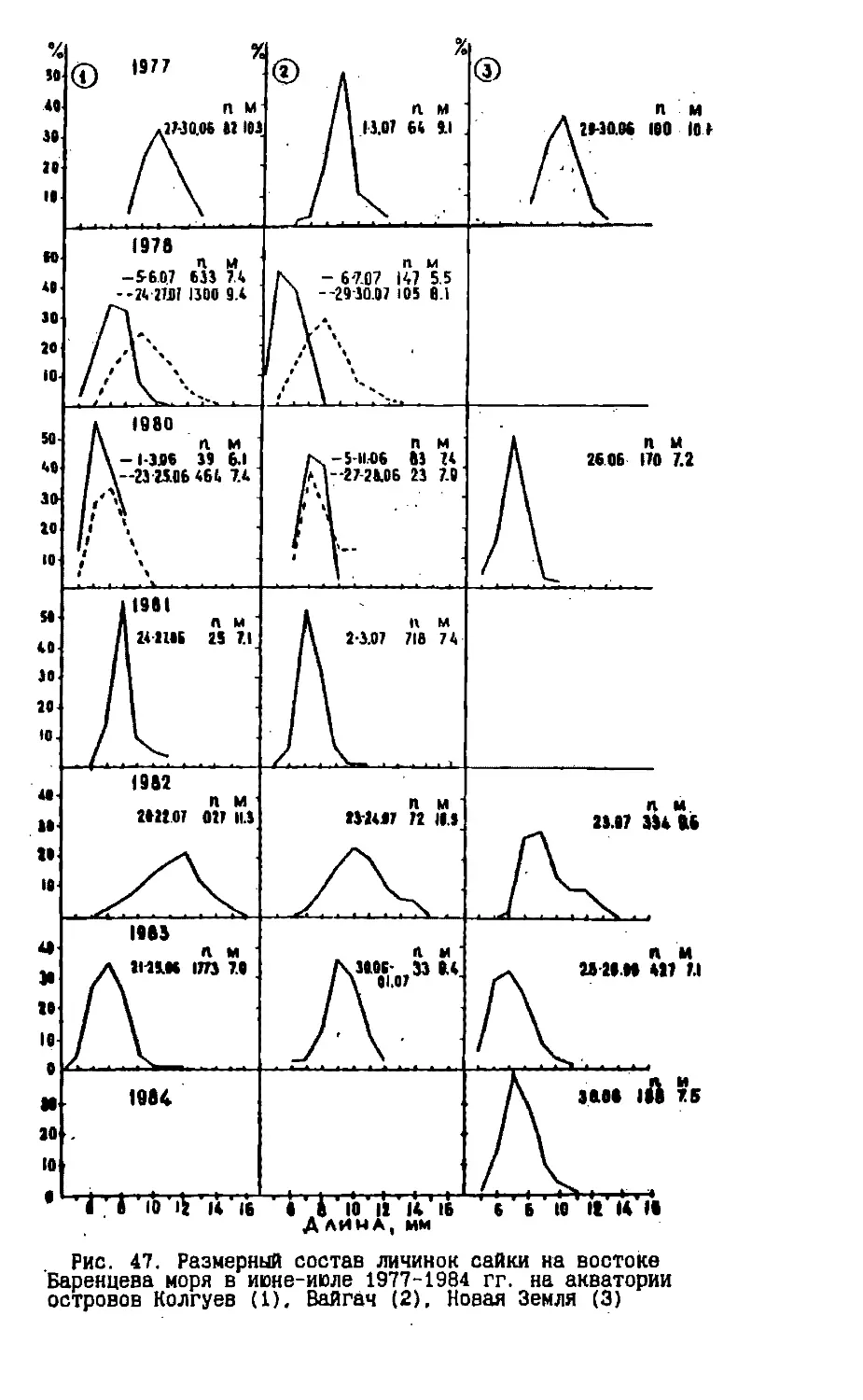
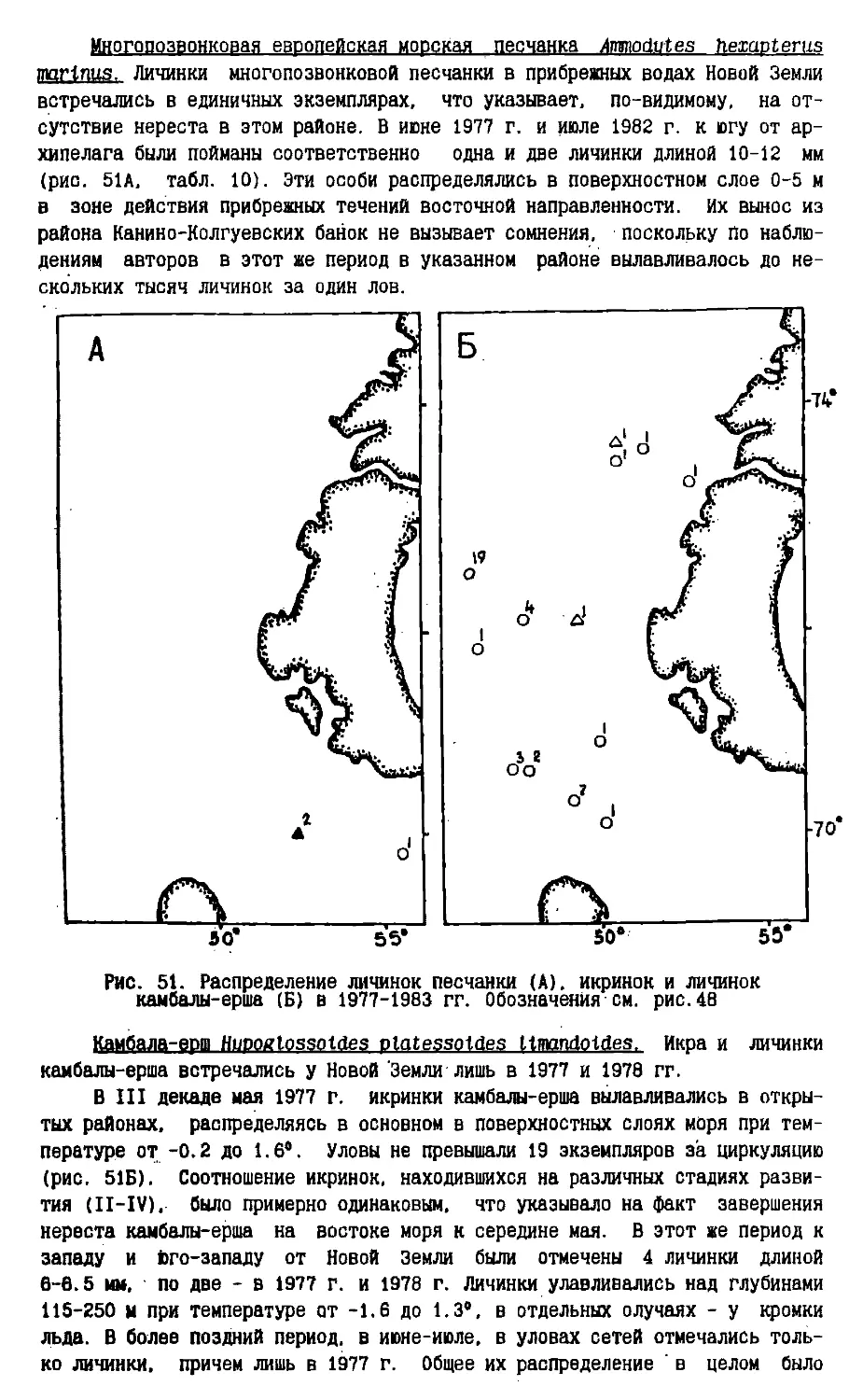
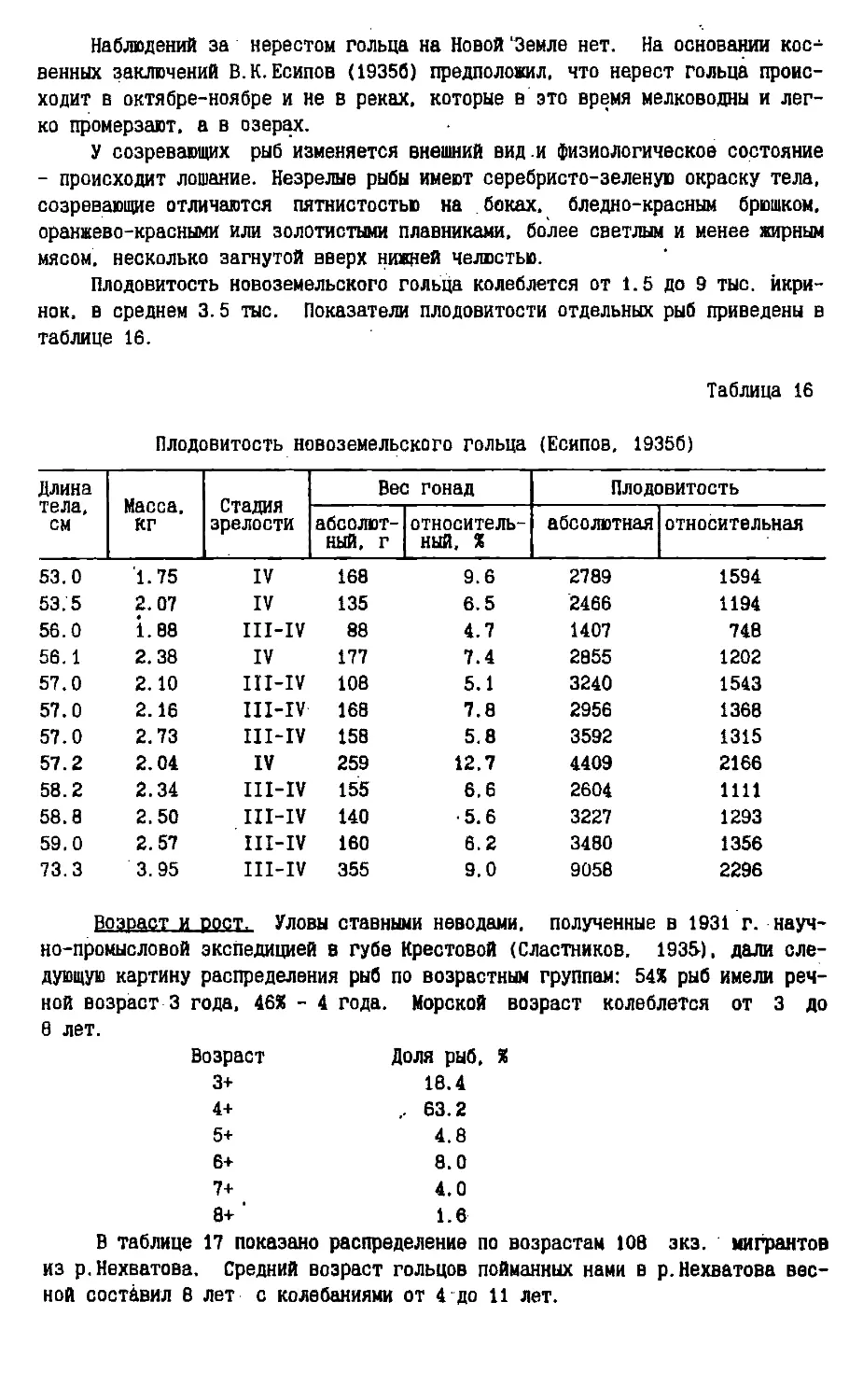
/



































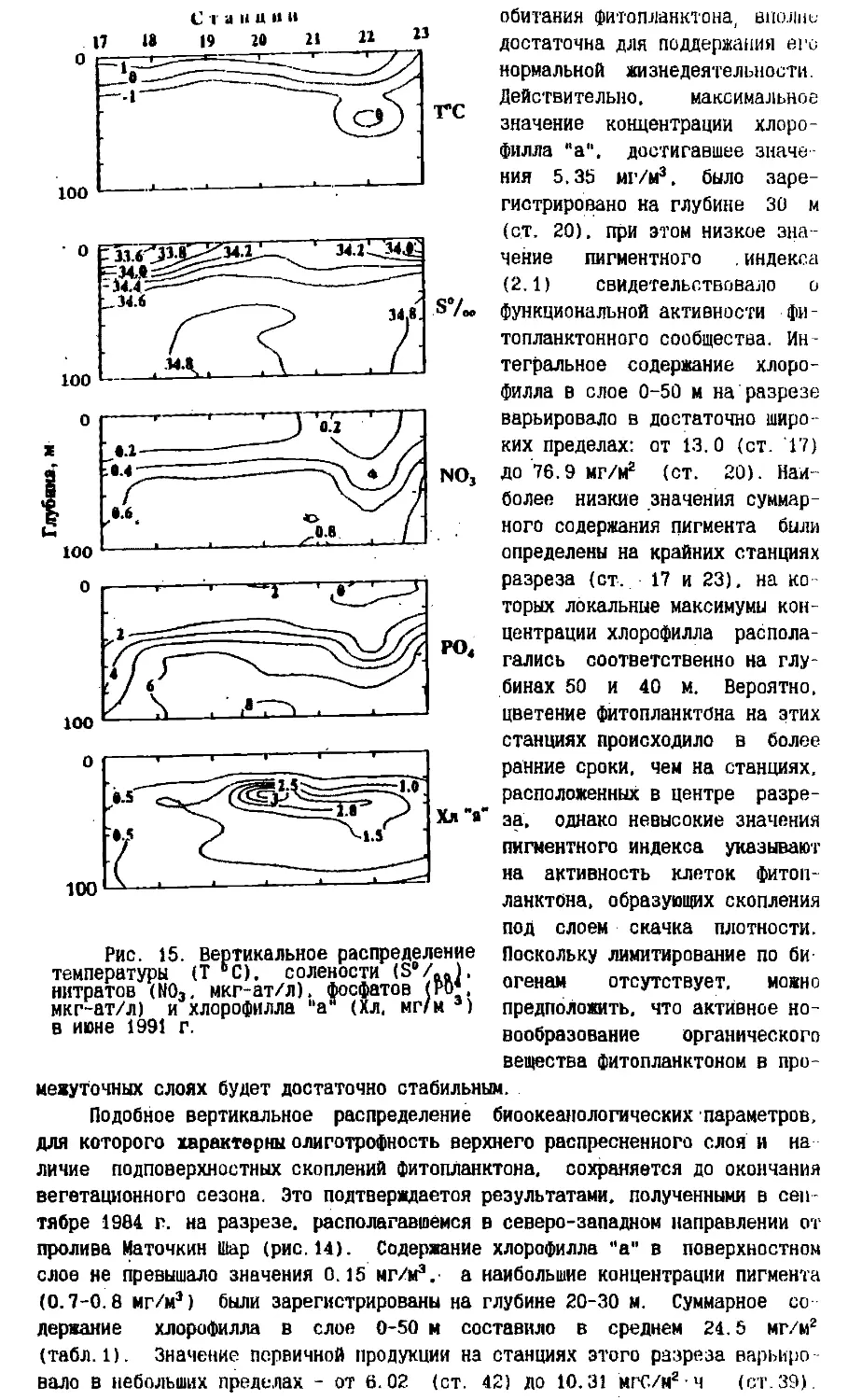



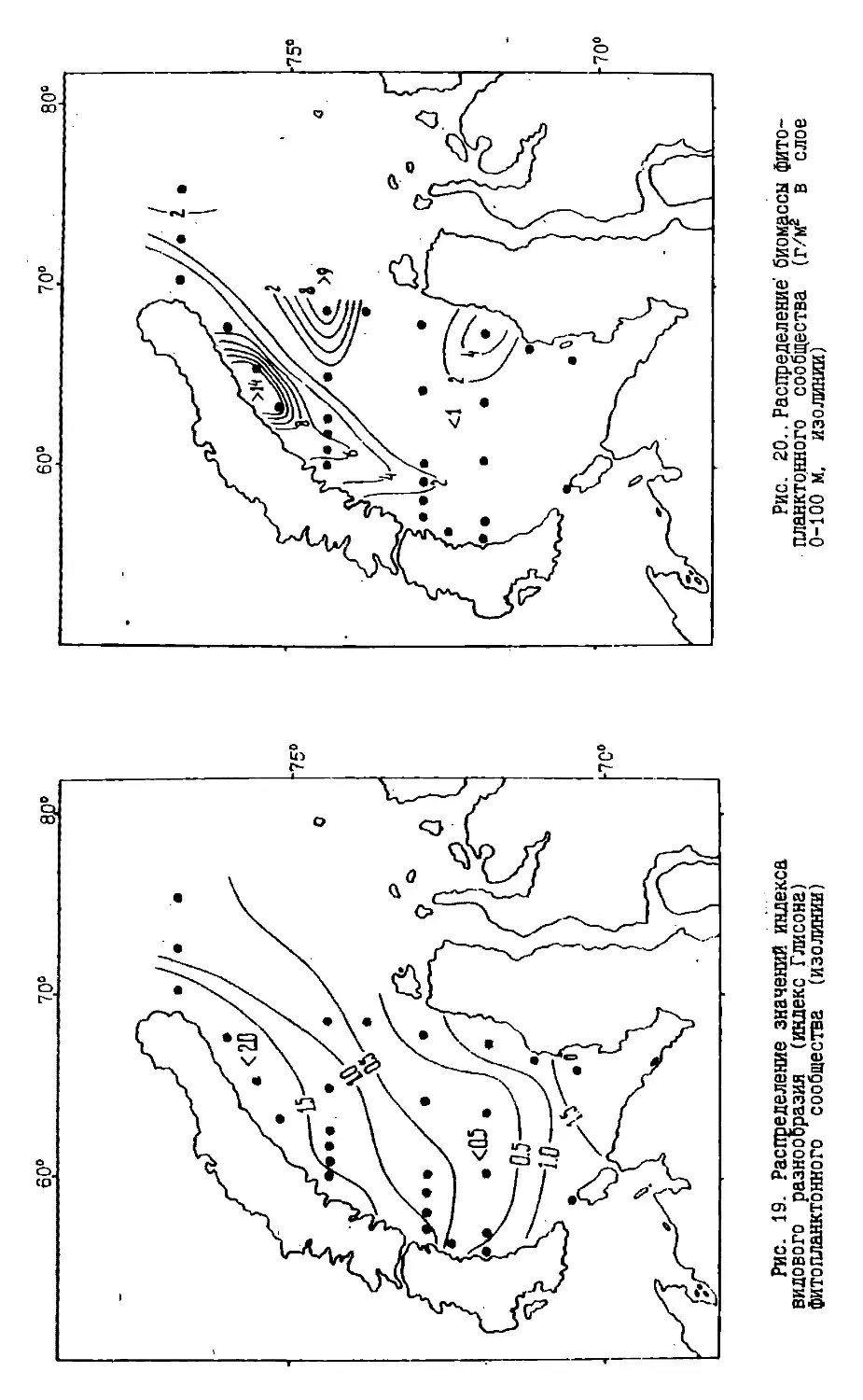
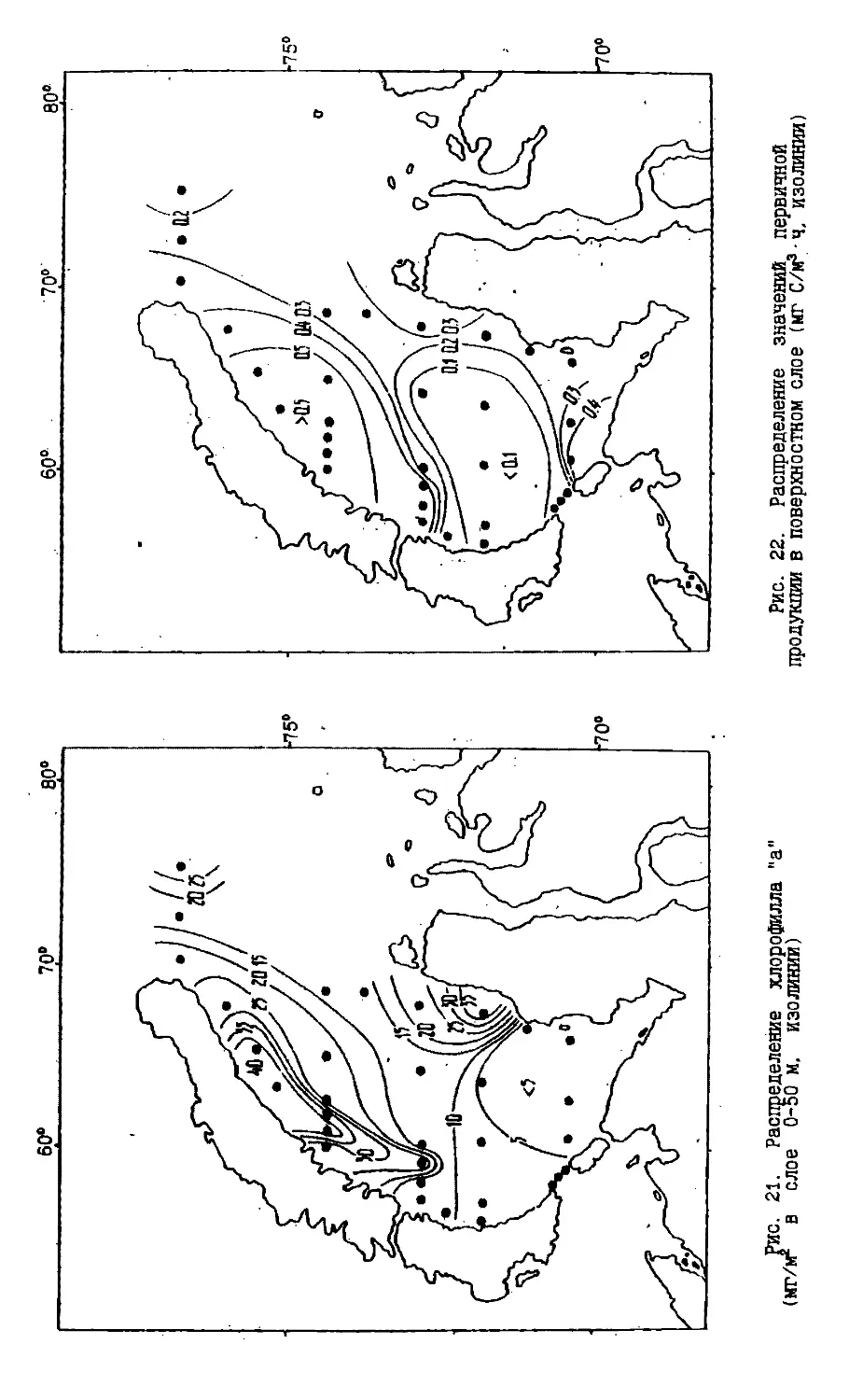

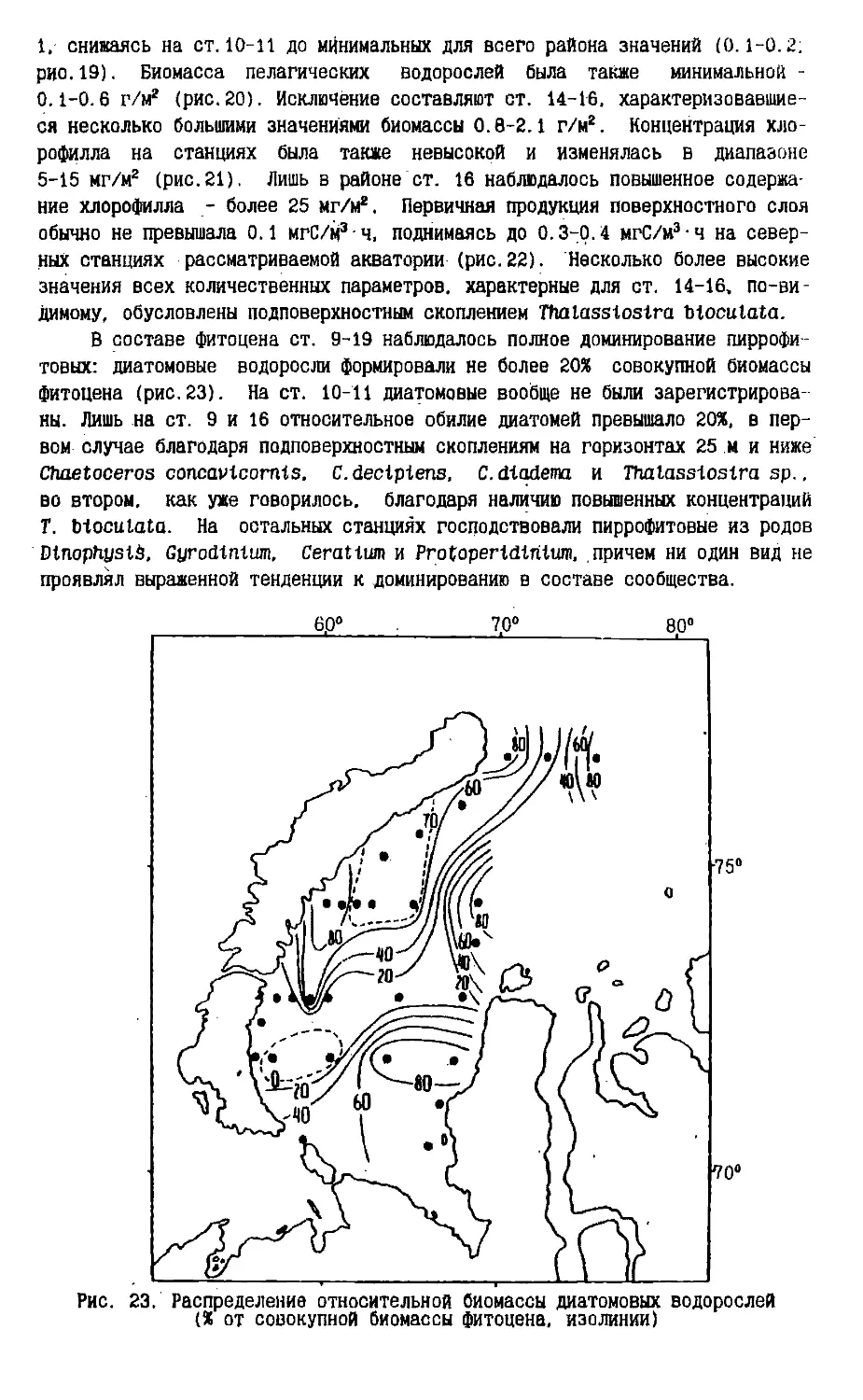








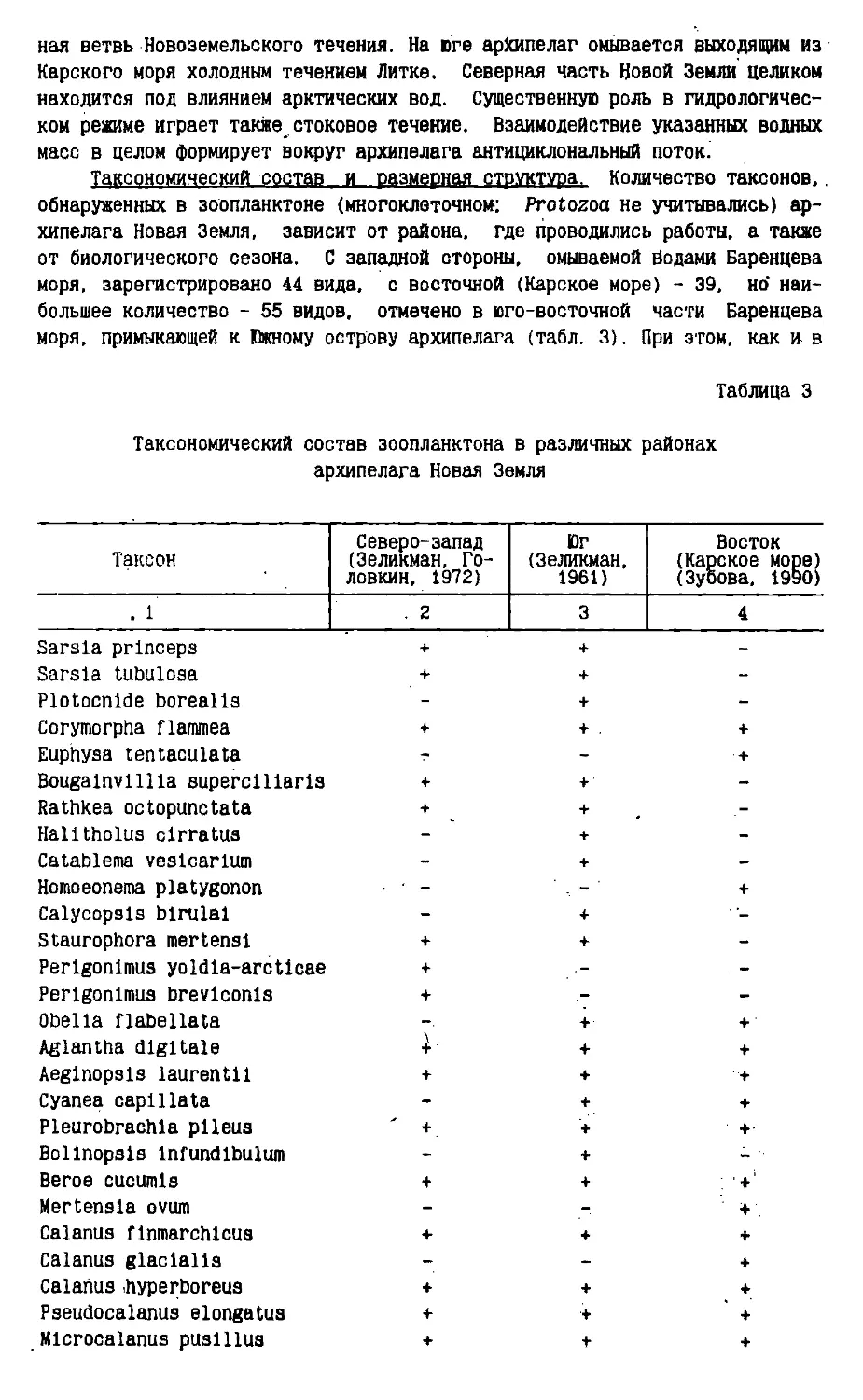

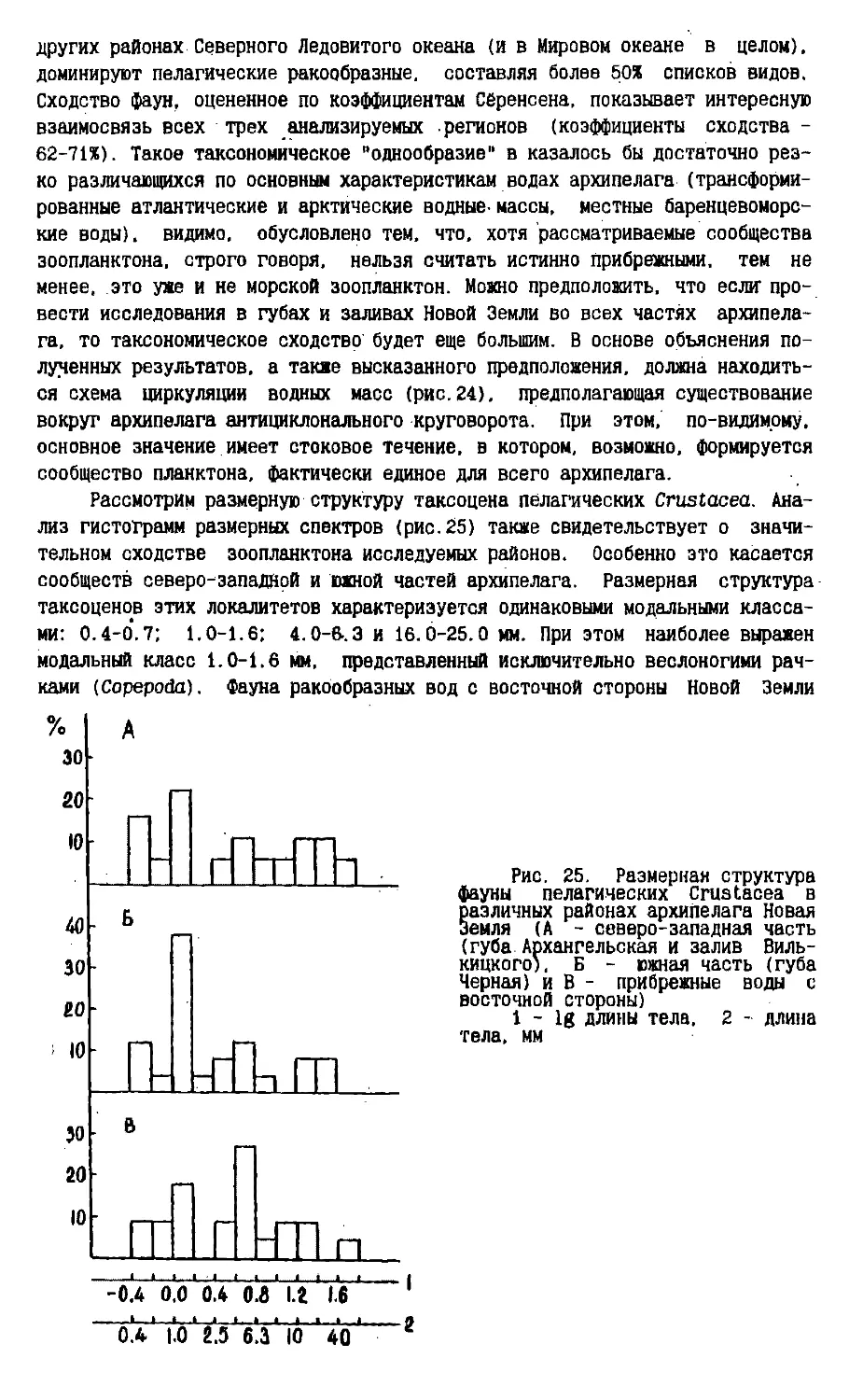





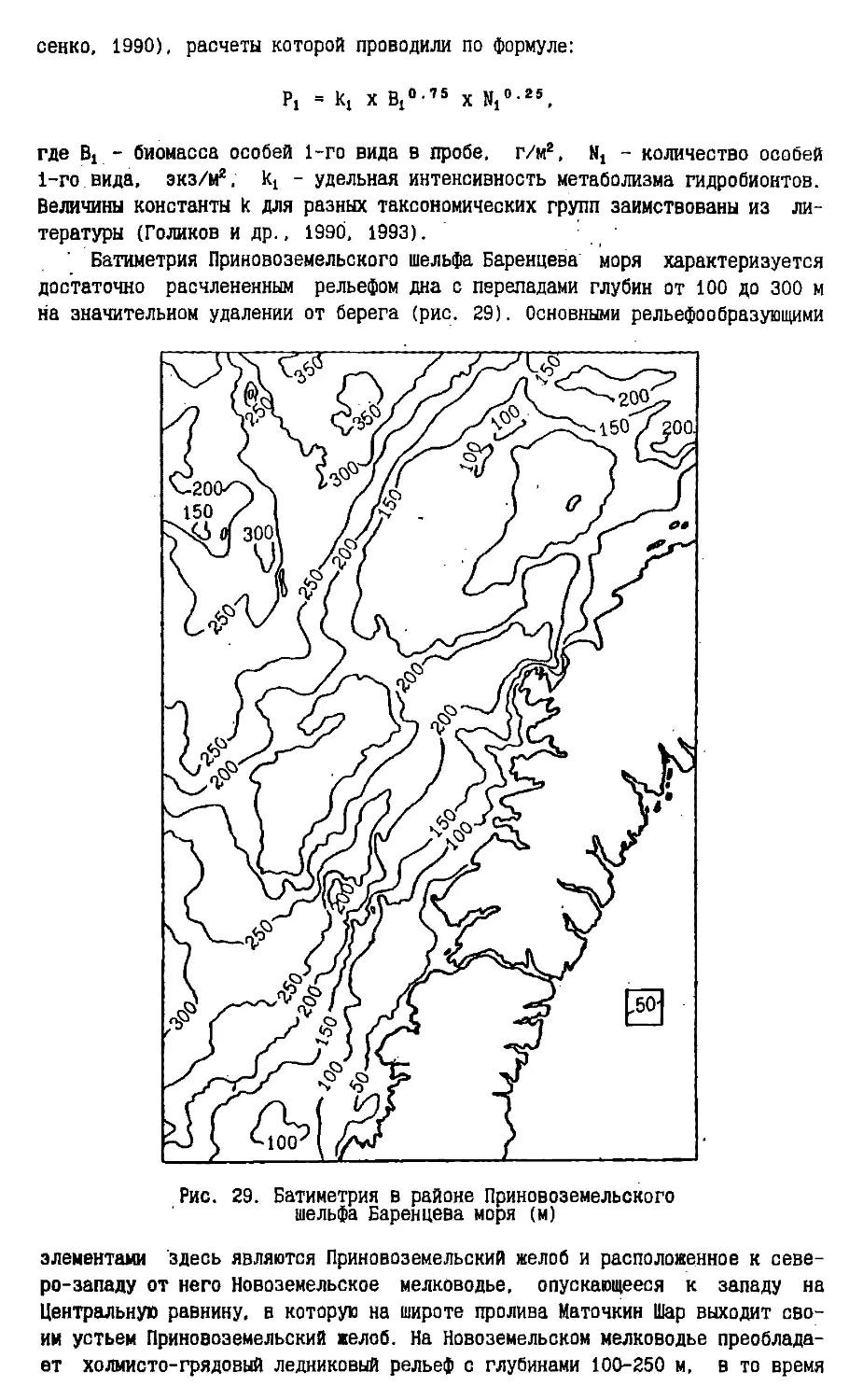


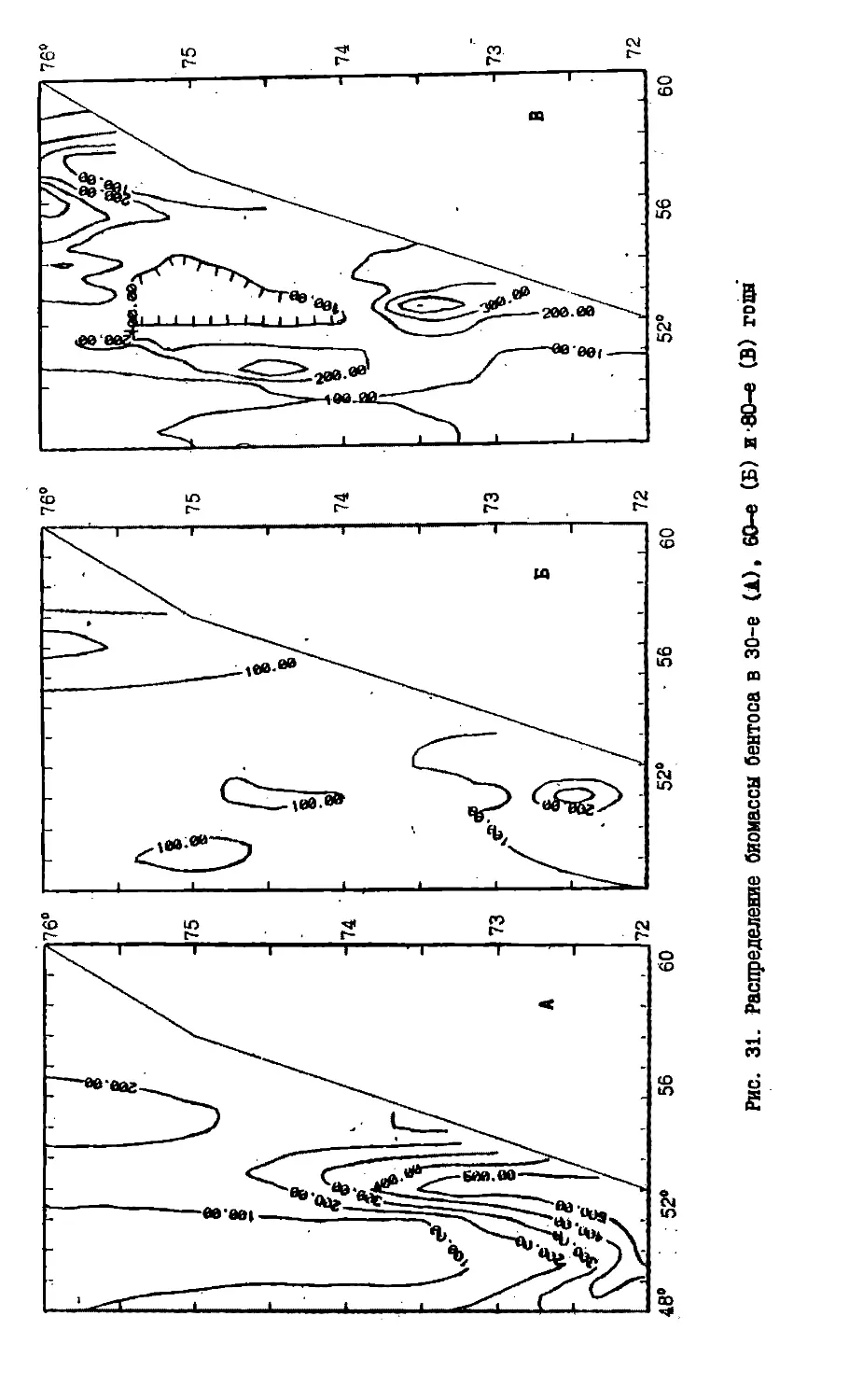

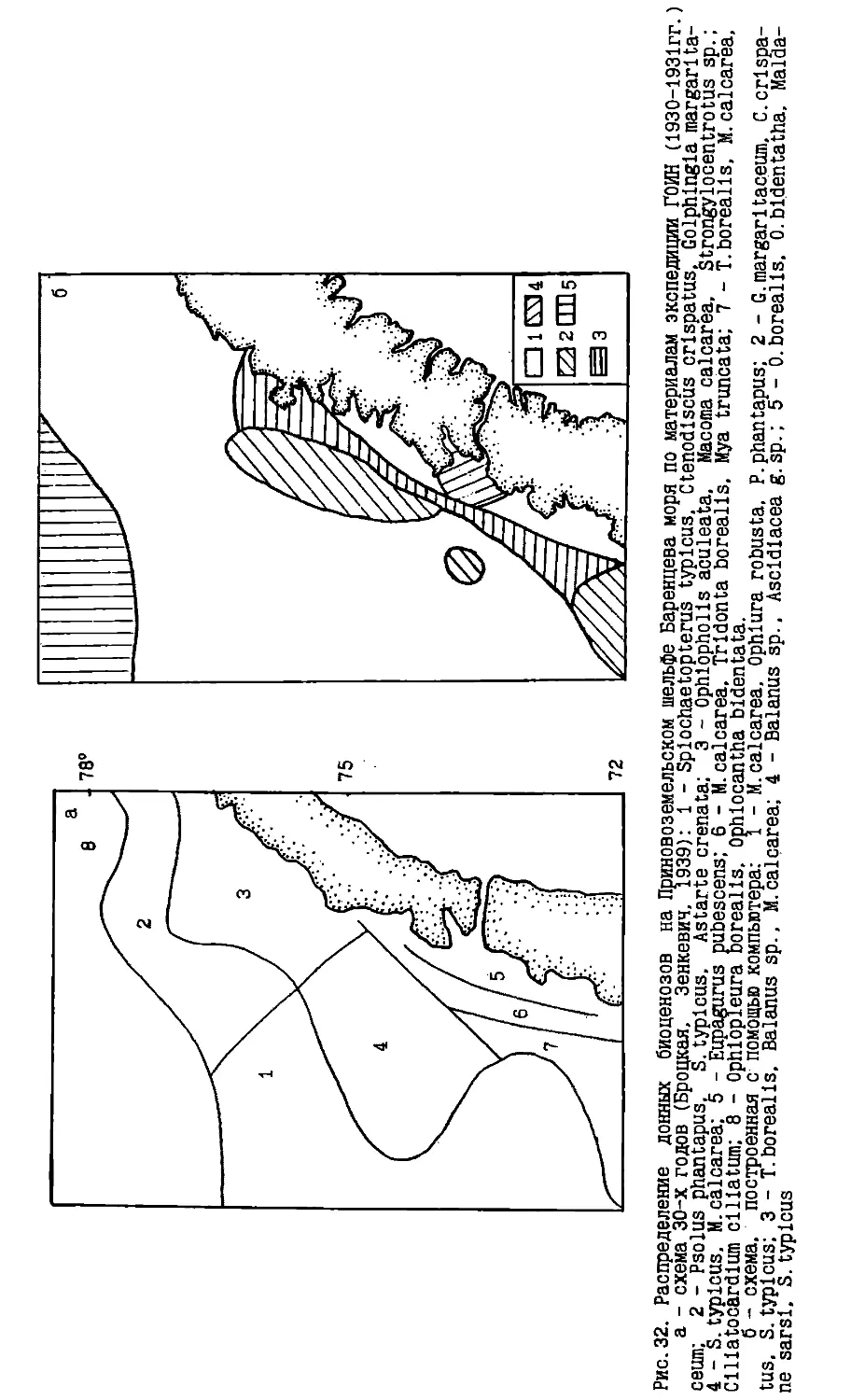


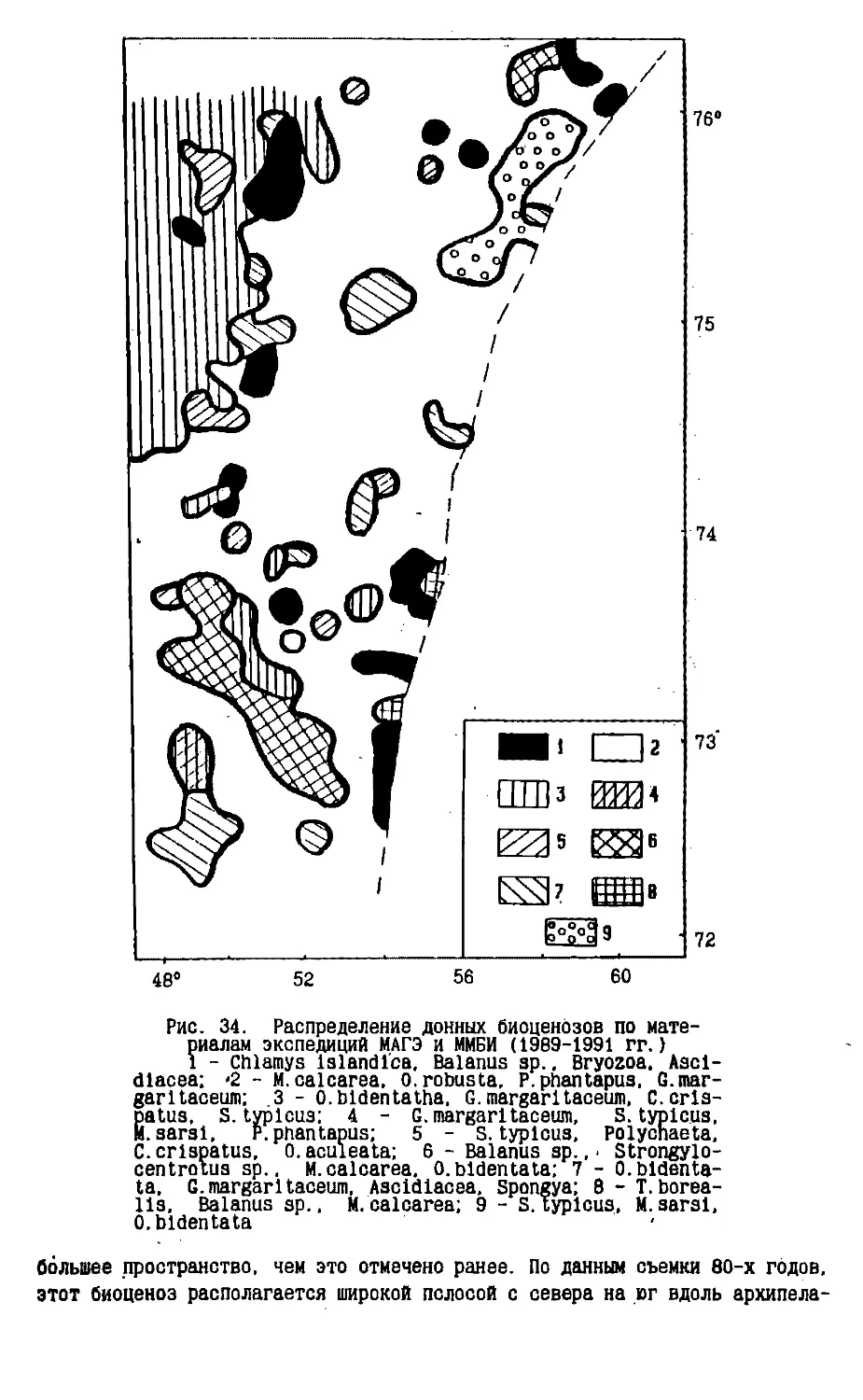









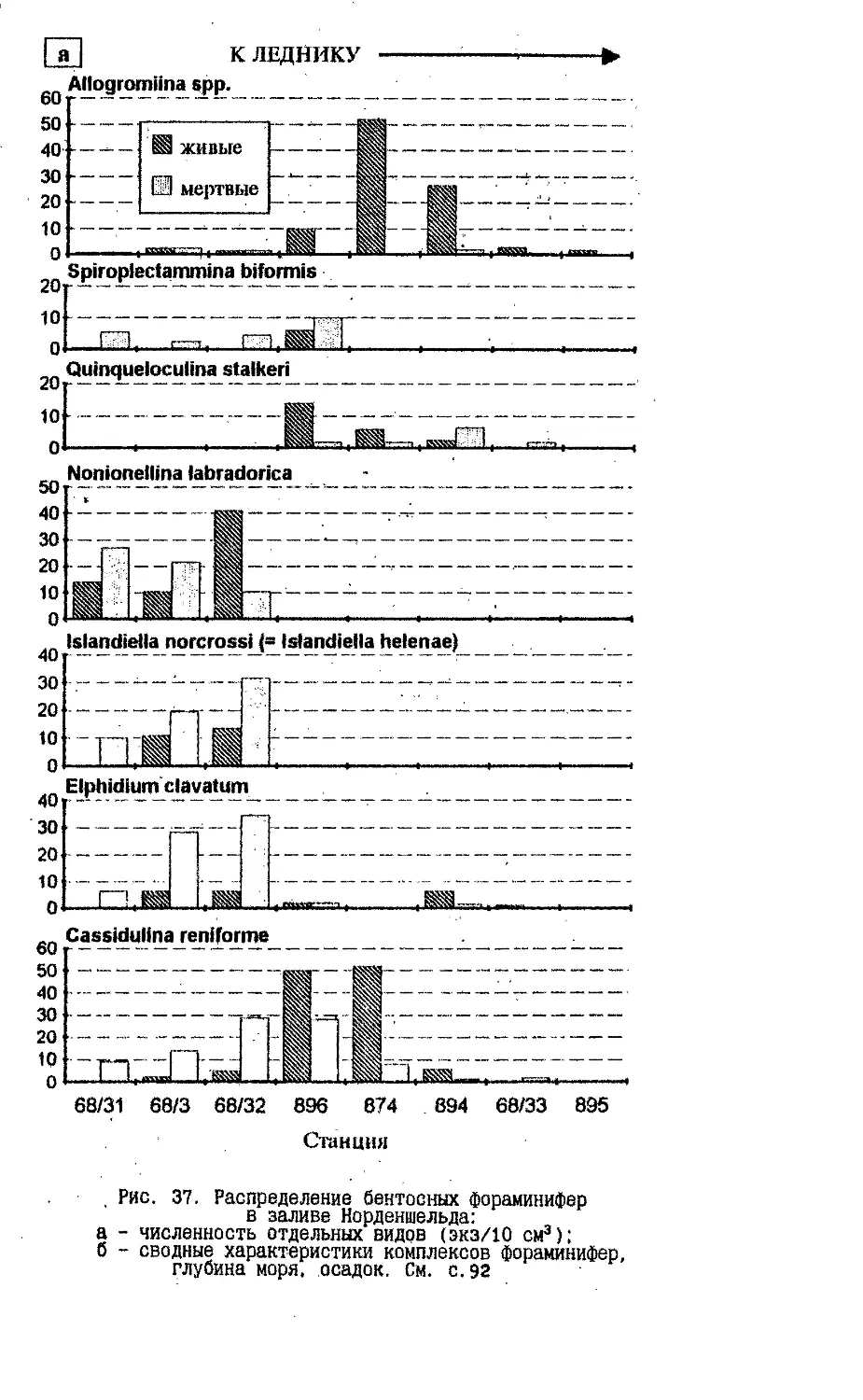
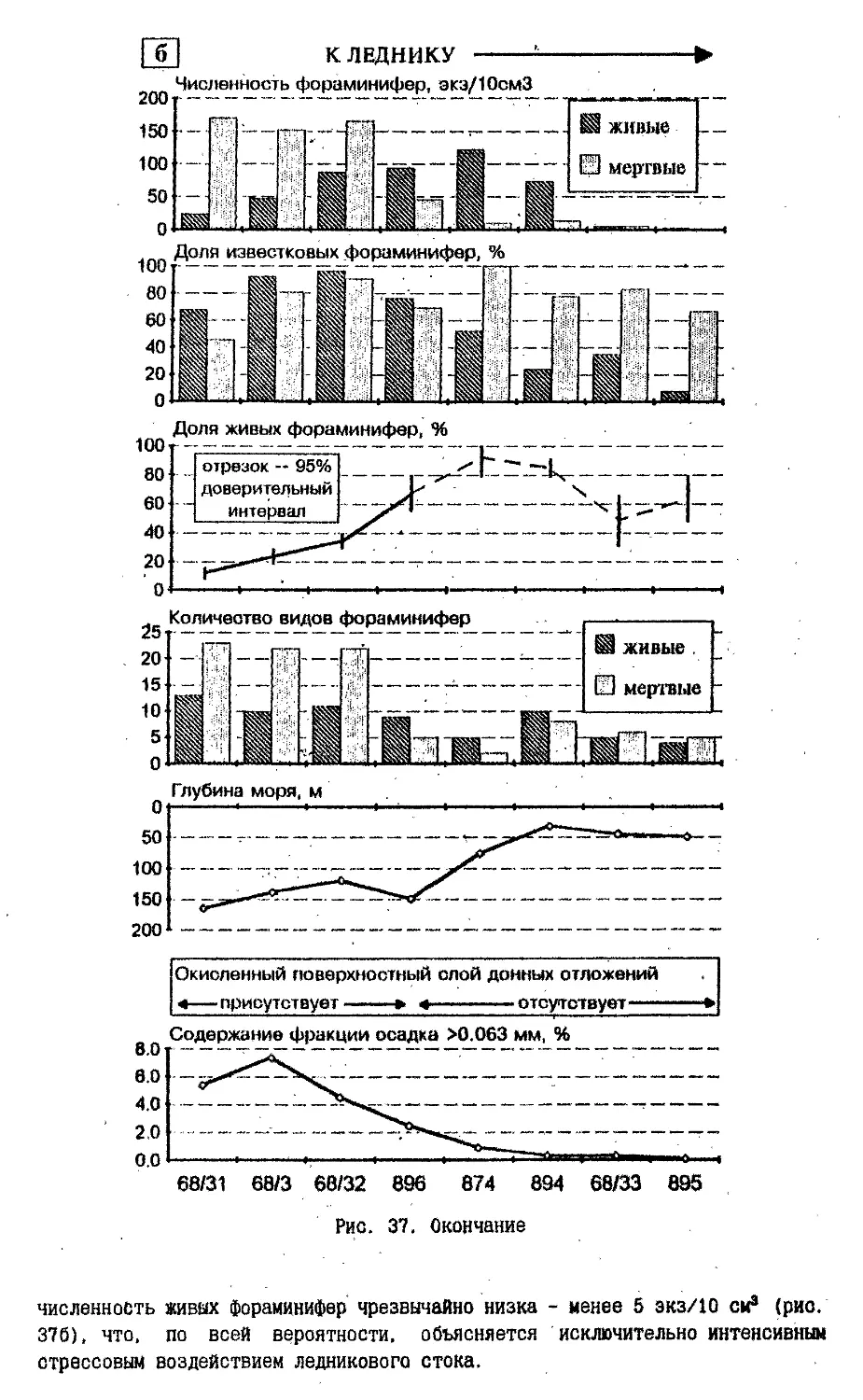

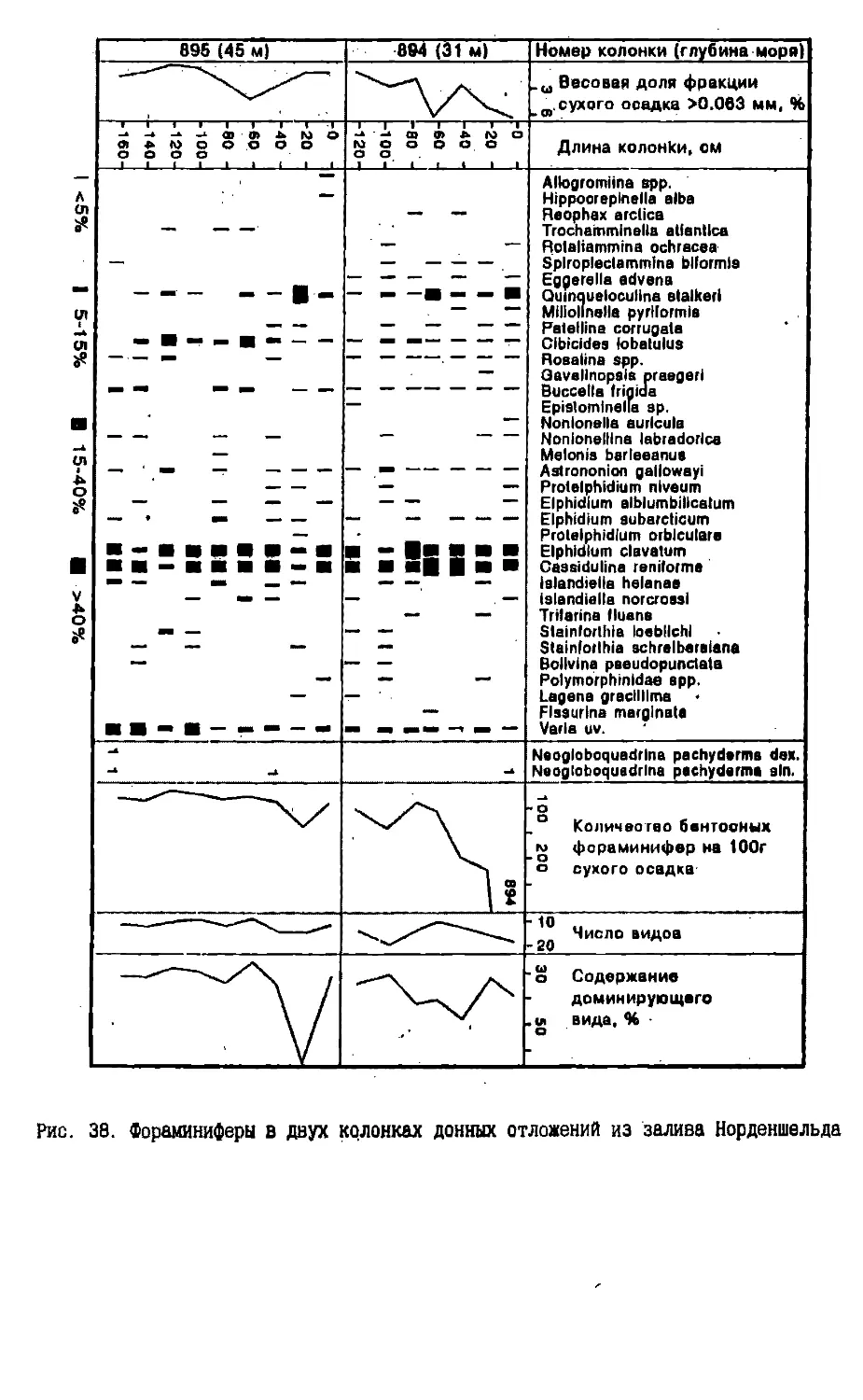


































































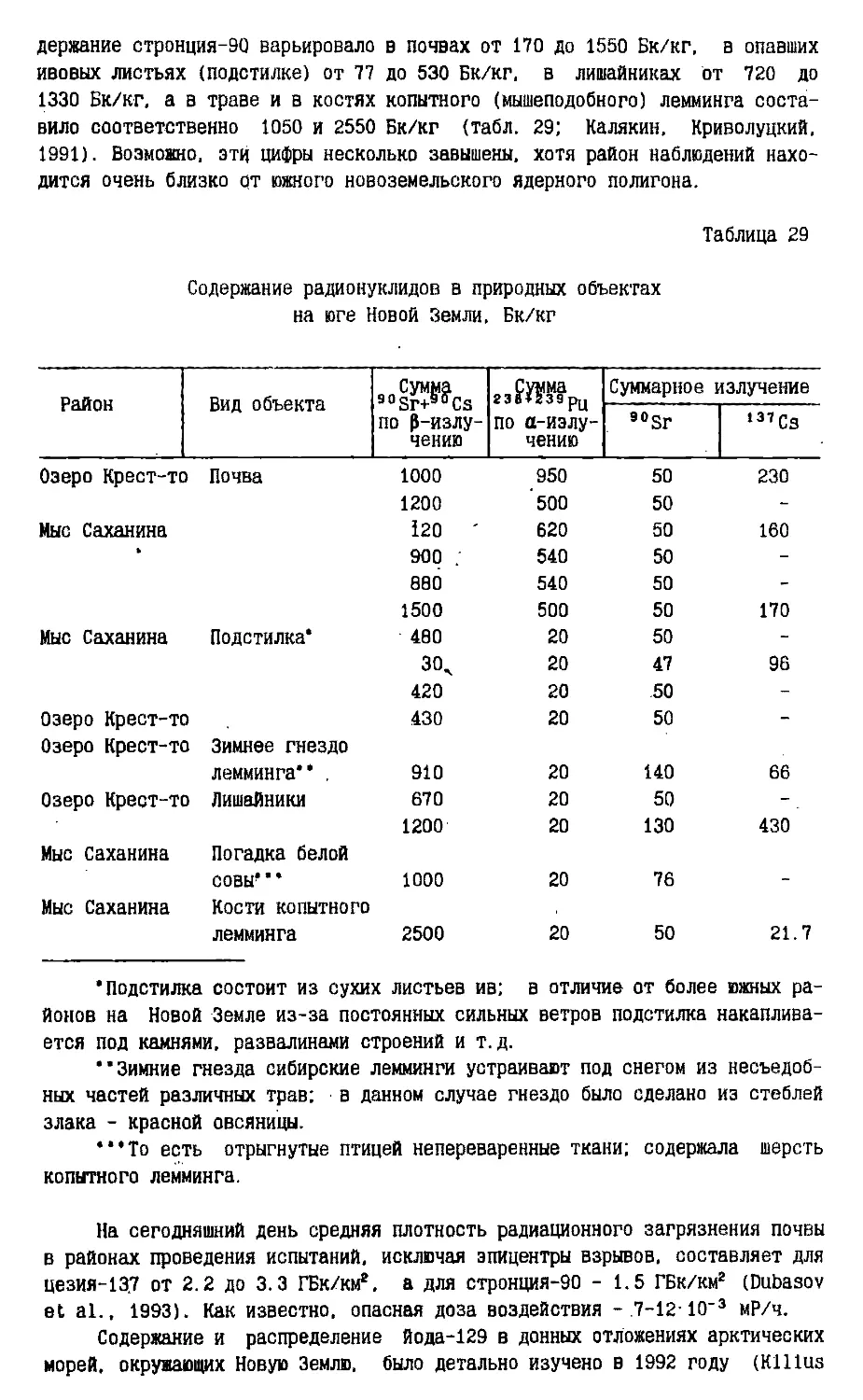
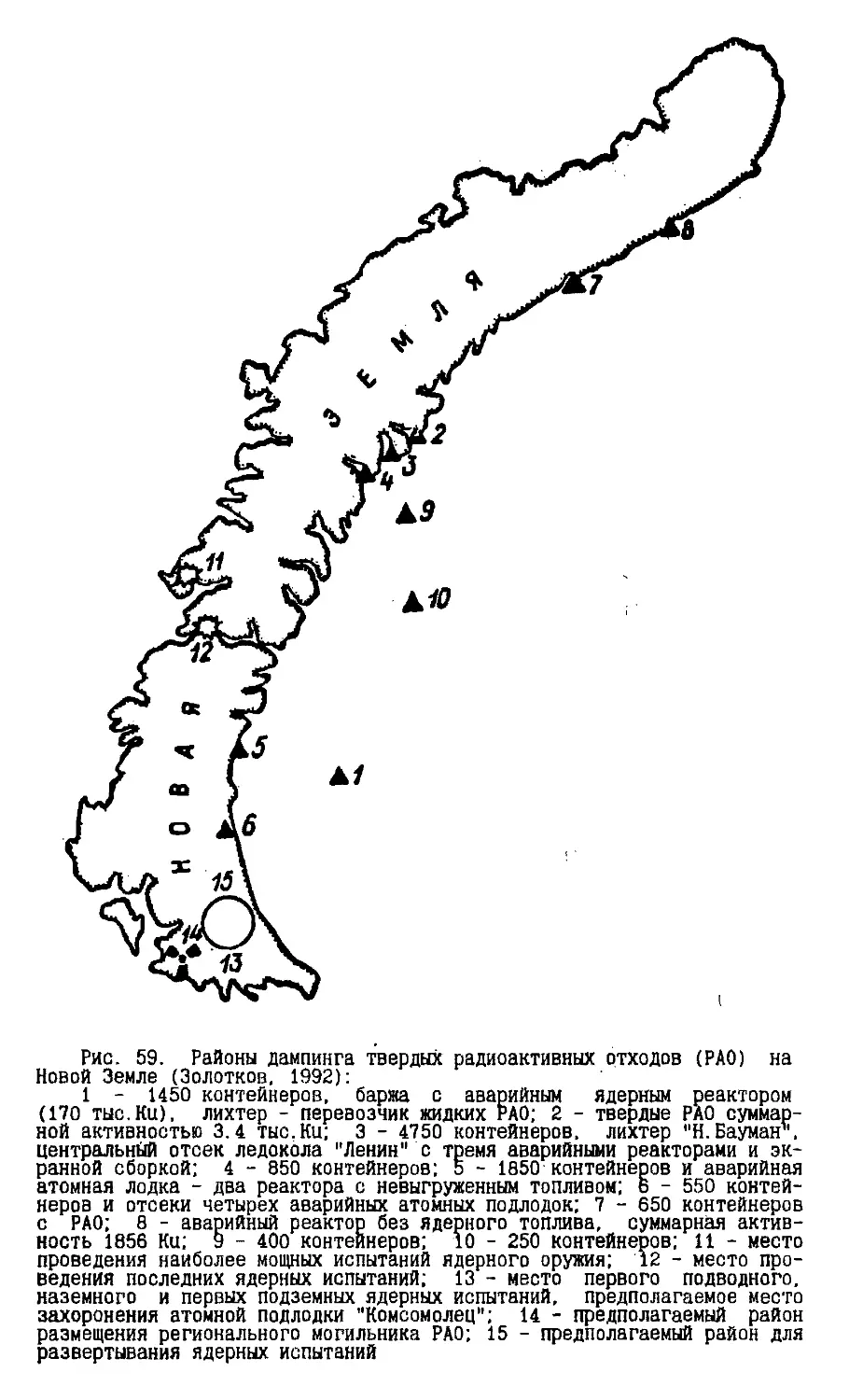


























Text
СРЕДА ОБИТАНИЯ
И ЭКОСИСТЕМЫ НОВОЙ ЗЕМЛИ
Архипелаг и шельф
ВВЕДЕНИЕ
Архипелаг Новая Земля является морским продолжением Уральского
хребта. Как Уральские горы разделяют два континента - Европу и Азию, так и
Новая Земля, отделяя Баренцево море от Карского, разделяет два океана -
Атлантический и Северный Ледовитый.
Такое положение архипелага определяет его барьерную роль во
взаимодействии водных и воздушных масс. Новая Земля как восточная граница
Баренцева моря является и восточным пределом распространения вод
Гольфстрима, точнее, северо-восточных его .ответвлений, находящихся за тысячи миль
от мест е,го рождения. Острова Новая земля долгое время привлекали внимание
географов. Наибольшую загадочность они представляли в период от Г.Меркато-
ра до В.Баренца. Большинство историков считает, что открытие Новой Земли
призошло в XIII-XIY веках. Некоторые специалисты по истории русского
Севера дбпускают, что поморы еще в XI веке посещали Железные Ворота, как
называли в древности пролив Карские ворота.
Новая Земля издавна привлекала своими биологическими ресурсами. Рус-
окие промышленники с Белого моря и Печоры посещали Новую Землю уже в
XV веке. Они добывали здесь морских зверей, птиц, арктического гольца. Во
второй половине XIX в. на Новую Землю устремились ученые - геологи,
ботаники, зоологи, ив начале XX в. началось настоящее паломничество членов
Общества любителей естествознания, антропологии и этнографии, художников.
. В предвоенные годы были организованы комплексные экспедиции
Арктического института, целью которых являлось всестороннее исследование природных
ресурсов, минеральных и биологических, морских и островных, их
использование в интересах индустриализации Севера и Советского Союза в целом.
Результатами этих работ явились систематизированные данные о геологическом
строении, климате, жизни моря и островов, предложения по оптимальному
использованию богатств архипелага.
Но в послевоенные годы приоритеты были отданы военному использованию
этого региона. На Северном и Южном островах, в прилегающих акваториях
начал функционировать северный ядерный полигон, здесь проводились наиболее
мощные наземные, подводные и подземные взрывы ядерного оружия. • Режим
секретности и многочисленные запреты на мореплавание и заходы судов в бухты
Новой Земли сделали архипелаг недоступным для исследователей, исключив его
на несколько десятилетий от отечественной и мировой науки.
По мере того, как Мурманский морской биологический институт усиливал
конплекснооть своих работ, расширял область своих интересов, исследование
Новой Земли становилось все более необходимым. Первые попытки получить
океанологические и биологические данные основывались на редких, трудно
разрешаемых подходах к берегам, на партизанских высадках на острова. Лишь
в 1986 г. при содействии Главкома Военно-Морского Флота СССР и командовав
ния Краснознаменного Северного флота Институт получил разрешение на
проведение первой в послевоенные годы экспедиции на Южный остров для изучения
новоземельского гольца. В последующие годы оказалось возмоиным выполнить
ряд более комплексных исследований на архипелаге и прилегающих акваториях.
Геологическое развитие Новой Земли охватывает палеозойский,
мезозойский и кайнозойский периоды. Особый .геоморфологический и экологический
след на архипелаге и прилегающем шельфе оставило четвертичное материковое
оледенение. Современный ледниковый щит Новой Земли, мощность которого
около 700 м. в несколько раз тоньше, чем древний покров льда. Сейчас в
западной части архипелага к морю спускается 25 ледниковых потоков.
Современная динамика экосистем Новой Земли определяется главным
образом колебанием климата и островного оледенения. Жизнедеятельность птичьих
базаров, популяций арктического гольца и полярной тресочки (сайки) во
многом происходит под влиянием теплых вод ледников. В 30-40-е годы животный
мир архипелага подвергался промысловому прессу рыбаков и охотников.
Особенно пострадали популяции кайры и гаги из-за массового сбора яиц и пуха.
