издании «Общей физиологии» значительно выросла библиография,
увеличился и общий объем книги. Но общий характер книги как
теоретического и практического руководства, объединяющие ее идеи и
целенаправленность остались неизменными. Книга имеет в виду дать
возможность молодым специалистам-физиологам получить современное
представление по всем основным проблемам мышечной и нервной
физиологии и еще больше углубить его по цитируемым
первоисточникам, приучить критически относиться ко всем фактическим
положениям, из каких бы лабораторий они ни исходили, и теоретическим
толкованиям, кем бы они ни высказывались. Наконец, изложением
всех основных физиологических процессов в их динамике и развитии,
а также описанием методов исследования этих процессов и постановки
самих опытов эта книга должна помочь молодым специалистам-
физиологам приобрести диалектическое понимание физиологических
явлений.
Переработка этой книги происходила в два приема: в 1938—1940 гг.
ив 1944—1945 гг. После первой переработки книга была просмотрена
моим другом, ныне покойным профессором Иосифом Львовичем Каном.
Иосиф Львович, как и в отношении первого издания, оказал мне
существенную услугу, просмотрев рукопись и дав множество ценных
указаний.
Я. Беритов
Тбилиси. 5/IX 1945 г.
ПРЕДИСЛОВИЕ К П ЕРВОМ У ИЗДАНИЮ
Как показывает название книги, данное руководство рассматривает
только физиологию двух органических систем — мышечнсш и'нервной.
Изучение основных функций мышечной и нервной систем дает
представление об основных свойствах живой возбудимой системы и о тех
закономерностях, которые характеризуют биологические системы в
отличие о? физико-химических.
Мышцы 'и нервы играют большую роль в деятельности каждого
органа. Для правильного понимания характерных особенностей
функциональных отправлений того или другого органа необходимо
предварительное знакомство с общими законами нервно-мышечной
деятельности. Без этого были бы совершенно непонятны, например,
особенности движения крови в кровеносных сосудах, роль- сердца и мозга
в этом движении и т. д. Отсюда вытекает необходимость знакомства
с общей физиологией мышечной и нервной "систем еще до изучения
физиологии отдельных органов. ~
Важное" 'значение общей физиологии мышечной и нервной систем
заключается также в том, что при ее прохождении изучаются основные
методы физиологического исследования, например методы
изолирования органов и тканей, их раздражения, регистрации вызываемых
реакций и т. д. Усвоение методов физиологического исследования
имеет чрезвычайное значение не только для специалиста-физиолога,- но
и вообще для каждого биолога и медика. Без этих методов немыслима
никакая экспериментальная работа как в физиологии, так ив любой
биологической и медицинской дисциплине.
Наконец, общая физиология мышечной и нервной систем
представляет собой некоторую базу для исследований в области психолотии и
науки о поведении, так как устанавливает общие законы деятельности
центральной нервной системы. Каждый исследователь психо-нервных
реакций и поведения животных и человека при установлении их
характерных особенностей и закономерностей должен руководствоваться
законами нервно-мышечной деятельности, многочисленные проявления
которой имеют место в каждой лсихо-нервной реакции и в каждом
акте поведения. Только при этом условии специфические
закономерности, выявляемые для психологии и науки о поведении, будут-
действительно научны.
Настоящее руководство является попыткой систематического
изложения существующих знаний о імышечной и нервной системах с точки
зрения диалектического материализма. Такое изложение общей
физиологии —'дело, конечно, не легкое. Нужно иметь в виду,_ что основные
понятия общей физиологии, т. е. понятия о живой возбудимой системе,
были созданы в прошлом столетии главным образом под влиянием
VIH
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
двух философских направлений — идеалистического и (механистически-
материалистического; а так как то и другое направления влияли на
физиологов, как и вообще на естествоиспытателей, более или менее
стихийно, то очень часто в изложении одного и того же автора одни
понятия носили идеалистический характер, а другие механистический.
Этот примитивный эклектизм препятствует точному изучению
явлений, установлению действительных взаимоотношений и проникновении*
в их историческое происхождение. Такое положение общей физиологии
хороша сознается не только у нас в СССР, но и за границей. Так,
например, Винтерштейн, критикуя идеалистические моменты в
физиологических понятиях, зовет к махизму для методологического
обновления общей физиологии; Эдвин Голът призывает к радикальному
эмпиризму; Вахгольдер, наоборот, считает телеологию
единственным выходом из создавшегося положения. Но, конечно, насколько эти
авторы правы в критике современного состояния общей физиологии,
настолько же беспомощен избранный ими путь для ее
методологического обновления.
Из современного критического положения можно выйти путем
совершенно новой переработки общей физиологии на основе
диалектического материализма. Однако для диалектического понимания
физиологических явлений недостаточно одной доброй воли и даже обладания
диалектическим мышлением. Для этого прежде всего необходимо,
чтобы каждое анализируемое явление было вновь изучено по
первоисточникам, а его природа выяснена путем постановки новых опытов
во всех тех случаях, где по существующим данным не совсем ясно
происхождение и историческое развитие анализируемого явления.
Составлением настоящего руководства мы были заняты около пяти
лет, между тем изучение физиологических явлений по первоисточникам
и выполнение новых и дополнительных исследований в целях
установления- диалектического развития требует гораздо больше времени.
Отсюда понятно, что в нашем руководстве должно быть описано немало
таких физиологических явлений, которые еще не получили требуемого
диалектическим материализмом освещения. Диалектическому анализу
были подвергнуты главным образом такие основные жизненные
явления, как основной биологический процесс живой системы, процесс
возбуждения, взаимоотношение структуры и функции, физико-химические
процессы в мышцах, динамика рефлекторной деятельности, динамика
корковой деятельности и некоторые другие. Частью этой работы явилась,
между прочим, статья, напечатанная в «Успехах современной
биологии» за 1933 г. под названием «Живая возбудимая система и основные
законы ее деятельности».
Настоящее руководство не удовлетворяло: бы своему назначению
подготовить теоретически и практически вооруженного
специалиста-биолога, специалиста-медика, если бы оно ограничилось только
изложением теоретических знаний. Оно одновременно должно служить
практическим руководством. Поэтому каждая теоретическая глава (за
немногими исключениями) снабжена практикумом, где приводится
постановка основных опытов с включением операционной техники и
указанием возможных методических ошибок. Как уже указывалось выше,
знание основных методов исследования жизненных процессов является
первейшей необходимостью для всякого биолога-экспериментатора, а
это знание приобретается только путем личной постановки опытов.
Практический курс данного руководства рассчитан как на студента-
биолога и студента-медика, так и на специалиста-физиолога.
Вследствие этого определенный ряд опытов должен быть исключен из про-
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
IX
граммы практических занятий со студентами и перенесен в
практический курс для специалистов. Студенты проходят лишь те опыты,
которые не сложны, не требуют много времени и могут быть легко
осуществлены при данном состоянии оборудования лаборатории.
Например, студенты могут не ставить опытов на теплокровных
животных, если такие опыты требуют операционного вмешательства, а также
опытов с такими сложными аппаратами, как струнный гальванометр
Эйнтховена и т. п., или таких опытов, которые требуют предварительно
хорошей сноровки и длительной работы, как опыты с условными
рефлексами по Павлову.
Большое внимание в этой книге уделено изложению данных о
структуре органических образований, особенно новейших сведений, имеющих
прямое отношение к функциональной деятельности этих образований.
Новейшие физиологические исследования мышечной и нервной систем
очень часто производятся параллельно с гистологическими. В
результате был установлен ряд чрезвычайно важных положений о взаимной
связи определенных физиологических явлений и структуры
органических образований. Эти 'положения выводились учеными независимо
о,т диалектического мировоззрения. Но, имея в виду одно из основных
положений этого мировоззрения — единство структуры и функции, мы,
понятно, не могли обойти ни одного явления без рассмотрения его
взаимоотношений со структурой.
В этой книге после каждой главы дается перечень тех работ,
которые цитируются в тексте. Это добавление имеет существенное
значение для самостоятельной разработки того или иного вопроса по
первоисточникам. Однако должен заметить, что литературная обработка
каждой главы по новейшим источникам не может быть признана
вполне удовлетворительной. Это было вызвано прежде всего отсутствием
исчерпывающей литературы, а затем отсутствием времени для
ознакомления со всей литературой по общей физиологии. Нужно иметь в виду,
что в главных физиологических журналах, как Pfluger's Archiv f. d.
gesammte Physiologie, Americ. Journal of Physiology, Journal of
Physiology, а также в других большая часть статей посвящена общей
физиологии мышечной и нервной систем. Поэтому приходилось выбирать
материал и несколько раз приступать к (переработке руководства на
основании накопившихся данных.
Мы снабдили данное руководство рисунками, в большинстве
случаев оригинальными. Эти рисунки иллюстрируют определенные
эксперименты и теоретические выводы или же представляют образцы
миографических и электрографических записей внешних реакций, которые
должны быть воспроизведены во время практических занятий. Кроме
того, много рисунков дано специально для практических занятий. Они
изображают более или менее схематично постановку опытов или
необходимые для этой цели препараты и аппаратуру. Громадное
большинство этих рисунков сделано по тем опытам, какие ставятся студентами
при практических занятиях и специалистами-физиологами при
прохождении специального курса на физиологическом отделении Тбилисского
(Тифлисского) университета.
Считаю своим приятным долгом поблагодарить профессоров Иосифа
Львовича Кана и Михаила Андреевича Киселева за просмотр
рукописи и ценные указания. Равно благодарю художника Александра
Федоровича Гветадзе за большую работу по изготовлению рисунков.
И. Берито?
Тбилиси (Тифлис), 25/ХИ 1935 г.
ОГЛАВЛЕНИЕ
Стр.
Предисловие ко второму изданию V
Предисловие к первому изданию VII
Введение • 1
Часть первая
МЕТОДИКА
Глава /. Необходимые сведения из анатомии лягушки 9
Практикум • . . . ¦ 9
Глава if. Методика раздражения 12
Глава ///. Методика регистрации нервно-мышечных реакций 24
A. Миографический метод 24
B. Электрографический метод 29
Практикум 34
Часть вторая
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Глава /. Общие сведения о строении мышечной системы 39
Глава If. Возбудимость как основное свойство мышечной ткани 48
Практикум 61
Глава fff. Сократительная функция мышцы 66
Практикум 116
Глава fV. Структура живой возбудимо системы 123
Глава V. Основной биологический процесс 120
Практикум 144
Глава VI. Закон возбуждения 146
Практикум 164
Г лава Vfl. Процесс возбуждения мышцы 167
Практикум 188
Г лава Vflf. Повышение возбудимости при раздражениях • . . 192
Практикум 202
Глава fX. Утомление 204
Практикум 217
Глава X. Работа мышцы 220
Практикум 234
Глава XI. Тепловой эффект мышцы 237
XII ОГЛАВЛЕНИЕ
Стр.
Глава XII. Общая физиология сердечной мышцы 247
Практикум ' 259
Глава XIII. Общая физиология гладкой мускулатуры 262
Практикум 288
Глава X}V. Смерть мышечной ткани 292
Практикум - ... 299
Часть третья,
ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ
Глава I. Нейронная и нейропильная теория нервной системы 303
Глава II. Возбудимость нервного ствола 328
Практикум 344
Глава III. Возбуждение и его проведение в нервном стволе 349
Практикум 386
Глава IV. Утомление нерва 392
Практикум 400
Глава V. Общая физиология концевой двигательной пластинки в
скелетной мышце 403
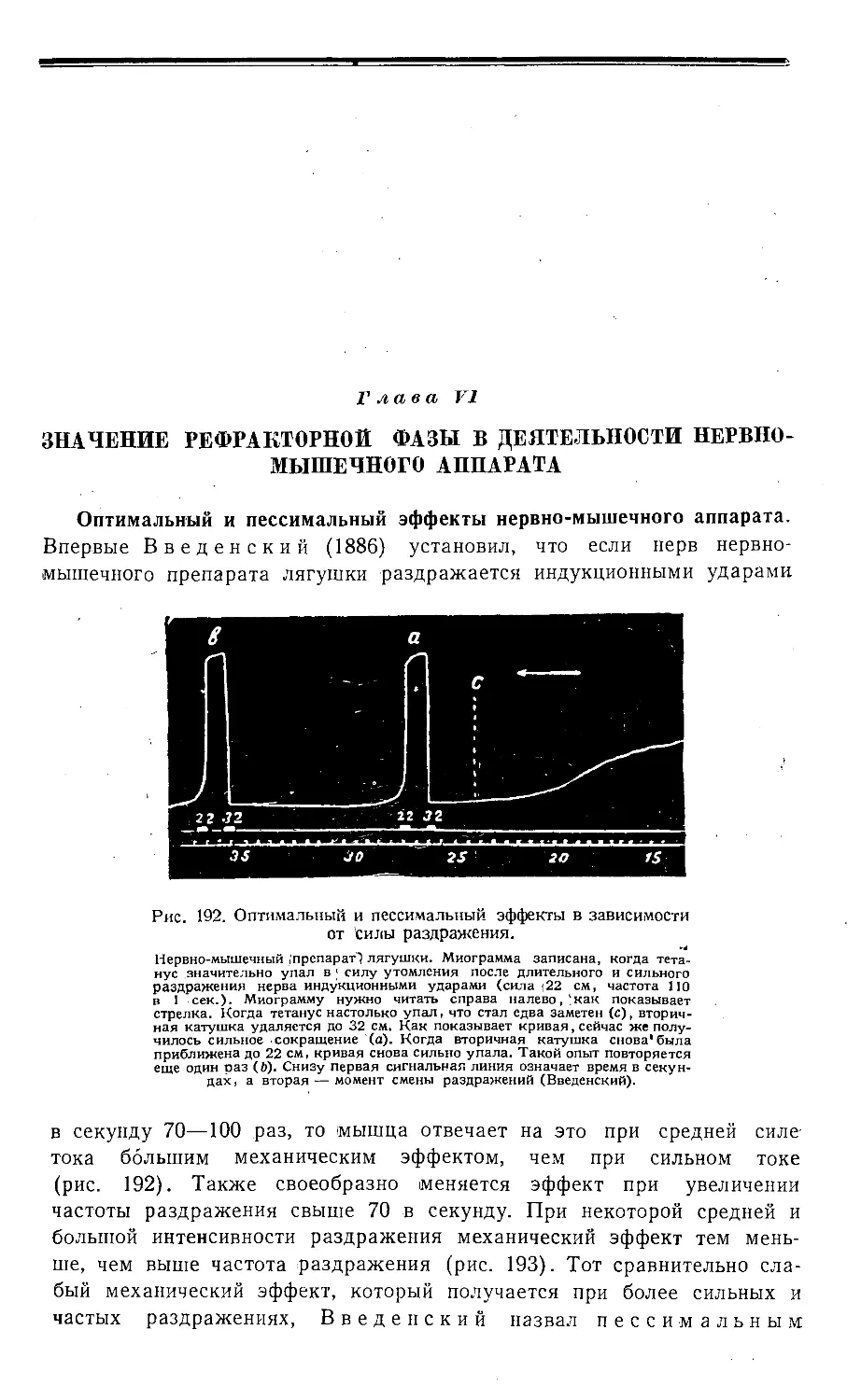
Глава VI Значение рефракторной фазы в деятельности нервно-мышечного
препарата 418
Практикум .' 428
Глава VII. Парабиоз 430
Практикум 446
Г л а в а VIII. Полярное действие электрического тока на нервно-мышечную
систему 449
Практикум 460
Ч асть четвертая
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ (АВТОНОМНОЙ) НЕРВНОЙ
СИСТЕМЫ
Глава I. Анатомические особенности вегетативной нервной системы. . . . 465
Глава II. Физиологические особенности вегетативной нервной системы . . 474
Глава ///.Антагонистическая деятельность симпатической и
парасимпатической систем 496
Глава IV. Роль вегетативной нервной системы в рефлекторной
деятельности внутренних орган ов 512
Глава V. Роль вегетативной нервной системы в деятельности скелетной
мускулатуры 519
Глава VI. Трофическое влияние симпатической нервной системы 528
Глава VII. Роль симпатических ганглиев в координации отдельных органов . 534
Глава VIII. Гомеостатическое значение симпатической системы 537
Практикум 540
Именной указатель 543
Предметный указатель - . . . • 551
ВВЕДЕНИЕ
ПРЕДМЕТ, ЗАДАЧИ И МЕТОДЫ ФИЗИОЛОГИИ И ЕЕ ОТНОШЕНИЕ
К ДРУГИМ БИОЛОГИЧЕСКИМ НАУКАМ
Предмет физиологии. Предметом физиологии является внутренняя
деятельность, (или такие внутренние функции организма, как
пищеварение, дыхание, кровообращение, движение конечностей и других
органов, выделение мочи, пота и других веществ, продукция тепла, нервное
л гуморальное взаимодействия органов, внутренняя секреция,
размножение и т. д. Каждая функция специфична для определенного рабочего
органа и в большинстве случаев для определенной ткани. Несмотря на
^то, ни одна из названных функций не осуществляется деятельностью
только одного органа. Напротив, можно утверждать, что каждая из
них выполняется при участии почти всех остальных органов, т. е.
целого организма. Так, например, координированное движение
конечности зависит не только от свойств нервно-мышечной системы и других
тканей, входящих в структуру конечности, но также от деятельности
центральной нервной системы, органов дыхания, кровообращения и т. д.
Под внутренней деятельностью, или внутренними функциями, мы
подразумеваем прежде всего деятельность отдельных органов и систем,
как, например, переваривание пищи в пищеварительном канале,
движение крови в кровеносных сосудах, газовый обмен в легких,
рефлекторное движение конечностей и т. д. Внутренними функциями
являются также те биологические процессы, которые протекают в каждой
ткани организма, а именно: сокращение мышц, производство и
выделение секрета железами, возбуждение рецепторов под влиянием
раздражений, распространение возбуждения по нервам и т. п. Далее, к
внутренним функциям относятся те физико-химические процессы,
которые возникают внутри клеток и в окружающей их лимфатической
¦:эеде, а также в кровеносных сосудах. Наконец, к внутренним
функциям относится взаимодействие отдельных органов, тканей и клеток,
. издающее физиологическое единство и целостность организма, вслед-
;твие чего деятельность каждой части определяется совокупной,
объединенной деятельностью всех составных частей целого.
II. С. Беритов
2
ВВЕДЕНИЕ
В первой половине прошлого столетия, -во времена Иоганна*
Мюллера, предметом физиологии являлась главным образам
деятельность отдельных органов. Во второй половине стало играть
крупную роль изучение биологических процессов в тканях, а также физико-
химических процессов во внутренней среде организма. В начале же
этого столетия и в особенности в настоящее время главным предметом
исследования являются, с одной стороны, биологические и
физико-химические процессы, протекающие непосредственно внутри клеток, и, с
другой — взаимодействие всех этих процессов в клетках, тканях,
органах и в целом организме.
Теоретические задачи физиологии. Главнейшими теоретическими
задачами физиологии являются: 1) изучение тех закономерностей,
согласно которым протекают жизненные явления в клетках и тканях,
сюда, например, относятся возникновение импульса в
нервно-мышечной ткани, его распространение внутри клетки и переход с одной клетки
на другие, его изменение под влиянием деятельности других тканей,
его воздействие на другие ткани и т. д.; 2) установление
закономерностей, определяющих деятельность каждого органа в отдельности и
взаимодействие между органами в целом организме; 3) изучение
законов, согласно которым в организме протекают такие процессы, как
усвоение питательных веществ клетками, электрические, химические и
тепловые явления в клетках и тканях, вызываемые действием
внешней среды, также те физико-химические процессы, которые возникают
внутри клеток и во внутренней среде в результате биологических
процессов: распад белков и углеводов с образованием разного рода
органических веществ, окисление последних или переход их в другие
соединения, изменение полупроницаемых перепонок, вызываемое тем
или другим путем, и т. д.; 4) изучение законов взаимодействия между
функцией и структурой, прежде всего в целях установления
изменений внутренней структуры клеток и тканей в связи с их
функциональной деятельностью; 5) наконец, изучение законов филогенеза и
онтогенеза всех жизненных функций организма и их
взаимоотношений со структурным развитием его.
Практические задачи физиологии. Практические задачи физиологии
многообразны. Основная задача заключается в служении
здоровью человека. Правильное понимание ненормального,
патологического состояния человека и искусственное восстановление нормы
путем лечения могут быть достигнуты только на основании точных
знаний о закономерностях протекания каждой функции и взаимодействия
функций. Немыслимо было бы, например, правильное лечение
патологических нарушений пищеварения, если бы эта функция не была нам
известна в норме. В частности, можно сказать, что только после того,
как была изучена ферментативная деятельность и иннервация
пищеварительной системы, стало возможным правильное терапевтическое и
хирургическое лечение заболеваний этой системы.
Второй основной практической задачей являются гиг иена и р а-
ВВЕДЕНИЕ
3
ционализация питания человека и сельскохозяйственных
животных: питательность, количество, пропорции разных пищевых
веществ, их отношение к работе и другие вопросы питания могут быть
научно разрешены только на основании изучения физиологической
деятельности организма.
Затем чрезвычайно важную практическую задачу представляют
гигиена и рационализация человеческого труда.
Прежде всего здесь важно установить такого рода взаимоотношения
между интенсивностью работы, ритмом ее, длительностью отдыха и
другими условиями, чтобы продуктивность работы была максимальна
и в то же время не имело места переутомление организма или
нарушение какой-либо внутренней функции организма. Ясно, что такой
рационализации труда должно -предшествовать изучение процесса труда,
а это изучение будет тем точнее и продуктивнее, чем лучше были
изучены все внутренние функции организма, чем точнее установлены
закономерности этих функций.
Дальнейшей практической задачей является внедрение
физиологических основ в физкультуру. Физкультура не может
способствовать здоровью человека, благоприятствовать его физическому
развитию, если она не будет всецело основываться на научных данных
о внутренних функциях организма. Так, например, необходимо, чтобы
для каждого типа физического упражнения были найдены свой ритм
и продолжительность соответственно возрастным, физическим и
половым особенностям физкультурника.
Наконец, ни одна наука кроме физиологии не в состоянии так ясно
и непосредственно путем наглядного опыта доказать вредное влияние
потребления алкоголя, курения табака, пользования морфием и
кокаином и другими подобными веществами на разные функции организма.
Следовательно, знание внутренних функций организма представляет
наилучшую основу для выработки правильных жизненных
привычек.
Методы физиологии. Так как физиология изучает внутренние
функции организма и проявление их в разных органах, тканях и клетках,
то главнейшим методом ее являются более или менее полное
изолирование органов, тканей и клеток и изучение их деятельности как
независимо от всего остального организма, так и в зависимости от той или
другой внутренней функции его или ряда функций. Так, например, для
изучения деятельности слюнных желез выводят наружу их проток.
Выходящую наружу слюну собирают и определяют ее качественный
и количественный состав, а также ее ферментативную деятельность.
Таким образом изучают собранную слюну при разных условиях:
например, при раздражении ротовой полости пищевыми или
отвергаемыми веществами, при нарушении кровообращения или иннервации
желез, при том или ином повреждении центральной нервной системы и т. д.
Первоначально, в первой половине и в середине прошлого столетия,
для изучения деятельности отдельных органов прибегали главным об*
4
ВВЕДЕНИЕ
разом к их изолированию. В конце прошлого столетия и в начале
нашего изучение изолированных тканей стало обычным явлением. В
настоящее время изолирование отдельных клеток, например нервных и
мышечных, и изучение их функций под микроскопом становится очень
распространенным и чрезвычайно плодотворным методом
физиологического исследования.
Как указывалось выше, в нормальном организме та или другая
функция осуществляется при участии всего организма. Это
обстоятельство делает очевидной недостаточность изучения функций на
изолированных частях организма. Деятельность организма всегда носит
целостный характер. Это значит, что функция того или другого органа
определяется прежде всего деятельностью всего организма.
Следовательно, физиология должна считать своей главной задачей изучение
той или иной функции в целом организме. Но, конечно, неизбежной
предварительной стадией является изучение той или другой функции
на изолированных частях. Оно должно предшествовать изучению ее в
целом организме.
Так как деятельность отдельных органов и систем сопровождается
освобождением той или другой энергии: механической, химической,
электрической и тепловой, то мы производим регистрацию этих энергий
специальными саморегистрирующими приборами. Автоматическая
регистрация дает возможность точно изучить всю динамику органических
процессов. Но так как чувствительность и точность приборов зависят
от общего состояния техники, то ясно, что с развитием техники вообще
происходит и усовершенствование техники физиологического
исследования. Современная физиологическая техника такова, что дает
возможность изучить в нервных волокнах электрический эффект, измеряемый
в миллиардных долях ампера, и тепловой эффект, измеряемый в
миллионных долях калории.
Физиология, биохимия и биофизика. Каждый биологический
процесс, протекающий в 'клетках, тканях и во всем организме, как рост,
размножение, возбуждение, рефлекс и т. д., с одной стороны,
обусловливается сложным комплексом физических и химических изменений, а
с другой, сопровождается возникновением целого ряда физических и
химических изменений в клетках и вне их во всей внутренней среде
организма. Эти физико-химические процессы протекают согласно
известным законам химии и физики. Соответственно этому исследование
их производится известными методами химии и физики. Тот отдел
физиологии, который изучает физические изменения в организме,
называется биофизикой, а тот, который изучает химические
изменения, — биохимией.
Изучение физико-химических процессов, протекающих в организме,
абсолютно необходимо для понимания условий возникновения и
протекания биологических процессов. Несмотря на это, ни в коем случае
нельзя думать, что путем такого исследования мы сведем
биологические процессы целиком к физико-химическим, что этого будет совер-
ВВЕДЕНИЕ 5
шенно достаточно для понимания самой природы биологических
процессов. Биологический процесс представляет качественно иное
явление, стоящее выше самых сложных физико-химических процессов. Он
управляется своими собственными закономерностями. Следовательно,
биофизика и биохимия должны служить физиологии главным образом
для успешного выявления специфических закономерностей
биологического процесса.
Физиология и морфологические науки. Физиология имеет
существенное значение для всех биологических и, в частности, для
морфологических наук. Давно уже прошло то время, когда зоологи, эмбриологи,
гистологи, анатомы изучали свой предмет — макроскопическое или
микроскопическое строение разного рода органических образований —
исключительно описательным путем, в одном определенном состоянии.
В настоящее время такого рода изучение даже нельзя считать
научным. Сегодня ту или другую отрасль морфологических знаний іможно
считать наукой только в том случае, если в ней господствует
исторический метод, т. е. явления изучаются не в одном статическом
состоянии, а в их постоянной изменяемости, обусловленной множеством
внутренних и внешних влияний. Сегодня в. биологию входят такого
рода отрасли знаний, как экспериментальная зоология,
экспериментальная гистология, экспериментальная эмбриология и др. Поэтому
для развития каждой из названных экспериментальных
морфологических наук безусловно необходимо знание физиологических
закономерностей и владение методами физиологического исследования.
Современный гистолог, например, исследуя ткани и клетку, сначала
изучает структурное образование при одном физиологическом
состоянии, а затем наблюдает изменение структуры при изменении
состояния тканей. Так были исследованы пищеварительные железы,
мышечные клетки, нервные волокна, нервные клетки и т. д. Для означенного
функционального изменения необходимо не только знать функции
изучаемого органического образования, но и владеть методами
воздействия на данную функцию и методами ее изучения. Ясно, что
гистология как экспериментальная и теоретическая наука не может
в настоящее время развиваться без физиологии. В таком положении
находится и всякая другая морфологическая дисциплина.
Физиология и психология. Научное изучение всякого рода
психонервной деятельности, в частности мышления, безусловно
подразумевает знание тех физиологических закономерностей, которыми
обусловливается выявление тех или других внешних реакций во время
психонервной деятельности. В этих реакциях одинаково принимают участие
рецепторы, эффекторы, центральная нервная система, а также другие
органы и системы, как кровообращение, пищеварение и т. д. Отсюда
следует, что психолог не может изучить психо-нервную деятельность
без ясного представления о некоторых внутренних функциях
организма, без знания некоторых методов физиологического исследования.
Однако это не означает, что психолог должен рассчитывать всецело на
6
ВВЕДЕНИЕ
физиологию при выяснении материальных процессов изучаемых им
психических явлений. Физиология не может дать тех закономерностей,
согласно которым протекают материальные процессы в субстрате мозга,
продуцирующем ощущения, представления и другие более сложные
субъективные переживания. Очевидно, изучение этих материальных
процессов должно стать первейшей задачей психологии.
Физиология и наука о поведении. Предметом науки о поведении
является та целостная деятельность организма, которая проявляется в
актах поведения. Этими актами организм, с одной стороны,
функционально приспособляется к среде, а с другой — так видоизменяет
среду, что она приспосабливается к потребностям организма. Эта
деятельность организма осуществляется посредством внутренних функций.
Ясно поэтому, что при изучении актов поведения физиология должна
играть крупнейшую роль. Значение физиологии при изучении
поведения настолько велико, что многие ученые считают акты поведения
предметом физиологии. Но такое представление неправильно не только
потому, что оно противоречит историческому развитию физиологии, но и
потому, что акты поведения нельзя считать внутренней функцией,
подобно пищеварению, дыханию и т. д. Поведение представляет
объединенную деятельность всех внутренних функций в виде актов
приспособления, причем в трудовом поведении человека сознание играет
главенствующую роль в объединении внутренних функций. Поэтому
основная задача науки о поведении состоит в изучении тех
закономерностей, согласно которым происходит как сознательное, так и
бессознательное объединение внутренних функций в актах поведения.
Сообразно с этим и методы исследования в науке о поведении являются
совершенно своеобразными: они должны быть направлены на изучение
взаимоотношений целого организма с внешней средой. Но, конечно,
при изучении поведения человека должны быть использованы и
психологические методы изучения субъективных переживаний. Итак,
физиология и психология должны быть положены в основу науки о
поведении, но, несмотря на это, наука о поведении — совершенно
самостоятельная наука, обладающая своим собственным предметом, задачами и
методами исследования.
Часть первая
МЕТОДИКА
Так как данная книга одновременно является и практическим
руководством, а описание опытов следует сейчас же после каждой
теоретической главы, імы сочли нужным предпослать всему курсу общие
методические указания, которыми следует руководствоваться при
постановке этих опытов. Здесь мы даем очень краткие сведения из анатомии
лягушки, сведения о методике электрического раздражения и
графической записи наблюдаемых явлений.
Глава I
НЕОБХОДИМЫЕ СВЕДЕНИЯ ИЗ АНАТОМИИ ЛЯГУШКИ
Практические занятия по общей физиологии животных происходили и
происходят главным образом на лягушках. Поэтому безусловно необходимо, чтобы каждый:
студент, прежде чем приступить к практическим занятиям по физиологии,
обязательно ознакомился с анатомией лягушки. Необходимо знать анатомию задних
конечностей, скелет и мускулатуру, 'нервы и сосуды; нужно иметь ясное
представление о расположении (внутренних органов, а также об отделах центральной нервной
системы и их положении. Вместо подробного описания анатомических соотношений
мы приводим (несколько подробных 'рисунков. Рис \ и 2 дают изображение
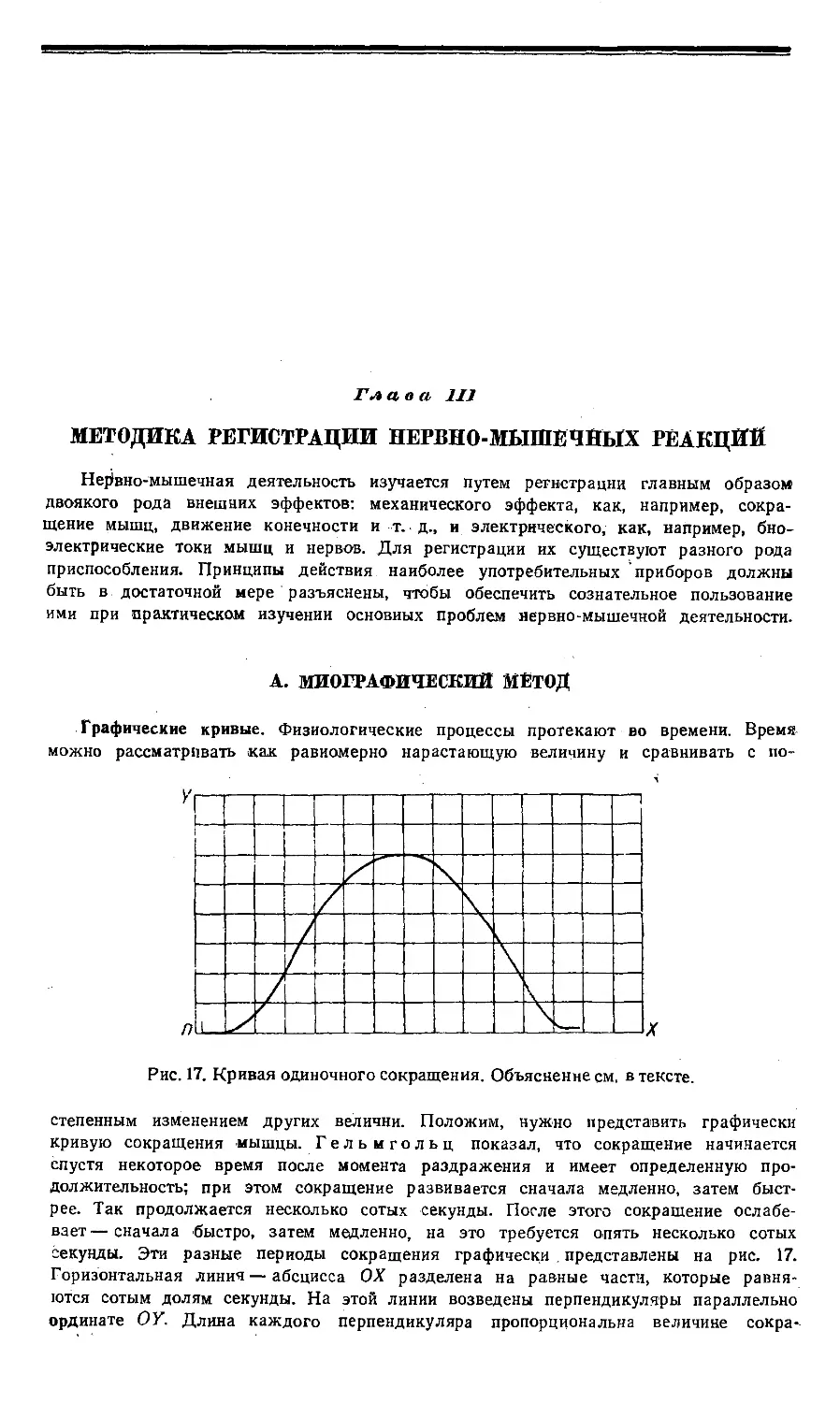
наружных мышц задней -ноги с задней іи передней сторон. Рис. 3 дает картину
(внутренних мышц бедра. На ірис. 4 представлена нервная система, центральная и
периферическая; іна рис. 5 — головной імоэг -сверху; наконец, рис. 6 дает представление
о расположении внутренних органов.
Практикум
1. Выделение главных мышц задней конечности. Берут полотенцем лягушку
левой рукой; мизіинцем и 'безыменным пальцами прижимают ее задние лапки
к ладони, большим и (средним пальцами фиксируют передние конечности, а
указательным — голову. Правой рукой производят ножницами іперерезку позвоночника
ровно іпо ісередине между атланто-затылочным сочленением и местом сочленения
тазовых костей; затем удаляют вое внутренности и отделяют заднюю половину
тела. Оинцетом и ножницами «снимают (кожу, держа пинцетом поіэвоночник.
Выделение мышц заключается в освобождении их по эсей длине от общих
фасций с другими мышцами. На бедре (выделяют следующие мышцы: triceps femoris
ileofibularis, sartorius, gracilis major, semitendinosus, semimembranosus, adductor tna-
gnas. На голени выделяют: gastrocnemius, peroneus, tibialis antialis, tibialis posticus..
10
МЕТОДИКА
Прослеживают, где каждая мышца начинается и где она кончается, заучивают
названия и зарисовывают расположение мышц.
2. Испытание характера действия мышц на суставы. Испытание действия мышц
на коленный и тазобедренный суставы производится следующим образом: берут
Рис. 1. Мышцы левой задней
ноги с задней (спинной) стороны
ляг/шки (Rana esculenta) no
Эккеру.
7 —m. coccygeo-iliacus; 2—т.
gluteus; 3— т. pyriformis; 4—т. vastus
externus; 5—т. trie, femoris; 6 —т.
tensor fasciae latae; 7 — т. gracilis
minor; 8 — m. semimembranosus;
9—m. ileofibularis; 10—m.
gastrocnemius; 11—m. tibialis anticus;
12 — m. peroneus.
Рис. 2. Мышцы левой задней ноги
с передней (брюшной) стороны
(Ranajesculenta) по Эккеру.
7 —m. sartorius; 2—т. adductor Iongus;
3— т. adductor brevis; 4—т. adductor
magnus; 5 — т. vastus internus trie,
femoris; б — т. gracilis major; 7 — m.
gracilis minor; 5 —m. gastrocnemius; 9—-
m. extensor cruris; 10—m. tibialis
anticus; 11—m. tibialis posticus; 12—m.
flexor tarsi.
пинцетом бедренную кость и поднимают лапку на воздух» затем другим пинцетом
потягивают ту или другую мышцу и 'наблюдают Движение суставов. Потягивание
m. triceps femoris, например, покажет, что эта мышца разгибает коленный сустав
и сгибает тазобедренный; m. semitendinosus при потягивании произведет обратное
действие: он сгибает коленный сустав и разгибает тазобедренный.
На голени аналогичное испытание происходит таким образом: берут лапку за
берцовую кость и поднимают ее на воздух. Потянув за m. gastrocnemius,
убеждаются (в том, что голеностопный сустав при этом разгибается, а коленный сгибается.
Обратное действие покажет гл. tibialis anticus. Таким же путем нужно
испытывать все другие мышцы и записать результаты.
СВЕДЕНИЯ ИЗ АНАТОМИЙ ЛЯГУШКИ
11
Некоторые мышцы Действуют на сустав разно: при одном положении сустава
«ни ее (разгибают, при другом сгибают. Например, m. ileofibularfs огибает
тазобедренный сустав, если угол при тазобедренном суставе меньше 90° и бедро находится
в одной (плоскости с корпусом, я, наоборот, разгибает его, если угол больше 90°.
3. Выделение нервов. Здесь
обращают внимание главным образом "на
нервы задней лапки: надо рассмотреть
образование седалищного сплетения из
трех корешков: VII, VIII и IX, образо-
-вание из него двух
нервов—седалищного (п. ischiadicus) и крестцового
(п. cruralis); наконец, деление
седалищного нерва на п. tibialis и п. рего-
neus; проследить на mm.
gastrocnemius, sartorius и нескольких других
мышцах места вхождения нерва.
Рис. 3. Глубоко лежащие мышцы левого
бедра (Rana esculenta) по Эккеру.
Перерезаны: m. tensor fasciae latae (7); т.
adductor longus (77); т. sartorius (72); т.
gracilis major (13); т. gracilis minor (74);
2 —m. adductor brevis; 3 —m. pectineus; 4—5
главы т. semitendinosus; б — пучки мышечных
волокон, которые отходят от semitendinosus к т.
adductor magnus; 7—т. ileopsoas; 8—т. adductor
-nagnus; 9—брюшко т. semitendinosus; 10—т.
vastus internus trie, femoris.
4. Расположение внутренних органов.
Для этих целей приготовляют
«спинномозговую лягушку» путем перерезки
«пинного мозга в области атланто-
затылочного сочленения. Затем
прикалывают лягушку булавками к
пробковой пластинке, протыкая их через
конечности и переднюю часть
головы. Разрезом кожи и мышц
живота обнажают внутренности.
Руководствуясь рисунком, знакомятся с их
г анатомией спинного и головного мозга
нервной системы.
Рис. 4. Спинной и головной мозг с
передней (брюшной) стороны (R. esculenta) по
Эккеру.
Не — большие полушария; L. ор — Iobi optlcl,
средний мозг; Sp т —спинной мозг;
обозначенная через М черта показывает пограничное место
между спинным и головным мозгом; /—п. olfac-
torius, обонятельный нерв; //—п. opticus,
зрительный нерв; ///—п. oculomotorius,
глазодвигательный нерв; IV — п. trochlears, блоковой
нерв; V — п. trigeminus (Va— г. ophtalmicus;
V& — г. palatinus; Vc—r. maxilaris sup.; Vd—r.
maxil. inf., Ve — r. tympanicus) — тройничный
нерв и его ветви; VI — п. abducens, отводящий
нерв; VII—п. facialis, лицевой нерв; X (вместе
с IX и X/) — п. vagus, блуждающий нерв;
Хг — г. communicans; Xz — п. glossopharyngeus,
языкоглоточный нерв; Х3— г. intenstinalis; Xt—г.
cutaneus; Mt—Мл0 — нервы спинного мозга от
1-го до 10-го; S —пограничный симпатический
ствол с симпатическими узлами S\—SJ0; SM—его
rami communicantes к спинномозговым нервам;
Sp —¦ верхний конец симпатического ствола
около ganglion Gasseri (G); No — n. obturatorius;
N — п. ischiadicus, седалищный нерв.
положением. Практическое знакомство
откладывается до изучения центральной
Г л а в а II
МЕТОДИКА РАЗДРАЖЕНИЯ
Преимущества индукционного тока (переменного) как раздражителя. Основные-
функции живой ткани можно изучать по ее деятельности. Последнюю же (можно
вызвать действием разнообразных раздражителей. При зксперименталыных
исследованиях большей частью пользуются электрическим током, а именно индукционными
ударами. 'Индукционный ток признан наилучшим раздражителем, во-первых, потому,
что он имеет высокое напряжение и проходит через ткани, обладающие большим
сопротивлением, сохраняя при ?том значительную силу; во-вторых, он протекает
кратковременными 'ударами, вследствие чего раздражающее действие продолжается
только в период (прохождения тока. Другие раздражители таким свойством или не
обладают совсем, или же обладают им не в такой степени. Химические раздражители,
например, постепенно проникают ів ткань 'и постепенно же выходят из нее, поэтому
действие их 'без нужды затягивается на продолжительное время. Точно так же
трудно быстро воздействовать температурным раздражителем и (быстро прекращать
это действие. Кроме того, температура очень сильно влияет на функциональное
состояние ткани. 'Механическое (раздражение вообще мало применимо для
раздражения живой ткани, (потому что сопровождается механическим повреждением ее
элементов. Гальванический (постоянный) ток .употребляется как раздражитель
в редких случаях, так как его трудно применять кратковременными ударами, а іпри
более или менее длительном действии іс ним связан значительный электролиз
тканевых соков и перенос продуктов электролиза с одного 'места на другое; это
обстоятельство сильно -меняет функциональное состояние раздражаемого участка іживой
ткани.
Отсюда очевидно, что приступающий к занятиям по 'физиологии должен иметь
совершенно ясное представление об индукционном токе, о гальваническом токе, об
электролизе и т. д.
Проводники и изоляторы. Все тела с точки зрения электрической проводимости
делятся на две группы. Тела, которые очень плохо или совсем не проводят
электрический ток, называются непроводниками или изоляторами. Такие суть: воздух,
сухое дерево, эбонит, стекло, резина, шелк, бумага и пр. Тела, которые хорошо
проводят ток, называются проводниками. Такие суть: все металлы, уголь, земля,
тело человека и животных, растворы. Причем последние, где прохождение тока
происходит с помощью ионов, называются проводниками второго рода (растворы
солей, кислот и щелочей). Металлы же будут проводниками первого рода.
Гальванические элементы. Если в проводник второго рода, иначе называемый
электролитом, опустить два проводника первого рода, например цинк и медь,
которые называются в данном случае электродами, и последние соединить между
собой медной проволокой, то ток пойдет через проволоку от меди к цинку.
Электрод, из которого ток выходит во внешнюю цепь, принято называть анодом;
второй электрод, к которому течет ток, называется катодом. Такое соединение пре-
МЕТОДИКА РАЗДРАЖЕНИЯ
13
водников первого и второго рода называется гальваническим элементом, а
получаемый при этом ток называется гальваническим током.
В элементе возникновение тока обусловлено разностью потенциалов на концах
опущенных в раствор
электродов. Электричество от
места большего потенциала
переходит к месту меньшего
потенциала. Отсюда понятно,
что если опустить в раствор
два совершенно одинаковых
по составу проводника и
-соединить их через гальвано-
Рис. 5. Головной мозг сверху
<Rana esculenta) по Эккеру.
Мо— medulla oblongata,
продолговатый мозг; S. гк. —fossa
rhomboidalis, ромбовидная ямка;
S—sulcus centralis, центральная
борозда; Li — limbus fossae
rhomboidalis; С— cerebellum,
мозжечок; L. op — lobus opticus,
средний мозг; Gp — gland, pinealis,
шишковидная железа; Th. о —
thalamus opticus, промежуточный
мозг; НС — большие полушария;
і — п. olfactorius, обонятельный
нерв; L. оі — lobus olfactorius,
обонятельные доли.
Рис. б. Внутренние органы самки лягушки (Rana
esculenta) no Эккеру.
И — сердце; L Ll, L* — печень; Lg, Lg*- — легкие; М —
желудок; Du — двенадцатиперстная кишка; D—тонкие кишки;
НВ —мочевой пузырь; Е,Е—яичник, который сильно разросся,
так как животное находится в периоде размножения.
метр, то последний не будет показывать тока, так
как на концах проводников, опущенных в раствор,
потенциал будет совершенно одинаков по величине,
и поэтому электричество не пойдет от одного
электрода к другому. Электрический ток
возникает от замыкания цепи при разности
потенциалов, что может случиться только при опускании в раствор разных проводников.
Сопротивление и проводимость. Гальванический ток при прохождении через
проводник испытывает сопротивление. Это сопротивление R прямо пропорционально
длине проводника / и обратно пропорционально его поперечному сечению $• Но,
кроме того, оно зависит от материала самого проводника р. Значит, R ==. п—
1 r s
Как івидно из формулы, если мы возьмем единицу длины (метр) и единицу
поперечного сечения {кв. мм), тогда сопротивление R будет равняться р\ р называется
удельным сопротивлением. Величина, обратная сопротивлению 1/R, называется
проводимостью.
Сопротивление живых тканей. Сопротивление живых тканей громадно. Оно
14
МЕТОДИКА
всегда превосходит (сопротивление металлов более чем /в миллион -раз; например, *
мышце оно в 2 500 000 раз больше, чем в ртути, а в нерве вдвое больше, чем в;
мышце. При проведении тока поперек нервных <или мышечных пучков
сопротивление оказывается гораздо большим, чем при продольном проведении. Это
обусловливается большой сопротивляемостью оболочек нервного и мышечного волокон.
Сопротивление небольшого участка нерва лягушки около 2 мм длиной к
0.7—1.0 мм толщиной (п. ischiadicus) равняется 20 000—40 000 омов. Сопротивления,
такого же участка более тонких -нервов (V іи VI спинальные нервы) диаметром
0.15—0.2 мм [равно 100 000—140 000 омов. Сопротивление нервов в сильной
степени зависит от их увлажнения. Указанные величины сопротивлений относятся іс
свежевырезанным увлажненным нервам. По мере подсыхания сопротивление растет.
Например, если подержать нерв на воздухе при комнатной температуре 19—22° в^
течение 10 минут, то сопротивление толстого нерва с 40 000 омов повышается до
80 000; сопротивление же тонкого нерва — с 130 000 ідо 380 000 омов. При этом
следует различать сопротивление наружное, которое зависит от степени увлажнения
наружной поверхности нерва, например физиологическим раствором, обычно
применяемым для предупреждения высыхания, и внутреннее сопротивление, которое
зависит от многих условий: от тканевой жидкости нервного ствола в прослойках между
нервными волокнами, от кровеносных сосудов, от нервных оболочек, нейроплазмьг
и нейрофибриллей нервных волокон и т. д. При высыхании сопротивление растет
сначала вследствие наружного высыхания, следовательно от увеличения наружного
сопротивления, а затем вследствие внутреннего высыхания — от увеличения
внутреннего сопротивления (Беритов, 10). Сопротивление нерва меняется также в
зависимости от температуры, >а именно с охлаждением сопротивление повышается.
Сопротивление отдельных мышечных или нервных волокон, конечно,
значительно превосходит сопротивление нервных стволов и целой мышцы. Так,
сопротивление мышечного волокна толщиной около 70 ц и длиной около 400 ц равняется
3 мега-омам. Это сопротивление в 30 раз больше сопротивления столбика
физиологического раствора такой же толщины и длины (Бухталь).
Сила тока. Силой тока называют количество электричества, проходящее через
любое сечение проводника в одну секунду. Сила тока / прямо пропорциональна
электродвижущей силе Е и обратно пропорциональна общему сопротивлению всей
Е
цепи R (закон Ома). Следовательно /= — •
Н
Единицы электричества. Сопротивление. За единицу сопротивления
принимается ом. Ом есть сопротивление ртутного столба сечением в 1 мм2 и длиной
106.3 см при температуре 0°.
Количество электричества. За единицу количества электричества
принимается кулон (К). Кулон есть такое количество электричества, которое,
проходя через концентрированный раствор медного купороса, выделяет на катоде
0.3294 г меди.
Сила тока. За единицу силы тока принят ампер (А). Это — такая сила тока,
при которой в единицу времени через поперечное сечение проводника проходит в
одну секунду один кулон.
Электродвижущая сила. За единицу электродвижущей силы (или
напряжения) принимается вольт (V). Вольт есть такая электродвижущая сила, под
влиянием которой по цепи с сопротивлением в 1 ом протекает ток в 1 ампер.
Ветвление тока. Если внешняя цепь на каком-либо участке делится на
несколько разветвлений, то общая сумма 'силы токов в разветвленном участке будет такая
же, как в неразветвленном. Если іветви неодинаковой длины, толщины или качествз,
тогда сила тока каждой ветви в отдельности будет обратно пропорциональна
величине ее сопротивления.
Реохорд. В физиологических лабораториях употреблялся специальный прибор
для отведения <в препарат тока желаемой силы путем образования добавочной.
цепи. Этот прибор называется реохордом Дюбуа-Реймона. В одну ветвь элемента
МЕТОДИКА РАЗДРАЖЕНИЯ
15
вводится живой препарат, а в другую — реохорд (рис. 7). Реохорд включает
большую систему проволок очень большого сопротивления (тонкая проволока из
никелина)» которая путем штепселей «^ S2 и т. д. и мостика VX может быть по частям
введена в цепь параллельно препарату. Изменение сопротивления в этой цепи
приводит к изменению силы тока в другой цепи, где находится препарат. Чем больше
сопротивление будет включено в реохорд, тем слабее будет ток в цепи реохорда и
тем сильнее он будет итти через препарат.
, 5. ? S, S, S% 7
\Ь \с
W
С
Рис. 7. Реохорд Дюбуа-Реймо-
на. Объяснение см. в тексте.
Рис. 9. Неполяриз^ющиеся
электроды Дюбуа-Реймона.
Объяснение см. в тексте.
Рис. 8. Компенсатор.
Между А и В натянута проволока большого сопротивления из никелина;
С — подвижной контакт.
Компенсатор. Так называется прибор, употребляемый ныне для ответвления токов
очень небольшой силы. Это — деревянная линейка, по которой протянута тонкая
проволока большого сопротивления длиной в 1 м (рис. 8). Линейка имеет два
неподвижных зажима на концах А В и один подвижной на мостике С. Через
неподвижные зажимы вся линейка вводится в главную цепь источника электричества, а
препарат отдельно соединяется с этой линейкой через подвижной С и один из
неподвижных зажимов А, т. е. он вводится в так называемую побочную цепь. Разность
потенциалов на концах линейки почти равна разности потенциалов источника тока.
Но если взять только часть линейки и измерить на концах этого участка
разность потенциалов, то она окажется меньше, а именно будет во столько раз меньше,
во сколько раз меньше сопротивление данного участка по отношению к общему
сопротивлению цепи.
16
МЕТОДИКА
Электролиз. Электрический ток, проходя через проводники второго рода, т. е.
растворы солей, кислот и щелочей, существенно изменяет их. Под влиянием
электрического тока в этих растворах происходит перенос ионов.
-+- +- +
При этом положительные ионы — катионы Н, К, Na и другие — направляются
к катоду и здесь отдают свой заряд, а освободившиеся химические продукты или
оседают на катоде, или вступают в реакцию с находящимися в растворе
(веществами. Отрицательные же ионы — анионы SO4 N03 С1 и т. д. — направляются к аноду,
здесь отдают -свой заряд и затем обычно (вступают в реакцию с веществом
растворителя или электродов. Например, разбавленная серная кислота содержит
электроположительные водородные ионы Н и электроотрицательные ионы S04. При
прохождении через нее электрического тока водородные ионы превращаются на
катоде в газообразный водород, а ионы SO4, теряя заряд на аноде, разлагают воду
с присоединением из ее состава водорода и освобождением кислорода.
Поляризация. Продукты разложения, собираясь на электродах, увеличивают
сопротивление для прохождения тока и, кроме того, изменяют ту самую
электродвижущую силу, которая вызывает разложение, так ікак между продуктами
разложения, в свою очередь, образуется электродвижущая сила, противоположная
электродвижущей силе основного тока. Поэтому действие последнего или совершенно
исчезает, или в значительной мере ослабевает. Это явление именуется поляризацией,
а вторично возникающая электродвижущая сила называется поляризационной.
Аккумуляторы. В качестве источника электричества употребляются главным
образом свинцовые и щелочные аккумуляторы. Свинцовый аккумулятор
состоит из двух пластинок, погруженных в разведенную серную кислоту. Если через 'них
проводить гальванический ток, то на анодной пластинке собирается перекись
свинца РЬОг, от которого пластинка становится коричневого цвета, катодная же
пластинка превращается в серый губчатый свинец. Это происходит вследствие того, что
из (первоначально образовавшегося сернокислого свинца на катоде выделяется РЬ
(в виде пористого свинца), а на аноде эта соль окисляется до образования РЬОг.
Окисление происходит потому, что здесь выделяется анион SO4, который разлагает
воду с образованием серной кислоты; освободившийся же кислород окисляет
анодную свинцовую пластинку. Если (после прекращения гальванического тока соединить
пористый свинец с пластинкой из окиси свинца проводником, то в последнем
возникает поляризационный электрический ток. Пластинка с пористым свинцом будет
катодом, а другая пластинка с окисью свинца—анодом. Поляризационный ток,
замыкаясь через проводник, приводит к восстановлению сернокислого свинца, и тем
самым ток постепенно уничтожается. Но и без соединения разнородных пластинок
проводником сернокислый свинец постепенно восстанавливается, и тем самым
электродвижущая сила поляризации уничтожается. Но она может быть снова создана,
если пропустить через аккумулятор гальванический поляризующий ток.
Для технических целей аккумуляторы обрабатываются поляризующим током до
тех пор, пока свинец одной пластинки на более или менее значительную глубину
не перейдет в РЬ02, а другая не станет пористой. В этом случае электродвижущая
сила обычно равняется 2.04 V. Во время зарядки она может доходить до 2.75 V,
но этот вольтаж быстро падает до 2.1 V.
Правила пользования аккумулятором:
1. Разряжение аккумуляторов должно происходить всегда через значительное
сопротивление. Нарушение этого условия ведет к порче того активного слоя
пластинок, который создает электродвижущую силу поляризации. Разумеется, чем больше
сопротивление в цепи, тем больше времени потребуется для разряжения.
2. При разряде аккумулятора вольтаж не должен падать ниже 1.8 V. Когда
напряжение достигает этого вольтажа, нужно прекратить пользование
аккумулятором -и поставить его на зарядку. Для полной зарядки пропускается ток в несколько
ампер в течение нескольких часов до «закипания» жидкости.
3. Перенос аккумулятора с места на место должен происходить с большей осто-
МЕТОДИКА РАЗДРАЖЕНИЯ
17
рожностыо, чтобы в силу сотрясения не отпал активный слой, дающий электричество,
и чтобы не выплеснуть кислоты, которая в таком случае сильно разъедает зажимы.
4. Аккумулятор разряжается и без работы, при этом пластинки покрываются
слоем сульфата свинца (PbSO*), отчего аккумулятор приходит в негодность.
Поэтому нельзя его оставлять на очень долгое время без зарядки. В случае
бездействия аккумуляторы нужно заряжать регулярно каждые один-два месяца или вылить
кислоту и наполнить их дестиллированной водой.
5. Для зарядки аккумуляторов должна применяться химически чистая серная
кислота, во всяком случае она не должна содержать железа и в особенности хлора.
Каждый раз вновь купленная серная кислота должна испытываться на содержание
хлора. Если имеются хотя бы следы хлора, кислота іне годится для аккумулятора.
6. Концентрация серной кислоты должна до зарядки соответствовать 1.14 по
удельному весу, после зарядки — от 1.18 до 1.2. При долгом стоянии аккумулятора
вода испаряется, и кислота становится более концентрированной. Поэтому нужно
доливать серную кислоту дестиллироЕанной водой до нужной концентрации.
Щелочным аккумулятор. Элемент щелочного аккумулятора состоит из двух
групп пластин, погруженных в1 ("раствор едкого калия или натрия (КОЦ или NaOH)
удельным весом 1.2. Анодом служит гидроокись никеля (N1 (ОН)з) и графит,
а катодом губчатое железо и кадмий (Fe + Cd). Рабочее напряжение одногр
элемента 1.25 V. Аккумулятор можно разряжать до! V на элемент^ после чего ста-.
вится на зарядку. В конце заряда напряжение доходит л^Г 1.8 V. По сравнению
с кислотными аккумуляторами Щелочные аккумуляторы более вынослибы,
нечувствительны к механическим сотрясениям, большим разрядным и зарядным токам
и легче свинцовых.
Неполяризующиеся электроды. Если в цепь гальванического тока вводится влдж-
ная ткань (нерв или мышца), то металлические проводники тока'не должны» прямо
касаться такой ткани. Во влажной ткани сейчас же начнется электролиз и
возникновение электродвижущей силы поляризации. Кроме того, соприкосновение металла
с тканевой жидкостью само может стать источником электричества, если
металлические электроды хоть немного не одинаковы. Во избежание всего этого пользуются
так называемыми неполяризующимися электродами. Такие электроды впервые были
сделаны Дюбуа-Реймоном (рис. 9). Провода от источника электричества
соединяются с цинковыми палочками Znt последние опущены в'стеклянные трубки Л,
наполненные концентрированным раствором сернокислого цинка. Противоположный
конец каждой трубки заткнут глиняной пробкой Г. Глина берется чистая, много раз
промытая яестиллираванной водой, и замешивается «а физиологическом растворе.
Цинковые палочки перед употреблением амальгамируются. Для этого палочки
сначала опускаются ів крепкую серную кислоту, а затем в ртуть или натираются ртутью.
После они промываются в проточной воде и так опускаются в стеклянные трубки.
Если такие электроды глиняными концами прикладываются к ткани, то
поляризация не будет иметь места, ибо при прохождении тока в местах соприкосновения
металла с жидкостью химических изменений не іможет быть. Под влиянием тока
сернокислый цинк будет разлагаться на Zn и S04. В анодном электроде S04 будет
стремиться растворить цинковую пластинку, а металлический цинк осядет на дно
трубки; в другом электроде цинк осядет на цинковой пластинке, a S04 обратно
растворит цинк. Таким образом, как в одном, так и в другом электроде химический
состав почти не изменяется, и поэтому поляризационный ток не сможет возникнуть.
В последнее время пользуются в качестве неполяризующихся электродов
серебряными электродами, покрытыми хлористым серебром. Приготовляют их для
каждого опыта следующим образом: обычные электроды из чистого серебра опускаются
з 1% раствор хлористого натрия. Оба полюса серебряных электродов соединяются
с анодом аккумулятора, а другой полюс аккумулятора соединяется с тем же
раствором хлористого натрия через угольный электрод. После пропускания тока
в течение нескольких минут серебряные полюсы ' электродов покрываются белым
налетом хлористого серебра. Такая комбинация Ag- — AgCl в соприкосновении
— И. С. Беритов
18
МЕТОДИКА
с нервом через физиологический раствор дает очень слабый поляризационный ток»
которым можно пренебречь при ряде физиологических опытов.
Чтобы серебряные проволоки покрылись тонким непроницаемым слоем
хлористого серебра, следует пропускать слабый ток, применив сопротивление в одну
электрическую лампу. До пропускания тока необходимо очистить проволоки от
жира промыванием ів щелочи. Наконец, следует несколько раз менять направление
тока, т. е. соединять серебряные іправолоки на одну минуту с анодом, затем на
минуту с катодом, затем вновь с анодом.
Индукция. Электрический ток, проходя через замкнутую цепь (первичную),
возбуждает во второй параллельной цепи вторичный, или индукционный, ток.
Причиной этого тока является пересечение вторичной цепи магнитными силовыми линиями,
расходящимися от первичной катушки. Поэтому индукционный ток возникает в
момент замыкания первичного тока или в момент его быстрого усиления
и ослабления, или же быстрого приближения и удаления первичной цели от
вторичной. При замыкании и усилении первичного тока, а также при сближении цепей
направление индукционного тока обратное индуктирующему току. Если замыкание
или размыкание, усиление или ослабление первичного тока и т. д. происходит
постепенно, без скачков, то возникающая электродвижущая сила индукции очень
мала, и тем меньше, чем медленнее происходит означенное изменение первичного
тока. Индукционный ток обычно протекает очень быстро в виде ударов. Для
большей интенсивности индукционного тока как первичной, так и вторичной цепи
придается форма спирали. Первичная спираль состоит из толстой медной проволоки
с небольшим количеством оборотов, вторичная — из тонкой проволоки с большим
количеством оборотов.
Санный индукционный аппарат Дюбуа-Реймона. Для физиологических целей
употребляется индукционный аппарат, сконструированный Дюбуа-Реймоном
(рис. 10). Из элемента D ток проходит через первичную спираль (1 мм в диаметре
и 200 оборотов) XX и через электромагнитный прерыватель тока df. В момент
замыкания и размыкания этой цепи во вторичной катушке КК (0.15—0.2 мм в
диаметре и 5 000—10 000 оборотов) возникают замыкательный и размыкательный
индукционные удары. Вторичная катушка передвигается по салазкам р—/?, и, смотря
по расстоянию между обеими катушками, интенсивность ударов меняется, а именно:
она тем больше, чем расстояние іменыне. Это изменение интенсивности в зависимости
от расстояния происходит по логарифмическому ряду. Для усиления тока вдвое
надо уменьшить расстояние между катушками тем больше, чем дальше они
отстоят одна от другой. Для усиления индукционных ударов внутри первичной
спирали помещается пучок железных проволок, так называемый сердечник, так как
присутствие сердечника усиливает магнитное поле первичной катушки. При одном
замыкании и размыкании первичной цепи (что производится рукой посредством
ключа Дюбуа-Реймона) ©о вторичной возникает по одному замыкательному и раз-
мыкательному удару. Если же хотят получить быстрый ряд этих ударов, то
заставляют работать электромагнитный прерыватель Вагнера, которым снабжен индукто-
рий Дюбуа-Реймона (рис. 10). Из элемента ток проходит по проволоке а к столбу S
и по пружине F, которая оканчивается якорем е, переходит на винт b и отсюда по
проволоке с идет в первичную спираль катушки XX, потом вокруг электромагнита Н
и, наконец, по проволоке fg возвращается в элемент. Во время замыкания тока
электромагнит намагничивается, и якорь е притягивается. Это прекращает
соприкосновение пружины F с винтом Ь, и ток размыкается. Электромагнит сейчас же теряет
магнетизм, и якорь отскакивает от него обратно, а ток снова замыкается. Это вновь
намагничивает электромагнит, за ним следует новое притяжение якоря, размыкание
тока и т. д. Чем меньше расстояние между якорем и полюсами электромагнита,
тем чаще происходят замыкание и размыкание первичного тока. Хорошими
электромагнитами можно легко вариировать частоту индукционных ударов от 15 до 80
в секунду. Определить частоту ударов можно по слуху, прислушиваясь к звуку
прерывателя, но, конечно, для точного определения ритма нужно ввести электро-
МЕТОДИКА РАЗДРАЖЕНИЯ
19
магнитный сигнал (рис.. 24) в. первичную цепь индуктория и записать на кимографе
колебания электромагнитного сигнала от замыкания и размыкания первичной цепи
вместе с хронографом Жаке или камертоном (рис. 23).
Рис. 10. Индукционный аппарат Дюб\а-Ре#мона (из Ландуа). Объяснение см. в тексте.
Характеристика замыкательного и размыкательного индукционных ударов.
Количество электричества при замыкании и размыкании совершенно одинаково. Но
амплитуда, в смысле вольтажа, а также временное течение индукционных ударов
разное, а именно: замыка-
тельный удар протекает с
меньшей амплитудой и
гораздо медленнее, чем размыка-
тельный (рис. 11).
Продолжительность
индукционных ударов, наиболее
распространенных в
физиологических лабораториях индукто-
риев, очень мала и сильно
меняется в зависимости от
расстояния вторичной катушки от
первичной. Так, по последним
исследованиям, при большом
расстоянии катушек (54 см),
достаточном для раздражения „ ^' L1' №6|?ия потен*иала в первичной (abed)
^ к и вторичной (з, Р) катушках индукционного аппарата,
нерва, длительность размыка-
тельных ударов около 0.25
сигмы (тысячных секунды), а
при значительном сближении
катушек (30 см) она
увеличивается в 2.5 раза — около
0.65 а (сигмы). При еще
большем сближении катушек (до
20—10 см) продолжительность
равняется нескольким сигмам. Продолжительность замыкательных ударов при
больших расстояниях катушек всегда значительно больше размыкательных. Так, при
тех же 54 и 30 см расстояния между катушками продолжительность, соответственно,
около 1.5 и 1.9 а (1>озенберг, .13).
Причина разного протекания замыкательных и размыкательных ударов заклю-
В момент ab — замыкание гальванического тока в
первичной цепи. В это время во вторичной цепи—индукционный
ток обратного направления — замыкательный удар (з).
Между моментами бис сила тока в первичной цепи не
меняется. Поэтому во вторичной никакого тока нет. В момент
cd первичная цепь размыкается, во вторичной цепи возникает
индукционный ток — размыкательный удар (Р) большого
потенциала, очень большой крутизны и того же
направления. Здесь же показано колебание тока в первичной и
вторичной катушках при наличности придатка Гельмгольца
прерывистой линией — се и р.
Т
20
МЕТОДИКА
чается в следующем. Когда гальванический ток замыкается через первичную спи-
раль, индукционный: ток противоположного направления возникает не только во
вторичной спирали, но также в первичной, что в физике называется, экстратоком,,
или самоиндукцией. Этот экстраток возникает вслед за замыканием и действует
на первичный ток в смысле задерживания постепенного нарастания
гальванического тока до максимума. Это ведет в свою очередь к более длительному протеканию
индукционного удара во вторичной спирали. При размыкании гальванического
тока в первичной спирали также возникает экстраток, на этот раз того же
направления, что и индуцирующий, но он продолжается значительно меньшее время
вследствие разрыва цепи. Поэтому падение
первичного тока до нуля не так сильно замедляетг
ся от экстратока. Это, конечно,
благоприятствует более быстрому протеканию
индукционного удара во вторичной цепи. Разница между за-
мыкательным и размыкательным ударами тем
сильнее, чем больше расстояние между
первичной и вторичной катушками. При сближении
катушек растет длительность индукционных
ударов, но длительность размыкательного
удара увеличивается быстрее, чем длительность за-
мыкательного, а поэтому при некотором
сближении разница между ними значительно
уменьшается.
Метроном с электрическими контактами.
Для раздражения одиночными индукционными
ударами через точно определенные промежутки
времени от К сек. до 2 сек. применяется
метроном. Он снабжен двумя эбонитовыми
чашечками с ртутью (рис. 12). Маятник метронома
В, при своем качании движет коромысло с
платиновыми наконечниками на концах. При
качании влево платиновый наконечник опускается в
левую чашечку Л/, а при качании вправо—'В
правую iV. Если ввести в первичную цепь инду-
ктория одну сторону, тогда при одном полном
качании маятника первичная цепь будет
замыкаться и размыкаться только один раз, если
же ввести обе стороны, то замыкание и
размыкание первичной цепи произойдет два раза.
При размыкании тока получается искра,
которая окисляет ртуть. Ртуть от этого покрывается черным слоем окислов, которые
являются дурными проводниками тока. Они покрывают не только ртутную
поверхность, но и платиновый наконечник. Этим самым затрудняется соприкосновение
наконечников с ртутью. А так как с каждым качанием положение и толщина
окислов меняются на ртутной поверхности, то каждый раз ток замыкается
с разной силой. Кроме того, при опускании контакта в ртуть происходит двойное
замыкание тока: сначала замыкается ток в момент соприкосновения с окислами,
а затем второй раз — при соприкосновении с чистой ртутью. Первый раз ток
нарастает от нуля до некоторой небольшой силы, а затем — от этой силы до
некоторой большей. При выхождении контакта из ртути также дважды происходит
падение силы тока. Сообразно этому возникают две пары замыкательных и размы-
кательных ударов. Во избежание загрязнения контактов поверхность ртути в чашке
нужно все время промывать проточной водой. Для этой цели чашки снабжены
двумя боковыми отверстиями. Ток воды уносит с собой образуемые окислы ртути.
Придаток ч Гельмгольца к индукторию Дюбуа-Реймона,, Иногда нужно, .чтобы
Рис.
12. Метроном с ртутными
[контактами.
В и С — зажимы; N и М — чашки
со ртутью: R — маятник.
МЕТОДИКА РАЗЧРАЖЕНИЯ
21
физиологическое действие замыкательного и размыкательного ударов было
одинаково. До некоторой степени; к этому можно приблизиться, если в первичной
спирали устроить добавочную, совершенно замкнутую спираль. В таком, случае экстра-
ток в момент размыкания цепи будет так же долго продолжаться в первичной
спирали, как при замыкании. Поэтому в первичной цепи падение тока до нуля
значительно задерживается. Это приведет к уравнению замыкательного и размыкательного
индукционных ударов во вторичной
спирали в отношении как
амплитуды, так и продолжительности
(см. рис. 11).
Униполярные индукционные
токи. Индукционные удары возникают
во вторичной цепи не только при
замкнутой вторичной спирали, но и
при ее разомкнутом состоянии —
униполярное действие
индукционного тока. Поэтому индукционные уда-
• ры действуют раздражающим
образом также и тогда, когда живая ткань
соединена только с одним концом
вторичной цепи; в этом случае для
ее раздражения потребуется
гораздо большее приближение катушек,
чем при биполярном соединении
вторичной катушки с живой тканью.
Ключ Дюбуа-Реймона. Для замыкания и размыкания гальванического тока
в первичной цепи, а также для того, чтобы регулировать действие индукционных
ударов на живую ткань, пользуются ключом Дюбуа-Реймона. Способ введения этого
ключа ©о /вторичной цепи показан на рис. 10. От вторичной цепи проволоки идут
к'Двум медным брускам у и z, а отсюда — к электродам для раздражения. Между
брусками помещен мостик г, который то соединяет их, то разъединяет. В первом
случае ток пойдет в значительной мере по мостику ввиду малого сопротивления
его в сравнении с сопротивлением живой ткани, которая введена в побочную цепь.
Ток, ответвляющийся в живую ткань, будет настолько мал, что не будет раздражать ее.
Во втором случае при разъединенных брусках весь ток направится через ткань.
КлЬч Дюбуа-Реймона в первичную цепь вводится иначе. Один брусок ключа
соединяется с гальваническим элементом, а другой — с катушкой, так что в
первичной цепи ток замыкается только п{ш соединении брусков мостиком. Для этой
цели употребляются также ртутные ключи. Чашечка с ртутью соединяется с
элементом, а мостик—с катушкой. Когда конец мостши погружается в ртуть, ток
замыкается.
Ключ Гельмгольца. Чтобы можно было записать:на кимографе (см. графический
метод) время раздражения индукционными ударами, пользуются ключом
Гельмгольца (рисМЗ). Это двухплечий ключ, где одно плечо вроде ключа Дюбуа-Реймона
служит для включения живой ткани ©о вторичную цепь, как на рис. 10, а другая
сторона включается в особую цепь с электромагнитным сигналом. Пишущее перо
сигнала пишет на уровне миографа (см. графический метод). Когда открывается
первое плечо для раздражения, сейчас же замыкается цепь через второе плечо
с сигналом. Когда же первое плечо закрывается и раздражение прекращается,
размыкается цепь через второе плечо. Значит, во время раздражения электромагнитный
сигнал пишет на кимографе свою кривую, точно отвечающую времени раздражения.
Релаксационный раздражитель. Индукционный аппарат Дюбуа-Реймона наряду
со ' своими преимуществами имеет целый ряд. существенных недостатков:
неустойчивость частоты, силы и продолжительности раздражения, а также невозможность
их точного определения. В последнее время вводятся в эксплоатацию такие элек-
Рис. 13. Двухплечий ключ Гельмгольца.
На эбонитовой или деревянной пластинке установлены
три медных столбика: средний для оси подвижного
плеча, а крайние для контактов. На одной стороне
сильная пружина притягивает плечо книзу и вызывает
контакт. Эта сторона служит для соединения с индук-
торием. Другая сторона открыта и. служит для
соединения с электромагнитным сигналом (треть
натуральной величины)"'.
')')
МЕТОДИКА
выход
трические раздражители, где в принципе лежит разряд конденсатора на ткань.
Здесь можно точно знать силу и продолжительность раздражающего тока. Для
получения друг за другом следующих раздражающих токов применяют
специальные устройства, где используются неоновые, тиратронцые или усилительные
лампы, Схема\>дного из таких раз-
Л2 JL^ § | I дражителей приведена на рис.
13а. Здесь первая лампа
работает как генератор
остроконечных импульсов, вторая же
лампа работает как усилитель
и для среза противоположных
волн импульсов, так чтсГ
импульс принимает
остроконечную форму одного
направления. Величину и частоту
импульсов можно менять
независимо друг от друга в
широких пределах. Все величины
частоты, силы и
продолжительности точно могут быть
определены и стабилизированы.
Металлические электроды
Индукционные удары могут
быть проведены через живую
ткань с помощью
металлических электродов:
платиновых серебряных, золотых.
Здесь не нужны неполяризующиеся электроды. Гак как замыкательные и раз-
мыкательные индукционные удары продолжаются очень короткое івремя и, кроме
того, они противоположны по направлению, поляризационный ток не успевает раз-
Рис. 13а. Схема релаксационного раздражителя.
Л,Л, — лампы У Б— 240, і?,—сопротивление ТО 0-1 mS, i?,—
сопротивление СС 10к&, і?,—реостат 2.5 м2,
/?„—потенциометр проволочный 1000 2, Rt — сопротивление типа ТО
0.8 м2, С, Ся — конденсаторы 0.05 v-F, С3 — конденсатор
0.015 (J-F, Г—трансформатор релаксатора, S2 —
выключатель питания, S,— кнопка раздражения, Бн— батарея 2V
2НКН—ю, БД—батарея 90 V типа БАС—80, Яс—батарея
сухая б V.
N
&
Рис. 14. Платиновые электроды для раздражения нервов и мышц лягушки.
А.—обычные двухполюсные электроды, укрепленные на вилке; В — двухполюсные элек-.
троды с кольцом Геринга; слева — вид спереди, а справа — сзади. Кольцо из
серебряной проволоки укрепляется на эбонитовой основе электродов; С— трехполюсные
электроды; D — двухполюсные электроды, в которых один полюс образует кольцо, а другой
находится в центре кольца.
биться в такой мере, чтобы повлиять на силу индукционных ударов. Так бывает
и в том случае, если индукционные удары следуют один за другим десятки и сотни
раз в секунду. В зависимости от назначения форма электродов бывает разная. На
рис. 14 показаны электроды для раздражения нервов и мышц лягушки. Электроды
фиксируются на вилке, которая воткнута в пробковую пластинку, служащую для
МЕТОДИКА РАЗДРАЖЕНИЯ
23
прикрепления препарата лягушки — А. Для ослабления внеполюсного
распространения тока, так называемых петель тока, биполярные электроды помещаются в
металлическом кольце из платины или серебра. Это кольцо называется кольцом
Геринга—В. Но можно прибегнуть и к другому способу: один полюс электрода
раздвоен и соединен с анодом, а другой, соединенный с катодом, помещается ровно
Ь
Рис. 15. Электроды Шеррингтона для раздражения нервов у теплокровных
животных.
А — для перерезанных; В — для целых нервов; а — стеклянная трубка; Ь —
платиновые электроды, которые путем пробки с укрепляются в боковом отростке стеклянной
трубки; d — пробка, которая защемляет нить, привязанную к нерву; е—боковая щель
для укладывания целого нерва в трубку. В электродах В — два боковых отверстия для
раздражения двух участков нерва (см. объяснение в тексте). (А —3/3 натуральной
величины; В—натуральная величина).
по середине между двумя анодными полюсами — С. Та же цель достигается, если
анод образует кольцо, а катод помещен в центре кольца — D.
Электроды для раздражения нервов у теплокровных. Для раздражения нервов у
теплокровных животных, как кошки, собаки, кролики, пользуются электродами
Шеррингтона. Электроды эти, употребляемые для раздражения перерезанных нервов,
представляют собой стеклянную трубку, диаметром з 5—8 мм, длиной в 5 см н
с боковым ответвлением (рис. 15, Л). К раздражаемому концу нерва привязывается
нитка. Нерв вводится в трубку ниткой;
на другом конце' нитка закрепляется
пробкой; через боковое отверстие под- ,+ •
п
водится под нерв пара платиновых
электродов. Затем электроды
укладываются глубоко в ранку, чтобы нерв Рис- 16- Электроды для раздражения непе-
п ререзанных нервов теплокровных животных,
не мог остыть. Снаружи остается толь- к f **« ^ п н v v
кп к-пнртт чттектпплпи г чяжимями Пття я — основная пластинка из эбонита, которая закру-
ко конец электродов с зажимами, аля чивается в конце. Провода проложены в толще
раздражения неперерезанных нервов этой пластинки и выведены наружу в закрученной
части. Нерв вкладывается в эту часть, а подвижная
стеклянная Трубка имеет сбоку Прорез пластинка (Ь) прикрывает ее; с — кусок резиновой
по йгрй л/ттинр Hprm иртяптшртгя r т-nvfi трубки. Эти электроды делаются разной величины:
по всей длине, перв вставляется в труо- для тонких и коротких нервов меньше, чем на
ку через эту щель (рис. 15, В). Можно рисунке; для толстых и. длинных — больше.
приделать к трубке два боковых
ответвления для раздражения нерва в двух участках, как это дано на рисунке.
Раздражение неперерезанных нервов теплокровных животных производится
также особыми электродами, изображенными на рис. 16, так называемыми погруженными
электродами. Это — эбонитовая пластинка с загнутым концом. На пластинке по
желобку проложены платиновые или серебряные проволоки, которые также
загибаются. Они служат полюсами для привода электрического тока. Основная
пластинка сверху прикрывается эбонитовой же пластинкой, которая удерживается
в этом положении резиновым кольцом с. Загнутый конец подводится под нерв и
прикрывается верхней пластинкой. Электроды закладываются в рану, и рана
зашивается. Снаружи выдаются противоположные концы электродов. Во избежание
значительного распространения петель тока лучше брать трехполюсные электроды.
Глава IIJ
МЕТОДИКА РЕГИСТРАЦИИ НЕРВНО-МЫШЕЧНЫХ РЕАКЦИЙ
Нервно-мышечная деятельность изучается путем регистрации главным образом
двоякого рода внешних эффектов: механического эффекта, как, например,
сокращение мышц, движение конечности и т. д., и электрического, как, например,
биоэлектрические токи мышц и нервов. Для регистрации их существуют разного рода
приспособления. Принципы действия наиболее употребительных 'приборов должны
быть в достаточной мере разъяснены, чтобы обеспечить сознательное пользование
ими при практическом изучении основных проблем нервно-мышечной деятельности.
А. МИОГРАФИЧЕСКИЙ МЕТОД
Графические кривые. Физиологические процессы протекают во времени. Время
можно рассматривать как равномерно нарастающую величину и сравнивать с по-
У
п
\
"*ч
kl
Л
Рис. 17. Кривая одиночного сокращения. Объяснение см. в тексте.
степенным изменением других величин. Положим, нужно представить графически
кривую сокращения мышцы. Гельмгольц показал, что сокращение начинается
спустя некоторое время после момента раздражения и имеет определенную
продолжительность; при этом сокращение развивается сначала медленно, затем
быстрее. Так продолжается несколько сотых секунды. После этого сокращение
ослабевает— сначала быстро, затем медленно, на это требуется опять несколько сотых
секунды. Эти разные периоды сокращения графически . представлены на рис. 17.
Горизонтальная линия—абсцисса ОХ разделена на равные части, которые
равняются сотым долям секунды. На этой линии возведены перпендикуляры параллельно
ординате OY. Длина каждого перпендикуляра пропорциональна величине сокра-
РЕГИСТРАЦИЯ НЕРВНО-МЫШЕЧНЫХ РЕАКЦИЙ
25
щения в данный момент; Та кривая, которая соединяет концы ординатных
перпендикуляров, представляет кривую сокращения. Всякие сложные изменения и
взаимоотношения можно выразить графически кривыми. Это освобождает от
подробных описаний и приведения длинного ряда цифр.
Саморегистрирующие аппараты. Впервые Гельмгольц применил
автоматическое записывание кривой сокращения (1850 г.). Эта запись носила очень
примитивный характер: к дистальному сухожилию мышцы привязывался длинный рычаг,
конец которого писал на установленной вертикально пластинке чернилами или же
царапал на закопченной бумаге. Если пластинка стоит, рычаг пишет вертикальную»
Рис. 18.^Изображение опыта с сердечным пинцетом и запись сердечных
движений на кимографе Марея.
Здесь идет запись сердечных движений особым прибором (Я), который
называется сердечным пинцетом. Лягушка приколота булавками к пробковой доске.
Последняя в горизонтальном положении держится зажимом (D) на штативе. Этот
штатив передвигается автоматически на салазках, когда кимограф вертится. Та
винтовая нарезка, которая передвигает штатив, приходит в движение вместе
с цилиндром путем бесконечного ремня (PFP).
линию; если же пластинка будет передвигаться с равномерной скоростью, рычаг
напишет кривую, как на рисунке 17. Знаіл скорость, легко вычислить все временные
изменения сокращения.
Пишущий рычаг принято называть миографом; он впервые был
применен Гельмгольцом. Аппарат, на котором записывается миографическая
кривая, называется кимографом. Первый кимограф был построен известным
физиологом Людвигом (1847 г.). Он построил его специально для исследования
изменений внутриартериального давления крови. Гельмгольц впервые
использовал этот кимограф для записи мышечных сокращений.
Кимограф Марея. Французскому физиологу М а р е ю принадлежит большая
заслуга в улучшении графического метода. Он дал прекрасный тип кимографа,
который получил во Франции большое распространение (рис. 18). Вращающийся
цилиндр кимографа приводится в движение часовым механизмом. Скорость
движения можно регулировать по желанию. Для. опыта цилиндр оклеивают бумагой,
которую покрывают тонким слоем копоти от керосиновой лампы или от коптящего
плаліени светильного газа, Пишущее перо миографа касается цилиндра и при
движении последнего пишет белую линию на черном фоне.
26 МЕТОДИКА
Кимограф Людвига. Людвиг ввел особый тип кимографа, который широко
распространен в германских лабораториях (рис. 19). Часовой механизм (ящик С)
приводит в движение диск D, который путем трения вращает ось цилиндра.
Скорость вращения может быть произвольно изменена. Для практических занятий со.
студентами употребляется несколько упрощенный кимограф, который отличается
тем, что на оси часового механизма установлено передвижное маленькое колесо,
а диск находится на оси цилиндра. На всех типах цилиндр может быть поднят
Рис. 19. Кимограф Людвига. Объяснение
см. в тексте.
Рис. 20. Изотонический миограф без
ts инерции в соединении с мышцей.
Я —. капсула, которая имеет впереди щель
(S). Внутри капсулы ходит пластинка Dr к
которой прикреплен через щель рычаг (Р).
Рычаг соединен с мышцей (М) нитью через
ролики (R, /?,). При сокращении пластинка
с рычагом поднимается вверх. Узкая щель
(S) оказывает сопротивление выхождению
воздуха, и этим путем устраняется
движение рычага по инерции (1/3 натуральной
величины).
и опущен винтом, и потому конец пишущего
пера легко можно установить на разной
высоте. Здесь также цилиндр оклеивается
закопченной бумагой. Барабану кимографа можно
придать как горизонтальное, так и
вертикальное положение.
Для сохранения кривых, написанных на закопченной бумаге, нужно
фиксировать их после записи. Для этого снимают бумагу с цилиндра и смачивают 5%
спиртовым раствором шеллака.
Кимограф с бесконечной лентой. Людвигу же принадлежит конструкция
такого кимографа, в котором бумага, накинутая на вращающийся цилиндр,
беспрерывно передвигается перед концом миографа. В этом случае пишущее перо
миографа пишет кривую чернилами. На конце миографа укреплен легкий
стеклянный резервуар с длинным выводным капилляром. 3 резервуар наливают чернила,
а конец капилляра прикладывают к бумаге.
Миографы. Пишущее перо миографа для регистрации мышечных сокращений
представляет обыкновенно легкий рычажок,^заканчивающийся- .:остртем - из перга
РЕГИСТРАЦИЯ НЕРННО-МЫШ^ЧНЫХ РЕАКЦИИ
27
ментной бумаги или обрезка тонкой алюминиевой пластинки. В одних случаях
двигательный орган действует непосредственно на рычажок, а в других он связан
с ним ниточкой через блоки или без них. Посредством блоков и нити движение
может быть передано на очень большое расстояние. В большом ходу миограф
Энгельмана (см. рис. 76). Но этот миограф при быстрых сокращениях дает
движение по инерции: он отклоняется больше, чем сокращается мышца. В некоторых
опытах движения по инерции искажают ход кривой сокращения, и потому эти
движения должны быть устранены. Для этой цели может быть применен скон-
Рис. 21. Капсула Марея. Объяснение см. в тексте.
струированный нами миограф, в котором воздушное сопротивление заглушает
движение рычага по инерции (см. рис. 20).
Передача путем воздушных капсул Марея. Воздушная передача мышечных
сокращений была введена в физиологическую практику и усовершенствована
Маре ем. Капсула, или барабан, Марея представляет круглую плоскую коробку
диаметром в 3—5 см, обтянутую с одной стороны резиноівой пленкой (рис. 21).
Эта капсула сообщается с ідругой такой же капсулой через резиновую трубку.
Каждое движение пленки в одной капсуле, вызванное сокращением мышцы, будет
менять давление воздуха во всей системе. Поэтому, если снаружи надавить на
s —
Рис. 22. Электромагнитный сигнал.
катушка электромагнита; А — якорь; F — пружина, которая натягивает якорь
(2 3 натуральной величины).
пленку одной капсулы, это давление передается на другую капсулу и производит
здесь выпячивание пленки. В последней капсуле пленка соединена подвижным
коленом с обычным миографам.
Передача электрическим током. Некоторые движения могут быть переданы
посредством электрического тока, действующего на электромагнит. К якорю А
последнего прикреплено пишущее перо, которое может записать на закопченной
бумаге движения якоря (рис. 22). Если каким-нибудь образом замыкается и
размыкается цепь, железный якорь то притягивается к электромагниту, то отходит от
него. Этим путем можно передать на далекое расстояние и записать момент
раздражения, начало и конец внешней реакции органа, время для контроля скорости
вращения цилиндра.
Миограф для регистрации напряжения мышцы. Мышца при сокращении
развивает определенную механическую силу — напряжение, если она встречает
препятствие к укорочению. При невозможности укорочения временное течение напряжения
в общем такое же, как и сокращения. Степень сокращения ненагруженной мышцы
28
МЕТОДИКА
определяется прежде всего длиной мышечных волокон и их сократительной
способностью. Степень же напряжения определяется не только этим, но еще количеством'
Рис. 23. Миограф для регистрации напряжения мышц.
М —мышца, которая ниткой привязана к оси миографа; у оси (Ach) начинаются два рычага под
прямым углом; один—горизонтальный—для регистрации кривой напряжения на кимографе, другой-
вертикальный — для отсчитывания величины напряжения по шкале (S/f); F, F. — пружины, которые
удерживают рычаги в определенном положении и создают препятствие для движения рычага;
Т — воздушный регулятор; это —цилиндр, в котором ходит поршень; последний подвижно связан
с горизонтальны* миографом. На дне цилиндра имеется отверстие, более или менее закрываемое
винтом. Просвет этого отверстия так регулируется этим винтом, чтобы сопротивление воздуха,
оказываемое движению поршня, устранило движение рычагов по инерции, но не влияло на
движение рычагов под влиянием напряжения^ мышцы.
сокращенных мышечных волокон. Следовательно, регистрация напряжения лучше
выражает степень деятельности всей мышцы, чем регистрация укорочения. Поэтому
Рис. 24. Хронограф Жаке.
во многих случаях для изучения мышечной деятельности лучше регистрировать
напряжение, чем укорочение мышц. Для регистрации напряжения употребляют разные
приборы, но принцип их один и тот же. Мышца действует на ось миографа,
который удерживается пружиной в одном положении. Под влиянием мышечного напря-
РЕГИСТРАЦИЯ НЕРВНО-МЫШЕЧНЫХ РЕАКЦИЙ
29
жения пружина немного растягивается: от этого ось чуть поворачивается, и
пишущий рычаг отклоняется в сторону мышцы. Чем сильнее напряжение, тем сильнее
деформация пружины и тем сильнее отклонение рычага. Конец рычага двигается по
шкале, где отмечено, какое напряжение в граммах соответствует данному положению
пишущего рычага. Можно рычаг направить на кимограф и регистрировать кривую
напряжения. На рис. 23 показан сконструированный нами миограф для регистрации
напряжения лягушечьих мышц.
Регистрация времени и контроль скорости вращения цилиндра. Для записи
скорости вращения цилиндра употребляют хронографы, или такие аппараты, кото-
\ Рис. 25. Диаграмма записывания времени.
Да^а электрическая цепь (К), которая включает электро-
магйитный сигнал (F), камертон-прерыватель (Е) и
гальванический элемент (Z). В камертоне буква а указывает место
контакта. Ток, проходящий в цепи, прерывается в этом
месте столько раз, сколько у камертона в секунду
колебаний (из Пембрея).
рые автоматически часовым механизмом или метрономом приводят в движение
пишущее перо через определенные промежутки времени. Таким прибором является
хронограф Жаке, который отмечает время в секундах или в пятых долях секунды
(рис. 24). Если требуется запись очень малых долей секунды (при быстрых
вращениях цилиндра), то прибегают к электромагнитным камертонам, дающим 25, 50,
100, 200... колебаний в секунду. Конец одного колена камертона снабжен пишущим
пером, которое прикладывается к закопченной бумаге и пишет при работе
камертона волнистую линию. Каждая волна будет соответствовать 1/г5, 'До и т. д.
секунды. Камертон приводится в постоянное движение электромагнитом,
прикрепленным к одному колену камертона (рис. 25).
В. ЭЛЕКТРОГРАФИЧЕСКИЙ МЕТОД
Струнный гальванометр Эйнтховена. Для регистрации электрических реакций
нервно-мышечной ткани до последнего времени был наиболее употребителен
струнный гальванометр Эйнтховена. 'Между полюсами сильного электромагнита натянута
тонкая платиновая или кварцевая посеребренная струна толщиной в несколько
микронов (рис. 26). При пропускании через струну электрического тока, смотря по
его направлению, струна отклонится вправо или влево.
Электромагнитное поле между полюсами создается путем пропускания
электрического тока через обмотку электромагнита. Для работы с биоэлектрическими
токами мышцы и нерва достаточно пропустить ток силой в 2—3 А.
Так как струна очень тонка и ее движения под влиянием нервно-мышечных
токов очень малы, то невозможно следить невооруженным глазом за движениями
струны. Необходимо, чтобы струна была освещена сильным светом и ее движения
увеличены в несколько сот раз. Для этой цели струна освещается с одной стороны
яри помощи осветителя GF (рис. 26), через который направляется свет от электри-
30
МЕТОДИКА
ческой вольтовой дуги (в 5—10 А). Движение струны наблюдается через
проекционный микроскоп DE (рис. 26) нецосредственно глазом или путем проекции ее теню
на экране. С помощью микроскопа и проекции движение струны увеличивается
в 500 раз и более.
Так как многие биоэлектрические токи протекают очень быстро, то нет
(возможности изучать их путем прямого наблюдения движений тени струны. Нужно их
регистрировать. Для регистрации прибегают к фотографии. Тень струны путем
проекционного микроскопа отбрасывается на щель фотографического аппарата. Перед
щелью движется фотографическая бумага, на ней получается изображение
движения струны. Колебание струны происходит настолько быстро, что оно может точно
регистрировать довольно быстро протекающие токи возбуждения, например сердеч-
Рис. 26. Схема струнного гальванометра Эйнтховена.
EF — полюсы электромагнита; СС — струна; GF —
осветительный микроскоп; АЕ — проекционный микроскоп;
D — проекционный окуляр.
ной мышцы. Но /в случае очень быстрого протекания биоэлектрического тока, как,
например, скелетной мышцы или цереброспинального нерва (2—4 сигмы, т. е.
тысячные доли секунды), колебание струны продолжается дольше этого тока, притом чем
слабее натянута струна, тем дольше длится ее колебание. Поэтому электрограмма
требует исправления. На рис. 27-1 дается схематическое изображение расположения
приборов при работе со струнным гальванометром.
Чувствительность струнного гальванометра очень велика. Она может быть
сделана большей или меньшей в зависимости от натяжения струны. При работе с
биоэлектрическими токами подбирают такое натяжение струны, что при двух амперах
в электромагнитном поле и увеличении колебания струны в 500 раз струна
отклоняется от одной тысячной вольта на 0.5—1.0 см. Но для улавливания очень слабых
токов можно расслабить струну настолько, что от одной стотысячной івольта струна
будет отклоняться на несколько миллиметров. Чувствительность струнного гальеано-
метра зависит также от силы магнитного поля и от массы струны. Следовательно,
чем меньше натяжение струны, чем больше сила магнитного поля и чем меньше
масса струны (т. е. чем она тоньше), тем выше чувствительность прибора.
Эти сведения о струнном гальванометре сообщаются не для того, чтобы можно
было работать на нем при практическом прохождении курса нервно-мышечной
физиологии. Струнный гальванометр — очень сложный инструмент; ознакомиться
РЕГИСТРАЦИЯ НЕРВНО-МЫШЕЧНЫХ РЕАКЦИЙ
зг
с работой на нем нужно под руководством специалиста. Приведенные сведения*
достаточны только для понимания многочисленных результатов, полученных с
помощью этого инструмента. Для желающих работать на струнном гальванометре-
в
^-<пи
-&5
-260
Рис. 27-1. Расположение приборов при работе со струнным гальванометром.
А — вольтова дуга проекционного фонаря; В — конденсор для света; С — стеклянный сосуд с водой*
для охлаждения световых лучей; D — осветительный микроскоп гальванометра; Е — струна; F —
проекционный микроскоп; G — вращающееся колесо Гартена для обозначения ординатных линий на
фотографической бумаге; И — передняя стенка фотографического аппарата с горизонтальной щелью.
Рис. 27-11. Электрограмма осциллографа катодных лучей и струнного гальванометра'1
для сравнения.
Л, В и С записаны осциллографом; а, Ь и с — струнным гальванометром; А, а — индукционные удары;.
В и b — токи возбуждения нерва лягушки; ОС и Ос— контрольные кривые от включения постоянного
тока в несколько милливольт. Регистрация тока возбуждения идет вслед за тем индукционным ударом,
который вызвал данное возбуждение. Осциллограф регистрирует их раздельно, а струнный
гальванометр — слитно: кривая тока возбуждения накладывается на кривую индукционного удара. Лучше всего1
преимущество осциллографа выступает из сравнения контрольных кривых. Осциллографическая
кривая поднимается сразу и потому образует прямой угол, а гальванометрическая—в течение четырех-пяти'
сигм (Эрлангер).
можно рекомендовать специальные статьи по методике, например работы нашего
выдающегося специалиста электрофизиолога проф. А. Самойлова (см.
библиографию).
Осциллографы. Струнный гальванометр ке является- идеальным прибором для*
регистрации быстро протекающих биоэлектрических токов. В последние годы в
физиологической практике вводятся такие аппараты, которые или совсем не
обладают инерцией, или инерция очень мала. Сейчас для исследования быстро
протекающих токов пользуются осциллографами. Для сравнительной характеристики
регистрирующей способности струнного гальванометра и осциллографа, дается рис. 27-11*.
32
МЕТОДИКА
Осциллографом называется регистрирующий .прибор, служащий для записи
мгновенных значений электрических величин. переменного тока. Осциллографы^
.делятся на две группы: инерционные и безинерционные. . - ¦¦>.-г.
К инерционным осциллографам относятся такие осциллографы, в которых
названная система является весомой и, таким образом, имеет инерцию. Такими
являются электромагнитные осциллографы. В силу инерции они могут быть
использованы для записи электрических колебаний до 20 000 в сек. К неинерционным
осциллографам относятся катодные осциллографы, где перемещение светового
пятна на флуоресцирующем экране производится пучком катодных лучей, котфрые
являются практически безинерционными. Эти осциллографы применимы для записи
колебаний до самых ультра-высоких частот.
3
Рис. 2S-A.
Схема
шлейфа.
N и
S—полюсы магнита,
Л— лента, р—
ролик, кк—
клеммы д/под-
водки токана,
п — пружина,
з—зеркальце.
Рис. 28-В. Оптическая схема хода луча шлейфного осциллографа:
И — источник света, А — фокусирующая линза, Щ — щель, О — преломляющая
призма, з — зеркальце на шлейфе, к — призма, р — развертывающий барабан,
ц — цилиндрическая линза, Б — кассета с светобумагой, Э — экран для визуального
наблюдения.
Электромагнитный осциллограф. Принцип работы
электромагнитного осциллографа основан на взаимодействии магнитного поля и
помещенного в нем проводника с исследуемым током. Основной частью
электромагнитного осциллографа является подвижная система, т. е. шлейф.
Поэтому электромагнитные осциллографы называются также шлейфными
осциллографами.
Шлейф представляет собой магнитоэлектрический гальванометр, где угол
отклонения подвижной системы в каждый данный момент прямо пропорционален
мгновенному значению тока, проходящего через него, как показано на ряс. 28-А.
Шлейф состоит из тонкой бронзовой ленты (<#), перекинутой через ролик (р).
Эта лента двумя концами закреплена на клемме (кк), через которую пропускается
исследуемый ток. Лента образует узкую петлю — шлейф. Посередине шлейфа укреп-
.лено маленькое зеркальце размером от 0.5X0-5 до 1 X 1 мм. Шлейф натянут
пружиной Я. Этим изменяются чувствительность и собственные колебания шлейфа. Вся
эта система помещена между полюсами магнита. При прохождения через шлейф
исследуемого тока, вокруг проводов шлейфа создается магнитное поле и
взаимодействует с полем постоянного магнита. Силы, действующие на провода шлейфа,
прямо противоположны, поэтому они вызывают поворачивание шлейфа с зеркалом.
Угол и направление поворота зависят от силы и направления тока через шлейф.
Чтобы сохранить пропорциональность отклонения в диапазоне исследуемых частот.
РЕГИСТРАЦИЯ НЕРВНО-МЫШЕЧНЫХ РЕАКЦИЙ
33
требуется, чтобы собственная резонансная частота шлейфа была немного выше
наивысшей исследуемой частоты. Кроме того, для уменьшения собственных
колебаний шлейфа нужно увеличить его затухание, для чего вся система погружается
в глицериновое или жидкое касторовое масло.
Обычно (шлейфные осциллографы делаются на 3—6 шлейфов, таким образом
можно записывать и наблюдать за несколькими одновременно протекающими лро-
цессами. Большое распространение получил шестишлейфный осциллограф 'Сименса,
изготовляемый уже в Союзе Харьковским электротехническим институтом. На
рис. 28-В дается схематическое расположение частей и хода луча в шлейфном
осциллографе.
Для упрощения на чертеже показана работа одного шлейфа. Здесь (И) —¦
источник «света, который может быть дуговой лампой или оке, что более практично,
Нить ,Х<г»0?
накала t /$нхн?
Ф/?цоресц экран
Отклоняющие
пластины
Ьм^-Шч і 1 ***** »а"р»ж
Анодная о?т
Рис. 29. Осциллограф катодных лучей (катодный осциллограф).
В броуновской трубке помещена спираль (катод), которая вследствие накаливания током испускает
электроны, и против катода находится металлическая диафрагма (анод), имеющая высокий потенциал
относительно катода. Поэтому излучаемые катодом электроны с большой быстротой устремляются
к аноду, проскакивают через отверстие и узким пучком направляются к флуоресцирующему экрану;
ударяясь об него, они вызывают зеленовато-голубое свечение. На пути электронного луча
расположены две металлические пластины в виде конденсатора (электронный луч проходит между этими
пластинами): Если к ним присоединить напряжение, например к нижней плюс, а к верхней минус, то,
под действием электрического поля на луч, последний меняет свое направление, отклоняясь к
положительной пластинке, прямо пропорционально напряжению, так что световое пятно на экране
перемещается вниз. При изменении направления тока в отклоняющих пластинках световое пятно
переместится кверху. При подаче на пластинки тока переменного напряжения световое пятно будет колебаться
вдоль вертикальной линии. Чтобы можно было визуально наблюдать периодические процессы, в
трубке заделывается другая пара отклоняющих пластин, перпендикулярно к первой. К этим пластинам
подводится пилообразное напряжение, которое равномерно отклоняет луч в горизонтальном
направлении слева направо (развертывает кривую) и затем очень быстро возвращает луч в исходное положение.
Если исследуемое напряжение периодически колеблется и подобрана соответственная частота
развертывающего напряжения, то на экране можно получить неподвижную кривую исследуемого напряжения.
лампой накаливания с короткой нитью (12V, 50W). Световой луч, пройдя
ахроматическую линзу (Л), диафрагмируется щелью («<)» преломляется призмой (о) и падает
на зеркальце шлейфа (з). Луч, отражаясь от зеркала, проходит на комбинированную
призму {К). Отсюда часть лучей падает на цилиндрическую линзу (ц) и
фокусируется ею в виде световой точки на фотобумагу, которая навернута на барабан
э кассете. При вращении барабана световой зайчик экспонирует в виде кривой,
которая становится видимою после соответствующей фотообработки. Часть луча,
выходящего из комбинированной призмы, падает на зеркало развертывающего барабана,
который, вращаясь, развертывает исследуемую кривую по времени; ее
визуально можно наблюдать на матовом экране. Вращение барабана происходит
электромотором со скоростью рулонной съемки от 3 до 125 см в сек.
Катодный осциллограф. Осциллограф катодных лучей, или катодный
осциллограф, впервые был применен для физиологических целей американским ученым
Эрлангером (1922). Он использовал трубку Брауна, в которой узкий поток
электронов направляется на флуоресцирующий экран (рис. 29). Электронный поток
в трубке может быть отклонен от своего прямого пути созданием электрического
поля поперек этого пути. В результате светящаяся точка на экране передвигается,
что можно наблюдать глазом или фотографировать. Масса подвижной части
механизма, именно потока электронов, практически равняется нулю. Следовательно,
записанная этим аппаратом кривая не требует никаких исправлений, как бы быстро
ни протекал регистрируемый ток.
3 И. С. Бернтов
34
МЕТОДИКА
Усилители. Биоэлектрические токи нерва или мышцы слишком слабы* чтобы.
можно было воздействовать на электронный луч в катодном осциллографе или
на шлейф в ^электромагнитном осциллографе. Так, например, для достаточного
отклонения луча катодного осциллографа требуется напряжение в 50—80 V, а для.
шлейфа ток '30—60 гпА. Поэтому при работе с ікатодными или со шлейфовьтми
осциллографами применяется усиление биотоков ламповым 'усилителем. Для
усиления пользуются трех- и (более электродными лампами. Применяется обычная схема
на сопротивлениях, как дающая наименьшие частотные и фазовые искажения.
Так как общее усиление должно быть очень большое; то ввиду экономических
соображений и устойчивости работы такое усиление достигается двумя усилителями:
предварительным и оконечным. Предварительный усилитель может быть 3-каскад-
ный, работающий на триодах 6Ф5 с питанием накала от 6 V аккумулятора и ано-::
дов от 90 V батарей. При изучении одного процесса усилитель может быть с одно-
тактным входом, т. <е. исследуемое колебание подается к сетке и земле первой
лампы. При одновременном 'изучении \двух и более процессов отведение их к двум
однотактным усилителям не является достаточно верным, ибо возможно пролезание
токов одного процесса на другой усилитель.
Во 'избежание этой помехи отведения вход усилителей должен являться
симметричным. Исследуемые колебания подводятся между сетками первого каскада
соответствующего усилителя. Общее усиление предварительного усилителя в 20 000 раз.
В усилителе имеется переключатель для уменьшения усиления в 2, 5, 10, 20 раз.
Предварительный усилитель может быть применен как для шлейфового, так и
для катодного усилителя. Оконечные же 'усилители должны быть разные: для
катодного с выходным напряжением, а для шлейфового с выходным током. Оконечный уси-.
литель для катодного осциллографа имеет коэффициент усиления 100, выходное
максимальное напряжение 100 V . Питается от выпрямителя. Оконечный усилитель для
д
шлейфового осциллографа имеет крутизну 100 пі—.. Максимальный выходной ток
30 тА. Питается от аккумуляторов. На рис. 30-А, В, С даны схемы усилителя с
электронными лампами советского производства.
Данные схемы усилителей построены для регистрации колеблющихся токов от
нескольких в секунду до многих тысяч. Для регистрации постоянных токов или
колеблющихся более 'редко, чем один іраз в секунду, применяются другого рода
усилители, так называемые усилители постоянного тока.
Практикум
Методика, электрического раздражения должна быть проработана в связи с по-
станковкой первого опыта с раздражением. Правда, студенты обвязаны иметь
основные понятия о постоянном и индукционном токах, но \в физиологической практике
употребляется целый ряд специфических электрических приборов, с которыми
физика их ее знакомит. Кроме того, в нашей методике даются подробные сведения
об индукционных ударах, об их физическом характере и физиологическом действии,
о чем физика упоминает только вскользь.
С графическим методом студенты знакомятся при первом опыте с кимотрафи-
ческой записью. Здесь они усваивают метод регистрации мышечных сокращений,
так называемый миографический метод. С регистрацией электрических эффектов
руководитель знакомит студентов на струнном гальванометре или на катодном
осциллографе. Здесь основная задача заключается в том, чтобы студент понял, как
вообще изучаются злектрическия явления в нервно-мышечной системе и как были
получены те многочисленные электрограммы, которые он встретит в данном руководстве.
Конечно, описание многочисленных приборов дано здесь не для заучивания.
Студент знакомится с тем или другим прибором во время практических занятий.
В методике же они приводятся для ориентации в случае надобности уяснить
принцип- прибора, его назначение и употребление.
Рис. З?-А. Предварительный усилитель с1- симметричным 'входом.
0 0
- 4 *
0 0 0 0
+ 160- -15 +
- f ¦
Рис. 30-В. Оконечный усилитель для шлейфного осциллографа..
?,Л77і
— ,і .+ і—4-
*:1
<f2Cr
OJfir \ ¦« |
— осцилогра
0ф9
б'дг
_ц ^>??g?> L
Рис. 30-С. Оконечный усилитель для катодного осциллографа (Квавилашвили).
36
МЕТОДИКА
Литература
Сведения относительно физиологической методики можно встретить во многих
руководствах по физиологии. Но существуют специальные практические руководства
по физиологии. Среди них следует упомянуть следующие:
1. Бурдон-Сандерсон, Фостер и Бринтон, Практический курс
физиологии, пер. с англ., 1886. Эта книга содержит много интересных сведений
по электрофизиологии, но в общем она сильно устарела.
2. Pembrey М., Practical Physiology. London, 1910.
3. Verworn M., Physiologisches Practicum. Jena, 1912.
4. As her L., Praktische Uebungen in-der Physiologie. Berlin, 1916.
5. Tigerstedt R., Physiologische Uebungen unci Demonstrationenfur Studierende.
Leipzig, 1913.
6. Tigerstedt R., Handbuch der physiologischen Methodik, 1910—19 П.
Многотомное руководство, где крупные специалисты поместили статьи по своей
специальности. Там есть, например, очень хорошие статьи С. Гартена об
электрографии, о фотографии; О. Франка — о принципах регистрации и т д.
7. Abderhalden Е., Physiologisches Praktikum. Berlin, 1922.
8. Самойлов A., Praktische Notizen zur Handhabung des Saitengalvanometers etc.
Arch. Anat. Physiol., Physiol. Abt., 1910. S. 477.
9. Воронцов Д., О способности струнного гальванометра регистрировать быстро,
протекающие токи, Русск. врач, 1915.
10. Б е р и т о в И., Методика электрического раздражения, Русск. физ. журн., т. 13,
стр. 422 и 442, 1930.
11. При изучении анатомии лягушки пользуются большим двухтомным труцом на
немецком языке: Eckeru. Wiedersheim, Anatomie des Frosches.
Braunschweig, 1896.
На других языках нет столь подробного труда по анатомии лягушки.
12. Р о ж а н с к и й Н. А., Практические занятия по физиологии животных, 1932.
13. Rosenberg Н., Pfl. Arch., Bd. 223, S. 582, 1929.
14. Erl anger J., Analysis of the Action Potential in Nerve. The Harvey Lectures, ser.
XXII, 1926—1927.
15. Гинецинский А. Г. и Лейбсон Л. Г., Практический курс
физиологии. ОГИЗ, 1933.
16. Ж ю л ь е н А., Практические занятия по физиологии животных и человека.
Москва, 1940.
17. Harris, Experimental Physiology. London, 1934.
18. Иордан Г., Практикум сравнительной физиологии. Биомедгиз, 1934.
Об осциллографической методике и об усилителях см.:
19. А р д е н н е М., Электроннолучевые трубки и их применение в технике слабых
токов Москва, 1936.
20. Holzer W., Kathodenstrahloszillographie in Biologie und Medizin. Wien, 1936.
21. Квавилашвили Ш., Методика осциллографического исследования
биоэлектрических явлений. Тр. Института физиологии им. Еериташвили, т. 6, стр. 435, 1945.
Часть вторая
ОБЩАЯ ФИЗИОЛОГИЯ
МЫШЕЧНОЙ СИСТЕМЫ
Глава 1
ОБЩИЕ СВЕДЕНИЯ О СТРОЕНИИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
Классификация мышц. Мышцы можно разделить на две основные
группы*, поперечнополосатых и гладких мышц. Поперечнополосатые
мышцы служат главным образом для движения скелета, поэтому они
иначе называются скелетными. К ним же относятся мышца сердца и
передняя часть пищевода. Поперечнополосатые мышцы называются
также еще произвольными, так как они могут приходить в действие
под влиянием нашей воли. Сердце и передняя часть пищевода
составляют в этом отношении исключение: они имеют поперечнополосатую
мускулатуру, однако не могут быть приведены в действие произвольно.
Гладкая мускулатура обслуживает все остальные органы; она
входит в состав стенок пищеварительного канала, кровеносных сосудов,
выводных протоков и резервуаров желез внешней секреции и
воздухоносных путей легких. От нее же зависит движение волос и перьев и
всего наружного покрова. Гладкая мускулатура иначе называется
непроизвольной, так как она не может сокращаться по нашей воле.
Микроскопическое строение поперечнополосатых мышечных
волокон. Основным морфологическим элементом поперечнополосатой мышцы
является мышечная клетка, или мышечное волокно. Оно имеет более
или менее вытянутую форму в виде нити толщиной 10—100 ji
(микронов,, тысячных миллиметра) и длиной до 5.3—12 см . (у человека), На
грбоих концах волокно заострено. Но бывают и такие волокна, у
которых один конец утолщен, а другой заострен. Таких утолщенных
концов особенно много в конечных сухожильных частях.мышц. Каждое
волокно заключено в очень тонкую, совершенно бесструктурную и
прозрачную эластичную оболочку, называемую сарколеммой
(рис. 31, А). При сокращении мышечного волокна она всецело следует
за изменением его формы. По мнению одних авторов, сарколемма
.является продуктом мышечной протоплазмы, а по імнению других —
продуктом соединительной ткани, связывающей мышечные волокна.
Внутри мышечной клетки содержатся протоплазматйческое
вещество — саркоплазм а, и тонкие ми офибрилли. В живой
мышечной клетке саркоплазма является однородной, бесструктурной средой,
40
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
которая, однако, на фиксированных и окрашенных препаратах
обнаруживает сетчатое строение. Протоплазматическая сеть саркоплазмы
очень густа и хорошо выражена в месте ее слияния с фибриллярной
сетью нервных окончаний. Концевые разветвления протоплазматиче-
ской сети плотно примыкают к анизотропному веществу миофибрил-
лей. По мере удаления от места вхождения нерва саркоплазматическая
сеть становится более редкой и контуры ее менее заметными (рис. 35),
но ее можно проследить даже между миофибриллями. Эта саркоплаз-
А — два мышечных волокна человека с окрашенными ядрами. Верхнее волокно посередине передае
лено,. и здесь видна сарколемма. Между волокнами видны соединительнотканные клетки. Увел, в J250
раз; В—диаграмма, показывающая связь между мышцей и костями; х — мышечные волокна,
которые начинаются и кончаются у костей в сухожилиях; у — оканчиваются у кости только одним
концом; между ними волокна, которые оканчиваются обоими концами в самой мышце; С — миофибрилли
мышечного волокна человека; D — пучок миофибриллей. Здесь темные широкие полоски (А) — двоя-
копреломляющее вещество; светлые полоски (У) — однопреломляющее; тонкие черные полоски,
делящие это вещество на две половины (Z), представляют пограничную линию между двумя соседними
сегментами миофибрилли. Увел, в 2 100 раз. Продольные темные полоски на границе миофибриллей
создают продольную исчерченность (по Леви из Максимова).
матическая сеть была впервые описана голландским гистологом Буке
(7) и названа им перитерминальной сетью. Впоследствии она была
описана и другими гистологами, например Лаврентьевым (12).
Затем в саркоплазме были найдены зернистые включения разного
характера, состоящие из гликогена, липоидов (жироподобные
вещества) и белков.
В мышечных волокнах высших позвоночных ядра, как правило,
располагаются под сарколеммой. Каждое из них окружено саркоплазмой
(рис. 34, 35). Ядра эти считаются центрами питания. У амфибий, птиц
и насекомых мышечные ядра обычно расположены по оси волокна
между миофибриллями. В области окончания двигательного нерва
наблюдается скопление ядер, несколько отличающихся от обычных.
Мышечные фибрилли. Миофибрилли располагаются по длине
мышечного волокна от одного конца до другого. Толщина их очень мала:
V—1.7 д, а расположенный между миофибриллями слой саркоплазмы
О СТРОЕНИИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
4L
еще тоньше — около 0.5у. . Это обусловливает продольную исчерчен-
ность мышечных волокон. Но в живых мышечных волокнах эта исчер-
ченность не так хорошо видна, как поперечная, которая
обусловливается сложной структурой миофибриллей (см. рис. 31, D и С).
Каждая из миофибриллей представляет колонку, состоящую из
многочисленных следующих один за другим коротких совершенно одинаковых,
цилиндриков. Каждый такой сегмент-цилиндрик миофибрилли является
призматическим телом. Центральный диск каждого сегмента темнеег
чем его крайние диски, и сильнее преломляет свет. Центральный слой
является анизотропным, обладающим свойством двойного
светопреломления — А, наружные же — изотропные, с простым
светопреломлением— J (рис. 31, D), Там, где светлый диск соприкасается с таким
же диском следующего сегмента, видна еще одна разделяющая их.
полоса в виде тонкой линии, именуемая полоской Z (рис. 31, D). Эта
полоска предполагается состоящей из коллагенного вещества и,
видимо, играет роль опоры и изолятора для двух пограничных
цилиндриков (Хоегквист, 27). Полоски, располагающиеся на всех фибриллях
мышечного волокна на одном уровне, делят волокно на столбики
высотой 2—2.5 jjl, числом около 25 000 в одном волокне длиной в 50 мм.
Чередованием темного слоя со светлым обусловливается наблюдаемая:
под (микроскопом характерная поперечная исчерченность
миофибриллей поперечнополосатых імышц (рис. 31, D и С). Эти составные части
миофибриллей хорошо видны как на мертвых, окрашенных мышцах,.
так и на живых (Бухталь и Линдгард, 22).
У некоторых животных, как, например, у насекомых, темный
центральный диск разделяется ровно пополам более светлым диском,
представляющим изотропное вещество (Энгельман, 2). Обычно миофиб-
рилли так располагаются в мышечном волокне, что светлые диски всех
фибриллей приходятся на одном уровне. Чередование их с темными
создает поперечную исчерченность всего мышечного волокна. Только
в так называемых красных мышцах не все светлые и темные диски
располагаются на одном уровне, поэтому здесь не получается полной:
поперечной исчерченности.
Высота анизотропного и изотропного дисков у различных животных
разная. У изученных до сих пор позвоночных оба диска вместе имеют
около 3 д; отношение между изотропным и анизотропным веществами
в мышцах лягушки равно около 1. В мышцах насекомого Hydrophilus
оба диска вместе составляют от 5 до 12 \х, а отношение между
изотропным и анизотропным менее 0.2 (Гюртле, 3).
Оригинальную точку зрения на поперечную полосатость скелетных
мышц высказывает Сцент-Гьорги (32). По его мнению мио-
фибрилля (представляет систему, состоящую из двух белков-—актина
и миозина. Актин образует длинные стержни, вокруг которых миозин
располагается в виде спирали. Это образование должно иметь <вид
штопора или винта. Спирали в миофибриллях плотно упакованы, так
что витки всех спиралей вкладываются друг в друга и находятся
42
ОВШАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
в одном положении. Это в свою очередь должно сообщить фибрйллям
поперечную полосатость, так как спирали должны иметь различные
оптические свойства в зависимости от расположения миозина по
отношению к актиновой нити. Отсюда делается вывод, что сегментарная
структура миофибриллей является чисто оптической иллюзией.
Доказывается это положение тем, что при вращении мышечной фибрилли
•поперечная полосатость смещается вдоль ее оси, на два периода при
одном полном обороте.
Мышечные фибрилли отсутствуют в эмбриональных мышечных
клетках—миобластах. Они возникают в них постепенно как продукт
Рис. 32. Развитие миофибриллей в миобласте зародыша цыпленка.
а — миобласт в 60-часовом зародыше; Ь ¦— миобласт в 76-часовом
зародыше; с — миобласт из 124-часового зародыша, когда впервые
появилась сегментация миофибриллей; due — последние стадии (по Дюсбергу
из Максимова).
деятельности саркоплазмы. Они появляются в виде коротких
неправильно расположенных ниточек без поперечной исчерченности. С
течением времени число их растет, самые волокна удлиняются,
располагаются правильными пучками и приобретают поперечную исчерчен-
ность (рис. 32). Скелетные мышцы содержат двоякого рода волокна.
Одни из них, обычно более тонкие и богатые саркоплазмой, имеют
больше ядер и мало миофибриллей. В них миофибрилли
распределяются группами из 3—10 фибриллей, между которыми находится более
толстый слой саркоплазмы. На поперечном разрезе эти группы
именуются полями Конгейма. В других волокнах, менее богатых
саркоплазмой « обычно более толстых, миофибриллей больше, и они однородно
распределяются по всему диаметру (Камерон, 23; Фюрлингер, 24).
Первые мышечные волокна, богатые саркоплазмой, имеют красный, цвет.
Мышцы, содержащие такие волокна в большом количестве, также
красного цвета и поэтому называются красными. Вторые
мышечные волокна являются бесцветными или окрашены в белый цвет.
Поэтому мышцы, лишенные красных волокон, или содержащие .их
в небольшом количестве, отличаются белой: окраской и называются
О СТРОЕНИИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
43
белыми. Красные волокна встречаются в імышцах всегда в
незначительном количестве, до 30%, и чаще всего располагаются на
внутренней стороне мышцы. Онтогенетически те «и другие волокна
развиваются самостоятельно (Фгорлингер, 24). К красным мышцам
относятся mm. ileofibularis, rectus abdomini, pectoralis pars abdominalis,
semitendinosus, gastrocnemius лягушки, semitendinosus кролика, soleus
кошки. 'К белым относятся mm. sartorius и semmimembranosus лягушки,
gastrocnemius кролика и кошки.
Строение мышцы. Мышечные волокна в одних мышцах
располагаются параллельно оси мышцы. Но они редко идут от одного конца
мышцы вплоть до другого. В sartorius лягушки, например, если один
конец начинается в сухожилии, то другой лежит в
соединительнотканной прослойке (perimisium) мышцы (рис. 31, В). Длина волокон в
общем равняется Чъ—2/з всей длины мышцы (Линдгард, 9). Но бывают
и очень короткие волокна. Так, например, в мышце sartorius лягушки
длиной 26 мм могут быть волокна длиной от 5.2— до 24.0 мм (Бух-
таль и Линдгард, 22). Толщина мышечных волокон также чрезвычайно
разнообразна. Она меняется от нескольких микронов до 100 и выше
(Бухталь и Линдгард, 22). С ростом животного толщина мышечных
волокон растет, а число тонких волокон уменьшается. На mm.
pectoralis pars abdominalis и sartorius лягушки было установлено, что с ростом
животного меняется и общее количество волокон. Но это наблюдается
до определенного возраста, когда лягушка достигает веса около 20 г.
При дальнейшем росте животного, до 100 г и выше, количество
волокон остается неизменным,'меняется только длина и толщина их. При
этом характерно, что мышечные волокна наружной поверхности,
прикрытые кожей, всегда наиболее тонки (Меписашвили, 26).
К мышцам с параллельными волокнами относятся: sartorius,
semimembranosus, gracilis major et minor, pectoralis pars abdominalis,
adductor lortgus и другие мышцы лягушки; -sartorius, semitendinosus,
а также мышцы живота и некоторые другие мышцы теплокровных
(Беритов, 10; Цхакая, 11).
В большинстве случаев, в особенности в крупных мышцах,
мышечные волокна не располагаются параллельно продольной оси мышцы.
От каждого конца мышцы заходят вглубь или тянутся по ее
поверхности сухожильные тяЖи, или пластинки. Мышечные же волокна
располагаются между ними наклонно к продольной оси, образуя с
'Последней углы разной величины. Только концы этих сухожильных частей
соединяются с противолежащим местом прикрепления мышцы прямыми
мышечными волокнами. В таких мышцах длина мышечных пучков
равняется одной трети или четверти всей длины імышцы. Такие мышцы
называются перистыми. К перистым мышцам относятся у лягушки,
например, mm. triceps femoris, ileofibularis, semitendinosus, gastrocnemius
tibialis anticus, peroneus :и другие мышцы. У теплокровных к перистым
относятся: rectus femoris, vastus externus et internus, gastrocnemius,
peroneus и некоторые другие.
44
ОБЩАЯ ФИЗИОЛОГИЯ МЫШНЧНОЙ СИСТЕМЫ
Рис. 33 дает точную схематическую картину расположения
мышечных пучков в ряде мышц лягушки.
Окончания двигательного нерва в мышечных клетках. Каждое
волокно двигательного нерва многократно разветвляется в мышце, и
каждое мышечное волокно получает по крайней мере по одной такой
ветке. Обычно в коротких мышечных волокнах, как, например, в икро-
f
*о
гглтп
Mlll'l
пит
П||11!
ниш
iiht'f
ими.
пищ
>>Иін
1111111
Кіііі!
іпіпі
ішіп
ММііі
IР III П
ШИН
іміщ
тми
(ниц
lltll;,
uin;
Mill
mil'/
WllD
A
*"&
Jf
— Hi-
fi
¦Ю
tli\» ;;;
a
—к
\A -4
¦b-~
p
Jp
Рис. 33. Строение мышц лягушки.
А — sartorius, В — m. semitendincsus, CD —¦ дорзальная глава; С. Y — вентральная, С —
gastrocnemius со спинной стороны, D — gastrocnemius с брюшной стороны, Е—gastrocnemius в продольном
разрезе.
Штриховые линии означают расположение мышечных пучков, сплошные же линии — положение
сухожилий. Цифры снаружи обозначают длину мышечных волокон и сухожилий, а внутри мышцы
рядом с сухожилием указывают на величину угла в градусах, который образуется в перистых
мышцах на месте прикрепления мышечных волокон к сухожилию. Р — проксимальный конец; D — дисталь-
ный. Как видно из рисунка, в m. ileofibularis длина мышечных волокон и величина углов меняются'
в меньшей мере, чем в gastrocnemius.
ножной мышце лягушки, имеется лишь по одному окончанию
двигательного нерва. Оно приходится как раз посредине мышечного волокна-
Но в длинных мышечных волокнах таких окончаний бывает несколько.
Так, в длинных волокнах m. sartorius лягушки оканчиваются по 2—3
нервных волокна (Лежава, 16). В большинстве мышечных волокон эти
нервные окончания 'принадлежат одному и тому же двигательному
корешку. Но в небольшой части мышечных волокон они принадлежат к
разным корешкам, т. е. соответствующие двигательные нервные волокна
выходят из разных сегментов мозга. В таких случаях говорят оплю-
рисегментальной (множественной) иннервации мышечных
волокон. В мышцах лягушки количество таких волокон очень изменчиво,
но не более 20% (Агдур, 4; Беритов, 8; Лаврентьев, 21). Каждое
нервное волокно, разветвляясь в мышце, снабжает несколько мышечных
волокон. Так, в глазных мышцах человека одно нервное волокно приходится
на три мышечных волокна. В других мышцах одно нервное волокно
приходится на значительно большее количество мышечных. Новейшие
авторы насчитывают у теплокровных животных до 160 мышечных
волокон на одно нервное (Кларк, 14), Аксон двигательной нервной
клетки и все мышечные волокна, которые получают окончания от
О СТРОЕНИИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ 45
этого аксона, являются в функциональном отношении элементарной
«единицей, которая носит название «двигательного элемента», или
«двигательной единицы» (Шеррингтон,
15). Вся центральная иннервация
скелетной імускулатуры
осуществляется этими элементами. У всех
позвоночных животных каждая
мышца состоит из множества таких
нервно-мышечных единиц. Но у
некоторых беспозвоночных многие
мышцы целиком иннервируются от
•одного двигательного и одного
тормозящего нервного волокна, т. е.
каждое мышечное волокно получает
по одной веточке от обоих
волокон. Таким образом, эти мышцы
представляют одну нервно-імышеч-
иую единицу. Таковы, например,
замыкатели и размыкатели клешни
речного рака и вообще мышцы
ракообразных (Мангольд, 25).
Рис. 35. Нормальное нервное окончание двигательного нерва
в мышечном "волокне летучей мыши.
N — нервное волокно, которое, гветвясь, образует нейрофибриллярную
сеть (т. п.). Эта сеть, окрашенная более темно, сливается с более бледно
окрашенной перитерминальной сетью (р. п.); an—анизотропное вещество;
is — изотропное вещество (Буке).
Каждое нервное волокно оканчивается в мышечных волокнах
сложно устроенным образованием, у высших позвоночных так называемой
концевой пластинкой, или двигательной бляшкой (рис. 34), а у
низших, как амфибии, концевым кустом.
Двигательное волокно, подходя к мышечному, теряет миэлин.
Соединительнотканная оболочка Генле переходит со своими ядрами на
^gtfggganaaiMa
і
!2ЗЕЗПШП^ U
ШЕН^ЯШЕШШ
Рис. 34. Схема двигательной пластинки.
GH — соединительная оболочка, подходящая
к пластинке нервного волокна; GS — шваннов-
ская оболочка; GM — миэлиновая оболочка;
С А — осевой цилиндр; В—место, где нервное
волокно теряет миэлин; С — концевая
пластинка; N V, N А, N F—три сорта ядер в ней;
D — мышечное еолокно (по Дювалю из
Максимова).
46
ОКЩАИ ФИЗИОЛОГИИ МЫШКгЧНОЙ-СИСТЕМЫ
поверхность, сарколеммы и. здесь исчезает так же,, как шванновская
оболочка. От нервного волокна входит-в : мышечное, волокно только
осевой цилиндр. В 'месте его вхождения саркоплазма образует под
сарколеммой скопление большей или меньшей величины (рис. 35, 3fl).
Некоторые авторы думают, что эта саркоплазма образована клетками,
шванновской оболочки нервного волокна (Максимов,, 31). ..
Осевой цилиндр разбивается, ц
ЩШіШШі^-і^ЖШЖ^ двигательной пластинке, на множе-
!Шіі|^ИіШЩ1ШШі^ЯЙі1ШЙі^ВШі ств0 конечных волоконец, нейро_
фибрилли которых образуют
тонкую сеть, По. данным Буке, эта
сеть непосредственно сливается
с протбплазматической сетью
саркоплазмы и через нее приходит
в связь с миофибриллями (рис.
35). (Буке, 1). Но другие, авторы
сомневаются в существовании
такой связи нейрофибриллярной сети
с миофибриллями. Это означало
бы сведение на-нет особой роли
концевой пластинки как
самостоятельного структурного
образования (Бухталь и Линдгард, 22).
Сконцентрированные в области
нервного окончания ядра резко от
личаются от остальных ядер
мышечного волокна. Этот ядерный
аппарат, видимо, имеет большое
функциональное значение, ибо он
резко реагирует на разногорода
воздействия. Так, например, при
перерезке нервного волокна количество-
ядер становится больше. Здесь же
в двигательной пластинке
капиллярная система развита значительна
сильнее, чем в других участках мышечного волокна (Буке, 17; Дюннинг
и Вольф, 18; Лаврентьев, Лежава, 28).
Рис. 36.щ Целая мышца (тп. sartorius)
маленькой лягушки, весом 1.6 г, окрашенная
прижизненно метиленовой синью.
Представлена проксимальная иннервационная зона
во всю ширину мышцы. Почти каждое
мышечное волокно получает здесь по
нервному окончанию.
А — проксимальная, В — дистальная сторона
мышцы, С — нервные стволы, подходящие к данной
зоне (Лежава).
Рис. 37. Распределение нервных окончаний в т. sartorius лягушки.
Дана конфигурация мышцы. Р — проксимальный конец; D — дистальный. В ней разветвляется wepF-
ный ствол, образуя четыре сегмента для нервных окончаний: а, Ь, с, d. Участки D, t и Р не содержат
нервных окончаний (Лежава).
В мышцах нервные окончания располагаются на определенных'
уровнях как бы сегментами (рис. 36). Поэтому в них определенные
участки совсем лишены концевых двигательных пластинок. Это так
О СТРОЕНИИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
47
называемые безнервные участки. В некоторых длинных мышцах такие
безнервные участки особенно велики на концах мышцы. Так, в гл. sar-
torius лягушки длиной 35—40 мм проксимальный конец лишен
нервных окончаний на расстоянии 6—9 мм, что составляет около Vs всей
длины мышцы (Майне, 20; Лежава, 16) (рис. 37). Длинные
безнервные участки имеются также в проксимальном и дистальном концах
m. pectoralis pars abdominalis лягушки (Лежава и Меписашвили, 30).
Литература
1. В о е 1< е J., Brain, 44, 1, 1921; Erg. Physiol., 19, 448, 1921.
2. Engelmann Th. W ., PH. Arch., 7, 155, 1873; 18. 1, 1878; 23, 571, 1880.
3. Hurtle K., Pfl. Arch., 126, Ь .1909»
4. Agduhr E., Anat. Anz., 52, 273, 1919,
5. Kultschizky N., Biol. Zbl., 7, 572, 1888.
6. Boheman, Anat. Anz., 10, № 10, 1894.
7. В о e k e J., Proc. Royal Acad. Sc. Amsterdam, Jan. 1915.
8. S e r i toff I., Pfl. Arch., 205, 455, 1924; Z. Biol., 85, 509 u. 521, 1927.
9. Lindhar d, Physiol, papers dedicated to prof. Krogh, 188, 1926.
10. Beritof.f I., Pfl. Arch., 209, 763, 1925.
11. Ц x a k а я M., Русск. физиол. журн., 10, 273, 1927.
12. Лаврентьев Б., Z. mikr.-anat. Forschung, 13, 289, 1928.
13. К a t z J., Pfl. Arch., 63, 1, 1896.
14. С 1 a r k D. A., J. Physiol., 70, XVIII, 1930; Amer. J. PhysioL, 96, 296, 1931.
15. Ш e p p и h г t о h 4. и сотрудники, Рефлекторная деятельность спинного мозга,
Биомедгиз, 1933.
16. Лежава А., Сборник «Проблемы нервной физиологии и поведения»,
посвященный проф. И. С. Бериташвили, изд. Груз, Фил. Ак. Наук СССР, стр. 329.
Тбилиси, 1936.
17. В о е k e J.. Penfieid Cytology of the nervous system. New Уогк, 1932.
18. D u n n i n g H. G. a. H. G. W о 1 f f, J. сотр. Neur., 67, 483, 1937.
19; Лаврентьев Б. И., Арх. биол. наук, 40, 194, 1938.
20. М і n e s G. R., J. Physiol., 37, 408, 1908.
21. Lawrentiew В. I., Zeitschr. mikr.-anat. Forsch., 14, 511, 1928.
22. В u с h t a 1 Fr. a. J. Lindhard, The Physiol, of striated muscle fibre-
Copenhagen, 1939.
23. С a m e г о n, J. Physiol., 68, 47, P., 1929.
24. F ii r 1 i n g e r Fr., Zool. Anz.. 90, 325, 1930; Kriiger, Duspivau.
Furling e r, Pfl. Arch., 231, 750, 1933.
25. Mangold E., Z. allg. Physiol., 5, 135, 1905.
26. Меписашвили И. С, Бюлл. эксп. биол. и мед., 10, 17, 1940.
27. Н 6 g g q u i s t G., Anat. Anz., 3, 273, 1920; Z. mikr.-anat. Forsch., 4, 605, 1925;
Handb. mikr. Anat., 2, Teil 3, 105, 1931.
28. Л e ж а в а А., Бюлл. экегк биол. и мед., 8, 40, 1939.
29. Лаврентьев Б. И., Арх. биол-. наук, 48, № 1—2, 194, 1937,
30. Лежава А. и М е п и с а ш в и л и И., Бюлл. эксп. биол. и мед., 7, № 1, 1939.
Зі. Max i mow A., a. Bloom W., A Text-book of Histologie, 1931.
32. Szent—Gy 6rgy i A., Studien on Muscle. Acta Physiol. Scand., 9, Suppl. XXV,.
1944; — Бюлл. эксп. биол. и мед., 20, 15, 145.
Глава II
ВОЗБУДИМОСТЬ
КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ
Общее понятие о живой возбудимой системе, возбудимости,
возбуждении и раздражении. Живой возбудимой системой называется такое
органическое структурное образование, которое на различного рода
внешнее воздействие отвечает определенного рода деятельностью, иначе
говоря, определенной реакцией. Например, живая імышечная ткань от
действия внешней силы или внутреннего нервного импульса производит
сокращение. Тот сложный биологический процесс, который возникает
в возбудимой системе імышцы под влиянием внешней силы или
нервного импульса и обусловливает сокращение, называется возбужде-
н и е м. Свойство же живой системы приходить в возбуждение при
изменении состояния внешней среды іили под влиянием нервного
импульса называется возбудимостью. 'А то изменение внешней
среды или та внешняя сила, которая вызывает возбуждение,
называется раздражителем, раздражающей силой. Внешняя
•сила производит возбуждение не прямо, (путем непосредственного
воздействия на возбудимую систему, а посредством определенного физико-
химического изменения в той жидкой среде, которая заполняет
межструктурные пространства саркоплазмы.
Такое физико-химическое изменение жидкой среды саркоплазмы,
от которого зависит возбуждение, называется раздражением. Но
часто понятие раздражения употребляется вместо раздражающей
•внешней силы (об этих понятиях см. подробно: Беритов, 2а; Мангольд, 1,
Фредерик, 24).
Не каждая живая система обладает возбудимостью. Последняя
является специфическим свойством мышечной и нервной систем. Это
свойство заключается в том, что когда в какой-либо части мышечной
или нервной системы возникает возбуждение, то оно само собой
распространяется по всей раздраженной структурной единице, а также
переходит на другие структурные части, если только с последними
имеются непосредственные протоплазматические связи. Можно,
следовательно, сказать, что когда под влиянием какой-либо внешней силы
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 49
где-либо в живой системе возникает возбуждение, то последнее само
становится источником раздражения других соседних частей.
В то же время в организмах с дифференцированными мышечной и
нервной системами очень много таких живых систем, которые не
обладают возбудимостью. Таковы прежде всего костная,
соединительнотканная и некоторые эпителиальные системы. Конечно, клеточные
элементы этих тканей не лишены способности реагировать на внешнее
воздействие. Мы знаем, что любая ткань, в частности эпителиальная,
сильно разрастается под влиянием длительных внешних воздействий.
Но это свойство уже не будет возбудимостью, ибо в этом случае
реакция ограничивается участком ткани, на который действует внешняя
сила. В отличие от возбудимости, это свойство живой системы можно
назвать раздражимостью или реактивностью.
Раздражимость есть общее свойство живой системы,
заключающееся в том, что живая система отвечает на определенного рода внешнее
воздействие усилением обмена веществ, в результате чего
происходят усиленный рост и размножение живой системы, усиленное
образование определенных продуктов и усиленное передвижение ее частичек.
Раздражимость свойственна всем живым клеткам и тканям.
Проявлением этого свойства являются амебоидное и всякое другое
движение одноклеточных; оно обусловливает все случаи размножения клеток
и рост тканей; от него зависят многочисленные явления морфогенеза
клеток и- их систем при онтогенезе, а также все явления
восстановления тканей и органов при полном или частичном удалении их во
взрослом организме. Проявлением его являются также все процессы
медленного морфологического 'Приспособления организма к длительным
изменениям внутренней или внешней среды.
В определенных границах раздражаемые клетки и ткани
реагируют тем сильнее, чем сильнее и длительнее внешнее или внутреннее
воздействие. Раздражимость клетки и ткани все время меняется, ибо
способность к реагированию зависит от состояния обмена веществ.
При всех тех условиях, когда обмен веществ повышается, например
при некотором повышении температуры, при некотором слабом
механическом, химическом или электрическом воздействии, усиливается
реактивная способность, значит, повышается раздражимость. Наоборот,
при ухудшении обмена веществ, как это, например, бывает от
повышения температуры выше некоторой границы, от некоторого сильного
механического, химического или электрического воздействия,
раздражимость понижается и исчезает совсем, когда обмен веществ
приостанавливается под влиянием некоторых более сильных воздействий.
Раздражимость есть свойство живой системы филогенетически
более древнего происхождения. Из него потом развилась возбудимость
нервной и мышечной тканей. Эта качественно новая особенность
живой системы, очевидно, произошла в связи с возникновением сложных
организмов в результате филогенеза. Когда одноклеточные
микроскопические организмы превратились в многоклеточные макроскопические,
4 И. С. Беритов
50 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
то на почве естественного отбора побеждали и сильнее развивались те
организмы, которые обладали свойством быстро реагировать на
внешние раздражения и передавать процесс возбуждения на большие
расстояния. Такая система обеспечивала организму как целому возможность
реагировать не только при влиянии внешней силы на весь организм,
но и при ее воздействии на отдельные части. Это имеет важное
значение для сохранения организма как неделимого целого.
Мы говорим «живая система», а не «живой белок», «живая
молекула», «живое или возбудимое вещество». В настоящее время
нельзя себе представить, чтобы іпростейшей или наименьшей частичкой
живого тела являлась одна очень сложная белковая молекула или
комплекс таких молекул. В живой системе белковое вещество играет
основную, руководящую роль в жизненных явлениях. Из него
построены основные структурные части живой системы. С ним связано
основное свойство живой системы — обмен веществ (Ф. Энгельс, 36).
Отсюда, однако, не следует, что белок может проявить это свойство вне
клетки, вне той сложной среды, которая имеется внутри клетки.
Только в клетке или, еще точнее, в протоплазме клетки даны основные
условия для жизненных явлений. Поэтому мы можем признать за
живое биологическое образование такую наименьшую частицу живого
тела, которая обладает всеми характерными признаками жизни, как
обмен веществ, рост, размножение, возбудимость и т. д. Таким
образованием в настоящее время является клетка со всеми своими
составными частями: протоплазмой, ядром, центрозомой, оболочкой и пр.
В многоклеточных животных и растительных организмах клетка
является единицей живой системы. Среди существующих простейших
также господствует клеточная организация, но у них весь организм
является неделимой единицей живой системы. Их нельзя разбить на;
части искусственно так, чтобы каждая часть сохранила способность к
росту и размножению. Но в более отдаленную от нас геологическую
эпоху живая система должна была иметь иную форму, вроде
современных бактерий и сине-зеленых водорослей, в которых ядерное
вещество не обособлено от протоплазмы. А до этой стадии развития
живая система должна была существовать в еще более простой форме,
в доклеточной, как голый комок протоплазмы, вроде фильтрующихся
вирусов, которые, вероятно, представляют собой живые, свободно
живущие мицеллы сложного строения (Немило-в, 44), или вроде коацер-
ватов — капелек, белковых коллоидных образований со сложными
протекающими внутри них ферментативными процессами распада и
синтеза (Опарин, 49). Вероятно, впоследствии, под влиянием
изменившихся геологических условий, эта первичная протоплазма в
результате эволюции приняла клеточное строение, которое нужно
рассматривать как один из важнейших актов приспособления живой протоплазмы
к новым геологическим условиям.
Одно время некоторые ученые думали, что каждая клетка состоит
из мельчайших живых единиц, подобно тому как какое-либо мертвое
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 51
тело состоит из молекул (Гейденгайн, 27). Теперь эта гипотеза почти
никем не защищается. В настоящее время господствует такое мнение,
что хотя каждая частичка клетки вовлечена в обмен веществ, растет,
участвует в размножении и обладает возбудимостью, но все эти
функции той или другой частички проявляются исключительно при их
совместном существовании в единой неделимой клетке (Гертвиг, 28).
Таким образом, между живым и мертвым образованиями существует
качественное различие в отношении внутренней структуры. Мертвое об-і
разование может быть доведено путем разложения до молекул или
комплекса их, так что образование не потеряет своих характерных
особенностей, своих качеств; живое же образование не может быть
разложено таким же образом до молекул без потери наименьшей
частичкой характерных признаков жизни.
Закон раздражения Дюбуа-Реймона. Не всякое внешнее
воздействие производит возбуждение, т. е. является раздражающим агентом.
Когда внешний агент, внешняя сила, действует на мышцу с большой
постепенностью, сначала очень слабо, а затем все сильней и сильней,
то мышца не раздражается. Необходимо, чтобы внешний агент в
момент действия на мышцу испытывал достаточно быстрое изменение в
интенсивности. При этом не имеет значения, является ли начальная
величина раздражителя нулевой или нет; точно так же не имеет
значения, в каком направлении меняется величина раздражителя: от
некоторой малой величины до некоторой большой или наоборот. Итак,
возбуждение вызывается не действием абсолютной силы раздражения,
а быстрым изменением ее от одной величины до другой; при этом
раздражающее действие тем сильнее, чем скорее происходит означенное
изменение. Это положение носит название закона раздражения Дюбуа-
Реймона (1845) (16). Для иллюстрации можно привести следующие
примеры:
1. Если на мышцу капнуть кипящим физиологическим раствором,
она возбудится и сократится. Но, если положить мышцу в холодный
физиологический раствор и затем медленно согревать его до кипячения,
мышца сварится без возбуждения.
2. Гальванический ток действует раздражающим образом только в
момент быстрого изменения его силы, или потенциала, например при
его замыкании и размыкании, когда разность потенциалов меняется
очень быстро от нуля до некоторой величины и обратно. Сила
раздражающего действия гальванического тока определяется высотой
изменения его вольтажа. Чем выше вольтаж, тем сильнее раздражение. Если
же сила или вольтаж гальванического тока остаются без изменения
или меняются очень медленно, то действие их на мышцу не является
раздражающим.
Однако в определенных случаях абсолютная сила имеет значение:
при гальваническом токе чрезмерно большой силы возбуждение
возникает также в период неизменного его действия, т. е. под влиянием
его абсолютной силы (Пфлюгер, 21). Это, вероятно, происходит
4*
'52
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
оттого, что прохождение сильного тока через мышцу сопряжено с
механическим повреждением живой системы, а также с ее термическими,
химическими и физическими изменениями. Все эти изменения в случае
их быстрого протекания являются источником новых раздражений,
вызывающих возбуждение и сокращение.
К закону раздражения Дюбуа-Реймона была сделана одна
существенная поправка. Еще Герман (18а), Бидерман (186), Г о о р-
в е г (46) и другие авторы указывали, что раздражающая сила
должна действовать некоторое определенное время на мышцу, чтобы
вызвать возбуждение последней. Чем ниже возбудимость мышцы,
тем дольше должно продолжаться внешнее воздействие.
Следовательно, для того чтобы внешняя сила подействовала на мышцу
раздражающим образом, недостаточно, чтобы эта сила быстро
изменилась от одной величины до другой; нужно еще, чтобы общее время
действия внешней силы не было меньше некоторого определенного
значения. Когда, например, возбудимость мышцы падает под влиянием
наркотического вещества, то, чтобы вызвать возбуждение, следует
увеличить не только силу внешнего агента, но и продолжительность его
действия. Минимальное время раздражающего действия зависит,
однако, и от абсолютной величины внешней силы. Чем выше абсолютная
сила, тем меньше это время. Время раздражения зависит также
от того, как меняется внешняя сила при своем воздействии на імышцу.
Чем быстрее меняется раздражающая сила, тем короче это время.
Поэтому при электрическом раздражении время раздражения прямо
зависит от скорости изменения раздражающего электрического тока.
Более подробное исследование минимального времени раздражающего
действия в зависимости от силы раздражения произведено В ей с сом
(47) и Гильдеімейстером (19).
Время, необходимое для того, чтобы минимальное раздражение
вызвало возбуждение, было названо Гильдемейстером
полезным временем (Nutzzeit). Так как продолжительность полезного
времени точно зависит от степени возбудимости, то, очевидно, все то,
что влияет на возбудимость, влияет и на полезное время.
Следовательно, полезное время зависит от обмена веществ, от рода животных,
от характера мышцы и т. д. Например, полезное время m. biceps
человека составляет 1—2 a, m. gastrocnemius лягушки — 3—7 а, гл.
gastrocnemius жабы—13 а> мышц клешни рака — 300 J (Лапик, 20).
Для измерения полезного времени пользуются главным образом двумя
способами: по одному из них, ©веденному Б е й с с о м (1901), определяется то время,
которое проходит от момента включения тока до -его выключения (47). В
настоящее .время чаще івсего пользуются для этой цели маятником Гельмгольца. По пути
движения маятника установлено два контакта. Один из них неподвижный, другой
подвижный, который можно установить на разных расстояниях от первого.
Маятник при своем качании размыкает сначала один, потом другой контакт.
Возбудимая ткань (нерв, мышца) через эти контакты вводится в цепь источника
электричества таким образом, что при размыкании маятником первого контакта она
включается te цепь постоянного тока, а при размыкании другого контакта, наобо-
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ
53
рот, — выключается. По другому способу, введенному Гоорвегом (1892),
полезное время измеряется путем применения конденсаторов (46). Как известно, ¦ при
разряде конденсатора через определенное сопротивление, чем меньше емкость
конденсатора, тем короче время его разряда. Пропуская через возбудимую
ткань разряды конденсаторов разной емкости, заряженной одним и тем же
пороговым вольтажем, легко установить тот разряд, который является пороговым.
Продолжительность этого разряда и будет полезным временем. Между этими
двумя способами существует характерная разница. При пользовании
маятником крутизна изменения тока в момент замыкания и размыкания
одинаково велика; в этом случае мы имеем, как говорят,
прямоугольные токи. При конденсаторных разрядах напряжение тока в ткани нарастает почти
мгновенно, как в прямоугольном токе, но падает сейчас же по достижении максимума,
н это падение происходит сначала быстро, а потом постепенно замедляясь, причем
время падения тем больше, чем больше емкость конденсатора.
Кривая силы-времени. Мы видели, что полезное время меняется
в зависимости от характера возбудимой ткани. Более возбудимая ткань
01^02 Ой 06 081 2 4 В.8 1 2 4 8 НО 20 W8080WQ6
Рис. 38. Кривая силы-времени m. sartorius лягушки, отравленной
кураре. На ординате вапряжение тока в вольтах, на абсциссе— общее
время в сигмах.
Кривая А — от зоны двигательных пластинок, В — от безнервного участка
мышцы (Цкипуридзе).
имеет маленькое полезное время, менее возбудимая — большое. Но
какова бы ни была ткань, время раздражения определенным образом
меняется в зависимости от изменения вольтажа, а именно — с
повышением вольтажа это время уменьшается. Если на абсциссе отложить
время действия в сигмах, а на ординате —¦ напряжение в вольтах, то
получится кривая силы-времени, которая хотя имеет одинаковый
характер у разных возбудимых тканей, но в деталях, а именно в
отношении длительности времени действия и величины вольтажа, будет
более или менее различаться. Так, на рис. 38 даются кривые силы-
времени для безнервного участка мышцы и для зоны двигательных
54
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
пластинок. Из этого рисунка видно, что для каждой возбудимой
системы с данной степенью возбудимости существует своя
характерная кривая силы-времени. Поэтому составление кривой силы-
времени является одним из хороших средств для характеристики
возбудимости ткани.
Закон электрического раздражения Нернста. Н е р н с т (25) в
определенной формуле связал силу раздражения и полезное время его
действия. Эта формула по отношению к замыканию постоянного тока
такова: *y7=const = &l где і—сила раздражения, t —полезное
время, a k — постоянная. Следовательно, по этой формуле,
произведение раздражающей силы постоянного тока на квадратный корень из
времени действия должно оставаться постоянным. Отсюда следует, что
для получения физиологического эффекта — возбуждения от
применения постоянного тока, действие тока на возбудимую ткань должно быть
тем дольше, чем ниже его напряжение. Если для одного тока с
известной силой найти полезное время и определить постоянную по
формуле Нернста, то легко можно вычислить приблизительную
величину полезного времени для каждой другой силы. По Н е р н с т у, для
мышц одного вида лягушки — R. esculenta эта постоянная составляет
335, а для другого вида — R. temporaria — 145. Ввиду того, что
формула Нернста определяет внешнее условие для вызова возбуждения, ее
называют еще законом электрического раздражения Нернста. Автор
исходит при этом из созданной им теории электрического раздражения,
согласно которой при пропускании электрического тока через живую
ткань изменение концентрации ионов іможет происходить только на
так называемых полупроницаемых перепонках, которые являются
естественным препятствием для перемещения ионов. Здесь происходит
повышение концентрации определенных іионов, которые своим
раздражающим действием вызывают возбуждение возбудимой системы.
Очевидно, чем сильнее электрический ток, тем скорее достигается
пороговая концентрация ионов, достаточная для того изменения
полупроницаемых перепонок, которое обусловливает процесс возбуждения.
Существует немало теорий физико-химических условий возбуждения.
Одной из наиболее разработанных является теория акад. П. Л а-
зарева, который исходит из известных исследований Леба и
Нернста — о действии ионов на живые клетки, а также из своих
собственных многочисленных исследований (22). По учению Лазарева, при
пропускании через ткани электрического тока на полупроницаемых
перепонках скопляется большое количество ионов, и тогда последние
действуют здесь на белковые вещества, вызывая их коагуляцию или
свертывание. Это изменение аггрегатного состояния Лазарев
называет «первой стадией процесса возбуждения». Аналогичные
изменения происходят в живой системе при действии ультрафиолетовых
лучей и при других раздражениях.
Раздражители мышечной ткани. Нормально мышечная ткань
возбуждается путем передачи возбуждения из центральной нервной си-
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧЛОЙ ТКАНИ 55
стемы через двигательный нерв. Но она может возбуждаться также
.при непосредственном воздействии любой внешней силы — химической,
физической, тепловой, электрической и т. д.
Химическое вещество іможет вызвать возбуждение, если оно
достаточно быстро проникает в мышечную клетку и здесь производит
быстрое изменение во внутренней жидкой среде протоплазмы. Таким
образом действуют многие неорганические и органические кислоты,
щелочи, соли, газы, спирты, алкалоиды, ацетилхолин и другие
соединения. Растворы веществ, которые совершенно не меняют химического
состава и физико-химического состояния мышцы, не влияют на мышцу
раздражающим образом. Таким является, между прочим, так
называемый физиологический раствор 0.65—0.9% хлористого натрия, который
употребляется в физиологических лабораториях для защиты
вырезанной или обнаженной живой ткани от высыхания. Для сохранения
мышечной ткани лягушки пользуются 0.65—0.7% раствором хлористого
натрия, а мышечной ткани теплокровных — 0.9% раствором. Подобные
растворы берутся потому, что они обладают таким же осмотическим
давлением, как и растворы солей в крови и лимфе соответствующих
животных. Кроме того, ионы натрия необходимы для сохранения
нормальной жизнедеятельности мышечной ткани (Овертон, 2Ь; Леб, 26).
Однако все эти искусственные растворы по своему физиологическому
действию на мышцы оказываются много хуже изоосмотической
кровяной сыворотки: работоспособность мышц поддерживается в ней и
дольше и на более высоком уровне (Купалов и сотр., 38). Прибавление
к искусственным растворам небольшого количества кровяной
сыворотки значительно улучшает их физиологическое действие на живые ткани.
Изотоничные растворы других веществ, например 6 % раствор
тростникового сахара, вредят мышечной ткани не столько своим
присутствием, сколько тем, что отнимают по законам диффузии ионы
натрия. Стоит только прибавить к изотоническому раствору тростникового
сахара немного соли натрия (0.1 %), чтобы такой раствор оказался
'индифферентным, подобно физиологическому раствору (Овертон, 2Ь).
Мышечная ткань и вообще живая ткань лучше сохраняет свои
жизненные отправления в таком растворе, который приближается к
етлазме крови не только по своим осмотическим свойствам, но и по
своему химическому составу. Такова, например, жидкость,
приготовленная впервые Р и н г е р о м, которая содержит все те соли и в том
количестве, что и кровь холоднокровных животных. Раствор Рингера
содержит 6.5 г NaCl, 0.14 г КС1, 0.12 г СаС12 и 0.2 г NaHC03 на
литр дестиллированной воды. Для сохранения живой ткани
теплокровных животных употребляют аналогичный раствор Локка; он содержит:
•9.0 г NaCl, 0.42 г КО, 0.24 г СаС12, 0.1—0.3 г NaHC03 и еще глюкозу
1—2 г на литр дестиллированной воды. Пользуются и другими
растворами, которые еще больше приближаются к кровяной плазме. Они
•содержат, кроме вышеуказанных соединений, MgCb и NaHP04 (Гедон,
Тироде).
56
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Воздействие температуры производит возбуждение мышцы. Но, как
уже указывалось выше, для этого нужно, чтобы температурное
изменение в мышечных клетках происходило быстро.
Точно так же вызывает возбуждение и механическое воздействие,.
как, например, удар, быстрое растяжение и т. д. Постепенное
растяжение или давление не производит возбуждения мышцы.
Электрический ток, как гальванический, так и индукционный, также
действует на мышечную ткань раздражающим образом не своей
абсолютной силой, а быстрым изменением последней. Действие
электрического тока изучалось давно и очень подробно. Дюбуа-Реймон и
Пфлюгер в 1840—1860 гг. создали целый отдел физиологии,
именуемый электрофизиологией. Этому отделу будет впоследствии
уделено особое место, поэтому мы не будем касаться здесь действия
электрического тока.
Во всех случаях раздражения, приведенных выше, наступление
возбуждения связано с одним и тем же физико-химическим изменением.
Оно заключается в изменении концентрации ионов на
полупроницаемых перепонках внутри клетки, иначе говоря, на поверхности
структурных элементов, которые целиком или в определенной своей части
составляют возбудимую систему. От количества и характера этих ионов,,
а также от быстроты, их скопления зависит возникновение
возбуждения в возбудимой системе. Значит, все внешние воздействия только
тогда производят раздражение, когда они, перемещая ионы внутри
клетки, создают определенное физико-химическое изменение на
полупроницаемых перепонках, притом достаточно быстро, в течение
определенного, так называемого полезного времени.
Процесс раздражения как основное физико-химическое условие
возбуждения имеет место не только при искусственном воздействии
какой-либо внешней силы на возбудимую систему, но и при ее
естественном возбуждении, т. е. когда последнее возникает путем
распространения самого процесса возбуждения. Процесс возбуждения, возникший
в каком-либо участке живой клетки, действует на соседний участок и
вызывает в нем возбуждение через то же самое физико-химическое
изменение раздражения, а именно: это изменение создается изменениями
электрического потенциала, которые возникают в живой системе —
клетке, в результате процесса возбуждения. Таким образом, при
естественном течении процессов внутри организма процесс раздражения,
е одной стороны, является условием — причиной процесса возбуждения,,
а с другой, — результатом — следствием этого процесса.
Определение степени возбудимости. Мы указали, что возбудимость
есть свойство возбудимой системы отвечать на внешние воздействия
процессом возбуждения. Но это свойство не всегда одинаково.
Возбудимость мышцы разная у представителей разных классов животных;:
она не одинакова в разных мышцах одного животного и е одной и
той же мышце в ее различных функциональных состояниях. Например,
мышца холоднокровных менее возбудима, чем теплокровных; отдох-
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 57'
нувшая мышца всегда обладает большей возбудимостью, чем
утомленная, и т. д. Но и одна и та же мышца не обладает одинаковой
возбудимостью во всех своих частях. Так, например, возбудимость m. sar-
torius лягушки на проксимальном конце, где нет двигательных
'Пластинок, ниже, чем в середине мышцы, в области двигательных
пластинок. Эта разница наблюдается и на мышце, отравленной кураре. После
такого отравления двигательные нервные импульсы перестают
возбуждать імышцу, но сама мышца реагирует на непосредственное
раздражение, причем возбудимость ее в нервных областях выше (Беритов, 2а, 15).
Химические вещества в малых концентрациях повышают
возбудимость, а в несколько больших дозах понижают ее. Так действуют соли,
кислоты, щелочи, алкалоиды. Температура также сильно влияет на
возбудимость. В лягушечьей мышце с повышением температуры от
комнатной до 40—42° возбудимость повышается, а с охлаждением
понижается.
Определение степени возбудимости производится следующим
образом. Какой-нибудь раздражитель слабой интенсивности, положим,
слабый индукционный удар, пробуется в отношении действия его на
мышцу. Если он не вызывает возбуждения, увеличивают
интенсивность и вновь пробуют. Таким путем находят ту минимальную силу
раздражения, которая производит минимальный эффект. Это
минимальное раздражение называется пороговым.
Интенсивность порогового раздражения является мерилом степени-
возбудимости. Когда возбудимость почему-либо понижается,
интенсивность порогового раздражения соответственно возрастает или, как.
говорят, порог раздражения становится выше. Наоборот, когда
возбудимость повышается, пороговое раздражение соответственно
уменьшается или, как говорят, порог раздражения становится ниже.
Хронаксия как мера возбудимости. Точное определение полезного
времени сильно затруднено необходимостью пользоваться токами
пороговой силы, ибо малейшее отклонение в сторону повышения пороговой'
силы ведет к значительному уменьшению времени раздражения.
Поэтому полезное время не может служить точным показателем
функционального состояния мышцы. Это обстоятельство побудило французского^
ученого Л а п и к а (20) выработать такой способ раздражения, с
помощью которого можно получить более точную характеристику
возбудимости как функцию времени. Согласно этому способу, сначала
находят минимальную силу постоянного электрического тока, которая в
состоянии вызвать возбуждение мышцы, независимо от длительности
действия тока. Эту силу тока Л а п и к назвал реобазой. Удвоив ее,
находят соответствующее минимальное время раздражения. Время, в<
течение которого ток силы удвоенной реобазы должен действовать на
ткань, чтобы вызвать минимальный эффект возбуждения, Л а п и к
назвал хронаксией. На кривой силы-времени хронаксия приходит-*
ся на ту часть кривой, где время раздражения сравнительно меньше
меняется в зависимости от силы раздражения (рис. 39).
58
0БЩЛ.Я ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
03
А'войн л
р?ас/аза и
реогіаза ?
О
Хронаксия представляет количественное выражение возбудимости,
ее временную характеристику. Она отображает скорость и
продолжительность жизненных процессов живой системы (Лапик, 31).
Вследствие этого хронаксиметрия является более точным методом измерения
возбудимости, чем определение последней по расстоянию
индукционных катушек.
Лапик и его ученики изучили хронаксию на ряде тканей у
многих животных при разных функциональных состояниях. У каждой
мышцы при определенных условиях была обнаружена хронаксия
определенной величины. Так, человеческие мышцы имеют хронаксию 0.1 —
!0.7 , причем хронаксия сгибателей вдвое меньше, чем хронаксия
разгибателей; точно так же как
хронаксия мышц
проксимальных суставов вдвое короче,
чем мышц дистальных суста-
эв. Хронаксия мышц
лягушки (gastrocnemius, ileofibu-
laris) около 0.3—0.56 а.
Хронаксию целой
нормальной мышцы не всегда можно
отнести к мышечным
волокнам. Мышцы содержат
нервы, а возбудимость нервов
высокая. Поэтому при
раздражении таких мышц
первым долгом должен
раздражаться нерв. Следовательно,
таким путем трудно выяснить
хронаксию мышечных волокон. Для определения хронаксии
нормальных мышечных волокон нужно брать их в изолированном виде. Такие
исследования были произведены, и оказалось, что изолированные
волокна имеют разнообразную хронаксию: она тем меньше, чем толще
волокно. Кроме того, хронаксия не одинакова вдоль по длине волокна.
Юна больше всего на заостренных концах волокна и меньше всего в
центральных участках, где имеются нервные окончания (Моор и
Брюкке, 30).
Точно так же, если изучать хронаксию нормальной мышцы в разных
участках ее, то она окажется весьма различной. В совершенно без-
-нервных участках хронаксия будет самой значительной. В тех зонах
•мышцы, где расположены двигательные пластинки, она будет вдвое-
втрое меньше. Но еще меньше она будет там, где проходят веточки
двигательного нерва в данной мышце. Так, в га. sartorhis лягушки на
проксимальном безнервном участке хронаксия равняется 0.92—1.60 а
в зоне двигательных пластинок — 0.40—0.70 j , а в области нервного
ствола —0.15—0.25 а (Цкшіуридзе, 43).
Хронаксия настолько характерна для функционального состояния
JpdHa/гсия
V*
Полезное время
С
j
Рис. 39. Кривая силы-времени с указанием отно-
¦шений реобазы неполезного времени к хронаксии
и двойной реобазе.
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 59
мышц, что теперь ею пользуются во многих клиниках для
установления степени дееспособности мышц при их патологическом состоянии.
Чем хуже функциональное состояние, тем больше хронаксия.
Например, в состоянии дегенерации после перерезывания двигательного
нерва хронаксия мышц сильно удлиняется, именно до 10—70 а (Бур-
тиньон, 23). От функционального состояния зависит, как это мы
увидим ниже, длительность и интенсивность процесса возбуждения,
скрытый период его, скорость проведения и т. д. И все эти явления стоят
е прямой связи с хронакоией: чем меньше хронаксия, тем сильнее
процесс возбуждения, тем меньше скрытый период, тем больше скорость
проведения. Очевидно, если с ухудшением функционального состояния
хронаксия удлиняется, это будет указывать и на то, что процесс
возбуждения ослабел, скорость проведения уменьшилась, скрытый период
увеличился и т. д.
Хронаксия мышц претерпевает определенное изменение в связи с
онтогенезом индивида. У новорожденных детей хронаксия мышц
значительно больше, чем у взрослых. Так, хронаксия двуглавой мышцы
(сгибателя локтевого сустава) в течение нескольких часов после
рождения в 6 раз больше, чем у взрослых: в среднем она равна 0.60 j , в
то время как у взрослых она в среднем равна 0.10 з. Кроме того, у
новорожденных хронаксия одной и той же мышцы меняется от опыта
к опыту и от одного младенца к другому в гораздо ббльших пределах,
чем у взрослых. Значит, с возрастом хронаксия уменьшается и
становится более постоянной. Наконец, у новорожденных отсутствует
указанное выше соотношение между хронаксиями сгибателей и разгибате-.
лей, а также імежду хронаксиями мышіц дистальных и проксимальных
суставов. Это соотношение устанавливается впоследствии, и весь
процесс заканчивается к 10 годам жизни ребенка (Вул, 40).
Определение изменения возбудимости путем хронаксии не всегда
дает точное представление об этом изменении. Дело ов том, что
величина хронаксии зависит от силы применяемого тока. При ухудшении
функционального состояния приходится сильно увеличивать реобазу, и
в таких случаях удвоенная реобаза может дать не увеличенную хронак-
сию, как это следовало бы ожидать, а, наоборот, уменьшенную.
Следовательно, в таких случаях хронаксия показывает вместо ухудшения
функционального состояния улучшение. Это, например, хорошо видно
в опытах Магницкого и Мужеева (33) при исследовании
хронаксии нерва при постепенном ухудшении его функционального
состояния под влиянием ядов: при постепенном нарастании реобазы
хронаксия сначала уменьшается, а затем только начинает расти.
Подробно о недостатках хронаксиметрического измерения
возбудимости будет сказано ниже, в отделе нервной физиологии.
Собственная возбудимость мышцы. Нормальная мышца, наряду
с мышечными волокнами, содержит двигательные нервные волокна.
Последние обладают возбудимостью, а потому непосредственное
раздражение нормальной мышцы может производить в мышце возбуждение
60
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
через двигательные нервные волокна. Но есть много способов
обнаружить собственную возбудимость мышцы.
В некоторых мышцах лягушки имеются такие участки, где нет
двигательных окончаний. Несмотря на это, раздражение таких участков
производит возбуждение. Например, в m. sartorius лягушки дисталь-
ный и проксимальный концы лишены нервных окончаний, однако
электрическое или механическое раздражение их вызывает сокращение.
Прямыми опытами можно доказать, что при раздражений
проксимального участка портняжной мышцы действительно возбуждается
непосредственно возбудимая система мышечных волокон. Именно,
берется самый конец проксимального участка, менее одной пятой всей
длины мышцы, и лосле физиологического обследования подвергается
обычной гистологической обработке для обнаружения нервных волокон
и окончаний. Оказывается, что и такие участки мышцы, которые
абсолютно лишены нервных окончаний, обладают возбудимостью (Беритов,.
45). Но © безнервных участках возбудимость імышцы ниже, чем в
нервных. Это хорошо видно на рис. 38.
Возбудимость мышцы лучше всего изучать на таких мышцах, где
почему-либо парализованы нервные окончания. Это бывает, например,
при дегенерации двигательного нерва. Если перерезать какой-либо
двигательный нерв у животного, то спустя две недели раздражение
периферического отрезка нерва не будет давать эффекта в мышцах.
Нерв, как говорят, перерождается, теряет все свои жизненные
функции. Но в это время прямое раздражение такой денервиро-
ванной мышцы будет вызывать ее сокращение (Броун-Секар, 4; Бид-
дер, 5). Кроме того, существуют вещества, действующие на мышцу
так, что раздражение двигательного нерва, так называемое
непрямое раздражение, не дает эффекта в мышце, но в то же время
прямое раздражение мышцы производит сокращение. Так действуют,
например, ионы магния (Вихман, 6). Типичным представителем таких
веществ является кураре. Этот яд из ряда алкалоидов оказывает такое же
действие на мышцы, как магний. После отравления кураре мышца более
не возбуждается при непрямом раздражении, но сохраняет способность
к возбуждению от прямого раздражения (Кюне, 7; Клод-Бернар, 8).
И на таких денервированных мышцах не трудно удостовериться, что
возбудимая система мышцы в нервных участках, т. е. в области
нервных окончаний, заметно более возбудима, чем в безнервных участках
(Беритов и Яшвили, 15).
Физиологическое действие кураре впервые было изучено Бернаром (1857),
а потом Кюне. Они нашли, что мураре не действует ни непосредственно на
мышечную ткань, ни на нервное волокно. Если приложить раствор кураре непосредственно
к нервному волокну, последнее совершенно не пострадает от этого. Оно будет
раздражаться после отравления точно так же, как и до него. Отсюда и было сделано
заключение, что если при отравлении мышцы нервное раздражение не вызывает
мышечного сокращения, то это явление должно быть обусловлено параличом
нервных окончаний, лишением их свойства передавать возбуждение.
Но уже давно были установлены факты, противоречащие этому первоначаль-
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 61
ному взгляду. Впервые английский ученый Лэ<нглій (13) показал (1906), что
кураре оказывает характерное действие и на денервированную мышцу. Л э н г л и доказал
это таким о-бразом. Он брал никотин и смазывал им денервированную мышцу.
Отравленный участок мышцы возбуждался и сокращался. Бели в момент сокращения
он смазывал ту же мышцу раствором кураре, то сокращение сейчас же исчезало.
Ясно, что кураре и никотин действуют на одну и ту же возбудимую систему мышцы.
Кроме того, по более поздним исследованиям оказалось, что очень малая доза
кураре повышает возбудимость мышцы как на прямое, так и на непрямое
раздражение (Гепферт и Шефер, 39). При несколько большей дозе кураре понижает
возбудимость ,в том участке мышцы, где оканчивается двигательный нерв, т. е. в зоне
двигательных пластинок. В других участках возбудимость не меняется (Лэнглк, I,
-Люкас, 10). Это действие кураре хорошо видно при изучении электрического
потенциала концевой пластинки на изолированных мышечных волокнах до и после кура-
ризации. До кураризации потенциал концевой пластинки мышечного волокна раз
в десять больше, чем в мышечном волокне ,вне концевой пластинки. После
кураризации прежде всего исчезает эта большая разность потенциалов между концевой
пластинкой и остальным мышечным волокном (Бухталь и Линдгард, 35). Только
при более высокой концентрации кураре возбудимость понижается во всех участках
мышечного волокна, и тем сильнее, чем больше доза кураре (Политцер, 11; Эдшт-
рем, 12). А при очень большой дозе мышца совершенно теряет возбудимость на
прямое раздражение. Итак, при слабом отравлении кураре возбудимость мышечного
волокна понижается прежде всего в области нервных окончаний. Вследствие этого
мышца теряет способность к возбуждению через двигательный нерв. В это время
¦возбудимость других участков мышечного волокна остается нормальной. При
несколько большей дозе кураре возбудимость понижается во всем мышечном волокне, а
при еще большем она исчезает здесь совершенно.
Но кураре безусловно действует и на нервное ©олокно, и прежде всего на голые
нервные окончании. Это видно из того, что если подвергнуть нерв задушению, а
петом восстановить в кислороде, то кураре произведет на такое нервное волокно
отравляющее действие. В результате произойдет резкое ослабление процессов возбуждения
в нерве (Фромгерц, 29). Очевидно, нечувствительность нормального нерва к кураре
зависит от того, что оболочки нервных волокон не проницаемы для кураре.
Невидимому, задушение и последующее 'восстановление так меняют состояние оболочки,
что кураре проникает внутрь волокна.
Практикум
НАБЛЮДЕНИЕ СОБСТВЕННОЙ ВОЗБУДИМОСТИ НА НОРМАЛЬНОЙ МЫШЦЕ
1. Опыт с m. sartorius. Для этого наблюдения следует брать m. sartorius
лягушки (у этой мышцы проксимальный конец на расстоянии б—9 mm лишен нервных
окончаний). У лягушки перерезают спинной мозг на границе с продолговатым,
разрушают его длинной иглой или проволокой и прикалывают неподвижную
лягушку к пробковой пластинке брюхом вверх. После этого производится операция с
-мышцей: снимают кожу с бедра, освобождают m. sartorius от других мышц,
навязывают на ее сухожильные концы нитки и затем вырезают.
Нитку с проксимального конца мышцы закрепляют в лапке на штативе, а
другую нитку перекидывают через блок, привязывая к ней гирьку в 10 г (рис. 41). На
проксимальный конец накладывают платиновые или серебряные электроды с
межполюсным расстоянием в 2 мм и раздражают мышцу отдельными индукционными
ударами. При нормальной возбудимости мышца дает сокращение от размыкательных
ударов при 20—15 ом расстояния между катушками, от замыкательных — при
10—15 см.
2. Опыт с магнием. Для этого опыта приготовляют нервно-мышечный препарат
лягушки. Перерезают позвоночник лягушки в грудной области и отделяют заднюю
62 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
часть, разрушают в ней остаток спинного мозга введением булавки в позвоночный
канал и затем удаляют все внутренности. Потом снимают кожу с обеих задних
лалок, удаляют копчиковую кость целиком, перерезают позвоночник продольно, а
вслед за ним весь таз по симфизу.
В одной половине обнажают и выделяют седалищное сплетение и продолжение
его на бедре, т. е. седалищный нерв вплоть до коленного сустава. Остатки таза и
мышцы бедра удаляются, остаются бедреная кость и тот кусок позвоночника,
который связан с седалищным сплетением. Таким
путем получается так называемый
нервно-мышечный препарат лягушки (рис. 40).
В 7ю нормальный раствор хлористого магния
(MgCl) погружается весь нервно-мышечный
препарат на несколько часов. Ионы магния убивают
нервные окончания много раньше мышцы. Поэтому,
.п, ._, .__ _ если раздражать нерв отдельными индукционными
ушг^^^ " Щ ' ударами, то нетрудно уловить тот момент, когда
раздражение нерва не будет производить
сокращения мышцы, тогда как раздражение самой мышцы
будет давать эффект.
3. Опыт Клод Бернара с отравлением
лягушки кураре. Неповрежденную лягушку прикалывают
булавками к пробочной пластинке спиной вверх.
На одной ноге перевязывают бедреную артерию
и затем приподнимают из раны седалищный нерв,.
f а все бедро под нервом перевязывают ниткой.
Этим достигается прекращение циркуляции крови и лимфы в оперированной ноге.
Впрыскивают в спинной лимфатический мешок 0,5—1 см3 1% раствора кураре.
Спустя несколько минут лягушка парализуется. Только оперированная нога,
куда не проник яд, сохраняет способность к движениям. Если раздражать где-либо
кожу (щипание, индукционный удар) и этим вызвать деятельность в центральной
нервной системе, то ответная реакция наступает только на оперированной ноге. Из
этого видно, что под влиянием «кураре как нервные стволы, так и центральная
нервная система не теряют возбудимости. Если теперь раздражать седалищный
нерв на другой ноге, то сокращения на ней при этом не получается. Но в то же
время непосредственно раздражение мышц током вызовет сокращение их.
Рис. 40. Нервно-мышечный
препарат лягушки.
Зажим держит нервно-мышечный"
препарат за отрезок бедреной кости (К).
С, Н. — спинномозговые корешки;
С. И. —- седалищный нерв; М —
икроножная мышца; А. С. — ахиллово
сухожилие, к которому приделан
крючок для соединения с миографом
(Лучиани).
ПРОВЕРКА ЗАКОНА РАЗДРАЖЕНИЯ ДЮБУА-РЕЙМОНА
1. Опыты с нагреванием. На задней лапке лягушки удаляют все мышцы бедра.
Остаются только бедреная кость и голень со стопой. Если такой препарат,
закрепив его за бедреную кость, опустить в химический стакан с холодным
физиологическим раствором и нагревать раствор с большой постепенностью до 40° и выше»
то можно сварить мышцу без единого сокращения. Если нагревание происходит
на большом огне и температура раствора поднимается очень быстро, то можно
наблюдать движение лапки от возбуждения мышц. Также получается движение, если
на препарат капнуть кипящим раствором.
2. Опыт с реономом Флейшля (рис. 41). Реономом можно менять интенсивность
гальванического тока то быстро, то постепенно. Реоном Флейшля представляет
эбонитовую четырехугольную доску. На поверхности ее имеется круговой
желобок, который во время опыта наполняется насыщенным раствором
сернокислого цинка или медного купороса. Желобок этот в двух противоположных местах
связан медными пластинками с борнами Л и В для зажима прозодов, которые идут
от аккумулятора. В центре круга вертикальная эбонитовая ось, к которой
прикреплены две изогнутые дугообразно медные пластинки, концы которых опущены в же-
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ 63
лобок. На этих пластинках имеются зажимы а и Ь, от которых провода идут
к электродам, наложенным на мышцу.
Если прибор установить, как показано на рисунке (изогнутые пластинки
находятся на равном расстоянии от зажимов А и В), тогда замыкание и размыкание
цепи ключом Дюбуа-Реймона не будет вызывать сокращения, ибо от зажима,
присоединенного к положительному
полюсу аккумулятора, потенциал падает рав- v***1***^ !\
номерно на обеих сторонах желобка до y^^^f _г\ А
другого зажима, присоединенного к отрица- yfrr'^^^^-^^=^^=t
Рис. 41.^Постановка опыта с реономом Флейшля.
р. Ф. — реоном Флейшля; Л и В — зажимы ' для соединения с аккумулятором
(Ак); а и b — зажимы для соединения с электродами (3); К — ключ Дюбуа-
Реймона; М — мышца, один конец которой фиксирован лапкой.
тельному полюсу. Вследствие этого пластинки будут находиться в точках
одинаковых потенциалов, и ток совершенно пе пойдет к препарату. Если изогнутые
пластинки приблизить к зажимам А и В, тогда между ними будет создаваться
максимальная разность потенциалов; и сила тока настолько увеличится, что замыкание
и размыкание цепи будет давать сокращение. Если же при замкнутой цепи тока
разность потенциалов постепенно меняется от минимальной величины до
максимальной путем медленного поворота изогнутых пластинок к борнам Л и В и обратно,
то мышца не дает сокращения. Мышца сократится только при быстром вращении
оси, т. е. при быстром изменении разности потенциалов между пластинками,
которое ведет к быстрому изменению силы гальванического тока.
С помощью реонома Флейшля легко обнаружить действие на мышцу большой
абсолютной силы раздражителя. Если при разности потенциалов в 6 вольт
усиливать ток даже с большой постепенностью, то мышца непременно начнет
сокращаться, когда сила тока достигнет большой величины.
ОПРЕДЕЛЕНИЕ СТЕПЕНИ ВОЗБУДИМОСТИ МЫШЦЫ
1. Определение порогов в раздражении мышцы. Накладывают электроды на такое
место мышцы, где нет поблизости нервных стволов, например на проксимальный
конец т. sartorius на расстоянии не более одной пятой длины всей мышцы.
Электроды соединены со вторичной катушкой через ключ Дюбуа-Реймона. В первичную
цепь также введен ключ для прерывания гальванического тока. Все расположение
приборов для этого опыта показано на рис. 42. Сначала находят порог для размы-
кательных ударов, т. е. то наибольшее расстояние индукционных катушек, при
котором получается сокращение мышцы. Для этого начинают действовать
индукционными ударами с 30—35 см расстояния между катушками, при котором мышца не
возбуждается. Затем сближают катушки до тех пор, пока не получится сокращение
мышцы. После определения порога для размыкательных ударов находят его и для
Ы ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
замыкательных. В последнем случае нужно поступить так, чтобы действие размыка-
тельных ударов было исключено. Для этой цели ключ во вторичной цепи остается
открытым только в момент замыкательного удара. Эту предосторожность нужно
строго выполнять во избежание изменения 'возбудимости под влиянием раздражения
размыкательными ударами. Необходимо, чтобы после каждого действующего
индукционного удара мышца отдыхала около 1—2 минут. Это правило необходимо
соблюдать вообще при определении порогов раздражения.
Рис. 42. Схема расположения и соединения приборов для изучения порогов раздражения.
А — аккумулятор; lull —¦ первичная и вторичная катушки индуктория Дюбуа-Реймона; В —-ваг-
неровский прерыватель к нему; К1 и К1і —¦ ключи Дюбуа-Реймона; О— провода, соединяющие
аккумулятор с первичной цепью индуктория для получения отдельных индукционных ударов; Т —-провода для
-тетанического раздражения. От ключа Дюбуа-Реймона во вторичной цепи идут провода к
электродам (3), которые положены на мышцу (М). .J
2. Изменение возбудимости под влиянием температуры. После того как
определены пороги раздражения m. sartorius, к мышце в области электродов
прикладывают минуты на две смоченный теплым рингеровским раствором (до 38°) кусочек
ваты. Затем удаляют вату, быстро очиідают лигнином капельки жидкости между
полюсами электродов и вновь определяют пороги раздражения. При этих условиях
опыта пороги понизятся на 2-—3 см. Чтобы опыт с согреванием прошел удачно,
требуется довольно низкая комнатная температура: 10—18°С.
3. Установление повышенной возбудимости в области нервных окончаний.
М. sartorius устанавливают, как на рис. 41. Сначала определяют пороги
раздражения на проксимальном участке. Затем передвигают электроды на полсантиметра
и вновь определяют пороги. Снова передвигают к середине мышцы на
полсантиметра и снова определяют пороги. Так исследуют всю мышцу. Затем составляют
диаграмму изменения возбудимости по всей длине мышцы.
Далее определяют также пороги раздражения на разных участках m. sartorius,
который пролежал в растворе хлористого магния до потери реакции на нервное
раздражение. В обоих случаях в средних частях мышцы пороги раздражения
окажутся ниже, чем в проксимальном участке.
Литература
(.Mangold Е., Erg. Physiol., 21, 360, 1923.
2а. В е г і t о f f J., Erg. Physiol., 23, 33, 1924.
26. Owerton E., Pfl. Arch., 92, 115, 146, 1902; 105, 176, 1904.
ВОЗБУДИМОСТЬ КАК ОСНОВНОЕ СВОЙСТВО МЫШЕЧНОЙ ТКАНИ
65
3. Kiihne W., Arch. Anat. Physiol., стр. 564, 1859.
4. Brown-Sequard, С. г. soc. biol., стр. 195, 1849.
5. Bidder F., Arch. Anat. Physiol., стр. 67, 1865.
6. Wiechmann E., Pfl. Arch., 182, 74, 1920.
7. К n h n e W., Arch. Anat. Physiol., стр. 542, 1860.
8. Bernard CL, Lemons sur les effets des substances toxiques, Paris, 1857.
9. L a n g 1 e у J. N., J. Physiol., 36, 347,{1907; Das auton. Nervensystem, Berl., 1922.
10. Keith LucasJ. Physiol., 35, 103, 1906.
11. P о li t z e r, J. Physiol., 7, 274, 1886.
12. E d s trom, Skand. Arch. Physiol., 41, 101, 1921.
13. Langley, Proc. Roy. soc, 78, 186, 1906.
14. В о e к e J., Int. Monatsschr. Anat. Physiol., 28, 1911; Erg. Physiol., 19. 448,1921.
15. Б e p и т о в И. и Я ш в и ли, Чвени мецниереба, на грузинск. яз., № 3, 1923.
16. D u Bois-Reimond Е., Unters. ub. d. tier. Elektr., 1, 258, 1871.
17. F г е у von, Arch. Anat. Physiol., стр. 43, 1883.
18a. Hermann L., Handb. Physiol., Bd. 1, Abt. 1, 1879.
186. Biederman h, Elektrophysiologie, стр. 157, Jena, 1895.
19. G i 1 d em e i s t er M., Z. Biol., 62, 358,, 1913.
20. L a p i с q u e L., С r. soc. biol., 67, 208, 1909; L'excitabilite en fonction du temps
Paris, 1926. ' .
21. Pfluger E., Unters. lib. d. Physiol. U Elektrotohus, стр. 446, Berlin, 1850.
22. Лазарев П., Исследования по ионной теории возбуждения; 1. ч. Изд.Моск-. на-
учно-иссл. инст., 1916; Ионная теория возбуждения, 1923; Функция нервной
системы и ее старение с физико-химической точки зрения. Доклад, 1932.
23. Bourgignon G., С. г. Acad. Sci., 178, 1452—1454, 1921.
24. F г ё d ё г і с q H., Physiol. Rev., 8, 501, 1928.
25. Nernst W., Pfl. Arch., 122, 276, 1908; Sb. K. Preuss. Ak. Wiss., 1,3, 1908.
26. L о e b J., Pfl. Arch., 116, 198, 1907.
27. H e i d e n h a i n, Plasma und Zelle, Jena, 1907.
28. H e r t w i g G., Handb. norm. path. Physiol., 1, 580, 1927.
29. FromherzJ. Physiol., 79, 67, 1933.
ЗО.Мооге A. R. a. Brucke E. T h., Pfl. Arch., 228, 619, 1931.
31. Л а п и к Л., Доклад на XV междунар. физиол. конгрессе, 1935.
32. Buchthal F г., Skand. Arch. Physiol., 70, 199, 1934.
33. М a g п і t z к у A. N. u. Mushejew W. A., Pfl. Arch., 232, 605, 1933.
34. Blair E. A. a. E r 1 a n g e r J., Am. J. Physiol., 106, 524, 1933.
35. Buchthal Fr. u. J. Lindhard, Сборник «Проблемы физиологии нервной
системы и поведения», поев. проф. Бериташвили, 1935.
36. Энгельс Фр., Диалектика природы. Арх. Маркса и Энгельса, кн. II,
стр. 49—50, 1925; Анти-Дюринг, 6-е изд., 1934.
37. Г и л л А. В., Физиол. журн. СССР, 19, 115, 1935.
38. Купалов П. С, Сб. докл. 6-го Всес. физиол. съезда в Тбилиси, стр. 110, 1937.
39. G 6 р f е г t Н. u. Schaefer H., Pfl. Arch., 239, 597, 1937.
40. В у л И. М., Невропат, и психиатрия, 6, вып. 11, 51, 1937.
