




























































































































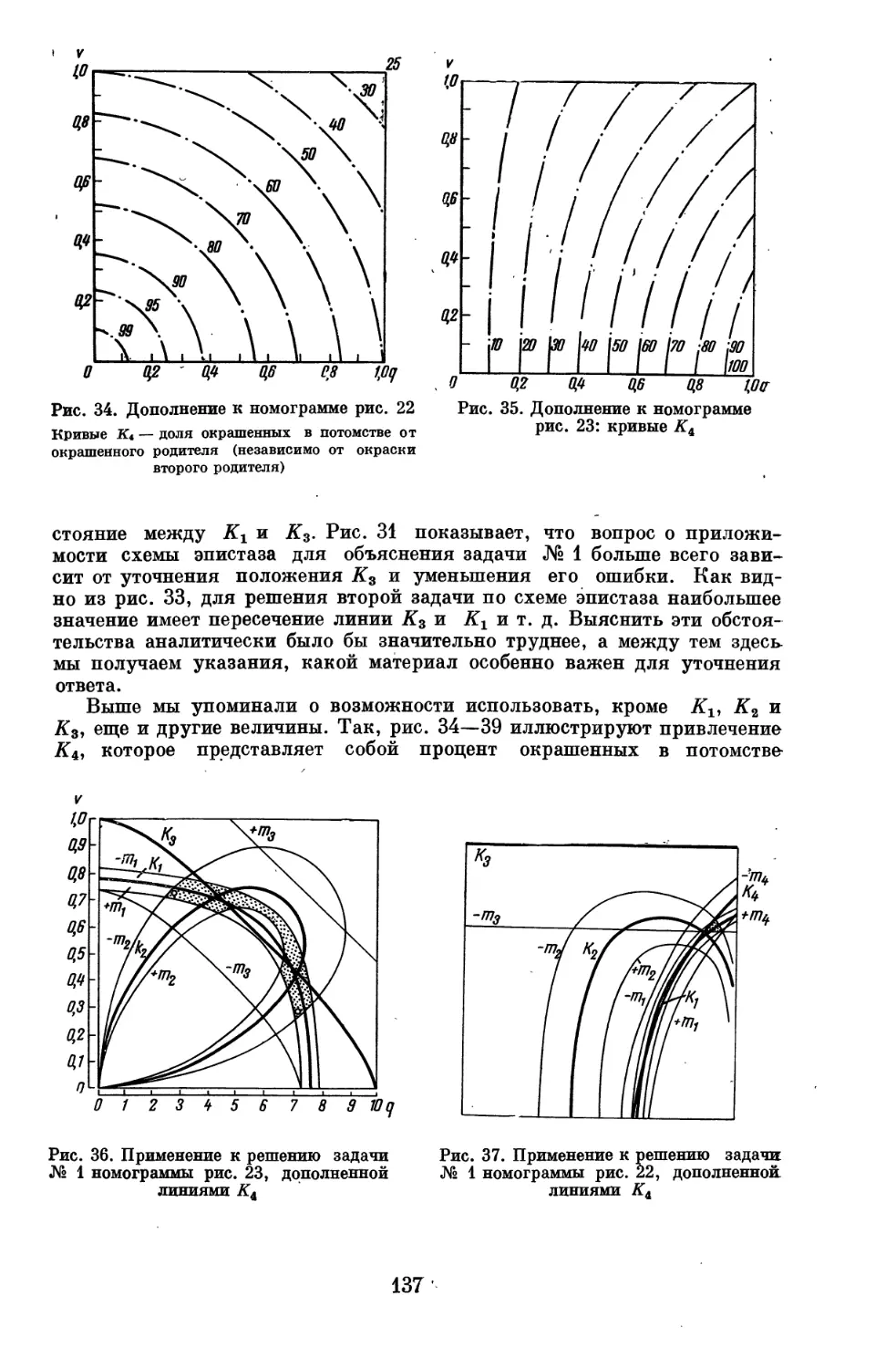






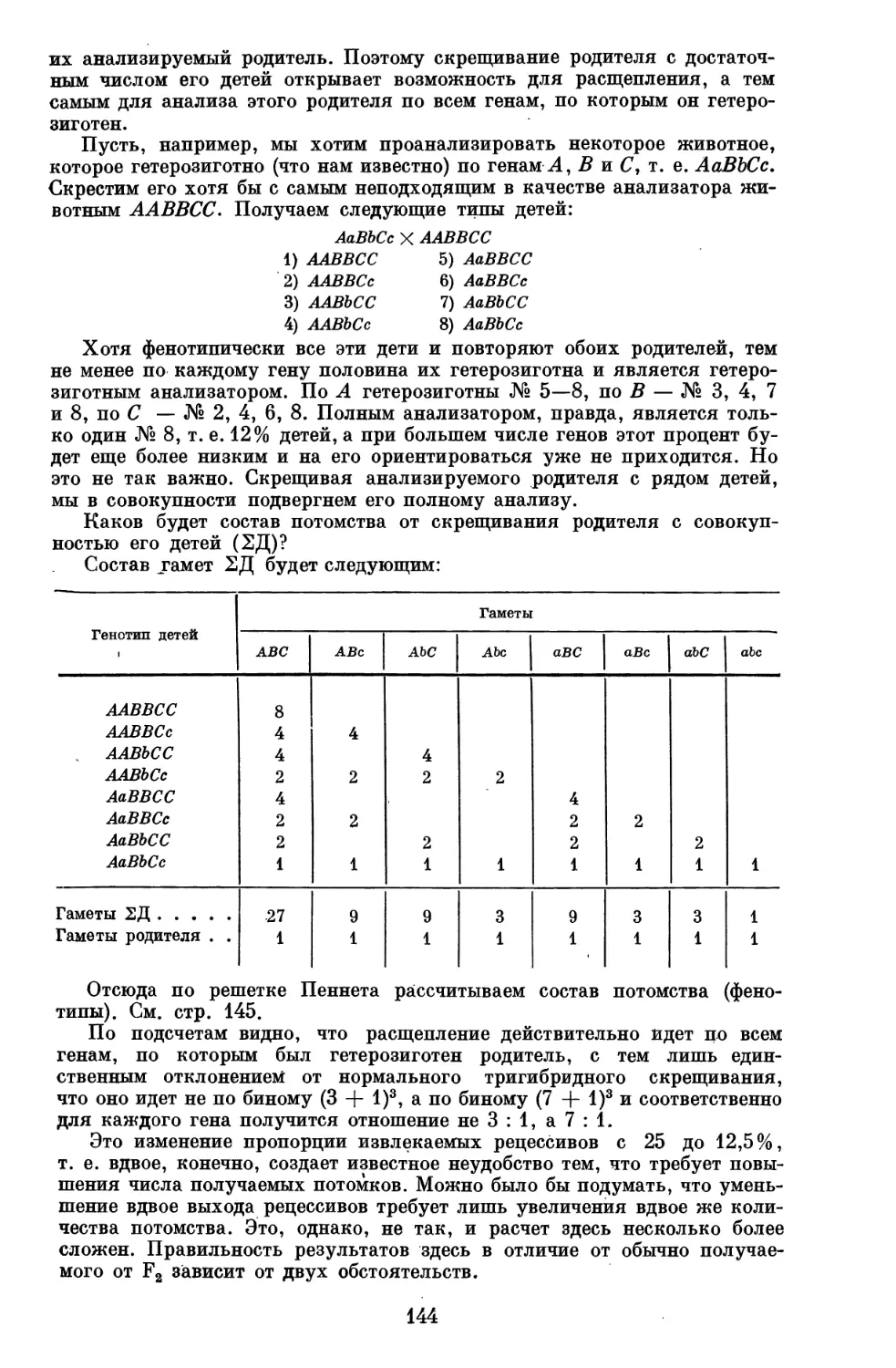





















































































































































































Author: Серебровский А.С.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез биология монография генетика селекция
Year: 1970




A
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ ОБЩЕЙ ГЕНЕТИКИ
А. С. СЕРЕБРОВШИЙ
ГЕНЕТИЧЕСКИЙ
АНАЛИЗ
в
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА1970
УДК 575 : 51
Генетический анализ. А. С. Серебровский. 1970 г.
Монография известного генетика А. С. Серебровского представляет
собой капитальный труд, над которым ученый работал много лет и
закончил его перед смертью. Не только в нашей, но и в зарубежной литературе
нет другой работы, где бы так всесторонне и оригинально были
рассмотрены сложные вопросы анализа наследственных особенностей
организмов. Ряд математических методов подобного анализа разработан А. С Се-
ребровским совершенно самостоятельно. Книга и теперь не утратила
значения фундаментального научного исследования.
Книга рассчитана на генетиков, селекционеров-исследователей, на
преподавателей генетики и селекции высших учебных заведений.
Иллюстр, 111, табл. 84.
Ответственный редактор
П. Ф.РОКИЦКИЙ
2-10-2
582-69 (I)
ПРЕДИСЛОВИЕ
Работая с 1925 г. над предлагаемой книгой, мы ставили перед собой
задачу дать более или менее систематическое изложение теории
генетического анализа, потребность в котором давно ощущается. Хотя
соответствующие материалы находятся почти в каждой генетической работе и
краткое изложение принципов генетического анализа дается во всех
курсах генетики, тем не менее систематического изложения теории
генетического анализа мы не имеем. К сожалению, это в значительной мере связано
и с тем, что в разработке самой этой теории имеются еще большие пробелы,
которые особенно велики в области анализа наиболее важных для
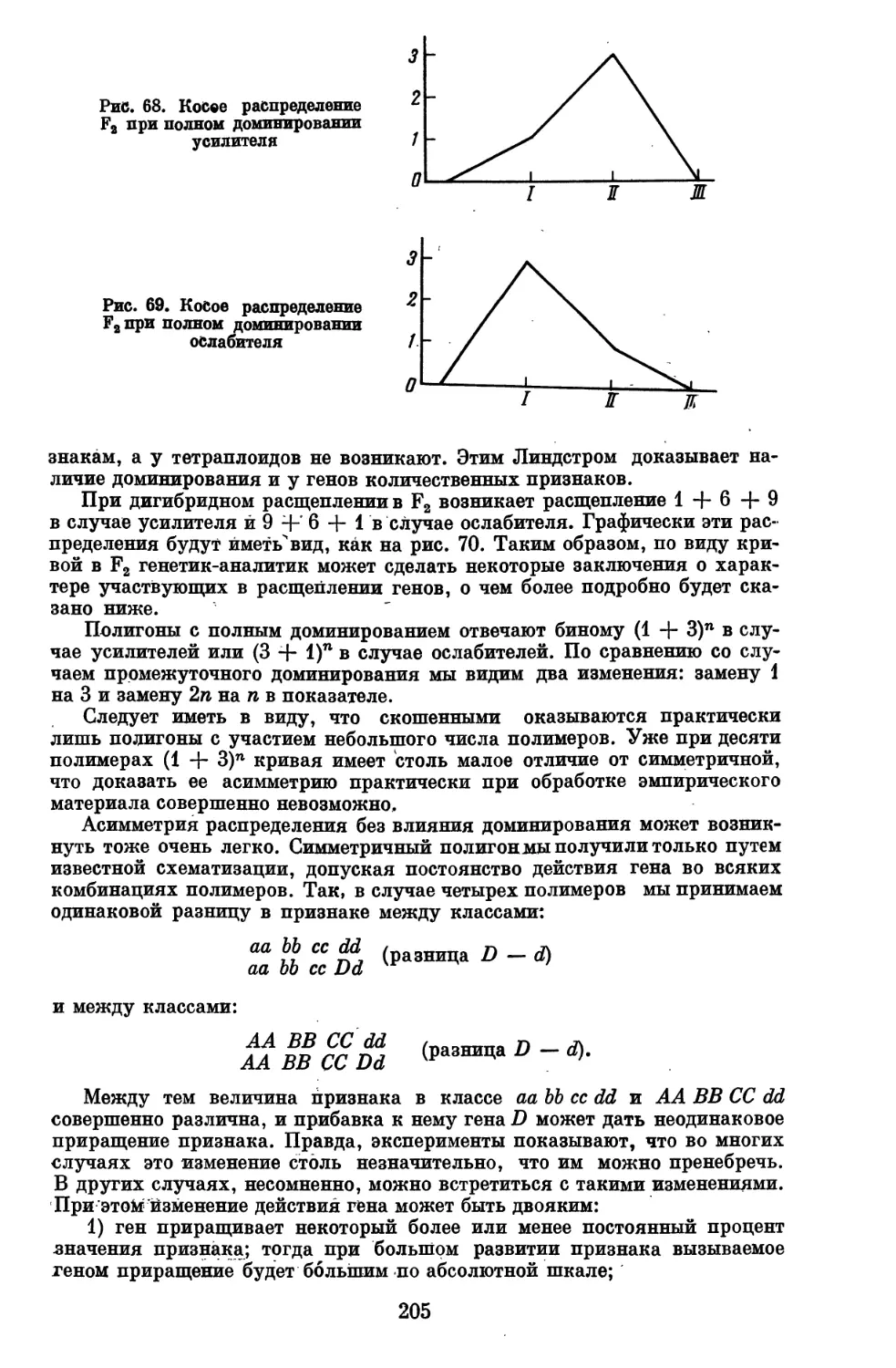
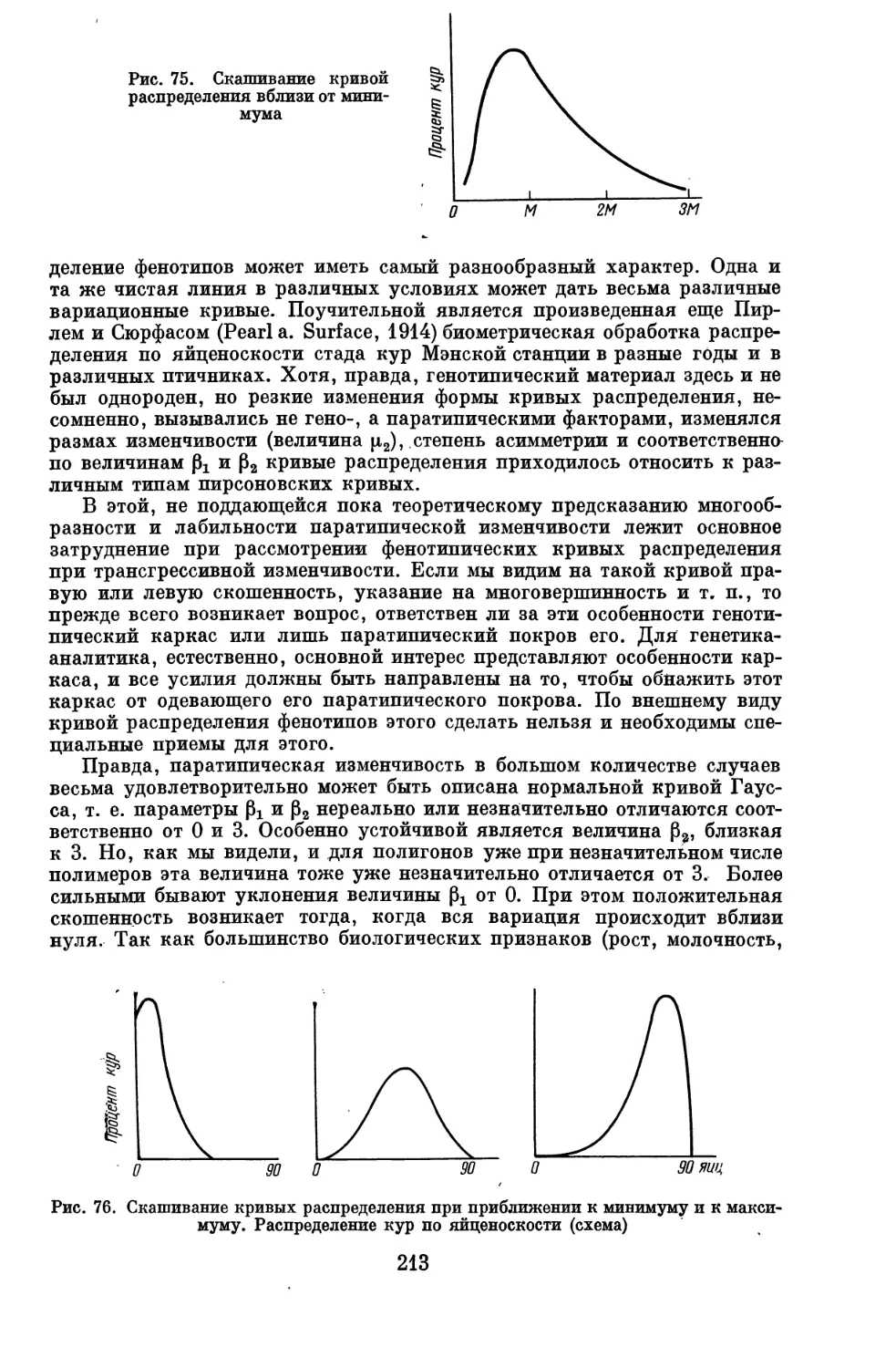
практики так называемых количественных (трансгрессивных) признаков.
Многие из этих вопросов нам пришлось разработать заново.
При построении курса генетического анализа можно было пойти по
пути составления компендиума и изложить конкретные результаты
генетического анализа различных животных и растений. От этого пути мы,
однако, отказались, так как он неизбежно привел бы к многотомной сводке
по частной генетике, в которой утонули бы принципиальные вопросы
генетического анализа. К тому же такие сводки по ряду животных и
растений уже имеются.
Мы избрали другой путь, на первый взгляд более трудный для
читателя, путь абстрагирования от конкретного материала частной генетики и
изложения теории генетического анализа как самостоятельной, в
значительной степени математической дисциплины, особенно во второй части книги.
При таком изложении становится яснее, что в данной дисциплине уже
сделано и что предстоит сделать, выявляются неразработанные участки и
прямые пробелы, из-за которых в ряде случаев мы еще не в состоянии решать
до конца задачи, особенно задачи анализа количественных и даже
качественных признаков в условиях популяций, с чем в основном и приходится
иметь дело генетику и селекционеру животных. Возможно, что у части
читателей это вызовет чувство неудовлетворенности. Но таково положение
дела и лучше заранее предостеречь от излишних надежд, чем
разочаровывать после.
Вместе с тем мы глубоко убеждены, что для развития генетического
анализа недостаточно одного накопления экспериментальных данных,
необходимо и абстрагирование от этого эмпирического материала, вплоть до
развития соответствующей математической теории. Только подобное
развитие генетического анализа позволит выработать приемы решения и
теории тех сложных задач, перед которыми мы сейчас вынуждены
останавливаться, не понимая, почему мы их не можем решить, какие условия
необходимы для этого, сколько решений возможно при данных условиях,
какие приемы должны привести к наиболее быстрому решению и т. д.
Нам остается отметить, что, исходя из общего плана книги, мы
оставили почти или вовсе без рассмотрения такие вопросы, как метод анализа
3
близнецов, некоторые методы, предложенные Райтом, и все вопросы
оценки суммарного генотипа и изучения ненаследственной изменчивости, как
входящие уже непосредственно в теорию селекции и неоднократно
излагавшиеся на русском языке в сводках С. Г. Давыдова (1936) и др. Мы
также почти не излагали основ генетики и биометрии, считая, что
интересующиеся вопросами генетического анализа уже знают эти дисциплины, по
крайней мере в объеме общих курсов.
Наиболее сложные (математические) методы генетического анализа
даются в виде дополнения, в которое включены: «Некоторые приемы
анализа геносвязи» М. И. Игнатьева, «Менделистическая алгебра» В. Гливен-
ко, «Метод треугольника» А. С. Серебровского.
Предлагаемые в приложении методы до широкого их использования в
практике генетического анализа нуждаются еще в дальнейшей разработке.
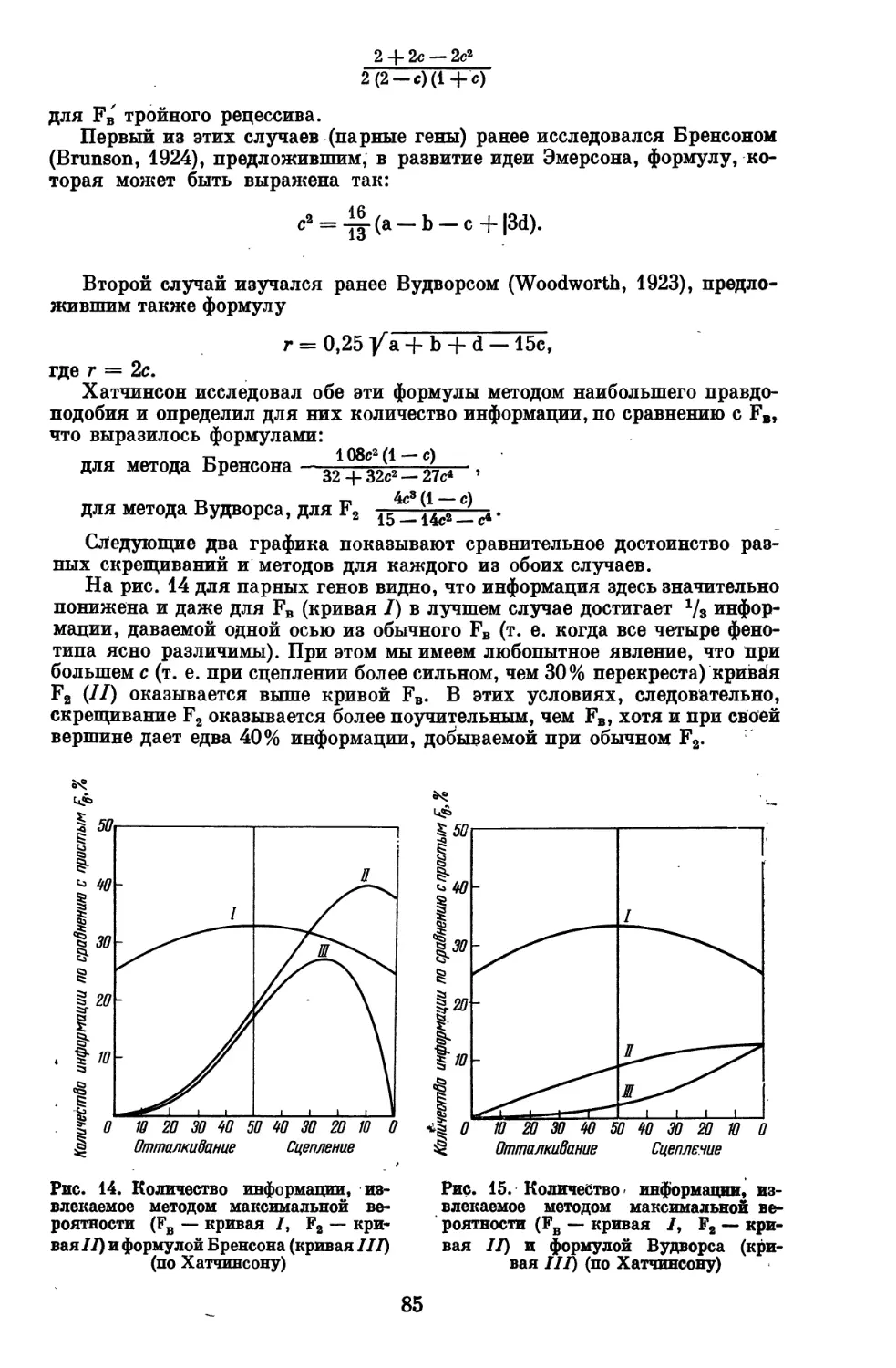
Выражаю глубокую благодарность товарищам, помогавшим мне в
подготовке рукописи к печати, в первую очередь Р. И. Серебровской,
М. В. Игнатьеву, А. А. Малиновскому и М. М. Местергази.
1 мая 1948 г. '
Автор
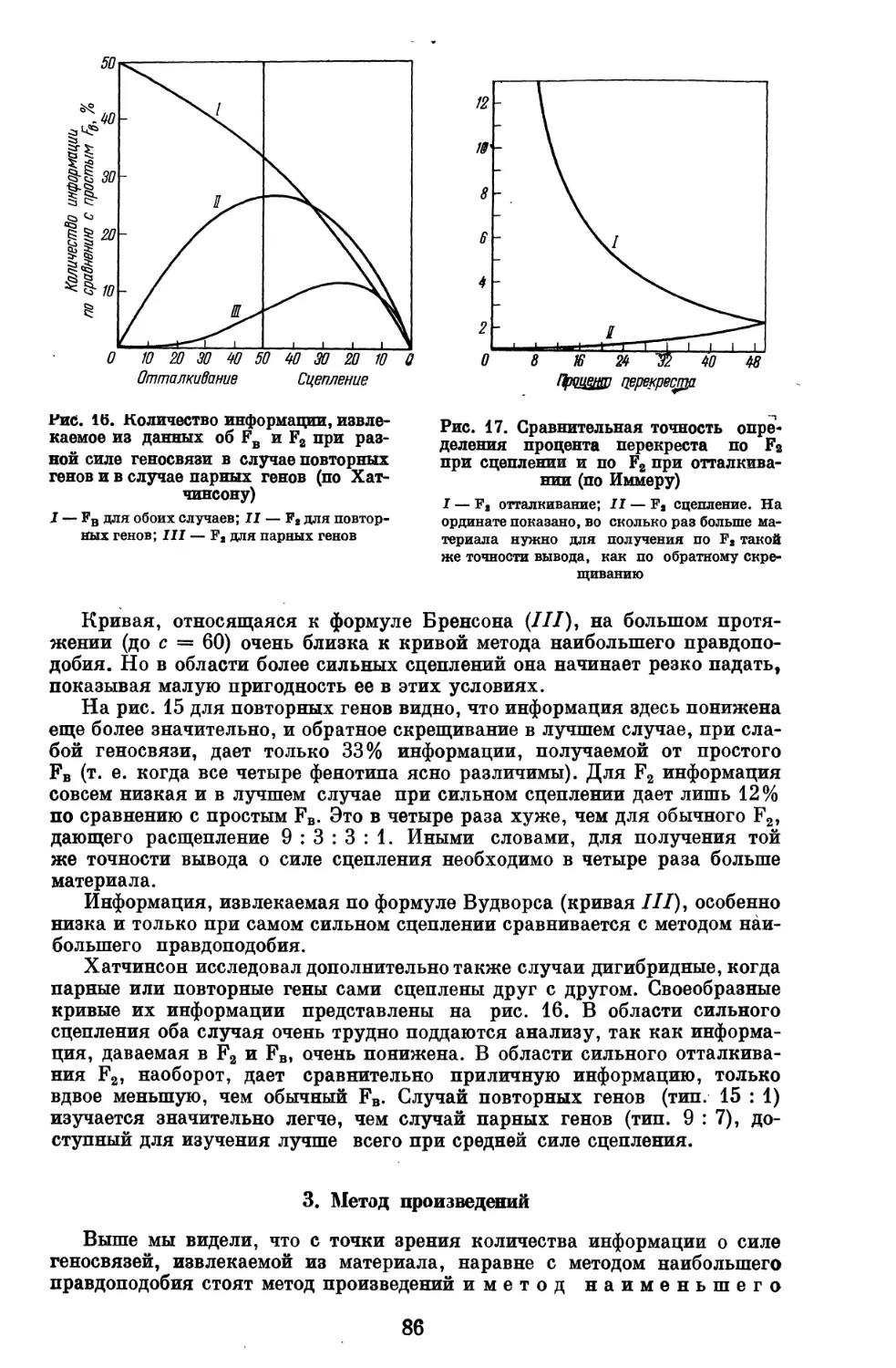
ВВЕДЕНИЕ
Генетическим анализом мы называем систему опытов, наблюдений и
вычислений, имеющих целью разложение свойств (признаков) организма
на отдельные наследственные элементы, «отдельные признаки», и изучение
свойств соответствующих им генов.
Генетический анализ начал разрабатываться после вторичного
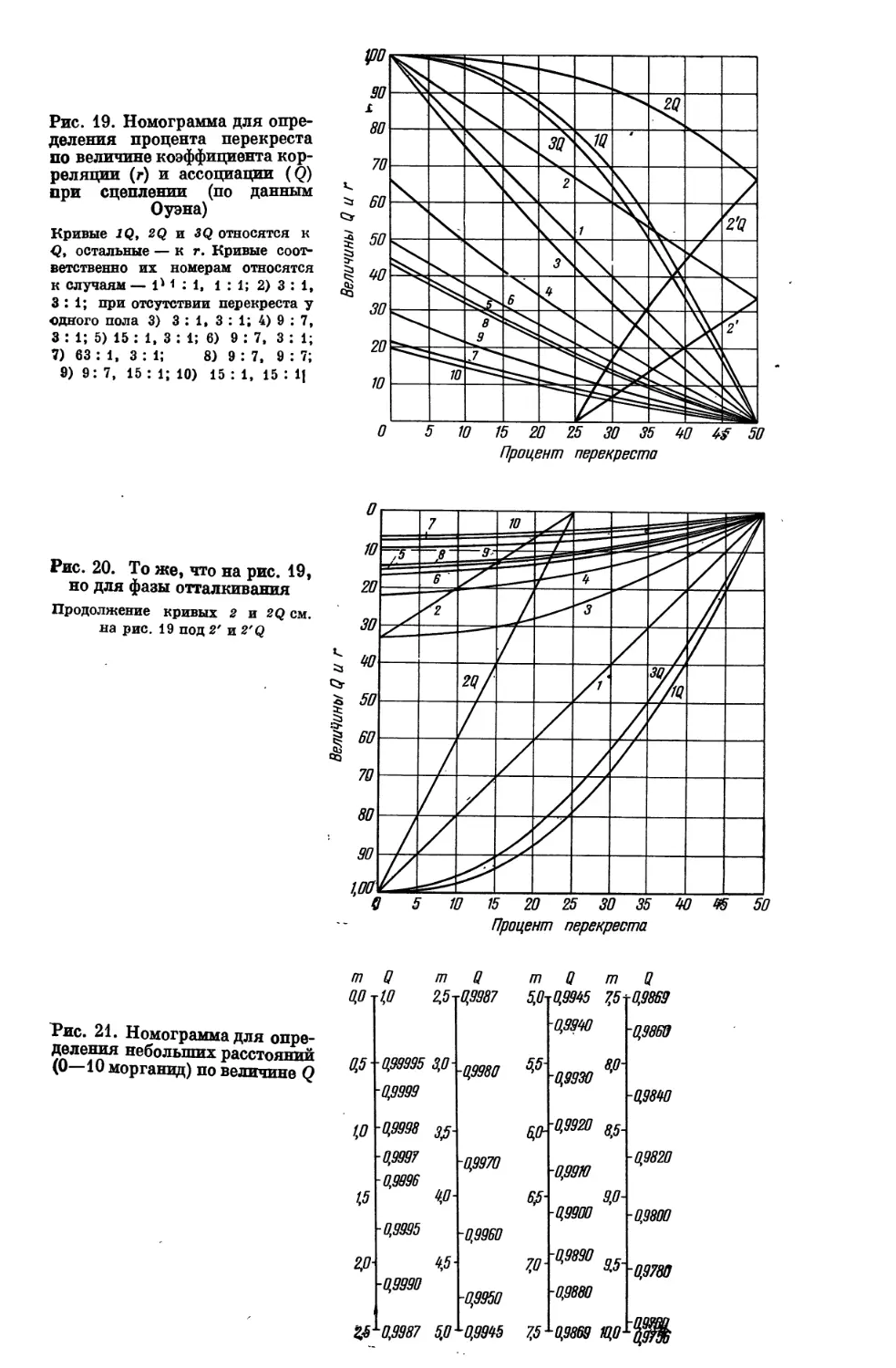
открытия менделизма и первыми генетиками-аналитиками надо считать у нас
Н. А. Юрасова и Е. А. Богданова, в Англии Бэтсона и Пеннета, во
Франции Кэно, в Германии Э. Баура и некоторых других исследователей
генетики кур, грызунов, львиного зева. Эпохальными фактами для биологии
явились такие открытия, что обычный «зоологический» признак — «серая
окраска мыши» — разлагается в новой, генетической плоскости на ряд
отдельных признаков: «наличие окраски» вообще, «наличие желтого
пигмента», «наличие пигмента, способного чернеть», «наличие усилителя
окраски», «наличие сплошного характера окраски» и т. д.
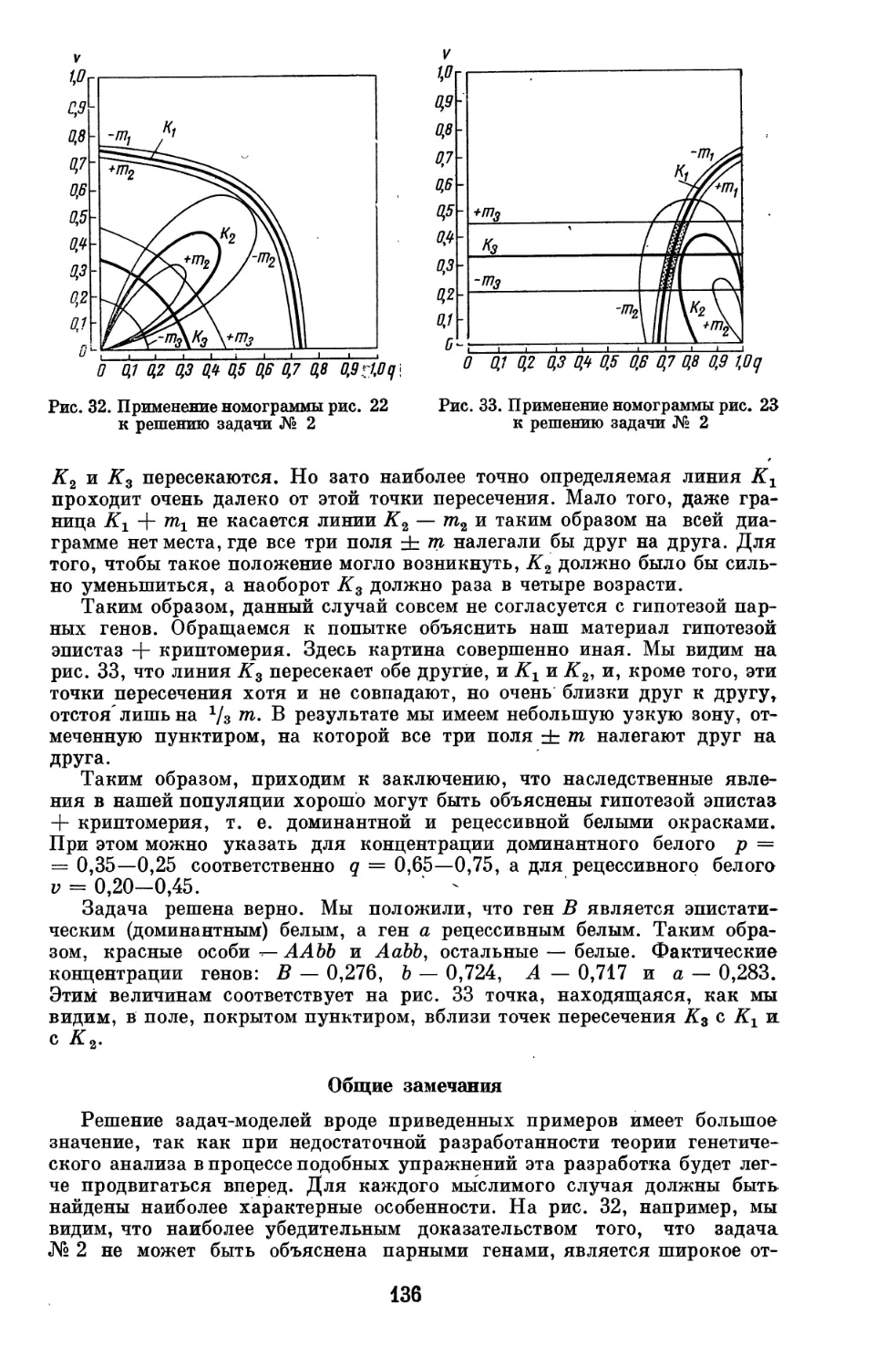
Подобно тому, как основой наших химических знаний являются данные,
добываемые химическим анализом, разлагающим вещества на составляющие
их простые элементы, так и в основе наших генетических знаний лежит
генетический анализ, разлагающий генотип организма на составляющие
его наследственные элементы. Как химик изучает свойства веществ и по
ним заключает об атомах, составляющих данное вещество, так и генетик,
наблюдая за наследованием признаков организма, заключает о
находящихся в его хромосомах генах. Каждому наследственному
различию в признаках соответствует различие хотя бы в одном гене, почему
часто упрощенно говорят, что признак вызывается геном. Наследственные
различия, соответствующие гену, мы называем феном. Однако связь
между феном и геном довольно сложна и обычно передается схемой
(схема I) *.
Схема I
Эта схема изображает зависимость фена не только от данного гена, но
и от среды, которая в свою очередь может быть разделена на среду
внешнюю и среду генотипическую, т. е. на совокупность всех остальных генов
организма, которые так или иначе влияют на признак. Например, мы
говорим, что длинные волосы кролика зависят от специального гена (v). Но
для того чтобы волосы выросли длинными, необходимо еще множество
* Объяснения ко всем схемам даются в тексте.
5
генов, без которых волосы вообще не могли бы расти и даже организм не
мог бы существовать. Кроме того, и внешние условия — холод, пища
и т. д.— способствуют или мешают росту.
Усвоение этой схемы полезно для того, чтобы избавиться от
метафизического представления о существовании строго постоянного фена,
отвечающего данному гену. Такого постоянства не только нет, но сплошь и
рядом один и тот же ген в различных условиях может играть совершенно
различную роль.
Ярким]примером этого может служить tifa-теяу кур. Введение этого
гена в генотип рыжей или глинистой курицы может превратить ее в черную.
Поэтому этот ген обычно называют «меланистическим». Однако тот же
ген, прибавленный * к генотипу особого рода золотистой курицы,
вызовет появление на каждом пере белого ободка, а прибавленный к генотипу
глинистой курицы с белыми пятнами — может дать сплошную белую
окраску.
rrv ГЧ ГЧ
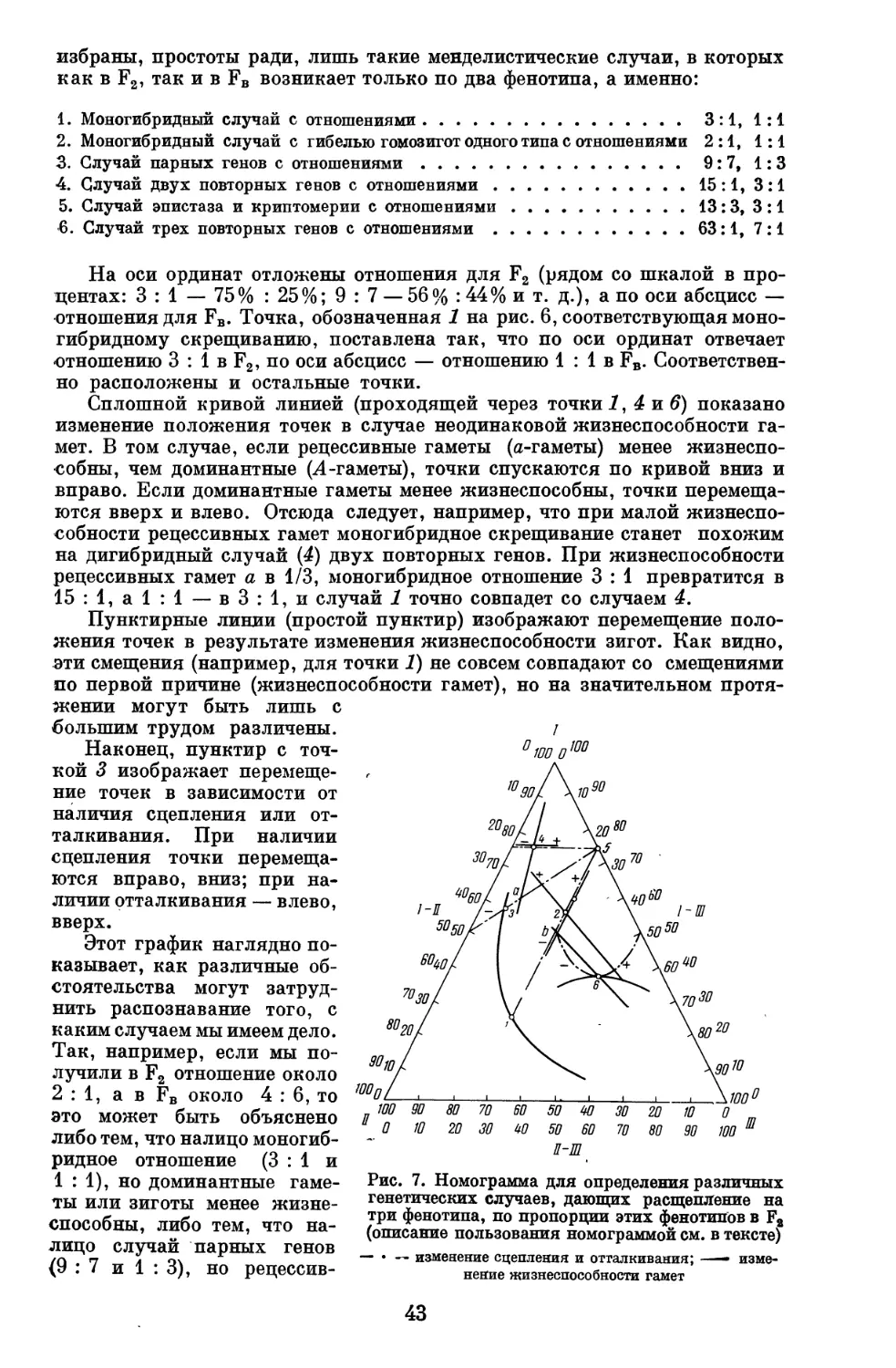
Рис. 1. Схема взаимоотношений фенотипа (q>),
генотипа (у), паратипа (я), голотипа (ri) и экзотипа (е)
Приведенная выше схема взаимоотношения между генотипом и
фенотипом несколько упрощена. В ней отсутствует существенный элемент,
который мы предложили назвать голотип ом (от 8Хоа — целый) **. Этим
термином мы обозначили структуру оплодотворенного яйца минус
качественный состав заключенного в ней генотипа. Таким образом в голотип
входит форма и размер яйца, относительное расположение в нем
различных частей, в том числе и положение ядра, количество, состав и
размещение питательных веществ в яйце и т. д. Голотип не может быть
противопоставлен генотипу, как цитоплазма — ядру, так как яйцо в целом
противостоит набору генотипов как части этого целого. Голотип важен тем, что
он является той отправной точкой, с которой начинается развитие
организма. Развитие организма и состоит в последовательном изменении голотипа
под действием внутренних и внешних факторов, которые соответственно
назовем эндотипом и эктотипом. Эндотип организма,
очевидно, состоит из (1) самого голотипа, являющегося производной фенотипа
матери, и (2) генотипа — набора генов, полученных как со стороны
матери, так и со стороны отца. Эктотип состоит из всех тех внешних
факторов, в которых проходит жизнь организма от начала его развития до того
момента, в который мы изучаем его фенотип. Наконец, генотип
противопоставляется паратипу, в который входят все остальные, кроме
генотипа, факторы онтогенеза.
Описанные взаимоотношения могут быть переданы схемой,
изображенной на рис. 1.
Отца можно рассматривать тоже как часть е, внесшего свой f в яйцо.
Для целей генетического анализа и теории селекции приходится часто
объединять голотип + эктотип под именем паратипа. С этой точки зрения
* «Прибавить ген» можно, конечно, не у данной курицы, а у ее потомства путем
соответствующего скрещивания и замены противоположного аллеля.
"■♦Предложенная А. С. Серебровским довольно сложная терминология не привилась.
В термине «голотип» нет надобности, так как свойства оплодотворенного яйца, как и
особенности всех последуюпщх стадий развития организма,— тот же фенотип.
Термины «паратип», «паратипические различия» теперь употребляют довольно редко.
Чаще говорят «средовые различия».— Прим. ред.
6
в фенотипических отличиях двух организмов мы различаем генотипиче-
ские и паратипические в зависимости от того, вызваны ли они гено- или
паратипическими факторами. В других случаях мы можем отмечать эндо- и
эктотипические различия.
Если оставить в стороне голотип, то генотип совпадает с эндотипом, а
паратип с эктотипом. Если это в целях упрощения иногда и допустимо, то
в таком случае необходимо помнить, что в паратипические факторы входит
и генотип предшествующего поколения, являющийся внешним фактором
по отношению к данному организму, а под именем генотипа данного
организма разумеется только часть генотипических факторов, действующих на
его развитие, именно та часть, которая заключена в самом данном
организме. Сказанное поясним примерами.
1. Два растения из чистой линии (или две морские свинки из линии,
проведенной через много поколений инбридинга) имеют одинаковый генотип.
Однако поставленные в различные условия (одно в хорошие, другое в
плохие) они развивают различные фенотипы, например, одно оказывается
крупным, другое — мелким. Это различие, таким образом, вызвано не
генотипом, и поэтому мы называем его паратипически обусловленным, или
паратипическим различием. Точнее, его следует назвать эктоти-
пическим, так как оно вызвано внешними факторами.
2. Две группы телят одинакового веса при рождении, выращенные на
ручной выпойке и в одинаковых остальных условиях, оказываются через
полгода различной величины. Крупные телята принадлежат к украинской
породе, более мелкие — к ярославской. Это различие, очевидно, зависит
от различий между генотипами этих пород, и мы называем его генотипи-
чески обусловленным, или генотипическим различием. Это
короткое выражение, однако, несколько двусмысленно, так как
генотипическим называют часто и различие между генотипами. Мы предлагаем
различие между генотипами называть генотипным различием (см.
ниже). Различие же между фенотипами — всегда фенотипическое
различие. Это фенотипическое различие может быть генотипически или
паратипически обусловленным (или обусловленным и тем и другим
вместе). Генотипически обусловленное мы называем обычно просто
генотипическим, прилагая при этом термин к различиям между генотипами лишь
благодаря отсутствию общепринятого термина, что часто ведет к
недоразумениям. Ниже мы дадим проект строгой терминологии, которая устраняла
бы возможность таких неясных выражений.
3. Две курицы с одинаковым генотипом находились: одна — за
стеклом, другая пользовалась прямым солнечным светом. Яйца их
инкубируются в одинаковых условиях, в одинаковых же условиях воспитываются
и цыплята. Однако цыплята первой курицы оказываются слабее, хуже
развиваются, чем цыплята второй. Между тем их генотипы и эктотипы
одинаковы. Разница объясняется тем, что эктотипы их матерей были различны,
и различными оказались голотипы — яйца первой курицы лишены
витаминов группы D. При сходстве эктотипов паратипы различны. Разница
в развитии цыплят является паратипической, а точнее, голотипической.
4. Завернутые вправо и влево моллюски являются наследственными
формами, но наследование это довольно своеобразно. Все дети одной
матери завернуты в одну сторону, даже если идет расщепление и генотипы
одних детей «левые», других — «правые». Это зависит, очевидно, от того,
что заворот определяется уже строением яйца, голотипом, и только в
следующем поколении устанавливается соответствие между фенотипом и
генотипом.
5. В некоторых скрещиваниях у дрозофилы потомство оказывается
нежизнеспособным, несмотря на то, что в генотипе самого этого
погибающего потомства ничего летального нет. Дело объясняется тем, что в
генотипе матери и в зародышевых клетках редукционного деления имелись гены,
7
повреждающие яйца, и хотя в результате редукционного деления эти гены
из яйца ушли,— яйцо оказывается нежизнеспособным. Таким образом,
эта гибель оказываетея голотипической природы, восходящей к генотипу
матери, и не имеет отношения к генотипу самого погибающего организма.
Терминология. Выше мы упомянули, что современная генетическая
терминология в части, касающейся генотипических, фенотипичеСких и
тому подобных различий, недостаточно выработана. Чтобы не затруднять
чтения книги лишней терминологией, мы воспользуемся возможностью
образования в русском языке двух форм окончания (генотипный и геноти-
пический и т. д.) и будем пользоваться следующей терминологией:
Фенотипические различия — различия между
фенотипами.
Генотипные различия — различия между генотипами.
Генотипические различия — различия между
фенотипами, обусловленные генотипными различиями, иначе, генотипически
обусловленные различия.
Паратипные различия — различия между паратипами (не
генотипными факторами онтогенеза).
Паратипические различия — различия между
фенотипами, обусловленные паратипными различиями.
Эктотипные различия — различия между эктотипами
(внешними средами).
Эктотипические различия — различия между
фенотипами, обусловленные различиями во внешней среде, и т. д,
В тех случаях, когда мы хотим обозначить различие между факторами
онтогенеза, мы будем употреблять окончание «типный», а в том случае,
когда захотим обозначать разницу между признаками, между эффектами
факторов,— окончание «типический». Генотипные различия выражаются
в генах, например, А и а, в числе генов-усилителей и т. п. Генотипические
различия, наоборот, выражаются в литрах молока, килограммах веса,
в окраске. В этих же мерах выражаются, очевидно, и всякие другие
«типические» различия: паратипические, эктотипические и т. д. Наоборот,
эктотипные факторы выражаются в градусах тепла, в силе света и т. д. и
несоизмеримы сами по себе ни друг с другом (температура и свет), ни с
генотипными факторами, но могут быть сопоставлены лишь через «типический»
эффект.
Поскольку предлагаемые нами термины «голотип» и «эктотип» еще не
стали употребительными, мы от их употребления в настоящей книге будем
по возможности воздерживаться, оперируя лишь терминами генотип,
паратип и фенотип. Повторим для ясности, что под генотипом мы разумеем
совокупность генов организма или клетки, или гаметы, под паратипом —
совокупность действующих на организм факторов онтогенеза, кроме гено-
типных (т. е. голотип + эктотип), под фенотипом — совокупность свойств
и признаков целого организма, взрослого или развивающегося.
Изучение гена в генетическом анализе прежде всего состоит в том,
чтобы указать, как проявляет себя данный ген, находясь в организме, т. е.
какое различие в признаках обусловливают его аллеломорфы. Из
предыдущего следует, однако, что фен гена может в разных случаях быть
весьма различным. При этих условиях наше знание о фене всегда имеет
несколько частный и условный характер, но мы с каждым новым
скрещиванием прибавляем что-нибудь к нашему знанию о гене. Часто оно
подвергается существенному пересмотру и уточнению, хотя дать исчерпывающее
описание действия гена мы, конечно, не можем, так как для этого его
необходимо изучить во множестве различных комбинаций и условий.
Приведем иллюстрацию из генетики морских свинок. Рыжие морские
свинки бывают двух сортов: с черными и с красными глазами. Легко
установить, что черноглазость доминирует над красноглазостью, и что здесь
8
имеется один ген, который обозначим через Е. Когда Е имеется, глаза
черные, в отсутствие 2?, т. е. у форм с генотипом ееч глаза красные. Мы можем
записать таким образом, что «ген Е вызывает черную окраску глаз».
Однако мы можем найти и серых морских свинок с красными глазами. Скрестив
их с рыжими черноглазыми, мы получим в F2 четыре типа свинок:
1) черные черноглазые (В, Е)\
2)][серые красноглазые (В, е);
3) рыжие черноглазые (6, Е)\
4) рыжие красноглазые (6, е).
Отсюда мы убеждаемся, сравнивая 1-й и 2-й типы, что ген Е влияет не
только на глаза, но и на окраску их шерсти, превращая серую в черную.
Эта окраска вызывается геном J9, так что Z?, Е — черная, J5, е — серая;
в отсутствие гена В ген Е влияет только-на глаза. Однако бывают свинки
рыжие с черными пятнами. Эти пятна вызываются геном V и легко
получить таких свинок с черными и красными глазами:
1) черноглазые, шерсть рыжая с черными пятнами;
2) красноглазые, шерсть рыжая^с черными£пятнами.
Отсюда заключаем, что ген Е действует не только при наличии i?, но и
при Ъ\ т. е. влияет на черный пигмент вообще, независимо от того,
покрывает ли он все тело или наблюдается только в виде пятен. Далее, скрестив
серую красноглазую свинку с коричневой черноглазой, мы сможем в F2:
получить четыре типа:
1) черные, черноглазые (Е, С)]
2) коричневые, черноглазые (Е, с);
3) серые, красноглазые (е, С);
4) светло-серые, красноглазые (е, с).
Отсюда мы заключаем, что ген Е не только превращает серую свинку в
черную (3-й тип в 1-й), но и светло-серую в коричневую (4-й во 2-й), т. е.
он вовсе не вызывает обязательно черную окраску, а только усиливает
окраску светло-серую до коричневой, серую до черной.
Каждое новое скрещивание прибавляет что-либо к нашему пониманию
действия гена, и теперь вместо того чтобы сказать — «ген Е вызывает
черную окраску глаз», мы должны сделать довольно длинное описание
гена Е как усилителя одного из типов окрасок, именно вызываемого тем
пигментом, который способен чернеть и который вызывается генами Bub'*
Но, конечно, появление какой-нибудь новой мутации потребует, может
быть, дальнейших дополнений к этому описанию.
Признаки качественные и количественные. Все признаки и различия
организмов генетики делят на качественные и
количественные. Различия между черной и рыжей коровой, между рысаком и
иноходцем, между рогатым и комолым скотом являются примерами
качественных различий. Различия между курами по яйценоскости, по росту
и др.— различия количественные.
Это разделение исторически вызвано тем, что генетический анализ
качественных различий, как правило, несравненно легче. Анализ
количественных различий, наоборот, как правило, настолько труден, что генетики
долгое время спорили о том, подчиняются ли количественные признаки
тем же менделевским законам, как и качественные, или нет. В настоящее
время установлено, что различий между теми и другими в способе
наследования не существует; как те, так и другие могут быть разложены на
генетические составные части (фены) с отвечающими им генами. Однако изучение
количественных генов технически несравненно труднее и требует
специальной методики, почему мы будем рассматривать генетический анализ
качественных и количественных признаков отдельно.
9
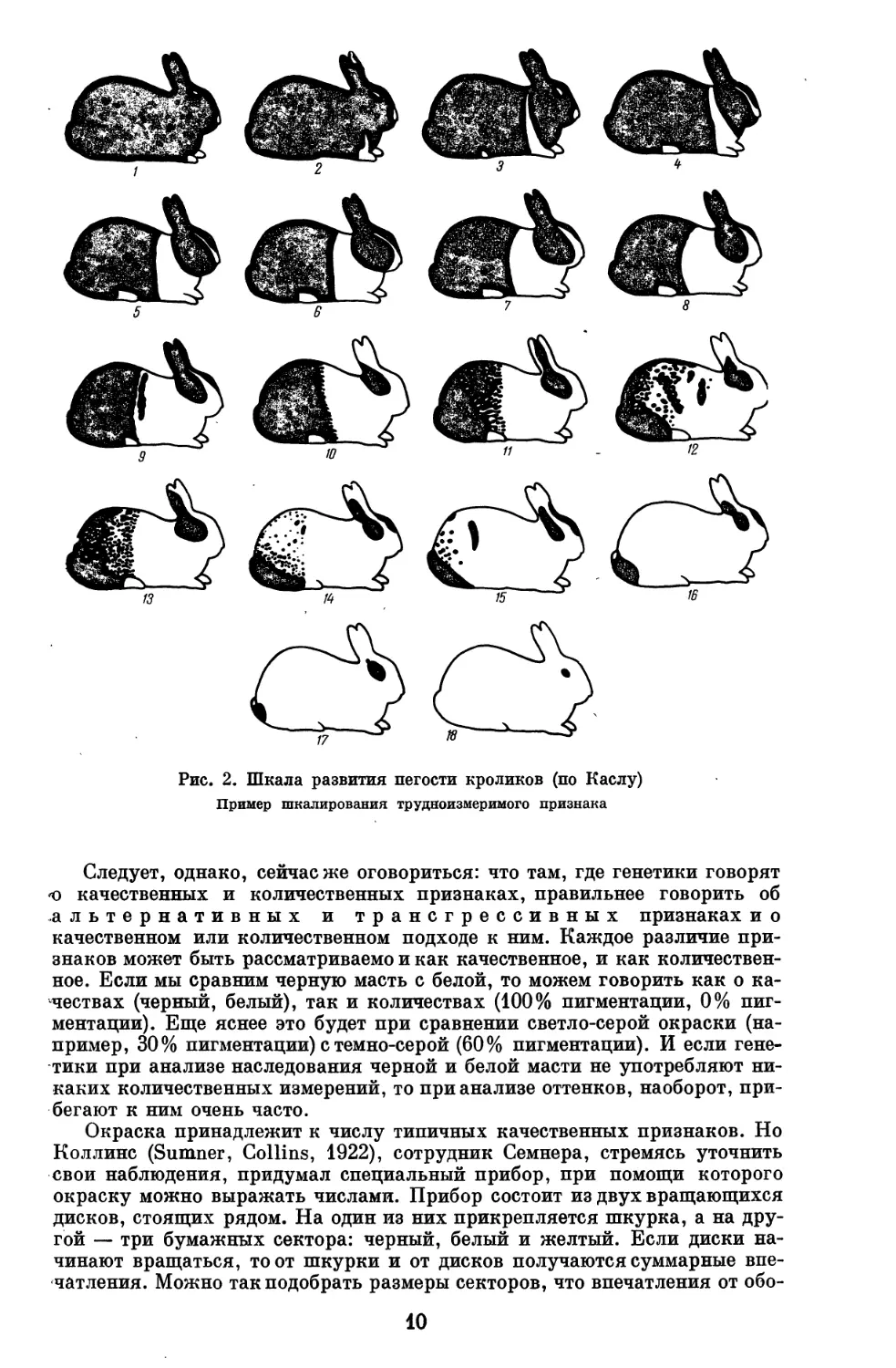
Рис. 2. Шкала развития пегости кроликов (по Каслу)
Пример шкалирования трудноизмеримого признака
Следует, однако, сейчас же оговориться: что там, где генетики говорят
ю качественных и количественных признаках, правильнее говорить об
альтернативных и трансгрессивных признаках и о
качественном или количественном подходе к ним. Каждое различие
признаков может быть рассматриваемо и как качественное, и как
количественное. Если мы сравним черную масть с белой, то можем говорить как о
качествах (черный, белый), так и количествах (100% пигментации, 0%
пигментации). Еще яснее это будет при сравнении светло-серой окраски
(например, 30% пигментации) с темно-серой (60% пигментации). И если
генетики при анализе наследования черной и белой масти не употребляют
никаких количественных измерений, то при анализе оттенков, наоборот,
прибегают к ним очень часто.
Окраска принадлежит к числу типичных качественных признаков. Но
Коллинс (Sumner, Collins, 1922), сотрудник Семнера, стремясь уточнить
свои наблюдения, придумал специальный прибор, при помощи которого
окраску можно выражать числами. Прибор состоит из двух вращающихся
дисков, стоящих рядом. На один из них прикрепляется шкурка, а на
другой — три бумажных сектора: черный, белый и желтый. Если диски
начинают вращаться, то от шкурки и от дисков получаются суммарные
впечатления. Можно так подобрать размеры секторов, что впечатления от обо-
10
их дисков будут одинаковыми. Тогда окраска шкурки выразится тремя
цифрами, выражающими в процентах участие трех цветов, например, 65%
черного, 20% белого, 15% желтого. Недостатком этого метода является
трудоемкость его. Семнер поэтому перешел к использованию фотометра.
Райт (Wright, 1917) употреблял шкалу из набора шкурок от белой через
серые до черной. Касл (Castle, 1919) составил шкалу развития пегости
кроликов (рис. 2).
С другой стороны, такой типичный количественный признак, как рост,
может в удобных случаях выступать как качественный признак («высокий
горох» и «низкий горох» в известном опыте Менделя). В значительной
степени от самого исследователя зависит, рассматривать ли признак
качественно или количественно и пользоваться одним или другим методом
анализа *.
Наиболее существенным различием признаков является, таким
образом, не «качество» или «количество», а наличие более или менее резкой
границы между ступенями развития признаков. Разница между черным и
белым, высоким и низким столь ясна, что никакого измерения не нужно
для того, чтобы характеризовать признак. В этих случаях генетический
-анализ идет легко и требуется только придумать необходимые
скрещивания для того, чтобы получить точный вывод. Если же признаки
непостоянные, трансгрессивно-изменчивы (например, разная степень молочности,
яйценоскости), то признаки приходится рассматривать как
количественные и прибегать в их анализе к соответствующей методике.
Задачи генетического анализа. Задачи генетического анализа могут
быть различными. Прежде всего может быть поставлена задача
подвергнуть генетическому анализу различие между двумя особями или породами,
выяснить, сколькими генами обусловлено это различие, и изучить
свойства этих генов. Здесь можно различить следующие частные задачи:
1. Установление гена.
2. Изучение его свойств путем изучения его действия на признаки в
различных комбинациях с другими генами.
3. Установление сцепления или отталкивания гена с другими генами,
ранее установленными.
4. Определение расположения гена среди других сцепленных с ним.
Первая задача может быть разрешена наблюдениями за наследованием
одного лишь данного гена.
Вторая и третья задачи требуют одновременного наблюдения за
наследованием не менее двух генов в одном и том же скрещивании.
Третья задача решается наиболее совершенно одновременным
наблюдением не менее как за тремя генами, специально подобранными для
этого.
Кроме различения и изучения гена, перед генетическим анализом
ставится задача анализа генотипа данной особи (например, генотипа данного
барана производителя) или данной породы (анализ леггорнов), или даже
вида (например, генетический анализ дрозофилы), или какой-либо
популяции (например, популяции кур Кавказа). Каждая из этих задач имеет
«вой специфические особенности и требует особых приемов, о чем будет
подробно говориться в соответствующих местах книги.
Необходимо подчеркнуть важность применения генетического анализа
в селекционной практике. Так, советскими селекционерами
животноводами разработан метод разведения серых смушковых овец путем
генетического анализа серой окраски (Глембоцкий, 1935; Васин, 1933). Было
установлено, что серая окраска вызывается доминантным геном, обладающим
плейотропным действием: в гомозиготном состоянии этот ген влияет на
* Сейчас во всех сводках по генетике количественных признаков разбирается вопрос
о выборе необходимой шкалы для каждого признака (иногда это может быть
логарифмическая шкала).— Прим. ред.
11
перистальтику желудка, что приводит к тяжелому заболеванию и смерти*
Выявление этого факта не только объяснило причину невозможности
закрепления серой окраски у смушковых овец, но и причину огромного
падежа ягнят, наблюдающегося в течение многих лет в хозяйствах,
разводящих этих овец и скрещивающих серых с серыми. При скрещивании же
гетерозиготных серых маток с черными баранами и, наоборот, черных
маток с серыми гетерозиготными баранами рождается только 50% серых
ягнят, но гибель ягнят полностью ликвидируется.
Селекция на тот или другой тон серой окраски смушки основывается на
наличии ряда генов-модификаторов и достигается путем использования
для скрещивания с серыми таких черных животных, которые сами
произошли от серого родителя желательного оттенка. Изложенный метод
разведения *серь$х смушковых овец, целиком основанный на использовании
данных генетического анализа, применяется в настоящее время во всех
хозяйствах, разводящих этих овец.
Другим примером использования генетического анализа для селекции
может служить борьба с крипторхизмом у тонкорунных овец породы
прекос (Глембоцкий, 1941). В племенных совхозах, разводивших эту породу,
ежегодно рождалось от 10 до 25% бесплодных крипторхов, что наносило
большие убытки племенным хозяйствам и снижало темпы метизации.
Генетический анализ установил, что крипторхизм у прекосов вызывается
рецессивным геном, тесно сцепленным (перекрест 4%) с доминантным геном
комолости.
Селекция на комолость, проводившаяся в течение многих лет у
прекосов, повлекла за собой сильнейшее распространение крипторхизма, но не
достигла своей основной цели — закрепления комолости, так как почти
все гомозиготные по комолости бараны оказывались крипторхами, т. е.
бесплодными, а все или почти все плодовитые комолые бараны были гете-
розиготами.
Я. Л. Глембоцким был предложен метод борьбы с крипторхизмом у
прекосов, исходя из данных генетического анализа этого признака.
Рогатые производители скрещивались с матками, имеющими роговые зачатки
(гомозиготы по гену рогатости), а рогатые производители — с матками, не
имеющими роговых зачатков. При подобном методе подбора рождаются
в равном количестве ненормальные и рогатые бараны, крипторхизм же
полностью исчезает.
Приведенные примеры с достаточной убедительностью показывают
исключительную важность генетического анализа для животноводческой
практики.
Необходимо иметь, однако, в виду, что для разных
сельскохозяйственных животных могут быть использованы отдельные элементы этого
анализа. Другими словами, не все излагаемые нами приемы генетического
анализа в равной степени применимы для различных объектов. Так,
несомненно, что топографический метод генетического анализа имеет больше шансов
быть плодотворно использованным, скажем, у шелкопряда, чем у каких-
либо крупных животных. То же можно сказать и о хромосомном анализе
и т. д.
С этой оговоркой именно и нужно давать оценку перспективности
различных методов генетического анализа.
Метод генетического анализа. Основным приемом химического анализа
является введение испытуемого вещества в реакцию, во время которой
молекулы вещества разлагаются на отдельные радикалы, группы атомов
и т. д. Подобным же образом генетик, желая узнать генетический состав
данного организма, вводит его в скрещивания и получает от него потомство.
При известных условиях признаки производителя как бы рассыпаются на
отдельные составляющие их элементы: одни потомки получают одни
признаки, другие потомки —- другие признаки. Изучение этого потомства, под-
12
•счет числа получивших данный признак и не получивших его позволяют
ответить на вопросы, интересующие генетика-аналитика.
Несмотря на принципиальное отличие организма от химического
соединения, механизмы редукционного деления, перекреста и
оплодотворения создают почву для большого сходства метода работы химика и
генетика. Это сходство особенно велико в логике построения тех выводов, ко-
тррые делают оба эти исследователя. Ввиду этого формального сходства
полезно подчеркнуть некоторые различия, которые делают работу генетика-
аналитика несравненно более трудной. Различие это состоит прежде всего
в том, что химик имеет возможность выделить свои элементы в чистом виде
и изучить свойство этих выделенных элементов -=- кислорода, водорода,
меди и т. п. Генетик лишен этой возможности, так как организм способен
жить, только обладая более или менее полным набором разнообразных
тенов, и немыслимо получить, например, морскую свинку, имеющую в
своем генотипе одни только гены Е. Поэтому мы находимся в положении
такого гипотетического химика, который вынужден был бы изучать свои
элементы, только сравнивая между собой различные, очень сложные вещества,
не имея возможности разложить их на простые или хотя бы значительно
их упростить.
Впрочем, на заре органической химии и химики находились в известной
мере в таком положении и вынуждены были оперировать многочисленными
радикалами, не зная их строения.
С этим связано и второе отличие наших работ от работ химика — мы не
имеем «чистых реактивов». В самом деле, если нужно подвергнуть
генетическому анализу курицу породы леггорн, то следует скрещивать ее с
другими породами — род-айленд, фавероль, бентамкой, дикой курицей и т. д.
Но генотипы этих пород тоже не известны, или известны лишь частично.
Поэтому в процессе анализа породы леггорн приходится анализировать
генотипы и всех других пород, которые используются в качестве
«реактивов». Ясно, что это чрезвычайно осложняет работу.
ЧАСТЬ ПЕРВАЯ
ГЕНЕТИЧЕСКИЙ АНАЛИЗ
КАЧЕСТВЕННЫХ (АЛЬТЕРНАТИВНЫХ) РАЗЛИЧНЕЕ
ГЛАВА I
МОНОГИБРИДНОЕ РАСЩЕПЛЕНИЕ.
УСТАНОВЛЕНИЕ ГЕНА
ЧИСЛО ГЕНОВ В РАСЩЕПЛЕНИИ
Изучая какое-либо скрещивание, мы наблюдаем, что потомство рас-
падается на различные фенотипные классы, или классы фенотипов,— одни
оказываются серыми, другие черными или белыми и т. д. На большом
материале выясняется, что численности этих классов находятся в более или
менее простых кратных отношениях друг к другу, установление чего и
составило величайшее открытие Менделя, по своему значению для биологии
сопоставимое с открытием Дальтоном закона кратных отношений в химии.
Так, потомство может распасться, например, на серых и черных в
отношении 3 : 1 или 1 : 1 или на серых, черных и белых в отношении
9 : 3 : 4 и т. д.
На основании этих менделевских отношений мы делаем заключение о
том, сколькими генами различались взятые для скрещивания исходные
родители. Часто говорят при этом, например, что в скрещивании участвует
один ген, два гена, и что нам удалось определить число генов,
участвующих в расщеплении. Что разумеется под этими словами? При каждом
наследовании от родителей к детям переходит громадное количество генов.
Однако, если мы скрещиваем две особи
AABBCCDDEE... X aabbCCDDEE...,
то гены С, J9, 2?..., присутствующие у обоих родителей в гомозиготном
состоянии, перейдут к 100% потомства. О таких генах говорят, что они в
данном скрещивании не менделируют или не принимают участия в
расщеплении. Наоборот, А и В в дальнейших поколениях будут менделироватьг
так как по этим генам скрещиваемые родители различаются.
Если исходные родители различаются по одному гену, то в дальнейшем
потомство будет расщепляться по так называемой моногибридной формуле.
При различии родителей по двум генам расщепление пойдет по дигибрид-
ной формуле, при трех генах — по тригибридной и т. д. Каждая из этих
формул имеет свое приложение в генетическом анализе, и мы должны
начать с подробного изложения моногибридной и дигибридной формул.
ИЗУЧЕНИЕ ОТДЕЛЬНОГО ГЕНА
В этой первой задаче изучение отдельных генов скрещивания нужно
вести так, чтобы в результате получить по возможности картину
моногибридного скрещивания. При скрещивании двух особей, различающихся па
14
одному признаку, скрещивание идет по схеме:
Р ЛАх ла
Fi Аа
F2 AA + 2Аа + аа
Как только это достигнуто и различные проверки показывают, что
перед нами действительно моногибридное скрещивание, наша
задача решена, и мы можем говорить, что в нашем опыте менделирует один
ген, относящийся к изучаемому различию. После этого мы можем
описать рецессивную форму, доминантную и гетерозигот. Однако нужна
иметь в виду, что абсолютного доказательства моногибридного характера
наследования быть не может. Всегда возможно сомнение, что перед нами не
один, а два гена, сильно сцепленных. Увеличением числа изученных
потомков вероятность обнаружения моногибридности непрерывно
повышается.
Как потомство в моногибридном скрещивании распадается на классы
фенотипов, зависит от того, будем ли мы заранее, априори, знать, что
родители гетерозиготны или же узнаем об их гетерозиготности апостериорно,
по их потомству. Конечно, наше знание не может повлиять на процесс
расщепления. Но так как для суждения о скрещивании мы часто собираем
статистический материал, то, как сейчас увидим, наше знание отражается
на выборе материала. Поэтому мы должны рассмотреть как априорные
моногибридные формулы, так и апостериорные. Последние особенно часто
находят себе применение при анализе домашних животных.
АПРИОРНЫЕ МОНОГИБРИДНЫЕ ФОРМУЛЫ
Наиболее популярной является моногибридная формула второго
поколения, по которой при скрещивании особей Аа X Аа их потомство
распадается на два фенотипа, А и а, в отношении ЗА : 1а в случае полного
доминирования, или на три фенотипа в отношении 1АА : 2Аа : \аа в случае
неполного доминирования, при котором удается различить по фенотипу вс&
три генотипа.
Однако часто значительно более удобным является получение не второго
поколения (F2) с отношением 3 : 1, а так называемого возвратного, или
обратного скрещивания (FB), с рецессивным родителем по схеме:
Аа X аа-+- Аа : аа.
Здесь возникай^ две категории А а и аа в равных количествах,
благодаря этому получается точная статистическая оценка результата (рис. 3)..
Этот тип скрещивания удобен тем, что его легко продолжать из
поколения в поколение, скрещивая или обе категории потомства одну с другой,
или доминантную форму с рецессивным родителем.
При таком способе потомство каждый раз распадается на две численно-
разные группы, и этот результат должен повториться из поколения в
поколение. Если через несколько поколений мы замечаем, что результат
скрещивания все тот же, и тем самым убеждаемся, что перед нами
действительно моногибридное скрещивание, мы можем суммировать результат
всех поколений и получить более или менее обширное итоговое число
для дальнейшей оценки, о которой скажем ниже.
Употребление второго поколения в этом отношении невыгодно,
особенно в случае полного доминирования. Из числа получающихся в F2 3i4 + la
особей с фенотипом А мы уже не можем брать для дальнейшего опыта, так
как не можем априори знать, будет ли взятая особь иметь генотип А А или
Аа. Решение же вопроса о гомо- или гетерозиготности особи по ее
потомству часто может быть затруднительно, особенно при малой плодовитости
15
Aa
-act Aa-
Aa-
Aa-j-aa
итЛ
-aa
—i aa Aa
-aa
A
/\
aa Aa-
-aa
итЛ
Рис. З. Две схемы обратных
скрещиваний типа А а X ад на
протяжении ряда поколений
крупных животных. Для них такое апостериорное решение относительно их
гетерозиготности достижимо только для самцов, от которых получается
«более 8—10 потомков. Если же, скрещивая черную корову с рыжим
быком, мы получаем трех черных телят, то это еще не доказывает гомозигот-
ность коровы по черной окраске. В самом деле, гетерозиготная черная
корова при скрещивании с рыжим*быком может дать трех телят следующими
равновероятными способами:
-й теленок
черный
черный
черный
черный
2-й теленок
черный
черный
рыжий
рыжий
3-й теленок
черный
рыжий
черный
рыжий
1-й теленок
рыжий
рыжий
рыжий
рыжий
2-й теленок
черный
черный
рыжий
рыжий
3-й теленок
черный
рыжий
черный
рыжий
Каждый из этих способов равновероятен; следовательно, из каждых
восьми гетерозиготных коров одна даст подряд трех черных телят, и мы,
же имея указаний на гетерозиготность, отнесем ее к числу гомозиготных.
Мы, таким образом, в 12,5% случаев сделаем ошибку. Этот процент
достаточно велик и с ним нельзя не считаться.
АПОСТЕРИОРНЫЕ МОНОГИБРИДНЫЕ ФОРМУЛЫ
Так как все-таки мы часто оказываемся вынужденными пользоваться
особями из F2 (или особями с-неизвестным происхождением), относительно
которых вопрос об их гомо- или гетерозиготности решается только апосте-
риорно, то необходимо иметь в виду, что при этом менделевские ожидания
будут искажаться. В разобранном нами случае скрещивание шло по
формуле:
черные коровы рыжий бык
Аа X <м>
и по априорной формуле менделизма мы должны были ожидать 50%
черных телят и 50% рыжих. Но, выбирая гетерозиготных коров по их
потомству, мы выбросили из восьми коров одну, случайно давшую трех черных
телят подряд, и признали гетерозиготными только остальных семь коров
(87,5%). Подсчитав их потомство, мы получаем: 9 черных (43%) + 12
рыжих (57%), т. е. рыжих значительно больше, чем черных.
При большом количестве исследованных коров мы можем усмотреть
в такой систематической нехватке черных телят намек на их меньшую
жизнеспособность и т. п., что, очевидно, будет ошибкой.
Поэтому при апостериорном определении гетерозиготности мы можем
заменить менделевское отношение 1 : 1 таким:
(*-*)=
1 или, иначе
' \ 2n-i "г"-!/'1
16
где п есть число потомков, получаемых от каждой из данных особей, а N —
общее число изученных потомков.
В данном случае п = 3 и 2п — 8, откуда ожидаемое отношение 3/4 : 1
или по второй формуле 9 : 12.
Если число потомков у разных особей различно, то средним числом
потомков пользоваться нельзя, расчет будет не совсем точным.
Правильнее разбить всех производителей на отдельные группы по числу потомков
(1, 2, 3 и т. д.), для каждой группы по указанной формуле вывести
ожидаемое отношение и затем полученные отношения суммировать (с учетом
весов).
Пример. От четырех подозреваемых в гетерозиготности черных коров
получено по два теленка и от восьми — по три. Вычислить ожидаемое
отношение черных и рыжих телят в потомстве коров, которые подтвердят
свою гетерозиготность, при гипотезе участия 1 гена.
Вычисляем отдельно:
1. Для четырех коров по два теленка.
Из четырех гетерозиготных коров при двух телятах будет забраковано
за недоказанностью их гетерозиготности 4-1/22 = 1 корова с двумя
телятами. Останется 4-2 — 1-2 = 6 телят. Среди них ожидается отношение:
или
6[1/3 черн. + 2/3 рыж.] = 2 черн. + 4 рыж.
2. Для восьми коронно три теленка.
Из восьми коров забракуется 8-1/23 = 1 корова с тремя телятами.
Останется 8-3 — 1*3 = 21 теленок.
Среди них ожидается отношение:
0, Г22 —1 22 134 oi Г 3 ,4 1
21 [23-ZTT: 2^1 J = —: —» или 21 [—черн. + -у-рыж.] =
= 9 черн. + 12 рыж.
Суммируя, получаем:
2 черн. + 4 рыж.
9 черн. +12 рыж.
11 черн. +18 рыж.
Аналогично может быть выведена формула и для апостериорного мен-
делевского расчета вместо априорной формулы 3:1. Приведем пример
такого расчета для случая рождения двух потомков. Задача,
следовательно, такова. Скрещивается большое число черных коров и черных быков,
некоторые из них гетерозиготны. Эту гетерозиготность ляожно установить
только по рождению рыжих телят. От каждой коровы получено по два
теленка. Каким должно быть моногибридное отношение черных и рыжих
телят в потомстве тех коров, от которых получены рыжие телята?
В потомстве от скрещивания Аа X Аа возникают четыре типа: АА, Аа,
аА и аа. Назовем АА черным первым, Аа — черным вторым, аА —черным
третьим и аа — рыжим. Тогда два теленка могут родиться следующими
равновероятными способами:
1) 4i4i 5) 4241 9) Ч3Ч1 13) p4i
2) Ч№ 6) Ч2Ч2 10) чзЧ2 14) рч2
3) 4i43 7) Ч2Ч3 11) чаЧз 15) рч3
4) 4ip 8) ч>р 12) ч3р 16) рр
2 А. С. Серебровский 17
Гетерозиготными родители смогут быть признаны только в тех случаях т
когда родился хотя бы один рыжий, теленок, т.е. в случаях 4,8,12,13, 14,
15 и 16. В этих случаях всего родится черных — 6, рыжих — 8. Вместо
отношения Зч : 1р мы должны ожидать Зч : 4р, т. е. отношение, сильно
уклоняющееся от априорного 3:1. Тем не менее, получив среди телят Зч : 4рг
мы должны будем сказать, что это отлично реализованное
моногибридное отношение для F2 (но только апостериорное!), и не пугаться, что оно
не похоже на знакомое 3:1.
ПЕРЕХОД ОТ АПОСТЕРИОРНЫХ РАСЧЕТОВ К АПРИОРНЫМ
Полезно иметь в виду, что априорное знание гетерозиготности
достигается не только знанием родителей данной особи. Так, если черная
корова родила уже рыжего теленка, то, очевидно, она гетерозиготна, и для
предсказания относительно следующих телят мы располагаем уже
априорным знанием ее гетерозиготности. В разобранном только что примере
в случаях 13, 14, 15 и 16 уже первый теленок оказался рыжим.
Следовательно, мы устанавливаем, что родители этих телят гетерозиготны. И для1
следующих тех же пар родителей мы должны ожидать обычное менделев-
ское отношение 3 : 1. И действительно, мы видим, что вторые телята в трех
случаях (13, 14 и 15) черные, а в одном (16) случае — рыжий. Необходима
только помнить, что телят, при помощи которых мы установили гетерози-
готность родителей и всех предыдущих, нужно исключить из дальнейшего
рассмотрения.
В случае крупных животных (коров, лошадей и пр.) методом
апостериорных расчетов менделевских ожиданий приходится пользоваться
особенно часто.
ПОПРАВКА ВЕЙНБЕРГА
Уже давно Вейнберг (Weinberg, 1912) заметил описанные искажения
при сборе статистического материала и предложил способ вычисления
действительной пропорции изучаемого признака. Этот способ известен под
именем метода братьев и сестер (Geschwistermethode). По этому способу
действительный процент признака определяется по формуле:
<? =
Ъкхх(х — 1)
2 кхх (/>-!)
•100 о/0.
Здесь/? — число детей в семье, х — число носителей признаков в семье,
а к — число семей с данным числом х носителей признака. Не вдаваясь
в изложение выводов этой формулы, покажем, как можно высчитать
правильный процент рецессивов в нашем предыдущем примере.
От скрещивания двух черных родителей мы получили «семьи» по два
теленка: в шести семьях 1 черный + 1 рыжий и в одной семье — 2 рыжих.
Здесь, следовательно, р = 2 и кх = 6, к2 = 1. Составим таблицу:
X
1
2
**
6
1
(*-1)
0
1
k^ix-i)
6-1.0 = 0
1-2.1=2
2
р
2
2
(р-1)
1
1
*^*<Р-1)
6-1.1 = 6
1.2-1 = 2
8
<?=-|-.100о/0=25о/0.
18
Как мы знаем, при скрещивании двух гетерозигот действительно
извлекается 25% рецессивов (критические замечания к методу Вейнберга см..
у К. М. Лютикова, 1931) *.
ТАБЛИЦЫ И ПОЛЬЗОВАНИЕ ИМИ
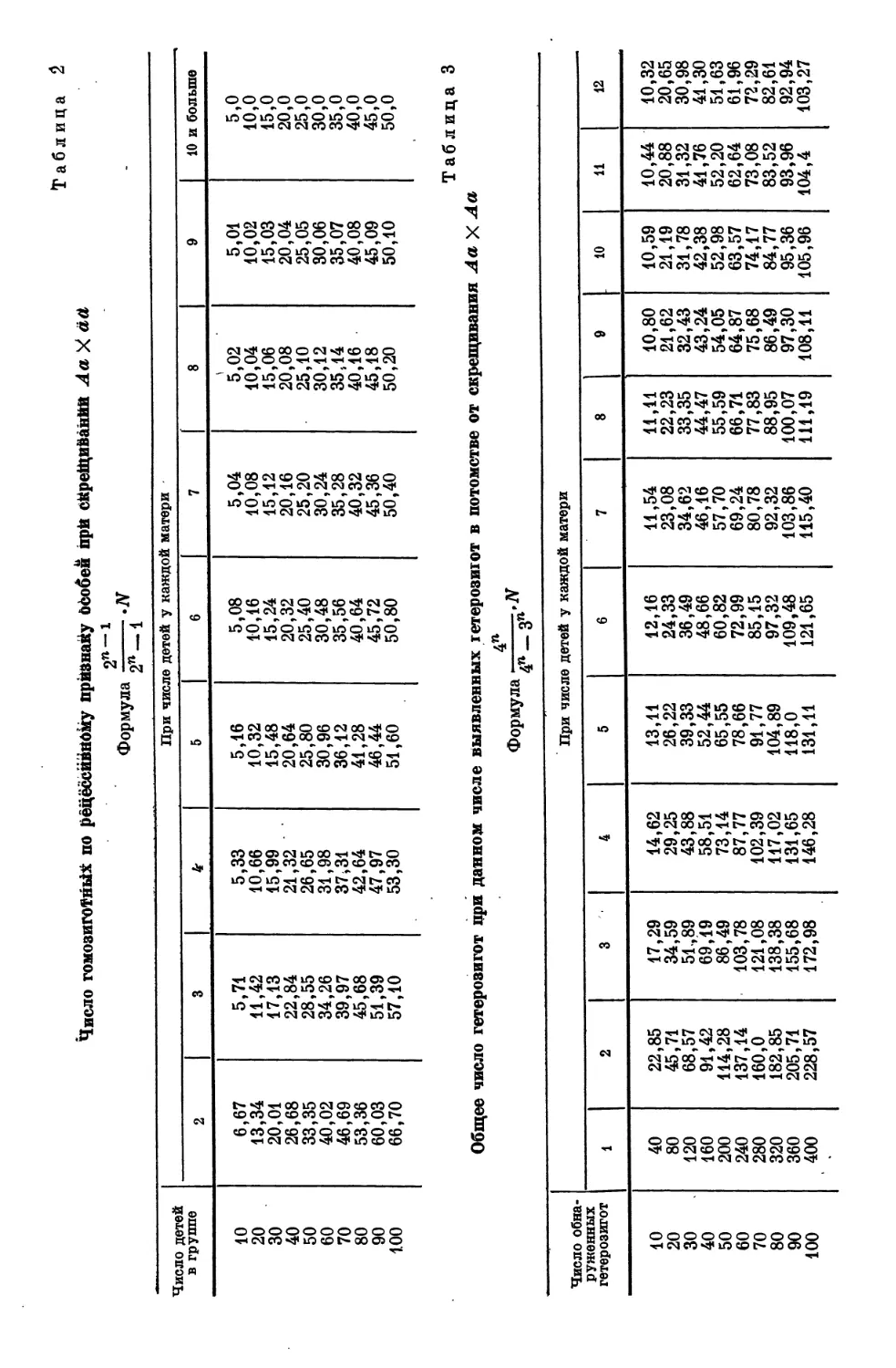
К. М. Лютиковым (1931) высчитаны таблицы, облегчающие быстрый
расчет при применении апостериорных формул (табл. 1).
Таблица 1
Число гомозиготных рецессивных особей в потомстве при скрещивании А а X Аа,
4П~ i
Формула 4n_3n -N
Число
детей
в
группе
10
20
30
40
50
60
70
80
90
100
При числе детей у каждой матери
2
5,714
11,43
17,14
22,86
28,57
34,29
40,00
45,71
51,43
57,14
3
4,324
8,649
12,97
17,30
21,62
25,94
30,27
34,59
38,92
43,245
4
3,657
7,314
10,97
14,63
18,33
21,94
25,60
29,26
32,84
36,57
5
3,278
6,556
9,834
13,11
16,39
19,67
22,94
26,23
29,50
32,78
6
3,041
6,083
9,124
12,16
15,21
18,25
21,29
24,33
27,31
30,41
7
2,885
5,770
8,655
11,54
14,425
17,31
20,19
23,08
25,97
28,85
8
2,778
5,582
8,354
11,11
13,89
16,67
19,45
22,22
24,94
27,78
9
2,702
5,405
8,108
10,81
13,51
16,22
18,92
21,62
24,27
27,026
10
2,649
2,298
7,948
10,597
13,24
15,89
18,54
21,19
23,79
26,49
11
2,61
5,221
7,831
10,441
13,05
15,66
18,27
20,88
23,49
26,10
12
2,582:
5,16В
7,745
10,327
12,94
15,4$
18,07
20,6S
23,23.
25,32.
Решим следующую задачу: пусть мы получили 138 телят от 23 черных
коров, скрещенных с черными быками, причем каждая из 23 коров дала по
6 телят и среди них хотя бы одного рыжего. Надо рассчитать по таблице*
сколько всего должно было быть рыжих, если случай моногибридный.
Соответственно 6 телятам на корову выбираем в табл. 1 столбец с числом
6 в заголовке. 138 телят разбиваем на 100 + 30 + 8. Эти числа смотрим
в левом столбце. Здесь имеется 100 и 30, но нет 8. Зато имеется 80 и
соответствующее 80 число в столбце 6 делим на 10. Выписываем из таблицы:
100 ..... . 30,41
30 9,124
8 2,433
138 41,967
Среди 138 телят надо ожидать 41,967 рыжих.
Применение этой таблицы полезно при тех расчетах, когда нужно
сопоставить фактически наблюдаемый материал по извлеченным рецессивам
с ожидаемым по моногибридной гипотезе.
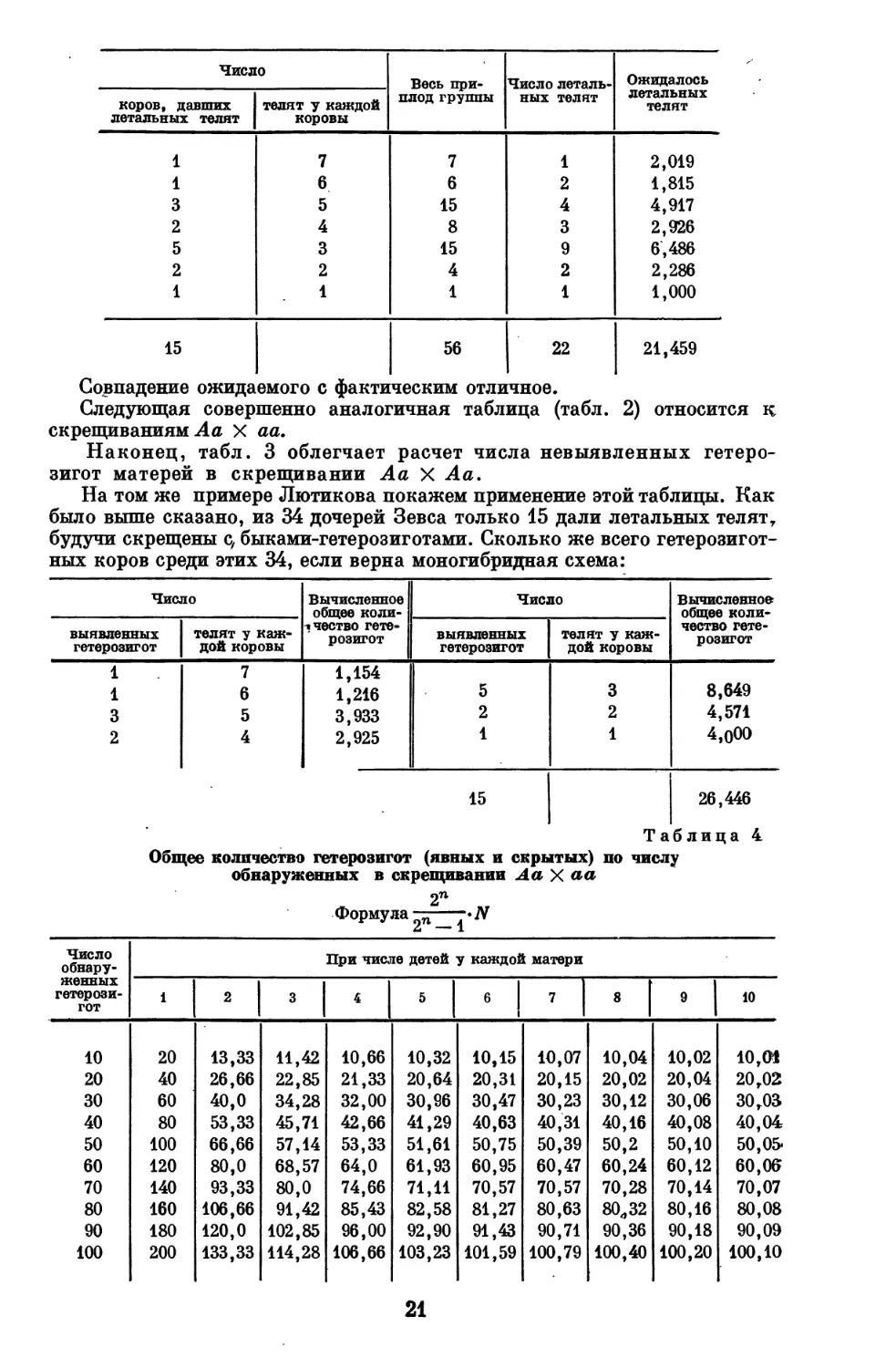
Лютиков приводит следующий пример. На Безенчукской станции
осталось в хозяйстве 34 дочери (из 60) быка Зевса, оказавшегося
гетерозиготным по летальному гену. Из числа этих коров 15, будучи скрещены с бы-
ками-гетерозиготами Зевсом и Примусом, дали хотя бы по одному
летальному теленку, а остальные 19 дали от тех же быков нормальных телят.
Обработка по табл.1 дает следующий результат (см. стр. 21):
* Методы Вейнберга подверглись уточнению и углублению рядом авторов (см.
соответствующую главу в книге Weber E. Mathematische Grundlagen der Genetik,
Fischer, Jena, 1967).— Прим. ред.
19 2*
ей
S
ю
а
X
f
а
&
"О
I
§
-з
г
g
о
ч
в
ев
а
е
й матери
При числе детей у каждо
<
<
1 t
05 1
со 1
*"■
со
**
1 °°
см
1
в группе
oooooooooo
1ЛОЮОЮОЮОЮО
-чЧ-чЧСМСМООСО^ЧРЮ
*ечСМОО**ЮсОЬ^ОООЭО
О О О ОО OOjO^O^
юоюоюоюоюо
««гЧ^СМСМСОСО^-^Ю
см*#соооосм*#соооо
*#оосмсоо*#оосмсоо
ОО'чн-гнСМСМСМООСОчИ
юоюоюоюоюо
tHtHCMCMCOOO^NFiO
00сОчНСМО00СО*#СМО
0-«-iCMCOvl»vl»iOCDt»00
юоюоюоюоюо
^■чнСМСМООСО^ЧРт
со см оо *# о со см оо чи о
-*-• со *а* со оо а> -*-• см чН со
lOClOOiOOCDrHCOrH
^ -чч СМ СМ СО СО *# ^ Ю
со со а см ю оо -«н *# t>- о
СО СО О) СО СО Од СО СО О) СО
Ю О Ю -чч CD -<Н !>• CM С» СО
1 -*н -чн СМ СМ СО СО *# 4t* Ю
1 •*■< СМ 00 *# Ю СО t^ 00 С* О
1 с^ *#^ оо ю см^о со со -*-•
1 Ю •** t* CM 00 чИ Oi \£^t^
I чН -^ СМ СМ СО 00 ** Ю Ю
1 г^*#-^ооюсмоэс©ооо
1 СО 00 О СО СО О СО^СО О t>-
1 CDCOOCDCOQCDCOOCD
1 ^ч СМ СМ СО чН *# Ю CD CD
I OOOOOOOOOO
1 -гч CM 00 *# Ю CO t* 00 О) О
1 чг£
СМЮ00ОСОсОСЭ^*#1>
ООСООЭООСОСЭСМСОСЭСМ
о*оо~^^T^cvTсм см со
«^CM00**iOCDt^0005O
*# 00 СМ СО О -* 00 СМ СО _^
*#00001>СМСООЮОЭЧ1<
оо^^смсмсосоео^
^4CMCO**iOcDt^OOOO
OO000000t^t»t>.CDC0
Ю-г-<С--СООЭЮ«ччС--00СЭ
«^CMCO^lOCOt^OOOJO
OCMC0**i0t*00CaO^4
OOcDvPCMOOOCO^OO'*-»
O-r^CMC
««HCM CO*
> »tf ** Ю CD t^ CO
•lOCOC^OOOO
^00i0t*O<^00i0t^05
rHMCO^iOt*OOCJOr4
ч-iCMCO^lOCDt^OOO^
^CMCO**iOCOt^OOO-H
«^C0**CDt-05OCMC0l0
«^СМОО^ЮСООООЭО'Н
СОООСЭСОСМСЭЮСМООЮ
^4 CO 4|^CD 00 05 -гч 00 ЧН СО
CM**C000OCMl0t*05-rH
^4CMC04t*CDt^00OOCM
«4-1 см со *# ю со с- a •«■<
тНСМСО*Ч,ЮСОС*00О'^
■«СМСОЮСО С-СЭС
СМЮ00-*-<*#С^ОЭСМЮ00
СОСМООЮ-^t^OOOcOCM
^OOOOOCOt*CMt^«rHCD
«*HCM*#iOt^OOO«*-«00*#
СЭОЭОЭСЭСЭОООООООООО
CM Ю ОО.чН 4ft*O00cDC5
r^CClOCDC
• OO-^OO ЮСМ
)OMCOiON
Ю -«H C- CM 00 4f lO«^t^
00 t>- Ю *# CM -r^O 00 fr- ifl
CM^COOJ^OOCOOOOCM
«чН -*-4 -^H <^H CM CM
OOOOOOOOOO
ЧИООСМСООЧРООСМСОО
•^•rHCMCMCMCOCOVt*
OOOOOOOOQ
«^CMCO**iOCOt*00Ob
Число
коров, давших
детальных телят
1
1
3
2
5
2
1
15
телят у каждой
коровы
7
6
5
4
3
2
1
Весь
приплод группы
7
6
15
8
15
4
1
56
Число
летальных телят
1
2
4
3
9
2
1
I 22
Ожидалось
детальных
телят
2,019
1,815
4,917
2,926
6,486
2,286
1,000
21,459
Совпадение ожидаемого с фактическим отличное.
Следующая совершенно аналогичная таблица (табл. 2) относится к
скрещиваниям Л а X аа.
Наконец, табл. 3 облегчает расчет числа невыявленных
гетерозигот матерей в скрещивании Аа X Аа.
На том же примере Лютикова покажем применение этой таблицы. Как
было выше сказано, из 34 дочерей Зевса только 15 дали летальных телят,
будучи скрещены с, быками-гетерозиготами. Сколько же всего гетерозигот-
ных коров среди этих 34, если верна моногибридная схема:
Число
выявленных
гетерозигот
1
1
3
2
телят у
каждой коровы
7
6
5
4
Вычисленное
общее
количество
гетерозигот
1,154
1,216
3,933
2,925
Число
выявленных
гетерозигот
5
2
1
15
телят у
каждой коровы
3
2
1
Вычисленное
общее
количество
гетерозигот
8,649
4,571
4,000
26,446
Таблица 4
Общее количество гетерозигот (явных и скрытых) по числу
обнаруженных в скрещивании Аа X аа
2п
Формула 2п_±-Я
Число
обнаруженных
гетерозигот
10
20
30
40
50
60
70
80
90
100
1
20
40
60
80
100
120
140
160
180
200
2
13,33
26,66
40,0
53,33
66,66
80,0
93,33
106,66
120,0
133,33
3
11,42
22,85
34,28
45,71
57,14
68,57
80,0
91,42
102,85
114,28
При числе детей
4
10,66
21,33
! 32,00
42,66
53,33
64,0
74,66
85,43
96,00
106,66
5
10,32
20,64
30,96
41,29
51,61
61,93
71,11
82,58
92,90
103,23
у каждой матери
6
10,15
20,31
30,47
40,63
50,75
60,95
70,57
81,27
, 91,43
101,59
7
10,07
| 20,15
! 30,23
40,31
50,39
60,47
70,57
80,63
90,71
100,79
»
8
10,04
20,02
30,12
40,16
50,2
60,24
70,28
80„32
90,36
100,40
9
10,02
20,04
30,06
40,08
50,10
60,12
70,14
80,16
90,18
100,20
10
10,01
20,02
30,03
40,04
50,05.
60,06
70,07
80,08
90,09
100,10
21
Таким образом, из 34 коров (дочерей Зевса) согласно вычислению 26,5
оказывается гетерозиготными. Если бы летальный ген передавался им
только от отца, их должно было бы быть лишь 17. Эта неувязка может
быть объяснена либо тем, что деталь попала к дочерям Зевса не только от
него, но и от их матерей, либо тем, что гипотеза недостаточно
правильна и т. д.,
Таблица 4, имеющая такое же значение, относится к скрещиванию
Аа X аа.
ЗНАЧЕНИЕ FB и F2
Прием обратного скрещивания более выгоден в отношении количества
использованного материала. Приведенный выше расчет подчеркивает эту
выгодность, так как в случае получения поколения F2 при апостериорном
изучении расщепления очень высок процент «брака». Как видно из 16
случаев 9 (1, 2, 3, 5, 6, 7, 9, 10 и 11), в которых рыжие телята вовсе не
родились, не могут быть использованы, хотя родители этих телят и были гете-
розиготами. При обратном же скрещивании пропадает только 1/4 часть
коров, у которых родилось по два черных потомка, т. е. 25% вместо 56%.
Однако для установления доминантности обратным скрещиванием в
некоторых случаях удовлетвориться нельзя, особенно в случае крупных
животных, где нельзя поручиться за гомозиготность родителей. Если при
скрещивании взятых из популяции черного быка и рыжей коровы
получается один или два черных теленка, то это может быть равно объяснено и
тем, что черный цвет доминирует над рыжим, и тем, что доминирует
рыжий, но рыжий родитель, был гетерозиготен. Тогда полезно подыскать
скрещивания, в которых оба родителя были бы черными или оба рыжими.
В этом случае получение F2 почти незаменимо, так как если у двух черных
родится рыжий теленок, это сразу доказывает доминантность черного
{в случае моногибридности).
Точно так же F2 необходимо для решения вопроса о влиянии гена на
жизнеспособность, так как жизнеспособность гомозигот часто резко
отличается от таковой гетерозигот.
ИСКАЖЕНИЯ МОНОГИБРИДНЫХ ОТНОШЕНИИ
На практике, конечно, почти никогда точного отношения 1 : 1 не
получается. Поэтому необходимо тщательно изучить всевозможные причины
искажений этого основного отношения.
Главнейшие из них_ следующие.
1. Случайность (рассматриваемая в теории вероятностей).
2. Различная жизнеспособность категорий Аа и аа.
* 3. Недостаточно резкое различие между фенотипами Аа и аа, потому
часть А а может быть причислена к аа и обратно (трансгрессия).
4. Участие других генов.
5. Недостаточно строгий выбор материала, ввиду чего среди родителей,
предполагавшихся как А а, могут оказаться и АА.
6. Фиктивные менделевские явления.
1. Случайность
Этот источник искажения отношений 1:1, как и всяких других, столь
неизбежен, что об искажении только тогда и можно серьезно говорить,
когда доказано, что искажение не случайно. Пределы случайных или случай-
до .возможных уклонений определяются при помощи формулы квадрати-
22
ческой ошибки *. Если мы ожидаем отношения 1 : 1 и получили всего п
потомков, то квадратическая опшбка т вычислится при достаточно большом
п (не менее 25) так:
т
-i/II-dy-i-itf
Поэтому всегда следует теоретически предполагаемый результат писать
<с указанием возможных уклонений.
Если ожидаемое отношение не 1 : 1, а а : Ь, то квадратическая ошибка
вычисляется по формуле: т = + 1/ , , .
Пример: получено 100 телят, среди которых предполагается
моногибридное расщепление 1 : 1 на черных и рыжих. Сколько ожидается тех и
других?
Отношение 1 : 1 получится при 50 черных и 50 рыжих телятах.
Квадратическая ошибка лг = ±-^~— =±5. Следовательно, ожидается
50 ± 5 черных и 50 ± 5 рыжих.
Теория вероятностей учит, что примерно в 2/3 случаев уклонение не
должно выходить из этих пределов. Однако нет ничего невероятного, что
фактический материал случайно уклонится и до двух квадратических
ошибок (в данном примере 50 ± 10) и в виде большей редкости достигнет
тройного квадратического уклонения.
О несомненно не случайном искажении отношения 1 : 1 можно
говорить только при уклонении фактических результатов от теоретически
вычисляемых за три квадратические ошибки. Однако уже при уклонении
больше чем за две квадратические ошибки можно предполагать
достоверность этого уклонения (т. е. не только случайный его характер) и
принимать меры к проверке этой достоверности. Лучше всего проверку
осуществить сбором второй порции такого же материала. Если и в ней снова
окажется уклонение в ту же сторону и примерно такой же величины,
это может служить уже довольно веским доказательством неслучайности
этого уклонения.
Иногда пользуются не квадратическим, а вероятным уклонением **.
Если обозначить его через Е, то Е = 0,6475 т.
В данном случае, т. е. для отношения 1:1,
Е = 0,6475. &- = 0,3237 У п.
Вероятная опшбка является таким уклонением, что фактически
получаемые числа в половине случаев уклоняются от ожидаемых на величину
меньшую, а в половине случаев — на величину большую, чем она. Это
свойство делает ее иногда очень удобным приемом оценки. Пусть,
например, мы имеем 20 стад ив каждом из них мы ожидаем отношения 1:1.
Вычислив для каждого стада 1) теоретически ожидаемые числа, 2)
фактически наблюденные, 3) вероятные уклонения и 4) уклонение
фактических чисел от теоретических, мы должны получить (если наша гипотеза
верна), что примерно в половине случаев (в 10 стадах) фактические
уклонения окажутся больше вероятных, а в половине — меньше (однако
средние из вероятных и из фактических уклонений не должны совпасть). Двум
* Достоверность фактических отклонений от теоретически ожидаемых отношений
при расщеплении теперь чаще всего проверяют методом «%-квадрат».— Прим. ред.
^♦Вместо «квадратических уклонений» теперь пишут «квадратических отклонений».—
Прим. ред.
23
квадратическим уклонениям примерно отвечают три вероятных. Поэтому
уклонение можно считать достоверным, когда оно больше чем в 3 раза
превышает вероятное.
2. Различная жизнеспособность
Это самая обычная причина искажений отношений 1 : 1, и для очень
многих случаев они возникают именно вследствие разницы в
жизнеспособности тех или других классов. Здесь нужно различать два близких
явления. Во-первых, могут быть различны жизнеспособности (или способности
к оплодотворению) уже гамет, из-за чего в образовании зигот участвует
неравное количество гамет Л и гамет а. Или же сами зиготы Л а и аа
оказываются неодинаково жизнеспособными и часть их вымирает. Первый
случай является редким, второй — очень частым.
Оба эти типа искажения можно отличить от очень важного четвертого
(участия других генов) по безрезультативности отбора. Какие бы группы
особей мы ни рассматривали, у них всегда будет получаться не 1:1,
а искаженное отношение, например 1 : 0,7.
В том случае, если искажение получается благодаря ослабленной
жизнеспособности зигот, установить наличие этой причины можно в
некоторых случаях по способу, предложенному Харрисом (Harris, 1918).
Способ этот основан на следующем. Пусть мы имеем группу семейств,
в которых потомство распадается на две категории: Аа и аа. Так как
рождение тех или других основано на случайности, то между рождением Аа
или аа не должно существовать никакой корреляции. Точно так же не
должно существовать корреляции между общим числом детей в данной семье
и процентом А а и аа в этой семье.
Не то получается, если, например, аа будут погибать, т. е. погибать
после зачатия, в состоянии зигот. Гибель их, во-первых, уменьшит
соответствующую семью и в то же время исказит отношение Аа и аа в пользу
первых. Благодаря этому между отношением А а и аа и величиной семьи
возникнет корреляция.
Открыть эту корреляцию можно вычислением ее между числом аа
форм и числом потомков в семье, но для того чтобы не впасть в ошибку и
не понять неизбежную при этом корреляцию между величиной помета и
числом аа или Аа в помете за искомую нами, необходимо пользоваться
специальной формулой:
|/l-?a+(%-^)2 f
где г является коэффициентом корреляции между общим числом членов
семьи и числом членов семьи категории аа, очищенной от упомянутой
корреляции *; г1а — тот же коэффициент, вычисленный обычными способами,
и, следовательно, еще не очищенный от участия другой корреляции;
vi = °ilMi — отношение конкретного числа членов семьи (аг) к
среднему числу членов семьи (Мг); иа = oJMa •— отношение конкретного
числа аа членов в семье (aa) к среднему числу аа членов в семье (Ма).
Пример (Harris, 1918). Дербишайр, изучая наследование альбинизма
у мышей, вместо отношения 3 : 1 получил отношение 3,21 : 0,79. Получи-,
лось ли это искажение в результате гибели альбиносов в стадии зигот или
нет?
* В данном случае применена модификация формулы для коэффициента частной
корреляции.— Прим. ред.
24
Харрис по данным Дербишайра вычислил следующие величины:
Afj = 4,5863 (среднее число мышат в пометах);
Afa = 1,1322 (среднее число альбиносов в помете);
<5j = 1,7518 (квадратическое уклонение числа мышат в пометах);
<50 = 1,0752 (квадратическое уклонение числа альбиносов в помете);
1,7518
vi = 38,19396 (= gj/Mj.IOOo/o = 4^5868,100%);
1,0752
г?а= 94,963о/0 (= ва/Ма.100% = ^дд^-ЮОо/о);
г1а = 0,3756 ±0,527.
г = — 0,0287 ± 0,0612, т. е. коэффициент корреляции оказывается
меньше своей ошибки, и поэтому никакой корреляции не обнаруживается.
Следовательно, недостаток альбиносов в опыте Дербишайра вовсе не
зависит от их меньшей жизнеспособности в утробной жизни.
С другой стороны, различная жизнеспособность гамет может быть
исследована другим приемом, а именно двумя обратными скрещиваниями:
°&АаХ Яаа
$ЛаХ с?яя
С их помощью мы определяем предполагаемые различные
жизнеспособности гамет, которые у мужских и женских гамет могут оказаться,
конечно, различными. Это будет служить подтверждением сделанной
гипотезы разной жизнеспособности гамет, а не зигот. Но мы можем получить и
дальнейшее подтверждение. Пусть в первом случае мы определяем
отношение т : 1, а во втором / : 1. Тогда, очевидно, при скрещивании с? Аа с 9 Аа.
мы должны получить
(тА + la) (fA + la) = mfAA + mAa + fAa + laa
или, распределяя по фенотипам,
A (mf + т + /) + la.
Например, если т = 1, 2; / = 1,1, то искомое отношение в F2 будет равно»
3,62 : 1 или 3 : 0,83. Если предсказание оправдывается, то мы можем
предполагать именно в неравной жизнеспособности гамет источник искажения
отношения 1:1.
3. Трансгрессия
Самым трудным для анализа случаем является такой, когда
исследуемые категории Аа и аа недостаточно резко разграничиваются, и мы при
классификации потомства делаем систематическую ошибку, зачисляя
часть Аа форм с не вполне проявившимся признаком в категорию аа (или
обратно).
Этот случай не редок при изучении, например, рецессивных пегостей.
В этом случае формы аа — пегие, формы Аа — сплошь окрашенные. Но
у таких гетерозигот иногда проскальзывают отдельные отметины, и па
ним можно Аа формы зачислить в аа категорию.
Подобный типичный случай мы встречаем при анализе куриной лохмо-
ногости, в которой гетерозиготные голоногие птицы имеют часто
незначительную лохмоногость. Произведенное исследование показало, что
поэтому формальному признаку отделить гомозиготных рецессивов от
гетерозигот нельзя. Однако был найден признак — указатель, позволяющий
теперь это разделение проводить почти с абсолютной точностью. Было
найдено, что у гетерозигот А а перышки располагаются исключительно на вне-
25
шней стороне цевок ног, а у гомозигот аа распространяются на переднюю и
внутреннюю стороны.
Однако возможны случаи, когда таких указателей оказывается
невозможно найти, и Аа сливаются с аа без ясной границы. Такой случай мож-
но|уже рассматривать как количественный и оперировать специальными
приемами, о которых речь будет идти дальше.
В том случае, однако, если процент ошибок незначителен и если в
большинстве случаев мы можем определенно относить особь в А а или аа
группу, целесообразнее не прибегать к сложным методам анализа
количественных признаков. Здесь, очевидно, должен быть выяснен вопрос, как часто
мы делаем ошибку. Предположим, что вместо отношения 1А : 1а мы
получили отношение 0,9Л : 1,1а, но подозреваем наличие моногибридного
расщепления. Если эта гипотеза верна, то, следовательно, 0,1 часть попала
из А в а-фенотип, так как тогда, вернув эту 0,1 к Л, мы получим
отношение 1 : 1.
Действительно ли 0,1 из 1,1 или 9% попали сюда ошибочно из класса
А, надо проверить исследованием дальнейшего поведения а-форм. Если
при скрещивании части из них с аа примерно у 9% появятся в потомстве
Jia-формы, то, следовательно, наша гипотеза была верна. При этом,
очевидно, незачем проверять всех а, а только некоторое количество,
достаточное для того, чтобы доказать, что около 9% а-формы генотипически
оказываются Ла-формами, что, действительно, подобная ошибка возможна.
Хороший пример подобного искажения — игрековидная форма гребня
у кур. Часто раздвоение оказывается настолько незначительным, что
гребень можно принять за нераздвоенный. Однако при скрещиваниях
некоторых кур с простым гребнем в потомстве появляется снова около 50% иг-
рековидных гребней.
Так как решение вопроса о том, имеем ли мы дело с влиянием данного
гена на жизнеспособность или только ошибаемся при классификации,
очень важно, то прием и должен быть всегда рекомендуем.
4. Участие других генов
Участие других генов в данном скрещивании ведет к искажению
отношения 1 : 1 в том, например, случае, если этот добавочный ген мешает
проявлению признака А или, как говорят, тормозит его развитие. Если это
торможение происходит обязательно, то мы будем иметь случай уже дигиб-
ридного или полигибридного скрещивания, о чем рень ниже. Но если это
тормозящее действие гена не сильно, то только часть особей Аа не разовьет
признака А, и мы получим некоторое искажение отношения 1 : 1 в сторону
А <а.
В этом случае полезно построить вариационную кривую по проценту
форм А а в различных семействах. Участие более или менее сильного
тормозящего гена даст соответственно более или менее заметное
двухвершинное распределение. Необходимо отметить, однако, что двухвершинность
может достоверно быть обнаружена лишь при большом (не менее 200) числе
наблюдений. В одну группу войдут все семьи, свободные от тормозящего
гена, и для этой группы вершина придется против 50%, в другую группу
войдут все семьи с участием тормозящего гена, и эта вершина будет лежать
где-нибудь между 0 и 50.