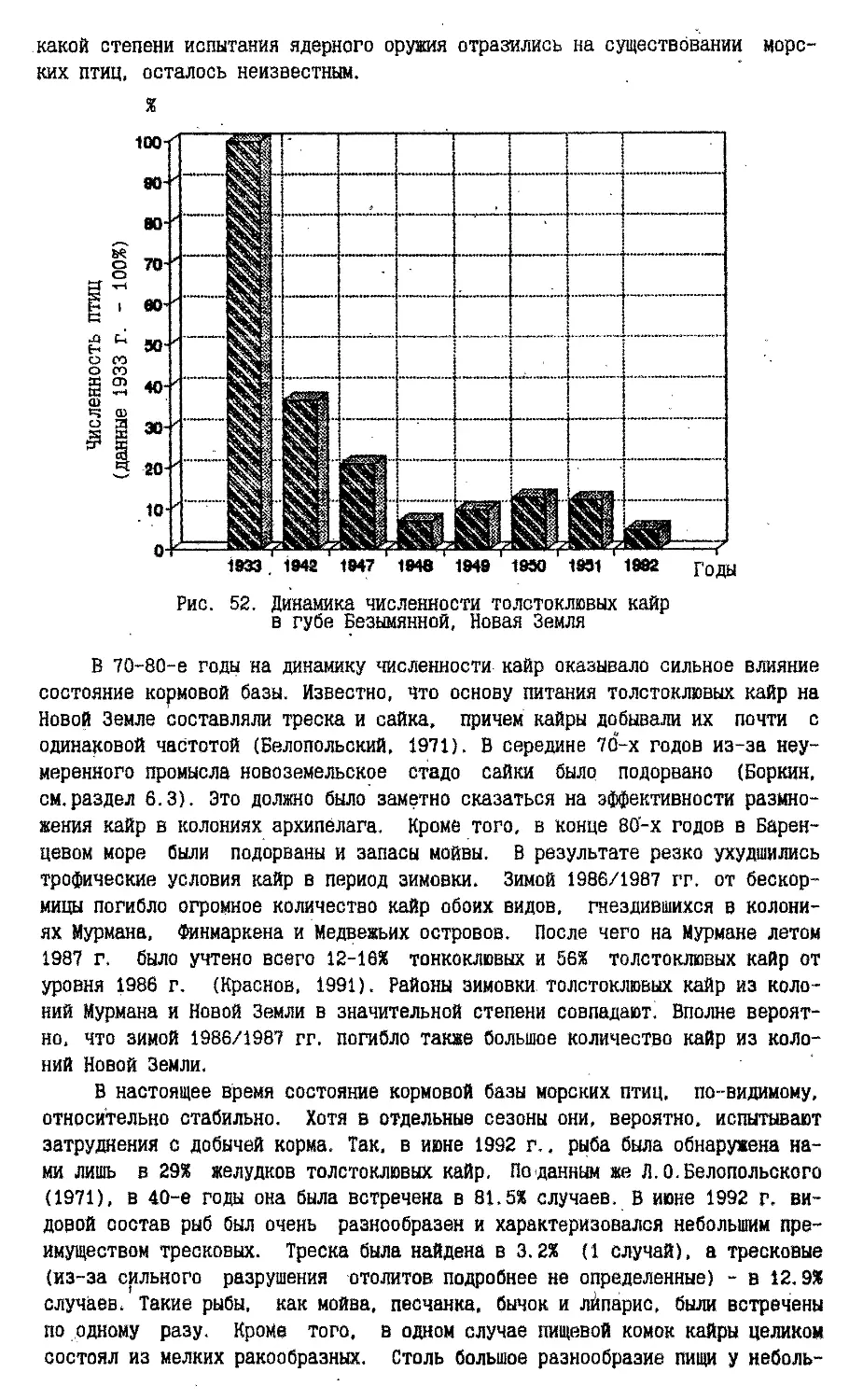
Начиная с середины 50-х годов и до .1963 года экосистемы архипелага,
подвергались мощному воздействию радиоактивного загрязнения во время испытаний
атомного оружия.
Данная книга продолжает серию публикаций о жизни арктических
архипелагов, начатую монографией "Среда обитания и экосистемы Земли
Франца-Иосифа (архипелаг и шельф)" Апатиты, 1994. В ней обобщены собственные данные
сотрудников ММБИ и литературный материал об абиотических условиях и
обитателях Новой Земли, о ее биологических ресурсах и о ситуации, сложившейся
после многих лет ядерных испытаний.
Являясь первой современной сводкой по экологической ситуации на Новой
Земле и прилегающих акваториях, книге, естественно, не может претендовать
на полноту и всесторонность всего накопленного материала. Ее цель .-
подведение некоторых итогов в изучении высокоширотных экосистем, определение
перспективы для более углубленных исследований, привлечение внимания
широкого круга специалистов к проблеме охраны уникальной природы Русской
Арктики.
Глава 1. ОБЩАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
Острова Новая Земля и Вайгач, как и расположенные поблизости
многочисленные мелкие острова и островки, образуют один из самых крупных
архипелагов Русской Арктики. Он имеет вид дуги, .выгнутой на северо-запад и
протягивающейся от мыса Гребень на юге острова Вайгач (69°38' с.ш.) до
мыса Карлсен - крайней северной точки Новой Земли (77°0Г с.ш.). Остров Вай-.
гач вытянут с юго-востока на северо-запад на 106 км (при ширине 44 км);
Новая Земля простирается с юго-запада на северо-восток на 925 км при
наибольшей ширине Южного острова 143 км, а Северного - 123 км. Таким образом,
общая протяженность архипелага в субмеридиональном направлении превышает
1000 км, что определяет отчетливые проявления широтной зональности
ландшафтов в его пределах. Площадь о-ва Вайгач - 3383 кмг, о-ва Южный -
33275 кмг, о-ва Северный - 48904 кмг. Новой Земли в целом - 82179 кмг,
мелких островов - около 1000 кмг (Черняева, 1957).
Остров Вайгач отделен от материка проливом Югорский Шар. а от Новой
Земли - проливом Карские Ворота. Северный и Южный острова разделены
проливом Маточкин Шар; ' от юго-западного побережья Южного острова про.лив Костин
Шар отделяет самый крупный из мелких островов - о-в Междушарский.
Основные черты литогенной основы ландшафта - геологического строения
и рельефа - определяются принадлежностью архипелага к Урало-Новоземельской
складчатой области позднегерцинского-раниекиммерийского (на севере)
возраста. Интенсивно дислоцированные породы палеозоя образуют ряд структур-
но-фациальных зон. вытянутых субпараллельно оои архипелага; в этом же
направлении протягиваются основные орографические линии.
Развитая вдоль побережий прибрежно-морская равнина с высотами до
80-100 м на о. Вайгач и юге Новой Земли сменяется холмистым и пластообраз-
ным рельефом с отметками до 170 м (губа Болванская) на о-ве Вайгач. 200 м
на юге Новой Земли. Постепенно повышаясь к северу, поверхность внутреннего
плато Южного острова на широте реки Безымянной приобретает среднегорный
характер с максимальными отметками (1342 м) в массиве Черный камень на
северном окончании Южного острова. Большая часть Северного острова имеет
горный рельеф с отметками 700-1300 м и выше. На юге этого острова
наибольшие высоты приурочены к восточному побережью, севернее залива Медвежьего
высоты смещаются в центральную часть и далее следуют вдоль западного
побережья, где в районе залива Норденшельда находится высшая точка архипелага
с отметкой 1590 м. Свободный от ледника северо-восточный край Северного
острова представляет собой холмистую равнину.
Береговая линия островов за исключением южной части восточного
побережья сильно изрезана. Преобладает расчленение фьордового и фиардового
типа, в целом более интенсивное на западном побережье, а также местами
шхерное. Такой характер побережья обуславливает исключительное разнообразие в
пространстве (при относительно небольшом наборе типологических элементов)
местообитаний и определяет сложную пространственную структуру прибрежных
экосистем как моря, так и суши.
Своеобразие климата Новой Земли помимо основных климатообразующих
факторов - географичеокого положения, количества поступающей радиации и
циркуляции атмосферы - определяется рельефом островов и резкими различиями
теплового режима Баренцева и Карского морей. С ноября по март архипелаг
находится под влиянием барической ложбины исландского минимума, с чем
связана интенсивная циклоническая деятельность. Летом давление повышается.
Различия теплового режима Баренцева и Карского морей определяют величину и
направление барического градиента - разницы атмосферного давления на
восточном и западном побережьях архипелага. С этим связана характерная-
особенность климата Новой Земли - частая повторяемость сильных ветров, в том
числе знаменитой боры. Скорости ветра при боре достигают ураганной силы:, в
Малых Кармакулах - 58, в Русской Гавани - 50, на мысе Желания - 40 м/с.
Средняя скорость ветра за год составляет на мысе Желания 8.2, в Русской
Гавани - 7.2, в Малых Кармакулах - 8.6 м/с (Советская Арктика, 1970).
Значение ветрового режима для функционирования экосистем суши в данном
регионе в основном заключается в перераспределении снежного покрова. Для
морских экосистем существенными являются сильное волнение, движения воды и
плавучих льдов, вызванные ветром.
Особенности температурного режима Новой Земли связаны с отепляющим
влиянием' атлантических вод. заходящих в Баренцево море. Это влияние
определяет сравнительно высокие, несмотря на северное положение архипелага,
зимние температуры, а также резкие различия температур западного и
восточного побережий. Самый холодный месяц - март, средняя температура которого
на западном побережье -14, -15 °С„ на восточном - -19, -22 °С. Абсолютный
минимум температуры воздуха составил на мысе Желания -50 °С, в Маточкином
Шаре -49 °С. Самый теплый месяц - август, со средней температурой в Малых
Кармакулах - 6.6, на мысе Желания - 2.4. в Русской Гавани - 3.8 "С
Абсолютный максимум составил на мысе Желания 21, в Малых Кармакулах - 24 °С.
На продолжительности безморозного периода сказывается большая
субмеридиональная протяженность островов: так, на мысе Желания он длится в
среднем 27 дней, в Маточкином Шаре - 48, в Малых Кармакулах - 84 дня.
Вместе с тем характерна контрастность годового хода температур: в любом
месяце лета возможны заморозки, а зимы - оттепели (Советская Арктика,
1970).
Обычная высокая относительная влажность воздуха способствует
образованию облаков и туманов в основном в летнее время. Число дней с туманами
в году на мысе Желания - 69, в районе Маточкина Шара - 25. В связи с
высокой облачностью число ясных дней в году не превышает 29. Тем не менее.
осадков выпадает мало: годовая их сумма на мысе Выходном 300 мм. на мысе
Желания - 189 мм. На леднике осадков выпадает больше - до 400-600 мм.
Наибольшее их количество приходится на летнее время и выпадают они
преимущественно в твердом виде.
Высота снежного покрова невелика: на мысе Желания максимальная из
декадных высот достигала 29 см.. в Малых Кармакулах - 38, у мыса Выходной -
46 см. Количество дней с метелями достигает 100. Из-за сильных ветров,
особенно на западном побережье, снежный покров залегает крайне
неравномерно (Советская Арктика. 1970).
Отмеченные особенности климата существенны для экосистем островов.
Большая облачность и частые туманы уменьшают поступающее во время вегета-
ционного периода на земную поверхность количество прямой солнечной
радиации. Малые мощности снежного покрова или его отсутствие на плакорных
поверхностях способствуют вымерзанию растительности зимой. Помимо низких
температур, лимитирующим фактором для развития растительности является
недостаток свободной влаги в почве. Особенно это видно в северной части
Северного острова, относимой рядом исследователей к зоне полярных пустынь
(см. ниже). Здесь на низких прибрежных поверхностях растительность
сосредоточена почти исключительно' на более или менее влажных местах: вблизи
снежников, ручьев, в увлажненных западинах, в микроложбинах делювиального
смыва на пологих склоновых шельфах. На остальной поверхности сосудистые
растения отсутствуют.
Недостаток влаги в грунте несколько компенсируется высокой
относительной влажностью (в среднем 80%, летом на мысе Желания до 92%). Кроме
того, разумеется, положительный фактор для развития растительности -
длинный полярный день.
Климат Новой Земли определяет существование разнообразных форм
современного оледенения, включая покровное. Ледники разных типов группируются в
4 областях (Оледенение..., 1968; Долгушин, Осипова, 1989). Область
покровного оледенения занимает большую часть Северного острова к северу от губы
Машигиной'на западном побережье и залива Цивольки на восточном. Обширный
ледниковый покров с мощностью льда до 500 м имеет в длину 413 км и в
ширину до 95 км; площадь его вместе с выводными ледниками 19330 кмг. Высота
ледораздела составляет 900-950 м в центральной и южной частях и постепенно
понижается к северу до 500-600 м.
Глубокое понижение коренного ложа на продолжении залива Иностранцева
отделяет от основного покрова его часть - Северный ледниковый купол. На
большей части покрова рельеф коренного ложа не проявляется, и его
морфология определяется свободным растеканием льда. На периферии покрова
появляются выводные ледники, контролируемые рельефом ложа, количество которых
превышает 60, а длина достигает 40 км и более (ледник Норденшельда -
62 км). Большая часть выводных ледников достигает моря и продуцирует
айсберги. Скорость движения льда была измерена на леднике Шокальского и
составила от 15-20 м/год у границы питания до 150 м/год у обрывающегося в
море фронта ледника.
Область горно-покровного оледенения лежит в виде неширокой полосы
южнее ледникового щита. Здесь сильнее проявляется влияние интенсивно
расчлененного рельефа коренного ложа, и оледенение имеет сетчатый характер с
многочисленными горными гребнями и нунатаками. В длину ледники достигают
20 км и более. За исключением нескольких крупных ледников, большинство из
них не доходит до моря и оканчивается на суше. Общая площадь ледников
1852 км*.
Область горного оледенения расположена севернее и южнее пролива Ма-
точкин Шар. протягиваясь с севера на юг на 160 км. Здесь преобладают
каровые и долинные ледники, самый крупный из которых имеет длину 9 км.
Область малых ледниковых форм находится в южной части Южного острова.
Настоящих ледников с признаками активного движения льда здесь нет, но
много навеянных снежно-ледовых образований в понижениях рельефа.
В период с 1933 по 1952 гг. Новоземельский ледниковый покров сокра-
0(ался ежегодно на 3.6 км3 льда или 3.2 км3 воды. При этом отекание льда
в море и откалывание айсбергов составляет 15-20% величины стаивания
(Оледенение..., 196В).
Для понимания влияния ледникового покрова на морские экосистемы важно
учитывать то обстоятельство, что динамика покрова подчиняется многолетней
разнопорядковой цикличности и периоды деградации ледников сменяются фазами
их наращивания.
Благодаря большой меридиональной протяженности Новой Земли граница
питания ледников меняет свое положение от .700 м у южного предела
оледенения до 300 м в районе мыса Желания, то есть понижается на 400-600 м (Гово-
руха. 19В8).
Поскольку большинство выводных ледников выходит в море, покровное
оледенение Новой Земли следует рассматривать как феномен островной суши,
оказывающий очень важное, хотя и недостаточно изученное влияние на морские
экосистемы прибрежных вод. Оно проявляется через продуцируемые ледником
айсберги и ледяные глыбы, сток пресных и холодных талых ледниковых вод.
вынос твердого материала - как включенного в лед, так и в виде взвеси -
"ледникового молока" (а в подледных потоках также и влекомого). Как
показали исследования, выполненные в 9 губах западного побережья Северного
острова, содержание взвеси на поверхности колебалось от 33-16 до
90-113 мг/л. у дна - от 4-12 до 41 иг/л, а мощности современных леднико-
во-морских илов во фьордах достигают 40-50 м (Арктический шельф..., 1987).
Ископаемые льды • на Новой Земле принадлежат к двум основным категори-.
ям: погребенный под моренными, флювиогляциальными и морскими отложениями
глетчерный лед (в основном .на Северном острове, в районе губ Крестовой,
Южной Сульменевой, Глазовой, Машигиной и в других местах) и
повторно-жильные льды в низменной части Южного острова, где развита многолетняя
мерзлота. Глубина летнего протаивания мерзлых грунтов на прибрежных равнинах
изменяется в пределах 30-45 см, на холмистых плато достигает 80-100 см
(Советская Арктика, 1970).
Речная сеть наиболее широко развита на Южном острове и в южной части
Северного острова. Длина самых крупных рек (Абросимова,. Безымянной и
других) не превышает 50-60 км. Главный водораздел протягивается вдоль оси
островов и несколько смещен к востоку; в результате на Южном острове
водосборный бассейн Баренцева моря занимает 2/3 его площади. Наиболее
крупные реки Северного острова расположены к югу от губы Северная Сульменева -
Гусиная. Митюшиха, Промысловая и Южная Крестовая; основные реки Южного
острова - Саханина, Рогачева, Пуховая, Безымянная. Абросимова. Савина.
Большинство рек течет в субширотном направлении, пересекая геологические
структуры вкрест простирания, и имеет полугорный и горный характер со
значительными уклонами русла (более 0.005); лишь в низовьях, выходя на
прибрежную равнину, реки меандрируют. Исключение составляют реки юго-западной,
равнинной части Южного острова, имеющие выработанный продольный профиль и
извилистое русло.
В устьях рек, впадающих в бухты и заливы, довольно обычны дельты,
часто несоизмеримо большие по сравнению с длиной рек.
Реки Северного острова - короткие, быстрые, с большим количеством
водопадов и порогов.
В целом реки Новой Земли относятся к снежно-ледниковому типу питания
о небольшой долей дождевого; на реках с преимущественно ледниковым
питанием максимальные расходы наблюдаются в июле и августе, когда у рек со
снеговым питанием самый низкий уровень. Паводок у этих рек происходит раньше.
во время довольно интенсивного весеннего снеготаяния, когда расходы но
сравнению с меженными увеличиваются в 20-50 раз, а уровень от десяткоь
сантиметров до 1-2.5 м (Советская Арктика. 1970).
Замерзают реки обычно в середине октября; в декабре промерзают до
дна, за исключением углублений в руслах крупных рек (глубже 1 м в межень),
а вскрываются в мае. Температура воды в ледниковых водотоках низкая - от
0.1-0.2 до 1.8 °С, в неледниковых - 8-9 °С.
Озера на Новой земле довольно многочисленны и достигают 1.5 км в
длину. Они принадлежат нескольким типам: реликтовые прибрежные (бывшие лагу-
ньь потерявшие связь с морем в результате, современного поднятия островов),
прибрежной равнины, ледниковых долин, горные, а также термокарстовые
(Советская Арктика, 1970). Питание озер - то же, что и рек; они обычно
проточные или имеют сток. Преобладают неглубокие озера (до Ю-11 м); наиболее
глубоки оз.Дерюгина (90 м) и оз,Нехватова (74 М). Толщина зимнего ледяного
покрова на озерах достигает 2 м, и условия для зимовки рыбы существуют
лишь в глубоких озерах.
Условия для формирования почвенного покрова на Новой Земле крайне
неблагоприятны. Большие площади заняты или каменистыми россыпями, или
территориями с примитивными скелетными" почвами со слабыми признаками
почвообразования'. Накоплению гумуса препятствуют низкие температуры,
ограничивающие деятельность микрофлоры, а также небольшой прирост биомассы основных
компонентов растительного покрова - мхов и лишайников за короткий
вегетационный период.
На большей части Северного острова преобладают арктические типичные
почвы - зональный тип почв арктических пустынь; на прибрежных равнинах
Южного острова и в крайней юго-западной части Северного острова - тундровые
глеевые почвы. На юге встречаются также дерново-арктические, болотные тор-
фяно-глеевые. аллювиально-маршевые солончаковые (на илистых осушках вдоль
побережья), редко - дерново-луговые почвы тундровых луговин (Советская
Арктика, 1970).
Во взглядах на положение Новой Земли в системе физико-географических
(ландшафтных) зон и регионов Арктики единства у исследователей нет. Так,
Е.С.Короткевич. автор капитальной комплексной монографии о полярных
пустынях (1972), при проведении районирования оставил Новую Землю за пределами
этой природной зоны. В.Д.Александрова (1983) по геоботаническим признакам
включила в зону полярных пустынь крайнюю северную часть Северного острова;
крайний юг о-ва Вайгача отнесен ею к субарктическим тундрам, а остальная
часть архипелага - к арктическим тундрам. В монографии "Советская Арктика"
(1970) на Новой Земле выделяются-две зоны: арктических пустынь и тундровая
зона в ее северном варианте - подзоны арктических тундр. Прибрежные
участки Северного острова по характеру растительности отнесены к арктической
пустыне, южнее широты Машигиной губы низменные и прибрежные участки
островов представлены арктической тундрой; горные районы 'занимает горная
полярная пустыня,' переходящая на юге в горную арктическую тундру.
Глава 2. ГЕОЛОГО-ГЕОМОРФОЛОГНЧЕСКАЯ ХАРАКТЕРИСТИКА
Разрез коренных пород островов Новой Земли и Вайгача представлен
почти исключительно палеозойскими осадочными и частично вулканогенными
образованиями, отражающими развитие герцинской геосинклинали. Представлены все
системы палеозойской группы. Встречаются также переотложенные породы
мезозоя, отложения неогена и антропогена (Геология СССР, 1970; Красножен и
др., 1986).
Породы кембрий (сланцы, известняки, конгломераты, туфогенные
песчаники и порфирита) обнажаются в ядрах антиклиналей на побережьях пролива
Карские Ворота и на западе Новой Земли. Отложения ордовика, силура, девона
и карбона развиты на юге, в центральной и северной частях архипелага. Они
представлены смятыми в складки терригенными и органогенными породами
(сланцы, • песчаники, конгломераты, известняки, доломиты и переходные
разности), изверженными интрузивными и эффузивными породами основного и
кислого состава. .
Пермские отложения на Вайгаче протягиваются узкой полосой вдоль
северо-восточного берега, а на Новой Земле распространены очень широко, слагая
большую часть Южного острова до р.Безымянной на севере, а на восточном
побережье продолжаясь к северу от Маточкиного Шара и уходя под ледниковый
покров, а также выходя отдельными пятнами на западном и восточном
побережьях.
Мощность этих отложений до 2000-3000 м, а состав - терригенный; они
трансгрессивно залегают на породах от верхнего карбона до девона.
Пермский, а также более поздний (триас - начало юры) возраст имеют интрузии
гранитоидов. развитые в ряде участков побережья Новой Земли. В целом для
проявлений магматизма характерна определенная последовательность:
извержения основной магмы происходили- в раннем палеозое, силуре и наиболее
интенсивно в начале девона; интрузии гранитоидов развивались в раннем девоне,
на границе перми и триаса и в конце триаса - начале юры.
Характер дислокаций пород палеозоя позволяет выделить три структурных
комплекса; байкальский (более развитый на юге Пайхойско-Новоземельской
системы), каледонский и герцинскЬ-раннекиммерийский (Хаин. 1977).
Каледонский комплекс охватывает отложения от кембрийских до среднедевонских,
слагающие две формации - нижнюю терригенную и верхнюю карбонатную
мощностью 3-3.5 км. Герцинский комплеко отделен несогласием от каледонского и
также состоит из двух формаций - нижней миогеосинклинальной карбонатной
мощностью 1.5-2.5 км и верхней раннеорогенной - пермской морской молаосы.
Описанные комплексы слагают три основные тектонические структуры
архипелага: Южный Новоземельский антиклинорий, протягивающийся от Пай-Хоя на
Вайгач и в южную часть Новой Земли, Кармакульский синклинорий, в основном
занимающий область сплошного распространения пермских пород на Южном
острове, и Северный Новоземельский антиклинорий, начинающийся в северной
части Южного острова и занимающий почти весь Северный остров. По северо-за-
падному крылу антиклинория проходит крушое тектоническое нарушение -
Главный Новоземельекий разлом. Кроме описанных структур, вдоль западного и
восточного побережий протягиваются подводные части соответственно
Баренцева и Карского синклинориев (Ковалева и др., 1982). Из менее крупных
структур следует отметить Литкенскую брахиантиклиналь с ядром из девонских
отложений, прорванных интрузиями диабазов, осложняющую строение Кармакуль-
ского синклинория в районе залива Литке, и огромную (длина более 300 км,
ширина 25-30 км) синклиналь залива Медвежьего в восточной части Северного
Новоземельского антиклинория, сложенную умеренно дислоцированными
пермскими отложениями. В пределах антиклинориев-на. дневную поверхность выходят в
основном допермские отложения.
На протяжении герцинского тектонического цикла район Вайгача и Новой
Земли развивался как миогеосинклинальная зона, для которой в первой
половине цикла было характерно чередование неглубоких морских, прибрежно-морс-
ких и отчасти континентальных обстановок осадконакопления. Начиная с
середины каменноугольного времени, проявляются складчатые тектонические
движения, поднятия и орогенный магматизм, максимум которых пришелся на перм-
ское-раннетриасовое время. С простиранием складок связаны не только
особенности орографии, но и образование напоминающего далматинский исходного
типа расчленения береговой линии на юго-западе архипелага, а также
некоторые черты конфигураций береговой линии к северу от реки Безымянная.
В конце триаса - начале юры происходили интенсивные блоковые
движения! сопровождающиеся интрузивным гранитоидным магматизмом. Эти и более
поздние циэъюнкгивныб дислокации сыграли большую роль в формирований
блоковой структуры островов с многочисленными продольными, позже поперечными
и диагональными разломами, впоследствии предопределившими направления
речной и ледниковой эрозии. Особенно интенсивно раздробленной оказалась
внутренняя часть Северного Новоземельского антиклинория - так называемая
Мозаичная область на юго-западе Северного острова и северо-западе Южного. В
результате в триасе регион представлял собой, вероятно, область с горным
рельефом, который к середине юры постепенно был разрушен.
По мнению Н.Г.Загорской (Горные страны.,., 1974), возможно с этим
временем связано образование четко выраженного в рельефе уступа вдоль
р. Безымянной, ограничивающего с юга Мозаичную область и отделяющего
выходящие на поверхность складки Северного антиклинория от расположенного
южнее поля развития ранней перми в пределах Кармакульского синклинория.
Уступ сохранил выраженность в современном рельефе благодаря неоднократному
тектоническому подновлению, абразионной и денудационной обработке.
Предполагается также, что плоские вершины горных возвышенностей в
области расчлененного альпинотипного рельефа севернее уступа представляют
собой остатки самой верхней и самой древней (четвертой) поверхности
выравнивания, возможно, имеющей допозднетриасовый возраст, т.е.
сформировавшейся раньше уступа. Это предположение кажется маловероятным, поскольку
интенсивные движения не способствовали развитию процессов^егионального
выравнивания. Возможно, плосковершинный характер гор связан, с более поздними
процессами типа педиментации и гольцовой планации.
Ниже уступа располагается третья поверхность выравнивания, в
современном рельефе имеющая вид денудационного плато на высотах 300-600 м и
занимающая в основном центральную часть Южного острова в области развития
перми. Данных о возрасте этой поверхности нет; одним из оснований для ее
выделений является то, что на Южном острове-она четким, более чем
50-метровым уступом отделена от нижележащей пологохолмистой равнины - второй
поверхности выравнивания. На Северном острове узкие прибрежные полосы этой
поверхности глубоко расчленены эрозионными врезами (Горные страны
1974).
Судя по характеру переотложенных мезозойских пород (коренные
отложения этого возраста неизвестны), со средней юры до конца раннего мела
западная и южная части Новой Земли были заняты морем, а побережье
характеризовалось низким слабо расчлененным рельефом и сильно изрезанной береговой
линией (Демокидов и др., 1957). Возможно в это время сформировалась вторая
поверхность выравнивания - пологохолмистая равнина с преобладающими
высотами 150-250 м, широко развитая на Саном острове и местами в
периферических частях Северного острова (Горные страны..., 1974),
Отсутствие коренных отложений позднего мезозоя и палеогена не
позволяет проследить историю развития территории на этом этапе (см. ниже.),
датируется миоценом.
По существующим представлениям, новейшие поднятия в основном
наследовали простирания древнего структурного плана и оживили многие разломы.
Наибольшая - амплитуда поднятий отмечается в пределах Мозаичной области, а
также вдоль западного побережья северной части архипелага, где были
сформированы горы Эдуарда Толля. Менделеева и Ломоносова. В целом для морфо-
структуры гор характерно наличие нескольких субпараллельных цепей, от 2-3
на север'е Северного острова до 8-10 на широте залива Норденшельда и
полуострова Адмиралтейства (Оледенение..., 1968). ширина разделяющих горные
цепи продольных долин невелика, 2-3 км; поперечные долины делят горные
хребты на короткие отрезки.
Для флювиальных долин Новой Земли характерны большая глубина, невыра-
ботанность продольного профиля, в случае ледниковой обработки -
расширенное днище. Местами долины, врезанные даже в низкие поверхности рельефа,
имеют вид каньонов и ущелий. Своеобразная черта долинного расчленения -
наличие сквозных долин, пересекающих горную страну от восточного побережья
до западного. Так, крупная долина Русанова протягивается от губы Крестовой
до залива Незнаемого. Севернее располагаются еще три сквозных долины.
Долина Самойловича соединяет губу Южную Сульменеву с бухтой Романова в
заливе Медвежьем; ответвление этой долины протягивается в губу Крестовую;
долина Ермолаева соединяет губу Северную сульменеву и залив Медвежий: долина
Хольтедаля идет от губы Машигиной к заливу Ога. а долина Пересечений
связывает верховья губ Митвшихи и Белушьей. Крупнейшей сквозной долиной
.является пролив Маточкин Шар (Оледенение..., 1968).
По данным экспедиций М.М.Ермолаева, ледниковый покров Северного
острова скрывает расчлененный рельеф с амплитудой до 500 м; некоторые его
участки лежат ниже уровня моря (Оледенение.... 1968).
К южной оконечности архипелага и на его периферии амплитуда поднятия
уменьшалась; здесь сформировалась самая нижняя, первая, денудационная
поверхность, срезающая на острове Вайгач и в юго-западной части Южного
острова складки Южного антиклинория. Считают (Каплин, 1962: Оледенение...,
1968). что это стрендфлет. сформированный совокупным действием абразии,
льдов и 'морозного выветривания. Поверхность перекрыта морскими и
ледниковыми отложениями и в современном рельефе представляет грядово-холмистую
равнину с высотой внутреннего края 80-100 и. В настоящее время она припоя-
нята и местами ограничена со стороны моря абразионными уступами высотой до
50 м.
Поскольку главными событиями четвертичного этапа, помимо сводово-бло-
кового поднятия, были оледенение и морские трансгрессии, на островах
широко распространены ледниковые и морские формы. Известное представление о
Новоземельском ледниковом покрове как реликте плейотоценового оледенения в
настоящее время оставлено; вместе с тем многие данные говорят о
существовании в прошлом на архипелаге оледенения значительных масштабов, чем
современное (Оледенение..., 1968). Наиболее отчетливые свидетельства этого
древние моренные гряды и валы, расположенные на некотором расстоянии от
края современных ледников. В области покровного оледенения Северного
острова прослеживаются две самостоятельные гряды конечных морен; на Южном
острове выраженные в рельефе конечно-моренные гряды встречаются до
73° с.ш.
Эти образования состоят в сложных взаимоотношениях с морскими
абразионными и аккумулятивными террасами: древние морены носят следы воздействия
моря, более молодые перекрывают морские уровни. А.Б.Бажев и В.Я.Бажева
(Оледенение..., 1968) выделяют три комплекса морских террас: верхний на
высотах 270-420 м, средний на 90-240 м. нижний на отметках 10-80 м, а
также современные береговые валы. Наиболее высокие террасы в виде плоских,
почти горизонтальных поверхностей с хорошо окатанной галькой, раковинами
моллюсков и плавником обнаружены в группе нунатаков в районе залива
Русская Гавань. Они связаны с бореальной трансгрессией, за которой
последовало максимальное и самое раннее из геологически документированных (в
пределах островов) оледенение. Толщина ледникового покрова достигала
700-1000 м. а поверхность располагалась на высоте 1100-1200 м. Вероятно,
максимум оледенения пришелся на заключительные этапы зырянского
оледенения.
Последовавшая трансгрессия способствовала сокращению оледенения и
сформировала уровни террас второго комплекса, из которых наиболее
выраженный на высотах 200-230 м. На северо-востоке Северного острова
сформировалась широкая морская поверхность, протягивающаяся вдоль всего восточного
побережья; трансгрессия перекрывала большую часть Южного острова. Когда в
ходе последующей регрессии уровень моря опустился до отметок около 120 м
над современным, произошло освобождение пролива Маточкин Шар от ледника,
разьединение ледникового покрова и его значительное сокращение.
Новое продвижение ледников (кармакульокая стадия, по А. Б. Бажеву и
В.Я.Бажевой) развилось на фоне продолжающейся регрессии; размеры
оледенения превосходили современные, достигая 73° с.ш. Краевые части ледников
этой стадии в районах губ Крестовой, Машигиной, Грибовой позднее оказались
перекрыты чехлом моренных и морских отложений и в погребенном состоянии
сохранились до настоящего времени. ""
Отступание ледников, связанное со среднеголоценовым потеплением
климата, совпало'с новой трансгрессией моря, в результате чего в моренах были
выработаны террасы нижнего комплекса высотой до 80 м. Во время
климатического оптимума оледенение, вероятно, исчезло полностью, и современный
ледниковый покров - молодое голоценовое образование.
В позднем голоцене имели место еще два эпизода активизации оледенения
- стадия Седова около 2.5 тыс. лет назад и стадия Адмиралтейства после ХУ1
века (малый ледниковый период), когда на побережье Северного острова
морена была надвинута на 8-10-метровую морскую террасу (Оледенение..., 1968).
Новые материалы по истории Новой Земли1 были получены в ходе
геологической съемки масштаба 1:200000 в конце 70-х - начале 80-х годов. Они
позволили существенно уточнить и дополнить более ранние представления, а
также высказать принципиально отличную от. общепринятой концепцию формирования
морфоструктуры архипелага (Красножен. 1982; Красножен и др., 1982. 1986).
Согласно этим взглядам, к концу палеогена в регионе сформировался
пенеплен, фиксированный корой выветривания, астатки которой в виде оранжевых
глин и суглинков мощностью 0.1-0.3 м, содержащих пыльцу палеогенового и
мезозойского возраста, обнаружены на поверхностях высотой 800-1100 м.
В миоцене на Карском шельфе сформировалось обширное сводово-блоковое
поднятие. Составлявшая его западную периферию территория будущего
архипелага была раочленена сетью долин, приуроченных к радиально расходящимся от
гипотетического центра поднятия разломам и врезанных на глубину 100-200 м.
Заполняющие их миоценовые осадки представлены песками, гравийно-галечными
отложениями и конгломерато-брекчиями мощностью 4-8 м. перекрытыми в районе
озера Невзорова морскими плиоценовыми глинами.
В конце миоцена произошло обрушение сводового поднятия и обособление
морфоструктуры Новой Земли по сиотеме разломов, с которыми связана находка
вулканитов на восточном побережье Северного острова с возрастом по
калий-аргону менее 10 млн. лет.
В плиоцене началась трансгрессия, продолжавшаяся (возможно, с
перерывом) в раннем плейстоцене; с ее началом связано формирсвание педиплена на
современных высотах 600-800 м. В среднем плейстоцене район испытал
неглубокое эрозионное расчленение, оледенение, сопровождавшееся выработкой пе-
диментов на высотах 260-360 м и сменившееоя бореальной (казанцевской, ми-
кулинской) трансгрессией. По определению абсолютного возраста раковин,
начальная фаза трансгрессии датируется временем около 146 тыс. лет назад. С
трансгрессией связано формирование осадков кумжинской толщи мощностью до
30 м, пологонаклонной морской равнины на высотах 200-300 м серии террас (в
частности, на абсолютной отметке (а.о) .220-240 м). Результаты изучения фо-
раминифер указывают на двукратное повышение уровня моря в бореальное
время.
В конце зырянского времени море отступило, произошло врезание долин
на глубину 50-200 м и развилось оледенение (мощность ледниковых отложений
2-5 м). В каргинское (молого-шекснинское; абсолютная датировка древесины
на а.о. 120 м 40330+940 лет) время произошла трансгрессия до отметок около
200 м, сопровождавшаяся накоплением морских, ледово-морских, аллювиаль-
но-морских отложений крестяхинской толщи и формированием морской
террасированной равнины на высотах 80-200 м. Наиболее выразительна абразионно-ак-
кумулятивная терраса на а.о. 180-200 м.
В сартанское время произошла глубокая (до 100 м ?) регрессия,
сопровождавшаяся возникновением горно-долинного оледенения, заложением
современной гидросети и ее врезанием на 30-40 м; накапливались ледниковые, вод-
ноледниковые и озерные осадки абросимовской толщи мощностью до 12 м.
Абсолютный возраст торфа из озерных отложений - 15310±650 лет. Признаки
покровного оледенения в позднем плейстоцене в рельефе и осадках Вкного
острова Новой Земли отсутствуют.
В голоцене возникло современное оледенение, в долинах сформировалась
серия речных террас (16-20, 5-8, 3 м, два уровня поймы - 1.5 и 0.3-0.5 м),
а на побережьях серия морских террас: 42-60. 22-40. 12-20, 6-10 м и совре-
менная - 2-3 м (Красножея и др., 1982, 1986).
Сделанный на основании материалов геологической съемки вывод о
существенно меньших, чем предполагалось ранее, масштабах оледенения Новой
Земли в позднем плейстоцене находит свое подтверждение в результатах
геолого-геоморфологических исследований заливов западного побережья
архипелага (Дунаев и др., 1990; Павлидис и др., 1990). Эти исследования показали,
что коренное ложе дна фьордовых заливов, расположенных севернее 75° с.ш.,
перекрыто маломощными ледниковыми отложениями позднего плейстоцена и голо-
ценовыми приледниковыми илами с примесью материала ледового разноса.
В заливах центральной части побережья, между 75° и 73° с.ш., мощность
выполняющей толщи намного больше и строение ее сложнее. Как показало сейс-
мопрофилирование в одном из заливов этой толщи, мощность рыхлых отложений
здесь достигает 150-200 м, и она состоит из шести горизонтов, из которых
нижние разделены стратиграфическими перерывами, а верхние накапливались
без существенных перерывов: По-видимому, в позднем плейстоцене выводные
ледники в основном не перекрывали заливы, и в них происходило накопление в
подледных условиях водно-ледниковых отложений, постепенно в голоцене
сменившееся осаждением темно-серых илов с относительно небольшой примесью
материала ледового разноса. В этом районе на местных водоразделах
сохранились в рельефе уступы морских террас.
В заливах, расположенных южнее 73° с.ш., морфологически не являющихся
фьордами, обогащенные органическим углеродом голоценовые илы подстилаются
ледниково-морскими отложениями позднего плейстоцена - плотными черными
глинами со значительной примесью щебнистого материала.
Судя по этим данным, в позднем плейстоцене на шельф выходили в
основном ледники Северного острова: в центральной части побережья архипелага
выводные ледники преимущественно останавливались в вершинах фьордов.
Вышедшие на шельф ледники сливались в Западно-Новоземельском желобе и
образовывали мощный выводной поток, двигавшийся на юг: в районе мыса Сухой Нос
этот образовал крупный конечно-моренный комплекс. В разрезе отложений,
заполняющих Южно-Новоземельский желоб, признаки воздействия ледников на дно
отсутствуют (Павлидис и др., 1990; Дунаев и др.. 1990).
Судя по немногочисленным абсолютным датировкам, для современного
этапа развития характерны очень высокие скорости поднятия: в районе
полуострова Адмиралтейства за последние 200 лет скорость поднятия составила
15 мм/год, за 1400-1800 лет - 4-5 мм/год (Ковалева, 1974). Судя по
интервалам высот казанцевских и каргинских террас, на регрессивных фазах
развития этих бассейнов скорости поднятия островов также были очень высоки.
Многие черты современной морфологии речных долин и побережий также
определяются этим явлением.
В историческом плане такие интенсивные вертикальные движения играют
большую роль в эволюции прибрежных экосистем, обуславливая миграцию
батиметрических зон по вертикали и латерали, а также характер морфолитодинами-
ческих процессов.
Высокие скорости свидетельствуют в пользу изостатичеокой природы
движений, однако, в свете приведенных выше данных об отсутствии следов
покровного оледенения на Южном острове в позднем плейстоцене и голоценовом
возрасте современного ледникового покрова Северного острова проблема
соотношения гляцио-. гидроизостазии, собственно тектонических движений,
оледенений и трансгрессий далека от разрешения и требует дальнейшей разработки.
Особенно важным представляется получение необходимого количества
абсолютных датировок. Это дало бы, в частности, возможность провести детальный
анализ следующей предполагаемой последовательности событий: относительно
ранняя деградация крупных низкоширотных ледниковых покровов - эвстатичес-
кое повышение уровня океана - увеличение мощности слоя воды в чаше Барен-
цевоморского шельфа - прогибание шельфа - трансгрессия на островной суше -
ее дегляциация - гляциоизостатический и гидроизостатический
(компенсационное воздымание периферии погружающегося шельфа) подъем островов -
регрессия в их пределах - новое оледенение.
По данным А.С.Красножен и соавторов (1986), нами были составлена
графики высотного положения разновозрастных морских террас Новой Земли и
вертикальных движений архипелага в позднем плейстоцене с учетом изменений
уровня Мирового океана (рис.1 А,Б). При построении графиков были сделаны
следующие основные допущения: принимая проведенное указанными
исследователями на основании фациального и палеоэкологического анализа отнесение
конкретных террас к определенным фазам трансгрессии - восходящей,
максимальной, нисходящей - положение их на временной оси показано условно,
поскольку абсолютные датировки доголоценовых отложений отсутствуют (за
исключением террасы с высотой 120 м и возрастом около 40 тыс. лет; для
начальных стадий казанцевской трансгрессии указан возраст - около 146 тыс.лет.
но отсутствует высота); изменения уровня Баренцева моря приняты
аналогичными таковым в позднем плейстоцене Мирового океана: трансгрессия
последнего межледниковья несколько превышала современный уровень (до +10 м).
повышение уровня в среднем вюрме было несколько ниже современного (до -10 м).
величина двух гляциоэвстатических регрессий принята равной - около 100 м.
В действительности регрессия раннего вюрма была, по-видимому, несколько
меньшей - до 50 м и состояла из нескольких стадий, а позднего вюрма -
несколько большей (Каплин. 1976; Мысливец и др., 1976; Chappel, Shackleton,
1986). Возрастные рубежи зырянского и сартанского оледенений приняты по
В.М.Макееву (1989).
Нижняя кривая показывает изменение высотного положения условной
точки, соответствующей современной береговой линии. Ее глубина определялась,
исходя из высоты соответствующей террасы с поправкой на положение уровня
океана в это время.
Несмотря на вполне очевидную гипотетичность, построенный график
вертикальных движений архипелага позволяет отметить некоторые особенности.
Так. общее тектоническое поднятие Новоземельского орогена в позднем
плейстоцене происходило с интегральной скоростью около 2 мм/год; скорость
поднятия в голоцене намного выше и достигает 10-13 мм/год. На это поднятие
накладывались пульсации изостатической природы, хотя не исключены и
собственно тектонические движения возвратно-поступательного характера. Во
время пульсаций скорости существенно превышали среднюю для позднего
плейстоцена.
На фоне общего поднятия довольно отчетливо выделяются два цикла,
каждый из которых охватывает трансгрессию, оледенение и позднее послеледни-
ковье. Максимумы обеих трансгрессий отмечены относительными опусканиями,
причем для казанцевского времени величина опускания Северного острова
может значительно - на 100 м - превышать показанную на графике величину,
если подтвердится соответствующий возраст террас на высотах 400-420 м. Как
отмечалось выше, возможно, эти опускания имеют гидроизостатический харак-
тер и обусловлены вовлечением архипелага, в общее погружение Баренце
во-Карского шельфа.
ВЫСОТА,*
300
200
ЮО
300
2W
200
Л
КАЗАНЦЕВСКАЯ
ТРАНСГРЕССИЯ
150
М
120
£9
80
ЗЫРЯНСКОЕ
ОЛЕДЕНЕНИЕ
КАРГИНСКАЯ
ТРАНСГРЕССИЯ яп
САРТАНСКОЕ 5Р
40
ОЛЕДЕНЕНИЕ -^
I I /I /\х
А
Л
-шо l ВРЕМЯ, тыс. лет
150 W 130 120 НО 100 90 80 70 60 50 40 30 20 10 0
ГЛУБИНА, м
390
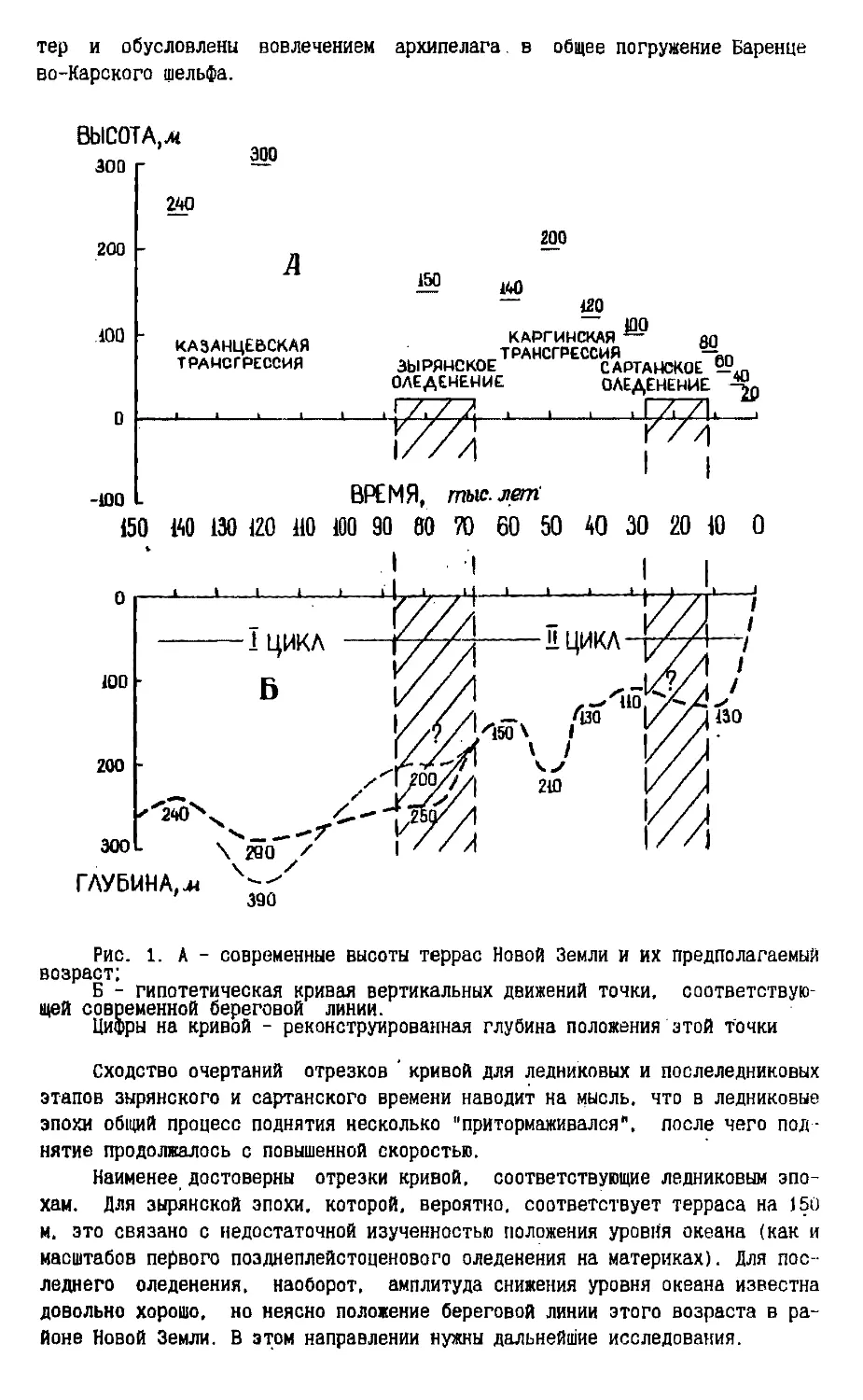
Рис. 1. А - современные высоты террас Новой Земли и их предполагаемый
возраст;
Б - гипотетическая кривая вертикальных движений точки,
соответствующей современной береговой линии.
Цифры на кривой - реконструированная глубина положения этой точки
Сходство очертаний отрезков ' кривой для ледниковых и послеледниковых
этапов зырянского и сартанского времени наводит на мысль, что в ледниковые
эпохи общий процесс поднятия несколько "притормаживался", после чего
поднятие продолжалось с повышенной скоростью.
Наименее достоверны отрезки кривой, соответствующие ледниковым
эпохам. Для зырянской эпохи, которой, вероятно, соответствует терраса на 150
м. это связано с недостаточной изученностью положения уровня океана (как и
масштабов первого позднеплейстоценового оледенения на материках). Для
последнего оледенения, наоборот, амплитуда снижения уровня океана известна
довольно хорошо, но неясно положение береговой линии этого возраста в
районе Новой Земли. В этом направлении нужны дальнейшие исследования.
Глава 3. АБИОТИЧЕСКИЕ УСЛОВИЯ МОРЯ
3.1. Исторические и современные представления
о морских водах района Новой Земли
Новая Земля считается не только условной границей между
субарктическим Баренцевым и арктическим Карским морями, но и настоящим барьером для
взаимодействия водных и воздушных масс. Казалось бы. для последних острова
архипелага не представляют серьезного препятствия, потому что интенсивные
воздушные течения в силу их масштабности преодолевают гораздо более
высокие и протяженные препятствия, существующие на материках. Однако в океане
существуют специфические условия энерго- и массообмена с атмосферой. Здесь
на гидрометеорологический режим влияет не столько орография' земной
поверхности, сколько фактор разделения архипелагами водных масс. В данном случае
имеются в виду водные массы, обладающие высокой способностью отеплять
атмосферу и поступающие из Атлантики, и ледовые воды, не обладающие такой
способностью и формирующиеся в результате действия трех факторов:
интенсивного охлаждения поверхности океана, выпадения твердых атмосферных
осадков и стока рек. в основном, Печоры. Оби и Енисея. Эти опресненные воды,
распространяющиеся на значительной акватории, замерзают, не только
вследствие осенне-зимнего охлаждения, но и при соприкосновении с высокосолеными
водами, имеющими отрицательную температуру круглый год.
К западу от Новой земли противоборство между атлантическими и
ледовыми водами заканчивается в пользу первых, а к востоку от нее - в пользу
вторых. Это стало известным еще в прошлом веке после морских экспедиций
вдоль берегов Новой Земли, которые провели гидрометеорологические и
океанографические измерения в водах Баренцева и Карского морей.
Острова Новая Земля долгое время были центром внимания географов,
наибольшую загадочность они представляли в период от Г.Меркатора до
В.Баренца. Большинство иоториков считает, что открытие Новой Земли призошло в
XIII-XIV веках. Некоторые специалисты по истории русского Севера
допускают, что поморы еще в XI в. посещали Железные Ворота, как называли в
древности пролив Карские ворота.
В XVI-XVIII вв. представления географов о гигантской Новой Земле были
положены в основу картографии наравне с другими более изведанными
материками. В те далекие времена люди еще не знали очертаний берегов Новой
Земли, бытовало даже мнение о том, что она соединена с Северной Америкой.
Другая крайняя точка зрения заключалась в том, что Новая Земля - это
только скопление льдов, которое время от времени собирается в одно место. По
мнению Герарда Меркатора. Новая Земля простиралась далеко на восток и
отделялась от Америки Проливом Аниан.
Впервые обследовать Новую Землю по всей протяженности удалось с моря
в 1596 г.. когда экспедиция Виллема Баренца обогнула северную точку, мыс
Желания, и в попытке пройти на юг вдоль восточного берега архипелага
21 августа судно было зажато плавучими льдами в небольшой бухте на
северо-восточном берегу Новой Земли, названной Ледяной гаванью. Это
историческое место стало первым источником информации о климате Арктики.
В материалах экспедиций В.Баренца 1594-1597 гг. описана природа Но -
вой Земли и очерчен западный берег архипелага. В 1598 г. в Амстердаме была
издана первая карта Новой Земли. Судя по русским названиям почти всех
географических объектов на ней, российские поморы обжили западное побережье
южного острова и делали попытки проникновения на северный остров
архипелага. На карте 1598 г. береговая черта обрывается на параллели Ледяной
гавани и далее на восток обозначена суша вплоть до мифического пролива,
разделяющего Новую Землю и Америку.
Все дальнейшие наблюдения ученых, их попытки представить теплые
морские течения, прорывающиеся сквозь ледовые арктические заграждения, были
связаны с глобальной системой течений, начинающейся от Флориды, где
зарождается Гольфстрим. Система Гольфстрима, включающая в себя течения
Северо-Атлантическое, Ирмингера. Восточно-Гренландское, Норвежское.
Шпицбергенское, Нордкапское. Мурманское и Новбземельское, снабжает Арктику теплыми
высокосолеными водами, которые ооздают специфические благоприятные условия
для существования живых организмов от бактерий.. фито- и зоопланктона до
китов и других млекопитающих.
Познание заполярных морских районов успешнее всего могло
осуществляться через климатические исследования. Немецкий метеоролог Генрих Лове
составил первые карты изотерм земного шара для каждого месяца. С 1848 года
он высказывает мысль о проникновении Гольфстрима до Новой Земли.
Основанием для такого заключения служили аномально.высокие температуры воздуха в
проливе маточкин Шар на Новой Земле.
Новая Земля, таким образом, стала привлекать наибольшее внимание не
только как восточная граница Баренцева моря, но и как восточный предел
распространения вод Гольфстрима, а вернее северо-восточных его
ответвлений, находящихся за тысячи миль от мест его рождения.
Август Петерман с 1852 года развивал мысль о теплом свободном ото
льда Северном Полярном море. В 1870 году до 60 норвежских судов ходили к
Новой Земле на звериный промысел. Один из норвежских капитанов,. Иоганне-
сен, беспрепятственно обогнул Новую Землю с севера и вблизи мыса Нассау
встречал предметы из обихода норвежских рыбаков, которые свидетельствовали
о переносе их с Лофотенских островов и из Финмаркена. Подобные факты
укрепляли Петермана в том, что течения северных ветвей системы Гольфстрима
распространяются повсеместно в • Северном Ледовитом океане, препятствуя
сплошному оледенению северных морей. На основании наблюдений и записей
вахтенных журналов норвежских судов им были построены карты изотерм
Баренцева и Карского морей.
В 1870 году опубликована обобщающая все материалы по Северному
Ледовитом океану работа, где Петерман (Petermann, 1870) выражает убеждение и
том. что ветви Гольфстрима доходят до Новой Земли и проникают в Полярный
бассейн.
В 60-е годы предположения о влиянии Гольфстрима на воды Мурмана
высказывались русскими исследователями А.миддендорфом, Ф. Яржинским и
Н.Данилевским. Особенно большое значение имело участие авторитетного ученого,
известного независимостью взглядов и бескомпромисностью. академика Мидден-
дорфа. принявшего активное участие в проверив гипотезы Петермана на
паровом корвете "Варяг", совершившим ; под командованием вице-адмирала К.Пось-
ета рейс по маршруту Архангельск - Новая Земля - Исландия в 1870 г. Уае 29
сентября 1870 г. Миддендорф выступил в Петербурге с докладом на тему
"Гольфстрим на востоке от Нордкапа", в котором не только подтвердил
истинность гипотезы Петермана, но и поведал о главном теплом течении Баренцева
моря, названном им Нордкапским, и о его огромной роли в смягчении климата
заполярных широт. Отдавая должное блестящим предвидениям Петермана, автор
доклада назвал петермановские утверждения вопреки считавшимся в то время
слишком смелыми "лишь осторожными".
Следует отметить, что работавший в те годы на Мурмане русский
исследователь Э.Майдель разделял точку зрения Миддендорфа.
Идеи глубокого проникновения теплых атлантических вод в Ледовитый
океан стали приобретать не только многих сторонников среди ученых, но и
овладевать умами мореплавателей. В 1869 г.. вслед за Скорессби и- Парри
теплые воды восточнее о. Медвежий были обнаружены Бессельсом.
К тому же обилие метеорологического материала в истории исследования
геосфер способствовало более высокому уровню изучения океана, позволяя
проводить' статистические расчеты и оперировать языком цифр: Экхольм
оценил влияние Гольфстрима на климат Швеции и всей Европы (Ekholm, 1899). Он
сделал вывод о том; что влияние тепла Гольфстрима распространяется на
большую часть севера Европейской России. По картам январских изаномал
температуры воздуха, Мурманский- берег характеризуется величинами от +20 до
+13°. летом аномалии на Кольском п-ове близки к нулю: от -1 до +4°. По
этим очень скупым цифровым данным был сделан важный вывод о мощном
поступлении тепла в Баренцево море именно зимой, когда активизируется
энергообмен между океаном и атмосферой в полярных районах.
Известный норвежский исследователь Х.Мон, базируясь на материалах
температуры воды, атмосферного давления и преобладающих ветров,
наблюдаемых в экспедициях 1876-1877 гг., пришел к выводу о том, что воды Нордкап-
ского течения, следующие вдоль Норвежских и Мурманских берегов, изменяют
направление в районе юго-восточных мелководий Баренцева моря. На севере
под действием восточных ветров теплое течение принимает западное
направление. Часть теплых вод уходит в Полярный бассейн, огибая с севера Новую
землю. Холодные полярные воды, считал Мон, поступают из Арктики в
поверхностном слое вместе с плавучим льдом. В то же время автор допускал
формирование холодного промежуточного слоя в процессе зимней конвекции
(Mohn,1885).
Во второй половине XIX в. на Новую Землю устремились геологи,
ботаники, зоологи, и в начале XX в. началось настоящее паломничество членов
Общества любителей естествознания,. антропологии и этнографии, художников.
Безусловно, участники экспедиций были знакомы с гипотезами влияния
Гольфстрима на западный берег Ижного острова Новой Земли и на все северное
побережье Кольского п-ова. Биологи Общества смогли убедиться в обитании
неарктических Форм морских растений и животных в водах, омывающих архипелаг.
Сравнительная доступность для экспедиций и большое разнообразие
неведомых ранее арктических природных комплексов привлекли внимание многих
исследователей, й безусловно, всех интересовали характеристики климата и
механизм его изменчивости. После того, как географическое положение Новой
Земли было установлено и открыты острова, расположенные к северу и востоку
от нее, образ естественной подводной стены между Баренцевым и Карским
морями был принят в качестве основополагающего для моделирования морских
течений. Не случайно самый большой пролив между архипелагом и континентом
был назван Карскими воротами. И до сих пор принцип сообщающихся сосудов
лежит в основе оценок водообмена через проливы, главным образом, через
Карские ворота.
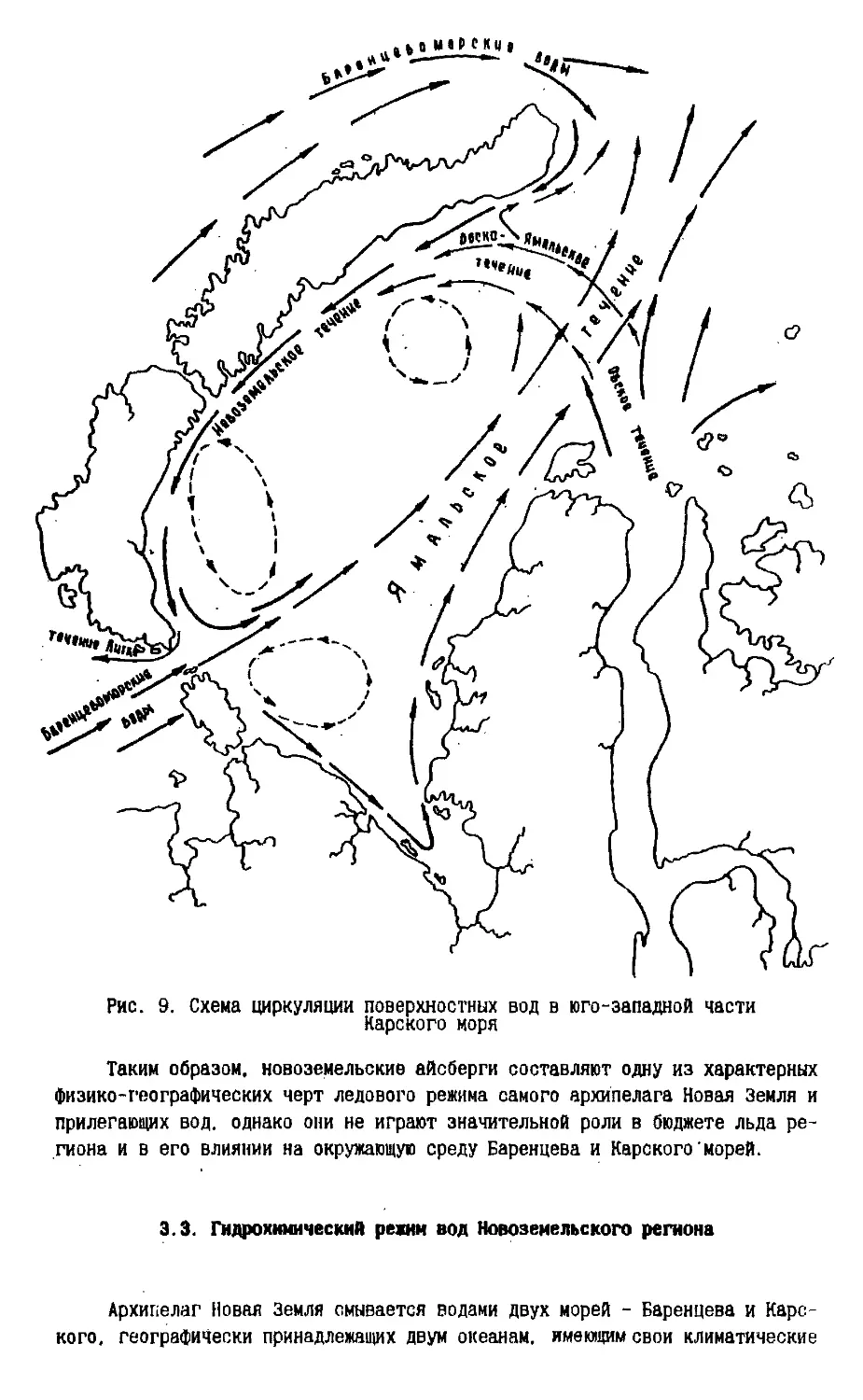
Наиболее общей чертой циркуляции вод Баренцева и Карского морей,
рассчитываемой с помощью прочно вошедшего в обиход океанологов
динамического метода вычисления течений, была циклоничность движения вод,
предполагаемая еще X.Моном и подтвержденная Ф.Нансеном. Вслед за Ф.Нансеном такое
представление о схеме циркуляции подтвердили В.А.Березкин (1938),
Н.Н.Зубов (1944). А.В.Соколов (1932) и А.И.Танцюра (1959).
Если следовать схемам течений Баренцева и Карского морей, вокруг
Новой Земли существует антициклонический круговорот вод, то есть западный
берег омывается течением, следующим с юго-запада на северо-восток, а
восточный берег - течением, направленным с северо-востока на юго-запад. Но
обычно предпочитают говорить о циклонических круговоротах Баренцева и
Карского морей в целом, поскольку такое представление служит аргументом в
пользу существования конвективного опускания вод в центральных районах
морей. Существуют подробные описания расходов вод,. тепловых и ледовых
характеристик различных районов (Чаплыгин, 1961; Панов, 1961; Никифоров и др.,
1974; Степанов, 1979). которые мы не приводим, потому что они вызывают
гораздо больше вопросов, чем дают ответов.
Учитывая неравномерность обеспеченности исследуемой акватории
измерениями и отсутствие истинных представлений о механизме трансформации вод,
мы будем использовать только самые надежные статистические материалы
наблюдений для получения самых общих выводов о свойствах водных масс,
омывающих Новую Землю.
Наилучшим показателем притока теплых вод служит положение границы
плавучего льда (ГПЛ). Можно отрицать воздействие на ледовитость вод,
характеризующихся высокой температурой, но нельзя не согласиться с тем, что
свободная ото льда акватория занята именно теми водами, которые своей
способностью обогревать огромные массы воздуха смягчают климат Арктики. Эти
воды принадлежат системе Гольфстрима.
Достаточно длительное наблюдение за положением ГПЛ дает объективное
представление о динамике фронта ледовых арктических вод. Нет необходимости
рассматривать круглогодичную изменчивость ледовитости исследуемого района,
потому что в летний период только на крайнем севере наблюдаются ледовые
поля. Начало летнего периода, судя по максимальной площади открытой воды,
наступает в июле, а окончание - в октябре. На рис. 2 представлены положения
ГПЛ с ноября по июнь за И-летний период наблюдений.
Если сопоставить карту циркуляции Баренцева моря и динамику ледового
покрова, то можно заметить ряд противоречий в представлении течений,
которые заключаются в следующем.
Согласно принятой специалистами схеме течений, вдоль-Новой Земли
существует постоянная струя теплых вод, навстречу которой движется холодное
Центральное течение, отделяющееся от теплого Западного Новоэемельского
течения зоной смешения. Холодные воды располагаются над подводными
возвышенностями, а теплые - над Центральной впадиной. Ежемесячная междгодовая
динамика ГПЛ свидетельствует об отсутствии постоянного выноса ледовых вод
40 45 50 55 6D 65 70 40 '45 SO 55 60 66 70
—l ^т- I Г ' " I 1 1 1 1 1. ' |
Рис. 2. Положение границы плавучего льда с ноября по июнь 1977-1978 гг.
Центральным течением и таким же отсутствием ежегодного притока теплых вод,
способным противостоять зимнему оледенению акватории, расположенной к
западу от Новой Земли. Таким образом, понятия теплых и холодных вод. а также
представления о постоянной схеме циркуляции вызывают большие сомнения.
Эти представления имеют глубокие корни, •■ связанные с разработкой
первых карт морских течений на основе эпизодических измерений температуры
воды и предположений о том, что изотермы оконтуривают потоки глобальных
систем течений. При этом физическая природа циркуляции не содержит конкретных
физических обоснований. Впоследствии для расчетов течений стали
предлагаться гидродинамические методы, которые не принесли положительных
результатов в оилу целого ряда причин, в основном из-за невозможности проведения
вычислительных экспериментов, направленных на доказательство правомерности
применения предлагаемых методов для расчета тех физических процессов,
которые вызывают наблюдаемые изменения гидрофизических свойств морских вод.
В связи с этим возникает вопрос о хотя бы приблизительном, но
физически обоснованном подходе к анализу наблюдаемых изменений показателей
водных масс. Прямыми показателями циркуляции вод или тепло- и влагообмена
между океаном и атмосферой мы не располагаем, а о косвенных
характеристиках этих процессов можно судить по материалам измерений температуры,
солености и растворенного кислорода. Два последних параметра имеют особенно
важное значение, потому что соленость является показателем влагообмена
между океаном, атмосферой, ледовым покрытием криосферы и речным стоком
гидросферы, а кислород - показателем" конвекции, наиболее интенсивно
проявляющейся в' полярных районах.
Отсутствие большого дефицита кислорода в Северном Ледовитом океане
свидетельствует о максимальной конвекции, благодаря которой вся водная
толща Арктического и Северо-Европейского бассейнов аэрирована до самого
дна до величин не менее 8058-ной относительной концентрации растворенного
кислорода. Лишь прибрежные мелководные районы арктических морей,
находящиеся под влиянием, обильного стока рек летом и интенсивного
льдообразования зимой, характеризуются дефицитом кислорода, доходящим, например, в
южной части Карского моря до 6058.
Ограничение действия конвекции обусловлено резким пикноклином за счет
высокой разности между соленостью вод поверхностных и подповерхностных
слоев. Разности температуры воды в этих же слоях не создают больших
эффектов расслоения, что определяет специфические условия формирования
стратификации водной толщи. Более южные районы Мирового океана обладают высокими
диапазонами изменения температуры.. поэтому в системах циркуляции открытого
океана наблюдается корреляция между температурой и соленостью.
Примером такой системы циркуляции является система Гольфстрима, воды
которой омывают подводные склоны Новой Земли. Вплотную атлантические воды
подходят к западному берегу архипелага зимой, во время наиболее
интенсивного взаимодействия между океаном1 и атмосферой (Адров и др., 1993).
К востоку от Новой Земли даже в холодные периоды года наблюдается
акватория, свободная ото льда. Она располагается вдоль подводного склона
желоба Св.Анны. Поскольку мы не располагаем данными зимних съемок в Карском
море и не можем утверждать.. что пространство открытой воды имеет отношение
к атлантическим водам, то выскажем лишь предположение об этом. Аргументом,
подтверждающим это предположение, является сходство термогалинных свойств
основной масс'ы вод Центральной впадины и желоба Св. Анны.
Из.наблюдений известно, что глубоководные районы полярных морей, под-
аерженных адвекции вод систем циркуляции открытого океана, гораздо дольше
сохраняют способность сопротивления арктическому холоду. Это объясняется
действием механизма конвекции, охватывающей атлантические воды до самого
дна, В мелководных районах тепло атлантических вод расходуется быстрее,
поэтому поверхность моря здесь покрывается льдом раньше.. К западу от Новой
Земли таким районом является Центральная возвышенность, а к востоку - весь
юго-восточный склон Карского моря,
В 1929-1932 гг. Н.Н.Зубов разработал теорию образования холодного
промежуточного слоя и показал, что холодные воды Центральной и других
возвышенностей образуются в результате зимней^ конвекции. Автор назвал
Центральную возвышенность лабораторией холодных придонных вод (Зубов, 1929,
1944).
Акватории, свободные от плавучего льда зимой, можно рассматривать как
полыньи, существование которых имеет важное значение в формировании водных
масс и гидрометеорологическом режиме Арктики. Великие полыньи Северного
Ледовитого океана могут сохранять свою сопротивляемость арктическому
холоду только вследствие работы механизма конвекции в водах, имеющих высокую
соленость на поверхности. Пример по; )бных вод мы имеем в Баренцевом мрре,
в районах Центральной впадины и Центральной возвышенности. Сходство
механизмов взаимодействия океана и атмосферы во всех районах океана позволяет
сделать заключение о единстве природы трансформации незамерзающих вод по
обе стороны от Новой Земли.
Действительно, статистические термогалинные диаграммы вод Западного и
Восточного Новоземельских течений отличаются друг от друга лишь
незначительно диапазонами температуры незамерзающих (атлантических) и солености
ледовых (неатлантических) водных массЛрис.З).
Теоретические положения расчета адвективной и конвективной
составляющих термогалинной и термооксигенной трансформации водных масс содержатся в
работе, посвященной системе Гольфстрима (Адров, 1993). Линейные
зависимости между температурой и соленостью, которые были положены в основу расчета
уравнений трансформации водных масс, присутствуют и на рис.3, только в
S%o
Рис. 3, Экстремальные T.S-профили атлантических (сплошная линия)
и неатлантических (пунктир) вод к западу (а)
и востоку (б) от новой Земли
сильно "урезанном" виде. Для сравнения приведем статистические точечные
диаграммы, построенные по данным температуры и солености, • измеренным аа
много лет в 10е-ных квадратах, расположенных западнее района Новой Земли
(рио.4а)' и отражающих термогалинную структуру вод Баренцева. Норвежского
и Гренландского морей. Другую крайность представляют диаграммы вод
Арктического бассейна (рис.46), на которых атлантические воды представлены
едва заметными отрезками прямых, характеризующих воды с почти одинаковой,
максимальной для каждого рассматриваемого квадрата соленостью.
Чо
S3
о..
UO
о
*е
il
:Г
П
: Щ
п
■Лая
:Ш
•'.'•»•,'*'•.!
I Г I ""I 1 ' » Т*
ег
-|—i—i—i—г~1—i—i—г
ег £i и с I-
бв сг а и
I- ег сг а и
Расчет границ водных масс и вычислительные эксперименты на материалах
Баренцева моря (Адров, Смоляр, 1988; Адров и др., 1992; Адров, Феопентов.
1994) позволят сделать некоторые выводы о пространственной и временной
изменчивости водных масс, окружающих Новую Землю.
Атлантические водные массы, отдавая в атмосферу тепло и влагу,
создают- аномальные условия циркуляции поверхностных вод и дрейфа ледовых полей.
Зимой; то есть в период максимального энергообмена между океаном и
атмосферой, наблюдается значительное увеличение количества атлантических вод,
Летом на поверхности моря атлантических,вод не наблюдается.
Центральная впадина Баренцева моря и желоб Св. Анны Карского моря
всегда заполнены атлантическими водами, почти полностью исчерпавшими запас
тепла и имеющими поэтому температуру, близкую к минимальной.
Для прогнозирования тепловых характеристик сезонов значительно более
показательными, чем температура воды, характеристиками являются величины
солености. Карское море, как самое обильное среди ледовитых морей
пресноводным стоком, имеет обширную акваторию с постоянным опресненным верхним
слоем. Этот слой служит субстратом для ледового покрова только в
мелководной юго-восточной половине моря. В северо-западной части атлантические
воды преодолевают влияние пресноводного отока, холодных арктических
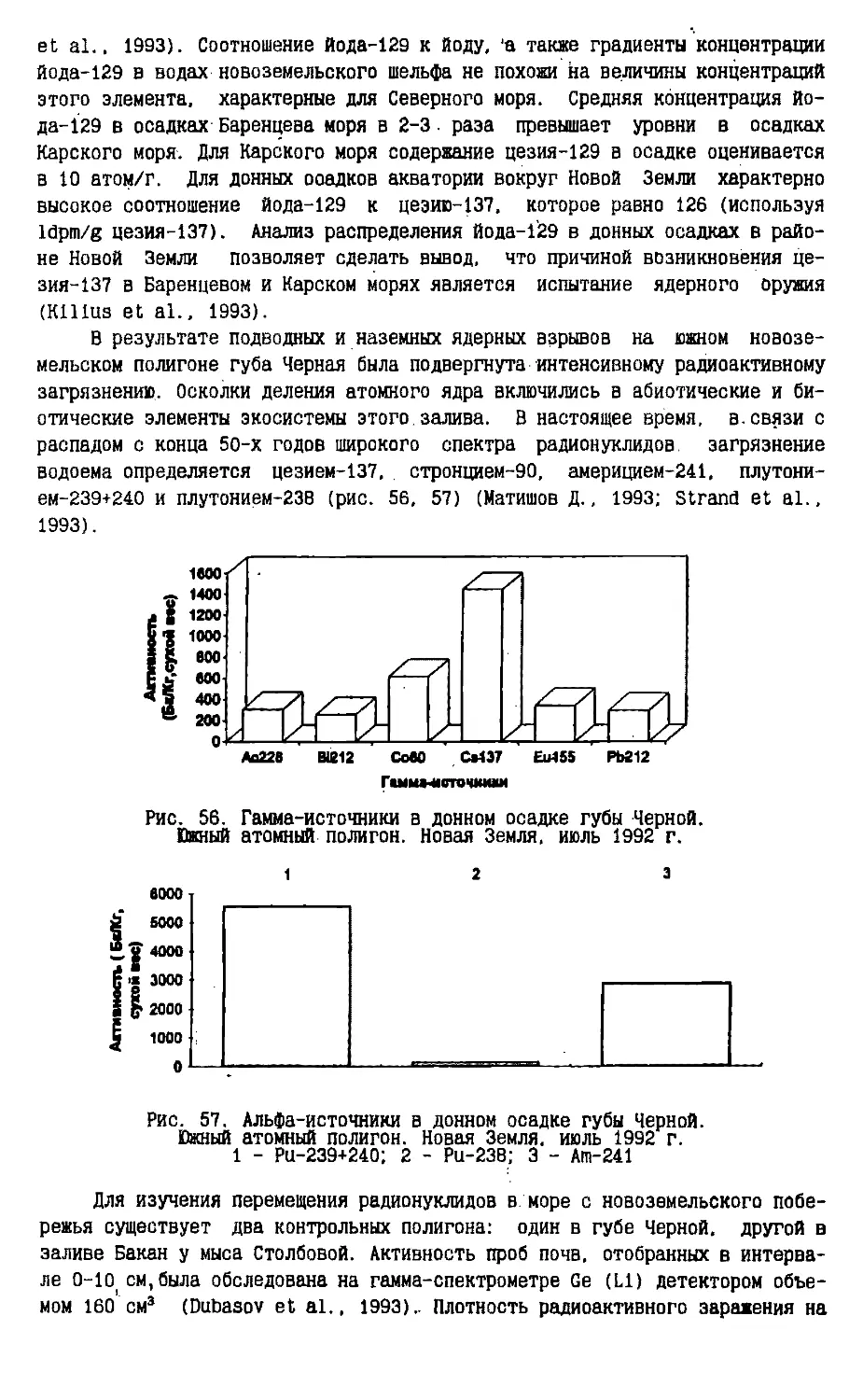
воздушных масс й атмосферных осадков. Там, где слой атлантических вод обладает
достаточной мощностью, формируются пространства воды, свободные ото льда,
которые называются арктическими полыньями.