41. М і n e s G. R., J. Physiol., 37, 408, 1908.
42. Lapique L., Cold Spr. Harb. Symp. Quant. Biol., 4, 98, 1936.
43. Цкипуридзе Л., Тр. Инст. Физиол. им. Бериташвили, 4, 460, 1941.
44. Немилов А. В., Успехи соврем, биол., 9, 331, 1938.
45. Б е р и т о в И., Физиол. журн. СССР, 27, 147, 1939.
46. Hoorweg L., Pfl. Arch., 52, 87, 1892.
Al. Weiss G., С. г. soc. biol., 53, 253, 1901.
48. У ф л я н д Ю. M.j Теория и практика хронаксиметрии, Ленинград, 1941.
49. Опарин А. И., Новый мир, № 7, стр. 233, 1940.
^ И. С. Беритов
Глава III
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
Изменения формы мышцы при сокращении. Когда мышца приходит
в возбуждение, она претерпевает следующее механическое изменение:
длина мышцы уменьшается, а толщина увеличивается. Это проявление1
мышечной деятельности называется сокращением. Сокращение
наступает в каждом мышечном волокне, и изменение формы всей
мышцы является суммой изменений всех сокращенных мышечных
волокон. Объем мышцы при этом почти не меняется, если не считать
очень незначительного уменьшения. Но это уменьшение не связано
непосредственно с самим процессом сокращения. Оно обусловливается
главным образом химическими процессами расщепления, которые имеют
место как во время возбуждения, так и во время сокращения (Эрнст,
74; Мейергоф, 40, 78).
Изменение миофибриллей во время сокращения. Укорочение и
утолщение мышечных волокон во время сокращения является результатом
аналогичных изменений імиофибриллей: новейшими микроскопическими1
исследованиями на живых мышцах в покое и в сокращенном состоянии
было установлено, что при сокращении существенно изменяется
анизотропное вещество, а не изотропное. Так, Гюртле (2), исследуя
микроскопическую картину живой мышцы насекомого {Hydrophilus)y ясно
показал,.что высота анизотропного вещества уменьшается почти вдвое.
Изотропное же вещество остается почти без изменения или даже
увеличивается. При этом во время сокращения объем анизотропного
вещества остается без изменения (рис. 43). Что же касается
преломляемости, она оказалась во время сокращения более или менее одинаковой
в изотропном и анизотропном веществах вследствие уменьшения ее в
анизотропном веществе.
Такие же результаты были получены на живых мышцах лягушки
Гольцем (58). Он определил, что в покое отношение изотропного
вещества к анизотропному составляет 0.9—1.0. При изотоническом
сокращении это отношение нарастает вследствие уменьшения высоты
анизотропного вещества до 1.32. С помощью более
усовершенствованной техники было установлено, что это отношение всегда больше-
единицы. Так, в пучке волокон m. semitendinosus лягушки в покойномі
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ ?7
и нерастянутом состоянии высота анизотропного вещества равняется
1.37 ;і, а изотропного — 0.87 ]і. При сокращении от воздействия
индукционного удара уменьшение длины миофибриллей происходит
исключительно за счет анизотропного
вещества, причем уменьшение высоты
анизотропного диска тем
значительнее, чем сильнее сокращение. Высота
же изотропного вещества не меняется
(Бухталь и Линдгард, 97).
Если мышца растянута и концы
се так фиксированы, что она не
может укоротиться, то наблюдаются
несколько иные изменения. При
растяжении анизотропное вещество
удлиняется в два раза сильнее
изотропного. При сокращении такой мышцы
укорачивается опять-таки
анизотропное вещество; изотропное же
вещество, наоборот, удлиняется (Бухталь
и Линдгард, 97) (рис. 44).
При сокращении скелетной мышцы
позвоночных также меняется
преломляемость анизотропного вещества.
Она уменьшается при сокращении
(Муральт, 59). При точном измерении
двоякой преломляемости живых
изолированных волокон было
обнаружено, что во время сокращения
двойная преломляемость уменьшается на
20—30% (Бухталь, 86). Вследствие
этого поперечная исчерченность во
время сокращения выступает хуже,—
она становится расплывчатой. Все
эти исследования на живых мышцах привели авторов к утверждению,
что сокращение мышцы осуществляется путем изменений в двоякопре-
ломляющем анизотропном веществе миофибриллей.
Первоначально строение миофибриллей изучалось на мертвых мышцах в условиях
фиксирования и окрашивания. Таким путем Энгельман (1) изучил мышцы того
же насекомого Hydrophilus. Подвергая гистологической обработке покоящиеся и
сокращенные мышцы, он обнаружил, что высота обоих веществ миофибриллей —
анизотропного и изотропного — во время сокращения уменьшается, но много
сильнее в изотропном вещестзе, чем в анизотропном. Общий объем того и другого
вещества при сокращении не меняется, но объем одного вещества —
анизотропного— увеличивался за счет изотропного. Преломляемость анизотропного вещества
при сокращении уменьшалась, а изотропного вещества, наоборот, увеличивалась.
Поэтому при обычном свете почти все части элемента миофибриллей оказывались
более или менее однородными.
Рис. 43. Волна сокращения в свежем
изолированном мышечном волокне из
разгибателя задней ноги Hydrophilus
в поляризованном свете.
Увеличение в 250 раз. В средней части волна
сокращения: светлые поперечные полоски —
изотропное вещество, темные полоски —
анизотропное. В покойных участках темные полоски
являются двойными. В сокращенном участке
темные полоски становятся более темными и
много тоньше, чем в покойных участках.
Мышечное волокно снабжено двумя двигательными
нервными окончаниями (N) (из Гюртле).
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Основные изменения мышечных элементов, умерщвленных во время сокращения,
хорошо видны на рис. 45. Здесь приводится боковая волна сокращения на мышце
Cassida equestris. На поверхности мышечного волокна оканчивается нервное
волокно. Как раз в этом месте сокращение максимально; темные полосы
представляют изотропное .вещество, а серые — анизотропное. Слева мышечное волокно
находится в покое. Очень хорошо выступают все переходные стадии сокращения
фибрилл ей.
Аналогичные результаты получил Франк (Frank, 29а), изучая гистологическое
отроение замороженной и уже мертвой мышцы лягушки (m. sartorius), а именно:
л замороженной, мышце в покойном состоянии отношение высоты изотропного ве-
I
I
ГШ
Ш
1
7
Рис. 44. Схематическое изображение отношения
анизотропного вещества к изотропному в живой мышце:
покойной, растянутой и сокращенной.
а — покой,, b — сокращение при фиксированных концах (изолет-
рическое сокращение).
7 — волокна в покойном состоянии; 2 — растянутые на 25°/0;
3—на 40°/0; 4—7—волокна в покое, растянутее на О 45% длины
в покойном состоянии (Бухталь, Кнаппейс и Линдгард).
щества к анизотропному равнялось 1.1—1.3. Если же мышца замораживалась в
момент изометрического сокращения, это отношение высот уменьшалось до 0.65
вследствие увеличения анизотропного вещества за счет изотропного. При замораживании
же мышцы в период изотонического сокращения оно уменьшалось до 0.5—0.45.
Как видно, структурные отношения между анизотропным и изотропным
веществами на мышцах, умерщвленных во время сокращения, оказываются совершенно
иными и даже противоположными тому, что наблюдается на живых мышцах во
время их сокращения.
О природе миофибриллей. На основании некоторых фактов нужно
считать миофибрилли неживым органическим образованием. Это было
предположено нами первоначально на основании некоторых косвенных
соображений (11), Теперь оно нашло подтверждение в ряде фактов.
Доказано, например, что мышца, окончательно лишенная возбудимости
под влиянием таких сильных наркотических веществ, как уретан,
хлороформ, не теряет еще сократительной способности. Она отвечает на
воздействие слабых растворов кислоты контрактурой такой же высоты
и длительности, как и нормальная мышца (рис. 66). В лаборатории
Бете это было показано в отношении как скелетной, так и гладкой
мышцы (13, 14, 15). Очевидно, миофибрилли не принадлежат к
живой системе, в противном случае сократительная способность -мышцы
должна была бы исчезнуть под влиянием наркоза.
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
69
Чрезвычайно характерно, что при микроскопическом исследовании
тренированных -мышц, утолщенных благодаря упражнению,
обнаруживается сильное увеличение саркоплазмы, т. е. той части мышечного
волокна, которую імы считаем живой, в то время как толщина миофи-
бриллей остается без изменения. Некоторыми авторами отмечается
увеличение числа миофибриллей, что, вероятно, обусловлено
образованием новых фибриллей из деятельной саркоплазмы. Следовательно,
сами миофибрилли, как неживая система, не способны к росту под
Рис. 45. Боковая волна сокращения в мышечном волокне Cassida
equestris. Объяснения в тексте (рисунок Роллета из гистологии
Максимова).
влиянием упражнения (Иамин, 37; Штейнгауз, 45; Морпурго, 46;
Зиберт, 47).
Самое лучшее доказательство того, что миофибрилли не являются
живым образованием, было дано Be б ером (133). Он показал, что
нити белка миозина, анизотропного сократительного вещества
миофибриллей, образуемые при впрыскивании в дестиллированную воду
раствора миозина, воспроизводят с большой полнотой оптические и
молекулярно-структурные свойства миофибриллей мышечного волокна:
двойное лучепреломление и типичную рентгеновскую диаграмму
мышечного волокна, которая свидетельствует о резко выраженном
удлиненном строении молекул миозина. Кроме того, эти нити миозина
обладают определенным ферментативным свойством: расщепляют аде-
нозинтрифосфорную кислоту, т. е. являются ферментом — аденозинтри-
фосфатазой. Все эти свойства миозин легко теряет одновременно с
исчезновением оптических свойств, например, при согревании выше
40° С. Вся эта характеристика 'искусственно приготовленных миозино-
вых нитей чрезвычайно уподобляет их анизотропному веществу
миофибриллей (Энгельгардт, 134). Итак, следует считать экспериментально
установленным, что сократительное вещество миофибриллей, подобно
70
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
сухожильным, соединительнотканным и другим фибриллям, назначение
которых состоит в выполнении чисто механической задачи,
представляет собой неживой продукт протоплазмы.
Многочисленные теории сокращения рассматривают сократительное
вещество как неживой продукт. Мы ниже приводим ряд теорий
сокращения: Энгелыана, Бернштейна, Гилла, Гарнера,
К. Мей'ера, С ц е н т - Г ь о р г и, исходящих из таких
физико-химических положений, которым подчиняются простые соединения
неорганического или органического характера.
Происхождение сокращения. Энгельман первый дал
своеобразно обоснованную теорию сокращения (1880). По ?нгельману (1),
сокращение імышцы происходит вследствие перехода воды из
изотропного вещества в анизотропное, причем процесс перехода воды
объясняется Энгельманом следующим образом. Мышца при
сокращении развивает тепло; под влиянием тепла анизотропное вещество
подвергается разбуханию, подобно другим двоякопреломляющим
волокнам (сухожильные нити, хрящ, волос), при этом оно отвлекает воду
из изотропного вещества и разбухает в поперечном направлении.
Теория Энгельмана в свое время пользовалась большим
распространением. Сомнению подвергали только причину разбухания.
До самого последнего времени многочисленные авторы признавали
ближайшей причиной сокращения те кислоты, которые образуются
в імышце в связи с ее возбуждением: молочную и фосфорную.
Образование кислот, в частности молочной, было известно уже давно. Еще
Энгельман знал об этом. Но он предпочел остановиться на
тепловом происхождении мышечного сокращения. Впервые Флетчер и
Гопкинс (12) в начале этого столетия (1907) указали на молочную
кислоту, как на причину сокращения миофибриллей. Именно они
нашли, что мышца может сокращаться и в бескислородной среде,
причем образовывается молочная кислота из гликогена. Они же
установили, что кислород нужен для восстановления молочной кислоты
в гликоген. Если после утомления мышцы в бескислородной среде дать
ей отдых в такой же среде, то это не ведет к восстановлению
сократительной функции мышцы. Но если отдых производить в кислородной
среде, то восстановление сократительной функции произойдет; при этом
молочная кислота обратно перейдет в гликоген. Вот тот основной
факт, который послужил Флетчеру и Г о п к -и -н с у для гипотезы
о значении молочной кислоты для сократительного процесса.
Благодаря работам Мейергофа (5) и Гилла (30) это
воззрение было сильно подкреплено. Мейергоф произвел
количественное определение молочной кислоты при разных условиях
мышечной работы. Так, например, во время покоя мышца лягушки (г. tern-
poraria) образует в течение одного часа 0.02% молочной кислоты. При
одиночных (изометрических) сокращениях образуется за 7 минут
0.226% молочной кислоты; при коротком тетаническом (изотоническом)
сокращении до одной минуты она достигает 0.13%; при длительном
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ 71
тетаническом раздражении до 'полного утомления образуется 0.23%.
Зти количественные определения производились в условиях,
исключающих возможность окисления молочной кислоты, т. е. в анаэробных
условиях.
Одновременно Мейергоф изучал роль фосфорной кислоты. На
основании этих исследований он представлял себе роль молочной и
фосфорной кислот в тако:м виде: (молочная кислота действует прямо
на сократительное вещество; фосфорная же кислота участвует ;в
образовании молочной кислоты из гликогена, который представляет
главный источник энергии (ом. в X главе).
Это воззрение Мейергоф а скоро пришлось оставить. Работами
Э м б д е н а было доказано, что в мышце при раздражении отдельными
индукционными ударами прежде всего образуется фосфорная кислота.
Отсюда он делает заключение, что фосфорная кислота является
необходимым веществом для начала сократительного процесса (6).
Вскоре после Эмбдена П. Эгглетон и Г. Эгглетон (1926)
(41) показали, что в период возбуждения в мышце происходит
расщепление находящегося в ней фосфагена на креатин и фосфорную
кислоту и что это расщепление происходит раньше образования молочной
кислоты. Отсюда эти авторы заключили, что начало сокращения мышцы
должно быть обусловлено расщеплением фосфагена на креатин и
фосфорную кислоту.
Это предположение нашло фактическую поддержку в последующих
исследованиях Мейергоф а (28) и его сотрудников. Они
установили, что в мышце три четверти фосфорной кислоты связаны с
креатином в виде креатинофосфорной кислоты. Некоторые авторы называют
ее фосфагеном.
Формула фосфагена такова:
/ОН
/NH —Р^О
C = NH \)H
I
N — СНоСООН
сн3
В период деятельности это вещество распадается на креатин и
фосфорную кислоту и сейчас же после сокращения восстанавливается
обратно, частью анаэробно, частью аэробно. Чем скорее протекает
процесс, чем меньше хронаксия, тем больше этого вещества распадается.
Несколько позднее Лундсгаард (42) доказал прямыми опытами,
что мышца действительно может сокращаться без образования
молочной кислоты, а именно: он нашел, что в мышце, отравленной моноиод-
уксусной кислотой, при сокращении гликоген расщепляется только
до стадии гексозофосфатов, являющихся промежуточным звеном при
72
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
образовании (молочной кислоты. В такой мышце в связи с сокращением
усиленно расщепляется фосфаген, и сокращение вызывается все то
время, пока в мышце происходит образование фосфорной кислоты
вследствие распада фосфагена. После 100—120 одиночных
вздрагиваний мышца перестает сокращаться, но в то же время она перестает
образовывать и фосфорную кислоту вследствие исчерпания всего
запаса фосфагена.
В нормальной мышце, при тех же анаэробных условиях, когда
имеет место образование молочной кислоты, полное истощение
наступает іпосле 500 сокращений, и освобождается в 6 раз больше тепла,
чем в отравленной імышце. Если при этом образуемая молочная кислота
устраняется из мышцы путем растворения в окружающем растворе,
полное .истощение наступает еще позднее, после 1500 сокращений; при
этом выделяется тепла в 15 раз больше, чем в том случае, когда
мышца лишена возможности образования молочной кислоты.
Характерно, что если в рингеровскую жидкость, в которой находится мышца,
прибавляется глюкоза, то работоспособность мышцы удерживается
еще дольше: она может дать до 2000 сокращений; при этом выделяется
больше молочной кислоты, чем в предыдущем случае, и освобождается
в 20 раз больше тепла, чем в отравленной мышце. Ясно поэтому, что
образование молочной кислоты играет существенную роль в работе
мышцы в анаэробных условиях (Гилл, 104).
Несмотря на то, что с точки зрения энергетики распад гликогена
до молочной кислоты и фосфагена до фосфорной играет существенную
роль в мышечном сокращении, в настоящее время ни молочная, ни
фосфорная кислота не считаются теми веществами, которые
обусловливают начало сократительного процесса. В 1929 г. Ломан (105)
открыл в мышце аденилпирофосфат, представляющий собой соединение
адениловой и пирофосфорной кислот. Формула аденилпирофосфорной
кислоты такова:
N = C-NHo
I I
НС С—N
I
N
сн он он он он он
— C — N —СН —СН —СН —СН—СНГ—О—Р —О-Р —О —Р-ОН
0 0 0 0
По новейшим исследованиям Парнаса и сотрудников (99), М е й е р-
г о ф а и Ломана (112) и др., распад этого аденилпирофосфата
является одним из первичных процессов, дающим энергию для
сокращения. Правда, сам аденилтрифосфат едва ли вызывает сокращение,
ибо, как показал Энгельгардт (134), под влиянием этого
вещества сократительное вещество миофибриллей — миозин — не
сокращается, а расслабевает. Но при расщеплении, его происходит осво-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
7.Ч"
бождение большого количества энергии, с образованием адениловой-
и пирофосфорной кислот. А это влечет за собой сложный
биохимический процесс, который и должен привести к сокращению. Именно,,
при распаде образуется адениловая и пирофосфорная кислоты..
Фосфорная кислота частично передается гликогену и принимает
участие в его распаде путем образования промежуточных соединений —
фосфорных сложных эфиров. Сейчас же после расщепления
происходит восстановление аденилпирофосфата. При этом исчезают
адениловая и фосфорная кислоты. Для этого процесса фосфорная кислота
берется также от фосфагена с освобождением креатина.
Полное восстановление аденилпирофосфата происходит за счет-
энергии, освобождаемой как при гликогенолизе, так и при окислении
молочной кислоты.
Связанная с углеводами фосфорная кислота в конце концов
освобождается в результате расщепления этих углеводов и частью
соединяется с адениловой кислотой, а частью с креатином — для
восстановления фосфагена.
При работе мышц происходит усиленный распад и синтез
фосфагена и аденилпирофосфорной кислоты. Это — постоянный процесс,,
сопровождающий мышечную деятельность. При этом существует
определенная связь между величиной произведенной работы, распадом
фосфагена и аденилпирофосфорной кислоты и образованием фосфорной
кислоты (Мейргоф и Ломан, 112).
На основании новейших исследований роль аденилпирофосфата,
фосфагена и гликогена в сократительном процессе можно представить,
следующим образом.
Под влиянием процесса возбуждения происходит распад
аденилпирофосфата. Это ведет в присутствии гликогена к образованию
фосфорных эфиров. Этот распад аденилпирофосфата, может быть, и
обусловливает начало сократительного процесса.
В дальнейшем происходит восстановление аденилпирофосфата за
счет распада фосфагена и гликолитического процесса. Происходящий
при этом распад фосфагена и углеводов должен, в свою очередь,
действовать на миофибрилли, усиливая и удлиняя тот сократительный"
процесс, который был вызван в самом начале распадом
аденилпирофосфата. Предполагается при этом, что главную роль играют
фосфорная и молочная кислоты. Именно их непосредственному действию на
миофибрилли приписывается сокращение последних. Почти все
известные теории сокращения исходят из этой точки зрения. Однако это
предположение в настоящее время у некоторых ученых вызывает
сомнение (Эрнст, 108).
Фосфаген был найден во всех мышцах позвоночных животных. Но-
количество его заметно вариирует: в красных мышцах он содержится
в заметно меньшем количестве, чем в белых (Крюгер, Дуспива
и Фюрлингер, 96). Фосфаген появляется в определенный период
онтогенеза, уже после того как эмбриональные мышцы приобрели-
74
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
способность сокращаться. У кроликов он возникает впервые на 20—
22-й день эмбрионального развития, именно в тот период, когда
кролики начинают давать на локальное раздражение локальные
рефлекторные реакции.
Количество фосфагена затем нарастает, но особенно сильно оно
возрастает после рождения, в период прозревания, когда вообще
функциональное состояние скелетных мышц сильно приближается
к взрослому состоянию (Коштоянц, 106).
Поперечнополосатые мышцы насекомых и других беспозвоночных
не содержат фосфагена ів виде креатинофосфорной кислоты. В них был
найден другого рода фосфаген, представляющий собой неустойчивое
соединение фосфорной кислоты и аргинина (Мейергоф и
Ломан, 53). В мышцах беспозвоночных существует аденилпирофосфат.
В поперечнополосатых мышцах речного рака аденилпирофосфат
расщепляется гфи работе на адениловую и пирофосфорную кислоты. При
восстановлении он синтезируется из этих продуктов распада (Ферд-
ман, 100).
Как указывалось выше, сокращение миофибриллей, по мнению
некоторых ученых, обусловливается, между прочим, действием молочной
и фосфорной кислот на миофибрилли. Поэтому много работ было
посвящено изучению искусственного действия этих и других кислот на
мышцы.
Копылов (7), Швенкер (8) и другие испытали действие многих веществ
на живую мышцу. Оказывается, что кислоты, проникшие внутрь мышцы путем
диффузии, вызывают сокращение.
Так действуют не только те кислоты, которые встречаются в мышце, —
молочная, фосфорная и угольная, — но и всякого рода органические и неорганические
кислоты, например, валериановая, соляная, серная.
Так же действуют щелочи и спирты. Значит, сократительное действие молочной
или фосфорной кислоты іне является специфической особенностью этих кислот. Но
нормальное сокращение, повидимому, связано с действием именно вышеуказанных
кислот.
В последнее время было выяснено, что напряжение, развиваемое мышцей под
действием кислоты, очень мало. Оно почти в пять раз меньше, чем при тетаниче-
ском сокращении, которое вызывается частыми индукционными ударами (Бете, 10).
Из этого, а также из других наблюдений Бете заключает, что тетаническое
сокращение нельзя считать результатом влияния молочной или фосфорной кислоты.
Таким результатом может быть признана только контрактура, которая наступает от
сильных и частых раздражений и во время которой развивается точно такое же
малое напряжение, как от прямого действия молочной кислоты. Тетаническое
сокращение должно вызываться, по мнению Бете, под влиянием какого-то другого
продукта обмена веществ, а не кислоты. Но едва ли можно сравнить действие
кислоты, проникшей ,в мышцу путем диффузии, с тем действием, которое кислота
производит, когда она возникает в самих мышечных волокнах. По этому поведу
Г ил л, один из выдающихся физиологов нашего времени, делает такое остроумное
замечание: «Мышца представляет собой высокоорганизованную структуру, поэтому
сравнивать 'влияние кислоты, омывающей целую мышцу, с действием ее в живом
механизме, примерно настолько же логично, как сравнение влияния бензина,
вылитого на автомобиль, с его действием в моторе» (30).
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
75
Происхождение расслабления сокращенной мышцы. Если
предположить, что сокращение основывается, с одной стороны, на
расщеплении аденилпирофосфата, фосфагена, гликогена, а с другой — на
появлении адениловой, фосфорной и молочной кислот, то прекращение
сокращения должно обусловливаться - прекращением расщепления
названных веществ и исчезновением названных кислот. Наблюдения
показывают, что при нормальном кровообращении кислоты исчезают
вслед за образованием. Во время коротких тетанусов нельзя даже
открыть повышения концентрации водородных ионов, которое
указывало бы на накопление той или другой кислоты. Даже при крайнем
утомлении повышение концентрации водородных ионов в мышце
невелико. Повидимому, это происходит оттого, что сейчас же по
прекращении возбуждения аденилпирофосфат и фосфаген перестают
расщепляться. В связи с этим прекращается и расщепление гликогена.
Далее очевидно, что еще во время сокращения, сейчас же после
образования кислот, адениловая кислота восстанавливается обратно в
аденилпирофосфат, фосфорная кислота также сейчас же исчезает,
главным образом путем восстановления обратно в фосфаген и в аде-
нилопирофосфат; наконец, молочная кислота исчезает путем
восстановления в гликоген и 'нейтрализации.
По Мейергофу общее количество щелочных солей
(бикарбонатов и фосфатов) недостаточно, чтобы объяснить нейтрализацию
больших количеств кислоты, наблюдаемую при крайнем утомлении.
Поэтому было сделано предположение, что в мышце имеется сильный
буфер из натриевого (или калийного) соединения белка, обладающего
способностью нейтрализовать кислоту. Образуемые при этом фосфаты
и лактаты уже не способны вызывать сокращение. В изолированной
мышце при отсутствии кровообращения запасы буферных веществ
мышцы быстро приходят к концу. Вследствие этого при длительном
раздражении в мышце накопляется большое количество свободной
молочной кислоты; она остается и после раздражения; поэтому
сокращение не должно прекращаться вместе с раздражением. Кроме того,
по Э м б д е н у, усиленное расщепление гликогена до молочной кислоты
происходит также некоторое время после раздражения. Вследствие
этого мышца может сокращаться, пока в ней происходит образование
молочной кислоты.
По окончании сокращения мышца достигает длины покойного
состояния. Это происходит и в том случае, если на мышцу не
действует никакая растягивающая сила, даже собственный вес, т. е., если
она лежит в горизонтальном положении. Значит, удлинение
сокращенной мышцы в связи с прекращением раздражения является активным
процессом. Удлинение не наступает только в том случае, если
мышечные волокна сократились значительно более, чем на одну треть. Тогда
наступает определенное структурное изменение внутри волокна,
вследствие чего мышца застывает в состоянии укорочения (Рамзей и Стрит,
118). Об активной природе расслабления сокращенной мышцы говорят
76
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
также наблюдения над белком миозином, которому приписывается
сократительная функция. Под влиянием аденозинтрифосфорной кислоты
миозиновые нити удлиняются почти вдвое (Энгельгардт). Повидимому,
когда ослабевает .процесс расщепления аденилпирофосфата и верх
берет его синтез, тогда проявляется его расслабляющее действие на
анизотропное вещество миофибриллей.
Объемніые изменения мышцы в связи с химическими процессами.
Мейергоф и его сотрудники изучали объемные изменения мышцы
при сокращении с точки зрения участвующих в них химических
процессов. Мышца при одиночном сокращении претерпевает небольшое
объемное изменение: объем ее уменьшается. Это уменьшение при
одиночных изометрических сокращениях составляет от 1.5ХЮ"5 до
2.5 X 10 "5 см3 на 1 г веса мышцы. При изотонических сокращениях
изменение объема несколько меньше. При ряде следующих друг за
другом сокращений объем мышцы уменьшается еще больше, ибо
каждое последующее уменьшение объема накладывается на
предыдущее. Но при длительных тетанических раздражениях происходит
сначала уменьшение, а потом, наоборот, увеличение объема: вначале
объем возвращается к исходному, а затем превышает его (Гарт-
ман, 54).
В лаборатории Мейергофа подробно изучали причину
объемных изменений мышцы при сокращениях. Было установлено, что
уменьшение объема обусловлено распадом фосфагена и аденилпиро-
фосфорной кислоты, ибо появление продуктов их ферментативного
расщепления в жидкости производит уменьшение объема жидкости..
Увеличение же объема обусловлено появлением молочной кислоты
в результате ферментативного расщепления гликогена и
восстановленного фосфагена. Если устранить появление молочной кислоты
отравлением моноиодуксусной кислотой, то уменьшение объема остается и
в промежутках между раздражениями (40, 78).
Теории мышечного сокращения. По мнению Бернштейна (9) и
других авторов, кислота действует на миоф-ибрилли путем изменения
поверхностного натяжения между анизотропным веществом и
саркоплазмой. Под влиянием кислоты это натяжение возрастает; поэтому
высота анизотропных слоев должна уменьшаться, и сообразно
увеличивается толщина их, причем объем данных слоев остается без
изменения. По этой теории изотропные слои не должны меняться при
сокращении. Это представление хорошо согласуется с гистологическими
наблюдениями на живых мышцах.
Г и л л (30) высказывается против роли поверхностного натяжения
в сокращении мышцы. Он рассматривает мышцу как вязкое и упругое
тело, аналогичное резиновой трубке, наполненной густой патокой.
Г и л л находит, что еще раньше механического эффекта,
регистрируемого миографом, в мышце происходит внезапное и быстро проходящее
остудневание определенных составных частей мышечного волокна
в результате действия кислоты. Это внутреннее изменение мышечного
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
77
вещества внешне проявляется с некоторой задержкой, ибо нужно время
на то, чтобы внутреннее напряжение, произведенное остудневанием,
преодолело сопротивление вязкости и произвело необходимое изменение
формы мышечного волокна: укорочение одних участков, удлинение
других. Г и л л подробно развивает свою гипотезу и находит, что многие
характерные особенности теплообразования імышцы во время пассивных
и активных изменений хорошо объясняются именно с его точки зрения,
по которой мышца является упруго-вязкой системой.
О природе самого сократительного процесса существует много
гипотез. Хорошо разработанная гипотеза принадлежит Гарнеру (39); Он
кладет в основу ее то положение, что сократительный процесс связан
с двойным светопреломлением определенных сегментов миофибриллей,
т. е. анизотропного вещества. Двойное преломление свидетельствует об
определенном расположении молекул в этих сегментах, характерном
для жидких кристаллов. Г а р н е р утверждает, что определенные
сегменты в действительности состоят из жидких кристаллов протеина,
жирной кислоты, лецитина, холестерина и других веществ этого рода.
Он обращает внимание на недавние исследования структуры жидких
кристаллов, в которых оказалось, что молекулы кристаллического
вещества располагаются линейно правильными рядами, причем каждая
молекула кристалла граничит с жидкостью. В этих жидких кристаллах
расстояние между молекулами кристаллического вещества много
больше, чем в твердых кристаллах того же вещества. Отсюда Г а р н е р
заключает, что, когда концентрация кислоты нарастает на поверхности
анизотропного вещества, имеющего линейное кристаллическое
строение, отдельные молекулы кристалла становятся ближе одна к другой,,
¦и таким образом происходит укорочение самих кристаллов, а значит,
всего анизотропного вещества. Автор.приводит разные доводы в пользу
своей теории.
Как указывает Фултон (38), многие факты сократительного
процесса мышцы хорошо согласуются именно с этой гипотезой Г а р н е р а.
Но мне кажется, что более новая теория сокращения, которая
принадлежит К- М е й е р у (57), является в настоящее время наиболее
обоснованной. Изучая тонкое строение, крепость ц сократимость
животных тканей, Мей ер пришел к заключению, что белковые сократи-.
тельные элементы миофибриллей, подобно другим сложным
органическим образованиям, составлены из вытянутых в длину молекул. Этим
белковым телом, очевидно, является миозин, который обладает
двоякой преломляемостью, подобно анизотропному веществу миофибриллей.
На том основании, что степень двоякой преломляемости уменьшается
при сокращении, а также по характеру рентгенограмм сокращенной
мышцы, М е й е р заключил, что мицеллы сократительного вещества
(объединенный комплекс молекул) в расслабленной фибрилле
растянуты и располагаются параллельными пучками, а при сокращении бнй
сгибаются и теряют параллельное расположение. Этим сгибанием,
собственно говоря, и обусловливается, по мнению М е й е р а, сокращение
78
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
-/ЯГ-
С/ *#№¦}
/ — \ _
щ%сі -г
* * л. '
tStS\
4
СОвН
о** Л.
Н-ис-яая среда
мышцы. Это скрючивание мицелл М е й е р объясняет следующим
образом. Известно, что в водном растворе аминокислот существуют так
называемые цвиттерионы, в
которых конечные аминные
и карбоксильные группы,
будучи заряжены
противоположным электричеством,
притягиваются друг к другу, образуя
внутримолекулярную соль. В
связи с [этим притяжением
конечных групп происходит
сгибание всей молекулы
аминокислоты. Благодаря
присоединению кислоты к аминно
группе или щелочи к
карбоксильной группе, цвиттерионы
превращаются в простые
ионы. Мейер полагает, что
аналогичным изменениям
должны подвергнуться двоя-
к опреломляющие мицеллы
миофибрилли, так как они
состоят из аминокислот. При
покойном состоянии эти
мицеллы вытянуты благодаря
ионизированному,
заряженному состоянию конечных
частей вследствие соединения
с кислотой или щелочью. При
этом двоякопреломляемссть
максимальна, Во время
сокращения мицеллы согнуты
благодаря образованию цвитте-
рионов путем * притягивания
друг к другу противоположна
заряженных конечных групп.
В связи с этим двоякопре-
ломляемость понижена. На
рис. 46 дается схематическое
изображение изменения
мицеллы белка по К. Meй[е'р!у.
Этому изменению может быть
уподоблен процесс сокраще-
^щ
Изоэяеятричесяая точке
Щ
слои
щелочная ере fa
Рис. 46. Схема изменения длины белковой
мицеллы в зависимости от изменения концентрации
водородных ионов среды. ¦¦¦¦
В кислой среде расстояние между функциональными груп"
нами белковой молекулы (аминной и карбоксильной)
равно 15.5 А (ангстрем), а в изоэлектрической точке—4 А-
Кроме того, схема показывает, что в кислой среде
ионизируется аминная группа, а в щелочной—карбоксильная.
Вокруг ионизированных групп образуется водная оболочка
(сольвэтаідионный слой). В изоэлектрической точке, т. е.
в нейтральном состоянии, в результате образования цвит-
терионов (внутримолекулярной соли) белок уже не имеет
ионизированных групп, равно как и слоев сольватации
(К. Мейер).
нйя анизотропного -вещества миофябрнялй. Согласно такому
представлению сокращение должно возникать каждый раз при нейтрализации
раствора, в который погружены мицеллы, а расслабление — вытя-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
79-
гивание — при подкислении или подщелачивании этого раствора. Как.
известно, внутри мышечных волокон содержится много больше калия,
чем в окружающей лимфе (Нейшлосс, 61; Гёбер, 62). Возможно, что
в покойном состоянии каждая імицелла защищена от скрючивания
присутствием ионов калия. С появлением фосфорной и імолочной кислот
происходит нейтрализация раствора, и імицеллы скрючиваются. В связи
же с устранением, этих кислот под влиянием буферных веществ
внутриклеточной среды імицелла вновь вытягивается.. Этим объясняется,,
новидимому, активное удлинение мышечного волокна после
сокращения.
Совершенно новая и оригинальная теория сокращения дается в
последнее время Сцент-Гьорги (135). Этот автор, в результате
тщательного изучения поведения імиозиновых нитей в растворах разного
электролитного состава и при действии ряда веществ, пришел к
выводу, что сложная белковая система миофибриллей должна состоять, по
крайней мере, из трех компонентов: актина, миозина и аденозинтрифос-
форной кислоты. Актин представляет добавочный белковый компонент-
миофибриллей, открытый в лаборатории Сцент-Гьорги. Этот
белок при определенной концентрации солей в результате ассоциации
глобулярных частиц переходит в фибриллярную форму, образуя
нити.
Физико-химические свойства комплекса, состоящего из актина и
миозина, называемого автором для краткости актомиозином, находятся в
исключительной зависимости от электролитного состава внешней среды..
При физиологических концентрациях калия, магния и кальция комплекс -
актомиозин находится в диссоциированном состоянии, в виде гидрати-
рованного геля. Малейшие сдвиги концентрации калия вызывают
быстрые изменения в физико-химическом состоянии актомиозина.
Понижение концентрации калия на 10% во внешней среде обусловливает
переход актина в фибриллярную форму, которая связывается с імиозином.
При этом нити актомиозина претерпевают сильные объемные
изменения, переходя в «сверхсокращенное состояние». Этот процесс является
обратимым. Повышение концентрации калия до первоначальной
величины обусловливает расслабление нити с одновременной
диссоциацией. Аденозинтрифосфорная кислота в процессе «сокращения» нитей
играет исключительную роль. В присутствии этого соединения
актомиозин не только коагулирует, но он отдает гидратационную воду,
сжимаясь до малого объема.
Роль актина по представлениям Сцент-Гьорги заключается в
том, что этот белок, благодаря легкому переходу из глобулярного
состояния в фибриллярное, дает с миозином такую систему двух
коллоидов, где особенно резко выявляются сократительные функции
миофибриллей. По мнению Сцент-Гьорги, актин образует длинные нити,
к которым только с одной стороны присоединяется імиозин. Получается
система, состоящая из двух стержней, скрепленных бок о бок друг с
другом. Если допустить, что в системе, образованной из двух стерж—
.-so'
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
.ней, один сжимается или оба они сжимаются, но не одинаково, то
малейшее изменение относительно объеліа одного из них вызывает
скручивание системы в полную окружность, с уменьшением длины
на 2/з- Это явление можно уподобить искривлению фанеры после
увлажнения.
Рис. 47. Одиночное сокращение m. gastrocnemius, pectoralis pars abdominalis и sar-
.tonus лягушки. Звезды обозначают добавочное поднятие от сокращения красных
волокон
, Таким образом, по Сцент-Гьорги, нужно допустить, что
сокращение миофибриллей обусловливается изменением концентрации
электролитов в саркоплазме. При этом аденозинтрифосфорной кислоте
приписывается специфическая роль в увеличении чувствительности белков
миофибриллей к сдвигам концентрации солей.
Одиночное сокращение. Когда на мышцу действует какая-либо
внешняя сила своим однократным изменением, как, например, одно
замыкание или размыкание электрического тока, мышца сокращается
.на определенную величину. Это сокращение в своем течении не
зависит от свойств раздражителя. Укорочение мышцы сначала происходит
медленно, затем быстрее и, наконец, при приближении к максимуму,
снова медленно.
Весь период от начала сокращения до момента максимального
укорочения у лягушечьей мышцы равняется 0.05—0.07 сек. Когда мышца
достигает максимума укорочения, она сейчас же начинает расслаб-
.ляться, т. е. удлиняться. Это удлинение также сначала происходит
медленно, затем быстрее и, наконец, снова медленно.
Продолжительность и высота одиночного сокращения очень сильно
меняются в связи с функциональным состоянием імышц. Мышца
сокращается тем скорее и тем интенсивнее, чем она больше отдохнула,
чем выше температура мышцы. Вообще говоря, когда течение физико-
.химических процессов в мышце затруднено, продолжительность со-
СОКРАТИТЕЛЬНАЯ ФУНКЦИИ МЫШЦЫ
81
кращения увеличивается, а высота его уменьшается. Обратная картина
получается при создании таких условий, которые благоприятствуют
течению физико-химических процессов в мышце.
На рис. 51 дается характерная картина изменений сокращения
m. gastrocnemius лягушки в зависимости от температуры. Например,
при температуре в 18° сокращение мышцы длится 0.16 сек., а при
38° —всего 0,09 сек.
Продолжительность одиночного сокращения у теплокровных
животных короче, чем у холоднокровных, но у одного и того же вида
животных в одних мышцах она больше, чем в
других. В красных мышцах, как m. rect.
abdom. лягушки, soleus кролика,
продолжительность одиночного сокращения много
больше, чем в белых мышцах, как m. sar-
torius лягушки, т. gastrocnemius кролика,
а именно, за счет удлинения нисходящей
фазы, на которой намечается новое
поднятие, отвечающее медленней
сокращающимся красным волокнам (рис. 47).
У беспозвоночных продолжительность
одиночного сокращения поперечнополосатой
мышцы такая же, как у позвоночных; из
жуков, например, у Hydrophilus в мышцах
конечности продолжительность 0.350 сек.,
у Dytiscus — 0.112 сек. (Роллет, 113). Но
она может быть гораздо длительнее, как,
например, у некоторых мышц клешни
ракообразных, а также много короче, как
например, у мышц, производящих движение
крыльев у насекомых. Продолжительность
одного сокращения у последних может
равняться 0.0033 сек. В клешне
ракообразных находят двоякого рода мышечные
единицы. Одни из них производят медленные
сокращения, а другие — быстрые, причем
порог раздражения медленных мышц ниже;
ими производятся все обычные движения
клешни. Порог раздражения быстрых выше;
быстрыми движениями осуществляется
раздавливание предметов, требующее большого напряжения (Пэйнтин, 109).
Продолжительность сокращения сильно іменяется в одной и той же
мышце в связи с онтогенезом мышц. В эмбриональном периоде и
в первое время после рождения длительность одиночного сокращения
всегда больше, чем в более позднем возрасте (Коштоянц и Рябиноз-
ская, 95) (рис. 48).
Как указывалось выше, мышца при сокращении не только укорачи-
6 и. С. Бернтов
Рис. 48. Кривая одиночного
сокращения m. gastrocnemius
кролика от непрямого
раздражения.
А — на 2-й день после рождения,
В—на 7-й, С—нз П-й, D — на
14-й и К —на 45-й день. Время —
в пятых секунды (Коштоянц и
Рябиновская).
82
ОБЩАЯ ФИЗИОЛ01ИН МЫШЬЧНОИ СИСТЬМЫ
вается, но и утолщается. Утолщение мышцы точно так же может быть
записано миографическим путем. Во время одиночного сокращения
кривая утолщения мало чем отличается от кривой укорочения.
Мышца как эластическое образование обладает упругостью.
Напряжение мышцы тем больше, чем больше нагрузка или вообще
растягивающая ее сила. Если концы мышцы не фиксированы и мышца
может укоротиться, то начальное напряжение мышцы, зависящее от
нагрузки, во время сокращения остается без изменения. Такое
сокращение называется изотоническим (рис. 49).
Рис. 49. Изотоническое сокращение мышцы.
М — мышца во время сокращения поднимает пишущий рычаг а и тяжесть Р,
которая привязана у самой оси. Рычаг записывает на кимографе изотоническое
сокращение (Бидерман).
Если же укорочение мышцы встречает препятствие в виде сильной
пружины или большого груза, которого она не может преодолеть,
тогда укорочения во время сокращения не происходит; меняется лишь
напряжение мышцы: оно увеличивается и проходит параллельно с
сокращением. Такое сокращение мышцы, при котором длина ее остается
без изменения, называется изометрическим (рис. 50).
Как чистое изометрическое, так и чистое изотоническое
сокращение — искусственные явления. В нормальном, неповрежденном
организме ни то, ни другое сокращения не имеют места. Каждый сустав
бывает со всех сторон прикрыт антагонистическими мышцами или их
сухожилиями. Поэтому каждая мышца представляет значительное
препятствие для свободной деятельности антагониста. Кроме того, когда
мышца сокращается, она производит известную работу, так как
приводит в движение какую-либо часть организма: поднимает ее или
переносит с одного места на другое. Таким образом, в нормальных
условиях сокращение мышцы не является чисто изотоническим, так
как мышца во время укорочения преодолевает известное
сопротивление; оно не может быть и чисто изометрическим, так как, несмотря на
сопротивление, мышца все-таки укорачивается.
СОКРАТИТКЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
«й
Скрытый период сокращения. Сокращение мышцы всегда
начинается спустя некоторое время после раздражения. То время, которое
проходит от момента раздражения до начала сокращения, называется
скрытым периодом сокращения. В этот период возникает
процесс возбуждения, а под влиянием его происходит усиление распада
аденилпирофосфата, фосфагена и гликогена, от чего начинается
сокращение. Так как сокращение записывается миографическим путем,
Рис. 50. Изометрическое сокращение мышцы.
М — мышца поднимает только короткое плечо рычага (erf), которое
вращается вокруг оси (с); е—-спиральная пружина препятствует
укорочению. Развиваемое при сокращении напряжение сдвигает
рычаг ей на небольшую величину. Но это движение записывается
на кимографе другим рычагом (а) в сильно увеличенном виде.
Выше приведены кривые изотонического (/. t.) и изометрического
(і. т.) сокращения. Первая дает ход укорочения, а вторая —ход
напряжения при почти неизменной длине. Волнистая линия
отмечает время в сотых долях секунды (Видерман).
то в скрытый период входит не только физиологический момент, но и
физический, как время, которое нужно для преодоления инерции
миографа и нити, связывающей миограф с мышцей.
Само собой разумеется, что физический момент скрытого периода
изменчив. Например, когда механический эффект мышцы записывается
капсулами Марея, скрытый период гораздо больше, чем при
регистрации его прямым миографом, т. е. путем нитей, блоков и рычагов, а при
фотографической регистрации сокращения мышцы этот момент
сводится к нулю.
Но и физиологический, истинный латентный период не постоянен.
Он сильно меняется в зависимости от функционального состояния
мышцы. Все условия, благоприятствующие обмену веществ, ускоряют
все жизненные процессы мышечной ткани. Так, повышение температуры
благоприятствует обмену веществ и этим ускоряет как возникновение
возбуждения и его проведение вдоль імышцы, так и обусловливаемый
им физико-химический процесс сокращения. Поэтому с повышением
6*
84
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
температуры скрытый период сокращения уменьшается. Утомление
сопровождается ослаблением обмена веществ в возбудимой системе, и
в связи с этим скрытый период возрастает. Особенно сильно влияет
обеднение мышцы водой: оно значительно удлиняет скрытый период
(Дюриг, 70).
Скрытый период реакции на икроножной мышце лягушки
составляет около 0.015 сек., если запись ведется прямым миографом при
комнатной температуре 18°. При температуре 38° скрытый период —
около 0.009 сек. (рис. 51). У красных, медленно сокращающихся
Рис. 51. Одиночное сокращение m. gastrocnemius лягушки.
Верхняя кривая записана при температуре мышцы в 38", а нижняя —
при 18'. Сокращение продолжается в первом случае 0.09 сек., а во
втором— 0.16 сек. Вертикальная линия обозначает момент раздражения.
Сокращение начинается в первом случае спустя 0.009 сек. от момента
раздражения, а во втором—-спустя 0.017 сек., внизу каждой кривой
волнообразная линия обозначает время: каждая волна равняется 0.01 сек.
мышц скрытый период больше, например, у m. hypoglossus он равня-
няется 0.3 сек. У теплокровных скрытый период сокращения короче, чем
у холоднокровных. Так, скрытый период m. gastrocnemius кошки равен
0.004 сек., а красной мышцы soleus — 0.01 сек. (Купер и Экклс, 55).
Скрытый период сокращения не одинаков даже в пределах одной
мышцы. В нервных участках он короче, чем в безнервных. Это
наблюдается даже в том случае, если мышца кураризована. Значит, в тех
участках імышцы, где возбудимость выше, где процесс возбуждения
протекает интенсивнее, там сокращение начинается раньше. Так, у
m. sartorius лягушки в нервных участках скрытый период равнялся
0.02 сек., а в проксимальном безнервном участке он составлял
0.025 сек. (Картозия, 79).
Во всех вышеприведенных случаях сокращение записывалось
миографическим путем на кимографе.
Если изучать одиночное сокращение мышцы фотографическим путем,
фотографируя самую мышцу во время сокращения, и этим путем
совершенно исключить физический момент, тогда скрытый период
оказывается значительно меньше. Так, на мышце лягушки он оказался
около 2.5 с (Pay, 49; Бете и Гаппель, 50). Но возможно, что в таких
случаях авторы имели дело не с сокращением мышцы, а с изменением
ее объема, которое наступает, в связи с возбуждением, еще раньше
сокращения (см. об этом явлении в гл. VI).
Тетаническое сокращение. Как было отмечено, одиночное
сокращение вызывается быстрым однократным изменением раздражающей
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
85
силы. Если это изменение повторится настолько быстро, что произойдет
во время сокращения, вызванного первым раздражением, то оно
вызовет прирост начавшегося сокращения. Если же за вторым
раздражением быстро последуют третье, четвертое и т. д., сокращение будет
увеличиваться с каждым новым раздражением до тех пор, пока
укорочение мышцы не достигнет предельной высоты; после этого
дальнейшие раздражения не прибавят ничего к высоте сокращения, они только
удлинят существующий эффект. Такое длительно протекающее
сокращение называется тетанусом, или столбняком. Тетанус бывает
полный и зубчатый. Если раздражающий агент повторяется с таким
интервалом, что каждое последующее раздражение наступает после
достижения вершины предыдущего сокращения, то каждое
раздражение вызывает новый подъем сокращения. Такой эффект называется
неполным или зубчатым тетанусом (рис. 52). В том же случае,
Рис. 52. Сокращение мышцы при разной частоте раздражения.
Сначала даны три одиночных сокращения, затем зубчатый тетанус, наконец,
полный тетанус. Внизу мышечной кривой идет линия, обозначающая .моменты
раздражений (Ферворн).
когда интервал настолько мал, что каждое новое раздражение
приходится раньше достижения вершины предыдущего сокращения,
последнее имеет сплошной характер, — оно протекает без зубцов. Такой
тетанус называется полным (рис. 52).
Как указывалось выше, продолжительность одиночного сокращения
зависит от температуры мышцы и вообще от ее функционального
состояния. Поэтому чем ниже будет температура мышцы или чем более
мышца будет утомлена, тем более редкое раздражение вызовет полный
тетанус. Необходимый для полного тетануса ритм раздражения
зависит также от физиологических особенностей мышцы. М.
gastrocnemius лягушки, например, дает полный гладкий тетанус при частоте
раздражений до 20—-30 в секунду, в то время как m. myoglossus
лягушки дает гладкий тетанус при 12 раздражениях в секунду. У
теплокровных животных и у человека полный тетанус получается при
частоте раздражений свыше 30 в 1 секунду, если только условия
обмена веществ были особенно благоприятны. Но и в самых благоприят-
86
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ных условиях полный тетанус получается при разных частотах. Так,
например, красная мышца soleus кошки дает сплошной изометрический
тетанус при 35 импульсах в секунду, а белая мышца gastrocnemius —
при 90 в секунду (Купер и Экклс, 55).
Тетаническое сокращение характерно отличается от одиночного тем,
что при одном и том же функциональном состоянии мышцы амплитуда
тетанического сокращения всегда больше одиночного. Более того, при
одиночном сокращении укорочение імышцы никогда не является
максимальным. Даже самое сильное однократное раздражение произведет
меньшее укорочение, чем слабое тетаническое раздражение.
Укорочение некоторых белых мышц во время тетануса в 2—3 раза больше
одиночного укорочения, а тетаническое укорочение красных — в
8—9 раз сильнее одиночного. Нельзя думать, что эта разница в
укорочении зависит от участия іразного количества мышечных волокон.
Индукционные удары большой интенсивности охватывают всю толщу
мышцы при однократном применении так же полно, как и при
многократном.
Точно так же напряжение мышцы при изометрическом тетанусе
значительно превосходит таковое при одиночном сокращении.
Напряжение нарастает с учащениемі раздражения. У теплокровных
животных оно достигает максимума при частоте около 60 о секунду.
В этом случае напряжение оказывается в 4 раза больше, чем при
одиночном сокращении (Купер и Экклс, 55).
Вопрос о происхождении разницы в высоте между одиночным
сокращением и тетанусом интересовал физиологов еще со времени
первой миографической записи мышечных сокращений. Гельмгольц,
открывший это явление, предположил, что при тетанусе имеет место
чисто механическое накладывание одного одиночного сокращения
на другое, что предыдущее сокращение является основанием, на
которое опирается последующее сокращение и потому может поднять его
еще выше. Это предположение было подтверждено опытами Фр'ея
(68), который нашел, что если создать препятствие расслаблению
мышцы, то рядом последовательных одиночных раздражений можно
довести высоту ее сокращения до той, которой она достигает при
тетанусе.
Однако этой внутренней или внешней поддержкой можно объяснить
только превосходство высоты сокращения при зубчатом тетанусе. При
частоте раздражения, когда тетанус является полным, высота
мышечного сокращения становится значительно выше, чем это могло бы быть
достигнуто только путем внешней поддержки (Вахгольдер, 69)
(рис. 53).
Для понимания этого явления нужно исходить из того факта, что
при полном тетанусе внутренние физико-химические процессы,
обусловливающие сокращение, протекают интенсивнее, чем при одиночном
сокращении или даже чем при зубчатом тетанусе. Нужно иметь
в виду, что при некоторых частых раздражениях процесс возбуждения
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ Щ
усиливается во время самого раздражения (Введенский, 101). Это
бывает, как мы увидим ниже, когда каждый последующий
индукционный удар попадает после предыдущего возбуждения в фазе повышенной
возбудимости. На этом основании іможно утверждать, что
сократительный процесс в миофибриллях под влиянием ряда быстро
следующих друг за другом импульсов возбуждения протекает интенсивнее,
чем от одного импульса или ряда редких импульсов. Вследствие этого
как высота мышечного укорочения, так и величина мышечного
напряжения при частых тетанических раздражениях должны быть выше, чем
от одиночного раздражения или ряда редких раздражений. Понятно,
что, когда после ряда раздражений сократительный процесс достигнет
Рис. 53. Зубчатый и полный тетанус при учащении
раздражения.
М. coracoradialts лягушки. В опыте А производится ряд одиночных
раздражений при постоянной повышающей поддержке, т. е. при
предварительном все большем и большем поднятии рычага миографа
леред каждым раздражением. В опыте В производится ряд
раздражений с нарастающей частотой. При редких раздражениях
тетанус достигает той высоты сокращения, какая наблюдается при
внешней поддержке. Но при дальнейшем учащении раздражения,
когда наступает полный тетанус, высота сокращения становится
значительно выше (Вахгольдер).
максимальной интенсивности, дальнейшие раздражения ничего не
-прибавят к высоте сокращения или напряжения, ибо последующие
импульсы возбуждения будут лишь в состоянии поддерживать
существующий максимальный сократительный процесс (рис. 52).
Однако если взять очень слабое, околопороговое тетаническое
раздражение, то можно получать небольшие тетанические эффекты, даже
ниже, чем при сильных одиночных сокращениях. Это особенно легко
наблюдать на толстых мышцах, как икроножная. При некоторых
слабых раздражениях, когда сокращается только один небольшой > пучок
мышцы, он не может укоротиться в полной мере, ибо он не в
состоянии потянуть за собой главную массу несокращенных мышечных
волокон в силу их тяжести и спаянности между собой.
Растяжимость и эластичность скелетной мышцы.
Поперечнополосатые скелетные мышцы являются упругими образованиями. Под
упругостью понимают способность деформированного тела возвращаться
в большей или меньшей степени к первоначальному своему состоянию
по удалении силы, вызвавшей деформацию. Упругость мышцы удобнее
всего изучать путем ее растяжения. Кривая растяжения мышцы с на-
88
ОБЩАЯ ФИЗИОЛОГИИ МЫШЕЧНОЙ СИСТЬМЫ
растающим грузом показывает, что при растяжении ее требуется все
большая и большая сила для достижения одинакового прироста длины.
Кривая имеет вид гиперболы с вогнутостью в сторону оси
напряжения (рис. 54) (Блике, 51).
Степень растяжения мышцы зависит также от времени действия
груза. Когда привешивают груз к мышце, последняя сначала
удлиняется сразу на определенную величину, но и потом она продолжает
удлиняться долгое время, только все меньше и меньше,—
асимптотически приближаясь к некоторой конечной длине, соответствующей
данному грузу. Чем больше груз, тем больше конечное удлинение
мышцы. Быстрая фаза удлинения длится 1—2 сек., медленная же фаза
может длиться несколько часов (рис. 56).
Рис. 54. Кривая растяжения и установка для изучения растяжения
с нарастающим грузом по Бликсу.
Между двумя стрйками (*?/?) горизонтально передвигается угольник (АВО). На вер
тикальной части последнего свободно висит мышца (М), прикрепленная к пишущему
рычагу (SS), который шарниром связан с горизонтальной частью (недалеко от ( ). По
пишущему рычагу передвигается стержень (rftf) с грузом на нижнем конце. Если
угольник (АВО) передвигать в направлении стрелки, то нагруженный стержень (dd) будет
придвигаться ближе к мышце, вследствие чего растяжение мышцы будет постоянно
возрастать. Сначала записывают, передвигая соответственно груз, кривую растяжения
покоящейся мышцы о, а, Ь> с, е, затем повторяют то же, приведя мышцу в состояние
тетануса посредством электрического раздражения, и получают кривую; Л, '", к (по
Бликсу из Ландуа).
Растяжимость разных мышц не одинакова. Красные мышцы
растягиваются больше белых, мышца с параллельными волокнами ¦—
больше мышц с перистым строением.
Общий характер кривой растяжения отдельных мышечных пучков
совершенно таков же, как у целой мышцы (Бухталь и Линдгард, 97).
Но в изолированных мышечных волокнах, если они не повреждены и
находятся в хорошем функциональном состоянии, кривая растяжения
при нарастающем грузе носит прямолинейный характер, т. е.
растяжение происходит по закону Г у к а (Зихель, 111). Отсюда следует,
что особенности кривой растяжения целой мышцы, должно быть,
обусловливаются особенностями строения целой адышцы: неодинаковой
длиной мышечных волокон, расположением их под углом друг к другу
и существованием соединительнотканных прослоек.
На рис. 55 даются кривые растяжения пучка мышечных волокон
при разных условиях. Сравнивая кривые 1 и «?, ясно видно, что при
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
89'
одной и той же нагрузке длина мышцы іменыііе при сокращении, чем
в покойном состоянии. Так, при нагрузке 200 г длина сокращенной
мышцы 13 мм, а покойной — около 17 мм.
По удалении нагрузки мышца как упругое или эластическое тело
имеет тенденцию возвратиться к исходной длине. Сначала это
происходит быстро-—в течение 1—2 сек., а потом все медленнее и медленнее —
в течение многих минут. Это последнее явление известно как
эластическое последействие. Но путем
эластического последействия мышца может не
всегда достигнуть исходной длины. В
определенных условиях она остается на
долгое время более или менее
удлиненной. Значит, мышца не обладает
совершенной упругостью. В существовании
остаточной деформации выражается
другое свойство мышцы — ее пластичность.
Эта остаточная деформация почти
отсутствует при кратковременных малых
действиях груза небольшой величины.
Она бывает выражена тем сильнее, чем
больше растягивающая сила и чем
дольше ее действие. Само начальное быстрое
укорочение выражено тем слабее, чем
дольше действие растягивающей силы
(рис. 56). Вот эти особенности
мышечной деформации указывают, что
состояние структурных элементов,
обусловливающих упругость мышцы,
существенным образом меняется от
продолжительности растяжения.
Некоторые авторы приписывают
эластические особенности мышечных
волокон миофибриллям (Зихель, 111). При этом в миофибриллях
эластичность анизотропного вещества считается большей по сравнению
с изотропным. Это заключение было сделано на том основании, что
в покойном состоянии анизотропное вещество каждого сегмента мио-
фибрилли от растяжения удлиняется вдвое больше, чем изотропное. Во
время сокращения растяжимость того и другого вещества почти
уравнивается. Предполагается, что это происходит благодаря повышению
эластичности изотропного вещества (Бухталь и Линдгард, 97). Видимо,
при сокращении изотропное вещество играет как бы роль буфера
против мгновенных изменений длины анизотропного вещества (Бухталь).
По исследованиям других авторов, эластичность мышцы должна
зависеть главным образом от сарколеммы и івообще соединительнотка-
ных элементов мышцы. Это доказывается таким образом. Раздавливают
изолированные мышечные волокна между двумя гладкими стеклян-
чг
Ш
?Оя*
0% 2і % 50% 75% ЮО%
Рис. 55. Кривые длины —
напряжения небольшого пучка из 12мышеч-
ных волокон m. semitendinosus
лягушки.
На абсциссе — длина мышцы в
миллиметрах и удлинение в процентах длины
покойного состояния, на ординате —
напряжение в миллиграммах.
1 — кривая длины — напряжения от
растяжения во время сокращения;
2 — мышца сначала растягивается, а
потом раздражается; 3 — растяжение
в покойном состоянии (Асмуссен).
'90 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ными пластинками; из них вытекает свернувшееся содержимое во-
.локна. Такие волокна, состоящие из одной сарколеммы, наполненной
прозрачной жидкостью, показывают почти такую же кривую
изменения напряжения в зависимости от растяжения, как нормальное
мышечное волокно или целая мышца (Раімзей и Стрит, 118).
Против такого утверждения говорит факт изменения эластичности
три функциональном изменении мышцы. Так, например, известно, что
* Л
J>art
1=&мм
/
I
Se
?" *' я' ю' 7г' /J' гг' isf г? гз* гУ г?1 гэ1 зг'
,:rfn^,H,^ іПіііііііі «¦t.ttH«».,t«^lilllllHIHltlHniffllMfrH4lllMMff/tMI»H»fltHil
PpLO.
іміиіііііиіі
L ~ 39aim
10' 12' 15' гі' гз' 25' 27' & 31' 33' 35' 37' 40'
fMniifiniiii!iiiti4HMiniiiiMiMiii4iiniM4fnyfiiitiittiji "luuii імшііііпіишііііміияіцчиіц
Рис. 56. Пластическое удлинение мышц при коротком и длительном растяжениях на
m. sartorius (верхняя кривая) и p. abdominalis (нижняя кривая).
Обе мышцы растягиваются сначала в течение 3—4 сек., а потом в течение 10 мин. В последнем случае.
после начального быстрого уцпичечия, мычгда продолжает удлиняться на всем протяжении этого периода.
По удалении тяжести первоначальное быстрое укорочение после длительного растяжения меньше.
чем после короткого. Наоборот, пластическое удлинение после длительного растяжения больше, чем
после короткого. В конце каждого опыта миэграф поднимается на одну минуту (звезда!), дабы
устранить и его собственную тяжесть (около 0.5 г). Восстановление длины ускоряется, но все же исходная
длина не достигается.
при окоченении мышцы теплокровного животного эластическая
растяжимость падает, и восстановление исходной длины после растяжения
становится более полным. То же бывает при контрактурах,
вызываемых под влиянием калия или тетано-токсина: в начальной стадии
развития контрактуры эластическая растяжимость несколько
уменьшается, а позднее — в стадии максимальной контрактуры — она
значительно падает, но исходная длина по удалении растягивающей
силы восстанавливается лучше (Рэнсон, 120). Утомленные мышцы
растягиваются сильнее свежих. Тетанически сокращенная мышца
растягивается заметно больше, чем несокращенная, и притом тем больше,
«чем сильнее сокращение (Блике, 51). Когда імышца сокращена от воз-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЙ МЫШЦ 91
действия кислоты и хлороформа, она растягивается сильнее, чем в
нормальном состоянии (Верзар, 89; Шлейер, 90).
Все эти факты указывают, что эластические свойства мышцы имеют
сложное происхождение: они определяются как эластическими
свойствами сарколеммы и соединительнотканных элементов, так и
эластичностью саркоплазмы и миофибриллей. Изменение эластических свойств
при функциональных изменениях должно зависеть главным образом от
изменения физико-химических свойств саркоплазмы и миофибриллей.
Растяжение мышцы, как и всякого другого упругого тела,
сопровождается необратимым выделением тепла. То же бывает при
расслаблении растянутой мышцы. Это, очевидно, обусловлено
превращением механической энергии в тепло вследствие трения в вязкой системе
мышцы. На этом основании Г и л л предполагает, что и при настоящем
сокращении мышцы затрата энергии вызывается причинами,
связанными с трением (30).
Активное напряжение без сокращения. В нормальном организме
всякая нагрузка, всякое растяжение конечности вызывает в мышцах
¦напряжение. Но это напряжение не обусловлено всецело действием
растягивающей силы или сократительным процессом, как это бывает на
изолированных мышцах. Установлено, что у теплокровных напряжение
мышцы обусловливается также действием центральной нервной
системы. Далее известно, что человек в состоянии произвольно
напрягать свои імышцы без заметного сокращения. Однако это произвольное
напряжение не вызывается в одной какой-либо мышце или в одной
синергетической группе их. Произвольное напряжение наступает сразу
во всех мышцах одного сустава или нескольких.
На человеке это явление было изучено экспериментальным путем.
У человека была ампутирована рука в области локтевого сустава по
способу Зауербруха. Сухожилия мышц предплечья были выведены
наружу для протеза. Пользуясь этими сухожильными концами, іможно
было регистрировать мышечное сокращение на кимографе. Когда
оперированному предлагали согнуть руку в предплечье, сгибатели плеча
сокращались, но при этом оставались мягкими, без признаков
напряжения; когда предлагали разогнуть руку, то сокращались разгибатели
также без затвердения. Но если предлагали напрягать мышцы плеча,
то все мышцы затвердевали без сокращения (Плаут, 16).
На нормальных людях не всегда удается наблюдать произвольное затвердение
'без сокращения. Можно вызвать изолированное сокращение без затвердения, но
затвердение обычно наступает вместе с сокращением (Зюснер и Гейнрих, 73).
И к с к ю л ю (29Ь) удавалось наблюдать произвольное затвердение после некоторой
тренировки; именно, он вызывал затвердение — активное напряжение в двуглавой
мышце плеча без сокращения ее или ее антагониста. Вообще легко удается
наблюдать непроизвольное затвердение без сокращения, которое вызывается раздражением
ироприоцепторов, т. е. мышечных чувствительных органов, в особенности на больных
людях (Гейнрих, 72; Зюснер и Гейнрих, 73). Икскюль и его
сотрудники,исследовавшие это активное напряжение, полагают, что в мышцах существуют двоякого
рода мышечные воломна: одни для движения, а другие для активного напряжения
92
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
или, как они называют, для залирательного действия (Sperrung). Между этими-
двумя системами признается определенная связанность функций: запирательная
часть мышцы активно напрягается в том положении, на которое установится
двигательная часть мышцы посредством центральных нервных импульсов. Пока это
запирательное действие произвольной мускулатуры наблюдалось на человеке в виде
центральных нервных реакций. На позвоночных животных это еще никому не
удалось наблюдать іни под влиянием центральной иннервации, ни путем раздражения.
двигательного нерва.
Активное напряжение было изучено также на поперечнополосатой мышце
sphincter ani. Эта мышца обычно находится в состоянии большого напряжения.
Лишь моментами под влиянием центральных нервных импульсов напряжение то
усиливается, то ослабевает. Процесс возбуждения, поскольку он проявляется в
соответствующих биоэлектрических токах, наблюдается только в эти переходные моменты
(Бек, 56). Когда же после определенного изменения напряжения центральная инг
нервация прекращается, достигнутое напряжение остается на долгое время. Только
много позднее оно постепенно меняется, более или менее возвращаясь к исходной
величине. Это постоянное напряжение, очевидно, является функцией самой мышцы.
Итак, наряду с пассивным напряжением, возникающим в мышце-
от растяжения или в связи с сокращением, когда мышца не в
состоянии укоротиться (изометрическое сокращение, большая нагрузка),
существует совершенно особое активное напряжение, возникающее
независимо от нагрузки и сокращения.
Об изменении сократительной способности в зависимости от
растяжения. Сократительная'способность мышцы в сильной степени зависит
от величины растяжения. При небольших растяжениях с помощью
небольших грузов сократительная способность нарастает. При больших
растяжениях, производимых большой нагрузкой, сократительная
способность, наоборот, уменьшается и при некоторых очень больших
нагрузках сходит на-нет. Это изменение сократительной способности в
зависимости от растяжения является законом для всех
поперечнополосатых мышц. Эту зависимость лучше всего можно исследовать путем
изучения напряжения мышцы изометрическим миографом. Так,
например, при регистрации напряжения m. sartorius лягушки можно
заметить, что если мышца не растянута и начальное напряжение ее близко'
к нулю, она при тетаническом раздражении самой мышцы или
соответствующего нерва развивает максимум напряжения в 100—150 г.
Если же мышцу предварительно растянуть силой до 100 г, т. е.
заранее создать в ней напряжение до 100 г, то такое же тетаническое
раздражение может развить напряжение только в 20—30 г, т. е. на 80—¦
70%' меньше (рис. 57).
Означенное изменение сократительного процесса в связи с
растяжением не зависит от аналогичного изменения процесса возбуждения.
Правда, первоначально с усилением растяжения мышцы процесс
возбуждения нарастает, как и сократительный процесс. Но, достигнув
определенной интенсивности, он с дальнейшим увеличением
растяжения не меняется и остается интенсивным даже при том максимальном
растяжении, когда сократительный процесс отсутствует (Беритов, 32).
Это падение сократительной способности не зависит также от
сократительная функции мышцы
УЗ
каких-либо изменений энергетических процессов: и при сильном
растяжении вслед за возбуждением происходит обычный распад аденилпиро-
фосфата, фосфагена и гликогена. Правда, известно, что в связи с
растяжением усиливается теплопродукция и потребление кислорода, но
это увеличение происходит
всего в два-три раза (Фенг, 127).
Известно также, что от
растяжения меняется рН, в смысле
сильного сдвига в щелочную
сторону. Но это изменение
совершенно обратимо, исчезая
немедленно после разгрузки.
Это явление скорее всего
нужно приписать изменению
физико-химического состояния
полупроницаемых мембран (Мар-
гариа, 128). На этом
основании следует признать, что
уменьшение и даже
исчезновение сократительной
способности в связи с растяжением не
может зависеть от каких-либо
химических процессов,
обусловленных растяжением.
Маргариа, как и Гил л,
считает наиболее вероятным,
что от растяжения меняются
физико-химические свойства
самих мышечных протеинов.
Можно предположить, что в
сильно растянутых мышцах
обычные энергетические
процессы, возникающие при
раздражении, не могут привести
в действие двигательную
машину миофибриллей
вследствие тех физико-химических
изменений, какие наступают в
протеинах миофибриллей вследствие сильного растяжения.
С точки зрения теории сокращения К. М е й е р а можно допустить,
что в связи с растяжением миофибриллей растягиваются как мицеллы,
так и интермицеллярное пространство сократительного вещества —
миозина. Вследствие этого карбоксильные и аминные группы
значительно удаляются друг от друга. Это затрудняет образование цвит-
терионов. При возбуждении мышцы аминные и карбоксильные группы
в белке ионизируются, но их притягательная сила недостаточна для
Рис. 57. Влияние начального напряжения на
сократительный процесс.
М. sartorius лягушки. Проксимальный конец
фиксирован. Дистальный конец соединен с изометрическим
миографом. Кроме того, проксимальная треть мышцы
соединена с изотоническим миографом. Верхняя
кривая записывает изменение длины проксимального
участка, нижняя кривая—напряжение всей мышцы.
В опыте Л начальное напряжение небольшое — 5 г.
Здесь комбинируется тетаническое раздражение обоих
участков. Первый раз сначала идет раздражение
проксимального участка (нижний сигнал); оно вызывает
укорочение проксимального участка и довольно сильное
напряжение всей мышцы (70 г); к нему присоединяется
раздражение дистального участка. Это раздражение дает
очень слабые эффекты, не меняя характера
существующей реакции. Второй раз сначала идет
раздражение дистального участка; при этом проксимальный
учссток удлиняется, и в общем развивается такое же
напряжение, как в первом случае; затем
присоединяется раздражение проксимального участка. Но и
теперь второе раздражение не вносит никаких
значительных изменений в существующую реакцию.
В опыте В мышца с самого начала сильно растянута
напряжением в 70 г. Раздражается проксимальный
участок; при этом он не укорачивается, напряжение же
нарастает всего только на 16 г, т. е. на 77"/,, меньше,
чем ¦& предыдущем опыте.
94 ОБЩАЯ ФИЗИОЛОіИМ МЫЩЬЧНОЙ СИСТЕМЫ
сгибания мицелл и образования цвиттерионов. Вследствие этого
сокращение не может наступить или происходит только частично, ибо цвит-
терионы могут образоваться между некоторыми наименее отдаленными
аминными и карбоксильными группами.
Механическое взаимодействие между различными участками
мышцы. Мышца при раздражении более или менее укорачивается во всех
своих участках только в том случае, если ей дается полная свобода в
укорочении. Если же оба конца мышцы зафиксированы или мышца
нагружена, то при сокращении одни участки укорачиваются, а другие-
Рис. 58. Механическое взаимодействие разных участков
сокращенной мышцы.
М. sartorius лягушки. Дистальный конец связан с изометрическим миографом,
а проксимальный конец, длиной в 10 мм — с изотоническим миграфом. Верхняя
кривая записывает изменение длины проксимального участка, а нижняя —
напряжение всей мышцы изометрическим миографом. В опыте А производится тета-
ническое раздражение дистального участка при разных силах раздражения (20,
15 и 12 см расстояния катушек). В опыте В тот же дистальный участок
раздражается отдельными индукционными ударами. В опыте С проксимальный участок
раздражается отдельными индукционными ударами. В первых двух опытах, когда
раздражается дистальный участок, проксимальный участок удлиняется, а в третьем
опыте при непосредственном раздражении проксимального участка последний
укорачивается (верхняя кривая).
удлиняются или остаются без изменения (рис. 58). Для понимания
этого явления нужно иметь в виду, что при возбуждении мышцы через нерв
сократительный процесс возникает не одновременно ид, всей длине
мышцы, а сначала в нервном экваторе, в области окончаний
двигательного нерва, и отсюда уже постепенно распространяется к концам
мышцы при прямом раздражении сокращение наступает сначала в
раздражаемом участке, а уже потом в других. В обоих случаях, вследствие
фиксации концов или наличия нагрузки, первоначально сокращенные
участки, укорачиваясь, растягивают другие участки, т. е. происходит
укорочение первых за счет удлинения других (Бете, 31; Беритов, 32).
Но даже если сократительный процесс происходит во всей мышце
одновременно, как это, например, имеет место при раздражении всей
мышцы (например, при пропускании индукционных ударов через всю мышцу),
то и тогда происходит укорочение одних участков за счет удлинения
других. Наиболее богатые нервными окончаниями участки мышцы со-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
95-
кращаются сильнее, чем другие. Поэтому они развивают большее
напряжение, чем другие, вследствие чего другие участки растягиваются,
несмотря на то, что они также возбуждены. То же самое бывает при<
тетаническом раздражении через нерв. Волна возбуждения все время
доходит до обоих концов мышцы. Энергетические превращения
происходят в большей или меньшей степени во всех участках, но вследствие
того, что сократительный процесс в нервном участке імного сильнее,.
тем в других участках, последние не укорачиваются, а удлиняются.
В последнее время такие же изменения длины при изометрическом'
сокращении были найдены на изолированных мышечных волокнах.
В мышечных волокнах конической формы было установлено, что при
по-ібуждении от прямого раздражения толстая часть, где
сократительный процесс должен быть сильнее всего, укорачивается, а другой,
тонкий конец, удлиняется. Между ними находится участок, длина которого*
не меняется (Бухталь и Линдгард, 97).
Механическое взаимодействие разных участков мышцы при сокращении
изучается следующим образом: один конец мышцы фиксируется, а другой соединяется
с изометрическим миографом. Кроме того, какая-либо часть мышцы вблизи
фиксированного конца соединяется с изотоническим миографом (рис. 77). Оба миографа.
пишут на одном кимографе. Изометрический миограф записывает кривую
напряжения, а изотонический — кривую изменения длины той части мышцы, которая лежит
между фиксированным концом и тем местом мышцы, с которым связан
изотонический миограф.
Во всех вышеприведенных случаях, когда в одних участках мышцьр
происходит сильное сокращение, другие участки не сокращаются, а
удлиняются. Это происходит главным образом потому, что волна
напряжения, возникшая в первично возбужденном участке,
распространяется вдоль мышцы быстрее возбуждения (Фишер, 33). Поэтому
когда волна возбуждения переходит в конечные участки мышцы,
последние уже оказываются растянутыми. Вследствие этого возбуждение не-
может вызвать здесь более или менее значительный сократительный'
процесс. Это легко обнаружить, если приложить тетаническое
раздражение к удлиненному участку мышцы; оно очень слабо повлияет на
существующий характер реакции: удлиненный участок останется
удлиненным, а общее напряжение мышцы изменится чрезвычайно слабо'
(рис. 57). Очевидно, в условиях растяжения мышцы второе
раздражение не вызывает того сократительного процесса, который оно обычно»
производит в отсутствии предварительного растяжения. Отсюда понятно,
что если тетанизирующее раздражение прикладывается не к нервным
участкам, а к какому-либо концу мышцы, то и тогда раздраженный,
сравнительно бедный нервами участок дает более сильное
сокращение, развивает более сильное напряжение, чем другие участки,
которые в момент возбуждения оказываются сколько-нибудь
растянутыми (Беритов, 32).
Затем очень важно отметить, что напряжение распространяется в*
мышце с некоторым ослаблением. Сообразно с этим напряжение,
возникшее в сокращенном участке, действует на суставы тем слабее, чем*
'96
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
.дальше от них находится первично сокращенный участок. Так, m. sar-
torius длиной 40 мм, при раздражении одного конца, например
проксимального, на отдаленный дистальный сустав действует на 33—37%
^слабее, чем на проксимальный (Беритов, 32).
Указанное механическое и физиологическое взаимодействие разных
участков мышцы, очевидно, имеет место и при нормальных условиях
деятельности мышцы в организме. Как указывалось выше, в организме
мышечное сокращение является в значительной мере изометрическим.
'Чтобы мышцы укоротились, они должны преодолеть определенное
сопротивление со стороны других антагонистических мышц, а также со
.стороны тяжести самой конечности. В условиях работы очень большое
.значение имеет то сопротивление укорочению, какое оказывает
нагрузка. Очевидно, при нормальной работе в каждой деятельной мышце
одни, а именно наиболее богатые нервными элементами, участки
укорачиваются, а другие удлиняются. Чем больше будет таких нервных
участков и чем равномернее они будут распределены по всей длине
мышцы, тем меньше будет растянутых участков, а следовательно, тем
полнее будет сократительный процесс в разных участках мышцы, тем
выше будет общее напряжение мышцы. И, что особенно характерно,
как раз те мышцы, на долю которых выпадает наиболее тяжелая
работа, как gastrocnemius, quadriceps femoris, triceps brachii, biceps bra-
chii, имеют такое строение, что в них длина мышечных волокон не
превышает одной трети всей длины мышцы, и располагаются они в косом
[Направлении между сухожильными тяжами, проникающими с концов
до половины и даже двух третей всей длины. Вследствие такого
строения множество мышечных *волокон сокращается сразу по всей длине
мышцы, и, таким образом, растяжение каких-либо участков мышцы
становится невозможным.
В. мышцах же с параллельными волокнами механическое действие
на суставы, очевидно, будет зависеть от местонахождения главной
массы окончаний двигательного нерва. При центральном положении
йіннервационной области точки прикрепления мышцы будут испытывать
•одинаковой силы напряжение. Если эта область лежит ближе к дис-
тальному прикреплению, то мышца будет действовать при нормальной
деятельности сильнее на дистальный сустав, чем на проксимальный, и
наоборот. И понятно, чем длиннее будет участок мышцы от иннерва-
ционной области до места прикрепления к суставу, тем слабее будет
действие мышцы на этот сустав (Беритов, 32).
Пластичность скелетной мышцы. В физике пластичностью
называется свойство твердых тел удерживать раз данную им длину и вообще
раз данную форму, которая была им придана внешней силой. Она
проявляется у всех тел с несовершенной упругостью. Деформация,
вызванная внешней силой, по удалении этой силы проходит не
полностью, а частично. Энергия, приобретенная телом при деформации,
отдается назад на устранение деформации только частично, сейчас же
после удаления внешней силы. Другая часть энергии остается, она
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
97
удерживает тело в более или менее измененном состоянии. Это и
называется пластичностью. Обычно, чем больше внешняя деформирующая
сила, тем больше эта остаточная энергия, тем больше пластическое
изменение. С другой стороны, эта остаточная энергия или
пластическое изменение тем больше, чем дольше времени действовала внешняя
сила.
Скелетные мышцы владеют всеми этими свойствами несовершенна
упругих тел. Если растянуть мышцу действием тяжести на несколько
процентов первоначальной длины, то после удаления тяжести
первоначальная длина не восстановится сейчас же полностью; длина ее
укорачивается с большой постепенностью, поэтому она в течение
продолжительного времени превосходит исходную величину. То же самое
произойдет, если немного укоротить мышцу действием какого-либо
раздражения; по удалении раздражителя мышца может остаться более
или менее укороченной на долгое время. При значительных
растяжениях и укорочениях імышца сначала стремится возвратиться к исходной
длине в силу своей упругости. В силу же пластичности нормальная
длина не восстанавливается сразу же полностью; в течение
значительного времени, иногда измеряемого минутами, мышца остается более
или менее укороченной или удлиненной (Блике, 51; Вахгольдер, 119;
Беритов, 103) (рис. 56 и 59).
Для полного выявления пластического укорочения или удлинения
нужно некоторое время. После очень кратковременного укорочения
или растяжения, измеряемого долями секунды, а иногда и
секундами, мышца может не дать пластических явлений. По імыеди
Бликса (51), разделяемой в последнее время (многими физиологами
(Гилл, 68; Рэнсон, 120), мышца представляет собой «упруго-вязкую»
систему. Отсюда вытекает, что каждой длине мышцы должно
соответствовать свое особое распределение или расположение структурных
элементов мышцы, а для того чтобы імышца могла, в результате
распределения этріх элементов, принять новую постоянную длину,
нужно время.
Так, если т. pect. pars abdom. или т. rect. abdom. лягушки
нагрузить 5—20 г на несколько секунд, то после снятия груза мышца сейчас
же возвращается почти к первоначальной длине в течение одной — двух
минут. При большей растягивающей силе імышца удлиняется больше и
не возвращается к исходной длине после снятия груза. Если действие
растягивающей силы продолжается несколько секунд, мышца остается
удлиненной всего на несколько мм. Если же растягивающая сила
действует в течение нескольких минут, остаточное удлинение
увеличивается тем сильней, чем дольше действует внешняя сила, т. е. чем больше
она удлинилась. В таких случаях она может оставаться удлиненной на
много миллиметров (рис. 56). Очевидно, при длительном растяжении
упругие свойства мышцы ослабевают (Беритов, 103).
Пластическое укорочение легко вызвать длительным тетаническим
раздражением, после которого мышца остается более или менее укоро-
7 И. С. Беритов
$8 оьшдя физиология мышечной системы
іенной. Если спустя некоторое время после раздражения мышцу рас-
тянуть небольшим грузом в 5—10 г, то длина ее превысит исходную
(рис. 59). При полном удалении нагрузки (поднятием миографа) мышца
укорачивается почти до исходной длины. На это нужно не много
времени— 1—2 мин. Этот опыт с пассивным растяжением и укорочением
можно повторять много раз; однако с каждым разом величина
пластического укорочения после сближения концов мышцы будет уменьшаться.
Отсюда следует, что сократительный процесс повышает пластичность
мышцы на некоторое время.
Не ©се скелетные мышцы пластичны в одинаковой степени. У
красных мышц, которым приходится очень долго удерживать определенное
Рис.^59. Пластическое укорочение после сокращения и пластическое удлинение от рас-'
тяжения. М. pect. pars abdom.
Раздражается тетанически проксимальный конец. В опыте А пороговое раздражение вызывает
небольшое пластическое последействие, которое устраняется под действием падающего миографа весом около
0.5 г: миограф был поднят немного и затем отпущен (звезда!). В опыте В, после сильного тетанического
раздражения, пластическое укорочение продолжается более минуты, и затем производится
пластическое удлинение действием тяжести в 5 г. В опыте С такое же углинекие производится спустя 10 сек.
после раздражения. В обоих случаях в момент, указанный стрелкол, тяжесть удаляется, и после
удаления мышца остается удлиненной. Внизу — время в секундах.
положение тела, пластичность проявляется в значительно большей
мере, чем у белых (Вахгольдер, 119). Таковы, например, сгибатели
бедра задней конечности, как m. semitendinosus, которым приходится
постоянно удерживать задние конечности в согнутом положении. Таковы
сгибатели передней конечности у самцов лягушки, которыми
осуществляется обнимательный рефлекс в период оплодотворения; таковы мышцы
хвоста, шеи и конечностей черепахи, которыми эти органы затягиваются
внутрь панцыря; таковы же сгибатели задних конечностей кролика,
которые удерживают обычное согнутое положение ноги. Особенно большой
пластичностью обладают мышцы живота. Они растягиваются под
влиянием поступившей в пищеварительный канал пищи, а также
развивающихся в матке зародышей, без того чтобы мышцы живота оказывали
обратное давление на внутренности. Растяжение происходит под
давлением изнутри, и растянутое положение удерживается до тех пор, пока
объем внутренностей не уменьшится. Так, например, если ввести в
брюшную полость лягушки шприцем несколько десятков кубиков рас-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
99
твора Рингера, живот соответственно растягивается, но при этом
давление жидкости нарастает только в самый момент введения жидкости,
затем сейчас же падает до нуля. Об этом можно судить, если соединить
живот с небольшим водяным манометром (Беритов, 103).
Пластические свойства скелетных мышц совмещаются с
эластическими, как этого нужно ожидать согласно представлению о том, чтэ
мышца предстазляет собой упруго-вязкую систему* Но каким
элементам мышечного волокна следует приписать то или другое свойство по
преимуществу? Прежде всего надо указать, что сократительное
вещество миофибрилли должно обладать обоими свойствами, так как, когда
мышца растягивается изометрическим миографом, то по мере
растяжения, параллельно с увеличением напряжения, сократительная
способность уменьшается. Но при данном растяжении с течением времени
напряжение в мышце падает в связи с пластическим изменением
(Беритов, 103).
За пластические свойства миофибриллей говорят также факты
изменения пластичности в связи с такими функциональными
изменениями, которые приводят к мышечному укорочению. Так, например,
под влиянием ацетилхолина пластичность сильно возрастает (Вахголь-
дер, 119); при окоченении теплокровной мышцы, в связи с
прекращением кровообращения, пластичность сначала повышается, затем
понижается, а в конце опять повышается. То же бывает при
контрактуре от тетано-токсина (Рэнсон, 120). При окоченении мышцы лягушки
от согревания до 30—33° С пластичность сначала понижается, а затем
сильно повышается (Беритов и Нивинская, 121).
Наиболее важным и бесспорным пластическим веществом является
саркоплазма. Обладая большой вискозностью и текучестью, она,
очевидно, абсолютно лишена эластических свойств.
Пластичность должна быть свойственна сарколемме и другим
соединительнотканным элементам мышцы. Эти образования проявляют
пластические свойства наравне с эластическими, подобно целой мышце.
На этом основании можно утверждать, что мышечное волокно
представляет очень сложное упруго-пластическое тело, где сарколеммы и
миофибрилли одинаково обладают эластическими и пластическими
свойствами, саркоплазма же представляет собой чисто пластическое
образование.
Как известно, многие органические образования обладают упруго-
пластическими свойствами. Например, полоски невулканизированного
каучука дают такие же кривые растяжения и стягивания, как живая
мышца. На этом основании явления растягивания мышцы под
влиянием внешней силы и восстановления исходной длины после
растяжения приписываются некоторыми авторами всецело физическим
особенностям мышцы (Маас, 117). Но мы ниже увидим, что это не
совсем так. Как кривая растяжения, так и в особенности кривая
восстановления после растяжения, по существу, зависят и от
физиологических процессов, протекающих внутри мышечных волокон.
7*
100
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Собственный тонус мышцы. Пластическое удлинение мышцы не
удерживается на все время. После удаления той силы, которая
произвела изменение длины, при хорошем функциональном состоянии мышца
через некоторый промежуток времени сама возвращается к
минимальной исходной длине. Так, если вырезать мышцу живота m. pect. pars
abdom. у лягушки и затем положить в раствор Рингера, то она сама в
течение нескольких минут может укоротиться более чем на 50'% той
длины, какую она имела в момент выпрепарирования. Если такую
мышцу растягивать действием какой-либо тяжести, то, как уже
указывалось выше, после удаления груза мышца сначала быстро
укорачивается на определенную длину благодаря своей эластичности, затем
продолжает укорачиваться очень медленно и этим путем в течение
многих минут іможет достигнуть первоначальной длины (рис. 56).
Способность к укорочению как после отпрепарирования, так и -после
растяжения в значительно большей мере свойственна красным,
наиболее пластическим мышцам, вроде m. pector. pars abdominalis
лягушки, чем белым, менее пластическим мышцам, как m. sartorius лягушки.
Означенное медленное укорочение мышцы, доводящее мышцу до
некоторой минимальной длины, помимо возбуждения и без особого
напряжения, не является всецело эластическим последействием, оно в
значительной мере зависит и от жизнедеятельности мышечной клетки,
а потому можно его называть собственным мышечным тонусом.
Очевидно, некоторую часть означенного укорочения по удалении
растягивающей силы нужно считать проявлением упругого последействия,
свойственного всем вообще не совершенно упругим телам. Наряду с
этим физическим процессом, укорочение должно вызываться теми
физико-химическими процессами, которые протекали в покойной мышце в
яиде постоянного расщепления аденилпирофосфата, фосфагена,
гликогена и других веществ. Как известно, и при покойном состоянии в
мышце образуются также продукты расщепления, как во время
сокращения, вызванного возбуждением. Между прочим, в нем образуются
при покойном состоянии фосфорная и молочная кислоты, хотя и в
очень малом количестве. Эти продукты расщепления должны
действовать на миофибрилли и тем самым обусловливать укорочение мышцы
независимо от возбуждения. Такое укорочение, собственно говоря, и
представляет собой подлинное выражение собственного тонуса мышцы.
Это положение подтверждается тем фактом, что чем лучше
функциональное состояние мышцы, тем полнее и быстрее мышца
восстанавливает свою исходную длину после растяжения. Например, мышца,
пролежавшая один или два дня в физиологическом растворе, не
восстанавливает совсем исходной длины. Она тем больше остается
удлиненной после растяжения, чем дольше она пролежала в
физиологическом растворе, чем хуже стало ее функциональное состояние (Бери-
тов, 103).
Такой же результат получается, если в течение нескольких часов
нагревать мышцу лягушки при температуре 31—33° С в физиологиче-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ Ю1
ском растворе. Нагревание ускоряет распад энергетического
материала. От этого собственный тонус мышцы усиливается. Но так как при
этом происходит обеднение мышцы энергетическим материалом, то
после такого согревания укорочение вслед за растяжением проявляется
слабее, мышца остается более или менее удлиненной. При этом по той
же причине ослабевает и тетаническое сокращение (Беритов и Нивин-
ская, 121). Все это хорошо видно на рис. 60.
Собственный тонус как выражение некоторого очень слабого
сократительного процесса протекает с очень небольшим напряжением.
Рис. 60. Влияние согревания на собственный тонус m. pect. pars abdom. лягушки.
А и В — до согревания. Сначала определяется порог тетанического раздражения (20 см расстояния
между катушками) и вызывается максимальное тетаническое сокращение при 18 см. Затем происходит
растяжение мышцы грузом в 10 г в течение 4 мин. Мышца удлиняется на 14 мм. После удаления груза
исходная длина — 31 мм — почти восстанавливается в течение 8 мин. Затем мышца согревается в
физиологическом растворе при 33°С. В результате нагревания в течение 2 ч. 30 м. мышца укоротилась на
3 мм (теперь длина мышцы равна 28 мм). В опыте С после согревания мышца вновь растягивается
грузом в 10 г: она удлиняется на 12 мм. После снятия груза мышца укорачивается в течение 10 мин. всего
на 4 мм; до исходной длины нехватает 8 мм. Проба электрического раздражения показывает низкую
возбудимость (порог около 5 см) и небольшое тетаническое сокращение при 2 см. В обоих случаях
растяжения в период восстановления миограф приподнимается несколько раз на несколько минут, чтобы
устранить растягивающее действие его. Эти моменты обозначены звездочкой.
Для m. pect. pars abdom. это напряжение измеряется 2—5 г, в то время
как при тетаническом сокращении оно достигает 100 г. Конечно, чем
слабее выражен собственный тонус, чем слабее тот основной обмен
энергетического материала, который обусловливает сокращение, тем
быстрее и сильнее растягивается мышца под влиянием столь малых
грузов, как 2—5 г. Так, на рис. 61 ясно видно, что при согревании
m. pect. pars abdom. напряжение в собственном тонусе больше 5 г.
Сократительный процесс, вызванный раздражением, сильно
способствует быстрому восстановлению длины мышцы до исходной величины.
Это может произойти менее, чем в одну минуту. Нужно только, чтобы
раздражение было достаточно сильным и длительным. Так, на рис. 62
полное восстановление исходной длины получается после
тетанического раздражения в течение 6 сек. Эта быстрое восстановление
исходной длины после тетанического сокращения должно происходить глав-
1?2
ОВЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ным образом в результате прямого воздействия определенных
химических веществ, образующихся в мышце во время сокращения, на
сократительное вещество, на імиофибрилли. Очевидно, при этом уничтожается
?Lsb
Pp.
а.
Рис. 61. Собственный тонус т. pect. pars abdom. при согревании.
Мышца была опущена в раствор Рингера, нагретый до 32° С, и сей-
час же нагружена 5 г. В первые 2 минуты мышца растягивается,
а затем постепенно укорачивается. Через час прибавляется груз
в 5 г, и с того момента мышца вновь начинает растягиваться.
то; смещение частичек сократительной субстанции, какое было вызвано
до тетанического раздражения растяжением.
Характерно, что красные мышцы, которые обнаруживают больше
пластичности и более способны к собственному тонусу, содержат
больше фосфагена. В этих мышцах в покойном состоянии распад фосфа-
В
ю
&
г" /' г' е- го' п' ір'
nilMHlllUnillllltiMMIIMNUIillllUliniUllllllllllllMiLtMMIIHinillMnilinilllllllNMltHWfgfl.iffrillf'iltji
Рис. 62. Восстановление собственного тонуса m. pect. pars abdom.
сокращением.
Мышца растягивается грузом 4 г в опыте Л б течение 2—3 сек., а в опыте В—10 мин.
После снятия груза (стрелка) через несколько секунд вызывается тетаническое
сокращение мышцы. В обоих опытах после тетануса исходная длина оказывается
восстановленной.]
гена и его последующее восстановление должны происходить в более
интенсивной форме, чем в белых мышцах. Повидимому, это
обусловливает то явление, что красные мышцы обладают способностью
удерживать укороченную длину, а также собственным тонусом в большей
мере, чем белые.
Итак, собственный тонус скелетной мышцы, поскольку он
выявляется в самопроизвольном достижении исходной длины по удалении
растягивающей силы или же в удержании длительного укороченного
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
103
состояния после сокращения, представляет собой небольшой
сократительный процесс, обусловленный тем спонтанным расщеплением аденилпи-
рофосфата, фосфагена и гликогена, какое имеет место все время в
покойном состоянии мышцы.
Контрактура. Существует своеобразная форма сокращения, которая
протекает очень длительно при малом напряжении. Такое сокращение
называется контрактурой. Контрактура вызывается сильным и длнг
тельным раздражением. Когда применяют такое раздражение,
сокращение не прекращается сейчас же вслед за прекращеним раздражения.
На это явление впервые обратил внимание Тигель (17); он же
назвал его контрактурой (рис. 63).
'.Рис. 63. Контрактура после тетаническсго раздражения на мышце зимней лугушки.
М. gastrocnemius. Раздражается седалищный нерв. В опыте 7 применяется слабое тетаническое
раздражение (а), затем два отдельных размыкательных удара (Ь, с); в опыте 2 — дважды
сильные индукционные удары. В обоих опытах нет контрактуры. В опыте 3 сначала слабое
тетаническое раздражение (/), затем сильное, но короткое тетаническое раздражение (g). В обоих
случаях последействие маленькое. Вслед за тем дается сильное и длительное раздражение около
¦4 сек. (л); оно производит продолжительную контрактуру. Во время этой контрактуры отдельные
индукционные удары (/", /) существенно не влияют на контрактуру. На сигнальной линии цифры
означают силу раздражения в сантиметрах раст. инд. катушек. Время — в секундах.
Чем хуже функциональное состояние мышцы, тем сильнее и
длительнее контрактурное последействие. Поэтому контрактура
наблюдается лучше всего при утомлении, при понижении температуры
мышцы, при воздействии кислот, а также различных ядовитых веществ, как
алкоголь, хлороформ и т. д. (Швенкер, 8). Если обмен веществ очень
ослаблен, то контрактуру можно вызвать и отдельными индукционными
ударами. Так происходит, например, с мышцами сильно охлажденной
лягушки (Беритов, 18) (рис. 64).
Контрактура Тигеля выступает не в одинаковой степени у
красных и белых мышц. Она всегда сильней выражена у красных, где она
наступает при более редких и слабых раздражениях, чем у белых.
Так, например, mm. semitendinosus, gastrocnemius и pect. pars abdom.
лягушки, содержащие больше красных волокон, реагируют
контрактурой сильнее и длительнее, чем m. sartorius или semimembranosus
лягушки. У красных мышц контрактурное последействие переходит в
пластический тонус: мышца на долгое время, иногда на много минут,
застывает в укороченном состоянии. Этот пластический тонус легка
•бнаружить, если мышцу растянуть до нормальной длины: при налич-
104
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ности контрактуры растянутая мышца вновь сейчас же укорачивается
соразмерно тому, какова была степень контрактурного сокращения.
Если же состояние укорочения было исключительно пластического
характера, тогда после растяжения мышца остается растянутой (рис.59).
Очевидно, во время тетанического сокращения мышца приходит в
пластическое укорочение определенной высоты, которое длится дольше, чем
контрактурное последействие. Пластическое укорочение перекрывает
контрактуру. Поэтому, если растянуть мышцу до нормальной длины в
то время, пока еще есть контрактура, мышца после растяжения может
Рис. 64. Одиночное сокращение охлажденной до 0°
икроножной мышцы лягушки.
Раздражение 4 раза отдельными индукционными ударами. Второй
удар спустя 12 сек. после первого; третий также спустя 12 сек.
после второго; четвертый же спустя 2 мин. после третьего. Каждый
раз раздражение вызывает сначала быстро протекающее
сокращение, затем медленно протекающее. При повторениях через 12 сек.
медленное сокращение ослабевает и снова вырастает после отдыха
в 2 мин. Вторая линия указывает опусканием вниз моменты
раздражения. Цифры обозначают интенсивность раздражения в
сантиметрах. Третья линия обозначает время в секундах.
укоротиться почти до вйсоты контрактуры. Если же растянуть ее по
прекращении контрактуры, мышца после растяжения не укоротится
совсем. Так, на рис. 65 брюшная мышца лягушки растягивается
несколько раз после тетанического раздражения в течение трех минут
(оп. В), и каждый раз после снятия груза мышца укорачивалась.
Контрактура вызывается независимо от раздражения под влиянием
разнообразных химических веществ. Если, например, вырезанную
мышцу погрузить в деци- или сантинормальный раствор какой-либо кислоты
или щелочи, то мышца впадает в длительное сокращение. Высота
такого сокращения может достигнуть высоты самого полного тетануса
(Копылов, 7; Бете, Френкель и Вильмерс, 14). Это сокращение обычно
протекает в двух фазах: в первую фазу сокращение очень быстро
достигает максимума, и затем быстро же происходит частичное
расслабление. Вслед за этим начинается новая фаза, когда сокращение достигает
максимума очень медленно, спустя несколько минут. Первая, короткая
фаза, производится через возбудимую систему, т. е. возбуждением, ибо
*
она исчезнет при наркозе мышцы, после потери ею возбудимости. Вторая,
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
105
длительная фаза остается при этом без изменения (Тауер, 71):. (рис. 66)*
Отсюда следует, что вторая фаза контрактуры всецело обусловлена ШШ;
действием кислоты прямо на миофибрилли, которые, очевидно, не под*
вергаются, как неживое образование, наркотическому действию, или же
повышением собственного тонуса, т. е. ускорением тех химических
Рис. 65. М. pect. pars abdom. В опыте А испытывается в покойном состоянии действие*
нагрузки в 2 и 5 г.
В опыте В производится сильное тетаническое раздражение. Сейчас же после раздражения нагрузка,
в 5 г удлиняет мышцу значительно, но не до той длины, какая получилась в опыте А при той же
нагрузке. Это указывает, что мышца сокращена, находится в контрактуре. Повторная нагрузка через 1 и
3 мин. вызывает большее удлинение, но контрактура все еще остается. В опыте С повторяется то ж«
тетаническое сокращение. После раздражения мышца не нагружается; она остается сильно
укороченной в течение 7 мин.
процессов, которые ведут к сокращению імиофибриллей и которые в.
небольшой степени протекают в мышце и помимо возбуждения.
Было установлено, что кислота вызывает укорочение и
соединительнотканных элементов — фасций, сухожилий. Она даже может дать-
б них, первую, круто начинающуюся фазу. Но это кислотное укорочение
соединительнотканных элементов развивается медленнее, чем таковое
скелетной мышцы, и не достигает такой высоты (рис. 67) [Тауер, 71;
Гетт (Hett) 93]. Это дает основание утверждать, что кривая кислотного
сокращения мышцы является результирующей из кривых контрактуры
мышечных волокон и укорочения коллагенных соединительнотканных
элементов. При этом мышечные волокна приходят в действие вначале
через возбуждение, а затем непосредственно путем воздействия на
миофибрилли или на физико-химические процессы, ведущие к
сокращению.
Контрактура, вызванная сильным тетаническим раздражением,
зависит от действия определенных продуктов обмена веществ на
сократительное вещество. Среди продуктов обмена веществ, повидимому,
большее значение должна иметь прежде всего імолочная кислота. Эта
кислота не успевает сейчас же после своего возникновения целиком
нейтрализоваться или расщепиться до угольной кислоты или
синтезироваться в гликоген. При определенных условиях она накапливается
в мышце во время раздражения, а потому ее действие на сократитель-
106
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ное вещество продолжается и после раздражения. Естественно, это
последействие будет тем сильнее, чем больше накопилось молочной
кислоты. По Эмбдену (27) повышенное образование молочной
кислоты происходит не только во время тетаническога раздражения, но и в
течение 5—10 мин. после него. Следовательно, даже если молочная
кислота будет нейтрализоваться во время раздражения, контрактура
может вызываться той молочной кислотой, которая выделяется после
раздражения. Таким образом, вызванная тетаническим раздражением
Рис. 66. Кислотная контрактура m. sartorius без
предварительного наркоза 20% метилалкоголем (а)
и после наркоза (&). В последнем случае отсутствует
первая фаза. Вторая же наступает почти без
изменения. Время в полминутах (Тауер).
.контрактура может происходить под
влиянием кислот, образуе ных в мышце
во время раздражения и вскоре поем
него.
Как указывалось, при плохом функциональном состоянии даже
отдельные индукционные удары вызывают двойной эффект: сначала оди-
-ночное вздрагивание, а затем контрактурное сокращение. Исходя из
того, что при ряде раздражений отдельными индукционными ударами
длительное сокращение наблюдается в самом начале и быстро исчезает
при повторении раздражения, Б р е м е р заключает, что оно не может
^обусловливаться, подобно контрактуре Тигеля, накоплением кислых
•продуктов. Б р е м е р предполагает, что при плохом функциональном
• состоянии происхождение контрактуры точно такое же, как при
отравлении вератрином. Предполагается, что то вещество, которое
обусловливает длительное сокращение при одиночном раздражении, в мышце
• существует еще до раздражения. Известно, что очень слабые
концентрации кислот, в частности молочной (0.005 норм, раствора), не
вызывают или вызывают очень слабую контрактуру. Но если одновременно
-с действием кислоты раздражать мышцу индукционными ударами, т?
сейчас же наступает сильная контрактура (Швенкер, 8). Аналогичное
явление имеет место, вероятно, в опыте Б р е м е р а. Но только таким
^веществом, предрасполагающим к контрактуре, здесь мог явиться
один из других продуктов обмена веществ — аммиак (Бюттнер и Гейм-
Рис. 67. Кислотная
контрактура т. sartorius (а) и
изолированной полоски
фасции из т. gastrocnemius
(b). Действие п '200 HCI.
На обеих кривых первая стрелка
указывает момент наливания
раствора Рингера, а вторая —
кислоты. Контрактура мышцы
много выше и достигает обоих
максимумов намного раньше, чем
,.- укорочение фасции (Тауер).
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
107
брехт, 67) или ацетилхолин (Рештейнер, 65). В определенных случаях
контрактура в связи с раздражением наступает заведомо при полном
отсутствии молочной кислоты. Это бывает при отравлении мышцы емо-
ноиодуксусной кислотой, когда распад гликогена происходит, но
молочная кислота не образуется. Характерно, что это совпадает с
расходованием фосфагена, с распадом которого связывается сокращение такой
мышцы (Кларк, Эгглетон и Эгглетон, 64). Очевидно, контрактура от
раздражения имеет сложное происхождение.
Существуют такие химические вещества, которые вызывают
контрактуру через возбудимую систему. К таким веществам следует
отнести кофеин, никотин, вератрін, ацетилхолин и др. Это видно из того,
что они не действуют на невозбудимую мышцу, а в возбудимой мышце
вызывают возбуждение вместе с контрактурой (Риссер, 19; Беритов,
102). Поэтому такую контрактуру принято называть контрактурой
возбуждения (Риссер, 19). Эта контрактура, повидимому,
обусловливается наступлением тех химических процессов, какие обычно имеют
¦место в результате возбуждения. Они сопровождаются замедленным
образованием молочной кислоты сообразно с замедленным характером
сокращения (Гилл). Некоторые же вещества, как КС1, НС1 и NaOH
вызывают контрактуру обоими путями: прямым действием на
сократительное вещество и, кроме того, раздражающим действием на
возбудимую систему (Фишер, 34).
Некоторые физиологи считают контрактуру функцией особого сократительного
•механизма, который отличается от механизма, обусловливающего одиночное
сокращение и тетанус. Например, Б о т а ц ц и С122) предполагает, что контрактура Тигеля,
как вообще длительные сокращения, вызываемые химическими веществами,
обусловливается сокращением саркоплазмы. Однако такое предположение опровергается
.разного рода опытами. Известно, например, что при длительных раздражениях
способность к контрактуре ослабевает параллельно с ослаблением способности к
одиночным сокращениям и к тетанусу.
Предполагается также, что контрактура свойственна особым мышечным
волокнам или особого рода возбудимой системе (Бремер, 123). В подтверждение
последнего предположения приводится тот факт, что очень часто на изолированной мышце
или на нервно-мышечном препарате контрактура іне наступает от одного нервного
импульса, а только от двух-трех, и при том только в том случае, если они следуют
друг за другом через 2—3 сигмы, т. е. при частоте выше 125 в секунду. Однако
против такого предположения говорит, во-первых, то, что на мышцах лягушки
можно получить контрактуру при любой частоте раздражения в большей или
меньшей степени, и, 'во-вторых, то, что при медленных укорочениях в мышце протекают
такие же химические процессы, как и при тетанусе (Гартри и Гилл, 124; Фурусава
и Гартри, 125; Мейергоф и Ломан, 126).
Медленное сокращение контрактурного характера производится наряду с
быстрыми сокращениями и поперечнополосатыми мышцамл высших беспозвоночных. Так,
¦у ракообразных раздражением двигательных нервных солокон можно вызвать как
•быстрое, так и медленное сокращение, причем быстрое вызывается через наиболее
толстые нервные волокна, а медленное — через сравнительно тонкие (Гарревельд и
Вирсма, 129). Было установлено, что в обоих эффектах участвует один и тот же
сократительный субстрат. Такое заключение основывается на ряде наблюдений.
Например, известно, что быстрое сокращение благоприятствует медленному, если
первое предшествует последнему (Вирсма к Гарревельд, 129, 131); при быстром и
108
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
медленном сокращении одинаковой высоты развивается одно и то же напряжение
(Вирсма и Гарревельд, 130). Также одинаковыми оказались количественные
.изменения фосфа-гена и '.молочной кислоты при медленном и быстром сокращениях
одинаковой высоты (Берген и Виромл, 132). На этом основании всю разницу в характере
сокращения авторы приписывают своеобразию процессов, протекающих в
двигательных пластинках толстых и тонких двигательных волокон (Вирсма и Гарревельд, 130).
О действии ацетилхолина на скелетную мышцу. Действие
ацетилхолина на мышцу подвергалось особенно тщательному исследованию, ибо
ацетилхолин содержится во многих тканях организма, а также в
мышцах- (Платтнер и Краних, 66). Ему приписывается определенное
функциональное значение в отношении мускулатуры. Именно,
предполагается, что ацетилхолин выделяется двигательными нервными окончаниями,
в мышечных волокнах при их возбуждении и затем, действуя
раздражающим образом на возбудимую систему мышцы, обусловливает
возбуждение последней (Дэйл, Фельдберг и Фогт, 80). Поэтому многие
авторы называют ацетилхолин передатчиком возбуждения с нерва на
мышцу, иначе говоря — медиатором или посредником между нервом и
мышцей. Мы подробно коснемся фактических оснований этой гипотезы
в связи, с рассмотрением физиологии двигательной нервной пластинки.
Сейчас же рассмотрим физиологическое действие ацетилхолина при
его искусственном приложении к скелетной мышце.
По исследованиям Риссера (19), Вахгольдера (69) и др., ацетилхолин
вызывает контрактуру при очень слабых концентрациях, как 1 :50 000 и даже
1:100 000. Но авторы находили, что при слабых концентрациях контрактуру дают
не все мышцы, а только некоторые. Так, например, у лягушки т. rectus abdom.,
т. coracoradialis обычно дают сильную контрактуру, m. sartorius часто дает только^
начальное вздрагивание без контрактуры, mm. ileofibularis и gastrocnemius дают
сначала одиночное вздрагивание, а затем длительную контрактуру. Первые мышцы
поэтому были названы тоническими, а 'вторые — нетоническими. Третья группа мышц,
представлялась состоящей из тех и других элементов. Некоторые авторы находят-
даже возможным изолировать в них тонические волокна от нетонических (Зоммер-
кампф, 110).
По последующим исследованиям оказалось, что ацетилхолин
производит как быстрое, так и медленное сокращение во всех скелетных,
мышцах лягушки. Вся разница заключается в том, что длительное
сокращение одних імышц, которые содержат больше красных волокон,.
как, например, m. pect. pars abdom., продолжается много минут, а
других, которые содержат очень мало таких волокон, как m. sartorius,
весь эффект продолжается менее одной минуты (рис. 68). Поэтому
в отношении к ацетилхолину мышцы лягушки можно разделить только
на две группы: 1) мышцы, дающие длительное ацетилхолиновое
укорочение, и 2) мышцы с кратковременным укорочением. В определенных
случаях, когда мышцы вырезаны и возбудимость их понижена,
ацетилхолин не производит начального быстрого сокращения, а только
медленное. Это случается со веемой мышцами лягушки, однако с красными
чаще, чем с белыми (Беритов, 102).
В мышцах теплокровных ацетилхоллин производит в малых д?зах
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
109
только вздрагивание и короткие тетанусы. При больших дозах сначала
наступает быстрое сокращение, а потом более или (менее длительная
контрактура (Броун, 81; Розенблют и Луко, 83). Сокращение можно
вызвать в мышцах лягушки и такими малыми дозами, как 0.005—0.28 у
(тысячных миллиграмма), если вводить в мышцу эту дозу через ее же
артерию. Такая доза производит только медленное сокращение. Если
Рис. 68. Действие ацетилхолина на мышцы лягушки.
Верхняя кривая — от m. pect. pars abctom., а нижняя — от m. sartorius. В опытах А и В тетанические
сокращения от прямого раздражения той и другой мышцы. В опыте С ацетилхолин в растворе 1: 20000
приливается по нескольку капель, сначала на m. sartorius, а затем на m. pect. pars abdom. На т. sar
torius сначала быстрые вздрагивания, затем контрактура продолжительностью около минуты. Ham.
pect. pars abdom. —только контрактура продолжительностью свыше 7 мин. Стрелки указывают
остановку кимографа на одну минуту. Звезда — момент небольшого случайного растяжения мышцы.
же брать ацетилхолин в количестве больше 0.5—10 у, то медленному
сокращению предшествует быстрое (Равентос, 82).
Ацетилхолин не только производит сокращение, но и влияет на
возбудимость. В малых дозах он повышает возбудимость, а в больших,
когда он дает сильное контрактурное сокращение, он угнетает ее
(Броун, 84; Розенблют и Луко, 83; Беритов, 103). Опыт с угнетением
показан на рис. 69. Здесь применялось локальное ацетилхолиновое
отравление в средней трети m. sartorius, как показано там же на схеме.
Дистальный конец раздражается тетанически с разной силой (18—15—
12 см расстояния между катушками). Во время полной контрактуры
эти раздражения почти не дают сокращений в проксимальной трети.
Здесь сокращения наступают только при более сильных
раздражениях — 10 см.
Первое быстрое сокращение носит всецело тетанический характер.
Это видно прежде всего из того, что при локальном отравлении одной
части мышцы с параллельными волокнами, как m. sartorius,
сокращение наступает во всей мышце. Медленное же сокращение не является
тетанусом, ибо при локальном отравлении в определенных случаях оно
возникает только в отравленном участке (Риссер, 19; Нарикашвили, 48)
(рис. 69). То обстоятельстве, что в отравленном участке во время
медленного сокращения возбудимость падает и проведение возбуждения
1 К) ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
затрудняется, свидетельствует, что это медленное сокращение
наступает также через возбудимую систему. Предполагается, что ацетилхо-
лин в малых дозах или при длительном действии производит частичное
расщепление возбудимой системы, подобно субминимальному
раздражению (см. ниже о действии субминимальных раздражений — гл. VIII)..
Рис. 69. Локальное отравление m. pect. pars abdom. ацетилхолпном.
Сбоку миограммы дается схематическое изображение опыта: D — свободный дистальный
конец, соединенный с миографом; этим способом записывается нижняя кривая; Р —
проксимальный конец, фиксированный. Проксимальная треть также записывается отдельно
на кимографе — верхняя кривая. Ff В — электроды для раздражения. Платиновые
электроды связаны с мышцей посредством нитки, смоченной в физиологическом растворе.
': При сокращении нитки передвигаются вместе с мышцей. Ас .. — место приложения
кусочка фильтровальной бумажки, смоченной в растворе 1 : 10 000 ацетилхолина. В
опыте А эффекты раздражения дистального конца при 18, 15 и 12 см расстояния между ка-
: тушками; при 15 и 12 см сокращается и проксимальный конец. В опыте В производится
ацетилхолиновое отравление; во время контрактуры, которая не распространяется на
проксимальную треть, то же раздражение дистального конца не дает сокращения в
проксимальном конце. Только более сильное раздражение — 10 см — дает сокращение
и в этой части. В опыте С ацетилхолиновая контрактура на 3-й минуте после
приложения ацетилхолина устраняется действием груза в 5 г; по удалении груза (стрелка)
мышца немного укорачивается, но полная контрактура восстанавливается после
электрического раздражения. Теперь проксимальный эффект значительно сильнее, чем в опыте В.
Благодаря этому здесь, в отравленном участке локально усиливается
тот физико-химический процесс, который ведет к сокращению. Большая
же разница в длительности медленного сокращения зависит от того,
что в красных мышцах ацетилхолиновое локальное сокращение
усложняется наступающим при этом пластическим укорочением, которое
может удерживаться в течение многих минут. То обстоятельство, что
спустя 3—5 минут ацетилхолиновое укорочение легко устраняется
небольшой тяжестью, как 5 г, свидетельствует о том, что здесь
действительно имеется чисто пластическое укорочение (см. опыт С на рис. 69).
В белых мышцах при этом же сокращении пластический тонус
значительно слабее или совсем отсутствует, а потому сокращение
продолжается только, пока действует ацетилхолин (Вахгольдер, 119; Бери-
тов, 102).
Мышечные шумы и тоны. Как указывалось выше, каждое
одиночное сокращение в благоприятных условиях продолжается меньше одной
десятой секунды, а его восходящее колено на мышце лягушки не
превышает 0.05 сек. Отсюда вытекает, что эта мышца может производить
зубчатый тетанус при раздражении с ритмом до 20 в секунду. В
мышце теплокровных животных одиночное сокращение протекает еще быст-
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
111
рее, поэтому мышца этих животных может показать еще более частую
зубчатость. При зубчатом тетанусе мышца издает шум, который можно
услышать или непосредственно, приложением уха к мышце, или, еще
лучше, через стетоскоп.
Однако и во время полного тетануса мышцы издают шум. Это
объясняется в одних случаях тем, что и во время тетануса сокращение всех
мышечных волокон не протекает одновременно. Начало и конец
сокращений разных мышечных волокон происходят в разное время, в
разной последовательности, а потому мышца и во время полного тетануса
представляет колеблющееся тело. Но в определенных случаях звук адо-
Рис. 70, Мышечный шум сгибателя [flexor carpi ulnaris при
произвольном сгибании руки. Конденсаторный микрофон приложен к мышце. Токи
из микрофона регистрируются осциллографом (Тренделенбург и Шютц).
жет получиться оттого, что при полном тетанусе и одновременном,
сокращении всех мышечных волокон в мышце получается некоторое
изменение напряжения под влиянием каждого импульса возбуждения.
На мышцах лягушки удается регистрировать столь высокий ритм
напряжения, как 50—70 в секунду (см. Введенский, 101; Фультон, 38), а.
при очень чувствительной методике и еще выше — до 120 в секунду —
(мышцу приводят в соприкосновение с мембраной микрофона и ведут
регистрацию колебаний мембраны через телефон; Шефер и Гепферт..
84). На теплокровных наблюдался ритм механического эффекта до 100*
в секунду. Следовательно, мышца может издавать звуки, отвечающие
50—120 в секунду во время полного тетануса. Кроме того, звук может
получиться от изменения объема мышцы в связи с возбуждением. Как.
будет указано ниже, по исследованиям Эрнста, мышца лягушки
испытывает объемные изменения по ритму возбуждения до 150 в
секунду. Следовательно, она может отвечать звуком и в 150 колебаний я-
секунду. В действительности было замечено, что между высотой
мышечного тона и частотой импульсов возбуждения сокращенной мышцы
существует близкое соотношение, ибо они протекают почти в одном \г
и том же ритме. Так, например, на рис. 70 дается мышечный шум
сгибателя руки человека при произвольной иннервации, зарегистрирован-
ный с помощью конденсаторного микрофона, который прикладывался
к мышце. Изменения тока в микрофоне регистрировались
осциллографом. Частота механического эффекта достигает 156 в секунду и
равняется ритму произвольной иннервации этих мышц (Тренделенбург и
Шютц, 91; Брандштетер, 92).
Заслуживает внимания то обстоятельство, что, когда мышца
человека или животных сокращается произвольно, она издает низкий тон,.
1 |2 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
который соответствует 19.5—20 колебаниям в секунду (Гельмгольц, 20).
Такой же низкий тон слышится при рефлекторном возбуждении мышцы,
например при раздражении чувствительного нерва. Во всех этих
случаях слышимый тон выражает собой ритм действия наиболее сильных
механических колебаний, а не ритм возбуждения. Как потом увидим,
ритм возбуждения во время произвольного и рефлекторного
сокращений гораздо выше.
Когда мышца раздражается непосредственно индукционными ударами,
мышечный тон вполне следует за ритмом раздражения. С учащением последнего мышечный
гон также повышается. Слышен мышечный тон в 704 колебания в секунду (Лёвен,
21) и в 1000 колебаний (Бернштейн, 22). Такие же высокие тоны, но только более
слабые, были получены и при непрямом раздражении мышцы, т. е. через нерв.
Эти высокие мышечные тоны не выражают собой ни ритма мышечного сокращения,
ни ритма мышечного возбуждения. Последний ритм даже при наиболее
благоприятных условиях не превосходит 400 колебаний в секунду. Если же можно
стетоскопом услышать тон в 704—1000 колебаний, то этому должна быть другая причина.
Известно, что всякое тело, проводящее индукционный (переменный) ток, издает звук.
При раздражении мышцы л нерва индукционными ударами мышца становится
звучащей по этой же причине, так как авторы употребляли индукционные удары столь
большой интенсивности, что физическое действие последних должно было
распространиться на весь препарат (см. подробно о мышечных шумах в статье Боумана н
Рпиберка, 107).
Градация механического эффекта. Величина мышечного сокращения
зависит от многих условий. Прежде всего существенное значение
Ри. 71. Высота мышечного сокращения в зависимости от силы
раздражения.
Мышца раздражается прямо индукционными ударами. М — замыкатель-
ный удар; ь — размыкательный. Внизу цифры отмечают расстояние
между индукционными катушками в сантиметрах (Пембрей).
имеет сила раздражения. При отсутствии нагрузки или при очень
небольшой нагрузке градация механического эффекта очень мала: уже
при некоторых слабых раздражениях нервного участка или самого
нерва можно получить максимальное укорочение. Это происходит
оттого, что для максимального укорочения всей мышцы достаточно
возбуждения лишь части мышечных волокон. Но когда мышца сильно
нагружена, то величина укорочения в значительных пределах меняется в
зависимости от силы раздражения: чем сильнее раздражение, тем
больше мышечных волокон возбуждается и тем выше поднимается груз
(рис. 71).
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
ИЗ
Раздражение, которое значительно выше порогов, но еще не
достаточно для вызова максимального эффекта, называется субмакси-
.мальныім. То раздражение, которое дает максимальный эффект,
называется (максимальным. Раздражение сильнее максимального
принято называть сверхмаксимальн ььм. Так, на рис. 71 для
размыкательных ударов 16-сантиметровое расстояние между катушками
является пороговым, от 15 до 9 см — субмаксимальным, 8
см—максимальным, а 7—5 см — сверхмаксимальным.
При постепенном усилении прямого раздражения безнервного
участка мышцы, когда раздражение нарастает без скачков,
механический эффект тоже нарастает постепенно. Для этой цели лучше всего
брать кураризованную мышцу. Если же імышца раздражается через
нерв или ток пропускается через всю імышцу и тем раздражаются
нервные волокна, заключенные в мышце, тогда с постепенным усилением
раздражения механический эффект нарастает скачками. Иногда этих
скачков бывает несколько. Это зависит от толщины раздражаемого
нерва и толщины, імышцы. Чем они толще, тем больше могут быть
градации (рис. 72). Отсутствие скачков при прямом раздражении
безнервного участка объясняется прежде всего тем, что с усилением
раздражения все более и более значительный участок мышцы
раздражается одновременно как в поперечном, так и в продольном
направлении, а это имеет существенное значение для силы механического
эффекта. Чем больше раздраженных волокон, тем больше
механический эффект. Далее: когда один участок сокращается, другой
растягивается. А, как указывалось выше, растяжение ослабляет
сократительный процесс, который вызывается потом распространением
возбуждения. С усилением раздражения первоначально сокращающийся участок
увеличивается и сообразно уменьшается другой, растягиваемый участок.
Этими двумя обстоятельствами и обусловливается постепенное
нарастание механического эффекта всей імышцы при прямом раздражении.
При раздражении мышцы через нерв эффект нарастает скачками
прежде всего потому, что самое вступление нервных волокон в реакцию
происходит скачками. Во-первых, нервные волокна имеют разную
возбудимость, поэтому одни из них возбуждаются при более слабых
раздражениях, другие — при более сильных; во-вторых, нервные волокна
данной мышцы не располагаются в нервном стволе или в самой мышце
совместно, а пространственно разделены друг от друга. Поэтому при
электрическом раздражении нерва сначала приходят в возбуждение
одни волокна, а потом другие. Но, кроме того, вступление в действие
каждого нового нервного волокна приводит к сокращению целой
группы связанных с ним мышечных волокон, а потому механический эффект
всей мышцы должен нарастать скачками.
Разная сократительная способность у мышц одного и того же
животного. Сократительная способность мышц различна для разных мышц.
Одни мышцы укорачиваются больше, другие меньше. Так, из мышц
лягушки tibialis ant. и peroneus укорачиваются на 20—30% нормальной
8 И. С Беритов
114
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
/
длины — это при наилучшем функциональном состоянии мышцы,
sartorius и semitendinosus укорачиваются на 45—70%; чем лучше
функциональное состояние, тем сократительная способность больше. Но и
при равных условиях одни імышцы укорачиваются сильнее других.
Следовательно, означенная разница в степени укорочения зависит не от
разного функционального состояния, а от природы самой мышцы
(рис. 73) (Беритов и Яшвили, 35). Такая же разница в сократитель-
Tfb.a
Sem\ \VS.s:
1 =27мм
1*33 мм
і \го.з% иех.\ ш%
I-3?мм 1-ЗОмм
Qast
1=30 мм
Vm. \
1~?7мм
1-28мгл
Ш.зЯ г
Pect </г?%
30.4% 1=45мм
2Э% Sar \J12Z
Js/fOMfA
Рис. 72. Одиночные сокращения при
разных условиях раздражения.
М. gastrocnemius лягушки. В опыте 1
раздражается свежая мышца пропусканием раз
рядов конденсатора по длине мышцы с
разной восходящей интенсивностью. В опыте
2 такое же раздражение двигательного
нерва. В опыте 3 раздражается кураризованная
мышца, как в опыте 7 (Асмуссен).
Рис. 73. Высота максимальных
укорочений /разных мышц весенней лягушки.
Прямое раздражение через всю мышцу. Для
тонких мышц, как sartorius, сила раздражения
8 см; для толстых мышц, как gastrocnemius,.
7 см (пороги раздражения были 14—16 см).
Частота раздражения 40 ударов в 1 сек. Длина
и степень укорочения мышцы в процентах
мышечной длины даны на кривых: tib. с. — tibialis
anticus; Per — peroneus; gast. —. gastrocnemius;
V. in. — Vastus int. trie, fern.; il — ileofibularis;
Sem — semitendinous; V. ex. — Vastus ext.;
Pect — pectoralis; Sar — sartorius.
ной способности была обнаружена у мышц кошки. Здесь sartorius
укорачивается в два раза сильнее, чем tibialis anticus и rectus fem.
(Цхакая, 26).
Разная сократительная способность различных мышц вполне
соответствует разнице расстояний между точками прикрепления при
пассивном максимальном сгибании и разгибании суставов. С увеличением
этой разницы сократительная способность увеличивается; значит,
каждая мышца производит приблизительно такое максимальное
укорочение, какое требуется для максимального движения суставов. Отсюда
можно заключить, что сократительная способность каждой мышцы
филогенетически развивалась так, как этого требовало ее
анатомическое положение (Беритов и Яшвили, 25). ,„... _
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
115
Сократительная способность в разных участках одной мышцы.
Путем регистрации отдельных участков мышц с параллельными
волокнами было установлено, что даже в пределах одной мышцы
сократительная способность не одинакова. Она больше там, где оканчивается
больше всего нервных волокон. Это выражается как в степени
укорочения, так и в степени напряжения. Так, в мышце sartorius лягушки
сократительная способность средних нервных участков почти вдвое
больше, чем в конечных, сравнительно безнервных участках. Например,
в одной серии опытов мышца укорачивалась в нервной части на
Рис. 74. Напряжение m. sartorius целой мышцы (1) и отдельных частей.
Мышца мысленно разделена на 5 одинаковых частей, и каждая часть записывается
отдельно. Сначала записывается напряжение всей мышцы — 52 г, затем идут записи
напряжения от частей мышцы, начиная с проксимального конца. Первая проксимальная
часть показывает напряжение 10 г (2), следующая—16 г (3); центральные части (4 и б*)
показывают 18 и 19 г, а дистальная часть (6)—10 г.
40—65%, в среднем — на 54.8%. Вся же мышца в среднем
укорачивалась на 41.7%. Проксимальный и дистальный концы показывали в
среднем 28.2% и 38.2% укорочения (рис. 74) (Беритов и Яшвили, 35;
Картозия, 75). Эта разная сократительная способность не зависит от
самих миофибриллей, т. е. от особенностей самого сократительного
вещества. Она, повидимому, прежде всего обусловливается тем, что
в наиболее нервных участках процесс возбуждения протекает сильнее,
чем в сравнительно безнервных (рис. 98). Вследствие этого в нервных
участках должен происходить более интенсивный обмен энергетического
материала — фосфагена и гликогена, чем в безнервных. Изучение
биохимических процессов в области нервных и безнервных участков
показало, что ферментативные процессы расщепления и синтеза в
нервных участках протекают интенсивнее, чем в безнервных. В
результате этого сокращение в нервных участках должно происходить
сильнее, чем в безнервных (Кометиани, 76).
Это обстоятельство имеет важное значение в нормальной работе
мышц. Чем больше будет нервных окончаний в мышечном волокне, тем
сильнее будет сократительная способность волокна, тем выше будет
развиваемое напряжение. Следовательно, множественная двигательная
иннервация мышечных волокон имеет большое биологическое значение.
Мб
ОБЩАЯ ФИЗИОЛОГИ» МЫШЕЧНОЙ СИСТЕМЫ
Практикум
1. Укорочение и утолщение мышцы во время сокращения. У мышц semitendinosus
(одной головки ее) перевязывают оба конца ниткой. Один конец прикрепляют
к штативу, а к другому концу привязывают гирьку в 5 г и перебрасывают
через блок, как на рис. 41. Мышца раздражается тетанзирующими токами.
Циркулем измеряют длину и толщину-
мышцы до и во время сокращения.
2. Одиночное сокращение и
скрытый период. Берут лягушку
и перерезают мозг в атланто-за-
тылочном сочленении. Разрез
приходится в области calamus scrip
tortus продолговатого мозга.
Такой препарат называется
спинномозговым. Препарат укладывают
на пробочную пластинку дорзаль-
ной поверхностью кверху и
прикалывают булавками. Перерезают
седалищный нерв в области
бедра, чтобы избежать
рефлекторного возбуждения мышц. Затем
обнажают гл. gastrocnemius,
перерезают ахиллово сухожилие, на
которое предварительно
навязывается нитка для соединения с
миографом.: На проксимальный конец
мышцы накладываются электроды,
соединенные со вторичной
катушкой. В этом опыте употребляется
кимограф Криса (рис. 75). Он
устроен так, что цилиндр делает
только один быстрый поворот и
при этом повороте размыкает
первичную цепь индуктория.
Вызванное этим раздражением одиночное
сокращение записывается на
цилиндре во время поворота. При
отсутствии кимографа Криса
можно употребить любой быстро
вращающийся кимограф. В этом
случае кимограф останавливается
рукой. Кимограф имеет
приспособление, с помощью которого
цилиндр при своем вращении
размыкает первичную цепь (рис. 76).
Сначала отмечают на
неподвижном цилиндре то положение
миографа, в котором он должен
находиться в момент
раздражения; замыкают цилиндром цепь и
записывают миографом
вертикальную линию. Затем пускают
кимограф в ход. Сокращение
Рис. 75. Полусхематический рисунок кимографа
Криса.
D—диск вращается вместе с цилиндром всего один раз.
Вращается же он под действием тяжести Р, после того как
отпустят клин Ь. Сделав один оборот, цилиндр снова
задевает за клин и останавливается. На диске имеется два
штифтика. Один открывает контакт Hlt отодвинув С,
который прижимается пружиной е; а другой открывает таким
же образом другой контакт Ка с противоположной стороны.
Один штифтик неподвижный, он находится на том месте,
где стоит (Xj), а другой подвижный, он виден спереди (Хв).
Если подвижной штифтик поставить на 0 делений на диске,
тогда интервал между размыканием контактов будет
равняться нулю. После передвижения штифтика в ту или
другую сторону размыкание одного контакта будет
происходить раньше другого. Эти контакты вводятся в первичную
цепь индуктория. При записи одиночного сокращения
вводится в цепь только один контакт.
С0КРА1И.ТЕЛЫ1АЯ- ФУНКЦИЯ МЫШЦЫ
іі7
начинается после вертикальной линии. Во время опыта записывается колебание
электромагнитного сигнала в 50—100 колебаний в секунду. Этим пользуются для
определения длительности скрытого периода и сокращения.
3. Влияние температуры на одиночное сокращение и на скрытый период его.
Сначала записывают сокращение описанным выше способом при камнатаой
температуре, а затем согревают мышцу накладыванием ваты, смоченной теплым
физиологическим раствором (38°). Новая запись происходит при этой температуре. Конечно,
во время записи вата и термометр удаляются.
Необходимо, чтобы кривая сокращения — миограмма— в точности
воспроизводила акт сокращешя. Для этого нужно избегать сильного трения пера о цилиндр.
Р из. 76. Постановка опыта с записью одиночного сокращения на быстро вращающемся
кимографе. Объяснения' см. в тексте.
Такое трение может совершенно исказить кривую сокращения, сильно удлиняя ее
нисходящее колено. Во избежание искажения кривой сокращения в силу трения
пишущее перо должно лишь слегка касаться закопченной бумаги, и, кроме того, к
миографу должна быть привешена небольшая тяжесть возможно близко к оси
миографа.
4. Зубчатый и полный тетанус. Тот же препарат лягушки. Теперь берут
m. gastrocnemius другой стороны. Для получения полного тетануса пользуются
электромагнитным прерывателем, который имеется при индуктории Дюбуа-Реймона.
Для получения зубчатого тетануса необходимы раздражения не реже 5—10 в
секунду. Это можно сделать рукой. Для получения максимального тетануса
вторичная катушка ставится приблизительно на 10 см выше порогов.
5. Контрактура. Тот же препарат и та же мышца. К мышце прикладывается
сильное тетаническое раздражение продолжительностью в 10 сек. После сокращения
мышца расслабляется с большой постепенностью. Это остаточное сокращение и есть
контрактура. Она будет тем длительнее и интенсивнее, чем сильнее и длительнее
раздражение и чем ниже температура.
6. Эластические и пластические свойства; собственный тонус. У лягушки
вырезывают m. sartorius осторожно, чтобы не растянуть сильно. Один конец укрепяют в шта-
118
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
тиве, как на рис. 76, а другой соединяют с миографом без груза. Последний пишет
на кимографе. Во время медленного хода кимографа привешивают груз, в 20 г.
Мышца растягивается на определенную длину. Если сейчас удалить груз,
мышца укортится почти на ту же длину. Это будет проявлением эластических
свойств мышцы.
В следующем опыте оставить груз в 20 г висеть полчаса. После начального
быстрого удлинения мышца будет удлиняться понемногу в течение всего получаса.
Затем удалить груз. Мышца тотчас же укоротится до определенной длины, а
затем будет укорачиваться дальше очень медленно в течение получаса. Это
медленное удлинение при постоянном грузе и медленное укорочение по удалении
груза выявляют пластические свойства. Еще лучше наблюдать пластические
свойства на pect. pars abdom или на rectius abdom. На этих мышцах можно поставить
еще такой опыт. Вырезать ту или другую мышцу, измерить длину и положить в
раствор Рингера. Через 20—30 минут мышца окажется значительно короче. Можно
растянуть мышцу и вновь положить в раствор Рингера. Она .вновь укоротится.
В этом выражается активный характер собственного тонуса мышцы.
7* Действие ацетилхолина на мышцы. Взять m. sartorius, проксимальный конец
зафиксировать, а дистальный соединить с изотоническим миографом. Кроме того, с
другим изотоническим миографом соединить проксимальную треть, как на рис. 69.
Оба миографа нагрузить по 5 г. Затем приложить к середине дистальной трети
кусочек фильтровальной бумаги или лигнина величиной в 3—4 мм2, пропитанной раствором
ацетилхолина в Рингере 1 : 10 000—30 000. Ацетилхолин вызовет сначала быстрое, а
потом медленное сокращение. Быстрое сокращение 'будет наблюдаться и на
проксимальной трети, а медленное — только на дистальной части. Для удачи этого опыта
надо, чтобы функциональное состояние мышцы было хорошим, т. е. возбудимость
была высокой. На мышцах, долго пролежавших в физиологическом растворе,
утомленных или взятых от истощенных лягушек, быстрого сокращения не будет, но
может отсутствовать и .медленное сокращение.
Если вместо гл. sartorius взять т. pect. pars abdom. и поставить на ней
такой же опыт, результат будет такой же, как (и іна т. sartorius, с той разницей,
что медленное сокращение будет продолжаться не около минуты, как в случае
m. sartorius, а несколько минут, благодаря повышенной -пластичности этой мышцы.
Если через 2—4 минуты мышцу растянуть до нормальной длины, которая была
до отравления, она сохранит эту длину. Это доказывает, что данное укорочение
через 2—4 минуты является чисто пластическим.
8. Сократительная способность разных мышц лягушки. В спинномозговом
препарате освобождают ряд мышц по всей длине, оставляя их в связи только
с одним суставом. К свободным концам привязывают нитки. Затем одну за другой
записывают іна 'кимографе кривые максимального укорочения мышц. Для этого
опыта -удобно, чтобы оба плеча миографа были одинаковой длины. В этом случае
высота кривой точно выражает величину укорочения. Один электрод накладывается
на один конец мышцы, а другой — на другой конец, чтобы ток проходил через всю
мышцу. Испытываются следующие мышцы: sartorius, semitendinosus, gastrocnemius,
peroneus. M. peroneus освобождается с проксимального конца, и нитка
привязывается к этому концу. Остальные мышцы освобождаются с дистального конца.
Определить процент укорочения.
9. Сократительная способность разных участков одной и той же мышцы. Из
мышцы sartorius вырезают экваториальную часть ее, т. е. тот участок, куда входит
нерв, длиной 'В 10—15 імм. Затем берут проксимальный участок такой же длины.
На оба конца навязывают до нитке. Одна нитка закрепляется в зажиме, другая
соединяется с миографом. іРаздражают тетаиически обычными электродами. Эффект
записывается на кимографе. Определяется процент укорочения.
10. Напряжение мышцы и его изменение в зависимости от начального
напряжения. Для регистрации напряжения мышцы взять изометрический миограф с
воздушным регулятором (рис. 23). Сначала установить воздушный регулятор так, чтобы
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
U9
рычажок миографа не давал собственных колебаний по инерции. Это достигается
поворачиванием винта регулятора на определенное расстояние от нуля. Затем
проверяют действие миографа: на какое расстояние отклоняется конец вертикального
миографа от груза в 50, 100, 500 и'1000 г. После проверки соединяют миограф • с
m. gastrocnemius спинномозгового препарата лягушки. Раздражают седалищный
нерв максимальными отдельными индукционными ударами при разных начальных
напряжениях. Сначала регистрируют напряжение, вызываемое раздражением при
отсутствии растяжения мышцы, когда (вертикальный рычаг стоит на 0. Записывают
кривую напряжения на кимографе. Затем- мышцу растягивают так, чтобы
вертикальный рычаг ^передвинулся по шкале на расстояние, соответствующее 50 г.
523Л5^ч>
Рис. 77. Схематическое изображение одновременной регистрации
напряжения всей мышцы с помощью изометрического миографа и изменения длины
одного участка мышцы .изотоническим миографом.
Здесь на рисунке проксимальный конец (Р) связан с изометрическим миографом, а ди-
стальная треть—с изотоническим. El обозначает участок раздражения.
Снова раздражают нерв той же силой и записывают кривую напряжения. Вновь
увеличивают начальное напряжение до 100 г и вновь записывают кривую
напряжения. Также записывают напряжение от сокращения при начальном напряжении
в 200, 500 г и выше, пока добавочное напряжение от сокращения мышцы не станет
равным нулю.
Аналогичные опыты производятся с тетаническим раздражением.
11. Механическое взаимодействие разных участков m. sartorius при сокращении.
Сначала устанавливают изометрический миограф так, чтобы при изменении
напряжения в пределах до 100 т 'вертикальный рычаг передвигался более или менее
чувствительно. Для этого (вставляют более слабые пружины, затем устанавливают
шкалу путем действия грузов в 10, 20, 50 и 100 г. После этого фиксируют один
конец изолированной мышцы sartorius, а другой соединяют с
изометрическим миографом. 'Кроме того, соединяют ту же мышцу с изотоническим
миографом, как .показано на рис. 77. Оба миографа записывают на одном /кимографе.
На проксимальном и дистальном участках мышцы расположены электроды для
раздражения. Каждая пара электродов связана с особой индукционной катушкой.
Производят следующие опыты: а) раздражают фиксированный конец сначала
отдельными индукционными ударами, затем тетанизирующими токами; б) раздражают
другой конец мышцы таким же образом; в) сочетают тетаническое раздражение
одного конца с раздражением другого отдельными ударами или тетанизирующими
токами. Получают эффекты, подобные изображенным на рис. 57 и 58.
120
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Литература
1. Engelmanft Th., Pfl. Arch., 23, 571, 1880.
2. H firth le К., РП. Arch., 126, 129, 1909.
3. Stubel H., Pfl. Arch., 180, 209, 1920.
4. В r ti с к e E., Denkschr, W. Akad. Wiss., 15, 1858.
5. Meyerhoff O., Erg. Physiol., 22, 301, 1923.
6. Embde n G., Z. physiol. Chem.,93, 1914; 113, 1921; 118, 1922; 165, 1927;Handb-
norm. path. Physiol., Ill, 1, 369-, 1925.
7. Ко py 1 off G., Pfl. Arch., 157, 218, 1913.
8. Schwenker G., Pfl. Arch., 157, 443, 1914.
9. Bernstein J., Pfl. Arch., 92, 54, 1902.
10. Bethe A., Pfl. Arch., 199, 491, 1923.
11. Бериташвили И., Общая физиол. мыш. и нервн. систем, Тифлис, 1922.
12. F 1 е t с h е г W. и. Н о р к і n s F., J. Physiol.,, 35,247, 1907; Proc. Roy. Soc,
В, 89,444, 1917.
13. S t а и b e W ., Pfl. Arch., 194, 574, 1922.
14. Bethe A., F r a e n к e 1 M. u. Wilmers J., Pfl. Arch., 194, 45, 1922.
15. Schott A., Pfl.Arch., 194, 271, 1922.-
16. Plaut K., Pfl.Arch., 202, 410, 1924.
17. T i e g e 1 E., Pfl. Arch., 13, 71, 1876.
18. В е г i t о f f J., Pfl. Arch., 198, 590, 1923.
19. Riesser Ot., Arch. exp. Path. Pharm., 91, 342, 1921; 92, 254, 1922.
20. 4 e I m h о I t z H., Wissensch. Abh., 2, 924, 1883.
21. Loven С h., Arch. Anat. Phys., стр. 368, 1881.
22. В e r n s t e i n J ., Pfl. Arch., 11, 191, 1875.
23. Grutzner, Bresl. Aerztl. Ztg., № 18, 1881; № 1, 1887.
24. Rosner A., Pfl. Arch., 81, 105, 1900.
25. В er it off J. u. Jasch w i 1 i D., Pfl. Arch., 205, 465, 1924.
26. Z с h а к а у а M., Pfl. Arch., 209, 753, 1925.
27. E.mbden G., Klin. Wschr., 3, 1393, 1924.
28. MeyerhofO., Naturwiss., H. 18—19, 1929.
29a. Frank G., Pfl. Arch., 218, 37, 1927.
29b. UexkullJ. u. Stromberger K., Pfl. Arch., 212, 625, 1926.
30. H i 1 1 A. V., Muscular Activity, 1926. Есть русск. перевод.
31. Bethe A., Pfl. Arch., 19Э, 491, 1923.
32. Ber itoff J., Z. Biol., 87, 574, 1928.
33. Fischer E., Pfl. Arch., 203, 580, 1924.
34. Fischer E., Pfl. Arch., 213, 352, 1926.
35. Beritoff J.u. Jaschwili D., Pfl, Arch., 205, 475, 1924.
36. A z и m a R., Proc. Roy. Soc, B, 96, 338, 1924.
37. Jamin Fr„ Handb. norm. path. Physiol., Bd. 8, 540, 1925.
38. F и 11 о n J., Muscular Contraction etc., London, 1926.
39. Garner W., Proc. Roy. Soc, B, 97, 40, 1925.
40. Meyerhoff O., Naturwiss., Dezemb., стр. 977, 1932.
41. E g g 1 e t о n P. и. E g g 1 e t о n G. P., Bioch. J., 21, 190, 1927.
42. L и n d s g a ar d, Bioch. Z., 217, 162; 220, 1, 8; 227$ 51, 1930; 230, 10; 233, 322, 103!*
43. H i 1 1 A. V., Physiol. Rev., 12, 56, 1932.
44. Bremer F г., J. Physiol., 76, 65, 1932.
45. S t e i n h а и s A. H., Physiol. Rev., 13, 103, 1933.
46. M о r p и г g о В., Virchow's Arch., 90, 522, 1897.
47. Siebert W. W., Z. klin. Med., 99, 350, 1928.
48. НарикашвилиС. Бюлл. эксп. биол. и мед., 7, 142, 1939.
49. R a u h F г., Z. Biol., 76, 25, 1922.
50. Bethe А. и. Н а р р е 1 P., Pfl. Arch., 201, 157, 1923.
СОКРАТИТЕЛЬНАЯ ФУНКЦИЯ МЫШЦЫ
ш
51. В 1 і х М., Skand. Arch. Physiol., 5, 150, 1895; ibid., 3, 295, 1893; ib., 4, 399, 1894-
52. Fenn W. Q., Handb. norm. path. Physiol., Bd. 8, H. II, 5, 146, 1925.
53. M e у e r h о f O.u. LohmannK., Bioch. Z., 196, 22, 49, 1928.
54. Hartmann, Bioch. Z., 270, 164, 1934; Hartmann u.Meyerhoff, Pfli
Arch., 234, 722, 1934.
55. Cooper S. a. E с с 1 e s J. C, J. I. Physiol., 69, 377, 1930.
56. Beck A., Pfl. Arch., 224. 278. T^O.
57. Meyer Kurt H., Bioci.. Z., 214, 253, 1929.
58. Holz Bern, Pfl. Arch., 23U, 246, 1932.
59. v. Muralt A., Pfl. Arch., 230, 299, 1932.
60. Мужеев В. А.иБауэр Ф.С., Биолог, журн., 3, вып. 2—3, 292, 1933.
61. Ncuschloss, Pfl. Arch., 199, 410, 1923; 204, 374, 1924; 207, 27, 37, 1925; 213,,
19, 40, 47, 1926.
62. Hober R., Pfl. Arch., 221, 479, 1929.
63. Hurthle K., Pfl. Arch., 223, 685, 1930.
64. Clark, Egg le ton. a. E g g 1 e t о n, J. Physiol., 74, 1, 1934.
65. Rehsteiner R., Pfl. Arch., 217, 430, 1927.
66. PlattnerF. u. Kranich E., Pfl. Arch., 229, 730, 1932; 230, 356, 1932..
67. Btittner H. E. u. Heimbrecht В., Pfl. Arch., 221, 43, 1928.
§8. v. F r e y, Festschrift fur Ludwig, 55, 1887.
69. Wachholder K., Pfl. Arch., 233, 683, 1934; Wachholder u. v.Lede-
bur J. Fr., Pfl. Arch., 229, 657, 1932.
70. D ii r i g A., Pfl. Arch., 87, 42, 1901.
71. Thauer R., Pfl. Arch., 234, 276, 1934.
72. Herinrich A., Pfl. Arch., 230, 596, 1932.
73. Stissner H. u. Heinrich A.,Pfl. Arch., 234, 521, 1935.
74. Ernst E., Pfl. Arch., 209, 613, 1925; 213, 144, 1926; 235, 562, 1935.
75. К a p t о 3 и я А., Тр. Физиол. инст. Тифл. унив. им. Сталина, I, 1936.
76. Кометиани П., Тр. Физиол. инст. Тифл. унив., 1.. 1936.
77. Meyerhof О., Die chemischen Vorgange im Muskel, S. 105—110, Berlin," 1930..
78. Meyerhof O.u Wolf W., Pfl. Arch., 236, 533, 1935.
79. К a p t о 3 и я А., Тр. Физиол. инст. Тбил. унив. им. Сталина, 3, 245, 1937.
80. Dale Н. Н., F е 1 d b е г g W. а. V о g t M., J. Physiol., 86, 353, 1936.
81. Brown G. L., J. Physiol., 89, 12P, 1937; 90, 438, 1937.
82. R a v e n t о s J., J. Physiol., 90, 9 P, 1937.
83i Rosenblueth A. a. Luc о J. V., Am. J. Physiol., 120, 181, 1937.
84. S с h a e f e r H. a. G 6 p f e r t H., Pfl. Arch., 239, 579, 1937.
85. Nakanishi S., L. Chosen med. assoc, 27, № 8, 128, 1937.
86. В u с h t h a 1 Fr., J. Physiol., 90, 37P, 1937.
87. Feng T. P., Chin. J. Physiol., 10, 417, 1936.
88. Feng T. P. a. Shen S. C, Chin. J. Physiol., 11, 417, 1937.
89. Verzar Fr., Bioch. Z., 132, 67, 1922.
90. Schleier JM Pfl. Arch., 197, 543, 1922.
91. Trendelenburg W. u. Schotz E., Z. Biol., 89, 41, 1930.
92. Brandstater P., Pfl. Arch., 240, 348, 1938.
93. Hett O., Pfl. Arch., 240, 588, 1938.
94. Cooper a. E с с 1 e s, J. Physiol., 69, 377, 1930.
95. Коштоянц и Рябиновская, Pfl. Arch., 235, 418, 1939; Коштоянц,-
Физиол. журн. СССР, 24, 221, 1938.
96. KrugerP., DuspivaF. u. Fur linger F г., Pfl. Arch., 231,?; 250, 1932.
97. В u с h t h a 1 F. a. L i n d h a r d J., The physiol. of striated muscle fibre,
Copenhagen, 1939.
98. Fischer E., Cold Spr. Harb. Symp. Quant. Biol., IV, 214, 1936.
99. P a r n a s J. О., О s t e r n P. u. Mann Т., Bioch. Z., 272, 64, 1934; 279, 24^,
1935. Parnas et Baranowski Т., С. г. soc. biol., 120, 307, Paris, 1935..
322 общая физиология мышечной системы
00f Ф<ердман Д., Сб. докл. VI Всес. физиол. съезда, стр. 432,
01. Введенский Н., О соотношении между возбуждением и раздражением при
тетанусе, П., 1886.
02. Верит о в И., Физиол. журн. СССР, 27, вып. б, 1939; Докл. Ак. Наук СССР,
23, 597, 1939.
03. Б е р и т о в И» Сборн. трудов, посвящ. 50-летию научно-педагогич.
деятельности проф. В. Воронина, стр. 66, Тбилиси, 1941.
04. Г и л л А., Эпизоды из области биофизики, Биомедгиз, 1935.
05. Lohmann, Naturwiss., 17, 624, 1929.
06. Коштоянц X. С, Физиол. журн. СССР, 24, 221, 1939.
07. Bouman Н. D. u. Rijnberk G. v., Neerl. de Physiol, de l'homme et
des animaux, 23, 441, 1938.
08. Ernst E., KHn. Wschr., № 45, 1936, Jahrg. 15.
09. Pant in С F. A., J. Exp. Biol., 13, 148, 1930.
10. S о m m e r k a m p f H., Arch. exp. Path. Pharm., 128, 107, 1928.
11. S i с h e I F. S. AL, J. Cell. Сотр. Physiol., 5, 21, 1934.
12. Lohmann K., Der Stoffwechsel des Muskels. Handb. d. Biochem. Erg.-Werk.,
Bd. 3, 1936.
13. R о 1 1 e t, по Бидерману, Elektrophysiologie, стр. 55, 1895.
14. F u 1 t о n F., Proc. Roy. Soc, 97—B, 406, 1925.
15. Biedermann W., Elektrophysiologie, SS. 62—65. Jena, 1895.
16. Nachmanson D. et Marnay A., C. r. soc. biol., 121, 1311, Paris, 1936.
17. M a a s J. A., Die visco-elast. Eigenschaften von Kautschuk und glatten Muskeln
usw. Dissrt. Naturw. Fak. Univ. Utrecht, 1938; Arch. Neerl. Physiol., 23, 141, 1938.
18. R a m s e у R. W. a. Street S. F., J. Cell. Сотр. Physiol., 15, 111, 1940.
19. Wachholder K., Pfl. Arch., 226, 255, 1930.
20. R a n s о n S. W., Am. J. Physiol., 86, 302, 313, 1928.
21. Берйтов И. и H и в и н с к а я О., Тр. Инст. физиол. Тбил. унив.
им. Сталина, 4, 1941.
22. Bottazi, Arch. Anat. Physiol., стр.377, 1901.
23. Bremer F., J. Physiol., 76, 65, 1932.
24. Hartr ее W. a. H i 1 1 A. V., J. Physiol., 56,239, 1922; ibid., 58, 441, 1924
Proc. Roy. Soc, B, 103, 207, 1928.
25. F u r u s a w a K. a. Hartree W., Pfl. Arch., 211, 644, 1928.
26. M e у e r h о f f O. u. Lohmann K., Pfl. Arch., 210, 790, 1925.
27. Feng T. P., J. Physiol., 74, 441, 1932; ibid., 74, 455, 1932.
28. Margaria R., J. Physiol., 82, 496, 1934.
29. H a r r e w e 1 d A. v. a. Wiersma С A. G., J. Exp. Biol., 14, 448, 1937.
30. W i e r s m a С A. G. a. H a r r e w e 1 d A. v., Physiol. Zool., 12, 43, 1939.
31. H a r r e w e 1 d A. v. a. Wiersma С A. G., J. Physiol., 88, 78, 1936.
32. В e r g e n W. R. a. Wiersma C. A. G., J. Gen. Physiol., 22, 193, 1938.
33. Weber H. H., Ergebn. Physiol., 36, 109, 1935; Pfl. Arch. 235, 205, 1939.
34. Энгельгардт В. А., Усп. совр. биол., 14, 177, 1941.
35. Szent-Gy orgy i A. N., Studien on Muscle, Acta Physiol. Scand., 9, Suffl,
XXV, 1944; Бюлл. эксп. биол. и мед., 20, 15, 1945.
Глава IV
СТРУКТУРА ЖИВОЙ ВОЗБУДИМОЙ СИСТЕМЫ
Саркоплазматическая сеть и ее значение. Как указывалось выше,
плотная часть мертвой окрашенной саркоплазмы, имеющая в области
нервных окончаний ясно выраженное сетчатое строение, является
возбудимой системой. Гистолог Буке (1) так характеризует эту сеть*
названную им перитерминальной: «Ч^сто іможно видеть более или
менее сетчатое строение в той саркоплазме, где лежит нейрофибрилляр-
ное сплетение двигательного нервного окончания. Саркоплазматическая
сеть представляет как бы прямое продолжение этого сплетения. Здесь
она развита сильнее всего. Подальше от нервного окончания эта сеть
постепенно уменьшается: сетчатость реже и видна хуже. Еще дальше
она редко заметна. Эта сеть, с одной стороны, сливается с нейрофиб-
риллярной сетью, а с другой — стоит в связи с миофибриллями,
а именно с их анизотропным веществом» (рис. 35), По исследованиям
Буке, здесь в анизотропном веществе перитерминальная сеть
кончается тончайшим разветвлением. Все это сетчатое образование, по
мнению Буке, представляет как раз то вещество, которое Лэнглй
называет воспринимающим веществом, ибо нейрофибриллярная сеть
непосредственно связана только с этим сетчатым образованием. Мы
же, как указывали выше, называем это образование возбудимой
системой. Следовательно, мышечное волокно получает нервные импульсы
возбуждения через эту сетчатую структуру, через нее же возбуждение
распространяется вдоль по волокну. Возбуждение мышцы при прямом
раздражении также должно быть функцией этой саркоплазматической
сетчатой структуры. С этим хорошо согласуется отмеченное выше
действие кураре и никотина на денерв'ированную мышцу, а также то
явление, что в области нервных окончаний, где саркоплазматическая
сеть больше развита, возбудимость выше, чем в безнервных участках
(Беритов и Яшвили, 2), а процесс возбуждения протекает значительно
сильнее (Беритов, 3).
Ячеистое строение саркоплазмы Буке установил на мертвых
мышцах, обработанных гистологическими методами. Ячейки возникают, пови-
д'имюму, в результате коагуляции — свертывания белка, составляющего
ш
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
основную массу протоплазмы. В живой мышце нет ячеистого строения.
Саркоплазма, как и всякая другая протоплазма, находится в
коллоидальном состоянии; она состоит главным образом из гидрофильных,,
т. е. богатых водой, коллоидальных белковых частичек. Но, безусловно,,
наличие определенных ячеистых структур после гистологической
обработки свидетельствует, что и в живой клетке протоплазма организована,
определенным образом, что основная белковая імасса ее в каждой части
(мышечной клетки составляется из мицелл, распределенных в
определенном порядке.
Коллоидальное состояние живой возбудимой системы. Протоплазма и
ядро мышечной клетки образованы из сложных коллоидальных систем.
Из этих систем составляются плотные структурные образования,
которые по своим свойствам являются гелеобразным гидрофобным плотнымг
коллоидом. Эти более твердые образования находятся в равновесии
с гидрофильными, жидкими коллоидами, которые заполняют все
свободное от плотных образований пространство клетки. Различие свойств*
плотных и жидких коллоидов протоплазмы обусловлено особым физико-
химическим состоянием белковых веществ и связанных с ними липоидов.
Частички белка в гидрофильном коллоиде содержат воду в виде
водной оболочки, где растворены соли и органические вещества. Но эта
водная оболочка не сплошная. Известный специалист Лепешки н-
(14) утверждает, что по многим признакам она находится в петлях
боковых углеводных цепочек, которые имеются на поверхности
белковых мицелл. Вода, кроме того, заполняет все интермицеллярные
пространства. В этой интермицеллярной фазе также растворены соли и
органические соединения. Как эта вода, так и находящаяся в
гидрофильном коллоиде не является простым растЕзрителем, ибо она содержит
липоиды — сложные жироподобные вещества в коллоидальном
состоянии. Поэтому протоплазма, вытекающая из пораненной клетки, не
смешивается с водой.
Протоплазма в целом представляет единое целостное образование
из коллоидальных частичек и воды, где вода удерживается в
определенном пространственном расположении взаимодействием
электростатических сил воды и коллоидальных частичек. Когда эти силы
почему-либо становятся меньше, молекулы воды освобождаются и, сливаясь,,
образуют вакуоли.
Гидрофильные коллоидальные частички легко переходят в
гидрофобные, бедные водой; при этом они коагулируют, что связано с
потерей водной оболочки. Все повреждающие воздействия солей, кислот и
других ядов, а также высыхание и механическое влияние вызывают
постепенную коагуляцию живой системы. В нормальной живой клетке
также существуют гидрофобные коллоидальные частички. Из этих
частичек составляются такие плотные образования, как нейрофибрилли,.
миофибрилли, оболочки ядра, хондриозомы, митохондрии и др.
По Лепешкину, гидрофильные и гидрофобные коллоидальные
частички представляют химические соединения белков и липоидов.
СТРУКТУРА, ЖИВОЙ ВОЗБУДИМОЙ СИСТЕМЫ
125
Этим объясняется,. что протоплазма хорошо растворяет в себе все те
вещества, которые растворяются в воде и липоидах. Липоиды имеются
также в интермицеллярной фазе. Это способствует проникновению и
распространению в клетке тех веществ, которые растворяются в
липоидах. Липоидобелок, или липопротеид, и представляет тот плотный
коллоид, из которого строятся все внутренние структурные образования
живой протоплазмы и в котором протекает обмен веществ. Он не
окрашивается органическими красками. Наоборот, оболочки протоплазмы и
ядра, состоящие из одного белка, окрашиваются. Протоплазма в целом
дает диффузную окраску только при повреждении клетки, когда уже
происходит расщепление липопротеидов на белок и линоид.
При повреждении, ©идимо, происходит не только расщепление
липопротеида на белок и липоид, но и денатурация белка. С денатурацией
же происходит повышение активности свободных аминокарбоксильных
групп или же увеличение их, вследствие чего повышается их
способность связывать как основные, так и кислые краски (Александров и
Насонов, 21).
Липопротеиды, находясь в коллоидальном состоянии, являются
местом ферментативных процессов, протекающих в живой клетке.
Вещества, которые адсорбируются коллоидальными частичками липопро-
теида, приходят в такое тесное соприкосновение с другими, и притом
в такой концентрации, что очень легко и быстро осуществляются
разного рода сложнейшие синтетические процессы. Липопротеиды могут
связать химически как металлы, так и разного рода органические
соединения.
Липопротеиды — очень мало устойчивые соединения. При разного
рода внешних воздействиях, а также без них они легко расщепляются
при участии тех же ферментов, переходя в более устойчивые
соединения, как белок, липоиды,.органические кислоты, энзимы, гормоны и т.д.
При этом освобождается энергия в .виде тепла, лучистой энергии,
электричества. После расщепления данной частички липопротеида
происходит восстановление другой такой частички липопротеида с
помощью тех же ферментов и перед тем освобожденной энергии (Лепеш-
хин, 14).
Нужно думать, что и помимо внешних влияний все время происхо*
дит не только расщепление липопротеида, но и более глубокое
изменение — денатурация белка. Но в живой клетке даны все условия, чтобы
денатурированная часть белка сейчас же восстановилась обратно в ту
форму белка, которая вступает в связь с липоидом и образует
липопротеид. Осуществляется же этот процесс, очевидно, путем обмена
веществ, той энергией, которая освобождается ів связи с обменом
веществ (Александров и Насонов, 21).
Данное выше представление о структуре живой системы
существенно отличается от господствующего до сих пор воззрения на структуру
протоплазмы как на смесь коллоидов, где в одном коллоиде, более
жидком, внешняя фаза является водой, а в другом, более плот-
126
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ном, внешней фазой являются белок и липоид. На границе между
этими коллоидами предполагалось существование полупроницаемых
осадочных перепонок, которым приписывалось регулирование
проникновения и распространения разного рода веществ внутри клетки (Ге-
бер, 4; Лилли, 13 и др.). Но в настоящее время Лепешкин и другие
авторы находят, что полупроницаемые перепонки не всегда существуют
на поверхности протоплазмы или составных частей ее, а там, где они
существуют, они не обусловливают в полной мере проникновение
веществ внутрь протоплазмы.
Обе составные части живой системы, как коллоидные липопротеид-
ные частички, так и промежуточная, интермицеллярная фаза, содержат
соли, жиры, углеводы, белки, ферменты и др. Однако характер их
существования в обеих средах не одинаков. В интермицеллярной фазе
они, повидимому, находятся, подобно тому как в плазме крови или
лимфе, в виде самостоятельных простых соединений; в липопротеидных
частичках они, повидимому, образуют очень сложные химические
соединения, объединенные вокруг белковой основы и притом имеющие
очень непостоянный характер, так как эти соединения все время
претерпевают процессы расщепления и восстановления.
Некоторые ученые придерживаются того мнения, что протоплазма
нормальной клетки не имеет структуры, так как в микроскопе
протоплазма клетки выглядит бесструктурной. Но в последние годы многие
исследователи стали на ту точку зрения, что гомогенный вид
протоплазмы является одной из форм ее жизнедеятельности. Известно,
например, что протоплазма клетки при кариокинетическом делении
становится структурной. Некоторые же нормальные клетки всегда являются
структурными; например, нервные клетки показывают нейрофибрилляр-
ное строение после прижизненной окраски метиленовой синькой (Боц-
лер, 11). Цитоплазма свежих нервных клеток (чувствительных и
двигательных) амфибий, при изучении с помощью поляризационного
оптического прибора, дает доказательство присутствия ориентированных
частичек протеина; они должны 'представлять материал для
производства видимых на глаз нейрофибриллей (Чинн, 17).
Установлено, что в живых нервных волокнах головастика лягушки
нейрофибрилли становятся видимыми в результате всякого рода
раздражений: химического, механического, температурного и т. д. (Шпей-
дель, 18). Далее было замечено, что вообще структура нервной клетки
меняется в связи с функциональными изменениями. Например, на
живом сердце лягушки цитоплазма нервных клеток внутрисердечных
ганглиев в покойном состоянии не обнаруживает структуры. Но при
переходе возбуждения с волокон блуждающего нерва на эти клетки
цитоплазма меняется: в ней появляется грануляция, контуры ядра лучше
видны, концевые бляшки синапсов становятся более выраженными
(рис. 155-П). Очевидно, оптические свойства протоплазмы изменились
в связи с изменением структуры (Степанова и Крохина, 25). Такое же
изменение наблюдается при действии на сердце переменных токов
СТРУКТУРА ЖИВОЙ ВОЗБУДИМОЙ СИСТЕМЫ
\2Т
ультравысокой частоты (Лаврентьевой Федоров, 12). Наконец,
известно, что нейрофибрилли нервных клеток ящериц в активном состояния
становятся тоньше и количество их неизмеримо возрастает. С другой
стороны, во время зимней спячки в тех же клетках замечается только»
несколько толстых нейрофибриллей (Ариенс Кдпперс, 20).
Но и тот факт, что почти все клетки при гистологической обработке-
обнаруживают структуру, причем всегда одного и того же типа для
каждого рода клеток, должен свидетельствовать, что эта структура не
является всецело искусственным продуктом. Она безусловно должна,
выражать форму изменения функциональной деятельности клетки в
период ее умирания. Подробные исследования установили, что всякое
повреждающее действие — химическое, механическое, электрическое,
термическое — производит во всех живых клетках одно и то же
структурное изменение, которое при небольшой степени наносимого
повреждения является обратимым, т. е. структура клетки возвращается к норме-
по удалении повреждающего агента. Это структурное изменение
заключается в денатурации белков внутри клетки, в связанном с нею
понижении дисперсности клеточных коллоидов и в повышении сорбцион-
ного уровня белкового состава к прижизненным красителям (Насонов,.
16; Макаров, 15).
Итак, можно утверждать, что структура является основой для
функциональных отправлений клетки. При этом, с одной стороны, рост и
развитие структуры обусловливают развитие функций, с другой —
усиленная функциональная деятельность ведет к усиленному росту и
развитию структуры. Это положение было выдвинуто еще Фр.
Энгельсом (19).
Впоследствии оно много раз доказывалось прямыми опытами. Так,.
В а р б у р г (7) обнаружил, что газовый обмен растет
пропорционально количественному росту структуры. Мы выше указывали, что липо-
протеид является основой обмена веществ. Поэтому, когда внутренняя*
структура клетки, строящаяся из липопротеида, растет и развивается,
безусловно должен расти и обмен веществ. Это же утверждают в
настоящее время выдающиеся биологи, как Гертвиг (5), Чайльд
(6) и др. Вот, например, как формулирует Чайльд взаимоотношение-
между структурой и функцией: «Развитие есть процесс
функциональной конструкции, т. е. если начать с какой-либо структуры и функции,.
продолжение функционирования вызовет изменение структурного
субстрата, а это в свою очередь изменит функцию и т. д.» (6).
Итак, живая возбудимая система представляет коллоидное
образование, в котором необходимо отличить гидрофильно-коллоидные и
дисперсионные (гидрофобные) частички, с одной стороны, и интермицел-
лярную фазу — с другой. Из коллоидных гидрофильных и
гидрофобных частичек составляется вся структурная часть живой протоплазмы,,
интермицеллярная фаза заполняет свободные от этой структуры
пространства, причем эта структура живой системы чрезвычайно
изменчива, так как она всецело определяется функциональной деятельностью»
128 ОБЩАЯ ФИЗИОЛОГИИ МЫШЕЧНГОЙ СИСТЕМЫ
которая все время меняется под влиянием внешних и внутренних условий.
О взаимодействии между коллоидными частичками и
интермицеллярной фазой. Все вещества, проникающие в клетку из внешней среды,
как кислород, белок, жир, углеводы, соли, сначала распространяются
в водяной среде, а потом, путем адсорбции, втягиваются коллоидными
частичками внутрь и здесь под влиянием ферментов (энзимов)
претерпевают синтез, образуя сложное белковое тело, которое ©след за тем,
под влиянием тех же энзимов, претерпевает сложный распад (Вар-
бург, 7; Рона 8). Как известно, энзимы действуют на вещества как
катализаторы, т. е. ими обусловливаются все синтетические и
аналитические процессы в клетке. По новейшим исследованиям Опарина (23),
синтезирующее действие фермента непосредственно связано со
структурой. Находясь в клетке в адсорбированном, связанном состоянии,
фермент оказывает синтезирующее действие, очевидно, на все другие
вещества, адсорбированные той же структурой. Когда же фермент
находится в водной среде в свободном, растворенном состоянии, тогда он
оказывает только гидролизирующее действие. По Опарину, в
каждой живой клетке фермент находится в том и другом состояниях: гид-
ролизирующем — в интермицеллярной фазе и синтезирующем — в
мицеллах, в структурах. Соотношением этих состояний и определяется
направленность ферментативного действия в данной клетке.
По імнению многих авторов, ферменты, или энзимы, играли главную
роль в происхождении живой системы и во всей ее эволюции (Деркум
9; Опарин, 24). Итак, нужно думать, что внутри клетки разного рода
вещества то объединяются под влиянием энзимов вокруг липопротеида
в одно целое сложное белковое образование, то расщепляются на
простые соединения, и эти два процесса в каждой частичке клетки, чпови-
димому, непрерывно следуют один за другим.
В результате этого ферментативного процесса в интермицеллярной
фазе живой системы возникают разного рода продукты распада, как
креатин, аммиак, фосфорная кислота, угольная кислота и др. Отсюда
уже часть этих продуктов выходит из клетки наружу, другая же часть
принимает участие в синтетическом процессе, который происходит в
коллоидных частичках после их адсорбции. Таким образом, интерми-
целлярная фаза выполняет еще одну специальную роль: она является
передаточной средой между внешней средой и коллоидными мицеллами,
где протекают синтетические ферментативные процессы.
Изменение 'интермицеллярной фазы со стороны содержания воды
сильно отзывается на процессах, протекающих в мицеллах.
Достаточно, например, несколько уменьшить содержание воды в
интермицеллярной фазе, как живая система видоизменяется до такой степени, что
теряет все свои функции не только на время уменьшенного
содержания воды, но навсегда, т. е. даже после того как вновь возвратить ей
обычное содержание воды. Так, например, если высушить сердце
лягушки в эксикаторе не более чем на 25%, оно не умирает:
последующее увлажнение возвращает ему обычную для него спонтанную ритми-
СТРУКТУРА ЖИВОЙ ВОЗБУДИМОЙ СИСТЕМЫ
129
ческую деятельность. Дальнейшее же высыхание вызывает смерть
(Морозов, 10). Отсюда вытекает, что при сравнительно сильном высыхании
живая система существенно меняется. Очевидно, меняются при этом
обе фазы. Это ©едет к прекращению обычного обмена веществ в
клетке. По Лепешкину, уменьшение воды ів интермицеллярной фазе
ведет к повышению концентрации солей, которая в первую очередь
обусловливает -расщепление и коагуляцию липопротеидов, а затем еще
денатурирование, т. е. свертывание белка. При некоторой высокой
степени концентрации солей это изменение захватывает всю массу
протоплазмы и потому становится необратимым процессом, производящим
умерщвление живой системы.
Итак, интермицеллярная фаза и липопротеидная структура живой
системы находятся в таком взаимодействии или обусловленности, что
обе фазы составляют одно целое образование.
Литература
1. В о е k e J., Erg. Physiol., 19, 448, 1921.
2. Б е р и т о в И. и Я ш в и л и Д., «Чвени мецниереба», № 2—3, 1923.
3. В е г і t о f f J., Pfl. Arch., 213, 206, 1926.
4. H 6 b e r R., Physikalische Chemie d. Zelle u. Gewebe, 2. H., 1924.
5. H e r t w i g G.f Handb. norm. path. Physiol., Bd. 1, 580, 1927.
6. С h i 1 d С h. M., The Origin and Development of the Nervous System, Chicago, 1921.
7. Warburg O., Erg. Physiol., 14, 253, 1914.
8. Rona P., Handb. norm. path. Physiol., 1, 68, 1927.
9. D ere urn F г., Arch. Neurol. Psychol., 19, 1133, 1928.
10. Морозов Б., Журн. эксп. биол. мед., 6, № 16, стр. 63, 1927.
П. В о z 1 е г Е., Z. Zellforsch., 5, 1927; Z. vergl. Physiol., 6, 1927.
12. Лаврентьев Б. И. и Федоров Б. Г., Биологическое действие
ультравысокой частоты, стр. 145, ВИЭМ, 1937.
13. L і 1 1 і е R. SM Protoplasmic Action of the Nervous System, Chicago, 1932.
14. L e p e s с k i n W. W., Kolloidchemie des Protoplasmas, II AufL, Dresden u.
Leipzig, 1938.
15. Макаров П. В., Арх. анат., гистол. и эмбриол., 19, 55, 1938.
16. Насонов Д., Тр. Физиол. инст. ЛГУ, 14, 1933; Насонов Д. и
Александров, В., Арх. биол. наук, сер. А, 36, 1934; Биол. журн., 6, 1937.
17. С h i n n Р г., J. Cell. а. Сотр. Physiol., 12, 1, 1938.
18. Speidel С. С, Cold Spr. Harb. Symp. Quant. Biol., IV, 13, 1936.
19. Э h г e л ь с Ф р., Диалектика природы, стр. 20—21, 1925.
20. Ariens Kappers, The Сотр. Anat. of the Nervous System of Vertebr. inc.
Mann., vol, I, 1936.
21. Александров В. Я. иД.Н. Насонов, Арх. анат., гистол. и эмбриол., 22,
11, 1939.
22. Парнас Я. О., Усп. совр. биол., 12, 393, 1940.
23. Опарин А. И., Синтез органич. вещества и роль витаминов в растен ях,
изд. АН СССР, стр. 5, 1940.
24. Опарин А. И., Новый мир, № 7, стр. 233, 1940.
25. Степанова С.С. и Крохина Е. М., Арх. биол. наук. 61, 107, 1941.
9 и. С. Беритов
Глава V
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
Понятие об основном биологическом процессе. Процесс
расщепления и восстановления, который беспрерывно происходит в плотном
коллоиде, является основным признаком жизнедеятельности живой
системы. В этом заключается та особая форма движения, которая
свойственна живой системе в отличие от мертвой (Ф. Энгельс, 12). Этот
процесс не зависит по существу от изменений внешней среды. Он
протекает даже при полном отсутствии каких-либо внешних влияний, как
говорят, «самопроизвольно» или «спонтанно»: живая возбудимая
система в каждой своей части постоянно претерпевает расщепление
и восстановление. При расщеплении освобождается энергия, которая
может потребляться на-восстановление той же возбудимой системы; так,,
во всяком случае, 'бывает в зародышевой ткани зерна, в тканях
голодающих животных при совершенно неизменном состоянии внешней.
среды. Это значит, что расщепление и восстановление живой
возбудимой системы происходит и без поступления извне каких-либо
питательных веществ и без каких-либо изменений во внешней среде, В этом
характерном для живой системы процессе расщепления и
восстановления белок играет первенствующую роль. Вернее, этот -процесс является
свойством белка (Энгельс, 12). Но это свойство белка обнаруживается
исключительно в условиях существования его в живой системе,
вероятно, в виде лшіопротеидов в коллоидальном состоянии. Вне этой
системы белок-липопротеид не может расщепляться и восстановляться
беспрерывно.
О процессе расщепления и восстановления живой системы мы
прежде всего судим по обусловленному им обмену веществ. Благодаря
означенному «спонтанному» процессу живая возбудимая
система вступает в определенные обменные отношения с внешней
средой. Поступающие через кровь и лимфу питательные вещества: белки,
углеводы, жиры, разнообразные соли и другие вещества,
ассоциируются белковым образованием в период восстановления и являются
главным источником для производства энергии в период расщепления.
В этот период, который в конечном счете приводит к окислению
некоторых продуктов свободным кислородом, образуются разного рода
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
131
простые соединений, как углекислота, вода, аммиак, холин, креатин,,
мочевина и разные аминокислоты, разного рода соли, как фосфаты,
карбонаты, хлориды, и другие продукты. Эти продукты частью
выделяются наружу, частью вновь ассоциируются белковым образованием
вместе с другими веществами, поступившими из внешней среды. Это
восстановление происходит за счет той энергии, которая выделяется
при расщеплении и окислении.
Так' как в обмене веществ живой системы процессы расщепления
и восстановления липопротеида играют центральную, руководящую
роль, то в силу этого организация клеточной структуры и ее развитие
всецело находятся в зависимости от этого белка. Это он создает
клеточную структуру в результате обусловливаемого им обмена
веществ. Процесс беспрерывного расщепления И восстановления следует
считать свойством не всех белков, а исключительно сложных белковых
образований — липопротеидов. Эти образования должны находиться
только в структурных элементах живой клетки. Например, белки в ин-
термицеллярной фазе, подобно белкам лимфы или плазмы крови,
должны быть лишены этого свойства. Нет никаких указаний на то, чтобы
белки лимфы или плазмы крови были способны к обмену веществ и:
росту. То же самое можно сказать о белках интермицеллярной фазы.
Очевидно, условия существования белковых образований в мицеллах
совершенно своеобразны, и это своеобразие обусловливает их
чрезвычайную динамику в смысле беспрерывного расщепления и
восстановления.
Весьма вероятно, что ферментативное действие осуществляется в
живой протоплазме, в основном, входящими в нее белковыми телами.
Можно представить, что мицеллы белка содержат в своей структуре
на своей поверхности каталитически активные группы разного
характера. Они обусловливают как клеточное дыхание, так и другие
ферментативные процессы (Энгельгардт, 22).
Когда в какой-либо частичке живой системы происходит процесс
расщепления сложных белковых образований и возникают обычные
продукты распада, то для данной частички мы имеем фазу
диссимиляции, или катаболическую фазу. За этой фазой следует
восстановление тех же образований. Это будет фаза
ассимиляции, или анаболическая фаза. Оба эти процесса, как
указывалось выше, должны протекать при участии ферментов. Клетки
содержат множество чрезвычайно активных веществ с ферментативным
действием, с помощью которых поступившие из внешней среды
вещества, а также продукты внутриклеточного распада веществ принимают
участие в восстановлении сложных соединений белка и вообще в
организации клеточной структуры; таким путем происходит усвоение
веществ, или анаболический процесс. Но при участии тех же ферментов
этот белок все время претерпевает расщепление, «ли катаболический
процесс. Катаболическая фаза имеет существенное значение для
следующей анаболической, ибо во время расщепления освобождается'
9*
132
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
энергия, необходимая для анаболического процесса. Но безусловно,
как уже указывалось выше, энергия, освобождаемая в результате
ферментативного окисления некоторых продуктов белкового
расщепления, также должна итти на восстановление возбудимой системы.
Из вышеприведенного следует, что катаболический процесс
неизбежно является основным условием для обмена веществ, роста и
размножения живой системы, так как свободная энергия, необходимая
для усвоения питательных веществ во время анаболического процесса,
берется прежде всего из энергии, освобождающейся во время катабо-
лического процесса. Отсюда становится понятным, почему за катаболи-
ческим процессом всегда следует анаболический, и чем чаще будет
происходить катаболический процесс, тем сильнее будет развиваться
структура живой системы.
Беспрерывный процесс расщепления и восстановления живой
системы безусловно является результатом ее долгого исторического
развития. В этом развитии первостепенную роль должны играть
историческое возникновение и дальнейшее развитие ферментов — этих
высокоорганизованных катализаторов. Только при активном участии таких
ферментов могли возникнуть сложнейшие белковые образования и
осуществиться характерные для живой системы процессы обмена веществ,
роста и размножения. Правда, при всех этих процессах точно так же
происходят физико-химические процессы, как возникновение
электролитов, изменение проницаемости, потребление кислорода, образование
углекислоты и т. д. Но все эти известные физико-химические процессы
являются следствием того сложного и своеобразного ферментативного
процесса, который претерпевает липопротеид в живой системе. Так
как этот ферментативный процесс іразложения и восстановления
характеризует только живую систему, то мы предлагаем называть его
биологическим процессом; а так как он лежит в основе, всех
жизненных отправлений, как, например, обмена веществ, роста и
размножения, то мы считаем еще более подходящим называть его
основным биологическим процессом.
Характеристика основного биологического процесса. В обычных
условиях существования организма во внешней среде живая
возбудимая система все время испытывает очень сложное воздействие внешней
среды, главным образом через нервную систему. Но и внутренняя
среда, которой окружена каждая частичка живой системы, все время
меняется благодаря поступлению в кровь и лимфу многочисленных и
разнообразных продуктов расщепления, происходящего в живых
тканях организма. Все это сильно влияет на динамику живой
системы, на ее основной биологический процесс в смысле усиления или
ослабления этого процесса. Но нельзя себе представить, чтобы живая
система во всех своих частях испытывала вполне одинаковое внешнее
влияние. Поэтому процесс восстановления и распада должен
происходить в разных частичках живой системы в разное время. Скорость
того и другого процессов так же может быть не одна и та же в раз-
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
133
ных частичках. А так как влияние среды на данный участок живой
системы меняется с течением времени, то безусловно с течением
времени меняется также скорость процессов расщепления и
восстановления в одном и том же участке. Эта гетерохронность процессов
расщепления и восстановления в разных частичках живой системы
является основным свойством, характеризующим основной
биологический процесс (рис. 78).
Норма Согревание
Иолич
бозб 45 5$ SS 67 59 57 60 60 43 50 $8 5/ 50 52 65 б! 82 56 70 60 70 67 63 63 77
?: lg1 0 і ± ¦ * * л i і і і і —і „і 1— і 1 1 1 1—_j 1 1 і 1 1 1 1 (
^<§ 0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 50 54 68 72 76 80 84 88 92 96 104108
Рис. 78. Основной биологический процесс.
Волнообразные линии а, Ь, с, d, е, f, g, h и / выражают процесс расщепления и восстановления разных
частичек возбудимой системы. Волны этих частичек начинаются и кончаются в разное время.
Нисходящее колено каждой волны обозначает фазу расщепления, а восходящее — фазу восстановления.
Эти процессы сначала даны для некоторого нормального состояния, а. потом при изменении его под
влиянием согревания и вообще при условии, благоприятствующем обмену веществ.
Под волнообразными линиями основного биологического процесса цифры означают количество
возбудимой системы. Эти цифры представляют общую сумму вертикалей всех волн в условной единице,
которая равняется десятой части полного восстановления при нормальном состоянии.
В самом низу кривая означает степень возбудимости сообразно с количеством возбудимой системы.
Здесь абсцисса обозначает время (в сигмах), а ордината — количество возбудимой системы. При
согревании степень возбудимости повышается вместе с увеличением общего количества.
Мы можем вполне определенно указать, сколько 'Именно времени
должен продолжаться тот или другой процесс в нормальных условиях
клетки. Мы при этом исходим из наших знаний о процессе
возбуждения нерва и мышцы, который по своему характеру является обычным
катаболическим процессом. В отношении процесса возбуждения в
настоящее время известно, что он продолжается в нервно-мышечной
системе всего несколько сигм и немного меняется под влиянием
внешней среды. Указанные благоприятные для обмена веществ условия
уменьшают эту продолжительность процесса возбуждения,
неблагоприятные же увеличивают ее. Но эти изменения происходят в
пределах нескольких сигм. Точно так же хорошо изучено течение процесса
134
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
восстановления после возбуждения. Этот анаболический процесс
обычно продолжается много дольше катаболического и подвергается
изменениям под влиянием внешней среды значительно сильнее. Период
восстановления в нервно-мышечной системе даже в самых лучших
условиях обмена веществ продолжается в два-три раза дольше
процесса возбуждения; но уже малейшее ухудшение производит заметное
удлинение, а более значительное ухудшение может замедлить процесс
восстановления до нескольких секунд и даже іминут (Беритов, 1). По
нашему мнению, в нервно-імышечной системе обычный катаболический
и анаболический процессы протекают в каждой частичке с точно
такою же скоростью и интенсивностью, как при возбуждении.
Следовательно, и іво время основного биологического процесса катаболическая
фаза в каждой живой частичке протекает в течение нескольких сигм,
я эта продолжительность мало меняется под влиянием внешних
условий. Анаболическая фаза в каждой частичке продолжается при
некоторых средних условиях несколько дольше; при неблагоприятных же
условиях восстановление может продолжаться несколько секунд и даже
минуты. Можно утверждать, что невозбудимость возбудимой системы
после сильного воздействия внешней среды должна зависеть прежде
всего от чрезмерного удлинения фазы восстановления или от его
полного прекращения.
Далее, в отношении фазы восстановления после возбуждения
известно, что в начале этой фазы возбудимость живой системы равняется
нулю, и она постепенно повышается до нормы. В связи с этим, тотчас
же после возбуждения, когда еще нет более или менее значительного
восстановления возбудимой системы, даже сильное раздражение не
может дать возбуждения. В дальнейшем раздражение дает новый
процесс возбуждения, но для этого требуется более сильное раздражение,
чем в норме, и притом тем более сильное, чем раньше приходится
раздражение. Наконец, в период восстановления новое раздражение
дает более слабое возбуждение, чем в норме, и притом тем слабее,
чем раньше приходится раздражение в этот период восстановления.
Отсюда мы заключаем, что как возбудимость, так и интенсивность
возбуждения являются функцией количества возбудимой системы
(подробное фактическое обоснование см. ниже в главе о возбуждении).
Все эти фактические данные насчет анаболической фазы после
возбуждения мы переносим и на весь тот период, когда возбуждения нет.
Именно в этот период степень возбудимости возбудимой системы
должна определяться во всякое время количеством восстановленной
части возбудимой системы. Чем больше ее в данный момент в живой
системе, тем выше должна быть возбудимость, тем слабее нужно
раздражение для возбуждения такой системы. Поэтому, если под влия-
' нием тех или иных благоприятных условий замечается повышение
возбудимости, то это должно указывать, что в каждой частичке
возбудимой системы восстановление липопротеидных образований происходит
в большей мере и в связи с этим общее количество возбудимой си-
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
135
стемы больше, чем обычно. Если же при неблагоприятных условиях
возбудимость, наоборот, падает, то это должно указывать прежде
всего на более слабое восстановление липоиротеида возбудимой
системы, а значит уменьшение общего количества восстановленной
возбудимой системы.
Итак, живая возбудимая система «самопроизвольно», т. е. помимо
воздействия внешней среды, беспрерывно разрушается и
восстанавливается. Однако в каждый данный момент различные образующие
живую систему частицы находятся на различных стадиях распада и
восстановления, причем в зависимости от внешних и внутренних условий
эти процессы протекают в разных частицах с разной интенсивностью
и скоростью. Эти противоположные процессы зависят один от другого:
после восстановления следует расщепление; расщепление каждой
частички связано с освобождением энергии; за счет этой энергии
происходит последующее восстановление. Оба процесса так неразрывно
связаны один с другим, что их нужно считать за единый неделимый
процесс. Это динамическое состояние, как наиболее характерное для
живой системы, лежащее в основе обмена веществ, роста,
размножения и т. д., является основным биологическим процессом,
представляющим собою по существу объединенную, но неустойчивую систему
ферментативных процессов.