5. Ошибки в классификации родителей
Искажения, возникающие на почве недостаточно строгого выбора
материала, вследствие чего в число родителей наряду с гетерозиготами
попадает часть гомозигот, не поддаются учету. Некоторый учет может быть
26
сделан только тогда, когда можно предположить, что эта примесь основана
на некоторой статистической закономерности: например, если в данной
местности некоторый ген А довольно редок, то можно предположить, что
все А-особи суть гетерозиготы. На самом деле, некоторый процент особей
окажется гомозиготами. С обработкой таких случаев мы познакомимся
жиже, в главе о популяциях.
6. Фиктивные менделевские отношения
Наконец, следует иметь в виду возможность возникновения
отношений, фиктивно напоминающих менделевские. Если, например, мы будем
рассматривать скрещивание лошадей, имеющих гриву направо, с
лошадьми, имеющими гриву налево, то нередко у нас могут получиться, например,
отношения 1:1, как бы указывающие на менделевское расщепление.
Фиктивность его, однако, вскрывается легко рассмотрением такого
скрещивания, в котором должно было бы получиться иное отношение. Если,
например, мы получили, как в только что упомянутом случае, отношение 1:1,
то в случае скрещивания двух «правогривых» лошадей друг с другом мы
должны были бы получить уже не 1 : 1, а 3 : 1 (если «правая грива»
доминирует) или 0 : 6 (если «левая» доминирует). В случае гривы мы, однако,
получаем одно и то же отношение в потомстве (со случайными
уклонениями, конечно), независимо от того, какой тип скрещивания
рассматривается. Это указывает на то, что положение гривы не имеет никакого
отношения к менделизму и к наследственности вообще, а определяется иными
факторами.
Наиболее важным приемом для борьбы с разного рода искажениями
менделевских отношений (благодаря ошибкам классификации потомков
и родителей, примеси других генов и пр.) нужно признать подыскание
таких генотипов, в которых изучаемый ген давал бы наилучшее
проявление. Этим приемом можно для многих генов совершенно устранить
возможность ошибок.
Возьмем в качестве примера ген, вызывающий у кур поперечную
волосатость оперения, который мы называем trage-теяом. Обычно он вызывает
поперечную полосатость. Но если к генотипу, соответствующему обычной
рыжей курице, прибавить этот ген, разница получится столь
незначительной, что 1) у цыплят в пуху ее, обычно, не удастся заметить, а 2) во
взрослом оперении ее удастся заметить с значительной определенностью только
у петухов, у кур же при недостаточной опытности процент ошибок может
быть значителен.
Однако стоит только ввести в скрещивание ген Ufa, который вызывает
развитие у цыплят черного пуха, как ген trage с полной достоверностью
может быть установлен уже у едва вылупившихся цыплят обоих полов.
Этот же самый ген Ufa, являющийся одним из лучших генов окраски
кур, в некоторых генотипах (у рецессивных форм atrasi) можно заметить
с большим трудом, и часто возможны ошибки.
Изучение генов пегости выгоднее вести на генотипах с яркими
цветными пятнами, а не бледными, при изучении одной пегости следует
устранить другие гены пегости и т. д.
В этом выборе подходящего генотипа в значительной степени лежит
залог успешности решения задачи. К сожалению, здесь трудно дать какие-
либо подробные указания и приходится ограничиться общими указаниями,
подобными только что сделанным. В этом отношении генетический анализ
«оказывается не только наукой, но и искусством, предоставляя
исследователю широкий простор в выборе приемов работы.
27
ИДЕНТИФИКАЦИЯ ГЕНОВ
При помощи моногибридного скрещивания мы можем только
установить в данном скрещивании наличие одного менделирующего гена, но о
самих свойствах гена получаем только предварительное понятие.
Пользуясь им, мы можем установить много липших генов, т. е. таких, которые
уже были ранее установлены в несколько другом проявлении или у других
пород. Например, скрещивая черных и кофейных собак, мы устанавливаем
моногибридный характер этого различия и говорим о гене, который
отличает черных собак от кофейных. Точно так же, скрещивая рыжих собак
с желтыми, мы находим и там один ген. Но, пользуясь только
моногибридным скрещиванием, мы не можем решить вопроса о том, имеем ли мы
дело с двумя разными генами — одним, вызывающим черный цвет, и
другими, вызывающими рыжий цвет, или перед нами только один ген,
который в разных случаях несколько различно проявляется, но который
может быть назван геном-усилителем,— кофейную окраску он усиливает
до черной, палевую (желтую) — до рыжей.
Решение этого вопроса достигается вторым основным приемом
генетического анализа, именно дигибридным скрещиванием.
ГЛАВА II
ДИГИБРИДНОЕ РАСЩЕПЛЕНИЕ
ЗНАЧЕНИЕ ДИГИБРИДНОГО РАСЩЕПЛЕНИЯ
Поскольку моногибридное скрещивание является основой изучения
отдельно рассматриваемого гена, постольку дигибридное скрещивание
оказывается основой генетического анализа в широком смысле слова.
Только владея этим скрещиванием, мы получаем возможность подвергать
анализу наследственные признаки, т. е. разлагать их на составные
генетические элементы, узнавать число генов, необходимых для возникновения
данного наследственного различия и для идентификации генов.
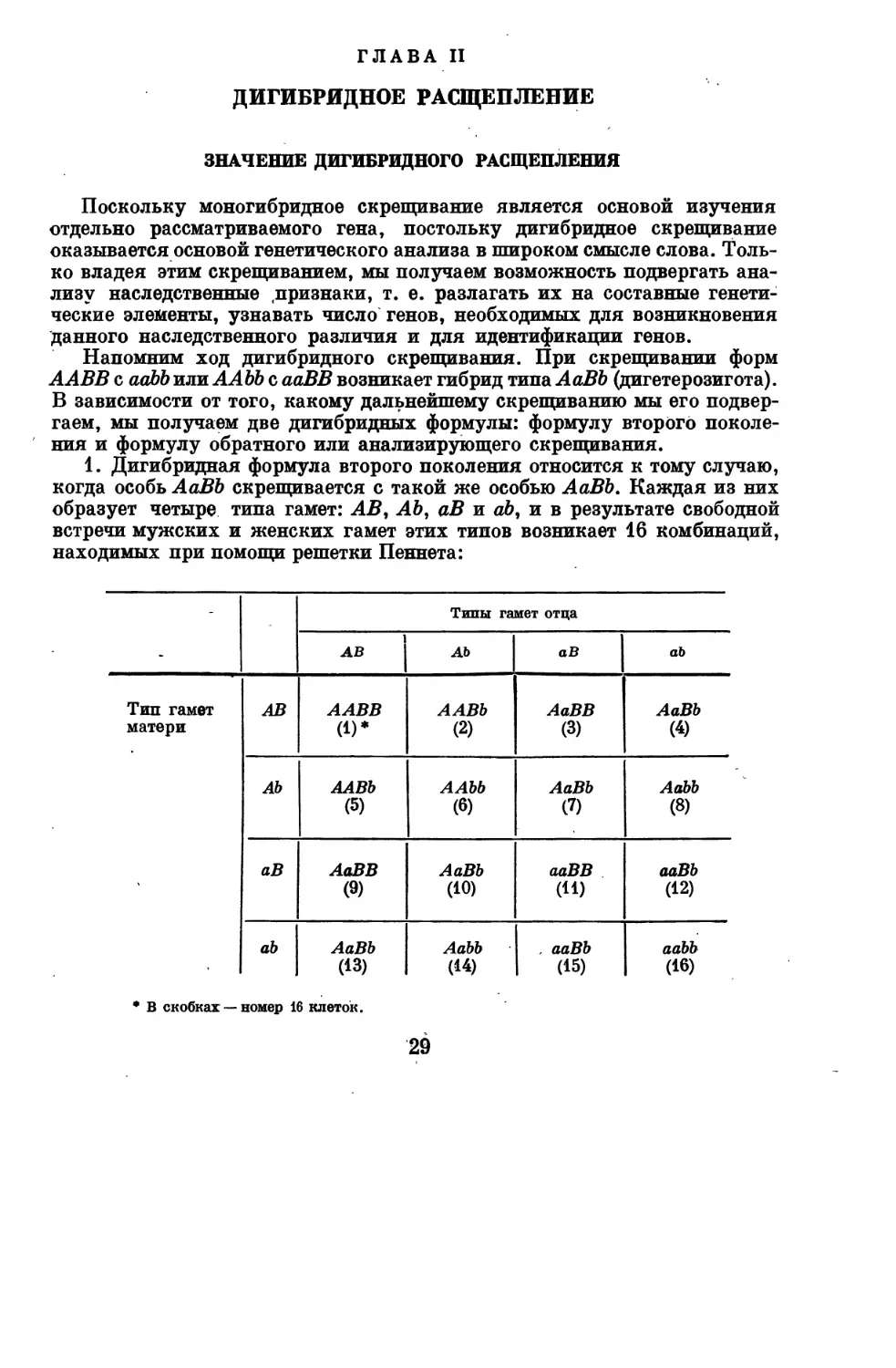
Напомним ход дигибридного скрещивания. При скрещивании форм
А АВВ с aabb или A Abb с ааВВ возникает гибрид типа АаВЬ (дигетерозигота).
В зависимости от того, какому дальнейшему скрещиванию мы его
подвергаем, мы получаем две дигибридных формулы: формулу второго
поколения и формулу обратного или анализирующего скрещивания.
1. Дигибридная формула второго поколения относится к тому случаю,
когда особь АаВЬ скрещивается с такой же особью АаВЬ. Каждая из них
образует четыре типа гамет: АВ, АЬ, аВ и аЬ, и в результате свободной
встречи мужских и женских гамет этих типов возникает 16 комбинаций,
находимых при помощи решетки Пеннета:
-
-
Тип гамет
матери
■
1
АВ
АЬ
аВ
аЬ
Типы гамет отца
АВ
ААВВ
(1)*
ААВЬ
(5)
АаВВ
(9)
АаВЬ
(13)
АЬ
ААВЬ
(2)
ААЪЬ
1 (6)
АаВЬ
(10)
АаЬЪ
(14)
аВ
АаВВ
(3)
АаВЬ
(7)
ааВВ
(11)
, ааВЬ
\ (15)
аЬ
АаВЬ
(4)
АаЬЪ
(8)
ааВЬ
(12)
ааЪЪ
\ (16)
* В скобках — номер 16 клеток.
29
В случае полного доминирования А над а и В над Ъ второе поколение
(F2) по внешности распадается на четыре фенотипа в отношении 9 : 3 : 3 : 1,
а именно:
_ Всего
Фенотип случаев
а, В —11, 12, 15 3
а, Ь — 16 1
2. Гораздо выгоднее во многих отношениях дигибридная формула
обратного или анализирующего скрещивания, т. е. скрещивание форм АаВЬ с
ааЬЪ. В том случае, когда гибрид АаВЬ был получен скрещиванием форм
ААВВ с aabb, разбираемое скрещивание будет действительно обратным
(FB) скрещиванием гибрида с родительской формой ааЬЪ (не обязательно,,
конечно, чтоб с самим родителем). Если же гибрид АаВЬ получен от
скрещивания ААЬЬ с ааВВ, скрещивание АаВЬ с ааЬЪ уже не будет обратным
с родителем. Форма ааЪЬ будет, как мы называем, «рецессивным
анализатором», а самое скрещивание — скрещиванием с рецессивным анализатором
или анализирующим скрещиванием.
При скрещивании с анализатором гибрид образует, конечно, те же
четыре типа гамет Ab, ab, aB, ab, а так как гаметы анализатора все ab, то
фенотипов возникает четыре (правый столбец или нижняя строка) и в том же
отношении, как и гаметы, т. е. 1-4, В : 14, 6; 1а,# : 1а,Ь, причем для этих
фенотипов сразу же известен и генотип, т. е. мы имеем:
lAaBb : lAabb : laaBb : \aabb.
Эти формулы принадлежат, очевидно, к числу априорных.
Апостериорная формула анализирующего скрещивания будет иметь вместо 1 тМ :
: 1 : 1 вид:
(i—^rj AaBb:(i--^)Aabb:(l~-±r>jaaBb:laabb,
где п — число потомков.
ДОСТОИНСТВА И НЕДОСТАТКИ ДВУХ ДИГИБРИДНЫХ СКРЕЩИВАНИЙ
Для практики генетического анализа дигибридные скрещивания
второго поколения (F2) и анализирующего скрещивания (FB) имеют свои
достоинства и недостатки, которые необходимо иметь в виду, ставя то или
другое скрещивание. Проведем вкратце сравнение их.
1. Как увидим ниже, при анализе приходится решать основной
вопрос, сколько разных категорий потомков возникает. При отношении
1:1:1:1 уже при 32 потомках можно быть уверенным в том, что все
категории уже возникли. Между тем при отношении 9:3:3:1
необходимо получить в 4 раза больше потомков, чтобы гарантировать себя от
случайного невозникновения последней категории потомков — laabb,
возникающих в числе 1/16.
2. Так же важно решить вопрос, какая категория потомков
возникает в большем количестве, какая в меньшем. При отношении, например,
9:3:4 решить, где мы имеем дело с 3/16 и где с 4/16, можно лишь на
большом материале. В соответствующем анализирующем скрещивании
отношению 9:3:4 отвечает отношение 1 : 1 : 2, и здесь установить,
какая категория составляет 50%, а какая 25%, очень легко. Это
соображение особенно важно при наличии искажений и других причин, о которых
мы говорили в предыдущей главе.
3. Важно, чтобы при расщеплении количество разных категорий было
возможно меньшим, иначе дигибридное расщепление трудно отличить от
Л Всего
Фенотип случаев
А, В —1, 2, 3, 4, 5, 7, 9, 10, 13 9
Л, Ь — 6, 8, 14 3
30
тригибридного и трудно установить, в каких числовых отношениях идет
расщепление. При полном доминировании во втором поколении возникает
столько же категорий, сколько и в анализирующем скрещивании. Но при
неполном доминировании в F2 может возникнуть до 9 разных категорий
потомков (а при наличии полового диморфизма даже до 18), т. е. такая
пестрота, разобраться в которой будет совершенно невозможно. Между тем
при анализирующем скрещивании число возникающих категорий всегда
одно и то же, независимо от степени доминирования.
4. Если в анализирующем скрещивании возникает больше четырех
категорий, это сразу указывает на участие в расщеплении больше чем двух
генов. По второму поколению судить об участии третьего гена трудно, так
как пестрота расщепления может зависеть от неполного доминирования.
5. Потомство, возникающее в F2, неудобно для дальнейшего изучения,
так как при полном доминировании гомозиготы и гетерозиготы будут
неразличимы. При анализирующем скрещивании возникают только
гетерозиготы (по доминантному признаку), которые могут использоваться для
дальнейших скрещиваний.
6. Однако получение F2 оказывается незаменимым, если анализатор,
т. е. aabb, нам неизвестен. Тогда необходимо прибегнуть к получению F2r
хотя бы уже для того, чтобы получить этот анализатор, так как в F2 он
возникает в количестве 1/16 потомства.
7. Наконец, F2 незаменимо как прием извлечения неожиданных рецес-
сивов, что составляет одно из звеньев генетического анализа пород.
Под термином «неожидаемые рецессивы» мы разумеем следующее.
Генетика дала нам уже ряд случаев, когда при скрещивании внешне очень
сходных форм во втором поколении начинают извлекаться такие
рецессивы, ожидать извлечения которых не было никаких оснований. Наиболее
эффективный пример этого мы имеем в работе Хагедорнов (HagedoornA. L.
a. A. G., 1922), посвященной скрещиванию двух близких видов крыс. Те
различия между ними в окраске, которые видны по внешности, давали
возможность ожидать в F2 расщепление по каким-либо слабым усилителям,,
модификаторам и пр. Результат, однако, оказался совершенно
неожиданным, и в F2 появились пегие, желтые и иные особи. Правда, мы здесь
имеем дело с межвидовым скрещиванием, нам неизвестна цитологическая база
этого явления, но методическое значение примера от этого не
изменяется.
Но и при скрещивании пород аналогичные явления могут встретиться.
Если мы скрещиваем кур с розовидным гребнем с курами со стручковым
гребнем, то (не зная того, что мы теперь уже знаем об их генетике) мы
никогда не могли бы ожидать получить в потомстве листовидные гребни, и
при обратных скрещиваниях этих листовидных гребней мы никогда бы
неполучили, как видно из прилагаемых схем (рис. 4).
Получение F2 особенно ценно тогда, когда мы скрещиваем породы,
бывшие раньше в изоляции (например, географической), или при скрещивании,
видов. Но главнейшим назначением этого скрещивания должно быть
извлечение неожиданных рецессивов или, что то же самое, открытие новых:
генов.
При обратном скрещивании гибрида с анализатором выясняются сразу
два вопроса: вопрос о числе генов, участвующих в скрещивании, и вопрос
о «взаимодействии генов» (по проявлению), т. е. о том, как проявление
одного гена влияет на проявление другого гена. Обе эти задачи удобно
решаются только тогда, когда в скрещивании участвует два гена. В случае
участия трех, четырех и больше генов получается слишком большое
разнообразие, даже при скрещивании с анализаторами. Чтобы в таком
скрещивании разобраться, нужно обширное число потомков, доступное толька
ботаникам или работающим с мелкими животными, или, наконец, лишь,
при очень простых взаимодействиях фенов.
31
стр \
/ч
стр.
орех.
1:1
орех.
орех.
А р
пч
/
'\
орех.
стр.
роз.
1:1
роз.
...
\лис-\
1 тов. i
I 1 -у
Моногибридная гипотеза подходит
Поколение F2 опровергает моно -
гибридную гипотезу
вместо 1:2:1 получено 9:3 • <?:/
стр.-
Рис. 4. Роль FB и F2 в анализе форм гребня кур
- стручковый гребень; роз.— розовидыый; орех. — ореховидный; листов,-
листовидный
Поэтому в случае, когда при скрещивании с анализаторами получается
больше четырех категорий потомков, часто приходится считать такое
скрещивание предварительным, ориентировочным, и различных
представителей полученного расщепления снова скрестить с анализатором. При
этом некоторые из них дадут желаемое дигибридное расщепление.
Итак, основным приемом генетического анализа является получение
дигетерозиготы и скрещивание ее с анализатором. В случае невозможности
почему-либо это сделать, например при отсутствии анализатора,
приходится прибегать к получению F2 поколения. К этому мы вернемся.
Говоря о трудности анализа в том случае, когда в расщеплении
участвует больше двух генов, мы, конечно, имели в виду случай, когда более
чем два гена влияют на один и тот же признак, например, на окраску.
Если же мы имеем ряд генов, влияющих на различные признаки, то можно
изучать скрещивание и с большим числом одновременно менделирующих
генов.
Так, у курицы можно без труда наблюдать одновременное мендели-
рование формы гребня, хохла, бак, голошеести, скорости оперения,
строения пера, окраски, числа пальцев, оперения ног, окраски яиц и пр. и по
каждому из этих признаков наблюдать по паре генов. Но одновременное
наблюдение за генами различных признаков имеет интерес
преимущественно для топографического анализа, о котором будет речь ниже.
РАЗЛИЧНЫЕ СЛУЧАИ ДИГИБРИДНЫХ РАСЩЕПЛЕНИЙ
Только в самом простом случае скрещивание с анализатором дает
четыре категории потомков в равных количествах, а в F2 отношение
9:3:3: 1. Большей частью получается измененное отношение, и
нужно знать всевозможные изменения дигибридного отношения, чтобы
сразу понять, с чем оно имеет дело.
Разберем на конкретных примерах наиболее обычные случаи
видоизменения дигибридной формулы, с которыми приходится встречаться
генетику-аналитику.
1. Простая кооперация
Анализируется разница между черным пойнтером и желтым. В первом
поколении доминирует черная окраска, поэтому в качестве анализатора
используем желтого пойнтера. От скрещивания черного Fi с желтым ана-
32
лизатором получаем:
25% черных 25% рыжих
25% кофейных 25% желтых
Обсуждение. Наличие четырех фенотипов в равных
количествах говорит о том, что мы имеем дело с дигибридным расщеплением
АаВЪ X ааЪЪ, дающим 1А,В : L4, Ъ : 1а, В : la, b. Моногибридным это
расщепление быть не может, так как при нем возможны лишь две категории
потомков, а не четыре. Тригибридвгое расщепление в некоторых случаях
может дать четыре фенотипа, но не в равных количествах. Следовательно,
наш случай дигибридный, и четыре класса фенотипов соответствуют
четырем возможным в таком скрещивании генотипам АаВЪ, АаЪЪ, ааВЪ и ааЪЪ.
Возникает вопрос, какой из этих генотипов соответствует каждому из
четырех фенотипов: черному, кофейному и т. д.
Рассуждаем так: черному соответствует генотип АаВЪ, так как генотип
АаВЪ должен быть у F1? а это ¥г было черным; следовательно, генотип
черных из FB тоже АаВЪ. Желтому фенотипу должен отвечать генотип ааЪЪ.
В самом деле, если один из родителей, давших FB, был черным и имел
генотип АаВЪ, то для того, чтобы в FB появилось отношение 1:1:1:1,
генотип второго родителя должен быть ааЪЪ. Действительно, если бы этот
второй родитель имел генотип ААЪЪ или ааВВ, то в FB получилось бы лишь
два фенотипа: в первом случае А, В + А, Ъ, во втором случае А, В + а, В.
Если бы генотип второго партнера был бы АаЪЪ или ааВЪ, то ¥г состояло
бы либо из ААВВ + АаВЪ, либо из ААВЪ + АаВЪ и в FB не могло бы
получиться отношение 1 : 1 : 1 : 1, а получилось 3 : 1 : 3 : 1 и т. п.
Следовательно, второй родитель FB поколения не может быть иным, как ааЪЪ.
Этот родитель — желтый. Следовательно, фенотипу желтый отвечает
генотип ааЪЪ.
Двум фенотипам мы, таким образом, с уверенностью приписали
генотипы АаВЪ и ааЪЪ. Остающимся двум фенотипам кофейный и рыжий,
следовательно, отвечают генотипы АаЪЪ и ааВЪ, причем безразлично какой, так
как с символами А и В пока не связано ничего больше, как только то, что
А ж В вместе дают черный, а без А и без В получается желтый.
Поэтому пишем:
А, В —черный а, 2? — рыжий
А, Ъ — кофейный _ а, Ъ — желтый
Всем фенотипам приписаны генотипы. Вторая часть задачи выполнена.
Остается третья часть — сформулировать действие генов.
По первым двум фенотипам — черному и кофейному, различающимся
лишь геном В,— устанавливаем, что «ген В превращает кофейную
окраску в черную».
По второй паре фенотипов — рыжий и желтый — таким же образом
устанавливаем, что «ген В превращает желтую окраску в рыжую». Отсюда
можно описать действие гена В так: «ген В превращает кофейную окраску
в черную, а желтую в рыжую». Это будет верно, но слишком примитивно.
Генетик-аналитик должен здесь не просто соединить, но обобщить
частные определения. Это наиболее трудно, так как нужно так сформулировать
разницу между кофейным и черным, чтобы она оказалась приложимой и к
паре желтый — рыжий. В данном случае, например, подобной общей
формулировкой может явиться такая: «черный несколько темнее кофейного».
Очевидно, будет также правильно написать и «рыжий несколько темнее
желтого». Найдя такую общую формулировку, подставляем вместо
фенотипов генотипы. Получаем:
\АаВЪ\ \АаЬЪ\
«Фенотипы, отвечающие | _, \, несколько темнее, чем \ ,, \, или,
отбрасывая А-тея: «ВЪ несколько темнее, чем ЪЬ».
3 А. С. Серебровский
33
Сформулировав разницу между ВЪ и ЪЬ, можем, перефразируя, дать
обобщенную формулировку действия гена В, например, так: «ген В
несколько затемняет окраску, например, от желтого до рыжего, от
коричневого до черного».
Переходя к гену А, сравниваем пару фенотипов черный — рыжий
(АаВЬж ааВЪ) и кофейный — желтый {АаЪЪ и aabb). Находим две частные
формулировки:
1) «ген А превращает рыжую окраску в черную»;
2) «ген А превращает желтую окраску в коричневую».
Что общего в этих двух частных формулировках? В обоих случаях под
действием гена А окраска резко изменяется. По сравнению с этим разница
между рыжим и желтым или между черным и коричневым (кофейным)
в значительной степени стушевывается. Например, коричневый можно
назвать «почти черным». С другой стороны, ни рыжий, ни желтый
«нисколько не черны». Поэтому можно написать, например, так:
3) «ген А превращает светлую (нисколько не черную) окраску в
более или менее черную».
4) «ген А переводит окраску из группы рыжих в группу черных».
Заключение. Разница между черной окраской и желтой у
пойнтеров зависит от двух полнодоминирующих и независимо наследующихся
генов А и В, из которых сильно действующий ген А переводит окраску из
группы рыжих в группу черных, а ген В несколько затемняет окраску, со
степени коричневой до черной или со степени желтой до рыжей.
Совместное действие обоих генов является простейшей формой кооперации.
Для проверки полученных выводов хорошо поставить, например,
скрещивание кофейных (А, Ъ) с рыжими (а, В), так как [при [этом должна}
возникнуть в потомстве черная окраска (А, В).
2.* Взаимная модификация
Анализируется разница между ореховидным и листовидным гребнями
кур.
Первое поколение оказывается с ореховидным гребнем, поэтому в
качестве анализатора выбираем форму с листовидным гребнем.
Скрещивание «ореховидного» гибрида с анализатором дает потомство:
25% с ореховидными (гребнями (1) 25% со стручковыми гребнями (1)
25% с розовидными » [(1) 25% с листовидными - [(1)
Обсуждение. На основании тех же соображений, как и в первом
примере, заключаем, что имеем дело с дигибридным обратным
скрещиванием, идущим по формуле 1 : 1 : 1 : 1, и что четырем фенотипам отвечают
генотипы AaBb, Aabb, ааВЪ и aabb.
Фенотипу «ореховидный», повторяющему фенотип Fx, очевидно,
отвечает генотип АаВЬ, а листовидному — aabb. Поэтому пишем опять, как и:
в первом случае:
ореховидный — АаВЪ стручковый — ааВЪ
розовидный — АаЪЪ листовидный — ааЪЪ
Сравнивая попарно фенотипы, находим частные формулировки:
1) «ген В превращает розовидный гребень в ореховидный»;
2) «ген В превращает листовидный гребень в стручковый».
Все четыре формы гребня столь резко различны, что найти обобщенную
формулировку для 1 и 2 очень трудно, комбинация А, В является
новообразованием.
34
Можно удовлетвориться поэтому, например, такой формулировкой:
«ген В превращает листовидный гребень в стручковый, а в присутствии
гена А превращает розовидный гребень в ореховидный».
Однако при более внимательном попарном сравнении гребней
листовидного со стручковым и розовидного с ореховидным, можно заметить то
общее, что стручковый гребень ниже листовидного, а ореховидный —
ниже розовидного. Поэтому можно обобщить действие гена В так:
«ген В делает гребень более низким».
Аналогичным образом сравнивая листовидный с розовидным, а
стручковый с ореховидным, можно сформулировать действие гена А так:
«ген А делает гребень более широким».
Заключение. Разница между листовидным гребнем и
ореховидным вызывается двумя генами, А и В, из которых ген А делает гребень
более широким, а ген В делает гребень более низким. Однако совместное
действие генов А ж В ведет к некоторым новообразованиям, например,
прорастанию гребня мелкими перышками в виде полосы поперек гребня и т. п.
Этот случай поэтому можно назвать случаем взаимной модификации.
В F2 от скрещивания двух ¥г с ореховидными гребнями возникло бы:
56% с ореховидным гребнем (9) 19% со стручковым »ребнем (3)
19% с розовидным » (3) 6% с листовидным » (1)
Такое скрещивание может быть поставлено для проверки сделанной
гипотезы.
3. Криптомерия
Анализируется разница между гнедой лошадью и рыжей. Первое
поколение оказывается гнедым, так что в качестве анализатора выбирается
рыжая лошадь. От этого скрещивания гнедого гибрида с рыжим анализатором
получается потомство:
25% гнедых (1)
25% вороных (1)
50% рыжих (2)
Обсуждение. Разница между гнедой лошадью и рыжей зависит
не менее чем от двух генов (так как расщепление не моногибридное).
Предположим, что только от двух. При этом должны возникнуть четыре
фенотипа А, В; А, Ъ\ а, В и а,6. Возникло же только три, очевидно, вследствие
того, что какие-либо два фенотипа неразличимы. Это подтверждается
тем, что категория рыжих возникла в количестве 50%; можно думать, что
здесь суммированы два класса генотипов (25% + 25%). Какие же это
генотипы? Так как рыжая окраска рецессивна, то, очевидно, 25% а, Ъ
фенотипа входят сюда; А, В фенотип входить не может, так как он, судя по Flt
должен быть гнедым. Следовательно, суммировались АЬ + ab или аВ +
+ ab, что конечно, безразлично. Мы имеем
Л, В—гнедые а, В — рыжие
А, Ь — вороные а, Ь — рыжие
Заключение. Разница между рыжей лошадью и гнедой, взятыми
для скрещивания, оказалась зависящей от двух генов: ген А имеется у
гнедой лошади и у вороной, отличая их от рыжей (аа). Второй ген В при а
не проявляется, т. е. а, В и а, Ъ неотличимы, а при А ген В вызывает
ослабление вороной масти до гнедой. Ген В является модификатором
по отношению к А, в частности модификатором-о слабителем,
причем у рыжих лошадей он может иметься вкриптомерном
состоянии (т. е. скрытом, непроявленном).
35:
з *
В'F« возникло бы
56% гаедых
19% вороных
25% рыжих
В таком скрещивании выяснить и доказать, что именно рыжих
получилось больше, чем вороных, а не наоборот, было бы трудно, а малейшие
ошибки в регистрации повели бы к совершенно превратным выводам.
4. Криптомерия с новообразованием
Анализируется разница между окраской белых минорок и бурых
леггорнов.
Первое поколение неожиданно оказывается черным. Это
«новообразование» мейделизм объяснил возникновением такого сочетания генов,
которого не было ни у одного из родителей. В простейшем случае
предполагаем, что один родитель был ААЪЪ, другой ааВВ, а в гибриде АаВЪ гены А
и В встретились и дали черную окраску. Проверим это объяснение двумя
обратными скрещиваниями с родителями:
1. Скрещивание черного гибрида Fx с белой миноркой дает:
50% белых
50% черных
Результат, очевидно, моногибридный, по схеме АаВЪ X ааВВ, где мен-
делирование по В незаметно. Обозначим менделирующий ген через А, тогда
черные формы — Л, белые — а.
2. Скрещивание черного гибрида Fx с «бурым» леггорном дает:
50% бурых
50% черных
Как и предполагалось, мы имеем снова моногибридное расщепление,
но уже по гену В по схеме АаВЪ х ААЪЪ, где незаметно расщепление по А.
Заключение. Различие окраски белых-минорок от окраски
бурых леггорнов зависит от двух генов А и В. В изменяет бурую
окраску в черную, при аВ окраски нет, птицы белые. Форма aabb, т. е.
анализатор, остается пока неизвестной. Такую форму можно извлечь из F2. Она
возникает в таком составе:
56% черных (9 частей)
19% бурых (3 части)
25% белых (4 части)
Это отношение является измененным дигибридным 9:3:3:1,
указывая, что два каких-то фенотипа неразличимы. Фенотипы А, В
(черные), А, Ъ (бурые) и а, В (белые) нам уже известны, следовательно,
последний фенотип а, Ъ неотличим от какого-то из них.
В зависимости от того, к какой из этих категорий присоединится
неизвестный фенотип, мы будем иметь такие возможности:
1(А, В + а* Ь) П(А, Ъ + а, Ъ) Ш (а, В + а, Ъ)
черных (10) 62% (9) 56% (9) 56%
бурых (3)19% (4)25% (3)19%
белых (3) 19% (3) 19% (4) 25%
Если наш экспериментальный материал окажется достаточным, для
того чтобы с уверенностью сделать выбор из этих трех гипотез, то задача
будет решена.
36
5. Эпистаз
Анализируется разница между
окраской белого и бурого леггорнов.
Первое поколение оказывается белым
с отдельными черными перьями (рис. 5).
Используем в качестве анализатора
бурого леггорна и получаем (цыплят):
50% белых
25% черных
25% «бурых» или «диких», т. е. с
коричневыми, черными и светлыми продольными
полосами.
При расположении полученных
фенотипов лучше всего усвоить такой
порядок, чтобы вверху располагался
фенотип, подобный первому поколению
гибридов, а внизу — фенотип, подобный
анализатору. Тогда верхнему фенотипу
будет отвечать генотип А ,5, а нижнему—
a, fc, что облегчает понимание.
Обсуждение. Возникновение
трех фенотипов в FB говорит за участие
по крайней мере двух генов. Если это так, то в группу белых,
возникших в числе 50% и повторяющих по внешности первое поколение,
должно, очевидно, войти 25% А, В + 25% каких-либо других: А, Ъ или
а, 6, что безразлично. Тогда будем иметь
25% Л, В белые 25% а, В черные
25% А, Ь белые 25% а, Ь «дикие»
Заключение. Разница между окраской белых и бурых леггорнов
оказывается зависящей от двух генов. Из них один — А — вызывает
белую окраску или, иначе, тормозит развитие окраски. А — неокрашенные,
аа — цветные. Ген В заметен только на аа цыплятах, вызывая черную
окраску в противоположность ab1 бурой или дикой окраске. В присутствии
А ген В незаметен *, так как А эпистатирует над В.
В F2 мы имели бы:
75% белых (12)
19% черных (3)
6% бурых (1)
6. Эпистаз и криптомерия
Анализируется скрещивание белого леггорна и белой минорки.
Гибриды ¥г оказываются тоже белыми, но с некоторым числом чисто
черных перьев. Подобно предыдущему, мы имеем явное новообразование.
Скрещиваем с леггорном (1-е обратное) и получаем
50% белых
50% белых с отдельными черными перьями •
Скрещиваем с минорками (2-е обратное) и получаем
25% белых с черными перьями
25% черных
50% белых
* Во взрослом состоянии фенотипы А, В и А, Ь различимы, так как А, Ъ рыжеют.
Кроме того, и в цыплячьем возрасте часть А, В имеют отдельные черные пушинки или
пятна, отсутствующие у Л, Ь,
Рис. 5. Цыпленок ¥г леггорн X ланг-
шан
Белая окраска с черными
исключительными перьями благодаря гетерозиготности
эпистатического белого (todix) при наличии
меланизма (Ufa)
37
Обсуждение. Наиболее интересным результатом оказывается
совершенно неожиданное появление в FB черных, хотя оба родителя этих
черных цыплят — белая минорка и гибрид Fx — были белыми. То
обстоятельство, что черные цыплята возникли в количестве 25%, указывает на
дигибридность этого обратного скрещивания, так что мы можем написать
для Fx формулу АаВЪ и для белой минорки — aabb. Фенотип Fx был
«белый с черными перьями», следовательно, этому фенотипу и отвечает
генотип АаВЪ. Белые минорки были чисто белыми, и раз мы приняли для них
генотип aabb, то, следовательно, этот генотип отвечает белой окраске.
При этом следует добавить, что, поскольку у нас при скрещивании двух
белых пород в Fx произошло новообразование, постольку необходимо
принять, что генотипы белых могут быть различны. Для большей
определенности поэтому подчеркнем, что генотип aabb отвечает фенотипу белых
минорок.
Возвращаемся к истолкованию второго обратного скрещивания, где
мы получили 25% с черными пятнами, 25% черных и 50% белых. Мы
располагаем их в таком именно порядке по принятому выше правилу,
начиная с фенотипа, повторяющего фенотип F1? и кончая фенотипом,
повторяющим тип анализатора. Осложнение в данном случае состоит в том, что
вместо четырех фенотипов в отношении 1:1:1:1 имеются три фенотипа
в отношении 1:1:2. Это легко истолковывается как 1 : 1 : (1 + 1), т. е.
тем, что фенотип анализатора (аа ЪЪ) и фенотип, отвечающий генотипу
ааВЪ, неразличимы по внешности. Теперь мы можл:: приписать генотипы
всем трем фенотипам:
белые с черными перьями . . . . АЬ ВЬ
черные Аа ЬЬ
белые аа ВЬ
белые аа ЬЬ
Переходим к формулировке действия генов. Сравнивая первую пару
фенотипов, генотипы которых различаются лишь геном В, находим:
1) ген В превращает черную окраску в белую, кроме отдельных перьев,
остающихся черными.
• Судя по второй паре фенотипов, никакого действия гена В нет, т. е.
2) ген В оставляет белую окраску без изменения.
Сравнивая теперь первый фенотип с третьим и второй с четвертым,
находим для гена А:
1) ген А превращает белую окраску в белую с черными перьями;
2) ген А превращает белую^окраску в черную.
Очевидно, общей формулировкой действия гена А будет:
3) ген А вызывает появление черных перьев.
Итак, мы заключаем, что в скрещивании участвуют два гена, А и В,
из которых ген А вызывает появление черных перьев, а ген В мешает этому.
Поэтому при аа — окраска белая, при Л, Ъ — черная и при А, В — снова
почти белая, кроме отдельных перьев.
Таким образом, обратное скрещивание ¥г X Р минорка мы
благополучно истолковали. Переходим к истолкованию Fx X Р леггорн. Наличие
расщепления здесь несколько неожиданно, так как при скрещивании
АаВв х АА ВВ все фенотипы должны относиться к классу Л, В.
Раз мы имеем в этом обратном скрещивании расщепление по фенотипам,
то можно предположить:
а) участие еще третьего гена (тогда для нашего скрещивания будем
иметь такое истолкование ¥г: (ААВВее X аа ЬЬ СС -+ Аа ВЬ Се);
б) фенотипическое различие между гомо- и гетерозиготами. Пока не
доказана непригодность дигибридного объяснения, не следует переходить
к тригибридному объяснению. Поэтому останавливаемся сначала на
втором предположении.
38
В обсуждаемом обратном скрещивании АаВЪ X А А ВВ должны быть
следующие генотипы:
АЛ ВВ
АЛ ВЬ
Аа ВВ
Аа ВЬ
(здесь пишем генотип, отвечающий Fx, последним, а генотип А А ВВ —
первым). Фенотип, отвечающий генотипу ААВВ, нам известен, это фенотип
белых леггорнов. Известен и фенотип генотипа A abb — это фенотип белых
с черными перьями. Фенотипы, отвечающие генотипам ААВЪ и АаВВ, нам
неизвестны. Можно сделать две гипотезы (так как по предыдущему АаВВ
ж ААВЪ для нас уже не безразличны):
i п
1) А А В В —белые А А В В— белые
АА ВЬ — белые АА ВЬ — белые с черными перьями
2) АаВЪ— белые с черными перьями Аа [ВВ — белые
АаВЪ—белые с черными перьями Аа ВЬ — белые с черными перьями
Обсудим, какая из этих гипотез вероятнее.
По первой гипотезе из попарного сравнения фенотипов для гена В
не находим никакого действия, а для гена А в обеих парах находим, что
ген А (1 и 2) в гетерозиготном состоянии дает белых с черными перьями,
в гомозиготном — чисто белых, следовательно, ген А подавляет развитие
черного.
Этот вывод, очевидно, совершенно противоречит тому выводу о
действии гена А, который мы делали раньше («ген А вызывает развитие черных
перьев»). Это нас не удовлетворяет. Поэтому исследуем вторую гипотезу.
Здесь из сравнения первой пары и второй пары фенотипов получаем
для гена В (1 и 2): ген В в гетерозиготном состоянии дает белых с черными
перьями, а в гомозиготном — чисто белых. Следовательно, ген В
подавляет развитие черного.
Этот вывод оказывается в полном согласии с тем, который мы сделали
для гена В по второму обратному скрещиванию. Это нас может
удовлетворить. Для гена А не находим никакого различия между действием в гете-
ро- и гомозиготном состояниях.
Заключение. Данный своеобразный случай объясняется
участием двух генов, А и В, из которых А вызывает развитие окраски (черной) *
перьев и доминирует полно, а В подавляет развитие окраски (черной)
перьев в гомозиготном состоянии вовсе, а в гетерозиготном оставляет
некоторые перья окрашенными.
В F2 в таком скрещивании должны возникнуть:
37% белых с черными перьями (6 частей)
19% черных (3 части)
44% белых (7 частей)
7. Парные или дополнительные гены
Анализируется разница между белым розовоглазым и белым
голубоглазым кроликами.
* В том случае, когда участвует белая окраска, правильнее ей противопоставлять не
какую-либо частную окраску (например, черная), а вообще «окраску», признавая
«белую» не за окраску, а за отсутствие окраски, т. е. делая противопоставление не
«белый—черный», а «окрашенный—неокрашенный».
39
Скрещивание между этими белыми кроликами дает 100 % окрашенных
с темными глазами. Мы, очевидно, опять имеем новообразование типа
ааВВ X ЛАЪЬ -> АаВЪ.
Скрещивая с каждым из родителей, мы получаем по 50% белых и 50%
окрашенных, т. е. опять нечто иное, чем в предыдущих случаях, где одно
обратное скрещивание давало 100% белых, а другое — 75% белых.
Настоящий случай легче для понимания, так как если мы правильно поняли
происхождение новообразования, то в FB должны получить:
АаВЪ X ааВВ-*50% А, В + 50% а, В
АаВЪ X ААЪЪ ^ 50о^ А, В + 50% А, Ъ
В каждом из обратных скрещиваний в половинном количестве получится
А, .В-фенотиц и в половинном — не Л, В, и соответственно половина
оказывается цветной, половина — белой.
Заключение. Разница между двумя белыми кроликами зависит
от двух генов А и В. Если они имеются одновременно, то возникают
окрашенные формы, если какого-либо нет, то получается белая. Можно думать,
что, если не будет ни а, ни Ь, форма вряд ли будет окрашенной. Если так,
то в F2 можно ожидать:
56% цветных (9)
19% )
19% I белых (7)
6% J
8. Повторные гены
Анализируется различие между голоногими и оперенноногими курами
(один из изученных случаев).
При скрещивании лохмоногих брама с голоногими виандотами
потомство оказывается лохмоногим. Для анализа Fx скрещиваем с виандотами и
в потомстве возникает:
75% лохмоногих
25% голоногих
Возникновение всего лишь двух категорий говорит как бы за моногиб-
ридность скрещивания. Но полученное здесь отношение 3 : 1 при
моногибридном скрещивании возникает в F2, а вовсе не при обратном скрещивании.
Следовательно, толкование здесь должно быть иным. Появление 25%
голоногих, сходных с анализаторами, говорит о том, что перед нами фенотип
а, Ъ. Тогда надо предположить, что 75% =25% А, В-\- 25% А,Ь +
+ 25% а, Ь.
Заключение. Разница между лохмоногими брама и голоногими
виандотами зависит от двух генов А и В, каждый из которых вызывает
оперение ног и только при их одновременном отсутствии ноги остаются
голыми.
В F2 при этом случае надо ожидать:
94% лохмоногих (15)
6% голоногих (1)
Отличить по второму поколению этот случай от обычного
моногибридного можно только на большом материале. По обратному же скрещиванию
(75% : 25% вместо 50% : 50%) они отличаются резко.
* Фактически обычно окрашенные бывают [пегими, что зависит от специальных
генов пегости, часто имеющихся в генотипе кроликов-альбиносов.
40
9. Димерия, однозначные гены
Последний случай, который мы рассмотрим, встречается, по-видимому,
особенно часто при наследовании количественных признаков, но довольна
редок в случае признаков качественных. Он непосредственно примыкает
к предыдущему случаю, в котором наследовались два повторных
одинаковых гена А и В, каждый из которых вызывал развитие перьев на ногах.
Классифицируя потомство, мы разделили его только на «лохмоногих» ж
«голоногих». Тогда вышло, что А, Ь; а, В и А, В все лохмоноги. Но если
бы мы подошли более внимательно к этому случаю, разделив лохмоногих
на категорию «сильно» и «слабо лохмоногих», то у нас категория А, В
отделилась бы в самостоятельную группу «сильно лохмоногих» и мы имели
бы:
25% сильно лохмоногих (А, В)
50% слабо лохмоногих (А, Ь + а, В)
25% голоногих (а, Ь)
в F2 имели бы:
56% сильно лохмоногих (9 А, В)
38% слабо лохмоногих [6 (А, Ь + а, В)]
6% голоногих (1 а, Ъ)
Таблица 5
Возможные типы взаимодействия фенов1 и формулы расщепления
Характер взаимодействия фенов
Типичный дигибридный
случай, простая кооперация . . .
Взаимная модификация ...
Криптомерия
Эпистаз + криптомерия . . .
Парные или дополнительные
гены
Однозначные гены с полным
доминированием
Однозначные гены с
неполным доминированием . . . .
Криптомерия и эпистаз с
неполным доминированием ...
Простая кооперация;
неполное доминирование одного гена
Простая кооперация;
неполное доминирование обоих генов
Формула расщепления
bFb
1:1:1:1
1:1:1:1
1:1:2
3:1
3:1
1:3
1:2:1
1:2:1
1:1:2
1:1:1:1
1:1:1:1
в F2
9:3:3:1
9:3:3:1
9:3:4
12:3:1
13:3
15:1
9:7
9:6:1
1:4:6:4:15
6:3:7
6:3:3:2:1:1
4:2:2:2:2:1:1:1:1
примерный объект
Окраска пойнтеров
Гребни кур
Гнедые лошади
Белые леггорны, бурые-
леггорны3
Белые леггорны, белая
минорка
Лохмоногость брама8
Формы коробочки у
пастушьей сумки
Белые куры иди белые-
кролики
Лохмоногость брама4
Белые леггорны 2
Белая минорка*
1 Автор говорит о взаимодействии фенов, что принципиально более правильно, нежели обычно-
фигурирующее в генетической литературе обозначение случаев как результата генных
взаимоотношений.— Прим. ред.
2 Если исследовать цыплят и не обращать внимание на различие между чисто белыми и
белыми с черными отметинами.
3 Если делить на лохмоногих и голоногих.
4 Если лохмоногих делить на сильно и слабо лохмоногих.
5 Наблюдаются классы фенотипов в Р2, отсутствующие в Рв #
6 Если различать цыплят чисто белых и белых G черными отметинами.
41
о
wo
% Rat
a wo
10 90
20 80
30 70
W 60\
fz50 50
60 40|
70 30
80 20
90 10
90
Ь9
20 3D
80 70
MPS 1-2
ltd
50
50
60 70 80
hO 30 20
ЫЫ 3-1M
SO
10
9-1
/C0%
0
~7/?
100
в
1
\
\
\
\
*M^
I
^
\
\ 4
\
4
V
>
4f
^
v
^
^J
'^
?^
•<
^£
4
Как правило, однако,
разделить ясно категории с
двумя генами (А,В) от категории
Л, Ъ + а, В бывает очень
затруднительно и получается
более или менее
значительный процент ошибки.
Приведенные случаи и
некоторые другие
видоизменения дигибридной формулы
в результате взаимодействия
фенов полезно сопоставить
в табл. 5.
Из сказанного ясно, сколь
разнообразной может
оказаться картина дигибридного
скрещивания, несмотря на
его теоретическую простоту.
Одни лишь явления эпистаза,
криптомерии и неполного
доминирования могут дать
самые различные числовые
отношения между различными
I — изменение жизнеспособности гамет; II — изменение
жизнеспособности зигот (для DD и Dd одинаково);
III — изменение сцепления и отталкивания; "D —5до-
минанты; R — рецессивы; Rat — отношение
Рис. 6. Номограмма для определения различных
генетических случаев, дающих расщепление на
два!феномена, по пропорциям этих феноменов в F2 -^
hj*F*i (описание пользования номограммой см. Щкатегориями потомков. Если
~в gB тексте)! » же мы вспомним, что было
сказано в предыдущей главе
об искажениях менделевских
отношений, возникающих на
почве различной
жизнеспособности, недостаточной
проявляемое™ признаков,
благодаря случайности и т. д., то станет ясным, что разнообразие здесь
поистине безгранично. Кроме того, и в дигибридных скрещиваниях нам
придется иметь дело и с априорными, и с апостериорными формулами.
Поэтому сказать, глядя на получившиеся цифры разных категорий
потомков, с каким случаем имеешь дело — на практике оказывается делом
часто чрезвычайно трудным.
Генетические задачи решаются легко только тогда, когда они...
предварительно уже решены другими. Поэтому необходимо предостеречь тех, кто
впервые приступает к генетическому анализу, от уныния и пессимизма,
если их первые опыты окажутся неудачными. Генетик должен
предварительно в совершенстве постичь азбуку менделизма, чтобы всевозможные
менделистические выкладки не затрудняли бы его. Затем он должен
держать в памяти возможно большее число разных «случаев», часть которых
мы привели выше, а еще лучше поупражняться в решении подобных задач
самостоятельно. Получив конкретный материал, нужно придумать
несколько возможных менделистических гипотез для объяснения его и из
них выбирать наиболее подходящую и простую. При этом особенно
важно представлять себе пределы возможных случайных уклонений и не
стараться педантично подгонять менделевские отношения до полного
совпадения их с конкретными цифрами — это часто приводит к нагромождению
многих гипотез, статистически не обоснованных.
На рис. 6 представлено графически, как некоторые случаи моно-, ди-
и тригибридных скрещиваний приобретают сходство друг с другом под
влиянием некоторых факторов: различной жизнеспособности гамет,
различной жизнеспособности зигот и сцепления и отталкивания, о которых
речь будет в следующей главе. Рисунок построен следующим образом:
42
избраны, простоты ради, лишь такие менделистические случаи, в которых
как в F2, так и в FB возникает только по два фенотипа, а именно:
1. Моногибридный случай с отношениями 3:1,1:1
2. Моногибридный случай с гибелью гомозигот одного типа с отношениями 2:1, 1:1
3. Случай парных генов с отношениями 9: 7, 1: 3
4. Случай двух повторных генов с отношениями 15:1, 3:1
5. Случай эпистаза и криптомерии с отношениями 13:3, 3 :1
6. Случай трех повторных генов с отношениями 63:1, 7:1
На оси ординат отложены отношения для F2 (рядом со шкалой в
процентах: 3:1 — 75% : 25%; 9 : 7 — 56% : 44% и т. д.), а по оси абсцисс —
отношения для FB. Точка, обозначенная 1 на рис. 6, соответствующая
моногибридному скрещиванию, поставлена так, что по оси ординат отвечает
отношению 3 : 1 в F2, по оси абсцисс — отношению 1 : 1 в FB.
Соответственно расположены и остальные точки.
Сплошной кривой линией (проходящей через точки 1, 4 и 6) показано
изменение положения точек в случае неодинаковой жизнеспособности
гамет. В том случае, если рецессивные гаметы (а-гаметы) менее
жизнеспособны, чем доминантные (4-гаметы), точки спускаются по кривой вниз и
вправо. Если доминантные гаметы менее жизнеспособны, точки
перемещаются вверх и влево. Отсюда следует, например, что при малой
жизнеспособности рецессивных гамет моногибридное скрещивание станет похожим
на дигибридный случай (4) двух повторных генов. При жизнеспособности
рецессивных гамет а в 1/3, моногибридное отношение 3 : 1 превратится в
15 : 1, а 1 : 1 — в 3 : 1, и случай 1 точно совпадет со случаем 4.
Пунктирные линии (простой пунктир) изображают перемещение
положения точек в результате изменения жизнеспособности зигот. Как видно,
эти смещения (например, для точки 1) не совсем совпадают со смещениями
по первой причине (жизнеспособности гамет), но на значительном
протяжении могут быть лишь с
большим трудом различены.
Наконец, пунктир с
точкой 3 изображает
перемещение точек в зависимости от
наличия сцепления или
отталкивания. При наличии
сцепления точки
перемещаются вправо, вниз; при
наличии отталкивания — влево,
вверх.
Этот график наглядно
показывает, как различные
обстоятельства могут
затруднить распознавание того, с
каким случаем мы имеем дело.
Так, например, если мы
получили в F2 отношение около
2 : 1, а в FB около 4 : 6, то
это может быть объяснено
либо тем, что налицо
моногибридное отношение (3 : 1 и
1 : 1), но доминантные
гаметы или зиготы менее
жизнеспособны, либо тем, что
налицо случай парных генов
{9 : 7 и 1 : 3), но рецессив-
Е-Ш
Рис. 7. Номограмма для определения различных
генетических случаев, дающих расщепление на
три фенотипа, по пропорции этих фенотипов в F2
(описание пользования номограммой см. в тексте)
— • — изменение сцепления и отталкивания; —
изменение жизнеспособности гамет
43
ные зиготы менее жизнеспособны, или парные гены несколько
сцеплены друг с другом.
Еще труднее будет поставить диагноз, если, например, мы получим в
F2 отношение около 6 : 1, а в FB около 2:1, так как близкая картина
может получиться при смещениях точек 1 и 2 под влиянием
жизнеспособности рецессивных зигот или точек 4 и 5 под влиянием сцепления и т. д.
Следующая диаграмма (рис. 7) также показывает, как под влиянием
некоторых причин различные менделевские случаи приобретают сходство
друг с другом. Здесь сопоставлено несколько случаев, при которых в F2
возникают три фенотипа, а именно:
1. Моногибридный с неполным доминированивхМ 1:2:1
2. Случай криптомерии 9:3:4
3. Случай димерии 9:6:1
4. Случай эпистаза 12:3:1
5. Случай полного сцепления 9:0:3
(=3:1)
6. Случай криптомерии и эпистаза с неполным доминированием 6:3:7
Диаграмма построена по способу треугольника. Например, точка i,
отвечающая отношению 1:2:1 или 25 : 50 : 25, помещена в данном
месте треугольника потому, что: 1) проходящая через нее горизонталь
(параллельно основанию, т. е. ребру II—III) отсекает на двух других
ребрах 25% (I фенотип), 2) линия, проходящая параллельно ребру 1-г-Ш,
отсекает на двух других ребрах 50% (II фенотип), 3) линия, проходящая
параллельно ребру I—II, отсекает на двух остальных ребрах снова по
25% (III фенотип). Другие точки расположены по этому же правилу.
Сплошные линии показывают перемещение точек в случае изменения
жизнеспособности гамет. Так как точки 2, 4 и 6 отвечают дигибридным
случаям, в которых изменения в жизнеспособности гамет, зависящие от
одного или другого гена, по-разному влияют на положение точки, то через
точки 2, 4 и 6 проходят по две сплошных линии.
Линии пунктирные с точкой указывают перемещение точек в
результате сцепления (отмечено «+») или отталкивания (отмечено «—»).^Например,
точка 2 (случай 9:3:4) в случае сцепления приближается к точке 5
(9:0:3 = 3:1), а в случае отталкивания приближается к точке 1
(1 : 2 : 1). Наличие перекрещивающихся линий указывает на то, что
некоторые отношения могут возникнуть из искажения различных случаев. Так,
через точку а (пересечение 60-й горизонтали и 60-й вертикали) проходят
две линии: сплошная — идущая через точку 4, и пунктирная —
проходящая через точки 3 и 5. Это значит, что отвечающая точке пропорция трех
фенотипов 60 : 30 : 10, или 6:3:1, может возникнуть либо из пропорции
9:6:1 (точка <?), благодаря слабому сцеплению генов, либо из
пропорции 12 : 3 : 1 (точка 4), благодаря понижению жизнеспособности гамет,
несущих доминантную аллель эпистатирующего гена. Очевидно, что если
пополнить эту диаграмму линиями перемещения точек под влиянием
различной жизнеспособности зигот, ошибок при классификации и т. д., то
можно будет получать при помощи такого чертежа указания о возможных
для каждого данного случая гипотезах.
Пусть, например, мы получили в F2 три фенотипа в отношении 50% :
: 25% : 25%, или 2:1:1 (точка Ъ). Точка эта лежит вблизи пересечений
четырех различных линий, и поэтому возможны для ее объяснения
следующие гипотезы:
1) имеется случай 9:3:4 (точка 2), слегка искаженный
отталкиванием;
2) или тот же случай, но искаженный неравной жизнеспособностью
гамет;
44
3) или случай 6:3:7 (точка б), сильно искаженный отталкиванием;
4) или, наконец, этот же случай, но сильно искаженный неравной
жизнеспособностью гамет.
Возможны будут и другие гипотезы, более сложные, например, что
точка Ъ есть случай 9:6:1 (точка <?), дважды искаженная сцеплением
(передвижение точки 3 по направлению к точке 5) и жизнеспособностью
(продвижение точки 3 по дуге вниз к точке 1) и т. д. В соответствии с этим
должно быть построено дальнейшее изучение данного случая. Однако при
надлежащих дополнительных опытах все эти возможности, по крайней
мере в простейших, а может быть в гибридных случаях, ясно вскрываются.
ГЛАВА III
СОСТАВЛЕНИЕ И РЕШЕНИЕ ЗАДАЧ
ПО ГЕНЕТИЧЕСКОМУ АНАЛИЗУ
Для овладения методом генетического анализа и приобретения
навыков в построении гипотез и их проверки необходимы: 1)
экспериментальные работы с дрозофилой, лабораторными грызунами и пр. и 2) решение
задач. Задачи могут быть двух типов — «ограниченные» и
«неограниченные». Под первыми мы разумеем такие задачи, когда сразу дается весь
необходимый материал и упражняющимся предлагается найти решение в
рамках этого материала. Примером такой задачи может служить
следующая.
Условие. Скрещены кролики А черный и В белый. Получено Fx в
составе двух черных и трех серых. Черные ¥г при скрещивании друг с другом
дали в F2 28 черных и 10 белых. Серые ¥г при скрещивании друг с другом
дали 35 серых, 11 черных и 15 белых. При скрещивании серых из этого
F2 с исходным черным родителем (А) получено от различных самок: а) 8
серых; в) 3 серых и 2 черных; с) 4 серых и 5 черных; d) 3 серых и 5
черных; е) 7 серых.
Найти:
1) число генов, участвующих в расщеплении;
2) сформулировать действие каждого из генов;
3) дать формулу исходных родителей А и В.
Решение. Наличие в расщеплениях трех фенотипов возможно:
1) при моногибридном, если доминирование не полно; 2) при дигиб-
ридном, если два из четырех фенотипов неотличимы друг от друга (эпи-
стаз, где в F2 12 : 3 : 1; криптомерия, где в F2 9 : 3 : 4; димерия, где в
F2 9 : 6 : 1).
Первая гипотеза моногибридного расщепления отпадает, так как при
ней в Fx должен был бы возникнуть лишь один фенотип (АА X аа -*■ Аа)
или если два, то повторяющие родителей (Аа X аа-+Аа X аа).
При дигибридной гипотезе полученное в F2 от ее серых Fx отношение
35 сер. : 11 черн. : 15 бел. близко отвечает отношению 9:3:4, так как
точное ожидание будет 34,4 + 11,4 + 15,2. Обе другие гипотезы эпистаза
и димерии не подходят ни по числовым отношениям, ни по характеру Fx.
Наоборот, случай криптомерии подтверждается характером Fx, если
принять для белого родителя (В) формулу ааВЬ, а для черного (А) — ААЪЪ.
ААЬЬ X ааВЬ -» АаВЪ + АаЪЪ
черн. бел. сер. черн.
Удовлетворяют гипотезе и все остальные скрещивания, так как
черные Б\ inter se должны дать 3 : 1 черных и белых (aabb), а серые из F2 по
гену В должны быть гомо- и гетерозиготами в отношении 1 : 2, почему при
46
скрещивании серых F2 с исходным черным родителем (-4) в одних случаях
должно получиться лишь серое потомство:
ААЬЬ X ААВВ -» ААВЬ
ААЬЬ X АаВ В -» ААВЬ + АаВЬ,
черн. сер. сер.
а в других случаях — серые и черные в отношении 1 : 1
ААЪЬ X ААВЬ -» ААВЬ + ААЬЬ
ААЬЬ X АаВЬ -» ААВЬ + АаВЬ + ААЬЬ + АаЬЬ
черн. сер. сер. черн.
Так как гипотеза дигибридности и криптомерии является простейшей
из возможных и вполне удовлетворяет всем заданным условиям, то она
может быть принята в качестве наиболее вероятного решения. «Наиболее
вероятным» это решение является потому, что можно составить и ряд
дальнейших, более сложных гипотез (тригибридных, тетрагибридных, со
сцеплением, летальностью и т. д.), которые также будут удовлетворять
заданным условиям и на ограниченном материале задачи не могут быть
опровергнуты.
Ограниченные задачи могут быть проработаны в большом числе
занимающимися. Такие задачи приведены в руководствах Синнота и Денна
(1934) и Рокицкого (1932, 1934) *. Очень удобно, если упражнения
проводятся вдвоем и занимающиеся по очереди задают друг другу
самостоятельно составленные задачи. Для составления задачи сначала намечается
число генов и их свойства, выписываются все возможные генотипы и
соответствующие им фенотипы, затем задаются генотипы исходных родителей,
рассчитываются в буквенных символах (т. е. генотипах) несколько
нисходящих скрещиваний, затем принимаются «фактически полученные»
частоты, более или менее отвечающие теоретическим расчетам, и, наконец, все
переводится в фенотипы. В этом переведенном в фенотипы виде
скрещивания и предлагаются второму занимающемуся для решения.
Составление приведенной выше задачи выразится в следующих этапах.
1. Выбор условий. Принимаем участие двух генов: А дает окраску,
а — неокрашенный, белый; В изменяет окраску при наличии А в серую,,
при ааВ не действует (криптомерия); у А и В — доминирование полное.
2. Составляем список генотипов (9) и фенотипов:
ААВВ — серый АаВ В — серый ааВВ — белый
ААВЬ — серый АаВЬ — серый ааВЬ — белый
ААЬЬ — черный АаЬЬ — черный ааЬЬ — белый
3. Для начального скрещивания выбираем, например:
ААЬЬ — черный
ааВЬ — серый
4. Расчет первого скрещивания:
ААЬЬ X ааВЬ -♦ АаВЬ + АаЬЬ
5. Принимаем частоты, приблизительно отвечающие теоретическому
расчету; вместо точного 1 АаВЬ : lAabb даем ЪАаВЪ + 2ЛаЬЬ.
6. Переводим в фенотипы: 3 серых + 2 черных и т. д.
Достоинствами ограниченных задач является простота их составления
и то, что они могут быть издаваемы в качестве сборников, даваться для
решения студенческим группам в размноженном виде и т. д. Однако эти за-
* Набор задач по генетике приведен также в более новых руководствах: А. М ю н т-
ц и г. Генетика. М., изд-во £«Мир», 1967; Щж. Брюбейкер.
Сельскохозяйственная генетика». М., изд-во «Колос», 1966.— Прим. ред.
47
дачи имеют очень крупные недостатки: 1) они лишают решающего задачу
инициативы в выборе тех или иных материалов, в «постановке» нужных ему
скрещиваний, 2) они открывают решающим задачу сразу весь материал,
тогда как при действительном решении задачи в исследовательской работе
материал раскрывается постепенно, по мере развития исследования.
Между тем для овладения методом генетического анализа особенно важна
выработка инициативы. Необходимо ставить скрещивания только такие,
которые вытекают из сделанных гипотез, должны дать ответ на
поставленный вопрос. Генетик-аналитик должен вырабатывать в себе умение решать
задачу с наиболее экономным расходованием материала и делать свои
доказательства предельно убедительными. Развитие инициативы поэтому
является для него очень важным.
НЕОГРАНИЧЕННЫЕ ЗАДАЧИ
Этому требованию удовлетворяет предлагаемый нами метод
«неограниченных» задач. Он состоит в своего рода диалоге между решающим
задачу и тем, кто задает ее. Составив свое условие, дающий задачу сообщает
решающему лишь данные по одному-двум первым скрещиваниям.
Например, в нашем случае сообщается:
«черный кролик № 1, скрещенный с белым № 2, дал два черных (№ 3
и 4) и три серых (№ 5, 6, 7)», причем заранее определяется пол каждого
номера. Получивший задачу должен сам заявить, какие дальнейшие
скрещивания он желает поставить, и дающий задачу в дальнейшем лишь
пассивно отвечает: «получено в результате то-то и то-то». При этом
обязательно, чтобы все «возникающие» животные получали определенные номера,
так как при сходстве фенотипов они могут иметь различные генотипы.
Соответственно и решающий задачу назначает «скрещивание № 15 с
№ 22» и т. п.
Решение задачи должно вестись в письменной форме с
формулированием сделанных гипотез и мотивов, заставляющих решающего ставить
то или иное скрещивание. После окончания решения дающий задачу
совместно с решающим или с несколькими решающими обсуждают
правильность различных решений, стройность доказательств, целесообразность и
нужность отдельных скрещиваний и т. д.
В неограниченных задачах требуется правильная работа дающего
задачу, т. е. сообщающего результаты поставленных скрещиваний.
Для того чтобы эти ответы подчинялись законам вероятности, мы
применяем следующую методику. Берется урна с шарами (карточками)
четырех цветов (например, белый, желтый, синий и зеленый), по 10 шаров
каждого цвета. Шары (карточки) перенумерованы от 1 до 9*. Набор шаров
дает возможность моделировать самые разнообразные «скрещивания»
путем последовательных тиражей. Методику эту мы с успехом применяли при
практических занятиях по геноанализу в МГУ, и, как увидим дальше, для
исследовательских целей при разработке теории геноанализа.
1. Пол
Назначаем: светлые — белые и желтые шары (светлые карточки,
красные карты) — самцы, темные (синие и зеленые) — самки. Если нам нужно
«получить» 7 потомков, выбираем по очереди 7 шаров и записываем по
порядку кто «родился». Например, № 103 — самец, № 104 — самец,
№ 105 — самка и т. д. Если мы хотим, чтобы отношение полов было не
Вместо карточек можно брать простые карты четырех мастей от туза до десятки.
48
1 : 1, а, например, 105 : 100, можно один синий шар удалить или лучше
просто не считать, тогда состав урны будет 20 светлых к 19 темным, или
105,1 : 100*.
2. Расщепление 1:1
Так же точно можно моделировать расщепление. Примем светлые
шары — А, темные — а. Если мы должны получить «потомство» от
скрещивания типа Аа X аау-^Аа + аа, выбрасываем гаметы гетерозиготы.
Если выпадает светлый шар, записываем Аа, если темный, записываем аа.
Аналогично поступаем и при других скрещиваниях, например Аа X АА.
Так как гомозигота АА дает все гаметы Л, то при выпадении светлого шара
записываем АА, темного —Аа.
3. Расщепление 1:2:1
В этом случае, при скрепщванииЛа X Аа, берем сразу два шара, в
результате чего выпадают четыре возможные комбинации: если выпадают
два светлых шара, записываем АА, если выпадают светлый + темный,
записываем А а (и аА), при двух темных записываем аа.
4. Ди- и полигибридные расщепления
Ди- и полигибридные расщепления обрабатываем таким же образом
отдельно для каждого гена. Пусть нам нужно получить «потомство» от
скрещивания A abb X АаВЬ. Сначала вынимаем два шара для А, так как здесь
скрещивание типа А а X Аа, затем один шар для В, так как здесь
скрещивание (по В) типа ВВ X ВЪ. Чтобы не ошибиться, удобнее сначала
моделировать для всех потомков «пол», потом «расщепление по-4», «расщепление
по В» и т. д.
В качестве примера приведем расчет одного скрещивания:
АаВЬ X АаВЬ
Тиражи (к — краен.,
ч —черн.)
Пол.
К
К
Ч
К
А
ЧЧ
кк
чк
чк
в
ч
к
ч
к
Записи
№ 103 cf ааВЬ
№ 104 cf ААВВ
№ 105 9 АаВЬ
№ 106 сГ АаВВ
АаВЬ X АаВЬ
Тиражи (к — краен.,
ч —черн.)
Под.
к
ч
к
А
кч
кч
ЧЧ
В
AAA
Записи
-
№ 107 cf АаВЬ
№ 108 ? АаВЬ
№ 109 с? ааВЬ
Вместо красный — черный можно с таким же успехом брать чет —
нечет. Еще удобнее оперировать сразу тем и другим, так как при этом один
шар дает сразу два независимых тиража. Например, светлый шар с
цифрой 10 дает сразу АА, так как он и чет и светлый.
Покажем, как можно подобным образом моделировать и всевозможные
осложнения.
* Если мы имеем дело с карточками или картами, то после каждого тиража стопка
карточек немного тасуется.
4 А. С. Серебровский 49
5. Неполное проявление
Допустим, мы хотим моделировать скрещивание при условии, что гете-
розиготы Аа в 20% случаев не проявляют действия Л и фенотипически
попадают в класс аа. Для этого назначаем, что те гетерозиготы, для которых
выпадает светлый шар с цифрой 1 и светлый шар с цифрой 2 или красная
двойка (два шара из десяти, т. е. 20%), будут такими «непроявленными»
гетерозиготами. Таких записываем, например, в скобках (Аа) и при
переводе в фенотипы приравниваем их к аа. Для случая непроявления в 10%
назначаем в «непроявленцы» только получивших светлый шар с цифрой 1»
Для 5% непроявления назначаем из светлых — желтый шар с цифрой 1
и т. д.
6, Нежизнеспособность гамет
Для моделирования, например, 10% гибели гамет назначаем в
«погибающие» темные шары с цифрой 1. Если выпадает темный шар с цифрой 1,
то тираж не записывается и вынимается новый шар.
7. Нежизнеспособность зигот
Пусть, например, мы задаемся условием, что 20% погибает. Если мы
приняли, что за аа отвечает более двух темных шаров, то можно назначить
в погибающие такие зиготы, для которых выпадает два темных шара
равного достоинства — с цифрой 1, с цифрой 2, с цифрой 3 и т. д.
Таких парных сочетаний 10, а всего возможно парных сочетаний темных
шаров 100. Таким образом «погибать» будет 10%. Отмирание в 25% может
быть обеспечено назначением на отмирание двух синих или двух зеленых
шаров. Сочетание темных шаров, в состав которых входит хотя бы один
шар с цифрой 1, будет 19% и т. д.
Раз овладев этой методикой моделирования, можно легко придумать
наиболее простые и удобные модели для всевозможных случаев, с
которыми можно встретиться в генетике. Создаваемые при этом модели (если
соблюдены необходимые правила возврата шаров и перемешивания) будуг
настолько строго отвечать стохастическим закономерностям, что материал
может подвергаться биометрической обработке и может быть определена
надежность сделанных гипотез и выводов и т. д. Исключительная ценность
этих материалов состоит в том, что в нужный момент могут быть вскрыты
все записи генотипов и проверены все шаги решавшего задачу.
В 1935/36 учебном году мы провели подобные упражнения по генана-
лизу со студентами-генетиками МГУ. Студенты были разделены на группы
по четыре человека. В каждой группе одна пара придумывала задачу,
которую решала вторая пара и обратно, вторая пара придумывала задачу
для первой пары. После утверждения задач руководителем началось их
решение и в конце решение было представлено в письменной форме с
изложением всего хода решения: всех делавшихся гипотез, их подтверждений
или опровержений. Большинство задач были дигибридными с тем или
иным взаимодействием генов и большей частью с каким-либо одним
осложняющим обстоятельством — или сцеплением, или нарушением
жизнеспособности, или с неполным проявлением.
Все задачи были решены и при этом обнаружилось очень ценное
обстоятельство — почти каждое решение несло черты оригинальности в подходе
к задачам. Так, одни студенты сосредоточили свое внимание на отыскании
рецессивных анализаторов и оперировали дальше при помощи этих
анализаторов. Другие старались наметить гипотезу, охватывающую сразу всю
картину в целом. Третьи обратились к изучению в первую очередь скрещи-
50
вания, дававшего наиболее разнообразное потомство. Четвертые
применили метод инбридирования и т. д. Хотя в настоящеее время далеко не всегда
мы можем сказать, какие из этих приемов более правильны и какие менее
правильны, но несомненно, что анализ ряда таких решений позволит осве<-
тить эту важную часть теории генанализа.
Объективным показателем правильности или неправильности приемов
является количество материалов, которое потребовалось для решения
данной задачи. Если при решении одной и той же задачи, но разными
приемами, одному генетику-аналитику потребовалось получить 1000 «потомков»,
а другому лишь 50 или 100, то надо будет заключить, что приемы второго
генетика более правильны, более рациональны, идут к цели более
прямыми путями, более логичны.
Необходимо подчеркнуть, что логичность построения гипотез и их
проверки является важнейшим моментом в работе генетика-аналитика.
Изучая указанные решения задач, можно было отчетливо видеть, как
решавший попадал на ложный путь, допустив недостаточно обоснованную
гипотезу или не заметив, что из данного факта можно сделать не один вывод, а
два, или предположив более сложную гипотезу там, где была возможна
простая и т. д. В этом отношении решение задач по генетическому
анализу является отличной школой логики.
Призер решения задачи. Мы даем образец решения подобной
неограниченной задачи.
Полную запись скрещиваний см. ниже, в табл. 6. Эта таблица является
как бы журналом вивария (отсутствуют лишь даты рождения). Первые
4 столбца не требуют объяснения. Последний столбец — номера
нисходящих скрещиваний — указывает, в каких скрещиваниях данное животное
участвовало. Например, против 1 указано 1, 2 и 8. Действительно,
отыскивая эти скрещивания по левому столбцу, в соседнем столбце (№ отца) при
этих скрещиваниях видим запись «№ 1». 71—80, 81—89, 128—151
записаны в строчку лишь ради экономии места для печати.
Даны три грызуна: темно-серый № 1 и два серых № 2 и № 3.
Скрещивание 1 с? X 92 дало:
темно-серых ... 4
серых 5
Скрещивание 1 с? X $ 3 дало:
темно-серых ... 4 светло-серых ... 1
серых 3 черных 2
Найти. Сколько генов участвует в расщеплении? Как действует
каждый из генов? Каковы формулы исходных животных?
Решение. Первое скрещивание дало результат, очень близкий к 1 : 1,
что указывает на возможность моногибридного расщепления типа АаХаа*
Отсюда, естественно, вытекала бы постановка скрещивания темно-серых
друг с другом и серых друг с другом для выяснения того, кто именно —
темно-серые или серые — являются гетерозиготными.
Однако уже скрещивание № 2 (1 с? X $ 3) осложняет картину, так как
мы получаем:
темно-серых ... 4 светло-серых ... 1
серых 3 черных 2
Здесь мы видим появление сразу двух новых, неожиданных
фенотипов — светло-серых и черных. Мы получили, следовательно, четыре
фенотипа, что невозможно при моногибридном различии между серыми
и темно-серыми. Здесь несомненно участие не менее двух генов. При этом
условии легко объяснимо и то, что $ 2и 9 3 при сходстве фенотипов дали
51
4*
различное потомство. Следовательно, суммировать результаты обоих
скрещиваний рискованно и даже недопустимо, и придется вести дальнейший
анализ отдельно для семей 1 X 2 и 1 X 3.
Как следует провести дальнейшее скрещивание?
а. Первое расщепление проще, с него следует начать. Поставим
скрещивание темно-серых с темно-серыми (7 (J X 5 §, 8 $), а серых с
серыми (4 rf X 9 $, 10 $).
б. Из скрещивания № 2 обратим внимание на новый фенотип —
черный. К сожалению, оба черных — самцы. Поэтому придется скрестить
одного из них с матерью, так как при этом можно рассчитывать на получение
-большего процента черных (14 с? X 3 £)•
Также используем единственную светло-серую № 13, скрестив с отцом
<1 с? х 13 $).
Использование темно-серых и серых из скрещивания № 2 пока ничем
не диктуется.
Ставим намеченные скрещивания (№ 3 и № 4). Оба скрещивания
темно-серых друг с другом (3 и 4) дают сходную картину:
Скрещивание Темно-серые Серые Черные
2. 7 с? X 5 9 3 1 3
4. 1 d X8 9 3 2 4
(Г 3 7
Несомненно, что темно-серые являются гетерозиготами. Наличие трех
«фенотипов говорит о моногибридном характере с неполным
доминированием или о дигибридном с недостатком одного из фенотипов. По первой
моногибридной гипотезе следовало бы ожидать 1:2:1. При этом, так
как темно-серые не могут быть гомозиготами, следовало ожидать:
1 серый : 2 темно-серых: 1 черный
или 4±1,7 8±2 4±1,7
Получено 3 6 ^7
Фактическая картина не очень хорошо отвечает ожидаемой (избыток
черных), но при таком небольшом числе это противоречие невелико.
Посмотрим, что дали остальные скрещивания.
Серые X серые (скрещивание № 5 и скрещивание № 6) дали 15 серых.
Это делает очень вероятным, что серые являются гомозиготами, что
соответствует и данным только что рассмотренных скрещиваний темно-серых.
Однако если серые — гомозиготы, то странно, как могли одинаковые серые
самки № 2 и 3 оказаться различными генотипически (ср. скрещивания
1и2)?
Скрещивание (№ 7) черного самца (14) с его матерью (3) дало:
серых 3
черных 3
коричневых •) 2
Этот результат совершенно неожиданный: появился еще новый фено^
тип — коричневый. Мы имеем, следовательно, уже пять фенотипов:
темно-серый, серый, светло-серый, черный и коричневый. Число фенотипов
более четырех указывает либо на участие более двух генов, либо на
неполное доминирование одного или обоих генов, что при дигибридном
скрещивании может дать шесть или девять фенотипов.
Наконец, последнее из поставленных скрещиваний (№ 8): темно-серый
1 (51 Х13 9 ~ светло-серая дало:
темно-серых 3
серых 3
светло-серых 4
52
Результат малопонятный, но ценный тем, что дал нам группу
светлосерых.
Подводя итог поставленным скрещиваниям, мы переходим к выводам.
1. Скрещивание по меньшей мере дигибридное, а если дигибридное,
то с неполным доминированием хотя бы одного гена (так как число
фенотипов более четырех).
2. Скрещивание в семье 1x2 более простое и возможно, что лишь
моногибридное, хотя это требует подтверждения.
3. Темно-серые несомненно — гетерозиготы, так как во всех
скрещиваниях (скрепщвания 1, 2, 3, 4 и 5) дают расщепление.
4. Серые — более гомозиготы, так как в скрещиваниях 5 и 6 не дали
никакого расщепления. Однако генотип серых, по-видимому, может быть
различным (ср. 2 $ и 3 9)-
Начнем также скрещивания:
а. Исследование новых фенотипов:
1) черных с черными (14 с? X 54 $ и 14 tf X 60 §);
2) коричневых с коричневыми (57 с? X 55 9)1
3) светло-серых со светло-серыми (63 с? X 62 $ и 63 с? X 70 $).
б. Проверка отношения 1:2:1 в семье 1 с? X 2 $ (повторение
скрещивания 3 и 4).
Так как эти скрещивания должны дать несомненные новости, то
подождем их результатов.
Результаты:
а. Черные X черные (скрещивание 9 и 10) дали 19 черных без всякого
расщепления, что подтверждает сделанное выше предположение о
гомозиготном характере черных.
б. Коричневые X коричневые (скрещивание И) дали:
коричневых 4
черных 4
белых 1
Опять новый фенотип — белый! Таким образом, мы имеем уже шесть
фенотипов. Очевидно, что белые являются извлеченным рецессивом аа по
сравнению с окрашенными. Правда, отношение 8 + 1 не вполне отвечает
3 : 1 или 6 : 2, но при малом числе возможно истолковать его как
моногибридное. Тогда коричневые — Аа и черные — АА, что совпадает с данными
предыдущих скрещиваний о гомозиготности черных.
в. Светло-серые X светло-серые (скрещивание 12 и 13) дали:
светло-серые серые белые
12. 63 в X 62 9 5 1 2
13. 63 в X 70 9 3 2 2
8 3 4
Снова извлеклись белые! Оба скрещивания очень сходные и их можно
суммировать. При этом получаем опять отношение, очень близкое к 1 : 2 :
: 1, именно 3 серых : 8 светло-серых : 4 белых. Невольно напрашивается
сопоставление с предыдущим скрещиванием коричневых — в обоих
скрещиваниях извлекались белые, в обоих скрещиваниях имеется по три
фенотипа, в обоих скрещиваниях отношение может быть принято за
моногибридное 1:2:1. При этом в обоих случаях фенотип скрещенных родителей
стоит посредине между двумя появившимися в потомстве новыми
фенотипами:
коричневый — между белым и черным
светло-серый — между белым и серым
53
Отсюда мы можем построить гипотезу:
аа — белые
Аа— светло окрашенные
АА— ярко окрашенные
При этом имеем два ряда — серый и черный:
Серый
аа 4 (бел.)
Аа 8 (светло-сер.)
АА 3 (сер.)
Черный
1 (бел.)
4 (кор.)
4 (черн.)
Всего
5
12
7
Теоретически
6±2,12
12±2,45
6±2,12
Картина начинает проясняться. Гипотеза, очевидно, очень подходит
к полученным результатам и объясняет пять фенотипов: белый,
светло-серый, серый, коричневый и черный. Остается сделать гипотезу об
отношении черного и серого рядов друг к другу. Так как мы начали наши
скрещивания с темно-серого с? и серых $ $ ,' а черные извлекались, то можно
принять черных за рецессивных, а серых за доминантных. Обозначив
серый через 5, получим такую гипотезу:
Я-ряд Ь-ряд .
ааВЬ — белые ааЬЬ — белые
АаВЬ — светло-серые АаЬЬ — коричневые
ААВЬ — серые ААЬЬ — черные
Без гипотезы остается лишь фенотип темно-серых. Прежде чем его
истолковывать! суммируем новые скрещивания темно-серых друг с
другом. Всего получаем:
Скрещивания
3. 7 сГ Х5 9
14. 7 сГ X 5 9
4. 7 сГ Х8 9
15. 7 сГ Х8 9
Ожидалось при 1:2:1
Потомство
серые
1
4
2
0
7
7,5
темно-
серые'
3
3
3
4
13
15
черные
3
1
4
2
10
7,5
Новые данные о темно-серых качественно не отличаются от прежних
(иных фенотипов не появилось). По-прежнему чаще всего встречаются
темно-серые, а серые и черные примерно одинаково часты. Картина еще более
приблизилась к моногибридной с промежуточным доминированием. Если
принять такую схему, то черные, темно-серые и серые, очевидно, можно
будет передать через ЬЬ, ВЪ и ВВ. Так как белые {аа) не извлекаются, то,
следовательно, все они гомозиготны по А А.
Гипотеза будет такова:
серые ААВВ
темно-серые ААВЬ
черные ААЬЬ
Если это правильно, то сопоставленный выше серый ряд (5-ряд) и
черный ряд (Ь-ряд) надо написать как 55-ряд и ЬЬ-ряд, а отсюда вытекает
возможность и 5Ь-ряда, из которого мы знаем член ААВЬ.
54
Сделаем сопоставление этих рядов:
Я#-ряд БЬ-ряд 6&-ряд
ааВВ (белый) ааВЬ ааЬЬ (белый)
АаВВ (светло-серый) АаВЬ АаЬЬ (коричневый)
ААВВ (серый) ААВЬ (темно-серый) ААЬЬ, черный)
Остаются неизвестными два фенотипа, отвечающие генотипам ааВЬ и
АаВЬ. Первый из них аа, и по аналогии с ааВВ и ааЪЪ, между которыми он
стоит, должен быть белым. Фенотип АаВЬ — неизвестен, по аналогии
между светло-серыми и коричневыми он должен быть светлее ниже стоящего
члена АА, т. е. светлее темно-серого, очевидно каким-то серым. Но для
серого мы уже дали формулу ААВВ. Этот серый стоит, однако, в другом
ряду, 55-ряду,, а серый (АаВЬ) должен стоять в 5Ь-ряду. Чем же он
отличается от ААВВ серого? Необходимо получить этот фенотип
скрещиванием. Очевидно, что генотип АаВЬ можно получить двумя способами:
1) ААВВ X ааЬЬ -» АаВЬ 2) ААЬЬ X ааВВ -» АаВЬ
(серый) (белый) (какой?) (черный) (белый) (какой?)
Для осуществления этих скрещиваний необходимо иметь белых ааЬЬ
и ааВВ. Такие белые в нашем распоряжении уже имеются: ааЬЬ есть белый,
возникший от скрещивания коричневых друг с другом (белая 97 $ из
скрещивания 11), а белые ааВВ должны были возникать в скрещивании
светлосерых друг с другом (белые 99 $ , 101 $ , 109 $ и 110 §). Мы,
следовательно, можем проверить нашу гипотезу, и она будет верна, если в обоих
скрещиваниях возникнут одинаковые фенотипы, при этом в 100%, и будут
какого-то серого оттенка, светлее темно-серых.
Назначим следующие скрещивания с предполагаемыми генотипами:
Скрещивание № 16: $97 ааЪЪ (белая) X 6 Мб ААВВ (серый)
» № 17: $99 ааВВ (белая) X d 14 ААЬЬ (черный)
» № 18: $ 107 ааВВ (белая) X 6 98 ААЬЬ (черный)
Результаты оказываются следующими.
Во всех скрещиваниях получились исключительно серые потомки.
Ожидавшийся одинаковый результат оправдался. Отсутствие расщепления
также оправдалось. Полученный фенотип оказался более светлым, чем
темно-серый, и тоже оправдал ожидание. Несколько неожиданным
оказалось лишь то, что появившийся фенотип оказался уже знакомым ранее
«серым», ничем не отличающимся от серого ААВВ. Однако это прекрасно
отвечает тому заключению, которое нам пришлось сделать в самом начале
анализа, что серые $ 2 и $ 3, несмотря на внешнее сходство, генетически
различны. Очевидно, одна из них была ААВВ, другая АаВЬ. Можно даже
установить, какая из самок несла какой генотип. Так как темно-серому
самцу должен быть дан генотип ААВЬ, то его скрещивание с генотипом
ААВВ и АаВЬ должно дать такие результаты:
ААВЬ X ААВВ -* ААВВ + ААВЬ
(темно-серый) (серый) (серый) (темно-серый)
ААВЬ ХАаВЪ-*ААВВ +2ААВЬ +ААЪЬ
(темно-серый) (серый) (серый) (темно-серый) черный
+АаВВ +АаВЬ +2АаЬЬ
(светло-серый) (серый) (коричневый)
Очевидно, скрещивание 1 tf X 2 J, давшее 5 серых и 4 темно-серых,
близко совпадает с первым расчетом, а скрещивание 1с? X 3 $, давшее
3 серых, 4 темно-серых, 1 светло-серый и 2 черных, совпадает со вторым
расчетом. Следовательно, 2 $ имеет генотип ААВВ, а 3 $ — АаВЬ.
55
Таблица 6
Скрещивание и их результаты
(модельная задача)
щивания
1
2
3
'
4
-
5
JVft животного, фенотип и генотип
отец
1
темно-серый
ААВЬ
1
темно-серый
ААВЬ
7
темно-серый
ААВЬ
7
темно-серый
ААВЬ
4
серый
ААВВ
мать
2
серая
-4ЛВЯ
3
серая
АаВЬ
5
темно-серая
ЛАВ&
8
темно-серая
1 ААВЬ
9
серая
ААВВ
№ животного, пол, фенотип
и генотип
1 с? темно-серый ААВЬ
2 9 серая ААВВ
3 9 серая <4а#6
4 с? серый А4ЯЯ
5 9 темно-серая ААВЬ
6 с? серый ААВВ
7 с? темно-серый ААВЬ
8 9 темно-серая AAZu
9 9 серая ААВВ
10 9 серая ААВВ
11 9 темно-серая ААВЬ
12 с? серый ААВВ
13 9 светло-серая Ла£2?
14 с? черный ЛЛ66
15 с? темно-серый А42?6
16 с? серый ААВЬ
17 с? серый АаВЬ
18 9 темно-серая ЛАЗЬ
19 9 темно-серая ААВЬ
20 с? черный ЛЛ66
21 с? темно-серый ААВЬ
22 9 серая АаВЬ
23 с? черный АА&Ь
24 9 черная ААЬЬ
25 9 серая ААВВ
26 9 черная ААЬЬ
27 9 темно-серая ААВЬ
28 9 темно-серая ААЯЬ
29 9 темно-серая ААВЬ
30 9 черная ААЬЬ
31 с? серый ААВВ
32 с? серый ААВВ
33 с? черный ЛЛ66
34 9 темно-серая ААВЬ
35 с? черный ЛЛ66
36 с? темно-серый ААВЬ
37 9 черная ААЬЬ
38 9 темно-серая ААЯЬ
39 9 серая ААВВ
40 $ серый А452?
41 9 серая ААВВ
42 с? серый ААВВ
43 с? серый AA&S
44 с? серый А42?2?
45 <$ серый А4ДЯ
! 46 9 серая ААВВ
№
нисходящих скрещи-J
взний
1, 2, 8
1
2, 7
5, 6
3, 14
3, 4, 14, i5
4, 15
5
6
8
7, 9, 10, 17
56
Таблица 6 (продолжение)
№
щива-
ния
6
7
8
9
10
11
12
№ животного, фенотип и генотип
отец
4
серый
ААВВ
14
черный
А<Ш
1
темно-серый
ААВЬ
14 черный
ААЬЬ
14 черный
57
коричневый
АаЬЬ
63
светло-серый
4аЯ£
мать
10
серая
ААВВ
3
серая
АаВЬ
13
светло-серая
54 черная
ААЬЬ
60 черная
ААЬЬ
55
коричневая
Лабб
62
светло-серая
АаВВ
Ка животного, пол, фенотип
и генотип
47 9 серая ААВВ
48 с? серый ААВВ
49 9 серая ААВВ
50 9 серая А4ЯЯ
51 9 серая ААВВ
52 9 серая ААВВ
53 с? серый ААВВ
54 9 черная ААЬЬ
55 9 коричневая Лабб
56 с? черный ААЬЬ
57 с? коричневый Ла2%
58 9 серая АаВЬ
59 9 серая АаВЬ
60 9 черная А466
61 с? серый АаВЬ
62 9 светло-серая АаВВ
63 с? светло-серый АаВВ
64 9 серая АаВЬ
65 с? темно-серый A4Zft
66 с? серый Ла£6
67 с? темно-серый ААВЬ
68 с? светло-серый АаВВ
69 9 темно-серая ААВЬ
70 9 светло-серая Ла2?2?
71—80 черные ААЬЬ
81—89 черные ААЬЬ
90 с? черный ЛЛ66
91 с? черный 4466
92 с? черный Л466
93 9 коричневая АаЬЬ
94 с? коричневый АаЬЬ
95 9 коричневая Ла&6
96 с? серый ААЬЬ
97 9 белая ааЬЬ
98 с? черный ААЬЬ
99 9 белая ааВВ
100 с? светло-серый Ла55
101 9 белая ааВВ
102 с? светло-серый АаВВ
103 с? светло-серый АаВВ
104 9 светло-серая ЛаЯб
105 с? светло-серый ЛаЯЯ
106 с? серый ААВВ
J4&
нисходящих
скрещивании
9
11
11
10
12
12, 13>
IS
57
Таблица 6 (окончание)
№
скре»
щива-
ния
13
14
15 .
16
17
18
Jsis животного, фенотип и генотип
отец
63
светло-серый
АаВВ
7
темно-серый
ААВЬ
7
темно-серый
ААВЬ
106 серый
ААВВ
14 черный
AAbb
98
черный
AAbb
.
мать
70
светло-серая
АаВВ
5
темно-серая
ААВЬ
8
темно-серая
ААВЬ
97 белая
aabb
99 белая
аа££
101
белая
ааВВ
№ животного, пол, фенотип
и генотип
107 с? серый ААВВ
108 9 светло-серая АаВВ
109 9 белая ♦ ааВВ
110 9 белая ааВВ
111 9 светло-серая АаВВ
112 9 светло-серая АаВВ
113 с? серый ААВВ
114 9 серая ААВВ
115 с? темно-серый ЛАВ6
116 & серый 4АВЯ
117 9 черный AAbb
118 с? черный AAbb
119 9 серая ААВВ
120 9 серая А4&В
121 б темно-серая ААВЬ
122 9 темно-серая ААВЬ
123 с? темно-серый ААВЬ
124 9 черная- AAbb
125 9 темно-серая ЛЛ#6
126 (J темно-серый ААВЬ
127 сГ черный AAbb
128—136 серые АаВЬ
137—143 серые АаВЬ
144—151 серые АаВЬ
.№
нисходящих
скрещиваний
Анализ может быть закончен постановкой еще некоторых добавочных
шроверочных скрещиваний, например, получение коричневых в
скрещиваниях черных с белыми типа aabb и т. д.
ГЛАВА IV
ЛОКАЛИЗАЦИЯ ГЕНА
ПОСТАНОВКА ВОПРОСА
После того, как существование гена установлено, дальнейшее его
изучение идет по двум направлениям. Во-первых, путем комбинирования с
другими генами и изменений внешних условий подробно изучаются
свойства гена как фактора онтогенеза, о чем мы уже говорили выше. Во-вторых,
не дожидаясь окончания этого изучения, а параллельно с ним, должен
ставиться вопрос о локализации гена. Под этим разумеется установление:
1) в какой из хромосом локализован ген; 2) на каких расстояниях он
расположен от других генов, ранее локализованных в этой хромосоме;
3) в каком порядке эти гены расположены, или конкретно — между
какими из ранее известных генов расположен вновь изучаемый ген.
Довести изучение гена до получения ответа на эти вопросы важно по
следующим причинам. Во-первых, суждение о гене по его фенотипическо-
му проявлению не всегда надежно. Мы видели, хотя бы на примере tifa-
гена курицы, что это проявление может меняться до неузнаваемости. С
другой стороны, фенотипическое проявление различных генов может быть до
неразличимости сходным. Так, например, у кролика мы знаем уже три
различных гена, вызывающих укорочение ости шерсти, характерное для
«короткошерстных» кроликов реке. Эти три типа рексов не различимы по
<фенам, но их гены различно локализованы.
У кур есть два сходных типа альбинизма. У дрозофилы почти не
различимы алые окраски глаз vermilion, scarlet и cinnabar, вызываемые
совершенно различными генами, и т. д. И поэтому окончательная
характеристика гена должна содержать указание на то, что он участвует, например,
в вызывании «алой окраски глаза», и на то, что он расположен во //
хромосоме на 57,7 морганиде. В этом случае ясно, что речь идет о гене
cinnabar, а не о каком-либо ином.
С другой стороны, точная локализация гена важна и для целей самого
генетического анализа. Установив, что ген Л локализован в таком-то
пункте такой-то хромосомы, мы получаем хорошее орудие для дальнейшего
изучения свойств прилегающих участков этой хромосомы. Ген А
становится «сигнальным» и позволяет решать целый ряд дальнейших задач
генетического анализа, в частности в области количественных признаков.
ЛОКАЛИЗАЦИЯ В ПОЛОВОЙ ХРОМОСОМЕ
Первым вопросом, на который приходится отвечать при подходе к
локализации гена, это вопрос о том, не локализован ли ген в
половой хромосоме. Вопрос этот следует ставить первым потому,
59
что он может быть разрешен даже в том случае, когда у данного вида
животных ни одного другого гена неизвестно, и, кроме того, потому, что оя
решается очень легко. Лишь по получении отрицательного ответа на этот
вопрос целесообразно планировать длительные и обширные исследования,
необходимые для установления локализации генов аутосомных, т. е.
локализованных в одной из аутосом, неполовых хромосом.
Классическое разрешение вопроса о том, локализован ли ген в половой
хромосоме или в аутосоме, сводится к постановке двух реципрокных
скрещиваний. Реципрокными скрещиваниями называются такие, когда
изучаемый признак в одном случае имеется у самца (и отсутствует у самки),
в другом случае имеется у самки (и отсутствует у самца).
Если реципрокные скрещивания (сохраняем первичный термин)
дают одинаковый результат, то изучаемый ген локализован в аутосоме.
Если же реципрокные скрещивания дают различный результат и в
частности в одном из реципрокных скрещиваний отцовский признак переходит
только к дочерям или только к сыновьям, а материнский — только к
сыновьям или только к дочерям, то изучаемый ген локализован в одной из
половых хромосом.
В этих случаях говорят, что признак сцеплен с полом. Возможны
следующие варианты этого явления (см. схемы ниже).
1. В одном реципрокном скрещивании (схема II, 1) изучаемый
признак (зачернен), имевшийся у самца, исчезает в Fl9 а в другом
реципрокном — переходит от матери к сыновьям. Это служит указанием на то, что
признак рецессивен, и его ген расположен в Х-хромосоме. Подобные
случаи наблюдаются у млекопитающих, рыб, мух, жуков, у которых мужской
пол имеет половые хромосомы X и Г, а женский — X и X, и могут быть
пояснены схемой II, 2 с половыми хромосомами.
сП>
D-1-O Р-гС
а _
XY
Г"
А
Фено- XY
типы: А
-АА
XX
I
Аа
XX
А
А _
XY
а
XY
_ аа
XX
I
Аа
XX
А
Схема II
а _
XY
_Аа
XX
1 1
А а Аа аа
XY XY XX XX
А а А а
Этот случай осложняется существованием самок А а, дающих как
сыновей, так и дочерей с А и с а (на схеме II, 3 справа).
2. В одном реципрокном скрещивании (схема III, 1) изучаемый
признак (черный) переходит от отца к дочери, в другом — от матери ко всему
потомству. Это указывает на то, что изучаемый признак доминантен и
локализован в Х-хромосоме, что поясняется схемой III, 2 (слева и в
середине).
Этот случай осложняется существованием самок Аа, дающих как
сыновей, так и дочерей с А и с а (на схеме III, 3 справа).
3. В одном реципрокном скрещивании (схем$ IV) признак отца ослаб-
ленно переходит к дочерям, а признак матери более или менее точно
повторяется у сына. Во втором реципрокном — признак матери опять точно
переходит к сыну и ослабленно — к дочери. Заметим, что здесь оба
скрещивания сильно похожи в том отношении, что сын повторяет точно мать»
а дочь оказывается промежуточной между отцом и матерью. Различие
60
■о
п-
-о
ЛЬ £б £Ъ5Ъ
АА.
ZZ
Аа
$i%'ZA
_а аа_
ZW ZZ
а
ZW
А
Аа
ZW
А
_А
ZW
а
ZW
Аа.
ZZ
ZW
Аа
ZW
А
1 1 1
аа А а
ZZ ZW ZW
а А а
Схема III
скрещиваний в том, что во втором реципрокном скрещивании средняя для
всего потомства степень признака значительно выше, чем в первом. Эти
различия указывают на то, что в Х-хромосоме имеется не полно
доминирующий ген.
■о о
Схема IV
4. Этот случай, похожий на предыдущий, отличается тем, что
промежуточный характер дочерей выражается в пятнистом проявлении признака.
Примером такого случая являются реципрокные скрещивания черных и
рыжих котов и кошек, где возникающие дочери получают пятнистую чер-
до-рыжую окраску, отражающую гетерозиготное состояние (схема V).
B-r-OD-r-O
Сх'ема V
5. В первом из реципрокных скрещиваний (схема VI, 1) признак отца
передается его сыновьям, во втором — признак матери передается всему
потомству. Это указывает на то, что в первом скрещивании ген изучаемого
признака локализован в половой У-хромосоме, а во втором случае — в
половой Х-хромосоме (схема VI, 2). При этом, однако, возможно отыскание
ж третьего типа скрещиваний, подобного тому, который мы видели в
случае № 2 (стр. 60) — признак переходит от отца к дочери.
Этот случай осложняется возможностью существования самцов XY
(АА), дающих все потомство с А, и существования самок XX (Аа), дающих
как сыновей, так и дочерей с А и с а.
Хромосомные схемы этих пяти скрещиваний, расшифровывающих
характер наследования, показаны на схеме VI, 2.
Примеры таких скрещиваний мы находим в опытах с жуком-листоедом
Phytodecta variabilis и у дрозофилы с участием гена bobbed,
локализованного как в Х-, так и в У-хромосомах.
61
aA.
XY
-О D-
■O
■O O-
П i4 £* Л Щ*
XX XY
.AA Aa_
XX XY
_ aa AA_
XX XY
XX
aa_
XY
_Aa
XX
Г- I I ■ I I ' 1 | 1 | f—y ,
j£ aa Aa Aa aa Aa a A Aa Aa Aa Aa aa
Фено-Хлу XX XY XX XY v XX XY XX XY XY XX XX
типы: A a A A * A A „.-. A A A A a
Схема VI
6, В шестом случае признак в одном скрещивании переходит от отца
к дочери, а в другом скрещивании исчезает в потомстве (схема VII, 1).
Случай этот похож на случай № 1 (стр. 60), с той лишь разницей, что
всюду самцы заменены самками, и самки — самцами. Объяснение этому
случаю сходное же (схема VII, 2): изучаемый признак рецессивен и его гевг
локализован в половой хромосоме, но все строение хромосомного полооп-
ределяющего аппарата иное — такое, как у птиц и бабочек. Здесь
строение самца ZZ и самки ZW, или, если не делать различия в употреблениях
букв, самца XX, а самки XY, в противоположность млекопитающим и
мухам, у которых самцы были XY, а самки XX. По такой схеме у кур
наследуется рецессивная зеленая окраска ног в скрещиваниях с желтоногими
курами. Этот случай тоже осложняется наличием самцов Аа, дающих как
сыновей, так и дочерей с А и с а.
»
6
аа
ZZ
2
г°
о
А
ZW
Аа
Фено-2.г
miinu: A
1
а
ZW
а
О-
АА
ZZ
I—
Аа
ZZ
А
-Ъ
ZW
D-
■Ъ
6^ Й"16^>
А
ZW
А
Аа_
ZZ
ZW
I ' 1—I
Аа аа А а
ZZ ZZ ZW ZW
А а А 4
Схема VII
7. Этот случай (схема VIII, 1) идет в сравнении с № 2 (стр. 60), но
также с заменой самцов на самок, а самок на самцов. Он указывает b&ZW —
■о
D-
D-
6ё hi £b¥b
А _
XY
Феио-ХУ
типы: а
_аа
XX
Аа
XX
А
а _
XY
А
XY
А
АА
XX
—I
Аа
XX
А
а _
XY
-Аа
XX
П
I 1 Г-
А а Аа аа
XY XY XX XX
А а А а
Схема VIII
62
тип полоопределяющего аппарата и на доминантный характер изучаемого-
признака (схема VIII, 2). Так наследуется, например,
поперечнополосатый рисунок пера кур плимутрок в скрещивании с черными.
8. Случай сравнивается с № 3 и соответствует неполному
доминированию гена, локализованного в Z-хромосоме. Схема его такая же, как в>
№ 7, но гетерозиготные Аа сыновья не полно проявляют признак.
Подобные случаи могут быть встречены у птиц и бабочек (схема IX).
ЮО
-0
mm 6
Схема IX
9. Случай, параллельный четвертому — с мозаикой, но строение
самцов А а.
ЪЪ
10. Могут быть ожидаемы также и случай наследования через FF-xpo-
мосому в параллель тому, что изображено для ХГ-системы в случае пятом
(схема X). ' , ' • .
-о
ОП-г-О П-п-О D
П Л) ЛбЪ сГб hh
Схема X
Схема, показанная на рис. 8, позволяет определить, с каким случаем^
сцепленной с полом наследственности мы имеем дело. Построение схемы
следующее. Приравняем изучаемый нами признак единице (1), а аллело-
морфный ему — 0. Тогда скрещивание 1 (J X 0 § будет обозначать
скрещивание, в котором самец имеет изучаемый нами признак, а самка —
нет. Такие скрещивания отнесены в левую половину чертежа, а реципрок-
ные им (0 (J X 1 $ ) - в( правую. Если сыновья все получили|данныж
1**0%
0 1/2
М дочерей .
0**1%
л
•
г
ТС/
i, . „
i i
9&\
У S^-'
У У
>/>Ч. <У
y*L/ \6jr *
1 - . .1 ll
1 1 T "
"JZsK J© ®@"
*^^ /^ / S 1
у ч / / ,
уУ \ / / '
уУ Ч / / 1
- ^У^У \ !
^^^ y' N '
'^У^ у N 1
Ц&С Й -
^fwK. - - irnh
' i i i
</2
'I
Y/2 I
Рис. 8. Схема для определения различных случаев сцепления- с полом
по результатам репипрокных скрещиваний
Объяснение в тексте
63
признак, их М = 1, если все не получили, то М = 0, если идет
расщепление, то их М = 0,5. То же для дочерей. Результат скрещивания
изображен кружком, поставленным так, что он отвечает величинам М
сыновей и М дочерей. Кружки, относящиеся к реципрокным скрещиваниям,
соединены линиями, сплошными для типа XY и пунктирными для типа ZW.
Номер в кружке указывает на номер случая, описанного выше.
Пользование схемой можно разобрать на следующих примерах.
Пример первый. Пусть мы изучаем наследование серебристой окраски
кур. Приравниваем серебристый к 1 и золотистый к 0. Скрестив
серебристого петуха с золотистой курицей (1 (J X 0 £ ), получаем в потомстве
всех сыновей и дочерей серебристых (Мс = 1, Мд = 1). В левой половине
рис. 8 соответственно Мс = Мд = 1 видим кружки с №№ 5, 7 и 10.
Следовательно, наш случай относится либо к 7-му, либо к 10-му типа ZW, либо
к 5-му типа XY.
Случай 10 отличается, впрочем, от 7-го тем, что в 10-м ген изучаемого
признака может перейти в W-хромосому и передаваться только по женской
линии. Но раз мы имеем дело с серебристым петухом, то, следовательно,
искомый признак наследуется, как в случае 7, и серебристый цвет вызван
доминантным геном, локализованным в Z-хромосоме, либо как в случае 5.
Для того чтобы доказать, что мы имеем дело со сцепленным с полом
случаем, необходимо сравнение с реципрокным скрещиванием. Здесь
получаем всех сыновей серебристых (Мс = 1), дочерей золотистых (Мд = 0).
Разница между результатами реципрокных скрещиваний налицо,
следовательно, мы в праве пользоваться таблицей. В первой ее половине,
соответственно Мс = 1 или Мд = 0, находим три кружка (1,7 и 10). Случай 1
отпадает, так как для него невозможен результат Мс = 1 и Мд = 1,
полученный в первом реципрокном скрещивании. Остаются те же 7 и 10, между
которыми мы уже сделали выбор.
Пример второй. Исследуем признак «узкие глаза» (М = 1) у
дрозофилы. В скрещивании 1 с? X 0 9 (0 — круглые глаза) получаем: все
сыновья круглоглазые (Мс = 0), а все дочери с глазами промежуточными
(Мд = 1/2). В левой половине рис. 8 соответственно Мс = 0 и Мд = 1/2
находим кружок с № 3 и 4. Следовательно, мы имеем дело с типом XY и со
случаями либо 3, либо 4; но так как случай 4 относится к мозаикам, то
остается лишь случай 3 — тип XY с промежуточным доминированием. Для
проверки ставим второе реципрокное скрещивание (0 с? X 1 $ ) и
получаем сыновей с узкими глазами (Мс = 1) и дочерей с промежуточными
(Мд = 1/2). Смотрим правую половину схемы и, действительно, находим
в соответствующем месте слева кружок с № 3 и 4, что подтверждает
правильность диагноза.
От наследования, сцепленного с полом, очень важно отличать
наследование, ограниченное полом. Последний термин относится к тем
случаям, когда признак проявляется только у одного пола
(борода мужчин, вымя коровы, черная грудь у петухов дикой окраски). При
ограничении полом нельзя, вообще говоря, поставить реципрокные
скрещивания, так как нельзя, например, скрестить быка, имеющего развитое вымя,
с коровой, вымени не имеющей. Однако следует иметь в виду, что в
некоторых случаях невозможно отличить ограниченную полом наследственность
от сцепленной с полом, когда доминантный признак локализован в У- или
WP-хромосоме. В самом деле, если все особи вида имеют, например, в W-
хромосоме данный ген, а в Z-хромосоме он не встречается, то реципрокные
скрещивания поставить нельзя и нельзя этот случай отличить от
ограниченности полом. Так, например, как упомянуто, у кур грудь «светлая»
(оставляя в стороне некоторые окраски, например, черную или белую и т. п.),
а у петухов «черная» (также оставляя в стороне некоторые окраски —
белую, рыжую, уничтожающие половой диформизм). Обычно это
трактуется как пример ограничения полом, тем более, что при кастрации самки
64
она развивает петушиный окрас, чем доказывается зависимость «светлой»
груди от гормона, вырабатываемого яичником. Иными словами,
допускается, что половой диморфизм вызывается только генами, определяющими
пол.
Однако не исключена возможность и того, что в W-хромосоме курицы
локализован специальный ген, влияющий хотя бы и через половую
железу на окраску. В том случае, если в этом гене произойдет мутация и
появятся черногрудые же куры (самки), этот вопрос будет выяснен, а пока
остаются обе гипотезы,
11. Ограничение полом иногда может сильно затруднить генетический
анализ. Пример этого мы встречаем у живородки Lebistes, в У-хромосоме
которой изучен ряд генов, вызывающих красивую цветную раскраску
самцов. У самок же эти раскраски не проявляются вовсе (кроме случаев
превращения самок в самцов, т. е. когда они, собственно говоря, перестают
быть самками). Поэтому реципрокные скрещивания поставить нельзя, а
возможны типы скрещиваний, показанные на схемах (стр. 61), так как
здесь возможно наличие гена как в Х-, так и в Y-хромосоме (см. случай 5).
12. Два из этих скрещиваний, типа 1 tf X О 9» показаны на рис. 8
под № 5. Они оказываются тоже характерно расположенными, не
повторяя ни одного другого случая. Соответствующий случай для типа ZW
показан на правой части рисунка, под № 10. Его два кружка
расположены тоже характерно.
ДОКАЗАТЕЛЬСТВА СЦЕПЛЕННОСТИ С ПОЛОМ
Если на основании сравнения реципрокных скрещиваний возникает
возможность считать, что изучаемый ген локализован в половой
хромосоме, то вопрос еще не должен считаться решенным, а полученный вывод
должен рассматриваться как гипотеза, которую необходимо проверить.
Однако факта различия в результатах реципрокных скрещиваний еще
недостаточно для того, чтобы считать гипотезу доказанной, хотя во многих
случаях вероятность ее и оказывается очень высокой. Однако следует
иметь в виду, что различие в реципрокных скрещиваниях может быть
вызвано и иными причинами. Так, сходная картина может получиться при
наличии так называемой материнской наследственности, установленной,
например, у тутового шелкопряда, у которого некоторые признаки
окраски развивающихся яиц определяются непосредственно материнским
организмом. При реципрокных скрещиваниях крупных пород кур с мелкими
Flf по-видимому, может оказаться несколько различным по величине и
жизнеспособности. Это может стаять в связи с тем, что в одном
скрещивании цыплята первого поколения развиваются из мелких яиц матери
мелкой породы, а в другом — из крупных яиц крупной породы *.
Поэтому для окончательного доказательства локализации гена в
половой хромосоме необходимо поставить серию скрещиваний,
подобранных так, чтобы в различных скрещиваниях ожидаемый результат был
также различным. Гипотеза может считаться доказанной лишь в том случае,
если все предсказания, сделанные для этих проверочных скрещиваний,
оправдаются. Проверочными скрещиваниями явятся F2, оба F и некоторые
F3 и т. п. Останавливаться здесь на подробном рассмотрении всех этих
случаев мы не будем, так как сцепленная с полом наследственность доста-
* Отметим, впрочем, что хороших исследований на эту тему еще нет из-за трудно»
сти'скрещивания крупных кур с мелкими петухами. В настоящее время, когда
искусственное осеменение для птиц разработано, открылась возможность проведения
подобных исследований.
При скрещивании крупных и мелких пород млекопитающих условия для развития
плода в реципрокных скрещиваниях будут также различными.
5 А, С. Серебровский
65
точно детально излагается во всех курсах генетики. Ограничимся одним
примером.
Пусть мы поставили два реципрокных скрещивания между курами
серебристыми и золотистыми с такими результатами:
с? золотистый х 9 серебристая -* <5 с? серебристые + $ $ золотистые
<J серебристый X $ золотистая -> с? с? и $ 9 серебристые
Находим выше случай 7-й и изображаем напш скрещивания так, как там
(схема XI).
аа
ZZ
\
Аа
ZZ ^
С
А АА
ZW ZZ
и
1 1
а Аа
ZW ZZ
х е м а XI
а
ZW
1
А
ZW
Если наша гипотеза верна, то F2 от /7 скрещивания, где все Fx
серебристые, должно пойти по схеме XII.
III
АА
ZZ
ееребр.
Аа
ZZ
Аа
ZZ
ееребр.
Схем
А
ZW
1
А
ZW
ееребр.
а XII
а
ZW
золот,
? 2
Здесь, следовательно, возникает отношение 3 ееребр.: 1 золот. и
притом так, что все извлекшиеся золотистые будут курочками.
Для сравнения можно поставить скрещивание этого же самого петуха
¥г не с сестрой Fx, а с чистопородной серебристой курицей. Так как обе
эти серебристые курицы имеют (если гипотеза верна!) одинаковую струк^
туру 7W » то и результат скрещивания должен быть одинаковым,
несмотря на различное происхождение кур.
Совершенно такой же результат (если гипотеза верна!) должен полу-
А
читься от скрещивания чистопородной курицы 7W с ее сыном ¥г
(/скрещивание).
Наоборот, F2 от / скрещивания должно дать иной результат, так как
(если гипотеза верна!) это скрещивание пойдет по схеме XIII.
Аа
ZZ
а
ZW
IV
\\
Аа аа
ZZ ZZ
ееребр. золот.
—г~
А
ZW
ееребр.
а
ZW
эолош.
Схема XIII
66
Здесь должно возникнуть отношение 1 серебр. : 1 золот. как среди
петухов, так и среди кур.