Судя по масштабности и скорости изменений ледовитости и термогалинных
характеристик морских вод к востоку и западу от Новой Земли, можно
предположить, что эти изменения вызваны не горизонтальным притоком вод, который
имеет слишком малую скорость для того, чтобы вызвать быстрые перемены в
соотношении ледовых и свободных ото льда акваторий, а вертикальным
изменением термогалинной структуры водной толщи вследствие регуляторной
деятельности системы океан-атмосфера. Суть механизма регулирования энерго- и мае-
сообмена между водными и воздушными массами в полярных районах заключается
в том, что максимальное наступление холодных арктических воздушных масс
вызывает максимальные тепло- и влагоотдачу атлантических водных масс и тем
самым удерживается климатическое равновесие. Экстремальные нарушения
режима тепла и осадков не бывают продолжительными, а эпохи похолодания всегда
бывают перманентными и сменяются стсшь же непрерывными периодами
потепления.
3.2. Льды и айсберги
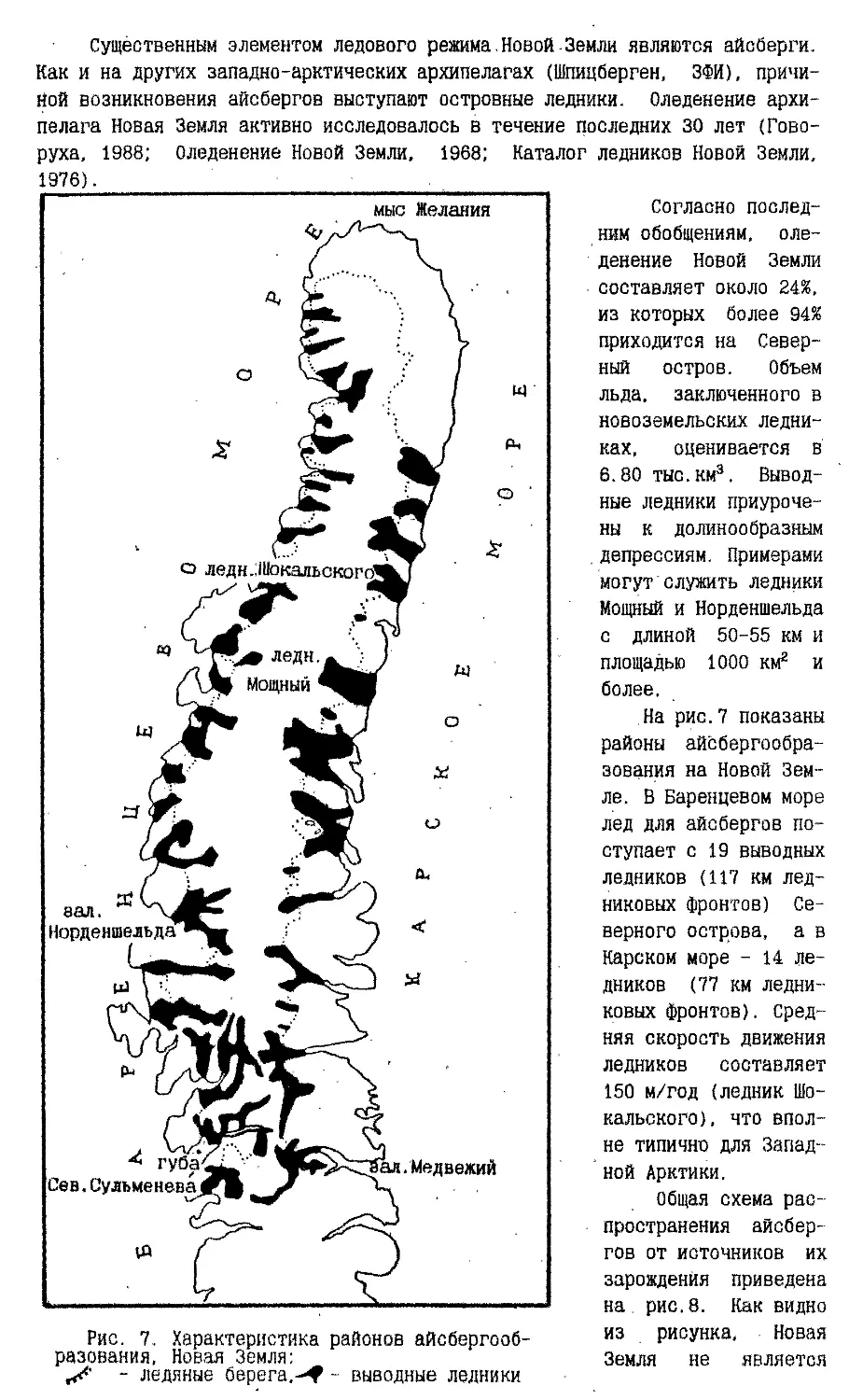
Архипелаг Новая Земля является .естественной протяженной (более
900 км) границей между;двумя морями Западной Арктики - Баренцевым и
Карским.
Главное физико-географическое следствие существования такого
природного барьера - принципиальное различие между морями, омывающими архипелаг.
Одно из них, Баренцево, никогда полностью не замерзает и может
рассматриваться как морской бассейн со свободной) кромкой. Другое, Карское,
общепризнанно как "истинно" арктическое море, которое полностью покрывается
льдом на многие месяцы. Причина таких различий - "экранирование" Новой
Землей мощной адвекции атлантического тепла, поступающего в Арктику с
воздушными (атмосферные циклоны) и морскими (квазипостоянные течения)
потоками (Советкая Арктика, 1970). В результате западные и восточные берега и
прибрежные воды архипелага Новая Земля относятся к различным районам
Атлантической климатической области: Баренцево (включая Печорское) море -
это южный район, а Карское море - восточный район (Атлас Арктики, 1985).
Такое подразделение в пределах одной климатической области хорошо
показывает не только существенные физико-географические различия
рассматриваемого региона, но и некоторые общие черты сходства в развитии
гидрометеорологических и ледовых процессов в районе Новой Земли. Особенно ярко ота
двойственность выступает при рассмотрении ледового режима, подчеркивая
общепризнанное мнение о ледовом покрове как наиболее чувствительном
комплексном показателе взаимодействия атмосферы и гидросферы.
Общими чертами ледового режима зоны вокруг архипелага являются начало
льдообразования, как правило, на чистой воде и полное окружение Новой
Земли дрейфующими льдами в годы, когда гидрометеорологическая обстановка
складывается по типу среднемноголетней. Главными же различиями в этих
принципиально сходных для обоих морей процессах выступают существенные
сдвиги в темпах и сроках протекания процессов льдообразования и таяния.
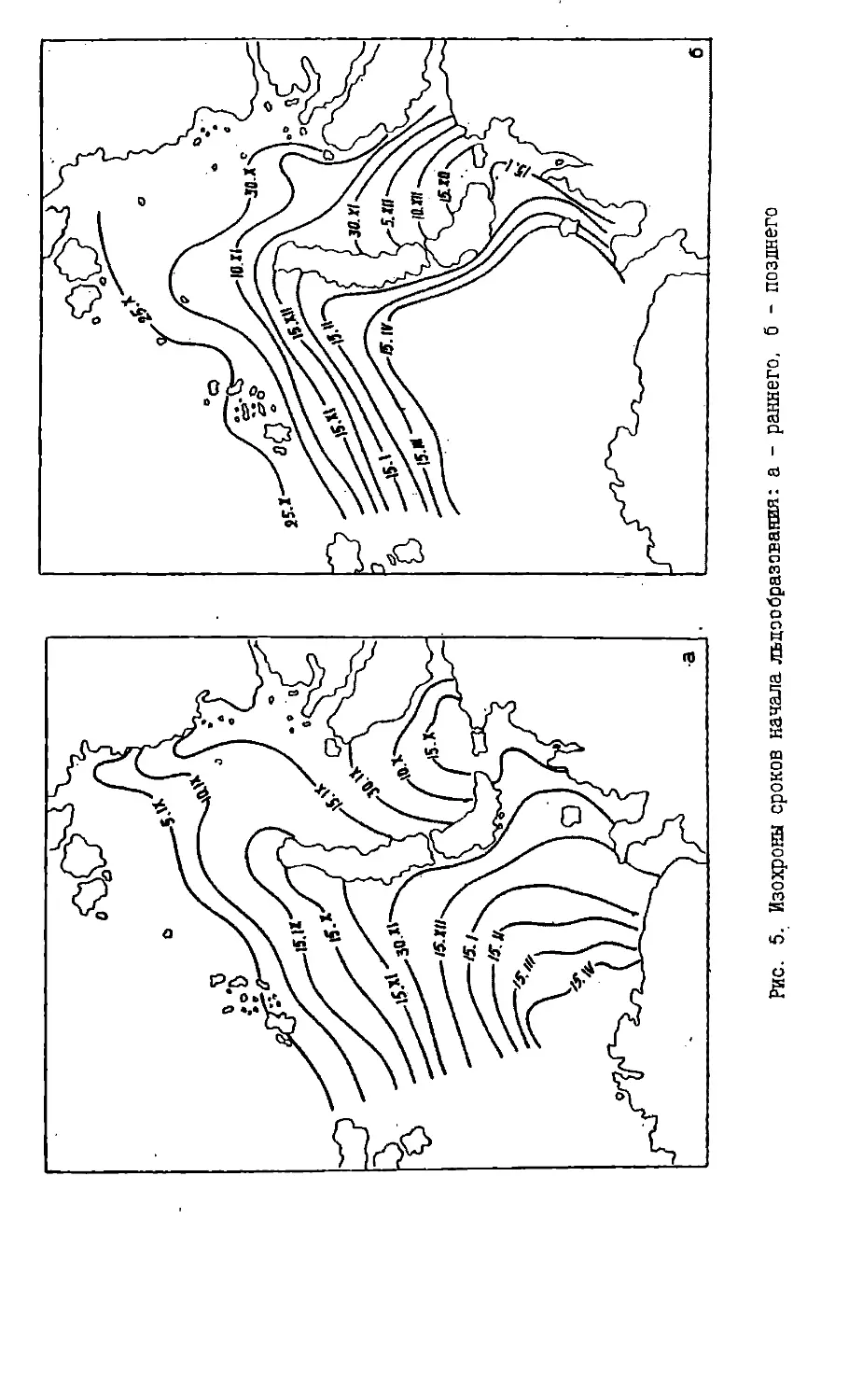
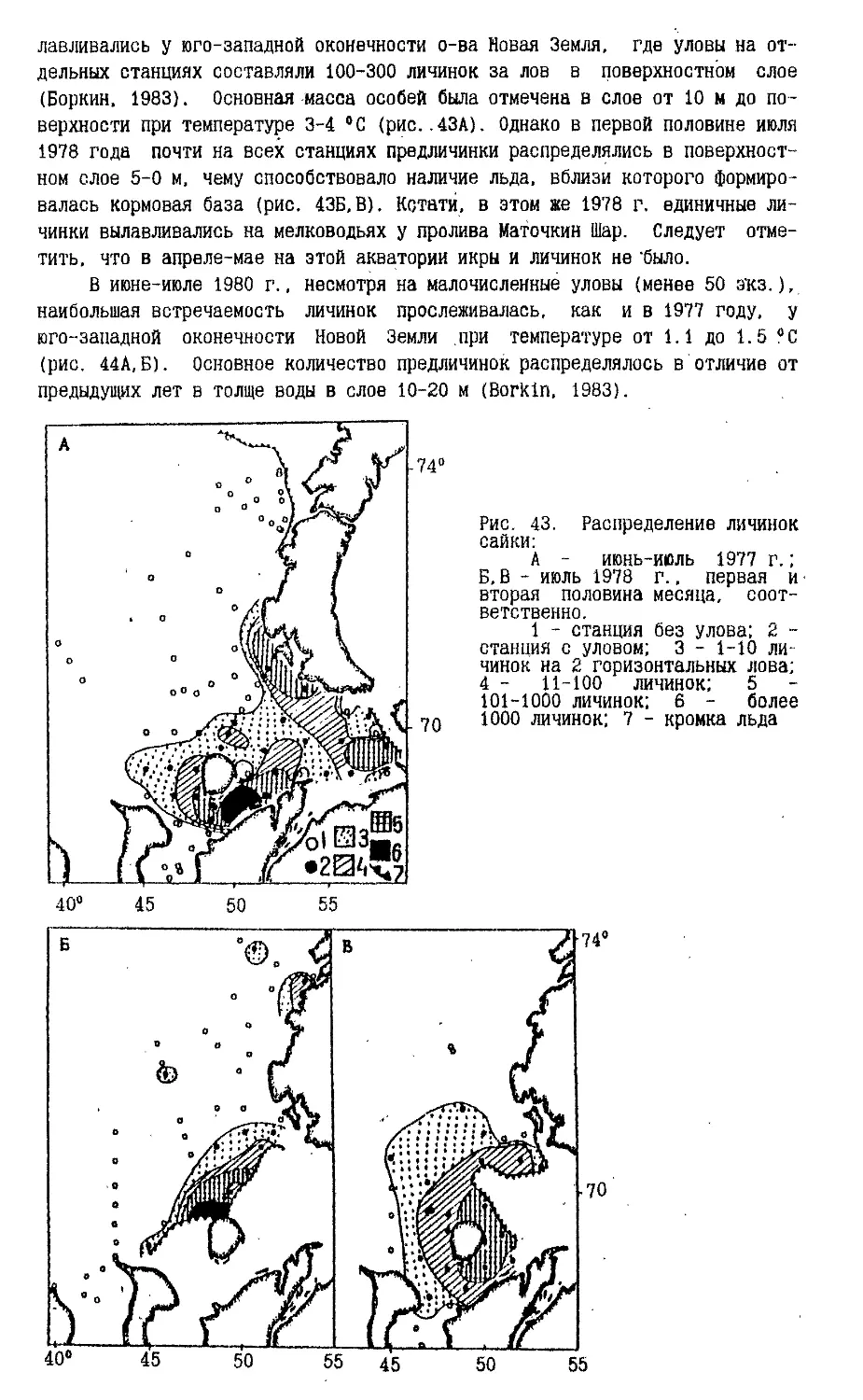
На рис.5 (а,б) показаны изохроны начала льдообразования в Баренцевом
и юго-западной части Карского морей для раннего и позднего (аномалии
среднего режима) наступления льда (Чилингаров, 1976). Хорошо видно, что
архипелаг Новая Земля разрывает изохроны, создавая зональный сдвиг в сроках
замерзания окружающих вод на расстояние до 900 км. Особенно хорошо это
можно проследить на рис.5, где, например, южное положение изохроны 15
октября в Карском море означает практически полное замерзание прибрежных вод
у восточных берегов Новой Земли на всем их протяжении, но лишь только
самое начало льдообразования в Баренцевом море у западных берегов в районе
мыса Желания. Приведенный пример иллюстрирует ту основу климатических
различий к западу и востоку от архипелага Новая Земля, которые проявляются во
всем годовом цикле развития ледовых условий на стыке двух морей.
На рис.6, где показано положение кромки льда для одного из зимних
месяцев, наглядно видно, что только в аномально теплые годы западные берега
архипелага могут быть свободными от льда. В среднемноголетних условиях со
стороны Баренцева моря кромка льда занимает меридиональное положение и
закрывает подходы к новоземельскому прибрежью на всем его протяжении, как
бы смыкаясь с более суровыми Печорским и Карским морями (Доронин. 1986).
Таким образом, архипелаг Новая Земля можно считать по ледовым условиям
частью единого ледового района, западной границей которого является Барен-
цевоморский новоземельский шельф.
Распределение и количество льдов в районе островов Новая Земля
зависит от режима ветров, течений, температуры воздуха. Неподвижный ледяной
покров образуется только в губах, заливах и проливах. У открытых побережий
наблюдается преимущественно однолетний дрейфующий лед (Гидрометеорология и
гидрохимия морей СССР, 1990). Западные берега Новой Земли являются районом
наибольшей сплоченности (до 4-5 баллов) молодых льдов до 30 см толщиной.
По мере замерзания толщина льда увеличивается, и к апрелю у западных
берегов Северного острова возникает наибольшая площадь однолетних тонких
(30-70 см) льдов сплоченностью 3-4 балла. В баренцевоморском секторе
архипелага однолетние толстые льды О 70 см) зимой представляют собой язык
редких и разреженных льдов, генетически связанных с массивом льдов
Карского моря (Зубакин. 1987), К востоку от Новой Земли толщина льда к концу
зилы выше, чем к западу.
Сильная динамичность
ледового покрова
рассматриваемого района
реализуется в повышенной торо-
систости и в формировании
и поддержании новоземель-
ских полыней. Так; при
западных преобладающих
ветрах чаще всего
обрадуется заприпайная полынья
вдоль всего восточного
побережья архипелага
Новая Земля с максимальной
шириной до 60-70 миль
(Добровольский, Залогам,
1965). Очевидно, что
полыньи важны не только для
морских животных, но
имеют существенное
прикладное значение для морских
транспортных операций.
В годовом цикле
существования ледяного
покрова важно отметить факт
сезонного очищения ново-
земельского района от
льдов во второй половине
лета, однако у восточных
берегов архипелага обычно
сравнительно длительное
время сохраняются льды
Новоземельского ледового
массива, полное
разрушение которого происходит в
третьей декаде августа, а
окончательное очищение
акватории - в конце
августа или в начале
сентября. Однако в аномально тяжелые по ледовым условиям годы окончательного
и полного очищения ото льда может и не произойти. Сохранность небольших
скоплений остаточных льдов вблизи архипелага до начала нового
льдообразования при этом не превышает обычно 556 от площади массива.
Припайные льды, особенно в Карском секторе, как правило, толще
дрейфующих. К апрелю они достигают 140-150 см с максимумом до 180-200 см.
Взлом и ^разрушение припая, начинается, соответственно в среднем, на 20-ЗО-е
оутки после начала таяния при толще 70-120 см и протекает в северном
направлении; от южных новоземельских проливов и далее вдоль побережий (Чилин-
гаров, 1976).
Рис. 6. Положение кромки льда для
наиболее ледовитых месяцев трех типов навигаций:
легкой (1), нормальной (2) и тяжелой (3)
мыс Желания
w
А*
о
Существенным элементом ледового режима,Новой-Земли являются айсберги.
Как и на других западно-арктических архипелагах (Шпицберген, ЗФИ),
причиной возникновения айсбергов выступают островные ледники. Оледенение
архипелага Новая Земля активно исследовалось в течение последних 30 лет (Гово-
руха, 1988; Оледенение Новой Земли, 1968; Каталог ледников Новой Земли,
1976). "
Согласно
последним обобщениям,
оледенение Новой Земли
составляет около 24%,
из которых более 94%
приходится на
Северный остров. Объем
льда, заключенного в
новоземельских
ледниках, оценивается в
6.80 тыс.км3.
Выводные ледники
приурочены к долинообразным
. депрессиям. Примерами
могут служить ледники
Мощный и Норденшельда
с длиной 50-55 км и
площадью 1000 км2 и
более.
На рис.7 показаны
районы айсбергообра-
зования на Новой
Земле. В Баренцевом море
лед для айсбергов
поступает с 19 выводных
ледников (117 км
ледниковых фронтов)
Северного острова, а в
Карском море - 14
ледников (77 км
ледниковых фронтов).
Средняя скорость движения
ледников составляет
150 м/год (ледник
Шокальского), что
вполне типично для
Западной Арктики.
Общая схема
распространения
айсбергов от источников их
зарождения приведена
на рис.8. Как видно
из рисунка. Новая
Земля не является
вал,
Норденшельда
4 губа
Сев.Сульменева
.Медвежий
Рис. 7. Характеристика районов айсбергооб-
разования, Новая Земля;
**' - ледяные берега.--* - выводные ледники
Рис. 8. Пути распространения айсбергов в Баренцевом
море от районов их образования
ключевым очагом продуцирования и распространения айсбергов в отличие от
более северных архипелагов Шпицберген. ЗФИ и Северная Земля. Это вполне
закономерно, если сравнивать число вьюодных ледников на ЗФИ (488) и Новой
Земле (33) или общую длину ледяных берегов на ЗФИ (2650 км), Шпицбергене
(1030 км) и Новой Земле (290 км). Расход льда на айсберги для Новой Земли
оценивается всего в 2.0 км3 (Каталог ледников Новой Земли, 1976). Внутри-
годовое распределение стока айсбергов неравномерно, с максимумом в летний
период (в августе 14» или 0.28 км стока). Можно предположить, что межгодо-
вые колебания числа айсбергов (расхода льда) должны быть значительными, но
подтверждающих материалов нет. Помимо значительно меньшей мощности
источников продуцирования, на распространение новоземельских айсбергоа влияют
местные физико-географические особенности. Так. еще Н.Н.Зубов отмечал, что
как со стороны Баренцева, так и со стороны Карского моря, айсберги у Новой
Земли очень редки, хотя почти в каждом фьорде Северного острова имеется
спускающийся в море ледник (Зубов, 1944). Причины состоят в системе ветров
и течений, доминирующих вдоль архипелага: у обоих побережий (западного и
восточного) айсберги прижимаются к берегу, отклоняясь вправо от
направления преобладающих новоземельских течений (рис.9). Кроме того, мелководные
пороги не выпускают айсберги из фьордов. По этой причине размеры
новоземельских айсбергов не могут соперничать максимальными размерами айсбергов,
достигающих в длину около 200 м, в ширину 150 м и в высоту 30 м. с
характерным соотношением надводной и подводной частей как 1:7. Дрейф айсбергов
происходит в южном направлении, особенно в Карском море, что способствует
их быотрому таянию.
Рис. 9. Схема циркуляции поверхностных вод в юго-западной части
Карского моря
Таким образом, новоземельские айсберги составляют одну из характерных
физико-географических черт ледового режима самого архипелага Новая Земля и
прилегающих вод. однако они не играют значительной роли в бюджете льда
региона и в его влиянии на окружающую среду Баренцева и Карского морей.
3.3. Гидрохимический реши вод Новозеиельского региона
Архипелаг Новая Земля омывается водами двух морей - Баренцева и
Карского, географически принадлежащих двум океанам, имеющим свои климатические
особенности и определенные различия гидрологического и гидрохимического
режимов.
Наиболее изучен гидрохимический режим вод юго-западного сектора
региона. Связано это прежде всего с большей хозяйственной освоенностью этих
вод и наличием свободных от льда акваторий, на протяжении большей части
года доступных для экспедиционных исследований.
Химическая структура вод Новоземельского региона определяется в
значительной мере циркуляцией вод и типом водных масс, поступающих в регион.
Большую роль в формировании гидрологического и гидрохимического режима в
регионе играют атлантические воды. Наибольшее влияние атлантические' воды
оказывают на западный сектор. Поступая с ветвями Новоземельского течения,
атлантические воды вследствие повышенной солености и соответствующей
платности опускаются в глубинные слои и распространяются системой течений
практически на всей акватории.
Особенности циркуляции и типы формирующихся водных масс позволяют
выделить в Новоземельском регионе три крупных района с однотипной структурой
вод, которые можно обозначить как западный, восточный и южный (Никифоров,
Шпайхер, 1980). В западном районе верхний слой занимают поверхностные
арктические • и трансформированные в течение зимы атлантические воды. Южный
(Печорский) район представлен в весенне-летний период прибрежной водной
массой. В нижних слоях располагаются баренцевоморские воды. В восточном
районе (Карское море) структуру определяют арктические и баренцевоморские
воды, поступающие между мысом Желания и Землей Франца-Иосифа, а также
через новоземельские проливы. Характерной чертой этого района является
присутствие в глубоководных желобах атлантических вод. поступающих из
Арктического басоейна. В придонных слоях скапливаются местные зимние воды.
Значительное влияние на гидрохимический режим восточной части региона В
весенне-летний сезон оказывают стоковые воды новоземельских ледников и, в
какой-то мере, веды материкового стока.
Динамика элементов гидрохимического состава вед в регионе имеет
четкий годовой цикл, обусловленный климатическими факторами и циклом
сукцессии фитопланктонных сообществ в этих водах. В весенне-летний период
северная часть региона характеризуется максимальными концентрациями кислорода.
Насыщенность достигает 117-12558, а иногда и больше. Высокое содержание
кислорода вызвано поступлением сюда холодных поверхностных.вод из
Евразийского суббассейна и интенсивным фотосинтезом. Бурное цветение
фитопланктона наблюдается в зоне таяния льдов. В. Ю. Визе (1943) показал, что
содержание кислорода в активном слое разреженных льдов превышает его
содержание в зоне сплошных льдов и на открытых акваториях. Наиболее ярко это
проявляется в северной части региона, слабо подверженной влиянию вод
материкового стока. Концентрация растворенного кислорода составляет
8.5-9.0 мл/л и имеет тенденцию снижения по направлению на юг.
Для западного района характерно высокое содержание кислорода на всех
глубинах с максимумом в слое фотосинтеза. Средняя его концентрация летом в
верхних слоях изменяется в интервале от 8.50 мл/л - у мыса Желания до
7.25 мл/л - на границе Печорского района, и от 7.25 до 7.00 мл/л - в водах
Печорского района. В придонных горизонтах концентрация кислорода
выравнивается по акватории и варьирует от 7.50 до 7.25 мл/л (рис.Ю). Хорошая
аэрация придонных вод на уровне 96-9258 определяется, главным образом,
динамическими процессами. Даже в конце зимы насыщенность не снижается ниже
Рис. 10. Концентрация растворенного
кислорода в поверхностном слое Новоземельского
региона летом, 0г мл/л
86-905J. Повышенное содер
жание 0г в придонном
горизонте наблюдается в
годы с хорошо развитым Се-
веро-новоземельским тече-
нием и снижается в период
усиления воздействия
ветвей атлантических течений
(Норина, 1965)
Между западной и
восточной частями региона
отмечены некоторые
различия в распределении
кислорода. В восточном
секторе региона концентрация
этого элемента может
превышать 8-9 мл/л.
Тенденция к снижению выражена
слабо (Широколобое, 1982).
Благоприятные условия для
развития фитопланктона
здесь создаются за счет
стока талых вод ледников,
расположенных вдоль
восточного берега
архипелага. К тому же гораздо
более низкая температура
воды поверхностных слоев
Соответственно, в областях
имеющих высо-
обусловливает большую растворимость кислорода,
притока баренцевоморских вод (Маточкин Шар, Карские Ворота),
кую температуру и соленость как летом, так и зимой, формируются зоны
относительного снижения концентраций кислорода. За пределами эвфотической зоны
(ниже 25-40 м) происходит быстрое снижение концентраций и насыщенности
вод. В придонных горизонтах относительное и абсолютное содержание
растворенного кислорода составляет соответственно около 8058 и 6.8 мл/л.
С началом зимы сильное влияние на формирование кислородного режима
оказывает быстрое выхолаживание вод и установление ледового покрова.
Разрушение летней стратификации водных слоев приводит к обеднению этим
элементом поверхностных слоев. На ■ севере региона отчетливо прослеживается
влияние арктических, хорошо аэрированных вод (около 8.0 мл 0г/л). На юге
заметно ощущается дефицит кислорода в придонном слое вследствие окисления
органического детрита. Происходит снижение его концентрации до 4.8 мл/л. В
водах Карского моря дефицит в отдельные годы превышает 405J (Русанов и др.,
1979; Гидрометеорологические условия, 1985).
Из-многочисленных микрокомпонентов химического состава морской воды
особо важное значение имеют соединения биогенных элементов: фосфора:
азота; кремния. Участие биогенных элементов на всех стадиях производства и
деструкции органического вещества приводит к отчетливо выраженным
межсезонным изменениям их содержания во всем слое вод от поверхности до Alia.
Особенно отчетливо эти изменения проявляются в поверхностных водах. Рас-
иределение этих веществ испытывает ряд характерных изменений, которые
служат показателями гидрологических, физико-химических процессов. Из-за
значительной широтной протяженности акватории весенне-летний период наступает
здесь не одновременно, что обусловливает различия физиологического
состояния биологических сообществ, и соответственно, различия биохимических
процессов в водных массах и пространственно-временного распределения
биогенных веществ.
Летом в поверхностных водах фосфаты практически на всей акватории
интенсивно ассимилируются развивающимся фитопланктоном. Поскольку развитие
продукционных процессов происходит неравномерно, то в зависимости от
момента наблюдений их концентрация варьирует от аналитического нуля до
0.2-0.5 мкг-ат/л (рис.11), что особенно характерно для восточной части
региона (Русанов и др.. 1979; Ильин, 1980; Широколобое 1982). Значительный
рост концентраций минерального фосфора происходит ниже слоя пикноклина, с
горизонта 40-50 м. В восточной части содержание фосфатов в глубинных и
придонных слоях увеличивается до 1.1 мкг-ат/л. в западном районе - до
0.90-1.0, а в Печорском районе от 0.5 до 1.1 мкг-ат/л в зависимости от
характера донной водной массы (атлантическая или зимняя баренцевоморская). В
мелководных районах южного сектора эвфотическая зона простирается до дна, и
концентрация фосфатов в таких районах удерживается на уровне поверхностных
вод.
Рис. И. Распределение минерального фосфора в
поверхностном слое летом. Р-Р04 мкг-ат/л
Распределение неорганических форм азота на акватории также
неравномерно. Отмечается обеднение эвфотического слоя нитратами и нитритами.
Однако некоторые различия между районами, вызванные главным образом
особенностями сукцессии фитопланктонных комплексов,
ранственного распределения нитратов (рис.12).
заметны на картах прост-
Нитриты обнаруживаются в
следовых количествах. За
пределами звфотической
зоны, обнаруживается
резкий рост концентрации
нитратов: в
баренцевоморских водах - до
8-10 мкг-ат/л; в водах
Карского моря - до
10-12 мкг-ат/л.
Содержание нитритов также
увеличивается за счет
погружения в придонные
горизонты детрита до
0.15-0.20 мкг-ат/л в
юго-западном секторе, до
0.55 мкг-ат/л - в
юго-восточном секторе. На
севере максимум нитритов
отмечается не в придонном
горизонте, а в слое пик-
ноклина - 35-75 м.
Определение
органических форм фосфора и
азота проводились лишь в
водах южного сектора - в
Печорском море. Здесь
концентрация Рорг в
летний период удерживается в
интервале 0.30-0.40 мкг-ат/л, a Nopr - в интервале от 18 до 30 мкг-ат/л.
Воды восточного и отчасти южного секторов региона характеризуются
повышенным содержанием кремния, что связано с притоком в эти районы
материковых вод. Поэтому минимальные концентрации силикатов отмечаются в
северной части , занятой поверхностными арктическими водами. В областях,
подверженных влиянию материкового стока, содержание этого биогенного элемента
более 10 мкг-ат/л (рис.13). В областях проникновения баренцевоморских вод
кремния в летний период содержится менее 5.0 мкг-ат/л (Мусина. 1960; Белы-
шева. 1970). И хотя весь весенне-летний период происходит интенсивная
ассимиляция кремния диатомовым фитопланктоном, в прибрежной зоне в области
стока ледниковых вод с Новой Земли удерживается сравнительно высокая
концентрация силикатов: от 1 до 5 мкг-ат/л. Межгодовое и отчасти межсезонное
перераспределение кремния в поверхностных водах восточного сектора
определяется гидрометеорологической ситуацией (Шпайхбр, Русанов, 1972). При
ослаблении Западно-Таймырского течения и усилении течений Святой Анны и
Ямальского обогащенные кремнием и распресненные воды проникают до мыса
Нелиния.
Рис. 12. Распределение нитратов
верхнего слоя летом, N-N03 мкг-ат/л
в водах
В западном районе в
водах Баренцева моря
концентрация кремния также
редко падает до
аналитического нуля. С глубиной
содержание кремния
возрастает в. арктических
водах до 6, а в
атлантических " водах до -
8-9 мкг-ат/л. В придонных
горизонтах глубоководных
участков концентрация
силикатов составляет для
атлантических вод около
24 мкг-ат/л. для
арктических вод - около
13 мкг-ат/л.
Таким образом,
отличительной чертой
восточного сектора (Карское
море) по сравнению с
западным (Баренцево море)
является более короткий
период интенсивного
функционирования экосистем.
Более суровые климатические
условия сокращают период
вегетации фитопланктона в
Карском море до 4 месяцев, что почти вдвое короче вегетационного сезона в
Баренцевом море (Романкевич и др., 1982). Естественно,- этот параметр во
многом определяет цикл основных гидрохимических параметров вод и их
пространственное распределение, а также определяет продукционные показатели
экосистем, которые в Карском море резко снижаются .относительно Баренцева
моря. Гидрохимический режим деятельного слоя вод восточного сектора
формируется адекватно влиянию вод Баренцева моря и пресноводного стока.
Значительная плотностная стратификация вод летом и отсутствие притока хорошо
аэрированных вод зимой с установлением ледового покрова приводит к
заметному дефициту кислорода в придонных слоях вод восточного сектора. В то хе
время в западном и южном районах аэрация вод остается высокой во всей
толще до дна.
Рис. 13. Распределение кремния летом в
поверхностных водах Новоземельского региона,
S1-S10, мкг-ат/л
Глава 4. ПЛАНКТОН
4.1. Хлорофилл и первичная продукция
Вопросы, связанные с изучением уровня и распределения первичной про-
дукции и концентрации хлорофилла в прибрежных водах архипелага Новая Земля
освещены в гидробиологической литературе крайне скудно. Имеющийся к
настоящему времени материал представляет собой лишь результаты небольшого числа
океанологических экспедиций, проведенных в разные годы в Баренцевом и
Карском морях, в процессе выполнения которых некоторые разрезы
располагались вблизи побережий архипелага. К их числу можно отнести исследования,
проведенные в Баренцевом море в августе-сентябре 1984 г. в июне 1991 г. а
также в Кароком море в августе-сентябре 1981 г. (рис.14). Материалы этих
экспедиций, частично опубликованные ранее (Бобров и др.. 1989; Савинов.
Бобров. 1990), и легли в основу данного раздела.
На формирование первичной
продукции в районах,
прилегающих к архипелагу Новая Земля,
определяющее влияние оказывают
гидрологические условия и
ледовый режим омывающих его
водоемов. Главной особенностью
прибрежных районов Новой Земли
является наличие ледового
покрова, который присутствует
здесь большую часть года. Меа-
годовые колебания ледовитости
могут быть весьма существенны.
Так. например, в аномально
холодные годы западное побережье
Новой Земли полностью
очищается ото льда лишь в
июле-августе, а с восточной - ее стороны
дрейфующий лед может
сохраняться на протяжении всего
летнего периода. В аномально теплые годы баренцевоморское,побережье Новой
Земли очищается ото льда уже в апреле-мае, а со стороны Карокого моря
ледовая кромка отступает от берегов архипелага в мае-июне (Vlnje, 1982).
Поэтому продолжительность сезона вегетации фитопланктона, а следовательно и
годовая величина первичной продукции варьирует из года в год в зависимости
от ледовых условий Карского и Баренцева морей, цикличность изменений
которых составляет 3-5 лет (Lunde, 1965).
Весенне-летнее таяние льда начинается у южных и юго-западных берегов
архипелага под воздействием атлантических вод Колгуево-Печорского и Ново-
Рис. 14. Схема расположения станций,
выполненных в 1981 г. (1), в 1984 г. (2)
И 1991 г. (3)
земельского течений Баренцева моря, а затем' в результате радиационного
прогрева охватывает все баренцевоморское побережье архипелага. Со стороны
Карского моря таяние льда происходит под влиянием прогретых баренцевомор-
ских вод, поступающих через новоэемельские проливы Югорский Шар. Карские-
Ворота и Маточкин Шар, а также подповерхностных атлантических вод Восточ-
но-Новоземельского течения, оледующего вдоль восточных берегов архипелага
(Петров и др.. 1989).
Известно, что таяние льда и возникновение в связи с этим
стратификации водной толщи создают предпосылки для эффективного продуцирования
органического вещества фитопланктоном. Изучению процессов, протекающих в прик-
ромочных областях центральной части Баренцева моря, посвящен целый ряд
работ (Rey, Loeng, 1985; Rey et al., 1987; Skjoldal et al., 1987 и др.). Как
показали исследования, проведенные в весенне-летний период,
продолжительность стадии от начала развития фитопланктона до наступления пика цветения
в водах, подверженных распреснению. варьирует от 3 до 5 недель (Skjoldal
etal., 1987). В период непосредственно предшествующий цветению, когда
уровень хлорофилла "а" примерно равен 0.5 мг/м э. первичная продукция фо-
тического слоя находится в пределах 100-400 мгС/м г в день, а затем резко
возрастает до величины 2 гС/м "й в день (Rey et al., 1987). Несмотря на то,
что приведенные выше результаты исследований были получены на материале,
собранной в центральной части моря, они несомненно справедливы для всей
акватории Баренцева моря, подверженной распреснению талыми водами, .в том
числе и для районов, примыкающих к архипелагу. Еще большая интенсивность
весеннего цветения фитопланктона может наблюдаться в районах мелководий,
например, в юго-восточной чаоти моря. Так исследования, проведенные в
Печорском море летом 1978 г. показали, что в период прикромочного цветения
фитопланктона, охватившего всю водную толщу, величина первичной
продукции в мелководном Вайгачско-Печорском районе достигала значения
354 мгС/м гч (Хромов, 1981), т.е. оуточная продукция превышала величину
6 гС/м г.
Бурное весеннее цветение фитопланктона быстро приводит к истощению
зимнего запаса биогенных элементов в верхнем слое водной толщи, а мощный
пикноклин, образованный талыми водами, препятствует проникновению
необходимых элементов питания из нижележащих слоев. Поэтому после завершения
очень кратковременной кульминации цветения происходит снижение первичной
продукции, а затем ее стабилизация. Основная часть первичной продукция
создаетоя за счет фитопланктона, образующего подповерхностные скопления,
что можно проиллюстрировать на примере данных, полученных в июне 1991 г.
на разрезе, проходящем вдоль западного побережья Северного острова
архипелага (рис. 15). Как видно из рис.15, верхний слой водной толщи занимают
распресненные воды, происхождение которых связано с таянием зимних
дрейфующих льдов. Низкие значения концентраций биогенных элементов (по данным
Г.В.Ильина, личное сообщение) и хлорофилла в этом слое свидетельствуют о
том, что к началу исследований весеннее цветение миновало период своей
кульминации. Обеднение поверхностных слоев минеральными элементами питания
приводит к снижению первичной продукции и погружению клеток фитопланктона
в более глубокие слои. Здесь они находят среду, где не чувствуется
недостатка й биогенных элементах, вследствие чего и происходит концентрация
клеток в промежуточных слоях. Благодаря тому, что верхний олиготрофный
слой обладает высокой прозрачностью, доля света, проникающая на глубину
€ i a ii u ii и
19 20 21
100
100
^•.2 -— —
:M ^
* *
-> XJ,
0.8 /
scZZ: j ■ /
NO,
Рис.
температуры
нитратов ""
мкг-ат/л)
в июне 1991
хлорофилла
обитания Фитопланктона, вполни
достаточна для поддержания его
нормальной жизнедеятельности.
Действительно, максимальное
значение концентрации
хлорофилла "а", достигавшее
значения 5.35 мг/м3. было
зарегистрировано на глубине 30 м
(ст. 20), при этом низкое
значение пигментного .индекса
(2.1) свидетельствовало о
функциональной активности фи-
топланктонного сообщества.
Интегральное содержание
хлорофилла в слое 0-50 м на разрезе
варьировало в достаточно
широких пределах: от 13.0 (ст. 17)
до 76.9 мг/м2 (ст. 20).
Наиболее низкие значения
суммарного содержания пигмента били
определены на крайних станциях
разреза (ст. 17 и 23), на ко
торых локальные максимумы
концентрации хлорофилла
располагались соответственно на
глубинах 50 и 40 м. Вероятно,
цветение фитопланктона на этих
станциях происходило в более
ранние сроки, чем на станциях,
расположенных в центре
разреза, однако невысокие значения
пигментного индекса указывают
на активность клеток
фитопланктона, образующих скопления
под слоем скачка плотности.
Поскольку лимитирование по би
огенам отсутствует, можно
предположить, что активное
новообразование органического
вещества фитопланктоном в
промежуточных слоях будет достаточно стабильным.
Подобное вертикальное распределение биоокеанологических параметров
для которого характерны олиготрофность верхнего распресненного слоя и
наличие подповерхностных скоплений фитопланктона, сохраняется до окончания
вегетационного сезона. Это подтверадаетоя результатами, полученными в
сентябре 1984 г. на разрезе, располагавшемся в северо-западном направлении от
пролива Маточкин Шар (рис.14). Содержание хлорофилла «а» в поверхностном
слое не превышало значения 0.15 мг/м3. а наибольшие концентрации пигмента
(0.7-0.8 мг/м3) были зарегистрированы на глубине 20-30 м. Суммарное со
держание хлорофилла в слое 0-50 м составило в среднем 24.5 мг/м2
(табл.1). Значение первичной продукции на станциях этого разреза
варьировало в небольших пределах - от 6.02 (ст. 42) до 10.31 мгС/м2ч (ст 39)
Таблица 1
Суммарное значение хлорофилла "а" и первичной продукции в слое 0-50 м
(Баренцево море, август-сентябрь 1984 г.)
Станция
Хлорофилл "а", мг/мг Первичная продукция, мгс/м^ч
9
И
12
13
14
15
16
17
18
19
39
40
42
43
44
45
46
47
48
24.1
27.3
22.8
20.2
26.6
26.8
14.9
16.2
13.0
13.1
14.5
27.
29.
26.
24.
14.
22.
27.5
25.5
1.61
3.10
1.88
4.90
10.12
2.61
2.19
3.28
4.60
1.77
10.31
7.75
6.68
6.02
7.70
4.00
2.64
3.00
1.12
В юго-восточной части Баренцева моря в этот же период исследований
характер вертикального распределения хлорофилла "а" во многом определялся
особенностями гидрологического режима. Так. в северной части разреза
мыс Русский Заворот - губа Черная наблюдалась переслоенность водной толщи
водными маосами различного генезиса (рис. 16). Распресненные воды,
образованные талыми водами и береговым стоком, подстилались холодными водами
течения Литке, поступающими сюда из глубинных слоев Карского моря.
Следствием мощного поступления этих вод явилось уменьшение глубины залегания
пикноклина, а наличие необходимых элементов питания предопределило
образование на границе стыка различных водных масс скопления фитопланктона, на
что указывают достаточно высокие для этого периода вегетации значения
хлорофилла "а". Так. на ст.14, расположенной в прибрежье Новой Земли,
концентрация хлорофилла для слоя 5-20 м в среднем составляла 0.9 мг/мэ.
тогда как в поверхности этот показатель был равен 0.17 мг/мг. На этой же
станции были определены наибольшие для этого разреза интегральные значения
содержания этого пигмента (26.6 мг/м2) (табл.1). Величина первичной
продукции в слое 0-50 м. измеренная на станциях этого разреза, закономерно
уменьшалась с ослаблением влияния карских вод в направлении с севера на
юг. варьируя в интервале от 10.12 (ст. 14) до 1.77 мгс/м* ч (ст. 19),
составляя в среднем 4.1мгС/м*ч. Аналогичная картина вертикального
распределения океанологических характеристик наблюдалась и на разрезе,
расположенном между островами Междушарским и Колгуев. Хотя значения
первичной продукции на станциях этого разреза были несколько ниже, однако
сохранилась тенденция приуроченности максимума к прибрежью Новой Земли
(табл.1).
Станции
17 16
100
— ГС
100
0.8 у
-hu" '
<^wl
о.в~
-0.4-—
-о, /
Хл"а"
В целом, средняя
величина первичной
продукции прибрежных
вод архипелага,
измеренной в августе-
сентябре 1984 г.
(4.5 мгС/м гч ),
была значительно ниже по
сравнению с западной и
центральной частью
Баренцева моря, где
она составляла
10-20 мгС/м гч
(Савинов. Бобров, 1990).
Вертикальное
распределение
гидролого-гидрохимических
характеристик водных
маео и концентрации
хлорофилла в сентябре
1981 г. в Карском море
достаточно подробно
описано ранее (Петров
и др., 1989; Петров,
Ильин. 1989; Бобров и
др., 1989). Позволим
себе лишь кратко
остановиться на
особенностях распределения
интегральных значений
содержания хлорофилла
Рис. 16. Вертикальное распределение
температуры (Т °С), солености (S°Ae), нитратов
1К0,Г мкг-ат/л), фосфатов, (Р0\ мкг-ат/л)
и хлорофилла "а" (Хл. мг/м 3) в августе 1984 г.
по разрезу,
му вдоль
побережья
Новая Земля
Как видно,
наименьшие
проходяще-
восточного
архипелага
(рис. 17).
из рис.17,
значения
концентрации
хлорофилла были
зарегистрированы на южных станциях
разреза. Суммарное содержание этого пигмента в слое 0-50 м, рассчитанного
для станций 12-14, варьировало от 3.6 до 5.6 мг/м *. Значение этого
показателя увеличивалось далее по направлению на север, достигало своего
максимума (23.5 мг/м е) на станции 27. а затем происходило его уменьшение до
значения 11.3 мг/м е на станции 30. Среднее содержание хлорофилла в водах,
омывающих восточное побережье Новой Земли, составляло 11.8 мг/м г, что в
два раза меньше, чем значение этого показателя (22 мг/м г. см. табл.1),
рассчитанного для акватории западного и юго-западного прибрежья архипелага
в такой же период вегетации. Локальные максимумы концентрации хлорофилла
на станциях этого разреза располагались, как правило, под слоем скачка
Ст, 12 Ст. 13 Ст. 14 Ст. 2в Ст. 27 Ст. 28 Ст. 29 Ст. 30
Рйо. 17. Распределение суммарного содержания хлорофилла "а"
в слое 0-50 м (мг/м г) в сентябре 1981 г.
плотности, а нижняя граница распространения повышенных концентраций
пигмента располагалась вдоль изопикнической поверхности со значением условной
плотности,равной 27.7 отн.ед. (Бобров и др., 1989), ниже которой
располагались холодные глубинные воды (Петров и др., 1989). Оценка летней
биомассы мезозоопланктона Карского моря, по данным 1981 г., показала, что она
лишь в три раза уотупает летней баренцевоморской (Фомин. 1989). а
пространственное распределение биомассы, в основном, определяется глубиной
залегания плотных собственно карскоморских вод, в частности, нижняя граница
распределения 7555-го уровня , биомассы мезозоопланктона повторяет границу
слоя вод, обладающих плотноотью 27.7-27.8 отн. ед. (Фомин и др., 1984).
Пространственное совпадение повышенных концентраций хлорофилла и 75Х-го
уровня биомассы мезозоопланктона говорит о важном экологическом значении
подповерхностных скоплений фитопланктона. Отсутствие лимитирования по
биогенам, приводящее к стабильности подобных скоплений, и жесткая
пространственная закрепленность определяют их особое значение для питания
растительноядного зоопланктона в силу своей гарантированности.
Таким образом, в летний период уровень первичной продукции прибрежных
районов архипелага Новая Земля относительно невысок, в среднее он
составляет лишь 25-40% от уровня первичной продукции, измеренной в западном и
центральном районах Баренцева моря. Основная доля новообразованного
органического вещества формируетоя за счет фитопланктона, обитающего в
промежуточных слоях водной толщи под слоем скачка плотности и имеюцего важное
значение для питания растительноядного зоопланктона.
4.2. Фитопланктон прибрежной зоны Карского моря
Работ, посвященных изучению фитрпланктонного сообщества Новоземель-
ского прибрежья Карского моря, немного. Наиболее интенсивные исследования
количественных характеристик микроводорослей данного района проводилиоь в
30-40-х гг., их результаты обобщены в работе П.И.Усачева (1968). В 80-х
годах широкомасштабные исследования акватории были возобновлены в ММБИ,
основные итоги данных изысканий были отражены в коллективной монографии
(Экология— 1989).
В основу данного раздела положен материал, собранный в рейсе НИС
•'Дальние Зеленцы" в августе-сентябре 1981 г. (рис.18). Отбор и обработка
проб фитопланктона велись стандартными методами (Федоров, 1979;
Руководство..., 1980; Макаревич, 1989). Для удобства анализа количественных
характеристик фитоцена, исследуемая акватория была разделена на три района
(рис.18), выделенных в результате флористического районирования (Дружков,
Макаревич, 1992) с применением методов корреляционно-плеядного анализа
(Вёльдре. 1964; Выханду. 1964; Шмидт, 1984),, с использованием полных
матриц коэффициентов сходства Серенсена-Чекановсксго (Schandelmeier,
Alexander, 1981; Шмидт, 1984). Для оценки видового разнообразия вычислялся
индекс Глисона (Margalef, 1960).
Рис. 18. Карта-схема расположения станций отбора проб
и флористические районы юго-западной части
Карского моря (объяснения в тексте)
район I: ст. 1-7. Фитопланктонное сообщество, занимающее пелагическую
зону на от. 1-7, несмотря на достаточно высокие значения индекса видового
разнообразия (d до 1.5; рис. 19), характеризовалось незначительным
количественным развитием: биомасса микроводорослей составляла менее I г/мг
(рис.20), а концентрация хлорофилла "а" - менее 5 мг/мг (рис.21).
Первичная продукция поверхностного горизонта находилась на среднем уровне,
изменяясь в диапазоне 0.2-0.4 мгС/м3ч (рис. 22).
Рис. 19. Распределение значений индекса
видового разнообразия (индекс Глисона)
фитопланктонного сообщества (изолинии)
Рис. 20.. Распределение' биомассы
фитопланктонного сообщества (г/мг в слое
0-100 м. изолинии)
ас. 21. Распределение хлорофилла "а"
(мг/м2 в слое 0-50 м. изолинии)
(-75°
h?o°
75°
Г706
Рис. 22. Распределение значений первичной
продукции в поверхностном слое (мг С/м3^, изолинии)
Необходимо отметить, что значительные различия в интегральных
количественных параметрах меаду ст. 1-7 района I и рассмотренными ниже
станциями района И могут быть интерпретированы не только в терминах абсолютных
уровней количественного развития фитопланктона на отдельных горизонтах, но
также могут объясняться и мелководностью района I (18-80 м) в сравнении с
районом II (78-370 м). Столь значительное "сжимание" жизненного
пространства планктонного сообщества на ст. 1-7. несомненно, сказывается на
значениях интегральных количественных характеристик пелагических водорослей. В
частности, именно мелководностью можно объяснить отсутствие в районе I
подповерхностных пиков фитопланктона, отмеченных на ряде станций"
района II.
В структурном отношении фитопланктонное сообщество на ст. 1-7 было
очень вариабельно: доля диатомовых в его составе изменялась от 21 до 77%
общей биомассы фитоцена, и ни одна популяция микроводорослей не достигла
лидирующего положения в количественном отношении.
Район II: ст. 25-30. Фитопланктонное сообщество в данной области пе-
лагиали характеризовалось наибольшим на исследуемой акватории
количественным развитием. Биомасса пелагических водорослей в слое 0-100 м достигала
14.3 г/ма> (ст. 28), и фактически весь район (кроме ст. 26) располагался в
зоне, очерчиваемой изопланктой 6 г/ма (рис.20). Соответственно здесь же
отмечены наибольшие значения концентрации хлорофилла (до 40 мг/м2;
рис.21), первичной продукции поверхностного слоя (более 0.5 мгС/м^ч:
рис. 22) "и видового разнообразия (d до 1.9; до 37 таксонов в пробе;
рис.19).
В структурном отношении в этом районе наблюдалось доминирование
диатомовых, составляющих от 75 до 84% совокупной биомассы фитопланктона
(рис.23). Исключение составляет ст. 29 (46%), где на горизонтах 0-10 м
значительного развития достигают популяции пирофитовых водорослей Gyrodt-
ntum lachryma (до 51 мкг/л) и Ceratiw arcttcum (до 47 мкг/л).
Количественная основа фитоцена на других станциях формировалась за счет двух
видов: Cnaetoceros dectptens и Thalasstostra с/, dectptens (на ст. 30 - в
основном второй вид). Популяция С. dectptens в слое 0-100 м образовывала
15-47% совокупной биомассы фитопланктона. Исключение составляют ст. 29 и
30, где биомасса этой водоросли была незначительной. Еще больший вклад в
структуру сообщества вносила Г. cf. dectptens - 2В-70%. Из других видов
только Jtfitzosoienta deltcattsstra достигала заметного развития на ст. 28 -
18% общей биомассы фитоцена. "
Фитопланктонное сообщество ст. ; 8 (глубина 134 м) по своим
качественным и количественным характеристикам было идентично фитоцену ст. 25-30
района II за исключением умеренной фотосинтетической активности.
Район III: ст. 9-19. 22-24. Станции этого района сходны по общему
таксономическому составу фитопланктона, заметно отличаются лишь некоторые
специфические черты, в основном,количественного характера. Поэтому мы
посчитали нужным разделить станции этого района на 2 группы: ст. 9-19 и
22-24, и рассмотреть их по отдельности.
Ст. 9-19. Фитопланктонное сообщество этой области пелагиали
характеризовалось наименьшими для района исследований уровнями развития
микроводорослей и носило явные черты деградации большинства популяций. Видовое
разнообразие фитоцена (d) на этих станциях принимало значения меньше
1, снижаясь на ст.10-11 до минимальных для всего района значений (0.1-0.2;
рио.19). Биомасса пелагических водорослей была также минимальной -
0.1-0.6 г/м2 (рис.20). Исключение составляют ст. 14-16.
характеризовавшиеся несколько большими значениями биомассы 0.8-2.1 г/м2. Концентрация
хлорофилла на станциях была также невысокой и изменялась в диапазоне
5-15 мг/м2 (рис.21). Лишь в районе ст. 16 наблюдалось повышенное
содержание хлорофилла - более 25 мг/м2. Первичная продукция поверхностного слоя
обычно не превышала 0.1 мгС/м3ч, поднимаясь до 0.3-0.4 мгС/м3-ч на
северных станциях рассматриваемой акватории (рис.22). Несколько более высокие
значения всех количественных параметров, характерные для ст. 14-16,
по-видимому, обусловлены подповерхностным скоплением Thalasstostra btoculata.
В составе фитоцена ст. 9-19 наблюдалось полное доминирование пиррофи-
товых: диатомовые водоросли формировали не более 20Х совокупной биомассы
фитоцена (рис.23). На ст. 10-11 диатомовые вообще не были
зарегистрированы. Лишь на ст. 9 и 16 относительное обилие диатомей превышало 20%, в
первом случае благодаря подповерхностным скоплениям на горизонтах 25 м и ниже
Chaetoceros concavicornis. С.declptens, С.dtadera и Thalasstostra sp.,
во втором, как уже говорилось, благодаря наличию повышенных концентраций
Г. btoculata. На остальных станциях господствовали пиррофитовые из родов
Dtnophysts, Gyrodinium, Cerattum и Protopertdlrilum. причем ни один вид не
проявлял выраженной тенденции к доминированию в составе сообщества.
Рис. 23. Распределение относительной биомассы диатомовых водорослей
(% от совокупной биомассы фитоцена. изолинии)
Ст. 22-24, Состав сообщества здесь характеризовался средними
значениями индекса видового разнообразия (d=i.i-1.2; рис.19) и биомассы
(1.0-5.5 г/м2; рис.20), а также высокими концентрациями хлорофилла
(20-35 мг/м2; рис.21) и интенсивными процессами фотосинтеза (более
0.5 мгС/м3-ч; рис.22).
Биомасса фитопланктонного сообщества формировалась в основном за счет
одного вида диатомей - С. Oecipiens, составляющего 47, 20 и 66% совокупной
биомассы фитоцена на ст. 22, 23 и 24 соответственно. Значительный вклад в
биомассу сообщества на отдельных станциях и горизонтах вносят также и
некоторые другие представители рода Ctaetoceros, популяции которых
характеризовались средними (Ctaetoceros borealis, Ctaetoceros concavtcornts) и
максимальными (Chaetoceros convolutus) значениями количественных
параметров. Из многочисленных видов пиррофитовых, зарегистрированных на ст.
22-24, можно выделить всего несколько форм: Dinophysis arc t tea. Gontauiax
catenata. Protopertdtntum breve, Protopertdtntum btpes, Protoperidinlum
brevipes и Protopertdtntum ovatum, - достигающих заметного количественного
развития в верхних горизонтах пелагиали (на уровне средних значений
биомассы) .
Таким образом, на исследуемой акватории количественные параметры
фитоцена варьировали в широких диапазонах значений в зависимости от стадий
сезонного развития. Характерной особенностью распределения биомассы
фитопланктона в данной области является ее периферийная концентрация в
прибрежных районах.Эта тенденция хорошо прослеживается как по распределению
биомассы, так и концентрации хлорофилла и первичной продукции.
Наибольшие биомассы фитопланктона (5-15 г/м2 в слое 0-100 м). концентрации
хлорофилла (20-40 мг/мг в слое 0-50 м) и значения первичной продукции
(0.3-0.5 мгС/м3-ч в поверхностном слое) обнаруживались у побережья
северо-восточной части о.Северного архипелага Новой Земли.
Несмотря на очень ярко выраженный характер сезонной стратификации,
отчетливой тенденции к сосредоточению микроводорослей в области пикноюииа
с образованием подповерхностного максимума не обнаружено. На некоторых
глубоководных станциях эти пики образовывались за счет оседания клеток,
формировавших ранневесеннее цветение, но в большинстве случаев выраженной
закономерности в локализации пиков (одного или двух) не прослеживалось -
они могли обнаруживаться во всем диапазоне глубин от 0 до 50 м. В целом
верхний 100-метровый слой пелагиали содержал 92-9856 биомассы
фитопланктонного сообщества.
4.3. Фитопланктон прибрежной зоны Баренцева моря
Описывая планктонную флору микроводорослей новоземельского прибрежья
Карского и Баренцева морей, мы имеем дело с двумя различными ситуациями,
обусловленными географическим положением этих бассейнов и связанным с ним
комплексом природных факторов. При этом, если в первом из них фактически
всю юго-западную часть моря и по гидрологии, и по составу пелагического
фитоценоза можно рассматривать как единый приновоземельский сектор (см.
Иакаревич. наст. изд., предыдущий раздел), то во втором выделение области
с аналогичными водами будет очень условным. Вообще фитогеографическое рай-
онирование Баренцева моря до сей поры не проводилось вследствие большой
акватории с широким диапазоном изменчивости гидрологических характеристик
и отчасти связанных с этим технических трудностей; однако,
преимущественная обусловленность пространственных различий в составе баренцевоморского
пелагического фитоценоза разновременностью наступления в нем сезонных
изменений в разных частях бассейна (Ларионов, 1993) ставит под вопрос смысл
проведения такого районирования в принципе. Принимая во внимание
последнее, иначе воспринимается заключение одной из работ П. Р.Макаревича и
Н.В.Дружкова (1994) о принадлетаости юго-западной части Карского моря и
юго-восточной Баренцева к одному фитогеографическому району: очевидно, что
на приведенном уровне сходства видового состава планктонной альгофлоры
(60-7055) к одному району можно отнести и все Баренцево, и Карское моря, а
то и весь арктический бассейн. Отметим, кстати, что. в частности, так и
было сделано Л.А.Виноградовой и Л.В.Грузовым (1990), - более того, и все
формы микроводорослей, встречающиеся в нем, они также отнесли к одной, ле-
дово-неритической группе.
В свете вышеизложенного логично будет ограничиться в настоящем
разделе рассмотрением лишь баренцевоморских вод, расположенных вблизи
архипелага Новая Земля. Кроме того, следует исключить крайнюю северную область, в
течение вйего года покрытую льдами.и особенно - южную, входящую в т.н.
Печорское море. - часть Баренцева, испытывающую на себе влияние 80S? всего
речного стока баренцевоморского побережья (Елшин..1979), и вследствие
этого и некоторых других климато-географических факторов являющую собой
совершенно особый водоем со специфическим фитоценологическим обликом.
Исследователи-фитопланктонологи не очень жаловали вниманием
очерченный нами район Баренцева моря. Из трудов Мурманской научно-промысловой
экспедиции лишь в одном отчете мы находим единичные сборы отсюда (Линко,
1915). Также отдельные станции обследовалиоь здесь в последние годы (Васю-
тина, 1991; Larlonov. 1992). Гораздо тщательнее этот сектор
непосредственно у Новой Земли был изучен экспедицией герцога Орлеанского, материалы
которой по микроводорослям представлены в знаменитой книге А.Менье (Meunler,
1910). Но, к сожалению, данный автор ни для одного фитопланктера не
указывает точные места находок, и нам остается лишь гадать, какие из
баренцевоморских видов обнаружены именно в этом районе. Целенаправленным
исследованием новоземельских прибрежных вод явилась экспедиция, предпринятая"
Мурманским морским биологическим институтом в августе 1967 года, в задачу
которой входило изучение влияния на продуктивность расположенных на берегах
архипелага птичьих базаров. Изданная по ее'итогам монография "Особенности
биологической продуктивности вод близ птичьих базаров севера Новой Земли"
(1972 г.. ред. а!Н.Головкин) содержит теоретические выкладки и описания
распределения биогенных элементов в исследованном районе, полностью
подтверждающие высокую степень такого влияния: все это опровергается, однако,
материалами по фитопланктону, представленными в одной главе данной книги
С.А.Соколовой (1972): на станциях расположенного "по-соседству" разреза
через юго-восточную часть Баренцева моря, специально изученного для
сравнения, численность микроводорослей была минимум на порядок больше, чем в
прибрежье у птичьих базаров, да и видовое разнообразие их выше. Впрочем, и
другие публикации А.Н.Головкина (1967; Головкин, Позднякова, 1964),
посвященные данному вопросу, грешат отдельными натяжками и несколько вольной
интерпретацией представленных рисунков, но обсуждение этой темы выходит за
рамки настоящего издания.
Материалом для данного раздела послужила часть сборов фитопланктона
из 60 рейса НИС "Дальние Зеленцы"; рассмотрен разрез, расположенный вдоль
западного берега о-ва Северный (Новая .Земля) (см. рис.14); исследования
проводились в июне 1991' года. Отбор проб осуществлялся 3-литровым
пластиковым батометром с 5 горизонтов, выбираемых, исходя из графиков
вертикального распределения температуры и солености .воды, полученных с помощью
гидрологического зонда ADM Elektronlk GmbH. Пробы'обрабатывались стандартным
способом с фиксацией 4%-и формалином и концентрированием отстойным методом
(Руководство.... 1980). Таксономическая идентификация и подсчет клеток
микроводорослей проводился под световым микроскопом МББ-lA в камере Ножот-
та по стандартной методике (Федоров. 1979). Картины распределения значений
отдельных гидролого-гидрохимических показателей, а также содержания
хлорофилла "а" - основного фотосинтетического пигмента фитопланктона и важной
функциональной характеристики состояния фитоценоза - на тех же станциях
представлены В.М.Савиновым выше в настоящем издании.
Очевидно, что подробный количественный анализ результатов
исследования короткого разреза из 7 станций в данном случае лишен смысла. Подобный
материал может служить лишь для предварительного ознакомления с
планктонной альгофлорой ранее не изучавшейся области баренцевоморского бассейна, в
целом, достаточно хорошо исследованного фитопланктонологами. В таблице 2
представлен список всех обнаруженных в пробах микроводорослей с указанием
их относительного обилия. Анализируя эти данные, сразу можно отметить, что
большинство видов имеют единичную встречаемость (не более нескольких
десятков клеток в литре воды), и биомасса фитопланктона в большей части проб
не превышает 0.5-1 мкг/л. Впрочем, те организмы, которые достигают
наибольшей численности (до нескольких десятков тысяч клеток на литр) - Nitz-
schia grunowii, Chaetoceros /urcellatus, С. /ragilis, Phaeocystis pouche-
ttt. Dinobryon balttcum, - имеют столь малые размеры, что общая биомасса
их в пробе составляет максимум около 10 мкг/л. Четко прослеживается
закономерность в вертикальном распределении микроводорослей: основная масса их
сконцентрирована в среднем слое воды, в области скачка температуры и соле-
нооти (есть также сгущения Chaetoceros /urcellatus и С. soctatts на
глубине 100 м, но там до 90% клеток представлено спорами). Наличие непрерывных
профилей распределения этих показателей, полученных с помощью зонда,
позволяет более тщательно изучить наблюдаемую зависимость. Так. на станциях 2
и 4 максимальные концентрации Wttzschta grunowtt, Chaetoceros fragilis и
С. /urcellatus отмечены на 30 м, а на ст. 3 - несколько глубже, на
40. Сравнив положение термоклина на этих станциях, убедимся, что он на
третьей продолжается также до несколько большей глубины. Аналогично
различным горизонтам, наибольшего обилия Dlnobryon balttcum на ст. 6 и 7
соответствуют разные положения скачка температуры.
Таким образом, проанализировав распределение только общих
количественных показателей развития сообщества микроводорослей у западного
побережья Новой Земли, мы можем заключить, что, достигая в целом крайне
невысоких значений, они тем не менее, обнаруживают неоднородность,
свойственную летнему сезону, а именно, увеличение в зоне пикноклина. Рассмотрев
однако изменения в видовом составе фитопланктона, мы увидим гораздо более
сложную картину развития. Нагляднее всего пронаблюдать их, передвигаясь от
одной отанции к другой по направлению разреза. На 1-й станции, северной.
Таблица 2
Встречаемость различных видов, микроводорослей на станциях исследуемого разреза
Вид
Станция N
> 1 *
3
4 | 5 '
6 J 7
Горизонт, м
0
5
22
30
100
0
27
30
35 100 0
..'
15 30 40
100
0 20
30
50
100
0
12 24 30
100 0
18 24
30
100
0 22 36 60J10C
Navlcula
vanhceffenii Gran
N. peiaglca CI.
Nitzschia
longlsslma (Breb.) Ralfs
N. seriate CI.
N. grunowii Hasle
Thalassloslra gravlfla CI.
T. nordenskloeldll CI.
Cosclnoslra
polyciiorfla Grim.
Skeletonema
costatum (Grev.) ci.
Chaetoceros
fragllls Meunler
С furcellatus Ball.
С soclalls Lauder.
+ X
+
X
+
X
X X +
+ 4
+ +
•444 4
X +
X X +
+ +
+
+ .+ X +
+
a.
4 4 + 4 4
4 4
t
0 4
4 0 4
■ 4 4
4
+ ■
4 4 4
4 4 •
4'
x ;.+
0 X X
4 4
'4.
'•*' +
4 . 4
4 X'
OXO
4 X
4 4 4
4
4 4 4 4
4
4
X
- + +
• x
X
4 4
4
X
4 4 4
0
0
4
4
X +
■ 4 4
4
+
0 X
A.
Продолжение таблицы 2
Вид
Станция N
Горизонт, м
О 5 22 30 100 0 27 30 35 100 0 15 30 40 100 0 20 30 50 100 0 12 24 30 100 0 18 24 30 100 0 22 36 60 100
С. septentrlonalis Oestr.
С. clnctus Gran
С; declplens CI.
С. borealls Ball.
Tnalasslonema
nltzschloldes Grim.
Tnalasslothrix
longlsslma CI. & Grim.
Eukanpla zodiakus Ehr.
Phaeocystls
pouchetll (Harlot) Lager. +
Dlnobryon
baltlcum Lenm.
Protoperldinlum
pellucldum Bergh +
P. quarnerense
(Schrod.) Balech
P. brevlpes
(Paula.) Balech + +
+ . +
+
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+
+ + +
x x +
о ■ + x
X X + +•■■+■ '+ 0; +
+ X
+■■■■■+ +
Окончание таблицы 2.
Вид
Станция N
1
2
• 3
4
5
6
7
Горизонт, м
0
5
22 30
100
0 27 30 35
.100
0
15
30 40
100 0 20 30 50
100 б 12 24 30 100 0
18 24 30
100 0 22 36 60
100
P. lslandlcum ,
(Pauls'.) Balech + .'+ + +
P. depressum
(Ball.) Balech + + • +
Ceratlum
arctlcm (Ehr.) CI. + +
ПРИМЕЧАНИЕ, о - доминирующие виды: х - виды со средней встречаемостью: + - единичные находки.
организмов очень мало, и лишь ранневесенняя Hitzstihia grunowil кое-где
встречается в значительном количестве. На 2-й станции к ней
"присоединяются" такие виды, как Ctoetoceros fragilis и С. /urcellatus. в меньшем
количестве Thatassloslra gravida, Т. nordensktoeldtt и Coscinisira polychorda
- все также типичные представители весеннего комплекса диатомовых.
Появляется и Phaeocystls pouchetii - золотистая водоросль, стоящая особняком
из-за своей жизненной стратегии: обычно, следуя за указанным комплексом в
"классическом" сукцессионном цикле, она в первую очередь реагирует бурным
развитием на избыток в воде биогенных элементов (Кашкин,' 1964). На 3-й
станции ранневесеннее сообщество в расцвете (правда, "зажатое" в один
исследованный горизонт), феоцистиса нет. На 4-й станции - более поздняя
весна, максимальное видовое разнообразие, появление многочисленных фитопланк-
теров и в нижних слоях воды. Станция 5 отличается резко уменьшившимся
количеством микроводорослей, при этом в верхних горизонтах возросла
численность летнего представителя Dtnobryon baLUcum. Наконец, на станции.6 он
слегка смещается вниз к облаоти скачка плотности, а пространство под ним
занимают оставшиеся весенние виды (некоторые образуют споры). Такая же, но
несколько более четко выраженная картина наблюдается и на последней, 7-й
станции.'
Безусловно, все это описание - не более чем приближенная схема
пространственной структуры пелагического фитоценоза, к тому же усложненная
присутствием в пробах большого числа единично встреченных видов
микроводорослей, несущим в себе вследствие этого значительный элемент случайности.
Так. дополнительным подтверждением наличия именно летнего комплекса в
верхних слоях воды на последней станции можно считать находки там
нескольких видов p.p. Protopertdtntum и Ceratium, однако они встречаются в
отдельных пробах и до этого. Тем не менее, сам факт наблюдения в плоскости
исследованного разреза нескольких сезонных комплексов фитопланктеров,
иными словами, упоминавшейся выше "развертки" сезонного хода развития
фитоценоза в пространстве (и не только по горизонтали, но и по вертикали!),
представляется совершенно очевидным.
В формировании именно такой структуры, по-видимому, играет роль не
только последовательность наступления климатических изменений (увеличение
инсоляции, таяние льда и т.п.). но и сама кратковременность вегетационного
периода фитопланктона в этом районе моря. Ведь всю описанную
последовательность событий (сукцессионный цикл!) мы наблюдаем в почти идентичной
гидрологической обстановке: при летней стратификации столба воды. Нужно
учесть, что с момента "запуска" цветения микроводорослей (в прибрежье
таким толчком оказывается минимальное значение температуры воды) до его
непосредственного начала проходит определенный достаточно длинный период
(Бардан и др., 1989). Очевидно, он необходим и организмам в
рассматриваемом приновоземельском районе, но сезонные гидрологические перестройки там
происходят быстрее, и сроки самих сезонов сокращаются (Бойцов, 1980). В
результате биологический сезон несколько "сдвигается" по отношении к
гидрологическому, и мы наблюдаем и отсутствие цветения при сформировавшемся
термо- и галоклине, и продолжение его одновременно с появлением летнего
комплекса миксотрофных и даже гетеротрофных планктонных простейших. При
этом, однако, зти процессы разделяются в пространстве по вертикали:
происходит "опускание" цветения - процесс, механизм которого пока совершенно
неясен.