Рис. 78 дает схематическое изображение основного биологического
процесса и его изменения под влиянием температуры.
Основной биоэлектрический ток и его происхождение. Основной
биологический процесс каждый раз в период расщепления производит
ряд ионизированных івеществ. Вследствие этого все время происходит
передвижение ионов из гидрофильно-коллоидно-дисперсионной фазы в
интермицеллярную и обратно. Особое значение придается иону калия,
который в мышечных волокнах, а также в нервных волокнах и
клетках находится в большом количестве и связан со сложными
органическими образованиями, вероятно, липопротеидами. По новейшим
исследованиям, содержание калия внутри поперечнополосатых мышечных
клеток равно в среднем 0.28% при исчислении во влажном состоянии
и 0.98%—на сухое вещество (Кумингс, 21). В плазме и лимфе, т. е. в
наружной среде мышечных клеток, калия во много раз меньше. Поэтому
при расщеплении получается избыток свободных ионов калия.
Возникающие при этом крупные органические анионы малоподвижны и
непроницаемы для перепонок. Этим создается условие для разности
потенциалов, для возникновения электродвижущих сил, связанных
главным образом с калием (Фенн и сотр., 17; Эмберсон, 18).
По представлениям Мейе р а (13), характером белковых коллоидных
частичек обусловливается направление движущихся ионов. Эти частички
обладают боковыми цепочками, которые заканчиваются кислотными и
основными группами. Если в цепочке преобладают основные группы,
тогда коллоидные частички являются по преимуществу основными
белками; если же — кислотные группы, тогда белки носят по преиму-
136 ОБЩАЯ ФИЗИОЛОГИЯ МЫШНЧНОИ СИСТЕМЫ
ществу кислотный характер. Кислотные цепочки присоединяют и
проводят сквозь себя главным образом катионы, а основные цепочки —-
главным образом анионы. В соответствии с этим, если в живой
системе преобладают основные белки, то возникшие в результате
химического процесса ионы должны сделать эти участки живой системы
источником отрицательного заряда; в случае же преобладания
кислотных белков — источником положительного заряда. Скорость процесса
расщепления и восстановления, как указывалось выше, не может быть
совершенно одинаковой в разных участках
возбудимой системы. Вследствие этого
количество подвижных ионов и степень
проницаемости для тех и других ионов в одних
участках больше, чем в других. Поэтому если
соединить живую нервно-мышечную ткань с
гальванометром, то можно получить
отклонение гальванометра в случае более или менее
значительной разницы в количестве ионов
на разных полюсах отводимого участка,
т. е. імы обнаружим электрический ток.
Судя по направлению и величине
отклонения гальванометра, мы можем решить, в
каком участке живой ткани сильнее всего
протекает основной биологический процесс,
а значит и обусловленный им процесс обмена
веществ.
При отведении от двух безнервных
участков изолированного мышечного волокна
получается небольшая разность потенциалов, да
одного милливольта, причем направление
тока меняется от участка к участку. Особенна
большая разность наблюдается, когда
отводятся безнервный и нервный участки одного
мышечного волокна. В этом случае она почти
в 10 раз превышает разность при отведении от двух безнервных
участков (рис. 79) (Бухталь и Линдгард, 19). Большая разность получается
также, если взять пучок мышечных волокон с нервной заной
посередине. Если отводить безнервные участки, разделяемые промежутком в
0.5 см, то разность потенциалов будет составлять не более 1 mV; если
же отводить нервную зону и безнервный участок, разделяемые таким
же расстоянием в 0.5 см, то разность потенциалов будет равна 2—3mV.
Особенно в больших пределах меняется разность потенциалов в том
случае, если один из отводящих электродов передвигается по
мышечному пучку, а другой соединен с окружающей средой. Когда в одном из
опытов отводящий электрод находился на нервном участке, разность
потенциалов равнялась 4—5 mV, когда же его прикладывали к
остальным участкам, она колебалась в пределах 0—2 mV. (Кометиани, 20).
Рис. 79. Основной
биоэлектрический ток .мышечного
волокна m. semitendinosus.
Продольное отведение 'в
струнный, гальванометр через
небольшой усилитель. Верхняя кривая
при отведении участка нервной
пластинки и безнервного участка.
Цепь гальванометра замыкается
несколько раз. Разность
потенциалов около 10 mV. Нижняя
кривая при отведении двух
безнервных участков; межполюсное
расстояние такое же. Разность
потенциалов 0.5—1.0 mV
(Бухталь и Линдгард).
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
137
Вышеприведенные наблюдения указывают на то, что в нервных,
участках биологические процессы протекают во много раз интенсивнее,.
чем в безнервных. Но, что характерно, и в этом случае направление
тока не одинаково. В большинстве случаев нервный участок является
электроотрицательным, но в других он становится
электроположительным. С точки зрения изложенных выще представлений М е й е р а эта
разница в направлениях тока может быть объяснена тем, что в нервных
участках коллоидальные липопротеидные частички в одних случаях
являются основными и тогда они при расщеплении заряжаются
отрицательно, а в других — кислотными, тогда они при-^расщеплении
становятся Положительно заряженными.
Во всех тех случаях, когда основной биологический процесс
почему-либо ослабевает, ослабевает и основной . электрический ток. Так,
под влиянием наркотического вещества — хлороформа разность
потенциалов исчезает параллельно с понижением возбудимости как в
нервных, так и в безнервных участках мышечного волокна (Бухталь и
Линдгард, 19). Так же действует понижение температуры. С
понижением температуры понижаются ферментативное расщепление и
восстановление; вместе с тем понижается и количество свободных ионов.
Соответственно ослабевает основной биоэлектрический ток, т. е.
разность потенциалов імежду двумя неповрежденными участками. Так,
если при данной температуре в 25° С имеется известная разность
потенциалов імежду двумя неповрежденными участками мышечного
волокна, то с охлаждением до 12° С эта разность постепенно падает,
пока не установится на одном уровне, отвечающем данной
температуре, что бывает спустя несколько минут от начала охлаждения.
После согревания до первоначальной температуры разность потенциалов^
быстро восстанавливается в течение нескольких секунд (Бухталь и
Линдгард, 19).
Из этих фактов ясно следует, что существование разности
потенциалов в изолированных мышечных волокнах указывает на разную
интенсивность основного биологического процесса в различных участках
волокна.
Биоэлектрический ток возникает также при локальном повреждении
живой системы, как, например, при прижигании, при разрезе и т. д.
Этот ток принято обозначать током повреждения, а также
демаркационным током. Если мы соединим через гальванометр
поврежденный участок с каким-либо другим неповрежденным, то
гальванометр обнаружит существование отрицательного электричества
в поврежденном участке. Но в нормальном организме такой ток не
встречается, и поэтому нельзя приписать ему какое-либо
биологическое значение.
Величина демаркационного тока зависит от многих условий. Так,
например, охлаждение уменьшает его, а согревание, наоборот,
увеличивает. В лягушечьей мышце он равняется 0.035—0.075 V. Эта
электродвижущая сила появляется вскоре после повреждения и длится
138
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
„иногда целыми часами, постепенно уменьшаясь. Она настолько
значительна, что ею можно пользоваться для раздражения
нервно-мышечного препарата.
Этот электрический ток физиологами называется также «т о к о т
покоя» (2). Тер>мин указывает на связь этого тока с таким
состоянием возбудимой системы, когда нет возбуждения. Но такое
представление неправильно. В живой возбудимой системе, как указывалось
выше, все время протекает процесс расщепления и восстановления,
даже при полном отсутствии изменений во (внешней среде.
'Вследствие этого нельзя говорить о действительном покое живой системы.
Одно время был большой спор между физиологами о
происхождении демаркационного тока. В полемике принимали участие Дюбуа-
Ре.ймон (6), Герман (2), Геринг (7) и др. В конце концов
восторжествовала точка зрения Германа, кратко выраженная в
следующем положении: умирающее вещество относится к живому
электроотрицательно. Значит, когда под 'влиянием повреждающих агентов
живое вещество разрушается, то в нем появляется отрицательное
электричество в силу возникновения химических процессов, производящих
'большое количество катионов, способных к диффузии.
Обычно для изучения демаркационного тока повреждают дисталь-
ный конец мышцы. Поэтому направление тока повреждения по
условиям опыта совершенно противоположно тому электрическому току,
который возникает при возбуждении в неповрежденном участке.
Стрелка в гальванометре должна отклоняться от тока повреждения в одну
сторону, а от тока возбуждения — в другую. Поэтому Д ю б уа - Р е й-
м о н назвал ток повреждения положительным колебанием тока, а ток
возбуждения — отрицательным.
Позднейшие авторы также пользовались этими терминами, хотя
они не отвечали действительности, потому что кажущаяся разница в
направлении этих токов обусловлена тем, что при возбуждении
разность потенциалов между возбужденной отводимой точкой и
поврежденным концом уменьшается вследствие возникновения
отрицательного потенциала также в возбужденной точке. На рис. 80 дается
направление токов в мышце после ее повреждения разрезом.
Между током повреждения и биоэлектрическим током возбуждения
существует большая принципиальная разница. Ток возбуждения
распространяется во всей імышце, следуя за возбуждением; ток же
повреждения ограничивается местом повреждения. Ток возбуждения от
однократного раздражения всегда имеет, как известно, очень короткую
продолжительность, измеряемую в каждом участке мышцы
небольшими долями секунды, а при тетаническом раздражении весь
электрический эффект состоит из ряда таких быстро протекающих токов; ток
же повреждения всегда имеет сплошной характер и длится очень
долго, иногда по нескольку часов.
Теории происхождения электродвижущих сил в живой ткани. Мы
выше рассмотрели происхождение электродвижущих сил с точки зре-
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС 139
с водой,
ния новейших фактических оснований и предположений. Но
существуют и другие теории. Одна теория предполагает, что
биоэлектрические потенциалы по существу обусловлены различной скоростью
диффузии ионов. Другая теория предполагает, что биоэлектрические
потенциалы суть фазовые потенциалы живых мембран. Они
возникают в результате разной растворимости ионов, с одной стороны,
в водной фазе, а с другой — в фазе, не смешивающейся
иными словами — в белке-липопротеиде, из
которого состоят в основном перепонки живой
системы. Эта избирательная растворимость
или проницаемость обусловливается как
характером электрического заряда перепонок,
так и их другими физическими
особенностями.
По современному состоянию знаний всем
означенным моментам происхождения
электромоторных сил нужно приписать
определенное значение в происхождении основного
биоэлектрического тока. Значение это не
одинаково. Так, электрическому заряду и
физическим особенностям живых перепонок и вообще
пограничным потенциалам следует приписать
только небольшую роль в происхождении
электродвижущих сил живой ткани. Более
важную роль должны играть диффузионные
потенциалы. Но при физиологических условиях
те и другие вместе никогда не в состоянии
достигнуть более чем нескольких милливольт,
а потому в происхождении биоэлектрических
потенциалов должны играть роль также
другие процессы, а именно освобождение ионов,
в особенности калия, внутри клетки в
результате распада сложных коллоидальных
систем, о чем уже говорилось выше (см.
Эмберсон, 18). Весьма вероятно, что в мышечных клетках ни
сарколемма, ни прилегающая к ней пограничная с саркоплазмой перепонка
существенно не влияют на происхождение разности потенциалов
живого мышечного волокна. При отведении с поверхности сарколеммы
разность потенциалов меньше, чем при отведении изнутри волокна
(Бухталь и Линдгард, 19). Если бы означенные поверхностные
оболочки имели значение для происхождения токов живого мышечного
волокна, то такое отведение изнутри должно было бы дать более
слабый ток, а получается наоборот. Это происходит оттого, что основной
источник разности потенциалов лежит внутри клетки, он 'возникает при
расщеплении коллоидных частичек липопротеида. Поэтому при
отведении токов мышечной клетки через сарколемму, представляющую
Рис. 80. Схема направления
токов повреждения в
перерезанной мышце.
На схеме электроды и
гальванометр не приведены. Стрелки на
линиях показывают направление
токов. Толстые линии
изображают возникновение сильных
токов в означенных направлениях
(Ландуа).
140
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
большое сопротивление, электродвижущая сила должна быть меньше,,
чем при отведении их изнутри клетки.
Влияние внешней силы на основной биологический процесс.
Каждая внешняя сила может -повлиять на основной биологический
процесс, если только она может произвести какое-либо изменение в
жидком коллоиде живой системы. При проникновении химических веществ
в клетку происходит изменение в составе интермицеллярной фазы, и
в зависимости от этого изменяются и коллоидно-дисперсионные
частички. Механическое действие также должно влиять на основной
биологический процесс через интермицеллярную среду. По всей
вероятности, прикосновение или сдавливание вызывает в ней повышение
давления, а это, в свою очередь, іможет привести к усиленному
расщеплению липопротеидов. Под влиянием электрического тока также должна
измениться сначала, интермицеллярная среда. Вызванное здесь
электрическим током передвижение ионов может влиять на расщепление и
восстановление липопротеидов как в одном, так и в другом
направлении. Также должны действовать изменение температуры, лучистой
энергии, высыхание и. т. д. Во всех вышеприведенных случаях
действие внешнего агента выражается сначала в (Повышении, затем и
понижении возбудимости. Но при некоторых сильных воздействиях тога
или другого агента возбудимая система совершенно лишается
возбудимости, а потом умирает.
Если принять во внимание, что внешние агенты действуют в смысле
изменения проницаемости не только на пограничные перепонки, но и
на всю протоплазму в целом, -и далее, если принять во внимание, что
из внешней среды самые сложные вещества, подобные декстрину и
яичному альбумину, могут проникать в возбудимую систему вопреки,
мембранной гипотезе (Насонов, 15, 16) и без изменения
проницаемости, наконец, если учесть и то обстоятельство, что химические
вещества или ионы проникают и накапливаются в том и другом участках
живой системы не только путем адсорбции, но и путем установления*
химических связей с ней (Лепешкин, 14), то действие внешних
агентов на возбудимую систему может быть представлено в следующем
виде. Рассмотрим для примера действие постоянного электрического
тока. Электрический ток производит катафорез — перенос составных
частей протоплазмы в интермицеллярной среде. Одновременно он
производит электролиз составных частей. Вследствие этого на катоде
собираются водородные ионы, а также другие катионы, в особенности (в
первое время) одновалентные, а на аноде кислотные остатки — анионы.
В дальнейшем здесь также получают преобладающее значение
двувалентные ионы вследствие медленного передвижения их по сравнению со
скоростью движения одновалентных. Это химическое изменение в
интермицеллярной среде оказывает на коллоидно-дисперсионные частички, а
значит и на возбудимую систему в целом, определенное действие.
Сначала оно производит повышение вязкости благодаря усиленному
расщеплению липопротеидов и затем коагуляцию белков. При этом мо-
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
141
жет также происходить изменение проницаемости. Но некоторые
авторы считают изменение проницаемости необратимым явлением,
возникающим в случае сильного тока в результате повреждения клетки под
влиянием катафоретического переноса дисперсных частичек (Лепеш-
кин, 14). При таком понимании действия электрического тока
повышение возбудимости на катоде и понижение ее на аноде нужно
объяснить действием химических веществ на возбудимую систему. Вещества,
¦собирающиеся на катоде, своими химическими связями с липопротеи-
дом, а частью своим влиянием на ферменты, благоприятствуют
ферментативным процессам расщепления и восстановления липопротеидов, в
результате чего происходит увеличение количества возбудимой системы
и, следовательно, повышение возбудимости. На аноде, наоборот,
собираются такие ионы и в таком количестве, что они тем же путем
препятствуют ферментативным процессам расщепления и восстановления
липопротеидов. Это уменьшает возбудимую систему и соответственно
понижает возбудимость.
При некотором сильном и длительном действии электрического тока
как на аноде, так и катоде возбудимость сходит на-нет. Это, должно
быть, объясняется в отношении катода расщеплением всей массы
липопротеидов и коагуляцией белка без последующего восстановления, а
в отношении анода — чрезвычайным уплотнением перепонок и
прекращением ферментативных процессов. Если по прекращении тока
возбудимость не восстанавливается, тогда, должно быть, имеет место уже
денатурирование белка, а денатурированный белок не в состоянии
участвовать в ферментативном синтезе липопротеидов.
Итак, действие каждого внешнего агента на возбудимую систему
•обусловливается изменением всей протоплазмы в целом. Внешний
агент производит внутри клетки в интермицеллярной среде такие
физико-химические изменения, которые оказывают на коллоидо-дисперс-
яую фазу, а значит и на возбудимую систему, определенные
воздействия в смысле изменения химических связей и ферментативных
процессов, а тем самым в смысле изменения обмена веществ и
возбудимости.
С точки зрения мембранной гипотезы, действие внешнего агента
на живую ткань влияет на возбудимость живой ткани путем изменения
проницаемости полупроницаемых поверхностных перепонок. Когда
внешний агент (одновалентные ионы, катод электрического тока)
действует на эти перепонки разрыхляющим образом, тогда в возбудимую
систему должно проникать больше водородных ионов, а также воды и
других веществ. Все это сильно благоприятствует ферментативным
процессам расщепления и восстановления; этим самым основной
биологический процесс должен усилиться. Соответственно этому возбудимая
система должна увеличиться и возбудимость должна повыситься.
Когда же под влиянием внешнего агента (двувалентные ионы, анод)
происходит уплотнение поверхностных перепонок, проницаемость
перепонок уменьшается, и тогда в возбудимую систему должно проникать
142
ОЫЦАЯ ФИЗИОЛОГИЯ МЫіШіЧМОЙ СИСТЕМЫ
меньше водородных ионов и других веществ, чем обычно; поэтому-
основной биологический процесс протекает медленнее. Сообразно с
этим количество возбудимой системы уменьшается. В этом случае
возбуждение должно возникать при более сильном раздражении, и
потому это состояние будет представлять стадию пониженной
возбудимости.
Как указывалось выше, лри некотором длительном или сильном
воздействии одновалентных ионов, независимо от того, будет ли это
вызвано непосредственным воздействием соответствующих растворов
или пропусканием электрического тока, живая ткань теряет
возбудимость. С точки зрения мембранной теории это явление следует
признать результатом сверхоптимального нарастания проницаемости
перепонок. Оно должно создавать чрезвычайно неблагоприятные условия
для восстановительного процесса, и живая система должна перестать,
возбуждаться от внешних раздражений. Так получается, между
прочим, под влиянием <ионов калия. Многие ученые приписывают
невозбудимость нервно-мышечной ткани под влиянием калия именно этому
обстоятельству (Гёбер, 3). Ему же приписывают понижение
возбудимости в катодном участке, так называемую катодную депрессию
(Макут, 9). Точно так же под влиянием длительного или
кратковременного, но интенсивного действия двувалентных ионов или анода,
постоянного тока живая возбудимая система теряет возбудимость.
Явление это приписывается резкому уменьшению проницаемости. Это
обстоятельство должно сильно задерживать переход ряда веществ-
через перепонки. Вследствие этого основной биологический процесс
¦ в возбудимой системе должен чрезвычайно ослабнуть. В таком
состоянии живая система не может реагировать возбуждением (Гёбер,.
3; Васильев, 4; Макут, 9).
Итак, с точки зрения мембранной теории, при повышении
проницаемости пограничных перепонок под влиянием одновалентных ионов
биологический процесс сначала усиливается, а затем чрезвычайно
ослабевает вследствие затруднений восстановительного процесса. При
понижении проницаемости тех же перепонок под влиянием двувалентных:
ионов биологический процесс постепенно ослабевает до чрезвычайно
малых пределов благодаря затруднению обмена веществ между интер-
мицеллярной и коллоидно-дисперсной фазами.
Среда и основной биологический процесс. Влияние среды на живую
систему никогда не бывает простым, действующим только в одном
направлении. Даже в искусственных лабораторных условиях нельзя себе
представить, чтобы живая система испытывала воздействие только
одного внешнего агента, приложенного экспериментатором.
Одновременно с этим живая система всегда подвергается целому ряду других
воздействий. Представьте, например, что экспериментатор действует на
нервно-мышечную ткань электрическим током. Для этой цели он
пользуется металлическим или иными электродами, которыми прикасается
к ткани. Повидимому, это обстоятельство будет сопряжено с механиче-
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС 143-'-
гким раздражением. Опыт всегда происходит при какой-либо
определенной температуре: очевидно, характер реакции будет зависеть и or
этой температуры. При -прохождении электрического тока произойдет"
передвижение ионов не только внутри ткани, но и на поверхности, так.
как эта поверхность обычно увлажняется солевым раствором, как
раствор Рингера. Поэтому живая система будет испытывать также
действие этих наружных ионов, т. е. химическое влияние. Кроме
того, живая система все время находится в чрезвычайно изменчивой
среде, будет ли то искусственная среда, когда система взята в
изолированном 'виде, или внутри организма. Отсюда ясно, что в тот момент,,
когда экспериментатор пользуется одним каким-либо воздействием на
живую систему, последняя испытывает всегда очень сложное влияние-
со стороны внешней среды, и самый агент, избранный
экспериментатором, действует «а'живую систему очень сложно.
Далее, то изменение, которое производит среда -в живой системег..
нельзя считать суммой изменений, імогущих возникнуть от отдельных
компонентов. Правда, каждый компонент производит в водной интер-
мицеллярной фазе определенное физико-химическое действие:
температура "действует на скорость передвижения іионов; механическое
влияние—на количество воды, а следовательно и на концентрацию ионов;
химическое влияние — на состав ионов; электрический ток — на
направление передвижения ионов. Но если принять во внимание, что
каждое действие зависит от другого: движение ионов зависит от
концентрации ионов, концентрация от передвижения .ионов и т. д., то
следует сказать, что то изменение, которое возникает в интермицел-
лярной фазе живой системы под влиянием среды, ни в коем случае
нельзя считать суммой компонентных действий. Это изменение
является своеобразным целостным процессом.
Сообразно с этим основной биологический процесс под влиянием:
внешней среды в каждый момент испытывает одно целостное
изменение: он или усиливается, или ослабевает. Всякое усиление или
ослабление основного биологического процесса ограничивается главным
образом тем участком, на который действует данная внешняя среда. Оно
не распространяется на более или менее отдаленные участки, где не
было воздействия данной среды.
Таково действие наркотических веществ, а также электрическое,,
химическое, механическое .и температурное. Это, конечно, касается
такого воздействия внешней среды, которое не вызывает возбуждения,,
а только меняет существующий основной биологический процесс
в смысле усиления или ослабления. Что это изменение биологического*
процесса действительно локальное, не распространяющееся и вообще
не влияющее на более отдаленные участки, на которые данный
внешний агент не действует непосредственно, вытекает из определенных
фактов, установленных по отношению к возбудимой системе нервной-
ткани. Вследствие этого речь об этих фактах будет итти в отделе-
нервной системы (см. главы о парабиозе и электрофизиологии).
544 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Итак, действие среды на живую систему, если даже оно не
вызывает процесса возбуждения, всегда является более или менее
сложным, а вызванное средой физико-химическое изменение живой
системы представляет собой одно целостное изменение, которое нельзя
считать суммой изменений, вызываемых компонентами среды в
отдельности. Это физико-химическое изменение является местным процессом:
оно происходит главным образом там, где данная среда действует.
Сообразно с этим, связанное с означенным физико-химическим
процессом изменение основного биологического процесса всегда локально и
не распространяется на более или менее отдаленные участки.
Практикум
Наблюдение демаркационного тока. Сделав надрез на т. gastrocnemius или
triceps лягушки, накладывают на разрез седалищный нерв нервно-мышечной лапки.
Предварительно этот нерв обрезают около позвоночника. Если при
накладывании нерва образуется цепь, то ток повреждения замкнется через нерв и
последний будет раздражаться, как от замыкания гальванического тока; при этом
мышцы препарата сокращаются. Сокращение последует и при поднятии нерва, как
от размыкания гальванического тока.
Регистрация основного биоэлектрического тока гальванометром. Для отведения
этих таков <в гальванометр пользуются ншоляризующими электродами. Описание
этих электродов, а также гальванометров ом. в методике.
На m. sartorius лягушки накладывают отводящие электроды; один полюс
по середине мышцы, а другой на сухожильном конце ее, соединяют с
гальванометром и отмечают отклонение струны. Затем согревают весь препарат путем
прикосновения іваты, смоченной согретым до 40° раствором Рингера. Вгновь соединяют
с гальванометром и следят за отклонением стрелки. Оно будет значительно сильнее,
чем 'в первом случае.
Наблюдение основного биоэлектрического тока в нервных и безнервных участках
пучка мышечных волокон. Взять от m. semitendinosus лягушки пучок мышечных
волокон длиной около 2—2,5 см. В этом пучке нервная зона находится по середине.
Если отводить (в какой-либо чувствительный гальванометр безнервный конец мышцы
е межполюсным расстоянием около 0.5 см, то получим небольшое отклонение
гальванометра в ту 'или другую сторону. Если же соединить так, чтобы один полюс
был на нервной зоне, а другой на безнервном (участке, отклонение гальванометра
будет в несколько раз сильнее.
Если подействовать на мышцу парами эфира так, чтобы она потеряла
возбудимость, то такое же отведение лучка к гальванометру не даст означенной (большой
разницы потенциалов.
Литература
1. Beritoff J., Z. Biol., 78, 331, 1923.
2. Hermann L., Handb. Physiol., Bd. 1, T. I, S. 235, 1879.
3. H б b er R., Physiol. Chemie d. Zelle u. d. Gewebe, 2. Halite, 1924.
4. Васильев Л., Новое в рефлекс, и физиол. нервной системы, т. 1, стр. 1, 1925.
5. Лапицкий Д., Новое в рефлекс, и физиол. нервной системы, т. 3, 56, 1929.
6. Du Bois-Raymond E., Untersuch. uber tier. Elektrizitat, Bd. I, 671, 1878.
7. H e r i n g E., Lotos, N9,36, 1889.
8. Лазарев П., Исслед. по ионной теории возбуждения. Изд. Моск. научн. инст.
1916.
ОСНОВНОЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС
145
9. Маски th Е., Pfl. Arch., 214, 612, 1926.
10. Win ter st e in H., Naturwiss., S. 247, 1931; Bioch. Z., 232, 196, 1931.
11. Бауэр Э. С, Теоретическая биология, изд. ВИЭМ, 1935.
12. Э н г е л ь с Ф р., Диалектика природы. Архив Маркса и Энгельса. Кн. II, стр.
49—50, 1925. Анти-Дюринг, б-е изд., стр. 257—259, 1934.
13. М е у е г К u r t Н., Schw. med. Wschr., II, 826, 1937.
14. L е р е s с h k i n W. W., Kolloidchemie des Protoplasmas, II Aufl., Dresden u.
Leipzig, 1938.
15. H а с о н о в Д. Н. Тр. VI Всес. физиол. съезда, стр. 273, 1937.
16. Насонов Д. Н. и Александров В. Я., Биол. журн., т. 6, вып. 1, 117,
1937.
17. Fenn W. О., Cobb D. М., HeynauerA. Н. а. М а г s h В. S., Am. J.
Physiol., 110, 74, 1934.
18. Amber son W. R., Cold Spr. Harb. Symp. Quant. Biol., IV, 53, 1936.
19. Buchthal Fr. a. LindhardJ. The Physiol, of str. muscl. fibre, Copenhagen,
1939.
20. К о m e t и а н и П., Тр. Инст. физиол. им. Бериташвили, 4, 593, 1942.
21. Cumings J. N., Biochem. J., 33, 642. 1939.
22. Энгельгардт В. А., Усп. совр. биол. 14, № 2, 1941.
Ю и. С. Беритов
Глава VI
ЗАКОН ВОЗБУЖДЕНИЯ
Общее понятие о законе возбуждения. Для живой возбудимой
системы чрезвычайно характерно то своеобразное изменение, которое
возникает под влиянием раздражения и именуется возбуждением. Это
изменение само собой распространяется в живой клетке, а также
переходит из одной клетки в другие, если она имеет с ними
непосредственные протоплазматические связи. Иначе говоря, возбуждение,
вызванное в определенном участке возбудимой системы, переходит на
соседние участки, даже если последние не подвергались воздействию'
внешнего раздражения. Следовательно, при раздражении одного-
участка в пучке мышечных волокон возбуждение распространяется па
всей длине пучка, а при раздражении одного участка двигательного
нерва возбуждение распространяется по всему нерву и затем
переходит на мышцу.
Процесс возбуждения имеет основное значение в
жизнедеятельности организма. Благодаря ему происходят все быстро протекающие
акты приспособления организма как единого целого к внешней среде,,
а также все быстрые функциональные приспособления одной части
организма к другим частям или же к изменениям внутренней среды.
Процесс возбуждения нервно-мышечной системы позвоночных
животных характеризуется одним чрезвычайно своеобразным признаком:
он протекает в каждом участке живой возбудимой системы всегда
с максимальной интенсивностью, т. е. в действие приходит всегда вся
возбудимая система целиком, разряжается, так сказать, вся ее
потенциальная энергия. Отсюда следует, что в данной возбудимой системе
процесс возбуждения не градуируется. При данном функциональном
состоянии интенсивность возбуждения будет одна при всех силах
раздражения. Только при изменении функционального состояния
может измениться интенсивность возбуждения. Так, например, при
утомлении, наркозе интенсивность возбуждения уменьшается, но и в этом
случае она будет одинаковой при всех силах раздражения. Значит,,
в каждый данный імомент интенсивность процесса возбуждения
мышечного волокна зависит от его функционального состояния и не зависит
ЗАКОН ВОЗБУЖДЕНИЯ
147
от силы раздражения. Точно так же при распространении процесса
возбуждения в каждом последующем участке мышечного волокна
интенсивность процесса зависит от функционального состояния этого
участка. Эта особенность процесса возбуждения, что он возникает
в раздраженном участке при участии всей возбудимой системы и затем
при своем распространении охватывает в каждом последующем
участке всю возбудимую систему полностью, является законом для
Рис. 81. Сокращение мышечного волокна при разных силах прямого
раздражения.
Кинематографический снимок. Снимается движение одного конца волокна.
Другой конец раздражается непосредственно. В каждом опыте первый снимок при
покойном состоянии, второй — вовремя раздражения. Опыт 7 —пороговое
раздражение 140 мм расст. кат. Нет сокращения. Опыт 2—сила раздражения 13« мм.
Волокно укорачивается на 10 мм. Опыт 3 — раздражение 130 мм. Укорочение •- ,
то же самое. Опыт 4—раздражение 100 мм. Укорочение опять на 10 мм (Като).
возбудимой системы мышечной и нервной тканей всех позвоночных
эюивотных, а также высших беспозвоночных.
Наиболее убедительное доказательство закона возбуждения для
возбудимой системы мышцы было дано японским ученым Като (25).
Он изолировал одно мышечное волокно и поместил его в узкой
ванночке с рингеровским раствором. Один конец волокна раздражался,
а передвижение другого конца регистрировалось посредством
киносъемки. Оказалось, что укорочение волокна происходит при разных
силах раздражения всегда на одну и ту же величину (рис. 81). Если
ухудшить функциональное состояние імышцы, например влиянием
наркотика, величина укорочения мышцы уменьшается. Но и в этом случае
мышца будет укорачиваться на одну и ту же величину при всех силах
раздражения (рис. 82). . •
Чтобы получить сокращение всего мышечного волокна в ответ на
раздражение одного какого-либо участка, необходимо, чтобы
мышечное волокно было совершенно свежим, неутомленным и
неповрежденным, т. е. чтобы возбудимость его была достаточно высокая. Если же
ю*
148
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
мышечное волокно почему-либо не обладает достаточно высокой
возбудимостью, тогда сокращение, вызванное в одном участке, не
распространяется (Гинтер, 19; Стейман, 39).. Такое локальное сокращение по
существу ничем не отличается от сокращения всего волокна. Но длина
сокращенного участка градуируется в зависимости от силы
раздражения, т. е. чем сильнее раздражение, тем более длинный участок
приходит, в сокращение. Соответственно и суммация наблюдается при
мм
w -
tfOpMQ
Наркоз
Ю'
*з-
'іжші
/if1 Ш 90 /3S 90 /28 90 /20 SO /О/ 70
?2 ; /Si ; /30 . /20 і ffO i 90
{ J -—._ » J I
Пороги
Рис. 82. Величина укорочения мышечного волокна в
нормальном состоянии и при наркозе .
В каждой группе по три сокращения при разных силах раздражения. Под
горизонтальной линией цифры обозначают расстояние между
индукционными катушками в миллиметрах: первый ряд для применяемых
раздражений, а второй — для порогов. На вертикальной линии дается масштаб
укорочения в миллиметрах. Как в норме, так и во время наркоза в каждой
группе сокращений величина укорочений одна и та же (Като).
любом интервале раздражений, особенно в том случае, если
последовательные раздражения прикладываются к двум разным участкам
мышечного волокна (Зихель и Проссар, 40). Эти наблюдения ни
в какой мере не противоречат закону возбуждения. Дело в том, что
при значительно пониженной возбудимости сильные электрические
удары в состоянии возбудить мышечное волокно в раздраженном
участке и тем вызвать здесь сокращение. Понятно, этот сокращенный
участок будет тем больше, чем сильнее раздражение, ибо с усилением
приложенного электрического удара будет увеличиваться и
раздражаемый участок. Но ток возбуждения, который возникнет здесь, будет
настолько слаб, что не сможет возбудить соседний участок и тем
обусловить распространение сокращения вдоль ©сего волокна.
В условиях опыта Като изолированное мышечное волокно сокращалось
изотонически— волокно іничем не было нагружено. Фотографируется дзижение свободного
конца іволокша, как показывает рис. 81. Но если волокно нагружено, тогда
сокращение мышечного волокна будет тем больше, чем сильнее раздражение, т. е. высота
сокращения будет точно градуироваться ъ зависимости от силы раздражения. Это
объясняется тем, что, когда сокращается один раздражаемый участок, другие
участки растягиваются, а в растянутых участках сократительная способность падает,
как об этом уже говорилось выше. Но, благодаря распространению тока,
раздражаемый участок становится тем больше, чем сильнее ток. Это увеличивает область
максимально сокращенного участка и уменьшает область растянутого участка, где
сократительный процесс протекает слабее. Вследствие этого с усилением
раздражения увеличивается общее укорочение волокна. Так было в опытах Б р о у н а и
ЗАКОН ВОЗБУЖДЕНИЯ
149
3 и х е л я (34), а также Асмуссена (35), у которых изолированное мышечное
волокно действовало на пружину из тонкой стеклянной палочки. Так было и в
опытах С е р к о в а (36) и Макарова (37), которые держали волокно в растворе
Рингера и нагружали его несколькими миллиграммами. Но у последних авторов один
раздражающий электрод был соединен с наружным концом мышцы, а другой — с
раствором, куда было опущено волокно. Небольшой участок волокна выдавался над
рингером, и ток пропускался через «его. В этих опытах условия распространения
тока по мышечному волокну были особенно благоприятны, и потому с усилением
раздражения все больший и больший участок волокна возбуждался непосредственно
от тока и, соответственно, мышечный эффект мог «градуироваться особенно хорошо.
Кроме того, своеобразие процесса возбуждения проявляется в
следующем: если во время возбуждения, вызванного каким-либо
раздражением, подействовать снова раздражителем, то второе іраздражение
не дает никакого эффекта. Оно не усилит и не удлинит процесса,
вызванного первым раздражением. Только после того, как этот
процесс закончится, второе раздражение в состоянии вызвать эффект,
причем этот второй эффект в. течение некоторого короткого времени
будет слабее первого. Чем меньше времени проходит после окончания
возбуждения, тем слабее возбуждение от второго раздражения.
Аналогично меняется .и степень возбудимости. Во время возбуждения
возбудимости нет; в этот период новое раздражение, какой бы силы
оно ни было, не дает эффекта. Вслед за окончанием процесса
возбуждения новое раздражение дает эффект, но при очень большой
интенсивности. Пороги раздражения постепенно возвращаются к норме. Период
абсолютной невозбудимости называют абсолютной
рефракторной фазой, а последующий за ним период
относительной невозбудимости — относительной рефракторной
фазой (рис. 85) (Люкас, 1; Беритов, 2; Дитлер и Оинума, 3).
На основании того факта, что возбудимая система дает
возбуждение всей своей потенциальной энергией, физиологи называют эту
закономерность законом «все или ничего». Под этим термином
подразумевают, что если возбудимая система вообще реагирует
возбуждением, то реакция непременно будет максимальной, она непременно
будет потреблять всю потенциальную энергию. Но в то же время
термин «все или ничего» неправильно указывает и на то, что если
возбудимая система не реагирует на раздражение возбуждением, то
она никак не реагирует на него. Такое понимание в настоящее время
едва ли кто разделяет. Известно, что слабое подпороговое
раздражение, которое не в состоянии вызвать возбуждения, не остается совсем
без влияния на возбудимую систему. Если такое раздражение
повторить быстро одно за другим, оно дает возбуждение. Очевидно, слабое
подпороговое раздражение и при разовом применении производит
определенное изменение, которое продолжается некоторое время после
раздражения. Вследствие этого последующее раздражение производит
еще большее влияние на возбудимую систему, еще больше изменяет
ее. Это изменение еще более нарастает под влиянии дальнейших
раздражений. Оно проявляется в повышении возбудимости, которая и
150
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
может' стать настолько большой, что повторяемое подпороговое
раздражение становится пороговым.
Указанный нами факт, что основной биологический процесс
претерпевает под влиянием внешней силы постоянное изменение в виде
усиления и ослабления, хорошо подтверждает положение, что внешнее
воздействие производит реакцию в живой системе не только в виде
возбуждения. А потому известный термин — закон «все или ничего»
для обозначения закономерности, лежащей в основе возбуждения, не
отвечает действительности. Наиболее подходящим термином для
обозначения этой закономерности следует считать просто закон
возбуждения.
Итак, под влиянием раздражения возбудимая система приходит
в особое состояние, известное под названием возбуждения, которое
затем быстро распространяется по всей живой единице, охватывая всю
целиком возбудимую систему. В этот момент вся живая система не
может быть вновь возбуждена новым раздражением. Вслед за этим
процессом начинается восстановление возбудимой системы, причем по
мере восстановления возбудимость постепенно возвращается к норме.
О внешних условиях, вызывающих возбуждение. Всякое внешнее
воздействие: химическое, термическое, механическое, электрическое,
которое может повлиять на обмен веществ возбудимой системы, на ее
основной биологический процесс, в определенных условиях вызывает
также процесс возбуждения.
Мы уже указывали выше, что внешняя сила вызывает возбуждение
только в том случае, если она быстро меняется от одной величины
к другой, и притом если время ее действия превосходит определенный
минимум, который тем меньше, чем быстрее происходит изменение
внешней силы и чем лучше внутреннее состояние возбудимой системы
в отношении физико-химических процессов.
О внутренних условиях, вызывающих возбуждение. С точки зрения
мембранной теории возникновение возбуждения следует приписать
передвижению одновалентных ионов в живой системе и быстрому
накоплению их на поверхности полупроницаемых перепонок. Под
влиянием этих ионов происходит сильное увеличение проницаемости
перепонок. Об этом свидетельствует прохождение таких веществ, которые
обычно не проходят.
Известно, например, по Эмбдену (4), что при раздражениях,
производящих возбуждение, из мышечных волокон выходит наружу
больше ^фосфорной кислоты, калия и холестерина, а внутрь мышечных
волокон проникает 'больше хлора, чем обычно.
Означенные физико-химические процессы еще не являются
возбуждением. Возбуждение наступает под влиянием этих процессов и
выражается в превращении основного биологического процесса в
другой вид биологического -процесса.
С точки зрения другой, более новой теории главное значение
придается вообще проникновению и накоплению определенных химиче-
ЗАКОН ВОЗБУЖДЕНИЯ
151
ских веществ в живой системе. Это определяется прежде всего
растворимостью этих веществ в воде и липоидах и затем их химическим
действием на липопротеиды и ферменты. При определенной
концентрации определенных веществ происходит расщепление липопротеидов
всей возбудимой системы. Вследствие этого основной биологический
процесс существенно меняется: он из гетерохронного состояния
переходит в изохронное.
Ниже мы подробно остановимся на этой особенности
биологического процесса возбуждения, а сейчас рассмотрим, отчего именно он
должен возникать.
Известно по исследованиям Михаэлиса (5) и других, что
действие ферментов в значительной мере усиливается при определенной
концентрации водородных ионов. Если концентрация водородных ионов
больше или меньше этой величины, ферментативное действие
ослабевает. По утверждению же Бете (6), водородные ионы имеют
исключительное значение при вызове возбуждения; где концентрация их
растет, там возбудимость повышается; значит, усиливается основной
биологический процесс, и там же возникает возбуждение. Автор
подкрепляет это положение рядом опытных данных. Нужно полагать, что
раздражающая сила производит в живой системе передвижение
водородных ионов вместе с другими ионами. Водородные ионы влияют на
ферменты и тем ускоряют как расщепление, ^ак и последующее
восстановление возбудимой системы.
Как указывалось выше, согласно закону раздражения Дюбуа-
Р е й ім о н а, возбуждение не вызывается медленным изменением
внешней силы. Лазарев (7) объясняет это следующим образом. При
медленном изменении внешней силы одновалентные ионы не могут
накопиться на поверхности перепонок в такой концентрации, чтобы
вызвать возбуждение, так как значительная доля их должна успевать
пройти сквозь перепонки, не задерживаясь на поверхности. С точки
зрения строения живой протоплазмы по Лелешкину это явление
объясняется тем, что одновалентные ионы не накапливаются в водной
оболочке коллоидных частичек в такой концентрации, чтобы вызвать
усиленный ферментативный процесс, необходимый для возникновения
процесса возбуждения.
Необходимость определенного времени действия раздражающего
агента для вызова эффекта возбуждения также вполне понятна
с точки зрения последней концепции. Чтобы в возбудимой системе
произошел полный разряд потенциональной энергии, накопление ионов
в мицеллах должно достигнуть определенной величины. Понятно, чем
быстрее будет происходить передвижение ионов вследствие изменения
внешней силы, тем скорее будет достигаться эта предельная
величина. Очевидно также, что чем медленнее протекает
ферментативный процесс, тем дольше должно продолжаться раздражающее
действие, чтобы одновалентные ионы собрались в достаточном
количестве в водяной оболочке, а водородные ионы проникли в доста-
152
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
точном количестве внутрь мицелл для вызова процесса
возбуждения.
Существует более или менее прямое фактическое подтверждение
того положения, что возбуждение действительно возникает под
влиянием одновалентных ионов. Воронцов (8) действовал на
возбуждаемый участок нерва в одних опытах катодом и анодом
электрического тока, а в.других одновалентными и двувалентными ионами и
получил следующие чрезвычайно важные результаты: 1) во время
возбуждения состояние нерва совершенно сходно с тем состоянием,
которое вызывается действием катода. Это видно из того, что эффект
возбуждения нервно-мышечного препарата уменьшается, если на
возбужденный участок подействовать анодом, который по своему
физиологическому влиянию противоположен действию катода; 2)
предварительная обработка нерва одновалентными ионами (К, Na, а также
МНз) очень ослабляет отрицательное действие анода на
возбужденный участок; предварительная же обработка двувалентными ионами
(Са, Mg, Ba) сильно благоприятствует отрицательному действию
анода на этот участок. Такой эффект наблюдается в начале и в
середине абсолютной рефракторной фазы. Эти факты представляют
наилучшее подтверждение того положения, что возбуждение действительно
обусловливается перегруппировкой одновалентных ионов и что
вызванное ими физико-химическое изменение в живой системе составляет
неизбежное условие для получения возбуждения.
Таким образом, более или менее быстрое изменение
раздражающей силы вызывает в возбудимой системе более или менее быстрое
физико-химическое изменение, прежде всего в смысле передвижения и
накопления ионов. Только когда это изменение достигает
определенной величины, наступает процесс возбуждения.
Общее понятие о биоэлектрическом токе возбуждения. С
возникновением процесса возбуждения в возбудимой системе возбужденный
участок становится источником электричества. Поэтому, если этот
участок отводится в гальванометр, соединенный другим полюсом
с каким-либо невозбужденным участком, то в силу разности
электрических потенциалов возникает электрический ток. Направление тока,
указывает, что возбужденный участок относится к невозбужденному
электроотрицательно.
Электрический ток в каждом возбужденном участке продолжается
вообще столько же времени, сколько длится абсолютная рефракторная
фаза. При охлаждении, при наркозе, при утомлении время протекания
этого тока удлиняется подобно абсолютной рефракторной фазе, а при
согревании, отдыхе, наоборот, укорачивается. На основании этих
фактов принято считать, что электрический эффект соответствует процессу
возбуждения, стоит в определенной связи с возбуждением (рис. 85).
Означенный электрический эффект в физиологии называется «током
действия». Этот термин, введенный лет 70 назад Германом (9),
основан на представлении, что возбудимая система якобы испытывает
ЗАКОН ВОЗБУЖДЕНИЯ
153
деятельное состояние только в период возбуждения, тогда как на
самом деле живая возбудимая система находится в деятельном
состоянии постоянно. Протекающий в ней основной биологический процесс
есть не что иное как деятельное состояние. Поэтому в настоящее
время совершенно неправильно называть электрический ток
возбуждения «током действия». Правильнее всего называть его термином
«биоэлектрический ток возбуждения», как впервые назвал его видный
чешский физиолог Чермак (22).
Происхождение биоэлектрического тока возбуждения. Во время
возбуждения внутри мышечного или нервного волокна освобождается
некоторое количество свободных ионов как результат -расщепления
сложнейших белковых образований липопротеидов возбудимой
системы. Многочисленные факты говорят за такое новообразование ионов.
Прежде всего об этом свидетельствует факт теплообразования при
распространении возбуждения в нерве. Гилл высчитал, что только 1 %
начального теплообразования в нерве может быть обусловлен волной
деполяризации, распространяющейся по нервному волокну. Остальное
тепло должно быть обусловлено происходящими внутри волокна
восстановительными химическими (процессами (Гилл, 26). Очевидно, то
же самое следует сказать по поводу возбуждения мышечных
волокон. Оно должно быть сопряжено с физико-химическими процессами,
освобождающими значительное количество ионов. Это предположение
подтверждается наблюдением Эрнста (27) над ритмическим
уменьшением объема мышцы по ритму раздражения до 100—150 в секунду.
Это ритмическое колебание объема мышцы может быть обусловлено'
только освобождением некоторого количества ионов в связи
с каждым импульсом возбуждения, ибо известно, что при растворении
некоторых кислот, как, например, Н3РО4, а также при растворении
некоторых солей, как, например, солей калия, наблюдается уменьшение
объема раствора. От нейтрализации этих кислот и ионов, наоборот,
объем увеличивается (Оствальд, 28; Нернст, 29). Растворение
неэлектролитов также сопровождается уменьшением объема растворителя
(Фрике, 30). Затем Эрнст показал, что ритмическое изменение
объема мышцы в определенных условиях раздражения имеет свой
собственный ритм 50—200 в секунду, не зависящий от ритма
раздражения, подобно собственному ритму возбуждения (см. об этом в
следующей главе). Наименьшая продолжительность этого колебания
составляла 3—5 а (рис. 83) (34). На основании всех этих фактов Эрнст
(33) полагает, что уменьшение объема мышцы обусловливается
повышением ионизации внутри мышечных волокон, которое имеет место
при возбуждении и тесно с ним связано. Эрнст полагает, что при
непрямом ^возбуждении мышцы изменения проницаемости мышечных
волокон, видимо, совсем не происходит, ибо при этом не выделяются из
мышцы ионы калия. Увеличение проницаемости и выхождение ионов,
калия имеют место при прямом раздражении. Следовательно, в этом
случае под влиянием внешнего поляризующего тока происходит новы-
154 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
дгение проницаемости оболочек для ионов. Однако возникновение
биоэлектрических токов при возбуждении должно быть обусловлено
главным образом возникновением разного рода ионов в результате
ферментативного расщепления липопротеидов.
В самое последнее время эта химическая природа происхождения
биоэлектрического тока возбуждения находит большую поддержку в
лице М е й е р а (44). Он утверждает, что в основе тока возбуждения
лежит новообразование ионов, в виде продуктов расщепления
сложных органических соединений. Из этих ионов анионы связываются с
белковыми цепочками, которые носят в большинстве случаев основной
характер и потому возбужденный участок должен стать
отрицательным по отношению к соседним невозбужденным участкам. Автор
находит, что ток возбуждения должен вполне соответствовать означенному
химическому процессу.
Рис. 83. Изменение объема мышцы при изотоническом
сокращении. Две икроножные мышцы лягушки.
В опыте А они раздражаются тетанически 50 раз в 1 сек. Внизу
сигнальная линия (S) отмечает моменты раздражения. Объем
мышцы (М) увеличивается и во время раздражения колеблется по
ритму раздражения. По прекращении раздражения увеличенный
объем остается еще некоторое время. В опыте В дается короткий
тетанус от размыкания постоянного тока. Объем увеличился, и
в самом начале — пять быстрых колебаний (Эрнст).
По мнению Лун да (45), источником биотока является реакция
окисления, и величина биотока определяется интенсивностью
окислительных процессов. Известно, что внутри клетки на поверхности
сложных коллоидных структур-перепонок все время протекают процессы
окисления и восстановления. Эти процессы должны обусловить
возникновение электрического тока благодаря электронной проводимости
пограничных 'поверхностей (Кометиани, 46).
Со времени Германа до настоящего времени господствует
другая теория, основанная на поляризации наружных полупроницаемых
ЗАКОН ВОЗБУЖДЕНИЯ
155
оболочек мышечного и нервного волокон. Предполагается,, что эти
оболочки являются поляризованными ввиду того, что они одни вещества
пропускают, а другие нет. Например, известно, что концентрация ионов
калия внутри мышечных и нервных клеток превышает таковую в
окружающей тканевой жидкости. От этого мембраны являются сильно
поляризованными, причем на наружной их поверхности накапливаются
положительные ионы, а на внутренней — отрицательные. Многие авторы
полагают, что в случае временного увеличения проницаемости в
каком-либо участке мышцы или нерва (например, под влиянием
индукционного удара) произойдет лередвижение ионов снаружи внутрь и
тем самым получится электрический разряд. Далее предполагается,
что этот электрический разряд повлияет на находящуюся здесь
возбудимую систему и вызовет в ней определенные биохимические процессы.
Но этот же электрический разряд повлияет на мебраны
соседнего участка мышечного волокна и вызовет в них увеличение
проницаемости. Это обусловит деполяризацию ближайших участков, новый
электрический разряд и т. д. Эта гипотетическая волна деполяризации,
распространяющаяся по мышечному волокну, и признается многими
авторами за биоэлектрический ток возбуждения (Герман, 9; Лилли,
18; Гилл, 26). Более подробно теории возникновения и
распространения биотока возбуждения будут изложены в главе о нервной
физиологии.
Однако мало вероятно, что деполяризация наружных мембран
играет такую исключительную роль в происхождении биоэлектрического
тока возбуждения и распространении процесса возбуждения. Прежде
всего, ни в мышце, ни в нерве не найдено анатомической структуры,
которую можно было бы отождествлять с теми пограничными
перепонками, которые фигурируют в мембранной теории возбуждения.
Сама поверхностная оболочка — сарколемма, как и прилегающий
изнутри слой саркоплазмы, безусловно, не играют такой роли, ибо эти
поверхностные перепонки не являются поляризованными и не
участвуют в происхождении электростатических токов, как на это уже
указывалось выше.
Отмечаемое некоторыми авторами уменьшение проницаемости
оболочек при возбуждении настолько незначительно, что этим нельзя
объяснить происхождение биоэлектрического тока возбуждения (Дюбюис-
сон, 31). Вообще, наблюдения говорят против такого мгновенного
изменения проницаемости мышечных перепонок, которое могло бы
объяснить факт возникновения возбуждения (Фенн, 32). Далее, против
мембранной теории говорит также тот факт, что как в мякотном
нервном волокне, так и в мышечном, эти мембраны-оболочки
обнаруживают колоссальное сопротивление. Так, сопротивление мышечного
волокна толщиной 70^ и длиной 400 р. равняется 3 млн. омов. Оно
в 30 раз больше сопротивления такого же цилиндрического тела,
наполненного 0.6% раствором поваренной соли. При возбуждении
сопротивление уменьшается, но сравнительно слабо — всего на 50—80%
156
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
(Бухталь, 24). Это сопротивление мышечного волокна обусловливается
не только поверхностными мембранами волокна, т. е. сарколеммой, на
и пограничными мембранами, лежащими внутри волокна
перпендикулярно к поверхности (Бухталь, 24). В силу таких больших
сопротивлений імне кажется, что интенсивность поляризационного тока должна
быть очень слабой, не достаточной для раздражения невозбужденного-
участка.
Наконец, против гипотезы происхождения биотоков мышцы
вследствие деполяризации наружных мембран указывают новейшие
наблюдения относительно изменения концентрации калия внутри мышечной
клетки в связи с возбуждением. Оказывается, что даже в состоянии
значительного утомления в результате периодического тетанического
сокращения концентрация калия внутри мышечных клеток не
меняется, несмотря на выход калия из клетки во внеклеточное
пространство. Калий выходит наружу вместе с водой, и потому
концентрация его внутри клетки, а также отношение концентрации калия внутри
и вне клетки остаются теми же, что были до утомления (Кометиани и
Гоголашвили 41). Абсолютное количество калия больше в свежей
мышце, чем в утомленной, и так как, кроме того, в нервных участках
калия оказалось больше, чем в безнервных, все это дает право
утверждать, что калий находится внутри клетки в химической связи с
протоплазмой, т. е. с возбудимой системой мышцы. Он освобождается и.
выходит' из мышечной клетки наружу при расщеплении возбудимой
системы в связи с возбуждением. Отсюда следует, что роль калия в
поляризации мембран ничтожная и ему нельзя приписать существенное
значение в происхождении биотоков возбуждения.
О биологическом значении биоэлектрического тока возбуждения-
Расщепление белковых образований во время возбуждения протекает
очень быстро — всего в течение нескольких сигм, поэтому
биоэлектрический ток развивается с большой быстротой. В возбужденном
участке происходит быстрое нарастание и убывание электрического
потенциала. Это изменение биоэлектрического тока настолько значительно,
что оно не может не повлиять на соседние невозбужденные участки
раздражающим образом. Можно предположить, что биоэлектрический
ток, распространяясь внутри мышечного волокна, замыкается обратно
в значительной мере через внутреннюю же среду волокна, а
небольшой частью — через наружную. Этим самым в отношении соседних
невозбужденных участков ?тот ток будет служить катодом, а в отноше^
нии возбужденного участка — анодом. Под влиянием катода
биоэлектрического тока соседние невозбужденные участки возбудятся.
Одновременно с этим, в результате возникновения <и передвижения ионов, здесь
возникнет новый биоэлектрический ток, который, в свою очередь,
должен подействовать раздражающим образом на дальнейший
невозбужденный участок. Теперь этот участок возбудится и даст
биоэлектрический ток. Таким образом, биологический процесс, который возникает
первоначально в раздраженном участке, распространяется по всей
ЗАКОН ВОЗБУЖДЕНИЯ 157
живой единице при помощи .биоэлектрического тока, и (вся живая
единица постепенно приходит в состояние возбуждения.
¦ На рис. 84 дается схематическое изображение тока возбуждения и
$го распространения через соседние участки.
Это чрезвычайно.важное значение биоэлектрического тока, ©первые
указанное Германом (9) ів 1872 г., было признано потом целым
рядом - физиологов (Кремер, 10; Гильдетенстер, 11; Лнлли, 12
и др.).
Итак, биоэлектрический, ток, с одной стороны, является следствием
возбуждения раздраженного участка, а с другой — сам является
источником раздражения для соседних невозбужденных участков.
Рис. 84. Диаграмма распространения волны
возбуждения по возбудимой системе.
Область В—F занята волной возбуждения. Ее длина в мышечном
волокне скелетной мышцы лягушки, принимая во внимание
скорость распространения в 2.5 м при 20°С и продолжительность
волны—-0.003 сек., будет равняться 7.5 мм. Волна возбуждения
началась в участке В, достигла максимума в участке D и
подлежит распространению в участке F—G. Наверху дана волна
биотока возбуждения, а стрелки указывают на распространение
этого тока по волокну и в окружающей среде.
Характеристика биологического процесса возбуждения. Известно,
Что при наличии возбуждения в мышце возникают в общем такие же
продукты распада, как и при его отсутствии во время основного
биологического процесса. Отсюда следует, что в мицеллах при
возбуждении должны протекать такие же ферментативные процессы, как во
время основного биологического процесса. Однако при возбуждении
должно иметь место такое изменение биологического процесса, какого
не бывает в возбудимой системе под обычным влиянием среды. По
нашему мнению, это изменение должно заключаться в том, что в
раздраженном участке вся возбудимая система одновременно
Подвергается расщеплению,- катаболическому процессу, и здесь не остается ни
одной частички, которая бы не подверглась этому процессу вследствие
раздражения. Значит, во время основного биологического процесса
ферментативный процесс расщепления и восстановления протекает в
разных частичках разновременно, т. е. гетерохронно. При
раздражении же, когда возникает возбуждение, этот процесс расщепления и
восстановления происходит одновременно, т. е. изохронно. Это
изохронное состояние не остается в одном раздраженном участке. Оно
быстро распространяется по всей возбудимой единице, а также от
одной единицы к другим, с которыми возбудимая система имеет непо-
158
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
средственные протаплазматические связи. Поэтому сначала гетеро-
хронное состояние переходит ів изохронное в раздраженном участке, а
затем это изменение быстро распространяется по всей возбудимой
системе.
То обстоятельство, что изохронное расщепление происходит
одновременно во всех частичках данного участка возбудимой системы и
тем обусловливает возникновение большого количества ионов в
данном возбужденном участке, должно быть признано причиной
происхождения биоэлектрического тока возбуждения. Очевидно,
продолжительность, интенсивность и скорость распространения биоэлектрического
тока должны выражать продолжительность, интенсивность и скорость
распространения изохронного процесса расщепления.
Изохронное состояние, выражающееся в одновременном
расщеплении и восстановлении всей возбудимой системы, сменяется гетерохрон-
ным, как только прекращается раздражение. Это обусловливается
разным физико-химическим окружением частичек живой системы.
Восстановление частичек заканчивается в течение разного времени, и
потому последующее расщепление не может произойти точно в одно
время. Однако переход изохронного состояния в гетерохронное
происходит не сразу. Первоначально некоторая часть возбудимой системы
будет расщепляться и восстанавливаться более или менее
одновременно, как это и видно на рисунке 85. Вследствие этого количество
возбудимой системы, а также степень возбудимости должны колебаться
некоторое время после возбуждения. Сообразно с этим и возбужденный
участок должен давать вскоре после возбуждения более или менее
длительный и колеблющийся ток, выражающий собой основной
биологический процесс в этот переходный период от изохронного состояния
в гетерохронное.
На рис. 85 дается схематическое изображение перехода основного-
биологического процесса в процесс возбуждения и обратное
восстановление основного биологического процесса. При этом даются также все
последствия в смысле изменения количества возбудимой системы и
степени возбудимости.
В настоящее время данное положение, вытекающее из нашей
концепции насчет ритмического изменения возбудимости и основного
биоэлектрического тока, получило подтверждение в последних
исследованиях Эрлангера и Гассера, произведенных- над возбудимой
системой нервных волокон (см. гл. III в отделе нервной системы).
Локальное изменение возбудимой системы и локальное сокращение.
Из нашей концепции возникновения возбуждения вытекает, что, если
Енешняя сила не вызовет изохронного расщепления и не даст
возбуждения, это еще не означает, что она вообще не влияет на возбудимую
систему. Если сила раздражения достаточно близка к порогам, она
должна повлиять на основной биологический процесс 'и изменить его.
Чтобы раздражение вызвало возбуждение, произведенное им
изменение должно превзойти определенную величину. Это та -величина, за
ЗАКОН ВОЗБУЖДЕНИЯ
159
которой должно последовать возникновение такого биоэлектрического
тока, который был бы достаточен для раздражения соседних
невозбужденных участков. Прекрасная фактическая иллюстрация этого-
представления была дана в исследованиях американского ученого
Гетещжронное
Изохронно?
Гетерохронн&е
а
f
I
1
J
КояичестЗ?
возб. системы
$1 (Л
4в <Ш 5/ 4$
Я §2 60 «# 78 0 3884 71 & 55 46 50 47 60
Ъл
-L L
0 4 8 12 W Ю 24 28 323&Ы U 43 5? 5§ SO 64 68 7273 80 8488 92 №№Ш
Рис. 85. Схематическое изображение биологического процесса возбуждения.
Здесь же показан биоэлектрический ток возбуждения и изменение возбудимости. Волнообразные іли-
нии а, Ь, с, d, е, f,g,hn і изображают биологический процесс в разных частичках живой системы,
сначала при гетерохронном состоянии, затем при изохронном, а в конце—вновь при гетерохронном,
которое устанавливается постепенно после возбуждения. Во время гетерохронного процесса каждая
волна начинается не только в разные моменты, но и продолжается разное время. Во время
возбуждения расщепление начинается во всех частичках одновременно, и все они одновременно начинают
восстанавливаться после расщепления. Цифры под волнообразными линиями обозначают количество
возбудимой системы, как на рис. 78. Это количество немного меняется от момента к моменту во время
основного биологического процесса; во время же возбуждения, которое вызывалось дважды,
сначала оно падает до нуля, а потом постепенно нарастает. Каждый раз в конце гериода восстановления
количество возбудимой системы превосходит то, которое было до возбуждения.
Выше волнообразных линий дается кривая биоэлектрического тока от двух возбуждений. Первый раз
возбуждение приходится во время гетерохронного состояния, и в этот момент общее количество
возбудимой системы равняется 60 условным единицам. Второй раз возбуждение наступает в конце периода
изохронного восстановления после первого возбуждения. Здесь количество возбудимой системы
равняется 76 единицам. Соответственно интенсивность биоэлектрического тока в этом случае выше, чем в гервом.
В нижнем ряду приведена кривая изменения возбудимости. Абсцисса отмечает время (в сигмах), а
ордината — количество возбудимой системы. Степень возбудимости немного колеблется во время основ_
ного биологического процесса. Во время возбуждения возбудимость исчезает (абсолютная
рефракторная фаза), после возбуждения она постепенно восстанавливается сначала до нормы (относительная
рефракторная фаза), а потом становится выше нормы (фаза сверхнормальной возбудимости).
Гельфана (13). Он раздражал электрическим током под
микроскопом отдельные мышечные волокна при посредстве тончайших
электродов с наконечниками диаметром в 5—16 jx. Один электрод
прикладывался с помощью микроманипулятора к поверхности волокна. Так
как толщина 'мышечного волокна составляет около 100 р., то электрод
покрывал только небольшую часть его диаметра. Другой электрод
вводился в раствор Рингера, в котором находилось волокно. Гельфан
обнаружил, что при раздражении электрическим током слабой
интенсивности мышечное волокно приходило в действие только в раздра-
160
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
женном участке, именно сокращался только этот участок, причем
сокращение было тем сильнее, чем больше было раздражение. Но когда
сила раздражения достигала определенной большой величины и
вследствие этого раздражающее действие простиралось на более
значительную часть мышечного Диаметра, все волокно приходило в действие,
т. е. волна сокращения распространялась по всей длине мышечного
волокна. Дальнейшее усиление раздражения не сопровождалось
усилением этого эффекта. Он всегда был максимальным. Очевидно, что
при сравнительно слабых раздражениях локальный и градуированный
эффект был обусловлен (малыми размерами электродов. Когда Гель-
фа н, а также другой исследователь Пратт (14) пользовались более
толстыми электродами, в 36—50 jjl, которые прикрывали значительную
часть диаметра мышечного волокна, тогда даже пороговое
раздражение производило с самого начала
только максимальный эффект,
распространявшийся по всему волокну.
Локальная реакция возбудимой
системы мышцы сопровождается
соответствующим биотоком. Этот биоток
отводится от раздражаемого участка
и на расстоянии нескольких
миллиметров от него. Причем интенсивность
тока круто падает с удалением от
раздражаемого участка. При
пороговых электрических раздражениях на
вершине кривой локального
потенциала наблюдается биоток
возбуждения (Шефер и сотрудники, 42;
Эдриан и Гельфан, 43). Локальные
биотоки возникают в каждом возбудимом образовании и лучше всего
изучены на нервном волокне. Поэтому природа этих биотоков и их
отношение к биотоку возбуждения будут рассмотрены в отделе
физиологии нервного ствола.
Локальные эффекты были получены К а т о и его сотрудниками при
раздражении изолированного мышечного волокна с помощью
капиллярных электродов (7—10 р.), когда они плотно касались мышечного
волокна. Если между электродами и поверхностью мышцы оставалось
небольшое пространство, мышца отвечала согласно основному закону
возбуждения и при пороговых раздражениях. В опытах К а т о
локальное сокращение получалось только при подпороговых раздражениях в
пределах изменения расстояния катушек на 3—4 мм, и оно
распространялось не больше, чем на 0.5 мм. Особенно легко было
демонстрировать его, если волокно долго пролежало в рингере. Так, на рис. 86
локальное сокращение наступало при слабых раздражениях (110—
108 мм расст. кат.), а при более сильных раздражениях (107—50 мм)
наступали сокращения одинаковой максимальной высоты во всем во-
/// 110 109 108 107 100 50
Рис. 86. Эффект раздражения
мышечного волокна капиллярными
электродами.
При слабых раздражениях (110—108 мм
расст. кат.)—только локальные сокращения,
а при сильных (107—50 мм) наступают такие
сокращения, которые охватывают все
волокно (Като).
ЗАКОН ВОЗБУЖДЕНИЯ
161
локне (Като, 25). Это локальное градуируемое и нераспространяющееся
изменение возбудимой системы, конечно, не сопровождается
рефракторными фазами (Зихель и Проссар, 40).
Из вышеизложенного ясно видно, что, когда внешнее раздражение
производит локальное небольшое изменение в небольшом участке
возбудимой системы и не по всему диаметру мышечного волокна, тогда
наступает локальное сокращение. Нужно предположить, что при таком
раздражении только незначительная часть возбудимой системы
приходит в изохронное расщепление. Основной биологический процесс не
изменяется по существу, поэтому здесь не развивается такой
биоэлектрический ток, который был бы в состоянии произвести раздражение
соседнего участка, вызвать волну возбуждения и тем самым обусловить
сокращение всего мышечного волокна.
Заслуживает внимания тот факт, что Като находит полное сходство между
локальным сокращением мышечного волокна и «постнаркотическим сокращением»,
т. е. таким сокращением, которое вызывается локально в области действия
наркотического вещества и іне дает биоэлектрических токов возбуждения. Оба эти
явления были исследованы в лаборатории Като с разных сторон, и авторы
пришли к заключению, что локальное сокращение и «постнаркотическое сокращение»
суть явления одной и той же природы и оба они характерно отличаются от рас-
нространяющегося (сокращения (Като, 25).
Чтобы понять, почему именно происходит локальное сокращение при слабых
раздражениях через тончайшие электроды, нужно принять во внимание, что
сокращение является функцией миофибршілей, а не возбудимой системы. Если где-либо
на одном уровне мышечного іволонна наступило сокращение, оно не
распространится само собой по волокну. Миофибрилли приходят в сокращение под влиянием
определенных физико-химических процессов при непременном участии адениловой,
фосфорной и молочной кислот. Если часть мышц опустить в раствор таких кислот,
то сокращение наступит только в этой части (Вильмерс, 16). Эти кислоты
возникают не только под влиянием возбуждения, но и помимо него, когда нет
возбуждения, т. е. во время основного биологического процесса. Известно, что и во время
основного биологического процесса фосфорная и молочная кислоты могут достигнуть
такого количества, что мышца сократится. Так происходит, например, при
согревании мышцы.
Если принять все это во внимание, то не трудно будет объяснить, почему
именно при применении очень тонких электродов сокращение может быть
локальным, ограничивающимся в участке раздражения. Согласно данной выше концепции,
во время основного биологического процесса в возбудимой системе мышцы
расщепление и восстановление происходят в разных ее частичках гетерохронно. Под
влиянием этого процесса в мышце все время образуется фосфорная и молочная
кислоты, но только в незначительном количестве. Действием этих кислот
обусловливается происхождение собственного тонуса мышцы. При раздражении через
тончайшие микроэлектроды некоторая более или менее значительная часть
возбудимой системы сразу придет в изохронное состояние. От этого может
образоваться так много фосфорной и молочной кислот, что в области ее образования
может получиться небольшое сокращение. Като и ученики исследовали
метаболический процесс в участках локального сокращения. Они не нашли здесь увеличения
молочной кислоты, но они открыли в участке «постнаркотического сокращения»
усиленное расщепление фосфагена без последующего восстановления. Значит, локальное
сокращение может быть обязано сильному распаду фосфагена, который обычно
происходит во время основного биологического процесса. Но как только
раздражение усиливается в такой мере, что в раздраженном участке почти вся возбудимая
11 И. С. Беритов
162 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
система претерпевает изохронное расщепление, должно последовать возбуждение
с распространением его во всем волокне. А от этого физико-химические процессы
разовьются во всем волокне в такой интенсивности, что все миофибрилли
мышечного волокна придут в сокращение.
Итак, биологический процесс возбуждения характеризуется тем,
что в период возбуждения все частички возбудимой системы
претерпевают ферментативное расщепление в одно и то же время, г. е.
изохронно, в то время как при основном биологическом процессе
расщепление происходит в разных частичках разновременно, г. е. гетерохрон-
но. Переход гетерохронного [биологического процесса в изохронный
происходит при такой тле раздражения, которая в состоянии вызвать
изохронное расщепление в значительной части возбудимой системы и
тем произвести такой биоэлектрический ток, который может
раздражать соседние невозбужденные части возбудимой единицы.
По поводу методики исследования закона возбуждения на мышце. Закон
возбуждения нельзя изучить на целой мышце, пользуясь минимальным раздражением.
Рис. 87. Аппарат для изолирования волокон.
1 — эбонитовая платформа; 2 — резиновые полоски; 3 — железная подставка; 4 —
вогнутое зеркало; «5—стеклянная пластинка; б— пробка;.7 — парафиновые валики <Като).
Мышца состоит из многих пучков мышечных волокон, из которых только часть
приходит в состояние возбуждения при минимальных раздражениях. Раздражение
должно быть настолько сильным, чтобы охватывать всю толщу мышцы, тогда
только второе раздражение действительно не даст никакого эффекта во время
существующего возбуждения. Если же первое раздражение не достаточно сильно и
вызывает возбуждение только одной части мышечных волокон, то новое
раздражение может вызвать сокращение других іволокон, и этот новый эффект даст
усиление существующего сокращения. Этим объясняется -большое расхождение
авторов по вопросу о продолжительности абсолютной рефракторной фазы. Например,
по Самойлову (17)4 абсолютная рефракторная фаза мышцы лягушки равна
0-0011 сек. В последнее время такую же величину нашел Фос (18) при комнатной
температуре (19°). Если бы это было так, то нужно было ожидать, что ритм токов
возбуждения мышцы уже на лягушке превышал бы 500 в 1 сек., между тем этот
ритм не превосходит даже на летних лягушках 300 в 1 сек.
ЗАКОН ВОЗБУЖДЕНИЯ
163
Можно хорошо выявить закон .возбуждения при исследовании небольшого
пучка мышечных волокон. Первоначально было показано Люка сом (21) на
тонкой полоске m. cutaneus dorsi из 15—20 волокон, что при постеленном усилении
раздражения сократительный эффект нарастает скачками, причем каждая «овая
ступень сокращения соответствует действию новых волокон или новой группы их.
Эти опыты также убеждают в том, что (процесс возбуждения, а значит и
вызванное им сокращение отдельных волокон, не меняются от силы раздражения.
/й
h
?
U
В
и
Рис. 88. Инструментарий для изолирования мышечныхл*
нервных волокон.
А — Zn — Си пара: 7 — цинк, 2 — медь; В — препаровальные иглы;
С — скальпели; D — стальной пинцет (Като).
В самое последнее время закон возбуждения был доказан на отдельных
мышечных волокнах. Именно Гинтер (19) исследовал под микроскопом отдельные
мышечные волокна из membrana basihyoidea лягушки и нашел, что как на, прямое
раздражение мышечных волокон, так и іна раздражение соответствующего
двигательного нерва свежие препараты отвечают каждый раз сокращением одной
величины. Следовательно, возбуждение отдельных волокон не усиливается с усилением
раздражения, а это свидетельствует, что отдельные волокна реагируют согласно
закону возбуждения.
До Гинтера исследовали волокна этой же мышцы Фишль и Кан (20).
Они, наоборот, наблюдали, что интенсивность сокращения этих волокон зависит
от силы раздражения, что, следовательно, на них не оправдывается закон возбуж
дення. Но Гинтер показал, что такое явление наблюдается только на повреж
11*
164
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
денных мышечных волокнах, которые вследствие повреждения местами с большим
трудом проводят возбуждение.
Методика исследования закона возбуждения на изолированных мышечных
волокнах была лучше всего разработана К а то. Она изложена ниже в практикуме.
Практикум
Методика изолирования мышечных волокон. Изолирование производится на
специальном аппарате, который был придуман одним из сотрудников К а то
{рис. 87). Главные части состоят из стеклянной пластинки 5 около 10 см длины и
7 см ширины, покоящейся на двух резиновых полосах 2, прикрепленных к
эбонитовой платформе /, и из вогнутого зеркала для отражения света от электрической
лампы. По краям стеклянной пластинки имеется парафиновый валик 7, а на одном
из концов ее парафином же прикреплен небольшой кусок пробки 6. Длинная
эбонитовая платформа служит для опоры рук во время операции.
Мышечный или нервный препарат укладывают «а стеклянную пластинку, куда
предварительно был налит раствор Рингера, и сильный пучок света зеркалом на-
Рис. 89. Расположение мышечного волокна
для раздражения и для наблюдения его
укорочения.
В — боковые расширения желобка для вставления
электродов; М — выступы желобка для укрепления
волокна хлопковыми ниточками; Р—конец
волокна для наблюдения (Като).
Рис.90. Неполяризующийся электрод.
Стеклянная трубка, которая внизу заткнута
каолином, пропитанным раствором Рингера.
Наверху сернокислый цинк, куда погружена
палочка цинка {Zri) с зажимом (Като).
правляют на препарат. Изоляция одного волокна после расщепления нерва ведется
под микроскопом, но после некоторой практики она может быть произведена и
без него.
Для изолирования требуются специальные инструменты (рис. 88): тонкие
стальные иглы В с острием около 15 щ скальпели с очень острым лезвием С,
тонкий пинцет D и гальванический пинцет Zn — Си (А) для раздражения волокна.
Для изолирования мышечных волокон лучше всего пользоваться мышцами
semitendinosus, sartorius и hypoglossus лягушки. Один конец мышцы перевязывают
шелковой ниткой и прикалывают булавкой к пробке на стеклянной пластинке.
Затем мышцу освобождают от фасций острым скальпелем, по возможности по всей ее
длине. После этого мышцу расшепляют продольно иглами на две части, затем на
ЗАКОН ВОЗБУЖДЕНИЯ
165
четыре и так далее, пока не получатся пучки, состоящие из 10—20 мышечных
волокон. От одного из пучков отделяют одно волокно, начиная с периферического'
свободного конца по всей длине. Под микроокопом при слабом увеличении
неповрежденное волокно имеет совершенно одинаковый вид и диаметр по всей длине.
При большем же увеличении может быть наблюдаема поперечная и продольная
исчерченность. На таких препаратах можно ставить опыты в течение 10 часов.
Если волокно где-либо повреждено, оно имеет там угловатое утолщение. Такое
волокно не годится для опыта.
Доказательство закона возбуждения на изолированном волокне.
Неповрежденное изолированное волокно длиной около 3 см помещают в парафиновом желобке,
который имеет на одном конце с каждой стороны по два выступа, как это указано
на рис. 89. Первые конусообразные выступы В служат для присоединения неполя-
ризующихся сапожкоЕЫх электродов (рис. 90); вторые маленькие выступы М —
для закрепления волокна путем накладывания поперек него нескольких ниточек.
Обведенный кружком участок Р служит для наблюдения под микроскопом или для
каносъемки. Желобок и все выступы наполнены раствором Рингера. Волокно
раздражают в участке В индукционными ударами, а укорочение его
наблюдают в участке Р под микроскопом с помощью окуляр-микрометра. Сначала
определяют порог путем отдельных индукционных ударов, затем производят
раздражение индукционными ударами разной интенсивности. При всех силах
раздражения укорочение будет одинаковым, как видно на рис. 81 и 8G.
Закон возбуждения на наркотизированном мышечном волокне. Вместо раствора
Рингера ів желобок наливают тот или другой раствор наркотика. Берут такой
раствор, который привел бы к глубокому наркозу через 30 минут. Опыты с
раздражением отдельными индукционными ударами производят через каждые 5 минут.
Определяют каждый раз порог, и затем испытываются три раздражения разной
интенсивности. В каждом испытании укорочение будет одинаковым при любой силе
раздражения, как на рис. 82.
Литература
1. Keith Lucas, J. Physiol., 41, 368, 1910.
2. В er it off I., Z. Biol., 62, 125, 1913.
3. D i 111 e r u. Oinuma, Pfl. Arch., 139, 279, 1911.
4. Em b den G. u. Lange H., Klin. Wschr., S. 129, 1924.
5. M i с h a e 1 i s. Die Wasserstoffionenkonzentration, Berlin.
6. Be the A., Pfl. Arch., 163, 147, 1916.
7. Лазарев П., Исследования по ионной теории возбуждения, ч. 1, изд. Моск.
научн. инст., 1916.
8. Woronzow D., Pfl. Arch., 218, 148, 716, 1927.
9. Hermann L., Handb. Physiol., Bd. 2, Т., I, 194, 1889; Grundr. d. Physiol.,
4. Aufl., 323, 1872.
10. Kremer M., Nagel's Handb. Physiol., 4, 793, 1909.
11. Gildemeister M., Z. Biol., 83, 145, 1925.
12. Li Hie В., J. Gener. Physiol., 7, 473, 1925.
13. G elf an S., Am. J. Physiol., 93, I, 1930; Gelfan a. Gerard, Am. J. Physiol.
95, 412, 1930; Gelfan, Am. J. Physiol., 96, 16, 1931.
14. Pratt Fr., Am. J. Physiol., 93, 9, 1930.
15. Gelfan a. Bishop, Am. J. Physiol., 101, 679, 1932.
16. Wilmers J., Pfl. Arch., 178, 193, 1920.
17. Samojloff A., Arch. Anat. Physiol., S. 39, 331, 1909.
18. Voss O., Z. Biol., 87, 608, 1928.
19. H inter H., Pfl. Arch., 224, 608, 1930.
20. Fischl H. u. Kahn R., Pfl. Arch., 219, 33, 1928.
21. Keith Lucas, J. Physiol., 33, 125, 1905—1906.
166
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
22. Ts с her та к A. v., PH. Arch., 175, 165, 1919.
23. Hill A. V., Adventures in Biophysics, 1931; Русск. перев.: Эпизоды из области
биофизики, Биомедгиз, 1935:
24. Вис h thai F г., Skand. Arch. Physiol., 70, 199, 1934.
25. Kato G., The microphysiology of nerve, Tokyo, 1934.
26. H i 1 1 A. V., Chemical wave transmission in the nerve, 1932.
327. Ernst E. и. К о s z k a s, J., Pfl. Arch., 235, 389, 1935.
28. Ostwa Id, Lehrbuch d. allg. Chemie, Bd. 1, S. 788, 1910; Bd. 2, S. 798, 1911.
29. Nernst, Theoretische Chemie, 5. Aufl., S. 382, 1907.
30. Fricke, Kolloid. Z„ 35, 264, 1925.
31. D и b и i s s о n M., Arch, intern, de Physiol., 38, 85, 1934; 41, 177, 1935.
32. Fenn W. O., Physiol. Rev., 16, 450, 1936.
33. E r n s t E. a. Koszkas, Pfl. Arch., 239, 691, 1938.
34. В г о w n D. J. S. a. S i с h e 1, F. J. M., Science, 72, 17, 1930.
35. Asmussen E., Pfl. Arch., 230, 263, 1932.
36. С e p к о в Ф. H., Докл. на Укр. физиол. съезде в Днепропетровске, 1939.
37. М а к а р о в П. О., Тр. Ленингр. общ. естествоисп., LXVII, вып. 1, 1939.
38. Cybulski N., Anzeig. d. Akad. Wissensch., Krakau math.-natur. KL, Reihe B;
Biol. Wissensch., 1912.
39. Steimann S. E., Am. J. Physiol., 118, 492, 1937.
40. Sichel F. J. M. a. Prossar С L., Am. J. Physiol., 128, 203, 1940.
41. Кометиани П. А. и Гоголашвили Ш., Сообщ. Акад. Наук Груз. ССР,
2, № 10, 1941.
42. Schaefer H., Scholmerich P. u. Haass P., Pfl. Arch., 241, 310,
1938.
43. A dr і а п Е. D. a. G е 1 f a n S., J. Physiol., 78, 271, 1933.
44. Meyer К. Н., Schw. med. Wschr., II, 826, 1937.
45. Lund, Exp. Zool., 51, 265, 1928.
46. Кометиани П., Соо?ц. Ак. Наук Груз. ССР, 4, 261, 1943.
Г лае а VII
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
Об электрическом эффекте мышцы. Наши знания относительно
процесса возбуждения основываются главным образом на изучении
биоэлектрических токов возбуждения. По характеру и изменениям токов
возбуждения заключают о характере и изменении процесса
возбуждения.
При раздражении двигательного нерва ток возбуждения сначала
возникает в том участке мышцы, где оканчиваются нервные волокна.
При прямом же раздражении імышцы он появляется прежде всего в
участке раздражения. Если этот участок соединить с каким-либо
другим невозбужденным участком через гальванометр, то окажется, что
возбужденный участок мышцы относится к любому невозбужденному
участку электроотрицательно (Герман, 1). Если с гальванометром
соединить два участка мышцы и подвергнуть последнюю раздражению,
тогда гальванометр будет дважды находиться под влиянием тока
возбуждения. Возбуждение возникает в одном какоім-либо участке, но,
распространяясь отсюда, оно вызывает возбуждение сначала под одним
электродом, отводящим биоток к гальванометру, а потом под другим.
Сообразно с этим подвижная часть гальванометра сначала
отклоняется в одну сторону, затем —в другую. Вследствие этого электрический
эффект является двойным. Такой эффект называется двуфазным
током возбуждения (рис. 91). Но регистрация двуфазных
токов часто мешает точному изучению электрического эффекта. Для
избежания этого повреждают каким-либо образом конечный участок
мышцы (например, обжиганием) и затем соединяют с гальванометром
этот поврежденный участок вместе с каким-либо неповрежденным
участком. Вследствие этого возбуждение отводится только одним
электродом, т. е. в этом случае ток возбуждения является однофазным
(Герман, 1). Такой же однофазный ток можно получить, когда один
отводящий электрод лежит на сухожилии, а другой — на мышце
(рис. 92).
При раздражении отдельными индукционными ударами ток
возбуждения в каждом участке мышцы проявляется в виде одного быстро
168
ОБЩАЯ ФИЗИОЛОГИЯ МЬШПЧНОЙ СИСТЕМЫ
протекающего колебания электрического тока. При повторных же
раздражениях колебание электрического тока возникает повторно: когда
раздражение тетаническое, т. е. вызываемый им механический эффект
проявляется сплошным тетанусом, электрический эффект протекает в.
виде быстро следующих одно за другим ритмических колебаний элект-
/Vjj^c
Ф
-^у—1
т
п Злектрогрсмлтя
і
Ф
1
о
} \
D
к
<2>
3
о
А
і К.
Рис. 91. Схематическое изображение распространения
тока возбуждения ,'в мышце с параллельными волокнами
m. sartor ius.
Мышца раздражается в участке Р —¦ проксимальный конец. Волна
возбуждения обозначается поперечными линиями. Мышца
соединена с гальванометром. Рис. А представляет момент раздражения;
волна возбуждения находится на проксимальном конце вне
отводимого участка. Рис. В изображает момент, когда волна
возбуждения достигает ближайшего отводящего электрода. Гальванометр
отклоняется в одну сторону, при этом получается первая фаза
электрограммы. Рис. С представляет момент, когда волна
возбуждения находится между отводимыми участками. В этот момент
ток отсутствует. Рис. D дает момент волны возбуждения под ди-
стальным электродом. Теперь гальванометр отклоняется'в другую
сторону, при этом получается вторая фаза электрограммы
(по Фультону).
рического тока (рис. 93), причем частота колебаний электрического
тока в секунду, т. е. его ритм, зависит как от частоты и интенсивности
раздражения, так и от функционального состояния мышцы: чем чаще
и сильнее раздражение, чем лучше функциональное состояние, тем
выше ритм тока возбуждения. Итак, электрический эффект мышечной
деятельности протекает прерывисто, т. е. ритмически, причем каждое
колебание тока соответствует одному импульсу возбуждения, вызван-
ному однократным раздражением.
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ 16>
Напряжение тока возбуждения мышцы довольно велико. Так,
сравнительно тонкая мышца — m. sartorius лягушки — развивает напряжение
до 20 mV при возбуждении всей мышцы одним индукционным ударом.
Рис. 92. Токи возбуждения m. sartorius лягушки. Мышца
кураризована. Раздражается одним размыкательным
индукционным ударом. Отводится в катодный осциллограф
на расстоянии |5 мм от раздражающих электродов.
Межполюсное расстояние отводящих электродов 9 мм.
А—двуфазный ток возбуждения при продольном отведении
нормального участка мышцы. В — однофазный ток возбуждения при^по-
перечно-продольном отведении, т. е. когда один отводящий электрод
лежит на нормальной продольной поверхности, а другой — на
поврежденном участке. В первом случае ток усиливался почти вдвое
меньше, и скорость фотографирования была почти вдвое меньшей,
чем во втором случае (Шефер).
а небольшой пучок или одно волокно ее дает напряжение при том же
раздражении, или при спонтанном возбуждении в связи с
повреждением, минимум 0.16 mV (Шефер, 56). Это напряжение тока возбуж-
Рис; 93. Электрический эффект во время тетанического сокращения m. semitendinosus
кошки.
Раздражается 300 раз в секунду двигательный нерв. Регистрация электрического эффекта происходит
струнным гальванометром. Быстро колеблющаяся кривая принадлежит этому эффекту, а сплошная
кривая—механическому эффекту мышцы. Нижняя линия отмечает время по одной пятой секунды. Ритм
колебаний тока следует по половинному ритму раздражения, т. е. по 150 кол. в 1 сек., местами же отвечает
ритму раздражения, т. е. по 300 в 1 сек. (участки X)
дения целой мышцы настолько велико, что можно пользоваться этим
током для раздражения нерва, если нерв приложить к мышце и затем
возбуждать ее через собственный нерв. При раздражении нерва
одного нервно-мышечного препарата можно получить как одиночные,
170 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
так и тетанические сокращения мышцы другого такого же препарата,
приложив нерв последнего к мышце первого. Это так называемое
вторичное сокращение впервые получено Матеучи в 1842 г.
В определенных условиях можно іполучить вторичное сокращение и от
раздражающего действия токов возбуждения одной мышцы
непосредственно на другую. Так бывает на мышцах немного подсушенных или
при сильном сжатии. Если такие мышцы плотно соприкасаются в
одной какой-либо точке, то механическое раздражение одной мышцы
вызывает сокращение обеих. Получение вторичного раздражения путем
раздражающего действия биотоков одной мышцы прямо на мышечные
волокна другой доказывается тем, что описанное явление наблюдается
и на кураризованных мышцах. Оно не наблюдается, если между
мышцами находится тонкий изолятор (Кюне, 65; Бидерман, 66).
Подсушивание или сжатие имеет то значение, что при этом возбудимость
мышечных волокон повышается, а в межклеточной жидкости
электропроводность понижается вследствие уменьшения межклеточной
жидкости. Это создает хорошие условия для распространения биотоков одной
мышцы на мышечные волокна другой.
Ритмический электрический эффект в мышце возникает не только
при ритмическом раздражении, но и при разного рода неритмических
раздражениях, например при пропускании постоянного электрического
тока, при сдавливании, при действии таких активных химических
веществ, как ацетилхолин. При этом не имеет значения, где
прикладывается ацетилхолин — в нервном или безнервном участке. Так,
например, на рис. 94 показан электрический эффект безнервного участка
мышцы от ацетилхолина.
В определенных условиях можно получить ритмический эффект и
от одиночного индукционного удара или от одного нервного импульса.
Это, например, бывает на подсушенных мышцах (Кюне, 65; Бидерман,
66; Дюриг, 58). В этих случаях дело идет о возбуждении одних
мышечных волокон под влиянием токов возбуждения других.
Но в определенных случаях высокой возбудимости мышечной ткани
один нервный импульс может вызвать ритмический разряд в каждом
волокне, независимо от соседних волокон. Это, например, наблюдали
Экклс и О'Коннор (70) на мышце теплокровного животного.
На рис. 95 дается хороший пример такого рода ритмического
разряда мышцы: на один нервный импульс мышца отвечает 10
импульсами. В определенных случаях, когда один импульс не дает
ритмического разряда, два быстро следующих друг за другом импульса
производят его (рис. 95). Такие ритмические разряды должны быть связаны
с высокой возбудимостью мышечных волокон, с хорошими условиями
восстановления возбудимой системы после каждого расщепления в
связи с возбуждением. Поэтому ритмические разряды особенно хорошо
наблюдаются после искусственного повышения возбудимости,
например под влиянием малых доз эзерина (Броун, Дейл и Фельдберг, 71;
Броун, 72).
процесс возбуждения мышцы
171
Происхождение ритмических разрядов мышцы в ответ на один
нервный импульс хорошо согласуется с данной выше концепцией о
процессе возбуждения. Как показано на рис. 95, импульсы возникают в
ритмическом разряде мышцы теплокровных с большой правильностью
через каждые 0.005 сек. Это указывает, что возникновение каждого
.¦последующего импульса в наиболее возбудимых волокнах происходит
после исчезновения рефракторной фазы, в фазе сверхнормальной
возбудимости. На этом основании мы должны предположить, что в
мышечных волокнах с повышенной возбудимостью изохронное
восстановление после первого возбуждения заканчивается более или
менее одновременно в значительной части возбудимой системы. Вследствие
Рис. 94. Токи возбуждения безнервного участка m. sartorius от ацетилхолина.
Свежевырезанная мышца. Ацетилхолин в растворе 1 : 50 000 фильтровальной бумажкой локально
прикладывается в проксимальном участке на расстоянии 3—5 мм от конца. Токи возбуждения отводятся
от проксимальной трети в струнный гальванометр через небольшой усилитель (усиление в 5 раз). Мышца
фиксируется на границе средней и дистальной третей путем зажима: возбуждение свободно может перейти
в этог участок, но мышца здесь не передвигается. Движение концов мышцы записываются
фотоаппаратом. Э — электрический эффект; Р —механический эффект с проксимального конца мышцы; D —
механический эффект с дистального конца. Между опытами А и В интервал в 2 сек. Ритм токов
возбуждения около 150 в 1 сек. Значительный механический эффект наступает на проксимальной стороне.
Дистальная треть дает лишь незначительное укорочение.
этого последующее спонтанное или «самопроизвольное» расщепление ее
произойдет в одно и то же время в, этой значительной части
возбудимой системы, и тем самым вновь может возникнуть
распространяющийся процесс возбуждения. Это іможет повторяться многократно,
пока не утомится возбудимая система и тем самым не 'ухудшатся
условия ее. восстановления.
Ток возбуждения и сокращение. Обычно ток возбуждения,
вызванный одним индукционным ударом, (Прекращается почти до начала
механического эффекта (рис. 96). Это с очевидностью показывает, что
означенный электрический ток не зависит от сократительного вещества
мышцы. Кроме того, при утомлении мышц от многократного
повторного раздражения отдельными индукционными ударами ослабление
механического и электрического эффектов не протекает параллельно:
механический эффект может уменьшиться и даже исчезнуть раньше
электрического (Брюкке, 2; Беритов, 3). В период длительной
мышечной работы электрический эффект продолжает нарастать в то время,
когда механический уже начинает ослабевать (рис. 108) (Беритов, 4).
Далее, если в стадии сильного утомления от длительного раздражения
отдельными индукционными ударами возобновить раздражение после
небольшого перерыва, то одиночные сокращения вначале бывают более
172
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
сильными, чем до перерыва, затем они быстро падают и только потом
начинают нарастать. Токи же возбуждения после перерыва вначале
более слабы, чем до перерыва, и они с самого же начала быстра
нарастают (Беритов, 4). Итак, изменение токов возбуждения
при продолжительной деятельности не идет параллельно
изменению (механического эффекта: усиление токов возбуждения может иметь
место при одновременном ослаблении механического эффекта. Из всех
приведенных фактов прежде всего нужно заключить, что в происхож-
Г'.'/: ,'7,'Л.'-'.'/г-ггтТ77і
Рис. ;95. Ритмический * разряд
биотоков возбуждения мышцы
на один нервный импульс.
М.tibialis ant. кошки.
В опыте 1 раздражается m. tibialis
одним индукционным ударом. В ответ
наступает разряд из 10 импульсов.
В опыте 2 раздражение происходит
двумя ударами через разные
интервалы. При большом интервале —
0.04 сек. (нижняя кривая) — каждое
отдельное раздражение дает по
одному импульсу; при интервалах
менее 0.01 сек. получается ритмическое
последействие, которое тем
длительнее, чем меньше интервал. Стрелки
указывают моменты нанесения
индукционных ударов. Время — 0.01 сек.
(Экклс и О'Коннор).
Рис. 96. Скрытый период электрического
и механического эффекта мышцы лягушки
(m. gastrocnemius). Раздражается
двигательный нерв одним индукционным
ударом. Верхняя кривая—от электрического
эффекта, нижняя — от механического.
1 — время .проведения возбуждения по нерву
от момента раздражения; 2 — время около
3 а для передачи возбуждения от нерва
к мышце через двигательную пластинку;
5—время от начала биотока возбуждения мышцы до
начала механического эффекта около 1.5 з;
4 — время около 4 а для медленно
нарастающего колена до начала быстро нарастающего
эффекта (Фультон).
дении тока возбуждения мышцы
сократительный процесс не участвует; затем из
этих же фактов видно, что
сократительный процесс не определяется одним
только возбуждением. Очевидно, ток
является прямым, непосредственным
следствием возбуждения, и потому он всегда
предшествует сократительному процессу (Эрнст, 42).
При тетанических раздражениях, прежде чем сокращение мышцы
достигнет своего максимума, мышца должна возбудиться несколько
раз. Соответственно в момент нарастания механического эффекта
мышца дает ряд биоэлектрических токов возбуждения; с каждым
новым импульсом, следовательно, с каждым новым током возбуждения,
сокращение повышается, пока оно не достигнет максимума.
Последующие токи возбуждения возникают уже на фоне существующего
максимального сокращения. При этом токи возбуждения всегда
протекают ритмически, в то время как сокращение имеет сплошной, не-
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
173
прерывный характер (рис. 93, 97). Эта характерная разница между
механическим и электрическим эффектами опять-таки указывает на
разное происхождение того и другого эффекта.
Эйнтховен (5) путем особого усовершенствованного прибора
для регистрации механического эффекта заметил, что начало
(механического эффекта почти совпадает с началом электрического, а иногда
даже предшествует последнему. Отсюда автор заключает, что
«отрицательное колебание тока» вызывается не волной возбуждения. Он
предполагает, что оно связано с самим сократительным процессом. Но
в самое последнее время сопоставили токи возбуждения с изменением
-светопреломления в сокращенной мышце. Преломление регистриро-
Рис. 97. Взаимоотношения между возбуждением и сокращением во время
коротких тетанусов. М. semitendinosus кошки.
Дается іэлектро грамма (/) и механограмма (//) этой мышцы от раздражения
двигательного нерва индукционными ударами с интервалом в 0.01 сек. При двух импульсах (токи
возбуждения b и d) сокращение сильнее, чем при одном импульсе (с); при четырех (а)
еще больше, ;а при шести (е, /) получается максимальное сокращение.
івалось путем фотоэлемента, куда попадали лучи света по выходе из
мышцы. Возникший при этом ток регистрировался осциллографом
катодных лучей. Оказалось, что сокращение мышцы начинается не
раньше, чем через 4 сигмы после начала тока возбуждения (Шефер и
Гёпферт, 50). Эти наблюдения ясно свидетельствуют, что ток
возбуждения ни в какой мере не связан с сократительным процессом.
Последний процесс также сопровождается током, но он, как это будет
видно ниже, совершенно другого характера. В опытах Эйнтховен а,
весьма возможно, регистрировалось не укорочение, а уменьшение
объема мышцы; это изменение могло зависеть и от возбуждения, как
об этом уже говорилось выше.
В последнее время выставляется и такое положение, что в мышце
биоэлектрический ток возбуждения возникает только в сеязи с .возбуждением нервных
окончаний, что возбуждение самого мышечного волокна лишено этого тока. Нервное
¦скончание в мышце или концевая двигательная пластинка признается источником
электричества, подобно электрической пластинке электрического органа рыб. Как
известно, двигательная и электрическая пластинки являются гомологами. Первое
сообщение было сделано в 1920 г. (Генрик и Линдгард, 33), и затем появилась
серия исследований в подтверждение этой гипотезы. Авторы нашли, что кураризо-
174
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ванная мышца не дает биоэлектрических токов возбуждения, хотя при
непосредственном раздражении мышца (сокращается (Линдгард, 34). То -же наблюдали в>
последнее время и в другой лаборатории (Гёфер, 39). Аналогичное явление
наблюдалось при раздражениях мышцы после дегенерации нерва, а также при
раздражении безнервных участков нормальной мышцы (Линдгард, 34). Далее, авторы
находили в некоторых случаях то же самое и на изолированных мышечных волокнах
при прямом раздражении (Бухталь, 32). Кроме того, авторы отмечают, что сокраще
кие кураризованной мышцы или изолированных мышечных волокон находится в
полной зависимости от силы раздражения: с усилением раздражения сокращение
нарастает с большой постепенностью. Наоборот, если на нормальной мышце
раздражается двигательный нерв или непосредственно концевая пластинка, сокращение
растет скачками при постепенном усилении раздражения, и, кроме того, каждый раз=
появляется биоэлектрический ток возбуждения (Асмуссен, 35; Бухталь, 32).
На этом основании заключают, что
мышечное волокно независимо от нерва и
нервных окончаний не подчиняется
закону «все или ничего», т. е. закону
возбуждения. Эти данные противоречат
наблюдениям старых и новых авторов.
Другим авторам приходилось
наблюдать на кураризованных мышцах
биоэлектрические токи возбуждения (рис. 98)
(Борутау, 44; Беретов, 37; Шефер, 56, 52).
Наблюдались также токи возбуждения
при пороговых раздражениях безнервных
участков m. sartorius, именно при
раздражении нескольких волокон на расстоянии,
2—3 мм от проксимального конца. При
этом в случае продольного отведения
возбужденных волокон токи возбуждения
получались двуфазные с большим
интервалом покоя между ними (Бишоп и Гиль-
сон, 38). Разряды токов
возбуждения получаются на денервированных
мышцах при введении ацетилхолина, а также
спонтанно в связи с фибриллярным
сокращением денервированной мышцы теплокровных (Броун, 51). Такие же разряды
с очень высоким ритмом можно наблюдать на безнервных участках мышцы лягушки^
под влиянием ацетилхолина, как это дано выше на рис. 94 (Беритов, 63).
Если сотрудники Лиіндгарда не наблюдали токов возбуждения на безнервной
мышце, то это должно зависеть от их метода исследования. Первое наблюдение-
они сделали на кураризованной икроножной мышце. Раздражали проксимальный
конец, а отводили дистальную треть (Бухталь и Линдгард, 62). При этих условиях,,
конечно, они не могли наблюдать токов возбуждения, ибо ввиду перистого строе
ния этой мышцы мышечные волокна, находящиеся в проксимальном конце, не
доходят совсем до дистальной трети (рис. 33). На кураризованной мышце с
параллельными волокнами, как m. sartorius, они не наблюдали биотоков возбуждения,
только в некоторых случаях. Вероятно, это случалось тогда, когда раздражался
один конец мышцы іи отводился другой на большом отдалении, когда все
раздраженные волокна (прерывались еще до отзодимого участка. Далее, они не
наблюдали токов возбуждения на изолированных мышечных волокнах в случае
механического раздражения безнервных участков, но наблюдали его в случае-
раздражения им нервного участка. Очевидно, применявшееся ими механическое
раздражение было достаточно для возбуждения концевой пластинки, где
возбудимость высока, но оно было недостаточно для возбуждения безнервного участка*
'J—lX-L-L
Рис. 98. Токи возбуждения' от нервного
и безнервного участков мышцы. Курари-
зованная мышца лягушки m. sartorius
отводится в струнный гальванометр
Эйнтховена.
Электрограмма А получена при отведении
среднего нервного участка, один отводящий электрод
лежит на нервном участке, а другой — на
безнервном проксимальном конце мышцы.
Раздражается дистальный конец на расстоянии 10 мм
от отводящих электродов.
Электрограмма В получена при отведении
проксимального участка, бедного нервами; оба
отводящие электрода находятся на этом участке.
Мышца раздражается с дистальной стороны на
расстоянии 10 мм от отводящих электродов.
Верхняя линия отмечает время в пятых секунды.
процесс возвуждения мышцы
175
где возбудимость значительно ниже. Здесь наступало только локальное
сокращение в 'месте раздражения, как в опытах К а то при пороговых раздражениях
(рис. 86). Характерно, что те же авторы получали полное сокращение волокна путем
электрического раздражения одного конца при таких силах раздражения, которые
были не далеки от порогов. Этот факт они рассматривали как результат
распространения токов раздражения вдоль всего волокна, но это предположение ничем
не было обосновано. Таким образом, все основные факты, выставленные Бухта-
лем иЛиндгардом против появления токов возбуждения в безнервных
участках, а значит, против существования вообще возбудимой системы в мышце,
одинаково не выдерживают критики.
Ток сокращения. Путем исследования биоэлектрических токов
мышцы наиболее совершенным прибором, осциллографом катодных
лучей, было установлено, что при одиночных раздражениях
монофазный электрический эффект состоит из быстро нарастающего
восходящего колена (и зубца) продолжительностью около 3 с, которое
предшествует сокращению, и последующего, медленно протекающего
нисходящего колена, продолжительностью в 50—70 а с очень малой
амплитудой, которое совпадает с сокращением (Бишоп и сотр., 6;
Шефер, 56). Так, на рис. 92 приводятся двуфазный и однофазный токи
возбуждения кураризованной мышцы m. sartorius, где при
однофазном отведении нисходящее колено продолжается около 50 з. Это
[медленное низковольтовое колебание тока при одиночных раздражениях
может продолжаться дольше одиночного сокращения -и может
кончиться гораздо раньше него (Серков, 55; Шефер, 56). Такой же ток
получается во время сокращения отдельных мышечных волокон, если он
отводится к осциллографу с помощью івоткнутых через сарколемму
микроэлектродов. Обнаруженный при этом ток возникает сейчас же
после раздражения и удерживается некоторое время после
сокращения в течение нескольких секунд (Бухталь и Петерфи, 31, 32). Было
установлено, что імедленное электрическое колебание определенным
образом связано с сократительным процессом. Во-первых, при
обработке імышцы водой она теряет сократительную способность наравне
с длительной частью электрического эффекта, в то время как
проведение возбуждения наравне с быстро протекающим эффектом
сохраняется (Бишоп и сотр., 6). Во-вторых, при отравлении моноиодуксус-
ной кислотой это медленное колебание тока 'исчезает или сильно
редуцируется, хотя при этом ни токи возбуждения, ни одиночное
сокращение не уменьшаются (Шефер и Шелмерих, 61) (рис. 99). На этом
основании можно высказать предположение, что низковольтовый
потенциал зависит от какого-то биохимического процесса, который
протекает в імышце в нормальных условиях в связи с сокращением и на
который отравление моноиодуксусной кислотой действует
расстраивающим образом.
Биоэлектрический ток, выражающий сократительный процесс,
проявляется еще с большей интенсивностью во время тетануса и
контрактуры. При тетаническом сокращении мышц лягушки сократительный
ток нарастает во время тетануса и удерживается после него во время
176
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
контрактурного последействия. Он уменьшается параллельно с
контрактурой и исчезает почти одновременно с ней. Этот ток протекает
без колебаний, подобно механическому эффекту. Подобно контрактуре
он становится все слабее и слабее при многократном повторении
раздражения и перестает вызываться при полном утомлении мышцы.
Этот сплошной электрический ток во время контрактурного
сокращения был назван контрактурным током (Беритов и Воронцов, 10).
о оі аг ш —тЬ—?^" обсек
Рис. 99. Биоэлектрический эффект сокращения m. sarto-
rius лягушки до (а) и после (Ь) отравления моноиодуксус-
ной кислотой.
Регистрация1 катоднолучевым осциллографом до отравления и
через 10 минут после введения 0.1% раствора этой кислоты в
лимфатический мешок. Мышца не вырезана. От начального, быстро
протекающего тока возбуждения видно только нисходящее колено.
Длительное добавочное колечко от сокращения целиком
устраняется моноиодуксусной кислотой (Шефер и Шелмерих).
Многие авторы наблюдали возникновение постоянного
биоэлектрического тока также при контрактурах, вызываемых химическими
агентами, как, например, ацетилхолином и вератрином (Риссер и Штейн-
таузен, 7), лактатом калия (Бухталь, 74), и вообще при воздействии
на мышцу веществ, вызывающих контрактуру, как К, НС1, СНС13
(Фишер, 8; Нерлицка, 9).
Продолжительность процесса возбуждения. Судя по токам
возбуждения, продолжительность процесса возбуждения, вызванного
однократным раздражением, например одним индукционным ударом,
зависит прежде всего от рода животного. У теплокровных она меньше, чем
у холоднокровных. В мышце лягушки, например, один ток
возбуждения продолжается 3—4з, у теплокровных — 2.5—Зл В изолированных
мышечных волокнах лягушки продолжительность процесса возбужде-
ПРОЦЕСС ВОЗБУЖДенИЯ МЫШЦЫ
177
ния 1.5—2 J. Следовательно, продолжительность 3—4а на целой мышце
является результатом неодновременного наступления возбуждения
в отводимом участке (Шефер, 56; Экклс и О'Коннор, 69). Повышение
температуры и отдых уменьшают, а понижение температуры и
утомление увеличивают это время (Беритов, 3). Вообще, можно сказать, что
во всех случаях, когда условия благоприятствуют обмену веществ, ток
возбуждения, а следовательно и процесс возбуждения, протекают
более быстро.
В связи с возбуждением наступает не только означенный выше
быстро протекающий ток, но также следующий за ним длительный
ток небольшой интенсивности. Он проявляется в виде медленного
опускания нисходящего колена кривой тока возбуждения. Мы выше
рассмотрели сократительный ток, который сопровождает сокращение.
Но едва ли всякое медленное последействие тока можно поставить
в связь с сократительным процессом.
Вероятно, это медленное опускание нисходящего колена тока
возбуждения, а также то низковольтовое колебание, которое
прекращается еще на высоте одиночного сокращения портняжной мышцы
(Шефер, 56) и продолжается несколько долей секунды после тетаниче-
ского раздражения (Беритов, 30), имеют такое же происхождение, как
аналогичное низковольтовое колебание в нервном стволе. Это
электрическое последействие более подробно изучено на нерве в связи с
нервным возбуждением. Поэтому некоторые гипотетические соображения
о его происхождении даются в отделе нервной системы.
Интенсивность процесса возбуждения. Интенсивность возбуждения
в каждой точке возбужденного волокна зависит, во-первых, от
функционального состояния данной точки и, во-вторых, от морфологических
особенностей самого волокна в данном участке. Чем большее, по той
или другой причине, количество возбудимой системы участвует в
возбуждении в данный момент, тем сильнее процесс возбуждения. Об
интенсивности судят по биоэлектрическим токам возбуждения. Правда,
к гальванометру обычно отводится не одно мышечное волокно, а вся
мышца, но и при этих условиях ясно івидна правильность данного
положения. Интенсивность тока возбуждения данного участка* мышцы
при наркозе, при длительной деятельности, а значит, при определенном
функциональном изменении, сначала увеличивается, а затем
уменьшается (Беритов, 4, 12). Очевидно, это является результатом
аналогичных изменений тока возбуждения в каждом мышечном волокне.
Однако интенсивность тока возбуждения разных участков не
одинакова даже при одинаковом функциональном состоянии. При изучении
токов возбуждения мышц с параллельными волокнами, и притом
более или менее одинаковой толщины по всей длине (m. pect. pars
abdom., т. sartorius), было обнаружено, что интенсивность токов
возбуждения значительно больше в тех участках мышцы, где
оканчивается больше всего двигательных нервных волокон. Например, на
концах m. sartorius, где микроскопическим «исследованием не было об-
12 И. С. Беритов
178 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСЕМЫ
наружено окончаний двигательного нерва, ток возбуждения в 2—3 раза
слабее, чем в центральном нервном участке. Эта разница в токах
возбуждения наблюдается как при непосредственном раздражении
мышцы, так и при возбуждении ее раздражением двигательного нерва.
Опытами установлено, что нервные окончания сам'и по себе не играют
существенной роли в происхождении большой интенсивности токов
возбуждения нервных участков. Та же разница наблюдается на кура-
ризованных імышцах при прямом раздражении (рис. 98). Отсюда было^
сделано заключение, что в мышечных волокнах процесс возбуждения
наступает сильнее всего в области нервных окончаний, т. е там, где
находится больше всего возбудимой системы мышцы. Этим, между
прочим, обусловливается то явление, что «нервные» участки при
сокращении укорачиваются сильнее и развивают больше напряжения,,
чем «безнервные» (см. гл. III) (Беритов, 37).
Ритмическое течение возбуждения. Мы выше указывали, что
электрический эффект при тетаническом сокращении мышцы всегда
протекает ритмически. На этом основании признают, что и
возбуждение всегда протекает ритмически. Лучше 'всего ритмическая природа
проявляется при сплошном неколеблющемся раздражении, как,
например, при раздражении постоянным электрическим током, ацетил-
холином и т. д. В таких случаях імышечные волокна отвечают
повторными импульсами почти предельно высокого ритма (рис. 94).
Вначале высокий, ритм потом постепенно уменьшается до 10—-
20 в 1 сек. (Гартен, 60; Эдриан и Гельфан 57; Беритов, 63). При
раздражении фарадическим током, если частота раздражения не слишком
велика, а интенсивность не слишком імала, ток возбуждения точно-
следует ритму раздражения. При малой интенсивности возбуждение
тоже протекает ритмически, но уже не соответствуя ритму
раздражения. Ритм возбуждения будет ниже, и притом тем ниже, чем слабее
раздражение. При некоторой высокой частоте раздражения ритм
возбуждения не совпадает с ритмом раздражения ни при какой силе. Он
всегда будет ниже ритма раздражения. Однако при любой высокой"
частоте и интенсивности раздражения этот эффект будет ритмическим
и будет слагаться из отдельных токов возбуждения. Так как ток
возбуждения сопровождает процесс возбуждения, то следует заключить,,
что процесс возбуждения мышцы всегда протекает ритмически. На
основании изучения токов возбуждения было установлено, что ритм
возбуждения тем выше, чем чаще и сильнее раздражение. Но, если
постепенно увеличивать частоту й- интенсивность тетанического
раздражения, ритм возбуждения достигнет предельной высоты, выше
которой он не будет подниматься, как бы ни усиливать и ни учащать
дальше раздражение. Этот предельный ритм возбуждения называется
собственным ритмом мышцы, так как он зависит прежде всего от
функционального состояния ее. В охлажденной мышце зимней лягушки
предельный ритм мышечного возбуждения не превышает 150—200 в
секунду, <в мышце летней лягушки он достигает 300, а в мышце те-
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ 179
плокровных ритм возбуждения доходит до 400 (рис. 100) (Беритов,
13, 14).
Лабильность. Высший предельный ритм возбуждения является
наилучшим показателем функционального состояния мышцы и вообще
возбудимой ткани. Этот ритм точно отображает функциональное
состояние: с улучшением функционального состояния, иначе говоря, с
повышением подвижности жизненных процессов или с ускорением
Рис. 100. Высший предельный ритм возбуждения m. semitendi-
nosus летней лягушки.
Верхняя электрограмма (А) получена при 200 раздражениях в 1 сек.,
нижняя (В)—при 300 раздражениях. В обоих случаях сила раздражения
максимальная—на 10 см сильнее порогового раздражения. На верхней
электрограмме вначале ритм возбуждения 200 в 1 сек., т. е. отвечает ритму
раздражения, потом он местами становится вдвое меньше. На нижней
ритм возбуждения только небольшими участками (X) совпадает с ритмом
раздражения, в остальных участках он вдвое меньше. Внизу—время в
сотых долях секунды. По этим электрограммам ясно, что высший ритм
возбуждения в данной мышце в летнем состоянии около 300 в 1 сек.
основного биологического процесса, высший предельный ритм
возбуждения становится выше. Введенский придавал подвижности тех
жизненных процессов, или элементарных реакций, которыми
сопровождается физиологическая деятельность возбудимой системы,
большое значение и считал ее характеристикой возбудимой ткани. Он
ввел специальный термин — функциональная подвижность, или л а-
бильность — для обозначения этого явления и считал мерилом
лабильности высший предельный ритм возбуждения (Введенский,
25, 40).
Мы выше указывали, как на показатель функционального
состояния, на хронаксию и пороговую силу раздражения. Этими
показателями мы определяем возбудимость, иначе говоря, функциональное со*
стояние возбудимой системы в определенный момент времени. Высший
ритм возбуждения же, который является мерилом лабильности, ото-
12*
180
ОБЩАЯ ФИЗИОЛОГИЯ МЫ'НЕЧНОЙ СИСТЕМЫ
бражает всю динамику возбудимой системы за определенный
промежуток времени, т. е. — весь характер изменения функционального
состояния во время деятельности. Но лабильность данной системы
зависит не только от исходного функционального состояния или от
тех или других внешних условий, но и от воздействия самих
импульсов, «меняющихся на ходу реакций», как говорит Ухтомский (41).
Поэтому лабильность 'меняется ©се время в связи с деятельностью.
В определенных условиях предельный ритм возбуждения нарастает во
время деятельности, значит, лабильность повышается, в других
условиях он падает, значит, лабильность понижается на ходу реакции.
Изменение лабильности в связи с деятельностью, вследствие
чего возбудимая система отвечает по новому, более высокому или
более низкому ритму, Ухтомский фигурально называет усвоением
ритма. Это повышение лабильности, несомненно, связано с действием
продуктов усиленного обмена веществ, вызванного распространением
импульса возбуждения в возбудимой системе (Ухтомский, 68).
В мышцах такую роль должны играть метаболиты, связанные не
только с процессом расщепления возбудимой системы, но и с распадом
аденилпирофосфата, фосфагена и гликогена, ибо обычно повышенное
функциональное состояние в мышцах наблюдается очень долго, в
течение многих минут после короткого раздражения.
Судя по предельному ритму возбуждения, лабильность мышц не
одинакова у холоднокровных и теплокровных животных. Она ниже
у первых, чем у последних. Но и в мышцах одного животного она не
одинакова. В красных мышцах она ниже, чем в белых.
Лабильность мышечных волокон вообще ниже, чем
соответствующих двигательных нервов. Вследствие этого предельный мышечный
ритм возбуждения значительно ниже предельного ритма двигательных
волокон. В отношении же хронаксии мышечное волокно в области
нервных окончаний довольно близко стоит к соответствующему нерву.
Этим и объясняет Ляп я к возможность передачи возбуждения
с нерва на імышцу. При этом сопоставлении хронаксии и лабильности
лучше всего проявляется преимущество установления предельного
ритма возбуждения для характеристики функционального состояния
ткани сравнительно с хронаксией. Хронаксия отображает лишь
скорость наступления одного импульса, предельный же ритм — течение
этих импульсов, их динамику за определенный промежуток
времени.
Продолжительность абсолютной и относительной рефракторных фаз.
Продолжительность абсолютной фазы почти такова же, как и
возбуждения, и меняется параллельно с возбуждением. Однако всякий раз
она чуть длительнее процесса возбуждения. Для мышцы лягушки про-
должительность ее равняется 3—4 $? а для теплокровной мышцы —
2.5—3 а. В отдельных мышечных волокнах теплокровных, в условиях
ненарушенного кровообращения, по утверждению Экклса и сотр.
(69), абсолютная рефракторная фаза не превышает 1.9. з.
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
181
Абсолютная рефракторная фаза не одинакова в пределах самой
мышцы. Она больше в безнервном участке. Так, в проксимальной
безнервной части портняжной мышцы лягушки абсолютная рефракторная
фаза равняется 4—5j, в то время как в нервном участке она
составляет 2—3j. Это не зависит от присутствия нервных окончаний в
нервном участке, ибо та же разница наблюдается после кураризации
(Картозия, 48). Очевидно, абсолютная рефракторная фаза тем короче,
чем выше возбудимость, чем больше возбудимая система.
Продолжительность абсолютной рефракторной фазы изменяется подобно
возбуждению. При. охлаждении абсолютная рефракторная фаза
удлиняется, то же происходит при утомлении. Наоборот, повышение
температуры укорачивает эту фазу (Бромвель и Кейзс Люкас, 17; Бери-
тов, 13).
Относительная рефракторная фаза более длительна, чем
абсолютная. При комнатной температуре и нормальном обмене веществ она
в два-три раза превышает абсолютную, но с ухудшением обмена
веществ процесс восстановления возбудимой системы затрудняется, и,
соответственно этому, относительная рефракторая фаза удлиняется
еще более. Так, на рис. 101 абсолютная рефракторная фаза мышцы
кошки продолжается свыше 1.6 о,, а относительная — около 8 а.
Рефракторная фаза и ритмическая природа возбуждения. Выше
было отмечено, что процесс возбуждения всегда протекает ритмически.
Объяснение этому явлению таково. При возбуждении возбудимая
система реагирует на раздражение всем своим составом, всем запасом
своей потенциальной энергии. Это лежит в основе абсолютной
рефракторной фазы. Каждое новое раздражение вызовет новое возбуждение,
после того как произошло некоторое восстановление возбудимой
системы; следовательно, между двумя следующими одно за другим
возбуждениями всегда должна существовать некоторая пауза покоя. Этим
объясняется, почему процесс возбуждения не іможет быть сплошным,
почему он должен всегда, при всех условиях протекать ритмически,
а также почему с изменением функционального состояния мышечной
ткани изменяется предельный ритм мышечного возбуждения. Так,
например, с улучшением функционального состояния под влиянием
отдыха, согревания и т. д. предельный ритм возбуждения увеличивается.
Это находится в связи с уменьшением продолжительности процесса
возбуждения, которое, в свою очередь, обусловливает укорочение
абсолютной рефракторной фазы, а чем последняя короче, тем чаще может
повторяться возбуждение, тем выше будет предельный ритм возбужден
ния. Если, например, абсолютная рефракторная фаза равняется 2.5 з,
то предельный ритм достигает 400; значит, в каждом определенном
случае предельный ритм мышечного возбуждения зависит от
продолжительности абсолютной рефракторной фазы.
Рефракторная фаза и механический эффект. Мы выше указали, что
при некоторой малой частоте тетанического раздражения, когда
каждое новое раздражение, каждый новый индукционный удар приходится
182 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
во время фазы сверхнормальной возбудимости, последующие импульсы
возбуждения становятся сильнее начального импульса. При большой
же частоте раздражения, когда каждый удар раздражения
приходится во время относительной рефракторной фазы, импульсы
возбуждения становятся слабее начального. Это обстоятельство, конечно,
влияет на механический эффект. В первом случае сокращение
максимальное, оптимально большое, а во
втором случае, наоборот, ниже
максимального, и тем ниже, чем чаще
раздражение. Известный русский ученый
Введенский (25), изучавший это
явление на нервно-мышечном
препарате, назвал тот большой
механический эффект, который получается при
сравнительно малой частоте
раздражения, оптимальным, а тот слабый
механический эффект, который
получается при более частых раздражениях,
пессимальным.
Оптимальный эффект мышцы
характерно отличается от пессимального.
После начального быстрого подъема
он поднимается еще дальше, но уже
с большой постепенностью или после
начального быстрого подъема сначала
опускается довольно быстро, а потом
вновь поднимается с большой
постепенностью. При пессимальном же
раздражении он сначала также быстро
поднимается, как и оптимальный
эффект, но сейчас же вслед за этим
наступает безостановочное падение.
Быстрота падения тем больше, чем
чаще раздражение (рис. 102) (Кро-
некер и Гоч, 64; Введенский, 25).
В наиболее убедительной форме оптимальное и пессимальное
проявления механического эффекта мышцы іможно наблюдать, если менять
частоту во время самого раздражения. Нужно начать раздражение
с большой частоты, 80—100 в секунду, приложив его к 'безнервному
проксимальному участку мышцы. При этом индукторий следует
установить на такой силе раздражения, которая дает максимальное
сокращение. Если в тот момент, когда сокращение начнет падать, сделать
раздражение редким, 30—50 в секунду, то сокращение усилится. Если
перевести раздражение вновь на большую частоту, сокращение опять
ослабеет, и т. д. (рис. 103).
Нарастание (механического эффекта при некоторой малой частоте
HU->V
Рис. 101. Токи возбуждения,
возникающие в нервной области т. рего-
neus tertius кошки.
раздражается двигательный нерв
индукционными ударами через разные интервалы.
Моменты раздражения обозначены течками.
Первое небольшое колебание от петли
индукционного удара. При интервале менее
чем 1.6 о второе раздражение не дает токов
возбуждения (опыты В и С). При интервале
около 2.2 з (on. D) второе раздражение дало
эффект. Значит, абсолютная рефракторная
фаза продолжалась около 2 з. Через 3 и
9 а второе раздражение дает ослабленный
эффект (опыты Е и F), а через 14 з — без
ослабления (опыт G). Значит, абсолютная
и относительная рефракторные фазы вместе
продолжались около 10 з. На долю
относительной приходится около 8 а (Экклс и
О'Коннор).
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
183
возбуждения, вероятно, обусловливается преимущественно усилением
процесса возбуждения под влиянием сверхнормальной возбудимости.
Рис. 102. Пессимальный и оптимальный эффекты мышцы.
Проксимальный участок m. sartorius длиной в 7 мм. В опыте
А частота раздражения 150 в 1 сек.: пессимальные сокращения;
в опыте В — 25—30 в 1 сек.: оптимальные сокращения. На
сигнальных линиях цифры обозначают силу раздражения в
сантиметрах расстояния между индукционными катушками.
Ослабление же механического эффекта мышцы в связи с учащением
раздражения объясняется тем обстоятельством, что каждый новый
удар раздражения приходится на время рефракторной фазы. Это,
конечно, должно обусловливать ослабление импульсов возбуждения.
Рис. 103. Явления оптимума и пессимума на безнервном
участке мышцы.
Проксимальный участок m. sartorius длиной 8 мм. Раздражается
тетанически на расстоянии 2—4 мм от проксимального конца.
Сила раздражения — 22 см расстояние между катушками (порог
27 см). Частота раздражения вначале 40 в 1 сек., затем три раза,
на короткое время, раздражение переводится на высокую
частоту — 60—80 в 1 сек. Каждый раз при этой частоте сокращение
ослабевает. Внизу — время в секундах. Гистологическое
исследование данного участка мышцы не обнаружило двигательных
нервных окончаний.
Кроме того, с большой частотой возбуждения связано и более быстрое
утомление возбудимой системы, особенно в мышцах вырезанных и
обескровленных. Вследствие этого рефракторные фазы удлиняются.
Это вызывает быстрое падение интенсивности процесса возбуждения
в течение тетанического раздражения. Ослабление процессов
возбуждения, в свою очередь, обусловливает ослабление тех биохимических
процессов, которые ведут к сокращению. В результате получается
184
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
пессимальный механический эффект: быстрое падение сокращения
после начального быстрого подъема.
Проведение возбуждения в мышечных волокнах. Как уже
отмечалось выше, при раздражении одного участка мышечного волокна
возбуждение не ограничивается этим участком: оно проводится в обе
стороны по мышечному волокну. Это свойство мышечной ткани
называется проводимостью. В скелетной поперечнополосатой
мускулатуре каждое мышечное волокно изолировано от других волокон
сарколеммой, которая не является живым образованием мышцы.
Поэтому возбуждение не может переходить от одного мышечного волокна
к другому. На рис. 91 дается схематическое изображение
распространения волны возбуждения по мышце с параллельными волокнами.
Двустороннее и изолированное проведение возбуждения является законом
для всех мышечных волокон скелетной мышцы. Но в определенных
искусственных условиях биотоки одних мышечных волокон могут
раздражать прилегающие к ним другие мышечные волокна, и тем самым
возбуждение может переходить от одних мышечных волокон на другие.
Это бывает, как уже указывалось выше, на сдавленных или
подсушенных мышцах, где возбудимость повышена, а межклеточная жидкость
заметно уменьшена.
Методика исследования проведения возбуждения. Так как распространение
возбуждения от раздраженного участка к нераздраженному требует тем больше
времени, чем дальше отстоит последний участок от первого, то, естественно, что как
возбуждение, так и сокращение сначала возникают в соседстве с
раздраженным участком, а потом race дальше и дальше от него. Отсюда ясно, что скорость.
распространения возбуждения легко может быть определена, если установить то
время, какое нужно для распространения возбуждения или сокращения на
известное расстояние. Первоначально скорость проведения возбуждения изучалась,
путем регистрации механического эффекта — утолщение мышцы /в двух разных
участках ее. Впоследствии стали изучать эту скорость регистрацией тока
возбуждения гальванометром. Понятно, что если к гальванометру отведены два
неповрежденных участка, то при раздражении одного конца імышцы гальванометр
отклонится сначала от тока возбуждения под одним электродом, а затем под другим.
Скорость проведения, т. е. величина расстояния (в метрах), на которое может
распространиться возбуждение в единицу времени—1 сек., высчитывается по фор-
г ,
муле х ——' гДе Г есть расстояние между электродами в сантиметрах, а г —
время в секундах, в течение которого происходит распространение возбуждения
между ними.
Скорость проведения возбуждения по мышечному волокну.
Мышечная проводимость зависит от многих условий и прежде всего от рода
животного. В мышцах холоднокровных скорость проведения значительно
меньше, чем у теплокровных. В мышце лягушки эта скорость
возбуждения при комнатной температуре равна 3—4 ім в секунду (Бернштейн,
20); в мышце человека она достигает 12—13 м (Герман, 21). Скорость
проведения также зависит от свойства самих імышц; например, в белой
мышце кролика возбуждение проходит 6—11 м в секунду, а в красной
3—3.4 м (Кольрауш, 22).
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
185
Скорость проведения меняется в связи с функциональными
изменениями. При температуре 6.5° лягушечья мышца проводит возбуждение
со скоростью 1.4 ім. Если же утомить імышцу, то скорость проведения
становится еще меньше — около 0,8 м (Кольрауш, 22). В портняжной
мышце лягушки при ненарушенном кровообращении скорость
проведения в среднем составляет 4.3 м в секунду, а в вырезанной и
обескровленной мышце она в среднем равна 2.1 м в секунду (Энгельман, 67).
Следовательно, можно сказать, что в мышце при условиях,
благоприятных для обмена веществ, и вообще при большем количестве в
мышечных волокнах возбудимой системы скорость проведения возбуждения
больше, чем при неблагоприятных условиях и вообще при меньшем
количестве возбудимой системы.
Проведение возбуждения и распространение сократительного
процесса. Возникшее в раздраженном участке возбуждение волнообразно
распространяется вдоль мышечного волокна по возбудимой системе.
Деятельность же сократительного вещества точно следует за волной
возбуждения: в каждый данный момент начинает сокращаться тот
участок миофибрилли, которого достигает волна возбуждения.
Сокращение же, вызванное в данном участке миофибрилли, само не
переходит на другие участки.
Это заключение было сделано нами на основании такого опыта:
если опустить половину m. sartorius в раствор Рингера при 34—37°,
то с течением времени согреваемый конец приходит в контрактуру,
вероятно под влиянием усиленного расщепления фосфагена и
гликогена на фосфорную и молочную кислоты, производящие сокращение —
контрактуру. Эта контрактура, вызванная под влиянием тепла в
согреваемом участке, не распространяется на другой несогреваемый
участок (Беритов, 23). То же 'бывает с химической контрактурой: если
погрузить половину мышцы в раствор кислоты, который вызывает в
погруженной части контрактуру, последняя не распространяется на
непогруженную половину (Вильмерс, 24). Кроме того, мы выше указывали,
что локальное сокращение, вызванное электрическим раздражением
путем микроэлектродов, или контрактурное сокращение, вызванное
локальным приложением ацетилхолина, не распространяется. Отсюда
следует, что сократительный процесс сам по себе не распространяется
от одного участка мышцы к другому, что наблюдаемая в мышце волна
сокращения в точности следует за волной возбуждения. Этим же
объясняется то характерное явление, что волна сокращения точно
с такой же скоростью распространяется вдоль по мышце, как и
процесс возбуждения.
Но, конечно, для того чтобы волна возбуждения вызвала
сокращение, необходимы определенные физико-химические условия. Если
этих условий нет, возбуждение может возникнуть и распространиться
без сокращения. Это, например, случается, если опустить мышцу
в теплый раствор Рингера на несколько часов. Вследствие усиленного
расщепления веществ, потребных для сокращения и других физико-
186
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
химических изменений, раздражение імыінцы не будет вызывать
сокращения, но может вызвать биоэлектрический ток возбуждения
сравнительно с нормой почти без ослабления (Беритов и Волынский, 45). То
же самое случится, если у двигательного нерва m. sartorius перерезать
ту веточку, которая распространяется в проксимальном участке
мышцы, и затем абсолютно утомить мышцу длительным раздражением
двигательного нерва. В этом случае в дистальной части мышцы
происходят такие физико-химические изменения, что прямое раздражение
дистального участка не произведет здесь сокращения, но вызовет его
в проксимальном участке (Беритов, 46). Очевидно, в дистальном
участке возбуждение вызывается и проводится, но оно не в состоянии
вызвать здесь .сокращение из-за отсутствия тех условий, которые
требуются для сократительного процесса.
О зависимости между количеством возбудимой системы,
возбудимостью, возбуждением и проведением. Выше мы видели, что степень
возбудимости, интенсивность возбуждения и скорость проведения
возбуждения определенным образом зависят от количества возбудимой
системы. Чем больше эта система, тем выше возбудимость, тем
интенсивнее процесс возбуждения, тем больше скорость проведения
(возбуждения. Значит, количество возбудимой системы представляет
главный фактор, от которого зависят основные функциональные явления
живой возбудимой системы.
В определенных случаях количество живой возбудимой системы
мышцы меняется в силу различных превращений самой живой
системы, например в связи с возбуждением, утомлением и наркозом. Во
время абсолютной рефракторной фазы количество возбудимой системы
сразу сходит на-нет, а при утомлении и при наркозе она уменьшается
постепенно вследствие превращения ее в систему, не способную к
возбуждению. В относительной рефракторной фазе, во время отдыха,
после удаления наркотического вещества, возбудимая система
постепенно увеличивается; вследствие этого живая система становится все
более и более возбудимой.
В других случаях количество возбудимой системы мышцы меняется
в связи с изменением обмена веществ. Так, при условиях, не
благоприятствующих обмену веществ, возбудимая система уменьшается
в силу общего уменьшения живой системы. Это имеет іместо при
денервации, при недостаточном питании, при отсутствии
упражнения. Наоборот, когда условия благоприятствуют обмену веществ, как,
например, при упражнении, при хорошем питании, а также в период
эмбрионального роста мышечной ткани, возбудимая система
увеличивается, ибо растет живая система мышцы. В нервных участках мышцы
возбудимая система потому белее развита, что
окислительно-восстановительные процессы здесь протекают сильнее, чем в безнервных участках,
да и вообще условия обмена веществ здесь лучше благодаря
усиленному развитию капиллярной системы в области двигательных пластинок.
Закон возбуждения по отношению к поперечнополосатым мышцам
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ 187
беспозвоночных животных. Закон возбуждения был установлен по
работам на мышцах позвоночных животных. По отношению к мышцам
беспозвоночных прямых исследований не имеется. Существуют лишь
работы, выполнявшиеся на нервно-мышечных препаратах, Гофман
(27) нашел, что імышцы клешни рака, в которых все мышечные во-
локна получают двигательные нервные окончания из одного
двигательного нервного волокна, при рефлекторном возбуждении
показывают градацию токов возбуждения не только в отношении частоты их,
но и интенсивности.
То же самое было обнаружено Серков ым (47), одним -из
сотрудников Воронцова, на нервно-мышечном препарате клешни рака.
Токи возбуждения показывали не только градацию интенсивности, но
и явление суперпозиции, характерное для сократительного процесса.
При двух раздражениях с -интервалом 5—7 а получался более сильный
однофазный электрический эффект, чем от одного раздражения, а при
больших интервалах наступал эффект с двумя зубцами, причем второй
зубец оказывался выше первого. Но эти токи возбуждения не
(выражают сократительного процесса, ибо, во-первых, даже при
значительных токах возбуждения сократительный процесс может отсутствовать,
во-вторых, если он и наступает, то начинается много позднее тока
возбуждения, спустя 20—40 а. Градация и накладывание токов
возбуждения должны указывать на то, что мышечные клетки не
реагируют согласно закону возбуждения всем запасом потенциальной
энергии возбудимой системы.
Однако на том же нервно-мышечном препарате рака Белоусов
(29), из лаборатории Введенского, получал при сильных и частых
раздражениях более слабый механический эффект (пессимальный),
чем при редких и слабых раздражениях (оптимальный). Серков
установил, что при некоторых малых интервалах, как 2.4 а, второе
раздражение нерва не дает мышечного эффекта. По этим данным
видно, что и в нервно-мышечном препарате рака при возбуждении
получается рефракторное состояние, а значит, он отвечает согласно
закону возбуждения. Весьма возможно, однако, что эта рефрактор-
ность проявляется в нерве, а не в мышце.
Опытом на клешне Cambarus darkii в последнее время было
показано, что в мышечных волокнах этой клешни возбуждение не
распространяется. Каждое мышечное волокно получает множество нервных
окончаний, которые выходят из окружающего его густого нервного
сплетения. Одно мышечное волокно длиной 3.5 мм получает до 20—
40 нервных окончаний. Поэтому при раздражении двигательного
нерва каждое мышечное волокно получает импульсы во всех своих
участках, вследствие чего мышца сокращается сразу и целиком. Но,
если мышцу повредить так, чтобы нервное сплетение перестало
возбуждаться, как это, например, бывает при механическом повреждении
мышцы в связи с операцией изолирования ее, тогда раздражение
нерва не дает вовсе сокращения, а прямое электрическое раздражение
188
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
мышцы дает сокращение, но только в области электродов (Гарре-
вельд, 79).
Эти факты показывают, что імышцы некоторых ракообразных не
производят и не проводят возбуждения по закону возбуждения.
Ира ктипум
Наблюдение тока возбуждения вторичным сокращением. Два нервно-мышечных
препарата помещены на пробковой пластинке; под препараты лодкладывают
смоченную физиологическим раствором вату или фильтровальную бумагу. Нерів одного
препарата обрезают около позвоночника и накладывают на m. gastrocnemius
другого препарата. Нерв последнего препарпта .раздражают тетанически «ли
отдельными индукционными ударами. Токи возбуждения, возникшие в m. gastrocnemius,
раздражают нерв другого препарата, отчего сокращаются мышцы последнего. Это
сокращение известно в физиологии под именем вторичного сокращения и считается
классическим опытом, подтверждающим возникновение биоэлектрических токов во
время возбуждения мышц. Если раздражение умеренной силы, то легко заметить,
что сначала возбуждается первично раздражаемая лапка, а затем вторично
раздражаемая.
Регистрация токов возбуждения струнным гальванометром. Вырезают m. sarto-
rius и соединяют его с изометрическим миографом или подвешивают груз в 70—
100 г, чтобы мышца сильно не укорачивалась при сокращении. На дистальный
конец накладывают 'пару платиновых электродов на расстоянии 5 мм от
сухожилия и раздражают индукционными ударами. Другую пару платиновых электродов
связывают с проксимальной частью мышцы нитью, 'окрученной из ваты и смоченной
физиологическим раствором. Она служит для отведения. Это важно сделать для
того, чтобы при сокращении отводящие электроды не передвигались на мышце.
Для отведения токов возбуждения мы не предлагаем пользоваться неполяризую-
щимися электродами, так как для быстро протекающих токов они не нужны.
Расстояние между полюсами отводящих электродов должно быть не меньше 10 мм.
Они должны находиться на некотором расстоянии от раздражающих электродов, не
ближе, чем на 10 мм, чтобы петли раздражающего тока не попадали в
гальванометр. Колебания струны от токов возбуждения будут получаться уже при
пороговых раздражениях: по одному колебанию от каждого удара. С усилением
раздражения колебания усилятся. При тетанических раздражениях получится ряд
колебаний по ритму раздражения, если частота последнего не очень высока, не
выше 100 в секунду. При длительных раздражениях с утомлением мышцы
амплитуда и частота колебаний струны (уменьшаются.
Если отводить в гальванометр то центральный, то проксимальный участки
m. sartorius, легко заметить, что при отведении проксимального участка колебания
струны будут значительно меньше, чем при отведении центрального. В этих опытах
раздражающие электроды накладываются на мышцу каждый раз на расстоянии
одного сантиметра от отводящих. Этот опыт показывает, что процесс возбуждения
в центральном нервном участке мышцы протекает интенсивнее, чем в проксимальном.
Наблюдение оптимального и пессимального сокращения. Берут от m. sartorius
лягушки проксимальный участок длиной в Vs всей мышцы. Проксимальный конец
не отделен от тела. Перерезанный конец соединяется с миографом при пяти- или
десятикратном увеличении (запись производится на кимографе). Мышца
загружается умеренно, 5—10 г. Электроды для раздражения накладываются на
расстоянии 2—5 мм от проксимального конца. Прерывают ток вагнеровским молоточком,
установив его на частоту 100—80 в секунду. Сила раздражения такова, что
дает максимальное укорочение. Начать раздражение при частоте 100—80 в секунду,
а когда сокращение будет падать, перевести частоту на 30—50 в секунду. Сейчас
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ
189
же -кривая сокращения поднимется — оптимальный эффект. Вновь перевести
раздражение ;ка большую частоту, и кривая вновь «опустится — пессимальный эффект. Этот
опыт легко удается на свежих, мало подвергавшихся раздражению препаратах.
Если мышца утомлена раздражением или истощена от долгого лежания без
кровоснабжения, то падение механического эффекта с учащением раздражения будет
происходить, ио последующее уменьшение частоты не приведет к усилению эффекта
вследствие развития сильного утомления за время раздражения.
Определение скорости проведения возбуждения в мышце. Берут m. gracilis
лягушки с параллельными мышечными 'волокнами и прикрепляют к пробковой
пластинке 'булавками, чтобы мышца .но могла укоротиться во время сокращения.
Регистрируется утолщение мышцы \ъ двух возможно отдаленных участках. Для этого
устанавливают связь их с прямыми миографами Энгельмана при посредстве особых
вертикальных ножек — пелотов. Раздражается один конец мышцы отдельными
индукционными ударами. Замыкание и размыкание тока производятся автоматически
вращающимся цилиндром кимографа. Для этой цели употребляют кимограф Криса
или вообще быстро вращающийся кимограф с контактами. В ответ на раздражение
сначала сокращается мышца под 'ближайшим пелотом, и от утолщения пелот
поднимается вверх, а затем иод другим пелотом. Разность скрытых периодов
записываемых при этом кривых означает время, которое требуется для прохождения
возбуждения от одного регистрируемого участка до другого. Время определяется
путем одновременной регистрации колебаний электромагнитного камертона, дающего
100 колебаний в секунду (о регистрации времени см. методику).
Опыт с непроводимостью возбуждения от одного мышечного волокна к другому
в скелетной мускулатуре. Крупную мышцу, как m. triceps или т. gracilis,
разрезают зигзагообразно, растягивают г. виде ленты и раздражают один конец.
Сокращение наступит только ів этом конце. Понятно, что если бы возбуждение
передавалось от волокна к волокну, то сократилась бы вся лента мышцы.
Измерение продолжительности рефракторных фаз. Если раздражать нерв нервно-
мышечного препарата или один конец длинной мышцы, как m. sartorius, двумя
следующими один за другим индукционными ударами, .можно найти такой небольшой
. интервал, при котором второе раздражение не производит эффекта. Тот
наибольший интервал, во время которого второе раздражение не дает эффекта, равняется
продолжительности абсолютной рефракторной фазы. В этих опытах эффект
записывается миотрафически на кимографе Криса. Раздражение наносится автоматически
посредством кимографа. Последний снабжен двумя контактами для двух
раздражений. Перемещением одного из этих контактов можно мекять по желанию
интервал между раздражениями.
При отсутствии кимографа Криса можно пользоваться маятником Гельмгольца
с двумя контактами для двух раздражений. На стоячем же кимографе записывается
высота сокращений. В обоих случаях один контакт неподвижен и служит для
размыкания постоянного тока в первичной цепи одного индуктория; этим наносится
первое раздражение. Другой контакт подвижен и служит для размыкания
первичной цепи в другом индуктории, чтобы нанести второе раздражение. Этот опыт
удается только в том случае, если первое раздражение настолько сильно, что
возбуждает все волокна мышцы.
Миографическим путем можно хорошо изучать абсолютную рефракторную
фазу. Для относительной рефракторной фазы нужно обратиться к
гальванометрической регистрации токов возбуждения. Так как каждый ток, выражающий один
процесс возбуждения, протекает совершенно самостоятельно, можно установить - тот
наименьший интервал, при котором второе раздражение дает совершенно такой же
эффект, как и первое. Этот интервал будет разняться абсолютной и относительной
фазам вместе. Отсюда вычитанием абсолютной фазы определяется
продолжительность относительной рефракторной фазы.
190
овіцая физиология мышечной системы
Литература
1. Hermann L., Pfl., 16, 191, 410, 1878; 24, 294, 1881.
2. Br иске Е. Т h., Pfl. Arch., 124, 215, 1908.
3. Beritoff J., Z. Biol., 82, 119, 1924.
4. Beritoff J., Z. Biol., 85, 15, 1926.
5. Einthoven W., Handb. north, path. Physiol., 8, Abt. 2, 785, 1928.
6. Bishop H. u. Gilson A., Am. J. Physiol., 82, 478, 1927.
7. Riesser u. Steinhauser, Pfl. Arch., 197, 288, 1922.
8. Fischer E., Pfl. Arch., 203, 580, 1924.
9. Herlitzka A., Arch, di Physiol., 14, 157, 317, 1916.
10. Beritoff J. u. Woronzow D., Z. Biol., 84, 418, 1926.
11. Ernst E., Pfl. Arch., 209, 613, 1925; 213, 144, 1926; 218, 137, 1927.
12. Beritoff J., Z. Biol, 85, 509, 1927.
13. Beritoff J., Z. Biol., 62, 125, 1913.
14. Beritoff J., Z. Biol., 80, 171, 1924.
15. Buchanan F L, J. Physiol., 27, 95, 1901.
16. Dittler R. u. Tichomirof f M., Pfl. Arch., 125, 112, 1908.
17. Br am we 11 С r. u.Keith Lucas, J. Physiol., 42, 495, 1911.
18. Snyder, Am. J. Physiol., 17, 350, 1906.
19. Bazett G., J. Physiol., 36, 414, 1907.
20. Bernstein J., Ueber d. Erregungsvorgang im Nerven u. Muskelsystem, 1871.
21. Hermann L., Pfl. Arch., 16, 420, 1873.
22. Kohlrausch A., Arch. Anat. Physiol., S. 283, 1912.
23. Б e p и t о в И., Неопубликованная работа.
24. Wilmers J., Pfl. Arch., 178, 193, 1920.
25. Введенский В., О соотношении между возбуждением и. раздражением при
тетанусе. П., 1886. .
26. Беріато'В И., Руоск. физиол. журн., 1, 1917.
27. Hoffmann P., Z. Biol., 64, 247. 1914.
28. Вето хин И., Русек. физиол. журн., 12, 85, 1929.
29. Белоусов М., Часть результатов была опубликована в монографии И. Бе-
ритова: Учение об основных элементах центр, коорд., П., 1916.
30. Beritoff J., Z. Biol., 85, 521, 1927.
31. Buchthal Fr.u. P e t e r f i Т., Pfl. Arch., 234, 527, 1934.
32. Buchthal F г., Skand. Arch. Physiol., 68, 187, 1934.
33. Henri ques u. L i n d h а г d, Pfl. Arch., 183, 1, 1920.
34. Lindhard J., Skand. Arch. Physiol., 64, 299, 1932; ib., 69, 59, 1934.
35. Asmussen E г., Skand. Arch. Physiol., 65, 261, 1933; ib., 70, 233, 1934.
36. К a t о G., The microphysiology of nerve, 1934.
37. Beritoff J., Pfl. Arch., 213, 206, 1926.
38. Bishop G. a. Gilson A., Am. J. Physiol., 89, 135, 1928.
39. Hoefer P., Z. Biol., 94, 247, 1933.
40. Введенский H. E., Возбуждение, торможение и наркоз, 1930.
41. Ухтомский А. А., Парабиоз и доминанта. Изд. Комм, академии, Москва,
1927.
42. Ernst Е. u. Koszkas J., Pfl. Arch., 23,5, 562, 1935.
43. Ernst E. u. Fricker, Pfl. Arch., 234, 399, 1934.
44. Boruttau H„ Pfl. Arch., 63, 158, 1896.
45. Б e p к t о в и В о л ы и с к и й, Неопубликованная работа.
46. Beritoff J., Z. Biol., 82, 119, 1924.
47. Serkoff Ph. N., Pfl. Arch., 236, 481, 1935.
48. іКартозия А., Тр. Физиол. инст. Тбил. уинв., 3, 245, 1937.
49. Hodgkin A. L., Proc. Roy. Soc, В., 123, 403, 1937.
50. Schaefer H. u. Gopfert H., Pfl. Arch., 238, 685, 1937.
ПРОЦЕСС ВОЗБУЖДЕНИЯ МЫШЦЫ 191
51. Brown G. L., J. Physiol., 89, 438, 1937.
52. G о р f е г t Н. u. S с h а е f е г Н., Pfl. Arch., 239, 597, 1937.
53. Meyer К. Н., Schw. med. Wschr., 11, 826, 1937.
54. S te im a n n S. E. a. Pratt F. H., Am. J. Physiol., 122, 27, 1938.
55. С e p к о в Ф. H., Учен. зап. Каз. унив., вып. 3, стр. 10, 1934.
56. Schaefer Н., Pfl. Arch., 237, 329, 1936.
57. A d r i a n E. D. a. G е 1 f a n S., J. Physiol., 76, 271, 1933.
58. D ц г і g A., Pfl. Arch., 97, 457, 1903.
59. Pfl tiger E., Untersuch. tib. d. Physiol, d. Elektr., S. 133, Berlin, 1859.
60. GartenS., Abh. d. Mathem.-phys. Kl. d. Konigl. Sachs. Gesellsch. d. Wiss.,
26, 331, Leipzig, 1901.
61. Schaefer H. u. Scholmerich P., Pfl. Arch., 240, 542, 1938.
62. Buchthal a. Lindhard, The Physiol, of str. muscle fibre, 1939.
63. Б e p ит о в И., Об эленлрич. яЕлениях мышцы при ацетилхолин. отправлении.
Сообш. Акад. Наук Груз. ССР, т. 1, 1940.
64. Kronecker Н. u. Gotch F., Arch. Physiol., S. 438, 1880.
65. Kuhne W., Z. Biol., 24, 383, 1888; 26, 203, I860.
66. В iederma nn W., Sitztmgsber. Wien. Akad., 97 (3), 145, 1S88.
67. Engelmann T h. W., Pfl. Arch., 66, 574, 1897.
68. Ухтомский А. А., Физиол. жури. СССР, 25, 767, 1938.
69. E с с 1 e s I. С. а. О ' С о n n о r W. I., J. Physiol., 97, 44, 1939.
70. E с с 1 e s J. С a.O'Connor W. I., J. Physiol., 94, 78, 1938.
71. BrownG.L, Dale H. H. a. F e 1 d b e r g W., J. Physiol., 87, 394, 1936.
72. Brown G. L., J. Physiol., 89, 220, 1937.
73. В u с h t h a 1 F г., Acta Psych, et Neurol., 15, 43, 1939.
74. H a r r e v e 1 d A. v., J. Сотр. Neurol., 70, 267, 1939.
Глава Г III
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
Понятие о повышении возбудимости. В возбудимой системе мышцы,
кроме возбуждения, раздражение вызывает повышение возбудимости.
Последнее происходит неизбежно при каждом деятельном
раздражении как однократном, так и многократном (Введенский, 1; Самойлов,
2, и др.).
Если сравнить пороги раздражения до и после возбуждения, то
окажется, что через некоторый небольшой промежуток времени после
раздражения потребуется меньшая раздражающая сила для (вызова
порогового эффекта, чем это было до того. Повышение возбудимости
проявляется еще в том, что при быстром следовании раздражений
одного за другим интенсивность импульсов возбуждения івначале
возрастает (Введенский, 3) (рис. 104). Повышенная или сверхнормальная
возбудимость наблюдается после рефракторных фаз, когда
совершенно закончилось восстановление системы, возбужденной
предыдущим раздражением. Значит, возбудимость, сведенная на-нет во время
возбуждения, сначала постепенно нарастает до нормы, а затем
превышает ее. В состоянии сверхнормальной возбудимости мышца
остается некоторое время, измеряемое долями секунды и даже
секундами, затем повышенная возбудимость постепенно проходит, возбуди-
імость возвращается к норме (рис. 85).
Следует отметить, что в данном случае возбудимость повышается
во всей мышце вследствие распространения возбуждения, вызванного
раздражением. Полное понимание этого явления само собой вытекает
из концепции, согласно которой восстановление происходит сейчас же
после возбуждения, одновременно в каждой частичке возбужденного
участка мышечного волокна. Вследствие этого в период изохронного
восстановления в мышце присутствует больше возбудимой системы,
чем в какое-либо другое время. Поэтому, когда происходят два
последовательных раздражения, первое раздражение попадает в гетеро-
хронное состояние, а второе может случиться в тот момент, когда уже
закончилось изохронное восстановление. В силу этого как процесс
расщепления, так и биоэлектрический ток возбуждения будут сильнее
в ответ на второе раздражение, чем на первое.
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
193
Сверхнормальная возбудимость проявляется не только в усилении
процесса возбуждения, но еще -в уменьшении порогов раздражения.
Мы здесь имеем дело с одной особенностью возбудимой системы:
ферментативное расщепление возбудимой системы происходит тем
легче, чем больше ее количество. Многие факты подтверждают это
положение: всякий раз, когда по какой-либо причине уменьшается
возбудимая система, ослабевает процесс возбуждения; тогда и степень
возбудимости меньше. Так происходит во время наркоза, при охлаждении,
во время относительной рефракторной фазы. Поэтому можно с полным
правом утверждать, что величина порогового раздражения зависит
прежде всего от количества возбудимой системы (Беритов, 17).
Рис. 1U4. Повышение возбудимости при тетаническом раздражении
небольшой частоты.
Регистрируются токи возбуждения т. semitendinosus лягушки гальванометром
Эйнтховена. В секунду производится по 20 замыкательных и размыкательных
ударов. Первые удары дают ничтожные эффекты (А); с повторением ударов
эффекты нарастают и в конце концов достигают максимальной величины (В).
Электрограмма В представляет продолжение Л с небольшим пропуском.
Время — в сотых долях секунды.
Как уже отмечалось выше, изохронный процесс постепенно
сменяется гетерохронным. Вследствие этого в каждый последующий
момент общее количество восстановленной возбудимой системы
постепенно уменьшается. Пока существует изохронное восстановление в
значительной части возбудимой системы, пока оно не заменится вполне
гетерохронным состоянием, степень возбудимости должна быть
больше, и процесс возбуждения, вызванный в это время, должен быть
сильнее, чем при гетерохронном состоянии.
В определенных условиях сверхнормальная возбудимость імышцы
продолжается многие секунды и даже многие іминуты. Причина этого
явления — не изохронное восстановление возбудимой системы, а, по
всей вероятности, действие продуктов обмена веществ, которые
образуются в мышце под влиянием возбуждения.
Спрашивается, почему степень возбудимости во время гетерохрон-
ного состояния должна быть меньше, чем при изохронном восстанов-
13 И. С. Беритов
194
ОБЩАЙ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
лении, если в первом случае часть возбудимой системы всегда
находится в состоянии полного восстановления. Дело в том, что мы судим
о степени возбудимости по порогам раздражения. Возбуждение
вызывается только в том случае, если раздражение производит расщепление
во всей возбудимой системе, когда получается такая разница
электрического потенциала, которая достаточна для раздражения
соседних невозбужденных участков. Безусловно, во время гетерохрон-
ного состояния реактивность вполне восстановленных частичек больше,,
чем еще не вполне восстановленных, которые находятся в разных:
стадиях восстановления. Поэтому пороговое раздражение для тех и
других частичек не должно быть одно и то же. Совершенно
восстановленные частички легче и скорее будут расщепляться в ответ на
более слабое раздражение, чем другие. Очевидно, при некоторых:
слабых раздражениях только наиболее восстановленные частички
будут расщепляться. Это и случается фактически, как об этом уже
говорилось выше (стр. 158). Но только возникший при этом
биоэлектрический ток не будет достаточен для раздражения соседних:
частичек. Чтобы это произошло, нужно применить такую силу
раздражения, которая произвела бы расщепление в большом количестве
частичек, не только вполне восстановленных, но также еще не вполне
восстановленных. Лишь при этом условии может получиться такаяг
большая разность электрического потенциала, которая будет в
состоянии раздражать соседние участки. Отсюда понятно, почему при гете-
рохронном состоянии следует применять для возбуждения более
сильное раздражение, чем во время полного изохронного восстановления..
Повышение возбудимости при субминимальных раздражениях.
Раздражение производит повышение возбудимости не только ів случае
возбуждения, но. и тогда, когда оно является субминимальным,.
т. е. не вызывает (возбуждения. При быстром следовании таких
раздражений одного за другим возбудимость повышается настолько, что
бывшее раньше субминимальное раздражение становится пороговым,
т. е. начинает вызывать возбуждение (Штейнах, 4; Крис и Сьюал, 13).
Можно взять такое субминимальное раздражение, что при
некотором малом интервале уже одного повторения его будет достаточно для
вызова эффекта. При этом, чем сильнее это субмйнимальное
раздражение, тем больше может быть этот интервал. Последний зависит
также от функционального состояния мышцы и от рода мышцы. Так,,
если субминимальное раздражение на 5% меньше порогового, то
интервал суммации для m. sartorius при 13° составляет 1—2 а; после
утомления интервал удлиняется до 100 а (Люкас, 14).
Повышение возбудимости, вызванное субминимальным
раздражением, существенно отличается от повышения при наличии
возбуждения. В последнем случае повышенная возбудимость устанавливается
по всему волокну вслед за возбуждением. При субминимальном же
раздражении возбудимость повышается лишь в раздраженном участке,
причем это происходит немедленно вслед за раздражением. Возникно-
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
195
вение возбуждения после ряда субминимальных раздражений
называется явлением суммации.
Явление суммации при электрическом раздражении мышечных
клеток зависит от силы раздражения, от интервала (между
раздражениями, от продолжительности каждого удара раздражения и от числа
раздражений. Л а пи к впервые показал это путем опыта (21). Он
установил, что определенная подпороговая сила раздражения может
вызвать возбуждение через определенное число повторений в
зависимости от интервала и продолжительности каждого раздражения при
данной подпороговой силе. Чем больше интервал и чем короче
каждое раздражение, тем чаще должно повторяться раздражение, чтобы
вызвать возбуждение. При постоянных величинах интервала и
продолжительности раздражения подпороговое раздражение тем скорее
вызывает возбуждение, чем больше сила раздражения. Явление суммации
свойственно каждой возбудимой
системе как мышечной, так нервной
и железистой тканей. Поэтому
подпороговая суммация
является всеобщим
законом суммации для всех
воз б уд имых тканей (Шри-
вер, 22).
Явление суммации в мышцах
может наблюдаться при интервале
до 1 секунды и больше (рис. 105).
Оно, видимо, обусловливается
физико-химическими изменениями, производимыми подпороговым
раздражением. Известно, что и при подпороговом раздражении происходит
усиленный распад фосфагена (в раздраженном участке и даже может
наступить локальное сокращение. Это, очевидно, связано с
расщеплением наиболее возбудимых частичек возбудимой системы, как уже
указывалось выше. В связи с этим расщеплением в возбудимой системе
освобождаются одновалентные ионы, прежде всего водородные. С
повторением раздражения эти ионы імогут накопиться в возбудимой
системе и повысить возбудимость до такой степени, что данное
раздражение окажется <в состоянии вызвать расщепление ©о всех частичках
возбудимой системы. Это вызовет появление биоэлектрического тока
такой интенсивности, которая обусловит возбуждение соседних
невозбужденных участков. Если же раздражение настолько слабо, что оно
не в состоянии произвести расщепление даже в наиболее
восстановленных частичках, то оно не будет давать возбуждения и при
многократном повторении; тогда мы не будем иметь субминимального
раздражения.
Итак, явление суммации должно основываться на активном
действии субминимального раздражения на наиболее восстановленные
частички возбудимой системы. Но возникшие в результате расщепле-
13*
МШИ
«инйиийввшнтнмсшввитямняиники! іінявіт(ііііитіт»ш гол ін і
Рис. 105. Явление суммации в мышце. М.
gastrocnemius лягушки раздражается
непосредственно следующими друг за
другом субминимальными индукционными
ударами с интервалом по 5 сек. Точки
указывают моменты тех ударов, которые
не произвели возбуждения (Пембрей).
196
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ния этих частичек ионы не в состоянии дать такой биоэлектрический
ток, который смог бы раздражать другие соседние частички. Такой
ток возникает только при повторных раздражениях, когда в
результате накопления водородных ионов и вызванного им повышения
возбудимости в раздраженном участке произойдет полное расщепление
всей возбудимой системы.
Явление лестницы. Явлением лестницы называется нарастание
эффекта мышцы при редких повторных раздражениях, когда каждое
последующее раздражение приходится на фазу повышенной
возбудимости от предыдущего эффекта. Это явление имеет место даже в том
Рис. 106. Явление ]лестницы и'повышенная возбудимость.
Спинномозговая лягушка. Верхняя кривая — от m. semitendinosus, нижняя — от
m. vastus externus. Раздражается седалищное сплетение. Сначала происходит
одно пробное раздражение, затем около 80 раздражений через полсекунды. По
прекращении этого раздражения прикладываются пробные раздражения через
различные промежутки времени в течение 16 минут. Даже последние пробы
вызывали все еще более сильные одиночные вздрагивания, чем в начале опыта.
случае, если раздражение максимально. Следовательно, при
повторных раздражениях нормальный максимальный эффект становится еще
больше, делается сверхнормальным (рис. 106). Нарастание
эффекта происходит до некоторой предельной величины, и эта
величина достигается тем скорее, чем чаще раздражение. Иной раз
нарастание максимального эффекта происходит не с самого начала: сперва
амплитуда одиночных сокращений падает, а уже потом начинает
нарастать (рис. ИЗ).
Если в момент нарастания раздражение прекратить, то при
возобновлении его спустя несколько минут новое сокращение окажется
ниже, чем было последнее сокращение до перерыва, но будет выше
первого сокращения предыдущего ряда. Так может случиться даже
спустя 10 минут после раздражения (рис. 106). Значит, то
функциональное изменение, которое создается в мышце в результате
повторных сокращений, удерживается после раздражений в течение многих
минут.
Явление лестницы и вообще длительное повышение возбудимости в
ответ на деятельное раздражение наблюдается в мышце во всех ее
возбужденных участках. В вышеприведенном.примере (рис. 106) мыш-
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
ца приходит в активное состояние через нерв. Можно было подумать,
что это обусловлено повышением возбудимости исключительно в
двигательных пластинках и вообще в зоне нервных окончаний. Но то же
явление наблюдается при изучении совершенно безнервного участка
Рис. 107. Повышение возбудимости от одиночного и тетанического
сокращений.
Проксимальный безнервный участок т. sartcrius длиной в 9 мм. На расстоянии
1—3 мм от конца раздражение производится тетанически (верхний сигнал), на
расстоянии 5—7 мм — одиночными индукционными ударами (нижний сигнал).
При ряде раздражений отдельными ударами (опыт Л) и после тетануса (опыт В)
одиночные вздрагивания повышаются.
мышцы. И здесь после одного непрямого или прямого раздражения в-
течение нескольких секунд, а после ряда раздражений в течение
многих минут возбудимость является повышенной, как это видно, например*
на рис. 107 (Беритов, 20).
Явление нарастания механического эффекта мышцы было изучено
многими авторами еще в 80-х годах прошлого столетия (Кронекер, 6;
ИВЕ
Рис. 108. Явление лестницы токов возбуждения m. semitendinosus лягушки.
Отводится дистальная треть в гальванометр ЭЙнтховена. Раздражение происходит через
2 сек.: сначала даются электрограммы по одной в разные стадии раздражения, затем — ме-
ханограммы из тех же стадий. Электрограмма Л — первый ток возбуждения; В — десятый
ток; С — через 5 минут в период максимального повышения возбудимости; D — через 25 минут
в стадии значительного утомления. Механограмма 7 соответствует электрограмме А; меха-
н о грамм а 2 — электрограмме В; механограмма 3 соответствует электрограмме С;
механограмма 4 — электрограмме D. Остальные объяснения см. в тексте.
Тигель, 8; Функе, 7; Россбах, 9, и др.)- Изучение нарастания
электрического эффекта производилось только в последнее время. Точного па*
раллелизма между нарастанием механического и электрического
эффектов нет. Например, в некоторых случаях электрический эффект
нарастает много сильнее, чем 'механический. Это видно на рис. 108, где
при раздражениях с интервалом в 2 сек. электрический эффект
нарастает на 37%, а механический изменяется только чуть заметно; через
25 минут, в период значительного утомления, когда імеханический
эффект ослаб почти втрое (механограммы 3 и 4)у электрический эффект
198
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
оказался таким же большим, каким был в конце периода нарастания
(ср. электрограммы С и D) (Беритов, 10).
По поводу происхождения явления лестницы одни авторы полагают,
что начальное повышение ее происходит под влиянием образуемых в
мышце кислых продуктов, а именно молочной и угольной кислот (Ли,
15; Грубер, 16). Как известно, эти кислоты в малых концентрациях
вызывают повышение возбудимости. Такую же роль должны играть
образуемые при возбуждении адениловая и фосфорная кислоты. По
новейшим исследованиям, как при повторных одиночных сокращениях,
так и при повторных коротких тетанических сокращениях в мышце
первоначально нарастает рН, т. е. реакция мышцы изменяется в
сторону щелочности. Лишь спустя много времени величина рН начинает
уменьшаться, и реакция переходит в сторону кислотности.
Первоначальная щелочность приписывается образованию основания в
результате расщепления фосфагена (Дюбюиссон, 23). Следовательно,
длительное повышение возбудимости в результате длительной мышечной
деятельности, должно быть, обусловливается повышением
концентрации водородных ионов, а также ионов калия вследствие расщепления
липопротеидов самой возбудимой системы. Нужно думать, что ионы
водорода и калия благоприятствуют восстановлению частичек
возбудимой системы после каждого их «самопроизвольного» расщепления,
а значит, обусловливают увеличение возбудимой системы. Это же
должно привести к усилению процесса возбуждения и вызываемого им
сократительного процесса. Однако на вырезанных мышцах и вообще
на мышцах с расстроенным кровоснабжением может случиться, что,
несмотря на усиление процесса возбуждения, сокращение не усилится.
Это будет происходить всякий раз, когда в мышце окажется
недостаток аденилпирофосфата и фосфагена, вследствие недостаточно
быстрого восстановления их после каждого расщепления.
Но все своеобразие изменения сократительного процесса нельзя
объяснить влиянием возбуждения и тех физико-химических дроцессов,
которые ведут к сокращению миофибриллей, ибо известно, что.
теплопродукция при одиночных сокращениях во время лестницы может не
усилиться; она может остаться на одном уровне или даже
уменьшиться. А этот факт указывает на то, что усиление одиночных сокращений
(может происходить без усиления тех физико-химических процессов,
которые приводят к сокращению. Очевидно, есть еще другой
существенный фактор. Мы знаем, что при растяжении мышцы меняется
сократительная способность миофибриллей: при малом растяжении
она увеличивается, а при большом падает. Далее, известно, что
молочная кислота меняет анизотропное вещество, уменьшая его двупрелом-
ляемость (Бухталь, 24). На этом основании можно предположить, что
при длительном раздражении под влиянием продуктов расщепления
сократительное вещество претерпевает обратимое коллоидохимическое
изменение, которое проходит спустя некоторое время после
сокращения. Это коллоидное изменение может быть такого рода, что оно бу-
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
199
дет повышать чувствительность миофибриллей к тем химическим
веществам, которые 'производят сокращение, т. е. повышать сократительную
способность імышцы. Весьма возможно, что такое изменение
миофибриллей играет некоторую роль в явлениях лестницы (Фишер, 18).
С точки зрения мембранной теории для явления лестницы было
предложено следующее объяснение. По представлению Эббеке (11)
а других, процесс возбуждения характеризуется разрыхлением прото-
ялазматических перепонок, из которых почти целиком состоит
возбудимая система. После возбуждения происходит постепенное
восстановление нормального состояния этих перепонок. Это обусловливает, по
мнению Эббеке, относительную рефракторную фазу. Но после
восстановления имеется еще некоторое последействие функционального
характера, которое проявляется, между прочим, в том, что разрыхление
перепонок на новое раздражение наступает в более глубоком и более
растянутом во времени виде. В этих случаях авторы имеют в виду
процесс возбуждения, а не сократительный процесс. Нарастание
последнего прежде всего, конечно, обусловлено нарастанием процесса
возбуждения. Но своеобразное изменение сократительного процесса, о
котором говорилось-выше, нельзя относить всецело за счет возбуждения.
Повышение возбудимости при тетаническом раздражении. При те-
танических раздражениях повышение возбудимости наблюдается после
коротких раздражений. Чем свежее мышечный препарат, чем лучше
его функциональное состояние, тем лучше выступает повышение
возбудимости, тем дольше оно удерживается. Если тетаническое
раздражение длится долго, функциональное состояние мышцы, наоборот,
ухудшается, возбудимость падает. Чтобы вызвать повышение
возбудимости в свеже вырезанных мышцах лягушки, тетаническое
раздражение должно длиться не более нескольких десятков секунды. Но для
этого достаточно тетаническое раздражение и в несколько секунд.
И в этом случае присутствие нервных пластинок не имеет
существенного значения в повышении возбудимости. И на проксимальных
безнервных участках после короткого тетанического раздражения
возбудимость повышается на много секунд (Беритов, 20) (рис. 107). Это
наблюдается также на мышцах денервированных и кураризованных
(Броун и Эйлер, 25). Следовательно, нервные окончания не играют
существенной роли в явлении повышения одиночных сокращений
после короткого тетанического раздражения.
Повышение возбудимости наблюдается и при больших частотах
раздражения, когда каждый последующий импульс приходится на
время относительной рефракторной фазы: повышение возбудимости
ясно наступает по прекращении раздражения, и в таких случаях
повышенная возбудимость может держаться очень долго. В этом легко
убедиться, если раздражение прекратить и спустя некоторое время
начать вновь. При повторении первые удары раздражения дадут более
интенсивные эффекты, чем в начале первого раздражения (рис. 109)
{Введенский, 1; Беритов, 5).
200 общая физиология мышечной системы
В определенных случаях можно наблюдать, что при таких частотах
раздражения, когда каждое раздражение попадает во время
относительной рефракторной фазы, все-таки происходит нарастание
биоэлектрических токов возбуждения. Так, на рис. 109 каждый новый эффект
наступает через 0.008 сек. и нарастает, хотя несомненно приходится
на время относительной рефракторной фазы (Беритов, 5). Усиление
процесса возбуждения, несмотря на рефракторную фазу, по всей
вероятности, объясняется тем, что при некоторых слабых раздражениях
первоначально возбуждается некоторая часть мышечных волокон,
другие же, хотя и не возбуждаются, но все же испытывают влияние раз-
Рис. 109. Повышение возбудимости при тетаническом раздражении большой частоты.
Регистрируются [токи возбуждения m. semi tend inosus лягушки гальванометром Эйнтховена. Частота
раздражения 250 в сек. Мышца отвечает 125 раз. Электрограмма В есть продолжение А с небольшим
пропуском. Во время второго раздражения первые токи возбуждения сильнее, чем в начале первого
раздражения. Во время первого 'раздражения токи возбуждения нарастают постепенно. Время —
, ¦ ; [в toTbix долях секунды.
дражения и претерпевают повышение возбудимости, как от
субминимального раздражения. В результате, при повторном раздражении и
эти волокна начинают возбуждаться. Поэтому кривая внешнего
мышечного эффекта должна нарастать, невзирая на то, что каждое новое
раздражение приходится на время рефракторной фазы от
предыдущего возбуждения.
Повышение возбудимости наблюдается даже в том случае, когда
мышца непосредственно раздражается индукционными ударами такой
высокой частоты, которая производит слабый, т. е. пессимальный,
механический эффект, несмотря на большую силу раздражения. В таких
случаях повышение возбудимости обнаруживается после раздражения.
Во время раздражения сокращение падает и может даже сойти на-нет.
Несмотря на это, возбудимость оказывается повышенной после
раздражения (рис. 110). Из этих фактов ясно вытекает, что длительное
повышение возбудимости после тетанического раздражения по существу
определяется продуктами химического обмена веществ, а не самим
возбуждением и что этот процесс по существу того же рода, что и при
оптимальном сокращении (Беритов, 20).
Повышение возбудимости после тетанического сокращения вновь
стало в последнее время предметом подробного исследования. Одни
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ 201
авторы пришли к заключению, что оно обусловлено присутствием
химического медиатора в концевых пластинках; при этом имелось в виду
накопление ацетилхолина во время тетанического раздражения
(Гутман и сотр., 26). Но это предположение противоречит тому факту, что
данное явление наблюдается и на безнервных препаратах, в которых
образование ацетилхолина при возбуждении не было доказано.
Поэтому другие авторы отрицают роль ацетилхолина в этом явлении и
приписывают его накоплению калия за время тетанического раздражения
в результате химических изменений, сопровождающих сокращение..
Было замечено, что в мышцах, частично отравленных курарином,
после искусственного введения калия наступает совершенно такое же
повышение сократительного и электрического эффектов, как после тета-
г
Рис. ПО. Повышение возбудимости после пессимального
сокращения.
Безнервный 'проксимальный участок m. ^arteritis длиной в 9 мм
раздражается на расстоянии 1—3 мм от проксимального конца тетанически
(частота 150 в ,1 сек.) (нижний сигнал), а на расстоянии 5—7 мм —
отдельными индукционными ударами (верхний сигнал). Пссле сильного
тетанического раздражения происходит значительное повышение одиночных
сокращений (опыты В и С),- а после слабого' раздражения ¦— едва заметное
повышение (опыт А).
нического раздражения (Браун и Эйлер, 25; Розенблют и Морисон,,
28). Но и против этого предположения было сделано основательное
возражение. Именно, было указано, что комбинация тетануса и
действия калия не влияют на величину и продолжительность іпоеттетани-
ческого усиления одиночных сокращений, что излишек калия
подавляет сократительный процесс. Кроме того, при частом и длительном-
раздражении двигательного нерва, когда возбудимость нерва
определенно понижается, в двигательных пластинках не наблюдается
понижения возбудимости, что должно было бы непременно наступить, если-
бы в них происходило накопление калия (Грумбах и Виллер, 27).
Поэтому названные авторы склонны рассматривать данное явление как
результат того электрического изменения, которое сопровождает
местное изменение основного биологического процесса. Кроме того, в
противоположность другим авторам, они признают способность к пост-
тетаническому усилению только за нервным участком, двигательными
пластинками, следовательно, связывают ее с возникновением местного
процесса в двигательных пластинках.
202 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Практикум
1. Наблюдение суммации. На свежем спинномозговом препарате определяют
пороги раздражения мышцы по отношению к отдельным индукционным ударам,
прерывая ток от *руки через электромагнитный прерыватель. Если препарат свежий,
не подвергался перед этим сильным и длительным раздражениям, то порог для
одиночных раздражений окажется много выше порога тетанического раздражения.
Приведенные опыты производятся на некураризозанных препаратах. Поэтому можно
приписать означенные случаи повышения эффектов повышению возбудимости не
мышечных, а нервных 'волокон или нервных окончаний. Однако такой же эффект
можно получить на безнервном проксимальном участке m. sartorius.
2. Наблюдение явления лестницы. Берут такой же препарат, как и выше;
m. gastrocnemius соединяют с миографом. Раздражают перерезанный седалищный
нерв сильными индукционными ударами через каждые 2 сек. посредством метронома.
Когда кривая одиночных сокращений начнет нарастать, следует на несколько секунд
участить удары метронома вдзое и затем івиовь перейти на прежний ритм. Когда
высота одиночных сокращения достигнет максимума, прекратить раздражение и
через каждые 2—3 минуты производить по одному индукционному удару, чтобы
проследить, когда возбудимость возвратится к норме. Определить процент оверх-
нормального сокращения.
Наблюдение повышения возбудимости. Приготовляют спинномозговой препарат
лягушки, как в опыте с одиночным сокращением. Вызывают в записываемой
икроножной мышце тетаническое сокращение раздражением двигательного нерва, а
повышение возбудимости прослеживают с помощью одиночных индукционных
ударов, приложенных к самой мышце в ^е проксимальном участке. Раздражение
производится метрономом регулярно через интервалы в 1—2 сек. (Ом. о пользовании
метрономом в методике.) Одиночное сокращение от этих ударов после тетанического
сокращения может оказаться сильнее, чем до. него. Этот опыт хорошо удается на
совершенно свежем и неутомленном препарате.
Если на том же препарате раздражать мышцу несильным тетаническим
раздражением и затем прекратить последнее на секунду и более, то окажется, что
повторное раздражение дает вначале более сильное сокращение, чем было до этого.
В предыдущем опыте мышца возбуждалась через нервные окончания. Но такой
же результат получится, если раздражать овеже вырезанный проксимальный без-
иервный участок m. sartorius. Постановка опыта на этом участке мышцы уже
описана выше. Препарат готовится и соединяется с миографом и электродами, как
в опыте с пессимальным сокращением (см. стр. 188). Сначала записывают
пороговый тетанический эффект, потом производят сильное тетаническое раздражение
продолжительностью © 2—4 сек. Спустя несколько секунд вновь пробуется то же
пороговое раздражение. Теперь пороговый эффект будет много сильнее, чем в
первом случае.
Литература
1? Введенский Н., О соотношении между раздражением и возбуждением
при тетанусе, стр. 278, 1886.
2. Samojloff A,, Arch. Anat. Physiol., SuppL, S. 1, 1908.
3. Введенский H., Тр. физиол. лаб. Петерб. унив., стр. 1, 1906.
4. Steinach Е., Pfl. Arch., 125, 318, 1908.
5. Beritoff J., Z. Biol., 62, 125, 1913.
6. Kronecker, Arb. Physiol. Anst. Leipzig, S. 177, 1871.
7. Funke J., Pfl. Arch., 8, 213, 1875.
8. T i e g e 1, Arb. Physiol. Anst. Leipzig, 1875, S. 1.
9. Rossbach M. u. Harteneck K., Pfl. Arch., 15, I, 1877.
10. Beritoff J., Z. Biol., 85, 1, 1926; ib., 85, 15. 1926.
ПОВЫШЕНИЕ ВОЗБУДИМОСТИ ПРИ РАЗДРАЖЕНИЯХ
203
11. Ebbecke U., Pfl. Arch., 195, 555, 1922.
12. Л а з а р е в П., Ионная теория возбуждения, Москва, 1923.
13. К г і е s v. u, S e w а 1 I, Arch. Anat. Physiol., S. 66, 1881.
14. К e i t h L u с a s, J. Physiol., 39, 461, 1910; The conduction of th& nervous
impulses, London, 1917.
15. Lee F. S.f Pfl. Arch., 110, 400, 1905.
16. Gruber С. М., Am. J. Physiol., 63, 338, 1923.
17. В er it off J., Erg. Physiol., 23, 33, 1924.
18. Fischer E., Pfl. Arch., 225, 532, Tokyo, 1930.
19. С a t о G., The microphysiology of nerve, 1934.
¦20. Б e p и t о в И., Флзиол. журн. СССР, 27, 147, 1939.
21. La pique L., С. г. Acad. Sci., Paris, 155, 70, 1912; Melanges biologiques, vol.
jubil. dedie a Ch. Richet, 1912; С. г. Acad. Sci, 189, 77, 1934.
-22. Schr iever H., Erg. Physiol., 38, 878, 1936.
23. D u b u i s s о n M., Pfl. Arch., 239, 314, 1939; Dubuisson u. Schulz, Pfl.
Arch., 239, 776, 1939.
24. В u с h t h a 1 F. a. L i n d h a r d J., The physiol. of striated muscle fibre, 1939-
25. Brown G. L. a. v. E u 1 e r U. S., J. Physiol., 93, 39, 1938.
26. Guttman S. A., Horton A. G. a. Wilber В. Т., Am. J. Physiol,, 119,
463, 1937.
27. Grumbach L. a. Wilier D. Т., Am. J. Physiol., 130, 433, 1940.
28. R о s e n b 1 u e t h A. a. M о г i s о n R. S., Am. J. Physiol., 119, 236, 1937.
Г л а в a JTX
УТОМЛЕНИЕ
Общее понятие. Мышца не может работать беспрерывно. С
течением времени деятельность мышцы постепенно ослабевает. Это
ослабление или полное прекращение мышечной деятельности в результате-
самой деятельности называется утомлением. В мышце во время
деятельности протекают два основных процесса: процесс
возбуждения, который выражает своеобразное изменение возбудимой
системы, и процесс сокращения, в котором видную роль играют
расщепление и восстановление фосфагена и гликогена. Как один, так и
другой процессы не могут протекать бесконечно. Если запасы кислорода
и энергетического материала в мышечных клетках исчерпались, а кровь,
не может снабдить мышечные клетки достаточным количеством
кислорода и энергетического материала, то возбуждение должно ослабевать,,
так как в этом случае восстановление возбудимой системы после
каждого возбуждения не достигает нормы. Если же исчерпались запасы
фосфагена и гликогена, а кровь недостаточно снабжает ими мышечные
клетки, тогда должно происходить ослабление сокращения. Но
недостаток этих веществ не должен отозваться на течении возбуждения..
Отсюда следует, что ослабление мышечной деятельности при ее
большой длительности нельзя (всегда приписывать утомлению возбудимой-
системы. Оно может быть вызвано в определенных условиях
исчерпанием запасов фосфагена и гликогена.
Заслуживает внимания тот факт, что в покоящейся мышие
потребление кислорода, т. е. так называемый основной метаболизм,
поразительно слабо. Например, мышца лягушки при 22° С на 1 г веса в час
поглощает всего 30—50 мм3 кислорода. Доказано, что эта энергия
дыхания в покое идет не только на восстановление возбудимой системы-
мышцы, но и на восстановление фосфагена (Мейергоф, 40),
расщепление jco.Top.oro происходит все время и в состоянии покоя (Белицер, 47).
Этим объясняется, что вырезанная мышца сохраняет возбудимость и*
сократительную способность в покоящемся состоянии в течение многих
дней не только на воздухе, но и в бескислородном растворе Рингера
(Гилл, 33). Очевидно, в бескислородной среде обмен веществ
возбудимой системы мышцы поддерживается за счет энергии расщепления*
УТОМЛЕНИЕ
205
фосфагена, аденилпирофосфата и гликогена. Этот процесс, как уже
указывалось выше, протекает в імышце івсе время в бескислородной
-среде. Следовательно, возбудимая система может черпать
необходимую энергию для своего /восстановления также и анаэробным путем.
Потребление кислорода, наоборот, чрезвычайно увеличивается при
работе імышцы. Работающая (мышца лягушки, например, потребляет на
1 г ткани в 1 час примерно 1000 мм3 кислорода (Гилл и Купалов,
46). Это избыточное потребление кислорода стимулируется распадом
фосфагена, вернее — продуктами его распада — креатином и фосфорной
кислотой, с одной стороны, и продуктом распада гликогена —
молочной кислотой, с другой (Белицер, 47).
По наблюдениям некоторых авторов, в условиях кровообращения в
-организме утомление імышцы при длительной работе не зависит от
количества произведенной работы. Как это мы увидим ниже (см.
главу о работе імышцы), при разных нагрузках количество работы
различное: с увеличением нагрузки в довольно больших пределах
количество работы нарастает. Несмотря «а это, утомление наступает через
-одинаковое число раздражений — одинаковое число рабочих движений
(Риссер и Миура, 35). Так, например, при нагрузке m. gastrocnemius
-лягушки в 20—60—-100 г утомление наступает через одно и то же
количество рабочих движений. То же бывает при нагрузке в 40, 100 и
200 г. В опытах Р и с с е р а на одном препарате количество работы
''при 20 г составляло 2165 гсм, при 60 г—5692 гсм, а при 100 г —
7108 гсм. Если при работе в 2165 гсм и при работе, втрое большей —
7 108 гсм утомление наступает после одного и того же числа
раздражений, то отсюда ясно следует, что в данных условиях утомление
зависит не от количества произведенной работы, а от числа возбуждений,
т. е. от изменения функционального состояния возбудимой системы
мышцы, которое наступает в связи с повторной деятельностью
мышцы (35).
Однако это положение нельзя распространить на все случаи
мышечной деятельности. Например, если мышца сокращается без нагрузки,
а значит, без напряжения, утомление наступает значительно позднее,
<іем при нагрузке или изометрическом сокращении (Топсое-Иенсен,
43). Это, очевидно, связано с тем обстоятельством, что при
растяжении грузом дыхание значительно повышается. Кроме того, на невыре-
занных мышцах с кровообращением растяжение безусловно влияет на
-кровообращение в смысле затруднения его, а потому чем сильнее
'будет растяжение мышцы от нагрузки, тем скорее должно наступить
утомление. Но в общем можно заключить, что действительное
утомление мышцы представляет только функцию возбудимой системы и
наступает вследствие длительной деятельности последней благодаря
ослаблению процесса возбуждения (Беритов, 1).
Основные факторы утомления. Отмеченное выше замедление или
даже задержка в восстановлении возбудимой системы после
многократного возбуждения при недостатке кислорода и питательного імате-
206 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
риала является не единственным и даже не главным фактором
утомления мышцы в нормальном организме. Указанная причина утомления
характерна прежде всего для вырезанных мышц, лишенных
кровоснабжения. При наличии же нормального кровоснабжения мышц с
нормальным дыханием и питанием организма не может возникнуть
такое положение, когда для восстановления возбудимой системы не было-
бы кислорода или других веществ. В нормальном организме
при нормальном дыхании и питании не могут также создаться
условия, при которых мышцы потеряли бы способность сокращаться из-за
недостатка аденилпирофосфата, углеводов или фосфагена. Известно*
например, что в утомленной мышце при нормальном кровоснабжении
содержание гликогена не отличается от нормы (Кобаяши, 18). Между
тем любая мышца в нормальных условиях кровоснабжения
утомляется при более или менее длительной »и интенсивной деятельности.
Причина этого явления лежит в том, что в результате мышечной
деятельности в мышце возникают разного рода ядовитые продукты
обмена веществ. К таким продуктам прежде всего нужно отнести
фосфорную и молочную кислоты. Эти вещества при больших
концентрациях действуют отравляюще на возбудимую систему. Вследствие этого»
затрудняется восстановление возбудимой системы после каждого
процесса возбуждения, сообразно с этим падает и ее возбудимость.
Насчет молочной кислоты существует прямое наблюдение, что
накопление ее понижает мышечную деятельность: в бескислородной атмосфере
мышца утомляется скорее, чем в бескислородном рингеровском
растворе, когда молочная кислота имеет возможность
диффундировать из мышцы в раствор. При прибавлении карбонатов к ірингеров-
скому раствору увеличивается выход молочной кислоты, и в то же
время повышается работоспособность мышцы (Матсуока, 5). Также
относительно фосфорной кислоты было установлено, что она
благоприятствует утомлению. Именно, введение ее в неутомленную мышцу
в очень малой дозе производит ослабление деятельности со всеми
обычными признаками утомления (Ранке, 6; Ли, 9).
Однако по количеству накопившихся молочной и фосфорной кислот
нельзя еще судить о степени утомления. Как указывалось выше, при
работе с разной нагрузкой как при нормальном кровообращении, так
и в анаэробных условиях мышцы утомляются через одно и то же
число сокращений. Между тем накопление фосфорной и молочной кислот
не одинаково при утомлении в условиях разной нагрузки. С
увеличением нагрузки количество этих кислот сначала растет, а потом падает.
Так, например, в вырезанной икроножной мышце лягушки при
нагрузке в 100 г образуется на 100% больше фосфорной и молочной
кислот, чем при нагрузке в 10 г. С дальнейшим увеличением нагрузки
это количество уменьшается и при нагрузке, близкой к абсолютной
силе, достигает такого минимума, какой наблюдается при отсутствии
всякой нагрузки (Нагана, 38). Очевидно, при разных нагрузках
условия расщепления и восстановления фосфагена и гликогена не одина-
УТОМЛЕНИЕ 207
ковы, в силу чего количество накопляющихся кислот различно. Но
отравляющее действие их на возбудимую систему определяется
главным образом не количеством накопившейся кислоты, а количеством
образуемой, ибо отравление должно происходить преимущественно
в момент образования кислоты.