Далее, хорошим доказательством правильности гипотезы может явиться
скрещивание золотистой курочки из / скрещивания с ее отцом. Если
гипотеза правильна, все потомство от этого скрещивания должно быть
золотистым, как и в чистой породе, хотя курица и относится к Fv
Серия подобных скрещиваний, если предсказания оправдаются,
может сделать нашу гипотезу доказанной. Следует, однако, иметь в виду,
что не всегда сделанное предсказание будет оправдываться точно.
Например, там, где ожидаются отношения 3 : 1 или 1:1, может получиться
искажение. В первой главе мы подробно обсуждали вопрос о том, каковы
источники этих искажений и в каких случаях наличие этих искажений не
мешает признанию того, что сделанное предсказание оправдалось.
В данном случае могут быть и иные источники искажений. Например,
„ а
наличие той же самой хромосомы летального гена может привести к от-
//
миранию части золотистых дочерей в скрещивании III или TV, С
аналогичным случаем столкнулся Денн (Dunn, 1922) при изучении наследования
гена альбинизма, локализованного в аутосоме, у белых виандотов. Но
такие искажения не являются решающими для того, чтобы отвергнуть
сделанную гипотезу. Решающими будут такие несоответствия, которые
вступают в противоречие с основами принятой гипотезы. Так, если бы,
например, в указанном выше скрещивании III появились золотистые
петухи или от скрещивания золотистого отца с его золотистой дочерью из
скрещивания / появились бы серебристые и т. п., т. е. возникли бы
явления, не укладывающиеся качественно в гипотезу,— эти явления пришлось
бы признать существенными для отказа от гипотезы сцепления с полом.
Следует, однако, помнить, что механизм сцепленной с полом
наследственности допускает некоторые исключения из обычных правил
наследования. Так, в скрещивании типа / изредка, в виде исключения, можно
ожидать появления наряду с серебристыми сыновьями и золотистых, а
наряду с золотистыми дочерями и серебристых. Сходный случай встретил
Кру при опытах с курами (Кру, 1931), на большом количестве объектов он
изучен у дрозофилы. Эти исключения, на первый взгляд противоречащие
гипотезе сцепления с полом, оказались одним из наиболее убедительных
доказательств сцепления, так как они связаны с «нерасхождением»
половых хромосом или с перекрестом между Х- или У-хромосомами (рыбки
Lebistes).
Однако как нерасхождение, так и перекрест между X и У являются
достаточно редкими явлениями и их можно привлекать к объяснению
встретившихся противоречий при локализации гена в половой хромосоме с
большой осторожностью, и необходимо немедленно ставить дальнейшие
скрещивания для доказательства того, что нерасхождение действительно
имеет здесь место.
Локализация гена в половой хромосоме может быть установлена и в
том случае, если специальных скрещиваний для этого поставить нельзя
или вообще приходится пользоваться популяционным материалом (см.
следующую главу).
ОПРЕДЕЛЕНИЕ НАЛИЧИЯ ГЕНОСВЯЗИ
После того, как установлено, что ген не локализован в половой
хромосоме, можно приступить к отысканию геносвязи (т. е. сцепления или
отталкивания) его с другими уже известными аутосомными генами или, если
у данного вида уже намечены группы сцепленных генов, то к определению
того, к какой группе сцепленных генов относится данный новый ген.
67
5*
Основным приемом этой работы является использование ддгибридной
формулы обратного скрещивания, о которой мы говорили выше. Для того
чтобы определить, имеется ли геносвязь между геном А и изученным ранее
геном 2?, нужно получить гибрид АаВЬ. В целях анализа это достигается
параллельно двумя способами:
1) ААВВ Х\ааЬЪ -+АаВЪ
2) ААЬЬ X ааВВ *+ АаВЬ
При анализе полученных гибридов АаВЬ скрещиванием с
анализатором ааЪЬ в случае наличия связи между А ж В получаются различные
результаты. В первом случае («фаза сцепления») А ж В обнаружат сцепление
и мы получим:
АаВВ {п)
АаЬЬ (1)
АаВЬХааЬЬг+ааВЬ (1) /г > 1
ааЬЬ (л)
Во втором случае («фаза отталкивания») А и В обнаружат
отталкивание:
АаВЬ (1)
АаЬЬ (п)
АаВЬХааЬЬ^ааВЬ ^ тг>1
ааЪЬ (1)
Для каждой пары генов в обоих случаях величина п должна быть
одинаковой. Чем ближе расположёны друг к другу гены А и В в хромосоме,
тем больше величина п. Наоборот, чем меньше величина л, тем дальше
гены друг от друга.
/. Задача изучения геносвязи распадается на две: 1) установить,
наследуется ли ген А независимо от генов B,C>D,E... или обнаруживает
геносвязь, и 2) определить расстояние гена А от тех генов, с которыми он об-
наруяшвает геносвязь.
Итак, для решения вопроса о наличии геносвязи, получаем потомство
от скрещивания АаВЬ X ааЪЬ, зная наперед по происхождению гибрида,
что именно мы можем ожидать — сцепление или отталкивание. Пусть мы
ожидаем получить сцепление, тогда фенотипы Л, В и а, Ъ должны
возникнуть без перекреста, фенотипы Л, Ь и а, В — с перекрестом.
В случае полной независимости генов А и В мы должны получить:
А,В +а,Ь =А,Ь + а,В. (1)
В случае наличия геносвязи (в данном случае в форме сцепления):
А,В + а,Ь>А,Ь + а,В. (2)
Так как мы имеем доло со статистическим явлением, то даже в случае
полной независимости генов А и В равенство (1) обычно точно не
осуществляется. С другой стороны, если гены А и В локализованы в хромосоме не
очень близко, морганид на 30 и более, то неравенство (2) будет выражено не
слишком явно. Поэтому практически задача сводится к вопросу о том,
можно ли для полученного результата
доказать его достоверное уклонение от равенства.
/ Достигается это при помощи вычисления ошибок.
Пример. 1. Получено А, В •— 38; А, Ь — 34; а, В — 32 и а, Ь — 37.
Ожидалось сцепление или независимость (не отталкивание!). 4,2? +
+ а, Ъ = 75; А, Ъ -+- а, В =?=: 66; всего 141. В случае независимости
ожидалось 70,5 + 70^5.
68
Квадратическая ошибка
"» = ]/ Щ£^ = У35Г25 = ± 5,95.
Следовательно, ожидалось 70, 50 ± 5,95 : 70,5 ± 5,95. Фактическое
уклонение, 75—70,5 = 4,5 ± 5,95, оказывается меньше своей квадратической
ошибки, и наличие сцепления в этом скрещивании остается не доказанным
и даже маловероятным.
2. Получено: А, В — 6; 4, Ъ — 25; А% В — 21; а, Ъ — 8. Ожидалось
или отталкивание или независимость (не сцепление!).
А, В + а, 6 = 14; Л, В + я» В= 46; всего 60. В случае независимости
ожидалось 30 : 30
/~3(Г
■у = ± 3,9.
Фактическое уклонение 16 ± 3,9, т. е. наличие геносвязи несомненно.
Иногда картина сцепления или отталкивания затемняется
привходящими обстоятельствами. Разные фенотипы могут оказываться или
неодинаково жизнеспособными, или гены А и В не всегда хорошо проявляются.
Так, например, при изучении наследования игрековидной формы
гребня курицы и полидактилии мы встречаемся с таким случаем: игрековидный
гребень проявляется примерно в 80% случаев, так что вместо отношения
1 : 1 мы получаем 4:6. Примерно то же получаем и у полидактилии
4 : 6. Тогда вместо отношения 14, В : 14, Ъ : 1а, В : 1а, Ъ мы получаем
16 4, В: 244, Ъ : 24а, В : 36а, Ъ.
Сумма А, В + а, Ъ оказывается 52, сумма 4, Ь +А, В = 48.
Ложность этой геносвязи можно установить также вычислением так
называемого коэффициента ассоциации (Yule, 1900):
" ~~ А,В-ауЬ + А,Ъа,В '
устраняющего влияния различий в жизнеспособности. Для данного
примера:
rt_ 16-36 — 24-24 _ft
^ — 16-36 + 24-24 ~"U-
Однако ложность геносвязи вскрывается вычислением ассоциации
только в простейшем случае, вроде приведенного. Нередко встречается
осложнение в виде различного проявления гена В на фоне 4 и а: например,
в присутствии 4 гены2?и Ъ ясно проявляются, а при а ген В проявляется
не всегда. Возможны и другие искажения, при которых и коэффициент
ассоциации не равен 0. Такие случаи приходится проверять путем
постановки скрещивания типа отталкивания. Если перед нами геносвязь, то
сумма средних членов (4, Ъ + а, В) должна быть больше суммы крайних
членов (-4, В + а, Ь).
Но если геносвязи нет, а картина, напоминающая сцепление, вызвана
искажающими факторами (непроявление, разная жизнепособность и т. п.),
то и в скрещивании, рассчитанном на получение отталкивания, вместо
отталкивания мы можем получить опять картину сцепления. Поэтому
наличие геносвязи только тогда может считаться доказанным, если оно будет
доказано и для типа сцепления, и для типа отталкивания.
ЗНАЧЕНИЕ FB и F2
Постановка обратного и анализирующего скрещивания для уловления
геносвязи является наиболее выгодной. Однако она упирается часто в
отсутствие анализатора ааЪЬ. В этом случае анализатор приходится созда-
69
вать при помощи извлечения дирецессива aabb в F2. Но здесь следует иметь
в виду, что как раз наличие геносвязи между а и b затрудняет получение
от скрещивания ааВВ X AAbb в F2 комбинации aabb. Поэтому, ставя это
скрещивание, нужно установить за ним тщательное наблюдение, тем более,
что наличие геносвязи может быть обнаружено и при изучении поколения
F2. При наличии геносвязи происходит изменение дигибридной формулы,
различное в случаях сцепления и в случаях отталкивания.
Состав F2 дигибридного скрещивания вычисляется по следующим
формулам:
Фаза
Сцепление
Отталкивание
А, В
|(3-2/>+Л
|(2 + Л
А, Ь и а, В
|(2/>-р°)
\Vp~p1
а, Ь
1"
где р — величина перекреста, т. е. относительное число случаев, в
которых принадлежащие к локусам А и В гены расцепились.
Результаты, к которым приводят эти формулы при разных значениях р,
даются в табл. 7.
Таблица 7
Структура F2 дигибридного скрещивания (в %) при разном проценте перекреста
Процент
перекреста
0
5
10
15
20
25
30
35
40
45
50
Отношение при сцеплении
А, В
75,00
72,58
70,25
63,06
66,00
64,06
62,25
60,56
59,00
57,56
56,25
А, Ь
0,00
2,44
4,75
6,94
9,00
10,94
12,75
14,44
16,00
17,44
18,75
а, В
0,00
2,44
4,75
6,94
9,00
10,94
12,75
14,44
16,00
17,44
18,75
| а, Ъ
25,00
22,56
20,25
18,06
16,00
14,06
12,25
10,55
9,00
7,56
6,25
Отношение при
А, В
50,00
50,06
50,25
50,56
51,00
51,56
52,25
53,06
54,00
55,06
56,25
А, Ь
25,00
24,94
24,75
24,44
24,00
23,44
22,75
21,94
21,00
19,94
18,75
отталкивании
| а, В
25,00
24,94
24,75
24,44
24,00
23,44
22,75
21,94
21,00
19,94
18,75
а, Ъ
0,00
0,06
0,25
0,56
1,00
1,56
2,25
3,06
4,00
5,06
6,25
В случае наличия сцепления группы А, Ь и а, В, входящие в дигибрид-
ную формулу в виде 3 : 3, начинают уменьшаться и при полном сцеплении
исчезают вовсе. Равенство между категориями А, Ь и а, В сохраняется. Что
касается отношения фенотипов А, В : а, Ь, то вместо 9 : 1 оно превращается
в 3 : 1. При наличии отталкивания, наоборот, фенотип a, b
уменьшается в числе, а при полном отталкивании исчезает вовсе. Таким образом, ди-
гибридная формула при полном сцеплении или отталкивании принимает
вид:
Полное сцепление Независимость Полное отталкивание
3 (75%) 9 (5бо/0) , 2 (50%)
0 (0%) 3 (19%) 1 (25%)
0 (0%) 3 (19%) 1 (25%)
1 (25%) 1 (6%) 0 (0%)
70
Рис. 9. Номограмма для
определения процента
перекреста по проценту фенотипов
(А, В + а,Ь) или (А, Ь + а,
В) в F2
Объяснение в тексте^
Формула сцепления резко отличается от формулы независимости и при
сравнительно небольшом материале даже не очень сильные сцепления
могут быть статистически уверенно обнаружены.
В отличие от формулы сцепления формула отталкивания в F2 может
быть с уверенностью отличима лишь при довольно большом количестве
потомства по отсутствию фенотипа а, Ъ и несколько повышенному числу
А, Ъ + а, В. При неполном отталкивании это оказывается очень
затруднительным, что видно из очень пологого хода кривой «отталкивание» на
рис. 9. При не совсем правильном проявлении некоторых генов или иных
источников искажения здесь может получиться ложная картина
геносвязи. В то время как при анализирующем скрещивании для того., чтобы
создалась такая картина, необходимо отклонение от отношения 1 : 1 у обоих
генов, в F2 для того чтобы возникло отношение, похожее на геносвязь,
достаточно неполного проявления даже одного гена.
Цример. Бели ген А вместо отношения 3 : 1 даст отношение 3 : 2, а В
даст правильно 3 : 1, то в F2 получим
(ЗА + 2а) (35 + Щ = 9А, В + ЗА, Ь + 6а, В + 2а, Ъ.
3 4-3
В то время как при независимости процент средних членов —^— • 100 % =
о I a
= 37,5%, здесь мы имеем —^—100% =45%, т. е.
отношение,характерное для случая отталкивания с 32% перекреста.
Вычисляя ассоциацию, получаем:
9-2 — 3-6
9.2 + 3-6
0,
т. е. и здесь вскрываем ложность полученного впечатления об
отталкивании. Отсутствие геносвязи вскроется коэффициентом ассоциации и при
независимом неправильном распределении обоих генов. Например:
(ЗА + 2а) (45 + Щ = 12Л, В + ЗА, Ь + 8а, В + 2а, Ь,
откуда числитель коэффициента ассоциации
12-2-3.8 = 0.
ЛОКАЛИЗАЦИЯ ПРИ ОТСУТСТВИИ ПЕРЕКРЕСТА
У ОДНОГО ИЗ ПОЛОВ
У некоторых животных задача локализации аутосомных генов сильно
упрощается отсутствием перекреста у одного из полов. У всех изученных
видов дрозофил перекрест отсутствует у самцов, у тутового шелкопря-
71
да — у самок. В этом случае для локализации ставят скрещивание так,
чтобы ди-гетерозиготным был пол, не дающий перекреста, у дрозофилы,
следовательно, им будет самец. Тогда скрещивание идет по одной из схем
(схема XIV, I, II).
йв
X О—
+ па
1
ав
ц<? * гш
ии
и
1
ав
ав
Схема
Ав
ав
XIV
1
аВ
ав
Потомство оказывается только двух типов, а не четырех. В I
скрещивании все дети, получившие ген А, получают и ген В (полное сцепление), а не
получившие А, не получают и В. Во II скрещивании все дети,
получившие А, не получают В, а не получившие А, наоборот, получают В (полное
отталкивание). Этим сразу доказывается, что изучаемый ген В
локализован в той же хромосоме, как и А, и дальнейшее исследование должно лишь
выяснить расстояние между А и В, так как проведенные скрещивания не
дают по этому вопросу никаких указаний: гены А и В могут лежать и
рядом, и по разным концам хромосомы, но благодаря отсутствию здесь
перекреста их сцепление или отталкивание будет все равно полным.
Для дрозофилы с ее небольшим числом аутосом (3 пары) выработан
следующий типический прием локализации нового рецессивного гена.
В лаборатории должна иметься линия с двумя доминантными генами,
одним из 77 хромосомы, другим из III. В зависимости от характера
признака, вызываемого новым геном, выбирают такие доминанты, чтобы они не
слишком мешали наблюдению за новым геном. Удобной линией является,
например, LH (Lobe — уменьшение глаза, II хромосома; Hairless —
отсутствие некоторых щетинок на голове и др., III хромосома). Пусть мы
локализуем новый ген п (nova). Ставим скрещивание гомозиготной пп <j? с LH tff
идущее по одной из трех схем в зависимости от того, локализован п во
//, /// или IV хромосоме.
// $
п++ LH+
ш $ п
П+ +
+П+
X
Lff+ , L++ , +Н+ , +++
X
LH+
п++ '
LH+
iv $
+71+
++П
++п Л+++"
П++
L+ +
/1++
п++
+++
X
+71+
LH+
+п+
+п+
тЬ#+
+71+
++ +
++п -г ++п "Г ++п "Г ++п
Если сыновья не проявляют признака nova, то, следовательно, ген
локализован в аутосоме и можно его исследовать дальше. Для этого, отобрав
из ¥г самца LH (рецессивный п внешне не проявляется), скрещиваем его
обратно с $ $ nova.
В зависимости от того, в какой хромосоме локализован новый ген,
получается потомство одного из следующих трех<сосгавов:
IV
LH +
LHn
L++
L+n
и
LH +
—
L++
—
Ill
LH +
—
—
L + n
IV
+ # +
+ Нп
+++
++п
II
—
+ Нп
—
++П
III
+ я +
—
—
++п
В случае локализации пв IV хромосоме (обозначено IV) возникают все
возможные восемь комбинаций из трех параллелей. Это указывает на то,
72
что nova свободно комбинируется как с L, так и с if и, следовательно, ло*
кализована не во II и не в III хромосоме. В случае дрозофилы это
указывает на локализацию в IV хромосоме.
В случае локализации п во II хромосоме возникает только четыре
комбинации, а не восемь. Не возникают те четыре комбинации, в которых
имелись бы комбинации Ьп и Н—К т. е. в которых возникли бы новые
комбинации геновЬжп против комбинаций, бывших в исходных мухах (L + и
+ п). Отсутствие перекомбинировки Ьжп указывает на то,
что оба гена локализованы в одной хромосоме. Иначе — п локализован
во II хромосоме.
В случае локализации п в III хромосоме возникает также только четыре
комбинации. Не возникают те, в которых имелась бы перекомбинировка
генов Н и п (Нп и Н—\-). Это указывает на локализацию п в той же паре
хромосом, как и Я", т. е. в третьей.
Иногда в таком скрещивании могут наблюдаться даже не четыре,
а лишь две комбинации ЬН-\- и Н—Ьи, указывая на то, что п не вступает
в перекомбинировку ни с L , ни с Н и как бы находится одновременно и
во 7/, и в 777 хромосомах. Такой результат указывает на то, что nova
связана с транслокацией между II и III хромосомами (о чем см. ниже).
Когда хромосома установлена, выбирают самок, гетерозиготных по и,
и доминанту в той же хромосоме и, скрещивая с самцами тг+, ставят опыт
на определение процента перекреста.
ЛОКАЛИЗАЦИЯ ПРИ ПЕРЕКРЕСТЕ У ОБОИХ ПОЛОВ
Локализация при перекресте у обоих полов осложнена этим
обстоятельством. При близком расстоянии между генами (до 20—25%
перекреста) это осложнение еще не так велико, но все же требуется уже
достаточное число потомков, чтобы ответить на вопрос определенно. При более
далеких расстояниях, когда ожидаемое расщепление мало отличается от
1:1:1:1, разные побочные искажающие обстоятельства делают
определение малонадежным или требуют большого потомства. Поэтому прямое
значение имеет лишь положительный ответ — геносвязь имеется.
Отрицательный же ответ — геносвязи нет — правильнее заменить другим:
геносвязь не доказана. Иными словами, отрицательный ответ правильнее
формулировать так: «ген А лежит либо в другой хромосоме, чем ген В, либо
в той же, но далеко от него».
Поэтому, если мы уже имеем у данного вида установленную группу
сцепления, например, из генов В — С — -D, то, получив отрицательный ответ
относительно недоказанности геносвязи между А и 5, следует поставить
опыт с участием А и D; D — выбирается потому, что он лежит на другом
конце сцепленной группы В — С — D и может оказаться ближе к А, если
А локализован в той же хромосоме, но далеко от В:
В -С -D -А.
Такое исследование целесообразно ставить, конечно, лишь в том случае,,
если расстояние В — D достаточно велико, иначе расстояния В — А и
D — А будут почти одинаковыми и ждать ясной геносвязи между D и А
при ее неясности между В и А не будет оснований.
У млекопитающих, по крайней мере у грызунов, установлено уже, чта
перекрест у различных полов идет с различной легкостью. У самцов
процент перекреста примерно на 7з ниже чем у самок. При локализации
поэтому выгоднее использовать вначале гетерозиготных самцов, так как
пониженный перекрест облегчает уловление геносвязи и получение более
определенных данных.
73
ОПРЕДЕЛЕНИЕ РАССТОЯНИЙ МЕЖДУ ГЕНАМИ
После того как наличие геносвязи установлено, т. е. установлено, что
два исследуемых гена наследуются не независимо друг от друга, возникает
задача более точной локализации гена. Наличие геносвязи между двумя
генами означает локализацию их в одной и той же хромосоме или
в одной и той же паре хромосом. Если гены отталкиваются, то один из
них локализован в одной из парных (аллеломорфных) хромосом, а
другой ген — в другой. Если гены обнаруживают сцепление, то
они локализованы в общей хромосоме.
А
(!)
а
В
b
А
а
b
(2)
В
Таково расположение генов при сцеплении (1) и при отталкивании (2).
Из прилагаемой схемы видно, что и сцепление, и отталкивание суть
две стороны одного и того же явления, так как если А отталкивается от J5,
то значит, что А сцеплено с Ь, и наоборот, если А сцеплено с JB, то А будет
отталкиваться от Ь.
Но если два гена расположены в одной хромосоме, то возникает вопрос,
каково расстояние между ними. В настоящее время мы не умеем
измерять это расстояние в абсолютных единицах и за меру его принимаем
ту легкость, с которой сцепленные гены расщепляются путем перекреста.
Расстояние между генами, выраженное таким способом, по формулировке
Моргана, приблизительно так же отражает действительность, как
отражает географию железнодорожная карта, на которой расстояние между
станциями выражено не в километрах, а в часах и минутах.
Если в анализируемом скрещивании, в котором гены А и В
обнаруживают сцепление, возникли четыре категории потомков —А, В; А, Ъ; а, В
и а, Ь, то расстояние между А и В вычисляется в простейшем случае по
формуле:
со/л - А,Ъ + а,В .юно/
С/о - А,В + А,Ъ + а,В + а,Ъ 1W/°-
Величина с% при небольших расстояниях между генами будет числом
единиц расстояния, или числом морганид. Она является числом процентов
случаев, в которых гены А и В расцепились так, что с% = 100% р.
Если в подобном же скрещивании гены А ж В обнаружили отталкивание,
то расстояние между ними вычисляется по формуле:
с% = ЛВъ+*'Ьп, ,'100%.
А, В + А, b + a,B + a,b /u
В обеих этих формулах А, В; А, Ь; а, В; а, Ь означают количество особей,
имеющих фенотип А, В; А, Ъ и т. д.
Расстояния, определяемые по этим формулам, имеют, однако, два
источника ошибок (помимо случайных). Во-первых, в тех случаях, когда
гены А и В расположены далеко друг от друга, далее 10—20 морганид,
между ними возможны «двойные перекресты», т. е. возможны два
перекреста в разных пунктах участка между А и В. После первого перекреста
вместо порядка АВ возникает А, Ь, а после второго АЪ снова перейдет в
ЛВ, и мы, таким образом, не заметим вовсе никакого перекреста. При
расстоянии в 30 морганид это произойдет примерно в 2% случаев
(несколько менее j^ ^ .-у^ = 2,25%), и мы заметим поэтому только
-30% — 2% = 28% случаев разрыва А я В. Чем дальше расстояние А от В,
тем двойные перекресты чаще и тем сильнее будет неувязка между дейст-
74
вительным расстоянием и вычисленным по указанным формулам.
Процент перекреста не бывает больше 50, если наблюдаются только 2 гена,
<а расстояние в морганидах может быть и больше 100. Избежать этой ошибки
нельзя, нельзя вычислить и точной поправки. Благодаря наличию так
называемого явления интерференции, зависящего от своего рода
«упругости» хромосом, которая в различных хромосомах и участках
хромосом различна, вычислить ожидаемое число двойных перекрестов можно
лишь приблизительно: так, при расстоянии в п морганид число
двойных перекрестов будет не больше, чем п2/400; но при увеличении п
возникают не только двойные, но и тройные, и четверные кроссоверы,
поэтому и этот расчет оказывается неточным.
Для дрозофилы составлены уже специальные таблицы, позволяющие
вносить соответствующие поправки для любой пары генов (для половой
хромосомы см. «Курс генетики». Синнот и Денн, изд. Биомедгиза, 1934) *.
Второй источник ошибок в вычислении расстояний между А и В
зависит от тех причин, которые искажают моногибридное отношение: неполная
проявляемость, различная жизнеспособность и т. д.
Цример. При анализирующем скрещивании получены генотипы в
таком отношении: 40 АаВЬ : 10 АаЪЪ : ЮааВЬ : 4Qaabb. Но из-за плохого
проявления гена В 10% от АаВЬ зачислено в фенотип AAbb и 10% от ааВЬ
в aabb. Отношение фенотипов поэтому таково:
36Л, В'ЛЬА, Ъ:9а, 5:41а, Ь
•откуда
44. 4- Q
с% = 36 + 14 + 9 + 41 -1000/0=230/0,
вместо правильного
С% = 40+ 10 +10°+40 •10°°/о = 20%-
Для устранения этой неправильности поступаем таким образом. Если
с% — наш искомый процент перекреста, то фенотипы 4,В;4,5 и т. д.
должны были бы возникнуть в количествах:
4,
А,
а,
а,
В =
ъ =
в=
ъ =
:50-
с
'~2Г
е
:50~
с
с
~9~
Но из-за того, что В в некоторой части случаев (к) не проявляется,
фенотипы фактически возникают в количествах:
1) А,В = (50—1-)_*(50—J-)
2) A,b=-t- + k(50--f)
3) а,В=\ A-J-
4) а,Ъ= (50 |-}+ £.-!-
♦См. также Н.Н.Медведев. Практическая генетика. Мм Изд-во «Наука»,
1966.— Прим. ред.
75
Складывая почленно 2 с 4 и 1 с 3, получим:
А,Ъ + а,Ъ = ^- + 50к-^- + 50-~ + 4г==50-Ь50&;
А,В + а,В = 50--% 50А+^- + ^-—у- = 50-50Л.
Вычитая из первого равенства второе, получаем:
(А, Ъ + а,Ъ) — (А,В + а, В) = 100&
к = (Л,Ь + а, Ь) - (Л, БЧ-^, В±
100
Если мы хотим выразить к в процентах, то
к% = (i4, Ь + а, Ь) - (Л, 5 + а, 6).
Если количество фенотипов А, В, А, Ь, а, В ж а,Ъ было выражено не
в процентах, а в абсолютных единицах, то
, (А,Ъ + а,Ъ)-(А,В + а,В)
К~~ А,В + А,Ъ + а,В + а,Ъ •
Зная к, нетрудно найти и значение с. По формуле, применявшейся ранее,
мы находим приближенное его значение (обозначим с'), а затем и истинное
значение:
, „ с' — 50*
В нашем примере мы имеем:
(14+41)-(36+ 9) _j!0_n,.
К 36 + 14 + 9 + 41 100— и,х'
<* = 36 + Ц+1 + 41 -*00% «23%;
со/о=!^0_ОИ_ = 20о/о.
Можно и непосредственно соединить эти вычисления в одну формулу,
имеющую следующий вид:
А,В + А,Ъ\+а,ВА-ащЬ
° А,В + а,В
Второй случай искажения может возникнуть на почве различной
жизнеспособности зигот или гамет. Например, ген «а» может понизить
жизнеспособность в I раз; а ген «Ь» в /' раз. Тогда задача может быть решена,
если мы допустим гипотезу, что особи аЪ будут иметь жизнеспособность,
пониженную в IV раз. Эта гипотеза, хотя и не совсем правильна, но
вычисления, произведенные на ее основе, все же дают достаточно хорошие
результаты.
Мы допускаем, следовательно, что вместо теоретических отношении
фенотипов
(50 J-) А, В:-£- A, 6:-J-e, В : (б0—f)a, Ь
в живых останется
Л,В-(50—f)
a,B = ±-V
а, Ь = (50 - -|Л W
76
Освободиться от искажающего влияния I и V мы можем, произведя вы-
'числение коэффициента ассоциации:
П _ А,В-а,Ъ — А,Ъ-а,В
^"~ А,В-а,Ъ + А,Ъ-а,В
Подставляя вместо Л, В,А,Ь и т. д. их значения, мы получаем:
(»--^)(»—j-)<r—j-.-fir (50-^-)2-(-f)2
(»-■f)(so—J-)w + -r-f «* (5°-Т-)а+(-гГ '
Между коэффициентом Q и морганидами существует довольно
сложная связь, поэтому для определения по Q числа морганид можно
воспользоваться номограммой (рис. 10).
Очень удобно определить с в том случае, если имеются два
противоположных скрещивания с сцеплением и отталкиванием. Тогда имеем:
«Фенотип
А, В
А,Ъ
При
сцеплении j
50—т
—А
- О СМ
Фенотип j
\а9в
А,Ъ
При
отталкива ии
с
' ("-*)'
Фенотип
а9В
а, Ъ
При
сцеплении
2
("Ч-)"
Фенотип
а, В
а, Ь
При
отталкивании
(50-т)''
Л w
2
Расположим фактический материал (для сопоставления формул)
следующим удобным образом:
Генотип
АаВЬ
АаЬЬ
ааВЬ
ааЬЪ
1 Классы
сцепления
{А, В)
(А,Ъ)
(«. В)
(«,*) 1
отталкивания
{А, В)'
(А, Ъ)'
(а, В)'
К ъу
Формулы
сцепления
50-'
2
2
2
О®-!)"'
отталкивания
с
~2~
("-т)'
("-т)1.
2
Легко показать, что
с =
Ц.В) + (Л,Ь) + (д,В) + (а, 6)
(Л,5) + (А, Ь) + (а, 5) + (а, 6) + (Л, В)' + (А, Ъ)' + (а, Б)' + (а,6)'
В самом деле, подставляя вместо {А,В), (А, В)' ж т.д. их формулы,
получаем:
-|-(1+* + /' + //') -с
: ЮОо/о -г
-J--100
50
= с.
(бО--J^(i +Z +/' + //') + -|-(1 +| + /' + /Г)
Величины (4,5), (4, 6)... должны быть выражены здесь в процентах
шли долях единицы.
77
ПРОБЛЕМА ТОЧНОГО ОПРЕДЕЛЕНИЯ ВЕЛИЧИНЫ
ПЕРЕКРЕСТА
Мы изложили очень кратко простейшие
приемы определения расстояния между генами. Но-
в более широкой постановке вопрос этот
оказывается значительно более сложным, так как на
сцену выступают следующие два сложнейших
фактора. Во-первых, не всегда возможно
пользоваться обратными скрещиваниями.
Приходится иметь дело и с F2 или с менее стандарт*
ными случаями, при которых, например, один
ген расщепляется в отношении 1 : 1, а другой
в отношении 3:1.
Возникающие при этом частоты четырех
фенотипов имеют уже более сложную
зависимость от величины перекреста.
Второе осложнение может зависеть от того,,
что в скрещиваниях будет участвовать не два, а
более генов. Например, может быть поставлена
задача определить силу сцепления между
генами А и JB, причем ген В вместе с геном С дает
расщепление по формуле 9 : 7 и т. д. При этом,
конечно, может иметь место дифференциальная
жизнеспособность и пр.
Над вопросами наилучшего определения
величины перекреста работало много авторов,
стремившихся найти формулы достаточно простые, достаточно точные и
допускающие определение статистической ошибки. Бэтсон и Пеннет
(1911) дали первый метод на основе своей теории редупликации гамет,
Коллинс (Collins, 1912, 1924), Бридже (Bridges, 1914) и мы (Serebrov-
sky, Ivanova, Ferry, 1929) использовали коэффициент ассоциации Юла.
В 1917 г. иную формулу предложил Эмерсон (Emerson, 1917), а Вудворс
(Woodworth, 1923) развил ее для более сложных случаев. Холдэйн (Hal-
Рис. 10. Номограмма для
приблизительного
определения процента перекреста
(с) по коэффициенту
ассоциации Q
Таблица 8
Влияние различных факторов, искажающих соотношение классов,
на определенные разными методами проценты перекреста
Искажающие факторы
Нормальная популяция
Гибнет 25% гам*ет а
Гибнет 25% гамет аЪ
Гибнет 25% аВ зигот
Гибнет 25% аа зигот
Гибнет 25% аа и ЪЪ зигот
Гибнет 25% аа и ЪЪ зигот
25% аа отнесены к А
Двойные рецессивы не
обнаружены
Процент классов зигот Fa
3
51,56
54,06
57,82
49,22
55,00
58,86
50,57
57,42
52,38
23,44
27,55
20,66
35,26
25,00
20,07
30,65
23,83
23,81
•о
23,44
17,22
20,66
14,08
18,75
20,07
17,24
17,58
23,81
'О
1,56
1,15
0,86
1,47
1,25
1,00
1,53
1,17
0,00
Процент перекреста,
определенного различными
методами
■
р.
а>
S
ГО >»
л
О о
Ко
25,00
32,34
41,61
11,74
35,36
44,42
20,53
41,46
21,82
>>
8
о
К
25,00
23,77
22,63
24,96
25,00
25,00
25,00
26,00
0,00
о i»S
25,00
23,35
23,48
22,87
25,00
26,18
24,14
26,40
0,00
о
и
25,00
27,10
28,74
26,11
28,20
30,83-
25,41
30,65
12,50
78
done, 1919) предложил свою формулу для вычисления перекреста по F^
и для вычисления вероятной ошибки. Различные варианты предлагали
Хор (Ног, 1924), Бэбкок и Клаузен (Babcock a. Clausen, 1927), Альберте
(Alberts, 1926), Капперт (Kappert, 1927), Мазер (Mather, 1938) и др.
Известный английский статистик Фишер (Fischer a. Balmakund, 1928)
писал, что нет ничего легче, как придумывать различные методы для
вычисления показателей связи, которые могут быть использованы для
вычисления перекреста, но что необходимо поиски этих методов подчинить
строгой теории, добиваясь того, чтобы возможно полнее исключить
различные искажающие факторы и до минимума снизить статистические ошибки.
Так, Фишер показал, что формула Эмерсона годится только в случаях
тесного сцепления, но в случае сильного отталкивания или при слабой
геносвязи дает очень незначительные, ненадежные результаты. Приведем^
здесь одну из таблиц, рассчитанных Оуэном (Owen, 1928), которой он
иллюстрирует, как отражаются на результатах, полученных различными
методами, искажающие факторы. В качестве примера взят случай
отталкивания двух генов с 25% перекреста в F2 (табл. 8).
Таблица показывает, что из-за различных искажений пропорция
четырех фенотипов в F2 вместо правильных 25 % перекреста по формуле Эмер-
сона * исчисляется от 11,74 до 44,42; по формуле Юла (коэффициент
ассоциации) —- от 0 до 26, по методу Фишера —- от 0 до 26,40 и при помощи
вычисления коэффициента корреляции — от 12,50 до 30,83. Если
исключить последний случай полного отсутствия aabb, то колеблемость ответов,
по трем последним методам резко снижается:
по Эмерсону . . 44,42—11,74 амплитуда ... 32,68
по Юлу 26,00—22,63 » ... 3,37
по Фишеру . . . 26,40—22,87 » ... 3,53
по г 30,83—25,00 » ... 5,83
Такой эмпирический путь сравнительной оценки различных методов,,
конечно, слишком груб — в разных типах скрещиваний при различных
иных искажениях амплитуды могут оказаться иными.
Фишером предложена теория оценки различных методов и произведена
сравнение пяти различных методов. Эти методы следующие.
1. Метод сумм, представителем которого является метод Эмерсона
п# = а — Ь -— c + d.
Здесь и в следующих формулах х — квадрат перекреста, выраженнога
в долях единицы [например, 10% перекреста: х = (ОД)2 = 0,01]; а, Ь,
с, d — частоты четырех классов фенотипов в F2, п — число особей, т. е.
n = a + b + c + d.
2. Метод взвешенных средних — тот же метод сумм, но с учетом
удельного веса а, Ь, с, d. В основу его кладется выражение а + ЗЬ + Зс + 9d,
которое приводит к уравнению
4пх = 2а — 2Ь — 2с — 10d.
3. Метод произведений, родственный коэффициенту ассоциаций, на
ограничивающийся отношением bc/ad при сцеплении и ad/bc при отталки-
Формулы Эмерсона следующие: если при геносвязи гаметы А, В А, Ъ, а, В, а,Ъ
образуются в пропорции г : s : s : г, то г = 0,5 Уе — М и s = 0,5 Ve + M — г*
где Е — сумма особей А, В + а, Ь (крайние члены дигибридного расщепления)у
а М — сумма А, Ъ + а, Ъ (средние члены).
Иначе те же формулы могут быть записаны так:
Г= -i/ (А,В + а,Ъ-А,Ь + а,вГ и 5 = -i/ A,B+ А,Ъ + а,В+ а,Ъ .
79
©ании, вместо л , .с и ,с , ал , по Юлу. Здесь выражение имеет вид:
ad + be be + ad
z(2 + z) __ ad_
(1 —ж)2""" be *
4. Метод наибольшего правдоподобия (Фишера), о котором подробнее
«скажем ниже и который приводит к уравнению
пх2 — (а — 2Ь — 2с — d) — 2d = 0.
5. Метод минимума %2, приводящий к уравнению 4-й степени.
На одном конкретном примере дигибридного расщепления] Фишер
сравнил эти методы и сравнил также квадратическую ошибку каждого
метода. Следующая таблица показывает это сопоставление:
Найдено
Вычислено:
1-й ^етод
2 »
3 »
4 »
5 » .
Классы фенотипов
Л, В
1997
1954,25
1972,87
1953,71
1953,77
1953,84
А, Ъ
906
905
916,37
925,54
925,47
925,40
а, В
904
905
916,37
925,54
925,47
925,40
-
а, Ъ
32
54,75
43,37
34,21
34,27
31,34
Процент
перекрестка
23,88
21,26
18,88
18,90
18,92
±т
4,268
2,348
1,454
1,545
1,545
Хз
9,7170
3,8600
2,0158
2,0153
2,0153
Чтобы дать сравнительную оценку методам, Фишер уделил особое
внимание анализу квадратических ошибок. Формула ошибки, как известно,
или
/?•
Подкоренное выражение Фишер называет вариансой искомой
величины х. При разных методах формула вариансы искомого х
имеет следующий вид:
1-й метод их «
2-й метод v% =
п »
1+6а:-~ i
4я
2*(1— х)(2 + х)
3-й метод "з= \(i + 2s) I
4-й метод ]
5-й метод) какуз-
В специальном исследовании Фишер доказал, что метод наибольшего
правдоподобия дает наименьшую из возможных варианс (при больших
выборках).
Поэтому можно сказать, что методы, имеющие минимальную ошибку,
извлекают всю доступную информацию из фактов и могут
быть названы эффективными (efficient) методами. Другие методы
извлекают из фактов только часть доступной информации и их эффективность
ниже. Эта эффективность может быть выражена отношением варианс, де-
80
Рис. 11. Сравнительная
точность пяти способов
вычисления процента перекреста при
различной силе геносвязи (по
Фишеру и Бальмуканду)
L
It
||
I
too
80
60
40
20
ъш
- 5/
^Г \
Ту/
1 1
V
1 1 1 1 J
10 20 SO 40 50 40 30 20 10 О
Оттагяидание Сцепление
Рекомбинации, %
лением варианс метода наибольшего правдоподобия на вариансу данного
метода.
Для первого и второго методов степень (процент) эффективности
определяется выражениями:
Efh
2х (1 — х) (2 + х) (1 — х2)
п (1 + 2х) Г п
рИ _ 2а;(1— х)(2 + х) . 1+в
^"2 — п(1+2х) " А»
4п
2х (1 — х) (2 + х)
2х (2 + х)
(1 + 2х) (1 — х2) (1 + 2х) (1 + х)
4а:2 _ 8ж(1 — ж) (2 + ж)
(1 + 2х) (1 + 6а: — 4а:2)
Эти формулы ценны тем, что позволяют судить о достоинствах равных
методов не вообще, а конкретно в различных областях значения х. Так,
например, видя в числителе обеих формул х в качестве множителя, мы
сразу заключаем, что при х = 0 оба метода совершенно неэффективны
(х = 0 соответствует полному отталкиванию, х = 1/4 — независимости
и х = 1 — полному сцеплению), при х = 1 числитель второго выражения,
благодаря множителю (1 — х), тоже превращается в 0. Следовательно, и
для исследования сильных сцеплений второй метод непригоден. Наоборот,
формула эффективности первого метода при х = 1 превращается в 1, т. е.
этот метод, при исследовании случаев сильного сцепления, оказывается
столь же эффективным, как и три последних метода.
На рис. 11 эти отношения показаны графически. График обнаруживает
неожиданно, что второй метод непригоден при х = 0 и при х = 1 и
достигает полной эффективности при х = 1/4, поэтому может быть вполне
использован при исследовании слабых случаев сцепления и
отталкивания.
В связи с этими сопоставлениями Фишер вводит в употребление
понятие «количество информации», являющееся не чем иным, как величиной,
обратной вариансе (1/у), или более интересную величину, получаемую
делением на п. Получаемая величина является «количеством
информации, получаемой при помощи одного растения (или другого объекта)».
Эта последняя величина удобна тем, что сразу показывает, во сколько
раз следует увеличить (или уменьшить) материал для получения желаемой
точности. Если, например, «количество информации на одну вариансу
(особь)» по одному методу в пять раз меньше, чем по другому, то для
получения одинаковой точности вывода нужно по первому методу увеличить
материал в пять раз.
Аналогичным способом можно исследовать и более сложные
положения. Одним из таких является случай криптомерии.
1. Криптомерия
Для случая F2 с криптомерией, дающей отношение 9ь: 3 : 4, расчет
начинают, исходя из фазы отталкивания, дающей более простое выражение.
6 А. С. Серебровский
81
лвЛ.
Abi^l
<,Р*-е
2
*т
АВ
с
2
С2
4
с (1-е)
4
i с(1-с)
1 4
с2
4
ль '
1-е 1
2
С(1-с)
4
с(1—с)2
4
(1-е)2
4
с (1-е)
1 4
аВ
1-е
2
С (1-0)
4
(1-е)2
4
(1— су
4
с (1-е)
4
аЬ
с
2
С2
4
с (1-е)
4
с(1-с)
4
1 с*
4
Из таблицы для четырех различных фенотипов F2 находим:
4
с)2^ 2с (1— с) _ 1 — с2
4 i 4"
J Ю Зс* л_ *?(* —<0 I 2(l-~'c)2 2 + с2
А»^ т""1 4—' 4—^ ~*—;
J Ь (1-<02 , 2с(1-с) _ 1-е2.
А%и 7 I 7 4— «>
ЛЯ (1-е)2 2(1-с)__1-с2 .
*»* 4 + 4 ""Г-'
В случае криптомерии два последних класса неразличимы и мы имеем
лишь три класса:
2 + с2
4 с а, В] i
1-е2
а А, В
Ь АУЪ
4
а'В\±
о,Ь1 4
Очевидно, ожидание последнего класса не зависит от с. По методу
максимальной вероятности пишем:
a log (2 + с2) + Ь log (1 - с2) + (с + d) log 1 = max.
Максимализируя это выражение, получаем первую производную и
приравниваем ее к 0:
2ас 2Ъс __ ft
откуда
1-е2
2 + с2
и задача решена.
Количество информации и квадратическое уклонение величины с
вычисляется из того же выражения для первой производной:
2ас 2Ьс
2 + с2 ~~~ 1 —с2 *
Дифференцируя это выражение еще раз по с, получаем, заменяя а и Ь на
их выражение через с и меняя знаки, количество
информации:
Зтгс2
(2 + с2) (1-е2) в
Обратная величина 3!гс2 ~~ бУДет случайной вариансой с и
квадратный корень из нее — квадратической ошибкой.
82
Рис. 12. Количество]! информа-
. ции, даваемое одной особью F2
в различных генетических
случаях (по Иммеру)
If
L
^\ Ч 1 1
s^htt**-.
~j0^
ttii
10 20 30 W 50 40 30 20 Ш Г
Отталкивание Сцепление
Делением вариансы нормального F2 на вариансу данного случая
находится эффективность определения геносвязи в данном случае:
(1 — с2)(2 + с2) . (1-е2) (2 +с2) Зс2
Eff-
In (1 + 2с2)
(1-е2) (2 +с2) _
Зле2 2 (1 +2с2)'
При приближении с к 0 данная относительная эффективность падает
до 0. При с = 0,50 эффективность 0,25 и при с = 1 становится равной г/2.
На рис. 12 показано сравнительное количество информации, даваемой
одной особью F2, в процентах от количества информации, даваемой одной
особью FB (когда все фенотипы отвечают генотипам). Верхняя линия
касается обычного F2, дающего отношения 9:3:3:1. Из графика видно,
что в фазе сцепления информация выше, требуется меньше особей для
достижения той же точности вывода, которая достигается относительно фазы
отталкивания. Чем сильнее сцепление, тем выше информация.
Кривая для F2 криптомерии идет гораздо ниже и только при сильном
сцеплении достигает 50% информации, даваемой FB. Особенно плоха
информация при отталкивании, даже слабом.
Причина столь низкой информации F2 зависит от того, что фенотипы F2
вообще, а при криптомерии особенно мало соответствуют генотипам.
2. Использование F3
Иммером (Immer, 1930) продолжено исследование криптомерии и
выяснен вопрос о возможности использования F3. Если каждое растение F2
(кроме ааЪЪ) самоопылять и высевать, например, по 25 особей на F3, то
можно выявить генотипы этих растений F2. При этом можно выявить:
1) гетерозиготность и 2) для дигетерозигот АаВЬ их строение АВ/ab или
Ab/аВ. Последнее труднее и при слабых геносвязях требует большого
числа особей F3 от каждой особи F2. Мы не будем приводить всех очень
интересных формул, выведенных Иммером, и дадим лишь графическое
сопоставление результатов, прочитать которые мы теперь сумеем.
На графике (рис. 13) даны три кривых. Кривая / относится к случаю,
когда при помощи F3 полностью проанализированы двойные доминанты,
вскрыты их гомо- и гетерозиготность и отделены АВ/ab от Ab/аВ. Кривая
III относится к тому же, но вскрыта только гетерозиготность, без
подразделения на структуры особей АаВЬ. Кривая 77 относится к разделению
особей Л, Ъ из F2 я&ААЬЬ ж АаЪЪ.
Интересным здесь оказывается следующее.
1. Прежде всего то, что проявление генотипа двойных доминантов
(А, В) резко повышает информацию и при с — 0 до 200% по сравнению
с FB. Это понятно из того, что каждая такая особь дает информацию о
перекресте двух ее родителей, а при FB — только о перекресте одного
гетерозиготного родителя.
2. Сравнение кривых I и III показывает, какую решающую роль
играет проявление у АаВЬ особей F их структуры АВ/ab или АЪ/аВ.
83
б*
О Ю 20 30 40 50 40 30 20 Ю
Отталкибание Сцепление
Рис. 13. Количество информации,
даваемое одной особью F2,
проанализированной при помощи F3 (по Иммеру)
I—полная классификация А, В доминантой;
II — одиночные доминанты, разделенные
на АА и Аа; III — двойные доминанты,
разделенные на ди- и моногетерозиготы
3. Из сравнения кривых II и 777
видно, что при с ^> 18% ис<^69%
(при этих точках кривые пересекают
друг друга) кривая II идет выше
кривой III. Это указывает, что
экспериментатору выгодно при слабых гено-
связах проявлять, при помощи F3
особей, не А, В, а4,6 особей F2. При
с <] 18%, т. е. при сильном
отталкивании, и то и другое примерно
одинаково ценно. Наоборот, при с ^> 69%
(т. е. при сцеплении, более сильном,
чем 31%) значительно выгоднее
проявлять генотип .особей Л, В.
4. При сильном отталкивании
информация, даваемая одной особью
F2, очень мала и в то же время
информация, даваемая каждой
проявленной особью, более чем в 25 раз выше.
Соответствующая формула получается
делением информации при
проявлении особи А, В на информацию F2
и имеет вид:
2 +с2
с (1 + с) (1 + 2с2)
При с <[ 0,75 эта величина более 25. При этих условиях оказывается
выгоднее затрачивать силы на то, чтобы вместо 25 особей F2 вырастить
25 особей F3, так как они дают больше информации (при другом числе
особей F3, выращиваемых вместо F2, будут и другие условия выгодности).
Мы видим, таким образом, что развитие идеи Фишера дает чрезвычайно
интересную теорию всей работы генетика г определяющего силу
сцепления, и целый ряд конкретных указаний о том, как и каким образом
организовать свое исследование. Необходимо расширить приложение этого
метода, распространив его на другие вопросы геноанализа.
Иммер исследовал значение F3 и для случая F2 криптомерии, дающей
в F2 отношение 9:3:4 [например, когда «криптомерия» — сходство всех
аа (я В и Ъ) обусловлено тем, что аа-формы рано погибают и не позволяют
отличить аВ от аЪ\. При этом выгода оказывается еще большей и может
быть использована при еще более широких рамках значения с.
Другой ученик Фишера — Хатчинсон (Hutchinson, 1929) исследовал
два более сложных тригибридных случая, с которыми сталкивались
ботаники. Один случай таков. Сцеплены гены А и В, причем ген В вместе
с третьим геном С являются парными генами, давая в F2 9:7. Второй
случай аналогичен, но гены В ж С .являются повторными генами и дают в F2
15:1. ]
Количество информации, даваемых одной особью F2, по сравнению
с одной особью обычного FB, выражается для первого случая формулами:
Зс3(5 + 2с2 —4с«)
(2 + с2) (1 + с) (2 — с2) (1 + Зс2)
для F
2»
1+2 — с2
2(2-с)(1 + с)
для FB тройного рецессива aabbcc.
Для второго случая формулы имеют вид:
с (11+2с2 — 4с2) „
Ч4. + с2) (4-с2) (1 + с)ДЛЯ *2:
84
2 + 2c — 2c2
2(2-c)(l+c)
для FBy тройного рецессива.
Первый из этих случаев (парные гены) ранее исследовался Бренсоном
(Branson, 1924), предложившим, в развитие идеи Эмерсона, формулу,
которая может быть выражена так:
c2 = ^.(a-b-c+|3d).
Второй случай изучался
жившим также формулу
ранее Вудворсом (Woodworth, 1923), предло-
г = 0,25 /а + Ь + d — 15с,
где г = 2с.
Хатчинсон исследовал обе эти формулы методом наибольшего
правдоподобия и определил для них количество информации, по сравнению с FB,
что выразилось формулами:
т? 108с2 (1 — с)
для метода Бренсона 32 + 32с*-27с* '
4с8 (1 с)
для метода Вудворса, для F2 1Ъ_Ш2_^.
Следующие два графика показывают сравнительное достоинство
разных скрещиваний и методов для каждого из обоих случаев.
На рис. 14 для парных генов видно, что информация здесь значительно
понижена и даже для FB (кривая /) в лучшем случае достигает 7з
информации, даваемой одной осью из обычного FB (т. е. когда все четыре
фенотипа ясно различимы). При этом мы имеем любопытное явление, что при
большем с (т. е. при сцеплении более сильном, чем 30% перекреста) криваЕя
F2 (II) оказывается выше кривой FB. В этих условиях, следовательно,
скрепщвание F2 оказывается более поучительным, чем FB, хотя и при своей
вершине дает едва 40% информации, добываемой при обычном F2.
5^
i
f
i
ии
30
го
10
i
—*ff^j I i
/я*
1 t I
1
*
I 1
0 10 20 30 40 50 40 30 20 10 0
Отталкивание Сцепление
Щ 0 10 20 30 40 50 40 30 20
i§ Отталкивание Сцепление
Ю О
Рис. 14. Количество информации,
извлекаемое методом максимальной
вероятности (FB — кривая /, F2 —
кривая II) и формулой Бренсона (кривая III)
(по Хатчинсону)
Рис. 15. Количество < информации,
извлекаемое методом максимальной
вероятности (FB — кривая /, F2 —
кривая II) и формулой Вудворса
(кривая IIГ) (по Хатчинсону)
85
DU
Л0
*Ч*
^
t^
§•§ 30
■S-^
5;^
§ ^
& <o
S> 3=
3= &
^л
lie
8
^s/
l^
/
/ 5^
L—i н^-т 1
1 1 ' ' ^
0 10 20 30 40 50 40 30 20 10 Q
Отталкивание Сцепление
Рис. 16. Количество информации,
извлекаемое из данных об FB и F2 при
разной силе геносвязи в случае повторных
генов и в случае парных генов (по
Хатчинсону)
1 — Рв для обоих случаев; II — Р2 для
повторных генов; III — Fa для парных генов
8 W 24 12 40
Проищ«п перенресгрд
Рис. 17. Сравнительная точность
определения процента перекреста по F2
при сцеплении и по F2 при
отталкивании (по Иммеру)
I — Fa отталкивание; II — Fa сцепление. На
ординате показано, во сколько раз больше
материала нужно для получения по Fa такой
же точности вывода, как по обратному
скрещиванию
Кривая, относящаяся к формуле Бренсона (Ш), на большом
протяжении (до с = 60) очень близка к кривой метода наибольшего
правдоподобия. Но в области более сильных сцеплений она начинает резко падать,
показывая малую пригодность ее в этих условиях.
На рис. 15 для повторных генов видно, что информация здесь понижена
еще более значительно, и обратное скрещивание в лучшем случае, при
слабой геносвязи, дает только 33% информации, получаемой от простого
FB (т. е. когда все четыре фенотипа ясно различимы). Для F2 информация
совсем низкая и в лучшем случае при сильном сцеплении дает лишь 12%
по сравнению с простым FB. Это в четыре раза хуже, чем для обычного F2,
дающего расщепление 9:3:3:1. Иными словами, для получения той
же точности вывода о силе сцепления необходимо в четыре раза больше
материала.
Информация, извлекаемая по формуле Вудворса (кривая ///), особенно
низка и только при самом сильном сцеплении сравнивается с методом
наибольшего правдоподобия.
Хатчинсон исследовал дополнительно также случаи дигибридные, когда
парные или повторные гены сами сцеплены друг с другом. Своеобразные
кривые их информации представлены на рис. 16. В области сильного
сцепления оба случая очень трудно поддаются анализу, так как
информация, даваемая в F2 и FB, очень понижена. В области сильного
отталкивания F2, наоборот, дает сравнительно приличную информацию, только
вдвое меньшую, чем обычный FB. Случай повторных генов (тип. 15 : 1)
изучается значительно легче, чем случай парных генов (тип. 9:7),
доступный для изучения лучше всего при средней силе сцепления.
3. Метод произведений
Выше мы видели, что с точки зрения количества информации о силе
геносвязеи, извлекаемой из материала, наравне с методом наибольшего
правдоподобия стоят метод произведений иметод наименьшего
86
5j2. Последний метод, связанный с решением уравнений 4-й степени,
слишком трудоемок и не представляет больших преимуществ в
других отношениях, поэтому он и не имеет перспектив для применения и
развития. Наоборот, метод произведений имеет крупное достоинство в том,
что легко может быть снабжен таблицами, почти устраняющими какую-
либо техническую вычислительную работу. Этот метод предложен и
разработан Фишером и Бальмакундом (Fisher, Balmakund, 1928) и особенно
Иммером (Immer, 1930).
Метод произведений сводится к тому, что для разных скрещиваний
рассчитывается теоретическая вероятность четырех классов фенотипов
(как и для всякого другого метода) — а, Ь, с, d и затем выводятся
соответствующие формулы для отношения двух произведений ad/bc. Применение
этого метода возможно, очевидно, только тогда, когда возникают четыре
класса фенотипов, так как если их три, то образование такого отношения
уже невозможно.
Следующая таблица (табл. 9) сопоставляет формулы для вычисления
величины с в разных генетических случаях, а также формулы вероятных
Таблица 9
Сопоставление различных формул для вычисления величины перекреста с
Фенотипиче-
ские
отношения
Формула для вычисления
Р
Вероятная ошибка
3:1 и 1:1
ad
1+р*
be 2 — Zp + p2
0,6745
Y4i
(2^р)(1-^)
(1 + 2^ — 2^)
3:1иЗ:1
ad _ 2^+jp*
be 1 — 2/>2 + р*
0,6745^ ~/Н2+/>
V 2N(14- 2юЦ
9:7 и 3:1
ad
2 + 7^ + Зр*
be
6 — V + 3/>4
o,6745i/(lziMHiz^!L(iz^)
*27: 37 и 3 :1
ad _14+25jp* + 9jp*
be 30 — 3 V + 9/>4
) H7AK1Г IS - P2) (2 + P*) (*Q - 3/»') (7 + V)
У 27NP* (9 + 2/>2 - 4/>«)
15 :1 и 3 :1
ad _ iip^ + p*
be 4-5/>2 + j&*
У N(11 + 2p* — 4j*«)
63:1 и 3:1
ad_
be
Zip* + p*
16 -r 17/>* + p*
1ЯадЛ1"^)(Ц-Л(47-Л
У tf(47 + 2/>2 — ЬрР)
9:7 и 9:7
ad _ 20 + 28jp2 + 9/>*
be 36 — 36/>2 + 2/>*
),6745]/"(10 + 9*2H4-/*)
18iV>2(3 + 2/>2)
9: 7 и 15:1
ad _11 + 34jp2 + 3^
be 27 — 30/>2 + 3/>4
0 ft74?l/(1 + 3>*> <9 - **> <* - *2) <A1 + ^)
Г * 6W/>2(13 — p2-2p*)
15:1 и 15 :1
ad _ 56jp0 + />4
be
16 —8/>2 + />4
0,6745]/(4"^(56 + ^)
2ЛГ(7 + 2р2)
87
Рис. 18. Сравнительная
точность определения процента
перекреста по F2 для
различных генетических случаев
На ординате показано, во сколько
раз больше требуется материала для
получения той же точности вывода
о силе сцепления, как в F2
простейшего случая 3 : 1 и 3:1 (по Иммеру)
П 8 ~ 16 24 Я Л 48 0 8 16 24 32 40 48
Процент перекреста
ошибок. Для формулы квадратических ошибок^следует всюду отбросить
коэффициент 0,6745.
Эффективность этого метода в различных случаях также различна.
Рис. 17 сравнивает для случая простой кооперации (3 : 1 и 1 : 1)
количества материала, потребные для получения той же точности ответа в
скрещиваниях FB, F2 со сцеплением и F2 с отталкиванием. Этот и следующий
графики можно было бы пересчитать в показателях информации, но мы
оставляем их так, как они построены Иммером. Здесь на ординате прямо
указано, во сколько раз больше материала требуется в разных случаях по
сравнению с FB. Числа по оси ординат представляют собой квадрат отношения
вероятных ошибок, т. е. величины, обратные количеству информации.
Из рис. видно, что F2 со сцеплением почти также хорошо использует
материал, какиГв, и лишь при слабых сцеплениях требует в 2,25 раз
больше материала. При слабых отталкиваниях требуется несколько больше
материала, зато при сильных отталкиваниях использование F2 очень
малоэффективно. При отталкивании с 8% перекреста требуется в 13 раз больше
материала, а при еще меньшем перекресте необходим огромный материал.
На рис. 18 тем же способом дано сравнение FB, F2 со сцеплением и F2
с отталкиванием при различных генетических случаях со случаем простой
кооперации. Можно заметить, что при повторных генах в фазе
отталкивания разница менее сильна, чем в фазе сцепления.
Формула для вычисления величины перекреста и их вероятные ошибки
по методу произведений представлены в табл. 9. При отталкивании
с% = 100%, т. е. совпадает с процентом перекреста, выраженным в
десятичных дробях; при сцеплении с% = (1 — р) 100%.
Иммером вычислено несколько числовых таблиц (табл. 10), по которым
на основании определяемой из опытов величины отношения ad/bc или
bc/ad непосредственно дается величина перекреста с и величина, при
делении которой на получается вероятная ошибка.
Например, в скрещиваниях, когда А и В дают отношение 3 : 1, в фазе
отталкивания получено на основании 100 особей ad/bc = 0,06218.
Отыскав эту величину во втором столбце, находим в первом столбце цифру
0,170, т. е. 17% перекреста между А и В. В четвертом столбце находим
величину 0,6509 и, деля ее на /100, получаем вероятную ошибку 0,06509,
т. е. ± 6,5%. Квадратическая ошибка будет больше в 1 : 0,6745 раза, т. е.
±10%.
4. Метод коэффициента ассоциаций Q
Коэффициент ассоциаций Юла был впервые применен Коллинсом для
определения величины перекреста (Collins, 1924). Оперируя
произведениями ad и be, видно, что этот метод близко примыкает к методу произведений.
С другой стороны, коэффициент ассоциаций относится к числу показателей
88
oooooooooooooooooooooooooooo
^(JN^^^rfN^COCOCOCOCOCOWCOCOCOlsalNSlNaiSDlNS^tO
oooooooooooooooooooooooooooo
CnOCn005tOOOCnisbc005COl^0005^tOOCO^OidSCOtOOCOOO*4
tOH^COOOOJCbCOC^COba^^tOCO-^lOi^OOH^.C^OOiCOl-^COOOOOCO
СЛ rfN H*
OOOOOOOOOOOOOOOOOOOOOOOOOOOO
С^<Й|^О05|А00|^|Л(»СЛМО^СЛС0Нк«5<10>^С15Н'ОсО00«ас5
i^g0C0C0OC0t0QC0t0CtiCnO0)0)N>00|^^H>0)N)C0^0i0}0)<l
O^rf^^C0cOH*^CnC0|SPCPC0C00000lS300^a)-^000000O5tO^400
CO 00 1* -^
OOOOOOOOOOOOOOOOOOOOOOOOOOOO
1^СЛС^СЯСЯС^С^№СЯСЛСЛСЛСЯСП|^ФФО)ФО)0)С))С)0)0)0)0)№
^cooococoi^co^cocoooto^tocr>^cn<oco^^cnootocnco^cn
OOOOOOOOOOOOOOOOOOOOOOOOOOOO
^l(S^^ijS^i^|N^^i|S|^^^WCtf00CCWCOWWWCOWO5WM
ОО^^Ь&СЛ|^|^Й5ММ1АООСООС100<10505СЯ^СООЗЬ5И-ОСО
IS3^b^cncOCO<^0»^OOH*CnOOtOCnOOIS3CnOOOC005COl-*'COCn~4cO
(O^WMtO^CO^tn^OOWOOMCnOOOOO^^i^O^^CDCDOO
OOOOOOOOOOOOOOOOOOOOOOOOOOOO
COCOCOCOCOe0COCOCOCOCOCOGOCOCOCOCOCO'O9№№№
COCOUUOdO}COMb9N}^IS3lA|A^M>OQO(OCOCOOOOO<l«JQa
Фl^l^ci}^9иocoчcяWl^ф^^lA(OФ^9coc/(|^:oocccO|jseO|^
^CTsoococo*J^p»fsc7apo^cn|N595cpi^H^coc/«cpH^H^co^^ooc^
о о о
00 *J О)
0,05462
0,06218
0,07033
о о о
04540
,05225
05973
о о о
СЭ ОЭ ОЭ
JS СП СЛ
оо о со
to со сп
о о о
2741
2829
2914
о о о
to to to
СП СЛ rfs
со со ^j
о о о
СЛ ** СО
0,03527
0,04118
0,04763
о о о
02832
03347
03915
о о о
6605
6583
6560
о о о
2463
2558
2651
о о о
to to to
(^ СО tO
0,12
0,02986
о
02369
о
6625
о
2363
о
to
со
|S3
0,11
0,02495
о
,01954
о
6644
о
2260
о
2110
оооооооооо
Ocooo^Scnrf^cotOb*.
оооооооооо
ооооооооо'о
tOh^h^-OOOOOOO
OOJCOCD^CnWi^OO
cnenocotootopoooto
H*COh*>tSPC&COh*»OQO
И* СЛ »-»> СО £* О О
00 О
СЛ
ОООООООООО
ОООООООООО
СЛ1-*00О0ОООО
ootoco-^cncotoH^oo
0>C75«<lCOCOOatOtOCni-»>
tOCOOit^tOOOOiCnCO
COOiOOCOCOlSPl-^Cn
Oi 05
оооооооооо
OJslCDpi^MWW^i^
Ю^5Н*£чСЛ£чНь«<1£0|£ч
оооооооооо
HpCD^OJCnCOhkCOQ
WONOJM'tn^^tnCn
СП h*>
оооооооооо
Величина
перекреста
с
ad/Ьс
отталкивание
bc/ad
сцепление
Fa
отталкивание
F2
сцепление
Отношение произведений
Величины, дающие при делении на
y~N вероятную ошибку
Я
I
•о
я?
£
я
аков
м
09
3
§
м
оше
и
н
ерекр
8
и
09
л
н
о
мет
о
S»
о *<
00
•*
ш
2
3
Я
1
а
н
м
о
»
е
я
Щ
3
о
а
со
I
3
я
№
tt
©
Ь
Л
Р
Й
^*
Таблица 10 (окончание)
Величина
перекреста
с
0,47
0,48
0,49
0,50
0,51
0,52
0,53
0,54
0,55
0,56
0,57
0,58
0,59
0,60
Отношение произведений.
ad/bc
отталкивание
0,8082
0,8676
0,9314
1,0000
1,0738
1,1533
1,2390
1,3316
1,4317
1,5400
1,6574
1,7848
1,9234
2,0742
bc/ad
сцепление
0,8071
0,8661
0,9313
1,0000
1,0736
1,1526
1,2373
1,3282
1,4260
• 1,5312
1,6446
1,7668
1,8989
2,0417
Величины, дающие при делении на
y~N вероятную ошибку
Fa
отталкивание
0,5225
0,5170
0,5115
0,5059
0,5002
0,4945
0,4888
0,4829
0,4771
0,4712
0,4652 •
0,4592
j 0,4531
! 0,4469
F*
сцепление
0,4888
0,4945
0,5002
0,5059
0,5115
0,5170
0,5225
0,5279
0,5333
0,5386
0,5438
0,5490
0,5541
0,5592
F»
х в
0,3366
0,3370
0,3372
0,3372
0,3372
0,3370
0,3366
0,3362
0,3356
0,3348
0,3339
0,3329
0,3317
0,3304
тсорреляции и именно для измерения корреляции был предложен Юлом.
Пирсон и Херон (Pearson a. Heron, 1913) показали малую пригодность его
для этой цели, так как он не может конкурировать с коэффициентом
корреляции. Однако некоторые недостатки Q как показателя корреляции делают
его как раз более пригодным для целей определения геносвязи. Этой
особенностью является то, что, как было продемонстрировано выше, величина
Q не изменяется от различной жизнеспособности классов фенотипов, если
влияние на нее генов А и В кооперируется по закону умножения.
Связь величины Q с величиной перекреста довольно сложна и, к
сожалению, не очень удобна для алгебраических операций. Однако Оуэном
(Owen, 1928) были выведены формулы Q для различных скрещиваний и
генетических случаев и для некоторых из них даны таблицы (через 5 морга-
яид). Впрочем, интерполяция при таких больших интервалах и криволи-
нейности зависимости далеко не точна. На рис, 19 и 20 представлены
графики изменения Q по проценту перекреста для FB и F2 и для F2 с отсутствием
перекреста у одного пола. О. А. Ивановой были рассчитаны таблицы
значений Q для FB для малых расстояний (0—10% перекреста) с интервалом
в 0,1 морганиды. Эту таблицу мы приводим здесь в графическом
изображении (рис. 21).
Формулы для Q следующие:
1. Для FB
Q- 2с-1
2. Для F2
<? =
2С2 — 2с + Г
4с2—1
2с* +1 *
3. Для F2 при отсутствии перекреста у одного пола
V 2с* + 1'
Ошибка Q вычисляется по формуле:
2 Ka^b^c^d-
90
Рис. 19. Номограмма для
определения процента перекреста
по величине коэффициента
корреляции (г) и ассоциации (Q)
ори сцеплении (по данным
Оуэна)
Кривые 1Q, 2Q и 3Q относятся к
Q, остальные — кг. Кривые
соответственно их номерам относятся
к случаям —1*1:1, 1 : 1; 2) 3 : 1,
3:1; при отсутствии перекреста у
одного пола 3) 3 : 1, 3 : 1; 4) 9 : 7,
3 : 1; 5) 15 : 1, 3 : 1; 6) 9:7, 3 : 1;
7) 63 : 1, 3 : 1; 8) 9 : 7, 9 : 7;
9) 9: 7, 15 : 1; 10) 15 : 1, 15 : 1|
80
70\
60
50
30
20
10
гч
10
5^
7^
5
Ф
2
з\
^
^
/
ч^
2'Q
2'
5 10 15 20 25 30 35 UO 4f 50
Процент перекрести
Рис. 20. То же, что на рис. 19,
но для фазы отталкивания
Продолжение кривых 2 и 2Q см.
на рис. 19 под 2' ш 2'Q
С2г
I
10
2о\
30\
40
50\
60
70
90
/,00\
пи
/
к
7
~.8~
yf
/
/
/
^
;—3-
2Q/
/
\*
10
— J^
/'
ч
^3
/1
//
V
3Q//
У/щ
/ >
/
//
Q 5 10 15 20 25 30 35 40
Процент перекреста
Щ 50
т Q /77 Q m Q m U
0,0
Рис. 21. Номограмма для
определения небольших расстояний п5л.аш95 30
(0—10 морганид) по величине Q Ц'° Т^шуо d>u
[09999
W
1,5
Щ
&
W 2,5Т
[0,9996
4,0
4,5
0,9987 5,0т0,9945 7,5
Ш970
[-09960
0t9950
6J5-
70
10,9987 5,0 L 0,9945 7,5 L 0,9869 Щ01
8,0-
Ш840
0,9920 85.
\0,99Ю
9,0\
Я5-
[0,9890
0,9880
5. Метод коэффициента корреляции
Возможно также определение величины перекреста по величине
коэффициента корреляции г при четырех полях.
г легко вычисляется по следующей формуле Келли:
d-
— 919» *
VpiPrtiQt
а
(а, В)
с
(а, В)
ъ |
(А,Ъ)
d
. (а,Ь) 1
где ръ=
а + Ь
а + с
«1 =
c + d
?2 =
b + d
Формулы, связывающие г с с, в различных генотипических случаях
следующие:
1:1 г = 2с — 1
4с2 —1
1) 1:1
2) 3:1
3:1 г =
3) 3:1 3:1 при отсутствии перекреста у одного пола
4с —1
г =
4) 9:7
5) 15:1
6) 27:37
7) 63:1
8) 9:7
9) 9:7
10) 15:1
3:1
3:1
31
3:1
3:1
15:1
15:1
г =
г =
г ,-—
г =
г =
г =
г =
3
4с2 —1
4с2—1
3 VW
4с2—1
V37
4с2 —1
ЗУ"21
4с2—1
7
4с2 —1
105
4с2 —1
15
На рис. 19 и 20 изменение г в зависимости от с показано графически.
6. Дальнейшие проблемы
Во всех изложенных выше методах имеется то общее, что все они
приводятся к уравнению с одним неизвестным — величиной перекреста.
Благодаря этому в рассчитываемых теоретических пропорциях для F*
оба средних члена (Л, Ь и а, В) всегда равны, а для FB обакрайние члены
(А,В ж а>Ъ) равны друг другу. Наиболее частое искажающее обстоятель-
* Более подробно о различных формулах вычисления корреляции при четырхпольнои
решетке см. в кн.: Ю л Дж., К е н л э л М. Теория статистики. М., Госстатиздат,
I960,— Прим. ред.
92
ство, вызываемое разной жизнеспособностью, стремятся исключить
методом произведений и другими методами. Между тем в разных генетических
исследованиях нередко не соблюдается по разным причинам равенство
членов А, Ъ и а, В, и для решения задачи следовало бы сводить ее к
отысканию большего, чем одно, числа неизвестных. Так как это трудно, то
возможно вместо теоретического 9:3:3:1 брать иное, учитывающее
уклонение каждого гена от формулы 3:1. Например, если ген А дает
фактическое расщепление 2,5 : 1, а ген В — 3,2 : 1, то дигибридное отноше-
жие при независимости должно быть 8 : 3,2 : 2,5 : 1.
Поэтому полученное в опыте фактическое отношение надо сравнивать
не с отношением 9:3:3:1, ас аномальным (см. Kappert, 1927). Это
может иметь особое значение при оценке совпадения фактического с
теоретическим по методу %2, так как отсутствие хорошего совпадения с
нормальным отклонением может вовсе не указывать на то, что сила сцепления
определена неправильно. Наоборот, попытка найти, например, методом
«минимального %Ч наилучшее совпадение может, по-видимому, привести
к отысканию неверной величины сцепления. К сожалению, этот вопрос еще
не освещен теоретически в достаточной степени.
Мы изложили вопрос об определении величины перекреста подробнее,
так как эволюция взглядов и углубление знаний в этом вопросе являются
очень поучительными. Начиная с довольно элементарных попыток
решения вопроса Бэтсоном и Пеннетом и Эмерсоном, мы пришли к настоящему
времени к очень стройным идеям в этой частной области, пример которой
показывает, как должны будут в дальнейшем развиваться и другие главы
генетического анализа, чтобы последний мог вырасти в теоретическую
дисциплину, освещающую практику работы генетика-аналитика.
7. Уточнение локализации
Описанными выше приемами мы узнаем, что изучаемый нами ген
обнаруживает сцепление или отталкивание с некоторым ранее изученным геном
ж дает с ним такой-то процент перекреста (с%). Этим самым мы узнаем, что
наш ген А локализован в той же хромосоме, что и ген В, и на расстоянии
с% от него. С некоторыми приблизительными поправками мы узнаем это
расстояние и в морганидах — число морганид есть тоже процент
перекрестов, если предположить, что все перекресты замечены и учтены.
Однако на этом локализация еще не является законченной. Мы знаем
лишь расстояние от А до В, но не знаем, в какую сторону от В расположен
ген А. Принято условно один конец хромосомы считать правым, другой —
левым. Следовательно, надо узнать, вправо или влево от В находится ген А.
Если в хромосоме только и известны гены А и В, то этот вопрос
решается произвольно — произвольно мы можем принять, что А находится левее
или наоборот. Но если в хромосоме известно более двух генов, т. е. по
крайней мере гены В и С, помимо А, то возникает задача узнать, находится
ли ген А от гена В в сторону С или в противоположную сторону.
Для решения этой задачи необходимо знание всех трех расстояний
А — В, А — С ж В — С. Самое большое расстояние, очевидно, разделяет
крайние гены из этой тройки. Если расстояние В — С самое большое, то,
следовательно, А расположен между В и С, и порядок генов В — А — С.
Если А — В самое большое, то, следовательно, ген А расположен от гена
В за геном С, и порядок генов В — С — А. Наконец, если самое большое
расстояние А — С, то порядок генов А — В — С. В зависимости от того,
какой ген, В или С, считается расположенным правее, узнаем, где
расположен А — вправо или влево от В и от С.
При небольших расстояниях между этими тремя генами большое
расстояние равно сумме двух меньших. Но если расстояния более или менее
93
значительны, то сумма двух меньших несколько превышает большее
расстояние и можно построить некоторый треугольник (схема XV).
В
A^L ±С
Схема XV
Так как наименее правильным является самое большое расстояние,
наиболее уменьшаемое двойными перекрестами, то его увеличивают так,
чтобы оно равнялось сумме двух меньших, и при помощи этих двух
меньших расстояний укладывают три гена в одну линию.
Для получения наилучших данных ставят, наконец, «трехгенный»
опыт, выбирая такие два гена В и С, кроме А, чтобы А было посредине.
Ставят скрещивания
ВАС Ъас ВаС Ъас
Ъас Ъас ЪАс Ъас *
при этом на одном и том же материале определяются расстояния В — А7
А — С, что и дает наиболее точные результаты.
8. Установление и локализация леталей
Леталями, или летальными генами, называют гены, приводящие
организм к гибели, т. е. лишающие его жизнеспособности. Эта гибель
может происходить на самых различных стадиях развития. В тех случаях,
когда эмбрион погибает незадолго до рождения или вылупления или
вскоре после него, установление и локализация леталей не представляет
больших затруднений. Невозможным оказывается лишь применение
анализирующего скрещивания, так как особи аа не жизнеспособны.
Однако имеется, судя по дрозофиле, очень распространенная
категория леталей (так называемых зиготических леталей), которые убивают
организм на самой ранней стадии развития эмбриона, вскоре после
оплодотворения. При такой ранней гибели заподозрить существование летали
оказывается обычно невозможным. Если, например, у коровы и быка будет
в гетерозиготном состоянии находиться такая лета ль, не проявляясь в их
фенотипе, то 25% зачатий будут как бы безрезультатными. Можно будет
думать, что просто не произошло оплодотворения, в то время как оно на
самом деле произошло, но эмбрион рано погиб и рассосался, дав только
яловость, причиной которой могут быть, как известно, и иные факторы.
Только после развития топографической генетики и открытия у
дрозофилы летальных генов удалось дать метод для их изучения.
Наиболее просто устанавливают летали в половой хромосоме. Здесь
показателем наличия летали служит ненормальное отношение полов в
потомстве самок, несущих леталь. Скрещивание с такой самкой идет по схеме
XVI.
В итоге потомство оказывается состоящим из 67% самок и 33% самцов,
или 2 ^ : 1 с? против нормального отношения полов, близкого к 1 9 :1с?-
По одному скрещиванию, конечно, не всегда удается уверенно
утверждать, что доля с? в 33% достоверно отличается от доли в 45—50%. Для
проверки ставят новые скрещивания, используя полученных дочерей
(с какими самцами — безразлично, так как все самцы имеют L). Как вид-
94
LI
LL
XX
?
ID
\ y
XX
1
LI
XX
?
ft)
Ч(аеут-ъ—
XY
L
XY
I
XY
Гибнет
Схема XVI
но из схемы, половина дочерей (тип. 2) будет иметь генотип матери,
гетерозиготные по летали, они снова должны дать «летальное отношение
полов», т. е. 2$ : 1(5*, и так исследование можно вести из поколения в
поколение, пока картина не станет вполне убедительной. У животных
малоплодных такое исследование, конечно, очень трудно, так как отделить
дочерей, дающих отношение 29 : 1 с?^ от их сестер, дающих 1$ : 1с?»
практически невозможно. Приходится суммировать их потомство, в
котором, следовательно, отношение полов должно быть 4 9 : Зс?» или 57% § :
: 43%с?, что довольно близко к 50%.
Установление аутосомной летали еще затруднительнее и должно
использовать какой-либо сигнальный ген, локализованный в той же
хромосоме. В случае дрозофилы такое скрещивание идет обычно по следующей:
схеме:
Гаметы
отца
la
La
l±xI±
La La *
Гаметы матери
Без перекреста 1
ин-г!
1А 100 —с
La 4
\lA 100 — с
\LA 4
r 1C0 — с
La—2~
LA 100 — с
IA 4
La 100 — с
\La 4
С перекрестом
*г
la с
TaT
la с
La~T
"Г
LA e
IA 4
i LA с
La 4
* с — процент перекреста между А и L.
Распределяя по фенотипам, получаем:
LA 2 100А~С + 2 4- =50 (в итоге 6?о/0)
1,А
4
100 — с
4
= 25 (в итоге 33%)
4 ' 4
"I—т- = ^5 (гибнут)
Из-за гибели 25% (L4-oco6h) среди оставшихся в живых получается*
отношение 2А : la вместо нормального 3:1, которое получилось бы, если
бы 1А особи не погибли.
Именно по тому признаку, что ген А, который ранее давал совершенно
нормальное отношение 3:1, вдруг стал давать отношение 2:1, мы и
подозреваем участие летали в этом скрещивании. О самой летали мы узнаем
пока лишь, что она расположена в той же хромосоме, как и сигналь А. Но*
95
о расстоянии L —А мы судить еще не можем, так как из формул видно, что
величина с не отражается на окончательных процентах фенотипов.
Определение места летали приходится вести, используя возникающих
в описанном скрещивании самцов la/La. Скрестив их с самками lAILa,
получаем:
Гаметы
отца
la
La
■ 1
Гаметы матери
без перекреста
1А 2
1Л 100 — с
la 4
1А 100 — с
La 4
, 100—с
La 100 — с
la 4
La 100 —с
La 4
с перекрестом
. с
1ат
la с
1а~Т
la с
Та~1Г
"г
LA с
la 4
LA с
La 4
Распределяя по фенотипам, получаем:
L,A 2
100-
4
.+ 2-|- = 25 + -f
L,a 2i^ + -|- = 50-^-
l,A -i^L-25—f
, С С
гибнут
Вместо отношения 1^4 : la мы получили снова Л в недостатке,
указывающем на участие в скрещивании летали в той же паре хромосом. Но
здесь уклонение от нормы зависит уже от величины с. При с = 0
отношение А : 2а, при с = 50% отношение 1:1, при промежуточных величинах
с получается и отношение А : а, промежуточное между 1 : 2 и 1 : 1.
Следовательно, по величине этого отношения можно заключить о расстоянии
между L и А. Для этого могут послужить формулы:
с = 200% - За%
с = ЗА% -100%,
где А% и а% обозначают процент полученных в скрещивании фенотипов
А или а.
Приведем небольшую таблицу для определения величины с по
процентам фенотипов А и а в этом типе скрещиваний.
Процент фенотипов
a
67(66,7)
65
60
А
33
35
40
с%
0
5
20
| Процент фенотипов
a
55
50
А
\ъ
50
с%
35
50
Определение расстояния L — А можно вести и в других скрещиваниях,
например,
la
96
Здесь имеем:
Гаметы
отца
А
la
без перекреста
LA<V
LA 100 —с
LA 4
LA 100— с
la 4
. 100—с
la 100 — с
LA 4
la 100 — с
la 4
с перекрестом
ч-
La с
LA IT
La с
la~T
"г
1А с
LA 4
1А с
La 4
Распределяя по фенотипам, получаем:
100 — с
L,A 2
L, а
JL — ^о !L • 67 —
100 —с
^— + 2-^ = 25 + ^-;
, 100 —с
I, a
4
25 %-
гибнут
33 +
Отношения здесь точно обратны предыдущему скрещиванию. Точная
локализация летали производится тоже в опыте с двумя сигналями —
одной, расположенной влево от летали, и другой — расположенной вправо.
Ограничимся рассмотрением случая с деталью в Х-хромосоме. Имея самку
А1В „ ,
строения —ут— f получаем от нее сыновей следующих генотипов (сх —
процент перекреста А — L, с2 — процент перекреста L — В):
Перекресты
0
1
2
1,2
Генотип
А1В
abb
ALb
alB
Alb
aLB
ALB
alB
Частоты зигот
50 c% °2 __-ClC2
2 2 200
50 °x ^2 4-ClC2
2 2 ^200
T 200
Ci С1С2
"2" 200
C2 ClC2
T" 200
c% C1C2
"2" 200
CiC2
200
C1C2
200
Частоты типов,
оставшихся в живых
Гибнут
Гибнут
Гибнут
Гибнут
т-«-«+т
ci-—
100
С2-Т
ClC2
100
£i£2
100
7 А. С. Серебровский
97
Подсчитываем отношения фенотипов А : а и В : Ъ (в процентах;:
. _ £l£2 £l£2 _ _ ..
Л - Cl 100+100 - Сь Cl — Лу
[отсюда сх = 100% — а.
а = 100%— сь
СЮъ , С1С2
loo"1" loo
100% — с2, отсюда с2 = 100% — Ь.
Я — Г CiC2 C1C2 _ с — В'
** - с* ~~ loo ' юо ~ °2' °2 — °*
Иными словами, процент фенотипов А и В дает непосредственно
процент перекреста сг и с2. Чем меньше самцов Л, тем ближе к нему
расположена деталь I.
При локализации аутосомных деталей обычно (у дрозофилы)
применяют такой способ. Иметь самцов, наиболее удобных для анализа alb/alb,
невозможно по их нежизнеспособности. Но можно использовать гетеро-
зигот, имеющих одну хромосому строения alb и снабдив другую каким-
либо сигнальным доминантом, например Су (для изучения деталей во //
хромосоме). Таким образом ставится скрещивание, например:
MB Q Су
"25" * Х о^Гг
Из полученного потомства отбрасываем всех мух, имеющих признак
Су. Остаются мухи, получившие от отца лишь хромосому alb. Мы как бы
использовали все же гомозиготного самца alb. Остальные расчеты
аналогичны тем, какие мы только что проделали для половой хромосомы,
только там мы разбирали структуры ALB/alb, а сейчас разбираем структуру
A IB/abb.
Соответственно получим:
сг = а% или сг = 100% —.4%;
с2 = Ь% или с2 = 100% —В%.
АНАЛИЗ АЛЛЕЛИЗМА
Установление множественного аллелизма
Многие гены, давая многократно или повторно мутации, способны
изменяться при этом различными способами. В результате из одного
нормального аллеля (базигена) возникает несколько мутантных аллелей
(трансгенов), составляющих вместе с базигеном серию аллеломорфов,
или аллелей, аллосерию*. Аллосерии известны теперь уже
у большинства достаточно изученных организмов и имеются все основания
считать это явление настолько распространенным, что можно ожидать
наличие серий для всех повторно мутировавших генов даже тогда, когда
вновь возникающие трансгены чрезвычайно похожи друг на друга. Во
всяком случае установление аллелизма является важным элементом
генетического анализа.
Метод установления аллелизма мало чем отличается от установления
сцепления, так как аллелизм во многих случаях не отличим от абсолютно--"
го сцепления, а открытие ступенчатого аллелизма
устанавливает, как мы полагаем, и по существу переход между сцеплением и
* Термины базиген и трансген очень удобны для определения гена до мутации и нового
гена, появившегося после мутации. Очень жаль, что в большинстве руководств по
генетике они почти не употребляются.— Прим. ред.
98
аллелизмом через частичный аллелизм, о котором скажем ниже. После
того, как абсолютное сцепление доказано, остается некоторыми
добавочными соображениями установить, с чем мы имеем дело: со сцеплением
двух негомологичных генов или с аллелизмом.
Следующий пример покажет ход этого анализа.
При скрещивании окрашенного кролика (например, черного) с
альбиносом получаем типичную моногибридную картину: черный доминирует
над белым, в F2 получаем 3 черных: 1 белый, в FB — 1 черный: 1 белый и
т. д. Отсюда устанавливаем ген окраски А и ген альбинизма а. Черный
кролик-гомозигот будет А А, альбинос — аа. Имеется еще одна окраска
кроликов, так называемая горностаевая, при которой белая
окраска туловища и красные глаза соединяются с черной окраской ушей,
носа и лап. При скрещивании горностаевого кролика с черным получаем
тоже моногибридную картину: черный доминирует над горностаевым, а
в F2 возникает расщепление на 3 черных: 1 горностаевый. Отсюда
устанавливаем еще аллопару: N — сплошь окрашенный и п — горностаевый
или акромеланистический (акры — конечности) *. В результате этого
черный кролик получает уже дигибридную формулу AANN, а
горностаевый A Ann. Так как скрещивание черного с альбиносом шло по
моногибридной формуле, надо предположить для альбиноса формулу aaNN, так
как при формуле аапп в F2 от скрещивания черного с альбиносом
расщепление должно было бы идти по дигибридной формуле и дать горностаевых,
чего не было:
9.4, N — черных
ЗА, п — горностаевых
За, ЛГ—белых
la, n —белых
Отсюда следует, что при скрещивании горностаевых {A Ann) с
альбиносами (aaNN) мы должны получить в Fx черных (AaNn) и в F2 9 черных : 3
горностаевых : 4 белых, как только что указано.
Однако поставленные Кастлом (Castle, 1919, 1925, 1930) и др. скрещи-,
вания не оправдали ожидания. Странным образом скрещивание
горностаевых с белыми пошло тоже по моногибридной схеме: в Fx доминирует
горностаевый над белым, в F2 возникает 3 горностаевых: 1 белый и т.д.
Сколько раз это скрещивание ни ставилось, результат всегда оставался один
и тот же— кроликов структуры aaNN не удалось найти.
Мы имеем таким образом случай, противоречащий элементарному
менделизму: три различные фенотипа (черный, горностаевый, белый)
наследуются во всех скрещиваниях как моногибриды. Правда, при неполном
доминировании одна аллопара может дать три фенотипа, отвечающие трем
генотипам А А, Аа, аа, но в данном случае это не объясняет дела, так как
горностаевый кролик не всегда является гетерозиготным, может быть и
гомозиготным.
Возникают две возможных гипотезы: или (1) генотипы aaNn
нежизнеспособны, или (2) гены А и N сцеплены друг с другом.
По первой гипотезе в Е2-скрещивании черного AANN с белым (аапп)
должно быть:
9Л, N — черных
ЗА, п —горностаевых
За, N —(гибнут)
la, n —белый
Полное изложение вопроса о серии аллелей основного гена окраски у кроликов см.
в кн.: Д у б и н и н Н. П., Г е п т н е р М. А. Руководство по генетике и селекции
кроликов. М.— Л., Сельхозгиз, 1932.— Прим. ред.
99
7*
т. е. 9 черных : 3 горностаевых : 1 белый, что совершенно не отвечает
данным опыта.
По второй гипотезе мы должны получить в F2 какое-либо из отношений,
отвечающих сцеплению той или иной силы, например:
22
5
5
4
41
7
7
9
342
21
21 и т.
100
Д.]
3#+4& + 2
2& + 1
2А? + 1
где к есть величина > 1, показывающая,
АВ и аЬ возникают чаще, чем АЪ и аВ:
кАВ : 1АЪ : 1аВ : каЪ.
во сколько раз гаметы
Большой опыт показал, что в F2 горностаевых не возникает вовсе, как
не возникает и при обратных скрещиваниях ¥г с альбиносами по
предлагаемой схеме AaNn X аапп. Мы, таким образом, вынуждены заключить,
что Ли.ЛГочень тесно или даже абсолютно
сцеплены друг с другом. Обозначив это обстоятельство скобками,
получим следующую схему наших скрещиваний:
черный
горностаевый
черн. X бел.
(AN)(AN) X (an) (an)
Fi
черя.
(An) (an)
F2
черн. черн.
(AN) (AN)+2(AN) (an)+
бел.
+ (an) (an)
(AN) (AN)
(An) (An)
альбинос
не найдено
черн. X горност.
(AN) (AN) X (An) (An)
Fi
черн.
(AN) (An)
F2
черн. черн. горност.
(AN) (An) + 2 (AN) (An) + (An)(An) +
горност.
+ (An)(An)
(an) (an)
(aN) (aN)
горност. X бел.
(An) (An) X (an) (an)
Fi
горност.
(An) (an)
F2
горност.
2 (An) (an) +
бел.
+ (an) (an)
Эти схемы, как видно, вполне удовлетворяют наблюдаемым в
экспериментах фактам. Таким образом, по данным скрещивания можно принять
гипотезу абсолютного сцепления А ж N генов.
Однако кое-что в этой гипотезе нас не должно удовлетворить. В самом
деле, исходный генотип дикого кролика был AANN. Когда возникла
мутация горностаевого — из A ANN возникло A Ann изменением одного гена.
При возникновении альбинотической мутации следовало бы ожидать
мутации тоже лишь в одном гене, на этот раз в Л-гене, в результате чего
получился бы генотип aaNN, а для возникновения альбинизма аапп
должны были бы возникнуть две мутации сразу, в Л-гене и в iV-гене. Между тем
генотип aaNN не существует. Поэтому приходится признать, что при
возникновении альбинотической мутации возникла сразу двойная мутация:
(AN) -> (an).
Это необходимое добавление к принятой выше гипотезе абсолютного
сцепления. Мы имеем, очевидно, такую историю мутаций:
(AN)
Базиген (полная окраска)
an
Альбинизм
An
Акромеланузм
100
При этом условии, очевидно, более вероятной становится гипотеза
аллосерии, т. е. того, что AN есть единый базиген, давший два трансгена,
качественно или количественно различных. Поэтому мы можем отбросить
аллопару N — га, введя обозначение для третьей аллели Л-серии,
например, а':
А-полная окраска
а . а1
Альбинизм Горностаевая окраска
Характерна для аллосерии возможность уложения членов серии в ряд
по степени доминирования:
А>а'>а,
т. е. аллель А доминирует и над а, и над а', в то время как а' доминирует
над а, но рецессирует под А.
Таким образом, установление аллосерии (в случае трех членов)
сводится:
1. К доказательству, что три фенотипа находятся между собой в
моногибридных отношениях, не сводимых к различию между АА, Л а, аа.
2. К доказательству, что эти отношения не могут быть сведены к
нежизнеспособности одной из четырех дигибридных комбинаций.
3. К аргументации в пользу того, что доказанное пунктами 1 и 2
абсолютное сцепление может быть более вероятно объяснено серией из трех
аллелей Л, а', а благодаря тому, что
а) имеется ряд доминирования А > а' > а,
б) имеется известное единство всех затронутых признаков по их
характеру,
в) гипотеза абсолютного сцепления требует дополнения гипотезой
двойной мутации.
В том случае, если аргументы (а, б и в) отсутствуют, гипотезы
абсолютного сцепления и сериальности оказываются равновероятными или даже
первая гипотеза становится более вероятной.
Особенно трудно довести анализ до окончательного вывода в том
случае, когда изученные гены не укладываются в ряд по доминантности. Такие
случаи имеются, например, у кузнечиков Paratettix, Apotettix и других,
у которых известен целый ряд окрасок, наследующихся по
моногибридной схеме, позволяющей установить ряд аллопар Л— а, Z? — 6, С — с
и т. д. Однако в комбинации Л5, АС, ВС... нет доминирования или эписта-
за, и действие каждого гена отчетливо видно. Здесь можно поэтому
построить две схемы:
(Abe) (aBc) (abfl А' А" Х*'\
Обе схемы одинаково удовлетворяют данным скрещивания и для
объяснения возникновения групп (Лбе), (aBc), (abc) нет надобности требовать
добавочной гипотезы одновременных соседних или цепных мутаций. В
отличие от известных в большом числе аллосерии, группы (Лбе), (aJ5c), (abC)...
не образуют никакое ряда. Это обстоятельство затрудняет признание
№
101
в них аллосерии а, А\ А", А'", т. е. второй схемы. Поэтому более
вероятной является первая схема абсолютного спепления, а не аллелизм.
Доказательство правильности гипотезы абсолютного сцепления добавочно
дает сравнительное изучение 1) нескольких других генов, у которых
сцепление оказывается уже не абсолютным и 2) других видов, у которых очень
сходные с данными гены уже явно дают перекрест или даже свободно
комбинируются.
Частичный, или ступенчатый, аллелизм
Частичный, или ступенчатый, аллелизм (Дубинин, 1929, 1930, 1931;
Серебровский, Дубинин и др., 1929) устанавливается по признакам,
совмещающим признаки сериального аллелизма и абсолютного сцепления.
С абсолютным сцеплением он сходен тем, что члены лестницы не
укладываются в ряд по доминантности, но отличается тем, что компаунды из
двух рецессивных аллелей дают частичный или полный возврат к норме.
На серию аллелей лестница аллелей похожа тем, что члены ее хотя и
не укладываются в ряд, все же дают явно однородные видоизменения, но
отличаются от серии тоже тем, что компаунды дают возврат к норме. Так,
в наиболее изученном примере гена scute у Drosophila melanogaster мы
имеем, например, такие члены: scute —achaete —scute6—scuteb—-scute3 и
нормальный базиген, по отношению к которому все остальные являются
рецессивными. Наоборот, scute3 по отношению ко всем остальным является
рецессивным.
Однако компаунды achaete/scuteQ имеют фенотип нормальных мух, а
scutejscutes едва можно отличить от нормальных и только при
биометрическом исследовании. Мы получаем таким образом картину, сводимую
к схеме:
Базиген
(ABCDE)
(aBCDE) (AbcDE) (ABcdE) abode
achaete scute6 scute5 scute3
Компаунды, составленные из этих ступенчатых аллелей, будут
проявлять те или другие признаки, в зависимости от того, по каким из малых
букв получится гомозиготность, как показывают следующие примеры:
achaete aBCDE achaete aBCDE
scute, = AbcDE = норм' Фенотип^ Scutez = abcde = «А^И&внотнп;
$cute5 ABcdE с scute, AbcDE
scute, = AbcDE =~ "*внотип; scute, = abcde = «^-фенотип.
В данном случае нам приходится допустить цепные мутации, особенно
при возникновении scute3 других подобных. Но так как все отмеченные
буквами Л, Z?, С, Z>, E элементы очень близки по своему действию, то их
приходится считать уже не «абсолютно сцепленными генами», а
отдельными частями единого гена, «центрами». Линейное расположение этих
центров приходится устанавливать сопоставлением отдельных членов лестницы
друг с другом.
Следующая таблица сопоставляет рассмотренные случаи:
102
Абсолютное
сцепление
двух
—нескольких
генов
Лестница
аллелей
Гены не
укладываются в ряд по
доминированию
Отдельные члены
дают фены,
неродственные или мало
родственные
У компаундов
фены обоих членов
выражены, или
имеет место эпистаз!
Вообще не
укладываются, но по
отдельным элементам
фенов
укладываются
Фены всех генов
обнаруживают
явное родство —
влияют на один и тот
же признак, более
или менее
различно количественно
Компаунды обнару
живают частичный
или даже полный
возврат к доминант-]
ному признаку
Различия между
генами не поддаются
количественному
объяснению
Серия
аллелей
Укладываются в ряд
у младших членов
доминирование
неполное
У компаундов
фенотип либо
промежуточный, либо с
доминированием,
промежуточным у
младпгах членов
Допустимо
объяснение количественное
(более слабое и
более сильное
изменение базигена)
Теория ступенчатого аллелизма, исходящая из представления о
сложности строения гена, разработанная автором этой книги, не является в
настоящее время общепризнанной*. Необходимо указать, однако, что в
последнее время начинают появляться новые факты, обосновывающие
правомочность высказанных в этой теории представлений (Рапопорт И. А.,
1947).
Не вдаваясь в обсуждение самого вопроса по существу, мы считаем эту
главу важной, как рассматривающую метод установления самого явления,
давшего основание для развития теории ступенчатого аллелизма.
Составление задач
Для освоения решения задач с участием геносвязи полезно также
составление и решение неограниченных задач, задач-моделей. Явление
сцепления и отталкивания можно достаточно легко моделировать при
помощи тех же шаров, как было описано выше, со следующей особенностью.
В этом случае тиражи производятся сразу для обоих сцепленных
генов. Пусть, например, мы должны получить потомство от особи ABlab
при условии, что гены А и 2? дают 20% перекреста. Такая особь должна
дать 4 типа гамет в пропорции: 44S : 1АЪ : 1аВ : iab.
Делаем такое назначение:
Шары
12 3 4
5
6
7 8 9 10
Соотв.
А,
А,
я,
я,
гаметы
В
ъ
в
ь
То же для "отталкивания
1 5
2345 1234
6 7 8 9 ИЛИ 7 8 9 10
10 6
При таком условии при «сцеплении» вероятность тиража «гаметы А, В»
в четыре раза больше, чем вероятность тиража «гаметы А, Ь» или «гаметы
а, В», что отвечает условию задачи. В одном опыте мы вынули с возвра-
После открытия ступенчатого аллеломорфизма Н. П.Дубиниными А. С. С е-
ребровскимв 1929—1930 гг. проблема структуры гена прошла сложную и
длинную историю. В настоящее время сложная структура гена считается
установленным фактом (см. обзор вопроса в статье: Н. П. Дубинин. Теория гена,
история и современные проблемы.—Бюлл. Моск. об-ва испытат. природы, т. 69, вып.
1, 1964).
Для доказательства аллелизма или псевдоаллелизма сейчас применяют
предложенный Льюисом так называемый цис-транс-тест. В принципе он очень близок к
функциональному тесту на аллелизм Т. Моргана (см. об этом М. Е.Лобашев.
Генетика. Изд-во ЛГУ, 1962).— Прим. ред.
103
тон следующие 20 шаров:
718735513 063 824 4 62 60.
Распределив эти цифры по принятому значению, получаем
13 13 3 2 4 4 2 = 9А,В
5 5 = 24, 6
6 6 6 = За, В с% = 5/20-100% = 25% ± 9,7%
7 8 7 0 8 0 = 6 а, Ъ
Приводим образцы моделей для различных величин перекреста.
Указано, что соответствует гаметам типа АЬ и аВ
перекреста, %
40
30
20
10
. 5
А, Ь
А, Ь
Л, Ь
А, Ь
А% Ь
4 и 5 всякие
светлые 4, всякий 5
5 всякий
5 светлый
5 белый
а, В
а, В
а, В
а, В
а, В
Шары
6 и 7 всякие
6 всякий, 7 светлый
6 всякий
6 светлый
6 белый
" Можно использовать также иной принцщп, который будет описан ниже
(в главе XI), для моделирования паратипической изменчивости.
Для случая отталкивания частоты соответственно перемещаются, как
показано выше для примера 20% перекреста.
АВ
Для получения потомства от скрещивания, например; типа —-г с? X
я г^ АЬ „
X V "if • r^e ОД™ родитель имеет структуру сцепления, а другой —
отталкивания, для каждого потомка сначала надо выбросить отцовскую гамету
(по типу сцепления), а затем материнскую по типу отталкивания. Так,
выбросив два шара 6 и 2, читаем: От отца получена гамета аВ (= 6 по
таблице сцепления), от матери гамета АЬ (= 2 по таблице отталкивания).
Поэтому генотип потомка aBIAb.
ГЛАВА V
ГЕНЕТИЧЕСКИЙ АНАЛИЗ В ПОПУЛЯЦИЯХ
ГЕНЕТИЧЕСКИЙ ЭКСПЕРИМЕНТ И СТИХИЙНЫЕ СКРЕЩИВАНИЯ*
Нет сомнения, что главным орудием генетического анализа является
эксперимент. Только путем планомерных скрещиваний, каждое из
которых должно давать краткий и ясный ответ на заранее поставленный вопрос,
подтвердить или отвергнуть рабочую гипотезу,— только этим путем
генетический анализ организма может быть проведен достаточно глубоко.
Однако в ряде случаев мы либо вовсе не в состоянии ставить специальных
экспериментов (например, при генетическом анализе человека), либо
сталкиваемся с рядом технических или экономических препятствий. Было бы,
например, совершенно недопустимым ставить многочисленные
специальные скрещивания, чтобы изучить генетику пегости крупного скота, так как
они могли бы нарушить интересы работы с этими дорогостоящими
животными. С другой стороны, на такую работу потребовались бы десятки лет
и она не могла бы быть доведена до конца, даже если бы представляла
очень большой интерес.
Однако генетический эксперимент характеризуется счастливой
особенностью — он широко и непрерывно ставится самой жизнью. Можно
без преувеличения сказать, особенно по отношению к домашним
животным, что здесь почти на любую тему, интересующую генетика,
скрещивания уже ставились раньше и продолжают ставиться и теперь людьми, даже
не знающими слова Генетика. И перед генетиком-аналитиком поэтому
возникает задача не только ставить нужный ему эксперимент, но и
отыскивать уже поставленный и подробно регистрировать и изучать его. Можно
думать, что для решения более простых вопросов этот второй метод
окажется едва ли не более продуктивным, и, несомненно, более быстрым.
Такие скрещивания на разнообразные темы могут быть найдены в
любом стаде, где они пропадают не зарегистрированными. Регистрирование
и изучение их не может быть названо экспериментом в точном смысле
слова. Точно так же не может быть назван экспериментом и анализ уже
зарегистрированных скрещиваний в том^виде, в каком мы находим их в
различных родословных записях, в племенных книгах и пр. Однако из этого
материала генетик может извлечь ценные данные для генетического
анализа, если только сумеет наладить работу в достаточно широком масштабе
и подойти к обработке собранных данных с некоторыми специальными
приемами.
Основным отличием собранного таким образом материала от
получаемого в планомерных экспериментах является отрывистость и неполнота
* Вместо названия «стихийные скрещивания» лучше было бы писать «свободные» или
«случайные» скрещивания.— Прим. ред.
105
генеалогических данных. Там, где в эксперименте мы можем вести какое-
либо исследование непрерывно на протяжении десятка поколений, мы
вынуждены использовать десятки родословных протяжением в одно-два
поколения, редко более, или даже располагать данными об одном родителе, не
зная другого, и т. д. Mill \ | В § i
ЗАКОНОМЕРНОСТИ СТРОЕНИЯ СВОБОДНЫХ ПОПУЛЯЦИИ
Возможность проведения генетического анализа в популяции (хотя,
правда, и ограниченная) основана на возникновении в популяции
стохастических закономерностей, благодаря которым она оказывается не каким-
то беспорядочным собранием различных особей, но более или менее
стройным целым, части которого определенным образом связаны друг с другом.
Хорошую иллюстрацию этого мы наблюдали при наших геногеографиче-
ских исследованиях кур. Во многих селениях мы производили
взвешивание кур и измерение яиц. При этом в большинстве случаев яйца
принадлежали не тем именно курам, которые подвергались взвешиванию. Между
тем оба ряда чисел обнаруживали всегда большой параллелизм: в тех
селениях, где взвешенные куры показывали высокий вес, яйца тоже
оказывались крупными и наоборот. Иными словами, взвесив некоторую выборку
кур, можно было достаточно точно представить и величину яиц, которые
будут снесены другими, не взвешенными курами, единственной
особенностью которых являлось лишь то, что они были членами той же куриной
популяции, как и куры взвешенные. Точно так же определив, например,
процент кур с листовидным гребнем, можно было сказать, каков будет
процент таких же гребней у петухов в этом селении, опять-таки потому
только, что эти петухи являются частью той же популяции.
Эти закономерности в общем крайне просты и заключаются в том, что
различные генотипы возникают в популяции по законам вероятности.
Для этого, однако, популяция должна быть «свободной».
Свободной популяцией называется такая, в которой спаривание самцов
с самками происходит стихийно, причем имеется равная вероятность
скрещивания любого самца с любой самкой в популяции, а не регулируется
сознательным вмешательством человека. Если, например, в какой-либо
популяции имеется 70% черных животных и 30% рыжих, то «свободной»
популяция будет в том случае, если черные самцы будут встречаться с
черными самками в 70%, ас рыжими — в 30% и рыжие самцы точно так же
будут встречаться с 70% черных самок и 30% рыжих. Иными словами, эти
всевозможные типы встреч черных и рыжих животных для размножения
будут осуществляться в такой пропорции:
черн. <5Х9 черн. 70-70% =49%
черн. вХ 9 рыж. 70.30о/о=21%
рыж. в X 9 черн. 30-70% =21%
рыж. с? X 9 рыж. 30-30% = 9%
100%
То же можно записать в общей форме:
™ „ Вероятность _ „ Вероятность
Тип скрещиваний скрещиваний Тип скрещивании скрещиваний
АвХЯА Р(А).Р(А) ВвХЯА Р(В)-Р(А)
AdXQB P(A).P(B) В 6X9 В Р(В)-Р(В),
где Р — процент встреч.
Очень важно подчеркнуть, что популяция может быть свободной по
одним признакам и несвободной по другим. Например, мы можем вести
106
в каком-либо стаде внимательный отбор животных по молочности, но при
назначении в случку не обращать внимания на подбор по окраске, и
окраска останется в популяции свободной.
Ряд исследований на домашних животных (куры, коровы, овцы)
показывает, что «свобода» этих популяций по множеству признаков
действительно имеет место. Даже там, где человек вмешивается в размножение
животных, это вмешательство может быть не систематическим, не
согласованным и не нарушать заметным образом свободу популяции. Одним,
например, может больше нравиться вороная масть лошадей, другим —
рыжая, третьим — гнедая и в отдельных заводах могут подбираться
сознательно линии и по мастям. Но в более или менее обширной популяции
подбор по масти может не отозваться на свободе скрещиваний лошадей
различного размера или различного темперамента, поскольку в пределах
одной масти распределение этих признаков может быть таким же, как
в другой.
Познакомимся с некоторыми следствиями из этого общего свойства
свободной популяции.
1. Биномиальное распределение гомо- и гетерозигот
(закон Харди — Пирсона)
В свободной популяции генотипы АА, А а и аа распределяются по
биномиальной формуле:
р2АА + 2pqAa + q2aa,
где р — доля гамет, несущих аллель А, и q — доля гамет, несущих аллель
а. При этом р + q равняется 1.
Представим себе множество сперматозоидов, среди которых рА и qa
сперматозоидов, и множество яиц, среди которых тоже рА и qa яиц. Если
мы смешаем между собой сперматозоиды и яйца, то произойдет свободная
встреча гамет в процессе оплодотворения, что можно записать так:
(рА + qa)*(pA + qa) = p2AA + 2pqAa + q2aa.
Математические исследования показывают, что какую бы мы смесь
особей ни взяли, уже через одно поколение популяция придет к такому
строению *. Скрестив, например, $ АА X $аа, получим первое
поколение А а.
Скрестив их друг с другом, получим, как известно, в F2: 1 АА-\-2Аа+
+ 1аа. Это характерное для F2 распределение генотипов является не чем
иным, как частным случаем общего распределения указанного выше при
р = q = 1/2, что можно увидеть, написав его применительно к условию
р + q = 1, как 1/4 АА + 2/4 А а + 1/4 аа. Легко видеть, что, скрещивая
($Аа и QAa, мы ввели поровну гаметы с А и с а.
Возьмем произвольный случай — 20с? С? АА + 30(5* с?## и 30$ $ АА +
+ 1009 9 Аа+100 9 9аа- Пустив их в свободное скрещивание и получив
от каждой самки хотя бы по одному потомку, будем иметь:
* В популяционной генетике различают закон Харди — Вейнберга, т. е.
математическое выражение структуры панмиктической популяции в соответствии с формулой
Харди — Вейнберга, и закон стабилизирующего скрещивания Пирсона. Согласно
последнему в случае, если популяция не находится в равновесном состоянии, иначе
говоря, если соотношение генотипов и фенотипов в популяции не соответствует
формуле Харди — Вейнберга, свободное скрещивание обязательно приведет популяцию
к новому соотношению генотипов и фенотипов по формуле Харди — Вейнберга,—
Прим. ред.
107
/
от 30 $ $ AA
от 100 $ $ Aa
от loo 9 9 аа
2 = 230
с 20 d Aa
6 АА + Ь Аа
10 АА + 20 Аа + 10 аа
20 Аа + 20 аа
16 АА+ 46 Ла + 30 аа *
с 30 с? аа
18 Ла
30 Ла + 30 аа
60 аа
48 Аа + 90 аа
Итого 16 ЛЛ + 94 Ла+ 120 аа,
или (в %) 7,0% АА + 40,8% Ла + 52,2% аа
Это распределение еще не вполне свободно, но уже сильно
приблизилось к нему. В исходном материале было 30 АА + 1204а + 130аа, что
в пересчете на проценты дает 10,7%АА + 42,8%Аа + 46,5%аа.
Определим, каково будет распределение этой популяции, когда она
достигнет свободного состояния? В образовании F2 участвуют гаметы:
А ... (2x16) + 94 = 126 или р == 0,274
а... 94+ (2x120) = 334 илид = 0,726
460 1,000
Соответственно р2 = 0,075, 2pq = 0,398 ид2 = 0,527, а биномиальное
распределение, выраженное в процентах, будет
7,5%АА.+ 39,8°/Ua + 52,7%aa.
Легко показать, что уже в F2 наступит такое распределение, если в ¥г
будут отобраны $ $? и с? с?» имеющие генотипы, отвечающие составу
популяции *.
2. Устойчивость популяции
Если популяция достаточно велика и свободна и генотипы АА, Аа ж
аа не подвержены отбору в различной степени, то раз достигнутое
биномиальное распределение сохраняется и в следующих поколениях.
Пусть мы имеем популяцию в составе р2АА + 2pqAa + фаа, причем
таково распределение как среди самцов, так и среди самок. При свободной
встрече полов возникнут следующие типы скрещиваний и соответственно
результаты их:
Типы
скрещиваний
ААХАА
ААХАа
ААХаа
АаХАа
АаХаа
ааХаа
Частоты
скрещиваний
р*
ip*q
2pV
. 4pV
4р$»
г4
Потомство
АЛ
Р*
2p*q
—
pV
—
Аа
2p*q
2pV
2pV
i 2р?»
aa
.
—
—
PV
2p?8
**
* Необходима оговорка, что стационарное состояние достигается в популяции сразу
после первого же свободного скрещивания только по аутосомным генам.— Прим. ред.
108
Суммируя и помня, что р + q = 1, получим частоты генотипов
АА, Аа и аа в потомстве:
АА. . . р4 + 2p*q + p*q* =р2(р2 + 2pq + g2) = р2 (р + g)2 = р2
Ла . . . 2р3? + 4/>V + 2pq* = 2pq (р2 + 2pg + g2) = 2pg (p + g)2= 2pg
аа . . p2g2 + 2p?3 + g4 = g2 (p2 + 2p<? + <?2) = ?2 (p + qf = 92
Таким образом, популяция состава р2ЛЛ + 2рдЛа + фаа при
свободном размножении дала популяцию такого же состава. Строгий
теоретический анализ показывает, что подобная устойчивость свойственна
бесконечно большой популяции. Так как каждое поколение является лишь
известной выборкой из популяции, происходящей не от всех членов предыдущего
поколения, то каждое новое поколение возникает с некоторым
статистическим уклонением. Эти уклонения в ряде поколений могут
суммироваться и небольшая популяция приходит через некоторое количество поколений
к изогаметации (Ромашев и Дубинин, 1931), состоящей в том, что либо
Л-аллель, либо а-аллель выцадают вовсе. Однако по ряду причин, в
обсуждение которых здесь входить не будем, в действительных природных
популяциях этот процесс идет медленно. Для эволюционного учения тем
не менее он представляет большой интерес. Для генетического же анализа
важно лишь установить, что в огромном большинстве случаев в популяции
мы сталкиваемся с распределением генотипов, чрезвычайно близким к
биномиальному, что позволяет опираться на эти закономерности в геноана-
литической работе.