Конечно же, невозможно на столь ограниченном материале одной съемки
пытаться создать всеобъемлющую теорию функционирования фитопланктонного
сообщества в приновоземельских водах Баренцева моря, и даже представленные
краткие рассуждения в значительной мере спекулятивны. Тем более не следует
делать этого, не имея данных по развитию в данном районе нанопланктонных
организмов, роль которых может оказаться тем важнее, чем менее обилен
микрофитопланктон. Этот вопрос, как и многие другие, - тема дальнейших
кропотливых исследований сложных биопродукционных процессов, протекающих в
пелагиали такой интересной части Баренцевоморской акватории, как район
архипелага Новая Земля.
4.4. Зоопланктон прибрежных вод
Зоопланктон прибрежных вод Новой Земли - слабоизученная область
гидробиологии Баренцева и Карского морей. Существует всего три публикации, в
которых рассмотрен зоопланктон пролива Маточкин Шар и более северных вод с
западной стороны Новой Земли (Хмызнйкова, 1931). губы Архангельской и
залива Вилышцкого (северо-западная часть архипелага) .(Зеликман, Головкин,
1972) и губы Черной (южная оконечность Новой Земли) (Тимофеев, 1992).
Кроме того, имеется некоторое количество информации, рассеянной в
статьях, посвященных зоопланктону более мористых вод.
Ниже представлен обзор опубликованных сведений по зоопланктону
прибрежных вод Новой Земли. Привлечены также данные, полученные в экспедициях
ММБИ КНЦ РАН в различных районах архипелага.
Анализ планктона немыслим без учета структуры и динамики водных масс,
поэтому очень кратко остановимся на этом вопросе. Южная и западная стороны
архипелага находятся под влиянием теплых, несущих трансформированные
атлантические воды, потоков (рис.24): Колгуево-Печорское течение и Прибреж-
Рис, 24. Течения в районе
архипелага Новая Земля (по: Атлас .... 1971)
(цифры в кружках - номера течений: 1 -
Колгуево-Печорское течение. 2 - Прибреж-
, ная и 3 - Восточная ветви Новоземельско-
го течения, 4 - течение Макарова, 5 -
течение Литке. 6 - Печорское течение)
1 - атлантические, 2 - арктические
и 3 - местные опресненные водные массы
ная ветвь Новоземельского течения. На юге архипелаг омывается выходящим из
Карского моря холодным течением Литке. Северная часть Новой Земли целиком
находится под влиянием арктических вод. Существенную роль в
гидрологическом режиме играет также стоковое течение. Взаимодействие указанных водных
масс в целом формирует вокруг архипелага антициклональный поток.
Таксономический состав и размерная структура. Количество таксонов..
обнаруженных в зоопланктоне (многоклеточном; Protozoa не учитывались)
архипелага Новая Земля, зависит от района, где проводились работы, а также
от биологического сезона. С западной стороны, омываемой Водами Баренцева
моря, зарегистрировано 44 вида, с восточной (Карское море) - 39, но*
наибольшее количество - 55 видов, отмечено в юго-восточной части Баренцева
моря, примыкающей к Пкному острову архипелага (табл. 3). При этом, как и в
Таблица 3
Таксономический состав зоопланктона в различных районах
архипелага Новая Земля
Таксон
. 1
Sarsla prlnceps
Sarsla tubulosa
Plotocnlde borealls
Corymorpha flammea
Euphysa tentaculata
Bougalnvlllla superciliaria
Rathkea octopunctata
Halltholus clrratua
Catablema veslcarlum
Horaoeonema platygonon
Calycopsls blrulal
Staurophora mertensl
Perlgonlmua yoldla-arcticae
Perlgonlmua breviconls
Obella flabellata
Aglantha dlgltale
Aeglnopsls laurentll
Cyanea caplllata
Pleurobrachla plleua
Bollnopsls lnfundlbulum
Beroe cucumls
Mertensla ovum
Calanus flnmarchlcua
Calanus glaclalla
Calanus .hyperboreua
Pseudocalanus elongatua
Mlcrocalanus puslllua
Северо-запад
(Зеликман,
Головкин, 1972)
. 2
+
+
-
+
-
+
+
-
-
• -
-
+
+
+
-.
;•
+
-
+
-
+
-
+
-
+
+
+
Юг
(Зеликман.
1961)
3
+
+
+
+ .
-
+
+
+
+
' , - '
+
+
.•-
-
+ ■
+
+
+
+
+
+
-
+
-
+
+
+
Восток
(Карское море)
(Зубова, 1990)
4
-
-
-
+
■ +
-
.-
-
-
+
-
-
-
+ ■
+
■■+
+
+
: ' + '
' ♦'.
+
+
+
+
+
Окончание таблицы э
1
Mlcrocalanus plgmaeua
Centropages hamatus
Limnocalanus grlmaldll
Temora longicornls
Eurytemora hlrundoldes
Drepanopus bungei
Metrldla longa
Metrldia lucens
Acartla longlremls
Acartia clausl
Acartla biflloaa
Heterocope appendlculata
Chirldius obtuslfrons
Gaidlus brevlsplnus
Heterorhabdua norveglcus
Olthona slmllis
Olthona afoantlca
Oncaea borealIs
Mlcrosetella norvegica
Verruoa stroemli
Balanus balanus
Balanus balanoldea
Balanus crenatus .
Mysls oculata
Hyperla galba
Hyperoche inedusaruni
Parathemlsto abyssorum
Parathetnlsto llbellula
Thysanoessa longlcaudata
Thysanoessa lnermls
Thysanoessa raschl
Crangon sp.
Sclerocrangon sp.
Sablnea зр.
Panda1из sp.
Pagurus sp.
Hyas sp.
Llmaclna hellclna
Llmaclna retroversa
CUons llmaclna
Sagltta elegans
Olkopleura labradorlensla
Olkopleura vanhoffenl
Frltlllarla borealls
Frltlllarla polarls
2
-
+
+
+
-■
-.
+
-'
. +
-
-
-
!•-.
- - •
-
"+.
+
+ ,
-
_.
t
-
+ . . ' '
+
-
+
+
t
+ '
-
+
+
+
_
+ r
>■'
*■•'•.
+
+
+
■ +
+
+
+
3
-
+
+
+
■'+,'_
+ .
i +
+
. +
t
.+
+
-
+
-
+
+.
+
+
+
+
+
+
+
+'
+
.-
,' -
t
+
-
-.
+
t'
+
"t
■" '■. -
•■'•''+'■ ■ .
: + ..' ■
■"<■*.'
+
-
4
+
-
+
+
-
-
+
+
+
_
-.
-
+
+
.+
+
+
■ +
-■
-
-
+
-
-
+
+
..+
+
+
-
-
-
-
-
-
+
'. .-
+
+
■+'
+
+
-
других районах Северного Ледовитого океана (и в Мировом океане в целом),
доминируют пелагические ракообразные, составляя более 50% списков видов.
Сходство фаун, оцененное по коэффициентам Серенсена, показывает интересную
взаимосвязь всех трех анализируемых .регионов (коэффициенты сходства -
62-71%). Такое таксономическое "однообразие" в казалось бы достаточно
резко различающихся по основным характеристикам водах архипелага
(трансформированные атлантические и арктические водные- массы, местные баренцевоморс-
кие воды). видимо, обусловлено тем, что. хотя рассматриваемые сообщества
зоопланктона, строго говоря, нельзя считать истинно прибрежными, тем не
менее, это уже и не морской зоопланктон. Можно предположить, что если
провести исследования в губах и заливах Новой Земли во всех частях
архипелага, то таксономическое сходство будет еще большим. В основе объяснения
полученных результатов, а такие высказанного предположения, должна
находиться схема циркуляции водных масс (рис.24). предполагающая существование
вокруг архипелага антициклонального круговорота. При этом, по-видимому,
основное значение имеет стоковое течение, в котором, возможно, формируется
сообщество планктона, фактически единое для всего архипелага.
Рассмотрим размерную структуру таксоцена пелагических Crustacea.
Анализ гистограмм размерных спектров (рис.25) также свидетельствует о
значительном сходстве зоопланктона исследуемых районов. Особенно это касается
сообществ северо-западной и южной частей архипелага. Размерная структура
таксоценов этих локалитетов характеризуется одинаковыми модальными
классами: 0.4-о'л; 1,0-1.6; 4.0-а.3 и 16.0-25.0 мм. При этом наиболее выражен
модальный класс 1.0-1.6 мм, представленный исключительно веслоногими
рачками (Copepoda). Фауна ракообразных вод с восточной стороны Новой Земли
Рис. 25. Размерная структура
фауны пелагических Crustacea в
различных районах архипелага Новая
Земля (А - северо-западная часть
(губа Архангельская и залив Виль-
кицкого), Б - южная часть (губа
Черная) и В - прибрежные воды с
восточной стороны)
1 - lg длины тела. 2 - длина
тела, мм
0.4 1,0 2.5 6.3 10 40
имеет некоторые отличия, выражающиеся в том. что здесь не четыре, а три
модальных класса (1.0-1.6; 4.0-6.3 и 10.0-25.0 мм), а доминирующий смещен
вправо, т.е. в сторону больших размеров (4.0-6.3 мм), несмотря на то. что,
как и в других районах, его образуют представители Copepoda. Такой
своеобразный "гигантизм" таксоцена Crustacea в Карском море, по-видимому, есть
проявление общих закономерностей изменчивости размеров тел животных,
известных как "экологическое правило Бергмана". Баренцевоморские сообщества
развиваются в условиях достаточно заметного влияния- теплых вод
атлантического происхождения, тогда как в Карском море воздействие этого потока
имеет локальное значение и не может выступать в качестве одного из .ведущих
факторов, определяющих условия существования прибрежных сообществ Новой
Земли. Исключение составляют, видимо, лишь районы проливов, через которые
в Карское море поступают водные массы из Баренцева моря (Югорский Шар,
Карские Ворота, Маточкин Шар). Основным природным явлением, которое
оказывает наибольшее влияние на жизнь прибрежных вод, скорее всего выступает
таяние морских льдов и наземного снежного и ледового покрова.
Биомасса. Данные, характеризующие биомассу зоопланктона вокруг
архипелага в течение одного сезона, отсутствуют..В связи с этим представленный
рисунок 26 является в значительной мере приблизительным, поскольку в его
основе лежат сведения, полученные в различные месяцы и годы (Зеликман,
Головкин, 1972; Коптев, Нестерова. 1983; Фомин, Петров, 1985; Тимофеев.
1992).
Биомасса зоопланктона в водах Новой Земли может достигать
500-700 мгм"3 (для слоя дно-0 м), но ее распределение неравномерное.
Больше всего масса зоопланктона в водах Колгуево-Печорского течения, а
также в районе птичьих базаров в северо-западной части архипелага. Особое
внимание следует обратить на губы и заливы. Так, в губе Черной '(южное
побережье Южного острова Новой Земли) в июле 1992 г. биомасса составляла
161 мгм"3 (Тимофеев, 1992), тогда как в губе Архангельской и заливе Виль-
кицкого (северо-западная сторона Северного острова Новой Земли) в августе
1967 г. она превышала 1000 мгм"3 (Зеликман, Головкин. 1972). Такие
различия могут быть обусловлены многими факторами, среди которых основными,
видимо, являются следующие: 1) сезонная и межгодовая изменчивость биомассы
зоопланктона, 2) поступление в воду губы Архангельской и залива Вилькицко-
го значительных количеств биогенных веществ с расположенных здесь больших
колоний морских птиц, что ведет к бурному и обильному развитию
фитопланктона и формированию благоприятных кормовых условий для пелагических
фитофагов (Головкин и др.. 1972; Соколова, 1972; Зеликман, Головкин, 1972).
В губе Черной такого дополнительного источника биогенов в виде
экскрементов птиц нет (либо очень мало).
Сезонная компонента пространственной вариабельности биомассы
зоопланктона состоит в том, что на севере планктон развивается позднее, чем на
юге. Например, в 1978 г. около южной оконечности Новой Земли весенние
процессы в пелагиали отмечались в июне, тогда как на северных участках
(севернее 75° с.ш.) - в июле. Соответотвенно. наблюдалась месячная задержка и
в формировании максимума биомассы зоопланктона (Коптев, Нестерова, 1983).
Однако следует учитывать также то, что северо-западная часть Новой Земли -
район, где в течение всего зимнего периода существует квазистационарная
полынья (Martin. Cavallerl, 1989). Данный феномен может оказывать
существенное влияние на сроки начала развития планктона, сдвигая их на более
ранние даты. Такие ситуации известны, например, для Карского моря
(Численно. 1972) и. по-видимому, роль полыней, как своеобразных сезонных "оазисов
жизни" в пелагиали арктических морей, должна стать предметом специальных
исследований.
В Карском море, вдоль восточного
побережья Новой Земли, биомассы
зоопланктона невелики (рис. 26) и не
превышают 200 мг м"э. При этом наибольшие
концентрации животных приурочены к
районам, находящимся под влиянием барейцево-
морских вод: проливы Карские Ворота и
Маточкин Шар, северная оконечность
Северного острова архипелага.
В целом, распределение биомассы
зоопланктона может быть объяснено с
привлечением концепции "зргоклина" (Legend-
re, Demera. 1985), адаптированной к
арктическим морям (Тимофеев. 1988).
Согласно этой концепции, для формирования
повышенной биологической продукции в
регионе необходим приток добавочной энергии
(тепловой, химической и т.д.).
Максимальное обилие планктона вокруг Новой
Земли обнаружено как раз в таких
районах, где наблюдается либо повышенное
теплосодержание водных масс (потоки
атлантических водных масс), либо
поступление больших количеств биогенных веществ
(районы птичьих базаров).
Рис. 26. Распределение
биомассы зоопланктона
(дно-поверхность) в водах архипелага Новая
Земля 1 - более 501. 2 - 301-500.
3 - 201-300, 4 - 101-200,
и 6 - 0-50, мгм"3
51-100
Таблица 4
Биомасоа зоопланктона (мг м . дно-поверхность)
в Карском море вдоль побережья Новой Земли
Станция*
2
3
4
12
13
14
26
27
28
29
Средняя
1981 г.
114
62
65
50
147
138
114
88
62
200
106.0
1982 г.
122
76
171
88
133
426
134
178
22
177
152.7
* Расположение станций на акватории приведено в работах В. Н. Широколо-
бова (1982) и С.Ф.Тимофеева (1985).
Данные, характеризующие многолетнюю динамику биомассы зоопланктона
прибрежных вод Новой Земли, отсутствуют. Имеются лишь сведения но обилию
зоопланктона вдоль восточного берега архипелага для 1981 и 1982 гг. Пробы
отбирались фактически в одни и те же календарные сроки и в одних и тех же
местах. Анализ данных (табл. 4) свидетельствует о том, что количественное
развитие планктона в 1981 и 1982 гг. было сходным, несмотря на некоторые
отклонения на отдельных станциях. Видимо, это обусловлено одинаковыми гид-
рологичеокйми условиями, имевшими место в 1981-1982 гг: (оба года являлись
холодными) (Бойцов и др.. 1986).
Глава 5. БЕНТОС
5.1. Количественное распределение бентоса н структура донных
сообществ на Лринрвоаемельском шельфе Баренцева моря
Систематические комплексные исследования бентоса в прибрежье Новой
Зенли начались после 1880 года, когда были проведены датская и русская
экспедиции у юго-западных берегов архипелага и в проливе Маточкин Шар
(Галкин, 1979). В 1893 и 1894 гг. Н.М.Книповичем на крейсере "Наездник"
были взяты пробы на нескольких станциях в южной части Приновоземельского
шельфа. В 1889-1906 гг. с организацией Экспедиции научно-промысловых
исследований (ЭНПИ) у берегов Мурмана началось изучение южной части моря до
76е с.ш. Работы выполнялись под руководством Н. М. Книповича и охватывали
почти всю.акваторию, называемую Приновоземельским шельфом. В 1901 г. на
первом русском ледоколе "Ермак" были проведены исследования донной фауны у
северной оконечности архипелага, позже в 1910 г. здесь было выполнено еще
10 станций. Юго-западное прибрежье архипелага было охвачено сборами
бентоса в 1914 г.. но уже с борта ледокола "Вайгач".
В 20-х годах, после окончания гражданский войны, исследования донной
фауны Баренцева моря были возобновлены. С целью уточнения имеющихся данных
по гидрографии, гидрологии и гидробиологии прибрежья Новой Земли в районе
архипелага работали экспедиции под руководством авторитетных российских
гидробиологов (Дерюгин, 1927: Крепе, 1927; Гурьянова, Ушаков, 1928;
Ушаков, 1930). Последующая организация Плавучего морского научного института
позволила начать систематические бентосные работы по всей акватории
Баренцева моря, включая и прибрежье Новой Земли. Они осуществлялись на ледоколе
"Малыгин" под руководством И. И. Месяцева. В 1924 г. тем же Инотитутом на
судне "Персей" впервые в практике отечественной гидробиологии было
положено начало систематическому применению количественных орудий сбора бентоса
- дночерпателей.
Известные русские гидробиологи на протяжении более чем 15 лет
участвовали в различных экспедициях по изучению бентоса моря с помощью
количественных орудий лова. Благодаря исследованиям Л.А.Зенкевича, Г. И.
Горбунова, В.А.Броцкой, Е.М.Крепоа, П.В.Ушакова и других, появилась возможность
анализа не только качественного состава донной фауны, но и ее
количественной представленности в разных районах, обусловленной факторами окружающей
среды (температурой, соленостью, рельефом дна. течениями и связанным с
ними характером донных осадков). . Результаты этих работ были
проанализированы, обобщены и опубликованы в основном в конце 30-х годов (Броцкая, Зение-
вич, 1939). Долгое время они оставались единственными в своем роде и
послужили основой для анализа трофической отруктуры донного населения
Баренцева моря (Кузнецов, 1970; 1980).
К изучению донной фауны Приновоземельского района гидробиологи
вернулись только спустя 30 лет, в конце 60-х годов, когда Полярным институтом
рыбного хозяйства и океанографии (ПИНРО) был организован ряд морских
экспедиций, охватывающих почти всю акваторию Баренцева моря, включая и Прино-
воземельский шельф. Собранные материалы были использованы для составления
общей карты распределения биомассы бентоса (Антипова, 1975), а также для
анализа распределения в водоеме животных отдельных таксономических групп
(Антипова. 1978; Анисимова, 1984; 1989; Галкин, 1989; Денисенко, 1986;
1990; Сикорский. 1991).
В конце 80-х - начале 90-х годов Мурманской арктической
геологоразведочной экспедицией (МАГЭ) совместно с Мурманским морским биологическим
инотитутом было выполнено несколько экспедиций по изучению Приновоземель-
ского шельфа, которое включало исследование видовой и биоценотической
структуры донного населения.
Рис. 27. Сетка станций, выполненных экспедициями
МАГЭ совместно с ММБИ в 1989-1991 ГГ.
* - дночерпатель. • - фотосъемка
Основу настоящего раздела составили материалы, полученные в течение
летних сезонов 1989-1991 гг. экспедициями на судах "Березкин" и "Карача-
евск". Всего было выполнено более 650 станций для исследования
гранулометрического состава донных осадков и подводной фотосъемки. Расстояние между
соседними станциями составляло не более 15 миль (рис.27). На каждой
станции по стандартной методике (Гуревич, 1990} отбирались пробы донных
осадков для двенадцатифракционного гранулометрического анализа.
Фотосъемка осуществлялась малосерийной глубоководной фотокамерой
ГФК-6, укрепленной на дночерпателе "Океан-50". Фотографирование дна
производилось на расстоянии 1.5 м от грунта до момента опускания на него дно-
черпателя. фотографируемая площадь при этом составляла 1 ме. Камеральная
обработка фотоснимков включала идентификацию зарегистрированных на них
донных осадков, представителей ин- и эпифауны, а также некоторых придонных
животных.
Параллельно с фотографированием более чем на 110 станциях были
отобраны количественные пробы бентоса, животных отмывали от грунта на
капроновом сите с размером ячеи 1.0 мм.и фиксировали 5Х-м раствором формалина.
Камеральная обработка, проб включала первичную разборку по крупным
таксонам, с последующей перефиксацией животных 75°-м спиртом и идентификацию
видов различных таксономических групп специалистами.
Помимо фаунистических матери- •
алов 1989-1991 гг.. нами
использованы карточки видовых определений
количественных бентосных проб,
собранных в районе Приновоземель-
ского шельфа в 1930-1932 годах
экспедициями Государственного
океанографического института (ГОИН)
(рис. 28 ) на судне "Персей" (на
26 станциях отобрано по 1-2 пробы
дночерпателем Петерсена с площадью
захвата 0.25 м2) и карточки
видовых определений количественных
бентосных проб, собранных в
экспедициях ПИНРО (рис. 28 ) на судах
"Маслсв" и "Водник" в 1970 г. (на
32 станциях отобрано по 2 пробы
дночерпателем "Океан-50" с
площадью захвата 0.25 ме).
Результаты таксономической
обработки использовали для
определения биогеографической
принадлежности, трофической и биоценотичес-
кой структуры донного населения.
Кластеризация донных сообществ
выполнена по оригинальной программе
с помощью компьютера. Основу для
выделения бентосных сообществ
составляли относительные величины
продукции (Р) каждого вида (Дени-
•А
till 4
Рио. 28. Сетка бентосных
станций, выполненных экспедициями
ГОИНа и ПИНРО • в разные
периоды исследования:
* - оборы ГОИН (1930-1931 гг).
а - сборыПИНРО (1968-1970 гг.)
сенко, 1990), расчеты которой проводили по формуле:
Р, = kj х Bt0-75 х Щ0-2*.
где Вх - биомасса особей 1-го вида в пробе. г/мг, Щ - количество особей
1-го вида, экз/м2. kt - удельная интенсивность метаболизма гидробионтов.
Величины константы к для разных таксономических групп заимствованы из
литературы (Голиков и др., 1990, 1993).
Батиметрия Приновоземельского шельфа Баренцева моря характеризуется
достаточно расчлененным рельефом дна с перепадами глубин от 100 до 300 м
на значительном удалении от берега (рис. 29). Основными рельефообразующими
Рис. 29. Батиметрия в районе Приновоземельского
шельфа Баренцева моря (м)
элементами здесь являются Приновоземельский желоб и расположенное к
северо-западу от него Новоземельское мелководье, опускающееся к западу на
Центральную равнину, в которую на широте пролива Маточкин Шар выходит
своим устьем Приновоземельский желоб. На Новоземельоком мелководье
преобладает холмисто-грядовый ледниковый рельеф с глубинами 100-250 м, в то время
как поверхность равнины имеет пологоволнистый характер с глубинами над
поднятиями от 170 до 250 м (Матишов, 1986).
Выполненная нами фотосъемка дна показала, что результаты
гранулометрического анализа неудовлетворительно характеризуют донные осадки в точках
опробования и без привлечения фотоматериалов не могут использоваться для
объяснения закономерностей распределения бентосных организмов. Так
оказалось, что наряду с глиной, алевритом и супесью, практически вся площадь
Новоземельской прибрежной равнины, Новоземельского мелководья,-. Поднятия
Кленовой и Плато Моллера в большей или меньшей степени покрыты крупной
галькой и разноразмерными валунами. Присутствие последних на дне хорошо
объяоняет факт частого развития в зонах преобладания мягких мелкозернистых
осадков не только многочисленных детритофагов. но и прикрепленных сестоно-
фагов, использующих в качестве субстрата крупнообломочный материал.
Обсуждение вопроса о происхождении этих крупнообломочных включений осадков не
входит в задачи настоящей работы и является скорее предметом специальных
литодинамических исследований. Тем не менее необходимо отметить, что
присутствие галек и валунов на поверхности мелкозернистых осадков может
маркировать как зоны с повышенной гидродинамикой у дна (зоны размыва), так и
избирательно-постоянные зоны вытаивания обломочного материала из льдин и
айсбергов, выносящих камни и гальки с литорали и верхней сублиторали.
Помимо дополнительной информации о характере донных осадков,
подводная фотосъемка позволила нам уточнить видовую структуру ряда бентосных
сообществ, 'обитающих на мелкозернистых осадках. В частности, благодаря
большей площади опробования фотосъемка регистрирует такие немногочисленные, но
крупные формы беопозвоночных, как морские ежи, асцидии, роговые кораллы,
губки и десятиногие ракообразные. Кроме того, отмечено, что на
фотографиях, как правило, насчитывается на 1.5-2.0 порядка больше мелких офиур, чем
в дночерпательной пробе в пересчете на 1 м2 (рис. 30). Соответствующая
коррекция биомассы часто выводит эту группу иглокожих в ранг доминирующих
форм. . -
Согласно проанализированным материалам, средние для Приновоземельско-
го шельфа Баренцева моря значения общей биомассы бентоса в начале 30-х
годов составляли . 200 г/мг (данные ГОИНа), в 1968-1970 гг. „ 90 г/мг
(данные ПИНРО) и в 1989-1991 гг.. . 240 г/мг (наши собственные данные).
Наблюдаемые расхождения в оценке средней величины биомассы могут, объясняться
двумя причинами: различиями в методике сбора первичного материала и
возможными климатическими флуктуациями, повлекшими за собой снижение обилия
бентоса за счет деградации поселений относительно теплолюбивых форм в
период наблюдавшегося в 60-е годы похолодания.
Вместе с тем, в распределении общей биомассы бентоса на Приновозе-
мельском шельфе Баренцева моря для разных лет исследований наблюдается
общая картина. Во всех трех случаях выделяются два основных района с
повышенной биомассой бентоса (рис. 31). Первый из них располагается на
прибрежных мелководьях, простирающихся вдоль Новой Земли на глубинах до 100 м
с галечно-гравийными и гравийно-песчаными донными отложениями. По данным
В.А.Броцкой, Л.А.Зенкевича (1939) и нашим материалам, биомасса бентоса
здесь достигает 200-300 г/м2, а в отдельных местах превышает S00 г/м*. По
данным ПИНРО, в этом районе также отмечены высокие по сравнению с другими
районами значения биомассы, но величины ее значительно ниже и не превышают
300 г/м2.
^<
;*?£
ЫЪ*
•£$'■
•^--
/w
:*v;
Рис. 30. Подводные фотоснимки участков дна, занятых биоценозами с доминированием офиур
CO
-
»
F^
-
-
"
1Г>
1 1
^\V».«i~~
L _1
1 ■
-два*"
' —\&.
->-
t-
'
(
\
1 . II
TO
1 1
^-—
^>
\
■ L_
1
U
<и£и£
^^7~~
C\J
r-
-
-
► ^ -
*
о
CO
s
о
U
N
ш
l
о
CO
m
cd
о
о
Ш
о
а
о
о
<и
S
о
IS
о
ш
S
I
со
о
£
Другой участок с повышенными значениями биомассы отмечен на
северо-востоке района исследования, где величины ее нередко достигают
400-500 г/мг. Глубины здесь составляют 100-150 м. а донные отложения
представлены алевритами с галькой или с отдельно лежащий крупными
валунами. В центральной части района на глубине 100 м и алевритовых, с редкой
галькой, осадках также отмечается повышение биомассы бентоса до величии
200-300 г/мг, по результатам исследований 30-х и 80-х годов, и до
200 г/мг, по материалам 60-х годов.
Понижение биомассы бентоса отмечено в глубоководном желобе, идущем
вдоль побережья Новой Земли, на глубине 200-300 м с пелитовыми донными
отложениями. Величины ее здесь только на отдельных редких участках
составляют 200 г/мг, тогда как в основном.не превышают 100 г/мг. Минимальные
значения биомассы - менее 100 г/мг - отмечены в северо-восточной части района
исследования на глубинах от 150 до 250 м песчано-алевро-пелитовых донных
отложений.
В целом сходная для разных периодов исследований картина в
распределении биомассы бентоса на Приновоземельском шельфе не противоречит
выдвигаемому предположению о возможном снижении ее в результате имевшего место
похолодания, поскольку участки с пов'ышенными значениями маркируют лишь
наиболее высокопродуктивные зоны, которые остаются таковыми, по нашему
мнению, вне зависимости от мелкомасштабных климатических флуктуации. В то же
время деградация поселений относительно теплолюбивых форм без последующей
замены их поселениями относительно холодолюбивых видов представляется
маловероятной. Таким образом, однозначного вывода о причинах низких
зарегистрированных значений биомассы бентоса на Приновоземельском шельфе в
конце 60-х годов сделать сейчас невозможно, поскольку для этого необходимо
проанализировать имеющуюся информацию о распределении и величинах биомассы
бентоса в других районах Баренцева моря в различные периоды исследований.
Выделение сообществ по единой методике с использованием материалов
трех указанных выше съемок дало ряд расхождений при сопоставлении бентони-
ческих группировок,описанных В.А.Броцкой и Л.Д.Зенкевичем (1939), с
выделенными нами (рис. 32а,б). Последнее связано не только с иными подходами к
кластеризации доминирующих форм, но и с самим понятием доминанты в
биологическом сообществе. Так, например, в сообществе Eupagurus pubescens,
расположенном в непосредственной близости от архипелага Новая Земля,
практически напротив пролива Маточкин Шар, к доминантам нами отнесены
прикрепленные сестонофаги - балянусы и асцидии (табл.5), а не всеядные и
подвижные раки-отшельники (Броцкая, Зенкевич, 1939). И, по-видимому, не случайно
для этого района П.В.Ушаков (1930) описал сообщество Stegophiura nodosa, в
котором, помимо названной офиуры, преобладали также фильтраторы-сестонофа-
ги.
Границы распространения комплекса Ophiopleura boreatts -• Ophtocanta
btdentata (no: Броцкая, Зенкевич. 1939) совпадают с биоценозом, выделенном
нами по данным этих авторов. Биоценоз расположен на глубинах свыше 200 м и
илистых донных осадках с подстилающей глиной. К доминирующим видам, помимо
двух указанных форм, отнесены Maldane sarst, Spiochaetopterus typtcus
(рис. 326). Аналогичное сообщество было выделено в северной части Приново-
земельского шельфа по результатам экспедиций ПИНРО (рис. 33). Наряду с уже
отмеченными видами в этом сообществе доминируют Htatetta arettca и Ophto-
photts aculeata.
Рис.32. Распределение донных биоценозов на Приновоземельском шельфе Баренцева моря по материалам экспедиции ГОИН (1930-1931гг.)
а - схема 30-х годов (Броцкая, Зенкевич, 1939): 1 - Splochaetopterus typlcus, Ctenodlscus crlspatus, Golphlngla margarlta-
ceum; 2 - Psolus phantapus, s.typlcus, Astarte crenata; 3 - Ophlopholls aculeata, Macoma calcarea, Strongylocentrotus sp.;
4 - S.typlcus, M. calcarea; 5 - Eupagurus pubescens; 6 - M. calcarea, Trldonta borealis, Mya truncata; 7 - T.borealls, M.calcarea,
Clllatocardlum clllatum; 8 - Ophlopleura borealis, Ophlocantha bldentata.
6 - схема, построенная с помощью компьютера: 1 - М.calcarea, Ophlura robusta, P.phantapus; 2 - G.margarltaceum,
Сcrlspatus, S. typlcus; 3 - T.borealls, Balanus sp., M.calcarea; 4 - Balanus sp., Ascldlacea g.sp.; 5 - 0.borealis. O.Mdentatha, Malda-
ne sarsl. S.typlcus
Таблица 5
Средняя биомасса (В, г/мг) и плотность поселений (N. экэ/м2)
донных сообществ Приновоземельскогс шельфа
Сообщество
1
гоин
1930-е годы
В
2
N
3
ПИНРО
1968-70 ГОДЫ
В
4
N
. 5
ИМБИ
1989-90 ГОДЫ
В
6
N
7
Balanus sp. 61 88
Ascldlacea
Stegophlura nodosa
Maldane sarsl 27 116
Splochaetopterus typlcus
Ophlacantha bldentata
Macoma calcarea 315 609
Ophiura robusta
Psolus phantapus
Golflngla margarltaceum
66 60
91 141
197 483
Ophlacantha bldentata
Golflngla margarltaceum
Ctenodlscus crlspatus
Splochaetopterus typlcus
150 1284 143 459 141 557
Ophlacantha bldentata
Golflngla margarltaceum
Ascldlacae
Spongya.
9 49
Golflngla margarltaceum
Splochaetopterus typlcus
Maldane sarsl
Psolus phantapus
Splochaetopterus typlcus
Maldane sarsl
Ophlacantha bldentata
78 50 252 1125
85 337
Trldonta borealls
Balanus spp.
Maooma calcarea
330 776
118 231
Nephthys spp.
Holothuroldea
Pectlnarla hyperborea
Splochaetopterus typlcus
13 36
Окончание таблицы 5
1
4 5
6 7
Chlamys islandlua
Balanus spp.
Bryozoa
Ascldlacea
108 275
Balanua app.
Strongylocentrctus spp.
Macoma calcarea
Ophlacantha bidentata
87 183
Splochaetopterus typlcus
Polychaeta
Ctenodlscus crlspatus
Ophicpholls aculeata
33 156
Ж
^
У X&t:
-г*
I ■ S
^ 3^4t
^ # §
^ V
*4$
ПШ2БЭ'
EZ33
Рис. 33. Распределение донных
сообществ по материалам бентосной
съемки ПЙНРО (1968-1970 гг.)
1 - М. calcarea. 0. robusta,
P.phantapua; 2 - O.borealls, O.bl-
dentatha, M.saral, S. typlcus; 3-
И. aarsi, G.margarltaceum, S.
typlcus; 4 - Nephtnys sp., Hcloturol-
dea. Pectinaria hyperborea,
S.typlcus; 5 - 0. bldentatha, G.marga-
rltaceum. С crlspatus. S. typlcus
Биоценоз Macoma calcarea - Ophlura robusta - Psolua phantapus
занимает на Приновоземельском шельфе, согласно нашим построениям, наибольшие по
площади участки дна (рис. 34) и располагается на илистых склонах Приново-
земельокой впадины. Границы его распространения в разные годы значительно
изменяются. Так. по материалам В.А.Броцкой и Л.А.Зенкевича (1939), он
располагается в юго-вссточной части района исследования, а на северо-воотоке
замещается сходным сообществом Psolus phantapus - Splochaetopterus typlcus
- /starts crenata. По материалам ПИНРО, первый из указанных биоценозов у
Южного острова смещается ближе к берегу, а у Северного острова - занимает
—I • ' • ■ ■ ■—
48° 52 56 60
Рас. 34. Распределение донных биоценозов по
материалам экспедиций МАГЭ и ММБИ (1989-1991 ГГ.)
1 - Chlamys lslandlca. Balanus зр., Bryozoa. Asel-
dlacea; <2 - M.calcarea. O.robusta, P.phantapus.
G.margarltaceum; 3 - O.bldentatha, G.margarltaceum, C.crls-
Batua, S. typlcus; 4 - G.margarltaceum, S. typlcus,
.sarsi. P.phantapus; 5 - S. typlcus, Polychaeta,
С crispatus, 0.acuieata; 6 - Balanus зр.,• Strongylo-
centrotus sp.. M.calcarea. O.bldentata; 7 - O.Mdenta-
ta, G.margarltaceum, Ascldlacea, Spongya; 8 - T.borea-
11з, Balanus sp., M.calcarea; 9 - S. typlcus. M.sarsi,
O.bldentata
большее пространство, чем это отмечено ранее. По данным съемки 80-х годов,
этот биоценоз располагается широкой полосой с севера на юг вдоль архипела-
га до 73°30' с.ш. В северной части границы его ареала совпадают с таксвыми
по материалам ПИНРО. На юго-востоке района исследования на основе съемки
ММБИ-МАГЭ выделяется близкое по видовому составу локально расположенное
сообщество Gol/tngta margaritaceum - S. typicus - Maldane sarst - Psolus
plumtapus, которое при* некотором отличии в видовом составе, располагается
в пределах основного сообщества, описанного в 30-х годах.
Второе из наиболее типичных сообществ,. выделенных для
рассматриваемого региона Баренцева моря. - Golphlngia margaritaceum - Ctenodtscus crts-
patus - Spiocbaetopterus typicus - оопряжено с глубинами более 200 м и
илистыми донными осадками с подстилающей глиной. По материалам ГОИНа, его
расположение на дне дискретно и приурочено к желобу, идущему вдоль
побережья Новой Земли ст Центральной впадины, сообщество со сходным набором
доминирующих видов - М. sarst - Golphlngia margaritaceum - Ctenodtscus
crtspatus -выделено нами по материалам съемки 60-х годов. Однако
расположение его значительно отличается от нашей сводки по данным ГОИНа. Оно
занимает участки дна, где в 30-е годы доминировали Macoma calcarea и Opntura
robusta. По данным ММБИ, рассматриваемое сообщество располагается в
северо-западной части района исследований, частично захватывая склоны желоба,
ведущего к Центральной впадине.
В пограничных участках между двумя рассмотренными крупными
сообществами выделено три переходных биоценоза, располагающихся мозаично. В первом
из них доминирует Ophtacantha btdentata и в большом количестве
присутствуют Gotpnt'ngta margaritaceum. . Ctenodtscus crtspatus, кроме того, весьма
часто встречаются фильтраторы (асцидии. губки). Доминирующими видами во
втором переходном сообществе являются Sptochaetopterus typicus, Maldane
sarst, Opntocanta btdentata, Opntopleura borealts. В последнем доминируют
Spiocbaetopterus typicus, Cerrianthus lloydt, С crtspatus, Ophturotdea.
Эти сообщества приурочены к значительным глубинам и преимущественно мягким
илистым донным отложениям с небольшим включением грубообломочного
материала.
Методика, используемая нами для идентификации сообществ, выявила в
биоценозе, описанном В.А.Броцкой и Л.А.Зенкевичем (1939), как Мдо truncata
- Trtdonta boreatts, другой набор доминирующих Форм: Trtdonta borealts -
Batanus sp. - Macoma calcarea. При этом границы его ареала также
изменились. Аналогичное сообщество было выявлено и по материалам 80тх годов,
однако ближе к берегу, на глубинах, не превышающих 100 м на мягких
илисто-песчаных донных осадках.
По данным съемки 80-х годов, в южной части района исследований и на
крайнем северо-востоке на илистых донных осадках с добавлением отдельных
камней и гальки на глубинах 100-150 м обитает сообщество Macoma calcarea -
Strongylocentrotus sp. - Balanus sp.. по расположению оно совпадает с
комплексами "8" на юге и "2" на севере (рис. 32а). но имеет другой набор
доминирующих форм.
Кроме того, по материалам съемки 80-х годов, в районе исследований
выделено незарегистрированное здесь раньше сообщество сестонофагов. Оно
располагается дискретно на различных участках Приновоземельского шельфа и
сопряжено о высоким содержанием грубообломочного материала (камней,
гальки) в дойных.осадках (рис. 34). К доминирующими формам в этом сообществе
относятся Chlamus tstandtea. Balanus sp., различные виды Brj/ozoa и Hsctdt-
acea.
Анализируя существующие различия в количественной представленности
бентоса, видовой структуре и пространственном распределении донных
сообществ в различные периоды текущего столетия, можно разделить возможно
вызвавшие их причины на две категории. Первая категория - естественные
флуктуации, вероятно связанные с климатическими изменениями. Вторая
категория - методические, связанные с отбором, промывкой, сортировкой и
хранением бентосных проб, определением местоположения судна.
Возможную роль естественных флуктуации мы уже частично обсуждали вы-,
ше. По-видимому, она может быть весьма значительной, но лишь в плане
казуальности пространственной локализации и структурных изменений донных
сообществ на таксономическом уровне. Способность климатических изменений
существенно сказываться на суммарной .биомассе бентоса, по нашему
мнению,маловероятна.
Относительно неточное определение местоположений судов вдали от
берегов (в основном по счислению), практиковавшееся почти до середины 80-х
годов, также может быть причиной некоторых расхождений в локализации
сообществ согласно материалам разных лет. По мнению специалистов, ошибки в
определении местонахождения судна за 2-3 дня без контроля по секстанту,
системе спутниковой навигации или системе радиомаяков могли достигать 20-30
миль. Поэтому смещение локализации тех или иных сообществ по материалам
разных лет наблюдений на указанное расстояние может быть признано как
следствие методической погрешности.
Таким образом, при анализе возможных изменений в донной фауне
Баренцева моря вследствие климатических или других естественных причин по
материалам различных периодов наблюдений, следует особое внимание обращать на
качество исходного.материала, которым и будет в основном определяться
объективность тех или иных сопоставлений.
Относительно вышерассмотренной' ситуации можно констатировать, что
при оценке каких-либо прямых изменений в количественных характеристиках
зоббентоса проанализированный материал должен использоваться весьма
осторожно. Более смелс сн может применяться для анализа структурных изменений
в сообществах на таксономическом уровне, особенно если ограничиться
группами доминирующих и характерных видов. Достоверность оценки изменений в
локализации сообществ будет в данном случае определяться лишь
пространственной дискретностью наблюдений и точностью их местоположений.
5.2. Бентосные фораминиферы в заливах Новой Земли >
Активно тающие в летний период ледники Новой Земли сбрасывают в
заливы большое количество минеральной взвеси с концентрацией до 88 мг/л
(Аксенов, 1987). Насыщенные взвесью талые веды формируют поверхностный слой
водного столба в заливах и выходят в открытое море (Taraaov et al., 1993).
Осаждаясь, взвесь формирует глинистые ледниково-морские фации (Гершанович.
1974). Современные процессы в заливах, находящихся под воздействием талых
ледниковых вод, представляют собой модель ситуации, существовавшей в
регионе в момент распада плейстоценовых ледниковых щитов (Матишов, 1984).
Летопись осадков несет информацию о палеообстановках. Фораминиферовый анализ
является одним из наиболее эффективных методов палеоэкологии. Чтобы ин-
терпретировать смену состава четвертичных комплексов фораминифер,
необходимы сведения о распределении современной фауны. Распределение фораминифер
в заливах Новой Земли до настоящего времени не изучалось. Настоящая
публикация' имеет целью частично восполнить этот пробел.
Пробы, послужившие материалом для исследования, были собраны автором
в 61-м (1991) и 68-м (1992 г.) рейсах НИС "Дальние Зеленцы" (табл. 6,
Таблица 6
Список станций
Станция
Координаты
N
Е
Дата
отбора
Глубина,
м
Грунт
Отбор
проб
Губа Безымянная
68/1
Бз-лит
Гл-лит
68/3
68/31
.68/32
68/33
874
894
895
896
Но-Зм
Но-ЛИТ
68/4
Вц-лит
Вз-лит
72?58.8'
72 59.0
75 07.0
'75 28.0
75 33,3
' 75 28.5
,75 21.5
75 22.0
75 19.7
75 20.0
75 24.7
75 20.5
. 75 20.2
75 35.6
75 35.0
76 31.0
53»03.4'
53 04.0
56 25.0
56 44.2
56 27.7
57 10.0
57 35.8
57 29.0
57 30.2
57 45.0
57 06.0
57 40.0
57 44.0
. 57 58.9
57 59.2
65 15.0
18.08.92
18.08; 92
Бухта Глазова
31.08.91
28.5
0
0
Залив Норденшельда
19.08.92
30.08.92
30. 08.92
30.08.92
• 18,08.91
29.08.91
29.08.91
30.08.91
30.08.92
30.08,92
139
165
120
45
76
31
49
149
3
0
Залив Вилышцкого
20.08.92'
20,08.92
Залив Визе
20.08.92
56
0
0
П
П
Гр
И
И
и
и
и
и
и
и
п
. Гр
И'
гр
Гр
Ван Bi
Box
Box
Box
Box
Океан
Океан
Океан
Океан
В
Box
ПРИМЕЧАНИЕ. Грунт: Гр - гравий, П - песок, И -ил. Отбор проб; Океан -
дночерпатель "Океан", Ван Вин - дночерпатель Ван-Вина, Box - коробчатый
дночерпатель, В - водолазные сборы.
рис. 35), Пробы жесткого субстрата (камни, ракуша, плотные покровы донной
фауны), на котором обитают прикрепленные фораминиферы, просматривались под
бинокуляром в течение рейса. Поверхностные пробы мягкого осадка объемом
20-100 см3 отобраны из верхнего сантиметра грунта, фиксированы и хранились
в 70°-м спирте; окрашены бенгальским розовым для разделения раковин о
цитоплазмой ("живых") и без цитоплазмы ("мертвых"); промыты на почвенном
сите о ячеей > 0,063 мм. Живые и мертвые фораминиферы подсчитаны во фракции
0.125 мм (табл. 7). . Две колонки донных отложений (ст. 894 и 895) отобраны
ударной трубкой. Образцы толщиной 5 см высушивались и затем отмывались на
сите с ячеей > 0.063 мм; для выделения раковин фораминифер использована
тяжелая вддкооть (СС14); раковины подсчитаны во фракции > 0.063 мм. Нате-
52°
56е
60е
64е
68°
Пробы:
• дночер пател ыше
• литоральные
Рис. 35. Места отбора проб для анализа распределения фораминифер
риал составил 7 проб жесткого субстрата, 16' поверхностных проб мягкого
осадка и 16 образцов из грунтовых колонок. В пробах отмечено 60 видов фо-
раминифер:
Adercotryma glomerata (Brady), 1878
Allogromllna зрр.
Alveolophragmlum crasslmargo (Norman). 1892
Ammodlscus зр.
Ammotlum cassis (Parker), 1970
Astacolus hyalacrulus Loebllch & Tappan. 1953
Astrcnonlon gallowayi Loebllch & Tappan, 1953
Bollvlna pseudopunctata Hoeglund, 1947
Buccella frlglda (Cushman). 1922
Buccella tenerrlma (Bandy). 1950
Cassldullna renlforme Norvang. 1945
Clblcldes lobatulus (Walker J. Jacob), 1798
Cornusplra sp.
Crlbrostorooldes Jeffreysl (Williamson), 1858
Dentallna baggl Galloway & Wlssler, 1927
Eggerella advena (Cushman). 1922
Elphldlum alblumbllicatum (Weiss), 1954
Elphldlum bartlettl Cushman. 1933
Elphldlum clavatum cushman. 1930
Elphldlum subarctlcum Cushman. 1944
Eplstomlnella sp.
Flssurlna marglnata (Montagu), 1803
Gavellnopsls praegerl (Heron Allen & Earland). 1913
Globobullmlna turgida (Bailey), 1851
Hyperammlna subnodosa Brady. 1884
Hlppocreplnella alba Heron-Allen & Earland, 1932
Islandlella helenae Feyllng-Hanssen & Buzaa. 1976
Islandlella norcroasl (Cushman). 1933
Lagena gracllllma (Seguenza). 1862
Lagena semlllneata Wright, 1886
Melonls barleeanus (Willamson), 1858
Mlllollnella pyrlformls (Schlumberger), 1894
MUlollnella spp.
Neogloboquadrlna pachyderma (Ehrenberg). 1894
Nonionella auricula Heron-Allen & Earland, 1930
Nonlonelllna labradorlca (Dawson), 1960
Oolina borealis Loebllch & Tappan. 1953
Patelllna corrugata Williamson. 1858
Peloslna variabilis Brady, 1879
Polymorphlnldae зрр.
Proteiphldlum nlveum (Lafrens). 1863
Protelphldium orblculare, (Brady), 1881
Proteonlna sp.
Psammosphaera sp. 2
Qulnqueloculina semlnula (Llnne), 1758
Qulnqueloqullna stalkerl Loebllch & Tappan. 1953
Recurvoldes turblnatus Brady. 1881
Reophax arctica Brady, 1881
Reophax atlantlca (Cushman). 1944
= Proteonlna atlantlca Cushman, 1944
Reophax scorpiurus Montfort, 1808
Reophax scottll Chaster, 1892
Robertlna arctica d'Orbigny, 1846
Rosalina spp.
Rotaliammina ochracea (Williamson), 1858
Spiroplectammlna blformls (Parker fc Jone), 1865
Stalnforthla loebllchl (Feyllng-Hanssen), 1954
Stalnforthla schrelberslana Czjzek, 1848
Textularla earlandl Phleger, 1952
Trlfarlna fluens (Todd). 1947
Trochammlna nana (Brady), 1881
= Trochammlna karlca Stschedrlna, 1946
Trochammlnella atlantlca F.Parker. 1952
Trochammlnella bullata Hoeglund, 1947.
Литоральные пробы собраны в пяти заливах, расположенных на западном
побережье Новой Земли (рис. 35). Ни в одном из заливов не встречены
илистые отложения в межприливной зоне. В пяти пробах литорального песка и
гравия живые фораминиферы не обнаружены. Исключение составляет самая южная
станция (губа Безымянная), где найден один живой экземпляр Polymorphinidae
gen. sp., предположительно вынесенный на литораль прибоем. На валунах и
гальке в пяти исследованных заливах не встречены макроводсросли;
прикрепленные фораминиферы отсутствуют. Немногочисленные пустые раковины,
обнаруженные в песке и гравии на литорали трех, из пяти исследованных заливов,
абрадированы и имеют крупный размер. Среди шести отмеченных видов только
один - Ctbtctdes tobatulus - входит в фауну фораминифер литорали Кольского
полуострова, где этот вид встречается как прикрепленный на камнях и
макроводорослях, остальные виды в Баренцевом море встречаются только ниже уреза
воды (Корсун и др., 1993). Таким образом, все встреченные раковины,
по-видимому, переотложены. Интенсивное выпахивание осушной зоны льдом,
вероятно, является основной причиной, обуславливающей отсутствие живых
фораминифер на литорали Новой Земли.
В верхней сублиторали была собрана только одна проба - на песчаном
грунте (глубина 3 м) в заливе Норденшельда (рис. 36). Живые фораминиферы в
этой пробе.не обнаружены, пустые раковины, вероятно, также переотложены.
Все дночерпательные пробы отобраны на небольшом удалении от берега в
зоне ледниксвс-морского осадконакопления, где донные отложения
представлены глинистым илом с выраженной слоистостью (Tarasov et al., 1993).
Немногочисленные гальки ледового разноса оказались целиком погруженными в
чрезвычайно разжиженный осадок, таким образом, в пробах отсутствует жесткий
субстрат, на' котором обитают прикрепленные фораминиферы. Как живые, так и
мертвые, неприкрепленные фораминиферы в мягком осадке обнаружены на всех
станциях (табл. 6).
Наиболее подробно исследован залив Норденшельда (рис. 36). Леднико-
вс-морские отложения во внутренней части залива (ст. 874. 894, 68/33 и
895) имеют слоистость с шагом от 1 до 10 см, которая, по мнению Тарасова с
соавторами (Tarasov et al., 1993), определяется сезонной цикличностью
режима седиментации. Соответственно, скорость осадконакопления составляет
Таблица 7
Численность живых и мертвьк фораминифер, экз/10 см3 .(не включены станции Но-лит и Вз-лит
фораминиферы отсутствуют)
Вид
1
Станция . .
68/1
2
68/31
3
68/3
4-
68/32
5
896
6
874
7
894
8
68/33
9
895
10
68/4
■ и
Бз-лит
12
Гл-лит
13
Но-Зм
.' 14"',
Вц-лит
15
Allogromllna spp.
Hlppocreplnella alba
Psansnosphaera sp.2
Proteonlna sp.
Pelosina variabilis
Нурегапп1ла subnodosa
Ammodlscus sp.
Reophax atlantlca
Reophax scorplurus
.^
0.5
L&
1.0
0Л
0.5
2JL '
3.5
-2.5
2.5
iTs
1.-0
£L5
L£
1.5
^ ■
. 0T5
Го
JJU
52Л
О
2&Л
2.0
10.5
0.5 .
0.5
1B.0
0?5
Ш
. 1.0
' 1Д
0.3
0.2
U>
0.8
0~3
3.0
1.5
Продолжение таблицы 7 .
10
и
12
13«
14
15
Reophax" arctica
Reophax scottll
Ammotlum cassis
Alveolophragmlum
crasslmargo
Crlbrostomoldes
Jeffreysl
Adercotryma glomerata
Recurvoldes turblnatus
Trochammlnella
bullata
Trochanmlna nana
Splroplectammlna
Textularla earlandl
Eggerella advena
Cornusplra sp.
2.5
■
2.5
12.0
55.0
8.5
0.5
33.5
i
1.0
1.0
15.5
6.0
0~5
- LA
10.5
JLS
5.5
3~0
0.5
2.0
8.5
m^^^
2.0
2.5
' ■
2.0
2.5
2.5
0.5
0.5
1.0
2.5
^m^_
0.5
Г75
,^^_
2.5
. 4.5
0.5
4.0
2.0
&JL
10.0
QJ.
0.3
0.3
0.3
1.3
0.8
0.3
6.3
0.5
с
о
l«J.
Id
Iю
1 с!
В
о
«
о
п.
is-
id
аз -a i
• о
12
is-
э
э
15
3
а
о
(71
п
inj о ю|
«н in d
1°
Icvt
I*
in
о
t-
CM
Ю
as i
I™
! H I
3 & Д
О i-l
О О О)
.-I в) <-* iH С
«и о) ьн
я а я о> r-i
& С & Ai О
си вни
и В н а) П
I Is я э и я
РЦ
3
В
В
S i
п
О гН
Я К
»-t О
О р.
8
ш
о
о
S
5
б
о
чн ой Э 9
t. Вше ri ' »
«-■ Я о о -от
о»
о
I
с
о о -от
гн ч-i щ-t чн ц a
■rj Jj S Л ш 3
О) О О Q, г-| тЧ
СО С гН Э в
о а о о) о *н
н (, (< ИнД
о S и СТ,н
^ ■<
рц о ы
Продолжение таблицы 7
10
11
12
13 14 15
Elphidium clavatum
Cassldullna renlforae
Islandlella norcrosll
- I.helenae
Bollvlna pseudoptmctata
Trlfarina fluens
Stalnforthia loebllchi
2.5
0.5
5.5
йЛ
1.0
•
Stalnforthla schrelberslana '
Globobullmlna turglda
Robertlna arctlca
Polymorphlnidae spp.
Astacolus hyalacrulus
Dentallna baggl
1Л
0.5
0.5
6.5
9.0
ioTo
0~5
0~5
JL5
0.5
6.5
28.5
2^5
14.0
ИЛ
19.5
0.5
0.5
<L5_
1.0
&Л
34.5
5J>
29.0
13.5
31.5
1.5
0.5
2^5
3.0
-
67?
2.0
2.0
50.0
28.0
52J2
8.0
6.0
&Л
1.5
ЁЛ
1.5
M'-
l.Q
0.3
йЛ
2.0
„ ш
0.4
SLL
0.5
24
1.3
0.8
Li
16.3
0.5
0.3
fi^
0:-3
SLA
1.9 0.5
0.2 0.6 0.2
Lagena semlllneata
0.5
окончание таблицы 7
1
Oollna borealls
Численность,
ЭКЗ/10 СИ3
Известковые
фораминиферы, %
Выборка, экз.
Количество видов.
всего
Количество видов
известковых
Живые особи, %
Планктон (HeogloboquacIrinE
2
20.0
97.5
40.0
37.9
39
193
7
16
_£
12
17
i pachj
3
22,5
170
68.1
45.9
47
320
12
23
_§ ■
10
.12
4
шл
152
92.6
81.2
Л2_
264
10
22
7
11
24
5
SLS.
166
3§Л
90.9
Ш-
268
Л
22'
__в
12
35
ruerma sin), экз.
6
94-0
48.0
ж&
69.6
47
23
_1
5
5
3
67
7
122
10.0
Ь2|Т Q
100
-Ж
5
5
2
__2
2
92
8
73.5
13.5
24,5
77.8
147
27
10
8
6
5
84
9
'5J
6.0
3i2
83.3
17
18
_5_
6
з.
5
49
. 10
' гж
1.5
7.7
66.7
26
15
_4
5
2
3
63
11
Д,у. р
23.3
11Л
83.9
52
92
11
14
7
7
.37
12
0.4
0.8
ш
.100
' 1
2
1
2
1
2
wO
13
Р,
5,0
100
■ _р_
24
..„ ш „, I,
4
4 '
0
14
0.63
А
6.9
100
JL
10
——
6 '
6
0
15
й_
0.9
100
0
5
—
3
3 >
0
1
Фракшя осадка
> 0.063 Ш. * 97.1 5.4
7.4
4.5'
2.5 0.9 0.3 0.3 0.1 2.6
100
80.0 70.5 97.7
ПРИМЕЧАНИЕ. Числитель - живые, знаменатель - мертвые фораминиферы.
56e30*
57°00'
57"30'
S8"O0"
75034.
-7S°30*
Рис. 36. Схема расположения станций в заливе Норденшельда
1-10 см/год. При столь высокой скорости седиментации на поверхности осадка
не успевает формироваться окисленный слой, типичный для мягких отложений
открытых районов Баренцева моря; цвет осадка по всему разрезу -
светло-серый. На дистальных станциях {68/31, .68/3, 68/32) скорость седиментации
значительно ниже и на поверхности светло-серого глинистого ила
присутствует слой окисленного осадка светло-оливкового цвета толщиной около 2 см
(рис. 37). На станции 896 ситуация промежуточная - окисленный слой
(толщиной 2 см) присутствует, но цветовая дифференциация слаба. Макрозообентос
беден в заливе Норденшельда. В дночерпательных пробах встречены
малочисленные моллюски-детритофаги Porttandta arctica, Nucula tenuis и Yotdtetta
fraterna (определение Н.А.Анисимовой и Е.А.Фроловой, ММБИ).
Раздельный подсчет живых и мертвых фораминифер позволяет
охарактеризовать их распределение во временных срезах различного масштаба. Скорость
репродукции бентосных фораминифер в Баренцевом море составляет 2-3
поколения в год (Воронова, 1976; Корсун и др., 1993). Соответственно,
состав.живых комплексов фораминифер в заливе Норденшельда характеризует летний
период 1991 и 1992 гг.
Интерпретация комплексов мертвых фораминифер зависит от оценки
скорости осадконакопления и оценки интенсивности процесса посмертной
дезинтеграции раковин. По-видимому, в исследованном районе тафономические
преобразования комплексов фораминифер значительно слабее, чем в открытых
районах Баренцева моря, где пустые раковины остаются в поверхностном слое
осадка в течение десятков и сотен лет, подвергаясь биотурбации и разрушаясь
за счет химической коррозии (известковые раковины), а также бактериального
и механического воздействия. В заливе Норденшельда, благодаря высокой
скорости седиментации, пустые раковины находятся в поверхностном слое осадка
в течение непродолжительного времени (несколько месяцев - несколько лет) и
интенсивность посмертного разрушения раковин не велика. Видимо, лишь самые
неотойкие псевдохитиновые раковины аллогромиид разрушаются прежде чем
попадут в подповерхностные слои осадка. .
Соотношение численности живых и мертвых фораминифер (рис, 376)
подтверждает оценку скорости осадконакопления. сделанную Тарасовым с
соавторами (Tarasov et al., 1993). На самой удаленной от ледника станции 68/31
доля живых особей составляет 12%. то еоть интервал отбора проб (0-1 см)
содержит скелетные остатки приблизительно восьми генераций фораминифер и,
соответственно, отложился за период порядка четырех лет. На станциях
внутри залива доля живых превышает 50% (рис, 376). Следовательно, верхний
сантиметр осадка отложился за период менее года.
Число видов живых фораминифер несколько убывает по направлению к
леднику (рис. 376), что, видимо, объясняется усилением экологического
стресса, источником которого выступают, талые ледниковые воды. Разнообразие
мертвых фораминифер резко уменьшается на входе в залив, между станциями
68/32 и 896. Однако этот тренд вряд ли является достоверным, так как число
учтенных видов сильно зависит от объема выборки, а количество подсчитанных
пустых раковин в пробах из залива на порядок величины меньше, чем в
пробах, взятых на удалении от берега (табл. 7).
Численность живых фораминифер на всех станциях, за исключением двух
ближайших к леднику, составляет от 20 до 120 экз/10 см3, что не уступает
средним величинам численности, известным для открытых районов Баренцева
моря (Корсун и др., 1993). На двух проксимальных станциях (68/33 и 895)
К ЛЕДНИКУ
Allogromiina spp.
60
50
40 ■'----
30
20
живые
мертвые
20Г
10
Spiroplectammina biformis
_EZU
20r
10
Quinqueioculina stalker!
0
№rt<mdlinaJa_lHadOTica_^__-
Islandiella norcrossi (= Islandiella helenae)
40
30
20
10
0
Elphidium ctavatum
_□.
ш
60
Cassidulina reniforme
50
40
30
20
10
68/31 68/3 68/32 896 874 894 68/33 895
Станция
. Рис. 37. Распределение бентосных фораминифер
в заливе Норденшельда:
а - численность отдельных видов (экз/10 см3);
б - сводные характеристики комплексов фораминифер,
глубина моря, осадок. См. с.92
К ЛЕДНИКУ
200
150
100
50
Численность фораминифер, экэ/ЮсмЗ
д
живые
мертвые
Доля известковых фораминифер, %
^
Доля живых фораминифер, %
25
20
15
10
5
0
0
50
100
150
200
Количество видов фораминифер
живые .
мертвые
■<«■" i
Глубина моря, м
Окисленный поверхностный слой донных отложений
«—присутствует » « ■отсутствует -—»
Содержание фракции осадка >0.063 мм, %
68/31 68/3 68/32 896 874 894 68/33 895
Рис. 37. Окончание
численность швшх фораминифер чрезвычайно низка - менее 5 экз/10 си3 (рис.
376), что, по всей вероятности, объясняется исключительно интенсивным
стрессовым воздействием ледникового стока.
На илистых грунтах открытых районов Баренцева моря среди живых фора-
минифер попеременно доминируют Melonts barieeams, Reophax atlantica, Re-
opfmx scorpiurus и ^logromtiria spp. (Корсун и др., 1993). В заливе Нор-
деншельда все массовые формы за исключением аллогромиид. не входят в этот
список (рис. 37а), что говорит о качественном своеобразии фауны залива;
В Бар'енцевоморском регионе фиорды Шпицбергена сходны с заливами Новой
Земли в том, что в них тоже впадают выводные языки ледников. Накоплена
определенная информация о современной фауне фораминифер Шпицбергена. Во
фиордах, особенно во внутренних частях, обильны известковые фораминиферы
Elphtdim clavatum и Cassidultna reniforme- (Nagy, 1965; Elverhoi.et al.,
1980). Эти виды рассматриваются как характеризующие ледниково-морское
осадконакопление (Osterman, Andrews, 1983). Среди агглютинирующих форм в
качестве массовой отмечена Spiroplectamlna bl/ormts (Elverhed et al.,
1980).
В пробах из залива Норденшельда MonioneUtrta labrauorica и Istandtet-
la noerosst встречены только на удаленных от ледника станциях (68/31.
68/3, 68/32). первый вид доминирует по численности среди живых фораминифер
(рис. 37а). Ближе к леднику (ст. 896, 874, 894) отмечены максимумы
численности Cassidultna reniforme, Altogromiina spp. и Qutnqueloeultna staUcert.
На ближаййих к леднику станциях (68/33 и, 895) в составе фауны фораминифер
остаются почти исключительно одни аллогромииды. К сожалению, имеющийся
материал не дает возможности охарактеризовать распределение Elphiditm
clavatum, вида,, как и С. reniforme, часто доминирующего в стрессовых биотопах.
Можно заключить, что в заливе Норденшельда С. reniforme, Allogromiim spp.
и Q. stalkert характеризуют проксимально-ледниковые условия, а N. labrado-
rtca и I. nocrosst, - менее стрессовые дистально-ледниковые.
Примечательно обнаружение значительных величин численности "Quinquelo-
cuttna stalkert. С одной стороны, этот вид, в отличие от всех остальных
форм, встреченных в заливе Норденшельда, крайне редок в осадках открытой
части Баренцева моря (Корсун и др., 1993), с другой стороны, Q. stalkert -
типичный представитель фиордовой фауны бентосных фораминифер западного
Шпицбергена (Elvertwl et al., 1980). Можно ожидать, что этот вид имеет
палеоэкологическое значение как региональный маркер, ледниково-морского осад-
конакопления.
Две дночерпательных пробы взяты за пределами залива Норденшельда.
Одна из них 68/4 отобрана в следующем к северу заливе - заливе Вилькицкого
(рис. 35). Состав фораминифер в этой пробе такой же, как в пробах из
внутренней части залива Норденшельда. В пробе 68/1, отобранной на два с
половиной градуса южнее - в губе Безымянной, преобладает Eggerella advena. Этот
агглютинирующий вид доминирует в фауне бентосных фораминифер,
расположенного к югу Печорского мелководья (Дигас, 197.0).
Две колонки донных отложений, от.894 и 895, отобранные близ ледника
Норденшельда (рис. 36), имеют длину 125 и 165 см. При существующей
скорости седиментации такая мощность осадка могла накопиться менее, чем за
столетие. Каждый образец (толщиной 5 см) содержит раковины, отложившиеся за
период от 1 до 5 лет.
Планктонные фораминиферы представлены единичными раковинами самого
обычного для полярных регионов вида /feogloboquadrlna pachydermia. Малая
численность раковин планктонных фораминифер типична для донных отложений
всего северо-восточного сектора Баренцева моря (Дигас, 1970), и в этом
S96 (45 м|
694 (31 м)
Номер колонки (глубина моря)
,ш Весовая доля фракции
.сухого осадка >0.063 мм, %
-1—г~
О 01
о о
1—г-
О О
—1_
ш
о
-т-
« « и о
Длина колонки, ом
л
ш
СП
01
I
*.
о
а?
V
*■
о
-■■
Allogromiina spp.
Hippooreplnella alba
Heophax arclica
Trochammlnella atlanllca
Rolaliammina ochracea
Splropleclammina bilormla
Eggerella advene
Qmnquelocullne alalkerl
Mlllollnalla pyrllormie
Palellina corrugate
Cibicides lobalulus
Rosaline spp.
aavallnopsls praegerl
Buccella Irigida
Epislomlnella эр.
Nonlonelle auricula
Nonlonelllne labradorlca
Melonia barleeanue
Astrononion galloweyi
Prolelphidium nlveum
Elphidlum alblumbillcalum
Elphidium subarclicum
Prolelphidium orblculare
Elphidlum clavatum
Caasidulina renilorme
Islandiella helanae
lalandiella norcroasl
Trilarina lluene
Slainlorthia loebllchl
Slainlorlhia schralbaralana
Bollvlna paeudopunclata
Polymorphlnldae app.
Lagena gracllllma
Flsaurlne marglnata
Varla uv.
Neogloboquadrlna pachyderms dan.
Neogloboquadrlna pachydarma aln.
Количеогао бянтооных
ы фораминифор на ЮОг
о сухого осадка
10
Ь-во
Число видов
о Содержание
доминирующего
.и вида, % •
Рис. 38. Форамигаферы в двух колонках донных отложений из залива Норденшельда
смысле осадки залива Норденшельда не отличаются от осадков открытого
моря. Следует отметить, что раковины теплоаодных планктонных фораминифер,
обнаруженные в пробах бухтового льда из заливов Северного острова Новой
Земли, по мнению авторов цитируемой статьи, переотложены из береговых те-
расс (Потехина и др., 1991).
По всей длине изученных колонок массовыми формами являются Casstdult-
na rent forme и Elphiuitm clavattm - типичные доминанты ледниково-морских
местообитаний (рис. 38). В ряде интервалов ведущая роль переходит к Qutn-
quelocutina statkeri. Согласно анализу распределения фораминифер в
поверхностных пробах, этот вид тоже характеризует стрессовые
проксимально-ледниковые условия, наблюдающиеся ныне в заливе Норденшельда. Сочетание
перечисленных трех видов как доминирующих свидетельствует, что за время
накопления вскрытой разрезом толщи осадка условия в месте взятия колонок мало
отличались от нынешних. Наблюдаемые в колонках колебания процентного
содержания этих трех видов, причем, весьма значительные, видимо, вызваны
какими-то малыми по амплитуде сезонными или многолетними изменениями
климатической обстановки, выяснение природы которых требует дополнительных
исследований.
По всей длине колонок практически полностью отсутствуют dltogromUna
spp. Можно полагать, что аллогромииды давали значительную долю
представителей в исходных комплексах, но их чрезвычайно нестойкие псевдохитиновые
раковины разрушились в процессе захоронения, а также, возможно, при
хранении и. камеральной обработке образцов.
Nontonetltna labradorica и Istandtetta nocrosst не являются
доминирующими ни в одном из интервалов. Исходя из анализа поверхностных образцов,
можно заключить, .что за период накопления осадка стрессовое воздействие
ледника ни разу не ослаблялось наотолько. чтобы допустить массовое
развитие этих видов, аналогичное тому, которое наблюдается ныне на удаленных от
. ледника станциях 68/31, 68/3 и 68/32.
В результате проведенных исследований установлено:
в литоральной зоне пяти исследованных заливов Новой Земли фораминифе-
ры не обитают;
Casstdultna rent/orme, /lllogromttna spp. и Quinquelocultna stalker!
характеризуют проксимально-ледниковые, a Nontonelltna labradortca и Islan-
dtelta nocrosst - дистально-ледниковые условия в заливе Норденшельда;
в последнее отолетие экологические условия в заливе Норденшельда мало
отличались от нынешних;
в Баренцевом море Quinquelocultna stalkert - наиболее специфичный
маркер стрессовых условий в заливах, находящихся под воздействием активно
тающих ледников.
Глава 6. НХЛЮШНА
6.1. Ихтиофауна прибрежных вод
Биологические исследования архипелага Новая Земля и его прибрежных
вод начаты в 1768 году, когда среди основных целей первой
правительственной экспедиции, заключавшейся в поиске различных руд и минералов, .ее
участникам следовало выяснить наличие жемчужных раковин, зверей, птиц и
морских животных (Визе, 1939). В материалах этой и последующих экспедиций
конца 18 - начала 19 столетий (Розмыслова, Лудлова, Лазарева. Литке),
имелись сведения о наличии в реках, озерах, губах и заливах Новой Земли толь-,
ко арктического гольца, который успешно вылавливался русскими и
норвежскими промышленниками. Первыми сведениями о морских рыбах прибрежья Новой
Земли, по-видимому, следует считать черновые записи П.К.Пахтусова после
экспедиции 1832-33 гг., где указывалось на лов трески у губы Митюшихи
(Топорков, 1937). Впоследствии немногочисленные сведения об отдельных видах и
перечне видов, пойманных когда-либо около Новой Земли, были объединены в
сводке Е. В. Бурмакина (1957) "Рыбы островов Советской Арктики", которая
стала практически завершающей работой по ихтиофауне архипелага, поскольку
с начала 50-х годов исследования губ и заливов не проводились.
Список рыб, представленных в работе Е.В. Бурмакина, включает в себя 30
видов, которые встречались непосредственно у берегов Новой Земли. Следует
отметить, что литературные указания на наличие отдельных видов, основаны
на устных сообщениях опытных промышленников, хорошо знавших промысловых
рыб, и вероятность ошибки в этом случае минимальна.
В данной работе предотавлен список из 35 видов (табл. 8) в который,
кроме рыб. указанных в сводке Е.В. Бурмакина (1957). включены полярная
акула и чешско-печорская сельдь, упоминавшиеся в работе А.Н.Дубровского
(1941); увеличен и изменен видовой состав липарид (Чернова, 1991) и
лососевых рыб (Chernltsky, 1991).
Таблица 8
Перечень видов рыб. встречавшихся в прибрежных водах Новой Земли
Вид рыб
1
Западное побережье
(Баренцево море)
2
Восточное побережье
(Карское море)
3
Somnlosus microcephalia - +
Clupea harengus harengus +
Clupea palassl suworowl - +
Окончание таблицы 8
1
Oncorhynchua.gorbuaha
Salmo salar
Salvellnus alplnus
Coregonua autumnalla
Coregonus lavaretus pldachiari
Mallotua vllloaua villoaua
Scombereaox saurua
Boreogadua saida
Eleglnua navaga
Gadua morhua morhua
Melanogranvnua aegleflnua
Pollachlua vlrena
Pungltlus pungitiua
Gasteroateua aculeatus
Lumpenua fabricll
Gymnells vlrldia
Cyclopterus lumpua
Liparls tunlcatua
Llparls glbbus
Liparls fabricll
Ammodytes marlnua
Scomber scombrus
Trlglopsls quadricornls
Myoxocephalus acorplua
scorplus
Gymnocanthus trlcuspia
Artedlellua acaber
Icelua blcornls
Trlglops plngell
Ulclna olrlkl
Llopaetta glaolalls
Hlppog10330Ides platessoldes
llmandoldes
Hlppogloaaus hlppogloaaus
2
+
+
■+
+
'" -
+
+.-
+
+
+
+
+
+
+
+
+
■ +
+
+
+
''+
+
+
+
+
• +
+
+
+
+
+
+
3
+
+
+
+ ■
+
-
- ,
+
-
+
-
-
-
-'
-.
,
+
+
+
' +
+
•
: +
-
_
_
-
-
V
-
-'
ПРИМЕЧАНИЕ. + поимка зафиксирована, - поимка не зафиксирована.
Виды рыб. описанные для прибрежных вод Новой Земли по данным
экспедиций, а также на основании устных сообщений рыбаков и промышленников, были
отмечены в следующих районах:
Полярная акула - Somniosus ratcrocephatus (Schneider), Встречалась у
залива Кротова (72° с.ш.), на восточном берегу (Дубровский. 1941).
Атлантическая сельдь - Clupea harengus harengus L. Северная точка
поимки - в районе Крестовой губы (74е с.ш.), отмечена также в губах Грибовой
и Белушьей (Агапов. Топорков. 1937).