Далее, как показали Палладии и его сотрудники (37), в
тренированной мыщце усиливаются синтетические и аналитические процессы.
В связи с этим было замечено, что в утомленной тренированной імышце
не происходит накопления молочной кислоты. Следовательно,
утомление может наступить в мышце, несмотря на отсутствие накопления
молочной кислоты. Это явление, однако, также не противоречит
приведенному выше положению об отравляющем действии кислых
продуктов на возбудимую систему мышцы. Эти вещества все-таки образуются
в тренированной імышце, как и в нетренированной, во время работы.
В обоих случаях они должны действовать на возбудимую систему
отравляющим образом в момент образования. Но только в
тренированной мышце эти кислоты исчезают сейчас же после возникновения
вследствие чрезвычайного ускорения синтетических и окислительных
процессов.
Большое значение в развитии утомления приписывают солям калия.
Известно, что лактат калия в малых концентрациях сначала повышает
возбудимость мышцы, а затем действует на нее угнетающим образом
(Бурридж, 7).
Детальное исследование содержания калия, натрия, кальция и
магния в мышцах теплокровных (кошка, крыса) при покойном состоянии
и после многоминутной деятельности (за 30 мин.) показало, что в
результате деятельности содержание калия уменьшается и это
уменьшение балансируется одновременным повышением содержания натрия.
Содержание кальция и магния существенно не менялось (Фенн и
сотр., 41). Нужно думать, что это нарушение нормального соотношения
одновалентных ионов может иметь определенное значение для
жизнедеятельности мышцы и ее работоспособности.
Углекислота, которая также образуется в мышце в большом
количестве, не имеет существенного значения. При нормальном
кровообращении она не может накапливаться в мышцах в такой
концентрации, которая могла бы оказать на них отравляющее действие. При
задержке кровообращения угольная кислота ускоряет развитие
утомления.
Одно время утверждали, что в мышце при длительной деятельности
развивается особое ядовитое вещество, которое вызывает утомление.
Были даже попытки к экспериментальному доказательству (Вейхардт,
8). Но эта идея не оправдалась. Правда, если экстракт утомленной
мышцы впрыснуть здоровому неутомленному животному, последнее
обнаружит такой признак утомления, как повышение температуры.
Но теперь известно, что и экстракты неутомленных мышц оказывают
точно такое же действие (Ли и Аронович, 9).
208 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Что продукты обмена веществ действительно оказывают на мышцу
-отравляющее действие, можно доказать простым опытом. Если
сначала утомить вырезанную мышцу длительным раздражением до полной
недеятельности, а затем пропустить через кровеносные сосуды раствор
Рингера и этим в некоторой мере очистить мышцу от накопившихся
продуктов обмена веществ, то к ней вновь возвращается способность
-к усиленной деятельности, хотя при этом мышца не восстановила
своего запаса кислорода или углеводов (Ранке, 6).
Рис. 111. Изменение электрического эффекта при утомлении m. gastrocnemius лягушки.
•Раздражается 50 раз в секунду. В самом начале электрограммы восходящее колено первого
колебания длится 0.004 сек., в конце же ее —-0.015 сек. Гальванометр Эйнтховена.
Итак, одним из основных факторов утомления является
отравляющее действие продуктов обмена веществ, в особенности молочной и
фосфорной кислот, на возбудимую систему мышцы.
Изменение возбуждения во время утомления. Судя по токам
возбуждения, возбуждение при длительной деятельности претерпевает
следующие изменения: интенсивность его уменьшается, а
продолжительность растет (рис. 111) (Беритов). Так, в одном опыте на мышцах
лягушки длительное непрямое раздражение отдельными
индукционными ударами произвело следующие изіменения токов івозбуждения:
продолжительность восходящего колена до утомления равнялась 2—3 о,
лосле же некоторого числа раздражений удлинилась до 6—7 а, т. е. на
200—300%. При этом амплитуда токов возбуждения была понижена
всего только на 25% (Беритов, 26).
Но такое изменение (возбуждения происходит на свежей мышце не
с самого начала длительного раздражения. В начале деятельности
возбуждение усиливается, а только затем ослабевает. Следовательно, в
отношении возбуждения імы имеем точно такое же изменение, как в
отношении возбудимости, которая также при длительной
деятельности сначала повышается >и только затем ослабевает (Брюкке, 16;
Беритов, 15, 26).
Означенное ослабление возбуждения, ло всей вероятности,
находится в прямой зависимости от отравляющего действия больших
концентраций кислых продуктов обмена веществ на возбудимую систему,
ибо аналогичные изменения испытывают токи возбуждения при
отравлении вообще живой ткани наркотиками и кислотами, а также при
помещении в бескислородную среду.
Изменение сокращения во время утомления. Одиночные
сокращения изменяются при утомлении следующим образом: амплитуда их с
течением времени уменьшается, а длительность каждого сокращения
УТОМЛЕНИЕ
209
нарастает. На совершенно свежей мышце амплитуда уменьшается не с
самого начала раздражения. Первоначально амплитуда сокращений
нарастает, как это уже указывалось выше. На рис. 112 показано
увеличение длительности одиночного сокращения, а на рис. 113 —
изменение высоты одиночного сокращения; сначала видна стадия повышения
высот сокращений. Затем их падение. Но и на последнем рисунке ясно
видно также удлинение одиночного сокращения в связи с утомлением.
Вначале нисходящее колено сразу доходит до абсциссы и почти
сливается с восходящим, а потом, в особенности в конце записи,
оно не сливается с нисходящим коленом и не сразу доходит до
абсциссы.
Чем выше температура и вообще чем лучше обмен веществ, тем позже
и слабее проявляется удлинение сокращения в связи с утомлением.
Ритм мышечной деятельности также сильно влияет на развитие
этих изменений. Чем чаще следуют раздражения, тем скорее эти
сдвиги наступают. На .величину ритма, при которой развивается утомление,
сильно влияет температура. Если температура імышцы лягушки
довольно низка (10—15°), то мышца при нормальном кровообращении
утомляется довольно быстро, уже при интервале в 4 сек. между
раздражениями. Если же температура мышцы равна 28—30°, то развитие
утомления не наблюдается 1и при интервале раздражений в 1 сек.
(Аккерман, 14).
Характерно, что в стадии сильного утомления, когда высота
сокращений уменьшилась на 50—60%, одиночные сокращения перестают
протекать длительно (Беритов, 15). Удлинение мышечного сокращения
при утомлении, по существу говоря, обусловливается возникновением
контрактуры. Следовательно, появившееся при утомлении контрак-
турное сокращение перестает вызываться при глубоком утомлении
мышцы.
Тетаническое сокращение мышцы при длительной деятельности
изменяется следующим образом: высота сокращения постепенно
уменьшается и затем исчезает полностью; ів определенных условяих, а
именно при слабых тетайизирующих раздражениях, в момент утомления
сплошной тетанус становится зубчатым, клоническим. В период
утомления, (По прекращении тетанизирующего раздражения всегда следует
контрактурное последействие. Оно тем интенсивнее и длительнее, чем
ниже температура, чем хуже условия для обмена веществ
(Беритов, 15).
Это контрактурное последействие, очевидно, того же
происхождения, как и удлинение одиночного сокращения при утомлении. Об этом
свидетельствует и тот факт, что в период сильного утомления, когда
одиночное сокращение возвращается к нормальной длительности, тета-
низирующие раздражения производят сильные тетанусы без контрак-
турного последействия (Беритов, 15).
Происхождение характерных изменений механического эффекта.
Хотя степень сокращения мыщцы находится в определенной зависимо-
14 И. С. Беритов
210
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
сти от интенсивности возбуждения, однако характер сокращения не
всегда отражает предшествующий ему процесс возбуждения. Как уже
указывалось выше, на мышцах лягушки сокращение усиливается в
первое время длительного раздражения параллельно с усилением
возбуждения. Но в дальнейшем сокращение меняется более независимо:
оно начинает падать раньше ослабления процесса возбуждения и
прекращается раньше
прекращения этого
процесса (рис. 108) (Брюкке^
16; Беритов, 15).
Д алее,
специальными опытами было
установлено, что
прекращение механического
эффекта на вырезанной
мышце при длительном
раздражении не
обусловлено также
израсходованием гликогена, так
как и после такого
утомления в мышце
достаточно много гликогена
(Мейергоф, 40).
Так, после
утомления в результате
длительного раздражения
отдельными
индукционными ударами или
повторной короткой
фарадизацией (по 1 сек.
через 1 сек.) количество
гликогена падает только
до одной трети
первоначальной величины
(Беритов и Волынский,.
31; Кометиани, 48). Но это прекращение механического эффекта при
этих же условиях может быть обусловлено израсходованием фосфагенаг
ибо количество его падает более чем в 10 раз и даже исчезает
полностью (Кометиани, 48). В этих опытах мышца, потерявшая
сократительную способность, еще сохраняла возбудимость, ибо на раздражение
она еще отвечала токами возбуждения.
Итак, прекращение механического эффекта на вырезанной мышце
при длительном раздражении, может быть, обусловливается не
прекращением возбуждения и не израсходованием запасов гликогена, а
исчезновением фосфагена или еще какими-то изменениями, имеющими
ближайшее отношение к сократительному процессу.
Рис. 112. Изменение механического эффекта при
утомлении m. gastrocnemius лягушки.
/ группа дает одиночные сокращения мышцы до утомления. //,
III и другие группы представляют их при постепенном развитии
утомления, что проявляется в постепенном удлинении сокращения
и в повышении их амплитуды. Если бы запись продолжалась
дольше, высота сокращения стала бы меньше (Роллет).
УТОМЛЕНИЕ
211
При нормальном функциональном состоянии организма утомление
мышц при их непрямом раздражении через двигательный дерв вообще
не сопровождается уменьшением кюл'ичества гликогена (Кобаяши, 18)
или накоплением молочной кислоты (Бюрги, 19). Утомление в таких-
случаях безусловно является следствием весьма сложных процессов,
среди которых 'израсходование запасов фосфагена и «отравляющее
действие молочной 'и фосфорной кислот на возбудимую систему, по-
видимому, играют существенную роль. Можно думать, что молочная
и фосфорная кислоты действуют на возбудимую систему в момент их
возникновения. А при известной очень івысокой частоте раздражения
это может привести сначала к повышению возбудимости, а потом
к понижению ее, параллельно же с этим и к ослаблению проце?са>
возбуждения. --*
ъ связи с утомлением происходит также удлинение одиночных
сокращений. Это, разумеется, не может зависеть от удлинения процесса
возбуждения. Последний удлиняется только на несколько сигм, тбгда
как одиночное сокращение может удлиняться от десятой доли секунды
до многих секунд. Можно было бы думать, что удлинение одиночных
сокращений обусловливается действием кислых продуктов
обмена;-веществ на сократительное вещество — миофибрилли. Известно,
например, как это уже указывалось выше, что мышца, погруженная в
санти- или миллинормальный раствор кислоты, сокращается. А если
она даже при этом не сокращается, то изменяется функционально
под влиянием кислоты таким образом, что отдельные индукционные
удары производят чрезвычайно длительные эффекты (Швенкер, 20).
Аналогичное явление может иметь место при некотором накоплении
фосфорной и молочной кислот в мышечной клетке. Однако опытным
путем доказано, что удлинение одиночных сокращений получается
в связи с их повторением и при отсутствии образования молочной
кислоты, а именно, на мышцах, отравленных моноиодуксусной кислотой,
причем в последнем случае удлинение наступает еще скорее, чем на
нормальной мышце. Далее, удлинение сокращения ! не находится
в прямой связи с утомлением, ибо оно получается еще раньше
утомления, еще в той стадии, когда высота сокращений растет. Оно не
зависит также существенным образом от кровообращения и
температуры, ибо оно получается при всякой температуре и в условиях
кровообращения (Паркинсон, 36). Нужно думать, что удлинение
одиночного сокращения в связи с его повторением имеет чрезвычайно
сложное ^происхождение: кислые продукты, низкая температура,
отсутствие кровообращения, несомненно, «играют роль в происхождении
его. Но, повидимому, есть, и другие факторы, обусловливающие это
явление. Все эти факторы производят такое физико-химическое
изменение миофибриллей, что последние дают сильно растянутый
сократительный процесс.
Об изменении миофибриллей при утомлении, между прочим,
свидетельствует изменение двойной преломляемости. Она уменьшается при
14*
212 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
утомлении на 20—30%. В сокращенной мышце вообще двойная
преломляемость падает. При утомлении это изменение анизотропного
вещества удерживается некоторое время и после сокращения, т. е.
когда миофибрилли не находятся в сокращении. При этом оказывается,
что уменьшение двойной преломляемости обусловлено действием
молочной кислоты на сократительное вещество (Бухталь, 44).
Ослабление тетанического сокращения при длительном
раздражении в значительной мере должно обусловливаться ослаблением
процесса возбуждения. Об этом свидетельствуют специальные опыты
с одновременной регистрацией механического и электрического
эффектов. В определенных условиях оба эти эффекта ослабевают 'и пре-
Рис. ИЗ. Первая и вторая стадия утомления мышцы одиночными раздражениями.
Спинномозговая лягушка. Верхняя кривая от m. ileofibularis, средняя—от m. semitendinosus, а нижняя—
от т. vastus externus. Раздражается периферический отрезок седалищного сплетения; сила 40 см
расстояния между индукционными катушками (порог 48 см). Раздражение размыкательными
индукционными ударами через каждые 2 сек. продолжается 17 мин. За это время трижды производилось
учащение раздражения с интервалом в полсекунды: в самом начале, через 8 мин. и через 17 мин. Впервые
8 мин. (А, В, С и D) стадия повышения механического эффекта; Е — переходный момент к ослаблению;
F •— стадия падения механического эффекта. Сигнальная линия обозначает моменты раздражения
индукционными ударами.
кращаются одновременно (Фултон, 28). Это может произойти
независимо от изменения запасов аденилпирофосфата, фосфагена или
гликогена и помимо накопления молочной и фосфорной кислот, именно
в силу отравляющего действия кислых продуктов на возбудимую
систему в (момент их возникновения, еще раньше их нейтрализации или
превращения ів индифферентное соединение.
Утомление мышцы при раздражении двигательного нерва
одиночными индукционными ударами. При утомлении мышцы путем
повторных одиночных раздражений двигательного нерва наблюдается, между
прочим, одно очень характерное явление. Именно, в зависимости от
того, в какой стадии утомления мышца находится, последняя
реагирует на переход от одного ритма раздражения к другому то
усилением, то ослаблением одиночных сокращений. В начале длительного
раздражения, когда одиночные сокращения нарастают, учащение
раздражения благоприятствует их повышению: максимальная высота до-
УТОМЛЕНИЕ
213
^кШШ^шкшшиИМішш
..іл.ліиШііз.;и.
стигается быстро. В период убывания одиночных сокращений следует
различать две стадии: сначала учащение ведет к ослаблению
сокращений, а замедление, наоборот, к увеличению их; впоследствии же,
в более глубокой стадии утомления, происходит как раз обратное:
учащение ведет к значительному повышению одиночных сокращений
(Беритов, 27). Первая и вторая стадии показаны на рис. 113, третья —
на рис. 134.
Происхождение первой и второй форм изменения, а именно
повышение одиночных сокращений в начальной стадии длительного
раздражения, а затем падение их в связи с учащением раздражений, не
может быть поставлено в исключительную связь с аналогичным
изменением процесса
возбуждения, как на" это уже
указывалось выше. Точно так
же в третьей стадии
утомления нарастание
одиночных сокращений при
учащении раздражений не
находится в строгой связи
с нарастанием
[возбуждения: электрический эффект
нарастает значительно
слабее механического и часто
продолжает нарастать в то
время, когда механический
эффект начинает ослабевать
(Беритов, 27).
Специальное
исследование происхождения этого
явления показало, что все эти изменения механического эффекта
происходят в нервных участках мышцы и, повидимому, зависят от продуктов
метаболизма нервных окончаний. Можно думать, что эти продукты
вызывают в мышечной клетке локальные физико-химические изменения,
вследствие чего в этой области возрастают как возбудимость, так и
сократительная способность (Гедевани, 39). С точки зрения мембранной
теории, эти изменения могут касаться поверхностных перепонок
саркоплазмы. С этой точки зрения процесс возбуждения должен
сопровождаться разрыхлением перепонок, в силу чего они становятся более
проницаемыми для ионов, действующих, с одной стороны, на
возбудимую систему, а с другой — на сократительное вещество миофибрил-
лей. Можно предположить, что при утомлении разрыхление перепонок
становится сверхнормальным и потому сильно повышается действие
этих ионов на возбудимую систему, вызывая тем самым ухудшение ее
функционального состояния.
Далее, можно предположить, что при учащении раздражения
происходит усиленное образование таких веществ, которые, наоборот, ве-
Рис. 114. Третья стадия утомления мышцы.
Спинномозговой препарат лягушки. М. semimembranosus
(верхняя кривая) и vastus externus (нижняя кривая). 3 часа
после начала длительного раздражения. Раздражается
периферический отрезок седалищного сплетения размыкательными
индукционными ударами. Обычный интервал раздражения 1 сек.
Между опытами 1-м и 2-м кимограф был остановлен на
несколько минут, но раздражение не прекращалось. В обоих опытах
испытывалось уменьшение интервала до полусекунды, и
каждый раз это вызывало сильное нарастание одиночных
сокращений.
214
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
дут к уплотнению этих мембран. В связи с этим должно происходить
улучшение обмена веществ, вследствие чего будет усиливаться как
процесс возбуждения, так и сократительный процесс. О том, что
уплотнение полупроницаемых мембран в области нервных окончаний
может иметь именно такой результат, іможно было судить из
следующих фактов. Промывание мышцы, утомленной непрямым раздражением,
таким раствором Рингера, который содержит некоторый излишек
ионов кальция (0.06—0.2%), тоже производит увеличение
сократительного эффекта (Валидов, 34). Точно так же если мышцу,
утомленную непрямым раздражением, положить в рингер, содержащий
0.1% СаС12, то утомление проходит быстрее, чем в чистом рингере
(Серков, 42). Как известно, характерное действие ионов кальция
заключается в уплотнении полупроницаемых перепонок. Значит, на
основании этих опытов можно предположить, что утомление мышцы
сопровождается разрыхлением поверхностных оболочек в области
нервных окончаний, и это обусловливает ухудшение функционального
состояния двигательной пластинки. Но с этой точки зрения совершенно
непонятно, почему в состоянии утомления учащение раздражения
должно привести к избыточному выделению двувалентных ионов
и к улучшению функционального состояния двигательной пластинки.
Характерно, что если утомление производится прямым раздражением
нормальной или кураризованной ткани, такое же увеличение
концентрации кальция не приведет к усилению мышечных сокращений.
Большая же концентрация кальция производила угнетение сокращений
(Валидов, 34).
По мнению Воронцова (45), утомление при длительном непрямом
раздражении обусловливается теми локальными изменениями, которые вызываются в нервных
окончаниях иод, влиянием притекающих токов возбуждения нервных импульсов. Эти
локальные изменения заключаются в деполяризации нервных окончаний, в резуль ¦
тате чего подавляются их функции. Но с этой точки зрения также нельзя
объяснить, почему в стадии утомления учащение раздражения должно привести к
усилению возбуждения и сокращения. Ведь при учащении деполяризация еще более
должна была усилиться.
Утомление мышцы при тетаническом раздражении двигательного
нерва. Ослабление мышечной деятельности при длительном непрямом
раздражении не зависит от утомления нерва. Как это імы увидим в
отделе нервной физиологии, для утомления нерва требуется гораздо
более длительное раздражение, чем для утомления мышцы. Известно,
например, что 'Вырезанный нерв в атмосфере воздуха не утомляется
при тетанизирующем раздражении даже в течение восьми часов
(Введенский, 21). В тех же условиях раздражения імышца утомляется
абсолютно в течение нескольких минут. Но это абсолютное утомление
(мышцы через нерв не обусловливается абсолютным утомлением
возбудимой системы мышцы, так как, несмотря на абсолютное утомление
через нерв, імышца еще хорошо отвечает возбуждением и
сокращением на прямое раздражение. Но это возбуждение и сокращение бу-
УТОМЛЕНИЕ
215
дут ослаблены и будут носить все признаки утомления.
Следовательно, при абсолютном прекращении сокращения через нерв мышца
находится в состоянии относительного утомления.
Это состояние относительного утомления прежде всего наступает
в области нервных окончаний. Этим собственно и вызывается
прекращение, передачи возбуждения с нерва на імышцу. Здесь імы имеем яв-
.ление, подобное слабому отравлению кураре. Это доказывается целым
рядом фактов. Так, 'например, при утомлении через нерв
возбудимость падает прежде всего в нервной области мышцы (Беритов, 30).
Затем контрактурное сокращение, в результате нервного возбуждения,
наступает по преимуществу в нервной области (Беритов, 29).
Хронаксия также значительно меняется в нервной области: до утомления
хронаксия в нервном участке нормальной портняжной мышцы в 2—
3 раза короче, чем в безнервном проксимальном участке. После
утомления через нерв хронаксия увеличивается в нервном участке и
уравнивается с хронаксией безнервных участков, которая при этом совсем
не меняется (Цкипуридзе, 50). Очевидно, в нервной области, где
процесс возбуждения сильнее и где сократительная способность больше
всего, образование кислых продуктов, как фосфорной и молочной
кислот, а также ацетилхолина и калия, идет более интенсивно, чем
в других участках мышцы, и потому отравляющее действие их на
возбудимую систему должно происходить здесь в большей степени,
чем в других участках. Вследствие этого возбудимость в нервной
области падает значительно, и соответственно передача возбуждения
с нервного окончания на возбудимую систему мышцы становится
затруднительной. Повидимому, биоэлектрический ток возбуждения
нервного окончания не в состоянии произвести раздражение возбудимой
системы мышцы.
Можно указать еще на другой фактор, который должен
действовать в том же направлении. Голое нейрофибриллярное ветвление
нервного окончания также должно подвергнуться отравляющему
действию продуктов метаболизма мышечной клетки. Кроме того, нервное
окончание само должно утомиться при длительном тетаническом
раздражении, независимо от мышечного сокращения. Об этом
свидетельствует тот факт, что передача возбуждения с нерва на мышцу
прекращается тем быстрее, чем чаще раздражение, хотя сокращение само
при частых раздражениях бывает слабее, чем при редких, как об этом
говорилось выше.
Следовательно, при длительном тетаническом раздражении
двигательного нерва мышца перестает реагировать вследствие >
утомления как возбудимой системы мышщы, так и нервного окончания.
Этим обусловливается ослабление биоэлектрического тока
возбуждения и его раздражающего действия на возбудимую систему
мышцы.
Деятельность мышцы без утомления. В нормальных условиях, когда
мышца беспрерывно получает через кровь питательный материал и
216 ОБЩАЯ ФИЗИОЛОГИЯ МЫШКЧНОЙ СИСТЕМЫ
кислород, утомление наступает позже, чем ів условиях изоляции
мышцы от кровообращения. В последнем случае утомление мышцы
должно наступить значительно скорее не только потому, что она
имеет в своем распоряжении определенный запас и извне ничего не
может получить, но и оттого, что продукты обмена веществ целиком
накопляются в мышце. В нормальных условиях эти продукты частью
восстанавливаются в те самые высокоэнергетические материалы,
откуда они произошли (липопротеид, аденилпирофосфат, фосфаген,
гликоген), частью нейтрализуются путем соединения ионов
противоположного знака, частью вымываются из мышцы кровью. По этой
причине мышце і^ожно дать такую нагрузку и такой ритм работы, при
которых она сможет работать более или менее длительное время без
утомления. Если деятельность мышцы является умеренной, т. е.
мышца не сильно нагружена и не укорачивается максимально,
утомление іможет не наступить в течение многих часов. Субмаксимальное
сокращение и сравнительно небольшая нагрузка потому имеют
существенное значение, что в мышцах, сокращающихся максимально и
производящих максимально напряженную работу, кровообращение
сильно затрудняется благодаря сжатию сосудов между туго
натянутыми мышечными волокнами. При умеренном сокращении и
напряжении, наоборот, мышцы лучше снабжаются кровью благодаря
расширению сосудов (Баркрофт и Миллен, 49). В этом случае должно
происходить наилучшее обеспечение работающей мышцы всеми
материалами, необходимыми для сократительного процесса. Поэтому, когда
мышца сокращена не максимально и не напряжена очень сильно, она
может сокращаться в течение многих часов без утомления. Так,
в одном опыте мышцы кошки тетанически сокращались с
субмаксимальной интенсивностью в течение восьми часов и не давали ясных
признаков утомления. Сокращение вызывалось раздражением
соответствующих двигательных корешков. В этом опыте возбуждение мышцы
производилось ритмом 50—60 в секунду беспрерывно (Беритов). Тем
более мышца может сокращаться без утЪмления, если тетаническое
раздражение производится с лерерывами или применяется
раздражение отдельными индукционными ударами, как это было показано в
лаборатории Ашера Голлигером (2) иШмидом (3).
Заслуживают особого внимания следующие опыты из лаборатории
Ашера. Здесь изучали утомление мышц кролика в совершенно
неповрежденном организме. Вводили в грудную область спинного мозга
кокаин и тем вызывали паралич задних йог. Раздражался седалищный
нерв электрическим током в области бедра через булавки, воткнутые
глубоко в бедро по линии местоположения этого нерва. Эффект
раздражения—движение голеностопного сустава — записывался на
кимографе. Раздражение производилось так, чтобы мышцы возбуждались
периодически через определенные промежутки времени: например,
мышцу периодически раздражали в течение 1—4 сек. и после каждого
раздражения давали отдых 1—4 сек. Так поступали при тетаническом
УТОМЛЕНИЕ
217
раздражении. Когда же применяли отдельные индукционные удары, то
меняли только промежуток между ударами от одной секунды до
нескольких. Этими исследованиями было обнаружено, что при некоторых
малых паузах первоначальная высота сокращений удерживалась только
очень короткое время — несколько минут, после чего падала до
некоторого уровня, на котором удерживалась в течение 3—5 часов. Чем короче
была пауза отдыха, тем скорее происходило это начальное падение-
сокращений (Голлигер, 2); чем больше была нагрузка мышц, тем ниже
падала высота сокращений (Вичи* 23). Температура тоже оказывала
влияние: при некоторой высокой температуре деятельность мышцы1
без утомления наблюдалась при более коротких паузах отдыха, чем
при низкой температуре (Аккерман, 14). Аналогичные опыты были
поставлены Е. Закарая в лаборатории Беритова на собаках
(32). Он раздражал двигательный нерв тетанически беспрерывно или
с 'Короткими паузами: 1—2 сек. тетаническое раздражение и 1—2 сек.
пауза. Раздражение продолжалось 10—13 часов, но сокращение мышц,
ослабевало только в самом начале — в течение первого часа
раздражения; затем оно не менялось. В какой мере это изменение
механического эффекта при нормальном кровообращении действительно
отражает утомление возбудимой системы, трудно сказать, так как в этих
опытах не производилось изучения токов возбуждения.
Праптгтум
Наблюдение изменений одиночного сокращения при утомлении. Приготовляют
спинномозговой препарат лягушки. Обнажают икроножную мышцу и соединяют с
миографом- Седалищный нерв перерезают и раздражают через 1—2 сек.
автоматическим вращением кимографа, как в опыте с одиночным сокращением.
Одиночные сокращения записывают все время или же по временам с пропуском
значительного числа их, как на рис. 112. В этом, опыте обращают внимание на измене-
кие длительности, высоты и скрытого периода сокращения.
Три стадии утомления. Спинномозговой препарат готовится, как в предыдущем
опыте. Раздражается седалищный нерв через каждые 17г—2 сек. метрономом,
который введен в первичную цепь. Одиночное сокращение икроножной мышцы
регистрируют на медленно вращающемся кимографе. Как в стадии нарастания
одиночных сокращений, так и в последующей стадии их падения пробуют учащать раз»
дражение в течение нескольких секунд. Этим путем устанавливают вое три стадии
утомления. Обращают внимание «а появление и исчезновение контр актурного
сокращения. Когда отдельные индукционные удары совсем перестанут вызывать
сокращение, пробуют тетаническое раздражение. Последнее дает еще больший эффект.
Наблюдение зависимости утомления от груза. Опыт на спинномозговом
препарате. Раздражают тетанически периферический отрезок седалищного нерва.
Записывается икроножная мышца іна медленно вращающемся кимографе. Сначала мышцу
нагружают малым грузом в 10—20 г. Раздражение продолжается, пока высота
сокращения не упадет наполовину. После отдыха в 5—10 минут наігружают мышцу
грузом в 100—200 г и снова раздражают до падения мышечной кривой наполовину.
Опять дают отдых в 5—10 минут, меняют нагрузку на прежнюю в 10—20 г и опять
раздражают. В первом и третьем опытах продолжительность сокращения мышцы
до половинной высоты будет больше, чем во втором. Во избежание влияния одного-
опыта на другой можно поставить опыт с малым грузом на икроножной мышце
одной стороны, а с большим — на другой.
218
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Изменение возбудимости мышцы после полного утомления непрямого
раздражения. Опыт на спинномозговом препарате лілгушки. Длительное тетаническое
раздражение седалищного сплетения. Записывается m. sartorius. Бременами
определяют порог раздражения самой мышцы. Для этой цели мышцу соединяют с другим
иядукгорием, причем электроды накладывают на проксимальный конец. В период,
когда тетаничеокое раздражение перестанет вызывать эффект, прямое раздражение
будет вызывать эффект, но возбудимость мышцы будет несколько понижена.
Литература
1. В ег it off J., Z. Biol., 81, 119, 1924.
2. H о 1 1 i g e r M., Z. Biol., 77, 261, 1922.
3. Schmid E., Z. Biol., 77, 271, 1922.
4. Beritoff J., Erg. Physiol., 23, 33, 1924.
5. Kennosure Matsuoka, Pfl. Arch., 202, 573, 1924.
<6. Ranke J., Arch. Anat. Physiol., S. 422, 1863; S. 320, 1864.
7. Burridge W., J. Physiol., 41, 285, 1910; 42, 359, 1911.
8. Weichardt W., Munch, med. Wschr., ряд работ 1904—1906; Ueber Er-
mudungsstoffe, Stuttgart, 1912.
9. Lee Fr. S. a. Aronovitch. В., Am. J. Physiol., 69, 92, Ш, 1924.
10. Schoenlein X., Pfl. Arch., 45, 134, 1889.
11. В er it off J., Z. Biol., 62, 125, 1913.
12. R о 1 I ett A., Pfl. Arch., 71, 209, 1898; Zbl. Physiol., 13, 721, 1900; 14, 435, 1900.
13. L.ohmann A., Pfl. Arch,, 91, 338, 1902; 92, 387, 1902.
14. А с k e r m a n n M., Z. Biol., 78, 331, 1923.
15. Beritoff J.,'Pfl. Arch., 198, 590, 1923.
16. Br u eke Er. Th. v., Pfl. Arch., 124, 215, 1908.
17. Meyer h off O., Pfl. Arch., 182, 232, 1920; 182, 284, 1920; 188, 114, 1921.
H8. К о b а у a s h i Mag., Z. BioL, 81, 263, 1924.
19. Burgi Fr., Z. Biol., 81, 253, 1924.
20. Schwenker C, Pfl. Arch., 157, 371, 1914.
;21. Введенский'H. E., Телеф. исслед. над электр. явл. в мышц, и нервном
аппарате, СПб., 1884.
22. Беритов И. и Яшвили Д., Чвени Мецниереба, № 3, 153, 1923.
.23. Witschi Er., Z. Biol., 85, 1, 1927.
.24. EmbdenGu. LangeH., Klin. Wschr., 3. Jahrg., S. 123, 1924; Embden G.
u. Adler, Z. Physiol. Chem., 118, I, 1922.
25. Lee F. S., Am. J. Physiol., 20, 170, 1907.
26. Beritoff J., Z. Biol., 85, 521, 1927.
27. Beritoff J., Z. Biol., 85, 1, 1927.
28. F u 11 о n J. F., Muscular contraction and the reflex control of movement.
Baltimore a. London, 1926.
29. Beritoff J., Pfl. Arch., 205, 458, 1924.
30. Beritoff J., Z. Biol., 82, 126, 1924.
31. Беритов И. и Волынский А., О количестве гликогена и процессе
возбуждения в период абсолютного утомления вырезанной мышцы. (Не опубликовано).
32. 3 а к а р а я Е., Физиол. журн. СССР, 16, 675, 1933.
33. Hill A. V., Chemical wave transmission in nerve. Cambridge Univers. Press, 1932.
34. Walidow I., Pfl. Arch., 235, 147, 1934.
35. R i e s s e r O. u. R i с h i M i u r a, Pfl. Arch., 232, 513, 539, 1933.
36. P a r k i n s о n I. L., J. Physiol., 78, 106, 1933.
37. Палладии А. В., Физиол. журн. СССР, 19, 277, 1935.
38. Nagaya Т., Pfl. Arch., 221, 720, 1929.
39. Ге дева ни Д., Физиол. журн. СССР, 19, 1052, 1935.
40. М е у е г h о f О., Die chemischen Vorgange im Muskel, Berlin, 1930.
УТОМЛЕНИЕ
219
41. Fenn W. О. и сотр., Cold Spr. Harb. Symp. Quant. Biol., IV, 212, 1936; Physiol.
Rev., 16, 450, 1936; Am. J. Physiol., 121, 595, 1938.
42. Серков Ф. H. Учен. зап. Каз. унив., вып. 3, стр. 2, 1934.*]
43. Topsoe-Jensen А., цитир. noBuchthal u. Lindhard, The Physiol.
of str. muscle fibre, 1939.
44. В и с h t h a 1 Fr., J. Physiol., 90, 33P, 1937.
45. В о p о h цо в Д. С, Сб. докл. VI Всес. физиол. съезда, стр. 75, 1937.
46. Н і 11 А. а. К и р а 1 о w Р., Ргос. Roy. Soc. Lond., ser. В, 105, 313, 1929.
47. Б e л и ц e p Б. А., Успехи совр. биол., 6, 235, 1937.
48. К о м е т и а н и П., Об уменьшении содержания гликогена и фосфагена в
вырезанной мышце в связи с утомлением. 1938. (Не опубликовано.)
49. В а г с г о f t Н. а. Е. М і 1 1 е n J. L., J. Physiol., 97, 17, 1939.
50. Цкипуридзе Л., Сообщ. Ак. Наук Груз. ССР, 2, № 5, 1941.
Тлае а X
РАБОТА МЫШЦ'Ы
Общее понятие. Механическая работа мышцы так же, как работа
в механике, измеряется произведением поднятого груза на величину
укорочения мышцы. Но в отношении работы мышца обладает
замечательным свойством: при
постепенном
увеличении груза укорочение
мышцы от одного и
того же раздражения
сначала растет, а
затем уменьшается.
Между грузом и
укорочением мышцы нет
пропорционального
отношения: как нарастание со-
кращения,такиуменыне-
ние его менее
значительно, чем
соответствующее изменение нагрузки.
Сообразно с этим
работа мышцы в связи с нагрузкой меняется согласно следующему
закону: с прибавлением нагрузки работа сначала растет, потом постепенна
падает, а при некоторой большой нагрузке равняется нулю (Вебер, 2).
Отсюда следует, что для каждой мышцы максимальная работа
получается для некоторых средних нагрузок. Это называется законом
средних нагрузок. На рис. 115 дана диаграмма Уоллера,
выражающая соотношение величины груза, высоты укорочения и
произведенной работы для икроножной мышцы лягушки. Горизонтальные линии
первого столбца диаграммы обозначают величину нагрузки в граммах,
линии второго столбца — высоту сокращения в миллиметрах, а третьего
столбца —- работу мышцы в граммиллиметрах. Кроме того, мы
приводим также рисунок 116, где хорошо видно повышение кривой
одиночного сокращения, пока нагрузка икроножной мышцы лягушки меняется
в малых пределах, от 10 до 50 г.
9
О
50
100
150
200
250
300
35д
UO0
450
500
550 L
мм
г/мм
6
7
7
6
5
3
2.5
2
15
1
О
350
700
900
1000
WOO
900
875
800
675
500
О
Рис. 115. Диаграмма Уоллера, выражающая
соотношение нагрузки в граммах, сокращения в миллиметрах и
работы в граммиллиметрах икроножной мышцы лягушки.
РАБОТА МЫШЦЫ
221
Конечно, в основе изменения мышечной работы в связи с
увеличением нагрузки должны лежать определенные физиологические
процессы. Прежде всего под івлиянием растяжения, произведенного
нагрузкой, происходит характерное изменение возбудимости; с
повышением нагрузки возбудимость нарастает, а при очень больших нагруз-
Рнс. 116. Увеличение одиночного сокращения мышцы с увеличением груза.
М. gastrocnemius лягушки. Цифры на кривых отмечают нагрузку в граммах (Пембрей).
ках, когда уже происходит повреждение мышечной ткани, уменьшается
(Фик, 1; Гоциридзе, 21).
Это увеличение возбудимости, вероятно, связано с отмечаемым
резким возрастанием концентрации водородных ионов в імышце при ее
растягивании. Это изменение исчезает сейчас же после разгрузки.
Поэтому изменение концентрации водородных ионов нельзя считать
следствием химических реакций. Наиболее вероятным объяснением
считается изменение физико-химических свойств липопротеидов при
изменении напряжения мышцы (Маргариа, 31). Водородные ионы, как
известно, действуют на возбудимую систему, повышая возбудимость
ее (Бете, 32).
В связи же с повышением возбудимости должно находиться
усиление процесса возбуждения. И действительно, судя по
биоэлектрическим токам возбуждения, с увеличением растяжения імышцы под
влиянием груза, или, вернее, с нарастанием начального напряжения мышцы,
процесс возбуждения усиливается. При дальнейшем же увеличении
он перестает нарастать (Фультон, 3). В мышцах с параллельными
волокнами, как га. sartorius лягушки, ток возбуждения усиливается при
растяжении почти на 100% (Форбс и сотр., 43). В мышцах с пери-
•стым строением, как m. gastrocnemius лягушки, усиление может
произойти на 1000% (Шефер, 39). Но в икроножной мышце такое
большое усиление обусловлено не столько повышением процесса
возбуждения в отдельных мышечных волокнах, сколько изменением их
расположения, вследствие чего улучшаются условия отведения токов
возбуждения.
Функциональным изменением мышцы, наступающим в связи с
растяжением, можно объяснить нарастание как укорочения мышцы, так
222
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
и выполняемой ею работы. Но тот большой прирост работы, который
при этом происходит, не может зависеть от усиления процесса
возбуждения. По всей вероятности, он обусловлен изменениями физико-
химического характера в самих миофибриллях, т. е. увеличением того
фактора, который является, по Г и л л у, первопричиной механической
реакции (30). Как уже указывалось выше, растяжение до
определенного предела ведет к таким изменениям, которые обусловливают
повышение сократительной способности мышцы. При увеличении
нагрузки выше определенного предела работа мышцы, наоборот,
уменьшается. Это явление нельзя объяснить понижением возбудимости от
повреждения мышцы, так как оно происходит раньше возможного
повреждения, когда процесс возбуждения еще является оптимальным.
Падение работоспособности при больших грузах хорошо объясняется
с точки зрения уже известного нам факта уменьшения
сократительного .процесса при больших растяжениях мышцы, т. е. больших
начальных напряжениях. Именно, с увеличением растяжения мышцы
сократительный процесс сначала растет, а затем уменьшается и при
некоторых больших растяжениях сходит на-нет. Процесс возбуждения
в сильно растянутых мышцах возникает и распространяется
беспрепятственно и без ослабления, но он (не в состоянии вызвать
сокращение (Беритов, 22).
По новейшим исследованиям, от величины нагрузки зависит также
образование молочной и фосфорной кислот. С повышением нагрузки
образование молочной и фосфорной кислот при работе повышается.
Максимум достигается при некоторой средней нагрузке. С дальнейшим
увеличением нагрузки образование той и другой кислоты в связи с
работой параллельно уменьшается. Минимум образования кислот
соответствует нагрузке, близкой к абсолютной силе, когда мышца не в
состоянии поднять груз при сокращении (Риссер, 29; Нагайа, 34).
Итак, начальное повышение работы в связи с повышением нагрузки,,
повидимому, объясняется прежде всего повышением возбудимости и
увеличением процесса возбуждения, а также увеличением
сократительного процесса. Уменьшение же работы при увеличении нагрузки выше-
определенного предела обусловливается прежде всего ослаблением-
сократительного процесса под влиянием сильного растяжения мышцъи
Но безусловно заметную роль играет также то обстоятельство, чта
в первом случае расщепление фосфагена и гликогена нарастает, а во
втором случае, наоборот, оно уменьшается.
Мышца и машина. Мышца обладает чрезвычайно своеобразным
двигательным механизмом, так как она, во-первых, от частой работы
становится сильнее и выносливее и, во-вторых, утилизирует горючий
материал с наибольшим полезным эффектом. Ни в одной машине
химическая энергия не переходит целиком в работу, всегда значительная
часть ее превращается в тепловую. Чем больше потенциальной
химической энергии переходит в работу, тем больше полезный эффект
машины. В наиболее совершенной паровой машине из горючего материала
РАБОТА МЫШЦЫ
223-
утилизируется только 15% энергии, в бензинном двигателе 25%»
а в двигателе Дизеля 37—41|% (Реах, 4). Многочисленными же
исследованиями в лаборатории Ц у н ц а (6) на человеке были получены
такие результаты: человек <при восхождении на гору потребляет из
всей химической энергии питательного материала максимум 37*>/ov
в среднем ЗГ%, на механическую работу. По исследованиям Гилла
(7), полезный эффект мышцы следует признать еще выше. Если
нагрузка мала и раздражение слабо, полезный эффект равняется 50%;
при увеличении нагрузки и усилении раздражения полезный эффект
уменьшается до 35—30%. Гил л высчитывает полезный эффект мышцы
на основании выделяемого мышцей тепла.
Таким образом, полезный эффект мышцы при некоторых условиях
выше, чем наиболее совершенной машины. Но этот факт не должен
казаться сколько-нибудь странным, так как в мышце тепло не играет
такой роли, как в машине, где химическая энергия сначала должна
превратиться в тепло, а затем уже тепло производит работу.
Сократительные элементы мышцы — миофибрилли работают по принципу,
хемодинамической машины, в которой химическая энергия прямо
переходит в механическую.
Величина мышечной работы. 1. Величина мышечной работы прежде
всего зависит от количества мышечных волокон. Чем толще мышца,,
тем больший груз она может поднять. При прочих равных условиях
мышца производит тем больше работы, чем больше поперечное
сечение (Вебер, 2).
Мышечные волокна располагаются отнюдь не всегда параллельно»
продольной оси (мышцы. В большинстве случаев у .высших
позвоночных мышечные волокна располагаются под некоторым углом к
продольной оси мышцы (рис. 33). В таких мышцах перистого строения:
длина мышечных іволокон значительно меньше всей длины мышцы::
в некоторых случаях она равняется одной трети и даже одной четверти
всей длины. Но зато общее количество мышечных волокон очень
велико. В этих мышцах величина укорочения сравнительно мала, а
грузоподъемность ючень велика. Следовательно, в перистых мышцах
работоспособность не может быть пропорциональна поперечному сечению
(Сеченов, 8; Цхакая, 9).
2. Работа мыЩцы с параллельными волокнами зависит также от
длины ее. Работоспособность мышцы при всех прочих равных
условиях тем больше, чем длиннее мышца. Эта закономерность имеет
место только в том случае, если мышцы не очень длинны, и потому
возбуждение почти сразу охватывает мышечные юолокна по всей длине
их. В длинных мышцах, с соответственно длинными мышечными
волокнами, как m. sartorius лягушки, между работой и длиной мышцы
нет прямо пропорциональной зависимости. С удлинением мышцы
работоспособность растет, но значительно в меньшей степени. Причина
заключается в том, что возбуждение и сокращение всегда возникают
сперва в одних участках; в это время невозбужденные участки, растя-
224
ОВЩЛЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
л
'0-\
•'# г<?
гиваясь, теряют в значительной мере сократительную способность
(Беритов, 22).
В перистых \мышцах работа мышцы зависит уже не от длины всей
мышцы,, а от длины наибольшей массы мышечных пучков и затем от
углов, образуемых ими с центральным сухожилием. При сокращении
углы эти увеличиваются в несколько раз: мышечные пучки принимают
еще более косое положение. Вследствие этого мышечные пучки,
сокращаясь, укорачивают імышцу не на ту величину, на какую они
передвигаются сами, а на 15—30%
больше (Беритов, 19) (рис. 117).
3. Работа мышцы зависит также
от числа иннервационных
областей в мышце или, вернее, от
распределения в ней двигательных
нервных окончаний. Доказано, что
мышечное напряжение, а значит, и
работа мышцы возрастают, когда
мышечные волокна возбуждаются
сразу в двух местах. Когда
двигательные нервные окончания
распределены по всей длине, мышца не мо-
Рис. 117, Схематическое изображение жет растянуться ни в одном своем
m. vastus int лягушки в лохое (1)ъ во участке в такой мере, чтобы это по-
время сокращения (II).
/ влияло на его сократительную
способность; поэтому
работоспособность такой мышцы должна быть
больше, чем в тех мышцах, где
двигательные окончания
сосредоточены в одной области
(Беритов, 22).
4. Наконец, имеет большое
значение местоположение главной ин-
нервационной области. Если она
-лежит ближе к дистальному суставу, то механическое действие мышцы
на этот сустав будет много сильнее, чем на проксимальный, ибо волна
напряжения, распространяясь от сокращенной иннерБационной области,
достигнет дистального сустава с меньшим ослаблением, чем
проксимального (Беритов, 22).
Абсолютная сила мышцы. Каждая мышца при определенном
функциональном состоянии .способна поднять груз не больше определенной
величины. То максимальное напряжение или та «максимальная сила,
какую ыышца развивает при сокращения в случае, когда ода уже не
в состоянии поднять груз, называется абсолютной силой.
Абсолютную силу можно просто выразить величиной того наименьшего
груза, который мышцы не в состоянии поднять (Карвало и Вейсс, 41).
По более старому определению, мерой абсолютной силы является та
Дорзо-вентральный разрез: Р — проксимальный
конец; D — дистальный. Непрерывной линией
обозначены сухожилия; Z, Z , Z'' —центральное
^сухожилие, идущее с проксимального конца; А,
А', А /, А" —наружное сухожилие, идущее с
дистального конца. Прерывистой линией
обозначены мышечные пучки. Цифры обозначают дли-
™у сухожилий и мышечных пучков в
миллиметрах, а те цифры, которые стоят в углах,
образуемых мышечными пучками при центральном
сухожилии, указывают на размер
соответствующих углов в градусах. Как показывает рисунок,
при сокращении мышца утолщается и углы
растут, вследствие чего вся мышца укорачивается
гв большей мере (8 мм), чем наиболее длинные
мышечные пучки {5.6—6 мм).
РАБОТА МЫШЦЫ
225
тяжесть, которая растягивает сокращенную мышцу до начальной
длины, т. е. до длины покойной ненагруженной мышцы (Вебер, 2).
Это определение абсолютной силы лучше івсего производить так:
тяжесть привешивается к покойной імышце, но так, чтобы не
растягивать імышцу. С этой целью тяжесть или 'миограф заранее подпирается.
Затем подвергают мышцу, максимальному тетаническому
раздражению. Находят тяжесть, которую мышца едва приподнимает. Она будет
выражать абсолютную силу. Абсолютная сила, измеренная по
последнему способу, всегда значительно меньше находимой по первому
способу. Оба способа имеют существенные недостатки. В первом 'случае
мышца растягивается, что меняет физиологические свойства мышечных
волокон. Кроме того, при некоторой большой нагрузке происходят
разрывы в мышечных волокнах (Карвало и Вейсс, 41). При втором
способе определения сократительная способность мышцы понижена
благодаря отсутствию предварительного растяжения.
Абсолютная сила икроножной мышцы лягушки средней величины
равняется в среднем 1000 г. Абсолютная сила мышц с параллельными
волокнами тем больше, чем больше их диаметр и длина. В перистых
мышцах она зависит главным образом от количества мышечных
волокон и угла прикрепления мышечных волокон к продольному
сухожилию (Вейсс, 10а; Цхакая, 9).
Для определения абсолютной силы нужно пользоваться тетаниче-
ским раздражением. При одиночном сокращении мышца не сможет
развить максимальное напряжение. Пользуясь тетаническим
раздражением, нужно брать раздражение такой силы, чтобы возбудились все
мышечные волокна данной мышцы; раздражение должно быть такой
частоты, чтобы каждый последующий стимул приходился после
окончания рефракторных фаз. В противном случае мы будем иметь дело
с пессимальньим сокращением, которое не способно развить
максимальную силу.
Влияние упражнения на мышечную работу. Частая напряженная
работа влияет определенным образом на анатомо-ги(стологичес!кое
строение мышцы. Это выражается макроскопически в утолщении
мышцы, микроскопически же в значительном увеличении диаметра
мышечных волокон. У человека в наиболее интенсивно работающих мышцах
мышечные волокна толще. Так, например, в верхней конечности
сильнее всего работают сгибатели локтевого сустава, и в них мышечные
волокна толще, чем в разгибателях, а в нижней конечности они толще
в разгибателях, которые здесь работают сильнее сгибателей. Тоньше
всего мышечные волокна в глазных и других наименее работающих
мышцах (Кохаши, 38). В связи с упражнением происходит
увеличение веса мышцы. Увеличивается при этом совершенно
пропорционально и абсолютная сократительная сила мышцы. Вследствие этого
сократительная сила на единицу веса мышцы остается неизменной. Точно
так же при атрофии денервированной мышцы в первое время
сократительная сила мышцы падает параллельно <с уменьшением веса мыш-
15 И. С. Беритов.
-26
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ды. При этом двоякая преломляемость мышцы остается без
изменения. Последняя начинает падать при дальнейшей атрофии мышцы, и
тогда только начинает уменьшаться сократительная сила на единицу
веса (Фишер, 47).
При гистологическом исследовании тренированной мышцы было
найдено, что увеличение імышцы происходит не за счет появления,
новых волокон, а за счет разрастания существующих. При этом не
меняется ни длина волокон, ни количество ядер, ни количество фиб-
риллей или их толщина. Увеличивается в них только саркоплазма
(Морпурго, 25; Зиберт, 49).
Эти и другие наблюдения единогласно показывают, что в
тренированных мышцах сократительная способность повышается, не вследствие
увеличения сократительных свойств миофибриллей или же увеличения
их толщины, а в результате разрастания саркоплазмы и изменения 'ее
физико-химических -свойств (Швальбе и Майеда, 24, 26; Морпурго, 25;
Фишер, 47; Штейнгаус, 48).
Изменение физико-химических свойств мышцы ів ісвязи с
упражнением было изучено впервые Эмбдеиом и сотрудниками (23). Они
исследовали сухой остаток тренированной мышцы и затем химические
процессы в кашице из тренированной мышцы. Они пришли к выводу,
что в тренированной мышце должно быть больше таких ионов,
которые благоприятствуют сократительному процессу. Они же показали,,
что в тренированной мышце происходит увеличение содержания
гликогена — гексодомонофосфата, играющего определенную роль
промежуточного продукта на пути расщепления гликогена до молочной
кислоты (23). Кроме того, тренированная мышца при работе теряет
меньше гликогена, выделяет меньше сахара, образует меньше
молочной кислоты, чем нетренированная (Феррари, 40). Отсюда следует,
что в тренированной мышце работа производится много эконрмнеег
чем в ^тренированных.
Очень подробное исследование изменения биохимических
процессов под влиянием тренировки произведено П а л л а д и н ы м и
его сотрудниками (33). Они установили, что при тренировке в мышцах
увеличивается количество таких веществ, как креатин, фосфорная
кислота, карнозин, которые играют важную роль в динамике
мышечной деятельности. Эти вещества обусловливают возможность работы
мышцы, а потому с увеличением их повышается работоспособность
мышцы. Кроме того, они же установили, что в результате тренировки
изменяются окислительно-восстановительные системы мышцы. Именно
в связи с тренировкой происходит увеличение количества каталазы и
ускорение обесцвечивания метиленовой синьки. Это свидетельствует,,
что в тренированных мышцах окислительные процессы протекают
быстрее и энергичнее, чем в мышцах нетренированных.
Наконец, ими же было установлено, что тренировка влияет на
синтетические процессы; например, она ускоряет синтез сложных
фосфорных соединений мышцы. Вероятно, в связи с этим находится то
РАБОТА МЫШЦЫ
227
явление, что в тренированных мышцах во время утомительной работы
отсутствует накопление молочной кислоты или она накопляется
в меньшей мере, чем в нетренированных мышцах.
Итак, упражнение, тренировка увеличивают работоспособность
мышцы вследствие определенного морфологического и
физико-химического изменения саркоплазмы.
Работа при трудовых движениях. При определении
работоспособности человека следует принимать ,во внимание не только /количество
работы в данный короткий промежуток времени или при одном
трудовом движении, но и количество движений, которые могут повториться
в назначенное рабочее время. Средняя величина дневной работы
мужчины при 8-часовом рабочем дне равняется 6.3—10 (максимум 10.5—-
11.0) кг/м в 1 сек. Значит, максимальный дневной полезный эффект
равняется 288 000 кг/м. Работа лошади »в 1 сек. равняется 75 кг/м
(лошадиная сила).
Работоспособность зависит прежде всего от величины самой
мышечной деятельности. Когда мышцы сокращаются максимально, они
утомляются скорее, чем в случае субмаксимальных сокращений. При
субмаксимальной деятельности мышцы могут работать
продолжительное время без утомления. Поэтому количество работы больше при
субмаксимальных сокращениях.
Количество работы зависит еще от величины груза. При некоторой
большой нагрузке утомление мышц происходит очень быстро, а потому
при такой нагрузке общий полезный эффект не может быть
оптимальным. Оптимальное количество груза, при котором імышца выполняет
максимальную работу, зависит от мощности мышцы. Работа мышц.
максимальна, когда подобран соответствующий груз, не слишком
большой и не слишком малый.
Далее, количество работы зависит от характера чередования
рабочих движений с отдыхом. Работа всегда протекает ритмически, но
характер ритма, т. е. оптимальное число движений в одну минуту или в
час, подвергается индивидуальным колебаниям. Он зависит как от
нервной системы рабочего, так и от тренировки, т. е. от индивидуального3
опыта. В условиях коллективного фабрично-заводского труда
вырабатывается некоторый средний ритм работы, который легко дается
наибольшему количеству рабочих. Путем тренировки этот ритм может
быть значительно повышен. Итак, количество работы мышц в
значительной мере зависит и от ритма работы.
Наконец, работоспособность мышцы зависит от питания и вообще
от всех жизненных условий. Основным условием для оптимальной
ежедневной работы мышц является ежедневное питание организма
соответствующим количеством энергетического материала, каким являются
в первую очередь углеводы. Человеческий организм обладает некоторым
запасом углеводов в самих мышцах и в печени; поэтому он может
работать некоторое время после прекращения питания. Но за счет
этих запасов оптимальная работа человека не может быть поддержана
15*
228 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
даже в течение одного дня. Следовательно, оптимальная
работоспособность мышц требует ежедневного снабжения организма питательными
веществаіми.
, Биохимия мышечной деятельности. Выше было указано, что
сокращение імышц сопровождается превращениями ряда сложных веществ,
играющих в мышце энергетическую роль, как, например, гликогена,
фоофагена и аденилпирофосфата. В процессе обмена энергетического
материала образуются киісло'ты: адениловая, фосфорная, молочная,
а также основания: аммиак, креатин, холин и т. д. Сейчас мы опишем
сложный путь расщепления и новообразования означенных веществ.
Основным материалом для сократительного процесса нужно
считать гликоген. По новейшим представлениям самое близкое участие
в превращениях гликогена принимают: аденозинтрифосфорная кисло'та
(аденилпирофосфат), фосфокреатин (фосфаген), трифосфопиридин-
нуклео'тид (коэнзим), а также ряд других веществ, играющих роль
или ферментов, или гормонов, или же активаторов биохимических
реакций (Парнас, 45).
Колоссальная работа, которая была проведена главным образом
в лабораториях Эмбдена, Мейергофа, Парнаса,
Вар-бурга, а также ряда других исследователей, дала возможность выяснить
в деталях .те биохимические реакции, которые сопровождают
сократительный пріоцесс (мышц.
В первую очередь гликоген претерпевает расщепление в результате
так называемого фосфоролиза. Сущность этой реакции заключается
в том, что при воздействии фосфорной кислоты из гликогена
получается г?ксозомюнофоофат. Последний в присутствии аден'озинтри-
фосфорной кислоты превращается в гексозодифосфат:
Гексозомонофосфат (+] аденозинтрифосфорная кислота =
гексозодифосфат !+ адениловая кислота.
К этой реакции сейчас же присоединяется другая,
восстановительная реакция:
2 фосфокреатин + адениловая кислота = 2 креатин -f- аденозино-
трифосфорная кислота.
Затем происходит расщепление гексозодифосфата с образованием
фосфоглицериновой и фосфопировиноградной кислот.
В этом начальном процессе еще нет молочной кислоты. За этой
начальной стадией следует другая стадия с образованием /молочной
кислоты:
Гексозодифосфат + 2 пировиноградная кислота = 2 фосфопирови-
ноградная кислота г+! 2 молочная кислота.
Гексозодифосфат \+' адениловая кислота = аденозинотрифосфор-
пая кислота ¦+ 2 молочная кислота.
Гексозодифосфат Ц-\ 2 креатин = 2 молочная кислота и 2
фосфокреатин.
В этой стадии параллельно с образованием молочной кислоты
происходит ресинтез других веществ, участвующих в реакции.
РАБОТА МЫШЦЫ
229
В заключительной стадии окончательно устраняются те йарушения,
которые были вызваны в предыдущих стадиях! Например, фосфопи-
ровиноградная кислота реагирует с креатином, и образуются фосфо-
креатин и пировиноградная кислота. Пировиноградная затем идет ни
образование аденилпирофосфата и молочной кислоты.
По новейшим исследованиям ведущими ферментами в
биохимических процессах мышцы являются основные белки мышцы. Так,
белок миозин обладает свойством расщеплять аденйлпирофосфат, т/ е.
является аденилтрифосфатазой. Белок миоген, главная составная часть
саркоплазмы, служит ферментом, под влиянием которого шестиугле-
родная цепь глюкозы расщепляется на две трехуглеродные молекулы,
"из которых потом образуется молочная кислота. Значит, белок миоген
отожествляется с ферментом альдолазой. Один из воднорастворимых
белков мышцы служит лактикодегидразой, ферментом, окисляющим
мелочную кислоту до пировиноградной (Энгельгардт, 50).
Вышеописанный сложнейший ферментативный процесс мышцы
может протекать одинаково хорошо в анаэробных и в аэробных
условиях, т. е. не требует присутствия кислорода. Так, расщепление
аденилпирофосфата происходит Гидролитически, а восстановление его
нормально осуществляется главным образом за счет энергии распада
фосфагена, который при этом расщепляется на креатин и фосфорную
кислоту. Аденйлпирофосфат при своем расщеплении в присутствии
гликогена ведет к образованию из гликогена гексозодифосфата, из
которого образуется молочная кислота. Этим самым он стимулирует
расщепле'ние гликогена до молочной кислоты.
Расщепление фосфагена является второй основной реакцией. Оно
также происходит в результате процесса, не требующего кислорода.
Его восстановление из креатина и фосфорной кислоты также может
происходить без кислорода, но только частично. Полное
восстановление его требует кислорода. При синтезе фосфагена, как и при
синтезе аденилпирофосфата, поглощается энергия. Необходимая энергия
доставляется в первую очередь расщеплением аденилпирофосфата, но
главным образом расщеплением гликогена . и окислением молочной
кислоты. Но есть основание предположить, что восстановление
фосфагена происходит частично за счёт энергии окислительного процесса,
который усиливается под влиянием креатина, выделяемого «из
фосфагена. Этот окислительный процесс не связан с окислением молочной
кислоты, ибо такое же усиленное потребление кислорода наблюдается
при отравлении мышцы моноиодуксусной кислотой, когда образование
молочной кислоты отсутствует (Белицер, 37). Повидимому, в этом
случае мы имеем дело с усиленным дыханием самой возбудимой
системы, т. е. с усилением основного 'биологического процесса. В пользу
этого говорит между прочим и очень небольшой размер избыточного
«фосфагенного» дыхания. Количество поглощенного кислорода не
превышает 100 мм3 на грамм мышцы за час. Таким образом,
усиленный распад фосфагена ведет к усиленному дыханию возбудимой си-
230 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
стемы, а благодаря этому ускоряется обратный процесс — ресинтез
фосфагена. Конечно, этот источник энергии приобретает большое
значение в таких аэробных условиях, когда ігач?му-либо образование
(молочной кислоты отсутствует. Восстановление фосфагена в аэробных
условиях при наличии молочной кислоты должно происходить
главным .'образом за счет энергии окисления молочной кислоты.
Расщепление гликогена до молочной кислоты также является
результатом ферментативного процесса. Оно может протекать в
бескислородной среде. Правда, во время работы імышца выделяет С02
даже в том случае, если сокращение изюлнрованной мышцы
происходит в бескислородной среде. Но при этом условии происходит не
новообразование С02 в результате какого-либо окислительного
процесса, а освобождение С02 из карбоксильных групп воздействием
специфичных ферментов, а также из карбонатов от действия на них
вновь возникших фосфорной и молочной кислот. В бескислородной
среде не происходит новообразования угольной кислоты во время
сокращения (Флетчер, 11).
Все химические процессы мышечной деятельности распадаются на
две фазы: 1) рабочая фаза, включающая процессы расщепления
аденилпирофосфата, фосфагена и гликогена с образованием аденило-
вой, фосфорной и молочной кислот; эта фаза протекает без участия
кислорода и потому называется анаэробной; 2) восстановительная фаза,
заключающаяся в процессах восстановления из названных кислот
аденилпирофосфата, фосфагена и гликогена. Эта фаза в полной мере
протекает при наличии кислорода и потому называется аэробной, или
окислительной, фазой (Мейергоф, 15; Гилл, 14; Лундсгаард, 27).
Однако в нормальных условиях работа мышцы безусловно зависит
от «окислительных процессов. Утомление мышцы в бескислородной
среде происходит гораздо раньше, чем в атмосфере воздуха или
чистого кислорода. Если, например, в іатмосфере азота утомить мышцу
абсолютно и з^т?м перенести в атмосферу кислорода, то мышце
возвращается способность к работе. При этом было замечено, что
в мышце исчезает определенное количество молочной кислоты и
выделяется такое количество углекислоты, которое точно соответствует
количеству поглощенного мышцей кислорода (Флетчер и Гоп-
кинс, 12). Но даже в нормальном организме кислород потребляется
мышцей главным образом не во время работы, а после '(Верзар, 13).
Химический процесс образования и ресиитеза молочной кислоты впервые был
изучен на мышечной кашице. Кашица сама собой с течением времени образует
молочную кислоту, как нормальная мышца в период покоя. Бели к такой кашице
прибавить гликоген или тростниковый сахар, это не повлияет на образование
молочной кислоты. Наоборот, прибавление гексозофосфорной кислоты увеличивает
образование 'молочной кислоты (Эмбден, 16). Отсюда было сделано заключение, что
молочная кислота происходит из гликогена не непосредственно, а через определенное
промежуточное вещество, которое содержит фосфорную кислоту. По мнению
Э м б д е н а, это должно быть такое же соединение углевода с фосфорной кислотой,
какое впервые было открыто Ивановым и Гарднером при спиртовом бро-
РАБОТА МЫШЦЫ
231
жении сахара как переходное к спирту промежуточное вещество. Эмбден
действительно получил из мышечного экстракта такое вещество и назвал его лакта-
ци до геном. Впоследствии обнаружили его и в нормальной мышце. Быстро
сокращающиеся белые мышцы дают его в большем количестве» чем красные,
медленно сокращающиеся. После интенсивной работы количество лактацидогена в
мышцах уменьшается и одновременно увеличивается количество фосфорной кислоты
б моче (Эмбден, 23). Лактацидоген был идентифицирован как гексозодифосфат, и
в данный момент ему приписывается роль исходного вещества при распаде
углеводов (Эмбден и сотр., 46).
Восстановление гликогена происходит за счет окисления продуктов
анаэробного расщепления гликогена. Принимают, что субстратом
окисления служит главным образом молочная кислота. Как
указывалось, вырезанная мышца в бескислородной среде утомляется очень
быстро. При ?том образуется как раз столько молочной кислоты,
сколько исчезает гликогена. Возникающая в мышце молочная кислота
остается без изменения. Она исчезает только в том случае, если
перенести мышцу в среду кислорода. По исследованиям Мейергофа
(17), исчезновение молочной кислоты не обусловливается окислением
всей молочной кислоты. Окисляется только небольшая часть ее, от
7в До ги всего количества молочной кислоты. Количество образуемой
при этом углекислоты и потребленного кислорода как раз
соответствует XU—1/е молочной кислоты. Остальная молочная кислота
превращается обратно в гликоген. Это восстановление происходит за
счет той энергии, которая выделяется при окислении части молочной
кислоты. По первоначальному мнению Эмбден а, Мейергофа
и др., и это восстановление должно было происходить посредством
лактацидогена. В последнее время Мейергоф (35) и др.
сомневаются в значении лактацидогена в процессе расщепления и
восстановления гликогена. Предполагают, что пр*и сокращениях весь фосфат
образуется из фосфагена. Правда, лактацидоген, или гексозодифосфат,
в мышце имеется, но он возникает как побочный продукт, и его
динамика не подтверждает приписываемую ему роль. Например, при
длительных и частых раздражениях количество гексозодифосфата растет,
а не уменьшается. Его изменения не имеют ничего общего с
начальным теплообразованием мышцы, хотя оно может быть связано с
замедленным теплообразованием после тетануса (Гилл, 30).
Итак, в фазе восстановления молочная кислота не уничтожается
целиком путем окисления. Окисляется лишь небольшая часть ее для
освобождения той энергии, которая необходима для обратного
превращения молбчной кислоты в гликоген. Вследствие этого последний
вновь используется в целях работы. Еще П а с т е р у было известно,
что брожение сахара в аэробных условиях дает меньше выхода спирта,
чем в анаэробных. Мейергоф выяснил сущность этого явления.
И здесь, в дрожжевой клетке, в аэробных условиях происходит ресин-
тез гликогена за счет окисления спирта. Этот ресинтез углеводов
принято называть реакцией Пастер — Мейергофа.
По исследованиям Мейергофа, во время покоя в мышце про-
232
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
текают такие же процессы, как во время работы. В бескислородной
среде происходит лишь расщепление гликогена до молочной кислоты
под влиянием мышечных ферментов. В кислородной же среде проис:
ходит, на ряду с этим, полное окисление части імолочной кислоты и
превращение остальной,' большей части ее, обратно в гликоген.
Количество окисленной молочной кислоты зависит от условий обмена
веществ. Чем лучше эти условия, тем меньше імолочной кислоты
окисляется, следовательно, тем больше молочной кислоты переходит в
гликоген,. Когда мышца сильно утомлена, то окисляется больше одной
четверти молочной кислоты. Многие яды и травматическое
повреждение, также сильно увеличивают окисляемую часть молочной кислоты.
Разумеется, работа мышцы протекает тем экономнее, чем меньшая
часть імолочной кислоты окисляется. Когда окисляется V4, по расчетам
Мейергофа (15, 17), полезный эффект гликогена составляет 40%,
в наиболее же благоприятных условиях в неповрежденной мышце,
когда окисляется Ve імолочной кислоты, полезный эффект Достигает
50—60%.
По исследованиям Змбдена (16, 23), окисленная часть
молочной кислоты достигает в редких случаях- у6 всей молочной кислоты.
Она сильно колеблется от этой величины до Vi2 части.
Также по опытам Маргариа и его сотрудников (44) известно,
чііо.при отдыхе человека после напряженной работы отношение
исчезнувшей молочной кислоты к окисленной равно 10. Следовательно,
полезный эффект гликогена, по этим исследованиям, должен быть
признан много выше — до 90%.
Фаза восстановления, по Мейергофу, продолжается много
часов после сокращения; после утомления тетаническим раздражением
она продолжается, например, 10—15 часов. Даже после
кратковременного тетануса восстановление гликогена достигает максимума через
40 сек. Наиболее короткий период восстановления наблюдается после
одиночного сокращения. Именно, если .при 28—30° мышца лягушки
раздражается отдельными индукционными ударами через каждые две
секунды, утомления не происходит (Аккерман, 18).
Скорость восстановления сильно зависит от температуры: после
утомлен'ия одиночными индукционными ударами при комнатной
температуре восстановление продолжается 20 часов, а при 7° — 30 часов.
При 0° восстановление замедляется гораздо больше (Мейергоф, 17).
Биохимические процессы в мышцах человека во время работы
протекают более или менее сходно с таковыми в изолированных
мышцах животных только в определенных условиях: если работа
человека не длительна — около 10 минут — и достаточно напряжена.
При более продолжительной и менее напряженной работе
биохимические процессы принимают иной характер, вероятно, вследствие
регулирующего влияния центральной нервной системы (Маргариа и сотр., 44).
Нужно отметить, что полный отрыв фазы восстановления от фазы
рабочей, фазы окисления от фа'зы анаэробной, является чисто искус-
РАБОТА МЫШЦЫ
233
ственным представлением, продуктом лабораторного опыта. В
действительности оба процесса должны протекать параллельно как ^во
время работы, так и в покое. Если только мышца снабжается
кислородом, окисление молочной кислоты, а такж<е восстановление в
гликоген не могут не происходить и во ъремя работы. Точно так же распад
адениліпирофосфата и фосфагена и их обратное восстановление
должны происходить в течение всей мышечной работы. Поэтому нельзя
говорить, что фаза восстановления начинается .после рабочей фазы,
после*ра'-боты. Эти противоположные процессы расщепления и
восстановления протекают все время и в каждый отрезок времени как
единый неразрывный процесс. Едиными же являются эти процессы не
только потому, что они протекают одновременно, а и потому, что
каждый из этих процессов хотя и противоположен другому, но при
совместном течении они поддерживают друг друга. Расщепление аденил-
пирофосфата и -фосфагена и образование фосфорной кислоты
благоприятствуют расщеплению гликогена до молочной кислоты; процесс
сжигания молочной кислоты, наоборот, ведет к новообразованию
гликогена и фосфагена.
Если работа не интенсивна и промежутки отдыха імежду
отдельными моментами работы достаточно велики, то окисление и
восстановление молочной кислоты смогут и-тти в уровень с образованием ее
из гликогена. Точно так же распад и восстановление аденилпирофос-
фата и фосфагена могут итти с одинаковой интенсивностью. В таком
случае в мышце не произойдет накопления ни той, ни другой кислоты.
Так, очевидно, и было в определенных опытах в лаборатории Ашер а,,
а также в опыте Беритова, где мышца тетанически сокращалась,
часами без утомления. Только в случае более или менее интенсивной
работы окисление и восстановление не успевают за образованием
фосфорной и молочной кислот, что вызывает накопление последних
вовремя работы. Окисление же и восстановление накопившихся кислот
происходят постепенно после сокращения, лосле работы, растягиваясь
на более или менее- значительный промежуток времени.
Биохимические процессы в нервных и безнервных участках мышцы.
Как указывалось выше, в нервных участках сократительная способ-
кость больше, чем в безнервных. Это выражается в том, что в нервных
участках процент укорочения мышцы, сравнительно с нормальной
длиной, и величина напряжения на единицу длины больше, чем в
безнервных участках; скрытый же период сокращения в нервных участках
меньше, чем в последних. Соответственно с этими функциональными
особенностями все химические процессы, ведущие к сокращению, в
нервных участках протекают гораздо интенсивнее, чем в безнервных. Такг
Кометиани (42) установил, что содержание фосфагена и аденилпи-
рофосфата в нервных и безнервных участках совершенно одинаково, но
и их распад и обратный синтез в нервных участках происходят
интенсивнее, чем в безнервных. Точно так же распад гликогена до молочной
кислоты и его окислительный ресинтез в нервных участках протекают
234 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
«интенсивнее, чем в безнервных. Этим объясняется то характерное
явление, что при аэробных условиях молочная кислота в безнервных
участках накапливается больше, чем в нервных. В анаэробных же
условиях, когда окислительный процесс выключен, конечно, происходит
.наоборот: молочная кислота накапливается больше в нервных участках,
вследствие чего яри длительном раздражении контрактурное
последействие выражено сильнее в нервном участке, чем в безнервном, как на
это уже указывалось выше.
Вообще окислительно-восстановительный процесс в нервных
участках протекает интенсивнее, чем в безнервных. Соответственно с
этим восстановление не только гликогена, но и фосфорных соединений
при аэробных условиях в нервных участках протекает интенсивнее.
Для мышц, отравленных моноиодацетатом, работа которых
выполняется за счет распада фосфорных соединений, также было доказано, что
возбуждение вызывает более интенсивный распад фосфорных
соединений там, где больше нервных окончаний. Вследствие этого в
анаэробных условиях нервные участки сокращаются сильнее безнервных, но
утомляются быстрее их. Таким образом, было установлено, что в
нервных участках мышцы, находящихся все время под непосредственным
¦влиянием нервных импульсов, скорость и интенсивность химических,
превращений, ведущих к сокращению, значительно выше, чем в
безнервных участках. Этим 'обусловливается более высокая
совратительная способность и более быстрое наступление сокращения в нервных
участках сравнительно с безнервными.
Практикум
Наблюдение высоты сокращения и работы в зависимости от груза. В
спинномозговом препарате лягушки перерезывается седалищный нерв. Икроножная
мышца соединяется с миографом «репкой нитью. Записывается только высота
сокращений в івиде (вертикальных линий. Сокращение мышцы вызывают максимальным
тетаническим раздражением седалищного нерва. При каждой нагрузке раздражение
продолжается 5—10 сек. После каждого раздражения дают отдых в 2—3 минуты.
Сначала записывают сокращение мышцы без нагрузки. Затем рукой поворачивают
цилиндр кимографа на полсантиметра и снова записывают при нагрузке в 10 г;
в том же порядке записывают кривые при нагрузках в 20, 50, 100, 200, 500 г. На
основании записанных линий легко высчитать истинное укорочение мышцы по
уравнению х=— )Где /—длина вертикальной линии, т — длина плеча, к которому
привязана нить от мышцы, а п—длина пишущего плеча рычага. Определив укорочение
и помножив на груз, получим работу. На основании результатов построить
диаграмму Уоллера.
Определение общего количества работы при разных нагрузках. Спинномозговой
препарат лягушки. Сначала записывают на кимографе сокращение икроножной
мышцы одной стороны, нагрузив ее сравнительно небольшим грузом в 100—200 г,
и раздражают седалищный нерв тетанизирующими индукционными ударами через
1—2 сек. Это делается с помощью метронома. Метроном включается во вторичную
цепь таким образом, <что, когда контакт подвижного плеча опускается в ртуть,
индукционные удары раздражают нерв; когда же контакт выходит из ртути,
раздражение прекращается. Раздражение продолжается, пока в силу утомления мышцы
РАБОТА МЫШЦЫ
2-35
высота сокращения не упадет более или менее значительно, например до
половины. Затем производят аналогичный опыт на икроножной мышце другой стороны.
Здесь мышцу нагружают сравнительно большим грузом >в 300—500 г и раздражают
не-рв столько же времени, как в перпом случае. Определяют общее количество
произведенной работы © том и другом случаях. Груз нужно подбирать сообразно
с толщиной мышцы. Чем она толще, тем больший нужен груз для той и другой
сторон. Если грузы оказались хорошо подобранными, то количество работы прн
меньшем грузе 'будет больше, чем при большем.
Проще ©сего определение общего количества работы производится таким
образом. Измеряют величину поднятия груза © начале и конце утомления и определяют
среднее поднятие груза. Затем подсчитывают, сколько раз мышца поднимала груз.
Помножив величину среднего поднятия в миллиметрах на количество поднятий и
затем на величину груза в граммах, мы получим общее количество работы в грам-
миллиметрах.
Литера ту р а
1. F і с k A., Beitrage z. vgl. Physiol., стр. 53, Braunschweig, 1863.
2. Weber E, Wagners Handworterb. d. Physiol., 3. Abt., 11, стр. 81, 184G.
3. Fulton S. R, Proc. Roy. Soc, B, 97, 406, 1925.
4. Reach R, Fortschritte naturw. Forschung, 10, 97, 1914.
5. Zunz N., Pfl. Arch., 68, 191, 1897.
6. Zunz NM Hagemann O., Lehmann C, Frentzel J., Landwirtsch.
Jahresber., 18, 1889; Erganzungsband, III, 27, 1898.
7. H i II A. W., J. Physiol., 46, 435, 1914.
8. Сеченов И., Очерки рабочих движений, 1903.
9. ZchakaiaNV., Pfl. Arch., 209, 753, 1925.
10. Цхакая M., Русск. физиол. журн., 8, 273, 1926.
10а. Weiss G., Traite de physique biologique, 1, 90—99, 1903.
П. F 1 e t с h e r W. M., J. Physiol., 22, 410, 1898; 28, N 5, 1902.
12. F 1 e t с h e r u. Hopkins F. G., J. Physiol., 35, 247, 1907.
13. Verzar R, J. Physiol., 44, 243, 1912; Erg. Physiol., 15, 1, 1916.
14. H i 1 i A. V., Erg. Physiol., 22, 299, 1923.
15. Meyer hoff O., Erg. Physiol., 22, 299, 1923.
16. Embden G. u. L a q u er R, Z. physiol. Chem., 93, 60, 1915; ibid., 93, 94, 1915
98, 681, 1916.
17. Meyer h of f O., Pfl. Arch., 182, 232, 1920; 182, 284, 1920; 188, 114, 1921.
18. Acker ma nn M., Z. Biol., 78, 331, 1923.
19. В er it off J., Pfl. Arch., 209, 763, 1925.
20. H i 11 A. V., Muscular activity, Baltimore, 1925. (Есть русский перевод.)
21. Г о ц и p и д з е. А., Мед. биол. журн., вып. 6, 1930.
22. В er it off J.,Z. Biol., 87, 513, 1928.
23. Embden G., Handb. norm. path. Physiol., 8, H. 1, 369, 1925.
24. S с h w a I b e G. u. Mayeda R., Z. Biol., 27, 482, 1890.
25. Мог pur go В., Virchow's Arch. path. Anat. Physiol., 150, 522, 1897.
26. Mayeda R., Ueber d. Kaliberverhaltnisse d. querstreift. Muskelfasern. Inaug.
Diss., Strassburg., 1890.
27. Lundsgaard, Bm:h. Z, 217, 162, 193Э; 220, 1, 8, 1930; 227, 51, 1930; 230, 10
1931; 233, 222, 1931.
28. Мейергоф О., Успехи совр. биол., т. 1, вып. 1—2, стр. 40, 1932.
29. RiesserO. u. Riich ipl., Pfl. Arch., 232, 513, 1933.
30. H i 1 1 A. V., Adventures in biophysics, 1931. Русский перевод: «Эпизоды из области
биофизики», Биомедгиз, 1935.
31. М а г g а г і a R., J. Physiol., 74, 496, 1934.
236
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
32. В е th e A., Pfl. Arch., 163, 147, 1916.
33. Палладии А. В., Физиол. журн. СССР, 19, 277, 1935. Докл. наVI Всес. фи-
зиол. съезде, 1937.
34. N a gay а Т., Pfl. Arch., 221, 720, 1929.
35. Meyer h off O., Die chemischen Vorgange im Muskel, Berlin, 1930.
36. Фердман Д. Сб. докл. VI Всес. съезда физиол., стр. 432, 1937.
37. Б е л и ц е р В. А., Биохимия, 2, 332, 1937. Сб. докл. VI Всес. физиол. съезда, стр.
449, 1937.
38. К о h a s h i G., Okajikas Fol. anat. jap., 15, 775, 1937.
39. Schaefer H., Pfl. Arch., 237, 339, 1936.
40. Ferrari R., Pfl. Arch., 230, 639, 1932.
41. С a r v a I 1 о u. WeissG., Pfl. Arch., 75, 591, 1899.
42. Кометиани П., Сб. докл. VI Всес. физиол. съезда, стр. 436, 1937.
43. F о г b e.s, Hay a. Hopkins, Am. J. Physiol., 65, 300, 1923.
44. Margaria R., Edward H. T. a. D i 1 1 D. В., Am. J. Physiol., 106, 689„
1933.
45. Parnas J. К., Физиол. журн. СССР, 24, 277, 1938.
46. Em b den, Deutike u. Kraft, KHn. Wochenschr., S. 213, 1933.
47. Fischer E., Am. J. Physiol., 131, 156, 1940.
48. Steinhaus.A. H., Physiol. Rev., 13, 103, 1928.
49. S i e b e r t W. W., Z. f. klin. Med., 109, 350, 1928.
50. Энгелыардт В. А., Усп. совр. биол., 14, №'2, 1941.
Глава XI
ТЕПЛОВОЙ ЭФФЕКТ МЫШЦЫ
Общее понятие. Физико-химические процессы, протекающие в
мышце, сопровождаются определенным тепловым эффектом. Некоторые из
этих процессов являются экзотермическими, т. е. сопровождаются
выделением тепла. К таким процессам относятся прежде всего окисление
молочной кислоты до угольной кислоты, расщепление гликогена до
молочной кислоты, аденилпирофосфата и фосфагена до фосфорной
кислоты, растворение этих кислот в мышечной плазме, нейтрализация
их и другие процессы. К эндотермическим процессам относятся
восстановление молочной кислоты в гликоген, ресинтез фосфагена и
аденилпирофосфата.
Уже давно стало известно, что мышцы при сокращении выделяют
тепло (Гейденгайн, 1; Фик, 2). Отсюда и было сделано заключение
3 н г е л ым а н о м, что імышечная работа, подобно работе паровой
машины, обусловлена переходом теоіла ів механическую энергию: что
химическая энергия сначала превращается в тепловую, а уже потом
тепловая энергия переходит в механическую. Но <в настоящее время
точно установлено, что выделяемая во время мышечной деятельности
теплота не играет никакой роли в происхождении мышечного
сокращения. Теплота образуется как во время сокращения, во время
работы, так и после сокращения, и даже больше после сокращения, чем
во время него.
Детальное исследование течения теплообразования было
произведено Гйллоім (3, 4) и его учениками. Он точно установил
количество тепла, выделяемого мышцей при сокращении. Он, например,
показал, что при одиночном сокращении изолированной мышцы
температура последней повышается на 0.003°, а при тетаническом — на 0.008°.
Он же впервые заметил, что во время сокращения при коротких тета-
нусах теплота выделяется сразу в большом количестве, а после
сокращения медленно в течение многих минут и даже часа. Первая быстрая
часть теплообразования называется начальной фазой, а последующее
медленное теплообразование — восстановительной фазой. Это деление
на фазы вызвано тем обстоятельством, что источники их происхожде-
238
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
кия совершенно различны. В настоящее время работами Г ил л а и его
сотрудников точно установлен весь ход теплообразования в імышце
яри разных условиях сокращения.
Начальная фаза теплообразования. Начальная фаза отличается от
восстановительной тем, что она главным образом определяется
химическими процессами, которые ведут к сокращению. Эта теплота
выделяется в анаэробных условиях. При одиночных сокращениях
начальная фаза теплообразования (Происходит не целиком во время
сокращения, т. е. в рабочей фазе, но в некоторой імере и в период
расслабления. 30—35'% всего начального теплообразования приходится на этот
период расслабления (рис. 118). В анаэробных условиях тепло обра-
>
^)
>г
-1-
ft
fc
к
Сг
<to
5
•о
Б
<J
с>
8-
^
<0
05
0U
0.2
02
01
•
/ >
*¦* \
¦***"
тт^-ть.^
^^ «
•ч. V
^^
* ——>
t^^^a
0 0.2 0A 0,6 QS 10 I?
Секучдьі
Рис. 118. Ход теплообразования при анаэробных условиях во время начальной фазы.
Сплошная линия изображает результаты 'анализа скорости 'теплообразования при Одиночных
сокращениях m. sartorius лягушки при 0°. Пунктирная линия показывает напряжение при изометрическом
сокращении (Гилл).
зуется и после сокращения. Эта часть анаэробного теплообразования
называется запаздывающим анаэробным
теплообразованием. На долю ее при коротких тетанусах приходится около 20'%'
всей анаэробной теплоты. Она продолжается всего несколько минут
(Гилл, 9).
В атмосфере кислорода в начальной фазе тепла образуется то же
количество, как и в отсутствие его. Всего на начальную анаэробную
фазу теплообразования приходится, по Гилл у, 40% всей теплоты,
выделяемой мышцей при наличии кислорода. При этом считается, что-
в анаэробной фазе имеют место исключительно экзотермические
процессы.
Происхождение начальной ф'азы теплообразования. Пока еще точно-
не установлено, какие именно химические экзотермические процессы
участвуют в происхождении начальной фазы теплообразования.
Известно, что в этой фазе не 'Принимает заметного участия расщепление
гликогена до молочной кислоты, ибо в мышцах, отравленных моноиод-
уксусной кислотой, начальное теплообразование в анаэробных уело-
тепловой эффект мышцы
2Ш
виях протекает почти так же, как в нормальной мышце (Гартри, 12)..
Поэтому начальная теплота должна быть обязана своим
происхождением прежде всего тем химическим превращениям, какие претерпевают
в мышце аденшширофосфат и фосфаген, а затем обмену веществ
возбудимой системы імышцы. Следует отметить, что распад фосфагена,
возможно, также не играет существенной роли в начальной
теплопродукции, ибо сколько тепла выделяется яри его распаде, столько
поглощается сейчас же для восстановления аденилпирофосфата.
Одновременно, конечно, происходят и другие экзотермические реакции, как,.
например, расщепление гликогена на промежуточные ступени. Но и
они, возможно, не играют существенной роли в происхождении
начальной теплоты, ибо одновременно происходят и другие реакции
эндотермического характера (Каттел, 16).
В происхождении начальной теплоты должны участвовать, імежду
прочим, эластические и пластические особенности імышцы. Как об этом:
будет сказано ниже, при пассивном укорочении растянутой -мышцы
всегда происходит выделение тепла, то же происходит при
растяжении. Мы ,знаем, что при сокращении, когда одна часть імышцы
сокращается, другая удлиняется; когда анизотропное вещество миофибрил-
ли сокращается, изотропное удлиняется. Надо полагать поэтому, что*
определенная доля начального теплообразования обусловлена
эластическими и пластическими изменениями внутри імышцы (Каттел, 16).
Как указывалось выше, в начальную теплоту входит 20%
замедленного теплообразования, которое возникает после сокращения..
Гил л относит его за счет замедленного образования молочной кис-
лоты. По Мейергофу, (при коротких тетанусах почти 30—50%
молочной кислоты образуется после сокращения. Но замедленное тепло
меньше того, которое должно соответствовать этой молочной кислоте,
что объясняется происходящим в это время восстановлением
фосфагена. Замедленное теплообразование можно считать балансом между
экзотермической реакцией образования [полочной кислоты и
эндотермической реакцией восстановления фосфагена (Гилл, 10).
Одно івремя Г'ллл и Мей ер го ф связывали начальное теплообразование
з мышце главным образом с образованием молочной кислоты.
Гилл определил, что при образовании в мышце молочной кислоты в
анаэробных условиях выделяется 370 кал. тепла. Отсюда, по его вычислению, только частьг
235 кал., следует отнести на расщепление гликогена до молочной кислоты. Гил я
исходил из теплоты сгорания молочной кислоты и гликогена. Одному грамму
молочной кислоты соответствует 0.9 г гликогена. При сжигании 1 г гликогена
развивается, по С л е т е р у, 3836 кал., а 1 г молочной кислоты по Бенедикту и
Мейергофу — 3601 кал. Разница составляет 235 кал. Но фактически в мышце
на 1 г молочной кислоты выделяется 370 кал. Разницу >в 135 кал. Гилл объяснял
нейтрализацией молочной кислоты щелочными соединениями белка.
Мей е pro ф определял 385 кал. для анаэробной фазы на 1 г молочной
кислоты. Но на образование молочной кислоты из гликогена он относил всего 180 кал..
Такое расхождение получалось потому, что Мейергоф исходит не из данных
Слетера о теплоте сгорания гликогена, а из собственных данных. Он же
определил эту теплоту равной 3782 кал. Остальное количество теплоты из начальной
240
общ^я физиология мышечной системы
¦фазы Мейергоф относил: 110 кал. на реакцию молочной кислоты с щелочными
белками, бифосфатами и бикарбонатами, 14 кал. на растзорение молочной кислоты
в мышечной плазме и, наконец, 80—40 кал. ,на изменение мышечного белка после
.воздействия .на него молочной кислоты.
Эти расчеты Гилла и Мейеіргофа были произведены в то івремя, когда
.еще не была известна роль аденилпирофосфата и фосфагена в мышечной
деятельности. Как указывалось выше, во время мышечной деятельности происходит
также расщепление большого количества аденилпирофосфата и фосфагена.
А этот процесс выделяет значительное количество тепла. Именно, при
¦ образовании 1 г фосфорной кислоты выделяется 120 кал. {Тилл, 6). Отсюда следует,
что в происхождении теплоты в анаэробных условиях значительную роль должно
.играть расщепление аденилпирофосфата 'и фосфагена.
Восстановительная фаза теплообразования. При сокращении
вырезанной мышцы при свободном доступе кислорода образуется гораздо
больше тепла, чем в анаэробных условиях, причем наибольшая часть
тепла образуется в фазе восстановления, т. е. в фазе окисления
молочной кислоты до угольной кислоты. Как уже указывалось выше, в
этот период происходит сжигание от У4 до Уб молочной кислоты и
восстановление остальной части ее в гликоген. В общем, по Г и л л у,
60'% всей теплоты приходится на фазу восстановления. Именно в этой
фазе теплообразование превышает в \х/% іраза все начальное тепло. Но
.здесь 25% приходится на ту запаздывающую теплоту, которая
наблюдается и в анаэробных условиях. Следовательно, теплообразование в
.связи с окислением превосходит начальную теплоту в 1.25 раза. При
сжигании Уб г гликогена во время восстановительного периода в
атмосфере кислорода фактически выделяется імышцей по Г и л л у 379 кал.
Между тем при лабораторном сжигании У5 г гликогена образуется
757 кал. Следовательно, разность 757 — 379 = 378 кал. ^поглощается»
мышцей во время восстановления. Очевидно, это та самая теплота,
которая нужна для превращения Уб г молочной кислоты в гликоген. Но,
как указывалось, в одних случаях исчезает в мышце не У5, а *Д или
Уб. Соответственно и в фазе восстановления выделяется большее или
меньшее количество тепла.
Но и эти расчеты Гилла и Мейергофа не вполне
соответствуют действительности. Известно, что при тетаническом сокращении
часть імолочной кислоты образуется после сокращения. В связи с этим
часть тепла, образуемого после сокращения, следует отнести за счет
расщепления гликогена, а не за счет сжигания молочной кислоты.
Далее, тепло поглощается не только при восстановлении молочной
кислоты в гликоген, но и при восстановлении фосфагена и
аденилпирофосфата. '
На основании сказанного можно было бы себе представить, что во
время нормальной мышечной деятельности не происходит никакой
другой теплопродукции, кроме той, которая характерна для рабочей
фазы, т. е. кроме экзотермических реакций распада гликогена,
аденилпирофосфата и фосфагена. Между тем не может быть сомнения в
том, что при нормальных условиях мышечной работы, при ее длитель-
ТЕПЛОВОЙ ЭФФЕКТ МЫШЦЫ
241
ном протекании в продолжение многих минут и часов, сгорание
молочной кислоты и восстановление гликогена, адецилпирофосфата и
фосфагена происходят одновременно. Теплообразование начинается
начальной фазой в связи с образованием первых порций фосфорной и
молочной кислот; но сейчас же вслед за ней должна начаться фаза
восстановления как аденилпирофосфата, фосфагена, так и гликогена.
И то обстоятельство, что при нормальном кровообращении даже
длительная утомительная работа не сопровождается уменьшением глико-
во
¦о
«о
50
йО
30
20
0
О
чі^г:—
/
/,..
-.?,....•
с
¦,,.....
с
А
^9
О
8
Ш
ft
12 3 4 5 6?
Рис. 119-1. Примерный ход теплообразования во время работы и после
нее в присутствии кислорода.
Ордината обозначает высоту теплообразования, а абсцисса — время в минутах. В течение
первых четырех минут производится работа —73 одиночных изометрических вздрагивания.
Во время раздражения сильное теплообразование — кривая А. По прекращении
раздражения кривая А сразу падает до некоторого уровня, а затем продолжает
падать очень медленно в течение многих минут. Прерывистая кривая С дает
вычисленный ход начального теплообразования. Кривая В — высота изометрических сокращений.
Излишек А по отношению к С выражает теплопродукцию во время восстановления.
Этот излишек имеет место уже во время раздражения, т. е. в рабочей фазе (Гилл).
гена и накоплением молочной кислоты, прямо указывает на наличие
восстановительных процессов с неизбежной теплопродукцией
окисления еще во время работы (рис. 119—1). Характерно, что даже в
анаэробных условиях тепловой эффект начальной фазы не ограничивается
только экзотермическими реакциями. Восстановление также имеет
место в этой фазе, а именно восстанавливается аденилпирофосфат за
счет энергии, выделенной при расщеплении фосфагена, и фосфаген —
за счет энергии образования молочной кислоты. Это видно из того,
что лри исключении образования (молочной кислоты, как это бывает
при моноиодуксусном отравлении, распад фосфагена значительно
более силен, чем обычно. Очевидно, в нормальных условиях фосфаген
восстанавливается обратно за счет энергии образования молочной
кислоты и ее сжигания.
Теплообразование при пассивном растяжении и укорочении мышцы.
Как указывалось выше, мышца представляет собой упруго-вязкую сн-
16 и. С. Беритов
242
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
стему. Отсюда следует, что в мышце как упругом теле должно
происходить при растяжении выделение тепла, а при последующем
укорочении — поглощение тепла. Но как вязкое тело мышца должна
выделять тепло как при растяжении, так и при укорочении. В первом
случае імы имеем дело с обратимым процессом, во втором — с
необратимым. Экспериментальное исследование теплообразования в мышце
при ее пассивном растяжении и укорочении дает именно такую кар-
тину выделения тепла. Как показано на рисунке 119-НЛ при рас-
ч
' \ А
^1 'v4y
Рис. 119-И.
А — ход теплообразования при пассивном растяжении живой мышцы m. sartorius
при постоянной нагрузке в 155 г; В — ход теплообразований при пассивном
укорочении после удаления растягивающей силы — в 155 г; С — ход
теплопродукции мертвой мышцы (m. sartorius) при многократном растяжении и
укорочении: 163 г «нагружаются» и «снимаются» с промежутком в 1 сек. (Гартри и Гилл).
тяжении выделяется тепло, которое представляет сочетание тепла^.
выделяемого упругими и вязкими элементами мышцы. При укорочении,.
после удаления растягивающей силы, сначала теплопродукция падает
вследствие поглощения тепла эластическими элементами в момент их
укорочения. Затем она усиливается, что обусловливается выделением*:
тепла вязкими элементами (рис. 119-115). Такие же изменения
теплопродукции происходят при растяжении мертвых мышц (рис. 119-И С)..
Характерно, что пассивное растяжение сопряжено с выделением
тепла не при всех условиях. Мышца нагревается от растяжения
только при некоторых імалых начальных нагрузках. Так, в одном опыте
m. sartorius при начальной нагрузке до 20 г от растяжения нагревается;
а при увеличении нагрузки до 100 г та же мышца от растяжения
охлаждается, т. е. происходит извращение теплового эффекта. Выделение
тепла от растяжения при малых начальных нагрузках находится в
связи с изменением мышечных элементов. А извращение при высоких
нагрузках находится в зависимости от изменения
соединительнотканных элементов, растяжение которых сопровождается поглощением
тепла. При этом поглощается больше тепла, чем его образуется от
мышечных элементов, поэтому последнее теплообразование
маскируется (Фенг, 13).
Теплопродукция при растяжении живой мышцы в значительной
мере зависит также от функционального состояния: чем лучше функ-
ТЕПЛОВОЙ ЭФФЕКТ МЫШЦЫ
243
циональное состояние, тем больше выделяется тепла при растяжении.
Так, например, в атмосфере кислорода выделяется вдвое больше
тепла, чем в азоте. Это сопряжено с большим потреблением кислорода
при растяжении в атмосфере кислорода. На теплопродукцию при
растяжении сильно влияет и предыдущая активность: так, например,,
если в атмосфере азота вызвать 50 одиночных вздрагиваний, то это»
уменьшает теплопродукцию при растяжении вдвое или даже еще
больше. После восстановления імышцы в кислороде теплопродукция при.
растяжении вновь достигает первоначальной величины (Фенг, 14).
После отравления моноиодуксусной кислотой теплопродукция прет
растяжении такова же, как в норме. Но если длительным
раздражением вызвать утомление мышцы, сопряженное с расходованием
запасов фосфагена, то растяжение не производит усиленного выделения
тепла. Характерно, что после этого и перевод імышцы в атмосферу
кислорода не благоприятствует выделению тепла при растяжении. На
основании этих наблюдений можно считать вероятным, что высокое
содержание фосфагена необходимо для повышенной теплопродукции
при растяжении и что растяжение каким-то образом стимулирует
экзотермические химические процессы (Фенг, 14). Очевидно,
теплопродукция при растяжении по своему происхождению представляет очень
сложное явление: в нем участвуют не только выделение тепла эла*
стическими и пластическими элементами мышечных волокон, не
только поглощение тепла соединительнотканными элементами мышц, но
и экзотермические химические процессы с повышенныім потреблением
кислорода и расщеплением фосфагена.
Теплообразование в разных условиях мышечного сокращения.
Теплообразование при изометрическом и изотоническом сокращениях не
одинаково. При изотоническом сокращении тепла выделяется больше,
чем при изометрическом . сокращении той же продолжительности
(Фенн, 7; Азума, 8). При изотоническом сокращении
теплообразование на 41% больше, чем при изометрическом.
Этот избыток тепла хорошо проявляется также, если изометрически
сокращенную мышцу сразу освободить и дать ей возможность
свободно сократиться. С момента освобождения теплопродукция повышается,
и притом тем больше, чем больше укорачивается мышца (Гилл, 18).
Во время расслабления изометрически сокращенной мышцы
теплообразование сейчас же падает почти на 15%. Это обстоятельство
указывает, что при расслаблении происходит прекращение или замедление
тех химических энергетических процессов, которые обусловливают
теплопродукцию. Это уменьшение теплопродукции более значительно, чем
то увеличение ее, которое должно иметь место в связи с пассивным
растяжением эластических и пластических элементов.
Количество выделяемой теплоты при изометрическом сокращении
зависит от степени развиваемого при этом мышечного напряжения:
чем больше напряжения, тем больше образуется тепла. А при
изотоническом сокращении теплообразование зависит от величины подай-
is*
244
ОБ'ЦАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
маемого груза. Чем больше груз, тем больше образуется теплоты
(Фенн, Гилл и Гартри). Если же мышца сокращается совершенно без
нагрузки, т. е. без напряжения, и не поднимает даже своего
собственного веса, например, если во время опыта мышца находится в
растворе Рингера, она развивает особенно мало тепла, даже меньше, чем
при изометрическом сокращении. Соответственно и потребление
кислорода минимально, когда мышца сокращается без напряжения. В
этом случае при тетаническом раздражении продолжительностью в
15 сек. потребление кислорода увеличивается сравнительно с покоем
всего на 10—30%, в то время как лри изометрическом тетаническом
сокращении той же продолжительности потребление кислорода
нарастает на 100—150% и выше (Асмуссен, 15).
Скорость увеличения тепла и общей энергии (тепло Ч-
механическая работа) при изотоническом сокращении зависит от поднимаемой
тяжести. Чем больше груз, тем медленнее сокращается мышца и тем
меньше скорость (выделения энергии. Эта особенность не зависит от
вискозных свойств мышц, от условий внутреннего трения. Это хорошо
видно, между прочим, из того факта, что если тетанически
сокращенную мышцу удлинить путем приложения тяжести, которая
превосходит величину напряжения, развиваемого при ее изометрическом
сокращении, то теплопродукция сокращенной мышцы падает, и вообще
энергия, выделяемая мышцей, уменьшается. Следовательно,
энергетические отношения при удлинении обратны тем, что наблюдаются при
укорочении в результате сокращения. Поэтому предполагается, что
химические процессы, обусловливающие выделение энергии,
регулируются действующей извне силой: большая сила обусловливает более
медленные энергетические реакции, вследствие чего и скорость
выделения энергии является пониженной (Гилл, 18).
Во время контрактур, вызванных химическими веществами, как ве-
ратрин и кофеин, тепло выделяется за время сокращения
пропорционально напряжению мышцы. Всякое замедление механического
эффекта, по Гиллу (3), сопровождается замедленным- теплообразованием,
так же как и замедленным выделением молочной кислоты.
Отношение общего теплообразования (включая окислительное) к
начальному, т. е. анаэробному, теплообразованию значительно
меняется в зависимости от условий сокращения. Это отношение при
одиночных сокращениях в среднем равняется 1.69. Во время серии одиночных
сокращений (30—40 в 1 мин.) оно доходит до 2.4. При тетанических
сокращениях оно достигает 2.5. Таким образом, можно сказать, что
с увеличением мышечной деятельности на одно и то же начальное
анаэробное теплообразование приходится все больше и больше
теплообразования в окислительной фазе, т. е. мышечная работа протекает
все экономнее (Бюньяр, 11).
При повторных раздражениях отдельными индукционными ударами
или короткими тетанусами теплообразование происходит следующим
образом. При работе на свежих мышцах, когда в отношении сокра-
ТЕПЛОВОЙ ЭФФЕКТ МЫШЦЫ
245
щения наблюдается явление лестницы, начальное теплообразование
или не меняется вообще, или же оно вначале івелико, а потам падает
до некоторого постоянного уровня. Таким образом, общая сумма
химических процессов, обусловливающих теплообразование, не является
первопричиной явления лестницы (Фишер, 13).
Методика исследования теплопродукции. Для исследования мышечной
теплопродукции применяют легкую термоэлектрическую батарею, состоящую из ста пар
спаев, образуемых тонкими проволоками из константана (сплав) и железа. Каждая
пара дает 52 микровольта при разнице температуры в 1 градус. Применением
подходящего гальванометра можно регистрировать тысячные доли градуса. Спаивание
пар первоначально производилось посредством обычного способа спаивания
проволоки. В виду тонкости проволок это было сопряжено
с затруднениями. Недавно Гамильтон Уильсон
изобрел более легкий способ изготовления термоэлектрической
батареи: сплошная константановая проволока на
отдельных участках гальванопластически покрывается серебром.
Каждая пара переходов между посеребренными отрезками
проволоки и непосеребренными составляет
термоэлектрическую пару. В последнее время Г и л л и его ученики поль- /?
зуются такой термоэлектрической батареей, на ряду с
изготовляемой путем сплавления разнородных металлов.
Термоэлектрическая проволока натягивается на
эбонитовую раму в виде спирали, имеющей несколько десятков Рис* 120. Термоэлек-
оборотов. В каждом полном обороте четыре спая, из кото- ТРИЧ ская оатарея со
^ к ^ спаянными элемен -
рых два располагаются внутри рамки так, чтобы поверхность , тами
мышцы соприкасалась со всеми внутренними спаями. Сюда ^Тешіые)> спаи в середи-
же внутрь рамки подводятся электроды для прямого раз- не: холодные— по бокам;
/ 1Г1~Ч ™. _, ЕЕ—платиновые, раздра-
дражения (рис. 120). Термоэлектрическая батарея с мышцей жающие электроды; GG—
помещается в специальной камере, и применяются все меры, провода к гальванометру
чтобы наружная температура батареи оставалась все время
постоянной.
При сокращении согреваются внутренние спаи, и, благодаря существованию
снаружи спаев с неизменной температурой, в замкнутой цепи этой спирали
устанавливается ток. Если концы спирали отвести в гальванометр, последний даст
отклонение. Г и л л пользуется для этой цели очень чувствительным гальванометром
Доунинга или Зернике.
Несмотря на довольно простой принцип, термоэлектрическое исследование
связано с очень большими трудностями, так как для получения точного представления
о ходе теплообразования нужно учесть целый ряд факторов: инерцию самой
термоэлектрической батареи, теплоемкость которой слишком велика, в то время как
проведение тепла происходит слишком медленно, число спаев в батарее, температурное
равновесие в мышечной камере, соотношение между электродвижущей силой и
температурой, особенности гальванометра и др.
Литература
1. Heidenhain, Mechanische Arbeit, Warmeentwicklung und Stoffumsatz
bei d. Muskeltatigkeit, Leipzig, 1864.
2. F i с k A., Mechanische Arbeit u. Warmeentwicklung bei d. Muskeltatigkeit,
Leipzig. 1882.
3. Hill A., Muscular activity, London, 1924. Русский перевод: «Работа мышц».
Госиздат, 1929.
246
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
4. Н і II А. V. и. М е у е г h о f О., Erg. Physiol., 22, 299 u. 329, 1923.
5. Mey.etfhof О., Handb. norm. path. Physiol., VIII/I, 500, 1925.
•6. Hill A. V., The Revolution in the Muscul. Physiology, Physiol. Rev., 12, 56,
1932.
7. Fenn J„ J. Physiol., 58, 175, 1923; 58, 373, 1923.
8. Azuma R., Proc. Roy. Soc, B, 96, 338, 1924.
•9. Hill A. V., Adventures in Biophysics, 1931. Русский перевод: «Эпизоды из
области биофизики», Биомедгиз, 1936.
10. Bugnard, J. Physiol., 82, 509, 1934.
11. Bugnard, J. Physiol., 80, 441, 1933.
12. Fischer E., Pfl. Arch., 225, 532, 1930.
13. Feng T. P., J. Physiol., 74, 455, 1932.
14. Feng T. P., J. Psysiol., 74, 441, 1932.
15. Asmussen, цитир. по Buchthal u. Lindhard, The physiol. of str,»
muscle fibre, 1939.
16. С a 11 e 1 M с К е е n, Cold Spr. Harb. Symp. Quant. Biol., IV, 227, 1936.
17. E n g e I m a n n T h., Pfl. Arch., 23, 1880.
18. H i 1 1 A. V., Proc. Roy. Soc:, B, 126, 136, 1938.
19. Hart г ее W., J. Physiol., 60, 269Д1925; 72, 1, 1931.
Iлав а XII
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
Анатомические и физиологические особенности сердечной мышцы.
Сердечные мышечные волокна, подобно скелетным, являются
поперечно-полосатыми, но в сердце каждое волокно связано с соседними
волокнами протоплазматическими мостиками (рис. 121). На этой анато-
Рис. 121. Гистологическое строение мышечной ткани сердца.
Лг\— мышечная ткань желудочка (из Гейденгайна); J3— разрез через атриовентрикулярный узел
<Тавара) овцы; х — поперечный разрез волокон; у — сердечная мышечная клетка из
предсердия (по Тавара из Максимова).
мической особенности основывается следующее физиологическое
свойство: при раздражении асе сердце реагирует, как одно мышечное ёо-
локно. Раздражение каждого участка сердца, если только оно
вызывает возбуждение, производит его во всей сердечной мышце, так как
возбуждение от раздраженного участка посредством протоплазматиче-
ских мостиков обходит все сердце.
Сердечная мышца характерно отличается от скелетной еще тем,
что в ней все физиологические процессы происходят гораздо
медленнее. Процесс возбуждения в каждой точке сердца удерживается
настолько долго, что почти уравнивается в длительности с сокращением
данной точки. Поэтому, если регистрировать одновременно
биоэлектрический ток возбуждения и сокращение определенного участка сердца,
то получается такая картина: сначала появляется ток возбуждения,
затем наступает сокращение, потом оба эффекта протекают параллель-
248
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
но, только ток возбуждения не удерживается до конца сокращения,.
он прекращается немного раньше последнего. Такая
последовательность электрического и механического эффектов должна быть
совершенно понятна. Электрический эффект выражает возбуждение, поэтому
он должен наступить раньше сокращения, которое возникает под
влиянием возбуждения; точно так же ток возбуждения должен окончиться
раньше сокращения, так как возбуждение не может продолжаться
дольше вызываемого и
сокращения. На рис. 122 дается
хорошая иллюстрация указанного
соотношения электрического и.
механического эффектов
предсердия и желудочка
лягушки.
Такое же длительное
возбуждение и такое же
соотношение его с механическими
эффектами наблюдается в сердечной
мышце теплокровных животных
и человека. Характер кривой
электрического эффекта у
человека и лягушки по существу
одинаков. На рис. 123
изображена электрограмма сердца
человека, полученная струнным
гальванометром. На людях
электрограмму сердца отводят через
руки и ноги. Электрический ток
Рис. 122. мшнограша и электрограмма ля- в03буждения сердца распростра-
Самая верхняя, быстро колеблющаяся криваяТуказы- няетСЯ ПО ВСему телу. ЭТО рЭС-
вает время в 0.05 сек. Вторая кривая принадлежит пппгтпаириир аярт/тпыирг^пгп тл-
механическому эффекту предсердия; третья механи- UpULipdHCHMC ЗЛСКіричсскши ш
ч1^эЙ^пр^^& )TSSSUa0^r" ка происходит в таком виде, как.
это дано на рис. 124. Поэтому
при отведении обеих рук и ног в гальванометр получается совершенно'
такой же электрический эффект, как если бы отведение производилось
непосредственно от сердца.
Электрическая активность сердца. Нормальная электрограмма
сердца человека и всех позвоночных животных имеет три
характерных зубца: один всегда маленький зубец, отвечающий предсердию
(рис. 123, Р), затем высокий и острый зубец (R), выражающий
начало деятельности желудочка и наступающий еще в скрытом периоде
систолы. Этот зубец очень постоянен. Наконец, третий зубец (Г),
отмечающий период диастолы желудочка. Этот зубец легко меняется
в зависимости от разных условий как в своем направлении, ибо он
может итти и вверх, и вниз от абсциссы, так и в своей
интенсивности.
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
249
Многозубчатый характер электрограммы желудочка объясняется
сложным строением проводящей системы сердца. В каждом участке
^ийтай
л^ч^ЯВГтчФ?
Рис. 123. Электрограмма сердца человека.
Наверху кривая 'пульса от сонной артерии (С). Внизу
электрограмма сердца (Я). Колебание Р от тока возбуждения
предсердия; Q, R, S, хТ—от тока возбуждения желудочка
(Эйнтховен). - Чі ч
Рис. 124. Распространение
электрических напряжений в теле,
вызванных возбуждением сердца (по
Уоллеру).
сердца пробегающая волна
возбуждения должна дать, как
в скелетной мышце, одно
колебание экектрического тока.
И действительно, если
соединить с гальванометром один
Рис. 125. Анализ электрограммы сердца.
Даны две формы электрограммы —С R,T, Р и О, J?i» Т,,
Р при отведении основания и верхушки желудочка.
Кривые, изображенные прерывистой линией, обозначают
изменения потенциалов под отводящими электродами: а, Ь,
с, d — под электродом на основании желудочка, п, I, і,
h, f, е—под электродом на верхушке, п, т, к, і, ft, g, et —
когда верхушечный электрод лежит посередине
желудочка. Электрограмма О, R, Т, Р является результирующей
из кривых а, Ь, с, d и п, I, /, Л, /, е, а электрограмма
О, Rlf Ти Р —из кривых a,b c,d ип. m, k, i, h, g, е. Когда
верхушечный отрицательный потенциал достигает
максимума позднее базального и проходит позднее него,—
получается первая форма электрограммы; когда же
верхушечный потенциал достигает максимума раньше
базального и оканчивается раньше него, —получается вторая
форма (Воронцов).
участок нормальный, а другой
поврежденный, то получится одно колебание электрического тока. Но,
в отличие от скелетной мышцы, этот ток не только предшествует
сокращению, но и сопровождает его (Воронцов, 3). Если желудочек
250
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
дает во время систолы несколько колебаний или несколько зубцов, и
лритом разного характера, то это должно являться результирующим
электрическим эффектом волны возбуждения імногих таких участков
(Эйнтховен, 22).
По Воронцову (3), возникновение острого зубца (R)
зависит от тока возбуждения основания желудочка, куда возбуждение
поступает из проводящих пучков Гисса раньше, чем в другие участки.
Последующее быстрое ослабление данного зубца происходит от
возникновения возбуждения в других отделах желудочка, в результате
алгебраической суммации токов. Возникновение же зубца (Т) зависит
от того, что возбуждение кончается на основании и верхушке
желудочка не одновременно. Если возбуждение кончается раньше на
основании, тогда зубец Т имеет направление, обратное зубцу R. Если
же возбуждение кончается раньше на верхушке желудочка,
направление зубца Т будет такое же, как и зубца R (рис. 125).
При нормальной автоматической деятельности сердца лягушки в
области возникновения спонтанных импульсов, значит, в венозных
синусах, каждому току возбуждения предшествует колебание
потенциала небольшой амплитуды (0,2 mV и меньше) с медленным постепенным
нарастанием. На вершине этого потенциала насаживается ток
возбуждения с быстрым нарастанием. Этот предварительный потенциал
локален и не распространяется, как это бывает при локальных изменениях
основного биологического процесса мышцы и нерва в результате суб-
мишшального раздражения. Этот локальный потенциал тем сильнее,
чем ближе к месту возникновения спонтанных распространяемых
импульсов. В поврежденной сердечной ткани локальные потенциалы
могут повторяться много раз с большой регулярностью без
возникновения распространяемых потенциалов возбуждения (Боцлер, 34).
Низковольтные потенциалы возникают в мышечной ткани сердца точно
также в связи с раздражением в виде длительного положительного
последействия (Боцлер, 34). Это электрическое последействие
сопровождается одновременным изменением напряжения мышцы. После сокращения
напряжение, как и потенциал, падает ниже уровня, существующего
до сокращения. Это указывает на существование тонического
сокращения в «покойном» состоянии, т. е. собственного тонуса мышцы
(Боцлер, 35).
В каждой точке сердца лягушки процесс возбуждения
продолжается десятые доли секунды, а иногда и целые секунды. Он вообще
широко (меняется в связи с изменением возбудимости сердечной мышцы.
Так, например, при повышении температуры продолжительность
возбуждения уменьшается, при понижении ее, наоборот, она
увеличивается. Так же действуют наркотические вещества, как эфир, хлороформ:
в малых концентрациях и при кратковременном действии они
повышают возбудимость, тем самым укорачивая процесс возбуждения, а в
больших концентрациях удлиняют его. В сердечной мышце зимней
лягушки процесс возбуждения продолжается Р/г—2 сек. При слабом
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
251
отравлении эфиром он укорачивается до 0.8 сек., т. е. почти вдвое
(Беритов и Цкиманаури, 1).
Интенсивность процесса возбуждения зависит от степени
возбудимости: она тем больше, чем выше возбудимость. Соответственно
меняется и сократительный процесс, так как он тем сильнее, чем сильнее
возбуждение (Беритов и Цкиманаури, 1).
Проведение возбуждения в сердечной (мышце также происходит
чрезвычайно медленно. По Энгельману (2), в полоске
лягушечьего сердца скорость распространения возбуждения равна 10—55 мм.
Замедление особенно велико в соединительных пучках Г и с а и Та-
в а р а, которые связывают мышечный узел синуса с мускулатурой
предсердия и желудочка. По определению Воронцова (3), в
соединительных пучках лягушечьего сердца скорость проведения
возбуждения равна приблизительно 12 мм в сек.
Означенные физиологические особенности — переход возбуждения
тэт одного мышечного элемента к другому и большая длительность
процесса возбуждения — делают сердечную мускулатуру чрезвычайно
хорошим объектом для изучения рефракторных фаз. Благодаря тому,
что возбуждение продолжается почти столько же времени, сколько и
сокращение, легко изучить рефракторную фазу прямо на механическом
эффекте мышцы. И действительно, рефракторная фаза была открыта
и изучена на сердечной мышце миографическим путем (Марей, 4;
Кронекер, 5).
Закон возбуждения в сердечной мыщце. Сердечная мышца
подчиняется закону возбуждения. Соответственно, при возбуждении ее
сначала наступает абсолютная, а потом относительная рефракторная
фаза. Абсолютная рефракторная фаза, сопровождающая возбуждение,
продолжается почти в течение всего времени сокращения сердечной
мышцы, т. е. во все время систолы, (потому что, как указывалось выше,
возбуждение удерживается в каждой точке сердца почти столько же
времени, сколько и сокращение. Однако это не значит, что абсолютная
рефракторная фаза продолжается во время сокращения всего сердца. Во-
первых, предсердие сокращается раньше желудочка, следовательно,
когда абсолютная рефракторная фаза в последнем начинается, она
кончается в предсердиях. Во-вторых, и в пределах одного желудочка
абсолютная рефракторная фаза не может проявиться одновременно во
всех его частях. Как ни медленно протекает процесс возбуждения в
каждом мышечном волокне, все-таки он начинается и заканчивается во
время систолы в одних частях желудочка раньше, в других позже.
Поэтому абсолютная рефракторная фаза продолжается не все время
сокращения желудочка. Когда сокращение начинает ослабевать, т. е. во
•время диастолы, абсолютная рефракторная фаза всегда сменяется
относительной, так как к этому времени в первоначально возбужденных
участках сердца возбуждение заканчивается -и начинается процесс
восстановления, т. е. относительная рефракторная фаза. В отношении
электрограммы эта смена происходит перед наступлением зубца Г. На
252 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
рис. 126 (видно, что индукционный удар не вызывает эффекта, если он
падает за 0.08 сек. до вершины зубца 7\ Но он вызвал его, когда
упал за 0.04 сек. до нее, причем этот эффект сильно уменьшен
сравнительно с нормальным.
На бьющемся желудочке сердца раздражение не вызывает
эффекта в момент систолы; эффект получается только в период диастолы,
т. е. с того момента, когда систолическое сокращение начинает
ослабевать. Этот добавочный эффект называется экстрасистолой.
Последняя всегда бывает значительно меньше нормальной систолы как па
Рис. 126. Рефракторная фаза и экстрасистола желудочка на остановленном сердце
лягушки.
Струнный гальванометр. Даны три записи. На каждой записи сверху вниз идут: время по 0.04 сек.-
момент одного раздражения желудочка, момент другого раздражения желудочка, электрограмма
желудочка и кривая механического эффекта. На первой записи дан нормальный эффект от второго
раздражения, на третьей — нормальный эффект от первого раздражения, а на средней записи дана
комбинация обоих раздражений. При этом второе раздражение дало небольшой эффект, сильно отличный
от нормального. В третьей записи также было произведено второе раздражение, но оно пришлось в
абсолютную рефракторную фазу и не дало эффекта. Везде моменты раздражения отмечаются и на
электрограмме небольшим колебанием благодаря забеганию в цепь гальванометра петли раздражающе го-
тока.
интенсивности, так и по длительности (рис. 127). Это происходит
главным образом оттого, что во время экстрасистолы возбуждается и
сокращается не все сердце, а только та часть его, которая
освободилась к моменту раздражения от предшествующего возбуждения. Ноу
конечно, имеет значение и то обстоятельство, что экстрараздражение
падает в імомент относительной рефракторной фазы: ів силу неполного
восстановления возбудимой системы эффект не может оказаться
полным. Чем ближе к концу систолы падает новое раздражение, тем
меньше должна быть вызванная им экстрасистола, и тем больше
должна быть пороговая сила раздражения, требуемая для вызова
экстрасистолы.
Таким образом, на сердечной мышце чрезвычайно удобно изучить
миографическим путем все характерные особенности той и другой
рефракторных фаз (рис. 127).
Следующая за экстрасистолой желудочка пауза до нового
систолического сокращения значительно удлиняется сравнительно с
нормальной паузой (рис. 127). Это происходит оттого, что экстрасистола
создает новую рефракторную фазу в том отделе желудочка, который
раньше всех освободился от систолического возбуждения. Поэтому
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ 253
тотчас же после экстрасистолы первый возбуждающий импульс со
стороны предсердия приходит в желудочек во время этой
рефракторной фазы и не возбуждает более его, не вызывает нормальной
систолы. Последняя наступает только от последующего второго
импульса» когда рефракторная фаза кончилась или достаточно ослабла.
Значит, пауза удлиняется после экстрасистолы в силу выпадения одной
нормальной систолы. Эта пауза
именуется компенсаторной. Такое
название нужно считать
устаревшим, не отвечающим
действительности, так как пауза
удлиняется не потому, что
вследствие усиленной деятельности
сердца потребовался усиленный
отдых, компенсирующий эту
деятельность, а в силу
рефракторной фазы от экстрасистолы.
Если бы после одной систолы
экстрасистола вызывалась без
перерыва несколько раз одна за
другой, то и тогда пауза после
последней экстрасистолы
удлинилась бы в такой мере, как
после однократно вызванной
экстрасистолы (Энгельман, 29).
Если охлаждением синусов
сильно удлинить интервал между
систолами, тогда за
экстрасистолой желудочка не следует
компенсаторная пауза, ибо
последующие импульсы приходят
в желудочек после окончания
рефракторной фазы от
экстрасистолы (Тренделенбург, 30).
На сердце лягушки
аналогичное явление наблюдается также
при раздражении предсердия. И
здесь за экстрасистолой
предсердия следует компенсаторная
пауза. В этом случае выпадает один импульс, идущий от синусов.
Но при раздражении синусов во время их деятельности за
экстрасистолой синусов следует точно такая же пауза, как после нормальной
систолы (Энгельман, 29). Это совершенно понятно, ибо в нормальном
сердце в синусах возникают периодические импульсы, и каждая новая
систола наступает через один и тот же интервал после начала
предыдущей.
Рис. 127. Миограмма лягушечьего сердца,
демонстрирующая экстрасистолу,
рефракторную іфазу и компенсаторную паузу.
Порядок записывания снизу вверх. Линия 00
указывает на те систолы, во время которых раздражается
сердце. Момент раздражения везде обозначен
буквой х. На 1-й кривой раздражение приходится в
самом начале систолы и не дает эффекта. На 2-й и 3-Й
кривых раздражение падает несколько позднее и
опять не дает эффекта. На 4-й раздражение
приходится очень близко к вершине систолы и дает
эффект. На 5—8-й кривых раздражение падает все
позднее и позднее, и соответственно вызываемая ими
экстрасистола становится все сильнее и сильнее;
с, с — компенсаторная пауза. На 1-3-й кривых
раздражение падает во время абсолютной
рефракторной фазы, а на 4-7-й во время относительной
рефракторной фазы (Марей).
254
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Это сохранение нормальной периодичности сердца при разных
условиях раздражения было обозначено Энгельманом как закон-
сохранения физиологического периода раздражения в сердце.
Повышение возбудимости в сердечной мышце. Сердечная мышца
подобно скелетной отвечает на раздражение не только возбуждением,.
А
МцрщРНЯ/щйЯШ^^
ШЛ
Рис. 128. Действие сердечного гормона на остановленное сердце.
Сердце стоит несколько дней. Через канюлю внутрь вводится рингеровский раствор, в котором
лежало автоматически бьющееся основание, желудочка. Появление и постепенное нарастание
пульсации обусловлены действием гормона, вышедшего из основания желудочка (Д аоерландт).
но и повышением возбудимости сверх нормы. Эта сверхнормальная:
возбудимость после каждого деятельного раздражения удерживается
несколько секунд. Для постановки соответствующих опытов выгодна
пользоваться остановленным сердцем. Остановку сердца можно
вызвать крепкой перевязкой сердца на границе предсердий и венозных
синусов (первая лигатура или перевязка Станниуса, рис. 129). На
таком остановленном желудочке обнаруживается повышение
возбудимости после каждой систолы, вызванной
раздражением; например, если сердце
раздражается повторно
индукционными ударами, следующими один за
другим, то второй эффект будет
сильнее первого, третий сильнее
второго и т. д., пока не установится
максимальный эффект (лестница Боу-
дича).
Повышение возбудимости можно
наблюдать и на автоматически
бьющемся сердце. Первая систола
после компенсаторной паузы часто-
бывает сильнее, чем остальные;
видимо, при нормальной пульсации-
сердца относительная рефракторная фаза продолжается от одной
систолы до другой. Удлинение паузы, естественно, благоприятствует
повышению возбудимости. Возбудимость повышается в сердечной мышце
и при субминимальном раздражении: при многократном повторении
такого раздражения на остановленном сердце возбуждение наступает,
сердце сокращается (явление суммации) (Баш, 7).
Тетаническое сокращение сердечной мышцы. Совершенно
нормальное сердце не отвечает ни на какие раздражения тетанусом. При-
быстром следовании отдельных индукционных ударов получается кло-
нический, т. е. прерывистый, эффект, где каждый зубец не выше
нормальной систолы. Это, разумеется, происходит оттого, что в каждом:
Рис. 129. [Опыт Станниуса /на сердце
лягушки.
А — первая перевязка (лигатура); В —
вторая перевязка; 1 — предсердия; 2 —
желудочек; 3 — синусы; С — граница между
желудочком и предсердиями; L — нить, которой
произведена первая перевязка (по Гедону).
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
25S
данном участке сердечной мышцы возбуждение, а значит, и
абсолютная рефракторная фаза продолжаются во время всей систолы. По*
этому второе раздражение произведет новый сократительный эффект
только во время диастолы. Этот эффект будет представлять собой
совершенно самостоятельное сокращение тех элементов сердца, где уж?-
окончились возбуждение и сокращение. Но если почему-либо
параллельное течение сокращения и возбуждения нарушено, например при
согревании, после длительного раздражения, при отравлении
хлороформом, хлористым натрием и др. веществами, под влиянием
раздражения блуждающего нерва и при многих других условиях, когда
возбуждение, а значит, и абсолютная рефракторная фаза значительна
короче сокращения, полный тетанус может наблюдаться и на
сердечной мышце. В этом случае второе возбуждение данного участка может
наступить еще во время его сокращения, и механический эффект
может оказаться не только сплошным, но и выше обычной систолы, т. е
обнаружить все характерные черты тетануса (Вальтер, 8).
Раз сердечная мышца реагирует по закону возбуждения сначала1
абсолютной, а затем относительной рефракторной фазой, она может
ддть пессимальный тетанус при некотором высоком ритме
раздражения. Но здесь характерно, что абсолютная рефракторная
фаза на нормальном желудочке продолжается несколько десятых
секунды. В соответствии с этим, нужно было бы ожидать пессималь-
ного эффекта от раздражения в ритме 10 раз в секунду при
оптимальном ритме 3 в секунду. Фактически пессимальный эффект на
лягушачьем сердце получается при ритме 60—100 в секунду (Шерешев-
ский и Шошина, 33). Но известно, что сердце обладает очень большой
изменчивостью своей лабильности и вследствие этого длительность
абсолютной рефракторной фазы может меняться в очень широких:
пределах.
Автоматическая деятельность сердечной мышцы. В отличие от
скелетной мускулатуры, сердечная мышца обладает свойством
автоматической деятельности, т. е. деятельности помимо внешних раздражений
и каких-либо нервных влияний. Это доказывается разнообразными
опытами, прежде всего тем, что сердце обнаруживает автоматическую-
деятельность в такой стадии эмбрионального развития, когда в нем
нет еще нервных элементов (Льюис, 20) или когда сердце денерви-
ровано вследствие перерезки обоих блуждающих и симпатических
нервов (Габерланд, 16). Ритмические импульсы возбуждения
возникают у холоднокровных, например у лягушки, в мышечном узле
венозных расширений, так называемых синусов, а у теплокровных тамг
где полые вены входят в предсердие, в так называемых узлах Кейт-
Флака. Здесь в мышечных элементах этого узла заложен тот
механизм, который обусловливает правильное периодическое возникновение
импульсов, — пэйсмекер, как говорят англичане (Экклс и Гофф, 26). Из
этих узлов возбуждение переходит на предсердия к атриовентрику-
лярным мышечным узлам Тавара, а затем по соединительным пучкам
256 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Гиса в желудочки. Узлы синуса содержат такие же мышечные
элементы, как пучки Тавара и Гиса: бедные фибриллями и 'богатые
саркоплазмой. Такие мышечные элементы вообще, видимо, представляют
раннюю стадию эмбрионального развития мышечной ткани сердца
(рис. 121—5).
Что автоматическая деятельность сердца впервые возникает в
мышечном узле синуса, это видно, между прочим, из того, что повышение
температуры в этом месте учащает биение сердца. Отделение его на
лягушечьем сердце путем перевязывания на границе предсердия и
полых вен останавливает биение сердца (первая перевязка Станниуса).
Спустя некоторое время остановленное сердце вновь начинает
сокращаться, особенно если раздражать его каким-либо агентом. Это
связывают с автоматической деятельностью мышечных узлов атрковен-
трикулярной системы (Эйстер, 12).
Автоматическая деятельность не является функцией одних синусов.
Атриовентрикулярная система точно так же владеет этой
способностью. Если отрезать желудочек, то последний через некоторое время
начинает пульсировать. Нужно только раздражать основание
желудочка на границе с предсердием, где лежат узлы этой системы,
электрическом током или механически (укол булавкой). Автоматическая
деятельность свойственна также корню аорты (Скрамлик, 13).
Если остановленное после первой перевязки Станниуса сердце
перевязать еще на границе предсердий и желудочка (вторая перевязка
Станниуса) (рис. 129), то возникает на некоторое время ритмическое
сокращение как предсердий, так и желудочка. Это следует считать
результатом раздражения гисовского пучка атриовентрикулярной
системы. Такое же раздражение на скелетной мышце вызвало бы только
одно сокращение.
Таким образом, мышечные узлы синусов, атриовентрикулярной
системы и корня аорты являются центрами автоматической деятельности.
Но автоматическая деятельность синусов выступает сильнее других
элементов. Это хорошо видно, если сравнить автоматическую
деятельность вырезанных кусков всех этих отделов. Отрезки синусов
показывают наиболее длительную автоматическую деятельность и наиболее
высокий ритм ее. Поэтому в нормальных условиях синусы играют
ведущую роль в автоматической деятельности всего сердца.
В некоторой малой степени (автоматическая деятельность
свойственна и остальным отделам сердца. Отжатая верхушечка желудочка
(2/3 последнего) без раздражения лежит неподвижно. Но в ответ на
однократное механическое раздражение (укол булавкой) или на
пропускание гальванического тока, или на повышение давления путем
соединения со столбом дефибринированной крови она отвечает
ритмической деятельностью. Отсюда видно, что автоматическая деятельность
есть общее свойство сердечной мышцы; она только ів большей мере
свойственна мышечным узлам синуса, атриовентрикулярной системе и
корню аорты, чем другим отделам сердца.
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
257
В эмбриональном сердце еще раньше* чем начинает в нем
циркулировать кровь, все отделы обладают свойствоім пульсации, и
каждый отдел обладает своим собственным внутренним ритмом.
Зачатки желудочка начинают пульсировать даже раньше предсердия.
Только когда разовьется предсердие, замкнется полость сердца и
кровь начинает циркулировать через него, ритм предсердия становится
господствующим: желудочки теряют свой ритм и подчиняются
предсердию (Копенгавер, 36, Госс, 37).
Происхождение автоматической деятельности сердца. В сердце
процессы возбуждения возникают под влиянием внутренних
раздражений. Известно, что вырезанное сердце продолжает автоматическую
деятельность. Нужно только пропускать через сердце изотоничные
растворы некоторых солей. В этих условиях вырезанное сердце
лягушки может работать в течение двух дней. Но и сердце теплокровных
животных может биться очень долго — в течение нескольких часов,
если только пропускается через него под определенным давлением
изотонический раствор Локка, согретый до 37°.
До последнего времени господствовало мнение Л ё б а (9), что
автоматическая деятельность синусных узлов происходит вследствие
чередования преимущественного (влияния одновалентных и
двувалентных ионов.
Лёб основывался при этом как на своих наблюдениях над
скелетной мышцей, так и на наблюдениях других авторов (Самойлов, 28;
Майне, 27). Этими авторами было установлено, что одновалентные
ионы повышают возбудимость и вызывают периодически повторяемые
вздрагивания, которые иногда протекают правильным ритмом.
Двувалентные ионы, наоборот, понижают возбудимость и подавляют
спонтанные сокращения, вызванные одновалентными ионами.
Теория Л ё б а поддерживается Лазаревым. Один из его
учеников подобрал такой раствор, под влиянием которого скелетная
мышца обнаруживает очень регулярную периодическую деятельность
подобно сердцу (Яуре, 21).
В последующих исследованиях особенно большое значение
придается ионам калия, которые оказывают на ритм сердечной
деятельности ускоряющее действие (Киш, 25; Риглер и Зингер, 24). Но и
другие составные части крови признаются существенно необходимыми
для нормальной автоматической деятельности сердца. Так, Манс-
фельд (10) недавно открыл важное значение углекислоты. Он
наблюдал, что автоматическая деятельность сердца прекращается, если
пропускать через сердце раствор Рингера, который содержит какую-
либо щелочь и тем отнимает у сердца запас углекислоты. Если потом
прибавить С02 к раствору Рингера, сердце вновь начинает
действовать автоматически.
Таким образом, по Мансфельду, следует считать углекислоту,
этот конечный продукт обмена веществ, естественным раздражителем
сердца.
17 И. С Бержтов
258 общая физиология мышечной системы
Сердечный гормон. В настоящее время целый ряд исследователей
приписывает автоматическую деятельность особому гормону,
вырабатываемому в мышечных клетках самого сердца. По Габерланду
(23), он производится в мышечных узлах синусов и во всей
проводящей системе. Сердечный гормон легко извлечь из сердца. Нужно
вырезать синусы из одного или нескольких сердец крупных лягушек и>
дать им автоматически сокращаться в 1 см3 раствора Рингера в
течение 15 мин. и более. Если потом этим раствором подействовать на
изолированный желудочек, то последний начинает биться сильнее и
чаще. Если он был в покое, то начинает 'биться. Очевидно, во время-
сокращения из синусов вымывается какое-то вещество, которое
производит возбуждение мышечных клеток сердца. Такое возбуждающее
вещество можно получить из основания желудочка, где имеются
соединительные пучки Гиса. На рис. 128 (см. стр. 254) дана
иллюстрация действия сердечного гормона.
Сердечный гормон автоматической деятельности был открыт также
в сердцах теплокровных (Демур, 17). Его находят в водных и
алкогольных экстрактах предсердечных узлов сердца разных животных,
а' также субэндокардиальных конечных разветвлений пучков Пур-
кинье (Демур и Рилан, 18).
Состав сердечного гормона еще не известен. Одни считают era
гистаминолодобным веществом (Риглер, 24), другие отрицают это
(Габерланд, 23), но происхождение его в некоторой степени известно.
'По исследованиям Цваардемакера, он существует в проводящей
системе сердца в виде недеятельного гормона и переходит в активный
гормон под влиянием радиоактивного излучения калия, которым
богато сердце. В недеятельном виде гормон находится и в скелетной
мускулатуре. Он адсорбируется (извлекается) тальком (talcum vene-
tum), его можно растворить в воде, алкоголе. Путем действия
радиоактивных веществ его легко можно превратить в активный гормон
сердца (Цваардемакер, 19).
Многие авторы высказываются против значения радиоактивности
в автоматической деятельности сердца на том основании, что обычный
калий нельзя заменить радиоактивным натрием или другим
радиоактивным веществом. Очевидно, ионы калия важны сами по себе для'
автоматии сердца (Глацко и Гринберг, 32).
Новейшие исследования на сердцах теплокровных указывают на
еще более сложное происхождение нормальной автоматической
деятельности. Именно, в мышечных узлах предсердия образуется сердечный
гормон, обусловливающий автоматическую деятельность. Но эта
деятельность происходит нерегулярными и неравномерными периодами.
Регулярность в автоматии устанавливается другим активным
веществом из мышечных узлов Кейт-Флака и Тавара. Это последнее
вещество не обладает способностью вызывать 'появление
автоматической деятельности. Значит, оно только придает автоматической
деятельности регулярный характер (Демур, 31).
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ.
259
Исходя из данного фактического материала, надо полагать, что ни
ионы натрия, калия и т. п., ни углекислота, ни сердечный гормон и
«активное вещество» мышечных узлов отдельно не являются
единственными возбудителями сердца. Только совокупностью их всех
вместе со многими другими составными частями крови создаются
условия для автоматической деятельности. В нормальном сердце под
влиянием всего состава крови и вырабатываемого в синусах гормона
прежде всего приходят в ритмическое активное состояние синусы.
Возникающее здесь возбуждение далее передается другим отделам по
проводящей мышечной системе, вызывая активное состояние других
отделов сердца. Образуемый в синусах, а также в проводящей системе
гормон, со своей стороны, диффундирует в кровь и вследствие этого
действует на все отделы сердца. Это действие на двигательную
(мускулатуру сердца выражается в повышении возбудимости, вследствие чего
усиливается как процесс возбуждения, так и обусловливаемый им
сократительный процесс.
Практикум
Наблюдение абсолютной и относительной рефракторных фаз и компенсаторной
паузы. Для этого опыта пользуются большой лягушкой. Спинномозговой препарат
прикалывают к доске брюхом вверх. Разрезают кожу на груди, вырезают грудную
кость, снимают сердечную сорочку и перерезают уздечку, находящуюся на спинной
стороне сердца. Регистрация производится так называемым сердечным пинцетом.
Проще всего записать биение сердца простым миографом Энгельмана. На
верхушку сердца накладывают маленькую и очень легкую зацепку из тонкой проволоки
и потом нитью соединяют с миографом. Для раздражения один полюс вторичной
катушки индукториума соединяют тоненькой проволокой с означенной зацепкой.
Другой иолюс соединяется такой же проволокой и зацепкой с основанием
желудочка.
Если условия опыта с методической стороны безупречны, то раздражение
желудочка во время систолы, т. е. восходящего колена кривой сокращения, не дает
эффекта; значит, в это время раздражение попадает в абсолютную рефракторную
фазу желудочка. Если же раздражение приходится в период диастолы, на
нисходящем колене кривой или немного позднее, то эффект, так называемая экстраси-
стола, наступит, и тем сильнее, чем позднее произойдет раздражение. Следовательно,
мы здесь будем иметь дело с относительной рефракторной фазой. После экстраси-
столы наступает компенсаторная пауза; она длиннее нормальной паузы и притом
там длительнее, чем раньше произойдет экстрараэдражение. После компенсаторной
паузы первая нормальная систола может быть более сильной, чем остальные. Это
будет результатом повышения возбудимости от удлинения паузы.
Весьма важно, чтобы при опытах с раздражением между электродами
заключался только один желудочек. В противном случае раздражение будет вызывать
экстрасистолу желудочка даже при попаданий ъ самом начале систолы. Это
произойдет оттого, что в это время предсердия свободны от рефракторной фазы и.
возбуждаются: отсюда возбуждение передается желудочку в тот момент, когда
систола его кончается и потому вызывает экстраснетолу. Нельзя также употреблять
очень большую силу раздражения: такое раздражение действует на блуждающий
нерв, в результате чего получается остановка сердца. В таких случаях за экстра-
систолой следует не истинная компенсаторная пауза, а длительная остановка, или
замедление сердечной деятельности. Кроме того, очень сильное раздражение может
вызвать так называемое трепетание сердца, которое (выражается в том, что, сердде
17*
260
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
дает ряд экстрасистол быстро одну за другой. Это является результатом
раздражения атриовентрикулярной системы Гиса (Габ?рланд, 14).
Опыты Станниуса /для демонстрации іавтоматической деятельности разных
отделов сердца. Обнажают сердце лягушки; накладывают нить по белой пограничной
линии между предсердием *и синусами, затем крепко перевязывают (первая
перевязка Станниуса) (рис. 129). После этого синусы продолжают биться, а предсердия
и желудочек останавливаются. Таким препаратом пользуются для изучения
скрытого периода сокращения желудочка, повышения возбудимости в ответ на
раздражение, явления суммации.
Затем накладывают нить на границе предсердий и желудочка и опять крепко
перевязывают (вторая перевязка Станниуса) (рис. 129). За этим следует
возобновление автоматической деятельности предсердий и желудочка на некоторое время.
Наблюдение распространения возбуждения в сердечной мышце.
Зигзагообразным разрезом сердца приготовляют мышечную ленту и растягивают булавками.
Если синусы остаются ів связи « работают, тогда сокращается вся лента:
сокращение на-глаз распространяется по участкам предсердия, а потом желудочка. Если
сердце стоит, тогда искусственное раздражение на одном конце вызывает
сокращение, которое на-глаз распространяется до другого конца. Этим доказывается
переход возбуждения от одного мышечного ©олокна к другому.
Наблюдение возникновения импульсов возбуждения в синусах. Стеклянную
палочку согревают до 40° в физиологическом растворе и прикладывают к синусам.
Сейчас же ритм сердечного сокращения сильно увеличивается. Такое же согревание
одного ж<елудочка не повышает ритма, оно только усиливает сокращения желудочка
и уменьшает их длительность вследствие повышения возбудимости.
Литер ату ра
1. Beritoff J. u. Zkimanauri G., Z. BioL, 82, 21, 1924.
2. Engelmann, Pfl. Arch., 17, 68, 1878.
3. В о p о h ц о в Д. С, Электрограмма сердца лягушки. Одесса, 1917.
4. М а г еу, С. R. Acad. Sci., 82, 408, 1876; 89, 203, 1879; J. Anat. Physiol., 13, 60,
520, 1877.
5. Kronecker H., Beitr. Anat. Physiol, стр. 173, Leipzig, 1874.
6. Wast 1 H., Z. BioL, 75, 289, 1922.
7. В a s с h v., Wiener Sitzungsber., 79(3), 37, 1879;JArch. Anat. Physiol., стр. 283, 1880.
8. Walther A., Pfl. Arch., 78, 597, 1899.
9. Loeb J., Festschr. fur Fick, S. 99, 1899.
10. Mansfeld u. Szent-Gy or gy i v., Pfl. Arch., 184, 236, 1920.
11. S t a n n i u s, Arch. Anat. Physiol., стр. 85, 1852.
12. E у s t e r J. A. E. u. Meek W. J., Am. J. Physiol., 61, 117, 1922.
13. S с r a m 1 i k E. v., Pfl. Arch., 183, 109, 1920.
14. H a b er 1 a n d t L., Das Herzflimmern Samml. anat. u. physiol. Vertrage u.
Aufsatze, Jena, 1914; Z. Biol., 66, 407, 1916.
15. Haberlandt L, Z. Biol., 76, 49, 1922; Z. Biol., 81, 161, 1924.
16. Haberlandt L./Handb. BioL Arbeitsmethoden, Abt. 5, Bd. 8, стр. 237,1928.
17. Demo or J., Arch/int. Physiol., 20, стр. 29 и 446, 1922—1923.
18. D e m о о r J. et Rylant F., С r. Soc. BioL, 93, 814, 1925.
19. Zwaarder maker, H., Pfl. Arch., 218, 374, 1927.
20. Lewis Warren H., Bull. Hopkins Hosp., 35, 252, 1924.
21. ЯуреГ. Г., Журн. зксп. биол. мед., 8, № 19, стр. 43, 1927.
22. Е і n t h о v e n W., Handb. norm. path. Phys., Bd. 8, Teil 2, стр. 785, 1927.
23. Haberlandt L„ Pfl. Arch., 219, 279, 1928; 221, 576, 1929.
24. R i g i e r R. и сотрудники, Pfl. Arch., 220, 56, 1926; 221, 509, 1929; 222, 450, 1929.
25. К i sc h В., PfL Arch., 214, 662 u. 669, 1926; 215, 675, 1927.
ОБЩАЯ ФИЗИОЛОГИЯ СЕРДЕЧНОЙ МЫШЦЫ
¦¦261
26. Е с с 1 е s J. С. а. Н. Е. Н о f f, Proc. Roy. Soc, В., 115, 307, 1934.
27. M i n es G. R., J. Physiol., 37, 408, 1908.
28. Samojloff A., Arch. f. Anat u. Physiol., S. 145, 1907.
29. Engeimann W., Pfl. Arch., 59, 333, 1894; ib., 65, 137, 1897.
30. Trendelenburg, Arch. Physiol, (u. Anat.), S. 311, 1903.
31. Demo or L., Физиол. журн. СССР, 24, 126, 1938.
32. G 1 a z к о A. J. a. D. M. G r e e n b e rg, Am. J. Physiol., 125, 405, 1939.
33. Шерешевский Л. M. и Н. А. Ш о ш и н а, Учен, зап. Ленингр. унив., сер-
биол. н., вып. 10, стр. 45, 1939.
34. Bozler Е., Am. J. Physiol., 138, 273, 1943.
35. Bozler E. Am. J. Physiol., 139, 477, 1943.
36. Cope nh aver W. M., J. Exper. Zooh, 80, 193, 1939.
37. G oss C. M., Am. J. Physiol., 137, 146, 1942.
Гл а в а XIII
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
Строение гладкой мышечной ткани. Гладкие мышечные клетки
у позвоночных животных большей частью имеют характер длинных
веретенообразных волокон, в середине более или менее толстых, а по
концам постепенно суживающихся и заостряющихся. Длина клеток
обычно не велика — 0.12—0.5 мм, а диаметр ее в 5—10 раз меньше.
В живой клетке содержимое является гомогенным (бесструктурным).
Но при обработке посредством самых различных веществ и красок
в клетке гладкой мышцы всегда обнаруживается продольная исчер-
ченность, обусловленная присутствием тонких волоконцев — миофиб-
риллей. Следовательно, гладкие мышечные клетки, подобно
поперечнополосатым, содержат протоплазму и миофибрилли. Отсутствие видимой
продольной исчерченности в живой клетке объясняется тем, что
показатель преломления миофибрилли очень близок к таковому саркоплазмы.
Многочисленные миофибрилли тянутся по всей длине мышечной
клетки. В противоположность фибриллям поперечнополосатых мышц,
фибрилли гладких состоят только из анизотропного вещества.
Вследствие этого мышечные клетки не имеют поперечной исчерченности и
поэтому называются гладкими. Протоплазмы в клетке тем больше,
чем меньше фибриллей. Она заходит между фибриллями и окружает
более компактной массой ядро клетки. В некоторых мертвых гладких
мышцах (в двигателях игл ежа, в стенках кровеносных сосудов) после
обработки имеются фибрилли не сплошные, а состоящие из продольных
точек или черточек (Буке, 62) (рис. 130).
Протоплазма внутри гладких мышечных клеток позвоночных (кошки,
аксолотля) не однородна. По новейшим исследованиям Роскина (49),
следует различать межфибриллярную плазму, которая, повидимому,
играет определенную роль в сократительном процессе и потому
называется киноплазмой. Она обволакивает фибрилли іиногда
сплошным слоем, иногда распадаясь на отдельные капельки. Кроме того,
существует мелкозернистая плазма, так называемая саркоплазма.
Она более прозрачна, слабо окрашивается красками, лежит вокруг
ядра и, видимо, играет определенную роль в обмене веществ.
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
263
Клетки гладкой мускулатуры содержат по одному ядру, которое
занимает центральное положение. Ядра имеют довольно сложную
структуру: под оболочкой ядра лежит спиральное волоконце,
закрученное вокруг ядра много раз (Роскин, 49). Гладкая мышечная клетка
не имеет оболочки наподобие сарколеммы, которую можно
изолировать. Но она покрыта тонкой миолеммой (Роскин) и окружена
тончайшим чехлом из сплетения коллагенных волоконец и межклеточного
Рис. ИЗО.
А — изолированные гладкие мышечные клетки из стенки желудка кошки. Увел. 220. В —¦ гладкие мы.
шечные клетки из кишечника тритона. Миофибрилли хорошо видны. Увел. 1430 (по Леви из Максимова).
вещества. Этот коллагенный чехол исходит из соединительнотканных
прослоек мышечной стенки. Роскин рассматривает мышечную клетку
вместе с этим коллагенным чехлом как единое целостное образование
и называет его м и о н о м.
В гладких мышечных клетках позвоночных, обработанных по Буке,
нервное окончание образует нейрофибрильную сеть, как в
поперечнополосатых мышцах, или же кольцо или петлю. Вокруг него выступает
сетчатое строение протоплазмы. Иногда нервные окончания прилегают
к ядру (рис. 131) (Буке, 63; Лаврентьев, 33).
В гладкой мышце всегда имеется густое нервное сплетение, из
которого мышечные волокна получают свои нервные окончания. Раньше
эти окончания находили в немногих мышечных клетках. Но в
последнее время их обнаруживают во многих органах почти в каждой
мышечной клетке, например в ресничном теле, зрачке, в кровеносных
сосудах (Буке, 62,-63), в желудке (Штёр, 78) (рис. 131).
264
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Гладкие мышечные клетки у беспозвоночных животных имеют
разнообразное строение. По исследованиям Р о с к и н а, они всегда
имеют миолемму — тонкую оболочку, саркоплазму с ядром, причем
ядро занимает обычно центральное положение в клетке, затем особое
протоплазматическое вещество, участвующее в сокращении, так
называемую киноплазму, и фибрилли, которые расположены в клетках
одних животных поверхностно, в клетках других — центрально, треть-
Рис. 131. Нервное сплетение в мышце сфинктера радужной
оболочки у обезьяны.
д — продольный разрез. Ясно выраженный синцитий мышечных волокон
с ядрами и нервными окончаниями. В—две мышечные клетки из того же
объекта с нервными окончаниями (Буке).
их — равномерно по всей плазме. Киноплазма — более жидкое
вещество, чем саркоплазма, не содержит зерен и ячей. Оно сильно
преломляет свет, подобно миофибриллям поперечнополосатой мышцы. Мио-
лемма и фибрилли — эластические образования. Они составляют как
бы скелет мускульной клетки, т. е. придают ей определенную форму.
Длина некоторых мышечных волокон достигает 4 мм, а толщина —
до 100 ji (рис. 132) (Роскин, 49). В некоторых специальных
образованиях они могут быть гораздо длиннее; так, в переднем ретракторе
биссуса Mytilus edulis они тянутся от одного конца мышц до другого-
на расстоянии 4—б см в расслабленном состоянии (Флетчер, 39).
Гладкие мышечные волокна в некоторых случаях соединены между
собой протоплазматическими мостиками: две соседние клетки анасто-
мозируют своими концами. Но эти мостики не всегда хорошо
различимы. Хорошо заметны они, например, в сфинктере радужной оболочки
(рис. 131). Буке (62) находит,что там, где гладкие мышцы образуют
более или менее толстую стенку, как в артериях, кишечнике,
мочеточнике, очень трудно определить границы клеток, ибо они очень тесна
прилегают друг к другу. На продольных разрезах иногда даже не
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСК^ЛАТЧ'РЫ 265
видно продольных линий между клетками. Что же касается миофи-
бриллей, то они не ограничиваются пределами одной клетки, а часто
проходят е нескольких клеточных элементах. Следовательно, гладкие
мышечные клетки в таких случаях построены по синцитиальному типу.
У беспозвоночных иногда встречаются заметные анастомозы между
двумя соседними клетками. У некоторых моллюсков мышечные клетки
связаны даже с эпителиальными клетками: миофибрилли мышечных
клеток переходят в
эпителиальные элементы
(Максимов, 58).
Возбуждение и
возбудимость гладкой мускулатуры.
Возбудимость гладкой
мускулатуры значительно ниже
возбудимости
поперечнополосатой. Для возбуждения ее
требуется относительно
большая интенсивность и большая
продолжительность
раздражения. Для возбуждения гладкой
мускулатуры желудка
лягушки отдельными
индукционными ударами нужно сдвинуть
катушки до 3—5 см. Это
сближение катушек имеет значение
не только в отношении
усиления индукционных ударов, но
и главным образом в
отношении их удлинения. Лучше всего
применять длительные
разряды конденсаторов и гальванический ток. Так, для матки и
мочеточника кошки, морской свинки, кролика можно брать разряды
конденсаторов в 7 jiF или прямоугольные толчки постоянного тока
продолжительностью в 0.7—0.05 сек. (Боцлер, 60).
Стенки желудка обнаруживают большую чувствительность к
механическим раздражениям; даже легкое прикосновение к желудку
вызывает в нем сокращение, но эта чувствительность зависит от нервов.
Если на желудок подействовать атропином и тем парализовать
действие нервов, прикосновение к желудку не вызовет эффекта. Только
сильное механическое раздражение, вроде укола, вызовет эффект.
В случае денервации особенно сильно страдает возбудимость на
электрическое раздражение. Отдельные индукционные удары перестают
вызывать сокращение. Да и тетаническое раздражение не производит
такого большого сокращения, как при нервном возбуждении (Кэн-
нон, 36). Некоторые гладкие мышцы, расслабляющиеся при
раздражении нерва, как, например, мышцы беременной матки, на прямое элек-
Рис. 132.
А — эпителиальн о-м ышечные клетки
Hydra. Сократимый мышечный тяж представляет
собой жидкий плазматический столбик сократительной
плазмы (киноплазмы), окруженной тонкой
эластической мембраной. Внутри киноплазмы проходит твердая
эластическая нить. Подобные же нити проходят по телу
клетки сверху вниз. В — мышечная клетка
В е г о ё (ктенофора). Часть клетки с ядром; снаружи—
киноплазма, которая отделена от осевого слоя
саркоплазмы тоненькой мембраной (Роскин).
266
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
трическое раздражение вовсе не реагируют. На этом основании
К э н н о н совершенно отрицает электрическую возбудимость гладких
мышц. Но другие авторы не согласны с ним. Так, например, на
небеременной матке кролика или кошки электрическое раздражение
вызывает сокращение, которое потом само распространяется. Такое
сокращение наблюдается даже в полоске матки,. не содержащей нервного
сплетения (Боцлер, 60). Также на денервированной мышце
мигательной перепонки кошки доказано, что гладкая мышца хорошо
возбуждается электрическим раздражением. Даже при одиночных
раздражениях она сокращается. Сокращение наступает в ней и при химическом
раздражении, например от адреналина. При этом возникают токи
возбуждения. Все это указывает на существование возбудимой системы
в гладких мышечных клетках.
Особенно большую чувствительность обнаруживают некоторые
гладкие мышцы к ацетилхолину. При этом ацетилхолин вызывает в них
в большинстве случаев почти такое же сокращение, как электрическое
раздражение. Особенно чувствительны к ацетилхолину те мышцы,
которые вообще отвечают медленными сокращениями, так называемые
тонические мышцы, например, продольная мышца голотурии, мышца
моллюска, сокращающиеся при действии ацетилхолина в концентрации
1 : 100 000 (Риссер, 68).
Все физиологические процессы в гладкой мускулатуре протекают
очень медленно, даже значительно медленнее, чем в сердечной. Судя
по электрическим токам возбуждения, процесс возбуждения может
длиться несколько секунд. Так, на желудке собаки при раздражении
соответствующего двигательного нерва (п. vagus) ток возбуждения
продолжается почти все время сокращения. Если один электрод
находится на желудке, а другой где-либо на наружной поверхности тела,
то ток возбуждения является четырехфазным: 1-я и 2-я фазы —
протекают быстро, каждая длится от % до 1 сек., а 3-я и 4-я фазы —
протекают медленно, по 5—15 сек. Первые две фазы приходятся на время
сокращения продольных мышц, а последние — на время сокращения
кольцевых (Рихтер, 11). Как и в сердечной мышце, ток возбуждения
начинается еще в скрытый период сокращения, достигает максимума
раньше максимума сокращения и раньше него заканчивается. Так, по
крайней мере, наблюдалось при одновременной регистрации
механического и электрического эффектов желудка лягушки (Чермак, 4)
(рис. 133). При длительной тонической деятельности, как это бывает
в гладкой мускулатуре retractor penis, токи возбуждения следуют
ритмически. Чем выше «тонус» сокращения, тем больше частота и
амплитуда токов возбуждения (Брюке, 12) (рис. 134).
При спонтанных маятникообразных движениях, которые возникают
в кольцевых мышцах кишечника, каждой волне сокращения
соответствуют два-три импульса. При перистальтических же движениях,
происходящих от сокращения продольных мышц кишечника, возникает
длинный ряд импульсов (Боцлер, 60) (рис. 135). Итак, при всех почти
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
267
условиях возбуждение в гладкой мускулатуре протекает ритмически,
подобно тому как в скелетной мускулатуре и нервах.
По новейшим исследованиям, электрический эффект гладкой мышцы
на непрямое раздражение бывает двоякого типа. Одни гладкие мышцы
Рис. 133. Электрический и механический эффекты желудка лягушки.
Кольцо, вырезанное из средней части желудка. Ток возбуждения отводится в струнный гальванометр.
Верхняя кривая представляет электрограмму, нижняя — механограмму. В самом низу дается время:
каждая черточка соответствует одной секунде. Регистрируется автоматическая деятельность желудка:
две последовательные волны возбуждения и сокращения. Ясно видно, что соответствующий волне
возбуждения электрический ток начинается раньше и продолжается значительное время в течение
сокращения (Чермак).
реагируют на раздражение нерва индукционным ударом одиночным
сокращением, которое сопровождается многофазным электрическим
эффектом. Таковы пиломоторы (мышцы, заведующие движением
волос) , мочевой пузырь, мигательная перепонка. Число фаз достигает
семи, как, например, на пиломоторах и мигательной перепонке кошки
(Розенблют и Позо, 77). По данным Экклса и Магладери (38),
Рис. і134. Электрограмма т. retractor penis собаки в состоянии тонического
сокращения.
Токи возбуждения записаны струнным гальванометром. Мышца отводиіся неполяризу-
ющимися электродами. Токи возбуждения следуют один за другим правильным ритмом
через каждые 5 сек. Внизу — время te секундах (Брюке).
мигательная перепонка кошки отвечает на раздражение соответственного
постганглионарного симпатического нерва сначала одним начальным
комплексом, а затем рядом электрических волн, протекающих в ритме
от 3 до 1 в секунду. Они возникают в базальной части, где
оканчивается нерв, «и распространяются к ее верхушке. То же бывает при
интравенозной инъекции адреналина, причем каждое колебание
электрического потенциала сопровождается сократительным эффектом,
который начинается вслед за началом электрического эффекта. Чем
сильнее раздражение, тем дольше держится ритмический эффект; при
двух последовательных раздражениях он длится дольше, чем при
одном (см. рис 136).
268 общая физиология мышечной системы
Э к к л с (38) полагает, что начальный комплекс электрического эф*
фекта возникает под влиянием нервного тока возбуждения;
дальнейший же ритмический разряд производится активным веществом (симпа-
Рис. 135. Токи возбуждения тонкой кишки.
А — периодический разряд тонкой кишки кролика; В — периодические
перистальтические разряды тонкой кишки кошки; С и D — токи возбуждения при маятникообрао-
ных движениях тонкой кишки^ кошки (Боцлер).
тином), которое освобождается под влиянием нервных импульсов.
Такого же мнения придерживается Монние (50). Этот автор
находит, что вообще в гладкой мускулатуре длительные и тонические
сокращения, равно как отвечающие им медленные ритмические измене-
Рис. 136. Электрический и механический эффекты мигательной
перепонки кошки.
Раздражается постганглионарный нерв верхнего шейного ганглия
отдельным^индукционным ударом. В каждом опыте одна кривая
представляет запись механического эффекта, а другая (сильно колеблющаяся)—
от электрического. В опыте 3 нерв раздражается одним индукционным
ударом, а в опытах 7 и 2 — двумя ударами с интервалом 15 и 310 а.
А и В—волны начального комплекса, следующие за раздражением,
R —-ритмический разряд (Экклс и Магладери).
ния потенциала, вызываются химическим веществом — медиатором,
которое выделяется нервным окончанием, в то время как быстрые
вздрагивания мышцы и соответственно быстрые колебания электрического
потенциала возникают в мышце под влиянием нервных импульсов
непосредственно. Некоторые авторы находят, что два последних
медленных компонента из каждого многофазного эффекта на одиночное раз-
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ 269
5mV
дражение нерва связаны с сокращением, а не с возбуждением. Именно,
шестой по счету компонент, который совпадает с периодом развития
механического эффекта, вероятно, выражает химический процесс,
соответствующий этому развитию, а седьмой компонент, который следует
Ео время сокращения и некоторое время после него, должно быть,
выражает процесс восстановления (Розенблют и Позо, 77).
Другие гладкие мышцы, вовсе не отвечая на одиночное
раздражение нерва, реагируют только на ряд индукционных ударов или на
действие сильного постоянного тока. По Л а п и к у, эти мышцы являются
итеративными органами, т. е. требующими повторного
раздражения для вызова двигательного эффекта. Соответственно хронаксия их
очень велика. Обычно такие
сек
мышцы на длительное
раздражение отвечают серией
биоэлектрических токов, как и при
спонтанной автоматической
деятельности. Но каждое колебание
носит сложный характер и
включает в себя и быструю и
медленную фазы. Таковы мышцы
матки, retractor penis и др.
(Розенблют и сотр., 34).
Характерно, что при действии
адреналина наступает только
медленная фаза. На быстрые фазы он,
наоборот, действует
отрицательно. К э н н о н полагает, что
медленная фаза всецело имеет
химическое происхождение. Быстрые же колебания биотока он не
приписывает импульсам возбуждения, а считает результатом
деполяризующего действия медиатора на ту клетку, где он выделился, а
также на другие клетки. По его мнению, адреналин потому действует на
них отрицательно, что он сам производит аналогичную
деполяризацию (37).
Сложная форма токов возбуждения при отведении от гладкой
мышцы вовсе не обязательна. Она безусловно связана со сложным
строением мышечной ткани. В случае простого строения, с
параллельным течением мышечных волокон, токи возбуждения имеют такую же
простую форму однофазного и двуфазного тока, как в скелетных
мышцах с параллельными волокнами. Так, например, в переднем ретрак-
торе биссуса моллюска Mytilus edulis, при прямом раздражении, на
равных расстояниях от раздражаемого участка токи возбуждения
являются двуфазными при продольном отведении и однофазными при
поперечно-продольном (Флетчер, 39) (рис. 137).
Проведение возбуждения в гладкой мышце происходит очень
медленно. В гладкой мускулатуре желудка лягушки, например, возбужде-
\
Рис. 137. Токи возбуждения переднего рет-
рактора биссуса Mytilus edulis:
а — при"продольном отведении, b — при попере^но-
продольном. Сверху время в секундах, сбоку
напряжение токов возбуждения в милливольтах
(Флетчер).
270
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ние переходит с одного участка на соседний настолько медленно, что
приходится выжидать десятки секунд, чтобы заметить этот переход.
В гладкой мускулатуре теплокровных животных возбуждение
распространяется быстрее. Так, например, в мышце тонкой кишки человека
скорость распространения возбуждения равняется 1 см в секунду
(Геитер, 5). Такое медленное проведение возбуждения, повидимому,.
связано с замедленным переходом возбуждения от одной клетки на
другую. В желудке длина мышечных волокон менее 1 мм, а потому
замедленный переход возбуждения должен происходить часто. В
пределах мышечной клетки возбуждение, должно быть, распространяется
значительно быстрее. Этим и объясняется, видимо, тот факт, что в тех
органах, где гладкие мышечные волокна достаточно длинны, скорость
проведения возбуждения оказывается значительно больше. Так,
например, в мышечных клетках мигательной перепонки кошки возбуждение
распространяется со скоростью 50—80 см в секунду (Экклс и Магла-
дери, 38), в мочеточнике кролика со скоростью 18 см, в матке кошки —
7 см (Боцлер, 60).
В длинной полоске матки, в кишечнике и в мочеточнике кошки,
кролика возбуждение распространяется, даже если произвести в них
зигзагообразный разрез (Боцлер, 60). Это указывает, что в гладкой
мыіпце означенных органов возбуждение распространяется
непосредственно от одной мыщрной клетки к другой. Это зависит от того, что»
между мышечными клетками существуют протоплазматические
мостики, которые проводят возбуждение, или* же токи возбуждения
каждой мышечной клетки настолько сильны, что возбуждают соседние
мышечные клетки. Но весьма возможно, что в нормальном желудке и
тонкой кишке позвоночных животных возбуждение переходит с одной
части мышечного слоя на другие пр'и участии нервной системы,
а именно через ауэрбаховское сплетение. Кроме того, в определенных
случаях в распространении возбуждения существенную роль должны
играть активные вещестга, выделяемые нервными окончаниями.
В некоторых неповрежденных мышечных волокнах возбуждение-
распространяется без ослабления, т. е согласно закону возбуждения.
Это было показано с очевидностью на переднем ретракторе Mytikts
edulis. В этой мышце длиной 5—6 см возбуждение распространяется со
скоростью 13—22 см в секунду без всякого декремента (Флетчер, 39).
8 матке и мочеточнике теплокровных животных точно так же
возбуждение, повидимому, распространяется без ослабления (Боцлер, 60).
Механический эффект гладких мышц. Сокращение гладкой мышцы
характеризуется очень большой длительностью, особенно в
нисходящем колене. У лягушки все сокращение гладкой мышцы желудка
длится 80—100 сек., причем на восходящее колено приходится 15—
20 сек. Следовательно, период, ослабления почти в 5 раз больше
периода нарастания сокращения. Но сокращение гладкой мышцы может
протекать и значительно быстрее, например, полное сокращение memb-
гапа nictitans зайца продолжается всего 5 сек.
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
271
Сократительный процесс в гладкой мышце происходит настолько
медленно, что его волну можно наблюдать в одном мышечном
волокне, если раздражать мышечное волокно под микроскопом. Во время
сокращения веретенообразное волокно укорачивается и утолщается.
й*-*-*«* ™-^^f^w'>4^j*a^.-^w«^s?7W'
.а^ы
\
•ц, *. , - Wi
Рис. 138. Сокращенная клетка гладкой мышцы из желудка воробья.
* В ней видно шесть волн сокращения (рис. Соли из Максимова).
Можно даже видеть в одном волокне гистологически обработанного
фиксированного препарата несколько волн сокращения (рис. 138).
Гладкая мышца при раздражении отдельными индукционными ударами
укорачивается на 45%. При максимальном же тетаническом
сокращении укорочение доходит до 60—75% (Шюллер, 6; Шультц, 1).
Рис. 139. Сокращение спинной мышцы пиявки. Длина мышцы в покое около 2 см.
Ток пропускается через всю мышцу.
В опыте А раздражение производится одним индукционным ударом. ,Момент раздражения обозначен
на пинии а. В опыте В раздражение производится рядом индукционных ударов. Моменты раздражений
отмечены на линии Ь. Сокращение дает в обоих случаях длительное последействие, продолжающееся:
в первом случае около 13 мин., а во втором—свыше получаса. Во время последействия кимограф
останавливался много раз: в опыте А—по 1 мин., в опыте В—по нескольку минут. В конце опыта В миограф'
9 раз поднимался и опускался; только после этого последействие сошло на-нет (Гогава).
У беспозвоночных сокращение гладкой мышцы, вызванное
однократным раздражением, например, одним индукционным ударом,
может длиться еще больше — в течение нескольких минут. При частых
раздражениях получается тетаническое сокращение. Смотря по частоте
раздражения, оно может быть неполным клоническим тетанусом и
полным тетанусом (рис. 139).
Механический эффект мышцы беспозвоночных на одиночное
раздражение часто бывает двоякого рода: быстропротекающий, наподо-
272
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНЭЙ СИСТЕМЫ
бие одиночного сокращения скелетной мышцы лягушки, и медленно
протекающий, который может тянуться в течение многих десятков
секунд. Это медленное сокращение протекает с меньшим напряжением,
чем быстрое. Обычно медленное сокращение сливается с быстрым, но
в определенных случаях медленное сокращение отделяется от
быстрого: оно начинается после того, как быстрое сокращение мышцы
достигло максимума. При определенных условиях медленное
сокращение значительно ослабевает и даже исчезает без аналогичного
изменения быстрого сокращения. Например, это бывает при согревании до
30—40° С и при охлаждении до —2° С. При истощении препарата
вместе с выпадением собственного тонуса исчезают и длительные
последействия (Гогава, 52). Все это указывает на то, что субстрат быстрого
и медленного сокращения не один и тот же.
По мысли Иордона в мышце существуют две сократительные
системы: быстросокращающаяся и медленносокращающаяся. Весьма
вероятно, что миофибрилли являются сократительным субстратом
с функцией быстрого сокращения. Что же касается медленного
сокращения, то его можно считать функцией так называемой киношіазмы,
которая, по исследованиям Р о с к и н а (48), является
дифференцированной частью саркоплазмы и прилегает к миофибриллям. Кино-
плазма претерпевает такого рода морфологические изменения при
сокращении, что ей можно приписать сократительную функцию.
Гистологическое изучение живых мышечных клеток во время
сокращения ясно показывает, что в определенных случаях действительно
сокращаются фибрилли. Так, в последнее время Боцлер (7)
исследовал микроскопически отдельные мышечные волокна из личинки
кишечнополостных животных (Вегоё forskali). Эти волокна были
длиной 2—4 мм, а толщиной 5—10 jx'. При раздражении отдельными
индукционными ударами волокна сокращались до 7в первоначальной
длины. При этом укорачивались одни миофибрилли, а поверхностный
слой саркоплазмы образовывал неправильные складки. Отсюда
Боцлер заключил, что фибрилли являются единственным сократительным
элементом. Однако Роек и н (48), производивший детальное гисто-фи-
эиологическое исследование живых и мертвых объектов разных
беспозвоночных (черви, иглокожие, моллюски), приходит к заключению, что
сокращение гладкой мышцы производится при участии киноплазмы,
которая прилегает к миофибриллям. При сокращении киноплазма не
просто образует неправильные складки, как указывает Боцлер, а
изменяется структурно: из стекловидно-прозрачной массы переходит
в беловато-мутную. При этом в ней возникают сгущения, пучности,
узлы сокращения (рис. 140). Р о с к и н наблюдал сокращение
киноплазмы на таких объектах, где нельзя было обнаружить фибриллей.
Поэтому можно утверждать, что сокращение гладких мышц
беспозвоночных тесно связано с функцией киноплазмы, миофибрилли же в
некоторых клетках играют главным образом опорную роль и, как
эластические нити, должны принимать участие в распрямлении мышечной
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
273
клетки после сокращения, в возвращении ей нормальных формы и
длины (Роскин, 48).
Гисто-физиологическое исследование гладких мышц позвоночных
также указывает на сократительную функцию киноплазмы. Р о-
скин (49) исследовал, гисто-физиологически гладкие мышцы кошки
н аксолотля и пришел к заключению, что в гладких мышцах
позвоночных сокращение есть функция как миофибриллей, так и імежфибрил-
лярной плазмы,
представляющей собой киноплазму. При
сокращении межфибриллярная
плазма образует более
интенсивно окрашиваемые
некоторыми красками капли,
которые повисают на фибриллях;
с течением времени маленькие
капли сливаются в большие.
Словом, наблюдается та же
картина сокращения, что и в
гладкой клетке
беспозвоночных (рис. 141).
Итак, по новейшим
данным, сократительный
механизм гладких мышц вообще
качественно отличается от со-
кратителъного механизма
поперечнополосатых мышц. В
последнем случае сокращение
является исключительной
функцией миофибриллей. В
гладких же мышцах мио-
фибрилли играют главным образом опорную, формоопределяющую
роль Сокращение есть функция особой межфибриллярной плазмы —
киноплазмы. Но уже у некоторых беспозвоночных миофибрилли несут
также сократительную функцию, наравне с опорной. Такую
двойственную функцию, несут, вероятно, и миофибрилли гладких мышц
позвоночных.
Филогенетическое развитие сократительной функции следует
представить в следующем виде: первоначально протоплазма обладала
способностью к сокращению. Такова протоплазма амеб, лейкоцитов и тому
подобных одноклеточных животных с амебоидным движением. На
следующей ступени развития двигательной функции произошла
дифференциация протоплазмы: она разделилась на киноплазму с
сократительной функцией и плазму с трофической функцией (Роскин). На
более высокой ступени развития из протоплазмы выделились фибрилли,
которые стали ' играть сначала формообразующую роль, образуя как
бы скелет клетки, причем они еще не принимали определенного уча-
-9 И. С. Беритов
Рис. 140. Сокращение изолированных мышеч
ных клеток Bolina
А и В — сильное утолщение киноплазмы (кольцевого
слоя), сопровождающееся образованием особых вздутий
или узлов сокращения. С — сокращенная клетка
нарисована при более глубокой установке фокуса
объектива (Роскин).
274
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
стия в самом сократительном процессе. Таковы они у многих
одноклеточных и низших беспозвоночных. Затем фибрилли начинают
принимать определенное участие в сократительном процессе, например,,
в гладких мышцах некоторых высших беспозвоночных и всех
позвоночных животных. Наконец, они принимают на себя всю сократитель?
иую функцию. Такова их роль в поперечнополосатых мышцах
позвоночных и, видимо, беспозвоночных животных. Сократительная функция:
миофибриллей качественно другого рода и стоит много выше
сократительной функции киноплазмы. Характерными особенностями
сокращения миофибриллей являются: высокое напряжение, большая
быстрота укорочения, быстрое расслабление. Вероятно, и биохимические
Рис. 141, Волны сокращения в гладкой мышечной
клетке из кишечника кошки.
Тонкие линии от миофибрилл. В участках сокращения
межфибриллярная киноплазма уплотнена (Роскин).
процессы, обусловливающие сокращение миофибриллей, характерно
отличаются от таковых для сокращения киноплазмы.
Новейшие исследования Хлопина (69) и др. свидетельствуют
о разном эмбриональном происхождении мышечной ткани. Так,
гладкая мышца у позвоночных происходит из мезенхимы, а у многих
беспозвоночных из эктодермы или энтодермы. Поэтому едва ли можно
сказать, что гладкие мышцы позвоночных возникли из гладкой мышцы:
беспозвоночных в результате их филогенеза. Но это не противоречит
вообще намеченной тенденции филогенетического развития
сократительной функции. Поперечнополосатая мышечная ткань признается:
происходящей из элементов, очень сходных с миоэктодермальными
элементами кишечнополостных. Следовательно, наряду с развитием
сократительной функции из первоначального древнего источника по>
намеченному выше плану, на определенной стадии филогенеза
животного мира выдвинулся новый источник — мезенхима
(соединительнотканные элементы) для развития той же сократительной функции,,,
приблизительно тем же путем, как это было с эктодермальными
элементами у низших беспозвоночных.
Сокращение в гладких мышцах обусловливается такого же рода
химическими процессами, как в поперечнополосатых. В них
происходит распад фосфагена и гликогена; в результате возникает
сокращение. Но не во всех гладких мышцах находят фосфаген как креатино-
фосфат. У беспозвоночных животных находят фосфаген в виде арги-
нинфосфата. Но в мышцах некоторых низших беспозвоночных не на-
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ 275
ходят совсем фосфагена. Так, например, его нет в мускулатуре
медузы. Здесь есть другого рода фосфорные соединения, которые при
сокращении распадаются, образуя фосфаты. Ресинтез фосфорных
соединений связан главным образом с процессами окисления молочной
кнслоты. Гликоген в гладких мышцах как позвоночных, так и
беспозвоночных животных распадается до молочной кислоты анаэробно.
Затем происходят окисление части молочной кислоты и восстановление
асей остальной молочной кислоты в гликоген за счет энергии окисле-
а.ія (Мейергоф, 65; Болдуин, 66; Вержбинская, 67). Характерно, что
на известных ступенях онтогенеза у некоторых высших позвоночных
также не был найден фосфаген. Значит, у них сокращение происходит
без участия фосфагенного механизма. Следовательно, надо полагать,
что фосфагенный механизм, участвующий в производстве сокращения,
является одним из достижений эволюции (Коштоянц).
Скрытый период сокращения. В гладкой мышце желудка лягушки
скрытый период составляет около 0.25—1 сек. (Альварец, 9). В
мышцах мигательной перепонки скрытый период сокращения равен 0.2 сек.
(Экклс и Магладери, 37). Но в других гладких мышцах позвоночных
животных скрытый период длится дольше. Особенно длителен он
у мышц, которые реагируют только на тетаническое раздражение.
Здесь он может продолжаться до нескольких секунд. Длительность
скрытого периода зависит от силы раздражения: скрытый период тем
короче, чем сильнее раздражение (Будде и Гельхорн, 10). Степень
возбудимости, конечно, также имеет большое значение. Например, при
Ескусственном повышении возбудимости под влиянием малых доз
пплскарпина или адреналина скрытый период уменьшается. Наоборот,
при понижении возбудимости в случае отравления большими дозами
тех же веществ скрытый период увеличивается (Будде и Гельхорн, 10).
Когда груз велик, продолжительность сокращения уменьшается, -и
скрытый период удлиняется. При повышении температуры сокращение
усиливается, длительность его, а также скрытый период уменьшаются.
Это изменение достигает максимума при температуре 39°-,
Закон возбуждения и рефракторные фазы. В первое время
утверждали, что гладкая мышца не реагирует по закону возбуждения, не
знает рефракторных фаз. Такое заключение сначала было' сделано
Шульцем (1). Он исходил из того наблюдения, что сократительный
эффект от двух быстро следующих одно за другим раздражений всегда
больше, чем от одного. Однако, по более поздним исследованиям,
следует признать, что это заключение было сделано неправильно: Так,
Шюллер (6) установил, что -мышца прямой кишки лягушки
отвечает на раздражение сначала абсолютной рефракторной фазой,
которая приходится на время скрытого периода и в начале сокращения,
а затем относительной, которая приходится на время сокращения и
немного после него. Далее было найдено, что тонкая кишка
теплокровного животного на тетаническое раздражение отвечает не
тетанусом, а клоническим эффектом, подобно сердцу (Магнус, 13). Наиболеіе
18*
276
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
демонстративно это было доказано в последнее время на денервиро-
ванной мышце мигательной перепонки кошки. При двух максимальных
раздражениях второй удар не дает электрического эффекта только
при интервале менее 0.2 сек. (Экклс и Магладери, 37). При
спонтанных ритмических эффектах каждый комплекс волн сопровождается
рефракторностью, т. е. невозможностью вызвать в этот период новый
У
2-
электрический эффект посторонним
раздражением (Экклс и
Магладери, 38).
За абсолютной рефракторной
фазой следует относительная. На
мочеточнике кролика она длится
после абсолютной рефракторной
фазы около 1 сек. При этом
характерно, что в начале относительной
рефракторной фазы сокращение
возникает на катоде раздраженного
участка, но оно не
распространяется (Боцлер, 60). Повидимому,
возникающий при этом
биоэлектрический ток настолько слаб, что он не
в состоянии раздражать соседний
участок.
Прекрасную иллюстрацию
закона возбуждения дает Боцлер
(60) на гладких мышцах іматки и
мочеточника теплокровных
животных,. На мышечной полоске матки
и отрезках мочеточника величина
мышечного напряжения не зависит
от силы раздражения. Так, на полоске іматки кошки каждый
деятельный стимул производит максимальный эффект (рис. 142).
Точно так же у некоторых беспозвоночных гладкие мышцы
реагируют по закону возбуждения. Так, Боцлер (7) на гладкой мышце
Вегоё показал, что при микроскопическом наблюдении мышечные
волокна укорачиваются одинаково сильно, до Vs первоначальной длины,
как при умеренных, так и при очень сильных раздражениях
отдельными индукционными ударами. Легко выявляется закон возбуждения
и иа мышцах медузы. На небольшой полоске мышечного кольца,
расположенного на краю колокола медузы, каждая волна сокращения,
вызванная одним электрическим ударом, сопровождается абсолютной
рефракторной фазой, в течение которой нельзя вызвать
дополнительного сокращения каким-либо новым раздражением. Эта мышца,
подобно сердцу, в нормальном состоянии не дает тетануса. При
некоторых высоких частотах раздражения, как 50—100 в секунду, получается
пессимальный эффект, а при низких частотах мышца отвечает ритми-
Рис. 142. Полоска гладкой мышцы из
матки кошки для демонстрации закона
возбуждения.
Раздражается прямоугольными токами
(постоянный ток) разного вольтажа, продолжительностью
О.З сек. Верхняя кривая указывает на
изменения вольтажа, а нижняя — на напряжение
мышцы при изометрической записи. При каждом
сокращении развивается напряжение около 1 г.
Температура 33°С (Боцлер).
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
277
ческими эффектами по ритму 1.5—1.8 в секунду. Словом, мышечная
полоска медузы обнаруживает такие же физиологические свойства,
как сердце. Подобно сердцу, она обладает низкой лабильностью, а во
время активного, состояния лабильность сильно увеличивается, что
ведет к укорочению рефракторной фазы іи хронаксии (Жуков, 57). Из
=всех этих наблюдений, ясно следует, что возбудимая система гладкой
мышцы, подобно скелетной и сердечной мышцам, подчиняется закону
возбуждения.
При некоторых сильных раздражениях на гладкой мускулатуре
позвоночных можно получить тетанический эффект, как это было в опытах
Шульда. Однако это еще не означает, что в таких случаях
абсолютная рефракторная фаза отсутствует. Тетанический эффект должен
наблюдаться во всех тех случаях, когда процесс возбуждения
начинается раньше механического эффекта и кончается прежде, чем
наступает максимум сокращения. Итак, при всех условиях раздражения,
возбудимая система гладкой мышцы отвечает на раздражение
согласно закону возбуждения.
Пластичность и собственный тонус. Гладкие мышцы, подобно
поперечнополосатым мышцам, обладают упругими и пластическими
свойствами. Но только у гладких мышц эластические свойства
выражены слабее, а пластические, наоборот, сильнее, чем у
поперечнополосатых мышц. Гладкие мышцы растягиваются под действием
растягивающей силы сначала быстро, потом медленно и принимают разную
длину без заметного изменения напряжения. По удалении этой силы
они стягиваются обратно сначала быстро, потом медленно.
Особенно сильно выражены пластические свойства в мышцах
полостных органов. Как у позвоночных, так и у беспозвоночных
животных с увеличением содержимого полостных органов мышечные стенки
растягиваются, но при этом давление внутри органа не меняется. Так,
например, желудок собаки, заключая в себе 240 см3 жидкой пищи,
развивает такое же небольшое давление, как и при содержании
460 см3 жидкости, а именно около 7 см водяного столба (Келлинг).
Точно так же в сильно наполненном мочевом пузыре это давление
составляет 8—10 см водяного столба. Значит, сократительные элементы
желудка и мочевого пузыря при любой длине в известных пределах
могут пребывать в покое и обнаруживать лишь незначительное
напряжение. Изменение напряжения гладких мышц наблюдается только при
сокращении. Следовательно, в полостных органах давление
повышается только при сокращении мышечных элементов.
По удалении растягивающей силы полостной орган быстро
уменьшается до некоторого объема, но не остается в этом состоянии, а
продолжает постепенно уменьшаться до того состояния, какое было до
действия растягивающей силы. При этом напряжение в мышечных
стенках не нарастает: оно остается на одном и том же уровне. Если
вырезать кольцо из середины желудка лягушки или поперечную
полоску и подвесить небольшую тяжесть, то удлинение будет происхо-
278
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
дить очень долго, в течение многих минут безостановочно.. Если потом
снять груз, то мышца укоротится сейчас же, но очень, незначительно.
Она останется более или менее долго растянутой, ибо восстановление
исходной длины происходит с большой постепенностью. Обычно в
гладкой мышце также возникает напряжение в связи с растяжением, как
в скелетной мышце. Но это напряжение быстро проходит. Если
растянуть мышцу изометрическим миографом до определенной длины, то
в течение одной-двух минут это напряжение сильно ослабнет и даже
сойдет на-нет. Но есть и такая гладкая мышца, которая вообще не
развивает напряжения. Таков, например, кожиомышечный покров
голотурии. Он может быть растянут до любой длины без возникновения
какого-либо напряжения. Так, например, полоска покрова в 6 см
может быть растянута в течение нескольких часов до 38 см. Эта мышца
является чисто пластическим образованием. Она не способна к
сокращению. Внутри кожномышечного покрова имеются настоящие
сократительные волокна — кольцевые и продольные. Сокращение их
уменьшает объем полости водяного легкого, из которого выжимается вода.
При этом сжимается и масса пластического кожного покрова. По
прекращении сокращения сократительных волокон уменьшенный объем
полости сохраняется благодаря вязкости кожного покрова.
Отсюда следует, что гладкие мышцы способны к такому изменению
формы, при котором происходит прирост напряжения; но, кроме того,
в них происходит смещение частиц относительно друг друга без
изменения формы мышцы, без изменения напряжения (Иордан, 25)..
Подобно скелетным мышцам, гладкая мышца сама укорачивается
до некоторой минимальной длины, если только на нее не действует
какая-либо растягивающая сила. После любого растяжения гладкая
мышца сама может принять свою начальную длину. Так, на поперечных
отрезках желудка лягушки, которые вообще растягиваются сильно,
исходная длина восстановится в одну минуту, если растяжение
происходило в течение нескольких секунд; если же растяжение
происходило несколько минут, тогда исходная длина восстановится в
течение нескольких минут, как это показано на рис. 143 А и В. В этом
выражается собственный тонус гладкой мышцы. Он проявляется тем
сильнее, чем лучше функциональное состояние мышцы. При некоторых
длительных растяжениях полного восстановления не бывает, мышца
остается более или менее удлиненной, если только не сократится
спонтанно, автоматически или под влиянием какого-либо внешнего
раздражения. В этом случае восстановление может произойти, при
любом растяжении в первую же минуту после удаления растягивающей
силы. Поперечный отрезок желудка, взятый из кардиальной области,
обычно производит спонтанные ритмические сокращения. Такой
отрезок восстанавливает первоначальную длину сейчас же по удалении
растягивающей силы (рис. 143 С) (Беритов и Ннвинская, 70):.