Приведем пример. В одном из стад Средней Азии Б. Н. Васиным (1928)
была обследована отара, в которой имелись три типа овец: с нормальными
ушами, с укороченными и «безухие», имеющие ушную раковину в 1—2 см.
В отаре на 843 головы оказалось: длинноухих — 729, короткоухих — 111
и безухих — 3. Так как короткоухие являются гетерозиготами, то состав
гамет стада таков:
А. . .(2 х 729) + 111 = 1569 или р = 0,93
а . . 111 + (2 X 3) = 117 или q = 0,07
4686
Отсюда по формуле Харди мы должны иметь такое распределение
генотипов: р2 АА = 0,865; 2pqAa = 0,130 и фаа = 0,005, а для отары в 843
головы 729,3 АА + 109,6 Аа + 4,1 аа.
Таким образом, фактическое распределение почти совпадает с
биномиальным по формуле Харди.
3. Распределение генотипов в популяции,
гетерозиготной по серии аллелей
Если в популяции встречаются не два аллеля, а три: 4, а' и а с
частотами р4, qa' и га, то различные генотипы дадут триномиальное
распределение:
р2АА + 2pqAar + фа'а' + 2prAa + 2qra'a + гЧа.
Этот многочлен алгебраически получается простым перемножением
трехчленов:
(рА + qa' + ra)*{pA + qa' + га).
Отсюда легко перейти и к случаям четырех и более аллелей,
f Серия аллелей представляет собой достаточно частое явление в
генетике. В частности, наибольшую известность приобрел случай трех
аллелей, определяющих четыре группы крови у людей. В связи с распростра-
109
нившейся, особенно во время войны 1914—1918 гг., операцией
переливания крови было обнаружено, что в ряде случаев такое переливание
приводило к тяжелым последствиям — агглютинации (слипание кровяных
шариков). При изучении этого вопроса выяснилось, что имеется четыре группы
людей, различающихся особенностью крови. Различия эти показали ге-
нотипическую природу, причем было установлено три аллеля А, В и R*.
Одна группа является компаундом АВ, другая имеет генотип А А или AR,
третья В В или BR и четвертая наиболее рецессивная — RR. В
подавляющем большинстве людских популяций имеется распределение:
2pqAB + р2АА + 2prAR + q2BB + 2qrBR + r2RR
IV III II I
4. Распределение генотипов в популяции по гену,
сцепленному с полом
Если ген локализован в половой хромосоме, то в популяции имеет
место следующее распределение:
Гомогаметный пол Гетерогаметный пол
р2АА + 2pqAa + q2aa рА + qa.
Таким образом у гетерогаметного пола (самцы у млекопитающих, мух
и пр., самки у птиц, бабочек и др.) распределение генотипов прямо
отражает состав гамет, а у гомогаметного пола имеет место обычное
биномиальное распределение.
Покажем, что несмотря на разницу генотипов у полов, такая
популяция тоже обладает устойчивостью. Для удобства расчетов напишем гете-
рогаметных животных так: AY и aY. Тогда гаметы, образуемые самцами
и самками, будут иметь состав:
р2А А -> р2А
2pqAa -» pq A + pqa
q2aa —* q2a
P (P + Я) Л + q (p + q) a = pA + qa'
pAY^^-pA + ^-pY
1 1
qaY->-Tqa + -2-qY
~ I I I I 1
— PA +-g" да + -24 (P + 9) y="2"pA + Tqa + ~2~У
В результате свободной встречи самцов с самками возникает потомство:
(pA + qa) (^TpA + ^qa + ±-Y) = ~р*АА +
+-j- pqAa \+ -^ q2aa + -у pAY + -у- qaY
или -±- (р2АА + 2pqAa + q2aa) + -\ (pAY + qaY).
В потомстве таким образом возникла половина особей гомогаметного
пола и половина особей гетерогаметного пола и в пределах каждого из них
* Эту систему групп крови у человека сейчас обозначают как АВО. Известно также
несколько других систем групп крови (см. об этом O.Prokop. Die menschlichen
Blut- und Serumgruppen.— Ver. G. Fischer, Jena, 1963).— Прим. ред.
110
распределение генотипов оказалось таким же, как в популяции
родительского поколения.
Характерной особенностью данного случая является следующее. Если
сопоставить частоту особей, обнаруживших рецессивный признак среди
разных полов, т. е. частоту особей аа у гомогаметного пола и особей а у
гетерогаметного, то получим простое отношение.
Так как q <С 1 (дробь), то q2 < q, и, следовательно, у гомогаметного
пола рецессивный признак встречается реже, чем у гетерогаметного, в q
раз:
аа .
aY,
Г
Я
иначе ^— = д.
Соответственно у гетерогаметного пола признак встречается чаще
в IIq раз. Если q = 0,1, то IIq = 10, признак встречается у гетерогаметов
чаще в 10 раз. Если q = 0,01, то IIq = 100 и т. д.
Хорошим примером подобных отношений является дальтонизм
(цветовая слепота) у человека. Этот признак встречается примерно у 1/20
мужчин (около 5%). У женщин же он несравненно реже, спускаясь до долей
процента. Но при частоте у мужчин в 5%, или 1/20, дальтонизм должен
встречаться у женщин (гомогаметный пол) в 20 раз реже, т. е. в 1/4%,
что, как мы видим, и имеет место в действительности (Серебровская, 1930).
Среди птиц, где гетерогаметным является женский пол, отношения
должны быть обратными. Удобными для наблюдений здесь являются
гены окраски — серебристой и поперечнополосатой. Соответствующие им
рецессивные признаки действительно чаще у кур, чем у петухов, что видно
из следующих примеров (в %):
Популяция
Дагестана
Армении
Дагестана
Южвого Дагестана
Армении
Петухи.
серебр.
30,2
18,2
поперечнополосат.
41,0
21,3
13,2
30Л0Т.
69,8
81,8
равномерн.
59,0
74,7
86,8
Куры
серебр.
14,7
14,2
поперечнополосат.
| ' 18,3
14,7
8,5
30Л0Т.
85,3
85,8
равномерн.
81,7
85,3
91,5
Эти поучительные картины еще раз подтверждают возможность
рассматривать распределение генотипов в фактически изучаемых популяциях
как совпадающие с биномиальным, с различными его модификациями.
5. Распределение генотипов в дигибридной популяции
В свободной популяции, гетерозиготной по двум генам, распределение
генотипов приближается к свободной комбинации двух биномиальных
распределений:
(р2 АА + 2pqAa + q2aa) (r2BB + 2rvBB + г?ВВ),
111
[то равно девятичленному многочлену:
fr2AABB + 2p2rvAABb + p2v2AAbb + 2pqr2AuBB + kpqrvAaBb +
+ 2pqu2Aabb + q2r2aaBB + 2q2rvaaBb + q2v2aabb.
3 несколько другом виде это выражение нагляднее показывает свободное
комбинирование генотипов по гену А и В. Вынесем за скобки элементы
гена А:
р2АА (r2BB + 2rvBb + u2bb) + 2pqAa (r2BB + 2rvBb + u2bb) +
+ q2aa(r2BB + 4rvBb -f v2bb).
Мы видим, что генотипы А А, Аашаа совершенно одинаково
комбинируются с генотипами ВВ, ВЪ и bb.
При р = q = 1/2 и г = v = 1/2 многочлен принимает вид, знакомый
из элементарного менделизма в качестве строения:
^-ААВВ + ^ААВЬ+±ААЪЪ + ^АаВВ + ^АаВЬ +
+ А АаЪЪ + -^ааВВ + -^ ааВЪ + -^ ааЪЪ.
Для распределения девяти членов нашего многочлена по классам
Л, В; А, Ь; а, В и а, Ь получаем такую общую формулу:
ф2г2 + 2p2rv + 2pqr2 + kpqrv) А, В + (pV + 2pqv*) Л, Ъ +
+ {q2r2 + 2q2rv) а, В + q*v* а, 6,
* ли
(Р2 + 2pq) (г2 + 2rv)A,B + (р2 + 2pq) ФА, Ь +
+ ф (г2 + 2rv) а, В + фт?а, Ъ
или, так как
р2 + 2pq = 1— q2 и г2 + 2гг? = 1 — г?2,
то
(1 - д2) (1 - г;2) Л, 5 + (1 - q2) v2 A, b + q2 (1 - г;2) а, 5 + gVa, Ъ.
В частном случае при g = г; = 1/2 получаем опять знакомое нам из
менделизма отношение:
То, что отношение 9:3:3:1 есть лишь частный случай строения
популяции при условии q = г? = 1/2, иногда упускается из виду генетиками,
полагающими, что подобное отношение является вообще свойственным
популяции. На самом деле, конечно, в зависимости от величины q и v
(и соответственно риг) могут возникнуть всевозможные отношения, как,
например:
А, В А,Ь а, В а,Ь
9 = 10"» v=*Tq 90»1 8>9 °'9 М
9 = 15"» v = l0 90jl °'9 8'9 °'*
9 = *ЙГ. у==Тб- 4М 4,6 44,6 4,4
9 = Ж* V = ~W 9'7 41'3 9'3 39'7
Многочисленные наблюдения, собранные нами и другими авторами
при изучении куриных популяций, показали, что свободное комбиниро-
112
вание генов в популяции действительно имеет место и что, следовательно,
приводимые выше формулы освещают теоретически явления,
происходящие повседневно в популяциях. Следующие примеры показывают это:
Розовидная форма гребней кур (А)
и наличие
хохла (В) А, В А, Ь А, В а, Ъ
Найдено 130 277 119 232
Вычислено • 133,7 273,3 115,3 235,7
Приведенная формула комбинирования генотипов в популяции может
быть расширена применительно и к более сложным случаям к генов. Тогда
она будет иметь вид
(р2АА + 2pqAa + q2aa).. -.{р\КК + 2pkqkKk + q\kk).
Полезно пояснить, что любой член, который получается при
перемножении этих генотипов, можно написать без всякого труда, пользуясь
следующими правилами, которые просто видны из примера.
Пусть мы хотим написать, с какой частотой в популяции должен
встречаться генотип АаВВСс. Соответственно каждой букве, символизирующей
ген, запишем частоты:
Аа ВВ Сс
PiQi pi РзЯз1
где под большими буквами — р, под малыми — q. Коэффициент является
произведением стольких двоек, сколько гетерозиготных пар в генотипе —
в данном случае Аа С с : 2 X 2 = 4. Итак, в полном виде получим:
Арг qiptp3q3A аВВСс.
Несколько других примеров:
plptqlAABBcc, Ър^^р^ъР^АаВЬСс, 2qlqlp3q3aabbCc.
Также автоматически можно записывать частоту встречи самца с самкой
определенных генотипов. Например, Аа ВЬ X аа ВВ будет встречаться
с частотой
4Pi2lP2?2' ЯгР\ = ^PiViPlq*,
а каждая из четырех категорий потомства от этого скрещивания будет
встречаться с вероятностью:
Чтобы не путаться со значками, при дигибридном скрещивании удобнее
употреблять, как мы делали выше, для генов А — а буквы р и q, для
генов В — Ъ буквы г и г; и т. д.
РОДИТЕЛЬСКИЕ ГРУППЫ
Изложенные выше закономерности свободных популяций позволяют
нам ввести некоторые новые понятия, которыми оказывается очень полезно
оперировать генетику-аналитику. Такими понятиями прежде всего
являются «Родительские группы» — «отцовская и материнская».
Пусть мы имеем в большом стаде, состоящем из черных и рыжих коров,
рыжего быка. Если известно, что разница между черными и рыжими
зависит от одного гена, то оказывается возможным с достаточной точностью
рассчитать окраску потомства, которая получится от данного быка.
8 А. С. Серебровский
из
Если данная популяция свободна (хотя бы по данному гену), то
генотипы коров распределяются по формуле
р2АА + 2pqAA + q2aa.
При скрещивании их с быком аа мы получим:
Генотип
матерей
АА
Аа
аа
Гаметы
матерей
А
| 1 1
а
Всего . . .
Гаметы
отца
а
а
а
Генотипы
телят
Аа
1 л 1
~2 Аа + ~2~аа
аа
Частоты
р2
2рд
1 9Z
Популяция
телят
р*Аа
pqAa + pqaa
фаа
Р(р + Я)Ла + д(р-\-д)аа = рАа + qaa.
Следовательно, популяция телят от рыжего быка должна иметь
структуру рАа + qaa, и нам остается определить коэффициенты р и q. Пусть,
например, в нашем стаде (если оно — свободная популяция по окраске)
было 75% черных коров и 25% рыжих. Отсюда 0,25 = q2 и q = 0,50.
Следовательно, р = 1 — q = 0,50, и мы должны ожидать среди телят 0,5 Aa-f-
+ 0,5аа (1 черный : 1 рыжий).
Популяцию коров, участвующих в скрещивании с нашим быком,
можно рассматривать как материнскую группу, имеющую генотип популяции,
р2АА + 2pqAa + q2aa.
Соответственно наше скрещивание запишем так:
аа X (р2АА + 2pqAa + q2aa).
Очевидно, вместо нашего рыжего быка может быть группа быков,
сборная по признаку окраски. В качестве материнской группы мы тоже при
желании можем взять не всю совокупность коров стада, но какую-либо
группу, например, черных коров. Рассчитаем, что должно получиться от
такого скрещивания. Формула материнской группы черных коров будет
следующая:
рЫА + 2рдАа __ рАА + 2дАа __ рАА + 2дАа __ рАА 2q
p* + 2pq - p + 2q " 1+q ~T+?+T+T
Скрещивание при условии р + q = 1 запишем так:
-"(тт? А +-ГТ-, «) = ТТТЛ' + Т7Т°°-
Если, как в выбранном примере, р = q = 1/2, то будем иметь: рыжий
бык X материнская группа черных коров -> 2/3 черных + 1/3 рыжих.
Рассчитаем, наконец, скрещивание отцовской группы черных быкон
с материнской группой черных коров:
1 А А 2q A q*
_1T+WЛА + TTw а + Т+^аа'
— > _—\у < ,
черн. тел. рыж. тел.
114
При q = 1/2 отцовская группа черных быков X материнская группа
черных коров ->• 8/9 черных + 1/9 рыжих.
Мы видим, что, оперируя родительскими группами, мы весьма просто
производим расчеты для различных типов скрещивания и, в частности,
для моногибридного случая получаем следующие результаты:
Скрещивание
родительских
групп
Потомство
аа^аа
ааУ^Аа
АаХЛа
1 + д
1
Аа -
i+q
АА-
lq
(1+?)2 ™т (1 + 9)"
или по фенотипу
l + 2g ga
(1+ff)* A+ (1+?)г
• Аа
(1 + я)2
ПРИЛОЖЕНИЕ К АНАЛИЗУ СЛУЧАЕВ
С ДВУМЯ ФЕНОТИПАМИ
Имея эти формулы, мы, очевидно, можем их использовать для целей
генетического анализа, так же, как в свое время использовали
мендедиетические формулы 3 : 1, 1 : 2 : 1 и 1 : 1.
Пусть, например, мы нашли в популяции, имеющей 75% черных
животных и 25% рыжих, что
1) рыжие X рыжие дали 100% рыжих;
2) рыжие X черные дали 67% черных + 33% рыжих;
3) черные X черные дали 89% черных и 11% рыжих.
Спрашивается, с каким случаем мы имеем дело, т. е. является ли этот
случай моногибридным и какая окраска доминирует?
Решение. То обстоятельство, что рыжие X рыжие дали только
рыжих, указывает на вероятную рецессивность рыжих. Появление рыжих
от скрещивания двух черных подтверждает этот вывод. Если рыжие
действительно рецессивны и случай моногибридный, то 25% рыжих во всей
популяции отвечает группе q2aa, т. е. ф = 0,25 ж q = 0,50.
От скрещивания черных с рыжими должно ожидать q/(l + q) рыжих,
т. е. при q = 0,50 1/3 рыжих, или 33%, а от скрещивания черных с
черными должно ожидать q/(l + qf рыжих, т. е. 1/9, или 11% рыжих. Все
расчеты совпадают с фактически полученными результатами и позволяют
признать нашу гипотезу удовлетворительной и качественно и
количественно.
1. Парные (дополнительные) гены в популяции
Совершенно аналогичным образом, только более кропотливо, могут быть
рассчитаны и гибридные и более сложные скрещивания.
Для примера выведем формулу для случаев парных генов (например,
две различные рецессивные белые окраски кур).
115
8*
Дигибридная свободная популяция будет иметь структуру:
р2г2ААВВ окрашенные kpqrvAaBb окрашенные
2p2rvAABb » 2pqv2Aabb белые
q2r* aaBB »
рЧ2ААЪЬ белые 2q2rvaaBb »
2pqr2AaBB окрашенные qb?2aabb »
Белая родительская группа будет иметь формулу:
р2у2ААЪЪ + 2pqv2Aabb + q2r2aaBB + 2q2rvaaBb + q2v4abb
рЧ2 + 2pgv2 + gh-2 + 2g2rv + g2v2
Знаменатель упрощается до v2 — q2v2 + q2 = z.
Окрашенная родительская группа будет иметь формулу:
р2г2ААВВ + 2p2rvAABb + 2рдг2АаВВ + ApqrvAaBb
p2r2 + 2p2rv + 2pq i* + 4,рдгг7
или, выводя рг за скобки,
рг {ргААВВ + 2/>г?А4Д6 + 2дгАаВВ + ЬдуАаВЪ)
рг (рг + 2^г? + 2gr + kqv)
Знаменатель упрощается:
рг [р (г + 2v) + 2q(r + 2v)\ = рг [(р + 2q) (г + 2v)J =
= pr(l + g)(l +У) = (1 -g)(l + g)(l +i;)(l _») = (1 -g2)(l _*■) = Jfle
Эта величина (.Кд) является одновременно долей окрашенных во всей
популяции.
Гаметы белого родителя будут:
-j- (pV-A.il» + 2pgv24abfe + q2r2aaRB + 2q2rvaaBb + q2v2aabb) ->
-► 4" О?2^6 + PQv2Ab + pqv2a> + q2r2aB + q2rvaB + q2^ +
+ Q2v2ab) = -i- [(pV + pqv2) Ab + (q2r2 + q2rv) aB + (pqv2 +
+ q2ru + q2v2)ab] = -L \pv2Ab + q2raB + qv (pv + qr + qu)db].
Гаметы окрашенного родителя будут:
-g (ргААВВ + 2pvAABb + 2qrAaBB + iqvAaBb) -»
-> -|r- (prAB + pv AB + pvAb + qrAB + qraB + qVrAB +
+ qvAb + qvaB + qvab) = -J£ [AB + y^fe + gaB + qvab].
Теперь нетрудно рассчитать результаты скрещивания:
1) белые X белые, 2) окрашенные X окрашенные.
1. Белые X белые дадут:
-i- [p2v*AAbb + 2pv2q2rAABb + 2pv*q (pv + q) Aabb + q*r2aaBB +
+ 2q*rv (pv + q) aaBb + q2v2 (pv + q) aabb].
Из них окрашены только ААВЬ, доля которых
2pq2rv2 _ 2pq2rv2 __ к
116
а белых соответственно
. 2pqW
2. Окрашенные X окрашенные дадут:
(АВ + vAb + qaA + qvaVf = ААВВ + IvAABb + IqAaBB +
AqvAaBb + v*AAbb + фааВВ + 2qv*Aabb + IfvaaBb + q^aabb.
Из них белыми будут:
v2AAbb + q2aaBB + 2qv*Aabb + 2q2vaaBb + q*z?aabb,
что составит от всего этого потомства долю:
р« + д* + 2qv* + 2дЧ + gV __ . __ 1+2д 1 + 2» _
(1+v + q + gv)2 ~~ (i+q)2' (l + *)2-A*
Остальные, окрашенные, образуют долю
l+2v + 2q + kgv _ l+2g 1 + 2v
(i + v + q + qv)2 ~~ (i + q)2 ' (i + vf
Полученные формулы для расчетов доли белых, получаемых от
окрашенных, и доли белых, получаемых от белых, довольно сложны и для
прямого использования неудобны. Можно, однако, применить следующий
графический способ (рис. 22).
Вычисляем значения К% (доля окрашенного потомства от белых) для
различных значений (при построении рисунка были вычислены К2 при
значении ^иг;:0;0,1;0,2,..., 0,9;1). Полученные величины нанесем
на квадратную таблицу, на которой абсцисса — значение д, а
ордината — значение v. Затем соответственно нанесенным на таблицу величинам
проведем интерполяционные кривые. На нашем чертеже такие кривые
начерчены через каждые 5%. Линия 5% соединяет все точки поля,
отвечающие таким величинам, при которых от скрещивания белых с белыми
получается 5% окрашенного потомства. В рассматриваемом случае эта
кривая носит замкнутый характер с точкой перегиба при q = v = 0. Из этой
же точки выходят и все остальные замкнутые кривые для 10%, 15%
и т. д. (на рис. 22 — сплошные линии).
Таким же образом построены еще две системы кривых на том же
рисунке: кривые для процента белых среди потомства от скрещивания
окрашенных друг с другом К3 (пунктирные линии через каждые 5%) и кривые
Рис. 22. Номограмма для
анализа дигибридной популяции в
случае парных генов (две
рецессивных белых окраски)
I — К2, процент окрашенного
потомства от скрещивания бел. X
хбел.; II—Kit процент белого
потомства от скрещивания окраш. X
X окраш.; Ill— Ktt процент
окрашенных особей во всей популяции
0 0,1' 0,2 0,3 0,4 0,5 0,6 0J 0,8 0,9 Щ
117
для процента окрашенных особей во всей популяции К1 (тонкие сплошные
линии). По такому графику легко можем находить элементы,
характеризующие данную популяцию.
Возьмем, например, точку в центре таблицы, отвечающую q = 0,5 и
v = 0,5. По положению точки относительно тонких линий (между 60% и
50%) узнаем, что в такой популяции окрашенных особей несколько более
55% (точно 56,25%). По положению точки относительно замкнутой
кривой 15% узнаем, что в популяции от скрещивания белых особей друг с
другом должно возникать около 16% окрашенных, а по положению точки
относительно пунктирной кривой 20 % узнаем, что от скрещивания
окрашенных друг с другом должно возникать около 21 % белого потомства.
Пусть, например, мы изучили популяцию и установили, что в ней
имеется 70% окрашенных особей и 30% белых, причем окрашенные при
скрещивании друг с другом дают около 15% белого потомства, а белые —
около 15% окрашенного потомства. Находим по номограмме кривую
(тонкую) 70% и ведем по ней до пересечения ее со сплошной замкнутой
кривой 15 % и с пунктирной линией 15 %. Тонкая линия 70 % и
пунктирная линия 15% идут на значительном протяжении близко одна от другой,
а сплошная линия 15% круто сечет их в двух точках. Беря эти точки
сечения, находим отвечающие им v около 0,49 и q около 0,29 или v около 0,29
и q около 0,49.
Мы нашли таким образом такие значения q и v, при которых могут
возникнуть те проценты, которые мы наблюдали в популяции. Таких точек
в данном случае оказалось две, что вполне понятно,— мы разбираем
случай двух различных рецессивных белых окрасок аа и ЪЬ. Поскольку они
не различимы друг от друга и обе рецессивны, совершенно безразлично,
будет ли иметься 0,49 аа и 0,29 ЪЬ или, наоборот, 0,29 аа и 0,49 ЪЪ —
отношения белых и окрашенных будут одинаковыми.
Пусть мы имеем теперь популяцию, состоящую из 20% окрашенных К1
и 80% белых, причем от белых возникает около 9% окрашенного
потомства К2, а от окрашенных — около 16% белого К3. Какова структура этой
популяции? Находим тонкую линию 20% и ведем по ней до пересечения
с жирной линией 9%. Такого пересечения не оказывается вовсе, и даже
при наибольшем сближении (при пересечении большой диагонали)
расстояние между этими двумя линиями (сплошная линия в 9% не нанесена,
она идет несколько снаружи от сплошной линии в 10%) достаточно велико.
Что касается пунктирной линии в 16% (она проходит несколько
правее пунктирной линии 15%), то пересечения с ней тоже нигде не
происходит. В области наибольшего сближения линий (К1 = 20% и!2 = 9%)
проходит линия К3 = 32 %.
Если мы изучили нашу популяцию на достаточно большом материале
и сможем отличить для К3 16% от 32%, то мы сможем сказать, что на
нашей таблице не имеется точки, удовлетворяющей наблюдаемым данным.
Этот вывод придется истолковать так, что наша популяция не относится
к случаю двух рецессивных белых окрасок, разбираемому здесь.
Подобно тому, как мы начертили систему кривых для случаев парных
генов, могут быть вычислены и начерчены, в виде номограммы, системы
кривых для всяких иных генетических случаев, причем в каждом случае
мы получаем свою специфическую, не повторяющуюся систему.
2. Эпистаз + криптомерия в популяции
Для сравнения возьмем другой, внешне сходный случай, при котором
тоже имеются два фенотипа, белые и окрашенные, и также возможно
появление белых от окрашенных и окрашенных от белых, но иной
генетической природы. Таким случаем является эпистаз + криптомерия, на-
118
пример, случай эпистатической [и рецессивной белой окрасок у кур,
дающий в F2 отношение 13 : 3.
Подобная популяция будет иметь структуру:
р*г*ААВВ
2p3rvAABb
РЧ2ААЬЬ
2рог2АаВВ ' белые благодаря АА или Аа ,
ApqrvAaBb
2pqv°Aabb J
q*faaBB \
2q2rvaaBb ) окрашенные - аВ
qVaahb — белые bb
Белая родительская группа имеет формулу:
рФААВВ + 2p2rvAABb + p2v2AAbb + 2pqr2AaBB + kpqrvAaBb + 2pqv2Aabb + gVaabb
p2r2 + 2p°-rv + p*v* + 2pq r2 + Apq rv + 2pqv2 + q2v2
Знаменатель упрощается до
p2 + 2pq + q2v2 = 1 — q2 + q2v2 = z±.
Окрашенная родительская группа имеет формулу:
g2i°aaBB + 2g2rvaaBb
qb* + 2q2rv *
Знаменатель упрощается до q2 (1 — г?) = Кг.
Величина Кг есть доля окрашенных в популяции.
Гаметы белой родительской группы будут
— (p2r2AB + p2rv АВ + p2rv Ab + p2v2Ab + pqr2AB + pqr2aB + pqrvAB +
-j- pqrvAb + pqruab -f pqv2Ab + pqv2ab + q2v2ab) = — [(p2r2-\-p2rv +pqr2-\-
+ pqru) AB + (p*rv + p2v2 + pqrv + pqv2) Ab + (pqr2 -f- pqrv) аВ -\-
+ (pqrv + pqv2 + q2v2) ab] = — \pr = J—T— AB +
+ PV pr + pV+qr + qv Ab + pqr(r + v)aB + qv(pr + pv + qv)ab] =
« 1
= — [prЛВ -f- pvAB + pqraB + qv(p+ qv) ab].
Гаметы окрашенной родительской группы будут
к~ (q2r2aB + q2rvaB + q2rvab) = -^-1
-=г- (q2r2aB + q2rvaB + q2rvab) = -^г- [#2r (r + у) аВ + q2rvab] =
= -g-(q2raB + q2rvab).
Скрещивание двух белых родительских групп друг с другом даст
— [prAB + pvAb + pqraB -f qv (p + qv) ab]2 =
zi
= 4r [P2r2^^fiB + 2p2rvAABb + 2p2qr2AaBB + 2pqrv (p + qv) AaBb +
zl
+ p2v2AAbb + 2p2q2rvAaBb + 2pqv2 (p + qv) Aabb + p*q2r2aaBB +
+ 2pq2rv (p + qv) aaBb + q2v2 (p + qv)2 aabb].
119
Рис. 23. Номограмма для
анализа дигибридной популяции
в случае эпистаза и
криптомерии (эпистатическая белая и
рецессивная белая окраски)
Обозначения, как на рис. 22
О 0,1 0,2 ф Ц4 0,5 0,6 0,7 0,8 0,9 Щ
Среди них окрашенных (ааВ) будет:
р^дЪ* + 2pq9-rv (р + gv)
(1 — д2 + я2»2)2
рд2г(р+ру + 2ду°')
(1 — Я2 + №2)2
= к9
Скрещивание двух окрашенных родительских групп друг с другом даст
— (q2raB + q2rvab)2 = ~(аВ + vabf = ^(ааВВ + 2vaaBb + v2aabb).
Среди них белых будет
д*гг;3
g*rW
r'v1
к\
д* (1 — г;2)2
(1 — v)2 (1 + vf
(i+v?
V \2
1+v)
= к*
Рис. 23 представляет систему кривых (интерполированных) для этого
случая. Мы замечаем сразу ее резкое отличие от системы кривых рис. 22 и,
в частности, ее асимметрию, отвечающую тому, что здесь два белых гена
не равноценны; например, две популяции, одна из которых имеет v = 0,3
и q = 0,5, а вторая имеет v = 0,5, q = 0,3, будут уже резко различаться
по всем процентам белых и окрашенных.
Выше мы попытались найти в номограмме парных генов популяцию,
характеризующуюся наличием 20% окрашенных и 80% белых особей,
причем белые с белыми давали 9% окрашенных, а окрашенные около 16%
белых. Такой точки нам не удалось найти. Обратившись к рис. 23 и идя
по тонкой кривой 20%, мы видим, что она на значительном протяжении
очень близко подходит к сплошной линии 9%, а далее находим точку 16,
где обе эти линии пересекаются друг с другом и с пунктирной линией 16.
Таким образом, точка, отвечающая v = 0,67 и q = 0,56, удовлетворяет
всем нашим фактическим данным.
Для некоторых популяций диагноз устанавливается таким методом
с большой уверенностью. Например, популяция, характеризующаяся 10%
окрашенных особей и 90% белых с К2 = 5% и К3 = 20%, находится на
номограмме 23 в точке, для которой v = около 0,81 и q = 0,55, а на
номограмме 22 подобной точки совершенно нет. В других же случаях сходные
точки допустимы на обеих номограммах. Такова, например, популяция,
характеризующаяся отношением 80% окрашенных и 20% белых, причем
белые дают 20% окрашенных, а окрашенные — 7% белых. По номограмме
парных генов находим приближенно точку, отвещающую v = 0,25 и
q = 0,35 или v = 0,35 ж q = 0,25, по номограмме для эпистаза + крипто-
диетрия сходную точку находим при v = 0,35 и q = около 0,95.
120
Однако вопрос может быть освещен более детальным изучением
популяции. На наших таблицах мы ограничивались пока нанесением лишь
трех систем кривых. Аналогичным образом могут быть нанесены и другие
кривые.
Такие кривые могут изображать, например, результаты скрещиваний:
1) белые X окрашенные;
2) белый отец X белые дочери;
3) белый отец X окрашенные дочери;
4) окрашенный отец X окрашенные дочери;
5) окрашенный отец X белые дочери;
6) белые братья X белые сестры.
3. Повторные (полимерные) гены.
Особенности этого случая
Третий типовой случай, при котором возникают два фенотипа (будем
их и здесь обозначать как цветной и белый),— это повторные гены, два
одинаково действующих, но не аллеломорфных гена. В мендедиетическом
эксперименте этот случай характеризуется, как известно, своеобразным
отношением в F2, а именно 15 : 1, чем и отличается характерно как от
моногибридного, так и от других дигибридных. Подобно тому, как мы делаем
это для двух предыдущих случаев, выводятся следующие уравнения,
характеризующие популяцию в случае димерии:
Кг=1 — q2v*
#2 = 0,
#4 = 1
(! + *»)*
1 — qv
(К± — доля окрашенных от окрашенного родителя — см. ниже).
Качественное отличие этого случая от двух вышеразобранных состоит в
постоянном К2 = 0, т. е. от белых особей inter se не возникает окрашенных. Все же
остальные величины имеют чрезвычайно сходные уравнения, и кривые,
их изображающие на рис. 24, оказываются в интересующем нас поле
(0 < q < 1) почти параллельными гиперболами. Это значит, что,
определив из эмпирического материала любую одну или все три из этих величин^
мы не можем определить какой-либо точки на нашем поле, а находим лишь
линию, отвечающую некоторой величине qv (во всех формулах всюду
имеемся лишь множитель qv или q2i?). Самое интересное, что мы не можем
в таком случае сказать, не имеет ли место равенство q = 1 или v = 1,
при котором наш случай превращается из димерии в мономерию, т. е. в
обычный моногибридный случай. Иными словами, мы фактически лишены
возможности сказать, имеем ли мы дело с моно- или дигибридной
популяцией, даже в том случае, когда q = 1/2 и v = 1/2. Действительно,
соответствующие уравнения для моногибридной популяции имеют вид:
^ = 1-5»
#2 = 0
Я2
#3 =
(i+<7)2
(1+9)
Достаточно предположить, что q = д^, чтобы получить уравнение-
димерии.
121
Рис. 24. Номограмма для
анализа дигибридной популяции
в случае двух повторных генов
I — JBTi, процент окрашенных в
популяции: II — Кь, процент белого
потомства от скрещивания окраш.Х
X окраш.; III— К*, процент
белого потомства от окрашенного
родителя
О 0,1 0,2 0,3 ОМ 0,5 ОД 0,7 0,8 0,9 Щ
То же самое оказывается справедливым и для полимерии.
Уравнения ее будут иметь вид:
о
(1 + ятяз... qj-
и т. д. И здесь мы имеем сложный множитель ЯгЯ2Яз-~Яп или {ЯгЯъЯз-'-Яп)2-
Положив его равным q, перейдем к моногибридной популяции.
Следовательно, изложенными способами отличить моно-, ди- и
полимерные популяции оказывается невозможным. Необходимо дальнейшее
исследование этого обстоятельства. О некоторых перспективах мы скажем
ниже, в главе о методе сигнальных генов. Отметим, что это
парадоксальное явление стоит в связи с тем, что основное отличие димерии от мономе-
рии состоит в том, что участвующие в ней доминантные аллели не аллело-
морфны друг другу, не обнаруживают отталкивания. Но в популяции
сцепление становится незаметным, уравновешивается отталкиванием, поэтому
и исчезает основной критерий различения димерии от мономерии.
В приведенных примерах мы осветили три случая, дававшие по два
фенотипа (окрашенные и белые). Подобным же образом могут быть
исследованы и всякие другие, дающие три, четыре и так далее фенотипа. Для
них соответственно могут быть даны другие системы кривых, а также
указаны качественные отличия. Например, в разобранном примере, когда
белые дают в потомстве окрашенных и окрашенные — белых,
исключается как случай димерии моногибридный, такидигибридный, ибо при этом
появление окрашенных от белых невозможно. К сожалению, в настоящее
время необходимые для этого предварительный анализ и расчеты еще не
произведены. Мы обращаем на это внимание наших читателей, так как
обработка различных случаев является одновременно и отличными
упражнениями и темами небольших исследований. Если бы каждый генетик
потратил несколько дней на обработку одного — двух таких случаев, мы
могли бы иметь целую коллекцию таких номограмм, которыми можно было
бы пользоваться в практической работе по генетическому анализу попу-
ляционного материала.
Ограничимся здесь лишь в качестве иллюстрации метода выводом
формулы для скрещивания: окрашенный отец X окрашенные дочери в
моногибридной популяции, где А — окрашенные, аа — белые. Такие дочери
могут возникать в ряде скрещиваний с различными частотами.
U0 30 20 10
К3 =
122
Типы
скрещиваний
Отец Мать
ААХАА
ААХАа
ААХаа
АаХАА
АаХАа-
АаХаа
Дочери
АА
АА + Аа
Аа
АА+Аа
АА + 2Аа + аа
Аа + аа
Частоты
скрещиваний
Р*
2рЦ
рЧ%
Частоты дочерей
АА] Аа аа
Р*
p*q p*q
pV
2 или р; *
2p3q
4/>V
2pq*
p3q рЦ
/>V 2p9q* p*q*
pq* pq*
2 p*q pq pq2
окрашенные белые
или р/2 1/2 q/2
Всего скрещиваний отцов с их окрашенными дочерями будет:
Отец А А
рЧр + q) _ р2
p[p + q(p + V] p(i+pq)'
Отеп Аа ^(^ + 1) -/*(*» + *)-.
итец Aap[p + g(p + i)] - p{i+pq)-
Белое (аа) потомство появится только от скрещивания Аа отцов с Аа
дочерями
Отец Дочь Частота Дети
Р Л Л . 2P(I А . Pq
-%АА + -j- Аа + -J" а«.
Аа
Аа
pq
2M~2fi=Pq
Итого белого потомства в этих [скрещиваниях появится ^,а во всех
скрещиваниях А отцов с А дочерями будем иметь белых:
рд - q -100%.
своеобразное от-
моногибридного
Ьр (1 +рд) 4 + bpq
В случае р = q = 1/2 эта величина равна 10%, т. е.
ношение 9:1, характеризующее потомство обычного
скрещивания от с? (5*^4 (F2), каждый из которых скрещен со своими
дочерями того же типа А (из F3).
Выше мы приводили уже формулу расщепления при скрещивании до-
минантов друг с другом, причем при появлении генотипа аа нашли фор-
мулу (1 )а . В данном случае доля оказалась уже иной. Это изменение,
вызванное условиями родственного отношения отец — дочь, объясняется
тем, что в свободной популяции самцы А А и Аа имеют состав самок
одинаковый, а среди скрещиваний отец X дочь, как мы видим, отцы АА
скрещиваются с дочерьми состава рАА + qAa, а отцы Аа — состава р/2АА +
+ 1/2 Аа.
НОМОГРАММА ДЛЯ ОПРЕДЕЛЕНИЯ ТИПА СКРЕЩ ИВАНИИ ПО Кг и К9
Следующий график (рис. 25) позволяет примерно решить, имеем ли мы
дело со случаем эпистаза + криптомерия или со случаем
моногибридным или двух или нескольких одинаковых генов.
123
fOOx
40
20
IaV"?
-3£ X X
Г \ vs-t
Г Моногибридный д ^ >^.
Y случай и случай \ \^>
1 повторных ген од \ \г~
1 1 1 JL -L. 1
< .
СнН
Рис. 25. Номограмма для
определения различных
генетических случаев (для популяций,
состоящих из двух фенотипов)
по Кх и Кь
Объяснение в тексте
10
20
30
W
ЯК*
Определяя величину Кг и К3 из эмпирических данных, смотрим по
номограмме точку, отвечающую им, и в зависимости от ее положения
определяем тип расщепления, после чего уже проверяем сделанный вывод
более детальным методом.
УРАВНЕНИЯ КРИВЫХ НОМОГРАММ
ДЛЯ ОПРЕДЕЛЕНИЯ ТИПА СКРЕЩИВАНИЯ
Эти кривые связывают Кг с К3.
Кривая /. Для случая парных генов, полагая q = v (так как при этом
Кг наибольшее), имеем:
JfiT1 = (l-?2)(l-i;2) и #з = 1-
1) #1==(l-<jr2)2
2) #8=1
Из (1) имеем
1 + 2? 1 + 2»
(1+?)2
(l + 2g)2
(1 + vf •
(1+?)*'
q=Yl-YK1.
Подставляя в (2), получаем:
Л3 = 1 j- = •
Таково несколько сложное уравнение кривой (а).
Кривая //. Так как выражения для Кх и Kz симметричны относительна
q и v, то можно обобщать их:
#1=(1-</2)п
(1 + 2?)"
#я=1
(1 + Я)"
2"
где п — число генов.
Для моногибридного случая га = 1 и соответственно
#з = 1-
1 + 2 Vi — Ki
(i + yt^tkiY '
124
Последнее выражение преобразовываем так:
к = (1+ У\-Кх?-\ -2 |М- Ki _^ 1 + 2 VT^Kl + (l-Ki)- l-2YT=Ki
3 (i + VT=Kly (i + VT=T5)»
и получаем
3) Ав- (1 + /1_Ж)а.
Теперь покажем, что и для случая двух повторных генов имеет место та
же связь Кь и Кг (при условии q = v, дающем максимальное К3). Для этого
случая имеем (см. выше):
Кг = 1 - gV; ^ = 1 - <Д ^ = 1 - Ки
или, полагая q — v:
__ д2„2 __ gt
Л3 — (1 + qv)2 > Л3 — (1 + д2)2 '
Отсюда, подставляя значение д4, получаем
8 (l+Vl-ft)!1
т. е. та же формула, как и в (3). При условии q = v = 1... и для случая
с несколькими генами будем иметь ту же связь К$ и Кг.
Как мы видели выше, для случая нескольких повторных генов:
Кг = 1 — gVZ2...; К3 gW'" •
(1 + grf...)» ■
или, полагая g = v = I =
#х = 1 _ g2n?
Подставляя значение g*n, получаем
а,2п
* * 1» в (1+?П)2
*,- *-*
(1 + VI— АГх)2 '
т. е. опять формула не отличима от моногибридной.
Разберем теперь случай для двух генов при q =f= v. Пусть v = kg; тогда
X1 = l-g2(kg)2
[l + <7(k?)p-
^3==-^i^!
Из первого имеем:
9* (k# = 1 - Kiq (k?) = УГ^Ж-
Подставив это в выражение для К3, получаем:
#з =
(i+Vi-Ki)* '
т. е. опять то же выражение.
То же самое легко показать и для случаев нескольких одинаковых
(повторных) генов, положив v = Kxq = K%q... и т. д.
Наоборот, из формулы парных генов видно, что при v =f= q выражения
для Кх и К9 меняются. При одной и той же q + v (но q Ф v) Kx
уменьшается, а К3 растет.
125
В самом деле, например
Кг = 1 — q2 — v2 + q2v2.
Так как величина qv наибольшая при их равенстве (при q -\- v = const) r
то и Кг наибольшее при равенстве q = и.
Если в случае парных генов один ген становится очень редким (v ж 0),
то дигибридное распределение превращается в моногибридное, и связь
Кг с К3 идет по формуле (3). Поэтому линия II на графике рис. 25 является
нижним пределом значения К3 при данном Кг в случае парных генов.
С другой стороны, линия // является верхним пределом К3 при данном
Кг в случае эпистаза. Этот предел осуществляется, когда в скрещивании
с участием эпистаза этот эпистатический ген приближается к
исчезновению. Тогда Кг указывает на процент только рецессивных белых и
относится к моногибридному. Но меньший процент рецессивных белых при
постоянном проценте белых может зависеть и от наличия эпистатического
белого. В этом случае v может быть невелико, и поэтому К3 может быть меньше
предела, даваемого кривой //, даже может быть равно 0, если рецессивного
альбинизма нет вовсе, а К± определено лишь наличием эпистатически
белого. К случаю эпистаз + криптомерия относится все поле между // и
осью ординат.
Когда подобным образом аналитически будут изучены всякие другие
случаи (с тремя фенотипами и т. д.), можно будет рассчитывать получить
аналогичные простые номограммы и для определения этих типов
расщепления в популяциях.
ОПРЕДЕЛЕНИЕ ДОМИНИРОВАНИЯ
Нередко исследователи допускают грубую ошибку, полагая, что
определить доминантность или рецессивность признаков можно простым
подсчетом того, какой из признаков чаще встречается в популяции. Зная, что
при наличии доминирования доминантный фенотип встречается втрое
чаще, чем рецессивный, переносят это отношение и на популяцию, что,
конечно, совершенно неверно.
Как мы видели, рецессивный генотип в популяции встречается с
вероятностью q2, а доминантный с вероятностью р2 + 2pq = 1 — q2. При
величине q = У0,5 = 0,707 q2 — 1— g2, т. е. фенотип рецессивный и
доминантный встречаются одинаково часто. Для тех же случаев, когда q^> 0,707
ир< 0,293, рецессивный фенотип будет встречаться чаще, чем
доминантный. Между тем для огромного большинства доминантных мутаций
р < 0,293 (обычно даже гораздо меньше) и доминантный фенотип
соответственно встречается значительно реже рецессивного. Поэтому исходить
в суждении о доминантности из наблюдения за частотой того или иного
фенотипа невозможно.
Для решения этого вопроса необходимо знать какие-либо родственные
связи.
1. Если имеется родословная, в которой на протяжении ряда
поколений идет видимое наследование какого-либо признака, то это может
служить хорошим доказательством доминирования признака. При этом,
однако, нужно иметь в виду, что если признак не очень редок, то в порядке
случайности среди многих родословных может быть найдена такая, в
которой и рецессивный признак может быть наблюдаем в ряде
последовательных поколений. Чем реже данный признак, тем такие случайности
менее вероятны и тем вероятнее доминирование изучаемого признака.
Очень часто мы встречаемся, однако, с «проскоками». Проскоками
называют такие случаи, когда, например, признак, производящий
впечатление доминантности, имеется у деда и внука, а у отца отсутствует. Если та-
126
Рис. 26. Повышение
вероятности извлечения гомозиготных
рецессивов (Z, в логарифмах)
в потомстве от родственных
скрещиваний при уменьшении q
ких проскоков мало, то можно сделать заключение о доминантности
признака, допустив, что он не всегда проявляется. Но при наличии ряда
проскоков надо быть очень осторожным, чтобы не принять случайное
скопление в родословной рецессивных признаков за признаки, доминирующие
с непостоянным проявлением.
2. Важное основание для суждения о доминантности и рецессивности
признаков мы получаем из подсчета родственных отношений родителей.
Шансы для проявления рецессивных гомозигот резко повышаются при
родственных скрещиваниях. Мы видели выше, что в свободной популяции
генотип аа возникает от скрещивания Ах X Аг с вероятностью q .2 .
При величине q = 0,001 эта вероятность падает до 1 миллионной. Между
тем при скрещивании отцов с дочерями, как мы видели, эта вероятность
равна g,— , что при малом q близко к <?/4. При q = 0,001 эта величина
равна 1/4000, т. е. в 250 раз больше, чем при свободной встрече гетерози-
гот А а друг с другом.
Отношение вероятности извлечения рецессивов при тесном
инбридинге и в свободной популяции выразится величиной
4(1+^)
По мере убывания q множитель
(1 + Я)2 = (1 + <?)2
Я2 t+pq
(1 + qf
4?
1
4g
е.
1+pq
чрезвычайно быстро возрастает
стремится к 1, а все отношение
График этого отношения
представлен на рис. 26. Так как по отношению к рецессивным мутациям
q может быть очень мало, падая до миллионных долей, то такие мутации
могут перейти в гомозиготное состояние практически лишь при
инбридинге. Поэтому, если, исследуя какой-либо редкий признак, мы замечаем,
что носители его происходят преимущественно от родственных
скрещиваний, то есть основание видеть в этих носителях извлеченные рецессивы.
В 1934 г., например, у ярославской рекордистки Золотой родился теленок
урод, с укороченными ногами, с неразвитой нижней челюстью и т. д.
Выяснилось, что этот теленок Призрак произошел от покрытия Золотой ее
внуком Маем. Это дало основание заподозрить, что в данном случае мы имеем
не паратипическое уродство, а извлечение рецессива. Было предсказано
поэтому, что при дальнейших родственных скрещиваниях в этом семействе
должны появляться подобные же уроды. Предсказание это подтвердилось
рождением в 1934—1936 гг. еще двух таких же уродов от скрещивания
близких родственников Мая друг с другом. Наследственный характер
этого уродства был, таким образом, подтвержден (Лютиков, 1934, 1936)-
127
20 30 4ff 50 60 70 80 90
Процент признака А 6 популяции
Рис. 27. График для
определения доминантности в
популяции по проценту разнородных
двоен и троен (по Васину)
Следует, между прочим, иметь в виду, что для диагноза рецессивности
важно не то, что настоящий признак возникал при инбридинге, jVT0> что
в этих случаях частоты в инбридинге выше, чем в популяции. Поэтому,
«ели имеются сомнения, следует выбрать на удачу серию пробандов
нормальных и подсчитать, какой процент их происходит от родственных
скрещиваний.
3. Во многих случаях вопрос о доминантности может быть решен
сравнением признаков у братьев и сестер (сибсов). Б. Н. Васин (1928)
разработал этот вопрос применительно к двойням овец, считая, что его можно
использовать и в других случаях, когда мы можем без выбора взять
достаточное количество пар или троен полных братьев и сестер и определить
у них наличие или отсутствие изучаемого признака. Для свободной
популяции можно вывести формулы, показывающие, как среди двоен-троен
распределяются такие, у которых оба брата будут иметь доминантные
признаки, оба рецессивные, или, наконец, двойня будет разнородной:
один брат будет иметь доминантный признак, другой — рецессивный.
Формулы эти имеют такой вид:
Оба брата (сестры) несут
рецессивный признак
Один брат несет рецессивный,
другой — доминантный признак . . . .
РФ
Двойни
.(? + *>'
' 2
(3 + 0
Тройни
.(ЗУ + Р2
? 4
. (9 + 3g)
РФ 4
На графике (рис. 27), составленном по расчетам Васина с некоторыми
изменениями, по оси абсцисс процент изучаемого признака в популяции
А или5, по оси ординат слева отложен процент разнородных двоен, а
справа — процент разнородных троен. Совместимость графика для двоен и
троен на одном графике возможна потому, что уравнения кривых для
двоен и троен различаются лишь постоянным коэффициентом 3/2, который
и отражен разницей масштаба ординат справа и слева.
Уравнения кривых (у — процент разнородных двоен, у' — процент
разнородных троен) следующие.
Для двоен:
1)y = p2(3~2P~p2).100o/0, где/^l-g, 100^ = Д Я=100% -Л;
2) V,.*8-2,'-* .1004.
Для троен:
3 , (3-2Р-Я
1) !^= у Р 2
2) у'- т 1"'
(3-2g-gr)
•100%;
■100%.
128
Рис. 28. График для
определения доминантности по проценту
признака А в популяции и по
проценту потомства с А от
родителя с А (по Васину)
О 20 40 60 80 100
Процент признака А в популяции
Пример. В отаре 20% черных овец. Среди тысячи зарегистрированных
двоен 19% разнородных, точнее, 19 ± 1,2%. Узнать, доминантна или
рецессивна черная окраска?
По оси абсцисс находим 20% и по оси ординат слева (двойни) находим
19%. Найденная по ее двум координатам точка лежит вблизи кривой,
указывающей на рецессивность нашего признака, т. е. черный — рецес-
сив. Линия доминантности проходит несколько ниже, в точке 16,5%. Сле-
19 16 5
довательно, наша точка лежит от нее на ' = 2,1 квадратической
ошибки, т. е. определение довольно убедительное, хотя и недостаточное
для полной уверенности. При 50 % признака А в популяции обе кривые
пересекаются. Очевидно, что при этом становится невозможным решение
вопроса и надо искать стадо с другим процентом признака 4. Как видно
по ходу кривых, они наиболее удалены друг от друга при 15—25% и
75—85 % признака А в популяции. Такого состава стада наиболее выгодны
для определения доминантности. Но во всяком случае нужно иметь в виду,
что обе кривые проходят рядом и поэтому определение доминантности
возможно по этой величине (т. е. по проценту разнородных двоен) лишь
при большом материале. Например, при 36% признака А в популяции,
когда кривые еще заметно разделены, уверенный ответ можно получить
не менее как по 6000 двоен!
Кривые для троен более выгодны, так как все расстояние между
кривыми в полтора раза больше. Но зато, очевидно, сбор материала по
тройням труднее, что парализует эту выгодность.
Если известна связь: один родитель — потомок, то в некоторых
случаях вопрос о доминантности можно также решать. Для этого может
служить график рис. 28. Здесь по оси абсцисс отложен процент признака А
в популяции, а по оси ординат — процент приплода с признаком А,
полученный от родителей (например, матерей) с этим признаком.
Уравнение кривых имеет вид:
1) у = Ю /Л, или у = 100q
2> ' = т % -5tW • ™ '-11 - (ТТТЩГ)]ЛЖ1 *•
где А — процент данного признака в популяции, а Б —
противоположного признака, т. е. 100% — А = В.
Требуется узнать, которая из двух окрасок доминантна. Берем в
качестве Л-признака черный. Процент, черных в популяции — 20%,
процент черных в потомстве от черных матерей — 45%. Соответственно
J I I I I I I I L
9 А. С. Серебровский
129
20 W 60
Процент признака Л S популяции
Рис. 29. Номограмма для
определения доминантности по
проценту признаки А в популяции
и по проценту разнородных
Двоен и троен у родителя с А
(по Васину)
20% по абсциссе и 45% по ординате находим точку и видим, что возле нее
проходит кривая рецессивности, следовательно, черная окраска
рецессивна.
Пример. Пусть мы имеем стадо состава
Матка
Черные
Белые
Всего . .
Ягнята
п
200
800
1000
черные
90
110
200
белые
110
690
800
В процентах по группе
матерей
черные"
45
13,7
белые
55
86,3
г
Проверяем по белым. Берем в качестве ^4-признака белую окраску:
процент белых в популяции —80%;
процент белых в потомстве от белых — 86,3%.
Найдя соответствующую точку, видим, что возле нее проходит кривая,
указывающая на доминантность белых.
Из хода кривых видно, что при 50% они пересекаются, т. е. при этом
условии задача неразрешима. С большим трудом, конечно, разрешима
она и в области, близкой к А = 50% (40—60%), и вблизи 100%.
Значительно легче решается вопрос о доминантности, если известен
признак у матери и группы ее детей — у двоен, троен и т. д. Номограмма
рис. 29 составлена по Васину и имеет две кривые. Уравнения этих кривых
следующие:
1. Процент разнородных двоен (троен) у матерей, имеющих
доминантный признак:
У =
У =
2(1 + ?)
3g*(3-g)
4(1 + (?)
100%,' (для двоен)
•100% (для троен)
2. Процент разнородных двоен (троен) у матерей, имеющих
рецессивный признак:
У = (Я — Я2) • 100 % (для двоен)
3
У = у (Я — Я2) • Ю0% (для троен)
130
Для использования номограммы необходимо определение двух
величин: процент признака в популяции и процент разнородных двоен (троен)
у матерей (либо отцов), несущих этот признак.
Пример. В стаде около 4% черных овец, остальные белые. У черных
маток около 16% разнородных двоен (белых + черных). Доминантна
или рецессивна черная окраска?
По номограмме против точки абсциссы 4% и точки ординаты 16%
находим точку, возле которой проходит кривая рецессивности.
Следовательно, черный цвет рецессивен.
Эта номограмма удобна тем, что кривые ее не пересекаются нигде (кроме
А = 100%) и довольно широко расходятся, что позволяет решать задачу
при меньшем материале. Так как кривые сильно расходятся при малых q,
то можно думать, что удобнее выбирать для обсуждения именно признак,
встречающийся в популяции реже (например, имея 4% черных и 96%
белых, мы можем ставить вопрос и о редком черном и частом белом
признаке). Однако это не совсем так: для редкого признака мы можем меньше
собрать материала и этим затруднить решение. Следующий пример
иллюстрирует это соображение, относящееся и к предыдущей номограмме.
Пример. В стаде 100 маток: 96 черных и 4 белых, у всех имеются
двойни. У 96 черных — 2 разнородных и 94 однородных, у 4 белых — 2
разнородных и 2 однородных.
Отсюда величины, необходимые для решения вопроса о том,
доминирует ли черная окраска (А), будут:
1) процент признака А в популяции — 96%;
2) у маток А разнородных двоен -— 2,08 ± 1,46%.
По номограмме (рис. 29) против абсциссы 96% и ординаты 2,08%
находим точку, лежащую близко от кривой £, указывающей на
рецессивность черных. Однако недалеко проходит и кривая доминантности,
имеющая при абсциссе 96 ординату 4,67. На сколько ошибок отстоит эта точка:
4,67-2,08 = 2,59; -||§ = 1,77.
Мы видим, таким образом, что наш вывод о рецессивности черных не
очень надежен, так как обе эти кривые против этого пункта абсциссы очень
близки и обследование 100 овец недостаточно.
Но не поможет ли нам, если мы перевернем задачу и вместо вопроса о
черной окраске станем выяснять, доминирует ли белая?
Для этого у нас имеются следующие данные:
1) процент признака (в такой постановке вопроса тоже А) — 4%;
2) у маток А разнородных двоен — 50%.
На той же диаграмме, но уже против точки 4% и точки ординаты 50%
находим точку, вблизи которой проходит кривая доминирования. Кривая
рецессивности очень далеко внизу, и вывод кажется очень ненадежным.
Но так как нам пришлось оперировать всего с 4 двойнями от белых маток,
то ошибка очень велика
50% ±25%.
Кривая рецессивности при абсциссе 4% имеет ординату 16%.
Отсюда реальность разницы
50% - 16% = 34%; 34/25 = 1,36.
Таким образом, кривая рецессивности отстоит от нашей эмпирической
точки лишь на 1,36 квадратической ошибки. Вывод о доминантности
белого еще менее надежен, чем вывод о рецессивности черных (1,36 <С 1,77).
При таком неблагоприятном проценте цризнака А необходимо
дальнейшее увеличение материала в три-четыре раза или использование
131
9*
троен. Но нужно иметь в виду, что хотя тройни дают более ясный диагноз
доминантности, но сами тройни значительно реже двоен и это может
нейтрализовать выгодную сторону троен.
ПОСТРОЕНИЕ И РЕШЕНИЕ ЗАД4Ч-М0ДЕЛЕЙ
Задача 1. Для упражнений в решении задач на генетический анализ
популяционного материала можно применить тот же метод, который мы
описывали выше. Применительно к популяции он требует дополнения,
учитывающего, что различные аллели находятся в популяции в
различных концентрациях.
Пусть, например, мы хотим моделировать дигибридную популяцию,
в которой аллели А и а находятся в концентрациях р и q, а аллели В иЬ
в концентрациях гиг;, причем р = 0,7, q = 0,3, г = 0,3 и v = 0,7.
Условимся, что при тираже шаров для А и а будем придавать шарам такие
значения:
i .12 3 4 5 6 7 = 7/10;
а 8 9 10 = 3/10;
для В и Ъ такие:
В. . . .
ь. . . . ,
12 3 = 3/10;
456789 10 = 7/10.
Вынув четыре шара, например, 7, 2, 3, 5, записываем соответственно
ААВЬ, выбрав 1, 8; 8, 9, записываем АаЪЪ и т. д. В результате многих
тиражей получаем нужное количество генотипов, которые в достаточной
степени будут соответствовать заданному условию. Записав эти генотипы
попарно, получаем случайные скрещивания и от каждого из них получим
потомство. Запись будет выглядеть так (берем пример из одного нашего
опыта):
Родители
ааЪЪХааВЬ
ААВВхАаВЪ
ААЪЪХААЪЪ
Дети
1
ааВЪ
АаВВ
ААЪЪ
2
ааВЪ
АаВЪ
ААЪЪ
1 Родители
АаВЬхААВЬ
1 ААЪЪХААВЬ
Дети
1
АаЪЪ
ААВЪ
2
АаВЪ
ААЪЪ
После того как создана такая модель генотипов, можно переводить ее
в фенотипы. Положим, например, что мы имеем дело с парными генами
и формы А, В — черные, а4,лиВйЬ- белые.
Написанную выше таблицу генотипов переводим в фенотипы и
записываем: •
Родители
бел.
черн.
бел.
черн.
бел.
и
бел.
черн.
бел.
черн.
бел.
т. д.
1 *
бел.
черн.
бел.
бел.
черн.
и
2
бел.
черн.
бел.
черн
бел.
т. д.
132
Всего родителей и детей:
белых 95 (59,4%);
черных 65 (40,6%).
После первых 40 скрещиваний в этой задаче мы имеем такую картину:
Число
скрещиваний
14
18
8
Типы скрещиваний
Бел.Хбел.
Бел.Хчери.
Черн.хбел.
Черн.Хчерн.
Потомство
фенотип
Бел.
Черн.
Бел.
Черн.
Бел.
Черн.
количество
28
0
18
18
3
13
процент
по
группам
100
0
50
50
18,7
81,3
Для решения задачи по номограммам используем три величины:
процент черных в популяции 40,6 = Ai
процент черных от скрещивания белых с белыми . . 0,0 = #2
процент белых от скрещивания черных с черными . . 18,7 = Кз
По номограмме для случая парных генов эти данные приводят к
решению v = 0; q = 0,77 или v = 0,77; q = 0. По номограмме для эпистаэа +
+ криптомерия получаем ответ q = 1,0; v = 0,77. Иными словами, мы
получаем моногибридное решение. В первом случае все особи имеют генотип
АА и расщепляются лишь по .Вили, обратно, все—ВВ расщепляются лишь
по А, что для задач о парных генах безразлично. Во втором случае q = 1
и v = 0,77, это указывает, что все особи аа, т. е. что белой эпистатической
окраски вовсе нет и расщепление идет лишь по рецессивному альбинизму
в порядке моногибридной популяции.
Оба решения, будучи неверными, хорошо объясняют наблюдающуюся
картину, в которой благодаря условию задачи и случайности (от
скрещивания белых с белыми не появилось ни одного черного) К2 = 0. В
соответствии с условиями задачи такие черные должны были бы составить около
6% среди потомства белых X белых. На 28 детей это составило бы 1,4 ±
± 1,15 и в порядке случайности такой единственный черный не появился.
Всего в популяции
белых 278 (57,9%);
черных 202 (42,1%).
Для решения по номограмме используются три величины:
процент черных в популяции 42,1 ±2,25(^1 ± mi)
процент черных от скрещивания белых с белыми 7,9±2,9 (Къ ± тц)
процент белых от скрещивания черных с черными 25,0±8,17(.йГз ± тпь)
ш
После получения 120 скрещиваний и 240 детей мы имеем такую
картину:
Число
скрещиваний
33
■
68
14
Типы
скрещиваний
бел.Хбел.
-
бел. Хчерн.
черн.хбел.
черн. Хчерн.
Потомство
фенотипы
бел.
черн.
бел.
черн.
бел.
черн.
количество
70
6
57
79
7
21
проценты
по группам
92,1
7,9
41,9
58,1
25
75
Найдем на двух номограммах (рис. 22 и 23) линии, отвечающие Къ
К2, К3, а также линии, отвечающие Кг + т1 и Кг — mly К2 + т2 и
К2 — т2, К2 + т3 и К3 — иг3, и нанесем их на чертеже, как этом
сделано на рис. 30 и 31. Мы видим, что все линии (рис. 30) пересекают друг
друга, хотя и не в одной точке, но довольно близко к точкам q = 0,72 и
v = 0,44 (и обратно q = 0,44 и v = 0,72). Далее мы видим, что на
протяжении некоторой полосы, отмеченной пунктиром, все три полосы,
ограниченные ± т, набегают друг на друга. Поэтому мы можем прийти
к заключению, что где-то на этой полосе находятся две точки,
отвечающие искомому решению и что наиболее вероятное положение этих точек
должно быть недалеко от точек q = 0,72 и v = 0,44 или обратно.
Вместе с тем мы видим, что наше решение еще не очень ограничено.
Особо широки пределы К3 (процент белых от скрещивания черных с
черными), нисколько не ограничивающие решение, даваемое двумя другими
условиями: Кг ± Щ и К2 ± гп2. Это обстоятельство указывает, что
собранный нами материал по 120 скрещиваниям при данных условиях еще
недостаточен. Действительно, для вычисления величины К3 ± т3 мы
располагаем всего 28 особями (7 белых + 21 черных), и для того чтобы
уточнить решение, эта часть материала должна быть значительно
усилена.
Итак, судя по номограме рис. 30, наша задача может быть
объяснена по схеме парных генов. Никаких противоречий при таком
объяснении мы не встречаем, и вопрос может идти об уточнении этого решения.
Найденное решение q = 0,44, v = 0,72 (приблизительно) действительно
Довольно близко к истине. Несколько портит решение то обстоятельство,
что от скрещивания белых с белыми появилось на 76 детей 6 черных при
теоретическом ожидании 4,2, что, конечно, вполне в пределах
случайности. Задача точного определения концентрации — задача особая и
требует соответственно большого материала. Нами же сейчас решался в
первую очередь вопрос о характере расщепления, причем мы показали,
что, во-первых, моногибридное объяснение отпадает, во-вторых, дигйб-
ридное по схеме парных генов хорошо подходит.
Не окажется ли, однако, удовлетворительным объяснение и по схеме
эпистаз + криптомерия. Обратимся к рис. 31. Мы видим, что кривые
Кг и К2 пересекаются в точке, отвечающей q = 0,85 и v = 0,70. Но зато
линия К3 проходит очень далеко от этой точки. Правда, на небольшом
участке (показанном пунктиром) три поля ± т налегают друг на друга,
так что возможность объяснить наш материал по схеме эпистаза +
криптомерия еще не исключена. Но при увеличении материала для вычисления
К3 и уменьшения его ошибки скоро станет очевидным, что К3 проходит
134
Рис. 30. Применение номограммы рис. 22 Рис. 31. Применение номограммы рис. 23
к решению задачи № 1 к решению задачи № 1
в стороне от точки пересечения линий Кх и К2. Для этого потребуется
собрать по скрещиванию черные X черные около 100 потомков.
Задача 2. Пользуясь той же генетической основой популяции-модели,
но изменив в условии действие генов (как именно — увидим ниже),
получаем следующую картину. Популяция состоит из белых и красных
особей. Белых всего 249 или 51,9 ±2,8%. Красных 231 особь, или 48,1+
± 2,8. Потомство их распределяется следующим образом:
Число
скрещиваний
25
71
24
Типы
скрещиваний
бел.хбел.
бел. X краен. 1
краен. X бе л./
краен. X краен.
Потомство
фенотип
бел.
краен.
бел.
краен.
бел.
краен.
количество 1
40
10
85
57
3
45
проценты
по группам
80,0±5,6
20,0±5,6
59,9
41,1
6,25±3,5
93,75±3,5
Картина на первый взгляд мало чем отличается от той, которую мы
имели в первой задаче. Можно лишь отметить, что среди потомства белых
несколько выше процент окрашенных — 20 вместо 7,9, а среди
потомства красных процент белых несколько ниже — 6,25 вместо 25. Но так как
и общий процент окрашенных в популяции несколько изменился — 48,1%
вместо 42,1%, то должны были измениться и все другие проценты.
С каким случаем мы имеем здесь дело?
Воспользуемся опять теми же тремя величинами:
процент красных в популяции 48,l±2,8(lTi ± mi
процент красных от скрещивания белых с белыми 20,0±5,6 (Яг ± ш%
процент белых от скрещивания красных с красными • • • • 6,25±3,5(1Гз ± и»3
Нанесем, как и раньше, эти данные на номограммы (полученные
картины см. рис. 32 и 33). На схеме парных генов (рис. 32) видим, что линии
135
V
C,3[
Щ
Ц7\
o,s\
0,3
0,2\
0,1
=3-
~НП2~~
к.
"у-Щ^з \+тз 111
цд\
0,8
oj\
0,6
Щ
ofi
о,з\
Щ
0,1
+щ
Кз
■щ
J I I 1
0 0,1 0,2 0,3 4* 0,5 0,6 0,7 0,8 0,9 ^Og\ ° V' 02 0,3 0,4 0,5 0,6 0,7 0,8 0,9 f,Oq
Рис. 32. Применение номограммы рис. 22
к решению задачи № 2
Рис. 33. Применение номограммы рис. 23
к решению задачи № 2
Кg и Ks пересекаются. Но зато наиболее точно определяемая линия Кг
проходит очень далеко от этой точки пересечения. Мало того, даже
граница Кг + т1 не касается линии К2 — т2 и таким образом на всей
диаграмме нет места, где все три поля ± т налегали бы друг на друга. Для
того, чтобы такое положение могло возникнуть, К2 должно было бы
сильно уменьшиться, а наоборот Къ должно раза в четыре возрасти.
Таким образом, данный случай совсем не согласуется с гипотезой
парных генов. Обращаемся к попытке объяснить наш материал гипотезой
эпистаз + криптомерия. Здесь картина совершенно иная. Мы видим на
рис. 33, что линия К3 пересекает обе другие, жК1ж К2, и, кроме того, эти
точки пересечения хотя и не совпадают, но очень близки друг к другуг
отстоя'лишь на 7з т. В результате мы имеем небольшую узкую зону,
отмеченную пунктиром, на которой все три поля ± т налегают друг на
друга.
Таким образом, приходим к заключению, что наследственные
явления в нашей популяции хорошо могут быть объяснены гипотезой эпистаз
+ криптомерия, т. е. доминантной и рецессивной белыми окрасками.
При этом можно указать для концентрации доминантного белого р =
= 0,35—0,25 соответственно q = 0,65—0,75, а для рецессивного белого
v = 0,20-0,45.
Задача решена верно. Мы положили, что ген В является эпистати-
ческим (доминантным) белым, а ген а рецессивным белым. Таким
образом, красные особи ^-ААЪЪ и Aabb, остальные — белые. Фактические
концентрации генов: В — 0,276, Ъ — 0,724, А — 0,717 и а — 0,283.
Этим величинам соответствует на рис. 33 точка, находящаяся, как мы
видим, в поле, покрытом пунктиром, вблизи точек пересечения Kz с Кг и
с К2.
Общие замечания
Решение задач-моделей вроде приведенных примеров имеет большое
значение, так как при недостаточной разработанности теории
генетического анализа в процессе подобных упражнений эта разработка будет
легче продвигаться вперед. Для каждого мыслимого случая должны быть
найдены наиболее характерные особенности. На рис. 32, например, мы
видим, что наиболее убедительным доказательством того, что задача
№ 2 не может быть объяснена парными генами, является широкое от-
136
Рис. 34. Дополнение к номограмме рис. 22 Рис. 35. Дополнение к номограмме
Кривые К* — доля окрашенных в потомстве от Рис* ^' кРивые Л4
окрашенного родителя (независимо от окраски
второго родителя)
стояние между Кг и К3. Рис. 31 показывает, что вопрос о
приложимости схемы эпистаза для объяснения задачи № 1 больше всего
зависит от уточнения положения Ks и уменьшения его ошибки. Как
видно из рис. 33, для решения второй задачи по схеме эпистаза наибольшее
значение имеет пересечение линии Ks и К± и т. д. Выяснить эти
обстоятельства аналитически было бы значительно труднее, а между тем здесь
мы получаем указания, какой материал особенно важен для уточнения
ответа.
Выше мы упоминали о возможности использовать, кроме Кг, К2 и
Kz, еще и другие величины. Так, рис. 34—39 иллюстрируют привлечение
К±, которое представляет собой процент окрашенных в потомстве
Рис. 36. Применение к решению задачи Рис. 37. Применение к решению задачи
№ 1 номограммы рис. 23, дополненной № 1 номограммы рис. 22, дополненной,
линиями КА линиями Кл
137
ЗРис. 38. Применение к решению задачи
-№ 2 номограммы рис. 23, дополненной
линиями К±
Рис. 39. Применение к решению задачи
№ 2 номограммы рис. 22, дополненной
линиями Км
окрашенного родителя безотносительно к окраске второго родителя.
Для случая парных генов эта величина равна:
\ 1 + я) \ i + »/
Для случая эпистаза + криптомерия:
*•-*{*—&)'
Выгоды этих величин в том, что они могут быть получены обычно с
^большей точностью, чем К2 и К2. Невыгода в том, что эти кривые
пробегают сходно с кривыми Кг. На рис. 36 видно, что благодаря
использованию К± решение этой задачи несколько уточняется. Покрытая
пунктиром подоса, в которой налегают друг на друга все поля + лг, здесь, по
«равнению с рис. 30, распалась на два участка, что все более ограничивает
решение и приближает его к верному. Для исключения неверного решения
ло схеме эпистаза К± не дало ничего нового, близко совпадая с Кг (рис.
37). Тот же малый эффект К± виден на рис. 38, но здесь еще сильнее
подчеркивается непригодность схемы парных генов для решения задачи
№1.
Когда будут подобным образом обработаны различные ди- и тригиб-
ридные случаи, будет ясно, какими элементами эти случаи могут быть
наиболее безошибочно отличены друг от друга, и вместе с тем будет
освещен и важный вопрос о том, на какой именно материал при его сборе
должно быть обращено особое внимание (как в нашей задаче № 1 на сбор
материала для уточнения К3, т. е. процента белых в потомстве от
скрещивания окрашенные X окрашенные).
Вместе с тем решением подобной задачи может быть освещен и
важный вопрос о количестве необходимого для решения задачи материала.
Мы видели, например, что 120 скрещиваний и 240 потомков оказалось
вполне достаточным для решения задачи № 2 в тех пределах, в каких мы
задачу решали, т. е. чтобы отличить случай парных генов от случая
эпистаза + криптомерия., Но тот же материал оказался недостаточным
для решения задачи № 1, поскольку мы не могли исключить неверного
решения по схеме эпистаза, а сделали его только маловероятным. Без
выяснения того, какие цифры, характеризующие популяцию, являются
для данного случая решающими, невозможно ответить на вопрос о том,
138
какой материал для этого достаточен. Между тем львиная доля ошибок
<при генетическом анализе зависит именно от того, что недостаточный
материал принимается за достаточный и, наоборот, не делается вывод там,
тде он может быть сделан.
Особенности проблемы анализа в популяциях
Очень важно подчеркнуть, что при генетическом анализе популяцион-
ш.ото материала сама проблема подвергается существенным изменениям.
Выше, при решении задач обычным экспериментом или задач-моделей,
мы ставили целью исследования найти, во-первых, сколько генов участ-
^вyют в расщеплении, во-вторых, каково действие каждого из генов в
различных комбинациях и, в-третьих, каковы генотипы определенных
особей.
В популяционных задачах на первое место выходит второй вопрос и
одновременно появляется следующий — каковы концентрации генов.
Однако вопрос о числе генов ставится здесь оригинально. В самом деле,
если мы в только что решенную нами задачу-модель включим еще третий
ген С с концентрациями р3 = 0,99 и q3 = 0,01, то это нисколько не
повлияет на ответ.
Подавляющее большинство наших особей будет СС и лишь около де-
еятка из всех 480 особей будет Сс и, вероятно, ни одной сс, так как такие
особи встречаются в одном случае на 10 000. Мы установим,
следовательно, что в нашей популяции наследственные явления объясняются двумя
генами А с р = 0,30 и В с г = 0,70. С формальной стороны мы как будто
сделаем ошибку, проглядев ген С. Однако по существу наше решение
будет верным, так как аллопара Сс фактически почти не существует в
нашей популяции, состоящей почти дацело из особей СС.
Следовательно, при анализе популяционного материала вопрос
ставится не вообще о числе генов, менделирующих в популяции, а лишь о
том числе, которое определяет, так сказать, основное лицо ее, т. е. о
«наиболее менделирующих генах».
ГЛАВА VI
АНАЛИЗ ГЕНОТИПА ОСОБИ
ВТОРАЯ ЗАДАЧА ГЕНЕТИЧЕСКОГО АНАЛИЗА
Второй задачей генетического анализа является анализ генотипа,
особи. Правда, в практической работе генетиков решение первой задачиг
изучение свойств генов, и решение данной второй задачи теснейшим
образом переплетаются, ибо для анализа любого скрещивания бывает
необходимо знать как свойства тех генов, с которыми мы имеем дело, так
и генотипы скрещиваемых особей и их потомков.
Однако эти проблемы все же имеют совершенно различный характер.
При изучении генов мы можем обычно забывать про многие основные
свойства генотипов животных, интересуясь лишь теми генами, которые
нам либо мешают, либо облегчают работу по изучению особенно нас
интересующих генов. Наоборот, при изучении многих сторон генотипа
особи приходится часто работать с генами, уже достаточно изученными и с
этой точки зрения уже не интересными, используя их анализ лишь в
качестве методического приема для решения интересующей нас более
глубокой проблемы. Если в первом случае нас интересуют свойства данного
гена, его идентификация, отличие от других, его действие в различных
комбинациях с другими и местоположение в хромосомном аппарате, то
в настоящей проблеме нас будут интересовать следующие вопросы.
1) Какие гены входят в генотип данной особи (или в генотип
совокупности особей, породы, вида)?
2) Как устроен наследственный аппарат данной особи (породы, вида)„
являющийся носителем генотипа?
КАЧЕСТВЕННЫЙ АНАЛИЗ СОСТАВА ГЕНОТИПА
В этом анализе можно различить следующие части.
1. Выяснение наличия в генотипе уже изученных ранее генов:
а) доминантных и эпистатических;
б) рецессивных и гипостатических (в частности, летал ей),
находящихся в гетерозиготном состоянии.
2. Выяснение наличия в генотипах неизученных генов:
а) доминантных и эпистатических;
б) рецессивных и гипостатических (в частности, летал ей),
находящихся в гетерозиготном состоянии.
Часть известных уже по прежним исследованиям генов может быть
узнана при изучении фенотипа животного. По фенотипу черно-пестрого
ярославского быка узнаем о наличии в его составе гена черной окраски
140
и гена белоголовости. Это — совершенно очевидно, и требуется лишь
подчеркнуть, что в таких случаях следует взвесить следующие
обстоятельства:
а) возможность непроявления гена под влиянием генотипических
факторов (криптомерия, эпистаз и т. п.);
б) возможность существования аналогичных генов.
Так, при осмотре белой курицы, прежде чем записать ее генотип
по данным фенотипа, необходимо вспомнить:
а) что альбинизм изредка возникает как паратипическое изменение;
б) что чисто белая окраска может вызываться рецессивным геном
atedu и комбинацией двух доминантных генов todi — tifa;
в) что относительно всех остальных доминантных генов окраски
необходимо отметить невозможность их констатации, а не писать «нет».
Анализ рецессивных составных частей генотипа может быть
произведен:
а) по данным о генотипе родителей и других родственников;
б) по данным о потомстве.
Что касается данных о родителях, то они могут дать основания для
суждения о генотипе исследуемой особи либо вполне достоверные, либо
условные или вероятные. Последние тоже могут представлять
определенную ценность либо для практических выводов, либо для постановки
проверочных исследований.
Точные данные о генотипе данной особи по данным о родителях мы
лолучаем только тогда, когда один из родителей гомозиготен по
рецессивному признаку. Так, в скрещиваниях Аа X аа можно с уверенностью
говорить о том, что у Аа в генотипе будет иметься ген а, хотя бы он был
скрыт под доминантным А или не мог бы проявиться по другим причинам.
Так, если мы получаем в скрещивании кроликов
серый X черный
I
белый
то можно с уверенностью записать в генотип альбиноса ген «черной
окраски» или, точнее, ген отсутствия агути (зонарности), несмотря на то, что
кролик родился белым, очевидно, благодаря расщеплению обоих
родителей по гену цветности
СсА. X Ссаа
I
сс.а
Точкой обозначаем здесь аллеломорф, о котором мы не можем судить с
достоверностью: если у серого родителя на месте точки стоит А, то и у
белого на месте точки должен стоять А, а если у серого родителя на
месте точки стоит а, то у белого на месте точки с одинаковой вероятностью
может стоять либо Л, либо а.
Вероятное заключение приходится делать в том случае, если
скрещиваются особи Аа X Аа. Так, если известно, что оба родителя
гетерозиготны по гену агути и изучаемый нами их потомок тоже по окраске агути, то
о второй его аллели мы должны сказать, что она с вероятностью х/г
может быть А и с вероятностью 2/з — #
Аа X Аа
\
А.
Такой будет вероятность потому, что от скрещивания двух гетерози-
гот возникает среди особей, имеющих ген А, отношение АА : 2Аа.
Следовательно, с вероятностью 2/з можно утверждать, что изучаемая особь
имеет генотип Аа и, следовательно, на месте точки в ее формуле стоит а.
-141
Такие же вероятные заключения могут быть сделаны и по другим!
родственникам. Например, на основании родословной
№ 1 серый X № 2 серый
1
№ 3 серый X № 4 серый № 5 белый
№ 6 серый
мы заключаем
1) что серые № 1 и № 2 гетерозиготны по альбинизму (№ 5 — белый);
2) что, следовательно, с вероятностью 2/з серый № 4 тоже
гетерозиготен;
3) что с вероятностью 72 рецессивная аллель С от № 4 могла попасть
к № 6;
4) что, следовательно, вероятность попадания к № 6 аллели С через
№ 4 будет у • у = у;
5) и так как № 3 тоже с некоторой (зависящей от характера той
популяции, к которой он принадлеясит) вероятностью может быть
заподозрен в гетерозиготности, то
6) вероятность, что № 6 гетерозиготен, будет ^> Vs-
Приведем еще один пример:
№ 1 серый X № 2 серый X № 3 серый
№ 4 серый № 5 серый X № 6 серый,
№ 7 белый
Изучая родословную серого № 4, заключаем следующее:
1. Серые № 5 и 6 гетерозиготны по альбинизму, так как дали белого
№ 7.
2. Если серый № 5 Сс, то для № 2 и 3 возможны гипотезы:
а) Сс X СС;
б) СС X Сс,
в) Сс X Сс.
3. Так как при Сс X Сс № 5 мог бы оказаться (в одном из 4 случаев)
и альбиносом, то можно утверждать с вероятностью 1 против 3/4, что
имеет место тип скрещивания (а) или (б), а не (в).
С другой стороны, так как по гипотезе «а» и «б» вероятность
появления гетерозиготности у № 5 в качестве единственного серого потомка
равна 0,5, а при гипотезе «в» — 2/з» то полная вероятность гипотезы «а» +
«б», с одной стороны, и «в», с другой — такова:
а)+б) Т*Т = Т
ч 3 2 2
т. е. гипотезы «а» + «б» и «в» — равновероятны.
4. Отсюда вероятность каждой из этих гипотез такова:
а)^; 6>Т; в)4*
5. Так как по гипотезам «а» и «б» серый № 2 гетерозиготен, то
вероятность гетерозиготности № 2 равна г/4 + 2U == 3/4«
6. Следовательно, вероятность для № 4 унаследовать через № 2 ген
альбинизма равна: 3/4*7з = 3/8.
142
7. И полная вероятность для № 4 быть гетерозиготом (так как не*
исключена гетерозиготность № 1) ^> 3/8.
Очевидно, что при расчете вероятности должно быть в некоторых
случаях принято во внимание и число всех особей, получающихся при
скрещивании. Так, например, если в только что разобранный пример мы
внесем небольшое изменение, сказав, что у № 5 имеются еще четыре полных
брата и сестры и все они серые, то вероятность для № 4 оказаться гетеро-
зиготой уменьшится.
АНАЛИЗ ПО ПОТОМСТВУ
Анализ по потомству является наиболее плодотворным путем анализа^
генотипа особи. Преимущество его перед анализом по восходящей
родословной и ее боковым ветвям обнаруживается из того, что два полных.
брата имеют, очевидно, одинаковые родословные и по данным этой
родословной должны получить одинаковую оценку. Между тем благодаря
расщеплению они могут быть (и обычно бывают) генотипически различными.
И это различие, если оно не обнаруживается в фенотипе, может быть
исследовано только через потомство. Недостатком этого источника познания
генотипа является его длительность, измеряемая обычно несколькими,
поколениями.
Анализ генотипа особи, говоря вообще, ведется при помощи
анализаторов, т. е. путем скрещивания с другими особями, имеющими
такие генотипы, при помощи которых может быть получено расщепление
в потомстве генов анализируемой особи. Наиболее удобными являются
рецессивные анализаторы, т. е. особи, гомозиготные по
тем рецессивным аллелям, по которым мы хотим проанализировать нашу
особь. Так, чтобы подвергнуть анализу генотип особи, фенотип которой
ABCREF (т. е. обнаруживает наличие в генотипе генов ABCflEF, но
неизвестна его гомо- или гетерозиготность), наилучшим рецессивным
анализатором будет aabbccddeeff. В самом деле, при таком скрещивании
сразу обнаружится, по каким генам наша особь гетерозиготна и по каким
гомозиготна.
С несколько меньшим удобством возможно использование и
гетерозиготных анализаторов, т. е. использование вместо
анализатора строения аа ЪЪсс... анализаторов строения Аа ВЪ Сс Dd... При
этой замене мы проигрываем на том, что расщепление по генам,
гетерозиготным у анализируемой особи, будет теперь идти уже по формуле 3:1,
а не 1:1. Для получения точного ответа потребуется соответственно
больше потомков, а при невозможности увеличения потомства придется
удовлетвориться меньшей степенью уверенности в заключениях. С другой
стороны, гетерозиготные анализаторы имеют свою выгоду, так как
нередко такие гетерозиготы (например, у дрозофилы) оказываются более
жизнеспособными и плодовитыми, особенно потому, что в ряде случаев
гомозиготы по некоторым генам могут быть вообще нежизнеспособными или
бесплодными. Так, при анализе на летали, очевидно, нельзя иметь в
качестве анализатора гомозиготную по летали особь.
АНАЛИЗ ПУТЕМ СКРЕЩИВАНИЯ РОДИТЕЛЕЙ С ДЕТЬМИ
(ДЕТИ-АНАЛИЗАТОРЫ)
Теоретические основы анализа путем скрещивания родителей с детьми
состоят в том, что дети, полученные от любых скрещиваний, но только в
достаточном количестве будут, по крайней мере, в своей
совокупности гетерозиготными по всем тем генам, по которым гетерозиготен
143
их анализируемый родитель. Поэтому скрещивание родителя с
достаточным числом его детей открывает возможность для расщепления, а тем
самым для анализа этого родителя по всем генам, по которым он
гетерозиготен.
Пусть, например, мы хотим проанализировать некоторое животное,
которое гетерозиготно (что нам известно) по генам А, В и С, т. е. АаВЬСс.
Скрестим его хотя бы с самым неподходящим в качестве анализатора
животным ААВВСС. Получаем следующие типы детей:
АаВЬСс X ААВВСС
1) ААВВСС 5) АаВВСС
2) ААВВСс 6) АаВВСс
3) ААВЬСС 7) АаВЬСС
4) ААВЬСс 8) АаВЬСс
Хотя фенотипически все эти дети и повторяют обоих родителей, тем
не менее по каждому гену половина их гетерозиготна и является
гетерозиготным анализатором. По А гетерозиготны № 5—8, по В — № 3, 4, 7
и 8, по С — № 2, 4, 6, 8. Полным анализатором, правда, является
только один № 8, т. е. 12% детей, а при большем числе генов этот процент
будет еще более низким и на его ориентироваться уже не приходится. Но
это не так важно. Скрещивая анализируемого родителя с рядом детей,
мы в совокупности подвергнем его полному анализу.
Каков будет состав потомства от скрещивания родителя с
совокупностью его детей (2Д)?
Состав jaMeT 2Д будет следующим:
Генотип детей
'
ААВВСС
ААВВСс
. ААВЬСС
ААВЬСс
АаВВСС
АаВВСс
АаВЬСС
АаВЬСс
Гаметы родителя . .
Гаметы
ABC
\
8
4
4
1 2
4
2
2
1
-27
1
АВс
4
2
2
1
9
1
АЬС |
4
2
2
1
9
1
АЪс
2
1
3
1
аВС
4
2
2
1
9
1
аВс
2
1
3
1
аЪС
2
1
3
1
аЪс
1
1
1
Отсюда по решетке Пеннета рассчитываем состав потомства
(фенотипы). См. стр. 145.
По подсчетам видно, что расщепление действительно идет по всем
генам, по которым был гетерозиготен родитель, с тем лишь
единственным отклонением от нормального тригибридного скрещивания,
что оно идет не по биному (3 + I)3, а по биному (7 + I)3 и соответственно
для каждого гена получится отношение не 3 : 1, а 7 : 1.
Это изменение пропорции извлекаемых рецессивов с 25 до 12,5%,
т. е. вдвое, конечно, создает известное неудобство тем, что требует
повышения числа получаемых потомков. Можно было бы подумать, что
уменьшение вдвое выхода рецессивов требует лишь увеличения вдвое же
количества потомства. Это, однако, не так, и расчет здесь несколько более
сложен. Правильность результатов здесь в отличие от обычно
получаемого от Fa з^ависит от двух обстоятельств.
144
Гаметы ЗД
СС
)Г0 ОТЦ
1ируем<
анализ
аметы
и.
ЛВС
\АВс
АЬС
АЬс
аВС
аВС
аЬС
аЬс
27 АВС
27 А, В, С
(1)
27 А, В, С
\ (1)
27 А, В, с
| (1)
27 А, В, С
(1)
27 А, В, С
(1)
27 А, В, с
(1)
27 Л, В, с
(1) .
27 А, В, с-
(1)
9 АВс
9 А, В, С
(1)
9 А, В, с
1 (2)
9 А, В, С
(1)
9, Л, В, с
(2)
9 А, В, С
(1)
9 Л, В, с
(2)
9 Л, Я, с
(1)
9 Л, М
(2)
9 ЛЬС
9 Л, В, С
(1)
9 Л, В, С
(1)
|9Л,&,С
(3)
9 Л, Ъ,С
(3)
9 Л, В, С
(1)
9 Л, В, С
CD
9 Л, 6, С
(3)
9 Л, 6, с\
(3)
3 Лбе
3 Л, 5, С
(1)
г а, в, с
(2)
|зл,ь, с
1 О)
3 Л, 6, с
(4)
зл,я, с
(1)
ЗА, В, с
(2)
ЗЛ,Ь, С
(3)
ЗЛ,6,с
(4)
9 аВС
9 А, В, с
(1)
9 л, 5, с
(1)
9 Л, 5, С
<1>
9 Л, 5, С
(1)
9 а, 5, С
(5)
9 а, 5, С
(5)
9 а, 5, С
(5)
9 а, 5, с\
(5)
3 айс
ЗЛ,Я, С
(1)
ЗЛ,£, с
(2)
\ЪА,В,С
(1)
ЗЛ,#,с
(2)
3 а, 5, С
(5)
За, В, с
(6)
3 а, Bt С
(5)
За, 5, с
(6)
3 абС
ЗЛ,£,С
(1)
зл,я, с
(1)
|зл,ь, с
(3) '
зл,ь,с
(3)
За, В, С
(5)
3 а, 5, С
(5)
За, 6, С
(7)
За, Ъ, С
(7)
1 аЬс
1А,В,с
(1)
1 Л, 5, с
| (2)
1Л,Ь,С
(3)
1 Л, 6, с
(4)
1а, В, С
(5)
1 а, 5, с
(6)
1 а, 6, С
(7)
1 а, 6, с
(8)
Примечание. В скобках — номера фенотипов.
Итого фенотипов
(1) Л, В, С 343 (5) а, 5, С 49
(2) Л, 5, с 49 (6) а, В, с 7
(3) Л, Ь, С 49 (7) а,Ь,С 7
(4) Л, Ь, с 7 (8) а, Ь, с 1
Отношение по отдельным
генам
Л:а = 7:1
5:6 = 7:1
С:с = 7:1
1. Введение в скрещивание достаточного количества детей для того,
чтобы гарантировать наличие среди этих детей гетерозигот по всем типам.
2. Получение достаточного количества потомков от скрещивания с
гетерозиготными детьми для того, чтобы гарантировать выход 12,5% рецес-
сивов.
Ясно, что при несоблюдении первого требования мы можем не ввести
в скрещивание необходимых категорий детей, тогда даже бесконечное
количество потомства не поможет извлечению рецессивов.
Так как дети по каждому гену распадаются на гомо- и гетерозиготы в
отношении 1 : 1, то уже при 8 детях, введенных в скрещивание, с
вероятностью 127/128 можно считать, что хотя бы одна гетерозигота по данному
гену будет введена в скрещивание.
Эту цифру (8) детей следует принять как минимум, опускаться ниже
которого уже рискованно.
У"7*1
При отношении 7 : 1 ошибка т = —g— =
получить Зт<^12,5%, решаем:
0,936. Для того чтобы
100 о/о.±. |Л-^-< 12,5 %,
где х — число особей, откуда 8х ]> 63.
10 А. С. Серебровский
145
Действительно, уже получив от 8 детей (или большего числа) 64
потомка, мы будем иметь ожидание для рецессивов:
8 ± 2,64, или 8/2,64 = 3,02, т. е. > 3.
Таким образом, достаточную гарантию анализа особи по всем ее
моногибридным признакам мы имеем при скрещивании ее не менее как с
8 детьми и при получении от такого скрещивания не менее 64 потомков.
Только те свойства, которые дают расщепление по более сложным
отношениям (15 : 1, 63 : 1 и т. п.), останутся вскрытыми без достаточной
уверенности, и для их анализа число потомков должно быть увеличено еще
значительнее, до 200 и более.
Основными недостатками метода детей-анализаторов являются:
1) почти полная неприложимость этого метода к анализу
малоплодовитых самок или самок, не пригодных для повторного оплодотворения
(пчелы);
2) сравнительно большое количество требуемого для анализа
потомства;
3) необходимость двух поколений для анализа.
Достоинствами метода детей-анализаторов являются:
1) одинаковая приложимость его для анализа как по известным, так и
по неизвестным генам;
2) наилучшая приложимость для анализа полулеталей и леталей;
3) возможность использования для начала анализа (для получения
детей) любых животных.
ВЫБОР ПРИЕМОВ АНАЛИЗА
Из обзора разобранных приемов можно извлечь необходимые
указания по вопросу о том, чем должны отличаться приемы анализа по
известным и по неизвестным генам. В первом случае, при анализе по известным
генам, задача сводится к тому, чтобы подобрать нужные анализаторы.
В идеальном случае должен быть взят анализатор, рецессивный по всем
нужным нам генам. Если это невозможно сделать, то придется иметь
несколько частичных анализаторов: один для анализа по
одним, другой — для анализа по другим генам. В этом случае анализ
завершается уже получением Fl9 кроме каких-либо более сложных случаев,
которые могут потребовать добавочных скрещиваний.
При невозможности иметь необходимый набор рецессивных
анализаторов или при анализе на летали и полулетали придется воспользоваться
гетерозиготными анализаторами. Это увеличивает количество потребного
потомства, но не удлиняет анализа.
Наконец, при отсутствии анализаторов придется прибегнуть к методу
детей-анализаторов, что уже удлиняет анализ на одно поколение.
При анализе на неизвестные вообще гены или тогда, когда
неизвестно, по каким генам надо вести анализ (например, когда мы хотим проана-
л изировать особь на содержание леталей; леталей, даже изученных,
много, и мы не знаем, какая именно леталь может быть у данной особи),
основным методом становится метод детей-анализаторов, так как, получая
детей, мы тем самым приготовляем анализаторов именно по тем генам, по
которым необходимо вести анализ, хотя бы мы и не знали, по каким.
Осложнение анализа гетерозиготной части генотипа возникает,
например, тогда, когда приложение метода детей-анализаторов оказывается
невозможным. К сожалению, такие случаи могут встретиться очень
легко, хотя бы, например, тогда, когда интересующая нас особь погибнет
прежде, чем достигнут половой зрелости ее дети, или когда
скрещивание с детьми-анализаторами окажется технически невозможным: пчели-
146
ную матку невозможно осеменить ее сыном, корову, ушедшую в другое
хозяйство, трудно осеменить ее отцом и т. д.
В таких случаях анализ придется вести уже по частям по нисходящим
ветвям родословной.
Анализ по нисходящим ветвям сводится к тому, что мы анализируем
генотипы детей, внуков и т. д. анализируемой особи и, получив данные о
генотипах этих детей, умозаключаем о генотипе интересующей нас особи.
Простейшим примером такого анализа по Fa будет, например,
изображенный ниже:
серая № 3 X серый № IX серая № 2
I I
(1) Fi: серые № 4—10 (2) Fi серые № 11—20
I I
(1) Fi: 225 серых (2) F2 90 серых
15 черных 6 черных
16 белых
Анализируется генотип серого № 1.
Рассуждаем так. По обоим F2 мы видим появление черных в
пропорции, отвечающей 15 : 1. Это показывает, что Fx состоит из особей Аа и
АА в равных количествах, откуда возникает возможность 4 типов встреч:
АА X АА, АА X Аа, Аа X АА и Аа X Аа, в последней из которых в
отношении 3 : 1 извлекаются черные.
В (1) F2, кроме того, в той же пропорции извлекались и белые, а это
указывает на то, что (1) Fx состоит также из СС и Сс в отношении 1:1.
1. Возможны следующие четыре гипотезы о строении № 1:
a) ААСС с) АаСС
b) ААСс d) AaCe
2. Для того чтобы в Fa появилось наблюдавшееся потомство, самки
должны иметь генотипы:
№ 3 .
АаСс
АаСС
ААСс
ААСС
Н 1
ААСС
ААСс
АаСС
АаСс
№ 2
АаСС
АаСС
ААСС
ААСС
3. Гипотезы Ь) и d), предполагающие для № 1 Сс, исключаются, так
как по ним в (2) F2 должны возникнуть белые, так как (2) ¥г будет
состоять из С с и СС особей.
4. Гипотезы а) и с) обе возможны, но по гипотезе «с» появление
черных в обоих F2 зависит от № 1. Это несколько более вероятно, чем
гетерозиготность по А обеих самок № 2 и № 3.
5. Поэтому принимаем для № 1 генотип АаСС, оговариваясь, что
нисколько не исключена возможность и ААСС генотипа.
Если бы мы скрестили № 1 отца еще с 2—3 самками и всюду в F2
получили бы черных, то это значительно подкрепило бы наш вывод о АаСС
генотипе № 1.
Из разобранного примера следует, что основное осложнение,
связанное с применением метода анализа по нисходящим ветвям, состоит в
необходимости различить, что именно было привнесено в эти нисходящие
ветви анализируемым предком и что особями, с ним скрещенными. В том
случае, когда все нисходящие ветви восходят к одной паре родителей,
эта задача для независимо наследующихся генов становится
неразрешимой; обнаружив в нисходящих ветвях рецессивы а, Ъ, с..., мы можем
утверждать, что они имелись в генотипе родителей, но какого именно
родителя — неизвестно (заметим, что мы имеем здесь на двух родителей одну
нисходящую ветвь, хотя бы и разветвленную). При двух ветвях на трех
147
ю*
родителей, как в только что разобранном примере, уже появляется
возможность сделать заключение более уверенно.
Достоверное (однозначное) решение получаем в том случае, если
расщепление по данному признаку наступает лишь в одной из двух ветвей.
i— 2 3
сер.—j—сер.—j—сер.
сер. сер.
I I
сер. сер.
черн.
В прилагаемой схеме выщепление черных наступило лишь в одной
ветви. Судя по этому расщеплению, возможны две гипотезы:
a) 1 Аа 2 АА 3 АА
b) 1 АЛ 1 Аа Ъ АА
Но гипотеза (Ь) невозможна, так как при ее справедливости и во
второй ветви должно было бы обнаружиться расщепление. Следовательно,
справедлива лишь гипотеза (а).
При расщеплении в обеих ветвях, наоборот, возможны, как сказано
выше, две гипотезы:
сер.—j—сер. —г—сер.
сеР- сеР- а) Аа А А Аа
се|р. Jj>. b>^ Л* AA
черн. черн. ■ ч
И обе гипотезы справедливы.
При наличии четырех родителей и трех ветвей почти все решения
оказываются уже однозначными, кроме одного, когда расщепление
появляется опять-таки во всех ветвях.
Распространяя анализ на большее число родителей, получим вывод,
что достоверное однозначное решение будет в том случае, когда по
данному признаку хотя бы в одной из ветвей в F2 не наступает расщепления.
Это относится к случаям, когда в ¥х расщепления нет. В противном
случае возможны однозначные решения и при расщеплении во всех ветвях.
ПотомствЪ ¥г на всех схемах опущено, так как оно везде здесь серое.
сер.—j—сер. Решения два, и по А и по В:
Аа — ААВЪ — ВВ
сер.
черн.
бел.
АА — Аа ВВ—ВЬ
сер. Решение достоверное:
АаВЬ — ААВВ — ААВВ
сер.—г—сер.—j—сер. Решений два:
СЛ СЛ &) ААВВ —АаВЬ —ААВВ
черн. черн. g АаВЬ - ААВВ - АаВЬ.
бел. бел. Из ня^ "олее вероятно «а»
сер.—j—сер.-—j—сер. Решение [вероятное по черному
сер. сер. и Д°стсверное по'белому
черн. черн.
бел.
сер.—г—сер.—j—-сер.—j—сер. Решение достоверное:
сер. сер. сер. ААВВ - АаВЬ - ААВВ - ААВВ
черн. черн.
бел. бел.
148
сер.—|—сер.—j—сер.—j—сер. Решение достоверное:
eel сер. сер. ААВВ - АаВВ - ААВЪ - ААВВ
черн. черы. бел.
бел.
сер.—j—сер.—j—сер.—j—сер. Решение достоверное:
сер. сер. сер. МВБ - ААВЪ - ААВВ - АаВВ
черн. бел. черн.
бел.
сер.—j—сер.—j—сер.—-.—сер. Решение достоверное:
сер. сер. сер. Аа-АА~АА-Аа
черн. черн.
сер.—j—сер.—j—сер.—j—сер. Решение достоверное:
сер. сер. сер. АА — Аа — АА — АА
черн. черн.
сер.—j—сер.—j—сер.—j—сер. Два решения:
»Л пЛ ' а) Аа — АА — Аа — АА
%L «S; 2£» Ъ)АА-Аа-АА-Аа.
черн. черн. черн. '
Рассматривая случаи двузначных решений, а именно
a) АА — Аа; а) АА — Аа — АА; а) АА — АА — АА — Аа\
b) Аа — АА; Ъ) Аа — АА — Аа; Ъ) Аа — АА — Аа — АА и т. д.,
замечаем, что самцы и самки в этих рядах должны располагаться
попеременно. Поэтому всюду все самцы имеют один генотип, все самки —
другой. Этим исключается возможность постановки скрещивания особей с
одинаковыми генотипами и получение новых ветвей, которые дали бы
однозначные решения (у растений это возможно).
Обходящие ветви. В ряде случаев могут быть использованы
обходящие ветви, как можно назвать ветви, восходящие не к данному
анализируемому животному, а к его родителям.
1 X 2|
сер. | сер.
сер.
1
1
сер.
черн.
1
1
сер.
1
сер.
черн.
В данной схеме мы имеем от № 1 и № 2 нисходящую ветвь с выщеп-
лением в ней в F2 черных. Отсюда, как выше сказано, возможны две
гипотезы:
a) АА — Аа;
b) Аа — АА.
Но имеется еще обходящая ветвь, которая восходит не к самому № 2?
а к его родителям. Наличие в этой обходящей ветви тоже выщепления
черных указывает на то, что сибс * номера 2. (на схеме он не имеет
номера) гетерозиготен по А {Аа + АА) и, следовательно, кто-то из
родителей № 2 — тоже Аа. Следовательно, вероятность того, что серый № 2
гетерозиготен, возрастает, т. е. вероятность гипотезы (а) выше вероятности
гипотезы (Ь), хотя все же возможность гипотезы (Ь) не исключается.
* Термин «сибс» (братья и сестры) заимствован из английского языка (сокращенное
Sisters and Brothers).
149
Наличие двух обходящих ветвей делает эту вероятность близкой к
достоверности (см. схему слева) или снова возвращает к исходной
равновероятности (справа):
-?
, Г г , г г ,
I I II II
сер. сер.—j—сер. сер. сер. сер.—|—сер. сер.
II III I
сер. сер. сер. сер. сер. сер.
] черн. черн. | черн.
I I
сер. сер.
черн. черн.
в зависимости от того, появится ли или нет в этой второй обходной
ветви расщепление по этому же признаку.
ПРОБЛЕМА ТЕОРИИ АНАЛИЗА ПО ВЕТВЯМ
Присматриваясь к этим простым примерам, правильность решения
которых легко показать простыми рассуждениями, мы замечаем, однако,
что в одних случаях получаем одно вполне определенное решение, в
других два разновероятных решения и т. д. Естественно, возникает
вопрос, от каких условий зависит определенность или неопределенность
ответа? Очевидно, для генетика-аналитика чрезвычайно важно ответить на
этот вопрос. Если мы сможем найти общую теорию этого ответа, то мы
тем самым сделаем ясным, какие именно скрещивания и вообще данные
необходимы для того, чтобы иметь возможность отвечать совершенно
определенно на вопрос о том, каков генотип данного животного. Имея
такую теорию, мы будем в состоянии вести анализ по кратчайшей линии к
поставленной цели. В настоящее время, к сожалению, мы такой теории
еще не имеем и вряд ли кто сможет четко ответить, почему в одних из
приведенных примеров решение определенное, а в других нет, и что
именно характеризует обе эти категории примеров.
Между тем мы сейчас увидим, что анализ этого вопроса вполне
возможен и применительно к нашим простым примерам довольно легок.
Обратимся к приведенному выше примеру и применим такой способ:
будем обозначать неизвестные аллели буквами х, у, z ...У трех Серых
родителей одна аллель обязательно А, а другие неизвестны. Запишем это так:
1
сер.
Ах
2
сер.
Ау
3
сер
Az
От скрещивания Ах с Ау получена популяция детей и внуков.
Очевидно, эта популяция имеет состав из тех же аллелей, которые имели
данные родители, т. е. 2А + х + у.
Но состав F2 — серые и черные в отношении 15 : 1. Как показано
выше, F2 состоит из потомства следующих четырех типов скрещиваний:
Типы скрещиваний Потомство
ААХАА 4АА
2Аа X АА ЫА + Ыа
АА + 2Аа + аа
ЛаХАг 9AA + GAa + aa
В образовании этого F2 участвуют 244 + 8 а или ЗА + а. Таков же
состав, очевидно, и Fx, следовательно, и Р — родительского поколения.
150
Таким образом, для родительского поколения № 1 + № 2 мы имеем
две характеристики состава:
1) 2А + х + у9
2) ЗЛ + а.
Очевидно, что 2А-\-х + у = ЗА+а или х + у = А + а.
Для второй ветви, происходящей от № 2 и 3, получаем также две
характеристики:
1) 2A + y+z,
2) 44.
Последнее ясно из того, что все F2 во второй ветви серые, никакого
расщепления нет и все F2 АА, как и все Fx, а, следовательно, и Р.
На этом основании напишем
2А + у + z = 44 или у + z = А + А.
Мы имеем два уравнения с тремя неизвестными:
х+у=А+а
y+z=A+A
Особенности этого рода уравнений таковы, что решение их носит
качественный характер. А и а не могут дробиться и поэтому у = А% z = A.
Подставляя значение у в первое уравнение, получаем х = а.
Итак, все неизвестные найдены: # = а, у = Л, z = А, следовательно,
формулы особей родительского поколения таковы:
Аа — АА — АА.
Иначе обстоит дело со вторым случаем. Рассуждая совершенно так же,
получаем два уравнения:
х + у =А + а
у + z = A +a
отсюда х + у = у + z\ х = z.
Ничего большего мы, оказывается, узнать не можем и, следовательно,
не можем больше сузить значение неизвестных. Положив х = Л,
находим z = А и у = а — решение, не приводящее ни к каким
противоречиям. Положив х = а, находим z = а, у = А — тоже вполне
правильное решение.
Аналогичную картину мы получаем и в другом примере, а именно:
х + У = А + а
у + z = A +a
z + w = А + а
Вычитая из первого второе, получаем
х — я = 0; х =*= z.
Вычитая из второго третье, получаем
У — и> — 0; у =w.
Опять-таки значение неизвестных ограничивается лишь тем, что х == z
и у = w, но которое из них А и которое а — неизвестно. Если х = z = A%
то у = w = а, и наоборот.
Задача. Решим этим методом несколько .более сложную задачу.
Дано: с? № 1 был скрещен с $ $ № 2 и № 3. Полученная от этого скрещива-
151
ния большая группа дочерей (Fa) была покрыта самцом № 4, а
полученные от этого скрещивания самки и самцы (Fb) дали inter se Fc — 126
серых + 2 белых. Кроме того, самка № 3, скрещенная с самцом № 4, дала
самцов и самок (F3), которые inter se дали Fe — 120 серых + 8 белых.
Найти: генотипы и фенотипы самцов № 1 и № 4, самок № 2 и № 3
и фенотипы Fa, Fb и Fd.
d № 1 ; (? № 2+ $ № 3)
$ $ Fa j с? № 4X 2 № 3
>Fd
xy-
-(zu + vw)
-Г5-
cd
1С
po " [e (JA + aY (ЗЛ + а)2
126 серых 120 серых
2 белых 8 белых
Полагаем, что случай моногибридный, обозначаем буквами х, у, г, s,
у, я, и и w неизвестные аллели родоначальников № 1 — № 4, а по Fr
и Fe, последним поколениям (единственным, фенотип которых нам
известен), определяем состав гамет:
9 А 7 1
1) ^g = g2, q = -g-. Состав гамет Fc-g- Л + -д-я или 7 Л + la
2) ^ д2, q = i-. Состав гамет F*-|- Л + ^а или 34 + la.
Подсчитываем участие исходных гамет в определении Fc и Fe.
Соответственно числу аллелей слева приравниваем и число аллелей
справа от знака равенства: »
1) 2 (х + у) + [(z + и) + (v + w)} + 4 (г + в) = 2(7А + la),
2) (г + s) + (v + w) = ЗА + la.
Итак, имеем два уравнения:
1) 2х + 2у + z + и + v + w + 4г + 4s = 144 + 2a,
2) v + w + г + s = ЗА + la.
Из (1) непосредственно следует, что г =f= a и s =f= а, так как и г и s
входят в состав Fe с коэффициентом 4, а аллель a — с коэффициентом 2.
Следовательно, r = Ans = An генотип № 4 АА.
Подставляя значение г и s в (1) и (2) и уменьшая правые и левые части
на соответствующее число А, получаем:
1') 2х + 2у + z + и + и.+ w = 6Л + 2а;
2') I? + и; = Л + я.
Равенство (2') дает нам сразу генотип № 3 — А а.
Подставляя значение (v + w) из (2') в (1') и уменьшая на А + а,
получаем (1") 2х + 2у + z + и = 5А + а.
Отсюда непосредственно следует, что х =f= а и у =^= а, следовательно,
#=-4 иу = 4и генотип № 1 — АА.
Подставляя значение х и у в (1") и сокращая на 4 А, получаем:
1"') z + и = л + а,
т. е. непосредственно генотип № 2 — Аа.
Итак, нами найдены генотипы всех родоначальников и теперь могут
быть даны и их фенотипы:
№ 1 АА ...... серый № 3 Ла серая
№ 2 Аа . . . . . • серая № 4 ЛЛ серый.
152
Так как № 1 АА, то и все Fa серое. Точно так же, так как генотип № 4
АА, то и все F5 и F1 — серое. Задача решена и нам удалось осветить всю
родословную на три поколения вверх:
сер.
-(сер. + сер.)
Аа Аа
I
сер. X сер. х сер.
| АА [ Аа
I I
сер. сер.
I I
126 серых 120 серых
2 белых 8 белых
АНАЛИЗ ГОМОЗИГОТНОЙ ЧАСТИ ГЕНОТИПА
Анализ гомозиготной части генотипа является наиболее сложной
задачей и притом до конца никогда не может быть доведен. Поскольку весь
генетический анализ основывается на изучении расщеплений, постольку
подвергнутой анализу может оказаться лишь та часть генотипа, которая
сможет быть переведена в гетерозиготное состояние. Но для того чтобы
какую-либо гомозиготную часть генотипа АА (соответственно аа) можно
было перевести в гетерозиготное состояние А а, необходимо, чтобы в
природе существовали уже аллели А ж а (или в более общей форме А и А').
Это событие может осуществиться благодаря тому, что 1) ранее уже
возникла трансгенация * из А и а (соответственно из а в i), 2) мы такун>
трансгенацию можем получить искусственно, 3) у какого-либо соседнего
вида вместо А путем ли мутации или какого-нибудь иного процесса
возник и имеется ген М\ иной, чем А **. Если такая аллель в нашем
распоряжении имеется или может иметься, гомозиготная часть генотипа АА
(соответственно аа) может быть переведена в гетерозиготное состояние
соответственно путем 1) скрещивания с особью, аа того же вида, 2)
скрещивания с особью АА', например, с соседним видом, 3) превращением А А
в Аа действием мутагенных факторов, таких, как рентген, радий и др.
Как только перевод АА вАа осуществлен, возникает возможность
анализа этой части генотипа. Но, очевидно, что еще надолго в каждом виде
останется такая часть генотипа, которая не сможет быть переведена в А а,
потому что второй аллели нет в нашем распоряжении или, например,
потому, что она имеется у такого вида, с которым анализируемый^вид
при скрещивании не дает потомства. С этой точки зрения генотип всякого
животного, вводимого в скрещивание, может быть разделен на
следующие части:
1. Гетерозиготная часть.
2. Гомозиготная часть:
А) гомозиготная надставка
Б) гомозиготная база:
а) гомозиготная база породы
б) гомозиготная база вида
в) гомозиготная база рода и т. д.
* Автор употребляет термин «трансгенация» в смысле генной мутации, т. е. перехода
гена, бывшего до мутации («базигена»), в новый ген после мутации («трансген»).—
Прим. ред.
** Мы выделяем этот случай, может быть, как самостоятельный, так как не исключена
возможность того, что в эволюции генов, наряду с трансгенациями из Л в а или и*
А в А', есть мутации, отличные по природе от трансгенации.
153
/ ены видовых Гены родовы, Ге^Мы!ШеЖ1их
признаков признаков зПачешШ-
у а gA CCD DEEF F.P G HJtJJ.KKJ- LMM NNOO и т.д.
Гетероз. Гомозиг Гомозиг. Гомозиг. Уо~мозйг.