Чешско-печорская сельдь - Clupea palasst suworowt Robtnerson. Изредка
попадалась в гольцовые оети в районе залива Кротова (Дубровский, 1941).
Хотя автор сообщавг о случаях поимки такой оелыда, как Clupea ар.,
несомненно, это может быть только чешско-печорская сельдь, поскольку
других видов или подвидов сельдей в Кароком море не отмечено (Андрия-
шев, 1954).
Арктический голец - Salvellnus alplnus L. Его распространение по
западной стороне Новой Земли прослежено на север до Русской гавани (около
76° с.ш.), а по восточной до о.Пахтусова (около 74° с.ш.). Южнее этих
пунктов встречался у побережья повсеместно (Бурмакин. 1957). Этому виду
специально посвящен в данной книге раздел 6.4.
Омуль - Coreganus autumnalis (PallasJ. У западного побережья
вылавливались единичные экземпляры (Агапов. 1937). но. по-видимому, не далее
72° с.ш. - северной точки его распространения в реках, поскольку места
поимок в море не указаны. В прибрежье восточной стороны Новой Земли отмечен
в районе залива Кротова (Дубровский, 1941) и в районе Маточкина Шара у
мыса Выходного (около 73° с.ш.) (Бурмакин. 1957).
Сибирский сиг - Coregonus lovaretus pldschtan (Gmelln). Однажды был
встречен у побережья восточной стороны Новой Земли (Бурмакин. 1957).
Отмечен в устье р.Нехватова (Chernltsky. 1991).
Мойва - Mallotus villosus vilosus (Afuller). У западного побережья
Новой Земли встречается повсеместно, от южной оконечности острова до мыса.
Желания (79°30' с.ш.), где в августе-сентябре можно наблюдать ее нерест
(Лука и Др.. 1986). В Карское море не проникает (Солдатов. 1923).
Макрелещука - Scomberesox saurus (Itoloaum). Единственный случай
поимки имел место у западной оконечности Маточкина Шара (Берг, 1939).
Проникновение данного вида к Новой Земле можно считать случаем невероятным,
поскольку макрелещука крайне редка даже в западной части Баренцева моря. л
Сайка - Boreogadus solda (Lepechin). Встречается в прибрежье
западного и восточного берегов повсеместно, иногда в больших количествах
(Бурмакин. 1957). Этому виду специально посвящен в данной книге раздел 6.3.
Навага - Eleglnus navaga (Pallas). Распространена по западному
побережью Новой Земли. Поимки отмечены в районах губы Крестовая. Малых Карма-
кул, губы Белушья (Агапов. 1937, Бурмакин. 1957).
Атлантическая треска - Gadus morhua morhua L.. Распространение трески
вдоль западного берега Новой Земли к северу прослежено до губы
Архангельской и Горбовых островов (около 76° с.ш.) (Агапов, 1937). Основные
скопления встречаются на глубинах до 50 метров между 71°22' и 73°20' с.ш.
(Бурмакин, 1957). У восточного побережья самой северной точкой поимки является
залив Медвежий, где она попадалась в гольцовый ставной невод (Дубровский.
1941). Единичные экземпляры, выброшенные штормами на берег, найдены еще
севернее, у островов Пахтусова. Южнее Медвежьего залива треска встречалась
между мысами Журавлева и Узким, у мыса Вишневского, а в промысловых
количествах - в заливе Кротова. В заливе Кротова она появляется ежегодно
(Дубровский. 1941). В целом треска держится у Новой Земли с июля по октябрь.
Пикша - Melanograroius aeglefinus (L.). Отдельные особи встречаются
вдоль западного побережья Южного острова архипелага до Иаточкина Шара
(Андрияшев. 1954).
Сайда - Pollachtus vlrens (L.). В летний период держится у западного
побережья Новой Земли и встречается до губы Крестовой, заходит в Маточкин
Шар (Агапов, 1937. Андрияшев. 1954).
Девятииглая колюшка - Pungitius punglttus II.). Отмечена в губе Бе-
лушьей (Есипов, 1935а).
Трехиглая колюшка - Gasterasteus aculeatus L.. Есть сведения о
наличии данного вида на Новой Земле, но конкретных географических районов не
указано (Бурмакин, 1957).
Люмпенус Фабрициуса - Lumpenus fabricii Reinhardt. Встречается у
западной стороны Новой Земли на участке от губы Черной (Гурвич, 1937) до
губы Грибовой (Бурмакин, 1957).
Гимнелис обыкновенный - Gymnelts viridis (Fabrielus). Отмечен в губе
Черной (Гурвич. 1937).
Пинагор - Ci/clopterus lunipus L.. Распространен вдоль западного
побережья Новой Земли до п-ва Адмиралтейства (75° с.ш.), а также в проливе Ма-
точкин Шар (Агапов, 1937).
Арктический липарис - Liparis tunteatus Reinhardt. В сводке Е.В.Бур-
макина представлен как европейский липарис, ареал которого распространен
только до Мурмана (Чернова, 1991). Вполне возможно, что это были другие
виды липарисов представленных ниже, поскольку существенный пробел в
видовом составе и положений ареалов рода липарид в евроазиатской Арктике
устранен совсем недавно (Чернова, 1988). Нахождения у Новой Земли приурочены
к району от Карских ворот до Маточкина Шара и непосредственно в нем
(Бурмакин. 1957; Чернова, 1991).
Горбатый липарис - Liparis gibbus Bean. Описания встречаемости у
Новой Земли отсутствуют, но судя по карте местонахождений (Чернова, 1991),
обнаружен в районах губы Саханихи и Петуховского Шара, а также у мыса
Желания.
Чернобрюхий липарис - Liparis fabricii Kroyer. отмечен в районах губы
Саханихи, Крестовая, в проливе Маточкин Шар (Гурвич. 1937; Чернова, 1991).
Песчанка - Amodytes marlnus Raltt. В прибрежье западной стороны
Новой Земли встречалась от 71°32' с.ш. до губы Грибовой (Кириллов, 1936;
Агапов. 1937). В массовых количествах вылавливалась в губах Средней и
Долгой и в заливе Рогачева . У восточного побережья отмечена в заливе Брандта
(73° с.ш.) (Бурмакин, 1957).
Скумбрия - Scomber scombrus L.. Только однажды в 1936 г. обнаружена
выброшенной на берег в губе Нехватова (около 71°,с.ш.) (Берг. 1939).
Ледовитоморская рогатка - Triglopsis quadricornis L.. Распространена
у берегов Новой Земли повсеместно (Книпович, 1901; Андрияшев. 1954).
Керчак - Myoxocephalus scorplus scorpius (L.). По западному побережью
встречается от губы Черной до Маточкина Шара, по восточному - в заливе
Брандта (Бурмакин, 1957).
Арктический шлемоносный бычок -. Gymnocanthus tricuspis (Reinhardt).
Отмечен в основном у юго-западного побережья (Агапов, 1937; Андрияшев,
1954).
Шероховатый крючкорогий бычок - drtedteltus scaber Knlposvltsch. Места
поимок приурочены к западному побережью, от Петуховского шара до Крестовой
губы (Андрияшев. 1954; Бурмакин, 1957).
Атлантический двурогий бычок - Icetus btcornts (Reinhardt).
Зафиксирован у западного побережья Новой Земли, в районах губ Черная, Безымянная.
Крестовая (Гурвич, 1937).
Остроносый триглопс - Triglops ptngelt Reinhardt. Отмечен в заливе
Моллера и в губе Крестовой (Гурвич, 1937).
Ледовитоморская лисичка - Ulctna olrikt-fLutlcen). Встречалась в губах
Безымянной и Крестовой (Бурмакин, 1957).
Полярная камбала - Ltopsetta gtactalis (Pallas). Попадается в
западном прибрежье Новой Земли . Весной в больших количествах встречается в
губе Белушьей (Агапов. 1937).
Камбала-ерш - Htppoglossoides ptatessotdes Itmandotdes (Btoch). Места
поимок приурочены к районам губы Безымянной, и пролива Маточкин Шар (Гур-
вич, 1937).
Палтус - Hippogtossus hippogtossus (L.). Единичные осоВи обнаружены в
заливе Моллера (Агапов, 1937).
Данный список видов рыб нельзя считать полным, поскольку специальных
полномасштабных исследований ихтиофауны прибрежных районов Новой Земли не
проводилось, а имеющиеся сведения получены для относительно ограниченных
пространств побережья. Достаточно много видов, отмеченных в западных
прибрежных районах, могут быть реально найдены и по восточному берегу. Новой
Земли, особенно у Южного острова, поскольку такие виды,как навага, девяти-
иглая колюшка, люмпенус Фабрициуса, остроносый триглопс. керчак, ледовито-
морская лисичка, полярная камбала, камбала-ерш и другие являются
достаточно обычными в Карском море (Антонов, Чернова, 1989), особенно в
юго-западной его части.
6.2. Ихтиопланктон прибрежных вод
Сведения по ихтиопланктону прибрежных вод о-ва Новая Земля
фрагментарны и малочисленны. Практически отсутствуют данные об эколого-биологи-
ческих характеристиках личинок. Известные работы, выполненные в ЗО-40-е
годы в восточной части Баренцева моря, охватывают в основном акваторию
Чешской губы, прибрежье Печорского залива и Канино-Колгуевский район
(Расе, 1949; Алексеева, 1949; Пономарева, 1949; Казанова, 1949).
В 1958-1959 гг. А.А.Кашкиной в рейсах на экспедиционном судне
Мурманского морского биологического института АН СССР "Профессор Дерюгин"
частично были изучены размножение и развитие некоторых рыб. а также
видовой состав йхтиопланктона а открытой части Печорского залива я в районе
между материком и Новой Землей (Кашкина, 1962). В этот период у берегов
Новой Земли ею обнаружены 23 личинки трески длиной и.9-22.9 мм, 3 личинки
сайки и мальки ледовитоморской лисички от 10.8 до 22 мм.
По литературным данным известно, что у Новой Земли в небольшом
количестве происходит нерест мойвы (Поздняков, 1962). В уловах икорной сети в
1959 году в экспедициях на судах ММБИ и ПИНРО попадались личинки мойвы к
западу и юго-востоку от Новой Земли. В более поздний период исследований
личинки мойвы у Новой Земли встречены не были, что возможно было
обусловлено резким снижением численности популяции под влиянием перелова (Боркин,
1985).
Большинство научных работ, посвященных изучению раннего онтогенеза
рыб востока Баренцева моря, касаются преимущественно исследований сайки
(Rass, 1936; Клюге. 1936; Клумов, 1937; Мантейфель, 1943; Тамбовцев, 1952;
Никольский, 1954; Пономаренко. 1963. 1964. 1965; Ponomarenko. 1964). В
этих работах приводятся первые сведения о времени и районах нереста, рас-
пределении преднерестовых и нерестовых скоплений, встречаемости и биологии
личинок и мальков сайки. Наиболее полной сводкой, касающейся раннего
периода жизни сайки, является работа А.С. Бараненковой с соавторами (1966).
Весьма ограниченные сведения о видовом составе ихтиопланктона у
восточных берегов Новой Земли известны лишь из работ Л.А.Пономаревой (1949).
В. К.Есипова (1952) и Г.В.Норвилло с соавторами (1982). В этих работах
сообщается о поимке в 1935-1945 гг. у северной оконечности архипелага между
75°46'-78е07' с.ш. и 68е28"'-74°20' в:д. трех мальков камбалы-ерша длиной
4-8 см (Есипов. 1952; Пономарева, 1949). В экспедиции ММБИ в Карское море
в 1981 году в проливе Карские Ворота выловлен малек европейского керчака,
а две личинки люмпенуса Фабрициуса обнаружены в юго-востоку от Новой Земли
(Норвилло и др., 1982; Норвилло, 1989).
Сведения о размножении сайки вдоль восточного побережья Новой Земли
практически отсутствуют. В вышеуказанных работах сообщается лишь о
единичных случаях вылова поздних личинок и мальков этого вида.
Более детальные исследования по ихтиопланктону в восточной части
Баренцева моря предприняты Полярным научно-исследовательским институтом
(ПИНРО) В 1977-1984 ГГ. (Боркин. 1983. 1990;' ВогкШ. 1979. 1983. 1985.
1986). . '
Материалы, собранные одним из авторов в экспедициях 1977-1984 гг. на
НИС "Ф.Нансен". "Гумма", "Процидн" и "Мензулинск". послужили основой
данного раздела*. При облове личинок использовали ихтиопланктонную сеть
ИКС-80. изготовленную из капронового газа N 140 с диаметром входного
отверстия 80 см.
На каждой станции проводили вертикальный облов горизонтов 50-0 и
дно-0. а также горизонтальные ловы в слое 0-3 и 10-15 м при работе судна в
режиме циркуляции продолжительностью 10 мин. С целью изучения
вертикального распределения в районах высокой плотности скоплений личинок выполняли
серии вертикальных ловов в горизонтах 0-5, 0-10, 0-20, 0-30, 0-50 м и до
придонного слоя. Скорость движения сетей при вертикальном облове личинок
составляла 1-1.2 м/сек. Во время циркуляции ловушка перемещалась со
скоростью 2-2.5 узла. Личинок фиксировали 2%-м раствором формалина. Материал
обрабатывали по методике Т.С.Расса и И.И.Казановой (195В). Длину личинок
измеряли при помощи окуляр-микрометра под микроскопом МБС-l с точностью до
0.1 мм.
За время исследований на 744 станциях обработано 1742 пробы, учтено
3468 и измерено 2218 личинок. Положение ихтиопланктонных станций
представлено на рис. 39-42. Список видов личинок, встречающихся в ихтиопланктоне
прибрежья архипелага Новая Земля, < приведен в табл. 9. При построении карт
распределения личинок по плотностям использовали стандартный метод
изолиний (Аксютина. 1970) и разработанную в ПИНРО методику с использованием
данных горизонтальных и вертикальных обловов (Боркин и др.. 1987).
Характеристика гидрологического режима районов распределения личинок
приведена по материалам отдела промысловой океанологии ПИНРО. Помимо
этого, учитывали данные о температуре воды в слое 0-200 м на 3-7-й станциях
разреза "Кольский меридиан" как показатель тепловых условий южной части
Баренцева моря в целом.
'В 1979 г. в связи с тяжелыми ледовыми условиями в летний период ихтио-
планктонная съемка на востоке моря не выполнялась.
:. ^
40° 45 50 55
40° 45 50 55
Рис. 39. Положение ихтиопланктонных станций
в \Ш г-: £ ~ май> Б " ивнь-июль;
в 1978 г.: В - апрель, Г - май, Д - июль,
Е - июль-август
Рис. 40. Положение ихтиопланктонных станций
в 1980 г.: А,Б - первая и вторая половина июня, соответственно:
в 1981 г.: В - июнь, Г - июнь-июль
N
• • • • • •[ л* • •
Рис. 41. Положение ихтио-
планктонных станций а 1982 г.
40°
45 50 55
§
ас
ь"
о i
aw
3<n
om
• • • • ill?**
. • • • :*!»&i
• • • • • • . »g2
, • • • " » •••^
« ••••# • ••
ё
ой
si.
Ш
: :
Таблица 9
Список видов личинок, встречающихся в ихтиопланктоне
архипелага Новая Земля
Семейство, вид, подвид
Зоогеографическая
характеристика
(по Рассу, 1949)
Частота
встречаемости
в уловах, экз.
Gadidae
Boreogadus salda (Lepechln)
Сайка
Gadus morhua morhua L.
Треска
Cotttdae
Myoxocephalus scorplus (L.)
Керчак европейский
Gymnacanthus trlcuspia Relnhardt
Арктический шлемоносный бычок
Trlgllps plngell Relnhardt
Остроносый триглопс
Lumoentdae
Lumpenus medlus Relnhardt
Люмпенус средний
pteuronecttdae
Hyppoglossoldes platessoldes
llmandoldes (Bloch)
Камбала-ерш
Pleuronectes platessa (L.)
Морская камбала
Ulclna olrlkl (Lutken)
Ледовитоморская лисичка
АтюйШйде
Ammodytes hexapterus marlnus
Raltt
Многопозвонковая европейская
морская песчанка
Ltpqrtdae
Llparls koefoedl Parr
Чернобрюхий липарис
Llparla llparls (L.)
Европейский липарис
Арктический Тысячи
Северо-бореальный Единицы
Арктическо-бореальный Десятки
Арктический • Десятки
Арктический Десятки
Арктический
Десятки
С
Арктическо-бореальный Единицы
Бореальный Единицы
Высокоарктический Единицы
Северо-бореальный Единицы
Арктический Единицы
Арктическо-бореальный Единицы
При изложении материала использовано общепринятое промысловое
районирование (Труды ПИНРО, 1957).
Сайка Boreogadus satda1. Результаты ихтиопланктонных съемок,
выполненных у Новой Земли,показали, что вдоль западного побережья и южной
оконечности практически постоянно происходит массовый нерест сайки. Так, во
второй половине июня - начале июля 1977 г. личинки в большом количестве вы-
лавливались у юго-западной оконечности о-ва Новая Земля, где уловы на
отдельных станвдях составляли 100-300 личинок за лов в поверхностном слое
(Боркин, 1983). Основная масса особей была отмечена в слое от 10 м до
поверхности при температуре 3-4 °С (рис..43А). Однако в первой половине июля
1978 года почти на всех станциях предличинки распределялись в
поверхностном слое 5-0 м, чему способствовало наличие льда, вблизи которого
формировалась кормовая база (рис. 43Б.В). Кстати, в этом же 1978 г. единичные
личинки вылавливались на мелководьях у пролива Маточкин Шар. Следует
отметить, что в апреле-мае на этой акватории икры и личинок не 'было.
В июне-июле 1980 г., несмотря на малочисленные уловы (менее 50 экз.),
наибольшая встречаемость личинок прослеживалась, как и в 1977 году, у
юго-западной оконечности Новой Земли при температуре от 1.1 до 1.5 ?С
(рис. 44А,Б). Основное количество предличинок распределялось в отличив от
предыдущих лет в толще воды в слое 10-20 м (Borkln, 1983).
Рис. 43. Распределение личинок
сайки:
А - июнь-июль 1977 г.;
Б,В - июль 1978 г.. первая и
вторая половина месяца,
соответственно.
1 - станция без улова; 2 -
станция с уловом; 3 - 1-10
личинок на 2 горизонтальных лова;
4 - 11-100 личинок; 5
101-1000 личинок; 6 - более
1000 личинок; 7 - кромка льда
40° 45 50 55
Рис. 44. Распределение личинок сайки
в *§§? г-: я и 1 ~ первая и вторая половина июня, соответственно;
в 1981 г.; В и Г - июнь и июнь-июль, соответственно.
Обозначения см. рис.43
В первой половине июня 1981 г.. на обследуемой акватории была поймана
всего одна личинка длиной 7 мм у юго-западного побережья о-ва Новая Земля.
В конце июня - начале июля личинки концентрировались на небольшом участке
у западного побережья о-ва Новая Земля (рис. 44В,Г). Среди разрушающегося
льда уловы .достигали 500 экз. за 1о мин циркуляции в поверхностном слое
0-5 м при температуре 0-1 °с.
В июле 1982 года в условиях температурного режима вод,
характеризующегося тенденцией сохранения на востоке Баренцева моря отрицательных
аномалий, было отмечено необычайно широкое распространение личинок сайки в
западном и северном направлениях (рис. 45). Личинки в небольшом количестве
вотречались на значительной акватории западного побережья Новой Земли.
Рис. 45. Распределение личинок сайки в июле 1982 г.
Обозначения см. рис. 43
Ихтиопланктонной съемкой, выполненной в июне 1983 года, были учтены в
основном единичные особи, отмечаемые, как и в предыдущем году, вдоль
побережья Новой Земли до 75° с.ш. (рис. 46А). Определенной закономерности в
вертикальном распределении, обусловленной близостью льда, в этом году не
было отмечено, хотя наибольшие скопления личинок выявлялись именно у
ледового припая в слое 0-20 м.
В 1984 году лишь на двух отанциях у южной оконечности Новой Земли
было поймано несколько ранних личинок на горизонтах 10-15 м (рис. 46Б). Как
и в 1983 г., оформившиеся личинки держались, как правило, на глубинах
10-20 м.
Необходимо отМетить, что вертикальное распределение предличинок и
личинок в значительной мере зависит от местоположения ледовой кромки, о
которой связано их существование в период инкубации и после выклева. По мере
роста и перехода на внешнее питание предличинки из приповерхностного слоя
погружаются в толщу воды. Случаи опускания предличинок на глубину 10-20 м
в приледовых участках обусловлены, по-видимому, штормовыми условиями. Уход
в спокойные слои моря несомненно снижает их гибель.
80°-
76
72
68
Рис. 46. Распределение личинок сайки
в июне-июле 1983 г. (А) и 1984 г. (Б). Обозначения си. рис.43
Анализ распределения личинок сайки в июне-июле 1977-1984 гг.
позволяет предположить наличие обособленного нереста в прибрежье юго-западной
оконечности архипелага Новая Земля.
Размерный состав личинок, выловленных у Новой Земли, в разные годы
значительно варьировал в зависимости от района и времени выклева, а также
времени года и температурного режима вод (табл. 10,11). На рис. 47
показана длина сайки в прибрежье Новой Земли в сравнении с размерным составом
личинок у о-вов Колгуев и Вайгач.
Анализ темпа роста показывает, что он различен в разные годы и
зависит в основном от климатических условий, а, соответственно, и теплоеодер-
жания водных масс. Так, за 23 дня в июле 1978 г. средняя длина личинок
увеличилась на 2.62 мм. В июне 1980 г. в среднем за 20 дней прирост
составил 0.53 мм. Очевидно слабый прогрев воды (1.5-2° за 2.5-3 недели) в этом
районе был основной причиной столь низкого темпа роста. Кроме того,
участки моря у южной оконечности о-ва Новая Земля постоянно подвержены влиянию
холодного течения Литке, поэтому темп роста личинок в этом районе всегда
ниже и средняя длина их обычно меньше, чем на других акваториях, где
значительно теплее. Вместе с тем установлено, что на начальных стадиях
развития размеры личинок увеличиваются незначительно и составляли за период
исследований 1977-1984 гг. в июне в среднем 7.1-7.5 мм. В июле, по мере
роста, средние значения длины изменялись от 7.1-9.6 мм в первой половине
месяца до 9.3-11.1 мм - во второй. Самый высокий темп роста личинок
отмечен в августе.
Европейский керчак МиохосерПдШз seorotus. Личинки европейского кер-
чака постоянно встречаются в ихтиопланктоне востока и, особенно,
юго-востока Баренцева моря (рис. 48А). Особи длиной 6-10 мм вылавливались,
начиная с мая по мере разрушения льда. Уловы в июне-июле в среднем редко
превышают 6-10 особей за траление сетью. Максимальный вылов наблюдали в
1977 г., когда за циркуляцию в слое 0-5 м к юго-западу от о-ва Междушарс-
кий было поймано 34 личинки.
Необходимо отметить, что по мере удаления от Новой Земли количество
личинок в уловах уменьшается. На станциях, выполненных над глубинами 100 м
и более,редко отмечались уловы более 2-3 экземпляров на сеть. Основная
масса особей распределялась в поверхностном слое воды 0-5 м.
Длина личинок за период исследований варьировала от 6.до 21 мм, в
среднем 12 мм. Размерный состав особей представлен в таблицах 10,11. Темп
роста можно проследить по материалам 1977 г.. когда длина личинок в мае
составляла 6-10 мм, в июне - 7-16 мм (средняя 11.9 мм), в июле -7-20 мм
(средняя 13.9 мм).
Темп роста, как правило, зависит от теплосодержания водных масс и
динамики разрушения льда в весенний период. Личинки керчака встречались при
температуре от -0.7 до 7.2°.
Арктический шлемоносный бычок Gwnacanthus trtcuspts. Шлемоносный
бычок на востоке моря встречался в среднем вдвое реже личинок керчака,
однако, как и предыдущий вид, он является обычным в новоземельском
ихтиопланктоне. Наиболее часто личинки шлемоносного бычка вылавливались вдоль
юго-западного побережья архипелага, где уловы составляли 1-10 особей за
циркуляцию (рис. 48Б). Отмечена тенденция уменьшения количества личинок в
уловах по мере удаления от береговой линии. Максимальный улов (28 экз.)
был отмечен к югу от Новой земли над глубинами 150 м.
Таблица 10
Размерный состав личинок, выловленных в прибрежье Новой Земли
в весенне-летний период 1977-1984 гг., %
Длина,
им
1
4
5
6
7
8
9
10
И
12
13
14
15
16
17
Вид
Boreogadus salda
сайка
2
0.2
2.0
16.0
23.0
23.0
14.8
8.0
5.4
3.0
2.6
1.0
0.6
0.3
0.1
Hyppoglossoldes
piatessoldes
llmandoldes
камбала-ерш
. 3
,.
57.2
42.8
Myoxocephalus
scorpms
европейский
бычок
4
0.2
1.4
2.0
.е. 6
13.0
16.0
24.0
13.6
5.8
5.0
4.8
1.2 '
Gymnacanthus
trlcuspls
арктический
шлемоносный
бычок
5
Л-.'8
5.6
12.5
12.5
11.0
18.8
13.2
13.2
5.7
3.8
1.3
Trlglops
plngell
остроносый три-
глопс
6
20.0
20.0
20.0
20.0
20.0
Lumpenus
medlus
лгомпенус
средний
7
0.7
1.9
2.7
2.7
5.8
9.3
12.8
13.2
Liparls
koefoedl
чернобрюхий
липарис
8
2.4
5.0
7.2
8.4
14.4
28.0
16.0 -
13.4
4.0
1.2
Ulclna olrlkl
ледовитомор-
ская лисичка
9
16.7
-
25.0~
25.0
16.7
8.3
8.3
Ammoflytes hexap-
terus marlnus
песчанка
Ю .
-
33.3
33.4
33.3
э
id
Е-
Ж
о
о
ю
CM СМ
маппюиг'Пнпп'п
nOKDCOnrlONHHOIO'l 1
"; ■
t-
.' 1 . 1 °
'
со
о
со
CD ,
256
269
со
о
"8
о>
ю
■*!* СО ^* О
■** о *-3 -ri
со ■ •«*
00 О]
см
см
+ ■+■+ +
§
СП
о
см
СО С3> О чН
«"*< *~t СМ СМ
о
с-
а>
ЙЯ^ЙЙ^ЙЙ^ЙЙЙ^ЙЙЙЁЙЁ
Таблица 11
Размерный состав личинок наиболее массовых видов рыб по годам, шт.
Длина.
мм
1
Вид
Myoxocephalus scorplus
1977г.
2
1980г.
з
1981Г.
4
1982Г.
5
1983Г.
6
Gymnocanthus trlcuspls .
1977Г.
7
1980Г.
8
1981Г.
9
1982г.
10
1983г.
И
Lumpenus mefllus
1977г.
12
1978Г.
13
1980г.
14
1981Г.
15
1982Г.
16
1983Г.
17
6
7
8
9
10
11
12
13
14
15
16
17"
18
19
20
21
22
1
3
3
14
25
42
82
54
25
26
21
5
2
-
2
3
3
7
7
2
1
2
2-
6
19.
22
24
6
-
"-
1
1
-
2
-
2
1
1
2
,1. :
5
3
5
5
1
2
14
13
10
11
3
2
1
4
8
7
7
25
15
16
3
1
3
6
1 6
2
3
1
-
2
2
6
5
8 '1
2
5
5
6
5
2
1
2
6
. 2
1
7
2
6 ...
8
8
18
9
8
4
3
3
4:
1
3
9
13
2
_
2
1
1'
2
1
_
-
3
3-
- -8
18
21
17
5
1
1
-
1
.2
1
1
2
1
4
3
2
6
2
о
Cv»
О
ж
о
о
П W (О П ^ (D и
о
см
00
см
о
со
я
I
ЙЗЙЙЙЯЙ
ш
£ Ё!
У
SO
40
30
at
it
n
41
10
20
Ф
1Э77
n м
J7-J0.06 II 101]
50-
40
3»
10
10
»1
40
10
10
w-I
4t
»
10
4*
JO
to
le
м
10
i
1978
n м
-S607 633 7.4
-24 Щ? 1300 9Л
n м
- 6-7.07 147 5.5
--2910.07 IOS 8.1
1980
n и
-I-3JB6 J9 6.1
-23 Ш6 46b 7.4
%
п м
13.07 64 5.1
®
1981
n м
2411IS 2S 7.1
1982
n и
201107 01? II.J
lift}
n м
IMS.* 1773 7.0
1964
n м
S 1106 83 74
27 2106 23 7.0
n м
2-3.07 718 74
П M
IM4J7 72 IIS
1M,B.;.,»"■
n м
Zl-30.es 100 №
n ы
2606 170 7.2
n м
23.17 Ш W
ft И
ад zi.M ш м
■I i t. ii* л
зал lA те
' 4 . 8 10 12 |i 16 • A 10 IS 14. 16 6 6 10 It 14 »•
Длина, мм
Рис. 47. Размерный состав личинок сайки на востоке
Баренцева моря в июне-июле 1977-1984 гг. на акватории
островов Колгуев (1). Вайгач (2). Новая Земля (3)
Рис. 48. Распределение личинок европейского керчака (А)
и арктического шлемоносного бычка (Б) в 1977-1983 гг.:
О - 1977 г.; д - 1978 г.; а - 1980 Г.; + - 1981 г.;
▲ - 1982 Г.: • ■ - 1983 г.
Большинство личинок распределялось в поверхностном слое воды 0-5 м
при температуре от 2 до 7°. Размерный состав личинок за период наблюдений
колебался от 7 до 18 мм и был различным в разные годы в зависимости от
времени выполнения съемок (табл. 10,11).
Остроносый триглопс Trtglops pttu?etl. Остроносый триглопс весьма
редок в ихтиопланктоне Новой Земли. За период наших исследований учтено лишь
5 личинок, которые выловлены в основном к западу от архипелага (рис. 49А).
Одна личинка была поймана у самого берега напротив пролива Маточкин Шар.
Наиболее раннее появление триглопса в ихтиопланктоне отмечено во второй
половине мая 1977 г., когда особь длиной 9 мм была обнаружена в улове на
отанции, выполненной у кромки льда. Обычно же личинки вылавливались в
июне-июле в слое от 50 м до поверхности над глубинами 30-160 м.
Длина особей варьировала от 7 до 16 мм (табл. 10). Температура воды
на станциях, где были встречены личинки, варьировала от -0.7 до 4.2°.
Люмпенус средний Lumpenus medtus. Личинки люмпенуса среднего, как и
сайки, наиболее многочисленны в ихтиопланктоне прибрежных вод Новой Земли.
Они ежегодно вылавливались в количестве от нескольких экземпляров до сотни
и более особей за лов. Максимальные уловы отмечены в иине-июле 1977 г. у
южной оконечности архипелага (рис. 4ЭБ). Личинки распределялись как в
поверхностных слоях воды, так и в слое 0-50 м над глубинами 30-170 м.
Рис. 49. Распределение личинок остроносого триглопса (А) и
люмпенуса (Б) в 1977-1983 гг. Обозначения см. рис.48
Появление личинок люмпенуса в ихтиопланктоне наблюдали, начиная с
мая, на акватории открытых участков моря и у кромки льда при температуре
от -1.0 до 1.5°. В массе личинки улавливались в июне-июле. Как и у
большинства арктических видов, у люмпенуса среднего нерест растянут на
протяжении всего зимнего периода, а выклев приурочен к разрушению льда весной и
зависит от поступления в восточные районы моря тепла с водами Мурманского
течения.
Уловы в мае, как правило, единичны; в июне-июле, в основном, по
несколько экземпляров за лов, редко по 5-10 особей за траление. Максимальный
улов составил 129 личинок за 10 мин циркуляции в слое 0-5 м в июле 1977 г.
Длина особей в мае составляла 11-21 мм. в июне - 10-28 ми. в июле -
15-36 мм (табл. 10.11). Температура веды, при которой встречался люмпенус.
колебалась от -1.0 до 7.0°.
Ледовитоморская лисичка t/tctna olrtkt. Личинки ледовитоморской
лисички обычны, хотя и малочисленны в ихтиопланктоне Новой Земли. За период
наших исследований в общей сложности было учтено 12 особей длиной 9-16 мм
(табл. 10).
Географически личинки лисички встречаются, по-видимому, повсеместно,
поскольку уловы были отмечены от полуострова Адмиралтейства до южной
оконечности архипелага над глубинами от 20 до 180 м (рис. 50А). Большинство
Рис. 50. Распределение личинок лисички (А) и чернобрюхого
липариса (Б) в 1977-1983 гг. Обозначения см. рис.48
особей вылавливалось в поверхностном слое воды 0-5 м.
Температура, при которой встречались личинки, составляла 1-3.5°.
Чернобрюхий липарис Liparts koefoedi. В наших сборах личинки
чернобрюхого липариса наиболее часто вылавливались в 1977 г., когда было учтено
96% всех проанализированных особей. В остальные годы липарис встречался
единично или вообще не был отмечен в уловах.
Практически все личинки распределялись у юго-западной оконечности
Новой Земли над глубинами от 30 до 160 м. Лишь 2 экземпляра были встречены
значительно севернее - на станции в 25 милях от берега на 74°10' с.ш.
(рис. 50Б). Количество особей в уловах, в основном, не превышало 1-2 зкз.
на сеть. Однако на двух станциях меяду архипелагом и о-вом Колгуев было
поймано И и 76 экземпляров чернобрюхого липариса за 10 мин циркуляции в
поверхностном слое. Личинки удерживались на участках смешения вод
встречных течений - теплого Канино-Колгуевского и холодного течения Литке,
несущего свои воды из Карского моря. Длина личинок липариса составляла 7-16 мм
{табл. 10).
Европейский липарис Ltoarta tlparta, Единственная личинка
европейского липариса длиной 7.2 мм была обнаружена на станции, выполненной в начале
июля 1977 г. между островами Новая Земля и Колгуев в координатах
70°05' с,ш. и 50°00* в.д. при температуре поверхности воды 3.6°. Глубина
моря составила 85 м.
Многопозвонковая европейская морская песчанка Amount es hexapterus
martnus. Личинки многопозвонковой песчанки в прибрежных водах Новой Земли
встречались в единичных экземплярах, что указывает, по-видимому, на
отсутствие нереста в этом районе. В июне 1977 г. и июле 1982 г. к югу от
архипелага были пойманы соответственно одна и две личинки длиной 10-12 мм
(рис. 51А, табл. 10). Эти особи распределялись в поверхностном слое 0-5 м
в зоне действия прибрежных течений восточной направленности. Их вынос из
района Канино-Колгуевских банок не вызывает сомнения, поскольку По
наблюдениям авторов в этот же период в указанном районе вылавливалось до
нескольких тысяч личинок за один лов.
Рис. 51. Распределение личинок песчанки (А), икринок и личинок
камбалы-ерша (Б) в 1977-1983 гг. Обозначения см. рис.48
Камбала-ерш Huooalossoiues platessoldes ttmandotdes. Икра и личинки
камбалы-ерша встречались у Новой 'Земли лишь в 1977 и 1978 гг.
В III декаде мая 1977 г. икринки камбалы-ерша вылавливались в
открытых районах, распределяясь в основном в поверхностных слоях моря при
температуре от -0.2 до 1.6°. Уловы не превышали 19 экземпляров за циркуляцию
(рис. 51Б). Соотношение икринок, находившихся на различных стадиях
развития (II-IV). было примерно одинаковым, что указывало на факт завершения
нереста камбалы-ерша на востоке моря к середине мая. В этот же период к
западу и юго-западу от Новой Земли были отмечены 4 личинки длиной
в-в.5 мм. по две - в 1977 г. и 1978 г. Личинки улавливались над глубинами
115-250 н при температуре от -1.6 до 1.3е, в отдельных олучаях - у кромки
льда. В более поздний период, в июне-июле, в уловах сетей отмечались
только личинки, причем лишь в 1977 г. Общее их распределение ' в целом было
идентичным распределении в мае. Количество особей на станциях варьировало
от 1 до 7 экз. на сеть. Личинки вылавливались в слое СЬ50 м над глубинами
55-260 м в зоне действия теплого Новоземельского течения при температуре
от 0.6 до 4.4°. Длина их составляла 6-7 мм в июне и 9-14 мм в июле.
Морская камбала Platesaa platessa. За весь период ихтиопланктонных
исследований единственная икринка морской камбалы IV стадии развития была
поймана в мае 1977 г. в поверхностных слоях моря при температуре воды 0.2°
в координатах 70°38' с.ш. 47°15' в.д. Две личинки морской камбалы длиной
6 и 6.5 мм обнаружены к юго-западу от Новой Земли в июле 1978 г.
Поскольку нерест этого вида проходит преимущественно в прибрежье Мур-
мана с последующим дрейфом икринок и личинок в струях Мурманского течения
вплоть до Канинских банок, то появление личинок морской камбалы у берегов
Новой Земли необходимо расценивать как случайное или эпизодическое,
связанное с повышенной активностью прибрежных течений в отдельные годы.
Треска Gadus morhua mortma. Одна икринка II стадии развития была
выловлена 26 мая 197В г. в координатах 72°27' с.ш. 4В°4Г в.д. в северной
части Новоземельского мелководья при вертикальном облове слоя 50-0 м.
Глубина моря здесь составляла 150 м. температура поверхности воды О "С. В
улове на этой станции также были обнаружены 2 неопределенные икринки
бычковых и з личинки люмпенуса и триглопса.
Обнаружение икринок трески в этом участке моря в 20 милях от кромки
льда на первый взгляд несколько парадоксально. Однако, по наблюдению
одного из авторов настоящей работы, в отдельные годы в мае-июне на склонах
Гусиной банки (смежный с Новоземельской банкой район) отмечались редкие
случаи поимки в донный трал преднерестовых или текучих самок трески.
Несомненно, что факт нереста трески на востоке моря носит скорее аномальный
характер, вместе с тем не очитаться с этим нельзя. Следует отметить, что
ранее В.И.Владимировым (1937) высказывалось предположение о возможном
нересте трески на Гусиной банке. Однако данная гипотеза позже на основе
многочисленных наблюдений была подвергнута сомнению как несостоятельная (Расе,
1949).
Проведенные исследования показывают, что нерест большей части
описанных видов происходит в прибрежных водах Новой Земли.
Наибольшим разнообразием отличается ихтиопланктон западной и особенно
юго-западной части архипелага. Несмотря на то. что во время
ихтиопланктонных съемок были отмечены личинки лишь 12 видов, количество размножающихся
здесь рыб значительно больше (Андрияшев, 1954).
Личинки пойманных видов рыб принадлежат к 7 семействам и относятся к
следующим зоогеографическим формам:
высокоарктические - 1 вид (ледовитоморская лисичка);
арктические - 5 видов (сайка, арктический шлемоносный бычок,
остроносый триглопс, люмпенус средний, чернобрюхий липарис);
арктическо-бореальные - 3 вида (керчак европейский, камбала-ерш,
европейский липарис);
северо-бореальные - 2 вида (многопозвонковая европейская морская
песчанка, треска);
бореальные - 1 вид (морская камбала).
Наиболее многочисленным видом в ихтиолланктоне была сайка, в меньшей
степени встречались личинки люмпенуса среднего и европейского керчака. Еще
реже в уловах отмечались личинки арктического шдемоносного бычка.
Личинки песчанки и европейского липариса были представлены единичными
экземплярами.
Таким образом, в ихтиопланктоне присутствуют в основном личинки
арктических видов, размножающиеся преимущественно в восточных районах
Баренцева моря.
В то же время личинки арктическо-бореальных и субарктических видов
появляются в ихтиопланктоне в результате их выноса водами теплых
прибрежных течений с Запада. В процессе эволюции личинки арктических видов рыб
адаптировались к суровым температурным условиям и имеют больше преимуществ
для успешного выживания, нежели представители бореальных видов.
Таким образом, климатические и гидрологические особенности региона
обусловливают в значительной степени видовой состав ихтиопланктона и
принадлежность видов к фаунистическо-зоогеографическим формам.
6.3. Сайка
Сайка Boreogadus saida (Lepechin) - наиболее многочисленный
представитель арктической ихтиофауны, распространена циркумполярно и в Баренцевом
море встречается почти повсеместно (Книпович, 1926; Клумов, 1935. 1937;
Есипов, 1952; Световидов. 1935, 1948; Тамбовцев, 1952; Юданов, 1947,
1964). В пределах шельфовой зоны архипелага Новая Земля в процессе
эволюции она заняла чрезвычайно важное место в трофической цепи экосистемы
данного региона и стала его неотъемлемой частью (Гептнер, 1937; Мантейфель.
1943; Гринкевич. 1957; Пономаренко, 1962. 1963, 1964, 1968; Боркин и др.,
1987а, б, 198Ва,б; Боркин, 1990 ).
Исследования показывают, что в арктических морях существует множество
изолированных популяций (стад) сайки, что обусловлено расчлененностью
арктической области крупными островами и архипелагами на ряд географически
обособленных морей и. как следствие, появление достаточных условий для
изоляции и образования таксономических отдельностей (Солдатов. 1923;
Книпович. 1926; Световидов, 1935; Клумов, 1937; Киселев. 1940; Москаленко,
1964; Есипов, 1952; Кравчук. 1958; Иванова, 1959; Шепель, 1971; Печеник и
др.. 1973; Юданов. 1976). Однако наиболее многочисленные, к которым в
первую очередь относится восточнобаренцевоморская (или новоземельская) сайка,
приурочены к полярным фронтам, которые образуются в южных участках ареала
в результате активных горизонтальных и вертикальных перемещений
разнородных водных масс теплых и холодных течений. В таких участках моря
происходит обогащение водной толщи биогенными солями и на их базе развивается
богатый фито- и зоопланктон, составляющий основу питания сайки. Области
полярных фронтов являются не только акваториями откорма, но и районами
размножения. Весь жизненный цикл восточнобареицевоиорской сайки тесным образом
связан с системой Новоземельского и Колгуево-Печорского теплых течений и
холодных арктических течений Литке и Макарова.
Первые сведения о миграциях сайки в Баренцевом и Карском морях
приведены в .работе Б.К.Москаленко (1964) и в дальнейшем были дополнены и
уточнены исследованиями В.П.Пономаренко (1968) и В.Н.Шлейника (Shlelnlk,
1970).
Наиболее подробно изучены миграции восточнобаренцевоморской сайки, на
скоплениях которой в течение ряда лет базировался промысел. В'
январе-феврале на мелководьях юго-восточной части Баренцева моря, как правило, у
о-ва Колгуев, проходит нерест сайки. В марте-апреле посленерестовая рыба
постепенно мигрирует в северо-западном, и северном направлении, достигая к
лету центральных районов моря, распределяясь преимущественно в зоне
действия Новоземельского течения.
В июне-июле более крупные особи обычно продолжают двигаться на
северо-восток, тяготея в основном к шельфовым участкам Новой Земли. До конца
августа сайка достигает своих окраинных, северо-восточных участков ареала,
доходя до мыса Шелания. Возможно, заходит в Карское море. В холодные в
гидрологическом отношении годы миграции сайки в первом полугодии часто
имеют более западную ориентацию, миграции же северного направления
сглажены и не столь ярко выражены.
В сентябре-октябре совершаются преднерестовые миграции рыбы из
северо-восточных районов вдоль архипелага Новая Земля в юго-восточные. районы
моря. Впервые созревающая сайка, распределяющаяся в основном в центральных
районах (Центральный желоб, Новоземельская банка), сдвигаясь постепенно к
Новой Земле через склоны Гусиной банки и Новоземельского мелководья, также
устремляется в южном направлении к о-ву Колгуев и мелководным участкам
сопредельных вод.
В октябре-ноябре- преднерестовая рыба по мере созревания формирует
нерестовые стаи и совершает локальные перемещения на акватории от о-ва Вай-
гач до Нанинских банок. В холодные годы часть рыбы выходит к мурманским
берегам или в Белое море, где нерестится в губах и заливах. В эти же'
месяцы неоднократно прослеживалась миграция сайки от о-ва Вайгач в западном
направлении. О происхождении этой рыбы судить трудно, однако можно
предположить, что эта сайка из Карского моря.
С декабря по март сайка нерестится в тех районах, которых она
достигла к декабрю-январю, а после нереста совершает обратную миграцию в районы
откорма.
Рассмотренная схема миграций сайки во многом схожа с миграциями
других морских промысловых видов рыб. весь жизненный цикл которых связан с
определенными системами течений. Более того, у сайки хорошо прослеживаются
характерные для тресковых преднерестовые миграции против течений и после-
нерестовые миграции по течению.
Помимо сезонных миграций, сайка совершает суточные вертикальные
миграции, которые наиболее четко проявляются в весенний и осенний периоды
(Печеник и др., 1973). В светлое время суток рыба опускается на дно и
держится у грунта, ночью поднимается к поверхности. Во время полярной ночи и
полярного дня распределение сайки по слоям не зависит от освещенности, а
определяется в основном ее биологическим состоянием или кормовой базой.
Для новоземельской сайки характерно распределение рыбы в
весенне-летний период при отрицательной температуре в районах с наибольшими для этого
вида глубинами - до 300-370 м. Причем в июле-августе сайка предпочитает
наиболее низкие температуры (-1.0- -1.8°). В осенне-зимний период рыба
выходит на мелководные участки моря с глубинами 40-80 м. У о-ва Колгуев на
воотоке косяки часто в маосовом количеотве подходят близко к берегу и
держатся некоторое время на глубине 5-15 и (Шлейник, 1979). В этих случаях
нередки массовые выбросы сайки на берег (Пономаренко, 1968).
Во второй половине года сайка повсеместно распределяется, в основном,
при положительной температуре от 0 до 3.5°, а в октябре-ноябре иногда и
при более высокой температуре: 4-5°. В последние годы в весенний период
скопления рыбы, отмечавшиеся в сопредельных водах центральных и западных
районов Баренцева моря, удерживались при положительной температуре.
Сведения по размерному и возрастному составу сайки из различных
районов обитания приводятся в работах многих исоледователей (Покровский. 1936;
Мантейфель. 1943; Илумов. 1949; Тамбовцев, 1952; Пономаренко, 1964: Бор-
кин, 1983а, б; Borkln, 1979. 1983а; Borkln, Shlelnik; 1981, 1985). Освоение
промыслом этого объекта позволило вести регулярные наблюдения за
биологическими характеристиками сайки, начиная с 1969 г. Анализ имеющегося
материала показал, что в Баренцевом море встречаются особи длиной до 40 см,
среди которых при облове доминирующее значение имеют рыбы 12-24 см.
Размерный состав уловов зависит от районов и времени промысла. Анализ
изменений длины сайки по годам показал, что наиболее показательны эти изменения
в восточных районах моря, где промысел ведется с 1969 г. и. как правило,
во второй половине года. Неоднороден размерный состав сайки в различных
районах Баренцева моря в одни и те же сроки.
В западных и северо-западных районах моря, а также у Шпицбергена
преобладали особи длиной менее 16 см, в центральных и восточных районах
встречалась более крупная рыба преобладающих размеров. - 18-25 см (Шлейник,
.1973).
Значительные изменения размерно-возрастного . состава уловов сайки
прослеживаются в течение года в традиционных районах образования
промысловых скоплений, какими являются восток и юго-восток моря. На нерестилища в
октябре-ноябре, как правило, сначала подходит крупная рыба. Несколько
позже в скоплениях появляются особи младших возрастов меньшей длины. В летний
период более мелкая, впервые созревающая сайка пополняет промысловую
часть популяций,и средний размер рыб, составляющих такие скопления, вновь
снижается (Shlelnik, 1971, 1973, 1974). В августе-сентябре с началом
формирования преднерестовых косяков процент содержания крупных особей
постепенно увеличивается.
В процессе эксплуатации восточнобаренцевоморской сайки удалось
проследить изменение ее размеров в межгодовом аспекте,(табл. 12).
В целом в уловах встречались особи от 5 до 40 см, среди которых в
разные годы доминировала сайка различной длины. В 1969 и 1970 гг. в уловах
преобладала сайка размерами от 20 см и более. В 1971-1974 гг. основу
уловов составляли особи длиной 16-20 см. В последующие годы, вплоть до
1977 г., вершина размерного ряда приходилась на рыб длиной 16-19 см при
одновременном увеличении в уловах рыб размерами менее 15 см. В следующие два
года (1978-1979) количество мелкой сайки в уловах еще более увеличилось и
основная масса рыб имела длину 12-16 см. Наиболее мелкая сайка
вылавливалась в 1980 г., когда средний размер ее в уловах составлял 11.6 см. В стаде
преобладали рыбы длиной 6-7 см. С 1981 г. началось укрупнение рыб в уловах,
и в 1983 г. средний размер сайки в промысловых уловах составил 18.8 см.
Однако с 1985 г. размеры особей интенсивно эксплуатируемого запаса вновь
начали уменьшаться и снизились до средней величины 12.6-12.8 см в 1986-1987
гг., а в .1988 г. составили минимальную за весь период исследований величину
11.1 см.
В последние годы (1992-1993 гг.) с выходом запаса сайки из состояния
глубокой депрессии средний размер особей увеличился до 14-15 см.
Таблица 12
Состояние популяции сайки на востоке Баренцева моря
Год
1969
1970
1971
1972
1S73
1974
1975
1976
1977
1978
1979
1980
1981
1982 ,
1983
1984 .
1985
1986
1987
1988
1989
1990
1991
1992
1993
Обищй
запас,
тыс. т*
1096
961
697
361
261
172
86
22
17
8
4
63
859
540
172 '
180
219
93
208
82
288
190
317
915
931
Вылов,
тыс.т
136.4
243.4
348.1
166.8
82.2
123.7
63.1
12.2
8.0
5.1
0.3
0.1
9.2
90.1
37.3
5.6
В. 2
0.6
0.004
-
0.2
0-1
-•
20.6
Уроисайность
поколения, индекс
численности
191'
168
150
124
26
200
5
31
30
140
302
247
140
50
39
16
334
366
153
120
41
48
239
"■ 118
45. О" * 156
Средний
возраст,
лет
4.6
4.3
4.1
3.8
3.6
3.7
2.9
3.1
3.1
2.3
2.5
2.5
1.9
2.5
3.3
2.1
2.5
1.5
1.7
1.6
1.5
1.7
1.7
2.2
2.3
Средняя
длина,
см
21.7
21.0
20.0
19.0
18.4
18.5
17.8
18.6
17.7
15.4
15.9
16.3
14.5
16.9
18.8
15.9
17.5
12.6
12.8
11.1
12.2
14.4
15.9
14.3
15.2
•Запас 1969-1980 гг. рассчитан методом ВПА. с 1981 г. - акустическим
методом. "Данные ИКЕС. '"Предварительные данные.
Предельный возраст сайки редко превышает 7-8+ лет. За период
исследований нами отмечено лишь 5 экземпляров в возрасте 10+ лет. которые были
выловлены на востоке моря. Промысловая часть баренцевоморской сайки
состоит из особей нескольких поколений.
До освоения промыслом сайки в условах преобладали особи в возрасте 4+
- 5+ лет (Печеник. Пономаренко. Шепель. 1973). однако с момента
организации тралового промысла на востоке Баренцева моря наблюдалось постепенное
уменьшение количества крупной рыбы (Шлейник, 1979). Средний возраст
облавливаемой популяции снизился от 4.6+ в 1969 г.. до 3.1+ в 1977 г.. а уже в
1981г. эта величина составила 1.9+. Кроме того, в уловах стала исчезать
рыба старших возрастных групп - сначала в возрасте 8+. затем 7+ лет. С
1978 г. ,в уловах перестала попадаться семилетняя (6+) сайка, несколько
экземпляров которой было отмечено лишь в 1982 и 1983 гг. Причина этих явле-
ний в интенсификации промысла, который способствует уменьшению в популяции
численности старших возрастных групп.
В 1989 г. средний возраст сайки снизился до экстремально низкой
величины 1.5+ лет, когда в популяции 60% особей составляли годовики.
В первые годы жизни, т.е. до наступления половой зрелости, сайка
растет довольно быстро. Длина особей интенсивно увеличивается до возраста 2+.
Ежегодный линейный прирост в это время составляет 40-50Я. В дальнейшем
темп роста снижается, и к 5-7-летнему возрасту не превышает 10Я, имея
общую тенденцию к сокращению.
Иначе происходит рост массы тела. В первые два года прирост ее
незначительный, но после достижения половой зрелости нарастание массы
происходит более интенсивно, составляя около 10Ж ежегодно. На последних годах
жизни прирост достигает 20Х и более.
Наши исследования указывают на прямую зависимость между темпом роста
и температурными условиями. Хорошо эта зависимость прослеживается на
первом году жизни, когда сеголетки сайки особенно чутко реагируют на
изменение теплосодержания водных масс.
Имеются заметные различия в росте самцов и самок: на 2-3-м годах
самцы в среднем несколько крупнее вамок, а на 5-7-м - наоборот.
Сокращение численности сайки на востоке моря привело к ее более
раннему созреванию. В 80-х годах темп созревания в первые два года жизни
увеличился в 1.5-2 раза по отношению к 70-м годам. С 1987 г. в нерестовых
скоплениях стали появляться зрелые особи уже длиной 11 см (Боркин, 1990).
Соотношение полов в течение года и в преднерестовый период всегда
было близким 1:1 с незначительным преобладанием самок. В возрасте 1+ самцов
в популяции всегда было больше в 4 раза, у.трехлеток соотношение
выравнивается, однако у рыб старших возрастов самки постоянно преобладают.
Анализ отруктуры нерестовой популяции сайки в Баренцевом море показы-
, вает. что со временем пополнение стало постоянно доминировать над
остатком, в связи с чем согласно классификации Г.Н.Монастырского (1949)
нерестовая популяция относится ко второму типу.
Нерест сайки на юго-востоке моря происходит преимущественно в
январе-феврале в прибрежных участках от п-ова Канин до о-ва Вайгач, а также на
акватории вокруг о-ва Колгуев. Эпизодический нерест отмечается у берегов
о-ва Новая Земля и в центральных районах Баренцева моря.
В зависимости от гидрологических условий возможно колебание сроков
размножения от конца ноября до марта (Расе, 1934; Есипов, 1937, 1952; Пок-
ровокий, 1936; Сохнов. 1975; Borkln, 1979). Основные нерестилища сайки
располагаются в прибрежной зоне, • однако нерестующую рыбу находили и вдали
от берегов (Клумов, 1935: Клюге, 1936; Мантейфель, 1943; Пономаренко,
1963, 1964. 1965; Hogneatad, 1966, 1968а,Ь). Массовый нерест протекает
подо льдом или у кромки льда над глубинами 20-60 м, реже - на значительном
удалении от берега над глубинами до 200 м. Нерест у сайки единовременный.
В холодные годы наблюдаются массовые заходы преднерестовой рыбы в губы и
заливы Белого моря.
Половой цикл сайки изучали В.П.Пономаренко (1965), Л.Н. Печеник с
соавторами. (1973), В.В.Сохнов (1975). Более подробно он рассмотрен О.Л.Хрис-
тофоровым (1978а.б), которым было показано, что для . сайки характерно
асинхронное созревание ооцитов. Высокие значения максимального
коэффициента зрелости у самок связаны с единовременным типом икрометания.
Сведения об индивидуальной абсолютной плодовитости сайки (ИАП) весьма
немногочисленны (Расе. 1941; Мантейфель, 1943; Клумов. 1949). Более полный
анализ ИАП за период 1969-1976 гг. дан в работе В.Н.Шлейника (1978).
Поздние исследования показывают, что на протяжении 1969-1988 гг. ИАП новозе-
мельской сайки изменялась от 1.8 до 69.7 тыс. икринок, в среднем составляя
14.4 тыс. икринок (Боркин. 1990). С увеличением длины, массы и возраста
особей линейно увеличивается ИАП. Наименьшее, количество икринок (1.8 тыс.)
отмечено у самки длиной 13 ом в возрасте 1+, максимальное (69.7 тыс.
икринок) - у 8-летней особи длиной 30 см. Минимальная длина сайки с икрой (3.2
тыс. икринок) составила И см (Боркин. 1990; Боркин, Терещенко. 1993).-
В процессе уменьшения численности нерестового стада среднегодовые
значения ИАП уменьшались от 24.7 тыс. икринок в 1970 г. до 14.6 тыс.
икринок в 1987 г. Вместе с тем за период 1969-1980 гг. была отмечена обратная
зависимость между средней ИАП у одноразмерных особей и с одинаковой массой
тела и общей численностью популяции, что. видимо, является видовым
приспособлением, направленным на сохранение численности стада. Среди
представителей семейства тресковых у сайки наиболее крупная икра, размеры в
диаметре составляют 1.44-2.11 мм. в среднем 1.75 мм (Перцева, 1936; Расе. 1949;
Пономаренко, 1965; Махотин. 1982). Внешний вид и строение икринок
указывает на адаптацию к условиям Арктики - большие размеры, тонкая и непрочная
оболочка, позднее развитие и недостаточная пигментация, отсутствие жировой
капли и прозрачность.
Инкубационный период продолжительный и длится от 1.5 до 3.5 месяцев,
в зависимости от температуры и продолжительности ледостава может достигать
4-5 месяцев (Казанова, 1949; Бараненкова и др.. 1966; Алтухов, 1979;
Боркин, 1983а).
Выклев личинок происходит с апреля по июль, но может быть и в марте,
а иногда продолжается до сентября (Бараненкова и др., 1966; Боркин. 1983а.
1985, 1986). Массовый выклевало нашим наблюдениям,происходит во время
интенсивного разрушения льда в мае-июне. Сроки выклева личинок колеблются по
годам и в значительной степени зависят от температуры воды и
продолжительности ледостава (Арснович и др., 1974; Боркин, 1983а; Borkln, 1984а.Ь,
1988. 1993).
В первые дни жизни личинки распределяются у поверхности воды в слое
0.5 м. Через 2-3 недели основная их масса располагается на горизонтах
5-Ю м, а в целом от поверхности до 20 м. реже 30 м (Borkln. 1979. 1983Ь).
Минимальный размер предличинок в наших сборах составил 4.4 мм. Средние
размеры в июне составляют 6-8 мм. в июле 8-10 мм, однако различны по годам
и зависят от сроков выклева. Отмечена прямая связь между темпом роста
личинок и прогревом водной толщи на различных участках моря. Наблюдения за
распределением личинок сайки по Баренцеву морю, выполненные в последние
годы, указывают на их локальную обособленность у Новой Земли и у
Шпицбергена. Многолетние исследования по распределению мальков в августе-сентябре
на акватории Баренцева моря и сопредельных вод подтверждают разобщенность
скоплений сайки в диаметрально расположенных участках моря (Апоп. 1988).
Личиночная стадия заканчивается при длине 27-35 мм.
В августе-сентябре значительная часть личинок переходит в стадию
малька, распределяясь в толще воды. В этот период длина сеголеток
колеблется от 3.0-3.5 до 5.5-6.5 см. иногда и более.
Необходимо отметить,. что На формирование численности поколений сайки
в раннем онтогенезе значительное влияние оказывают такие факторы, как
температура, ветровой режим, лед, обеспеченность кормом (Боркин и др.,
1987а).
Проведенные в 1977-1989 гг. исследования на востоке моря показывают,
что многочисленные поколения сайки появляются в холодные годы, неурожайные
- в теплые. Применение метода корреляционного анализа между тепловым
состоянием моря и количественными показателями молоди позволило выявить
статистически значимые связи численности личинок и температуры воды на ГМС
"Северный Колгуев" в июле (г= -0.91). Тесная.обратная связь прослеживается
между индексами численности 0-группы сайки и теми же значениями
температуры в июле (г= -0.73).
Активное формирование в районах нереста ледового покрова в зимний
период и значительная продолжительность ледостава способствуют повышенному
выживании сайки на ранних этапах развития и формирования высокой
численности ее поколений. Зависимость между ледовитостью в юго-восточной части
моря в первом полугодии и численностью личинок оказалась довольно высокой
- коэффициент корреляции составил 0.86, для.периода январь-февраль на
стадии 0-группы эта величина оказалась'равной 0.68.
Личинки, распределяясь у поверхности моря, наиболее уязвимы и в массе
гибнут при активном волнении под влиянием штормовых ветров (при скорости
более 10 м/с). В годы с повышенной ветровой активностью в период выклева
личинок и их первых недель жизни появляются бедные поколения, при слабой
ветровой деятельности - многочисленные (г= -0.93). Аналогичный характер
влияния прослеживается и в формировании сайки на стадии 0-группы
(г» -0.63).
Концентрация пищевых организмов в летний период в районах* массового
распределения личинок составляет 0.6-1.6 экз/л. Принимая во внимание
высокую степень адаптации сайки в арктических условиях, на ранних этапах
развития (длительный период резорбции желтка, продолжительное время
смешанного питания, способность долго голодать, сохраняя жизнеспособность, слабая
интенсивность питания, быстрое расширение пищевого спектра, замедленный
процесс переваривания пищи), а также мелкомасштабность
сконцентрированности в естественной среде кормовых организмов (Wlynsen, 1977; Tilaeth, 1984)
и поведенческие характеристики личинок в период откорма (скорость
плавания, поисковая реакция, направленность движения (Михеев, Пакульская. 1988;
Hanterm, Thomas, 1974), можно судить о достаточной ее обеспеченности пищей
в исследуемые годы.
Сведения о питании крупной' сайки немногочисленны, но дают ясное
представление о качественном составе потребляемых организмов и
интенсивности питания (Клумов, 1935а, б. 1949; Мантейфель, 1943; Никольский, Рада-
ков. 1968; Белова. Тарвердиева, 1964; Пономаренко. 1967а). В течение жизни
характер питания сайки меняется весьма значительно. Основу пищи молодых
особей составляют фито- и зоопланктон. По мере роста в желудках начинают
встречаться мелкие рыбы и их молодь, моллюски, черви, крабы, медузы и
т. д.. однако в пищевом комке всегда преобладают низшие ракообразные - в
основном копеподы и эвфаузииды.
Перед нерестом (ноябрь-декабрь) средний балл наполнения желудков
составляет всего 0.5-0.8, т.е. интенсивность питания минимальная. Посленерес-
товая рыба питается более активно - в феврале-мае средний балл достигает
1.1-1.7. С июня по сентябрь активность питания сайки наибольшая, что сов-
падает с массовым развитием зоопланктона на востоке моря. В июле-августе
средний балл наполнения самый высокий - 2.3-2.5. с октября интенсивность
питания постепенно снижается. В осенне-зимние месяцы в желудках сайки доля
креветки и рыб (в основном мальков сайки и мойвы) увеличивается. Весной и
летом основу рациона соотавляют калянус и капшак.
Одним из важнейших показателей откорма сайки является ее жирность.
Наименьшая жирность наблюдается в марте-апреле (5.6%). Летом этот
показатель увеличивается и достигает наибольшей величины (14%) в
сентябре-октябре.
Являясь важным звеном в экосистеме Баренцева и других морей Северного
Ледовитого океана, сайка одновременно представляет интерес как объект
специализированного промысла (Печеник. Шепель, 1970; Шлейник, 1973; Shlelnik.
1971). Вопрос о необходимости изучения биологии сайки и возможности ее
промысла в Баренцевом и сопредельных морях ранее неоднократно ставили
советские и зарубежные исследователи (Покровский, 1936; Есипов. 1937;.Мас-
лов, 1957; Дмитриев, 1958; Кравчук, 1958; Пономаренко. 1962, 1965а). По
мнению И.Г.Юданова (1947) и С.К.Клумова (1949), промысел только в губах и
на мелководьях юго-восточной части Баренцева моря мог бы составить сотни
тысяч центнеров рыбы. Однако технический уровень промышленности и
отсутствие должного спроса в довоенный период не стимулировали соответствующего
промысла.
В 40-50-х годах в Чешской губе ежегодно вылавливали от нескольких
оотен тонн до 1-6.5 тыс.т сайки. С 1969 г. начался специализированный
траловый промысел объекта, чему в значительной степени способствовало
ухудшение состояния сырьевой базы донных рыб Баренцева моря (треска,
пикша) и атлантическо-скандинавской сельди в Норвежском море.
В апреле 1969 г. экспедиция в составе двух РТЛ и ПР "Советск"
положила начало отечественному морскому промыслу сайки. Вылавливали рыбу донными
и пелагическими тралами среднётоннажные и крупнотоннажные суда. Норвежские
суда промышляли ее пелагическими тралами и кошельковыми неводами. Кроме
Баренцева моря, промысел сайки ведется в Чукотском море.
В начале 70-х годов доля сайки достигла уже 40-50% общей добычи
рыбных объектов советскими судами в Баренцевом море.
Анализ динамики уловов показал, что промысел, начатый в конце 60-х
годов и основанный на преднерестовых и нагульных скоплениях, • быстро
возрастал и достиг максимума в 1971 г. - 348 тыс. т при запасе 1.1 млн.т
(табл. 12). причем до 1973 г. основу уловов составляла крупная рыба в
возрасте 5-6 лет. Но уже к 1973 г. средний возраст сайки из промысловых
уловов снизился до 3.6 против 4.6 в 1969 г. Одновременно почти вдвое
уменьшилась и средняя масса рыбы.
Высокая степень эксплуатации повлияла на снижение уловов, и уже в
1978-1980 гг. в традиционных районах распределения рыба практически не
ооздавала скоплений. В значительной мере этому способствовало снижение
нерестового запаса до критического уровня и, как следствие, появление в
1975-1978 гг. малочисленных поколений (см. табл. 12). Следует отметить,
что с 1974-1975 гг. основу уловов составляли рекруты в возрасте 2+ - 3+
лет, имеющие среднюю длину 15 см.
В 1982-1983 гг. наметилось восстановление численности стада аа счет
появления богатых поколений 1979-1980 гг. Однако интенсивный облов этих
поколений на стадии молоди и рекрута вновь поверг стадо в депрессивное
состояние (Bull. stat. 1966, 1994).
Через 5 лет повторно создавались благоприятные условия для появления
еще двух высокоурожайных поколений - 1985 и 1986 гг., индексы численности
которых по данным международных съемок 0-группы оказались на уровне самых
урожайных поколений (Anon, 1988). Однако значительного пополнения запаса
стада, которое ожидалось от этих поколений, не произошло, а наоборот,
продолжалось падение запаса.
Наметившееся в 1987 г. восстановление запаса было нейтрализовано
высокой степенью естественной смертности в.. результате хищничества. Это, в
свою очередь, объясняется возникшим в середине 80-х годов дисбалансом
трофических связей в экосистеме Баренцева моря. Резкое уменьшение численности
мойвы, креветки,.молоди сельди - традиционных объектов питания хищных гид-
робионтов (гренландский тюлень, треска, морские птицы) - способствовало
частичному переносу пресса хищничества на сайку. С возникшим в 1985- 1986
гг. дефицитом кормовой базы потребление сайки в осенне-зимний
период,прежде воего тюленями и треской резко увеличилось.
Расчеты показывали, что естественная смертность увеличилась в среднем
в 3-4 раза, а максимально в 5-10 раз. В межгодовом аспекте коэффициент
естественной смертности составлял 0.54 в среднем для периода 1969-1984 гг.,
а уже в 1988 г. этот показатель, достиг величины 4.25.
После продолжительного периода глубокой депрессии величина общего
запаса сайки на востоке моря в 1992-1993 гг. резко увеличилась (табл. 12). В
значительной мере данный факт обусловлен высокой степенью выживания рыбы в
связи с восстановлением стада мойвы и сельди. Необходимо также отметить,
что в эти годы многократно увеличился нерестовый запас популяции, величина
которого обеспечивает появление богатых и средних по урожайности поколений
даже при неблагоприятных условиях выживания. А это в свою очередь
благоприятствует высокому уровню пополнения, а следовательно - стабилизации
общего и промыслового запасов.
Таким образом, основной причиной уменьшения запаса в 70-80-х годах
следует считать перелов нерестовой части популяции и вылов молоди, что в
конечном итоге подорвало репродуктивную способность вида. В то же время
нельзя недооценивать влияние на запас естественных факторов среды, и
прежде всего, степени воздействия хищников, смертность от которых при
определенных условиях может многократно возрастать, а в отдельных случаях
намного превышать промысловую смертность даже в годы интенсивной эксплуатации
стада.
Исследования предшествующих, лет показали, что у сайки много врагов и
все они в большей или меньшей степени связаны между собой в один большой и
чрезвычайно интересный биологический комплекс, основным стержнем которого
является именно сайка (Боркин и др., 1992; Borkln, 1990; Borkin et al.,
1992). Экосистема восточной части Баренцева моря в процессе эволюции
сформировалась удивительно гармонично и рационально - основные" потребители
сайки откармливаются ею в строго определенный сезон года, запасая
энергетический потенциал перед размножением (табл. 13).
Так. например, с ноября по февраль на юго-востоке моря стада
гренландского тюленя интенсивно откармливаются нерестовой сайкой, после чего
устремляются в Белое море на щенные залежки. В эти же районы из Карского
моря выходит белуха и с января по апрель активно питается сайкой. Чуть
позже, в апреле-мае, у самок белух появляется потомство. С мая по август
длится гнездовой сезон кайр и других колониальных морских птиц западного
побережья Новой Земли, основу питания которых составляет посленерестовая
сайка и ее молодь-(Горбунов. 1925; Успенский, 1956; Головкин. 1972).
Откорм сайкой крупной и созревающей трески на банках и мелководьях востока в
осенний период был известен давно (Зацепин. Петрова, 1939; Гринкевич.
1957; Маслов. 1968). После нагула эта рыба мигрирует к Лофотенским
островам на нерест.
Таблица 13
Питание морских гидробионтов сайкой в различные сезоны года
Вид
Лето
VI
VII
VIII
Осень
IX
X
XI
Зима
XII
I
II
Весна
III
IV
V
Кайра + + + ■ • +
Треска + + + +
Гренландский
тюлень + + + +
Белуха + + + +.
Исследования показывают, что наиболее активными потребителями сайки в
последние.годы являются гренландский тюлень и треска. По данным норвежских
исследователей, в настоящее время в Баренцевом море обитает примерно 2 млн
особей гренландского тюленя, которые в год потребляют 4.6 млн т пищи, в
том числе 501 рыбы. Причем на мойву и сайку приходится, соответственно. 32
и 6% (т.е. 1.47 и 0.28 млн т) (Markussen, tfrltaland. 1985).
Однако нельзя не принимать во внимание того, что гренландский тюлень
беломорской популяции значительную часть своей жизни - проводит в районах
поотоянного местообитания сайки. Значение этой рыбы в питании тюленя
оказывается неоомненно более значительным (Клумов. 1936. 1937; Мантейфель,
1943). Расчеты показывают, что в период осенне-зимней миграции
(октябрь-январь) вдоль Новой Земли через юго-восточные районы Баренцева моря,
где в это время распределяются основные скопления преднерестовой сайки,
популяция гренландского тюленя численностью 1-0.8 млн особей поедает от
100 до 300 тыс.т сайки (табл. 14). Кроме гренландского тюленя, сайкой
питаются морской заяц (численность 50-60 тыс. особей), кольчатая нерпа
(около 50 тыс, особей), в меньшей степени хохлач, серый и обыкновенный тюлень,
морж (Клумов, 1935а,б; Клюге. 1936; Чапский, 1938; Рутилевский. 1939).
Из китообразных сайку потребляют белуха, малый полосатик (кит Минне),
нарвал и некоторые другие.
Значение сайки в жизни белухи особенно велико, поскольку жизнь этого
кита постоянно связана с присутствием ледовых полей. Являясь типичным
ихтиофагом, белуха у кромки льда в первую очередь поедает сайку, а ее
миграции часто связаны именно с распределением этой рыбы (Клумов. 1936;
Остроумов, 1929. 1935). Исследования показывают, что ежегодно в зимний период
распределение скоплений белухи и концентраций нерестовой сайки совпадают.
При суточном рационе 50 кг рыбы, в результате хищничества белухи в течение
6-7 месяцев (с ноября по май) сайки погибает от 50 до 100 тыс.т
(табл. 14).
Таблица 14
Годовое потребление сайки морскими млекопитающими,
птицами и треской на востоке Баренцева моря
Вид
Гренландский тюлень
Численность,
тыс.экз.
800
(беломорская популяция)
Морской заяц
Нерпа кольчатая
Белуха
Кайра
50-60
50
15-20
2000
Треска 500000-700000
Всего:
Средний
суточный
рацион,
кг
10-15
15-20
5-в
40-50
0.3-0.4
Период
откорма
сайкой,
месяц
ноябрь-
Доля сайки
в пищевом
рационе, %
25-50
-февраль
январь-
25-50
-декабрь
ноябрь-
-март
декабрь-
-май
июль-
-август
0.02-0. 04 октябрь-
-ноябрь
50
50-70
50-100
30-50
Количество
потребляемой
сайки в год,
тыс.т
100-300
50-100
15-30
50-100
70-80
100-200
385-810
В последние годы наиболее существенным потребителем сайки стала арк-
то-норвежская треска. С начала 80-х годов основу ее питания составляла
мойва и северная креветка. Ежегодное их потребление варьировало в пределах
2-3 и 0.3-0.7 млн т соответственно (Пономаренко и др., 1978; Bolstad,
1987). Но уже в 1986 г. треска выедала остатки запасов мойвы и креветки, а
в 1987 г. практически была лишена своей обычной кормовой базы. В'
результате жирность и упитанность уменьшились, темп роста замедлился.