Как указывалось выше, многие гладкие мышцы беспозвоночных
способны к двоякого рода сокращениям ів ответ на раздражения:
ОБЩАЙ ФИЗИОЛОГИИ ГЛАДКОЙ: МУСКУЛАТУРЫ §?9
к медленному и быстрому. Медленное сокращение оставляет за собою
длительное тоническое последействие. Это тоническое последействие
может продолжаться часами и даже днями. Так, например, по данным
Гогава, на мышце пиявки тоническое последействие проходит в
течение многих десятков минут (рис. 139), а по данным Коштоянца и
J. =г?мм
/піП1ііііііііііііііпііііміііііііі)Иінміпііііііііічіііііпіп tun мшит inn и mum і іппиіПіііііін
<ntnMMi\!niMMitiunUniiim.tninnnmtt^iMMMi.Mt^nillM'MMninnl'lUn,nnnMMlinlinMMnM1-
Рис. 143. Растяжение іи укорочение поперечного отрезка желудка лягушки.,
Л*—длина нерастянутого отрезка .желудка 22 мм. Растягивается грузом в 5 г, сначала на несколько
секунд, а потом на 10 мин. (опыт В). После растяжения мышца разгружается и миограф
приподнимается (звездочки!). После короткого растяжения, одноминутного поднятия миографа (кимограф
на это время остановлен) оказалось достаточным для восстановления исходной длины. После длительного
растяжения до 40 мм исходная длина не восстановилась и в течение 10 минут. С — другой препарат,
сокращавшийся спонтанно. Этот отрезок также растягивался в течение 10 минут грузом в 5 г. После
удаления этого груза исходная длина восстановилась в течение 2—3 мин., потом мышца укоротилась еще
сильней вследствие спонтанных сокращений, происходивших приблизительно через 1 мин. Звездочки
отмечают [момент остановки кимографа, после того как спонтанное сокращение достигало максимума.
М у ж е е в а, мускулатура ноги улитки приходит в полный покой
через четыре дня после раздражения. Это длительное последействие
сокращения, безусловно, пластического происхождения. Во время
сокращения происходит такое перераспределение молекул в вязкой
системе мышцы, что после сокращения мышца остается укороченной.
Зто пластическое последействие проходит очень (медленно под
влиянием тяжести миографа и собственной тяжести мышцы, если нет
спонтанного удлинения, происходящего под влиянием нервной системы.
280
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Пластическое происхождение последействия доказывается тем, чт®
оно всецело зависит от температуры. Лучше всего оно выступает при
некоторой средней температуре—16—20° С. При повышении
температуры оно выступает все хуже и хуже и уже при температуре 32G
мышца расслабевает быстро после сокращения, в течение нескольких
секунд, а при 40—45° С последействие почти отсутствует. Это
изменение тонического последействия с повышением температуры происходит
параллельно с изменением собственного тонуса. При повышении
температуры выше 16—20° С собственный тонус падает, мышца удли-
Рис. 144. Удлинение спинной мышцы пиявки при охлаждении.
Раствор Рингера комнатной температуры (19°С) заменяется охлажденным до 2°С Охлаждение мышцы
продолжается в течение 1 часа; когда мышца перестает удлиняться, холодный раствор заменяется
комнатным. Мышца укорачивается от согревания до комнатной температуры (Гогава).
няется, одновременно уменьшается пластическое последействие после
сокращения (Гогава, 52; Коштоянц и Мужеев, 26). Это объясняется
тем, что вязкость системы уменьшается, когда температура
повышается. Но если сильно и надолго охладить гладкую мышцу ниже
6° С, то и в этом случае собственный тонус ослабевает. Так бывает
при охлаждении ножки аплизии (Иордан, 25), а также спинной мышцы
пиявки (Гогава, 52) (см. рис. 144).
То обстоятельство, что собственный тонус и пластическое
последействие изменяются параллельно, указывает, между прочим, на то, чтз
субстрат медленного сокращения и пластического укорочения должен
быть одним и тем же. Этим субстратом по всей вероятности является
главным образом киноплазма, которой приписывается функция
медленного сокращения.
Собственный тонус гладкой мышцы, как он выявляется в
тоническом укорочении после сокращения, вызванного раздражением, илк
в спонтанном укорочении после удаления растягивающей силы,
рассматривается многими авторами как чисто физическое явление,
свойственное многим неживым образованиям, например, пластическому
каучуку (Иордан, 24; Боцлер, 43). Были проведены специальные
опыты в отношении обмена веществ при разных условиях состояния
гладкой мышцы, в частности, во время пассивного укорочения после
разгрузки, причем не было найдено никаких изменений в обмене
веществ. Обмен веществ изменялся только во время сокращения и
контрактуры под влиянием нервных импульсов (Маас, 55).
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
281
Но опытами Коштоянца и сотрудников было показано, что
тонические явления гладкой мышцы в определенной мере обусловлены
обменом веществ, ибо они зависят от окислительных процессов, с
ослаблением которых тонические явления выпадают (Коштоянц и Му-
жеев, 27; Кудрявина, 56). Надо полагать, что собственный тонус
гладкой мышцы обусловливается теми физико-химическими процессами,
которые обычно протекают в мышце в .состоянии покоя. Беспрерывное
расщепление гликогена и в особенности фосфагена, которое имеет место
все время и при полном покое, должно иметь особенно большое
значение в происхождении этого тонического состояния. Мышца после
растяжения сама укорачивается или удерживается в укороченном
состоянии после тетануса главным образом под влиянием, этих
процессов. Поскольку собственный тонус зависит, с одной стороны, от
вязкости мышечной структуры, а с другой — от энергетического обмена
веществ, то всякое изменение вязкости и обмена веществ должно
влиять и на степень собственного тонуса. У позвоночных, животных
большую роль играет в этом отношении вегетативная нервная система.
Она влияет как своими нервными импульсами, так и гормонами,
которые она выделяет, например, адреналином, ацетилхолином. У
беспозвоночных собственный тонус также регулируется как нервной системой,
так и гормонами. Удаление ножного ганглия улитки повышает
вязкость гладких мышц ноги: нога становится твердой наощупь и
оказывает большое сопротивление растяжению. Значит, нормально ножной
ганглий оказывает длительное тормозящее влияние на собственный
тонус (Иордан, 45). У осьминога тонус поддерживается гормоном
тирамином, который вырабатывается задней слюнной железой;
гормон бетаин, наоборот, уничтожает его (Серени, 47).
Многочисленными учеными, применявшими как старые, так и
новейшие методы биохимического исследования (Бете, 21; Парнас, 20;
Паркер, 28; Маас, 55), было доказано, что во. время собственного то:
нуса гладкой мускулатуры очень часто не происходит, повышения
обмена веществ. Но в определенных случаях ясно улавливается
некоторое усиление обмена веществ. Так, например, в гладкой мышце
ретрактора глотки виноградной улитки происходит небольшое, но
заметное повышение теплопродукции при длительном укорочении (Боц-
лер, 29). Также в мышцах пиявки было замечено небольшое
повышение потребления Ог и выделения С02 (Конгейм и Икскюль, ,54).
Однако считают возможным, что это повышение обмена веществ было
результатом повреждения мышцы, нанесенного операцией и самой
манипуляцией исследования. Известно, что всякое повреждение мышцы
повышает обмен веществ (Маас, 55).
В определенных случаях тоническое укорочение сочетается с
некоторым тоническим непрерывным напряжением. Это бывает при
сильных раздражениях. После тетанического сокращения остается еще
сокращение характера контрактуры, какая бывает в аналогичных
условиях у скелетной . мышцы. Такая контрактура наблюдается в ноге
*282
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
улитки, после сильного механического раздражения. При этом заметно
повышенное потребление кислорода. Предполагается, что этот эффект
¦обязан непрерывному нервному влиянию, вызванному раздражением
(Маас, 55)..
«Запирательное» действие гладкой мышцы. Механическая
деятельность гладкой мышцы выражается не только в быстрых и медленных
сокращениях ив собственном тонусе. Некоторые гладкие мышцы
обладают еще способностью затвердевать в одной приданной им длине;
при этом теряется свойство растяжимости. В первый раз это явление
было обнаружено в запирательной мышце двустворчатых моллюсков
(Pecten). Эта мышца состоит из двух пучков разного цвета. Один
пучок более серый или стекловидный. Он производит обычное
сокращение, закрывая раскрытую раковину. Но он не в состоянии удержать
раковину в этом состоянии. Эта функция выполняется другим, белым
пучком. Он следует за укорочением мышцы совершенно пассивно и
только по окончании движения приходит в такое состояние, что
раковина оказывается прочно закрытой на неопределенное время (Мар-
¦со, 42; Парнас, 20; Бете, 21). Похоже на то, как если бы мышца
в определенный момент перешла из мягкой упруго-вязкой массы
в твердую, прочную массу. Достигаемое при этом механическое
действие на определенных объектах всегда максимально: оно не меняется
¦от тяжести. Оно наступает под влиянием центральной нервной
системы, но может протекать независимо от нее, ибо, будучи вызвано,
удерживается и после перерезки нервов (Будденбрйк, 74; Икскюль, 71).
Явление это принято называть запирательным действием (Sperrang)
из-за сходства его с состоянием шестерни, в зубцы которой вставлена
•собачка, вследствие чего она может без всякой затраты энергии
противостоять силе, которая стремится ее повернуть.
В последнее время запирательное действие было изучено И к с к ю-
¦лём (14) на мышцах голотурии. Он нашел у них запирательную
мускулатуру в коже. Ее деятельность проявляется в затвердении кожи и
способности удерживать большие тяжести. И это тойическое
напряжение может происходить без участия нервов. Такой же эффект
наступает в мышце, если изолировать ее вместе с кожей. В теплой воде это
запирательное действие постепенно проходит без изменения самой
формы мышцы, но в норме «запирательное» действие регулируется
центральной нервной системой: под влиянием ее мышца быстро прихо-
.дит в состояние покоя. Всякое же повреждающее воздействие- на кожу
вызывает затвердевание, образование своего рода пйнцыря вокруг
тела; по миновании опасности центральная нервная система
прекращает это затвердение. Однако эта запирательная мускулатура
голотурии не лишена некоторой сократительности. Если на вполне
расслабленную мышцу слегка надавить, то она суживается без заметного
затвердения. При усилении давления сужение наступает одновременно
с затвердением. У млекопитающих к чисто запирательным мышцам
относится сфинктеры желудка, мочевого пузыря, заднего прохода. Их
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
283
состояние постоянно активно. Наблюдаемое временами расширение
.просвета этих сфинктеров обусловливается, временным прекращением
запирательного действия.
Итак, определенные гладкие мышцы все время находятся в
состоянии затвердевания, запирательного действия; для них это состояние —
норма. Лишь в определенные моменты, в них происходит временное
прекращение запирательного действия.
У некоторых беспозвоночных элементы запирающего механизма,
отличны от элементов, производящих быстрые движения. Классическим
объектом является указанная выше запирательная мышца
двухстворчатых моллюсков. Точно такие же мышцы были найдены у морского
ежа. Мышцы, которыми морской еж приводит в движение свои иглы,
состоят также из двоякого рода мышечных пучков: двигательного и
запирательного. Но запирательное действие этих мышц характерно
•отличается от вышеприведенного запирателя раковины. Последний
всегда, при всех условиях, развивает максимальное действие, в то
время как сила запирательного действия мышц иглокожих зависит от
величины сопротивления, которое она преодолевает. Сила
запирательного действия- нарастает до тех лор, пока не уравновесится
действующая сила тяжести (Икскюль, 71).
Изменчивое запирательное действие наблюдается также, у пиявки.
Если к пиявке, присосавшейся к предмету задней или передней при-
соской, привесить тяжесть 10 г в момент сокращения, то животное
держит тяжесть в течение многих минут в одном и том же
укороченном состоянии. Если тяжесть увеличить до 70—100 г, пиявка останется
в том же укороченном состоянии, но запирательное действие
соответственно нарастает. Это проявляется нарастанием затвердения с
увеличением тяжести. На данную тяжесть устанавливается запирательное
.действие такой степени, какая может уравновесить тяжесть (Икскюль,
71). Можно увеличить тяжесть до 100—— 150 г, но в таком случае
животное не в состоянии долго удержать ее на одной длине: с течением
времени оно удлиняется (Беритов, 72).
Запирательное действие проявляется при любой степени укорочения
животного, иначе говоря при любой степени сокращения продольных
мышц. Если раздражением усилить сокращение, то от этого
запирательное действие не меняется: пиявка может удержать без
удлинения тяжесть 50—100 г при разной степени сокращения.
Сократительная и запирательная функции у пиявки, видимо, принадлежат одним
¦¦а тем же мышечным элементам. Но при сокращении пиявка может
іподнять гораздо меньший груз (не более 50 г)„ чем она в состоянии
поддержать путем запирательной функции (Беритов, 72).
/ Как указывалось выше, в организме запирательное действие
регулируется нервной системой. Предполагалось существование
специальных нервов, которые прекращают запирательное действие. Так,
у двустворчатой раковины наблюдалось запирание раковины при
раздражении определенного нерва., идущего от, висцерального ганглия
284 оёщая физиология мышечной системы
к мышцам-запирателям, а при раздражении другого нерва это запи-
рательное действие прекращалось (Павлов, 46) .По новейшим,
наблюдениям, возникновение и прекращение запирательного действия
этой мышцы обусловливается не специфическим действием разных
нервов, а состоянием нервно-мышечного передаточного аппарата, т. е.
нервной пластинки в .мышечных волокн-ах, и физиологическим,
состоянием самой мышцы. Предполагается, что вязкость играет главную-
роль. Когда она велика, нервные импульсы не действуют. Когда она
мала и происходит расслабление мышцы, нервные импульсы начинаю*
оказывать влияние: они вызывают сокращение или же обусловливают
запирательное действие на новом уровне укорочения (Жирмунская, 75) ~
У: пиявки запирательное действие возникает рефлекторным путем.
Оно, безусловно, не зависит от головного ганглия, ибо наблюдается
и после удаления, головной части. Не зависит оно и от хвостовог©
ганглия или хвостовой присоски, ибо оно наблюдается и после
удаления этой присоски. Оно исчезает после удаления нервной цепочки.
Значит, запирательное действие у пиявки вызывается рефлекторным
путем, посредством брюшной цепочки (Беритов, 72).
Запирательное действие мышц сопровождается: чрезвычайно малым
повышением обмена веществ. Потребление кислорода-в запирательных
мышцах пресноводных моллюсков повышается всего на 15—60%
сравнительно с состоянием покоя (Жуков, 3). Это потребление кислорода
в 500 раз меньше по сравнению с. тем, что наблюдается при тетаниче-
ском сокращении скелетных мышц лягушки. На этом основании можн©
утверждать, что запирательный механизм качественно .отличается ог
сократительного процесса. Однако такое запирательное действие,
которое непосредственно связано с сокращением, как это, например,
бывает у пиявок, должно сопровождаться значительным повышением
обмена веществ. .В отношении пиявки это доказано Конгеймом ш-
Икскюлем (73) и Кометиани (76)..
Сущность запирательного действия мышцы не известна. Весьма
возможно, что запирательное действие в определенных мышечных
образованиях связано с таким физико-химическим изменением внутр»
мышечных клеток, которое делает мышечные клетки нерастяжимыми,
временно лишает их упруго-пластических свойств. Но, видимо,- зап»~
рательное действие продольной мышцы пиявки иного происхождения.
Это запирательное действие сопровождается усиленными биотоками
возбуждения мышцы. Осциллографическое исследование показало, чт©
токи возбуждения мускулатуры пиявки в состоянии максимального
укорочения, но без нагрузки, суть сравнительно большой длительности —
0,1—0,05 сек. и очень небольшой амплитуды—20—50 ji V. При
нагрузке до 80 г токи возбуждения усиливаются, доходя до'500р V*
и учащаются, достигая местами 70 в секунду. При дальшейшём
повышении до 150 г то?и возбуждения усиливаются еще больше, но частота
падает, местами до 10 ровных, очень крупных колебаний в секунду.
Зто наблюдается при сохранении одной и. той же степени укороче-
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
285
¦ния. Если- імышца при сокращении не удерживается в запертом
состоянии и она удлиняется при нагрузке, несмотря на сокращение,
тогда биотоки мышцы усиливаются лишь в незначительной степени, да
д то в самом начале действия груза (Беритов, 72).
На основании этих наблюдении можно предположить, что стеаень
запирательного действия мышцы-у пиявки определяется интенсивностью
активного состояния возбудимой системы мышцы. Она тем выше, чем
¦интенсивнее это; активное состояние. Последнее обусловливается
действием нервных импульсов из центральной нервной системы, которое,
в свою очередь, тем интенсивнее, чем больше груз, ибо с увеличением
груза усиливается раздражение кожно-мышечных рецепторов.
Автоматическая деятельность гладкой мускулатуры. Органы с
гладкими мышцами обладают способностью к автоматической ритмической
деятельности. Такие органы сокращаются периодически без внешних
раздражений не только в организме, но и будучи вырезаны из
организма. Каждое такое сокращение, подобно сердечной систоле,
представляется одиночным сокращением, которое вызывается одним
возбуждающим импульсом.
Характерно, что при автоматической деятельности каждое
сокращение максимально. Прямая кишка лягушки, например, сокращается
на 60—75%, что составляет максимум сокращения для этого органа
(Шюллер, 6). А так как абсолютная и относительная рефракторные
фазы продолжаются во время сокращения и некоторое время после
него, то ритм автоматической деятельности определяется
продолжительностью рефракторных фаз. Во всех тех условиях, когда
рефракторные ф.азы укорачиваются, автоматический ритм
увеличивается. Так бывает под влиянием температуры: с повышением
последней ритм повышается, а с понижением уменьшается (Магнус, 13).
Когда почему-либо автоматическая деятельность прекратилась, она '
может быть вновь вызвана на некоторое время одним коротким
раздражением. Так, па рис. 136 дается ритмическая деятельность
мигательной перепонки кошки после одного и двух индукционных ударов.
Происхождение автоматической деятельности. Прежде всего следует
©тметить, что для автоматической деятельности гладкой мускулатуры,
подобно сердечной, существенное значение имеет обычный солевой
состав крови, именно ионы натрия, калия и кальция. Изотонический
раствор хлористого натрия поддерживает автоматию долгое время.
Хлористый калий действует возбуждающим образом на многие органы
о гладкой мускулатурой, как кишечник, мочевой пузырь, бронхи; матку.
Он является антагонистом (многих парализующих ядов (Цондек, 15).
Хлористый кальций в малых дозах безусловно нужен для
автоматической деятельности. Если действуют раствором Рингера, свободным от
Са, Mg, то автоматическая деятельность останавливается (опыты на
матке кошки). Но Са действует возбуждающим образом на гладкие
мышцы одних'органов (кровеносные сосуды)1 и парализующим на
мышцы других (желудок, кишечник, мочевой пузырь) (Цондек, 15).
286 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Определенное соотношение всех этих ионов поддерживает
нормальную деятельность гладких мышц (Спиро, 16).
Углекислота также имеет существенное значение для
автоматической деятельности. Если петлю кишки поместить в раствор Рингера,.
где бикарбонат (NaHCOs) заменен эквимолекулярным количеством
какой-либо щелочи, которая связывает угольную кислоту, петля кишки
перестает сокращаться автоматически. Если вскоре после этого ее-
перенести обратно в обычный раствор Рингера, петля вновь начинает
сокращаться (Мансфельд, 17).
Однако ни металлические ионы, ни углекислоту нельзя признать
специфическими возбудителями автоматической деятельности.'Мы видели,.
Рис. 145. Действие холина и его эфира с пировиноградной кислотой. Петля
кишки кролика находится в растворе Тироде.
(Раствор Тироде кроме составных частей локковской жидкости включает еще хлористый
магний и фосфорнокислый натрий.) К нему прибавляют в опыте А сначала 1 мг холина,
затем 2 мг. В опыте В испытывается действие 1{10о мг холинового эфира пировиноградной
кислоты, а в опыте С '/«о мг того же эфира. Момент действия холина везде обозначен
стрелкой. Как показывают кривые, холиновый эфир производит в 75—100 раз более
сильное действие, чем один холин (леГо).
что аналогичное действие они оказывают на сердце и даже на
скелетную мышцу. Специфическим возбудителем гладкой мышцы всего ки*
шечника следует считать гормон холин и ацетилхолин, которые
выделяются в стенках кишечника (Le Heux, 18). Если петлю кишки
теплокровного животного опустить в раствор Локка температуры 37°, то-
петля будет сокращаться некоторое время, затем остановится. В этом
растворе был обнаружен холин, составляющий 3Д всего того вещества,
которое выделяется в растворе из петли. Если воздействовать холином
на неподвижный кишечник, например, после отравления хлороформом
или вследствие обнажения во время операции при перитоните, то это*
¦вызовет восстановление автоматической деятельности (Араи, 19).
Многие органические кислоты, а также их соли более или менее-
благоприятствуют автоматической деятельности кишечника. Так
действуют, например, уксусная, пировиноградная, пропионовая кислоты и
их соли. Есть основание для предположения, что действие их
происходит путем образования эфиров с холином, т. е. холинового эфира..
Этот эфир производит более сильное возбуждающее действие на
«мускулатуру кишечной стенки, чем один холин (рис. 145). Образова-
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
287
ние холинового эфира в самой кишке приписывается действию
специфических ферментов (Le Heux, 18).
В последнее время было обнаружено, что во многих случаях этим
веществом является ацетилхолин или ему подобное вещество. Так,
например, в стенках мочевого пузыря собаки, при его активном
сокращении, образуется вещество, которое, подобно ацетилхолину, угнетает
сердце лягушки (Гендерсон и Рэпке, 41). Ацетилхолин образуется
в большом количестве в стенках беременной матки, и ему
приписывается определенная роль в механизме родов (Си-Чун Чанг и Уонг, 53).
Он выделяется в кишечном соке и, видимо, вырабатывается в
железистых клетках (Блох и Нехелес, 61). По новейшим исследованиям,,
автоматическая деятельность гладкой мышцы кишечника не
обусловливается только ацетилхолином. Известно, что продольные и
циркулярные мышечные пучки кишечника производят автоматические
сокращения, причем характер сокращений того и другого пучка не одинаков..
Эти сокращения наступают при несомненном отсутствии ацетилхолина
и, видимо, происходят от каких-то специфических активных веществ,
образуемых в самом мышечном пучке. Ацетилхолин же, который, как
известно, выделяется нервными окончаниями парасимпатической
системы, только усиливает эту автоматию (Демур, 59).
Вышеозначенное активное действие металлических ионов,
углекислоты, а также холина и ацетилхолина на автоматическую
деятельность гладких мышц, по мнению одних авторов, происходит через,
нервную систему, а именно через то нервное сплетение, которое лежит
в самой стенке кишечника и называется ауэрбаховским сплетением;
(Магнус, 13). По мнению других, автоматическая ритмическая
деятельность не зависит вообще от нервов и, в частности, от ауэрбаховского*
сплетения. Это, например, заключают из того, что данная функция
сохраняется в желудке до 48—72 часов по прекращении
кровообращения. Считают невероятным, чтобы нервные клетки сохранили
способность к деятельности в данных условиях на столь длительный
промежуток времени, но существуют и более прямые доказательства. Так,.
например, денервированная мигательная перепонка, всецело лишенная
нервных элементов, может развивать спонтанную деятельность. Так
же ведут себя другие органы в отсутствии раздражения нервов.
(Морин, 51). Известно также, что полоска гладких мышц желудка,
сохраняет способность к автоматической деятельности при
несомненном отсутствии ауэрбаховского сплетения (Гунн и Андергил).
Однако, по гистологическим исследованиям Эсвельда (23), известно,,
что в гладкой мускулатуре кишечника, как и во всех других органах
с гладкой мускулатурой, существует богатая нервная сеть, или
синцитий, без нервных клеток. Функция этой сети еще не определена. Если:
кусок кишки без ауэрбаховского сплетения и вообще без ганглиозных
клеток показывает ритмическую деятельность, которая под влиянием"
холина усиливается, то это еще не говорит в пользу миогенного
происхождения этой деятельности, так как вышеназванный синцитий мог
288
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
бы обусловить эту ритмическую деятельность. По физиологическим же
исследованиям Эсвельда (22), холин и другие сложные
органические соединения, как пилокарпин, физостигмин, ацетилхолин, муска-
рин и гистаімин, действуют на препараты кишки без ауэрбаховского
сплетения в 100—1000 раз слабее, чем на препараты с таким.
сплетением; это свидетельствует скорее всего о том, что указанные вещества
действуют возбуждающим образом как на іауэрбаховское сплетение,
так и на гладкую мускулатуру непосредственно. По аналогии с
сердечной мышцей можно заключить, что автоматическая деятельность
есть функция гладких мышц, но эта деятельность поддерживается и
управляется все время ауэрбаховским сплетением. Повидимому,
нормально все химические воздействия влияют на кишечник прежде всего
через это сплетение. Например, по наблюдениям Э ев е льда, на
препаратах без ауэрбаховского сплетения названные вещества совсем не
влияют на ритм. Такое 'влияние наблюдается при наличии этого
сплетения.
Итак, надо полагать, что автоматическая деятельность нормальной
кишки — очень сложное явление, осуществляемое мышечными
элементами при активном участии всей нервной системы кишечника.
Практикум
Наблюдение перистальтического движения на кишечнике лягушки. У
спинномозговой лягушки обнажают кишечник и где-либо на желудке или яа топкой кишке
производят механическое раздражение: надавливание, легкий щипок пинцетом,
укол. В раздраженном месте в результате сокращения кольцевых мышц произойдет
сужение. В течение минуты это сужение перейдет на соседние участки.
Миографическая запись гладкой мышцы. Постановка опыта изображена на
рис. .146. Берут лягушечий желудок. Вырезают кольцо из середины желудка
шириной 0.5 см., которое затем перерезают в одном месте и привязывают нитки к
свободным концам. Один конец полоски укрепляют за нитку, а другой обычным порядком
соединяют с миографом. Миограф должен быть очень легким, и вся нагрузка' не
должна превосходить 1—0,5 г. Для раздражения электрод накладывается на
определенный участок мышцы, лак это показано на рис. 146, или ток пропускается
через всю мышцу, как на рис, 149.
Опыт с раздражением. Для вызова сокращения требуется очень большая сила
тока (10—5 см расстояния между катушками). Для записи механического эффекта
кимограф ставится на очень медленное вращение. Наблюдают одиночное сокращение
и скрытый период, продолжительность которых измеряется секундами.
Продолжительное тетаническое раздражение дает более длительное и более сильно.е
сокращение, чем одиночное раздражение.
Если желудок взят не во время зимней спячки, а весной, летом и ранней
осенью, то поперечный отрезок будет сокращаться ритмически спонтанно.
Действие ацетилхолина на гладкую мышцу. На поперечный отрезок желудка,
присоединенный к миографу, наносят несколько капель раствора ацетилхолина
1 : 10 000 или 1 : 1000. Мышца сокращается, как от электрического раздражения;
при этом сокращение от ацетилхолина 1 : 10 000 слабее, чем от 1 : 1000.
Опыт с растяжением поперечного отрезка желудка лягушки. Подвесить к
миографу груз в 5 г и через несколько секунд снять. Исходная длина восстанавливается
сама, если вес миографа не превышает 1 г. Начальное быстрое восстановление будет
представлять эластическое .укорочение. Если нет полного восстановления, нужно под-
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
289
нять миограф на минуту, т. е. разгрузить мышцу полностью. Тогда наступает полное
восстановление. Повторить опыт с длительной нагрузкой; іподвесить гирю на 5 минут
я передвигать кимограф по 5—-10 сек. через минуту. При хорошем функциональном
состоянии после удаления груза полное восстановление произойдет в несколько минут,
если миограф будет подниматься или если мышца будет производить
автоматические сокращения. Это восстановление в медленной фазе обусловливается собствен -
Рис. 146. Изображение постановки опыта с миографической записью сокращения
мышечной полоски из желудка лягушки. Необходимые объяснения даны в тексте.
ным тонусом мышцы. При плохом функциональном состоянии, а также если
растяжение производилось значительно дольше, 10—30 минут, то после удаления груза
и тяжести миографа мышца остается более или менее удлиненной. Это будет
пластическое удлинение.
Спинная мышца пиявки. На гладкой спинной мышце пиявки можно хорошо
изучить пластические и сократительные функции гладких мышц беспозвоночных
животных. Мышца вырезается вместе с кожей вдоль по спине, полоской в 3—4 см
длины и 0.5 см ширины, с внутренней стороны очищается и располагается в
вертикальном положении. Нижний конец фиксируется, верхний конец соединяется с
миографом. Миограф нагружен 2—3 г. Оба конца мышцы, кроме того, соединены
с индукторием, как показано на рис. 149. Можно поставить следующие опыты:
Л. Опыты с раздражением. Определяют пороги раздражения одиночными и те-
таническими индукционными ударами и записывают двигательные эффекты на
кимографе при разных силах раздражения.
B. Опыт с согреванием и. охлаждением. Опускают мышцу в раствор Рингера,
согретый до 40° С; мышца удлиняется. Снова определяют пороги и записывают
двигательные эффекты. Затем охлаждают до комнатной температуры и вновь
производят запись на кимографе. Наконец, мышцу охлаждают до 6° и зновь
определяют пороги и записывают двигательные эффекты.
C. Опыт с растяжением. Привешивают груз 10 г и записывают постепенное
удлинение в течение получаса. Затем снимают груз и наблюдают восстановление исход-
19 и. С. Беритов
290
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
ной длины: быструю эластическую фазу и медленную фазу, выражающую
постепенное исчезновение пластического удлинения под влиянием собственного тонуса.
Опыт с запирательным действием на пиявке. Целую пиявку перевязывают
толстой ниткой посередине и около задней присоски. Головную часть удаляют.
Когда животное успокоится, задний перевязанный участок оказывается сильно
укороченным. В этот момент осторожно берут за нитку, расположенную около задней
присоски, и укрепляют пиявку в вертикальном положении. Затем к другой нитке
привешивают гирьки 10—100 г. Если задняя часть животного находится в состоянии
запирательното действия, то при всех этих тяжестях удлинение ее будет одинаково
небольшим, причем оно не будет нарастать с течением времени. Если же
укорочение было обусловлено только сокращением без запирательното действия, то будет
происходить значительное удлинение, тем большее, чем больше тяжесть и чем
дольше она действует.
Литература
1. S с h u 11 z P., Arch. Anat. Physiol., 1895, S. 517; 1897, S. 322, 307; 19C3, Suppl., S. L
2. Grutzner P., Erg. Physiol., 3, 2, 1904.
3. Guth E., Pfl. Arch., 85, 118, 1901.
4. Tschermak A. v., Pfl. Arch., 175, 165, 1919.
5. Ganter G., Pfl. Arch., 201, 115, 1925.
6. Schuller J., Arch. exp. Path. Pharm., 90, 196, 1921.
7. Bozler E., Z. wiss. Biol., Abt. C, 6, 361, 1927.
8. Bottazzi P., Arch. Anat. Physiol., S. 377, 1901.
9. Alvarez W. C, Am. J. Physiol., 42, 423, 1917.
10. В u d d e W. u. G e 11 h о г n E., Pfl. Arch., 203, 170, 1924.
11. Rich ter C, Am. J. Physiol., 67, 612, 1924.
12. Brucke Fr. v., Pfl. Arch., 133, 313, 1910; 136, 502, 1910.
13. Magnus R., Pfl. Arch., 102, 349, 1904; 103, 515, 525, 1904; 108, 1, 1905; 111, 152,.
1906.
14. Uexkull v., Z. Biol., 39, 73, 1899; 44, 307, 1907.
15. Zondek S. u, Bioch. Z., 132, 302, 1922.
16. Spiro K., Schvv. med. Wschr., 51, 457, 1921.
17. M a n s f e 1 d G., Pfl. Arch., 188, 241, 1921.
18. Le H e u x, Pfl. Arch., 173, 8, 1919; 179, 177, 1920; 190, 280, 301, 1921; 191, 99, 192L
19. Arai K., Pfl. Arch., 193, 359, 1922.
20. P a r n a s J. K„ Pfl- Arch., 134, 441, 1910.
21. Bethe A., Pfl. Arch., 142, 291, 1911.
22. Es veld L. W., Arch. exp. Pathol. Pharm., 134, 347, 1928.
23. Es veld L. W., Z. mikr.-anat. Forsch., 15, 1, 1928.
24. Jordan H., Allgemeine vergl. Physiol, d. Tiere, S. 407, 1929.
25. Иордан Г., Физиол. журн. СССР., 19, 141, 1935; Юбил. сб., поев. проф. И. Бери-
ташвили, стр. 71, 1936.
26. Коштоянц X. С. и Мужеев В. А., Биол. журн., 2, вып. 6, 1933.
27. Коштоянц, X. С. и Мужеев, В. А., Сб. «Некоторые вопросы сравнит,
физиологии»*, 1934.
28. Parker G. Н., J. Gen. Physiol., 5, 45, 1923.
29. В о z 1 е г Е., J. Physiol., 69, 442, 1930.
30. Жуков, К., Физиол. журн. СССР, 19, 933, 1935.
31. F r a n k О., Erg. Physiol., 3, II Abt., 495, 1904.
32. Tiegs О. W., Austr. J. Exp. Biol. a. Med. Sci., 1, 131, 1924.
33. Lawrentiew B. J., Z. mikr. Forsch., 6, 467, 1926.
34. Rosenblueth А. и сотр., Am. J.Physiol., 116, 387, 1936; ib., 106; 103, 659,.
1933; ib. 114, 147, 1935.
35. M о n n i e r A. M. а. В а с q Z. M.. Arch. Intern. Physiol., 40, 485, 1935.
ОБЩАЯ ФИЗИОЛОГИЯ ГЛАДКОЙ МУСКУЛАТУРЫ
291
36. Cannon W. В. a. Rosenblueth A., Auton. neuro-effector system, 1937.
37. Е с с 1 е s J. С. а. М a g 1 a d е г у J. W., J. Physiol., 89, 45Р, 1937.
38. Е с с I е s J. С. а. М a g 1 a d е г у J. W., J. Physiol., 90, 31, 1937; ib., 90, 68,1937.
39. F I е t с h е г М., J. Physiol., 90, 233, 1937.
40. Eccles J. С, Erg. Physiol., 38, 389, 1936.
41. H e n d e r s о n V. E. a. R о e p к e M. H., J. Pharmacol., 51, 97, 1934.
42. Marceau, С. г. Soc. Biol., Paris, 141, 278, 1905; Arch. Anat, 17, 1909.
43. В о z 1 er E., Z. vergl. Physiol., 6, 361, 1927; 7, 379, 407, 1G08; 8, 371, 1928; 12, 579,
1930; 14, 429, 1931.
44. R i e s s e r O., Erg. Physol., 38, 133, 1936.
45. И о p д a h Г., Практикум сравнит, физиологии. Биомедгиз, 1934.
46. Павлов И., Pfl. Arch., 37, 6, 1885.
47. S е г е п і Е., Arch. Farmacol. sperim., 47, 1929.
48. Р о с к и н Г. И., Учен. зап. МГУ, вып. 13, стр. 227, 1937.
49. Роскин Г. И., Бюлл. эксп. биол. мед., 4, 152, 1937. Z. ZeUforsch. Mikr. Anat.,
24, 85, 1935.
50. М о n n i er A. M., Cold Spring Harb. Sympos. Quant. Biol., 4, 111, 1936.
51. M о r i n G., L'automatisme intenstinal des vertebres et la regulation, Lyon, 1935.
52. Гогава M., Сообщ. Ак. Наук Груз. ССР, т. V, 711 , 1944.
53. Н s i-C h u n Chang a. Amos Wong, Chin. J. Physiol., 7, № 2, 151, 1933.
54. С о n h e i m О., Z. physiol. Chem. 76, 298, 1912; ConheimO.u.Uexkiil 1 J. v;f
Z. Physiol. Chem., 76, 314, 1912.
55. Ma as J. A., Arch. Neerl. Physiol., 23, 1, 1938.
56. Кудрявина H. А., Физиол. журн. СССР, 26, 432, 1939.
57. Жуков. Е. К., Учен. зап. Ленингр. гос. унив., сер. биол. н., № 10, стр. 111, 1939.
58. М а х і m о w A. A., A text-book of Histology, 1931.
59. Demo or L., Физиол. журн. СССР, 24, 126, 1938.
6j. Bozler E., Am. J. Physiol., 122, 614, 1938; 124, 502, 1938; 127, 301, 1939.
61. В 1 о с h E. a. N e с h e 1 e s H., Am. J. Physiol., 122, 631, 1938.
62. Boeke J., J. Com. Neur., 56, 27, 1932.
63. Boeke J., W. Penfields Cytology of the nervous system, sect. VI, 243, 1932.
64. S t о e h r Ph., Z. ZeUforsch., 16, 1932; Z. mikr.-anat. Forsch., 33, 233, 1933.
65. M e у e r h о f O., Die chemischen Vorgenge im Muskel, 1630, Berlin.
66. В a 1 d w i n E., An introduction to ccmpar. biochemistry, Cambridge 1937.
67. В e p ж б и н с к а я Н. А., Физиол. журн. СССР, 2\і 415, 1936.
68. Riesser О., Arch. exp. Path. Pharm., 161, 34, 1931.
69. X л о п и н Н. Г., Арх. анат., гистол. и эмбриол., 15, 3, 1936.
70. Беритов И. и Нивинская О., О собственном тонусе гладкой
мускулатуры. 1940. (Не опубликовано).
71. U е х к іі 1 1 J. v., Umwelt und Innerwelt der Tiere, 1909, Berlin.
72. Б e p и t о в И., О «запирательном» действии пияеки. Сообщ. Ак. Наук Груз.
ССР, т. V, 723, 927, 1944.
73. Cohnheim О. u. J. v. U е х к іі 11. Цитир. по Jordan, Allg. vergl. Physiologie der
Tiere, S. 419, 1929.
74. Buddenbrock W., Grundriss d. vergleichenden Physiologie, Berlin, 1928.
75. Ж и p м у h с к а я E. А., Физиол. журн. СССР, 29, 536, 1940.
76. К о м e т и а н и П., Сообщ. Ак. Наук Груз. ССР, 5, 65, 1944.
77. Rosenb lueth А. а. Е. С. del Pozo, Am. J. Physiol. 137, 263, 1942.
78. Stohr Ir. Ph., Mikroskop. Anatcmie d. vegetat. Nervensystems, Berlin, 1928.
79. Har reveld v. A., J. E>:per. Biol., 16, 398, 1939.
19*
Глава XIV
СМЕРТЬ МЫШЕЧНОЙ ТКАНИ
Общее понятие. Когда в мышечной ткани обмен веществ
становится невозможным вследствие остановки кровообращения, она теряет
возбудимость и вообще жизненные свойства. Кроме того, она
укорачивается и утолщается, т. е. претерпевает точно такие изменения, как
при сокращении. Это изменение мышечной ткани называется
окоченением (rigor mortis). Окоченение сходно с сокращением не только
с внешней стороны, но и со стороны внутренних химических
процессов. В окоченевающей мышце развивается большое количество
молочной кислоты (Флетчер и Гопкннс, 1 и 2) и фосфорной кислоты
(Эмбден и сотр., 3).
Как мы указывали выше, молочная и фосфорная кислоты
развиваются в мышце при совершенно нормальных условиях не только во
время деятельности, но и в состоянии покоя. Только в этом последнем
случае обе кислоты образуются в очень малых количествах. После
остановки кровообращения эти кислоты продолжают образовываться как в
покое, тем же самым путем: из гликогена и фосфагена путем
ферментативного расщепления, причем процесс нейтрализации кислот в этом
случае сильно ограничен небольшими запасами буферных веществ.
Кроме того, превращение молочной кислоты или лактата в гликоген
ограничено небольшим запасом кислорода. По исчерпывании этих
запасов молочная и фосфорная кислота накапливаются в мышце в таком
количестве, как это бывает <в деятельной мышце под влиянием
возбуждения и даже в больших размерах. Вследствие этого мышца
сокращается и притом тем сильнее, чем больше в ней накапливается кислот.
В конце концов окоченевающая мышца достигает максимального
укорочения. Установлено, что в окоченевающей мышце молочной кислоты
накапливается даже больше, чем при сильном тетаническом
сокращении, именно в среднем до 0.6—0.7% (Фюрт, 4; Мейергоф, 20).
Одновременно с молочной кислотой из фосфагена образуется фосфорная
кислота в большем количестве, чем обычно. В нормальных условиях
значительная часть фосфорной кислоты обратно соединяется с
креатином, образуя вновь фосфагенГВ^ окоченевающей мышце этот процесс
СМЕРТЬ МЫШЕЧНОЙ ТКАНИ
293
восстановления сильно задерживается, и потому фосфорной кислоты
накапливается больше, чем в покое или даже во время деятельности
(Эмбден и Адлер, 5).
О роли молочной и фосфорной кислот в явлении окоченения.
Исходя из факта накопления молочной и фосфорной кислот в окоченевшей
мышце, окоченение приписывают действию этих кислот на
сократительное вещество. Особенно большую роль в этом явлении должна играть
молочная кислота, ибо известно, что вырезанные мышцы не
окоченевают, если поместить их в атмосферу кислорода, и в особенности, если
поместить в раствор Рингера, через который пропускается кислород.
Последний диффундирует внутрь мышцы, вследствие чего, во-первых,
часть молочной кислоты сжигается, а, во-вторых, за счет
освободившейся при этом энергии восстанавливается другая часть обратно в
гликоген (Флетчер, 1; Винтерштейн, 25).
На вырезанных мышцах раздражение благоприятствует
окоченению, ибо деятельное состояние ускоряет распад гликогена и фосфагена,
а значит, и накопление кислот. Но, опять-таки, в условиях снабжения
кислорода из окружающей среды окоченение раздражаемой мышцы
может быть сильно задержано, в особенности в тонких мышцах, и даже
отсутствовать. На мышцах теплокровных животных это удается
наблюдать гораздо реже (даже на очень тонких мышцах), чем на мышцах
холоднокровных. Это, повидимому, объясняется тем, что мышцы
теплокровных легко повреждаются от наркоза, применяемого при операции,
от самой операции, от высыхания и от последующих манипуляций в
связи с раздражением (Вольф, 26).
Однако, по новейшим наблюдениям, нельзя считать молочную
кислоту основным существенным условием окоченения мышцы, ибо стало
известно, что окоченевает также мышца, отравленная моноиодоуксусной
кислотой, в которой расщепление гликогена не доходит до образования;
молочной кислоты (Лундсгаард, 24). Поэтому нужно предположить, что
посмертное окоченение, подобно сокращению, зависит не от одной
кислоты, а от всей суммы биохимических процессов, протекающих в
мышце в покое и при деятельности. А, как известно, в этом сложном
процессе главную роль играет расщепление липопротеидов — основного
вещества возбудимой системы, затем происходит распад аденилпиро-
фосфата и фосфагена. Поэтому вероятно в отравленных моноиодуксус-
ной кислотой мышцах окоченение вызывается главным образом
кислыми продуктами распада липопротеида, аденилпирофосфата и фосфагена.
Во время окоченения выделяется также углекислота, но она в
данном случае іне является продуктом окисления (Вакер, 6). Она, вероятно,
возникает, как вообще в анаэробных условиях, в результате
расщепления кетокислот, образуемых при анаэробном распаде углеводов.
Условия, определяющие окоченение мышцы. Окоченение мышцы
предполагает одно существенное условие: образование и накопление
продуктов расщепления в мышце. Отсюда ясно, что если по какой-либо
причине затрудняется образование продуктов расщепления, окоченения
294 ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
не произойдет или оно будет очень слабым. Известно, что мышцы
лягушки окоченевают очень поздно, если их держать при низкой
температуре, около 0°; в этом случае окоченение начинается через 4—7 дней.
Исхудавшая, бедная гликогеном, мышца окоченевает много позднее,
чем упитанная, богатая гликогеном. Естественно, все те условия,
которые благоприятствуют образованию молочной кислоты,
благоприятствуют также окоченению. Работающая мышца, уже богатая продук-
Рис. 147.
А — положение быстро окоченевшего трупа после смерти от холеры. Все
суставы сильно согнуты, как при «каталептическом оцепенении». В — положение
того же трупа спустя некоторое время, когда окоченение прошло вследствие расслабления
окоченевших мышц. Предыдущая поза исчезла; все конечости упали (из Данилевского).
тами расщепления, окоченевает раньше, чем мышца, находящаяся в
покое. У лягушки разгибатели передних конечностей работают меньше,
¦чем сгибатели, и поэтому окоченевают позднее вторых. 'Мышца с
перерезанным двигательным нервом окоченевает позднее остальных
мышц (Эйзельсберг, 7; Нагель, 8). Мышцы теплокровных, где обмен
веществ происходит более интенсивно, где, следовательно, образование
кислых продуктов происходит быстрее, чем в мышцах холоднокровных,
окоченевают раньше последних. Часто случается, что какая-либо
группа мышц перед смертью, т. е. перед остановкой
кровообращения, работала особенно сильно и вследствие этого в них уже
накопилось большое количество кислот. Такие мышцы окоченевают раньше
других. Поэтому окоченевающий труп принимает ту самую позу,
какая была при жизни наиболее длительной. Так, например, на поле
сражения часто находят убитых в позе прицеливания из ружья.
Точно так же труп умершего от холеры принимает ту неестественную
позу, какая типична для холерных судорог (рис. 147).
Легко можно устранить окоченение, если каким-либо образом
повлиять на накопление продуктов расщепления. Например, если
маленькую мышцу поместить в атмосфере кислорода, она окоченевает
позднее, чем в бескислородной среде, или даже совсем не окоченевает
(Флетчер, 1; Винтерштейн, 10). Это зависит от того, что кислород
проникает в мышцу путем диффузии и производит обычный
восстановительный процесс: сжигает часть молочной кислоты и освободившейся
при этом энергией способствует превращению не только остальной
части молочной кислоты в гликоген, но и фосфорной кислоты в фосфаген.
СМЕРТЬ МЫШЕЧНОЙ ТКАНИ
295
Мышца, погруженная в раствор Рингера или в физиологический
раствор, окоченевает медленнее. Это происходит вследствие того, что часть
продуктов расщепления диффундирует из мышцы в раствор, и таким
образом задерживается накопление ее в мышце (Мангольд, 12).
Известно также, что мышца лягушки не окоченевает, если пропускать
через нее щелочной раствор и тем самым нейтрализовать в ней
кислоту. Лактат же и фосфаты не вызызают сокращения (Бауман, 11).
На ірис. 148 дается прекрасная иллюстрация окоченения (или
контрактуры) мышцы под влиянием искусственного воздействия молочной
кислоты и затем прекращения этого окоченения под влиянием ЫаНСОз,
который, нейтрализуя кислоту, производит расслабление мышцы.
Возбудимость и окоченение. Во время окоченения мышца обычно
теряет возбудимость, однако, между этими явлениями нет прямой
зависимости. Первоначально окоченевшая мышца не лишена возбудимости.
Если окоченение не максимально, раздражение производит
сокращение (Мангольд, 12). Если даже во время окоченения раздражение не
дает эффекта, то это не означает, что мышца мертва. Во-первых, это
может быть вследствие максимального укорочения окоченевшей мышцы,
которая поэтому не дает дополнительного сокращения на раздражение.
Во-вторых, возбудимая система мышцы может находиться в состоянии
парабиоза, в которое она впадает под влиянием накопившихся кислот.
Это такое состояние, когда мышца лишена возбудимости, но еще живет.
Если удалить кислоты, которыми было обусловлено данное состояние,
мышце возвращается возбудимость (см. подробно об этом явлении в
нервной физиологии). Что в окоченевшей мышце действительно
наступает парабиотическое состояние, хорошо видно из следующих опытов.
Если через окоченевшую и лишенную возбудимости мышцу пропустить
щелочную или насыщенную кислородом жидкость, то возбудимость
мышцы восстанавливается и одновременно исчезает окоченение, т. е.
мышца становится нормальной (Мангольд). Рис. 148 хорошо
иллюстрирует отношение мышцы во время искусственного окоченения к
электрическому раздражению. Окоченение мышцы постепенно достигает
высоты максимального тетануса. Отдельные индукционные удары все
время дают дополнительные эффекты, но только тем слабее, чем
сильнее окоченение. А на максимальной высоте и отдельные индукционные
удары и тетаническое раздражение производят ничтожный эффект.
Пороги раздражения сначала повышаются, затем возвращаются к норме.
Структурные изменения в саркоплазме при смерти. Функциональное
изменение при прекращении кровообращения мы поставили в связь с
накоплением кислых продуктов от расщепления тех веществ, которые
имеют непосредственное отношение к сокращению. Но такого рода
кислоты, а также другие химические вещества все время образуются при
расщеплении самого липопротеида. Все эти кислые продукты, действуя
на возбудимую систему, безусловно, должны производить такие
изменения, какие вообще вызываются, когда действуют на протоплазму
кислотами. Известно, что кислоты влияют на белок двояким образом:
296
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
во-первых, они вступают в связь с белками, которые играют в некоторой
мере роль слабых оснований; во-вторых, они ускоряют путем
воздействия своих водородных ионов денатурирование белковых
образований. По всей вероятности, в слабых концентрациях эти кислые
продукты ускоряют расщепление липопротеидов на белок и липоид;
в больших концентрациях, вероятно, происходит образование кислых
соединений белка и одновременно вызываются коагуляция и денатури-
Рис. 148. Кислотное окоченение мышцы.
Сначала мышца j опущена в раствор'Рингера и произведено раздражение отдельными
индукционными ударами іразной силы. Максимум одиночных сокращений получается при 9 см.
Производится также проба тетанического j раздражения (Т). Затем раствор Рингера заменяют молочной ,кисло-
той 0.02 норм, раствора. Время действия 5 мин. Мышца дает контрактурное сокращение. В это же
самое время вызывают одиночные сокращения. Наконец, мышцу переносят обратно в раствор Рингера
с прибавлением NaHC03. Окоченение в первое время усиливается, достигает высоты максимального
тетануса, а затем постепенно проходит в течение 6 часов. На кривых Т обозначает тетаническое
раздражение, ta цифры—силу раздражения в сантиметрах ^расстояния между катушками. Внизу (на
^ " линиях ..цифры означают время (по Швенкеру).
рование белка (Лепешкин, 23). Повидимому, понижение возбудимости
мышцы по прекращении кровообращения находится в прямой
зависимости от количества расщепленного липопротеида. Чем больше
расщепляется липопротеид под влиянием кислых продуктов, тем меньше
становится возбудимой системы и, сообразно, тем ниже будет возбудимость
мышечной ткани. Когда произойдет полное расщепление всего
липопротеида, наступит полная потеря возбудимости мышцы. Но это еще не
будет смерть, ибо с возобновлением кровообращения или удалением
кислых продуктов мышца вновь начинает возбуждаться. Смерть
должна наступить уже в результате накопления большого количества кислых
продуктов, когда произойдут коагуляция и денатурирование белка,
образованного из липопротеида. К этому моменту, вероятно, нарушается
вся сложная система саркоплазмы, обусловливающая обмен веществ.
СМЕРТЬ МЫШЕЧНОЙ ТКАНИ
297
Прекращение окоченения мышцы. Окоченение держится несколько
дней, затем оно проходит, мышца становится мягкой и удлиняется.
Если во время окоченения труп имел какую-либо неестественную позу„
то последняя, исчезает в силу размягчения мышц (рис. 147).
Все те условия, которые ускоряют обмен веществ, благоприятствуют
прекращению окоченения. Имеется подробное исследование влияния
температуры на окоченение и его прекращение. Оно произведено на
мышцах кролика и дало следующие результаты (Бирфрейнд, 19):
Начало окоченения Полное прекращение
При 4° через іу2"часа через 48 часов
. 20° , і ; , . . за .
. 25° „ 45 ' мин. , 4 .
I 4Г ; ИГ . ; 2Уз .
Размягчение окоченевших мышц не зависит от бактериальных
процессов. То же самое случается, если держать труп в совершенно асе-
птических условиях (Карпа, 13). Далее, размягчение наступает,
несмотря на то, что продолжается выделение той самой молочной кислоты,
которая вызвала окоченение. Вопросы о происхождении размягчения-
окоченевших мышц еще не вполне выяснены. Но все связывают его
с накоплением в мышце больших количеств кислоты. По мнению
некоторых ученых, окоченение, как и сокращение, является результатом
гидратации сократительного вещества миофибриллей. Эти ученые
объясняют размягчение окоченевших мышц прекращением способности к
гидратации, разбуханию. Так, по мнению Ф ю р т а (4), разбухание
наступает в результате присоединения воды к белковому веществу.
Прекращение разбухания Ф ю р т считает последствием выделения
воды из этого вещества. Последний процесс приписывается воздействию
кислоты в большой концентрации.
Существует и другое представление о размягчении окоченевших
мышц, согласно которому размягчение считается следствием
разрушения анизотропного вещества, причем из мышцы выходит не вода, а
водный раствор белка. Таково мнение В а к е р а (6). Но такое изменение
анизотропного вещества этот автор приписывает воздействию не
молочной кислоты, а угольной, которая освобождается молочной кислотой из
карбонатов. Согласно прежним воззрениям, прекращение окоченения
зависит от растворения мышечных белков под влиянием больших
количеств молочной кислоты (Меланби). Самое окоченение, по этому
воззрению, представляет свертывание белка от малых концентраций
молочной кислоты. Но в настоящее віремя доказано, что большая
концентрация молочной кислоты не может растворить свернутый белок.
Только минеральные кислоты в состоянии растворить его.
Существовало также мнение, что окоченевшая мышца размягчается вследствие
аутолиза, т. е. самопереваривания (Гедин и Роуланд, 14; Фогель, 15).
Но если бы это было так, в мышце должны были выделяться обычные
продукты белкового распада: альбумозы, пептоны, аминокислоты;
фактически же этих продуктов при прекращении окоченения не находят.
298
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
Искусственное окоченение мышцы. Достаточно согревать мышцу в
растворе Рингера или в физиологическом растворе при 33—40° в
течение одного часа, чтобы вызвать максимальное окоченение (Иенсен, 16).
При согревании в мышце образуется вдвое больше молочной
кислоты, чем при сильном утомлении от тетанического раздражения
(Флетчер и Гопкинс, 2). Поэтому многочисленные авторы приписали это
окоченение накоплению большого количества молочной кислоты (Фюрт,
4; Мейергоф, 20). Это окоченение исчезает, если переложить мышцу
в холодный физиологический раствор на несколько часов. По мнению
Ф ю р т а, это происходит потому, что, с одной стороны, часть
молочной кислоты выходит наружу, а с другой, нейтральные соли проникают
внутрь мышцы и вызывают дегидратацию белкового вещества, т. е.
вытеснение воды из него. В последнее время было установлено, что
отравление імоноиодуксусной кислотой хотя и препятствует
образованию молочной кислоты, но не устраняет теплового окоченения при
согревании до 39—40°. Более того, такое отравление благоприятствует
ему. Повидимому, здесь имеет место какое-то еще неизвестное действие
моноиодуксусной кислоты (Л'иппаи и Давид, 21).
Когда температура мышцы повышается выше 40—45°, мышца
укорачивается еще больше. Но это укорочение не похоже на естественное
•сокращение. Окоченение при температуре до 40°, как указывалось
выше, есть обратимый процесс, как и естественное сокращение. А то
укорочение, которое вызывается при согревании выше 40—45°, не
проходит в холодном растворе Рингера, сколько бы времени мышца ни
лежала в нем. Очевидно, в этом случае мы имеем дело с необратимым
процессом.
Искусственное окоченение можно вызвать также воздействием
химических веществ: хлороформа, эфира, уретана, кислот, щелочей и т. д.
При малых дозах кислоты и щелочи наступает обычная контрактура,
о которой говорилось уже выше. Каким образом действует хлороформ
и другие наркотики, сказать очень трудно. Раньше полагали, что
хлороформ благоприятствует расщеплению гликогена до молочной кислоты
и тем вызывает кислотное окоченение. Но сейчас установлено, что
хлороформ вызывает контрактуру и после отравления моноиодуксусной
кислотой, т. е. когда образование молочной кислоты не может иметь
места. Но это окоченение значительно слабее и развивает меньше
напряжения, чем в нормальной мышце. Поэтому можно думать, что
молочная кислота, если и не обусловливает всецело хлороформную
контрактуру, то является одним из факторов последней (Липпаи, 22). Но
все эти кислоты и наркотические вещества, а также те, которые
вызывают контрактуру через возбуждение, при больших дозах производят
такое же необратимое окоченение, как высокая температура. При
больших дозах происходит свертывание — коагуляция мышечного белка.
Вследствие этого окоченение не проходит ни от времени, ни при
помещении в такие условия, которые благоприятствуют выхождению наркотика
или продуктов обмена веществ (Росси, 17; Фюрт, 4; Швенкер, 18 и др.).
СМЕРТЬ МЫШЕЧНОЙ ТКАНИ
299
Практикум
Наблюдение обратимого и необратимого теплового окоченения. Заднюю лапку
лягушки опускают до колена в раствор Рингера или физиологический раствор,
медленно согревают до 37е и затем держат при
этой температуре 1 час (лапка без кожи). Спустя
5—15 минут m. gastrocnemius начинает
сокращаться и достигает максимума через час.
Вследствие этого голеностопный сустав разгибается.
Потом лапку кладут в холодный раствор Рингера.
В течение 2—3 часов разгибание проходит, ибо
длина m. gastrocnemius возвращается к норме.
После этого ту же или другую такую же
лапку согревают до 50—60°. В течение нескольких
минут мышца укорачивается на 80% своей
первоначальной длины. Если потом положить ее в
холодный раствор Рингера и оставить ее так до
следующего дня, укорочение не пройдет.
Наблюдение окоченения от кислоты.
Постановка опыта дана на рис. 149..Берут m. gastrocnemius
или т. sartorius лягушки и опускают сначала
б раствор Рингера; на кимографе записывают
максимальные сокращения. Затем вместо Рингера
добавляют 0.02% норм, раствор молочной кислоты.
Через каждую минуту испытывают раздражение
разной силы, вызывая максимальное сокращение. Ког-
ла кислотное укорочение достигнет такой
величины, что раздражения перестанут давать
сокращения, кислота заменяется раствором Рингера с
прибавлением NaHCCb. Окоченение достигнет макси-
чума в течение нескольких минут.
Повторить опыт с фосфорной кислотой: взять
0.02% на физиологическом растворе и
подействовать на m. sartorius. Через 20—30 сек. наступает
максимальное укорочение. Если сейчас же
заменить раствор фосфорной кислоты раствором
Рингера, сокращение начнет ослабевать, и мышца
достигнет исходной длины через 1—2 минуты.
Возбудимость мышцы после действия фосфорной
кислоты несколько падает.
Можно взять более слабый раствор фосфорной
кислоты, как 0.001%. Сокращение будет
происходить с большей постепенностью, чем в
предыдущем случае, и будет менее интенсивным. Возбу-
ZHMocTb мышцы почти не изменится.
Рис. 149. Видоизмененный
аппарат Копылова для
миографического изучения изолированной
мышцы.
Мышца находится в стеклянном
резервуаре (М) и соединена с
миографом (My) тонкой проволокой (К).
Эта же проволока служит для
подведения тока,к мышце от провода Е,
идущего от индуктория. Другой
конец мышцы привязан к проводу,
который проходит через стеклянную
трубку (А) и затем припаян к
металлическому стержню F. Этот
провод служит другим полюсом, по
которому направляется ток от провода Е1.
Эбонит X—X препятствует переходу
тока от Е' к Е помимо мышцы. Диск
N служит для движения
металлического стержня F со стеклянной
трубкой вниз и вверх. Краны Сі, Cs и С.ч
служат для перемены испытуемой
жидкости во внутреннем резервуаре.
S — винт, который давит на
пластинку Т, он служит для передвижения
миографа в горизонтальной
плоскости. L—задвижка для фиксации
миографа на разной высоте.
Литература
1. F 1 е t с h е г W. М., J. Physiol., 23, 10, 1898; 28, 354, 474, 1902; 43, 286; 1911; 44,
361, 1913.
2. Fletcher a. Hopkins, J. Physiol., 35, 247, 1907.
3. Е m b d е n G. и его сотрудники, Z. physiol. Chem., 93, стр. 46, 54, 84, 1915; 118,
1, 1922.
4. F u r t h Ot. v., Erg. Physiol., 17, 363, 1919.
5. Adler E. u. Em b den, Z. physiol. Chem., 118, 1, 1922.
a Wacker L., Pfl. Arch., 165, 452, 1916.
-300
ОБЩАЯ ФИЗИОЛОГИЯ МЫШЕЧНОЙ СИСТЕМЫ
7. Е і s е I s b е г g A. v., Pfl. Arch., 24, 229, 1881.
8. N age 1 W. A., Pfl. Arch., 58, 279, 1894.
9. Fletcher, J. Physiol., 28, 974, 1902.
10. Win ter stein H., Pfl. Arch., 190, 225, 1907.
11. BaumannW., Pfl. Arch., 167, 114, 1917.
12. Mangold E., Pfl. Arch., 96, 498, 1903.
13. Karpa P., Pfl. Arch., 119, 119, 1906.
14. Hedin u. R о 1 a n d, см. статью v. F ti г t h, Erg, Physiol., 17, 363, 1919,
15. Vogel R., Deut. Arch. klin. Med., 1902.
16. Jensen P., Pfl. Arch., 160, 333, 1915.
17. Rossi E., Z. Biol., 54, 299; 56, 253, 1911.
18. Schwenker G., Pfl. Arch., 157, 371, 1914.
19. Bierfreund M., Pfl. Arch., 43, 195, 1888.
20. Meyer h of Q., Die chemischen Vorgange im Muskel, S. 566, 1930.
21. L i p p а у F. u. David F ., Pfl. Arch., 230, 70, 1932.
22. Lippay F., Pfl. Arch., 229, 538, 1932.
23. Lepesch к i n W. W., Kolloid. d. Protoplasmas, 1938.
24. Lundsgaar d, Bioch. Z., 217, 162, 1930; 220, 18, 1930; 227, 51, 1930; 230, 10,
1931; 233, 322, 1932.
25. Win ter ste in H., Z. ges. gerichtl. Med., 2, 1, 1923.
26. Wolf H., Pfl. Arch., 217, 210, 1927.
Часть третья
ОБЩАЯ ФИЗИОЛОГИЯ
ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ
Глав a I
СТРОЕЯДЕ НЕРБНОЙ СИСТЕМЫ
Элементы нервной системы. Нервная система состоит из следующих
элементов: нервных клеток с заключенными в них ядрами,
многочисленных протоплазматических отростков клетки, так называемых ден-
Рис. 150, Гигантская нервная клетка из передней центральной извилины
головного мозга человека.
Справа виден нейрит и начало его миэлиновой оболочки. Остальные отростки
принадлежат дендритам. В дендритах и в теле клетки видна сеть нейрофибриллей, особенно
густая около ядра клетки. Отсюда нейрофибрилли переходят в нейрит.
дритов, и особых дифференцированных отростков, так называемых
нейритов, или осевых цилиндров (аксонов).
Величина нервной клетки колеблется в очень широких пределах:
у млекопитающих она вариирует от 4 до 130 jjl, у беспозвоночных,
например у моллюсков, достигает 300—500 ji. Форма нервной клетки
чрезвычайно разнообразна. Она зависит, между прочим, от числа,
места отхождения и толщины отростков (рис. 150).
Дендриты отходят от клетки в различном количестве, имеют разную
длину и форму. Толстые дендриты по своему строению не отличаются
от тела клетки. Они состоят из нейроплазмы и нейрофибриллей и, кро-
304 ОБЩ\Я ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНаЛЬНОЙ ПіРИФдРИЧіСКОЙ НЕРВНОЙ СИСТЕМЫ
л
Рис. 151. Схематическое
изображение миэлинового
нервного волокна.
Л—продольный;
В—поперечный разрез. В А налево ми-
элин растворен, и занятые им
места имеют более светлый вид.
В А направо миэлин показан
черным, каким он бывает
после обработки осмиевой
кислотой; ах — осевой цилиндр с
центральной частью, занятой
нейрофибриллями; ах'—
периферическая, свободная от фиб-
риллей часть осевого
цилиндра; х — слой свернувшегося
вещества, одевающий осевой
цилиндр; ра — периаксиаль-
ное пространство; у — прото-
плазматнческая, образованная
шванновскими клетками сеть,
пронизывающая миэлиновую
массу; SL — утолщенные
места сети в форме волокон,
обусловливающие существование
лантермановских насечек;
Si—шванновские клетки с их
ядрами; S—шванновская
оболочка; R — перехват Ранвье ¦
(по Немилову).
гконцевое разветвление
миэлиновая оболочка
ме того, содержат тигроиды, или тельца Ниссля.
Тонкие дендриты не содержат тигроидов (Ле-
ношек, 2). Тонкие дендриты возникают путем
ветвления толстых. Все дендриты начинаются
широким коническим основанием и затем
ветвятся подобно дереву, откуда и произошло их
название. Большинство авторов находит, что
дендриты оканчиваются свободно. Разветвления
дендритов нередко снабжены по длине шипами
или кругловатыми утолщениями. Существуют
нервные клетки, не имеющие дендритов.
Нейрит, или осевой цилиндр, характерно
отличается от дендритов: он никогда не содержит
тигр*оидов, а только нейрофибрилли и нейро-
плазму. Поверхность нейрита всегда гладкая.
Обычно от тела клетки отходит один нейрит,
начинаясь иногда на одном из дендритов.
Толщина нейрита по всей длине одинакова, и его
диаметр равняется нескольким микронам.
Длина нейрита может быть очень велика. У
крупных позвоночных животных, например, нейриты
двигательных нервных клеток бывают
протяжением больше одного метра. Некоторые нейриты
покрыты более или менее сложно устроенными
оболочками, образуя нервное волокно. Самая
наружная, очень тонкая оболочка —
соединительнотканного происхождения. Она называется
шванновской оболочкой или неврилеммой. Под
ней лежит другая, более толстая оболочка,
которая вследствие содержания в ней миэлина
называется миэлиновой, или мякотной. Волокна
с миэлиновой оболочкой называются миэлино-
выми (рис. 151). Другие нейриты не имеют
миэлиновой оболочки. Такие волокна называются
безмякотными.
Миэлиновая оболочка на нервных волокнах
бывает разной толщины. Но івообще, чем
меньше диаметр нервного волокна, тем эта
оболочка тоньше. Сообразно меняется и диаметр
осевого цилиндра. В тонких волокнах не только
миэлиновая оболочка, но и осевой цилиндр
меньше диаметром (Гассер и Грундфест, 56; Ме-
писашвили, 49). Миэлиновая оболочка не сплошь
покрывает осевой цилиндр. Начало нейрита и
лишены ее. Кроме того, через каждые 1—2 мм
прерывается на протяжении нескольких микро-
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
305,
нов. В этих участках осевой цилиндр покрывается лишь неврилеммой,
т. е. очень тонкой соединительнотканной оболочкой сетчатого строения
(Эттих и Иохимс, 29). Участки эти, именуемые перехватами Ранвье,,
располагаются в тонких волокнах чаще, чем в толстых (Зоттерман, 57;:
Такахаши, 60). Кроме того, на оболочке нервного волокна имеются так
называемые насечки Лантермана. Они углубляются почти до осевого
цилиндра, ко
захватывают только часть
[окружности. Эти насечки
располагаются много чаще
в толстых волокнах, чем
в тонких. Судя по
ультрамикроскопическим
изменениям насечек в
живом волокне под
влиянием электролитов, эти
насечки должны служить
участками для обмена
веществ между осевым
цилиндром и
окружающей внутренней средой
(Эттих и Иохимс, 29).
Нейриты на своем
пути могут отдавать
побочные ветки-коллатера-
ли. Как нейрит, так и
коллатерали
оканчиваются телодендриями.
Последние веточки тело-
дендрий оканчиваются
в мозгуГсвободно'на теле
клеток или дендритов,
не сливаясь с теми элементами, к которым они подходят (рис. 152).
Каждый нервный элемент включает нейрофибрилли и
межфибриллярное вещество, так называемую нейроплазму. Присутствие
тончайших нитей нейрофибриллей доказано во всех положительно нервных
элементах. Яснее всего они выступают в объемистых клетках передних
рогов спинного мозга. Но они видны и во всех отростках вплоть до
конечных разветвлений. В теле клетки фибрилли тянутся параллельно
более или менее густыми пучками, пробегая в различных направлениях
и располагаясь особенно густо вокруг ядра (рис. 150). По мнению
одних авторов, нейрофибрилли анастомозируют между собой и тем
образуют истинный сетчатый остов (Раман-Кахаль, 3). По (мнению других,
нейрофибрилли не образуют истинной сети, все фибрилли идут в
нейрите или в нервной іклетке изолированно друг от друга, но более или
менее значительными пучками. Иногда они распадаются на небольшое
20 И. С. Беритов
Рис. '152. Двигательная клетка переднего рога спинного
мозга собаки с многочисленными оканчивающимися на
ее поверхности в виде больших (а, Ь, с,і,е) и малых (df h)
концевых ножек нервными волоконцами; от больших
ножек g, f и др. отходят в виде продолжений тончайшие
фибрилли, оканчивающиеся маленькими петельками (по
Рамон-Кахалю),
305 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
число мелких пучков, анастомозирующих с соседними пучками, но без.
слияния самих фибриллей (Апати, 17; Бете, 7; іБоцлер, 21). Нейрофиб-
рилли признаются всеми современными авторами за
дифференцированные части протоплазмы. Их нужно считать наиболее характерной?:
структурной частью живой системы нервных элементов. Они замечены
Боцлером и де-Рени на некоторых живых объектах с полной
очевидностью (Максимов и Блюм, 45). Их находят в осевых цилиндрах
живых нервных волокон при
ультрамикроскопическом
исследовании (Эттих и Иохимс, 29).
В живых нервных волокнах
головастика лягушки нейрофиб-
риллярное строение осевого
цилиндра обнаруживается каждый
раз в результате раздражения
(химического, механического,
электрического). С отдыхом
нейрофибрилли перестают быть
видными (Шпейдель, 43). Нейрофиб-
рилли не являются постоянным
образованием, ибо форма и
количество их вариируют при разных
условиях. С повышением
функциональной деятельности они
становятся тоньше, количество их
при этом возрастает (Петерфіг,
по Жерарду, 53). Это, например.
происходит в двигательных
нервных клетках спинного мозга
вследствие длительной
деятельности во время стрихнинных
судорог. При гистологической
обработке спинного мозга лягушки,,
находящейся до этого в покойном состоянии, нейрофибрилли толще,,
сильнее окрашиваются, располагаются прямыми пучками по длине всей
клетки. Во время же стрихнинных судорог нейрофибрилли утончаются,
число их становится больше и они заполняют всю клетку, змеевидно*
изгибаясь (Гурвич, 59). То же наблюдается в двигательных клетках
спинного імозга при сравнении их во время спячки и после
пробуждения, как это видно на рис. 153 (Кахаль, 58). Быть может, с этим связан
также тот факт, что в чувствительных волокнах нейрофибрилли вообще
тоньше, чем в двигательных (Ариенс Каплерс, 50).
Нервные отростки у низших беспозвоночных не дифференцированы
на дендриты и нейриты. Они все являются однородными образованиями
и все они голые подобно дендритам (рис. 153а). Таковы отростки у всех
кишечнополостных, но они встречаются также у других беспозвоночных:
Рис. 153. Изменчивость нейрофибриллей
у ящерицы.
А, а — во время спячки; В, b — в бодрствующем
состоянии, после того как ящерица нагревалась в
течение нескольких часов при температуре 30е С
(Рамон-Кахаль).
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ 307
наряду с дифференцированными нервными отростками. У высших
беспозвоночных нервные отростки дифференцированы на нейриты и дендри-
ты. Как недифференцированные, так и дифференцированные отростки
большей частью имеют гладкую поверхность. Но в некоторых случаях
они покрыты утолщениями по всему ходу волокон и их разветвлений,,
так называемыми варикозными утолщениями (см. рис. 153а). Некоторые
авторы предполагают, что эти утолщения являются искусственными
Рис. 153а. Из эктодермального нервного сплетения медузы (Rhizostoma).
Мультиполярная клетка своими варикозными отростками
соприкасается с гладкими нервными отростками, которые принадлежат
биполярным нервным клеткам (по Боцлеру).
продуктами и возникают в результате гистологической обработки (Рах-
матуллин, рис. 65).
У беспозвоночных в случае дифференциации отростков нейриты
покрываются оболочками. Но у них не бывает типичной миэлиновой
оболочки с перехватами Ранвье и насечками Лантермана. Часто эти
оболочки более или менее толстые и перекрывают также клетки. Толщина
их іможет достигнуть значительной величины. Так, у ракообразных в
наиболее толстых волокнах осевой цилиндр равняется 15—20 ji, а весь
диаметр волокна достигает 80 . Оболочка содержит вещество, которое
окрашивается подобно коллагену соединительной ткани позвоночных.
Однако оболочка нервных волокон содержит также жироподобное
вещество, только оно не образует определенного слоя, как в миэлиновой
оболочке. Диаметр нервных волокон у беспозвоночных чрезвычайно
разнообразный: от самых тонких голых волоконец в несколько
микронов до видимых на-глаз, диаметром в 11 мм; таковы, например,
гигантские волокна у головоногих моллюсков (Юнг, 44). Чаще всего тонкие
волокна лишены оболочек; лишь когда эти волокна ложатся вместе в
большом количестве, они покрываются соединительно тканными
оболочками. Осевой цилиндр нервных волокон беспозвоночных имеет
полужидкий характер, принимающий фибриллярное строение при
раздражении (Шпейдель, 43).
Синапсы. Нервное волокно оканчивается в центральной нервной
системе как на теле нервных клеток, так и на их дендритах, а также
3D8 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧ6СКОЙ НЕРВНОЙ СИСТЕМЫ
свободно в межклеточном пространстве. У позвоночных эти окончания
имеют очень разнообразную форму и величину, но в общем они
напоминают пуговку, луковицу или петлю. На дендритах этот тип
окончаний наиболее распространен. Но на теле клетки окончания могут быть
более сложного строения. Они образуют более или менее правильную
спираль вокруг клетки или вокруг того холмика, где начинается
нейрит или ветвятся древовидно, и каждая последняя веточка :
многократно прикасается к телу клетки (рис. 154).
На гистологически обработанных препаратах обычно не находят
между конечной луковицей и телом клетки или дендритом никаких
протоплазматических связей. Находящуюся между ними физическую
среду Шеррингтон и Форстер назвали синапсами. Но
сейчас этим именем обычно обозначают конечные луковицы или петельки,
которыми заканчивается аксон. Основанием для суждения о раздельном
существовании синапсов служит разное окрашивание нейрофибрилляр-
ных элементов нервных окончаний и тела клетки или дендритов. Кроме
того, в конечных луковицах нейрофибрилли могут показывать
совершенно иное расположение и иную толщину, чем в том участке тела
клетки или дендритов, к которому они прилегают (Бодиан, 41).
Синаптические окончания >в одних случаях действительно являются
окончаниями нервного волокна —> конечной луковицей или петлей. Но в
других случаях голое ответвление аксона подходит к клетке илидендри-
там и ікасается их во многих местах, утолщаясь каждый раз на месте
прикосновения, а потом только оканчивается где-либо луковицей
(рис. 154). Такие 'боковые синапсы одни авторы называют мимоходными
почками (boutons en passant), а другие — коллатеральными (боковыми)
синапсами (Лоренте де-Но).
Число конечных синапсов на теле одной клетки может доходить до
100 и больше (Гофф, 37). Но на дендритах одной клетки количество
синапсов может достигать нескольких тысяч (Лоренте де-Но, 38; Бо-
дла-н, 41). Это особенно хорошо видно на рис. 155-1, где поверхность
дендритов густо • усеяна оинаптическими окончаниями; число их на
приведенной дендритной части около 300. Однако на некоторых
ганглиозных клетках число- синапсов много меньше, а иногда они даже
могут отсутствовать (Фален и Давенпорт, 39).
В спинном мозгу взрослого человека синаптические окончания,
величиной 0.5—5 jx, имеют форму пуговки, петли и луковицы. На
100 квадратных микронов поверхности промежуточной нервной клетки
приходится максимум 14.5 окончания, а на двигательной клетке —
'23.1 окончания. Вообще по всей поверхности маленьких промежуточных
жлеток находят 114 окончаний, а на теле двигательных клеток — до 1832.
Нервные окончания также очень густо располагаются на поверхности
дендритов. Поэтому общее число их на теле и дендритах одной клетки
достигает нескольких тысяч (Минклер, 61).
На живой нервной клетке позвоночных также удается ясно
распознать синаптические окончания. Так, на живой нервной клетке
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
309
¦і
f
S ы ^
^^жщ.
•~т'
л
J4i?
л/У X _ n
*ЧУ
Рис. 154. Синаптические окончания нейрофибриллей на теле двигательных клеток
(А—Е) и на большой промежуточной клетке (HS) спинного мозга 15— 16-дневной
кошки.
d, а — синаптические окончания на дендритах, последние не зарисованы. Одни и те же нейро-
фибрилли, Еетвясь, образуют многочисленные синапсы как на теле клеток, так и на дендритах.
Цифры указывают на нервные волокна (Лоренте де-Но).
сердца лягушки из узла Лрдвига в покойном состоянии трудно заметить
очертания синаптических окончаний. Но после воздействия на узел
токами ультравысокой частоты или при раздражении блуждающего нерва,
или при механическом раздражении самой клетки прикосновением ми-
310 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
кроиглой, синапсы становятся хорошо видны: они претерпевают так
называемую желатинизацию. Самая протоплазма клетки обнаруживает
большую структурность, благодаря изменениям преломляемости лучей
в некоторых составных частях протоплазмы (рис. 155-П) (Лаврентьев
и Федоров, 51; Смиттен, 55).
Несмотря на тесную связь синапсов с клеткой, дегенерация первых
не ведет к заметным изменениям клетки. По исследованиям
Лаврентьева (42), в ганглиях автономной нервной системы после перерезки
Фиг. 155-1. Двигательная клетка с дендритом из спинного мозга кошки.
Обработка по Кахалю. Видны синаптические окончания: на данной поверхности клетки находится
около 100 окончаний, а на дендритах — около 300 (Лаврентьев).
нервов соответствующие им синапсы перерождаются, но не изменяется
структура тех клеток, вокруг которых они оканчиваются. Если
регенерирующее нервное волокно подходит к телу клетки, то вокруг клеток
вновь возникают синапсы, причем тип этих синапсов будет тем же,
что и раньше.
Существование чисто физической среды между нервным окончанием
и телом клетки не признается всеми гистологами. Так, например,.
Буке (40) находит, что наблюдаемое при гистологической обработке
разное окрашивание нервных окончаний и тела клетки не может
служить доказательством перерыва между нейрофибриллями синапса и
тела клетки. В определенных случаях разное окрашивание получается
й при совершенно ясной протоплазматической непрерывности, как это,
например, имеется в нервной двигательной пластинке, где нейрофиб-
риллярная сеть нервного окончания сливается с протоплазматической
^етью мышечного волокна, несмотря на разное их окрашивание.
У беспозвоночных типичные синаптические окончания отсутствуют.
Нейриты обычно свободно оканчиваются заостренными волоконцами
в петлях нервных отростков или их конечные участки присоединяются
к другому нервному отростку. Если отростки варикозны, тогда они могут
закончиться одним из утолщений, но эти конечные утолщения ничем не
отличаются от тех, которые существуют по всему ходу волокна. Они
также свободно лежат в петлях или же прилегают к другим отросткам.
Боцлер (64) описал такие случаи у сцифомедуз и признал их за
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ 311
типичные синапсы (см. рис. 153а). Но едва ли эти утолщения,
существующие по всей длине отростка, являются синапсами. Характерно,
что на рис. ках Б о ц л с р а но •
¦еле утолщения еще тянется
небольшой участок последнего
волоконца без утолщения. Это
подтверждает, что мы здесь имеем
простой перерыв волоконца, а не
синаптические окончание.
У эмориона позвоночных на
ранних стадиях развития не на-
.ходят также конечных
луковиц. Нервные отростки
пересекают друг друга или идут
параллельно пучками и свободно
заканчиваются в петлях без
утолщения, как в нейропиле
беспозвоночных. Синапсы
возникают на поздних стадиях
эмбрионального развития. Это
было установлено на зародышах
разных млекопитающих (кошка,
крыса) и человека (Фален и
Давенпорт, 39; Уйндль, 54;
Минклер, 61).
Понятие о нейронной теории.
За единицу нервной системы
многие авторы признают
нервную клетку со всеми ее
отростками и со всеми окончаниями
.этих отростков (рис. 156). Этот
комплекс нервных элементов
Вальдейер (1) назвал
нейроном; отсюда самая теория
получила название
нейронной. Нейронная теория
предполагает, что в нервной системе
.всех животных каждый^нейрон,
каждая единица нервной'системы
существует: совершенно
изолированно от других нейронов и
что нейриты этих нейронов не
сливаются с другими нервными
элементами, а оканчиваются на поверхности клеточных тел или ден-
дритов многочисленными синапсами (рис. 152, 154, 155-1). В качестве
^примера укажем на центростремительный (афферентный) чувствитель-
Рис. 155- II. Синапсы в живой нервной клетке
сердца лягушки.
А — крупная нервная клетка людвигова узла в покое.
Синапсы не видны. В и С—две стадии изменения
клетки под влиянием раздражения блуждающего нерва.
Синапсы (S) стали видны благодаря их желатинизации,
меняется и самое содержимое клетки; N — ствол
блуждающего нерва (Лаврентьев и Федоров).
312 ОБЩАЯ ФИЗ !ОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
с
ріый нейрон позвоночных животных. Клетка его лежит в спинномозго^
бом ганглии; из клетки выходит один нейрит, который в ганглии же
делится на две ветви: одна идет на периферию и здесь оканчивается в.
хоже или в других тканях, в воспринимающих
нервных образованиях, так называемых
рецепторах; другая ветвь входит в мозг и здесь
делится на две веточки, из которых одна
направляется вперед, в краниальном направлении, а
другая — назад, в каудальном. Обе ветви
отдают множество коллатералей на разных
уровнях мозга. Все эти разветвления
чувствительного нейрона оканчиваются в соответствующей
симметричной половине мозга, частью около
клеток, лежащих целиком в головном и спинном
мозгу, частью около двигательных клеток и их
дендритов. Другой хороший пример
представляет центробежный (эфферентный)
двигательный нейрон позвоночных. Клетка и дендриты его
лежат в передних рогах спинного імозга или в
аналогичных образованиях головного мозга.
Нейрит выходит из мозга на уровне того же
сегмента, где находится и клетка, направляется
в периферические двигательные органы и
оканчивается здесь в волокнах поперечно полосатых
мышц, в так называемых эффекторах (рис. 157).
После Вальдейера нейронная теория была
признана рядом выдающихся гистологов,
например, Леношеком (2), Кахалем (3),
Догелем (47), и физиологов, например, Ф е р-
в о р н о м (4), Бидерманом (5), У о л л е-
ром (6), Шеррингтоном (48) и др.
Анатомические основания нейронной теории..
Нейронная теория основывается прежде всего;
на анатомических исследованиях позвоночных
животных.
Многие авторы находят, что у этих
животных каждая нервная клетка со всеми
отростками анатомически изолирована от других:
элементов нервной системы. Всякая связь
между этими элементами осуществляется путем
контакта или прикосновения (Кахаль, Лоренте
де-Но и др.).
Понятие о нейроне как о единице нервной системы вытекает также
из наблюдений, над эмбриональным развитием нервной системы.
Доказано, что каждая клетка со всеми своими отростками развивается из.
одной эмбриональной клещи — нейробласта. По мнению К а х а л я>.
е
156. Схематическое
изображение нейрона.
Z — ядро нервной клетки;
D — дендриты; ах — аксон
или нейрит; К — коллатера-
ли; S — шванновская
оболочка; Я — перехват Ранвье; а—
голая часть нейрита; b—часть
нейрита с миэлиновой
оболочкой; с — часть нейрита с
миэлиновой оболочкой и с
оболочкой Шванна; d — часть
аксона, покрытая только
оболочкой Шванна; е — голые
древовидные разветвления
окончания нейрита. Такие же
древовидные разветвления
имеются у коллатералей (по
Штёру).
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
313
Рис. 157. Взаимоотношения между двигательными и чувствительными нейронами.
Схематическое изображение: А — спинной мозг; В — двигательный (передний) корешок; С —
чувствительный (задний) корешок; D — спинномозговой (межпозвоночный) узел; Е — кожа; F — мышца
скелета; а — клетки чувствительных нейронов в спинномозговых узлах; b — клетки двигательных
нейронов в передних рогах; с — периферические отростки чувствительных нейронов, которые
оканчиваются в коже, образуя разного рода рецепторы; d — центральные отростки (нейриты) тех же нейронов;
е — их коллатерали, которые оканчиваются около клеток и дендритов интраспинального и
двигательного нейронов; /—¦ нейрит двигательных нейронов, оканчивающихся в скелетных мышцах (по Догелю).
это онтогенетическое развитие нервных элементов повторяет собой
развитие филогенетическое (рис. 158).
Далее, важное значение имеет в этом отношении дегенерация
синапсов в вегетативной нервной системе и в спинном мозгу после
перерезки
соответствующих нервов, а также ^ -і^М' (%? -^v t / s \\\ YY \\
последующая регенерация
их в связи с врастанием
в нервные узлы того же
или другого нерва.
Структурные изменения
пострадавшего нейрона не
распространяются на нейро-
фибриллярный аппарат и
клеточное тело
следующего нейрона
(Лаврентьев, 42).
Физиологическое
основание нейронной теории.
Физиологи также
выставляли одно важное
доказательство нейронной
теории. По их наблюдениям,
нервная клетка оказывает
на свои отростки трофи-:
Рис. 158, Филогенетическое и онтогенетическое
развитие нейрона по Рамон-Кахалю.
Верхний ряд дает филогенетическое развитие: А — лягушки;.
В — ящерицы; С — крысы; D— человека. Нижний ряд
показывает онтогенетическое развитие нейробласта в пяти следующих..
одна за другой стадиях а—е.
314 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
ческое, т. е. регулирующее обмен веществ, влияние. Так, например,
Уоллер показал, что если перерезать передний, т. е. двигательный,
корешок, то периферический отрезок отмирает в течение
нескольких дней вплоть до мышц; в этом отрезке меняется структура
нервных волокон, как говорят, они перерождаются, дегенерируют.
Одновременно они меняются и физиологически: они теряют
все жизненные свойства. В центральном же отрезке, где нервные
волокна имеют связь с клетками,
они сохраняют как нормальную
гистологическую структуру, так и все
свои физиологические свойства.
Если перерезать задний корешок
между ганглием и спинным мозгом,
то в центральном отрезке нервные
волокна перерождаются, а в
периферическом нервные волокна
сохраняются без изменения, так как
они остаются в связи с клетками
ганглия (рис. 159). Таким путем
Уоллер доказал, что нервные
волокна сохраняют способность к
жизненной деятельности только в
том случае, если их анатомическая
связь с клеткой остается в
полной сохранности. Отсюда вытекает,
что клетка дказывает на свой аксон
какое-то действие,
обусловливающее правильное питание,
необходимое для его существования.
После перерезки спинного
мозга перерождение происходит ниже
перерезанного места в пирамидных
и других нисходящих путях.
Перерождаются все нервные волокна,
отрезанные от своих клеток, а также
их окончания на клетках спинного імозга. На эти последние клетки
перерождение не распространяется.
Однако, по новейшим исследованиям, перерезка аксона не остается
совсем без влияния на жизнедеятельность клетки и на ту пасть аксона,
которая остается в связи с ней. Они также страдают от этого, и
притом тем сильнее, чем ближе к клетке произошла перерезка аксона.
Так, после перерезки двигательного корешка очень близко к мозгу
двигательные клетки претерпевают дегенеративное изменение. Тельца
Ниссля растворяются, ядро принимает эксцентрическое положение,
клетки впоследствии могут даже погибнуть (Гейст, 35). То же
происходит при перерезке задних корешков. Если перерезка прошла слиш>
Рис. 159. Корешки спинного мозга и их
перерождение после перерезки.
П. к. — передний корешок; з. к. — задний
корешок; уз — спинномозговой ганглий; о — место
перерезки корешка; зачерченный
отрезок—перерожденный участок. Схема 1 иллюстрирует
перерождение периферического нервного ствола
чуть ниже места соединения двигательного и
чувствительного корешков; схема 2
—перерождение периферического отрезка двигательного
корешка после перерезки; схема 3 — перерождение
периферических чувствительных волокон,
отрезанных от ганглия; схема 4 —перерождение
центрального отрезка чувствительного корешка
.©следствие перерезки между мозгом и ганглием
(по Данилевскому).
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
315
ком близко к спинальному ганглию, часть клеток в ганглии
перерождается (Гинзей и сотр., 36). Но, как правило, в большинстве клеток
дегенеративные процессы через некоторое время проходят и клетки
принимают свой обычный вид; в это же время начинает вырастать
новый аксон вместо отрезанного и переродившегося.
Эта последующая регенерация аксона и его окончания может также
служить некоторым основанием для нейронной теории. Но характерно,
что направление растущего аксона к иннервируемой ткани определяется
нейротропным действием самой иннервируемой ткани, точнее, каким-то
химическим действием ее. Иннервируемая ткань в определенных
случаях обусловливает и форму регенерируемого окончания. Это
случается, если иннервируемая ткань хорошо дифференцирована.
Следовательно, если можно говорить об организующей роли клетки на свой
гксон и его окончания, а также на иннервируемую им ткань, то можно
говорить и об обратном организующем действии иннервируемой ткани
на подходящий к ней аксон (Лаврентьев, 42).
Одно обстоятельство очень благоприятствует признанию и
распространению нейронной теории. Многие физиологи утверждают, что
нервная клетка является источником тех нервных процессов, которые
обусловливают целесообразную и координированную деятельность всех
органов. Например, общепризнано, что все импульсы координированного
произвольного или рефлекторного движения возникают в клеточной
массе нервных ядер. Имеется прямое физиологическое
доказательство, что нервное ядро служит тем местом, где возникают разряды
импульсов. Например, локальное отравление стрихнином клеточной
массы определенного центрального нервного ядра обусловливает
периодические разряды быстро протекающих импульсов. Эти
импульсы стрихнинного (Происхождения в определенных случаях
распространяются только в пределах отравленного нейрона. Они не возникают
при стрихнинном отравлении нервных волокон и окончаний. Этим
подтверждается, что стрихнин вызывает означенные разряды" своим
действием на клеточные тела (Дюссер де Баренн, 52). Таким
образом, по утверждению физиологов, клеточная масса представляет
самую главную составную часть нервной системы, волокна же служат
проводниками процессов, возникающих в клеточной массе.
Нейронное строение нервной системы было установлено
первоначально на позвоночных животных, но потом оно было признано
обязательным и для беспозвоночных, т. е. для всего животного мира,
универсальным (Заварзин, 66). Многие исследователи нашли
нейронное строение у червей, насекомых, ракообразных (Кахаль, 3;
Ретциус, 20; Леннош?к, 2; Заварзин, 66) и даже у кишечнополостных
(Бодлер, 65). По исследованиям этих авторов нервная система
беспозвоночных животных в основном также состоит из нейронов. И у них
нервные клетки вместе со своими отростками — дендритами и
нейритами — составляют нервную единицу, анатомически обособленную от
других таких элементов (рис. 159а). Но есть одно существенное
316 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
различие: у позвоночных нейрит одного нейрона своим конечным
разветвлением или боковыми ответвлениями заканчивается синапти-
чески на теле или на дендритах другого нейрона. У беспозвоночных
разветвления нейритов не заканчиваются такими синапсами. Она
Рис. 159а, Область двигательных нервов второго торакального
ганглия личинки стрекозы.
VWN, VWN, — перемычка между первым боковым нервом и коннективой. На
краях ганглия расположены двигательные нервные клетки, а в центре находится
нейропиль, который составлен из дендритных разветвлений этих клеток. Каждая
клетка дает один отросток, который на некотором расстоянии от клетки образует
расширение, а уже от него отходят многочисленные дендриты и рдин нейрит.
Нейриты выходят из узла через боковые нервы. Ясно видно, что каждая клетка
имеет свою систему дендритов и один нейрит, т. е. образует нейрон (по Заварзину).
вступают в связь с дендритами других нейронов путем пересекания:
или сопровождения их без специальных связей и большей частьк>
свободно заканчиваются утонченными гладкими или
варикозными волоконцами в петлях нервного сплетения (Заварзин, 66; Ганст-
рём, 25).
Нейропильная теория. Нейронная теория не получила всеобщего'
признания. На основании исследований Гольд ж и и его
последователей, затем Бете, Ни селя и в особенности Апати (17),
создалось отрицательное отношение к нейронной теории. Бете (7) объявил
эту теорию гипотезой, требующей основательных доказательств. Он,,
как и Апати, опирался при этом на наблюдения над
беспозвоночными животными. Вместе с Апати он утверждает, что между
воспринимающим раздражение чувствительным' нервом и производящим
внешний эффект двигательным нервом существует беспрерывная связь,,
осуществляемая нейрофйбрйллямй. Последние переходят из одного
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
317
нервного, элемента в другой без перерыва, Нейрофибрилли идут
е нервных, клетках в разных направлениях, пересекая одна другую под
разными углами; затем нейрофибрилли одного нервного отростка
непосредственно переходят в, другой отросток, помимо нервных клеток.
Ни начала, ни конца, ни'перерыва нейрофибриллей нигде не удается
Рис. 160._ Расположение .и направление інейрофибрилльй в
нервной системе.
Схематический рисунок. ' Rz — клетка чувствительного нерва; Cz —
клетка двигательного нерва;^т — мышцы (Бете).
найти (рис. 160). Так образуется своего рода сеть нейрофибриллей,
которую назвали нейропилем. Отсюда и теория получила название
пейропильной.
Бете сам не изучал строения нервной системы позвоночных.- Он
выступал против нейронной теории по другим соображениям. Бете
пытался выставить такие факты, которые должны были говорить
вообще против нейронной теории. Он приводит наблюдение над
крабом Carcmus maenas, у которого имеется несколько пар
конечностей и сообразно с этим несколько пар нервных узлов в брюшной
.нервной цепочке. Рефлекторная деятельность каждой конечности
происходит при посредстве соответствующего нервного узла. В этом узле
нервные клетки большей частью расположены с одной стороны.
Бете воспользовался этим обстоятельством. Он удалил клеточную
сторону. В узле осталась лишь сеть нервных отростков — нейропиль.
Однако после такой операции рефлекторная деятельность соответствуй
ющей конечности не прекратилась. Отсюда он делает заключение, что
нервные клетки не являются центром нервных процессов, что
координированная деятельность может протекать помимо них в сети
нейрофибриллей, следовательно, она является функцией нейропиля. .
Аналогичный опыт был проделан Ш т е й н а х о м (8) на
поясничном межпозвоночном узле лягушки. Он вырезал из узла клеточную
318 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
кассу; узел продолжал проводить возбуждение; при этом
рефлекторная деятельность задней лапки сохранилась без изменения.
Были попытки поколебать и эмбриональное основание нейронной
теории. Янсен (18) и Шульце (19) утверждают, что нейриг
вырастает не прямо из ганглиозной клетки; он сначала 'возникает из
других эмбриональных элементов, а уже потом соединяется с
ганглиозной клеткой. Бете и некоторые другие авторы отрицают также
исключительное трофическое влияние нервной клетки на отростки. По
наблюдениям Бете, у молодых іживотных периферический отрезок:
нерва, лишенный связи с клеткой, все-таки способен к регенерации;
такой отрезок, по Бете, сначала перерождается, но потом
регенерирует, восстанавливаясь не из соответствующей нервной клетки,,
а совершенно из других элементов.
Итак, защитники нейропильной теории, отрицая вообще нейронное
строение, признают синцитиальный нейропиль, как общий тип строения-
нервной системы всего животного мира.
Критическое рассмотрение нейронной и нейропильной теории.
Прежде всего нужно указать, что не все доводы Бете против
нейронной теории основательны. Так, например, опыты на Carcimis
maenas ни в каком случае не говорят против этой теории. У
ракообразных и вообще у членистоногих координированное движение
конечностей по существу не зависит от нервных узлов. Как мы узнаем
впоследствии, двигательная координация у этих животных составляет
функцию периферической нервной системы. У позвоночных же
животных координация обусловливается исключительно центральной нервной
системой. Вот почему в опытах Бете удаление клеточных элементов
нервного узла не должно было лишить конечность способности к
координации. Аналогичный опыт Штейнаха на позвоночных ничуть не
противоречит нейронной теории. В настоящее время ни один физиолог
не считает межпозвоночный узел за центральную нервную систему;
координация движений происходит помимо него. Вот почему удаление
клеточной части узла не должно было сопровождаться потерей
координирующей способности. Межпозвоночному узлу принадлежит только-
проводящая функция; если же с удалением клеточной части эта
функция не пострадала, то это щроисходит оттого, что возбуждение
из периферического отростка клетки может переходить в центральный,
непосредственно через общий нейрит.
Характерно еще то, что опыты Бете и Штейнаха,
выставленные против нейронной теории, с очевидностью подтверждают
доминирующее значение нервной клетки. В опытах обоих авторов
функциональная деятельность узла сохранялась только недели две. Позднее
эта деятельность расстраивалась и совершенно прекращалась. Этим
подтверждается одно из оснований нейронной теории, а именно
трофическое влияние клетки на свои отростки.
По более поздним гистологическим исследованиям, как уже.
указывалось выше, существование типичного синцитиального нейропил&
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
319
у высших беспозвоночных животных подвергается сомнению. Ряд
исследователей отвергает в настоящее время синцитиальный ней-
ролиль как основную форму нервной системы у червей и ракообразных
^Кахаль 3; Ретциус, 20; Ленношек 2; Боцлер, 21; Заварзин, 62). Эти
исследователи специально задавались целью изучить строение нейро-
пнля, но приходили к убеждению, что и у названных животных
нервная система состоит из самостоятельных нейронов, что нейропиль
представляет сплетение множества тончайших нервных волокон»
принадлежащих разным нейронам.
Кроме того, по новым исследованиям, периферический отрезок
нерва восстанавливается не по Б е т е, т. е. не независимо от
центрального отрезка (аутогенная регенерация), а путем разрастания
центральных, не отделенных от клеток нейритов (Гарриоон, 14, Бэрроус, 15,
Лаврентьев, 42). Только оболочки новообразованных участков нервных
волокон являются по существу чем-то вроде постороннего образования,
лишь вторичным образом соединенного с нейритом (Кахаль, Гейден-
гайн).
Однако неосновательность возражений Бете против нейронной
теории не іможет быть использована для утверждения универсальности
нейронного строения. Прежде всего следует указать, что у низших
беспозвоночных, у кишечнополостных нервная система не построена по
нейронному типу. Во-первых, у этих животных отростки нервных клеток не
дифференцированы на нейриты и дендриты; во-вторых, очень часто эти
отростки сливаются, образуя синцитий. Такое строение нервной системы
является первичньш в ее филогенетическом развитии.
Некоторые авторы в настоящее время считают такое синцитиальное
слияние нервных отростков искусственным продуктом, результатом
гистологической обработки (Заварзин, 66). В особенности большую
роль сыграли в этом отношении наблюдения Боцлер а (65) над сци-
фомедузамй (Rhizostoma). Он описал у них в экзодерме нервное
сплетение из двоякого рода нервных элементов. Одни элементы
представляют биполярные клетки с толстыми гладкими отростками, а другие —
мультиполярные клетки с сравнительно тонкими и варикозными
отростками. Эти варикозные отростки местами прилегают к гладким
отросткам и такие участки признаются за синаптическую связь (рис. 153а).
Но можно усомниться в синаптической природе этих связей. Мы здесь,
вероятно, имеем обычное явление соприкосновения двух отростков.
Наличие варикозности по всей длине отростка, а не только в области
прилегания к другому отростку, свидетельствует, что эти варикозности
не являются синапсами. Возможно даже, что эти варикозности
являются искусственным продуктом, как это бывает, например, при
окраске безмякотных нервных волокон метиленовой синькой (Рахма-
туллин, 67).
Нейропильное строение нервной системы у позвоночных отмечается
рядом авторов в отношении вегетативного отдела периферической
нервной системы. Нервные сплетения Ауэрбаха и 'Мейснера, а также сим-
320 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
патические нервные узлы рисуются, ими .как синцитиальный нейропиль
наподобие нервной сети низших беспозвоночных животных (Штёр, 22;
Буке и др.)- Но более поздними специальными исследованиями
Лаврентьева и его сотрудников .было доказано, что, в симпатических
узлах преганглионарные нервные волокна заканчиваются на теле и ден-
дритах посттанглионарных. клеток синоптически.
Однако, по мнению Лаврентьева, в ауербаховском сплетении
имеется точно так же подлинный синцитий нервных элементов; он
образуется благодаря слиянию дендритных . отростков . (64).
В отношении центральной нервной системы точно так же многие
авторы отмечали существование. в/ней- нейропиля наряду с нейронами.
Впервые Ретциус (20) описал нейропиль как широкопетлистое «ей-
рофибриллярное. образование, которое заполняет свободное от клеток
и нейритов пространство серого вещества мозга. Существование
.нейропиля в центральной нервной системе позвоночных признается
выдающимся .неврологом. Ар иен с; К.аипе.рео.М! (13).
Многими авторами отмечено нейропильное строение в некоторых
элементах головного мозга, например в слое нервных клеток Пуркинье
(Бильшовский, 10; Вольф, 11; Удендаль, 12).
Наконец, Буке (9), Т.игс (16) и,другие утверждают, что
двигательные окончания переходят в саркоплазматическую сеть мышечной
клетки без перерывов. Связь окончания чувствительного нервного
волокна с нервным элементрм рецепторов также осуществляется при
посредстве слияния нейрофибриллей. окончания с нейрофибриллями
рецепторного элемента.
Буке (26) даже полагает, что промежуточная среда на месте
перехода от одного нейрона к другому ,не может быть чисто физической,
неживой средой, каким-то цементом. И эта среда должна представлять
«живое вещество», т. е. живую систему, и иметь строение наподобие
перитерминальной сети, связывающей нейрофибриллярное окончание
с нейрофибриллярной сетью внутри ,клетки. Ввиду того что живая
система синапса, по Буке, характерно отличается от тех образований,
к которым она прилегает, она может обусловливать физиологическую
самостоятельность связываемых ею нейронов.
Но с точки зрения физиологии не имеет особого значения,
существует ли между нервным окончанием и клеткой или дендритом
физическая среда или протоплазматический субстрат. Главное
заключается в том, что здесь происходит перерыв нейрофибриллей, и это
обстоятельство не служит препятствием для передачи возбуждения от
нервного окончания на тело клетки или на дендриты.
Нейронно-нейропильная теория. Новейшие гистологические
исследования центральной нервной системы дают возможность составить
совершенно новое представление о строении мозга. Согласно.этим
исследованиям, нейропиль, как сеть нервных элементов, является основной
формой строения нервной системы у низших беспозвоночных животных,
как, например, у кишечнополостных. Нейропиль здесь распространяется
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
321
по всему организму и выступает как нервное сплетение без
дифференциации нервных отростков на нейриты и дендриты и без обособленного
скопления нервных клеток в виде нервных узлов. При этом нейрофиб-
рилли встречающихся нервных отростков местами сливаются одна
Рис. 161. Нервная сеть гидры (Hydra fusca). Она образуется из протоплазматических
отростков. Лежит на мышечном слое, но не видна связь между нервной сетью и мышечной
тканью (из книги Рожерса, 1927).
с другой. В результате образуется нейрофибриллярная сеть синцитиаль-
ного характера (рис. 161, 162). В связи с таким строением стоит
главная физиологическая особенность нейропиля: он диффузно проводит
возбуждение одинаково во все стороны, причем каждый элемент
участвует в проведении возбуждения как в одном, так и в другом
направлении.
Это строение нервной системы характерно для низших
беспозвоночных. Оно является древней формой строения нервной системы. Из
нее развиваются обособленные нейронные комплексы. Однако ней-
ропиль сохраняется и у позвоночных вплоть до человека (Паркер, 46).
У высших беспозвоночных в нервных узлах мы также находим
нервную сеть, которая обычно занимает центральное место. Она образуется
путем пересечения множества нервных отростков, отходящих от
чувствительных, двигательных и промежуточных клеток. В одних случаях
21 И. С Беритов
322 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
эти отростки дифференцированы на дендриты и нейриты, а в других —
нет, но в образованных ими сплетениях мы обычно не видим ни слияния
нервных отростков между собою, ни связывания анастомозами.
Нейриты, как и дендриты, свободно оканчиваются в петлях (рис. 159а).
Ято образование также называется нейропилем, в отличие от нервных
стволов, нервных путей и клеточных скоплений.
Рис. 162. Схематическая картина эктодермальной нервной системы кишечнополостных.
Наверху фигуры небольшой участок эктодермы (эпидермис;. Эпителий состоит из поддерживающих
клеток (SO, между которыми находятся чувствительные клетки (S). Базальный конец последних
распадается на тонкие волоконца, которые распространяются в подэпителиальном нервном сплетении.
Эпителий также содержит мышечные клетки (ет), базальные концы которых образуют сократительные
волоконца. Там же находится мышечная ткань, которая лежит под эпидермисом (т). Нервное
сплетение, которое простирается между чувствительными клетками эпителия и мышечными клетками,.
содержит нервные клетки (g) различного вида (Гертвиг).
Выдающийся невролог Джадсон Херрик (23) считает, что
также у круглоротых рыб и хвостатых амфибий нейропиль является
основной структурной формой центральной нервной системы. Этот
нейропиль представляет собой густое сплетение тончайших разветвлений
аксонов, переплетенных с многочисленными разветвлениями дендритов.
Но только, по Херрику, нейропиль позвоночных, в отличие от нейро-
пиля беспозвоночных представляет собой не синцитий или простое
сплетение нервных отростков. В нейропиле позвоночных аксонные
волоконца одних нейронов оканчиваются на теле и дендритах других
нейронов путем синапсов (рис. 163). Поэтому Херрик называет ней-
ропильные участки синаптическими полями, которые служат для
объединения всех более или менее дифференцированных нервных
образований, давно известных под названием нервных центров и .нервных
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ ' 323
путей. Нейропиль, как густой войлок, не только заполняет все
пространства между этими дифференцированными образованиями, но также
проникает во все эти образования, заполняя все межклеточные
пространства. В образовании нейропиля принимают главное участие как нервные
отростки дифференцированных нервных центров, их аксоны и дендриты,
так и нервные отростки других, недифференцированных образований,
как это имеет место в formatio reticularis и substantia gelatinosa
Rolando. Тончайшее строение центральной нервной системы высших
позвоночных особенно детально было изучено Лоренте де-Но. Он
описал тончайшие сплетения разветвлений дендритов и нервных
волокон, а также их синаптические связи. На рис. 163 представлена
схематически картина синаптического
нейропиля в области
двигательных клеток спинного мозга.
Итак, нервная система, по
существу говоря, как у
позвоночных, так и у высших
беспозвоночных животных состоит из
клеточных скоплений и нервных
путей, с одной стороны, и
нейропиля, с другой. Нервная система
в основном построена по
нейронному типу, но так как она
включает всегда ыейропильные
образования, мы называем эту новую
теорию строения нервной
системы нейронно-нейропилъной. При
этом мы не утверждаем, чтс
нейропиль позвоночных всецело
построен по синаптическому
типу. Возможно, что в
исключительных случаях происходит
слияние нейрофибриллярных
элементов, образуется нервная
сеть — синцитий, как например,
в ауербаховском сплетении. Мы находим эту теорию наиболее
приемлемой и будем придерживаться ее при объяснении физиологических яв^
ЛеНИЙ. :. .-;
Если нейропиль является постоянным образованием центральной сип
стемы, то он должен играть существенную роль в центральной деятель-;
ности. Об этом свидетельствует прежде всего чрезвычайно богатое
снабжение нейропиля капиллярами. Установлено, что в коре переднего
мозга млекопитающих, во всех ее слоях, а также и симпатическом
шейном ганглии, затем в области окончания VIII нерва в ядре Маут-
нера в головном мозгу головастика, капиллярная система развита
вдвое-втрое больше, чем в нервном стволе или белом веществе мозга;
21*
¦п
:ш:^:^ш
¦я
Рис. 163. Схематическое изображение
нейропиля в сером веществе спинного мозга.
В — клеточные тела, D —¦ дендриты, S — нервные
окончания с синаптическими утолщениями. Эти
утолщения более толсты на теле клетск, чем на
дендритах (Лоренте де-Но).
324 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
более того, установлено, что вообще в сером веществе повышенная
васкуляризация находится в связи с нейропилем — сплетением дендри-
тов и нервных окончаний, а не с клеточной массой (Дюннинг и Вольф,
33; Бартелмец и Гоер, 34). Безусловно, богатое кровоснабжение ней-
ропиля должно свидетельствовать об интенсивной функциональной
деятельности. Мы признаем за нейропилем сложные функции, которые
ни © коем случае не могут быть осуществляемы наличием нервных
центров и нервных путей.
Эмбриональное развитие нейропиля у позвоночных. Нейропиль как
сетевидное образование нервных отростков эмбрионально закладывается
в результате разрастания нервных отростков.
В курином зародыше нейропиль впервые намечается в возрасте
девяти дней: межклеточное пространство серого вещества спинного
мозга достаточно обогащается сагитально и поперечно лежащими
волоконцами. В последующие дни сплетение растет, оно богатеет как
разветвлениями дендритов, так и нейритов. На 13-й день уже
появляется типичная нейропильная сеть в виде сложного войлока. В
дальнейшем происходит еще большее уплотнение этого войлока (Зураба-
швили, 62).
Сравнительное физиологическое и структурное исследование спинного
мозга котенка на разных стадиях постэмбрионального развития
показывает, что нейропильное строение характерно для спинного мозга в
ранние стадии его развития. У котенка в первые дни после рождения, когда
он производит только общие целостные реакции на всякое
раздражение, в спинном мозгу имеются нервные клетки и сплетение их отростков,
но нет синаптических луковиц, нет перицеллюлярных нервных
сплетений, очень слабо развиты прямые нисходящие пути, связывающие
отдаленные сегменты мозга. С появлением быстрых локальных реакций,
нормального стояния на ногах и локомоции нервное сплетение, или
нейропиль, становится значительно гуще, нервные отростки
располагаются плотнее, а с другой стороны, появляются синапсы на
клетках, перицеллюлярные сплетения и ассоциативные длинные пути
(Уиндль, 54).
И у человеческого эмбриона нейропиль как сплетение нервных
отростков закладывается очень рано в спинном мозгу, еще на б-й
неделе, раньше возникновения способности к рефлекторным движениям,
т. е. раньше установления связей между чувствительными и
двигательными элементами и формирования мышечных клеток и нервных
двигательных пластинок. Уже потом с сетевидным образованием спинного
мозга связываются заднекорешковые волокна и нервные пучки,
нисходящие из продолговатого мозга и верхних отделов спинного мозга. При
этом устанавливается способность к недифференцированным
рефлекторным реакциям, простирающимся на весь организм (Уиндль, 32;
Минковский, 31).
Из этих наблюдений на эмбрионах следует, что в онтогенезе
нервной системы нейропильное строение и обусловленные им общие цело-
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
325
стные реакции предшествуют развитию нейронной системы с ее
изолированными местными реакциями и быстрым односторонним
проведением возбуждения, подобно тому, как это происходило в процессе
филогенеза.
Разделение нервной системы на периферическую и центральную.
При делении нервной системы на центральную и периферическую имеют
в виду чисто анатомическое основание. Все, что сгруппировано в
позвоночном канале и в черепной коробке, именуется центральной нервной
системой, а все, что лежит вне их, называется периферической. Так как
последняя система главным образом состоит из нервных волокон, то
под этим термином часто подразумевают лишь периферические нервные
стволы, состоящие из нервных волокон. Фактически это неправильно.
Обе нервные системы включают все нервные элементы — как нейропиль,
так и нейроны с их клетками, дендритами и нейритами. Так,
периферическая система включает целый ряд межпозвоночных ганглиев,
а один из отделов ее, так называемая вегетативная, или автономная,
нервная система, включает широко распространенный нейропиль и еще
целый ряд ганглиев.
Судя по морфологическому составу, периферическая и центральная
нервные системы существенно не отличаются одна от другой. Если
физиологи все-таки считают нужным рассматривать ту и другую систему
отдельно, то основанием этому послужило другое весьма существенное
обстоятельство. У позвоночных животных каждая координированная
реакция организма обусловливается функциональной деятельностью
спинного и головного мозга, т. е. той системой, которую принято
называть центральной.
Периферическая система не играет существенной роли в этом
отношении: она только проводит возбуждения с периферии в
центральную систему и обратно. Следовательно, основная функция
периферической системы — проведение возбуждения. Эта функция значительно
проще, чем координирующая функция центральной нервной системы,
и потому целесообразно сначала рассмотреть ее.
Периферическая нервная система позвоночных животных не является
функционально однородным образованием. Она как анатомически, так
н функционально делится на две системы: цереброспинальную или
соматическую, и вегетативную или автономную. Поэтому общая
физиология той и другой системы будет изложена отдельно.
Центральная нервная система, в свою очередь, также делится
анатомически и функционально на два отдела: на стволовую часть
головного мозга и спинной мозг, с одной стороны, и на большой мозг,
с другой. Сообразно с этим общая физиология их будет излагаться
отдельно: сначала будет изложена прирожденная нервная деятельность
спинного мозга и стволовой части головного мозга, а затем
индивидуально приобретенная деятельность коры больших полушарий.
Изложение общей физиологии нервной системы мы начнем с
периферической цереброспинальной системы.
326 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Литература
3. Waldeyer W., Dtsch. med. Wschr., 44, 1891.
2. Lenhossek M. v., Der feinere Bau d. Nervensystems im Lichte neuerer For-
schungen, 2. AufL, Berlin, 1895.
3. Ramon у С a j a 1, Revista de cientias] medicas de Barcelona, 16, 20, 23, 1892.
Structure et connections des neurones, Conf. Nobel faite a Stockholm, Dec. 1906.
4. Verworn M., Das Neuron in Anat u. Physiologie, Jena, 1900.
5. Biedermann W., Elektrophysiologie, Leipzig, 1895.
6. Waller А., С r.-soc. biol., 34, 1852 (шесть сообщений).
7. В e t h e A., Allgemeine Anat. u. Physiol, d. Nervensystems, Leipzig, 1903.
8. S t e i n а с h E., Pfl. Arch., 78, 291, 1899.
9. Boeke J., Erg. Physiol., 19, 448, 1921.
10. В i e 1 s с h о w s к y, J. Psych. Neur., 5, 1904.
11. Wolf, Anat. Anz., 33, 1903.
12. О u d e n d a 1, Psych, en Neurol. Bladen, Amsterdam, 1912.
13. A r i e n s К a pper s C. U., Die vergleichende Anatomie d. Nervensystems d. Wirbel-
tiere und Menschen, 1, 90, 52, 1910.
14. Harrison, J. exp. Zool., 8, 1900; Arch. Entw. Mech., 30, 1930.
15. Burrows, J. exp. Zool., 10, № 1, 1911.
16. Tiegs O. W., Austr. J. Exp. Biol. Med. Sci., 3, 46, 69, 161, 1926; 6; 111, 1929.
17. Apathy, Mitt. Zool., Stat. Neapel, 12, 1897.
¦18. J a n s e n, Die Entwicklungsmechanik d. Nervenbahnen im Embrio d. Saugetiere,
Kiel, 1903.
19. Schultze, Arch. mikr. Anat., 66, 1905.
,20. R e t z i u s, Proc. Roy. Soc, B, 80, 1908.
21. BozlerE., Z. Zellforscher., 5, 1927; Z. vergl. Physiol., 6, 1928.
22. Stohr Ph., Z. wiss. Biol., Abt. C, 5, 167, 1927; Mikroskopische Anat. d. vegetati-
ven Nervensystems, Berlin, 1928.
23. J u d s о n H err i с к С, Proc. Nat. Acad. Sci., 16, № 10, 643; J. сотр. Neur., 59,
93, 1934.
24. С о g h i 1 I F., Anatomy and the Problem of Behavior, Oxford, Univ. Press, 1929.
25. Hanstrom В., Vergleichende Anatomie d. Nervensystems d. wirbellosen Tiere,
Berlin, 1928.
26. Boeke J., Nerve endings, motor and sensory, в книге W. P e n f i e 1 d: Cytology
of the nervous system, New York, 1932.
27. Forbes A., Physiol. Rev., 2, 361, 1922.
28. Hill A. V., Proc. Roy. Soc, B, 105, 163, 1929.
29. E 11 i с h G. u. Jochims I., Pfl. Arch., 215, 518, 1926.
30. Auerbach L., Pfl. Arch., 222, 493, 1929.
31. M i n k о w s k у М., Zum gegenwartigen Stand d. Lehre von den Reflexen, Zurich-
Berlin, 1925.
32. W i n d 1 e W. F. a. Fitzgerald J. E., J. Сотр. Neur., 67, 493, 1937.
33. D u n n i n g H. S. a. Wolff H. G., J. Сотр. Neur., 67, 483, 1937.
34. Barteimez G. W.a. Hoerr N., J. Сотр. Neur., 57, 401, 1933.
35. Geist F. D., Arch. Neur. a. Psych., 29, 88, 1933.
36. H i n s e у J. С, К r u p p M. A. a. Lhomon W. Т., J. Сотр. Neur., 67, 205,
1937.
37. H off E. C, Proc. Roy. Soc, B, 111, 175, 226, 1932.
38. Lor en te de No, J. Psych. Neur., 45, 1933; ib., 46, 113, 1934.
39. P h a 1 e n G. S. a. D a v e n p о r t H. A., J. Сотр. Neur., 68, 67, 1937.
40. Boeke J., Сб., посвящ. И. Бериташвили, изд. Груз. фил. АН СССР, стр. 63, 1936.
41. В о d i a n D., J. Сотр. Neur., 68, 117, 1937,
42. Лаврентьев Б. И., Арх. биол. наук, 48, вып. 1—2, 195, 1938.
43. S р е і d е 1 С. С, Cold Spr. Harb. Symp. Quant. Biol, 4, 13, 1936.
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ
327
44. Young J. Z., Cold Spr. Harb. Symp. Quant. Biol., 4, 1, 1936.
45. M a x i m о v A.a.Bloom W., A Text-book of Histology, 1931, pp. 240, 253.
46. Parker G. M., The elementary nervous system, 1919.
47. D о g i e 1 A. S. Многочисленные исследования, напечатанные в Arch. mikr. Anat,
за 1890—1906 гг.
48. Sherrington Ch., The integrative action of the nervous system, 1906.
49. Меписашвили И., Сообщ. Ак. Наук Гр. ССР, 1, № 8. 1940.
50. Ariens Kappers, The^comp. anat. of thejnervous system of vertebr., 1936.
51. Лаврентьев Б. и Федоров Б., Биолог, действие ультравысоких частот
(ультракоротких волн). ВИЭМ, стр.195, 1937.
52. Dusser de Barrenne J. G, а. Мс С u II о с h W. Т., Am. J. Physiol. 127,
620, 1939.
53. Gerard R. W., Quart. Rev. Biol., 6, 59, 193.
54. Win die W. F., J. Сотр. Neur., 50, 439, 1930.
55. Смиттен H., Бюл. эксп. биол. мед. 8, 195, 1939.
56. Gasser H. a. Grundfest H., Am. J. Physiol., 127, 393, 1939.
57. Zottermann Y., Skand. Arch. Physiol., 77, 123, 1937.
.58. Ramon у Cajal, Trab. d. Lab. de invest. Univ. de Madrid, 3, fasc. 1, p, 9t 1904.
59. Gurwitsch L. F., Pfl. Arch., 187, 147, 1922.
60. Katashi Takahashi, J. сотр. Neur. Psychol., 18, 166, 1908.
61. Minckler J., The Anat. Rec, 77, 9, 1940.
62. Зурабашвили А. Д., Тр. Инст. физиол. им. Бериташвили, 5, 169, 1943.
63. Tiegs О. W., Austr. Exp. Biol. Med. Sci., 4, 79, 1927.
64. Лаврентьев Б. И., Морфология автономной нервной системы, Сб. работ по
ред. проф. Лаврентьева, стр. 5, 1939.
65. Bozler Е., Zeitschr. Zellforsch., 5, 1927 (цитир. по Заварзину).
¦66. Заварзин А. А., Очерки по эволюц. гистологии нервной системы. Медгиз,
1941, стр. 46.
Глава II
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
Возбудимость нерва. Нервное волокно, подобно мышечным
волокнам, обладает возбудимостью. Степень возбудимости нервных волокон
значительно выше возбудимости мышечных, поэтому для раздражения
чувствительных и двигательных нервных волокон требуется гораздо
меньшая сила электрического тока, чем для скелетных мышц.
Возбудимость различных нервных волокон, входящих в состав одного
нервного ствола, не одинакова. Исследованиями американских ученых
Эрлангера и Гассера установлено, что в седалищном нерве
холоднокровных и теплокровных животных существуют три рода
волокон, называемых Л-, Б-, С-волокнами. При этом возбудимость Л-воло-
кон больше, чем В, В-волокон больше, чем С. Если принять пороговую
силу индукционного удара, необходимую для возбуждения Л-волокон,
за единицу, то для В-волокон она будет равна 2.4, а для С — 15.7.
Но даже возбудимость группы волокон Л не одинакова. Поэтому
при слабых раздражениях возбуждается только часть их. То же самое
следует сказать для групп В и С. Для возбуждения всех волокон
группы Л требуется 2.1 единицы, для группы В— 6, а для группы
С—203. Если выразить силу индукционных ударов в сантиметрах
индукционных катушек и представить, что порог и максимум силы тока
для группы Л-волокон равняются 30 и 20 см, то для группы В они
будут равняться 18 и 10 см, а для С — 8 и 4 см.
Разная возбудимость нервных волокон находится в связи с
толщиной их. К группе А принадлежат исключительно толстые миэлиновые
волокна — свыше 15 jjl. По толщине эти миэлиновые нервные волокна
делятся на три группы, причем волокна наибольшей толщины дают
наиболее быстро протекающую волну возбуждения (альфа), волокна
средней толщины — более медленную волну (бета), а меньшей
толщины— наиболее медленную волну (гамма). Группа В-волокон
включает тонкие миэлиновые волокна, С-волокна признаются за немиэли-
новые волокна (рис. 164). Но к ним же относятся некоторые тонкие
миэлиновые волокна (Эрлангер и Гассер, 17). В передних и задних
корешках, т. е. среди двигательных и чувствительных волокон, нахо-
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА 329
дятся все три рода миэлиновых волокон группы А Волокна В проходят
через rami communicantes grisei, волокна С — через г. commun. gr.,
а также через задние корешки и г. commun. albi.
На рис. 164 дается поперечный разрез нервного ствола (п. saphenus
кошки), где ясно видно существование нервных миэлиновых волокон
разной толщины, а на рис. 165 — количественное распределение нервных
волокон разной толщины в п. peroneus жабы.
Возбудимость нервных волокон меняемся в очень больших пределах.
Такое изменение может происходить под влиянием наркотических
веществ, кислот, щелочей, солей.
.*?
в
8*::
Малая концентрация и
кратковременное действие производят
повышение возбудимости, а
большая концентрация и
продолжительное действие — понижение
возбудимости, сразу или же
после начального кратковременного
повышения возбудимости.
По новейшим исследованиям,
возбудимость нерва меняется
строго параллельно концентрации
водородных ионов. При
определенной концентрации нерв
сохраняет нормальную возбудимость
в течение многих часов. Для
нерва теплокровных животных при
температуре 37° С это бывает при
рН=7.4. Если рН повышается до
7.8, т. е. реакция нерва меняется
в сторону щелочности,
возбудимость повышается; при
дальнейшем повышении концентрации до 8.0 и выше происходит
ритмическое возбуждение нерва. Когда рН падает, возбудимость
понижается, а когда рН становится ниже 7.4 и реакция меняется в
сторону кислотности, возбудимость становится ниже нормы (Леман, 46).
Температурные изменения также оказывают большое влияние на
возбудимость нерва. Постепенное согревание до 40° ведет к повышению
возбудимости, постепенное охлаждение, наоборот, — к понижению ее.
Но возбудимость нервных волокон меняется и сама собой, так
сказать, спонтанно, при внешне одинаковых условиях. Это видно из
постоянной изменчивости порогов раздражения и скрытых периодов
реакций (Блэир и Эрлангер, 18). Это показывает, что основной
биологический процесс все время меняется: он то усиливается, то ослабевает,
соответственно меняется и количество вполне восстановленной
возбудимой системы с течением времени, вследствие чего меняются
возбудимость и скрытый период.
L
Рис. 164. Поперечный разрез пучка нервных
волокон из VII заднего корешка кошки.
А— толстые А-волокна диаметром 10—22 р.,
В—тонкие миэлиновые B-волокна диаметром I—6 ^ (Лежава
и Меписашвили).
330 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Возбудимость мякотного нервного волокна вдоль его поверхности не
одинакова. Оно чрезвычайно чувствительно к электрическому раздра-
Рис. 165. Кривая распределения волокон в п. peroneus жабы.
Результат гистологического исследования. На рисунке каждая точка отвечает одному волокну.
Абсцисса соответствует толщине волокон (в микронах), а ордината — количеству волокон.
Ступенчатой же линией показана общая толщина всех нервных волокон за соответствующие полмикрона.
Между обозначившимися тремя группами существует небольшое количество волокон переходной
толщины (Гассер и Эрлангер)
жению в области перехватов Ранвье, где нет миэлиновой оболочки.
Наоборот, там, где нервное волокно покрыто мякотной оболочкой, оно
не раздражается обычными индукционными ударами большой интенсив-
/Г
)(
ш
5
4 300
ч>
і
1
>гоо
§
^
ч>
^
%1М
$
$
^
<$
^Т ,g
і і ,
)(
_ _і_. л . і.
ности. Это открытие
принадлежит
лаборатории К а то и было
установлено на
изолированных нервных
волокнах. Между двумя
перехватами Ранвье
расстояние около 2ммш
Приизучении порогов
волокна
раздражались биполярными
микроэлектродами с
межполюсным
расстоянием около 20 ji или
один микроэлектрод
(катод)
прикладывался к изолированному
волокну, а другой (анод) — к центральному нервному стволу.
В обоих случаях при положении электродов на перехватах Ранвье
нервное волокно возбуждалось при расстоянии между катушками около
400 мм, в то время как при положении электродов посредине между
перехватами эффект получался при 200 мм (рис. 166). Путем матема-
О
0.3
06 09 12 /5
Расстояние от R б ям
/<7
Zf
Рис. 166. Кривая порогов электрического раздражения
нервного волокна в разных участках от одного перехвата
до другого.
На абсциссе — расстояние в миллиметрах от перехвата Ранвье, а
на ординате — расстояние между индукционными катушками в мм.
Электроды биполярные (Като)
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
331
тического анализа авторы заключают, что и этот последний эффект
вызывается не из среднего миэлинового участка, а путем распространения
тока на область перехватов Ранвье. Следовательно, миэлиновая нервная
оболочка является хорошим изолятором, достаточным для того,, чтобы
электрический ток употребляемых нами индукционных катушек не
действовал через нее на осевой цилиндр. Очевидно, и при раздражении
нервных стволов электрическим током возбуждение возникает в
нервных волокнах прежде всего в области перехватов Ранвье (Rato, 37).
Раздражение нервного ствола. Всякий действующий на нерв
внешний агент, интенсивность которого быстро меняется от одной величины
до другой, вызывает в нерве возбуждение. Следовательно,
деятельность нервного ствола подчиняется известному закону раздражений
Дюбуа-Реймона.
Возбуждение возникает в нерве не просто под влиянием абсолютной
силы действующего агента, а от быстрого изменения этой силы.
Как и в случае мышечной ткани, для вызова возбуждения в нерве
необходимо, чтобы раздражитель действовал определенное время;
слишком кратковременное действие электрического тока не оказывает
раздражающего действия. При этом имеет значение межполюсное
расстояние электродов. Так, при определенной силе тока время 0.45 з
достаточно для возбуждения, если расстояние между полюсами 2 мм. Если
же это расстояние равняется 16 мм, то ток должен действовать в
продолжение 1,2 j (Вейсс, 7). Разумеется, это время тем меньше, чем
лучше функциональное состояние нерва, чем выше его возбудимость.
О местных изменениях основного биологического процесса, или о
местных реакциях нерва. При некоторых подпороговых раздражениях
нервное волокно отвечает местным изменением основного
биологического процесса. Этот процесс сопровождается местным колебанием
электрического потенциала. В нем, должно быть, участвуют наиболее
возбудимые частички возбудимой системы раздражаемого участка. Это
электрическое колебание очень слабо, но в настоящее время оно хорошо
изучено с помощью осциллографов. Впервые занялись им Раштон
(38) и Катц (39). Они раздражали нерв подпороговыми
индукционными ударами и обнаружили, что в участке раздражения возникает та:
кой же ток, как при возбуждении, но только меньшей амплитуды.
Кроме того, этот ток локален, т. е. наблюдается только в области
раздражения, не дальше чем на расстоянии нескольких миллиметров от
раздраженного участка. Он усиливается с усилением подпорогового
раздражения. Достигнув определенной критической величины, он
переходит в ток возбуждения, который распространяется по всему нерву.
Эта необходимость определенного предварительного местного
потенциала для возникновения возбуждения была отмечена еще раньше,
40 лет назад, Чаговцем по чисто теоретическим соображениям (59).
Продолжительность локального потенциала значительно меньше
продолжительности распространяемого биотока. В нерве лягушки при
комнатной температуре локальный процесс продолжается до 0,18 а;
332 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
в безмякотном нерве ракообразной Carcinus maenas он продолжается
при той же температуре 0,25 о (Катц, 39). Как интенсивность, так »
продолжительность локального процесса меняются в зависимости от
разных условий. Они тем больше, чем сильнее раздражение. При низ*
кой температуре локальный процесс удлиняется. При наркозе
локальный процесс ослабевает, и требуется большая сила раздражения для
его наступления (Катц, 39).
При детальном исследовании локального процесса на изолированных
нервных волокнах рака оказалось, что
местный процесс состоит из двух
частей — поляризационного потенциала,,
обусловленного оболочками волокна
на месте электрического раздражения^
и местного электрического ответа. Они
характерно отличаются друг от друга,
Поляризационный ток возникает как'
на катоде, так и на аноде. Они
одинаковы по амплитуде, но
противоположно направлены. Эти эффекты
продолжаются все то время, пока действует
удар, и нарастают совершенно
пропорционально нарастанию силы
удара. Они не меняются ни от
рефракторной или сверхнормальной фаз,
которые оставляют за собой
возбуждение, ни от удара к удару. Местный
электрический ответ возникает только
на катоде, длится и после удара в.
течение полусигмы, нарастает с
усилением раздражения гораздо быстрее
по мере приближения к пороговому
раздражению; во время рефракторной
фазы ослаблевает и даже исчезает,
во время сверхнормальной фазы
увеличивается. Он меняется значительна
от удара к удару и распространяет*
ся дальше поляризационного тока;
(рис. 167). Вот этот ток и находится в определенном отношении к току
возбуждения; когда в нормальном нервном волокне местный ответ
достигнет одной пятой величины тока возбуждения, который в наиболее
возбудимых волокнах равняется 60—100 mV, тогда только начинается!
распространяющийся процесс возбуждения (Ходкин, 31). Все это
свидетельствует, что возникновение процесса возбуждения
обусловливается именно этим местным электрическим ответом, а не
поляризационным потенциалом. Область местного процесса в мякотных
нервах ограничивается несколькими миллиметрами, в пределах кото-
В
[
F
"V
V
V"*"
~v—
о
с
н
Рис. 167. Электрические явления
в нервном волокне ракообразных в
области приложения раздражающих
электродов при индукционных ударах
разной силы:
Электрограммы А и В — при силе 1.5
условных единиц: А — на катоде, В — на. анодв.
Электрограммы С и D—при силе 1.0 (С—на
катоде, D — на аноде); Е и F — при силе
1,0 через некоторое время при более низкой
возбудимости (Е—на катоде, F—на аноде);
G и И — при силе 0.01 (G — на катоде,
И — на аноде). На электрограмме С и В
ясно видно, что вслед за поляризационным
током возникает другой небольшой ток,
который продолжается значительно дольше
поляризационного. В электрограмме С этот
местный ток предшествует току возбуждения.
В электрограмме А он мало заметен из-за
быстрого наступления тока возбуждения.
В электрограмме G он отсутствует вследствие
слабости раздражающего тока. В
электрограммах В, ?>, F и И от анодной области
местный ток совершенно отсутствует, как и
ток возбуждения (Ходкин),
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
333
рых он распространяется с декрементом, т. е. с постепенным
уменьшением. Так, в седалищном нерве лягушки местный процесс от
подпорогового раздражения затухает на расстоянии 9 мм от места
приложения электродов (Караев, 51). Местный процесс качественно
отличается от процесса возбуждения. Он ограничивается одним
раздраженным участком, и интенсивность его градуируется по силе
раздражения, постепенно затухая по мере удаления от места раздражения.
Напротив, процесс возбуждения распространяется по всему нерву с
одинаковой интенсивностью и не зависит в своей интенсивности от силы
раздражения. Поэтому здесь, в переходе от местного процесса в
распространяющийся процесс возбуждения, нужно видеть проявление
одного из всеобщих законов развития: количественное изменение основ-
ного биологического процесса с определенного момента скачкообразно
переходит в качественное изменение — процесс возбуждения.
Характерно, что при очень частых подпороговых раздражениях, выше
1000 в секунду, локальная электрическая реакция носит длительный
характер, который удерживается без колебания в течение нескольких
минут (Катц, 39). Это указывает, что в участке раздражения при этом
условии не происходит изохронного расщепления и восстановления. По-
видимому, как при согревании или при химическоім раздражении,
ускоряется процесс расщепления и восстановления, но это происходит не
одновременно для всех частичек, а в самое различное время. Возможно,
что частый фарадический ток производит раздражение не толчками,
л сплошь, через те тепловые и химические реакции, которые при этом
вызываются.
Скрытый период возбуждения. Подобно тому как в мышце, в
нервном волокне возбуждение возникает не сразу, а через определенный
промежуток времени, который в нерве лягушки равняется 0.3—0.4 а
(рис. 171). Скрытый период возбуждения нерва при раздражении
отдельными индукционными ударами почти не зависит от силы
раздражения. Для наиболее возбудимых волокон он всегда один и тот же.
Но при действии постоянного тока скрытый период зависит от силы
раздражения. Он тем больше, чем слабее ток (Гёпферт, 30).
В основе скрытого периода лежит тот местный процесс, которым
начинается каждый эффект в раздраженном участке. Местный процесс
выражается в усиленном расщеплении наиболее возбудимых частичек
возбудимой системы. Он сопровождается, как уже указывалось выше,
возникновением электрического потенциала. С ним находится в связи
возникновение процесса возбуждения. Когда местный ток достигает
определенной критической величины, местный процесс переходит в
процесс возбуждения, который уже охватывает всю возбудимую систему.
Скрытый период возбуждения именно и равняется времени,
требующемуся для того, чтоб местный процесс достиг этой критической
величины.
Хронаксия как мера возбудимости нерва. Хронаксия нервных
волокон изучалась многосторонне как Лапиком и его сотрудниками, так
334 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
и другими учеными. Первоначально изучалась хронаксия целого нерва.
Этим путем было найдено, что хронаксия седалищного нерва лягушки
равняется 0.36 а (Лапик, 12). Хронаксия нервов ллечевого пояса
человека равна 0.06—0.16 а (Бургиньон, 41). Было установлено, что эта
хронаксия сильно меняется в зависимости от функционального
состояния. Так, например, при охлаждении от 8.5 до 3° хронаксия удлиняется
от 0.96 до 1.70 а (Дворкин и Флоркин, 3).
Хронаксия нервов сильно меняется в онтогенезе. У котят, например,,
в период внутриутробной жизни хронаксия нервов (п. cruralis, нервные
ветви к сгибателям колена) в 3—6 раз больше, чем у взрослых. После
рождения хронаксия несколько укорачивается уже в течение первых
часов. Но наиболее сильное укорочение происходит впоследствии,
в связи с прозреванием (Аршавский, 26).
Хронаксия нерва сильно зависит и от соединительнотканных
оболочек, покрывающих нервные волокна. С удалением наружной оболочки
нерва хронаксия укорачивается вдвое (Тасаки, 47). Она зависит и от
межполюсного расстояния электродов: хронаксия уменьшается, когда
уменьшается это расстояние (Раштон, 48).
По исследованиям хронаксии изолированных нервных волокон лягу*
шечьего нерва оказалось, что минимальная хронаксия 0.2 j характерна
для нервных волокон, проводящих со средней скоростью, —10 м в
секунду; наоборот, волокна, проводящие более быстро, 25—30 м в секунду,
имеют большую хронаксию — 0.3 j. Большая величина хронаксии
свойственна также нервным волокнам, которые проводят возбуждение со
скоростью меньше 10 м в секунду. Здесь она для наиболее тонких,
медленно проводящих волокон равна 7 j. Реобаза в этих случаях меняется
правильно, следуя изменениям скорости проведения (Блэир и Эрлангер„
18). Хронаксия расходится с остальными показателями
функционального состояния и в других случаях, например во время повышенной
возбудимости — супернормальной фазы, которая бывает после
рефракторных фаз. Именно, она удлиняется, вместо того чтобы укорачиваться
(Хайаси и Брюкке, 19). То же бывает при наркозе; при заведомом'
ухудшении функционального состояния, когда реобаза сильно
увеличивается, хронаксия уменьшается (Цкипуридзе, 55). Также при наркозе
изолированных двигательных волокон жабы с увеличением
концентрации наркотического вещества реобаза увеличилась от 38.15 до 80 mV,
хронаксия уменьшилась от 0.42 до 0.33 о. По мере приближения к
критической концентрации, когда нервное волокно перестает реагировать,
реобаза возрастает до 105 mV; хронаксия же теперь увеличивается да
0.45 а (Тасаки, 47). Таким образом, в определенных случаях изменений
возбудимости, именно когда возбудимость сильно падает или сильна
повышается, изменения хронаксии им не соответствуют: при падении
возбудимости хронаксия не увеличивается, а, наоборот, укорачивается;
при повышении не укорачивается, а, наоборот, удлиняется. Значит,
путем хронаксии нельзя проследить действительное функциональное
изменение возбудимой системы, действительное изменение возбудимости. Эта
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
335
явление объясняется тем, что время действия раздражителя зависит от
силы раздражения. Оно тем меньше, чем больше сила раздражения.
Когда реобаза сильно нарастает в связи с понижением возбудимости,
удвоенная реобаза оказывается настолько большой силой, что она
производит эффект при меньшем времени, чем было до понижения
возбудимости. Точно так же при сильном повышении возбудимости
реобаза настолько падает, что удвоенная реобаза производит эффект
при большем времени, чем до повышения.
Эта противоречивая зависимость между реобазой и хронаксией
в условиях значительного отклонения возбудимости нерва от нормы
побудила авторов к некоторой математической обработке величин реобазы
и хронаксии. Так, некоторые авторы, например Ренквист и
сотрудники (57), ;в таких случаях предпочитают судить о функциональном
состоянии нерва по произведению реобазы на хронаксию (RhXChr). Для
этой же цели Л а с с а л ь (56) предложил формулу . I Однако
(Rh)*-Chr
условные величины, получаемые при обработке реобазы и хронаксии
этими формулами, не дают прямого указания о состоянии
возбудимости нерва. Чтобы привести в соответствие показания хронаксии
с функциональным состоянием нерва, Цкипуридзе (54, 55)
применил способ обработки реобазы и хронаксии, при котором хронаксия
становится прямым показателем функционального состояния нерва. При
воздействии на нерв какого-либо наркотизирующего агента, когда
реобаза прогрессивно возрастает, а хронаксия укорачивается, он прекращал
действие наркотика и восстанавливал возбудимость нерва до нормы. На
таком нерве заново определялось необходимое время действия тока при
тех же повышенных реобазических силах, которые требовались
вовремя действия наркотика. Эти величины времени были значительно
короче хронаксии во время наркоза. Затем устанавливались отношения
между хронаксиями, которые получались во время действия наркотика,
и теми временами действия тока, которые были установлены от тех же
реобаз при восстановлении возбудимости, путем деления первых на
вторые. Эти отношения показывали, насколько дольше должен
действовать ток в разных стадиях наркоза, чтобы вызвать возбуждение.
Произведение наркотической хронаксии на соответствующее отношение
дает величину времени действия тока в сигмах, которая довольно верно
характеризует состояние нерва в данной стадии наркоза. Это время
действия тока дает уже прогрессивное увеличение с углублением
наркоза.
Подобную же картину мы можем получить, если использовать
формулу Л а с с а л я. Нужно ту условную величину, которая получается
по формуле Лассаля в отношении нормального нерва, разделить на
условные отношения, полученные для разных стадий наркоза.
Полученные отношения вполне совпадают с отношениями при вышеописанном
способе. Значит, они будут выражать время действия тока в сигмах,
которое будет нарастать с углублением наркоза.
336 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Таким образом, применение формулы Лассаля с установлением
отношений с нормальным нервом дает возможность легко и точно
следить за изменением возбудимости по принципу времени действия тока.
Означенный способ пригоден как для случая пониженной возбудимости,
когда хронаксии являются укороченными, так и для случая повышенной
возбудимости, когда хронаксии являются удлиненными.
Субординационная хронаксия нерва. Хронаксия двигательного нерва,
как и мышцы, в нормальном организме в значительной мере зависит от
центральной нервной системы. Было установлено, что при отделении
нерва от спинного мозга хронаксия удлиняется (Лапик, 33). Очевидно,
вследствие постоянного прохождения импульсов из центральной
нервной системы по двигательным нервным волокнам возбудимость
последних поддерживается на некотором высоком уровне. Это явление
подробно было исследовано Лапиком с сотрудниками. Оказалось, что
.все те условия, которые благоприятствуют рефлекторной деятельности
центральной нервной системы, укорачивают хронаксию. Так действует
растяжение мышцы на хронаксию ее двигательного нерва. Этим же
объясняется укорочение хронаксии нервов у котят в связи с
прогреванием. Те же условия, которые не благоприятствуют рефлекторной
деятельности, наоборот, удлиняют хронаксию. Так, например,
хлороформирование животного удлиняет ее и делает стабильной. Лапик (15)
называет хронаксию, зависящую от импульсов, идущих из
центральной нервной системы, «субординационной», подчиненной, а хронаксию
вне нервной зависимости, которая выражает свойство собственно
органа,— «конституциональной» (Брюкке, 19; Шошары, 20).
Такая же зависимость хронаксии от центральной нервной системы
•существует в отношении скелетной мускулатуры. Например, отмеченные
выше различные хронаксии сгибателей и разгибателей человека, а
также одинаковая хронаксия всех мышц, работающих одновременно,
зависят от центральной нервной системы, значит, указанные хронаксии
являются субординационными. Но эти субординационные хронаксии
мышц следует отнести за счет двигательных нервов мышц, ибо у
человека хронаксия мышц определяется через нерв.
Установлено, что субординационная хронаксия главным образом
сбусловливается деятельностью головного мозга, а именно средних
отделов его, в особенности лежащих здесь красных ядер, которые регулируют
тонус скелетных мышц. После перерезки или разрушения данного
отдела или после охлаждения его хронаксия мышц и периферических
двигательных нервов удлиняется. Аналогичное удлинение наблюдается при
общем рефлекторном торможении, которое вызывается сдавливанием
кожи на голове (Лапик, 32). (См. об этом общем торможении отдел
центральной нервной системы, том II.)
Поскольку деятельность центральной нервной системы, в частности
тонических и автоматически действующих центров головного мозга,
зависит от разного рода периферических раздражений, то, безусловно,
эти раздражения должны определять субординационную хронаксию.
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
337
Периферические раздражения не одинаково действуют на центральную
нервную систему и через нее на периферические органы. А потому,
в зависимости от того, что и как раздражается, субординационные
изменения хронаксии выражаются в уменьшении или в увеличении
хронаксии двигательного нерва или двигательных точек мышцы, т. е. тех
нервных участков, которые обычно раздражаются при определении
хронаксии імышцы (Уфлянд и сотр., 25).
Изменение возбудимости двигательных нервов под влиянием
центров— давно известный факт. Оно исследовалось на разных животных,
разными путями: в отношении порогов раздражения двигательного нерва
(Штарке, 42; Иедерхольм, 43; Гедевани, 44), в отношении
демаркационного тока и основного биологического тока (Резвяков, 45). Эти
изменения возбудимости двигательных нервов под влиянием центров одни
авторы считают результатом распространения разнообразных
центральных процессов на периферические органы и допускают существование
особого рода тормозящих волокон (Иедерхольм, 43); другие
предполагают поляризацию двигательных нервных волокон под влиянием тех
токов, которые возникают в спинном мозгу при его деятельности
(Моннье и Джаспер, 36, 35; Резвяков, 45).
Существование особых тормозящих волокон в цереброспинальной
системе, подобных блуждающему нерву для сердца, отрицается в
настоящее время всеми физиологами. Поляризационные токи
распространяются из спинного мозга по двигательным и по заднекорешковым
волокнам на очень небольшое расстояние — всего на несколько
миллиметров (Баррон и Мэттьюс, 34). Кроме того, мы выше указывали, что
яри подпороговых раздражениях нервного ствола возникающий
поляризационный ток распространяется на очень короткое расстояние,
измеряемое миллиметрами. А потому нет оснований для утверждения, что те
изменения возбудимости, какие устанавливаются по всему протяжению
двигательного нерва, обусловливаются распространением
поляризационного тока. Мы думаем, что уменьшение хронаксии или повышение
двигательного эффекта обусловливается небольшим повышением
возбудимости вследствие постоянной импульсации двигательных нервов из
тонических и автоматически действующих центров средних отделов
мозга. Увеличение же хронаксии . или уменьшение двигательного
эффекта должно происходить вследствие прекращения этой импульсации
з результате торможения деятельности тонических и автоматических
центров или повреждения означенных центров. С этим хорошо
согласуется тот факт, что в периферическом нерве, находящемся под
влиянием центральной нервной системы, аммиака выделяется больше, чем
з нерве, отделенном от центральной нервной системы (Гальтер, 40).
Это указывает,' что под влиянием центральной нервной системы
расщепление белковых соединений усилено, значит, активность возбудимой
системы нерва повышена, что может происходить только при
наличии постоянного притока импульсов из' центральной нервной
системы.
22 И. С Беритов
338 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Надо отметить, что некоторые исследователи не смогли подтвердить на
млекопитающих факта существования субординационной хронаксии. Так, Ф о р б с с сотрудн^
(60) нашли, что если нерв перевязывается ближе чем на 2 см от раздражающих,
электродов, тогда наблюдается понижение реобазы и удлинение хронаксии. Эти
изменения авторы приписывают местному процессу повреждения нерва. При
расположении раздражающих электродов на 3 и более см от места перерезки они не
обнаружили никаких изменений реобазы и хронаксии. Перерезка спинного мозга,
лроприоцептивные раздражения и наркоз тоже не давали заметных изменений
реобазы и хронаксии. На основании этих и других фактов Форбс и Девис (61)
отрицают самый факт существенных сдвигов в хронаксии. Непостоянство
явления субординации при проприоцептивных раздражениях отмечает и Б р юкке
(62). Вул и Коников (63) после перерезки седалищного нерва у кролика также
не получали никаких существенных сдвигов в хронаксии. Они, наоборот, отмечают,
что само оперативное вмешательство (разрез кожи, раздвигание мышц и т. ы.)-
вызывает в большинстве случаев заметные изменения реобазы и хронаксии нервной
системы. Если после такого вмешательства произвести перерезку нерва, то это
невызовет дальнейших существенных сдвигов в реобазе и хронаксии. Последние факты
свидетельствуют, что изменения реобазы и хронаксии нервно-мышечного аппарата,.
в связи с перерезкой нерва, обусловливаются не только выпадением центральных
тонических импульсов, но и разного рода влияниями, идущими от раневого поля
в виде многочисленных залпов нервных импульсов и физико-химических сдвигов*.
вызванных рефлекторно, т. е. раздражением многочисленных рецепторов и
чувствительных нервов в операционной области (Вул и Коников, 63).
Изохронизм. По данным Л а пи к а (12), хронаксия нерва и мышцы
з нормальных условиях более или менее одинакова. Во всех тех
случаях, когда возбуждение с нерва переходит на мышцу, хронаксия.
последней или точно такова же, как и нерва, или немного больше, но
не более чем в два раза. При еще большей разнице нервное
возбуждение перестает переходить на мышцу. Отсюда Лап и к заключает о*
физиологическом изохронизме мышцы и нерва, который является
основным условием передачи возбуждения с нерва на мышцу. Л а п и к
полагает, что нервный импульс, достигший нервных окончаний в мышце,.
своим биоэлектрическим током производит возбуждение эффекторногО'
органа благодаря тому, что последний координирован с нервом
в отношении временного течения процессов, в отношении хронаксии.-
В случае нарушения этой координации, например путем
увеличения хронаксии мышцы, для получения того же результата был бы:
необходим нервный импульс большей продолжительности и
интенсивности. Но если величина нервного импульса остается без
изменения, то удлинение хронаксии в мышце неизбежно поведет к
прекращению передачи возбуждения с нерва на мышцу (Лапик, 15). Так,,
например, по Л а пику кураризация прекращает непрямое
возбуждение мышцы, когда хронаксия мышцы становится вдвое больше нормы.
Было исследовано действие также других ядов на нервно-мышечный"
препарат. В одних случаях яд меняет хронаксию нерва; в других —
мышцы, — и тем создает гетерохронизм, препятствующий передаче
возбуждения с нерва на (мышцу. Стрихнин, например, уменьшает нервнуку
хронаксию; пилокарпин, физостигмин, никотин первоначально
уменьшают мышечную хронаксию. Значит, по Л а пику, выключение нерва:
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
339
происходит не только при удлинении хронаксии мышц, но и при
укорочении (21).
Однако некоторые авторы не подтвердили самого .факта изохронизма
в некоторых случаях для нормального нервно-мышечного препарата.
Так, например, у котят в возрасте до 10—12 дней хронаксии нерва
и сгибателей колена относятся к друг другу, как 1 : 3 или 1:2, т. е.
налицо подлинный гетерохронизм. Несмотря на это, возбуждение нерва
передается на мышцу (Аршавский, 26). Кроме того, Вахгольдер
и Леде бур (4) нашли, что хронаксия «нетонических» мышц лягушки
превышает таковую нерва гораздо больше, чем в три раза. Несмотря
на это, возбуждение передается с нерва на такие мышцы. Авторы
показали это как на вырезанных мышцах, так и на мышцах с
ненарушенным кровообращением. В проверочных опытах Л а п и к (22) подтвердил
результаты Вахгольдера и Ледебура. Но он объяснил это
изменениями мышцы в результате повреждения от долгого
соприкосновения с воздухом. Если держать вырезанную неповрежденную мышцу
в физиологическом растворе и оттуда вынимать только для
раздражения, то хронаксия оказывалась постоянно малой, около 0,35 а , как
з двигательном нерве.
Безусловно, в нормальных условиях для беспрепятственной передачи
возбуждения с нерва на мышцу возбудимость мышцы должна быть
более или менее подходящей, чтобы нервные импульсы могли возбудить
мышцу. Чем быстрее протекают нервные импульсы, чем короче
биоэлектрические токи возбуждения нервных окончаний, тем выше должна
быть возбудимость мышцы в области нервных окончаний. Этим
объясняется, повидимому, то явление, что возбудимость мышцы наиболее
высока в области нервных окончаний. Здесь, действительно, хронаксия
мышцы должна быть сходной с таковой нервных волокон. Но, ,как
известно, в других участках мышцы возбудимость гораздо ниже, сробраз-
но хронаксия должна быть длительнее. Это и видно по опытам Моорз
и Б р ю к к е (16), которые доказали на изолированных мышечных
волокнах языка и membranae basihyoidea, что хронаксия в середине их
значительно меньше, чем на краях. Возможно, что и в опытах
Вахгольдера и Ледебура, а также других авторов, большая
хронаксия мышцы получалась в связи с раздражением безнервных
участков ее.
Явление адаптации, или аккомодации. Впервые Нернст (52)
обратил внимание на то, что нерв обладает свойством приспосабливаться,
г. е. он тем труднее поддается раздражению, чем медленнее нарастает
ток. Это свойство возбудимой системы Нернст и Гилл называют
аккомодацией, а другие авторы — адаптацией.
В последнее время Гилл с сотрудниками изучали это явление
более детально (28). Оказалось, что время аккомодации при медленно
нарастающих токах не одинаково в разных тканях и в одной и той же
ткани при разных функциональных состояниях. Следовательно, для
характеристики скорости процесса раздражения не достаточно учитывать
12*
340 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ. ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
тот фактор времени, который известен как полезное время, или хро-
наксия.
Нужно также учитывать фактор времени «аккомодации».
Изменение порогов раздражения в связи с функциональным изменением нерва
зависит; между прочим, и от последнего фактора.
Время аккомодации по способу Г ил л а при экспоненциальном
нарастании тока для двигательного нерва лягушки в среднем составляет
35 а при комнатной температуре, для рыб оно сильно колеблется
между 11—20 а, для нерва крабов и омаров —между 800—1600 , для
человека — 58 а. При увеличении ионов калия и кальция в нерве
лягушки время аккомодации уменьшается. Уменьшение же ионов
кальция, наоборот, ведет к увеличению этого времени, а при отсутствии
кальция аккомодация становится бесконечной, т. е. нерв не
возбуждается (Соландт, 29).
Предполагается, что в случае медленного нарастания тока нерв
каким-то образом приспосабливается, аккомодируется к действующему
раздражителю и что время этой аккомодации тем больше, чем
медленнее нарастает ток. Время аккомодации^ в существенной мере зависит
также от особенностей возбудимой ткани. Чем ниже возбудимость, тем
больше время аккомодации. Мерой аккомодации, по Шриверу (27),
является время экспоненциально нарастающего тока, при котором
происходит возбуждение нерва, если пользоваться :t силой тока, равной
двойной реобазе. Г и л л (28) же принимает как меру аккомодации
время возвращения изменившегося в процессе раздражения'
порогового состояния к состоянию покоя. Г ил л исходит от возврата в
состояние покоя, а не наоборот, от перехода порога из состоянии покоя к
состоянию при раздражении; он считает, что возвращение к норме
лучше отображает свойства ткани, ибо оно не зависит от особенностей
применяемого тока. Между временем аккомодациии по Шриверу и
Г и л л у существует определенное соотношение. Время аккомодации по
Шриверу в 2.89 раз (меньше того времени, которое находят
экспериментальным путем по Гил л у.
Аккомодации следует приписать и то явление, что при пропускании
постоянного тока изменение возбудимости, в смысле повышения на катоде и понижения
на аноде, наблюдается больше -всего в -самом начале, а потом это изменение
довольно быстро проходит и достигает уровня, близкого к покойному состоянию.
Скорость возвращения возбудимости к норме за время прохождения тока является
мерой амплитуды аккомодации. Но характерно, что на невырезанных нервах с
ненарушенным кровообращением начальное изменение возбудимости остается на все
время прохождения тока. Обычно это изменение далее в некоторой мере
повышается. Если пропустить надпороговый ток, то на вырезанном нерве импульсы
возбуждения возникают только в начале пропускания тока. На нервах же с
сохранившимся кровообращением разряды импульсов возникают все время.
Отсюда .может быть сделано заключение, что по существу аккомодация к
действию постоянного тока свойственна вырезанным нервам, находящимся в
ненормальных Физиологических условиях, но она равняется нулю в нормальных нервах
с'. ненарушенным кровообращением (Паррак, 53).
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
341
Естественное возбуждение нерва. Нормально нерв возбуждается н.е
от непосредственного раздражения. Если он чувствительный, то он
возбуждается вследствие распространения возбуждения от. периферических
органов чувств, так называемых- рецепторов. Если нерв двигательный,
то он возбуждается в силу распространения возбуждения из
центральной нервной системы. Но нервный ствол, а значит и нервное волокно
обладают способностью возбуждаться от непосредственного
раздражения внешним агентом.
Раздражители нервного ствола. Химическое раздражение.
Химические вещества, только тогда вызывают возбуждение, - если они быстро
проникают внутрь нервного волокна и тем производят очень быстрые
физико-химические изменения в нервном волокне. Так как оболочки
нервного волокна состоят из жироподобных веществ, вроде
холестерина, лецитина, так называемых липоидов, то ясно, что только такие
вещества могут быстро проникнуть в нерв, которые хорошо
растворяются в этих липоидах; вещество, растворяющееся в воде, тоже проникает
в нерв, но гораздо медленнее.
К химическим раздражителям следует отнести быстрое высыхание,
концентрированные растворы кислот, также слабые растворы щелочей,
как 0.8%, растворы нейтральных солей, органические соединения, как
эфир, хлороформ и др.
Значение ионов натрия. Среди химических веществ особое место
занимают ионы натрия. Последние необходимы для сохранения всех
жизненных свойств нерва. Поэтому для долгого сохранения
вырезанного нерва следует его погрузить в физиологический раствор или
смачивать им. В физиологическом растворе температуры 0—2°
вырезанный нерв сохраняет возбудимость до 20 дней. В изотоничном же
растворе тростникового сахара (6%) нерв теряет возбудимость в течение
50 часов. Стоит только к такому раствору прибавить немного ионов
натрия — 0.1% хлористого натрия, чтобы возбудимость в нерве
удержалась так же долго, как в физиологическом растворе. Вместо
хлористого натрия можно взять другие соли натрия, а также соли лития. Но
нельзя пользоваться другими щелочами и солями (Овертон, 8).
Температурное раздражение. Быстрое изменение температуры
вызывает в нерве, как и в мышце, возбуждение. Так, быстрое повышение
температуры лягушечьего нерва до 35—50° или быстрое охлаждение до»
4Э действует как раздражение. Медленное согревание не вызывает
возбуждения. Оно может умертвить нерв, если температура дойдет до
45°. Быстрое согревание производит не одиночное возбуждение, а ряд
импульсов возбуждения.
Механическое раздражение. Быстрое механическое воздействие
также действует на нерв раздражающим образом: удар, давление, разрез,
гянпок и т. д. При таком раздражении центростремительного нерва
возникают различные ощущения, как, например, боль и мурашки-в
пальцах при раздражении п. ulnaris; при таком же раздражении
двигательного нерва наступает сокращение мышцы (Оинума, 9). Постепенные
34} ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
механические изменения могут убить нерв без возбуждения и изменения
возбудимости. При перерезке нерва очень тонкими, сильно
заостренными ножницами или скальпелем раздражение двигательных волокон
может не произойти, ибо в этом случае механическое раздражение
продолжается меньше полезного времени.
Раздражение индукционными ударами. Лучшим раздражителем
нерва считается индукционный ток. При раздражении индукционными
ударами возбуждение в нерве возникает от таких слабых ударов,
которые совершенно не действуют даже на наиболее чувствительные
поверхности, как, например, поверхность языка. Сильные индукционные
удары вызывают возбуждение не только в момент их возникновения,
но и в момент их исчезновения. Поэтому на один размыкательный или
на один замыкательный удар могут возникать по два импульса
возбуждения. Если сила удара особенно велика, то іможет получиться еще
больше импульсов. Это объясняется тем, что с приближением
индукционных катушек сильно увеличивается не только сила удара, но и
его продолжительность. Действие такого индукционного удара
уподобляется действию сильного гальванического тока, вызывающего тетанус
в силу теплового, химического и механического раздражений живой
системы. Конечно, такое сильное раздражение действует на
раздражаемый участок нерва умерщвляющим образом.
Электрический ток действует сильнее, когда он проходит по длине
нерва, а не поперек него. Это, видимо, зависит от того, что проведение
тока в поперечном направлении встречает на своем пути гораздо
меньше перехватов Ранвье, чем в продольном. Как подробно было изложено
выше, раздражающее действие электрического тока на нервные
волокна в основном происходит через перехваты Ранвье.
При раздражении нерва индукционными ударами возбуждение
возникает в участке приложения электродов только при пороговых силах
раздражения. Путем регистрации скрытого периода Мышечного
сокращения было установлено, что с увеличением силы раздражения нервно-
мышечного препарата возбуждение возникает сразу все на большем и
большем пространстве. От этого скрытый период возбуждения мышцы
уменьшается.
Если порог раздражения нерва размыкательными индукционными
ударами составляет 30—35 см расстояния между катушками, то уже
при расстоянии в 20 см возбуждение возникает на стороне катода на
пространстве около 20 мм. Длина такого одновременно возбужденного
участка зависит от многих условий, между прочим от толщины нерва,
от увлажнения нерва и т. д. Это происходит от распространения
раздражающего тока или, как говорят, от петель тока вдоль по нерву
(Като, 5).
Униполярное раздражение. Индукционные удары действуют раздражающим
образом и в том случае, если нерв соприкасается только с одним электродом, а
другой электрод свободно кончается в воздухе. При этом ток распространяется очень
далеко по препарату, и это может привести к большим методическим ошибкам.
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА 343
-Можно ослабить такое распространение тока, если нерв вблизи раздражаемого
участка соединен с землей хорошим проводником. Еще лучше поместить нерв на
кольце из серебряной или платиновой проволоки и раздражать нерв внутри этого
.кольца. Петли тока в значительной мере будут замыкаться через кольцо и не будут
распространяться далеко по нерву за пределы кольца (Гартен, 13; Беритов, 6).
Это кольцо носит название кольца Геринга по имени физиолога, который впервые
предложил это средство для ослабления униполярного действия электрического
тока (рис. 14). Униполярное раздражение электрическим током имеет место и при
двухполюсном действии электрического тока. Электрический ток распространяется
вдоль по нерву по обеим сторонам электродов: он идет на поверхности нерва по
тому же направлению, как и между электродами, а внутри нерва в обратном к
нему направлении.
Такое внеполюсное распространение тока происходит -и в том случае, когда
один или оба конца нерва подняты, т. е. при отсутствии ветвления тока. При
некоторых сильных раздражениях это внеполюсное распространение тока
бывает настолько значительно, что может произвести раздражение нерва
на большом расстоянии от раздражающих электродов. Например, если на
расстоянии 1 см от электродов к раздражаемому нерву приложить конец нерва другого
нервно-мышечного препарата, то при некоторых средних силах раздражения, 15—
20 см, реакция наступит на обоих препаратах. Это внеполюсное распространение
тока называется электротоническим, и о происхождении его будет сказано ниже,
з главе о полярном действии электрического тока.
Особенно сильно униполярное распространение тока наблюдается в том случае,
если один препарат соединен с двумя индукториями, например если один конец
нерва или один корешок соединен с одним индукторием, а другой конец или
другой корешок — с другим. При одновременном раздражении ток одного индуктория
переходит во вторичную катушку другого индуктория. Катушка действует как
заземление. Это обстоятельство может оказать существенное влияние на эффект
препарата, если применяемая сила раздражения достаточно велика или если обе пары
электродов прилегают к нерву достаточно близко друг от друга. В таких случаях
ток от одного индуктория может воздействовать на нерв раздражающим образом
через обе пары электродов (Введенский, 50).
Внеполюсное распространение тока является большой помехой для локального
раздражения нерва электрическим током. Можно в значительной степени
ограничить это распространение. Для этого нужно, чтобы нерів был свеж и хорошо
увлажнен и чтобы межполюсное расстояние было маленькое — около 1 мм.
Установлено, что с удлинением межполюсного расстояния до 6 мм и с высыханием нерва
внеполюсное распространение тока значительно усиливается (Беритов, 6). Кроме того,
знеполюсное распространение тока сильно ослабевает с применением трехполюсных
электродов (рис. 14-С). Еще в лаборатории Сеченова было принято
бороться с внеполюсным распространением тока путем применения трехполюсных
электродов с катодом внутри (Вериго, 49). С этой же целью можно применять и
электроды с кольцом Геринга.
Действие гальванического тока. Этот ток действует на нерв при его
замыкании и размыкании. Но если ток сильный, то раздражающее
действие продолжается и во время прохождения тока. Если
раздражается таким током двигательный нерв, то мышца отвечает тетанусом
{Пфлюгер, 58). Чувствительный нерв также возбуждается от прохождения
сильного тока. При раздражении кожной поверхности или нерва током
чрезмерной силы субъект чувствует сильную боль. При помощи
струнного гальванометра было доказано, что при замыкании и размыкании
сильного гальванического тока в нерве возникает не один импульс
возбуждения, а целый ряд их (Гартен, 10; Эббеке, 11). Причиной этого
344 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТемы
явления должны служить те сильные химические, температурные и
механические изменения, какие вызываются сильным электрическим
током в нерве.
Практикум
1. Проверка закона раздражений Дюбуа-Реймона на нерве реономом Флейшля.
Постановка этого опыта уже была описана (:на мышце). Для этой цели пользуются
нервно-мышечным препаратом лягушки. Раздражают седалищный нерв и наблюдают
эффект на лапке.
Рис. 168. Изображение постаовнки опыта на нервно-мышечном препарате лягушки.
Препарат помещен во влажной камере. Лапка висит, а нерв растянут горизонтально; его свободный!
конец с 'позвоночником лежит на особой подставке. Посередине нерва наложены электроды. Над
камерой изображена ее крышка, обтянутая фильтровальной бумагой.
2. Определение порогового раздражения путем отдельных индукционных ударов
и тетанического раздражения. Этот опыт должен быть произведен на
нервно-мышечном препарате лягушки. Определяя пороги раздражения через каждые 2—3
минуты, можно заметить чувствительное изменение порогов. Увлажнение нерва
физиологическим растворам или жидкостью Рингера возвратит первоначальный порог.
Это изменение порогов стоит в связи с высыханием нерва между полюсами
электродов. При незначительном высыхании порог понижается, а при значительном
повышается. Причина этого явления лежит в том, что первоначально нерв
высыхает снаружи, в связи с чем .растет наружное сопротивление неріва. Вследствие
этого электрический ток сильнее распространяется по нервным волокнам, лежащим;
внутри ствола. Когда же высыхание распространяется ів глубь нерва, то тогда
только нарастает и внутреннее сопротивление нерва. От этого раздражающее дей-
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА 345-
ствие тока ослабевает и, конечно, тем сильнее, чем больше высыхание. Но с
'высыханием связано также изменение физиологического состояния нерва: возбудимость,
сначала повышается, а затем понижается до полной потери возбудимости. Чтобы
избегнуть этого изменения физиологического и физического состояния нерва,
необходимо через каждые 5—10 минут смачивать нерв физиологическим 'раствором или
раствором Рингера.
Во избежание быстрого высыхания нерва на воздухе нервно-мышечный
препарат помещают во влажной камере (рис. 168). Влажная камера представляет
металлическую квадратную коробку со стеклянными стенками. Камера эта употребляется
для защиты препарата от высыхания. Влажный воздух в камере создается при
помощи обклеивания крышки с внутренней стороны мокрой фильтровальной
бумагой, а также опусканием нижнего края камеры в желобок, наполненный водой*
В одной из стенок камеры имеется отверстие для соединения мышцы с миографом.
Электрический ток приводится к наружным зажимам, которые через дно камеры
связаны с внутренними зажимами. Последние же соединяются с электродами.
Приготовление нервно-мышечного препарата уже было описано выше. Для
миографической записи икроножной мышцы удаляют всю ненужную часть лапки
ниже колена, перерезая голень около коленного сустава (см. рис. 40).
Помещение во влажной камере не гарантирует всецело препарат от
высыхания. При комнатной температуре нерв заметно высыхает в камере в течение 10—
20 минут. Тонкие нервы могут высыхать так же сильно, как на открытом воздухе.
Значит, порог раздражения меняется и во влажной камере. Но все же высыхание
в камере происходит несколько медленнее. Отсюда следует, что и во влажной
камере нерв необходимо увлажнять физиологическим раствором по крайней мере
через каждые 10 минут.
3. Изменение порогов в зависимости от толщины и межполюсного расстояния
нерва. Порог электрического раздражения нерва зависит очень сильно от толщины
нерва. Спинномозговую лягушку прикалывают спиной вниз и обнажают
седалищное сплетение. Затем определяют пороги раздражения того или другого корешка
отдельными индукционными ударами. При этом оказывается, что чем. тоньше нерв,
тем ниже порог, т. е. тем слабее пороговое раздражение. Тонкие нервы легче
раздражаются электрическим током главным образом потому, что наружное
межполюсное сопротивление при тонких нервах много больше, чем при толстых, а
потому на долю внутрилежащих нервных волокон в первом случае приходится больше
тока, чем во втором. Вообще всякое улучшение проводимости электрического тока
по поверхности нерва ведет к повышению порогов. Так действует, например,
накопление крови или лиімфы, капля физиологического раствора между полюсами
электродов, наличие 'в этом 'месте кровеносного сосуда или какой-либо рыхлой
ткани и т. д.
Порог электрического раздражения сильно зависит и от межполюсного
расстояния. Берут нервно-мышечный препарат и определяют порог при наименьшем
расстоянии между электродами, около 0.5 мм. Затем раздвигают электроды до 1, 2, 4
к 8 мм и вновь определяют пороги того же нерва. Пороги определяют каждый раз
после смачивания. При этом окажется, что с увеличением ¦межполюсного
расстояния до 2—3 мм порог понижается почти на 10 см, а при дальнейшем увеличении
порог более не изменится. Это зависит от изменения сопротивления в межполюсном
пространстве. При малом межполюсном расстоянии рингеровский раствор заполняет
все межполюсное пространство. От этого внешнее сопротивление очень мало
сравнительно с внутренним и электрический ток с большей силой идет по
внешней жидкости, а на нервные волокна действует слабо. Если межполюсное ¦
расстояние больше 2—3 мм, то эти слои жидкости около электродов очень малы
сравнительно с остальным участком и внешнее сопротивление значительно
вырастает. В данном случае на долю нервных волокон, т. е. внутреннего сопротивления,
приходится больше тока, чем в предыдущем, что и вызывает заметное снижение
порога.
346 ОБЩ.ЧЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
При еще большем раздвижении электродов пороги не меняются более или
менее значительно, так как при этом растет как внешнее, так и внутреннее
сопротивление.
4. Изменение порогов в зависимости от функционального изменения нерва.
Согревание нерва физиологическим раствором комнатной температуры до 37—40°
приведет (к значительному понижению порогов раздражения вследствие повышения
возбудимости нерва.
5. Определение максимальной силы электрического раздражения. Для этой цели
производится регистрация механического эффекта икроножной мышцы на кимографе.
При этом должно регистрироваться не укорочение мышцы, а напряжение ее, для
чего берут изометрический миограф. Наблюдение можно производить на обычном
нервно-мышечном препарате. Но можно взять и спинномозговую лягушку:
раздражать периферический отрезок седалищного неріва на бедре, а записывать
напряжение икроножной мышцы. Раздражается нерв отдельными индукционными ударами
или тетанически. Для получения оптимальной силы раздражения увеличивают его
до тех пор. пока не получится максимальное напряжение, т. е. такое напряжение,
которое не может быть увеличено дальнейшим усилением тока. Чем тоньше нерв, а
также чем тоньше мышца, тем скорее достигается максимум напряжения.
6. Повреждающее действие сильных индукционных ударов на нерв.
Накладывают на седалищный нерв нервно-мышечного препарата две пары электродов на
расстоянии не менее 2 см одна от другой. Определяют пороги раздражения через
ту и другую пары электродов. Затем пропускают через ту пару электродов, которая
дальше отстоит от мышцы, электрический ток при 8—12 см расстояния между
катушками в течение одной минуты. По прекращении этого раздражения вновь
определяют порог раздражения через ту же и через другую пару электродов. Порог
перераздраженного участка будет сильно повышен^ а порог другого участка
останется без изменения. Если раздражающая сила достаточно велика, нерв
умерщвляется при раздражении в течение одной минуты. Такой силой является для
седалищного нерза 8 и меньше сантиметров расстояния между катушками. Чем
тоньше нерв, тем меньше та сила тока, которая оказывает повреждающее действие
(Беритов, 6).
7. Униполярное действие электрического тока. Приготовляют спинномозговой
препарат лягушки и прикрепляют его к пробковой пластинке спиной вниз.
Обнажают седалищное сплетение. На один из карешков у выхода их из позвоночного
столба навязывается нитка; затем он перерезается выше перевязки. Другой конец
данного корешка также перерезается на месте соединения его с другими
корешками. Потом этот же конец привязывается ниткой к остальным корешкам.
Вырезанный корешок накладывается на электроды так, чтобы свободный конец корешка
висел в воздухе. Пропуская через электроды ток большой силы (10—5 см), можно
заметить движение лапки. Это получается от распространения тока по данному
корешку на другие неперерезанные корешки со стороны ближайшего электрода.
Чем ближе этот электрод к неповрежденным корешкам, тем сильнее будет
униполярное действие тока. При этом оно будет много сильнее в том случае,
когда катод находится на стороне этих корешков (см. постановку опыта на
рис. 169, В).
С іцелью ослабления (униполярных токов можно применить трехполюсные
электроды с катодом внутри (рис. 14, С) или электроды с кольцом Геринга (рис. 14, Б).
Нерв накладывается на эти электроды так, чтобы катод приходился ровно
посредине 'между анодами. Теперь для активного униполярного действия потребуются
значительно более сильные токи.
8. Внеполюсное распространение тока путем ветвления тока. Для демонстрации
прибегают к тому же препарату, что и в предыдущем опыте. Та же постановка
опыта, только теперь проксимальный конец опускается так, чтобы он прикасался
к седалищному сплетению (рис. 169, Л).
При раздражении корешка в 10—12 см произойдет движение лапки от
ветвления тока по замкнутому нервному кругу. Ветвление тока происходит неизбежно,
ВОЗБУДИМОСТЬ НЕРВНОГО СТВОЛА
347
¦когда раздражение нерва производится внутри организма. В таких случаях ток
ветвится по окружающей ткани. Это, конечно, мешает локальному раздражению
данного нерва. Для ограничения ветвления тока нужно применять трехполюсные
электроды с катодом внутри или электроды с кольцом Геринга. В то время как
Рис. 169. К постановке опыта для демонстрации ветвления
тока (А) и униполярного действия (В).
Дано изображение седалищного сплетения и обычных платиновых
электродов: на рис. А IX корешок перетянут с обеих сторон ниткой и поднят
на электроды. На рис. В тот же корешок поднят на электроды, но здесь
проксимальный конец перерезан и свободно висит в воздухе, а дистальный
перетянут ниткой. В первом случае ток пойдет как между полюсами, так
и по всему замкнутому кругу, вследствие чего произойдет раздражение
и других корешков. Во втором случае петли тока распространяются к не-
перерезанным корешкам по одной стороне поднятого на электроды корешка.
с обычными двухполюсным.и электродами при данной установке опыта деятельное
зетвл?ние тока получается при 13—10 см, с трехполюеными электродами или с
кольцом Геринга деятельного ветвления не будет и при 0 см (Бернтов, 6).
Литература
1. Erlanger J., The Harvey Lecture, Series XII, 1927.
2. G a s s e r H. S. a. Erlanger J., Am. J. Physiol., 80, 522, 1927.
3. D w о r k i n K. a. F 1 о r k i n M., Am. J. Physiol., 95, 139, 1930.
4. Wachholder u. Ledebur V., Pfl. Arch., 228, 183, 1931; 231, 77, 1932,
229, 126, 1931; 232, 708, 1933.
5. Kato G., The theory of decrementless conduction in narcotised region of nerve,
Tokyo, 1924; The further studies in decrementless conduction, Tokyo, 1936.
6. Беритов. И., Русск. физиол. журн., 13, 442, 1930.
7. W е і s s G., С. R. Soc. Fiol., 53, pp. 400, 440, 466, 543, 1901; J. Physiol., 5, 239, 1903;
С. R. Acad. Sci., 132, 999, 1901.
"8. Overton E., Studien iiber die Narcose, Jena, 1901.
9. О i n u m a S., Z. Biol., 53, 309, 1910.
30. Garten's., Z. Biol., 52, 534, 1909; Ber. Verh. Sachs. Ges. Wiss., Leipzig, Math
phys. KL, 60, 85, 1908.
11. E b b e с k e U., Pfl. Arch., 203, 336, 1924.
12. LapicqueL, L'excitabilite en fonction du temps, Paris, 1926.
13. Garten S., Tigerstedt's Handbuch d. Physiol. Methodik, 2, H. I, S. 338, 1911.
14. X и л л (Г и л л) А. В., Природа, № 12, 1933.
15. Л а п и к Л., Новейшие успехи в познании нервного механизма. Речь на 15-м
Международном конгрессе физиологов, 1935.
16. М о о г е А. и. В г й с к е Е., Th., Pfl. Arch., 228, 619, 1931.
17. Е г 1 a n g е г I. a. Gasser Н. S., Am. J. Physiol., 92, 43, 1930.
18. В 1 а і г А. а. Erlanger J., Am. J. Physiol., 106, 554, 1933.
348 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
19. Брюкке Е. Т., Физиол. журн. СССР., 19, стр. 53, 1935; — Hayasi К. u. Brucke
Е. Th., Pfl. Apch., 235, 31, 1935.
20. Ш о ш а р А. и В. и П. Ш о ш а р, Проблемы биологии и медицины. Сборн.,
посвященный проф. Л. Штерн, 1935, стр. 50.
21. Лапик Луи, Физиол. журн. СССР, 19, 227, 1935.
22. L а р і с q и е L. и. М., Pfl. Arch., 230, 381, 1932.
23. Магницкий А. Н. и МужеевВ. А., Тр. физиол. отд. Гос. Тимир. научно*
иссл. инст., 1930, стр. 77.
24. Макаров П. О., Сб. докл. VI Всес. физиол. съезда, стр. 88, 1937; Тр. Ленингр.
общ. ест., LXVII, вып. 1, 1939.
25. Уф ля н д Ю. М., Сб. докл. VI Всес. физиол. съезда, стр. 245, 1937.
26. А р ш а в с к и й И. А., Сб. докл. VI Всес. физиол. съезда, стр. 379, 1937.
27. Schriever Н., Z. Biol., 91, 173, 1930; ib. 93, 123, 1932.
28. Гил л А., Физиол. журн. СССР, 19, 115, 1935; Proc. Roy. Soc, В. 119, 705, 1936.
29. S о 1 a n d t D. Y., Proc. Roy Soc, B. 119, 355, 1936.
30. G 6 p f e r t. H., Pfl. Arch., 241, 538, 1938.
31. Hodgkin H. L., Proc. Roy Soc, B. 126, 87, 1938.
32. L a p i с q u e L., Cold Spr. Harb. Symp. Quant. Biol., 4, 98, 1936.
33. Lapicque L„ С r. soc. biol., 88, 46, 1923; 105, 848, 1930.
34. Barron D. H. a. Matthews B. H. C, J. Physiol., 86, 29P, 1935.
35. Monnier A. M., Arch, internat. physiol., 37, 337, 1933; Cold Spr. Harb. Symp.
Quant. Biol., 4, 111, 1936.
36. Monnier A. M. a. Jasper H. H., С. г., Acad. Sci., 194, 224Э, 193?..
37. К a t о Gen., The microphysioiogy of nerve, 1935; Cold Spr. Harb. Symp. Quant.
Biol., 4, 202, 1936.
38. Rush ton W. A. H., Proc. Roy. Soc, B. 124, 210, 1937.
39. Katz В., Proc. Roy, Soc, B. 124, 244, 1937.
40. Halter K., Bioch. Z., 257, 313, 1933.
41. В о u r g u i g n о n G., La chronaxie chez l'homme, Paris, 1923.
42. Starke Jon., Zbl. Physiol., S. 596, 1898.
43. Jaderholm G. A., Pfl. Arch., 114, 248, 1906.
44. Гедевани Д. M., Тр. Физиол. инст. Тбилис. унив., 3, 478, 1937.
45. Р е з в я к о в Н. П., Юб. сб. Тр. Ивановского мед. инст., 1935; Физиол. журн. СССР
19, 1021, 1936; Сб. докл. VI Всес. Физиол. съезда, стр. 90.
46. Lehmann J. Е., Am. J. Physiol., 118, 600, 613, 1937.
47. Т a s a k i I с h i j i, Am. J. Physiol., 125, 367, 1939.
48. R u s с h t о n W. A. K., J. Physiol., 63, 357, 1927.
49. Werigo Br., Pfl. Arch., 76, 517, 1899.
50. В в e д e н с к и й Н. Е., Тр. СПб. общ. естеств., 38, 1907; то же в т. 4. Собр. соч.
Введенского, стр. 69, 1938.
51. Караев. А., Бюлл. эксп. биол. мед., 6, № 1, 1938.
52. Nernst W., Pfl. Arch., 122, 276, 1908; Ab. K- Preuss. Akad. Wiss., 1, 3, 1908.
53. Par rack H. O., Am. J. Physiol., 130, 481, 1940. ;
54. Цкипуридзе Л., Сообщ. Акад. Наук Груз. ССР, 4, № 5, 1943.
55. Цкипуридзе Л., Сообщ. Акад. Наук Груз. ССР, 3, № 9, 1942.
56. L a s s а 1 е Н., Sunti della communicazioni XIV Congresso Intern, di Fisiologia^
152, Roma, 1932.
57. RenquistJ., Leskinen V. a. Parvianen S., Skand. Arch. Physiol.,
61, 113, 1931.
58. Pf I ii g e r E., Arch, pathol., Anat., 13, 437, 1858; Physiologie desElectrotonus, S. 446, 1859'
59. Чаговец В., электрические явления в живых тканях, Anat 2, 1906.
60. Lambert Е. F., Sk i nner В. F. a. F or bes A., Am. J. Physiol., 106, 721 „1.933.
61. Davis H. a. Forbes A., Physiol. Rev, 16, 407, 1936.
62. Брюкке E. Т., Физиол. журн.-СССР., 19, 53, 1935.
63. Вул И. М. и Коников А. Л., Бюлл. эксп. биол. и мед., 4, 347, 13971
Г л а в a III
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
Состояние возбудимой системы нервных волокон во время
основного биологического процесса и во время возбуждения. В возбудимой
системе нервных волокон, подобно тому как в мышечных волокнах,
процесс расщепления и восстановления протекает все время и при
отсутствии возбуждения. Это проявляется прежде всего в постоянной
теплопродукции. Но этот процесс не должен протекать совершенно
одинаково в разных частичках нервного волокна. Невозможно себе
представить, чтобы внешние и внутренние условия пространственно
разных частичек возбудимой системы были совершенно одинаковы, а
потому в разных частичках процесс расщепления и восстановления
должен протекать в течение разных промежутков времени. Значит,
возбудимая система нервного волокна при отсутствии раздражения пребывает
в гетерохронном состоянии, характерном для основного биологического
процесса.
При раздражении нерва, когда он приходит в возбуждение,
согласно нашей гипотезе, все частички возбудимой системы одновременно
расщепляются и затем одновременно же начинают восстанавливаться,
т. е. состояние нервных волокон становится изохронным.
Во время расщепления в возбудимой системе возникают разные
ионы. При гетерохронном состоянии нормального нерва эти ионы
должны появляться во всех участках его в более или менее одинаковом
количестве, а поэтому между ближайшими участками нерва не должно быть
разности электрического потенциала. Но после раздражения в каждом
возбужденном участке в связи с изохронным расщеплением появляется
значительно больше ионов, чем в других участках с гетерохронным
состоянием. Поэтому между участками устанавливается разность
электрических потенциалов. Вследствие этого между участком
возбуждения и ближайшим невбзбужденным участком возникает ток, так
называемый биоэлектрический ток возбуждения.
Интенсивность и продолжительность процесса возбуждения. Судя по
токам возбуждения, разного рода нервные волокна производят волну
возбуждения разной интенсивности и продолжительности. Волокна груп-
350 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
пы А дают наиболее сильные токи возбуждения. Электродвигательная
сила наиболее активного нервного волокна лягушки принимается окола
100 mV (Тасаки, 99; Гассер, 108). Наиболее слабые токи производятся
волокнами группы С. Если принять амплитуду отклонения С-волокон
за единицу, то в седалищном нерве лягушки отклонение
В-волокон будет равняться 5, а Л-волокон — 100.
В кожном нерве лягушки высота отклонений для тех
же волокон относится, как 1:3: 40, а в кожном нерве
собаки п. sapfienus — как 1 : 4 : 40 (Эрлангер и Гассер,
44) (рис. 170).
Продолжительность процесса возбуждения в
различных волокнах также варьирует очень сильно. В одной
группе Л седалищного нерва лягушки наименьшая
продолжительность— 1.2 а (альфа-волна), характерная для
наиболее толстых волокон. В волокнах средней
толщины она равняется 1.6 а (бета-волна), а в тонких
волокнах — 3.3 q (гамма-волна) (Эрлангер, Гассер и Бишоп,
58). На рис. 170 дана схематическая картина всех
75 /ОО /25 /50 /75 200 225 250 275 300
Рис. 170. Интенсивность и продолжительность процесса возбуждения в разных нервных
волокнах.
Седалищный нерв лягушки. Схематическое изображение токов возбуждения на расстоянии 90 мм от
раздражаемого участка. Регистрация токов возбуждения происходила катодным осциллографом.
Ордината отмечает амплитуду в условных единицах, а абсцисса — время в сигмах. Волны а, % у относятся
к А-нервным волокнам, дальше идет волна В-волокон а затем волна С-волокон. Большой скрытый
период последней волны обусловлен очень малой скоростью распространения возбуждения по С-волокна»
(Эрлангер, Гассер и Бишоп)
этих волн возбуждения седалищного нерва лягушки на расстояний
90 мм от раздражаемого участка.