часть > надет, особи надет. надет, надет рода
породы вида
База рода
База вида
База породы
Гомозиготная часть генотипа особи
Рис. 40. Схематическая структура генотипа особи
Гетерозиготная часть анализируется любым технически удобным
скрещиванием. Гомозиготная часть требует предварительного перевода в
гетерозиготное состояние (в потомстве). Гомозиготная надставка состоит
из генов, иные аллели которых имеются у других особей той же породы,
и поэтому надставка может быть проанализирована путем различных
внутрипородных скрещиваний. Но далее у нас остается гомозиготная
база породы, состоящая из генов, свойственных всем особям данной
породы в гомозиготном состоянии. Для анализа этой части генотипа
необходимы уже межпородные скрещивания. Подвергнув такими
скрещиваниями анализу гомозиготную породную надставку, мы добираемся уже
до гомозиготной базы вида. Путем межвидовых скрещиваний мы сможем
проанализировать видовую надставку и добраться до гомозиготной базы
рода и т. д. Впрочем, здесь мы уже близко подходим к пределам
возможных скрещиваний, и дальнейший анализ станет возможным лишь путем
получения мутаций соответствующих генов.
Не нужно, однако, думать, что это связано с каким-то разделением
генов на породные, видовые, родовые и т. п. Оставаясь в пределах
породных скрещиваний, мы можем, например, анализировать летальные гены,
которые по характеру действия своих нормальных аллелей относятся,
может быть, как раз к генам видовым и даже еще к более общим. Таким
образом, под термином гомозиготная база породы мы разумеем те гены,
которые имеются в гомозиготном состоянии у всех особей породы, и не
мутировали*, т. е. даже в порядке поисков мутаций мы не находим в
пределах породы особей, которые могут быть использованы в качестве
анализаторов. Сказанное о структуре генотипа иллюстрирует рис. 40.
Для проведения анализа гомозиготной части генотипа особи или
породы необходимо последовательное, а в возможных пределах и
одновременное скрещивание анализируемой особи с другими отличными от нее
особями той же породы, с представителями других пород и с другими
видами.
Так как нам заранее не могут быть вполне известны генотипические
различия анализируемой особи I от скрещиваемых с нею форм (II, III
* Необходимо сделать оговорку, что наличие генов в гомозиготном состоянии у всех
особей какой-либо породы еще не доказывает, что в прошлом они ни разу не
мутировали. Очень велика вероятность, что возникающие вследствие мутации новые гены
исчезнут из популяции по случайным причинам или вследствие отрицательного
отбора.— Прим. ред.
154
и т. д.), то выбор этих форм идет в значительной степени вслепую.
Скрещивание I X П, I X Шит.д. каждый раз анализирует лишь некоторую
часть генотипа I, именно ту часть, которой I отличается от партнера, с
ним скрещиваемого.
Каждое скрещивание (I X II, I X III и т. д.) следует проводить
возможно подробнее, получая Fx, F2, оба FB, а если скрещивание слишком
сложно, то и F3 и т. д., чтобы в результате можно было сказать, что та
часть генотипа, которой I отличается от II (соответственно!отIIIи т. д.),
действительно подвергнутая анализу, уяснена. Так как каждое такое
скрещивание анализирует только некоторую часть генотипа, то для
выполнения анализа в возможно большей степени, вообще говоря, надо
поставить и изучить скрещивания со всеми остальными породами и со
всеми остальными видами, с которыми это технически возможно. Ввиду
необъятности этой задачи полезно руководствоваться указаниями:
1. Удобнее начинать анализ со скрещивания с породами, не
слишком сильно отличающимися от анализируемой, так как при этом мы
будем получать не слишком сложные расщепления.
Более далекие скрещивания кажутся иногда более заманчивыми, но
расщепления в них могут быть такой сложности, что понять их окажется
делом слишком трудным. Лучше, начав с более простых скрещиваний,
изучить участвующие в них гены и после этого усложнить скрещивание.
2. Удобно с этой же точки зрения ставить сразу ряд скрещиваний
I X II, I X III и т. д. По своей сложности эти скрещивания всегда
окажутся различными, и анализ более простых позволит легче уяснить себе
и более сложные скрещивания.
3. Следует обратить прежде всего внимание на FB скрещивания,
всегда более простые, чем Fx X Б\. Расщепление в FB легче понять и, поняв
его, затем легче понять и картину расщепления в F2.
4. В том случае, если F2 дает слишком сложную картину,
целесообразно не задерживаться слишком на его изучении, но поставить ряд
скрещиваний на F3, подбирая для этих скрещиваний особей, наиболее
сходных по фенотипу и притом отличающихся от F1? так как именно при
таком подборе мы можем рассчитывать получить в F3 более простые и
наилегче понятные картины расщепления.
5. Систему скрещивания I. X II, II X III, и т. д. полезно
пополнять скрещиваниями II X III и т. д., в результате чего получаются
«кольца», облегчающие понимание расщепления.
/
#^— *Ш
В нашем личном опыте при анализе на Аниковской генетической
станции генотипа исчезнувшей ныне павловской породы кур мы работали с
такой системой скрещиваний, начатых Н. К. Кольцовым. Павловский
петух был скрещен с курами пород орловской, индобойцовой, фавероль и
бентамками. Наличие этой системы следующим образом облегчило,
например, нам анализ оперения ног. Из названных пород павловцы,
фавероль и бентамки имели оперенные ноги, орловцы и индобойцовые были
голоногими. Каждая из названных трех лохмоногих пород имела
несколько отличный характер лохмоногости.
155
Бентамки-
susuSt St
Генотипы
в F2
Фавероль-
SuSuStSt
-Павловские-
su su stst
I
I
SuSuStSt SuSuStSt
susuSt st SusuStSt
su su st st , susuStSt
SusuStst
susuStst
SuSustst
Susustst
susustst
Полные переходы Полные переходы
между типами между типами ро-
обоих родителей дителей и до
полной голоногости
-Индобойцовые
Su Su st st
-Орловские
Su Su st st
Su Su st st Su Su st st
Su su st st Su su
su su st st su su
ist st \
stst \
sfst J
Распадение на оба
родительских типа с некоторым чио
лом слаболохмоногих форм
на базе неполного
доминирования Su. Скрещивание
с орловцами осложнилось
модификатором, а
скрещивание с . индобойцами легче
всего удалось свести к
отношению 3: 1
Генотипы в F2
В результате исследования оказалось, что имеется в основном 2 гена
suso и susta (su и st), причем Su является геном голоногости (напротив,
su вызывает рецессивную лохмоногость) и St является геном
доминантной лохмоногости. Бентамки имели «двойную лохмоногость» (su su St St)t
павловцы — рецессивную (su su st st), фавероли — доминантную (Su Su
St St), а орловцы и индобойцовые имели генотип Su Su st st. Так как
каждый из этих генов доминировал неполно и гетерозиготы сильно
колебались по степени оперенности ног, то вначале картина наследования
была очень неясна, скрываясь за бесконечными переходами от голоногих
к лохмоногим во всех скрещиваниях. Однако, как видно из приложенной
схемы, скрещивание павловских с индобойцовыми оказалось самым
простым и с его помощью удалось открыть и установить аллопару Su — sur
после чего выяснились и остальные все скрещивания.
Наоборот, по окраске павловско-индобойцовое скрещивание
оказалось необыкновенно сложным, F2 расщеплялось не менее как по 6—7 генам,
и чуть не каждый цыпленок был на свой образец. Эту сложность не удалось
бы распутать, если бы не одновременное наличие скрещивания павловских
с фаверолями, где расщеплялись лишь три аллопары, в том числе одна
ранее неизвестная, которую удалось при этом изучить и осветить затем,
хотя и не до конца, павловско-индобойцовое скрещивание (Серебровский
А. С, 1926).
ГЛАВА VII
ТОПОГРАФИЧЕСКИЙ АНАЛИЗ ГЕНОТИПА
ЗАДАЧИ ТОПОГРАФИЧЕСКОГО АНАЛИЗА ГЕНОТИПА
По установлении аналитической формулы организма (фактически
одновременно) можно переходить к выяснению того, как эти гены
распределены по хромосомному аппарату данной особи (или породы, вида).
Здесь возникают три следующих вопроса.
1. Сколько групп сцепления образуют эти гены, а в конечном счете
между сколькими хромосомными парами распределен генотип.
2. Как распределены гены в каждой из групп сцепления (и
соответственно в каждой из хромосом), каковы порядок генов, взаимные
расстояния между ними и общая длина каждой из этих групп сцепления и
каковы некоторые другие свойства.
3. Нет ли в хромосомном аппарате данной особи каких-либо
особенностей, отличающих его от типа, свойственного виду (нехватки, инверсии,
трансклокации и т. п.).
ОПРЕДЕЛЕНИЕ ЧИСЛА ГРУПП СЦЕПЛЕНИЯ И ХРОМОСОМ
Для решения этого вопроса надо определить наличие сцеплений генов
друг с другом. В характере проявления генов никаких указаний на
наличие между ними сцеплений не имеется: сцепленными могут быть гены
как совершенно несходного проявления (форма гребня и длина ног,
окраска шерсти и длина шерсти и т. д.), так и очень сходные (два гена «реке»
у кроликов). Поэтому в начале работы с неизвестным животным
приходится идти совершенно вслепую и все установленные гены исследовать
попарно на наличие или отсутствие между ними геносвязи. Это
исследование может быть запланировано следующим образом. Пусть мы изучили
у данного вида гены Л, В, С,...ЛГ, тогда составляем квадратную матрицу:
Гены
А
*}~
С
D
Е
• • •
N
А
В
1
С
2
3
D
4
5
6
Е
7
8
9
10
N
79
80
81
82
83
и т.д.
157
Цифрами 1, 2, 3 ...обозначены номера опытов, которые нужно
поставить. В опыте № 1 исследуем наличие геносвязи между генами А и В, в
опыте № 2 — между А и С, в опыте № 3 — между В и С и т. д. Всего та-
п(п— 1)
ких опытов при тг-числе генов надо запланировать—у—=—- , что дает при
10 генах 45 опытов, при 20 генах — 190 и т. д. Число этих опытов, таким
образом, достаточно велико и необходимо установить рациональную
последовательность их, дабы не распылить внимания и сил. Приведенная
нумерация дает как раз одну из таких рациональных последовательностей.
Идея ее состоит в том, что, исследовав на геносвязь два гена А и В, мы
затем исследуем третий ген С как с А, так и с В. Затем исследуем ген D
как с А, так ис5исСит.д. Пусть, например, мы показали в опыте № 1г
что А и В независимы. Отсюда мы заключаем, что А и В относятся к
разным группам сцепления.
Получив отрицательные результаты так же в опытах № 2 и № 3, мы
заключаем соответственно, что и ген С относится к третьей группе, так
как он не относится ни к Л-группе, ни к Б-группе. Если бы мы, не
поставив опыта № 3, поставили бы опыт № 4, то (при отсутствии сцепления)
мы не могли бы утверждать не только то, что имеется четыре
независимых группы, А; В; С; D, но даже и три, так как у нас оставалась бы не-
опровергнутой возможность геносвязей В — С и В —D. Следовательног
мы могли бы утверждать лишь: 1) наличие двух групп А и В и 2) что гены
С и D не относятся к Л-группе. В итоге, поставив вместо опытов №1,2,
3, три же опыта, но № 1, 2, 4, мы получили бы меньше данных о числе
групп сцепления.
В пределах вертикального столбца порядок постановки опытов
безразличен (4, 5, 6 или 5, 6, 4, или 6, 5, 4 и т. д.). Но по возможности не
следует забегать вперед и переходить к опытам последующего
вертикального столбца, не добившись окончания всех опытов предыдущего столбца.
После того как какая-либо геносвязь найдена, в этот план работы
может быть введена некоторая поправка. Если, например, оказалось, что
С и D сильно сцеплены, то отпадает нужда обязательно ставить для гена
Е отдельно опыт ECnED. Следующий столбец D можно слить со столбцом
С. Но этого нельзя делать, если связь С и D не сильная, так как при
расположении, например, С — D— Е ген Е может обнаружить с D
уловимую связь, а с расположенным далее геном С связь Е будет очень слабой,
оставаться неуловленной.
Если далее мы уловим связь трех генов CDE, доказав такой их
порядок, то следующий ген F следует в первую очередь исследовать с генами
Е и С, а не с расположенными в середине. В самом деле, при
расположении гена F между С и Е (т. е. C—F—E) геносвязь F либо с С, либо с Е
мы обязательно уловим, так же как и при расположении F — С — Е и
С — Е — F, если F будет расположен не слишком далеко вне участка
С — Е. Если же мы сначала поставим опыт FD, то отрицательный
результат еще не будет говорить против принадлежности гена F к группе С —
— D —Е, так как это расстояние может быть слишком велико, чтобы
сцепление могло быть уловленным.
При подобном изучении очень важно возможно скорее узнать:
1) у обоих ли полов4 происходит перекрест;
2) если у обоих, то одинаково ли легко. Для облегчения
исследования, естественно, следует ставить опыт так, чтобы гетерозиготой в
скрещивании
АВ аЪ АЪ аЪ
явилась бы особь того пола, у которого перекрест отсутствует или по
крайней мере затруднен. У дрозофилы используется самец (перекреста
158
нет вовсе), у шелкопряда — самка по тем же причинам. У
млекопитающих выгоднее брать самца, так как, по данным о грызунах, перекрест у
самцов идет несколько слабее, гены как бы «сближены» и наличие гено-
связи у них легче обнаруживается.
Исследование этого вопроса не представляет принципиальных
трудностей; как только будут найдены первые геносвязи путем сравнения
реципрокных скрещиваний, например:
вопрос решается непосредственно. Однако с достаточной уверенностью
вопрос о наличии перекреста у данного пола может быть решен и до
отыскания сцепленных пар.генов. Этот вопрос нам пришлось впервые
решать применительно к курице. Ответ был найден следующим образом.
По цитологической картине было найдено, что число пар хромосом у
курицы около 16. При этом условии и при условии отсутствия у кур
перекреста у женского пола следовало бы ожидать, что при изучении
наугад взятых пар генов не реже как 1 : 16 было бы абсолютное сцепление
(или отталкивание) между взятыми генами. Нами было поставлено с
различными генами 79 опытов типа
$) АаВЬ X с? ааЪЪ
и ни в одном случае абсолютного сцепления не было найдено. Этим с
высокой вероятностью было доказано, что у кур (9 $) перекрест в ауто-
сомах должен иметь место, что позднее удалось подтвердить на генахг
сцепление которых было доказано прямым опытом.
УПРОЩЕНИЕ ИССЛЕДОВАНИЯ
Описываемая часть топографического анализа может быть
значительно упрощена совмещением ряда опытов в одном и том же
экспериментальном материале.'Например, поставив тетрагибридное скрещивание
AaBbCcDd X aabbccdd,
мы сразу ставим все 6 первых опытов приведенной выше таблицы: А — \В,
А— С, В — С, A —D, B—D, C—D. Пентагибридное скрещивание
дает материал сразу для 10 опытов и т. д. Райту (Wright S., 1927) у
морских свинок удалось поставить даже октагибридные скрещивания. При
этом следует лишь заботиться о том, чтобы изучаемые одновременно гены
не мешали проявлению друг друга.
У кур, например, могут быть поставлены скрещивания с участием
следующих генов:
розовидного гребня; полидактилии;
стручкового гребня; коротконогости;
хохла; лохмоногости;
бак, или голой шеи; скорости оперения;
2—3 гена окраски оперения; курчавоперости;
1—2 гена окраски ног; окраски яиц;
т. е. 13—15 генов одновременно. Трудности такого опыта сводятся:
1) к получению особей, гетерозиготных одновременно по всем этим
генам;
159
2) к невозможности продолжения этого опыта, так как в потомстве от
таких гетерозигот практически невозможно получить особь, снова
гетерозиготную по всем этим генам;
3) к трудности выбора партнера-анализатора, гомозиготного рецес-
сива по всем этим генам.
ЧИСЛО ГРУПП СЦЕПЛЕНИЯ И ЧИСЛСГПАР ХРОМОСОМ
Современная генетика доказала, что у всякого организма число групп
сцеплений должно совпадать с числом пар хромосом, так как именно
распределением генов по хромосомам и опричинено наличие групп
сцепленных генов. Однако нужно помнить, что в начальной стадии изучения
число групп сцеплений может не совпасть с числом хромосом. Это может
быть вызвано 1) тем, что не для всех хромосом найдены сцепленные
группы и 2) тем, что для некоторых более длинных или легко
перекрещивающихся хромосом может быть первоначально установлена более чем одна
группа сцепления. Так, установив, например, сцепленные пары А — В и
К — L и показав, что А и В не обнаруживают геносвязи ни с К. ни с L,
мы еще не можем быть убеждены в том, что А — В и К — L
локализованы в различных хромосомах.
В этом случае может оказаться, что все эти гены расположены в одной
хромосоме, но А и В на одном конце и К и L на другом, на расстоянии в
50—60 морганид от первой группы. Тогда В и К будут обнаруживать
лишь настолько слабую геносвязь, что ее с трудом можно будет доказать.
Так, например, в половой хромосоме курицы на одном конце
расположены гены trage (поперечнополосатый окрас пера) и trufege (желтые ноги)
с расстоянием между ними в 8 морганид, а на другом конце — гены tuge
(серебристый окрас) и Suke (медленное оперение) с расстоянием между
ними в 20 морганид. Но tuge и trage расположены друг от друга все же
так далеко, что дают 43—46% перекреста. Если бы по ходу сцепленной с
полом наследственности не было бы ясным, что все эти гены находятся в
половой хромосоме, то осталась бы очень сомнительной доказанность того,
что это не две разные группы, но лишь одна. Только при нахождении
нового гена, расположенного где-либо возле середины расстояния между
tuge и trage, который показал бы геносвязь с обоими этими генами в
25—30%, мы получили бы полную уверенность в единстве всей этой
группы (см. Mac Arthur, 1933).
СОСТАВЛЕНИЕ ПЛАНА ХРОМОСОМЫ
Составление плана хромосомы не представляет каких-либо
принципиальных трудностей, и к тому, что было нами изложено в разделе,
посвященном локализации гена, здесь особенно нечего прибавить. Пусть
мы показали, что гены А, В, С... относятся к единой группе сцепления.
Расстояние А — В равно 10 морганидам. При помощи трехгенного
эксперимента локализуем третий ген С и наносим его на план, например, так,
как показано на рис. 41 на второй схеме сверху. Затем такими же трех-
генными или более сложными экспериментами локализуем гены D и Н
(третья схема рис. 41), исправляя расстояние А — В и значительно
удлиняя схему вправо. Далее локализуем остальные гены, получив, например,
план хромосомы, как на нижней схеме рис. 41.
Исчерпав все относящиеся к группе гены при достаточном их числе,
получаем уверенность в том, что левее К и правее Н вряд ли имеются
еще большие участки (хотя, конечно, еще не исключена возможность
и того, что один из следующих генов придется локализовать левее К или
160
правее Н). Но для удобства работы один из концов, например К,
примем за начало хромосомы или за левый конец и к нему отнесем нулевую
морганиду, перенеся ее из А, где она находилась в начале исследования.
Мы получили, таким образом, план хромосомы, на котором указан
порядок генов, расстояния между ними и расстояния от начала. При
небольшом числе генов этот план будет еще очень приближенным и по мере
_4_а_4~_
О 10
* 10 В 5С
нч--
О 10 W
-Ч 1*1 * I l-f
О 9 11 1В 38
К on 0 а А п J9DnB с С лг Е in F 7 И
i н-ч—8 гг\ 5 i - 1—-—н^н—
О 20 24 323436 Ы 56 66 69
Рис. 41. Пример построения плана хромосомы с последовательным уточнением
внесения в него следующих генов будет совершенствоваться в следующих
двух направлениях:
1) более точное определение начала и конца путем изучения генов
левее К и правее Н;
2) более точное определение расстояний между генами путем
изучения промежуточных генов.
Пример (2) на приведенной схеме мы видим в расстоянии между
генами А и В. Первоначально оно было определено в 10 морганид, но после
отыскания промежуточных генов — J ж D — увеличилось до 12,
благодаря тому, что эти промежуточные гены позволили улавливать двойные
перекресты, ускользавпше от внимания ранее. Соответственно можно
полагать, что расстояния К — Gil С — Е пока несколько преуменьшены,
и когда будут найдены гены, локализованные между К и G и между С и
Е, эти расстояния (а вместе с тем и общая длина хромосомы) будут
несколько увеличены.
Дальнейшее изучение свойств хромосомы (или группы сцепления)
может быть направлено на выяснение ряда более глубоких свойств этой
хромосомы. Такими вопросами в свете современных генетических знаний
являются:
1) место прикрепления нити веретена;
2) наличие в хромосомах инертных, лишенных генов, участков;
3) «упругие» свойства хромосомы, выражающиеся в явлениях
интерференции (соответственно коинсиденции);
4) абсолютное значение (в долях микрона) одной морганиды в разных
местах хромосомы и т. д.
В эти вопросы здесь мы входить, однако, не будем, отсылая к
специальной литературе.
АНАЛИЗ ОСОБЕННОСТЕЙ ХРОМОСОМНОГО АППАРАТА
Среди типичных особей вида, имеющих хромосомный аппарат,
устроенный «нормально» — типично для вида, могут встретиться особи с
аномалиями в этом отношении. К таким аномалиям относятся:
1) нехватки — выпадения небольших участков хромосом, как правило,
больших, чем один ген;
11 А. С. Серебровский
161
2) делеции — очень крупные нехватки, когда выпавшая часть
хромосомы превышает (в морганидах) остающуюся;
3) транслокации — перемещение частей одной хромосомы в другую,
или, по-видимому, более правильно — обмен между хромосомами
негомологичными участками;
4) дупликации — удвоение в хромосомном аппарате отдельных
участков хромосом;
5) инверсии — поворот части хромосомы на 180° вокруг поперечной
оси, проходящей через эту часть хромосомы.
Хромосомные аберрации, или дислокации, как называют эти
видоизменения, в некоторых случаях (многие транслокации, делеции) легко
узнаются под микроскопом. Развитие пайнтеровской методики на
хромосомах слюнных желез дрозофил для этого объекта дает на пути
цитологического метода более легкие и точные результаты (в том числе и для
инверсий), чем данные, получаемые генетическим путем. Тем не менее
цитологическим методом (помимо пайнтеровского метода, приложимого пока
только к некоторым насекомым), мы можем усмотреть лишь достаточно
грубые изменения хромосом, а нехватки, удвойки и инверсии, кроме
редких исключений, остаются за пределами метода.
Поэтому генетический метод анализа хромосомных аберраций вполне
сохраняет и сохранит свое значение, и следует лишь стремиться к тому,
чтобы анализируемый объект одновременно подвергался бы и
цитологическому изучению, учитывая то, что оба метода отлично дополняют и
помогают друг другу. Это относится даже и к случаям нарушения числа
хромосом. Если, например, в хромосомном аппарате курицы окажется
лишняя хромосома из числа крупных, то цитологически будет
легче и проще обнаружить это или подтвердить предположение, сделанное
по данным скрещиваний. Но недостаток или излишек одной из мелких
хромосом курицы цитологически почти не удается подтвердить, так как
до сих пор даже не удается установить «нормальное» число этих мелких
хромосом, вследствие трудности наблюдений столь многочисленных
образований под микроскопом.
Основным принципом установления наличия хромосомных аберраций
является установление противоречия (несоответствия) хода наследования
каких-либо признаков с типичными для данного вида. Такими
несоответствиями могут быть следующие: рецессивный признак проявляется там,
где он нормально не должен был бы проявляться; рецессивный признак
исчезает там, где он должен был бы проявиться; независимо
наследующиеся в норме гены вдруг обнаруживают сцепление; сцепленные гены,
дававшие определенный процент перекреста, вдруг становятся как бы
абсолютно сцепленными и т. д.
Для каждого типа хромосомных аберраций характерно появление того
или иного типа аномалий в ходе наследования, и, установив эти
последние, мы можем заключать и о наличии определенных хромосомных
аберраций. После этого желательна и цитологическая проверка вывода.
АНАЛИЗ НЕХВАТОК
Наличие нехваток устанавливается на основании неожиданного
проявления рецессивных признаков.
Если мы имеем у гетерозиготы две близко расположенные
рецессивные аллели а и 6, то при нормальном строении противоположной
хромосомы имеющиеся в ней доминантные аллели А и В не дают возможности
рецессивным аллелям проявиться (схема 1 рис. 42).
Если же в хромосоме имеется нехватка и кусок хромосомы с А и В
выпал, то рецессивные аллели а и Ь получают возможность проявиться
(схема 2 рис. 42).
162
Рис. 42. Анализ нехватки
I — рецессивы а и Ь подавлены
АВ-хромосомой; 2 — благодаря
удачному расположению нехватки
а и & одновременно проявляются;
3 — нехватка расположена
«неудачно» и проявляется лишь Ь, что
неотличимо от 4 — рецессивной
трансгенации Ъ'-
Установить нехватку можно лишь в сравнительно редких
благоприятных случаях, когда в нашем распоряжении имеется две или более
достаточно близко расположенные рецессивные аллели, но притом
расположенные так, что обе они попадают в участок, захваченный нехваткой.
При несоблюдении этого условия, как на схеме 3 (рис. 42), сможет
проявиться лишь одна из рецессивных аллелей. В этом случае мы уже не
сможем отличить нехватку 'от трансгенации, а сможем сказать лишь, что
в противоположной хромосоме возникла рецессивная же аллель Ъ'
(схема 4 рис. 42), обычно более рецессивная, чем Ъ. Действительно, по
данным скрещиваний, случаи 3 ж 4 будут неразличимы. Поскольку в
определение нехватки мы ввели указание на выпадение из хромосомы более
чем одного рядом расположенного гена, для доказательства того, что
мы имеем дело именно с нехваткой, а не с трансгенацией, именно этот
пункт и требуется доказать. Поэтому ясно, что только в том случае, когда
мы располагаем рядом или близко расположенными рецессивными
генами, мы это доказательство и можем осуществить. Если же мы этого
доказать не можем, нам приходится остановиться на выводе, что мы имеем дело
либо с трансгенацией, либо с недоказуемой нехваткой. Несомненно, что
в целом ряде случаев, мутации, которые мы сегодня считаем за рецессивные
трансгенации, на самом деле являются недоказанными нехватками.
Самое исследование ведется следующим образом. Пусть, например,
мы обнаружили новую мутацию (nova), в гомозиготном состоянии
дающую летальный эффект. Поскольку нехватки, как правило, в
гомозиготном состоянии дают летальный эффект, то мы можем задаться вопросом,
не является ли наша мутация нехваткой. Для решения этого вопроса
определяем локализацию новой мутации. Определив с максимальной
точностью ее локус, смотрим по топографическому плану этой хромосомы,
нет ли в непосредственной близости к этому локусу других ранее
изученных и имеющихся в нашем распоряжении рецессивных мутаций.
Если мы такими рецессивами обладаем, то ставим скрещивание ее
с нашей nova. Если ни одна из этих соседних рецессивных аллелей в Fx
с nova не проявляется, следовательно, nova либо трансгенация, либо если
и нехватка, то такая, которая не захватывает исследуемые локусы. Если
одна из взятых для скрещивания рецессивных аллелей (например, а) в
Fx с nova проявляется, мы делаем вывод, что nova либо новая аллель в
той же самой серии, как и аллель а, либо если нехватка, то столь
короткая, что, захватывая локус а, не доходит до соседних известных нам ло-
кусов. И, наконец, если при скрещивании с nova проявляются две (три
и т. д.) рецессивные аллели а и Ъ (и с и т. д.), то мы можем уже с большой
уверенностью признать нашу nova за нехватку, захватившую все эти
локусы. Если между а и Ъ имеется еще какой-либо рецессив а, то
скрещивание с ним будет хорошей проверкой вывода. В самом деле, если мы
действительно имеем дело с нехваткой, захватившей локусы А и 2?, то
должны выпасть и все гены, расположенные между этими локусами.
\A\B\
Г7Щ—
Нехдатка
i i
\ч*\
-2
Нехватка
А\ I
—г^ш—
\A\V\
„ igm ..
163
11*
ОПРЕДЕЛЕНИЕ ДЛИНЫ НЕХВАТОК МЕТОДОМ ПЕРЕКРЕСТА
В случае, если мы не располагаем необходимыми рецессивами для
анализа нехватки (а это вполне обычно), то для определения ее длины
может быть привлечен метод перекреста. Этот метод основан на том, что
перекрест между двумя хромосомами осуществляется только на
гомологичных участках и на всем участке, захваченной нехваткой, перекрест
осуществиться не может. Пусть мы имеем нехватку в районе между ло
кусами генов А и В. Если построить гетерозиготную структуру так, как
показано на рис. 43,2, то, очевидно, что теперь перекрест между А ж В
будет осуществляться уже не с прежней частотой, но реже, так как ранее
он мог происходить в любом пункте между А и 2?, а теперь лишь на тех
участках, которые не захвачены нехваткой.
А В
Н—I И—
2
+ч ь-ь-
a b
Рис. 43. Расположение аллелей для измерения длины нехватки
1 и 2 ■— опыты
Для определения длины нехватки следует в одинаковых условиях
поставить два опыта для определения расстояния (в морганидах) между
А ж В. В первом опыте (рис. 43,1) ж А ж В расположены в нормальной
хромосоме, а во втором опыте (рис. 43, 2) в хромосоме, содержащей
нехватку. Пусть в первом случае расстояние между А и В определено в сг
морганид, во втором — в с2 морганид. Очевидно, что длина нехватки будет
равна (с± — с2). Опыты эти, конечно, следует повторять и в структурах
АЫаВ.
Впервые Бриджес этим методом определил длину нехватки forked-
Bar, оказавшейся равной 0,7 морганиды (Bridges, 1917), и затем еще с
большей убедительностью Mop (Mohr, 1923) при работе с Notch-8
нехваткой, длина которой могла быть достаточно точно определена и прямым
путем — скрещиванием с рецессивами, расположенными на участке,
захваченном нехваткой. Оба определения дали согласные результаты —
около 4 морганид. Следует, впрочем, иметь в виду, что данный метод
определения длины нехваток не очень надежен.
Во-первых, при длине нехватки, например, 0,7 морганиды, как в
случае f-B нехватки, требуется чрезвычайная точность опыта и большой
материал для того, чтобы сделать реальной разницу в величине
перекреста по сравнению с контролем. Для этого необходимо использовать гены,
наиболее близко расположенные к нехватке. В своем опыте Бриджес
использовал счастливо расположенные гены rudimentary ж fused,
разделенные участком в 4,4 морганиды.
С другой стороны, этот метод может дать слишком большую
величину, если возникновение нехватки будет сопровождаться изменениями в
хромосоме на соседних участках с понижением перекреста. Так именно
было со второй нехваткой, исследованной Бриджсом (Bridges, 1919).
По сравнению с нормой нехватка дала понижение перекреста на 5%,
но несомненно, что сама нехватка захватила гораздо менее 5 морганид и
остальное пало на долю затруднений перекреста в соседних местах.
164
АНАЛИЗ ДЕЛЕЦИЙ
Делеция является очень большой нехваткой, захватывающей всю
среднюю часть хромосомы и оставляющей нетронутыми лишь концы
хромосом. Поэтому делетированная хромосома состоит из
соединенных друг с другом концов исходной хромосомы. В таком виде
делетированная хромосома может сохраняться лшпь в качестве добавки к
нормальному хромосомному аппарату. В настоящее время делетирован-
яые хромосомы обнаружены у дрозофилы (в результате облучения
рентгеном) и лишь применительно к Х-хромосоме *.
Обычный прием улавливания делетированных хромосом, или деле-
ций, таков. Рентгенизуются самцы, имеющие нормальную Х-хромосому,
в частности, имеющие на левом конце нормальные аллели у+ sc+w+, а на
правом сг* ЬЬ+, и скрещиваются с самками, например, yv из линий со
сцепленными Х-хромосомами или, что хуже, со вторичным
нерасхождением. Такие самки-нерасхожденки имеют строение XyvXyvY. В случае
возникновения в сперматозоидах рентгенизированного самца делеций к
обычным категориям потомства в таком скрещивании (на схеме верхний
ряд) прибавляются единичные экземпляры новых категорий (нижний
ряд потомков) — схема XVII.
XyvXyvY*XY
Без деле- I I I j '
щий-.XyvXyvX XyvX XyvXY Xyv Y XyvYY YY
(гибнут) 9(1) 9(2) cr(3) o~(4) (гибнут)
С деле-
иией (д): XyvXyvb Xyv л XyvbY
$ (5) QT (6) cTfTf
Схема XVII
Фенотипически эти категории выглядят следующим образом:
Самки: (1) (2) — нормальные
(5) — vermilion (v)
Самцы: (3) и (4) — yellow-vermilion (yv)
(6) и (7) — vermilion (v)
Следовательно, все мухи vermilion несут делеций. Самцы (6)
бесплодны, так как лишены У-хромосомы, самки (5) и самцы (7) плодовиты и
могут быть использованы для дальнейшего размножения. Впрочем,
нередко мухи с делециями оказываются бесплодными благодаря
стерилизующим свойствам самой делеций (вернее, остатка делетированной
хромосомы).
Генетический анализ делеций сводится к тому, чтобы определить
границы или межи делеций, т. е. по возможности точно определить, какая
именно часть хромосомы оказалась выпавшей. Для этого вводят в
скрещивание с самцами, имеющими делецию, самок, гомозиготных по
рецессивным генам, локализованным вблизи предполагаемого местоположения
левой межи и правой межи. Часть сыновей такого скрещивания получит
Х-хромосому матери и делецию от отца. Оставшиеся в делеций
(делетированной хромосоме) доминантные аллели закроют соответствующие
рецессивы в материнской хромосоме, и мы увидим, таким образом,
какие именно аллели в делеций сохранились. Все же рецессивы,
находящиеся в районе делеций, будут беспрепятственно проявляться.
* Делеций и дефишенси обнаружены у многих организмов, в том числе и у растений.—
Прим. ред.
165
Пример. Самка, гомозиготная по генам у sew f cr bb, скрещена с
самцом, несущим делецию. Получены сыновья двух категорий:
1) у sc w f cr ЬЬ
2)wfcr
Сравнивая эти две категории и учитывая, что благодаря гомозигот-
ности матери различия между ее сыновьями не могут быть вызваны
перекрестом, заключаем, что делеция закрыла рецессивные аллели у, sc и ЬЬ.
Следовательно, состав делеции (остатка делетированной хромосомы)
у+ sc+ bb+. Левая межа прошла где-то между sc ц w (между нулевой и
1,5 морганидой), а правая межа между сг и ЬЬ. Вся часть хромосомы
между этими межами выпала.
Последняя часть заключения, вероятно, не вполне достоверна. Как
показали исследования Меллера (Muller, 1935), при образовании делеции
в оставшейся части хромосомы могут возникнуть мутации вблизи межи
или на некотором от нее расстоянии. Может случиться поэтому, что,
например, в случае прохождения левой межи немного правее локуса
white (w) сам этот локус даст рецессивную трансгенацию. Поэтому
строение делеции (остатка) будет у* sc? w bb+ и у мух, несущих делецию, но
потому, что он дал трансгенацию w. В нашем выводе будет ошибка —
межа прошла правее и?, а не левее. Вопрос может быть доисследован
введением в скрещивание рецессивов, ближайших к w справа, например,
facet. Если facet закроется делецией, следовательно, межа прошла правее
facet, а ген w+ дал трансгенацию w. В случае отрицательного результата
(facet проявился) все же остается сомнение, прошла ли левая межа левее
w или правее его, но между w и fa.
Совершенно убедительного генетического метода для решения этого
вопроса мы сейчас не имеем, но для дрозофилы этот вопрос точнее
решается методом Пайнтера — изучением хромосом слюнных желез.
АНАЛИЗ ИНВЕРСИЙ
Анализ инверсий сводится также к решению двух вопросов: 1)
установление наличия в хромосоме инверсий и 2) установление
местоположения меж. Характерным признаком возникновения в хромосоме инверсии
является резкое снижение в ней перекреста вплоть до полного
прекращения — по крайней мере того перекреста, который улавливается по
наблюдениям над появлением в скрещивании потомков-перекрестников
(кроссоверов).
у+ v+ 4+
Пример. Имеем самку строения -^—j- и ожидаем от нее сыновей
таких категорий:
без перекрестов О
перекрест на первом месте .... 1
перекрест на втором месте .... 2
перекрест двойной 1 . . 1—2
* А. С. Серебровский был склонен к замене иностранных терминов русскими. Термин
«перекрестник» не прижился. Обычно говорят: кроссоверные или некроссоверные
(классы особей или особи).— Прим. ред.
166
Г y+v+f+ \
X У v f ]
} У>/ \
Л yv+Г /
( У+vy \
\ У v Г /
\ У v+f f
около 51%
около 26%
около 16%
около 7%
около 49%
Рис, 44. Пара Х-хромосом
дрозофилы «Hjg
В нижнем ряду — порядок генов
нормальный (j/to+u/), в верхнем —
[инвертирован на участке,
показанном скобками
Вместо этого бесперекрестники возникают почти в 100% и лишь
изредка появляются самцы v и #/, т. е. двойные перекрестники. Перекрест
оказывается, таким образом, резко подавленным и притом весьма
оригинальным образом: в то время, как нормально двойные перекрестники
должны появляться гораздо реже одиночных, здесь одиночные исчезают
вовсе, а двойные все же появляются и иногда почти в нормальном
проценте и даже повышенном.
Объяснить это явление легко тем, что в хромосоме произошла
инверсия, и структура самки стала, например, такой, как показано на рис. 44.
При этих условиях перекрест оказывается возможным лишь при
специальном положении хромосом (рис. 45).
На рис. 45 дана упрощенная схема взаимного положения нормальной
и инвертированной хромосомы при перекресте. Фактически при этом
хромосомы находятся в более сложном положении, изгибаясь петлей, так
что конъюгируют все участки хромосом одновременно, как и точки
прикрепления нитей веретена. Подобные петли можно видеть в слюнных
железах дрозофилы. Для целей настоящего изложения достаточно, однако,
представить взаимное положение хромосом так, как на рис. 45.
При этом одиночные перекресты приводят к хромосомам совершенно
ненормального строения, например, при перекресте между v и /, как на
рис. 45 внизу.
В одной из таких перекрестных хромосом имеются лишние участки
(в районах у и у+), а в другой — большие нехватки (нет ни у, ни у*).
Кроме того, неправильно распределяются и нити веретена, и хромосомы
оказываются нежизнеспособными. Что касается двойных перекрестов
(например, левее и правее #), то они дают хромосомы столь же нормаль-
лого строения, как и до перекреста (рис. 46). Так как все одиночные
перекресты погибают, то среди оставшихся классов бесперекрестников и
двойных перекрестников процент последних может оказаться
(кажущимся образом) нормальным или даже повышенным.
Таким образом, на наличие инверсии указывает уменьшение
перекрестов, особенно за счет одиночных перекрестов. При постановке диагноза
приходится, однако, принимать во внимание следующее.
1. Уменьшение перекреста может вызываться не только инверсией,
по и другими генетическими факторами. Так, например, известен
открытый Гоуэном рецессивный трансген в 777 хромосоме, прекращающий
в гомозиготном состоянии перекрест во всех хромосомах. Присоединение
к хромосоме транслокации тоже иногда заметно понижает перекрест.
В этих случаях перекрест, очевидно, вообще затрудняется, почему
двойные перекресты становятся особенно редкими. Отношение двойных
перекрестов к одиночным является поэтому очень важным
диагностическим признаком.
2. При наличии инверсии одиночные перекресты или во всяком случае
особи, которых можно принять за одиночных перекрестников, не всегда
отсутствуют. Во-первых, перекрест идет и вне районов инверсии
!/scw
н+
9**
тг
wT
167
Рис. 45. Схема одиночного
перекреста с участием
инвертированной хромосомы;
возникают «негодные» хромосомы
Рис. 46. Схема двойного
перекреста с участием
инвертированной хромосомы; «негодные»
хромосомы не возникают^
Рис. 47. Схема взаимного
положения хромосом при
перекресте в районе инверсии
(вверху) и вне его (внизу)
Пунктиром отмечены районы &
возможностью перекреста
Рис. 48. Одиночный перекрест
между нормальной хромосомой
и инвертированной (перекрест
вне района инверсии)
У* U
У
y+ kf* y* i,
I
I I I I I I
-»—h-
+ f
r
l2
H *
н—h
I 1 1 —I—I— I 1 1 -H h-
Летальна Лщпальна
Летально Нелетальнп
*r-fc
<Г± v+ VT у
Рис. 49. Схема одиночного и двойного] перекреста с участием двух деталей,
имитирующих участие инверсии]
(рис. 47), и здесь одиночные перекресты дают хромосомы тоже нормаль
ного строения, как и двойные перекресты в инвертированной хромосоме.
3. В некоторых случаях картина, сходная с прекращением перекреста,
может получиться при возникновении в хромосоме (половой) сразу двух
язталей (рис. 48 и 49).
Любой одиночный перекрест между Zx — Z2 даст всегда хромосому,,
несущую леталь, и если мы изучаем перекрест в половой хромосоме,
наблюдая по самцам-сыновьям, то такие одиночные перекрестники не будут
появляться. Двойной перекрест в районе между леталями будет давать
жизнеспособных перекрестников. Таким образом, будем получать: 1) без-
перекрестников обычного типа (с l[ — Z2), 2) одиночных перекрестников
вне участка 1г — Z2 и 3) двойных перекрестников в районе lx — Z2 и
притом относительно в повышенном проценте. Это повышение процента
двойных перекрестов будет основано на том, что общее число потомков
будет уменьшено, благодаря гибели всех одиночных перекрестников. Таким
образом, картина будет в точности повторять то, что мы имеем при
инверсии, в частности, в том очень обычном случае, когда инверсия
сопровождается летальностью.
Отличить эту «псевдоинверсию» можно, во-первых, потому, что среди
гомозиготного пола (самок у дрозофилы), где летали не будут проявлять
ся, перекрестники одиночные и двойные будут в нормальном
соотношении, как и вообще все классы перекреста, во-вторых, потому, что дочери
этих самок, получившие при перекресте лишь одну из деталей, дадут
обычную для летали картину и позволят понять этот оригинальный
случай.
4. Очень трудны для диагноза случаи коротких инверсий, когда в
этом инвертированном участке двойные перекресты не имеют шансов
произойти. Тогда мы наблюдаем лишь уменьшение процента перекреста
с сохранением перекреста вне этого района. Если отсутствуют двойные
перекресты, отсутствует и характерный диагностический признак и
остается возможность сомневаться, произошло ли изменение в этом
участке благодаря наличию здесь транслоканта или по какой-либо иной
причине. Возможно, что известные нередко случаи изменения процента
перекреста являются случаями недоказанных инверсий.
Вторая задача анализа инверсии — установление местоположения
меж — решается генетическим анализом с большим трудом и с меньшей
точностью, чем по отношению к делециям. Аргументы для выводов
черпаются из тех же данных по перекресту, как и доказательства самого
наличия инверсий. Если некоторая аллель может войти в анализируемую
хромосому при одиночном перекресте, то локус этой аллели лежит вне
инверсии. Если это введение может быть осуществлено только двойным
перекрестом, то локус лежит в области инверсии.
169
Пример. При разведении известной инверсии ВС1 (или С1В) в такой
•структуре самок (на данной схеме расположение генов показано
нормальным)
у+ sc w+ v в
i i i 1 1
< i i i —-j
у 5С+ W У+ В
изредка наблюдается соединение с Ваг генов у, sc+ и w. Это соединение
происходит в порядке одиночного перекреста. Наоборот, v и v* меняются
местами лишь в порядке двойного перекреста. Отсюда заключаем, что
левая межа инверсии прошла между w и v, так что локус w остался вне
инверсии, а локус v вошел в инверсию.
Трудности установления точного местоположения межи инверсии
связаны с тем, что в окрестностях межи перекрест часто вообще очень
затруднен. Перекрестники появляются очень редко, и гены, близко
расположенные к меже, почти вовсе не вовлекаются в перекрест.
Тем более неточно определяется это положение межи, если
поблизости от нее нет удобных для экспериментов локусов. В случае дрозофилы,
однако, мы имеем теперь блестящий метод (Пайнтера) *, пользуясь
которым можно устанавливать положение меж с величайшей точностью
(порядка 1/2оо~~1/4со длины хромосомы). Обычным (не пайнтеровским)
цитологическим наблюдением инверсии могут быть установлены лишь в
исключительных случаях, когда в инверсии оказываются точки
прикрепления нити веретена и притом не в центре инверсии. Тогда при
сравнении инвертированной хромосомы с нормальной кажется, будто у нее
переместилась точка прикрепления нити веретена.
Значительно повысить точность определения положения меж инверсии
ложно в том случае, если в распоряжении генетика имеется несколько
различных инверсий. Располагая двумя такими различными инверсиями,
можно получить искусственные нехватки, что было одновременно
проделано Меллером (Muller, 1930) и нами при изучении инверсий sc9 и i/4
(не опубликовано).
Пусть мы имеем две различные инверсии с таким порядком генов:
ABCIHGFEDIJKL
ABIGFEDCIHJKL
Получив особь, гомозиготную по двум таким инверсиям, мы получаем
затем перекрестные хромосомы при помощи одиночного перекреста в
области инверсии (например, между F и Е):
AB/GFED/JKL
ABC/HGFEDC/HJKL
В верхней хромосоме имеются две нехватки, так как отсутствует Н и
отсутствует С, т. е. состав хромосомы будет:
АВ - DEFG — JKL
Нам остается подвергнуть анализу протяжение этих нехваток
описанным выше приемом проявлений рецессивов. Очевидно, что длина С-
,нехватки равна расстоянию между левыми межами обеих инверсий,
а длина //-нехватки — расстоянию между правыми межами. Таким
приемом можно определить местоположение меж довольно точно. Опасность
"* Имеется в виду изучение хромосомных нарушений с помощью цитологического
метода гигантских хромосом слюнных желез (на дрозофиле).— Прим. ред.
170
«сделать ошибку, приняв за область нехватки локус, захваченный
[одновременно возникшей вблизи межи мутацией, о чем шла речь при анализе
делеции, здесь уменьшена, так как нет шансов, чтобы в обеих несколько
различных инверсиях добавочно мутировали бы одни и те же локусы.
Этот метод, не получивший еще широкого распространения, имеет
большие перспективы, так как позволяет путем последовательного получения
искусственных нехваток подвергать анализу и такие
участки хромосом, которые еще не дали мутаций. Мы получаем возможность
тсак бы вырезать из хромосомы узкие «ломтики» и, изучая фенотипический
эффект таких заведомых отсутствий, определить физиологическое
действие этих «ломтиков» в нормальной хромосоме.
АНАЛИЗ ТРАНС ЛОКАЦИЙ
При анализе транслокаций приходится решать следующие вопросы:
1) установление наличия транслокации, 2) между какими хромосомами
произошел обмен участками, 3) местоположение разрывов (ыеж) и
прикреплений транслокаций.
Основным признаком происшедшей транслокации является изменение
геносвязей и притом, что наиболее характерно, таким образом, что гены,
бывшие ранее независимыми, начинают давать картину сцепления друг
с другом.
Этим феноменом пользуются при отыскании транслокаций в
новейших опытах по рентгеномутациям. Для опыта берут самцов, полученных
от рентгенизованного отца и являющихся гетерозиготами по генам,
расположенным в разных парах хромосом. Обычно пользуются для этой
цели двумя доминантными генами, например, в II ж III хромосомах.
В нормальном хромосомном аппарате расщепление идет следующим
образом:
W-
(2)-
(3)-
Гаметы самца
А
В результате образования таких четырех типов гамет возникают и
четыре типа потомков Л, В; А, Ъ\ а, В и а, Ь, примерно в равных
количествах, иллюстрируя независимое комбинирование генов А и J5,
расположенных в разных негомологичных хромосомах.
Пусть теперь хромосомы самца будут нести транслокацию таким
образом, что куски II и III хромосом обменялись участками аир (межа
показана черточкой):
/3 | А а I В
, _ _ _ __ ^
171
Тогда возникнут гаметы (тоже в числе четырех типов, благодаря
отсутствию у самцов дрозофил перекреста) такого состава:
(О
(2)
(3)
t
ОС
ОС
А
а
а
а
13
В
b
а | В
Л b
ocj
2/3
2сс
ос,р
Всматриваясь в эти типы гамет, замечаем, что типы (2) и (3)
отличаются нарушением баланса генов. В типе (2) нехватает куска а (гипопло-
идия), a (J представлен два раза (гиперплоидия). В типе (3), наоборот,
не хватает |3 и имеется лишняя а. Очень часто случается, что эта гипо-
гиперплоидность оказывается столь тяжелым нарушением генотипа, что
зиготы, получившие эти гаметы, погибают (замечается гибель 50% яиц)
и выживают только зиготы, возникшие от гамет 1 и 4-го типов, где
баланс генов не нарушен и произошло лишь перераспределение генов между
хромосомами. В результате этой дифференциальной смертности
возникают зиготы А, В и а, Ь, т. е. гены А и В как бы проявляют полное
сцепление друг с другом. Иными словами, возникает кажущаяся картина
того, что хромосомы II и III сцепились в одну. Сколько бы генов в этих
хромосомах мы не изучали,^ картина «полного сцепления» их друг с другом
будет одна и та же.
Если зиготы от гамет (2) и (3) гибнут не нацело или гибнет из них лишь
один тип (2) или (3), то мы получим картину неполного сцепления. Это
может служить указанием на то, что один из обменных участков очень
мал и гипоплоидность по нему отражается на жизнеспособности
подобно, например, рецессивному летальному гену. Если таков, например |3,
то мы будем иметь зиготы Л, В (33%); а, В (33%) и а, Ъ (33%). Вычисляя,
получим между А и В процент «перекреста» 33%.
Если в гетерозиготном состоянии транслокационные нарушения
(нехватки и удвойки) не нарушают заметно жизнеспособности, то это может
обнаружиться во втором поколении. Получив от особи (1) типа F2,
получим (оставляя временно без внимания возможность перекреста у
самок):
$AolB
рлр&
oluolB
<xa$b
3*АаБ
Л, В
«,РЗ
л, в
а, Р
Л, В
«. Р
а, в
а, Р
/ЗА/ЗЬ
А, В
а. р
А, Ъ
Р, Р.
А, В
«, Р
А, Ь
«. Р
аааВ
А В
а, р
А, В
а, р
а, В
а, а
а, В
а, Р
aajSb
А, В
а, Р
А, Ь
а, Р
я, В
а, Р
а, Ъ
а, р
Греческими буквами отмечена лишь гомо- или гетеровиготность зигот по нехватками
172
до транс- А в с #
локации '
п С BAD
После н—| 1 —Н ь
С L В А D
-и—| i —Н ь
ч-
а be
с . В А ,
н— £— --Н-
2 -Н 1 1 -Н Н
* b A x В
3 —l 1 1 1 I
a be d
'> —f 1 н h-
Рис. 50. Схема образования гамет при наличии обменной
транслокации А ж С
1—4 — типы гамет
В двух средних клетках большой диагонали мы имеем особей, уже
гомозиготных по нехваткам. Такие особи погибают почти наверное и мы
ммеем в потомстве
94, Я; 2А,Ь; 2а, В; 1а,Ь.
Процент иерекомбинированных А и В равен всего 4/i4-100% = 28,6%
вместо полагающихся для независимого расщепления 6/16 • 100 % = 37,5 %.
Следовательно, гены Л и В и здесь обнаруживают некоторое «сцепление».
Вычисление коэффициента ассоциации дает
п_ (9Х1)-(2Х2) _ 5 лоос
V - (9Х1) + (2Х2) - 13 - ">6ЬЬ*
что отвечает расстоянию от А до В приблизительно в 35 морганид.
Наличие перекреста у самок будет ослаблять эту картину сцепления
и при далеком расстоянии генов А и В от меж сделает его совсем
незаметным. Эта картина сцепления, конечно, ложная, отражающая лишь
дифференциальную жизнеспособность особей со сбалансированным и с
несбалансированным генотипом. Однако если в перенесенном участке
гимелся какой-либо сигнальный ген, то можно обнаружить уже настоящее
♦сцепление генов, принадлежавших ранее к различным хромосомам.
Следующая схема пояснит сказанное. Пусть первоначально имелись
хромосомы с двумя генами АяВ во II яСжБв III хромосомах. В
результате транслокации они переместились: А в хромосому /// и С в хромосому
// (рис. 50).
Рассчитывая гаметы, как ранее, получаем четыре типа (1 — 4 на
рис. 50).
При условии полной жизнеспособности всех типов имеем в 50% СВ и
J5, в 50% cb или Ь, и также в 50% AD и в 50% ad или d, т. е. гены С и В
оказываются сцепленными, как А и/), а бывшие ранее сцепленными А и
В или С vlD теперь наследуются независимо. При больших транслокациях
и нежизнеспособности картина еще осложняется, но возникновение
новых сцеплений всегда ясно.
Таким образом, транслокация оказывается установленной, и мы
получаем указание на то, какие именно хромосомы оказались связанными
транслокациями, хотя не всегда легко узнать, какиче концы оказались
перемещенными.
173
27++Я
b++B
Рис. 51. Схема
[расположения хромосом
при конъюгации с
наличием обменной
транслокации
ОПРЕДЕЛЕНИЕ МЕЖ ТРАНСЛОКАЦИЙ
В простейшем случае, как у дрозофилы и у
шелкопряда, когда у одного пола отсутствует перекрест,
исследование вопроса о том, какие именно локусы
попали в транслокацию, можно вести, вводя в
скрещивание различные рецессивы в той области,
которая ориентировочно определена, пока не будет
обнаружено, что из двух близко расположенных по
плану генов один изменил свое сцепление, а другой не
изменил его. Межа, следовательно, проходит где-то
между ними.
Этим методом можно вести наблюдение, однако,
лишь тогда, когда несбалансированные генотипы
не летальны. В частном случае их летальности,
когда по расщеплению кажется, что обе хромосомы
сцепились, этот метод не дает возможности решить,,
переместился ли ген из одной хромосомы в другую
или нет. В таком случае необходимо вести
наблюдение за перекрестом между генами,
расположенными в обеих связанных трансклокацией хромосомах,
вводя в опыт гены, локализованные в транслокациях
и вне ее.
Для понимания картин, наблюдающихся при расщеплениях с участием
транслоцированных хромосом, необходимо рассмотреть ход перекреста
в этих условиях. В настоящее время, к сожалению, детали этого вопроса
изучены еще недостаточно, но основные его особенности выяснены. В
гомозиготных особях, где обе гомологичные хромосомы имеют одинаковое-
строение, перекрест не имеет особенностей, но у гетерозигот, у которых,
следовательно, две хромосомы нормальные, а две связанные
транслокацией, перекрест может идти очень оригинально благодаря возникновению*
в мейозе «колец» из четырех хромосом. Такие кольца возникают потому,,
что хромосома, участвующая в транрлокации, составленная, например,
из части II и части III хромосомы, может одновременно конъюгировать
с гомологичными частями обеих Этих хромосом. Рис, 51 показывает
возможное расположение при конъюгации четырех хромосом в этих
условиях. Мы видим, что несмотря на произошедший обмен участками, все-
хромосомы лежат так, что гомологичные части их лежат рядом, могут
конъюгировать и давать перекрест.
При расстоянии от А до межи ЪА и от D до межи ЪСв (в долях единицы)
эти четыре типа хромосом возникнут с частотами, как показано в правой
графе, принимая для простоты отсутствие интерференции.
Если проследить, например, за хромосомой Л—D, то нетрудна
понять, что из конъюгации она может выйти с четырьмя различными
видоизменениями своего состава
С частотой
С а ~САС
AUD
CD — CACI>
b'Avl>
1. Без перекреста в составе . . . . Л — D
2. С перекрестом между А и межой а — D
3. С перекрестом между межой и D А — d
4. С обоими перекрестами • ... a — d cAc
Наблюдаемое расстояние А — D будет, следовательноу
[AD] = {cA +cD -2cAcD)A00t
или, считая 100 СА = тА и 100 cD = mD,
[AD] = mA + mD — —mAmD.
A*D
174
Также четыре типа будет после конъюгации и у других хромосом.
В том случае, когда сохраняются лишь сбалансированные генотипы*
совершенно такая же картина сцепления оказывается между всеми
четырьмя аллопарами и, например, «расстояние» между АиВ будет также
равно:
[АВ] = тА + тв — -gg- тА тв.
В результате мы получаем систему уравнений:
1) [АВ] = тА+тв — 0,02 тАтв\
2) [АС] = тА + тс — 0,02 тАтс;
3) [AD] = mA + mD — 0,02 mAmD;
4) [ВС] = тв+тс — 0,02 твтс;
5} [BD] = mB + mD — 0,02 mBmD*
6) [CD] = mc + mD-+ 0,02 mcmD.
Так как величины [АВ] = а; ]АС] = Ь; [AD] = d и т, д. находятся
из опыта, то остаются четыре неизвестных тА, тв, mD и тс,
соответственно х, у, z и и;, которые легко могут быть найдены.
Это решение, как и вывод формул, основано на предположениях, что-
перекрест идет без каких-либо местных изменений и интерференция
отсутствует. В действительности интерференция, особенно при малых
расстояниях, имеет немалое значение.
Практически проверяя эти формулы на одной из транслокаций,
полученных Стертевантом и Добржанским (Sturtevant a. Dobzhansky, 1931),
мы нашли, что гораздо более близкие результаты получаются при
использовании формул такого более простого вида:
а = х + у — ху
Ъ = х + z — xz
d = У + z — yz, ■
Таблица 11
Перекрест с участием транслокации Е и полученное потомство
Без перекреста между Е и Sb
{ Sb
\ al dp b pr с
Г al Sb
\ dp b pr с
( al dp Sb
2 \b pr с
(cSb
4 \ al dp b pr
J al с Sb
4'4\ dpb pr
t Г al dp с Sb
2Л\Ърг
566
зоз
13
8
9
9
203
133
4
3
5
6
1262
'
С перекрестом между Е
0 1 +
и \ al dp b ре е Sb
1 I al
\ dp b pr с Sb
J al dp
2* | b pr с Sb
4 I'
\ al dp b pr Sb
J al b pr с Sb
U2\dp
1,4 1 dp b pr Sb
, Г al dp с
2>*{bprSb
2
и Sb
164
87
2
4
5
6
47
41
2
0
3
5
4
0
370
= 1632
175
где все величины выражены не в процентах перекреста или морганидах,
«а в долях единицы, т. е. принимая 100% = 1. Решая эту систему
уравнений, приходим к такому квадратному уравнению:
я2 — 2х + R = 0;
тде
Отсюда
Я =
d — ab
1-d
x = l±Yl—R.
Стертевантом и Добржанским (Sturtevant a. Dobzhansky, 1931) при
изучении перекреста, с участием транслокации Е, связывающей // и /77
.хромосомы, были получены результаты, приведенные в табл. 11.
Подсчитывая перекресты между отдельными генами, получаем:
Варианты
1. al — dp
2. dp — Ъ
3. Ъ — pr
4. pr — с
5. dp— с
6. al — с
Число
перекрестов
44
46
0
454
470
480
с%
2,7
2,8
0
27,8
28,8
29,4
| с
0,027
0,028
0,000
0,278
0,288
0,294
Варианты
7. al — Sb
8. dp — Sb
9. b — Sb
1Q. c — Sb
11. a/ — b
Число
перекрестов
398
382
370
714
86
c%
24,4
23,4
22,7
43,7
5,3
с
0,244
0,234
0,227
0,437
0,053
Если расположить эти гены на плане хромосом (схема XVIII), то ера-
;зу~станет ясна непригодность линейной схемы. Например, гены dp, С л
■U-+
4-
af dp 2
+
4-»
+
а1 dp b pr
4-
Схема XVIII
Sb связаны расстояниями (в перекрестах) так: Sb— с (самое большое) =
= 43,7%, &dp — с = 28,8% ирг —Sb = 23,4%, как бы образуют такой
1>яд:
Sb-
dp
"~v—
—v—
47,7
28JB
JHo ген al также образует с dp ж с ряд
al . dp -
-V—
2J
28,8
29£
176
Отсюда следовало бы предположить порядок
Sb-
al
dp
•с.
при котором ген al должен давать с Sb примерно на 2,5% меньше
перекреста, чем dp — Sb, так как ближайшим с Sb геном оказывается al. На самом
деле, однако, расстояние al — Sb равняется 24,4, т. е. больше чем Sb — dp.
Еще ярче это противоречие видно на генах sb — Ъ — с ж al — Ъ — с.
Обе эти тройки также укладываются в ряды:
Sb-
22,7
-~v—
43J
218
al
5,3
v
29,4
218
Отсюда опять порядок генов должен быть Sb — al — b — с и
расстояние Sb — al должно быть примерно на 5,0 меньше, чем Sb — Ь. На самом
деле, как мы видели, Sb — al равняется 24,4, a Sb — Ъ = 22,7, т. е.
обратное отношение. Мы видим, следовательно, интервсное явление, что Sb
находится ближе всего к Ь, но не может быть расположен ни вправо от
него, ни влево.
Очевидно, должна быть применена крестообразная схема с
перекрестом в месте межи транслокации. Тогда все отношения становятся
понятными (рис. 52).
Расстояние al — Sb есть сумма расстояний: (al — межа) + (межа — Sb);
расстояние с — Sb есть сумма расстояний: (с — межа) 4- (межа — Sb)
и т. д.
j-Sh
ч—н
al dp
+Sb
t
*e
22,2
tfi 27j
H
Рис. 52. Построение крестообразного плана двух пар хромосом с обменной
транслокацией между ними
Фактически эти расстояния несколько меньше, чем указанные суммы,
благодаря двойным перекрестам.
Попробуем теперь решить такую задачу, пользуясь данными о
расстояниях dp — *УЬ, dp — с и Sb — с, найти место «пересечения» хромосом, как
будто нам неизвестно, что это пересечение близко к Ь.
12 А. С. Серебровский £77
Строим систему уравнений (а = 0,234; Ь = 0,288; d = 0,437), как
указано выше:
х + у — ху = 0,234;
х + z — xz = 0,288;
У + z — yz = 0,437.
Отсюда
D 0,234 + 0,288 — 0,437 - 0,234 — 0,288 n nQ9
R = i-0,437 = U'UdZ#
Наше уравнение приобретает вид:
я2 — 2х + 0,032 = 0;
откуда
х = 1 ± f\ -0,032
или
хг = 1,984,
х2 = 0,016;
хг — лишено смысла (198,4% перекреста), и, следовательно, х2 = 0,016,
или 1,6%. Подставляя значение х в остальные уравнения, находим:
у = 0,222 и z = 0,277 (рис. 52).
Так как межа расположена где-то около Ъ (перекрест в этой области
очень затруднен), то полученные решения очень близки к правильным,
особенно, если принять во внимание не очень большой материал,
положенный в основание экспериментального определения величин а, Ь и d.
К сожалению, как в данном случае, так и во многих других
возникновение транслокаций сильно понижает процент перекреста в соответствующей
половине хромосомы, и это положение делает невозможным достаточно
точно указать на нормальном плане хромосомы место, в котором находится
межа. Например, в разбираемом примере на огромном расстоянии между
dp — с перекрест составляет всего около 30 %. Если разделить нормальное
расстояние dp — с, равное 62 морганидам, пропорционально вычисленным
нами величинам хж у, то мы получим положение межи:
п 1,6
1,62,5 1,6 + 27,7'
1,6-62,5 о ,,
П= 29,3 =3'41'
т. е. положение межи на 3,41 морганиды правее dp, а всего на 15,13 морга-
ниды, т. е. очень далеко от места black (48,5). Эта невязка происходит
только потому, что благодаря транс локации вся половина хромосомы
al — рг в 54,5 морганиды длины дает всего лишь 5,3% перекреста, т. е.
раз в восемь меньше нормы. Если бы мы располагали данными лишь о трех
генах Sbdpc, то мы не могли бы знать, где именно понижен перекрест и
наше решение нельзя было бы переносить на нормальный план.
Детальным анализом транслокаций занимались еще очень мало,
особенно с интересующей нас здесь точки зрения. К тому же основным
объектом этого изучения служила и служит дрозофила, но для этого объекта мы
имеем в настоящее время метод Пайнтера, дающий возможность многие
вопросы решать быстрее и точнее цитологическим исследованием слюнных
желез. Поэтому более детально на методе анализа транслокации мы
останавливаться не будем.
178
+ Типы гамет Типы зигот Фетттипь/
1 1 —С 1 •
-*7
2 1 t
5
1 £_ ZLZ .Л
+
4
b
i
—rb
Рис. 53. Схема скрещивания с участием дупликации Ь+
Объяснение в тексте
АНАЛИЗ ДУПЛИКАЦИИ (УДВОЕНИЙ)
Дупликацией (удвоением) является участок хромосомы,
перенесенный при транслокации со своего нормального места в другое место той же
или другой хромосомы. Если транслокант не лишает организм
жизнеспособности, то после ряда скрещиваний хромосома, от которой транслокант
оторван и где получилась соответствующая нехватка, заменяется
нормальной хромосомой. В результате мы получаем в общем нормальный
хромосомный аппарат, но один участок его оказывается удвоенным и «удвойка»
сидит в качестве добавки к нормально расположенному участку, где-либо
в другом месте другой или той же хромосомы.
Характерным признаком удвоения является образование
«подавителя», т. е. гена, подавляющего действие известного раннее рецессивного
гена, или как бы перемещение известного ранее гена в другое место с
изменением доминирования. Если, например, кусок нормальной хромосомы
с нормальным аллеломорфом black (Ь+) переместился в 777" хромосому и
там будет находиться в следующих поколениях как удвойка, то обратное
скрещивание пойдет так, как показано на рис. 53.
Ген Ъ может проявиться лишь в четвертой категории зигот, так как
в (1) и (2) Ъ находится в гетерозиготном состоянии, а в (3) хотя он
гомозиготен (ЬЬ), но его подавит нормальный аллель, сидящий в III хромосоме.
Вместо 1 : 1 расщепление идет в отношении 3:1.
b b+ b с
. 1 —* J . ' 1 i_
i i + \ 1
b с b £
Типы гамет Типы зигот Фенотипы
Ь b+ Ь Ь+
5 "- —* ' —^ +
В*
УЬ £^
Г"' 1 г- **
b с
С
Рис. 54. Схема скрещивания с участием дупликации Ь+ и сигнали с
(продолжение рис. 53)
179
12*
b* b b
_i с v i
т ~s
Типы гамет Типы зигот Фенотипы
Ь* Ъ Ь+ b
-J —* ' < Ь
о й?
-L
а
—г
Ъ
Рис. 55. Схема скрещивания с участием дупликации &+ при
доминировании двух Ь над Ь+
Для того чтобы выяснить причину этого искажения менделевского
отношения, следует маркировать сигнальными генами вдобавок и другие
хромосомы и вести дальнейшее изучение (3) категории, отыскав ее среди
других последовательными скрещиваниями с этими сигналенными
линиями (добиваясь гомозиготности по 6). В потомстве тип (3) будет вести себя
следующим образом (рис. 54).
Тип (6) зигот проявит признак 6, а (5) не проявит. Вместе с тем (6)
проявит и признак с, а (5) не проявит его. Следовательно, мы получим картину
того, что ген Ъ как бы переместился в хромосому к С+, тогда как если в
хромосоме, действительно несущей 6, имеются другие гены, они не будут
обнаруживать сцепления с сигналью с+.
Взяв для исследования и тип (2), мы получаем уже иную картину
Ъ+ с+ Ь с
X
Ъ с Ъ с
Здесь, очевидно, будет обычное правильное расщепление по Ъ+ и 6, и
признак Ъ будет свободно комбинироваться с признаком с.
Таким образом, беря для исследования различных особей из
расщепления, будем получать локализацию гена то в одной хромосоме, то в
другой.
Локализация самого удвоения особых затруднений не представляет.
Совершенно такую же картину мы будем иметь, если в третьей
хромосоме возникнет просто аналогичный ген, тормозящий проявление данного
рецессива. Различение этих случаев с полной убедительностью почти
невозможно, кроме тех случаев, когда в удвоении сразу оказывается два,
а тем более два рядом лежащих гена. В этом случае сильно возрастает
вероятность того, что мы имеем дело с удвоением, а не с гомологичными
«тормозящими» генами (например, случаи Sable vermilion — удвоения,
изученного Бриджесом (Bridges, 1919).
Весьма оригинальной и трудной для анализа задачей является
случай, когда две рецессивные аллели не подавляются доминантом (например,
в случае vermilion, Bridges, 1919) и когда в удвоенном участке хромосомы
находится рецессивная аллель. В этом случае скрещивание пойдет по
схеме рис. 55.
Из скрещенных особей одна (b+ b+ Ъ) не проявляет признака 6, а
другая (bb) проявляет.
Из двух типов дочерних зигот один (1) проявит 6, так как Ь+ не
подавляет двух 6, а (2) не проявит. Полученное отношение 1 : 1 говорит за
гетерозиготность исходного родителя, не проявляющего признака 6,
так что ему можно дать формулу ВЪ как и (2), а (1) — формулу bb. Однако,
180
ъ*
\
ь
1_
b
L_
ь*
т
ь
Типы
>
гамет
Ъ
b
< _
ь*
~Ъ.
ПС
,ь
ь
1
ь
Типы зигот
Ъ
^
Ь
Фенотипы
0
+
Ь
" b
Рис. 56. То же, что на рис. 55, но иное скрещивание
если мы скрестим (2), хотя бы с ЪЪ родителем, то получим
противоречащий результат (рис. 56).
В результате скрещивания наблюдается совершенно неожиданное
расщепление и появятся нормальные особи, у которых при дальнейшем
исследовании не будет обнаружено ничего ненормального. Среди особей
^-фенотипа часть (1/3) снова даст такую же картину расщепления в
отношении 1 : 3. Наличие этого отношения указывает на дигибридность,
поэтому дальнейшее исследование надо вести как при дигибридном
расщеплении. В дальнейшие детали этого анализа мы входить не будем*
ГЛАВА VIII
ГЕНЕТИЧЕСКИЙ АНАЛИЗ ГЕНОФОНДА ПОПУЛЯЦИЙ
ТРЕТЬЯ ЗАДАЧА ГЕНЕТИЧЕСКОГО АНАЛИЗА
Третьей основной задачей генанализа является выяснение строения
генофонда, т. е. той совокупности генов, которая находится у особей,
составляющих некоторую популяцию. Такой популяцией может быть
отдельное стадо, группа особей, занимающая административную или
национальную территорию, порода, кряж пушных зверей, наконец, вид *.
В состав популяции входят всегда особи с различными генотипами, и
перед генетиком возникают задачи выяснить и учесть это многообразие,
охарактеризовать популяцию и ее генофонд в целом.
Отметим разницу в постановке этого вопроса и вопроса о составлении
топографического плана хотя бы хромосом курицы. В последнем случае
мы привлекаем к нашему исследованию, конечно, не одну птицу, но
тоже множество птиц, скрещиваем различные породы и т. д. Тем не менее
в результате нашей работы мы получаем план хромосом, который дает
нам характеристику любого отдельного животного. Мы узнаем, что в
половой хромосоме у него расположены четыре гена (в виде той или иной
аллели), нам известные. Один — влияющий на скорость оперения, два—
на окраску оперения, один — на окраску ног. Самое большое, что при
этом мы узнаем для характеристики совокупности кур, это то, что у
различных особей этой совокупнЬсти хромосомы могут различаться аллелями:
у одних имеются Suke, у других asuke и т. д., и что при наличии тг-генов
возможно существование 2П комбинаций. Мы узнаем, так сказать,
потенциальное многообразие этой совокупности, но мы ничего не узнаем еще о
фактическом многообразии. Зная, например, возможность возникновения
миллиона различных окрасок и существование, например, «ненасиживаю-
щего» гепотипа, мы остаемся в неведении того, сочетались ли когда-либо
эти окраски с ненасиживанием или нет.
Для характеристики генофонда и популяции, его несущей, нужно
узнать: 1) список генов, входящих в данный генофонд, что, конечно
совпадает со списком генов данного вида в целом, 2) концентрации всех аллелей
или относительные частоты, с которыми аллели входят в генофонд, 3)
характер комбинирования генов, осуществляющийся в данной популяции,
4) характер распределения концентраций аллелей по территории или по
другим элементам популяции, 5) общее строение популяции, разумея
например, ее однородность или ее возникновение в результате бывшего
ранее или идущего в настоящее время смешения из двух или нескольких
* Проводить знак равенства между популяцией и видом не следует. Популяция —
систематическая категория ниже вида, обычно состоящего из многих популяций.
Случаи, когда вид состоит из одной популяции, очень редки.— Прим. ред.
182
популяций, и 6) дополнительно — характер мутационного процесса,
идущего в популяции.
Первый вопрос в данном разделе не отличается от того, который мы
рассматривали при анализе генотипа особи.
Понятие о концентрации гена в популяции мы тоже рассматривали
выше и также познакомились с методами вычисления р я q я если нужно г
и др. Поэтому здесь нам необходимо остановиться лишь на методических
указаниях по сбору материала.
Сбор материала
Для получения правильных данных прежде всего необходимо
заботиться о том, чтобы обследованные животные правильно отражали популяцию.
В том случае, когда популяция может быть описана целиком, опасность
искажения отпадает. Но, как правило, мы делаем для описания выборки
из популяции и необходимо заботиться о том, чтобы выборки отражали
популяцию правильно. Это связано, во-первых, с вопросом о величине
выборки, и, во-вторых, с тем, чтобы выборки не сопровождались отбором,
хотя бы и бессознательным. Такой отбор может произойти в том случае,
если мы станем, например, искать «типичных животных», отбрасывая
«не типичных», или если станем обращать внимание на «интересных»
животных, или, наконец, не учтем того, что по особенностям признаков одни
животные легче попадают в поле нашего внимания, чем другие.
Рассматривая, например, издали кур, бродящих по поселку, мы будем легче
замечать белых и пропускать окрашенных в дикий покровительственный
окрас. В результате мы можем дать повышенную оценку процента белых
кур.
Точно так же редкие признаки привлекают наше внимание больше,
чем обычные. Мы постоянно встречались с совершенно преувеличенными
представлениями у местных наблюдателей о проценте кур голошеек или
бесхвосток: признаки эти производят впечатление часто встречающихся,
тогда как более правильное исследование обнаруживает большую
редкость этих генов по сравнению с другими, менее заметными.
Поэтому при анализе популяции необходимо, учтя особенности
данного вида животных, организовать осмотр особей так, чтобы исключить
возможность хотя бы бессознательного выбора. Ловя, например, кур для
описания, нужно вылавливать всю попавшую в поле зрения группу, но отнюдь
не допускать усиленного преследования брлее заинтересовавших. Если же
тем не менее некоторых особенно интересных животных приходится
добывать специальными усилиями, то их следует учитывать отдельно, не вводя
в основную выборку, взятую для характеристики популяции.
Несоблюдением этих требований были сильно испорчены выводы многих
исследований, которые в результате способны быть в большей степени материалом
для характеристики интересов и образа мыслей обследователя, чем для
объективных особенностей популяции. Лучше всего пользоваться для
производства выборки какими-либо объективными, случайными в
отношении признаков показателями, например, четными и нечетными номерами
животных, алфавитом фамилий владельцев, брать каждого третьего или
каждого десятого из случайно подводимых и тому подобными приемами.
Нужно помнить, что от продуманности этого вопроса зависит результат
всей работы.
Для тех генов, у которых разница р и q не слишком велика, можно
ограничиться простым описанием фенотипов достаточно большого
количества наудачу выбранных животных, так как здесь мы можем встретить
как особей с доминантной аллелью, так и q2 гомозиготных рецессивов.
Если q велико и р мало, т. е. доминантная аллель редко встречается, то
183
необходимо лишь увеличить описываемый материал. Гораздо хуже
обстоит дело в случаях, если q мало. В этом случае q2 вовсе мало, и
гомозиготные рецессивы почти не имеют шансов быть встреченными.
Следующая таблица иллюстрирует это различие между доминантами и
рецессивами.
я.
0,01
0,05
0,10
0,30
0,50
V
0,99
0,95
0,90
0,70
0,50
Процент в популяции II
фенотипов
рецессивных
0,01
0,25
1,00
9,00
25,00
доминантных
II
99,99 I
99,75
99,00
91,00
75,00 II
1
Q
0,70
0,90
0,95
0,99
Р
0,30
0,10
0,05
0,01
Процент в популяции
фенотипов
рецессивных
49,00
81,00
90,25
98,01
доминантных
51,00
19,00
9,75
| 1,99
При q = 0,05, или 5%, даже большая популяция может состоять из
одних доминантных фенотипов и производить впечатление гомозиготной.
При тех же 5% доминантной аллели (р = 0,05) в популяции почти 10%
доминантных фенотипов.
Так как многие рецессивные гены могут находиться в популяции в
концентрациях ниже 5%, то достаточно полный анализ популяции должен
быть дополнен экспериментами или специальным сбором дополнительного
материала, направленного к специальному анализу этих более редких ре-
цессивов.
Значение инбредного материала для анализа популяций
При родственных скрещиваниях (инбридинг, внутрибрачие)
вероятность возникновения рецессивных гомозиготных комбинаций резко
повышается, различно для различных степеней инбридинга. Так, например,
при скрещивании отца с дочерями вероятность обнаружения рецессива у
отца повышается до 1/8. Пусть, например, в популяции, где доля рецессива
q = 0,05 (= 5%), мы собрали данные о 100 телятах, полученных от
скрещивания быков с их дочерями. При 5% концентрации гена из №Q-2pq =
95 5
= ^"Ш" * "Ш" "^^ ^ ^ ^удут гетерозиготами и среди их телят V8 X
X 9,5 = 1,2 теленка будут гомозиготными рецессивами. Иными словами,
если раньше один рецессив возникал на 400 особей (0,05-0,05 =
= 0,0025 = 0,25%), то здесь он возникает уже один на 83, в 5 раз чаще.
Чем реже встречаются рецессивы, тем большую выгоду представляет
инбридинг для его проявления.
Таким образом, сбор дополнительных данных по инбредным животным
дает возможность
1) выявить гетерозиготность популяции по более редким рецессивам;
2) повысить уверенность в гомозиготности по доминантным аллелям
(так, например, если на 100 животных мы не нашли рецессивов, то можно
сказать, что 0^ q ^ 0,05, а при таком же числе инбредных, что 0 ^ q <
< 0,01) *;
3) повысить уверенность в том, что редко встречающиеся фенотипы
имеют генотипическую, а не паратипическую обусловленность.
*ЛПри каком q на 400 животных при инбридинге на отца появится один рецессивный
гомозигот? 400-2pgr-V8 = 1. Отсюда, зная, что р = 1 — д, имеем 100 (д — q2) = 1,
откуда q ^ 0,01. j
184
Рис. 57. Схема анализа путем
скрещивания отца с дочерьми
I — самец; II — самка; III — оба
пола; IV — гетерозигота Аа; V —
гомозиготы АА]и аа; I, 3,4 —
число особей
Ц-г-О
О
■о- □
о
-rOTbhVTQjO-^ О
<>5 4
□
/
//
///
IV
V
Эксперименты
В тех случаях, когда для анализа популяции можно применить
эксперимент, он ставится различными способами в зависимости от
биологических особенностей животного. Наилучшим методом является скрещивание
самца (гетерозиготного) с несколькими его дочерями. Как показывает
схема (рис. 57), дочери (безразлично, от одной или нескольких матерей)
оказываются в половине случаев гетерозиготными, почему в потомстве от
обратного скрещивания с отцом получается
От гомозиготных дочерей .
От гетерозиготных дочерей
АА
2
1
Аа
2
2
1
Итого
3 4 1
или 7:1
Как более подробно объяснялось выше (дети-анализаторы), для
анализа особи было необходимо стремиться к тому, чтобы от каждого
анализируемого отца было пущено в скрещивание достаточное (более 8)
число дочерей. Для анализа популяции это требование не имеет значения
и важно лишь общее число исследуемых отцов.
Поэтому для анализа популяции имеет такое же значение и
скрещивание сына с матерью, как и отца с дочерью. Ответ о концентрации рецесси-
ва мы получаем из общего процента рецессивов в этих инбредных
скрещиваниях, а не из числа подвергнутых анализу отцов.
При невозможности скрещивать детей с родителями приходится
прибегать к скрещиванию братьев и сестер и полубратьев и полусестер *.
В первом случае схема скрещивания такова (рис. 58).
Как видно из нижнего ряда рисунка, состав F2 получается следующим:
АА — 9; Аа — 6; аа — 1.
В итоге в F2 мы получаем отношение 15А : \а или вероятность
появления рецессовов 1/16 = 6,25%. В случае скрещивания полубратьев с
полусестрами (рис. 59) схема ничем почти не отличается и вероятность
появления аа равна 1/16.
Разница, однако, состоит в том, что при скрещивании братьев-сестер
(сибсов) мы одинаково анализируем из Р и самца и самку, так как каждая
особь F2 имеет одинаковых дедов и бабок. При скрещивании же полубратьев
и полусестер (полу-сибсов) особи F2 инбредированы только на деда, а
бабки по отцу и по матери у них различны. Следовательно, если среди
женских особей (Р) тоже будут встречаться гетерозиготы, то они в первом
случае обнаружатся, а во втором — нет.
* К этому приему прибегали исследователи популяций дрозофилы (С. С. Четвериков,
Н. П. Дубинин и др.).
185
I Н-т-О
U-r-6 И-гО Lb-0 Hbji
<i> <> <> <> <> <S> <!> 4 6 ф <!> 4 <^4 i
Рис. 58. Схема'анализа при помощи скрещиваний брат X сестра
Поэтому при скрещивании сибсов для получения процента гетерози-
готов среди Р надо полученный в F2 процент рецессивов умножить на 8,
а во втором случае на 16. В самом деле, получив в F2 от сибсов на 100
особей к рецессивов, мы имеем 16А пар родителей (дедов), из которых один
гетерозигот, так что 16А приходится на 200 особей (дедов) откуда имеем
2щр • 100 % = 8 к % г етерозиготов. При скрещивании полусибсов те же
16 к рецессивов приходятся на 100 дедов, а не пар дедов, и процент
гетерозигот будет:
^.100о/0 = 16А:о/0.
Так как эта величина равна 100 X 2pq, то в первом случае
2pq = 0,08А, или pq = 0,04 А,
а во втором
2pq = 0Д6А:, pq = 0,08 к.
Имея в виду, что р = 1 — д, получаем для сибсов
q = 0,5 — /0,25 — 0,04к = 0,5 — 0,1 У25 — 4&, для полусибсов
q = 0,5 - /0,25 — 0,08 к = 0,5 -[0,1 У 25 - 8 fc.
При 1 % рецессивов для q получаем соответственно 0,042 й 0,088, или
в процентах гамет 4,2 и 8,8.
Для изучения состава популяции по генам, имеющим плохое
проявление, необходимо заботиться о том, чтобы в F2 получить достаточно
большие группы потомков от 1 пары родителей (сибсов). Из вышесказанного
следует, что при этом из каждых четырех пар сибсов в F2 у одной пары
наступает расщепление в отношении 3:1. Если число потомков здесь
невелико и рецессивы извлекаются в числе 1—2, то относительно плохо проявляю-
От-Ч>г<>^Ч>На-гЧ>гЧ>т--С>тО
п-
■о
О D-r-® Н-г-0
<> 6 <> <> <> ф <!> 4 <J>4 <> i <> <i> 4 4
Рис. 59. Схема анализа при помощи скрещиваний полубрат X полусестра
186
щихся признаков будет трудно решать, являются ли они наследственными
или ненаследственными паратипическими уклонениями. Если же мы
получим большое потомство, то и рецессивов с заданным колеблющимся
видоизменением возникает целая группа, так что наследственный характер
этого изменения станет очень вероятным. В этом случае статистическую
ошибку определения концентрации q надо вычислять, принимая за число
вариант не число полученных потомков, а число особей (с? с? и § §),
взятых из популяции для анализа.
Для оценки величины вычисляемых р и q важно иметь характеристику
пипуляции в отношении ее «свободы», или, наоборот, степени ее инбредно-
сти в последнем поколении, т. е. степень родства, связывающего
скрещивающиеся популяции самцов и самок. В самом деле, имея, например,
к % особей аа, мы вычисляем q по формуле:
q- y ТОО f
достулируя, что в популяции имеет место распределение по формул
Харди. Но такое распределение имеет место лишь при отсутствии родствен
ных связей между родителями, давшими исследуемое поколение. Если же
изучаемое потомство произошло, например, исключительно от скрещивания
братьев с сестрами, то, произведя подобное вычисление, мы сделаем
ошибку.
Анализ структуры популяции
После того как концентрация генов в популяции изучена, может быть
начат дальнейший анализ популяции. Здесь могут быть поставлены на
разрешение следующие вопросы.
1. Распределение концентрации отдельного гена по различным частям
популяции: по территории (геногеография), по экологическим станциям,
по частям популяции, выделенным по другим принципам (например, часть
популяции скота, записанного в племенную книгу) и, наконец, по времени.
2. Соотношение в распределениях концентраций двух и более генов
в одной и той же популяции по ее частям, опять-таки территориальным,
временным и др.
По первому пункту особых методических указаний не требуется. При
изучении геногеографии концентрации гена могут быть нанесены на
карту, при изучении изменения концентраций во времени — начерчены
соответствующие таблицы и кривые и т. д. И поскольку основные
интересующие исследователя вопросы о причинах особенностей этого распределения
выходят уже за пределы компетенции генетического анализа, переходя
в область эволюции, истории и т. д., мы здесь останавливаться подробнее
на них не будем.
Что касается изучения соотношения в распределении концентрации
различных генов друг с другом, то это изучение является хорошим
методом изучения структуры популяции. В самом деле, пусть мы
нашли, что на каком-либо пути между географическими пунктами М и N
концентрация гена А систематически падает. На основании этого мы ничего
большего сказать не можем и в состоянии лишь поставить ряд вопросов
о причинах этого изменения. Но иное дело, если мы устанавливаем, что
на том же пути параллельно с падением концентрации гена А падает
концентрация генов 5, С и D и, наоборот, растут концентрации генов Е, F
и G. При этом мы, например, обнаруживаем, что от пункта М до речки N
процент изменения всех этих концентраций невелик, а при переходе на
другой берег речки соотношение концентраций всех этих генов резко
изменяется (подобную картину, например, мы наблюдали в популяции кур
187
Дагестана при переходе через речку Аварское Койсу). Наличие резкой
смены целого ряда концентраций может быть принято в качестве
доказательства того, что в данном месте мы перешли из одной популяции в
другую, из одной части генофонда в другую, а область, равномерно
обнаруживающая изменение ряда концентраций, может быть принята за область,
в которой идет диффузия одного генофонда в другой генофонд. В данном
случае генетический анализ подходит к вопросам о низших систематических
единицах (расы, кряжи, отродья и т. п.), к анализу процессов
дифференциации единого генофонда на его части, и обратно процессов
перемешивания этих частей при новых встречах, естественно, или под влиянием
хозяйственной деятельности человека (метизация и т. д.).
Установление различия между популяциями и его реальность
Для этих целей может быть применено вычисление средней или
средней квадратическои разницы между сравниваемыми популяциями по-
ряду генов.
Пусть мы имеем две популяции (I—II), изученные по нескольким
генам*.
Гены
А
В
С
D
Е
F
G
Н
I
К
Концентрации
I
0,088
0,095
0,165
0,000
0,000
0,372
0,111
0,151
0,019
0,280
II
0,110
0,112
0,315
0,025
0,000
0,324
0,063
0,266
0,048 *
0,422
Л
0,022
0,017
0,150
0,025
0,000
0,048
0,048
0,115
0,029
0,142
2А2
-J0~= 0,062736
V ю
Дз
0,000484
0,000289
0,022500
0,000625
0,000000
0,002304
0,002304
0,013225
0,000841
0,020164
: 10 = 0,006273
. = 0,079
Средний квадрат разницы (или корень квадратный из него) может
служить мерилом различия между сравниваемыми популяциями. Чем он
больше, тем больше различие. При 0 мы имеем полное сходство, при
1 — максимально возможное различие. Беря средний квадрат вместо
среднего из первых степеней (абсолютных значений), мы придаем несколь-
ко большее значение крупным различиям в концентрациях, что вполне
закономерно, так как мелкие разницы чаще могут быть случайного
характера.
Однако у этого коэффициента имеется недостаток в том, что он дает
большой вес генам, находящимся в популяции в концентрациях, близких
к 0,500. Естественно, что более редкие гены (или, обратно, более частые,
* Пример из работы А. С. Серебровского (1926, 1927). Речь идет о популяциях кур
двух ущелий Дагестана.
188
если брать другой их аллеломорф, никогда не смогут дать больших
слагаемых и заметно отразиться на итоге.
Это уравновешивание можно произвести, например, разделив разницу
между а и Ъ на их сумму, т. е. построив индекс h = а~}ъ • Так как для
каждого гена имеются две концентрации р и д, то в зависимости от того,
какие аллеломорфы мы возьмем, числитель будет оставаться одним и тем
же, а знаменатель (тем самым и весь индекс) меняться. Этого, однако,
легко избежать, условившись использовать в знаменателе всегда
концентрации менее 1/2.
Наше вычисление примет такой вид:
А
В
С
D
Е
F
G
Н
I
К
а
0,088
0,095
0,165
0,000
0,000
0,372
0,111
0,151
0,019
0,280
ъ
0,110
0,112
0,315
0,025
0,000
0,324
0,063
0,266
0,048
0,422
(а-Ъ)
—0,022
—0,017
—0,150
—0,025
0,000
0,048
0,048
—0,115
—0,029
—0,142
(а + Ь)
0,198
0,207
0,480
0,025
0,000
0,696
0,174
0,417
0,067
0,702
h
-0,111
—0,082
-0,313
-0,1000
+0,069
+0,276
—0,276
-0,433
-0,197
2Д = —1,037;
/12
0,01232
0,00674
0,09750
1,00000
0,00474
0,07617
0,07617
0,18749
0,03870
2Д2 = 1,49983
Недостатком этого способа является, наоборот, преувеличение малых
концентраций, где, как в случае гена D, разница в концентрациях 0,025
дала сразу h = 1,000.
Удобный метод анализа структуры популяции мы применяли при
изучении геногеографии кур. Метод этот состоит: 1) в составлении для каждой
отдельной популяции (селения, района) особого графика, наглядно
представляющего ее особенности, и 2) в сопоставлении друг с другом ряда
графиков, групп графиков соседних популяций, изображенных и
расположенных на листе бумаги в соответствии с географическим
расположением отдельных селений, районов и пр. Этот метод, не претендуя на
точную количественную оценку сходства или различия строения
сравниваемых популяций, дает возможность в удобной, простой и наглядной форме
выявить наблюдаемое сходство или различие.
Применительно к курам графики составляются следующим образом.
1. Выбираются наиболее удобные гены, лучше других наблюдаемые и
с не слишком малыми рид.
2. Для каждого гена величина концентрации мутантной аллели
(трансгена) откладывается в виде вектора, начинающегося из общей начальной
точки, причем вектор каждого гена имеет свое постоянное направление.
3. Концы вектора соединяются линиями сплошными или
пунктирными, и некоторые линии проводятся еще внутри графиков для придания
большей выразительности чертежу.
4. Удобно из центра (начальные точки вектора) описать круг радиусом
0,500, который бы служил масштабом при сравнении графиков,
изображенных в разном масштабе.
Мы в своих работах довольствуемся 8 векторами, так как при
независимой изменчивости этих векторов случайное совпадение такого числа
вариантов совершенно маловероятно.
189
Использованные нами гены * следующие:
Ген
sune — хохол
wene — розовидный гребень
wegue — стручковый гребень
trage —- «плимутроковый» рисунок
tuge — серебристый окрас
atedu — рецессивный альбинизм
at rase — «ситцевый» рисунок
tifa — меланизм
Вектор
величина
Р
Р
Р
Р
Р
Я
Я
Р
направлен.
" W
NW
N
NE
Е
SE
S
SW
Рис. 60—64 показывают графики некоторых популяций кур,
иллюстрируя наблюдавшееся разнообразие более или менее больших
популяций. При помощи построения и сравнения друг с другом подобных
графиков возможно освещение целого ряда вопросов строения и истории
популяции. Перечислим некоторые из таких вопросов.
^-7 /
i
i
i
w
'ш
-^ iv
Рис. 60. Графики популяций беспородных кур, обнаруживающие значительное
сходство
1 — Нежин;]11 — Ярославль; 111 — Кировск. южн.;]1У]— Полтава;}V— ^Московская обл.|
1. Установление ареала популяций. Если
сравнить графики некоторых популяций Украины, Московской и Ярославской
областей и южной части района Ветлуги (рис. 60), то видно замечательное
сходство, показывающее, что на всем этом огромном пространстве мы
имеем дело, несомненно, с единой популяцией.
2. Установление границ между популяциями.
Сравнение группы графиков из Северного Дагестана — районов,
населенных аварским и андийским племенами (рис. 61),— позволяет видеть,
наоборот, резкую границу, проходящую по реке Аварское Койсу и
разделяющую графики на два совершенно различных типа: аварский — на
востоке и андийский — на западе. Одновременно видно, что два андийских
графика довольно схожи друг с другом, а из трех аварских два тоже сходны
друг с другом, отличаясь несколько от третьего, но не так сильйб, как оба
они отличаются от андийских.
А. С. Серебровский обозначает гены курицы по разработанной им по предложению
Н. К. Кольцова еще в 1921 г. десятичной системе обозначения'генов. Первоначально
были разработаны таблицы числовых символов, которые для удобства были
переведены в буквенные. Так, основной ген цветности у кур, грызунов и других животных
имеет символ 8111, или буквами tede, тогда альбинизм надо обозначать как atede.
К сожалению, и до сих пор в обозначении генов у различных видов животных и
растений'царит большая путаница.
Предложенная Серебровским десятичная система^еще ждет своего претворения
в жизнь.— Прим. ред.
190
Рис. 61. Графики двух популяций кур,г-аварской и андийской [(Горный
Дагестан)
1
\ 3 \ 7 \ /
\ /
N /
N I
\1
\/
Рис. 62. Графики популяций кур из 4 селений Сисиана (Армения)
Правые графики обнаруживают сильную изогаметацию. График суммы четырех
этих популяций см. на рис. 63
i V в У ш\/
Рис. 63. Графики популяций кур Сисиана. (Ср. сходство шравого
графика с левым, являющимся суммой 4 различных популяций,
изображенных на рис. 62)
I — Сисиан без Ангехакота; II — Сисиан; III — Ангехакот
Рис. 64. Анализ популяций вычитанием. (Ср. сходство двух правых
графиков — фактического и разностного.)
I — Гидатль; II— Карах: III — Кусур; IV — 2 \ Кар-Гид
3. Установление распада популяции на
отдельные «диферент ы». Рис. 62 показывает группу графиков
популяций соседних деревень в Горной Армении (Сисиан). Легко видеть,
что все эти графики чрезвычайно резко отличаются друг от друга и даже
имеют довольно странный вид благодаря чрезвычайному развитию одних
векторов и, наоборот, недоразвитию других. Однако стоит лишь все эти
изолированные популяции (изоляты) сложить вместе, и мы получаем
популяцию, очень сходную с популяцией большого селения Ангехакота,
одного из центров этого района (рис. 63). Это сходство позволяет сделать
вывод, что популяции отдельных деревень являются частями единой
популяции, подвергшейся в результате изоляции процессу «изогаметации»
(Д. Д. Ромашов, 1931), и ставшими, таким образом,
дифференцированными элементами, «дифферентами» единой популяции.
4. Установление слияния популяции. Рис. 64
представляет собой графики для популяции трех аварских ущелий: Гидатля
(наиболее северо-западное), Караха и Кусура (наиболее южные). Можно
заметить, что эти три графика почти по всем элементам образуют как бы
ряд с Карахом посредине. В самом деле, если из графика Караха
вычесть график Гидатля, то мы получаем изображенный справа график,
чрезвычайно близкий к графику Кусура. Это служит указанием о том, что
Карах действительно можно рассматривать как промежуточное звено
между Гидатлем и Кусуром, возникшее, по-видимому, в результате слияния
популяции типа гидатлинской и кусурской. Гипотеза расхождения из
Караха двух других популяций маловероятна. В этом случае мы
наблюдали бы расхождение по одним генам и не наблюдали бы по другим. В
действительности же все почти гены дают в крайних популяциях
противоположные отклонения. Подобную же картину мы видим на рис. 65,
представляющем группу графиков популяций селений юго-восточной Балкарии.
Графики образуют довольно постепенный переход от округлого графика,
характерного для многих мест Горного Кавказа, в частности близкого
к андийским графикам (см. рис. 61), до очень вытянутых с W на Е
графиков Осетии.
.Можно поэтому попробовать решить хотя бы приблизительно задачу,
с какой именно популяцией слилась осетинская на Череке и в какой
именно пропорции произошло слияние двух популяций на Череке. Вопрос
решаем так.
Если черекская популяция является смесью чисто осетинской и
некоторой иной (X), то мы можем эту иную (Х-популяцию) восстановить,
вычитая из черекской популяции (Ч) осетинскую (О) и изучая оставшийся
остаток.
Вопрос приходится решать методом проб, так как у нас два
неизвестных — состав популяции X и пропорции смешения с ней осетинской.
/ ^' п ш ш
Рис. 65. Анализ популяции сложением — определение пропорции
смешения двух популяций. (Ср. сходство двух средних
графиков — фактического (Черек) и суммированного)
I — 24 — 0;\ II — Черек (Балкария Ср.); Ill] IP""1 IV—Осетинская
(Куртатинское ущ.)
192
Рис. 66. Анализ популяций вычитанием — определение
пропорции смешения двух популяций. (Ср. сходство правого
графика с левым — разностного и фактического)
I и II — яфетическая; III — 34 ^ 20
Сделаем два расчета. Предположим, что черекская популяция (Ч)
является смесью осетинской (О) ж третьей (X) в пропорциях 1 : 1 и 1 : 2.
В первом случае (пропорция 1 : 1) концентрации по каждому гену
популяции определяются из уравнения: '.";."
1 + 0_ = Ч; Х = 2Ч-0$
.9- ... .
где Ч и О — концентрации соответствующих генов черекской и осетинской
популяций.
Для пропорции смешения 1 : 2 уравнение будет иметь вид;
^ X + 2.Q _„
—з— -*<
откуда X = 3-5 — 2*0*
Рис. 65 и 66 представляют собой полученные графики Х-популяции по
обеим гипотезам. Как видно по гипотезе 1:2, мы извлекли Х-популяцию,
замечательно похожую на «яфетическую», изображенную на рис. 66, справа.
Это обстоятельство, конечно, не случайно и говорит о правильности нашей
гипотезы о том, что популяция Черека возникла из слияния двух
популяций — «яфетической» и осетинской, в пропорции 1:2.
Проверяем эту гипотезу слиянием фактически наблюденных
популяций осетинской (Куртатинское ущелье) и верхнебалкарской
(«яфетической»). Рис. 65 (посредине) показывает полное сходство
вычисленного, таким образом графика с фактически наблюдаемым для Черека.
13 А. С. Серебровский
ЧАСТЬ ВТОРАЯ
ГЕНЕТИЧЕСКИЙ АНАЛИЗ КОЛИЧЕСТВЕННЫХ
(ТРАНСГРЕССИВНЫХ) РАЗЛИЧИИ
ГЛАВА IX
ОБЩИЕ ОСНОВЫ. ГЕНЕТИЧЕСКАЯ КОМБИНАТОРИКА
ТРИ МЕТОДА ГЕНЕТИЧЕСКОГО АНАЛИЗА
КОЛИЧЕСТВЕННЫХ (ТРАНСГРЕССИВНЫХ) РАЗЛИЧИЙ
В настоящее время далеко не всякая задача по анализу
количественных признаков и не при всяких условиях может быть решена. Для
решения этих задач в зависимости от условий (характер признака, степень его
паратипической изменчивости, способ размножения организма,
численность потомства, число хромосом и особенности перекреста, степень
изученности генетики данного вида и т. д.) приходится прибегать к различным
приемам, пользоваться различными методами, комбинировать их. Поэтому
необходимо научиться владеть всеми предложенными методами, уяснить
их достоинства и недостатки и те условия, в которых каждый из этих
методов может быть с успехом применен.
В настоящее время можно говорить о трех методах анализа
количественных признаков. Эти методы можно обозначить тремя терминами: менде-
листический метод, алгебраический, или метод уравнений, и метод сигна-
лей. Здесь мы дадим сначала краткий обзор этих методов, а затем
перейдем к их более подробному изложению.
Менделистический метод состоит в приложении
обычных менделистических формул и наблюдений за происходящими
расщеплениями. Но так как при наличии трансгрессивной изменчивости
распределение потомства по классам расщепления по фенотипу оказывается
невозможным, то приходится прибегать к наблюдению за следующими
поколениями и к постановке специальных скрещиваний для того, чтобы
решить, к какому же классу расщепления следует отнести данную особь.
Таким образом, принцип этого метода состоит в замене особи группой
ее потомства с таким расчетом, чтобы при этом оказалась нейтрализован*
ной паратипическая изменчивость.
Наилучшие условием для применения этого метода является
способность организма к вегетативному размножению. Для иллюстрации этого
метода мы разберем следующий гипотетический пример. Пусть мы
скрещиваем две формы растений, желая проанализировать разницу в их росте.
Исходные формы имеют Мх = 20 см и М2 = 26 см. Для Fx M3 = 23 смг
а в F2 мы имеем вариацию с М4 = 23 см и с непрерывной изменчивостью от
17 до 29 см.
Сантиметры 17 18 19 20 21 22 23 24 25 26 27 28 29
Процент особей .... 0 2 5 8 10 14 16 14 10 8 5 2 0
194
Усмотреть в этом распределении какие-либо классы расщепления,
очевидно, совершенно невозможно, поэтому и нельзя сказать, вызывается ли
разница между скрещенными формами одним геном или несколькими
генами.
Возьмем, однако, из F2 20 особей из самых различных классов, но>
пропорционально частоте этих классов (или свободным тиражом из общей
массы) и получим от каждого растения вегетативным размножением по
100 дочерних растений. Заменим таким образом каждое из 20 растений
сотней его потомков-отводков, полученных вегетативно и, следовательно,,
обладающих тем же генотипом. Вырастив эти растения и измерив их,
вычислим для каждой сотни средние. Тогда окажется, что эти средние
распадаются на ясные три группы:
I
19,8
19,9
20,0
20,0
20,1
II
22,6
22,9
22,9
23,0
23,0
23,0
23,1
23,2
23,2
23,5
III
25,8
25,9
26,0
26,2
26,2
Мы видим, что между этими группами имеются большие различия:
20,1—22,6 и 23,5—25,8, тогда как между членами одной группы различия
ничтожны. ^
Особенно наглядно распадение на три группы выразится на схеме
XIX, где каждая точка отвечает отдельной средней.
J& .£*. At
■ i ' » i i » 1 1 « .
18 13 20 21 22 23 2+ 25 2f 27/f
Схема XIX
Вместе с тем мы замечаем: 1) что прбпорция отдельных групп
совпадает с менделевским моногибридным отношением 1 : 2 : 1 и 2) что величина
признаков в I и III группах совпадает с величиной признака у скрещенных
форм, а во II группе — с величиной признака в Fle Эти данные позволяют
считать, что родители аа X А А различаются одним геном, в ¥г мы имеем
А а с промежуточным развитием признака, а в F2 идет расщепление по
формуле аа : 2Аа : АА.
Распадение F2 на резко обособленные группы произошло потому, что
заменив особь ее 100 потомками, мы в /Too = io раз уменьшили транс-
грессивность изменчивости и этим выявили «качественный» характер
якобы «количественного» признака, или, вернее говоря, «альтернативный»
характер внешне трансгрессивной изменчивости.
Поясним структуру этого схематического примера. Было принято
условие, что одна раса растений имеет строение АА, другая аа, что имеет
место промежуточное доминирование и каждый аллель А увеличивает
признак на 3 см. Таким: образом генотипически скрещивание идет по такой
схеме:
195
13*
Поколения
, Pi
Р2
Fx
F2
20
aa
,
aa
21
22
23
Aa
2Aa
24
25
26
AA
AA
Но одновременно имеет место паратипическая (модификационная,
ненаследственная) изменчивость, благодаря которой каждый генотип
представлен разнообразными фенотипами. Схематически мы приняли
распределение этих фенотипов по биному (1 + 1) 6. Тогда все скрещивания
приобретают такой вид:
Поколение и его
генотипы
Pi(««)
Ра (АА)
¥i(Aa) (
Fa *
И
(ady
&Аа)
{ (АА)
Bee F2
В п]
роцентах
17
1
1
1
0,40
Классы растений по длине.
18
6
6
6
2,35
19
15
15
15
5,85
20
20
1
20
2
22 '
8,60
21
15
6
15
12
27
10,55
22
6
15
6
30
36
14,05
23
1
1
20
1
40
1
42
16,40
24
6
15
30
6
36
14,05
еде
25
15
6
12
15
27
10,55
26
20
1
2
20
22
8,60
27
15
15
15
5,85
28
6
6
6
2,35
29
1
1
1
0,40
Сильная трансгрессия трех генотипов, составляющих F2, не дает
возможности по внешнему виду кривой распределения F2 различить эти три
группы. Заменяя каждую особь средней из 100 ее потомков, мы уменьшаем
колеблемость средних величин в 10 раз, трансгрессия уменьшается, и
строение F2 снова становится ясным.
К построению таких схематических примеров мы будем прибегать и
в дальнейшем при разработке теории генанализа количественных
признаков, и научиться этому приему весьма полезно.
Меддёлистическим методом пользовались в своих работах Ист (East,
1916), Филидпченко (1926), Ленин (1928) в исследовании пшениц
и др.
Успешность применения рассматриваемого менделистического метода
в сильнейшей степени зависит от того, насколько правильно удается
произвести замену особи группой ее потомков. При вегетативном
размножении это наиболее просто, так как все потомки образуют однородный клон,
состоящий из особей с одинаковым генотипом. Однако вегетативным
размножением обладают почти исключительно растения, да и то далеко не
все. При получении «заменяющего потомства» половым путем дело сразу
осложняется, так как гетерозиготные особи начинают расщепляться,
и заменяющее потомство оказывается уже генотипически разнородным.
У самоопыляющихся растений это затруднение сравнительно мало
осложняет картину. У организмов аллогамных дело осложняется уже
значительно, так как скрещивать друг с другом приходится различные особи
196
F2, гетерозиготность которых может быть разной. Наконец, максимум
осложнений возникает тогда, когда организмы оказываются
малоплодными и для получения заменяющего потомства приходится скрещивать друг
с другом целые группы животных, например, быка с многими коровами
и т. п. В этих условиях метод оказывается уже почти безрезультатным и
его применение малоцелесообразным.
Второй метод алгебраический, или метод решения уравнений,,
основан на том, что различное число генов, участвующих в скрещивании,,
различная степень доминирования, различная сила генов и т. д.
определенным образом отражаются на характере кривых распределения в различных,
поколениях. Эти кривые распределения могут быть описаны при помощи
различных параметров — средней величины, моды, квадратического
уклонения, показателя асимметрии, показателя эксцессивности и т. д.—
и могут быть выведены многочисленные формулы, связывающие эти
параметры с генотипичёскими элементами. Генотипические элементы являются
здесь в качестве неизвестных, х, у, z..., а параметры могут быть
вычислены из данных наблюдений. Таким образом могут быть составлены
уравнения. Если удается составить столько уравнений, сколько имеется
неизвестных, то неизвестные могут быть найдены.
Если же наблюдения дают больше уравнений, чем число неизвестных,
то для такого случая математика предоставляет возможность найти
наиболее вероятные значения искомых величин.
В качестве простейшего примера приложения этого метода разберем
случай биномиального распределения. Хотя распределение не всегда
биномиальное, но это наиболее частый случай и дальнейшие относятся только
к нему.
Распределение, отвечающее биномиальному ряду (1 + 1)Л,
обладает тем замечательным свойством, что для него имеет место равенство
или
-У\<
9 П
V* = <з2 = т.
Пример: (1 + 1) 6
Д/ Д1/
1
6
15
20
15
6
1
64
|А2 =
—3
-2
—1
0
+1
+2
+3
= <52 =
— 3
-12
-15
—
+15
+12
+ з
96 6
64—4 =
9
24
15
—
15
24
9
96
п
= 4
Это свойство дает нам возможность найти показатель степени бинома,
если почему-либо эта степень нам неизвестна. Пусть, например, нам дан
ряд:
1.х; 4.4; 11.7; 20.6; 24.6; 20.б; 11.7; 4.4; 1в1
Требуется найти степень бинома (1 + 1) #, наиболее близко
отвечающего данному ряду.
197
Рещениез
/ Л Д/ А*/
1Д _4 — 4,4 17,6
4,4
11,7
20,5
24,6
20,5
11,7
4,4
1,1
100,0
1*2
—3
-2
—1
0
+1
+з
+2
+4
, = в«:
-13,2
-23,4
-20,5
+20,5
+ 23,4
+13,2
+ 4,4
249
-100 —2,5
39,6
46,8
20,5
20,5
46,8
39,6
17,6
249,0
>.
Так как <з2 в -|-, то # = 4а2. Подставляя значение <та = 2,5, находим:
х = 4<х2'= 10.
Следовательно, к данному распределению очень близко подходит бином
десятой степени (1 + 1)10, дающий ряд: 1; 10; 45; 120; 210; 252; 210; 120;
45; 10; 1. Действительно, этот ряд и был взят в приведенном выше
условии, но члены его были даны в виде процентов, и два самых крайних члена
присоединены к соседним.
К генетическому анализу эта примерная задача имеет то прямое
отношение, что расщепление с участием полимерных генов дает приближенно
именно биномиальное распределение, и задача о степени бинома есть
задача о числе аллелей, участвующих в расщеплении. В самом деле,
моногибридное отношение А А + 24 а + аа отвечает ряду 1 + 2 + 1 или биному
(1 + I)2, дигибридное расщепление дает ряд (при промежуточном
доминировании) 1+4 + 6 + 4+1, отвечающий биному (1 + I)4, и т. д.
Задачи, фактически встречающиеся в генетическом анализе
количественных признаков, всегда значительно сложнее. Здесь приходится
отыскивать несколько неизвестных — число генов, силу действия этих генов,
степень доминирования и т. д. Для решения задачи приходится строить
систему уравнений. Эти необходимые уравнения можно получать, используя
различные гибридные поколения, обратные скрещивания и т. д. или
вычисляя иные параметры — моменты высших степеней, моды и пр.
Теоретически часто можно построить необходимый ряд уравнений для решения
задачи (более подробно об этом см. ниже). Однако поскольку все параметры
вычисляются из эмпирического материала с неизбежными статистическими
ошибками, постепенное осложнение задачи приводит к накоплению этих
ошибок, и решение может в конце концов оказаться столь неточным, что
иногда теряет всякое значение. Правда, повышение точности может быть
достигнуто увеличением используемого материала, получением большого
количества потомства и пр., но это не всегда, конечно, возможно.
Возрастание неточности вызывает, к сожалению, довольно быстро предел
применению данного метода. К тому же и с математической стороны метод
уравнений еще очень мало разработан и поэтому в настоящее время почти
не применяется. Однако дальше мы увидим, что в целом ряде случаев
этот метод может и должен быть использован и заслуживает дальнейшей
теоретической разработки *.
Наиболее совершенным методом генетического анализа количественных
признаков, безусловно, является третий метод — метод сигнал ь-
* Алгебраический метод анализа количественных признаков сейчас приобрел
значительно большее значение в связи с разработкой теории наследуемости (см.
послесловие),— Прим. ред.
.198
ных генов (сигналей). Идея его состоит в том, чтобы, используя
локализацию генов в хромосомах и явление сцепления, исследовать влияние
на изучаемый признай того участка хромосомы, в котором расположен
данный сигнальный ген.
Приведем типовую задачу на применение этого метода. Пусть мы имеем
две инбредные линии дрозофилы, различающиеся числом щетинок на
стернитах брюшка и, кроме того, одним геном, например sepia (глаза
цвета сепии, символ se, локализован на 26 морганиде в /77 хромосоме).
Примем, что мухи сепия имеют 18 щетинок на стерните, мухи нормальной
линии — 20. Скрещиваем обе линии и получаем Fx с нормальными глазами
(sepia — признак рецессивный) и с 20 щетинками. Берем самца ¥г и
скрещиваем его обратно с чистыми самками сепия. При этом, естественно,
потомство получается двух типов — с глазами нормальными и с глазами
сепия, в пропорции 1 : 1 (по схеме А а X аа ->■ А а + аа). Подсчитав
щетинки у мух нормальных и мух сепия, получаем, например:
нормальные — в среднем 20 щетинок
\^ сепия —в среднем 18 щетинок
Этот результат можно объяснить двояко: или тем, что сам ген сепия
в порядке плейотропии влияет на число щетинок, понижая их с 20 до 18,
или тем, что в той же /// хромосоме локализован специальный ген,
влияющий на число щетинок, причем в хромосоме, несущей сигнальную аллель
нормальных глаз, расположен щетиночный ген, увеличивающий число
щетинок с 18 до 20 (а соответственно в хромосоме с сигнальной аллелью
сепия расположена рецессивная щетиночная аллель). Этот второй случай
можно пояснить схемой XX.
Se
А
t t
se
se+
1
\-se
4-fl
Se
A
+ +se
т -M
se+ 4-se
a+ +a
Схема XX
Какое из двух^объяснений правильно, можно исследовать
специальными приемами, о которых речь будет ниже. Если будет доказано, что сам
ген сепия на щетинки не влияет, то можно будет считать доказанным второе
объяснение, и тем самым разница между исходными линиями окажется
проанализированной: мы показали, что разница между инбредными
линиями в две щетинки зависит от гена (или генов), расположенного в III
хромосоме в линии с нормальными глазами, и что действие этого гена носит
доминантный (или адоминантный*) характер.
Обычно результат получается более сложный. Например, в подобном
скрещивании мы могли, бы получить такой результат среди потомков самца
¥г с самками сепия чистой линии:
нормальные — в среднем 19,5 щетинок
сепия —в среднем 18,5 щетинок
* В смысле неполного доминирования.