Оказавшись в 1987 г. на голодном пайке, треска в поисках пищи
вынуждена была в осенний период компенсировать энергетические затраты
интенсивным откормом сайкой на востоке моря, где долгое время распределялась в
водах с предельной для ее обитания отрицательной температурой от 0 до -1°.
По результатам анализа питания трески в сентябре-ноябре, доля сайки в
пищевом рационе составляла от 2 до 8935, частота встречаемости в желудках
25-401, суточные рационы 2.1-3.758. В районах совпадения ареалов сайки и
трески на востоке моря распределялось от 500 до 750 тыс.т трески (по
данным ПИНРО). Выполненные по методу Пономаренко и др. (1978) расчеты
показали, что потребление сайки треской в этот период составило порядка
100-200 тыс.т.
В 1988 г. условия откорма трески оказались более благоприятные, что
обусловлено высокой биомассой капшака. Тем не менее распределение трески и
сайки было близким к 1987 г., и по предварительным расчетам гибель
последней от хищничества составила не менее 100 тыс.т.
Кроме трески, сайку поедает и некоторые другие хищные рыбы, ареалы
которых на южных и западных окраинах в Баренцевом море частично совпадают:
палтус, зубатка, камбала-ерш. навага, арктическая треска.
Неотъемлемой частью трофоценоза на востоке моря являются
многочисленные морские птицы - чайки, кайры, гагарки и другие, которых на западном
побережье Новой Земли летом гнездится более 2 млн особей (Успенский. 1956,
19876). Кайры, составляющие основу птичьих базаров, наиболее приспособлены
для добывания рыбы на большой глубине - до 130 м (Vader, 1987). Являясь
типичным ихтиофагом, кайра питается преимущественно сайкой, причем
ежедневно потребляет около 300 г рыбы и в среднем 100 г приносит своим птенцам
(Головкин, 1963).
За гнездовой сезон, который длится у этих птиц 120 дней, выедание
сайки составляет 70-80 тыс.т. Следует отметить, что большинство кайр
остается зимовать в этих же районах и. распределяясь в течение всей полярной
зимы вблизи кромки льда или на полыньях, продолжает питаться, по-видимому,
также сайкой (табл. 14). Достоверные сведения по составу питания за' этот
период отсутствуют.
Таким образом, становится очевидным, что сайка в экосистеме Арктики
является гораздо более важным элементом, чем это представлялось ранее и, в
первую очередь, именно как объект питания многочисленных животных, птиц и
рыб.
Анализ динамики численности сайки показывает, что этот вид должен
крайне осторожно эксплуатироваться промыслом и лишь в те периоды, когда
наблюдается существенное увеличение популяции за счет появления урожайных
поколений: При этом обязательно должен учитываться возможный объем
потребления рыбы морскими гидробионтами.
6.4. Арктический голец
Арктический голец, являющийся самой северной пресноводной рыбой,
известен на Новой Земле с 15 столетия, где русские промышленники с берегов
Белого моря и из Печорского края занимались охотой на морского зверя и
птиц. Первые научные исследования биологии новоземельского гольца были
выполнены в 1931-1932 гг. во время экспедиции Арктического института и после
50-летнего перерыва пополнены материалом, собранным в экспедициях
Мурманского морского биологического института в 1986-1989 гг. Результаты
экспедиций, а также литературные данные об общей характеристике вида дают
лишь некоторые представления о гольце островов Новая Земля.
Систематическое положение и диагноз. Арктический голец Salvelims at-
ptnus L.: отряд сельдеобразных Ctupet/ormes. подотряд лососевидных Salmo-
notdei, семейство лососевых Salraontdae, род гольцов Salvettnus.
Д.С.Берг (1948) дает следующие характеристики данного вида: Д III-IV
36-37
8-11. A III-IV 7-9. 1.1.130-140, squ. 190 240. жаберных тычинок на 1-й
38
дуге 18-30, жаберных лучей 10-12, пилорических придатков (в р.Каре) 30-44.
В одной десятой части длины тела (по Сиитту) впереди Д находится 26-37
чешуи. Верхняя челюсть у молодых не заходит за вертикаль заднего края глаза.
у старых заходит, но не так далеко, как у S.ralma. Спина с
синевато-стальным отливом, темных пятен ни на плавниках, ни на теле нет; в период
нереста на спине появляются мелкие красные или оранжевые пятнышки. Длина
35-65 см', вес 1-3 кг, наибольшая длина до 88 см и вес до Ю-15 кг.
В целом арктический голец очень изменчив. Внешний вид, особенности
биологии, другие показатели сильно отличаются как у рыб. обитающих в раз-
ных водоемах, так и у рыб, населяющих один и тот же водоем. Даже такой
показатель, как цвет мяса, может варьировать у рыб разных популяций или у
рыб одной популяции в разные годы.
Жизненный цикл новоземельского гольца (Andrews, Lear, 1956)
определяется суровыми условиями его обитания: скудностью кормовой базы пресных вод
и низкими зимними температурами в море. Поэтому для откорма голец должен
мигрировать в море, а для зимовки - в непромерзающие глубокие озера
архипелага.
Молодь гольца первые 3-4 года жизни проводит в пресной воде. Данных
об этом периоде очень мало. Л.О.Ретовский (1935) приводит сведения о
нахождении мальков в мелких озерах и реках, соединяющих озера с морем.
Анализ питания показал, что в состав пищи входят пресноводные
ракообразные и личинки насекомых, а также взрослые комары и мухи. Мелкая молодь
питается придонными объектами, а воздушные насекомые встречаются лишь в
желудках крупных форм (Ретовский. 1935). За пресноводный период малек
вырастает до 11-27 см (Сластников, 1935). Начиная с 3-4-летнего возраста,
гольцы ежегодно мигрируют в море на нагул.
Мигрирует голец в море весной, но данных о конкретных сроках ската
молоди нет (Есипов, 19356). От первого ската в море до созревания проходит
не менее 4'лет, и все эти годы голец возвращается на зимовку.
В море голец растет быстро (Сластников, 1935). но данных о биологии
гольца в морской период жизни нет. В.К.Есипов (19356) считает, что
нагуливающийся голец не уходит далеко от устьев рек, но анализ паразитофауны
новоземельского гольца дает основания предполагать и длительные морские
миграции этих рыб (Митенев. Шульман. 1986).
Возвращается голец из моря в озера, начиная со второй декады августа.
В реки губы Крестовой идут те особи, которые будут нереститься *в данном
году. Их ход заканчивается в конце августа, в сентябре они попадаются уже
единично. Рыбы, пропускающие нерест в данном году, заходят в реки во
второй половине августа, максимум хода отмечен в первых числах сентября
(Есипов, 19356). К середине сентября ход заканчивается, завершают его мелкие
рыбы, проведшие в море один-два года. В других реках Новой Земли сроки
хода могут быть иными.
Наши наблюдения за ходом гольца в р. Нехватова, проведенные в первую
неделю сентября 1986 г., показали следующее распределение мигрирующих рыб
по полу и стадии зрелости (табл. 15).
Таблица 15
Распределение мигрантов гольца в р. Нехватова
по полу и стадии зрелости в 1986 г., %
Пол
Стадии зрелости
II
III
IV
Самцы (135 экз.) 87 8 5
Самки (155 экз.) 85 15 О
Наблюдений за нерестом гольца на Новой'Земле нет. На основании
косвенных заключений В.К.Есипов (19356) предположил, что нерест гольца
происходит в октябре-ноябре и не в реках, которые в это время мелководны и
легко промерзают, а в озерах.
У созревающих рыб изменяется внешний вид и физиологическое состояние
- происходит лошание. Незрелые рыбы имеют серебристо-зеленую окраску тела,
созревающие отличаются пятнистостью на боках, бледно-красным брюшком,
оранжево-красными или золотистыми плавниками, более светлым и менее жирным
мясом, несколько загнутой вверх нижней челюстью.
Плодовитость новоземельского гольца колеблется от 1.5 до 9 тыс.
икринок, в среднем 3.5 тыс. Показатели плодовитости отдельных рыб приведены в
таблице 16.
Таблица 16
Плодовитость новоземельского гольца (Есипов, 19356)
Длина
тела,
см
Масса,
кг
Стадия
зрелости
Вес гонад
абсолютный, г
относительный. %
Плодовитость
абсолютная относительная
53.
53.
56.
56.
57.
57.
57.0
57.2
58.2
58.8
59.0
73.3
1.75
2.07
1.88
2.38
10
16
73
04
34
50
57
95
IV
IV
III-IV
IV
III-IV
III-IV
III-IV
IV
III-IV
III-IV
III-IV
III-IV
168
135
88
177
108
168
158
259
155
140
160
355
9.6
6.5
4.7
7.4
5.1
7.8
5.8
12.7
6.6
•5.6
6.2
9.0
2789
2466
1407
2855
3240
2956
3592
4409
2604
3227
3480
9058
1594
1194
748
1202
1543
1368
1315
2166
1111
1293
1356
2296
Возраст и рост. Уловы ставными неводами, полученные в 1931 г.
научно-промысловой экспедицией в губе Крестовой (Сластников. 1935). дали
следующую картину распределения рыб по возрастным группам: 5455 рыб имели
речной возраст 3 года,
8 лет.
4635 - 4 года. Морской возраст колеблется от 3 до
Возраст
3+
4+
5+
6+
7+
8+*
Доля рыб, %
18.
63.
4.
8.
4.
1.6
ИЗ р
В таблице 17 показано распределение по возрастам 108 экз. мигрантов
Нехватова. Средний возраст гольцов пойманных нами в р.Нехватова
весной составил 8 лет с колебаниями от 4 до 11 лет.
Длина и вес рыб различного возраста из уловов в губе Крестовой и в
р.Нехватова представлены в таблицах 18-21 (Сластников, 1935).
Таблица 17
Возрастной состав уловов гольца р.Нехватова (Сластников, 1935)
Речной возраст
Количество, %
3 56
4 44
Морской возраст
0+
37
32
■ . 1+
14
10
2+
4
2
. 3+
1
0
Таблица 18
Размерно-возрастной состав уловов гольца р.Нехватова
и губы Крестовой, см
Речной возраст
3
4
Морской возраст .
3+
46.3
45.9
4+
52.7
52.6
5+
6+
52.0 58.8
57.6 60.8
7+
8+
61.8 64.7
73.3
Таблица 19
Вес гольца губы Крестовой, г
Речной возраст
3
4
Речной возраст
Морской возраст
3+
4+
5+
6+
1186 1726 1598
1124 1750 2207
Длина гольца р.Нехватова,
2142
2840
СМ
7+
2385
8+
2310
3945
Таблица 20
Морской возраст
0+
1+
2+
3+
3 24.3 30.9 40.6 54.0
4 25.0 30.8 38.5
Вес гольца р. Нехватова. г
Таблица 21
Речной возраст
Морской возраст
0+
1+
2+
3+
157
171
330
350
755
71В
1210*
*В оригинале - 210, очевидно,- опечатка.
Экспедиция ММБИ в 1986 г. исследовала гольцов в двух реках: Гусиная и
Нехватова (Черницкий и др. 19ВВ). Размерно-весовые показатели из наших
уловов представлены в табл. 22.
Таблица 22
Размерно-весовые показатели гольцов Новой Земли
(по данным ММБИ, 19В6)
Река
Количество,
экз.
Длина,
см
Вес.
кг
Упитанность
(по Фультону)
Гусиная
Нехватова'
25
88
49.6+1.1
47.4+0.7
1.57+0.10
1.31+0.07
1.28+0.03
1.16+0.02
Данных по размерно-возрастной характеристике гольца Карского
побережья Новой Земли нет, за исключением результатов исследования 76 рыб из
залива Брандта (Есипов. 1952). Длина рыб составила в среднем 53.4 см
(32-75 см), вес 1.В5 кг (0.4-5 кг), а возраст не превышал 14-15 лет. По
устным сообщениям, полученным нами от новоземельских рыбаков, вес гольца
из рек восточного побережья может достигать 15 кг.
Питание. В море голец питается главным образом рыбами, в меньшей
степени ракообразными (табл. 23). Наши наблюдения в сентябре 1986 г. показа-
Состав пищи гольца, пойманного в губе Крестовой (Есипов.
Таблица 23
19356)
Компонент
Встречаемость. %
Треска
Тресковые
Мойва
Пеочанка
Бычки и их мальки
Мальки рыб (не тресковых)
Рыбы неопределенные
Икринки рыб
Ракообразные
Бокоплавы
Черви
Прочее
2В
16
6
5
3
13
2
1
7
6
5
8
ли. что голец в устьевой зоне р.Нехватова питался очень интенсивно,
наполненность желудков 5 баллов, в содержимом преобладали молодь трески, сайда,
песчанка, амфиподы, мизиды. Весной у рыб,пойманных в этом районе, желудки
были пусты. В озерах системы реки Нехватова (Скальное. Питьевое) основной
пищей гольцов являются остатки высших растений, двустворчатые моллюски,
амфиподы, личинки насекомых, девятииглая колюшка, мелкие бычки и гольцы.
Максимальный улов новоземельского гольца (80 т) отмечен в конце
прошлого - начале нынешнего веков, в настоящее время промысел гольца имеет
лишь локальное значение, достигая по ориентировочным подсчетам до 10 т в
год.
Глава 7. МОРСКИЕ ПТИЦЫ
(РЕТРОСПЕКТИВНЫЙ АНАЛИЗ РАЗВИТИЯ ПОПУЛЯЦИЙ) ,
В Баренцевом море наиболее грандиозные колонии морских птиц
расположены на западном побережье островов Новой Земли. Тем не менее, период
систематического изучения орнитофауны Новой Земли был сравнительно короток.
Первые авифаунистические исследования и количественные учеты были начаты в
20-х годах, и их результаты были отражены в работах Г.П.Горбунова (1925,
1929), Л.А.Портенко (1931), С.К.Красовского (1937). В этот период новозе-
мельские колонии морских птиц представляли собой определенный интерес в
качестве промыслового объекта, способного иметь большое значение при.
дальнейшем хозяйственном освоении региона. Многоплановые и систематические
исследования морских птиц проводили только в период 1947-1951 гг. Их
начало связано с организацией по инициативе Л. 0. Белопольского в губах Грибовая
и Безымянная филиала заповедника "Семь островов". Основные результаты этих
работ были отражены в публикациях С.М.Успенского (1956) и
Л.0.Белопольского (1971). К сожалению, с момента ликвидации новоземельского филиала
заповедника (15 октября 1951) орнитологические наблюдения были здесь
исключительно редки. Так. в августе 196? г. в некоторых губах Северного острова
Новой Земли провел учетные работы А.Н.Головкин (1972).
Сведения о современном качественном и количественном составе авифауны
Новой Земли, экологии и гнездовой биологии даже наиболее массовых видов в
литературе почти полностью отсутствуют. Хотя именно сейчас существует
огромная потребность в сведениях такого рода, поскольку в последние
десятилетия экосистемы региона Новой Земли испытывали чрезвычайные антропогенные
нагрузки (включая проведение ядерных испытаний), последствия которых
трудно предсказуемы. Исходя из этого, основной целью нашего исследования было
выявление основных тенденций развития популяций фоновых видов морских птиц
Новой Земли. Поскольку в губе Безымянной находятся одни из наиболее
крупных колоний толстоклювой кайры (Urla lomtUa), подробно описанные в период
существования заповедника, то именно этот район и был выбран в качестве
полигона для работы комплексной экспедиции Мурманского морского
биологического института и Кандалакшского государственного природного заповедника
в июне 1992 года. Позднее, в июле этого же года губу Безымянную посетили
сотрудники ВНИИприрода С.М.Успенский и Г.В.Хахин (1993 г.). Поэтому
возникла исключительная ситуация, позволяющая сопоставить результаты
исследований, проведенных независимыми группами в одном районе. В августе 1993 г.
губы Безымянную и Грибовую посетил А.Б.Савинецкий. который любезно
поделился с нами своими наблюдениями.
В ходе работы нами были использованы материалы из архива заповедника
"Семь островов", хранящиеся в настоящее время в Кандалакшском
государственном природном заповеднике. В его основу были положены данные,
собранные по программе "Летописи природы" рядом сотрудников заповедника, и в
первую очередь Л.О.Белопольским в 1947 г., Т.В.Боголюбовой - в 1947 г..
Н.Н.ПУТЯТИНЫМ - В 1949-1950 ГГ.. В. А. Сушко - в 1947-1951 гг. И СМ. Успенс-
ким - в 1948-1950 гг. Информация, содержащаяся в годовых отчетах
руководителя работ Л.0.Белопольского. позволила получить подробное представление о
методике учета птиц-базарников, а значит,и о пределах достоверности
данных, полученных при ее использовании. Последнее особенно необходимо при
проведении ретроспективных анализов развития авифауны.
Основу методики, применявшейся в новоземельском филиале, составлял
подсчет взрослых особей кайр в колониях. Нередко конкретный исполнитель
предполагал, что на момент учета часть оообей кайр вместе с птенцами
покинула птичьи базары и ушла в море. Количество' таких особей определяли
произвольно и выражали в виде процента от числа учтенных птиц. После чего
полученные данные суммировали и умножали на два. так как предполагалось, что
в колониях присутствует только один партнер из каждой размножающейся пары.
Иногда исполнитель произвольно вводил поправку на птиц, ■ находившихся в
воде и в воздухе (в 1947 г. она была определена в 2055). после чего
суммировал полученные данные и делил их пополам. Подобные методы учета могли
приводить к искажению реальной картины, ибо количество кайр, покинувших
птичьи базары, определяли совершенно произвольно, причем не от количества
птиц, действительно размножавшихся. * а от количества особей, подсчитанных
на базарах'. Как показали результаты почти 60-летнего мониторинга кайр на
Семи островах, кроме насиживающих яйцо или обогревающих птенца птиц в
колониях всегда присутствует определенное количество их партнеров. Здесь же
держатся взрослые особи, по каким-либо причинам не размножавшиеся в данном
сезоне. В отдельные годы их количество бывает особенно велико. Например,
на Семи островах в 1987 г. не размножалось около 90% держащихся в колониях
кайр. В то же время, как показали результаты кольцевания и цветного мече-
ния, здесь же держатся молодые птицы, еще никогда не приступавшие к
размножению, но уже имеющие взрослый наряд. Конечные результаты подобных
учетов слабо отражают реальную численность размножавшихся птиц в сезоне и при
этом мало приемлемы при сравнении с данными других лет и других
исследователей. Поэтому на практике при изучении долговременной динамики
численности кайр мы использовали учеты количества особей, присутствующих в
колониях в конце периода насиживания. Но и эти результаты не подходят для
определения доли птиц, размножавшихся в конкретном сезоне, однако они в
меньшей степени зависят от субъективных качеств учетчика и вполне
объективно описывают реальные изменения численности птиц в популяциях. Так.
массовая гибель кайр во время зимовки в Баренцевом море в конце 80-х
годов, независимо друг от друга, была описана рядом исследователей на
основании учетов в колониях разных'районов региона (Краснов, 1991; Vader.
1987).
Чтобы выявить изменения, произошедшие в авифауне исследуемого района.
мы решили .за точку отсчета взять наиболее детально описанный й закартиро-
ванный учет'прошлых лет, выполненный в 1948 г. (архив заповедника).
Учитывая сложность поставленной задачи, в июне 1992 г. мы проводили
исследования только на Северных базарах губы Безымянной, при этом, кроме общего
подсчета птиц, проводили детальное сравнение карты-схемы птичьих базаров
1948 года с реально существующей обстановкой. Обследование колоний
проводили с суши, а на недоступных участках - с воды, с моторной лодки,
регистрируя кайр в колониях и гнезда моевок {Rissa trtdactyta) (расчетные
коэффициенты не использовали). Птиц других видов, таких как чистики (Ceppfms
grylle), тупики {Fratercula arctica), бургомистры (Larus hyperboreus),
учитывали путем подсчета особей, встреченных на маршруте.
Анализ литературных и архивных источников информации, а такие
собственных данных, полученных в июне 1992 г.. позволил нам составить общее
представление о характере развития поселений кайр в губе Безымянной за
весь период орнитологических наблюдений на Новой Земле. Оно основано на
материалах, представленных нами в табл. 24. . Как уже упоминалось нами
ранее, использование разными исполнителями своих специфических методических
подходов привело к трудно сопоставимым результатам, поэтому в табл. 24
приведены лишь первичные материалы (количество птиц, учтенных в колониях).
При сравнении карты-схемы Северных птичьих базаров 1948 г. с реально
существующими в 1992 г. колониями установлено, что количество отдельных
групп (субколоний) кайр уменьшилось на ЗОЖ, а их численность снизилась до
41% от уровня 1948 г. По сравнению же с 1950 г. кайр в колониях учтено еще
меньше - 12.5%. Несколько иная картина получается при детальном анализе
данных по Южным базарам губы Безымянной. По сообщению СМ. Успенского и
Г.В.Хахина (1993), В 1992 г. здесь было учтено по сравнению с 1948 г. 7055,
а по сравнению с 1950 г. - 58.3% кайр. Однако при анализе эти авторы
оперируют данными, для получения которых были использованы различные
поправки, упоминавшиеся при описании методик учета. В табл. 24 нами представлены
первичные' данные этих учетов из годовых отчетов С. М. Успенского и Л. 0. Бело-
польского (архивы заповедника). По ним в 1992 г. на Южных базарах кайр
было учтено'лишь на 13.656 меньше, чем в 1948 г. и на 41.7% меньше, чем в
1950 г. Строго говоря, ориентируясь только на данные по Южным базарам губы
Безымянной, действительно можно прийти к выводу лишь о "некотором
сокращении числа гнездящихся на птичьих базарах кайр" (Успенский. Хахин. 1993).
Тем не менее анализ всего комплекса доступных нам материалов
свидетельствует о реальном и значительном сокращении численности толстоклювых кайр в
районе исследования. Противоречивость же результатов по Южным базарам
объясняется, вероятнее всего, применением неадекватных методик учета.
Основные тенденции изменения численности толстоклювых кайр в разные
периоды времени в целом в губе Безымянной показаны на рис. 52. Из него
следует, что в губе Безымянной их численность была наивысшей в начале 30-х
годов и минимальной - в 1992 г, С нашей течки зрения,это вполне объяснимо.
Промысел морских птиц на Новой Земле, активно развивающийся в. 20-30-е
годы, к концу 30-х годов, вероятно, стал оказывать заметное влияние на
динамику численности кайры. Активная заготовительная деятельность велась во
многих губах Новой Земли, в том числе и в губе Безымянной. Не прекращалась
она даже в годы Великой Отечественной войны. О ее масштабах можно судить
по результатам экспедиции под руководством Л.О.Белопольского на Новую
Землю в 1942 г. Ее участниками (всего 50 человек) на птичьих базарах Новой
Земли с середины июня по середину августа было собрано 0.5 млн яиц кайр.
Кроме того, содержимым уже насиженных яиц кайр было заполнено 25 бочек
(оно использовалось как корм для зверей на зверофермах). За тот же срок
экспедицией было заготовлено 14 тыо. взрослых кайр (Белопольский, личное
сообщение). Именно активной эксплуатацией колоний мы объясняем снижение
численности кайр в 30-40-х годах. В пользу этого же предположения
свидетельствует и то, что во время существования в губе Безымянной заповедника
(1947-1951 гг.) численность кайр была относительно стабильной (рис. 52).
Информацией о последующем десятилетии мы не располагаем. Каким образом и в
Таблица 24
Численность толстоклювых кайр в губе. Безымянной (Новая Земля) в разные периоды времени (в особях)
Год
1933
1942
1947
Северные базары
Количество
116557
100000*
10В950
птиц
Источник информации
Красовский С.К. (1937)
Белопольский Л.О.
(по: Успенский, 1956)
Белопольский Л.0..
Боголюбова Т. В.
(архив заповедника)
Южные базары
Количество
1510946
500000*
235400
ПТИЦ
Источник информации
Красовский С.К. (1937)
Белопольский Л.О.
(по: Успенский. 1956)
Белопольский Л.0..
Боголюбова Т. В.
(архив заповедника)
Всего
Количество
1644503
600000*
344350
в губе Безымянной
птиц
Источник информации
Красовский С.К. (1937)
Белопольский Л.0.
(по: Успенский, 1956)
Белопольский Л.0.,
Боголюбова Т. В.
(архив заповедника)
30313
26113
49675
85000
10670
89300 Успенский СМ, • 124713 Успенский СМ.
(архив заповедника) (архив заповедника)
80750 Успенский СМ. (карто- 111967 Успенский СМ.
(картосхемы, архив заповедника) схемы, архив заповедника)
105375 Успенский СМ. 160550 Успенский СМ.
(архив заповедника) (архив заповедника)
120000 Белопольский Л.О. 210500 Белопольский Л.О.
(архив заповедника) (архив заповедника)
Рассчитано по Сушко В. А.
(архив заповедника)
70000** Успенский СМ., 81000** Рассчитано по нашим
данным и Успенскому,
Хахину (1993)
ПРИМЕЧАНИЕ. В таблице 'указана численность птиц, непосредственно учтенных в колониях, без применения расчетных
коэффициентов. Не приводятся сведения по о-ву Кутоаому. они включены в итоговые данные по губе Безымянной.
'Данные Белопольского Л.О. за 1942 г. приводятся по Успенскому СМ. (1956). В годовом отчете за 1947 г. Белопольский Л.О.
(архив заповедника) оценивает численность кайр в губах Грибовой и Безымянной а 2400000 экз.
**В таблице приведены данные Успенского СМ. и Хахина Г.В.. (1993), уменьшенные нами вдвое, так как они использовали
методику, по которой количество птиц, учтенное в колониях, увеличивалось минимум в два раза.
Успенский СМ.
(архив заповедника)
Успенский СМ.
(картосхемы, архив заповедника)
Успенский СМ.
(архив заповедника)
Белопольский Л.О.
(архив заповедника)
Наши данные
Успенский СМ.,
Хахин Г.В. (1993)
200000
какой степени испытания ядерного оружия отразились на существовании
морских птиц, осталось неизвестным.
%
1933 1942 1947 1948 1949 1930 1931 1992
Рис. 52. Динамика численности толстоклювых кайр
в губе Безымянной, Новая Земля
Года
В 70-80-е годы на динамику численности кайр оказывало сильное влияние
состояние кормовой базы. Известно, что основу питания толстоклювых кайр на
Новой Земле составляли треска и сайка, причем кайры добывали их почти с
одинаковой частотой (Белопольский, 1971). В середине 70-х годов из-за
неумеренного промысла новоземельское стадо сайки было подорвано (Боркин,
см.раздел 6.3). Это должно было заметно сказаться на эффективности
размножения кайр в колониях архипелага. Кроме того, в конце 80-х годов в
Баренцевом море были подорваны и запасы мойвы. В результате резко ухудшились
трофические условия кайр в период зимовки. Зимой 1986/1987 гг. от
бескормицы погибло огромное количество кайр обоих видов, гнездившихся в
колониях Мурмана, Финмаркена и Медвежьих островов. После чего на Мурмане летом
1987 г. было учтено всего 12-16SS тонкоклювых и 56Х толстоклювых кайр от
уровня 1986 г. (Краснов, 1991). Районы зимовки толстоклювых кайр из
колоний Мурмана и Новой Земли в значительной степени совпадают. Вполне
вероятно, что зимой 1986/1987 гг. погибло также большое количество кайр из
колоний Новой Земли.
В настоящее время состояние кормовой базы морских птиц, по-видимому,
относительно стабильно. Хотя в отдельные сезоны они, вероятно, испытывают
затруднения с добычей корма. Так. в июне 1992 г.. рыба была обнаружена
нами лишь в 29Х «елудков толстоклювых кайр, Поданным »е Л.О.Белопольского
(1971), в 40-е годы она была встречена в 81,55S случаев. В июне 1992 г.
видовой состав рыб был очень разнообразен и характеризовался небольшим
преимуществом тресковых. Треска была найдена в 3.2Х (1 случай), а тресковые
(из-за сильного разрушения отолитов подробнее не определенные) - в 12.956
случаев. Такие рыбы, как мойва, песчанка, бычок и липарис, были встречены
по одному разу. Кроме того, в одном случае пищевой комок кайры целиком
состоял из мелких ракообразных. Столь большое разнообразие пищи у неболь-
шого количества особей (9 экз.) свидетельствует о том, что в этот период в
прибрежной зоне крупного источника какого-либо массового корма не было,
Правда, в губе Безымянной непосредственно у самых птичьих базаров мы
наблюдали кормежку моевок и бургомистров на скоплении нерестящейся мойвы. Но,
судя по количеству собравшихся птиц, оно было небольшим. С борта оудна нам
удалось определить расстояние, на которое в этот период времени улетали
кайры в поисках корма. Как оказалось, основная масса птиц искала пищу не
далее 20 км от берегов губы Безымянной, и только отдельные кайры улетали в
поисках корма на большее расстояние (до 35-40 км). После окончания периода
размножения, с середины августа, кайры из.кодюний Новой Земли уходят в
центральные районы Баренцева моря. Во всяком случае при учете морских птиц
в 1993 г. от кромки льдов до берегов Мурмана, скопления толстоклювых кайр
были обнаружены нами 31 августа в районе с координатами 77°00' с.ш.,
43*00* в.д. и 73°26' с.ш., 44°38' в.д. Выводки кайр мы наблюдали в двух
участках маршрута: между 77 и 76° и в районе с координатами
73°30* с.ш., 43°18' в.д. - 73°28' с.ш., 43°13' в.д. Еонцентрация кайр в
этом районе объясняется, по-видимому, наличием здесь скоплений косяков
мойвы. Последняя к концу лета достигает оеверных участков своего ареала и
какое-то время распределена здесь стабильно (Боркин и др., 1992).
О зимнем размещении новоземельских кайр мы можем.оудить по встречам
окольцованных птиц. В нашем распоряжении имеется 15 возвратов. Сообщения о
декабрьских встречах окольцованных в губе Безымянной кайрах пришли из
районов, расположенных вдоль всего побережья Мурмана, от Горла Белого моря
до фиордов Норвегии. Какая-то часть новоземельских толстоклювых кайр
проникает и в Белое море. Так, 18 декабря 1955 г. толстоклювая кайра была
найдена на берегу. Колвицкого озера (Кандалакшский залив). У побережья
Гренландии новоземельские кайры были встречены в декабре и феврале (3
сообщения) . Мигрируют они, вероятно, по тем же путям, что и птицы из колоний
Мурмана, хотя сроки миграций, возможно, различаются. Во всяком случае,
новоземельские кайры были встречены у берегов Норвегии в марте (1
сообщение), а у берегов Западного Мурмана - в апреле и мае (по одному
сообщению).
Что касается второго массового вида - моевки» то ситуация с ним
несколько более сложная и запутанная (табл. 25). Безусловно, что в 1923 г.
моевки в губе Безымянной еще не гнездились (Самойлович, 1929). Не вызывают
сомнения и учеты СЕ.Красовского (1937). впервые зарегистрировавшего в
данном районе гнездовые колонии этого вида. Результаты же дальнейших
наблюдений могут трактоваться неоднозначно. Столь большие колебания
численности в 1947-1950 гг. могут быть 'объяснены недостоверными учетами, и в то
же время они вполне могут быть следствием естественных процессов в
популяциях моевки. Тем более, что некоторые архивные и опубликованные материалы
полностью противоречат друг другу. Так, в годовом отчете заповедника за
1950 г. (архив заповедника), Л.О.Белополыжий, ссылаясь на С.М.Успенского,
утверждает, что учет моевки в данном оезоне не проводился, и приводит чисто
глазомерные впечатления последнего. Но позднее С.М.Успенский (1956)
публикует количественные данные учета гнезд моевок, полученные в 1950 г.
Оценивая общуи численность моевок на Новой Земле в 15-20 тыс. пар, он приходит
к выводу, что колония в губе Безымянной наиболее крупная (в 1950 г. -
3830 пар). При этом С.М.Успенский (1956) совершенно игнорирует данные
Л. 0. Белопольского и Т.В.Боголюбовой за 1947 г., что, возможно, свидетель-
Количество гнезд моевок в губе Безьмянной (Новая Земля) в разные периоды времени
Таблица 25
Год
1923
1933
1947
1946
1949
1950
1992
Северные базары
Количество гнезд
0
?
9500
2085
1560
1020
4566
Источник информации
Успенский СМ.
Южные базары
Количество гнезд
0
9
12800
5550
(архив заповедника)
Успенский СИ.
6350
(архив заповедника)
Успенский, 1956
Наш данные
2810
6500
Источник информации
.-
Успенский СМ.
(архив заповедника)
Успенский СМ.
(архив заповедника)
Успенский, 1956
Успенский, Хахин.
1993
Всего в губе Безьмянной
Количество гнезд Источник информации
0
1800
22300
7635
7910
3830
11066
Самойлович Р.Л.
(1929)
Красовский С.К.
(1937)
Белопольский Л.0.
Боголюбова Т.В.
(архив заповедника)
Успенский СМ.
(архив заповедника)
Успенский СМ.
(архив заповедника)
Успенский. 1956
Рассчитано по нашим
данным и
Успенскому, Хахину. 1993
ствует о его недоверии к полученным ими результатам, поскольку эти авторы
в 1947 г. учли в губе Безымянной 22300 пар моевок, т.е.больше, чем по
оценке С.М.Успенского, их гнездилось во всех колониях Новой Земли. Но
столь высокая численность была зарегистрирована через 15 лет после
последнего учета в этом районе, выполненного С. К. Красовским, поэтому она вполне
может объясняться естественным увеличением количества гнездящихся птиц.
Лучшим тому подтверждением является детально отслеженный рост популявди
моевок в колониях Семи островов. В период с 1962 по 1976 гг. количество
гнездящихся здесь птиц возросло с 2270 до 23800 пар. К тому же для этого
вида характерны периодические резкие сокращения численности гнездящихся
птиц. Их также неоднократно наблюдали на Семи островах (Краснов, 1991).
Объясняются они соответствующими изменениями трофических условий.
На Новой Земле подобной связи отмечено не было. Судя по данным,
приведенным С.М.Успенским (1956). и материалам архива заповедника, птицы в
период гнездования не испытывали затруднений с кормом, а состояние
трофической базы в период зимовки оставалось неустановленным. Ситуация с
современной численностью моевок нам представляется более простой. Как уже
упоминалось выше, за точку отсчета в своих исследованиях нами был выбран учет
1948 г. как наиболее подробно описанный и детально закартированный. Учет,
проведенный нами в июне 1992 г. на Северных Безымянных базарах, показал
увеличение численности гнездящихся моевок примерно в 2.2 раза. Как видно
из табл. 25. рост численности моевок на Южных Безымянных базарах, по данным
С.М.Успенского и Г.В.Хахина (1993). был несколько меньшим. В материалах
учета по Южным Безымянным базарам за 1948 г. (архивы заповедника)
существует некоторое расхождение между первичными и итоговыми данными. На
карту-схему Южных базаров нанесено 5550 гнезд вместо 4500, указанных в
окончательных итогах и позднее приведенных в публикации С.М.Успенского и
Г.В.Хахина (1993), поэтому численность моевок в 1992 г. была лишь относи-
. тельно большей, чем в 1948 г., причем тысячная разница распределена на
11 км маршрута. Следует, видимо, признать, что такая величина различий
находится в пределах допустимой ошибки при учетах на таком протяжении и
сложном маршруте. Кроме того, по данным С.М.Успенского (архив
заповедника), в 1949 г. на Южных базарах гнездилось 6350 пар моевок. Таким образом,
если исходить из данных по Южным Безымянным базарам, необходимо сделать
вывод, что действительного увеличения гнездовий моевок в губе Безымянной
не произошло. Тенденция увеличения численности может быть выявлена лишь
при использовании материалов учета по Северным Безымянным базарам. Если же
признать численность моевки в губе Безымянной, зарегистрированную Л.О.Бе-
лопольским и Т. В.Боголюбовой, реальной. • то итоги учета в 1992 г. будут
свидетельствовать лишь о частичном восстановлении ее гнездовий.
Мы полагаем, тем не менее, что. несмотря на все колебания численности
моевок в колониях Новой Земли, существовала общая тенденция роста ее
гнездовий в течение всего периода наблюдений. Первый ее этап наблюдали в
начале 30-х годов. Возможно, он продолжался до середины 40-х годов. Второй
этап увеличения ее численности должен был наблюдаться в 70-80-е годы, так
же как и во всех колониях Мурмана (Краснов. 1991) и Финмаркена (Furnesa.
Barrett.. 1985). Объясняется он единой для всего региона причиной - ростом
общих, запасов мойвы, который наблюдался в Баренцевом море вплоть до 1975
года (Vader et al., 1989). Попытки не увязать рост колоний моевки с
освобождением "жилплощади" после сокращения гнездовий кайр (Успенский. Хахин.
1993) нам представляются неубедительными. Они основываются на взглядах
Л.О.Белопольского (1957), полагавшего, что в условиях птичьих базаров рост
численности моевок сдерживается нехваткой мест, пригодных для гнездовия. и
которые, как правило, заняты кайрами. В настоящее время эти взгляды
опровергнуты 40-летней историей развития колоний Мурмана, где в 60-80-е годы
наблюдался одновременный рост как количества кайр, так и количества
моевок. Число каждого вида возросло в несколько раз, что свидетельствует о
существовании значительного резерва мест, пригодных для гнездования этих
видов (Краснов, 1991).
Таким образом, развитие колоний толстоклювых кайр и моевок в
условиях, по крайней мере. Южного острова Новой Земли, как и в других частях
региона (Мурман, Финмаркен), определялось действием, по-видимому, двух
факторов. На первом этапе, в 20-40-е годы, определяющей была эксплуатация
колоний этих видов и добыча взрослых особей, а в последующем - состояние их
кормовой базы.
Что же касается современного состояния гнездовий других видов морских
птиц, то судить о них можно лишь приблизительно. Видимо, существенно
увеличилась численность тупиков. Так, на о-ве Шестакова (губа Грибоаая) в
августе 1993 г. А.Б.Савинецкий (личное сообщение) насчитал около 70 пар
тупиков. В годы существования здесь заповедника (1948-1950 гг.) она
колебалась в пределах 22-25 пар. На о-ве Крестоватике (губа Грибовая) количество
пар этого вида возросло с 3 в 1940 г. до 7 в 1993 г. В то же время на
Северных базарах губы Безымянной в июне 1992 г. мы обнаружили лишь 5 пар
тупиков (в 1948 г. - 22 пары). Вполне вероятно, что в этом случав мы могли
допустить значительный недоучет, так. как их гнезда располагались в
труднодоступных осыпях и расщелинах песчаника.
Из чайковых птиц, видимо, уменьшилось количество бургомистров,
гнездящихся в губе Безымянной. В июне 1992 г. этот вид крупных чаек гнездился
в районе птичьих базаров губы Безымянной с плотностью 1.8 пары на 1 км
береговой полосы, а в 1950 г.. по данным С.М.Успенского (1956). с плотностью
4.4 пары.
По всей видимости, численность гнездившихся обыкновенных гаг также
была ниже обычного для конца 40-х годов уровня, на о-ве Крестоватике она
изменялась от 243 в 1947 г. до 330 гнезд в 1949 г. (архив заповедника). Но
в 1993 г. А. Б.Савинецкий оценил количество гнездящихся здесь уток в
100-120 экземпляров (личное сообщение). Подобные изменения могут
объясняться не только воздействием антропогенного фактора (в частности, высоким
уровнем беспокойства в губе Грибовой), но и естественными колебаниями
численности, характерными для обыкновенных гаг и в других местах ареала в
Баренцевом море.
В то же время необходимо отметить, что в гнездовой авифауне мороких
птиц Новой Земли появились два новых вида. В июне 1992 г. в губе
Безымянной мы обнаружили гнездо большого поморника {Catharacta skua). В настоящий
момент это первые сведения о размножении больших поморников на архипелаге.
В последнее время этот вид активно колонизирует острова южного побережья
Баренцева моря. В России он был найден на гнездовании на Семи островах в
1988 г. (Краснов, 1990). и с тех пор количество размножающихся пар
постепенно растет. В связи с этим находка размножающихся больших поморников на
Новой Земле была вполне ожидаемой.
Другой вид - морская чайка (Larus imrtnus). В 1993 г. взрослых птиц
этого вида наблюдал в губе Грибовой А. Б. Савинецкий (личное сообщение).
Здесь же. на о-ве Крестоватике. им были найдены оперенные, но еще не
летающие птенцы морской чайки, вокруг которых беспокоились две пары взрослых
птиц этого вида. Рядом, на о-ве Топориков, он также отметал две пары
морских чаек.
Таким образом, за период, истекший с конца регулярных
орнитологических наблюдений (1951 г.), в авифауне морских птиц Новой Земли произошли
вполне очевидные изменения. По всей видимости, с , проведением здесь в
50-60-х годах испытаний ядерного оружия, они не связаны. Подобные
изменения в колониях морских птиц в этот период имели место на побережье.Мурмана
и Финмаркена и определялись единым для всего региона фактором -
трансформацией кормовой базы под воздействием интенсивного индустриального
промысла рыбы.
Глава 8. ЮРСКИЕ МЛЕКОПИТАЮЩИЕ
8.1. Китообразные
Вследствие большой протяженности островов Новая Земля о юга на север
видовой состав китообразных в прибрежных водах архипелага сильно
различается. Для северных районов характерны холоднолюбивые виды, в прибрежье Ш-
ного острова встречаются китообразные, свойственные умеренным водам. При
этом большое значение имеет гидрологический и температурный ренимы
конкретного года. Так, если летние температуры оказываются выше средних,
появляются не свойственные прибрежью китообразные, мигрирующие с Гольфстримом:
отмечены .подходы к берегам Новой Земли беломордых и белобоких дельфинов,
атлантической афалины и ряда других представителей умеренных вод
Атлантики.
В прибрежных водах архипелага Новая Земля встречаются представители
пяти семейств китообразных: DelpMntdae, Monodonttdae. ZiphUdae, Salae-
nopter-tdaen Balaenidae (табл. 26).
Таблица 26
Видовой состав и размерные характеристики китообразных,
встречающихся у берегов архипелага Новая Земля
(дана максимальная длина тела, в скобках - средние величины).
Семейство
Род
вид
1
Длина тела,
м
2
Размеры
новорожденных,
см
3
Семейство настоящих дельфинов (Delphlnldae)
Род афалин (Turslops)
Афалина (Т. truncatus) 3.3 (2.6)
Род короткоголовых дельфинов (Lagenorhynchua)
Беломордый дельфин (L.alblrostrls) 3.5 (3.0)
Белобокий дельфин (L.acutus) 3.0 (2.7)
Род косаток (Orclnus)
Косатка (О.огса) 10.0 (6.5)
Род гринд (Globicephala)
Обыкновенная гринда (G.melaena) 6.0 (4.0)
Род морских свиней (Phocoena)
Обыкновенная морская свинья (P.phocoena) 2.0 (1.6)
110
.1.10
100
260
175
75
Окончание таблицы 26
Семейство нарвалов (Monodontldae)
Род белух (Delphinapterua)
Белуха (D. leucas) 6.0 (3.9) 165
Род нарвалов (Monodon)
Нарвал (M.monoceros) 6.0 .,(5.0) 150
Семейство клюворылов (Ziphiidae)
Род бутылконосов (Hyperoodon) .
Высоколобый бутылконос (Н.ampuleatus) 9.0 (8.0) 250
Семейство полосатиков (Balaenopterldae)
Род настоящих полосатиков (Balaenoptera)
Финвал (B.physalua)
Гренландский кит (B.myatlcetus) .
Сейвал (В.borealla)
Малый полосатик (B.acufcorostrata)
Семейство гладких китов (Balaenldae)
Род южных гладких китов (Eubalaena)"
Южный гладкий кит (Е. glacial is) 20.0 (.15.0) 500
27.8 (18.8)
21.0 (17.0)
18.0 (14.0)
10.0 (8.5)
600
450
450
300
Из приведенного списка китообразных постоянными (или сезонными)
обитателями прибрежных вод, омывающих Новую Землю, можно считать белуху,
косатку, обыкновенную морскую свинью, финвала, гренландского кита и нарвала.
Появление у берегов Новой Земли животных других видов носит эпизодический
характер.
Афалина. В зоне действия Гольфстрима атлантическая афалина
заходит в Баренцево море, где может достигать Новой Земли. Дельфины этого
вида предпочитают прибрежные воды с относительно небольшими глубинами.
Афалины - стадные животные, но больших скоплений не образуют. Обычно
группы дельфинов состоят из десятка или нескольких десятков голов. Скорость
передвижения животных в среднем составляет 7-8 миль в час, максимальная -
15-18 узлов (Земский, 1980).
Обитая на малых глубинах, афалины питаются преимущественно бентосными
животными и придонными рыбами (камбала, пикша). Пелагические рыбы,
ракообразные и моллюски играют второстепенную роль в спектре питания этих
дельфинов.
Беломордый делвфин. Вид занесен в Красную книгу СССР.
В Баренцевом море беломордые дельфины встречаются у побережья Кольского
полуострова, могут заходить в прибрежные воды Южного острова архипелага
Новая Земля.
Обычно животные держатся мелкими группами или парами, но иногда
образуют крупные скопления в тысячи голов.
Беломордый дельфин питается преимущественно донными и придонными
рыбами. В желудках дельфинов находили навагу, мойву, сельдь, треску, а также
некоторых моллюсков, в том числе головоногих (Томилин, 1962).
Численность беломордого дельфина не установлена.
Белобокий дельфин. Дельфины этого вида охраняются с
1976 года, а сам вид занесен в Красную книгу СССР.
Белобокие дельфины встречаются главным образом в. юго-западной части
Баренцева моря, но иногда доходят до Воронки Белого моря и прибрежных вод
архипелага Новая Земля.
Животные держатся обычно небольшими группами (до 50 голов), в районах
скопления могут образовывать стада численностью до 1.5 тысяч животных.
Белобокие дельфины питаются главным образом пелагическими стайными
рыбами (сельдь, треска, навага, мойва). Кроме того, в спектр питания
входят брюхоногие и головоногие моллюски, ракообразные.
миграции и численность белобокого дельфина в арктических водах не
установлены.
Косатка. Подходы косатки к берегам Новой Земли отмечены
главным образом в летний период. Косатка является самым крупным
представителем семейства настоящих дельфинов с ясно выраженным половым диморфизмом (у
самцов спинной плавник намного больших размеров, чем у самок, и может
достигать 150 см в высоту). Косатки ведут стайный образ жизни как в открытом
море, так и у берегов. Обычно они.передвигаются небольшими группами, могут
заходить в крупные реки. В период пищевых кочевок косатки движутся со
скоростью 6-7 миль в час. однако могут развивать скорость до 17 узлов
(Виноградов, 1949).
Косатка - единственный вид китообразных, который можно отнести к сак-.
рофагам, так как наряду с другими объектами питания она поедает
теплокровных животных. В желудках косаток находили остатки тюленей, моржей,
дельфинов, морских птиц и даже крупных китов (малых полосатиков, сейвалов).
Основное же питание косаток составляют рыбы (лососевые, сельдь, треска,
мойва, палтусы и скаты) и головоногие моллюски (преимущественно кальмары).
Одной из поведенческих пищевых особенностей косаток является способность
организовывать стаи для охоты на дельфинов, тюленей и рыб (Смирнов. 1935).
В целом, косатка - малоизученный вид китообразных: не определены
пути миграции этих животных в северо-восточной Атлантике-, неизвестен
возраст, при котором происходит половое созревание, различаются данные о
сроках беременности, не определена даже приблизительно численность косаток,
хотя и считают этот вид одним из самых многочисленных в Арктике (Земский,
1980).
Г р и н д а. Атлантическая, или обыкновенная, гринда встречается в
водах Баренцева моря преимущественно в его юго-западной части. Однако в
теплые годы так же.как и афалина, может достигать берегов Новой Земли.
Гринды являются стадными животными, обычно держатся небольшими
группами, но в местах концентрации пищи образуют косяки в тысячи голов.
Нередко гринды встречаются в группах с другими видами дельфинов - афалинами,
короткоголовыми дельфинами и T.nv
Объектами питания гриндам служат почти исключительно несколько видов
крупных кальмаров из числа образующих скопления. При отсутствии кальмаров
гринда в небольших количествах поедает рыб (сельдь, треска, камбала).
Полагают (Томилин. 1962). что у гринд весь период воспроизводства
занимает 40 месяцев, поэтому темп пополнения стада очень медленный.
Гринда может быть отнесена к видам промысловой численности, так как
во многих местах ареала наблюдаются косяки гринды в 1000 и более голов.
Морская свинья. В Баренцевом море, у берегов Новой Земли
и в западной части Карского моря встречаются преимущественно представители
подвида североатлантической обыкновенной свиньи. В северной и восточной
частях ареала эти дельфины проводят обычно летние месяцы, покидая на зиму
холодные воды.
Морские свиньи являются стадными животными и обитают главным образом
в прибрежных мелководных районах. Численность стад может достигать
нескольких тысяч голов (Дубровский. 1938: Гептнер и др., 1976).
Спектр питания морских свиней представлен различными видами рыб
(сайка, треска, пикша, мойва, навага, песчанка, мелкие лососевые),
ракообразными и головоногими моллюсками (Чапский. 1941).
Белуха. Среди китообразных по количественному составу в водах,
омывающих Новую Землю, преобладает белуха. Животных этого вида можно
наблюдать практически в любом районе прибрежных вод как Северного, так и
Южного островов архипелага.
У берегов Южного острова белухи особенно многочисленны зимой. С
мая-июня начинается миграция этих дельфинов к Северному острову вдоль
западного и через Карские ворота и Югорский Шар восточного прибрежья
архипелага. Во время подхода сайки на нерест к островам Вайгач и Новая Земля
здесь наблюдаются большие концентрации зимующих белух (Буторин, 1964,
1966). • '
Белуха - стадное животное, возрастной и половой состав стад может
быть различным. В поисках пищи белухи обычно не заныривают глубже 8-10 м.
В то же время в экспериментах с обученными животными была показана
способность белух достигать глубин свыше 600 м. В среднем стадо животных
движется со скоростью 2-5 узлов. Напуганные дельфины могут ее увеличивать до 10
узлов (Клейненберг и др., 1964). Белухи не избегают льдов и опресненных
районов. Отмечены случаи захода полярных дельфинов в губы и реки
архипелага.
Спектр питания белух . представлен преимущественно рыбами (сайка,
сельдь, мойва, навага, треска, пинагор и т.д.) и ракообразными. Имеется
сезонная и возрастная смена объектов питания.
Ориентировочная численность белух в Белом море составляла 8-Ю тыс.
особей, в Карском море 40-50 тыс. (Клумов. 1939).
Нарвал. Вид внесен в Красную книгу СССР и РСФСР. Нарвал обитает
в арктических водах вблизи кромки плавучих льдов. В водах архипелага эти
китообразные встречаются преимущественно у берегов Северного острова,
образуя группы до 20 особей (Громов и др., 1963). В других частях
северо-восточной Атлантики стада нарвала достигают численности до сотни голов.
Крупные косяки представлены животными всех возрастных групп, мелкие - или
самками с детенышами, или самцами.' В прошлом веке отмечались стада
численностью в несколько тысяч голов. _^
Биология нарвала, как и косатки, изучена недостаточно. Считают
(Земский, 1980), что самки рождают одного детеныша с перерывом в 2-3' года.
Объектами питания нарвалов являются головоногие моллюски, а также
рыбы (камбала, палтус, сельдь, тресковые, реже - лососевые).
Общая численность вида оценивается в 20-30 тыс. особей. В водах
России численность нарвала неизвестна (Соколов, 1990).
Высоколобый бутылконос - редкий вид,
занесенный в Красную книгу СССР.
В водах России высоколобый бутылконос встречается у западного
побережья Новой Земли, в прибрежье Кольского полуострова, иногда дельфины это-
го вида заходят в Белое море. Животные обитают преимущественно в
пелагических районах, но нередко их можно встретить у берегов материка и даже в
устьях крупных рек (Громов и др.. 1963).
Высоколобые бутылконосы образуют небольшие группы, численностью до 20
голов, представленные обычно разновозрастными животными и одним крупным
самцом. Иногда встречаются группы животных, состоящие из одних самцов. Для
высоколобых бутылконосов характерны сезонные миграции: весной эти киты
появляются в северной части ареала, осенью отходят к югу.
Основу корковой базы высоколобых бутылконосов составляют головоногие
моллюски. Второстепенными объектами питания являются некоторые рыбы
(треска, сельдь).
В период с 1946 г. по 1972 г. было отмечено снижение численности вы-
соколобого бутылконоса, что явилось, по-видимому, результатом его
перепромысла. По состоянию на начало 70-х годов численность высоколобого
бутылконоса в восточной Атлантике оценена в 6-9 тыс. особей (Соколов. 1990).
Ф и н в а л. Эти киты чаще встречаются в северо-восточной части
Баренцева моря, реже - у берегов Новой Земли и в Воронке Белого моря,
эпизодически наблюдаются в Карском море.
Финвалы совершают регулярные сезонные миграции. В летний период они
заходят в Баренцево море из вод. омывающих северо-западную Африку и
.Испанию. Некоторые киты могут зимовать и севернее, в районах влияния
Гольфстрима (Смирнов, 1935).
Питаются финвалы в основном ракообразными и рыбой.
В 70-х годах численность этих китов в северной Атлантике была оценена
в 20 тыс. голов (Земский, 1980).
Сейвал- вид, занесенный в Красную книгу СССР. Промысел сейвала
запрещен с 1980 г. Киты спорадически встречаются в открытой части
Баренцева и Белого морей одиночно или парами. Лишь в местах скопления пищевых
объектов (планктонных беспозвоночных, ракообразных, кальмаров, рыбы)
сейвалы сосредотачиваются по нескольку десятков особей. У Новой Земли эти
киты могут встречаться к западу от Южного острова.
Форма фонтана у сейвала - обычная для полосатиков, но высота конуса
не превышает 5 м (Смирнов, 1935).
Китобои считают сейвалов самыми быстроходными китами, хотя скорость
их до сих пор точно не определена (Виноградов, 1949).
Основные объекты питания сейвала - мелкие планктонные беспозвоночные
(Calanus /trunarchtcua и др.). Реже в желудках этих китов обнаруживали
крупных планктонных ракообразных и рыб (морской окунь, макрель, мойва,
минтай).
Общая численность сейвала невысока. До запрета на его промысел в
водах северной Атлантики добывали не более 200 голов. Столь значительное
сокращение северо-атлантического стада сейвала связывают с перепромыслом
вида в целом (Соколов. 1990).
Малый полосатик. Представители этого вида китообразных
являются самыми мелкими китами в семействе полосатиков. Малый полосатик
встречается в Баренцевом море практически на всей его акватории, иногда
заходит в Карское море. Пути и сроки миграций животных до сих пор
недостаточно изучены.
В отличие от других видов полосатиков, этот кит питается в основном
рыбой (сельдь, навага, треска, мойва, морской окунь), в меньшей степени -
ракообразными и головоногими моллюсками.
Форма фонтанов у малых полосатиков соответствует таковой у других
китов этого рода, но высота фонтанов не превышает двух метров. Под водой
киты находятся не более 10-15 мин, а питаясь, выныривают через каждые
3-4 мин.
Полагают (Громов и др., 19635. что половой зрелости малые полосатики
достигают уже в двухлетнем возрасте, при длине самцов 7.5 м, самок -
8.0 м.
Численность малых полосатиков не определена, однако считают, что этот
вид наиболее многочисленный среди усатых китов, обитающих в Арктике.
Норвежские китобои до 70-х годов добывали ежегодно 2-3 тыс. этих китов, в
период 1980-85 гг. - до 1.5 тыс. животных (Потелов, 1986).
Гренландский кит. До XVII века этот вид был весьма
многочисленным в Субарктике и арктической зоне Атлантики. Китобойный
промысел существенно снизил количество гренландских китов, поэтому в 1935
году был введен запрет на их промысел. В настоящее время гренландский кит
внесен в Красные книги МСОП и СССР.
В Баренцевом море гренландский кит встречается преимущественно в его
северной части, а также у побережья Новой Земли и острова Вайгач.
Одиночные животные или немногочисленные группы китов обычно обитают вблизи
ледовой прикромочной зоны или среди сплоченных дрейфующих льдов.
В водах архипелага Новая Земля встречаются главным образом особи
шпицбергенского стада (популяции), которые зимуют восточнее юга Гренландии
(Дубровский. 1938).
Питаясь только в верхних горизонтах моря, гренландский кит не
совершает глубоководных погружений (апноэ не превышает 5-10, редко -
15-20 мин) . При вентиляции легких между погружениями киты выпускают от 4
до 9 двуструйных фонтанов, высота которых не превышает 5-6 м.
Скорость передвижения гренландских китов в период кочевок не
превышает 4 миль в час, максимальная - до 9 узлов.
Основу кормовой базы у гренландского кита составляет планктон (Cala-
nus fimarchicus, Limactna hettctna).
Общая численность вида в 70-х годах оценивалась в одну тысячу голов
(Земский, 1980).
Южный гладкий кит. Для этого вида усатых китов
характерны сезонные миграции: весной животные перемещаются с мест размножения и
зимовки из теплых районов ареала в умеренные и холодные воды, осенью
возвращаются обратно.
В летний период южные киты держатся в большинстве случаев поодиночке,
реже парами. В местах концентрации пищевых объектов животные могут
образовывать небольшие скопления по нескольку десятков голов.
Фонтан у китов ясно выражен, имеет две струи высотой не более 5 м. Во
время питания южные гладкие киты закыривают не более, чем на 10 мин.
предпочитая верхние горизонты моря.
Основными объектами питания китам служат различные виды калянусов;
иногда в желудках животных находили моллюсков (Громов и др.. 1963).
'Точные данные о численности гладких южных китов отсутствуют, но
считают (Земский. 1980), что в результате интенсивного промысла в конце
прошлого и в начале текущего столетия численность животных катастрофически
сократилась. В настоящее время добыча южных гладких китов запрещена во
всем Мировом океане.
Обсувдая вопросы хозяйственного значения китообразных, обитающих в
прибрешных водах архипелага Новая Земля, следует отметить, что до
последнего времени постоянным объектом промысла являлась лишь белуха. Лимиты
добычи этих китов в настоящее время составляют для Белого моря - 500, в
Баренцевом и Карском - 1500 голов в год. Однако из-за трудоемкости промысла
и небольшого количествам получаемого продукта, колхозы европейского Севера
проводили последний забой белух в 1990 г.
В связи с развитием российско-норвежских экономических отношений
рассматривается вопрос о возможном совместном промысле малого полосатика
(Научно-практическая конференция "Развитие прибрежного промысла и аква-
культуры в Баренцевом море", 15-16 декабря 1993 г., Мурманск).
8.2. Ластоногие
Ластоногие играют важную роль в функционировании экосистем прибрежья
Новой Земли. Оценить их реальное значение в настоящее время не
представляется возможным, так как имеющаяся современная информация о состоянии
популяций тюленей и моржей, их биологии в районе весьма скудна. Одним из
основных источников данных о морских млекопитающих архипелага были поморы,
промысловики. Но с сокращением или прекращением промысла получение
необходимой информации сократилось практически полностью. Дело было осложнено и
тем, что Новая Земля, как ядерный полигон находилась продолжительное
время за завесой секретности.
Поморский промысел на Новой Земле начал свою историю в XVI веке.
Первая российская научная экспедиция на архипелаге была проведена в
1768-69 гг. под руководством Ф. Розмыслова (Чапский, 1939). В этой и
большинстве последующих экспедиций исследования ластоногих проводились попутно
с другими работами в очень ограниченных объемах, достигнув апогея в период
активного промысла в довоенное время. Они были прекращены после Второй
мировой войны, когда архипелаг стал военным объектом. Поэтому имеющиеся
данные по биологии ластоногих этого района очень неполные, основаны в
большинстве своем на довоенных, и даже дореволюционных, источниках.
Уровень изученности биологии большинства арктических видов ластоногих
крайне низок. В настоящее время в некоторой степени изучаются лишь
эксплуатируемые человеком виды (гренландский тюлень; морской заяц и нерпа в
меньшей степени).
Некоторую информацию нам удалось собрать во время 62-го, 67-го, 68-го
и 75-го рейсов НИС "Дальние Зеленцы", которые проводились ММБИ в
июле-сентябре 1992, 1993 ГГ. (табл. 27).
Для западного района Российской Арктики и особенно Баренцевого моря
характерны молодые и тонкие однолетние льды, ледяной покров тут разделен
многочисленными каналами и трещинами. Кроме того, у Новой Земли припай
всторощен, а с продвижением на восток снежный покров на льдах
увеличивается (Беликов и др.. 19В9). Акватория архипелага в зимнее время покрывается
льдами. ' Ледовый режим оказывает большое влияние на распределение и образ
жизни ластоногих.
Берег Баренцева моря сильно изрезан, многочисленные губы имеют защи-
щенные от волновой деятельности места. У берегов часто встречается
значительная глубина. В губах и заливах лед образуется сравнительно поздно.
Таблица 27
Ластоногие, отмеченные у Новой Земли во время рейсов
НИС "Дальние Зеленцы" в 1992-93 гг.
Дата
17.08.91
18.08.91
18.08.91
18.08.91
16.08.92
18. 08. 92
20. 08. 92
21.08.92
21.08.92
21.08.92'
21.09.93
21.09.93
17.08.91
17.08.91
18.07.92
30. 08. 91
18.07.92
Координаты
73°51'
75 21
76 14
76 21
70 06
75 35
76 30
77 12
77 16
77 08
70 30
70 23
74 24
75 04
70 23
70 24
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
с.ш.
50" 08*
57 38
56 44
56 39
40 95
58 05
65 12
62 37
62 25
62.50
58 15
58 И
Количество
Морской
в. д.
в. д..
в. д.
в. д.
в. д.
в. д.
в. д.
в. д.
в. д.
в. д.
в'.д.
в. д.
Гренландский
52 06
54 30
55 27
55 23
в. д.
в. д.
в. д.
Кольчатая
в. д.
заяц .
2
1
тюлень
5
1
1
нерпа
1
1
' Примечание
С рыбой во рту
Зал. Норденшельда
Неперелинявший
Неперелинявший, гл. 75 м
Зал. Вилькицкого
Зал. Иностранцева
Неперелинявший
Неперелинявший
Глубина 100 м
Карские Ворота, гл. 85 м
Карские Ворота
До берега 8 миль
Карские Ворота
Зал. Глазова, гл. 34 м
Карские Ворота
Побережье же Карского моря имеет гораздо меньше защищенных мест.
Ледовый покров на этом участке держится значительно дольше, чем в первом, а
в отдельные годы не сходит оовсем (Антипин, 1938а.б).
В акватории архипелага Новая Земля обитают 4 вида ластоногих: морж,
кольчатая нерпа, морской заяц, гренландский тюлень. Обитание здесь серого,
обыкновенного тюленей и хохлача не доказано, но не исключены заходы этих
видов ластоногих.
В 1840 г. К.Е.Бэр в материалах экспедиции Литке, собранных на Новой
Земле, обнаружил череп пятнистого тюленя с западного побережья Новой
Земли, что может свидетельствовать о дальних миграциях этих животных
(Смирнов. 1903).
Имеются данные о наблюдениях серого тюленя вблизи архипелага: 12 и
14 августа 1976 г. наблюдали 3 тюленей в Карском море у поселка Амдерма
(Вишневская и др.. 1990; Соколов и др.. 1990). А в августе 1988 г. у
западного побережья о-ва Вайгач отмечены молодые тюлени этого вида (Калякин.
Мужчинкин. 1990).
В работе А.В.Яблокова и В.Н.Минеева (197В) указывается на
существование залежки тевяков на полуострове Гусиная Земля (Новая Земля), но эта ин-
формация, к сожалению, не конкретизируется. •
Моржи, обитающие на Новой Земле, относятся к номинальному подвиду -
атлантический морж (Odobenus. г. rosmrus). По сравнению с двумя другими
подвидами - лаптевским и тихоокеанским, это наиболее мелкая форма. Средняя
длина тела самца 345 см, самки - 293 см, максимальная соответственно 375 и
338 см. В Карском море первогодки достигают 133 см, годовалые - 181 см,
двухлетки - 218 ом (Чапский. 1936). Все размеры приведены по
горизонтальным промерам (Lev).
Численность атлантического моржа была значительно сокращена в
результате длительного бесконтрольного промысла. В настоящее время промысел
запрещен. Подвид занесен в Красные книги МСОП и (с 1978 г.) - СССР (II
категория). Биология и поведение этего подвида наименее изучены; вероятно, что
он в настоящее время самый малочисленный.
Об относительно большой численности моржей у побережья архипелага в
XIX в. можно судить по тому факту,. что каждой из 15 лодок, промышляющих в
этом районе в 1857 г., было добыто до 50 моржей (Чапский, 1939).
Во время экспедиции В.Баренца у Новой Земли моржи были отмечены у
о-ва Берха, мыса Нассау, п-ва Адмиралтейства и на Оранских островах.
Участники экспедиции Де-Фера в июне 1597 г. у о-ва Нассау наблюдали массу
этих ластоногих, залегающих на льдах. В конце XVIII в. И.Лепехин отмечал,
что на севере Новой Земли моржей больше, чем каких-либо животных (Чапский,
1939).
В заливе Норденшельда в августе 1991 г. нами было найдено 3 старых
черепа моржей с выбитыми клыками.
В 20-х годах нашего столетия у Новой Земли моржи стали большой
редкостью. У многочисленных в то время становищ и полярных станций они
практически не появлялись. Исключением была лишь губа Черная (Горбунов, 1929).
Тут они появлялись ежегодно в ноябре, на островах в губе их собиралось
значительное количество. Автор полагал, что животные заходят в этот район,
отходя из Карских Ворот через Петуховский Шар, однако отмечал при этом,
что моржи до губы Белушьей не доходят.
В 1934 г. численность моржей в Баренцевом и Карском морях оценивалась
не более 1200 голов, а годовой прирост популяции не более 12%
(Чапский. 1939).
В первой половине нашего зека моржи регулярно отмечались у Карских
Ворот, в районе мыс Спорый Наволок - мыс Желания и на Оранских островах. К
началу 40-х годов их численность у Новой Земли оценивалась в 2-3 тыс.
особей (Чапский. 1941).
К середине 70-х годов на Новой Земле функционировало лишь одно
лежбище на о-ве гемскерк, хотя в 1952 г. их отмечали на Оранских островах.'на
о-вах Бритвин и Пуховом, на побережье заливов Течений и Анны, а также на
мысе Константин (Белобородое, Тимошенко, 1974).
В 1978 г. численность карской популяции оценивалась в несколько сотен
голов. Для стадного животного, каким является морж, такой популяционный
уровень очень низок (Бычков, 1976). В связи с запретом промысла,
введенного в 1956 г., состояние атлантического моржа заметно не улучшилось, однако
имеются некоторые свидетельства, позволяющие допустить улучшение состояния
популяции.
На протяжении ряда лет в конце 80-х годов наблюдалось лежбище моржей
на западном побережье о-ва Вайгач (Калякин, йужчинкин. 1990). В 1992 г. до
30 моржей залегали на берегу в Маточкином Шаре вблизи становища Лагерное
(Успенский, Хахин, 1993).
Миграции и распределение моржей в районе Новой Земли тесно связаны с
ледовой обстановкой и изучены очень плохо. Зимовку и весенний период моржи
проводят в юго-восточной части Баренцева моря, затем со льдами
продвигаются на север. Летом моржи отмечаются на льдах Карского моря, включая и
прибрежную зону северной части Новой Земли, также могут отмечаться на льдах
севернее архипелага. В июне севернее Новой Земли могут встречаться
мигрирующие особи (Арсеньев, 1976).
В летнее время, в основному августе-сентябре, при отсутствии льдов
моржи образуют береговые лежбища, на которых держатся до 1.5 месяцев.
Самки с детенышами ложатся ближе к воде.
По наблюдениям 1937 г., на о-ве Гемскерк моржи появились 2 июля, все
они отходили в северном направлении. Залежки, состоящие из отдельных
самцов, появляются с первых чисел августа. Самки прибывают позже. В сентябре,
с отходом льдов, моржи концентрируются на побережье северо-восточной части
архипелага, образуя береговые залежки в районе от Оранских о-вов до мыса
Спорый Наволок. Залежки были на о-ве Гемскерк и на побережье от мыса
Спорый Наволок до мыса Флиссингского. "Со второй половины сентября отдельные
группы из'самок и детенышей следуют на юг, самцы встречаются редко.
Последний раз в году моржей наблюдали в середине ноября на шое Девеер
(Антипин, 1938а).
В октябре моржи могут образовывать лежбища в районе Карских ворот,
проходя от северной части архипелага вдоль берегов на места зимовки.
Зимне-весенний сезон животные проводят в юго-западной части Баренцева
моря на льдах. Моржи предпочитают залегать на молодых льдах, которые имеют
разводья, каналы и трещины (Беликов и др., 1989). Во время весенней
подвижки льдов часть моржей переходит в Карское море через Карские Ворота и
огибает Новую Землю с севера. К концу лета они откочевывают на запад опять
же двумя путями. Во второй половине лета животные образуют многочисленные
береговые залежки на побережье островов.
В юго-восточной части Баренцева моря зимуют не более 100-120 моржей
(Лукин. 1978). Зимой они наблюдаются редко, например, в 1937 г. отмечены
8 февраля и 21 мая (район мыса Константин), двигавшиеся с юга на север
вдоль кромки льдов.
Вопрос о миграции моржей от северной части Новой Земли до Земли
Франца-Иосифа и обратно остается открытым ввиду отсутствия каких-либо данных.
Основу питания моржей составляют донные беспозвоночные: моллюски,
ракообразные, реже черви, иглокожие, рыба. Может хищничать, используя в пищу
тюленей, реже птиц. В.М.Антипин (193Ва) отмечает хищничество по лахтаку,
указывая на то. что моржи-хищники более крупные и хорошо упитанные,
Питаются моржи в основном на глубинах до 50 м. но могут заныривать гораздо
глубже - до 180 м. В Северной Атлантике, как и в других районах
распространения моржей, в летний период в рационе животных преобладают моллюски. В
желудках обычно встречаются черви и ракообразные, изредка - представители
других групп. Из рыб в желудках была найдена сайка. Н.А.Смирнов (1908)
указывает на случаи нападения и поедания тюленей моржами, при случае они
могут поедать трупы китов и других животных. Имеются данные о нападении
моржей на гренландских тюленей, поедании подстреленной гаги (собственные.
наблюдения) и кайры (личное сообщение К.В.Галактионова. Ю.В.Краснова).
Щенка, по всей видимости, проходит с конца апреля до конца мая на
льдах. В этот период моржи держатся семейными группами по 3-6 животных,
состоящих из самки, самца и детенышей разного возраста. .Н.А.Смирнов (1908)
отмечал спаривание моржей в районе Канино-Колгуевского мелководья в конце
мая - начале июня.
Лактационный период у самок моржей длится до двух лет и по этой
причине они рождают детенышей не чаще 1 раза в 3-4 года (Попов, 1960).
Самки со щенками были отмечены на о-ве Гемскёрк. где полностью на
берег не выходили, а обычно лежали в прибойной полосе. Спаривание не
наблюдали. Кормление сосунков происходило в воде, его продолжительность
составила 20-45 мин. Щенок может находиться под водой 5-7 мин (Антипин, 1938а).
Морской заяц (Erlgnathua barbatus) - обычный вид прибрежья
архипелага, встречается практически по всему побережью. Пагетод, биологически
связан с малыми, до 100 м. глубинами. Со льдами может встречаться над
большими глубинами. Местом обитания часто выбирает губы и острова с изрезанной
береговой линией, хорошо защищенные от штормов и нагромождения льдов. Бен-
тофаг. основу его питания составляют ракообразные и моллюски, а в
осенне-зимний период - сайка. В течение большей части года ведет одиночный
образ жизни.
Общая численность морских зайцев оценивалась в 8 тыс. особей
(Доронина и др., 1970). Долгое время этот вид, как и в настоящее время, был-
объектом промысла. Промысел сетями велся с середины сентября, зимой - отстрел-
на льду (Антипин, 1938а,б). В 1933 г. на 4 зверобойных судна, работавших в
северо-восточной части Карского моря, добыча составила всего 8 голов (Еси-
пов, 1935а). Можно предположить, что прекращение судового промысла в этом
районе в годы войны и послевоенное время способствовало увеличению
численности.
В летний период лахтак наиболее часто встречается в Карском море, в
южной и восточной частях Баренцева моря, в Мезенском заливе и Горле Белого
моря (Потелов. 1969).
Весной лахтаки продвигаются на северо-восток вдоль Новоземельского
побережья, вплоть до мыса Желания. Концентрируясь в полосе рассеянных
льдов, они оказываются довольно далеко от берега, часть животных
рассеивается по побережью. Во второй половине лета и осенью распределяются в
прибрежной полосе на мелководьях, к этому времени к берегу подходят и звери с
кромки дрейфующих льдов (Чапский, 1976). Так. 20 августа 1992 г. лахтак
был отмечен в зал. Иностранцева, на воде в его южной части.
Осенняя миграция на юг проходит по кромке льда вдоль побережья Новой
Земли. В районе мыса Константин мигрирует одиночками с севера на юг с
конца августа. В конце февраля мигрирует на север (Антипин, 1938а).
С ноября-декабря и в течение всего января лахтаки держатся на льдах
в прибрежных водах архипелага. В феврале животные концентрируются у
разводий, а со второй половины марта распределяются по репродуктивным областям.