По опытам на изолированных нервных волокнах теплокровных
(кошка) продолжительность процесса возбуждения в Л-волокнах при
температуре 37° значительно меньше, чем в аналогичных нервных
волокнах лягушки, а именно около 0.5 а для спинальных корешков и 0.7S
для п. phrenicus (Гассер, 50) (рис. 171).
Во всех случаях одна треть или одна четверть всей
продолжительности волны возбуждения приходится на восходящее колено, а
остальное время на угасание возбуждения (Розенберг, 60; Эрлангер, 58).
Много раз исследовалось изменение токов возбуждения в
зависимости от изменения функционального состояния. Тепло, отдых уменьшают
продолжительность волны возбуждения, а охлаждение, утомление,
наркоз увеличивают ее (Воронцов, 3; Тигерштедт, 4).
Поскольку возбуждение есть функция возбудимой системы, которая:
в нервном волокне представлена осевым цилиндром, всей суммой ней*
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 35Е
роплазмы и нейрофибриллей, нельзя считать, что амплитуда и
продолжительность процесса возбуждения в миэлиновых волокнах зависят
вообще от диаметра нервного волокна. Диаметр осевого цилиндра
меняется параллельно с диаметром всего волокна, он приблизительно-
вдвое тоньше диаметра всего волокна (Гассер и Грундфрест, 101;
Рис. 171. Ток возбуждения одного нервного волокна
теплокровного животного.
Стрелка указывает момент раздражения одним индукционным
ударом. Ток возбуждения появляется после интервала, который
составляется из скрытого периода раздражения в области катода
и времени проведения от раздражаемой точки до отводимогоучастка.
Время — в десятитысячных секунды (Гассер).
'Меписашвили, 94). Сообразно с этим мы должны считать, что
интенсивность и продолжительность процесса возбуждения в миэлиновом
волокне определяются толщиной осевого цилиндра. Продолжительность
его тем меньше, а интенсивность тем выше, чем больше диаметр
осевого цилиндра,
Изучение биоэлектрических токов возбуждения отдельных нервных волокон
ведется в течение последних 15 лет. Оно стало возможным благодаря высоко
развитой технике усиления слабых токов в десятки и сотни тысяч раз и применению'
осциллографа катодных лучей и других приборов, которые с помощью усилителей
точно отображают изменение электрического тока во времени. Прежде
биоэлектрические токи возбуждения исследовались такими приборами, как струнный
гальванометр капиллярный электрометр, которые обладают инерцией, поэтому нельзя было
изучать быстро протекающие токи; кроме того, эти приборы настолько мало
чувствительны, что нельзя было регистрировать токи возбуждения отдельных нервных
волокон. Этими старыми приборами было установлено, что в общем нервном стволе
лягушки продолжительность биоэлектрического тока возбуждения составляет около
2 а (Воронцов, 1), а в нервном стволе теплокровных —около 1.5 j (Гартен, 2).
Эти токи, конечно, отображали всю сумму токов возбуждения всех нервных
волокон, а потому они оказывались более или менее удлиненными.
Продолжительность возбуждения наиболее возбудимых толстых
волокон изучается прежде всего путем раздражения нерва пороговыми
силами. В этом случае возбуждаются только Л-волокна, и осциллограф
будет регистрировать только токи возбуждения их. Для изучения
продолжительности других волн отводят нерв возможно далеко от раздра-
352 ОБАЯЩ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
жен.ного участка. Это делают потому, что нервные волокна разной
толщины проводят возбуждение с разной скоростью. Поэтому при
отведении тока возбуждения около раздраженного участка мы можем
получить одну волну электрического тока, выражающую сумму токов
возбуждения всех волокон. Но если отводить, на большое расстояние, тогда
мм
30
20
/0
0
30
20
10
0
20
Ю
0
30
20
10
0
20
W-
Рис. 172. Изображение токов возбуждения А-волокон седалищного нерва
жабы на разных расстояниях от раздраженного участка.
Токи возбуждения были зарегистрированы осциллографом катодных лучей. На абсциссе
обозначается время в сигмах, а на ординате — высота кривых в миллиметрах. Кривая
А дает электрограмму на расстоянии 12 мм от раздраженного участка. Здесь ток
возбуждения (а) имеет одну волну. Она только в области восходящего колена несколько
искажена действием петли раздражающего тока. Если бы этого действия не было,
восходящее колено имело бы характер К-волны. Кривая В —в 31.0 мм; здесь намечается
на нисходящем колене явный горб от бета-волны. Кривая С—в 46.5 мм; здесь бета-волна
выражена еще сильнее и намечается гамма-волна. Кривая D — в 62.5 мм и дает три
волны: альфа-, бета- и гамма-волну. Кривая Е на расстоянии 82 мм. Здесь те же волны,
но расхождение их еще сильнее (по Эрлангеру)
токи возбуждения разных волокон будут отводиться в разное время и
соответственно будет получаться двойная, тройная или четверная
электрограмма (рис. 172).
Можно поставить специальные опыты для .выявления в нервном стволе воли
возбуждения тех и других (волокон. Так, ^например, если раздражать седалищный
нерв пороговыми раздражениями, то мы получим альфа-волну возбуждения. При
некотором сильном раздражении получается двойной эффект: альфа- и бета-волны.
Но мож;НО отделить волны бета от волн альфа. Для этого нужно сначала
возбудить слабым раздражением нервные волокна, дающие альфа-волну, затем через
небольшой промежуток, до I а, произвести второе, более сильное раздражение. В©
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 353
время второго раздражения нервные волокна, производящие альфа-волну, уже
находятся в возбуждении, поэтому они более не возбудятся. Возбуждение может
получиться только в других волокнах, /производящих бета-волну возбуждения.
Налицо будет только альфа.возбуждение от первого раздражения и бета-возбуждение
от второго (рис. 173).
Продолжительность возбуждения в двигательных нервных волокнах
беспозвоночных значительно больше. Даже у высших
беспозвоночных — ракообразных — процесс возбуждения сравнительно длительнее,
чем у позвоночных. Так, в двигательном нерве Maja squinado наиболее
быстрая волна возбуждения длится свыше 3—4 а , а в чувствительных
Рис. 173. Осциллограммы седалищного нерва жаоЪі на
большом расстоянии от раздражаемого участка
д — двойной ток возбуждения — альфа- и бета-волна от первого
сильного раздражения; В — альфа-волна от пороговой силы
первого раздражения; С — двойной ток, как в А, от второго сильного
раздражения; D — результат присоединения второго сильного
раздражения к слабому первому раздражению через 0.85 а. Белые
черточки означают время в сигмах. Прочие объяснения см. в
тексте (Эрлангер)
волокнах — около 4 о (Боге и Розенберг, 80). У низших
беспозвоночных продолжительность процесса возбуждения должна быть еще больше.
Проведение возбуждения. Возбуждение возникает в нерве в том
месте, где действует раздражение, и отсюда быстро распространяется
в обе стороны от раздраженного участка. Так, например, в
двигательном нерве возбуждение распространяется от раздражаемого участка
не только в нормальном для данного нерва направлении, т. е. в сторону
мышцы, но и в необычном для него направлении, т. е. в сторону мозга.
Это свойство проводимости нерва Кюне (9) доказал на нерве
m. sartorius. Кюне обнаружил, что каждое нервное волокно названной
мышцы еще раньше вхождения нерва в мышцу делится в самом
нервном стволе на две ветви: одна идет к одному концу мышцы, а
другая — к другому. Он перерезал мышцу пополам в области вхождения
нерва в мышцу без повреждения нерва и раздражал один конец
мышцы. В результате сокращение получилось не только в раздражаемой
половине мышцы, но и в другой. Отсюда ясно, что в раздражаемом
участке возбудились нервные волокна; по ним возбуждение распро-
23 И. С. Беритов
354 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
странилось в обратном, необычном для двигательного нерва
направлении; затем, достигнув места разветвления, оно перешло по другой
ветви в другую половину мышцы.
Двустороннее проведение возбуждения является
общим законом нервной деятельности. Это было доказано для
каждой проводящей системы. Так, например, Б а б у х и н показал в
очень демонстративной форме двустороннее проведение на нерве
электрического сома (10). Весь нерв электрического органа этой рыбы
состоит из одного разветвленного нейрита. Раздражая одну из ветвей„
он наблюдал эффект во всем электрическом органе. Ясно, что и здесь
первоначально возбуждение распространилось в необычном для нерва
центральном направлении, а затем, перейдя на другие ветви, оно
пошло в обычном для них направлении. В центральной нервной системе
возбуждение точно так же распространяется в нервных волокнах по
обоим направлениям. Это было с очевидностью доказано в отношении,
заднекорешковых волокон Шеррингтоном и Фрелихом (11).
Они перерезали задние столбы в области продолговатого мозга
(funiculus gracilis), ло которым проходят заднекорешковые волокна,
начинающиеся в поясничной области спинного мозга. Эти волокна
нормально проводят возбуждение только в головном направлении. Но-
если искусственно возбудить их в области разреза, то возбуждение
распространяется *и в обратном направлении; оно, например, вызывает
сгибание задней ноги на раздражаемой стороне.
О механизме проведения возбуждения. Проведение возбуждения в
нервных волокнах производится, как в мышечных волокнах, с помощьк>
биоэлектрического тока возбуждения: после того как он возникает в.
одном участке в связи с процессом возбуждения, он становится
источником раздражения для соседнего невозбужденного участка. Об этом
'высказывался еще Герман (91) в 1879 году; затем (К ре мер (92)
математически доказывал это; в последнее время Л ил ли (93)
представил прекрасную модель распространения возбуждения по нервному
волокну. Но прямые фактические доказательства этого положения были
добыты в самое последнее время при изучении локальных процессов в
участке электрического раздражения.
Как мы указывали выше, при пороговом раздражении также
происходит местный физиологический процесс в раздражаемой области.
Этот процесс, безусловно, представляет собой местное изменение
основного биологического процесса. Он сопровождается электрическим
током такого же характера, как нервный ток возбуждения. Когда
местный ток достигнет определенной интенсивности и захватит
определенную длину, тогда наступает распространяющийся процесс
возбуждения (Раштон, 74; Катц, 75) (рис. 167).
Но іи до достижения этой интенсивности (местный ток действует
совершенно определенно на соседний участок, а именно, подобно под-
пороговому раздражению, он повышает возбудимость участка. В
седалищном нерве лягушки это влияние распространяется на расстоянии
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 355
не менее 9 мм, если интенсивность подпорогового раздражения на 5'%'
ниже порогового (Караев, 64).
Нужно предположить, что при подпороговых раздражениях пока
имеет место только локальный процесс, в раздражаемом участке
остается гетерохронное состояние. Под влиянием подпорогового
раздражения на некотором небольшом участке нервного волокна
расщепляются только наиболее возбудимые частички. Возникающий при этом
электрический ток настолько слаб, что, распространяясь электротонически по
нервным волокнам, он не в состоянии произвести изохронное
расщепление всей возбудимой системы и тем вызвать процесс возбуждения.
При пороговых и сверхпороговых раздражениях местный процесс
захватывает всю возбудимую систему на некотором участке нервного
волокна, измеряемом долями миллиметра: все частички ее испытывают
изохронное расщепление; при этом возникает электрическая реакция
такой амплитуды, что она в состоянии раздражить соседний участок и
вызвать здесь изохронное расщепление во всей возбудимой системе.
Соответственно и возникающий здесь электрический эффект настолько
значителен, что он также становится источником раздражения для
дальнейшего невозбужденного участка. Таким путем возбуждение доходит
до конца нервного волокна.
В последнее время явление распространения нервного импульса
посредством биотоков стало предметом детального исследования. Так,
например, X о д к и н (73) показал, что если путем передавливания или
охлаждения небольшого участка нерва уничтожить в нем возбудимость
и проводимость или, как говорят, создать блок, то, при возбуждении
нерва с одной стороны от блокированного участка, на другой стороне,
вблизи него, на протяжении, нескольких миллиметров, наблюдается
повышение возбудимости. Здесь каждый раз в это время возникает
небольшое колебание электрического потенциала, которое имеет такую
же конфигурацию, как биоток возбуждения. Оно только значительно»
слабее, чем обычный ток возбуждения, и сильно снижается по мере
удаления от блокированного участка, совершенно исчезая на
расстоянии 8 мм от него. X о д к и н доказал, что локальное повышение
возбудимости за блокированным участком обусловлено воздействием этого
локального тока, возникающего в результате электротонического
распространения тока возбуждения через блокированный участок. Отсюда
автор делает заключение о фактической возможности распространения
нервного импульса путем электротонического действия биотоков
возбуждения с одного возбужденного участка на соседние невозбужденные
участки.
О возникновении и распространении возбуждения с точки зрения мембранной
теории. Предполагается, что нервное волокно сильно поляризовано вследствие
неравномерного распределения некоторых ионов, в особенности ионов калия,
концентрация которого внутри волокон, по Фенну и сотр. (121), превышает таковую
снаружи почти в 65 раз. Определенного мнения относительно того, какие именно*
структурные образования обусловливают эту поляризацию, — миэлиновая оболочка,
23*
356 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
шванновская оболочка, аксиолемма или поверхность нейрофибриллей, — не
существует. Предполагается только существование поляризованных образований —
оболочек — й еще возможность изменения их проницаемости под влиянием внешних и
внутренних воздействий. Возникновение так называемого тока покоя или, вернее, тока
повреждения в связи с повреждением нерва (перерезка, обжигание) 'считается
наилучшим доказательством поляризации нервных оболочек, ибо предполагается, что
ток повреждения возникает в результате деполяризации поврежденного участка.
С мембранной точки зрения и биоток возбуждения возникает в результате
деполяризации нервных оболочек в участке раздражения. Предполагается, что под
влиянием любого раздражения (электрического, химического, механического)
проницаемость оболочек увеличивается в раздражаемом участке нерва и вследствие этого
здесь происходит деполяризация оболочек, возникает электрический разряд: поток
ионов врывается снаружи внутрь волокна или вообще проскакивает через
разрыхленные мембраны. Это обусловливает определенное изменение возбудимой системы,
так называемый процесс возбуждения. В результате возникает разность потенциалов
между этими активными и соседними неактивными участками. Происходит
передвижение ионов. (Возникают локальные токи, которые выходят через оболочки наружу
и возвращаются к возбужденному участку. Под влиянием этих локальных токов,
когда они достигают определенной интенсивности, в соседнем участке проницаемость
перепонок и оболочек также увеличивается. Здесь также происходит деполяризация,
возникают 'возбуждение и новые локальные токи. Последние опять замыкаются
через соседний невозбужденный участок и опять производят возбуждение его.
Таким образом, наряду с распространением возбуждения внутри волокон в осевом
цилиндре происходит распространение деполяризации в мембранах нервных волокон,
причем ток возбуждения признается выражением волны деполяризации.
Следовательно, по этой теории, возникновение и распространение тока возбуждения
представляют собой чисто физические явления.
На рис. 174—схематическое изображение возникновения биотоков
повреждения (А) и возбуждения (В) с точки зрения мембранной теории, как это дано было
крупным физиологом Гассером.
Ряд наблюдений приводится авторами для подтверждения этой теории.
Так, например, Коуэн (122), работая над нервами крабов, нашел, что содержание
калия внутри нервных волокон в 13 раз больше, чем в крови или морской воде.
Если нерв погрузить ів такой раствор, который содержит калия в пять раз больше,
чем кровь краба или морская вода, то как токи повреждения, так и биотоки
возбуждения сильно ослабевают; при этом нерв становится навоз б удимым. Процесс
этот полностью обратим: промывание обычной морской водой восстанавливает и
возбудимость и биоток. Угнетающее действие калия частично устраняется
прибавлением хлористого кальция,—очевидно, © связи с уплотнением мембран. Если
уравнять концентрации ионов калия внутри и снаружи нерва, то ток повреждения
падает от GO mV до 3.5 mV. При дальнейшем (повышении концентрации калия
снаружи ток повреждения сходит на-нет. Коуэн же наблюдал, что при тетаниче-
ском раздражении калий выходит наружу в большом количестве— в 20—30 раз
больше, чем в покойном состоянии, и накоплению его на поверхности нерва
приписывается ослабление активного состояния — утомление. Подобно калию действуют,
по Коуэн у, рубидий и цезий, и притом совершенно одинаково, если взять их в
эквивалентных количествах. Но рубидий и цезий действуют на токи возбуждения
угнетающим образом значительно сильнее, чем на токи повреждения. Автором
приводятся и другие наблюдения, которые также говорят за физическое происхождение
тока возбуждения и за .существенную роль ионов калия при этом.
Однако в покойном состоянии ионы ікалия диффундируют через оболочки
чрезвычайно слабо. Предполагается, что в покое калий связан с такими анионами, для
которых оболочка не проницаема. При раздражении образуются такие анионы,
которые -могут проникать через оболочку; івыходя, они выводят с собой и ионы
калия. Так как проницаемость мембран должна меняться под влиянием разных
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 357
факторов, то величина токов повреждения и (возбуждения может меняться з
широких пределах.
Изменение сопротивления нервных оболочек Коуэін (122) считает возможным
объяснить следующим образом: известно, что сопротивление системы «вода + масло»
меняется в сильной степени в зависимости от того, что является дисперсионной
средой. В системе, где такой средой служит вода, сопротивление меньше; в
системе «эмульсия воды в масле» сопротивление больше. Если предположить, что
нервная оболочка, смотря по функциональному состоянию нерва, меняет свои
дисперсионные свойства, то можно объяснить как моментальное возникновение токов,
так и их исчезновение.
Осциллограф
с,
Побр?жд
область
биоток &оз</джд.
Поверхность ос/олочни
Раздраж
электроды
т -*• -*¦
'*»f. ..I****** * "* * + "+¦ * ¦*¦ "*¦
— III I I I ¦ llf ^^^^¦¦¦¦H IM. I ¦
¦iftni'ififrmn
Лонал сойотом
Ьшхшшш
-¦¦
rff<mufi область
Рис. 174. Диаграмма нервного волокна для демонстрации
разницы потенциалов между наружной и внутренней
поверхностями оболочки.
Представлено вероятное происхождение биотоков от повреждения и
от возбуждения с мембранной точки зрения.
Наверху (А) изображено происхождение тока повреждения путем
выравнивания потенциалов через поврежденную оболочку. Внизу (В)
дано распространение тока возбуждения в виде распространения
быстропреходящего повреждения в оболочке, произведенного
раздражением (Гассер)
Эта конкретизация мембранной теории в отношении биотоков повреждения и
возбуждения нерва не согласуется го многими известными фактами. Например, эта
теория не может объяснить того наблюдения Фенна (123), что, в зависимости от
напряжения С02 и буферности окружающей среды, калий то івыходит из мышцы
наружу, то, наоборот, входит в мышцу. Если снаружи напряжение СОг больше и
рН меньше, то калий выходит из клетки; если же рН меньше внутри клетки, то
калий входит в клетку. Изменение рН в клетке зависит от интенсивности обмена
веществ в ней. Следовательно, перераспределение ионов калия и других ионов
существенно зависит от интенсивности и характера метаболизма внутри клеток (Бре-
мер, 112; 'Верзар, 113).
Затем Пондер (114) показал, что поврежденная поверхность мышцы может
стать электроположительной nG отношению к неповрежденной после обработки
мышцы таким веществом, которое вызывает контрактуру. Разностью концентраций
калия нельзя объяснить и того известного факта, что вода, свободная от
электролитов, вырабатывает в точке соприкосновения с нервом электрополо>жительноеть
порядка 100 mV. Если это явление чисто физического порядка, то оно вызывается
не нервными волокнами, а соединительнотканной оболочкой нерва, т. е. калий здесь
353 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
уже не піри чем. Бели же оно обусловлено физиологическими реакциями, подобно
тому как это наблюдается при контрактуре мышцы, то переходом калия из клетки
наружу нельзя объяснить ни величины, ни знака наблюдаемого потенциала (114).
Все эти факты, не согласные с гипотетической ролью калия в возникновении
.и распространении ібиотоков ів нерве, (наоборот, указывают на то, что в основе их
лежат сложные биохимические процессы, протекающие внутри нервного волокна.
Роль перехватов Ранвье в распространении возбуждения. Уже давно
русский ученый Верило (1899) доказал, что распространению
возбуждения по нервному волокну не препятствует понижение
возбудимости или даже исчезновение ее на маленьком участке до 5 мм (104).
В е р и г о вызывал 'невозбудимость маленьких участков длиной 2—5 мм
действием наркотических веществ или электрического тока, и
возбуждение перескакивало через такие участки. Согласно с теорией
Германа, Вериго предположил, что это происходит благодаря биотоку
возбуждения: биоток возбуждения, распространяясь электротонически
через невозбужденный участок, может произвести такое
электротоническое изменение за пределами его в возбудимом участке, которое может
вызвать в нем возбуждение. В новейшее время это явление
перескакивания возбуждения подверглось тщательному исследованию, и было
обнаружено, что возбуждение перескакивает от одного перехвата
Ранвье к другому. Так, Блэйр и Эрлангер (85) показали, что
если действием анода постоянного тока блокировать один или два
перехвата Ранвье, а в соседних сегментах нервных волокон повысить
возбудимость действием катода постоянного тока, то биоэлектрический
ток возбуждения, возникший с одной стороны анодного
блокированного участка, распространяясь по осевому цилиндру, может вызвать
возбуждение на другой стороне его, так сказать, перескочив через
блокированный участок. Биоэлектрический ток возбуждения в нерве
считается в 2.5 раза сильнее того порогового тока, который раздражает
нерв, а при распространении по волокну на 1—2 мм его интенсивность
ослабляется вдвое. Следовательно, можно предположить, что в
нормальных нервных волокнах биоток возбуждения свободно может
распространиться скачками от одного перехвата Ранвье к другому, перескочив
через 1—2 мм межперехватного участка. Этот фактический вывод
совершенно согласуется с тем, что нам известно сейчас относительно
электрической возбудимости миэлинового нервного волокна. Мы выше
указывали, что нервное волокно возбудимо электрическим путем только
в перехватах Ранвье. Поэтому биоэлектрический ток, распространяясь
по осевому цилиндру, может выйти наружу и замкнуться через
окружающую среду только в перехватах Ранвье. Этим самым он
произведет, раздражение возбудимой системы исключительно на перехватах
Ранвье. Ток возбуждения одного перехвата может оказать
раздражающее действие не только на один, но на два и три последующих
перехвата. При некоторой высокой возбудимости нервного волокна
возбуждение может перескакивать через два-три межперехватных участка.
Во всяком случае опыты показывают, что в лягушечьем нерве возбу-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 359
ждение может перескакивать через два невозбудимых перехвата, значит,
через три межперехватных участка общим протяжением около 6 мм
(Тасаки, 99). Понижение скорости проведения в связи с наркозом или
охлаждением нерва на большом расстоянии должно обусловливаться
главным образом уменьшением числа перескакиваемых волной
возбуждения межперехватных участков. На рис. 175 дается схематическое
изображение скачкообразного распространения возбуждения в нервном
волокне.
Ток возбуждения от данного перехвата нервного волокна должен
замкнуться через наружную среду, чтобы раздражать соседние
перехваты. С этим положением хорошо согласуется тот факт, что
перескакивание возбуждения можно затормозить и облегчить изменением
электрического сопротивления в наружной среде, а также пропусканием
Рис. 175. Распространение 'биотока возбуждения по мякотному нервному
волокну.
Дано 6 сегментов с перехватами Ранвье. На* уровне 2 и 3-го сегментов находится
волна возбуждения. Сверху приведена кривая биотока возбуждения. Там большая стрелка
указывает направление распространения .волны возбуждения. Стрелки внутри волокна и
на его поверхности указывают на физическое распространение биотоков возбуждения.
Они проходят по осевому цилиндру от возбужденного участка и, выйдя наружу через
перехваты Ранвье, возвращаются обратно по наружной оболочке к возбужденному
участку. Интенсивность этих токов сильно ослабевает после каждого ^перехвата, что
отображено в количестве и толщине стрелок.
¦подпороговых индукционных ударов восходящего направления
навстречу распространению тока возбуждения (Тасаки, 99).
Однако скачкообразное распространение возбуждения от одного
перехвата к другому не исключает возможности распространения
возбуждения по осевому цилиндру без перерыва. При возбуждении
должна устанавливаться разность потенциалов между возбужденным и
невозбужденным участками нейрофибриллей осевого цилиндра.
Вследствие этого между ними должен возникнуть ток, который будет
проходить главным образом по нейроплазме и может послужить
источником для раздражения ближайшего невозбужденного участка
нейрофибриллей. С возбуждением нового участка возникает новый местный ток,
который послужит для дальнейшего распространения возбуждения по
осевому цилиндру. Но, безусловно, эти местные токи играют
определенную роль и в скачкообразном распространении возбуждения.
К а то (90) показал, что если охладить межперехватный участок
изолированного нервного волокна и создать тем самым здесь
непроводимость возбуждения, последнее вообще перестает распространяться.
Повидимому, для того чтобы ближайший перехват Ранвье возбудился
путем перескакивания тока, нужно, чтобы этот ток развивался с
достаточной интенсивностью и продолжительностью. Весьма возможно, что
(процесс возбуждения одного перехвата Ранвье не достаточен для этой
360 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
дели. Но он может быть достаточным для возбуждения соседнего
участка, покрытого миэлиновой оболочкой. Возникший при этом местный
ток также будет действовать на ближайшие перехваты. Вследствие
этого электротоническое действие биотока возбуждения на -соседний
перехват Ранвье может усилиться и удлиниться в такой мере, что
произойдет его возбуждение. Отсюда следует, что если холодом понизить,,
блокировать реактивность межперехватного участка, то это должна
привести к ослаблению раздражающего действия биотока
возбужденного перехвата на ближайший перехват. Но если каким-либо образом
повысить возбудимость в ближайшем перехвате, т. е. укоротить era
полезное время, как это было в опытах Блэйра и Эрлангера, то
уничтожение возбудимости в межперехватном участке не ликвидирует
проведения возбуждения, ибо, несмотря на уменьшение длительности
биотока данного перехвата, последний окажется выше порога по
отношению к ближайшему перехвату с повышенной возбудимостью.
Это новое представление о механизме проведения возбуждения
вдоль ,по нервному волокну точно так же трудно связать с
мембранной гипотезой возбуждения. Едва ли можно утверждать, что в
области перехватов Ранвье на небольшом протяжении — около 2 р.—
имеются все условия для создания той большой поляризации, которая
приписывается нервному волокну и от нарушения которой должно
зависеть возникновение тока возбуждения. Наилучшим доводом, против
такоРі гипотезы является тот факт, что даже четырехчасовое
возбуждение нерва тетаническим раздражением не ведет к такому повышению
проницаемости нервных оболочек, которое обусловило бы заметное
выхождение <ионов калия или натрия изнутри нервных волокон (Нет-
тер, ПО). Поэтому імы считаем -более вероятным, что деполяризация
оболочек не играет роли в процессе возбуждения. Мы предполагаем,
что, когда в каком-либо перехвате возникает возбуждение,
освобожденные при этом катионы образуют локальные токи, которые
распространяются по осевому цилиндру и выходят наружу через соседние
перехваты. Выхождение локальных токов действует на перехваты Ранвье,
как катод постоянного тока. Поэтому здесь происходит раздражение
возбудимой системы и возникает процесс возбуждения, который в свою-
очередь освобождает катионы и производит новые локальные токи^
могущие послужить источником раздражения для следующих
перехватов, и т. д. Очевидно, в каждом перехвате до возникновения
возбуждения действуют петли тока возбуждения из близлежащих
возбужденных перехватов, как это и видно по опытам Ходкина и др.
и как это представлено схематически на рис. 175.
Скорость проведения. В цереброспинальном нерве позвоночных
животных возбуждение проводится много быстрее, чем в скелетных
мышцах. У теплокровных, например в нерве собаки, по определению
многих авторов, скорость проведения составляет максимум 100—80 м
в секунду (Мюрних, 15; Эрлангер, 5). У холоднокровных животных
скорость проведения значительно меньше. В нерве зимней лягушки она
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 362
равна приблизительно 25 м. В нервах беспозвоночных животных
проводимость еще более низка; например, в двигательном нерве
ракообразных, как Maja squinado, скорость проведения возбуждения
составляет в разных волокнах 5.3—1.4 мм в секунду (Боге и Розенберг,.
80). У низших беспозвоночных скорость проведения еще ниже;
например, в двигательном нерве беззубки (Ainodonta) скорость проведения
всего 1 см, в нерве пиявки—32—40 см (Бете, 19).
На проводимость нерва сильно влияет его функциональное
состояние. Во всех условиях, благоприятствующих обмену веществ
(согревание, отдых и т. д.), проводимость увеличивается. Наоборот, при
неблагоприятных условиях (охлаждение, утомление, наркоз и т. д.) она
уменьшается. Сила раздражения не влияет на скорость проведения.
При слабом и сильном раздражении возбуждение распространяется с
одинаковой скоростью (Пипер, 14; Дюбуа-Реймон, 16).
Указанные выше скорости проведения возбуждения максимальны,..
ибо они относятся к целым нервным стволам. При исследовании
отдельных нервных волокон оказывается, что скорость проведения в
отдельных волокнах не одинакова. В толстых волокнах большой
возбудимости скорость проведения больше всего. Чем тоньше нервное волокно,.
чем ниже возбудимость, тем меньше и скорость. Так, скорость
проведения для Л-волокон лягушки, производящих альфа-волну, при комнатной
температуре равняется 42 м в секунду, для волокон бета-волны — 25 м,
для волокон гамма-волны—17 м, для Б-волокон — 5.5—1.3 м, а для
С-волокон — 0.7 м (58). У теплокровных (собака, кошка) при
температуре тела 37° для Л-волокон скорость составляет около 90—30 м в
секунду, для 5-волокон — 20—10 м (Эрлангер, 5; Эрлангер и Гассер, 44).
Отсюда следует, что скорость проведения находится в связи с
толщиной волокон, подобно продолжительности и интенсивности
возбуждения и возбудимости. Именно, авторы находили, что скорость
проведения отдельных нервных волокон меняется пропорционально квадрату
диаметра волокна (Блэйр и Эрлангер, 107). Но, конечно, поскольку
ток возбуждения волокон тем сильнее, чем толще волокно, то»
между скоростью возбуждения и амплитудой токов возбуждения
также существует определенная зависимость, и эта зависимость
прямая, т. е. чем больше амплитуда тока возбуждения, тем выше скорость
проведения (Зоттерман, 86; Эрлангер, 58). Повидимому, скорость пра-
ведения возбуждения в нервных волокнах холоднокровных и
теплокровных животных совершенно одинакова при равной толщине их и равной
температуре (Гассер и Эрлангер, 7).
По новейшим исследованиям, скорость проведения меняется
пропорционально не общему диаметру волокон, как это раньше думали,-а
диаметру осевого цилиндра, как это и следовало ожидать, и?о в.
нервном волокне осевой цилиндр представляет возбудимую систему.
Отношение диаметра осевого цилиндра к диаметру волокон в
толстых волокнах, выше 10 д, составляет 1 : 2. В тонких же волокнах,,
тоньше 9 р., отношение растет, т. е. толщина миэлиновой оболочки
332 ОБЩДЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
уменьшается в большей мере, чем осевого цилиндра (Гассер и Грунд-
фест, 101).
Определенное соотношение существует также между диаметром
осевого цилиндра и длиной межперехватных участков. Чем толще
осевой цилиндр, тем длиннее сегменты волокна между перехватами Ранвье
(Зоттерман, 86; Меписашвили, 94). Отсюда следует наличие
определенного соотношения между толщиной осевого цилиндра, длиной меж-
лерехватных участков и скоростью распространения возбуждения
(Гарш, 102). Очевидно, чем толще осевой цилиндр, а значит чем
сильнее ток возбуждения в перехватах Ранвье, тем длиннее межперехват-
лый участок, через который должен перескочить ток возбуждения.
Чем быстрее протекает процесс возбуждения в каждой точке нерва,
тем быстрее, конечно, должно распространяться возбуждение. В
соответствии с этим произведение времени подъема кривой тока
возбуждения на скорость является постоянной величиной (Эрлангер, 58).
Скорость проведения возбуждения, безусловно, зависит в значитель-
лой мере и от интенсивности тока возбуждения, ибо чем он іинтенсив-
лее, тем большее число перехватов Ранвье он может возбудить сразу
л тем меньше будет скрытый период возбуждения перехвата. По
Тасаки (118), скрытый период возбуждения перехвата Ранвье ори
пороговом раздражении 'равняется 0.45 а, а при удвоении пороговой
силы он уменьшается до 0.2 а.
Скорость проведения в сильной мере меняется в постэмбриональном
развитии нервных волокон. Она очень низка в первые дни после
рождения и растет с ростом животного. Так, у четырехдневного котенка в
наиболее толстых волокнах п. saphenus скорость проведения составляет
10 мв секунду, затем она постепенно растет и в 78-дневном
возрасте достигает нормальной скорости, характерной для
взрослой кошки, — 60 м в секунду. Максимальный диаметр миэлиновых
,волокон п. sapheniis меньше в первые дни после рождения, а
лотом постепенно он достигает нормы взрослых животных. Вообще
характерно, что скорость проведения меняется в строго линейном
^отношении с ростом животного: чем длиннее становятся ноги, тем
быстрее происходит проведение. Вследствие этого в первые дни после
рождения движение дистальных суставов наступает точно через такое
.же время, как у взрослых животных (Гарш, 103). Характерно, что
это постэмбриональное изменение скорости проведения точно
совпадает с увеличением диаметра нервных волокон и в особенности с
удлинением межперехватных участков. Рост последних происходит
параллельно удлинению нерва в растущем организме (Бойкотт, 109;
Такахаши, sill). Это еще раз указывает, что скорость проведения
возбуждения в миэлиновых волокнах связана главным образом с длиной
межперехватных участков.
Скорость проведения в изолированных нервных волокнах зависит
также от внешних, чисто физических условий — от электропроводности
-окружающей среды. Так, на нервных волокнах краба было обнару-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 363
жено, что эти волокна, будучи- в морской воде, проводят возбуждение
на 14—40% быстрее, чем в масле. Скорость проведения в гигантских
волокнах, когда они находятся в морской воде, на 80—140% выше,
чем на воздухе или в масле. Скорость эта повышается, если соединить
нервное волокно с рядом металлических скобок (Ходкин, 88). Это
явление хорошо согласуется с представлениями о роли биотока в
распространении возбуждения. Чем меньше наружное сопротивление, тем
с большей силой ток возбуждения замыкается через эту среду, тем
скорее возбуждаются соседние участки, тем дальше простирается
раздражающее действие биотока. Этими опытами X о д к и н а
показывается, ЧТО У высших беспозвоночных, не обладающих миэлиновой
оболочкой, но имеющих соединительнотканную оболочку, возбуждение
распространяется при помощи биотока возбуждения.
Нервные волокна отличаются друг от друга в отношении
проведения и в связи с их функциональным назначением. Так, например,
различают афферентные нервные волокна тактильной и болевой
чувствительности. По Эрлангеру и Гассеру (44), у млекопитающих имеются
такие волокна, которые проводят возбуждения со скоростью 1.3—0.7 м
в секунду. Эти волокна очень тонки, частью безмякотны. Авторы
относят их к системе, связанной с болью. Однако среди таких «болевых»
волокон или волокон, возбуждаемых деформацией кожи, существуют
волокна, которые проводят возбуждение со значительно большей
скоростью. Например, у млекопитающих различные защитные рефлексы
вызываются по чувствительным волокнам, распространяющим
возбуждение со скоростью 30 ім в секунду (Экклс и Шеррингтон, 45; Эдри-
ан, 43). Тактильные волокна проводят возбуждение более быстро., Они
являются толстыми мякотными чувствительными волокнами. Скорость
проведения этих волокон у лягушки равна около 15 м в секунду,
а у млекопитающих около 50 м (Эдриан, 43).
Определение скорости проведения возбуждения в нерве
впервые было произведено Гельмгольцем (12). Он пользовался методом,
принцип которого заключается в том, ічто при коротком действии ггока отклонение
стрелки гальванометра зависит как от интенсивности тока, так и от его*
продолжительности. Данный ток, проходящий через гальванометр, замыкается в момент
раздражения, а размыкается ів момент сокращения; по степени отклонения стрелки
судят о промежутке времени между раздражением и сокращением. Если
раздражать сначала наиболее отдаленный участок от мышцы, а затем наиболее близкий,
то ів измерениях времени окажется разница: в первом случае промежуток времени
будет больше, чем во втором. Разница должна быть отнесена на счет времени,
необходимого для проведения возбуждения от дальнейшего участка к ближайшему.
Зная это и зная расстояние между данными участками, можно высчитать скорость
проведения. Скорость проведения была определена Гельмгольцем еще другим
способом, а именно миографически. Этот способ получил большое распространение.
Записываются два сокращения одно за другим: одно от раздражения дальнейшего
участка нерва, другое от ближайшего. Разность скрытых периодов сокращения
укажет на время, необходимое для проведения возбуждения между данными
участками.
В последнее время для измерения скорости проведения предпочитают пользо-
364 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
ваться биоэлектрическими токами возбуждения. Одновременно регистрируют момент
раздражения и биоэлектрический ток гальванометром. Биоэлектрический ток
отводится в гальванометр от двух наиболее отдаленных участков нерва. Здесь
вычисляют скорость проведения на основании разницы скрытых периодов тока
возбуждения (Пипер, 14).
Проведение возбуждения без декремента и с декрементом. У всех
позвоночных животных проведение возбуждения в нервной системе
происходит без ослабления, без декремента, на какое бы расстояние
оно ни распространилось (Гоч и Бурч, 20).
Наиболее простое и убедительное доказательство мы находим
в микрофизиологических исследованиях К а т о (49) іи его сотрудников.
Изолируется одно двигательное нервное волокно в середине п. tibialis
Рис. 176. Кимографическая запись одиночных сокращений
нервно-мышечного препарата (седалищный нерв — m. gastrocnemius) через одно
двигательное нервное волокно.
Изоляция нервного волокна по середине седалищного нерва. Проксимальный
конец нерва раздражается отдельными индукционными ударами. При расстоянии
между катушками в 382 мм эффекта нет. Впервые эффект получается при
расстоянии 381 мм. Этот эффект^ совершенно такой же, .как при сильных
раздражениях при расстоянии в 379—200 мм (Като).
лягушки, которое снабжает m. gastrocnemius. Все остальные волокна
перерезаются. Проксимальный конец нервного ствола раздражается
обычными индукционными ударами, а сокращение мышцы
регистрируется на кимографе. Одно нервное волокно снабжает 50—70
мышечных волокон, поэтому возбуждения одного нервного волокна достаточно,
чтобы вызвать сокращение значительной силы. Уже при пороговых
раздражениях нерва мышца отвечает максимальным укорочением.
Такое же укорочение получается при всех больших силах раздражения.
Отсюда следует, что возбуждение нервного волокна всегда обладает
одной и той же интенсивностью и что это возбуждение
распространяется и достигает мышцы всегда с постоянной интенсивностью, т. е.
без ослабления, без декремента (рис. 176).
Была произведена также регистрация биоэлектрических токов
возбуждения мышцы при вышеприведенных условиях опыта. На рис. 177
показано, что величина биоэлектрического тока возбуждения при всех
применяемых силах раздражения, от 334 до 150 мм расстояния между
катушками, была одной и той же, точно такой, как при пороговом
раздражении в 334 мм.
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
365
Однако у беспозвоночных животных, наряду с такой нервной
системой, где возбуждение распространяется без ослабления, существует
и такая, где возбуждение проводится с ослаблением, с затуханием, как
говорят — с декрементом. Повидимому, у низших беспозвоночных
эта своеобразная нервная система господствует. Так, у
кишечнополостных и иглокожих с нейропильным строением нервной системы
возбуждение на локальное раздражение не может охватить весь организм.
Ответная реакция всегда ограничивается определенным участком
организма, который тем больше, чем сильнее раздражение (Ганстрем, 21).
Рис. 177. Электрическая реакция мышцы при непрямом
раздражении через одно нервное волокно.
Раздражается проксимальный нерасщепленный конец седалищного нерва
и отводится m. gastrocnemius к гальванометру Эйнтховена. Возбуждение
проводится через средний расщепленный участок с одним нервным
волокном. В опыте 7 раздражение—размыкательный индукционный удар в 335 мм
расстояния между индукционными катушками — мышца не дает эффекта.
В опытах 2, 3 и 4 при более сильных раздражениях— 334, 250 и 150 мм —
получается ток возбуждения, и он при всех раздражениях одной
амплитуды. В опыте 4 первый эффект обязан замыкательному удару (Като).
Декрементное проведение возбуждения вообще свойственно и
низшим моллюскам, гастроподам. Так, было установлено, что в длинном
нерве ноги ашшзии возбуждение проводится с декрементом, ибо
мышечная реакция в ответ на раздражение нерва тем меньше, чем
дальше от нее раздражается нерв (Фрэлих, 120). Если наркотизировать
в этом нерве один участок, то интенсивность процесса возбуждения
падает при распространении как в нормальном, так и в
наркотизированном участках; кроме того, нормальный участок нерва проводит
возбуждение с той интенсивностью, с какой оно выходит из
наркотизированного участка (Иордан, 52; Люллис, 55).
Декрементное проведение возбуждения установлено было также по
токам возбуждения нерва. Они ослабевают при распространении
возбуждения по нерву (Кан и сотр., 56). При этом декремент испытывают
все нервные волокна, — как проводящие очень медленно, так и
проводящие более или менее быстро, при самом свежем состоянии (Крийгс-
ман, 119).
366 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ1
Декрементное проведение возбуждения в нервах беспозвоночных
должно быть связано в некоторой степени также с особенностями их
структуры. Так, например, известно, что нервные волокна в комиссуре
беззубки чрезвычайно тонки, в среднем 0,35 ji, не имеют оболочки,
слабо изолированы друг от друга и, повидимому, не содержат нейро-
фибриллей или эти нейрофибрилли настолько тонки и малочисленны.,.
что не поддаются современным методам исследования. Вследствие
этого продольный срез через нерв дает картину гомогенного вещества
с едва заметной продольной исчерченностью (Скадовская, из
лаборатории Роскина, 106). Эти структурные особенности проводящей системы-:
беззубки таковы, что едва ли нервные волоконца представляют
самостоятельные проводящие единицы. Вероятнее всего, что весь нерв
фигурирует как одно нервное волокно, в котором нет условий для
изохронного расщепления всей возбудимой системы. Повидимому, при
распространении возбуждения вдоль по комиссуре все меньшая и
меньшая часть нервных волоконец приходит в изохронное расщепление.
У других беспозвоночных нервные волокна проводят возбуждение
в нервном стволе совершенно независимо друг от друга. Таков,
например, кишечный нервный ствол улитки (п. intestina'lis, Helix pomatia).
В нем имеются нервные волокна разной толщины и разной
возбудимости. Наиболее возбудимые волокна проводят возбуждение со скоростью
40—48 см в секунду, наименее возбудимые — со скоростью 5.1 см. Во
всех нервных волокнах проведение происходит с декрементом. При
некотором пороговом раздражении процесс возбуждения
ограничивается почти участком раздражения. По всем признакам он
совершенно сходен с тем локальным процессом, какой обычно наблюдается
в мякотном нерве при подпороговом раздражении. С усилением
раздражения локальный процесс нарастает в интенсивности, и
одновременно нарастает область его распространения по нерву (Кан, 56;.
Крийгсман, 119). Декрементная нервная система признается и у
высших беспозвоночных. Так, известно, что в нервах конечности речного
рака и морских крабов (Maja squinado) большая часть нервных
волокон проводит возбуждение без декремента (Боге и Розенберг, 80;.
Ходкин, 84). Эти волокна — с соединительнотканными оболочками —-
обладают высокой возбудимостью и проводят возбуждение со
скоростью 1.1—3.6 м в секунду. Они очень напоминают волокна
позвоночных животных. Но наряду с ними некоторая часть очень тонких
волокон нервного ствола (одна треть), лишенных оболочек и наименее
возбудимых, проводит возбуждение очень медленно, от 0.1 до 0.5 м в
секунду, и, повидимому, с декрементом (Люллис, 55).
Низшие беспозвоночные не лишены того типа нервной системы,
который проводит возбуждение без декремента. Подобные нервные
образования имеются у них наряду с теми, которые проводят с
декрементом; например, среди кишечнополостных медузы владеют хорошо
развитой бездекрементной системой. Так, В е т о х и н (32) вырезал
кольцо из края колокола медузы и в зтом кольце вызывал беспрерыв-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 367
ное движение іволны возбуждения в одном направлении в течение
нескольких часов без ослабления. Та и другая системы характерно
различаются между собой не только физиологически, но также и
гистологически. Декрементный нейропиль у кишечнополостных состоит из-
более толстых нервных клеток и нервных волокон. Самая скорость
проведения у них не одинакова. В декрементной системе она равняется
1 см в секунду, в то время как в бездекрементной скорость достигает
24 см (Будденброк, 23; Боцлер, 24).
Иордан с Люллисом (55) находят также, что у высших
беспозвоночных ^декрементное проведение возбуждения свойственно
определенным волокнам, а именно наиболее тонким. Волокна с толстыми-
оболочками принадлежат к другому типу; они проводят возбуждение
без декремента. Авторы сравнивают последние волокна с группой
В-волокон позвоночных животных, а декрементные волокна — с
группой С-волокон. Правда, известны и такие факты, когда толстые волокна
при 'известных условиях дают градуированные эффекты. Так, если на
клешне рака раздражать нерв замыкателя клешни, который состоит
всего из одного нервного волокна, то реакция мышцы будет тем
сильнее, чем сильнее раздражение (Сегаар, 22). По-видимому, здесь
градация эффектов обусловливается не декрементным распространением
возбуждения, а тем, что с усилением раздражения в нерве возникает
не один, а два и более импульсов. Кроме того, при повторных
раздражениях каждое последующее раздражение дает повышенный эффект
благодаря повышению возбудимости под влиянием предыдущих
импульсов. Все это вместе обусловливает градацию механического*
эффекта мышцы в ответ на раздражение нерва отдельными
индукционными ударами (Пэнтин, 83).
Нервная система, проводящая возбуждение без декремента,
безусловно является более поздним филогенетическим образованием. Ее
возникновение должно1 быть связано с появлением крупных, свободно
подвижных организмов, так как целостная реакция таких организмов-
в борьбе за существование не могла быть обеспечена без системы,
проводящей возбуждение быстро и без декремента.
Бездекрементное проведение возбуждения настолько существенно
отличается от декрементного, что нужно считать их качественно
отличными друг от друга. Если же принять во внимание, что декрементное
проведение возбуждения у беспозвоночных сопряжено с более
высоким обменом веществ (см. ниже), чем бездекрементное проведение
в нерве лягушки, что, следовательно, бездекрементное проведение
возбуждения гораздо более экономно, чем декрементное, мы должны
признать, что бездекрементная нервная система представляет высший
и качественно новый продукт филогенетического развития нервной
системы (Кан, 56).
Закон изолированного проведения возбуждения в нервных
волокнах. В нормальном нервном стволе с дифференцированными нервными.
волокнами, как у позвоночных и высших беспозвоночных животных,
368 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
импульсы возбуждения распространяются по нервным волокнам
совершенно изолированно. В нервном стволе возбуждение не передается
-с одного нервного волокна на другое. Закон изолированного проведе-
дения возбуждения был установлен впервые Иоганном
Мюллером в 1834 г. На этом основывается возможность многообразия
движений іи ощущений в соответствии с многообразием раздражений.
Понятно, что при смешении нервов разнообразных двигательных и
рецепторных органов возможность изолированных движений и
ощущений была бы исключена, если бы возбуждение переходило от данного
волокна к соседним. Но в нервных стволах низших беспозвоночных,
где голые нервные волокна чрезвычайно тонки и не изолированы друг
от друга, едва ли возбуждение проводится по отдельным волокнам
совершенно изолированно. Как мы указывали выше, у этих животных
нервный ствол, должно быть, ведет себя так, как одно нервное
волокно.
Что в нервном стволе беспозвоночных совершенно возможен
переход возбуждения от одних нервных волокон на другие, прилегающие
к ним волокна, доказывается недавними опытами Джаспера и
Монние (125). Они брали из конечностей ракообразных (Cancer
pagurtis и Maja squinado) нервные стволики длиной 3—8 см и плотно
прижимали друг к Другу, перевязывая шелковой ниткой на расстоянии
одного сантиметра. При химическом или электрическом раздражении
свободного конца одного стволика они інаблюдали биотоки
возбуждения в свободном конце другого стволика. Значит, в месте
прикосновения двух пучков от первично возбужденного пучка возбуждение
передалось на другой нераздраженный пучок. Это осуществляется, как
показали те же авторы, путем биотоков возбуждения. Ракообразные
относятся к высшим беспозвоночным. Отсюда нужно заключить, что
переход возбуждения от одного нервного волокна к другим путем
биотоков возбуждения, невидимому, характерен вообще для всех
беспозвоночных. Такое заключение было сделано нами в отношении пиявок.
Эти животные реагируют сокращением мышц при раздражении
кожных рецепторов после удаления нервной цепочки. Так как
соответствующие чувствительные волокна не имеют никакой прямой связи с
мышцами, следует заключить, что возбуждение переходит от
чувствительных волокон к мышцам через двигательные волокна. Эти волокна
•проходят *в кольцевом нервном стволе вместе с чувствительными,
а потому они могут раздражаться биотоками возбуждения довольно
толстых чувствительных волокон (Беритов, 126).
Но, по последним исследованиям, и у высших животных, с
дифференцированными волокнами, возбуждение проводится в волокнах не
всегда изолированно. Так, например, при искусственной обработке
нерва высокими концентрациями NaCl, мочевины, сахара и других
веществ, когда обработанный участок еще обладает возбудимостью и
проводимостью, нарушается изолированное проведение возбуждения.
В измененном участке возбуждение переходит с одних волокон на дру-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
369
гие; Так, например, раздражением п. peroneus можно вызвать
сокращение m. gastrocnemius, если химическому воздействию подвергался
седалищный нерв. Отсюда следует, что в измененном участке
возбуждение нервных волокон п. peroneus переходит на нервные волокна
"П. tibialis (Квасов и Науменко, 95).
Аналогичное явление наблюдается при повышении возбудимости
нервных волокон в области анода, вслед за размыканием постоянного
тока (Квасов, 95). Это, конечно, происходит оттого, что в означенных
случаях ток возбуждения раздраженных нервных волокон,
распространяясь по межтканевой жидкости, оказывается достаточно сильным
для возбуждения соседних нервных волокон. Характерно, что при
раздражении отдельными индукционными ударами получаются не просто
одиночные вздрагивания, а тетанизированные сокращения. Это
указывает, что ток возбуждения одних волокон раздражает наиболее
близкие волокна, а ток возбуждения последних — следующие, близкие
к ним волокна; от тех, в свою очередь, могут снова возбудиться
первые. В результате получается тетанический эффект.
Роль нейрофибриллей в проведении возбуждения. По опытам
Бете (17), возбуждение проводится по нейрофибриллям, а нейро-
плазма, или перифибриллярное вещество, в этом не участвует. Опыты
Бете заключались в следующем: с помощью особого приспособления
нерв лягушки сдавливался в одном участке. Оказалось, что этим
путем можно очень сильно уменьшить перифибриллярное вещество (почти
в 218 раз сравнительно с нормой), іи все-таки нерв обнаружит в сдав
ленном месте способность к проведению. Сама фибриллярная масса
при этом не уменьшается. Значит, способность к проведению должна
принадлежать уцелевшим нейрофибриллям.
Другое доказательство Бете заключалось в следующем. У пиявки
в нервных волокнах нейрофибрилли располагаются в нейроплазме
в виде спирали. Если растянуть нерв, то нейрофибрилли
расправляются; при этом, конечно, длина нервных волокон значительно
увеличивается. Бете изучил время распространения возбуждения в
нормальных и растянутых нервных волокнах. Оно оказалось совершенно
одинаковым. Отсюда ясно, что возбуждение распространялось по
нейрофибриллям. Если бы оно распространялось по нейроплазме, в
растянутых нервах время распространения возбуждения должно было бы
увеличиться (19).
Еще одно основание для признания основной роли нейрофибриллей
дается гистологом Буке (18). Согласно его исследованиям, нервные
элементы связываются с периферическими органами, как мышечные
клетки, кожные рецепторы, посредством нейрофибриллей. Например,
в мышечных волокнах нейрофибриллярная сеть сливается
непосредственно с перитерминальной сетью возбудимой системы мышцы. Нейро-
плазма в этом связывании не принимает участия. Отсюда вытекает,
что проведение и передача возбуждения от нервных окончаний к
мышечным волокнам должны происходить через нейрофибрилли.
24 и. С. Беритов
370 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Нейрофибриллярное строение нервных волокон и нервных клеток:
характерно для позвоночных животных и высших беспозвоночных ..
У низших беспозвоночных в нервных элементах, проводящих
возбуждение с декрементом, существование нейрофибриллей не очевидно.
Поэтому можно предположить, что в этих волокнах возбуждение
возникает и распространяется по всей плазме, как в мышечных клетках.
Ритмическая природа возбуждения. Нервный ствол или изолиро-
ранное нервное волокно обычно отвечает на одно кратковременное
раздражение, например на один индукционный удар, одним импульсом
Рис. 178. Биоэлектрические токи нерва при тетаническом раздражении.
Седалищный нерв кошки вырезан и помещен во влажной камере при 37°. Один конец нерва раздражается
150 замыкательными и 150 размыкательными индукционными ударами, а другой отводится в
гальванометр Эйнтховена. В начале электрограммы сила раздражения небольшая (24 см расстояния между
катушками); потом во время самой регистрации она увеличивается приближением катушек до 10 см.
Токи возбуждения в начале электрограммы следуют по ритму размыкательных ударов—150 в секунду,,
а затем — по ритму размыкательных и замыкательных вместе — 300 в секунду. Линия наверху
обозначает время по 0.2 секунды. = *
возбуждения. Если раздражение повторяется или действие внешней,
силы длится долго, дольше, чем развивается один импульс
возбуждения, то вслед за первым импульсом возникает ряд других.
Возбуждение протекает ритмически при всяком прерывистом раздражении, как
фарадический ток. В этом случае ритм возбуждения может равняться
ритму раздражения, но может быть и меньше последнего. Величина.
ритма возбуждения в каждом данном случае зависит от интенсивности
и частоты раздражения, подобно тому как это мы видели на мышце..
Когда частота раздражения не слишком велика, ритм возбуждения
может вполне следовать за ритмом раздражения — полный ритм
возбуждения. В нерве летней лягушки возбуждение может протекать
по ритму раздражения до 500 в секунду, а в нерве зимней — до
300 (Введенский, 25). При этом, если частота раздражения
небольшая, до 100—150 в секунду, то при пороговых силах ритм возбуждения
совпадает с ритмом раздражения. При больших же силах может
получиться удвоенный ритм возбуждения, ибо возбуждение будет
возникать как от размыкательных, так и от замыкательных ударов,
(рис. 178). Если частота раздражения выше, то для получения
однородного ритма возбуждения требуется тем большая сила раздражения,
чем выше частота последнего. В противном случае ритм возбуждения
будет более редким: он может быть вдвое (половинный ритм) или
даже в три и четыре раза более редок. Иначе говоря, возбуждение,
будет возникать в ответ на каждое второе, третье или четвертое
раздражение. Если ритм раздражения нерва лягушки равен приблизи-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
371
тельно 300 в секунду, то при слабых раздражениях слабые импульсы
возбуждения чередуются с сильными. Такой ритм возбуждения
называется двойным (Беритов, 26).
Высший, или предельный, ритм возбуждения. При очень частых и
сильных раздражениях нерв отвечает одним и тем же высшим ритмом
возбуждения, на который он способен в данном функциональном
состоянии. На зимних препаратах лягушки высший ритм составляет 300
в секунду, на летних — 500 (Беритов, 26). Высший ритм возбуждения
нерва у теплокровных животных при температуре тела 38° больше,
чем у лягушки. По новейшим исследованиям, он может равняться 600
в секунду (Форбс, Рэй и Грифитс, 27) и даже превышать 1000 (Гас-
сер, 50) (рис. 188). Высший ритм, разумеется, зависит от
функционального состояния нерва. По опытам Гартена, например, высший
нервный ритм у кролика при температуре 38° достигал 500, а при
низких температурах (25—30°) ритм возбуждения составлял всего 100—
200 в секунду. В этих опытах нерв раздражался или индукционными
ударами частотой в 2200 в секунду или сильным постоянным током (2).
Высший предельный ритм возбуждения в двигательном и
чувствительном нервах позвоночных больше, чем в скелетных мышцах. Это
доказывает, что лабильность в наиболее возбудимых нервных волокнах
выше, чем в скелетных мышцах. На том же основании лабильность
в двигательных и чувствительных нервах теплокровных выше, чем
в нервах холоднокровных. Но лабильность одного и того же нерва не
всегда одинакова. Она меняется не только под влиянием внешних
условий (см. по этому поводу главу о парабиозе), но и в результате
самой деятельности. В свежем и нормальном нерве теплокровных
после некоторой интенсивной деятельности лабильность первое время
повышается (Гассер, 79). В нервах же с плохим функциональным
состоянием лабильность быстро падает во время деятельности (рис. 188).
Лабильность нервов у беспозвоночных различна, но в общем она
ниже, чем у позвоночных. Даже у высших беспозвоночных
(ракообразных) предельный ритм не превышает 200—400 в секунду
(Кац, 81).
Абсолютная и относительная рефракторные фазы. Как указано
выше, возбуждение нерва протекает ритмически. Эта ритмическая
природа возбуждения объясняется тем, что деятельность нервного
волокна, подобно мышечному, происходит согласно основному закону
возбуждения. Каждое деятельное раздражение производит лол'ное
изохронное расщепление живой системы нервного волокна, которое затем
распространяется по всему волокну. Отсюда понятно, что в момент
этого изохронного расщепления в нерве возбудимость исчезает и вновь
появляется после возбуждения, по мере восстановления возбудимой
системы. Значит, в нерве, как и в мышце, возбуждение сопровождается
абсолютной рефракторной фазой, а после него следует
относительная рефракторная фаза. Прямым следствием
этой закономерности является тот факт, что при одновременном раз-
24*
372 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
дражении нервного ствола на обоих концах иолна возбуждения,
распространяющаяся с одного конца, не достигает другого. Каждая из
них затухает в месте перехода в рефракторную область другой.
Поэтому по одному нервному волокну не могут итти сразу две волны
возбуждения.
Абсолютная рефракторная фаза продолжается столько же, сколько
длится процесс возбуждения, вернее — чуть больше: значит, в
смешанном нерве летней лягушки она длится около 2 а, а у зимней — 3—5 а.
Разумеется, у лягушки в нервных волокнах разной толщины и
возбудимости продолжительность абсолютной рефракторной фазы различна.
В наиболее толстых волокнах группы А она меньше—1.20 а; в
волокнах средней толщины группы А она равна 1.68 j, а в тонких
волокнах, дающих гамма-волну, продолжительность еще больше — 3.13 а
(Эрлангер, Гассер и Бишоп, 5). Найденная абсолютная рефракторная
фаза двигательного нервного ствола, очевидно, выражает абсолютную
фазу наименее возбудимых двигательных волокон. У теплокровных
животных абсолютная рефракторная фаза гораздо короче, в
особенности при хорошем функциональном состоянии. Для наиболее
возбудимых волокон, дающих альфа-волну возбуждения, абсолютная
рефракторная фаза равна приблизительно 0.41—0.44 а (Гассер). Конечно,
когда нервные волокна возбуждены, их повторное раздражение не
производит никакого изменения в процессе возбуждения ни в смысле
усиления, ни в смысле удлинения его. От продолжительности
абсолютной рефракторной фазы зависит высший, предельный ритм
возбуждения. Например, мы указывали выше, что предельный ритм
возбуждения в нервных стволах летних лягушек составляет 500 в секунду. Так
как абсолютная рефракторная фаза в нерве летних лягушек
продолжается около 2 а, то естественно, что возбуждение может повториться
в секунду не более 500 раз (Беритов, 26).
Продолжительность относительной рефракторной фазы в 3—4 раза
превышает продолжительность абсолютной фазы, но только при
отличном функциональном состоянии. Продолжительность относительной
рефракторной фазы в сильной мере зависит от функционального
состояния нерва. При условиях, неблагоприятных для обмена веществ,
период этот значительно удлиняется, может даже достигнуть
нескольких секунд, как это бывает при парабиозе (см. ниже) (Беритов, 28).
Во время относительной рефракторной фазы повторное
раздражение дает более слабый эффект, чем в норме. Этот эффект тем слабее,
чем раньше приходит раздражение во время относительной
рефракторной фазы. Нормальный эффект получается только в конце этой фазы.
Далее, во время относительной рефракторной фазы требуется для
вызова возбуждения более сильное раздражение, чем в норме; оно
должно быть тем сильнее, чем ближе к началу относительной фазы оно
приходится, т. е. чем меньше в момент раздражения количество
восстановленной возбудимой системы (Беритов, 26, 29; Люкас, 30).
Наконец, скорость проведения возбуждения во время относительной реф-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
373
ракторной фазы значительно ниже, чем
в норме при отсутствии возбуждения
(Гассер и Эрлангер, 31). Вследствие
этого при двух последовательных
раздражениях индукционными ударами,
когда второе раздражение приходится
на время относительной рефракторной
фазы, интервал между двумя эффектами
всегда заметно длиннее, чем интервал
между ударами. Это обусловлено
замедленным возникновением и
распространением второго эффекта (Эрлангер и
Гассер, 58). Для иллюстрации
рефракторных фаз приведен опыт на рис. 179. В
этом опыте производится одновременное
тетаническое раздражение нерва в двух
участках. Ввиду разницы в ритмах
раздражения индукционные удары того и
другого раздражения приходятся друг
за другом через разные промежутки
времени. Когда промежуток времени между
двумя ближайшими ударами не
превышает 3—4 j, второй эффект не
получается. При промежутке 5.4—9.8 а
получается слабый эффект, тем более
слабый, чем меньше этот промежуток.
Электрограмма получена на зимних
лягушках, и потому как абсолютная, так и
относительная рефракторные фазы
растянуты.
По новейшим исследованиям,
означенная выше разница между абсолютной
и относительной рефракторными фазами
наблюдается в отношении толстых
волокон, проводящих возбуждение с
большой скоростью. Так, в п. phrenicus
кошки абсолютная рефракторная фаза
составляет около 0.5 а, а относительная —
около 2 с, как это дано на рис. 180,/.
В отношении тонких волокон с малой
скоростью проведения известно, что у
них относительная рефракторная фаза
длится обычно не дольше абсолютной, а
при скорости около 1 м <в секунду она
бывает короче абсолютной (Эрлангер и
Гассер, 58). Это, повидимому, связано с
з
а
к-?«
я
о.
о
4)
03
о
X
X
3
СП
СЗ
-?*
3
х
о.
о
си
а>
Си
К
ез
X
Л
с:
н
?
о
о
X
о
X
н
г
о
о
о
о
Я
X
I
(в I
9"»
я*
Sis
Я I
с? I
н ч 5
«> 3* *
С о Й
Я _CQ
in
Я
о "g
«5
О O.S!
!*?
' гі а.
Sou
si
?a*
м^
CD
A
3
re p
° ir
я ^>
>¦%
я с
ь 9
я
я я
о *?
? e> °
G я
rt Я „
Ш
« 2 о
(О
«
<D
<о
к о
О f- rf
$ « О
(О т
5°
«в 1
О. I
Я в>
!§
к аз
8.8
I!
V О
о8
374 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
тем обстоятельством, что тонкие волокна находятся в лучших условиях
обмена веществ с окружающей их лимфатической средой, чем толстые
волокна с толстыми миэлиновыми оболочками.
Абсолютная рефракторная фаза обычно определяется путем применения двух
следующих один за другим индукционных ударов максимальной интенсивности,
т. е. такой интенсивности, которая достаточна для возбуждения всех нервных
волокон данного ¦смешанного ствола. Но
если второе раздражение
сверхмаксимальное (10—15 см расстояния между
индукционными катушками), т. е.
такое, которое производит сильное
физиологическое изменение катодного
или анодного характера, тогда при
некоторых определенных условиях второе
раздражение влияет на импульс
возбуждения от первого раздражения: при
одних условиях он усиливается и
удлиняется, при других, наоборот,
ослабевает и укорачивается (Воронцов и
сотр., 35). Этот факт Воронцов
выдвигает против известного закона «все
или ничего», а следовательно, против
закона возбуждения. Раз возбуждение
меняется под влиянием последующего
раздражения, то, очевидно, оно не
должно выражать собой разряжения
потенциальной энергии всей возбудимой
системы. Но дело в том, что сильные
индукционные удары производят в
нерве местные, довольно сильные
поляризованные явления, пр ічем на
катоде и на аноде они имеют
противоположное направление. Возникающие при
этом поляризационные токи могут
распространяться электротонически на
протяжении нескольких миллиметров (і^ац,
75; Ходкин, 73). Чтобы локальное
изменение, вызванное первым ударом,
могло перейти в процесс возбуждения,
должно пройти определенное время для
его развития, как на это уже
указывалось выше. При самых благоприятных условиях оно должно составлять не
менее 0.4—0.5 а (см. также Багно, 78). Поэтому, если второй удар следует быстро
за первым, поляризационное явление от первого удара должно измениться под
влиянием последующего второго удара. Оно или усилится, если оба удара одного
направления, или, наоборот, ослабеет в силу алгебраической суммацки, если второй
удар противоположного направления. Отсюда следует, что в опыте Воронцова
под влиянием второго удара меняются физические условия возникновения
возбуждения от первого удара, а не самое возбуждение непосредственно.
Отмеченное выше изменение ритма возбуждения, а именно
половинный и двойной ритмы, обусловливается действием относительной
рефракторной фазы. При слабых раздражениях большой частоты
получаемый половинный ритм возбуждения происходит оттого, что каж-
Рис. 180-1. Токи возбуждения п. phrenicus
кошки при 37.5°.
По два сверхминимальных индукционных удара с
разным интервалом. Возбужденные участки
отводятся в катодный осциллограф на расстоянии 1 см от
раздражающего катода. В 1-м опыте интервал около
2?, где второй эффект немного слабее первого; во 2-м
Интервал 1.2 з и второй эффект значительно слабее
первого; в 3-м интервал около 0.8 з, и второй эффект
еще меньше. В 4-м опыте второй удар пришелся
через 0.4 j и не дал эффекта (Гассер).
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 375
дое четное раздражение попадает в относительную рефракторную фазу
в начале ее и не вызывает возбуждения; каждое нечетное
раздражение производит его, так как к этому времени рефракторное состояние
¦вполне или же в значительной мере заканчивается. Двойной ритм
получается в тех случаях, когда каждое четное раздражение, попадая
в рефракторную фазу, вызывает возбуждение не во всех, а только
в некоторых наиболее поверхностных волокнах, на которые раздраже-
"ние действует сильнее, и еще, вероятно, в сравнительно толстых
волокнах, у которых относительная рефракторная фаза сравнительно
коротка. Вследствие этого первое раздражение возбуждает все
волокна, второе — только указанные выше волокна, третье — опять все
волокна, ибо к этому времени относительная рефракторная фаза
заканчивается и в тех волокнах, которые не возбуждались от второго
раздражения. Иначе говоря, одна часть волокон будет возбуждаться
лри каждом раздражении, а другая—-только через одно раздражение.
В случае попадания раздражений в относительную рефракторную
•фазу полный ритм возбуждения получается тогда, когда сила
раздражения настолько велика, что преодолевает рефракторное состояние во
всех возбужденных волокнах, но при этом, конечно, эффект будет
слабый, ибо в моменты раздражения возбудимая система во всех
волокнах восстановлена частично (Беритов, 26, 33). *
Хронаксия во время относительной рефракторной фазы. Несомненное
понижение возбудимости во время относительной рефракторной фазы своеобразно
отражается на хронаксии. При сильном нарастании 'реобазы, соответственно понижению
возбудимости, хронаіксия укорачивается. При этом количество электричества, потреб-
:ное для раздражения, почти не меняется, ибо с нарастанием реобазы
пропорционально уменьшается время действия тока (Макаров, 53). Исходя из того, ічто во
©ремя относительной рефракторной фазы реобаза и вообще пороговое раздражение
повышаются, а іпроцессы возбуждения и скорость проведения его ослабевают, нужно
искать, конечно, причину укорочения хронаксии не в улучшении функционального
состояния ткани во время этой фазы, а в самом методе хронаксиметрического
определения времени реакции. Как известно, время реакции зависит от силы
раздражения: оно тем короче, чем сильнее раздражение. При определении хронаксии
берется удвоенная реобаза. Реобаза же в рефракторную фазу растет очень сильно:
в 2—4 раза. Удвоение такой повышенной реобазы оказывается физиологически
более сильным агентом, требующим значительно более короткого времени для
раздражения, чем удвоение малой реобазы при покоящемся состоянии неріва.
Абсолютная и относительная рефракторные фазы нервов с декре-
ментным проведением возбуждения. Выше мы показали, что в нервах
низших беспозвоночных возбуждение проводится с декрементом. По
отношению к этим нервам раньше утверждали, что абсолютная
рефракторная фаза у них отсутствует и что их эффект всегда тем выше,
чем сильнее раздражение. Это положение было принято на основании
•наблюдений за механическими эффектами мышцы нервно-мышечного
препарата или за электрическими эффектами, регистрируемыми
баллистическим гальванометром (Иордан и Люллис, 52; Люллис, 55; Кан 56).
Но, по новейшим исследованиям нерва улитки (п. intestinalis от Helix
pomatia), оказалось, что вблизи раздражаемого участка ток возбужде-
376 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
шЫт
ния нерва с усилением раздражения достигает определенной высоты
и дальше не усиливается. При дальнейшем усилении раздражения
возникают новые волны возбуждения: во-первых, от возбуждения
других, менее возбудимых
групп нервных волокон и,
во-вторых, вследствие
возникновения новых повторных
волн возбуждения как в тех
же самых высоковозбудимых,
так и в других нервных
волокнах (ркс. 18Q-II). То и
другое обстоятельство, очевидно,
и обусловливает широкую
градацию механического
эффекта мышцы или
электрической реакции нерва,
наблюдаемой по
баллистическому гальванометру. Но при
регистрации электрической
реакции нерва струнным
гальванометром оказалось, что во
время начального колебания
биотока, отвечающего
возбуждению всех наиболее
возбудимых волокон, повторное
раздражение не дает эффекта.
Оно вызывает небольшую
новую волну возбуждения лишь
через 7—8 а. Полный эффект
от второго раздражения
получается через 25 а. Отсюда
следует, что абсолютная
рефракторная фаза
продолжается около 7 с, а относительная
длится до 25 а (yMaja—до 34 а)
(Крийгсман, 119). Это хорошо
видно на рис. 180-П, где второе
раздражение не дает эффекта
через 5 а, а через 10, 15 и 20 а
Рис. 180-И, л и В. Токи возбуждения п. mtestj.
nalis от Helix pomatia при нарастающей силе ра3
Дражения (размыкательные индукционные удары ч"
Регистрация струнным гальванометром Эйнтховена. Цифры
означают силу раздражения в единицах пороговой силы.
При усилении раздражения в 7 раз быстрый ток
возбуждения от наиболее возбудимых волокон достигает
максимума (А). При силе в 12—37 единиц дополнительно возникают
длительные колебания струны от токов возбуждения
нервных волокон низкой возбудимости (В). Определение
рефракторных фаз двумя последовательными индукционными
ударами (С и D): 1 — биоток возбуждения наиболее
возбудимых волокон от одного раздражения; 2 — от двух
раздражений, разделенных интервалом в 5а; 3—то Же при
интервале 10з, 4 — при интервале 15з, 5—при интервале 20j.
Внизу рремя — Юг (Крийгсман).
дает ослабленные эффекты-
Несмотря на то, что данный нерв улитки может возбудиться со всей
своей потенциальной энергией при некоторой силе раздражения,
возбуждение распространяется в нем с декрементом. Это доказывает, что
возникающий в раздраженном участке нерва биоток возбуждения не
настолько интенсивен, чтобы целиком возбудить соседний участок.
Ввиду этой особенности, чем дальше ток возбуждения распростра-
ВОЗБУЖДЕНИЙ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 377
няется, тем меньше становится его раздражающее действие на
последующий участок.
Рефракторные фазы были установлены и в других нервах
беспозвоночных. Так, в нерве конечности Maja абсолютная рефракторная
фаза для наиболее возбудимых волокон достигает 1.4—2.8 а, а для
менее возбудимых — 4—7 а, относительная фаза у Maja — 34 а (Люл-
лис, 55; Боге и Розенберг, 80). В нерве конечности Carcinus абсолютная
рефракторная фаза равняется 3—4 о (Пэнтин, 83). С большой длитель-
Рис. 181. Электрограмма седалищного нерва лягушки.
В секунду дается по 20 замыкательных и размыкательных ударов. Сила
раздражения—21 см расстояния между катушками (порог 25 см). Первый замыкательный
удар не дает эффекта, а второй, спустя 0.035. сек. после эффекта, вызванного
первым размыкательным ударом, дает заметный эффект; третий замыкательный
удар производит уже такой же эффект, как и размыкательный.
ностью абсолютной рефракторной фазы связан и более низкий
предельный ритм возбуждения этих нервов.
Сверхнормальная возбудимость нерва. Если нерв отвечает на
раздражение возбуждением, то вслед за тем имеет место повышение
возбудимости сравнительно с имевшейся до возбуждения. Эта сверхнормальная
возбудимость наблюдается после относительной рефракторной фазы,
т. е. к моменту полного восстановления возбудимой системы
(Введенский, 25; Беритов, 26; Люкас, 30). Сверхнормальная возбудимость
в нерве удерживается сравнительно с мышцей очень недолго, но и
здесь она длится гораздо дольше, чем возбуждение или рефракторные
фазы. Например, на рис. 181 продолжительность ее превышает
0.038 сек. По Люкас у, она продолжается до Vio сек. при 15°.
Повышение возбудимости в данном случае происходит во всем нерве, где
распространяется возбуждение, а не только в раздражаемом участке.
В этот период повышенной возбудимости процесс возбуждения,.
поскольку он выражается в электрическом эффекте, наступает сильнее;
кроме того, повышается скорость распространения возбуждения
(Грэим, 59).
По новейшим исследованиям, сверхнормальная возбудимость после
рефракторных фаз сменяется пониженной, т. е. становится ниже
нормы. Понижение возбудимости на нервах теплокровных начинается
спустя 0.02 сек. после возбуждения и может продолжаться до 0.06 сек.
Но на свежих препаратах пониженная возбудимость вновь сменяется
повышенной. В период пониженной возбудимости скорость проведения
уменьшается, но интенсивность тока возбуждения заметно не умень-
378 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
яіается сравнительно с нормой (Грэим, 59). Фаза пониженной
возбудимости проявляется сильнее на неповрежденном нерве в организме.
После тетанического раздражения сверхнормальная возбудимость
укорачивается и может совершенно сойти на-нет. Наоборот, пониженная
возбудимость, субнормальная, нарастает; при повторных возбуждениях
•она углубляется и удлиняется (Эрлангер и Гассер, 58).
Таким образом, после возбуждения и восстановления нерва от
рефракторных фаз в нерве наблюдается ритмическое изменение
возбудимости выше и ниже нормы.
Характерно, что длительные изменения возбудимости после
рефракторных фаз не в одинаковой мере свойственны всем нервным
волокнам. У волокон В теплокровных в нормальном состоянии почти не
наблюдается стадии длительного повышения возбудимости. После
импульса возбуждения возбудимость остается ниже нормы на долгое
время и постепенно, в течение 100—150а, возвращается к норме. В
волокнах А теплокровных обе фазы после одного импульса
продолжаются всего 100 а, в то время как в волокнах С (симпатических) они
длятся больше 700 о (Гассер, 97).
Как мы уже отметили в отделе мышечной физиологии,
сверхнормальное повышение возбудимости должно быть обусловлено
одновременным изохронным восстановлением всей возбудимой системы.
Вследствие этого после возбуждения наступает момент, когда масса
возбудимой системы бывает больше, чем в норме. Этим объясняется не
только сверхнормальное повышение возбудимости, но и наблюдаемое
при этом сверхнормальное усиление процесса возбуждения в ответ на
іновое раздражение.
Так как настоящее гетерохронное состояние наступает не сразу,
з. до этого определенная часть возбудимой системы разряжается и
восстанавливается более или менее изохронно, то возбудимость
неминуемо должна колебаться в связи с изменением количества
возбудимой системы после первоначального восстановления, как это ясно видно
«а рис. 85, изображающем переход изохронного состояния в
гетерохронное.
Сверхнормальная возбудимость после раздражения легко может
'быть выявлена, если один участок раздражать слабыми
индукционными ударами, а другой — максимальными. При комбинации их
окажется, что и слабые удары начинают давать максимальные эффекты,
>если они попадают вскоре после сильных ударов (рис. 181). В
отдельности слабые удары возбуждают только наиболее возбудимые и
наиболее доступные для раздражения наружные волокна. Действие этих
ударов на внутренние волокна должно значительно ослабевать ввиду
того, что им приходится преодолевать при прохождении поперек нерва
большое сопротивление в виде нервных оболочек. Если после сильных
ударов, вызвавших возбуждение всего ствола, и слабые удары
вызывают эффект, то это должно указывать на возбуждение менее
возбудимых и более глубоко лежащих волокон вследствие повышения в них
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ
379
возбудимости под влиянием волны возбуждения от предшествующих
сильных ударов.
Ритмический эффект после одного кратковременного раздражения.
В совершенно нормальном функциональном состоянии нерв отвечает
на одно короткое раздражение одним импульсом возбуждения. Но
в определенных случаях изменений внутреннего состояния нерва
однократное раздражение вызывает ряд быстро следующих друг за другом
импульсов. Это бывает, например, при накоплении активных ионов
внутри нервных волокон: при высыхании нерва, при увеличении
концентрации ионов калия, при уменьшении ионов кальция (Шривер и
Цебулла, 62).
Этот ритмический разряд возбуждения в ответ на одно
раздражение находится в связи с отмеченным выше ритмическим изменением
возбудимости или количества возбудимой системы после каждого
возбуждения. Накопившиеся в нервном волокне активные ионы
обусловливают изохронное расщепление той части возбудимой системы,
которая более или менее восстановилась после предыдущего общего
расщепления. Возникающий при этом биоток настолько велик, что он
в состоянии воздействовать на соседние частички возбудимой системы
и вызвать распространяющийся процесс возбуждения.
Ритмический разряд может возникать и в нормальном нерве после
чрезмерно сильного индукционного удара или после повреждения от
перерезки (Эдриан, 117). Это уже обусловливается теми более или
менее длительными физико-химическими .изменениями, какие вызываются
сильными индукционными ударами или повреждением живой системы
от перерезки.
Ритмический разряд импульсов после одного кратковременного
индукционного удара некоторые авторы считают фактом,
противоречащим закону «все или ничего», а значит и закону возбуждения (Бете,
124; Макаров, 116). Но, как видно, это не так. Повторный импульс
после одного раздражения возникает не за счет остатков той энергии,
которая была к моменту раздражения, а за счет той энергии, которая
успела восстановиться после предыдущего импульса возбуждения.
Следовая отрицательность или электрическое последействие
возбуждения. Вслед за быстро протекающим током возбуждения большой
интенсивности наступает другой ток, который проявляется в виде
замедленного опускания нисходящего колена кривой быстро
протекающего тока или же в виде нового дополнительного тока, который на
кривой может быть отделен от быстро протекающего некоторым
углублением. Этот ток был впервые описан Воронцовым (1926) (51),
а позднее подробно изучен рядом американских исследователей
(Эмберсон и Доунинг, 36; Гассер и Эрлангер, 37; Жерард, 35а, и др.)>
а также самим Воронцовым (38, 39).
Указанное электрическое последействие имеет такой же
отрицательный характер, как и ток возбуждения, но значительно слабее его:
амплитуда его в редких случаях достигает 10% высоты быстро проте-
380 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
кающего тока возбуждения. Но продолжительность его большая: она
может достигнуть многих сигм и даже десятков сигм (рис. 182
и 183). При тетаническом раздражении это отрицательное
последействие несколько усиливается и удлиняется. Аналогично влияют на него
повышение температуры, іионы кальция, вератрин. В обратном
направлении действуют охлаждение, цианистый калий, хлористый калий и
другие вещества. Это последействие возникает и распространяется
в нерве подобно току возбуждения (Воронцов, Эрлангер и Гассер, 58).
Отрицательное электрическое последействие затем сменяется
положительным, еще более слабым и продолжительным. После одного
в и
Рис. 182 и 183. Токи возбуждения седалищного нерва лягушки.
Струнный гальванометр. Чувствительность струны выражается
в отклонении на 40 см от 1 милливольта (без включения препарата
в цепь струны). Однофазное отведение тока возбуждения. Нерв
раздражается отдельными индукционными ударами. Левая
электрограмма (рис. 182) получена при комнатной температуре 14° без
предварительной тетанизации. Правая электрограмма (рис. 183)
получена при согревании от 14 до 16э и после тетанизации в течение
1 мин. В первом случае нисходящее колено продолжается около
0.09 сек., во втором—свыше 0.3 сек. Уже на первой электрограмме
нисходящее колено кривой'тока возбуждения в девять раз больше
в сравнении с контрольной кривой, когда постоянный ток
прекращается мгновенно (Воронцов).
импульса положительный потенциал не превосходит 5 микровольт
(5 миллионных вольта), а длительность его достигает многих десятых
секунды. На рис. 184 дана картина отношений интенсивности и
продолжительности тока возбуждения и электрических последействий.
Положительное последействие значительно усиливается после тетаниче-
ского раздражения: оно тем сильнее и длительнее, чем сильнее и чаще
раздражение (Эрлангер и Гассер, 58). Иногда положительное
последействие вновь может смениться отрицательным. Таким образом,
получается такая же ритмичность в колебании потенциала, как это
было указано выше относительно возбудимости после возбуждения. По
данным Г а с с е р а, оба эти .изменения точно совпадают во времени,
причем каждый раз повышение возбудимости сверх нормы совпадает
с отрицательным потенциалом, а понижение—-с положительным (50).
Разные нервные волокна показывают электрическое последействие
не в одинаковой степени. В волокнах А обе фазы выражены хорошо,
но длятся после одного импульса всего около 100 а, в нервных же
волокнах В в свежем состоянии почти отсутствует отрицательное
последействие. Ток возбуждения сразу переходит в длительное поло-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 381
жительное колебание, которое длится много дольше, чем в волокнах А
(Гассер, 97).
В симпатических волокнах С особенно хорошо выступает
положительное колебание. Оно быстро сменяет небольшую отрицательную
фазу сейчас же после быстро протекающего тока возбуждения (Эрлан-
гер и Гассер, 58).
Особенно длительно электрическое последействие протекает в
нервах беспозвоночных. В нервах рака оно после одного импульса
возбуждения продолжается несколько секунд, а после тетаниче-
ского раздражения — даже несколько минут (Левин, 42;
Боге и Розенберг, 80; Жуков, 105). Электрическое
последействие также сначала является отрицательным. Но при
определенных температурных условиях оно сменяется
положительным колебанием, которое продолжается много
минут. На нервах черноморского краба положительное
колебание появляется при температуре выше 20° С (Жуков, 105).
На межганглионарном нерве беззубки, при одиночном
раздражении после начального, сравнительно быстрого
колебания с продолжительностью около 1 секунды,
наступает сначала длительное отрицательное колебание, а затем
положительное, продолжающееся многие секунды. При
ряде раздражений получается сплошное отрицательное
колебание, продолжающееся все время раздражения. При
тетанических раздражениях последействие — сначала отри-
I
4
V
__ Отрицательное
последействие
Яоложитемюре
— последействие
/о
20
30
40
50
6?
70
00
Рис. 184. Течение всего цикла изменений электрического потенциала в нервном волокне
млекопитающих.
Диаграмма показывает точное отношение интенсивности и продолжительности между током
возбуждения и длительными электрическими последействиями (Гассер).
цательное, затем положительное — продолжается особенно долго, в
течение многих минут. На эти последействия сильное влияние оказывают
углекислота и замещение кислорода азотом. В первом случае
положительное колебание усиливается, а отрицательное исчезает, во втором
случае происходит наоборот (Кан и сотр., 56).
На вопрос о происхождении электрического последействия авторы
не дают единого ответа. Несомненно, что последействие происходит от
возбуждения, но еще точно не известно, что собственно оно выражает.
Воронцов (51) с самого начала утверждал, что остаточная
электроотрицательность нерва выражает состояние повышенной возбудимости,
ибо в этот период он всегда находил понижение порогов раздражения.
Он же полагал, что явление лестницы в нервном стволе всегда
совпадает с повышением остаточной электроотрицательности. В последнее
382 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
время Гас с ер и Эрлангер в общем утверждают то же самое.
Именно: они нашли, что ритмическое изменение остаточного
электрического потенциала точно совпадает с изменением возбудимости. Во
время отрицательного электрического- последействия возбудимость выше
нормы, а во время положительного, наоборот, ниже нормы. Точна
так же возбудимость меняется параллельно с электрическим
последействием под влиянием внешних агентов. Когда почему-либо
отрицательное последействие усиливается, возбудимость повышается; когда же,,
наоборот, усиливается положительное последействие, возбудимость
понижается (Леман, 98).
В настоящее время Воронцов и сотрудники полагают, что
следовая отрицательность выражает собой повышение проницаемости
поверхностных перепонок, которое обусловливает интенсивный обмен
веществ между волокном и внешней средой. Значит, следовая
электрическая отрицательность представляет физическую волну, возникающую
и распространяющуюся вслед за возбуждением (Воронцов и
Юденич, 39). Некоторые факты говорят против такого объяснения.
Известно, что запоздалая теплопродукция нерва после тетануса совпадает
во времени с продолжительностью электрического последействия. Кроме
того, существует точное соответствие между скоростью выделения
тепла и конфигурацией электрического последействия. А это
обстоятельство указывает на связь электрического последействия с обменом1
веществ. Это подтверждается и тем фактом, что при повышении обмена
веществ усиливается и электрическое последействие, причем это может
произойти без изменения быстро протекающего тока возбуждения
(Гассер, 108). Источником теплопродукции нерва, конечно, частично*
являются и биотоки нерва с их поляризационным действием. Но, по*
Гил л у, на эту фазу теплопродукции приходится не больше 1 % всей
теплопродукции (см. об этом ниже). На основании этих фактов нужно*
считать наиболее вероятным, что медленные электрические потенциалы
после возбуждения выражают изменение интенсивности основного
биологического процесса. После рефракторных фаз изохронное
восстановление удерживается на некоторое время в большинстве частичек
возбудимой системы. Это состояние связано с усиленным обменом веществ,
вследствие чего соответственный участок нерва является
электроотрицательным. Изохронно восстановленные частички постепенно
расщепляются и, прежде чем установится обычное гетерохронное состояние,„
наступает некоторое пониженное гетерохронное состояние, когда
значительная часть частичек находится в состоянии расщепления или
в начальных стадиях восстановления, т. е. когда нерв находится в со-
стоянии пониженного обмена веществ. В это время, конечно, данный
участок нерва будет электроположителен в отношении других
нормальных участков нерва и в то же время будет показывать пониженную
возбудимость. Все это ясно видно на рис. 85, где после рефракторных:
фаз общий уровень возбудимой системы сначала выше нормы, а потом
ниже.
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 38$
Данное выше объяснение годится только для первых десятых долей,
секунды медленных электрических колебаний. Что же касается таких
длительных электрических изменений, как секунда и больше, они
должны быть обусловлены действием метаболитов, возникших в
результате изохронного расщепления возбудимой системы, на возбудимую-
же систему, на ее основной биологический процесс. Этим, вероятно,,
объясняется, что длительные колебания потенциала более интенсивны
и продолжительны после тетанического раздражения, чем после
одиночных ударов. Кроме того, на это должен указывать .и тот факт, что-
углекислота и замещение кислорода азотом действуют на длительное
электрическое последействие, ибо это действие должно осуществляться,
путем влияния на основной биологический процесс.
Повышение возбудимости при субминимальных раздражениях.
Повышение возбудимости в нерве может быть наблюдаемо и при
раздражениях такими индукционными ударами, которые не производят
возбуждения, т. е. при субминимальных раздражениях. Это проявляется в том,
что при быстром повторении такого раздражения последнее становится
деятельным. Следовательно, нервным волокнам также свойственно-
явление суммации, как и мышце (Люкас, 48). Повышение
возбудимости в данном случае, конечно, происходит только в раздражаемом
участке.
Это повышение возбудимости от подпорогового раздражения в
лягушечьем нерве обнаруживается через 1—2 з и более. До этого
возбудимость, наоборот, понижена (Гильдемейстер, 67; Кан, 68). Очевидно,
частичное изохронное расщепление создает в расщепленных частичках
такое же состояние, как это бывает со всеми частичками при
сверхпороговых раздражениях: расщепленные частички сначала находятся
в состоянии рефракторной фазы, а затем, после восстановления,.
в состоянии повышенной возбудимости. Одновременно продукты
расщепления и возникший через них электрический потенциал должны
действовать на соседние участки и повышать их чувствительность к
последующим раздражениям.
При подпороговых раздражениях наблюдается еще один период,
в самом начале раздражения. Он продолжается очень короткое время,,
до 0.4 . Если повторное раздражение попадает в этот период, то
получается суммация — усиление эффекта с наступлением процесса.
возбуждения. Это бывает перед наступлением фазы пониженной
возбудимости. Очевидно, в этот период под влиянием двух воздействий
изохронное расщепление частичек настолько обширно, что оно дает-
биоток, способный распространить изохронное расщепление на всю
возбудимую систему.
Явление лестницы на нерве. На совершенно нормальном и свежем:
нерве нельзя наблюдать явление лестницы, но на нервах, измененных,
например, от вератрина, избытка кальция, асфиксии, явление лестницы
обычно наблюдается. На нервах теплокровных оно может быть
наблюдаемо и после некоторой интенсивной деятельности, именно если после:
384 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
такой активности приложить тетанизацию подпороговой силы. Во всех
этих случаях при недеятельных подпороговых раздражениях после
нескольких ударов недеятельное раздражение превращается в
деятельное, и вызываемые им эффекты усиливаются с каждым новым ударом.
При деятельном же раздражении электрический эффект растет с
каждым ударом с самого начала раздражения. Чем чаще раздражение,
тем выше поднимается вызываемый электрический эффект зозбужде-
? " А в *
Рис. 185. Токи возбуждения в нерве теплокровного. Изолиро
ванный нерв при t° — 37°С.
Раздражается ударами от тиратронного осциллатора. Токи возбуждения
регистрируются осциллографом. Частота раздражения в 1 сек. — 200 (7),
70(2), 30(3) и 15 (4). Все записи А сделаны после сильного тетанического
раздражения той же частоты. Все записи В—в состоянии отдыха. Внизу
время — Чм сек. Неодинаковая высота токов возбуждения обусловлена
спонтанными изменениями возбудимости нервных волокон (Гассер).
ния, а значит и процесс возбуждения. При некотором же ритме
явление суммации отсутствует; очевидно, каждый удар приходится после
периода повышенной возбудимости от предшествующего раздражения
(рис. 185).
Это лестницеобразное повышение зависит от двух обстоятельств:
1) каждый последующий удар возбуждения приходится на время фазы
повышенной возбудимости, на так называемый суммационный период,
вследствие чего возбуждаемые раздражением волокна производят
больший эффект в ответ на последующие раздражения, чем на первое;
2) последующие удары вовлекают в реакцию новые волокна, которые
не были возбуждены от первых раздражений, вследствие того же
суммационного периода.
Одиночное тетанизированное сокращение. Так называется явление,
которое получается при комбинации субминимального тетанического
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 385
раздражения одного участка нерва с максимальным одиночным
раздражением какого-либо другого участка. Отдельные индукционные удары
дают обычные одиночные сокращения, а при комбинации с
субминимальным тетаническим раздражением производят более или менее
длительные тетанизированные сокращения. Это явление было открыто
Рис. 186. Анализ одиночного тетанизированного сокращения.
А — электрограмма тока возбуждения седалищного нерва от раздражения одним
размыкательным индукционным ударом максимальной интенсивности; В —
электрограмма от комбинации того же одиночного раздражения с подпороговым
тетаническим раздражением, которое начинается до одиночного раздражения и
продолжается некоторое время после него. Ритм раздражения 50 ударов в 1 сек.;
С — механограмма и электрограмма m. gastrocnemius при комбинации тех же
раздражений, что и в электрограмме В. Под электрограммой сигнальная линия
поднятием обозначает момент одиночного раздражения. Внизу время в 0.2 сек.
Хорошо видно, что тетаническое раздражение само по себе не дает эффекта, а
после одной волны возбуждения производит ряд волн возбуждения по ритму тета-
нического раздражения сначала в нерве, а затем и в мышце (Киселев).
Введенским. Он же назвал его: одиночное тетанизиро-
ванное сокращение (46).
Введенский локализировал причину этого явления в нерве. Он
полагал, что волна возбуждения, распространяясь в нерве, производит
в нем определенное изменение функционального состояния в смысле
повышения возбудимости. Именно, мы должны здесь иметь в виду
фазу сверхнормальной возбудимости. Вследствие этого субминимальное
тетаническое раздражение становится на некоторое время
минимальным, производящим в нерве ряд импульсов. Это, в свою очередь,
обусловливает тетаническое сокращение. В последнее время многие
авторы занимались специальным исследованием биоэлектрических
явлений возбуждения нерва и мышцы; они нашли, что тетанизированное
сокращение, действительно, обусловлено возникновением в нерве ряда
импульсов от тетанического раздражения под влиянием волны
возбуждения от сильного электрического раздражения (Киселев, 47). Мы
здесь приводим рисунок Киселева, где дается прекрасный анализ
25 и. С Беритов
386 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
этого явления (рис. 186). На этом рисунке хорошо видно
происхождение одиночного тетанизированного сокращения. В опыте А дается
биоэлектрический ток возбуждения нерва от одного максимального
индукционного удара. В опыте В этот индукционный удар производится во
время субминимальной тетанизации нерва. Вслед за первым сильным
током возбуждения от отдельного индукционного удара следует ряд
постепенно ослабевающих токов возбуждения по ритму раздражения.
В опыте С дается тетанизированное сокращение мышцы и
биоэлектрические токи возбуждения мышцы. Эти токи протекают по ритму тета-
нического раздражения и притом так же постепенно ослабевают, как
в нерве. Ясно, что течение возбуждения в мышце во время одиночного,
тетанизированного сокращения всецело обусловливается притекающими
с нерва импульсами, которые возникают в нем от субминимального
тетаніического раздражения благодаря сверхнормальной возбудимости,
произведенной одиночным максимальным раздражением.
Ир а кпггіыум
1. Опыт Кюне с проводимостью возбуждения по нерву в обоих направлениях,
М. sartorius лягушки перерезается пополам в области вхождения нерва, на
так, чтобы не перерезать веточек нерва, идущих з ту и другую стороны.
Затем раздражается та или другая половина мышцы механически — путем щипания
или электрически — индукционными ударами. Каждый раз при сокращении одной
половины мышцы будет сокращаться и другая половина.
2. Наблюдение непроводимости возбуждения от одного нервного волокна к
другому. Заднюю половину лягушки освобождают от внутренностей и
привешивают ее на штативе так, чтобы ноги свободно висели в воздухе. Остаток спинного
мозга предварительно разрушают. Если теперь раздражать то один корешок, то
Другой из поясничного сплетения, движения ноги будут разные. Поясничное
сплетение лягушки состоит главным образом из трех корешков, двигательные волокна
которых потом 'большей частью вступают в общий ствол седалищного нерва.
Бели бы возбуждение переходило от одного нервного волокна к другому, то,
конечно, означенное разнообразие в движениях ног должно было бы отсутствовать.
3. Определение скорости проведения. Помещают нервно-мышечный препарат
лягушки во влажную камеру и записывают сокращение мышцы на кимографе
Криса или на другом, который вращается со скоростью около 100 см в секунду*
Раздражение и запись происходят, как в опыте с одиночным сокращением. Один
раз раздражается іближайший к мышце участок нерва, другой раз — дальнейший.
Возбуждение іво втором случае проходит всю длину нерва, а потому скрытый
период сокращения в этом случае будет больше, чем в первом. На основании
разницы времени и расстояния между раздраженными участками высчитывается
скорость проведения по нерву.
4. Наблюдение рефракторной фазы нерва. На нервно-мышечном препарате
нельзя точно изучить рефракторную фазу нормального нерва, пользуясь
механическим эффектом. Рефракторная фаза мышцы длиннее, чем рефракторная фаза
нерва, а потому если второе раздражение не дает эффекта или дает слабый
эффект, то это могло произойти от рефракторной фазы мышцы. Рефракторную фазу
нерва лучше всего изучить путем биоэлектрических токов возбуждения самого
нерва. Это можно сделать гальванометром Эйнтховена, капиллярным электрометром
Липмана или же осциллографом катодных лучей.
Постановка опыта может быть такой, как это было в опыте, данном на рис. 179»
но можно поступить и так,, как это было описано выше в отношении мышцы: да-
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 38Г
ются два быстро следующих одно за другим раздражения, интервал между
которыми может быть изменяем произвольно. Для этого пользуются маятником Гельм--
гольца с двумя электрическими контактами, которые введены в первичную цепь
индуктория. Контакты замкнуты. При качании маятника происходит размыкание
сначала одного контакта, затем быстро вслед за ним другого. Один из контактов-
является подвижным. Путем приближения его к другому, неподвижному контакту
время между размыкательными ударами можно менять по желанию.
Для определения продолжительности абсолютной рефракторной фазы оба
раздражения должны быть максимальной силы, т. е. такой, чтобы все нервные
волокна в раздражаемом участке нерва приходили в возбуждение. Для этого надо
приблизить вторичную катушку К первичной на 10—20 см, считая от порогов. В
случае, если первое раздражение не максимально, абсолютная рефракторная фаза
может быть сведена на-нет, так как после первого раздражения остались бы
невозбужденными некоторые волокна, которые поэтому могут быть возбуждены
вторым раздражением іпри любом 'интервале между раздражениями. Если же второе -
раздражение было слабо, не максимально, тогда оно 'Вызвало бы эффект только
з период более или менее значительного восстановления возбудимой системы, т. е.
в поздний период относительной рефракторной фазы, а потому регистрируемая фаза
невозбудимости была бы продолжительнее абсолютной рефракторной фазы. Для
определения относительной рефракторной фазы лучше брать также максимальные-
раздражения. Когда второе раздражение произведет такой биоэлектрический ток,.
как (первое, тогда, очевидно, наступил конец относительной рефракторной фазе.
При отсутствии чувствительных приборов для регистрации биоэлектрических.
токов нерва можно пользоваться для изучения рефракторных фаз механическим
эффектом мышцы, регистрируя его кимографом Криса. Кимограф снабжен двумя
контактами для размыкания электрического тока в первичной цепи индуктория.
Кимограф быстро вращается и при своем вращении размыкает сначала один, затем-
другой контакт. По івеличине механического эффекта можно судить о действии
второго раздражения. Этим путем можно определить только абсолютную рефракторную
фазу нерва и притом более или менее приблизительно.
5. Наблюдение биоэлектрических токов возбуждения нерва при тетаническом
раздражении с помощью гальванометра Эйнтховена. Берут два седалищных нерва
от крупной лягушки. Один конец івзятых нервов накладывается на платиновые
электроды для раздражения, а другой конец соединяется посредством платиновых
же электродов с гальванометром. іМежиолюсное расстояние в последних
электродах должно быть около 1 см, а между обеими парами электродов не менее
2 см. Находят пороговое раздражение, а затем усиливают его приближением
индукционных катушек. Наблюдают на экране колебание струны от возникающих в
нерве биоэлектрических токов возбуждения. Чем чаще раздражение, тем чаще
будет это колебание. Амплитуда же колебания будет расти с усилением раздражения.
При порогах около 35—40 см расстояния между катушками амплитуда будет
нарастать при усилении раздражения до 25—30 см и даже при еще большем усилении
его. Чем толще пучок нервов, тем больше нужно усиливать раздражение для
получения максимальных колебаний от биоэлектрических токов возбуждения.
Что наблюдаемое при этом колебание струны действительно вызывается
биоэлектрическими токами, в этом, можно удостовериться таким путем: нервы
умерщвляют аммиаком между раздражающими и отводящими электродами. После этого
раздражение в 25 см и более сильное не будет давать колебаний струны. Только
при 15—10 см и еще более сильном раздражении могут получаться колебания
струны. Но это уже будет не от биоэлектрических токов, а от распространения
петель раздражающего тока. В этом легко можно удостовериться при перемене
направления раздражающего тока. С переменой направления переменится и
направление колебаний струны. В случае биоэлектрических токов возбуждения
перемена направления раздражающего тока не повлияет на направление колебаний
струны.
25*
388 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
6. Наблюдение закона возбуждения на изолированном нервном волокне.
Изолирование нервного волокна по Методу Като заключается в следующем; берут гаерв-
но-мышечный препарат — седалищный нерв — m. gastrocnemius, помещают его на
стеклянную пластинку изоляционного аппарата, а центральный конец его
закрепляют на пробке булавкой. Затем удаляют соединительную ткань вокруг нервногй
ствола на небольшом протяжении. Первый этап операции состоит в расщеплении
соединительнотканной оболочки нерва, периневрия, острым скальпелем или иглой
в том месте, где должно производиться изолирование одного волокна. Оболочку
сначала снимают с одной стороны и затем перерезают поперек давлением иглы. Процесс
удаления оболочки очень существенен: он технически очень труден и требует
тренировки. Обнаженный нервный ствол перерезают с обеих сторон, оставляя только
небольшой (неповрежденный пучок посредине. Пучок расщепляют иглой, и
отдельные волокна из этого пучка подвергают раздражению прикладыванием
гальванического пинцета (рис. 88-Л). Если волокно двигательное и не повреждено, то разт
дражение даст сокращение мышцы. Бели такое волокно найдено, тогда все
остальные волокна перерезают. Операция происходит под микроскопом при небольшом
увеличении: до 50—100 раз. Но при некоторых навыках можно этого добиться и
без микроскопа. Микроокоп употребляется только для контроля. Препарат с
изолированным нервным волокном очень нежен, но может долго сохраняться после
операции в холодном месте (леднике). Нерасщепленный конец используется для
раздражения обычными платиновыми электродами. Коленный сустав фиксируется.
.Мышцу соединяют с миографом, и сокращение записывают на кимографе.
Опыт заключается в нахождении порогов раздражения отдельными
индукционными ударами и в записывании пороговых эффектов. Затем идет запись одиночных
сокращений при разных силах раздражения. Во всех случаях высота сокращений
будет одна и та же, как на рис. 176.
7. Наблюдение сверхнормальной возбудимости и одиночного тетанизированного
сокращения. Нервно-мышечный препарат лягушки располагают во влажной камере.
На нерв накладывают две пары электродов: одну пару близко к мышце, другую
на расстоянии 2 см от первой. Через близкую пару раздражают подпороговым те-
таническим током, а через дальнюю — одиночным индукционным ударом
максимальной силы. Сокращение записывают на медленно вращающемся кимографе. Сначала
испытывают то и другое раздражение в отдельности, а затем вместе. При
комбинации отдельный индукционный удар будет вызывать вместо одиночного
сокращения короткое тетаническое сокращение. Это будет указывать, что подпороговое
тетаническое раздражение стало пороговым вследствие повышения возбудимости от
возбуждения, вызванного отдельным индукционным ударом.
Литератур а
і. В о р о н ц о в Д., Изв. Петрогр. биол. лабор., 15, 45, 1915.
2. Garten S., Z. Biol., 52, 534, 1909.
3. Воронцов Д., Труды общ. естествоисп. в Петербурге, 43, кн. б, стр. 119, 1913
Учен. зап. Каз. унив., Физиология, кн. 2—3, вып. 1—3, стр. 56, 1932.
4. Tigerstedt К, Z. Biol., 58, 451, 1912.
5. Erlanger J., Gasser H. S. u. Bishop G. H., Am. J. Physiol., 81, 473,
1927,
6. Ros'enberg H., Pfl. Arch., 216, 300, 1927.
; 7. G a s s e r H. S. u. E r 1 a n g e r J., Am. J. Physiol., 80, 522, 1927.
8. E r 1 a n g e r J., Analysis of the action potential in nerve. The Harvey Lecture, ser.
XXII, 1926—1927.
9. К fl h n e W., Z. Biol., 22, 305, 1886.
10. Babuchin, Arch. Anat. Physiol., 1877, S. 262.
11. Sherrington С S. a. F г б 1 i с h F., J. Physiol, 28, 14, 1902.
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЕ В НЕРВНОМ СТВОЛЕ 389
12. Helmholtz Н. v., Monatsber. Berl. Akad., S. 14, 1850; Konigsberger naturw.
Abh., 2, 166, 1851; Arch. Anat, Physiol, wiss. Med., S. 71, 276, 1850; S. 199, 1852-
13. Helmholtz H. v. u. Baxt N., Monatsb. Berl. Akad., S. 228, 1867; S. 184'
1870.
14. Piper H., Elektrophysiologie menschl. Muskeln, Berlin, 1912.
15. Miirnich, Z. Biol., 66, 1, 1915.
16. Du Bois Reymond R., Arch. Anat. Physiol., Suppl., S. 68, 1900.
17. Be the A., Anat. Anz., 37, 129, 1910.
18. В о e к e J., Erg. Physiol., 19, 448, 1924.
19. В eth e A., Pfl. Arch., 122, 1, 1908.
20. G о t с h Fr. u. В u г с h G., J. Physiol., 24, 418, 1899.
21. Hanstrom В., Vergl. Anat. d. Nervensystems d. wirbellosen Tiere etc., Berlin,
1928.
22. Segaar J., Z. vergl.'PhysioI., 10, 120, 1930.
23. Buddenbrock W., Vergleich. Physiol, d. Nervensystems d. Wirbellosen, Handb.
norm. path. Physiol., 9, 8U5, 1929.
24. Bozler, Z. vergl. Physiol., 4, 70, 1926.
25. Введенский H. E., Тр. физиол. лаб. Петерб. унив., 3, 134, 1908.
26. Beritoff J., Z. Biol., 62, 125, 1913.
27. F о r b e s A., Ray a. G r i f f i t h F. В., Am. J. Physiol., 66, 553, 1923.
28. В e r i t о f f J., Z. Biol., 78, 231, 1923.
29. Б e p и t о d И., Русск. физиол. журн,, 1,1, 1917.
30. К e i t h Lucas, The conduction of the nervous impulse, London, 1, 1917.
31. Gasser H. S. a. Erl anger J., Am. J. Physio!., 73, 613, 1915.
'№. В e t о x и h И., Русск. физиол. журн., 12, 85, 1929.
33. Beritoff J., Erg. Physiol., 23, 33, 1924.
34. Barnes Т. С, Am. J. Physiol, 107, 447, 1934.
35. Woronzow D.S, Pfl. Arch., 218, 148, 716, 1927; 221, 725, 1928; 224, 490, 1920;
Журн. эксп. биол. мед., № 16, 101, 1927.
35a. Gerard R. W., Am. J. Physiol., 93, 337, 1930.
36. Amber son W. a. Dawning A., J. Physiol., 68, 19, 1929.
37. G a s s e r H. a. ErlangerJ., Am. J. Physiol., 94, 247, 1930.
38. Воронцов Д. и Шерешевский Л., Учен. зап. Каз. унив., вып. 1—2,
стр. 68, 1932.
39. Воронцов Д. и Юденич Н., Сб. работ Каз. мед. инст., IX—X»
стр. 108, 1933.
40. Beritoff J., Z. Biol., 85, 521, 1927.
41. В e r i t о f f J. u. Woronzow D., Z. Biol., 84, 417, 1926.
42. Lewin A., J. Physiol., 63, 113, 1927.
43. Э д p и a h E. Д., Учен. зап. Каз. унив., вып. 1—2, стр. 2, 1932.
44. Е г 1 a n g е г J. a. Gasser Н., Am. J. Physiol., 92, 43, 1930.
45. Е с с 1 е s J. С. a. Sherrington С. S., Proc. Roy. Soc, В. 107, 512. 193!-
46. В в е д е н с к и й Н. Е., О соотношении между раздражением и возбуждением при
тетанусе, П., 1886.
47. Kisseleff M., Pfl. Arch., 233, 469, 1933.
48. Keith Lucas, Proc. Roy. Soc, B, 85, 501, 1916.
49. К a t о G e n i с h i, The microphysiology of nerve, 1934.
50. G a s s e r H. S., Юбил. сб., посвящ. проф. И. Бериташвмли. Изд. Груз. филиала
АН СССР, 1936.
51. Воронцов Д. С, Научн. изв. Смоленск, унив., т. III, вып. II—Медицина,
стр. 46, 1926.
52. Иордан Г., Физиол. журн. СССР, 19, 141, 1935; Jordan u. bullies,
Z. vergl. Physiol., 19, 648, 1933.
53. Макаров П. О., Арх. биол. наук, 38, вып. 1, стр. 95, 1935; там же, вып. 3, стр.
683, 1935.
390 общая физиология цереброспинальной периферической нервной системы
54. Магницкий А. И. и Муже ев В., Pfl. Arch., 226, 1, 1933; 232, 604, 1933.
55. L u 1 1 і е s H., Pfl. Arch., 233, 584, 1933.
56. Кан И. Л., Сб. докл. VIBcec. физиол. съезда, стр. 82, 1937; Бюлл. эксп. биол.
мед., 6, № 1, 1938; Кан И. и Кузнецов Д., Бюлл. эксп. биол, мед., 6,
13, 1938.
57. S і m о п а г t A. a. Simonart Е., Arch, intern, pharm. ther., 49, 302, 1935-
58. E г 1 a n g e r J. a. Gasser H. S., Electr. signs of nerv. activity, 1937.
59. Graham H. t, Am. J. Physiol., 110, 225, 1934; 111, 456, 1935.
60. В ас q Z. M., Proc. Roy. Soc, B. 123, 418, 1937.
61. Eccles J. C, Proc. Roy. Soc, B. 123, 412, 1937.
62. К и б я к о в А., Сб. докл. VI Всес. физиол. съезда, стр. 198, 1937.
63. Гинецинский А. Г., Сб. докл. VI Всес. физиол. съезда, стр. 193, 1937.
64. Караев А., Бюлл. эксп. биол. мед., 6, № 1, 1938.
64а. Pant in С. F. A., Proc, Roy. Soc, В. 123, 397, 1937.
65. В и с h t h a 1 Fr., Proc. Roy. Soc, B. 123, 404, 1937.
66. Mar nay A. a. N а с h m a n s о h n D., C. r. Soc. biol., Paris, 124, 942, 1937.
67. Gi ldeme ister M., Pfl. Arch., 124, 447, 1908.
68. Кан И. Л., Журн. эксп. биол. мед., 4, № Ю—11, стр. 45, 1926.
69. Леонтович А., С. г. Acad. Sci., 127, 908, 1928; Биол. журн., 2, №213, 1932;
Физиол. журн. СССР, 22, 377, 1937.
70. Brown G. L., J. Physiol., 89, 459, 1937.
71. М а г п а у A. a. Nachmansohn D., J. Physiol., 89, 359, 1937.
72. В а с q Z. М. a. Nachmansohn D., J. Physiol., 89, 368, 1937.
73. Hodgkin A. L., J. Physiol., 90, 183, 211, 1937.
74. Rush ton W. A. H., Proc. Roy. Soc, B. 124, 210, 1937.
75. Katz В., Proc. Roy. Soc, B. 124, 294, 1937.
.76. Rosenberg H., Proc. Roy. Soc, B. 124, 308, 1937.
77. G 6 p f e r t H. u. S с h a e f e r H., Pfl. Arch., 239, 597, 1937.
78. В a g h n a u I t E., C. r. Soc. Biol., Paris, 125, 867, 1937.
79. Gasser H. S., Am. J. Physiol., 121, 193, 1938.
80. Bogue J. G. a. Rosenberg H., J. Physiol., 87, 158, 1936.
81. Katz В., J. Physiol., 87, 199, 1936.
82. S с h r i e v e r H. u. С e b u 1 1 a R., Pfl. Arch., 241, 1, 1938.
^3. P a n t i n С F. D., J. Exp. Biol., 13, 111, 1936.
84. H odgkin A. L., Proc Roy. Soc, B. 126, 87, 1938.
85. В 1 a i r E. A. a. E r 1 a n g e r J., Am. J. Physiol., 126, 97, 1939.
86. Zotterman Y., Skand. Arch. Physiol., 77, 123, 1937.
87. Graham H. T. a. Lorente deNo R., Am. J. Physiol., 123, 326, 1938.
88. Hodgkin A. L., J. Physiol., 94, 560, 1939.
89. Lapicque L., Cold Spr. Harb. Symp. Quant. Biol., 4, 98, 1936.
90. Kato G., Cold Spr. Harb. Symp. Quant. Biol., 4, 202, 1936.
91. Hermann L., Handb. Physiol., Bd. 2, T. 1, 194, 1879.
92. Cremer M., Nagel's Handb. Physiol., 4, 793, 1909.
93. Li Hie R., J. Gen. Physiol., 7, 473, 1925.
94. M e п и с а ш в и л и И., Сообщ. Ак. наук Гр. ССР, 1, № 8, 1940.
95. Квасов Д. Г. и Науменко А. И., Физиол. журн. СССР, 19, 1914, 1935;
Уч. зап. Ленингр. унив., сер. биол., № 10, 125, 1939.
96. М іі 1 I er J., Handb. Physiol., S. 1534, 4. Aufl., 1844.
97. Gasser H. S., J. Neurophysiol., 2, 361, 1939.
98. L e h m a n n I. E., Am. J. Physiol., 118, 600, 1937.
99. Та s a k i I с h i j i, Am. J. PhysioL, 127, 211, 1939.
100. Grundfest H., Am. J. Physiol., 127, 252, 1939.
101. Gasser H. a. Grundfest H., Am. J. Physiol., 127, 343, 1939.
102. Hursh J. В., Am. J. Physiol., 127, 131, 1939.
103. Hursh J. В., Am. J. Physiol., 127, 140, 1939.
ВОЗБУЖДЕНИЕ И ЕГО ПРОВЕДЕНИЙ В НЕРВНОМ СТВОЛ і 391
104. Werigo Br., Pfl. Arch., 76, 552, 1899.
105. Жуков E. К., Уч. зап. Ленингр. унив., сер. биол. н., № 10, стр. 99, 1939.
106. Скадовская Н. С, Арх. анат., гистол. и эмбриол., 16, 468, 1939.
107. Blair Е. A. a. Erl anger J., Am. J. Physiol., 106, 524, 1933.
1Э8. G a s s e r H. S., J. Appl. Physics, 9, № 2, p. 88, 1938; S с h m i 11 F. D. a. Gas-
ser H.,Am. J. Physiol., 104, 320, 1933.
109. Boycott A. E., J. Physiol., 30, ,371, 1904.
110. Netter H., Pfl. Arch., 215, 373, 1927.
111. Та kah as hi K, J. сотр. Neurol, a. Psychol., 18, 166, 1908..
112. Bremer Fr., Am. J. Physiol., 112, 41, 1935.
113. Verzar F., Verhandl. d. Vereins d. schweizer Physiol., 16 Tag. in Basel, 1940#
114. Ponder E., J. Physiol., 87, 67 P, 1936.
llo. Bishop G. H.. Выступление в прениях по докладу Amberson, Cold Spr.
Harb. Symp. Quant. Biol., 4, 53, 1936.
116. Макаров П., Физиол. журн. СССР, 28, 36, 1940.
117. Эдриан С. Д., Механизм нервной деятельности. Перев., Биомедгиз, 1935.
118. Tasaki I., Pfl. Arch., 244, 125, 1940.
119. К г і j g s m а п| В. I., Nederl. Akad. van Wetenschappen, 43, № 8, 3, 1940.
120. F r 6 h 1 i с h F. W., Z. allg. Physiol., 10, 418, 1910; 11, 121 и 141, 1910.
121. F e n n W. O., Cobb D. M.( Hegnauer A. H. a. Marsh B. S., Am. J.
Physiol., 110, 74, 1934.
122. Cowan S. L., Proc. Roy. Soc, B. 115, 216, 1934.
123. F e n n W. O. a. Cobb D. M., Am. J. Physiol., 129, 245, 1940.
124. Be the A., Pfl. Arch., 244, 43, 1940.
125. Jasper H. H. a. M о n n i e r A. M., J. Cellur. Сотр. Physiol., 11, 259, 1938.
126. Беритов И., Тр. Тбилис. гос. унив. им. Сталина, т. XXVII а, стр. 29, 1945.
127. Granit R., Leksell H. a. Skoglund С. R., Brain, 67, 125, 1944.
I л а, в а IV
УТОМЛЕНИЕ НЕРВА
Общие условия для утомления нерва. Если нерв, находящийся в
неблагоприятных условиях, возбуждается быстро повторяемым
раздражением, то он, подобно мышце, теряет способность к возбуждению,
т. е. утомляется. Но при более или менее благоприятных условиях
трудно утомить нерв до полной потери возбудимости. Введенский-
(1), например, показал, что изолированный нерв лягушки в
атмосфере воздуха может быть раздражаем в течение 8 часов без утомления.
Но если условия обмена значительно ухудшены, например, если нерв-
поместить в атмосферу азота и исключить возможность
проникновения кислорода снаружи, тогда, как показал Байер (2), нерв
утомляется при тетаническом раздражении в течение 3—5 часов. Это
утомление проходит полностью, если нерв перевести на 5—10 сек.
в атмосферу кислорода, что указывает на исчерпание запаса кислорода
как на причину утомления. При высокой температуре утомление-
в атмосфере азота наступает скорее.
Обмей веществ в нерве, несомненно, продолжается и при
отсутствии возбуждения. Поэтому изолированный нерв теряет возбудимость
и при отсутствии возбуждения, если его поместить в атмосферу азота.
Т э р н е р (3) показал, что если в атмосфере азота находятся два-
нерва, из которых один беспрерывно раздражается, а другой только по*
временам для пробы, то признаки ослабления нервного возбуждения'
обнаруживаются на обоих нервах, но на непрерывно раздражаемом.
это наблюдается раньше, чем на другом.
Относительное утомление нерва. В последнее время стало известно,,
что признаки утомления в нерве обнаруживаются уже в первые
минуты тетанического раздражения (Беритов, 4) и даже в первую секунду
(Весци, 5; Форбс и Райе, 6). Для иллюстрации приводится рис. 187,
где дается электрограмма от начала нервной деятельности и затем в
разные стадии утомления. Высота биоэлектрических токов возбуждения
постепенно ослабевает еще в течение первой секунды раздражения.
Судя по токам возбуждения на рис. 187 В и С, значительное
утомление наступает в течение первых 3—11 минут раздражения. Это прояв-
УТОМЛЕНИЕ ННРВЛ 39'}
ляется, с одной стороны, в уменьшении высоты токов возбуждения, а с
другой — в удлинении рефракторной фазы, т. е. замедлении процесса
восстановления. Как показывают электрограммы, при этом происходит
изменение ритма возбуждения, который становится двойным.
Последнее обстоятельство указывает на сильное ухудшение процессов вое-
Рис. 187. Утомление нерва.
Седалищный нерв лягушки. Частота раздражения 150. Сила раздражения 30 см,
порог 38 см. Электрограмма А дает эффект в начале длительного раздражения,
В — спустя 3 минуты, когда полный ритм стал двойным, а С—-спустя 11 минут,
когда этот двойной ритм обратно переходит в полный, а интенсивность токов
возбуждения становится в 3 раза слабее, чем было вначале. Время-—0.2 сек.
становления возбудимой системы. Здесь каждое раздражение следует
через 0.007 сек., и этот интервал оказывается недостаточным для
такого восстановления возбудимости, чтобы данное раздражение вызвало
возбуждение всех волокон нервного ствола. Таким образом, нервное
волокно обнаруживает способность к утомлению со всеми
характерными изменениями процессов возбуждения и восстановления
возбудимой системы.
Можно было думать, что уменьшение биоэлектрических токов
возбуждения при длительном раздражении обусловливается локальным
повреждением некоторых нервных волокон в раздраженном участке.
Но новейшие опыты показали, что при длительном раздражении одного
394 ОБЩАЯ ФИЗИОЛОГИЯ Ц'іРКБРОСПИНАЛЬНОЙ ПКРИФКРИЧКСКОЙ ПКРВИОЙ СИСТКМЫ
участка биоэлектрические токи оказываются ослабленными и в том
случае, если заменить раздражение этого участка подобным же
раздражением другого участка, конечно при том условии, если сила
раздражения не чрезмерно велика. Очевидно, ослабление
биоэлектрического тока, а значит возбуждения, обусловлено утомлением нерва по
всей длине распространения возбуждения (Форбс и Райе, 6).
В связи с утомлением нервных волокон наблюдается удлинение
рефракторной фазы. Об этом можно заключить прежде всего из факта
превращения обычного полного ритма биоэлектрических токов в
двойной (рис. 187) (Беритов, 4). Удлинение рефракторной фазы вообще
хорошо проявляется при смене частого утомляющего раздражения —
Рис. 188. Утомление п. pnrenicus кошю. Нерв помещен в атмосфере 5% С02 и 95%О.»
при іс 37.5° С
Токи "возбуждения регистрируются катодно-лучевым осциллографом. Наверху цифры означают время
в Секундах от начала раздражения, внизу — время в сигмах. Раздражение происходит по ритму 107о
в % сек. Токи возбуждения следуют этому ритму. Второй ток возбуждения несколько слабее первого;
это указывает, что уже второе возбуждение приходится ца время рефракторной фазы (Гассер).
200 в секунду раздражением частоты 16—35 в секунду. В последнем
случае токи возбуждения оказываются ослабленными сравнительно с
тем, что было до утомляющего раздражения. Это указывает на
удлинение всей рефракторной фазы (абсолютная+* относительная) до 30 о
и более (Форбс и Райе, 6).
Как указывалось, при ухудшении обмена веществ утомление нерва
от тетанического раздражения происходит много быстрее. Но и в этом
¦случае происходит такое же изменение возбуждения и рефракторных
фаз, как в нормальном нерве. Сначала происходит уменьшение
интенсивности возбуждения, затем ритм возбуждения становится двойным.
Точно так же происходит удлинение процесса возбуждения и
рефракторных фаз. Это, например, хорошо видно на рис. 188, где нерв
теплокровного животного (п. phrenicus) в условиях избытка С02 утомился от
тетанического раздражения в значительной степени в течение
полуминуты.
Специальными опытами было установлено, что это удлинение
происходит главным образом за счет относительной рефракторной фазы.
Абсолютная рефракторная фаза также удлиняется, но значительно
меньше. По исследованиям Воронцова, она увеличивается всего в
і1і2—3 раза. Так, в одном опыте продолжительность абсолютной рефрак-
УТОМЛЕНИЕ НЕРВА
395
торной фазы до утомления равнялась 1.8 а (седалищный нерв лягушки);
после тетанизации (по 100 индукционных ударов в секунду) в течение
3 час. 45 мин. это время возросло до 2.86 а ; в другом опыте до
утомления она равнялась 1.26 а, а после 2.5 час. тетанизации она
увеличилась до 3.4 о. По прекращении раздражения продолжительность
абсолютной рефракторной фазы сравнительно быстро (около 30 мин.)
возвращалась к норме (Воронцов, 23).
С утомлением нерва происходит также замедление распространения
возбуждения. Уже в течение первого часа скорость распространения
уменьшается в два-три раза. При дальнейшем утомлении она меняется
мало (Воронцов, 26).
Опыты с утомлением на нерве беспозвоночных, наоборот, показывают
их быструю утомляемость. Например, совершенно свежие нервы
клешни рака утомляются при фарадическом раздражении в течение 2 мин.
Восстановление происходит в 4—20 мин. Утомление получается сильнее
всего в раздраженном участке, но до известной степени утомляются и
другие участки в силу проведения возбуждения. Об изменении
возбуждения судили на основании токов возбуждения нерва (Левин, 7).
При раздражении кишечного нерва улитки (Helix pomatia)
отдельными индукционными ударами через каждую минуту утомление в нем
наступает через час (Крийгсман, 37). Но если импульсы возбуждения
следуют через большие интервалы, утомление, повидимому, не
наступает и в нервных волокнах беспозвоночных. Так, например, нервная
система медузы, как показал Ветохин (8), не утомляется в течение
нескольких часов, если одна волна возбуждения следует спустя 4—5 сек.
после другой.
Обмен веществ в нерве во время возбуждения и при покое. Мы
указали, что при условиях, неблагоприятных для обмена веществ, в
нерве наступает, как и в мышце, абсолютное утомление. При благоприятных
же условиях можно наблюдать только относительное утомление в са-
мом начале раздражения. Нервная деятельность сопровождается
определенными физико-химическими процессами, которые с давних пор
являлись предметом исследования. Прежде всего было установлено,
что нерв все время потребляет кислород и выделяет углекислоту (Тун-
берг, 9). По новейшим исследованиям, нервы лягушки R. esculenta
зимой потребляют 11—21 мм3 кислорода на грамм веса нерва в течение
1 часа, а нервы R. temporaria — 17—27 м>м 3. Весной потребление
кислорода больше; так, например, нервы R. esculenta потребляют 21—28 мм3
кислорода. Выделение углекислоты у зимних R. esculenta равно 12 мм3, а у
R. temporaria— 16 мм3 на 1 г веса за 1 час (Жерард, 10). В общем
оказывается, что нерв во время основного биологического процесса
потребляет кислород.так же, как мышца. При возбуждении многие авторы
отметили заметное увеличение потребления кислорода и выделения
углекислоты (Габерландт, 14; Тайтиро, 1'5, и др.). Новейшее
исследования ГиллаиМейергофа (16) с сотрудниками подтверждают
точными измерениями это первоначальное положение. Так, по Ж е р а р д у,
396 ОБЩАЯ ФЙЗИОЯОГЙЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
во время длительного фарадического раздражения нерва лягушки,
потребление кислорода в среднем повышается на 100%.
Обмен.веществ в нерве сопровождается выделением аммиака.
Образование аммиака связано с расщеплением азотистых соединений.
Поэтому нужно предположить, что он образуется в связи с основным
биологическим процессом возбудимой системы. По Винтерштейну
(29), в нерве лягушки за час возникает 83.1 X 10 ~7 мг NH3. Под.
влиянием наркоза выделение аммиака уменьшается до границы
изменяемости. В нормальном организме при ненарушенной связи со
спинным мозгом образуется больше аммиака, чем при нарушенной,
например, когда нерв перерезан. Так, седалищный нерв кролика, когда он
связан с мозгом, образует 13ХЮ~амг NH3 за час времени на 1 г
веса. Когда нерв отделен, образование аммиака падает до 4—5 X
[X Ю ~у мг (Гальтер, 30). Этот факт указывает, что при возбуждении
нерва со стороны центральной нервной системы образование аммиака
увеличивается. Зависимость образования аммиака от возбуждения была
доказана и прямым путем: раздражение нерва увеличивает продукцию-
аммиака почти втрое (Винтерштейн, 29; Гальтер, 30).
Во время основного биологического процесса происходит также
потребление глюкозы. Если положить пучок нервов в раствор Рингера,
содержащий глюкозу, то этот «свободный сахар» исчезает в растворе-
с течением времени. Глюкоза превращается в гексозофосфат. Этот
процесс протекает без участия кислорода, так как сахара исчезает больше,,
чем это 'могло быть вызвано окислением его за то же время. При-
анаэробном дыхании глюкоза превращается в молочную кислоту
(Винтерштейн, 12; Хольмс, Жерард и Соломон, 13).
Непосредственно после операции выделения нерв потребляет больше-
сахара, чем спустя некоторое время. Повышение потребления сахара
в начале опыта нужно считать последствием тех возбуждений, какие
испытал нерв при операции выделения.* Электрическое раздражение-
нерва также производит повышение потребления сахара. Это
наблюдается как в раздражаемом участке, так и во всем нерве в связи с
проведением возбуждения. В покойном состоянии при 37° 1 г нерва
потребляет 0.37 мг сахара за час времени. При раздражении это
потребление увеличивается на 100% (Винтерштейн и Френкель-Конрат, 12;
Хольмс, 13).
В нерве беспозвоночных, осуществляющем проведение с
декрементом, обмен веществ характерно отличается от обмена в нерве
позвоночных. Межганглионарная комиссура беззубки потребляет на 1 г веса-
в течение часа при покое (20—21° С) 150—200 мм3 кислорода, т. е..
в 5 раз больше, чем мякотный нерв лягушки при тех же условиях..
При раздражении в оптимальном ритме (120—150 в минуту)
потребление кислорода повышается на 40%. Так же велика разница в
отношении выделения аммиака. В покое на 1 г веса в течение 1 часа (при-
20—25° С) выделяется 0.091 мг — опять-таки в 5 раз больше, чем
в мякотном нерве лягушки. При раздражении в оптимальном ритме
УТОМЛЕНИЕ НЕРВА
397
-аммиака выделяется на 370% больше, чем прн покое (Кан и
сотр., 31).
Теплообразование в нерве. Теплообразование в нерве изучалось
А. В. Гиллом и его сотрудниками. Изучение стало возможным в
результате созданной ими чрезвычайно тонкой техники регистрации
миллионных долей калории. Небольшое количество тепла образуется
в нерве, подобно тому как и в мышце, еще при покое. В мякотном
нерве при 20° в покое образуется тепла приблизительно 41 X 10~3кал.
в минуту на 1 г. В атмосфере азота основной обмен веществ сильна
снижается, но полностью не прекращается. Известно, что в атмосфере
азота или водорода теплопродукция продолжается еще в течение
2—3 часов, и при этом нерв выделяет тепло в размере 1/5—У4 того
количества, которое выделяется в атмосфере кислорода (Березина, 32;
Жерард, 10).
Г и л л и его сотрудники изучили теплообразование в нерве при
разных условиях тетанического раздражения. Они установили, что
теплопродукция нерва при возбуждении усиливается. И эта продукция тепла
протекает в две фазы.- одна во время возбуждения — начальное
теплообразование, а другая после него — замедленное теплообразование.
Продолжительность последней фазы измеряется многими минутами.
Когда по нерву проходит один импульс, то теплообразование
начинается или во время возбуждения или непосредственно после
распространения его. Вслед за этим наступает позднее теплообразование,
которое сперва протекает быстро, а потом постепенно падает,
приблизительно в течение часа при 20°. Количество тепла, выделяемого в той
и другой фазах, ничтожно, но вполне измеримо.
'Максимальная величина начального теплообразования при
одиночном импульсе в імякотном нерве лягушки при 20° равна 7ХЮ~8кал.,
т. е. около 3 эргов на 1 г. При тетаническом раздражении при 20°
начальное теплообразование нерва в кислороде равно 7.5ХЮ"6 кал-
на 1 г, а теплообразование в период восстановления в 15 раз больше
(рис. 189).
По новейшим исследованиям, отношение всего тепла к начальной
фазе зависит от частоты раздражения: оно тем больше, чем реже
раздражение. Так, при частоте 40—50 в секунду оно составляет
6.9—9.2, а при частоте 1.8—6.4 <в секунду оно равняется 21.6—15.2
(Фенг, 33). Нарастание теплопродукции с учащением раздражения
происходит не пропорционально повышению ритма раздражения,
а несколько в меньшей степени. Отсюда следует, что с повышением
ритма возбуждения количество тепла, приходящегося на один импульс,
уменьшается. Когда частота раздражения превышает 600 в секунду,
общая теплопродукция нерва уменьшается, и притом тем больше, чем
чаще раздражение (Бюньяр и Гилл, 36).
Все изменение теплопродукции протекает параллельно с
изменением токов возбуждения. Поэтому можно утверждать, что
теплопродукция нерва связана с биологическим процессом возбуждения.
398 ОЫЦАИ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИС
Л'КМЫ
В анаэробных условиях обе фазы теплопродукции уменьшаются
в одинаковой степени. Это указывает, что, в отличие от мышцы, здесь
обе фазы обусловлены потреблением кислорода. В анаэробных условиях
абсолютная величина теплопродукции быстро падает с течением
времени, уменьшаясь спустя 100 мин. до 7% своей первоначальной
величины, а спустя 3 часа —даже до нуля (Жерард, 10). Потребляемый
'/
tff
I"
12
ST w
^
§
е -
г
о
у
і
/
/
Семцнды
W
го
?6
Рис. 189. Теплообразование нерва лягушки в кислороде во время и
после 12-секундного максимального тетанического раздражения при 20°.
На абсциссе — время в секундах, на ординате — скорость теплообразования»
полученная путем анализа отклонений гальванометра. В этом случае
теплопродукция равнялась 1410 ^Ю-1 кал. на 1 г, причем начальное теплообразование
составляло 90ХЮ—* кал на 1 г. Последнее равняется площади кривой
над пунктирной линией. Остальное тепло образуется при восстановлении,
которое закончилось через 25 минут. Отношение всего тепла к начальному теплу
равняется приблизительно 16 (Гилл, 28).
при этом кислород черпается из каких-то запасов, находящихся в нерве.
Наличие в нем таких запасов видно из того, что при переносе нерва
из анаэробных условий в аэробные он поглощает больше кислорода, чем
это соответствует увеличенному количеству выделяемой угольной
кислоты. Этот излишек кислорода, очевидно, идет на пополнение тех
запасов, которые были затрачены им при анаэробиозе. Но теплопродукция
при анаэробиозе может иметь место и без окислительного процесса,
в результате расщепления гликогена и фосфагена. Эти процессы
в нерве, безусловно, происходят, хотя и в очень слабых размерах
(Березина, 32; Шмит и Кори, 34; Жерард и Топиков, 35).
В нервах беспозвоночных теплообразование происходит гораздо
сильнее, чем в мякотных нервах позвоночных. Так, при 20° начальное
теплообразование в нерве краба при максимальном тетаническом
раздражении составляет около 70 X 10 ~6 кал. в секунду на 1 г, а при
одиночном импульсе — 8.5 X Ю-7 кал. Отношение всего тепла к
начальному значительно больше, чем в нерве позвоночных. Оно состав-
УТОМЛЕНИЕ НЕРВА
899
ляет почти 50. В отношении восстановления или второй фазы нерв
краба более подобен мышце, чем нерву лягушки.
Ход теплообразования в нерве, по Гиллу, характерно отличается
от наблюдаемого в мышце. Во-первых, теплообразование во второй
фазе, т. е. запаздывающее теплообразование, в нерве лягушки в 15 раз,
а в нерве краба в 50 раз больше начального. В мышце оно
больше начального только в 1.5 раза. Во-вторых, все запаздывающее
теплообразование происходит независимо от поступления кислорода
извне, в противоположность мышце, в которой часть запаздывающего
тепла выделяется анаэробно, а часть только в присутствии кислорода.
Предполагается, что в нерве имеется какое-то кислородное соединение,
которое обеспечивает возможность окисления, когда исключен доступ
молекулярного кислорода. Поэтому <в нерве нельзя отделить одну фазу
от другой путем помещения нерва в анаэробные условия. Как
указывалось выше, при 20° нерв лягушки сохраняет возбудимость в
течение 3—5 часов в атмосфере водорода или азота, а при 0° это может
длиться целый день. При истощении внутреннего запаса кислорода
в нерве весь процесс теплообразования прекращается. Однако нельзя
еще сказать, что обе фазы целиком обязаны своим происхождением
окислительным процессам, как это полагал Жерард. Гилл считает
этот вопрос более сложным и требующим дальнейшего исследования
(Гилл, 27, 28).
Источником теплопродукции при раздражении нерва можно было-
считать токи возбуждения, тем более, что теплопродукция в нерве
очень мала. Но Гилл доказал, что на долю токов возбуждения джоу-
лева тепла приходится всего 3.5—10 ~10 кал. на 1 г, т. е. меньше l%fc
начального тепла. Теплопродукция в нерве при раздражении,
безусловно, происходит от химических реакций. Но еще не вполне известно^
какие именно химические реакции участвуют в этой теплопродукции.
Пока доказано, что при раздражении не усиливается распад гликогена.
до молочной кислоты, но наблюдается некоторое увеличение распадам
фосфагена (Фенг, 33).
Возбуждение как окислительный процесс. Винтерштейн (17)
с сотрудниками находит, что положение О'б избыточном потреблении
кислорода при возбуждении верно только для самого раздраженного
участка нерва: здесь только происходит усиленное потребление
кислорода. Но при изучении газового обмена в возбужденном участке нерва
вне раздраженного участка самая тонкая современная микрометодика
не открывала никакого увеличения потребления кислорода или
выделения углекислоты. Отсюда Винтерштейн сделал заключение, что
проведение возбуждения не связано с усилением окислительного
процесса. Еще раньше Паркер не мог найти повышенного обмена веществ
вне раздраженного участка. Повышенный газовый обмен в
раздраженном участке Винтерштейн считает эффектом самого
электрического тока как раздражителя, а не физиологического процесса
возбуждения.
400 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Гилл изучал теплопродукцию нерва вне раздраженного участка,
и все-таки он и здесь нашел повышение ее. Затем, при пользовании
усовершенствованной методикой, было вновь показано, что потребление
02 увеличивается не только в раздраженном участке, но и вне его
(Харашима, 24). Отсюда было сделано заключение об усиленном
потреблении кислорода и при проведении возбуждения. Затем Фенн (19)
іпоказал, что существует параллелизм между биоэлектрическими токами
возбуждения и потреблением кислорода; этим самым была обнаружена
причинная зависимость между ними. Жерард (20) и Мейергоф
(21) произвели ряд других проверочных опытов, но всегда приходили
к заключению, что возбужденный: участок потребляет кислород.
Винтерштейн (18) не соглашается с этим. По его мнению»
в опытах Гилла и Мейергофа электрическое раздражение было
очень сильно, так как они брали по 5—8 нервов. А потому повышенная
теплопродукция или повышенное .потребление кислорода могли быть
обусловлены распространением.петель раздражающего тока или
поляризационного тока от раздраженного участка на исследуемый участок
перва.
Однако противники Винтер штейна отмечают в его опытах
некоторые методические недостатки, которые препятствовали наблюдению
избыточного потребления кислорода при проведении нервных
импульсов. Когда они устранили эти недостатки, избыточное потребление
кислорода при проведении нервного возбуждения оказалось таким же,
как в раздраженном участке (Харашима, 24).
Особенно демонстративны опыты Кана и его сотрудников на меж-
гаиглионарном нерве анодонты и на парабиотизированном нерве
лягушки. Первый опыт дает значительное избыточное потребление
кислорода спустя много минут после раздражения, а парабиотизированный
нерв лягушки, когда не дает возбуждения, то не производит и
избыточного потребления кислорода в раздраженном участке (25). На
основании этих фактов можно считать решенным в положительную
сторону вопрос об избыточном потреблении кислорода во время
возбуждения нерва.
Практикум
Опыт сравнительной неутомляемости нерва. Для наблюдения относительно
меньшей утомляемости нерва по сравнению с мышцей ставят опыт по Бернштейну.
Помещают нервно-мышечный препарат во влажной камере и определяют силу тета-
нического раздражения нерва, дающего на мышцах максимальный тетанус. Затем
в дистальной части нерва (ближе к мышце) пропускают слабый гальванический ток
(рис. 190). Этим вызывают уничтожение проводимости на анодном участке. Теперь
раздражение проксимальной части нерва не будет давать сокращения за
невозможностью 'проведения через анодный участок. Это раздражение продолжают десятки
минут и целые часы. Только по временам размыкается цепь постоянного тока, за
которым следует быстрое восстановление проводимости и наблюдается эффект тета-
нического раздражения на мышце. Опыт может [продолжаться несколько часов
только в том случае, если (постоянный ток не является сильным и тем 'самым не
умерщвляет анодного участка нерва. Во избежание этого сильным током пользу-
УТОМЛЕНИЕ НЕРВА
409
ются только вначале, шока не исчезнет проводимость. Затем сильно ослабляют его
с помощью компенсатора, так как раз вызванная непроводимость хорошо
поддерживается и очень слабым постоянным током. На другом препарате можно испьь
тать скорость утомления мышцы при таком же раздражении нерва в условиях
свободного доступа нервных импульсов к мышце.
Утомление нерва следует изучить с помощью токов 'Возбуждения самого нерва.
Берут седалищный нерів лягушки. Располагают его в хорошо увлажненной камере.
Рис. 190. Схематическое изображение опыта
сравнительной неутомляемости нерва.
Нервно-мышечный препарат. На проксимальную часть
наложены платиновые электроды (аа') для тетанического
раздражения. Средняя часть поляризуется с помощью неполя-
ризующихся глиняных электродов (&&')• Гальванический
ток проводят через эти электроды, ответвляя его с помощью
компенсатора (к).
Один конец нерва раздражают, а другой отводят в гальванометр или осциллограф.
Частота раздражения должна 'быть івысокая—150—200 в секунду. Интенсивность
раздражения (большая — на 10 см выше порогов. (Все время наблюдают за токами
возбуждения на экране или путем 'фотографии через определенные промежутки
времени. Во избежание ослабления нервного эффекта вследствие повреждения в
раздражаемом участке следует перемещать раздражающие электроды через каждые
30—60 минут.
Литература
1. Введенский Н., Телеф. исследование над электрическими явлениями в
мышце и нервном аппарате. 1884.
2. Bayer Н. V., Z. allg. Physiol., 8, 169 u. 180, 1903.
3. Thorner W., Z. allg. Physiol., 8, 530, 1908.
4. В er it off I., Z. Biol., 62, 125, 1913.
5. Veszi J., Z. allg. Physiol., 13, 320, 1912.
6. Forbes A. a. Rice L., Am. J. Physiol., 90, 119, 1929.
7. Lew in, Am. J. Physiol., 63, 113, 1927.
8. Ветохин И., Русск. физиол. журн., 12, 85, 1929.
9. Thunberg Т., Cbl. Physiol., 18, 553, 1904.
10. Gerard R. W., J. Physiol., 63, 280, 1927.
11. Gerard R. W., Am. J. Physiol., 82, 381, 1927.
12. Winter stein H. u. Frankel-Conrat, Bioch. Z., 247, 178, 132s
26 и. С. Беритов
402 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
13. Holmes Е. G., Gerard R. W. a. Solomon Е. I., Am. J. Physiol., 93„
342, 1930.
14. Haberlandt L., Arch. Anat. Physiol., S. 419, 1911.
15. Tashiro S.,J. Physiol., 32, 107, 1913.
16. Meyerhof O., Klin. Wochenschr., No 1, S. 1—10, 1929.
17. Winter stein H., Pfi. Arch., 224, 749, 1930.
18. Winter stein H., Bioch. Z., 232, 196, 1931.
19. Fen n W. O., J. Gen. Physiol., 10, 767, 1927.
20. G e r a r d a. Chang, Science, 72, 196, 1930.
21. Meyerhof O. u. Schulz, Bioch. Z., 228, 1, 1930.
22. H i 1 1 A. V., Proc. Roy. Soc, B. Ill, 106, 1932.
23. Воронцов Д., Учен. зап. Каз. унив., кн. 2—3, вып. 1—2, стр. 56, 1932.
24. Harashima S., J. Сотр. Physiol., 3, 419, 1933.
25. ГСан И. Л., Успехи coop, бнол., 3, 637, 1934.
26. Woronzow D., Pfl. Arch., 235, 96, 1934.
2Y. Н і I 1 А. V., Adventures in Biophysics, 1931. Русск. перевод: Эпизоды из области
биофизики, Биомедгиз, 1935.
28. Н і 11 А. V., Chemical Wave Transmission in Nerve, 1932.
29. Win ter stein H., Biochem. Z., 156, 138, 1925.
30. Halter K., Biochem. Z., 257, 313, 1933.
31. Кан И. Л., Сб. докл. VI Всес. физиол. съезда, стр. 82, 1937.
32. Beresina М., J. Physiol., 76, 170, 1932.
33. Feng Т. P., Erg. Physiol., 38, 73, 1936.
34. S с h m і 11 F. О. а. С о r i G. F., Am. J. Physiol., 106, 339, 1933.
35. Gerard a. T о p i k о w, Proc. Soc. Exp. Biol. Med., 27, 360, 1930.
3G. Bugnard L. a. Hill A., J. Physiol., 83, 383, 1936.
37. Krijgsman B. J., Z. vergl. Physiol., 28, 286, 1940.
Г л а в а V
ОБЩАЯ ФИЗИОЛОГИЯ К0>НЦЕВОИ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ
В СКЕЛЕТНОЙ МЫШЦЕ
Характеристика нервных участков скелетной мышцы. Тот небольшой
участок мышечного волокна, где находится окончание двигательного
нерва, так называемый нервный участок, как уже указывалось выше,
обладает рядом морфологических особенностей. Здесь имеется
скопление своеобразной цитоплазмы, которая включает по нескольку ядер;
здесь нейрофибриллярное разветвление нервного окончания приходит
в непосредственную связь с возбудимой системой этой цитоплазмы
(рис. 35). Таких специфических нервно-мышечных участков в
коротких мышечных волокнах длиной до 10—20 мм содержится не более чем
по одному на каждое. Но в длинных волокнах имеется по нескольку
таких участков. В скелетных мышцах лягушки они располагаются на
расстоянии около 10 мм друг от друга (Лежава, I). Нервное
окончание со всеми разветвлениями и связанное с ним саркоплазматическое
вещество со своими ядрами образуют более или менее обособленную
структуру, которая именуется концевым двигательным органом или
концевой двигательной пластинкой.
Нервные участки мышечного волокна, как указывалось выше,
отличаются от безнервных участков также и в физиологическом отношении.
Возбудимость в нервных участках значительно выше, чем в безнервных
участках. Соответственно хронаксия в нервных участках значительно
короче, чем в безнервных участках. Нервные участки характеризуются
более интенсивным биологическим процессом, чем безнервные участки,
и сообразно более высокой возбудимостью. Они развивают более
сильный и короткий процесс возбуждения по сравнению с безнервными
участками. Соответственно и биоэлектрический ток возбуждения в
нервном участке в несколько раз сильнее, чем в безнервном.
Сократительный процесс в нервном участке также в два-три раза сильнее, чем-
в безнервном. Все ферментативные процессы анализа и синтеза, обу^
словливающие сокращение, протекают в нервных участках гораздо
интенсивнее и быстрее по сравнению с безнервными.
Все эти характерные физиологические особенности нервного участка-
26*
404 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
следует рассматривать как особенности самой возбудимой системы
мышцы или саркоплазматического образования в области концевой
пластинки, ибо все они обнаруживаются и после того, как концевые
пластинки перестают возбуждаться через нервные окончания, как это,
например, бывает после кураризации или воздействия ионов магния.
Возбуждение концевых двигательных пластинок. Двигательная
пластинка, представляющая собой своеобразную модификацию возбудимой
системы нерва и мышцы, отвечает на раздражение по закону
возбуждения. Это лучше всего видно на изолированной нервно-мышечной
единице. При раздражении нервного волокна связанные с ним мышечные
волокна івсегда отвечают максимально. Все условия, повышающие или
понижающие функциональное состояние двигательной пластинки,
меняют ее физиологическое действие на мышцу, но передача возбуждения
всегда будет максимального характера: мышечное волокно будет
сокращаться одинаково при всех силах раздражения нерва. Точно
так же при химическом раздражении двигательной пластинки, например,
ацетилхолином, мышечное волокно отвечает рядом сокращений
одинаковой высоты (С т е й м а н, 52).
Возбуждение двигательной пластинки, как всякого другого
возбудимого образования, сопровождается колебанием электрического
потенциала. Этот биоток возбуждения обычно сливается с биотоком
возбуждения мышцы и потому не всегда удается заметить его при
регистрации электрических явлений мышцы. Но при определенных условиях
биоток концевой двигательной пластинки выступает с большой
отчетливостью. Это, например, бывает в m. sartorius лягушки в том случае,
если при непрямом раздражении биотоки отводятся точно от нервного
участка мышцы, где располагается большое количество нервных
окончаний. Тогда кривая мышечного биотока имеет на восходящем колене
перелом. До перелома кривая биотока представляет восходящее колено
биотока двигательной пластинки, а после перелома—-ток возбуждения
ближайшего участка мышцы. Вследствие этого с удалением отводящих
электродов от нервного участка эта начальная часть биотока уменьшается
и укорачивается. Она совершенно исчезает на расстоянии 1—2 мм от
него. Электродвигательная сила биотока двигательной пластинки на
свежих мышцах —0,7—1,4 mV. Ввиду побочного замыкания этого тока
через соседние мышечные волокна действительная величина ее
несомненно больше (Шефер и Хаас, 47).
У теплокровных животных при нормальном состоянии мышцы биоток
возбуждения двигательной пластинки обычно не выявляется. Так, по
опытам Э к к л с а и сотрудников (48 и 49) известно, что в m. soleus
кошюи этот биоток в редких случаях предшествует мышечному току
возбуждения в виде более или менее значительной предварительной
ступеньки высотой до 12—15% мышечного биотока. Обычно биоток
двигательной пластинки значительно слабее — до 2% высоты мышечного
биотока — или даже отсутствует.
Биоток двигательной пластинки, а значит и процесс возбуждения
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 405
ее может быть прослежен от начала до конца, если раздражать
двигательный нерв два раза с интервалом 0,8—2 а. В этом случае, благодаря
попаданию второго раздражения во время рефракторной фазы самой
мышцы от первого раздражения, мышца не будет реагировать вновь
возбуждением. Поэтому ток возбуждения двигательной пластинки имеет
возможность проявиться. Он в общем в 10 раз слабее мышечного тока
и продолжается 4—5 а (Экклс и О'К о н н о р, 32). Значит, процесс
возбуждения двигательной пластинки лягушки продолжается не больше
4—5 а. Но, безусловно, эта продолжительность тока возбуждения
двигательной пластинки должна 'быть намного больше норімального процесса
возбуждения, ибо он был вызван в двигательной пластинке во время ее
относительной рефракторной фазы, когда вообще процесс возбуждения
протекает более или менее замедленно.
На кураризованных мышцах лягушки легко изучить возбуждение
двигательных пластинок. В то время, когда мышца перестает
реагировать сокращением на непрямое раздражение, она производит очень
слабые биотоки в нервной области с продолжительностью до 20 а (Э к к л с
и О'К о н н о р, 37; Г и н е ц и н с к и й, 9). То же самое наблюдается на
кураризованной мышце кошки (Экклс и сотрудники, 48 и 49). Кура-
ризация, безусловно, ухудшает функциональное состояние двигательных
пластинок, производя ослабление и сильное удлинение процесса
возбуждения, в особенности его восстановительного периода. Видимо1, этим
объясняются малая интенсивность и большая продолжительность
(до 20 а) наблюдаемого на кураризованной мышце биотока
двигательных концевых пластинок. При слабой кураризации электрический
эффект начинается быстрым колебанием, как в норме, но сильно
удлиняется его нисходящее колено. Однако и в этом случае биоток
отводится только от нервной области (рис. 191). (Экклс и О'К о н н о р, 32).
Продолжительность процессов возбуждения и рефракторных фаз
двигательной пластинки можно определить также путем изучения
механического эффекта на нервно-мышечном препарате. Раздражая
двигательный нерв двумя следующими друг за другом индукционными
ударами и регистрируя механический эффект мышцы, можно
определить тот минимальный промежуток времени, когда второй удар не
влияет на механический эффект. Этот интервал времени обычно
значительно превышает абсолютную рефракторную фазу нерва. У взрослых
собак, например, он составляет 1.5—2а. Относительная рефракторная
фаза в этих же условиях также длится значительно дольше: 6—8 а.
С некоторой вероятностью эту абсолютную рефракторную фазу
можно отнести на счет мышечной возбудимой системы двигательных:
пластинок. *
Продолжительность рефракторных фаз гораздо больше у щенков;
в ранние стадии постэмбрионального развития; например,впервые дни'
после рождения абсолютная фаза составляет 6—8 и, а относительная —
40—60 j. С возрастом продолжительность той и другой фаз
уменьшается. Норма в отношении рефракторных фаз у взрослых животных
406 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
устанавливается через 1.5—2 месяца. Но наиболее резко, почти
скачкообразно, укорочение рефракторных фаз происходит на 12—14-й день,
тотчас после прозревания животных (Аршавский, 35). Так как
абсолютная рефракторная фаза лишь немного превышает продолжительность
процесса возбуждения, мы должны принять, что в двигательных
пластинках взрослых теплокровных животных процесс возбуждения
продолжается около 1.5—2 j, а в раннем
постэмбриональном периоде он гораздо
длительнее, как этого и следовало ожидать с
точки зрения эмбриогенеза функциональных
особенностей нервно-мышечного аппарата.
Утомление двигательных пластинок.
Нервный участок мышцы обладает также
такими свойствами, которые нужно отнести
исключительно к окончаниям двигательного
нерва. Прежде всего следует отметить
быструю утомляемость нервных участков при
непрямом раздражении. Как указывалось
выше, на тетаническое раздражение нерва
мышца перестает реагировать еще раньше
утомления нерва и самой мышцы. После
того как мышца перестала сокращаться под
влиянием нервных импульсов,
притекающих к ней через нервные окончания, она
может еще хорошо сокращаться в ответ на
непосредственное раздражение. Чем чаще
раздражение нерва, тем скорее мышца
перестает сокращаться и тем лучше она
продолжает реагировать на непосредственное
раздражение. Раз в этих опытах
возбудимая система нерва и мышцы не испытывает
абсолютного утомления, то можно
предположить с большой вероятностью, что
ослабление мышечного эффекта при непрямом
раздражении обусловлено утомлением
нервных окончаний, т. е. одного из переда-
нервом и мышцей. Иначе говоря,
предполагается, что в условиях нервно-мышечной деятельности в
первую очередь утомляется нервное окончание концевой пластинки.
Однако можно утверждать, что быстрая утомляемость нервного
окончания связана не со свойствами самого нервного окончания, а с
условиями пребывания нейрофибриллярной сети нервного окончания
в мышце. Голые тончайшие нейрофибрилли, представляющие
возбудимую систему нерва, легко подвергаются отравляющему действию
продуктов обмена веществ самой концевой пластинки и ближайших частей
мышечного волокна. Под влиянием продуктов обмена веществ, возни-
Рис. 101. Токи возбуждения
двигательной пластинки m. so-
leus кошки в ответ на
раздражение двигательного нерва одним
индукционным ударом. Биотоки
регистрируются осциллографом
через усилитель.
1 — при полной кураризации в
нервной области наступает одна
медленная волна; 2 — при неполной
кураризации: а — ток возбуждения и
последующая медленная волна в
нервной области, b — в безнервной
области, в 2 см от нервной, биотоки
отсутствуют, нет и сокращения; 3—до
кураризации: а—в нервной области,
b — в безнервной. Стрелки
указывают на момент раздражения. В опыте
3 усиление тока в 75 раз слабее, чем
в опыте 2, и в 50 раз слабее, чем в
опыте 7. Внизу время—0.01 сек.
(Экклс и О'Коннор).
точных звеньев между
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 407
кающих как при возбуждении двигательной пластинки, так и при
сокращении ближайшего участка мышечного волокна, должна меняться
прежде всего лабильность нейрофибриллей — она должна падать.
Сообразно должен уменьшаться предельный ритм возбуждения,
передаваемого с нерва на мышцу. А потом, в результате более длительной
и сильной мышечной деятельности, нервные окончания отравляются
в такой мере, что они перестают проводить нервные импульсы еще
раньше утомления самой возбудимой системы мышцы.
Итак, физиологические особенности двигательной нервной пластинки
определяются не особенностями нервного окончания, а исключительно
тем, что голые волоконца нервного окончания легко подвергаются
влиянию продуктов обмена веществ, который протекает в нервной области
особсппо иптенсивно.
Понижение лабильности нервных окончаний, вследствие
отравляющего действия продуктов обмена веществ мышцы на голые нейрофиб-
рилли нервного окончания, может стать также причиной
трансформации ритма возбуждения от нерва к мышце. При некотором высоком
ритме возбуждения нерва, как 100—300 в секунду, мышца вначале
реагирует по ритму раздражения, а затем довольно быстро ритм
возбуждения падает до половинного и ниже и на этом низком ритме
может держаться более или менее значительное время до полного
прекращения передачи нервных импульсов на мышцу.
Значительное накопление продуктов обмена веществ >в нервном
участке должно также повлиять на возбудимую систему мышцы в этом
участке. При длительном раздражении продукты обмена вещества
накапливаются в таком количестве, что вызывают локальную
контрактуру в нервном участке. Несомненно, количество их в нервном участке
достаточно велико, чтобы повлиять и на возбудимую систему,
обусловливая понижение возбудимости, падение лабильности ее. Это понижение
лабильности возбудимой системы мышцы в нервном участке играет
также определенную роль в происхождении утомления и
трансформации ритма возбуждения в нервно-мышечном препарате.
Утомление двигательной пластинки при длительном раздражении
нерва, видимо, связано с определенным изменением состава
ионов. Это видно из того, что при пропускании ионов Са через
кровеносные сосуды утомленной мышцы утомление проходит (Валидов, 2;
Воронцов, 3). По мнению Воронцова, утомление
обусловлено повышением проницаемости перепонок в области
двигательных пластинок, которое вызывается разрыхляющим действием на
них биоэлектрических токов нервных импульсов. Но, по нашему
мнению, утомление двигательной пластинки обусловлено действием
продуктов расщепления, которые возникают в концевой пластинке
и вообще в нервном участке в связи с процессом возбуждения
и сокращения мышечного волокна. Эти же продукты расщепления
должны обусловить, главным образом, увеличение проницаемости
перепонок. Ионы калия, освобождаемые при возбуждении, могут играть
408 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
особенно большую роль в этом явлении. Но, конечно, и токи
возбуждения со стороны нервного волокна и нервных окончаний также должны
играть в этом некоторую роль. На этом основании восстанавливающее
действие ионов кальция, вероятнее всего, базируется на уплотнении
полупроницаемых перепонок двигательной пластинки, вследствие чего
должно затрудняться прохождение ядовитых продуктов расщепления
от места их возникновения к нейрофибриллям и вообще в возбудимую
систему концевой лластинки.
Хронаксия двигательной пластинки. При изучении хронаксии
нормальной мышцы лягушки в разных ее участках было обнаружено три
рода хронаксии, три величины ее. В связи с этим было сделано
предположение о существовании трех возбудимых мышечных субстанций
(Люкас, 10). Но при параллельном морфологическом и хронаксиметри-
ческом исследовании участков мышцы (m. sartori'us, т. pect. pars abdom.
лягушки) было установлено, что наименьшая хронаксия мышцы, которая
равняется хронаксии нерва — 0.15—0.25 а, наблюдается как раз ів том
участке імышцы, где проходят веточки двигательного нерва, а
наибольшая хронаксия — 0.92—1.60 а—встречается в разных участках мышцы,
где нет вообще ни нервных веточек, ни двигательных пластинок. Отсюда
было сделано заключение, что наиболее короткая хронаксия в мышце
обусловлена возбуждением мышцы через нервные веточки и отвечает
нервным волокнам. Наибольшая хронаксия, обусловленная
раздражением безнервных участков мышцы, относится к мышечному волокну
непосредственно. Хронаксия средней величины, 0.4—0.7 а,
наблюдалась в нервных участках. Происхождение этой хронаксии
стало понятным после кураризации. На кураризованиой мышце исчезает
наиболее короткая хронаксия, отвечающая нервным волокнам, а
средняя и длинная хронаксии остаются, причем средняя хронаксия
наблюдается в нервных участках, а длинная — в безнервных. Отсюда было
сделано заключение, что средняя хронаксия отвечает двигательным
пластинкам, вернее, возбудимой системе мышцы в области
двигательных пластинок (Цкипуридзе, 11). Хронаксия возбудимой системы
мышцы в нервных участках очень близка к хронаксии нервных
волокон. Этим самым обеспечиваются наилучшие условия для передачи
возбуждения с нерва на мышцу.
При утомлении мышцы в результате длительного тетанического
раздражения нерва полностью исчезают хронаксии малой величины,.
отвечающие нервным участкам. Теперь в этих участках хронаксия
оказывается такой же большой, как в безнервных участках, в которых она
не меняется (Цкипуридзе, 51). Совершенно ясно, что наблюдаемое при
утомлении значительное удлинение хронаксии мышечной возбудимой*
системы в нервных участках может стать причиной прекращения
передачи возбуждения с нерва на мышцу.
Механизм передачи возбуждения в двигательной пластинке. Процесс
возбуждения в нервном окончании должен сопровождаться более
сильным электрическим током, чем процесс возбуждения в соответствую-
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 409
щем нервном волокне, ибо общая масса нейрофибриллей в нервном
окончании ввиду сильного ветвления и образования густой сети должна
быть больше, чем в нервном волокне. Этот ток должен раздражать
возбудимую систему мышцы в двигательной пластинке, тем более что
возбудимость ее здесь вообще достаточно высока.
Кроме того, как указывалось выше, хронаксия концевой
двигательной пластинки (нервного участка) довольно близко стоит к хронаксии
нервного волокна. Это обстоятельство, конечно, в сильной мере должно
благоприятствовать возбуждению двигательной пластинки биотоками
нервных импульсов. Более того, известно, что мышечные участки, даже
безнервные, возбуждаются электрическими ударами, продолжительность
которых равняется продолжительности нервного тока (Лапик, 12).
Наконец, установлено, что время передачи нервного импульса на мышцу
равняется продолжительности биотока нервного импульса. Очевидно,
полезное время для естественного возбуждения двигательной пластинки
н вообще нервного участка мышцы равняется продолжительности тока
возбуждения нервного окончания: около 1 а для скелетной мышцы
летней лягушки (Гёпферт и Шефер, 13) и около 0.6 с для глазных
мышц кошки (Лоренте де-Но, 15).
Биоток двигательной пластинки достигает такой
электродвигательной силы (0,7—1,4 mV и более), что он также должен являться
раздражителем в отношении ближайшего участка возбудимой системы
мышцы. И действительно, в тех случаях, когда биоток возбуждения
мышечного волокна следует после того, как биоток двигательной
пластинки достиг максимума, мы имеем возбуждение мышечного волокна
под влиянием двигательной пластинки.
На основании приводимых фактов можно утверждать, что в
нормальной мышце при отличном функциональном состоянии возбуждение
нервного волокна может передаться непосредственно возбудимой системе
мышцы, минуя двигательную пластинку. Эта пластинка возбуждается
также под влиянием того же нервного импульса, но отвечающий
возбуждению двигательной пластинки биоток не может проявиться, ибо
он наступает одновременно со значительна более сильным мышечным
биотоком возбуждения. Эта передача возбуждения в нервно-мышечном
аппарате, должно быть, осуществляется при посредстве биотока
нервного импульса, ибо этот биоток в состоянии раздражать возбудимую
систему как двигательной пластинки, так и окружающей ее области
мышечного волокна.
При некотором понижении возбудимости мышцы в области нервных
окончаний, как это, например, бывает во время рефракторного
состояния (Куфлер, 50), биоток возбуждения нервного волокна оказывается
не в состоянии воздействовать на возбудимую систему мышцы
непосредственно, но возбуждает двигательную пластинку, а через биоток
возбуждения ее производит возбуждение самой мышцы. В таких
случаях мышечному биотоку высокой амплитуды всегда предшествует
биоток двигательной пластинки небольшой высоты.
410 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
При кураризации, когда возбуждение нервного ствола дает лишь
.локальные биотоки двигательной пластинки, очевидно, функциональное
состояние нервно-мышечного аппарата оказывается в такой мере
пониженным, что биоток возбуждения нервного волокна возбуждает
двигательную пластинку мышцы, но не в состоянии возбудить ни
непосредственно, ни посредством биотока этой двигательной пластинки
•ближайший участок возбудимой системы імышцы.
При более глубокой кураризации возбуждение нерва не
вызывает возбуждения в соответствующих двигательных пластинках, но
мышца продолжает еще реагировать на прямое раздражение. Это
означает, что под влиянием кураре возбудимость в двигательных
пластинках понижается лишь в такой степени, что нервные биотоки
перестают возбуждать их.
Итак, возбуждение мышцы через нерв происходит путем
раздражающего действия биоэлектрического тока возбуждения нервного волокна
как на двигательную пластинку, и затем путем действия биотока
последней на возбудимую систему мышцы, так и непосредственно на
еозбудимую систему мышцы, иначе говоря, возбуждение переходит
с нерва на мышцу тем же путем, как в нервном волокне от
возбужденного участка на не возбужденный.
В определенных случаях возбудимость двигательной пластинки или
самой мышцы понижается в такой степени, что отдельные нервные
импульсы не производят возбуждения мышцы, но если они следуют
подряд друг за другом, мышца возбуждается. Так бывает при
утомлении мышцы и при неполной кураризации. В этих случаях отдельный
биоэлектрический ток нервного импульса производит в двигательной
пластинке локальное изменение основного биологического процесса,
сопровождаемое соответствующим биотоком. Происходит локальный
процесс подобно тому, как это бывает в мышце при субминимальном
раздражении. В связи с локальным физико-химическим изменением
двигательной пластинки здесь происходит локальное повышение
возбудимости, которое длится долгое время — до секунды и более.
Повторные нервные импульсы своими биоэлектрическими токами повышают
возбудимость еще более. В результате подпороговые импульсы
становятся пороговыми.
Характерно, что даже «в нормальной мышце определенные нервные
волокна возбуждают мышечные волокна ів результате суммации.
Таковы наиболее тонкие двигательные нервные -волокна, которые
оканчиваются в тонких же мышечных волокнах, образующих веретена для
проприоцепторов імышцы. Они не возбуждают отдельными импульсами
эти мышечные волокна. Требуется ряд нервных импульсов для
возбуждения последних (Дейк, 16).
Выделение ацетилхолина в нервных участках мышцы. Если мышцу
возбуждать тетанически через нерв, то при пропускании по кровеносным
сосудам мышцы искусственной питательной жидкости (Рингера, Локка)
в оттекающей от мышцы венозной жидкости появляется ацетилхолин,
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 411
или, вернее, ацетилхолинаподобное вещество. Этого не бывает, если
мышцу возбуждать тетанически с такой же интенсивностью прямым
путем. Более того, если тетанически раздражать денервированную
мышцу, ацетилхолин не будет выделяться. Наоборот, он будет
выделяться в кураризованной мышце при раздражении нерва, несмотря на
отсутствие сокращения (Дэйл и Фельдберг, 4; Дэйл, 5). На основании
такого рода фактов было сделано заключение, что нервные окончания
выделяют ацетилхолиноподобное вещество при их возбуждении
нервными импульсами. Но в последнее время было обнаружено, что
ацетилхолиноподобное вещество выделяется из мышцы и при покойном
состоянии б условиях перфузии искусственной жидкостью, но только
в меньшем количестве, чем при возбуждении (Кибяков, 6).
Ацетилхолиноподобное вещество содержится в скелетных мышцах
в разной концентрации. Его больше всего в тонических мышцах,
например, в сгибателях передних конечностей самца лягушки. Отношение
содержания его там к содержанию ацетилхолина разгибателей той
же конечности равно 7: 1. У самок лягушек нет почти разницы между
этими мышцами в отношении ацетилхолина. Особенно мало
ацетилхолина в мышцах бедра (Платнер и Крайних, 18).
Ацетилхолиноподобное вещество содержится в экстрактах не только нервных участков
мышцы, но и безнервных. Однако в последних участках количество
его в два-три раза меньше (Гоголашвили, 8).
Холинэстераза в нервных участках мышцы. В мышце находят
также фермент, расщепляющий ацетилхолин на холии и уксусную
.кислоту. Его называют холинэстеразой. Предполагается, что этот
фермент содержится на поверхности двигательных пластинок в особенно
большом количестве, а потому ацетилхолин, образуемый при
возбуждении, не может накопиться в двигательной пластинке. Он каждый
раз сейчас же после образования разлагается под влиянием холин-
зстеразы (Марнай и Нахмансон, 7, 20; Гинецинский, 27).
Гипотеза о химической передаче нервного возбуждения на мышцу.
В последнее время, в связи с установлением факта выделения ацетил-
холиноподобного вещества под влиянием нервного возбуждения, было
высказано предположение, что нервные импульсы вызывают в нервном
окончании выделение ацетилхолина, который затем раздражающим
образом действует на возбудимую систему мышцы. Таким образом,
.ацетилхолин считается передатчиком возбуждения —медиатором между
нервным окончанием и мышечными клетками. В основу этой гипотезы
была положена также отмеченная выше значительная чувствительность
скелетной мышцы к ацетилхолину. Можно даже получить эффект,
близко сходный с одиночным сокращением. Это бывает на мышцах
теплокровных, если быстро вводить маленькую дозу ацетилхолина через
мышечную артерию, предварительно опорожненную путем зажатия.
Большие же дозы производят тетанусы с таким же напряжением, как
это свойственно тетанусу, вызванному электрическим раздражением
нерва (Дейл, 5). Повторным введением ацетилхолина через артерию
412 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
можно вызывать сокращение многократно в течение нескольких часов
(Симонар, 21).
Далее, предполагается, что утомление мышцы при длительном
непрямом раздражении обусловливается уменьшением выделяемого
ацетилхолина. Когда количество его становится ниже порогового,
сокращение прекращается. Но возможны индивидуальные колебания в смысле
пороговой концентрации. В одних мышечных волокнах она будет выше,
чем в других. Соответственно раздражающее действие нервных
импульсов в одних імышечных волокнах может прекратиться раньше, чем
в других. С этим связывают то явление, что при длительной
деятельности сначала получается частичное утомление, а уже потом оно
становится полным (Розенблют и Морисон, 31).
Изучение биоэлектрических токов возбуждения мышцы при
действии ацетилхолина показало, что при этом возникает всегда более
или менее длительный ряд токов возбуждения (Броун, 17; Симонар,
21). От одного же нервного импульса обычно получается в мышце одно-
колебание биоэлектрического тока, один импульс возбуждения в мышце.
Защитники химической передачи возбуждения предполагают, что это
достигается действием холинэстеразы, которая, будто бы, все время
существует в двигательной пластинке или на ее поверхности в большой
концентрации. Благодаря ей ацетилхолин, освобожденный нервным
окончанием при возбуждении, сейчас же расщепляется, поэтому свое
активное действие ацетилхолин может оказать лишь в момент
образования (Дейл, 5; Броун, 17). Это предположение находит
подтверждение еще и в том, что на спинальных и вообще на
наркотизированных кошках, отравленных простигмином (эзерином), раздражение
двигательного нерва отдельными индукционными ударами производит
в мышцах не один импульс, а целый разряд импульсов.
Предполагается, что простигмин парализует действие холинэстеразы, и потому
выделяемый нервным окончанием ацетилхолин в состоянии длительно
действовать на мышцу и тем самым вызвать в ней разряд импульсов
(Дейл и сотр., 5, 37; Розенблют и сотр., 31, 45).
Гипотеза о химической передаче возбуждения с нерва на
скелетную мышцу в настоящее время находит целый ряд возражений
существенного значения. Против нее говорят прежде всего очень малый
скрытый период возбуждения и очень малая продолжительность
активного состояния после одного нервного импульса — всего в течение
нескольких сигм. Гипотеза допускает, что действие ацетилхолина и
холинэстеразы происходит моментально, ограничиваясь несколькими
сигмами. Известно, что при частых нервных импульсах мышца
теплокровных может воспроизвести ритм нервного возбуждения до 400)
в секунду. Отсюда следует, что освобожденный одним нервным им*
пульсом ацетилхолин сейчас же расщепляется холинэстеразой,
существующей до наступления нервного импульса или освобожденной
самими нервными импульсами, подобно ацетилхолину. Но вслед за
этим, почти вне времени, должна расщепляться и холинэстераза или
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 413
устраниться ее действие, ибо сейчас же последующий нервный импульс
в состоянии вызвать новое мышечное возбуждение.
Лабораторным путем была изучена скорость расщепляющего
действия холинзстеразы на ацетилхолин. Оказалось, что то количество
ацетилхолина, которое образуется в мышце лягушки от одного нервного
импульса (в условиях перфузии искусственного раствора), может
расщепиться тюд влиянием мышечной холинзстеразы до 50% в течение
5 мин., а ацетилхолин, образующийся в мышце теплокровных от
одного нервного импульса, — в течение 75 сек. (Марнай и Нахмансон,
20). Эти наблюдения в сильной мере обесценивают гипотетические
соображения относительно моментального действия холинзстеразы на
ацетилхолин.
Если все количество холинзстеразы, находимой в мышце, относить
на долю нервных окончаний, как это сделали впоследствии Марнай
и Нахмансон (20) и другие, тогда количество холинзстеразы могло
бы быть признано совершенно достаточным для расщепления
ацетилхолина за короткий срок абсолютной рефракторной фазы. Но нет
основания для такого утверждения. Во-первых, Кларк и его
сотрудники (43) показали, что концентрация холинзстеразы в нервных
участках мышцы такова, что скорость гидролитического расщепления
ацетилхолина должна быть значительно ниже, чем это приняли
Марнай и Нахмансон. При наивысшей концентрации холинзстеразы,
какая может быть в нервных окончаниях, количество ацетилхолина
может уменьшиться наполовину, по крайней мере, в 0.1 сек. Во-вторых,
холинэстеразу находят и в таких участках мышцы, где нет нервных
окончаний (Гинецинский, 27), а также и в таких тканях, где вообще
нет нервных окончаний, например, в белом веществе мозга
(Нахмансон, 39). В нервных участках мышцы холинзстеразы больше, чем
в безнервных, прежде всего потому, что в нервных участках все
ферментативные процессы протекают гораздо интенсивнее, и потому здесь
холинзстеразы как энзима должно быть гораздо больше, чем в
безнервных участках.
Указанный выше факт благоприятствования простигмина (эзерина)
тетаническим эффектам в ответ на один нервный импульс также не
говорит за ацетилхолиновое происхождение этого явления. Так бывает
и после отравления барием и гуанидином (Фенг и Шен, 46). Повиди-
мому, это связано с улучшением функционального состояния,
обусловливающим повышение возбудимости в двигательной пластинке и во
всем мышечном волокне под влиянием означенных веществ. Очевидно,
ферментативные процессы расщепления и синтеза возбудимой системы
усиливаются в такой мере, что двигательная пластинка уподобляется
рецептору (см. о рецепторах в отделе центральной нервной системы).
Возбудимая система ее претерпевает ряд изохронных расщеплений
и восстановлений и, сообразно с этим, производит ряд мышечных
импульсов в ответ на один нервный импульс.
Активное действие ацетилхолина на скелетные мышцы чрезвычайно
414 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
велико — оно обнаруживается в некоторых случаях при разведении
1 : 1 миллион. Но это действие такого рода, что трудно совместить era
с ролью передатчика возбуждения. Именно, к активному действию
ацетилхолина мышцы адаптируются после одного раза. Мышца
реагирует возбуждением и повышением возбудимости всего в течение около
1 минуты. В дальнейшем это действие исчезает, несмотря на наличие
ацетилхолина. По удалении ацетилхолина нужно подождать несколько
минут, чтобы повторное приложение ацетилхолина оказало активное
действие. Но в это время пороги прямого электрического раздражения
нормальны; нервные импульсы продолжают вызывать сокращения;
обычно и другие активные вещества, как КС1, производят активное
действие (Беритов, 25). То же самое было показано в отношении
изолированных мышечных волокон. Первое приложение ацетилхолина к
нервному участку мышечного волокна производит короткое тетаническое
сокращение, после чего повторное приложение ацетилхолина перестает
вызывать эффект. Но в этот момент прямое раздражение мышечного*
волокна, а также КС1 вызывают такое же быстрое тетаническое
сокращение нервного участка (Бухталь и Линдгард, 41). Отсюда вытекает,
что возбудимая система мышцы быстро адаптируется к такому
химическому раздражителю, как ацетилхолин, а потому нельзя допустить,
чтобы длительные повторные сокращения, какие вызываются с нерва
или прямым путем, происходили через ацетилхолин.
Против этой гипотезы говорит также тот факт, что при
нормальном кровообращении в мышце в ней не происходит выделения
ацетилхолина ни при каких условиях. Эти наблюдения были сделаны на
животных, которым был введен эзерин., Эзерин препятствует
расщепляющему действию холинэстеразы на ацетилхолин, и потому опыты,
с улавливанием ацетилхолина всегда ставятся на эзеринизированных
жіивотных. Оказалось, что при нормальной деятельности мышцы с
нормальным давлением крови в оттекающей из нее венозной крови
ацетилхолин не содержится (Фельдберг, 22; Дейл и сотрудники, 23;
Флейшль и сотрудники, 24). Ацетилхолин появляется при
определенном ненормальном функциональном состоянии мышцы, как это бывает
при пропускании искусственного раствора, а также при естественном
кровообращении, если только давление крови было сильно пониженным
(Флейшль и сотрудники, 24). На основании этих фактов следует
заключить, что выделение ацетилхолина при раздражении двигательного'
нерва связано с ненормальным течением, вернее ухудшением, обмена
веществ в нервных окончаниях и что оно совершенно не должно иметь
места при нормальном кровообращении и хорошем функциональном
состоянии мышцы.
В то же время ацетилхолин, выделяемый нервными окончаниями
или двигательной пластинкой при плохом функциональном состоянии
мышцы, безусловно должен оказывать определенное физиологическое
действие на двигательную пластинку и вообще на возбудимую систему
мышцы в близлежащем участке. Это действие должно заключаться
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 4tS
прежде всего в повышении возбудимости, ибо известно, что в малых
дозах ацетилхолин повышает возбудимость мышцы (Беритов, 25). На
этом изменении может основываться отмеченное выше повышение
одиночных сокращений утомленной мышцы лягушки в связи с
учащением одиночных раздражений. Это повышение происходит главным
образом в нервных участках, притом исключительно при
раздражении нерва (Гедевани, 26). Поэтому можно предположить, что оно
обусловливается повышением возбудимости под влиянием усиленного
выделения ацетилхолина нервными окончаниями в связи с учащением
нервных импульсов. Можно также допустить, что выделяемый
нервными окончаниями ацетилхолин благоприятствует раздражающему
действию биоэлектрических токов нервных окончаний на мышцу. Это,
например, может иметь место на слегка кураризованной мышце, когда
один нервный импульс не производит возбуждения в мышце, а ряд их
дает сокращение (Гёпферт и Шефер, 14).
Новейшие опыты ясно показывают, что образование ацетилхолина
в организме не связано с деятельностью нервных синапсов и вообще
с нервной деятельностью. Он образуется и в совершенно безнервной
мышечной системе. Так, в отношении m. sartorius лягушки было
доказано, что в проксимальном безнервном участке мышцы ацетилхолин
также присутствует, но только в концентрации в 3—5 раз меньшей, чем
в нервном участке из середины мышцы (Гоголашвили, 8). Далее,
установлено, что при развитии куриного эмбриона ацетилхолин
начинает возникать в организме на третий день инкубации, еще раньше
появления синапсов и вообще еще раньше появления более или менее
оформленных нервных элементов, когда вся нервная система
представлена просто агрегатом нейробластов. Содержание ацетилхолина на
грамм веса эмбриона достигает своей максимальной величины на
4—5-й день инкубации, в то время когда органогенез нервной и
мышечной систем только начинается. Несмотря на непрекращающееся
чрезвычайно интенсивное развитие нервно-мышечной системы во все
последующие дни инкубации, содержание ацетилхолина неизменно остается
одним и тем же (Цинг Ианг Кио, 40).
Характерно еще то, что при развитии эмбриона овцы, еще раньше
развития двигательных пластинок в мышечных волокнах, в нервном
участке мышцы образуется большее количество ацетилхолина, чем
в безнервном (Нахмансон, 44).
На основании этого рода фактов можно утверждать, что
ацетилхолин является метаболитом возбудимой системы вообще, как нервной,
так и мышечной, и что при активных процессах, как возбуждение,
некоторое количество ацетилхолина освобождается из сложного состава
липопротеидов наряду с другими продуктами обмена веществ.
По-видимому, его освобождается тем больше, чем более ненормальны
условия обмена веществ.
Но так как химические процессы возникновения и исчезновения
ацетилхолина должны определенным образом влиять на возбудимую
416 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
систему наряду с другими химическими превращениями, то
весьма вероятно, что превращения ацетилхолина в нервных
окончаниях играют определенную роль в процессе возникновения и
распространения возбуждения в двигательной пластинке. Только эта роль
ацетилхолина будет осуществляться не непосредственно, а
через биоток возбуждения. Более того, продукты
химических превращений, в частности ацетилхолина, если они почему-либо
не сразу исчезают в нервных окончаниях, должны определенным
образом действовать путем диффузии на возбудимую систему соседних
невозбужденных участков двигательной пластинки и самой
мышечной клетки, в первую очередь в смысле повышения возбудимости,
а также лабильности. На этом основании можно утверждать, что
химические продукты возбуждения нервных окончаний в мышце
играют определенную роль в передаче возбуждения с нерва на мышцу.
Именно, после первых нервных импульсов эти продукты должны
создавать более благоприятные условия для дальнейшего возбуждения
мышечной возбудимой системы, обусловливая, с одной 'стороны, более
высокий предельный ритм возбуждения, а с другой, — усиление
отдельных импульсов возбуждения. Аналогичные предположения
высказывались в последнее время многими физиологами (Экклс, Лапик,
Ухтомский, 36; Шефер, 47).
Л итература
1. Лежа в а А., Тр. Инст. физиол. Тбил. унив., 3, 275, 1937.
2: В а ли до в И. Г., Pfl. Arch., 235, 147, 1934; Физиол. журн. СССР, 17, 950, '934.
3. Воронцов] Д. С, Сб. докл. VI Всес. физиол. съезда, стр. 75, 1937.
4. D а 1 е Н. М. a. F е 1 d b е г g W., J. Physiol., 81, 39р, 1934.
5. D а I е Н. М., Cold Spr. Harb. Symp. Quant. Biol., 4, 193, 1936; Физиол. жу^и. СССР,
24, 116, 1938.
6. К и б я к о в А. В., Сб. докл. VI Всес. физиол. съезда, стр. 198, 1937. ^
7. Mar nay A. a. Nachmansohn D., J. Physiol., 89, 359, 1937; 92,37, 1938.
8. Гоголашвили Ш., Сообщ. Акад. Наук Груз. ССР, 2, № 9, 1941.
9. Г и н е ц и н с к и й А., Сб. докл. VI Всес. физиол. съезда, стр. 193, 1937.
10. Keith Lucas, J. Physiol., 34, 372, 1906; 35, 103, 1903; 35,; 310, 1907; 36, 113,
1907.
11. Цкипуридзе Л., Тр. Инст. физиол. анат. им. Бериташвили, 4, 460, 1941.
12. Lap ic que L., Le mechanisme physique et le mechanisme chimique de la
transmission nerveuse. Centre de documentation universitaire. Paris, 1936.
13. Schaefer M., Pfl. Arch., 237, 329, 1936.
14. Gopfert H. u. Schaefer H., Pfl. Arch., 239, 597s 1937.
15. Lor en te de N6, Am. J. Physiol., Ill, 272, 1935.
16. van Dij ik J. A., Arch, neerland. Physiol., 21, 203, 1936.
17. Brown G., J. Physiol., 80, 459, 1937.
18. P 1 a 11 n e r F. u. Krannich E., Pfl. Arch., 229, 730, 1932; 230, 705, 1932.
19. Беритов И., Физиол. журн. СССР, 27, 668, 1939.
20. Mar nay A. a. Nachmansohn D., С. г. Soc. Biol., Paris, 124, 942, 1937.
21. S i m о n a r t A. a. Simonart E. F., Arch, intern, pharm. ther., 49, 302, 494, 1935;
49, 381, 1935.
22. F eld berg W., Pfl. Arch., 232, 88, 1933.
ФИЗИОЛОГИЯ КОНЦЕВОЙ ДВИГАТЕЛЬНОЙ ПЛАСТИНКИ В СКЕЛЕТНОЙ МЫШЦЕ 417
23. D а I е Н. Н., F е 1 d b е г g W. а. V о g t M., J. Physiol, 86, 353, 1936.
24. Fleischl A., Sibul J. et К a e I i n, Arch, intern, physioL, 44, 24, 1937.
25. Беритов И., Физиол. журн. СССР, 27, 667, 1939.
26. Гедевани Д., Физиол. журн. СССР, 19, 1052, 1935.
27. Гинецинский А., Докл. на VI совещ. по физиол. проблемам в АН СССР
1939.
28. La р і с q u e L., Cold Spr. Harb. Symp. Quant. Biol., 4, 202, 1936.
29. Buch t a 1 W. a. Lindhard I., The physiology of str. mus. fibre, Copenhagen,
1939.
30. Кометиани П. А., Сб. докл. VI Всес. физиол. съезда, стр. 436, 1937.
31. Rosenb lueth A. a. Morison R. S., Am. J. Physiol., 119, 236, 1937.
32. E с с I e s J. С a. O'Connor W. I., J. Physiol., 95, 5P, 1939; 94, 8P, 1938.
33. Гинецинский А. Г., Физиол. журн. СССР, 19, 980, 1935.
34. We dens ky N., С. г. seances Acad. Sci., Paris, 24 Dec, 1894.
35. Аршавский И. А. и Розанова В. Д., Физиол. журн. СССР, 26, 629, 1930.
36. Ухтомский А. А., Физиол. журн. СССР, 25, 767, 1938.
37. Е с с 1 е s J. С. a. O'Connor W. I., J. Physiol., 97, 44, 1939.
38. Brown G. L., Dale H. H. a. Feldberg W., J. Physiol., 37, 394, 1936.
39. Nachmansohn D., Bull. Soc. chim. biob, 130, 1065, 1937.
'40. Zing Yang К у o, J. NeurophysioL, 2, 488, 1939.
41. В u с h t h a 1 F. a. Lindhard J., J. Physiol., 95, 59P, 1939.
42. В а с a Z. M. a. Nachmansohn D., J. Physiol., 89, 368, 1937.