199
Подобный результат можно истолковать так. В III хромосоме вместе
с геном нормальных глаз локализован щетиночный ген, прибавляющий
одну щетинку (19,5-^-18,5 = 1), но, кроме того, где-то в других хромосомах
локализованы гены, в совокупности прибавляющие еще одну щетинку.
В самом деле*, предположим, что такой второй ген локализован во II
хромосоме. Тогда все скрещивание пойснится схемой XXI.
в\\в
s\\S
АЛ\А
b++b
S++S
а++а
20щет. *'
+ +Ь
5++S
А+\а
b\\b
s+ +s
а++а
Линия сепия
1Вщет
в++ь
s\\s
Afia
b+\b
S\+s
A++a
B\\b
s-t+s
a++a
H+b
S++S
ff+43
Afi 20щет Kb 19щет.
Среднее Щ5щет.
afi 19щет. atb 18ш,ет^
Среднее 18,5 гцет.
Схема XXI
Вводя в анализ сигнали, расположенные в разных хромосомах и на
разных участках хромосом, мы можем подвергнуть анализу все эти
участки и выяснить влияние их на изучаемый признак (подробнее см. ниже
гл. XII).
Особое значение метод сигналей приобретает тем, что, раз установив
влияние участка на данный признак, мы получаем возможность не только
точно «инвентаризировать» гены, в нем расположенные, но и оперировать
с ними дальше почти так же, как мы оперируем с хорошо менделирующи-
ми генами — в скрещиваниях мы будем следить уже не за самим «щетиноч-
ным» или другим геном, а за сопровождающей его сигналью.
Таким образом, мы видим, что каждый из трех разобранных методов
имеет свои недостатки и свои достоинства. Поэтому в практической работе
геноаналитика приходится работать комбинированно всеми тремя
методами, применяя в подходящих условиях различные приемы, стремясь
довести решение задач до того уровня, который допускается особенностями
анализируемого вида животных и состоянием наших знаний как частной
генетики вида; так и теории самого генетического анализа.
Комбинаторика полимеров
Полимерными генами, или полимерами, будем, называть гены,
влияющие на один и тот же признак и в различных комбинациях способные
обусловливать при сходных внешних условиях различную стецень развития5
2ра
признака. В скрещивании организмов обычно принимает участие
несколько полимеров, более или менее свободно комбинируясь друг с
другом. Так как каждый полимер или усиливает развитие признака
(увеличивает рост, повышает молочность, усиливает яйценоскость, умножает
число волос и т. п.), или ослабляет признак, то полимеры называют либо
усилителями либо ослабителями.
Однако в этом словоупотреблении имеется некоторая
неопределенность. Иногда эти термины прилагают к аллелям. Если в паре аллелей
А и а А дает большее развитие признака, чем а, то говорят иногда, что
А есть усилитель, а а — ослабитель. Или же данный ген А вообще
называют усилителем, а про а говорят — отсутствие усилителя, называя
ослабителем только те гены, ослабляющее действие у которых имеет
доминантный аллель. В дрозофильной генетике, в которой принято, наоборот,
называть ген в соответствии с характером действия по преимуществу
рецессивных аллелей, усилителями обычно называются гены, рецессивные
аллели которых усиливают признак.
В дальнейшем усилителями (У) мы будем стремиться называть гены,
доминантные аллели которых усиливают признак, и ослабителями (О)—гены,
доминантные аллели которых ослабляют признак. Соответственно через
у мы будем обозначать отсутствие усилителя (т. е. наличие его
рецессивного аллеломорфа), а через о — отсутствие ослабителя. Эта
терминология, конечно, не имеет никакого отношения к теории присутствия —
отсутствия и сходна с нею лишь внешне, поскольку «отсутствие» здесь
означает лишь отсутствие одного аллеломорфа. В тех случаях, когда нам
придется говорить о генах, усиливающих признак, независимо от того,
являются ли они доминантными или рецессивными аллелями, мы так и
будем говорить: аллели-усилители, аллели-ослабители.
Помимо направления действия полимера, мы будем иметь в виду силу
действия, обозначая ее буквой £*. Говорят о генах, действующих сильно
или слабо, или просто о сильных генах и слабых генах. Эти термины
неопределенны. Более точно сказать, что сила полимера i = 2 щетинкам или
1 = 10%.
Это будет означать, что особи А а в среднем имеют на две щетинки, или
на 10%, больше, чем особи аа, т. е. аллель-4 прибавляет две щетинки, или
10% щетинок. К сожалению, вопрос о силе действия генов принадлежит
к числу очень слабо изученных. Можно, однако, уверенно сказать, что
выражать силу действия гена в абсолютных единицах можно лишь чисто
условно, подразумевая «при прочих равных условиях», так как в
зависимости от различных, хотя бы внешних, условий особи Аа ж аа будут
различаться неодинаково. По-видимому, выражение силы действия гена в
процентах признака тоже схематично, ив разных условиях этот процент
прибавки будет тоже меняться различно для разных признаков и, возможно,
для разных генов. Поэтому, говоря, например, что ген А усиливает
признак на 10%, мы имеем в виду «при некоторых средних условиях», при
которых сила гена определялась или при которых мы ведем расчет.
Под числом генов или числом полимеров,
принимающих участие в данном скрещивании, мы будем разуметь число пар
аллелей (число аллопар). По этому словоупотреблению в моногибридном
расщеплении принимает участие один ген А в его двух аллелях А ж а.
В дигибридном расщеплении принимают участие два гена А и В с
четырьмя аллелями А и а, В и Ъ и т. д. Остальные гены в данном скрещивании не
расщепляются или почему-либо не принимаются во внимание и в счет не
входят. Число генов мы будем обозначать обычно буквой п.
Для сознательного рассмотрения кривых распределения
потомства, возникающего в различных скрещиваниях, необходимо озна-
* Более подробно о смысле Гсм. ниже, гл. XII.
201
комиться с некоторыми свойствами генетической комбинаторики.
Ознакомление с ними позволит из рассмотрения кривых или
характеризующих их параметров извлекать материал для построения выводов и
гипотез, необходимых при аналитической работе»
ВОЗНИКНОВЕНИЕ БИНОМИАЛЬНОГО РАСПРЕДЕЛЕНИЯ ГЕНОТИПОВ
В скрещивании, в котором участвует несколько полимеров,
расположенных в разных хромосомах и вообще способных свободно
комбинироваться, возникает характерное распределение генотипов гамет, в
дальнейшем — зигот (потомства), отвечающее закономерностям коэффициента
■бинома Ньютона. Как отмечено, это распределение имеет место уже по
отношению к гаметам.
Пусть, например, мы имеем особь, гетерозиготную по двум генам
АаВЪ. Ее гаметы будут АВ, АЬ, аВ и ab. Если гены А и В сходны по своему
действию и равны более или менее по силе, которую примем за единицу, то
можно сказать, что генотипы гамет АЬ и аВ по силе действия, которую они
смогут в дальнейшем оказать на признак организма, равны.
Следовательно, гаметы будут трех классов.
I класс без усилителей аЪ 1
II ь с одним усилителем ... Ab + aB 2
III » с двумя усилителями . . АВ 1
Распределение 1 + 2 + 1 отвечает биному (1 + I)2, следовательно,
у дигибридной особи гаметы распределены по биному второй степени, т. е.
показатель степени бинома равен числу генов или аллопар.
У тригетерозиготы АаВЪСс распределение гамет будет следующим:
без усилителей аЪс • i
с 1 усилителем АЪс, аВс, аЬС ..... з
с 2 усилителями АВс, АЪС, аВС .... 3
с 3 усилителями АВС 1
Распределение 1+3 + 3 + 1 отвечает биному (1 + I)3, где
показатель степени также равен числу генов, по которым особь гетерозиготна.
При скрещивании двух дитетерозиготных особей друг с другом
возникает уже 16 комбинаций гамет, и по числу усилителей зиготы
распределяются так:
без усилителей ааЬЪ 1
с 1 усилителем 2 АаЬЪ, 2 ааВЬ 4
с 2 усилителями ААЬЪ, ааВВ, 4 АаВЬ .... 6
с 3 усилителями 2 ААВЬ, 2 АаВВ 4
с 4 усилителями ААВВ 1
Распределение зигот здесь (в отличие от гамет) 1 + 4 + 6 + 4 + 1
отвечает уже биному (1 + 1) 4, где показатель степени равен 2п, так как
в дигибридном скрещивании п = 2. Легко обнаруживается, что гаметы
распределяются в соответствии с биномом (1 + 1)п, а зиготы в F2: (1 + 1)2п.
Для большого числа генов получающиеся ряды легко вычислить при
помощи треугольника Паскаля, имеющего такой вид:
тг = 1 1 1
71 = 2 12 1
/г = 3 1 3 3 1
тг = 4 1 4 6 4 1
/г = 5 1 5 10 10 5 1
гс = 6 1 6 15 20 15 6 1
202
Рис. 67. Три изображения
одного и того же распределения
{1 + I)6 при различных
масштабах осей координат. (Наи-
^ более удачно № 3)
1 ~~~" Объяснение в тексте
Каждое число в этом треугольнике является суммой двух
расположенных над ним в предшествующем ряду.
Для нас важно отметить следующие свойства этих рядов: они
одновершинны (для четных степеней имеют один наибольший средний член,
для нечетных — два равных средних) и симметричны. Если
изобразить эти ряды графически *, то получим характерный «полигон», который
по мере повышения степени бинома будет все более приближаться к
плавной кривой, так называемой нормальной или гауссовой кривой.
Графическое изображение полигонов (и кривых) не дает еще
возможности полно судить об их характере. Так как единицы на абсциссе
(классовый промежуток и т. п.) и ординате (число вариант) совершенно
различны, то для каждой из них можно выбрать любой масштаб, который мы
сочтем удобным. Поэтому одну и ту же кривую или полигон можно
нарисовать совершенно различно: и высокой и сжатой, и низкой и растянутой
(рис. 67).
Однако полигоны могут быть действительно сжатые и высокие или
растянутые, если их сравнивать с нормальной, гауссовой кривой. Характер
кривой определяется вычислением так называемых «моментов» разных
степеней: момент второй степени ([а2)**> третьей (|х3) и четвертой (|х4).
Момент второй степени является средним квадратом уклонений вариант от
средней арифметической для всего распределения, момент третьей
степени — средним кубом уклонений от средней арифметической и т. д.
Способы вычисления моментов мы здесь не излагаем, отсылая к
соответствующим руководствам по математической статистике или биометрии (см.,
например, Лахтин, 1922, 1924; Романовский, 1938)***.
Пирсоном предложено характеризовать кривые двумя величинами
(помимо так называемого «критерия Пирсона», о котором мы говорить не
будем) — Pi и р2. Эти величины легко вычисляются из моментов по
следующим формулам:
**{|-".-£[-*]•
* График строится так: на оси абсцисс откладываются номера классов — I, II, III...
N, на оси ординат — соответствующие члены разложения бинома (1 + 1)^~х или
частоты /. Ветви ломаной линии, соединяющей полученные точки, сводятся вниз
к добавочным точкам (нулевому и N + 1 «классам»), которые имеют чисто условное
значение — их частоты равны 0.
** В генетико-селекционных работах момент второй степени обычно обозначают как
вариансу.— Прим. ред.
*** Указанные автором руководства — сейчас библиографическая редкость. Более
новые книги на русском языке: В. Ю. У р б а х. Биометрические методы. М.,
«Наука», 1964; П. Ф. Р о к и ц к и й. Биологическая статистика. 2-е изд.
Минск, изд-во «Вышэйшая школа», 1967; Дж. У. Снедекор. Статистические
методы в применении к исследованиям в сельском хозяйстве и биологии. М., Сель-
хозгиз, 1961; Н. А. Плохинский. Биометрия. Новосибирск, Изд-во Сиб. отд.
АН СССР, 1961.— Прим. ред.
0 IE Ш N 7 W
20[ 3
15
0 Гй
0 ГГШШ*Ч1ШУШ
203
Нормальная кривая Гаусса характеризуется величинами (Зх = О,
Р2 = 3. Величина рх не равна нулю, если кривая имеет несимметричный
вид. Для всех симметричных кривых \i3 = 0 иг следовательно, Pi = О тоже.
Если р2 > 3, то говорят об эксцессивной кривой, т. е. кривой выше
нормальной. Если р2 < 3, говорят о кривой ниже нормальной, о кривой,
имеющей отрицательный эксцесс. Соответственно величина |J2 имеет название
«показателя эксцесса», а рх — показателя асимметрии, или скошенности *•
ВЛИЯНИЕ РАЗЛИЧНЫХ ГЕНОТИПИЧЕСКИХ ЭЛЕМЕНТОВ
НА Pi и р2
Для симметричного биномиального распределения рх = 0. Конечно,
точное равенство получается лишь в теории (при неограниченном числе
наблюдений). Если мы вычислим ($х для эмпирического ряда, то равенство
рх = 0 может осуществиться лишь случайно, обычно же получается для
Рх какое-либо конечное значение. Однако полигон или кривую следует
признавать асимметричной лишь тогда, когда рх реально отличается от нуля,
превышая свою ошибку более чем втрое.
Асимметричный полигон возникает в случае участия в расщеплении
более или менее полно доминирующих генов в «извлекающем
скрещивании» (т. е. в F2 или вообще при встрече двух гетерозигот).
Пусть мы имеем простейшее моногибридное скрещивание с полно
доминирующим геном-усилителем А. В F2 имеем АА + 2Ла + аа. Так как
А А и Аа по фенотипу будут одинаковы, то мы получим лишь два класса:
без усилителя аа 1
с усилителем АА-\-2Аа 3
Графическое изображение покажет нам сильно асимметричный
полигон. Такой полигон называется скошенным влево в сторону
извлеченных рецессивов (рис. 68).
Обычно распределение называется скошенным влево, если средняя
арифметическая (М) лежит левее (или по своему значению меньше), чем
мода {Мб), т. е. наиболее частая варианса (вершина кривой). Например»
в данном случае, если фенотипу аа приписать значение 1, а фенотипу А А
и Аа значение 2, то средняя М равна (1 X 1 + 2 X 3) : 4 = 7/4 = 1,75,
тогда как мода Мо = 2,0, распределение скошено влево. Однако иногда
применяется и обратная терминология.
Скошенность вправо (рис. 69) получится, если
рецессивной формой окажутся формы с более сильным развитием признака, т. е*
если в скрещивании участвует ослабитель. Тогда в F2 получим:
без ослабителей оо 1
с ослабителем 00+2 Оо 3
Линдстром (Lindstrom, 1932) — один из немногих генетиков, уже ряд
лет работающий над исследованием количественных признаков,
воспользовался асимметрией очень интересным образом. Он поставил на инбридинг
ряд диплоидных и тетраплоидных томатов. У тетраплоидов, как известног
рецессивные гомозиготы извлекаются реже, либо вовсе не извлекаются
(например, среди АААа). Линдстром получил, что у ряда диплоидов в
потомстве возникают асимметричные распределения по количественным при-
* Методы характеристики кривых, разработанные Пирсоном, сейчас применяются
очень редко, за исключением эксцесса. Наиболее полно о них говорится в книге:
А. В. Л е о н т о в и ч, Г. А. Г р и г о р ь е в, А. И. М а н д з ю к.
Вариационная статистика. М., Сельхозгиз, 1935 (также ставшей библиографической
редкостью).— Прим. ред.
204
Рис. 68. Косое распределение
F2 при полном доминировании
усилителя
Рис. 69. Косое распределение
F2npn полном доминировании
ослабителя
знакам, а у тетраплоидов не возникают. Этим Линдстром доказывает
наличие доминирования и у генов количественных признаков.
При дигибридном расщеплении в F2 возникает расщепление 1 + 6+9
в случае усилителя и 9 + 6 + 1 в случае ослабителя. Графически эти
распределения будут йметь^вид, как на рис. 70. Таким образом, по виду
кривой в F2 генетик-аналитик может сделать некоторые заключения о
характере участвующих в расщеплении генов, о чем более подробно будет
сказано ниже.
Полигоны с полным доминированием отвечают биному (1 + 3)п в
случае усилителей или (3 + 1)п в случае ослабителей. По сравнению со
случаем промежуточного доминирования мы видим два изменения: замену 1
на 3 и замену 2п на п в показателе.
Следует иметь в виду, что скошенными оказываются практически
лишь полигоны с участием небольшого числа полимеров. Уже при десяти
полимерах (1 + 3)п кривая имеет столь малое отличие от симметричной,
что доказать ее асимметрию практически при обработке эмпирического
материала совершенно невозможно.
Асимметрия распределения без влияния доминирования может
возникнуть тоже очень легко. Симметричный полигон мы получили только путем
известной схематизации, допуская постоянство действия гена во всяких
комбинациях полимеров. Так, в случае четырех полимеров мы принимаем
одинаковой разницу в признаке между классами:
аа ЬЬ ее dd (разница D-d)
аа ЬЬ ее Dd
и между классами:
АА ВВ СС dd
АА ВВ СС Dd
(разница D — d).
Между тем величина признака в классе аа bb cc dd и АА ВВ СС dd
совершенно различна, и прибавка к нему гена D может дать неодинаковое
приращение признака. Правда, эксперименты показывают, что во многих
случаях это изменение столь незначительно, что им можно пренебречь.
В других случаях, несомненно, можно встретиться с такими изменениями.
При этой изменение действия гена может быть двояким:
1) ген приращивает некоторый более или менее постоянный процент
.значения признака; тогда при большом развитии признака вызываемое
геном приращение будет большим по абсолютной шкале;
205
8
6
k
2
0
L_
1
r"
l_
l\ Л
t \ / \
/ \ / \
/ Л \
' / x \
1 I \ \
1 / \ \
1 I \ \
1 ^y >ч 1
Рис. 70, Косые расдределения
F2 при полном доминировании
усилителя (сплошная кривая)
и ослабителя (пунктирная).
Дигибридное расщепление
1 I Ж Ш
2) по мере усиления признака дальнейший его рост будет совершаться
с большим трудом; действие гена в этих условиях будет ослабевать.
Изменение действия гена первого типа даст скошенность вправо,
изменение второго типа — скошенность влево.
Поясним сказанное частным примером.
Пусть мы имеем расщепление с двумя полимерами (с адоминировани-
ем), где каждый аллель дает прирост значения признака на 50%. Тогда
величина признака в каждом кларсе будет следующей:
I класс без усилителей ааЬЪ 100 1
II » с 1 усилителем 2 АаЬЪ + 2 ааВЪ 150 4
III » с 2 усилителями ААЬЪ + ааВВ + 4 АаВЪ 225 6
IV » с 3 усилителями 2 ААВЪ + 2 АаВВ 338 4
V » с 4 усилителями ААВВ 507 1
Графическое изображение обнаружит сильную скошенность вправо
(в плюс сторону), как на рис. 71.
В левой части мы видим классы более сближенными, в правой —
широко раздвинутыми. При ослаблении действия гена по мере усиления приз-
знака мы будем иметь противоположную картину — сближенными будут
классы в правой части, раздвинутыми в левой. К этой природе асимметрии
мы вернемся еще при учете паратипической изменчивости.
Генотипические факторы влияют также и на величину эксцесса-
Симметрическое биномиальное распределение (1 + 1)п имеет отрицатель,
ный эксцесс, т. е. р2 < 3. Так, для ряда 1 + 4 + 6 + 4 + 1 мы имеем:
/
1
4
6
4
1
16
А
-2
-1
0
+1
+2
А/
—2
-4
+4
+2
А*/
4
4
4
4
16
дз/
—8
-4
—4
-8
А«/
16
4
4
16
40
16 . 40 о г п И* 2,5 л г
В* = 16 = 1; Н* = J6 = 2,5; 32 = -^- = — = 2,5.
Полезно знать, что для биномиальных распределений имеют место
свойства:
п . п /Зл—"2\ о о %
Ъ = т, [*4 = т (-т-); Р2 = 3 - -.
206
Рис. 71. Косое распределение
благодаря неравенству классов
/ I
'I I
i i I
W
ш ш
Из этой формулы легко вычислить величину р2 Для биномов разных
степеней:
Степень
2
4
6
8
&
2,00
2,50
2,67
2,75
Степень
10
20
оо
&
2,80
2,90
3,00
Из этих данных видно, что по мере возрастания показателей степени
бинома р2 приближается к 3 и хотя достигает величины 3 лишь при оо-
степени, но уже при 10-й степени настолько близко подходит к 3, что
отличить по эмпирическому материалу реально эту величину от 3 будет трудно»
Нужно щсеть в виду при этом, что здесь показатель степени является не
числом генов, а числом аллелей (т = 2га) и бином 10-й степени возникает
при пяти полимерах: А, В, C,D, E.
Интересно узнать, что при одновременном участии усилителей и
ослабителей могут возникать более высокие кривые. Разберем случай
распределения с участием полно доминирующих усилителя и ослабителя. Ди-
гетерозиготы УуОо дадут в F2 такое распределение:
без усилителей yyOO + fcyyOo *
е 1 усилителем 4УуОо + 2УуОо+2УУОО+ууоо + УУоо . . Ю
с 2 усилителями УУоо +2 Ууоо 3
Это распределение 3 + 10 + 3 имеет тоже три класса, как и
распределение 1 + 2 + 1, но средний класс здесь не в 2 раза больше, как прежде,
а в 3,5, т. е. распределение более высокое.
р2 = 2,67, а не 2,0, как для распределения 1 + 2 + 1.
Рассчитав такое же скрещивание с участием четырех полимеров, двух
усилителей и двух ослабителей, получим распределение 9 + 60 + 118 +
+ 60 + 9, для которого р2 = 2,83, т. е выше, чем у бинома (1 + I)1
(рис. 72). Однако и здесь р2 останется все еще меньше 3.
Рис. 72. Высокое
распределение (а) 9 : 60 : 118 : 60 : 9 и
нормальное; \(б) 1:4:6:4:1
%50
to
30
W
10
/?1
г
-
/
//
//
//
0 Т F
а
л
А
1 i
Ш Ш
\
\\
\\
1 *"
V
W
207
Рис. 73. Ложная многовер-
пшндость
Распределение цыплят по строению
таза (из Промптова)
+/ +*2 ^классы
Распределение более высокое, чем нормальное, т. е. (J2 > 3, может
возникнуть лишь при некоторых особых случаях, например, при прогрессивном
отмирании крайних вариантов. Допустим, например, что в дигибридном
расщеплении 1 + 4 + 6 + 4+1 генотипы среднего класса
обнаруживают нормальную жизнеспособность, соседние с ними классы (4) отмирают
на 50% и крайние на 75%. Тогда распределение примет вид:
1/4 + 2 + 6 + 2 + 1/4или 1 + 8 + 24 + 8 + 1.
(*2 =
8
(i4 = т и р2 = 3,5,
Для этого ряда
т. е. наш ряд обнаруживает сильный положительный эксцесс. Легко
видеть, что если жизнеспособность плюс вариант иная, чем
жизнеспособность минус вариант, то в результате такой дифференциальной
жизнеспособности распределение примет асимметричный характер,
скопившись либо вправо, либо влево.
Пожалуй наибольший интерес для генетика-аналитика представляет
обнаружение отрицательного эксцесса, постепенно переходящего в д в у-
или многовершинность.
Следует постоянно иметь в виду, что в ряде случаев многовершинное
распределение может оказаться созданным искусственно и не иметь
никакого отношения к расщеплению. Это часто происходит при попытках
«троить кривые распределения для количественных различий, когда мы
искусственно назначаем границы классов. Легко понять, что если мы
разобьем какой-либо признак на классы разного интервала, то мы получим
вершины там, где интервал будет большим, д понижение там, где будет
интервал узкий. Распределяя в ряд признаки, не поддающиеся точному
измерению, например, вариации пегости, особенности устройства тазовой
кости и т. д., мы обязательно более или менее нарушаем равенство
классовых промежутков, не будучи в состоянии выразить их числами. Хороший
пример такой ложно многовершинной кривой мы заимствуем из интересной
работы Промптова (1926) о генетическом анализе строения таза у кур.
Разбивая кур по устройству таза на классы, он получил трехвершинную
кривую (рис. 73). Ее вид указывает лишь на то, что промежуточные классы
были установлены с очень узкими границами, так что немногие особи
попадали в эти границы. Хотя в данном случае расщепление имело место, но
«удить о нем по виду кривой невозможно.
Посмотрим, каким образом может возникнуть понижение величины |J2.
1. Влияние разной силы генов. Пусть мы имеем ди-
гибридное расщепление с участием генов разной силы и пусть
ген В вдвое сильнее, чем ген А. Для расчетов подобных примеров двойное
действие гена В можно символизировать через (ВВ), написав формулу ди-
гетерозиготной особи через АА (ВВ) Ъ.
208
Рис. 74. Трехверпшнное
распределение при дигибридном
скрещивании
В F2 мы будем иметь тогда следующее распределение генотипов:
без усилителей aabb -. . 1
с 1 усилителем 2 Aabb 2
с 2 усилителями ААЬЬ + 2 аа (ВВ) b 3
с 3 » 4 Аа (ВВ) Ъ 4
с 4 » аа(ВВ)(ВВ) + 2АА(ВВ)Ь 3
с 5 » 2 Аа (ВВ) (ВВ) 2
с 6 » ААВВВВ 1
Для этого сильно растянутого ряда
fi2 = 2,5; (х4 = 14,5; р2 = 2,32,
т. е. значительно ниже, чем |J2 = 2,5, свойственного биному (1 + I)4.
Рассчитав аналогичный пример, но с еще большей разницей в силе
действия (В в три раза сильнее -4), получим уже внешне заметную многбвер-
шинность (рис. 74).
Классы Генотипы Частоты Классы Генотипы Частоты
0 aabb 1 4 4 Аа (ВВВ) Ь 4
1 2 Aabb 2 5 2 АА (ВВВ) Ь 2
2 ААЬЬ 1 6 аа (ВВВ) (ВВВ) 1
3 2 аа (ВВВ) 6 2 7 2 Аа (ВВВ) (ВВВ) 2
8 АА (ВВВ) (ВВ) 1
Для этого ряда:
W = 5*> \*>* = 54,5; р2 = 2,18,
т. е. Р2 еще сильнее уменьшилась, приближаясь к 2, отвечающим ряду
1 + 2 + 1. Очевидно, если ген В будет бесконечно сильнее гена А, мы
получим именно ряд 1 + 2 + 1. Действительно, для двух генов разной силы
имеет место формула: N
fc-^i + ^y],
где А, есть число, показывающее, во сколько раз один ген действует сильнее
другого. При возрастании X второй член в» скобках будет стремиться
к нулю.
Сложение \i2 и ц,4. Воспользуемся данным примером, чтобы
ознакомиться с двумя формулами, которые позволяют при желании
рассчитывать [А2, |i4 и р2 теоретически, не прибегая каждый раз к вычислению
самого ряда. Эти формулы таковы:
I. \л2 (а + Ъ) = \i2 (a) + |х2(Ь)
II. [г4 (а + Ъ) = [г4(а) + \i4(b) + 6jx2 (a)\i2 (b).
14 А. С. Серебровский
209
Если признак варьирует под влиянием двух независимых причин а и Ь,
причем причина а отдельно вызывает вариацию с fx2 (#) и (л4 (я)» а причина
Ь отдельно вызывает вариацию с \i2 Ф) и pt4 (&)»т0 общая вариация
признака будет иметь \i2 (а + 6) и (л4 (а + 6), легко вычисляемые из моментов а и
Ь по приведенным формулам:
Приведем пример: один ген вызвал ряд 1 + 2 + 1 с р,2 = 2/4 иц4 = 2/4.
Два разных гена вызовут вариацию с
2,2 4 .
^2=4" + Т = Т==1;
V* ==Т + Т + 6Т' Т = 2'5;
Р2 = 2,5,
что мы видели выше для бинома (1 + I)4- Если один ген действует сильнее
другого в Я раз, то вызываемые им моменты будут соответственно:
(х2 = тЯ2 и [x4=TA,4.
Для гена тройной силы это дает:
18 162
1*2 =— И [Х4 =— .
Дигибридное расщепление с участием генов А и (ВВВ) даст поэтому
вариацию с
2 , 18 20 к 2 , 162 , « 2 18 к/ к
4 г 4 4
откуда
^ = 2,18,
что мы и видели в предыдущем примере.
2. Влияние сцепления генов. Сцепление генов
оказывает на характер распределения потомства подобное же влияние, как и
различная сила действия генов. Прием, который мы применили для
анализа влияния различной силы генов в предыдущих примерах, позволяет
нам легко понять и роль сцепления. В самом деле, обозначая ген двойной
силы символом (ВВ), мы можем этот же символ применить и для
обозначения двух генов ординарной силы, но абсолютно сцепленных друг с другом.
Для скрещивания с участием двух равносильных полимеров А и 2?, как
мы видим, характер распределения отвечает биномиальному ряду 1 + 4 +
+ 6 + 4 + 1 р (г2 = 1, [х4 = 2,5 и р2 = 2,5, Если гены А жВ будут
абсолютно сцеплены друг с другом, дигетерозиготы будут иметь строенце
(АВ) (ab) и потомство распределится уже в соответствии с рядом 1+2 + 1,
для которого
H>2=-2i И*=у и Р2 = 2.
Но так как действие этого комплексного полимера будет вдвое большим,
т. е. X = 2, то полные величины моментов будут:
\х2{АВ) = уЯ2=2 и М^)=4^4 = 8,
о
откуда все равно (J2 = — = 2*
210
Таким образом, та же пара генов без сцепления давала распределение
с р2 == 2,5, отвечающее биному (1 + I)4, а при полном сцеплении стала
давать распределение с |32 = 2, отвечающее биному (1 + I)2 —
отрицательный эксцесс возрос. Сцепление понижает величину |32.
3. Влияние типа скрещиваний. Различный тип
скрещиваний может тоже существенно отражаться на характере распределения
потомства и параметрах полигонов. Отношения здесь могут быть
чрезвычайно разнообразными. В качестве примера мы сравним распределение
потомства в F2 и в FB (АаВЬ X АаВЪ и АаВЬ X ааЪЪ).
Как разбиралось выше, при равной силе полимеров А и В и
промежуточном доминировании в F2 возникало распределение, отвечающее ряду
1 +4 + 6 + 4 + 1и биному (1 + I)4, в FB же возникает распределение,
отвечающее ряду 1 + 2 + 1 и биному (1 + I)2.
Класс
Без усилителя
1 усилитель
2 усилителя
3 »
4 »
F,
ааЪЪ
2 АаЪЪ + 2 ааВЬ
4 АаВЪ+ААЪЪ+ааВВ
2 ААВЬ + АаВВ
ААВВ
[А2 = 1
tk = 2,5
р2=2,5
/
1
4
6
4
1
(1+1)*
*в
ааЪЪ
АаЪЪ + ааВЪ
АаВЪ
1
1
^= —
ft = 2
/
1
2
1
(W
Замена F2 на FB при данных условиях снижает вдвое степень
соответствующего бинома, понижая соответственно и величину |$2, т- е- увеличивая
отрицательный эксцесс.
Эту закономерность легко уяснить, применив теоремы сложения
моментов. В самом деле, изменчивость потомства при промежуточном
доминировании является суммой изменчивости генотипов гамет обоих
родителей. Каждый из дигетерозиготных родителей АаВЬ дает гаметы АВ +
+ АЪ + аВ + ab, которые при равносилии генов А ж В отвечают ряду
1+2 + 1 или биному (1 + I)2, для которого моменты будут:
п 1 п «(Зп — 2) 1 ,
и« = т = т. и V*= т —~ = т ■
Для родителей ааЪЪ моменты, очевидно, равны нулю.
Соединяя в процессе оплодотворения гаметы обоих родителей, мы
получаем изменчивость зигот по теореме сложения:
для F2 для FB
0,М 1|1 А 1|Л 1
ш = 1*1 +1*1 _; T + Ts=zi; т + 0==т;
О , М , /? О М 1,1,/jl 1 ос
f*4 = i*« + f*f + m; Hi = у + у + 6у • у = 2->5^
где «О» означает отца, а «М» — мать.
Зная распределение генотипов гамет родителей, вводимых в
скрещивание, мы можем легко рассчитать распределение генотипов зигот потомства.
211
14*
В случае полного доминирования распределения F2 и FB могут
различаться по величине |3. Для свободно комбинирующихся генов
распределение генотипов в FB всегда симметрично (при
равной жизнеспособности). Наоборот, в F2, как мы видели, при
доминировании возникает асимметричный полигон с рх ^ 0. Сравним для примера
скрещивание АаВЬ X АаВЬ и АаВЬ X aabb, где гены А и В равносильны
и полно доминируют.
Класс
Без усилителя
1 усилитель
2 усилителя
F2
aabb 1
2 Aabb+ AAbb + 2 ааВЬ + aaBB 6
4 АаВЬ + 2АаВВ + 2ААВЪ + ЛЛЯЯ 9
(1+3)2
Fb
aabb 1
Aabb + ааВЬ 2
(l+l)2
Здесь в F2 мы видим резкую скошенность распределения, а в FB
совершенно симметричное, хотя в обоих случаях имеется по три класса
генотипов: без усилителя, с одним усилителем и с двумя.
4. Влияние паратипической изменчивости. Все
предыдущее изложение относилось лишь к генотипической комбинаторике.
Наблюдаемое в действительности распределение количественных
признаков имеет всегда существенно иной характер благодаря неизбежному
участию паратипической изменчивости. В этих реальных
распределениях фенотипов генотипическое распределение составляет то, что мы
предлагаем называть каркасом. В дигибридном расщеплении, например,
каркасом явится распределение 1+4 + 6 + 4 + 1. Каждое число в
этом ряду будет членом или классом каркаса.
Благодаря неизбежному наличию паратипической изменчивости
каждый член каркаса оказывается представленным уже не одним классом
фенотипов, а многообразной совокупностью, в которой каждая особь несет
свой собственный фенотип, если изменчивость непрерывна (вес, рост), или
несколькими классами, если изменчивость прерывиста (число щетинок,
число яиц и т. д.). При трансгрессивной паратипической изменчивости
изменчивость членов каркаса находит одна на другую и получается
некоторое единое распределение, особенности которого являются сложной
суммой особенностей генотипического каркаса и пар атипического
покрова *.
Закрепляя, как мы приняли выше за элементами фенотипа, генотипа и
паратипа, символы ф, уияв общей форме, мы будем иметь характеристики
фенотипического "распределения:
М^ср = JA2Y + \*>2п\
Если для генотипического каркаса мы имеем возможность дать
довольно стройную теорию, элементам которой было посвящено предыдущее
изложение, то для паратипического покрова такой теории дать почти
невозможно. Паратипическая изменчивость зависит от столь разнообразных,
часто трудно учитываемых причин, что возникающее на их почве распре-
* Вводя для удобства дальнейших рассуждений понятия генотипического каркаса и
паратипического покрова, мы ясно понимаем условность этого деления, так как
несомненно, что всякий организм представляет единство наследственного и
ненаследственного.
212
Рис. 75. Скашивание кривой
распределения вблизи от
минимума
О М 2М ЗМ
деление фенотипов может иметь самый разнообразный характер. Одна и
та же чистая линия в различных условиях может дать весьма различные
вариационные кривые. Поучительной является произведенная еще Пир-
лем и Сюрфасом (Pearl a. Surface, 1914) биометрическая обработка
распределения по яйценоскости стада кур Мэнской станции в разные гбды и в
различных птичниках. Хотя, правда, генотипический материал здесь и не
был однороден, но резкие изменения формы кривых распределения,
несомненно, вызывались не гено-, а паратипическими факторами, изменялся
размах изменчивости (величина р,2), степень асимметрии и соответственна
по величинам рх и р2 кривые распределения приходилось относить к
различным типам пирсоновских кривых.
В этой, не поддающейся пока теоретическому предсказанию
многообразности и лабильности паратипической изменчивости лежит основное
затруднение при рассмотрении фенотипических кривых распределения
при трансгрессивной изменчивости. Если мы видим на такой кривой
правую или левую скошенность, указание на многовершинность и т, п., то
прежде всего возникает вопрос, ответствен ли за эти особенности
генотипический каркас или лишь пар атипический покров его. Для генетика-
аналитика, естественно, основной интерес представляют особенности
каркаса, и все усилия должны быть направлены на то, чтобы обнажить этот
каркас от одевающего его паратипического покрова. По внешнему виду
кривой распределения фенотипов этого сделать нельзя и необходимы
специальные приемы для этого.
Правда, паратипическая изменчивость в большом количестве случаев
весьма удовлетворительно может быть описана нормальной кривой
Гаусса, т. е. параметры рх и р2 нереально или незначительно отличаются
соответственно от 0 и 3. Особенно устойчивой является величина рг, близкая
к 3. Но, как мы видели, и для полигонов уже при незначительном числе
полимеров эта величина тоже уже незначительно отличается от 3. Более
сильными бывают уклонения величины рх от 0. При этом положительная
скошенность возникает тогда, когда вся вариация происходит вблизи
нуля. Так как большинство биологических признаков (рост, молочность,
0 90 0 90 0 90 яиц
Рис. 76. Скашивание кривых распределения при приближении к минимуму и к
максимуму. Распределение кур по яйценоскости (схема)
213
плодовитость и пр.) не может принимать отрицательных значений, то
вариация от средней (М) в минус-сторону оказывается стесненной и не
может превысить величины М — 0 = М (рис. 75 и 76). Наоборот, в плюс-
сторону вариация может происходить свободнее и часть вариационной
кривой заходит за М + М = 2М.
Возможны и противоположные случаи, когда вариация происходит
далеко от нуля, приближаясь к некоторому верхнему пределу или
«потолку». Если, например, построить распределение кур по яйценоскости за
апрель — июнь, то может получиться в хороших стадах левоскошенная
кривая, т. е. многие куры подойдут по своей кладке к возможному пока
пределу 90 яиц за 90 дней, и вариация в плюс-сторону тем самым будет
стеснена. Наоборот, в минус-сторону будет более свободная возможность
варьировать до нуля (кривая 3, рис. 76).
В некоторых случаях несимметричную кривую удается
интерпретировать как симметричную, изменяя строение оси абсцисс, например,
разметив ее не на равнозначные классы, а так, чтобы каждый следующий класс
был в равное число раз больше предыдущего, например, не на 1, 2, 3, 4,...,
а на 1, 2, 4, 8, т. е. взяв за единицу масштаба оси абсцисс не абсолютную
величину признака, а ее логарифм. Обычно такая замена, впрочем,
оказывается слишком грубой, и правоскошенная кривая превращается не в
симметричную, а в левоскошенную. В этих случаях можно прибавить перед
логарифмированием какую-либо постоянную величину. Впрочем, такая
перестройка кривых является слишком кропотливым мероприятием и
редко удается вполне.
К счастью, как сказано, в большом числе случаев паратипическая
изменчивость вполне удовлетворительно передается нормальной кривой
Гаусса или биномом (1 + 1)п уже сравнительно невысокой степени
(тг = 10 и т. п.). Для разного рода теоретических выкладок вполне
возможно пользоваться подобной моделью, достаточно удобной, не требующей
интегрирования и достаточно близко моделирующей действительность.
Особенно удобным упрощением является при этом то, что распределение
генотипов и фенотипов характеризуется прерывистыми рядами (как
в случае прерывистой изменчивости), позволяя легче производить нужные
операции.
[[ВЛИЯНИЕ СООТНОШЕНИЯ ГЕНО- И П4РАТИЦИЧЕСК0Й ИЗМЕНЧИВОСТИ
НА ХАРАКТЕР РАСПРЕДЕЛЕНИЯ ^ФЕНОТИПОВ
Покажем на примерных расчетах, как изменение отношения размаха
паратипической и генотипической изменчивости отражается на характере
распределения фенотипов. Пусть мы имеем в F2 расщепление по одному
гену с & = 1, а паратипическую изменчивость схематично изобразим
биномом (1 + I)6. Следовательно, будем иметь каркас 1 + 2 + 1 с
интервалами между классами в единицу, а каждый член каркаса будет одет
паратипическим покровом в виде ряда 1+6 + 15 + 20 + 6 + 1. Общее
распределение фенотипов в F2 будет, следовательно:
1 1 6 15 20 15 6 1
2 2 12 30 40 30 12 2
1 1 6 15 20 15 6 1
1 8 28 56 70 56 28 8 1
Этот ряд является не чем иным, как разложением бинома (1 + I)5.
Обратим внимание на то, что суммация пара- и генотипической
изменчивости прошла по очень простому закону — из биномов (1+ I)2 и (1 + 1)в
214
Рис. 77. Появление трехвершинности в моногибридном F2 по мере увеличения силы
гена i и неизменной <з п
мы получили (1 + I)8, т. е. простым перемножением:
(1 +1)2-(1 +1)6 = (1 +1)8.
у п Ф
Распределение получилось даже с некоторым увеличением высоты
полигона (уменьшением отрицательного эксцесса), так как в этом примере
Par = 2; р2я = 2,67; р2, = 2,75,
Пусть теперь в аналогичном примере сила гена будет не i = 1, а
i = 2, i = 3 и т. д. Тогда соответствующие распределения фенотипов
примут вид, показанный в табл. 12.
Таблица 12
Распределение фенотипов по классам при разных значениях i
1 = 2
1
2 = 1
6
6
2
15
17
12
- 20
32
1
30
15
46
6
40
6
52
15
30
1
46
20
12
32
15
2
17
6
6
1
1
1
2 = 1
6
6
15
15
2
20
22
12
15
27
30
6
36
1
40
1
42
6
30
36
15
12
27
20
г
22
15
15
6
6
1
1
1 = 4
1
2 = 1
6
6
15
15
20
20
2
15
17
12
6
18
30
1
31
40
40
1 1
30
31
6
12
18
1 15
2
17
| 20
20
15
15
6
6
1
1
При графическом изображении этих распределений получим рис. 77.
Из рядов и особенно графиков наглядно видно, как по мере увеличения
силы действия гена распределение фенотипов из правильного
биномиального становится все более растянутым и неправильным и, наконец, в нем
обозначаются три ясные вершины, на которых сразу видно, что материал
генотипически неоднороден и в нем идет расщепление по моногибридной
схеме. Такой же самый результат, естественно, получился бы, если бы
215
абсолютная сила действия гена оставалась
бы равной единице, но уменьшился бы
размах паратипической изменчивости.
Отсюда ясно, что для характера кривой
распределения важнейшее значение имеет
именно соотношение степени (размаха)
паратипической и генотипической
изменчивости. Так как наилучшей мерой
изменчивости является ее квадратическое
уклонение, то можно сказать, что для
легкости понимания кривой распределения и
возможности заключать по ней о наличии
и характере расщепления основное
значение имеет отношение сгу/сгя.
Ту же роль имеет и отношение
, названное Райтом коэф-
V«
г+<
Рис. 78. Появление пятивершин-
ности в дигибридном F2 по мере
увеличения силы генов
(изображена лишь половина распределений)
фициентом корреляции между генотипом
и фенотипом.
В наших примерах величина <yY была
(с учетом силы действия i) равна J/ ^- -i2,
где п — степень бинома и i — сила гена.
В четырех разобранных случаях эти величины были, последовательно:
2 '
з yl
v±±-
2 У 2
2
4 У г
Величина <у„ всюду равна 1/ — = -^— , а отношение ау/оп было, по-
12 3 4
следовательно, тр=-, -у~, -г^=-, -у= или0,57; 1,15; 1,73 и 2,30. Приоу/ол =
=2,30 трехвершинность ясно видна, при 1,73—намечается, при 1,15 —
незаметна.
При расщеплении по двум генам при той же силе гена и aY = 4, F2
будет иметь такое распределение:
1 6 15 20 15 6 1
4 24 60 80 60 24 4
6 36 90 120 90 36 6
4 24 60 80 60 24 4
1 6 15 20 15 6 1
2=1 6 15 20 19 30 61 80 66 60 94 120 94 60 66 80 61 30 19 20 15 6 1
Здесь мы имеем aY = 4, а отношение ау/а„ = 8/^6 = 3,07. Пятивер-
шинность выражена, однако, здесь несколько менее резко, чем
трехвершинность при Оч/оп = 2,30. При i = 3, когда оу/в„ = 2,45, что близко
к 2,30, пятивершинность видна уже очень плохо (рис. 78, пунктир).
Таким образом, число генов, участвующих в распределении, тоже
влияет на ясность расщепления. При одном и том же отношеннии оу/оп
чем больше генов, тем каркас труднее уловим.
После этого краткого обзора генотипической комбинаторики перейдем
к более подробному изучению трех методов анализа количественных
(трансгрессивных) признаков.
216
ГЛАВА X
МЕТОД МЕНДЕЛИСТИЧЕСКИЙ ИЛИ ИЗУЧЕНИЕ РАСЩЕПЛЕНИЯ
В ПОТОМСТВЕ
УСЛОВИЯ ПРИМЕНЕНИЯ
Как мы видели выше, этот метод особенно успешно применим в том
случае, когда анализируемый организм может образовывать клоны
вегетативным размножением. При анализе животных эта возможность
практически отпадает, и об образовании клонов можно говорить лишь в
исключительных случаях, пока не использованных генетиками. Так, мыслимо,
например, применение культур тканей для генетического анализа
клеточных особенностей животных *. Мы можем ставить различные
скрещивания и от каждой особи брать штамм для культивирования его в различных,
но для разных особей в одинаковых условиях культуры. Таким образом,
вероятно, возможно будет подвергнуть детальному генетическому
анализу такие признаки, как величина клетки, скорость роста и деления,
особенности строения и физиологии, отношение к химическим воздействиям
и т. п. Попыток такого рода работ генетиками сделано не было, кроме
как для целей изучения кариотипа здоровых и больных тканей человека,
Можно думать, однако, что некоторые важные вопросы генанализа
сельскохозяйственных животных, связанные с проблемой конституции и роста,
могли бы быть интересно освещены этим приемом. При применении этого
приема внешняя характеристика особи замещалась бы цифрами,
получаемыми от культуры тканей. Так как эти культуры можно проводить через
ряд пассажей (пересадок), то паратипические особенности отдельной
культуры оказались бы элиминированы, и мы имели бы достаточно точную
характеристику клеток каждой особи, необходимую для выводов.
Как правило, однако, животные размножаются аллогамно, и
применение метода надо ориентировать на этот способ размножения.
АНАЛИЗ МЕЖРАСОВЫХ РАЗЛИЧИЙ
Мы начнем с анализа скрещивания двух рас или двух более или менее
кровных (инбредных) линий, предполагая, что несмотря на неизбежную
гетерозиготность, основное различие между этими расами все же более
или менее гомозиготировано. Значительная однородность Fx от
скрещивания, например, пород кур подтверждает это. Предположим, что мы
получили Fj и F2 и можем получить при желании оба возвратных (обратных)
скрещивания FB и F3.
* В культуре тканей животных и растений к настоящему времени достигнуты очень
большие успехи, так что мысли А. С. Серебровского по этому поводу отнюдь не были
фантастическими.— Прим. ред.
217
Какие выводы можно рассчитывать извлечь из такого материала и что
необходимо для решения различных задач?
Основной задачей является выяснение вопроса о характере
расщепления, происшедшего в F2, для чего требуется обнажить каркас F2 от пара-
типического покрова. Для этого возможно применение нескольких
приемов. Разберем их на простейших примерах.
Первый прием. Скрещивание осэбед F2 с чистыми расами
Теория этого приема следующая. Пусть, например, наши расы
различаются по одному гену А.
I раса II раса
аа Fi АА
Аа
F2
аа + 2Аа -f- АА
Так как благодаря трансгрессивной пар атипической изменчивости
генотипы F2 не различимы по фенотипу с достаточной точностью, то
следует взять особей F2 без какого-либо выбора и скрестить с одной из рас.
При неполном доминировании будет безразлично, скрещивать ли с I или
со II расой. При более или менее выраженном доминировании, узнаваемом
по приближении ¥г к одной из рас, надо выбрать рецессивную расу.
При моногибридном расщеплении скрещивание особей F2 с I расой
пойдет по следуюпщм трем типам:
(1) ааХаа > аа
(2) аа X Аа > Аа + аа
(3) аа X АА ► Аа.
Первый тип скрещивания дает потомство, подобное чистой расе аа.
Третий тип скрещивания дает потомство, подобное Fx, а второй тип дает
потомство строго промежуточное между I расой и III. При этом
возникнут такие закономерности:
1. Число скрещиваний (1) будет равно числу скрещиваний (3),
образуя по 25% всех скрещиваний.
2. Число скрещиваний (2) составит 50% всех скрещиваний.
3. Помимо величины М скрещивания будут характеризоваться и
величиной а (или |х2 = а2). В (1) квадратическое уклонение будет сходно
с I расой, в (3) — с Fx. Эти сигмы могут быть либо равными, либо в (3)
больше, чем в (1), так как гетерозиготы часто варьируют сильнее гомозигот.
4. В скрещивании (2) а будет увеличена.
Степень увеличения сигмы в скрещивании (2) можно рассчитать таким
образом. Разность между Fx и I расой равняется силе действия гена
А, т. е. L Отношение 1 : 1 в скрещивании (2) есть бином (1 + I)1,
следовательно, для него
Величину el находим по материалу ¥г или I расы, если они одинаковы.
Если они различны, берем среднюю из обеих. Отсюда для скрещивания
типа (2)
бср = <3Y + On = -j- + Gn
218
Успех этого приема будет зависеть от того, удастся ли от каждого
индивидуального скрещивания получить достаточное количество
потомства, чтобы скрещивания (1), (2), (3) отчетливо отличались бы друг от
друга. Потребное количество потомства будет зависеть от отношения силы
i гена и степени паратипической изменчивости. Для животных, имеющих
мало потомства и особенно рождающих одного детеныша, индивидуальные
скрещивания, конечно, не дадут результатов. В этом случае из F2 нужно
будет взять лишь самцов и каждого самца использовать на возможно
большем числе самок при помощи искусственного осеменения.
В случае дигибридного расщепления в F2 будет уже девять генотипов
и шесть типов скрещиваний с рецессивной расой (табл. 13).
Таблица 13
Расщепление в F2. Средние арифметические и а отдельных групп
Скрещивания и их потомство
/
М
(1) ааЪЪХааЪЬ -»
(2) ааЪЪхАаЬЪ-*
(3) ааЬЬХааВЬ^
(4) ааЪЪХЛАЪЪ^
(5) ааЪЪХааВВ-*
(6) ааЪЬхАаВЪ-*
+ aabb
(7) aabbXAABb-
(8) aabbXAaBB
(9) aabbXAABB
aabb
ааЬЬ + Aabb
ааЬЬ + ааВЬ
АаЬЬ
>ааВЬ
АаВЬ + АаЬЬ + ааВЬ-\-
-* АаВЬ + Aabb
-* АаВЬ + ааВЬ
-> АаВЬ
} М2 =
М3
Mi
Mi+ M3
/4-'
+ °l
Mi--
M3
M3+ M5
2
Ms
v_
2 о
+4
По величине средних арифметических (М) скрещивания распадаются
на 5 групп в пропорции:
Mi 1 Mi == Мр
М2 4 M2=Mp+0,5i
Мг 6 M3=Mp + i
М* 4 М* = Мр + 1,Ы
Ms 'l Ms = Mp+2i
где Мр — среднее арифметическое родительских рас.
Образовался тот же самый бином (1 + I)4, но только с классовым
промежутком вдвое меньшим, чем в F2. Такое распределение получится,
впрочем, лишь в том случае, если силы генов А ж В равны или более или
менее равны. В противном случае будет девять разных М, но более или
менее сгруппированных так:
Мг; M2MS; М^МЬМ%) М7МЪ; М9.
Что касается величины сигм, то они будут наименьшими у (1) и (9), т. е.
там, где средние равны Мг и М6. Эти сигмы будут равными соответственно
<т чистой породы и Fx. При (2), (3), (7) и (8) сигмы будут несколько
увеличены до
где i равно половине разности между расой и Fx.
В скрещиваниях (4), (5) и (6), имеющих одинаковую М3, величина а
будет очень различной. В двух случаях она будет близкой к ая и в четы-
219
рех случаях достигает наибольшей величины, равной ^/ -^-£2 + <з*. В том
случае, если а в чистой расе и в ¥г будет приблизительно одинаковой, среди
наших скрещиваний мы будем иметь три различных величины ап (удобна
сравнивать не а, а а2).
с2
<*
1 9
2 ..
/
4
8
4
Скрещивания
(1) (4) (5) (9)
(2) (3) (7) (3)
(6) -
Для того чтобы сделать данную картину распределения М и а
достаточно ясной, в дигибридном скрещивании потребуется, очевидно, еще
большее количество потомства, примерно в четыре раза большее, чем при
моногибридном, если разница между исходными расами и степень пара-
типической изменчивости а та же самая.
Использование FB. При применении этого приема нужно иметь
в виду возможность значительного выигрыша путем замены F2 на FB. При
этом скрещивание ведется так:
I раса X И раса
I
I раса X Fi
I
I раса X FB
Отметим, что такой ряд скрещиваний на зоотехническом языке носит
название грединга, или поглощения, в данном случае поглощение второй
породы первою. В процессе метизации, проводимой в СССР, такие
скрещивания встречаются часто.
В случае моногибридности это скрещивание изобразится так:
аа X АА
I
аа X Аа
I
аа[Ла
\ аа
Так же, как ранее, мы выбирали «случайно» особей из F2 и скрещивали
их с чистой расой, здесь мы выберем особей из FB.
Получим только два возможных типа скрещиваний
Скрещивания и потомство
(1) ааХ аа -» аа
(2) ааХ Аа-> аа-\- Аа
1
1
1
м
Mi
М2
о2
4
4.
По сравнению с представленным ранее анализом моногибридного F2
здесь картина будет значительно проще — ожидается не три типа
скрещиваний, а лишь два, соответственно чему будет лишь два типа средних
арифметических — Мг и М2, из которых Мг должно равняться М чистой
расы, а М2 = М для F|. Правда, так как интервал между Мг и М2 остается
прежним, в каждом скрещивании необходимо получить такое же количест-
220
во потомства, чтобы суметь отличить скрещивания типа (1) и (2) друг от
друга. Но зато нам надо было выявить распределение скрещиваний (1),
(2) и (3) по формуле 1 + 2 + 1, здесь же необходимо выявить
распределение типов скрещиваний (1) и (2) только по формуле 1+1. Ясно, что для
этого потребуется вдвое меньшее число скрещиваний, а это даст
возможность (например, при применении искусственного осеменения) вдвое
повышать число потомства в каждом скрещивании и в 1,4 раза (1,4 ^ }/~2)
повышать точность картины.
В дигибридном скрещивании аналогично мы получаем состав FB из
<aabb + Aabb + ааВЬ + АаВЬ и при скрещивании с расой aabb будем иметь
не 9, а 4 типа скрещиваний (табл. 14).
Таблица 14
Расщепление в FB. Средние арифметические Мне2 отдельных групп
Скрещивания и потомство
(1) ааЪЪХ аоЬЪ -» aabb
<2) aabbX Aabb -> Aabb -f aabb
(3) aabbX aaBb -» aaBb + aabb
<4) aabbXAaBb-* AaBb + Aabb + aaBb-\-
+ aabb
f
1
1
1
1
M
Mi
M2
M2
Mi+ Ms
2
>
(Mi +\)
(Mi + i)
0*
«S
1 ., 2
~fi +4
-§-'2 + °* = 4в
В случае равенства силы генов А и В в скрещивании будут лишь три
класса М: (1) — Aflf (2) и (3) — М2 и (4) — М3. В случае неравенства
(сила А = гг, сила В = i2) будем иметь четыре величины М, причем
такого строения:
Mi = М чистой расы; М2 = Mi + -«г;
Л/з = Mi + -у- и М4 = М! + ^-^- .
Определив из опыта величины Mv M2, М3, М4, мы сможем
непосредственно иметь величины ix и £2.
Выгода этого приема по сравнению с исследованием F2 также сводится
к возможности получения выводов на меньшем материале или получения
яа том же материале выводов более точных. В самом деле, каркас F2
имел строение 1+4 + 6 + 4 + 1, а здесь 1 +2 + 1. Для построения
картины потребуется в четыре раза меньше скрещиваний, а при том же
числе скрещиваний в некоторых случаях может быть получено в четыре
раза больше потомства в каждом, что даст в два раза более точный
результат. Кроме того, при неравенстве силы действия генов А ж В при анализе
F2 мы должны установить девять типов скрещивания, что уже очень
затруднительно. Здесь же лишь четыре. При таком малом числе типов
скрещиваний не представляет трудности решить, какую формулу дать тому или
иному скрещиванию и тем самым отличить (2) от (3). Отличить эти два
скрещивания друг от друга и от (1) очень важно, так как в (2) имеется ген
А без участия В и в (3) ген В без участия А. Различив скрещивания (2) от
(3), мы различаем ген А от гена В и можем в дальнейшем перевести его
в гомозиготное состояние и т. д. Оба гена окажутся таким образом «в
наших руках», что и представляет собой существенную цель генетического
анализа — не только познать гены, но и дать возможность овладеть ими.
При наличии в F2 9 генотипов внести такую же ясность в этот вопрос,
очевидно, значительно труднее.
221
Существенная положительная сторона использования FB для анализа
состоит в прямой возможности продолжения этого скрещивания. Мы
можем из наметившихся классов каркаса или из общей массы скрещиваний
выбрать представителей для дальнейшего скрещивания с исходной расой.
Ожидаемые при этом результаты легко рассчитать.
Недостаток первого приема. Изложенный прием, однако, даже при
достаточном количестве потомства не может дать несомненного ответа.
Причина этому следующая.
Мы всегда можем рассчитывать встретиться со скрещиванием такого
типа:
ааВВ X ААЪЬ
АаВЪ
где А — ген-усилитель, а В — ген-ослабитель (с полным
доминированием). Тогда картина скрещивания будет сходна с промежуточным
наследованием. Например, А = +1, В = —1, тогда:
ааВВ = — 2, ААЪЪ = + 2, АаВЪ = 0.
Но, благодаря доминированию, каркас в F2 будет таков:
без усилителя ааВВ -\~ 2 ааВЬ 3
с 1 усилителем 4 АаВЪ + 2АаВВ + 2 ААВЪ + ааЪЪ + ААВВ .... 10
с 2 усилителями ААЪЪ + 2 АаЪЪ 3
При обратном скрещивании F2 с расой ааВВ будем иметь следующие
величины Mug (табл. 15).
Таблица 15
Результаты скрещивания F2 с расой ааВВ
Тип скрещивания
(1) ааВВ X ааВВ
(2) ааВВ X ааВЪ
(3) ааВВ X ааЪЪ
(4) ааВВ X АаВВ
(5) ааВВ X АаВЪ
(6) ааВВ X АаЪЪ
(7) ааВВ X ААВВ
(8) ааВВ X ААВЪ
(9) ааВВ X ААЪЪ
Тип потомства
ааВВ
ааВВ + ааВЪ
ааВЪ
АаВВ + ааВВ
ааВВ + ааВЪ + АаВВ +
+ АаВЪ
ааВЪ + АаВЪ
АаВВ
АаВВ + АаВЪ
АаВЪ
Фенотип
) а, В
[а, В +
+ А, В
>
\
\ А, В
J
/
2 4
1
2
4 8
2
1
2 4"'
1
м
Мх
М2
М3
о
4
т-*+4
4
i
Здесь £ равно силе действия гена А и, с другой стороны,— половине
разности между скрещенными породами.
В результате мы обнаруживаем только три типа скрещивания:
a) дающее в потомстве лишь а, В
b) » » » » л, В + А, В
c) » з> » » Л, 5
Эти скрещивания встречаем в пропорции 4 + 8 + 4, т. е. иначе 1 + 2 +
+ 1. Сравнив с предыдущим, мы видим, что у нас получилась типичная
моногибридная картина, как будто породы ааВВ и ААЪЬ различаются
лишь одним геном и притом неполнодоминирующим, что, очевидно, не-
222
верно, так как в нашем примере участвовало два гена и притом полно
доминирующих. Получилось это потому, что раса, выбранная в качестве
анализатора, оказалась неправильно выбранной. Такой же
неверный ответ получился бы, если бы мы взяли в качестве
анализатора вторую расу ААЪЪ. Правильный ответ получился'бы, если бы
могли выбрать в качестве анализатора расу или генотип aabb, но хотя такие
генотипы среди F2 и существуют, найти их невозможно, не зная, как они
должны выглядеть. Заметить сделанную ошибку можно лишь исЬледова-
нием р2 для F2, распределение которого (3 + 10 + 3) дает значительно
большую Р2, чем распределение 1"+ 2 + 1, а именно 22/3 вместо 2.
Обнаружить, однако, это различие наблюдением очень трудно.
Второй прием. Скрещивание особей F2 или F^ с особями ¥г
Недостаток изложенного выше приема состоит в том, что мы вынужденны
были в последнем примере выбрать в качестве анализатора форму,
гомозиготную по одному из доминантных генов (ааВВ), и тем сделали
невозможным, при полном доминировании, обнаружение расщепления по гену В.
В качестве анализатора, однако, можно выбрать не чистую породу, а
особей ¥г. Эти особи ¥г заведомо гетерозиготны по всем
генам, участвующим в распределении, и
должны дать возможность обнаружения
расщепления по всем генам.
Разберем моногибридное скрещивание при этом приеме. При
скрещивании особей ¥г с особями F2 возникают такие типы скрещиваний (табл. 16).
Таблица 16
Результаты скрещивания особей F2 с особями Fi
Скрещивания и потомство
(1) Аа X аа-> аа + Аа
(2) Аа X Аа -» аа + 2 Аа + АА
(3) АаХАА^Аа + АЛ
f
1
2
1
м
Mi
м2
Ms
Qt
7Г? + °1
-г'г-К
—l +<£
При неполном доминировании возникнут три различные средние
величины потомства от этих скрещиваний:
М1==Мр+4; M2 = Mp+i = MFa и М3 = Мр+^.
Эти типы М возникают в отношении 1 : 2 : 1. Мы получим, следовательно,
такую же моногибридную картину, как и выше, только все величины М
будут несколько больше. Точно так же будут несколько больше (на V**2)
и величины а2.
Если с особями ¥г мы будем скрещивать не особей F2, а особей FB,
картина опять упростится. Мы будем иметь два типа скрещиваний:
Скрещивания и потомство
(1) АаХаа -» да + Аа
(2) Аа X Аа-* аа + 2 Аа + АА
f
1
1
м
Mi
м2
223
Выгоды такой замены F2 на FB те же самые, какие мы обсуждали выше.
Таким образом, мы видим, что и при замене в качестве анализатора
чистой породы особями ¥г мы получаем столь же четкую моногибридную
картину.
Некоторое отличие возникает при полном
доминировании гена А, которое уже будет заметно по Fx. В этом случае приведенные
три скрещивания дадут такие по фенотипам результаты (табл. 17).
Таблица 17
Результаты скрещивания F2 с Fi при полном доминировании А
Скрещивания и их потомство
(1) Аа X яа -* а\ + А\
(2) Аа X Аа -» <ц + ЪАг
(3) Аа X АА -* Аг
f
1
2
1
м
м1 = мр+^г
M2 = Mp + J^-i
M3 = Mp-{-i
о2
4-+**
О о о
— + а; = <5Fz
2
В этом случае распределение в скрещиваниях (2), наиболее частых,
будет несколько более асимметричным, чем в (1) и (2) (при симметрии пара-
типической изменчивости) или иметь скошенность в ту же сторону, куда
скошен и каркас. Если покров скошен в другую сторону, чем каркас, то
скошенность распределения в (2) будет, наоборот, меньшей, чем в (1) и (2).
В случае дигибридного распределения мы будем иметь при данном
приеме девять различныхскрещиваний (табл. 18).
Таблица 18
Тины скрещиваний на F2 при двух полнодоминантных генах с равным действием
Тип скрещивания
(1) AaBbXaabb
(2) АаВЪхАаЪЬ
(3) AaBbXaaBb
(4) АаВЪхААЪЪ
(5) АаВЬхааВВ
(6) АаВЪхАаВЪ
(7) АаВЪхААВЪ
(8) АаВЬхАаВВ
(9) АаВЪхААВВ
Тип потомства
ааЬЪ + АаЬЪ + ааВЬ + АаВЪ
ааЬЬ +2АаЪЬ + ААЬЬ+ааВЬ + 2AaBb+
+ ААВВ
ааЬЪ + 2ааВЪ + ааВВ+АаЬЬ+2АаВЬ+
+ АаВВ
АаЬЬ + ААЬЬ -f АаВЬ + ААВЬ
ааВЬ + АаВЬ + ааВВ + АаВВ
все как F2
АаЬЬ+2АаВЬ+АаВВ+ААЬЬ+2ААВЬ+
+ ААВВ
ааВЬ+2АаВЬ+ААВЬ+ааВВ+2АаВВ+
+ ААВВ
АаВЬ + ААВЬ + АаВВ + ААВВ
f
1
2
2
1
1
4
2
2
1
(iA = iB)
Mi = Mp + i
\
}
„ M2=Jfp + l,5i
. Me=Afp + 2i
J
k M4=Mp+2,5t.
M5 = Mp + 3£
Сравнивая с таблицей 13, мы видим лишь, что все М выше на i и, оче-
видао, также все а2 будут выше на 2/4 f2, даваемое дигетерозиготным
родителем. Все остальное оказывается результатом первого приема.
Посмотрим теперь, что дает этот прием при случае с полно до
минирующим усилителем и ослабителем, для которого мы получили первым
приемом неверный результат.
224
Пользуясь вышеописанной таблицей, перепишем скрещивание с
указанием получающихся фенотипов, (табл. 19).
Таблица 19
Типы скрещиваний на F2 при двух полнодоминантных генах, один из которых
усилитель, а другой ослабитель
Тип скрещивания
(1) AaBbXaabb
(2) AaBbXAabb
(3) AaBbXaaBb
(4) AaBbxAAbb
(5) AaBbXaaBB
(6) AaBbXAaBb
(7) AaBbXAAbb
(8) AaBbXAaBB
(9) AaBbxAABB
Распределение по классам
0
a, b
a, b
a, b
a, b
+1
4, b
34, b
A,b
24, b
ЗА, b
24, b
-1
a, B
a, B
3a, 5
2a, В
3a, 5
2a, В
0
Л, 5
34, В
ЗА, В
24, В
2АУ В
94, Я
64, В
64, В
4, В
/
1
2
2
1
1
4
2
2
1
—1
1
1
3
1
3
1
0
2
4
4
1
1
10
3
3
1
+1 j М
1
1
3
1
1
3
1
Mz
Мь
М2
Мъ
Мх
М3
м*
м2
Mz
Если ikfp = — 1 есть среднее для расы ааВВ, а i = 1 есть сила
действия гена А, то величины будут иметь следующие значения (при силе гена
В г2 = -1 = -*):
Afi = Mp + 0,50i 1
Af2 = Mp-;-0,75* ...... 4
Л*з = Мр + 1,00$ 6
Af4=Afp+lf25* 4
M5 = MP+1,50* 1
Мы видим, таким образом, что, несмотря на введенное осложнение,
каркас получился типично дигибридным. Таким образом, мы избежали
ошибки в определении числа генов. Зато, однако, от дигибридного случая двух
равных усилителей с неполным доминированием этот случай стал
отличаться уже заметно менее.
Эти отличия следующие.
1. Распределение М имеет а2 в 4 раза меньшую, а разность между Мь и
Мх в два раза меньшую, так как в случае двух усилителей с неполным
доминированием Мь — Мх = 2£, а в случае усилителя и ослабителя с
полным доминированием Мь — Мг = li. Это различие, однако, не
существенно, так как в обоих случаях эта разность равна половине разности между
скрещенными породами или равна разности между чистой породой и ¥г.
2. Значения J32 для распределений в F2 и в распределениях,
получившихся от суммирования скрещиваний ¥г X F2, заметно отличаются друг
от друга. Так, в случае двух усилителей каркас для S (Fx X F2) будет
1 + 4 + 6 + 4 + 1и|3 = 2,5, а в случае усилителя и ослабителя каркас
для 2 (Fx X F2) будет 3 + 10 + 3 и р2 = 2,67.
Это различие, однако, тоже стушевывается паратипическим покровом
и может быть обнаружено лишь методом решения уравнений.
3. В половине скрещиваний, а именно во всех, дающих М2 и М4
[скрещивания (2), (3), (7) и (8)], возникает асимметричное распределение
1+4 + 3, 1+3, 3 + 1,3 + 4 + 1. Это отличие очень интересно, так
как в предыдущем случае асимметрии не было вовсе.
Однако для проверки всех этих отличий наиболее убедительным
должно явиться сопоставление данных второго приема и первого приема, т. е.
15 А. С. Серебровский
225
скрещивания с ¥г и с чистой расой. В первом случае, при двух усилителях,
оба ответа одинаково говорят о дигибридности скрещивания, в случае же
доминантных усилителей и ослабителей ответы будут резко
противоречивыми.
Упрощение второго приема. И второй прием также может быть
значительно упрощен использованием для анализа не F2, a FB.
Сравним ход этих скрещиваний для двух последних дигибридных
случаев.
1. Случай двух усилителей при скрещивании FB X ¥г даст типы
скрещиваний:
Тип скрещивания
(1) АаВЬХааЬЬ -»
(2) АаВЪхАаЪЪ-*
(3) АаВЬХааВЪ -+
(4) АаВЬХАаВЬ
Тип потомства
ааЬЬ + АаЪЬ + ааВЬ + АаВЬ
ааЬЬ + 2АаЬЬ + АаЪЬ + ааВЬ +
+ 2АаВЬ + ААВЬ
ааЬЬ + 2ааВЬ + ааВВ + АаЬЬ +
+2АаВЬ + АаВВ
как в F2
/
1
1
1
1
м
, Л/р+. 1,5£=М2
Mv+2i = Mi
о*
2 . 9
3 -о 2
— *+<4
По сравнению с упрощением первого приема вся разница сводится
лишь к тому, что все М больше на i и все а2 больше на -г-*2, т. е. выгоды
этого упрощения и здесь те же самые, как и указанные выше.
2. В случае доминантных усилителя и ослабителя состав FB будет
таким:
АаВЬ X ааВВ -> ааВЬ + АаВЬ + ааВВ + АаВВ
и скрещивания ¥г X FB будут таких типов:
Тип скрещивания
(1) АаВЬХааВЬ-*
(2) АаВЬХАаВЬ -»
(3) АаВЪХааВВ-*
(4) АаВЬхАаВВ-*
Тип потомства
ааЬЬ+2ааВЪ + ааВВ+АаЬЬ+2АаВЬ+
+ АаВВ
как в F2
ааВЬ + АаВЬ + ааВВ + АаВВ
ааВЬ + 2АаВЬ + ААВЬ+ ааВВ +
+ 2АаВВ + ААВВ
0
а, Ь
1
1
+1
А%Ь
1
3
—i
о, В
3
3
1
1
0
А, В
3
9
1
3
или, объединяя классы a, b и А, В:
(1)
(2)
(3)
(4)
—i
4
10
1
3
+1
1
3
м
м2
Мг
Mt
Mi = Mp+-L.i
Ms^Mp+i
226
Таким образом, и здесь получился типичный дигибридный каркас,
в 1/4 случаев возникает высокое распределение каркаса (3 + 10 + 3) и
в 50% [в (1) и (4) скрещиваниях], дающих М2, распределение каркаса
асимметрично (если симметричен паратипический покров).
Отметим также, что если просуммировать результаты всех четырех
типов скрещиваний, то получим также сильно асимметричное
распределение каркаса (перед суммированием все отношения приводим к 16):
(3 + 4 + 1) X 2, (1 + 1) X 8 и т. д.:
(1) ,
(2)
(3)
(4)
—i
6
3
8
4
21
0
8
10
8
12
38
+2
2
3
5
В предыдущем случае при таком суммировании получился бы почти
симметричный каркас строения 9 + 24 + 22 + 8 + 1.
Эти каркасы по своей скошенности различны, но под покровом парати-
пической изменчивости это отличие обнаружить нелегко.
Третий прием. Инбридинг F2 или FB
Это? прием состоит в том, чтобы подвергнуть особей F2 или FB
инбридингу, хозяйственную сторону которого мы пока оставим. При продолжении
этого инбридинга на ряд поколений будут выделяться всевозможные
генотипы, и число таких генотипов явится анализом скрещивания. Но уже
первое инбредное поколение может быть использовано для выводов:
аа
2Аа
АА
аа
(1)*
1
<?
?
2 Аа
(2)
2
<?
(5)
2
АА
<?
(5)
2
<?
* В скобках — тип скрещивания, цифры под ними —
частота.
Здесь возможны два случая: а) в случае обильно плодовитых видов
инбридинги могут быть заложены от отдельных пар и б) в случае
малоплодовитых видов один самец может скрещиваться со многими самками.
Разберем на примере моногибридного скрещивания теорию этого при.
ема. В F2 моногибридного скрещивания имеем генотипы аа + 2Аа + АА.
Образуем всевозможные пары, скрещивая особей без подбора друг к другу-
Из таблицы видно, что возможны шесть типов встреч (табл. 20).
227
15*
Таблица 20
Шесть возможных типов скрещиваний при инбридинге из F2
Тип скрещивания и потомство
/
(1) аа X ла -»• аа
(2) аа X Ла -* аа + Ла
(3) аа X А4-> Ла
(4) Ла X Ла-> яя + 2Ла + ЛА
(5) Ла X ЛЛ-» Ла + ЛА
(6) ЛЛХ ЛЛ-> АА
f
1
4
2
4
4
1
М
при неполном
доминировании
Mi = Mp
iIf2 = Mp+0,5i
} Ms = Mp+l,0i
Af4==Mp+l,5i
M5 = Mp+2,0i
при полном
доминировании
Afi = Mp
M2=Mp-fO,5*
M4 = Mp+l,0i
Мз = Мр+0,751
М4=Мр+1,0*
Итак, распределение М будет иметь вид:
При неполном При [полном
доминировании доминировании
Мр 1 Мр 1
Мр + 0,5* 4 Мр+0,50* .... 4
Мр + 1,0*. . . . ? 6 Мр + 0,75г .... 4
Мр+1,5* 4 АГР +1,001 .... 7
Л/р + 2,0* 1
При промежуточном доминировании мы получим каркас дигибридного
типа, при полном — очень своеобразный четырехчленный, сильно
асимметричный, несколько напоминающий дигибридное отношение 9:3:3:1
с неодинаковыми интервалами.
Очевидно, и при этом приеме удобнее начинать инбридинг не с F2,
а с FB. Здесь отношение опять значительно упростится и примет такой
вид:
Тип скрещивания и потомство
(1) ааХаа-* аа
(2) аа X Аа -» Аа + аа
(3) АаХЛа^2 Аа + АЛ+аа
f
1
2
1
м
при промежуточном
доминировании
М2 = Мр+0,Ы
М8 = Л#Р+1,0*
при полном
доминировании
Мг^Мр
M2 = Mp+0,5i
M3=Mp+0,75i
При отсутствии доминирования будет симметричный каркас
моногибридного типа 1 + 2 + 1, при полном доминировании — асимметричный,
благодаря неравным интервалам между классами.
При инбридировании дигибридного FB будем иметь 10 типов
скрещивания.
ааЬЬ
ЛаЬЬ
ааЬЬ
1
2
АаЬЬ
2
5
ааВЬ
3
6
АаВЬ
4
8 I
ааВЬ
АаВЬ
ааЬЬ
3
4
АаЬЬ
6
8
ааВЬ
7
9
АаВЬ
9
10
Их результаты. представлены в табл. 21.
228
Таблица 21
Результаты инбридирования дигибридного FB
Типы скрещиваний и их потомство
М (неполное
доминирование)
(1) АаЬЬ X ааЪЪ
(2) aabb X Aabb
(3) aabb X ааВЪ
(4) aabb X АаВЪ
(5) ЛабЬ X 4аЬЬ
aabb
aabb + ЛаЬб
aabb + ааВЪ
aabb + Лдбб + АаВЪ + 4а#Ь
ааЬб + 2АаЪЬ + ААЬЬ
(6) ЛдбЬ X яа#& -» оабб + Aabb + <ш#6 -f АаВЪ
(7) ш*#Ь X яя#Ь -» <ш#Ь + 2аа£Ь + ааВВ
(8) Ла&Ь X 4л#Ь -* яа&Ь + 2Ла66 + ААЪЪ + ааВЪ +
+ 2АаВЪ + ААВЪ
(9) ааВЪ X 4аЯЬ -» aabb + 2аа#Ь + ааЯЯ + Aabb +
+ 2Ла#6 + АаВВ
(10) 4а#Ь X АаВЪ-* как в F2
Af2 = Mp+0,5i
M5 = Mp+l,0t
М4 = Мр+1,5/
М5 = Мр+2г
При промежуточном проявлении и при равной силе действия генов
получится дигибридное распределение М.
Сравнивая эти результаты с расчетами, полученными выше (табл. 18),
видим, что по распределению средних инбридирование FB и скрещивание
F2 с чистой расой дают совершенно одинаковый результат. Некоторые
отличия будут лишь в величинах и распределении а, так как в
скрещиваниях (5), (7), (8), (9) и (10) будут сигмы, не встречавшиеся там. На этом
вопросе мы останавливаться не будем.
Случай малоплодовитых животных. В случае
малоплодовитых животных, особенно млекопитающих, попарный
инбридинг, очевидно, не может дать необходимых данных для выведения
средних и определения характера распределения. В этом случае, однако,
возможно применение этого приема путем скрещивания самцов
из F2 со многими самками F2.
Теория этого приема следующая.
Цоложение. Достаточно большая выборка,из F2 в скрещивании рав-
ногаметна особям F1? т. е. совокупность многих особей F2 образует
таковой же состав гамет, как и отдельные особи ¥г.
Справедливость этого положения легко доказывается простым
подсчетом. Если взять достаточно большое число особей F2 без выбора, то их
генотипы образуют распределение, отвечающее теоретическому составу
F2. Подсчитаем состав гамет такой выборки особей F2 (табл. 22).
Таким образом, особи F2 образуют состав гамет аЪ + АЪ + аВ + АВ,
совершенно подобный составу гамет особей ¥г строения АаВв.
Если мы будем брать самцов F2 и скрещивать их с совокупностями
самок F2, то, очевидно, будем иметь девять различных типов скрещивания
соответственно девяти генотипам ¥г дигибридного скрещивания. Эти скре-
щивяция будут:
(1) aabb X 9 9 F2 = aabb X АаВЪ
(2) Aabb X 9 9 Fa = Aabb X АаВЪ и т. д.
Сравнив эту таблицу с табл. 18, мы увидим их полное сходство. Таким
образом, этот прием оказывается совершенно тождественным с приемом II
(скрещивание ¥г X F2).
229
Таблица 22
Состав выборки F* для дигибридного скрещивания
Генотипы в F2 и их
частоты
lAabb
Особь Fi (АаВЬ) . .
Гаметы
аЬ
1
1
1
1
4
1
АЬ
1
1
1
1
4
1
аВ
1
1
1
1
4
1
1
АВ
1
1
1
1
4
1
Вместе с тем легко понять, что скрещивание $ ¥± со многими £ $ F2
с данной точкц зрения не имеет смысла, так как оно совершенно подобно
скрещиванию Fx X F1? т. е. простому получению F2.
Четвертый прием. Скрещивание самцов F„ со многими самками FB
Закономерности состава гамет здесь будут следующими:
Тип скрещивания
Моногибридное
Дигибридное '
Тригибридное
Состав
аа
Аа
aabb
Aabb
ааВЬ
АаВЬ
aabbcc
Aabbcc
ааВЬсс
aabbCc 1
АаВЬсс
AabbCc
ааВЬСс
АаВЬСс
Состав гамет
а А
abc
8
4
4
4
2
2
2
1
27
Abc
4
2
2
1
9
аВс
4
2
2
1
9
аЬ
4
2
2
1
9
^
2
1
3
АЬ
2
1
3
&6С
4
2
2
1
9
1
1
аВ
2
1
3
АВс
2
1
3
АВ
1
1
46С
2
1
3
^
-
аВС АВС
2
1 1
3 1 .
230
Закономерность состава гамет в совокупности FB ясна и может быть
передана произведением биномов:
(А + За)-(Я + ЗЬ).(С + Зс)...
Это обобщение приложимо, однако, лишь к такому случаю, когда FB
получается из скрещивания Fx с рецессивной расой аа, aabb, ааЪЪсс и т. д.
Для FB на доминантнунГрасу будет иметь место обратное отношение:
(34 + а).(35 + Ъ)-(ЗС + с)...
В разбиравшемся нами неоднократно примере усилителя и ослаби- .
теля, когда FB идет на расу ааВВ, состав гамет FB, очевидно, по гену а
будет (А + За), а по гену В, наоборот, (32? + Ь), или, раскрывая скобки
(А + За)-(35 +Ъ) = ЗАВ + АЬ + 9аВ + Sab = 9аВ+ ЪАВ + За + 1АЬ.
Поэтому в зависимости от характера скрещивания инбридированное
Fb по IV приему может давать весьма различные картины, очень ценные
для генетика-аналитика. Вместе с тем подобное скрещивание явится
«разведением в себе» поколения FB, играющего в проводимой сейчас
в СССР метизации немаловажное значение и распространение.
Поэтому целесообразно подвергнуть это скрещивание более подробному
анализу.
1. Моногибридное скрещивание. Состав гамет FB :
А + За или а + ЗА.
Здесь будем иметь два типа скрещиваний, моногибридный каркас с
различными интервалами в зависимости от доминирования.
Тип скрещивания и потомство
(1) ааХ $$FB-*a(3a + i4) = 3 аа + Аа
(2) Аа X 9 9 FB-> (А+а) (3а+Л) ~Ъаа + 4Аа+
+ АА
1
1
1
Неполное
доминирование
Полное
доминирование
M^Mp + li
2. Дигибридное скрещивание FB от скре щи в а-
н и я aabb X F2 дает следующие результаты: L-
(1) aabb X 9 9 FB -> ab (9аЬ + ЪАЬ + ЪаВ +АВ) = 9 aabb + 3 Aabb + ЪааВЬ + АаВЬ;
(2) АаЬЬх 99 VB^(ab + Ab)(9ab + ZAb+3aB+AB) = 9aabb+l2Aabb +
+ ЪааВЬ + 4 АаВЬ + 3 ААЬЬ + ААВЬ;
<3) ааВЬХ 9 9 Ь*в -> (ab + AB) (9a6 + ЪАЬ + ЪаВ + АВ) = 9ааЬЬ+ ЪАаЬЬ +
+ 3 ааВЬ + 10 АаВЬ + ЪА аВВ + 3 ААВЬ+ ААВВ;
<4) АаВЬХ99 Яв->(аЬ + АЬ + аВ+АВ)(9аЬ + ЪАЬ + гаВ+АВ) = 9ааЬЬ +
+ 12 АаЬЬ + 12ааВЬ + 16 АаВЬ + ЪААЬЬ + 4ААВЬЛ-ЪааВВ +
+ ЬАаВВ+[ААВВ.
231
Возникающие в этом случае М в общей форме, если сила А = ix и
В = i*2i будут иметь следующий вид:
При неполном
доминировании
^"р+ТЬ+Т1»
M^lfp+^b+^b
^з=Мр+4-*1+-!г*2
м4=жр +_S1+_J2
1Г1=ЛГр+-*-,
д/2 = Л/р + i
Мз = Л/р + 1,5*
/
1
1
1
1
1
2
1
При полном
доминировании
M1 = Mv^-^-h + ±-h
л/а = л/р +_|_tl+_|_fa
*,в»ЛГр++,1+ .£.,,
или при $i = i2 = i:
Afi«lfp+4-<
Ма==Л/р+_^
M3 = Mp+^-i
/
1
1
1
1
1
2
1
Все каркасы будут иметь асимметричный характер и при равенстве
силы генов А и В образуют такие ряды:
(1) 9+6+1 (1) 9+6+1
((3)} 9+15+7+1 ((з) }<Н-«Ч-5
(4) 9+24+22+8+1* (4) 9+30+26
3. Дигибридное скрещивание. FB от скрещивания
ааВВ X Рх.
Состав гамет $ £ г FB = 9аВ + 34Я + ЗаЬ + АЪ.
Здесь четыре типа скрещиваний:
(1) ааВЬ X $ $ FB -> (а& + а#) (3 а& + 46+9 а£ +3 АВ) =3 ла$6 + АаЬЬ + 12 ааЯЬ +
+ 4 Ла#6 + 9 ааВВ + 3 4а£Я;
(2) АаВЬ X 9 9 FB г* № + Л6 +Л£+аЯ) (3 а&+ Л6 + 9 а£+3 АВ) = 3 ааЬЬ + 4 АаЬЬ +
+ 12 аа56 + 16 АаВЬ + А466 + 4 А4Я6 + 9 ааЯЯ + 12 АаВВ +
+ ЗЛАВВ;
(3) ааВВХ 9 9 FB~*a£(3a& + 46 + 9а# + 3 АВ) =ЪааВЬ + АаВЬ + 9ааВВ +
+ ЗАаВВ\
(4) АаВВХ 99 FB^(a£ + AB)(3a& + i46 + 9a£+3^B) = 3aaBb + 4^a£6 +
+ 9 аа#£ +12 АаВВ + А4Я6 + 3 ААВВ.
* Это семейство распределений легко получается по способу видоизмененного
треугольника Паскаля: ч
9 6 1
9 15 7 1
9 14 22 8 1
232
Возникающие в этих случаях М в общей форме имеют следующий
вид
При неполном доминировании _
(1)^1=^+4-^-4-*»
<2)ifta=ifP+4ii-4«i
®if.=jfp+4ii-4|»
(4)ж4=л/р+4^-4^
/
1
1
1
1
При полном доминировании
Ui=^P+4fc--4*2
U2=mp+4*1-4*2
ii/3=MP+4h
Nf4 = Mp+4$i
/
i
i
i
i
или при h = h = i*
м1 = мр—Lj
iif2 = Mp ...
M3 = Mp + -±-'$
M1=il/p+-l-J
j^3=MP+4«
л/2=мр + **
Л/4 = Мр+-^
В случае многократно разбиравшегося выше случая полнодоминиру
ющих А и В усилителя и ослабителя с ix = 1 и г2 = —1 будем иметь:
Мз = Мр+ | 1 М4 = Мр + ! 1
М1 = Мр+ §
M2 = Afp+ £
Каркасы при ^ = i2 будут иметь следующий вид:
(1) 3+13+13+3
(2) 3+16+26+16+3
(3) 3+10+3
(4) 3+13+13+3
(1) 3+22+7
(2) 3+26+35
(3) 12+4
(4) 12+20
Нетрудно видеть, что и при помощи этого приема различные случаи
расщепления, довольно легко отличаются друг от друга. Так, оба
разобранных дигибридных скрещивания (aabb X ААВВ и ааВВ х A Abb)
отличается друг от друга следующим.
При неполном доминировании:
а) косными каркасами отдельных скрещиваний в первом случае и
симметричными высокими — во втором;
б) величинами М\, М2, М3: во втором случае они все не меньше, чем
в первом, и к тому же Мг <С Мр и М2 = ilfp, тогда как в первом случае
все М > Мр.
При полном доминировании:
а) разным характером скошенности каркасов;
233
б) совершенно различными интервалами между М отдельных
скрещиваний, что особенно заметно при ix = i2, а также при £2 = —ix. В первом
случае всего три значения: Ми М2, Мъ на равных интервалах в -тр i, a
во втором четыре разных М с неравными интервалами между ними в 1/8,
2/8 и 1/8.
Приведенные на предыдущих страницах несколько утомительные и
схематические выкладки тем не менее необходимы, так как они могут
показать, что различные мыслимые типы скрещиваний открывают
совершенно различные возможности для получения нужных выводов. Без
таких выкладок трудно было бы, например, вскрыть, что одним из
наиболее перспективных для генетика-аналитика скрещиваний является
скрещивание с? Fb со многими самками FB и т. п. Возможно, что дальнейшим
анализом удастся найти и еще более интересные типы скрещиваний.
Очевидно, что значение свойств различных типов скрещиваний очень важно,
так как при той же затрате труда и материала удачно выбранный тип
скрещивания может дать ценные результаты, и немного иной тип — ис-
, портить всю работу.
Хотя фактические случаи всегда будут, конечно, более
сложными, чем моно- и дигибридные схемы, тем не менее вскрытые при
разборе этих более простых примеров особенности различных
скрещиваний будут иметь значение и в этих более сложных скрещиваниях.
АНАЛИ8 СЦЕПЛЕНИЯ
По каркасу расщепления можно делать выводы не только о числе
тенов и их доминировании и силе действия, но также и о наличии между
ними геносвязи (сцеплений или отталкиваний). Разберем на примере
простейшего случая дигибридного расщепления (скрещивание ааЪЪ X
X ААВВ с величиной перекреста между А и В в 25%) особенности
возникающей здесь картины.
Дигётерозиготные особи АВ/ab образуют при 25% перекреста гаметы
состава ЗАВ -f- Ab + аВ + ЗаЬ, в результате чего состав F2 будет:
3 АВ
Ab
аВ
3 ab
3 АВ
АВ
»-АВ
о Ab
6 АВ
аВ
ЪЧВ
ab
»Ив
АЪ
з^-
6 АЪ
А АЬ
i~W
, aB
i-w
л ab
аВ
6 аВ
А М
1 аВ
А аВ
1ж
л ab
Ъ1ПГ
3 аЬ
Я ab .
о АЬ
Ъ~сТ
л аВ
3~аТ
аЬ
91ЙГ
По первому приему (см. стр. 218) мы скрещивали особей F2 с чистой
рецессивной расой.
Наличие сцепления отразится а) на пропорции отдельных типов
скрещивания и б) на том, что 6-й тип ааЪЪ X АаВЪ здесь распадается на два
разных:
f^y^ n ab „ Ab
ab A abykL ab A аВ '
234
Рис. 79. Возникновение трех-
верпшнности дигибридного
каркаса благодаря сцеплению
1 — без сцепления; 2 — со
сцеплением
В результате мы будем иметь:
Тип скрещивания и потомство
<1) аЬаЬ X аЬаЬ -* аЬаЬ
(2) аЬаЬ X АЬаЬ -» abab + АЬаЬ
(3) аЬаЬ X аВаЬ -» аЬаЬ + аВаЪ
(4) аЬаЬ X АЬАЬ -> АЬаЬ
<5) аЬаЬ X аВаВ -» аВаЬ
<6а) аЬа& X -45а& -* dbdb + 4/?аЬ + АЬаЬ +
аВаЬ
<€Ь) а&аЬ X АЬаВ -* аЬаЬ + ЛбаЬ + аВаЬ + АВаЬ
<7) аЬаЬ X 4ВЛ& -> АВа& + АЬаЬ
(8) аЬаЬ X АВаВ -> аВа& + АЯаЬ
<9) abab XABAB-* АВаЬ
f
9
6
6
1
1
18
2
6
6
9
М (при неполном
доминировании)
Мр
ЛГр+4-Ь
"p+4-h
Mp+ii
Мр + *2
мр+4-^+—ia
il/p + -*-i1+-*-*2
Mv+b + ~Yh
Mv+-Lh + h
Mp + *i + *2
Итак, получаем каркас (£х = i2 = £):
^i = ^P 9 * Jtf1 = Afp + _|_| ...... 12 1,34
ж4=л/р+4-' 12 1«34 I
2 Мь = Мр + -%-1 9 1
M8 = Mp + -L i 22 2,45 *
Мы видим, таким образом, что наличие сцеплений в 25% изменило
весь характер каркаса (рис. 79), сильно растянув его, так что |52 вместо
2,5 для ряда 1+4 + 6 + 4 + 1 опустилось до 2,16.
Если сила сцепления отразилась на форме каркаса, и в частности на
величине (J2» T0 возникает вопрос, нельзя ли по этой величине найти силу
сцепления. Это оказывается в принципе вполне возможным, так как легко
установить функциональную связь между величиной сцепления (процент
перекреста) и величиной |J2.
Воспользуемся этим случаем, чтобы ознакомить читателя с
возможностью индуктивного метода отыскания различных связей в
данной области. Интересующую нас сейчас связь мы могли бы искать
чисто дедуктивным методом, установив, как образуются из элементов
кривой распределения величины ц,2 и щ и вычисляемая из них величина
235
p2=
_ P-4
2 . Однако этот путьчасто бывает слишком труден, требует серьезного
^2
математического аппарата и недоступен для начинающих. Однако, как
мы сейчас увидим, нередко мы можем решить задачу самым элементарным
путем и притом путем привычной натуралисту индукции.
Рассчитаем несколько каркасов для разной силы сцепления — 10%,
20%, 30%, 40% и 50% (независимость). Это нетрудно сделать хотя бы при
помощи приведенной на стр. 234 таблицы, если представить себе, что
гетерозиготная особь АВ/ab будет образовывать гаметы АВ, АЪ, аВ и ab в
зависимости от разной силы сцепления в различных пропорциях, а
именно:
Процент
перекреста
10
20
30
40
50
10 ТИПОВ С]
АВ
9
8
7
6
5
?рещивани]
Пропорция
АЬ
1
2
З1
4
5
Я В ЭТИХ CJ
гамет
аВ
1
2
3
4
5
гучаях воз]
аЬ
9
8
7
6
5
шкнут в
(обозначая частоты гамет АВ и ab через а, а частоты АЪ и аВ через Ь):
Тип
(1)
(2)
(3)
(4)
(5)
(6а)
(6Ь)
(7)
(8)
(9)
Частота
а3
2 ab
2 ab
Ь2
Ь2
2 а2
2 а2
2 ab
2 ab
а2
ю%
81
18
18
1
1
162
2
18
18
81
20%
64
32
32
4
4
128
8
32
32
64
30%
49
42
42
9
9
98
18
42
42
49
40%
36
48
48
16
16
72
32
48
48
36
50%
25
50
50
25
25
50
50
50
50
25
м
Mi
\ м2
,
> М3
1 М*
мъ
Так как величины М остаются независимыми от сцепления, такими
же, как и в разобранном выше случае 25% перекреста, то, соединяя
частоты / : (2) и (3); (4), (5) и (6) и (7) и (8), получаем такое распределение Mi
м
Мг
М*
Ms
Mi
Мъ
Частоты (/) при перекресте в
ю%
81
36
166
36
81
400
20%
64
64
114
64
64
460
30%
49
84
134
84
49
400
40%
36
96
136
96
36
400
50%
25
100
150
100
25
400
236
Высчитываем теперь для всех пяти рядов [л2 и (х4, однако не
производя никаких сокращений, так, чтобы видно было строение этих вели-
v, A2/ v. А4/
чин, т. е. в виде 2~Т~ и 2"л *
Тогда будем иметь:
720 2664
Ю 7ЙА 1ЛЛ 40
20 ^ ^i^k 50
30
Глядя на числители (х2, легко видеть их систематическое возрастание
на 80 по мере усиления сцепления, т. е. уменьшения величины перекреста
(с %) на 10%. Закономерность же изменения (х4 пока не ясна, кроме
возрастания.
Для упрощения рядов произведем сокращение всех величин \i2 и [i4
на 4.
Переписываем:
V»2
720
400
640
400
560
400
V*
2664
400
2176
400
1736
400
№
480
400
400
400
14
1342
400
1000
400
с%
10
20
30
100-с%
90 тлп
1*2
180
100
160
100
140
100
Нч
666
100
544
100
434
100
80 il АЛ Л П£\
70 ~
0%
40
50
100-с%
60 л лл л лл
1*2
120
100
111
И-4
336
100
250
100
50 * Af\f\ ЛП£\
Уже одного взгляда на ряд \i2 достаточно, чтобы понять закономерное
изменение \i2. Числитель является не чем иным, как удвоенной величиной
(100 — с%), т. е. удвоенным процентом неперекрестов.
Итак,
н _ 2(100-go/o)
V*- loo
9 2с%
^ — loo"
Этот успех позволяет нам надеяться найти закономерность строения
числителя и [i4. Попробуем сократить еще весь ряд на одну величину(оста-
вим пока знаменатель без внимания).
666 = 2-333
544 = 2-272
434 = 2-217
336 = 2-168
250 = 2-125
о%
10
20
30
40
50
100-с%
90
80
70
60
50
Дальнейшее общее сокращение ряда невозможно. Попробуем часто
удающийся прием выделения множителей,
возрастающих на единицу при переходе от числа к числу.
Нижнее число 125 делится на 5. Не делится ли 168 на 6? Делится. А 217
на 7? Делится!
Итак,
666 = 2-9.37
544 = 2-8-34
434 = 2-7-31
336 = 2.6-28,
250 = 2-5-25
с%
10
20
30
40
50
Ю0-с% ^
90
80
70
60
50
ю
1
2
3
4
5
Ю0-о%
10
9
8
7
6
5
237
Ряд множителей, 9, 8, 7, 6, 5, нам становится понятным из
сопоставления с рядом (100 — с%). Это есть (100 — с%)/10. Требует разъяснения
ряд множителей, 37, 34, 31, 28, 25. Нельзя, например, еще раз выделить
из них множители 9, 8, 7, 6, 5? Непосредственно это не удается.
Испробуем еще один прием разложения анализируемых чисел на сумму слагаемых,
из которых одно было бы понятно. После некоторого размышления
замечаем, что если написать 37 = 36 + 1, то 36 = 9-4, т. е. учетверению той
же самой знакомой нам девятки. Попробовав в следующем числе найти
4«8, видим, что 34 равняется 4-8 + 2. Изменение 1 и 2 оказывается
параллельным изменению с% от 10 к 20. Проверим уловленную
закономерность:
с% 100—с%
37 = 4-9+1 10 90
34 = 4-8+2 20 80
31 = 4-7+3 30 70
28 = 4-6+4 40 60
25 = 4.5+5 50 50
На этом мы можем анализ закончить, так как все 'стало ясным.
Заменяя 9, 8, 7, 6, 5 на ~g ° и 1,2, 3, 4, 5 на с%/10, можем написать:
2 100-с% r4(10°-g%) _i_ f%l - 2 (100 - с%) [4 (100-co/o)+g0/0]
1002
_2 100-*% r4(10Q-g%) . с%1 _
^ ~" 100 ' 10 ' L 10 "^ T0j ""
Теперь получим из выведенных нами формул:
В - В* -о 1 с%
PS ^2(100-c%) "
Проверка по имеющимся у нас рядам \i2 и [г4 показывает правильность
формулы.
Если считать не в процентах, а в долях единицы (10% составляют 0, l)t
то выведенная нами формула еще упрощается:
ц2 = 2-2с,
fi4 = 8 — 14с + 6с2,
В - 4~3с
Рз"" 2-2* •
Напомним, что эти закономерности выведены лишь для первого
приема (F2 X aabb). Для других скрещиваний надо вывести формулы заново,
что может явиться полезным упражнением, почему мы и не будем здесь
этим заниматься. Таким образом, индуктивным путем может быть
установлена весьма изящная связь между силой сцепления генов в дигибрид-
ном расщеплении и элементами, характеризующими каркас нашего
распределения. Нетрудно видеть, что теперь мы сможем решить и обратную
задачу нахождения с по величине |J2.
Преобразуя последнее выражение, находим:
232-4
6"~ 2(32-3 '
ВЫВОДЫ
Изложенное достаточно показывает, что имеются различные приемы,
позволяющие заменить альтернативные признаки отдельных
особей параметрами Af, о и пр., вычисляемыми по совокупности их
238
потомства, и этим из-под покрова паратипической изменчивости обнажить,
генотипический каркас распределения. В зависимости от того типа
скрещиваний, который мы примем, строение этого обнажившегося каркаса
будет весьма различным. И еще более различным этот каркас будет в
случаях, различных по самой природе анализируемого явления. Различное
число генов, различная их сила, элементы взаимодействия (эпистаз,
криптомерия), сцепление, различная жизнеспособность у различных членов
каркаса и т. д.— все это будет давать необозримое разнообразие
обнажившихся каркасов.
Поэтому расшифровка обнажившегося каркаса в свою очередь явится
делом достаточно трудным, и генетик-аналитик вовсе не должен думать,
что достаточно ему будет обнажить каркас и все расщепление станет ему
сразу понятным, даже в том случае, если ему удалось получить
достаточное потомство для преодоления затемняющего характера паратипическо-
го покрова. В некоторых случаях, конечно, обнажившийся каркас даст
сразу легко понятный ответ, например, четкое отношение 1 + 1; 1+ 2 +
+ 1 и т. д. Однако все те искажающие факторы, которые мы разбирали
в начале настоящей книги, будут действовать и здесь, и надо быть
готовым к тому, что для уяснения природы обнажившегося каркаса
потребуется постановка еще дополнительных исследований, применение для
контроля других приемов и т. д.
Конечно, в помощь генетику-аналитику можно было бы разобрать
большое число различных теоретически мыслимых примеров и по
полученным данным составить нечто вроде «определителя». Пример такого
определителя мы даем на следующих страницах.
Однако при почти бесконечном разнообразии возможных случаев,
даже в пределах моно-, ди- и тригибридных скрещиваний, эта задача очень
трудно выполнима. Поэтому важнее, чтобы генетик-аналитик овладел
достаточно хорошо самой идеей этого метода и теорией и техникой
необходимых расчетов для того, чтобы он мог по первым данным составить
себе ряд возможных гипотез, произвести расчеты и продумать
необходимые дополнительные или проверочные скрещивания. При этом нужно
еще раз подчеркнуть большое значение биологических особенностей
анализируемого вида. Анализ растений, шелкопряда, пчелы, курицы,
кролика, коровы благодаря этим особенностям должен вестись своими
собственными приемами. Исчерпать все возможные ухищрения, к которым
сможет прибегнуть аналитик, поможет лишь сама практика этой работы,
которая находится, к сожалению, пока в самом зачаточном состоянии.
ПРИМЕР ОПРЕДЕЛИТЕЛЯ ДЛЯ СКРЕЩИВАНИЙ с? F2 X F2 $ $ *
1. При скрещивании самцов F2 со многими самками -F2 обнажен
каркас:
трехчленный 2
пятичленный '.'.'. 4
семичленный 11
девятичленный 17
2. Каркас имеет строение 1 + 2 + 1 с равными интервалами
моногибридное расщепление 3
Каркас иного строения — какое-либо искажение более сложных
расщеплений.
МPi и Мр2 __ м родительских рас, с меньшим (Рх) и с большим (Р2) развитием
признака: il/F,» м¥2 — М поколений Fx и F2; М1У М2 —М различных членов каркас
са, как в тексте.
239
3. М2 (у среднего члена каркаса) = МFl:
2Мг = М Pl + Af Fl; 2M3 = Л/ р2 + A/Fl — моногибридное
расщепление с неполным доминированием.
М2 < AfPl; 4Af2 = AfPl + 3Mp2; M3 = AfFl = АГРа —
моногибридное расщепление с полным доминированием усилителя.
М2 > AfFl; 4М2 = 3MPt + Мр2; М3 = ATPl = MPl. —
моногибридное расщепление с полным доминированием ослабителя.
4. Каркас строения 1+4 + 6 + 4 + 1— дигибридное расщепление 5
Каркас иного строения — какое-либо искажение более сложного
расщепления.
5. Мь < МР2 6
М5>МР2 7
6. Мь = 1/4 (Afpt + ЗМр2) — скрещивание типа aabb x -4-452? с адо-
минированием.
Мъ = 1/8 (AfPl + 7Мр2) > 1/4 (AfPl + ЗМр8) - дигибридное
расщепление aabb x -4-455 g iB = 2iA. А доминирует
промежуточно, В — полно.
7. М5 = Мр2 — скрещивание типа aabb x ААВВ с полным
доминированием, 1г = i2.
Мь > Мр2 — скрещивание типа ааВВ X -4-466 8
8. MFl < Мр, 9
MFl>MPf . . 10
9. MFl = M?t = Afp2 — скрещивание типа ааВВ X -4-466, где А и
5 равносильные с неполным доминированием усилителя.
М = 1/2 (Mpt + MpJ; MPl < Мр2 — скрещивание типа ааВВ X
X -4-466 с полным доминированием обоих генов, А — усилитель,
В — ослабитель.
10. Мъ = AfFl — скрещивание типа ааВВ X -4.466, ix = £2, оба гена
с полным доминированием.
Мъ > MFl — скрещивание типа ааВВ < AAbb.
%в = 2^. В доминирует полно, А — неполно.
11. Каркас строения 1+6 + 15 + 20 + 15 + 6 + 1 — тригибрид-
ное скрещивание с ix = i2 — i3.
Каркас |строения 1+2 + 3 + 4 + 3 + 2 + 1— дигибридное
расщепление с ix = 2i2, т. е. один ген вдвое сильнее другого:
неполное доминирование. Или же ix — i2, но один ген доминирует
полно, другой неполно 12
12. ЛТ4 = MFl — адоминирование, дигибридное скрещивание
ix = 2i2 13
М4 < MFi 14
13. М1 <[ Afp2 — скрещивание типа аабб х ААВВ, ix = 2i2,
промежуточное доминирование.
М1 > Afp2 — скрещивание типа ааВВ X -4.466, i± = 2i2,
промежуточное доминирование.
14. М1 — ilfFl, i± = 2i2 — полное доминирование 15
М7 ]> AfPl, i± = i2, но А доминирует неполно, В — полно . 16
15. М7 = Mpt — скрещивание типа aabb X ААВВ, ix = 2i2, полное
доминирование.
240
М7 > ilfp, — скрещивание типа ааВВ х А466, i± = 2f2, полное
доминирование.
16. М7 <[ Л/р2 — скрещивание типа aabb x -4-466, ix = &а> -4
доминирует промежуточно, В — полно.
M7=ilfpt — скрещивание типа ааВВ х -4-466, ii—if, -^ доминирует
промежуточно, -В — полно.
17. Каркас строения 1+2 + 1 + 2 + 4 + 2 + 1 + 2 + 1 . . 18
Каркас строения 1+0 + 2 + 2 + 1+4 + 1+ 2 + 2 + 0 + 1
и т. д.
18. Интервалы между членами каркаса равные. Дигибридное
расщепление с ix = 3i2 ...• 19
Интервалы неравные 22
19. М9 = Мт?г — неполное доминирование . '. . 20
М9 = MFi — полное доминирование 21
20. М9 < И?* — скрепщвание типа аабб х ААВВ, ^ = 3ia,
неполное доминирование.
М9 = Afp, — скрепщвание типа ааВВ X -4-466, ^ = 3i2, неполное
доминирование.
21. Af9 = Mps — скрещивание типа аабб х ААВВ, ix = 3i2, полное
доминирование.
М9 > Мр, — скрещивание типа ааВВ х ААЪЪ, h = 3ia, полное
доминирование.
22. Интервалы между М8 и М4 и между Jlf ф и ДГ7 шире других, и в
частности:
вдвое — дигибридное расщепление с h = 4i2; с одинаковой
степенью доминирования А ж В или ^ = 2£2, если -4 доминирует
промежуточно иВ- полно;
втрое — дигибридное расщепление с ix = 5i2; с одинаковой сте
пенью доминирования А ж В;
вчетверо — дигибридное расщепление с ^ = 3i2, если Л домини
рует промежуточно, а -В — полно.
16 А. С. Серебровский
ГЛАВА XI
МЕТОД АЛГЕБРАИЧЕСКИЙ
Как оыло изложено кратко выше, идея алгебраического метода
сводится к составлению систем уравнений по числу требующихся для
отыскания неизвестных. Такими неизвестными являются: число генов,
участвующих в расщеплении (х), сила действия генов (£17 i27... ^),сила
сцепления (саЪ, сас...)у жизнеспособность различных генотипов (иг, i>2.--)> сте~
день доминирования или эпистаза (Zl7 Z2...) и т. п.
Что касается известных величин, то они могут браться из различных
поколений или из сравнения различных поколений друг с другом.
Таковы средние арифметические Af pt> Mp2, M-pt, M$2, М$9 и другие; разности
ikfp, — Мр2; Мрх — MVl или лучше а2 = \х2 разных поколений, (х4, Ца
и т. д.
Не всегда, однако, два уравнения могут дать возможность определить
два неизвестных. Так уравнения
х + у = 2; х = 2 — у;
2х + 2у = Ь; x=^L
являются по сути дела одним уравнением, так как одно просто
вытекает из другого и второе справедливо при любых значениях х ж у, при
которых справедливо первое. Для того чтобы решение было возможным,
необходимо, чтобы уравнения были не тождественными, а сопряженными.
Ниже мы увидим, что при применении «метода уравнений» существенною
трудностью является составление необходимого числа сопряженных
уравнений.
ВЫДЕЛЕНИЕ ГЕНОТИПИЧЕСКОГО КОМПОНЕНТА
Первой задачей, которую нам приходится решать, является
исключение паратипической изменчивости из уравнений. Идея решения этой
задачи, предложенная Райтом и разработанная нами, сводится к
следующему. Пусть мы имеем скрещивание двух рас, доведенное до F3. Во всех
поколениях, благодаря трансгрессивной изменчивости, мы увидим лишь
более или менее правильные кривые распределения без намека на какое-
либо явное распадение на классы расщепления (рис. 80).
Единственное, что мы замечаем, это увеличение разкаха изменчивости
в F2, заставляющее нас предполагать наличие расщепления. Но каково
это расщепление? Можно ли его выделить из-под покрова паратипической
изменчивости?
242
Рассуждаем так: в F2 изменчивость —
двойной природы, генотипической и
паратипической. По теореме сложения
моментов:
Ц
ъ
^
Мр
P»2F2 == \^2У + V>2n',
(1)*
бр^И^- (2)
Отсюда
Ply = V>2F2 — V>2*\ (3)
H4Y = \1&2 — \i& — 6ji2Yii2*. (4)
Какие элементы из правой части
равенств нам известны или могут быть
известны? [x2f2 и [i4F3 могут быть
непосредственно высчитаны из
распределения F2. Что касается ця, то эти
величины, характеризующие степень
паратипической изменчивости, могут быть
взяты из других поколений. Можно
приближенно принять, что паратипи-
ческая изменчивость, наблюдаемая нами в Fx и в Рх и Р2, имеет тот же
характер, как и в F2. Тогда мы можем принять, например, что
Рис. 80. Кривые распределения и со1
отношения М по в поколениях Fx и
F2 при скрещивании чистых линий
Pi.hP2
где п = 2, 4
И** = \hFt> или (хпя
И-4я — H'uFf
УпРг + H-nFi
И Т. П.,
Подставляя (5) в (3), находим:
P»2Y = H^Fa — H»2Fi»
(5)
(6)
(7)
т. е. (i2 генотипической изменчивости F2 равен разности между \i2 2-го и
1-го поколений.
Подставив теперь уравнения (6) и (7) в (4), находим и [i4Y:
M*4Y = H-uFa — f^uFi — 6 (jX2p2 — fX2Fi) H-2Fi •
(8)
Узнав (i2Y и |i4Y для F2, мы вычисляем важную для характеристики
генотипической изменчивости величину |52Y.
Каково качество полученного решения и от чего зависит правильность
его? Прежде всего мы допускаем некоторую условность в том, что в F2
паратипическая изменчивость равна изменчивости Fx. Это не совсем так.
Генотип особей ¥г одинаков, а в F2 — многообразен, поскольку
паратипическая изменчивость у разных генотипов может быть различной.
В случае моногибридного расщепления F2 имеет состав А А + 2Аа + аа,
и поэтому правильным будет принять вместо (5):
\lniz
M-nPi + V'nPz + ^nFi
(9)
где и = 2или4. В дигибридном расщеплении появляются новые генотипы:
ААЬЬ1 АаВВ, ААВЬ, ааВВ. Но так как по каждому гену отношения 1+2+1
сохраняются, то выражение (9) может быть принято и для дигибридного,
и для других расщеплений. На деле редко можно располагать достаточным
* Для современного читателя более привычно написание уравнения (1) как а]
=<5* + о%.— Прим. ред.
.2 _
243
16*
материалом по всем поколениям и приходится преимущественно опираться
либо на fjip, либо на fipt.
Далее нужно иметь в виду, что \in зависит от внешних условий и для
правильности решения (9) необходима увередность, что условия, в
которых находились все поколения, были одинаковы. Для этого, в частности,
желательно иметь все поколения одновременно, т. е. к моменту получения
F2 необходимо обеспечить дополнительное получение Fx и Р. Для этого,
однако, нужно, чтобы родители (Р), дававшие Fx, от которого получено
F2, и те, от которых получено новое Fx, были одними и теми же или
одинаковыми, что возможно только при гомогенном материале.
Предыдущее изложение велось нами применительно к скрещиванию
чистых линий, где в Р и ¥г все генотипы однородны и изменчивость
является целиком паратипической. Такие случаи при работе с животными,
очевидно, не встречаются. Возникает вопрос, приложимо ли сказанное к
скрещиванию животных, для которых в любой совокупности имеется
генотипическое расщепление наряду с паратипическим.
В этих случаях результаты операции вычитания [i2F* — fi2p,
оказываются действительно иными. Что остается в результате вычитания, легко
видеть из следующего. При наличии в ¥г расщепления благодаря него-
мозиготности родителей изменчивость уже в ¥± будет двоякой природы —
генотипическая плюс паратипическая, как и в F2. Записав
соответствующие уравнения и производя вычитание, получим:
Н«2Р2 = H-2P2Y + U2F3*
Р2Fi = MgEVr/Ь H»2F»*
H-2F8 ""P'SFi ==)12FaY "~ H'SFiY
т. е. разность между \i2$2 и fi2Fl есть уже разность генотипической
изменчивости F2 и Fx. Мы исключили не только паратипическую изменчивость,
ио и часть генотипической.
РЕШЕНИЕ ЗАДАЧИ О ЧИСЛЕ ГЕНОВ
И СИЛЕ ИХ ДЕЙСТВИЯ
Генотипическая изменчивость в F2, как мы видели, выражается
каркасом более или менее биномиального характера. Простейшим случаем
здесь будет такой, когда (1) все гены равны (i± = i2 = = i) и (2)
когда имеет место адоминирование А а = (А а + аа)]2.