Большие скопления морского зайца отмечались вблизи юго-западного
берега Южного о-ва Новой Земли, у пролива Маточкин Шар, п-ва Адмиралтейства,
мыса Желания (Чапский. 1976). Единичные экземпляры по побережью
встречаются круглый год. Зимой их численность меньше, чем осенью или весной.
Массовых залежек лахтаки не образуют. Весной отдельные особи лежат на небольших
льдинах или по кромке припая (Антипин,- 1938а,б). По наблюдениям в р-не
мыса Константин, зимой остаются в этом районе. С конца августа и в конце
февраля наблюдались мигрирующие одиночки (Антипин. 1938а).
Зимовка большей части новоземельского стада проходит на дрейфующих
льдах юго-восточной части Баренцева моря.
Морской заяц достигает максимальной численности в зимний период в
юго-западной части Баренцева моря (более 100 животных за 1 сутки
наблюдений), у мыса Желания (Потелов, 1969). У восточного побережья Новой Земли
их численность ниже (до 50 особей).
Период щенки - апрель-май. Основной район щенки - юго-восточная часть
Баренцева моря, по новоземельской окраине Баренцева моря, не севернее п-ва
Адмиралтейства. Небольшое количество самок щенится в западной части
Карского моря. В период щенки самки с детенышами .редко концентрируются.на
льдах на большом расстоянии друг от друга (Чапский. 1976). В районе мыса
Константин размножения не отмечено..В период гона издает "брачный крик'\
Интересно, что размеры новорожденного морского зайца составляют до
58.6% длины самки, в то время как у гренландского тюленя зтот показатель
достигает 51.1. а у моржа - 39%. Имеются сведения о наличии в приплоде
морских зайцев двойни (Чапский, 1938).
Линные залежки наблюдались с начала мая. Первыми залегают молодые
звери. В конце июня и в июле, когда морской лед уносит, заходят через
речки в пресные озера (оз. Реликтовое) и залегают здесь на льду. Позднее,
когда лед йа озере тает, лахтаки выходят.на берег и лежат на снегу или
льду ледника (Антипин, 1938а).
Период линьки значительно растянут - с середины апреля до ноября. В
это время много животных встречается в районе Канино-Колгуевского
мелководья, в Печорском море, на востоке Баренцева моря, у Земли Франца-Иосифа.
В авгуоте 1991 и 1992 гг. мы неоднократно наблюдали неперелинявших морских
зайцев у западного побережья Новой Земли (табл. 27).
В период щенки и линьки животные практически не питаются, 'они
малоподвижны и находятся в полусонном состоянии, осторожность их снижается.
Один из наиболее многочисленных видов настоящих тюленей. В северной
Атлантике в первой половине 80-х гг. норвежские и советские промысловики
добывали до 500 морских зайцев ежегодно (Потелов. 1986а).
Кольчатая нерпа (Pusa hlsplda) - пагетодный вид, обычный и
многочисленный в данном районе. Встречается по всему побережью Новой Земли,
единичные экземпляры - круглый год. Численность нерпы у берегов Новой Земли к
70-м годам оценивалась в 20 тыс. особей (Доронина и др.. 1970).
Нерпа, обитающая на карской стороне архипелага и его севере, крупнее
баренцевоморской в среднем на 10 см (Чапский, 1940).
Значительные скопления наблюдались в районе от Маточкина Шара до
о-вов Пахтусова. Во многих районах нерпа держится непостоянно, совершая
перемещения, вызванные изменениями ледовых и кормовых условий. Пути
миграции изучены мало, хотя мигрирующая - "ходовая" нерпа, на Новой Земле давно
была известна зверобоям. Отмечено, что с наступлением лета и исчезновением
льдов нерпа' мигрирует из прибрежной полосы южной части Новой Земли через
Карские ворота в Карское море, а осенью - в обратном направлении
(Чапский. 1976),
В летнее время, когда берега архипелага освобождаются ото льда, часть
популяции, держится у берегов, совершая небольшие пищевые кочевки. Другая
часть уходит на север вместе со льдами, предпочитая зону дрейфующих льдов
(Соколов и др., 1963).
По наблюдениям в районе мыса Константин, единичные особи отмечаются
круглый год. количество увеличивается в осенние месяцы и в феврале-марте
во время миграций. В осенние месяцы наблюдается движение зверей с севера
на юг, одиночками. В феврале-марте ход на север, полыньями в разреженном
льду (Антипин, 1938а).
У нерпы миграции слабо выражены; лишь осенью, в период подходов сайки
к берегам Новой Земли, отмечалось увеличение ее численности в прибрежных
районах (Чапский, 1941). Затем следует зимняя откочевка нерпы с карской
стороны архипелага. Второй раз в году нерпы образуют скопления в весенний
период (февраль-март) (Дубровский. 1938).
Для размножения выбирает неподвижные прибрежные льды, в частности, у
южного побережья Новой Земли. Береговых залежек в этот период не образует.
Основная масса нерпы, обитающей в Карском море и в восточной части
Баренцева моря, обычно щенится в период с конца последней декады марта по
начало второй декады апреля (Чапский, 1940). Размножение нерпы обычно
происходит на прочном льду припая у приглубых берегов, где самки устраивают
норы-логовища. Поздние щенки могут быть рождены на поверхности льда или
даже на берегу. Нередко среди щенков встречаются недокормленные, которых
называют "заморышами" или "кавадеями". Они чаще рождаются на берегу
(Смирнов. 1908). Считают, что "заморыши" - результат значительного недокорма
самкой. Причинами его могут служить беспокойство, гибель самки, разрушение
припайного льда штормом или ледоколом .' Наиболее благоприятные условия для
размножения нерпы и выкармливания потомства имеются в Карском море (Поте-
лов, 19866). В.М.Антипин (1938а) указывает, что нерпа залегает обычно на
ровном льду заливов и губ. вокруг лунок. Щенка весной, с середины марта по
середину апреля. В районе мыса Константин образует линные залежки в конце
мая - начале июня, ложится на ровный, более тонкий лед. У одной лунки от
1 до 10 шт. Ложится и на дрейфующий крупнобитый лед; торосистого льда, как
правило, избегает.
На о-ве Гемскерк были отмечены линяющие особи в конце апреля, у о-вов
Пахтусова - со второй половины мая по конец июня, в пелагических районах
юго-западной части Карского моря - в августе (Чапский. 1940). Иногда в
этот период нерпы могут образовывать линные залежки на льдах, на которых
залегают до 150-300 животных, но чаще менее многочисленные.
Гренландский тюлень (РакорШиз groenlandlca) встречается у Новой
Земли чаще в районе Карских ворот, у южной оконечности архипелага и по
западному побережью, по кромке льдов проникает далеко на север в
пелагической части Баренцева и Карского морей. Ярко выраженный обитатель холодных
еод, предпочитает прикромочную зону паковых и дрейфующих рассеянных льдов.
В неледовый период может придерживаться.прибрежной зоны, например, на юге
архипелага.
Распределение пагофильных тюленей в период размножения и линьки
связано с океанологическими и гидрометеорологическими факторами. У
гренландских тюленей эта связь наиболее ярко выражена (Тимошенко, 1987). В другие
периоды годового цикла определяющую роль в распределении этих ластоногих в
море играет пищевой фактор (Назаренко. 1986).
Общая схема миграций гренландских тюленей беломорской популяции
выяснена давно (Фрейман. 1939; Дорофеев, 1960). В различные годы по тем или
иным причинам миграционные пути их могут изменяться.
В прошлом гренландские тюлени появлялись у берегов архипелага в
октябре-январе, в летний период их встречи отмечались лишь на севере. На
открытой воде встречались, как правило, только серки. В.И.Антипин (1938а)
отмечает, что подходы гренландского тюленя к берегам Новой Земли не
постоянны, большей частью встречаются одиночные звери или небольшие стайки.
На карской стороне архипелага, впрочем, как и на западной, их
миграции в весенне-осенний период происходят в зависимости от изменения
положения кромки льдов (Дубровский, 1938). Нагульные животные могут залегать на
льду, обычно у самой кромки.
К концу мая льды, с расположенными на них линными залежками, обычно
выносит из Белого моря. По окончании линьки тюлени.рассредотачиваются по
Баренцеву и Карскому морям. Ранее они стадами шли на север вдоль берегов
Новой Земли, распределяясь от Шпицбергена на западе до островов Северной
Земли на востоке. В настоящее время в результате перелова основного
кормового объекта гренландских тюленей - сайки, пути их весенне-летних
миграций изменились. С конца 70-х годов они все чаще стали появляться у берегов
Мурмана в весенне-летний период, в настоящее время проводят тут большую
часть нагульного периода.
Зимой и весной тюлени наблюдаются в полосе льдов в районе Карских
ворот. К середине мая часть зверей мигрирует по кромке льдов до Гусиной
Земли, затем по кромке льдов движется на запад (к Земле Франца-Иосифа) и в
северо-западные районы Карского моря. . Севернее и восточнее Новой Земли
встречаются довольно рассеянно, небольшими группами, до начала сентября, к
зиме мигрируют на юг (Белое море, юго-восточная часть Баренева моря),
летом встречаются мелкими группами, к осени собираются в более крупные стаи.
Весенние миграции часто зависят от конфигурации ледовой кромки. В целом
тюлени следуют за отступающими льдами и держатся преимущественно около их
кромки.
Непосредственно к берегам Новой Земли подходят нечасто, в
июле-августе большое число зверей оказывается рассредоточенным на огромном
пространстве между Новой Землей и Землей Франца-Иосифа. Во время осенней
миграции на юг тюлени придерживаются берегов Новой Земли, питаясь сайкой. В
сентябре стаи гренландских тюленей регистрировались в районе мыса Желания,
а затем отмечались по всему побережью вплоть до южной оконечности
архипелага. Стаи передвигаются от мыса к мысу, в губы и заливы заходят редко.
Миграция вдоль карского побережья Новой Земли отмечена в очень небольших
размерах (Чапский, 19766). Например, В.М.Антипин (1938а) отмечает, что в
районе мыса Константин гренландского тюленя очень мало, встречается
главным образом в осенне-зимние месяцы.
Не исключена возможность размножения части беломорской популяции
гренландских тюленей на льдах у южной оконечности архипелага. Однако в
районах, где средняя температура в феврале-марте ниже -12°, регулярных
летных залежек не было найдено (Смирнов, 1927).
Новая Земля издавна являлась важным промысловым районом ho морскому
зверю. Основу новоземельского промысла составляли, главным образом,белуха и
голец, второстепенную, но не малую долю промысла составляли морж, тюлени и
белый медведь. В начале 30-х годов XIX века ежегодно в новоземельском
районе промышляли до 80 поморских зверобойных судов, а в 1835 г.- 137 судов
(Брейтфус, 1905). Русские промышленники отмечали, что помимо небольшого
количества местных зверей, во время миграций появляются большие скопления
ластоногих, так называемый "проходной", "ходовой" зверь, который в
основном и служил объектом промысла. Очевидно, массовое появление морского
зверя связано было с подходом косяков сайки: поэтому промысел тюленя был в
основном' оориентирован на добычу мигрирующих животных, сетями или путем
отстрела на льду, что при незнании особенностей миграционных путей не
могло обеспечить его доходность (Антипин, 1938а,б). Тем не менее промысел
существовал за счет обилия морского зверя и был сокращен, когда численность
моржей и тюленей существенно снизилась. В 1859 и 1860 гг. к Новой Земле
ходило не более 5-6 поморских судов, в 1898-1905 гг. тут работало 5-9
судов в год (Брейтфус, 1905).
Норвежский промысел в данном районе стал развиваться1 во второй
половине XIX века, в то же время поморский промысел у Новой Земли значительно
сократился. В 1871 г. тут промышляло 80-90 норвежских судов и только 8
русских. Иностранный промысел велся бесконтрольно и начинался он раньше,
чем туда успевали прийти поморские суда (после схода льда в Белом море)
(Нильсен, 1991). Размер добычи норвежскими судами в 1887 г., например,
составил 1200 моржей и до 14 000 тюленей (Брейтфус, 1905). В начале XX
века русским правительством были предприняты некоторые меры по прекращению
иностранного промысла на Новой Земле, но отдаленность района затрудняла
охрану и контроль в этих водах. Во время экспедиций В.А.Русанова в
1909-1910 гг. на северо-западе архипелага были обнаружены 5 действующих
промысловых пунктов норвежцев (Корякин, 1987).
До начала сороковых годов промысел ластоногих активно проводился в
районе губы Черной (фактория Красино). Помимо тюленей, тут добывались и
моржи. Лов тюленей производился в основном сетями длиной 50 м, которые
устанавливались на мысах до декабря включительно. Обычно к декабрю план по
добыче перевыполнялся. В марте после образования полыней начинали ледовый
промысел нерпы и морского зайца. Уровень промысла морского зайца на Новой
Земле (включая о-в Вайгач) по К.К.Чапскому (19415 приведен ниже: 1932 г. -
395, 1933 Г. - 386, 1934 Г. - 352, 1935 г. - 956, 1936 г. - 440, 1937 Г. -
840. 1938 Г. - 865.
В зимний период в районе Маточкина Шара велся сетной промысел нерпы и
морского зайца. После 30 апреля, когда начинался полярный день,
открывалась охота на нерпу, которая скапливалась у лунок (Рихтер, 1938). В 60-е
годы в Баренцевом и Карском морях среднегодовая добыча нерпы составляла
4.6 тыс.. а в 70-е - 2 тыс. особей (Тимошенко, 1982). С 1980 г. до 1985 г.
норвежскими и русскими зверобоями в северо-восточном промысловом районе
добывалось до 5 тыс. нерп ежегодно (Потелов. 19866).
Морж промышлялся в Русской Гавани, на косе Серебренникова, на о-вах
Гемскерк, Русанова (Антипин. 1938а.б). Имеющаяся у нас статистика промысла
очень неполная. Известно, например, что в 1933 г. четырьмя зверобойными
судами на северо-востоке Карского моря было добыто 172 особи (Есипов,
1935). Судовой промысел моржей у побережья Новой Земли, в Карском море
проводился в период 1929-1935 гг. Технология промысла допускала потерю
40-5056 убитых моржей, которые тонули. К.К.Чапский считал , что во время
проведения промысла в ареале ежегодно погибало 2.5-3 тыс. моржей. Судовой
промысел у Новой Земли и в западной части Карского моря велся с 1929 г. до
1935 г.; из-за нерентабельности был прекращен (Чапский, 1941).
Еще в дореволюционное время предпринимались попытки ограничения
иностранного промысла морского зверя на Новой Земле из политических
соображений. В дальнейшем пресс промысла снижали войны или экономические
спады. Лишь во второй половине XX века стали вводиться меры по охране морских
млекопитающих, обитающих на архипелаге. В 1947 г. на Шном острове был
организован филиал заповедника "Семь островов". Просуществовав три года, он
был закрыт с началом строительства ядерного полигона.
В конце 30-х годов К.К.Чапский указывал на необходимость введения
5-10-летнего моратория на промысел моржей, считая, что численность их
популяции, обитающей у Новой Земли, возрастет за десятилетний период от
1200-1300 особей до уровня 5000. В.М. Антипин, как и К.К.Чапский. предлагал
ввести временный запрет на промысел моржей, обитающих у Новой Земли, но он
был введен лишь в 1956 г. (Антипин, 1938а.б).
К 30-м годам XX столетия на Новой Земле, сохранились некоторые
береговые лежбища моржей, располагавшиеся на островах Оранских, Гемскерк. Брит-
вин и некоторых других в районе Карских Ворот, а также на участке от мыса
Бисмарк до мыса Спорый Наволок (Чапский, 1939).
Исследования, проведенные в 50-х годах, показали, что численность
мораей у архипелага крайне низка. Так, во время авиаучета на участке
мыс Спорый Наволок - мыс Желания были отмечены всего 4 моржа, залегающие
на удалении друг от друга, в 40-80 км от берега. Общая численность
популяции оценивалась в то время до 200 голов (Белькович, Хузин, 1960). К началу
70-х годов численность моржей, обитающих в Баренцевом и Карском морях, не
достигала и 1000 особей (Жирное и др.,; 1978). В этой связи предлагалось
заповедание Оранских островов и о-ва Гемскерк (Бычков, 1986).
Эти отрывочные данные свидетельствуют о том, что популяциям
ластоногих Новой Земли был нанесен колоссальный ущерб.
Несомненно, что свертывание ядерных испытаний на Новой Земле должно
благотворно повлиять на состояние популяций морских млекопитающих. Однако
снижение военной активности и контроля на архипелаге имеет и другую
сторону. Свободный доступ к побережью северных островов ведет к разграблению их
биологических богатств, усиливается фактор беспокойства. В данной ситуации
важнейшую роль должны сыграть вновь создаваемые заповедники и пропаганда
экологических знаний среди населения, экипажей судов и в воинских частях.
Несомненно, что необходима также разработка и внедрение новых нормативных
и законодательных актов, направленных на реальную охрану экосистем Севера
и регламентирующие хозяйственную и другую деятельность.
Как уже отмечалось, на распределение и численность ластоногих в водах
архипелага оказывают значительное влияние ледовая обстановка и другие
гидрометеорологические факторы. Длительное время популяции морских
млекопитающих испытывали пресс активного промысла. На фоне изменений, происходящих
в современных морских экосистемах, на восстановление запасов ластоногих на
архипелаге до первоначального уровня.раоочитывать не приходится даже при
условии введения строжайших мер охраны. Установлению тенденций изменений в
популяциях морских млекопитающих данного региона препятствуют, в первую
очередь, отсутствие информации о численности животных в популяциях, их
питания в настоящее время, степени загрязненности их организма
радионуклидами и характере воздействия этих загрязнителей на организм ластоногих.
8.3. Белый медведь
Режим секретности и многочисленных запретов на мореплавание и заходы
судов в бухты Новой Земли давно сделали весь архипелаг зоной
недоступности, фактически отключив его на несколько десятилетий от российской и
мировой науки. Не случайно, что различные стороны биологии одного из наиболее
интересных и ценных видов фауны Новой Земли - белого медведя - до сих пор
изучены явно недостаточно. Отрывочны сведения о его морфологии,' поведении,
характере миграций, недостаточно известна популяционная структура,
величина годового прироста, масштабы и причины.смертности. Почти не
исследованы адаптации зверя к обитанию в условиях низких температур и полярной
ночи, к длительному голоданию и крайне экономному расходованию энергии.
Вовсе не изучен механизм ориентации зверя во льдах. Все это, к сожалению,
касается не только новоземельского белого медведя, но и вида в целом.
Следует отметить, что в последние годы в исследованиях, связанных с белым
медведем, наибольшим вниманием пользуется вопрос о его динамике численности.
К его решению привлекаются данные учетов зимних убежищ самок белых
медведей, результаты авиаучетов животных, а также данные, получаемые от.
сотрудников полярных станций, охотников и других наблюдателей.
В Норвегии. Канаде. США. Гренландии исследования белого медведя более
разнообразны благодаря разработанным методикам обездвиживания и мечения
зверей, что позволяет изучать их морфологию, физиологию, биохимию,
генетику и даже проводить лабораторные эксперименты.
Систематическое положение. До настоящего
времени структура вида у белого медведя еще окончательно не установлена.
Неоднократно предпринимались попытки выделения различных географических рас
белого медведя. Различия в размерах тела и строении (пропорциях черепа)
зверей, встречающихся в отдельных районах, рассматриваются некоторыми
исследователями как доказательство существования обособленных географических
рас животных. Однако, как считает С.М.Успенский (1967). тут может быть и
другое объяснение: там, где белые медведи реже встречаются с охотниками,
они достигают более зрелого возраста, а следовательно, и большей величины.
С возрастом же не только увеличиваются, размеры тела животных, но и
изменяются пропорции их черепа.
Несмотря на то, что этот вид встречается в разных участках Полярного
бассейна, степень географической однородности вида довольно высока. Это.
очевидно, связано с однородностью условий его существования и, в
частности, с однообразием основных объектов питания. Обособлению отдельных
популяций препятствуют и особенности образа жизни белых медведей, связанные с
миграциями в поисках пищи и пассивными передвижениями с дрейфующими
льдами. Новоземельский белый медведь относится к западной популяции,
малочисленной по сравнению с центральной (море Лаптевых и западная часть
Восточно-Сибирского моря) и восточной (восточная часть Восточно-Сибирского и
Чукотское моря) популяциями (Беликов. 1990). Такое же деление белых медведей
на эколого-географические группировки предлагает С.Н.Успенский с
соавторами (1985). основываясь на данных геохимического анализа. Так. центральная
группировка достоверно отличается от двух других по содержанию в костной
ткани медведей калия, титана, никеля и алюминия. И все же вопрос о статусе
выделенных группировок остается открытым. Поскольку они не имеют
географической изоляции, к ядру группировок, т.е. особям в значительной степени
оседлым, периодически могут добавляться мигранты из других районов, не
задерживающиеся здесь на длительное время. В таком случае остаются
неизменными своеобразие и обособленность группировок по геохимическим критериям
(поскольку химический состав организмов связан с геохимией ландшафтов), но
осуществляется обмен генетической информацией. С.М.Успенский (1965)
считает, что геохимически выделенные группировки не могут рассматриваться как
локальные популяции белого медведя. Тем не менее, установленный факт
приуроченности медведей к определенным географическим районам вносит
коррективы не только в представление об экологических особенностях вида, но и в
стратегию его охраны. В частности, становятся необходимыми
дифференцированный подход к анализу динамики численности медведей в разных
группировках и дифференцированное определение оптимальной плотности животных и
квоты изъятия. .
Питание. Основная пища белого медведя состоит из ластоногих,
главным образом, нерп, реже - морских зайцев и моржей. Есть данные, что
иногда от белого медведя страдают и песцы, и олени, и мелкие грызуны -
лемминги (Адлерберг, 1935). Как случайную пищу, употребляемую
преимущественно в зимний период, отмечают морские водоросли (Антипин, 19386).
Описывая питание белого медведя, Г.П. Адлерберг (1935) замечает, что этот зверь
никогда не упустит случая разграбить встретившийся ему склад, оставленный
полярными путешественниками или промысловиками, поедая при этом не только
продовольствие (например, сухари, мясо, сахар, кофе), но и различные
предметы обихода; так приводятся, например, случаи, когда медведь поедал
табак, паруса и т.д.
Описаны случаи добычи белыми медведями белух (Lowry et al., 1987),
причем отмечается, что белухи являются обычным пищевым объектом медведя,
когда они попадают в ледяные ловушки, плавают в разводьях или выглядывают
из отдушин во льду.
Экспериментально установлено (Cushlng et al., 1988), что белый
медведь избирательно реагирует на подводные звуки различных потенциальных
пищевых объектов. На предъявление звуков кольчатой нерпы число
ориентировочных реакций белого медведя (настораживание ушей, принюхивание, движения
головой) оказалось достоверно большим, чем на предъявление звуков морского
зайца, гренландского тюленя, а также фоновых звуков.
Охотясь на нерпу, медведь затаивается около самой лунки и прикрывает
лапой морду, оставляя, однако, лунку в поле зрения (вероятно, маскирует
черный кончик носа). Показавшаяся нерпа попадает под удар лапы. В первую
очередь медведь сдирает с убитого тюленя кожу и жир и поедает их, лишь при
последующих неудачных охотах возвращаясь есть мясо. Медведь может поймать
нерпу в воде: учуяв нерпу по запаху с кромки льда (с подветренной
стороны), медведь тихо спускается в воду и плывет так. что наверху торчит
только кончик носа; изредка ныряет. Когда до нерпы остается небольшое
расстояние, медведь ныряет и хватает ее под водой. При этом наблюдалось, как
нерпа мгновенно исчезала в воде, а через несколько секунд медведь выбрасывал
ее на кромку льда (Марунин, Кищинский. 1969).
Пищеварительная система медведя, очевидно, приспособлена к
специфическому составу пищи. В частности, существует мнение о том, что
результатом адаптации к выоокому уровню витамина А в пище является большое число и
размеры клеток Ито в печени (крупные перисинусоидальные клетки, содержащие
в цитоплазме липидные капли).При атом оуммарное ооцержание ретинола и рети-
нолпальмитата в печени коррелирует со средней площадью.клеток Ито (Leigh-
ton et al., 1988).
Исследования эффективности переваривания белым медведем его основной
жертвы - кольчатой нерпы (Best, 1985) показали, что наиболее высокая пере-
вариваемость у пищи, состоящей из кош и жира нерпы, наиболее низкая - у
пищи, состоящей из мышц, внутренностей и скелета. Высекая степень
перевариваемое™ наблюдалась также у пищи, состоящей из рыбы. Основными
лимитирующими факторами потребления энергии для белого медведя является емкость
пищеварительного тракта (в основном, характеризующаяся объемом желудка) и
доля жира и мускулов в съеденной порции корма (Best.1985).
Годовой цикл, размножение. В ранних источниках,
например, в сводке Г.П.Адлерберга (1935) отмечается, что большинство белых
медведей (кроме беременных самок) бодрствует круглый гсд, хотя не
отрицается возможность залегания на зиму в берлогу и самцов. Современными
методами установлено (Bruce et al.,- 1989), что в сыворотке крови белых
медведей имеется фактор, индуцирующий зимнюю спячку: Природа этого фактора до
сих пор окончательно не установлена. Известно только, что он обладает опи-
оидным эффектом, не являясь, тем не менее, опиоидом.
Сон занимает летом 2536 бюджета времени, зимой - несколько больше.
Поза спящего белого мвдведя зависит от термических условий и весьма
изменчива - от свернувшегося клубком швотного с прикрытой лапой мордой до
лежащего навзничь, с распростертыми и поднятыми вверх лапами. Иногда белый
медведь спит, присыпанный снегом. На берегу часто пользуется укрытиями в
снегу или песчано-галечных наносах. В среднем сон продолжается 7ч 45мин и
приурочен, преимущественно, к дневному периоду, что. вероятно, связано с
преимущественно ночной активностью главной добычи белого медведя - тюленей
(Stirling. 1989). Есть данные экспериментов по изучению скорости
стандартного метаболизма у белого медведя в. искусственной берлоге (Watts et al.,
1987), показывающие, что ежедневно медведь теряет при спячке 0.7 кг веса.
Среднесуточный расход энергии составляет чуть больше 2000 ккал.
спаривание приходится у новоземельских белых медведей на
февраль-март. В это время нередко можно встретить нескольких самцов,
следующих за самкой, причем ближе всех к ней находится самый-крупный (Антипин.
19386). Первые роды у самок белых медведей бывают обычно в возрасте
4-5 лет (Ramsay. Stirling. 1988). Самки залегают в берлоги в сентябре, а в
конце января - в феврале, т.е. после 247-дневной беременности, родят 2.3 и
очень редко 4 медвежат, не.превышающих 700 г веса и 27 см длины (Мантей-
фель. 1938). Берлога у медведицы с новорожденными медвежатами имеет очень
маленькое отверстие. Если же медведица залегла с "лончаками" (медвежата-
ми-второгодками), то отверстие у берлоги большое и вблизи ее снежные
склоны бугров обкатаны и исцарапаны медвежатами (Антипин. 19386). В берлоге
новорожденные медвежата развиваются очень медленно: глаза открываются у
них только через 30 дней (Иантейфель. 1938). Выход самок с молодыми
медвежатами наблюдается в середине марта - начале апреля (Антипин, 19386;
Иантейфель. 1938). На основании личных наблюдений В.И.Антипин (19386)
заключает, что молодые медвежата остаются при матери, по крайней мере, на две
зимы. Медведицы приносят потомство один раз в два года (Мантейфель, 1938).
После 20-летнего возраста у белых медведей, как правило, плодовитость
снижается (Ramsay, Stirling. 1988).
Распределение и численность. Для сбора
сведений о распределении и численности белого медведя в Арктике, в том числе
и на Новой Земле, на ранних этапах исследований использовалась
разветвленная сеть полярных станций и гидробаз Главного управления Северного морско-
го пути. Сотрудниками Кандалакшского государственного заповедника с 1960
года во все населенные пункты Советской Арктики регулярно рассылались
специальные вопросники, на которые отвечали представители местных органов
власти, работники охотничьего хозяйства и зверобойного промысла, капитаны
кораблей и другие корреспонденты. Основными недостатками полученного таким
образом материала являются его территориальная и хронологическая
неравномерность и известная неравноценность данных, поступивших из различных
мест. Тем не менее, были получены сведения о распределении и численности
белого медведя за период с конца 30-х до 60-х годов. По данным
корреспондентской сети, на севере Новой Земли, как на восточных, так и на западных
ее берегах (до района Русской гавани включительно), в 30-40-х годах
медведи были многочисленны - 50-100 встреч за год (рис. 53). В следующем
десятилетии сходная встречаемость отмечалась только на крайней оконечности
острова, близ мыса Желания. Этот район звери посещали чаще, чем южные
части Новой Земли. Здесь и в настоящее время проходят их миграционные пути на
Землю Франца-Иосифа и Северную Землю. Сюда же, видимо, выходит часть
животных из центральных районов Карского моря при движении к Земле
Франца-Иосифа. В 60-х годах во всем этом районе встречаемость белых медведей
уменьшилась дс 10-40 встреч в год (Карпович. 1969).
Вдоль всего восточного побережья Новой Земли, вплоть до мыса
Меньшикова за описываемый период средняя встречаемость белого медведя почти не
изменялась, зверь был здесь обычен (10-40 встреч.в год). По опросным
данным, основные места залегания медведиц в берлоги также находятся на
восточном побережье Новой Земли (рис. 53) (Успенский, 1965). Р.Ш.Хузин (1960)
отмечает, что у Карского побережья Новой Земли в весенне-летнее время
часть медведей мигрирует не на север, как обычно в других частях Арктики,
а на юг. Автор связывает это с особенностями гидрологического режима
западной части Карского моря:, даже в теплые годы здесь до конца лета
сохраняются значительные массивы льда. К тому же этот район весьма благоприятен
ив кормовом отношении, так как летом здесь ежегодно держится много
морского зайца и нерпы. Следует отметить, что в начале 50-х годов в
юго-восточной части Новой Земли белые медведи в летнее время бывали редко. Тст же
Р.Ш.Хузин (1960) предполагает, что в этот период медведей отпугивал
проводившийся здесь береговой промысел морского зверя.
Качественно новый этап в изучении белого медведя, в частности, его
распределения, численности и ее динамики начался с применением
авиаметодов. В бывшем СССР систематически проводились ледовые авиаразведки на
больших площадях и по определенным маршрутам. Эти разведки осуществлялись
в течение всей светлой части года (с февраля по ноябрь) и равномерно
охватывали всю Советскую Арктику. В связи с тем. что во время таких разведок
наблюдатели-гидрологи и визуально оценивают ледовый покров и одновременно
имеют возможность регистрировать встречи животных, рекомендуется судить о
распределении белых медведей по сезонам года, по районам, а также в
зависимости от особенностей льда. Места встреч белых медведей,
зарегистрированные при ледовых авиаразведках в 1962, 1967-68 годах, показаны на
рис. 54. По сравнению с другими районами Арктики количество медведей у
Новой Земли неизменно было в эти годы большим. Во многом это связано с
благоприятными кормовыми условиями (Успенский, Шильников, 1969).
Самые близкие к нашему времени данные по учету белых медведей во
время ледовых авиаразведок, опубликованные в доступной литературе, относятся
193? - 1950 г.г. 19514960 г.г. 1961 - 19б7г.г.
Рис. 53. Распределение белого медведя в районе архипелага Новая Земля в 1937^1967 гг. ,по данным
корреспондентской сети (по: Карпович. 1969):
а - пункты корреспондентской сети; белый медведь многочислен; обычен; х - очень
редок; ? - сведений нет;*ч& - район, в котором в конце 50-х годов отмечено увеличение численности медведей;
-.• - места залеганий медведиц в берлоги (Успенский. 1965)
к периоду 1970-84 гг. (Горбунов и др.. 1987). У берегов Новой Земли в это
время медведи или их следы встречались относительно равномерно на
территории, покрытой льдом (рис. 54). В большинстве случаев наблюдались одиночные
звери. За период наблюдений тенденций к изменению численности зверей в
этом районе не прослеживалось.
60° 70 60° 70
Рис. 54. Распределение белого медведя в районе архипелага
Новая Земля, по данным авианаблюдений ледовой разведки:
1 - 1962, 1967-1968 гг. (по: Успенский, Шильников, 1969);
2 - 1970-1984 гг. (по: Горбунов. Беликов. Шильников. 1987);
■ - места встреч одиночных белых медведей; -• - места
наблюдений следов медведей; о - места встречи нерп
Данные о численности белых медведей на Новой Земле весьма
неопределенны. Есть сведения о их количестве в популяции, обитающей в Баренцевом и
Карском море - около 1000 экземпляров (Беликов. 1990). Однако следует
учитывать, что в это число входят и звери Земли Франца-Иосифа.
Белый медведь и человек. Белый медведь на
большей части его ареала всегда имел лишь второстепенное промысловое значение.
Существенная доля добытых зверей и их шкур использовалась охотниками для
своих нувд и обычно не фигурировала в торговой и промысловой статистике. В
нашей стране охота на белого медведя была полностью запрещена с 1956 года.
Промысел зверя на Новой Земле известен с XV века. По уровню
среднегодовой добычи белых медведей район Новой Земли занимал третье место после
Шпицбергена и побережья Чукотки. Среднегодовая добыча белых медведей на
Новой Земле (по: Успенский. 1969) приведена ниже: 1700-1799 гг. - 100.
1800-1849 ГГ. - 100. 1850-е гг. - 100. 1860-е гг. - 100. 1870-е ГГ. - 100,
1880-е ГГ. - 100, 1890-е ГГ. - 100. 1900-е ГГ. - 100. 1910-е гг. - 100,
1920-е гг. - 100, 1930-е гг. - 200, 1940-е гг. - 100, 1950-е гг. - 50.
Всего добыто с начала XVIII века 26500 зверей, т.е. 17% от общего числа
зверей, добытых на севере Евразии в 11 промысловых районах.
Учитывая, что, кроме русских поморов, белых медведей на Новой Земле и
в ее прибрежных водах добывали участники различных экспедиций (начиная с
Баренца в 1596г., а особенно часто в XVIII-XIX вв.), а также (с начала XIX
века) норвежские зверобои, можно считать, что в среднем за год в этот
период (особенно в XVIII-XIX вв.) здесь добывалось не менее, чем по 100,
медведей (Успенский, 1969).
Постоянное население появилось на Новой Земле с 1870 года, а в
1878 г. организовалась также первая полярная станция (Успенский, 1969).
Приведенные сведения о заготовке медведей факториями Новой Земли в
послереволюционный период не отражают полного объема добычи медведей, так
как значительное количество зверей добывалось норвежскими зверобоями; они
же вывозили шкуры медведей, добытых местным населением.
Увеличение добычи медведей в 20-х и 30-х годах связано, в основном, с
ростом промыслового населения, организацией промысловых поселков на севере
и востоке Новой Земли, деятельностью многочисленных экспедиций, ростом
сети полярных станций (табл. 28). С.Н.Успенский (1969) предполагает,.что
общий объем годовой добычи на островах в этот период превышал 200 медведей.
Начало 50-х годов характеризуется резким сокращением запасов зверей, а
также и объема их добычи.
Вопрос об охране белого медведя рассматривался еще в 1938 году, когда
вышло Постановление о запрещении в Советской Арктике охоты на зверей с
судов и без крайней необходимости на полярных станциях. В 1950 году было
введено повсеместное ограничение охоты, а в 1956 году вышло Постановление
"О мерах'по охране фауны Арктики", предусматривающее и полный запрет охоты
на белого медведя. Единственно допустимой формой хозяйственного
использования вида остается отлов медвежат для зоопарков по специальным лицензиям.
Таблица 28
Количество шкур и живых белых медведей, заготовленных
факториями Новой Земли (по: Успенский, 1969)
Год
1917
1919
1920
1921
1922
1923'
1925"
1926*
1927'
1928*
1929'
Шкур
медведей
34
78
35
33
81
22
33
103
74
48
53
Живых
медведей
0
0
0
0
0
2
3 '
- 0
1
0
2
Год
1930'
1931*
1932*
1933*
1934
1940
1941
1942
1943
1944
Шкур
медведей
36
73
45
44
67
72
73
108
92
29
ЖИВЫХ
медведей
4
2
1
4
0
0
0
0
0
0
* Заготовлено не только на Новой Земле, но и на островах Вайгач и.
Колгуев.
С 1968 года при отлове медвежат запрещено отстреливать сопровождающих их
медведиц.
В связи с большой популярностью белых медведей и спросом на этих
животных для зоопарков ежегодно в Арктике отлавливаются десятки, а то и
сотни медвежат, этот вид промысла осуществляется, как правило, в местах
массового залегания в берлоги медведиц (на Новой Земле - это восточное
побережье). Смертность.отловленных таким образом медвежат достигает 50Я (Ху-
зин, 1960). Одним из бескровных способов является отлов
медвежат-подростков в море с судов, предложенный и использовавшийся Р. III. Хузиным (1960).
В 1975 году вступило в силу международное Соглашение по охране белых
медведей между СССР, США, Канадой, Норвегией и Данией. Срок действия
Соглашения был продлен в 1981 году на встрече представителей стран-участниц.
Безусловно, перечисленные меры по охране белого медведя способствовали
некоторому увеличению численности зверя в Арктике. Однако естественный
ареал этого вида в российском секторе Арктики полностью еще не восстановился.
реальная численность зверей неизвестна (Саблина, Овсяников. 1988).
Вместе с ростом численности белого медведя и увеличением на Севере
населения стала более существенной проблема взаимоотношений человека и
белого медведя. Покровительственное отношение к медведю, запрещение его
добычи привели к полной потере некоторыми.белыми медведями боязни человека,
формированию "синантропных" групп, учащению конфликтов между медведями и
людьми. С.М.Успенский (1987а,б) отмечает: воли с 1930 по 1968 г. по
всей Советской Арктике погибли или получили увечья от белых медведей 10-12
человек, то всего за несколько лет с 1976 по 1979 г. - 6. Одним из
следствий участившихся контактов медведей с человеком и домашними животными
оказалось распространение среди медведей ряда новых заболеваний, в том
числе трихинеллеза (Успенский, 1965).
В связи с развитием добычи нефти и газа на шельфе возникает вопрос о
влиянии буровых вышек на обитающих в этих районах белых медведей.
Выдвинута даже гипотеза о "тяготении белых медведей, к морским буровым вышкам"
(Stirling, 1988). Вследствие постоянного перемещения льда вокруг вышек
образуются трещины, а по ходу потока - участки открытой воды разной площади,
которые являются местами концентрации тюленей - кольчатой нерпы и морского
зайца, что привлекает сюда медведей. Охотясь на тюленей, белые медведи
могут оставаться вблизи вышек в течение нескольких дней и дольше. Некоторые
медведи привыкают к присутствию людей и теряют страх перед ними, возникают
проблемы обеспечения безопасности персонала вышек. Бывают случаи отстрела
медведей. Таким образом, при увеличении числа вышек вполне реальна угроза
численности зверей на осваиваемых территориях.
В настоящее время существует реальная угроза снижения численности
белого медведя в Арктике под влиянием изменений среды обитания. Среди
негативных факторов, влиянвдх на численность белых медведей, отмечаются
следующие: загрязнение среды, глобальное потепление Арктики, влияющее на
паковый лед и затрудняющее добычу пищи, увеличение интенсивности
ультрафиолетового излучения, отрицательно влияющее на зрение, усиление фактора
беспокойства для зверей в связи с хозяйственным освоением Севера (01зеп,
1991-1992).
Глава 9. РАСТИТЕЛЬНОСТЬ
Растительность моря. Работы о водорослях Новой Земли немногочисленны,
большинство из них выполнено в конце прошлого начале текущего столетия.
Они дают общее представление о флоре водорослей и распределении сообществ
макрофитов вдоль берегов Новой Земли (Kjellman. 1878, 1883; Rosenvlnge.
1887; Зинова, 1925, 1929; Флеров, 1932; Флеров, Карсаков, 1932). Имеются
некоторые данные о биомассе и запасах ламинариевых у юго-западного
побережья (Гемп, Вызова, 1970: Сорокин. Пельтихина. 1991).
Флора бентосных водорослей Новой Земли - обедненная
североатлантическая. Всего для Новой Земли отмечается 158 видов (Зинова, 1962, 1974).
Низкая температура воды,, ограниченная солнечная радиация, широкое
распространение мягких грунтов, истирающее действие льда - основные факторы,
определяющие состав и распределение макрофитов у побережья Новой Земли. На
литорали из-за небольшой амплитуды приливов и отливов льда, влияние которого
прослеживается до 3-6 м глубины, водоросли редки и немногочисленны. В
сублиторали на скалиотых и каменистых грунтах с сильным течением или прибоем
отмечается разнообразная растительность.
На литорали местами наблюдается развитие сезонных водорослей - 1/го-
spora penicilliformis, Pilayella litoralls. Enteromorpha mlcrococca (=*Bll-
dtngto minima).
Пояс фукусов, представленных, главным образом, Fucus distlcfius,
опускается в сублитораль на глубину 0-2 м, иногда фукусы встречаются до 7-9 м
глубины. В этом поясе встречаются в основном однолетние, сезонные
водоросли Chordaria HagelU/ormts. ■ Dlctyoslphon /oentculaceua, Stlctyostphon
tortlUs, Monostroma, Enteromorpha, Chorda /Hum, С tomentosa, Spongo-
morpha, Ctadophora. PitayeUa Utoraiis, RhizocIonium, ffhodocnorton purpu-
reum. Глубже F.distichus встречаются иногда в южных районах Новой Земли
F.serratus и F.vesiculosus. Отмечается низкорослость фукусов и ряда других
видов этого пояса.
Практически повсеместно на глубине 2-20 м имеются обширные заросли
ламинариевых, наибольшего развития которые достигают на глубине 5-10 м.
Верхний ярус сообщества ламинариевых образуют Laminaria digitata. L.sac-
charlna, Ataria esculenta, Desmarestta acuteata, на севере, кроме того,
L. soltdungula. На юго-западном побережье в местах с интенсивным движением
воды в верхней части пояса ламинариевых обычно преобладает L. digitata. в
среднем - /.esculenta и в нижнем - L.sacchartna, L.digitata и A.esculenta
доминируют на открытых участках заливов, L. saccharine и D.acuteata - в
сравнительно защищенных местах. Широко распространены смешанные заросли
ламинариевых. Нижний ярус сообщества-ламинариевых формируют красные
многолетние водоросли Palmarla palmata, Halosaccton ramentaceum. Ptilota ptumo-
sa, Odonthatta dentata. Phycodrys rubens, Phyttophora brodlaei, Rhodometa
lycopodioides. На камнях и гальке встречаются корковые водоросли Lithoder-
ma, HUdenbrandtla и красные известковые водоросли. Отмечается, что при
довольно крупных размерах (Laminarta, Patmarta, Halosacclon) слоевище
водорослей тонкое. Максимальная длина и масса слоевища Laminarla saccharine.
может составлять 318 см и 0.7 кг, слоевища L.dtgttata - 255 см и 1.5 кг
соответственно (Сорокин. Пельтихина, 1991). Биомасса водорослей в
сообществе ламинариевых у юго-западных берегов составляет 0.5-10 (до
22) кг/м2. средняя биомасса ламинариевых невысокая - около 2-3 кг/м2.
На глубине 10-30 м развиваются красные водоросли, основными
компонентами сообщества являются Phycodrys rubens, Potystphpnta arcttca. Phyllo-
phora tnterrupta, Pttlota ptumosa. Odonthalta dentata. Местами широкое
распространение на глубине 20-50 м в районе с сильным течением имеют
красные корковые известковые водоросли (Ltthotnamnton, Phymatoltthon).
Общий характер растительности бентосных водорослей в целом одинаков в
разных районах Новой Земли. В северной части Новой Земли и у восточного
побережья сообщества беднеют в качественном и количественном отношении,
сообщество фукусов исчезает (на севере) или представлено отдельными
растениями. В районе Новой Земли по 75° с.ш. проводится морская фитогеографи-
ческая граница между двумя подобластями арктической области:
нижнеарктической и высокоарктической. Западное и восточное побережье Новой Земли
южнее 75° с.ш. входит в состав Печорско-Новоземельской провинции
нижнеарктической подобласти. Эндемичных родов и Видов бентосных водорослей в данном
районе не отмечается (Зинова, 1974).
Растительность суши. Ботанические исследования на Новой Земле были
выполнены в 20-30-е годы (Зубков, 1935; Толмачев, 1936, 1939;
Александрова, 1956. 1983). Остается много нерешенных вопросов о флоре данного района
и особенностях развития наземных растений в суровых условиях Арктики.
Остров Пиши, и южная часть Северного острова относится к арктическим
тундрам (Александрова, 1979). Растительность этого, района в" целом не
обильная, но довольно разнообразная (рис. 55).
Для Южного острова характерны ассоциации тундровых болот,
представленные осочниками и пушицо-дюпонтиево-моховыми группировками. В
ассоциациях тундровых болот большую роль играют виды пушицы (Ertophorum), дюпонтия
(Dupontta /tschert) и осока (Carex stans). Мхи нередко образуют сплошной
ковер, главным образом из гипновых мхов (Calttergon sarmentosum, С. stramt-
neum, Drepanocladus exannulatum). Роль мхов увеличивается по мере
нарастания торфа.
Широко распространены ассоциации пятнистой тундры на тяжелых
суглинках, представленные злаково-мохово-кустарничковыми или мохово-кустарничко-
выни группировками. Растительный покров пятнистой суглинистой тундры
довольно однообразен, хотя и богат' в видовом отношении. Хорошо дренированные
вершины холмов заняты ассоциацией щучки арктической (Deschapsla arcttca) с
полярной ивой (Saltx polarts), в напочвенном покрове развиты мхи. главным
образом - Hytocontum prolt/erum и Camptothectum trichotdes. Довольно часто
здесь встречаются из злаков: мятлики (Poa arcttca, P.alpigena), овсяница
(Festuca orevl/olta), из разнотравия: камнеломка (Saxt/raga Mrculus).
валериана (Valeriana capltata), лютик (Ranunculus ntvatts) и другие. По
верхним и средним частям склонов щучка остается главным компонентом
ассоциаций, . ива принимает меньшее участие в покрове, зато здесь хорошо
развивается камнеломка... Напочвенный покров образован мхами (Camptothectum
trtchotdes, Hytocomtum prolt/erum) и лишайниками (Cladonta sllvattca,
С. cocct/era, Cetraria tskandlca)... Нижние части склонов, где подолгу за-
леживается снег, заняты злаковой ассоциацией из мятликов (Роа alpigena,
P. alptna) с примесью щучки, овсяницы красной (Festuca rubra) и видов
разнотравий. Моховой покров здесь почти не развит (Зубков, 1935).
1
Рис. 55. Схема распределения растительных сообществ на
южных склонах возвышенностей в окрестностях радиостанции
Маточкин Шар (Новая Земля) (Толмачев, 1939)
а-а: зона снегового минимума, b-b: зона снегового макси- .
мума. 1(9) - весьма скудная растительность - наиболее
обедненные варианты полигональных тундр. Преобладает Saxlfraga
opposltlfolla, Papaver radlcatum, Potentilla marglnata, Poa
abbrevlata, Luzula confusa и др. Большая часть
щебнисто-глинистого грунта не покрыта растительностью; 2 - относительно
богатая растительность с почти сплошным или сплошным ковром
растений. Хорошо дренированная почва. Достаточное, но не
чрезмерное покрытие онегом в зимнее время. Silene- aculls,
Myositis alpestrls. Astragalus alplnus, Lazula confusa, Oxytro-
pls sordida. Polygonum vlvlparum, Saxlfraga hieracifolia, Poa
alpigena и др.; у нижнего края - Ranunculus borealIs; 3 -
щеористые, лишь отчасти заросшие участки; растительность
развивается поздно. Характерны Oxyrla dlgyna и Artemisia tlle-
sll; 4 - почти голый субстрат; 5 - снежник; 6 - почти голый
щебнистый субстрат. Отдельные кустики Ranunculus sulphureus,
Parrya nudlcaulls и др.; 7 - болотистая тундра. Значительное
покрытие снегом. Грунт глинисто-щебнистый с преобладанием
мелкоземлистых фракций. Сильное увлажнение. Почти сплошной
растительный покров. Относительно крупные злаки, осоки, Erio-
phorum, обильно развитые мхи. Растительность беднеет по
направлению к участку "6"; 8 - переход от болотистых тундр к
полигональным. Довольно густые заросли Sallx arctlca и Dryas
oc.topetala,. в пестрой смеси с Astragalus umbellatus и другими
растениями, в том числе с элементами болотистой тундры.
По направлению стрелок х и у мощность снегового покрова
постепенно увеличивается.
Большую площадь на острове Южном, особенно в центральных частях и на
восточном побережье, занимают ассоциации мохово-лишайниковой и
лишайниковой пятнистой тундры. Доминирующими видами в ассоциациях
мохово-лишайниковой тундры являются кустистые лишайники Cladonia stlvattca, Cetraria ts-
landica и мох Dtcranlum elongatum, цветковая растительность представлена
немногими видами - полярный мак, лапчатка альпийская,, мятлик арктический.
Растительность лишайниковой пятнистой тундры образована накипными
лишайниками,. Cladonia attwttca развита^слабее, иногда преобладает Sphaeroptwrus
globosus, большая площадь занята каменистыми пятнами, лишенными
растительности.
Самая северная часть Северного острова относится к полярным пустыням
(Александрова, 1983), Большая часть территории занята ледниками, свободна
ото льда узкая полоса около моря. Условия для жизни растений крайне
неблагоприятны. Растительность скудная, сформирована дерновинками разрозненных
растений или фрагментами фитоцекозов. Здесь растут накипные лишайники,:
реже встречаются кустистые лишайники (Cetrarta deltset), растут немногие мхи
(из рода Drepanocladus) с немногими видами цветковых растений - камнеломка
(Saxlfraga opposttUotta). ясколка (Cerasttum regeltt), мятлик (Poa alpt-
gena), кисловица (Oacyrta dlgyna), крупка (Draba alptna), щучка (Deschamp-
sta borealls). На 76° с.ш. собрано 70 видов цветковых растений и свыше 200
видов лишайников. На берегу Русской гавани (76°14' с.ш.) отмечено всего 25
видов цветковых растений (Толмачев, 1936, цит. по: Берг, 1955),
Фитогеографическая граница на суше между двумя зональными типами
наземной растительности тундровой и полярно-пустынной проводится около
76° с. ш. в районе Новой Земли. Растительность южной части Новой Земли
входит в состав Новоземельеко-Вайгачской подпровинции Новоземельско-Запад-
ио-сибирской-Центрально-сибирской провинции лодобласти арктической тундры.
Основными компонентами растительности Новой Земли являются лишайники
и мхи. Из кустарников встречается несколько видов ив - мелкие, прижатые к
земле кустики, растущие, главным образом, около моря. Отмечена на Новой
Земле карликовая береза (Betula nana). Из злаков широко распространены дюпон-
тия, щучка; много пушицы. Большие площади на Новой Земле заняты осоками.
Из разнотравия встречаются белокопытник, мятник, камнеломки, лютики,
астрагал, остролодка, незабудки, незабудочник, валериана, гречиха, наррия.
оердечник, синюха, полынь, кипрей, ясколка, куропаточья трава. Из ягод на
Новой Земле встречаются морошка и голубика. "Цветов на Новой Земле иногда
попадается очень много. Они все очень .ярки, и из них особенно красивы
незабудки, растущие густыми плотными кустиками, и желтые полярные маки. Но
цветы на Новой Земле производят очень странное впечатление" (Матусевич;
Соколов. 1927, с. 52).
Глава 10. СОДЕРЖАНИЕ РАДИОНУКЛИДОВ В СРЕДЕ И БИОТЕ
До последнего времени; радиоактивное загрязнение прибрежной зоны и
островных ландшафтов российских полярных архипелагов оставалось
практически не исследованным. Со ссылкой на военных специалистов в отечественной
литературе есть только самые общие сведения об уровнях искусственных
радиоизотопов и мощности новоземельских ядерных взрывов. Испытание ядерного
оружия проводилось на трех технологических площадках Новой Земли: зона А -
губа Черная, зона В - пролив Маточкин Шар и зона С - губа Сульменова.
После атмосферных испытаний уровни радиоактивных выпадений в различных
районах Арктики выросли в 200-3000 раз. После взрыва супербомбы в 54 Мт служба
Госгидромета в Амдерме зафиксировала плотность радиоактивных осадков,в 11
тыс. раз превышающую фоновую (Голубчиков, 1993; Матущенко. 1991).
Пс расчетам А.М.Матущенко (1991), за весь период новоземельских
испытаний в атмосферу попало до 5 тыс. Ки цезия-137. Суммарная мощность
энерговыделений от всех 132 испытаний превысила 273 Мт, что составляет 94% от
мощности всех советских ядерных взрывов.
На территории новоземельских полигонов есть три официальные
санитарно-защитные зоны с повышенными уровнями загрязнения искусственными
радионуклидами. В чаотности, от атмосферных взрывов до 1993 г. остались
северная (до 50 мкР/ч) и южная (до единиц мР/ч) санитарно-защитные зоны. Кроме
того, от подземного ядерного взрыва 2.08.87 г., когда газообразные
продукты попали в атмосферу, осталась также зона с фоном до 100 мкР/ч
(Матущенко, 1991). Вместе с тем, по данным аэрогаммасьемки южного острова
архипелага (Островская Э.Я., специалист бывшего Мингео СССР, личное сообщение)
существует несколько десяткой локальных участков с повышенным радиационным
фоном (до 2 мР/ч). Диаметр этих участков от нескольких сот метров до 5 км
(Экологические проблемы Арктики .... 1992).
Ориентировочно, в 1967 году концентрация цезия-137 в лишайнике и
мышцах оленя была до 10"' Ки/кг, а в новоземельском гольце - до Ю"9 Ки/кг. К
1990 году уровень цеэия-137 в указанных организмах снизился почти до
10"10 Ки/кг (Матущенко, 1991).
Содержание цезия и стронция'в различных объектах природной среды
Новой Земли по данным А.М.Матущенко (1991) колеблется от 30 до 2500 Бк/кг
(1 Ки = 3.7-Ю10 Бк). Уровень радиоактивности новоземельских рыб,
отловленных в 1985-1990 годах, не превышал 100-130 Бк/кг. Однако природный
радиоизотоп калия-40 обуславливает до 9035 этой активности в рыбах. Известно,
что комиссия Codex Allmentarlx Всемирного общества здравоохранения
рекомендовала (Бюлл. МАГАТЭ. Т. 31. N3.4. 1989) следующие допустимые уровни
радиоактивности загрязнения пищевых продуктов: по цеэию-137 - 1000 Бк/кг и
по стронцию-90 - 100 Бк/кг.
Концентрация радионуклидов в почвах и некоторых видах биоты на юге
Новой Земли в районах мыса Саханина и озера Крест-то в десятки и даже
сотни раз превышает фоновое значение (Калякин. Криволуцкий. 1991). Так, со-
держание стронция-ЭО варьировало в почвах от 170 до 1550 Бк/кг. в опавших
ивовых листьях (подстилке) от 77 до 530 Бк/кг, в лишайниках от 720 до
1330 Бк/кг, а в траве и в костях копытного (мышеподобного) лемминга
составило соответственно 1050 и 2550 Бк/кг (табл. 29; Калякин, Криволуцкий.
1991). Возможно, эти цифры несколько завышены, хотя район наблюдений
находится очень близко от южного новоземельского ядерного полигона.
Таблица 29
Содержание радионуклидов в природных объектах
на юге Новой Земли. Бк/кг
Район
Вид объекта
Сумма
90Sr+65Cs
по р-излу-
чению
238*239 рц
по а-иэлу-
чению
Суммарное излучение
90
sr
137
Сз
Озеро Крест-то Почва
Мыс Сахалина
Мыс Сахалина Подстилка*
Озеро Крест-то
Озеро Крест-то Зимнее гнездо
лемминга" .
Озеро Крест-то Лишайники
Мыс Саханина Погадка белой
совы' * *
Мыс Саханина Кости копытного
лемминга
1000
1200
120
900
880
1500
480
30,
420
430
910
670
1200
1000
2500
950
500
620
540
540
500
20
20
20
20
20
20
20
20
20
50
50
50
50
50
50
50
47
50
50
140
50
130
76
50
230
160
170
96
66
430
21.7
'Подстилка состоит из сухих листьев ив; в отличие от более южных
районов на Новой Земле из-за постоянных сильных ветров подстилка
накапливается под камнями, развалинами строений и т.д.
"Зимние гнезда сибирские лемминги устраивают под снегом из
несъедобных частей различных трав; в данном случае гнездо было сделано из стеблей
злака - красной овсяницы.
'"То есть отрыгнутые птицей непереваренные ткани; содержала шерсть
копытного лемминга.
На сегодняшний день средняя плотность радиационного загрязнения почвы
в районах проведения испытаний, исключая эпицентры взрывов, составляет для
цезия-13,7 от 2.2 до 3.3 ГБк/кмг. а для стронция-90 - 1.5 ГБк/кмг (Dubasov
et al., 1993). Как известно, опасная доза воздействия -7-1210"3 мР/ч.
Содержание и распределение йода-129 в донных отложениях арктических
морей, окружающих Новую Землю, было детально изучено в 1992 году (Klllus
et al.. 1993). Соотношение йода-129 к йоду, 'а также градиенты концентрации
йода-129 в водах новоземельского шельфа не похожи на величины концентраций
этого элемента, характерные для Северного моря, средняя концентрация
йода-129 в осадках Баренцева моря в 2-3. раза превьшает уровни в осадках
Карского моря. Для Карского моря содераание цезия-129 в осадке оценивается
в 10 атом/г. Для донных ооадков акватории вокруг Новой Земли характерно
высокое соотношение йода-129 к цезив-137. которое равно 126 (используя
ldpm/g цезия-137). Анализ распределения йода-129 в донных осадках в
районе Новой Земли позволяет сделать вывод, что причиной возникновения
цезия- 13? в Баренцевом и Карском морях является испытание ядерного оружия
(Klllus et al., 1993).
В результате подводных и наземных ядерных взрывов на южном новозе-
мельском полигоне губа Черная была подвергнута интенсивному радиоактивному
загрязнению. Осколки деления атомного ядра включились в абиотические и
биотические элементы экосистемы этого, залива. В настоящее время, в.связи с
распадом с конца 50-х годов широкого спектра радионуклидов, загрязнение
водоема определяется цезием-137, стронцием-90, америцием-241, плутони-
ем-239+240 и плутонием-238 (рис. 56, 57) (Матишов Д., 1993; Strand et al.,
1993).
1600
« моо
f 1200
1000
BOO
600
400
200
0
\ПЛ)\
mm
Ас22В ВШ2 СовО С*Ш EirfSS Pb212
Гаши-источшим
Рис. 56. Гамма-источники в донном осадке губы Черной.
Южный атомный полигон. Новая Земля, июль 1992 г.
Рис. 57. Альфа-источники в донном осадке губы Черной.
Южный атомный полигон. Новая Земля, июль 1992 г.
1 - PU-239+240; 2 - Ри-238; 3 - Ат-241
Для изучения перемещения радионуклидов в море с новоземельского
побережья существует два контрольных полигона: один в губе Черной, другой в
заливе Бакан у мыса Столбовой. Активность проб почв, отобранных в
интервале 0-Юсм,была обследована на гамма-спектрометре Ge (L1) детектором
объемом 160 см3 (Dubasov et al.. 1993).. Плотность радиоактивного заражения на
Рис. 59. Районы дампинга твердых радиоактивных отходов (РАО) на
Новой Земле (Золотков. 1992):
1 - 1450 контейнеров, баржа с аварийным ядерным реактором
(170 тыс.Юл), лихтер - перевозчик жидких РАО; 2 - твердые РАО
суммарной активностью 3.4 тыс.Ки; 3 - 4750 контейнеров, лихтер "Н.Бауман1',
центральный отсек ледокола "Ленин" с тремя аварийными реакторами и
экранной сборкой; 4 - 850 контейнеров; 5 - 1850 контейнеров и аварийная
атомная лодка - два реактора с невыгруженным топливом; 6 - 550
контейнеров и отсеки четырех аварийных атомных подлодок; 7 - 650 контейнеров
с РАО; 8 - аварийный реактор без ядерного топлива, суммарная
активность 1856 Ки; 9 - 400 контейнеров; 10 - 250 контейнеров; 11 - место
проведения наиболее мощных испытаний ядерного оружия; 12 - место
проведения последних ядерных испытаний; 13 - место первого подводного,
наземного и первых подземных ядерных испытаний, предполагаемое место
захоронения атомной подлодки "Комсомолец"; 14 - предполагаемый район
размещения регионального могильника РАО; 15 - предполагаемый район для
развертывания ядерных испытаний
побережье Черной губы изменяется от 0.4 кБк/м2 до 0.4 мБк/м2. Выброс от
взрывов 1957 года отразился в верхнем 5-сантиметровом слое, в котором
содержится до 7555 цезия-137. На дне Черной губы осадки имеют уровень
цезия-137 порядка 20-50 Бк/кг. Это в 2 раза выше содержания природных
радионуклидов Th-232 и U-238. На участке залива Бакан загрязнение почвы цези-
ем-137 варьирует от 0.4 кБк/м2 до 6 мБк/мг. В среднем эти величины
сопоставимы с уровнем глобального загрязнения в. северном полушарии.
В питьевой воде поселков Белушья, Рогачева и Северный на Новой Земле
концентрация цезия-137 и стронция-90 в 3-4 раза ниже, чем предельно
допустимая доза для населения. Однако уровни этих радионуклидов (11-20 Бк/м2)
вполне ощутимы.
В 1990-1991 годах концентрация цезия-137 в новоземельских мхах
местами составляла 200 Бк/кг (Dubasov et al.. 1993). В мышцах оленей, гусей,
уток и рыб накопилось радиоцезия до 40 Бк/кг. Содержание стронция-90 в
костях гольца, гусей и оленей достигало соответственно: 1-20. -10-20 и
20 Бк/кг.
Приведенные выше данные свидетельствуют о пестрой, мозаичной
радиоэкологической картине на новоземельском архипелаге. Ясно то. что
необходимо радиационное картирование островов и побережья, специальное
радиоэкологическое исследование почв, флоры и фауны.
Сегодня существует крайне трудная техническая проблема в связи с
утилизацией и захоронением радиоактивных отходов: Сегодня публикуются
различные картосхемы с указанием мест затопления в Карском и Баренцевом морях по
обе стороны Новой Земли более 11000 контейнеров с радиоактивными отходами,
а также 15 аварийных реакторов с атомных подводных лодок.
Загрязнение радиоизотопами морских акваторий при различного рода
захоронениях было достаточно мощным. В период 1963-1986 годов контейнеры с
твердыми радиоактивными отходами регулярно захоронялись в новоземельских
бухтах Карского моря (рис. 58), В заливах Абросимова, Степового, Цивольки,
Ога, Седова. Благополучия, Течений и других, погребено много тысяч
различных контейнеров (Золотков, 1991). В частности, в заливе Цивольки
затоплены: судно атомно-технологического обслуживания "Н.Бауман", бетонный
контейнер с ядерным реактором атомохода "Ленин". Жидкие отходы (в объеме
около 16 тыс.м3 и общей активностью 5500 Ки) были слиты в северо-восточном
секторе (74-77° с.ш.) Баренцева моря.
Имеющиеся материалы по радиоактивному загрязнении архипелага Новая
Земля позволяют сделать вывод о том, что ключевые элементы экосистемы
архипелагов; озерные и морские прибрежные воды (до 10-20 Бк/м3), донные
отложения (от 1 до 20-50 Бк/кг). мхи и лишайники (10-540 Бк/кг), арктические
гольцы, птицы (гуси) и северные олени (до 10-20 Бк/кг) - содержат
цезия-137. а также других радионуклидов на фоновом уровне. Исключение
составляют донные отложения и почвы локальных территорий (губа Черная и др.)
старых ядерных полигонов. В их пределах концентрация радиоизотопов в среде
и, возможно, биоте может быть от 0.5 до 5 тыс. Бк/кг и более. Для
восстановления прибрежных экосистем.и обеспечения охраны природы Новой Земли и
прилегающих морей, вероятно, следует ставить вопрос о создании системы
биосферных заповедников.
Как видно из материалов данной книги, архипелаг Новая Земля является
уникальным объектом с точки зрения многих естественных наук: географии,
геологии, биологии, экологии. Поэтому в будущем эти. острова должны
использоваться комплексно, с учетом интересов не только обороны и народного
хозяйства, но и науки. Было бы очень полезно провести исследования. влияния
прошлых испытаний ядерного оружия на экосистемы архипелага и шельфа.
Будем надеяться, что после .заключения международных соглашений о
прекращении испытаний ядерного оружия Новая Земля станет доступной для
международных исследований.
ЛИТЕРАТУРА
АГАПОВ И.Д. Биология трески (Gadua morhua L.) западного побережья
Новой Земли // Тр. / Арктического института.- 1937.- Т. 100.- С. 25-45. .
АГАПОВ И.Д., ТОПОРКОВ Г.Н. Некоторые данные о рыбах западного
побережья Новой Земли // Проблемы Арктики.- 193?.- N 2,- с. 11-25.
АДЛЕРБЕРГ Г.П. Хищные звери (Carnlvora flssipedla) Арктики.- Л.:
Изд-во Главсевморпути. 1935.- 316 с.
АДРОВ Н.И. Трансформация водных масс системы Гольфстрима.- Апатиты.
1993.- 171 с.
АДРОВ Н.М., СМОЛЯР И.В. Построение полей водных масс Баренцева моря и
их использование для решения задач гидрометеорологического и
рыбопромыслового прогнозирования,- Апатиты, 1988.- 44 с.
АДРОВ Н.М., ФЕОПЕНТОВ С.А. Автоматизированный анализ океанологической
информации.- Апатиты, 1994.- 54 с.
АДРОВ Н.М., ПРОЗОРОВ А. С., СЛОБОДИН В. П., ФЕОПЕНТОВ С. А. Варианты
формальных классификаций водных масс Баренцева моря для проведения
вычислительных экспериментов в целях экологического прогнозирования //
Теоретические подходы к изучению экосистем морей Арктики и Субарктики.- Апатиты,
1992.- С. 126-139.
АДРОВ Н.М., КОЛПАЧНИКОВ А. Н., СЛОБОДИН В.П. Атлас водных масс
Баренцева моря. Ч. 3. Классификация по принадлежности к глобальным системам
циркуляции.- Апатиты, 1993.- 52 с.
АКСЕНОВ А.А. (ред.) Арктический шельф Евразии в позднечетвертичное
время.- М.: Наука, 1987.- 278 с.
АКСЮТИНА З.М. Количественная оценка скоплений рыб методом изолиний //
Тр. / ВНИРО.- 1970.- Т. 71.- ВЫП. 2.- С. 302-308.
АЛЕКСАНДРОВА В.Д. Растительность Окного острова Новой Земли //
Растительность Крайнего Севера СССР и ее освоение.- 1956.- Вып. 2.- С. 187-306.
АЛЕКСАНДРОВА В.Д. Проект классификации растительности Арктики // Бот.
журн.- 1979.- Т. 64, N 12.- С. 1715-1730.
АЛЕКСАНДРОВА В.Д. Растительность полярных пустынь СССР.- Л.: Наука,
1983,- 144 С.
АЛЕКСЕЕВА СП. Икринки и мальки Печорского залива // Тр. / ВНИРО.-
1949.'- Т. 17.- С. 175-188.
АЛТУХОВ К.А. О размножении и развитии сайки Boreogadus salda (Lepe-
chln) в Белом море // Вопр. ихтиологии.- 1979.- Т. 19.- Вып. 5.-
С. 874-882.
АНДРИЯШЕВ А.П. Рыбы северных морей СССР.- М.-Л.: Изд-во АН СССР.
1954.- 566 с.
АНИСИМОВА Н.А. К фауне и количественному распределению иглокожих
Баренцева' моря // Бентос Баренцева моря. Распределение, экология и структура
популяций,- Апатиты, 1984,- С. 32-44.
АНИСИМОВА Н.А. Структура фауны, распределение и экология иглокожих
Баренцева моря и сопредельных районов Северного Ледовитого океана: Авто-
реф. дисс. ... канд. биол. наук. - Л., 1989. - 22 с.
АНТИПИН В.М. Морской зверь на Новой Земле // Сов. Арктика.- 1938а.-
N 7.- С. 98-101.
АНТИПИН В.М. Фауна позвоночных северо-востока Новой Земли // Проблемы
Арктики.- 19386.- N 2.- С. 153-171.
АНТИПОВА Т.В. Предварительные данные о видовом составе и
количественном распределении двустворчатых моллюсков Баренцева моря // Океанология.-
1975.- Т 15.- ВЫП. 2.- С. 330-332.
АНТИПОВА Т.В. Географическое распространение двустворчатых моллюсков
Баренцева и Карского морей.- М.: ВНИРО.- 1978.- 64 с. (Рук. деп. в ВИНИТИ.
АНТОНОВ С.Г., ЧЕРНОВА Н.В. Состав ихтиофауны Карского моря // Эколо
гия и биоресурсы Карского моря.- Апатиты, 1989.- С. 95-100.
Арктический шельф Евразии в позднечетвертичное время.- М.: Наука,
1987.- 278 с.
АРОНОВИЧ Т.М.. ДОРОШЕВ СИ.. СПЕКТРОВА Л.В. Биологические особенности
эмбрионального и личиночного развития беломорских рыб // Биология
промысловых рыб и беспозвоночных на ранних стадиях развития.- Мурманск: Изд-во
ПИНРО. 1974.- С. 9-11.
АРСЕНЬЕВ В. А. Морж // Млекопитающие' Советского Союза. Под
ред. В. Г. Гептнера. Т. 2, Ч. 3.- М.: Высшая школа, 1976.- С. 33-51.
Атлас Арктики.- М.: Изд-во ГУГК, 1985.- 204 с.
Атлас Мурманской области. - М.: Изд-во ГУГК-НИГЭИ, 1971.- 33 с.
Атлас океанов. Северный Ледовитый океан.- М.: Изд-во ВМФ СССР, 1980.-
235 с.
БАРАНЕНКОВА А.С.. ПОНОМАРЕНКО В. П., ХОХЛИНАН.С. Распределение,
размеры и рост личинок и мальков сайки в Баренцевом море // Вопр.
ихтиологии.- 1966.- Т. 6.- Вып. 3.- С. 498-518.
БАРДАН СИ., ДРУЖКОВ Н.В.. БОБРОВ В. А., БАЙТ A3 В. А. Комплексный
экологический мониторинг в губе Дальнезеленецкая (Баренцево море):
Зимне-весенний период 1987-1988 ГГ.- Апатиты, 1989.- 42 с.
БЕЛИКОВ СЕ. Белый медведь в Советокой Арктике // Тез. Всес. конф.
"Экол.проблемы охраны «ивой природы".- М.. 1990.- Ч. 1.- С 81-83.
БЕЛИКОВ СЕ., ГОРБУНОВ D.A., ШИЛЬНИКОВ В.И. Распространение
ластоногих и китообразных в морях советской Арктики и в Беринговом море зимой //
БИОЛ. моря. 1989.- N 4.- С. 33-41.
БЕЛ0Б0Р0Д0В А.Г.,ТИМОШЕНКО D.К. В защиту атлантического моржа //
Природа.- 1974.- N 3.- С. 97-99.
БЕЛОВА А.В.. ТАРВЕРДИЕВА М.И. Некоторые данные о питании сайки // Тр.
/ ММБИ.- 1964.- Т. 5.- С. 143-147.
БЕЛОПОЛЬСКИЙ Л.С. Состав кормов морских птиц Баренцева моря // Ученые
записки Калининградского ун-та.- 1971.- Вып. 6.- С. 41-67.
БЕЛЫЕВА Е. В. О распределении биогенных элементов и растворенного
кислорода в водах Арктического бассейна // Химические ресурсы морей и
океанов.- М.: Наука, 1970.- С. 35-42.
БЕЛЬКОВИЧ В.М., ХУЗИН Р.Ш. Атлантический морж. Сберечь и умножить
ценное животное Севера // Природа.- I960.- N 2.- С. 67-69.
БЕРГ Л.С. Появление бореальных рыб в Баренцевом море // Сб. посвящ.
научн. деят. Н.М. Книповича. - М.: Изд-во АН СССР, 1939.- С. 17-25.
БЕРГ Л.С. Рыбы пресных вод СССР и сопредельных стран. Т. 1.-М.-Л,:
ИЗД-ВО АН СССР, 1948.- 446 С.
БЕРГ Л. С Природа СССР.- М.: Гос. изд-во географ, лит., 1955.- 495 о.
БЕРЕЗКИН В. А. Динамика моря.- Л.: Изд-во АН СССР. 1938.- 542 с.
БЕРЕСТОВСКИЙ Е.Г., АНИСИМОВА Н.А., ДЕНИСЕНКО С.Г., ЛУППОВА Е. Н..
САВИНОВ В.М.. ТИМОФЕЕВ С.Ф. Зависимость между размерами и массой тела
некоторых беспозвоночных и рыб Северо-Восточной Атлантики.- Апатиты, 1989.-
29 с.
БОБРОВ Ю.А.. САВИНОВ В.М.. МАКАРЕВИЧ П.Р. Хлорофилл и первичная
продукция // Экология и биоресурсы Карского моря.- Апатиты. 1989.- С. 45-50.