43. Clark A. J., Raventos J., Stedman E. a. Stedman E., Quart.
J. Exp. Physiol., 28, 77, 1938.
44. Nachmansohn D., J. NeurophysioL, 3, 396, 1940.
45. Rosenb lueth A., Am. J. Physiol., 129, 22, 1940.
46. Feng T. P. a. Shen S. C, Chin. Physiol., 11, 51, 1937; Feng T. P., Chin.
Physiol., 12, 177, 1937; ib., 13, 119, 1938.
47. S с h a ef er H. u. H a a s P., Pfl. Arch., 242, 365, 1939.
48. Eccles J. C, B. Katz a. W. S. Kuf f I er J. NeurophysioL, v. 4, 402, 1941.
49. Eccles J. С a. S. W. Kuff ler J. NeurophysioL, 4, 402, 1941.
50. Kuff ler S. W., J. NeurophysioL, 5, 199, 1942.
51. Ц к и п у р и д з е Л., Сообщ. Акад. Наук Груз. ССР, 2, № 5, 1941.
52. Steiman S. Е., Am. J. Physiol., 140, 269, 1943.
27 И. С. Беритов
Глава VI
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ ФАЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-
МЫШЕЧНОГО АППАРАТА
Оптимальный и пессимальный эффекты нервно-мышечного аппарата.
Впервые Введенский (1886) установил, что если нерв нервно-
мышечного препарата лягушки раздражается индукционными ударами
Рис. 192. Оптимальный и пессимальный эффекты в зависимости
от силы раздражения.
Нервно-мышечный і'препарат"? лягушки. Миограмма записана, когда
тетанус значительно упал в ; силу утомления после длительного и сильного
раздражения нерва индукционными ударами (сила ,22 см, частота ПО
в 1 сек.)- Миограмму нужно читать справа налево,!как показывает
стрелка. Когда тетанус настолько упал, что стал едва заметен (с),
вторичная катушка удаляется до 32 см. Как показывает кривая, сейчас же
получилось сильное сокращение (а). Когда вторичная катушка снова* была
приближена до 22 см, кривая снова сильно упала. Такой опыт повторяется
еще один раз (&). Снизу первая сигнальная линия означает время в
секундах, а вторая — момент смены раздражений (Введенский).
в секунду 70—100 раз, то мышца отвечает на это при средней силе
тока большим механическим эффектом, чем при сильном токе
(рис. 192). Также своеобразно меняется эффект при увеличении
частоты раздражения свыше 70 в секунду. При некоторой средней и
большой интенсивности раздражения механический эффект тем
меньше, чем выше частота раздражения (рис. 193). Тот сравнительно
слабый механический эффект, который получается при более сильных и
частых раздражениях, Введенский назвал пессимальный
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ флЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-МЫШЕЧНОГО АППАРАТА 419
эффектом. Тот же большой эффект, который наступает от некоторых,
более слабых и редких раздражений, был обозначен им как
оптимальный (11).
Для свежего нервно-мышечного препарата лягушки с икроножной
мышцей оптимальный ритм лежит около 100 в секунду (Введенский,
17); для такого же препарата теплокровных — около 150 (Бриско, 13).
При более редких или более частых ритмах раздражения получается
более слабый механический эффект
мышцы. Оптимальный ритм нервно-
мышечного аппарата определяется
лабильностью нерва. Чем выше
лабильность, тем выше должен быть
оптимальный ритм и,
соответственно, тем выше должен быть тот
ритм, при котором оптимальное
состояние переходит в пессимальное.
Обычно пессимальный и
оптимальный эффекты изучаются на
нервно-мышечном препарате при
раздражении нерва. Но
Введенский (1) показал, что такие же
эффекты могут быть получены на
кураризованной мышце при прямом
раздражении. Выше, в мышечной
физиологии, мы познакомились с
пессимальным эффектом мышцы при
ее непосредственном раздражении.
Пессимальный эффект может
быть наблюдаем при самом свежем
состоянии препарата. Но утомление
благоприятствует его
наступлению. Однако Введенский установил, что это способствование
развитию песси'мального состояния не может быть признано прямым
результатом обычного утомления. Во-первых, при уменьшении частоты
или силы раздражения пессимальное состояние моментально может
смениться на оптимальное; во-вторых, при пессимальном состоянии
сократительная способность мышцы не снижается, в определенных
случаях она даже увеличивается, ибо тотчас по прекращении пессималь-
ного раздражения одиночные удары дают повышенные эффекты.
Введенский полагал, что при пессимальном состоянии имеет места
особого рода утомление в самой мышце и в нервных окончаниях,
возрастающее с частотой раздражения вследствие недостаточного
интервала. Но это утомление является не причиной пессимального
состояния, а только моментом, благоприятствующим ему.
Обычно оптимальный и пессимальный эффекты изучаются в
условиях изотонического сокращения мало нагруженной мышцы. В этих
Рис. 193. Оптимальный и пессимальный
эффекты в зависимости от частоты
раздражения.
Нервно-мышечный препарат лягушки
раздражается индукционными ударами: в одних
случаях с ритмом 20 в 1 сек. (а, Ь, с, е), в
других—с ритмом 100 (А, В, С, D, К). Одна
частота сменяется другой без перерыва раздражения.
Миограмма пишется слева направо, как покавы-
вает стрелка. В первое время высота сокращения'
при малой частоте меньше, чем при большой,,
но потом, когда мышца утомилась от
длительного (40 сек.) раздражения большой частоты (?>),
малая частота производит неизмеримо больший
эффект (е), чем большая частота (D и В}
(Введенский).
420 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
условиях смена оптимального эффекта пессимальным может произойти
только в том случае, если все мышечные волокна или главная імасса
их находятся в пессимальном состоянии. При пессимальном состоянии
одной части мышечных волокон мышца может оказаться укороченной
максимально под влиянием остальной части мышечных волокон.
Вследствие этого пессимальный механический эффект лучше всего изучать
в условиях изометрической записи. В этом случае регистрируется
напряжение мышцы, которое тем больше, чем больше сокращенных
мышечных волокон и чем они сильнее сокращены. Поэтому песси-
мальное состояние одной части мышечных волокон должно вызвать
уменьшение общего напряжения мышцы, уменьшение кривой
изометрического сокращения. Точно так же хорошо изучать пессимальное
сокращение мышцы при больших нагрузках. В этих случаях величина
укорочения мышцы или подъем тяжести всецело зависят от количества
и интенсивности сокращения возбужденных мышечных волокон.
Поэтому пессимальное состояние одной части мышечных волокон хорошо
отзывается на величине укорочения: чем больше таких волокон, тем
меньше укорочение, тем меньше подъем тяжести (Гоциридзе, 10).
Происхождение пессимального сокращения в связи с учащением
раздражения. Это явление не требует особых объяснений. Когда
частота раздражения нерва мала, каждое новое раздражение падает
после конца относительной рефракторной фазы и вызывает в мышце
максимальное возбуждение; отсюда тетаническое сокращение мышцы
должно быть максимальным и оптимальным. Но коль скоро, в силу
учащения раздражения, каждое новое раздражение падает во время
относительной рефракторной фазы, оно производит сравнительно
слабое возбуждение как нерва, так и мышцы. Поэтому тетаническое
сокращение становится слабым, пессимальным (рис. 193). Чем чаще
раздражение, тем слабее вызываемое им возбуждение нерва
и мышцы, а потому внешний эффект постепенно ослабевает (Эдриан, 3;
Беритов, 2). Очень хорошо выступают все соотношения между
возбуждением и сокращением, если одновременно с сокращением
регистрировать биоэлектрические токи возбуждения мышцы и нерва при
помощи струнного гальванометра. Соответственная запись дается на
рис. 194, 195. На этих электрограммах ясно видно, что как в мышце,
так и в нерве, чем чаще следуют биоэлектрические токи возбуждения,
тем они слабее и в то же время тем слабее механический эффект.
Это в свою очередь указывает на то, что малая интенсивность
импульсов возбуждения в связи с увеличением их частоты является главной
причиной пессимального механического эффекта, что, следовательно,
в таких случаях учащение возбуждения не благоприятствует
увеличению высоты тетанического сокращения.
Происхождение пессимального сокращения в связи с усилением
раздражения. Здесь мы должны иметь в виду два случая: когда
частота раздражения сравнительно невелика — 70—100 двойных ударов
в секунду — и когда она выше этого. В первом случае при некоторых
Значение рефракторной фазы в деятельности нервно-мышечного аппарата 421
слабых раздражениях возбуждение возникает в препарате только от
размыкательных ударов; значит, ритм возбуждения отвечает ритму
прерывателя тока. Но при усилении тока возбуждение возникает и от за-
мыкательных ударов, поэтому ритм возбуждения удваивается, следо-
^ШВ,?^.,^Ш^^^Ш^^^Ш^^^^^^^^^^^^Щ^^^Ш^^^^^^^^т^^^Ш^^Ш
Рис. 194. Течение биоэлектрических токов возбуждения мышцы при переходе от пес-
симального эффекта к оптимальному.
Регистрируются возбуждения m. semitendinosus кошки; раздражается п. ischiadicus очень близко
к тазобедренному суставу. Сначала частота раздражения 130 в 1 сек. Потом она постепенно уменьшается
до 70 в 1 сек. Верхняя сплошная линия принадлежит кривой мышечного сокращения, а вторая, сильно
колеблющаяся линия — токам возбуждения мышцы (гальванометр Эйнтховена1).
вательно он будет равняться 140—200 в секунду. Так как общая
продолжительность рефракторных фаз мышцы не меньше 10 з, то,
естественно, что при удвоенном ритме нервного возбуждения механический
эффект должен быть пессимальным, так как нервные импульсы будут
приходить в мышцу во время ее относительной рефракторной фазы.
&\':'у''Л'.':'у :* *¦• ..-¦ * .-.•¦ .. J% JPk -А». *•.,.. w ^.-иЯ^.
Рис. 195. Течение биоэлектрических токов возбуждения нерва при переходе от
оптимального эффекта к пессимальному.
Тот же препарат, условия опыта приблизительно те же, как на предыдущем рисунке. Только здесь
сильно колеблющаяся линия принадлежит токам возбуждения п. ischiadicus. Раздражается этот нерв в
области тазобедренного сустава, а отводится в гальванометр из области коленного сустава. Частота
раздражения в начале опыта 80 в 1 сек., затем она нарастает до 140 в 1 сек.
Соответственно, мышечное возбуждение будет ослаблено в сравнении
с нормой.
При усилении раздражения пессимальный эффект можно получить
и при меньшей частоте раздражения, например 30—50 в секунду.
Это наблюдается в тех случаях, когда мышца утомлена, ибо с
утомлением удлиняется относительная рефракторная фаза. Импульсы
раздражения, раньше попадавшие после этой фазы, теперь приходятся во время
нее и вызывают сравнительно слабые возбуждения.
При большой частоте раздражения, 150 в секунду и выше, на любом
нервно-мышечном препарате можно получить как оптимальный, так и
пессимальный эффекты. Когда раздражение слабо, не каждое после-
422 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
дующее раздражение производит возбуждение. Второе раздражение
попадает из ¦ относительную рефракторную фазу и вследствие малой
интенсивности не вызывает эффекта. Как указывалось выше, во время
относительной рефракторной фазы для вызова эффекта требуется тем
большая сила раздражения, чем оно приходится раньше. Третье
раздражение производит эффект, так как оно попадает ів конце или
после рефракторной фазы, причем эффект будет сравнительно
большой интенсивности. Четвертое раздражение снова не дает эффекта,
так как оно происходит очень рано во время относительной
рефракторной фазы, как и второе раздражение. Пятое раздражение вызывает
эффект подобно третьему и т. д. Следовательно, при большой частоте
раздражения слабое раздражение дает так называемый половинный
ритм возбуждения большой интенсивности, что обусловливает большой,
т. е. оптимальный механический эффект. Когда сила раздражения
настолько повышена, что каждое новое раздражение, попадая'' очень
рано в относительную рефракторную фазу, способно вызвать
возбуждение, то это возбуждение будет итти по ритму раздражения, но
интенсивность импульсов возбуждения будет значительно ниже, чем в случае
слабого раздражения. В связи с этим механический эффект нервно-
мышечного препарата должен стать слабым, пессимальным.
Итак, смена оптимального механического эффекта пессимальным
как в связи с учащением раздражения, так и в связи, с усилением
последнего зависит от учащения импульсов возбуждения и ослабления
их под влиянием относительной рефракторной фазы.
Наилучшим доказательством существенного значения
относительной рефракторной фазы в происхождении пессимального эффекта
является чрезвычайно быстрая смена пессимального состояния на
оптимальное. Достаточно сменить высокую частоту раздражения на малую,
чтобы сразу пессимальное состояние уступило место оптимальному
(Воронцов, 14). Так, например, на рис. 196-А пессимальная частота
100 в секунду была заменена редкой — 27 в секунду и уже первый
импульс редкой частоты дал усиленный оптимальный эффект; он
приходился вслед за последним ударом пессимального раздражения через
0.02 сек., т. е. сейчас же после рефракторной фазы от последнего
пессимального импульса. Вообще достаточно выпасть 1—3 импульсам
возбуждения из пессимального эффекта, чтобы в силу прекращения
влияния рефракторных фаз пессимальное состояние на время сменилось
оптимальным (как это представлено на рис. 196-В) (Воронцов, 14).
Отчего зависит пессимальное сокращение: от пессимального
состояния нерва или самой мышцы? Из предыдущего изложения еще не
совсем ясно, отчего именно возникает пессимальный механический
эффект: оттого ли, что создается пессимальное течение возбуждения
в нерве, или потому, что это течение имеет место © мышечных
волокнах, или от того и другого вместе. Любой из этих случаев может иметь
место. Выяснение же того, что именно происходит в каждом данном
случае, не представляет большой трудности. При анализе явления
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ ФАЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-МЫШЕЧНОГО АППАРАТА 423
нужно исходить из данных условий опыта и из функционального
состояния нервно-імышечного препарата. Возьмем для выяснения
несколько характерных случаев. Положим, частота раздражения равна
50—70 в секунду и при сильном раздражении получается пессималь-
лый эффект. Если функциональное состояние хорошее и температура
довольно высокая, тогда при сильном раздражении получаемый песси-
мальный механический эффект всецело должен зависеть от пессималь-
ного состояния возбудимой системы мышцы. При данных условиях не
будет пессимального состояния в нерве, несмотря на удвоение ритма
Рис. 196. Смена пессимального эффекта на оптимальный в икроножной мышце лягушки.
Регистрируются токи возбуждения мышцы. В обоих опытах пессимальное состояние вызывается
раздражением нерва в ритме 100 в 1 сек. В опыте А это пессимальное раздражение сменяется на оптимальное
в ритме по 27 в 1 сек. В опыте В пессимальное раздражение было прервано на короткий момент с
выпадением двух импульсов раздражения. Здесь кроме электрического эффекта (Эл^) даны механограмма
(М) и сигнальная линия (Сиг.), обозначающая перерыв раздражения. Остальные объяснения см. в тексте
(Воронцов).
возбуждения, так как в нерве каждое новое возбуждение будет
возникать после рефракторных фаз. В таком случае імышца получит от
нерва в секунду 100—140 одинаково сильных импульсов. Но
рефракторная фаза возбудимой системы мышцы более продолжительна, чем
в нерве, в особенности благодаря предшествующей оптимальной
деятельности, а потому каждый из нервных импульсов может действовать
на данную систему во время относительной рефракторной фазы.
Понятно, в этом случае імышца будет отвечать более слабыми
возбуждениями, чем при слабом раздражении, когда она получает только
50—70 импульсов. Таким образом, при частоте двойного раздражения
50—70 в секунду возбудимая система мышцы может впасть в
пессимальное состояние при оптимальном состоянии возбудимой системы
нерва. Следовательно, в рассмотренном случае пессимальный
механический эффект обусловливается пессимальным состоянием возбудимой
системы імышцы.
Как было указано выше, нервные импульсы должны действовать
на мышечную возбудимую систему раздражающим образом при
посредстве биоэлектрических токов возбуждения нервных окончаний.
Отсюда следует, что пессимальное состояние возникает в мышце под
влиянием прерывистого нервного биотока некоторой высокой частоты,
424 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
когда каждое электрическое колебание приходится еще во время
относительной рефракторной фазы и поэтому вызывает ослабленный
эффект — ослабленный процесс возбуждения. Этот теоретический вы*
вод хорошо, подтверждается путем прямого воздействия на безнервный
участок мышцы электрическим раздражением сравнительно высокой
частоты- Как указывалось выше (мышечная физиология), при
учащении раздражения до 80—-100 ів секунду мышца впадает в пессималь-
ное состояние подобно нервно-мышечному препарату.
При более высоких частотах раздражения, а именно 200—500 в
секунду, пессимальное состояние может наступить и в самом нерве.
При такой частоте каждое новое раздражение приходится во время
рефракторной фазы самого нерва. Если раздражение слабое, это
рефракторное состояние не будет преодолеваться им, а потому нерв
будет возбуждаться через один или два раздражения, причем каждое
возбуждение будет почти максимальным. Следовательно, мышца будет
получать в секунду 100—150 или еще меньше сильных нервных
импульсов. Если мышца будет находиться в совершенно свежем
состоянии, она может реагировать на это довольно сильными возбуждениями,,
и сообразно е этим, довольно сильным механическим эффектом.
Усиливая раздражение нерва, мы тем самым учащаем ритм возбуждения
в . нерве и уменьшаем интенсивность нервных импульсов под
влиянием рефракторной фазы. В этом случае мышца получает 200—500
очень слабых импульсов, на которые она должна отвечать слабой
деятельностью и соответственно слабым механическим эффектом, как
если бы непосредственно на мышцу падали слабые пороговые
раздражения. Следовательно, в момент пессимального состояния нерва мышца
дает слабые сокращения, с редким ритмом возбуждения, но эта
мышечная деятельность будет уже не пессимального характера, а
характера пороговых раздражений (Беритов, 2).
Итак, при высокой частоте раздражения — 200—500 в секунду — пес-
симальный механический эффект зависит от пессимального состояния
возбудимой системы нерва. Возбудимая система мышцы обнаруживает
в это время все признаки очень слабой деятельности, как это бывает
при непосредственном пороговом раздражении. Во время такого
пессимального сокращения мышца может дать еще сильное сокращение
при ее непосредственном раздражении. Значит, ів данных условиях
опыта сама мышца не находится в пессимальном состоянии*
Пессимальное состояние нервно-мышечного препарата может быть
обусловлено пессимальным состоянием не нерва или мышцы, а
окончаний двигательного нерва в мышечных волокнах. Окончания
двигательного нерва считаются легче утомимыми, чем нервное волокно и даже
чем мышечное волокно. Поэтому при некоторых высоких частотах и
силах раздражения двигательного нерва нервное окончание скорее будет
приходить в пессимальное состояние, чем мышца, а уже это, в свою
очередь, будет обусловливать уменьшение мышечного эффекта. Мы
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ ФАЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-МЫШЕЧНОГО АППАРАТА 425
выше показали, что нервное окончание в мышечном волокне при его
свежем состоянии не должно отличаться от обычного нервного
образования. Но так как нервное окончание состоит из голых тоненьких
нейрофибрилляриых волоконец, то оно легко> должно подвергаться
повреждающему действию продуктов обмена веществ в двигательной
пластинке. Поэтому, в связи с деятельностью двигательной пластинки,,
нервное окончание должно быстро подвергаться отравляющему
действию продуктов обмена и сообразно с этим быстро терять свою
лабильность. В результате этого пессимальное состояние нервно-
мышечного аппарата в определенных случаях действительно может
быть обусловлено пессимальным состоянием нервных окончаний.
О такой возможности свидетельствует наблюдение Трофимова-
(18), который нашел, что при пессимальном состоянии m. sartorius
нри высоких частотах раздражения нерва, как 160—600 в секунду^,
сильно удлиняется абсолютная рефракторная фаза нервных
окончаний, в то время как абсолютная рефракторная фаза проксимального*
безнервного участка імышцы увеличивается немного или даже не
меняется. Точно так же хронаксия безнервнсго участка мышцы
меняется слабо, в то время как в области нервных окончаний она меняется-
очень резко в смысле удлинения. Эти же наблюдения свидетельствуют
что пессимальное состояние нервных окончаний обусловливается
действием каких-то продуктов обмена веществ, ибо норма абсолютной
рефракторной фазы восстанавливается в течение многих минут после-
пессимума. Но, принимая во внимание, что в области нервных
окончаний капиллярная сеть сильно развита, надо полагать, что при
нормальном кровообращении пессимальное состояние в нервных
окончаниях не наступает раньше, чем в мышечных волокнах.
Пессимальное состояние нерва характеризуется понижением
обмена веществ, что, конечно, объясняется тем обстоятельством, что
процессы возбуждения, возникающие во время относительной
рефракторной фазы, сильно ослаблены. Это хорошо проявляется в отношении1
теплопродукции. Было установлено на нерве теплокровных животных,,
что при высоких частотах раздражения — 700—1500 в секунду —
теплообразование держится на высоком уровне некоторое время, а
затем падает. Это падение происходит тем скорее и резче, чем чаще
раздражение. Особенно резко проявляется падение теплообразования
при высоких частотах раздражения, если применять его на фоне
длительного редкого раздражения — около 100 в секунду. При смене
редкого раздражения частым теплообразование резко ослабевает. Если
потом опять перейти к редкому раздражению, теплообразование сразу
возвращается к повышенному уровню (Бюньяр, 6). Очевидно, при
пессимальном состоянии нерва теплообразование уменьшается, и тем
сильнее, чем глубже пессимум, чем слабее протекают процессы воз*
буждения в связи с их учащением (Бюньяр и Гилл, 11).
В определенных случаях нерв и мышца одновременно могут
находиться в пессимальном состоянии. Это должно иметь место, между
-426 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
.прочим, при средних частотах раздражения, как 100—200 в секунду, и
при плохом функциональном состоянии нерва, когда продолжительность
рефракторных фаз нерва больше, чем интервал между
раздражениями,— больше 0.01—0.005 сек. При некоторой малой силе
раздражения в нерве будут возникать импульсы возбуждения только от таких
ударов, которые будут попадать после рефракторных фаз. Эти
импульсы будут максимально интенсивны и с небольшим ритмом —
50—100 в секунду. На этот ритм сильных импульсов мышца будет
реагировать максимальным сокращением, оптимальным эффектом.
При некоторой же большей силе раздражения каждый стимул, попадая
в рефракторную фазу, будет вызывать ослабленный импульс
возбуждения, и тем самым нерв будет приходить в пессимальное состояние.
В это время мышца будет получать 100—200 нервных импульсов, хотя
и ослабленных сравнительно с оптимальным состоянием, но все-таки
довольно значительных. Возбудимая система мышцы может
реагировать на это полным ритмом возбуждений, но, конечно, еще более
ослабленных, так как каждое возбуждение будет приходиться здесь
в более раннюю стадию рефракторной фазы, чем в нерве. Это поведет
к ослаблению механического эффекта мышцы. Таким образом, при
.некоторой средней частоте раздражения — 100—200 в секунду — полу-
чаемое пессимальное сокращение мышцы может находиться в связи
с одновременным пессималъным состоянием возбудимой системы нерва
и мышцы.
Ослабление нервно-мышечного эффекта при некоторых сильных раздражениях
¦нерва может быть обусловлено локальными изменениями в раздражаемом участке.
Известно, что при некоторых сильных индукционных ударах раздражаемый участок
шерва впадает в состояние пониженной возбудимости, в парабиотическое состояние,
^как об этом (подробно будет 'изложено в следующей главе. Это явление хорошо
•было изучено еще Введенским. Впоследствии Гильдемейстер (7), а в
¦ последнее время Эрлаінгер с сотрудниками (8) нашли, что каждый
индукционный удар в области катода вызывает ів нерве сначала кратковременное повышение
возбудимости, а затем его более длительное понижение—катодную депрессию с
длительностью в несколько сигм. Кроме того, было' найдено, что второй
индукционный удар, попадая во время относительной рефракторной фазы, может не вызвать
возбуждения, но заметно меняет рефракторность, превращая относительную фазу в
абсолютную. На этом основании Като (9) и Эрлангер (8) полагают, что
угнетение нервно-мышечного эффекта при усилении фарадического раздражения может
быть обусловлено этими локальными изменениями в нерве. Конечно, это может
происходить, однако в условиях опыта Введенского пессимальное явление было
следствием не этих локальных изменений в участке раздражения. Пессимальное
состояние устанавливалось во всем нервно-мышечном препарате. Когда с одного нерв-
Tic го участка вызывался пессимальный эффект, тогда любое раздражение всякого
другого участка не устраняло эффекта, даже, наоборот, углубляло это.
Концепция Введенского о происхождении гтессимального состояния.
Первоначально Введенский полагал, что пессимальное состояние складывается в самой
мышце. Этот взгляд основывался на том, что нерв вообще не утомляем и что кура-
ризованная мышца легко дает пессимальный эффект в связи с учащением
раздражения (1). Впоследствии Введенский утверждал, что пессимальное явление
•создается исключительно в нервных окончаниях. Основанием служило то
наблюдение, что при пессимальном эффекте мышца сохраняет свою прямую возбудимость.
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ ФАЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-МЫШЕЧНОГО АППАРАТА 427
Далее, Введенский полагал, что в мышце вслед за возбуждением сначала
наступает период невозбудимости, т. е., по нынешним представлениям, рефракторная
•фаза, а затем период повышенной возбудимости, т. е. фаза сверхнормальной
возбудимости. По его мнению, при некоторой высокой частоте раздражения
возбуждение и сократительный эффект от каждого последующего удара угнетаются под
влиянием предыдущего, если каждый последующий попадает в период
невозбудимости предыдущего возбуждения. Когда же последующий удар попадает в
период повышенной возбудимости от предыдущего возбуждения, тогда, наоборот, по
мнению Введенского, получается повышение двигательного эффекта,
оптимальное сокращение. Это объяснение самого факта пессимального и оптимального
явлений очень сходно с изложенным выше.
Но пессимальное состояние мышцы в связи с усилением раздражения
Введенский объяснял несколько иначе. Он говорил, что, «когда по нерву к мышце идут
сильные возбуждения в неизменно' большом числе, то мышца, дошедшая
до известной стадии утомления, не способна отвечать на них сокращением; однако
она способна еще отвечать на возбуждения более слабые, которые достигают
до нее трансформированными в нервных окончаниях в небольшом числе
импульсов» (1). Эта формулировка в настоящее время не может быть признана правильной.
Во-первых, при пессимальном состоянии нервные импульсы не всегда сильны и
неизменны по своему ритму. Во-вторых, при оптимальном состоянии мышца получает
от нерва не слабые, а сильные импульсы. В-третьих, трансформация ритма
возбуждений происходит не только в нервных окончаниях, т. е. в мышце, но и в самом
нерве. Вследствие этих погрешностей, являвшихся результатом несовершенной
методики исследования биоэлектрических токов возбуждения нерва и мышцы, В в е-
де некий дает пессимальному явлению от сильных раздражений несколько иное
объяснение. А именно, он связывает пессимальное состояние с сильными нервными
импульсами, а оптимальное — со слабыми. Хотя и здесь роль фазы невоз-будоимости
подразумевается при создании пессимального состояния, ибо сильные импульсы идут
в «неизменно' большом числе», но все-таки в выводах главный акцент делается на
<снлу импульсов. Такая формулировка очень напоминает учение Введенского
о парабиозе, развитое им много позднее (4). С развитием же этого учения он
отказался совершенно от своей первоначальной концепции насчет фазы навозбудимости
вообще по отношению к пессимальному состоянию. Он тогда признал его за
проявление парабиоза нервных окончаний в мышце, наступающего благодаря учащению
и усилению притекающих сюда нервных импульсов.
Происхождение пессимального явления на нервно-мышечном препарате
послужило предметом исследования также с других теоретических позиций и получило
другое решение. Так, Воронцов, изучая действие электрического удара на нерв
в момент его возбуждения, заметил, что, если этот удар достаточно силен, он
задерживает, ослабляет существующий процесс возбуждения. На основании целого ряда
наблюдений и соображений он приходит к заключению, что и в окончаниях
двигательного нерва в мышце данный импульс возбуждения ослабевает, тормозится,
если вскоре вслед за ним поступает второй сильный импульс со стороны нервного
волокна. В этом случае биоэлектрический ток возбуждения нервного волокна играет
такую же роль по отношению к нервному окончанию, как и индукционные удары
к нервному волокну. Значит, по мнению Воронцова, токи возбуждения нервного
волокна вызывают локальное угнетение нервного окончания, производя его
деполяризацию. Этим самым они обусловливают пессимальное состояние мышцы.
Это понимание пессимального состояния предполагает, что ток возбуждения
нервного импульса при всех условиях таков, что он в состоянии деполяризовать
нервное окончание подобно длительному действию катода постоянного тока
'значительной силы. Но мы знаем, что пессимальное состояние получается тем лучше, чем
чаще раздражение, при частотах 250 в секунду и выше, когда нервные импульсы
ослабевают еще в нерве благодаря рефракторности нерва. А с ослаблением
импульсов ослабевают и токи возбуждения и их деполяризующее значение.
428 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОііПИНАЛЪНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Идея о роли тока возбуждения в происхождении п?ссимального состояния
ненова. Она была высказана еще Введенским. Но только ело исходная точка,
была другая. Он исходил из воззрения Германа, что ток действия,
сопровождающий волну возбуждения, должен поляризовать покоящиеся точки в сфере его
действия отрицательно, а возбужденные положительно. Поэтому Герман и вместе с
ним Введенский полагали, что возбужденное место,,как находящееся в анэлек-
тротоне, не возбудимо к новым импульсам в течение изівестного времени. Поэтому
при некоторых частых раздражениях импульсы возбуждения должны ослабнуть и
должен получиться пессимальный эффект. Если толковать пеосимальное явление с
точки зрения электротоничеакого действия тока возбуждения, то гипотетичеакое-
представление Введенского мне кажется более правильным, чем
Воронцова.
С точки зрения химической передачи возбуждения пессимальное явление
объясняется как результат накопления большого количества ацетилхолина в
двигательных пластинках. Как известно, избыток ацетилхолина угнетает возбудимую систему
(Коуен, 15; Гинецинокий, 16). Но против этого говорит, кроме всего того, что
сказано выше относительно химической передачи возбуждения, еще то обстоятельство,
что смена пессимального явления оптимальным происходит моментально, в
течение малой доли секунды. Это предполагает моментальное понижение концентрации,
ацетилхолина при переходе к оптимальному раздражению, что мало вероятно.
Против 'этого говорит и то обстоятельство, что при пессимальном явлении импульсы,
возбуждения нервных окончаний ослабевают; соответственно должно было бы ослаб-
иуТі> и гипотетическое 'выделение ацетилхолина нервными окончаниями, а потому
накопление его в избыточном количестве не должно ібыло бы иметь места.
Практикум
Наблюдение пессимального сокращения в зависимости от частоты раздражения..
Опыт с нервно-мышечным препаратом или спинномозговой лягушкой. Икроножную
мышцу соединяют с миографом, который пишет на кимографе. Мышца нагружается
большой тяжестью—200—300 г. Ток прерывается прерывателем Вагнера, которым,
снабжен индукторий. Заранее обращается внимание на то, чтобы прерыватель
хорошо работал при разной частоте. Для начала нужно брать частоту раздражения
30—50 в секунду. Затем, не прекращая раздражения, учащают прерывание тока до-
100 в секунду и выше. Это достигается путем приближения якоря к полюсам
электромагнита с помощью винта. Так раздражают нерв долго, до значительного
падения кривой в силу утомления, и затем (переводят раздражение обратно на
прежнюю малую частоту. При этом мышечное сокращение усиливается. Если после этого
раздражение снова перевести на большую частоту, то ослабление эффекта
произойдет значительно скорее.
Если функциональное состояние нервно-мышечного препарата очень хорошее, то-
может случиться, что взятое учащение раздражения до 100 в секунду недостаточно
для получения п?ссимального эффекта. В таком случае необходимо утомить
препарат длительным оптимальным раздражением, чтобы вызвать удлинение
относительной рефракторной фазы в мышце. После этого учащение до 100 ;в секунду
вызовет пессимальный эффект.
Наблюдение пессимального состояния в зависимости от силы раздражения^
Первый случай, когда частота раздражения мала: 70—100 двойных ударов ів
секунду. Сначала находят порог и оптимальную силу раздражения. Затем усиливают
раздражение еще больше приближением катушек на 10 см и более с таким
расчетом, чтобы и замыкательные удары вызывали возбуждение. В течение нескольких
секунд такого раздражения кривая мышечного сокращения падает вследствие
удвоения ритма (возбуждения в мышце и попадания каждого импульса в относительную-
рефракторную фазу. Если потом перевести вторичную катушку обратно на опти-
ЗНАЧЕНИЕ РЕФРАКТОРНОЙ ФАЗЫ В ДЕЯТЕЛЬНОСТИ НЕРВНО-МЫШіЧНОГО АППАРАТА 429
¦мальную силу, кривая сокращения поднимается. Опшг повторяется. Каждый раз с
приближением катушек кривая (Сокращения будет падать.
Второй случай — когда частота раздражения высокая: 200 двойных ударов и
выше. 'Сначала находят порог, затем оптимальную силу. Если после этого сблизить
катушки еще ;ка 3—5 см, то кривая сокращения сразу или медленно упадет.
Обратное удаление катушек на 3—5 ом вызовет восстановление оптимальной кривой.
Это пессимальное состояние наступает потому, что при оптимальной силе не все
размыкательные индукционные удары вызывают возбуждение в нерве, а только
каждый второй или третий удар вследствие попадания в рефракторную фазу. При
-пессимальной же силе все размыкательные удары или вообще большое количество
их вызывают возбуждение и притом во время рефракторной фазы.
Прл высоких частотах раздражения — 200—250 в секунду — можно получить два
¦пессимума. Первый будет иметь место ири приближении катушек после
оптимальной силы на 3—5 см, как было указано выше, а второй — при дальнейшем
сближении на 10 см и более. Второй пессимум обусловливается раздражающим
действием замыкательных ударов, которые начинают действовать наравне с размыка-
тельными, т. е. новым удвоением ритма возбуждения.
Литератур а
1. Введенский Н. Е., О соотношении между возбуждением и раздражением при
тетанусе. П., 1886.
2. Беритов И., Русск. физиол. журн., 1, 1, 1917.
3. Adrian Е., J. Physiol., 46, 38*4, 1914.
4. Введенский Н. Е., Pfl. Arch., 100, 1903; Возбуждение, торможение и наркоз,
П., 1901.
5. Keith Lucas, J. Physiol., 43, 46, 1912.
6. Bugnard L., J. Physiol., 80, 441, 1934.
7. Gildemeister M., Pfl. Arch., 124, 496, 1908.
18. Erlanger J. a. Blair E., Am. J. Physiol., 99, 108, 1931.
9. Kato и сотрудники, Am. J. Physiol., 89T 692, 1929.
10. Гоциридзе А., Физиол. журн. СССР, 19, 1031, 1935.
11. Bugnard L. a. Hill A. V., J. Physiol., 83, 383, 1935.
12. Briscoe Gr., J. Physiol., 90, 10P, 1937.
13. Воронцов Д. С, Сб. докл. VI Всес. физиол. съезда, стр. 75, 1937;
Arch, intern, physiol., 49, 273, 1939.
14. Cowan С. L., J. Physiol., 88, 3P, 1936; Proc. Roy. Soc, В 129, 337, 1940.
15. Гинецинский А. Г., Сб. докл. VI Всес. физиол. съезда, стр. 193, 1937.
16. Wedensky N., С. г. Seanc. Acad. Sci., Paris, 24, dec. 1894.
17. Трофимов Л. Г., Физиол. журн. СССР, 26, 231, 1939.
Г лае а VII
ПАРАБИОЗ
Характеристика процесса умирания. Живая возбудимая система под
влиянием разнообразных внешних агентов, которые так или иначе ме-
няют ее внутреннюю физико-химическую среду, теряет свои
жизненные свойства, т. е. умирает. К таким внешним агентам нужно отнести,
вообще все те вещества, которые не входят в состав кровяной плазмы
или отличаются от нее по концентрации растворенных составных
частей, и все то, что может вредить обычному течению обмена веществ.
В частности, к таким агентам следует отнести наркотические вещества,
как хлороформ, эфир, кокаин; растворы солей, кислот и щелочей;
высокую и низкую температуры, электрический ток и др. Под влиянием
всех этих агентов процесс умирания протекает более или менее
одинаково. Но начальные стадии процесса умирания обратимы: под
влиянием внешних агентов живая система постепенно теряет свои,
основные свойства; если же во-время удалить эти агенты, живая
система возвратит все свои свойства полностью.
Введенский (1) первый изучил это явление подробно (1901)..
Как он, так и другие после него изучали процесс умирания главным
образом на нервном стволе, вот почему мы сочли нужным произвести
здесь рассмотрение этого общего вопроса.
Процесс умирания следует разделить на три стадии. Первая ста-
дия —подготовительная (провизорная); в это время в
умирающем участке нерва меняется ритм возбуждения, высокий ритм
понижается, ровный становится неровным. Поэтому она называется еще
трансформирующей стадией. Обычно эти опыты
производились на нервно-мышечном препарате лягушки. Действию внешнего-
агента подвергался участок посредине нерва. Вторая стадия
характеризуется тем, что в умирающем участке возбудимость падает,
проводимость ослабевает. Затем, такое сильное тетаническое
раздражение умирающего участка или повыше его, в нормальной
проксимальной части, которое раньше давало оптимальное сокращение
мышцы, теперь производит пессимальное, т. е. дает слабый эффект в:
мышце. Слабое же раздражение попрежнему вызывает в мышце опти-
ПАРАБИОЗ
431:
малыши эффект (рис. 197). Во время этой стадии одновременное-
раздражение измененного и проксимального участков вызывает
гораздо меньшее сокращение, чем раздражение одного измененного-
участка. Это бывает даже в том случае, если проксимальное
раздражение не дает само по себе никакого эффекта. Так, например, если
раздражением измененного участка вызывается значительный
механический эффект, то с присоединением сильного тетанического раз-
197. Парадоксальная стадия.
Нервно-мышечный препарат лягушки. Первая кривая записана
спустя 22 мин. после смазывания нерва раствором кокаина.
Раздражается проксимальный участок 70 раз в 1 сек. Пороговое
раздражение при 41 см дает слабый тетанус, при 39—40 см вызывает
оптимальное сокращение, а при 38 см пессимальное (в виде
слабого тетануса); при 35—25 см производит гтакже пессимальный
эффект в виде начальных вздрагиваний (Введенский).
дражения в проксимальном нормальном участке существующий эффект
ослабевает или прекращается. Ввиду означенного неожиданного
отношения измененного участка нерва к раздражениям стадия эта была
обозначена как парадоксальная. Как показывают рис. 197 и
198, во время пессимального эффекта мышца дает только начальное
кратковременное сокращение. Судя по токам возбуждения, мышца в
это время получает от нерва один сильный импульс в самом начале,
а затем еще несколько очень слабых. Таким образом, сокращение это
по существу является одиночным (рис. 199) (Беритов, 2).
Тот последний период парадоксальной стадии, когда раздражение
нормального проксимального участка перестает давать эффект на
мышце, но устраняет эффект раздражения измененного парабиотиче-
ского участка, как это дано на рис. 200, принято выделять особо, как
тормозящую стадию. Эта последняя стадия вообще очень
кратковременна. Так, при развитии парабиоза в течение нескольких
часов тормозящая стадия длится всего несколько минут. Поэтому не
всегда удается ее наблюдать.
После тормозящей стадии возбуждение перестает проводиться
через измененный наркотизированный участок независимо от силы
раздражения проксимального участка. Одновременно и измененный
участок теряет возбудимость. Раздражение его не дает сокращения. Нерв
432 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
в этом участке как будто умирает, но фактически смерть наступает
только тогда, когда после тормозящей стадии действие
наркотизирующего агента продолжается еще долго. Если же скоро прекращают
Рис. 198. Парадоксальная стадия. Нервно-мышечный
препарат лягушки.
Кокаин 0.5°/0. В 1-м опыте нерв раздражается 20 раз в 1 сек., во
2-м—10 раз; в 3-м—70 раз. Интенсивность раздражения везде
одна — 25 см. В 3-м опыте раздражение вызывает пессимальный
эффект, если оно повторяется безостановочно. Но если раздражение
прекращается 3—4 раза в секунду, то эффект становится
оптимальным. Время раздражений указывается сигнальной линией
путем опускания ее вниз. Время — в секундах.
действие внешнего агента тем, что нерв переводят в чистый раствор
Рингера или физиологический раствор, он возвращает все свои
функциональные свойства. При восстановлении функциональных свойств
Шв Ш
Рис. 199. Биоэлектрические токи возбуждения нерва и мышцы во время
парадоксальной стадии.
Нервно-мышечный препарат лягушки спустя час после смазывания 0.57о раствором кокаина.
Регистрация струнным гальванометром Эйнтховена. В 1-м опыте (А) в гальванометр отводится проксимальный
край парабиотического участка на границе с нормальным, во 2-м (В)— наиболее парабиотизированный
участок, в 3-м (С)— мышца. Интенсивность раздражения одна и та же во всех опытах; частота тоже
одинакова — 20 индукционных ударов в секунду, действующих на нормальный проксимальный конец
нерва. На всех рисунках дана еще механическая кривая сокращения мышцы в виде одиночного
сокращения. В опыте А интенсивность токов возбуждения падает едва заметно, в опыте В она уменьшается
в течение 1 сек. почти в четыре раза, в опыте С мышца показывает всего два импульса. Время—0.2 сек.
нерв проходит те же стадии, только в обратном порядке: сначала
тормозящую, потом парадоксальную и, наконец, трансформирующую.
То состояние 'нерва, когда он потерял способность к
функциональной деятельности, но все еще жив, так как эта способность
возвращается к нему после удаления внешнего агента, было обозначено
Введенским как парабиоз.
ПАРАБИОЗ
433
Такое название само отмечает это характерное состояние живой
ткани; пребывание между смертью и жизнью. Но обычно этим
названием пользуются для обозначения всего процесса умирания, всех
его стадий.
Парабиоз как характерное изменение функционального состояния
живой системы свойствен всем возбудимым тканям, а не только
нервным волокнам. Парабиоз наблюдается также в мышцах, на железах
Рис. 200. Тормозящее действие со стороны нормального
проксимального участка на эффект раздражения измененного
парабиотического участка.
На нижней кривой верхний отметчик раздражения (.А) относится к
проксимальному участку, а нижний (В) к парабиотическому. В цифрах
выражено расстояние между индукционными катушками в сантиметрах.
Верхняя кривая дает опыт торможения одиночных сокращений, а нижняя —
тетанического сокращения, которые вызывались раздражением
парабиотического участка. Раздражение проксимального участка само по себе
не дает эффекта на мышце, а в комбинации оно тормозит тот и другой
эффекты '(Введенский).
и в центральной нервной системе. Наркотический сон, вызываемый
эфиром, хлороформом и другими наркотиками при операциях на
животных и на человеке, является парабиотическим состоянием
центральной нервной системы. Местная анестезия, производимая в коже или
в слизистых оболочках действием новокаина, конечно, является пара-
биотическим состоянием периферических чувствительных нервов и
рецепторов. Во всех случаях парабиоза імы имеем одно и то же
функциональное изменение, которое протекает в одном и том же виде
с прохождением одних и тех же указанных выше стадий. Поэтому
можно сказать, что парабиоз есть единая общая реакция живой воз-
будимой системы на всякое воздействие внешней среды, создающее
неблагоприятные физико-химические условия для жизни. Но
возникновение парабиоза может быть обусловлено самыми разными физико-
28 и. С. Беритов
434 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Химическими изменениями. Так, например, физико-химические
изменения в нервней ткани при воздействии одновалентных и двухвалентных
ионов будут не одинаковы, они скорее будут противоположного
характера. Этим объясняется то разнообразие явлений, ,какое получается
при воздействии тех и других ионов на живую ткань. Известно,
например, что парабиоз, вызванный воздействием двухвалентных ионов,
например кальция, можно снять, т. е. уничтожить воздействием катода
постоянного тока, и, наоборот, усилить воздействием анода, а
парабиоз, вызванный калием, можно снять приложением анода постоянного
тока и, наоборот, усилить приложением катода (Воронцов, 22;
Васильев, 16; Макут, 20). То же явление получается при воздействии
сильных индукционных ударов во время парабиоза. В случае парабиоза,
вызванного одновалентными ионами, перераздражение сильными
индукционными ударами углубляет ларабиоз, а в случае парабиоза от
воздействия двухвалентных ионов то же перераздражение ослабляет парабиоз
(Васильев и Иванов, 19), Очевидно, действие катода постоянного тока
или сильных индукционных ударов сходно по своим физико-химическим
последствиям в живой ткани с действием одновалентных ионов и
противоположно действию двухвалентных ионов. Но, несмотря на
противоположный характер вызываемых внешним воздействием
физико-химических процессов, результат будет один и тот же в отношении возбудимой
системы, если только эти физико-химические процессы обусловят
ухудшение обмена веществ. Поэтому и парабиоз будет протекать всегда
в одном и том же виде (см. Васильев, 16; Русинов, 23, и др.).
Основные функциональные изменения в парабиотическом нерве.
Главное изменение в функциональной деятельности живой системы под
влиянием необычного внешнего агента заключается в сильном
замедлении или ослаблении всех жизненных процессов.
Прежде всего это касается процесса возбуждения. Его
интенсивность падает. Об этом можно судить по биоэлектрическим токам
возбуждения, которые ослабевают, и, понятно, тем больше, чем сильнее
и длительнее действие внешнего агента. В отношении
продолжительности процесс возбуждения меняется довольно незначительно. По
опытам Воронцова (10), под влиянием кокаина продолжитесь^
ность возбуждения седалищного нерва лягушки от 3 J удлиняется до
4 а, т. е. всего на одну сигму. В парабиотическом участке меняется
равным образом и проведение возбуждения: уменьшается скорость прове-;
дения (Вериго, 5; Введенский, 6; Борутау, 7; Като, 12). В одном из
опытов Като при хлороформном парабиозе седалищного нерва
лягушки скорость уменьшилась с 18.4 м в секунду до 8.5 м. Но
интенсивность процесса возбуждения в связи с его проведением не меняется
во время парабиоза, именно она не ослабевает. В этом отношении
наркотизированный нерв не отличается от нормального, что было
доказано в лаборатории Като как для целого нерва, так и для
изолированных нервных волокон (18).
Определенным образом меняется и абсолютная рефракторная фаза::
ПАРАБИОЗ
¦435
она также удлиняется в соответствии с возбуждением. Можно было
думать, что с малым удлинением процесса возбуждения мало изме^
нится и абсолютная рефракторная фаза; но оказалось, что при
определенных условиях она может удлиниться значительно больше — от 2
до 8 а (Воронцов, 31; Като, 12). Повидимому, когда под влиянием
внешнего повреждающего агента процесс восстановления очень сильно
замедляется, повторное раздражение не может дать эффекта не
только во время возбуждения, но и некоторое время после него из-за
очень імалого количества восстановленной возбудимой системы и,
значит, из-за очень низкой возбудимости. По всей вероятности, если
раздражение следует вскоре за возбуждением, т. е. в самый ранний
период изохронного восстановления, получающаяся при этом разность
электрических потенциалов настолько мала, что она не в состоянии
раздражать соседний участок и служить причиной распространения
возбуждения.
В парабиотическом участке особенно сильно удлиняется
относительная рефракторная фаза, т. е. время восстановления (Борутау и
Фрэлих, 4). Это лучше всего видно из того, что в парадоксальной
стадии тот ритм раздражения, который раньше всегда производил
оптимальный эффект, например 20—30 раздражений в секунду, теперь
производит пессимальный эффект. Для иллюстрации этого явления
приводим рис. 198, где при частоте 20 в секунду наступает типичный
пессимальный эффект, только в меньшей степени, чем при частоте
70 в секунду. Этот опыт ясно указывает, что рефракторная фаза нерва
значительно удлинилась. Если при частоте 20 получается
пессимальный эффект, то это указывает на удлинение рефракторной фазы до
0.05 сек., в то время как в нормальном нерве она длится іменее
0.01 сек. Но в конце парадоксальной стадии относительная
рефракторная фаза удерживается еще дольше; она продолжается несколько
секунд и даже десятки секунд. Процесс восстановления замедляется
настолько сильно, что второе раздражение может вызвать заметный
эффект не раньше, чем спустя несколько секунд (Беритов, 2).
В связи с сильным замедлением процесса восстановления находится,
во-первых, ослабление процесса возбуждения и, во-вторых, понижение
возбудимости. Так как при значительном ухудшении обмена веществ
процесс восстановления заканчивается очень поздно, а то и совсем не
доходит до конца, то понятно, что порог раздражения в это время
должен быть много выше, а вызываемый эффект возбуждения много
меньше, чем при нормальном состоянии нерва.
Полный парабиоз в таком случае'должен обозначать такое
состояние, когда более или менее значительное восстановление возбудимой
системы становится невозможным вследствие глубокого расстройства
обмена веществ.
Парабиотический участок характеризуется еще очень быстрым
утомлением, иногда в течение какой-нибудь секунды. Вследствие этого
лабильность данного участка очень быстро понижается во время самой
28*
436 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
деятельности, ибо относительная рефракторная фаза во время тетани-
ческого раздражения значительно удлиняется и углубляется после
каждого импульса возбуждения. Это явление с очевидностью
выступает, если регистрировать токи возбуждения от середины парабиоти-
ческого участка. Интенсивность этих токов падает очень сильно уже
в течение одной секунды. Этим объясняется, что при пессимальном
раздражении с нерва передается на мышцу только один-два началь
ных импульса (рис. 199). Но стоит только значительно понизить ритм
раздражения до 10 в секунду, как уже та самая сила раздражения,
которая при высокой частоте, 20—70 в секунду, производила песси-
мальный эффект с одиночным сокращением мышцы, теперь будет
вызывать полный тетанус с передачей к мышце нервных импульсов по
ритму раздражения (рис. 198).
В парабиотическом нерве, судя по биотокам возбуждения,
происходит ослабление процесса возбуждения, с одной стороны, вследствие
ослабления этого процесса в отдельных нервных волокнах, а с
другой — ввиду неодновременного прекращения проведения возбуждения
в ряде нервных волокон. Последнее явление объясняется не только
неодновременным действием наркотика на все нервные волокна, ибо по
причине диффузии одни волокна будут наркотизироваться раньше
других, но и разной резистентностью их к данному яду, как об этом
будет сказано ниже. По выходе из наркотизированного участка
импульс возбуждения усиливается до нормы и распространяется по
нормальному участку, как при отсутствии наркотизации. Это и было
доказано наилучшим образом на изолированных нервных волокнах. Так,
К а т о с сотрудниками (33) исследовали наркотизированное
двигательное нервное волокно, раздражая при этом нерв отдельными
индукционными ударами и регистрируя мышечное сокращение. Оказалось, что
раздражение проксимального нормального участка все время вызывает
сокращение одной и той же максимальной высоты. Это продолжается,
пока не наступит парабиоз, когда раздражение сразу перестает
вызывать эффект. Точно так же биоэлектрический ток возбуждения
нервного волокна по выходе из парабиотического участка достигает в
нормальном участке всегда одной и той же интенсивности. Обе эти серии
опытов свидетельствуют, что характер импульса возбуждения' не
меняется при прохождении через наркотизированный участок.
В определенных стадиях наркоза было замечено, что тетанические
раздражения нерва с редким ритмом вызывают через парабиотический участок слитые
тетанусы небольшой высоты с длительным тоническим .последействием (Болдырев и
Квасов, 40). Наблюдаются и такие стадии, когда одиночные индукционные удары
перестают давать двигательные эффекты через наркотизированный участок, но те-
таническое раздражение через тот же участок производит его, причем этот эффект
развизается медленно, достигает очень небольшой высоты и очень медленно
ослабевает по прекращении раздражения. На этом основании некоторые авторы
утверждают, что в наркотизированном нерве импульс возбуждения «деформируется» и при
выходе его в нормальный участок остается таким же деформированным, способным
вызвать в мышце только медленное, или, как они говорят, «тонусоподобное», сокра-
ПАРАБИОЗ
437
щение (Макаров, 34; Свердлов, 35). При этом тонусоподобном сокращении не были
найдены токи возбуждения в мышце (Свердлов). Все эти опыты велись на нервно-
мышечном препарате икроножной мышцы лягушки. Указанные теоретические
выводы противоречат приведенным выше фактам К а то и других. При проверке
опытов Свердлова и Макарова фактическая сторона подтвердилась. Но
экспериментальный анализ опытов опроверг их оригинальные теоретические выводы.
Прежде всего оказалось, что медленное тонусоподобное сокращение вообще
свойственно всем нервно-мышечным препаратам с икроножной мышцей, если только они
возбуждаются через небольшой пучок нервных волокон, как, например, при
пороговом раздражении, расщеплении нерва до небольшого пучка, полном наркозе
значительной Чсісти нервных волокон и т. д. (Беритов и Цкипуридзе, 36). Это
объясняете* неспособностью икроножной мышцы укоротиться в полной мере, когда в ней
возбуждается небольшой пучок мышечных волокон. Этому мешают несокращенные
волокча и, в некоторой мере, также нагрузка мышцы. Если при этих условиях
сократительная способность мышцы в результате длительного опыта более или
менее ослаблена, то мышечные волокна вообще не будут в состоянии укоротиться от
одного импульса. Но ряд импульсов возбуждения в состоянии вызвать небольшое,
постепенно нарастающее укорочение.
Как уже говорилось выше, икроножная мышца содержит много красных
мышечных волокон с хорошо выраженными пластическими свойствами. Вследствие
этого сокращение переходит в пластическое укорочение и потому по прекращении
раздражения остается более или менее значительное пластическое последействие.
Этим объясняется медленное расслабление икроножной мышцы. Наконец, оказалось,
что при сокращении небольшого пучка мышечных волокон нельзя регистрировать
токи возбуждения струнным гальванометром с любого участка мышцы. Смотря по
тому, где лежит сокращенный пучок, с одних участков биотоки отводятся, а с
других нет. Если передвигать отводящие электроды по поверхности мышцы,
всегда можно найти такие участки, откуда токи возбуждения будут
регистрироваться хорошо (Беритов и Цкипуридзе, 37).
Парабиотический участок нерва является электроотрицательным в
отношении к нормальным участкам. Возникающий электрический ток
имеет сплошной характер, по которому он ничем не отличается от
обычного демаркационного тока (Введенский, 1). Этот возникающий
в парабиотическом участке ток представляет собою ток основного
биологического процесса, который обязан своим происхождением
процессу умирания живой системы, протекающему здесь под
влиянием внешнего агента.
Локализация наркотического действия. Парабиотические изменения
под влиянием того или другого химического агента наступают в
нервных волокнах разной толщины не с одинаковой скоростью. Раньше
всего парабиотизируются наиболее тонкие волокна—С. Более толстые
волокна А и В парабиотизируются впоследствии. Точно так же
восстановление после отмывания скорее всего происходит в более тонких
волокнах.
Так, например, эфир прежде всего наркотизирует С-волокна,
потом В и ів самом конце Л-волокна. Восстановление их по удалении
эфира происходит в обратном порядке (Гейнбекер и Бартлей, 42). На
клиническом материале, при применении кокаина к нерву, доказано,
что прежде всего исчезает чувство боли, потом тактильное и, наконец,
438 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
кинэстетическое. Обратное получается при сдавливании нерва (Же-
рард, 41). Есть исключение: например, цианистый калий быстрее
действует на волокна В, чем на С; волокна же А наиболее резистентны.
При отмывании цианистого калия возбудимость волокон С
восстанавливается позднее всех или даже вовсе не восстанавливается (Эско-
бар, 24). Такое разное отношение волокон к ядам зависит прежде
всего от структуры волокон. К а то (18, 33) находит, что, за редкими
исключениями, химические вещества проникают в осевой цилиндр
через перехват Ранвье. А так как чем толще нервные волокна, тем
толще осевой цилиндр, и тем дальше отстоят перехваты Ранвье, то,
соответственно, отравление толстых миэлиновых волокон должно происходить
позднее тонких. Относительно веществ, не содержащих солей, как
глюкоза, это было доказано с большой вероятностью (Эрлангер и Блэйр, 32).
Для солевых растворов большое значение должны иметь насечки Лантер-
мана, ибо ультрамикроскопическими исследованиями было обнаружено,
что при воздействии электролитов, как соли, щелочи и кислоты,
коагуляция белков происходит прежде всего в области насечек и перехватов
(Эттих и Иохимс, 30). Отсюда следует, что толстые волокна Л, в
которых насечек больше, чем в В-волокнах, должны наркотизироваться
электролитами раньше тонких.
Далее, изолированные нервные волокна наркотизируются очень
быстро. Полный наркоз достигается через несколько секунд, в то время
как тот же раствор наркотизирует нервный ствол в течение многих
минут. Было также испытано локальное отравление перехватов Ранвье
и межперехватного участка 0.1% раствором кокаина и 1.5-—2.5%
раствором уретана. В то время как через перехваты наркоз наступал почти
моментально в течение одной секунды, через миэлиновую оболочку
межперехватного участка наркоз наступал более чем через час
(Като, 33).
Отсюда следует, что большая длительность развития наркоза в
нервном стволе обусловливается медленной диффузией наркотика
через периневрий и другие образования, окружающие волокна. Но как
только наркотик доходит до волокон, последние сейчас же
наркотизируются через перехваты Ранвье и насечки Лантермана.
Характерно, что для каждого наркотического вещества существует
определенная пороговая концентрация, способная производить полное
прекращение проведения. Если взять более низкую концентрацию
наркотического вещества, полного наркоза не будет, как бы долго оно ни
действовало. Так, например, уретан в концентрации 1.5% производит
полный наркоз перехвата Ранвье через несколько секунд, а
концентрация 1.4% уже не производит его. Она только понижает возбудимость,
удлиняет абсолютную рефракторную фазу. При этом подпороговая
концентрация производит это функциональное изменение сразу в самом
начале воздействия и в дальнейшем не углубляет его. Иначе говоря,
степень функционального изменения зависит от концентрации
наркотика, а не от продолжительности его воздействия на нерв (^чато, ЪЪ).
ПАРАБИОЗ 439
О структурных (субстанционных) основаниях парабиоза. Указанг
ные выше физиологические, функциональные изменения, при действии
внешних повреждающих агентов, безусловно находятся в связи с
определенными изменениями внутри нервных волокон, в протоплазме их.
Еще Клод Бернар (1875) указывал, что наркоз есть проявление
первого физико-химического изменения протоплазмы клетки, которое
заключается в «полукоагуляции», т. е. такой коагуляции, которая
обратима (27). Затем Бинц (1891), исследуя влияние морфия и
хлороформа на нервные клетки мозга, установил в стадии наркоза
помутнение протоплазмы и появление резких границ на ядре и других
частях клетки и рассматривал это явление как начальную стадию
свертывания (28). Но детальное исследование изменения структуры под
влиянием наркотических веществ является делом последних лет. Так,
было показано, что наркотические вещества (не электролиты, как
этиловый спирт) производят определенные ультрамикроскопические
изменения нервных волокон, прежде всего в осевом цилиндре, в смысле
уменьшения дисперсности нейроплазмы, перевода ее из гидрофильного
состояния в гидрофобное и, наконец, коагуляции ее. Нейрофибрилли же
начинают более сильно преломлять свет (Эттих и Иохимс, 30).
Определенные ультрамикроскопические изменения наблюдаются в осевом
цилиндре и от электролитов, как соли, щелочи и кислоты. В іместах
проникновения их к осевому цилиндру, именно в области лантермановских
насечек и перехватов Ранвье происходит коагуляция нейроплазмы, а
нейрофибрилли становятся видны или хуже или лучше (Эттих и
Иохимс, 30). Наиболее детальные исследования в этом направлении были
проведены Насоновым с сотрудниками (25). Они установили, что в
протоплазме живых образований, при воздействии как наркотиков, так
и других іагентов, происходят одного и того же рода структурные
изменения— уменьшение дисперсности коллоидов протоплазмы, а также
увеличение ее вязкости. Авторы установили это, между прочим, путем
исследования отношения мышечных и нервных клеток к витальной, т.е.
прижизненной окраске такими индифферентными красками, как мети-
леновая синька и нейтральрот, в условиях наркоза разного рода
наркотическими веществами (спирт, эфир, хлороформ и т. д.). Они
нашли, что нормальная цитоплазма и кариоплазма не окрашиваются
вовсе, но в цитоплазме откладываются зернышки красок (гранулы),
в большинстве случаев около ядер, иногда же более или менее
значительно во всей клетке. Под влиянием малых концентраций
наркотического івещества отношение клеток к краскам таково же, как в норме.
Но под влиянием больших концентраций наркотических веществ
характерным образом меняется отношение клеток к краскам: цитоплазма
и кариоплазма окрашиваются более или менее значительно в
однородный цвет, в ядре появляются структурные образования, адсорбирующие
на себе краску; гранулообразование же красок в цитоплазме, наоборот,
прекращается. Этот процесс обратим. После переноса
наркотизированных клеток в нормальную среду, окрашенная протоплазма обесцвечи;
440 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
вается, структурные образования внутри ядра исчезают, появляются
зернышки красок. При еще более высоких концентрациях в клетках
возникают необратимые изменения.
Ультрамикроскопическое исследование также показывает
определенное изменение протоплазмы в связи с наркозом. В нормальной
нервной клетке, например, цитоплазма представляется черной с
небольшой опалесценцией, в особенности около ядра, ядро же оптически
пустым, бесструктурным, чернее цитоплазмы. Под влиянием наркотиков
цитоплазма начинает светиться равномерно молочно-белым светом.
Светится и ядро, но сильнее цитоплазмы, в особенности его оболочка.
Внутри ядра выступают светлые, неправильной формы глыбки, по всей
вероятности, хроматина. Иногда видно и ядрышко, которое темнее
остальной части ядра. Эти изменения обратимы. Они исчезают, если
перенести наркотизированную ткань в чистый рингеровский раствор
(Макаров, 29).
Эти обратимые структурные изменения в протоплазму,
наблюдаемые при наркозе, были названы Насоновым паранекротиче-
с к и м и, а самое состояние протоплазмы в это время —
паранекрозом. Эти структурные изменения предшествуют >в большинстве случаев
потере функциональной деятельности: простейшие перестают двигаться,
мышцы перестают сокращаться, останавливается мерцательное
движение эпителия, прекращаются рефлекторные реакции у низших
беспозвоночных— кишечнополостных, червей и моллюсков (Макаров, 26).
Таким образом, паранекроз есть структурная основа наркоза и
парабиоза (Насонов, 26). Нужно думать, что внешние агенты производят
прежде всего структурные изменения в смысле расщепления липопро-
теидов, коагуляции и денатурации белка. Это связано с нарушением
гидратационных оболочек вокруг коллоидных мицелл и укрупнением
этих мицелл, которые при этом переходят из гидрофильного состояния
з гидрофобное. В результате такого рода изменений в протоплазме
возникают ультрамикроскопические структуры. Коагуляция и
денатурация белка должны быть связаны со сложными изменениями в самой
структуре белка. По Эстбери (43), белковая цепочка при денатурации
выпрямляется, при этом происходит освобождение валентностей,
вследствие чего повышается химическая активность цепочки, в частности
ее окрашиваемость. Этим же, видимо, объясняется большая
адсорбционная способность денатурированного белка.
Итак, в основе парабиотического функционального изменения
лежит определенное структурное изменение возбудимой системы. В
результате этого изменения ухудшается обмен веществ, в частности,
ухудшаются условия для восстановления липопротеидов. Это ведет
к уменьшению возбудимой системы. Указанные же функциональные
изменения должны быть связаны главным образом с этим
уменьшением ее.
ь Восстановление нормальной структуры нервной протоплазмы и
вообще живой системы основывается, конечно, прежде всего на превра-
ПАРАБИОЗ
441
щении денатурированного белка в нормальный. Без этого не может
произойти восстановление липопротеида. Восстановление же
нормального белка, должно быть, происходит за счет энергии, доставляемой
обменом веществ (Александров и Насонов, 39). Так как расщепление
липопротеида и денатурация белка в некоторой ограниченной степени
происходят все время и независимо от внешних влияний, то
естественно ожидать, что всякое ухудшение обмена веществ должно
задерживать восстановление нормального белка, а значит должно привести, в
свою очередь, к структурным изменениям паранекроза. Так, например,
должна действовать асфиксия, т. е. нарушение обмена веществ
благодаря отсутствию кислорода. Отсюда следует, что не только явления
паранекроза могут стать причиной ухудшения обмена веществ, но и
самое ухудшение обмена веществ должно повлечь за собой
наступление паранекроза.
Все те агенты, которые вызывают наркоз живой ткани, производят
и умерщвление ее, если только действие агента более длительно или
более интенсивно. Смерть нервной ткани, конечно, также сопряжена с
определенным, более глубоким структурным изменением протоплазмы,
которое необратимо. Это состояние протоплазмы, ведущее к смерти,
известно под названием некробиоза. При окрашивании и
ультрамикроскопическом исследовании нервная ткань обнаруживает во время
некробиоза более значительные структурные изменения. Эти
изменения должны быть связаны с полной коагуляцией и денатурацией всего
липопротеида, а также с происшедшими от этого изменениями во всей
структуре нервного образования.
Явление декремента возбуждения в парабиотическом участке нервного ствола.
С давних пор было замечено, что при некоторых условиях наркоза одного участка
нерва проводимость в наркотизированном участке исчезает раньше исчезновения
возбудимости этого участка. Кроме того, было найдено, что в наркотизированном
участке интенсивность возбуждения ослабевает при его проведении и что вместе
с тем уменьшается и скорость распространения; вследствие этого возбуждение
затухает еще раньше, чем оно дойдет до другого конца наркотизированного участка.
Значит, в связи с парабиозом авторы отметили еще одно характерное явление, а
именно распространение возбуждения в парабиотическом участке с декрементом,
т. е. с ослаблением интенсивности (Борутау, 7; Фрэлих, 4; Люкас, 14).
Как было сказано выше, в нормальном нерве позвоночных животных
возбуждение распространяется без ослабления, без декремента. В каждой точке нервного
волокна каждый раз приходит в действие вся возбудимая система. Исходя же из
приведенных выше наблюдений на парабиотическом нерве, некоторые авторы
находят, что закон возбуждения не применим к ненормальному состоянию нерва. Однако
было бы совершенно неправильно допустить, что при распространении возбуждения
через парабиотический участок декремент происходит оттого, что возбуждается не
вся возбудимая система, а только часть ее. Данное явление можно объяснить
гораздо проще, без нарушения известных законов общей физиологии. Парабиоз, или
процесс умирания, протекает на всем протяжении парабиотического участка нерва,
чо не с одинаковой скоростью в различных его частях. В центральном месте, где
был приложен действующий агент, парабиоз должен развиться сильнее и скорее
всего. Но каков бы ни был этот агент — химическое вещество, электрический ток
или высокая температура, он действует в некоторой малой степени и на соседние
442 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
участки; последние тем сильнее будут подвергаться действию внешнего агента, чем
они ближе к месту его (приложения. Значит, парабиоз должен наступить >и в этих
участках, хотя и несколько позднее и слабее. Отсюда интенсивность тока
возбуждения, а значит и процесса возбуждения должна быть ослаблена в периферической'
части парабиотического участка значительно меньше, чем в центральной.
Следовательно, декремент возбуждения при его распространении через парабиотический
участок нерва происходит без нарушения основного закона возбуждения именно
потому, что количество более или менее восстановленной возбудимой системы по краям
этого участка больше, чем в середине (Беритов, 2).
Это положение нашло хорошее подтверждение в исследованиях К а то (12) и
его учеников. Они установили, что возбуждение распространяется с некоторым
ослаблением только в пограничной области парабиотического участка. Если
повреждающий агент действует на большой участок нерва, то на расстоянии нескольких
миллиметров от нормального участка возбуждение распространяется с ослаблением.
Это явление, очевидно, обусловливается тем, что, вследствие диффузии раствора
Рингера от нормального участка в наркотический, в пограничных частях образуется
область постепенно углубляющегося наркоза. В центральном нервном участке,
который находится в однородном пара-биотическом состоянии, возбуждение проводится
¦без ослабления и без изменения скорости проведения. Аналогичные результаты были
получены и другими авторами (Форбс и его сотрудники, 13; Кох, 15).
Повышение возбудимости под влиянием внешних агентов. Многие
внешние агенты вызывают понижение функциональных свойств нерва
не сразу, а после периода повышенной возбудимости. Так действуют
слабые растворы кислот и солей и наркотиков, слабый электрический
ток. Предварительное повышение возбудимости наблюдается точно так
же при расстройстве обмена веществ под влиянием внутренних причин:
мышечная ткань, нервная и даже железистая вскоре после перерезки
нервов сначала в течение нескольких дней испытывают повышение
возбудимости, а уже затем идет дегенерация с понижением возбудимости
вместе с ослаблением других функций (Введенский, 1; Хандворский и
Захариас).
О влиянии парабиоза на соседние непарабиотические участки нерва.
Введенский (1) при изучении парабиоза заметил, что в соседних непарабиотических
участках возбудимость изменяется в противоположном направлении. Когда под
влиянием химических агентов или сильных индукционных ударов в одной
соответствующей части нерва возбудимость падает, то в это время она повышается на
некотором расстоянии от этой части. При восстановлении же парабиотического участка
возбудимость в побочных участках возвращается к норме. Введенский
предположил, что это изменение возбудимости в побочных участках функционально
связано с изменением ее ів парабиотическом участке. Аналогичные опыты с
аналогичными (выводами были сделаны учениками Введенского Васильевым (16)
и другими.
При экспериментальном анализе этого рода явлений нами было обнаружено,
что изменение (возбудимости в побочных участках не находится ни в какой
функциональной связи с парабиотическим участком. Нами было установлено, что, во
время длительно протекающего химического парабиоза повышение возбудимости в
¦соседних участках на расстоянии 8—25 мм от парабиотического происходит
вследствие высыхания, из-за недостаточной влажности воздуха во влажной камере, а
понижение ее после удаления действующего агента происходит в связи со
смачиванием всего нерва физиологическим раствором. Если устранить высыхание путем
сильного увлажнения воздуха в камере или же путем укладывания
непарабиотических участков нерва в ванночку с физиологическим раствором, то при парабиозе
ПАРАБИОЗ
443
сдного участка в соседних участках возбудимость не меняется. Точно так !же
повышение возбудимости в соседних участках при парабиозе от сильных индукционных
ударов не находится ни ів какой функциональной связи с пар абиотическим
участком. Это поівышение происходит вследствие действия петель тока на соседние
участки нерва. Это видно из того, что при устранении петель тока путем кольца
Геринга, соединенного с землею, возбудимость в соседних участках не меняется
{Б-еритов и Цкипуридзе, 17).
* Происхождение характерных эффектов в разных стадиях парабио-
тического состояния. С точки зрения функциональных изменений легко
объяснить все стадии парабиоза со всеми характерными изменениями
механического и электрического эффектов. Трансформирующая стадия
характеризуется уменьшением ритма возбуждения и колебанием
интенсивности возбуждения. Это зависит от удлинения относительной
рефракторной фазы. Если в нормальном нерве 50—100 раздражений в
секунду производили 50—100 импульсов возбуждения одинаковой
интенсивности, то это происходило потому, что лабильность нерва была
нормальной и вследствие этого каждое раздражение приходилось после
рефракторных фаз во время нормальной или даже повышенной
возбудимости. В начале же парабиоза, когда лабильность поврежденного
участка начинает падать, каждое раздражение приходится во время
относительной рефракторной фазы. Поэтому при некоторых слабых
силах не все раздражения дают эффект, а каждое второе или третье.
Колебание интенсивности возбуждения характерно вообще для
пороговых раздражений; оно обусловлено главным образом изменчивостью
интенсивности самих индукционных ударов вследствие несовершенства
контакта в прерывателе п'ервичной цепи.
Характерное течение механического эффекта во время
трансформирующей стадии ¦— сначала быстрый подъем и сейчас же следующее
за ним постепенное опускание — зависит от удлинения относительной
рефракторной фазы. Вследствие быстрой утомляемости парабиотиче-
ского участка уже в течение первых секунд относительная
рефракторная фаза заметно удлиняется. От этого каждый последующий нервный
импульс возбуждения становится все слабее и слабее. А это <в свою
очередь производит ослабление механического эффекта сейчас же
вслед за первоначальным подъемом.
Особенности парадоксальной стадии, именно получение пессималь-
ного эффекта при той самой частоте раздражения, которая в норме
давала оптимальный эффект, объясняется удлинением относительной
рефракторной фазы, как об этом уже говорилось выше. Но в
парадоксальной стадии и усиление раздражения вызывает пессимальный
эффект гораздо легче, чем в норме. При усилении раздражения частота
возбуждения в парабиотическом участке увеличивается. Это
обусловливает быстрое уменьшение интенсивности возбуждения вследствие
попадания раздражений в относительную рефракторную фазу и быстрой
Утомляемости участка. В этом случае только один-два начальных
нервных импульса выходят из парабиотического участка и достигают
444 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
мышцы. Здесь они вызывают один-два импульса возбуждения и тем
самым обусловливают сокращение в виде одиночного вздрагивания.
Тормозящая стадия изучена очень імало ввиду трудности
наблюдения из-за кратковременности ее протекания. Кроме того, она
наблюдается не всегда. В тот период парабиоза, когда раздражение
проксимального участка не дает мышечного эффекта, а раздражение парабио-
тического дает его, очень часто при комбинации обоих раздражений
торможения нет. Поэтому трудно дать тормозящей стадии убедительное
объяснение.
По данным В'В еден с кого и других, для того чтобы получился тормозящий
эффект, опыт должен удовлетворять следующим условиям;
1. Проксимальный участок должен раздражаться сравнительно сильными
токами, а парабиотический, наоборот, сравнительно слабыми пороговыми токами, как
на рис. 200, где проксимальный участок раздражается очень сильно, при 20 см,
когда порог был 40 см, а парабиотический — при 31 см, когда порог был 34 см
(Введенский).
2. Частота раздражения проксимального участка должна быть большая, в то
время как парабиотический участок раздражается редкими раздражениями, которые
в состоянии вызвать оптимальный эффект (Русинов, 23).
3. В парабиотическсм участке раздражающие электроды должны лежать
ближе к верхнему краю, т. е. ближе к электродам проксимального нормального
участка (Русинов, 23).
Мы думаем, что в означенных условиях раздражение проксимального участка
действует на парабиотический участок не только импульсами возбуждения,
посылаемыми из нормального участка в парабиотический, но и чисто физически —
распространением самого тока на парабиотический участок. Эти петли тока, действуя
на парабиотический участок одновременно с токами, приложенными непосредственно
к парабиотическому участку, производят такое учащение раздражения, которое
обусловливает глубокий пессимальный эффект. Но, кроме того, петли тока, действуя
на парабиотический участок, могут повлиять на функциональное состояние его. В
определенных случаях они могут ухудшить это состояние и тем обусловить
ослабление возбуждения, вызываемого непосредственным раздражением.
Итак, процесс умирания нерва и вообще оюивой ткани следует
рассматривать как результат понижения всех жизненных функций
вследствие ухудшения обмена веществ. Все реакции умирающей живой
ткани свойственны и совершенно нормальному состоянию:
трансформация ритма возбуждения, явление оптимального и пессимального
эффектов, утомление и т. д. Вся разница заключается в том, что под
влиянием повреждающего агента функциональное состояние, вернее
лабильность, сильно ухудшается, и потому означенные явления в
нормальной ткани происходят при более частых и продолжительных
раздражениях, чем в умирающей, парабиотической.
Процесс умирания, или парабиоз, с точки зрения Введенского. Вопрос о
процессе умирания возник в связи с учением, которое рассматривало возбудимость и
проводимость как два по существу различные явления. Были произведены также
опыты, которые делали возможным полное разделение этих функций. Например,
некоторые авторы находили возможной потерю способности к возбуждению при
сохранении проводимости (Грюнгаген, 9; Герцен, 11). Введенский заинтересо-
ПАРАБИОЗ
445
вался этим ©опросом и стал изучалъ его, подобно своим предшественникам, на
нерве, действуя на него всевозможными повреждающими агентами. Он подробно
исследовал все стадии умирания и первый поставил вопрос о физиологической
природе этого процесса, т. е. парабиоза. Введенский выставлял такую гипотезу:
парабиотическое состояние есть состояние возбуждения, которое характерно
отличается от нормального возбуждения тем, что оно протекает непрерывно, неритмично,
или, как еще говорит В.веденский, парабиотическое состояние есть
своеобразное сплошное, неколеблющееся возбуждение. Сообразно с этим было объявлено, что
и рефракторное состояние, сопровождающее это возбуждение, является сплошным
непериодическим, что до полного парабиоза рефракторное состояние является
относительным, а іво время полного парабиоза абсолютным. Так как парабиотическое
состояние не распространяется на нормальные участки нерва, то Введенский
считал парабиотическое возбуждение лишенным способности к распространению.
Далее, он же полагал, что всякое раздражение как нормального участка, так и
парабиотического углубляет состояние парабиоза. Этому он приписывал все
характерные изменения эффекта .во время парадоксальной стадии. Так, например,
известное явление, что слабое тетаническое раздражение производит полный эффект, а
сильное—производит его в очень слабой форме, по Введенскому, зависит от
того, что сильное раздражение углубляет парабиотическое состояние и это
затрудняет проведение возбуждения через парабнотический участок. Если в этой же
стадии исходящие из нормального участка импульсы прекращают эффект,
вызываемый раздражением парабиотического участка, то это указывает, по
Введенскому, на то, что первые импульсы углубляют парабиоз, переводят
парабнотический участок из относительного рефракторного состояния в абсолютное.
Введенский пришел к этому гипотетическому представлению на основании
следующих наблюдений: под влиянием внешнего агента возбудимость нерва
первоначально повышается, потом понижается, а через некоторое время совершенно
исчезает вместе с проводимостью. Если во-воемя убрать внешний агент, возбудимость
и проводимость постепенно возвращаются к норме. Нечто подобное имеет место
при раздражении, которое вызывает возбуждение: во время возбуждения нет воз-
.будимости, затем она возвращается. Далее, измененный участок является источником
электричества. Он при соединении с неповрежденным участком дает ток, который
имеет неколеблющийся, сплошной характер. Опять некоторое сходство с обычным
возбуждением, которое также сопровождается электрическим током. Значит, между
обычным возбуждением и функциональным состоянием парабиотического участка
есть некоторое сходство.
Однако возбуждение, вызванное раздражением, характеризуется не только
пониженной возбудимостью и электрическим током. Это возбуждение протекает
ритмически в виде ряда быстро следующих один за другим кратковременных процессов,
и оно не ограничивается местом возникновения, а распространяется вдоль по нерву.
Затем возбудимая система реагирует на раздражение согласно основному закону
возбуждения — вовлечением в реакцию всей возбудимой системы. То, что наиболее
характерно для обычного возбуждения, не характерно для парабиоза, и само
гипотетическое определение природы парабиоза не включает в себе ничего другого,
кроме отрицания основных особенностей обычного возбуждения. Поэтому совершенно
недопустимо, чтобы в основу парабиотического состояния было положено
возбуждение.
В настоящее время гипотетическое представление Введенского пользуется
большой популярностью главным образом среди его учеников; среди других
физиологов оно не находит сочувствия. Благодаря исследованиям Люкаса (14),
Эдриана(3)и других, физиологи стали на совершенно иную точку зрения, а
именно они рассматривают характерные явления при парабиозе как результат удлинения
рефракторной фазы, декремента возбуждения и других наблюдаемых ими изменений
в функциональных свойствах нерва.
446 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Практикум
Постановка опыта с парабиозом. 1. Простой способ. В небольшой
стеклянной банке объемом около 500 смЗ располагают нервно-мышечный препарат
лягушки (рис. 201). Лапка свободно висит в воздухе, поддерживаемая за бедреную-
кость. Нерв лежит на электродах. Банка плотно
закрыта пробкой, через которую проходят провода от индук-
тория к электродам. Банка имеет сбоку небольшое
отверстие диаметром в 2—3 мм. На дне банки лежит
вата. Определяют порог раздражения. Затем через
отверстие вводят в банку градуированной пипеткой 1 см3
серного эфира, выливая его на вату. После этого
исследуют пороги раздражения через каждые 2—3 мин..
Вначале пороги понижаются, и, таким образом,
наблюдается первоначальная стадия повышения возбудимости.
Спустя 5—15 мин. пороги начинают повышаться. Через
20—40 мин. или еще позднее, в зависимости от
температуры, возбудимость исчезает совсем: 20—15 см
раздражения (при первоначальном пороге 40—50 см) не
дадут эффекта. В этот момент в нерве наступил
полный парабиоз. Если сейчас же открыть банку, вынуть
препарат и смазать физиологическим раствором, то через:
5—10 минут возбудимость начнет восстанавливаться
а через 10—20 мин. возбудимость нерва возвратится
к норме.
Этим способом хорошо и легко можно демонстри-
оовать изменение возбудимости в связи с парабиозом и
затем момент полного парабиоза. Отдельные
характерные стадии парабиоза не могут быть прослежены. Для
этого прибегают к другому способу, который был
применен Введенским для их изучения.
2. Сложный миографический способ.
Нервно-мышечный препарат лягушки помещают во
влажной камере и соединяют икроножную мышцу с
миографом для записи. На нерв накладывают две пары
электродов для раздражения: одну посредине нерва, а дру_
гую — на проксимальный участок. Каждый участок
раздражается отдельным индукционным аппаратом. Частота
раздражения 50—100 в секунду. Определяют пороги для
каждого {раздражаемого участка и записывают
оптимальное сокращение. Затем смазывают нерв посредине
на протяжении 1 см0.5°/0 раствором кокаина, пользуясь
для этого ватой (раствор кокаина должен быть
свежеприготовлен на физиологическом растворе). После этого про-буют раздражать нерв
одним и тем же оптимальным раздражением через 3—5 мин. Первое изменение
в механическом эффекте наблюдается спустя 15—20 мин. Если до отравления мышца
давала вначале быстрый подъем, а затем медленное нарастание сокращения, то
теперь этот медленный подъем сходит на-нет. После начального быстрого подъема
кривая, наоборот, медленно опускается вниз. Это является характерным признаком
первой подготовительной или трансформирующей стадии. Но сама трансформация
ритма возбуждения может быть наблюдаема путем соединения парабиотического
участка с гальванометром. Введенский изучал это явление путем соединения
этого участка с телефоном. Но иногда и миографической записью можно улозиггь
трансформацию ритма возбуждения. Это проявляется в зубчатом тетаническом со-
Рис. 201. Простая
постановка опыта с парабиозом.
Вся установка с электродами и
нервно-мышечным препаратом (М)
держится на деревянной
крышке К. Она свободно вынимается
из банки; препарат
устанавливается предварительно и затем
только вставляется в банку. На
дне банки под пробкой (Л) лежит
вата. Наркотическое вещество —
1 см3 эфира вносится через дырку
сбоку. Определяются пороги
раздражения нерва до внесения
эфира и после него через каждые
2—3 минуты. После полной
потери возбудимости препарат
вынимают и промывают
физиологическим раствором. Затем на
открытом воздухе определяют пороги
раздражения до восстановления
нормальной возбудимости. На
основании полученных цифр
составляют кривую изменения
возбудимости.
ПАРАБИОЗ
44Г
кращении, что указывает на уменьшение ритма и неравномерное течение импульсов,
возбуждения в нерве.
Следующая, парадоксальная стадия наступит через 25—35 мин., если комнатная
температура равна 17—20°. Если же она ниже, то парадоксальная стадия наступает
позднее. Наоборот, если согреть нерв до 30°, эта стадия начнется очень скоро. Для
наблюдения следующей, тормозящей стадии требуется соблюдение целого ряда
условий, как это указывалось выше: большая частота раздражений (100 в
секунду), положение нижних электродов в верхнем участке парабиотического участка..
Ввиду того что тормозящая стадия очень кратковременна, не нужно ждать полной
непроводимости; для сильных раздражений она наблюдается уже в конце
парадоксальной.
Полный парабиоз при комнатной температуре наступает через 40—50 мин. после-
отра.вления. Для наблюдения возвращения к нормальному состоянию сейчас же по
наступлении полного парабиоза нерв нужно вынуть из камеры и положить в
физиологический раствор на 15 мин. Затем его помещают вновь во влажную камеру,
пробуют раздражать и записывают эффекты. При удачном течении опыта
обнаруживается тот момент парадоксальной стадии, когда проксимальное раздражение
дает только начальное сокращение, но хорошо тормозит эффект, вызванный
раздражением парабиотического участка. Затем проксимальное раздражение также
начинает давать хорошие тетанусы, и наступает подготовительная стадия, а за ней и
нормальное состояние.
Литература
1. Wedensky N., Pfl. Arch., 100, 5, 1903; Возбуждение, торможение и наркоз,.
1901, Петербург.
2. Beritoff J., Z. Biol., 78, 231, 1923.
3. Adrian E., J. Physiol., 46, 384, 1913.
4. Boruttau H. u. Frolich F., Z. allg. Physiol., 4, 153, 1904.
5. Werigo В., Pfl. Arch., 76, 522, 1899.
6. Wedensky N., Pfl. Arch., 84, 134, 1900.
7. Boruttau H., Z. allg. Physiol., 4, 134, 1904.
8. Veszi J., Z. allg. Physiol., 13, 320, 1912.
9. Grunhagen, Pfl. Arch., 6, 180, 1876.
10. Воронцов Д., Тр. физиол. лаб. Петерб". унив., 1911—1913, т. 7—8.
11. Herzen, Zentr. Physiol., No 18, 1899.
12. Kato G., The theory of decrementless conduction in narcotised region of nerver.
Tokyo, 1924.
13. Davis H., Forbes A., Brungwick D. a. McHopkins M., Am. J.
Physiol., 76, 448, 1926.
14. Keith Lucas, The conduction of the nervous impluse, London, 1917.
15. Koch E., Z. Biol., 87, 249, 1928.
16. Васильев Л. Л., Новое в рефл. и физиол. нервной системы, 1, 1925.
17. Беритов И. и Цкипуридзе Л., Русск. физиол. журн., 18, 385, 1935.
18. Kato G., The Microphysiology of nerve, 1934.
19. Wassiliew L. L. u. IwanowK, Pfl. Arch., 235, 184, 1934.
20. Mackuth E., Pfl. Arch., 214, 612, 1926.
21. Hober R., Physikal. Chemie d. Zelle u. d. Gewebe, 2 Halite, 1924.
22. Woronzow D.S., Pfl. Arch., 203, 300, 1924; Pfl. Arch., 207, 279, 1925;
Трофимов Л. Г., Журн. эксп. биол. мед., 19, стр. 192, 1927.
23. Русинов В. С, Сбор. раб. физиол. лаб. Ленингр. унив., 1930, стр. 19.
24. Escobar R. A., Arch, intern, pharmacodynamic 56, 251, 1937.
25. Насонов Д., Тр. физиол. инст. ЛГУ, 14, 1933; Насонов Д. и
Александров В., Арх. биол. наук, сер. Л. 36, 1939; Биол. журн., 6, 1937; Учен. зап.
Ленингр. унив., 17, 1937.
26. Макаров П. В., Арх. анат., гистол., эмбриол., 19, 1, 1938.
448 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
27. Bernard CI., Lemons sur Ies anesthetiques etc., Paris, 1875.
28. Binz C, Vorlesung uber d. Pharmakologie, Berlin, 1891.
29. M а к a p о в П. В., Арх. анат., гистол., эу.бриол., 15, вып. 4, стр. 3, 1936.
30. Ettich G. u. Jochims I., Pfl. Arch., 215, 519, 675, 1927.
31. Воронцов Д. С, Учен. зап. Каз. унив., 92, 56, 1932.
32. Erlanger J. a. Blair E. A., Am. J. Physiol., 124, 341, 1938.
33. Kato G., Cold Spr. Herb. Symp. Quant. Biol., 4, 202, 1936; Физиол. журн. СССР,
24, 201, 1938.
34. Макаров П. О., Тр. Ленингр. общ. естествоисп., LXVII, вып. I, 1939.
35. Свердлов СМ., Учен. зап. Каз. унив., 1934.
36. Беритов И. и Цкипуридзе Л., Физиол. журн. СССР, 22, 575, 1937.
37. Беритов И. и Цкипуридзе Л., Работа не опубликована.
38. Трофимов Л. Г., Физиол. журн. СССР, 26, 231,- 1939.
39. Александров В. Я. и Насонов Д. Н., Арх. анат., гистол., эмбриол.
22, 11, 1939.
40. Болдырев В. Б. и Квасов Д. Г., Тр. физиол. инст. ЛГУ, 14, 112, 1934.
41. Gerard R. W., Quart. Rev. Biol., 6, 59, 1931.
42. Heinbecker P. a. Bar th ley S. H., J. Neurophysiol., 3, 219, 1940.
43. Ast bury W. Т., Fundrmentaly of fibre structure. Oxford, 1933; — Cold Spring
Harb. Symp. Quant. Biol., 2, 15, 1934.
Глава VIII
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА
НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ
Выше мы ознакомились с электрическими явлениями в живой
ткани. К ним относятся биоэлектрические токи возбуждения, следовые
потенциалы, или электрическое последействие возбуждения, локальные
потенциалы на месте раздражения, токи, связанные с повреждением
живой ткани, контрактурный ток мышцы. Теперь рассмотрим действие
самого электрического тока на живую ткань. С этой стороны мы
подробно коснемся лишь действия постоянного тока на нервно-мышечную
систему.
Полюсное действие постоянного тока. Гальванический ток во время
замыкания и размыкания действует раздражающим образом не на
весь межполюсной участок, а только на те места, куда ток входит и
откуда он выходит, т. е. где расположены катод и анод. В момент за-
мыкания раздражение происходит на катоде, а в момент размыкания —
на аноде. Это было доказано Д ю б у а - Р е й м о н ю ім (2)
следующим образом: анод помещался на умерщвленной части мышцы или на
сухожилии, а катод — на нормальном участке мышцы посередине. При
этом условии замыкание тока производило сокращение, а размыкание —
нет. Это, конечно, могло произойти только в том случае, если на катоде
раздражение происходит в момент замыкания, а в момент размыкания
не происходит. Если посередине мышцы помещался анод, а катод
переносился на нефункционирующий участок, то сокращение получалось
только при размыкании.
Сокращение, вызванное раздражением нерва или мышцы путем
размыкания и замыкания гальванического тока, имеет характер
одиночного сокращения, если сила раздражения сравнительно мала. Как
уже указывалось выше, при замыкании и размыкании сильного тока
сокращение имеет тетанический характер: сокращение продолжается
более или менее значительное время после замыкания и размыкания
тока. Ему отвечает, судя по биоэлектрическим токам, не один импульс
возбуждения, а целый ряд их.
Физиологический электротон. Действие постоянного тока на нерв
или мышцу вызывает, кроме возбуждения, повышение возбудимости.
29
И. С. Беритов
450 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Это явление подробно было изучено Пфлюгером (1). Было
установлено, что на катоде во время замыкания возбудимость повышается,,
а на аноде, наоборот,—понижается. Вместе с повышением возбудимо-
Рис. 202. Диаграмма физиологического электротона при
разных силах постоянного тока.
Nn — нерв (в то же время нулевая абсцисса); К — катод; А — анод
постоянного тока, цепь которого не приведена. Изменение возбудимости
в нерве изображено тремя волнообразными линиями; кривая а, Ь, с, d для
слабого тока; е, /,Н,к— для тока средней силы; тп, р, р, г -~ для тока
большой силы. Кривая над абсциссой показывает повышение
возбудимости, а под абсцжхой — понижение ее. Высота кривой на обеих
сторонах означает величину изменения; /'„, iit г показывают положение
индифферентного участка при данной силе раздражения. Стрелка показывает
направление тока в нерве (из Ландуа).
сти изменяется и проводимость. На катоде проводимость улучшается*
а на аноде — ухудшается. Это функциональное изменение на катоде-
называется катэлектротоном, а то же на аноде — анэлектротоном. Ознг-
ченное изменение происходит не только на полюсах, где лежат
электроды, но также по обеим сторонам электродов, на некотором
гЛ
+
с
Поп
2VA
а
W
тт
тт
Рис. 203. Схема расположения электродов на нерве при изучении электротонического-
и периэлектротонического действия.
Пол. — поляризующие электроды; р — проксимальные платиновые электроды в 3 мм от дистального
поляризующего электрода; d — дистальные платиновые электроды в 28 мм от него же. Обе пары-
служат для раздражения индукционными ударами. М — икроножная мышца.
расстоянии от них. Однако между полюсами существует также
индифферентный, неизмененный участок, который при слабых и
умеренных токах приближается к аноду, а при сильных токах — к катоду.
На рис. 202 дается подробная диаграмма означенного изменения
возбудимости.
В момент размыкания возбудимость и проводимость меняются в
обратном направлении около каждого полюса. При размыкании около
катода возбудимость понижается, а около анода она, наоборот,
повышается; проводимость же на катоде ухудшается, а на аноде
улучшается. Через несколько секунд возбудимость и проводимость на обоих
полюсах возвращаются к исходной величине.
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 451
¦ Электрическое действие постоянного тока очень хорошо можно
демонстрировать миографическим путем. Если раздражать нерв
отдельными индукционными ударами около поляризующего электрода,
ближайшего к мышце (электрод р на рис. 203), то при 'нисходящем
направлении поляризующего тока замыкание его, т. е. катод,
произведет усиление существующих одиночных вздрагиваний, а размыкание
SO*
SOv
Рис. 204. Электротоническое и периэлек-
тротоническое действия при нисходящем
поляризующем токе.
Нервно-мышечный препарат лягушки.
Проксимальная пара электродов находится у катода.
В опыте А нерв раздражается через
проксимальную пару электродов; во время вызываемых
отсюда одиночных сокращений производится
поляризация нерва. Под влиянием ее одиночные
сокращения усиливаются: явление катэлектро-
тона. В опыте В одиночные сокращения
вызываются через дистальную пару электродов.
Теперь под влиянием поляризующего тока
одиночные сокращения исчезают: явление пери-
электротона. Верхняя сигнальная линия
обозначает раздражение нерва индукционными
ударами. Цифры здесь указывают на силу
раздражения в сантиметрах. Нижняя сигнальная
линия обозначает время действия
поляризующего тока. Цифры здесь означают расстояние
в сантиметрах по шкале компенсатора, с
помощью которого происходит ответвление тока от
аккумулятора в препарат (Гедевани).
Рис. 205. Электротоническое и
периэлектротоническое действия
при восходящем поляризующем
токе.
Тот же препарат. Проксимальная
пара электродов у анода. Здесь в
опыте А одиночные сокращения
вызываются пороговым раздражением
отдаленного дистального нервного
участка. При действии поляризующего
тока (нижний сигнал) одиночные
сокращения усиливаются: явление пери-
электротона. В опыте В одиночные-
сокращения вызываются
раздражением ближайшего нервного участка^
Здесь поляризация нерва производит
исчезновение одиночных сокращений:
явление анэлектротона. Те же
сигнальные обозначения, что и на«
рис. 204 (Гедевани).
этого тока — ослабление их (рис. 204, А). При восходящем
направлении замыкание и размыкание поляризующего тока произведут
обратные эффекты: ослабление от замыкания и усиление от размыкания
(рис. 205, В).
При замыкании постоянного тока на аноде, а при размыкании —
на катоде возбудимость іможет исчезнуть совсем. Одновременно
исчезает и проводимость. Это имеет место при применении длительного и;
сильного постоянного тока (Вериго, 4). Но по прекращении
раздражения возбудимость и проводимость восстанавливаются на обоих
полюсах. Может случиться, что на аноде восстановление не произойдет
при пропускании чрезмерно длительного и интенсивного тока.
29*
452 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Эта закономерность в изменениях возбудимости под влиянием
постоянного тока наблюдается не при всех условиях опыта. Если действие
тока очень длительно или если интенсивность его очень велика, тогда
при замыкании возбудимость на катоде после первоначального
повышения быстро падает до исходной величины, а затем (может
спуститься и ниже, до полного исчезновения (катодная депрессия, Вериго, 4).
Одновременно проводимость на катоде ухудшается, иногда также до
полного исчезновения (Грюнгаген, 5; Пэрна, 6). В то же время
возбудимость анодного участка после начального падения возвращается к
исходной величине.
Физический электротон. Прохождение постоянного тока через нерв
производит, кроме электротонических изменений возбудимости, еще
следующее явление.^ По обеим сторонам полюсов в нерве возникает
электрический ток, направление которого одинаково с направлением
тока, протекающего в імежполюсном пространстве нерва. Этот ток тем
сильнее, чем ближе к электродам. Он распространяется по нервному
стволу быстрее возбуждения. Данный ток не выражает какого-либо
физиологического процесса. Он также имеет место при умерщвлении
нерва эфиром (Бидерман, 10). Кроме того, известно, что подобный
ток наблюдается в проволоке, покрытой влажной оболочкой (Матеучи,
12), но в то же время он отсутствует в таком нерве, который не имеет
миэлиновых оболочек. Отсюда видно, что этот электрический ток,
возникающий в нерве при прохождении постоянного тока, — чисто
физическое явление. Некоторые авторы рассматривают этот ток как
результат пограничной поляризации, т. е. накопления продуктов электролиза
между осевьщ цилиндром нервного волокна и імиэлиновой оболочкой
(Герман, 13). Предполагается, что на границе между осевым
цилиндром и миэлиновой оболочкой происходит в области анода накопление
анионов с положительным зарядом, а в области катода накопление
катионов с отрицательным зарядом. Эта поляризация вызывает в
ближайшей электролитической среде токи, которые имеют такое же
направление, как главный, поляризующий ток. Возникающие при этом
петли тока поляризуют ближайший участок нерва, вследствие чего
поляризация тем же путем может распространиться еще дальше.
Другие авторы думают, что данный физический ток возникает в
силу того, что миэлиновая оболочка является худшим проводником,
чем осевой цилиндр. Это создает условия для распространения тока
вдоль по нерву. Ток, так сказать, обходит плохо проводящие оболочки
и потому может распространиться далеко от электродов (Грюнгаген, 3).
По нашему мнению, при пропускании тока через нерв происходит как
поляризация нерва, так и ветвление тока далеко от полюсов. В
настоящее время известно, что сопротивление осевого цилиндра раз в
пять меньше сопротивления миэлиновых оболочек. По некоторым
данным, миэлиновые оболочки обладают такой высокой непроводимостью,
что едва ли применяемые для раздражения постоянный ток или
индукционные удары проникают к осевым цилиндрам через оболочки. Во
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 453
всяком случае, доказано, что они действуют раздражающим образом на
осевой цилиндр через перехваты Ранвье (Кат о). Следовательно,
можно утверждать, что ток проникает в осевой цилиндр главным
образом через эти перехваты. Поэтому внешний ток, приложенный к
нерву, должен сильно ветвиться по нерву, проникая в области анода в
одни и те же осевые цилиндры через разные перехваты Ранвье как в
интраполярных, так и в экстраполярных участках и затем выходят -из
осевого цилиндра в области катода через разные же перехваты
Ранвье.
Однако на перехватах Ранвье имеются соединительнотканная
оболочка Генле и оболочка Шванна. Да и самый осевой цилиндр, состоящий.
из нейроплазмы и нейрофибриллей, не должен представлять собой
простую электролитическую среду. Он весь состоит из полупроницаемых
мембран. А потому при пропускании тока происходит поляризация этих
оболочек и мембран, причем в области анода концентрируются анионы,
а в области катода катионы. Это создает поляризационный так,
проявляющийся по прекращении основного тока.
Описанный физический электротон производит определенное
функциональное изменение по длине почти всего нерва. Им обусловливается
физиологический электротон. Ему же обязан своим происхождением
описываемый ниже периэлектротон.
От рассмотренного нервного тока следует отличать другой
электрический ток, который также івозникает при прохождении постоянного
тока. Гальванизируемый участок является электроотрицательным по
отношению к каждому другому участку нерва. Этот ток тот же
биоэлектрический ток, который возникает при повреждении живой
нервной ткани и который характерен вообще для ларабиотического участка
(Введенский, 14). Прохождение постоянного тока, несомненно, вызывает
в нерве парабиотическое состояние, в связи с чем и возникает данный
ток чисто физиологического происхождения.
Происхождение физиологических электротонических явлений.
Наибольшей популярностью пользуется физико-химическое объяснение этих,
явлений, предложенное Л ё б о м (3). По его мнению, в нерве
прохождение электрического тока производит перенос ионов от одного полюса
к другому, как это происходит при электролизе, и воздействием этих
ионов на живую систему должно вызываться характерное изменение
возбудимости на полюсах. Л ё б первый отметил своеобразное
физиологическое действие ионов на живую ткань. По опытам Лёба (3),
при погружении лягушечьей мышцы в VV нормального раствора NaCl
приблизительно через час мышца начинает сокращаться. Если взять
более концентрированный раствор соли, %—% нормального раствора
при комнатной температуре, то сокращение начинается тотчас же.
Подобным же образом действуют соли калия; только калийное
сокращение не продолжается так долго. Наоборот, катионы Са и Mg
действуют угнетающим образом на сокращение, вызванное ионом натрия.
Отсюда Лёб заключает, что возбуждение при замыкании тока через
454 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
A* Na' К
нерв или мышцу может зависеть от повышения коэффициента с д. ¦.
От этого коэффициента должны зависеть также изменения возбуди-
«иости на полюсах, т. е. электротонические явления. Так, известно, что
действие ионов калия и натрия повышает возбудимость как в нерве,
так и в мышце. Действие же Са и Mg, наоборот, понижает ее.
С точки зрения приведенных наблюдений Лё б, Лазарев (8) и
другие авторы объясняют физиологическое действие постоянного тока
на возбудимую ткань следующим образом. Как известно, при
пропускании электрического тока через растворы электролитов перенос
одновалентных ионов происходит быстрее, чем двухвалентных. То же
происходит в нерве и імышце при пропускании тока. Вследствие этого ионы Na
н К скорее достигают катодного участка, чем Са и Mg, и поэтому на
катоде коэффициент повышается, а на аноде понижается. Этим
обусловливается повышение возбудимости на катоде и понижение ее на
аноде. Кроме того, принимается, что возбудимость на аноде понижается
под влиянием накопляемых здесь анионов (Чаговец, 7, и др.)» а на ка~
тоде повышается также под влиянием іводородных ионов, переносимых
электрическим током наравне с ионами калия и натрия (Бете, 16).
Раздражающее действие электрического тока в момент
размыкания гальванического тока считается проявлением обратного переноса
ионов от катода к аноду, т. е. поляризационного тока. Теперь в
области анода и катода отношение ионов должно измениться в обратном
направлении и, соответственно, функциональное состояние должно
измениться в противоположном смысле: происходит понижение
возбудимости на катоде и повышение ее на аноде вместе с возникновением
здесь возбуждения.
Недавние опыты Воронцова (15) наглядно показали
правильность господствующего взгляда на происхождение электротонических
явлений. Он обнаружил, что при обработке нерва одновалентными ка-
Na,K
тионами соотношение ъч значительно повышается в нерве срав-
Са, М г г
нительно с нормальным состоянием. Это соотношение неблагоприятно
для возбуждения, так как для возникновения возбуждения требуется
накопление одновалентных ионов до известной концентрации, а эта
концентрация уже достигнута в нерве предварительной обработкой. И
вот оказывается, что в этих условиях для вызова возбуждения
необходимо не действие катода, как обычно, а действие анода, т. е. не
дальнейшее повышение концентрации одновалентных ионов, а, наоборот,
. ее понижение. Поэтому возбуждение возникает при замыкании тока
не на катоде, а на аноде. После же обработки двухвалентными ионами,
например кальцием, анод быстро понижает проводимость, катод же
действует раздражающе.
Более того, Воронцов (21), Виноградов (22) и др. показали,
что парабиотическое состояние, вызванное в нерве двухвалентными
ионами, можно устранить действием катэлектротона, т. е. приложением
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 455
катода к парабиотическому участку. Обратно, парабиоз, вызванный
одновалентными ионами, уничтожается анодом.
Эти изменения возбудимости под влиянием одно- и двухвалентных
ионов, а также анода и катода постоянного тока, сочетаются с
определенными заметными на-глаз структурными изменениями. Действие К и
ОН, а также действие катода производит видимое в микроскопе
разрыхление вещества осевых цилиндров, действие же Са и Н, а также
.анода — уплотнение его. При этом разрыхление, вызванное К и ОН,
устраняется действием анода, а уплотнение, вызванное Са и Н,
устраняется катодом (Макут, 26).
Означенные структурные изменения происходят параллельно с
изменением возбудимости: с уплотнением нервных мембран возбудимость
падает, с разрыхлением их до определенной степени возбудимость
повышается, а при дальнейшем разрыхлении она падает, наступает так
называемая католическая депрессия (Эббеке, 27). В гистологических
наблюдениях насчет действия иона Н существуют противоречивые данные.
Водородный ион как будто уплотняет нервные мембраны, подобно
аноду. По Эббеке (28), ион Н в малых концентрациях уплотняет
мембраны, а в больших, наоборот, разрыхляет. По Бете и др., ион Н
действует подобно катоду: он повышает возбудимость и вызывает
возбуждение (16). Несмотря на эти противоречия, можно
утверждать, что явление катэлектротона вызывается действием
одновалентных ионов, а анода — действием двухвалентных.
С этим пониманием физиологического электротона хорошо
согласуется и явление католической депрессии. Как указывалось выше, на
катоде при замыкании сильного постоянного тока после
первоначального повышения возбудимости происходит понижение возбудимости.
Зто связано с накоплением большого количества катионов в области
катода. Как известно, под влиянием большой концентрации
одновалентных ионов происходит парабиотическое понижение возбудимости после
начального повышения. Поляризация нерва в связи с длительным
пропусканием постоянного тока связана с возникновением
поляризационного тока, которое имеет направление, противоположное
поляризующему току. Это, в свою очередь, должно повлиять на физиологическое
состояние на полюсах: на катоде возбудимость должна понизиться,
сравнительно с первоначальным повышением, до нормы, а на аноде —
повыситься, сравнительно с первоначальным понижением. С этим
явлением должно быть связано обратное повышение пониженной
возбудимости на аноде при длительном пропускании постоянного тока.
Явление Шеминского. Длительное электротоническое действие
постоянного тока проявляется также в постепенном ослаблении
вызываемого им нервно-мышечного эффекта благодаря локальному физико-
химическому изменению возбудимой системы в области катода. Это
наблюдается хорошо и при ритмическом раздражении, причем чем
меньше интервал, тем скорее получается означенное локальное
изменение. Если изменить направление тока после того, как произошло
456 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
значительное ослабление механического эффекта, то будут вновь
получаться сильные эффекты. Через некоторое время они тоже ослабнут, но
обратное изменение направления тока вновь поведет к восстановлению
эффектов (рис. 206); так можно повторить много раз. Это явление
впервые было изучено Ше минским и его сотрудниками (23) на
мышцах путем прямого раздражения. Он же анализировал это
явление и пришел к заключению, что падение эффекта при повторных
Рис. 206. Локальное действие катода постоянного тока.
Кураризованная мышца (т. gastrocnemius) лягушки. Раздражается
сверхмаксимальными разрядами конденсатора 8 pF. Цифры
означают силу тока по потенциометру, служащему для ответвления
тока. Порог при 8, максимум при 36, раздражается при 44; 7 —
первая кривая утомления; 2 — вторая кривая утомления, t — ток
восходящий, т.е. катод на проксимальном конце мышцы, анод на
дистальном сухожильном конце. 4> — ток нисходящий, т.е. на
проксимальном мышечном конце находится анод. Кривую читать справа
налево (Ф. Шеминский и Фр. Шеминский).
раздражениях током одного направления вызвано не утомлением
мышцы вообще, а действием постоянного тока на полупроницаемые
перепонки мышечных волокон. Именно, предполагается, что на катоде
суммируется католическое разрыхление, вследствие чего этот участок
становится более проницаемым, ухудшающим условия для тех физико-
химических процессов, которые обусловливают возбуждение. При
перемене направления анод попадает на разрыхленную область и
уплотняет ее; этим самым проницаемость восстанавливается, імышца вновь
начинает отвечать возбуждением на раздражение. Такое же явление
получается на нервно-мышечном препарате при раздражении нерва
постоянным током (Уфлянд, 24; Цкипуридзе, 25). Отсюда следует, что
при длительном применении постоянного тока наблюдаемое явление
ослабления эффектов в значительной мере следует отнести за счет
электротонического действия электрического тока на полюсах, а не
утомления возбудимой системы.
Периэлектротон Введенского. В последнее время было открыто еще
одно замечательное изменение возбудимости нерва при пропускании
через него электрического тока. За пределами электротонических
изменений функциональное состояние нерва не остается неизменным. На
стороне катода за пределом катэлектротона на расстоянии около 2 см
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 457
от поляризующих электродов возбудимость понижается, а на другой
стороне, за пределом анэлектротона, наоборот, повышается. Эти
изменения возбудимости были открыты Введенскшм (17) и названы
периэлектротоническими явлениями, короче — периэлектротоном. Эти
изменения возбудимости очень легко наблюдать на нервно-мышечном
препарате, если на стороне мышцы на расстоянии более 2 см от
поляризующих электродов нерв раздражать отдельными индукционными
Периэлектротон Натэлектротои Анолектротон Периэлектроток
Рис. 207. Диаграмма распределения петель в области электротона и перн-
электротона в нервном волокне.
О — осевой цилиндр; М — миэлиновая оболочка; Р — перехват Ранвье; Я — нервное
окончание. В середине нервного йолокна пропускается постоянный ток. Стрелки
указывают направление петель тока.
ударами и вызывать ими одиночные сокращения. При действии
поляризующего тока нисходящего направления, т. е. с катодом на стороне
мышцы, одиночные сокращения ослабевают и даже прекращаются
(рис. 204, В), а в случае восходящего тока, с анодом на стороне мышцы,
эти сокращения усиливаются (рис. 205, Л).
Происхождение этих изменений возбудимости Введенским не
было объяснено. Его выяснил Гедевани (18), который установил
появление особого тока за пределами эл?ктротонической области.
Этот ток по своему направлению противоположен поляризующему
току. Гедевани полагает, что поляризующий ток, распространяясь
по нервным волокнам в виде физического электротона, на стороне анода
заходит в осевые цилиндры на протяжении многих миллиметров от анода
и затем идет по осевым цилиндрам к электродам. Так как осевой
цилиндр является достаточно хорошим проводником, миэлиновая же
оболочка проводит ток чрезвычайно дурно, ток должен входить в
осевые цилиндры главным образом через перехваты Ранвье и затем
проводиться обратно к электродам опять главным образом через
осевой цилиндр. Но по этой же причине часть этого тока идет по
осевому цилиндру в противоположном направлении до голых нервных
окончаний в мышце. По всему пути, где миэлиновая оболочка
прерывается в перехватах Ранвье и нервных окончаниях, ток выходит
наружу. Возникает множество петель тока, замыкающихся через
соединительнотканную оболочку нервных волокон и поверхность нерва
с тем участком, откуда ответвляется осевой ток (рис. 207). Такое же
распространение осевого тока, конечно, имеет место и по другую
сторону от электродов, но там ток противоположного направления.
Это внеполюсное распространение тока принята также называть
электротоническим, ибо от него зависит электротоническое изменение
возбудимости. На стороне анода в электротонической области,
где электрический ток входит снаружи в осевые цилиндры, про-
458 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
исходит понижение возбудимости — анэлектротон, а дальше, в пери-
электротонической области, где ток выходит из осевого цилиндра наружу,
происходит повышение возбудимости, как на катоде, т. е. периэлектро-
тон. Обратное получается на стороне катода. Итак, в основе периэлект-
ротонических изменений лежит один и тот же физический электротон,
который возникает благодаря большому различию в электропроводности
осевого цилиндра и оболочек. Но только вблизи электродов он имеет
такое же направление, как поляризующий ток, а дальше, в области пе-
риэлектротона — противоположное направление. Этот периэлектротони-
ческий физический ток настолько силен, что им можно пользоваться
для раздражения и поляризации другого нерва. Если отвести в нерв
другого нервно-мышечного препарата ток из периэлектротонической
области исследуемого препарата, этот периэлектротонический ток будет
действовать на данный нерв раздражающим образом и вызывать в нем
электротонические явления. Кроме того, Гедевани показал, что
между электротоническими и периэлектротоническими функциональными
изменениями нерва не существует никакого взаимодействия. Периэлек-
тротонические изменения сохраняются, если умертвить нерв без грубых
нарушений его структуры на границе между электротонической и
периэлектротонической областями.
Взаимосвязь электрического и периэлектротонического участков
изучал также Цкипуридзе (29). Он установил, что при замыкании
восходящего слабого тока наблюдаемое в периэлектротонической
участке повышение возбудимости длится недолго, уже через 2—3 мин.
возбудимость возвращается к норме. В это самое время в
электротоническом участке пониженная возбудимость остается без изменения.
При замыкании слабого нисходящего тока в периэлектротонической
участке возбудимость оказывается пониженной часами, а в
электротоническом возбудимость повышается сразу, но скоро падает
наполовину в течение 3 мин., и затем только держится часами слабое
повышение. При замыкании относительно сильного постоянного тока
в нисходящем направлении в области катода развивается католическая
депрессия после начального повышения возбудимости, в области же
периэлектротона возбудимость понижается и остается пониженной
все время. Отсюда ясно следует, что изменение возбудимости в
периэлектротонической области не зависит от ее изменения в
электротонической; между ними нет функционального взаимодействия.
Закон сокращений Пфлюгера. На раздражение нерва замыканием и
размыканием постоянного тока мышца реагирует различно в
зависимости от направления и интенсивности тока.
1. Очень слабый ток вызывает сокращение только при замыкании.
Это происходит как при ^направлении тока в сторону мышцы, при так
называемом нисходящем направлении, так и при обратном,
восходящем направлении.
2. Ток средней силы вызывает сокращение как при замыкании, так
и при размыкании при обоих направлениях тока.
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 459
3. Ток очень большой силы производит сокращение только при
замыкании, если направление тока нисходящее, или только при
размыкании, если направление тока восходящее.
В основе этих явлений лежит полюсной закон действия
электрического тока и физиологический электротон. При раздражении слабым
током того и другого направления сокращение наступает только на
катоде при замыкании, так как замыкание является более сильным
раздражением, чем размыкание. Токи средней силы обоих направлений
вызывают сокращение при замыкании и размыкании, как того требует
полюсной закон, но, конечно, при замыкании возбуждение возникает
на катоде, а при размыкании на аноде. При пользовании сильными
токами большую роль в получаемых эффектах играют электротонические
изменения. В случае восходящего тока сокращение наступает только
при размыкании, но не при замыкании, ибо возбудимость и
проводимость на ближайшем к мышце полюсе, т. е. на аноде, настолько
понижаются в момент замыкания, что возбуждение, возникшее на катоде,
не может пройти через анодный участок и достигнуть мышцы. При
размыкании возбуждение возникает на аноде и свободно может
распространиться до мышцы. В случае нисходящего тока сокращение
получается только при замыкании, так как при замыкании возбуждение,
возникшее на ближайшем к мышце катодном полюсе, беспрепятственно
доходит до мышцы, а при размыкании возбуждение, возникшее на
.аноде, не может дойти до мышцы в силу непроводимости в катодном
участке.
Часто случается, что и при слабых токах возбуждение возникает не только при
замыкании, но и при размыкании, и даже скорее при размыкании, чем при
замыкании. Это происходит в том случае, если анод постоянного тока лежит на слегка
поврежденном участке нерва. Вследствие обусловленной повреждением повышенной
нозбудимости размыкание дает такой же или даже более сильный эффект, чем
замыкание. Бели повреждение настолько глубоко, что сопровождается понижением
возбудимости, то эффекты извращаются еще больше. В этом случае к
раздражающему действию постоянного тока присоединяется такое же действие тока
повреждения, т. е. его замыкание и размыкание, которое происходит одновременно с
замыканием и размыканием постоянного тока. Именно, если анод лежит на
поврежденном участке, то замыкание будет давать эффект, как обычно, а размыкание
произведет эффект только при сильном токе. Наоборот, если катод лежит на этом
участке, то замыкание всегда будет производить эффект при более сильном токе,
чем размыкание.
Электротоническое действие индукционных ударов. Так как действие
индукционных ударов очень кратковременно, то при пользовании ими
более или менее значительно выраженный электротон не имеет места.
Это же является причиной того, что возбуждение возникает только на
катоде, т. е. в момент нарастания силы тока. В момент же ослабления
или исчезновения тока возбуждение не возникает на аноде, так как
исчезновение тока приходится в период абсолютной рефракторной
фазы от предыдущего возбуждения. Но так бывает только в том
случае, если продолжительность индукционного удара меньше
продолжительности абсолютной рефракторной фазы. Если же она почему-либо
460 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
достаточно велика, как это бывает при большом приближении вторичной
катушки к первичной (продолжительность размыкательного удара
возрастает до 10 о и более), то возбуждение может возникать и на аноде,
т. е. в момент исчезновения индукционного удара. Таким образом, на
каждый индукционный удар получится по два импульса возбуждения.
От действия индукционных ударов получаются также изменения
возбудимости: в момент нарастания силы индукционного удара на катоде
возбудимость повышается, а на аноде понижается. Это, между прочим,
проявляется в том, что при сильном сближении катушек индукционные
удары восходящего направления производят сокращение все слабее
и слабее и даже могут перестать вызывать его совсем. Катодное
возбуждение перестает вызывать сокращение от исчезновения
возбудимости на аноде, как это бывает при пользовании сильным
постоянным током того же восходящего направления. При еще большем
сближении катушек мышца вновь производит сокращение, которое
постепенно нарастает с усилением ударов. Описанные пробелы в
реакциях нерва при постепенном усилении восходящего индукционного
удара были впервые замечены Ф и к о м, а потому называются
пробелами Фика (19).
Вышеописанное явление наблюдается не всегда. Оно не бывает на
совершенно свежих препаратах при пользовании быстро протекающими
размыкательными индукционными ударами. Легче всего его наблюдать
на препаратах в плохом функциональном состоянии и при пользовании
длительно протекающими замыкательнымн индукционными ударами.
Появление сокращений по другую сторону пробелов первоначально
считали результатом возникновения возбуждений на аноде вследствие
исчезновения анэлектротона. К этому заключению приходит в
настоящее время и Воронцов с сотрудниками (21). Но есть основание
предполагать, что в определенных случаях оно обязано своим
происхождением возникновению Еторичных катодов вблизи анода вследствие
распространения катодного возбуждения от катода к аноду. При
пропускании индукционных ударов возбуждение возникает в области
перехватов Ранвье. При пороговых раздражениях возбуждаются
ближайшие к катодному полюсу перехваты. Но с усилением раздражения
одновременно возбуждаются и дальнейшие перехваты. Это было
установлено путем изучения скрытого периода сокращения. С усилением
раздражения он укорачивается. При некоторых больших силах
катодное возбуждение настолько приближается к аноду, что оно как будто
перекрывает состояние анода, уменьшая его депрессирующее действие
(Бете и Гейнрих, 20).
Ир а ктипум
Наблюдение полюсного действия постоянного тока. Нервно-мышечный препарат.
Полюсы раздражающих электродов отстоят один от другого на несколько
сантиметров. В этом опыте пользуются постоянным током средней силы, когда сокращение
вызывается как замыканием, так и размыканием тока. Посередине между полюсами
ПОЛЯРНОЕ ДЕЙСТВИЕ ЭЛЕКТРИЧЕСКОГО ТОКА НА НЕРВНО-МЫШЕЧНУЮ СИСТЕМУ 461
раздражающих электродов нерв перетягивается ниткой или умерщвляется
аммиаком (смазывание аммиаком производят при помощи стеклянной палочки). При
восходящем токе сокращение наступает только от размыкания, а при нисходящем —
только от замыкания.
Наблюдение физиологического электротона. Нервно-мышечный препарат
помещен во влажной камере. Постоянный ток пропускают через нерв, пользуясь непо-
ляризующимися электродами. На расстоянии 1—2 мм от электрода, ближайшего
к мышце, к нерву прикладывается пара платиновых электродов для раздражения
фарадическим током с целью наблюдения за изменениями возбудимости (располо-
Рис. 208. Постановка опыта для наблюдения электротонических явлений.
Нервно-мышечный препарат помещен во влажной камере. На нерв наложена пара неполяризую-
щихся электродов, соединенных с аккумулятором через потенциометр, и пара платиновых, соединенных
с индукторием.
жение приборов и препарата см. «а рис. 208). Сначала находят порог и затем
замыкают постоянный ток средней силы. Если ток нисходящий, т. е. если
ближайший к мышце электрод является катодом, тогда в момент замыкания порог
раздражения значительно понизится, что укажет на 'повышение возбудимости. Тотчас после
размыкания тот же порог повысится, что укажет на понижение возбудимости. При
восходящем направлении тока ближайший к мышце электрод будет анодом.
Сообразно с этим после замыкания порог повысится, а по размыкании он, наоборот,
понизится.
Наблюдение периэлектротона. Та же установка, как в предыдущем опыте. Свежий
нервно-мышечный препарат. Платиновые электроды накладывают около самой мышцы
ка расстоянии около 2 см от неполяризующихся электродов для постоянного тока.
Через платиновые электроды определяют пороги раздражения отдельными
индукционными ударами или фарадическим током. Если постоянный ток нисходящий
(катод на стороне мыщцы), то пороги повысятся, если же он восходящий (анод на
стороне мышцы), то пороги понизятся. Эти явления наблюдаются уже при слабых
поляризующих токах.
Опыт с явлением Шеминского. Нервно-мышечный препарат лягушки во влажной
камере. Раздражается нерв постоянным током через ншоляризующиеся электроды.
462 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕРЕБРОСПИНАЛЬНОЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Ток прерывается метрономом. Сокращение икроножной мышцы регистрируется на
кимографе. После ряда раздражений, когда эффект ослабевает в значительной
мере, изменяют направление тока.
Опыт с законом сокращений Пфлюгера. Нервно-мышечный препарат во влажной.
камере.. Неполяризующкеся электроды. Сначала пробуют постоянный ток небольшой
интенсивности. Ослабление тока достигается применением компенсатора (рис. 208),.
которым ток небольшой силы ответвляется от аккумулятора к препарату. Затем
проводят ток средней силы. Для этой цели от аккумулятора ответвляют компенсатором-
ток большой силы или весь аккумуляторный ток пускают к препарату. Для
получения сильных токов к препарату подводят ток от двух или трех аккумуляторов.
Нельзя пробовать слабые токи после применения сильных токов. Возбудимость-
раздражаемого участка меняется в такой мере, что может получиться извращенный
эффект. Например, если на аноде возбудимость была повышена или на катоде она
была понижена предыдущим приложением сильного тока, то слабые токи скорее-
будут давать сокращение при размыкании, чем при замыкании. Точно такие же-
извращенные эффекты могут получиться в случае механического или химического-
повреждения нерва; например, если сернокислый цинк из ближайшего к мышце
неполяризующегося электрода почему-либо проникает к нерву через глиняную пробку,
то возбудимость в этом участке падает и потому при восходящем токе ни
замыкание, ни размыкание не будут давать эффекта, так как под ближайшим к мышце-
?лектродом возбудимость и проводимость окажутся сильно пониженными.
Литература
1. Pfl tiger Е., Physiologie des Elektrotonus, Berlin, 1859.
2. Du Bois ReymondE., Untersuch. ub. d. tier. Elektrizitat, 2, 383, 390.
3. Loeb J., University California Publication, 3, 9, 1905; Pfl. Arch., 116, 193, 1907;:
Динамика живого вещества. Русск. перевод, 1910.
4. Werigo В., Pfl. Arch., 84, 260, 1901; Pfl. Arch., 31, 417, 1883.
5. Griinhagen A., Elektromot. Wirkung lebend. Gewebe, Berlin, 1871.
6. Pearna N., Pfl. Arch., 100, 145, 1903.
7. Чаговец В., Электрические явления в живых тканях, ч. 1, 1903; ч. 2, 1906.
8. Лазарев П., Исследования по ионной теории возбуждения, Москва, 1916.
9. НбЬег R., Physikalische Chemie d. Zelle u. d. Gewebe, Leipzig, 1922.
10. В i e d e r m a n n W., Sitzber. K- Akad. Wien. Mat.-nat. KL, 97, Abt. 3, стр. 84, 1884..
11. Пэр на H., Тр. физиол. лаб. Петерб. унив., т. 6—7, 1911—1912.
12. Matteuchi Ch., С. г. Acad. Sci., 56, 760, 1863; 56, 151, 194, 884, 1867; 66, 580, 1864..
13. Hermann L., Pfl. Arch., 5, 264, 1872; 6, 312, 1872; 7, 301, 1873.
14. Wedensky N., Pfl. Arch., 100, 5, 1903.
15. Woronzow D., Pfl. Arch. 218, стр. 148, 716, 1927.
16. Be the A., Pfl. Arch., 163, 147, 1916.
17. Введенский H., Изв. Росс. акад. наук, стр. 333, 1920.
18. Ге дева ни Д., Русск. физиол. журн., 15, 395, 1932.
19. Fie k A., Gesammte Schriften, 3, стр. 159, 176, 203, Wtirzburg, 1904.
20. Be the A. u. Heinrich A., Pfl. Arch., 229, 787, 1932.
21. Woronzow D. S. u. Judeni tsch M., Pfl. Arch., 224, 490, 1930.
22. В и н о г р а д о в М., Работы физиол. лаб. Петрогр. унив., 1914; Pfl. Arch., 204, 1924..
23. Scheminsky Ferd. u. Scheminsky Fr., Pfl. Arch., 225, 145, 1930.
24. Уфлянд Ю. M. и Шошина H. А., Физиол. журн. СССР, 17, 7, 1934.
25. Цки пу ри дзе Л., Из лаборатории И. Беритова. Не опубликовано.
26. Macku th E., Pfl. Arch., 214, 612, 1926.
* 27. Ebbecke U., Pfl. Arch., 195, 555, 1922.
28. Ebbecke U., Pfl. Arch., 195, 300, 1922.
29. Цкипуридзе Л., Бюлл. экспер. биол. и мед., 4, № 2, 129, 1937.
Часть четвертая
ОБЩАЯ ФИЗИОЛОГИЯ
ВЕГЕТАТИВНОЙ (АВТОНОМНОЙ)
НЕРВНОЙ СИСТЕМЫ
¦лшгт*
Глава 1
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ
Общее понятие. Вегетативная нервная система позвоночных
включает ту периферическую нервную систему, которая обслуживает все
внутренние органы, кровеносные сосуды и железы. Центробежные пути
этой системы, выходящие из спинного и головного мозга, оканчиваются
в гладкой мускулатуре, <в железистой ткани и в поперечнополосатой
мышце сердца. Вегетативная нервная система отличается от
цереброспинальной тем, что центробежные пути ее не доходят прямо до своего
секреторного или двигательного органа. Они оканчиваются около
нервных клеток, которые находятся как в ганглиях симпатического
пограничного столба, так и во всевозможных других ганглиях,
разбросанных во внутренних органах и вообще по ходу волокон вегетативной
нервной системы. Здесь оканчиваются нервные волокна, берущие
начало в центральной нервной системе, а отсюда начинаются новые
нервные волокна, которые уже доходят до периферических органов.
Каждый центробежный путь прерывается около ганглиозных клеток только
один раз. Он может пройти через несколько ганглиев, но перерыв
произойдет около клеток только в одном из них. Единственным
исключением является центробежный путь к мозговой части надпочечников,
которая несет секреторную функцию. Этот путь не прерывается
(Эллиот, 9). Нервные волокна, берущие начало в центральной системе
и оканчивающиеся в ганглиях, называются преганглионарными;
те же волокна, которые начинаются в ганглиях и кончаются в гладкой
мышце или в железах, называются постганглионарными
(рис. 209). Этими фактическими данными мы обязаны видному
английскому физиологу Лэнгли (1).
Преганглионарные волокна по пути следования отдают коллатерали,
которые, в свою очередь, оканчиваются где-либо в ганглиозных узлах,
иногда в очень отдаленных. Количество коллатералей может быть
довольно значительно. Так, в шейном симпатикусе одно преганглионар-
ное волокно приходится на 32 постганглионарные клетки (Рэисон
и Биллингсли, 2).
30 И. С. Беритов
466
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
До последнего времени полагали, что все преганглионарные волокна,.
подобно обычному цереброспинальному волокну, имеют миэлиновук>
оболочку. Постгакглионарные же волокна лишены ее. По новейшим
исследованиям оказалось, что.некоторая небольшая часть преганглио-
[Ganglmestritj
ABC О Е
Рис. 209. Схематическое изображение строения автономной нервной системы.
А — кожная поверхность на спине; В — спинной мозг; С — симпатический пограничный столб; D —
симпатические ганглии во внутренних органах; Е — внутренние органы; Ь, ЬХ) Ь2—нервные клетки
спинного мозга; Dr. — потовые железы; At. — кожные артерии; Аг. — мышечные волокна, двигающие
волосы; Gangl. vert. — спинномозговые ганглии; R. comm. gr.—серые rami communicantes: R. comm.
alb. — белые rami communicantes; Periph. Gangl. — периферические ганглии вблизи внутренних
органов; Praev. Gangl. — крупные симпатические ганглии; е — артерии внутренностей; et—кишка; е2—
мочевой пузырь; преганглионарные нервные волокна изображены непрерывной линией, а постганглио-
нарные — пунктиром. Здесь только приведены центробежные пути: двигательные и секреторные (по
Шульцу).
нарных волокон также лишена миэлиновой оболочки. Наоборот, и среди
постганглионарных некоторая небольшая часть волокон обладает
миэлиновой оболочкой. Так, в отношении, симпатических волокон верхнего
шейного узла кролика известно, что около одной трети преганглионар-
ных волокон являются немиэлиновыми, а среди постганглионарных
около одной десятой являются імиэлиновыми. Миэлиновые
преганглионарные волокна всегда оканчиваются синапсами как на миэлиновых, так
и на немиэлиновых постганглионарных нейронах. Немиэлиновые преган-
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕГВНОЙ СИСТЕМЫ 467
глионарные волокна всегда оканчиваются на немиэлиновых постганглио-
нарных нейронах. То же самое было найдено в отношении верхнего
шейного узла у кошек и обезьян (Бишоп и Гейнбекер, 3).
Новейшие экспериментально-морфологические исследования
парасимпатической системы дают такую картину строения автономной
нервной системы. На основании изучения окончаний нервов
парасимпатической системы (п.vagus, п. erigens) в нервных сплетениях
пищеварительного канала (ауэрбаховское и мейснеровское сплетения) было
установлено, что волокна п. vagus и п. erigens прерываются около клеток
Рис. 210. Клетки из нервных узлов вегетативной системы.
А — клетка из ауэрбаховского сплетения в пищеводе. На ней перицеллюлярный аппарат с его преган-
глионарным волокном дегекерированы после перерезки блуждающих нервов (Колосов и Сабусов); В —
нервная клетка из верхнего шейного симпатического ганглия человека. Она окружена корзинковидным
сплетением пренглионарного волокна (Штёр мл.).
определенного типа, нейриты которых заканчиваются в мышечных
клетках. Эти клетки известны как двигательные клетки 1-го догелев-
ского типа. У этих клеток парасимпатические волокна образуют
сложный перицеллюлярный аппарат, который после перерезки
парасимпатических волокон перерождается вместе с волокнами (рис. 210)
(Лаврентьев, 4, 18; Колосов, 5). Этим путем было установлено, что
преганглионарные нейроны оканчиваются на теле клеток постганглио-
нарных нейронов. Синаптические окончания на постганглионарных
нейронах были открыты в последнее время и на живом объекте. Так,
на сердце лягушки при большом увеличении были замечены окончания
блуждающего нерва на теле нервных клеток в узле Людвига, а также
установлено их изменение в связи с возбуждением блуждающего
нерва. Они меняют свои оптические свойства: контуры делаются
отчетливыми, блестящими (см. рис. 155—II) (Лаврентьев и Федоров, 17).
Но наряду с нейронными элементами симпатические ганглии
содержат густое сплетение из тончайших нервных разветвлений преганглио-
нарных волокон, переплетенных с разветвлениями дендритов
постганглионарных нейронов (рис. 211). Эти нервные элементы, образуя
клубок, вступают между собой в многочисленные интимные связи.
В определенных случаях этот нейрофибриллярный клубок окружает
ганглионарные клетки в виде перицеллюлярных аппаратов, в других
30*
468 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
случаях он лежит в стороне от них; иногда он занимает в ганглии
центральное положение, а клетки окружают его, как это бывает
в нервных узлах беспозвоночных животных. Одни новейшие гистологи
утверждают, что в этом клубке нервные элементы преганглионарных
волокон сливаются с дендритами постганглионарных нейронов,
образуя первичный нейропиль синцитиального характера (Штёр, 6;
Тигс, 7). Другие отрицают это и находят строгое разграничение
между элементами того и другого нейронов. При перерезке
преганглионарных волокон дегенерирует не весь клубок, а только та часть
Рис. 211. Три нервных клетки, отростки которых образуют
клубок. Из верхнего шейного симпатического ганглия
человека. Увел, в 500 раз (Штёр мл.).
его, которая относится к разветвлениям преганглионарных волокон
(Лаврентьев, 4, 18; 'Колосов, 5; Рэнсон и Биллингсли, 2; Кастро, 8).
Это явление должно служить наилучшим доказательством того, что
преганглионарные волокна заканчиваются в означенном клубке нейро-
фибриллей путем синапсов. В самое последнее время хорошо были
изучены синапсы .преганглионарных волокон. Небольшими
округленными утолщениями диаметром 1—7 jji они располагаются как на теле
клеток, так и на поверхности дендритоз. Большинство из этих
синапсов боковые, причем одно конечное разветвление преганглионарного
волокна касается своими боковыми синапсами многих клеток и дендри-
тсв или одной и той же клетки во многих точках (Гибсон, 20). Итак,
в симпатическом ганглии мы имеем нейропиль, типичный для
центральной нервной системы позвоночных животных; он состоит из густого
сплетения дендритных отростков, в котором боковыми и конечными
синапсами оканчиваются разветвления преганглионарных нервных
волокон.
Единственное место, где синцитий, действительно, может быть
наблюдаем, это ганглии ауэрбаховского сплетения, где дендриты
соседних клеток 1-го догелевского типа сливаются между собой,
как показано на рис. 212. Такой синцитий наблюдали многие авторы,
между прочим Догель и Лаврентьев. Но Лаврентьев по-
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 469
лагает, что синцитиальные связи между клетками ауэрбаховского
сплетения составляют исключение и можно рассматривать их как не
завершившееся полностью отделение одного нейрона от другого в
момент развития (18).
Постганглионарные волокна той и другой систем, заканчиваясь
в гладких мышечных клетках, образуют небольшую нейрофибрилляр-
ную сеть или колечки, которые лежат в углублении ядра — или свободно
в протоплазме, или на поверхности клеток. Это было найдено в ресни-
чатой мышце человека (рис. 131) (Буке, 13), в стенке желудка чело-
Рис. 212. Ганглий ауэрбаховского сплетения из
области пищевода. Ясно видно синцитиальное строение.
Нервные клетки связаны между собой отростками
(Колосов).
века (Тигс, 7), в прямой кишке и в retractor penis кошки (Колосов, 12).
Аналогичное было найдено в мышечных волокнах сердца кролика и
человека. Здесь одни нервные волоконца пробегают в протоплазме и
могут заканчиваться даже в анастомозах между волокнами; другие
волоконца оканчиваются петельками вблизи ядер (Вуллард, 14). В
железах постганглионарные волокна образуют густое сплетение, откуда
отдельные веточки достигают клеток в виде многочисленных петелек
и утолщений на теле клеток. Так бывает, по крайней мере, в слюнных
и желудочных железах, а также в мозговой части надпочечников.
В печени волоконца из густого сплетения вступают внутрь печеночных
клеток и заканчиваются около ядер, образуя тонкую сетку, подобно
тому как это бывает в гладких мышечных клетках (Арнштейн, 15;
Ригель, 16).
Симпатическая и парасимпатическая вегетативные системы. Смотря
по тому, откуда выходят преганглионарные волокна, вегетативная
система делится на два отдела: на симпатическую и парасимпатическую
системы.
I. Симпатическая система включает в себя симпатический
пограничный столб. Преганглионарные волокна выходят из грудного и
поясничного отделов спинного мозга, начиная с первого грудного сегмента до
четвертого поясничного сегмента (рис. 213). Все преганглионарные
470 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
волокна проходят через пограничный столб. Эти волокна частью
прерываются здесь же в симпатических узлах столба, частью проходят
через один или несколько узлов и затем прерываются где-либо в
симпатических узлах внутренних органов.
II. Парасимпатическая система включает ту часть вегетативной
системы, преганглионарные волокна которой выходят из головного мозга
(краниальная система) и из крестцового отдела спинного мозга
(сакральная система). Головные вегетативные волокна выходят вместе
с глазодвигательными, лицевыми, языкоглоточными и блуждающими
нервами. Крестцовые же волокна выходят вместе с крестцовыми
нервами, участвуют в образовании plexus hypogastricus и здесь
распределяются в ветвях nervus pelvicus.
Область распространения симпатической и парасимпатической
систем не одна и та же. Симіпатическая система распространена во всем
организме, парасимпатическая же только в определенных областях.
Но эти области получают волокна и от симпатической системы.
Следовательно, одни органы получают волокна от обеих систем, а
другие— только от симпатической. К последним относятся: мигательная
перепонка кошки, селезенка, мочеточник, потовые железы, гладкие
мышцы кожи, а также стенки некоторых кровеносных сосудов во
внутренних органах. Значит, большая часть кровеносных сосудов и
внутренние органы получают нервные волокна из обеих вегетативных
систем (рис. 213, 214).
Некоторые авторы находят спинномозговую заднекорешковую парасимпатическую
систему. Полагают, что очень тонкие преганглионарные волокна ее начинаются
в спинном мозгу, выходят через задние корешки и синашически оканчиваются
в спинномозговом ганглии на клетках; здесь же начинаются постгангл.ионарные
волокна. Этим волокнам приписывается иннервация сосудистой системы и желез
внешней и внутренней секреции, гладких мышц желудка, кишечника, а также
скелетных поперечнополосатых мышц (Кюре и сотр., 10). Но существование такого
рода парасимпатических .волокон, которые прерываются в спинномозговом ганглии,
оспаривается и требует дальнейшего исследования (Капперс, 11; Лаврентьев, 18).
Основными сведениями о строении вегетативной нервной системы мы обязаны
главным образом упомянутому английскому физиологу Лэнгли (1). Он же дал ей
новое название автономной системы, имея в еиду ее анатомическое и
физиологическое своеобразие. Л э н г л и относит к автономной системе центробежные пути
симпатической и парасимпатической систем с их ганглиями, а также нервные
сплетения Ауэрбаха и Мейснера, которые находятся в стенках пищеварительного канала
и не имеют прямой связи с центральной нервной системой. Многие авторы находят,
что это название не соответствует действительности, что так называемая автономная
нервная система не является действительно автономной. Она, подобно остальной
периферической нервной системе, находится в очень тесной анатомической и
физиологической связи с центральной нервной системой. Поэтому данные авторы
предпочитают сохранить старое название вегетативной нервной системы как за
симпатическими нервами и узлами (в старом и узком смысле слова), так и за всеми
родственными им образованиями.
Никотинный метод исследования вегетативной нервной системы.
Гиршман (1863) первый показал, что никотин парализует у
кроликов симпатическую систему: например, после никотинного отравления
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 471
<Чі СО <Г
о и
? 2 о ' ^п
U ? Ь ют- fa
U~!
472 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
раздражение не вызывает расширения зрачка. Этим свойством нико-
тина широко воспользовался Лэнгли. Он отравлял им всё животное-
путем интравенозной инъекции никотина или смазывая определенные
ганглиозные узлы 1—2% раствором никотина. Действие никотина всегда
проявляется одинаковым образом: оно парализует передачу
возбуждения в ганглиозных узлах от преганглионарного волокна на клетки пост-
ганглионар'ных нейронов; но при этом совсем не влияет на преганглио-
нарные и постганглионарные волокна. Поэтому при отравлении всего
животного раздражение преганглионарных волокон совсем не
вызывает периферического эффекта, в то время как раздражение посттан-
глионарных волокон производит обычный эффект. В случае же
отравления одного узла раздражение соответствующих ему
преганглионарных волокон на периферии в одних местах вызывает обычный
эффект, в других—не вызывает. Значит,
некоторые волокна заканчиваются в
отравленном узле, а другие —¦ нет. Путем отравления
узлов, лежащих по пути следования
преганглионарных волокон, можно установить,,
где именно заканчивается каждое волокно.
Таким путем Лэнгли выяснил все
взаимоотношения нервных узлов и нервных
волокон вегетативной нервной системы.
По новейшим исследованиям,
характерное действие никотина, применяющегося в
некоторых малых концентрациях,
обусловливается параличом преганглионарных
окончаний, а не ганглиозных клеток
симпатического узла. Это видно из того, что после
никотинного отравления верхнего
симпатического ганглия раздражение
преганглионарных волокон не дает обычных для него
эффектоз, например расширения зрачка,
движения мигательной перепонки; введение
же ацетилхолина в ганглий через артерию
производит этот двигательный эффект,
очевидно, путем непосредственного
раздражения ацетилхолином ганглиозных клеток
(Бакурадзе, 19).
Литература
1. L a n g I е у J. N., Text book of Physiology, 2,
264, 1900; Brain, 26, 1, 1903; Erg. Physiol., 2, 813,
1903; Das autonome Nervensystem, Berlin, 1922.
2. R a n s о n S. W. а. В i 11 i n g s 1 e у P. R., J. comp,
Neur., 29, 313, 1918.
3. В i s h о p G. H. a. H e i n b e r k e r P., Am. J. Physiol., 100, 519, 1932.
4. Lawrentjew B. L, Z. mikr. Anat. Forsch., 23, 1931; 35, 1934.
8
9
Щ
Гт
11
Рис. 214. Диаграмма сердечных
нервов.
7—ганглиозная клетка блуждающего
нерва в продолговатом мозгу; 2 —¦
преганглионарное миэлиновое
волокно блуждающего нерва; 3—его
окончание при нервной клетке сердца (4);
5 — постганглион арное безмякотное
волокно с окончанием (б) в сердечной
мышце; 7 — клетка центробежного
ускоряющего нерва в мозгу; 8 — его
миэлиновое волокно, проходящее
через белый г. communicantes; 9 — его
окончание около нервных клеток gl.
stellatum (10); 11— постганглионар-
ное безмякотное волокно и его
окончание (72) в сердечной мышце (Пе-
фрей и Филиппе).
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 473-
5. Колосов Н. Г., Тр. Тат. инст. теор. и клин, мед., вып. 2, стр. 18, 1936.
6. Stonr Ph. jr., Mikroskop. Anat. d. vegetativen Nervensystems, Berlin, 1928.
7. Tiegs O. W., Austr. J. Exp. Biol. Med. Sci., 4, 79, 1927.
8. deCastro F., Cytology a. Cellular Pathology of the Nervous System, New York, 1932.
9. E 11 i о t T. R., J. Physiol., 46, 285, 1913.
10. Kure К- и сотр., Quart. J. Exp. Physiol., 18, 333, 1928; ib., 20, 51, 1930; ib.r
20, 1, 103, 1931; Z. Zellforsch. u. mikr. Anat, 22, 193, 1936.
11. Ariens Kappers, The сотр. anat. of the nervous syst. ttct p. 236, 1936.
12. Колосов п. Г. и Мещеряков A. M., Тр. Каз. мед- инст., вып. 2, стр. 229г
1938; Арх. анат., гистол., эмбриол., 20, 279, 1939.
13. ^oeke J., Kgl. Acad. Wetensch. Amsterdam. Proceed., 17, 1915.
14. Woo Hard H. H., J. Anat., 60, 345, 1926.
15. Arnste in C, Anat. Anz„ 10, 410, 1894.
16. Riegels L., Arch. mikr. Anat. Forsch., 14, 73, 1928.
17. Л ав d e нть ев Б. И. и Федоров Б. Г., Биол. действие ультравыс. частот
(ультракоротких волн), ВИЭМ, стр. 145, 1937.
18. Л а в р е нть е в Б. И., Морфология автономной нервной системы. Сб. работ под
ред. проф. Лаврентьева, стр. 5, 1939.
19. Бакурадзе А., О действии ацетилхолина и никотина на симпатический
ганглий. Не опубликовано.
20. Gibson С, J. Neurophysiol., 3, 235, 1940.
Г л а в а II
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ
Физиологические особенности преганглионарных волокон.
Возбудимость миэлиновых преганглионарных волокон, как это полагается
тонким миэлиновым волокнам из группы В> сравнительно низка. Чтобы
вызвать двигательный или секреторный эффект раздражением
преганглионарных волокон, необходимо приложить к ним более сильные
индукционные токи, чем к соматическому двигательному нерву. Так,
пороги раздражения преганглионарных волокон лягушки состазляют
15—20 см расстояния между катушками, в то время как для
двигательного нерва той же толщины пороги раздражения составляют
35—40 см.
Хронаксия преганглионарных волокон, судя по вызываемым ими
двигательным эффектам в гладкой мускулатуре, очень велика. Так,
например, по данным Уфлянда (44), хронаксия блуждающего нерва
по эффекту желудка лягушки равняется 1 j, а по эффекту сердца
лягушки — 2 а. Она оказывается даже длиннее, чем хронаксия более
тонких постганглионарных волокон (Шошары, 46). Но если определять
хронаксию преганглионарных волокон по токам возбуждения самих
преганглионарных волокон, то она оказывается значительно меньшей.
Так, например, хронаксия преганглионарных волокон верхнего шейного
симпатического ганглия кошки, судя по токам возбуждения этих же
волокон, равняется 0.4—0.5 а. Она в шесть раз меньше хронаксии
более тонких безмякотных постганглионарных волокон (Аршавский,
43). Эти противоречия объясняются тем, что преганглионарные волокна
вызывают периферический двигательный эффект через ганглий, а ган-
глионарные клетки при определенном функциональном состоянии не
возбуждаются одним нервным импульсом. Для этого требуется ряд
импульсов. Клетки возбуждаются в результате суммации. Поэтому при
определении реобазы преганглионарных волокон по эффектам мышцы
приходится значительно усилить постоянный ток, чтобы получить в
нерве ряд импульсов. По той же причине хронаксия получается
длительной, ибо она отвечает действию нескольких нервных импульсов на ган-
глионарные клетки. Правда, периферический эффект с преганглионар-
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 475
ных волокон получается и от одиночного стимула, но только в том
случае, если он длится долго — около 3 а (Аршавский, 43). В этом
случае в преганглионарных волокнах должен возникать не один
импульс, а ряд импульсов. Что действительно число и ритм импульсов
возбуждения преганглионарных волокон имеют значение для передачи
возбуждения через симпатический ганглий, хорошо видно из опытов
Б р ю к к е (47). Он наблюдал на кошках, что хронаксия
преганглионарных аолокон в отношении двигательного эффекта 3-го века или
расширения зрачка тем меньше, чем чаще следуют разряды
конденсатора. Она может быть снижена в 10 раз сравнительно с тем, что
бывает при одиночных разрядах.
Процесс возбуждения преганглионарных волокон, на основании
изучения биотоков возбуждения и рефракторных фаз, должен длиться
несколько сигм. Так, установлено, что у холоднокровных
продолжительность абсолютной рефракторной фазы равняется 3.5—7 а, а у
теплокровных — 2.2—4.5 J . Значит, продолжительность процесса
возбуждения преганглионарных волокон холоднокровных должна быть несколько
меньше: 3.5—7 j, а у теплокровных — несколько меньше 2.2—4.5 J
(Бишоп и Гейнбекер, 5). В некоторых случаях у теплокровных
животных продолжительность как тока возбуждения, так и абсолютной
рефракторной фазы уменьшается до 1.2—1.5 7 (Грундфест, 24).
Специальные исследования над биотоками возбуждения,
рефракторной фазой и скоростью проведения возбуждения в преганглионарных
волокнах выявили, что эти волокна не представляют вполне однородной
массы. По данным Э к к л с а (7), на основании изучения характера
биоэлектрических токов возбуждения, скорости распространения
возбуждения и других проявлений, преганглионарные волокна верхнего
симпатического ганглия кошки можно разбить на четыре группы,
причем первые три группы считаются состоящими из миэлиновых волокон,
а четвертая из немиэлиновых. Первая группа преганглионарных
волокон проводит возбуждение с наибольшей скоростью —- около 20 м в
секунду, вторая — около 12 м, третья — около 8 м, а четвертая — около
3 м. Первая группа волокон обладает несколько более высокой
возбудимостью, чем вторая, возбудимость третьей группы в 1!/г раза ниже,
а четвертой — в четыре раза ниже, чем первой.
После одиночного возбуждения и рефракторных фаз возбудимость
не возвращается сразу к норме и не устанавливается фаза сверхнор-
мзльной возбудимости. Относительная рефракторная фаза сильно
удлиняется, и возбудимость достигает нормы через 150 а , как это вообще
свойственно тонким миэлиновым волокнам. Соответственно, у этих
волокон после одиночного возбуждения обычно отсутствует фаза
отрицательного электрического последействия (рис. 215). После тока
возбуждения сразу устанавливается положительное последействие. Но
отрицательное электрическое последействие появляется при тетаниче-
ском раздражении. Оно возникает еще во время раздражения и
продолжается после него некоторое время (Грундфест, 24).
476 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Высший предельный ритм возбуждения преганглионарных волокон
должен быть высоким, поскольку абсолютная рефракторная фаза их
равняется нескольким сигмам; соответственно и лабильность их должна
быть сравнительно высокой. Специальных исследований по этому поводу
нет, но, исходя из явления рефракторных фаз, надо полагать, что
высший предельный ритм возбуждения должен быть у холоднокровных
150—300, а теплокровных — 200—450 в секунду.
Физиологические особенности постганглионарных волокон. К пост-
ганглионарным волокнам относятся главным образом тонкие безмиэли-
&%
110
200 300 - 400 50О бОО 700 ?ООб
Рис. 215. Кривая восстановления возбудимости после одного
импульса возбуждения в нервных волокнах.
На ординате цифры выражают степень возбудимости в числах, обратных
пороговой силе раздражения, взятой в процентах нормального порога. На
абсциссе время в сигмах. А — самые толстые миэлиновые волокна,
какие находятся в л. phrenicus кошки, В —ттонкие миэлиновые волокна и
С — миэлиновые волокна из 'симпатической системы ЧГассер).
новые волокна С, обладающие, как известно, низкой возбудимостью'.
Возбудимость постганглионарных волокон ниже преганглионарных
и вообще цереброспинальных. Это ясно видно при сравнении порогов
раздражения симпатических и цереброспинальных волокон: при
раздражении седалищного нерва для вызова симпатического эффекта на
кровеносных сосудах конечности требуется большая сила раздражения,
чем для вызова сокращения поперечнополосатых мышц конечности.
Хронаксия постганглионарных волокон, судя тю эффектам иннерви-
руемой ими гладкой мускулатуры, равняется нескольким сигмам. Так,
например, хронаксия постганглионарных волокон для 3-го века кошки
составляет 1.5—4.5 а, для вазоконстрикторов лягушки и собаки она
равна 2 а, для п. splanchnicus — 3.2 а (Уфлянд, 44; Аршавский, 43).
Процесс возбуждения в постганглионарных волокнах, как и должно
быть у волокон С, продолжается значительно дольше, чем в
преганглионарных, принадлежащих к волокнам В. Специальное изучение
физиологических свойств волокон п. splanchnicus показало, что в волокнах С
лягушки процесс возбуждения при 18—19° С продолжается около 8
Это заключение было сделано на основании определения продолжитель-
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 477
ности абсолютной рефракторной фазы и тока возбуждения. Затем
следует относительная рефракторная фаза, длящаяся около 30 а. Процесс
возбуждения сопровождается быстро протекающим током возбуждения.
Но за ним следует длительное положительное колебание
продолжительностью 1—2 сек. Отрицательное колебание наступает после одного
импульса при особых условиях, например после отравления вератрином.
У теплокровных в волокнах С из разных отделов вегетативной
системы процесс возбуждения продолжается 2 о. Сообразно с этим ток
возбуждения и абсолютная рефракторная фаза продолжаются 1.8—2 а.
Рис. 216. Токи возбуждения постганглионарных волокон кошки (при
ganglion stellatum) до и после тетанизации.
А — в покойном нерве; В — последний ток возбуждения с отрицательным
последействием после тетанического раздражения частотой 12 в 1 сек. и продолжительностью в 45
сек.; С — конец короткого 1-секундного тетанического раздражения свежего нерва; D —
конец 10-секундного тетанического раздражения в ритме 15 в 1 сек.; сильно,выраженное
положительное последействие (Бронк и сотр.).
Относительная рефракторная фаза длится около 20 а. Затем идет
сверхнормальная фаза, т. е. период повышенной возбудимости,
продолжительностью около 60 а. Затем она сменяется субнормальной фазой,
периодом пониженной возбудимости, длящимся свыше 600 а- Наряду
с изменениями возбудимости происходят длительные электрические
колебания. Непосредственно вслед за биотоком возбуждения
наблюдается длительное отрицательное отклонение. Оно совпадает с
периодом повышенной возбудимости. Затем следует положительное
колебание, которое совпадаете периодом пониженной возбудимости (рис.215).
Отрицательное, как и положительное последействия могут
совершенно отсутствовать. Оба появляются в связи с повторным
раздражением. Но первым, повидимому, возникает отрицательное колебание, а
потом оно постепенно уступает место положительному. Хорошая
иллюстрация токов возбуждения постганглионарных волокон и их
изменения в связи с повторным раздражением дана на рис. 216 (Бронк
в сотр., 18).
При сравнительном исследовании преганглионарных и
постганглионарных волокон теплокровных оказалось, что, в то время как в
преганглионарных волокнах абсолютная рефракторная фаза составляла
2.2—4.5 а , в постганглионарных волокнах она равнялась 4—5 а . У хо-
478 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
лоднокровных абсолютная рефракторная фаза постганглионарных
волокон длится больше — 4.5—10 г против 3.5—7 о в преганглионарных
(Бишоп и Гейнбекер, 5). Исходя из рефракторных фаз, высший ритм
возбуждения, а значит, и лабильность постганглионарных волокон,
несколько ниже, чем в преганглионарных. Для холоднокровных она
соответствует 100—220, а для теплокровных 200—250 в секунду.
Конечно, чтобы получить пессимальное явление на нервно-мышечном
препарате: постганглионарный нерв—гладкая мышца, можно не
повышать ритма раздражения до 100—250 в секунду. Так как нервные
импульсы ослабевают при гораздо меньших частотах раздражения
вследствие попадания их в относительную рефракторную фазу, то
пессимальное состояние можно получать при столь малой частоте
раздражения, как 60—120 б секунду. Так, например, при раздражении
постганглионарных волокон мигательной перепонки глаза кошки
(3-е веко) оптимальной частотой является 30 в секунду; 60
раздражений в секунду уже дают пессимальное сокращение мигательной
перепонки, а 160—только начальное вздрагивание (Аршавский, 43).
Проводимость возбуждения в постганглионарных волокнах также
оказывается значительно меньше, чем в цереброспинальных. Так,
например, в симпатических волокнах лягушки, проходящих через
седалищный нерв и оканчивающихся в стенках сосудов, скорость
проведения возбуждения равна 2 м (Фишер и Шильф, 6).
При сравнительном исследовании скорости проведения в
постганглионарных и преганглионарных1 волокнах симпатической системы
внутренних органов скорость проведения последних оказывается
значительно выше. У теплокровных в постганглионарных волокнах она
равняется 2—1 м в секунду, в преганглионарных— 15—10 м; у
холоднокровных в первых — 0.8—0.3 м, а в последних — 4.5—3 м (Бишоп
и Гейнбекер, 5).
Физиологические особенности разных постганглионарных волокон
неодинаковы. Волокна верхнего шейного ганглия млекопитающих делятся
на четыре группы, из которых первая группа относится к миэлиновым,
а остальные три — к немиэлиновым. Между ними была замечена
следующая физиологическая разница: первая группа проводит возбуждение
со скоростью около 5—8 м, а в остальных проведение происходит много
медленнее: 3—1 м в секунду (Экклс, 7).
Из всех приведенных іфактов вытекает, что в безмякотных
постганглионарных волокнах процесс возбуждения, его проведение и вообще все
физиологические процессы протекают более медленно, чем в
преганглионарных и цереброспинальных мякотных волокнах.
Физиологические особенности симпатических ганглиев. Деятельное
состояние в симпатическом ганглии нормально вызывается импульсами
возбуждения, притекающими из центральной нервной системы через
преганглионарные волокна. Искусственно оно может быть вызвано
электрическим раздражением преганглионарных волокон или
пропусканием через ганглий таких веществ, которые раздражают клеточные-
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
479'
элементы и не раздражают нервных волокон. Такими веществами
являются, например, определенные концентрации ацетилхолина или
калия. Вызванное этим путем деятельное состояние симпатического
ганглия изучается по токам самого ганглия или постганглионарных
волокон и по тем эффектам, которые наступают через постганглио-
нарные волокна в эффекторах.
При искусственном раздражении преганглионарных волокон
симпатический ганглий получает импульсы не в одно и то же время. Благо-
Рис. 217, Биоэлектрические
токи возбуждения верхнего
симпатического ганглия
кошки при разных силах
раздражения преганглионарного
нерва.
В 1-м опыте сила раздражения
равняется 3.3 условным
единицам, во 2-м — 5.0, в 3-м—20, в
4-м—10, в 5-м- 100, в 6-:д—50.
Цифры указывают нумерацию
волн. N обозначает
отрицательное длительное колебание
электрического последействия, а Р —
положительное длительное
колебание (Экклс).
Рис. 218. Биоэлектрические токи
возбуждения верхнего шейного ганглия кошки при
двух раздражениях преганглионарного нерва
во время длительного отрицательного
колебания.
Раздражение возбуждает только группу волокон,
производящих первую (волну. После тока
возбуждения длительное отрицательное колебание
электрического потенциала. Во время этого отрицательного
колебания повторное раздражение производит
усиленный ток возбуждения. В 1-м опыте второе
раздражение происходит через 35 а, во 2-м через 70s,
в 3-м через 110 з; в 4-м производится одно второе
раздражение, в 5-м через 150з, в 6-м через 90, в 7-м
через 55 з. Наверху время по 0.01 сек. (Экклс).
даря разной возбудимости и проводимости нервных волокон,
симпатический ганглий отвечает на одно раздражение нервного ствола рядом
импульсов. При слабом раздражении наступает только одна волна тока
возбуждения, соответствующая первой, наиболее возбудимой группе
волокон (рис. 217, оп. 1). За ней следует длительное отрицательное
колебание (/V), как это бывает и в цереброспинальных нервных
волокнах. При более сильном раздражении наступают первая и вторая
быстрые волны (оп. 2). За ними следует сна-чала длительное
отрицательное колебание, а потом длительное положительное. При еще более
сильных раздражениях получаются вначале три волны (оп. 3—4), а
затем и четыре волны (оп. 5—6). За ними следует сначала отрицательное
480 ОБЩАЯ ФИЗИОЛОРИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
колебание, а затем очень длительное положительное (Р).
Во время длительного отрицательного колебания возбудимость
ганглия повышена, а во время положительного она, наоборот, пони-
жена, как это известно в отношении цереброспинального нерва.
Это отчетливо выступает при повторении раздражения во время этих
длительных колебаний. На рис. 218, например, повторное раздражение
производится во время длительного отрицательного колебания и
вызывает почти вдвое более сильную волну возбуждения, чем первое
раздражение. На рис. 219, наоборот, повторное раздражение производится во
Рис. 219. Биоэлектрический ток возбуждения
верхнего шейного ганглия кошки при двух
раздражениях преганглионарного нерва во время
длительного положительного колебания.
Первое раздражение сильное, и оно возбуждает первые две
группы волокон, а второе раздражение разной силы.
Последнее в первых двух опытах слабое, в 3 и 4 — средней
силы, а в 5 и 6— большой силы. В каждой паре первый
опыт от одного второго раздражения. Интервал между
двумя раздражениями везде 213з. На всех осциллограммах
первое колебание от петли раздражающего тока.
Колеблющаяся кривая —время по 0.01 сек. (Экклс).
-время длительного положительного колебания и вызывает более
слабый эффект, чем первое раздражение. При этом второе раздражение
производит первую и вторую волны, но обе волны ослаблены.
Как уже указывалось выше, длительные электрические
последействия отводятся и от постганглионарных волокон. Но в ганглиях они
выражены сильней, — здесь при тетанических раздражениях они
достигают почти высоты токов возбуждения.
Во время тетануса отрицательное колебание сильно нарастает,
суммируется и продолжается еще некоторое время—до 2 сек. после
раздражения. Положительное колебание наступает после раздражения,
непосредственно вслед за отрицательным, и может продолжаться до
5 сек. (рис. 220) (Розенблют и Симеоне, 19).
Быстрые колебания электрического потенциала в симпатическом
ганглии безусловно выражают процесс возбуждения клеточных
элементов ганглия. Предполагается, что каждая клетка в ответ на
один залп импульсов со стороны преганглионарных волокон обычно
разряжается одним импульсом возбуждения, одним быстрым колебанием
электрического потенциала. Форбс и сотрудники (41) заключили об
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 481
этом на основании отведения токов возбуждения микроэлектродами,
введенными внутрь ганглия. Повидимому, при этом условии биотоки
отводятся от отдельных ганглиозных клеток. Эти авторы наблюдали, что
при активировании ганглия одним залпом импульсов отводимая клетка
разряжается одним быстрым колебанием потенциала. При частых же
раздражениях клетка отвечает своим собственным, очень низким
ритмом совершенно одинаковых быстрых волн по 0.4—2.4 в секунду. Это
Рис. 220. Биоэлектрические токи возбуждения верхнего шейного
симпатического ганглия кошки.
Раздражается преганглионарный нерв размыкательными и замыкательными
индукционными ударами; токи ганглия отводятся в осциллограф. Частота
раздражения: А—по 4 пары ударов в секунду, В—по 18 пар, С—по 36 пар. В опыте А
несколько мелких колебаний после первого большого принадлежат преганглионар-
ным волокнам с медленным проведением возбуждения. В опыте В токи
возбуждения следуют на замыкательные и размыкательные удары, причем токи
возбуждения размыкательных ударов несколько сильнее. В опыте С токи возбуждения
значительно слабее и следуют только по ритму размыкательных ударов (Розенблют
и Симеоне).
доказывает, что клеточная деятельность ганглия протекает по закону
возбуждения.
Однако в симпатическом ганглии разряды импульсов возникают не
только во время раздражения, но и после него. Это бывает после
частого раздражения и выражается в спонтанном возникновении
импульсов возбуждений довольно низкого ритма — сначала по 2 удара в
секунду, а затем постепенно урежающегося до 1 удара в секунду. Это,
например, было замечено Бронком на g. stellatum кошки (22). Нужно
думать, что здесь мы имеем проявление периодического изохронного
расщепления возбудимой системы после каждого изохронного
восстановления. Частое предварительное возбуждение, очевидно, создает в
ганглии соответствующие физико-химические условия для
одновременного расщепления и восстановления возбудимой системы.
В симпатических ганглиях на одних и тех же клеточных элементах
оканчиваются разные преганглионарные волокна. Вследствие этого
раздражение одних из них благоприятствует эффективности других волокон.
Мы здесь имеем, так сказать, пространственную суммацию
(Бронк, 22). При отведении от постганглионарных волокон простран-
31 И. С. Беритов
482 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
ственная суммация выявляется в повышении амплитуды тока
возбуждения, что указывает на увеличение числа возбужденных ганглиозных.
клеток, а также в появлении добавочных мелких колебаний вслед за
большим колебанием (см. рис. 220, Л).
Симпатическому ганглию свойственна также обычная временная
суммация, ибо возбуждение ганглиозных клеток зависит от частоты и.
числа притекающих к ним импульсов. Это выявляется при
ритмическом раздражении преганглионарных волокон в постепенном нараста-
Рис. 221. Явление нарастания тока возбуждения (явление лестницы)
постганглионарных волокон в ответ на ритмическое раздражение
преганглионарных. Время—0,5 сек. (Бронк).
нии тока возбуждения постганглионарных волокон (рис. 221).
Нарастание это свидетельствует об увеличении числа возбужденных ганглиозных
клеток: очевидно, некоторая часть ганглиозных клеток вступает в-
реакцию в силу повышения возбудимости под влиянием предыдущих
преганглионарных импульсов. Следовательно, мы имеем здесь явление-
лестницы (Бронк, 22).
На основании вышеприведенных фактов пространственной и
временной суммации ясно следует, что ганглиозные клетки симпатического*
ганглия при той степени возбудимости, какую они имеют в покое,
невозбуждаются одним импульсом через одно синаптическое окончание:.
Требуется повышение возбудимости до некоторого уровня, чтобы они
сами разрядились импульсами. возбуждения. Это повышение обычно*
достигается одновременным или разновременным действием импульсов
со стороны множества синапсов. Возбудимость такого образования,
которое нельзя возбуждать одиночным стимулом, Л а л и к (45) назвал*
итеративной (повторительной), а само возбудимое образование-
итеративным. Значит, для возбуждения итеративного образования
имеют существенное значение не только сила и длительность
раздражения, но и ритм и число действующих раздражений. Так как
вегетативная система в целом при раздражении преганглионарных волокон-
производит внешний эффект в том случае, если раздражение
многократно повторяется и в преганглионарных волокнах возникает ряд
импульсов, то Л а п и к всю вегетативную систему называет
итеративной. Но фактически симпатические волокна лишены этого свойства
(Аршавский, 43). Следовательно, итеративным свойством должны
обладать только ганглиозные клетки. Это же свойство присуще и ганглиоз-
ным клеткам центральной нервной системы, как будет видно ниже. Но
проявляется оно лишь в таких условиях, когда ганглиозные клетки
находятся в состоянии некоторой низкой возбудимости. Если под
влиянием ряда раздражений возбудимость повысилась и клетки возбудились
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ. НЕРВНОЙ СИСТЕМЫ 483
в результате суммации, то после этого уже каждое деятельное
раздражение может вызвать возбуждение, если только оно- не попадает
в рефракторную фазу.
Однако следует отметить, что итеративность как двойство,
основанное на суммации, не представляет чего-либо специфического только для
нервных клеток. Суммация свойственна всякой возбудимой системе в
большей или меньшей степени. Она проявляется каждый раз, когда,
действующий на возбудимую систему агент является подпороговым.
Как указывалось в мышечной и нервной физиологии, при действии
подпорогового раздражения в возбудимой системе происходит
локальное изменение основного биологического процесса, частичное
изохронное расщепление возбудимой системы, т. е. возникает локальный
процесс, который при многократном повторении раздражения
превращается в изохронное расщепление всей возбудимой системы," в
распространяющийся процесс возбуждения. Очевидно, в симпатическом
ганглии каждый синапс при своем возбуждении производит, вероятно,
через свой биоток возбуждения в нервной клетке только локальный
процесс, только частичное изохронное расщепление возбудимой
системы. Значит, его действие на нервную клетку является
подпороговым. Чтобы вся клетка пришла в состояние изохронного расщепления,
которое дало бы начало распространяющимся импульсам возбуждения,
нужно воздействие биотоков возбуждения со стороны многих синапти-
ческих окончаний в одно и то же время или в разное время. Только
в результате пространственной или. временной суммации. многих под-
пороговых влияний нервная клетка приходит в состояние возбуждения.
На этом основании мы не находим целесообразным приписывать
нервным клеткам симпатического ганглия так называемую итератив-:
ную возбудимость как особое свойство, в отличие от других нервных
образований. Поэтому мы не будем придерживаться этого термина'в
последующем изложении.
Химическое раздражение симпатического ганглия. Симпатический
ганглий очень хорошо возбуждается некоторыми химическими веществ
вами, как ацетилхолин и калий. В этом отношении он уподобляется
скелетной мышце: Пропускание через ганглий столь слабого раствора
ацетилхолина, как-25—ЮОу; [у (гамма) — миллионная часть грамма] на
1.0 ом3 перфузионной жидкости (Локка или Рингера),
вызывает-ритмические разряды импульсов, частота которых тем выше, чем выше
концентрация ацетилхолина. При более слабом растворе разряда импульсов не
происходит, а только повышается возбудимость. Пропускание растворов
кальция оказывает обратное действие: понижает возбудимость и
уменьшает ритм разрядов, вызываемых ацетилхолином (Бронк,, 22); Таким
же образом действует адреналин (Марраци, 25).
Если раздражать симпатический ганглий ацетилхолином, то на
периферии вызывается совершенно такой же эффект, как если бы
раздражался преганглионарный нерв электрическим током (рис. 222).
Это указывает, что преганглионарные нервные импульсы я ацетилхолин
31*
484 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЧРВНОЙ СИСТЕМЫ
действуют на одни и те же элементы. Доказано, что ацетилхолин
производит свой обычный эффект и в том случае, если преганглионарные
волокна и синапсы переродились вследствие перерезки преганглионар-
ного нерва. А это свидетельствует, что ацетилхолин действует на
клеточный состав ганглия подобно нервным импульсам.
Передача возбуждения в симпатическом ганглии. В симпатических
ганглиях преганглионарные волокна образуют около клеток очень слож-
Рис. 222. Віияние верхнего шейного симпатического ганглия
на третье веко кошки (membrana nictitans).
В опыте А реакция дается от электрического раздражения преганглионарных
волокон, а в опыте В —от введения ацетилхолина в симпатический ганглий]
(Бакурадзе).
ное перицеллюлярное образование, состоящее из нейрофибриллей,
которые касаются тела клетки множеством боковых и конечных синапсов.
Возбуждение этого образования, как указывалось выше, должно
сопровождаться биоэлектрическим током значительной интенсивности. Этот
ток в своей начальной фазе протекает так же быстро, как ток
возбуждения нервных волокон. А потому можно утверждать, что начальная
быстрая фаза биотока возбуждения преганглионарных нервных
окончаний должна являться раздражителем для постганглионарных клеток.
Следовательно, в симпатических ганглиях передача возбуждения через
синапсы должна происходить таким же физическим путем, как в
пределах нервного волокна от возбужденного участка к соседнему
невозбужденному участку. С этим представлением хорошо согласуется малый
скрытый период при передаче возбуждения в симпатическом ганглии —
всего 2—8 j . При этом и такое небольшое запаздывание следует
объяснить тем обстоятельством, что в пределах ганглия волна
возбуждения распространяется по очень тонким нервным волоконцам перицел-
люлярного аппарата с большим замедлением и что возбуждение пост-
ганглионарной клетки наступает в результате суммации множества
импульсов от одного и того же или многих преганглионарных нервных
волокон (Экклс, 27).
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 485
С указанной ролью синапсов в передаче возбуждения
хорошо согласуется также то явление, что при дегенерации преганглио-
нарных волокон после их перерезки биотоки возбуждения ганглия —
начальные быстрые колебания — ослабевают вместе с гистологическим
перерождением синапсов и исчезают совсем вместе с исчезновением
синапсов, т. е. передача возбуждения от преганглионарных волокон на
постганглионарные всецело зависит от структурной целости
преганглионарных волокон. Точно так же при регенерации преганглионарных
волокон передача возбуждения в ганглии восстанавливается с
появлением первых синапсов (Гибсон, 42).
Мы указывали выше, что симпатический ганглий на один преган-
глионарный импульс отвечает всего одним импульсом. При высокой
- частоте преганглионарных импульсов происходит трансформация их
в ганглии до очень низкого ритма — 2—3 в секунду. Однако в
определенных случаях передается без трансформации и довольно высокий
ритм, или же трансформация происходит, но только до некоторого
высокого ритма. Так, например, на рис. 220 при раздражении
преганглионарных волокон 4 и 18 двойными индукционными ударами в
секунду, когда эффекты наступают как от размыкательных, так и от
замыкательных ударов, верхний симпатический ганглий отвечает
импульсами по ритму раздражения (опыты А я В). Когда же частота
раздражения доходит до 36 двойных ударов в секунду, ганглий
отвечает правильно трансформированным ритмом по 36 в секунду, т. е.
только на размыкательные удары (Розенблют и Симеоне, 19).
Последний факт, по нашему мнению, был также одним из лучших
доводов в пользу физической передачи возбуждения в симпатических
ганглиях.
О происхождении длительного отрицательного колебания в
симпатическом ганглии. Медленные колебания электрического потенциала
симпатических ганглиев, которые наступают после возбуждения, не
могут быть обусловлены активным состоянием преганглионарных волокон
и их окончаний. Это видно, например, из того, что медленные
колебания симпатического ганглия возникают и при антидромном
возбуждении, т. е. в результате раздражения постганглионарного нерва. Они
наступают при антидромном возбуждении и после дегенерации
преганглионарных волокон.
Отсюда ясно следует, что медленные колебания потенциала
должны зависеть от тех элементов ганглия — клеток и дендритов, которые
возбуждаются обоими путями — преганглионарным и антидромным
(Экклс, 20; Ллойд, 26).
Как уже говорилось выше, длительное повышение возбудимости и
длительное отрицательное колебание после возбуждения наблюдаются
в соматических и вегетативных нервных волокнах. Аналогичное явление
должно иметь место в клетках симпатического ганглия при их
возбуждении. В случае возбуждения клеток ганглия, в результате
безразлично какого раздражения — преганглионарного или постганглионарного^
486 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
источник электричества, связанный с процессом возбуждения,
находится внутри клеток. Вследствие этого возникает ток, который, выйдя
из клеток наружу, создает катэлектротоническое изменение на
поверхности, вернее, в поверхностном слое клеток. Это должно обусловить
длительное отрицательное колебание потенциала всего ганглия, т. е.
колебание потенциала в том самом направлении, что и направление
тока возбуждения.
Что касается периода пониженной возбудимости и. положительного
колебания, который следует вслед за повышением возбудимости, то
он, повидимому, имеет несколько иное происхождение.
О происхождении положительного колебания и пониженной
возбудимости в симпатическом ганглии. Длительное положительное
колебание настолько продолжительно и настолько интенсивно, что его нельзя
целиком приписать тем низковольтным следовым электрическим
колебаниям, какие наблюдаются в нервных волокнах. Кроме того, при
возбуждении ганглия характерные медленные токи отводятся только от самого
ганглия и не отводятся от постганглионарных волокон, даже если они
в это время проводят импульсы возбуждения. В определенных случаях,
например при повреждающих раздражениях, положительные
колебания наступают без начальных быстрых колебаний (Форбс и сотр., 41).
На этом основании надо полагать, что медленное положительное
колебание возникает не в нервных волокнах и не является последействием
распространяющегося процесса возбуждения. Мы полагаем, что
положительное колебание возникает в основном от локальных процессов в ден-
дритах и клеточных телах на месте соприкосновения с синапсами. Как
указывалось выше, преганглионарные волокна, сильно ветвясь в ганглии,
заканчиваются как на клетках, так и на дендритах. Поэтому при
каждом возбуждении преганглионарных волокон должны активизироваться
как клетки, так и сплетение дендритов, т. е. нейропиль. Повидимому,
активное состояние нейропиля, а также локальный процесс в клетках
продолжаются значительно дольше распространяющегося процесса
возбуждения. Медленный ток, отводимый от ганглия, очевидно,
получается в результате суммации электрических токов, возникающих при
этих медленных процессах. Так как общая масса дендритов во много
раз превосходит общую массу клеточных тел, то, безусловно, в
происхождении медленного тока нейропиль играет главную роль. Поэтому
ради краткости мы будем называть этот ток нейропильным.
Длительное понижение возбудимости в клеточных элементах после
каждого начального импульса и небольшой фазы медленного
отрицательного колебания, вероятно, является прямым следствием медленного
нейропильного тока. Надо предполагать, что нейропильный ток,
распространяясь по межуточной жидкости, действует на близлежащие
клетки и их аксоны, подобно тому как это вообще свойственно
постоянному току. В местах проникновения нейропильного тока внутрь клеток
и аксонов должно происходить анэлектротоническое понижение
возбудимости. Подобного рода нейропильное угнетение нервных элементов мы,
физиологии зеки г особенности вегетативной нервной системы 487
тювидимому, имеем в центральной нервной системе, а потому более
подробное обоснование роли нейропиля в активном угнетении
клеточной деятельности будет дано в отделе центральной нервной системы.
Здесь укажем только на один факт, который хорошо согласуется с
означенной гипотезой относительно происхождения пониженной
возбудимости в симпатическом ганглии. Именно, было замечено, что
некоторые преганглионарные волокна (так называемые S2) не производят
повышения возбудимости, а только понижение, которое притом
развивается не сразу, а с некоторой постепенностью, достигая максимума
через 100 с (Экклс, 7; Ллойд, 27). Сверхнормальная возбудимость
после возбуждения обязательна для каждой возбудимой системы,
и тем более обязательна для всякой нервной клетки. И если в
симпатическом ганглии ее не наблюдается при раздражении определенных
преганглионарных волокон, то этому должна быть особая причина.
Вероятно, определенные преганглионарные волокна (S2) особенно
тесно связаны с нейропилем и особенно слабо с клеточными телами.
Поэтому при возбуждении их активное состояние нейропиля и его
угнетающее действие берут верх с самого начала. Клетки испытывают такое
анэлектротоническое действие со стороны нейропиля, что они не в
состоянии проявить свойственную им сверхнормальную возбудимость
после возбуждения.
С этим угнетающим действием нейропиля должно быть связано
¦явление трансформации ритма возбуждения в ганглии при некоторых
повышенных частотах раздражения. Так, например, было замечено,
что при раздражении преганглионарных волокон g. stellatum частотой
ниже 30—40 в секунду постганглионарные сердечные волокна
обнаруживают очень хорошее синхронное течение токов возбуждения.
Когда же частота раздражения доходит до 60 в секунду, токи
возбуждения постганглионарных волокон падают очень быстро почти до
исчезновения. Но такое быстрое падение не наблюдается в
постганглионарных волокнах, когда они раздражаются непосредственно. Его
не бывает и в преганглионарных нервных волокнах. Следовательно,
означенное быстрое падение токов возбуждения происходит в самом
ганглии (Бронк и Пумфри, 21). Но и в самом ганглии — в нервных
клетках постганглионарных нейронов — рефракторные фазы
продолжаются чуть дольше, чем в постганглионарных нервных волокнах. Так,
например, в шейном симпатическом ганглии кошки продолжительность
абсолютной рефракторной фазы около 2 а, а относительная — около
15 а (Броун, 36; Экклс, 23). Очевидно, оптимальный ритм возбуждения
клеток симпатического ганглия кошки 60—70 в сек. И если при
частоте преганглионарных импульсов еще до 60 в сек. токи возбуждения
постганглионарных волокон очень быстро падают до исчезновения, то
причиной тому должны быть не рефракторные фазы ганглиозных
клеток. Мы думаем, что эта трансформация ритма возбуждения
обусловлена усиленным анэлектротоническим действием нейропильного тока
на клетки ганглия, в силу чего лабильность их падает.
488 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Многие известные явления ослабленного периферического эффекта
при частых раздражениях преганглионарных волокон также должны
быть обусловлены этим обстоятельством. Известно, например, что
раздражение блуждающего нерва лягушки производит торможение
сердечной автоматической деятельности лучше всего при ритме 30
в секунду. Если частота больше или меньше, торможение ослабевает
(Воронцов, 2). Ослабление торможения с повышением частоты
раздражения преганглионарных волокон не может быть приписано пес-
симальному состоянию преганглионарных волокон. Частота
раздражения свыше 30 в секунду слишком мала для того, чтобы нормальные
нервные волокна цереброспинального происхождения пришли в песси-
мальное состояние. Значит, ослабление торможения в означенном
случае можно приписать активному состоянию нейропиля в сердечном
ганглии. Было найдено, что при такой малой частоте раздражения
блуждающего нерва, как одно раздражение через 1—2 сек., желудок
сокращается; если же частота превышает 4—5 в секунду, наступает
торможение, т. е. ослабление сокращения. Это явление рассматривалось
как результат рефракторной фазы, как пессимальное явление (Вич,
Шварц и Вейнштейн, 4). Но, по вышеприведенным соображениям, и это
ослабление сократительного эффекта должно быть обусловлено
активным состоянием нейропиля.
По новейшим наблюдениям Розенблюта (9), при раздражении
преганглионарных волокон оптимальный эффект получается при таких
низких частотах раздражения, как 20—30 в секунду. Так, для пило-
моторов он наступает при 20 в секунду, для мигательной перепонки
(3-е веко) — при 15, кишечника — при 20, беременной матки — при 20т
сердца — при 30, желудка — при 25, подчелюстных желез — при 35.
Но эти частоты, безусловно, не являются оптимальными ни для самого
раздражаемого нерва — преганглионарных волокон, ни для постганг-
лионарных волокон. Мы выше указывали, что предельный ритм
преганглионарных и постганглионарных волокон должен быть
сравнительно высоким — 100—300 в секунду. Если же, несмотря на это,
предельный оптимальный периферический эффект получается при
таком низком ритме, как 20—30 в секунду, то это указывает, что при
более высоком ритме раздражения происходят ослабление импульсов
возбуждения и трансформация их ритма .в самом ганглии (рис. 220).
Итак, явление трансформации ритма возбуждения в
симпатическом ганглии, а также угнетение его активной деятельности при таких
частотах раздражения, которые значительно ниже оптимального ритма
раздражения преганглионарных и постганглионарных волокон, должно
быть, обусловливается угнетающим действием нейропильного тока на
клетки и аксоны постганглионарных нейронов.
Симпатический ганглий можно возбудить также раздражением
постганглионарных волокон, так называемой антидромной волной
возбуждения. Возбуждаются, конечно, клетки симпатического ганглия.
Одновременно должны возбуждаться и наиболее толстые дендриты, выхо-
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 489
дящие из клетки. Но возбуждение не переходит на преганглионарные
волокна. Под влиянием антидромной волны после начального быстро
протекающего тока, отвечающего возбуждению клеточной массы
ганглия, происходит сначала сравнительно короткое отрицательное,
а потом длительное положительное электрическое последействие. При
этом, опять-таки, во время отрицательного последействия наблюдается
повышение возбудимости по отношению к преганглионарным
импульсам, а во время положительного, наоборот, понижение ее. Состояние
Рис. 223. Действие активных ве.цеств верхнего шейного симпа
тического ганглия.
В 1-м опыте дается реакция 3-го века кошки на раздражение преганглио-
нарных волокон, идущих к верхнемушейному симпатическому ганглию; во
2-м—реакция того же века после введения в данный ганглий локковской
жидкости, которая прошла этот же ганглий в период раздражения (Кибяков).
пониженной возбудимости длится довольно долго — иногда около
двух минут (Экклс, 23; Бронк, 22). Это длительное понижение
возбудимости после антидромного возбуждения, по всей вероятности, нужна
отнести насчет активирования клеток и некоторой части нейропиля.
Как известно, ганглиозные нервные клетки в симпатических ганглиях
обладают многочисленными дендритными отростками, образующими
местами значительные расширения — дендритические пластинки
(Лаврентьев, 48). Надо полагать, что при возбуждении ганглиозной
клетки в активное состояние приходят и ее дендриты на некотором
расстоянии от тела клетки. Вследствие этого сильно возрастает
разность потенциалов между клеточными телами и дендритами, с одной
стороны, и межуточной жидкостью — с другой. От этого возникает ток,
который, распространяясь через межуточную жидкость, должен оказать
на окончания преганглионарных волокон значительное анэлектротони-
ческое угнетающее действие.
О химической передаче возбуждения в симпатическом ганглии.
В настоящее время представители химической точки зрения на
передачу возбуждения в нервной системе утверждают, что в
симпатических ганглиях возбуждение передается исключительно посредством,
химического агента — ацетилхолина, выделяемого окончаниями пре-
490 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
ганглионарных волокон. Главным основанием для такого утверждения
служит то явление, что при определенных условиях в симпатических
ганглиях образуется ацетилхолин. Именно, К и б я к о в (29) нашел,
что питательный раствор, пропущенный через верхний шейный
симпатический ганглий (кошки) во время раздражения шейного
симпатического нерва, содержит вещество, которое, будучи (введено обратно
в верхний шейный симпатический ганглий, вызывает сокращение
мигательной перепонки (рис. 223). Это вещество, оказалось, обладает
характерными свойствами ацетилхолина (Фельдберг и Гаддум, 32).
Выделяется оно нервными окончаниями преганглионарных волокон. Было
даже определено количество ацетилхолина. От одного импульса при
одной клетке симпатического ганглия образуется 10—15 у
ацетилхолина, т. е. около 3 млн. молекул (Фельдберг и Вертиайнен, 31).
Аналогичное предполагается насчет действия преганглионарных волокон
блуждающего нерва на постганглионарные нейроны сердца. Это
действие будто бы осуществляется посредством того же передатчика —
•ацетилхолина (Дэйл, 33).
Кэннон и сотрудники (30) утверждают, что ацетилхолин в
симпатическом ганглии играет такую же роль, какую они приписывают
¦ему в эффекторах. Они полагают, что от него зависит переход
возбуждения в ганглии от преганглионарных нейронов на
постганглионарные. Они, между прочим, установили, что малая доза
ацетилхолина, введенная в ганглий через сосудистую систему, производит
возбуждение, а большая доза, наоборот, торможение. Если инъициро-
вать вещество, защищающее ацетилхолин от разрушения, как эзерин
(физостигмин), то в период ослабления ганглиозной деятельности
ацетилхолин усиливает ее, а в период максимальной деятельности,
наоборот, угнетает. Это явление объясняется в первом случае
увеличением количества ацетилхолина до оптимальной концентрации, а во
втором — увеличением ее до пессимальной.