В этом случае
I*2Y = -J-*'2f (1)
где |i2Y — момент второй степени биномиального ряда, п — степень
бинома и £ — сила действия гена. Чем сильнее действие гена, тем сильнее
будет, очевидно, размах изменчивости. Так как \i2 = а2 и есть средняя
сумма квадратов, то и i входит в формулу в квадрате. Иначе i равна
классовому промежутку каркаса.
Так как п есть показатель бинома, то он равен числу аллелей и,
следовательно, вдвое больше числа пар аллелей или, как обычно говорят,
числа генов. Обозначив число генов #, имеем х(2, следовательно,
H2Y=lH2, (2)
откуда
244
В этом уравнении два неизвестных х и i. Для нахождения их нужно
получить второе уравнение. Если мы скрещиваем две чистые расы ааЪЪ и
ААВВ, то разница в величине их признаков будет зависеть от совместного
действия всех четырех аллелей —А, а, В и Ъ. Для случая адоминирова-
ния и равенства генов можно считать, что D — М2 — Мх = ni или i = D/и,
т. е. сила действия одной аллели равна разности между средними
величинами признаков у скрещенных типов (М2 — Мг), деленной на число
аллелей. Так как п равно 2#, то
1 2х ' 1 4*2 ' w
Имея два уравнения (2) и (4) с двумя неизвестными х и i, находим и
значение х*. Поставив (4) в (2), получаем:
D2 х D2
*«2 - - —
откуда
# = -
D2
Так как
то
_ D2 /ft.
т. е. число генов (ж), участвующих в скрещивании, равно квадрату
разности между расами (D2), деленному на 8-кратную разность моментов
второй степени F2 и F1#
Это равенство справедливо при следующих условиях: 1) скрещиваются
гомозиготные расы, 2) сила генов одинакова, 3) адоминирование, 4) все
аллели-усилители находятся у одного родителя, все ослабители — у
другого, 5) гены не сцеплены друг с другом, 6) жизнеспособность различных
генотипов одинакова.
Мы видим, что применение формулы ограничено целым рядом
условий, притом таких, которые в действительности почти никогда не
осуществляются, особенно у животных: 1) животные всегда более или менее
гетерозиготны, 2) сила генов различна, 3) доминирование нередко
промежуточное, 4) гены между породами распределяются обычно
«неправильно», т. е. у каждой породы имеется часть усилителей, часть ослабителей,
5) нередко гены сцеплены и 6) жизнеспособность генотипов часто
различна. Этих элементов мы заранее не знаем. Построив, как мы сделали, три
уравнения, мы должны были ограничиться лишь тремя неизвестными,
допустив для остальных неизвестных некоторые условные значения.
Однако все 5 первых условий обладают тем свойством, что всякое
уклонение от них если и влияет на решение, то в одну сторону: число генов
(х) оказывается сниженным. Поэтому получающийся ответ является
минимальным ответом. Если мы получим ответ х = 2, то его можно с
полной уверенностью читать как «число генов, участвующих в скрещивании
не менее двух». Иного ответа, по сути дела, мы и не можем иметь, так как
в любом скрещивании принимает участие множество очень малых
(слабых) генов, учесть которые мы не имеем возможности.
Для генетика-аналитика минимальный ответ уже представляет
определенный интерес, и вопрос может состоять лишь в том, как обеспечить,
чтобы этот минимум не был слишком снижен.
* Собственно говоря, мы имели дело с тремя неизвестными: jx2y, x и i и нашли их из
трех уравнений: (2), (4) и (5).
245
Анализ показывает, что в этом отношении при применении этой
формулы особое значение имею* два условия: степень доминирования и
характер распределения генов между родительскими типами (3 и 4) *.
ДОМИНИРОВАНИЕ
Влияние доминирования может быть легко исключено двумя
способами. Во-первых, может быть использована более общая формула, вывод
которой мы здесь приводить не будем (Serebrovsky, 1928). Она имеет вид:
3D2 — 4DDi + 4D*
Здесь, помимо уже знакомых величин D и [г, участвует Dx, которое
равно разности между средней величиной признака у ¥х и у рецессивного
типа (т. е. отстоящего от Fx дальше). В зависимости от степени
доминирования положение |хр1 относительно родительских типов будет
различным. При неполном доминировании D равно 2Db и формула
превращается в приведенную ранее.
При полном доминировании D равно D1? и формула приобретает вид:
_ 3D2
Х~ 16 0i2Pa-!x2Fi) *
Влияние доминирования может быть исключено также и путем
использования не F2, a FB с рецессивным типом. В этом случае гомозиготы не
возникают вовсе. Возникают одни только гетерозиготы и, следовательно,
картина будет одинакова, независимо от степени доминирования. К
материалу FB должна быть применена формула:
= 3D2
Dx здесь стало вместо D, так как по нему правильнее определяется сила
действия гена.
ВЛИЯНИЕ РАСПРЕДЕЛЕНИЯ АЛЛЕЛЕЙ-УСИЛИТЕЛЕЙ
Наиболее сложно обстоит дело с влиянием распределения генов между
родительскими расами. В самом деле, при отсутствии между А и В
сцепления ¥г и F2 получаются совершенно одинаковыми в таких двух
скрещиваниях:
(1) ааЪЬ х ААВВ
(2) ааВВ X AAbb.
Но величина D при равных условиях будет меньше во втором
скрещивании, чем в первом, а в приведенном примере при условии ix = i2 во (2)
D = 0. Следовательно, и в формулах для определения х числитель будет
либо преуменьшен, либо сведен к нулю, и вместе с тем сведенным к нулю
окажется и х. Мы получаем ответ, что в скрещивании участвует «не менее
0 генов», т. е. ответ, не представляющий никакого интереса.
* Вопрос о методах определения числа генов, которыми различаются скрещиваемые
формы, подробно разбирается в монографии Мазера (Mather. Beiometrical
genetics). В ней есть ссылка на работу А. С. Серебровского (Serebrowsky, 1928).— Прим.
ред.
246
Если мы имеем скрещивания типа:
D х
(1) aabbccddeeff X AABBCCDDEEFF 12 6
(2) aabbccddeeFF X AABBCCDDEEff 8 2,67
(3) aabbccddEEFF X AABBCCDDeeff 4 0,67
(4) aabbccDDEEF.F X'AABBCCddeeff 0 0
к к кг
n
ряд решений х выражается формулой:
х_ (*-fa)»
п
Истинное решение по приведенной формуле дает нам ответ не о числе
генов, участвующих в расщеплении, а о характере их распределения.
Необходимо отыскать какое-либо второе уравнение, из которого
можно было бы произвести определение i, не прибегая к определению разности
между исходными родителями.
Исследуем для этой цели F3, например, полученное по второму
приему. Здесь в F3 мы получаем распределение М от отдельных скрещиваний:
М1 = Мр +1,00 1
М2 = Мр +1,50 4
Ms = ilfp + 2,00 6
М4 = Мр+2,50 4
М5 = Мр + 3,00 1
Это распределение М очень похоже на распределение отдельных
особей:
без усилителей 1
с 1 усилителем 4
с 2 усилителями 6
с 3 » 4
с 4 » 1
с той, однако, разницей, что между отдельными классами здесь интервал
равен £, а в вышеприведенном *-i.
Следовательно, для этого поколения [х2 каркаса будет в 4 раза
меньше. Поэтому можно написать
[A2F3 = -f-*'2 + H'2it.
К сожалению, новое уравнение не позволяет определить отдельно два
неизвестных х и i. Наоборот, \i21t отсюда может быть определено, как г
произведение x-i2. Уменьшение генотипической изменчивости будет в
четыре раза при всяком значении i и х.
Этот пример мы привели для того, чтобы показать наглядно, что не
всякое новое уравнение, которое мы можем составить на баёе еще нового
скрещивания, позволит нам найти одно лишнее неизвестное. Наоборот,
мы можем поставить много скрещиваний, подобных разобранным только
что, и на основе каждого составить новое уравнение. Но это будут лишь
«избыточные» уравнения, позволяющие лишь более точно определить
значение произведения xi2, входящего во все эти уравнения. Для того
чтобы найти и х и i отдельно, надо найти такое новое уравнение, где х и
i были бы представлены в какой-либо иной связи, чем xi2. Разность М?х —
—Мр2 = D тем и ценна, что там мы имеем связь x-i = 2)/2.
247
ИСПОЛЬЗОВАНИЕ р4
Такое новое уравнение может быть получено при помощи момента
более высоких степеней, чем обычно используемое [х2, а именно {л4* т. е.
момента четвертой степени. Для бинома (1 + 1)п имеем простую
закономерность:
*(3*-2)
1*4 = Jg •
Для каркаса расщепления при (1): равенстве генов и (2) адоминировании
будем иметь
1*4 J§* •
Здесь п = 2х и i связаны уже иначе, чем в jx2, что дает возможность
решить задачу. При отсутствии паратипической изменчивости решение
было бы просто. Достаточно было бы получить |32 делением jx4 на jut^.
о _ R _ #г (Зтг — 2)£а 16 __ о 2_
Рг ~~ »2 16 ггЧ* ~~ и '
^2
отсюда
2
з-р2 •
Так как здесь я — число аллелей, а число генов или мест вдвое меньше
(п == 2х), то
_ 1
*~ 3-р2 •
Это последнее выражение может быть использовано для обработки
каркаса, хорошо обнаженного от покрова по первому приему главы X.
При наличии же паратипической изменчивости дело осложняется. По
теореме сложения моментов имеем
И*ф = P>4y + R* + 6(1^ р»;
И»2ф = H-2Y +
Неоткуда определяем
( и«г = |»4ф — и*я — бщг и»; (1)
№* = И*ф — N*. (2)
И подставляя (2) в (1), получаем
|i4Y — (А4Ф — И»« — 6 (р,2ф — [ia„) jx2«;
В правой части имеем все величины, определяемые из эксперимента,
в левой — неизвестные. Получаем теперь p2Y = (WhIy-
где \1+ = fipt, |A« = fipt. Но, с другой стороны, как было, показано выше,
х = 1/(3 - Ы-
Из-за громоздкости подставлять значение для p2Y в это выражение
мы не будем, так как ясно, что задача решена. При этом мы обошлись без
вычисления разницы между родительскими расами и оперировали лишь
248
с F2 и Fx. Поэтому решение не будет зависеть от того, как распределены
между исходными породами гены, участвующие в расщеплении.
К сожалению, как мы видим, формула эта довольно сложна. Сложность
в данном случае вредит тем, что она влечет за собой многократные
вычитания, деления и умножения (шесть операций), при которых происходит
нарастание статистической ошибки. А так как само значение |32 уже при
трех или четырех генах отличается мало (2,67 и 2,75), то прием становится
не очень точным. Впрочем, если при его помощи удастся отличить дигиб-
ридное расщепление от тригибридного, то этого будет достаточно.
Теория этого вопроса требует дальнейшей разработки. Настоящая
формула применима при условиях: 1) скрещивание чистых пород, 2)
адоминирование, 3) равная жизнеспособность, 4) отсутствие сцепления.
Для полного доминирования ее легко переделать. Для разных степеней
доминирования перспектива осложняется, так как необходимо будет
ввести новое неизвестное — степень доминирования. Для
его определения можно воспользоваться разностью между ¥г и F2, так
как при промежуточном доминировании |ipa = [ip^ а при наличии
доминирования — неизбежен сдвиг F2 в сторону рецессивного фенотипа. Этот
вопрос может послужить темой специального исследования.
ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Мы изложили здесь «метод решения уравнения» как один из методов
генетического анализа количественных признаков. Однако более
внимательное рассмотрение вопроса показывает, что, идя именно этим путем,
мы должны будем дать стройную теорию генетическому анализу, который
пока все еще остается неприведенным в систему собранием эмпирических,
более или менее остроумных, правил. Легко показать, прежде всего,
что, например, приведенная выше формула для отыскания числа генов
3D2
п-
может быть приложена хотя бы к классическому менделевскому
моногибридному скрещиванию. Пусть, например, мы имеем скрещивание:
черный х белый
i
черные
3 черных: 1 белый
Положим белый = 0, черный = 1. Тогда D = 1; fji2Fl = 0; [х2р2 = 3/16*
Подставляя в формулу, будем иметь:
п - 3-1* __ 3 л
, "-»(£-.)-•-'•
т. е. получаем ответ, что мы имеем моногибридное скрещивание.
Если бы в F2 мы получили расщепление 9 черных, 6 серых, 1 белый,
то, полагая, что белые = 0, серые 1 и черный = 2, получили бы ответ:
„ 3-2* 9
i6-i6-
Это показывает, что и обычное моногибридное скрещивание можно было
бы решать алгебраически. Если мы этого не делаем, то потому, что не
имеем нужды, так как твердо знаем, что соотношение 3 : 1 в F2 отвечает
249
моногибридному, а отношение 9:6:1— дигибридному случаю. Иначе
говоря, мы решаем это уравнение, не формулируя, как решаем, не
формулируя тысячи других уравнений, предъявляемых нам жизнью.
Однако перед нами всегда остается основной закон, что мы можем
найти столько неизвестных, сколько сопряженных уравнений имеется в
нашем распоряжении, и в том случае, когда число неизвестных больше
числа уравнений, мы вынуждены давать некоторым неизвестным
условные, более или менее вероятные, значения. В данном примере отношение
3 : 1 может получиться, например, и при двух генах, если они
абсолютно или хотя бы очень сильно сцеплены друг с другом, или если процент
перекреста между ними с = 0. Имея отношение 3: 1 и говоря, что перед
нами моногибридное расщепление, мы принимаем некоторое вероятное
условие с ==/= 0. Для этого, конечно, имеются основания, так как с,
равное 0, явление редкое, маловероятное. Аналогично, отношение 3 : 1
может получиться, например, как искажение дигибридного отношения
9 : 7, если жизнеспособность белых (v) примерно 45%. Принимая, что
3 : 1 есть моногибридное отношение, мы допускаем условие, что v =/=45%
или что v — 100% и одновременно, что с ==/= 0 или с% = 50, так как
явления г; = 45% ис = 0 маловероятные.
Получая в наше распоряжение несколько скрещиваний, мы
распространяем наше суждение и на другие неизвестные. Например, имея
материал по F2 (3 черных : 1 белый) и по FB (1 черный : 1 белый), мы
получаем уверенность в том, что v =f= 45%, так как при парных генах и этом
условии в FB получилось бы отношение 1: 1,3, а отношение 1:1, очевидно,
было бы при v — 33%, при которых в F2 было бы отношение 3 : 0,75.
Иными словами, из дигибридного 9 : 7 для F2 и 1 : 3 для F в не может
получиться 3:1 и 1 : 1 с единой поправкой на жизнеспособность белых.
Какую бы задачу по генетическому анализу ни решали, мы всегда
имеем дело с несколькими уравнениями, из которых можем найти столько
неизвестных, сколько это можно сделать по правилам, вскрытым
математикой. Осознать это необходимо для того, чтобы, воспользовавшись
богатым опытом математики, получить возможность решать задачи
генетического анализа кратчайшим путем, сознательно намечая каждый
последующий шаг, не путаясь в паразитных уравнениях, выбирая наиболее
решающиеся скрещивания. К сожалению, сейчас мы еще далеки от
такого состояния генанализа. Если, например, мы составим задачу с
несколько осложненным условием (например, два гена А и В с 20% перекреста
между ними, Л в 30% случаев не проявляется, В эпистатирует над А и
лонижает жизнеспособность на 25%), то никто не сможет сказать, как
именно надо вести решение, чтобы получить возможно быстрее ответ,
сколько необходимо для этого поколений и сколько нужно получить
потомства.
В этом направлении генетики-аналитики только что начали
двигаться. Изложенный и в настоящей главе материал еще слишком
незначителен для утверждения, что метод решения уравнений может быть
практическим методом анализа количественных признаков. В качестве
подсобных приемов применение выведенных формул может быть
использовано при обработке материалов более или менее значительных
скрещиваний. При этом, однако, совершенно необходимо помнить о тех условиях,
для которых формулы выведены. Тем не менее дальнейшая разработка
теории в этом направлении, разработка, так сказать, алгебры
генетического анализа должна иметь для нашей дисциплины большое значение.
Может возникнуть вопрос, не являются ли задачи генетического
анализа, особенно взятые из жизни, безнадежно трудными для того, чтобы
они могли быть подвергнуты метематической обработке. Действительно, в
большинстве скрещиваний принимает участие много генов, каждый из
них имеет свою силу действия, по-своему влияет на жизнеспособность,
250
имеет свой процент проявления, обнаруживает с другими генами
специфическое взаимодействие, дает с ними различные проценты перекреста
и т. д. Все эти элементы являются неизвестными, и легко видеть, что их
можно насчитывать очень много. Мало того, задачи генетического
анализа принадлежат к таким своеобразным задачам, где неизвестным
оказывается самое число неизвестных.
Тем не менее уже эмпирически найдены некоторые приемы решения,
и задачи, очевидно, поддаются значительным упрощениям, которые лишь
ждут своей формулировки. Такое упрощение задач может заключаться,
например, в замене отдельных неизвестных сводными. Если, например,
в каких-либо скрещиваниях участвует х генов с различной силой
то перед нами задача с х неизвестными, требующая для решения х
уравнений. Но мы можем заменить их тремя сводными неизвестными х — число
генов, i — средняя сила действия генов иб{- характеристика
вариации силы действия. Для отыскания этих трех неизвестных требуется уже
ле х уравнений (число большое и к тому же неизвестное), а только 3.
Между тем, узнав х, i и аь мы получим довольно подробную
характеристику нашего случая. Получив такую ориентацию в нашем материале,
мы можем идти дальше, уточняя и конкретизируя сведения, например,
-о более сильных генах и т. д.
Таким образом, в зависимости от того, каким числом скрещиваний и
уравнений мы располагаем, должны составляться различные сводные
неизвестные, и структура этих сводных неизвестных должна нам дать ясную
картину того, до какой глубины мы сумели довести решение задачи, и что
осталось еще невыясненным.
ГЛАВА XII
МЕТОДЫ СИГНАЛЕН
Сигнальными генами, или сигналями, мы называем удобные для мен-
делистических наблюдений альтернативные гены с более или менее
известной локализацией, которые, не влияя на изучаемый трансгрессирующий
признак или влияя достаточно определенным образом, облегчают
генетический анализ этого признака, позволяя следить за наследованием того
участка хромосомы, в котором эти сигнали расположены. Так как на этом
участке хромосомы могут находиться некоторые из генов, участвующих
в определении полимерного признака, подвергаемого анализу, то при
помощи сигналей мы оказываемся в состоянии выделить из общей массы
полимеров эти гены и следить за их наследованием не по собственным их
фенам, а по фену расположенной вблизи от них сигнали.
Суть метода при этом сводится к тому, что можно называть «условной
плейотропией». Под плейотропией, как известно, разумеется свойство
многих генов влиять одновременно на несколько признаков, т. е. иметь
сложный фен. Одна часть этого фена может быть выражена очень
отчетливо, альтернативно наследуясь. Другая часть фена может сильно и
трансгрессивно варьировать. Многие гены дрозофилы влияют, например, на
жизнеспособность мух. Так, мухи с пурпуровыми глазами (purple, pr)
имеют жизнеспособность в среднем на 30—50% более низкую, чем мухи
с нормально красными глазами. Если бы мы захотели изучить этот ген,
наблюдая лишь жизнеспособность и не обращая внимания на окраску глаз,
то это было бы чрезвычайно трудной задачей благодаря сильно
трансгрессивной изменчивости жизнеспособности. Но если мы будем следить за
геном, наблюдая его действие на окраску глаза, задача решится
чрезвычайно легко.
В данном случае один и тот же ген действует и на трансгрессивный
признак и на альтернативный. Но что будет, если в хромосоме тесно
рядом будут находиться два самостоятельных гена, из которых один
будет влиять на трансгрессивный признак, а другой на альтернативный.
С точки зрения задач генетического анализа оба эти случая равноценны,
так как мы всегда сможем легко следить за интересующим нас геном
трансгрессивного действия по альтернативному фену его соседа. Этот
ген-сосед явится сигнальным геном.
Положение осложняется, если сигнальный ген в процессе перекреста
будет отрываться от изучаемого гена. В этом случае заключения, которые
мы будем делать о нем, наблюдая сигналь, будут носить менее точный,
условный характер, и это понижение точности будет тем сильнее, чем
дальше сигналь будет расположена от гена трансгрессивного действия,
или, точнее, чем легче будет между ними происходить перекрест.
В этих условиях положение может быть значительно улучшено, если
изучаемый ген удастся заключить между двумя сигналями, т. е. если уда-
252
стся найти два сигнальных гена, локализованных по длине хромосомы
один слева, другой справа от него. Все случаи, в которых обе сигнали
оторвутся друг от друга, будет легко отбросить. Вместе с тем в тех
случаях, когда обе сигнали остались соединенными, можно будет с большей
вероятностью утверждать, что изучаемый ген (ген трансгрессивного
действия) между ними сохранился. Нетрудно понять, что этот ген сможет
оторваться от сигнален лишь при помощи двойного перекреста. Даже при
расстоянии в 10—20 морганид между сигналями эти двойные перекресты
будут очень редкими событиями (соответственно менее 0,25 и 1%),
которыми во многих случаях можно будет пренебречь.
Методом сигналей могут решаться почти все задачи генетического
анализа — разложение признака на отдельные фены и тем самым
выделение отдельных генов, участвующих в определении полимерного
признака, исследование свойств этих генов; их локализация, т. е. основные
задачи глав I и IV. Так же может решаться и задача глав VI и VII,
анализ генотипа, изучение свойств хромосомы и отдельных участков. Почти
не могут быть, наоборот, решаемы задачи популяционные, так как здесь
нарушается сцепление.
Недостатком сигнального метода, как было сказано выше, является
то, что он может быть приложен лишь к тем видам животных, для
которых уже более или менее изучен топографический план хромосомы или
у которых, по крайней мере, известно достаточно большое число генов,
которые годятся для использования в качестве сигнальных генов.
Метод сигналей развился из работ по локализации генов у дрозофилы.
Еще в ранних работах Моргана, Меллера при изучении таких генов, как
vortex, Notch, eosin, было замечено, что их проявление сильно меняется
при комбинации с другими. Опыты показали, что не сами эти гены
влияют на проявление vortex, eosin и других, но какие-то иные, лишь
локализованные в тех же хромосомах. Таким образом, стали улавливаться
модификаторы, усилители или ослабители, которые можно было отличить
сразу от паратипических факторов, потому что сильное или слабое
проявление признаков оказывалось как-то связанным с сигнальными
генами.
К числу первых работ, совершенно ясно поставивших вопрос о
применении сигнальных генов к анализу количественных признаков,
относятся исследования, посвященные анализу величины яиц у дрозофилы.
Этот признак, несмотря на некоторую кропотливость работы с ним, очень
ценен для исследований такого рода, так как почти не отражает на себе
тех резких колебаний величины мух, которые вызываются различной
влажностью корма и степенью населенности культур. Эти колебания очень
осложняют эксперименты подобного рода с дрозофилой и относят почти
все ее количественные признаки к числу колеблемых паратипически.
Сравнительные исследования показали, что почти каждая
лабораторная линия дрозофилы дает особую среднюю величину яиц. Ставя
различные скрещивания с участием сигнальных генов, можно показать влияние
различных хромосом на величину яиц.
Несмотря на то что различие величины яиц, особенно М, исчисляется
сотыми долями миллиметра, метод сигналей сделал возможым установить
и здесь влияние различных хромосом, т. е. наличие многих генов,
локализованных в разных хромосомах. В 1926 г. этим методом Купченко в
нашей лаборатории определил влияние отдельных хромосом на
яйценоскость дрозофил, не ставя своей задачей более точно локализовать эти
гены. В нашей лаборатории генетики МГУ была подвергнута более
детальному изучению одна вторая хромосома, осигналенная геном Lobe (Коган,
Женихова). Вторым партнером была хромосома, так называемая дупль,
яесущая гены dumpy, black, purple, curved, plaexus, speck. Даже только
.в одной этой паре хромосом удалось открыть не менее четырех генов,
253
влияющих на величину яиц: один ослабляющий аллель, вблизи локуса
dumpy, и три усиливающих аллеля — в правой части хромосомы Lobe*
Ряд авторов пользовались методом сигналей для генетического анализа
и других организмов. Пеннет (Punnett, 1927—1930; Серебовская Р. И.,
Серебровский А. С, 1926) пробовали найти ген, влияющий на
яйценоскость кур, по предположению Пирля и Серфейса (Pearl. Surface, 1914),
локализованный в половой хромосоме. Касл (Castle, 1929) получил
отрицательный результат при попытке уловить гены роста кролика,
используя в качестве сигналей некоторые гены окраски. Наоборот, Грину (Green,
1932) удалось доказать, что у мышей имеется ген или гены, влияющие
на скорость роста в той же хромосоме, где локализованы аллели D — d
(голубая окраска). Линдстром использовал гены окраски томата для
анализа величины плода (Lindstrom, 1932) и т. д.
От изучения сигнальных генов следует отличать изучение плейотро-
пии генов. О. А. Иванова (1926) изучала наследование полимастии у
коров и показала, что на ее материале полимастные коровы давали несколько
более высокие удои. В данном случае предполагается, что тот же самый
ген, который влияет на развитие сосков, одновременно повышает
деятельность вымени. Правда, как было упомянуто, при сильном сцеплении
сигнального гена и гена трансгрессивного действия отличить этот случай
от плейотропии не всегда будет легко или даже возможно, но тем не менее
следует обе проблемы различать.
Например, плейотропию можно и даже особенно удобно изучать в
популяции, как, например, делала это О. А. Иванова (1931), а сигналями
можно пользоваться лишь в родословных. В самом деле, если какой-либо
ген Р имеет плейотропное действие, то все части его сложного фена будут
всегда более или менее связаны друг с другом. Но в некоторых
родословных, где в одной хромосоме с геном Р окажется локализованным какой-
либо модификатор, плейотропия окажется нарушенной. Если, например,
ген полимастии действительно несколько повышает удой, то в некоторых
родословных, наоборот, полимастные коровы могут оказаться генетически
менее молочными, если в той же хромосоме окажется локализованным
ген, понижающий молочность.
В популяции, где случаи сцепления и случаи отталкивания
встречаются равно часто, возможность такого искажения отпадает. Наоборот,
связь сигнального гена с геном трансгрессивного действия можно
наблюдать только в родословной —в популяции эта связь постепенно
исчезает. Поэтому в зависимости от того, что именно мы хотим изучать,
плейотропию или связь сигналей с генами трансгрессивного действия, мы
должны пользоваться различным материалом. Выводы о наличии плейотропии,
сделанные на основании изучения родословной, могут оказаться
неверными.
К сожалению, приходится констатировать, что применение метода
сигналей до последнего времени остается еще на очень примитивном
уровне. Фактически дело сводится лишь к тому, что если при расщеплении
по сигнали (S) обнаруживается разница в изученном признаке между
группами S и 5, то констатируется, что «в хромосоме S имеются гены,
влияющие на изучаемый признак». Если такого различия не
обнаруживается, делается вывод, что хромосома S «нейтральна». Первый вывод
правилен, но в таком виде мало интересен, так как заранее можно
сказать, что при тщательном изучении можно будет показать, что почти
каждая хромосома несет те или другие гены, влияющие на любой
количественный признак. Наоборот, вывод о «нейтральности» хромосомы большей
частью будет неверным. Как мы увидим ниже, хромосома может казаться
«нейтральной» и когда в ней локализованы сильные гены, но действующие
противоположно и расположенные определенным образом относительно
сигнали.
254
Одной из причин такого неудовлетворительного положения,
несомненно, является то, что до сего времени не уделялось внимания
разработке теории вопроса, не выяснялось, при каких условиях и какие именно
выводы можно и нельзя делать из наблюдений над опытами с участием
сигналей. Общей теорией здесь опять является вопрос о числе
неизвестных и числе и характере уравнений, необходимых для отыскания этих
неизвестных. Поэтому в настоящей главе мы должны уделить
необходимое место освещению вопросов теории сигналей.
СТРОЕНИЕ ХРОМОСОМЫ КАК НОСИТЕЛЯ
ПОЛИМЕРНЫХ ГЕНОВ
Сила гена и баланс хромосомы
Для того чтобы не запутаться в тех достаточно сложных явлениях, с
которыми мы имеем дело при анализе генотипа хромосомы с помощью
сигнальных генов, необходимо более внимательно обсудить стоящие здесь
перед аналитиком проблемы, так как недостаточная ясность постановки
проблемы не позволяет получить и ясные выводы. Поэтому мы должны
здесь остановиться на строении хромосомы как объекта нашего анализа,
как носителя тех генов-полимеров *, усилителей и ослабителей, которые
мы должны изучать.
Хромосома является удлиненным телом, состоящим из
расположенных в ряд генов. Вопрос о наличии межуточных элементов, связывающих
гены, сейчас для нас неинтересен. Можно оставить в стороне и вопрос о
природе и строении генов — для нас они сейчас интересны лишь как
источники воздействия на онтогенез. G этой точки зрения для нас важно, что
ген имеет некоторую силу действия, которую обозначим буквой /.
Различные гены обладают различным действием. Если даже взять гены,
влияющие на один и тот же признак, например, на величину яйца дрозофилы,,
где качественные различия действия генов трудно уловимы, то остаются
количественные различия — один ген действует сильно, другой слабо;
и различия в направлении — один ген усиливает, увеличивает признак,
другой ослабляет, уменьшает. Таким образом, у разных генов / будет
различное и иметь либо положительный, либо отрицательный знак.
В каждой хромосоме генов данного признака (например, величины
яйца) ^обыкновенно бывает несколько, различных по силе и по
направлению. Отсюда возникает новое качество — баланс хромосомы:
(Б), причем
Б = 11 + 12 + 13+ ...1п1
т. е. баланс хромосомы, ее итоговая сила действия на данный признак
может быть схематично выражен алгебраической суммой сил генов,**.
При наличии взаимодействия между отдельными генами эта формула
может осложняться, но для начала вполне достаточно (что в известных
пределах подтверждено экспериментально) принять для баланса такую
простейшую формулировку.
Аллосила и аллобаланс
Определение как баланса хромосомы, так и силы гена / является
достаточно трудной задачей. Гораздо чаще мы имеем дело не с этой полной
силой гена, а лишь с разницей в силе действия между двумя аллелями.
* Словом полимеры автор сокращенно называет полимерные гены.— Прим. ред.
** Развитые на этих страницах А. С. Серебровским взгляды на хромосому и на общий
баланс ее влияния на количественный признак несколько напоминают
представления Мазера о полигенах. Приоритет принадлежит Серебровскому.— Прим. ред*.
255
s
jfd
s
€1
±
net
S
\u iB |4r \k
iP С
Рис. 81. Строение баланса
S и ^(вверху) и аллобаланса
S (внизу)
Сравнивая действие на признак
двух аллелей, мы говорим,
например, «в хромосоме S расположен
слабый усилитель величины
яйца, его сила равна +0,02 мм».
Это значит, что в гомологичных
хромосомах S и s, различаемых
нами, например, при помопщ
сигнальных аллелей S ж s, находятся
два различных аллеля, из которых
аллель в хромосоме S является
более сильным в смысле
увеличения яйца, чем аллель в
хромосоме s.
Обозначая силу действия этих
аллелей символами Is и 18,
получаем
i == /fl —19. j
хромосом
хромосомы
Эту величину i назовем аллель-
ной силой гена, расположенного в
хромосоме, или его аллосияой.
Именно ее мы обозначаем здесь и ранее буквой г. В данном примере
i = +0,02. Аллосила другого аллеля, локализованного в хромосоме s,
будет
i = 18 — /s«
В данном случае она равна —0,02 мм. Если нужно, то можно
обозначить эти i полнее, is — *«, но каждый раз для сравнения сила второго
аллеля принимается за 0. Иначе говоря, is = — is*
Если сравниваемые хромосомы различаются несколькими генами,
то возникает новое качество, параллельное балансу,— аллобаланс
хромосомы S по сравнению с хромосомой s. Обозначая его через D,
пишем с тем же основанием, как и для Б:
D = i1 + *2+ ... + *«•
Можно для ясности написать Ds, т. е. аллобаланс 5-хромосомы.
Очевидно, что
Ds = Bs-B8, D3 = B8-Bs и Ds--D3.
Для лучшего пояснения этих отношений обратимся к рис. 81. Он
изображает пару хромосом гомологов, различаемых по сигналям S и s.
Стрелками на ней изображены полные силы (/) генов А... Направление
стрелки вверх показывает, что данный ген усиливает, направление стрелки
вниз — что он ослабляет. Длина стрелок указывает на силу генов
(полную); пометка def—на наличие в хромосоме нехватки; жирная часть
стрелок — на разницу в силе действия аллелей. Так, сила 1а = +3,
сила 1а = +1. Жирно отмечена аллосила:
iA = lA—Ia= +2-
На основании верхней части рис. 81 представлено строение Ds
аллобаланса хромосомы S (рис. 81, внизу). Направление стрелок указывает
направление действия генов, локализованных в хромосоме £, по
сравнению с их аллелями в s-хромосоме. Длина стрелок пропорциональна
величинам аллосил i.
256
Подчеркнем, что аллосилы и полные силы являются существенно
иными вещами. Так, аллель G в обеих хромосомах S и s одинаков, поэтому
аллосила этого гена в хромосоме S равна 0, хотя сам ген имеет I* = +2.
Далее, ген С сильнее гена А: 1С = 4, 1А = 3, 1С = 3,5, 1а = 1. Но
аллосила 1а = 2, a ic = +0,5, т. е. аллосила гена С меньше, благодаря
большому сходству обоих аллелей 1С и 1С.
Еще важнее, что 1В > 0 и 1Ъ > 0, т. е. оба аллеля усиливают признак,
ген является усилителем. Но аллосила гена В (в — —0,5, т. е. в
сопоставлении с аллелем Ъ ген В является ослабителем. Обратная картина
имеет место у гена D. Оба аллеля Dud, как показывают стрелки, являются
ослабителями признака, но iD = +2 и/)в сопоставлении с d фигурирует
как усилитель. Гены Е и F служат примером того, как благодаря
наличию нехваток здесь численно аллосила и полная сила совпадают и 1Е =
= 1е = +3, if = If =+1, но зато нехватка def F приобретает аллосилу
h = —1, h = —3, хотя, конечно, /F и 1г = 0, как нехватки, как
реальное отсутствие генного вещества.
Сигнальные аллосила и аллобаланс
Теперь необходимо ознакомиться с третьей характеристикой
гена, именно с силой гена, которую мы определяем, пользуясь сигнальным
методом.
Эту сигнальную аллосилу гена обозначим символом /.
Ниже будет показано, что
7 = i (1 — 2с) или / == i — 2ic,
где i — аллосила гена, ас — расстояние в долях единицы перекреста
(с = процент перекреста/100) между сигналью и данным геном. Если
полимер расположен рядом с сигналью, то разделяя в потомстве
сигнальные классы, например Ss и ss, мы будем иметь у всех особей класса
наш полимер и ни у одной особи ss. Разница в величине признака, т. е.
Ms — Ms будет непосредственно давать аллосилу полимера i. Но если
полимер будет отрываться в процессе перекреста от сигнали и попадать в
группу 55-потомков, разница в величине признака в классе Ss и в классе
ss будет уменьшаться и при очень далеком расстоянии (при с, близком к
0,5) исчезнет вовсе. Таким образом, аллосила полимера, определяемая
методом сигналей, благодаря перекресту будет меньше действительной
аллосилы на величину 2ic (на удвоенный момент гена относительно
сигнали).
При определении аллобаланса при помощи сигнали мы получим
также сигнальный аллобаланс (d), выражаемый формулами:
d = /i + /2 + .../n;
d = 2i(l-2c).
Список условных обозначений
полная сила гена;
сила гена, локализованного в хромосоме S;
сила гена, локализованного в хромосоме s\
Is — Is — аллельная сила гена, локализованного в хромосоме S;
I's + Is ... 1 л
\ баланс хромосомы;
1\ + h + ~- J
17 А. С, Серебровскм* 257
I -
18-
i =
Bs =
Б =
D = ix + г2 + ---аллосила хромосомы, или разность балансов;
D = Bs — Б8 — аллосила хромосомной пары;
Ds = Bs — Б8; Ds = —Ds — аллобаланс хромосомы S;
т — расстояние в морганидах вправо;
— т — то же, но влево;
|т| — расстояние в морганидах безотносительно направления;
с% — расстояние в процентах перекреста (всегда положительно);
7 — сигнальная аллосила гена;
г— сигнальная аллосила хромосомы;
\— сигнальный аллобаланс;
с — расстояние в долях единицы перекреста между сигналью и данным
числом.
ПРИЕМЫ АНАЛИЗА ПРИ ПОМОЩИ СИГНАЛЕЙ
После этих вводных соображений перейдем к более детальному
анализу применения сигналей для генетического анализа количественных
признаков и к выводу некоторых формул.
ОДНА СИГНАЛЬ В ХРОМОСОМЕ
Наиболее часто встречающимся случаем является такой, когда мы
располагаем для нашего анализа признаков сигналями,
расположенными по одной в разных хромосомах. В этих случаях перед нами могут стоять
две различные задачи:
1) анализ признака,
2) анализ влияния хромосом на признак и ее строение по генам
данного признака.
Полный анализ признака будет произведен, очевидно, когда будут
выполнены обе эти частные задачи, когда станет известным, какие
хромосомы влияют на признак, как в этих хромосомах расположены гены
данной категории и как каждый из них действует.
Сначала мы рассмотрим возможность освещения вопроса о строении
хромосомы с одним сигнальным геном. Типовая задача такого анализа
сводится к получению Fx, гетерозиготного по сигнали Ss и скрещиванию
F1? с рецессивом ss. В случае, если хромосома S (будем обозначать так
кратко хромосому, несущую аллель S) отличается от хромосомы s по
генам, влияющим на анализируемый признак, полученное потомство будет
состоять из двух классов, Ss и ss, различных по величине данного
признака.
ОДИН ПОЛИМЕР В ХРОМОСОМЕ
В простейшем случае, если в осигналенной хромосоме имеется лишь
один полимер, влияющий на анализируемый признак, то это скрещивание
изображается схемой XXII.
Здесь S и s обозначают аллели сигнали, А и а — гены, влияющие на
данный признак.
Что касается величин признака, то они будут таковы: если для
генотипа характерна при данных условиях средняя величина Мх, а
аллосила гена А будет г, то для генотипа Аа М2 = Мг + D, где D = г,
поскольку в скрещивании участвует лишь один ген.
258
s
s
5
S
Схема XXII
-4-
5 Л
•*———■—
i Ь-
$ a
S a
В потомстве группа ss будет состоять из 100 — с% форм sajsa и с%
форм sA/sa. Следовательно, средняя величина признака в этой группе
бурет:
м _ (Ш-с)%-Д/1 + со/0.Л/2
т*~ 100
или удобнее, используя с вместо с%:
М8=(1-с)Мг + сМ2, (1)
а для группы Ss соответственно наоборот:
М8 = сМг + (1^с)М2, (2)
откуда
d = Ms — Ms = сМг + (1 — с)М2 — (1 — c)Mx — cikf2 = (1 — 2с)(М2 — М х);
d = (l-2c)D,
откуда
или
<--И*-4).
со/0 =50(1-4)
(3)
Если изобразить эти скрещивания в виде вариационных кривых, то
мы будем иметь следующий рис. 82, из которого видна структура
вариационной кривой и положение М. Вместе с тем видно, что кривые потомства
являются скошенными в сторону перекрестных групп, что открывает
возможность привлечения для их изучения момента третьей степени ((л3) и
показателя асимметрии. На этом вопросе здесь мы, однако,
останавливаться не будем.
Определение величины D прямо из эксперимента возможно только у
тех видов, у которых один пол не дает перекреста (мухи, бабочки).
В этом случае, взяв в качестве гетерозиготы особь пила, не дающего
перекрест, получаем
SA ja_ SA , sa
sa sa . sa sa
Первая группа потомков имеет величину М2 = Mt + D, вторая Mlf й
D = i = М 2 — Мг.
259
t7*
F««-tf-»4
Рис. 82. Схема скрещивания с участием
сигнала и полимера (объяснение в
тексте)
Подставляя величину D в
формулу (3), находим с.
В этом простейшем случае
результат скрещивания дает нам
возможность узнать почти все, что нам
нужно]— аллосилу гена i = D и
расстояние А — 5 по формуле (3).
В частности, в случае d= D, с = О
ген А абсолютно сцеплен с S (или
сам ген S влияет на данный признак).
В случае d = 0, с = 0,5 или с% =
= 50 ген А свободно отрывается от 5,
т. е. расположен либо в другой
хромосоме, либо очень далеко от S
(порядка 100 морганид).
Вопрос о том, расположен ли ген
А вправо или влево от 5, здесь решен
быть не может, или может быть
решен лишь в том случае, когда
известно расстояние С гена от какого-
либо конца хромосомы, причем С<^ с.
В этом случае, очевидно, ген А не может лежать между S и
ближайшим концом хромосомы. В случае С ^> с это возможно и становится,
наоборот, невозможным сказать, локализован ли ген А правее или левее S.
Но в том случае, когда в хромосоме известен только один ген, от
нашего произвола зависит, назвать ли правой или левой ту сторону от S,
в которой лежит А.
Приведем иллюстрацию. Нами был взят в качестве сигнали ген
spineless, расположенный в /// хромосоме Drosophila melanogaster на 54 мор-
ганиде. Его признак — укорочение больших щетинок; на число
щетинок он влияния не оказывает. Кроме того, был взят доминантный ген
delta, который имеет плейотропное проявление: на концах жилок крыла
он дает треугольные расширения, послужившие основанием для названия,
а кроме того, он увеличивает число стерно-плевральных щетинок. Бели
у нормальной мухи (и spineless) число этих щетинок на одной стороне
груди 8—9, то у delta мух 10—11. Однако благодаря большой
трансгрессии у многих мух по числу щетинок не удается установить, являются ли
они delta или non-delta.
Была поставлена задача — определить положение усилителя числа
щетинок, пользуясь только сигналью и числом щетинок.
Неизвестных у нас два — сила действия гена и расстояние гена от
сигнали. Первое уравнение имеет следующий вид:
i = Ма — М а.
Для этого поставлено было такое скрещивание (А = delta):
SA * ч„ г* *sa SA . sa
sa ^ + sa sa l sa
Так как для скрещивания был взят самец, не дающий перекреста, то среди
потомства мы получаем две категории мух, различимых по сигнали spi-
nelless. При этом все мухи S должны нести «усилитель числа щетинок» и
следовательно:
Ms — М8 = МА — М'а = i.
В нашем опыте было получено:
Ms = 15,056; М8 = 11,000; i = 4,056.
260
%
60\
50
40\
30\
20\
10
0
[ Л
L ssl \/ \Ss
1 i ■ ■ i i fi '^» » V^ ■ t N>*—i
5. 6 7 8 9 10 11 12 13 П 15 16
-*—.#—»-
1
5
6
7
ss/
1 у
/ /
/ /
8 3
10
/4
\/ \ss
\ \
1 1 Г4^ ^^.-1
// 12 13 14 15 1Б
d-+
Рис 83. Сильно трансгредирующее распределение мух Ss и ss по числу щетинок
Из разности средних вычислены d и D и определено местоположение гена А (материал иной, чем
в тексте)
Вторым уравнением является
с*-50(1-4-)-
Для определения d было поставлено скрещивание:
-S-d-X?f-(100-co/o)^ + co/o^ + co/o^-f(10O-c0/o)^-.
Откуда найдено
d = М8 - М8 = 14,179 - 10,780 = 3,399.
Подставляя величину d и i в формулу, находим
*%-»(*--от)-8'15-
Следовательно, искомый ген, «усилитель числа щетинок», имеет силу
действия i = 4,056 и расположен в с% или в 8,15 морганиды от spineless
(58,50 морганиды). Так как знак расстояния неизвестен, то это либо
50, 35-я морганида, либо 66, 65-я.
В действительности ген delta расположен на 66,2-морганиде, f. е. наше
определение оказалось достаточно точным (Серебровский и
Бессмертная — не опубл.). Рис. 83 иллюстрирует один из опытов. %
Для определения направления («вправо» или «влево» от сигнален)
необходимо еще одно уравнение. В этом отношении «количественные»
гены ничем, конечно, не отличаются от «качественных». В приведенном
нами примере локализации гена delta мы использовали также вторую
сигналь, ген glass (стекловидные глаза), расположенный на 63,0 йорга-
ниде. Для этого гена было получено d = 3,694 и по-прежнему D = 4,056.
Отсюда с = 4,5%, Так как усилитель числа щетинок оказался
расположенным ближе к glass, чем к spineless, и так как расстояние от spineless
оказалось больше, чем оба другие расстояния, то очевидно, что порядок
генов таков: spineless — glass — усилитель. Таким образом, место
усилителя 66, 65 или 67,50, а в среднем 67,1 морганиды, правее обеих сиг-
налей, там, где действительно, локализован ген delta.
В процессе этого исследования нам удалось далее открыть еще один
слабый ген левее spineless. Благодаря тому, что ген delta имеет качест-
261
венное проявление на жилках крыла, мы могли отделить среди потомства
от скрещивания
^п w -. sa: sa л sA
— 9 х с? — МУХ — от —г •
sa + w sa J sa sa
При этом оказалось, что эти мухи имели меньшее число щетинок,
чем мухи sa/sa, полученные от реципрокного скрещивания (без
перекреста), а именно 14,560 против 14,986. Это обстоятельство естественно
можно объяснить тем, что анализируемые хромсомы имели такое
строение:
Ь S А В s a
X
В s а В s a '
где В обозначает усиливающий аллеломорф в хромосоме s. Для проверки
этого факта было поставлено дальнейшее исследование без участия гена
A (delta). Наличие гена В подтвердилось. Так как действие его было
незначительно, по сравнению с действием delta, то участие в скрещивании
гена В не отразилось сильно на вычислении положения А.
МНОГО ПОЛИМЕРОВ И ОДНА СИГНАЛЬ
Исследование генотипов хромосомы с одной сигналью имеет, как
видно из предыдущего, малое основание для успеха (кроме видов, у которых
один пол не дает перекреста, о чем скажем ниже). Причиной этого
является то, что в подобных условиях мы вынуждены ограничиваться
определением почти лишь одного сигнального аллобаланса d. Уже определение
аллобаланса D сопряжено с большими трудностями, так как для этого,
как мы видели, необходимо повторное скрещивание с изменением
величины перекреста. У аллогамных животных такое повторение практически
почти не осуществимо, так как мы еще не знаем методов изменения
перекреста, например, у баранов или петухов, а замена одного пола другим
теоретически возможна лишь тогда, когда мы располагаем
гомозиготными линиями. Скрестив две такие линии, например, инбредные линии
мышей, мы можем из Fx выбрать самцов и самок и, скрестив их с
соответствующего пола гомозиготными рецессивами, получить необходимые два
уравнения (и третье для определения разницы в перекресте). Вероятно,
этого значительно легче будет достичь у растений, особенно у способных
размножаться вегетативно. Одна часть клона может быть поставлена в
условия, дающие высокий перекрест, другая — в условия, дающие
низкий перекрест.
У баранов, овец, коров, быков, у которых можно проводить
искусственное осеменение, большое потомство благоприятно тем, что может дать
больший материал для достаточно точного определения.
Изучая величины d в каждом поколении у отца, сыновей, внуков,
можно будет сделать некоторые выводы о строении хромосомы и по тому,
как именно изменяются величины d. Например, при одном усилителе,
близко расположенном к сигнали, изменения в последовательных
поколениях будут иные, чем при усилителе далеком, при наличии усилителя
и ослабителя изменения будут опять иными и т. д. К сожалению, эти
заключения будут нередко очень неопределенными, так как нам будут
оставаться неизвестными или почти неизвестными хромосомы, приходящие
со стороны матери.
Использование одной сигнали более перспективно при анализе видов,
у которых перекрест происходит только у одного пола. Таким видами
являются, кроме лабораторной дрозофилы, важный
сельскохозяйственный объект тутовый шелкопряд, как, вероятно, и другие
бабочки. У мух перекреста не происходит у самцов, а у бабочек — у са-
262
мок. Отсутствие перекреста открывает возможность долго сохранять
хромосому одного и того же состава.
Разберем схему скрещиваний XXIII. Она показывает, что
скрещивание даже сильно гетерозиготного самца Drosophila с гомозиготной самкой
дает только два типа потомства, из которых один тип повторяет самца
(по данной хромосоме S).
г
а\ \а
bl lb
с\ \с
е\ \е
/
а\\А
ь\\в
с\\С
d\\D
1
Схема XXIII
а\ la
с\ \с
tf+ \d
//<
п
s\\s
al\A
b\\B
с\\с
d\\D
Е\ \е
44
XI
Так как этот отцовский тип представлен и самцами и самками, то из
потомства можно выбрать опять скрещивание, повторяющее
родительское, и хромосому SABCDE можно вести неизменной (пока в ней не
возникнут незамеченные мутации).
Это, в свою очередь, позволяет, если нужно, ставить грединг на
анализаторную линию по схеме ss ,x Ss и получить генотип, в котором все
остальные хромосомы будут, как у анализатора, кроме данной осигнален-
ной хромосомы. Поставив тогда реципрокное скрещивание
гетерозиготной самки с самцом-анализатором, мы будем иметь потомство, в котором все
расщепление будет зависеть только от изучаемой хромосомы и ее
перекрестов.
Разница М2 — Мх между двумя осигналенными классами $х и Sx в
потомстве от скрещивания без перекреста дает аллобаланс хромосом:
М2 - Мх = Б2 - Бх
D
(в данном примере разность ABCDe — abcdE).
Поставив скрещивание с перекрестом (реципрокное), мы получим уже
многообразное по полимерам потомство и можем определить для него
сигнальный аллобаланс.
Среди этих Ss потомков мы выбираем несколько самцов и от них ведем
снова линии без перекреста с самками анализаторов, сохраняя таким
образом в неизменности хромосомы нового строения. Поставив
скрещивание без перекреста (нааример, как на схеме XXIV), мы находим
новое.значение аллобалансa d.
263
i
h
с
s
я-;- f A
c\ ic
d\ \d
lU/mfyl
-s
3+
1 f
с
л1
е\ -К
//
XI
а\- -Iff
rfl }i7
//
П
a\ \A
ft tf
//
1%
Схема XXIV
S+ 45
a+ \a
b\ \b
c\ Iе
Д \d
£t tf
//.
t%
Таким образом, используя попеременно скрещивание с перекрестом и
без него, можно (теоретически говоря) исследовать строение хромосомы
с любой степенью точности и даже при довольно большом числе
полимеров в хромосоме. Приложение этого приема, как видим, возможно лишь к
видам, не дающим перекреста у одного из полов. К счастью, среди этих
видов мы имеем дрозофилу, важнейший генетический лабораторный объект,
и тутового шелкопряда, у которого уже целый ряд хромосом осигнален
1—2 генами.
Приблизительно по такой схеме Р. И. Серебровской и нами (1930)
было выполнено исследование III хромосомы дрозофилы по числу
щетинок.
Была взята хромосома с геном sepia (обозначим ее далее как
нормальную + + ) и исследовалось скрещивание:
+ +
st ca
X
st ca
st ca
На основании реципрокных скрещиваний были определены d и D.
Получено D = 3,31, ds, = 1,82, dca = 0,60.
Из скрещивания, где гетерозиготной была $, т. е. скрещивания с
перекрестом, были отобраны 10 самцов
+ +
(по другим генам они были
st ca
несколько различны, это подтвердило, что хромосома + + подверглась
перекрестам) и скрещены с самкой для «размножения» новых
хромосом + -f. Так как в качестве гетерозигот были взяты самцы, а самки из
чистой линии, то хромосомы нового строения (претерпевшие перекрест)
были размножены неизменными. После этого в каждой из новых линий
были поставлены снова по два реципрокных скрещивания и определены
те же величины. В большинстве случаев они оказались новыми, хотя за
реальность этих изменений не всегда можно было поручиться. Для полу-
264
чения реальности одна линия была исследована на большом материале и
дала довольно ясно ds = 1 и D = 1 щетинка. Эта картина повторялась и
в следующих поколениях. Очевидно, все полимеры оказались удаленными
из хромосомы + + и остался лишь один, тесно связанный с
нормальным аллелем гена scarlet, если только это действие (+ 1 щетинка) не
свойственно самому этому сигнальному аллелю.
В этом опыте, таким образом, показано, что первоначальное строение
хромосомы по полимерам, влияющим на щетинки, было отличным от
конечного строения, полученного путем перекреста.
ДВЕ И БОЛЕЕ СИГНАЛИ
Две сигнали в хромосоме позволяют продвигать анализ дальше. Чем
большим числом сигналей мы можем располагать, тем глубже может быть
изучена хромосома,
1. Две сигнали при возможности изменения величины перекреста
(полом, возрастом и другими условиями) позволяют, как мы видели,
определить d и D у видов, имеющих перекрест у обоих полов.
2. При одном полимере можно определить его положение. Задача
решается так: от каждой сигнали Sx и S2 возможно определение
сигнальной аллосилы dx и d2. Тогда имеем два уравнения:
d2 = h = * (1 - 2с,).
Здесь неизвестных три: г, сг и с2. Но сг и с2 могут быть сведены к одному
неизвестному. В самом деле, возможны три расположения полимера и
двух сигналей, где / — расстояние между сигналями.
Деля dx на d2, исключаем г и получаем -т-= узГгГ» откуда,
подставляя вместо с2 его значение, получаем три возможных гипотезы (схема
XXV):
Сг'С}+1
1 dil
гипотеза I ci= -J+di_^
1 cU-di <Ы
гипотеза II « = у • ^+^ + 5+И*
гипотеза III ci = -g"— д __ £ •
Схема XXV
Если в нашем скрещивании имеется один полимер, то только одна
гипотеза даст правильный ответ, а другие дадут абсурдные решения.
Найдя с, находим и i.
Пример: имеем й± = +6; d2 = +3; / = 10% или ОД; найти с
и *. Находим:
гипотеза / сх= 0,3, или 30%, с2 = 40%,
гипотеза // ci=* — 0,1, или —10% —абсурд,
гипотеза /// d= 0,7, или 70% —абсурд.
265
Подходящее решение дает только первая гипотеза, указывающая на
положение А — Sx —S2. Подставляя полученные значения ев
выражение d, находим £=+15 (схема XXVI). Легко проверить, что при таком
Схема XXVI
i и таких расстояниях сигнальной аллосилой будет (при принятом
схематическом расчете):
]\ = 15 [1 - (2 X 0,30)] = + 6.
U = 15 [1 - (2 х 0,40)] = + 3.
При наличии нескольких полимеров правильное решение получится
лишь в том случае, если обе сигнали будут находиться вне зоны
полимеров. Конечно, при этом будут определены не г и не D. Если хотя бы
одна из сигналей окажется между полимерами, решение будет неверным.
Эту неправильность решений в ряде случаев можно будет заметить уже
потому, что подходящие решения будут получаться не по одной гипотезе,
а по двум или по всем трем. Ошибка сможет быть замеченной и при
помощи применения метода грединга (см. дальше).
Но если один из полимеров будет значительно сильнее других,
решение даже и в таком случае будет довольно близким к правильному.
На прилагаемой схеме XXVII представлен случай с двумя
полимерами А и В, одним сильным (i2 = + 10) и одним слабым (гг = — 1),
расстояние между сигналями 10%. Сильный полимер расположен
посредине между сигналями, слабый — левее sx на 2%. При этих условиях
dt = 8,04, d2 = 8,24.
+Л7
77
+
ВЦ
+
0 12 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Схема XXVII
Определяя сх по dx и d2 при I = 0,1, находим по трем гипотезам:
гипотеза / ci = 4,5— абсурд,
гипотеза // с\ = 0,056, или 5,6%,
гипотеза /// а = 3,5 — абсурд. /
Очевидно, правильна лишь вторая гипотеза Sx — А —. S2. Определяя
D, находим:
п — * 804 — 9 04
U — 1 —0,112 ~у>и4-
Итак, искомый ответ сх 5,6% от S± и i = 9,04.
266
Так как в задаче участвуют два полимера, то мы определили не iy
a D. В действительности D = 10 — 1 = 9.
Следовательно, мы определили очень точно D и аллобаланс
хромосомы. В то же время мы довольно точно определили и местонахождение
сильного полимера В.
3. Два сигнальных гена могут быть использованы и иначе, с еще
большими перспективами для генанализа:
Si S%
I II III
Сигнали разбивают хромосому на три отрезка — два внешних
отрезка I и III, и II внутренний, ограниченный справа и слева сигналями.
Этот отрезок лежит между двумя сигналями и представляет собой то
качественно новое для анализа, что приносят две сигнали по сравнению
с одной. В самом деле, поставим скрещивание:
Si 52 X Si 52 *
В потомстве возникнут четыре категории особей, различных по сигналям:
1 \ *Si ^2 . <у\ Sl S2 '* Ч\ <^1 $2- • 4Л si S*
' 51 52 ' ' S\ 52 ' 51 52 ' ' Sl 52
Если возьмем первые два класса, то мы выберем хромосомы, не
претерпевшие перекреста на участке Sx — S2. Если расстояние S± — S2
невелико, это будут преимущественно хромосомы без перекреста, с
небольшой примесью хромосом, потерпевших двойной перекрест между
S± и 52. Этими, сравнительно редкими двойными перекрестами (макси
мальное количество которых легко вычислить) можно пока пренебречь
Выбрав из потомства особь —-—— и повторяя ее скрещивание с
51 52
анализатором, т. е. ведя «грединг на анализатора», мы получаем в конце
концов (если избежим двойного перекреста между S± и S2) особи, в
которых весь генотип будет от анализатора, кроме участка хромосомы между
St и S2 и небольших участков вправо и влево.
Мы приходим таким образом к типу нашей прежней задачи
(дрозофилы и шелкопряды), так как имеем возможность для анализа отрезка
^i — #2 располагать материалом и с перекрестом и без него. Весь необ-
' ходимый материал мы будем получать уже в одном скрещивании. Но зато
анализировать будем не всю хромосому, а лишь отрезок.
Пусть между 51 и S2 имеется модификатор А на расстоянии сх и с2 от
сигналей, действующих с силой i
Si A iS*2
Cl Сч
Состав четырех категорий потомства этих скрещиваний будет:
sx$2 aa
SXS2 Аа
s1S2 сгАа + с2аа
S1s2 c2Aa -f- с^аа
Отсюда средние арифметические будут:
МХ = М* (1)
М2 = М1+ i; (2)
М = ClMl + Ci (Ml + ** : (3)
3 Ci + Ci * '
м = g2Mi + ci (Mi + i) . ^v
4 C1 + C2 K '
267
Здесь Мх обозначает величину признака у форм Аа. Мы имеем здесь
четыре неизвестных и четыре уравнения или, если считать М известным,
так как оно непосредственно равно Ми то три неизвестных i, сг и с2 и
три уравнения. К сожалению, одно из последних уравнений является
паразитным, так как из третьего определяем:
Ms — Mi /c\
С»ДС1ЛГ,-Л#..' (Ь)
а из четвертого
М^ — М* /£\
c* = ci--m=m (6)
и исключить по отдельности ни сх, ни с2 не удается.
Поэтому для решения необходимо найти еще одно уравнение. Таким
будет
с = сх + с2, (7)
где величина, являющаяся частотой перекреста между Бг и £2,
определяется из наблюдения за двумя сигналями.
Из (7) и (5) находим:
ИЛИ
Cl~~C M2 — M1 '
а затем находим и с2.
Задача, таким образом, оказывается решенной.
Мы видим здесь совершенно те же отношения, как и в случае
дрозофилы и шелкопряда при работе с одним геном, т. е. введение второй
сигнали (на расстоянии не очень большом) искупает неудобства, вызываемые
наличием перекреста у обоих полов. Сами сигнали оказываются здесь
терминальными сигналями, т. е. сидящими по концам (в данном случае —
по концам участка), о преимуществах чего говорилось выше.
ДАЛЬНЕЙШЕЕ ИЗУЧЕНИЕ УЧАСТКА РАЗРЫВАМИ
Получив перекрестные формы S& и SxSg, можно получить форму
siS%
Si 1У2
и в обратном скрещивании с анализатором восстановить строение
В процессе двух последовательных перекрестов (сначала отрыв S± от 52,
а затем воссоединение их) строение отрезков между "5Х и S2 может
изменяться, что обнаружится перемещением фокуса. Ставя последовательно
разрывы и соединения сигналей, можно будет изучить строение отрезка
с любой точностью, как мы об этом говорили раньше, анализируя случай
с одной сигналью (Серебровский, Волкова, 1940).
Введение третьей и дальнейшей сигналей, не изменяя дело по
существу, позволит охватить изучением новые отрезки хромосомы. Это еще раз
подчеркивает то большое значение, которое имеют для генетического
анализа количественных признаков хорошие сигнальные гены, почему
отыскание, изучение и сохранение их должно быть отмечено как важное
дело.
268
ВАРИАНТ М. Л. КАРПА
Интересный вариант сигнального метода был применен в нашей
лаборатории Карпом (Карп, 1935) при анализе /// хромосомы дрозофилы по
генам стернитальных щетинок. Первоначально была выделена одна
хромосома, которую мы будем обозначать знаком + + + + + +» и по
размножению через гетерозиготного самца была подвергнута анализу при
помощи тоже единственной выделенной при помощи запирателей
хромосомы из линии «га си са». Эта хромосома имела следующие шесть
рецессивных генов: га (rughoid — грубые глаза), th (thread — щетинка усиков
голая), st (scarlet — алые глаза), си (curled — загнутые крылья), sr
(striped — темная полоса вдоль спинки груди) и са (dared — глаза винного
цвета). Был определен аллобаланс исследуемой хромосомы, оказавшийся
равным около + 6 щетинок.
После этого было поставлено скрещивание с перекрестом и из
потомства выделены 46 самцов с различными комбинациями сигналей.
Анализируемая хромосома таким образом была разбита на участки,
соединившиеся с участками анализаторной хромосомы. Через самцов без
перекреста эти новообразованные хромосомы были далее размножены и
сохранены.
Вариант Карпа состоит в оригинальном использовании этих
новообразованных хромосом. Пусть, например, в результате перекреста был
получен с? фенотипа га, имеющий строение, представленное на схеме
XXVIII.
1Ъ ++ + + +
1 1 I I I I
и »
ru tii St Си Sr Ca
Схема XXVIII
Здесь жирной чертой изображен отрезок хромосомы исходной,
анализируемой, а тонкой — хромосомы анализаторной. Если на участке
ги — th имелись гены, влияющие на щетинки, то, очевидно, аллобаланс
хромосомы ru +H—|—Ь будет отличаться от аллобаланса прежней
хромосомы + Н—Ь++ и это изменение можно будет учесть по потомству с
любой желаемой точностью.
Но перекрест на участке ги — th, дающий хромосому ги + + + + >
может произойти на любой точке между этими генами, поэтому и
хромосомы ги + + + + могут иметь различной длины участок, пришедший
с геном rut как видно из схемы XXIX.
ги ++ + + +
i I I i i I
ги ++ + + +
1 nil 1
ru tt t +
1 III)
ru ++ + + +
) I I I I I
Схема XXIX
Если на участке ru — th нет генов, влияющих на число щетинок, то
хромосомы + + + + + ига -\—К++ не будут отличаться по своему влия-
269
нию на щетинки. Если между ги и th имеется один такой ген (а), то
хромосомы будут двоякого типа:
1)то++++,
2) гиа ++++.
Первый тип не будет отличаться от +Н—\--\—Ь хромосомы, второй будет
отличаться на величину, равную силе действия гена а. Если между ги —
th имеются два модификатора а и 6,^ то хромосомы ги -\—|—Ь+ будут
трех типов:
' i)ru++++,
2) гиа -\—М—(-,
3) гиаЪ ++++.
Если мы получим в результате перекреста много различных,
независимо возникших самцов с хромосомами ги -\—Ь + + и изучим силу
влияния каждой из этих хромосом на щетинки (опять-таки с желаемой
точностью) и узнаем, сколько возможно различных типов хро-
МОСОМГИ + + Н—he точки зрения их влияния на щетинки, то отсюда
сможем заключить и о числе генов-модификаторов на этом участке и даже
приблизительно об их расположении. Правда, если, например, гены а и
Ъ расположены близко друг к другу, то тип (2) хромосом может не
возникнуть, и мы будем полагать, что имеем лишь один ген с суммарным
действием (ia + ib). Точно так же, если ген расположен близко к th, то
будет трудно доказать: расположен ли он левее th, т. е. на участке
ги — th, или правее th, т. е. на участке Sr — th. Но эти трудности
возникают при изучении и хорошо проявляющихся генов. Основная
трудность в данном случае состоит в том, что для того чтобы с уверенностью
сказать, сколько типов хромосом строения ги + + Н—\- возможно,
нужно изучить большое число таких хромосом, а так как для изучения
каждой хромосомы нужно исследовать большое число мух, то работа
получается весьма трудоемкой. С точки зрения методической мы, очевидно,
имеем здесь дело с соединением метода сигнал ей и метода проявления
каркаса.
В исследовании Карпа было изучено, например, семь различных
хромосом ги + + Н—Ь (рис. 84).
Балансы новообразованных хромосом довольно ясно распадаются на
три группы I, II, III, почему можно заключить, что на участке имеется
не менее двух генов, влияющих на число щетинок.
Рис. 84. Сопоставление семи
хромосом ги и + по их
действию на щетинки
Намечается распределение на три
группы: I — сильное ослабление;
II — слабое ослабление; III —
усиление (по М. Л. Карпу)
*2\
1 2 3*56
I ш
270
Этим, однако, исследование еще не заканчивается. Если даже
допустить, что имеются только два гена, то для трех классов будем иметь три
«, „ к п, thst n uCUn „ hsr „ А _ са
~i~~i—I / \ \ \\ к \ <
-ЗМ +6,28 -UJ2 -2,16+5,36-4,88-3,10*5,54 -6,52+3,64 -2,66
ги п h h thst. „ си _ sr . _ „ са
\ \ \ l Г ill
-3,04+2,16 +4,12 -5,36 -4,08 +6,52 -2,66-4,88
Рис. 85. Две гипотезы расположения полимеров в 771 хромосоме,
обнаруженных в исследовании М. Л. Карпа
генотипа: ++, «+, ab, но сказать, какой из этих генотипов какому типу
хромосом соответствует, еще невозможно и можно сделать любые
гипотезы. Для того чтобы узнать, какая из гипотез верна, необходимо эти
хромосомы пустить снова в перекрест, рассуждая следующим образом:
если I тип имеет генотип + + , то после скрещивания с перекрестом в
потомстве от него снова могут получиться все три типа -\—К 0+ и ab;
если I тип имеет генотип а+, то после перекреста в его потомстве
смогут получиться только два типа а-\- и ab, а тип ++ не сможет
возникнуть;
если I тип имеет генотип ab, то после скрещивания с перекрестом в его
потомстве может получиться лишь один тип ab, но ни.++, ни а+;
если, наконец, на участке ru — th имелось не два, а больше генов-
модификаторов, то смогут возникнуть еще новые типы.
Таким образом далее смогут быть изучены и все другие участки
хромосом.
Этим путем Карпом было показано наличие в изученной им хромосоме
не менее 6—7 генов, влияющих на число щетинок, частью усилителей,
частью ослабителей, расположенных более или менее вперемежку по всей
длине хромосомы. Окончательно порядок генов установлен автором не
был, но следующая схема показывает возможный порядок (рис. 85).
АНАЛИЗ НЕСКОЛЬКИХ КОЛИЧЕСТВЕННЫХ
ПРИЗНАКОВ ОДНОВРЕМЕННО
Интересное развитие метода возникает, если мы будем изучать не-
сколько различных количественных признаков одновременно.
sab S А
н з1 н- х н 1-
Схема XXX
Разберем здесь простейший пример (схема XXX), когда мы имеем
одну сигналь s и два гена А и В, влияющие на разные признаки, не
связанные друг с другом корреляцией.
271
генотип
9 (SAb
* \saB
л (Sab
1 \sAB
л о fSaB
частота 1 признак Л
(1 — ci)(l—ft)
с2 (1 — ci)
ci (1 — с2)
cica
М2
Mi
Ma
Mi
Mi
M2
Mi
M2
признак Б
Ni
Ni
Ni
Ni
Ni
Ni
Ni
Ni
Пусть расположение генов таково, тогда М — средние для
признака А; N — средние для признака В; Мг — при аа; Мг — при Аа; N± —
при 66; N2 — при ВЬ.
Исследуем корреляцию между обоими признаками в S-rpynne.
Мх М2 Если вычислять, например,
коэффициент ассоциации, т. е.
Ni
N2
ei (1 — с2)
cic2
с2 (1 — с2)
(1 —С!)(1-с2)
Q =
ad —be
ad + bc
то получим
с = с2 (1
b = cica
ci)
cb = ci (1 — ci) c22
1— 2ca
a = ci (1 — c2)
d = (l-ci)(l--ca)
ad = ci(l —ci)(l —d2)2
После простых выкладок получаем
Так как с2<^-^с2<^1 и числитель всегда больше 0, то мы имеем
положительную ассоциацию (несколько иное выражение корреляции).
Правда, благодаря наличию трансгрессивной изменчивости,
фактическая'формула ассоциации оказывается более сложной, величина ее
пониженной и зависящей не только от с2, но и от сг. Поэтому и определение
сг из этого уравнения] оказывается невозможным без дополнительных
уравнений. Но наличие положительной ассоциации может быть
использовано как указание на данный порядок генов.
Действительно, пусть теперь расположение генов таково:
с2 ci
В потомстве имеем хромосомы
генотип
»а™
л (BSa
1 \bsA
.. }bSA
* \Bsa
U* \BsA
частота
4(i-ft)(l~C)
ci (1 — c2)
C2 (1 — Cl)
ClCz
Характеристика
признак A
M2
Mi
Mi
M2
Ma
Mi
Mi
M2
признак В
Nt
Ni
Ni
Ni
Ni
Ni
Ni
Ni
272
Для группы S будет
ClC2
Cl (1 — С2)
С2 (1 — Cl)
(l__Cl)(l__C2)
Если вычислить коэффициент ассоциации
^ __ ad —be
^ "" ad + be 3
то увидим, что здесь он равен нулю.
В самом деле,
а = cic2 с = с2 (1 — ci)
d = (l-ci)(l~C2) b = ci (1 — c2)
ad = ci(l — c2)c2(l — ci) cb = ci(l — Сг)сг(1 — ci)
Следовательно, ad = be и ad — be = 0, a Q = 0.
При малых расстояниях число двойных перекрестов будет меньше,
чем сгс2, и возникает Q <^ 0.
Итак, в зависимости от расположения генов вокруг сигнал ей мы либо
получим корреляцию (ассоциацию) между признаками, либо не получим.
Если же мы вышеразобранными способами определим сг и с2 для этих
разных признаков, то сможем расположить гены вполне правильно, т. е.
дать план хромосомы в области сигнали. Между тем, если бы мы
исследовали каждый признак отдельно, то даже определив сг и с2, взаимного
расположения генов мы не узнали бы.
Очевидно, чем большее количество признаков мы будем изучать, тем
более интересными будут полученные результаты. При работе с
несколькими признаками может быть использован метод исчисления частичных
корреляций. Подробно эти вопросы здесь можно не разбирать.
Методом сигналей могут быть разрешены и трудные задачи
генетического анализа. Поэтому он является мощным оружием
генетика-аналитика. В настоящее время этот метод может иметь пока очень скромное
применение, лишь для немногих видов животных. Для того чтобы метод
сигналей использовать, необходимо, чтобы у данного животного было
изучено большое число генов, пригодных для использования в качестве
сигналей. Число таких генов должно быть значительным. Например, у
коровы мы имеем не менее 30 пар хромосом. Для того чтобы располагать
хотя бы одним геном в хромосоме, нужно не менее 30 сигналей. По ряду
причин их должно быть значительно больше, может быть до сотни и
более. Для того чтобы располагать таким числом сигналей, нужно
изучить частную генетику вида примерно так же хорошо, как мы изучили
генетику дрозофилы или кукурузы.
Даже у таких наиболее изученных животных, как кролик, курица и
шелкопряд, число имеющихся в нашем распоряжении сигналей еще
недостаточно, и самое применение метода у них сильно ограничивается.
Поэтому дальнейшее отыскание генов, пригодных для использования в
качестве сигналей, и применение уже известных должно явиться
существенной исследовательской задачей и генетиков, и селекционеров.
18 А. С. Серебровский
ЛИТЕРАТУРА
Васин Б. Н. 1928. Генетика овец. Наследование окраски и пежин. Изд. Центр.
генетической станции НКЗема РСФСР.
Васин Б. Н. 1928. Возможность анализа признака в популяции.— Журн.
эксперт*, биол., т. 4, вып. 1.
В а с и н Б. Н. 1933. Анализ окраски ширази и ее летального действия. Селекция
каракульских овец. Сельхозгиз.
Глембоцкий Я. Л. 1935. Новые данные о летальности ширази.— Проблемы
животноводства, № 2.
Глембоцкий Я.Л. 1941. Борьба с крипторхизмом у прекосов.— Советская
зоотехния, № 3.
Давыдов С. Г. 1936. Селекция сельскохозяйственных животных. М.— Л.,
Сельхозгиз.
Дубинин Н. П. 1929. Исследование ступенчатого аллеломорфизма у Drosophila
melanogaster.— Журн. эксперим. биол., т. 5, вып. 2.
Дубинин Н.П. 1930. Ступенчатый аллеломорфизм у Drosophila melanogaster.—
Журн. эксперим. биол., т. 6, вып. 4.
Дубинин Н. П. 1931. Центровая теория гена achaete-scute.— Труды IV Всес.
съезда анатомов, гистологов и зоологов.
Дубинин Н. П., Ромашов Д. Д. 1932. Генетическое строение вида и его
эволюция.— Биол. журн. т. 1.
Иванова О. А. 1926. О наследовании многососковости у крупного рогатого ско
та.— Журн. эксперим. биол., т. 3, вып. 4.
Иванова О. А. 1931. Генетика крупного рогатого скота. М.— Л.
Кру Ф. А. 1931. Генетика животных. М.— Л., Сельхозгиз.
Л а х т и н Л. К. 1922. Кривые распределения. М., ГИЗ.
Лахтин Л. К., 1924. Курс теории вероятностей. М., ГИЗ.
Л е п и н Т. К. 1928. Изменчивость количественных признаков у твердых пшениц.—
Изв. бюро генетики, № 6.
Лютиков К. М. 1931. Генетический анализ в отношении медленно плодящихся?
животных.— Журн. эксперим. биол., т. 7.
Лютиков К. М. 1934а. О племенной работе с семействами рекордистки
«Золотой».— Проблемы животноводства, № 2.
Лютиков К. М. 19346. Еще о семействе «Золотой».— Проблемы животноводства,
№ 3.
Петров С. Г. 1928. Генетический анализ популяции кур Ветлужского района.—
В сб. «Генетика курицы», II. Труды центр, ст. по генетике с.-х. животных, № 3.
Промптов А. Н. 1926. Генетика строения таза кур.— В сб. «Генетика домашнейг
курицы». М., изд-во «Новая деревня».
Рапопорт И. А. 1947. Химическая реакция с аминогруппой протеина в
структуре генов.— Журн. общей биол., т. 8, № 5.
Рокицкий П. Ф. 1932. Генетика. М.— Л., Сельхозгиз.
Рокицкий П. Ф. 1934. Практическое пособие к курсу генетики. М.— Л.,
Сельхозгиз.
Романовский В. И. 1938. Математическая статистика. М., ГИЗ.
Ромашов Д. Д. 1931. Об условиях равновесия в популяциях.— Журн.
эксперим. биол., т. 7.
Серебровская Р. И. 1930. Генетика дальтонизма.— Медико-биол. журн.,
вып. 4—5.
Серебровская Р. И., Серебровский А. С. 1926. Генетика яйца и
яйценоскости.— В сб. «Генетика домашней курицы». М., изд-во «Новая деревня».
Серебровская Р. И., Серебровский А. С. 1940. Опыт исследования
аллобаланса пары хромосом двумя сигналями.— Изв. АН СССР, серия биол. ,№1»
Серебровский А. С. 1926. Исследование по генетике курицы.— В сб.
«Генетика домашней курицы». М., изд-во « Новая деревня».
274
Серебровский А. С. 1926—1927. Генетический анализ популяций домашних
кур горцев Дагестана.— Журн. эксперим. биол., т. 3, вып. 1—4.
Серебровский А. С. 1934. О свойствах менделистических равенств.— Докл.
АН СССР, т. 2, № 1.
Серебровский А. С. 1935а. Геногеография домашних кур Кабарды п Балка-
рии.— Успехи зоотехн. наук, т. 1, вып. 1.
Серебровский А. С. 19356. Геногеография кур Армении.— Успехи зоотехц.
наук, т. 1, вып. 3.
Серебровский А. С. 1936. Опыт нового метода генетического анализа
количественных признаков.—Докл. АН СССР, т. 2(11), № 1 (87).
Серебровский А. С. 1936. Второй вариант метода треугольника,— Докл.
АН СССР, т. 4 (13), № 1 (105).
Серебровский А. С. 1937. Третий вариант метода треугольника.— Докл.
АН СССР, т. 14, № 9.
Серебровский А. С, Волкова К. В. 1940. Опыт анализа хромосомы
двумя сигналями.— Изв. АН СССР, серия биол., № 1.
Серебровский А. С,Дубинин Н. П. 1929. Искусственное получение му>
таций и проблема гена.— Журн. эксперим. биол., М., серия Б, вып. 4.
Синнотт Е. В., ДеннЛ. 1934. Курс генетики. M.t Биомедгпз.
Филипченко Ю. А. 1926. Изменчивость количественных признаков у мягких
пшениц.— Изв. бюро генетики, № 4.
Четвериков С. С. 1927. О некоторых моментах эволюционного процесса с точки
зрения современной генетики.— Журн. эксперим. биол., серия А, т. 2, вып. 1,4,
А 1 Ь е г t s H. W. 1926. A Method for Calculating Linkage Values.— Genetics, 11.
Babcock E. В., Clausen R. 1927. Genetics in Relation to Agriculture. N. Y.
BatesonW., PunnettR. С 1906. Experimental Studies in the Physiology of
Heredity.— Poultry Rept Evol. Com., 3.
Bateson W., PunnettR. C. 1911. The Inheritance of the Peculiar Pigmentation
of the Silky Fowl.— J. Genetics, 1.
Bridges С. В. 1914. Direct Proof Through Non-disjunction that the Sex-linked Genes
of Drosophila are borne by the X-chromosome.— Science, 40, 81.
Bridges С. В. 1917. Deficiency.— Genetics, 2.
Bridges СВ., 1919. Vermilion Deficiency.— J. Gen. Physiol., 1.
Brunson A. N. 1924. The Inheritance of a Lethal Pale Green Seedling Character
in Maize. Mem. Cornel Agric. Exptl. Sta, 71—72.
Castle W. E. 1919. Studies on Heredity in Rabbits, Rats and Mice.— Publ.,
Carnegie Inst. Wash., N 288.
Castle W. E., 1925. Heredity in Rabbits and Guinea Pigs.— Bibl. Genetics, 1.
Castle W. E., 1929. A Further Study of Size Inheritance in Rabbits with Special
Reference to the Existence of Genes for Size Characters.— J. Exptl. Zool., 5.
Castle W. E. 1930. The Genetics of Domestic Rabbits. Cambridge, Univ. Press.
Collins S. N. 1912. Genetic Coupling as a Cause of Correlation.—Amer. Naturalist, 46.
Collins S. N. 1924. Measurement of Linkage Values.— J. Agric. Rev., 27.
D u n n L. С 1922. Inheritance of Plumage Colour in Crosses of Buff and Columbian
Fowls.—Amer. Naturalist, 56.
East E. M. 1916. Studies on Size Inheritance on Nicotiana.— Genetics, 1.
Emerson R. A., 1917. Genetical Studies on Variagates Pericarp in Maize.— Genetics, 2.
Fisher R. 1930. Inverse Probability.— Proc. Cambridge Physiol. Soc, 26.
Fischer R.A., Balmakund B. 1928. The Estimation of Linkage from the
Offspring of Selfed Heterozygotes.— J. Genetics, 20.
G r e e n С V. 1932. Genetic Linkage in Size Inheritance.— Amer. Naturalist, 66.
G о w e n J. W. 1919. A Biometrical Study of Crossing-Over.— Genetics, 4.
H a g e d о о r n A. L. a. A. G. 1922. Species Crosses in Rat.— Z. indukt. Abtrammungs-
und Vererbungs lehre, 29.
Haldane J. B. S. 1919. The Probable Errors of Calculated Linkages Values and the
Most Accurate Method of Determining Gametic from Certain Zygotic Errors.— J.
Genetics, 8.
Haldane J.B. S. 1921. Linkage in Poultry.— Science, 54.
Hardy G. H. 1908. Mendelian Proportions in a Mixed Population.— Science, 28.
Harris J. A., 1918. Further Studies on the Interrelationschip of Morphological and
Physiological Characters in Seedling of Phaseolus.— Mem. Brooklyn. Bot. Garden, 1.
H о г К. S., 1924. Interrelation of Genetic Fastors in Barley.— Genetics, 9.
Hutchinson С. В. 1929. The Linkage of Certain Aleurone and Endosperm Factors
in Maize and Their Relation to Other Linkage Groups.— Mem. Cornel. Agric. Exptl.
Sta., 60.
I m m e r F. R. 1930. Formulae and Tables for Calculating Intensities.— Genetics, 15.
KappertH. 1927. Uber die Auswertung dihibrides Spaltungsreihe bei Koppelungs-
studien.— Z. indukt. Abstammungs- und Verbungslehre, 44.
Lindstrom E. W. 1932. First Chromosome Genes in the Tomato.— Genetics, 17.
MacArthur J. W. 1933. Sex-linked Genes in the Fowl.— Genetica, 18.
Mather K. 1938a. Crossing-over.— Biol. Rev., 13.
275 1&*
Mather K. 1938b. The Measurement of Linkage in Heredity. London, Methuen.
Mohr 0. 1923. A Genetic and Gytological Analysis of a Section Deficiency Involwing
Four Units of the X-chromosome in Drosophila melanogaster.— Z. indukt. Abstain-
mungs- und Vererbungslehre, 32.
M u 11 e r H. J. 1930. Types of Visible Variation Induced by X-rays in Drosophila.—
J. Genetics, 22. ,
M u 11 e r H. J. 1935. A Viable Two-gene Deficiency.— J. Heredity, 26.
Owen J. V. 1928. Calculating Linkage Extensities by Product Moment Correlation.—
Cenetics, 13. y
Pearl R., Surface F. M. 1914. A Biometrical Study of Egg Production on the
Characters of the Egg.— Bull. Bur. Animal. Ind. U. S. Dept Agric. 110.
Pearson K., Heron D. 1913. On Theories of Association.— Biometrics, 9.
PunnettB. С 1927—1930. Genetic Studies in Poultry.— J. Genetics, 11.
SerebrowskyA. S. 1928. An Analysis of the Inheritance of Quantitative Transgres-
sive Characters.— Zeitschrift fur induktive Abstammungs- und Vererbungslehre,
Bd. XLVIII, Heft 2.
Serebrowsky A. S., Ivanova 0. A., Ferry L. 1929. On the Influence
of Genesy, 1 and N on the Crossing-over close to their loci in the Sexchromosome of
Drosophila melanogaster.— J. of Genetics, vol. XXI, N 2.
Sturtevant A. N., Dobzhansky F. 1931. Contribution to the Genetics of
Certain Chromosome Anomalies in Drosophila melanogaster. Publ. Carnegie Inst.,
421.
Sumner F. В., С о 11 i n s H. N. 1922..Color Mutations in Peromyseus.— J. Exptl.
Zool., v. 36.
Weinberg W. 1912. Methode und Fehlerquellen der Untersuchung auf Mendelsche
Zahlen beim Menschen.—Arch. rass. und ges. Biol., 9.
WoodworthC. M. 1923. Calculation of Linkage Intensities Where Duplicate Factors
are Concerned.— Genetics, 8.
Wright S. 1917. Color Inheritance in Mammals.— J. Heredity, 8.
Wright S. 1927. The Effects in Combination of the Major Color-Factors in the Guinea
Pig.— Genetics, 12.
Yule G. W. 1900. On the Association of Attributes in Statistics.— Philos. Trans. Roy.
Soc, 194. 7
ДОПОЛНЕНИЯ
НЕКОТОРЫЕ ПРИЕМЫ АНАЛИЗА ГЕНОСВЯЗИ
М. В. ИГНАТЬЕВ
Вычисление ошибок применимо при сравнении численностей двух
классов. Но при скрещиваниях, производимых для выявления геносвя-
зи, обычно получается не менее четырех классов, и можно поставить
вопросы о том, насколько вся система численностей четырех классов
отклоняется от численностей, ожидаемых при отсутствий геносвязи. Для решения
этого вопроса чаще всего применяется функция %2.
Величина %2 является общей мерой отклонения наблюдаемых значений
от теоретических, определяемых по какой-либо гипотезе. Ее формула
лмеет вид:
где а — наблюдаемые численности классов, тп — ожидаемые, п —
численность всей совокупности, J — обозначает суммирование по всем
классам. Для суждения о том, укладывается ли полученная величина %2 в
пределах значений, допускаемых действием случайностей наблюдения,
служит таблица вероятностей значений %2, которая и прилагается здесь
(табл. 23) *. Вероятности значений %2 зависят от числа степеней
свободы. Под степенями свободы подразумевается число классов, которым
можно придавать произвольные значения. Так, например, если даны два
класса, то произвольно можно давать численности лишь какому-нибудь
одному из них, численность же другого определяется общей суммой и
численностью первого класса, так что при двух классах — лишь одна
степень свободы. Если же даны четыре класса, как например, при обрат-
лом скрещивании АаВЪ х aabb, то число степеней свободы равно трем.
Однако число степеней свободы не всегда равно числу классов без
одного. Ниже мы встретимся с несколько более сложными расчетами.
Если метод %2 применить к случаю двух классов, то он приводит к
тому же результату, что и вычисление ошибок, поэтому его можно
рассматривать как обобщение метода ошибок.
Примеры. Применим метод %2 к приведенным данным. Сумма А, В +
+а, В образует численность одного класса и равна 75, тогда как сумма
А, Ъ + а, В, входящая в состав другого класса, равна 66.
Совместимы ли эти числа с гипотезой независимости генов, согласно которой
численность каждого класса должна быть равна 70,5? Исчисление %2, которое
даст ответ на поставленный вопрос, производится по следующей схеме:
* Для лучшего освоения метода и возможности более широкого его использования мы
включили в статью полную таблицу распределения х2-квадрат из книги П. Ф. Рокиц-
кого («Биологическая статистика». Минск, 1967) вместо приведенных в статье
отдельных выдержек из этой таблицы.— Прим. ред.
279
eft
<м
се
tr
tsl
I©
cd
Ф
H
S3
I
ев
ft
Вероятности значения x2, превышающего табличное
0,01
0,0025
0,05
0,10
0,25
0,50
0,75
0,90
0,95
0,99
^ И" |
* cOr44^oOO^^oO<^t^^Hc>a(^<^^oOO^<^^i^cOO^v^oO'rHvt,e^
ФC^rtNOOOsf,OФCSII>(^^cOт^ЮO^J^OOrнЮQ(NЮQfOCOlCЛ(NЮOOCOr^CO^COHoO
. С0Фтн«ЮС0<ЙОгчС0^С0"^0)ОМС0^С0^то^М^ЮЮа>О5ОС0С000О01^Ю
"Ч-l "Ч-Н ТН чгН
ОсОСОгНОО^ОЮО^ФСО^тНЧГООтнЮООн^^ОСОСООгн^ЬО^СО^СООСО^Ю
Ю^Qr^^I^*•C0^05OнC0sl^CDI>CЯOн^|^*lnCO0005O^r^C0^1,ЮCDO>rнC0^ЛCD0005
rHTHrHT^^rHlNWNWMNiNWCOCOWMCOWCOstt^sli^st'slilO^OOQOTHN
■«Н чгН -чН
000100^0ЮОЮ05СОС1ЭОС5СООСОЮООгН^Ф05п^СОООнСОЮМ*ЮОЮООпСО
•^« ■«н ^н
THHW00\fvfWC00005a)W^CDH\tl>(5iOHWrH^O00C0st*(NCTiC0O^Of000CDO
^co(^l^lNcDOcocoQ^lюoooмюt>Ql^lsl^coмoNcoю^050(Nooтн^юююlC
ЧТИ ЧГН
(N^ri05C0sJi^iNQWOW00iNWN05ONC0C04f,4fsl,^MC0NriOiNC00000f0v't1^
СОЬ^С^(©ОООМСОЮ^(»0)тнМСО^С0^00050тН|^СО'^ЮС01>ООФСОО)ЮтнсО'Н
тНСЧ|^10С01Г^О^©^С^ГО^ЮГ>ООО^О^СЧеО^О«>-00050^С^
^MCOWMMPOCOCOmWCQCOCOWCOCOMOOCOCOWOOCOCOCOCOCOCQCOCOWCOCOCOCOCO
0^01СО^ЮФ^ООООгнММ^ЮСО^(Х>СЛОтнС\1СО^ЮФ^ООО^ФО}О^ФО^О^а)
ООгн^МС0^ЮЮС0^0005ОнгН01С0^ЮС0^00а105ОтнМ00^С0ММтНгнОО
ОМЮООМОО^гнООЮСООЬЮСООООСО^С^ОООСО^М^ОЗЬООСО^СС^^МСО
ОООтНтнММСО^^ЮСО^Ь000500НММ^^ЮСО^ООЭ005005^СОЮ^ССС<1
OlOrHlO^NCOCO^NCOQt'-COCO^OSNWOl^Qin^OOlOCOrHQrHCDQ^OJCOCO
гНС0^т^ЮгН^С90)ЮМ001ЙМ05С0С0тН00ЮССО00С0С0^051>^|Ю^,г1^'ССтн05
ОООНтн^МСОИ^ЮЮСО^^ОО^ООнМООСО^ЮСОСО^ООЮ^СОгнО^^
Мп01Л^^Ю05СОЮ1чгНСОСОтНгн^СОСОО^ОС01МОООСОСОЮСО'НОО^^ЮСО
ОгНООЮООМСООЮ01ЛНСОС^ОО^ОСОМО)ЮМоОЮС100ЮМО>-н^^^Ю1>0
OOOOOrH^N(NlWWslt4fiOlOCOt>'t>OOOOQOOrHWWOO^^NO>t*WOO'HO
rHMC0^l0C0^0005O^NC0^mc0N00O>OTH(MC0vl<lfi«0N0005OOOOOOOO
■чН
Число степеней свободы 1.
Получилось х2 = 0,574. Надлежит определить вероятность такого х2-
Для этого обращаемся к таблице его распределения, входя в нее с одной
степенью свободы, т. е. пользуясь первой строкой. Мы видим, что х2 =
= 0,574 лежит между значениями, вероятности которых равны 0,25 и
0,50, так что
0,25 <Р(%2)< 0,50.
Такого рода отклонения вполне могут вызываться случайностями
наблюдения, так что приведенные числа, несомненно, совместимы с
гипотезой отсутствия геносвязи.
Согласно принятой практике, они вообще были бы совместимы при
Р (х2) > 0,05, но не были бы совместимы при Р (%2) <[ 0,01.
Если же вероятность х2 меньше 0,05, но больше 0,01, то получается
сомнительная зона; во всяком случае, утверждать наличие геносвязи при
таком х2 было бы неосторожно.
Второй из приведенных примеров приводит к следующей таблице:
Класс
I
II
Всего
Наблюдаемое
число (а)
14
46
60
Ожидаемое
число (тп)
30
30
60
Отклонение
(а — тп)
-16
+16
0
тп
8,533
8,533
17,066
Степеней свободы 1.
Р(Х2)<0,01.
Данные несовместимы с гипотезой независимости генов; наличие
геносвязи можно считать несомненным.
Но метод х2 предоставляет также возможность для более полного
анализа данных, если применить его к оценке всей системы численностеи
четырех классов. В таком случае, кроме гипотезы отсутствия геносвязи,
можно также испытать соответствие наблюдений с менделевскими
расщеплениями по каждому из двух локусов — А и В. Так, в приведенных
примерах можно проверить, осуществляются ли расщепления Аа и ВЪ в
пропорции 1 : 1 или существует реальные от нее уклонения, которые
могут вызываться неполной проявляемостью аллелей, различием в их
жизнеспособности и т. д. Для решения этой задачи вычисляются три
функции х2» а именно Ха> %в> %ав ; первая — для уклонений расщепления Аа
от пропорции 1:1, вторая — для таких же отклонений по ВЬ, третья — для
уклонений от независимости расщеплений по А я В. Вычисление может
быть произведено по общей схеме, но удобнее воспользоваться
специальными формулами, применяемыми для рассматриваемой задачи. Они имеют
вид:
2 (gi + 02 — аз — ^4>2
__ (ai — Д2 + Дз — а*)2
*В— п
2 _ (fll — 02 + A3 + Д4)2
где ах, а2, ад и а4 соответственно равны численности классов А, В; А, Ь;
а, В я а, Ъ.
281
Примеры.
Для первого из разобранных примеров ах = 38, а2 = 34, а3 = 32 и
а4 = 37.
Вычисления располагаются так:
Вид уклонения
Расщепление по Аа
Расщепление по ВЪ
Совместное расщепление А я В
Всего . . .
х»
0,064
0,007
0,574
0,645
Степеней
свободы
1
1
1
3
Р(Х2)
0,80
0,95-0,98
0,25—0,50
0,80-0,90
На основании данных таблицы можно сделать вывод, что
расщепления по А и по В согласуются с менделевскими ожиданиями, так же как и
совместное расщепление согласуется с гипотезой независимости генов А и
В. Поэтому и вся система отклонений находится в соответствии с
теоретическими числами. В самом деле, для нее х2 == 0,645 при трех степенях
свободы, поэтому в таблице распределения %2 приходится пользоваться
третьей строчкой; в ней значится, что Р (%2) = 0,645, лежит между 0,75
и 0,9. Отметим, что %\в имеет естественное одинаковое значение с
полученными ранее в 1-м примере. Это отображает аддитивное свойство
функции х2» которое в свою очередь дает возможность определять его
косвенным путем; а именно можно сосчитать общее %2 по четырем классам
(по общей формуле), а из итога вычесть значения у?А и %%- Этот путь,
который иногда целесообразнее, иллюстрируется на материале
следующего 2-го примера. Для него дано:
ах = 6, а2 = 25, а3 = 21 и а4 = 8.
Ради иллюстративных целей мы поведем расчет иным путем, а
именно сначала рассчитаем общий х2- Это значит, мы проверим, насколько
приведенные данные согласуются одновременно с тремя гипотезами:
двумя — о менделевских расщеплениях по А — а и В — Ь (в соотношениях
1 : 1) и одной — о независимости расщеплений по А и В. При этих
предположениях численности классов АВ, АЬ, аВ и аЪ должны быть друг
другу равны. Так как общая численность равна 60, то на каждый класс
приходится по 15 особей. Вычисления располагаются следующам
образом: ч
Класс
I
II
III
IV
Всего
Наблюдаемое
число (а)
6
25
21
8
60
Ожидаемое
число (тп)
15
15
15
15
60
Отклонение
(а — тп)
—9
+10
+6
-7
0
тп
5,40.
6,667
2,400
3,267
17,734
Степеней свободы 3. Р (%2) 0,01.
Вывод получается отрицательный. Данные несовместимы с гипотезой,
требующей равные численности классов. Для того чтобы выяснить ис-
282
точник расхождений, произведем операцию, называемую разложением
X2 по степеням свободы. Эга операция основана на упомянутом выше
аддитивном свойстве функции %2, согласно которому возможно сложение и
вычитание %2.
В этой связи достаточно вычислить по предыдущему способу %2А и
у?в. Первое из них равно 0,067, второе 0,60, каждое имеет по одной
степени свободы. Вычитая их из итога, получаем у?АВ = 17,734 — 0,067 —
— 0,600 = 17,067 при одной степени свободы. (Можно проверить, что тот
же результат получится и при непосредственном вычислении по формуле
%2АВ.) Сводя итоги, строим следующую таблицу:
Вид уклонения
Расщепление по А
Расщепление по В
Совместное расщепление
х*
0,067
0,600
17,067
17,734
Степеней
свободы
1
1
1
3
Р(Х«)
0,25-0,50
0,25-0,50
0,01
0,01
Из таблицы видно, как разложилось %2. При соответствии
расщеплений Аа и ВЬ получилось явное несоответствие с гипотезой
независимости генов, причем это расхождение привело к нарушению
соответствия всей системы значений с испытываемыми гипотезами.
Следовательно, все дело в наличии геносвязи АВ.
Может показаться, что разница несущественна. Однако она будет
нереальной лишь при очень малом числе наблюдений. Если же
численности равны хотя бы тем величинам, которые здесь приведены, то метод
X2 вполне в состоянии вскрыть и отклонения от менделевских пропорций,
и отсутствие геносвязи. Действительно, в этом случае ах = 16, а2 = 24,
а3 = 24, а4 = 36.
Исчисление по формулам %2 для обратного скрещивания дает следующее.
Вид уклонения
Расщепление по Аа
Расщепление по ВЬ
Совместное расщепление АЪ
X*
4,00
4,00
0,16
8,16
Степеней
свободы
1
1
1
3
Р (X*)
0,02-0,05
0,02-0,05
0,50-0,75
0,02—0,05
Из данных таблицы видно, как плохое соответствие по двум менделев-
ским отношениям сочетается с хорошим соответствием по независимости
генов А и В.
В результате же совместного действия трех условий получилось
впечатление, что приведенные цифры будто бы отображают наличие связи,
которой на самом деле нет.
Для оценки реальности уклонений от гипотезы независимости генов
применяется тот же метод %2. Но если ожидаемые в F2 пропорции четырех
классов (Л, В; А, Ь; а, В и а, Ъ) составляют 9 : 3 : 3 : 1, то %2 вычисляются
283
по следующим формулам:
2 _ (ai + Д2 — 3 g3 — 3 а4)2
(ai — 3 02 + Лз — 3 а&)2
Зй
2 __ (ai — 3 д2 — За3 + &к)2
х| =
Пример. Допустим, что численности классов в F2 при скрещивании
АаВЪ X АаВЪ составляют: А, В = 151; А, Ъ = 67; а,В = 66 и а, Ъ = 16.
Спрашивается, совместимы ли эти числа с обычным дигибридным
расщеплением?
На первый взгляд может показаться, что перед нами типично дигиб-
ридное расщепление. Но произведем расчет %2, принимая аг = 151, а2 =
= 67, ав = 66 и а4 = 16.
Вид уклонения
Расщепление Аа
Расщепление ВЬ
Совместное расщепление АВ
Всего . . .
X*
0,871
1,138
4,006
6,015
Степеней
свободы
1
1 '
1
3
Р(Х2)
0,25-0,50
0,25-0,50
0,02-0,05
0,25-0,10
Таблица показывает, что расщепления Аа и ВЬ вполне соответствуют
менделевским, совместное же расщепление заставляет сомневаться в
независимости генов. В то же время нарушение пропорции в совместном
расщеплении перекрывается хорошим соответствием с менделевскими
расщеплениями по генам Л и В, так что в общем получается достаточно хорошее
соответствие. Для того чтобы рассеять сомнения, достаточно увеличить
число наблюдений. Так, например, если при сохранении наблюдаемых
пропорций численности классов были бы вдвое больше, то геносвязь
можно считать доказанной. В самом деле, в таком случае:
аг = 301, а2 = 134, az = 132 и а4 = 32
*tAB
будет равен 8,012, так как общий %2 = 12,030. Вероятности
каждого из них меньше 0,01, и наличие геносвязи становится несомненным;
расщепления же А а ж ВЬ соответствуют менделевским ожиданиям.
Приведем еще некоторые примеры, которые понадобятся в
дальнейшем. Допустим, что одна из аллелей локуса А, так же как и одна из
аллелей локуса В, проявляет неполную доминантность и поэтому в
результате скрещивания АаВЪ х АаВЪ в F2 обнаружилось девять классов
фенотипов. Их численности следующие:
Класс
ААВВ
ААВЪ
ААЬЬ
АаВВ
АаВЪ
Численность
94
109
47
121
247
Класс
АаЪЪ
ааВВ
ааВЪ
ааЬЬ
Чис
ленность
124
39
117
102
Итого
1000
Спрашивается, соответствуют ли эти численности ожидаемым при
дигибридном расщеплении и независимости генов А и В?
Для решения вопроса производим вычисления 9%2. Они
располагаются следующим образом:
284
Класс
(1) AABB
(2) ААВЬ
<3) ААЬЬ
(4) АаВВ
(5) АаВЬ ,
<6) АаЬЬ
(7) ааВВ
<8) ааВЬ
(9) аа&Ь
Всего . . .
Наблюдаемое
число (а)
94
109
47
121
247
124
37
117
102
1000
Ожидаемое
число (тп)
62,5
125,0
62,5
125
250
125
62,5
125
62,5
1000
Отклонение
(а — тп)
+31,5
-16,0
-15,5
-4,0
— 3,0
- i,o
-23,5
- 8,0
+39,5
0
v, (a — тп)
тп
15,876
2,048
3,844
0,128
0,036
0,008
8,836
0,512
24,964
56,252
Число степеней свободы рассчитывается так.
Расщепление по А дает две степени, так как оно производит три
класса. Расщепление по В дает тоже две степени и, наконец, связь между А и
В еще одну степень, как и ранее, так что всего получается пять степеней.
Это значит, что если нам представлено пять данных, допустим, например,
численности особей А В [ т. е. сумма (1) + (2) + (3)] А, а, [т. е. сумма
(1) + (4) + (5)] В, Ъ [(2) -F (5) + (8)], ЪЪ [(3) + (6) + (9)] и величина
перекреста, то мы можем рассчитать численности всех девяти классов.
Входим в таблицу %2 с 5 степенями свободы или смотрим 5-ю строчку.
Но ни в 5-й строчке, ни во всей таблице мы не видим числа 56,252, так
что соответствия теоретических и эмпирических значений не получилось.
Но надо решить вопрос, за счет чего происходит это нарушение
соответствия, за счет ли наличия геносвязи или за счет отступления от менде-
левских пропорций, вызываемых неполной проявляемостью или
дифференциальной выживаемостью и. т. п. Этот вопрос решается
разложением %2. Расчеты производятся следующим способом:
Вид уклонения
Расщепление А—а
АА
Аа
аа
Всего . .
Расщепление В—Ь
ВВ
ВЪ
Всего . .
Совместное
расщепление А и В
Всего . .
1 Наблюдаемое
число
250
492
258
1000
254
473
273
1000
—
—
Ожидаемое
число
250
500
250
1000
250
500
250
1000
—
Отклонение
0
-8
! +8
0
4
-27
+23
0
°~"
X*
0
0,128
0,256
0,384
0,064
1,458
2,116
3,638
52,230
56,252
| Степеней
свободы
—
—
—
2
—
—
—
2
1
5
Р (*>*
—
—
—
0,75-0,90
—
—
—
0,10-0,25
0,01
0,01
285
В'Этой таблице %2 для расщепления А—а и &—Ь произведены
непосредственно, а эс2 для совместного расщепления получено вычитанием %2А и
%з из найденного ранее общего %2.
Расчеты показывают, что расщепления по А и по В соответствуют
менделевским пропорциям, а все расхождение идет за счет совместного-
расщепления, что и обнаруживает наличие геносвязи.
Следующий пример относится к операциям сложения %2. ' v
Допустим, что произведены испытания на сцепление или отталкивание»
в двух экспериментах: в обратном скрещивании и в F2. Получились
результаты:
Класс
А, В
А, Ь
а, В
а, Ъ
Всего . .
Численность
обратное
скрещивание
310
208
189
293
1000
Ft
571
171
156
102
1000
Соответствует ли совокупность наблюдений генотипов в двух опытах:
независимости генов А и В? Для ответа на поставленный вопрос сначала
производится вычисление %2 отдельно по каждому скрещиванию и затем
складываются как величины %2, так и степени свободы.
Вычисления располагаются так:
Вид уклонения
Обратное
скрещивание: А, В
А, Ь
а, В
а, Ь
Всего . .
F2
А, В
Л, Ь
а, В
а, Ь
Be его . .
Общий итог .
Наблюдаемое
число
310
208
189
293 ч
1000
471
171
, 156
102
1000
—
Ожидаемое
число
250
250
250
250.
1000
562г5
Ш,5
187,5
62г5
1000
—
! Отклонение
+60
-42
-61
+43
0
+8,5
-16,5
-31,5
+39,5
0
—
Х2
14,400
7,056
14,884
7,396
43,736
0,128
1,452
5,292
24,964
31,836
75,572
Степеней
свободы
—
3
3
6
Р(Х2)
—
0,01
0,01
0,01
Из таблицы видно, что оба эксперимента, а следовательно и общи!?
итог, свидетельствуют о несомненном наличии геносвязи.
Заслуживает внимания итог по F2 в сравнении с предыдущим
примером.
286
В обоих случаях фигурируют те же данные, но во втором 9 классов
объединены в 4, как бывает при полной доминантности. Это приводит к
значительному увеличению соответствия с гипотезой независимости:
X2 с 56,252 снизилось до 31,836. Хотя оно и осталось, но самый факт
снижения очень показателен. Он иллюстрирует положение о том, что
уточнение классификации имеет весьма существенное значение, поскольку
сопровождается увеличением извлекаемых о геносвязи сведений.
Применение метода %2 и в данном случае освещает положение. Для
того чтобы показать это, придадим предложенным числам некоторые укло-.
нения за счет случайностей наблюдения (иначе %2 по В и по АВ окажется
равным 0, что свидетельствовало бы о специальном неслучайном подбора
материала). Пусть поэтому численности классов равны аг = А, В = 92,
а2 = А, Ъ = 30, а3 = а, В = 57 и а4 = а, Ъ = 21, при общем итоге п =
= 200.
В таком случае применение формул для дигибридного скрещивания
приводит к таблице:
Вид уклонения
Расщепление по А
Расщепление по В
Совместное расщепление АВ
> Всего .. .
X*
V
20,807
0,027
0,222
21,156
Степеней
свободы
1
1
1
. 3
Р(Х2)
0,01
0,75-0,90
0,50-0,75
0,01
Вычисления показывают, что гипотезу геносвязи приходится, несом-..
ненно, отвергнуть вследствие малого %|в; нарушение же менделевских
пропорций идет за счет гена А. Вопрос может быть решен и с помощью,
коэффициента ассоциации.
МЕТОД МАКСИМАЛЬНОЙ ВЕРОЯТНОСТИ
(MAXIMUM LIKELIHOOD) *
М. В. ИГНАТЬЕВ
Метод максимальной вероятности * имеет большое значение как для
задачи определения тесноты сцепления, так и далеко за ее пределами.
Поэтому как общий прием отыскания значений, получаемых в опыте
величин, он заслуживает подробного рассмотрения.
Его техника может иногда показаться сложной, но в действительности
он основан на простых положениях. Удобнее всего иллюстрировать их на
каком-либо конкретном материале. Для этой цели допустим, что имеются
наблюдения, относящиеся к обратному скрещиванию, а именно:
Класс Численность
А, В 310
А, Ь 208
а, В 189
а, Ъ 293
Всего 1000
Выше эти данные послужили примером для решения задачи о наличии
геносвязи. Геносвязь была с несомненностью установлена, и теперь
возникает вопрос, как оценить величину перекреста. Обозначим ее через р,
так что в принятых выше символах
4 р = с/100.
В таком случае, исходя из предпосылки сцепления, получим, что
вероятность появления особи, относящейся к классу А, В, выразится че-
рез (1/2)-(1—р), к классам А, Ъ и а, В — через -^-р и к классу а, Ъ
опять через (1/2) «(1—р). Вероятность получения проведенного сочетания
численностей, т. е. того, что при 1000 испытаний особь первого класса
появится 310 раз, второго 208, третьего 189 и четвертого 293 ра за может
быть найдена из разложения многочлена:
(4-(»-й+4-р+-гр+4-(»-йГ-
* Термин maximum likelihood принято переводить как максимальное правдоподобие
(см. «Англо-русский словарь математических терминов». М., ИЛ, 1962). Метод был
разработан Фишером (Fisher, 1930) и им же предложено его название,— Прим. ред.
288
Она равна:
п 1000! Г 1 ,, чзг0 / 1 \208 / 1 \189 1 /, ч2м
310! 208! 189! 293! [_
1000!
3101 208! 189! 293!
B*-*»F(T»r-
Метод максимальной вероятности требует, чтобы эта вероятность
была наибольшей.
Определение значения Р, обращающего приведенное выражение в
максимум, не представляет труда. Прежде всего отметим, что в задаче
отыскания максимума цифровые коэффициенты не играют роли, и
поэтому она сводится к определению максимума выражения:
(1 _ p)*o*pW.
Но максимум этого выражения достигается при том же значении р,
при котором достигает максимума его логарифм, т. е. величина
а = 603 log (1 — р) + 397 log p.
Теперь задача становится уже совершенно простой. Она сводится к
максимализации функции а. Для ее решения достаточно взять
производную а по р и, приравняв ее к нулю, решить полученное уравнение
относительно р. Как известно, производная логарифма равна:
dlogx _ 1
dx x
Следовательно,
jfa = _ 603 . 397
dp 1 — Р Р '
Остается решить уравнение
603 • «Ю7 _. Q
Из него вытекает
1—J» Р
-603 р + 397 - 397 р = 0
р = 0,397.
Чтобы довести решение задачи до конца, нужно определить ошибку
найденной оценки.
Для этого сначала отыскивается .ее варианса, представляющая квад-,
рат ошибки. Правило ее нахождения очень несложно, а именно в
выражении da/dp следует заменить 603 через аг и 397 через а2, так что
dCt щ &1 , 412
1р^~Т^р ~*~ р *
и продифференцировать его, получаемся:
<ЙХ fll Ог
*Р*~ l(1-P>2 P%
и затем заменить аг и а2 их теоретическими значениями, т. е. ожидаемыми
численностями суммы двух крайних и двух средних классов. Эта замена
дает
аг = 1000 (1 — р); а2 = 1000 р
и приводит к результату:
dp* ~
19 А. С. Серебровский
1000
_ 1000 _ _
\Pi "*" "
289
1000
Р (!-/>)
Искомая варианса находится из равенства
1 сРа
а = тп.
р - --
vp ~~dp*>
Пользуясь равенством, находим:
^р юоо
и ошибка
тр- V юоо
В данном случае при р ==0,397, i?p = 0,00024 и /wp = 0,015, так
что 'окончательный ответ имеет вид:
р ± тр = 0,397 ± 0,015.
Значение произведенного вычисления заключается в том, что
определение р по методу максимальной вероятности дает наименьшую ва-
риансу и, следовательно, наименьшую ошибку оценки, чем всякий
другой прием. Поэтому метод максимальной вероятности приводит к
наиболее точной из всех возможных оценок и обладает наибольшей
эффективностью.
Теперь не представляет труда переход к формулировкам в общем виде.
Пусть р — величина перекреста (относительное число рекомендаций),
причем она принимается одинаковой для обоих полов; т%, т?г2,..., mt —
ожидаемые пропорциональные численности индивидуумов в классах
1, 2..Л. Все они выражаютсй известным образом через р (например, в
1 1
рассмотренном примере тг = -у-(1 — р), ^2 = урит. д).
Пусть далее a1? a2... at — наблюдаемые численности в тех же классах,
.причем их сумма:
% + а2 + ...+ at
ъ систему значени]
шиях из разложеш
(т1 + т2+ ...+ mt)n
Вероятность получить систему значений р(аи а2,..., at)
находится при этих условиях из разложения многочлена
и равна
р (аи а2,..., at) = ^ ^ ^ (т^ (ти2)%..., {mt)at.
Метод максимальной вероятности требует нахождения р, приводящего
это выражение к максимуму.
Задача сводится к нахождению максимума функции
a = c^log//*! + a2logm2 + ...+ at logmt
или, что то же, к решению уравнения:
da, dlogmi . dlogrm , , dlognii Л
Варианса же определенного таким образом значения находится из
выражения
где J обозначает суммирование по всем t классам.
Применим полученные формулы к некоторым наиболее часто
производимым скрещиваниям.
290
1. ОБРАТНОЕ СКРЕЩИВАНИЕ: АаВЬ к ааЬЬ
а. Фаза сцепления. В этом случае численности классов выражаются
следующим образом:
Класс
к
А, В
а, Ъ
Наблюдаемая
численность
01
а2
Ожидаемая тс™*.
численность Класс
1
1
птщ,=. —«— пр
| а, В
а, &
Наблюдаемая
численность
«3
04
Ожидаемая
численность
1
пщ»=—пр
1
727714= "J" Л (1 — р)
Уравнение для отыскания р имеет вид
fll i 02 I 03 04
!-/>
^_ «4_=0
р 1—р •
откуда
Р =
02 + 03
В принятых выше символах эта формула обращается в
А,Ъ + а,В
с%
Варианса равна
и ошибка
А,В + А9Ъ + а,В + а,Ъ
L.*li-p)
•100%.
... ™„=±><^.
Читатель, несомненно, узнает обычную формулу ошибки, которая
применялась раньше для обдаружения геносвяэи.
б. Фаза отталкивания. Нетрудно видеть, что в этом случае
п
а ошибка остается той же.