БОЙЦОВ В.Д. Структура гидрологических сезонов в прибрежной зоне Мур-
мана // Физико-химические условия формирования биологической продукции в
прибрежной зоне Мурмана.- Апатиты,. 1980.- С. 18-25.
БОЙЦОВ В.Д.. БОЧКОВ D.А., ЕРГАКОВА Л.Р. и др. Баренцево. Норвежское,
Гренландское моря, Северо-Восточная и Северо-Западная Атлантика //
Промысловая океанография.- М.: Агропромиздат. 1986.- С. 248-268.
БОРКИН И.В. Эффективность нереста сайки Баренцева моря // Биология и
промысел пелагических рыб Северного бассейна.- Мурманск: Изд-во ПИНРО,
1983а.- С. 52-64.
БОРКИН И.В. Результаты исследований ихтиофауны в районе Земли
Франца-Иосифа и к северу от Шпицбергена // Исследования биологии, морфологии и
физиологии гидробионтов.- Апатиты, 19836.- С 34-42.
БОРКИН И.В. О биологии сайки района Шпицбергена // Географические
проблемы' изучения и освоения арктических морей.- Л.: Изд-во ГО СССР,
1985.- С. 152-153.
БОРКИН И.В. Распределение и особенности биологии сайки района
Шпицбергена в раннем онтогенезе // Экология и биологическая продуктивность
Баренцева моря.- Мурманск, 1986.- С. 193-195.
БОРКИН И.В. Размножение и ранний онтогенез сайки Boreogadus salda
(Lepechln) Баренцева-моря. Автореф. дис. ... канд. биол. наук. - М.. 1990.-
21 с.
БОРКИН И.В., ОЖИГИН В.К.. ШЛЕЙНИК В.Н. Влияние океанологических
факторов на урожайность поколений сайки Баренцева моря // Влияние
океанологических условий на распределение и динамику популяций промысловых рыб
Баренцева моря.- Мурманск: Изд-во ПИНРО. 1987а.- С. 227-243.
БОРКИН И. В.. ПОНОМАРЕНКО В. П.. ТРЕТЬЯК В. Л., ОШЕЙНИК В. Н. Сайка
Boreogadus salda (Lepechln) - рыба полярных морей (запасы и использование)
// Биологические ресурсы Арктики и Антарктики. - М.: Наука, 19876.-
С. 183-207.
БОРКИН И.В., БЕЗДЕНЕЖНЫХ А.В. .Анализ взаимоотношений типа
"хищник-жертва" между треской и сайкой в Баренцевом море // Тез. докл. 3 Все-
союз. конф. по морской биологии.- Киев, 1988а.- Т. I.- С. 258-259.
БОРКИН И.В., НЕСТЕРОВА В.Н. Рель кормовых условий в питании и
распределении личинок сайки // Питание морских рыб и использование кормовой базы
как элементы промыслового прогнозирования.- Мурманск: Изд-во ПИНРО,
19886.- С. 79.
БОРКИН И.В., ТЕРЕЩЕНКО Е. С. Взаимосвязь репродуктивного потенциала
сайки Баренцева моря с ее биологическим состоянием // Материалы отчетной
сессии по итогам НИР ПИНРО 1992 года.- Мурманск: Изд-во ПИНРО. 1993.-
С. 133-139.
БОРКИН И. В., ЧЕРНООК В. И., ПОНОМАРЕВ Я. И.. БОГОМОЛОВ В. D. Результаты
авиасъемки морских птиц Баренцева моря // Материалы отчетной сессии по
итогам НИР ПИНРО 1991 года.- Мурманск: Изд-во ПИНРО, 1992.- С, 277-290.
БРЕЙТФУС Л. Морской звериный промысел в Белом море и Северном
Ледовитом океане. Экспедиция для научно-промысловых исследований Мурмана.- СПб.:
Тип. И. Усманова, 1905.- 24 с.
БРОЦКАЯ В. А., ЗЕНКЕВИЧ Л.А. Количественный учет донной фауны
Баренцева моря 7/ Тр. / ВНИРО.- 1939.- Т. .4.- С. 5-126.
БУРМАКИН Е.В. Рыбы островов Советской Арктики // Тр. / Арктического
института.- 1957.- Т. 205.- С. 127-150.
БУТОРИН Д.А. Особенности промысла белухи и некоторые закономерности
ее распределения в Арктике // Рыбное хозяйство.- 1964.- N 5.- С. 19-21.
БУТОРИН Д.А. Где зимует белуха // Рыбное хозяйство.- 1966.- N 1.-
С. 28-29.
БЫЧКОВ В.А. Охрана и рациональное использование ластоногих.- М.:
ИЗД-В0 ВНИИТЭИСХ. 1976.- 62 С.
БЫЧКОВ В.А. Сохранение редких видов ластоногих и хищных млекопитающих
методом заповедания // Тез. докл. 9 Всес, совещ. по изуч., охране и рац.
использ. морск. млек. - Архангельск, 1986.- с. 79-80;'
Бюллетень МАГАТЭ.- 1989.- Т. 31, N 3-4..
ВАСЮТИНА И.П. Фитопланктон юго-восточной части Баренцева моря в
июле-августе 1977 г. // Исследования фитопланктона в системе мониторинга
Балтийского моря и других морей СССР.- М.: Гидрометеоиздат, 1991.-
ВЕЛЬДРЕ С. Р. О корреляционной структуре внешних морфологических
признаков ушастой круглоголовки Phrynocephalus mystaceus (Pallas,1776) //
Применение математических методов в биологии.- л.. 1964.- Вып. 3.- С. 75-85.
ВИЗЕ В.Ю. Моря Советской Арктики,- Л.: Изд-во Главсевморпути. 1939.-
ВИЗЕ В. Ю. Гидрохимические условия в области кромки льдов арктических
морей //Проблемы Арктики.- 1943.- К 2..- С. 13-21.
ВИНОГРАДОВ Н.П. Морские млекопитающие Арктики // Тр. / Арктического
института,- 1949.- Т. 202.- С. 76-98.
ВИНОГРАДОВА Л.А., ГРУЗОВ Л. В. О биоценологическом районировании эпи-
пелагиали Северной Атлантики и сопредельных районов // Тр. / ГиИН.- 1990. -
Т. 182.- С. 112-133.
ВЛАДИМИРОВ В. И. К биологии трески у Новой Земли // Учен. зап.
Пермского ун-та.- 1937.- Т. 3.- ВЫП. 1.-.С. 121-128.
ВОРОНОВА Н.Н. Норфо-экологические особенности баренцевоморских форм
фораминиферы Trucatullna lobata d'Orblgny // Биология Баренцева и Белого
морей.- Апатиты, 1976.- С. 81-86.
ВЫХАНДУ Л. К. Об исследовании многопризнаковых биологических систем //
Применение математических методов в биологии.- Л., 1964.- Вып. 3.-
С. 19-22.
ВИШНЕВСКАЯ Т.О., БЫЧКОВ В. А.. КОНДАКОВ А. А.. МИШИН В. Л. Серый тюлень.
Биология и современное состояние популяций,- Апатиты, 1990.- 46 с.
ГАЛКИН СИ. История исследований донной фауны Баренцева моря //
Экология донного населения шельфовой зоны.- М.: Наука, 1979.- С. 112-127.
ГАЛКИН Ю.И. Многолетние изменения донной фауны в переходных
биогеографических районах на примере моллюсков Баренцева моря // Жизнь и среда
полярных морей.- Л.: Наука. 1989,- С. 157-164.
ТЕМП К.П., ВЫЗОВА Л.Н. Водоросли пролива Карские Ворота (южное
побережье Новой Земли) // Материалы рыбохоз. исслед. Северного бассейна.-
1970.- Вып. 13.- С. 102-110.
ГЕПТНЕР В.Г. Материалы по млекопитающим острова Диксон, прилегающей
части Северо-Западного Таймыра и Карского моря // Сб. трудов Гос. музея
при МГУ.- Т937.- Т. 3.- С. 3-24.
ГЕПТНЕР В.Г.. ЧАПСКИЙ К.К.. АРСЕНЬЕВ В.А.. СОКОЛОВ В. Е. Млекопитающие
Советского Союза.- М.: Высшая школа, 1976.- 718 с.
Геологическое строение островов Новой Земли и острова Вайгач // Тр. /
НИИГА.- 1957.- Т. 81.- 216 С.
Геология СССР. Т. XXYI. Острова Советской Арктики. Ч. 1.
Геологическое описание.- М.: Недра, 1970.- 548 с.
ГЕРШАНОВИЧ Д.Е. Новые данные по стратиграфии позднечетвертичных
отложений Берингова моря и залива Аляска // Геология моря.- 1974.- N 3,-
С. 68-77.
Гидрометеорологические условия шельфовой зоны морей СССР.- Л.: Гидро-
метеоиздат, 1985.- Т. 6.- Вып. 1-2.- С. 92-118.
Гидрометеорология и гидрохимия морей СССР. Т. 1. Баренцево море.
Вып. 1. - Л.: Гидрометеоиздат. 1990.- 310 с.
ГОВОРУХА Л.С. Современное наземное оледенение Советской Арктики.- Л.:
Гидрометеоиздат, 1988.- 256 с.
ГОЛИКОВ А. Н.. СКАРЛАТО 0. А.. АВЕРИНДЕВ В. Г.. МЕНШУТКИНА Т. В.. •
НОВИКОВ O.K., ШЕРЕНЕТЕВСКИЙ A.M. Экосистемы Новосибирского мелководья и
некоторые закономерности их распределения и функционирования // Экосистемы
Новосибирского мелководья и фауна моря Лаптевых и сопредельных вод. - Л.:
Наука, 1990.- С. 4-79.
ГОЛИКОВ А.Н., АНИСИМОВА Н. А., ГОЛИКОВ А. А., ДЕНИСЕНКО Н. В., КАПТИЛИНА
Т. В., НЕНШУТКИН В. В., НОВИКОВ O.K.. ПАНТЕЛЕЕВА Н. Н., ФРОЛОВА Е. А. Донные
сообщества и биоценозы губы Ярнышной Баренцева моря и их сезонная •
динамика. - Апатиты, 1993. - 54 с.
ГОЛОВКИН А.Н. О выедании рыбы кайрами и моевками в гнездовий период в
Баренцевом море // Зоол. яурн. - 1963.- Т. 42.- Вып. 3.- С. 408-416.
ГОЛОВКИН А.Н. Влияние морских колониальных птиц на развитие
фитопланктона // Океанология.- 1967.- Т. 7.- Вып. 4.- С 672-682.
ГОЛОВКИН А.Н. Птичьи базары севера Новой Земли // Особенности биоло- .
гической продуктивности вод близ птичьих базаров севера Новой Земли.- Л.:
Наука, 1672,- С. 84-91.
ГОЛОВКИН А.Н. Метаболиты морских птиц: особый тип связи птиц с
морскими экосистемами // Тез. докл. 1В международного орнитологического
конгресса. - Н.: Наука, 1982. - С. 103.
ГОЛОВКИН А.Н.. ПОЗДНЯКОВА Л. Е. Влияние морских колониальных птиц на
режим биогенных элементов в прибрежных водах Восточного Нурмана // Новые
исследования планктона и бентоса Баренцева моря.- М.-Л.: Наука. 1964.-
С. 88-98.
ГОЛОВКИН А.Н., ШИРОКОЛ0Б0В В.Н.,. ГАРКАВАЯ Г. П. Особенности
распределения биогенных • элементов в районе птичьих базаров севера Новой Земли //
Особенности биологической продуктивности вод близ птичьих базаров севера
Новой Земли.- Л.: Наука, 1972.- С. 46-62.
ГОЛУБЧИКОВ С. Кому нужна Новая Земля? // Энергия: Экономика. Техника,
ЭКОЛОГИЯ.- 1993,- N 4.- С. 37-45.
ГОРБУНОВ Т.П. Птичьи базары Новой Земли // Тр. / Ин-та по изучению
Севера.- 1925.- Т. 26.- С. 1-4вГ
ГОРБУНОВ Г.П. Материалы по фауне млекопитающих и птиц Новой Земли //
Тр. / Ин-та по изуч. Севера.- 1929.- Вып. 40.- С. 169-239.
ГОРБУНОВ Ю.А.. БЕЛИКОВ С. Е., ШИЛЬНИКОВ В. И. Влияние ледовых условий
на распределение и численность белого медведя в морях Советской Арктики //
• БЮЛЛ. МОИП.- 1987.- ОТД. биол. Т. 92.- Вып. 5.- С. 19-28.
Горные страны Европейской' части СССР и Кавказ.- М.: Наука, 1974.-
ГРИНКЕВИЧ Н.с. Годовые изменения в питании трески Баренцева моря //
Тр. / ПИНРО.- 1957.- ВЫП. 10.- С. 88-105.
ГРОМОВ И. М., ГУРЕЕВ А. А.. НОВИКОВ Г. А., СОКОЛОВ И. И., СТРЕЛКОВ П. П.,
ЧАПСКИЙ К.К. Млекопитающие фауны СССР. Ч. 2.- М.-Л.: Изд-во АН СССР,
1983.- 923 с.
1
ГУРВИЧ Г.С. Донное население паотбищ трески Новоземельского
промыслового района //Тр. / Арктического института.- 1937.- Т. 100.- С. 77-91.
ГУРЕВИЧ В.И. Прикладная седиментология и геоэкология.- Л.: Недра,
1990.- 63 с. н
ГУРЬЯНОВА Е.Ф.. УШАКОВ П.В. К фауне Черной губы на Новой Земле //
Исследования морей СССР,- 1928.- Вып. 6.- С. 5-71.
ДЕНИСЕНКО Н.В. Количественное распределение мшанок в Баренцевом море
// Тез. докл. 7 Всесоюзн. коллоквиума по ископаемым и современным
мшанкам. - Н.. 1986.- С. 22-24.
ДЕНИСЕНКО Н.В. Распределение и экология мшанок Баренцева моря.-
Апатиты. 1990.- 156 с.
ДЕНИСЕНКО С.Г. Соотношение между биомассой, плотностью поселений и
продукцией двустворчатых моллюсков Chlamys Islandlca в Баренцевом'море //
Тез. докл. 5 Всесоюзн. конф. по промысловым беспозвоночным. - М., 1990.-
С. 108-110.
ДЕРЮГИН К.М. Гидробиологические исследования у Новой Земли //
Исследования морей СССР.- 1927.- ВЫП. 4.- С. 5-16.
ДИГАС Л.А. Зоогеографическое районирование Баренцева моря по форами-
ниферам // Вопросы геологии Южного Урала и Поволжья.- Саратов. 1970.-
С. 127-142.
ДМИТРИЕВ Н.А. О промысле сайки // Рыбное хозяйство.- 1958.- N 11.-
С. 12-14.
ДОБРОВОЛЬСКИЙ А. Д.. ЗАЛОГИН B.C. Моря СССР.- М.: Мысль. 1965.-350 с.
ДОБРОВОЛЬСКИЙ А.Д.. ЗАЛОГИН Б.С. Моря СССР.- М.: Изд-во МГУ. 1982.-
192 с.
ДОЛГУШИН Л. Д.. ОСИПОВА Г. Б. Ледники.- М.: Мысль, 1989.- 44В с.
ДОРОНИН Ю.П. Региональная океанология. - Л.: Гидрометеоиздат, 1986.-
302 с.
ДОРОНИНА Н.А.. ЖАДРИНСКАЯ н.Г. Новая Земля и Вайгач // Советская
Арктика. Моря и острова Северного Ледовитого океана,- М., 1970.- С. 363-388.
ДОРОФЕЕВ СВ. Основные черты годового цикла лизни беломорского лысуна
//Сов. рыбохоз. иссл. в морях Европейского Севера.-М.: ВНИРО, 1960.-
G. 443-456.
ДРУЖКОВ Н.В., МАКАРЕВИЧ П.Р. Пространственно-временная структура мик-
рофитопланктонного сообщества арктических морей (на примере юго-западной
части Карского моря) // Теоретические подходы к изучению экосистем морей
Арктики и Субарктики.- Апатиты, 1992.- С 30-38.-
ДУБРОВСКИЙ А.Н. Новая Земля // Природа.- 1938.- N 3.- С. 21-37.
ДУБРОВСКИЙ А. Н. Треска в Карском море //Природа. - 1941.- N 6.-
С. 74-75.
ДУНАЕВ Н.Н.. ИОНИНА. С. НИКИФОРОВ С. Л., ПАВЛИДИС С. А. Строение и
развитие западно-новоземельеких бухт в связи с проблемой поздневалдайского
оледенения // Современные процессы осадконакопления на шельфах Мирового
океана.- М.: Наука. 1990.- С. 94-103.
ЕЛШИН Ю.А. Приток речных вод в Баренцево и Белое моря и его колебания
внутри года и в многолетнем разрезе//Водные ресурсы. -■1979,- N 2.-
С. 65-69. -■
ЕСИПОВ В.К. Земля Франца-Иосифа.- Архангельск: Севкрайгиз. 1935а.-
75 с.
ЕСИПОВ В.К. Материалы по биологии и промыслу новоземельского гольца
// Тр. / Арктического института.- 19356.- Т. 17.- С. 1-70.
ЕСИПОВ В.К. Промысловые рыбы Баренцева моря.-М.: Пищепромиздат.
1937.- 110 с.
ЕСИПОВ В.К. Рыбы Карского моря. М.: Изд-во АН СССР. 1952. 145 с.
1ИРН0В Л.В.. ВИНОКУРОВ А.А.. БЫЧКОВ В.А. Редкие и исчезающие животные
СССР. Млекопитающие и птицы.- М.: Лесная промышленность. 1978,- 303 с.
ЗАЦЕПИН В.И.. ПЕТРОВ Н.с. Питание промысловых косяков трески в южной
части Баренцева моря // Тр. / ПИНРО.-.1939.- Вып. 5.- С. 1-170.
ЗЕЛИКМАН Э.А. К планктической характеристике юго-восточного сектора
Баренцева моря // Гидрологические и биологические особенности прибрежных
вод Мурмана. - Мурманск: Кн. изд-во. 1961,- С. 39-58.
ЗЕЛИКМАН Э.А.. ГОЛОВКИН А.Н. Особенности распределения и
продуктивности зоопланктона близ птичьих базаров севера Новой Земли // Особенности
биологической продуктивности вод близ птичьих базаров севера Новой Земли.-
Л.. 1972.- С. 92-114.
ЗЕМСКИЙ В.А. Атлас морских млекопитающих СССР. - М.: Пищевая
промышленность. 1980.- 184 с.
ЗИНОВА А.Д. Современное состояние и дальнейшие перспективы
флористических и биологических исследований по морским водорослям в СССР // Тр. /
совещ. работников водорослевой промышленности СССР.- 1962.- Т. 1.- С.
ЗИНОВА А.Д. Состав и фитогеографическое деление арктической флоры //
Гидробиология и биогеография шельфов холодных и умеренных вод Мирового
океана.- Л., 1974.- С. 12-13.
ЗИНОВА Е.С. Водоросли Карского моря // Тр. / Ленингр. об-ва
естествоиспытателей. - 1925.- Т. 55.- Вып. 3.- С. 1-116.
ЗИНОВА Е.С. Водоросли Новой Земли // Исследования морей СССР.- 1929.-
Вып. 10.- С. 41-128.
30Л0ТК0В А.А. Схема северного испытательного полигона (Новая
Земля) // Полярная правда.- Мурманск, 1991.- 25 сентября.
ЗУБАКИН Г.К. Крупномасштабная изменчивость состояния ледяного покрова
морей Северо-Западного бассейна.- Л.: Гидрометеоиздат, 1987.- 160 с.
ЗУБКОВ А.И. Оленеводство и оленьи пастбища на Новой Земле // Тр. /
Арктического института.- 1935.- Т. 22.- С. 7-34.
ЗУБОВА E.D. Видовой состав и распределение массовых видов
зоопланктона Карского моря // Структурно-функциональная организация экосистем
Баренцева моря.- Мурманск: Изд-во ММБИ КНЦ АН СССР. 1990.- С. 103-120. (Деп. в
ВИНИТИ. N 5272-В90).
ЗУБОВ Н.Н. К происхождению промежуточного холодного слоя в Полярном
бассейне // Тр. /Гос. Океанограф, ин-та.- 1929,- Т. 4.- Вып. 2.-
С. 47-68.
ЗУБОВ Н.Н. Льды Арктики.- М.: Главсевморпуть, 1944.- 360 с.
ИВАНОВА Е.И. Ихтиофауна и рыбный промысел Карской губы // Проблемы
Севера.- 1959.- Вып. 3.- С. 79-82.
ИЛЬИН Г.В. Биогенные вещества в водах юго-восточной части Баренцева
моря // Физико-химические условия формирования биологической продукции
Баренцева моря.- Апатиты. 1980.- С. 41-50.
КАЗАНОВА Н.И. Нерест, икринки и мальки рыб из юго-восточной части
Баренцева моря // Тр. / ВНИРО.- 1949.- Т. 17.- С. 157-174.
КАЛЯКИН В. Н., МУЖЧИНКИН В.Ф. Моржовые залежки острова Вайгач и
перспективы их сохранения // Тез. докл. 10 Всес. сов. по изуч., охр. и рац.
исп. мор. млекопитающих.- М., 1990.- С. 131-133.
КАЛЯКИН В.Н., КРИВОЛУЦКИЙ Д. А. Новая Земля: следствие'продолжается //
Энергия: Экономика, Техника. Экология.- 1991.- N 4.- С. 12-16.
КАПЛИН П.А. Фиордовые побережья Советского Союза. - М.: Изд-во АН
СССР. 1962.- 188 с.
КАПЛИН П.А. Изменения уровня Мирового океана в плейстоцене по данным
определений абсолютного возраста древних береговых линий // Проблемы па-
леогидрологии. - М.: Наука, 1976.- с. 95-101.
КАРПОВИЧ В.Н. Размещение белого медведя в Советской Арктике по данным
корреспондентской сети // Белый медведь и его охрана в Советской Арктике.-
Л.: Гидрометеоиздат, 1969.- С. 68-88.
Каталог ледников Новой Земли.- Л.: Гидрометеоиздат, 1976.- 79 с.
КАШКИН Н.И. Материалы к экологии Phaeocystls pouchetii (Harlot) La-
gerheim, 1893 (Chrysophyceae) в окраинных морях Северной Атлантики // Тр.
/ ММБИ.- 1964.- Т. 5.- С. 16-36.
КАШКИНА А.А. Ихтиопланктон юго-восточной части Баренцева моря // Тр.
/ ММБИ.- 1962.- Вып. 4.- С. 97-133. -•
КИРИЛЛОВ Ф.Н. О новоземельской песчанке Ammodytes marlnus Raltt //
Докл. АН СССР. Новая сер.- 1936.- Т. 4.- N 5.'- С. 225-227.
КИСЕЛЕВА Е.В. Сайка из Обской губы // Тр. / НИИ полярного земледеле-
ния, животноводства и промыслового хозяйства.- 1940.- Вып. 10.-
С. 103-110.
КЛЕЙНЕНБЕРГ С. Е., ЯБЛОКОВА.В.. БЕЛЬКОВИЧ В.М.-. ТАРАСЕВИЧ М. Н. Белу
ха. - М.: Наука. 1964. - 456 с.
КЛЕНОВА М.В. Геология Баренцева моря.-М.: Изд-во АН СССР, 1960.-
367 с.
КЛУМОВ С.А. Сайка - арктическая рыба // Рыбное хоз-во. - 1935а.- N 5.-
С. 18-19. '
КЛУМОВ С.А. Проблемы - сайки в Арктике//Рыбное хоз-во.- 19356. -
N П.- С. 95-96.
КЛУМОВ С. А. Распределение белухи на Северо-Азиатском Севере // Тр. /
Полярной КОМИССИИ АН СССР.- 1936.- ВЫП. 27.- С. 1-71.
КЛУМОВ С.А. Сайка и ее значение для некоторых жизненных процессов в
Арктике // Изв. АН СССР. Сер. биол. - 1937,- N 1.- С. 175-195.
КЛУМОВ С. А. Сайка // Промысловые рыбы СССР.- М., 1949.- С. .519-521.
КЛУМОВ С.К. Белуха Советского Севера (сырьевая база и промысел) //
Тр. / ВНИРО.- 1939.- N 12.-С. 23-31.
КЛЮГЕ Г. А. Биологические работы на полярной станции Мыса Желания в
1934-1935 гг. // Бюлл. Арктического ин-та.- Л., 1936.- N 4.- С. 170-172.
КНИПОВИЧ Н.М. Зоологические исследования на ледоколе "Ермак" летом
1901 г. // Ежегодник Зоол. музея Акад. наук.- 1901.- Т. 6.- С. 1-120.
КНИПОВИЧ Н.М. Определитель рыб морей Баренцева. Белого и Карского //
Тр. / Ин-та по изучению Севера.- 1926.- Вып. 27.- С. 157-158.
КОВАЛЕВА Г.А. Современные движения п-ва Адмиралтейства (Северный
остров Новой Земли) // Геотектонические предпосылки поисков полезных
ископаемых на шельфе Северного Ледовитого океана.- Л.: Недра, 1974.- С. 73-81.
КОВАЛЕВА Г.Н., ТИМОФЕЕВ О.П.. ТРУФАНОВ Г. В. Складчатые структуры
Южного острова Новой Земли // Геология Южного острова Новой Земли.- Л.:
Недра. 1982.- С. 88-99.
КОПТЕВ А.В.. НЕСТЕРОВА В.Н. Особенности широтного распределения
летнего кормового планктона в восточных районах Баренцева моря //
Исследования биологии, морфологии и физиологии гидробионтов. - Апатиты, 1983.-
С. 22-28.
КОРОТКЕВИЧ Е.С. Полярные пустыни.- Л.: Гидрометеоиздат, 1972.- 420 с.
КОРСУН.С.А.. ПОГОДИНА И. А.. ТАРАСОВ Г. А., МАТИШОВ Г.Г. Фораминиферы
Баренцева моря ( гидробиология и четвертичная палеоэкология).- Апатиты,
1993.- 220 с.
КОРЯКИН B.C. Владимир Александрович Русанов. Науч.-биогр. сер.- М.:
Наука. 1987.- 128 с.
КРАВЧУК В.А. Полярная треска - сайка Boreogadus saida (Lepechln) //
Изв. ВНИОРХ.- 1958.- Т. 44.- С. 207-214.
КРАСНОВ Ю. В. Гнездование большого поморника в СССР // Изучение
морских колониальных птиц в СССР.- Магадан. 1990.- С. 47-48.
КРАСНОВ Ю.В. Основные тенденции развития популяций и охрана морских
птиц Баренцева моря // Экологическая ситуация и охрана флоры и фауны
Баренцева моря.- Апатиты.- 1991.- С. 55-б2.
КРАСНОЖЕН А.С. История новейшего развития южной части Новой Земли //
Геология Юкного острова Новой Земли.- Л., 1982.- С 100-108.
КРАСНОЖЕН А. С. БАРАНОВСКИЙ О.Ф., ЗАРХИДЗЕ В. С.. МАЛЯСОВА Е. С.. ЛЕВ
О.Н. Верхнечетвертичные отложения Южного острова Новой Земли // Геология
Юкного острова Новой Земли.- Л... 1982.- С. 87-99.
КРАСНОЖЕН А. С.. БАРАНОВСКИЙ О.Ф.. ЗАРХИДЗЕ В. С.. МАЛЯСОВА Е. С.
Стратиграфия и основные этапы геологического развития архипелага Новая Земля в
кайнозое // Кайнозой шельфа и островов Советской Арктики.- Л.. 1986.-
С. 23-26.
КРАСОВСКИЙ С.К. Биологические основы промыслового использования
птичьих базаров. Этюды по биологии толстоклювой кайры // Тр. /
Арктического института.- 1937.- Т. 77.- С. 33-92.
КРЕПС Е.М. Гидрологический очерк Черной губы на Новой Земле и
реликтовых озер, к ней примыкающих // Исследования морей СССР.- 1927,- Вып. 5.-
С. 3-80.
КУЗНЕЦОВ А.П. Закономерности распределения пищевых группировок донных
беспозвоночных в Баренцевом море // Тр. / ИОАН СССР.- 1970.- Т. 88.-
С. 3-128.
КУЗНЕЦОВ А.П. Экология донных сообществ шельфовых зон Мирового океана
(трофическая структура морской донной фауны).- М.: Наука. 1980.- 244 с.
ЛАРИОНОВ В.В. Пространственное распределение и сукцессионные смены
видовых комплексов фитопланктона Баренцева моря. Автореф. дис канд.
биол. наук. - М., 1993. - 32 с.
ЛИНКО А.К. Планктон у побережья Западного Нурмана по отношению к
рыбному промыслу в 1906 г. // Тр. / Мурм. научно-промысл. эксп. Отчет
Л.Л.Брейтфуса за 1906 год.- С.-По.. 1915.- С. 137-151.
ЛУКА Г.И.. ОЖИГИН В.К., ПАНАСЕНКО А.Д. Мойва // Ихтиофауна и условия
ее существования в Баренцевом море.- Апатиты. 1986.- С. .7-12.
ЛУКИН Л.Р. О сроках и районах щенки атлантического моржа //
Экология.- 1978,- N 5.- С. 100-101.
МАКАРЕВИЧ П.Р. Микрофитопланктон // Методические рекомендации по
анализу количественных и функциональных характеристик морских биоценозов
северных морей. Ч. 1. Фитопланктон, зоопланктон, взвешенное органическое
вещество.- Апатиты. 1989.- С. 4-40.
МАКАРЕВИЧ П.Р., ДРУЖКОВ Н.В. Сравнительная характеристика
фитопланктона юго-восточной части Баренцева и юго-западной части карского морей //
Альгология.- 1994.- Т. 4, N 1.- С. 15-23.
МАКЕЕВ В.М. Основные особенности развития природной среды
Евроазиатской высокоширотной Арктики в позднем плейстоцене и голоцене // Проблемы
кайнозойской палеоэкологии и палеогеографии морей Северного Ледовитого
океана.- Апатиты, 1969.- С. 105-106.
МАНТЕЙФЕЛЬ П. Акклиматизация животных на Крайнем Севере // Сов.
Арктика.- 1938.- N. 7.- С. 74-81.
МАНТЕЙФЕЛЬ Б.Н. Сайка и ее промысел. - Архангельск: Севгиз, 1943.-
МАРУНИН В.Н.. КИЩИНСКИЙ А. А. Белый медведь в восточной части Карского
моря // Белый медведь и его охрана в Советской Арктике. - Л.: Гидрометеоиз-
дат. 1969.- С. 114-121.
МАСЛОВ Н.А. Миграции трески Баренцева моря // Тр. / ПИНРО.- 1968.-
Вып. 23,- С. 21-67.
МАСЛОВ Н.А. Об уточнении методики долгосрочных прогнозов состояния
запасов трески и пикши // Тр. / ПИНРО,- 1957.- Т. 10.- С. 30-53.
МАТЙШОВ Д.Г. Радионуклиды в донных осадках, биоте шельфа и побережий
Баренцева моря. Результаты радиоэкологических наблюдений, проведенных ММБИ
в 1991-1992 гг.- Апатиты, 1993.- 34 с.
МАТИШОВ Г. Г. Дно океана в ледниковый период.-Л.: Наука, 1984.-
176 с.
МАТИШОВ Г.Г. Геоморфология дна в экосистеме моря // Жизнь и условия
ее существования в бентали Баренцева моря.- Апатиты,' 1986.- С. 5-26.
МАТУСЕВИЧ Н. Н., СОКОЛОВ А. В. Новая. Земля. - Вологда: Изд-во Северный
печатник, 1927.- 101 с. '
МАТУШЕНКО A.M. Новая Земля: следствие заканчивается // Энергия:
Экономика. Техника, Экология.- 1991.- N 12.- С. 2-10.
МАХОТИН В.В. Особенности раннего онтогенеза тресковых рыб Белого моря
// Автореф. дис. ... канд. биол. наук. - М., 1982, - 25 с.
МИХЕЕВ В.Н., ПАКУЛЬСКАЯ Д.С. Пищевое поведение личинок рыб в среде с
неоднородным распределением корма // Питание морских рыб и использование
кормовой базы как элемента промыслового прогнозирования: Тез. докл.-
Мурманск, 1988.- С. 64-66.
МИТЕНЕВ В.К.. ШУЛЬМАН Б.С. Паразитбфауна рыб Баренцева моря.
Проходные рыбы>// Ихтиофауна и условия ее существования в Баренцевом море.-
Апатиты; 1986.- С. 151-160.
МОСКАЛЕНКО Б.К. О биологии полярной трески - сайки // Вопр.
ихтиологии.- 1964.- Т. 4.- ВЫП. 3.- С. 433-443.
МОНАСТЫРСКИЙ Г.Н. О типах нерестовых популяций рыб // Зоол. журнал.-
1949.- Т. 28.- Вып. 6.- С. 535-545.
МУСИНА А.А. Гидрохимическая характеристика Арктического бассейна //
Тр. / ААНЖ- I960.- Т. 218.- С. 5-64.
МЫСЛИВЕЦ В.И.. КАЛИНИНА Л.И., СОЛОВЬЕВА Г.Д. Опыт расчета изменения
объема Мирового океана в плейстоцене // Проблемы палеогидрологии.- М.:
Наука, 1976.- С. 102-112.
НАЗАРЕНКО Ю.И. Исследования гренландского тюленя в связи с проблемой
его питания // Тез. докл. 9 Всес. совещ. по изуч., охр. и рад. исп.
морских млекопитающих. - Архангельск. 1986.- С, 289-291.
НИКИФОРОВ Е.Г., ШПАЙХЕР А.О. Закономерности формирования
крупномасштабных колебаний гидрологического режима Северного Ледовитого океана.- Л.:
Гидрометеоиздат, 1980.- 265 с.
НИКИФОРОВ Е.Г., ПАНОВ В.В., ШПАЙХЕР А.О. Карское море //
Океанографическая энциклопедия.- Л.: Гидрометеоиздат. 1974.- С. 222-225.
НИКОЛЬСКИЙ Г.В. Частная ихтиология.- М.: Советская наука. 1954.-
458 с.
НИКОЛЬСКИЙ Г.В., РАДАКОВ Д. В. О пищевых отношениях пелагических рыб
северных морей // Материалы по экологии, трески Северной Атлантики. - М.,
1968.- С, 5-18.
НИЛЬСЕН Й.П. Русская опасность для Норвегии? // Оттар.- 1991.-
N 192.- С. 33-40.
НОРВИЛЛО Г.В. Ихтиопланктон // Экология и биоресурсы Карского моря.-
Апатиты, 1989.- С. 100-104.
НОРВИЛЛО Г.В., АНТОНОВ С.Г., ПЕТРОВ А.А. Некоторые результаты ихтиоп-
ланктонных работ в Карском море // Комплексные исследования природы
северных морей.- Апатиты, 1982.- С. 47-52. ,
НОРИНА A.M. Гидрохимическая характеристика северной части Баренцева
моря // Тр. / ГОИН.- 1965.- Вып. 83.- С. 243-271.
Оледенение Новой Земли.- М.: Наука, 1968.- 340 с.
ОСТРОУМОВ Н.К. К изучению белухи и промысел ее в Енисейском заливе У/
Тр. / Сибир. рыбохоз. научн. станции.- Красноярск. 1929.- Т. 4.- Вып. 1.-
38 с.
ОСТРОУМОВ Н.К. К систематике и размножению белухи Европейского Севера
и Карского моря // За рыбную индустрию Севера.- 1935.- N 11.- С. 28-35.
ПАВЛИДИС Ю.А., ДУНАЕВ Н.Н.. ЩЕРБАКОВ Ф. А. Актуальные проблемы
четвертичной геологии Баренцева моря // Современные процессы осадконакопления на
шельфах Мирового океана,- М.: Наука. 1990,- С. 76-93.
ПАНОВ В.В. Роль атлантических вод в гидрологическом и ледовом режиме
Арктических морей (на примере Карского моря) // Проблемы Арктики и
Антарктики. - 1961.- Вып. 8.- С. 75-77.
ПЕРЦЕВА Т.А. Определитель пелагических икринок рыб Баренцева моря.-
М.: Пищепромиздат, 1936. - 36 с,
ПЕТРОВ В.С., ШИР0К0Л0Б0В В.Н.. КРЫМСКИЙ А.В. Основные черты
гидрологического режима // Экология и биоресурсы Карского моря.- Апатиты. . 1989.-
С. 11-16.
ПЕТРОВ B.C., ИЛЬИН Г.В. Гидрохимические условия // экология и
биоресурсы Карского моря.- Апатиты, 1989.- С. 17-20.
ПЕЧЕНИК Л.Н.. ПОНОМАРЕНКО В.П., ШЕПЕЛЬ Л.И. Биология и промысел сайки
Баренцева моря. - М., 1973. - 65 с.
ПЕЧЕНИК Л.Н.. ШЕПЕЛЬ Л. И. Некоторые черты биологии полярной тресочки
Баренцева моря // Рыбное хоз-во. - 1970.- N 12. - С. 13-14.
ПОЗДНЯКОВ Ю.Ф. Распространение личинок мойвы в южной и юго-восточной
частях Баренцева моря.- Тр. / ММБИ, 1962.- Т. 4(8).- С. 134-145.
ПОКРОВСКИЙ СИ. Полярная треска - сайка // За рыбную индустрию
Севера. - 1936. - N 3. - С. 28-ЗоГ
ПОНОМАРЕВА Л.А. Икринки и мальки рыб Карского моря // Тр. / ВНИРО.-
1949.- ВЫП. 17.- С. 185-208.
ПОНОМАРЕНКО В.П. О промысле сайки и его возможностях у берегов
Баренцева моря // Промысл.-эконом, бюл. - 1962.- N 4.- С 39-43.
ПОНОМАРЕНКО В.П. Осенне-зимнее распределение преднерестовых и
нерестовых скоплений сайки в Баренцевом море // Тр. / ПИНРО.- 1963.- Вып. 15.-
С. 177-197.
ПОНОМАРЕНКО В.П. Районы и условия нереста сайки в Баренцевом море //
Материалы Сессии Учен. Совета ПИНРО за 1962-1963 гг.- 1964.- Вып. 3.-
С. 220-226.
ПОНОМАРЕНКО В.П. Развитие гонад и сроки нереста сайки в Баренцевом
море // Докл. АН СССР.- 1965а.- Т. 161.- Вып. 3.- С. 697-700.
ПОНОМАРЕНКО В.П. Распределение сайки в Баренцевом море // Рыбное
хоз-во.- 19656.- N 1.- С. 10-11.
ПОНОМАРЕНКО В.П. Питание личинок и мальков сайки в Баренцевом и
Карском морях // Материалы рыбохоз. исслед. Северного бассейна.- Мурманск,
1967а,- Вып. 10.- С. 20-27.
ПОНОМАРЕНКО В.П. Районы осенне-зимнего распределения сайки в южной
части Баренцева моря как индикатор весеннего распределения мойвы и мой-
венной трески у берегов // Вопр. ихтиологии.- 19676.- Т. 7.- Вып. 6.-
С. 1073-1079.
ПОНОМАРЕНКО В.П. Миграции сайки в советском секторе Арктики // Тр. /
ПИНРО.- 1968.- Вып. 23.- СГ 500-512.
ПОНОМАРЕНКО В.П.. ПОНОМАРЕНКО И. Я.. ЯРАГИНА Н.А. Потребление баренце-
воморской мойвы трескою и пикшей в 1974-1976 ГГ. // Тр. / ПИНРО.- 1978.-
Вып. 41,- С. 53-66.
ПОПОВ Л.А. Материалы по биологии размножения моржа моря Лаптевых //
. Бюл. МОИП, отд. биол.- I960.- Т. 65.- N 2.- С. 25-30.
ПОТЕЛОВ В.А. Распределение и миграции морского зайца в Белом,
Баренцевом и Карском морях // Морские млекопитающие.- М.: Наука. 1969 -
С. 245-250.
ПОТЕЛОВ В.А. Результаты научных исследований состояния запасов
морских млекопитающих северного бассейна // Тез. докл. 9 Всес. совещ. по
морским млекопитающим.- Архангельск, 1986а.- С. 327-329.
ПОТЕЛОВ В.А. Сравнительная характеристика районов щенки кольчатой
нерпы Белого, Баренцева и Карского морей // Тез. докл. 9 Всес. совещ. по
морским млекопитающим.- Архангельск, 19860.-С. 325-326.
ПОРТЕНКО Л.А. Производительные силы орнитофауны Новой Земли // Тр. /
биогеохим. лабор. АН СССР.- 1931.-Т. 2.-Приложение 2.- С. 3-52.
ПОТЕХИНА Е.М.. ХУСЙДТ.А.. БЕЛЯЕВА Н. В. Фораминиферы и минеральные
включения бухтовых льдов Северного острова Новой Земли // Бюл. МОИП., отд.
геология.- 1991.- Т. 66.- ВЫП. г.- С. 126-127.
ПРЕОБРАЖЕНСКИЙ Ю.В. Гидрометеорологическая характеристика Баренцева
моря.- Л.: Гидрометеоиздат, 1947.- 303 с.
РАСС Т.С. Нерест, икра и мальки промысловых рыб Баренцева моря //
Карело-Мурманский край.- 1934.- N 3-4,- С. 58-60.
РАСС*Т.С. Географические параллелизмы в строении и развитии костистых
рыб.- И.: МОИП, 1941.- 60 с.
РАСС Т.С. Состав ихтиофауны Баренцева моря и систематические признаки
икринок и личинок рыб этого водоема // Тр. / ВНИРО.- 1949.- Т. 17.-
С. 7-65. ■
РАСС Т.е.. КАЗАНОВА И.И. Инструкция по сбору икринок и личинок рыб.-
М.: ВНИРО, 1958.- 43 с. '■ ■
РЕТОВСКИЙ Л. 0. Питание молоди новоземельского гольца //Тр. /
Арктического института.- 1935.- Т. 17.- С. 89-109.
РОМАНКЕВИЧ Е.А.. ДАНЮШЕВСКАЯ А. И.. БЕЛЯЕВА А.Н., РУСАНОВ В. П. Биогео-
• химия органического вещества арктических морей.- М.: Наука. 1982.- 239 с.
Руководство по методам биологического анализа морской воды и донных
отложений.- Л.: Гидрометеоиздат. 1980.- 191 с.
РУСАНОВ В.П., ЯКОВЛЕВ Н.И., БУЙНЕВИЧ А.Г. Гидрохимический режим
Северного Ледовитого океана // Тр. / ААНИИ. - 197Э. - Т. 355. - С. 1-140.
РУТИЛЕВСКИЙ Г.Л. Промысловые млекопитающее полуострова Челюскина и
пролива Вилькицкого // Тр. / Ин-та полярного земледелия. Сер. Промысловое
хозяйство.- Л., 1939.- Вып. 8.- 48 с.
РИХТЕР Б. Маточкин Шар // советская Арктика.- 1938.- N 5.- с. 81-88.
САБЛИНА Т.Б., ОВСЯНИКОВ Н.Г. Охрана млекопитающих, требующих для
своего обитания больших территорий // Актуальные проблемы морфология и
экологии высших позвоночных.- М.: Наука, 1988.- С. 631-642.
САВИНОВ В.М., БОБРОВ D. А. Пространственные неоднородности
распределения первичной продукции Баренцева моря // Экология и биологическая
продуктивность Баренцева моря.- М.: Наука, 1990.- С. 9-17.
САМОЙЛОВИЧ Р.Л. Краткие сведения об экспедициях на Новую Землю в
1921-1927 гг. // Тр. / НИИ по изучению Севера.- 1929.- Т. 40.- С. 5-118.
СВЕТОВИДОВ А.Н. Второй вид трески из рода (Boreogadus agalls Rein-
hard t. ) // ДОКЛ. АН СССР.- 1935.- Т. 1. N 6.- С. 427-431.
СВЕТОВИДОВ А.Н. Трескообразные // Фауна СССР. Рыбы. Т.9.- Вып. 4.-
М.-Л.: Изд-во АН СССР, 1948.- С. 1-222.
СИКОРСКИЙ А.В. Полихеты сем. Splonidae Баренцева моря. Автореф. дис
...канд. биол. наук.- М.. 1991.- 24 с.
СЛАСТНИКОВ Г.С. О возрасте и темпе роста новоземельского гольца //
Тр. / Арктического института.- 1935.- Т. 17.- С. 71-88.
СМИРНОВ Н.А. о морских и звериных промыслах // Экспедиция для
научно-промысловых исследований Нурмана.- СПб., 1903.- 158 с.
СМИРНОВ Н.А. Очерк русских ластоногих // Зап. Имп. Академии Наук.-
СПб., 1908.- сер. 8.- Т. 23. N 4.- 75 с.
СМИРНОВ Н.А. Биологические отношения некоторых ластоногих ко льдам //
Сб. в честь проф. Н.М.Книповича. - М.: Изд. Наркомата Земледелия РСФСР,
1927.- С. 89-107.
СМИРНОВ Н.А. Морские звери арктических морей (ластоногие и
китообразные) // Звери Арктики.- М.: Изд-во АН СССР, 1935.- С. 455-570.
Советская Арктика (Моря и острова Северного Ледовитого океана). - М.:
Наука, 1970.- 528 с.
СОКОЛОВ А.В. Динамическая карта Баренцева моря // Тр. / океаногр.
ин-та.- 1932.- Т. П.- Вып. 2.- С. 33-48.
СОКОЛОВ В. Е. Редкие животные нашей страны. - Л.: Наука, 1990. - 235 'с.
СОКОЛОВ И. И. (ред.). Млекопитающие фауны СССР.- М.-Л.: Изд. АН СССР.-
Ч. 2.- 1963.- 962 с.
СОКОЛОВ В.Е., ВИШНЕВСКАЯ Т.Ю. .БЫЧКОВ В.А. Серый тюлень // Редкие и
исчезающие виды млекопитающих СССР.- М.: Наука. 1990.- с 7-43.
СОКОЛОВА С.А. Фитопланктон в районе птичьих базаров севера Новой
Земли // особенности биопродуктивности вод в районе птичьих базаров севера
Новой Земли.- Л.: Наука. 1972.- с 63-73.
СОЛДАТОВ В.К. Материалы по ихтиофауне Карского и восточной части
Баренцева морей по оборам экспедиции Института в 1921 г. // Тр. / Плов,
морск. научн. института.- 1923.- Вып. 3.- С. 1-80.
СОРОКИН А.Л., ПЕЛЬТИХИНА Т.с. Ламинариевые водоросли Баренцева моря.-
Мурманск: Изд-во ПИНРО, 1991.- 188 с.
СОХНОВ В. В. Характеристика нерестовой популяции сайки Баренцева моря
// Тр. / ПИНРО.- 1975.- ВЫП. 35.- С. 209-217.
СТЕПАНОВ СИ. Пространственно-временная изменчивость полей солености
в Карском море в навигационный период // Тр. / Аркт. и Антаркт. НИИ.-
1979.- Т. 361.- С. 59-61.
ТАМБОВЦЕВ Б.М. Сайка // Промысловые рыбы Баренцева и Белого морей.-
Л., 1952.- С. 157-161. i .
ТАНЦЮРА А.И. О течениях Баренцева моря // Тр. / ПИНРО.- 1959. Вып.
П.- С. 35-53.
ТИМОФЕЕВ С.Ф. К фауне мизид Карского моря // Зоол. «урн.- 1985.-
Т. 64, N П.- С. 1739-1741.
ТИМОФЕЕВ С.Ф. Трофодинамический анализ экосистем Баренцева, Белого и
Карского морей // Современные проблемы гидробиологии Баренцева моря.-
Апатиты, 1988.- С. 29-34.
ТИМОФЕЕВ С.Ф. зоопланктон // Международная
(американо-норвежско-российская) экологическая экспедиция в Печорское море, на Новую Землю.
Колгуев, Вайгач и Долгий.- Апатиты, 1992.- С. 14-21.
ТИМОШЕНКО Ю.К. Изучение питания и перспективы промысла кольчатой
нерпы в Белом, Баренцевом и Карском морях // Тез. докл. 8 Всес.совещ. по
изуч., охране и рац. исп. мор. млекопитающих.- Астрахань, 1982.-
С. 360-362.
ТИМОШЕНКО Ю.К. Влияние ледовых условий на распределение
гренландского тюленя в Белом море // 3 Съезд сов. океанол. Секц. Биол. океана. Ч. 3.-
Л., 1987.- С. 119-120.
ТОЛМАЧЕВ А.И. Обзор флоры Новой Земли // Arctlca.- 1936.- N 4.-
С. 143-178.
ТОЛМАЧЕВ А.И. О некоторых закономерностях распределения растительных
сообществ В Арктике // Бот. журн.- 1939.- N 5-6.- С. 504-517.
ТОМШШН А. Г. Китообразные фауны морей СССР. - М.: Изд-во АН СССР,
l^Db. — tCld С.
ТОПОРКОВ Г.Н. Новоземельский тресковый промысел// Тр. / Арктического
института,- 1937.- Т. 100.- С. 7-23Г
Труды ПИНРО.- 1957.- Т. 10.- 281 в. ' '
УСАЧЕВ П.И. Фитопланктон Карского моря // Планктон Тихого океана.-
М.: Наука. 1968.- С. 6-28.
УСПЕНСКИЙ СМ. Птичьи базары Новой Земли.- М.: Изд-во АН СССР, 1956.-
179 с.
УСПЕНСКИЙ СМ. Распределение, численность и охрана белого медведя в
Арктике // Бюлл. МОИП. Отд. биол. - 1965.- Т. 70.- Вып. 2.- С. 18-24.
УСПЕНСКИЙ СМ. Белый медведь должен жить'// Природа.- 1967.- N. 2.-
С 81-84.
УСПЕНСКИЙ СМ. Добыча белого медведя на севере Евразии в 18-20 веках
// Белый медведь и его охрана в Советский Арктике. - Л.: Гидрометеоиздат, -
1969.- С. 122-141.
УСПЕНСКИЙ СМ. Человек и медведь: условия мирного сосуществования //
Охота и охот, хоз-во.- 1987а.- N. 5.- С. 12-14.
УСПЕНСКИЙ СМ. Шнвая Арктика.- М.: Мысль, 19876.- 272 с.
УСПЕНСКИЙ С, ХАХИН Г. Новая Земля сегодня // охота и охот, хоз-во.-
1993.- N 1.- С. 1-3. ■ .
УСПЕНСКИЙ С. М., ШИЛЬНИКОВ В. И. Распределение и численность белых
медведей в Арктике по данным авианаблюдений ледовой разведки // Белый медведь
и его охрана в Советской Арктике. - Л., 1969. - С. 89-102.
УСПЕНСКИЙ С.М.. ГОЛОВКИН А.Н., ГУРЕВИЧ В.И., ЧЕЛИНЦЕВ Н.Г.
Географические группировки белого медведя в Советской Арктике, выделенные на
основе геохимических данных // Зоол. журн.- 1985.- Т. 64.- Вып. 4.-
С. 600-605.
УШАКОВ П. В. Бентонические группировки Маточкина Шара (Новая Земля) //
Исследования морей СССР,- 1930.- Вып. П.- С. 5-128.
ФЕДОРОВ В.Д. О методах изучения фитопланктона и его активности.- М.:
Изд-во МГУ, 1979.- 168 с.
ФЛЕРОВ Б.К. Распределение водорослей у берегов Новой Земли // Тр. /
Гос. океанограф, ин-та.- 1932.- Т. П.- Вып. 1,- С. 7-45.
ФЛЕРОВ Б.К.. КАРСАКОВ Н.В. Список водорослей Новой Земли // Тр. /
Гос. океанограф, ин-та.- 1932.- Т. П.- Вып. 1.- С. 46-73.
ФОМИН O.K. Некоторые структурные характеристики зоопланктона //
Экология и биоресурсы Карского моря.- Апатиты, 1989.- С. 65-85.
ФОМИН O.K.. ПЕТРОВ B.C. Роль природных факторов в распределении
биомассы планктона в Карском море // Природа и хозяйство Севера.- 1985.- Вып.
13.- С. 35-45.
ФОМИН 6.К.. САВИНОВ В.М., БОБРОВ D. А. Пространственное распределение
биомасоы.планктона в Карском море // Биологические науки.- N 11,- 1984.-
С 54-59.
ФРЕЙМАН CD. Q миграциях гренландского тюленя // Сб. посвящ. научн.
деят. Н. М. Книповича. - М.-Л.. 1939.- С 353-368.
ХАИН В.Е. Региональная геотектоника. Внеальпийская Европа и Западная
Азия.- М.: Недра, 1977.- 359 с.
ХМЫЗНИКОВА В.Л. Материалы к изученив зоопланктона Маточкина Шара и
прибрежных районов Новой Земли // Тр. / ГОИН.- 1931.- Т. 12.- С. 131-150.
ХРИСТОФОРОВ о.Л. Гаметогенез и половой цикл сайки (Boreogadus 3alda)
Баренцева моря // Тр. / ВНИРО.- 1978а.- Т. 130.- С. 33-46.
ХРИСТОФОРОВ О.Л. Особенности функции воспроизводительной системы
сайки Boreogadus salda L. Баренцева моря в связи с экологией и возрастом //
Автореф, дис. ... канд. биол. наук.-Л., 19786. - 28 с.
ХРОМОВ В.М. Первичная продукция органического вещества в восточной
части Баренцева моря //Тез. докл. 4 съезда ВГБО. Ч. 1.-Киев. 1981. -
С. 163И64.
ХУЗИН Р.Ш. Отлов и охрана белого медведя // Природа.- I960.- N 10.-
С. 54-56.
ЧАПЛЫГИН Е.И. О тепловой напряженности течений Карского моря //
Проблемы Арктики и Антарктики.- 1961.- Вып. 8.- С. 19-28.
ЧАПСКИЙ К.К. Морж Карского моря. Результаты исследования биологии,
географического распределения и запасов мораа Карокого моря и Новой Земли
// Тр. / Арктического института.- 1936.- Т. 67.- Вып. 17.- С. 10-62.
ЧАПСКИЙ К.К. Морской заяц Карского и Баренцева морей (биология и
промысел) // Тр. / Арктического института.- 1938.- Т. 123.- С. 7-36.
ЧАПСКИЙ К.К. Краткий исторический анализ современного состояния
запасов моржа в Баренцевом и Карском морях // Проблемы Арктики.- 1939.- N 3.-
С. 62-63.
ЧАПСКИЙ К.К. Нерпа западных морей Советской Арктики // Тр. /
Арктического института,- 1940.- Т. 145.- С. 1-72.
ЧАПСКИЙ К.К. Морские звери Советской Арктики. - М.: Главсевморпуть,
1941.- 365 с.
ЧАПСКИЙ К.К. Отряд китообразных, отряд ластоногих // Млекопитающие
фауны СССР.- М.-Л., 1963.- Ч. 2.- С. 641-740.
ЧАПСКИЙ К.К. Ластоногие и зубатые киты // Млекопитающие Советского
Союза.- М.: Высшая школа, 1976.- Т. 2.- Ч. 3.- С. 131-328.
ЧЕРНИЦКИЙ А.Г.. МАТИШОВ Г. Г., ЕРМОЛАЕВ В.В. Возможность использования
арктического гольца для товарного лососеводства в Баренцевом море.-
Апатиты, 1987.- 38 с.
ЧЕРНЯЕВА Ф.А. Площади северных арктических морей и их водосборных
бассейнов // Тр. ААНИИ, 1957.- Т. 208.- С, 32-48.
ЧЕРНОВА Н.В. Рыбы семейства липаровых (Scorpaenlformea: Llparldldae)
Баренцева моря и сопредельных вод. / Автореф. дис. ... канд. биол. наук.-
Л., 1988.- 16 с.
ЧЕРНОВА Н.В. Липаровые рыбы Евроазиатской Арктики.- Апатиты. 1991.-
ЧИЛИНГАРОВ А.Н. Ледовый режим и научно-методическое обеспечение моро-
ких транспортных операций в Карском море. Дис. ... канд. геогр. наук.- Л.:
Изд-во ААНИИ, 1976.- 187 С.
ЧИСЛЕНКО Л.Л. Зоопланктон бухты Диксон (Карское море) //
Географическая и сезонная изменчивость морского планктона. - л.: Наука. 1972.-
С. 228-238.
ШЕПЕЛЬ Л.И. Размерно-возрастной состав сайки в юго-западной части
Карского моря осенью 1969 г. // Вопр. ихтиологии.- 1971.- Т. П.- Вып. 1.-
С. 161-164.
ШЕПЕЛЬ Л.И. Методы и особенности поиска сайки Баренцева моря //
Материалы рыбохоз. исслед. Северного бассейна. Вып. 21.- Мурманск: Изд-во ПИН-
РО, 1973.- С. 230-238.
ШИРОКОЛОБОВ В.Н. Гидролого-гидрохимические условия юго-западной части
Карского моря // Комплексные исследования природы северных морей.- Апати-
ШЛЕЙНИК В.Н. Состояние запасов и перспективы промысла сайки (Boreoga-
dua salda) в Баренцевом море // Тр. / ПИНРО.- 1973.- Вып. 33.- С. 225-236.
1Ш1ЕЙНЖ В.Н. Плодовитооть сайки Баренцева моря // Тр. / ПИНРО.-
1978.- Вып. 41.- С. 156-166.
ПШЕННИК В.Н. Биологические основы рационального использования сайки
Баренцева моря // Автореф. дис. ... канд. биол. наук:- М., 1979,- 24 с.
ШМИДТ В. М. Математические методы в .ботанике. - Л.: Изд-во Ленинград.
ун-та, 1984.- 288 С
ШПАЙХЕР А. 0., РУСАНОВ В.П. Распределение кремния как индикатора
водных масс морей сибирского шельфа // Проблемы Арктики и Антарктики.- 1972.-
Вып. 40.- С. 64-70.
Экологические проблемы Арктики и перспективы ядерного разоружения //
Волна.- N 118.- 1992.- 15 окт., Архангельск.
Экология и биоресурсы Карского моря.- Апатиты, 1989.- 181 с.
ЮДАНОВ И.Г. Технологическая характеристика сайки // Рыбное хоз-во.-
1947.- N 1.- С. 39-41.
ПЛАНОВ И. Г. Пищевая ценность сайки // Рыбное хоз-во.- 1964.- N 1.-
С. 39-41.
ЮДАНОВ И.Г. Зоогеография сайки в Северном Ледовитом океане // Природа
и хозяйство Севера.- Апатиты, 1976,- Вып. 4.- С. 111-113.
ЯБЛОКОВ А.В., МИНЕЕВ В.Н. Заповедники для морских млекопитающих //
Природа.- 1978.- N 8.- С. 30-41.
ANDREWS C.W., LEAR E. The biology of Arctic Char (Salvellnus alplnus)
In Northern Labrador.- J. Fish. Res. Bd; Can.. 1956. Vol.13. N 6.-
P. 843-860.
ANON. Preliminary report of the International 0-group fish survey In
the Barents Sea and adjacent waters In August-September 1988 // ICES, CM.
1988/G:45,- 32 p.
BEST R.С Digestibility of ringed seals by the polar bear // Can. J.
Zool- 1985.- Vol. 63. N 5.- P. 1033-1036.
BOLSTAD H. A. 3.8 mill, torn flsk rett 1 torskemagane // Flskaren. -
1987.- N 76.
BORKIN I.V. Distribution and length composition of the polar cod
larvae In the Barentss Sea // ICES. CM. 1979/G:16.- 8 p.
BORKIN I.V. Distribution and age-length composition of the Barents
Sea polar cod stocks in 1980 // Annla blol.. Copenh.- 1983a (1980).-
V. 37.- P. 160-161.
BORKIN I.V. Distribution and age-length composition of the polar cod
larvae in the Barents Sea In 1980 // Annla blol., Copenh.- 1983b (1980).-
V. 37.- P. 161-162.
BORKIN'I.V. Soviet investigations on Polar cod larvae in the Barents
Sea In 1981 // Annla blol.. Copenh.- 1984a (1981).- V. 38.- P. 134-135.
BORKIN I.V. Distribution of polar cod larvae off the West
Spitsbergen // ICES. CM. 1984b/L:5.- 7 p.
BORKIN I.V. Soviet Investigations on Polar ood In the Barents Sea in
1984 // Annla blol., Copenh.- 1986.- V. 41.- P. 111-112.
BORKIN I.V.Ichthyoplankton in the coastal waters off West Spitsbergen
// ICES. 1988. ELHS.- V. 4.- Posters 74.- 16 p.
BORHIN I.V. Polar cof (Boreogadus salda Lepechln) in Barents Sea
ecosystem // Arctic research: asdvances and prospecrts.- Moskow: 1990.-
Pt. 2.- P. 230-231.
BORKIN I.V.. Fishes inhabiting coastal waters // Environment and
ecosystems of the Frantz Josef Land (Archipelago and shelf).- Apatity. 1993.-
P. 188-194.
BORKIN I.V., SHLEINIK V.N. Results of investigations on the Barents
Sea polar cod in 1977-1980 // ICES. СМ.- 1981/G:24.- 12 p.
BORKIN I.V.. SHLEINIK V.N. Results of investigations on the Barents
Sea polar cod In 1981-1984 //ICES. СМ.- 1985/G:8.- V. 4.- 12 p.
BORKIN I.V. CHERNOOK V. I., PONOMAREV Ya.T., B0G0M0L0V V. Yu.. GAVRILO
M. V. Results of the air survey on Sea birds In the Barents Sea In Autumn
1991 // Interactions between fish populations In the Barents Sea.-
Murmansk: PINRO, 1992.- P. 205-216.
BRUCE D.S.. DARLING N.K.. SEELANDK.J., OELTGEN P. R., AMSTRUP S. С
Does the polar bear (Ursus maritimus) hibernate? Further studies on
opioids and hibernation // FASEB J.- 1989.- Vol. 3. N 3.- P. 396.
Bulletin Statlstlque des Peches Marl times.- 1966.- V. 55.- 188 p.
Bulletin Statlstlque des Peches Marltlmes. - 1994.- V. 77.- 156 p.
CHAPPEL J., SHACKLETON N.J. Oxygen Isotopes and sea level // Nature.-
1986.- N 324.- P. 137-140.
CHERNITSKY A. G. 'Arctic char in Novaya Zemlla: Ecological observations
of some anadromous and landlocked populations // ISACF Inform. Ser.-
1991,- Vol. 5.- Inst, of Freshw. Res., Drottnlngholm. - P. 37-43.
CUSHING B.S., CUSHING N.L.. JONKEL С Polar bear responses to the
underwater vocalizations of ringed seals // Polar Biol.- 1988.- V. 9, N 2.~
P. 123-124.
DUBASOV YU.V., KRIVOKHATSKIJ A.S.. MALAKHOV P. B. . SAFRONOVV.G. Ra-
dloecologlcal monitoring on Novaja Zemlya nuclear test site in.1990-1992
// Proceedings of the International Conference on Environmental
Radioactivity.- Klrkenes, 1993.- P. 147-151.
DZUS В.Е. Blostereometrlsche Unterwasserausrustung Selenezkaja //
DDR. Blld und Ton.- 1981.- N 7.- P. 214-215.
EKHOLM N. Sverlges temperaturforhallanden jamforda med det ofrlga Eu-
ropas. Ymer.- 1899.- 37 p.
ELVERHOI A., LIESTOL 0.. NAGY J. Glacial erosion, sedimentation and
mlcrofauna in the Inner part of KongsfJorden,. Spitsbergen // Norsk Pola-
rlnst. Skrlfter.- 1980.- V. 172.- P. 33-61.
FURNESS R.W., BARRETT R. T. The food requirements and ecological
relationships of a seablrd community In North Norway // Ornis scand.- 1985.-
V. 16.- P. 305-313.
HANTER J., THOMAS С L. Effects of prey distribution and feeding
behaviour of larval anchovy Engraulls mordax G. // The early life history of
fish. Springer.- 1974.- P. 64-78.
HOGNESTAD P.T. Observations on Polar Cod in the Barents Sea // ICES.
CM. Symposium "The Ecology"- 1966. N 5.- 14 p.
HOGNESTAD P.T. Observations on Polar cod in the Barents Sea // Rap.
Process Verb. Reun.- 1968a,- Bd. 158.- S. 126-130.
HOGNESTAD P.T. Polar cod Boreogadus salda Lepechln In Norwegian
waters // Astarte by the Zoological Department Tromso Museum.- 19686.-
N 31.- P. 1-3.
KJELLMAN F.R. Uber die Algenvegetatlon des Murmanlshen Meers an der
Westkuste von Nowaja Semlla und Wajlgatsch // Nova Acta Regla Soc. Sclen-
tlar Upsallensls.- Upsala: 1878.- 32 p.
KJELLMAN F.K. The Algae of the Arctic Sea // Kongl. Svenska Vetensk.
Akad. Handl.- 1BB3. - Bd. 20.- N 5.- P. 1-350.
KILIUS L.R., RA1SBECK G.M., Ylon F. et al. A measurement of the 1Z9I
Distribution in Sediments sampled from the Arctic seas surrounding Novaja
Zemlya // Intern. Conf. on Environmental Radioactivity In the Arctic and
Antarctic. Klrkenes, August 1993.- 1993.- P. 333-336.
LARIONOV V.v. Spatial structure of the phytoplankton community of the
open sea // Phytoplankton of the Barents Sea.- Apatity. 1992.- P. 64-73.
LEGENDRE L.. DEMERS S. Auxiliary energy, ergocllnes and aquatic
biological production // Natur. can.- 1985.- Vol. 112, К 1.- P. 5-14.
LEIGHTON F.A.. CATTET M., NORSTROM B.." TRUDEAU S. A cellular basis
for high levels of vitamin A In livers of polar bears (Ursus marltlmus):
the Ito cell // Can. J. Zool.- 1988.- Vol. 66.- N 2.- P. 480-482.
LOWRY L.F., BURNS J.J., NELSON R.R. Polar bear, Ursus marltlmus, pre-
datlon on belugas Delphlnapterus leucas In the Bering and Chukchi seas //
Can. Field.-Natur.- 1987.- Vol. 101.- N 2.- P. 141-146.
LUNDE. T. Ice conditions at Svalbard 1946-1963 // Norsk Polarlnst.
Arb..- 1965.- P. 61-80.
MARGALEF R. Temporal succession and spatial heterogeneity In
phytoplankton // Perspectives In marine biology. Berkeley - Los Angeles: Univ.
of California Press.- I960.- P. 323-349. .
MARKUSSEN N.M.. ORITSLAND N. A. Granlandselens Matbehov III // Biolo-
glsk lnstitutt Avd. for general flslologl universltetett in Oslo.- 1985.-
61 p.
MARTIN S.. CAVALIERI D. С Contributions of the Siberian shelf polyny-
as to the Arctic ocean Intermediate and deep water // J. Geophys. Res.-
1989.- V. 94.- N 9.- P. 12725-12738.
MATISHOV D.. SZSZYPA E. Radionuclides In the bottom sediments and
biota of the shelf and of Barents Sea coasts // Proceedings of the
International Conference on Environmental Radioactivity.- Klrkenes: 1993.-
P. 205-216.
MEUNIER A. Mlcroplancton des Mers de Barents et de Kara. Due d'Orle-
ans Campange Arctlque de 1907.- Bruxelles: 1910.
(JOHN H. Stromungen des Europalschen Nordmeeres // Petermans Geogr.
Mitt.. Erganzungsh. - 1885.- N. 79.- P. 3-60.
NAGY J. Foraminlfera In some bottom samples from shallow waters in
Vestspltsbergen // Norsk Polarlnstltutt-Arbok 1963.- Oslo: 1965,-
P. 109-128.
OLSEN L. What's killing the polar bear? Toxins, tourism, greenhouse
overheating, UV radiation, top politicians // Bear news.- 1991-1992.- Vol.
8-9.- N 1-2.- P. 10.
OSTERMAN L.E., ANDREWS J.Т., Changes in glacial-marine sedimentation
in core HU77-159, Frobisher Bay, Baffin Islands, N.W.T. // Glacial Marine
sedimentation.- NY.: Plenum, 1983.- P. 451-494.
PETERMANN A. Der Golfstrom und Standpunkt der thermometrlschen Kenn-
tnlss des Nord-Atlantischen Oceans und Landgeblets 1m Jahre 1870 // Peter-
mann's Geographylsche Mittheilungen.- 1870.- Bd 16.- P. 1-56.
PONOMARENKO W.P. Results of Polar cod study In the Barents Sea //
Ann. Blolog.- 1964.- V. 19.- P. 93-96.
RAMSAY M.A.. STIRLING J. Reproductive biology and ecology of female
polar bears (Ursus marltlmus) // J. Zool.- 1988.- V. 214.- 8 4.-
P. 601-633.
RASS T.S. 1936. Spawning, eggs, fry of the foodflshes of the Barents
sea // Rev. gesamm Hydroblol. und Hydrographle.- 1936.- Bd. 33.- S. 11-19.
REY F.. LOENG H. The Influence of ice hydrographlc conditions on the
development of phytoplankton In the Barents Sea // Marine Biology of Polar
Region and Effects of Stress on Marine Organisms (eds. J.S.Gray and
M.E.Christiansen) J.Wiley and Sons. Ltd..- 1985.- P. 49-63.
REY F., SKJOLDAL-H.. SLAGSTAD 6. Primary production In relation to
climatic changes in the Barents Sea // The effect of oceanographic
conditions on distribution and population dynamics of commercial fish stocks in
the Barents Sea.- Bergen: Inst. Mar. Biol., 1987.- P. 29-46.
ROSENVINGE L.K. Alger fra Novala Semlla Dg Kara - Hevet, samluende
paa Dljmphna Expedition 1882-1883 // Dljmphn. Togtes, Zopl.. Bot. Udbyt-
te.- Kobenhavn: 1887.- 78 p.
SCHANDELMEIER L., ALEXANDER V. An analysis of the influence of ice on
spring phytoplankton population structure In the south-east Bering Sea //
LlmnoT. Dceanogr.- 1981.- V. 26.- К 5.- P. 935-943.
SHANNON C.E.. WEAVER W. The mathematical theory of communication.-
Urbana. 1963.- 177 p.
SHLEINIK V.N. Soviet investigation an age-size composition of Polar
cod In the Barents sea In 1970 // Ann. Biol.- 1971.- V. 27.- P. 113-114.
SHLEINIK V.N. Distribution and age-size composition of Polar cod'In
the Barents sea in 1971 // Ann. Biol.- 1973.- V. 28.- P. 131-132.
SHLEINIK V.N. Distribution and age-size composition of Polar cod In
the Barents sea in 1972-1973 // ICES. СМ.- 1974.- 6 p.
SKJOLDAL H.R., HASSEL A.. REY F.. LOENG H. Spring phytoplankton deve- .
lopment and zooplankton reproduction in the central Barents Sea In the
period 1979-1984 // The effect of oceanographic conditions on distribution
and population dynamics of commercial fish stocks In the Barents Sea.-
Bergen: Inst. Mar. Biol.- 1987.- P. 59-89.
SORENSEN T.A. A new method of establlishing - groups of equal
amplitude in plant sociology based on similarity of a species content and its
application to analysis of the vegetation on Danish commons // Kgl. Dan.
Vld. Selsk. blol. ser.- 1948.- Bd. 5.- N 4.- P. 1-34.
STRAND P., RUDIJORD A.L.. SALBU B. et al. Survey of artificial
radionuclides In the Kara Sea // Internation Conference on Environmental.- Kir-
klenes.- 1993.- P. 53-65.
STIRLING J. Affraction of polar bears Ursus marltlmus to off shore
drilling sites in the eastern Beaufort Sea // Polar Rec- 1988.- V. 24.-
N 148.- P. 1-8.
STIRLING J. Sleeping giants // Natur. Hist.- 1989.- Vol. 98.- N 1.-
P. 37-38.
TARASOV G.. NURENBERG D., GROT E.. CHASANKAEV N. The bottom sediments
// On the lntrenatlonal multl-dlsclpllnary cruise to high-latitude
archipelagoes of the Barents Sea (Frants Josef Land and Novaja Zemlya).-
Apatity, 1993.- P. 22-26.
TILSETH S. The distribution of cod larvae and prey organlsme In the
Lofoten area releted to critical prey concentrations // Reproduction and
recruitment of Arctic Cod. Proc. Soviet-Norwegian Symp. on Reproduction
and Recruitment of Cod,- Bergen: Л984.- P. 36-71.
VADER W. What is wrong with the gullemots of northern Norway // Fo-
redreg Nordisk kuvs, Reyklavik. - 1987.- V. 7.- P. 7-9.
VADER V., ANKER-NILSSEN T.. BAKKEN V., BARRETT R.T., STRANN К.В.
Regional and temporal differences In breeding success and population
development of flsheating seablrds In Norway after collapses of herring and
capellne stocks // Trans. 19 IUGB Congress.- Trondhelm: 1989.- P. 29-40.
VINJE Т.Е. Sea ice condition in 1982 // Norsk Polarinst. Arb.. 1982.-
P. 49-53.
WATTS P.D.. ORITSLAND N. A.. HURST R.J.-Standard metabolic rate of
polar bears under simulated denning conditions // Physiol. Zool.- 1987.-
V. 60.- N 6.- P. 687-691.
WLYNEN W.I. A mathematical model of the relationship between larval
anchovy (Engranlls mordax) growth, prey mlcrodlstrbutlon, and larval
behaviour // Envlronm. Biol. Fishes.- 1977.- N 2.- P. 211-233.