В последнее время находят, что в симпатических ганглиях
определенными волокнами выделяется не возбуждающее вещество —
ацетилхолин, а тормозящее вещество — адреналин. Доводом для такого
предположения служит легкая тормозимость активного состояния
ганглия адреналином, также искусственно введенным в очень малых
количествах, как 5 у на кошку весом 3—4 кг (Марраци, 25).
Против химической передачи возбуждения в симпатическом ганглии
говорит прежде всего то обстоятельство, что в нормально
функционирующем симпатическом ганглии при неповрежденном кровообращении
ацетилхолин не образуется или, точнее, не выделяется из ганглия при
его возбуждении. То же наблюдается и на эзеринизированных животных,
т. е. в условиях, когда ацетилхолин не должен расщепляться от холин-
эстеразы. Ацетилхолин появляется только при ухудшении
функционального состояния ганглия, и притом даже независимо от
возбуждения (Лоренте де-Но, 34). Локковская жидкость, которой обычно
Пользуются для перфузии симпатического ганглия, чтобы уловить аце-
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 491
тилхолин, не может заменить кровь. Даже прибавление небольшой
порции сыворотки крови (1 часть ее на 10 частей локковской
жидкости) , видимо, значительно улучшает функциональное состояние
ганглия, ибо при этом почти втрое уменьшается количество
выделяемого ацетилхолина (Мак Интош, 35). На этом основании нужно считать,
что ацетилхолин образуется при ненормально протекающем
метаболизме нервных элементов ганглия.
Против химической передачи возбуждения говорят также
небольшой скрытый период передачи возбуждения, а также незначительная
продолжительность рефракторной фазы. Рефракторный период верх-
не-шейного симпатического ганглия равняется 2 а (Броун, 36). Значит,
расщепление ацетилхолина холинэстеразой должно происходить с такой
быстротой, чтобы уже в течение 2 а ацетилхолин перестал действовать.
Последние исследования скорости расщепления ацетилхолина эстера-
зой из ганглия показали, что максимальная активность эстеразы очень
велика, ибо она в состоянии расщепить то количество ацетилхолина,
какое выделяется в ганглии от одного преганглионарного нервного
импульса, в течение 15 а. Но это время во много раз больше
рефракторной фазы. Поэтому едва ли холинэстераза в состоянии уничтожить
действие ацетилхолина в течение 2 а (Глик, 37). Кроме того,
приходится допустить, что холинэстераза возникает в ганглии одновременно
с ацетилхолином под влиянием каждого нервного импульса.
Характерно, что если пропускать через симпатический ганглий эзеринизиро-
ванную жидкость, то это не удлиняет продолжительности возбуждения
или рефракторной фазы. Понятно, если бы возбуждение клетки
постганглионарного нейрона зависело от ацетилхолина, а прекращение
возбуждения — от действия холинэстеразы, то эзерин должен был бы
удлинить процесс возбуждения и рефракторную фазу (Экклс, 27) или
обусловить возникновение длинного ряда колебаний потенциала от
одного нервного импульса, как это бывает при введении ацетилхолина
через артерию (Экклс, 38).
Исследования Кибякова, который впервые нашел активное
вещество в оттекающей из вены симпатического ганглия перфузионной
жидкости — растворе Рингер-Локка (14), подтвердили наблюдения
ЛорентедеНо. В условиях нормального кровообращения он также
не смог открыть образование ацетилхолина при раздражении преган-
глионарных волокон. Но он находит, что как при перфузии раствором
Рингер-Локка, так и при нормальном кровообращении в симпатическом
ганглии появляется другое активное вещество, которое по своим
физиологическим свойствам близко стоит к симпатину. Это вещество
вызывает повышение кровяного давления и усиление сердечной
деятельности; кроме того, оно возбуждает нервные клетки в самом
симпатическом ганглии (49). На этом основании Кибяков заключает, что
передача импульса с преганглионарных волокон на постганглионарные
нейроны происходит при участии активного вещества, имеющего
сходство с симпатином. Но автор не мыслит механизма передачи возбужде-
492 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
ния в симпатическом ганглии без участия биотока возбуждения
нервных окончаний преганглионарных волокон. По его мнению, ток
возбуждения нервных окончаний вызывает возбуждение ганглионарных
клеток, а активное вещество повышает возбудимость клеток, улучшает
их восприимчивость к приходящим импульсам (49).
Что касается медленных колебаний потенциала, следующих за
быстрым током возбуждения, многие авторы считают возможным
приписать их действию ацетилхолина (Экклс, 27; Монние, 39). Вообще
известны также факты, которые говорят за различное происхождение
начальных быстрых колебаний потенциала и последующих медленных.
Так, например, известно, что под влиянием малых доз никотина
длительное отрицательное колебание исчезает, а передача импульсов
с преганглионарных волокон на постганглионарные сохраняется
(Экклс, 77 27). Но если принять во внимание, что, во-первых, при
нормальном функционировании симпатического ганглия нет даже
следов ацетилхолина в оттекающей от ганглия крови и что, во-вторых,
эти медленные колебания потенциала не являются спецификой
симпатического ганглия, а наблюдаются в большей или меньшей степени во
всех нервных элементах, едва ли можно согласиться с тем, что аце-
тилхолин играет какую-либо роль в нормально функционирующем
ганглии и, в частности, в происхождении медленных изменений
потенциала.
Конечно, в тех случаях, когда в ганглии освобождается ацетилхо-
лин в значительном количестве как один из побочных продуктов
нарушенного обмена веществ, он должен играть определенную роль
в функциональных изменениях симпатического ганглия. Ему можно
приписать, например, уменьшение скрытого периода после ряда
раздражений до 1 7 а (Экклс, 7; Бишоп и Гейнбекер, 40), явление
лестницы на обескровленном ганглии на ряд раздражений (Бронк и
сотр., 18). Но в этом действии ацетилхолина нет ничего специфического.
Также должны влиять и многие другие продукты метаболизма
нервного процесса (Штерн, 28).
Явление суммации в эффекторах вегетативной системы.
Вегетативная нервная система производит периферический эффект главным
образом при повторном раздражении. Активное состояние того или
другого органа предполагает воздействие ряда нервных импульсов.
Так, вазоконстрикторы сокращаются после ряда раздражений
симпатической системы, матка сокращается после ряда раздражений
п. hypogastricus, железы выделяют секрет после длительного
раздражения секреторного нерва, полная остановка сердца получается после
ряда раздражений блуждающего нерва и т. д. Отсюда следует, что
для изучения роли вегетативной системы в той или другой функции
нужно пользоваться ритмическим раздражением, причем частота
раздражения имеет существенное значение.
В первое время Лапик (11) объяснял явление суммации в эф-1
фекторах тем, что хронаксия эффектора много длительнее тока
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 493
возбуждения нервного импульса. Поэтому эффект может наступить
только в результате повышения возбудимости в эффекторах под
влиянием ряда нервных имиульсов. При этом хронаксия эффектора
укорачивается в такой мере, что устанавливается изохронизм между ним и
эффектором. В настоящее время Лап и к допускает, что это
повышение возбудимости в эффекторе происходит путем выделения нервными
окончаниями физиологически активных веществ (12).
Об оптимальном и пессимальном эффектах вегетативной системы.
Впервые Введенский (14) наблюдал явление оптимального
и пессимального состояния на вегетативной системе (1893). Именно,
он установил их для подчелюстной железы. При раздражении chorda
tympani в ритме 40 в секунду наблюдалось наиболее обильное
отделение слюны, а при большей частоте раздражения секреция
ослабевала, наступало пессимальное состояние. Впоследствии эти явления
были изучены на многих других эффекторах вегетативной системы.
При этом было обнаружено, что оптимальный эффект получается при
очень разных частотах раздражения соответствующего двигательного
или секреторного нерва. Так, например, известно, что преганглионар-
ные волокна верхнего шейного узла вызывают оптимальное
сокращение — тетанус membrana nictitans кошки при частоте раздражения
40—200 в секунду. В то же время раздражение постганглионарных
волокон производит оптимальное сокращение при частоте не свыше
120 в секунду; при частоте раздражения 180 в секунду сокращения не
наступает вовсе. Очевидно, постганглионарные волокна хорошо
воспринимают ритм раздражения до 120, а преганглионарные — до 200 в се-
кунду. Но высокий ритм преганглионарных волокон трансформируется
в ганглии до 100 в секунду и ниже, и потому только и вызывается
максимум сокращений (Кверидо, 1).
В этой трансформации ритма возбуждений внутри ганглия
существенную роль должен играть нейропильный ток, как уже указывалось
выше.
Обмен веществ в симпатических ганглиях и постганглионарных
волокнах характерно отличается от соматической периферической
системы. В них потребление кислорода и выделение углекислоты
гораздо значительнее. Так, в изолированном симпатическом безмякотном
нерве селезенки быка потребление кислорода при комнатной
температуре в среднем составляет 65.3 мм3 на грамм нерва в час, в таком
же нерве теленка — 83.1 мм3; при температуре тела это потребление
становится гораздо значительнее, достигая нескольких сот кубических
миллиметров. Выделение С02 не соответствует количеству
поглощенного кислорода, — оно меньше. Это объясняется тем, что часть
кислорода идет на процессы дегидратации. Точно так же потребление
сахара значительно выше в безмякотных нервах, чем в мякотных того
же животного. Так, в безмякотном нерве селезенки быка на час
времени и на грамм веса потребляется в первый час опыта 1.462 мг,
а во второй — 0.724 мг. Аммиак в симпатических безмякотных во-
494 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
локнах выделяется в таком же размере, как и в мякотных; в среднем
на грамм веса в час выделяется при комнатной температуре 4.78 у.
Но при температуре тела аммиака выделяется на 70% больше (Розен-
баум, 15).
В изолированном симпатическом узле кошки обмен веществ
протекает несколько интенсивнее, чем в безмякотных волокнах. Так, за час
опыта на грамм веса при температуре тела кислород потребляется
в среднем 220 мм3. В атмосфере кислорода потребление его
усиливается, но никогда не достигает той величины, как это бывает в опыте
с мозгом кошки (Гольденберг, 8).
В связи с этим находится сравнительно малая утомляемость
симпатических ганглиев. Они утомляются много быстрее, чем нервы, но
сравнительно с мозгом они оказывают сильное противодействие
утомлению. Например, у кошки после полного прекращения
кровообращения верхний симпатический ганглий способен работать в течение
10—15 минут и вызывать сокращение мигательной перепонки в ответ
на тетаническое раздражение преганглионарного нерва. Даже после
70 мин. отсутствия кровообращения ганглий способен к полному
восстановлению функциональной деятельности (Б а р ж е т о н, 10).
Но если нерв не вырезан и не перерезан, а находится в условиях
нормального кровоснабжения, утомления не наступает и после многих
часов деятельности. Так, Бек (13) показал, что длительное
тетаническое раздражение шейного симпатического ствола производит
максимальное расширение зрачка у кошки в течение 9—17 часов. Если
нерв перерезывался, то ослабление эффекта наблюдалось уже через
час. Предполагается, что в этих опытах возбуждение передавалось
помимо ганглиозных клеток, которые вообще должны утомляться
скорее нервных волокон. Так, например, известно, что у
холоднокровных длительное, тетаническое раздражение блуждающего нерва
перестает тормозить сердце через несколько минут. Значит, передаточный
нервный механизм в сердце лягушки утомляется быстро в течение
нескольких минут. Но без такого длительного раздражения
передаточный механизм вырезанного сердца, если через него пропускать
раствор Рингера при определенном давлении, сохраняет способность
к работе в течение многих часов. Блуждающий нерв является преган-
глионарным, ганглий лежит в сердце. Очевидно, обмен веществ.
в ганглиозных клетках лягушки протекает на очень низком уровне,
если они сохраняют возбудимость в отсутствии кровообращения в
течение многих часов.
Л ите ратур а
1. Querido A., Am. J. Physiol., 70, 29, 1924.
2. Воронцов Д., Тр. Петерб. общ. естествоисп., 43, вып. 6, 1913'.
3. Trendelenburg W., Arch. Anat. Physiol., Supplb., стр. 294, 1902.
4. Veach H. О., Schwartz L. L. a. Weinstein M., Am. J. Physiol., 91,
451, 1930.
5. Bishop G. H. a. Heinbecker R., Am. J. Physiol., 94, 171, 1930.
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 495
6. Fischer u. Schilf Е., Pfl. Arch., 200, 227, 1923.
7. Eccles J. С, J. Physiol, 85, 179, 1935; ib., 95, 207, 1935; 85, 464, 1935.
8. Гольденберг E. Э., Сборы. «Опыт исслед. нервно-гумор. связей»), под ред~
Быкова, III, стр. 51, 1937.
9. Rosen blue th A., Am. J. Physiol., 101, 149, 1932.
іа В a r g e t о n D., Am. J. Physiol., 121, 261, 1938.
11. L a p i с q u e L., L excit. en fonction du temps, Paris, 1927.
12. Lapicque L., Cold Spr. Harb. Symp. Quant. Biol., 4, 98, 1938.
13. Beck A., Pfl. Arch., 124, 585, 1909.
14. Введенский H. E., Врач, 14, № 3, стр. 89, 1893.
15. Rosenbaum H., Bioch. Z„ 247, 189, 1932; ib., 257, 307, 1933; Halter K*
Bioch. Z., 257, 325, 1933.
16. Gasser H. S., J. Neurophysiol., 2, 361, 1939.
17. Grundfest M. a. Gasser. H. S., Am. J. Physiol., 123, 307, 1938.
18. Bronk, Tower, Solandt a. Lurabee, Am. J. Physiol., 122, 1, 1938.
19. Rosenb lueth A. a. Simeone F. A., Am. J. Physiol., 122, 688, 1938.
20. Eccles J. C, J. Physiol., 89, 41P, 1937.
21. Bronk D. W. a. Pumphrey R- J-, Proc. Soc. Exp. Biol. Med., 32, 1661, 1935.
22. Bronk D. W., J. Neurophysiol., 2, 380, 1939.
23. Eccles J. C, J. Physiol., 88, 1, 1936.
24. Grundfest H., Am. J. Physiol., 127, 252, 1939.
25. Mar razz i A. S., Am. J. Physiol., 127, 738, 1939.
26. Lloyd D. J., J. Physiol., 5, 464, 1939; 96, 118, 1939.
27. Eccles J. C, Ergebn. Physiol., 3 , 339, 1936.
28. Штерн Л., Физиол. журн. СССР, 19, 377, 1935.
29. Kibiakow A., Pfl. Arch., 232, 432, 1934.
30. Cannon W. В. a. Rosenblueth A., Auton. nervo-effector system, 1937.
31. Feldberg W. a. Vartiainen A., J. Physiol., 83, 105, 1934.
32. Feldberg W. a. Gaddum J., J. Physiol., 80, 12, 1933.
33. Dale H. H., Reiziiberragung d. chemisch. Mittel im periph. Nervensysteiru
Urban u. Schwarzenberg, 1935.
34. Lorente de N 6, Am. J. Physiol., 121, 321, 1938.
35. M с In tosh F. C, J. Physiol., 94,. 155, 1939.
36. Brown G. L., J. Physiol., 81, 228, 1939.
37. Glik D., J. Gen. Physiol., 21, 431, 1939.
38. Eccles J. С a. Magladery J. W., J. Physiol., 90, 31a, 68, 1937.
39. Monnier A. M., Cold Spr. Harb. Symp. Quant, biol., 4, 111, 1936.
40. Bishop G. H. a. Heinbecker P., Am. J. Physiol., 100, 519, 1932.
41. Therman P. O., Forbes A. a. Galambos R., J. Neurophysiol., 31,.
192, 1940.
42. Gibson C, J. Neurophysiol., 3, 238, 1940.
43. A p ш а в с к и й И. А., Арх. биол. наук, 46, 106,1937; Бюлл. эксп. биол. мед.г
3, 369, 1937.
44. Уфлянд Ю. М., Теория и практика хронаксиметрии, Ленинград, 1938.
45. Lapicque L., С. г. Acad. Sci., Paris, 155, 70, 1912; Cold Spr. Harb. Symp. Quant.
Biol., 4, 98, 1938.
46. С h a u с h a r d A. et P. et M-me В. С h a u с h a r d, С r. Acad. Sci., 201, 293„
1935; Физиол. журн. СССР, 22, 561, 1937.
47. В r u с k e Fr., Pfl. Arch., 233, 777, 1934.
48. Лаврентьев Б. И., Морфология автономной нервной системы, Москва, 1939.
49. Кибяков А. В., Тр. Казанск. гос. мед. инст., 1941, вып. 1, стр. 1.
Глава ill
АНТАГОНИСТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ СИМПАТИЧЕСКОЙ
И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ
Общее понятие. Центробежные пути симпатической и
парасимпатической систем, обслуживающие один и тот же рабочий орган, так
называемый эффектор, обнаруживают характерное противоположное
действие. Если нерв одной системы производит возбуждение данного
органа, то нерв другой системы задерживает его деятельность,
тормозит его. Например, краниальная система через глазодвигательный нерв
вызывает сужение зрачка, а идущий из грудного отдела симпатический
нерв производит расширение зрачка. Краниальный блуждающий нерв
тормозит сердце в смысле замедления и прекращения его
автоматической деятельности, а симпатический нерв, наоборот, учащает и
усиливает его автоматическую деятельность. У лягушки блуждающий
нерв смешан с постганглионарными волокнами симпатической системы.
Обычно раздражение его дает тормозящий эффект, так как
возбудимость иреганглионарных волокон блуждающего нерва выше постганг-
лионарных волокон симпатической системы. Но в некоторых, редких
случаях можно вызвать также ускорение сердечной деятельности.
Это, повидимому, бывает при поверхностном положении симпатических
волокон в раздражаемом участке нерва. При этом условии слабое
раздражение произведет ускорение сердечной деятельности,
замедление же ее получится при сравнительно сильном раздражении. При
некоторых сильных раздражениях возбуждение симпатикуса
проявляется после раздражения в виде усиления и учащения последующей
сердечной деятельности (рис. 224 и 225).
Краниальная система (п. vagus) усиливает автоматическое
движение кишечника, так называемую перистальтику, а симпатическая
(п. splanchnicus), наоборот, ослабляет и прекращает ее (рис. 226
и 227), Парасимпатическая система усиливает сокращение мочевого
пузыря (п. errigens), а симпатическая тормозит его (nn. hypoga-
strici).
Функциональное состояние каждого вегетативного органа зависит
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 497
от взаимоотношений между симпатической и парасимпатической
системами. Эти две системы действуют на органы в высшей мере
согласованно: усиление действия одной системы сопровождается
ослаблением действия другой. Так, например, при выключении симпатической
224. Тормозящее действие блуждающего нерва лягушки на сердце.
Нерв раздражается индукционными ударами большой интенсивности. Во время раздражения и
некоторое время после него сердце не бьется. После раздражения происходит учащение и усиление сердечных
сокращений (Пембрей и Филиппе).
системы путем отравления эрготамином значительно усиливается
действие блуждающего нерва на сердце, а при повышении тонуса
симпатической системы воздействием адреналина ослабевает или
совершенно уничтожается действие блуждающего нерва (Штерн, 1).
Рис. 225. Ускоряющее действие блуждающего
нерва лягушки на сердце.
Нерв раздражается индукционными ударами небольшой
интенсивности. Продолжительность раздражения обозначена
внизу линией. Во время раздражения происходит сильное
ускорение и усиление сердечной деятельности вследствие
преобладающего действия симпатических волокон
(Пембрей и Филиппе).
Антагонистическое отношение вегетативных органов к некоторым
веществам. Существуют такие вещества, которые на одни вегетативные
органы действуют, а на другие — нет. Адреналин, секрет надпочечных
желез, например, действует на органы, иннервируемые вегетативной
системой, подобно симпатической системе. При этом если раздражение
симпатических волокон повышает деятельность периферического
органа, то и адреналин повышает ее (сосуды, сердце, слюнные
железы) ; наоборот, если раздражение симпатической системы тормозит
деятельность органа (кишки, мочевой пузырь), тогда адреналин также
тормозит ее. Исключение составляют только потовые железы. Они
получают нервы из симпатической системы, но на них адреналин не
действует. Следует указать, что железистые клетки надпочечников
получают непосредственно преганглионарную симпатическую иннерва-
32 И. С. Беритов
498 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
цию. А выделяемый здесь адреналин, который потом через кровь
действует на весь организм, подобно симпатикусу, выполняет роль
постганглионарных волокон. Отсюда ясно, что надпочечники предста-
Рис. 226. Тормозящее влияние симпатической
системы на движение кишечника.
Внутрь кишечника введена резиновая груша, которая
соединена с пишущей капсюлей Марея. Сужение кишки
записывается поднятием кривой. Раздражается индукционными
ударами п. splanchnicus. Белая линия показывает время
раздражения. Автоматическое движение кишечника
приостанавливается на некоторое время (Старлинг).
вляют интегральную часть симпатической системы, так как они
производят широко распространенный симпатический эффект, когда вся
симпатическая система приходит в активное действие (Эллиот, 6).
К симпатическим ядам относится также эрготамин, который парализует
Рис. 227. Возбуждающее влияние
парасимпатической системы на движение кишечника.
Движение кишки записывается тем же способом, что и выше.
Раздражается п. vagus индукционными ударами. Время
раздражения обозначается поднятием сигнальной линии.
Вначале происходит кратковременная остановка движения, а
вслед за остановкой — усиленное движение. Максимум
движения приходится после раздражения. Полагают, что
п. vagus содержит также и тормозящие центробежные пути
к кишечнику (Старлинг).
в большинстве случаев действие симпатического нерва. После
воздействия этого яда раздражение симпатического нерва не дает эффекта.
Существуют и такие яды, которые действуют на вегетативные
органы подобно парасимпатической системе. Так, физостишин, муска-
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 499
рин, пилокарпин, холин возбуждают эффекторы подобно
парасимпатической системе. Здесь потовые железы точно так же составляют
исключение. Они получают нервы из симпатической системы, и, несмотря на
это, пилокарпин, мускарин и холин возбуждают их. Есть и такое
вещество, которое парализует парасимпатическое действие на эффекторы.
Так действует атропин. Например, после атропинизации сердца
блуждающий нерв перестает тормозить сердце; одновременно и ацетилхолин,
введенный в сердце, не тормозит его. Значит, атропин парализует
в сердце ту возбудимую систему, которая воспринимает активное
вещество — ацетилхолин.
Местом действия вегетативных ядов неЛьзя считать окончания
вегетативных нервов, ибо после денервации, когда перерождаются
нервные окончания, эти яды оказывают свое действие на эффекторы
в смысле понижения или повышения возбудимости, а также в смысле
возбуждения и угнетения 'возбуждения. Поэтому надо полагать, что
эти вегетативные яды действуют непосредственно на возбудимую
систему эффекторных клеток.
Антагонистическое действие симпатической и парасимпатической
систем на вегетативные органы не всегда наблюдается в той чистой
форме, как это указывалось выше. Известно, например, что
раздражением п. vagus вызывается сокращение кишечника, если мускулатура
или, вернее, вся периферическая структура находится в состоянии
низкой возбудимости. Если мускулатура находится в состоянии
высокой активности, раздражение п. vagus производит торможение (Мак
Эрла, Мак Суиней и Стопфорд, 63).
В определенных случаях через симпатическую систему происходит
не торможение кишечника, но, наоборот, повышение его
автоматических движений; например, если оба vagi перерезаны, а п. splanchnicus
цел, раздражение центрального конца одного из блуждающих нервов
производит через п. splanchnicus не торможение, а, наоборот, усиление
(Ходе, 64). По исследованиям Боцлера (65), раздражение
симпатической системы (п. hypogastricus), а также инъекция адреналина
производит в отношении тонкой кишки и матки небеременной кошки и
других животных в норме двухфазный эффект: сначала возбуждение —
усиление автоматических сокращений, а затем торможение —
угнетение последних. Путем анализа наблюдаемых фактов Б о ц л е р
приходит к заключению, что вообще симпатические импульсы способны
вызывать как возбуждение, распространяющееся в мышечных волокнах,,
так и одновременно понижение возбудимости их. Первоначально
нервные импульсы повышают активность мышц. Но с течением времени
мышца перестает возбуждаться под влиянием тех же симпатических
импульсов вследствие вызываемого ими гуморальным путем
понижения мышечной возбудимости и блокирования мышечного проведения-
Если эта блокада наступает быстро, то фаза возбуждения
маскируется, она отпадает, и поэтому реакция является с самого начала
тормозной.
32*
500 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Химическая природа возбуждающей и тормозящей деятельности
вегетативной нервной системы. Прежде всего нужно решить вопрос,
где именно возникает торможение, функцией какого нервного элемента
оно является. Преганглионарные волокна только проводят
возбуждение из центральной нервной системы, подобно цереброспинальным
нервам. Даже если раздражение преганглионарных волокон дает
торможение, то это не зависит непосредственно от них. Это видно прежде
всего из того, что раздражение соответствующих постганглионарных
волокон также вызывает торможение периферического органа.
Существуют ли среди постганглионарных волокон специфически
тормозящие волокна, или все дело в том органе, на который они
действуют? Имеются определенные факты, которые говорят в пользу
второго предположения. Известно, что адреналин действует на
гладкую мускулатуру и железы аналогично симпатическому нерву, а
пилокарпин — аналогично парасимпатическому. Было доказано, что оба
упомянутых вещества действуют на мышечную ткань и в том случае,
если влияние нервов уничтожено вследствие их дегенерации после
.перерезки (Левандовский, 2; Лэнгли, 3; Эллиот, 4; Андерсон, 5).
Следовательно, симпатическое и парасимпатическое действие этих ядов
происходит не при посредстве нервов. Яды действуют прямо на
двигательные и железистые клетки. На основании этого рода фактов
Лэнгли заключил, что в мышечных и железистых клетках, которые
иннервируются из обоих отделов вегетативной системы, существуют
двоякого рода «воспринимающие вещества»: одно производит
возбуждение, а другое — торможение. Характер симпатического или
парасимпатического эффекта будет зависеть от того, где заканчивается данный
нерв: в возбуждающем или тормозящем воспринимающем веществе.
Под воспринимающим веществом Лэнгли нужно понимать живую
систему. По мысли Лэнгли, в каждой мышечной и железистой
клетке должны существовать две живые системы: одна для
возбуждения, а другая для торможения.
Это представление Лэнгли сильно видоизменилось в свете
новейших фактов. Именно, в последнее івремя стало известно, что
возбуждение, блуждающего и симпатического нервов продуцирует в сердце
лягушки такое вещество, которое действует на сердце подобно самому
нерву. Если, например, долго тормозить сердце раздражением
блуждающего нерва и собрать всю кровь или жидкость Рингера, которая
успела пройти через сердце за это время, то собранная жидкость
окажет на бьющееся сердце тормозящее действие. Если же собрать
жидкость во время ускоренной сердечной деятельности, вызванной
ш сердце раздражением симпатического нерва, и затем воздействовать
ею на другое сердце, то в этом случае, наоборот, работа последнего
сердца ускорится (рис. 228) (Лёви, 7). За гормональную природу
действия нервных импульсов на сердце говорит и тот факт, что от одной
волны возбуждения блуждающего нерва торможение сердца наступает
с большим скрытым периодом — более 0.1 сек. и продолжается около
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 501
2 сек. Это проявляется в замедлении нескольких ритмических
импульсов, наступающих после раздражения (Броун и Экклс, 42).
Можно было думать, что тормозящее вещество возникает в сердце
в связи с прекращением сердечных сокращений и вообще от гиподи-
намического состояния сердца, а возбуждающее, наоборот, в связи
с усилением сердечной деятельности (Ашер и Шейнфинкель, 17). Но
было выяснено, что то и другое вещества не возникают в результате
процессов, имеющих отношение к механической деятельности сердца.
Рис. 228. Действие активных веществ сердца.
Сердце г. esculenta. Через сердце пропускается
чистый раствор Рингера. В момент 2 он заменяется таким
раствором Рингера, который прошел сердце в период
20-минутного раздражения п. vagus. Сейчас же высота
систолических сокращений падает. В момент 3
вводится 0.01 mg атропина. Вследствие паралича
тормозящего действия блуждающего нерва
раздражение его (момент 4) производит^ усиление
систолических сокращений от возбуждения
симпатических волокон сердца, проходящих через
блуждающий нерв. Собранный в этот период раствор Рингера
вновь вводится в сердце в момент 6, от чего высота
систол вновь нарастает (Лёви).
Именно, в некоторых случаях жидкость, собранная во время
торможения, вызванного раздражением п. vagus, вернее п. vagosympathicus,
лягушки, производит ускорение сердечной деятельности (Лёви, 7).
Этот опыт показывает, что при раздражении симпатикуса в стволе
блуждающего нерва в сердце выделяется возбуждающее вещество,
несмотря на остановку сердца, и что в определенных случаях
физиологическое действие этого вещества, конечно, при переносе жидкости из
данного сердца в другое, оказывается сильнее одновременно выделяемого
под влиянием волокон блуждающего нерва тормозящего вещества.
На основании этого рода фактов в настоящее время многие
физиологи высказываются в том смысле, что вообще действие вегетативной
системы на мускулатуру имеет чисто гормональную природу; постган-
глионарное волокно через свое окончание выделяет в иннервируемой
клетке вещество, которое производит или возбуждение, или
торможение возбудимой системы этой клетки.
Положение о гормональной регуляции деятельности гладкой
мускулатуры как будто подтверждается тем фактом, что в некоторых
органах большинство мышечных волокон не снабжено нервными
окончаниями. По Штёру (8), нервные волоконца образуют густое
сплетение вдоль мышечных клеток, но из 100 мышечных клеток не
более одной снабжается нервным окончанием.
502 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Кроме того, очень редко удается наблюдать в одной мышечной
клетке два нервных окончания, соответствующих симпатической и
парасимпатической системам. Буке всего на двух препаратах удалось
наблюдать по два окончания, да и то было не известно, относятся ли
они к разным системам (Буке, 27; Тигс, 28). Поэтому для нервной
регуляции мышечной деятельности может иметь существенное значение
выделение таких химических агентов, которые, распространяясь через
кровь, или путем диффузии через межуточную жидкость, будут
способствовать сокращению или расслаблению в безнервных мышечных
волокнах.
По отношению к симпатической системе таким активным гормоном
признается вещество, подобное адреналину. Близкое сходство этого
вещества с адреналином доказывается их синэргетическим действием
на разные органы, например на 3-е веко (membrana nictitans) кошки.
Подпороговое действие его на эту перепонку повышает ответ
последней на адреналин. Сходство адреналина и вещества, выделяемого
симпатической системой, доказывается еще следующим фактом.
Существуют такие вещества, как, например, эрготоксин, которые снимают
в эффекторе действие адреналина и одновременно действие
симпатической системы.
Однако адреналин в своем действии на эффекторы несколько
отличается от вещества, выделяемого окончаниями симпатических
волокон. Так, например, после инъекции эрготоксина адреналиноподобное
вещество вызывает повышение кровяного давления, а адреналин
производит его падение после короткого начального повышения.
Ввиду такой разницы Кэннон и сотрудники предложили для
вещества, выделяемого окончаниями симпатических волокон, особое
название симпатина (9). Но известно, что симпатикус в одних
органах производит возбуждение, а в друлих — торможение. Кэннон
и сотрудники полагают, что в органах, где оканчиваются
симпатические волокна, образуются двоякого рода вещества: один симпатии
для возбуждения, а другой для торможения. Кэннон полагает, что
симпатии в сущности везде один и тот же, но в эффекторных клетках
существует двоякого рода вещество, вроде тормозящей и/
^возбуждающей субстанции Л э н г л и. Симпатии, соединившись с одним из
них, производит возбуждение — сокращение, а соединившись с другим,
вызывает торможение — расслабление сокращения (30).
Симпатии содержится в экстрактах всех нервов, которые
производят адреналиноподобное действие на эффекторы, и всех тех органов,
где они оканчиваются. Но, нормально, он освобождается нервными
окончаниями при их возбуждении (Кэннон и Лиссак, 45).
Однако не все симпатические нервы выделяют своими окончаниями
адреналиноподобное вещество. В гладких мышцах некоторых
кровеносных сосудов, а также в потовых железах некоторых животных
симпатические нервы производят ацетилхолиноподобное вещество, подобно
парасимпатической системе.
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 503
По отношению к парасимпатической системе активным
гормональным продуктом ее деятельности, повидимому, является вещество,
схожее с ацетилхолином. Этот гормональный продукт, подобно ацетил-
лолину, не устойчив в щелочи и устойчив в кислотных растворах.
Оба растворяются в алкоголе и не растворяются в эфире. Они
разрушаются при согревании до 56° С. Человеческая кровь разрушает их
и при 40° С за 15 сек. Эзерин предохраняет оба вещества от
разрушения. Ацетилхолин или ему подобное вещество содержится в
экстрактах всех тех нервов и органов, которые производят ацетилхолинопо-
добное действие. Так, например, ацетилхолиноподобное вещество
образуется в сердце при раздражении блуждающего нерва (Дэйл, 10),
в матке при ее (возбуждении раздражением п. hypogastricus
(Шериф, 11), в кишечнике при его возбуждении раздражением блуждающего
нерва (Бантинг, Миик и Мааске, 20). В одном случае
ацетилхолин является тормозящим веществом (сердце), в других —
возбуждающим (матка, кишечник). Вследствие этого кровь, вытекающая из
летли сокращенного кишечника при раздражении блуждающего нерва
и содержащая активное вещество, на неподвижную денервированную
петлю кишечника действует возбуждающим образом, а на бьющееся
-сердце — тормозящим (20).
Раздражением нервно-сосудистого пучка петли кишечника,
содержащего как симпатические, так и парасимпатические нервные волокна,
в петле выделяется вещество как парасимпатического, так и
симпатического характера (Виталь, 57).
Аналогичное явление гормонального действия вегетативных нервов
<было установлено по отношению к железам. Так, например, известно,
что денервированная подчелюстная железа может выделять слюну
при раздражении chorda tympani противоположной стороны (Бабкин
и сотр.; 12). ' Известно также, что при раздражении симпатических
нервов, иннервирующих железу, сокращается мигательная перепонка.
Нарушение связи через кровь приводило к исчезновению этих эффектов
(Каттел, Вольф и Кларк, 13). Точно так же п. splanchnicus действует
на надпочечники, на железу внутренней секреции и вызывает секрецию
адреналина гормональным путем: преганглионарные волокна производят
вещество, подобное ацетилхолину, а уже это последнее вещество
благоприятствует секреции адреналина из клеток мозгового вещества
(Фельдберг, Минц и Тсудзимура, 32). Даже слюна, вытекающая при
раздражении chorda tympani, содержит активные вещества, которые
действуют на слюнную железу подобно раздражению
означенного нерва. Именно, если пропускать локковскую жидкость через
слюнную железу, железа останется в покое. Но если к локковской
жидкости прибавить вытяжку из деятельной слюнной железы или же
слюну, то секреция слюны наступает без раздражения нерва (Де-
мур, 58).
Точно так же установлено прямое действие ацетилхолина на
железу. Ничтожная доза ацетилхолина, как 0.003—0.1 у, введенная
504
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
в железу через артерию, производит секрецию, которая нарастает
с повышением дозы ацетилхолина ассимптотически. При этом было
замечено, что ацетилхолин не только вызывает секрецию слюны, но
делает железу чувствительной к повторному введению ацетилхолина
(Бецнак и Фаркас, 61).
О распространении активных веществ через кровь. Активные
вещества, выделяемые в мышечных и железистых клетках нервными
окончаниями вегетативной системы, не локализируются в этих
клетках. Они диффундируют из клетки наружу, поступают в кровь и через
кровь оказывают действие на весь организм, проявляясь в
активировании разных органов. Так, например, у кошки с денервированным
сердцем раздражение нижнего поясничного симпатического ствола,
который был изолирован от спинного мозга, приводило через-
30—60 сек. к заметному повышению кровяного давления, а через
3 мин. — к учащению сердцебиения. Так как связь между сердцем
и раздражаемой симпатической системой была только через кровь, то
очевидно, что симпатии или адреналиноподобное вещество, выделяемое
раздраженной симпатической системой, через кровь действовало на
сердце (Кэннон и Розенблют, 30). Аналогичное явление наблюдается
при активировании парасимпатической системы. Если, например, взять
кровь из сонной артерии одного кролика во время раздражения
блуждающего нерва и внести ее в ушную вену другого кролика, то
наблюдается торможение сердечного ритма, следовательно и тормозящее
вещество ¦— ацетилхолин передается через кровь (Бринкман и Вельде,
31). Демонстративно обнаруживается перенос ацетилхолина кровьк>
у беременных животных (кошки, кролика, свинки). Раздражение
блуждающего нерва матери вызывает остановку сердца не только
у матери, но и у плода (Гинецинскиц и Шамарина, 56). Особенно
хорошо получаются опыты с перенесением ацетилхолина при
предварительном введении в кровь эзерина (физостигмина), который
предохраняет ацетилхолин от расщепления. Один такой опыт приводим
здесь: раздражение chorda tympani на одной стороне вызывает
сокращение мигательной перепонки, секрецию подчелюстной железы
другой стороны, учащение дыхания и т. д. Все это получалось при
эзеринизации особенно сильно (Бабкин, Аллен и Ставраки, 12).
Каждый раз при возбуждении симпатической или
парасимпатической системы в крови человека появляются активные вещества,
подобные адреналину и ацетилхолину. Альперн и сотрудники (24)
установили, что при разных вегетативных диссимметриях кровь, оттекающая
от тканевого участка с нарушенной иннервацией, содержит ацетилхо-
линоподобное вещество, которое действует тормозяще на сердце
лягушки. То же бывает при феномене Ашера: во время замедления
сердца при надавливании на глазное яблоко кровь получает активные
свойства, как от раздражения блуждающего нерва. Такие активные
свойства приобретает кровь при разных патологических состояних,
сопровождающихся повышением функции парасимпатической или симпати-
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 505
ческой нервной системы, как язва желудка, гипертония, бронхиальная
астма и т. д.
О механизме действия активных веществ. Из приведенных выше
наблюдений ясно, что в эффекторах вегетативной нервной системы
при их возбуждении или торможении под влиянием нервных импульсов
образуются активные вещества, подобные адреналину и ацетилхолину.
Эти вещества оказывают на эффекторы, непосредственно или путем
распространения через кровь, определенное физиологическое действие,
а именно: адреналиноподобное вещество оказывает на данный
эффектор такое действие, как раздражение соответствующих симпатических
нервов, а ацетилхолиноподобное вещество действует на них подобно
раздражению соответствующих парасимпатических нервов.
Отсюда авторами было сделано заключение, что адреналин и аце-
тилхолин или подобные им вещества являются передатчиками —
медиаторами возбуждения и торможения с нерва на эффектор в
области вегетативной нервной системы. Каждый раз, когда вегетативный
нерв приходит в активное состояние, его окончания в гладких мышцах
и железах производят то или другое химическое вещество, а уже это
последнее непосредственно или через кровь возбуждает или тормозит
мышечные и железистые клетки.
Были произведены специальные опыты для установления
количества того или другого вещества, потребного для активирования
эффектора. Так, ацетилхолин действует на сердце лягушки
тормозящим образом в дозе 0,02 ^ IY =0.001 мг) на грамм свежей ткани.
Адреналин действует на один грамм желудка в дозе 0.06 v.
Концентрации ацетилхолииа и адреналина, необходимые для активирования
эффекторов, не выходят за пределы тех концентраций, в каких они
выделяются эффекторами при возбуждении вегетативной нервной
системы (Кэннон, 30).
Образование ацетилхолиноподобных и адрекалиноподобных
веществ в нервной ткани обычно связывают с обменом веществ, в
частности с углеводным обменом. Именно, предполагается, что
ацетилхолин образуется в нервной клетке из холина и из продуктов распада
углеводов (Манн, Тенненбаум - и Квостел, 46). Были поставлены
специальные опыты для проверки этого предположения. Было
испытано действие таких веществ, как фтористый натр, глицериновый
альдегид и другие, которые прекращают в той или другой стадии распад
углеводов и тем самым делают невозможным образование
ацетилхолииа. Оказалось, что влияние глицеринового альдегида на сердце
лягушки проявляется в прекращении тормозящего действия
блуждающего нерва (Коштоянц и Митрополитанская, 47).
Адреналиноподобное вещество оказалось таким продуктом, которое под влиянием
оксидаз, как фенолаза, тирозиназа, легко окисляется и тогда теряет
свое симпатикомиметическое действие на сердце. Так же действует
медь, которая принимает участие в окислительных процессах.
Некоторые оксидазы представляют комплексное соединение меди с белком.
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Поэтому после обработки сердца солями меди симпатический нерв
перестает действовать на сердце (Коштоянц и Митрополитанская, 47).
Критическое рассмотрение гипотезы о химической или гуморальной
передаче возбуждения в эффекторах вегетативной нервной системы.
Прежде всего следует указать на ряд фактов, которые не могут быть
всецело согласованы с этой гипотезой. В определенных случаях
химическая передача возбуждения не имеет места. Так, парасимпатическая
•система (п. vagus, n. pelvicus) действует возбуждающе на тонкие
кишки, мочевой пузырь и некоторые другие органы. При этом
выделяется ацетилхолин. Но атропин, который устраняет действие ацетил-
холина, не устраняет действия блуждающего нерва на тонкие кишки
и п. pelvicus на мочевой пузырь. В отношении желудка известно, что
атропин только ослабляет в нем двигательный эффект блуждающего
нерва (Кэннон, 30).
Есть и другие вещества — яды, которые понижают чувствительность
мышечных клеток к признанному химическому медиатору, но не
влияют на передачу возбуждения с нерва на мышцу. Это бывает,
например, при действии вещества, известного под цифровым
обозначением 933 F, на мигательную перепонку (35). На мигательной перепонке
кошки Бак и Бове (48) показали, что после отравления 933 F
меняется последействие сокращения, вызванного нервным импульсом,
.а не само начальное сокращение, — именно, чем больше доза 933 F,
тем меньше это последействие. Отсюда следует, что 933 F устраняет
только последействие, а на начальное сокращение не действует.
Значит, в этом сложном механическом эффекте только последействие
обязано своим происхождением химическому медиатору.
Наконец, известно, что чувствительность и реактивность гладкой
"мускулатуры к ацетилхолину и адреналину уменьшается при его
повторном действии, подобно тому как это наблюдается в отношении
действия ацетилхолина на скелетную мускулатуру. Это было замечено
в отношении разных гладких мышц, а также в отношении сердца
(Герман и сотр., 66; Тифено, 67). Это явление адаптации к активным
химическим веществам решительно говорит также против
исключительно гуморальной передачи возбуждения.
На основании всей суммы известного фактического материала
•можно допустить следующую форму передачи возбуждения. В
физиологии гладкой мускулатуры мы отметили, что под влиянием нервных
импульсов гладкая імышца отвечает сложным механическим и
электрическим эффектом. Первое быстрое сокращение с первым быстрым
электрическим колебанием и, вероятно, также второе сравнительно
•быстрое колебание обусловлены непосредственно обычным импульсом
¦возбуждения. Это доказывается тем, что абсолютная рефракторная
•фаза совпадает именно с этими колебаниями. Далее, там же было
показано, что под влиянием адреналина гладкая мышца дает
медленные сокращения и медленные электрические колебания, подобно
третьей медленной фазе электрического колебания, вызванного оди-
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПЯРАСИМПАТИЧЕСКОЙ СИСТЕМ 507
ночным нервным импульсом. Отсюда можно заключить, что токи
возбуждения нервных окончаний производят возбуждение возбудимой
системы, вследствие чего наступают быстрые сокращения и колебание
электрического потенциала. В результате процесса возбуждения
в нервных окончаниях или в двигательных пластинках мышцы
выделяется активное вещество — адреналин или ацетилхолин, который и
производит дальнейшее медленное сокращение с медленным
электрическим колебанием. Таким образом, сократительный эффект
начинается под влиянием процесса возбуждения, а под влиянием активного
вещества происходит дальнейшее продолжение его. Такого же мнения
в общем держатся Экклс, Монние и другие.
Тормозящее действие вегетативной системы на эффекторы должно
быть всегда химического происхождения. Так, например, можно
предполагать, что под влиянием нервных импульсов блуждающего нерва
в постганглионарных нейронах сердца образуется такое вещество,
подобное ацетилхолину, которое парализует возбудимую систему
проводящей мышечной ткани в синусных узлах, в пучках Гисса и т. д., от
которых зависит возникновение и проведение периодических импульсов
сердечной деятельности. Это химическое вещество едва ли образуется
непосредственно ів мышечных клетках, как это предполагает школа
К э н н о н а. Против этого предположения говорит прежде всего факт
-отсутствия двоякого рода и вообще двух нервных окончаний в
гладких мышечных клетках. Более вероятно, что торможение гладкой
мускулатуры происходит через нервное сплетение. Мы выше
указывали, что гладкие мышечные клетки получают нервные окончания из
периферического нервного сплетения, находящегося в стенках данного
органа. Это нервное сплетение посылает только импульсы возбуждения
к гладким мышцам. Торможение же, повидимому, заключается в
прекращении этих импульсов. Активное тормозящее вещество выделяется
здесь в нервном сплетении, и оно, вероятно, парализует, с одной
стороны, нервные элементы, через которые возбуждающие импульсы
передаются мышцам, а с другой — наиболее близко лежащие и наиболее
чувствительные мышечные клетки.
Итак, тормозящее действие вегетативной системы на эффекторы
происходит исключительно химическим путем и разыгрывается в том
нервном участке, где оканчиваются постганглионарные волокна.
Выделяемое здесь постганглионарными нейронами тормозящее вещество
тормозит как нервные элементы, передающие возбуждение к мышечным
клеткам, так и близлежащие мышечные клетки.
Тормозящее действие активного вещества имеет свое
электрическое выражение. Оно было изучено на предсердиях сердца черепахи
в связи с торможением при раздражении блуждающего нерва.
Электрический эффект возникает в виде сплошного положительного
колебания в самом начале раздражения, достигает максимума в течение
нескольких секунд и удерживается на много секунд после
раздражения, падая с большой постепенностью. . Автоматическая деятельность
508 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
начинается еще во время этого колебания (Гаскел, 53; Самойлов, 54)
(рис. 229). Такой длительный ток может возникать в скелетной мышце,
когда на нее действует ацетилхолин (Риссер и Штейнгаузен, 55). На
основании этих наблюдений можно утверждать, что медленное
электрическое колебание сердца под влиянием раздражения блуждающего нерва
обусловлено появлением активного тормозящего вещества — главным
образом ацетилхолина — в сердце.
Если же автоматия восстанавливается, несмотря на существование
положительного колебания, а, значит, и несмотря на действие активного
Рис. 229. Положительное электрическое колебание предсердия черепахи при
раздражении блуждающего нерва.
(а). Отводятся основание и верхушка остановленного правого предсердия в струнный гальванометр.
Зубцы на записи обусловлены забегающими токами возбуждения от других бьющихся отделов сердца.
При раздражении блуждающего нерва они прекращаются, и вместе с тем в отводимом предсердии
возникает медленное положительное колебание. Сигнальная линия Ъ отмечает время раздражения
блуждающего нерва; линия с — время в секундах (Самойлов).
вещества, то это хорошо согласуется с тем фактом, что и при
искусственном действии ацетилхолина автоматическая деятельность
прекращается в самом начале его действия, а потом она восстанавливается,
несмотря на продолжающееся действие ацетилхолина.
Однако едва ли тормозящие нервы способны производить только
торможение мышечной системы гуморальным путем и совсем не
способны вызывать в ней возбуждение своими возбуждающими
импульсами. Как указывалось выше, новейшие опыты Боцлера и других
свидетельствуют, что тормозящие нервы могут производить и
возбуждение мышцы: это бывает в самом начале раздражения, пока еще не
проявилось гуморальное действие тормозящего характера. Это дает
нам право утверждать, что любой нерв, как возбуждающий, так и
тормозящий, оказывает биотоками возбуждения своих нервных
окончаний раздражающее действие на периферический орган — мышечную
систему; одновременно он выделяет или благоприятствует выделению
активных веществ, производящих повышение или понижение
возбудимости мышечной системы и этим часто маскирует возбуждающее
действие начальных нервных импульсов на эту систему.
В отношении желез также имеются указания, что секреция не
обусловливается целиком действием активных веществ, выделяемых
нервами. По отношению к железам нервный механизм — нервные
импульсы должны играть главную роль. Разенков (25) считает
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 509
нервный механизм пусковым в отношении желез, полагая, что начало
секреции обусловливается нервными импульсами непосредственно.
При этом, конечно, образуется и активное вещество, действие
которого может быть обнаружено. Так, в отношении подчелюстной железы
chorda tympani безусловно является пусковым нервом. Это видно
из того, что чем реже частота раздражения, тем меньше содержится
в слюне плотного остатка, тем меньше слюны. Максимальный процент
плотного остатка получается при некоторых средних частотах. При
высоких частотах, как 100—¦ 150 индукционных ударов, получается
ослабленный, пессимальный эффект. При этом освобождается ацетил-
холиноподобное вещество. Его действие, видимо, проявляется в
длительном последействии слюнной секреции. В период последействия
состав слюны характерно отличается от слюны, получаемой во время
раздражения: она содержит плотного остатка почти в 2х/г раза меньше,
а органического вещества почти в 3Va раза меньше, чем во время
раздражения. Кроме того, на это последействие сильное влияние
оказывает эзерин: при малых частотах раздражения, когда выделяется
меньше ацетилхолина, последействие усиливается и удлиняется, а при
более частых раздражениях, когда ацетилхолин должен выделяться
больше, наоборот, последействие тормозится. Эзерин усиливает
активное действие ацетилхолина. А ацетилхолин, как известно, при малых
концентрациях возбуждает возбудимую систему, а при больших
угнетает ее. Вследствие этого при редких раздражениях эзерин усиливает
последействие,, а при частых угнетает его. И прямыми опытами было
показано, что введение в слюнную железу больших доз ацетилхолина
угнетает слюнную секрецию (Приходькова, 26). Эти данные ясно
показывают, что ацетилхолин действительно является таким активным
веществом, которое обусловливает слюнную секрецию; но и нервные
импульсы играют первенствующую- роль в этой секреции: они
начинают секрецию и гонят слюну, более богатую органическими
соединениями, а, значит, и ферментами.
Когда орган находится все время в состоянии автоматической
деятельности, как сердце, тогда вегетативные нервы, регулирующие
автоматик), могут проявить свое действие только в виде повышения или
понижения автоматии. В этих случаях, безусловно, действие
вегетативных нервов может быть исключительно гуморальным: нервы,
понижающие автоматию, должны производить это исключительно через
химическое вещество, угнетающее деятельность органа; нервы,
повышающие автоматию, — через вещество, облегчающее возбуждение.
Однако это действие нервов едва ли проявляется непосредственно
в эффекторах. Скорее всего их влияние осуществляется через то
нервное сплетение или через то образование, откуда исходит автоматия.
Ацетилхолин, например, тормозит автоматию сердца лягушки, если он
действует на синусы и предсердие, где оканчивается блуждающий нерв,
но не тормозит автоматию сокращений желудочка при локальном
приложении к желудочку (Беритов, 62). Отсюда следует, что тормозя-
510 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
щее действие медиатора на сердце не распространяется на
двигательные мышечные клетки. Оно, вероятно, ограничивается нервными узлами,,
синусными мышечными узлами и проводящей мышечной системой.
Литература
1. ШтернЛ., Физиол. журн. СССР, 19, 377, 1935.
2. Lewandowsky, Z. ges. Neur. Psychiat., 14, 281, 1913; Zentr. Physiol., 12, 599,.
1898; Arch. Anat. Physiol., 1899, стр. 360.
3. Lang ley J. N., J. Physiol., 33, 374, 1905; Proc. Roy. Soc. В., 78, 170, 1906.
4. E 1 1 i о t , J. Physiol., 32, 401, 1905; 35, 367, 1907.
5. Anderson, J. Physiol., 33, 414, 1905.
6. Elliot, J. Physiol., 31, 1901.
7. Loewi O., Pfl. Areh., 189, 239, 1921; 193, 201, 1921.
8. S t 6 h r Ph. Jr., Mikroskop. Anat. d. vegetativen Nervensystems, Berlin, 1928.
9. К э и и о h В., Некоторые выводы из факта химической передачи нервных
импульсов. Докл. на 15 Междунар. физиол. конгрессе, 1935.
10. Dale Н. Н., Reiziibertragung durch chemische Mittel im peripheren Nervensystem r
Urban u. Schwarzenberg, 1935.
11. Sherif M. F., J. Physiol., 85, 298, 1935.
12. В a b k i n В., Allen A. a. S t a v г а с k у С, Trans. Roy. Soc, Canada, Sec.
V., стр. 89, 1932.
13. С a 11 e 1 JVL, W о 11 H. С a. Clark D., Am. J. Physiol., 109, 375, 1934.
14. К i b i a k о w A., Pfl. Arch., 232, 432, 1934.
15. F e 1 d b e r g W. a. G a d d u m J., J. Physiol., 80, 12, 1933.
16. F e I d b e r g W. a. V a r t i a i n e n A., J. Physiol., 83, 103, 1934.
17. A s h e r L. u. S с h e i n f i n k e 1 N.. Z. Biol., 92, 263—275, 1932.
18. Eccles J. C, J. Physiol., 85, 179, 1935; 85, 207, 1935; 85, 464, 1935.
19. В i s h о p G. H. a. H e i n b e с k e r P., Am. J. Physiol., 100, 519, 1932.
20. Bunting H., Meek W. J. a. Maaske С A., Am. J. Physiol., 114, 100r.
1935.
21. F e 1 d b e r g W., M i n z B. a. Tsudzimura H., J. Physiol., 81, 286, 1934..
21a. Про п пер H. И. Сб. докл. VI Всес. физиол. съезда, стр. 60, 1937.
22. Бабский Е. Б., Сб. докл. VI Всес. физиол. съезда, стр. 189, 1937.
23. Р о с и н Я. А., Сб. докл. VI Всес. физиол. съезда, стр. 200, 1937.
24. А л ь п е р н Д. Е., Сб. докл. VI Всес. физиол. съезда, стр. 211, Химическая
природа нервного возбуждения в организме человека, УИЭМ, 1939.
25. Р а з е н к о в И. П., Сб. докл. VI Всес. физиол. съезда, стр. 11, 1937.
26. П р и х о д ь к о в а Е. К., Сб. докл. VI Всес. физиол. съезда, стр. 253.
27. Во eke J., Brain, 44, 1, 1921.
28. Tiegs О. W., Austr. J. Exp. Biol. Med. Sci., 1, 131, 1927.
29. Witanowski W. R., Pfl. Arch., 208, 694, 1925.
30. CannonW. B. a. Rosenblueth A., Auton. nervo-effector system, 1937, New-
York.
31. В r i n k m a n n R. u. v. d. V e I d e J., Pfl. Arch., 207, 492, 1926.
32. Feldberg W., Minz B. a. Tsudzimura H., Arch. Exp. Path. Pharm^
163, 66, 1931; Pfl. Arch., 233 657, 1933; J. Physiol., 80, 15—16P, 1934.
33. В a r s о u о G. S., G a d d u m J. H. а. К h a g у a 1 M. A., J. Physiol., 82, 9—10P,..
1934.
34. Rosenblueth A. a. McRioch D., Am. J. Physiol., 106, 365, 1933.
35. Monnier A. M., Proc. Roy. Soc. B, 123, 409, 1937.
36. E с с 1 e s J. С a. Magladery J. M., J. Physiol., 90, 31 a. 68, 1937.
37. С a n n о n W. B. a Rosenblueth A., Am. J. Physiol., 119, 221, 1937..
38. E с с 1 e s J. C, Erg. d. Physiol., 38, 339, 1936.
АНТАГОНИЗМ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ СИСТЕМ 51В
39. Brown G. L., Physiol. Rev., 17, 485, 1937.
40. Cannon W. В. а. В а с q Z. M., Am. J. Physiol., 96, 322, 1931.
41. Eccles J. C, Physiol. Rev., 17, 538, 1937.
42. Brown G. L. a. E с с 1 e s J. C, J. Physiol., 82, 211 a. 242, 1934.
43. Brown G. L., J. Physiol., 81, 228, 1934.
44. Glick D., J. gen. Physiol., 21, 431, 1938.
45. С a n n о n W. a. L i s s а к К., Am. J. Physiol., 125, 265, 1939.
46. Mann, Tennenbaum a. Quastel, Bioch. Journ., 32, 243, 1938.
47. Коштоянц X. С. и Митрополитанская Р. П., ДАН, 23, № 9, 1939_
48. В а с q Z. М. а. В о v e D., Arch, inter, pharmacol., 50, 315, 1935.
49. Monnier A. M., Cold Spr. Harb. Symp. Quant Biol., 4, 111, 1936.
50. Lorente de No, Am. J. Physiol., 121, 321, 1938.
51. Mcintosh F. C, J. Physiol., 94, 155, 1938.
52. Mcintosh, J. Physiol., 93, 46P, 1938.
53. G a s k e 1 W. H., Beitrage zur Physiologie C. Ludwigs zu s. 70. Geburtstage gewidmet,.
Leipzig, 1887, S. 114; J. Physiol., 7, 451.
54. Самойлов А. Ф., Изв. Росс. Акад. Наук, стр. 451, 1259, 1917.
55. R і е s s е г О. u. Steinhausen W., Pfl. Arch., 191, 288, 1928.
56. Гинецинский А. Г. и ШамаринаН. М., Физиол. журн. СССР, 25, 656,.
1938.
57. Вита ль Д. А., Физиол. журн. СССР, 27, 49, 1939.
58. De moor L., Физиол. журн. СССР, 24, 126, 1938.
59. Bronk, Tower, Solandt a. Lurabee, Am. J. Physiol., 122, 1, 1938.
60. Mar razz i A. S., J. Pharm.[Exp. Ther., 65, 395, 1939; Am. J. Physiol., 127, 738..
1939.
61. BeznikM. a. Farkas E., Quart. J. Exp. Physiol., 26, 265, 1937.
62. Беритов И., О локальном действии ацетилхолина на отдельные участки сердца..
Не опубликовано.
63. М с Е г 1 а Е. D., McSwiney В. A. a. S t о р f о г d J. S. В., Quart. J. Exp.
Physiol., 15, 211, 1925.
64. Hodes R., Amer. J. Physiol., 130, 642, 1940.
65. Bozler E., Am. J.Physiol., 130, 627, 1940.
66. Hermann K., Jordan F., M о r i n G. u. W i a I J., С r. Soc. biol., 129,
pp. 593, 595, 952, 1938; 131, 282, 1939. Цит. no Annual Rev. of Physiol., 2, 265, 1940..
67. Tiffeneau M. a. Scheiner M., С r. Soc. biol., 130, pp. 448, 627, 1939;
131, 1939. Цит. по Annual Rev. of Physiol., 2, 265, 1940.
Глава IV
РОЛЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ В РЕФЛЕКТОРНОЙ
ДЕЯТЕЛЬНОСТИ ВНУТРЕННИХ ОРГАНОВ
Центростремительные пути вегетативной нервной системы.
Вегетативная система включает, наравне с центробежными, также
центростремительные или чувствительные пути. По исследованиям Л э н г л и
(1), между симпатической и парасимпатической системами существует
в этом отношении некоторое различие. Краниальная и сакральная
вегетативные системы содержат в себе также чувствительные волокна,
симпатическая же большею частью состоит из центробежных путей.
Только в отношении внутренностей она содержит также и
центростремительные волокна, которые по своему строению не отличаются
от других центростремительных волокон: они снабжены миэлиновой
оболочкой и проходят всю систему без перерывов; клетки этих
волокон лежат в спинномозговых узлах. Скорость проведения возбуждения
у них так же велика, как в цереброспинальном нерве. Например,
в п. splanchnicus скорость проведения возбуждения достигает 40 м
в секунду (Деннинг, 2).
При изучении биоэлектрических токов симпатических стволов
новейшей техникой, в связи с раздражением внутренних органов, были
обнаружены двоякого рода центростремительные импульсы
возбуждения — быстрые и медленные и, кроме того, еще длительные волны.
Быстрые центростремительные импульсы возникали при изменении
напряжения внутренних органов. Медленные импульсы отвечали
повреждающим раздражениям, вызывающим рефлекторно внешние
реакции. Длительные же волны, обычно протекающие с
продолжительностью около 0.03 сек., видимо, возникали в результате раздражений
эфферентных симпатических волокон (Тоуэр, 3).
Чувствительные волокна вегетативной нервной системы оказывают
определенное действие на центральную нервную систему, вызывая
рефлекторную реакцию как во внутренних органах, так и в скелетной
мускулатуре.
Преганглионарные псевдорефлексы. Уже на основании
существования центростремительных путей, оканчивающихся в центральной
РОЛЬ ВЕГЕТАТИВНОЙ НЕЯВНОЙ СИСТЕМЫ В РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 513
нервной системе, можно утверждать, что сама по себе вегетативная
система неспособна к самостоятельной рефлекторной деятельности,
что последняя может совершаться только через центральную нервную
систему. Несмотря на это, некоторые авторы находят возможным
рефлекторное действие вегетативной системы. Так, например, после
разрушения спинного мозга раздражение центрального отрезка
п. splanchnicus вызывает повышение кровяного давления. Это
действие прекращается с 'перерезкой симпатического пограничного столба.
Р о ж а н с к и й (4) утверждает, что здесь мы имеем настоящий рефлекс.
Аналогичное наблюдение было сделано в лаборатории Р а з е н-
к о в а. Так, после удаления симпатической цепочки и разрушения
продолговатого мозга при раздражении центрального конца
перерезанного п. splanchnicus было замечено торможение перистальтики
кишечника. Этот эффект исчезал после общего или локального
отравления пограничного столба никотином. Отсюда было сделано
заключение, что мы здесь имеем рефлекс, который вызывается путем
передачи возбуждения от п. splanchnicus major к п. splanchnicus minor
через симпатические узлы (Аршавский, 5).
Но, по исследованиям Л э н г л и (1), эти реакции раздражения
симпатической системы не могут быть названы рефлексами. Они,
безусловно, происходят оттого, что каждое центробежное преганглионарное
волокно дает коллатерали в нескольких узлах. Когда раздражается
центральный отрезок преганглионарного волокна, то возбуждение,
распространяясь в обратном направлении, переходит через
коллатерали на ряд нервных узлов. Это, конечно, в свою очередь,
вызывает деятельность того периферического органа, в котором
распространяются нейриты узловых клеток. Следовательно, здесь мы имеем
дело не с переходом возбуждения от чувствительного нерва на
двигательный, а с распространением возбуждения в центробежном нерве
сначала в обратном, т. е. центростремительном, направлении, а затем
по коллатералям в обычном для нерва центробежном направлении. Это
явление Л э н г л и обозначил как «псевдорефлекс» или «преганглио-
нарный аксонный рефлекс». При дегенерации преганглионарных
волокон, в связи с их перерезкой, аксон-рефлексы с соответствующего
симпатического узла исчезают. Этот факт подтверждает правильность
положения Лэнгли об отсутствии нервных связей в симпатических
узлах наподобие рефлекторного центра спинного мозга
(Лаврентьев, 14).
Можно привести еще один пример «псевдорефлекса», который
также подробно был разобран Лэнгли. Соковкин (6) перерезал
у кошки все те нервы, которые соединяют g. mesentericum inferior со
спинным мозгом. Потом перерезал один из nn. hypogalstrici, который
направляется к plexus hypogastrics, и раздражал электрическим током
его центральный отрезок. В результате он получил через pi.
hypogastricus сокращение мочевого пузыря, sphincter ani int., и сужение
кровеносных сосудов в нижнем отделе mucosa recti. Лэнгли (7) дока-
3*3 и, с. Беритов
514 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
зал, что в данном опыте раздражается не чувствительный, а центро.-
бежный путь. Путем дегенерации он выяснил, что перерезанный
п. hypogastricus не содержит совсем чувствительных волокон.
Псевдорефлексы постганглионарных волокон. Наряду с прегангли-
онарными псевдорефлексами наблюдаются и постганглионарные
псевдорефлексы. Такие аксон-рефлексы наступают в очень ограниченной
области, в пределах распространения одного постганглионарного
волокна при раздражении одного из его отростков. Возможно, что в этих
постганглионарных рефлексах принимает участие существующая везде
в тканях нервная сеть. Согласно гистологическим исследованиям Л е-
о н т о в и ч а (9), эта сеть включает даже нервные клетки.
Очень легко демонстрируемый аксон-рефлекс наблюдается на
коже. А именно, если царапнуть или капнуть на нее горчичным
маслом, то в области раздражения происходит покраснение кожи, что
свидетельствует о расширении кровеносных сосудов. Это явление
не пропадает после перерезки соответствующих спинальных нервов,,
но оно исчезает после дегенерации их (Бр?слауер, 8). Это явление
можно рассматривать как постганглионарный аксон-рефлекс (Шильф, 10).
Но есть и другая точка зрения, которая относит означенную
реакцию к афферентной системе, к афферентному аксон-рефлексу
(Крог, 17). В настоящее время эта точка зрения наиболее
приемлема, ибо доказано, что заднекорешковые чувствительные
волокна на периферии наряду с проведением нервных импульсов от
рецепторов в центральную нервную систему своими боковыми
веточками производят расширение сосудов. Поэтому можно принять, что*
при раздражении кожных рецепторов наблюдаемое местное
расширение сосудов является афферентным аксон-рефлексом, протекающим
совершенно независимо от центральной нервной системы, а также от
симпатической.
О рефлекторных реакциях вегетативной системы. Все рефлексы
вегетативной нервной системы протекают через центральную нервную*
систему. Рефлекторная дуга начинается или во внутренних органах,,
или в периферических. Хорошо известно, что раздражением почти
любого периферического чувствительного нерва можно вызвать
изменение кровяного давления и, кроме того, задержку биений сердца.
Точно так же раздражением внутренних органов вызываются
рефлекторные реакции. По мнению одних, эти рецепторы исключительно
болевого характера; прикосновение и вообще слабые раздражения будто
бы не деятельны. Однако известно, что при некоторых патологических
процессах чувствительность увеличивается, и тогда слабые, неповреж*
дающие раздражения также производят рефлекс. На это указывает
известный опыт Гольца на лягушках. Этот опыт заключается в том,
что удары тупым предметом по внутренностям производят остановку
сердца через блуждающий нерв. Эта остановка вызывается много
легче, если внутренности предварительно подвергались длительному
воздействию воздуха. Кроме того, кровяное давление, несомненно,.
РОЛЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ В РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 515
является естественным раздражителем рецепторов кровеносных
сосудов. Так, например, полагают, что п. depressor, содержащий в себе
чувствительные волокна от корня аорты и от сердца, нормально
возбуждается вследствие изменения давления крови. Именно при
усилении давления происходит возбуждение депрессора. Это вызывает через
центральную нервную систему расширение кровеносного русла, что, со
своей стороны, обусловливает падение кровяного давления. Что вообще
кровяное давление может стать раздражителем, ясно видно из того,,
что пачиньевы тельца брыжейки в определенных случаях
разряжаются импульсами синхронно с пульсацией кровеносных сосудов
(Гаммой и Бронк, 12).
Не только давление в кровеносных сосудах, но и давление в
любом полостном органе (мочевой пузырь, прямая кишка, желудок,,
желчный пузырь, двенадцатиперстная кишка, петли толстой и тонкой
кишки), вызванное одним и тем же способом путем растяжения
раздуванием введенного в них тонкостенного резинового мешочка,
производит раздражение рецепторов этих органов. При этом наступает
рефлекторная реакция через центральную нервную систему при таких
давлениях, которые ни в коем случае нельзя считать болезненными,
напр., в мочевом пузыре кошки достаточно было растяжение в 10 мм
Hg. Ответная реакция заключалась, как при сдавлении кожной
поверхности, в развитии общего торможения, которое, между прочим,
выявлялось в торможении рефлекторных реакций скелетной
мускулатуры (Беритов и Бакурадзе, 15).
Основным и наиболее распространенным раздражением внутренних
рецепторов должны служить химические вещества. В таких органах,
как селезенка, почка, кишечник, присутствие СОг или отсутствие О?
в растворе, питающем орган, является раздражителем рецепторов этих
органов. За этим следует повышение кровяного давления в сонной
артерии, учащение и углубление дыхания. Введение в кровеносные
сосуды этих органов никотина, ацетилхолина, гистамина, адреналина
также вызывает рефлекторное повышение кровяного давления в
сонной артерии, учащение и углубление дыхания. Эти рефлекторные
реакции наступают быстро, с коротким скрытым периодом, но дают
длительные последействия. Новокаин, введенный в означенный орган,
снижает рефлексогенное действие химических веществ. Этим
доказывается химическая природа раздражения данных интероцепторов
(Черниговский, из лаборатории проф. Быкова, 16).
Тоническая деятельность вегетативной нервной системы. Известно,
что определенная часть преганглионарных волокон активно действует
на разнообразные органы для удержания их в состоянии некоторой
постоянной деятельности. Это тоническое влияние вегетативной
системы по временам то усиливается, то ослабевает. Так, известно, что
блуждающий нерв все время действует тормозящим образом на
сердце. Это видно прежде всего из того, что после перерезки
блуждающих нервов ритм сердечной деятельности повышается. Точно так
33 *
516 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
же симпатические волокна сердца все время посылают свои
ускоряющие импульсы к сердцу (Бронк и сотр., 13). Перерезка их ведет
к понижению ритма сердечной деятельности. Такое тоническое
влияние парасимпатическая и симпатическая системы оказывают и на
другие органы. Так, например, тоническое расширение зрачка зависит от
симпатической системы, что видно из того, что зрачок суживается
после перерезки симпатической системы. Точно так же тоническое
сужение кровеносных сосудов зависит от симпатической системы, потому что
с перерезкой последней сосуды расширяются; тоническая
двигательная деятельность пищеварительного тракта обусловливается
центральными влияниями через блуждающие нервы парасимпатической
системы.
Тоническая деятельность вегетативной нервной системы возникает
вследствие определенных периферических влияний. Так, тоническая
иннервация, ускоряющая сердечный ритм, обусловливается
раздражением рецепторов в корне аорты и в стенках сонных артерий от
периодического изменения кровяного давления, а также чувствительных
окончаний блуждающего нерва в легких при растяжении во время
вдоха и спадении их во время выдоха. Возникающие при этом
афферентные, т. е. центростремительные, импульсы оказывают через
симпатические центры імозга на ритм сердечного симпатического нерва как
возбуждающее, так и тормозящее влияния (Бронк и сотр., 13). Далее,
тоническая деятельность пищеварительного тракта зависит от
раздражения разного рода рецепторов, между прочим статического
рецептора в полукружных каналах. При раздражении их происходит
усиление тонуса мускулатуры пищеварительного тракта, а при разрушении
их, наоборот, временное ослабление этого тонуса (Василенко, 11).
Но есть и такие органы, которые свободны от тонического влияния
вегетативной нервной системы. Например, потовые железы, гладкие
мышцы кожи в норме не находятся в тоническом состоянии.
Тоническая деятельность вегетативной нервной системы прежде
всего зависит от центральной нервной системы. В продолговатом
імозгу существуют специальные центры для расширения и сужения
кровеносных сосудов, для ускорения и торможения сердца, откуда
нервные импульсы идут непрерывно (рис. 230). Там же, в верхнем
отделе, находится центр для симпатико-адреналовой системы и для
мускулатуры пищеварительного тракта. В промежуточном мозгу лежат
центры сложных эмоциональных реакций, которые наступают через
вегетативную систему. Например, у кошки при гневе через
симпатическую систему происходит выпрямление волос, появление пота на
пальцевых подушках, повышение артериального давления, выделение
адреналина. Таким образом, центральная нервная система
обусловливает как тоническую деятельность, так и всякую рефлекторную
деятельность вегетативной нервной системы.
После перерезки симпатической системы рефлекторная
деятельность через центральную нервную систему исчезает совсем. Тоническая
РОЛЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ В РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 517
же деятельность исчезает временно и частично. Поэтому в первое
время думали, что симпатические узлы обладают способностью
тонической иннервации. Но дело в том, что тонус периферических органов
помимо центральной нервной системы зависит еще от разнообразных
гормонов, выделяемых из желез внутренней секреции, а также от
определенных продуктов обмена веществ. Эти гормоны и продукты
разносятся кровью по ©сему организму и тем самым производят на
ткани определенное действие. Так, например, известно, что адреналин,,
гормон надпочечных желез, вызывает тоническое сужение кровенос*
Рис. 230. Биоэлектрические токи возбуждения^
симпатических нервов при тонической иннервации вазомоторов.
Запись осциллографом Мэтьюса. А—нерв^из солнечного сплтения
кролика; В — левый шейный симпатикус кролика; С — левый п.
hypogastrics кошки. Отметчик времени на верхнем краю каждой кривой — по
0.25 сек. Сильные токи обусловлены синхронным возбуждением
целого ряда ,постганглионарных волокон (Эдриан).
ных сосудов, тоническое ускорение сердечного ритма и т. д.; или,
например, известно, что при мышечной работе выделяется какое-то
вещество, которое через кровь производит расширение кровеносных
сосудов как в том же рабочем органе, так и в других. Затем известно,
что в стенках кишечника образуется вещество холин, который
действует на стенки кишечника, вызывая усиленную перистальтику,
подобно парасимпатической системе.
Ли терату р а
1. Lang ley N., Das autonome Nervensystem, Berlin, 1922.
2. Denning H., Z. Biol., , 395, 1929.
3. TowerS., J. Physiol., 78, 225, 1933.
4. Рожа некий H., К учению об отношении спинного мозга и симпатических
узлов к сосуд, системе, дисс, Казань, 1889.
5. Аршавский И., Труды физиол. отдел. Гос. Тимиряз. инст., стр. 95, 1930.
6. Sokowkin N., Pfl. Arch., 8, 600, 1874.
7. Lang ley a. Anderson, J. Physiol., 16, 435, 1894; La n g 1 e y, J. Physiol,
25, 468, 1900.
8. Breslauer, Dtsch. Ztsch. f. Chir., 150, 50, 1919.
9. Леонтович А., Журн. зкеп. биол. мед., 9, стр. 5, 1926.
518 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
10. Schilf Е., Dtsch. Z. Nervenheilkunde, 106, 196, 1928.
11. Василенко Ф., Некот. вопросы сравн. физиол. Сборн. раб. лабор. сравн.
физиол. жив. Биол. инст. им. Тимирязева, стр. 102, 1934.
12. Gammon G. D. a. BronkD. W., Am. J. Physiol., 114, 77, 1935.
13. Bronk D. W., Ferguson L. K., Margaria R. a. Solandt D. G.,
Am. J. Physiol., 117, 237, 1936.
14. Лаврентьев Б., Морфол. авт. нервн. системы. Сборн. работ ВИЭМ, стр, 5,
1939.
15. Беритов И. и Бакурадзе А., Тр. Инст. физиол. им. Бериташвили,
т. 5, 125, 1943.
16. Черниговский В. Н., Исследование рецепторов некоторых внутренних
органов, дисс, Ленинград, 1941.
17. Krogh A., Anatomie u. Physiologie d. Kapillaren, Berlin, 1924.
Г л а в а V
РОЛЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ В ДЕЯТЕЛЬНОСТИ
СКЕЛЕТНОЙ МУСКУЛАТУРЫ
Историческая справка. В 1884 г. Бремер (1) открыл, что в
мышечных волокнах конечностей и языка оканчиваются, наравне с мя-
котными, миэлиновыми нервными волокнами, безмякотные, голые
Болокна. Эти последние волокна были изучены голландским гистологом
Буке (2) и другими в его лаборатории (Агдур, 3). Нервные
окончания безмякотных волокон лмеют совершенно такое же строение, как
окончания обычных імиэлиновых волокон. Авторы заметили
дегенерацию окончаний голых волокон после перерезки симпатической системы
и не обнаружили ее после перерезки передних двигательных
корешков. Отсюда они сделали заключение о симпатическом происхождении
этих волокон.
Исходя из этих гистологических наблюдений, голландский ученый
д е Б у р (4) пытался обнаружить физиологическую роль
симпатической системы в деятельности скелетных мышц. Он разъединял
симпатическую систему от центральной системы путем перерезки rr. com-
municantes или удаления симпатического пограничного столба на
лягушках и кошках и потом изучал тонус задних конечностей. Он
заметил ослабление тонуса. Отсюда он заключил о зависимости тонуса
конечностей от симпатической системы.
Другие голландские же ученые нашли характерное изменение
обмена веществ в мышцах во время их тонической деятельности,
я именно повышенное выделение креатина, который является
продуктом белкового обмена. Углеводный обмен, наоборот, по данным этих
авторов, не усиливался во время тонических сокращений (Пекелха-
ринг и Гоугенгуйц, 5).
Приняв во внимание эти химические исследования, де Бур
предположил, что симпатическая система вызывает распад белковых тел
в саркоплазме и следствием этого процесса является сокращение
саркоплазматического вещества. Согласно этому предположению,
миофибрилли не играют никакой роли в производстве тонических
сокращений.
520 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Опыты де Бура ввиду их большого теоретического значения
были повторены во многих физиологических лабораториях. Но в
громадном большинстве случаев исследователи не получили тех
результатов, какие сообщались д е Б у р о ім. Если даже они получались,
авторы не соглашались с теоретическим пониманием д е Бура. Так,
например, утверждают, что разница в тонусе конечностей у лягушкиі
после перерезки симпатикуса на одной стороне обусловливается
рефлекторным выделением кислой слизи на нормальной стороне. Это
выделение раздражающим образом действует на кожу и вызывает сги-
бательный рефлекс (Гедевани, 6). Известно, что при раздражении
перерезанного симпатического нерва тонус усиливается на
соответствующей стороне и даже наступают изолированные движения
пальцев (Лангелаан, 7а). Это явление как будто подтверждает гипотезу*
де Бура. Но оказывается, что и это явление опять-таки
обусловливается выделением кислой слизи, которая вызывает рефлекторные
движения раздражением того же самого участка кожи, где она
выделяется (Гедевани, 6).
Кроме того, были поставлены другие опыты специально для
выяснения роли симпатической системы в деятельности скелетной
мускулатуры. Прежде всего было доказано, что рефлекторный тонус
сгибателей задних ног лягушки, который являлся предметом исследования
де Бура, ни в какой мере не зависит от симпатической системы.
И после удаления симпатической цепочки, проводящей возбуждение
из центральной нервной системы, означенный рефлекторный тонус
наступает без ослабления (Беритов, 8). Далее было показано, что и тот
нерефлекторный тонус мышц, который наступает в виде контрактуры
после сильного и длительного раздражения двигательного нерва, не
зависит от симпатической системы (Беритов, 9). Против д е Б у р а-
говорит тот общепризнанный факт, что прямое раздражение
симпатического нерва никогда не вызывает сокращения скелетных мышц, хотя
оно может производить сильные сокращения гладкой мускулатуры.
Многие авторы испытывали также действие адреналина на
скелетную мускулатуру. Они предполагали, что, если симпатическая система-
действует непосредственно на мышечные волокна, адреналин должен
был вызвать сокращение, как в гладкой мускулатуре. Однако и эти>
опыты не оправдали гипотезы де Бура. На нормальную скелетную
мышцу адреналин не оказывает никакого действия (Панелла, 10). Он
действует только на утомленную мышцу в нормальных условиях
кровообращения; при малой концентрации он усиливает сокращение, а при
большой — ослабляет его (Десси и Грандис, 11; Панелла, 10;
Грубер, 12). Но эти эффекты на утомленной мышце авторы объясняют
действием адреналина на кровеносные сосуды, сужением сосудистой
системы (Лэнгли, 13; Кэннон и Найс, 14; Вастль, 15). При малой
концентрации адреналина, когда происходит небольшое сужение сосудов,
кровообращение усиливается в силу ускорения тока крови. Это ведет
к лучшему снабжению мышечной системы кислородом и буферными
РОЛЬ ВЕГЕТАТИВНОЙ СИСТЕМЫ В ДЕЯТЕЛЬНОСТИ МУСКУЛАТУРЫ 52*
соединениями и лучшему вымыванию из нее продуктов обмена
веществ. Вследствие этого сокращение усиливается, утомление частично'
проходит. При большой концентрации адреналина, когда происходит
более сильное сужение сосудов, кровоснабжение мышцы сильно
ослабевает; это обусловливает уменьшение кислорода и буферных
соединений и затем накопление продуктов обмена веществ в мышце.
Ухудшаются условия мышечной деятельности, вследствие чего утомление
быстро углубляется.
Опыты на кошках также не оправдали предположения д е Б у р а~
Так, например, было установлено, что, хотя тонус десимпатизирован-
ной конечности и ослабевает сравнительно с нормальной конечностью,
но это ослабление временное. Спустя несколько дней или недель тонус-
восстанавливается — возвращается к норме (Дюссер де Баренн, 16).
Весьма возможно, как полагает Л э н г л и, что это временное
ослабление тонуса зависит от уменьшения в мышцах кислых
продуктов обмена веществ вследствие расширения кровеносных сосудов.
Это расширение наступает после перерезки симпатической системы,
так как тоническое сужение сосудов обусловливается, между прочим,.
центральными влияниями через симпатическую систему. Но
расширение является временным. Спустя некоторое время сосуды вновь
суживаются под влиянием других факторов, обусловливающих тонус сосудов-
Затем нужно отметить, что наблюдаемое Пекелхарингом и
Гоугенгуйцоім усиленное выделение креатина при тонической
иннервации не нашло подтверждения. Из лаборатории самого П е-
келхаринга впоследствии вышли такие работы, которые
указывают на неизменяемость содержания креатина при тонической
иннервации (Дюссер де Баренн и Терверт, 17). іКроме того, не была*
замечено изменения в образовании креатина в связи с симпатической
иннервацией (Шмид, 18; Ринберк, 40).
Существует большая литература по поводу опытов д е Б у р а^
Резюмируя эту литературу, многие авторы категорически отрицают
роль симпатической системы в тонической деятельности скелетной
мускулатуры (Кобб, 19; Шпигель, 20; Форбс, 21; Беритов, 22). Многие
гистологи также высказываются против утверждения Б у к е о
симпатическом происхождении окончаний немиэлиновых нервных волокон
в мышечных клетках (Телло, 23; Бильшовский, 24; Штёр мл., 25;
Лаврентьев, 30 и др.). В последнее время результаты специальных
гистологических исследований, поставленных с целью проверить Буке,
совсем не оправдывают его выводов. Они находят, что те голые
нервные волокна, на которые указывает Буке, или являются обычными
двигательными волокнами в известной стадии дегенерации, или же
заканчиваются не внутри мышечных клеток, а лежат снаружи ихг
служа для иннервации кровеносных сосудов (Тигс, 26; Гинзей, 27;:
Вилькинсон, 28; Рэнсон, 32; Тушнова, 48).
Эффект Гинецинского — Орбели на утомленной мышце. В
последние 20 лет О р б е л и со своими сотрудниками (29) утверждает, что
.522 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
симпатическая система иннервирует все возбудимые ткани, как
мышечную, так и нервную (мозг, рецептор), оканчиваясь внутри их
клеточных образований. Она регулирует в них обмен веществ, производит
какие-то химические изменения и тем самым лучше приспосабливает
их к функциональной деятельности. Эта адаптационно-трофическая
гипотеза симпатикуса была выведена авторами из многочисленных
разнообразных опытов на разных животных. Исходным же фактом
является наблюдение Гинецинского (31), что утомленная рядом
одиночных раздражений мышца до некоторой степени восстанавливает
¦свою сократительную способность под влиянием тетанического
раздражения симпатикуса.
Благоприятствующее влияние раздражения симпатической системы
на утомленную мышцу было подтверждено у нас и за границей во
многих лабораториях. Было также подробно изучено, при каких
именно раздражениях симпатической системы лучше всего проявляется
эффект Гинецинского. Так, например, К и р з о н (из лаборатории
Ухтомского, 54) установил, что этот эффект лучше всего получается
при частоте электрического раздражения симпатической цепочки 10—20
в секунду, опускающейся в отдельных случаях до 4 в секунду. Но
если препарат согреть и вообще повысить его лабильность, оптимум
частоты при раздражении прямоугольными токами можно поднять до
70 в секунду. Все это свидетельствует, что в эффекте
Гинецинского мы имеем настоящий симпатический эффект. Однако насчет
природы действия симпатической системы на скелетную мышцу
мнения разошлись. В то время как О р бели и его сотрудники
утверждают прямое, непосредственное влияние симпатических импульсов на
обмен веществ внутри мышечных волокон, другие авторы не согласны
с этим и дают другие объяснения. Так, в лаборатории Беритова
Гедевани подробно изучил эффект Гинецинского и пришел
к заключению, что он обусловливается передвижением остатков крови
в кровеносных сосудах изолированной мышцы. Путем наблюдения
мышцы под микроскопом было установлено, что и в условиях
остановки кровообращения на изолированной лапке раздражение
симпатического нерва производит более или менее значительное передвижение
остатков крови из артерий в капилляры, а отсюда в вены,
вследствие сужения артериальной системы. Из этого было сделано
заключение, что кровь, оставшаяся после прекращения кровообращения
в капиллярах мышцы, во время утомления, с одной стороны, лишается
запасов кислорода, а с другой — насыщается кислыми продуктами
обмена веществ. Это неизбежно должно обусловливать ослабление
процессов возбуждения и сокращения. При раздражении симпатического
ствола эта кровь заменяется свежей порцией из артериальной
системы, которая содержит некоторый запас кислорода и симпатина,
а также разного рода энергетические вещества, потребляемые мышцей,
и свободна от кислых продуктов обмена веществ. Вследствие этого
должно получиться восстановление мышечной деятельности. Наилучшим
РОЛЬ ВЕГЕТАТИВНОЙ СИСТЕМЫ В ДЕЯТЕЛЬНОСТИ МУСКУЛАТУРЫ 523
доказательством этого объяснения служит тот факт, что совершенно
такая же картина усиления одиночных сокращений на лишенных
кровообращения мышцах получается в том случае, если в период
утомления через сосуды мышцы пропускается свежая порция крови путем
открытия зажатых сосудов всего на несколько секунд (рис. 231) (Геде-
вани, 33). Хорошее доказательство значения свежих порций крови для
восстановления утомленной
мышцы дается также в
опытах Некрасовых (50),
показавших, что утомленная
мышца после обливания
сывороткой крови
восстанавливает свою работоспособность
и что в этом отношении
артериальная кровь
обнаруживает более мощное
восстанавливающее действие на
утомленную мышцу, чем венозная.
Над вопросом о природе
трофического действия
нервов на ткани много работал
И. П. Павлов. Он еще в
своей докторской диссертации
(„Центробежные нервы
сердца", 1883) привел веские
доказательства существования
таких нервных волокон в
системе сердечных нервов,
которые усиливают или
ослабляют сердечные сокращения
без изменения ритма. В
течение последующих пяти лет он
упорно продолжал работать
над тем же вопросом и
пришел к убеждению, что эти нервы не являются особыми трофическими
нервами, а представляют собой сосудорасширяющие или
сосудосуживающие нервы венечных сосудов сердца. В работе, опубликованной
з 1888 г., он характеризует «усиливающий нерв как такой, который
повышает вообще все жизненные свойства желудочкового мускула...»
При раздражении его «увеличиваются как его (желудочка) возбудимость,
так и проводимость». Таким образом, Павлов считал, что это истинно
трофическое влияние особых нервных волокон на сердце
осуществляется не путем прямой иннервации мышечных клеток, а посредством
действия на сосудистую систему, путем расширения венечных сосудов
сердца. «С признанием сосудистой натуры новых сердечных нервов, —
говорит Павлов, — вполне и без малейшей натяжки объясняются
Рис. 231. Влияние передвижения крови на кривую
утомления ,-мышцы.
Спинномозговая лягушка. Записываются одиночные
сокращения икроножной мышцы от раздражения седалищного
нерва. Верхняя кривая от препарата, на котором
кровообращение было остановлено путем перерезки сосудов. Вначале
дается серия сокращений до утомления, затем период
сильного утомления. Здесь два раза производилось раздражение
симпатического нерва при 20 и 18 см расстояния между
катушками. Оба раза оно вызывает усиление одиночных
сокращений и одновременно передвижение в капиллярах остатков
коови (наверху кривой указываетя это обстоятельство).
Нижняя кривая записана на препарате, на котором
кровообращение было остановлено путем сдавливания артерии.
Мышцы находятся в средней стадии утомления. Сперва на
короткое время перестали сдавливать артерию; вследствие
этого кровь начинает передвигаться в мышце; это сейчас же
вызывает повышение одиночных сокращений. После нового
сдавливания артерии мышечные сокращения опять убывают.
После этого раздражается симпатический ствол на
короткое время при 25 см расстояния между катушками. В
последнем случае та же картина нарастания и убывания
одиночных сокращений и одновременно передвижение крови.
Моменты прекращения сдавливания артерии и раздражения
симпатикуса даются на сигнальной линии (Гедевани).
524 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
все стороны действия этих нервов» (55). Характерно, что Павлов
обнаружил благоприятствующее действие усиливающих нервов и на
вырезанном сердце «и при минимальном количестве крови,
находящейся в сосудах вырезанного1 сердца», как замечает автор. Даже эта
обстоятельство не помейіало ему отрицательно относиться к прямому
действию усиливающих нервов на сердечную мускулатуру.
Опыты Гедевани подтверждают этот взгляд Павлова на
значение кровеносной системы в трофике ткани. Его опыты также
доказывают сосудистую природу благоприятствующего действия
вегетативной системы на ткани, а именно сосудистую природу
симпатического действия на утомленную мускулатуру.
Действие симпатикуса на утомленную мышцу в последнее время
подробно изучалось в лаборатории А ш е р а в Швейцарии.
Первоначально сотрудники этой лаборатории высказывались, на основании
своих исследований, за прямое действие симпатикуса на скелетную
мускулатуру, а именно непосредственно на сократительный механизм
мышцы (Майбах, 34; Лабгарт, 35; Харлет, 36; Михол, 37). Но в
последнее время, в связи с возникновением учения о гормональной
природе передачи возбуждения, с одной стороны, и в связи с новыми
гистологическими исследованиями, опровергающими симпатическую'
иннервацию скелетных мышц, —¦ с другой, лаборатория А ш е р а
провела новые исследования с целью обнаружения активных веществ в-
перфузионной рингеровской жидкости утомленной мышцы во время
раздражения симпатической системы. Было открыто вещество со
свойствами адреналина, ибо эта перфузионная жидкость производила
учащение сердцебиения (Ломан, 38). Аналогичный результат был
получен у нас Синицы ныім (39). У него перфузионная жидкость из
одной утомленной конечности проходила как через сердце, так и через
другую утомляемую конечность. При раздражении симпатикуса одной;
стороны усиление сокращений происходило как на сердце, так и на
мышцах другой конечности. Определенными опытами
устанавливается, что симпатический эффект на утомленной мышце совершенно-
таков же, каким он получается от адреналина в концентрации
1 : 1 000 000. Именно, адреналин, подобно симпатину, повышает
одиночные сокращения утомленной мышцы, когда эти сокращения
вызываются раздражением двигательного нерва, и не влияет совершенно
на неутомленную мышцу; далее, подобно симіпатикусу, он перестает
действовать после отравления эрготоксином, который, как известно,
парализует двигательный эффект симпатических нервов (Коркилл и
Тигс, 47).
Такое же двигательное влияние оказывает раздражение
парасимпатической системы на мышцу языка. Известно, что после перерезки
и дегенерации подъязычного нерва раздражение язычного нерва
производит длительную контрактуру языка (Вульпиан, 51). Это явление
обусловливается возбуждением chorda tympani, проходящего через
язычный нерв к кровеносным сосудам языка. Предполагается, что*
РОЛЬ ВЕГЕТАТИВНОЙ СИСТЕМЫ В ДЕЯТЕЛЬНОСТИ МУСКУЛАТУРЫ 525
окончания этого нерва выделяют ацетилхолин, который через кровь
действует на мышечные волокна, чувствительность которых к ацетил-
хслину в сильной мере повышается пЬсле перерезки двигательного
нерва (Дэйл и Гассер, 52; Фельдберг, 53).
Аналогичные факты улучшения сокращений утомленной мышцы
-были отмечены даже под влиянием раздражения симпатических
нервов противоположной стороны (Бетьер, 43).
Итак, не может быть сомнений в том, что симпатическая система
-действует на скелетную мускулатуру также гуморальным путем:
нервные окончания симпатической системы выделяют в гладких мышцах
артерий и артериол активные вещества, которые затем
распространяются путем диффузии и кровообращения; достигнув утомленной мыш-
ttbL они действуют на нее благоприятствующим образом, подобно
адреналину (Кэннон, 44; Бремер, 45).
Однако, новейшие наблюдения Кибякова говорят и за то, что
вообще перфузионная жидкость Рингера, протекающая по сосудам
свежих, неутомленных задних конечностей, приобретает свойство
снимать утомление нервно-мышечного препарата, которое было вызвано
раздражением нерва отдельными индукционными ударами. Такая пер-
"фузионная жидкость повышает и сердечную деятельность в случае ее
ослабления. По мере утомления задних конечностей протекающая
через них жидкость Рингера теряет это свойство. Далее, оказалось, что
это вещество испаряется вместе с водой, а потому оно не идентично
с симпатином. Наконец, К и б я к о в показал, что это вещество не
специфично, ибо оно содержится и в экстрактах печени, мозга, даже
периферических нервных стволов. Отсюда следует, что активное
вещество, действующее подобно симпатину, всегда должно находиться
в свежей крови. На это указывают и наблюдения Ковалевой и
Некрасова (42), обнаруживших наличие таких активных веществ,
способных воспроизвести «симпатический эффект» и без раздражения
симпатикуса. Они приписывают этим веществам частью гематогенное
происхождение, частью кожно-железистое и мышечное, а также
считают их обязанными сосудистой системе. Следовательно,
первоначальное заключение Гедевани о роли передвижения крови в капиллярах
в эффекте Гинецинского хорошо согласуется с новейшими
исследованиями, устанавливающими гормональный характер этого
эффекта, ибо вовстанавливающее действие всех этих активных веществ
должно происходить главным образом путем передвижения крови из
артерий в капилляры и в самых капиллярах в области двигательных
пластинок. В настоящее время мы должны признать, что улучшение
сокращений утомленной мышцы под влиянием симпатической системы
сложно по своему происхождению. Именно, оно должно
обусловливаться тем, что вследствие передвижения крови в капиллярах
утомленные мышечные клетки получают из нее активные вещества вместе
с кислородом и буферными веществами, а накопившиеся в них кислые
продукты обмана частью переходят в кровь, частью нейтрализуются.
526
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Итак, как физиологические, так и гистологические исследования
последнего времени опровергают гипотезу прямой непосредственной
симпатической иннервации мышечных клеток скелетной мускулатуры,
но в то же время физиологические исследования ясно указывают на
важную роль симпатической системы в мышечной работе, с одной
стороны, путем изменения тока крови в капиллярной системе мышц,
а с другой — путем выделения окончаниями симпатических нервных
волокон в стенках кровеносной системы гормонов, активно
действующих через кровь на мышечные клетки.
Л итература
1. Bremer, Arch. mikr. Ana т., 21, 165, 1882.
2. Boecke J., Int. Mschr. Anat. Physiol., 28, 377, 1911; Verh. Anat. Ges., Miinchen,
April, 1912; Anat. Anz., 44, 1913.
3. Agdhur E., Verh. K. Akad. v. Wet. Amsterdam, sect. 2, 20, № 6, 1920.
4. De Boer, Fol. NeurobioL, 7, 358, 837, 1913.
5. Pekelharing u. Hoogenhuyze v., Z. physiol. Chem., 64, 252, 1910.
6. Гедевани Д., Физиол. журн. СССР, 19, 1042, 1935.
7а, Langelaan J. W., Brain, 45, 134, 1922.
7b. Langelaatt J. W., Verh. K. Akad. v. Wet, Amsterdam, Afdeel. Naturkunde,
XXIV, № 1, 19, 1925; Arch, neerl. Physiol., 16, 253, 1931*
8. В er it off J., Fol. NeurobioL, 8, 421, 1914.
9. Beritoff J., Pfl. Arch., 198, 590, 1923.
10. P a n e 1 1 a, Arch. Hal. Biol., 48, 430, 1907.
11. Dessy u. Grand is, Arch. Ital. Biol., 41, 225, 1904.
12. Gruber Ch., J. Pharm. Exp. Ther., 23, 235, 1924.
13. Lang ley J. N., Das autonome Nervensystem., Berlin, 1922.
14. Cannon a. Nice, Am. J. Physiol., 32, 44, 1913.
15. Wastl H., Pfl. Arch., 219, 337, 1928.
16. Dusser de Barenne, Pfl. Arch., 166, 145, 1916.
17. Dusser de Barenne u. Tervaert Cohen, Pfl. Arch., 195, [370, 1922.
18. Schmid W., Bioch. Z., 201, 126, 1928.
19. Cobb S., Physiol. Rev., 5, 518, 1925.
20. Spiegel E., Handb. norm. path. Physiol., 9, 711, 1929.
21. Forbes A., Arch. Neur. Psych., 22, 264, 1929.
22. Б e p и t о в И., Доклад на Всес. съезде физиологов в Харькове, 1930.
23. Теі 1о J. F., Traw. Lab. invest, biol., Univ. Madrid, 15, 101, 1917.
24. В i e 1 s с h о w s k у M., По его личному сообщению Коббу, см. № 19.
25. St or Ph. Jr., Mikroskop. Anat. d. vegetativen Nervensystem s, Berlin, 1928.
26. Ti egs O. W., Austr. J. Exp. Biol. Med. Sci., 9, 151, 1932.
27. H insey Jos., Proc. of the Association of Research in Nervous a. Mental Disease,
9, 153, 1930; J. Сотр. Neur. Psych., 44, 87, 1927; J. Сотр. Neur. Psych., 47,23,
1928.
28. Wilkinson H., J. Сотр. Neurol., 50, 133, 1930; J. Сотр. Neurol., 51, 129, 1930,
29. Op бе ли Л., Изв. научн. инст. им. Лесгафта, т. 6, 1923; Юбилейный сборник,
посвященный 75-летию со дня рождения акад. Павлова, стр. 386, 1925; Врач, газета у
№ 3, 192.
30. Лаврентьев Б. И., Арх. биол. наук, 33, 605, 1932. Доклад на научной
конференции сектора морфологии ВИЭМ120 февр. 1933 г.
31. Гинецинский А., Русск. физиол. журн., 6, 139, 1924.
32. Ranson S. W., Ann. int. Med., 6, 1013, 1933.
33. Гедевани Д., Русс, физиол. журн., 13, 28, 1930; там же, 16, 484, 1933.
РОЛЬ ВЕГЕТАТИВНОЙ СИСТЕМЫ В ДЕЯТЕЛЬНОСТИ МУСКУЛАТУРЫ 527
34. Maibach Ch., Z. Biol., 88, 207, 1928.
35. Labhard Er., Z. Biol., 89, 218, 1929.
36. С h a r 1 e t G., Z. Biol., 90, 299, 1930.
37. Mi chol E., Z. Biol., 90, 313, 1930.
38. L ohm an G., Z. Biol., 92, 391, 1932.
39. Синицы h H. П., Физиол. журн. СССР, 19, 1060, 1935.
43. Rynberk G. v., Arch, neerl. Physiol., 1, 727, 1917.
41. К и б я к о в А. В., Сб. докл. VI Всес. физиол. съезда, стр. 198.
42. Ковалева Г. и Некрасов П., Опыты исследования нервно-гумор. связи^
под ред. Быкова, стр. 75, 1937.
43. Baet jer А. М., Am. J. Physiol., 93, 41, 1930.
44. Cannon u. Rosenblueth, Auton. Neuro-Effect., 1937.
45. Bremer F., Egr. Physiol., 34, 678, 1932.
46. Rosenb lueth A. a. Luco J. V., Am. J. Physiol., 120, 781, 1937.
47. С о r k i 11 A. B. a. T i e g s O. W., J. Physiol., 78, 761, 1933.
48. T у ш n о a a B. M., Бюлл. эксп. Оиол. мед., 5, вып. 1, стр. 29 и 31, 1938.
49. Е с с 1 е s J. С, Egr. Physiol., 38, 339, 1937.
50. Некрасов П. Н. и Некрасова Н. В., Физиол. журн. СССР, 21, 519г.
1936; 25, 5, 1938.
51. Vulpian А., С. г. Acad. Sci., 76, 146, 1873.
52. D а 1 е u. G a s s е г, J. Pharmacol., 29, 53, 1926.
53. Fe Id berg W., Pfl. Arch., 232, 88, 1933.
54. К и p 3 о h M. В., Тр. физиол. научно-иссл. инст. Ленингр. гос. унив., 14, 31, 1934.
55. Павлов И. П., Центробежные нервы сердца, дисс. 1883; Arch. f. Anat. Physiol.,
Physiol. Abt., 1887, S. 498; Еженед. клинич. газ., 1888. Приведенные в тексте
цитаты взяты из отд. оттиска последней статьи, стр. 27—29.
56. Черниговский В. Н., Физиол. журн. СССР, 29, стр. 1, 15 и 526, 1940.
Г л а в а VI
ТРОФИЧЕСКОЕ ВЛИЯНИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Понятие о трофике. Выше было указано, что основной
биологический процесс, обусловливающий обмен веществ живой системы, лежит
в основе питания клетки, а значит ее роста и размножения. В
условиях, благоприятных для обмена веществ, происходит усиленное
питание, в силу чего клетки быстро растут и размножаются. При
неблагоприятных условиях для обмена веществ питание ухудшается. От
этого происходит остановка роста и размножения клеток. Это свойство
живых клеток питаться путем обмена веществ иначе называется
трофическим свойством.
От клеточного обмена веществ зависят нормальный рост и
развитие тканей и органов. 'Как известно, функциональная деятельность
-сильно благоприятствует росту и развитию их, а она зависит от
внешних раздражений. Однако, установлено, что как эмбрионально, так и
постэмбрионально рост и развитие тканей не всецело определяется
¦функциональной деятельностью, вызванной внешними раздражениями.
Если на растущем организме изолировать заднюю часть спинного
мозга — крестцово-поясничную область — от всяких внешних
влияний, путем перерезки всех задних корешков, а также от
межцентральных влияний, путем перерезки спинного мозга на границе поясничного,
то рост изолированного мозга, а также связанной с ним мускулатуры,
будет итти таким же темпом, как в нормальных условиях (Тоуэр, 12).
•Отсюда следует, что все условия для нормального обмена веществ,
нормального питания тканей даны в самой ткани, с одной стороны,
в виде внутриклеточной деятельности, а с другой — в виде межуточной
жидкости, которая омывает тканевые клетки.
Понятие о внутренней среде. Межуточная жидкость является той
¦средой, в которой живет клетка, а потому иначе она называется
внутренней средой. На эту среду, на ее химический состав,
прежде всего, влияет кровь, затем самые клетки данной ткани
продуктами своего метаболизма и, наконец, нервная система, которая
через свои окончания выделяет продукты своей активной
деятельности. Кровь, в свою очередь, содержит в себе как вообще продукты
ТРОФИЧЕСКОЕ ВЛИЯНИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ 529
клеточного метаболизма всего организма, так и, в частности, разного
рода гормоны желез внутренней секреции и метаболиты нервного
происхождения. Следовательно, на питание клеток или тканей могут
оказывать так называемое трофическое влияние метаболиты и гормоны
всего организма (Штерн, 8; Разенков, 7).
Межклеточная жидкость каждой ткани по своему химическому
составу отличается присущими ей особенностями, которые
обусловливаются своеобразием гисто-гематического барьера, существующего на
грани между кровью и непосредственно питательной средой
изучаемого органа. Этим барьером является эндотелий капилляров. В силу
его структурных и функциональных особенностей из крови в
непосредственную питательную среду данного органа переходят только
определенные вещества, притом лишь в определенных концентрациях и
с определенной скоростью. Этим путем поддерживается определенный
состав межтканевой жидкости, определенный уровень питания клеток
(Штерн, 8).
Всякое изменение нормального состава внутренней среды, конечно,
ведет к изменению уровня обмена веществ в клетках, к повышению
или понижению уровня питания тканей, изменению трофики.
Нарушение это может быть вызвано как разного рода физическими и
химическими агентами, так и патологическими влияниями. Процесс нарушения
внутренней среды и его физиологическое значение для тканей
особенно подробно изучены на гемато-энцефалическом барьере, т. е. на
барьере между центральной нервной системой и кровью. Этот барьер,
по данным Штерн и ее школы, регулирует нормальный состав
спинномозговой жидкости. Спинномозговая жидкость представляет собой
единую межтканевую жидкость центральной нервной системы, не
только проникающую во все отделы этой системы, но и
сообщающуюся с подоболочечными пространствами и заходящую во все
периферические нервные проводники (Сперанский, 10; Закарая, 9).
Всякое вещество, попадая через барьер в эту жидкость, имеет тенденцию
распространиться по всей нервной системе. Направление, в котором
распространяется вещество в этой среде, видимо, определяется прежде
всего разностью давления в отдельных участках внутри самой
жидкости. Когда, например, химический или бактериальный агент попадает
в периферический нервный ствол, он распространяется по нервному
стволу в большинстве случаев в центральном направлении, т. е. к
мозгу, но в редких случаях он распространяется и в периферическом
направлении (Закарая, 9). Аналогичное движение веществ происходит
в пределах самой центральной нервной системы. При нормальных
условиях циркуляции ток цереброспинальной жидкости в подоболо-
чечных пространствах спинного и головного мозга происходит главным
©бразом в головном направлении. Поэтому при введении какого-либо
красящего вещества, например туши, в хвостовый отдел мозга оно
обнаруживается в цистернах основания мозга. Внутри же центрального
канала, наоборот, жидкость движется в каудальном направлении. Так,
34 и. С. Беритов.
530 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
например, тушь, введенная в полость 4-го желудочка, скопляется
через некоторое время в хвостовой части спинного" мозга
(Сперанский, 10),
Штерн со своими сотрудниками подробно изучила свойство гема-
то-энцефалического барьера, его роль в проникновении в спинномозговую
жидкость разного рода веществ, введенных в кровь, и вообще в
регулировании нормального состава этой жидкости (8).
Влияние удаления симпатической системы у взрослых животных.
С давних пор вегетативной системе приписывается трофическая роль
з отношении всех других тканей. И. П. Павлов первый высказался
в этом смысле в отношении вегетативной системы сердца (14). При
этом предполагается, что вегетативная система, в данном случае
п. yago-syrapathicus, содержит в себе такие волокна, которые
усиливают или ослабляют сердечные сокращения. Но, как уже указывалось,
выше, по первоначальному учению Павлова, это. трофическое
влияние сердечных нервов осуществляется не непосредственно путем иннерт
вацни тканевых клеток, а через сосудистую систему. Именно, усиление-
сердечной работы происходит путем расширения венечных сосудов-
сердца, а ослабление — путем сужения их. Но в последнее время Пав*
лов и некоторые другие ученые предположили прямое и
непосредственное трофическое влияние симпатической системы -на все ткани: железы,,
гладкие и поперечнополосатые мышцы, рецепторы и нервные центры.
В наиболее последовательном виде эта гипотеза разделяется акад. О р-
б е л и и его сотрудниками, как об этом уже говорилось выше.
Прямое трофическое влияние симпатической системы в последнее
время подвергалось основательному изучению во многих
лабораториях. В основном этого рода исследование заключалось в следующем:
тот или другой орган или ткань, кожа и мышцы одной конечности
лишались симпатической иннервации путем перерезки всех
соответствующих нервов. Каждый раз такое исследование кончалось
утверждением, что симпатическая система абсолютно не влияет на питание
тканей, на их обмен веществ. У некоторых млекопитающих (морские
свинки, кошки, собаки, козы) удаляли симпатический пограничный
столб на одной стороне и затем в течение многих месяцев изучали
гистологическое строение, вес и рост тканей, а именно мышц, костей
и кожи. И, несмотря на многократные тщательные физиологические
исследования, не было замечено на оперированной стороне никаких
отклонений от нормы. Животные жили долго, сохраняя и развивая
все двигательные функции на оперированной стороне, как на
нормальной (Такахаши, 1; Коатс и Тигс, 3; Кэинон, 4).
Влияние удаления симпатической системы растущих организмов.
Опыты с удалением симпатической системы были произведены также
на растущих организмах (котята, щенки). Так, у щенков удалялась
симпатическая система одной задней ноги — вся поясничная и
крестцовая цепочка симпатических узлов. Животные жили в лаборатории
годами. Задняя нога, лишенная симпатической иннервации, росла со-
ТРОФИЧЕСКОЕ ВЛИЯНИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ 531
вершенио так же, как и другая задняя нога. Никаких расстройств в
обмене веществ не было замечено: мышцы, кости, кожа с волосами
росли на обеих ногах одинаково. Только в течение первых месяцев
был замечен более быстрый рост десимпатизированной ноги, что,
видимо, стояло в связи с более усиленным током крови в сосудах,
лишенных симпатического тонического сужения (Закарая, 5).
Аналогичные опыты были произведены на котятах в Америке в
лаборатории очень крупного специалиста по симпатической .
иннервации — Кэннона (4). Здесь в некоторых опытах удалялась вся
симпатическая система с обеих сторон, начиная от шейных узлов до
крестцового. Здесь также пришли к полному убеждению, что
симпатическая система не несет трофических функций в отношении
мускулатуры или кожи,
Трофическая роль симпатической системы легче всего могла быть
обнаружена на. двигательных органах. Опять-таки оказалось, .что де-
симпатизированная задняя нога в своей работоспособности не уступает
другой, нормальной ноге. Даже, наоборот, обычно она развивает
больдне напряжения и утомляется позднее нормальной (Закарая, 5).
Итак, по новейшим исследованиям, симпатическая система не несет
никакой непосредственной тррфической функции по отношению к
тканям, как во взрослых, так и & растущих организмах.
Влияние симпатической системы на регенерацию нервных
элементов. Как известно, нервная клетка и центральная нервная система
действуют на рост аксонов, выходящих из мозга, .и на их окончания
трофически. Если перерезать аксон, отрезанный от нейрона, участок
аксона и его окончание дегенерируют. Но центральный отрезок будет
расти, и, когда дойдет до иннервируемой ткани, произойдет
регенерация окончания. При этом регенерируется точно такое окончание, какое
свойственно данной ткани. Это бывает как с цереброспинальной
двигательной и чувствительной, так. и с симпатической нервной системой.
Доказано специальными опытами, что эта регенерация зависит от
импульсов, исходящих от мозга по регенерируемому волокну, что в
регенерации данного волокна и нервного окончания и вообще в
пределах данной системы не играют никакой роли другие волокна, другая
система. В частности, симпатическая система не играет никакой роли
ни в дегенерации, ни в регенерации двигательных окончаний в
скелетной мышце или окончаний чувствительного нерва в рецепторах
(Лаврентьев, 13).
Влияние вегетативной системы на химические процессы в тканях.
Несмотря на отсутствие прямого влияния на физиологические
особенности тканей, раздражение или удаление симпатической системы,
безусловно, оказывает влияние на химизм тканей. Мы ниже увидим, в чем
заключается назначение вызываемых при .этом изменений. Сейчас мы
коснемся только одного вопроса: в чем именно заключаются
означенные изменения в химизме тканей? Прежде всего известно, что как при
действии вегетативных ядов, так и при раздражении вегетативных
34*
532 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
нервов меняется тканевое дыхание. Кроме того, было обнаружено, что
тканевое дыхание в мышцах конечности меняется спонтанно, в сроки
не более 2—3 часов, и это явление исчезает после полной десимпати-
зации конечности (Шатенштейн и сотр., 11). Затем было установлено
изменение в содержании электролитов, азотистых, липоидных и
углеводных соединений под влиянием симпатической системы. Так,
например, известно, что перерезка одного или обоих стволов симпатической
системы в брюшной полости меняет на определенное время
проницаемость так называемого гемато-энцефалического барьера в отношении
сахара, молочной кислоты и фосфора. Далее, известно, что в
оттекающей крови конечности при повышенной симпатической иннервации
наблюдается более или менее выраженное уменьшение ионного
коэффициента, уменьшение сахара и жира. Наоборот, после десимпатиза-
ции получается на протяжении известного времени в оттекающей
крови конечности (ообаки) повышение сахара. Если к этой операции
присоединить перерезку задних корешков, количество сахара
становится нормальным. Таким образом, было установлено, что в
определенных условиях и задние корешки влияют на химизм тканей на
периферии. Эти факты были установлены Альперном и его
сотрудниками (6) наравне с другими фактами, и на основании их было
сделано заключение о гуморальном действии симпатической
системы, регулирующем химические процессы в тканях на периферии
и в центрах.
Эти же авторы нашли, что парасимпатическая система лишена
означенного действия, она рассматривается исключительно как
моторная система. Однако в отношении некоторых желез пищеварительного
канала доказано трофическое влияние парасимпатической системы,
а именно блуждающего нерва на поджелудочную железу и частично
также на желудочные железы (Разенков, 7).
Далее, симпатическая система осуществляет свою химическую
функцию еще тем, что она иннервирует всю кровеносную систему,
регулируя кровоснабжение всего организма в целом и каждого
органа, каждой ткани в частности.
Наконец, симпатическая система иннервирует важнейшие
внутренние органы, имеющие отношение к составу и движению крови, как
надпочечник, селезенку, печень. Поэтому симпатическая система путем
иннервации кровеносной системы и других органов может производить
целый ряд изменений во внутренней среде — в крови и лимфе — в
отношении химического состава и физических особенностей. И все
химические процессы, которые определяются этой деятельностью
симпатической системы, имеют существенное значение для трофики
тканей. Отсюда следует, что мы не можем говорить о
непосредственной трофической функции симпатической системы, т. е. о
непосредственных трофических импульсах, как бы посылаемых через
симпатическую систему к тем или другим тканевым элементам. Симпатическая
система осуществляет свою трофическую функцию путем тех физика-
ТРОФИЧЕСКОЕ ВЛИЯНИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ 533
химических изменений в составе крови и лимфы, которые
обусловливаются ее гуморальным действием, а также деятельностью иннерви-
руемых симпатической системой органов кровеносной системы,
надпочечника, селезенки и печени.
Литература
1. Takahaschi V. N., Pfi. Arch., 193, 322, 1922.
2. Lang ley J. N.. J. Physiol., 31, 244, 1904.
3. С о a t e s A. E. a. T i e g s С W., J. Exp. Biol. Med. Sci., 7, 37, 1930.
4. С a n n о n W. В., Newton H.( Bright, Men kin V. a. Moore R.,
Am. J. Physiol., 89, 84, 1929.
5. 3 а к a p а я E., Вестник хирургии и псгран. сблгстей, № €8—С9, 1931; Русск.
фнонол. журіь, 16, 675, ?1033.
6. Аль пер н Д. Е., Физиол. журн. СССР, 19, И, 1935.
7. Разенков И. П., Сб. докл. VI Всес. физиол. съезда, стр. 15, 1037.
8. Штерн Л. С, Сб. докл. VI Всес. физиол. съезда, стр. 15, 1937.
9. 3 а К а р а я Е., Тр. Физиол. инст. Тбил. унив., 3, 309, 1937.
10. Сперанский А. Д., Элементы построения теории медицины, 1935.
11. Ш ат е н ш т е й н Д. И., Сб. докл. VI Всес. физиол. съезда, стр. 346, 1937.
12. Tower S. S., J. Сотр. Neurol., 67, 109, 1937.
13. Лаврентьев Б. И., Арх. биол. наук, 48, вып. 1—2, 1941.
14 Павлов И. П., Еженед. клинич. газета, 1888; Arch. Anat, Physiol, Physio!.
Abt., S. 498, 1887.
Глава Til
РОЛЬ СИМПАТИЧЕСКИХ ГАНГЛИЕВ В КООРДИНАЦИИ
ОТДЕЛЬНЫХ ОРГАНОВ
Общее понятие. Из симпатических ганглиев выходят как
возбуждающие, так и тормозящие нервы. Кроме того, преганглионарные волокна
оканчиваются своими коллатералями во многих симпатических ганглиях.
Да и в одном ганглии данное преганглионарное волокно оканчивается на
ряде постганглионарных клеток. Так, в верхнем шейном симпатическом
ганглии одно преганглионарное волокно приходится на 32
постганглионарных волокна (Рэнсон и Биллингсли, 6). На этом основании
можно было признать, что ганглии симпатической системы играют
определенную роль в координации движений и секреции
обслуживаемых органов. Первое физиологическое доказательство координирующей
деятельности симпатической системы было дано Гофманом
(1). Он нашел, что при раздражении первого или второго
грудного нерва, содержащего преганглионарные волокна, всегда получается
общее расширение зрачка, а при раздражении отдельных ветвей
п. ciliaris, т. е. постганглионарных волокон, вызывается только местное
расширение одной части зрачка. Отсюда он заключил, что ганглиозные
клетки, откуда происходят постганглионарные волокна, соединены
между собой комиссуральными волокнами для образования
координирующего центра. Но Лэнгли (2) на основании собственных
исследований опроверг это заключение. Именно, он наблюдал, что при
раздражении одной маленькой Еетви одного из трех грудных нервов
сокращение наступает не во всем расширителе зрачка, а только
в одной части его. Далее, Лэнгли заметил, что раздражение пре-
ганглионарных и постганглионарных волокон одного узла дает
совершенно одинаковые эффекты. Было замечено, что необходимо
раздражать некоторое число преганглионарных или постганглионарных
волокон, чтобы получить максимальное и полное расширение зрачка.
Если же число раздражаемых волокон очень невелико, то в обоих
случаях раздражения эффект является частичным. Отсюда Лэнгли
заключил, что в узлах вегетативной системы клетки не соединены
между собой комиссуральными волокнами и не образуют
координирующих центров.
РОЛЬ СИМПАТИЧЕСКИХ ГАНГЛИЕВ В КООРДИНАЦИИ ОРГАНОВ 535
По гистологическим исследованиям, соседние ганглиозные клетки
узлов соединены между собой путем нейрофибриллярных клубков или,
как это уже указывалось выше, путем нейропиля (рис. 211, 212).
Если же, несмотря на это, при раздражении небольшого числа пре-
ганглионарных волокон зрачок не давал в опыте Лэнгли
максимального расширения, то это должно было быть обусловлено тем, что
ганглии не работают как единое целое. Это, очевидно, зависит от
того, что все связи ганглионарных клеток между собой
осуществляются исключительно дендритами. А тонкие и голые разветвления
последних должны проводить возбуждение с большим декрементом,
вследствие чего возбуждение одной клетки не может переходить на
другую, а также непосредственное возбуждение какого-либо дендрита
не будет передаваться по дендритам же на клетки. Отсюда понятно,
что чем больше будет число возбужденных преганглионарных волокон,
тем больше элементов ганглия должны приходить в деятельное
состояние. Узловые нервные клетки вегетативной системы могут быть
уподоблены в функциональном отношении двигательным нейронам
скелетной мускулатуры, но с той разницей, что по одним из них
центральное возбуждение проводится к периферическим органам без
изменения, а по другим оно трансформируется в торможение. Преган-
глионарные нейроны должны быть уподоблены интраспинальным
координирующим нейронам обычной рефлекторной дуги. Они проводят
возбуждение, исходящее из координирующих центров мозга к
определенным узлам вегетативной системы для вызова возбуждений и
торможений в отношении определенных периферических органов.
Основная функция симпатических ганглиев. Согласно Лэнгли,
симпатические ганглии можно рассматривать прежде івсего как усилители
импульсов, исходящих из центральной нервной системы. Так как каждое
преганглионарное волокно отдает коллатерали во многие ганглии, то
функция ганглиев должна состоять в увеличении области
распространения возбуждений, приходящих через преганглионарные волокна.
Отсюда следует, что симпатическая система имеет некоторое сходство с
центральной нервной системой, которая производит широкое
распространение возбуждения. Но в вегетативной нервной системе границы
распространения возбуждения очень ограничены, так как эти границы
в значительной мере зависят от того, какие преганглионарные волокна
возбуждены.
В симпатических ганглиях существуют не только проводники
импульсов возбуждения в виде клеток и их аксонов, но и нейропиль
в виде густого сплетения разветвлений дендритов с синаптическими
окончаниями на них разветвлений преганглионарных волокон. Этот
нейропиль, как указывалось выше, выполняет функции угнетения
проводящей системы в случае более или менее сильного воздействия на
него, т. е. на нейропиль, преганглионарной системы. Нейропиль
спинного мозга также обладает функцией угнетения или торможения,
й через зту функцию он играет существенную роль в координации
535
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
спинномозговой деятельности. На этом основании мы можем
утверждать, что и симпатический ганглий должен играть определенную роль
в координации внутренних органов. За координирующую роль
симпатических ганглиев говорят и другие физиологические особенности этих
ганглиев, о которых уже говорилось выше, как временная и
пространственная суммация, большой скрытый период, длительное
отрицательное и положительное колебания потенциала после каждого
импульса возбуждения с повышением возбудимости во время
отрицательного колебания и с понижением ее во время положительного,
возможность ряда ритмических импульсов после тетанического
раздражения.
Роль центральной нервной системы. По всем тем физиологическим
и гистологическим фактам, какие нам известны сейчас, симпатическому
ганглию нельзя приписать решающей роли в координированной
деятельности вегетативных органов (Уиттеридж, 5). Центральная нервная
система определяет посредством преганглионарных нейронов характер
участия каждого ганглия в координации этих органов. Симпатические
ганглии поэтому должны играть только подчиненную роль: они должны
принимать определенное участие в передаче сложных
координированных иннервации из центральной нервной системы рабочим органам.
Они, с одной стороны, усиливают или ослабляют притекающие
центральные импульсы, а с другой — расширяют или суживают поле их
действия. Наилучшим доказательством этого положения можно считать
то явление, что симпатический ганглий не обладает спонтанной
деятельностью, в то время как центральные органы владеют этим
свойством. Характерно, что при нормальных связях с центральной нервной
системой ganglion stellatura кошки дает все время быстро протекающие
биоэлектрические токи возбуждения. Но оказалось, что это
возбуждение зависит от центральных влияний, ибо после отделения
пограничного симпатического столба спонтанные токи возбуждения исчезают
(Фишер и Левенбах, 4).
Итак, координированная деятельность внутренних органов по
существу обусловливается центральной нервной системой. Симпатические
ганглии же играют лишь подчиненную роль в этой координации,
участвуя в ней, с одной стороны, для усиления или ослабления притекающих
из центральной нервной системы импульсов, а с другой — для
расширения или суживания области их распространения.
Литератур а
1. Hofmann., Schmid. Jahrb. ges. Med., 281, 113, 1904.
2. L a n g 1 e у J. N., J. Physiol., 31, 244, 1904.
3. Eccles J. C, J. Physiol., 85, 179, 1935; 85, 464, 1935.
4. F i s с h e г M. N. u. Lowenbach H., Pfl. Arch., 233, 722, 1934.
5. Witteridge D., J. Physiol., 89, 99, 1937.
6. R a n s о n S. W. а. В i 11 i n g s 1 e у P. R., J. Сотр. Neurol., 29, 316, 1918.
Глава VIII
ГОМЕОСТАТИЧЕСКОЕ ЗНАЧЕНИЕ СИМПАТИЧЕСКОЙ СИСТЕМЫ*
Основная функция симпатического отдела вегетативной системы.
Мы выше рассмотрели физиологические особенности отдельных
составных частей вегетативной системы. Теперь мы коснемся роли
вегетативной системы в целом в жизнедеятельности всего организма.
Основная функция симпатического отдела вегетативной системы
заключается в удержании постоянства внутренней среды, в которой
живут живые части тела: например, в удержании постоянной
температуры тела, постоянного водного баланса тела, постоянного кровяного
давления и т. п. Эта основная функция была названа Кэнноном (1)
гомеостазисом. Цереброспинальная система, реагируя на
внешний мир, потребляет кислород и разного рода материалы (углеводы,,
жиры, белки), дающие энергию. Вследствие этого происходят
изменения в жидкой среде тела. Симпатическая же часть вегетативной
нервной системы обладает функцией регулировать внутренние
процессы, мобилизовать определенные силы для устранения наступающих
изменений. Кэннон подробно выясняет гомеостатическую роль
симпатической системы. Он, например, анализирует все нарушения во
внутренней среде при физической работе и указывает те пути, какими
симпатическая система осуществляет свою гомеостатическую функцию
в отношении каждого нарушения. Как известно, мышечная работа
сопряжена с потреблением углеводов, в частности сахара. При
двухчасовом беге собаки декстроза крови падает от 90 до 66 мг на 100 см3
крови. Это происходит несмотря на постоянный приток декстрозы из
печени. И вот, когда содержание декстрозы понижается до 70—80 мг
па 100 см3 крови, приходит в действие симпатико-надпочечная система.
Чем ниже падает сахарный уровень в крови, тем больше активность
этой системы. Именно усиленно выделяемый адреналин освобождает
сахар из запасов печени в большем количестве, чем обычно, и этим
самым устраняет быстрое падение сахара в крови. Кэннон показал,
что не только тяжелая физическая работа, но и хождение на коротком
расстоянии сопровождается повышением деятельности симпатико-над-
почечной системы.
538 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
D связи с мышечной деятельностью стоит теплообразование. В
покое человек весом 68 кг потребляет 250 см3 кислорода в минуту,
производя при этом 1.25 калорий тепла. Если бы при этом тепло не
уменьшилось, кроме излучения еще каким-то иным путем, то в
каждые 50 мин. температура тела увеличивалась бы на 1°. При тяжелой
работе потребляется в 18 раз больше кислорода; в этом случае
температура тела повышалась бы много быстрее: на 6° в каждые 16 мин.
Но высокая температура — серьезная опасность, особенно для нервных
клеток мозга. В устранении этой опасности существенную роль играет
симпатическая система. Путем торможения центрального
симпатического тонуса сосудистой системы, производящего постоянное сужение
периферических сосудов, происходит расширение последних. Это ведет
к усиленному кровообращению и через него к усиленному охлаждению
крови путем излучения и контакта кожи с более холодной средой.
Кроме того, усиливается деятельность симпатического центра
потоотделения; происходящее при этом усиленное выделение пота также
производит охлаждение тела.
Далее, в связи с мышечной работой происходит увеличение
кислотности вследствие образования большого количества угольной и
молочной кислот. Это изменение также не безопасно. Под влиянием
симпатической системы приходят в деятельное состояние бронхиальные
мышцы как через нерв, так и путем воздействия адреналина. Бронхи
расширяются, и через это облегчается вхождение воздуха в легкие
и выхождение его из легких. Усиленная вентиляция легких приводит
к уменьшению кислотности.
Наконец, при мышечной работе увеличивается потребность в
кислороде. Сосуды работающих мышц расширяются в 9 раз, а потребление
кислорода увеличивается в 18 раз. Очевидно, возникает «кислородная
недостаточность». Как же устраняется эта недостаточность? Под
влиянием симпатической системы увеличивается сердечный ритм; затем
значительное количество крови переносится из внутренних органов
в "рабочие органы в силу симпатического сужения сосудов внутренних
органов. Эти изменения увеличивают давление крови на 50 мм и даже
более, а вследствие этого повышается скорость передвижения крови
в работающих мышцах. К этому нужно добавить еще одно
приспособление, найденное Баркрофтом (2): в покое кровь селезенки
содержит красные кровяные шарики на 50—100% больше, чем
циркулирующая кровь. При многих обстоятельствах, в частности при тяжелой
физической работе, селезенка сокращается под влиянием симпатической
системы и выбрасывает свое содержимое в общее кровяное русло.
Вследствие этого число красных шариков в крови увеличивается на
25%. В результате, конечно, не только устраняется кислородный
недостаток в тканях, но усиленным темпом выносится главный продукт
сгорания— угольная кислота — из тканей в легкие.
Все изменения, наблюдаемые при мышечной работе, имеют место
?и при эмоциональных возбуждениях: страхе, ярости, гневе и вообще
ГОМЕОСТАТИЧЕСКОЗ ЗНАЧЕНИЕ СИМПАТИЧЕСКОЙ СИСТЕМЫ 539
при более или менее значительных двигательных реакциях. Во всех
этих случаях происходят более или менее значительные изменения во
внутренней среде тела, и во всех этих случаях симпатическая система
выступает в защиту нормального уровня этой среды своим гомеоста-
тическим действием.
Основная функция парасимпатического отдела вегетативной системы.
Парасимпатическая система не владеет гомеостатической функцией.
Функции ее скорее защитного или охранительного характера. Так,
третий головной нерв, так называемый глазодвигательный, защищает
сетчатку от света путем сужения зрачка; блуждающий нерв тормозит
деятельность сердца, когда нет надобности в частом ритме; он же
вызывает выделение желудочного, а также панкреатического' Сока
для переваривания пищи; 7-й и 9-й головные нервы вызывают
секрецию слюны также для переваривания пищи. Все.эти. рефлексы
краниальной вегетативной системы существенно отличаются от рефлексов
симпатической. И, что характерно, обе системы действуют
антагонистически: действие парасимпатической системы прекращается, как
только начинается действие симпатической системы.
Итак, симпатическая система играет чрезвычайно важную роль
в организме, удерживая сложный состав внутренней среды на одном
определенном уровне. Она возвращает ее к норме после всякого рода
нарушений, производимых воздействием внешней среды и работой
самих органов.
Эта концепция Кэннона о роли симпатической нервной системы
в жизнедеятельности организма точно определяет ту особо важную
функцию, какую несет симпатическая система в отличие от
цереброспинальной, а также и парасимпатической систем.
Новейшие исследования на десимпатизированных животных,,
уточнили гомеостатическое действие симпатической системы. Так, например,
в холодной среде у собак, совершенно лишенных симпатической
системы путем удаления симпатической цепочки с обеих сторон,
температура тела падает намного больше, а вызываемое охлаждением
дрожание тела начинается значительно раньше и протекает более
ускоренным темпом, чем у нормальных. Но при высокой внешней
температуре у оперированных животных наблюдается такое же повышение
температуры тела и одышка, как у нормальных.
Более значительная разница наблюдается в отношении
чувствительности к инъекции инсулина. Содержание сахара в крови падает
более низко после инъекции инсулина у оперированных собак, чем
у нормальных. Заметная разница наблюдается в отношении
сопротивляемости к низкому давлению кислорода: она понижена у
оперированных собак ('Мак Донаф, 4).
При напряженной мышечной работе, например при беге, утомление
наступает раньше, чем у нормальных животных. Но способность
к регулированию содержания сахара в крови как во время работы, так
и после нее у десимпатизированных животных точно такова, как и у
540
ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
нормальных. Также повышение содержания лактата в крови во время
работы одинаково у тех и у других животных (Кэннон и сотр., 5).
Значит, у животных, лишенных симпатической системы, имеется
добавочный механизм для устранения тех нарушений, которые возникают
во внутренней среде в связи с работой.
Практикум
1. Наблюдение тормозящего действия блуждающего нерва на сердце лягушки.
Находят блуждающий нерв на одной или на обеих сторонах и освобождают от
окружающих тканей. На один из них накладывают электроды. Биение сердца
записывается на кимографе. Расположение блуждающего нерва дано на рис. 232..
а) Раздражают нерв сравнительно слабыми индукционными ударами (15—20 см
расст. кат.) 20—30 раз в секунду. Биение сердца замедляется и ослабевает, но
может случиться ускорение н усиление сердцебиения под влиянием ускоряющих
шміпатическіих волокон при их более білашпрмятном для раздражения раевдложешиг
в нервном стволе.
б) Усиливают раздражающий ток, сближая катушки на 2—8 см. Сердцебиение
останавливается. После раздражения остановка продолжается еще некоторое время.
Если раздражение продолжается сравнительно долго, то пульсация сердца
начинается после остановки еще во время раздражения. После остановки нередко
следует более частая и более сильная пульсация сердца. Это — влияние симпатических
волокон.
2. Наблюдение действия никотина на тормозящие сердечные ганглии. Капнуть
0.1—0.5% раствором никотина на сердце. Деятельность сердца не меняется. Н?
теперь раздражение блуждающего нерва не остановит сердца вследствие паралича
ганглнозных клеток сердца, около которых оканчиваются преганглионарные волокна
блуждающего нерва. Наоборот, оно вызовет ускорение пульсации вследствие
одновременного раздражения постганглионарных ускоряющих волокон симпатической
системы. Если раствор слабее, то ів результате раздражающего действия никотина
на ганглиозные клетки сердцебиение замедлится, даже может остановиться на
некоторое время. В таких случаях раздражение блуждающего нерва произведет
остановку сердца при более слабых силах, чем до отравления. Сильные раздражения
производят остановку сердца, а потом учащение.
3. Наблюдение сужения кровеносных сосудов при раздражении симпатического
нерва. На спинномозговом препарате лягушки освобождается симпатическая цепочка
на уровне седалищного сплетения от прилегающих тканей. Центральный конец выше
6-го симпатического ганглия перерезан. Электроды взять трехполюсные или с
кольцом Геринга (рис. 14). Они подводятся под симпатический нерв на уровне 7—6 сим-
патических узлов. Конец нерва поднят на воздух во избежание ветвления тока.
Наблюдается под микроскопом кровообращение в плавательной перепонке.
Определяют пороги раздражения симпатического нерва. При пороговых раздражениях
кровообращение сначала замедляется, через 15—20 сек., а затем, через 30 сек. и более,
происходит полная остановка. При сильных раздражениях замедление начинается
раньше и полная остановка происходит раньше. Если симпатический нерв хорошо
освобожден от прилегающих кровеносных сосудов и соединительнотканных
элементов, порог раздражения довольно низкий; при обычном индуктории в 5000 оборотов
во вторичной цепи заметный эффект замедления получится уже при 40 см расст.
инд. кат., а полная остановка — при 30—35 см. В таких случаях более или менее
сильное раздражение, 15—20 см, убивает нерв, а потому нельзя применять его.
Сильные раздражения должны применяться только в том случае, если симпатический
нерв не освобожден от сосудистых и других пленок.
Если хорошо присматриваться к просвету артериол до и во время раздражения
симпатикуса, можно ясно заметить сужение просвета в связи с раздражением.
ГОМЕОСТАТИЧЕСКОЕ ЗНАЧЕНИЕ СИМПАТИЧЕСКОЙ СИСТЕМЫ
541
4. Действие ацетилхолина и адреналина на сердце. Ритмическая деятельность
обнаженного сердца спинномозговой лягушки записывается на кимографе. 1. Капают
на сердце раствором ацетилхолина 1 : 1 000 000,—происходит замедление сердечного
ритма. Если же каонуть на него более сильным раствором, как 1 : 100 000 и
1:10 000, сердце остановится на некоторое время. 2. Капают раствором адреналина
1 : 10 000, — происходит усиление и учащение сердечной деятельности. 3.
Прикладывают локально фильтровальную бумажку в 2—3 мм2, пропитанную ацетилхолином
Рис. 232. Положение нервов в грудной области"1 лягушки.
7 — языко-глоточный нерв; 2 — блуждающий; 3 — подъязычный; 4 — левое
предсердие; 5 — желудочек; б — левое легкое; 7— гортанный нерв; 8 —
луковица аорты; 9 — правое предсердие; 10—правое легкое (по Морату и ДоЙонуа).
е растворе 1 : 100 000—1:1000 000, на верхушку или середину желудочка
обескровленного сердца лягушки, — происходит усиление систолических сокращений
желудочка или же не оказывается никакого влияния. Если приложить туда же
ацетилхолин в более сильной концентрации, то это вызовет кратковременную контрактуру
с кратковременным ослаблением систолических сокращений. Если приложить локально
ацетилхолин в тех же концентрациях к синусам или к предсердиям, сейчас же
наступает более или менее длительная остановка сердца.
Результаты ясно демонстрируют, что ацетилхолин, выделяемый в результате
раздражения блуждающего нерва, должен действовать тормозящим образом на
проводящую систему сердечной мышцы, а не на двигательную.
5. Влияние раздражения симпатического нерва на кривую утомления
поперечнополосатых мышц. На одной стороне спинномозгового препарата лягушки на
<5едренную артерию накладывается зажимчик, или артерия так высоко
приподнимается стеклянной палочкой, что в конечности останавливается кровообращение.
Икроножная мышца соединена с кимографом. Седалищное сплетение перерезано повыше
присоединения гг. communicantes. Симпатический н?рв выпрепарирован, как в
вышеприведенном случае. Седалищный нерів раздражается отдельными индукционными
ударами через 1—2 секунды. В момент сильного ослабления одиночных сокращений
производится тетаническое раздражение симпатического нерва в течение 20—30 сек.
Под влиянием последнего раздражения одиночные сокращения сначала усиливаются
(скрытый период 5—10 сек.), а затем вновь начинают падать. В следующем опыте
вместо (раздражения снимают зажимчик с артерии на 10—20 сек. Спустя 3—5 сек.
одиночные сокращения начинают нарастать, как в предыдущем случае; достигнув
некоторой большой высоты, они начинают ослабевать. Можно повторить тот и
Другой опыт несколько раз.
542 ОБЩАЯ ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
6. Опыт Лёви с гуморальной передачей торможения по усовершенствованной-'
методике Бака. В сердце лягушки (Dj через аорту вставлена двойная канюля,
одна трубочка которой соединена с резервуаром раствора Рингера, а другая
с другим сердцем (R). При систолическом сокращении сердца D, а также под
давлением, создаваемым резервуаром, жидкость будет переходить из сердца D
в сердце R. Систолические сокращения обоих сердец должны записываться на
одном кимографе. Сердце D должно быть изолировано вместе с блуждающим нер-
Рис. 233. Опыт с гуморальной передачей торможения
Верхняя кривая — от сердца R, а нижняя — от сердца D. При
раздражении блуждающего нерва (сигнал 5) сначала
останавливается сердце -LD, а затем сердце R. Время —5 сек.
вом. Если этот нерв раздражать фарадическим током до полной остановки сердца»
то через некоторое время остановится и второе сердце: тормозящие продукты,
выделяемые в сердце блуждающим нервом, перейдут в раствор благодаря диф-
фуэии^ а после перехода в другое сердце повлияют на него также тормозящим
образом (рис. 233).
Литература
1. Cannon W. В., Arch. Neur. Psychiatr., 22, 282, 1929.
2. Barer of t, J. Physiol., 64, 1927.
3. Rosenblueth A., Annual Rev. of Physiol., 2, 263, 1940.
4. McDonough F. K., Am. J. Physiol., 125, 530, 1939.
5. В г о u h a L., Cannon W. B. a. Dill D. В., J. Physiol., 95, 431, 1939.
6. Edwards H. Т., Brouha L. a. J о h n s о n R. E., Am. J. Physiol., 124„
254, 1938.
*
ИМЕННОЙ УКАЗАТЕЛЬ
А
Александров В. Я. 125, 441
Альперн Д. Е. 504, 532
Аршавский И. А. 334, 406, 474, 476, 478,
482, 513
Б
Бабкин Б. 503, 504
Бабухин 354
Бакурадзе А. 472, 484, 515
Белицер Б. А. 204, 205, 229
Белоусов М. 187
Березина М. 397, 398
Болдырев В. Б. 436
В
Валидов И. 214
Василенко Ф. 516
Васильев Л. Л. 142, 434, 442
Введенский Н. Е. 87, 111, 179, 182, 192,
214, 343, 370, 377, 385, 392, 418, 426,
427, 430, 431, 433, 442, 444, 445, 453,
457, 493
Вержбицкая Н. А. 273, 323
Вериго Б. 343, 358, 434, 451, 452
Ветохин И. 366, 395
Виноградов М. 454
биталь Д. А. 503
Волынский А. 210
Воронцов Д. С. 152, 176, 187, 214, 249,
250, 251, 350, 351, 374, 379, 380, 381,
382, 394, 395, 407, 422, 423, 427, 428,
434, 435, 454, 450, 488
Вул И. М. 59, 338
Г
Гедевани Д. М. 213, 337, 415, 451, 457,
458, 520, 522, 523, 524, 525
Гинецинскій А. 405, 411, 413, 428, 504,
522, 525
Гогава М. 271, 272, 279, 280
Гоголашвили Ш. 156, 411, 415
Гольденберг Е. 494
Гоциридзе А. 221, 420
Гурвич Л. 306
Д
Данилевский В. 294, 314
Догель А. С. 312, 313, 468
Ж
Жуков Е. К. 277, 284, 381
3
Заварзин А. А. 315, 316, 318
Закарая Е. Ф. 217, 529, 531
Зурабашвили А. Д. 324
И
Иванов К. 230, 447
К
Кан И. Л. 365, 366, 367, 375, 381, 383,
397, 400
Караев А. 333, 355
Картозия А. 84, 181
Квавилашвили Ш. 35
Квасов Д. Г. 369, 436
Кибяков А. В. 411, 489, 4?0, 491, 525
Кирзон М. В. 522
Киселев М. А. 385
Ковалева Г. 525
Колосов Н. Г. 467, 468, 469
Комехиани П. А. 115, 136, 156, 210, 233.
284
Коников А. Л. 338
Копылов Г. 74, 104
Коштоялц X. С. 74, 81, 275, 279,280, 281,
505
Крохина Е. М. 26
Кудрявина Н. А. 281
Кузнецов Д. 365
Купалов П. С. 55, 205
544
ИМЕННОЙ УКАЗАТЕЛЬ
Л
Лаврентьев Б. И. 40, 44, 127, 263, 310.
313, 315, 320, 467, 468, 470, 471, 489,
500, 513, 521, 531
Лазарев П. П. 54, 151, 454
Лежава А. 44, 46, 47, 329, 403
Леонтович А. 514
Лепешкин В. В. 124, 126, 129, 140, 141,
151, 296
М
Магницкий А. Н. 59
Макаров П. В. 127, 440
Макаров П. О. 149, 375, 379, 437
Максимов А. А. 42, 45, 265, 306
Меписаштшли И. 43, 47, 304, 362
Мещеряков А. М. 469
Митрополитанская Р. П. 505
Морозов Б. 129
Мужсев В. А. 59, 279, 280, 281
Н
Нарикашвили С. П. 109
Насонов Д. 125, 127, 140, 439, 440
Науменко А. И. 369
Некрасов П. Н. 523, 525
Некрасова Н. В. 523
Немилов А. В. 50, 304
Нявинская О. А/99, 101, 278
О
Опарин А. И. 50, 128
Орбели Л. А. 521, 522, 530
П
Павлов И. П. VII, 284, 523, 524, 530
Палладии А. 207, 226
Парнас Я. О. 72, 228, 281, 282
Приходькова Е. К. 509
Пэрна Н. 452
Р
Разенков И. П. 508, 529, 532
Рахматулин 319
Резвяков Н. П. 337
Рожанскяй Н. А 512
Роскин Г. И. 262, 264, 272, 273, 274
Русинов В. С. 434, 444
Рябиновская А. М. 81
С
Самойлов А. 31, 162, 192, 508
Свердлов С. М. 437
Серков Ф. Н. 149, 175, 187, 214
Сеченоз И. М. 223
Синицын Н. П. 524
Скадовская Н. С. 366
Соковкин Н. 513
Сперанский А. Д. 529, 530
Степанова С. С. 126
Т
Трофимов Л. Г. 425
Тушнова В. М. 521
У
Уфлянд Ю. М. 337, 456, 473, 476
Ухтомский А. 180, 416
Ф
Феодоров Б. 127, 310, 467
Фердман Д. 74
Франк Г. 68
X
Хлопин Н. Г. 274
ц
Цкиманаури Г. 251
Цкипуридзе Л. Р. 53, 58, 215, 334, 335,
408, 437, 443, 456, 458
Цхакая М. 43, 114, 223, 225
Ч
Чаговец В. 331, 454
Черниговский В. Н. 515
Ш
Шамарина Н. 504
Шатенштейн Д. И. 532
Шерешевский Л. М. 255
Шошина Н. А. 255
Штерн Л. С. 492, 497, 529, 530
Э
Энгельгардт В. А. 69, J2
Энгельс Ф. 130
Ю
Юденич Н. 382
Я
Яуре Г. Г. 257
Яшвили Д. 60, 114, 115, 123
А
Ackermann М. (Аккерман) 208, 217, 232
Adler E. (Адлер) 293
Adrian Е. D. (Эдриан) 160,477, 363, 379,
420, 445, 517
Agdhur Е. (Агдур) 44, 519
Allen А. (Аллен) 503, 504
Alvarez W. С. (Альварец) 275
Amberson W. R. (Эмберсон), 135, 139, 379
ИМЕННОЙ УКАЗАТЕЛЬ
545
Anderson (Андерсон) 500, 513, 517
Apathy (Апати) 306, 316
Агаі К. (Араи) 286 -:
Aronovitch В. (Аронович) 207
Arnstein С. (Арнштейц) 469
Asher L. (Ашер) 233, 501
Asmussen Е. (Асмуссен) 89, 114, 149,174
244
Astbury W. Т. (Эстбери) 440
Azuma R. (Ацума) 243
В
В acq Z. М. (Бак) 506
Baetjer А. М. (Бетьер) 525
Baghnault Е. (Багно) 374
Baldwin E. (Болдуин) 273
Barcroft Н. (Баркрофт) 216, 538
Bargcton D. (Баржетон) 494
Barron D. Н. (Баррон) 337
Bartelrnez G. W. (Бартелмец) 324
Barthley S. Н. (Бартлей) 437
Basch V. (Баш) 254
Baumann W. (Бауман) 295
Bayer Н. V. (Байер) 392
Beck А. (Бек) 92, 494
Bergen W. R. (Берген) 108
Bernard С. (Бернар) 60, 61, 62, 439
Bernstein J. (Бер іштейл) 70, 76, 112, 184
Bethe А. (Бете) 63, 74, 84, 94, 104, 151,
221, 281, 282, 305, 316, 319, 361, 369,
379, 454, 455, 460
Beznak А. (Бецнак) 504
Bidder F. (Биддер) 60
Biedermann W. і(Бидерман) 52, 82, 83, 170,
312, 452
Bielschowsky М. (Билыповский) 319, 521
Bierfreund М. (Бирфрейнд) 297
Billingley Р. К. (Биллингли) 465, 468,
534
Bins? С. (Бинц) 439
Bishop G. Н. (Бишоп) 174, 175, 350, 372.
467, 475, 478, 492
Blair А. (Блэир) 329, 334, 358, 360, 361,
438
Вііх М. (Блике) 88, 90, 97
Bloch Е. (Блох) 287
Bodian D. (Бодиан) 308
Boeke J. (Буке) 40, 46, 123, 262, 263, 264,
310, 320, 369, 469, 502> 519, 521
de Boer (де-Бур) 519, 520, 521
Bogue J. G. (Боге) 353, 361, 366, 377,
.381
Boruttau H. (Bopyxav) 174, 434, 435, 441,
- . 508
Bottazzi: (Боттацци) 107
5 И. С. Беритов
Bouman H. D.. (Боуман) 112
Bourguignon G. (Бургиньон) 59, 334
Boycott А. Е. (Бойкот) 362
Bove D. (Бове) 505
Bozler E. (Боцлер) 126, 250, 265,266,268,
270, 272, 276, 280, 281, 306, 307, 310,
. 311, 319, 367, 499
Brandstater Р. (Брандштетер). Ill ; ¦
Bremer F. (Бремер) 106, 107, 357, 525
Bremer 519
Breslauer (Бреслауер) 514
Brinkmann R. (Бринкман) 504
BromwellC. (Бромвель) 181
Bronk D. W. (Бронк) 478, 481, 482, 483,
489, 492, 515
Brouha L. 540, 542
Brown G. L. (Броун) 109, 148, 170, 174,
201, 412, 487, 491
Brown-Seqard (Броун-Секар) 60
Brucke E. Th. (Бркжке) 58, 208, 210, 266,
267, 334, 335, 338, 339
Brucke Fr. 475
Brungwick D. (Брунгвик) 442
Bruchthal F. (Бухталь) 14, 41, 43, 46, 61,
67, 88, 89, 95, 136, 137, 139, 156, 174,
175, 198, 212, 414 . .
Budde W. (Будде) 275
Buddenbrock W. (Будденброк) 282, 367
Bugnard L. (Бюньяр) 244, 397, 429
Bunting H. (Бантинг) 503
Burch G. (Бурч) 364 ... .
Biirgi Fr. (Бюрги) 211
Burridge W. (Буридж) 207
Burrows (Бэрроус) 319
Buttner H. E. (Бюттнер) 106
С
Cajal R. (Кахаль) 305, 306, 312, 313, 319
Cameron J. (Камерон) 42
Cannon W. В. (Кэннон) 265, 266, 269,490,
502, 504, 505, 506, 507, 520, 525, 530,
531, 537, 539, 540
Carvallo J. (Карвалло) 225
Cattel M. (Каттел) 239, 503
de Castro F. (Кастро) 468
Cebulla R. (Цебулла) 379
Chauchard A. et P. (Шошар) 336, 474
Chauchard В. (Шошар) 336, 474
Charlet G. (Харлет) 524
Child Ch. M. (Чайльд) 127
Chinn Pr. (Чинн) 126
Clark А. (Кларк) 44, 413
Clark D. 44, 107, 503
Coates A. E. (Коотс) 530
Cobb S. (Кобб) 521
546
ИМЕННОЙ УКАЗАТЕЛЬ
Conheim О. (Конгейм) 281, 284
Cooper S. (Купер) 84, 86
Cori G. F. (Кори) 398
Corkill А. В. (Коркилл) 524
Copenhaver W. М. (Копенговер) 257
Cowan S. L. (Коуэн) 356, 357, 428
Сгетег М. (Кремер) 157, 354
Cumminds J. N. (Куминг) 135
D
Dale Н. Н. (Дейл) 108, 170, 411, 412, 414,
503, 525
David F. (Давид) 298
Davenport Н. А. (Давенпирт) 308, 311
Davis Н. (Девис) 338, 442
Demoor J. (Демур) 258, 287
Demoor L. 503
Denning H. (Деннинг) 512
Dercum Fr. (Деркум) 128
Dessy (Десси) 520
v. Dijik J. А. (Дейк) 410
Dill D. В. (Дилл) 540, 542
Dittler (Диттлер) 149
Downing А. (Доунішг) 379
Du Bois Raymond E. (Дюбуа Реймон)
17 — 18, 20—21, 51, 56, 138, 361, 450
Dubuisson M. (Дюбюиссон) 155, 198
Dunning H. S. (Дюнтшнг) 46, 323
Diirig А. (Дюриг) 170
Dusser de Barrenne (Дюссер де Баренн)
315, 521
Dworkin К. (Дворкин) 394
Е
Ebbecke U. (Эббеке) 199, 343, 455
Eccles J. С. (Экклс) 84, 86, 170, 176, 180,
255, 267, 268, 270, 275, 276, 363, 404,
405, 406, 416, 475, 478, 479, 480, 484,
485, 487, 488, 491, 492, 507
Ecker (Эккер) 10 — 13
Edstrom (Эдштрем) 61
Edwars H. Т. (Эдварс) 540, 542
Eggleton G. Р. (Эгглетон) 71, 107
Eggleton P. 71, 107
Einthoven W. (Эйнтховен) 29, 173, 249
250
Eiselsberg А. (Эйзельсберг) 294
Elliot T. R. (Эллиот) 465, 498, 500
Embden G. (Эмбден) 71, 75, 106, 150, 226
228, 230, 231, 292, 293
Engelmann T. W. (Энгельман) 41, 67, 70
185, 237, 251, 253, 254
Erlanger J. (Эрлангер) 33, 158, 328, 329,
330, 334, 350, 352, 353, 358, 360, 361,
362, 363, 372, 373, 378, 379, 380, 381,
382, 427, 438
Ernst E. (Эрнст) 66, 111, 153, 154, 172
Escobar R. А. (Эскобар) 438
Esveld L. W. (Эсвельд) 287, 288
Ettich G. (Эттих) 305, 306, 438, 439
Euler U. S. (Эйлер) 201
Eyster J. A. E. (Эйстер) 256
F
Parkas E. (Фаркас) 503
Feldberg W. (Фельдберг) 108, 170, 411,
414, 490, 503, 525
Feng Т. Р. (Фенг) 93, 242, 243, 397, 399,
413
Fenn W. О. (Фенн) 135, 155, 207, 355,
357, 400
Fenn J. 243
Ferrari R. (Феррари) 226
Fick А. (Фик) 221, 236, 460
Fischl H. (Фишль) 163
Fischer E. (Фишер) 95, 107, 176, 226, 478,
536
Fleischl А. (Флайшль) 414
Fletcher W. M. (Флетгер) 70, 230, 264,
269, 270, 292, 293, 294, 298
Florkin M. (Флоркин)'334
Fogt M. (Фогт) 108
Forbes А. (Форбс) 221, 338, 371, 392, 394,
442, 480, 486, 521
Foss О. (Фосс) 162
Fraenkel M. (Френкель) 104
Frankel-Conrat (Френкель-Конрат) 396
F redericq H, (Фредерик) 48
Frey (Фрей) 86
Frolich F. (Фрблих) 354, 365, 435, 441
Fromherz (Фромгерц) 61
Fulton J. F. (Фултон) 77, 111, 172, 218,
221
Funke J. (Функе) 197
Furlinger Fr. (Фюрлингер) 42, 43
Furguson L. К. (Фергюсон) 516, 518
Furusawa К. (Фурусава) 107
Fiirth О. (Фюрт) 292, 297, 298
G
Gaddum J. (Гаддум) 490
Gammon G. D. (Гаммон) 515
Ganter G. (Гентер) 270
Garner W. (Гарнер) 70, 77
Garten S. (Гартеі) 177, 343, 351, 371
Gaskel W. H. (Гаскел) 507
Gasser H. (Гассер) 158, 304, 328, 330, 350,
351, 356, 361, 362, 363, 371, 372, 373,
374, 378, 379, 380, 381, 382, 384, 394,
476, 525
ИМЕННОЙ УКАЗАТЕЛЬ
547
Geist F. D. (Гейст) 315
Gelfan S. (Гельфан) 159, 160, 177
Gellhorn E. (Гельхорн) 275
Gerard R. W. (Жерард) 379, 395, 396, 397,
398, 399, 438
Gibson С. (Джибсон) 468, 485
Gildemeister M. (Гильдемейстер) 52, 157,
383, 427
Gilson А. (Джильсон) 174
Glazko A. J. (Гладко) 258
Glick D. (Гликк) 491
Gopfert H. (Гепферт) 111/173, 333, 409,
415
Goliz Ber. (Гольц) 66
Goss С. М. (Госс) 257
Gotch Fr. (Гоч) 182, 364
Gottlieb (Готлиб) 471
Graham H. Т. (Грэим) 377, 378
Grandis (Грандис) 520
Greenberg D. M. (Гринберг) 258
Griffith F. В. (Грифит) 371
Gruber Ch. (Грубер) 520
Grumbach Z. (Грумбах) 201
Grunhagen (Грюнгаген) 444, 452
Grundfest H. (Грундфест) 304, 351, 362,
475
Guttman S. А. (Гутман) 201
H
Haas P. (Xaac) 404
Haberlandt L. (Габерландт) 255, 258
Halter R. (Гальтер) 337, 396
Hanstrom В. (Ганстрём) 316, 365
Happel P. (Гаппаль) 84
Harashima S. (Харашима) 400
Harreveld A. V. (Гарревельд) 107, 108,
188
Harrison (Гаррисон) 319
Hartree W. (Гартри) 107, 242, 244
Hay (Гэй) 221, 236
Hayasi К. (Хайаси) 334
Heidenhain (Гайденгаин) 51, 237, 319
Heimbrecht В. (Геймбрехт) 107
Heinbecker P. (Гейнбекер) 437, 467, 475,
478, 492
Heinrrich А. (Гейнрих) 91, 460
Helmholtz H. (Гельмгольц) 24, 25, 86,
112, 363
Henderson V. E. (Гендерсон) 287
Henriques (Генрик) 173
Hering E. (Геринг) 23, 138
Hermann К.(Герман) 506
Hermann L. 52, 138, 152, 155, 157, 167,
184, 354, 428, 452
Herrick C. Judson (Херрик) 322
Hertwig G. (Гертвиг) 51,- 127
Herzen (Герцен) 444
Hett О. (Гетт) 105
Le Heux (ле-Го) 286, 287
Hill A. V. (Гилл) 70, 74, 76, 91, 97, 107,
153, 155, 204, 205, 222, 230, 231, 237,
238, 239, 240, 243, 244, 245, 339, 340,
395, 397, 399, 400, 425
Hinsey J. С. (Гинзей) 315, 521
Hinter H. (Гинтер) 148, 163
Hodes R. (Ходе) 499
Hodkin H. L. (Ходкин) 332, 354, 360, 363,
366, 374
Hoerr N. (Гоер) 324
Hoefer P. (Гефер) 174
Hoff E. С. (Гофф) 308
Hoff H. E. (Гофф) 255
Hofmann F. В. (Гофманн) 534
Hoffmann P. (Гофманн) 187
НбЬег R. (Гёбер) 79, 142
Hogquist G. (Хогквист) 41
Holmes E. G. (Холмс) 396
Holt E. (Гольт) VI
Holliger M. (Голлигер) 216
Hoogenhuyze (Гугенгуйц) 519, 521
Hoorweg L. (Гоорвег) 52, 53
Hopkins F. G. (Гопкинс) 70, 230, 292,
298
Horton А. (Хортон) 201, 203
Hsi-Chun Chang (Си-Чун Чанг) 287
Hursh J. В. (Гарш) 362
Hurtle К. (Гюртле) 41, 66
J
Jaderholm G. А. (Иедерхольм) 337
Jamin Fr. (Иамин) 69
Jansen (Янсен) 318
Jasper H. H. (Джаспер) 337, 368
Jensen P. (Иенсен) 298
Jochims J. (Иохимс) 305, 306, 438, 439
Johnson R. E. (Джонсон) 542
Jordan H. (Иордан) 272, 278, 280, 281,
365, 367, 375
Jordan F. 506, 511
К
Kaelin E. (Кэлин) 414, 417
Kahn R. (Keh) 163
Kappers Ar. (Kannepc) 127, 306, 320, 470
Karpa P. (Карпа) 297
Kato G. (Като) 147, 148, 160, 161, 175,
330, 331, 342, 359, 364, 427, 434, 435,
436, 438, 442, 453
Katz В. (Катц) 331, 332, 333, 354, 371,
374
И. С. Беритов
548
ИМЕННОЙ УКАЗАТЕЛЬ
Kisch В. (Киш)-257
Kobayashi М. (Кобаяши) 206, 211
Kochashi G. (Кохаши) 225
Koch Е. (Кох) 442
Kohlrausch А. (Кольрауш) 184, 185
Krannich Е. (Краних) 108, 411
Kries V. (Крис) 194
Krijgsman В. J. (Крийгсман) 365, 366, 376,
395
Krogh А. (Крог) 514
Кгопескег Н. (Кронекер) 182, 196, 251
Krupp М. А. (Круп) 315, 326
Kuffler S. W. (Кафлер) 410
Kiihne W. (Кюне) 60, 170, 353
Кигё К- (Кюре) 470
L
Labhard Er. (Лабгарт) 524
Lambert E. F. (Ламберт) 338
Landois (Ландуа) 19, 139, "450
Langley J. (Лэнгли) 61, 123, 465, 470, 472,
500, 502, 512, 513, 520, 521, 534, 535
Lapicque L. (Лапик) 52, 57, 58, 180, 195,
269, 333, 334, 338, 409, 416, 482, 493
Lassale Н. (Лассаль) 335
Ledebur V. (Ледебур) 339
Lee F. S. (Ли) 208, 207
Lehmann J. E. (Леман) 329
Lenhossek M. v. (Ленношек) 312, 319
Leskinen V. 335
Lewin А. (Левин) 381, 395
Lewis Warren (Льюис) 255
Lillie R. S. (Лилли) 126, 155, 157, 354
Lindhard J. (Линдгард) 41, 43, 46, 61, 67,
88, 89, 95, 136, 137, 139, 173, 174, 414
Lippay F. (Липпаи) 298
Lissak К. (Лиссак) 502
Lhomon W. T. 315
Lloyd D. J. (Ллойд) 485, 487
Loeb J. (Лёб) 54, 55, 257, 453, 454
Lohmann К. (Ломан) 72 — 74, 107
Lohman G. 524
Loewi О'. (Лёви) 500
Loven Ch. (Левей) 112
Lorente de N6 R. (Лоренте де Но) 308,
309, 312, 323, 409, 490, 491
Lowenbach H. (Ловенбах) 536
Lucas К. (Лкжас) 61, 149, 163, 181, 194,
372, 377, 383, 408, 441, 445
Luco J. V. (Луко) 109
Lullies Н. (Люллис) 365, 366, 367, 375,
377
Lund (Лунд) 154
Lundsgaard (Лундсгаард) 71, 230, 293
Lurabee (Лураби) 492, 495
М
Maas J.-А. (Маас) 99, 250, 281, 282
Maaske С. А. (Мааске) 503
Mackuth Е. (Макут) 142, 434, 455
Magnus R. (Магнус) 275, 285, 287
Magladery J. W. (Мьгладери) 267, 270,
275
Maibach Ch. (Майбах) 524
Mangold E. (Мангольд) 45, 48, 295
Mann (Манн) 505
Mansfeld G. (Мансфельд) 257, 286
Marceau (Mapco) 282
Marey (Марей) 25, 27, 251
Margaria R. (Маргариа) 93, 221, 232, 516,
518
Marnay А. (Марнай) 411, 413
Marrazzi A. S. (Марраци) 483, 490
Matsuoka Kenn. (Матсуока) 206
Matteuchi Ch. (Матеучи) 70, 452
Matthews В. Н. С. (Мэтьюс) 337
Mayeda R. (Майеда) 226
Mc Donough F. К. (Мак Донаф) 539
Mc Erla E. D. (Мак Эрла) 499
Mc Intosh F. С. (Мак Интош) 491
Mc Swiney В. А. (Мак Суини) 499
Mc HopKins M. (Мак Гопкинс) 442
Meek W. I. (Миик) 260, 503
Meyer К. (Мейер) 70, 77, 78, 93, 135,
154
Meyerhof О. (Мейергоф) 66, 70, 72, 76,
107, 204, 210, 228, 230, 231, 232, 239,
240, 273, 292, 298, 395, 400
Michaelis (Михаэлис) 151
Michol E. (Михоль) 524
Millen J. L. (Миллен) 216
Minckler J. (Минклер) 308, 311
Mines G. R. (Майне) 47
Minkowsky M. (Минковский) 324
Minz В. (Минц) 503
Miura R. (Миура) 205
Monnier A. M. (Монние) 268, 337, 368,
492, 507
Moore A. (Moop) 58, 339
Morine G. (Морин) 287, 506, 511
Morison R. S. (Морисон) 201, 412
Morpurgo В. (Морпурго) 69, 226
Miiller J. (Мюллер) 368
Muralt А. (Муральт) 67
Miirnich (Мюрних) 360
Nachmansohn D. (Нахмансон) 411, 413,
415
Nagaya Т. (Нагайа) 206, 222
Nagel W. А. (Нагель) 294
ИМЕННОЙ УКАЗАТЕЛЬ
549
Necheles Н. (Нехелес) 287
Nernst W. (Нернст) 54, 154, 339
Netter Н. (Неттер) 360
Neuschloss (Нейшлосс) 79
Nice (Найс) 520
О
Oinuma S. (Оинума) 149, 341
O'Connor W. J. (О'Коннор) 170, 176, 450,
406
Ostwald W. (Оствальд) 154
Oudendal (Удендэль) 320
Owerton E, (Овертон) 55, 341
P
Panella (Панелла) 520
Pantin G. F. (Пэнтин) 81, 367, 377
Parker G. H. (Паркер) 281, 321, 399
Parkinson I. L. (Паркинсон) 211
Parrack H. О. (Паррак) 340
Pawianen S. (Павианен) 335, 348
Pekelharing (Пекельхаринг) 519, 521
Pembrey M. (Пембри) 29, 36, 112, 195,
221, 497
Peterfi Т. (Петерфи) 175, 306
Pfluger E. (Пфлюгер) 51, 56, 343, 450,
458
Phalen G. S. (Фален) 308, 311-
Piper H. (Пипер) 361, 364
Plattner P. (Платтнер) 108, 411
Plaut К- (Плаут) 91
Ponder E. (Пондер) 357
Politzer (Политцер) 61
del Pozo E. С. (Позо) 267, 269
Pratt Fr. (Пратт) 160
Prosser C. Z. (Проссер) 148, 161
Pumphrey R. J. (Пэмфри) 477, 495
Q
Quastel (Квостел) 505
R
Ramsey R. W. (Рамзей) 75, 90
Ranke J. (Ранке) 206, 208
Ranson S. W. (Рэлсон) 90, 97, 99, 465,
468, 521, 534
Raventos J. (Равентос) 109, 413
Rauh Fr. (Pay) 84, 371
Rehsteiner R. (Рештейнер) 107
Renquist J. (Ренквист) 335
Retzius (Ретциус) 319, 320
Rice L. (Райе) 392, 394
Richter С. (Рихтер) 266
Riesser О. (Риссер) 107, 109, 176,205, 222,
265, 508
Riegels L. (Ригельс) 469
Rigler R. (Риглер) 257
Rijnberk G. (Ринберк) 112, 521
Roepke M. H. (Рэпке) 287
Rogers (Рожерс) 321
Rollet (Роллет) 81
Rona P. (Рона) 128
Rosenbaum (Розенбаум) 494
Rosenberg H. (Розенберг) 353, 361, 366,
377, 381
Rosenblueth А. (Розенблют) 109, 201, 267v
269, 350, 412, 480, 481, 485, 488, 504
Rossbach M. (Россбах) 197
Rossi E. (Росси) 298
Rushton W. A. H. (Руштон) 331, 334,
354
Rylant F. (Рилан) 257
S
Czent-Gyorgyi (Сцент-Гьорги) 41, 79,
, 80
Schaefer H. (Шефер) 61, 111, 160, 169,
173, 174, 175, 176, 178, 221, 404, 409,
415, 416
Scheiner M. С. (Шеинер) 506
Seheinfinkel N. (Шейнфинкель) 501
Scheminsky Fer. (Шеминский) 456
Scheminsky Fr. 456
Schilf E. (Шильф) 478, 514
Schleier (Шлейер) 91
Schmid E. (Шмид) 216
Schmid W. 521
Schmitd F. О. (Шмит) 398
Scholmerich P. (Шёлмерих) 175, 178
Schotz E. (Шютц) 111
Schriever H. (Шривер) 195, 340, 379
Schuller J. (Шюллер) 271, 275, 285
Schultz P. (Шультц) 271, 275, 277, 466
Schultze (Шульце) 318
Schwalbe G. (Швальбе) 226
Schwarz L L (Шварц) 488
Schwenker С. (Швенкер) 103, 106, 211,
296, 298
SchwenKer G. (Швенкер) 74
Scramlik E. (Скрамлик) 256
Segaar J. (Ceraap) 367
Sereni E. (Серени) 281
Sewall (Сьюал) 195
Shen S. С. (Шен) 413
Sherif M. F. (Шериф) 503
Sherrington Ch. S. (Шеррингтон) 23, 45,
308, 312, 354, 363
Sibui J. 414, 417
Sichel F. S. M. (Зихел) 88, 89,. 148, 149, 161
Siebert W. W. (Зиберт) 69, 226
Simeone F. А. (Симеоне) 480, 481, 485
Siraonart A. a. Simonart E. (Симонар) 412
550
ИМЕННОЙ УКАЗАТЕЛЬ
Skinner В. F. (Скиннер) 338
Smitten N. (Смиттен) 310
Sokowkin N. (Соковкин) 513
Solandt D. Y. (Соландт) 340, 477, 495,
516
SolomDn Е. I. (Соломон) 396
Sommerkampf Н. (Зоммеркампф) 108
Speidel С. С. (Шпейдель) 126, 305, 307
Spiegel E. (Шпигель) 521
Spiro К- (Спиро) 286
Starke J. (Штарке) 337
Starling (Старлинг) 498
Stavracky С. (Ставрацки) 503, 504.
Stedman Е.(Штедман) 413, 417
Steiman S. Е. (Стейман) 148, 404
Steinach Е. (Штейнах) 194, 317, .318
Steinhaus А. Н. (Штейнгауз) 69, 226
Steinhausen W. (Штейнгаузен) 176, 508
Stohr Ph. (Штёр) 263, 320, 467, 468, 501>
521
Stopford J. S. В. (Стопфорд) 499
Street S. F. (Стрит) 75, 90
Siissner H. (Зюснер) 91
Т
Takahashi К- (Такагаши) 305, 362, 530
Tasaki Teh. (Тасаки) 334, 350, 359, 362
Tello J. F. (Тулло) 521
Tennenbaum (Тенненбаум) 505
Tervaert Cahen (Тэрверт) 521
Thauer R. (Tayep) 105
Therman P. О. (Тэрман) 486, 495
Thorner W.'(TspHep) 392
Thunberg Т. (Тунберг) 395
Tiegel (Тигель) 103, 197
Tiegerstedt К- (Тигерштет) 350
Tiegs O. W. (Тигс) 320, 468, 469, 502, 521,
524, 530
Tiffeneau M. (Тифено) 505
Topikow 398
Topsoe-Jensen А. (Топсё-Иенсен) 205
Tower S. (Тоуэр) 477, 495, 512, 528
Trendelenburg W. (Тренделенбург) 111,
253
Tschermak A. V. (Чермак) 153, ['266, 267
Tsudzimura H. (Тсудзимура) 503
U
Uexkull v. J. (Икскюль) 91, 281, 282, 283,
284
T
Vartiainen А. (Вартианен) 490
Veach H. О. (Виг) 488
v. d. Velde J. (Вельде) 504
Verzar F. (Верзар) 91, 230, 357
Verworn M. (Ферворн) 312
Veszi J. (Весци) 392
Vulpian А. (Вульпиан) 524
W
Wachholder К. (Вахгольдер) VI, , 86, 87,
97, 98, 99, 108, 110, 399 J
Wacker L. (Вакер) 293, 297
Waldeyer W. (Вальдейер) 311
Waller А. (Уоллер) 220, 249, 312, 314
Walther А. (Вальтер) 255
Warburg О. (Варбург) 127, 128, 228
Wastl H. (Вастль) 520
Weber E. (Вебер) 223
Weber H. H. 69
Weichardt W. (Вейхардт) 207
Weinstein M. (Вейнштейн) 488
Weiss G. (Вейсс) 52, 225, 331
Wial J. (Виал) 506, 511.
Wiechmann E. (Вихман) 60
Wiersma C. A. G. (Виргма) 107, 108
Wilber В. Т. (Вильбер) 201
Wilier D. Т. (Виллер) 201
Wilkinson H. (Вилкинсон) 521
Wilmers J. (Вильмерс) 104, 161, 184
Windle W. F. (Уйндль) 311, 324
Winterstein H. (Винтерштейн) VI. 293,
294, 396, 399, 400
Witteridge D. (Виттеридж) 536
Wolf H. (Вольф) 46, 293, 320, 503
Wong Amos (Уонг) 287
Woollard H. H. (Вуллард) 469
Z
Zing Jang Kyo (Цинг Ианг Кио) 415
Zondek S. G. (Цондек) 285
Zotterrnan Y. (Цоттерман) 305, 361, 362
Zwaardemaker H. (Цваардемакер) 258
Zunz N. (Цунц) 223
Y
Young J. Z. (Юнг) 307
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А
Абсолютная сила мышцы 224
Автоматическая деятельность сердца 255J
—¦ гладкой мышцы 285
Автономная нервная система см.
вегетативная н. сист.
Адаптация в нервном стволе 339
Аденилпирофосфат — расщепление и вос-
f становление 228 — 230
Аккумуляторы — кислотные 16,
щелочные 17, правила пользования 16
Активные вещества 500, симпатии 502,
ацетилхолин 503, распространение 504,
механизм действия 505;
Актин 79, актомиозин 79
Анаболическая фаза 131
Анатомия мышечной системы 9, 399
нервной системы 11, 303
Антагонистическая деятельность
вегетативной нервной системы 494,
происхождение ;500
Ассимилятивная фаза 131
В
Биоток возбуждения мышцы — понятие
153, происхождение 153,
биологическое значение 156, однофазный и дву-
фазный 167, ритмическое течение 170.
и сокращение 171, опыт 188
Биоток возбуждения сердца 248, нерва
349, преганглионарного волокня 475,
постганглионарного — 477,
симпатических ганглиев 476, двигательной
пластинки 409, опыты 387
Биоток основного биологического
процесса мышцы 135
Биоток повреждения 137,
демаркационный^, теория 138
Биоток сокращения 175
Биохимия скелетной мышцы 70, 228,
гладкой мышцы 274
В
Вегетативная нервная система —
анатомия 465, симпатическая и
парасимпатическая системы 469, никотинный
метод 470, антагонистическая
деятельность 496, рефлекторные реакции 514,
тоническая деятельность 515, действие
на скелетную мускулатуру 519, тро*
фическое влияние 528, гомеостатиче-
ское значение 537, опыты 540
Внутренняя среда — понятие 528
Возбудимая система — понятие 48, тете-
рохронное состояние 132,
изохронное — 157
Возбудимость мышцы—понятие 48,
определение степени 56, собственная
возбудимость мышцы. 59, 61, как
функция количества возбудимой системы
134, повышение ее 192, опыты 61, 63
Возбудимость нервного ствола 328,
нервных волокон 328, сверхнормальная
377, преганглионарных волокон 474,
постганглионарных 476
Возбуждение мышцы — понятие 48,
характеристика 146, 157, условия 150 —
151, биоток 153, продолжительность
его 176, интенсивность 177,
ритмическое течение 177
Возбуждение нерва 349,
продолжительность его 350, интенсивность 350,
проведение 353, ритмическая природа
370, высший или предельный ритм
371, многократное возбуждение на
одно раздражение 379,
преганглионарных волокон 475, постганглионарных
— 476, как окислительный процесс
399
Вторичное сокращение 170, опыт 188
Г
Гальванические элементы 12
Гальванометр Эйнтховена 29
552
ИМЕННОЙ УКАЗАТЕЛЬ
Гладкая мускулатура — строение 262,
возбудимость и возбуждение 265,
биоток 266, сокращение 271, скрытый
период 275, рефракторные фазы 275
пластичность 277, собственный тонус
277, запирательное действие 282,
автоматическая деятельность 285,
опыты 288
Гликоген — расщепление в v мышце 228,
восстановление 231, в гладкой мышце
274
Гомеостатическая функция вегетативной
нервной системы 538
д
Двигательная единица 45
Двигательная пластинка 44, возбуждение
ее 404, утомление 406, хронаксия 408,
механизм передачи возбуждения 408,
выделение ацетилхолина 410, значение
его 411
Декремент возбуждения 364, 441
Демаркационный ток — понятие 137,
мембранная теория 141, опыт 144
Диаграмма Уоллера 220
Диссимиляция 131
. Е
Единицы электричества 15
ль
Живая возбудимая система — понятие 48
50, структура 123, обмен веществ
130
3
Закон ["возбуждения — понятие 146, 150,
методика исследования 162, по
отношению к мышцам беспозвоночных
187, в сердце 251, в гладкой мышце
275, опыты 165, 259
Закон „все или ничего" 149
Закон изолированного ^проведения
возбуждения 367, 368, опыт 388
Закон сокращений Пфлюгера 458
Закон средних нагрузок 220
Закон раздражения Дюбуа-Раймона 51,
опыт 62
Закон электрического раздражения Нерн-
ста 54
Запирательное действие гладкой мышцы
282
И
Изохронизм нерва и мышцы 338
Индукционные удары 19
Индукционный аппарат 18, придаток
Гельмгольца 20
Индукционный ток — понятие 12,
униполярный 21, электротоническое
действие 459
Итеративные органы 269, возбудимость
их 482
К
Камертон 29
Капсула Марея 27
Катаболическая фаза 131
Кимограф Марея 25, Людвига 26, Криса
: 116
Киноплазма 262
Ключ [Дюбуа-Раймона 21, Гельмгольца
21
Кольцо Геринга 23
Компенсатор 15
Компенсаторная пауза 253
Контрактура 103, происхождение 105,
опыт 117
Кураре, физиологическое действие 60
Л
Лабильность мышцы 179
Липопротеиды 125
;Локальное сокращение 159
Локальный процесс в мышце — понятие
158, в нервных волокнах 331, в
сердце 250
Ж
Мембранная теория возбуждения 154,
355
Методика изолирования волокон 164
Метроном 20
Миографический метод 24
Миографы 26, для напряжения 27
Мион 263
Миофибрилли 40, сокращение 66,
природа 68
Мышечные шумы и тоны 111
Мышцы поперечно-полосатые 39,
строение 43
Н
Напряжение активное 91, от сокращения
92, 118
Нейронная теория 311, физиологические
основания 313, критика 318
Нейронно-нейропильная теория 320
ИМЕННОЙ УКАЗАТЕЛЬ
553
Нейропильная теория 316, критика 318
Нервная система —элементы 303,
синапсы 307, теории строения 311 — 323,
эмбриональное развитие 324
Нервное окончание в мышце —
структура 44, физиология 403
Нервные участки мышцы — структура 46,
возбудимость 60, сократительная
способность 115, биоток 135,
биохимические процессы 233, характеристика 403
Нервный ствол — возбудимость 328,
раздражение 381, локальный процесс 331,
скрытый период — 333, хронаксия 333,
раздражители 341
О
Обмен веществ в нерве 395,
симпатических ганглиях 493
Обмен веществ живой системы 130,
анаболическая и катаболическая фазы
131, самопроизвольный — спонтанный
130
Одиночное сокращение 80, скрытый
период 83, 116, продолжительность 81
Одиночное тетанизированное сокращение
384, опыт 388
Окоченение 292, условия 293,
прекращение 297, искусственное 298, опыты
299
Окончания двигательного нерва 44
Оптимальный эффект мышцы 182, внерв-
номышечном препарате 419, опыты
188, 428
Основной биологический процесс
мышцы— понятие 130, характеристика 132
биоток 135, влияние внешней силы 140,
влияние среды 142, опыт 144
Основной биологический процесс нерва
349
Осциллографы 31, электромагнитные 32,
катодный 33
II
Парабиоз — понятие 429,
подготовительная (трансформирующая) стадия 430,
парадоксальная 431, тормозящая 431,
происхождение их 443, локализация 437,
функциональные изменения 434, 442,
структурные изменения 439, концепция
Введенского 444, опыты 446
Парасимпатическая нервная система —
анатомия 406, основная функция 539
Передача возбуждения в мышце —
электрическая 408, гуморальная 410, в
симпатическом ганглии 484, в
эффектах вегетат. системы 500, механизм
передачи 500, критика 506
Периэлектрон 456, опыты 461
Пессимальный эффект мышцы 182, опыт
188: — в нервномышечном препарате
418, происхождение 420, — в нерве
424, — двигательных пластинках 424,
концепция Введенского — 426, в
вегетативной системе 493, опыты 428
Пластичность скелетной мышцы — 89, 96,
гладкой мышцы 277
Повышение возбудимости 192, при
субмаксимальных раздражениях 194, при
редких раздражениях 196, при тета-
нических раздраж. 199, опыты 202, от
растяжения 221, в сердце 254, в
нерве 377, 383
Полезное время 52
Поляризация 16
Полярное действие электрического тока
449, физиологические изменения 449,
456, физические 452, происхождение
их 453, опыты 460
Полярное действие индукционных ударов
459
Пороговое раздражение — понятие 56,
определение 63, 344
Постганглионарные нейроны — анатомия
465, физиология 476
Преганглионарные нейроны — анатомия
465, физиология 474
Предельный (высший) ритм возбуждения
мышцы 178, нерва 371, преганглио-
нарных волокон 476, постганглионар-
ных 478, симпатических ганглиев 487
Проведение возбуждения в не ве 355,
механизм 354, роль перехватов Ранвье
358, скорость 340, методика 363,
проведение с декрементом 364, —
изолированное 367, роль нейрофибриллей
369, опыты 386
Проведение возбуждения в парабиотйче-
ском участке 441, в преганглионар-
ных волокнах 475, в постганглионар-
ных 478
Проведение возбуждения в скелетной
мышце 184, методика 184, скорость
184, опыт 189
Проведение возбуждения в сердце 251, —
гладкой мышце 269
Продолжительность возбуждения в
скелетной мышце 176, в сердце 250, в
нерве 349
Псевдорефлексы преганглионарные 512,
постганглионарные 514
554
ИМЕННОЙ УКАЗАТЕЛЬ
Р
Рабочая фаза в мышце 230
Работа мышцы 220, 222, величина ее
223, влияние упражнения 225, влияние
нагрузки^227, опыты 234
Раздражение — понятие 48, пороговое 57,
субминимальное 194, униполярное
342
Раздражимость 49
Раздражитель — понятие 48, мышцы 54,
нерва 341
Раствор Рингера 55, — Локка55, — Тиро-
де 55
Растяжимость скелетной мышцы 87,
изменение сократительной способности
92, взаимодействие различ. участков
94, 119
Релаксационный раздражитель 21
Реобаза 57
Реохорд 14
Рефракторные фазы — понятие 149,
продолжительность их в скелетной мышце
181, — ритмическая природа 181, —
механический эффект 181, — нервных
участков 181, в сердце 251,259, в
нерве 371, — при декрементном
проведении 375, значение в нервно-мышечной
деятельности 418, — преганглионарн.
волокон 475, постганглионарных
волокон 477", опыты 189, 259, 386
С
Сарколемма 39
Саркоплазма 39
Сверхнормальная возбудимость мышцы
192, нерва 377
Сверхнормальный эффект 196
Сердечная мышца —^особенности 247,
биоток 248, рефракторные фазы 251,
автоматическая деятельность 255,
происхождение 257, сердечный гормон
258, опыты 259
Симпатическая нервная система —
анатомия 466, гомеостатическое значение
537, влияние на химические процессы
531
Симпатический ганглий — анатомия 467,
физиология 478, химическое
раздражение 483, передача возбуждения 484,
биотоки 479, происхождение их 485,
486, координирующая деятельность 534
Синапсы 307, 468
Скрытый период сокращения скелетной
мышцы 83, гладкой мышцы 275
Скрытый период возбуждения нервного
ствола 333
Следовая отрицательность 379
Смерть мышцы 292, окоченение 293, —
возбудимость 295, структурные
изменения 295
Собственная возбудимость мышцы 59, 61
Собственный тонус скелетной мышцы
100, гладкой мышцы 277
Сокращение — понятие 66,
происхождение 70, теории Бернштейна 76, Гилла
76, Гарнерг 77, Мейера 77, Сценц-
Гьорги 79, — одиночное 80, —
изотоническое 82, — изометрическое 82, — те-
таническое 84, градация 112, разных
мышц 113, 118, разных участков 115,
118, вторичное 170
Сопротивление живых тканей 13
Структура и функция 127
Субминимальное раздражение 194
Субординационная Х[онаксия 336
Суммация в мышце 194, в нерве 383,
симпатическом ганглии 481, опыты
202
Т
Теплообразование в мышце 237,
начальная фаза 238, восстановительная 240,
при растяжении и укорочении 241, при
разных условиях 243, методика 245
Теплообразование в нерве 397
Тетанус полный 85, зубчатый 85,
происхождение 86, опыт 117 ¦
Тетанус сердца 254, гладкой мышцы 277
Ток —покоя 138, —
повреждения—демаркационный 137,— действия 153 —
сокращения 175
Торможение вегетатирной н. системы
496, природа 500, 506
Тормозящая стадия парабиоза 431,
происхождение 444
Трофическое действие вегетативной н.
системы 528, роль симпатической
системы 530
У
Униполярное раздражение 342, 346
Усилители тока 34
Утомление двигательной пластинки 406
Утомление мышцы 204, факторы 205;
изменение сокращения 208 и
возбуждения 208, происхождение этих
изменений 209
ИМЕННОЙ УКАЗАТЕЛЬ
555
Утомление при одиночных раздражениях
212, при тетанических раздражениях
214, деятельность без утомления 215,
опыты 217
Утомление нерва 391, относительное 391,
опыты 400
Ф
Физиология — предмет 1, задачи 2,
методы 3, отношение к другим наукам
4-6
Фосфаген 71, 228
X
Холинэстераза в двигательных
пластинках 411
Хронаксия — понятие 57, — скелетной
мышцы 58, — нервных стволов и
волокон 333, — субординационная 336, —
двигательной пластинки 408, — преган-
глионарных волокон 474, — иостган
глионарных 476
Хронографы 28
Э
Экстрасистола 252
Эластичность скелетной мышцы 87, 117
Электроды неполяризующиеся 17,
металлические 23, Шеррингтона 23
Электрографический метод 29
Электролиз 16
Электроток физиологический 449,
физический 452, опыты 461
Электротоническое действие
индукционных ударов 459
Элементы гальванические 12
Эффект Гинецинского-Орбели 521,
происхождение 522
Я
Явление адаптации или аккомодации 339
Явление Гинецинского-Орбели 521
Явление лестницы в мышцах 196,
происхождение 198, в нерве 383, опыт 202
Явление нарастания сокращения мышцы
при учащении раздражения 212
Явление суммации в мышце 195, в нерве
383, в эффекторах вегетативной
системы 492
Явление Шеминского 455
Печатается по постановлению
Рёдакционно-издательского совета
Академии Наук СССР
*
Технический редактор Е. Н. Симкина
*
РИСО АН СССР № 2550. А05776. Тип. заказа
№ 70. Подп. к печ. 19/ХП 1946 г. Формат бум.
70ХЮ8і/і6. Печ. л. 343'4. Уч.-издат. 44,5.
Тираж 5000.
Набрано в 1-й Образцовой тип. ОГИЗа треста
«Полиграфкнига» при Совете Министров РСФСР,
Москва, Валовая, 28.
Отпечатано во 2 тип. Издат. Академии Наук СССР
Москва, Шубинский, 10