2. СКРЕЩИВАНИЕ F2. ПОЛНАЯ ДОМИНАНТНОСТЬ А ж В
а. Фаза сцепления. Если кроссинговер в обоих полах протекает
одинаково, то вероятности появления особей классов А, В; Л, Ъ; а, В и а, &
соответственно равны: ь
4-<3-2p + pV -*-(2i>-p2), -^-Р2)*-^1-2^^)-
Определение величины р по способу максимальной вероятности
становится в таком случае затруднительным. Поэтому вводится другой
параметр Р = (1 — р)2. Этот параметр име#г еще и то преимущество, что
он применим к случаям, когда кроссинговер протекает в обоих полах
по-разному. Тогда Р=. (1 — рг) (1 — pj, где pi и р2 „относятся к разным
291 .-
19*
полам. Численности классов три этих обозначениях выражаются
следующим образом:
Класс
Л, Ъ
А, Ь
Наблюдаемая
численность
«1
Ожидаемая
численность*
Класс
а, В
а, &
Наблюдаемая
численность*
04
Ожидаемая^
численность*
-TXi-P)
р
Функция а равна
а = аг1оё(2 + Р) + ^^(1 — Р) + M<>g(l -Р)'+ aJogP.
Отсюда уравнение для нахождения Р имеет вид:
dot
dP
ei
ог
о3
2+Р 1 —Р
- + *1 = 0
Р ^ Р и#
Оно приводится к виду: тгР +,(% — 2аа — 2а3 + а^Р — 2а4 = 0.
Решение этого уравнения дает искомое Р.
Величина же перекреста равна
Варианса оценки р находится из вариансы Р. Чтобы найти последнюю,
достаточно взять (Pa/dp2:
d*a __ (1+2Р)
dp* "2P(1— Р)(2 + Р)'
Так что, согласно общему правилу,
Отсюда
1 ,_ п(1+2РУ
vv 2P(1 —Р)(2+Р)#
_ 2Р(1~Р)(2 + Р)
"р-г" ■ ' * л(1+2Р)
Что касается вариансы величинк перекреста -Р, то можно показать, что
она равна
После подстановки Р = (1 — р)2 получается:
_ р(2-р)(г^-2р + р*)
иР
2л (3 — 4р + 2р2)
р e ± 1^^(2-^(3-2^ + ^)
2л (3 — 4/> + 2/>2)
Пример. Допустим, как в одном из предыдущих примеров, что в F2
получилось следующее распределение 1000 особей по классам: Л, В =
=571; А, 4=ч? 171; я, В — 156 ^ а, 6=102. Наличие геносвязи было дока-
292
зано ранее. Полагая фазу сцепления, требуется найти величину
перекреста. Мы икеем: аг == 571, а2 = 171, а9 = 156 и аА = 102.
Выражение: аг — 2а2 — 2а3 + а4 = 185 и уравнение для
нахождения Р имеют вид:
1000 Р2 + 185Р _ 204 = О: :
сОтсюда Р = 0,3685 и р = 1 — У 0,3685 = 0,3930, а в процентах с «
= 39,30.
Подставляя это значение в формулы вариансы, найдем vp = 0,000431
и щр = 0,021.
Итак, с = 39,30 ± 0,021%.
б. Фаза отталкивания. В этом случае, при одинаковом перекресте в
обоих полах, численности классов выражаются следующим образом:
Класе
АВ
АЪ
Наблюдаемая
численность
аг
Ожидаемая
численность лдасс
аВ
аЪ
Наблюдаемая
численность
«3
«4
Ожидаемая
численность
т (•-"•)
т"
Для удобства вычислений принимается х = р2 (или, если величина
перекреста в обоих полах разная, х = р1р2). Эта замена приводит
данный случай к только что рассмотренному с заменою Р на х.
В результате для нахождения х получается уравнение
па? + (аг — 2а2 — 2а8 + аА)х — 2а4 = 0.
Величина перекреста получается р = j^a?.
Варианса о; равна:
_ 2*(1-д).(2 + «)
*~~" п,(1+2а?) . '
Она приведена выше, как выражение vp. Переходя же от о? к р, получим
вариансу р, которая равна
Ур ~~ 2тг (1 + 2рР) » .
а ошибка
mP-±V 2*(1+V) •
Пример. Допустим* что мы имеем: а х = 1997, а2 = 906, а3 = 904,
а4 = 32 и п = 3839.
Уравнение для нахождения я имеет вид:
3839л8 + 1655а: - 64 = Q.
Отсюда х = 0,035712, величина перекреста р = )Л),035712 = 0,1890,
или в процентах с,= 18,90%; vp = 0,0002387 и игр= 0,01545,а в
процентах 1,545%. Эти величины и приведены в таблице.
3. F2 ПРИ НЕПОЛНОЙ ДОМИНАНТНОСТИ
Фаза сцепления. Одинаковая частота перекреста в обоих полах. В этом
случае фенотипически можно различать девяти классов. Их численности
приводятся в следующей таблице. ,
293
Класс
ААВВ
ААВЬ
ААЬЬ
АаВВ
АаВЬ
Наблюдаемая
численность
01
ог
0з
04
«5
Ожидаемая
численность
lv-py
Ipa-p)
п
IP2
\p(l-p)
/>a+f(l-p)2
Класс
АаВЬ
ааВВ
ааВЬ
ааЬЬ
Наблюдаемая
численность
0в
07
08
«9
Ожидаемая
численность
ра-р)
р2
p(i-P)
Ja -p)2
Функция а в этом случае равна:
а = (<*! + e9)log~-(l — р)2 + (в, + «в + в, + a8)log^-p(l — р) +
+(оз + a7) log-i-i*2 + % log 4 IP2 + (1 - Р)2]-
Уравнение для нахождения р имеет вид:
Д*& П — 01 02'— 03 — 07 — 05 205 (1 — 2р) Д + 01 + Д9 + 03 '+ 07 — 05
о> ~~ /> 1 —2/> + 2/>2~~ 1—/>
Это условие приводит к кубическому уравнению:
Р*-
•D]
2п
Р* +
2ъ— D — 0б
2а
-Р —
N — D^-аь
4/г
О,
где Z? = ах + а9 — а3 — a7. \
Таким образом, определение р может показаться несколько сложным,
хотя трудности решения кубических уравнений не следует
преувеличивать *. Варианса полученной оценки находится проще, а именно
обозначим
В таком случае
dL
dp
А
Р
В(1-2р)*
1— 2р + 2р*'
4Вр(1-р)
Р
С
£L_ А_ ,
dp2 ~~ Р2 ~*~ 1 — 2р + 2/>2 (1 — pf'
Теперь, согласно установленному правилу, следует произвести в
выражениях А, В и С замену а на их теоретические значения тп.
Следовательно,
А = п+ 07-^-09-^ = *+ J- (1 — р)2 5-(1 —/>)2 —
—*-(1-рГ = пРр-р).
Точно так же найдем:
В = -£-(1 -2р + 2р*) и С = п(1 -р2).
Подставляя эти значения в -^ , получаем:
d*L___ _ пр(2 — р) а(1-р2) , 2пр(1 — р){1-2р + 2р*)
1-2р + 2р*
dp2
(1-Р2)
* Обычно сначала находят приближенное значение уравнения: ж8 + ах2 — Ьх + с =
= 0. Его можно найти графическим путем, вычертив ход функции fix), даваемой
в левой части уравнения, и определив точку пересечения ее с осью абсцисс. Пусть
это значение равно зд. Уточняющее слагаемое равно п = / (#0)//i (*•)* так что
х = жо + л.
294
что приводится к виду
d*L__ _ 2/i (1 — 3/? + Зр2)
<*/>*- р{1-р)(1-2р + $Р)*
Поэтому варианса равна
„ _ p(l-j?)(l-;2p + 2^)
^Р "~ 2л (1 — Зр + Ър*) •
а ошибка — квадратному корню из этой величины.
Пример. Пусть в F2 наблюдается следующее распределение 1000
особей по классам: ААВВ = 94, ААВЪ = 109, ААЪЪ = 47, АаВВ = 121,
АаВЪ = 247, АаЪЪ = 124, ааВВ = 39, ааВЪ = 117 и ааЪЬ = 102.
Наличность геносвязи была доказана. Считая, что она выражается фазой
сцепления, требуется определить величину перекреста.
Решение. В этом случае ах = 94, а2 = 109, а3 = 47 и т. д. Производя
подстановку приведенных данных в уравнение для нахождения, р,
получаем:
р3 - 1,356856р2 + 0,658781р - 0,128909 = 0.
Это уравнение удовлетворяется при р = 0,400. Подставляя это
значение в формулу вариансы, получаем vp = 0,000223. Следовательно, тр =
= 0,015 и поэтому: р = 0,400 ± 0,015.
Ограничивая описание метода максимальной вероятности этими
задачами, сделаем некоторые сопоставления. В приведенных примерах при
одном и том же объеме наблюдений мы получили несколько оценок
величины перекреста, их вариансы и ошибки. Сводя результаты, строим
следующую таблицу.
Эксперимент
1. Обратное скрещивание
2. F^ полная доминантность
3. Fs неполная доминантность
Оценка
перекреста
(Р)
0,397
0,393
0,400
Варианса
оценки
(V
0,000240
0,000440
0,000223
Ошибка
оценки
(тр>
0,015
0,021
0,015
Мы видим, что все три эксперимента практически приводят к одной и -
той же величине перекреста, именно к 0,4. Однако из этого еще не
следует, что все они статистически, т. е. в смысле точности определения
искомой величины, равноценны; наоборот, видно, что второй эксперимент
дает значительно меньшую точность. Недостаток его точности, по
сравнению с обратным скрещиванием, может быть измерен отношением ва-
рианс. Оно равно 24 : 44 = 0,55. Можно сказать, что во втором
эксперименте извлекается 54% количества информации, доступной при первом
эксперименте. Это значит, что для достижения во втором эксперименте
точности, одинаковой с первым, потребуется значительно больше
наблюдений, а именно в 44/24 = 1,8 раза больше. Отсюда вывод, имеющий
практическое значение: оценка перекреста по обратному скрещиванию в
случае полной доминантности статистически предпочтительнее, чем по
F2. Однако, если доминантность неполная, так что можно различить 9
классов фенотипов, то обратное скрещивание теряет свое преимущество.
Оно даже становится несколько менее выгодным. Разумеется, вопрос
рассматривается здесь с чисто статистической точки зрения, так что
техническая сторона постановки экспериментов не учитывается, а
принимается, что оба типа скрещиваний одинаково осуществимы. Вопрос о выборе
295
типа скрещиваний на практике будет, конечно, решаться на основе
взвешивания всех обстоятельств. Однако было бы ошибочно среди этих
обстоятельств упускать из виду статистическую ценность эксперимента.
Приведенные примеры показывают, как ставится вопрос об
относительных преимуществах различных испытаний. Его рассмотрение
является предметом так называемой теории планирования экспериментов.
Она базируется на применении метода максимальной вероятности. Этот
метод, как указано выше, первоначально был применен при оценке
точности различных формул, служащих для определения величины
перекреста. Позднее он же был привлечен и для оценки точности результатов
разных скрещиваний. Причем под наиболее точным определением
искомой величины подразумевается то, которое дает наименьшую
статистическую ошибку и, следовательно, допускает наименьшее количество
испытаний.
Планирование эксперимента
В основе теории планирования эксперимента лежит представление
о количестве информации относительно величины перекреста при
разных генетических ситуациях. Чем больше количество информации, тем
точность оценки выше, а ее варианса меньше. Поэтому для измерения
количества извлекаемой в эксперименте информации применяется
величина, обратная вариансе оценки, полученной по методу максимальной
вероятности, как методу, дающему наименьшую ошибку оценки. Если
количество информации о величине перекреста р обозначить через /р,
то 1Р = 1/Vp. От величины информации, извлекаемой из всей
совокупности наблюдаемых особей, следует отличать среднее количество
информации на одну особь или количество, доставляемое в среднем одной
особью. Оно обозначается через ip, причем Ip = nip, где п — число
наблюдаемых особей. Сравнение различных экспериментов и различных
генетических положений удобнее производить при помощи количества
информации на одну особь.
В противоположность общему количеству информации оно не
зависит от объема совокупности. Величина ip является мерой статистической
ценности отдельного индивидуума в условиях данного эксперимента.
Так как варианса вычисляется по формуле
то
Для практических же целей часто удобнее пользоваться другой, но
идентичной формулой:
Оценка обратного скрещивания и F2
Применим эту формулу для оценки основных типов экспериментов,
производимых с целью определения величины перекреста: сюда
относятся обратные скрещивания {АаВЪ х ааЪЪ) и F2 (АаВЪ х АаВЪ) (в фазах
сцепления и отталкивания). В дальнейшем будем считать, что величина
перекреста в обоих полах одна и та же.
296
1. Обратное скрещивание. Относящиеся к обратному скрещиванию
расчеты приводятся в следующей таблице.
Класс
АВ/аЬ
Ab/ab
aB/ab
ab/ab
Итого» •
Сцепление
та
1
2'
1
5 (4-Л
1
dm
dp .
1
"2
1
2
1
2
1
~2
0
ip
1
2(1-Р)
1
2?
1
2*
1
2(1-.р)
1
Р(1-Р)
Отталкивание
m
1
2?
1
2?
! 1
dm
dp
1
1
""2
1
~2
1
2
0
*Р
1
2?
1
2(1-р)
1
2(1-р)
1
2р
1
| Р(1-р)
Класс
Сцепление
dm
dp
Отталкивание
dm
dp
AB/AB
АВ/АЬ
AB/aB
AB/ab
Ab/ab
Ab/Ab
Ab/aB
aB/aB
aB/ab
ab/ab
2<1-P)e
2?(1-P)
\p(i-P)
ad-*)*
^p{i-pf
2>(l-p)
J(l-P)2
-|(1-р)
|(l-2/>)
|(1 —2^)
-(1-P)
P
1
2*
|(l-2p)
1
IP
|(l-2p)
^
2(1-2P)S
P (1-/0
|(l-2p)2
P(i-P)
2
2
1
|(l-2p)*
P(l-P)
1
2(1-2*)*
P(l-P)
1
2P(l-i»)
2i>(l-P)
1
2^
5(l-p)2
|(1-P)S
|(1-P)
|(i-P)4
5P(l-p)'
l
IP
|(l-2p)
|(l-2p)
P
-(1-P)
-2(1-/0
g(l-2p)
-|(1-/»)
ld-2p)
1
IP
г(1"-2р)«.
P(l-P)
Jd-2p)»
P(l-P)
2
2
1
|(l-2p)»
P(l-P)
1
|(l-2j>)*
P(l-P)
1
Итого
P(l-P)
297
p(i-p)
Таблица содержит знакомые читателю элементы, в частности, среднее
количество информации на особь, как и следовало ожидать, равняется
уменьшенной в п раз величине обратной вариансы, как она была выведена
ранее. Информации одно и то же количество как в фазе отталкивания,
так и в фазе сцепления. ^
2. F2. Значения ip в F2 зависят от того, насколько детальна
классификация особей в изучаемой совокупности. Наиболее полная
классификация возможна при наибольшем объеме сведений о генотипах
наблюдаемых особей. Она включает в себя 10 классов, в состав которых входят
9 фенотипов, а двойные гетерозиготы АаВЪ разбиты после испытания их
потомства на два класса соответственно фазам сцепления и отталкивания —
АВ/ab и АЬ/аВ. Определение ip приводится в таблице на стр. 297.
Итог вычислений показывает, что полная классификация в F2 дает в
2 раза больше информации, чем обратное скрещивание. Это вызывается
тем, что F2 содержит сведения относительно рекомендаций в гаметогенезе
обоих полов.
Другой тип возможной классификации относится к условию неполной
доминантности, когда в обоих локусах можно различить по три фенотипа.
Общее количество классов равно теперь 9. Расчет ip для них отличается
от предыдущего лишь объединением двух классов АВ/аЬиАЦаВ, которые
ранее брались врозь. Этот класс имеет ожидание -т>-(1—2р + 2р2)
а • 2(1 — 2р)2
и b ip вносит долю, равную 1 * ,£ 2, тогда как прежде каждый из сос_
тавляющих его классов вносил по величине, равной 2. Отсюда получается
^изменение итога:
Sp- p(i-p)
-4
2(1— 2р)2
2(1~3р + 3р*)
1-2р + 2р* p(l-p)(l-2p+W
Обратная величина, помноженная на тг, была получена ранее как
значение вариансы для соответствующих условий.
Наконец, прежний план относится к случаю полной доминантности.
Расчеты для него приводятся ниже:
Класс
А, В
•я, В
•а, Ь
Итого
Сцепление
т
\(2р-р*)
|(l-2p + j»«)
1
dm
dp
(i-p)
~ 2
\-p
(1-/0
~~ 2
0 ,
*p
(i-p)*
3-2p + p*
2 (1 - p)*
2P-p*
1
2(3 —4p + 2p2)
lP(2-p)(Z-2p+p*)
Отталкивание
m
|(2+P2)
1
dm
dp
1
>
0
ip
P*
2 + p*
1— p*
1
2(1 + 2^)
|(2 + ^)(l-p2)
Этим заканчивается анализ отдельных простейших экспериментов.
Теперь представляется возможность сделать их сравнительную оценку.
Для этого количество информации, извлекаемой при обратном скрещива-
жии, принимается за единицу и определяется относительно значения для
других типов данных. Получаются следующие результаты:
298
_ Относительное количество
Эксперимент информации
Обратное скрещивание . . 1
F2, полная классификация 2
2(1 —Зр + 3/?2
F2, неполная доминантность 1 ~ 2р 4- 2в*
F2, полная доминантность
2(1-/»)(3-4р + 2р«)
сцепления (2 -/>) (3 — 2р + jj»)
отталкивания
2(1+2р*)
(2 + />2)(1+р)
На основании этой таблицы может быть составлен график зависимости
относительных значений количества информации от величины перекреста
р. Он наглядно показывает условия, при которых скрещивание F2 имеет
преимущество перед обратным скрещиванием и при которых оно
уступает ему. F2 имеет преимущество при неполной доминантности, особенно
при тесном сцеплении или отталкивании. При полной же доминантности
получается обратное положение: F2 не уступает в таком случае обратному
скрещиванию лишь при тесном сцеплении, при отталкивании же его
статистическая ценность всегда невелика.
Те сведения и выводы, которые были получены или намечались ранее
при рассмотрении конкретных примеров с р = 0,4, получают теперь
полное и обоснованное обобщение. Поэтому представляется возможным
решить вопрос о лучшем типе скрещивания в зависимости от генетических
условий, что имеет несомненное практическое значение.
Теоретически F2 содерягат вдвое больше информации, чем обратное
скрещивание. Однако невозможность отличить друг от друга различные
генотипы ведет к большому понижению познавательной ценности,
заключающейся в этом скрещивании. Отсюда получается вывод о том, насколько
важно стремиться к полноте сведений о генетическом строении
подвергаемых наблюдению особей.
Аналогичным способом можно произвести анализ относительной
ценности других типов скрещивания, в частности F3 и т. д. и вообще
исследовать вопросы, связанные с планированием экспериментов в различных и
более сложных изложениях.
МЕНДЕЛИСТИЧЕСКАЯ АЛГЕБРА
В. ГЛИВЕНКО
1. Отношения между генотипом двух последовательных поколений
обычно описываются в виде символических равенств, как, например:
, Аа X Аа = АА + 2Аа + аа9
АаВВ х АаЬЪ = ЫАВЬ + ЪАаВЪ +, ААаВв%
и т. п. Чтобы получить правую часть такого равенства иэ левой, иногда
требуются утомительные подсчеты. А. С. Серебровский (1934) предложил
автоматизировать эти подсчеты, применяя абстрактную алгебру с
операциями, символизирующими скрещивания, оплодотворение и т. д. Ему
это полностью удалось, по крайней мере для случая, когда отсутствует
сцепление генов. Но алгебра, предложенная А. С. Серебровским,
несколько сложна. Упрощение этой алгебры (для диплоидов) и является
предметом моего сообщения.
2. Мы называем менделистической алгеброй множество Я элементов
х, у, z, £..., удовлетворяющее следующим четырем условиям:
1. Каждой паре элементов х, у множества Я соответствует вполне
определенный элемент х + у множества Я, такой, что
х + у = у + х,
(х + у) + z= х+ (у+ z).
2. Каждой паре элементов л?, у множества Я соответствует вполне
определенный элемент л?, у множества 91, такой, что
ху = ух,
(х + y)z = xz + yz.
3. Каждой паре элементов, х, у множества Я соответствует вполне
определенный элемент х, у множества 91 такой, что
х-у = у-х,
(ar.y)-z= я- (y.z),
(x + y)-z = x-z + y -z,
(xy)-(zt) = (x-z)(y-t).
4. Множество Я содержит подмножество 93 (базис множества Я)
такое, что:
1. Каждый элемент множества Я может быть получен из
элементов базиса Я путем повторного применения операций х + у, ху и
ху;
300
2. Каков бы ни был элемент множества и каковы бы ни были
элементы |, т), базиса 95
(еч+ г\)х = (ец)х.
Как обычно, мы будем обозначать сумму одинаковых слагаемых х+х+
+...+ х через пх и произведение п одинаковых множителей хх....х через
хп. Нетрудно видеть, что
л (х ■+- у) = тг# + пу
ж что
> + y)2 = ^ + 2ay + ,y2.
3. В генетике мы составим множество Я из следующих элементов:
1) из генов, например, ^,а> &v
2) из гамет, например А, В; а.В...
3) из зигот, например Аа, АВ\ВВ =* (А *В) (а-В)...
* 4) из генных, гаметных, зиготных популяций, например,
24 + а, 42+4а,...
5) из элементов, которые получаются формальным применением
операций х + у, ху, х-у к элементам 1); 2); 3); 4) независимо от того
имеет результат биологической смысл, как, например (А + а)А =
=s Л2 + 4а,или не имеет, как например А + Аа, Аа,...
Базис здесь состоит из генов.
Чтобы рассчитать состав популяции, получающейся от скрещивания
популяции х и у, надлежит составить произведение ху.
Примеры:
<4а)(4а) = (А + а)(4а) = (А + а)(4 + а) = АА + 2Аа + аа; (Аа +
+ АА)(Аа) == (Аа)(Аа) + (44)(4а) = (АА + 2Аа +"аа) + 2(АА + 4а)=
— 344 + 4ка + аа; (4а.ЯЯ)(4а\В6)=(4а)(4а)".(ЙВ) (66) == (44 +2Аа +
+ аа).(4ВЬ) = 444 -ВЪ + 84а 56 + 4аа.£6.
МЕТОД ТРЕУГОЛЬНИКА
А. С. СЕРЕБРОВСКИЙ
В главе XII монографии были намечены приемы, позволяющие по*
дойти к анализу количественных различий в том случае, если в
распоряжении исследователя имеются более или менее гомозиготные расы,
позволяющие ставить достаточно обширные скрещивания с получением
типичных поколений гибридов: Flf F2, FB и т. д. К сожалению, в огромном
большинстве случаев в животноводстве эти условия не даны. Приходится
иметь дело с большим количеством отдельных скрещиваний, к которым
схема Fl9 F2, FB и т. д. неприменима. В поисках решения задачи в таких
условиях мы разработали метод, который можно назвать «методом
треугольника». Идея его сводится к следующему.
В моногибридной популяции все возможные типы скрещиваний могут
быть сведены к следующим шести типам, характеризующимся, с одной
стороны, различной частотой, а с другой, различным распределением
потомства. Эти типы следующие:
Тип
скрещивания
ааХаа
ааХАа
ааХАА
АаХЛа
АаХАА
ААХАА
Частота скрещивания
я*(я2-я2)
f 92-P2
4p2q2(2pq-2pq)
*pq\p*-2pq
p4p2-p2)
Состав
потомства
aa
aa-\- Aa
Aa
aa + 2Aa]+ AA
Aa + AA
AA
M сибса
потомства
Ма
Ma + \l
Ma + i
Ma+i
3
Ma + 2i
4
0
'"
0
i-
1*
0
4
°i
г*+Л
«S
iM-i
WeS
•S
Нанесем шесть типов сибсов ( = братья + сестры) в виде точек на
четырехугольном поле с ординатой М и абсциссой а2.
Мы получаем симметричный треугольник из шести точек (рис. 86).
В этом треугольнике будем различать основание, или бордюр,
обращенный к оси М, минус-угол при Ма и оI = ol, плюс-угол при Ma + 2i ш
ol = ol и вершину треугольника при Ма + i и аф = ал + -j- i .
После того, как мы будем иметь большое количество подобных сиб
сов, взятых без выбора из моногибридной популяции, определим для
302
Рис. 86. Частоты точек геноти-
пического каркаса моногибрид-
ного М— ^-треугольника {при
моногибридных скрещиваниях
в популяции)
6
м
ма+о
Ma^i
Ma+\i
%<§i
Mtt.fy
г л 1 1,2 г :2
i II
- .?♦
• bpq3
•2p2qz .ApY
• bp3q
- -P*
Рис. 87. Схема распределения
эмпирических точек в
моногибридном М — а2-треугольнике
М
.у
• • %•• •
Рис. 88. Примеры различных
шшо- и дигибридных
треугольников
1 — полное доминирование
-усилителя; 2 — неполное доминирование
усилителя; з — полное
доминирование ослабителя; 4 — неполное
доминирование ослабителя; 5 и б —
деталь; 7—два гена равной силы,
адоминирование; 8 — то же, у пчел;
9 — два гена, один вдвое сильнее
второго, адоминирование; 10 — два
гена, один — с адоминированием,
другой*— с полным
доминированием; т— два гена с полным домини
роваяием, оба усилителя; 12 — то
же, один усилитель, другой
ослабитель
•
• •
/
•
•
3
• -
• •
•* г
•
•
• *
•
•
• • •
•
•
5
•
• в
•
•
• • •
•
••
* *
• •
•
10
•
•
•
•
•
•
• •
• •
■ 'А
•
9
•
•
• - •
•
• •
•
• •
•
8\
•
•
••
• «
••
•
•
• |
•
• • •
• • • . • •
• • • • • •
• • • • • •
• • • • • •
• • •
каждого из них М жа% и затем
нанесем эти «эмпирические [точки» на
М — (Тф поле. Эти точки должны
будут расположиться, образуя
более или менее выраженные
скопления около наших шести
«теоретических точек» каркаса, как
показывает рис, '87,
, Бели паратипическая
изменчивость не слишком сильна и число
особей в каждом сибсе достаточно
велико, чтобы сделать
статистическую ошибку незначительной,
то при рассматривании всего
треугольника в целом мы сможем
заметить лежащий в его основе,
рисунок каркаса из шести точек или
центров скопления эмпирических
точек.
Легко показать на ряде
примеров, что различным генотипи-
ческим случаям соответствуют
весьма различные
треугольники, примеры которых даны на
рис. 88.
Таким образом, идея метода
треугольника сводится к тому, что
каждый генотипический случай
эмпирических точек вскрывается
на М — а2 поле, и открывается
возможность по числу и расположению этих центров* сгущения
распознать, с каким именно генотипическим случаем мы имеем дело.
Разработка теории этого метода в свою очередь сводится к тому, чтобы
для всех мыслимых генетических случаев дать предварительно строение
треугольника, исследовать, как различные генетические моменты
(доминирование, сцепление, число генов и их сила, летальность, сцепление
с полом и т. д.) отражаются на особенностях строения «треугольника».
Так как число мыслимых генетических случаев бесконечно велико и все
их учесть невозможно, то такой анализ ряда отдельных генетических
моментов должен будет внести некоторую классификацию в разнообразие
треугольников, расположив их в ряды. Следующий рисунок дает
предоставление о таких рядах (рис. 89).
Отметим влияние числа генов и доминирования на характер
треугольника.
ч Рис. 89. Ряды треугольников
I — изменение треугольника при
числа генов; II — изменение треугольника по
мере усиления доминирования; * in — изменение
треугольника по мере усиления разницы сил генов
ЧИСЛО ГЕНОВ
При неполном доминировании каждый новый равносильный ген
увеличивает число точек, причем по основанию, как и по другим сторонам,
прибавляются две липшие точки. Ниже мы увидим, что обстоятельство
это играет важную роль в понимании картины. В случае полного
доминирования новый ген прибавляет по основанию лишь одну новую точку,
так что два доминирующих гена прибавляют столько точек, сколько один
неполно доминирующий. Введение нового гена иной силы прибавляет
больше точек, до шести. Если новый ген равен прежнему,—
прибавляются, как мы видим, две точки. Если вдвое сильнее или слабее — прибав-
304
ляются четыре точки. Если более чем вдвое сильнее или слабее — шесть
точек. Шесть точек прибавляются и в том случае, если сила действия гена
не кратна силе действия прежнего.
ДОМИНИРОВАНИЕ
Доминирование полное приводит к' асимметричному треугольнику —
эта асимметричная точка соответствует типу скрещивания Аа X Аа.
Вершиной треугольника оказывается точка скрещивания Аа х аа. Если
асимметричная точка лежит от вершины в сторону минус угла — случай
доминантного ослабителя, если в сторону плюс угла — случай
усилителя. При неполном доминировании треугольник приобретает очень
неправильный вид, причиной чего является то, что по мере усиления
степени доминирования три точки перемещаются к остальным трем,
остающимся неподвижными, как показано на рис. 90.
Точка, лежащая в треугольнике для полного доминирования
асимметрично, интересна тем, что она отвечает скрещиванию Аа X Аа, дающему
при доминировании асимметричное распределение потомства,
расщепляющегося в отношении 3:1. Остальные три точки этого треугольника
дают распределение симметричное. Это обстоятельство открывает
интересную перспективу развития метода треугольника, перехода от
треугольника к тетраэдру. Если в качестве третьей оси взять р,3, то можно будет
располагать наши точки уже в пространстве. При адоминировании в этом
нужды нет, так как все fi3 = 0, кроме случайных уклонений в ту или
другую сторону. При участии доминирующих генов некоторые точки
выйдут из плоскости, либо подымаясь вверх при \i ^> О, либо опускаясь ниже.
Например, дигибридный треугольник с полным доминированием примет
вид тетраэдра с тремя слоями точек. Если вышерасположенные точки
обозначить более крупными кружками, мы будем иметь такой рисунок
(рис. 91).
Подробнее на этом вопросе пока можно не останавливаться. Но при
обработке эмпирических точек нужно высчитать для них и fi3. Если по
величине обнаружится косое распределение, можно будет полагать
участие доминирования. Однако, если fi3 ^ 0 будет у большинства сиб-
сов, это будет указывать лишь на скошенность паратипического покрова.
Рис. 90. Перемещение трех
точек моногибридного
треугольника по мере перехода от адо-
минированного к полному
доминированию
Рис. 91. Дигибридный
тетраэдр. Случай двух
полнодоминирующих генов
20 А. С, Серебровский
305
ЧАСТОТЫ ТОЧЕК КАРКАСА
Так как различные типы скрещивания встречаются и в популяции и
в различных поколениях с различной частотой, то и точки каркаса М —
а2-треугольника будут встречаться тоже различно часто. В
моногибридном случае распределение частот будет такдм: ч-
аа X аа (f Аа X Аа 4p2q2
аа X Аа 4p<f Аа X АА 4p3q
аа X АА 2pif АА х АА р*
Для различных значений р и q мы будем иметь такое распределение
частот:
1 (все частоты) _1_ 2 (все частоты)
Р = 9= 2 умножены на 16) Р —з » Я— з умножены на 81)
g4 4pq3
аа х аа 1 аа X Аа 4 16 *
32
2/>V 4/?V
аа X ЛЫ 2 Аа х Ла 4 8 16
/?4 4/fy 8
AAxAAi AaXAA 4 1
1 3_ (частоты умно- JU 9 (частоты умноже-
Р — 4 » ^ ~" 4 жены на 256) Р= до * ^ ^ То" ны на 16 тыс.)
В цифрах частоты выразятся так:
81 6561
108 2916
18 36 162 324
12 36
1 1
По мере усиления разницы р -г- q треугольник все сильнее
скашивается: при р > q — в сторону плюс угла, при р < q — в сторону минус
угла. Уже при р = 1/4 и q = 3/4 соответственно, а тем более при р =
= 1/10, частота угловой точки становится столь малой, что ее можно не
принимать во внимание — эмпирических точек здесь почти не будет, а
они будут располагаться косой полоской от вершины к минус-углу или
обратно (полосы к ^длюс-углу постоянно наблюдались, например, при
изучении сосков свиней — Серебровский, Жирнов, неопубл.). По тому,
направлена ли эта точка к^ минус- или плюс-углу, можно судить об
отношении ри д.
. •" /
12
б 60
1 60 ^~~160 ""^\
1 15 ,'240 240 \
'~ "\ 120 у'Л~п N 799
/24 32 \ ,ho H80 J J92.
6 f ' 4S U6 15 \240 —ojn /
i \24 32 / :«>v» ,/.,
*—' \ 12
I.. ._
Схема XXXI
306
Распределение частот в дигибридном скрещивании с равносильными
генами имеет несколько более сложный вид:
gV1
1 •
2p*q2v*
| +
2q*rW
•
рЧ*
<74Г2
4p202r2^2
•
2p9gV
+
2р4г2г?2
p*r*
4?4гг?3
ApqW
•
8р?д2гг?3
4p4rV
£p*qv*
Spq^v2
•
4р4гг?3
ЪрЦЧЧ
4p?3r4
8р*гг?3
4р4г3г;
+
4?W
16рд3гг;3
+
4р???г?4
•
16p9?W*
16р3дгг?3
lbpq*r*v
ш
16pg3rV
16р2?2гг?3
16р3дт?г?2
16pVr2tf2
По существу, однако, сложность этого распределения не так уж
велика. Мы имеем здесь наложение шести моногибридных треугольников
друг на друга по следующему простому закону: к шести точкам
моногибридного треугольника гена А приложены своими минус-углами шесть
моногибридных треугольников второго гена, с соответствующим
умножением частот.
Точками в нижнем углу квадратиков указан, например, треугольник
гена 2?, приложенный к точке треугольника ф гена А, ж частоты,
относящиеся к этому треугольнику, подчеркнуты. Во всех них участвует
д4, который может быть вынесен за скобки, после чего останется
моногибридный треугольник гена В. Для случая q = р — г = v = 1/2,
отвечающего например F2, распределение частот имеет вид, показанный на
схеме XXXI.
Здесь важно отметить, что основное большинство частот
сосредоточивается в шестиугольнике в средней части треугольника, а частоты вершин
начинают относительно убывать, эмпирические точки э этом случае будут
располагаться в виде более или менее округляющегося поля. Например,
точки при минус- и плюс-углах составляют лишь по 0,4% -общей суммы
частот и даже вместе с примыкающими двумя (1+4 + 8) составляют
лишь по 5%.
Это округленное поле будет еще более выражено при увеличении числа
генов до 3,4 и т. д., как показано пунктиррм на тригибрйдном
треугольнике. ; :
При условии р ф q и г =j= q дигибридщш треугольник по
распределению частот будет принимать разнообразные формы, в основном
сводящиеся к двум. Если р > q ж г ]> v или р < q ж г <^v, то треугольник
307
20*
Таблица 24
Общий обзор всех возможных типов скрещиваний в популяции при сцеплении генов А и В (объяснение в тексте)
АВ „АВ „АВ „ЛВ 2А6 АЬ аВ А£ ^В аЬ
АВ АЬ аВ fob аВ АЬ аВ jzb jab ab_
1 1 41
0
1
1 2 Зу,
1
4
I4
И 3
2
4 '
1 4
1 3 3»/2
1
4
14
1 12 3
2
4
1 8
20 3
2
т
1 4
/
I 4 3
(1-е)
1 4
I 13 2»/*
-§-<1-с>+г(1 + 0)
|218 1 2Vl
|-(i-c) + -i(H-c)
8
28 2
2(1-с)
4
5 3
с
4
1 14 2У,
22
т<2~с)+тс
29 2
1
8
35 2
2с
4
,
1 6 3
0
1 2
1 15 2у,
1 ~
1 4
1 23 2i/2
1
4
1 4
30 2
(1-е)
4
36 2
с
4
41 2
0
2
7 '- 3
0
2
1 16 2V2
1
4
1 4
1 24 2V2
1 4
31 2
(1-е)
4
37 2
с
4
42 2
0
2
46 2
1
8 ^ 21/,
1
4
1 17 2
2
4
1 8
25 2
2
4
1 8
32 IV*
8
38 li/t
т<а-°> + т0
8
43 1у, |
4 1/4
47 IV,
4 1А . ,
50 1
2
4
4
9 2«/«
4
| 18 2
2
4
1 8
26 2
2
4
1 8
33 IV,
-|<l-c)+i-<i-c>
8
39 IV,
т<2~с) + тс
8
44 IV,
4 1/4
48 IV,
4 Щ
51 1
2
4
8
53 1
2
4
4
10 2
0
2
19 ^ IV,
1 4
27 IV, 1
4 1
34 1
(1-е)
4
40 1
с
4
45 1 1
2
49 1
0
2
52 1/2
1/4
4
54 1/2
1/4
4
55 0
1 1
будет превращаться в косую полосу, идущую от вершины к
соответствующему углу — плюс-углу в случае р > q, r > v и к минус-углу в случае
Р <С 9» г <^ v. В том случае, если р^> q, r <^v или, наоборот, р <^ q,
г < v, то распределение частот принимает характер обращенного
треугольника, максимум частот находится у средней точки основания:
Частоты в процентах Частоты в процентах
6 2
17 20 1
3
17 43 9
6 1
8 20
1
Косая полоса Обращенный треугольник
р = 0,25 ? = 0,75 /> = 0,1 ' д = 0,9
г = 0,25 г? = 0,75 г = 0,9 г? = 0,1
Таким образом,- изменение концентрации гена в популяции
существенно и характерно отражается на форме каркаса треугольника.
СТРОЕНИЕ ТРЕУГОЛЬНИКА ПРИ НАЛИЧИИ ГЕНОСВЯЗИ
В свободной дигибридной популяции, в которой гены А и В имеют
геносвязь, нарушаемую в С случаях перекрестом [0 < с < 1/2), и в
которой достаточно установилось равенство случаев сцепления и
отталкивания, распределение генотипов, как мы видели выше, таково:
АВ , 0 АВ . су АВ . 9 АВ . су АЪ . АЪ , аВ , 0 АЪ , 9 аВ , аЬ
АВ+ *АЬ + *ЮВ + *lfi + *Ч5 + Ж + ЮВ + ^W + *13Г + ~аЬ
(р и q, r и и отброшены, чтобы не осложнять расчетов. При желании они
могут быть вставлены. Данный случай отвечает р = q и г = и).
Табл. 24 дает обзор всех скрещиваний, возможных в такой
популяции, их относительные частоты, М и сг^ сибсов.
В каждой клеточке:
в левом верхнем углу — номера типов скрещиваний от 1 до 55;.
в левом нижнем углу — частота данного типа скрещиваний (при р =
= Ф\
в правом верхнем углу — М потомства (опущены всюду i);
в центре — aY2 (опущены всюду i2).
Жирной чертой обведена, часть скрещиваний, у которых в формулы а2
входит переменное с.
Эта таблица может быть использована и для случаев без геносвязи,
являясь более общей. В случае независимости генов АВ с = 1/2 и графы
АВ/ab и Ab/аВ могут быть объединены.
Проставляя различные значения с в эту таблицу и нанося полученные
точки на М — а2-поле, получаем каркасы треугольников для различной
силы геносвязи. При сравнении с дигибридным треугольником без
сцепления в треугольнике со сцеплением можно выделить постоянную часть,
309
А У*' fy *A % */4 7A 8Ai*
-7
| (&
l (m—
i $7>v
^Jy"
L
Г a "■
<§)<$)
\ i
44
^ ^
44
—jl*
4 4
l \
, / / /
! / / /
X A' '
\88*
x88'
44..—
K88s ~~~^
\88^
,\ \ \
\ \ \
\ \ \
(5)(t) @@W®
. -
"©
~ш)
^^
Рис. 92.
Перемещение точек
переменной части ди-
гибридного треуго»
льника при
возникновении гено-
связи
а — независимые
гены; б — сильная
геносвязь
г Ь
i
г®/
й--;;
Ь 4
\-,&
1 1
8 .
—i—i—i—i—
1 \
? / 1
1 '
-| г
4--@
в которой aY не зависит от величины с, и переменную часть, в которой
0y меняются при изменении. Во всех случаях, как видно из следующей
таблицы, величины Ж от с не зависят:
о*2
0
1
2
3
4
о
1
4
6
4
1
1
4
8
24
24
8
2
4
16+| 8
32+J16
16+!_L.
Перс
3
4
32
32
шенная
±е
4
1
8Н
I
часть
Более детальное строение переменной части показано на рис. 92, а,
верхняя: схема (в кружках номера скрещиваний из табл. 30).
При с <[ 1/2 в каждой группе половина точек отодвигается вправо,
половина влево. Так, при с, близком к нулю, переменная часть
располагается так, как на нижней схеме рис. 92, б.
Вместе с постоянной частью мы получаем такое распределение точек:
2 п-L-LJL-i-JL 6 JL 8
a ° - 4 4 4 4 4 "Т 4 Т
М 0 1
1 8
18 16
40
2 18 32
40
3 8 16
8
4 1
16
16
16
310
При с, равном 1/4, получим еще более оригинальное «полосатое»
распределение £очек:
JLJLJLJlJLJLJL-2
0 4 4 4 4 44 4*
м
0
1
2
3
1
4
6
4
8
4
24
8
24
4
8
16
16
36
16
16
4
8
4
16
8
16
В качестве наиболее характерной особенности треугольника для
случаев сцепления надо отметить их высоту. При с, близком к 0, вершина
лежит уже при а? = 2г2, т. е. треугольник вдвое выше, чем при незави-
симых генах. При с = 0,25 вершина лежит при а2 = 1 -о~*2, т-е« в полтора
раза выше. Эти случаи сцепления напоминают случаи с неравными
генами, но там повышение треугольника сопровождается увеличением числа
точек по основанию, здесь же число точек основания остается равным
пяти.
СТРОЕНИЕ ТРЕУГОЛЬНИКОВ ДЛЯ МАЛЫХ СИБСОВ
Все приведенные выше треугольники относятся к случаю достаточно
больших сибсов (большое число братьев'+ сестер). В подобном случае
действительно можно полагать, что, например, скрещивание А а X аа
дает сибс состава 50% Аа + 50 %аа, для которого характерно
1 1
М = Ма + -у i и а2 — £2 jj0 возьмем маленький сибс, например, из
двух особей. Тогда скрещивание Аа X аа реализуется в трех различных
типах, давая сибсов либо Аа + Аа (25 % случаев), либо Аа + аа (50%),
либо аа + аа (25% случаев). Эти сибсы будут иметь соответственно М =
1 1
=Ma+i и а^ = 0, Мв + -я-г и о* = — г2, Мй и crY2 = 0. Иными словами,
только 50% сибсов окажется в точке моногибридного каркаса, отвечающей
Аа X аа; 25% займут место возле точки каркаса аа X аа я 25% около
точки АА X аа. Что касается скрещивания Аа X Аа, то при двух
потомках оно может дать сибсы:
АА + АА (6) Аа + АА (5) аА + АА (5) аа + АА (3)
АА + Аа (5) Аа + Аа (3) аА + Аа (3) аа + Аа (2)
АА + аА (5) Аа + аЛ (3) аА + аА (3) аа + аА (2)
Ла + аа (3) Ла + аа (2) аА + аа (1) аа + аа (1)
По своим М и а2, эти сибсы отнесутся к самым различным (всем шести)
точкам моногибридного треугольника, как показывают цифры в
скобках. .
В результате мы получим (считая для упрощения, что от каждого
типа скрещивания получено пропорциональное число сибсов) для сибсов
из двух особей в каждом такое их распределение.
Моногибридный треугольник примет следующий вид. Вершинная
точка исчезает, так как при двух потомках отношение АА + 2Аа + аа
311
не может быть реализовано в отдельном сибсе. Сильно возрастают
частоты трех точек по основанию и особенно усиливается середина
основания:
22
9
12
12
Скрещивание
1) аа X ял
2) Аа X ол
3) ААХаа
4) Аа X Аа
5) Аа X АА
6) ААхАа
Частоты
1
4
2
4
4
1
Число
сибсов
4
16
8
16
16
4
Распределение сибсов по точкам
1
4
4
1
9
2
8
4
12
3
4
8
6
4
22
1
4 1 5
4
8
[ 12 1
6
1
4
4
9
При трех членах в сибсе отношение Аа : аа реализуется уже 4
следующими способами:
м
Ъаа + ОАа Ма
2aa + iAa Ma + \i
2
iaa + 2Aa Ma + ^i
Oaa + ЪАа Ma + i
0
s«
4
о
Частота
1
3
3
1
Мы видим, что здесь окажется невозможной точная реализация
отношения 1:1 и, следовательно, исчезает точка (2). Но вместо нее
возникают две новых точки с новыми М и оД Вместо точки (2), таким
образом, получится группа точек, расположенных по параболе.
При четырех членах в сибсе точка (2) реализуется следующим
образом (схема XXXII и XXXIII).
На
Ма+Цз\
Ma+2/3i
Ma+i
0 2/9=8/361А =.
1 i i i i i i i i
Схема XXXII
Ма
Ма+1/Ы
Ma*2lki
Ma+3/ti
Man
0 f/4
1 I 1 1 Г
- '
-.""*"
Схема XXXIII
По мере увеличения числа членов в сибсе, рассеивание точки (2)
будет происходить следующим образом: по параболе, опирающейся на
точки (1) и (3), с вершиной, проходящей через точку (2) будет
располагаться биномиальный ряд частот. Основная масса этих частот, образуя
подобие серпа луны, будет все более и более концентрироваться у точки (2),
312
Рис. 93. Схема каркаса точек
А а X А а при малых сибсах
Рис. 94. Точечная схема рис. 93,
но для сибсов при п = 6
Каждая точка равна 1% всего материала
Рис. 95. То же, что на рис. 94, но для
сибсов при п = 10
в пределе превращаясь в единственную точку. Уже при сибсе в 20
членов концентрация рассеянных точек каркаса (не эмпирических) достигнет
удовлетворительной степени.
Очень сложен закон рассеивания при малых сибсах точек (4) Аа х Аа
(рис. 93). Схема состоит из системы парабол, на пересечениях которых
располагаются ряды биномиальных коэффициентов, вычисляемых по
формуле
/
с^<*
•2Ь,
п п-Ъ
где Ъ = Аа, с = аа, а = АА, а + Ь + с = 10 различных степеней.
31&
Следующие рисунки (рис. 94 и 95) дают каркасы точек Аа х Аа для
величины сибсов 6 и 10. Чтобы не оперировать большими числами,
рисунки даны в удобном и достаточно точном виде — в виде
«однопроцентных точек», таких точек, каждая из которых равна 1/100 всей суммы
частот треугольника.
На полной схеме треугольника при п — 10 были бы видны три ясно
выраженные «точки» основания (1), (3) и (6), две столь же ясно
выраженные, но серпообразной формы точки (2) и (5). Что же касается «точки»
(4), то она еще распылена в целый елкообразный рисунок, в центре
которого намечается несколько сгущений.
Лишь при сибсе в 20 особей точка (4) принимает уже достаточно
оформленный вид и устанавливается типичный для моногибридного случая
треугольник из б точек каркаса, оказывающихся все еще своего рода
созвездиями, а не точками.
При дигибридном расщеплении рассеивание при малых сибсах будет
давать еще более сложный рисунок.
Однако наличие паратипической изменчивости скроет все эти рисунки
и даст обычную картину скопления эмпирических точек. Эти скопления,
однако, будут тем слабее выражены, чем меньше будут сибсы.
ТРЕУГОЛЬНИКИ ДЛЯ СВОДНЫХ СКРЕЩИВАНИЙ
В предыдущем изложении мы имели дело с сибсами, т. е. с детьми
одной пары родителей. Для животных малоплодных получение таких
сибсов становится невозможным, зато для полигамных животных
возникает возможность использовать полусибсов, т. е. детей одного отца и
многих матерей. Посмотрим, какой вид примет треугольник в этом
случае. Как мы видели, при скрещивании самца с большим числом самок
возникают при моногибридном наследовании три типа скрещиваний:
Частота
я2
2pq
Р2
Родители
аа (р2АА + 2pqAa + q2aa)\
Aa(p4A + 2pqAa + qe>aa)
AA(p2AA + 2pqAa + q*aa)
Состав потомства
qaa + pAa
q Аа р
2^ + -2+^AA
qAa + pAA
М 1
ма + р^
1
(Ma+pi)+^i
(Ma + pi) + i
о2
РЯР
\pqi*+lV
pq.i*
Следовательно, мы получаем треугольник из трех точек (рис. 96). Весь
этот треугольник сдвинут по беи вправо на pq, так что основание
лежит на уровне о* = pq, а не о* = 0. При р = q положение
треугольника отвечает положению точек (2), (3) и (5) в обычном моногибридном
скрещивании для сибсов, так как pq = 4. Но там в качестве самки
участвует всюду особь Аа, а здесь целая популяция с составом гамет
рА + qa.
О pq 1/Ь 1/b+pq 1/2
1 1 1 1 п-
Рис. 96. Моногибридный
треугольник в случае скрещива- „
ния самца со многими самками
3*4,
{Ма*р1щ
Н/%
Н
Нетрудно показать, что для дигибридного скрещивания мы будем
иметь шесть точек в виде треугольника, сходного с моногибридным для
сибсов, но опять-таки сдвинутого на pqrv. Таким образом, замена сибсов
на полусибсов оказывается вполне возможной и даже очень выгодной,
так как, во-первых, строение треугольника упрощается, во-вторых,
точность эмпирических точек может быть значительно повышена
увеличением числа детей в полусибсе, что компенсирует понижение точности,
происходящее от замены одной матери на многих.
РАЗРЕШАЮЩАЯ СПОСОБНОСТЬ МЕТОДА ТРЕУГОЛЬНИКА
Всякий метод должен быть бценен с точки зрения того предела, после
которого он уже не может быть применен. В нашем случае важно
составить себе ясное представление о том, какой силы гены еще могут
улавливаться данным методом. К сожалению, ответить на этот вопрос довольно
трудно, так как суть метода состоит в том, чтобы сквозь «беспорядок»
эмпирических точек заметить «порядок», т. е. схему расположения центров
скопления точек и понять, с каким именно «рисунком» мы имеем дело.
Тем не менее важно посмотреть, при каких условиях в данном
«беспорядке» точек становятся заметными отдельные центры скопления.
Прилагаемые два рисунка поясняют проблему. На рис. 97 мы видим в
беспорядке разбросанные 35 точек. Каких-либо ясных скоплений здесь не
видно, хотя в трех местах (в центре и две вверху) скопления как будто
намечаются. На рис. 98, несомненно, видно три скопления. Самое
обильное вверху и бедное (из пяти точек) внизу. Не совсем ясно, ^вляется ли
среднее скопление двойным? Оба рисунка относятся к одной задаче и
представляют собой 35 полусибсов моногибридной популяции. Разница
между рисунками в том, что для рисунка 97 были взяты полусибсы из
10 особей, а для рисунка 98—из 20. В данном случае при величине полу-
сибса где-то между 10 и 20 уже наступили условия, при которых рисунок
стал ясен.
Мы имеем такие условия:
i = 3; о% + pqi2 = 4,7; п = 10 (рисунок каркаса незаметен),
i = 3; о% + pqi2 = 4,7; п = 20 (рисунок каркаса ясен).
Чем больше inn, тем легче распознавание рисунка каркаса, чем больше
<з2, тем труднее. Приблизительно можно положить, что S = —7= , тогда
i У п
• / ..
:. • •
• • • •
Рис. 97. Распределение
эмпирических точек для случая
рис. 96 при условии i = 3,
п = 10 и а* + pqi* = 4 г
Сгущения не выражены
! • *
Рис. 98. То же, что и на
рис. 97, но п = 20
Три сгущения выражены
315
можно характеризовать условия видимости рисунка. В данном случае
4,7
0,5 и £2
ступила видимость.
4,7
—-^ = 0,35. Где-то около S = 0,40—0,45 на-
2 5
На рис. 99 представлена та же задача, но при i = 1. Здесь 5 = —т= =
1 У 20
= 0,56, и рисунок опя!ъ исчез. Не виден он и на рис. 100 еще и при
п = 30, где S = А= = 0,49.
1 /30 N
Если действительно этот показатель может характеризовать суть
дела, то разрешительная способность метода примерно такова:
1
Величина полусибса
10
20
40
100
1000
_~02
0,4Х Уп *
0,79
0,56
0,40
0,25
0,09
• •••
• • •
• • •• .
Рис. 99. То же, что и на рис. 97t
но i = 1
Сгущения слились
Рис. 100. То же, что на рис. 98,
но п = 30
Сгущения едва намечены
Результат, как видно, довольно неутешительный, так как еще при
сибсе в 100 особей индивидуально улавливаются только гены, сила
действия которых не менее 0,25 Оф.
Поэтому возникает проблема дальнейшего уточнения метода.
В этом отношении оказываются следующие перспективы.
УТОЧНЕНИЕ МЕТОДА ТРЕУГОЛЬНИКА. СЛИЯНИЕ СИБСОВ
Первый прием. Слияние инбредного сибса (брат X сестра) с
родительским. Прием «слияние сибсов» состоит в следующем.
Пусть, например, мы имеем скрещивание типа Аа X аа. В потомстве
от такого скрещивания возникают особи Аа и аа. Если мы будем
скрещивать это потомство друг с другом, то при этом возможно образование
«дочерних скрещиваний» типа Аа X Аа, Аа X аа, аа X Аа я аа X аа>
потомство которых будет характеризоваться своими М и а2.
Ha M — о2-треугольнике моногибридного скрещивания, имеющем,
как было показано выше, 6 точек соответственно 6 генотипически
различным типам скрещиваний, упомянутые только что дочерние скрещивания
займут только три точки из 6. Это показано сплошными кружками.
316
От скрещивания аа X аа потомство получается лишь аа, и потому и
дочерние скрещивания займут только одну точку (верхний угол),
отвечающую типу аа X аа. Наоборот, дочерние скрещивания от Аа X Аа
займут все 6 точек и т. д.
Таким образом, каждый тип скрещивания характеризуется не только
тем, что возникающее от него потомство (сибс) имеет свою М и а2, но и
тем' что образуемые из этого сибса новые скрещивания, дочерние
скрещивания, занимают тоже определенные (одну или несколько) точки на М— а2-
треугольнике.
Чтобы отразить это явление графически, можно произвести слияние
дочернего сибса с родительским сибсом и охарактеризовать полученный
слитой сибс соответствующей М и а2. Так, например, если от
родительского скрещивания Аа X аа мы получили родительский сибс
состава Аа 4- аа и отсюда получили дочерние сибсы АА + 2Аа + аа; Аа +
+ аа; аа^ то, сливая каждый из них с родительским, получим следующие
три точки слитых сибсов:
1) 2Аа + 2аа (родительский сибс) 2) Аа + аа
АА + 2Аа + аа (дочерний сибс) Аа + аа
АА _[- 44а + Ъаа (слитой сибс)
3) Аа + аа
2аа
2Аа + 2аа Аа + Ъаа
М=Г
M=zi
*2=4*2 а2==16*2
Такие же простые расчеты, произведенные для всех 6 типов
родительских скрещиваний, дают все возможные типы слитых скрещиваний для
моногибридного случая с адоминированием. Таких типов,
сопоставленных в табл. 25, оказывается 12.
При нанесении этих точек с соответствующими им частотами на М —
сг2-поле, получаем рис. 101.
В левой части рисунка представлено распределение слитых сибсов, а
в правой, для сравнения,— простой моногибридный треугольник. Как
видно, от слияния сибсов характер моногибридного рисунка довольно
резко изменился. Наиболее интересным является исчезновение точки в
ма+о
+2/4i
+фг
+6/4i
+2i
0
1
••
••
~
-
- »
/А
1
п
•3
'>
•г.
я
&
1
«*
п
•1:
•
о
SS
&
/А
—1—
л.
Рис. 101. Моногибридный треугольник, измененный I приемом
(слева) и неизмененный, приравненные по числу точек (справа)
317
Результаты слияния сибсов с их родителями
Таблица 25
Типы родительского
скрещивания
аа X а<*>
Аа X аа
АаХАа
аа ХАА
АаХАА
ААХАА
Всего . . .
Относительная
1
частота при рд = к
Частота
<74
ЬрЯ*
2pV
£p*q
1
М, о8 и частоты родительско-дочерних слитых сибсов
М с*
0 0
4?4
V
4
АГ о8
1/4 3/16
&pq9
Apq*
4
М о«
2/4 4/16
8pq*
8pq*
8,. ,
ДГ о»
2/4 8/16
lpV
PV
1
M o«
3/4 1/16
4p03
4p9gr2
4p?*
8
M a»
1 4/16
2p^
I0i>v
10
A! o»
1 8/16
4p?tf2
4pV
4
Af C°"
5/4 7/16
4p^2
4p8g
4p°?
< 8
6/4 4/16
$p*q
1 /
8p*q
8
Af o«
. 6/4 8/16
w
PV
1
Af o»
7/4 3/16
V?
4p8g
4
1
А/ о8
2 0
4p4
4p4
I 4
Рис. 102. Моногибридный
треугольник, измененный II
приемом
ма+о
1/21
Hi
+3l2i
+2L
€
I
— •
-
_ •
//«•
i
•
•
ь
2h
1—
•
•
центре основания треугольника (при М = 1 и aj = 0). Благодаря этому
вершины треугольника 0,0 и 2,0 оказываются разделенными обширным
пустым полем, треугольник опирается на ось М как бы двумя широко-
расставленными (на интервал в 2 i) ножками. Это обстоятельство очень
полезно с методической точки зрения, так как облегчает счет числа генов,
участвующих в расщеплении. При простом построении треугольника в
моногибридном случае мы имеем 3 точки вдоль линии с$ = 0, в дигиб-
ридном — 5 точек и т. д. Счет этих точек является одним из наиболее
надежных признаков, по которым можно судить о числе генов,
участвующих в расщеплении. При слиянии сибсов мы получаем на том же
интервале уже меньше точек: при моногибридном — вместо 3 лишь 2, при
дигибридном (равносильные гены) — вместо 5 лишь 3 и т. д. Благодаря
увеличению интервала между точками с i до 2£ счет их облегчается.
Кроме того, он облегчается и тем, что благодаря слиянию сибсов получается
увеличение объема сибса и уменьшение рассеивания, положение
эмпирических точек приближается к теоретическим местам.
Правда, в результате слияния сибсов рисунок в целом осложняется.
Но упрощение одной детали рисунка, в данном случае «бордюра» вдоль
линии о* = 0, должно принести определенное облегчение в работе
аналитика.
Второй прием. Уничтожение той же средней точки основания (АА X
Xaa,M=l, о* = 0) может быть достигнуто и другими приемами. Очень
интересный рисунок возникает при использовании материала
скрещиваний отец X дочь и мать X сын (рис. 102). В этих скрещиваниях тиц
АА X аа невозможен, так как если, например, отец имеет генотип АА,
то у него не может ^>ыть дочери аа. Остальные типы скрещиваний
возможны, но частоты их также удачно изменяются в сторону уравнения (на
рис. 102 изображен случай р = q = 1/2).
4-^
2L
ис. 103. Дигибридные треугольники, упрощенные II приемом
319
Расчет частот этих 5 точек виден из следующей таблицы:
Родительские
скрещивания
тип
отец X
X мать)
аа X аа
аа X Аа
аа X АА
Аа X аа
АаХ Аа
АаХАА
ААХ аа
ААХАа
ААХАА
частота
я*
2pq*
Р2Я2
2рд*
крЧ* .
2р*д
p*q*
2рЧ
Р'
Относительные ч*<
при р =
1
-9 = 2
Тип дочерей
аа
аа + Аа
Аа
аа-\- Аа
аа + 2Аа + АА
Аа + АА
Аа
АА + Аа
АА
1СТОТЫ
Тип скрещивания отец X дочь и их частот
аа X аа
Я*
РЯ*
Я3
1
Аа X аа 1
аа X Аа \
РЯ*
РЧ2
РЯ*
Р2Я2
2pq*
2
АаХ Аа
РЯ*
2р*Я2
p*q
ря
2
Аа X АА
ААХ Аа
Р2Я2
РЪЯ
РЧ2
p*q
2р*Я
2
ААХАА
р*я
Р*
Р*
1
При дигибридном скрещивании в зависимости от равенства или неравенства
силы генов М — сг2-треугольники, видоизмененные этим приемом,
принимают следующий вид (рис. 103).
Третий прием. Слияние сибсов от скрещивания отца с несколькими
его дочерями (от одной матери).
Этот прием особого пояснения не требует. Если отец скрещивается с
несколькими своими дочерями (полными сестрами), то и в этом случае
аа X АА оказывается невозможным.
Расстояние между точками по линии <з\ = 0 получается равное 2L
В то же время, благодаря слиянию сибсов, точность определения
положения точки может быть значительно поднята, например, в
гомозиготных скрещиваниях при 4 дочерях точность возрастает вдвое, а так как
точки основания раздвинуты вдвое,
• . то общее повышение точности дости-
*• # гается 4-кратное, при 9 дочерях —
* • 6-кратное и т. д.
. • ' Не вдаваясь в подробности рас-
• • * четов, в принципе достаточно про-
/ дочь * jдочери стых (Серебровский, 1936), приведем
рис. 104, показывающий вид
моногибридного треугольника (при /? = q),
измененного этим приемом, при раз*
, ^ ном числе дочерей.
2 дочери
Много дочереи
Рис. 104. Моногибридные треугольники,
измененные по III приему, при разном
числе дочерей
ПРИМЕР ПРИМЕНЕНИЯ МЕТОДА
ТРЕУГОЛЬНИКА
На задних крыльях у пчел, как и
многих других перепончатокрылых,
имеется ряд крючочков, которыми
заднее крыло сцепляется с передним.
320
Число этих зацепок у домашней пчелы сильно варьирует, и есть
основание предполагать роль и генотипических факторов в этой изменчивости.
В 1926 и следующих годах С. Г. Петров, разрабатывавший методику
менделистического анализа пчел, собрал громадный материал по сотне
пчелиных семей, беря из каждой семьи молодых пчел и трутней и
подсчитывая у них число зацепок. Однако, несмотря на большой материал,
Петрову не удалось выяснить генетику числа зацепок из-за сильнейшей
паратипической колеблемости. Ряд М, высчитанных для отдельных семей,
оказывается почти непрерывным, не обнаруживая какого-либо
распадения на группы и не образуя даже ясных вершин, которые указывали бы
на обнажающийся каркас генотипов.
Попробуем применить к анализу этого ценного материала метод
треугольника. Однако посмотрим прежде всего, какой тип рисунка возникает
у популяции пчел с их оригинальным способом размножения
гаплоидными самцами. В моногибридной популяции пчел возможны следующие 6
типов скрещиваний:
Частота
q*.q = q*
Я2Р = PQ2
2pq.q = 2pq*
2pq-p=2p2q
Р2Я = P2'q
/?2./? = р3
Тип
скрещивания
аа X а
аа X А
АаХ о.
АаХА
ААХ а
ААХА
Потомство QQ
аа
Аа
аа-\- Аа
АаА-АА
Аа
АА
М
адоминирование
М
Mi
Mi+i
1
Afi + 2*
Mi+ 1 2 *
Mi+i
Mi + 2i
<>
0
0
1
l
- /2
4*
i °
0
и о2
полное
доминирование
M
Mi
Mi+ i
1
M i + 2 г
Mi+i
Mi+i
Mi + t
*r
0
0
1
V2
0
0
0
Рисунки этих типов будут следующие (см. рис. 105).
Для трутней, происходящих от матерей без оплодотворения и поэтому
гаплоидных, будем иметь независимо от доминирования:
Частота
?2
2рд
Ра-
Матери
аа
Аа
АА
Сыновья
а
а+А
А
М
Mi
1
Mi + 2 i2
Mi+i
*
0
1
I'1
0
Рис. 106 изображает треугольник для трутней.
Для дигибридного с адоминированием при различых ix и i2, полагая
для простоты р = £, будем иметь треугольники рис. 107.
Для самок (рабочих пчел), таким образом, является характерным
усеченный треугольник (или, вернее, трапеция) без вершины,
отвечающей у других животных типам скрещивания Аа X Аа, АаВЬ X АаВЬ,
невозможным здесь из-за гаплоидности самцов. Не входя пока в
дальнейшие подробности, посмотрим, какой вид имеет расположение
эмпирических точек по материалам, собранным Петровым.
21 А. С. Серебровский
321
Ч,+1/21
Mr+2/2i
M^3/2t
M,+*/2i\-
.pq
f/ti2
Mt
• 2pg2 MrH/2i\
.2р*Ч
О
1/W
•2pq2
•p+p*q
Рис. 105. Моногибридные треугольники для рабочих
пчел при адоминировании (слева) и при полном
доминировании (справа)
(
м,
4
0 tfct
t I
.2)
Рис. 106.
Моногибридный треугольник для
трутней
Рис. 108 и 109 представляют этот эмпирический материал для рабочих
пчел и для трутней.
Если обратить внимание на основную группу точек в районе М=19,5—
—21,5 то довольно легко заметить рисунок именно такой, какой можно
ожидать по рис. 107 и напоминающий цифру 8, где центры двух отверстий
приходятся: первого на М = 19,9 и о% = 2,3—2,4, а второго на М = 20,7
и (уJ = 2,4. Этот рисунок является частью дигибридного при более или
менее равных генах. Так как рисунок несколько асимметричен, можно
думать, что р > q. Положив, примерно, q — 3/5, v = 1/2, будем иметь
теоретическое распределение частот (в процентах), показанное на схеме
XXXIV. Основная часть точек (83%) образует 8.
V
v\ )V-V>
2К \.7-8,0
\ /
\ У
\ /■
#7
2,7
V-2J
W
Схема XXXIV
Две других схемы представляют собой распределение частот при
несколько иных значениях q и v. Фактическое распределение, конечно,
можно отнести к разным скоплениям очень условно, но, проделав с такой
оговоркой подсчет, находим, что теоретические частоты достаточцо
близко совпадают с характером рисунка в основном скоплении эмпирических
точек (схема XXXV).
Если мы примем одно из подобных объяснений распределения точек
для рабочих пчел, то у трутней из тех же семей должны увидеть рисунок
несколько иной, а именно: вместо «8» только один параллелограмм.
Нечто подобное действительно усматривается (на рис. 109) при М =
= 25,5 иа2 = 2,8, где достаточно ясно виден просвет, окруженный довольно
густым, более или менее ромбическим скоплением точек. К сожалению,
изменчивость у трутней значительно выше, чем у рабочих пчел, — у
322
?? lrh
</<?1г=12
$? hm2i2
£? Lr2lt
Рис. 107. Дигибридные треугольники
для пчел (вверху) и трутней (внизу) при
адоминировании
Слева — гены равносильны, справа — разно-
сильны (it = 2i2).
2§\
21
22
/% 'Ч
56*
т-
Рис. 108. Распределение [по М — а2
полю .эмпирических точек — семей пчел
($ 9) по числу зацепок (по материалам
Петрова)
1 1 9 4 5 В 7 8 9
~1 I 1 1 Г
// ш
т 1 1 1 1 1 1 г
19
20
21
22
23
24
25
2б\
27\
1— •
Г • *• • *
•*••••••
- •: .•
• •
_ *
W
•
Рис. 109. То же, что на рис. 108, но для трутней
21-
V ffi tJ9
Я 13 " Ю
/К
•
• \
Afi
# V # 2,8 W 19
V* 3,2 2,1
0,8 Qi,
9 W 7tt
X X X
q=Bjb v=l/2 q=3/u v=i/3 <j=*/5 V3S//3
ь
Схема XXXV
рабочих минимальные а2 в основном располагаются на уровне в 1,8—2»
у трутней при 2—2,5. Максимальные а2 трутней достигают величины 12
против 5,4 рабочих. Возможна, что в связи с этим улавливать какой-
либо рисунок по материалу трутней оказывается труднее.
Несомненно, однако, что гипотеза двух более или менее
равносильных генов приложима лишь к основному скоплению точек. Но имеется,
кроме того, немало точек и при М > 20,2 и а2 > 3. Особенно интересны 4
группы точек, идущих на таблице рабочих пчел (рис. 109) на уровне
а2 = 3,5 (при М = 20,6; 21,6; 22,3; 23,4). Возможно, что это —
указание на еще одну 8-образную группу, указывающую на участие третьего
гена, примерно вдвое более сильного, но редкого. Данных для того чтобы
делать это предположение более определенным, у нас недостаточно.
Конечно, при такой высокой паратипической изменчивости (з% ^
^ 2), при i —' 1 (т. е. ап в полтора раза выше силы гена) одного
рассмотрения точек недостаточно. Оно дает только возможность для
построения гипотезы. Наличие достаточно ясно видимых «окошечек»,
окруженных скоплением точек, позволяет сделанную дигибридную гипотезу
считать более или менее обоснованной. Для полной убедительности,
впрочем, необходимо было бы провести некоторые дополнительные
эксперименты. Поэтому, чтобы отчетливее пояснить метод, в следующем
параграфе мы продемонстрируем решение задачи-модели.
ЗАДАЧНМЮОДЕЛИ ПО АНАЛИЗУ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ
Методику составления задач-моделей мы можем распространить и на
наиболее важную и трудную область — область количественных признаков.
Здесь нам приходится моделировать две части явления — генотипическую
и паратипическую. Первая часть не представляет каких-либо
особенностей по сравнению с тем, что нам уже известно. Нужно лишь каждому
гену придать некоторую силу действия и степень доминирования. Пусть,
324
например, мы задаем дигибридную задачу. Положим силу действия гена
А : iA = 3, и силу действия гена В: iB = 1. Кроме того, зададим
некоторую «базу», т. е. величину признака, соответствующую генотипу aabb.
Пусть база Ъ = 10. При промежуточном доминировании и полном
доминировании различным генотипам будут соответствовать следующие
величины признака:
Генотип
aabb
ааВЪ
ааВВ
Aabb
АаВЪ
Y |
при
промежуточном
доминировании
10
11
12
13
14
при полном
доминировании
10
11
11
13
14
Генотип
АаВВ
ААЬЬ
ААВЪ
ААВВ
\
У
при
промежуточном
доминировании
15
16
17
18
при полном
доминировании
14
13
14
14
Но для количественных признаков характерно постоянное
«искажающее» влияние паратипических факторов, благодаря которому один и тот
же генотип дает различные фенотипы. В простейшем случае паратипичес-
кую изменчивость можно моделировать при помощи бинома (1 + 1)п,
меняя в различных задачах показатель п. На урнах-моделях это
моделирование можно осуществить таким образом:
1) бином (1 + I)2 = 1 + 2 + 1 ... частоты
т— 1, 0, + 1 ... уклонения.
я
= -1,
= 0.
два шара красных
Выбросив два шара черных, долагаем я =
= + 1; шары разные, черный и красный,.я
2) бином (1 + I)4 = 1+4 + 6 + 4 + 1 частоты,
— 2, —- 1,0, + 1, + 2 уклонения.
3) бином (1 + I)6 = 1 + 6 + 15 + 20 + 15 + 6 + 1... частоты,
— 3,— 2,—1, 0, 1, 2, 3 .... уклонения.
Можно выбрасывать три шара и либо считать на них (черный и чет),
положив 6ч = —3,5ч = —2, 4ч = 1,3ч = 0 и т. д., либо считать эти
три шара как трехзначное число. Например, I) 1, II) 10, III) 7 = 107.
В этом случае можно пользоваться следующей таблицей, где частоты
умножены на 15: /
1 15 001-015 = —3
6 90 016-105 = —2
15 225 106-330 = -1
20 300 331—630 = 0
15 225 631-855 =1
6 90 856-945 = 2
1 15 946-960 =3 ~
Выпадение шаров 0000 и 961 — 999 во внимание не принимается.
Можно ограничиться тиражом двух шаров, но покрасив их в 4 цвета
и написав на них предварительно следующие цифры:
На шарах
красный нечет,
красный чет .
желтый нечет
желтый чет
Цифры
3
2
2
1
На шарах
черный нечет
черный чет .
синий нечет
синий чет
Цифры
2
1
1
0
325
Можно выбрасывать четыре шара (4 черных = —2; 3 черных + 1г
красный = — 1; 2 черных + 2 красных = 0 и т. д.) или удобнее
выбрасывать два шара, но читать на них окраску (красную или черную) как
чет или нечет. Считая «черный» за «чет», запишем:
4ч = -2; Зч = —1; 2ч = 0; 1ч = + 1; Оч = + 2.
Можно, наконец, выбрасывать два шара, предварительно пометив их
цифрами 1—10, и читать первый — за десятки, второй — за единицу.
Например: 1) 1 + II) 5 = 15; I) 5 + II) 1 = 51 и т. д.
Для перевода чисел в величину паратипического уклонения составим
следующую таблицу, умножив каждую частоту, т. е. член бинома (1 + 1)г
на 6:
1 6 01 —06=-2
4 24 07 — 30 = — 1
6 36 31—66= 0
4 24 67 — 90= 1
1 6
16 96 91 —96= 2
Выпадение шаров 00, 97, 98 и 99 во внимание не принимается.
Мы умножили частоты на 6 потому, что 16x6 = 96, что близко к 100
комбинациям, возможным при выбрасывании двух шаров по данной
схеме. Лишними остаются лишь пары марок 00, 97, 98 и 99, выпадение
которых не принимается вовсе во внимание.
Так как вероятность каждой цифры будет 1/8 (5 шаров из сорока),
то выпадение двух троек будет 1/64, двух нулей 1/64 и т. д., а суммы двух
цифр распределятся как раз по биному:
Слагаемые
0,0
од
0,2
1,1
0,3
1,2
1,3
2,2
2,3
3,3
Частота л
1 = -3
15 = —1
20 = 0
15 =+ 1
6 = + 2
1 = + 3
Теперь мы можем приступить к моделированию наследования
количественных признаков. Получив путем тиража потомство и его генотип,
делаем тираж для определения паратипического уклонения и,
просуммировав базу, у и я, получим и фенотип. Полная запись опытов будет
иметь такой вид (условия: iA = 3, is = 1, промежуточное
доминирование, база = 10, я по (1 + I)6.
Пример записи:
№
скрещивания
1
№ и генотип
отца
3
АаВЪ
№ и гонотип
матери
6
ааВЪ
№, пол и генотип
потомства
11 d АаЬЬ
12 $ АаЬЬ
13 9 АаВЪ
14 d ааВВ
15 9 АаВЪ
16 с? ааВЪ
17 9 aabb
Ъ -J- Y
10+3
10+3
10+4
10+2
10+4
10+1
10+0
п
—1
—2
+1
0
-1
+2
+1
ф
12
11
15
12
13
13
11
326
и
т. д.
a— ■—
*
Рис. 110, Решение задачи.
Положение точек после первых //
25 скрещиваний
12
13
1$
4
При желании могут быть вводимы дальнейшие осложнения, например,
З1г -*- для всего помета в целом, я2 — для отдельной особи, я3 — половые
коэффициенты и т. д.
Решающий задачу получает только номер и пол потомков и их
фенотип (ф), а у и я остаются известными лишь дающему задачу.
Варьируя число генов, силу их действия, степень паратипической
изменчивости и т. д., можно делать задачи более легкими или постепенно
повышать их трудность. Необходимо добиться, чтобы генетики-аналитики
научились справляться с решением подобных задач и в такой степени
отточили бы орудие своего анализа, чтобы иметь возможность решать
задачи и на биологическом материале, как правило, еще более трудные и
пока недоступные для нас, кроме отдельных объектов и простейших
случаев.
Пример решения задачи. Получены два исходных скрещивания.
Исходные особи: № 1 tf, <р = 15 и $ $ № 2, ф = 9 и № 3, ср = 12.
Для первоначальной ориентации в материале мы поставили несколько
скрещиваний для освещения следующих вопросов:
1. Дает ли результат отбор, т. е. не является ли материал чистой
линией?
Так как в первых скрещиваниях все самые мелкие особи оказались
самками, то был поставлен вопрос:
2. Нет ли полового диморфизма?
На второй вопрос был получен отрицательный ответ, поэтому в
дальнейшем оба пола обрабатывались вместе.
Результат отбора начал быстро сказываться, показав наличие
различных генотипов. Например, в скрещивании № 15 (отбор в плюс-сторону)
были получены от двух родителей с ф = 17 особи только с ф = 14, 15 и
16 и М = 15,4, в то время как при минус-отборе в скрещивании № 11 от
родителей с ф 9 X 10 были получены особи с ф = 8—12 и с М = 10,2.
Вначале мы сделали несколько попыток построить гипотезы обычными
методами «менделистического анализа», но ввиду быстро выяснившейся
почти всюду значительной изменчивости, эти попытки пришлось оставить
и применить метод треугольника, ставя различные скрещивания и
вычисляя для каждого сибса М и а2.
После первых 25 скрещиваний мы получили такую картину (рис. 110).
На рисунке начинает ясно обозначаться треугольник. Минус-вершины
его находится около М = 9 и а2 = 1, плюс-вершины около М = 16 и
а2 = 1, а третья вершина при М = 13 и а2 = 5. В этом месте, очевидно,
скрещивались сильные гетерозиготы, а в минус- и плюс-вершинах
отобраны минус- и плюс-гомозиготы, чему соответствует малая изменчивость
потомства в этих «угловых» скрещиваниях. О числе и силе действия
генов при таком малом числе точек судить еще рано, но самый факт
обозначения треугольника говорит за то, что число генов невелико.
327
О 1 г2
7 8 6Z
17V
Рис. 111. Решение задачи.
Распределение точек после 167 скрещиваний
Для того чтобы проверить,
действительно ли минус-вершины и плюс-
вершины нельзя раздвинуть
дальнейшей селекцией, был поставлен ряд
дальнейших скрещиваний особей из
этих углов, но получить М больше
16 и меньше 10 не удалось. На
основании около 150 скрещиваний мы
получим следующую картину (рис. 111).
Положение вершин плюс и минус
обозначалось достаточно ясно. Очень
большое скопление точек у этих
вершин вызвано просто тем, что в
попытках раздвинуть эти вершины
было поставлено много скрещиваний.
Однако мы видим ясно скопление
точек и при М = 15 и при М = 12.
Это говорит за то, что вдоль по
основанию треугольника (обращенному
к линии М), несомненно, имеются
не только три скопления {аа X аа; аа X А А; АА X 4i), но и еще
некоторые точки. Если считать, что мы имеем дело с симметричным
треугольником, то скоплению при М = 16 должно соответствовать скопление
при М = 10. Наметившемуся ясному скоплению при М = 15 должно
отвечать скопление, при М = 11 (что действительно усматривается), а
скоплению при М = 12 должно отвечать отсутствующее скопление при
М = 14 (последнее легко объясняется тем, что в этой средней области
вообще ставилось очень мало скрещиваний). Таким образом, по
основанию треугольника можно ожидать скоплений при М, равном 10, 11, 12,
13, 14, 15, 16 —всего 7. Это отвечает 1) тригибридному скрещиванию, если
гены равны, 2) дигибридному, если один ген сильнее другого.
Этим случаям отвечают следующие две схемы расположения точек
(рис. 112).
Эти распределения точек, имея сходство в числе точек по основанию
(7), отличаются высотой. Вершина треугольника (точка максимальной
изменчивости) в первом случае лежит на уровне o2Y = 3 X 2/4 = 1,5,
во втором — на уровне а2, = 2,5. Наш фактический! треугольник еще
выше, его основание лежит на уровне примерно в* = 1, а вершина меж-
ду ог^ = 4 и о* = 5, а отдельные точки достигают 6,2 и 7,5. Так как
в каждом сибсе у нас лишь 10 особей, то ошибка о% при этом очень
велика, вследствие чего наши точки, несомненно, слишком рассеяны по оси
о2 и фактически треугольник должен быть ниже. Тем не менее, исходя
м
10
11
12
13
1$
15\
А
10
11/4 21/ь 31/ь Ф 1 2 Зб2
Рис. 112. Дигибридный каркас
при iA — 2iB (слева) и триги-
бридный
при iA
(справа)
328
П 1 2 3 Ь 5 'об'
-т 1 1 1 1 г
0 1
?A/
И-&..->*-
13
*§\
m
Рис. 113. Решение задачи.
Распределение 167 точек при сиб-
сах п = 20
5а*
м
W
11
Рис. 114. Решение задачи.
Распределение точек при сибсах
п = 30
• *-'"$&!• ' » * *
из этих соображений, приходится считаться с той гипотезой, которая
предполагает более высокий треугольник (два гена, один в два раза
сильнее другого).
Судя по нашему треугольнику, у которого основное скопление точек
располагается при а2 = 1—1,5, такой величины должен быть кадрат
паратипической сигмы, что отвечает биному (1 + I)4 или (1 + I)6.
Наметившиеся семь скоплений располагаются с интервалом в 1. Таким
образом, сила действия самого слабого гена в нашей задаче не может быть
больше 1, так как интервал между скоплениями определяется как раз
силой самого слабого гена. Отсюда можно заключить, что нам приходится
решать задачу, в которой сила действия слабого гена не превышает и
даже, может быть, меньше о£
Это обстоятельство заставило нас сосредоточить внимание на
уточнении положения наших эмпирических точек. Для этого мы пошли двумя
путями*
1. Начали повторять все поставленные скрещивания, сначала по два,
потом по три раза с тем, чтобы в каждом сибсе стало не по 10, а по 20 и
30 особей. Этим приемом мы уменьшили случайное уклонение наших
эмпирических точек от теоретического их положения.
2. Изменили типы скрещивания. Чтобы сделать теоретический
рисунок более простым, применили инбрвдинг.
На рис. ИЗ и 114 показано изменение эмпирического треугольника
при переходе к сибсам в 20 и 30 особей.
Общая картина изменений треугольника сводится к следующему:^
1. Основание треугольника передвинулось, так что на линии о — 1
осталось значительно меньше точек. Следовательно, а® должно быть > 1,
примерно, 1,25—1,50. 2
2. Резко снизилась вершина треугольника — примерно до <Хф == 3,5-—
—4, так что общая высота треугольника по оси а2 определилась в 2,5—2,75.
Следовательно, для максимальной изменчивости oY = 2,5—2,75, что
значительно выше, чем а? = 1,5, величины, соответствующей гипотезе
трех равных генов. ^
3. Вместе с тем в области М = 14—16 начал намечаться
моногибридный рисунок (рис. 113). /
4. Такой же рисунок начал намечаться в области М = 10—1^ (рис.
114).
5. Структура треугольника начала принимать несколько ячеистый
характер, что тоже соответствует больше схеме разносильных генов, в
22 А. С. Серебровский
329 ,
0
м\
to
11
12
13
п
15
16
17
1 2 3 4 5(
1 1 1 1 г
•
••
;•• •
•%■ •
- •.//• •
• • • . .
•• •
•
;•• • • •
г *•
I-
Рис. 115. Решение задачи.
Уничтожение средних сгущений
применением слияния сибсов
по I приему
М
10
11
12
13
15
17\
56*
.V.
Рис. 116. Решение I задачи.
Применение III приема]
которой, как видно из рис. 112 (слева), имеется неравномерное
распределение точек по площади треугольника, в противоположность схеме трех
равных генов (эта ячеистость более заметна в рис. 113 при сибсах в
20 особей).
Таким образом, внимательное рассмотрение треугольника позволяет
наметить следующие выводы:
1. а*; = 1,25-1,50.
2. Более подходяща схема двух генов, из которых один в два раза
сильнее другого.
Для изменения и упрощения теоретического рисунка мы применили
два приема: во-первых, выбрали все имевшиеся скрещивания братьев^с
сестрами и слили их с родительскими сибсами. Как показано выше, при
этом исчезает в основании треугольника точка аа X АА, и при
моногибридном расщеплении остаются по основанию (т. е. при о\ = 0) лишь две
точки, разделенные интервалом в 2i.
Как видно на рис. 115, середина основания треугольника
действительно расчистилась и осталось два болыпих*скопления точек — при М = 11
и М = 15,5. Расстояние между ними равно 4,5, и так как эта величина
равна 2£, то отсюда определяется
• т i — 2. Такова должна быть сила
\ I. действия сильного гена в нашем
т\1 . • # скрещивании.
•• • *•# • В качестве еще одного приема
.• /д # • мы использовали скрещивание от-
• •• • *•*•**• цов с их дочерями. Отцы были
д* #» •• "• * *• взяты из различных мест тре-
• •* • *•*•* ^ #. *• •• * угольника. Всего было получено
f **m • # *! * '' 30 таких точек. На рис. 116 вид-
• *•• *•*•*•' но, что по основанию действитель-
/•9 • Ч * но расположились лишь два ос-
• • *•# # новных скопления с интервалом,
J' # примерно, в 4—4,5.
На этом мы закончили решение
задачи, придя к правильному
выводу:
1) ol скорееJl ,25, чем 1;
Рис. 117. «Полосатые» треугольники при
наличии доминирования двух слабых
генов — двух [ослабителей (справа), усили-
теля и ослабителя (слева)
330
2) 1 ген с i = 2 при адоминировании;
3) несомненно наличие одного адоминирующего гена с i = 1.
При этом мы должны были отметить, что сходные картины могли
получиться при наличии вместо одного неполно доминирующего гена двух
полно доминирующих (рис. 117). Различение этих случаев по своеобразной
полосатости довольно затруднительно. Проведенные нами вычисления fi8
для ряда первых скрещиваний дали в среднем |х3 —- около 0, почему мы
пришли к заключению, что имеем дело с адоминированием.
Таким образом, в этом отношении решение тоже оказалось
правильным.
Следует подчеркнуть, что данная задача является достаточно трудной,
ввиду сильной паратипической изменчивости. Действительно, при такой
паратипической изменчивости, например, <р = 13 может быть у всех 9
генотипов, возможных в данной задаче. При этих условиях довольно
безнадежны попытки разобраться в отдельных скрещиваниях обычными
методами, которые мы применяем при анализе качественных различий.
Метод треугольника, однако, привел нас к правильному решению
задачи.
22*
ПОСЛЕСЛОВИЕ
Автор монографии Александр Сергеевич Серебровский (1892—1948)
принадлежал к группе тех ученых-биологов, которые оказали громадное
влияние на развитие генетики и селекции в СССР. Исследовательская
работа А. С. Серебровского началась еще в первые годы после Великой
Октябрьской социалистической революции и продолжалась вплоть до
самой его преждевременной смерти. Кроме 120 напечатанных работ, в
его научном архиве осталось еще около 30 неопубликованных, в том числе
несколько крупных монографий. Среди них находилась и рукопись
публикуемого сейчас «Генетического анализа». Первый его вариант был
набран еще перед Великой Отечественной войной, вторично монография была
принята к печати в 1946 г., но также не увидела света.
Круг интересов А. С. Серебровского как исследователя был очень
широким — от вопросов общей биологии и эволюционного учения и до
конкретных вопросов селекции отдельных видов сельскохозяйственных
животных.
В то же время он был очень сильным аналитиком и математиком.
Математический склад мысли А. С. Серебровского выявился еще в первых
его работах, например, в статьях «Опыт статистического анализа пола»
(1921), «Полигоны с фокусами и их значение для биометрики» (1925),
«Статистический метод в биологии» (1925) и др. Начав разработку генетики
домашней курицы, он неизбежно столкцулся с необходимостью
разработки теории генетического анализа, тех вопросов, которые сейчас входят
в так называемую математическую или статистическую генетику. Работ
в этой области было тогда очень мало, и А. С. Серебровскому пришлось
идти в значительной степени собственными, оригинальными путями.
Итоги длительной работы А. С. Серебровского по разработке теории
генетического анализа и обобщены в данной монографии. На ее страницах
автор многократно подчеркивает сложность задач генетического анализа,
при котором приходится иметь дело со многими неизвестными, а также и
неразработанность его методов. Он постоянно призывает читателя к
самостоятельной их разработке. Очень интересно сравнение работы лица,
проводящего генетический анализ, с работой химика-аналитика,
разлагающего молекулы вещества на составляющие их группы атомов,
радикалы и т. д. В обоих случаях одинакова логика анализа. Но перед
генетиком стоят значительно большие трудности, нежели перед химиком,
ибо первый не может непосредственно выделить из генотипа отдельные
гены. Это выделение может быть сделано только теоретически, на основе
анализа результатов скрещиваний. Химик же может разложить
вещество на составляющие его компоненты и в ряде случаев оперировать с
этими отдельными компонентами. Сопоставляя законы Менделя с законом
кратных отношений Дальтона, А. С. Серебровский также приходит к
выводу об общности методических подходов генетики и химии к вопросам
анализа.
332
План монографии очень прост и строго логичен.
Первая часть посвящена генетическому анализу качественных
(альтернативных) различий, вторая — количественных (трансгрессивных).
Анализ качественных различий начинается с простейшего случая —
моногибридного расщепления, затем автор переходит к дигибридному,
подробно разбирает различные осложнения дигибридного расщепления и
методы локализации гена в хромосоме. В последующих главах первой
части книги даны основы анализа популяций (также по качественным
признакам), методы анализа генотипа особи, методы топографического
анализа генотипа и генофонда популяций в целом. Во второй части
книги, где речь идет об анализе количественных различий, существенно
выделение трех методов: менделистического, алгебраического и метода
сигналей.
Главы, посвященные моногибридному и дигибридному скрещиваниям,
кажутся элементарными, но они будут очень полезными для целей
преподавания, особенно учитывая, что до сих пор нет сборника генетических
задач. Особое место в этом отношении занимает глава о составлении и
решении задач по генетическому анализу..
В то же время приводимые в книге детальные расчеты при
моногибридном и дигибридном наследовании демонстрируют, что по многим
причинам возможны многочисленные отклонения от классических
отношений, выводимых на основе закономерностей элементарного менделизма.
Для тех, кто работает с сельскохозяйственными животными,
существенно влияние на получаемые при расщеплении отношения малого числа
потомков в отдельных семьях.
Очень хорошо показано, какое значение имеет тщательный анализ
отдельных скрещиваний в популяциях и какие возможности открывает
популяционный анализ. Несмотря на то, что сельскохозяйственных
животных разводят с применением определенных методов отбора и подбора,
по очень многим признакам, по которым отбор не проводится,
скрещивания в популяции носят случайный характер. Поэтому к ним вполне при-
ложимы расчеты по формуле Харди — Вейнберга.
Материал главы VII, посвященной топографическому анализу
генотипа, может быть использован только в специальных исследованиях,
например, на дрозофиле. Но поразительна тонкость аналитического
подхода автора, например, когда знакомишься с предлагаемым им методом
анализа нехваток. Даже в этом узко специальном вопросе А. С. Сереб-
ровский верен себе. Он тщательно продумывает такую схему опытов,
которая позволила бы получать искусственные нехватки и их
анализировать.
В 30-х годах А. С. Серебровский пропагандировал идеи так
называемой геногеографии, разработал ее методы и сам провел несколько гено-
географических исследований. К сожалению, эти методы сейчас
основательно забыты. Будем надеяться, что прочтение разделов книги, где
говорится о геногеографии, вновь возбудит интерес к ней, и что будут
использованы многие предложенные А. С. Серебровским методы, в том числе
графические.
Большой интерес вызывают главы IX и последующие, в которых идет
речь о методах анализа количественных признаков, или, как совершенно
правильно пишет А. С. Серебровский, количественных различий.
Известно, что именно к этим вопросам перешло сейчас внимание генетиков, что
и отразилось в появлении монотрафихг таких авторов, как Фолконер,
Манер, Лернер и других (см. список литературы). Нельзя не отметить
оригинальности подхода А. С. Серебровского, в частности, в разработанном
им алгебраическом методе.
Очень ценны в познавательном отношении схемы анализа
наследования количественных признаков при различных скрещиваниях, в том
333
числе при разведении в себе первого или второго поколений или
проведении обратных скрещиваний на одну из исходных форм. Такие
скрещивания реально осуществляются в животноводческой практике при
создании различных гибридных форм животных или при выведении новых
пород.
С интересом прочтет современный читатель и раздел об определении
числа генов, которыми различаются скрещиваемые формы, хотя при
этом неизбежны некоторые упрощения. Так, очевидно, что разные
гены обладают разной силой действия на количественные признаки, но
можно оперировать со средней силой действия генов, введя при этом
новый параметр — характеристику вариации силы действия генов.
Интересно разобраны автором в 9 главе вопросы шкалирования
количественных признаков. Наметить правильную шкалу вариации
изучаемого признака совсем не так просто, как это может казаться с самого
начала, и этому вопросу уделяется особое внимание (см. Falconer, 1961).
Лица, лично знавшие А. С. Серебровского, хорошо помнят его любовь
к созданию терминов, а также к переводу их на русский язык. Нет
сомнения, что в любой науке надо создавать такие термины, с помощью
которых можно достаточно точно выделять определенные явления или
предметы и отличать их друг от друга. Надо сказать, что в некоторых частях
книги терминология несколько сложновата. Без некоторых терминов,
особенно из числа приведенных во вводных главах, можно было бы
обойтись. Но за новыми терминами всегда скрываются ценные и интересные
мысли, например, что генотип предшествующего поколения, через
фенотип матери, является как бы внешним фактором по отношению к данному
поколению. Ценно также, вне зависимости от того, надо ли вводить
новые термины или нет, указание, что надо различать генотипные различия,
то есть различия между генотипами, и генотипические, т. е. различия
между фенотипами, обусловленные генотипными различиями. В
современной литературе употребляют термины «генетические различия» и
«генотипические различия». Иногда также говорят о «генотипически
обусловленных различиях».
Хотя А. С. Серебровский писал свою монографию в расчете на то,
что ею будет пользоваться широкий круг читателей — генетиков и
селекционеров, но нельзя сказать, что все ее части написаны одинаково
.доступно и просто. Особенно трудны расчёты уравнений кривых
номограмм для определения типов скрещиваний (гл. 5), однако мы не сочли
возможным изменять или упрощать текст автора. В некоторых случаях
автор пропускает промежуточные звенья при расчетах, что также
затрудняет чтение. Однако единственный совет читателю, желающему осно-
»вательно разобраться в материале книги и, наконец, его усвоить — это
5итать книгу только с карандашом в руках, самому выполнять все
расчеты и таким образом следить за ходом мысли автора. Очень многие
методические приемы, примененные А. С. Серебровским, например,
использование номограмм, графиков, структура некоторых таблиц, могут
быть широко использованы в практике генетической и селекционной
работы.
В дополнении помещены 3 статьи математиков М. В. Игнатьева и
В. Гливенко и работа А. С. Серебровского «Метод треугольника».
В статье «Некоторые приемы анализа геносвязи» М. В. Игнатьев показал
значение критерия хи - квадрат для оценки степени соответствия
фактически получаемых частот теоретически ожидаемым при сцепленном
наследовании. В настоящее время метод хи-квадрат общепринят и
излагается в любой сводке по генетике и селекции, а также в книгах по
статистике (см. например Рокицкий П. Ф. Биологическая статистика. Изд-во
«Вышэйшая школа». Минск, 1987). Двадцать лет тому назад это было
новшеством. Мы Поместили статью М. В. Игнатьева имея в виду, что
334
приведенный в ней разбор применения критерия хи-квадрат к нескольким
случаям расщепления будет полезен при прохождении курса генетики.
Несколько особняком стоит вторая статья М. В. Игнатьева. В ней
говорится о применении разработанного Фишером метода максимального
правдоподобия для анализа сцепленного наследования. Примерно в том
же виде изложен этот вопрос и в монографии по математической генетике
Э. Вебер (Weber E., 1967).
Небольшая заметка математика В. Гливенко — попытка выразить
алгебраически некоторые генетические закономерности. В более широком
плане подошли к этому А. А. Ляпунов и А. Г. Маленков в статье
«Логический анализ строения наследственной информации» («Проблемы
кибернетики», 8, 1962).
Метод треугольника разрабатывался А. С. Серебровским в течение
ряда лет. Хотя он прямо относится к анализу количественных признаков,
но автор не счел возможным включать статью об этом методе
непосредственно в текст монографии ввиду его своеобразия и самостоятельного
значения. Основное в нем заключается в том, что результаты многих
скрещиваний наносятся на поле, в котором ось абсцисс обозначает значения
варианс, а ось ординат — значения средних арифметических. По
расположению эмпирических точек, каждая из которых соответствует
определенному значению обоих величин для результатов скрещивания, можно
сделать ряд выводов о числе и о характере действия (сила,
доминирование) генов, которыми различаются скрещиваемые формы.
Первоначальная схема кажется очень простой и ясной. Однако детальный анализ
многочисленных случаев наследования с участием разного количества генов,
различий в их доминировании и силе действия показал, что
распределение точек в поле может быть очень причудливым и в нем нелегко
разобраться. Может быть, последнее обстоятельство и явилось причиной того,
что метод треугольника применяли очень немногие исследователи,
преимущественно из числа лиц, связанных с А. С. Серебровским. Будем
надеяться, что после опубликования монографии этот интересный метод
станет достоянием многих генетиков и селекционеров, тем более, что поле
х — а2 может быть использовано и для других целей, например для
изучения процесса отбора, при анализе межгрупповых, межсортовых или
межпородных различий и т. д. Интересно также применение А. С.
Серебровским метода моделирования.
Монография была написана А. С. Серебровским почти 30 лет тому
назад. За это время по многим вопросам статистической генетики
достигнут немалый успех, что отразилось в опубликовании значительного
числа ценных монографий, список которых дан ниже. Из их числа надо
особо назвать большие монографии Дубинина Н. П. иТлембоцкого Я. Л.
(1967), Фолконера (Falconer, 1960), Кемпсорна (Kempthorn, 1957), Мазера
(Mather, 1964), и, наконец, Э. Вебер (Weber, 1967). Последняя книга,
носящая многозначительное название «Математические основы генетики»,
представляет собой наиболее полную современную сводку по
математической или статистической генетике. Последнее на наш взгляд более
правильно, так как основную~долю1в1кей занимают именно статистические
~1шдходйГ и методы. В большинстве указанных в списке книг отражены
и те вопросы, о которых говорится в «Генетическом анализе».
Подробное сравнение их содержания с содержанием монографии
А. С. Серебровского заняло бы слишком много места, поэтому
ограничимся лишь кратким разбором того, в каких направлениях протекало
развитие вопросов генетического анализа после работы А. С. Серебровского.
jjpежд,е_всего изменились акценты —. многое жз того, что казалось л
прежде важным и даже основным, отошло на задний план, наоборот,
выдвинулись на первое место другие вопросы. Наконец, появились новые
подходы и методы. В монографии А. С. Серебровского очень большое
335
внимание уделено генетическому анализу моногибридного и дигибридного
расщеплений и локализации генов. Само собой разумеется, конечно, что
это — арифметика генетики, которую надо знать каждому генетику и
селекционеру, однако удельный вес этих вопросов в конкретных
исследованиях сравнительно невелик. Кроме того, многие из задач по анализу
расщеплений решаются в более общем виде, на основе вероятностных
расчетов. |^^Шнную_и хорошо развитую область генетики превратилась
^ популяционная генетика^ что, в частности, выразилось в появлении
нескольких самостоятельных руководств (Дубинин Н. П., Глембоцкий
Я. Л., 1967; Lerner, 1950, 1955; Rasmuson, 1961). Кроме того, во всех
генетических и селекционных сводках популяционной генетике всегда
отводится большое место.
Еще большего развития достигла генетика количественных
признаков, причем на путях, несколько отличных от того, о чем писал А. С.
Серебровский. Общая часть анализа количественных признаков, а именно
характеристика обусловленности количественных различий многими
генами, осталась примерно той же, как она изложена, при этом
достаточно полно, в монографии. Однако произошла переоценка роли отдельных
методов. А. С. Серебровский считал наиболее совершенным методом
анализа количественных признаков_метод сигнальных генов.
Приведенные в монографии примеры такого анализа, действительно, очень
изящны и красивы. JHo^^eTo^ сигнал ей все же не нашел широкого применения,
прежде всего потому, что у большинства видов животных и растений,
количественные признаки которых имеют хозяйственное значение, как
правило, неизвестны маркированные хромосомы. Впрочем, попытки
установить "связь между наследованием отдельных качественных и
количественных признаков по-прежнему продолжаются. Большие надежды в этом
отношении возлагаются на иммуногенетику. Так как открыто много
независимых друг от друга генов, определяющих химические свойства
крови (а также и другие химические свойства) у животных, есть надежда,
что некоторые из них окажутся связанными (в порядке ли плейотропного
действия или тесного сцепления двух генов) с определенными
количественными хозяйственно-полезными признаками. Некоторые факты этого
рода получены на курах, свиньях.
Существенное значение для анализа вариации количественных
признаков в популяциях и установления ее причин сыграло использование
нового генетического параметра — коэффициента наследуемости. Идею
этого параметра можно найти в ранних работах американского генетика
jC.JPaHTa, большинство из которых было опубликовано еще при жизни
А. С. Серебровского. Странным образом А. С. Серебровский не уделил
им внимания. В монографии он упоминает и то вскользь, только о
величине а7/6ф, названной Райтом коэффициентом корреляции между
генотипом и фенотипом.
Ни слова нет и о т. н. путевых коэффициентах Райта. По-видимому,
логика исследовательской мысли А. С. Серебровского шла иными
путями, поэтому построения Райта были ему чуждыми. В настоящее время
коэффициент наследуемости, обозначаемый символом h2 и выражающий
долю генотипической вариансы Он в общей фенотипической вариансе ар
2
данного количественного признака (т.е. h2 = -^ |, занимает значительное
V 4 /
место во всех сводках и монографиях по генетике, в т. ч. и указанных в
прилагаемом списке литературы. Конкретные же Есследсванвя с
использованием коэффициента наследуемости, проведенные не только на
животных, но и на растениях, исчисляются многими сотнями.
Разработаны далее математические методы, с помсшью ксто{ых в
генотипической вариации признака, оцениваемой ая» могут быть выделены
336
отдельные компоненты, зависящие от аддитивного действия генов (а^),
от аллельного взаимодействия, т. е. от эффекта доминирования (a2D) и,
наконец, от взаимодействия неаллельных генов, получившего общее
наименование эпистатического взаимодействия (of).
Были разработаны и некоторые другие генетические параметры,
например, для установления генетической связи между разными признаками
(коэффициенты генетической корреляции в дополнение к уже известным
ранее коэффициентам фенотипической корреляции). Теоретически и
экспериментально была поставлена проблема т. н. взаимодействия
генотипа и среды, иначе говоря зависимости между различиями во внешней
среде и фенотипическими свойствами генетически отличающихся
организмов.
Сказанное выше отнюдь не умаляет значения монографии А. С. Се-
ребровского как важного этапа в разработке методов генетического
анализа, этапа в очень большой степени оригинального и самобытного.
Ознакомление с творческой лабораторией А. С. Серебровского,
овладение его подходами и методами будет очень полезно для повышения
теоретического и методического уровня современного читателя —
генетика и селекционера.
Академик АН БССР П. Ф. Рокицкий
15 ноября 1968 г.
НОВЕЙШАЯ МОНОГРАФИЧЕСКАЯ ЛИТЕРАТУРА
Бр юбейкер Дж. Л. 1966. Сельскохозяйственная генетика. М., изд-во «Колос».
Дубинин Н. П., Глембоцкий Я. Л. 1967. Генетика популяций и селекция.
М., изд-во «Наука».
Л е п е р П. Р., Н и к о р о 3. С. 1966. Генетико-математические основы различных
методов оценки племенных качеств животных. Новосибирск, изд-во «Наука».
Плохинский Н. А. 1964. Наследуемость. Новосибирск, изд-во «Наука».
Руководство по разведению животных. 1963. Т. II. Генетические основы
продуктивности и селекции. М., Сельхозгиз.
Теоретическая и математическая биология. 1968. М., Изд-во «Мир».
Фишер Р. А. 1958. Статистические методы для исследователей. М., Госстатиздат.
Biometrical Genetics. 1960. Ed. by 0. Kempthorne. London, Oxford, N. Y., Paris, Per-
gamon Press.
Falconer D. S. I960.. Introduction to Quantitative Genetics. Edinburgh and
London, Oliver a. Boyd.
Fisher R. A. 1965, The Theory of Inbreeding. 2nd ed. Edinburgh and London, Oliver
a. Boyd.
Hogben, Lancelot, 1946. An Introduction to Mathematical Genetics. N. Y.
Norton <& Co.
Kempthorne 0. 1957. An Introduction to Genetic Statistics. N. Y., Wiley & Sons.
Lerner J. M. 1950. Population Genetics and Animal Improvement. Cambridge
Univ. Press.
Lerner J. M. 1954. Genetic Homeostasis. N. Y., Wiley & Sons.
Lerner J. M. 1958. The Genetic Basis of Selection. N. Y., Wiley & Sons.
Li С. С 1955. Population Genetics. Univ. Chicago Press.
Lush J. L. 1945. Animal Breeding Plans. Ames, Iowa. Iowa State Coll. Press.
Malecot G. 1947. Les Mathematiques de THeredite. Paris.
Mather K. 1949. Biometrical Genetics. London, Methuen.
Mather W. B. 1964. Principles of Quantitative Genetics. Minn., USA, Burgess Publ.
Col.
Mather A., 1966. Statistical Analysis in Biology. 5th ed. London, Methuen.
Rasmuson M. 1961. Genetics on the Population Level. Stockholm, Swensk Bokfur-
laget.
Statistics and Mathematics in Biology. 1954. Ed. by Kempthorne 0. and others. Ames,
Iowa. Iowa State College.
Weber E. 1967. Mathematische Grundlagen der Genetic. Jena, G. Fischer Verlag.
ОГ ЛАВ ЛЕНИ Е
Предисловие 3
Введение 5
ЧАСТЬ ПЕРВАЯ!
ГЕНЕТИЧЕСКИЙ АНАЛИЗ КАЧЕСТВЕННЫХ
(АЛЬТЕРНАТИВНЫХ) РАЗЛИЧИЙ
Г л а в а I. Моногибридное расщепление. Установление гена 14
Число генов в расщеплении 14
Изучение отдельного гена 14
Априорные моногибридные формулы 15
Апостериорные моногибридные формулы 16
Переход от апостериорных расчетов к априорным 18
Поправка Вейнберга 18
Таблицы и пользование ими 19
Значение FB и F2 22
Искаженна моногибридных отношений 22
1. Случайность 22
2. Различная жизнеспособность 24
3. Трансгрессия . . : 25
4. Участие других генов 26
5. Ошибки в классификации родителей 26
6. Фиктивные менделевские отношения 27
Идентификация генов 28
Глава II. Дигибридное расщепление 29
Значение дигибридного расщепления 29
Достоинства и недостатки двух дигибридных скрещиваний . . ^ 30
Различные случаи дигибридных расщеплений 32
1. Простая кооперация 32
2. Взаимная модификация 34
3. Криптомерия 35
4. Криптомерия с новообразованием 36
5. Эпистаз 37
6. Эпистаз и криптомерия . . 37
7. Парные или дополнительные гены 39
8. Повторные гены 40
9. Димерия, однозначные гены 41
Глава III. Составление и решение задач по генетическому анализу 46
Неограниченные задачи 48
1. Пол „ - 48
2. Расщепление 1:1.., 49
3. Расщепление 1:2:1 49
339
4. Ди- и полигибридные расщепления 49
5. Неполное проявление ! 50
6. Нежизнеспособность гамет 50
7. Нежизнеспособность зигот . . * 50
Глава IV. Локализация гена 59
Постановка вопроса 59
Локализация в половой хромосоме 59
Доказательства сцепленности с полом 65
Определение наличия геносвязи 67
Значение FB и F2 69
Локализация при отсутствии перекреста у одного из полов 71
Локализация при перекресте у обоих полов 73
Определение расстояний между генами 74
Проблема точного определения величины перекреста 78
1. Криптомерия . . 81
2. Использование Fs . . 83-
3. Метод произведений 86°
4. Метод коэффициента ассоциаций 88
5. Метод коэффициента корреляции 92
6. Дальнейшие проблемы ......: 92
7. Уточнение локализации 93
8. Установление и локализация деталей . . . ., г 94
Анализ аллелизма . 98
Установление множественного аллелизма 98
Частичный, или ступенчатый, аллелизм 102
Составление задач 103
Глава V. Генетический анализ в популяциях 105
Генетический эксперимент и стихийные скрещивания 105
Закономерности строения свободных популяций 106
1. Биномиальное распределение гомо- и гетерозигот (закон Харди —
Пирсона) 107
2. Устойчивость популяции . 108
3. Распределение генотипов в популяции, гетерозиготной по серии аллелей 109
4. Распределение генотипов в популяции по гену, сцепленному с полом., . . 110
5. Распределение генотипов в дигибридной популяции 111
Родительские группы 113
Приложение к анализу случаев с двумя фенотипами 115
1. Парные (дополнительные) гены в популяции 115
2. Эпистаз + криптомерия в популяции 118
3. Повторные (полимерные) гены. Особенности этого случая 121
Номограмма для определения типа скрещиваний по Кг и К3 123
Уравнения кривых номограмм для определения типа скрещивания 124
Определение доминирования 126
Построение и решение задач-моделей 132
Общие замечания 136
Особенности проблемы анализа в популяциях 139
Глава VI. Анализ генотипа особи 140
Вторая задача генетического анализа , . . . . 140
Качественный анализ состава генотипа 140
Анализ по потомству 143
Анализ путем скрещивания^ родителей с детьми (дети-анализаторы) " 143
Выбор приемов анализа 146
Проблема теории анализа по ветвям 150
Анализ гомозиготной части генотипа 153
340
Глава VII. Топографический анализ генотипа 157
Задачи топографического анализа генотипа 157
Определение числа групп сцепления и хромосом 157
Упрощение исследования • 159
Число групп сцепления и число пар хромосом 160
Составление плана хромосомы . . . . 16^
Анализ особенностей хромосомного аппарата 161
Анализ нехваток 162
Определение длины нехваток методом перекреста 164
Анализ делеций 165
Анализ инверсий 166
Анализ транслокаций 171
Определение меж транслокаций 174
Анализ дупликаций (удвоений) 179
Глава VIII. Генетический анализ генофонда популяций 182
Третья задача генетического анализа 182
Сбор материала 183
Значение инбредного материала для анализа популяций 184
Эксперименты 185
Анализ структуры популяции 187
Установление различия между популяциями и его реальность ...... 188
ЧАСТЬ ВТОРАЯ
ГЕНЕТИЧЕСКИЙ АНАЛИЗ КОЛИЧЕСТВЕННЫХ
(ТРАНСГРЕССИВНЫХ) РАЗЛИЧИЙ
Глава IX. Общие основы. Генетическая комбинаторика 194
Три метода генетического анализа количественных (трансгрессивных) различий 194
Комбинаторика полимеров 200
Возникновение биномиального распределения генотипов 202
Влияние различных генотипических элементов на Рх и р2 204
Влияние соотношения гено- и паратипической изменчивости на характер
распределения фенотипов 214
Г л а*в а X. Метод менделистический или изучение расщепления в потомстве 217
Условия применения 217
Анализ межрасовых различий 217
Первый прием. Скрещивание особей F2 с чистыми расами 218
Второй прием. Скрещивание особей F2 или FB с особями Fx ...... . 223
Третий прием. Инбридинг Fa или FB 227
Четвертый прием. Скрещивание самцов FB со многими самками F в . . . . 230
Анализ сцепления 234
Выводы '. 238
Пример определителя для скрещиваний с}¥2 X 9 9F2 239
Глава XL Метод алгебраический 242
Выделение генотипического компонента 242
Решения задачи о числе генов и силе их действия 244
Доминирование 246
Влияние распределения аллелей-усилителей 246
Использование у4 • • • 248
Заключительные замечания 249
Глава XII. Методы сигналей 252
Строение хромосомы как носителя полимерных генов 255
Сила гена и баланс хромосомы . 255
Аллосила и аллобаланс 255
Сигнальные аллосила и аллобаланс 257
341
Список условных обозначений 257
Приемы анализа при помощи сигналей .- . 258
Одна сигналь в хромосоме 258
Один полимер в хромосоме 258
Много полимеров и одна сигналь . . . ., 262
Две и более сигнали 265
Дальнейшее изучение участка разрывами . 268
Вариант М. Л. Карпа 269
Анализ нескольких количественных признаков одновременно 271
Литература , __. . . 274
ДОПОЛНЕНИЯ
Игнатьев М. В. Некоторые приемы анализа геносвязи 279
Игнатьев М. В. Метод максимальной вероятности 288
Гливенко В. Менделистическая алгебра 300
Серебровский А. С. Метод треугольника 302
Послесловие 332
Новейшая монографическая литература 338
Александр Сергеевич Серебровский
Генетический анализ
Утверждено к печати
Институтом общей генетики
Редакторы ДО. С. Демин, В. А. Волков
Редактор издательства 9. А. Фролот
Художник Г.' А. Астафьева
Технический редактор С. Г. Тихомирова
Сдано в набор 28/1 1970 г. Подписано к печати 22/VI 1970 г.
Формат 70Xl08Vit Бумага № 2 Усл. пёч. л. 30,1 Уч.-изд. л. 26,1
Тираж 5000. Т-10219 Тип. зак. 173
Цена 1 р. 75 к.
Издательство «Наука»
Москва, К-62, Подсосенский пер., 21
2-я типография иад-ва «Наука»
Москва Г-99, Шубинский пер., 10
ИСПРАВЛЕНИЯ
И ОПЕЧАТКИ
Страница
15
33
39
97
114
145
174
210
225
232
236
243
268
318
Строка
12 сн.
23 св.
16 св.
1 сн.
3 св.
4 сн.
Рис. 51
20 сн.
Табл. 19,
1 гр. ел., 3 сн.
1 сн.
Нижняя табл., 1
3 гр. ел. /
12 св.
1 св.
Табл. 25, 1 сн.
Напечатано
разные
ААВВ
АаВЬ
alB
2pqAA
утл
8
Da
54,5
2
ААЬЬ
14
114
460
Л
Аа
РЯ
Должно быть
равные
АаВВ
АаВВ
alb
2pqAa
Y4
<- D d
54,5
25
ААВЬ
2A
144
400
F*
ал
p=q
А. С. Серебровский